/












































































































































































































































































































































































Автор: Фоули Р.
Теги: палеонтология экология эволюция эволюционное учение
ISBN: 5-03-001433-0
Год: 1990




Текст
Р Фоули
ЕЩЕ ОДИН
НЕПОВТОРИМЫЙ
вид •
Экологические Ли
аспекты
эволюции
человека *' & х
Издательство «Мир»
ANOTHER UNIQUE SPECIES
Patterns in human evolutionary ecology
ROBERT FOLEY
I.ohpman
Scientific and Technical
Cnpiibllahrd In the United Stales with John Wiley and Sons, Inc. New
Р Фоули
Еще один
неповторимый
вид
Экологические, аспекты эволюции человека
Перевод с английского
канд. биол. наук
Е. 3. ГОДИНОЙ, В. В. СВЕЧНИКОВА
под редакцией
д-ра ист. наук
Т. И. АЛЕКСЕЕВОЙ
Москва «Мир» 1990
ББК 28.1
Ф81
УДК 56
Фоули Р.
Еще один неповторимый вид. Экологические аспекты
Ф81 эволюции человека: Пер. с англ. — М.: Мир, 1990. — 368 с.,
ил.
ISBN 5-03-001433-0
В книге английского автора рассмотрены экологические аспекты проблемы
происхождения человека. Проведен анализ тех экологических фактов, которые
определили направление естественного отбора среди древних гоминид и их эво-
люцию. Книга написана строго научно, ио легко и доступно; благодаря оригиналь-
ному подходу она не дублирует имеющиеся издания по палеоантропологии.
Для студентов и специалистов в области антропологии, эволюционной био-
логии, зоологии и всех интересующихся проблемой происхождения человека.
1908000000—416
*041(01)—90 ПО — 90
ББК 28.1
Редакция литературы по биологии
ISBN 5-03-001433-0 (русск.)
ISBN 0-582-44690-2 (англ.)
© Longman Group UK Limited 1987.
This book was originally published
in the English language by Long-
man Group UK Limited of London
© перевод на русский язык, Годи-
на Е. 3., Свечников В. В., 1990
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
Автор этой книги читает лекции по разным аспектам био-
логической антропологии в Кембридже (Англия). Первые его
работы появились десять-двенадцать лет тому назад и сразу
же обратили на себя внимание модным сейчас и во многих
случаях действительно эффективным междисциплинарным под-
ходом. Формы и темпы эволюции ранних гоминид и древнейших
популяций человека современного типа исследовались и трак-
товались им под углом зрения приспособления к среде обита-
ния, коэволюции ,с соответствующими таксонами млекопитаю-
щих, поведенческих факторов, реконструкции среды обитания
в широком смысле слова, в том числе и характера питания.
За это время Р. Фоули опубликовал около десятка крупных
статей, которые постоянно цитируются в современной антропо-
генетической и палеоантропологической литературе.
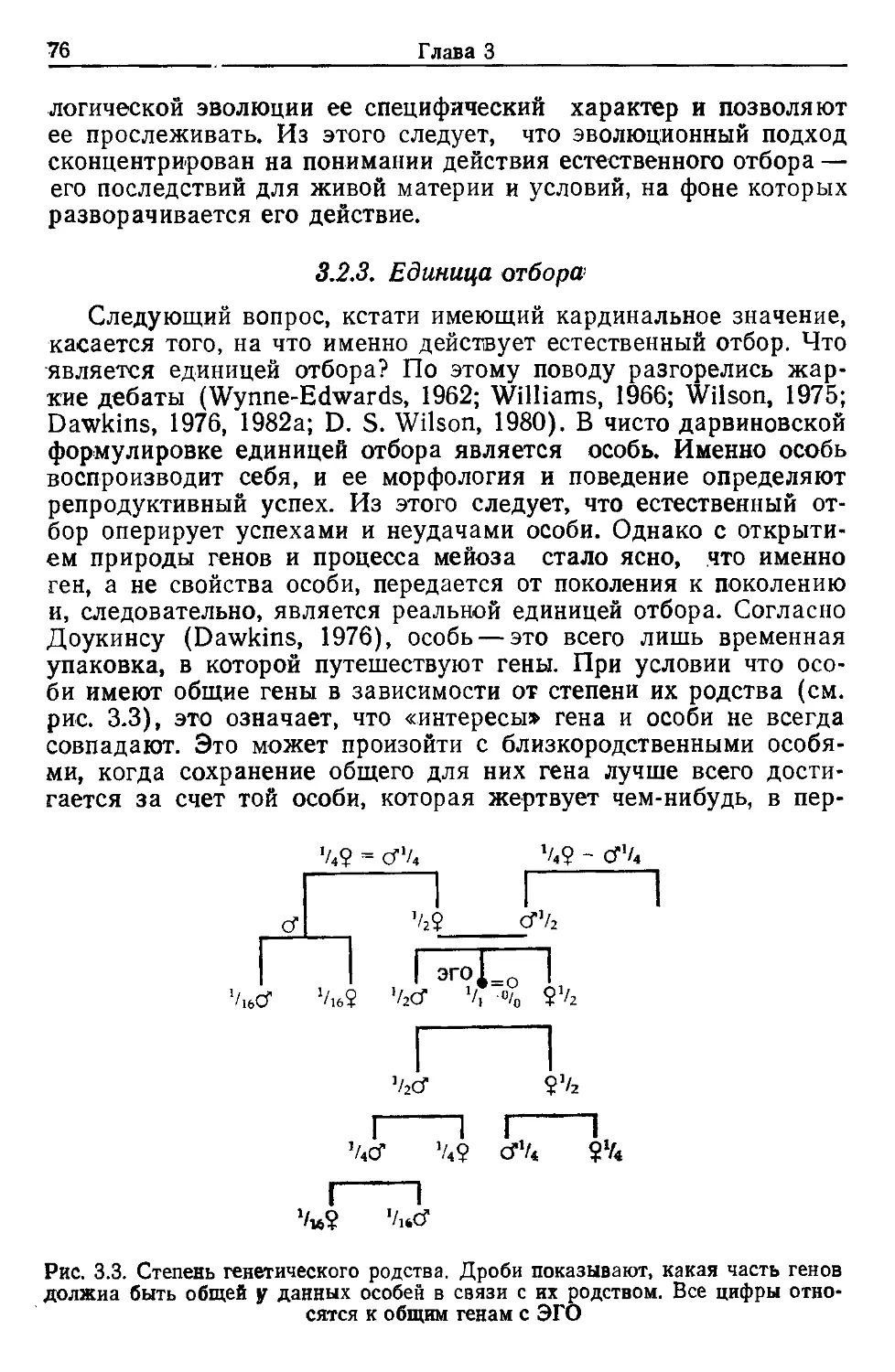
Но значение его деятельности не исчерпывается его само-
стоятельным вкладом в антропологию. Будучи прекрасным ор-
ганизатором, он провел два симпозиума, материалы которых
были им изданы и составили заметную веху в палеоантрополо-
гической литературе. Труды первого из этих симпозиумов вы-
шли в свет в 1984 г. под заглавием «Эволюция гоминид и эко-
логия сообществ: доисторическая адаптация человеческих по-
пуляций в биологической перспективе»1.
Как организатор данного симпозиума и издатель его трудов
Р. Фаули объединил усилия исследователей разных направле-
ний — геологов, палеонтологов, специалистов по теории эволю-
ции, палеоантропологов и археологов, — чтобы попытаться по-
лучить целостную картину эволюции ранних гоминид на фоне
эволюции других млекопитающих плиоцена и плейстоцена, вос-
создать картину древних способов охоты и вообще проследить
взаимозависимость между микродинамикой среды и микроэво-
люционными сдвигами в древнейших человеческих коллективах.
Изданный том ввел в науку много новых конкретных наблю-
дений, основанных на локальных разработках, а также много
интересных реконструкций реальных взаимосвязей, существо-
вавших в системе антропогеоценоз — биогеоценоз — человече-
ская популяция.
1 R. Foley (ed.) Hominid Evolution and Community Ecology: Prehistoric
Human Adaptation in Biological Perspective, New York—London, Academic
Press, 1984.
6 Предисловие редактора перевода
Тема второго симпозиума, труды которого были изданы
Р. Фоули вместе с Е. Круис в 1986 г., — «Зубы и антрополо-
гия»1. Эта тема также находится на стыке нескольких дисцип-
лин и весьма актуальна сейчас, когда наблюдается большой
интерес к развитию зубной системы человека и млекопитающих
и идет острая дискуссия о факторах эволюции зубов. На фоне
распространенного в кругах специалистов, в том числе и в
Советском Союзе, мнения об исключительной стабильности
зубной системы в ходе времени материалы этого симпозиума,
напротив, указывают на ее лабильность и, в частности, зависи-
мость от средовых факторов, в первую очередь от пищи. Роль
генетического аппарата в этом случае никоим образом не за-
малчивается, но он рассматривается как одна из составляющих
в общей картине эволюции, что в общем типично и для других
систем человеческого тела.
Книга Р. Фоули «Еще один неповторимый вид» своими ис-
токами восходит к тем исследованиям автора и его коллег,
которые получили отражение на симпозиуме, посвященном эво-
люции гоминид и экологическим сообществам. Вопросы клима-
тической адаптации древних гоминид, взаимоотношений с раз-
нообразными млекопитающими в процессе эволюции, роли миг-
раций в становлении адаптивных особенностей древнейших
предков современного человека, теснейшей связи экологии и
эволюции — эти вопросы, поднятые на симпозиуме, в представ-
ленной вниманию читателя книге изложены более углубленно,
и трактовка их представляется достаточно убедительной. В то
же время автор отдает себе отчет в трудностях анализа станов-
ления человека как вида и выбирает, по-моему, единственно
правильный путь — анализирует этот вид на фоне прошлых со-
бытий и процессов, которые обусловили эволюцию гоминид.
Рассматривая филогенетическое наследие человека и окру-
жающую его среду как соответственно внутреннюю и внешнюю
движущие силы эволюции, автор глубоко проникает в сущность
причин, способствующих формированию вида Homo sapiens.
«Эволюционная история нашего вида — это рассказ об адаптив-
ных изменениях его морфологии и поведения» — этими словами
автора обозначается замысел книги. И в самом деле, вся книга
читается как увлекательный рассказ об эволюционной экологии
гоминид, однако рассказ этот основан не на умозрительных за-
ключениях, столь обычных для темы, связанной с древнейшими
этапами человеческой истории, а на строгом, тщательном и ло-
гически глубоко продуманном анализе фактов и событий отда-
ленного прошлого. Т. Алексеева
1 R. Foley. Е. Cruwys (eds.). Teeth and Anthropology, В. A. R. Interna-
tional Series No. 227, Oxford, B. A. R., 1986.
Посвящается памяти
Дэвида Кларка и Глинна Айзека
ПРЕДИСЛОВИЕ
Одна из первых книг об эволюции человека была написана
Чарлзом Дарвином. «Происхождение человека» по-прежнему
остается одной из лучших книг на эту тему: в ней поставлены
многие из тех вопросов, которые и поныне стоят в центре вни-
мания, а ведь она была написана почти без ссылок на какие-
либо ископаемые материалы и притом в то время, когда окаме-
нелости гоминид были практически неизвестны. Тем не менее
по общему убеждению прогресс в изучении происхождения че-
ловека осуществляется лишь благодаря открытию новых иско-
паемых остатков. С появлением из земли нового «недостающего
звена» наука делает очередной шаг вперед, а к родословному
древу человека добавляется еще одна ветвь. Но не менее важ-
ны, чем сами находки, те теоретические концепции, на фоне
которых освещается рассказ об эволюции человека. Книга Дар-
вина сохраняет свое значение, потому что автор рассматривает,
как и почему происходит эволюция человека. Другие книги на
эту тему также не утратили своей актуальности, если обеспечи-
вали прочную и в конечном итоге «полезную» теоретическую
основу, которую после каждой новой находки не нужно было
отбрасывать, как отслужившую обертку от бритвенных лезвий.
Эта книга также посвящена основам изучения эволюции
человека. Если и существует какой-либо фон для изучения эво-
люции любого вида, в том числе и человека, то это, конечно,
экология. Достижения эволюционной биологии за последние
10 лет показывают, что особенности эволюционного процесса
теснейшим образом связаны с теми экологическими принципа-
ми, которые определяют взаимоотношения между особями, по-
пуляциями и видами, с одной стороны, и их окружением —
с другой. Эволюционная экология, оказывается, имеет большее
значение для эволюционной биологии, чем подчас бесплодные
споры о классификации и филогении. Как утверждает Майкл
Гизелин, она гораздо больше соответствует духу дарвиновской
оригинальной трактовки теории эволюции. Кроме того, эколо-
гические основы эволюционных моделей находят свое подтверж-
дение не только в морфологических и физиологических призна-
ках, но и аспектах поведения. В этой книге я попытался про-
"8
Предисловие
анализировать характер эволюционного процесса у одной
группы животных — гоминид — с точки зрения их экологии.
Идеи, вдохновившие меня на эту работу, по существу не
являются антропологическими. Изучение эволюции человека
было традиционно отделено от других разделов эволюционной
биологии. Несмотря на революционную мысль Дарвина о том,
что человек — это всего лишь еще одно животное, эволюция
человека рассматривалась как довольно специальный предмет.
То обстоятельство, что мы сильно отличаемся от большинства
других населяющих Землю видов, сильно повлияло на подход
к проблемам эволюции человека. В этой книге сделана попытка
избежать такой трактовки человека и его предков, которая рас-
сматривала бы их как нечто большее, нежели просто еще один
биологический вид, эволюционировавший на протяжении по-
следних нескольких миллионов лет. Озаглавив книгу «Еще один
неповторимый вид», я надеялся показать, что уникальность,
как это ни парадоксально, есть свойство всех видов, а не толь-
ко человека. Поэтому научную основу книги составляют в мень-
шей степени антропология и археология и в большей — эволю-
ционная экология. В этой связи на идеи и модели, представлен-
ные в книге, оказали сильное влияние работы таких биологов,
как Джон Мэйнард Смит, Эрик Пианка, Роберт Мартин, Пол
Харви и Тим Клаттон-Брок. Еще один биолог-эволюционист,
воздействие которого ощущается в книге, — это Джон Айзен-
берг, и, вероятно, стоит отметить, что во введении к своей
книге «Радиации млекопитающих» он ссылается на работу ант-
рополога Деррила Форда «Среда, хозяйство и общество», под-
сказавшую ему, в каком направлении следует рассматривать
взаимоотношения между окружающей средой и поведением.
Эти взаимоотношения явно недооценивались во многих обла-
стях антропологии и после выхода книги Форда; в настоящее
же время идеи об эволюционной и поведенческой экологии че-
ловека должны быть заимствованы антропологами из зоологии.
Хотя эта книга посвящена эволюции человека, она не содер-
жит исчерпывающего перечня всех исторических событий. Эво-
люцию человека можно представить как историю, разворачи-
вающуюся во времени, с сильным акцентом на хронологию.
Этот путь не был использован здесь. Мне хотелось найти под-
ход к изучению эволюции человека, проанализировать и в кон-
це концов объяснить ее, а не просто сообщить читателю основ-
ные факты в хронологическом порядке. В первой главе дается
общая характеристика процесса эволюции человека на языке
простой систематики, но в дальнейшем основной упор делается
на анализ одределенных проблем эволюции человека с точки
зрения экологии. Вот почему эту книгу следует рассматривать
не как простой учебник, а скорее как книгу для чтения по эко-
Предисловие
9
логии и эволюции, дополняющую общий очерк об эволюции
человека. Для реализации этого тематического подхода при-
шлось опустить немалую часть того сырого, описательного ма-
териала, который лежит в основе многих учебников по эволю-
ции человека. Я надеюсь, что книга представит интерес для тех
читателей, кто специально интересуется вопросами эволюции
человека, — антропологов и археологов, а также для биологов
общего профиля. При написании текста я пытался по возмож-
ности избегать употребления слишком специальных выражений,
чтобы сделать книгу доступной для более широкого читателя.
С этой целью многие разделы открываются вступлением и об-
суждением ряда основных понятий, терминов и затрагиваемых
проблем. Надеюсь, что это не оттолкнет от книги тех читателей,
которые уже знакомы с принципами эволюционной экологии.
Следует сделать и еще одну оговорку: я сконцентрировал все
внимание на ранних стадиях эволюции гоминид, т. е. на перио-
де времени от 5 до 1 млн. лет назад, когда линия гоминид воз-
никла в Африке и развивалась только в пределах этого мате-
рика. Это произошло вовсе не потому, что данный период не-
пременно является наиболее важным или интересным, но
потому, что он дает много примеров, иллюстрирующих ряд про-
цессов и механизмов эволюционной экологии. Тем не менее есть
все основания, чтобы сходным образом рассматривать и более
поздние периоды эволюции человека.
В создании этой книги участвовало, осознанно и неосознан-
но, большое количество людей, оказавших мне самую разнооб-
разную помощь. Я писал ее, будучи в Университете Дарема,
в промежутках между исполнением довольно-таки изнуритель-
ных обязанностей по отработке отведенных мне часов занятий
со студентами и попыткой спасти от банкротства наш буфет,
где я регулярно пил кофе. Мои коллеги в Дареме представляли
идеальную университетскую среду — дружественную, но гото-
вую поспорить. Так, можно было всегда рассчитывать на Гил-
берта Мэнли по части оживленных и острых дискуссий. Курсы,
которые я вел совместно с Робертом Лейтоном и Малькольмом
Смитом, стимулировали развитие идей, лежащих в основе этой
книги. Майкл Карризерс, хотя и не склонный всерьез воспри-
нимать название этой книги, ободрял меня как дружеской под-
держкой, так и вином, за что я ему бесконечно благодарен.
Остальные мои коллеги в Дареме, в особенности Мартин Джо-
унс, высказали много полезных замечаний и мыслей. Но прежде
всего я выражаю признательность нескольким поколениям сту-
дентов за то, что они терпеливо сносили все мои интеллекту-
альные выкрутасы, хотя в свое время и мечтал частенько о том,
чтобы их сочинения на заданные мною темы были покороче.
Я должен также поблагодарить студентов и сотрудников ка-
10
Предисловие
федры антропологии Университета Нью-Мексико за дружелю-
бие и интерес, проявленные к моей особе, когда я работал у
них в качестве приглашенного сотрудника.
Роберт Мартин прочел первоначальный вариант рукописи и
сделал много замечаний; те согласованность и смысл, которые
в ней, возможно, есть, книга в значительной степени приобрела
благодаря его усилиям. Целиком или по частям рукопись чи-
тали Клайв Гэмбл, Джон Спет, П. К. Ли и Элизабет Круис.
Все они высказали много ценных предложений (частью из ко-
торых я воспользовался!). Все ошибки остаются полностью на
моей совести. Сара Банни немало сделала для того, чтобы
улучшить стиль и научное содержание книги. Я выражаю так-
же особую признательность Полли и Джайлсу Куртисам за
доброту и гостеприимство, оказанные мне в Кембридже. Всем
этим людям, а также многим другим, особенно Джил Гуди,
я остаюсь бесконечно Обязанным.
И наконец, последнее. В студенческие годы, а также в на-
чале своей научной карьеры я имел счастье учиться, получать
поддержку и дружеское расположение у двух крупнейших ар-
хеологов, слишком рано ушедших из жизни. Эта книга посвя-
щена памяти Дэвида Кларка и Глинна Айзека, без вдохновляю-
щих идей которых я едва ли попытался бы взяться за подоб-
ную работу.
Роберт Фоули
Кембридж
Июнь 1986
ВВЕДЕНИЕ
«А почему утка?» — таков был один из занятнейших вопро-
сов, который задавал Гручо Маркс1. И хотя он оказал на ан-
тропологию гораздо меньшее влияние, чем его однофамилец
Карл Маркс, вопрос «А почему человек?» также чрезвычайно
интересен. Традиционные ответы на этот вопрос носят философ-
ский или теологический характер. Однако действительная за-
слуга совершенной Дарвином революции состоит в том. что
этот вопрос становится по существу практической проблемой в
рамках эволюционной биологии и его решение зависит в боль-
шей степени от эмпирических исследований, чем метафизиче-
ских, этических и телеологических дискуссий. Ответ, предла-
гаемый неодарвинизмом, рассматривает эволюцию человека как
следствие того, что человеческие (и гоминидные) особенности
позволили их носителям получить преимущество в отношении
размножения над теми, кто не обладал такими чертами. По-
этому проблема заключается в том, чтобы определить, каковы
эти благоприятные характеристики, и показать, при каких ус-
ловиях они становятся выгодными. Таким образом, вопрос
«А почему люди?» требует определения типа окружающей сре-
ды, как биологической, так и социальной, в которой преимуще-
ства человеческого бытия превышали бы затраченные усилия.
Это — вопрос локализации происхождения человека во времени
и пространстве.
Такой подход может быть не столь грандиозным, как более
философский подход, и не столь романтичным, как те теории,
согласно которым возникновение человека объясняется прибы-
тием на землю космических пришельцев или смешением разных
видов человекообразных обезьян, но тем не менее только он
способен принести ощутимые результаты. Задавая вопрос
«А почему люди?», мы выходим за пределы круга ископаемых
остатков и вступаем в область эволюционной экологии, которая
и обусловливает появление этих окаменелостей.
Ископаемые остатки составляют основу изучения эволюции
человека. Палеоантропологов, т. е. тех, кто исследует проис-
1 Американский комик, один из трех братьев — популярнейших киноакте-
ров в 1930-х гг. — Прим, перев.
12
Введение
хождение человека, легче всего представить ползающими на
загорелых коленях в какой-либо негостеприимной части света
в поисках мелких фрагментов окаменелостей, которые могут
послужить ключом к разгадке тайны нашей эволюции или, более
того, оказаться недостающим звеном; в обыденном сознании
это можно приравнять к тому, что найден Святой Грааль. Каж-
дая новая находка ископаемых костей человека становится сен-
сацией на страницах газет всего мира, получает остроумное
прозвище, а также научное название, которое гораздо труднее
произнести, и занимает свое место как составная часть загадоч-
ной мозаичной картинки. По мере того как появляется все
большее количество таких фрагментов нашего прошлого, рас-
сказ об эволюции человека теряет свою таинственность: чем
больше ископаемых находок, тем полнее наше знание.
Хотя характер эволюции человека сегодня, без сомнения,
более понятен, чем во времена, когда Дарвин писал первую
книгу на эту тему-—-«Происхождение человека», большинство
вопросов, поставленных в ней великим ученым, сохраняет свой
смысл и остается столь же дискуссионными, как и прежде.
Накопление ископаемых остатков дает ответы на многие вопро-
сы эволюции человека: как выглядели древние гоминиды?;
когда они возникли?; где они возникли?; как они эволюциони-
ровали?; но основной вопрос почему? остается по-прежнему
спорным.
В этой книге я высказываю предположение, что становление
человека обеспечило решение проблем, с которыми столкнулись
предковые формы в далеком прошлом. «Стать гоминидом» —
это решение оказалось лучшим с точки зрения адаптации по
сравнению с различными альтернативными решениями в рамках
доступного в то время генетического разнообразия, а также
теми альтернативами, которые были использованы другими ви-
дами, ныне вымершими. Причина заключается в том, что эво-
люция — это процесс решения проблем. Естественный отбор
благоприятствует таким «решениям», которые лучше справля-
ются с задачами, поставленными окружающей средой; эти ре-
шения распространяются в популяции, потому что их носители
характеризуются более высокой степенью репродуктивного ус-
пеха. Так популяции и виды приспосабливаются к условиям
своего обитания. Поэтому не существует никакой общей схемы
эволюции, никакого руководящего плана, никакого заранее
установленного ее направления. Относительная черта процесса
эволюции состоит, скорее, в разнообразии альтернативных ре-
шений экологических проблем, которые имеются в генетическом
и фенотипическом репертуаре у множества организмов разных
систематических групп; из этого множества решений естествен-
ный отбор благоприятствует «лучшим».
Введение
13
Этот процесс происходил с гоминидами и людьми, так же
как и с любыми другими организмами. Характер эволюции че-
ловека таков, что его черты можно обнаружить и в ископаемых
остатках, и в археологических находках. Он запечатлен также
в особенностях нашей сегодняшней биологии и поведения. Эти
черты, однако, сформировались под влиянием тех проблем,
с которыми сталкивались первые гоминиды. Чтобы ответить на
вопрос «А почему гоминиды?», мы должны понять, в чем за-
ключались эти проблемы и почему свойства гоминид обеспечи-
ли лучшее их решение.
Откуда, однако, возникают эти проблемы? И как они могут
быть истолкованы? Их происхождение связано с двумя при-
чинами. Это, во-первых, наша принадлежность к определенной
группе животных — млекопитающим, приматам, животным с
наземным образом жизни и т. д. — и, во-вторых, среда, в кото-
рой мы существуем. В конечном счете, следовательно, они воз-
никают в результате взаимодействия двух этих источников.
Цель настоящей книги заключается в том, чтобы определить те
проблемы, с которыми сталкивались первые гоминиды, и соот-
нести их с эволюцией адаптивной стратегии гоминид. Решение
этой задачи зависит от трех обстоятельств: от тех подходов,
которые используются при изучении прошлого как арены, где
разворачивались события эволюции древнейших гоминид; от
изучения специфических особенностей гоминид, что является
прерогативой антропологии; от биологической, в особенности
эволюционной и экологической, теории как основы, позволяю-
щей соединить отличительные особенности животного с усло-
виями, в которых протекали его жизнь и развитие.
Эти предпосылки определили структуру книги. Беглый об-
зор современного состояния исследований древнейших гоминид
дается в гл. 2. В гл. 3 очерчены принципы эволюционной эко-
логии— аналитическая основа книги. Трудности ее применения
к археологическим материалам и находкам ископаемых остат-
ков обсуждаются в гл. 4. После установления теоретических и
методологических рамок можно перейти к анализу действитель-
ных адаптативных проблем (гл. 5—10). До некоторой степени
выделение этих проблем должно носить произвольный, искусст-
венный характер, так как в «реальном мире», где речь идет о
выживании особей, существуют не дискретные категории, а не-
прерывное давление отбора. Однако для того, чтобы исследо-
вание стало возможным, я по очереди обсуждаю проблемы,
встающие перед гоминидами, в связи с тем, что они являются
тропическими животными, крупными млекопитающими, назем-
ными приматами, обитающими в условиях смены времен года,
сталкивающимися с межвидовой борьбой и осваивающими но-
вые экологические ниши. Каждая из этих проблем, как будет
14 Введение
показано ниже, сыграла свою роль в формировании направле-
ния эволюции гоминид.
Центральная задача этой книги состояла в том, чтобы при-
близить палеоантропологию к основному руслу эволюционной
биологии. Решению этой задачи препятствовал давно устано-
вившийся взгляд на человека как на уникальный, совершенно
новый тип организма, для которого тем самым требуется совер-
шенно особый способ объяснения, отличный от всех других,
используемых для понимания остального биологического мира.
Развитие эволюционной экологии человека зависит поэтому от
решения проблемы человеческой уникальности.
Глава 1
ПРОБЛЕМА ЧЕЛОВЕЧЕСКОЙ УНИКАЛЬНОСТИ
1.1. Отличаются ли люди от других существ?
1.1.1. Степень уникальности
Современный человек — это, без сомнения, совершенно осо-
бый вид животных. Речь, беспрецедентная пластичность пове-
дения, своеобразное выпрямленное положение тела, мозг, слиш-
ком большой для целей простого удобства, сложные техноло-
гические навыки — все эти черты буквально кричат о челове-
ческой уникальности. Хотя Дарвин показал, что мы происходим
от более ниэкораэвитых форм и шимпанзе, возможно, предпри-
нимают весьма серьезные попытки приблизиться к нам, овладе-
вая языком, изготовлением орудий, а также другими менее
приятными аспектами человеческого поведения, такими, как
каннибализм и стычки с себе подобными, в глубине души мы
знаем, что человек надежно защищен от нашествия орд живот-
ных непроходимой пропастью. Стены бастиона, именуемого
человеческой уникальностью, все еще достаточно прочны сегод-
ня, когда прошло более ста лет после ураганной дарвиновской
атаки.
В истории исследований об эволюции человека можно об-
наружить две противоположные тенденции. С одной стороны,
это позиция тех ученых, которые стремятся подчеркнуть уни-
кальность человека, концентрируя внимание на прерывистости
эволюции и отыскивая уникальные человеческие характеристи-
ки в ископаемых остатках, дошедших из далеких эпох. С другой
стороны, существуют и те, кто видит в приматах зеркальное
отражение человека и вследствие этого всячески преуменьшает
различия в поведении и строении тела между ними, доказывая
недавнее эволюционное расхождение ветвей, ведущих к чело-
веку и его ближайшим родственникам по эволюционному дре-
ву. Настоящая книга основывается на этой второй стратегии,
но вместе с тем ее автор понимает, что, несмотря на все на-
блюдения и аргументы, нельзя игнорировать тот факт, что че-
ловек — это совершенно особый вид. Трудность поэтому заклю-
чается в самом определении уникальности.
Современный человек — Homo sapiens sapiens — это уни-
кальный подвид. Уникальность становится проблемой, когда мы
пытаемся соотнести уникальное явление с общим классом яв-
лений, а затем стремимся объяснить характер выявленной для
16
Глава 1
этого класса изменчивости, исходя из общих принципов и за-
кономерностей. Необходимо, однако, сделать два замечания о
приложимости общих биологических принципов в отношении
нашего собственного подвида. Во-первых, термин «подвид» сам
по себе биологический, и если мы помещаем человека в био-
логические рамки, пользуясь такими терминами, как «вид» или
«подвид», то это означает необходимость использования биоло-
гических понятий при изучении обнаруженных характеристик.
Во-вторых, все биологические таксоны уникальны. Это основа
биологической таксономии и разнообразия жизни. Все виды
уникальны, потому что они составляют «группы действительно
или потенциально скрещивающихся между собой естественных
популяций, которые репродуктивно изолированы от других та-
ких групп» (Мауг, 1963, р. 120). Они являются, таким образом,
генетически уникальными. Генетическая изоляция может быть
результатом географической изоляции, но практически во всех
случаях она включает также фенотипические и, следовательно,
адаптивные различия. Все виды не менее уникальны в физио-
логическом, экологическом и поведенческом отношении, чем со-
временный человек. Экологи определяют эту адаптивную осо-
бенность с помощью концепции «экологической ниши», харак-
теризующей уникальное многомерное биологическое простран-
ство, занимаемое видом (Hutchinson, 1957). Хотя Homo sapiens
sapiens уникален, но столь же уникальны и все остальные под-
виды. Это справедливо и по отношению к прошлому, так что
Homo erectus также уникален и отличен от других видов го-
минид.
Из этого следует, что неприменимость биологических прин-
ципов к современному человеку нельзя объяснить его уникаль-
ностью. Можно использовать, однако, еще два разграничитель-
ных критерия. Либо различия между гоминидами и представи-
телями других таксономических единиц должны быть гораздо
больше, чем те, которые существуют для других видов, либо
человеческий вид возник в результате уникального процесса,
которому не подвержены другие организмы.
Обратимся к первому из этих критериев: насколько отлича-
ются люди от других организмов? Нашими ближайшими род-
ственниками являются крупные человекообразные обезьяны.
Физиологические основы нашего родства были давно продемон-
стрированы Дарвином (1871) и Гексли (1863); генетические
различия на удивление малы и составляют менее 1% (Sarich,
1971). Поведенческие различия значительны, хотя, как показал
Пассингем (Passinghem, 1982), человек и здесь укладывается
в рамки высших приматов. Однако, как указывает Дарвин
(Darwin, 1871, р. 152), расстояния между живущими видами
могут ввести исследователя в заблуждение:
Проблема человеческой уникальности 17
«Значительный пробел в органической цепи между челове-
ком и его ближайшими родичами, который не может быть за-
полнен ни одним из вымерших или живущих видов, часто вы-
двигался как серьезное возражение против мнения, что человек
произошел от какой-либо низшей формы. Но это возражение
не будет иметь особенного значения для тех, кто в силу общих
соображений, верит в общий принцип эволюции. Пробелы
встречаются постоянно во всех частях органической цепи, не-
которые из них значительны, резки и определенны, другие же
в различной степени, но менее значительны... Но все эти пробе-
лы зависят только от числа вымерших родственных форм.
В какой-нибудь из будущих периодов — и даже не слишком
отдаленный, если мерить столетиями —- цивилизованные чело-
веческие расы почти наверное уничтожат и вытеснят в целом
мире дикие расы. К тому же времени... человекообразные обезь-
яны будут без всякого сомнения уничтожены. Пробел между
человеком и его ближайшими родичами в этом случае сделает-
ся еще больше, потому что он будет лежать между человеком,
можно надеяться еще более цивилизованным... и какими-то
обезьянами, настолько низкоорганизованными, как павианы,
тогда как теперь этот пробел идет от негра или австралийца к
горилле»1.
Другими словами, если не принимать в расчет несколько
драматизированный и отдающий расизмом тон дарвиновского
изложения, степень уникальности организма зависит от того,
насколько сходен с ним ближайший родственник из числа ныне
живущих организмов, а это в свою очередь зависит от того,
какое количество родственных форм вымерло ранее. Поэтому,
подобно многим другим эволюционным спорным вопросам, эта
проблема переходит в историческую плоскость и подвержена
лишь относительным оценкам. Человеческая уникальность, как
мы теперь знаем, благодаря значительно более обширной кол-
лекции ископаемых остатков по сравнению с тем, что было в
распоряжении Дарвина, была чрезмерно преувеличена за счет
исчезновения австралопитеков, Homo erectus и неандертальцев,
не говоря уж о возможном исчезновении древних человекооб-
разных обезьян, которые состояли в более близком родстве с
гоминидами, чем сегодняшние шимпанзе и горилла. Даже если
взять поведенческие аспекты, археологические реконструкции
развития техники изготовления орудий свидетельствуют о су-
ществовании впоследствии исчезнувших промежуточных техно-
логических приемов. При рассмотрении ранних гоминид види-
мый разрыв между ними и другими организмами не должен
1 Цитируется по изданию: Ч. Дарвин. Сочинения, том 5, М., Из-во АН
СССР, 1953, стр. 170. — Прим, перев.
2—367
18
Глава 1
мешать более тесному союзу палеоантропологии с эволюцион-
ной биологией.
Второй критерий, который может быть использован для оп-
ределения уникальности человека, состоит не только в том, что
исключителен продукт развития (люди), но и в том, что про-
цессы и механизмы, участвовавшие в достижении конечного
результата, также уникальны. Так, хотя все организмы от
птиц-носорогов до лошадей исключительны в том смысле, как
это было описано выше, процессы, с помощью которых они эво-
люционировали, одинаковы для всех: естественный отбор ведет
к адаптации, генетической изоляции и характерному набору
поведенческих и морфологических особенностей. Здесь необхо-
димо рассмотреть вопрос о том, может ли этот процесс сохра-
нять свою силу и в ходе эволюции человека.
1.1.2. Неадекватность термина «культура»
Если и существует новый эволюционный процесс, который
можно обнаружить у человека, то это культурная эволюция.
Культура — это ключевое понятие для антропологии, вмещаю-
щее в себе большую часть того, что, по нашему мнению, явля-
ется отчетливо или исключительно человеческим. К сожалению,
существует почти столько же определений культуры, сколько
ученых-антропологов, но в широком смысле под культурой по-
нимаются небиологические аспекты поведения человеческого
вида, включая речь, изготовление орудий, возросшую пластич-
ность поведения, способность к символическому мышлению и
самовыражению с помощью символов; культура передается не
через систему генетических механизмов, а посредством обуче-
ния и усвоения. Хотя способность к культуре может быть видо-
специфической и генетически детерминированной, поведение,
которое ее определяет, таковым не является. С развитием этих
процессов эволюция человека оказалась отделенной от основ-
ного русла биологической эволюции. Для человека естествен-
ный отбор по меньшей мере дополняется параллельным процес-
сом культурного отбора, если не полностью заменяется им.
В последние годы были разработаны модели генно-культурной
коэволюции (Lumsden, Wilson, 1980; Cavalli Sforza, Feldman,
1981).
Однако является ли понятие «культура» действительно по-
лезным при изучении происхождения человека? Я попытаюсь
доказать, что нет. Культура — сложный термин, суммирующий
целый ряд характеристик, имеющих важное значение в жизни
современного человека. Но при изучении происхождения этих
культурных особенностей едва ли нужно связывать их всех во-
едино. Мы не знаем — и на самом деле именно это и хотим
Проблема человеческой уникальности
19
выяснить, — когда та или иная из них впервые появилась в ря-
ду гоминид. Каждая из них — будь то возросшая способность
к обучению, речь, изготовление орудий и т. д. — возможно, раз-
вивалась отдельно от остальных под влиянием независимых сил,
связанных с отбором, а поэтому смешивать их всех в понятии
«культура» значит отметать саму возможность, что в прошлом
гоминиды, вероятно, владели лишь частью своего современного
поведенческого репертуара или что этот репертуар состоял из
других комбинаций. Я не вижу поэтому, в чем состоят преиму-
щества использования термина, хотя и весьма краткого в на-
писании, но игнорирующего сам вопрос, на который мы пыта-
емся найти ответ, и теряющего вследствие этого аналитиче-
скую точность.
Многие черты человеческой культуры, если рассматривать
их в отдельности, .можно обнаружить в рудиментарной форме
у других животных. Шимпанзе, как известно, используют и из-
готовляют орудия; использование орудий, наряду с существен-
ной обработкой имеющихся в природе материалов (например,
при гнездостроительстве) встречается и у других представите-
лей животного царства. Шимпанзе обнаруживают способности
к систематическому использованию языка жестов (вроде аме-
риканского языка глухонемых), что свидетельствует о понима-
нии ими символов и грамматических структур. Более того,
в своем естественном состоянии приматы пользуются широким
набором коммуникативных систем. Сейфарт и др. (Seyfarth,
Cheyney, Marler, 1980), например, показали, что верветки
(Cercopithecus aethiops) используют голосовые сигналы в ка-
честве «слов». Типы поведения, сформировавшиеся под влияни-
ем обучения, конечно, также широко распространены среди
животных: от голубых синичек, научившихся открывать молоч-
ные бутылки, до таких птиц, как седлистая гуйя, подражающая
песням соседей, и японских макаков, научившихся смывать
песок со своей еды. В каждом из этих случаев имело место не
только обучение, но также быстрая передача информации и
развитие «традиций» в популяции.
Поэтому большая часть тех особенностей, которые известны
под названием «культура», если их рассматривать по отдель-
ности, встречаются в той или иной форме в царстве животных.
Использование термина «культура» в палеоантропологии за-
трудняет рассмотрение этой непрерывности. При исследовании
истоков поведения современного человека представляется более
плодотворным принять редукционистский подход, что позволит
иметь дело с доведенными до минимума категориями поведе-
ния, а не предполагать, пользуясь на априорных основаниях
термином «культура», что мы имеем дело с чем-то совершенно
от них отличным.
2»
20
Глава 1
Я не хочу сказать, что после того как эти характеристики
появились, они не оказывали влияния на дальнейший ход эво-
люции. Однако на данном этапе лучше рассматривать их как
следствия, а не причины поведенческой эволюции, т. е. как
факторы, изменяющие природу воздействующего на гоминид
давления отбора, а не механизмы, посредством которых это
давление действует. Признать наличие культурной эволюции
или коэволюции в самом начале анализа эволюции человека,
значит заранее предопределить неадекватность естественного
отбора. Задача всякого эволюционного экологического анализа
состоит в том, чтобы доказать, как обстоит дело, а не предуга-
дывать возможные решения. Соответственно в этой книге автор
избегает употреблять термин «культура» на том основании,
что он не является необходимым и полезным для нашего ана-
лиза. Поведенческая, а не культурная эволюция — вот адекват-
ный термин, обладающий при минимальных допущениях макси-
мальной гибкостью. Использование этого термина не устраняет
характеристик, делающих людей уникальными, таких, как рас-
тущие способности к обучению, изобретательство и подражание,
сложная коммуникация и беспрецедентная пластичность пове-
дения, но позволяет рассматривать их так, чтобы обеспечить
возможность межвидовых сопоставлений.
1.1.3. Методологический редукционизм
В данной книге используется редукционистский метод ис-
следования, и, возможно, примененный в антропологическом
контексте он потребует некоторых комментариев. «Редукцио-
нист — это тот, кто стремится объяснить даже самые сложные
явления, исходя из элементарных взаимодействий между более
или менее элементарными частями» (Anon, 1983, р. 301). В на-
шем случае закономерности сложной группы организмов, каки-
ми являются гоминиды, объясняются, исходя из элементарных
экологических и эволюционных процессов: естественного отбора
и адаптации. Как я уже пытался показать, такой подход оправ-
дан, с одной стороны, общей эволюционной историей гоминид
и, с другой, методическими трудностями, возникающими при
определении того, в какой степени люди освободились от дей-
ствия естественного отбора. «Значение редукционизма состоит
в том,... что он помогает более точно очертить постоянно изме-
няющуюся границу между тем, что можно определить как «из-
вестное», и «остальным» (там же). Под «известным» в данном
случае понимается общая биологическая характеристика и эво-
люционная история людей, под «остальным», или «неизвест-
ным»,— природа процессов и механизмов, приведших к отделе-
нию линии человека от его ближайших биологических родст-
Проблема человеческой уникальности 21
венников. Использование редукционистского подхода оправды-
вается тем, что, несмотря на очевидность возникших интригую-
щих особенностей, их сущность и механизм передачи остаются
совершенно неясными. Одна только попытка объяснить макси-
мум изменчивости, исходя из минимума частей и их взаимодей-
ствий, может привести к обнаружению свойств любого нарож-
дающегося явления.
Подведем итоги. Мы утверждали, что элементарные прин-
ципы эволюционной экологии подходят для анализа гоминид.
Они представляют собой наиболее удобный способ описания и
объяснения различий и сходства между человеком и другими
организмами. Наша очевидная уникальность не является доста-
точной причиной для использования специальных методических
приемов и концепций. Уникальность — характерная особенность
всех видов, а степень различий между видами — функция диф-
ференциальной смертности, но не абсолютное качество. Кроме
того, с методической точки зрения было бы совершенно непри-
емлемым устанавливать для гоминид еще до анализа особый
цабор правил и ограничений. Они могут возникнуть только в
результате анализа. Таковы в общих чертах предпосылки для
использования принятого в этой книге метода.
Итак, гоминиды представляют собой уникальную группу
животных. Пытаясь применить в ходе анализа общие методиче-
ские приемы и концепции эволюционной биологии, я вовсе не
хочу покончить с их уникальностью, а желаю, скорее, объяс-
нить ее. Цель этой книги состоит не в том, чтобы проследить
истоки человеческих характеристик вплоть до доисторического
«болота», но, скорее, в том, чтобы привлечь к рассмотрению
отдельных эволюционных событий мощный набор аналитиче-
ских и концептуальных приемов. Мы действительно являемся
уникальным видом, но нельзя забывать при этом, что мы всего
лишь еще один уникальный вид.
1.2. Наше место в природе
1.2.1. Человек как животное
Мысль о том, что процесс происхождения человека можно
объяснить с помощью эволюционных и экологических принци-
пов, базируется на одном ключевом предположении, состоящем
в том, что наш вид может рассматриваться как биологический
организм, а каждый из нас — как, по сути дела, животное.
Утверждение, что людей можно рассматривать как биологиче-
ские организмы, вытекает из того факта, что они эволюциони-
ровали путем естественного отбора. Это утверждение базирует-
ся на сопоставлении черт сходства и различия между современ-
22
Глава 1
ным человеком и другими видами. Несмотря на ограниченность
доступных в его время данных, Дарвин в «Происхождении че-
ловека» (1871) сумел показать, что наши анатомические, фи-
зиологические и онтогенетические характеристики весьма сход-
ны с таковыми представителей отряда приматов и что некото-
рые поведенческие особенности, такие, как выражение лица,
также можно успешно сравнивать у людей и других приматов.
Выводы, которые смог сделать Дарвин, послужили основой для
дальнейших исследований в течение последующих более чем
100 лет и составляют ключевую предпосылку, на которой бази-
руется настоящая книга. Археология и физическая антрополо-
гия были заняты установлением точного характера взаимосвя-
зей между человеком и другими животными (Le Gros Clark,
1964). До выхода в свет «Происхождения видов» природа этой
взаимосвязи не казалась хоть сколько-нибудь проблематичной,
так как между человеком — венцом творения — и всеми други-
ми организмами, казалось, лежала непреодолимая пропасть
(Foley, 1984а). Однако, после того как утвердилось мнение, что
мы характеризуемся достаточно установленным родством с
другими животными, на повестке дня встал вопрос о взаимо-
связях между человеком и приматами.
Нынешняя ситуация была критически проанализирована
Пассингемом (Passingham, 1982) в детальной монографии о
строении и поведении приматов. Признавая значимость измене-
ний формы, таких, как бипедия, манипуляторные способности
и т. д., он особенно подчеркивает важность изменений мозга.
Однако вывод, к которому приходит Пассингем (Passingham,
1982, р. 332—333), касается самой сути взаимоотношений меж-
ду человеком и биологическим миром. Мы можем сильно отли-
чаться даже от обезьян, но эти различия можно объяснить с по-
мощью элементарных биологических принципов. Он выделяет
три таких принципа, посредством которых достигается транс-
формация. Во-первых, количественные изменения могут иметь
последствия, которые будут носить качественный характер.
Прибегая к аналогии с компьютером, он подчеркивает, что
компьютер большего размера по сравнению с небольшим не
просто делает те же операции, но в более широком масштабе,
а может решать совершенно иные задачи и работает с большей
эффективностью. Во-вторых, радикальная реконструкция не яв-
ляется необходимой предпосылкой основных функциональных
изменений, а небольшие изменения могут иметь фундаменталь-
ные последствия. И наконец, в-третьих, «освоение нового спосо-
ба переработки информации может иметь революционные по-
следствия». Способ, о котором идет речь, это язык; хотя зачат-
ки этого способа (коммуникации) опять-таки имеются и у
других организмов, у человека его освоение связано с транс-
Проблема человеческой уникальности
23
формацией и овладением новыми функциями, в первую очередь
такими, как мыслительная деятельность.
Общий вывод, к которому можно прийти в результате всех
этих рассуждений, состоит в том, что современный человек во
многих отношениях сопоставим с другими видами животных, но
в процессе эволюции претерпел некоторые существенные и
скачкообразные изменения. Однако в центре внимания должна
находиться не просто уникальность человека, а возможность
объяснения как дискретной, так и непрерывной уникальности
с помощью основных биологических принципов.
1.2.2. Приматы
Приматы — это группа животных, которая по большинству
биологических характеристик не отличается от современных
человеческих популяций, а ее эволюционные пути разошлись с
путями человека позже, чем у всех остальных животных. Общие
характеристики человека и приматов служат как доказательст-
вом эволюции человека, так и источником идей и сомнений.
Приматы создают контекст для изучения эволюции человека.
Приматы представляют собой отряд класса млекопитающих,
характеризующийся сохранением некоторых примитивных черт
и прогрессивным развитием более специализированных призна-
ков, связанных с древесным образом жизни и высокой степенью
социализации. Некоторые тенденции выявляются в процессе
эволюции приматов (Le Gros Clark, 1949; Napier, 1971)
(рис. 1.1). Приматы сохраняют основной план строения тела
Увеличение
размеров тела
Увеличение мозга
Сохранение пяти пальцев
на кисти и стопе
Фронтально
расположенные глаза,
бинокулярное зрение
Укорочение морды
и потеря обонятельной
чувствительности
Полная подвижность
конечностей
Цепкая, способная
к манипулированию кисть
Рис. 1.1. Основные свойства приматов (по Napier, 1971)
24
Глава 1
Приматы
Рис. 1.2. Связь между основными таксономическими подразделениями приматов
млекопитающих, например пять пальцев -на кисти и стопе. Эти
сегменты конечностей стали гораздо более подвижными, за счет
чего приматы приобрели возможность умелого захвата предме-
тов и манипулирования кистями и стопами. Глаза сближаются
во фронтальной плоскости, что обеспечивает развитие биноку-
лярного зрения, морда укорачивается в связи с утратой обоня-
тельных способностей. Мозг увеличивается по отношению к
размерам тела, многие виды живут в сложных социальных и
природных условиях обитания. Существует также тенденция
к увеличению размеров тела с изменениями в абсолютной и от-
носительной продолжительности как отдельных фаз жизненно-
го цикла, так и продолжительности жизни в целом. В отноше-
нии пищи приматы оказываются на редкость легко приспосаб-
ливающимися животными, но в целом в рамках отряда отмеча-
ется тенденция к переходу от насекомоядности к питанию фрук-
тами и всеядности. Следует подчеркнуть, однако, что здесь
перечислены основные закономерности, а какая-либо отдельно
взятая группа приматов может либо усилить их, либо поменять
на противоположно направленные.
Ныне живущие приматы делятся на две основные группы
(рис. 1.2)—стрепсириновых и гаплориновых (Szalay, Delson,
1979). Первые — наиболее древние; к ним относятся полуобезь-
яны (Prosimii): небольшие по размерам, ведущие в основном
ночной образ жизни, насекомоядные формы — лемуры, лори и
родственные им виды. Другая группа — гаплориновые прима-
ты— состоит из трех основных таксонов—долгопятов, широ-
коносых и узконосых обезьян. Долгопяты — это реликтовая,
локализованная в пределах Юго-Восточной Азии, группа по
Проблема человеческой уникальности
25
Рис. 1.3. Относительное положение материалов в эоцене, около 55—35 млн. лет
назад, с изображением альтернативных путей распространения широконосых
обезьян: 1—из Северной Америки до образования связи между Северной
и Южной Америкой; 2 — из Африки через резко уменьшившийся в размерах
Атлантический океан (см. обсуждение Ciochon, Chiarelli, 1980; карта по Eisen-
berg, 1981)
преимуществу сходных с полуобезьянами форм, которые вновь
адаптировались к ночному образу жизни. Вследствие этого
многие из их характеристик сходны с таковыми стрепсириновых
приматов. Широконосые и узконосые обезьяны — это группы
человекоподобных высших приматов (Anthropoidea) или собст-
венно обезьян, Нового и Старого Света соответственно. Обе
Америки отделились от Евразии и Африки по меньшей мере
35 млн. лет назад (рис. 1.3), что позволило человекоподобным
приматам развиваться там в изоляции. Таким образом, эти две
группы представляют собой классический пример параллельной
эволюции, в ходе которой под действием одинакового давления
отбора образуются сходные эволюционные модели. Широконо-
сые остались целиком древесной группой, обитающей в тропи-
ческих дождевых лесах Центральной и Южной Америки. В чис-
ле широконосых — самые мелкие представители приматов
(мармозетки и тамарины), которые по крайней мере отчасти
заняли некоторые из свободных в Новом Свете экологических
ниш полуобезьян, а также более крупные обезьяны, которые
представлены формами, превосходно приспособленными к ла-
26
Глава 1
занью по деревьям, включая необычайно ловкую паукообраз-
ную обезьяну.
В Старом Свете, с другой стороны, адаптивная радиация
обезьян шла намного более интенсивно и в плане приспособле-
ния к наземному образу жизни. Узконосые обезьяны разделя-
ются на две группы: Cercopithecoidea (собакоподобные, или
низшие узконосные обезьяны) и Hominoidea (гоминоиды), куда
входят человекообразные обезьяны и человек. Низшие узконо-
сые обезьяны — наиболее пестрая группа среди современных
приматов. В нее входят тонкотелые обезьяны, специализирую-
щиеся на поедании листьев, и мартышковые, среди которых
много наземных форм, таких, как павианы, мандрилы, макаки
и патасы. В противоположность тонкотелым обезьянам мартыш-
ковые всеядны; обычно они питаются плодами растений, но в
ряде случаев употребляют в пищу и позвоночных животных.
К гоминоидам относятся современные люди и их ближайшие
родственники, человекообразные обезьяны, которые, по тради-
ции, делятся на крупных и малых человекообразных. Послед-
ние представлены гиббонами (Hylobatidae), которые ведут
исключительно древесный образ жизни и являются превосход-
ными и быстрыми брахиаторами, что позволяет им свободно
передвигаться в верхнем ярусе тропического леса. Эти виды в
настоящее время сохранились лишь в Юго-Восточной Азии.
Крупные человекообразные обезьяны фактически делятся на
две группы: азиатские формы, к которым относится орангутан,
и африканские, куда входят шимпанзе (два вида: обычный и
карликовый) и горилла, самый крупный из ныне живущих при-
матов. Люди, как мы увидим далее, находятся в наиболее
близком родстве с африканскими человекообразными обезья-
нами.
Именно с ныне живущими видами последних человека свя-
зывает теснейшее генетическое, морфологическое и эволюцион-
ное родство. Как не замедлили отметить биологи-эволюционис-
ты, люди — это настоящие приматы, получившие многие свои
особенности в наследство от общего предка всех приматов,
а также приспособившие удачные черты приматов для своих
собственных целей. В масштабе палеоантропологии основные
адаптивные особенности, приматов — хватательная конечность
(кисть или стопа), высокоразвитое зрение, редукция обоняния,
высокий уровень общественных отношений — представляют как
бы точку отсчета для изучения любых аспектов эволюции чело-
века, в то время как отклонения от характерной для приматов
формы — например, бипедия — должны рассматриваться как
особо важные с точки зрения эволюции.
Несмотря на многолетние исследования, таксономия прима-
тов все еще находится в стадии разработки и в настоящее
Проблема человеческой уникальности
27
Таблица 1.1. Классификация приматов (Kavanagh, 1984, с изменениями)
Отряд
Подотряд
Надсемейство
Семейство
Подсемейство
Род Обычное название
Primates
Strepsirhini! >
Lemuroidea
Cheirogaleidae
Microcebus
Cheirogaleus
Phanerinae
Phaner
Lemuridae
Lemur
Varecia
Hapalemur
Lepilemuridae
Lepilemur
Indriidae
A vahi
Propithecus
Indri
Daubentonioidea
Daubentoniidae
Daubentonia
Lorisoidea
Lorisidae
Loristnae
Loris
Nyciicebus
Arctocebus
Perodicticus
Galiginae
Galago
Haplorhini1'
Tarsioidea
Tarsiidae
Tarsius
(Platyrrhini)2'
Ceboidea
Callitrichidae
Cebuella
Приматы
Полуобезьяны
Лемуроморфные
Мышиные лемуры
Карликовые лемуры
Лемур фанер
Обыкновенные лемуры
Лемур вареция
Гапалемур
Изящные лемуры
Мохнатые индри, или
авагисы
Сифаки
Черные короткохво-
стые индри
Руконожки, или айе-
айе
Лоризиморфные
Лориевые
Тонкие лори
Толстые медленные
лори
Ангвантибо
Обыкновенный потто
Обыкновенный галаго
Долгопяты
Широконосые обезьяны
Карликовая игрунка
28
Глава I
Продолжение
Отряд
Подотряд
Надсемейство
Семейство
Подсемейство
Род Обычное название
Callithrix Saguinus Leontopithecus Callimiconidae Callimico Cebidae Saimiriinae Saimiri Aotinae Aotus Callicebinae Callicebus Alouattinae Alouatta Cebinae Cebus Pitheciinae Pithecia Chiropotes Cacajao Atelinae Lagothrix A teles Brachyteles (Catarrhini)2) Cercopithecoidea Cercopithecidae Colobinae Colobus Presbytis Nasalis Simias Rhinopithecus Обыкновенные игрун* ки, или мармозетки Тамарины Золотистые львиные тамарины Гельдиевая мармозетка Беличьи обезьяны, или саймири Ночные обезьяны Прыгуны, или тити Ревуны Капуцины Саки Мохнатые саки, или хиропоты Уакари Шерстистые обезьяны Паукообразные обезь- яны Брахителесы Узконосые обезьяны Собакоподобные, или низ- шие узконосые обезь- яны Тонкотелые обезьяны Гверецы Лангуры Носатые обезьяны Симиасы, или коротко- хвостые носатые обезь- яны Курносые обезьяны
Проблема человеческой уникальности
29
Продолжение
Отряд
Подотряд
Надсемейство
Семейство
Подсемейство
Род Обычное название
Pygathrix Пигатрикс, или обезь-
яна-герцог
Cercopithecinae Мартышковые
Cercopithecus Мартышки
(MiopithecusY') Карликовая мартышка.
или талапойн
(Erythrocebus)^ Мартышки патас
(Cercocebus)* 2 3 4^ Мангобеи
Pap io Павианы
(Mandrillus)4^ Мандрилы
Theropithecus Ге лады
Macaca Макаки
Hominoidea Гоминоиды
Hylobatidae Гиббоновые
Hylobates Настоящие гиббоны
(Symphalangus)4') Сиаманги
Pongidae3’ Понгиды
Pongo Орангутан
Африканские человекообразные обезь-
яны+люди(?)
Gorilla Горилла
Pan Шимпанзе
Homo Человек (люди)
*) Альтернативная классификация делит приматов иа два подотряда: полуобезьян
и обезьян. Принципиальное различие между двумя классификациям1и касается положе-
ния долгопятов (которых во втором варианте относят к полуобезьянам). Термином «по-
луобезьяны» иногда пользуются для обозначения приматов более низкого по сравнению
с обезьянами ранга.
2) Инфраотряд Platyrrhini (широконосые) — это обезьяны Нового Света; Catarrhini
(узконосые) — это люди, человекообразные обезьяны и низшие узконосые обезьяны
Старого Света.
3) Пересмотр таксономии гомииоидов приводит к терминологической путанице. Тра-
диционная классификация включает три семейства: Hylobatidae (гнббоны н сиаманги),
Pongidae (орангутан, горилла и шимпанзе) и Hominidae (люди). Ныне считается обще-
признанным, однако, что люди находятся в более близком родстве с шимпанзе и горил-
лой (африканскими человекообразными), чем кто-либо нз них с азиатской человеко-
образной обезьяной — орангутаном. В формальной классификации это должно означать,
что африканские человекообразные относятся к семейству Hominidae. Однако это при-
вело бы к значительной путанице, так как термин «гоминиды» используется преимуще-
ственно для обозначения человека и его непосредственных предков. Поскольку эта проб-
лема далека от разрешения, в приведенной выше схеме таксон «гомиииды» отсутствует,
4) Некоторые авторы рассматривают эти таксоны как подроды.
30
Глава 1
Таблица 1.2. Таксономия гоминид: место человека и гоминид в мире живого
(по Le Gros Clark, 1949; Szalay, Delson, 1979)
Таксон Лнннеевское название Обычное название Состав данной группы животных
Класс Mammalia Млекопитающие Все теплокровные живот- ные. покрытые шерстью, которые рождают живых детенышей
Отряд Primates Приматы Полуобезьяны, обезьяны человекообразные обезь- яны и человек
Подотряд Haplorhini Гаплориновые Долгопяты, обезьяны, человекообразные обезь- яны и человек
Инфраотряд Catarrhini Узконосые обезьяны Обезьяны Старого Света, человекообразные обезь- яны и человек
Надсемейство Hominoidea Гоминоиды Человекообразные обезь- яны и человек
Семейство Hominidae Гоминиды Человек и его предки
Род Вид Подвид Homo H. sapiens H. s. sapiens Собственно люди Человек Неоантроп Анатомически современ- ный человек (люди)
время претерпевает существенные изменения в связи с разви-
тием новых биохимических классификационных подходов. Это
справедливо как в отношении крупных таксонов — например,
изучается вопрос о включении долгопятов в группу Anthropoi-
dea (обезьян) или Prosimii (полуобезьян), а также о степени
родства обезьян Старого и Нового Света, — так и мелких: на-
пример, проблема выделения отдельных видов в роде Papio
(павианы). Ниже приводится основная классификация ныне жи-
вущих представителей отряда приматов (табл. 1.1).
1.2.3. Гоминиды и люди
Изменчивость ряда характеристик составляет основу для
классификации, в соответствии с которой приматы разделяются
на более крупные или более мелкие таксономические единицы
в зависимости от общности признаков. Точная оценка того,
о какой именно группе идет речь, имеет большое значение в
эволюционном анализе. Для нас особенно важно уточнить
смысл понятий гоминиды и люди.
Раньше часто считалось, что оба термина взаимозаменяемы.
Однако, поскольку в настоящее время мы до конца не знаем,
какова степень различий между исчезнувшими и сохранивши-
Проблема человеческой уникальности
31
мися таксонами гоминид, игнорирование различий приведет
лишь к излишней путанице. Термин «гоминиды» (происходящий
от названия семейства Hominidae) должен использоваться для
обозначения всех популяций и видов, с которыми нас объеди-
няет общая эволюционная история, отличная от таковой осталь-
ных приматов. В настоящей книге термин «ранние гоминиды»
относится к стадии развития гоминид до начала эволюции Но-
то sapiens. Термин «человек» («люди») должен быть сохранен
исключительно для обозначения представителей единственного
ныне живущего подвида гоминид — Homo sapiens sapiens — так
же, как термин «человеческий» для обозначения характеристик,
свойственных представителям ныне живущих популяций чело-
века. Для вымерших или анатомически отличных форм лучше
пользоваться общим термином «гоминиды», так как именно
различия между вымершими и живыми формами дают ключ к
эволюционной экологии гоминид. В табл. 1.2 суммируются
представления о таксономическом месте человека « гоминид в
биологическом мире.
Глава 2
РАССКАЗ ОБ ЭВОЛЮЦИИ ЧЕЛОВЕКА
2.1. Шкала эволюции человека
Эволюция Homo sapiens — это эволюция всего-навсего од-
ного из многих миллионов видов, живших на нашей планете.
Чтобы понять этот процесс, а также эволюцию гоминид в це-
лом, нужно взглянуть на них с точки зрения эволюции всей
органической жизни.
2.1.1. Геологическое время
Возраст Земли в настоящее время исчисляется в 4600 млн.
лет. Весь этот промежуток времени — геологическое время —•
разделен на ряд периодов, характеризующих эволюцию клима-
та Земли, ее географии и биологии (рис. 2.1). Каждому перио-
ду соответствуют определенные геологические отложения и ис-
копаемые остатки живых организмов, находящихся в
установленных географических и стратиграфических соотно-
шениях.
Протяженность геологического времени осознается с тру-
дом, и здесь могут возникать проблемы в связи с определением
понятий. Большинство людей, привыкших измерять время в ча-
сах, днях и годах в пределах одного поколения в ту или дру-
гую сторону от своего собственного бытия как точки отсчета,
не испытывают затруднений, исчисляя события прошлого, со-
отнесенные с этими доступными для их понимания единицами.
Большая часть исторических событий, измеренных в месяцах
и годах, также воспринимается без затруднений. Однако пере-
шагнуть за пределы письменной истории — это значит перейти
на новую шкалу, в которой годы жизни отдельных людей и от-
дельных поколений становятся незаметными, а поддающиеся
наблюдению единицы времени — тысячи и миллионы лет — не
имеют видимой связи с тем временным периодом, в котором мы
живем сейчас.
Для понимания эволюционных процессов нужно знать про-
тяженность тех промежутков времени, в которые они происхо-
дили. Это означает необходимость учитывать несколько различ-
ных аспектов геологического времени. Один из них состоит
лишь в том, что периоды геологического времени настолько
Рассказ об эволюции человека
33
Рис. 2.1. Геологическая временная шкала
Четвертичный
Тысячи
Миллионы
лет
продолжительны, что это выходит за рамки наших обычных
представлений, даже если речь идет об относительно недавних
эпохах. Например, самая последняя часть последней закончен-
ной геологической эпохи — поздний плейстоцен — это период
протяженностью 115000 лет, что более чем в 50 раз длиннее
всей нашей письменной истории и эквивалентно по протяжен-
ности жизни 4600 поколений современных людей. Значение
этих огромных промежутков времени становится еще более
ощутимым, если речь идет о более ранних эпохах, таких, как
плейстоцен в целом — почти 2 млн. лет, плиоцен — 3 млн. лет
и миоцен — 20 млн. лет. На первый взгляд необъятная протя-
женность времени не поддается объяснению, так как трудно
представить процессы, происходившие в масштабах такой шка-
лы. Однако парадокс геологического прошлого заключается в
том, что при всей грандиозности промежутков времени процес-
сы жизни, происходившие внутри них, действовали на основе
повседневных закономерностей. Иными словами, хотя геологи-
ческое время характеризуется долговременными изменениями
и непрерывностью, биологические процессы, действующие во
3-367
34
Глава 2
временных рамках одного индивидуального жизненного цикла,
по-прежнему применимы к нему. Именно это и позволяет нам
охватить масштабы эволюционных изменений.
Это не означает, разумеется, что мы отрицаем наличие про-
цессов, происходящих в масштабах долговременной шкал^ь
Суть в том, что такие процессы не отвергают существования
более коротких по протяженности. Методический подход, по-
зволяющий охватить длительные отрезки прошлого, состоит в
признании разнообразия процессов, происходящих в различных
временных шкалах. Существуют огромные по протяженности
процессы тектоники плит и континентального дрейфа, 20 000—
40000- и 100 000-летние циклы изменчивости орбиты Земли и,
следовательно, глобальных температурных вариаций, 10000-лет-
ние межледниковые циклы и циклы продолжительности жизни
всех видов, которые могут варьировать от нескольких тысяч лет
для калифорнийской остистой сосны до части суток для микро-
скопических организмов (Imbrie, Imbrie, 1979; Butzer, 1982).
Сложность жизни заключается не в господстве одной какой-ли-
бо временной шкалы, а в интеграции многих.
Возникают, однако, проблемы идентификации этих шкал,
поскольку они зависят от наших способностей измерять время.
Очень часто наше восприятие происходящих во времени процес-
сов зависит не столько от реальных биологических или физиче-
ских процессов, сколько от разрешающих возможностей спосо-
бов датировки. Например, в отдаленном прошлом виды возни-
кали с пугающей быстротой относительно протяженности того
времени, когда это происходило. Однако очень часто разреша-
ющая способность геологических датировок не выходит за пре-
делы миллиона лет. Следовательно, то, что в один геологиче-
ский период можно охарактеризовать как быстротечное, соста-
вит всю протяженность другого периода, в рамках которого
может произойти целый ряд новых событий. Поэтому возмож-
ность познать прошлое зависит от наших способностей к уста-
новлению градуированной шкалы.
2.1.2. Способы датировки
Способы датировки делятся на две большие группы: отно-
сительные, т. е. те, которые определяют возраст предметов или
событий относительно друг друга, и абсолютные, когда можно
вычислить возраст в реальном времени (ряд лет) (Oakley,
1966). История развития способов датировки заключается в
том, что абсолютные методы постепенно брали верх над отно-
сительными.
Правда, относительные способы все еще сохраняют свое
значение. Основной метод относительного датирования — это
Рассказ об эволюции человека
35
Рис, 2.2. Схематическое изображение основных методов датирования, корре-
ляции и калибровки
стратиграфия, или геологическое наложение (рис. 2.2). В боль-
шинстве случаев там, где геологическое отложение наклады-
вается на другое, оно относится к более позднему или более
раннему периоду. С помощью стратиграфического анализа и
корреляции устанавливается последовательность событий в
геологическом прошлом. Другой важный способ относительной
хронологии — это биостратиграфическая корреляции. В этом
случае ископаемые остатки исчезнувших животных и растений
используются для упорядочения геологических отложений.
Слои, которые содержат одни и те же виды или сходные сово-
купности видов, могут в эволюционном контексте расценивать-
ся как современные. Исходя из сходства и различий местона-
хождений ископаемых остатков можно построить относитель-
ную хронологию. В основе этого метода лежит предположение,
что существа, сохранившиеся в виде морфологически сходных
окаменелостей, жили на Земле в одно и то же время, и поэтому
слои со сходными типами ископаемых остатков отложились
почти одновременно. Если известна эволюционная последова-
тельность генеалогических линий ископаемых существ, биостра-
тиграфия может быть использована для построения обширных
хронологий, скоррелированных от одного местонахождения к
другому в пределах обширных территории.
Для повышения точности относительных методов датировки
был разработан ряд способов, основанных на достижениях
точных наук. Наиболее известно датирование по содержанию
фтора (Oakley, Montagu, 1949). Ископаемые остатки, как из-
вестно, поглощают фтор и другие микроэлементы из окружаю-
щих их отложений, поэтому концентрации этих элементов ука-
3*
36
Глава 2
зывают на синхронность или относительный возраст образцов.
Однако, поскольку содержание фтора сильно варьирует на раз-
ных территориях, эта методика применяется только в пределах
одной местности.
В большинстве случаев способы, основанные на достижени-
ях точных наук, использовались для получения абсолютных
датировок. Главным принципом, лежащим в основе большинст-
ва абсолютных датировок, является принцип радиоактивного
распада (Tite, 1972). Некоторые химические элементы встреча-
ются в нестабильной форме, т. е. в виде радиоактивных изото-
пов, которые распадаются с образованием других элементов
(стабильных изотопов). Этот радиоактивный распад происходит
с определенной скоростью, что позволяет использовать соотно-
шение между стабильными и нестабильными изотопами для
датировки прошлых событий. Скорость распада измеряется «пе-
риодами полураспада» — постоянными по длительности проме-
жутками времени, в течение которых половина данного коли-
чества нестабильного изотопа переходит в стабильную форму.
Такие «изотопные часы» позволяют хорошо датировать событие
в тех случаях, когда можно установить, в какой момент в про-
шлом они начали «тикать». Это справедливо для каждого из
известных методов.
Наиболее ценный метод датировки процесса антропогене-
за — калий-аргоновый. Калий (К) превращается в аргон (Аг)
с известной скоростью (период полураспада = 1,3 X Ю9 года;
Birch, 1951). Этот метод является достоверным лишь тогда,
когда известно время образования исходного уровня калия, и,
следовательно, наиболее подходит для скальных пород вулка-
нического происхождения. Однако благодаря очень большому
периоду полураспада датировка по соотношению 40К/40Аг (так
же как и с использованием родственного метода, по соотноше-
нию 40Аг/39Аг) годятся только для очень древних скальных
пород и имеет нижнюю границу своих возможностей не менее
500 000 лет.
На противоположном конце находится один из наиболее
известных радиометрических, или изотопных, методов датиров-
ки— радиоуглеродный (14С). Всякая живая материя поглоща-
ет из атмосферы содержащийся в ней в незначительных коли-
чествах нестабильный изотоп углерода (14С). Однако после
смерти организма это поглощение прекращается, и поскольку
14С — нестабильный изотоп, он превращается в азот. Скорость
радиоактивного распада опять-таки известна (период полурас-
пада = 5730 лет; Tite, 1972), и поэтому соотношение нестабиль-
ного и стабильного изотопов углерода (14С/12С) можно исполь-
зовать для определения времени смерти. Радиоуглеродный спо-
соб используется для определения возраста ископаемых
Рассказ об эволюции человека
37
остатков и поэтому зависит от наличия незагрязненного орга-
нического материала. Кроме того, короткий период полураспа-
да означает, что точность метода ограничена верхним пределом
в 40000 лет, хотя с помощью недавно разработанного атомно-
го ускорителя эта цифра может быть отодвинута до 100 000 лет
назад (Gillespie, Gowlett et al., 1984).
Промежуточное положение между границами точности, до-
ступными радиоуглеродному и калий-аргоновому методам, за-
нимают некоторые менее известные изотопные способы дати-
рования (например, по соотношению гз^/тть) (см. Bishop,
Miller, 1972). Плохо датированным остается, однако, важный
период от 500 000 до 40 000 лет назад. До некоторой степени
заполнить этот пробел можно с помощью других способов да-
тировки.
Т ермолюминесцентный метод (ТЛ) основан на том, что при
распаде урана в кристаллических веществах электроны застре-
вают в кристаллической решетке, и это используется для да-
тировки. При нагревании вещества происходит высвобождение
электронов в виде пучка света, который можно измерить, и на
основе скорости эмиссии вычислить возраст образца. Методом
электронного парамагнитного резонанса (ЭПР) измеряют то
же явление, но без разрушения образца. Датировка следов
расщепления также основана на распаде урана, но в этом слу-
чае протяженность отрезка времени, в течение которого распад
имел место, измеряется путем подсчета числа «следов», остав-
ленных эмиссией электронов на поверхности минералов и сте-
кол (Fleischer, Hart, 1972). Хотя все эти методы в принципе
могут помочь заполнить хронологический разрыв между време-
нами, надежно датированными с помощью двух основных спо-
собов, все же они еще недостаточно широко используются.
Еще один метод, на который опирается определение геоло-
гического времени, — это палеомагнитный. Направление маг-
нитного поля Земли не остается стабильным с течением време-
ни и в прошлые эпохи многократно менялось (Stacey, 1969).
Перемены полярности часто наблюдались в стратиграфических
отложениях. Палеомагнитные последовательности можно ис-
пользовать для стратиграфических корреляций, а отдельные на-
блюдения — как ограниченный тест верности других способов
датировки.
Палеомагнетизм — это фактически один из серии методов,
которые не дают реальных цифр, но позволяют установить кор-
реляции между различными территориями. Сходный способ,
часто используемый в наши дни, — анализ стабильных изотопов
кислорода в микроокаменелостях глубоководных морских отло-
жений. Анализ соотношения двух стабильных изотопов кисло-
рода (16О/18О) в материалах, извлеченных из морских глубин,
38
Глава 2
позволяет построить температурные кривые для многих различ-
ных регионов мира и разделить их на несколько серий (Shack-
leton, Opdyke, 1976). Были получены некоторые корреляции
этих кривых с наземной стратиграфией, особенно для лёссовых
районов Европы (Kukla, 1975). С помощью этих и других мето-
дов были установлены хронологические рамки эволюции чело-
века.
2.1.3. Хронологические рамки эволюции человека
Перед исследователями прошла длинная процессия ископае-
мых существ, которых считали самыми ранними стадиями эво-
люции человека, претендовавшими на роль легендарного «не-
достающего звена». Многие из этих ископаемых остатков ныне
не принимаются в расчет, поскольку относятся к другим груп-
пам животных. Например, миоценовая человекообразная обезь-
яна плиопитек некогда считалась ранним гоминидом на том
основании, что имела плоское (ортогнатное) лицо — характер-
ный признак гоминид. Ныне эта особенность признана резуль-
татом параллельной эволюции в пределах родственных линий.
Другие ископаемые остатки нашли свое место в различных
частях эволюционной истории человека.
Эволюция — это непрерывный процесс, в котором нет ес-
тественных разрывов, и поэтому эволюция человека не может
иметь одной-единственной исходной точки. Хронологическая
шкала может быть разработана только применительно к так-
сономической. Таксономия — это классификация предметов и
явлений, в нашем случае — растений и животных. Классифика-
ционная схема, используемая в биологии, известна как таксо-
номическая система Линнея по имени шведского натуралиста
Карла Линнея, разработавшего ее в XVIII столетии. Используя
степень сходства или различий в морфологической структуре
организмов, можно построить иерархическую классификацию,
группирующую животных во все более крупные единицы. Вслед-
ствие этого хронологическая шкала эволюции человека зави-
сит от таксономической шкалы.
Существуют значительные разногласия относительно време-
ни первого появления различных таксонов (Andrews, 1983;
Ciochon, 1983; Pilbeam, 1984). Источник этих разногласий по
большей части коренится в методах, используемых для рекон-
струкции филогенетических соотношений (Sarich, 1971; Good-
man, 1976; Elderedge, Cracraft, 1980). Традиционный подход
заключается в анализе морфологии ископаемых остатков и да-
тировке расхождений эволюционных линий на основе измерения
возраста геологических отложений. Например, обнаружение
окаменелостей с признаками, уникальными для семейства го-
Рассказ об эволюции человека
39
минид на рубеже в 12 млн. лет, будет означать, что линии го-
минид и человекообразных обезьян существенно разошлись по
крайней мере до этого времени.
Альтернативный подход состоит в использовании так назы-
ваемых «молекулярных часов» (Sarich, Wilson, 1976; Friday,
1981). В этом случае сравниваются биохимические особенности
ныне живущих видов и на основании степени их сходства ус-
танавливается время, прошедшее с момента отделения от ис-
ходной линии в эволюционной истории. Используются три
основных источника данных: белки, подобные тем, которые со-
держатся в сыворотке крови; «паразитические», или вирусные,
гены, связанные с определенными видами, и реальное картиро-
вание ДНК- Предполагается, что скорость изменения этих при-
знаков постоянна в течение длительных периодов времени и,
следовательно, количественная оценка различий может служить
основой исчисления возраста эволюционных событий. Конечно,
в какой-то момент «молекулярные часы» должны быть откалиб-
рованы по хорошо датированным меткам шкалы ископаемых
остатков, в противном случае построенные на их основе фило-
генетические схемы будут давать лишь относительную инфор-
мацию.
Когда Сарич и Уилсон (Sarich, Wilson, 1967) впервые вы-
ступили с идеей молекулярного подхода, они столкнулись с
резкой критикой: новый метод давал значительно более позд-
ние датировки по сравнению с основанными на ископаемых
остатках. Сегодня, однако, палеоантропологи и биохимики до-
стигли большего согласия относительно возраста основных со-
бытий в эволюции приматов и гоминид (Ciochon, 1983; Good-
man, Baba, Darga, 1983; Pilbeam, 1984). Это произошло благо-
даря пересмотру молекулярных датировок, более точной оценке
возраста ископаемых остатков и, что самое важное, благодаря
пониманию палеонтологами того факта, что не все ископаемые
формы должны быть прямо связаны с современными, представ-
ляя собой разнообразные варианты, существовавшие в про-
шлом, но не сохранившиеся до настоящего времени. В резуль-
тате возникло более сложное прочтение эволюционной истории
приматов.
Приматы, как считают в настоящее время, отделились от
ранних насекомоядных примерно 70 млн. лет назад (см. рис.
2.3) (Szalay, Delson, 1979; см. также подробное обсуждение
современных взглядов на эту проблему в статьях Ciochon, Сог-
ruchini, 1983). Ранние ископаемые остатки гаплориновых и
стрепсириновых приматов мало известны, но считается, что
группа предков узконосых обезьян отчетливо отделилась от
широконосых форм Нового Света по меньшей мере 35 млн. лет
назад. Время, когда разошлись линии предков человекообраз-
40
Глава 2
Рис, 2.3. Эволюционные связи основных групп современных приматов (см.
Szalay, Delson, 1979; Pilbeam, 1984)
ных и низших узконосых обезьян, плохо определяется по иско-
паемым остаткам, но составляет, вероятно, около 23 млн. лет.
Ветви африканских и азиатских гоминоидов разошлись, веро-
ятно, около 12 млн. лет назад. Специалисты по молекулярной
биологии датируют отделение ветви гоминид от остальных
африканских человекообразных обезьян приблизительно 6 млн.
лет назад; лучшими примерами весьма раннего появления го-
минид в ископаемых материалах служат остатки возрастом
примерно в 5 млн. лет из Табарина (Hill, 1985) и Лотагама
(Patterson, Behrensmeyer, Sill, 1970) в Кении (рис. 2.4).
В промежутке между 5 и 1,2 млн. лет назад гоминид можно
разделить на две большие группы: австралопитеков и предста-
вителей рода Homo (Howell, 1978). Первые представляют са-
мых ранних из известных гоминид, а последние появились на
сцене, по-видимому, около 2 млн. лет назад. С этого момента
и до настоящего времени род Ното претерпел ряд существен-
ных эволюционных изменений, главными из которых были эво-
люция Ното sapiens, начавшаяся около 200 000 лет назад,
и возникновение человека современного типа (Н. sapiens sapi-
ens) в последние 100 000 лет.
Итак, эволюция человека — это многомерный процесс. Как
отдельный вид и подвид человек характеризуется историей, ко-
торая насчитывает лишь сотни тысячелетий; род, к которому
мы принадлежим, эволюционировал на протяжении около 2 млн.
Рассказ об эволюции человека
41
Расстояние
ДНК-ДНК
От
2-
4-
6-
8-
10-*
Рис. 2.4. Таксономические расстояния для некоторых групп узконосых обезьян,
полученные методом гибридизации ДНК—ДНК. Шкала справа указывает на
подсчитанное время дивергенции (в млн. лет) и датировку самых ранних из
известных находок ископаемых гоминид (по Sibley, Ahlquist, 1984; Pilbeam,
1986)
лет, а наше семейство — Hominidae — имеет древность по
меньшей мере в 6 млн. лет. Понимание этих временных рамок
играет важную роль для определения самого типа вопросов и
объяснений, которые мы желаем задать и получить относитель-
но процесса эволюции человека. Далее, эволюция человека не
может быть ограничена появлением чисто человеческих таксо-
нов. Каждое эволюционное событие означает отделение друг от
друга различных групп животных. Чтобы понять, как происхо-
дило расхождение ветвей, необходимо знать предковые группы.
Так, многие факторы, определяющие ход эволюции человека,
лежат в основе процесса эволюции гоминоидов и узконосых
обезьян в целом. Хорошее знание шкалы эволюции человека —
это тот фундамент, на котором должен основываться эволюци-
онный и экологический анализ.
2.2. Ископаемые гоминиды
2.2.1. Происхождение гоминид
Предположение, что первые гоминиды появились в самом
конце миоцена, т. е. около 6 млн. лет назад, противоречит как
прежним взглядам на эволюцию человека, согласно которым
42
Глава 2
она происходила преимущественно в плейстоцене, так и более
поздним воззрениям, согласно которым обособление гоминид и
человекообразных обезьян (понгид) имело место не позднее
15 млн. лет назад (см., например, Pilbeam, 1972). Подобное
суждение основывалось на том, что рамапитек рассматривался
в качестве гоминида или существа, близкого по происхождению
к гоминидам (Simons, 1961; Pilbeam, 1968). Ramapithecus рип-
jabicus был впервые обнаружен в 1930-х гг. на Индийском
субконтиненте (Lewis, 1934). С тех пор большое количество
остатков рамапитека было обнаружено в Греции, Китае, К&нии
и Турции (Leakey, 1967; Pilbeam, 1968; Andrews, Tobien, 1977);
близкородственные формы найдены также в Венгрии (Kretzoi,
1975).
Причины, по которым эти ископаемые существа были отне-
сены к гоминидам, связаны с особенностями строения их зуб-
ной системы, единственными хорошо изученными анатомически-
ми признаками (Conroy, Pilbeam, 1975). Рамапитек имел ре-
дуцированные клыки, утолщенную зубную эмаль, увеличенные
ложнокоренные и коренные зубы и, как первоначально предпо-
лагалось, зубную дугу параболической формы. Все эти призна-
ки, встречающиеся и у более поздних гоминид, считались
характерными для всего семейства. На основе этого диагноза
генеалогия гоминид прослеживалась по меньшей мере вплоть
до среднего миоцена (рис. 2.5), а миоценовые гоминоиды рас-
сматривались в свете мнения об очень древнем обособлении
ветви, ведущей к современным человекообразным обезьянам.
Однако эта точка зрения на происхождение гоминид стано-
вилась объектом все более острой критики. Отчасти это было
связано с применением методов молекулярной биологии в эво-
люционной систематике, отчасти — с успехами в нахождении
костных остатков миоценовых гоминид как в отношении их ко-
личества, так и качества и наконец — с пересмотром таксоно-
мического значения ряда признаков, обнаруженных у рамапи-
тека и, как считали, свойственных исключительно гоминидам.
Так, например, оказалось, что толстая зубная эмаль — это
сложный признак, который встречается не только у людей, но
и у орангутана (Кау, 1981). Сходные характеристики обнару-
живаются и у других миоценовых гоминоидов, в частности Gi-
gantopithecus и Sivapithecus (Kay, 1981; Gannt, 1983). Как по-
казали недавно Бойд и Мартин (Boyde, Martin, 1984), характер
развития эмали у высших приматов исключительно сложен. По
мере накопления новых данных по ископаемым миоценовым
гоминидам становилось ясно, что те особенности, которые рань-
ше считались несомненной принадлежностью этого семейства,
на самом деле являются частью более общего типа, характери-
зующего эволюцию гоминоидов. Пилбим (Pilbeam, 1984; см.
Рассказ об эволюции человека
43
Узконосые
Рис. 2.5. «Классическая» модель эволюционного древа узконосых обезьян,
показывающая ранее отделение гоминид от остальных человекообразных и ра-
мапитека в качестве прямого предка более поздних гоминид
подробный обзор у Ciochon, Corruchini, 1983) убедительно
доказывает, что рамапитека нужно рассматривать как неотъ-
емлемую часть миоценовой модели эволюционной дивергенции,
не пытаясь связать его с более поздними событиями.
Рамапитек, после того как ему было отказано в принадлеж-
ности к гоминидам, пополнил ряды избранного общества не-
состоявшихся предков человека, каждый из членов которого
обладал, как поначалу предполагали, какой-нибудь уникальной
чертой гоминид. Однако дальнейший анализ каждый раз пока-
зывал, что многие из этих черт вовсе не обязательно принадле-
жат исключительно гоминидам, а связаны с решением общих
для гоминоидов адаптивных проблем.
Трактовка эволюции гоминоидов, увязывающая как палео-
антропологические, так и биохимические данные, состоит в том,
что из событий миоцена основное внимание уделяется раннему
расхождению различных ветвей гоминоидов в Африке, распро-
странению их оттуда в другие части Старого Света и (лишь в
самом конце этого периода) появлению гоминид. Следователь-
но, отдельные образцы или экземпляры, которых относили к
таким таксонам, как Kenyapithecus, Sivapithecus, Ouranopithe-
cus и т. д. (см. табл. 2.1), вовсе не обязательно родственны бо-
лее поздним или ныне живущим гоминоидам и гоминидам, но
44
Глава 2
Таблица 2.1. Характерные особенности, а также хронологические
и географические рамки основных групп миоценовых гоминоидов
Хроиологиче-
Географине- ские рамки
Род ское раопро- (млн. лет Описание
с гра нение1) до настоящего
времени)
Pliopithecus ЗЮЕП; ВЦЕП 15—9 Небольшая примитивная евро- пейская форма, которую часто связывают с гиббонами, но, ве- роятно, представляющая при- мер параллельной эволюции
Dryopithecus ЗЮЕП 12,5—9 Более крупный европейский примитивный гоминоид, вероят- но, ведущий древесный образ жизни и обитающий в лесу
Sivapithecus ВЦЕП; ПСП; СиП; ВАП? 15—7 Широко распространенная и весьма изменчивая миоценовая человекообразная обезьяна, приспособленная к обитанию на более открытых или лесистых пространствах; возможный пре- док орангутана
Ramapithecus ПСП; СиП; ВЦЕП? +11—7 Прежде рассматривался как са- мый ранний гоминид, ныне счи- тается азиатской человекооб- разной обезьяной, близко род- ственной сивапитеку (возмож- но, что это две различающиеся по полу формы одного таксона)
Gigantopithecus СиП 9,5—7 Самый крупный из известных гоминоидов, вероятно, наземная форма с очень крупными спе- циализированными коренными зубами; дожил до среднего плейстоцена
Ouranopithecus ПСП 12—10 Крупный гоминоид из Греции, вероятный предок всех иыне живущих крупных гоминоидов
Kenyaplthecus ВАП; ПСП 14—12 Довольно крупный африкан- ский гоминоид, прежде связы- вающийся с рамапитеком, но, вероятно, представляющий от- дельную группу; возможный предок более поздних африкан- ских гоминоидов
Рассказ об эволюции человека
45
Продолжение
Род Географиче- ское распро- странение1) Хронологиче- ские рамки (млн], лет до настоящего времени) Описание
Proconsul ВАП 22—12 Долго существовавшая группа африканских человекообразных обезьян, ранние формы кото- рой, вероятно, были предками всех современных гоминоидов; древесные и лесные жители с уникальной системой передви- жения
Limnopithecus ВАП 22—12 Более мелкий африканский го- мииоид
’) Географическое распространение определяется по зоогеографическим зонам Берно- ра (Вегпог, 1983): ЗЮЕП — западная и южная европейские провинции; ВЦЕП — восточ- ная и центральная европейские провинции; ПСП — провинция Субпаратетис», СиП — си- валикская провинция; ВАП — восточно-африканская провинция. Обсуждение и основные источники см. в тексте.
относятся к числу чисто миоценовых эволюционных явлений.
Этот вывод имеет важное значение, которое мы будем более
подробно обсуждать в гл. 7.
2.2.2. Плиоценовые гоминиды
Как уже указывалось выше (см. разд. 2.1.3), костные остат-
ки гоминид в основном начинают появляться около 5 млн. лет
назад (географическое распространение упоминаемых в тексте
местонахождений см. на рис. 2.6). Самые ранние местонахож-
дения— это Табарин возле озера Баринго в Кении, где была
найдена челюсть, принадлежащая, по-видимому, ископаемому
гоминиду, жившему 4,9 млн. лет назад (Hill, 1985), и Лотагам
на западном берегу озера Туркана; однако наиболее известные
и богатые местонахождения — район Хадар в Эфиопии и Ле-
толи в Танзании.
В Летели найдено большое количество зубов и фрагментов
челюстей, а также отпечатки стоп прямоходящего примата,
предположительно гоминида (М. D. Leakey, Hay et al., 1976;
М. D. Leakey, 1978). В Хадаре Джохансон и др. нашли сни-
скавшую ныне шумную популярность «Люси» — фрагментарный
скелет небольшого по размерам гоминида, а также множество
других окаменелостей, включая группу гоминид, очевидно, по-
гибших одновременно в результате наводнения (см. American
Journal of Physical Anthropology, 1982, vol. 57, № 4). Первое
местонахождение имеет возраст 3,6 млн. лет, второе — около
2,9 млн. лет.
46
Глава 2
Рис. 2.6. Места наиболее важных находок ранних гоминид (австралопитеко-
вых, Homo habilis и ранних представителей Н. erectus) в восточной и южной
Африке
Несмотря на большую изменчивость найденных ископаемых
костей, Джохансон и Уайт (Johanson, White, 1979) утверждают,
что все они принадлежат одному виду Australopithecus afaren-
sis (рис. 2.8). Представители этого вида как истинные гомини-
ды характеризуются длинной и узкой зубной дугой с прямыми
параллельными ветвями и крупными коренными зубами, но по
сравнению с другими гоминидами резцы и клыки у них также
более крупные. Лицо является прогнатным, т. е. выступает впе-
ред (особенно .несущая зубы область). Черепная коробка не-
большая с отчетливо выраженным мышечным рельефом и пост-
глазничным сужением. Таким образом, Australopithecus afaren-
Рассказ об эволюции человека
47
sis обладает чертами, которыми могли бы отличаться лишь
древнейшие гоминиды. Наиболее характерная посткраниальная
особенность это, наверное, то, что афарокий австралопитек
был явно способен к прямохождению, хотя некоторые признаки,
например изогнутые фаланги (кости пальцев), могут рассмат-
риваться как унаследованная приспособленность к древесному
образу жизни (Jungers, Stern, 1983; Stern, Susman, 1983). Еще
одна интересная особенность A. afarensis — это сильная измен-
чивость размеров тела, которая, возможно, означает половой
диморфизм данного вида (Johanson, White, 1979; Johanson,
Edey, 1981), но некоторые специалисты истолковывали ее как
наличие на этой ранней стадии по крайней мере двух видов
гоминид.
К середине плиоцена находки ископаемых гоминид встреча-
ются намного чаще. Хотя датировка южноафриканских мате-
риалов вызывает некоторые затруднения (см. разд. 2.1.2), кост-
ные остатки гоминид, относящиеся к периоду от 3 до 1,8 млн.
лет, обнаружены на территории Восточной (Кооби-Фора, Омо,
Канапои) и Южной Африки (Макапансгат и Стеркфонтейн)
(Howell, 1978). Эти, а также более поздние важные местона-
хождения, такие, как Олдувай, Пенинж, Чесованья, Сварткранс
и Крамдраай, переходящие из плиоцена в плейстоцен, свиде-
тельствуют, что в промежутке от двух до одного млн. лет назад
гоминиды разделились на несколько видов и в связи с этой ди-
вергенцией прослеживается два различных направления в эво-
люции человека.
Первое из них представлено грацильными и массивными
австралопитеками. Грацильные формы относятся к виду Aust-
ralopithecus africanus (рис. 2.8), который, по мнению Тобайеса
(Tobias, 1980), включает и более ранний вид Australopithecus
afarensis', это изящно сложенные гоминиды с округлым шаро-
образным черепом, прогнатным, блюдцеобразным лицом, уве-
личенными коренными и ложнокоренными зубами. Клыки ре-
дуцированы по сравнению с тем, что найдено у A. afarensis.
Массивные формы австралопитеков датируются несколько бо-
лее поздним временем; они представлены Australopithecus boi-
sei в Восточной Африке и Australopithecus robustus в Южной
Африке (рис. 2.8). Некоторые авторы (Howell, 1978) считают,
что в Южной Африке существует третий вид массивных авст-
ралопитеков A. crassidens. Все эти гоминиды имеют массивное
сложение с отчетливо выраженными выступами и гребнями,
служащими для прикрепления мускулов, с резко выраженным
сужением черепа в постглазничной области. Лицо крупное, ча-
сто широкое и квадратное в максиллярной области. Зубы явля-
ются самой характерной чертой этих гоминид: сильно увели-
ченные коренные зубы и внешне не отличающиеся от них лож-
48
Глава 2
Рис. 2.7. Геологические формации и датировки основных плиоценовых и ран-
неплейстоцеиовых ископаемых гоминид. Объяснения и ссылки см. в тексте.
Г обозначает положение ископаемых гоминид
покоренные сочетаются с редуцированными и тесно располо-
женными передними зубами — резцами и клыками. Нижняя
челюсть, как можно ожидать при таких крупных коренных зу-
бах, очень массивная с выраженным рельефом. Однако, подоб-
но всем остальным известным гоминидам, массивные австрало-
питеки приспособлены к прямохождению как обычному способу
передвижения. Другая линия — род Ното,впервые представлен-
ный Homo habilis (рис. 2.9). В отличие от массивных австрало-
питеков ранние представители рода Ното (гоминины, собст-
венно люди —см. табл. 1.2) характеризуются увеличением ем-
кости черепа вплоть до 650 см3. Зубы и челюсти, хотя и
превышают по своим размерам таковые современного человека,
тем не менее меньше, чем у австралопитековых. Остатки Ното
habilis обнаружены в различных местах Восточной Африки —
в первую очередь в Олдувае и Кооби-Фора, а также в Южной
Африке.
2.2.3. Плейстоценовые гоминиды
В начале плейстоцена гоминиды характеризовались наиболь-
шим разнообразием, чем когда-либо до или после этого: в Аф-
рике сосуществовали массивные австралопитеки и ранние пред-
Рассказ об эволюции человека
49
ставители рода Ното. Однако в раннем плейстоцене произошли
два важных события. Во-первых, около 1,6 млн. лет назад на
смену Homo habilis пришел Homo erectus (Howell, 1978) и, во-
вторых, вымерли австралопитеки. Великолепные экземпляры
Homo erectus были найдены как на восточном (Кооби-Фора)^
так и западном (Нариокотоме) берегах озера Туркана в Кении;
их возраст составил примерно 1,6 млн. лет. В частности был
найден почти полный скелет (KNM-WT 15 000) Homo erectus,
принадлежавший мальчику-подростку и указывающий на очень
массивное телосложение этого вида. Самые поздние из извест-
ных находок австралопитековых датируются временем чуть бо-
лее 1 млн. лет назад.
Homo erectus был первым гоминидом, вышедшим за преде-
лы нынешней суб-Сахары. Остатки представителей этого вида
обнаружены в Северной Африке, на Ближнем Востоке, в Китае
и на Яве; сходные, но, вероятно, более поздние формы найдены
и в Европе (рис. 2.9). Homo erectus характеризуется увеличе-
нием размеров тела, очень крупным лицом и зубами по сравне-
нию с современным человеком, резко выраженными надглаз-
ничными валиками (надбровные дуги), длинным низким чере-
пом с выраженными местами прикрепления мышц вдоль
теменных и затылочной костей, утолщением костей черепа и
4—367
50
Глава 2
Homo hobihs
Australopithecus robust us
Australopithecus africamis
Australopithecus afarenfis
Рис. 2.8. Самые ранние гоминиды (приведено к одному масштабу): A. Austra-
lopithecus afarensis; Б. Australopithecus africanus; В. Australopithecus robus-
tus; Г. Homo habilis (White, Johanson, Kimbel, 1983)
более высоким сводом последнего. В отношении посткраниаль-
ного скелета Homo erectus очень похож на современного чело-
века, хотя, по мнению некоторых ученых (Day, 1971; Day, Mol-
leson, 1973; Kennedy, 1984), в строении бедренных костей и та-
за между ними имеются сильные различия.
Согласно давно установившемуся мнению, может быть, са-
мое существенное отличие Homo erectus — это его постоянство
во времени (Pilbeam, 1975; Rightmire, 1981), а также эволюци-
онная стабильность, которая, правда, иногда оспаривалась
(Wolpoff, 1983). В период от 1,6 до примерно 0,4 млн. лет
назад Homo erectus представлял собой единственный вид рода
Ното и сравнительно мало изменился за этот промежуток вре-
мени, несмотря на большую протяженность занимаемого им
ареала. Однако примерно с 0,4 млн. лет назад этот морфоло-
Рассказ об эволюции человека
51
Рис. 2.9. Места наиболее важных находок более поздних ископаемых гоминид
(поздние Homo erectus и ранние Н. sapiens): 1 — устье реки Клазиес; 2 — Сал-
данья (Флорисбад); 3 — пещера Бордер; 4—Кабве (Брокен-Хилл); 5 — Олду-
вай и Летоли; 6 — Ндуту; 7-—Омо; 8 — Бодо; 9 — Тернифина; 10 — Сале;
И — Ирхуд; 12 — Убейдия; 13 — Гора Кармел; 14—Кафзех; 15 — Шанидар;
16 — Петралона; 17 — Вертешселлеш; 18 — Бильцингслебен; 19—Штейнгейм;
20 — Мауер; 21 —позднеплейстоцеиовые местонахождения юго-западной Фран-
ции (включая Ла-Феррасси и Ла-Шаппель-о-Сен); 22 — Торральба/Амброна;
23 — Исерния; 24 — Монте-Чирчео; 25 — Сванскомб; 26 — Понтневид; 27 —
местонахождения ранних гоминид на о. Ява; 28 — Иингку; 29 — Лантянь; 30 —
Чжоукоудянь
гический застой прекращается и начинается переход к Ното
sapiens. На самых ранних стадиях он отмечен очень крупными
экземплярами [Брокен-Хилл (Кабве), Бодо, Петралона], увели-
чением черепа, дальнейшей редукцией относительного размера
лица и мускулатуры головы. Пока не ясно, был ли этот переход
приурочен к определенной местности, но многие из ранних ис-
копаемых представителей Homo sapiens найдены в Европе
(Штейнгейм, Сванскомб, Мауер, Вертешселлеш и т. д.), что
свидетельствует о дальнейшей географической экспансии гоми-
нид. Эти архаические формы Homo sapiens достигают вершины
своего развития в неандертальцах — морфологически обособ-
ленной группе гоминид, обитавших, судя по находкам, в Европе
4
52
Глава 2
и некоторых частях Азии в период от 100 000 до 50000 лет на-
зад. Неандертальцы обладали массивным скелетом, выражен-
ными надбровными дугами, крупным прогнатным лицом, круп-
ными носовыми костями; они отличались и рядом других уни-
кальных характеристик, таких, например, как диастема между
третьим коренным зубом и восходящей ветвью нижней челюсти
/Trinkaus, Howells, 1979). По недавнему утверждению Трин-
кауса (Trinkaus, 1983), неандертальцы сильно отличались от
людей современного типа по анатомическим особенностям таза.
Переход от архаических форм Homo sapiens к анатомически
современному типу Homo sapiens sapiens — одна из давнишних
загадок палеоантропологии. В Европе процесс замещения не-
андертальцев неоантропами был, по общему мнению, относи-
тельно быстрым и, по мнению ряда ученых, две эволюционные
формы гоминид в течение некоторого времени сосуществовали
.(Stringer, 1984). В других местах из-за недостаточного количе-
ства находок и менее четкой их хронологии обнаружить одно-
временное существование неандертальцев и Homo sapiens sa-
piens не удается. Кроме того, как показывают найденные в Аф-
рике костные остатки, человек современного типа рано появля-
ется на этом континенте; возможно, именно здесь следует
искать истоки происхождения современного человечества (Bau-
er, 1984; Rightmire, 1984), и это событие имело место не позд-
нее 100 000 лет назад. Наверняка известно, что к 30 000 лет
назад человек современного типа оказался единственным пред-
ставителем гоминид, распространившимся по всему миру, хотя
австралийская находка Коу-Свомп, возраст которой составляет
лишь 10 000 лет, обнаруживает некоторые признаки, напомина-
ющие архаические формы Н. sapiens (Thorne, Macumber, 1972).
Представители нашего собственного подвида характеризуются
гораздо более грациальным скелетом, утончением костей череп-
ного свода, более высоким и округлым черепом, уменьшением
размеров лица и зубов, развитием выступающего вперед подбо-
родка (Howell, 1978).
2.2.4. Филогения гоминид
Значительное увеличение числа ископаемых находок, а так-
же улучшение их сохранности позволяют нам с некоторой точ-
ностью реконструировать процесс филогении гоминид. Ушли в
прошлое представления об однолинейном характере эволюции,
где, как по лестнице, осуществляется переход к высшим фор-
умам. На смену им пришла более разнообразная и сложная
эволюционная история. Однако даже сейчас наука не пришла
к полному согласию относительно всех деталей этой филогении
(рис. 2.11 и 2.12).
Рассказ об эволюции человека
53
Рис. 2.10. Более поздние ископаемые гоминиды (приведены к одному масшта-
бу): A. Homo erectus (1—KNM-ER3733; 2 — реконструкция черепа с о. Ява
S4; 3 — Чжоукоудянь); Б. Архаический Н. sapiens (1—Штейигейм; 2 — Каб-
ве; 3 — Шанидар; 4 — Моите-Чирчео); В. Homo sapiens sapiens
Australopithecus afarensis — самый ранний из известных
видов гоминид, — возможно, является предком всех более позд-
них форм. Однако даже на этой ранней стадии, возможно, уже
имела место некоторая дивергенция популяций (рис. 2.11).
Афарский австралопитек, вероятно, был предком Australopithe-
cus africanus, по крайней мере отчасти. Последний, по мнению
некоторых ученых (Tobias, 1980; Skelton, McHenry, Drawhorn,
1986), в свою очередь считается предком как массивных авст-
ралопитеков, так и рода Ното (рис. 2.11, В). Джохансон с сотр.
(Johanson, White, 1979; Johanson, Taieb, Coppens, 1982; White,
Johanson, Kimbel, 1983), однако, полагают, что расхождение
ветвей австралопитеков и рода Ното произошло до появления
Australopithecus afarensis (рис. 2.11,Б).
По всеобщему мнению, массивные австралопитеки отдели-
лись от ветки, ведущей к современным людям, хотя среди уче-
ных существуют некоторые разногласия по поводу того, явля-
ются ли они монофилетической группой или образуют всего
54
Глава 2
Миллионы лет назад Миллионы лет назад
Н. erectus
Н. habilis
A, boisei/robustus
A. africanus
Н. erectus "|"д. boisei/robustus
Н. habilis A. africanus
О
Рис. 2.11. Альтернативные филогенетические схемы эволюции ранних гоминид:
A. Walker, Leakey, 1978; Б. Johanson, White, 1979; В. Skelton, McHenry, Draw-
horn, 1986; Г. Модель, показывающая раннюю дивергенцию массивных австра-
лопитековых и видообразование в пределах рода Ното
9
о
1
2
Н. sapiens sapiens
I
H, sapiens neanderthalensis
H. sapiens (архаический)
H. erectus
H, erectus
H. habiiis Австралопитековые
A
H. sapiens sapiens
I H sapiens
I——^neanderthalensis
H. sapiens (архаический)
H. erectus
H. erectus
Ц. habiiis Австралопитековые
В в
Рис. 2.12. Альтернативные филогенетические схемы эволюции более поздних гоминид: А. Простая однолинейная
модель; Б. Классический взгляд на неандертальцев как боковую ветвь; В. По Andrews, 1984а
56
Глава 2
один вид (Howell, 1978; Rak, 1983). Общепризнано, что Ното
habilis относится к самым ранним из известных представителей
рода Ното, но в последнее время высказываются предположе-
ния о существовании более чем одного таксона Ното до появ-
ления Homo erectus (Wood, 1984) (рис. 2.11,7"). Далее, по
утверждению Эндрюса (Andrews, 1984b), Homo erectus, кото-
рый обычно понимают как гоминид среднего плейстоцена, мо-
жет быть в действительности представлен двумя отдельными
линиями, лишь одна из которых —• африканская — дала впо-
следствии начало Homo sapiens. Таким образом, и в отношении
более поздних стадий эволюционного развития человека ученые
не пришли к полному согласию в выборе моделей эволюцион-
ного процесса (рис. 2.12).
Период с конца миоцена до позднего плейстоцена охватыва-
ет более 5 млн. лет. Сегодня мы можем восстановить эволюци-
онную историю гоминид по ископаемым остаткам — начиная от
их появления в виде небольшой прямоходящей африканской
формы, через период видового разнообразия на территории
Африки, а также длительный период существования Ното
erectus до появления Homo sapiens и возникновения человека
современного типа около 30000 лет назад. Меняющаяся мор-
фология костных остатков гоминид, однако, рассказывает нам
лишь часть эволюционной истории гоминид.
2.3. Поведение гоминид
Эволюционная история нашего вида — это рассказ об адап-
тивных изменениях морфологии и поведения. Характер измене-
ний внешних, экологических условий жизни и поведенческих
реакций является, следовательно, частью эволюционной истории
человека. Если первое зафиксировано в геологической и пале-
онтологической летописи, то второе — в остатках материальной
культуры, которые дают материал для археологии.
2.3.1. Палеоэкология
Климат Земли никоим образом не остается постоянным, но
колеблется в связи с как внутренними, так и внешними факто-
рами. В то время когда происходила эволюция гоминид, климат
был более прохладным (рис. 2.13). В миоцене и плиоцене это
привело к исчезновению обширных массивов тропического дож-
девого леса и распространению более сухих тропических биото-
пов с выраженной сменой сезонов, а также установлению более
умеренного климата, который является характерной частью со-
временного мирового биома.
Рассказ об эволюции человека
57
Рис. 2.13. Климатические сдвиги позднего кайнозоя — температурные характе-
ристики субарктического дна (по Kenneth, 1977)
За прошедшие 2 млн. лет похолодание климата Земли до-
стигло своего апогея во время ледникового периода, когда ле-
довый панцирь распространился на обширные территории и
покрыл большую часть материковых массивов Северного полу-
шария. В это время значительно понизилась температура, про-
изошло падение уровня мирового океана, вследствие чего мно-
гие регионы оказались соединенными материковыми мостами,
а многие части тропиков были более сухими, чем сегодня. Пе-
риоды наступления ледника чередовались с более теплыми пе-
риодами, подобными тому, в котором мы живем сейчас. Во
время плейстоцена насчитывалось около 20 таких глациально-
интерглациальных циклов, вероятно, связанных с изменением
орбиты Земли по отношению к Солнцу (Roberts, 1984)
(рис. 2.14).
Эти климатические и средовые изменения, несомненно, игра-
ли важную роль в эволюции гоминид. Более сухой климат и,
как следствие, меньшее развитие лесного покрова в миоцене
обеспечили условия для эволюционирования многих видов жи-
вотных, заселявших мир, который сам находился в процессе
становления. Гоминиды наряду с другими наземными примата-
ми были одной из таких групп животных. Многие характеристи-
ки ранних гоминид, в том числе и бипедия, могли возникнуть в
ответ на требования со стороны этих новых условий.
Кроме того, полное оледенение эпохи плейстоцена, начавше-
еся в то время, когда гоминиды стали распространяться за пре-
делы тропического пояса, действовало одновременно и как
Стадии 1 3 5 7 9 11 13 15 17 19 21 23 Теплая
кислорода [246 8 10 12 14 16 18 20 22 Холодная
Межледниковый
Ледниковый
Тихоокеанский керн V28-239
180/’6О (на млн.)
Возраст, । j
,млн. лет | Г"
О
Поздний i
плейстоцен |
—I—I—I—I-
0,5
Средний плейстоцен
1,0
Ранний плейстоцен
2,0
Плиоцен
Рис. 2.14. Температуры в плейстоценовом океане, соотношения изотопов кислорода и геохронология плейстоцена
(по Roberts, 1984)
Рассказ об эволюции человека
59
движущая сила, и как мощный тормоз на поведение и адапта-
цию ранних гоминид. Например, понижение уровня моря в пе-
риоды оледенения привело к появлению материковых мостов,
ныне не существующих (рис. 2.15); в то же время понижение
температуры могло стать серьезным барьером на пути распро-
странения животного, которое связано своим происхождением
с тропиками. Однако к концу плейстоцена гоминиды заняли все
основные биотопы земного шара, оставив незаселенными лишь
отдаленные острова.
2.3.2. Технология
Основное различие между гоминидами и другими примата-
ми состоит в степени зависимости их существования от изготов-
ленных ими орудий, а также от того, насколько они могут воз-
действовать на окружающие предметы, видоизменяя их в своих
целях.
Хотя шимпанзе используют орудия, изготовленные из прути-
ков или травинок, для таких целей, как «ужение термитов»,
никто никогда не наблюдал, чтобы они употребляли каменные
орудия (van Lawick Goodall, 1970). Гоминиды, с другой сторо-
ны, изготовляли орудия в течение по меньшей мере 2 млн. лет
(см. обзор последних данных на эту тему Harris, 1983; Isaac,
1984). В Кооби-Фора (Кения), Омо (Эфиопия) и Олдувае
(Танзания) были обнаружены примитивно обработанные ка-
менные орудия в геологических отложениях, которые не моложе
1,8 млн. лет; в Када-Гона (Эфиопия) орудия найдены в слоях
древностью от 2,5 до 2,7 млн. лет (Roche, 1980; Roche, Tierce-
lin, 1977), Харрис (Harris, 1986) недавно сообщил об орудиях
возрастом в 2 млн. лет из слоев Семлики (Заир), что значи-
тельно расширяет границы географического распространения
этих древнейших изделий рук человеческих. Обычно считается,
что гоминиды пользовались ветками и палками в течение како-
го-то периода, предшествующего началу изготовления каменных
изделий.
Каменные орудия, образующие первичный материал ранней
археологической летописи, по-разному классифицируются уче-
ными в соответствии со способом их производства, различиями
по характеру выполнения и функциональной значимостью. Одна
из таких классификаций предложена Дж. Г. Д. Кларком
(Clark, 1968), выделившим пять основных типов изготовления
орудий (табл. 2.2). Эти типы нельзя рассматривать как хроно-
логически приуроченные или эволюционно преемственные; про-
стые способы производства встречаются на протяжении всей
первобытной истории, а более сложные сильно варьируют во
времени и пространстве. Тем не менее в таблице представлены
Рис. 2.15. Распространение в плейстоцене ледниковых щитов и расширение прибрежных областей суши в свя-
зи с понижением уровня моря (по Roberts, 1984)
Рассказ об эволюции человека
61
Таблица 2.2. Способы изготовления каменных орудий в первобытной истории
(по классификации J. G. D. Clark, 1968)
Способ производства Характеристика Название культуры Распространение
Способ 1 Простое прямое воздействие путем удара с получени- ем отщепов и чоп- перов Олдувайская Начиная с раннего плей- стоцена (особенно в Аф- рике, но встречается по- всеместно)
Способ 2 Производство крупных отщепов с обильной ретушью; ручные рубила Ашельская (ше.ть- Ранний и средний плей- ская) стоцен, широко распрост- (аббевильская) ранен в Африке, Европе (карари) и некоторых частях Азии (нижний палеолит)
Способ 3 Систематическая подготовка нуклеу- сов до производст- ва отщепов Мустьерская, ле- Начальные стадии позд- валлуазская (сред- него плейстоцена в Ев- ний палеолит) ропе и Африке
Способ 4 Производство но- жевидных пластин с уменьшенной пло- щадкой Ориньякская, гра- На протяжении позднего ветт (верхний па- плейстоцена во многих леолит) частях земного шара, особенно в Европе; раз- личные культуры с раз- личным распростране- нием
Способ 5 Производство мик- ролитических от- щепов и пластин с ретушью Вилтон, натуфий- Глобальное постплейсто- ская, маглемозий- ценовое распространение ская (мезолит и и ограниченное присут- неолит) ствие в позднем плейсто- цене
основные, хотя и связанные друг с другом способы произ-
водства.
Основная форма обработки камня заключалась в оббивке
кусков камня с получением чопперов и отщепов с простым, но
эффективным режущим краем (способ 1). Второй способ прин-
ципиально отличается от первого тем, что в процессе производ-
ства от нуклеусов отделяются значительно более крупные от-
щепы, которые затем подвергаются обработке ретушью.
Преимущественная форма орудий — ручные рубила с характер-
ной вторичной оббивкой по обеим сторонам. Технологический
прогресс третьего способа состоит в подготовке нуклеусов до
отделения от них отщепов. Этим достигалось получение отще-
пов определенного размера и формы, а также более эффектив-
ное использование сырья. Способ 3 в значительной мере сходен
со способом 4, продолжающим традицию обработки нуклеусов,
но производящим не отщепы, а длинные ножевидные пластины
62
Глава 2
с редуцированными, почти точечными ударными площадками.
Способ 5 также продолжает традицию миниатюризации камен-
ных орудий, включая производство большого числа очень мел-
ких пластин, как правило, отретушированных.
Следует подчеркнуть, что это технологическая, а не типо-
логическая или хронологическая классификация, которая пред-
назначена лишь для описания способов производства каменных
орудий. Во многих археологических памятниках наблюдаются
признаки использования не менее двух способов изготовления
орудий, причем изменения могли происходить не обязательно
в одном направлении. Однако, несмотря на все сложности,
можно все же выделить отчетливые типы изменчивости техно-
логических приемов.
Существует общая хронологическая тенденция в отношении
указанных способов производства (рис. 2.16). Ранний плейсто-
цен и большая часть среднего характеризуются изготовлением
простых чопперов, отщепов и ручных рубил, хотя распростра-
нение этих последних ограничено Африкой, Средним Востоком
и Западной Европой. К концу среднего плейстоцена начинают
изготовлять нуклеусы, причем по преимуществу в тех регионах,
где раньше существовали ручные рубила. Приблизительно
40 000 лет назад в некоторых частях земного шара, в первую
очередь в Европе и на Среднем Востоке, появляются ножевид-
ные пластины. Микролиты — это преимущественно постплей-
стоценовое явление, хотя в Африке в районе суб-Сахары их
появление отмечено уже около 40 000 лет назад (Klein, 1983).
При этом, однако, следует помнить, что в некоторых частях
света, таких, как Юго-Восточная Азия, технологических изме-
нений на протяжении плейстоцена практически не наблюдается.
Изменение технологических приемов у ранних гоминид
обусловлено несколькими факторами. Отчасти это отражение
их повышенных манипуляторных способностей. Но в некоторой
мере возрастающее усложнение процесса изготовления орудий
отражает изменения познавательных способностей гоминид. Как
показал Гоулетт (Gowlett, 1984), переход от чопперов и отще-
пов к стадии нуклеусов предполагает увеличение числа поня-
тийных категорий, которыми могли овладеть гоминиды. В этом
случае характер технологического процесса свидетельствует не
только о физических и экологических затруднениях или воз-
можностях гоминид, но также об их способности задумать и
спланировать такие действия, которые выходят за рамки сию-
минутных потребностей. Действительно, судя по новейшим ра-
ботам, можно предположить, что основной фактор, определяю-
щий сложность изготовления каменных орудий и, следователь-
но, особенности соответствующих археологических находок, —
это уровень планирования, необходимого для добычи пищи. По-
Рассказ об эволюции человека
63
Рис. 2.16. Классификация ранних фаз первобытной истории в связи с геологи-
ческой и временной шкалой и технологическими приемами (см. табл. 2.2)
(Isaac, 1972, р. 384, с изменениями)
64
Глава 2
этому обитание в более изменчивых и менее безопасных усло-
виях с выраженной сменой сезонов влекло за собой усложнение
технологических операций (Binford, 1980; Torrence, 1983). Этим
объясняется также локализация и нестабильность способов из-
готовления орудий у гоминид в плейстоцене, на которую обра-
щал внимание Айзек (Isaac, 1972).
2.3.3. Древнейшая адаптация гоминид
Изменения морфологических особенностей и технологиче-
ских приемов, несомненно, создали основу для определенного
образа жизни, свойственного гоминидам в плиоцене и плейсто-
цене. Этот образ жизни формировался под влиянием требований
окружающей среды. Адаптивная изменчивость затрагивала так-
же и поведение гоминид — стратегию размножения, социальные
связи, поиск пищи. Эти аспекты можно реконструировать на
основе ископаемых остатков гоминид и археологических данных.
Современные человеческие популяции отличаются огромным
разнообразием социальных связей, форм и пластичности. Одна-
ко в основе социального поведения людей лежат некоторые
фундаментальные характеристики, которые позволяют отличать
это поведение от такового у других видов животных. Можно
предположить, что именно эти особенности сыграли важную
роль в эволюции гоминид.
Один комплекс подобных поведенческих характеристик свя-
зан с заботой родителей о потомстве, с ростом и развитием
детей. Потомство человека зависит от других людей, в первую
очередь своих родителей, намного дольше, чем детеныши лю-
бого другого примата. Процесс роста замедлен, стадия зрело-
сти достигается относительно поздно. Отсюда вытекают раз-
личные последствия. Одно из них — продолжительный период
обучения и, следовательно, большие возможности для накопле-
ния информации и приобретения жизненного опыта. Другое
состоит в том, что человеческие индивидуумы оказываются в
большой степени взаимозависимыми; это справедливо как в
отношении самих детей, так и взрослых, которых связывает
присутствие детей. Кооперация между индивидуумами состав-
ляет обычный элемент человеческого поведения.
Доказательством замедленного периода созревания у доис-
торических популяций гоминид может, вероятно, служить ха-
рактер прорезывания зубов. Замедленные скорости роста могут
проявиться большими различиями в степени стертости разных
категорий зубов по мере того, как будет увеличиваться проме-
жуток времени между их прорезыванием. Ранние гоминиды
действительно обнаруживают это свойство. Хотя нельзя полно-
Рассказ об эволюции человека
65
стью исключить возможности увеличения скорости стирания
зубов, эти данные могут свидетельствовать о сходном типе по-
ведения ранних гоминид, а именно о таких его аспектах, как
забота о детях и взаимозависимость.
Одна из форм кооперации в человеческом обществе — это
способность делиться пищей. Поскольку этот тип поведения
встречается во всех человеческих обществах, но крайне редок
среди остальных приматов, он считается одной из ключевых
человеческих характеристик. Айзек (IsaaK, 1978а) на основе
археологических данных утверждал, что дележ пищи наблю-
дался у ранних гоминид плио-плейстоцена. Он предположил,
что способность делиться пищей возникает в результате раз-
деления труда, которое в свою очередь обусловлено включени-
ем мяса в пищевой рацион гоминид. Поскольку вследствие
этого самцы и самки по отдельности занимаются поисками пи-
щи, становится необходимым дележ пищи. В дальнейшем воз-
никает необходимость формирования специального места, где
этот дележ происходит, — иными словами, жилища. В качестве
доказательства приведены данные о наличии каменных изделий
вместе с остатками крупных млекопитающих, скопления кото-
рых были найдены в одной и той же местности. Такие материа-
лы получены со стоянки KBS (FxJiI) в Кооби-Фора (Кения)
возрастом 1,8 млн. лет и со сходных местонахождений в Олду-
вае (Танзания) (М. D. Leakey, 1972). Согласно этой точке зре-
ния, жилище, разделение труда и дележ пищи появляются на
ранних стадиях эволюции человека.
Полагают, что дележ пищи связан с употреблением в пищу
мяса. Как правило, обезьяны питаются растительной пищей,
лишь некоторые виды занимаются активными поисками пищи
мясной (Harding, 1981). Только у павианов (Papio spp.) и
шимпанзе наблюдались случаи охоты на млекопитающих (De
Vore, Washburn, 1963; Goodall, 1963; Harding, Teleki, 1981).
С другой стороны, современные люди часто включают в свой
рацион большое количество мяса (Hill, 1982). Возникновение
охоты, по утверждению некоторых ученых, представляет собой
критическую стадию эволюции, потребовавшую для своего до-
стижения (Новых навыков и способностей и открывающую новые
возможности (Washburn, Lancaster, 1968). Например, высказы-
вались предположения о том, что для охотничьей деятельности
нужен более высокий уровень кооперации между индивидуума-
ми, чем при сборе растительной пищи, поэтому охота выступала
в роли стимула для развития языка и социальных связей меж-
ду самцами. Следовательно, данные о ранних случаях поедания
мяса, развитии охотничьих навыков и способов ведения охоты
будут иметь важное значение для понимания эволюции пове-
дения гоминид.
5—367
66
Глава 2
Доказательствами в пользу поедания мяса служат кости
животных, найденные вместе с каменными орудиями, а также
следы, оставленные на костях этими орудиями (Isaac, Crader,
1981). Как уже говорилось, согласно археологическим данным,
такие скопления появляются, чуть позже рубежа в 2 млн. лет
назад, а многие примеры, относящиеся к эпохе плейстоцена,
показывают, что гоминиды имели доступ к источникам мясной
пищи в течение довольно длительного времени. Конечно, в позд-
нем плейстоцене наряду с возникновением человека современ-
ного типа появляются и археологические стоянки, содержащие
огромное количество костей животных; они красноречиво сви-
детельствуют о хищнических тенденциях гоминид.
Еще одна ключевая характеристика современного челове-
ка — масштабы социальных связей, которые он может и дол-
жен образовывать. Хотя это привело к радикальным преобра-
зованиям по сравнению с исходной системой социальных взаи-
моотношений в сообществе приматов, в основе этой человече-
ской характеристики лежит один элемент, имеющий важное
биологическое и эволюционное значение. Это свойство узнавать
и различать индивидуумов'среди большого их числа, а затем
корректировать в соответствии с этим свое поведение. Это до-
стигается с помощью улыбок, смеха, нахмуренных взглядов,
которые делают человеческое лицо таким эффективным источ-
ником информации. Человеческое лицо уникально по своей
изменчивости, поразительному контролю над выражением эмо-
ций и способности передавать огромный набор сообщений
(Darwin, 1871). Основу для этого образует сложная система
мышц, которая придает человеческому лицу определенные
очертания и выражения и служит доказательством эволюцион-
ного происхождения значительной части человеческих эмоций
и поведения. Не будь у человека способности узнавать множе-
ство различных индивидуумов, невозможны были бы и сложные
родственные связи, которые являются структурой социальной
жизни человека.
И все же, несмотря на выразительность человеческого лица,
основной способ коммуникации людей — это язык. Это свойст-
во опять-таки имеет отчетливое биологическое и эволюционное
происхождение, так как лингвистические способности человека
твердо базируются на строении гортани, умелой артикуляции
языка и губ, усложнении и увеличении мозга. Язык, обобщен-
ное использование звука в качестве символа, отражается в дру-
гих способах человеческого общения — музыке, живописи, ли-
тературе — и составляет фундаментальную основу человече-
ского поведения. Способность создавать символы в голове и в
предметном мире, которые моделировали бы прошлый опыт,
текущие понятия и будущие решения, образует основание по-
Рассказ об эволюции человека
67
ведения и культуры современного человека. Эта способность
должна была играть существенную роль в стратегии выжива-
ния ранних гоминид.
2.4. За рамками повествования
2.4.1. Еще раз об эволюции гоминид (резюме)
Гоминиды как отдельное семейство появляются в конце
миоцена около 5 млн. лет назад. Самые первые из них были
небольшими прямоходящими существами с небольшим мозгом
и, по-видимому, развитым половым диморфизмом. Вскоре, од-
нако, происходит их разделение на самостоятельные ветви —
этот процесс лучше всего характеризуется термином «адаптив-
ная радиация». Одна линия ведет к более поздним австралопи-
текам— животным с крупными зубами и лицом, часто массив-
ным скелетом и небольшим мозгом; они жили в Африке, ве-
роятно, на протяжении длительного периода от 3 до 1,2 млн.
лет назад. Другая линия — с более крупным мозгом и менее
мощной мускулатурой — привела к возникновению более позд-
них видов рода Ното — Н. habiiis, Н. erectus и Н. sapiens.
Африканские гоминиды позднего плиоцена и раннего плейсто-
цена, вероятнее всего Homo habiiis, по-видимому, ответственны
за создание самых первых каменных орудий, за начальные по-
пытки использования животной пищи в масштабах, сравнимых
с таковыми современного человека, и, возможно, за истоки со-
циальной жизни человечества — язык, кооперацию, дележ пи-
щи, — которые в конечном итоге привели к расцвету изменчи-
вых и сложных социальных взаимосвязей, характерных ныне
для нашего вида.
Мы намеренно упростили рассказ об эволюции гоминид и
первобытной истории, оставив в стороне многие важные проб-
лемы и изложив некоторые гипотезы с большей уверенностью,
чем это позволяют имеющиеся данные. Например, в последние
годы ведутся острые дискуссии о роли охоты в противополож-
ность собирательству, а также о том, насколько убедительные
доказательства об использовании животных ресурсов можно
получить из археологических материалов раннего и среднего
плейстоцена (Binford, 1980,1983; Isaac, 1983; Shipman, 1983; Potts,
1984a). Специалисты найдут здесь много таких мест, которые
им покажутся весьма спорными. Однако наша цель состояла
в том, чтобы дать краткий обзор современных палеоантрополо-
гических данных для тех, кто с ними не знаком, а также под-
черкнуть повествовательный элемент в истории эволюции гоми-
нид. Наше эволюционное прошлое можно трактовать как рас-
сказ, сюжет которого прослеживается в ископаемых остатках,
5»
68
- Глава 2
археологических материалах, данных по биологии и экологии
наших ближайших ныне живущих родственников — приматов.
Это рассказ, в котором персонажи сменяют друг друга с доста-
точной регулярностью, а главному герою сопутствует неизбеж-
ный адаптивный успех, благодаря его прямохождению и круп-
ному мозгу.
2.4.2. Окаменелости молчат
Не считая краткости и простоты, наш обзор далек от со-
вершенства, так как он является скорее повествованием, чем
объяснением, хотя основные события в нем обозначены и поме-
щены во временные и пространственные рамки, он мало спо-
собствует объяснению эволюции человека. Доминирующий под-
ход, используемый для истолкования событий, если он и есть,
состоит в ретроспективном взгляде, при котором трактовка со-
бытий прошлого основывается на современных характеристиках
представителей человеческого вида и условий их жизни. Это
все равно, что читать детективный роман, когда заранее зна-
ешь, кто убийца. Это знание, несомненно, накладывает отпе-
чаток на прочтение первых глав книги. Например, поскольку
интеллект и техническое производство являются основными сла-
гаемыми нашего сегодняшнего успеха, их часто рассматривают
в качестве наиболее важных элементов нашего поведения в
прошлом. Эта идея наверняка лежит в основе многих прежних
гипотез о том, что увеличение мозга предшествовало другим
изменениям в эволюции человека (Reader, 1981).
Причина такого преобладания ретроспективного подхода
кроется, возможно, в убеждении, будто ископаемые остатки,
если расположить их в правильном хронологическом порядке,
могут сами рассказать свою историю. Это, однако, далеко не
так. Ископаемые остатки, так же как и любые другие источ-
ники информации, зависят в своем истолковании от теоретиче-
ской основы. Другими словами, если что и отсутствует в нашем
рассказе об эволюции человека, так это изложение причин про-
исходящих изменений, а также механизмов, посредством кото-
рых они осуществлялись. На этом уровне лишь ретроспектив-
ный взгляд сообщает нам, что крупный мозг можно считать
успешной адаптацией, а бипедию — наиболее подходящей фор-
мой передвижения. Необходима теоретическая основа, которая
позволила бы не просто рассматривать характеристики ранних
гоминид в свете нашего собственного, гораздо более позднего
поведения, а сравнить их с альтернативами, встававшими в
свое время перед первыми гоминидами. Для этого нужно не
просто послушать, о чем, по нашему мнению, рассказывают
Рассказ об эволюции человека 69
ископаемые остатки, но и подумать, какие еще окаменелости
мы могли бы найти.
Чтобы перейти от простого повествования об эволюции че-
ловека к анализу, который реально показал бы, почему гоми-
нидам сопутствовал успех, надо располагать ископаемыми ма-
териалами, которые могут рассказать нам, что же произошло
в далеком прошлом, и соединить их с представлениями, кото-
рые развиваются в эволюционной экологии и позволят нам по-
нять, что могло произойти в этом далеком прошлом. Сравнив
наблюдаемое и ожидаемое, можно выйти за рамки ретроспек-
тивного взгляда на эволюцию человека и перейти к ее понима-
нию на основе здравых экологический идей.
Глава 3
ЭВОЛЮЦИОННАЯ экология
Эволюционная история гоминид представляет собой уни-
кальную картину биологических изменений. Центральная мысль
этой книги, однако, состоит в том, что уникальные биологиче-
ские формы могут складываться в результате обычных биоло-
гических процессов и механизмов и что это приложимо к го-
минидам в той же мере, как и к любой другой группе живот-
ных. Поэтому основные принципы теории эволюции и экологии
являются подходящей основой для понимания эволюции челове-
ка. Задача состоит не в создании какой-либо специальной тео-
рии, а в четком применении тех известных теорий, которые уже
доказали свою эффективность в более широкой области эволю-
ционной биологии. В этой главе будут описаны некоторые из
наиболее важных идей эволюционной экологии, а именно связь
между эволюцией и экологией, характер естественного отбора,
пути адаптации, использование эволюционных принципов при
анализе поведения.
3.1. Эволюция и экология
Экологи изучают взаимоотношения между организмом и
окружающей средой с тем, чтобы выявить принципы, их конт-
ролирующие. Однако сами эти взаимоотношения могут быть
крайне разнообразными. Как указывает Пианка (Pianka, 1978,
р. 2), организм и среда как составные части не остаются по-
стоянными: то, что представляет собой организм на одном уров-
не, на другом является частью окружающей среды. Пианка пред-
ложил термин «организменный уровень» как способ избежать
этой проблемы и выделил несколько таких уровней (рис. 3.1).
В экологии уровень, или единица анализа, может варьировать
от отдельного организма до целого сообщества, составленного
из нескольких видов. Каждый уровень может быть проанализи-
рован исходя из его взаимоотношений с внешней средой.
Среда — это в равной степени изменчивое понятие. Она
включает основные физические характеристики, такие, как тем-
пература, влажность, особенности почв и т. д., а также биоти-
ческие факторы, например присутствие симпатрических видов,
Эволюционная экология
71
Рис. 3.1. Взаимосвязь между организменным уровнем и окружающей средой
в экологии. Каждый расположенный выше уровень является частью окружаю-
щей среды по отношению к нижерасположенному
которые выступают как хищники, конкуренты или, напротив,
источники пищи. Кроме того, следует подчеркнуть, что окру-
жающая среда любого организма включает также других пред-
ставителей той же популяции и вида, с которыми организм на-
ходится в генетических и социальных взаимоотношениях. С этой
точки зрения сущность окружающей среды не исчерпывается
узким типологическим перечнем географических и биотических
факторов; к этой среде относится все, что действует на организ-
менном уровне, будь то физические, биологические или соци-
альные воздействия. Важно не столько то, каковы взаимоотно-
шения между ними, сколько механизмы, регулирующие эти
взаимоотношения.
Существует несколько категорий экологических взаимоотно-
шений. Многие взаимодействия между разными организмами
включают передачу энергии, например когда один организм по-
глощает другой частично или целиком (травоядные, хищники,
паразиты и т. д.). Характер энергии, описываемый законами
термодинамики, образует структуру этих энергетически зависи-
мых взаимоотношений. Эта структура определяется экологами
с помощью таких концепций, как трофическая пирамида — мо-
дель снижения популяционного и энергетического уровней по
мере продвижения вверх по пищевой цепи в результате неэф-
фективности преобразования энергии (рис. 3.2) — и экосистема.
Различия в структуре энергетических взаимоотношений между
организмами помогают объяснить изменчивость в распределе-
Глава 3
Высшие хищники
Третичный трофический уровень
Хищники
Вторичный трофический уровень Травоядные
Первичный трофический уровень Растения
Рис. 3.2. Трофическая пирамида: потеря энергии иа каждой стадии трансфор-
мации придает пирамидальную форму экологическим сообществам
нии растений и животных. Другие взаимоотношения основаны
на потоке веществ. Сюда относятся такие процессы, как круго-
ворот углерода или азота, без которых немыслима жизнь (пути
превращения и перемещения этих элементов в экосистеме в ос-
новном известны).
Менее заметными, но не менее значимыми являются конку-
рентные взаимодействия между организменными уровнями в от-
ношении таких ресурсов, которые приходится делить (пища,
брачные партнеры или жизненное пространство). Способность к
конкуренции или ее избежанию является основным фактором,
который определяет поведение, распределение и обилие расте-
ний и животных.
Наряду с этими основными взаимоотношениями существуют
такие, как социальные, пространственные и генетические связи,
но структура биологического мира, который служит объектом
наших эмпирических наблюдений, складывается в результате
изменчивого характера всех этих зависимостей. Задача эколо-
гической науки состоит в том, чтобы объяснить характер этих
соотношений и сформулировать принципы, управляющие ими.
Принципы экологии теснейшим образом связаны с теорией
эволюции, потому что именно естественный отбор в конечном
итоге формирует экологические взаимоотношения посредством
дифференциального репродуктивного успеха. Именно эта взаи-
мосвязь и составляет суть эволюционной экологии.
3.2. Естественный отбор
3.2.1. Дифференциальное воспроизведение
Неодарвинистская теория эволюции путем естественного от-
бора занимает центральное положение в современной биологии.
Биологическая эволюция — сложное явление, состоящее из
многих процессов, но в основе их лежит механизм естественного
отбора. В наиболее простом виде теория эволюции утверждает,
Эволюционная экология
73
что те особи, которые оставляют больше потомков по сравне-
нию с другими, будут генетически лучше представлены в следу-
ющих поколениях и, следовательно, последние будут особенно
схожи с этими, успешно размножавшимися организмами. Меха-
низм, с помощью которого осуществляется дифференциальный
репродуктивный успех, относится к естественному отбору. От-
бор есть не что иное, как избирательное выживание биологиче-
ских единиц (Futuyma, 1979).
Тогда естественный отбор — это механизм, определяющий
форму и частоту особей в каждом поколении, для того чтобы,
с одной стороны, сохранять в популяциях определенный фено-
тип (стабилизирующий отбор), а с другой —развивать их в но-
вом направлении, иными словами, стимулировать эволюцион-
ные изменения. Однако, хотя естественный отбор занимает
центральное положение в современной биологической теории, он
действует не изолированно, и, следовательно, для полного пони-
мания процесса биологической эволюции необходимо учиты-
вать другие биологические процессы, направляющие и сдержи-
вающие действие естественного отбора, т. е. рассматривать весы
контекст, в рамках которого происходит естественный отбор.
3.2.2. Факторы, ограничивающие отбор
Первое ограничение состоит в том, что отбор оперирует из-
менчивостью, имеющейся в популяциях. Дарвин вначале стре-
мился не к созданию теории эволюции, а к тому, чтобы ответить
на вопрос, почему существует внутри- и межвидовая изменчи-
вость животных и растений. Возможно, поэтому первые две гла-
вы «Происхождения видов» озаглавлены «Изменчивость в при-
роде» и «Изменчивость при одомашнивании», а сам термин
«эволюция» в книге фактически не фигурирует. В результате
естественного отбора те разновидности, которые выживают и
размножаются лучше других разновидностей, становятся доми-
нирующей формой в популяции или виде внутри сообщества.
Отбор определяет размах и частоту изменчивости в биологиче-
ских системах.
Однако отбор может эффективно действовать только тогда,
когда изменчивость существует и имеет адаптивное значение.
Сила отбора, а следовательно, направление и скорость эволю-
ции ограничиваются степенью и природой изменчивости внутри
популяции. Понимание механизма дифференциального воспро-
изведения зависит от существующего размаха изменчивости
формы как морфологической, так и поведенческой.
Однако не все вариации дают материал для эволюции; сле-
дующее ограничение — это генетические основы жизни. Отбор
оперирует фенотипами, т. е. реальным морфологическим, физио-
74
Глава 3
логическим, биохимическим и поведенческим проявлением ор-
ганизма (Futuyma, 1979, р. 506). Именно приспособленность фе-
нотипа определяет успех выживания и размножения. Однако
отбор может действовать лишь в том случае, если существует
способ, посредством которого фенотипические признаки могут
наследоваться, т. е. передаваться потомкам, и, следовательно,
продолжаться в ряду поколений. Без этого фенотипическая при-
способленность не имела бы эволюционного смысла. Именно от-
сутствие механизма, с помощью которого признаки могут на-
следоваться, привело к тому, что была отвергнута предложен-
ная Ламарком теория эволюции путем наследования приобре-
тенных признаков. В современной биологии признаются те за-
коны, управляющие наследственностью, которые были открыты
Менделем. Генетические основы жизни сдерживают силу естест-
венного отбора. Особенно важное значение имеют две генетиче-
ские характеристики. Первая состоит в том, что ген не меняется
в течение жизни. Информация может идти только в одном на-
правлении— от генотипа к фенотипу, но не наоборот. Вторая —
в том, что именно ген, в составе гаплоидной гаметы, передается
от родителей детям. Эти два обстоятельства, объединенные у
людей и большинства животных в процессе полового размноже-
ния, определяют, какие из особей выиграют, а какие постра-
дают в результате репродуктивного поведения других особей.
Именно ген сохраняет непрерывную нить эволюции. Ограничен-
ное число способов, посредством которых гены могут переда-
ваться от одного поколения к другому, действует как ограниче-
ние естественного отбора.
Новые гены появляются в популяции главным образом в ре-
зультате мутаций. Именно мутации поддерживают и увеличива-
ют уровень генетической изменчивости, создавая тем самым еще
один фактор, ограничивающий силу естественного отбора. Мута-
ция— это ошибка в репликации гена во время мейоза. Ее нали-
чие может привести к появлению нового фенотипа (т. е. вариан-
та), который будет конкурировать с уже имеющимися в гено-
фонде. Хотя существуют и другие источники новаций (напри-
мер, генный поток между популяциями, рекомбинации, гетеро-
зиготность), без мутаций отбор мог бы лишь поддерживать име-
ющуюся структуру или форму или, если она становилась нежиз-
неспособной, приводил бы к вымиранию популяции. Эволюцион-
ный анализ должен поэтому учитывать скорость и природу му-
таций или, другими словами, источник наследуемых изменений,
составляющих основу естественного отбора. Это положение име-
ет важные последствия. Одно из них состоит в том, что, хотя
процесс мутирования носит случайный характер (здесь имеются
в виду генные ошибки в процессе репликации), форма мутации
таковой не является; это означает, что особенности фенотипа,
Эволюционная экология
75
полученного в результате мутации, будут зависеть от природы
исходного фенотипа (Dawkins, 1976). У амебы не может про-
изойти мутация, которая привела бы к появлению человеческо-
го интеллекта: все ее мутации не выйдут за пределы амебоид-
ной организации. Именно это свойство может обеспечивать по-
степенный характер эволюции. Еще одно важное обстоятельст-
во заключается в том, что последствия мутации могут не все
проявляться немедленно и одновременно. Например, в случае
мутации, обеспечивающей повышение способностей к обучению,
некоторый эффект может наблюдаться сразу, однако полностью
значение мутации обнаружится лишь через несколько поколений
(при этом большую роль будет играть вид усваиваемой инфор-
мации).
Еще один фактор, оказывающий влияние на естественный
отбор,— это борьба за существование. Разрабатывая теорию эво-
люции, Дарвин заимствовал концепцию конкурентной борьбы у
политэкономов XIX в. .в целом и особенно из демографиче-
ского учения Мальтуса. Конечная численность имеющихся ре-
сурсов в сравнении с потенциально высокой скоростью воспро-
изводства и ростом населения позволила Дарвину увидеть ос-
новной фактор, который делает необходимым естественный от-
бор,— перевес выселения над ресурсами. Конкуренция — прямая
или непрямая — между особями за общие источники пищи — та-
кова обязательная предпосылка естественного отбора. Именно в
свете ограниченности ресурсов те из особей, которые лучше при-
способлены к овладению ими, получают преимущества в отно-
шении размножения, а следовательно, и преимущества в про-
цессе естественного отбора.
Это приводит нас к рассмотрению последнего ограничения
действия естественного отбора и одновременно возвращает к
ключевому понятию репродуктивных различий. Для того чтобы
какой-либо признак попал под действие естественного отбора,
нужно, чтобы этот признак влиял на способность особи к ус-
пешному размножению. При отсутствии взаимосвязи между
природой фенотипа и различиями в репродуктивном успехе есте-
ственный отбор не может действовать. Различия в фенотипах,
не оказывающие существенного воздействия на шансы выжи-
вания особи, не могут играть важной роли в эволюции.
Итак, стержнем эволюционной теории является принцип
естественного отбора, т. е. дифференциальный репродуктивный
успех биологических существ. Строго говоря, все остальные
черты эволюционного процесса носят вторичный характер. Но
поскольку естественный отбор оперирует в биологическом и фи-
зическом мире, он ограничен другими факторами, а именно мен-
делевским наследованием, генетической изменчивостью, мута-
циями, борьбой за существование и т. д., которые придают био-
76
Глава 3
логической эволюции ее специфический характер и позволяют
ее прослеживать. Из этого следует, что эволюционный подход
сконцентрирован на понимании действия естественного отбора —
его последствий для живой материи и условий, на фоне которых
разворачивается его действие.
3.2.3. Единица отбора
Следующий вопрос, кстати имеющий кардинальное значение,
касается того, на что именно действует естественный отбор. Что
является единицей отбора? По этому поводу разгорелись жар-
кие дебаты (Wynne-Edwards, 1962; Williams, 1966; Wilson, 1975;
Dawkins, 1976, 1982a; D. S. Wilson, 1980). В чисто дарвиновской
формулировке единицей отбора является особь. Именно особь
воспроизводит себя, и ее морфология и поведение определяют
репродуктивный успех. Из этого следует, что естественный от-
бор оперирует успехами и неудачами особи. Однако с открыти-
ем природы генов и процесса мейоза стало ясно, что именно
ген, а не свойства особи, передается от поколения к поколению
и, следовательно, является реальной единицей отбора. Согласно
Доукинсу (Dawkins, 1976), особь — это всего лишь временная
упаковка, в которой путешествуют гены. При условии что осо-
би имеют общие гены в зависимости от степени их родства (см.
рис. 3.3), это означает, что «интересы» гена и особи не всегда
совпадают. Это может произойти с близкородственными особя-
ми, когда сохранение общего для них гена лучше всего дости-
гается за счет той особи, которая жертвует чем-нибудь, в пер-
= d"/4 %? - с?"/4
________d”/2
I I I 471
W */2cT % $’/2
’/2cT ?’/2
’/4cT V4? cTV* $‘4
I---------1
Рис. 3.3. Степень генетического родства. Дроби показывают, какая часть генов
должна быть общей у данных особен в связи с их родством. Все цифры отно-
сятся к общим генам с ЭГО
Эволюционная экология
77
вую очередь репродуктивными усилиями, в пользу своих бли-
жайших родственников. Этот принцип получил название «род-
ственный отбор» и был сформулирован Хамилтоном (Hamilton,
1964). Логически действие отбора перемещается на уровень ге-
на, а не особи, хотя в большинстве случаев репродуктивные ин-
тересы гена и особи совпадают.
Существует, однако, поведение, при котором эти интересы
расходятся; оно называется альтруистическим. Альтруизм — это
акт, в результате которого увеличивается репродуктивная при-
способленность одной особи за счет другой. Если единицей от-
бора является особь, то, следовательно, должны эволюциони-
ровать те типы поведения, которые ей благоприятствуют, т. е.
повышают ее собственный репродуктивный успех. Поведение,
понижающее репродуктивные возможности особи (в частности,
альтруизм), должно быть элиминировано отбором, так как, по
определению, оно будет встречаться все реже, а не чаще в каж-
дом последующем поколении. Если такое поведение все же
существует, особь не может быть единицей отбора. В этом слу-
чае оказывается полезной концепция «эгоистичного гена» (Daw-
kins, 1976). Поскольку особи имеют общие гены со своими бли-
жайшими родственниками (рис. 3.3), репродуктивный успех (со-
хранение генов) каждой особи тесно связан с размножением ее
родственников, обладающих определенной частью тех же генов.
Так составляется совокупная приспособленность1 особи. При не-
которых обстоятельствах, например, в случае родителей и их по-
томства, братьев и сестер и т. д., приспособленность может уси-
ливаться не в результате размножения конкретной особи, а в
результате такого ее поведения, которое повышает репродуктив-
ный успех ее близких родственников, пусть даже и за счет са-
мой этой особи.
Таким образом, чисто технически ген, а не особь является
единицей отбора. Доукинс несколько прояснил этот вопрос, раз-
граничив два понятия: выживание репликаторов и отбор транс-
портных средств.
«Естественный отбор можно охарактеризовать двумя спосо-
бами. Оба они верны, но концентрируют внимание на различ-
ных аспектах одного и того же процесса. Эволюция происходит
в результате неодинакового выживания репликаторов. Гены
представляют собой репликаторы; организмы и группы организ-
мов таковыми не являются, это лишь средства для транспорти-
ровки репликаторов. Отбор транспортных средств — это про-
цесс, при котором некоторые транспортные средства лучше дру-
гих обеспечивают сохранение своих репликаторов» (Dawkins»
1982b, р. 46).
1 Этот термин также предложил У- Д. Хамилтон. — Прим, перев.
78
Глава 3
Хотя учитывать разницу между особями и генами, может
быть, и важно, оба уровня не являются взаимоисключающими в.
интерпретации теории естественного отбора, а скорее дополня-
ют друг друга. С точки зрения эволюционной экологии наибо-
лее интересной аналитической единицей является, конечно, особь
как целостный и организованный организм. Именно особи долж-
ны взаимодействовать с окружающей средой. Однако придет
время, когда поведение особи можно будет объяснить только
исходя из его генетической основы.
Совсем иной характер носят различия между отдельным ор-
ганизмом и группой их как единицей отбора. Здесь, если пользо-
ваться терминологией Доукинса (Dawkins, 1982), речь идет не
о дополняющих друг друга отношениях между репликаторами
и транспортными средствами, а о конкуренции между двумя
транспортными средствами. Групповой отбор — это отбор при-
знаков, которые благоприятствуют группе организмов в целом,,
за счет отдельных носителей этих признаков. Это опять-таки
альтруизм, но такой, который нельзя объяснить общими генами
и родством. По мнению некоторых ученых (Wynne-Edwards,.
1962; Wilson, 1980), вид с таким поведением будет находиться
в состоянии большего равновесия по отношению к своим ресур-
сам и, следовательно, будет иметь больше шансов выжить.
Проблема с групповым отбором, однако, заключается в том,,
что с точки зрения теории он не может осуществляться. Пред-
ставьте себе организм, который ведет себя таким образом, что-
бы уменьшить свои шансы на выживание: например, издает
сигнал тревоги, заметив хищника, и тем самым попадает в поле
зрения этого хищника, но зато повышает шансы на выживае-
мость группы в целом. Конечно, такие группы будут иметь пре-
имущества перед теми, в которых альтруисты отсутствуют, а на-
падения хищников происходят еще чаще. Однако этот тип пове-
дения не может сохраниться, так как (об этом уже говорилось в
отношении родственного отбора) в каждом последующем поко-
лении будет все меньше и меньше альтруистически настроенных
особей, издающих сигнал тревоги, пока в конце концов, в ре-
зультате более высокой смертности, они не исчезнут совсем. Рав-
новесие, следовательно, будет поддерживаться за счет группы
особей, не издающих сигнала тревоги. В этом случае, как мы
видим, групповой отбор слабее индивидуального.
Индивидуальный отбор обладает большей силой по сравне-
нию с групповым, потому что размножаются именно особи и
они же осуществляют транспорт генов. Хотя при снятии огра-
ничений отбор теоретически может происходить на любом чис-
ле уровней, на практике в силу того, что он ограничен законами
наследственности, единственным уровнем является индивиду-
альный. Это остается справедливым и в случае использования
Эволюционная экология
79
моделей, основанных на структурированных группах. Модель
группового отбора Д. С. Уилсона (Wilson, 1980), хотя и произ-
водит впечатление своим математическим изяществом, пред-
ставляет собой пример индивидуального отбора, действующего
при очень больших ограничениях (Harvey, 1985).
Этот вывод имеет важные последствия для анализа моде-
лей эволюционной экологии. Принципиальным среди них явля-
ется то, что при рассмотрении адаптивных преимуществ следует
исходить из уровня отдельных особей, а не их групп или видов,
если только не существует альтернативы гипотезе совокупной
приспособленности. Особи являются основным материалом для
эволюции и поэтому должны рассматриваться как аналитиче-
ская единица адаптивного поведения.
3.3. Отбор и адаптация
3.3.1. Адаптация
Результат естественного отбора — дифференцированное вы-
живание биологических существ — способствует развитию адап-
тации. Адаптация — это термин, который хотя и часто исполь-
зуется в экологии, но имеет несколько значений или оттенков
(Dunbar, 1982). Во-первых, существует адаптация как процесс,
т. е. процесс, с помощью которого организм меняется и приспо-
сабливается к условиям окружающей среды. В этом смысле
адаптация может быть продуктом естественного отбора в тече-
ние ряда поколений, а может быть и ответной реакцией, приоб-
ретенной или усвоенной в течение более короткого времени.
Второе значение термина «адаптация» касается действитель-
ных взаимоотношений между организмом и средой его обита-
ния. В этом случае термин показывает или пытается описать,
какие особенности организма связаны с определенными факто-
рами окружающей среды. Например, густой мех — это адапта-
ция млекопитающих к низким температурам внешней среды.
Здесь уместно подчеркнуть, что такое использование термина
правомерно лишь тогда, когда рассматривается как адаптивный
признак организма, так и обусловливающее его свойство окру-
жающей среды. Третье значение термина «адаптация» отража-
ет степень соответствия между организмом и средой. Причина,
вследствие которой термин приобретает количественный отте-
нок, состоит в том, что не существует адаптивно совершенных
организмов так же, как и идеальной адаптации. Оценка адапта-
ции может быть только относительной.
Эти терминологические оттенки могут привести к важным с
точки зрения экологии выводам. Один из них состоит в том, что
адаптация достигается посредством изменения целого ряда био-
80
Глава 3
логических характеристик: биохимических, физиологических,
морфологических и поведенческих. Все это способы приспособ-
ления организма к требованиям окружающей среды. Например,
животное может адаптироваться к высоким температурам за
счет уменьшения вязкости крови (биохимический способ), повы-
шения легочной вентиляции или потоотделения (физиологиче-
ский способ), увеличения относительной поверхности тела (мор-
фологический способ) или, наконец, оставаясь в тени и снизив
двигательную активность (поведенческий способ). Современный
человек раздвигает рамки адаптивных систем еще шире — путем
изобретения кондиционеров. При рассмотрении проблем адап-
тации нужно учитывать относительную стоимость и выгоды
каждого способа адаптации.
Еще один вывод состоит в том, что адаптация может быть
генетически детерминированным процессом, возникающим в от-
вет на требования естественного отбора, или фенотипической ре-
акцией особи, возникающей в течение ее жизни в ответ на неко-
торые средовые факторы. Например, шимпанзе как вид челове-
кообразных обезьян на протяжении нескольких миллионов лет
приспособились к питанию в основном плодами растений, о чем
можно судить по строению их зубов и желудка. Это пример
адаптации путем естественного отбора. С другой стороны, в по-
пуляции шимпанзе, ныне живущей в резервате Гомбе в Танза-
нии, обезьяны привыкли получать бананы от исследователей,
которые прикармливали их (van Lawick Goodall, 1971). Про-
изошла адаптация к новому источнику пищи путем обучения.
В первом случае имеет место изменение генных частот, во вто-
ром—фенотипические изменения, при которых геном не затра-
гивается. Однако между этими двумя типами адаптации нельзя
провести четкое различие. Фенотипические изменения могут быть
не только поведенческими. Например, у людей в условиях вы-
сокогорья происходят фенотипические изменения дыхательной
системы, в которых геном не участвует. В то же время поведен-
ческие изменения могут быть генетически обусловленными.
Вместе с тем как тот, так и другой тип адаптации нельзя ото-
рвать от естественного отбора. Если речь идет о поведении,
в основе которого лежит обучение, то оно может и не находить-
ся под прямым генетическим контролем, однако способности к
усвоению этих типов поведения могут варьировать и зависеть
от генетических различий. Следовательно, поведение хотя бы
косвенно является продуктом естественного отбора. Естествен-
ный отбор и адаптация прочно связаны друг с другом, хотя пу-
ти их могут изменяться.
Еще одно важное замечание в связи с рассмотрением терми-
на «адаптация» состоит в том, что при всех спорах о генах или
особях как единицах отбора, адаптируются к окружающей сре*
Эволюционная экология
81
де именно отдельные особи, а не группы их или гены (Williams,
1966), не культуры или общества (Foley, 1981). С описательной
точки зрения не будет ошибкой сказать, что благородный олень
приспосабливается к среде путем сезонных миграций, а племя
нуер адаптируется с помощью разведения крупного рогатого
скота, но это будут лишь статистические обобщения, указываю-
щие на поведение всех членов данной популяции или большей
ее части. Хотя эти утверждения и имеют описательную цен-
ность, они не основываются на каких-либо теоретических пред-
посылках. Это особенно справедливо для гоминид, где из-за
связанной с социальной организацией сложности поведения час-
то приходится слышать об адаптации культуры. Однако это не
так (Ellen, 1982). Особи создают, используют и отбрасывают
различные институты и формы культуры как средства адапта-
ции к социальной и биологической среде. Такой взгляд более
плодотворен, так как позволяет непосредственно связать тип по-
ведения с той самой особью, репродуктивный успех которой яв-
ляется предметом пристального внимания, а также позволяет
обнаружить и проанализировать изменчивость адаптивного по-
ведения.
Итак, мы рассмотрели три значения термина «адаптация»
и показали, что адаптация может быть достигнута различными
способами, по-разному связанными с естественным отбором —
от прямого генетического контроля до способностей к обучению.
Однако истинный характер адаптации пока неясен.
3.3.2. Пределы совершенства
Одна из точек зрения на адаптацию состоит в том, что она
характеризует идеальные взаимоотношения между организмом
и средой обитания или по крайней мере тенденцию к такому
совершенству (Cain, 1964). Этот взгляд, принятый в качестве
экологической и эволюционной аксиомы, привел к неправильно-
му пониманию теории Дарвина, согласно которому все, что дела-
ет животное, следует рассматривать как наилучшее из возмож-
ного в данных условиях. Это означает, что организмы не просто
приспособлены, а идеально приспособлены. Это представление
о совершенстве адаптационного процесса, или «программа адап-
тационистов», стало объектом серьезной критики со стороны
ряда ученых (Lewontin, 1978; Gould, Lewontin, 1979), критики,
которая содержала в себе существенное умаление аналитичес-
кой ценности концепции адаптации и действия естественного от-
бора как биологического процесса. Возражения этих авторов в
основном сводились к трем пунктам: 1) многие особенности ор-
ганизма являются следствием ограничений в процессе разви-
тия; 2) многие признаки связаны с аллометрией, т. е. опреде-
6—867
82
Глава 3
ляются размерами тела, а не прямой реакцией организма на
условия окружающей среды; 3) подавляющая часть генетиче-
ского материала нейтральна, как доказывают в настоящее вре-
мя специалисты по молекулярной биологии. Совсем недавно
Гулд и Врба (Gould, Vrba, 1983) предложили термин «экзапта-
ция» для обозначения тех признаков, которые не являются
следствием адаптивного процесса.
Однако их критика была направлена против двух положе-
ний, которые не обязательно являются частью программы адап-
тационистов. Первое состоит в том, что естественный отбор —
это свободный процесс, не имеющий пределов и обладающий
безграничной силой. Ранее было показано (см. разд. 3.3.1), что
это не так. Естественный отбор действует на фоне других био-
логических процессов, таких, как развитие и аллометрия. Это не
означает, что данные процессы не подвержены действию естест-
венного отбора или не зависят от давления отбора, а свидетель-
ствует о скупых средствах, с помощью которых отбор функцио-
нирует (нередко посредством генов, программирующих онтоге-
нез). Второе положение состоит в том, что адаптация — в рам-
ках дарвиновской теории — должна быть идеальной. Это, одна-
ко, противоречит тому определению адаптации, которое -мы дали
в предыдущем подразделе: адаптация есть мера соответствия
организма условиям его обитания и степень адаптации может
меняться от полной приспособленности к среде обитания до
плохой приспособленности. Весь этот размах адаптивной измен-
чивости должен быть представлен в мире живого.
Гулд и Левонтин (Gould, Lewontin, 1979) правильно подчер-
кнули опасность допущения возможности идеальной адаптации,
но были неправы, предположив, что нереальность существова-
ния идеальной адаптации и методические трудности, связанные
с изучением адаптационного процесса, будто бы подрывают ло-
гическую стройность теории естественного отбора. Если естест-
венный отбор действует в рамках некоторых ограничений, то
так же функционирует и его продукт — адаптация. Разные авто-
ры пытались дать перечень факторов, ограничивающих процесс
адаптации (Williams, 1966; Curio, 1973; Maynard Smith, 1978;
Oster, Wilson, 1978; Dawkins, 1982a), чтобы, как сказал Доу-
кинс (Dowkins, 1982а, р. 35), «знать, чего можно ожидать от
теории естественного отбора». Ниже перечислены основные
ограничения до Доукинсу (Dawkins, 1982а).
1. Временной разрыв между действием условий окружающей
среды и результатами отбора. Отбор оперирует в сжатые сро-
ки. Организмы отбираются на основе непосредственных требо-
ваний окружающей среды. Если меняются условия среды, то
признак, ранее бывший адаптивным, может потерять это свой-
ство. В зависимости от интенсивности отбора, может, однако,
Эволюционная экология
83
пройти несколько поколений, прежде чем организм вновь при-
способится к условиям среды, если это вообще произойдет. Дей-
ствительно, окружающая среда может характеризоваться неста-
бильностью, так что полная адаптация постоянно оказывается
недостижимой для вида. В соответствии с этим адаптации сле-
дует рассматривать в связи с историей тех биотопов, в которых
они возникли. Хотя отбор может действовать на протяжении
короткого отрезка времени, изучение адаптации предполагает
рассмотрение более длительной временной перспективы. Следует
учитывать также и миграции, когда животное переходит в но-
вые условия обитания, к которым оно не полностью приспо-
соблено.
2. Исторические ограничения. Отбор действует только на
живые организмы. Он не может начинать заново с каждым но-
вым поколением, но должен модифицировать и поддерживать
уже существующие адаптивные изменения. Следовательно, раз-
витие новых адаптаций происходит на фоне прежних. Следова-
тельно, тенденцию к совершенству можно оценить с учетом то-
го, что ей предшествовало. Например, вопрос о том, какая си-
стема передвижения является наилучшей — на двух или на че-
тырех ногах — не имеет никакого смысла. Логично спросить, яв-
ляется ли прямохождение более эффективным способом пере-
движения по сравнению с той локомоторной системой, на осно-
ве которой оно развивалось. Именно благодаря историческим
ограничениям сохраняются эволюционный консерватизм и адап-
тивное несовершенство.
3. Генетическая изменчивость. Подобно тому, как историче-
ские факторы влияют на направление адаптивных изменений,
наличие генетических вариантов определяет возникновение но-
вых адаптивных черт. Можно представить себе более совершен-
ную и даже идеальную адаптацию, но лишь при наличии гене-
тического материала.
4. Компромиссные решения. Давление отбора на организмы
никогда не определяется только одним фактором. Напротив, ор-
ганизмы постоянно подвергаются действию бесчисленного мно-
жества факторов, влияющих многими способами на их репродук-
тивный успех. Многие из этих давлений отбора противодейству-
ют друг другу. Например, для животного может быть выгодно
максимально увеличить количество потребляемой пищи, однако
время, затраченное на кормежку, могло быть также использова-
но на размножение или заботу о потомстве — процессы, интен-
сификация которых также дает ряд преимуществ. Выход из этих
противоречий — компромиссное, наиболее подходящее решение,
которое, не отдавая предпочтения какому-нибудь одному факто-
ру, обусловливающему давление отбора, благоприятствует об-
щей приспособленности и, следовательно, затрудняет усиление
6*
84
Глава 3
любой отдельной адаптивной черты. Можно посмотреть на эту
ситуацию и по-другому: хотя каждая адаптация сулит некото-
рые выгоды, она влечет за собой и расходы. Стоимость адапта-
ции может выражаться в энергетических или временных едини-
цах, а может оцениваться и по тем последствиям, которые обна-
руживаются на другом направлении адаптивной стратегии.
5. Единица отбора. Обсуждение вопроса о единице или уров-
не отбора (разд. 3.2.3) имеет важное значение и для понимания
процесса адаптации. Поскольку адаптация есть следствие отбо-
ра, она должна происходить на разных уровнях. Отсюда в свою
очередь можно заключить, что то, что имеет приспособительное
значение на одном уровне, может играть отрицательную роль
на другом. Например, адаптация особи может выражаться в
том, чтобы максимально увеличить репродуктивные показатели
и оставить в следующих поколениях как можно больше своих
потомков. Однако, если каждый из членов популяции будет
успешно придерживаться подобной стратегии, это приведет к
полной деградации среды обитания и гибели всей популяции.
Конечно, это крайний пример и до исчезновения популяции в
действие должны вступить другие механизмы. Однако он пока-
зывает те противоречивые факторы, которые препятствуют раз-
витию идеальной адаптации.
6. Непредсказуемость. Животные обитают в сложных усло-
виях окружающей среды, эксплуатируя ресурсы, неравномерно
распределенные во времени и пространстве. В соответствии с
принципами естественного отбора, сформулированными в нача-
ле главы, животные должны быть как можно лучше приспо-
соблены к условиям среды обитания. Однако эта среда состоит
из других организмов, каждый из которых также стремится
адаптироваться к ней. Процессы адаптации многих организмов
нередко противостоят друг другу (хищник и добыча, конкурент-
ные виды и т. д.). Кроме того, все сообщество может быть под-
вержено действию внешних факторов, таких, как изменения кли-
мата. Все это обусловливает нестабильность системы, в которой
критерий отбора постоянно меняется, удерживая тем самым
процесс адаптации от чрезмерной настройки на определенную
волну и, следовательно, от неизбежного стремления к совер-
шенству.
Адаптация — это экологическое проявление естественного от-
бора. Ограничивающие ее факторы перекрываются с теми, ко-
торые воздействуют на механизмы эволюции. Если адаптация
представляет собой «ожидаемый результат» естественного отбо-
ра, то достижение совершенства таковым не является.
Эволюционная экология
85
3.4. Адаптация как процесс решения задач
Вероятно, вернее и полезнее рассматривать адаптацию как
процесс решения задач (Maynard Smith, 1978; Dunbar, 1982).
Адаптивные признаки — это средства решения (в рамках тех
ограничений, которые обсуждались выше) проблем, поставлен-
ных перед организмом окружающей средой.
«Любой ныне живущий вид, безусловно, располагает готовым
набором решений некоторых основных экологических и биоло-
гических проблем. Он должен иметь гарантированный источник
пищи для пополнения запасов энергии, место для жилья, спо-
соб размножения... Обладание реальным набором таких реше-
ний называется адаптацией» (Dobzhansky, 1974, р. 323, цит. по
Jochim, 1981).
Эта точка зрения на адаптацию совпадает с реалиями био-
логического мира. Она объединяет адаптацию и естественный
отбор, создает практическую основу для эволюционно-экологи-
ческого подхода. Окружающая среда ставит проблему выжива-
ния перед организмом. Естественный отбор воздействует на по-
ведение, морфологию, физиологию и биохимию организма с тем,
чтобы, повышая его репродуктивный успех, решить эту пробле-
му или максимально ослабить ее остроту. Возможные решения
должны быть ограничены всеми упомянутыми выше фактора-
ми, но конечный продукт процесса — это адаптация. Роль эво-
люционно-экологического анализа должна поэтому заключаться
в том, чтобы определить задачи, стоящие перед организмом,
и оценить те решения, которым отбор благоприятствует или ко-
торые он отвергает.
3.4.1. Опасности, которые несет антропоморфизм
Согласно этой точке зрения, адаптация — это стратегия вы-
живания, причем под стратегией понимается набор действий
или признаков, способствующих лучшему решению задач, по-
ставленных окружающей средой. Описание адаптации как стра-
тегии решения задач создает полезную основу для соотнесения
характеристик организма (его адаптивных особенностей) со
свойствами окружающей среды (задачами, стоящими перед ор-
ганизмом).
К сожалению, оба термина — и «стратегия», и «решение за-
дач»— имеют отчетливое антропоморфное звучание. При обыч-
ном употреблении они подразумевают сознательное принятие
решения и выбор цели со стороны особи. Ни один из этих
смысловых оттенков не совместим с той точкой зрения, что ес-
тественный отбор — это слепая сила, а адаптация — результат
случайного взаимодействия с обычными и безликими биологи-
86
Глава 3
ческими силами. Было бы, конечно, неразумно ожидать, что пау-
ки «сознательно преследуют выбранную стратегию» или олень
«пытается решить задачу, как избежать давления со стороны
хищников». Даже если речь идет о гоминидах, было бы неверно
полагать, что они могли на любой конкретной стадии их эволю-
ции осознать поставленные перед ними проблемы, не говоря уж
о принятии подходящего решения. Современные люди с их не-
превзойденными познавательными и аналитическими способно-
стями могут осознать задачу, поставленную перед ними окру-
жающей средой, и выбрать соответствующую стратегию, ио и
это ни в коей мере не означает, что цели, которые они хотят
достичь с помощью своей стратегии, совпадают с «целью», под-
разумеваемой в эволюционной биологии и означающей репро-
дуктивный успех и дарвиновскую приспособленность. То, что их
стратегии оказывают воздействие на процесс размножения,
а следовательно, имеют эволюционный подтекст, должно быть
предметом особого разговора.
Использование терминов, которые четко обозначают наме-
ренное достижение цели и имеют смысловую нагрузку, слишком
много значащую для современного человека, на протяжении ря-
да лет служило объектом пристального внимания и критики
(см., например, Alden et aL, 1976). Следует, однако, заявить со
всей определенностью, что употребление таких терминов, как
«стратегия» и «решение задач», вполне допустимо в рамках
эволюционной биологии, так как они дают хорошие аналогии
тем способам, посредством которых действует естественный от-
бор и достигается адаптация. Кроме того, в этих терминах за-
ключена очень выразительная аналогия, и главное, они пред-
ставляют собой чрезвычайно полезные стенографические обозна-
чения для краткой характеристики конечного результата дея-
тельности всех факторов, вносящих свой вклад в эволюцию и
рассмотренных в этой главе.
Взгляд на эти термины как введенные по аналогии и пред-
ставляющие собой удобные стенографические символы был хо-
рошо обоснован Даямондом (Diamond, 1977, р. 296—297) для
понятия «стратегия».
«Стратегия состоит из трех элементов: действия мыслитель-
ных процессов, заранее поставленной цели и взвешивания аль-
тернативных способов достижения этой цели с тем, чтобы решить,
какой из них самый подходящий. В последнее время экологи и
биогеографы сочли полезным говорить и писать о животных и
растительных видах таким образом, как если бы те тоже при-
держивались определенных стратегий. Например, современный
эколог, нимало не смущаясь, рассуждает: «Вид птиц в условиях
нестабильных биотопов должен придерживаться стратегии быст-
рого размножения». Или: «Высокая величина индекса, опреде-
Эволюционная экологии
87
ляемого как время поиска, отнесенного ко времени преследова-
ния, характеризует оптимальную стратегию добычи пищи для
хищника, специализирующегося в процессе охоты на крупной
добыче, которую трудно поймать».
Вплоть до недавнего времени такие антропоморфные рас-
суждения о растениях и животных считались наивными и даже
вредными. Сейчас ясно, что использование термина «стратегия»
представляет собой удобный стенографический прием для опи-
сания как эволюционных, так и поведенческих явлений. На эво-
люционном уровне виды и особи различаются по своей адаптив-
ности; адаптация некоторых видов (или особей) позволяет им
оставить больше потомков по сравнению с другими в сходной
ситуации, поэтому первые в этой ситуации превзойдут по чис-
ленности последних. В связи с этим можно сказать, что эволю-
ция имеет цели, такие, как долговременное выживание или боль-
шое число выживших потомков. Вероятность достижения этих
целей есть функция биологических адаптаций, которые являются
аналогами альтернативных способов достижения цели. Сравни-
тельная оценка альтернативных стратегий осуществляется есте-
ственным отбором, а не за счет процессов мышления. Когда мы
говорим, что «вид птиц в условиях нестабильных биотопов дол-
жен придерживаться стратегии быстрого размножения», полу-
чается как бы стенограмма текста вроде следующего: «Виды и
особи различаются по скорости размножения. Чем менее стаби-
лен биотоп, тем выше вероятность локального исчезновения или
смерти организма вследствие случайных факторов. Чем меньше
популяция, тем также выше вероятность локального исчезнове-
ния вследствие случайных факторов. Чем выше скорость раз-
множения, тем короче промежуток времени между прибытием
первых поселенцев в вакантную экологическую нишу и дости-
жением популяций крупного размера, при котором эта ниша
была бы насыщена. Вероятность исчезновения организмов в этот
промежуток времени выше, чем по достижении популяцией
большого размера. Чем длиннее этот интервал и чем меньше
число доступных в данный момент экологических ниш, тем вы-
ше вероятность того, что вид может случайно исчезнуть одно-
временно из всех экологических ниш. Поэтому в нестабильном
биотопе естественный отбор ведет к предпочтительному выжи-
ванию видов с высокой скоростью размножения». (Курсив
мой. — Р. Ф.)
Если нам ясна логическая суть этих рассуждений и извест-
но, что такие термины, как «решение задач» и «стратегия»,
употребляются в точном, специальном значении и по аналогии,
тогда эти понятия могут оказаться исключительно ценными.
Действительно, концепция «решения задач» представляет собой
основу анализа, проведенного в последующих главах книги,
88
Глава 3
а определение стратегий выживания — важную цель этого ана-
лиза. В каждой главе сделана попытка выделить экологические
и средовые проблемы, стоящие перед ранними гоминидами и го-
миноидами, затем путем изучения всего размаха ответных реак-
ций в широком сравнительном аспекте идентифицировать и объ-
яснить избранные адаптивные стратегии.
3.4.2. Оптимальность
Рассмотрение адаптации как процесса решения задач имеет
еще и то преимущество, что нуждается в минимальном числе
допущений. Здесь не нужно оговаривать такие понятия, как ста-
бильность или единица более крупного масштаба, чем особь, не
нужно и ожидать совершенства в природе. И все-таки некото-
рые допущения лежат в основе этого подхода. Они согласуются
с принципами, получившими в эволюционной биологии название
принципов оптимальности.
Принцип оптимальности в биологии можно сформулировать
следующим образом:
«Организмы внутри популяции неодинаковы в отношении их
физиологии, морфологии и поведения. Из-за ограниченности ре-
сурсов— пищи, жизненного пространства, брачных партнеров и
т. д. — отбор будет благоприятствовать тем организмам, поведе-
ние и морфология которых повышают их возможности доступа
к ресурсам по сравнению с другими организмами и, следова-
тельно, увеличивают свою производительность по сравнению с
другими. Таким образом, результатом естественного отбора, дей-
ствующего в изменчивой популяции, является увеличение отно-
сительного количества тех особей, поведение которых увеличи-
вает доступ к ресурсам. Иначе говоря, тех, чье поведение отно-
сительно ресурсов может быть названо оптимальным. Поэтому
оптимизация есть ожидаемый результат естественного отбора,
действующего в мире, где ресурсы конечны и выступают в роли
сдерживающего фактора» (Foley, 1985; по Pyke, Pulliam, Char-
nov, 1977).
Отсюда следует, что адаптация — это тенденция к оптимиза-
ции соответствия между поведением организма и средой его оби-
тания. Отбор благоприятствует «оптимальному решению» про-
блем, с которыми сталкивается организм. При анализе эволю-
ции гоминид мы пытаемся понять, почему некоторые их харак-
теристики стали оптимальными решениями специфических про-
блем выживания, стоявших перед ранними гоминоидами и гоми-
нидами. В других своих работах (Foley, 1985) я подробно рас-
смотрел важность и ценность принципов оптимальности для ан-
тропологии. По большей части речь шла о факторах, ограничи-
вающих силу, масштабы и применение принципов оптимально-
Эволюционная экология
89
сти. Эти ограничения аналогичны тем, которые стоят перед
естественным отбором и адаптацией и уже обсуждались выше.
Кроме того, я останавливался на вопросе о том, в какой мере
современные люди вышли из-под давления естественного отбо-
ра и, следовательно, за рамки принципа оптимальности (кото-
рый базируется на дарвиновских предположениях). Поскольку
основное внимание в этой книге сконцентрировано на анализе
гоминоидов и самых первых гоминид, этот вопрос вряд ли име-
ет большое значение при обсуждении полученных результатов.
Стоит подчеркнуть, однако, два аспекта моделей оптималь-
ности. Первое — это понятие о фенотипическом наборе, и вто-
рое — использование допущений оптимальности как основы для
проведения сравнений между альтернативными типами пове-
дения.
Стратегия означает выбор одной из многих альтернатив в
качестве руководства к действию. Как мы видели, в эволюцион-
ной биологии «выбор делает» естественный отбор. Поэтому что-
бы анализ был продуктивным, необходимо знать не просто ре-
альный результат, а все разнообразие возможных альтернатив.
В эволюционной биологии под фенотипическим набором как раз
и понимается разнообразие альтернатив (Maynard Smith, 1978).
Оптимальный фенотип есть тот, которому благоприятствует от-
бор и который сохраняется во времени. Например, при анализе
эволюционного значения внутривидовой борьбы Мейнард Смит
(Maynard Smith, 1976b) охарактеризовал возможное разнооб-
разие альтернативных типов поведения, хотя далеко не все из
них встречались в природе. Следует помнить, например, что
эволюция ритуальных форм схваток, во время которых сопер-
ники имеют минимальный шанс получить серьезное увечье,
происходила на фоне гораздо более опасных альтернативных
типов поведения. Жизнеспособность и стабильность любой
стратегии должна оцениваться в свете возможных альтернатив-
ных стратегий, т. е. на основе фенотипического набора.
Однако недостаточно было бы просто констатировать, что в
этой связи понятие «оптимальный» лишь тогда отражает прин-
цип оптимальности, если при этом рассматриваются другие
альтернативы, включенные в фенотипический набор, а не просто
типы поведения. Из этого следует также, что качество анализа
стратегий зависит от адекватности фенотипического набора. Ес-
ли мы пытаемся объяснить эволюцию адаптивных черт у ран-
них гоминид, нужно, чтобы мы не забыли рассмотреть все раз-
нообразие открывавшихся перед ними альтернатив. Так как, по
определению, они не могли быть развиты и почти наверняка не
оставили следов в ископаемых остатках, возникают серьезные
методологические затруднения. Нам нужно, чтобы мы сумели
оценить, какие еще возможности выбора открывались перед
90
Глава 3
ранними гоминидами. Это можно сделать не с помощью прямых
наблюдений, а путем тщательного использования экологических
и этологических принципов, а также сравнительных данных.
Эволюционный анализ хорош лишь тогда, когда рассматрива-
ются альтернативы эволюционных путей (Foley, 1985). В этой
книге сделана попытка проанализировать те особенности гоми-
нид, которые развились в результате их эволюции, на фоне тех,
которые могли бы развиться.
При использовании принципов оптимальности делается осо-
бый упор на сравнительный метод, оценивается сходство и раз-
личия поведенческих и анатомических особенностей, исходя из
их энергетических затрат, преимуществ и сравнительной цены
выживания. На практике это делается таким образом, чтобы
определить проблемы, стоящие перед организмом, а затем пред-
ложить оптимальное решение — это единственное в ряду многих
альтернатив, которое максимально повышает преимущества и
снижает до минимума затраты. Это чем-то напоминает прогноз,
предсказывающий, что именно должно было получиться в ре-
зультате эволюции. Такой анализ предполагает наличие неко-
торого образца — «идеальной адаптации», — по которому мы
могли бы оценить то, что получилось на самом деле. На самом
деле это способ моделирования действия отбора на организм.
Сравнивая реальный результат с ожидаемым, мы можем оце-
нить, насколько точно были определены разновидности давления
отбора (проблемы, выдвигаемые средой), воздействующие на
организм. Принципы оптимальности являются инструментом для
измерений адаптации и адаптивных стратегий. Вооружившись
этим инструментом, мы вряд ли найдем в раннем плейстоцене
сверхэффективных гоминид, которые принимали бы эволюцион-
но совершенные решения с энергией, достойной спортивных тре-
неров, но зато сможем определить, почему эволюция пошла по
тому, а не иному пути.
3.4.3. Поведенческая экология
Хотя понятия решения задач и стратегии в соответствии с
заключенной в них подходящей смысловой аналогией можно ис-
пользовать для всех аспектов адаптации, они, вероятно, наибо-
лее уместны в сфере поведенческой экологии. Поведение жи-
вотных, их социальная организация, деятельность, связанная с
добыванием пищи, активность при размножении, реагирование
на хищников — все эти особенности в такой же мере подверже-
ны действию естественного отбора, как и морфологические при-
знаки. Поведение животных с большей очевидностью, чем струк-
тура, является стратегией выживания. Кроме того, в случае та-
ких сложных высших организмов, как приматы, прямой ответ
Эволюционная экология
91
на требования окружающей среды дает скорее поведение с его
возможностями к усилению' пластичности, чем структурные осо-
бенности, развивающиеся путем естественного отбора в ряду
поколений. Сюда могут быть включены и процессы принятия ре-
шений, что позволяет рассматривать понятие стратегии как неч-
то большее, чем просто модель-аналог.
Конечно, многие адаптивные изменения, возникшие в про-
цессе эволюции человека, являются поведенческими — развитие
склонности к питанию мясом, поиски пищи вокруг некоторого
центра, основанная на родственных связях социальная система
и т. д. Многие из тех задач, которые встают в процессе анализа
перед палеоантропологами, лежат в сфере поведенческой эко-
логии.
Кребс и Дэвис (Krebs, Davies, 1981) определяют поведенче-
скую экологию как науку о значении поведения для выжива-
ния. Они связывают поведение с экологическими условиями,
в которых обитает то или иное животное, и рассматривают по-
ведение как реакцию на ограничения и требования, которые
создаются этими условиями. Поведенческая экология, взятая в
таком аспекте, в основном касается функций поведения, т. е.
степени повышения уровней выживаемости и воспроизводства
за счет поведения. Именно это связывает поведенческую эколо-
гию с эволюционной биологией в целом. Прямая зависимость
между поведением, экологией и эволюцией подчеркивает в пер-
вую очеред функциональные, а не механистические причинно-
следственные связи (Tinbergen, 1963; Krebs, Davies, 1981).
Например, популяция приматов может образовывать круп-
ные социальные группы. Причины, лежащие в основе этого яв-
ления, можно рассматривать как в функциональном, так и ме-
ханистическом плане. Функциональная причина образования
групп состоит в том, что при таком образе жизни особь имеет
больше шансов не стать добычей хищников. Это иадеет сущест-
венное значение для выживания. Механистическая причина оп-
ределяется тем, как особи узнают друг друга и взаимодейству-
ют между собой, устанавливая социальные связи, которые и
объединяют их. Функциональная интерпретация зависит от эво-
люционных принципов, таких, как уже рассмотренный принцип
оптимальности, а также от экологических принципов, определя-
ющих характер среды, в которой живет организм. В анализе,
которому посвящены следующие главы этой книги, основное
внимание уделяется функциональному аспекту эволюционной
биологии.
Поиски причин развития, связанных с отбором признака, из-
давна занимали центральное место в эволюционной биологии.
В той области знания, где изучается поведение животных, эта
проблема выступила на первый план прежде всего в связи с
92
Глава 3
ростом социобиологии, а сравнительно недавно — в связи с рас-
ширенным использованием принципов оптимальности в эволю-
ционной биологии.
Как направление в биологии, социобиология возникла на ос-
нове классической этологии в связи с необходимостью более
строгого применения теории Дарвина (Wilson, 1975). В ней бо-
лее подробно рассматриваются взаимоотношения между родст-
венными особями (родственный отбор) и объясняются причины
альтруизма. Непосредственная реакция на книгу Уилсона сра-
зу после ее публикации состояла в распространении принципов
поведенческой экологии на людей и обсуждении природы гене-
тического детерминизма. Эти дискуссии часто приобретали по-
литическую и полемическую окраску. В дальнейшем, когда оп-
поненты перешли к детальному рассмотрению вопросов научной
значимости, дебаты потеряли свою остроту.
Многие из этих вопросов уже обсуждались в данной главе:
мы говорили о том, является ли адаптация ожидаемым резуль-
татом естественного отбора, а также об уровне адаптации и о
том, могут ли другие процессы, помимо естественного отбора,
приводить к адаптивным изменениям. Изложенные выше аргу-
менты вполне можно использовать при анализе поведенческих
особенностей, а не только физиологических. Однако что касает-
ся поведения, то здесь есть одна проблема, вызывающая неко-
торые затруднения, а именно связь между обучением и генети-
ческими основами поведения.
Как уже было показано, для того чтобы признак попал под
действие отбора, он должен оказывать некоторое воздействие
на репродуктивный успех и иметь генетическую основу. То, что
поведение влияет на размножение, почти аксиома, но вопрос об
уровне генетического контроля представляется гораздо более
спорным, особенно в отношении высших животных и человека.
Одна из крайних точек зрения состоит в том, что каждый тип
поведения контролируется одним геном, осуществляющим пря-
мой контроль. Например, Ротенбюлер (Rothenbuhler, 1964) су-
мел показать, что у пчел (Apis mellifera), которые соблюдали
«гигиену» (т. е. удаляли мертных личинок из улья), эта дея-
тельность находилась под двойным генетическим контролем и
включала два типа поведения — откупоривание ячейки и удале-
ние личинки. Проведя ряд экспериментов по скрещиванию, Ро-
тенбюлер (там же) доказал, что каждый из этих типов поведе-
ния находится под особым генетическим контролем. В данном
случае генетический контроль был весьма прямым и очень спе-
цифическим; сложные типы поведения возникали при добавле-
нии новых генов к поведенческому набору.
Другая крайность — это признание того, что в основе пове-
дения лежит научение и что гены им не управляют. Такое объ-
Эволюционная экология
93
яснение, как нередко считают, вполне подходит для большин-
ства видов человеческой деятельности (см., например, обсужде-
ние этого вопроса в работе Kaplan, 1978). Доказательством из-
менения поведения в результате научения служат нередко на-
блюдаемые примеры того, как поведение животных одного и то-
го же вида может меняться в зависимости от тех конкретных
условий, в которые попадает особь. В качестве классического
примера приматологи часто приводят привычку мыть клубни
батата перед едой, получившую распространенность в популяции
японских макаков (Масаса fuscata). Это поведение, заключав-
шееся в отмывании клубней от песка, было «изобретено» одной
особью, а затем распространилось путем подражания среди всей
популяции (Kawai, 1965).
Между этими двумя крайностями, однако, лежит целый ряд
других возможностей. Маловероятно, чтобы научение играло
большую роль у животных с быстрой сменой поколений, живу-
щих в неизменных и прогнозируемых условиях. С другой сторо-
ны, маловероятно, чтобы у животных с социальным поведением
и крупными размерами тела, каковыми являются большинство
млекопитающих, генетический контроль сам по себе был бы до-
статочным для реагирования на то широкое разнообразие усло-
вий, в которых может оказаться особь. Обучение и генетический
контроль должны, как можно ожидать, взаимодействовать друг
с другом. Лучшая модель подобной взаимосвязи — это, навер-
ное, игровая ситуация. Даже самые простые игры содержат
большое количество возможных решений и действий или ходов.
Было бы чрезвычайно утомительно рассказывать человеку, как
играть в ту или иную игру, объясняя каждый ход при всех воз-
можных обстоятельствах. С другой стороны, чтобы человек на-
учился играть сам путем одного экспериментирования, потребу-
ется очень много времени. Лучший способ обучения новой игре
состоит в том, чтобы дать краткое описание основной задачи
игры и взаимоотношений между игроками, изложив его в виде
нескольких правил. Зная эти правила, каждый игрок может вы-
работать оптимальное решение для каждой ситуации. Связь
между обучением и генами у высших организмов, в том числе и
у людей, носит, вероятно, сходный характер. Наследственность,
несомненно, поставляет основной набор правил, с помощью ко-
торых вырабатываются поведенческие стратегии. Эти правила,
однако, являются не шаблонами для копирования, а критерия-
ми, на основе которых принимаются решения. Зная их, особь в
состоянии начать «играть», но может также обучаться на основе
своего жизненного опыта (Passingham, 1982; Layton, Foley в
печати).
Эта игровая модель удовлетворяет основным условиям дей-
ствия отбора (в смысле наличия генетического компонента),
94 Глава 3
признавая в то же время пластичность поведения как одну из
его обычных характеристик. Отбор будет действовать не на са-
мо поведение, а на те правила, которыми оно определяется. Та-
кое решение имеет преимущество в том отношении, что избега-
ет генетического детерминизма, усложнившего развитие социо-
биологии, не отказываясь от представлений о той важной роли,
которую может играть поведение в эволюции организма и его
адаптации. Это особенно важно при изучении эволюции гоми-
нид, где, как мы можем без тени сомнения утверждать, пове-
денческие адаптации имели огромное значение, а типы поведе-
ния вряд ли находились под простым и прямым генетическим
контролем.
Глава 4
ПУТИ В ПРОШЛОЕ
Предыдущая глава, вероятно, поразила читателя почти пол-
ным отсутствием ссылок на гоминид и стремлением избежать
обсуждения проблем, связанных с ископаемыми остатками ор-
ганизмов и археологическими данными — единственными пря-
мыми свидетельствами древнейших эволюционных стадий. Это
выглядит весьма странно для книги об эволюции человека, но
было сделано сознательно, поскольку одно из центральных по-
ложений автора состоит в том, что процесс эволюции человека
нуждается в проверке с помощью общих эволюционных и спе-
цифических предположений о природе человека как вида.
Важно, однако, чтобы принципы, сформулированные в пре-
дыдущей главе, можно было проверить и воспользоваться ими
при анализе имеющихся данных. Для этого необходимо исполь-
зовать весь арсенал применяемых в палеоантропологии подхо-
дов. В связи с этим в данной главе обсуждаются некоторые во-
просы, относящиеся к реконструкции прошлого.
4.1. Палеоантропология
Можно считать, что палеоантропология как таковая возник-
ла в результате слияния двух отдельных ветвей антропологии:
физической или, биологической, особенно той ее части, которая
посвящена изучению ископаемых гоминид, и археологии, изуча-
ющей поведение человека на основе остатков материальной
культуры. Несмотря на очевидную общую цель — познание ран-
них стадий эволюции гоминид, — обе эти дисциплины были ис-
торически разобщены, различаясь как предметом исследования,
так и теоретической ориентацией. В последнее время, однако,
в палеоантропологии набирает силу комплексный междисципли-
нарный подход. Это главным образом связано с более широким
проникновением биологических и в первую очередь экологиче-
ских концепций в сферу изучения ранних гоминид. Хотя совре-
менные палеоантропологические исследования в большинстве
своем характеризуются общими целями и предпосылками, в их
основе лежат разные подходы (рис. 4.1).
96
Глава 4
4.1.1. Эволюция и ископаемые остатки
Хотя первые попытки построить модель эволюции человека
были предприняты Дарвином (Darwin, 1871) и Геккелем (Haec-
kel, 1876) еще до открытия каких-либо ископаемых гоминид и,
следовательно, независимо от них, остатки ранних представите-
лей этого семейства дают основной материал при изучении эво-
люции человека. Одна за другой находки ранних гоминид в
Европе, Юго-Восточной Азии, Китае и Африке становились
предметом ожесточенных споров (Reader, 1981). Окаменелости
(ископаемые остатки организмов)—это, строго говоря, мине-
рализованные остатки тканей растений и животных геологиче-
ского прошлого. В случае животных они представлены главным
образом костями и зубами, которые претерпели различные физи-
ческие и химические изменения после смерти их владельцев. Ин-
формация, получаемая при изучении ископаемых остатков, в ос-
новном анатомического характера. Большая часть ранних работ
по морфологии первых гоминид носила типологический харак-
тер; скелетные признаки интересовали исследователей как мар-
керы в процессе эволюции человека, а не как факторы, повыша-
ющие или понижающие вероятность выживания изучаемых осо-
бей. Этот подход можно назвать эволюционным, подразумевая
под эволюцией филогению или реконструкцию таксономических
соотношений, а не продукт естественного отбора или адаптив-
ные модели ранних гоминид. При таком анализе всех гоминои-
дов можно расположить в виде прямой линии (рис. 4.2), на од-
ном конце которой находятся человекообразные обезьяны, на
другом — современные люди, а между ними вдоль всей линии
располагаются ископаемые существа в соответствии с тем чис-
лом «человеческих баллов» (т. е. черт анатомического сходства
с современными людьми), которые им удалось собрать. Споры
между исследователями касались в основном добавления или
вычитания баллов с последующим перемещением соискателя в
ту или другую сторону по линии эволюции. Как неандертальцы,
Физическая антропология + Археология палеолита
Палеоантропология
Изучение
ископаемых
остатков
Сравнительная
биология
(приматы)
Сравнительное исследование
современных
человеческих популяций
Изучение
археологических
материалов
Рис. 4.1. Схематическое изображение составных частей палеоантропологии
Пути в прошлое
97
Homo sapiens sapiens-----►
Архаический ----►
Homo sapiens
Неандертальцы------------►
Homo erectus-------------►
Homo habilis-------------►
Australopithecus---------►
Ramapithecus------—------►
Шимпанзе ----------------►
Г орилла-----------------►
(Человекоподобный
Прогрессивный
Обезьяноподобный
Примитивный
Рис. 4.2. «Классическая» однолинейная модель эволюции человека, представ-
ляющая собой простейший путь от человекообразных обезьян к человеку
так и австралопитеки то выигрывали, то проигрывали в резуль-
тате частых перетасовок в недрах антропологических музеев.
Ограничения в интерпретации ископаемого материала были,
вероятно, вызваны двумя главными факторами. Первый из
них — неопределенность датировок и связанная с этим необхо-
димость использования самих ископаемых остатков в качестве
хронологических маркеров, вследствие чего наблюдать незави-
симое от времени варьирование морфологических особенностей
гоминид по методическим причинам становилось возможным
лишь на самом поверхностном уровне. Таким образом, исследо-
вания часто просто подтверждали существовавшие ранее пред-
положения. Второй фактор состоял в представлении об одноли-
нейном характере эволюции. В недавнем прошлом ученые на ос-
новании различий в поведении и морфологии между современ-
ными людьми и всеми остальными видами ныне живущих при-
матов приходили к выводу, что такое расхождение существо-
вало всегда. Уникальность современного человека приводила к
убеждению в уникальности его предков. В то время как в дру-
гих таксонах отмечалась адаптивная радиация многих форм от
близкородственных предков, гоминиды, казалось, эволюциониро*
7—367
98
Глава 4
вали в одном ряду форм, связанных между собой прямыми от-
ношениями типа предок — потомок. Представления об одноли-
нейном характере эволюции и гипотеза одного вида долго тор-
мозили аналитические исследования в палеоантропологии.
В последние годы благодаря прогрессу науки, эти препятст-
вия были преодолены. Развитие новых методов датировки, о ко-
торых говорилось в гл. 2, дало возможность датировать иско-
паемые остатки независимо от их морфологии. Хотя датировка
многих находок, таких, например, как южноафриканские авст-
ралопитековые, все еще оставляют желать лучшего, сейчас уже
ясно, что различные формы гоминид жили одновременно (Ho-
well, 1978). Следовательно, необходимо учитывать не только
временные связи между предками и потомками, но и взаимоот-
ношения иного типа. При наличии хорошо датированных иско-
паемых остатков стало возможно использовать аналитические
методы, а не только типологические. Ископаемые материалы
можно было теперь связать с другими организмами, с функци-
ей и средой обитания, а не только с тем или иным периодом
времени. Современные исследования по эволюции человека, ос-
нованные на изучении окаменелостей, пользуются теперь самы-
ми разнообразными методами, начиная от традиционной био-
метрии до микроскопического и химического анализа, чтобы
найти связь между формой гоминид и всем, что могло наложить
отпечаток на их кости — будь то мягкие ткани особи или попу-
ляционные отношения, в которых она жила, или же биотические
и физические условия существования популяции.
Непосредственное изучение ископаемых остатков гоминид
должно всегда оставаться в центре эволюционных исследований
по той простой причине, то только эти остатки позволяют прямо-
судить о действии отбора в прошлом на примере такой формы,
у которой можно точно определить время существования.
4.1.2. Эволюция и приматы
Если ископаемые материалы позволили создать классиче-
скую базу данных для исследований ранних гоминид, то совре-
менные приматы обеспечили ученым соответствующий материал
для сравнительных исследований. Основываясь на чертах сход-
ства и различия между человеком и другими современными
приматами, Карвин и др. предприняли свои исследования по
эволюции человека, также ставшие классическими. Однако пе-
реход от типологического анализа к функциональному, о кото-
ром уже упоминалось выше, еще более повысил роль примато-
логии в палеоантропологии. Модели эволюции человека теперь
основывались либо на прямых аналогиях с приматами, особен-
но павианами и шимпанзе (см., например, De Vore, Washburn,
Пути в прошлое
99
1963), либо на экстраполяциях общих закономерностей, кото-
рым подчиняется изменчивость морфологии и поведения прима-
тов (см., например, Martin, Мау, 1981). Значительное расшире-
ние масштабов полевых исследований поведения приматов за
последние два десятилетия позволяет намного полнее предста-
вить поведенческие возможности ранних гоминид.
Первоначально приматология, находясь на службе у палео-
антропологии, занималась «оживлением» ископаемого материа-
ла, используя для этого многочисленные и часто противоречи-
вые по своим результатам наблюдения на приматах. Теоретиче-
ской базы, позволявшей определить, какие особенности поведе-
ния современных обезьян можно связать с гоминидами, а ка-
кие— нет, фактически не существовало. Обезьяны часто слу-
жили всего лишь зеркалом, в котором палеоантропологи могли
найти отражение своих представлений о ранних гоминидах. Рас-
ширение рамок сравнительных исследований в приматологии
(см., например, Clutton-Broch, Harvey, 1977, а, b) наряду с бо-
лее четким использованием теории эволюции привело к сущест-
венному улучшению ситуации. Приматологические экстраполя-
ции, как теперь утверждают, должны быть связаны с отбором.
Тип поведения, скажем дележ пищи, обнаруженный у некото-
рых обезьян, нельзя положить в основу модели поведения ран-
них гоминид, если не учитывать той ситуации, в которой он ны-
не встречается и приобретает адаптивный смысл. Понимание
того, что поведение зависит от ситуации, существенно проясни-
ло роль сравнительноприматологических исследований в палео-
антропологии. Особенно важное значение в этой связи имело
развитие и более четкое использование теоретических исследо-
ваний в такой области знания, как изучение поведения живот-
ных (см., например, Wilson, 1975; Dawkins, 1976; Maynard
Smith, 1976, a, b, 1983; Krebs, Davies, 1981, 1984; King’s College
Sociobiology Group, 1982).
Можно выделить три основных приема при использовании
приматологических данных в исследованиях ранних гоминид.
Первый из них —это «видовой модельный подход». Вид обезь-
ян берется в качестве модели и с некоторыми модификациями
используется для реконструкции жизни ранних гоминид. Основ-
ные «модельные» виды — шимпанзе и павианы (Jolly, 1970;
Dunbar, 1976; Tanner, 1981; Zihlman, Cronin et al., 1978; Johnson
1981; МмНепгу, Corruccini, 1981; Zihlman, Lowenstein, 1983). Вы-
бор определенного вида современных приматов в качестве моде-
ли объясняется либо филогенетическим сходством с ранними
гоминидами (в случае шимпанзе), либо предполагаемым сходст-
вом условий жизни (в случае павианов), либо и тем и другим
(в случае некоторых популяций шимпанзе, живущих в саван-
не). Возможен и другой вариант — выделение ключевой особен-
100
Глава 4
Рис. 4.3. Относительные размеры зубов у представителей двух современных
родов павианов (коренные зубы приведены к одному и тому же масштабу).
Гелада (Theropithecus) (Л), имеет относительно крупные коренные и ложно-
коренные зубы и небольшие резцы и клыки. В противоположность этому Papio
(Б) имеет относительно крупные передние зубы. По утверждению Джолли
(Jolly, 1970), подобное строение зубов у гелады есть результат питания «мел-
кими пищевыми объектами»
ности, общей как для «модельного» вида современных обезьян,
так и для ископаемых гоминид. Например, по утверждению
Джолли (Jolly, 1970), гелады сближаются с ранними гоминида-
ми, особенно австралопитековыми, за счет редукции передних
зубов и увеличения задних (рис. 4.3). Он показал, что у гелады
это означает адаптацию к питанию мелкими пищевыми объек-
тами, в частности семенами. Отсюда Джолли делает вывод, что
ранние гоминиды, возможно, имели сходный пищевой рацион.
Такие модели при всей их информативности имеют один серь-
езный недостаток: не учитывают того, что два вида теоретиче-
ски не могут быть одинаковыми, иначе они не были бы двумя
видами. Модель должна принимать во внимание не только чер-
ты сходства, но и различия.
Второй, возможно, более обычный подход состоит в анали-
зе свойств. Обычно это заключается в том, что перечисляют ха-
рактерные особенности человека и затем констатируют разли-
чие— или отсутствие — их аналогов у приматов (например, та-
кие типы кооперации, как альтруизм и дележ пищи, а также
поедание мяса, язык и коммуникации). Если они встречаются у
обезьян, то помогают представить, как и в какой ситуации функ-
ционировали некоторые характеристики у гоминид. С другой
стороны, их отсутствие должно указывать на какую-то ключе-
вую особенность, отличающую человека от остальных приматов.
В течение многих лет такие ключевые особенности одна за дру-
гой предлагались на роль рубежа, непреодолимого для обычных
Пути в прошлое
101
приматов (использование орудий, охота, язык и т. д.), но при
проведении более детальных исследований чаще всего оказыва-
лось, что по крайней мере шимпанзе способны этот рубеж пре-
одолеть (например, Goodall, 1963). Тот факт, что уникальность
для современного человека многих его характеристик оказыва-
ется эфемерной перед лицом бурно развивающихся полевых ис-
следований поведения обезьян, существенно дискредитирует
этот подход. Его можно критиковать и за те несерьезные объяс-
нения, с помощью которых пытаются «почем зря» охватить все
возможные проблемы. Так, наличие какой-либо особенности го-
минид у других приматов рассматривают как аргумент в пользу
того, что она имелась и у ранних гоминид; отсутствие же этой
особенности у современных приматов якобы может служить до-
казательством ее первостепенной важности для ранних гоминид.
Третий подход также носит сравнительный характер. Широ-
кое сопоставление различных признаков у приматов приводит к
выявлению общих закономерностей. Гораздо плодотворнее экст-
раполировать не отдельные свойства, а всю модель на основе
некоторых переменных. Так, Клаттон-Брок и Харви (Clutton-
Brock, Harvey, 1977 а, Ь) на основе систематических обобщений
большого количества данных об адаптации приматов построили
интересные модели взаимосвязей между важными экологически-
ми переменными. Вслед за ранними работами Милтона и Мея
(Milton, Мау, 1975) они, например, показали соотношение раз-
меров тела приматов с величиной занимаемого ими ареала.
С помощью этой корреляции можно оценить величину ареалов
ранних гоминид на основе размеров их тела и сопоставить полу-
ченные результаты с другими источниками информации (Foley,
1984b). Этот подход отличается особой продуктивностью, так
как он зависит не от специфических характеристик «модельно-
го» вида, а от эмпирических закономерностей, установленных с
помощью наблюдений. Таким образом, здесь мы имеем дело с
моделью высшего порядка (Clarke, 1968) и примером правиль-
ного использования аналогий, когда экстраполируются во вре-
мени не отдельные типы поведения, а регулирующие их законо-
мерности.
Несколько иной пример использования данных по ныне жи-
вущим приматам для изучения эволюции гоминид дают молеку-
лярные или биохимические часы. Как мы уже объяснили в гл. 2
(разд. 2.1.3), сходство и различие биохимических признаков,
относящихся к белкам, иммунным реакциям и последовательно-
сти ДНК, можно использовать для измерения филогенетических
расстояний точно так же, как используются для этого морфоло-
гические признаки. Этот метод широко применяется для изуче-
ния эволюции приматов и уже оказал сильное влияние на со-
временную трактовку ископаемых приматов и эволюции челове-
102
Глава 4
ка (Sarick, 1983; Pilbeam, 1984). Он основывается на двух
главных предпосылках. Первая состоит в том, что сходство на
биохимическом уровне отражает последовательность диверген-
ции ныне живущих таксонов в процессе эволюции. Например,
•степень иммунологических и биохимических различий между
всеми гоминоидами и низшими узконосыми обезьянами боль-
ше, чем между видами внутри каждой группы (см. рис. 2.4).
Это означает, что гоминоиды и низшие узконосые разошлись
раньше, чем обособился хоть один из видов, входящих в состав
этих групп. Это предположение ныне является общепризнан-
ным. Другая предпосылка заключается в том, что скорость био-
химических изменений остается постоянной, поскольку рассмат-
риваемые признаки не имеют адаптивного значения, а значит,
не находятся под контролем отбора и зависят лишь от скоро-
сти мутаций. Если это справедливо, то можно измерить молеку-
лярные различия, составив шкалу для определения не только
последовательности дивергенций, но и их датировки. Данные о
молекулярной и биохимической эволюции могут быть поэтому
использованы в качестве часов. Это предположение сначала
подвергалось серьезной критике (Uzell, Pilbeam, 1971). Многие
из первых оценок времени дивергенции, давших очень поздние
цифры, были впоследствии пересмотрены с учетом таких фак-
торов, как время поколения (Sarich, Wilson, 1973), В настоя-
щее время молекулярные часы широко применяются и получили
всеобщее признание как ориентир, дающий представление о по-
следовательности эволюционных событий. Однако они ничего не
могут сказать нам ни о предшествующих эволюционных стади-
ях (Pilbeam, 1984), ни о тех генеалогических линиях, от кото-
рых не осталось живых потомков.
Ныне живущие приматы — важный источник информации о
ранних гоминидах; основные этологические закономерности и
наблюдения становятся все более серьезным элементом всякого
исследования в этой области. Однако в любом случае при ис-
пользовании модели-аналога, необходимо знать истинную при-
роду того, что лежит в основе аналогии, чтобы оценить ее до-
стоверность.
4.1.3. Современные человеческие популяции
Поскольку продуктом эволюции гоминид является современ-
ное человечество, есть основания утверждать, что на основе дан-
ных о популяциях ныне живущих людей можно реконструиро-
вать более ранние фазы эволюции. Основной источник такого
рода данных — это те популяции, в которых люди живут за
счет охоты и собирательства. Последние два вида деятельно-
сти— это такая адаптация, при которой популяции существуют
Пути в прошлое
103.
за счет использования имеющихся в природе ресурсов, практи-
чески не контролируя их воспроизводство, поведение или рас-
пределение (Foley, 1984). Именно сходство этой ситуации с те-
ми условиями, в которых жили ранние гоминиды, и составляет
основу для сравнения.
Аргументы в пользу этнографических аналогий или моделей
неоднократно приводились в археологической литературе. Не-
которые из них были наполнены утонченным философским
смыслом и касались природы аналогий в науке вообще (Binford,
1981; Hodder, 1982), другие концентрировали внимание на тех-
нических проблемах, связанных с природой археологических дан-
ных (Binford, 1967; Yellen, 1977). Рассматривался также вопрос
об исторической специфике сохранившихся групп охотников-
собирателей (Lee, DeVorce, 1968, рр. 4—5; Hayden, 1981). Со-
временные человеческие популяции обитают исключительно на
таких территориях, которые в настоящее время малопригодны
для сельского хозяйства. Более того, их жизнь далека от жизни
настоящих охотников и собирателей, где царит конкурентная
борьба человека с другими видами; они живут в мире, где
условия диктуются производителями пищи. Это отражается во
многих чертах их поведения и экологии.
При переходе к рассмотрению ранних гоминид, однако, воз-
никает гораздо более серьезная проблема: в то время как охот-
ники-собиратели позднечетвертичного1 времени относятся к то-
му же подвиду, что и современные люди, об их более древних
предшественниках этого сказать нельзя. Использование данных
по современным охотникам и собирателям в качестве модели
для реконструкции образа жизни ранних гоминид предполага-
ет нарушение таксономических границ и вследствие этого под-
падает под те же ограничения, что и уже обсуждавшиеся при-
матологические модели. Следовательно, должны быть разрабо-
таны приемы, которые позволили бы определить достоверность
той или иной аналогии, а также возможность распространения
на другие виды гоминид.
Обычно применяется следующая методика: сначала находят
основные или универсальные характеристики охотников-соби-
рателей, устанавливают их ключевую роль в адаптации и пове-
дении людей, а затем ищут следы их в ископаемых остатках и
археологических материалах. К подобным важнейшим характе-
ристикам относятся использование в пищу мяса, дележ пищи,
мобильность и т. д. Эта процедура дополняет те приемы, кото-
рые используются в приматологических моделях. Однако и в
этом случае может иметь место самореализация. Во-первых,
1 В настоящее время четвертичный период чаще называют антропогено-
вым. — Прим, перев.
104
Глава 4
здесь фактически не рассматриваются адаптивные и связанные
с отбором альтернативы — обязательный элемент любого эволю-
ционного анализа. Если в прошлом обнаруживались характери-
стики, аналогичные тем, которые имеются у нынешних охотни-
ков и собирателей, то делался вывод о сходстве поведения в
древних и современных популяциях; при этом практически не
анализировались другие причины, которые могли привести к
сходным типам поведения (Hill, 1984). При таком подходе мо-
жет показаться, что в далеком прошлом Земля была заселена
охотниками и собирателями, которые мало чем отличались от
своих современных собратьев. Такой взгляд вряд ли приблизит
нас к решению проблемы гоминид, которые отличались по пове-
дению и от современных обезьян, и от современных людей.
Чрезмерное увлечение моделями охотников-собирателей приве-
ло к представлению об эволюционной и экологической стабиль-
ности в первобытной истории человечества. Это сильно повлия-
ло на палеоантропологические исследования последних лет. По-
пытки доказать существование древнейших охотников-собирате-
лей в плиоцене и плейстоцене Восточной Африки (Isaac,
1978 а, Ь) привели к тому, что утвердилось представление о го-
минидах среднего и позднего плейстоцена как уже вполне сло-
жившихся охотниках и собирателях; это в свою очередь серьез-
но подкрепляло точку зрения, согласно которой неандертальцы
существенно не отличались от современного человека (см. глав-
ным образом Brace, 1964; Brose, Wolpoff, 1971). Сегодняшние
споры вокруг интерпретации плиоценового и плейстоценового
археологического материала (Binford, 1981, 1984; Bunn, 1981;
Gamble, 1981; Isaac, Crader, 1981; Potts, Shipman, 1981; Isaac,
1983; Pottts, 1984a), а также недавний анализ скелетных остат-
ков неандертальцев, проведенный Тринкаусом (Trinkaus, 1982),
вновь подняли вопрос о том, насколько древние гоминиды, осо-
бенно предшествующие Homo sapiens, отличались от современ-
ных охотников-собирателей и встречались ли у них такие фор-
мы поведения и организации, которые не имеют современных
аналогий.
Несмотря на все проблемы, связанные с использованием
данных по современным охотникам и собирателям в качестве
средства для понимания прошлого, это направление по-прежне-
му остается неотъемлемой частью палеоантропологических ис-
следований. При правильном использовании оно позволяет про-
никнуть в глубь изучаемых процессов, и, кроме того, эти охот-
ники и собиратели представляют один из полюсов эволюцион-
ной изменчивости, с которой антропологам приходится иметь
дело. Поскольку ныне живущие человеческие популяции оста-
ются единственным источником сведений об адаптации, вклю-
чая данные по технологии и материальной культуре, они долж-
Пути в прошлое
105
ны сохранить свою ведущую роль. Свидетельства в пользу адап-
тации древнейших людей черпаются в основном из остатков их
изделий, поэтому так важно понять связь между отработанным
материалом, технологическим процессом и поведением. Эта
связь лежит в основе этноархеологии, в которой изучение охот-
ников и собирателей играет главную роль (Jellen, 1977; Bin-
ford, 1978 а, b, 1979, 1980; R. Gould, 1980). Более того, охотни-
ки и собиратели уже давно занимают важное место в дискусси-
ях о роли окружающей среды в формировании организационной
структуры человеческого общества (Forde, 1934; Lee, DeVorce,
1968) и таким образом выполняют роль связующего звена меж-
ду биологически ориентированными аспектами палеоантрополо-
гии и другими разделами антропологии.
4.1.4. Археология ранних гоминид
Археология ранних гоминид во время плейстоцена — это ар-
хеология древнего каменного века, или палеолита. С тех пор
как каменные орудия были впервые признаны остатками прош-
лых популяций, их распространение и морфологические особен-
ности стали использоваться в качестве свидетельств о поведе-
нии древнейших людей. Благодаря своим способностям к изго-
товлению орудий, гоминиды создали собственную палеонтологи-
ческую летопись, независимую от их телесных остатков. Из на-
блюдений за ее записями — в виде морфологии изделий, их про-
странственного распространения, смены технологических прие-
мов с течением времени — возникают факты, на которых основа-
ны представления об адаптации и поведении ранних гоминид.
Вообще говоря, археологические исследования ранних гоми-
нид характеризуются двумя подходами: типологическим и пове-
денческим. В прежних исследованиях по археологии плейстоце-
на преобладала типология каменных орудий. Исходя из предпо-
ложения, что форма изделия отражает стилистические вкусы
тех, кто это изделие изготовил, их мастерство или намерения,
что в свою очередь обусловлено культурными пристрастиями
или способностями гоминид, утверждалось (хотя и не прямо),
будто типологическая изменчивость связана с пространствен-
ными и временными особенностями этно-культурных популяций.
Эти популяции можно было в свою очередь классифицировать в
соответствии с различными эволюционными схемами. Реально
использованные методы реконструкции указанных особенностей
сильно варьировали, начиная от использования «типовых изде-
лий» и до изощренного метрического и статистического анализа
(Bordes, 1961; Clarke, 1968). Реальные цели, которые ставили
перед собой исследователи, были, однако, довольно однообраз-
ными: характеристика древнейших культур в пространстве и
106
Глава 4
времени, анализ изменений орудий с учетом миграции, распро-
странения и внутренней эволюции. Были разработаны и новые
методы, но основная цель по-прежнему сводилась к описанию
культур различных групп гоминид (Isaac, 1971 а). Внутри этих
групп адаптивное поведение, по мнению исследователей, почти
ле менялось.
Более четкий интерес к поведению гоминид, выходящий за
пределы повествовательного описания или дополняющий этот
подход, возникает в связи с развитием в 1960-х гг. анализа ка-
менных орудий. В СССР Семенов (Semenov, 1964) разработал
функциональную интерпретацию каменных орудий путем изу-
чения их микростертости, в то время как в США Л. Р. Бинфорд
и С. Р. Бинфорд (Binford, Binford, 1966) пересмотроели трактов-
ку традиционных типологических частот, исходя из функции,
а не стиля орудий. Значение этих исследований заключалось в
том, что каменные орудия рассматривались в первую очередь
в связи с их предполагаемым использованием, а не просто как
случайные носители культурной информации. Каменные орудия
должны были способствовать выживанию гоминид, и поэтому
их следует рассматривать с точки зрения того, насколько они
удовлетворяли своему предназначению. Помимо упомянутых из-
менений, в изучении каменных орудий наметились и другие про-
цессы, отражающие общую тенденцию к тому, чтобы сконцент-
рировать все внимание на поведении и сделать анализ каменных
орудий одним из средств к его познанию.
Эту тенденцию отражает, в частности, все возрастающая
роль экологии в интерпретации археологических данных. Цент-
ральное направление археологического анализа сместилось на
изучение условий археологической стоянки на микроуровне,
в локальном и региональном масштабах (Vita-Finzi, Higgs,
1972), а также на изучение органических остатков, обнаружен-
ных вместе с каменными орудиями. С увеличением базы данных
расширялся и круг вопросов, на которые пытались найти от-
вет; теперь сюда включались вопросы о связях между древней-
шими популяциями и окружающей средой — другими словами,
вопросы по эволюционной экологии ранних гоминид. Нужно осо-
бенно отметить работы Глинна Айзека и Льюиса Бинфорда с
сотрудниками, которые разработали и использовали целый на-
бор технических методов, чтобы ответить на ряд специфических
вопросов о поведении ранних гоминид, не удовлетворяясь такой
ситуацией, когда круг вопросов определяется только техниче-
скими приемами (Isaac, 1971,а, b, 1976, 1983; Isaac, Crader,
1981; Binford, 1981, 1984).
Современные исследования по палеоантропологии включают
в себя четыре упомянутых подхода — изучение ископаемых
остатков гоминид, ныне живущих приматов, этнографические и
Пути в прошлое
107
археологические исследования. Эклектическое использование
широкого диапазона методических приемов, данных и идей,
склонность к междисциплинарным подходам создают основу
для нынешнего бурного развития исследований в области эво-
люции гоминид.
4.2. Природа прошлого
Л. П. Хартли писал, что «прошлое — это чужеземное госу-
дарство, где все делается по-другому». Для палеоантропологов
задача первостепенной важности понять эту страну, и потому
был разработан ряд идей и концепций, позволяющих отправить-
ся туда. Археологи и палеонтологи потратили немало времени,
пытаясь ответить на вопросы, как достигается знание прошло-
го и что ответить из наблюдений за событиями прошлого (см.
Binford, 1977, 1981). Все умозаключения по поводу истории на-
шего вида зависят от того, сколь адекватны будут наши зна-
ния о путях, по которым к нам приходит информация о прош-
лом. Следовательно, мы должны вкратце охарактеризовать
принципы и методы, используемые палеоантропологами для
изучения прошлого.
4.2.1. Прямое и косвенное наблюдение
Изучение прошлого и настоящего часто противопоставляют
друг другу, говоря, что первое зависит от непрямого наблюде-
ния, тогда как второе можно наблюдать прямо. Специалисты по
социальной антропологии, этологи и экологи — все они могут
изучать ныне живущие популяции и отдельных особей, делая
выводы из непосредственных наблюдений. Палеоантропологи,
с другой стороны, должны реконструировать системы прошлого
на основе косвенных наблюдений, по остаткам ископаемых ор-
ганизмов, орудий и т. д. Это должно означать, что «актуалисти-
ческие науки» располагают более убедительной суммой знаний,
чем «палеонауки».
Это противопоставление, по-видимому, не столь уж и оправ-
дано, как может показаться на первый взгляд (Mellor, 1974).
Любое знание, относится ли оно к настоящему или к прошлому,
основывается на выводах (Binford, 1981, р. 26), т. е. структури-
рованных заключениях, в основе которых лежат принципы, свя-
зывающие явления между собой, причем эта связь часто носит
причинно-следственный характер. Поэтому, когда специалист по
социальной антропологии заявляет, что такая-то группа людей
характеризуется определенной социальной структурой, этот вы-
вод основан на следующих принципах: «то, что говорит инфор-
матор, соответствует действительной структуре общества» или
108
Глава 4
«частота взаимосвязей между индивидуумами отражает соци-
альные хитросплетения». Эти принципы подвергаются провер-
ке, контролю и уточнению, но в любом случае они лежат в ос-
нове наших знаний о настоящем времени.
Информация о прошлом строится на основе положений, вы-
текающих из тех же принципов. Например, заключение о струк-
туре сообщества в один из прошлых периодов можно получить,
исходя из анализа относительных частот различных видов в ос-
татках фауны. Принцип, лежащий в основе этого заключения,
состоит в том, что частота ископаемых экземпляров отражает
соотношение численности живших в прошлом животных. Тогда
разница между информацией о прошлом и настоящем состоит
не столько в различии между прямым и косвенным наблюдени-
ем, сколько в разной длине цепочки умозаключений и силе
сцепления составляющих ее звеньев. Вполне возможно, что лю-
бая информация о прошлом должна базироваться на более
длинной и более труднодоступной цепочке выводов, чем инфор-
мация о настоящем, но эпистемология остается по сути той же
самой. В любом случае прогресс той или иной научной дисцип-
лины зависит от того, в какой мере цепочка выводов может
быть подвергнута проверке и уточнению.
4.2.2. Принцип единообразия
Главный принцип, лежащий в основе изучения событий и яв-
лений прошлого, — это принцип единообразия. Принято считать,
что разработка этого принципа Чарлзом Лайеллом в «Принци-
пах геологии» (Lyell, 1830) заложила основы современной гео-
логии и создала необходимые концептуальные предпосылки для
теории эволюции Дарвина и Уоллеса. В простейшем виде прин-
цип единообразия можно свести к следующему: события прош-
лого можно объяснить только с помощью процессов и механиз-
мов, действующих сегодня. Однако, как указывает С. Дж. Гулд
(Gould, 1977, рр. 150—151), принцип единообразия состоит из
четырех элементов:
1. Научные правила, принципы и законы единообразны в
пространстве и времени.
2. Процессы, лежащие в основе геологических событий, еди-
нообразны во времени.
3. Скорость геологических изменений постоянна.
4. Характер (рельеф) Земли единообразен во времени.
Гулд утверждает, что лишь первые два элемента остаются в
силе сегодня. Нам достаточно известно о движении материков
и изменении биологических сообществ в разные геологические
эпохи, чтобы не сомневаться в том, что мир в процессе своей
истории изменялся, и хотя эти изменения нельзя свести к не-
Пути в прошлое
109
скольким обширным и непродолжительным катастрофам, вроде
библейского потопа, нельзя сказать также, чтобы они проходи-
ли постепенно и гладко. Однако первые два положения принци-
па единообразия сохраняют свое значение при любом направ-
лении исследования прошлого.
Наиболее важная и полезная сторона принципа единообра-
зия состоит в том, что единообразны во времени процессы, фор-
мирующие мир, а не он сам. Единообразие — мощная и весьма
ценная концепция при анализе механизма или процесса, одна-
ко при анализе явления это — смирительная рубашка. Напри-
мер, сегодня можно провести интенсивные наблюдения за ка-
ким-нибудь видом обезьян, например шимпанзе, и детально
описать их поведение. Это современное исследование может
оказаться полезным для интерпретации поведения шимпанзе в
прошлом, реконструированного на основании ископаемых остат-
ков. Это, однако, произойдет лишь в том случае, если будут
экстраполированы принципы, лежащие в основе поведения по-
пуляций шимпанзе, а не сам тип поведения. Другими словами,
нельзя утверждать, что если шимпанзе живут сейчас стадами,
в которые входит по многу самцов, то так они жили всегда.
Это утверждение, однако, может оказаться справедливым, если,
исходя из теперешних наблюдений, будут уточнены условия об-
разования многосамцовых групп и будет показано, что подоб-
ные условия имелись в прошлом.
Подобное ограниченное использование принципа единообра-
зия, при котором тщательно контролируется правильность ис-
ходных допущений, позволяет неразрывно связать прошлое с
настоящим. В обоих случаях все зависит от выводов, сделан-
ных на основе наблюдений, и от фундаментальных предпосы-
лок, что принципы, управляющие окружающим нас миром, не
меняются в пространстве и во времени, даже если меняются
конечные продукты их действия. Поэтому методы, используе-
мые для изучения прошлого, должны быть тесно связаны с тем
миром, в котором мы живем сегодня. Это единственный путь,
по которому можно проникнуть в прошлое, спроецировав на не-
го наши сегодняшне знания, но не окрашивая их своим субъек-
тивным восприятием.
4.3. Установление связей между прошлым и настоящим
4.3.1. Теория средних значений
Бинфорд (Binford, 1977, 1981) сформулировал принципы,
связывающие прошлое и настоящее, в виде теории средних зна-
чений. Существует целый ряд теорий, в которых наблюдаемые
аспекты прошлого соотносятся с процессами, действующими се-
по
Глава 4
Рис. 4.4. Связь между прошлым и настоящим. Выводы о жизни в прошлом (Г)
зависят от изучения ископаемых остатков (В), однако при интерпретации
необходимо учитывать изменения, происходящие в процессе фоссилизации (Г,
В). Это в свою очередь зависит от изучения процесса образования ископаемых
остатков (В), а также от того, каким образом поведение, наблюдаемое в на-
стоящее время (А) может быть прослежено по этим остаткам в прошлом
годня, что позволяет сделать вывод о происходивших в прош-
лом изменениях. Подобные связующие аргументы издавна ис-
пользовались в геологии, палеонтологии и археологии, но в по-
следние годы они сделались более четкими и формальными,
так как стало очевидно, что одного только здравого смысла не-
достаточно для интерпретации ископаемых остатков. Вследствие
этого был выполнен ряд экспериментов по изучению таких раз-
личных явлений, как разрушение кости (Behrensmeyer, 1978),
разложение туши (Hill, 1975), стертость каменных орудий Ke-
eley, 1978; Keeley, Toth, 1981). Общей чертой всех этих экспе-
риментов была попытка смоделировать те условия, в которых
были образованы материальные остатки прошлого.
Бинфорд (Binford, 1981, р. 29) резюмирует основные поло-
жения своей теории следующим образом: «Все наши заключе-
ния о прошлом — это выводы из наблюдений, сделанных на со-
временном археологическом материале» (т. е. наблюдений, сде-
ланных в настоящее время на материале, который может быть
изучен методами археологии или палеонтологии), «точность,
с которой могут быть связаны прошлое и настоящее, зависит от
того, насколько они сопоставимы». Итак, наблюдения в на-
Пути в прошлое
111
стоящем должны быть сделаны в такой форме, которая допус-
кала бы аналогичные наблюдения в прошлом (рис. 4.4). Други-
ми словами, не имеет смысла, анализируя поведение, скажем,
сегодняшней антилопы, переносить его в прошлое в качестве
модели, если эти наблюдения никак не связаны с ископаемыми
данными — например, размерами тела или некоторыми анато-
мическими особенностями.
Теория средних значений — это направление палеоантропо-
логии, изучающее и описывающее процессы, с помощью кото-
рых информация о мире живой природы переносится на мир ис-
копаемых организмов, тем самым позволяя нам делать выводы
о мире живой природы в геологическом прошлом, исходя из
материала об окаменелостях, относящихся к этому прошлому.
4.3.2. Тафономические процессы
Практическое применение теории средних значений в основ-
ном связано с изучением того, как поведение человека или жи-
вотных оставляет определенные следы в виде археологических
-объектов или ископаемых остатков организмов. Это область та-
фономии, раздела палеонтологии, исследующего процесс захо-
ронения организмов или перехода из биосферы в литосферу (Ef-
remow, 1940; Behrensmeyer, Hill, 1980; Shipman, 1983). Или
иными словами, тафономия в основном пытается ответить на
вопрос, насколько убедительное представление о жизни в прош-
лом можно получить из ископаемых материалов?
Поскольку до сих пор не было разработано какой-либо об-
щей теории о том, как образуются ископаемые остатки организ-
мов и археологические материалы, общий ответ на этот вопрос
едва ли может быть получен. Факторы, оказывающие влияние
на структуру этих процессов, различны; проведен ряд исследо-
ваний, посвященных изучению изолированного действия отдель-
ных компонентов. Среди них можно назвать серьезные работы
о состоянии костных остатков при питании падалью (Hill, 1978;
Brain, 1981), в зависимости от условий погребения (Isaac,
1967), от различий в условиях обитания и размерах биотопа
(Behrensmeyer, Dechant Boaz, 1980), от размеров животного
(Behrensmeyer, 1983), от утаптывания почвы (Gifford-Gonzales,
Damrosch et al., 1985; Behrensmeyer, Gordon, Janagi, 1986) и от
размывания ее водой (Hanson, 1980). Изучались также отдель-
ные поведенческие процессы, такие, как разделывание добычи
(Binford, 1981), стирание зубов (Walker, 1981), шлифовка ору-
дий (Keeley, 1978), образование и распад поселений (Yellen,
1977). Однако общая природа тафономических процессов может
быть лучше всего продемонстрирована с помощью ряда проти-
вопоставлений.
112
Глава 4
Наиболее важным из них является контраст между теми
компонентами ископаемых остатков, которые отражают опреде-
ленные виды поведения, и геологическими компонентами. Про-
исхождение ископаемых остатков связано не только с биологи-
ческими факторами, действующими в течение жизни животного,
равно как и не только с геологическими процессами, действую-
щими после его смерти. Здесь скорее смешивается то и другое,
что необходимо учитывать при всяком анализе. В силу двойст-
венности происхождения ископаемых остатков изучение их при-
роды затрагивает как факторы, действовавшие при жизни орга-
низмов, так и факторы, вступающие в действие после их смерти
(факторы геологического характера). Изучение первых в основ-
ном относится к области археологии (этноархеологии) и сфоку-
сировано на том воздействии, которое определенный тип пове-
дения оказывает на структуру археологического материала. На-
пример, Бинфорд (Binford, 1978а) показал, что пространствен-
ное распределение находок орудий на эскимосской стоянке оп-
ределяется тем, как эскимосы выбрасывали (или просто роня-
ли) ненужные предметы, т. е. одна из характеристик стоянки
связана с определенным типом человеческого поведения. С дру-
гой стороны, Беренсмейер (Behrensmeyer, 1983) проводила дли-
тельное исследование процессов распада и разрушения биомас-
сы на популяции крупных восточноафриканских млекопитающих
с целью изучения судьбы костных остатков после смерти живот-
ного.
Еще одно противоречие касается изучения тафономических
процессов,действующих сегодня, на современном костном и ар-
хеологическом материале (это — неотафономия, или «актуалисти-
ческие исследования») и прямых исследований древнейших усло-
вий жизни. Первое обычно бывает основано на уже упомянутой
теории средних значений и нацелено на создание моделей про-
цесса фоссилизации, которые могут быть в дальнейшем исполь-
зованы при анализе ископаемого материала. Последние, напро-
тив, пытаются на месте определить, кому принадлежат опреде-
ленные ископаемые остатки или конкретный археологический
инвентарь. Первое направление представлено исследованиями
Хилла, проводившего анализ посмертных изменений костей жи-
вотных в условиях современного восточноафриканского ланд-
шафта (Hill, 1975) и предложившего модель расчленения скеле-
та (Hill, 1979), второе включает детальную реконструкцию до-
исторической среды обитания в Олдувае (Нау, 1976), а также
анализ фаунистических остатков в важном среднемиоценовом
месторождении Форт-Тернан (Shipman, 1982) —единственном
месте в Африке, где встречаются костные остатки гоминоидов
этой эпохи, включая кениапитека, которого одно время относили
к тому же виду, что и рамапитека, и считали поэтому гомини-
Пути в прошлое
ИЗ
дом. В целом исследования по палеотафономии представляют
собой закономерный итог тафономического анализа в том смыс-
ле, что они приходят к выводу о природе прошлого, исходя из
непосредственного изучения процессов, происходящих сегодня.
И наконец, заключительное противоречие вытекает из раз-
личий между археологией и палеонтологией. Палеонтологи, име-
ющие дело со всевозможными организмами, исключая человека,
в первую очередь интересуются факторами, действующими пос-
ле геологических отложений, функциональной анатомией и сре-
довыми аспектами адаптации. Археологи, не игнорируя значе-
ния этих факторов, должны учитывать важную роль социаль-
ных и культурных воздействий в образовании археологических
материалов, а также тот факт, что их информация базируется
не на костных остатках самого организма, а на материальных
проявлениях его деятельности. Следовательно, масштабы неота-
фономических исследований в археологии существенно расши-
ряются и включают многие методы и понятия социальной антро-
пологии, а также экологии.
Хотя и существуют другие подходы к изучению прошлого
без непосредственной интерпретации ископаемых и археологи-
ческих материалов (например, биохимические исследования и
модели, построенные на аналогиях с современными охотниками
и собирателями), изучение костных и материальных остатков
остается важнейшим и центральным аспектом подобных иссле-
дований. Именно они прокладывают главную дорогу к прошло-
му, но их интерпретация целиком зависит от нашего понимания
процессов их формирования и трансформации. Поэтому для
лучшего познания того, что происходило в прошлом, необходи-
мо дальнейшее развитие тафономических исследований и ис-
пользование теории средних значений.
4.4. Методы изучения прошлого
В связи с тем, что было сказано выше о том, какими спосо-
бами приобретаются наши знания о прошлом, можно предло-
жить ряд методов, которые послужат своеобразным руководст-
вом при изучении эволюционной экологии любой группы живот-
ных, в том числе и гоминид. Применение этих методов обеспе-
чит проведение палеоантропологического анализа на основе
прочных поведенческих и биологических принципов, а это в
свою очередь приведет к тому, что поведенческие теории, обра-
щенные в прошлое, будут подвергаться проверке.
4.4.1. Теория и модели
В гл. 1 было установлено, что окаменелости не могут рас-
сказать нам свою историю, поэтому необходима теория для
8—367
114
Глава 4
исследования событий прошлого, а также для того, чтобы от
простого описания эволюции гоминид перейти к ее анализу и
объяснению. На практике это означает, что должны быть по-
ставлены специфические вопросы. Роль теории состоит в том,
чтобы сформулировать эти вопросы. Как будет показано в сле-
дующей главе, для постановки вопросов об эволюции гоминид в
качестве теоретической основы может быть использована эво-
люционная экология. Основная идея заключается в том, что
теоретические выкладки могут непосредственно стимулировать
вопросы и прогнозы. Например, теория экологии утверждает,
что первичная продуктивность экосистемы в тропиках зависит
от количества осадков (Murphy, 1974). Прогнозируемый харак-
тер изменчивости можно использовать для определения потен-
циальных реакций животного. Прогнозируемая реакция затем
может быть использована для построения адаптационной моде-
ли, которую можно сравнить с имеющимися эмпирическими
данными.
Однако теория (т. е. логические умозаключения и прогнозы,
сделанные на основе нескольких аксиом) —не единственный ис-
точник вопросов о прошлом. Наблюдения за ископаемыми ос-
татками организмов, а также за современными биологическими
формами могут (при использовании в рамках какой-то теорети-
ческой основы) привести к выделению важных направлений ис-
следования в данной области. Например, бимодальное распре-
деление некоторых морфологических характеристик у ископае-
мых гоминид может навести на мысль о половом диморфизме. Эти
же вопросы могут возникнуть и при рассмотрении теоретических
моделей, освещающих те факторы, которые способствуют прояв-
лению полового диморфизма или тормозят его. В этом случае
наблюдения заставляют обратиться к теории, которая может
объяснить их результаты. Использование данных подобным об-
разом составляет суть научного метода, основанного на «рас-
познавании специфики данных» (Binford, 1983, р. 97).
Другой метод того же характера состоит в поисках анома-
лий или отклонений (Gould, 1980). Явные несоответствия меж-
му самими данными и их интерпретацией могут нередко ис-
пользоваться для формулировки новых вопросов относительно
источника информации и, следовательно, разработки новых под-
ходов и новых трактовок. Классическим примером в этом плане
из области физической антропологии может служить разоблаче-
ние пильтдаунской находки, явно аномальное положение кото-
рой на эволюционном древе человека привело к разработке но-
вых методов исследования. Опять-таки ведущую роль сыграла
теория, так как именно несоответствие между теорией эволюции
человека и особенностями пильтдаунского черепа привело к
постановке исследования.
Пути в прошлое
115
Однако наличия общей теории или фона еще недостаточно
для интерпретации фактов. Теория должна быть выражена в
такой форме, которая позволила бы провести прямой анализ
данных, доступных для ее проверки. Эту функцию берет на себя
модель, выступающая в роли посредника между теорией и фак-
тами. Модель описывает и прогнозирует структуру и изменчи-
вость явлений, исходя из теоретических закономерностей. Так,
например, теория может констатировать, что половой димор-
физм «обусловлен» соперничеством между самцами и что усло-
вия, при которых это соперничество будет наиболее выражено,
прямо связаны с распределением пищевых ресурсов, обеспечи-
вающим поиск пищи в составе больших групп. Отсюда вытека-
ет модель, характеризующая особенности изменчивости при по-
ловом диморфизме: отчетливые различия между самцами и сам-
ками должны быть отмечены там, где пищевые ресурсы распре-
делены в среде обитания неравномерно и кормежка проходит в
составе больших групп, тогда как минимальный половой димор-
физм характерен для тех групп, которые живут в условиях рав-
номерного распределения ресурсов. С использованием этой мо-
дели теория принимает такую форму, в которой ее можно при-
менять для изучения имеющихся данных, характеризующих по-
ловой диморфизм и распределение пищевых ресурсов.
Для изучения прошлого необходим, однако, второй уровень
построения модели. Это связано с тем, что, как уже говорилось
выше, ископаемые остатки и археологические материалы явля-
ются результатом действия не только поведенческих, прижиз-
ненных факторов, но и геологических, посмертных. Следователь-
но, предложенная модель полового диморфизма должна быть
преобразована в другую форму, которая даст возможность про-
верить ее применимость для прошлых геологических эпох. Это
означает, что нужно рассмотреть, как отражается явление поло-
вого диморфизма в ископаемых остатках и как можно реконст-
руировать изменения условий в давно исчезнувших экосисте-
мах. Это вновь возвращает нас к теории средних значений и та-
фономическим исследованиям.
Однако эту идеальную процедуру могут осложнить некото-
рые факторы. Из-за того что ископаемые остатки и археологи-
ческие материалы могут принимать ограниченные по своей при-
роде формы, различные поведенческие модели, будучи преобра-
зованы в такие модели, которые прогнозируют структуру этих
остатков, возможно, окажутся идентичными (Hill, 1984). К со-
жалению, трансформация различных поведенческих моделей в
их археологические или палеонтологические воплощения может
дать одинаковый результат: иначе говоря, различные типы по-
ведения приводят к появлению одинаковых ископаемых остат-
ков. Следовательно, едва ли достаточно предсказать, как будут
8*
116
Глава 4
выглядеть археологические и палеонтологические материалы,
если модель верна. Нужно рассмотреть возможные совпадения
моделей, а также четко обозначить черты несходства.
4.4.2. Четыре пути в прошлое
Ни теории, ни модели сами по себе не могут считаться до-
статочными для палеоантропологии, так как все они в той или
иной мере стремятся наложить настоящее на прошлое. Посколь-
ку теория должна, по крайней мере в первоначальном виде, ис-
ходить из нашего понимания мира, каким мы его видим сегод-
ня, она является отражением современных структур и их измен-
чивости. Чтобы избежать простого перенесения этой структуры
в прошлое, нужны независимые методы определения того, что
на самом деле тогда происходило. И если ископаемые остатки
оказываются бессловесными при отсутствии теоретической базы,
то и теория утверждает только самое себя, если не подвергается
эмпирическим проверкам. Можно предложить четыре способа
таких проверок.
1. Способ логических выводов. Можно сделать вывод о ха-
рактере ископаемого материала. Классический вывод (хотя и
почти наверняка неправильный) состоит в том, что сходство в
морфологии орудий свидетельствует о сходстве социальных и
культурных условий жизни их изготовителей. Такова основа
культурологических интерпретаций археологических данных.
Если каждое звено в логической цепи выводов (а их может быть
несколько) тщательно выковано и подкреплено эксперименталь-
ными и актуалистическими исследованиями, этот подход оказы-
вается очень действенным (см. критический обзор Binford, Sab-
loff, 1982). Справедливость вывода может быть доказана с по-
мощью этнографических или экспериментальных данных, а так-
же посредством моделей.
2. Изолирующие методы. Большинство методов, реконструи-
рующих прошлое, рассчитано на извлечение информации из ши-
рого спектра данных. Бинфорд (Binford, 1981) впервые предло-
жил альтернативный метод, который с большой долей скепти-
цизма относится к рассмотрению многочисленных и разнообраз-
ных данных, сосредоточиваясь вместо этого на небольших дета-
лях; они могут показаться довольно незначительными, но на са-
мом деле необычайно чувствительны к поведенческим вариаци-
ям, вследствие чего их можно использовать для разграничения
гипотез. В качестве одного из примеров приводится характер
разделки туш (Binford, 1981), в качестве другого — расчленение
скелета (Hill, 1975). Еще одним, чрезвычайно изящным приме-
ром использования этого метода служит изучение стертости зу-
бов с помощью сканирующего электронного микроскопа (Wal-
ker, 1981).
Пути в прошлое
117
3. Вероятностный метод. Не все методы позволяют разгра-
ничить типы поведения или гипотезы; вместо этого они нередко
предлагают статистический подход для оценки вероятности. Ма-
тематическая основа этого подхода обсуждается в работе Пил-
бима и Вайсниса (Pilbeam, Vaisnys, 1975). На практике здесь
речь идет о выделении процессов, которые могут привести к
сходным результатам, и экспериментальной или теоретической
попытке оценить вероятности различных исходов. Этот метод
редко оперирует единичными результатами и их повторяемость
является необходимым условием его применения.
4.Сравнительно-биологический метод. Проверить представле-
ния о прошлом до некоторой степени можно и без прямого об-
ращения к ископаемым материалам. Основным способом изуче-
ния ранних гоминид остается сравнительно-биологический. Ха-
рактер межвидовой и внутривидовой изменчивости среди рас-
тений и животных может быть использован для корреляции
биологических переменных. Были изучены — теоретически или
эмпирически — многие такие коррелятивные связи, например
между размерами тела и скоростью основного обмена (Kleiber,
1961). Если эти корреляции достоверны, они позволяют прямо
судить о ранних гоминидах, ограничиваясь лишь частичным об-
ращением к деталям строения ископаемых остатков. Конечно,
необходимо учитывать условия, в которых устанавливается лю-
бая корреляция. Однако при наличии в прошлом одной или не-
скольких из установленных переменных можно использовать
корреляции для того, чтобы на их основе прийти к выводам об
организмах, живших в далеком прошлом. В качестве классиче-
ского примера этого подхода приведем работу по биохимии бел-
ков, которая оказала такое большое влияние на палеоантропо-
логию и реконструкцию филогенеза (см. разд. 2.1.3).
Все эти методы изучения прошлого весьма ценны и использу-
ются в этой книге. В каждом из них может найтись место и для
традиционных подходов, таких, как изучение типологии орудий,
и для более изощренных средств, опирающихся на достижения
точных наук. Для всех них, однако, характерно то, что они ис-
ходят из представления о том, что процесс фоссилизации у раз-
ных организмов может протекать по-разному. Поэтому они
должны учитывать разнообразие типов поведения у представи-
телей широкого спектра видов. Во многих работах показано
(см., например, литературу о процессах фоссилизации в Восточ-
ной Африке — Behrensmeyer, Hill, 1980; Shipman, 1983), что ис-
копаемые остатки и археологические свидетельства существо-
вания ранних гоминид — это результат действия многих процес-
сов. При попытке определить относительную роль этих процес-
сов можно высказать лишь некоторые предположения.
118
Глава 4
4.4.3. Пример
Названные методы представляют собой взаимосвязанные
способы решения палеоантропологических проблем, сформули-
рованных на основе теоретических предпосылок, и проверки
соответствующих гипотез (рис. 4.5). Это можно проиллюстриро-
вать на примере, который ниже будет рассмотрен гораздо более
подробно, а именно на процессе включения мяса в рацион го-
минид.
Первый этап исследования заключается в осознании того,
что переход к мясоедению представляет собой важную палео-
антропологическую проблему. Это подтверждается разным уров-
нем плотоядности у современных приматов и человека (К. Hill,
1982), что свидетельствует о значительных адаптивных сдвигах,
происшедших в процессах эволюции гоминид. Подобные разли-
чия создают основу для рассмотрения этого вопроса в эволю-
ционном контексте.
Чтобы изучить эту палеоантропологическую проблему, необ-
ходимо соотнести ее с теоретической основой. Наиболее подхо-
дит экологическая теория, т. е. та, которая связывает трофиче-
Контропирующая Рабочая теория
теория
Общие Эволюционная
эволюционные экология
принципы
Естественный
отбор
ведет
к адаптации
к Адаптации
ф решают
проблемы
выживания
Биологические модели
Модели, основанные
на биологических
принципах
Определение адаптивных
проблем и прогнозирование
решений, основанных
на общих биологических
принципах
Пути в прошлое:
эмпирические тесты
Поиски моделей
в прошлом
Теория средних значений
Археологические
и палеонтологические
модели
Выводы
Изолирующие способы
Вероятностные методы
ГравнительнО'биопогический
подход
Модели, демонстрирующие
справедливость прогнозов
при изучении ископаемых
остатков
Рис. 4.5. Пути в прошлое: итоги способов изучения эволюции гоминид, обсуж-
давшиеся в этой главе
Пути в прошлое
119
-скую структуру с размерами ниши. Проблема поедания мяса у
гоминид в действительности представляет собой частный случай
из разряда более общих экологических вопросов: при каких
условиях организм меняет свое положение в трофической пира-
миде, переходя от растительной пищи к мясной? При каких
условиях организм расширяет сферу используемых им пищевых
ресурсов?
Сформулировав проблему в более общем виде, можно по-
строить экологическую модель, основанную на общих принци-
пах, а не на специфических особенностях какого-либо одного
типа организмов (в данном случае гоминид). Эта общая модель
предполагает использование широкого сравнительного подхода.
Например, животное расширяет границы ниши, когда затраты,
связанные с использованием новых источников пищи, ниже
средних затрат при использовании пищевых ресурсов в целом
(Mac Artnur, Pianka, 1966). Далее можно построить модель, оп-
ределяющую те условия, при которых это может произойти,—
например, истощение существующих источников пищи, расшире-
ние новых, развитие новой стратегии добывания пищи или
уменьшение конкурентной борьбы. Это позволит прийти к пред-
варительному заключению об условиях перехода к частичному
питанию мясом у гоминид.
Однако для проверки выдвинутого предположения необходи-
мо разработать модель с учетом «ископаемых доказательств».
Иными словами, нужно, чтобы были обнаружены следы усло-
вий, при которых возможно мясоедение и его реальные проявле-
ния. Подобное требование включает в себя тщательное опреде-
ление условий среды и последствий питания мясом. Например,
если условия предполагают снижение численности других хищ-
ников или всеядных организмов и, следовательно, уменьшение
конкурентной борьбы, то в ископаемых материалах должно
быть отмечено изменение частоты встречаемости и разнообра-
зия среди этих видов.
Чтобы удалось извлечь из ископаемых остатков такого рода
информацию, необходимо применить на практике те четыре
принципа, которые служат для постижения прошлого (см. пре-
дыдущий раздел). Можно сделать ряд выводов на основе тео-
рии средних значений: например, констатировать, что тесная
пространственная связь между орудиями гоминид и костями
животных, за которыми гоминиды охотились, указывает на то,
что осуществлялась разделка туш животных. Можно использо-
вать изолирующие методы. Например, по расположению надре-
зов на костях животных относительно мяса (мышц) можно
отделить те кости, на которых оставили следы исключительно
гоминиды, от костей животных, съеденных гоминидами, но
умерщвленных другими хищниками (Potts, 1984а). Может быть
120
Глава 4
использован и вероятностный метод. Например, можно вычис-
лить вероятность того, что кости животных и обработанные че-
ловеком орудия будут встречаться вместе при определенных по-
казателях смертности животных и частоты выбрасывания ору-
дий. И наконец, можно провести сопоставление между харак-
терными особенностями ныне живущих человеческих популяций
и других животных, в особенности приматов, чтобы определить,
существуют ли какие-нибудь физиологические особенности, ука-
зывающие на древность употребления в пищу мяса (например,
в анатомии пищеварительного тракта).
Следует отметить, что в этом примере использованы основ-
ные подходы к палеоантропологическому материалу, описанные
в данной главе, — изучение ископаемых остатков, современных
приматов, ныне живущих человеческих популяций и археологи-
ческих материалов, что свидетельствует о необходимости раз-
вивать аналитическую методологию на как можно более широ-
ком материале. Хотя этим не гарантируется решение любой па-
леоантропологической проблемы, в частности из-за обычной не-
хватки доказательств, привлечение самых разнообразных дан-
ных является необходимым залогом успешного анализа.
4.5. Аналитическая палеоэкология
В этой и предыдущих главах была сделана попытка описать
то, чем следует руководствоваться при изучении процесса эво-
люции гоминид. В первой главе подобную первичную основу
составляли ископаемые остатки и археологические материа-
лы — фактические данные, на которых должны базироваться ин-
терпретация и анализ. Однако, как уже говорилось, эти доказа-
тельства сами по себе недостаточны — ископаемые остатки не
могут рассказать нам свою историю. Отчасти это происходит из-
за фрагментарности ископаемых остатков или из-за косвенного
характера сравнений с современными организмами, что ослож-
няет их интерпретацию. Отчасти это возникает из-за отсутствия
подходящей теоретической основы, которая помогла бы связать
наши знания о прошлом с нашим пониманием того, как рабо-
тают биологические и экологические системы.
Такая основа была представлена в гл. 2. Было показано, что
эволюционные механизмы и процессы связаны с принципами
эволюции через центральное понятие адаптации. Особенное зна-
чение имеет высказанное предположение о том, что адаптация
есть процесс решения задач, точнее решение проблемы выжи-
вания, обеспечивающее индивидууму наибольший репродуктив-
ный успех. Это аналогично выбору стратегии, сделанному игро-
ком. Выявление проблем, с которыми сталкивались ранние го-
Пути в прошлое
121
Рис. 4.6. Диаграмма Венна, демонстрирующая уникальность человека и его
адаптивной стратегии (заштрихованная область) в виде пересечения различ-
ных биологических общностей, к которым принадлежат гоминиды. НП — на-
земные приматы; СС — сообщество саванны; ТО — тропические организмы;
КМ — крупные млекопитающие; Г — гоминиды
миниды, признается одной из важных исходных задач, постав-
ленных перед палеоантропологией.
Однако, как мы пытались показать в этой главе, хотя и важ-
но исходить из здравых эволюционных и экологических прин-
ципов и моделей, необходимо найти способы соотнести их с име-
ющимися данными. Для практического использования этих прин-
ципов требуются методы, позволяющие изучать прошлое. Здесь
подчеркивалось, что методы эти разнообразны, что существует
целый спектр возможных подходов, но что все они должны ис-
ходить из принципов единообразия и использовать теорию
средних значений.
Цель этих глав заключалась в том, чтобы показать, что па-
леоантропология приобретает более аналитический характер,
развиваясь в рамках эволюционной биологии. Этот факт имеет
далеко идущие последствия. Если адаптация есть решение стоя-
щих перед организмом проблем, тогда мы вправе спросить, ка-
кие проблемы стояли перед первыми гоминидами? Затем мы
должны рассмотреть эти проблемы в контексте общих биологи-
ческих принципов, что позволит изучать эволюцию гоминид на
более широкой сравнительной основе. Необходимо выделить и
обсудить факторы, определяющие потенциальные решения,
122
Глава 4
с тем, чтобы понять причины, ведущие к конкретным «решени-
ям гоминид». И наконец, эти эволюционные и экологические мо-
дели должны быть соотнесены с данными по ископаемым остат-
кам организмов и археологическим материалам.
Оставшиеся главы этой книги посвящены именно этой цели.
Выделен целый ряд обширных биологических и экологических
проблем, которые могли иметь существенное значение в эволю-
ции гоминид, таких, как обитание в тропическом климате, раз-
меры тела, наземный образ жизни, сезонность и межвидовая
конкуренция. Сделана попытка связать их с особенностями эво-
люционной экологии ранних гоминид. Каждая из этих проблем
стояла не только перед гоминидами, но и перед многими дру-
гими организмами, что создает основу для плодотворных срав-
нений. В итоге то, что можно назвать уникальностью человека,,
есть результат не какой-то одной адаптации, а комбинации це-
лого ряда адаптаций (рис. 4.6).
Глава 5
ГОМИНИДЫ КАК ТРОПИЧЕСКИЕ ЖИВОТНЫЕ
Гоминиды относятся к приматам, а этот отряд млекопитаю-
щих появился и до сих пор обитает главным образом в тропиче-
ском поясе. Можно, стало быть, утверждать, что гоминиды —
это исходно тропические организмы, которые вторично адапти-
ровались к широкому многообразию других климатических ус-
ловий и типов экосистем. И можно, следовательно, думать, что
многими из своих характерных признаков гоминиды обязаны
тем ограничениям и возможностям, с которыми связана жизнь
в низких широтах. В настоящей главе обосновывается мнение,
в соответствии с которым гоминид можно рассматривать как
тропических животных, а также обсуждаются особенности тро-
пической экологии и проблемы, связанные с жизнью в тропиках.
5Л. Эволюция гоминид и тропики
Возможность рассматривать ранних гоминид как порожде-
ние тропической среды основывается на данных, полученных из
трех независимых источников — по географическому распрост-
ранению современных приматов, распространению ископаемых
приматов (исключая человека) и изменениям распространения
ископаемых гоминид в процессе эволюции.
5.1.1. Распространение приматов
Тропическое происхождение приматов подтверждается гео-
графическим распространением их современных видов. Если
определять тропики, как географическую зону, заключенную
между тропиками Рака и Козерога, можно видеть, что в этой
области обитает большинство видов приматов, отличных от че-
ловека, и что практически ни один вид приматов не заходит на
севере дальше 25° северной широты, а на юге — дальше 30° юж-
ной широты (рис. 5.1). Исключениями из этого правила (не счи-
тая современных людей) являются лишь несколько видов рода
Масаса (Napier, Napier, 1967, р. 378). Масаса mulatto заходит
на север вплоть до Биджана в Китае, однако не исключено, что
этот вид был завезен сюда человеком уже в исторические вре-
мена. Японский макак, Масаса fuscata, обитает на острове Хон-
Рис, 5,1. Географическое распространение современных приматов. (Napier, Napier, 1967.)
Гоминиды как тропические животные
125
Рис. 5.2. Распределение современных приматов по тропическим биотопам..
(Napier, Napier, 1967.)
сю на 41° северной широты. Обычны макаки и в северной Афри-
ке, где распространены вплоть до Гибралтара (Масаса sylva-
nus), хотя современные популяции этих животных сформирова-
лись в результате их интродукции сюда лишь в 17-м столетии
(Napier, Napier, 1967, р. 378). Географическое распространение
приматов может быть связано с такими экологическими ограни-
чениями, как, например, низкая температура воздуха, меньшая
долгота дня, меньшая продолжительность сезона вегетации и
меньшая доступность пищи.
В тропиках приматы, отличные от человека, занимают самые
различные биотопы (рис. 5.2). Главным образом они встреча-
ются во влажном тропическом лесу, но живут и в высокогорных
районах, в лесах с выраженной сезонностью, лесистой и травя-
ной саванне, отчасти заходя даже в полупустынные области
(например, мартышка гусар, Erythrocebus patas) (Hall, 1962).
Тот факт, что в тропиках приматы буквально вездесущи, а в
других поясах практически отсутствуют, по всей видимости, под-
126
Глава 5
тверждает предположение о важной роли, которую сыграли в
эволюции приматов особенности экологических процессов в тро-
пиках.
Вполне правдоподобно, однако, звучит и то предположение,
что со временем произошло сужение ареала распространения
приматов под влиянием хозяйственной деятельности человека,
которая, судя по плотности населения и изменениям окружаю-
щей среды, была интенсивнее именно в более высоких широтах.
Для изучения такой возможности целесообразно изучить гео-
графическое распространение ископаемых приматов.
5.1.2. Распространение ископаемых приматов
Хотя находки ископаемых приматов случаются довольно ред-
ко, они все-таки дают некоторое представление о распростране-
нии этих животных в прошлом. Доисторический ареал обитания
приматов, безусловно, был шире, чем современный. Как отмеча-
лось в гл. 2, самые ранние приматы датируются концом мело-
вого периода (около 65 млн. лет назад) (Szalay, Delson, 1979).
Ранние представители группы Plesiadapiformes (примитивные
формы приматов), найденные в Северной Америке и Европе, от-
носятся к интервалу с конца мелового периода до позднего
эоцена. Strepsikhinae (сохранившийся подотряд приматов) рас-
пространяется по Северной Америке, Европе, Азии и Африке,
начиная с раннего эоцена. В раннем эоцене появляется и под-
отряд Haplorhinae, представители которого широко распростра-
няются по всем континентам. Широкое распространение пред-
ставителей этого подотряда сохраняется в олигоцене и миоцене.
Однако около 36 млн. лет назад в Северной Америке приматы
вымирают. В Европе приматы обычны в миоцене и полиоцене,
а в плейстоцене здесь уже встречаются только представители
рода Масаса. В последние 10 млн. лет распространение прима-
тов ограничивается преимущественно Южиой и Центральной
Америкой, Африкой и тропическими регионами Азии.
Поскольку на протяжении рассмотренного отрезка времени
происходили значительные климатические изменения, описан-
ный характер распространения ископаемых приматов полностью
•согласуется с «тропической природой» этого отряда. В начале
кайнозойской эры было гораздо теплее, чем теперь, и тропиче-
ские области, в особенности тропические леса, занимали гораз-
до больше места. Однако с середины миоцена начало происхо-
дить быстрое снижение температуры и значительное изменение
дрейфа материков, что привело к сокращению площади тропи-
ческой зоны (подробнее об этом см. разд. 5.4.1). В соответствии
с этими климатическими сдвигами произошло и сужение ареала
обитания приматов, что подтверждает предположение о сильной
зависимости их распространения от температурных условий.
Гоминиды как тропические животные
127
Рис. 5.3. Географическое распространение ископаемых гоминид в зависимости
от широты места находки. (Oakley, Campbell, Mollison, 1977.)
Современные люди и некоторые ранние гоминиды—единст-
венные приматы, которым удалось избежать этих ограничений.
Однако, хотя современный человек распространен повсеместно,
ископаемые находки указывают на то, что освоение высоких
широт шло постепенно и сравнительно медленно. На рис. 5.3
изображена связь между широтой, на которой были сделаны
находки ископаемых гоминид, и приблизительным возрастом
этих находок. Из рисунка видно, что миллион лет назад и ранее
распространение гоминид ограничивалось тропиками и субтро-
пиками, что к этому времени они освоили границы зоны умерен-
ного климата, и что только в позднем плейстоцене началось их
проникновение в высокоширотные области. Критические же со-
бытия в эволюции гоминид, связанные с их отходом от других
приматов и приобретением таких отличительных признаков, как
бипедия и способность к изготовлению орудий труда, происхо-
дили в тропических областях.
5.2. Особенности тропической экологии
5.2.1. Факторы, определяющие структуру среды
Приняв версию о «тропическом происхождении» гоминид.,
уместно спросить, чем тропические среды отличаются от уме-
ренных, и в чем разница между протекающими в них экологиче-
128
Глава 5
скими процессами? Особенности тропической экологии опреде-
ляются двумя главными причинами: высокой и практически по-
стоянной на протяжении круглого года температурой воздуха и
количеством осадков как основным фактором разнообразия тро-
пических биотопов.
С этими двумя главными характеристиками связаны и неко-
торые другие особенности тропической зоны (Еуге, 1968; Cloud-
sley-Thompson, 1975; Harris, 1980):
1. Долгота дня на протяжении круглого года остается по-
стоянной и примерно равной продолжительности ночи. Это
обстоятельство имеет важное значение для понимания суточной
организации активности животных.
2. Суточные колебания температуры выражены сильнее, чем
сезонные.
3. Скорость ветра в низких широтах незначительна, что мо-
жет приводить к низкой интенсивности транспирации.
4. Высокий уровень солнечной радиации, обусловливающий
не только высокую температуру воздуха (превышающую как
правило 25 °C), но и большую интенсивность ультрафиолетово-
го облучения, которое может оказывать на организм вредное
действие и вместе с тем положительно влиять на протекание в
нем метаболических процессов (например, синтез витамина D).
5. Если выпадают дожди, то они зачастую имеют резко вы-
раженный сезонный характер и могут быть циклоническими или
ливневыми. Одним из следствий этого обстоятельства может
быть то, что средние годовые показатели осадков не всегда на-
дежно отражают истинную доступность влаги.
6. Многие тропические области отличаются обилием и видо-
вым разнообразием растений и животных. В то время как в
экосистемах умеренного климата нередко доминируют много-
численные особи только нескольких видов, тропические экосис-
темы часто состоят из многих видов, каждый из которых пред-
ставлен небольшим числом особей. Именно так обстоит дело во
влажном тропическом лесу.
7. Поскольку температура в тропиках изменяется незначи-
тельно, характер среды в большей степени определяют другие
факторы. В тропических средах тип растительности нередко
сильно зависит от типа почвы. Тип же почвы определяется
местными геологическими факторами.
8. Из сказанного следует, что в пространственном отношении
организация биотопов в тропиках очень разнообразна. Биотопы,
занимающие большие пространства, подобные тем, которые
имеются в Евразии и Северной Америке, в тропиках встречают-
ся сравнительно редко. Структуру экосистем в тропиках часто
можно назвать мозаичной.
Гоминиды как тропические животные
129
3000
Влажный
тропический
лес
Сезонный
тропичес-
кий лес
Широколиственный
тропический лес ——
Колючее
редколесье
Пустыня
Саванна
----Полупустыня
12 И 10 987654 3210
Число месяцев в году без дождя
Рис. 5.4. Типы тропических биотопов в зависимости от среднего годового уров-
ня осадков и сезонности. (Fowley, 1982.)
Все эти характерные особенности тропиков должны сильно
сказываться (и это действительно имеет место) на растениях
и животных в связи с их приспособлением к условиям окружа-
ющей среды. Решающим же фактором являются дожди. В об-
щем структуру и тип биотопа в тропиках можно предсказать по
характеру выпадения осадков. Важное значение здесь имеют
три взаимосвязанных показателя. Первый из них — общее или
среднегодовое количество осадков. Из рис. 5.4 видно, что тип
биотопа прямо зависит от этого показателя. Как эмпирические
работы (Strugnell, Pigot, 1978), так и теоретические расчеты
(Murphy, 1974; Rosensweig, 1968) показали, что это обстоятель-
ство связано с увеличением первичной продуктивности и его
влиянием на структуру биотопа. С возрастанием количества
осадков происходит увеличение растительного покрова, биомас-
сы, растительной продукции и вторичной биомассы (рис. 5.5).
Вторым показателем является сезонное распределение дождей.
9—367
Число
сухих месяцев
Среда
10 9 8 7 6 5 4 3 2
Пустыня Открытая __ - Лесистая _ ___ Сезонный - Влажный
и полупустыня саванна саванна лес лес
Доминирующие
животные
Доминирующая
растительность
Небольшие
млекопитающие;
несколько
видов копытных,
мало зависящих
от воды
Стадные
травоядные
копытные,
зависящие
от воды
Копытные, питающиеся
травой и древесными
побегами;
очень крупные
травоядные
млекопитающие
Смешанная фауна —
несколько видов копытных,
многочисленные
мелкие млекопитающие,
приматы
Приматы,
мелкие
млекопитающие
и птицы
Ксерофитные
суккуленты,
немногочислен-
ные виды трав
и деревьев
Трава
и редко
рассеянные
деревья
и кустарники
Трава
и колючие
деревья
Немногочисленные виды трав;
листоладные мелколиственные
деревья; несколько видов
кустарников
и полукустарников
Многочисленные
виды деревьев,
лиан и эпифитов
Рис. 5.5. Экологические и энергетические характеристики тропических сред обитания
Гоминиды как тропические животные 131
Для увеличения продуктивности на протяжении круглого года
требуется влага, а следовательно, чем длительнее сухое время
года, тем беднее и менее разнообразнее растительное сообщест-
во. Тип биотопа зависит от продолжительности сухого сезона
столь же сильно, как и от среднегодового количества осадков
(из рис. 5.4 можно видеть, насколько тесно связаны эти два по-
казателя). Третьим фактором является предсказуемость дождя.
За величиной среднегодовых количеств осадков зачастую скры-
ваются сильные межгодовые колебания этого показателя. А это
обстоятельство может иметь решающее значение, так как часто
для адаптации и эволюции животных важным является не сред-
негодовое количество осадков, а частота, за счет которой дости-
гается их некоторый минимальный уровень.
Несмотря на всю сложность этого вопроса, в качестве опре-
деляющей переменной нередко используют среднегодовое коли-
чество осадков. Такое упрощение, связанное со скудостью кли-
матологических данных (в особенности касающихся прошлых
лет), все-таки отчасти оправданно, так как все три рассмотренные
переменные тесно взаимосвязаны (Harris, 1980). Чем меньше
среднегодовое количество осадков, тем сильнее выражен их се-
зонный характер и ниже предсказуемость. Засушливые экосис-
темы нередко характеризуются сильно выраженной «сезонно-
стью» и подвержены локальным и периодическим колебаниям
выпадения осадков; в более влажных биогеоценозах продолжи-
тельность засушливых сезонов незначительна (или таковых
здесь вообще не бывает), а ее годовые колебания выражены
слабо. Как будет показано в гл. 8, эта сезонность играла важ-
ную роль в эволюции ранних гоминид.
5.2.2. Биотопы
Рассмотренные выше три взаимосвязанные переменные оп-
ределяют структуру биотопов в тропиках. «Влажным экстрему-
мом» тропических биотопов являются влажные тропические ле-
са, «сухим» — пустыни. Между этими крайними случаями рас-
полагается непрерывный ряд биотопов, классифицировать кото-
рые можно по-разному и с разной степенью детализации. Общее
•описание основных типов тропических биотопов выглядит, од-
нако, довольно просто (Eyre, 1968; Cloudsley-Thompson, 1975;
Harris, 1980):
1. Влажный тропический лес характеризуется полностью
сомкнутым пологом и включает 3—5 ярусов. Деревья высокие,
покрыты лианами и эпифитами и на небольшой площади пред-
ставлены многочисленными видами. Для такого биотопа требу-
ется, как правило, 3000 мм осадков ежегодно с минимальной
•9*
132
Глава 5
продолжительностью периода засухи. Первичная продукция и
биомасса характеризуются высокими показателями; очень бога-
та фауна беспозвоночных, мелких млекопитающих и птиц. Круп-
ных млекопитающих, которые сумели приспособиться к услови-
ям влажного тропического леса, здесь мало, причем основную
их группу составляют приматы.
2. Тропический лес с выраженной сезонностью встречается
во влажных тропиках, где значительную часть года занимает
период засухи. Лесной покров сплошной, но тоньше, чем во
влажном тропическом лесу. Преобладают древесные формы,
представленные непрерывным рядом от полувечнозеленых до
сезонно листопадных видов.
3. Широколиственный тропический лес постепенно переходит
в редколесье, лишенное непрерывного древесного покрова. Де-
ревья в широколиственном тропическом лесу полувечнозеленые,
с крупными стволами, покрытыми толстой огнестойкой корой.
В зависимости от количества выпадающих осадков биотопы это-
го типа, встречающиеся главным образом на глубоких песчаных
или кислых почвах, могут частично перекрываться с соседними
биотопами других типов.
4. Колючий лес доминирует в зонах с более продолжитель-
ным засушливым сезоном. Деревья снова представлены вечно-
зелеными формами, приспособленными к сезонным изменениям
количества влаги, с мелкими, защищенными обильными колюч-
ками листьями. Деревья этих видов засухоустойчивы и огне-
стойки. Наличие в колючем лесу значительных открытых про-
странств привлекает в него крупных млекопитающих.
5. Саванна (см. также гл. 8)—термин, который широко ис-
пользуется для обозначения разнообразных и постепенно пере-
ходящих друг в друга типов тропических биотопов. Некоторый
порядок в терминологическую путаницу внес недавно Хэррис
.(Harris, 1980), охарактеризовавший саванну как тип тропиче-
ского биотопа, в котором преобладающей растительной формой
является трава. Хэррис различает три определенных, хотя и пе-
реходящих друг в друга, типа саванны: 1. лесистую саванну,
в которой основными формами нетравянистых растений являют-
ся деревья (обычно Acacia и Commiphora); 2. кустарниковую
саванну, по которой рассеяны низкорослые травянистые кустар-
ники; 3. травяную саванну, в которой преобладает непрерывный
травяной покров. Следует подчеркнуть, однако, что биотопы пе-
речисленных типов располагаются мозаично и что в каждом из
них преобладает трава. Тип биотопа зависит от количества
осадков, дренажа, типа почвы и частоты пожаров. В саванных
€иотопах всех типов изобилуют представленные разнообразны-
ми видами крупные травоядные млекопитающие и охотящиеся
на них хищники.
Гоминиды как тропические животные 133
6. Полупустыня и пустыня. Настоящих пустынь в тропиках
нет, но нередко встречаются здесь чрезвычайно засушливые об-
ласти с низким годовым уровнем осадков в виде непредсказуе-
мых, имеющих сильно выраженный сезонный характер, дождей.
Условия для роста травы в таких областях слишком суровы,
и главными формами растительности здесь являются устойчивые
к экстремальным ситуациям ксерофиты, способные запасать зна-
чительные количества воды и питательных веществ и быстро
реагировать на выпадение дождя. Очевидно, показатели пер-
вичной и вторичной биомассы и продуктивности здесь характе-
ризуются низкими значениями.
7. Высокогорные биотопы плохо вписываются в любую клас-
сификацию тропических сред, поскольку из-за деформации
условий существования, обусловленной снижением температуры
при большой высоте над уровнем моря, этим биотопам могут
быть присущи и черты биотопов умеренного и даже арктическо-
го климата.
Мы показали, что гоминиды относятся к отряду животных,
который возник и эволюционировал в тропическом климате,
а следовательно, и приспособлен к условиям этого климата.
В связи с этим мы попытались охарактеризовать особенности
тропических экосистем и тропической экологии. Эти особенно-
сти в первую очередь определяются влажностью — фактором,
от которого зависит и многообразие условий среды в тропиках,
п структура тропических биотопов разных типов.
Остается рассмотреть, каким образом эти экологические осо-
бенности могли отразиться на эволюции гоминид.
5.3. Ранние гоминиды в условиях тропиков
5.3.1. Терморегуляция
Из сказанного выше ясно, что адаптация к экваториальному
климату должна предполагать способность переносить высокую
температуру воздуха. Следовательно, можно думать, что раз-
витие этой способности было одной из тех адаптивных проблем,
с которыми столкнулись гоминиды в процессе эволюции. Это
предположение подтверждается и поистине замечательным
совершенством механизмов терморегуляции у человека. Совре-
менные люди, как и все высшие млекопитающие, являются
гомойотермными организмами, т. е. способны поддерживать
постоянную и высокую температуру тела за счет внутренней
теплопродукции и ее регуляции. У большинства млекопитаю-
щих независимость температуры тела от температуры окружаю-
щей среды обеспечивается за счет шерсти или волосяного по-
крова, который выполняет функцию изоляционного материала
134
Глава 5
(Me Nab, 1974). Так обстоит дело у большинства представите-
лей отряда приматов. Современный человек, однако, отличает-
ся от них отсутствием сколь-либо значительного волосяного
покрова, и регуляция температуры тела у него сильно зависит
от потоотделения (Eichna, Bean и др., 1945). В отличие от
большинства млекопитающих человек обладает лишь ограни-
ченной способностью регулировать температуру тела за счет
учащенного дыхания, однако при той же плотности и размере
потовых желез, что и у других приматов, интенсивность выде-
ления пота у человека выше, чем у какого-либо другого из
известных млекопитающих. Современные люди знают об этом
по поездкам в метро, где интенсивность потоотделения на про-
тяжении короткого времени может достигать двух и более лит-
ров в час и способна сохраняться на уровне 1 л-ч-1 более
длительное время (Weiner, Hellman, 1960; Ladell, 1964).
Тот факт, что в процессе эволюции у гоминид произошла
столь радикальная модификация системы терморегуляции и бы-
ла достигнута столь высокая ее эффективность, заставляет
предполагать, что тепловой стресс играл у них немаловажное
значение в отборе. Следовательно, необходимо определить усло-
вия, в которых может развиться тепловой стресс, и выяснить,
каким образом ему можно противостоять.
Температура воздуха в тропиках нередко близка к 30 °C,
однако фактически тепловой стресс зависит главным образом
от уровня солнечной радиации, который в свою очередь опреде-
ляется типом биотопа. Самый высокий уровень солнечной ра-
диации отмечается в пустыне, где он достигает 200 кал • м-2-ч~!.
В саванных биотопах этот показатель составляет обычно
100 кал-м~2-ч-1 (Ingram, Mount, 1975; Mount, 1979). Оказыва-
ют влияние на тепловой стресс и такие факторы, как отража-
тельная способность среды, влажность воздуха и ветер. Харак-
тер терморегуляции у животных, помимо прочего, связан с его
активностью в зависимости от суточных колебаний температу-
ры воздуха.
Что в этом отношении можно сказать о ранних гоминидах?
Насколько известно, вплоть до позднего плейстоцена или голо-
цена гоминиды в пустынях не обитали (см. разд. 10.1.1). Что
же касается ранних африканских гоминид, то, как свидетель-
ствуют палеоэкологические данные, самой сухой областью, где
находили их останки, была расположенная в Восточной Африке
местность Пениндж, хотя и она скорее всего представляла собой
травяную саванну (Isaac, 1965; 1967; 1984). Целый ряд данных,
однако, указывает на то, что гоминиды были сосредоточены
главным образом в саванном биоме (подробнее см. гл. 8) и,
следовательно, подвергались здесь достаточно высокой солнеч-
ной радиации. Районы саванн, как правило, характеризуются
Гоминиды как тропические животные 135
слабыми ветрами и низкой влажностью; первый из этих факто-
ров способствует развитию теплового стресса, второй препят-
ствует ему. Другими словами, хотя ранние гоминиды и обитали
в экосистемах со значительной солнечной радиацией, ее уро-
вень, по-видимому, не достигал тех экстремальных значений,
которые, как считается, могли бы вызвать развитие нового типа
терморегуляции.
5.3.2, Характер изменения активности
Многие авторы пытались объяснить высокую эффективность
терморегуляции у человека характером его активности (Monta-
gu, 1964; Campbell, 1974). Интенсивная мышечная деятельность
в жару ставит организм в очень сложное положение. Наиболее
широкое признание получило предположение, что развитие теп-
лового стресса связано с эволюцией охоты (Campbell, 1974).
Интенсивная двигательная активность, которой сопровождалось
преследование животных по саванне, по всей вероятности долж-
на была вызвать развитие новых механизмов охлаждения орга-
низма. Это объяснение, однако, страдает целым рядом недостат-
ков. Главный из них состоит в том, что потоотделение в ответ
на тепловой стресс развивается, как правило, с задержкой, и,
таким образом, оно вряд ли может принести организму какое-
либо облегчение непосредственно во время охоты (Newman,
1970). Кроме того, крупные хищники, по-видимому, от теплово-
го стресса не страдают. Противоречит приведенному выше
объяснению и тот факт, что в результате усиленного потоотде-
ления резко увеличивается потребность организма в воде, что
противоречит общей тенденции гоминид к расширению ареалов
пищевого поиска в связи с переходом на более высокий трофи-
ческий уровень (см. разд. 6.3.2).
Альтернативное объяснение также основывается на харак-
тере активности человека, но не в связи с причиной этой актив-
ности, а в связи со временем суток, когда она разворачивается.
Большинство животных, обитающих в тропиках, обнаруживают
четкие циклы активности, причем периоды минимальной актив-
ности («отдых») тесно коррелируют с периодами наиболее
высокой температуры воздуха и еще значительнее — с периода-
ми наиболее интенсивной солнечной радиации (рис. 5.6). Таким
образом, тепловой стресс, вероятнее всего, должен возникать у
тех животных, адаптация которых предполагает высокий уро-
вень активности в середине дня. Приуроченность активности к
дневным часам суток может быть связана с двумя обстоятель-
ствами. Первое заключается в том, что если пищи мало, живот-
ные должны затрачивать на ее поиск много времени. Так, в
сухое время года гелады тратят на поиски пищи гораздо больше
A. Г епада
В. Горилла
Рис. 5.6. Характер суточной активности у различных африканских млекопитаю-
щих. (Я. Dunbar, 1977; Б. Altmann, Altmann, 1970; В. Fossey, Harcourt, 1977;
Г. Wrangham, 1977; Д. Kruuk, 1972; Е. Schaller, 1972; Ж. Jarman, Jarman,
1973.)
Гоминиды как тропические животные 137
времени, а днем заметно активнее, чем в дождливый сезон
(Dunbar, 1977). Вторая причина может заключаться в том, что
полуденная активность животного позволяет ему избежать пря-
мой конкуренции с другими видами. Крайне важное значение
это обстоятельство может иметь для плотоядных и в особенно-
сти питающихся падалью животных. Острая конкуренция меж-
ду такими хищниками может разгореться, например, за туши
мертвых животных, и любая возможность избежать столкнове-
ния с более крупными хищниками этим плотоядным крайне
выгодна. Рассмотренные аспекты поведения при поисках пищи
будут обсуждаться и в последующих главах.
Таким образом, очевидно, существовали особые причины
для того, чтобы гоминиды проявляли активность, еще более
осложняющую им нелегкую жизнь в условиях жаркого климата,
причем естественный отбор позволил найти гоминидам из сло-
жившейся ситуации радикальный, хотя и «дорогостоящий» вы-
ход, чего совсем не потребовалось делать другим видам, спо-
собным избегать изнурительную жару.
Почему, однако, решение общей для всех млекопитающих
проблемы терморегуляции у человека произошло за счет исчез-
новения волосяного покрова и усиления потоотделения, т. е;
процессов, которые среди животных отмечаются крайне редко?
Прежде чем ответить на этот вопрос, необходимо выяснить, как
по-другому могла быть разрешена проблема терморегуляции
и в чем заключаются преимущества и недостатки этих альтер-
нативных решений.
Можно предложить несколько общих решений проблемы.
Во-первых, за счет увеличения площади поверхности тела от-
носительно его объема либо в результате относительного удли-
нения дистальных частей конечностей, либо вследствие умень-
шения размеров тела (правило Аллена и правило Бергмана);
Во-вторых, за счет расширения периферических кровеносных
сосудов, что могло бы сопровождаться более быстрым охлаж-
дением крови. В-третьих, путем избегания солнечной радиации
либо за счет снижения дневной активности, либо в результате
перехода к исключительно ночному образу жизни. В-четвертых,
за счет сокращения площади поверхности тела, подвергающей-
ся воздействию солнечной радиации. И наконец, благодаря
увеличению интенсивности дыхания. Таковы основные возмож-
ности млекопитающих справиться с проблемой терморегуляции
при перегреве. Каждый из этих способов решения проблемы,
однако, имеет свои недостатки.
В табл. 5.1 приводятся показатели длины конечностей у го-
минид и других приматов относительно массы тела. Представ-
ленные данные указывают на то, что у современного человека
произошло относительное удлинение конечностей, что, по-види-*
138
Глава 5
Таблица 5.1. Отношение общей длины конечностей к массе тела
у некоторых приматов
Виды приматов1)
Длина конеч-
ностей/масса
тела1)
(Х100)
Рыжий мышиный лемур (Microcebus rufus) 24,5
Крысиный маки (Cheirolagus major) 25,5
Серый гаполемур (Hapolemur griseus) 32,5
Лемур-вари (Verecia vaiegata) 30,8
Мохнатый индри (Avahi laniger) 39,1
-Короткохвостый индри (Indri indri) 35,2
Галаго Демидова (Galago demidoff) 32,9
Золотой потто (Arctocebus calabarensis) 38,9
Обыкновенный потто (Periodictus potto) 29,4
Карликовая игрунка (Cebuella pygmaea) 27,3
Золотистая игрунка (Leontopithecus rosalid) 30,2
Беличий саймири (Saimiri sciureus) 32,2
Шерстистая обезьяна Гумбольдта (Lagothrix lagotricha) 31,3
Зеленый колобус (Colobus verus) 31,7
Обыкновенный носач (Nasalis lavatus) 33,3
Мартышка талапойн (Micropithecus talapoin) 31,4
Павиан анубис (Papio anubis) 31,8
Белорукий гиббон (Hylobates lar) 50,0
Карликовый шимпанзе (Pan paniscus) 33,5
Горилла (Gorilla gorilla) 26,9
Современный человек (Homo sapiens sapiens) 42,7
*) Для всех приматов (за исключением человека) показатели рассчитывались по дан-
ным Джангерса (.lungers, 1985; табл. 1 и 2).
’) Приводимые показатели представляют собой сумму показателей Джангерса для
передних и задних конечностей, т. е. их можно рассчитать, по формуле (длина передних
конечностей+длина задних конечностей/мм)/маеса тела (г)0,33.
мому, сопровождалось и некоторым изменением терморегуля-
ции. С другой стороны, размеры тела человека в процессе
эволюции обнаруживали общую тенденцию к увеличению (под-
робнее см. гл. 6). Известно, что под влиянием теплового стресса
у современного человека происходит и расширение перифери-
ческих сосудов (Mount, 1979, р. 165—167). Этот процесс, одна-
ко, играет в терморегуляции лишь второстепенную роль, а кро-
ме того, он препятствует столь важному для человека поддер-
жанию постоянного и высокого уровня притока крови к мозгу.
Как уже говорилось ранее, отбор в связи с дневной активно-
стью, по всей видимости, происходит независимо, а следователь-
но, проблема терморегуляции вряд ли могла быть разрешена
за счет снижения уровня дневной активности (или активизации
ночного поведения). Четвертая возможность, связанная с умень-
шением у гоминид площади поверхности тела, подвергающейся
воздействию солнечных лучей, использовалась для объяснения
бипедии (Mount, 1970); соответствующая модель недавно обое-
Гоминиды как тропические животные 139
нована математически Уилером (Wheeler, 1984). Вертикально
стоящий человек получает лишь 2/3 той энергии, идущей непос-
редственно от солнца, которая достается четвероногому живот-
ному (этот показатель увеличивается по мере приближения
солнца к зениту). Хотя развитие бипедии вряд ли можно
объяснить одним только тепловым стрессом, последний, воз-
можно, является в данном случае важным, хотя и вторичным
фактором. Кроме того, как уже ранее подчеркивалось, усиление
теплового стресса прямо связано с увеличением уровня физи-
ческой активности (а не просто с увеличением расстояний во
время поиска пищи); во многих случаях активность эта застав-
ляет наклоняться, что уменьшает терморегуляторные преиму-
щества бипедии. И наконец, возможность решить проблему
терморегуляции за счет учащенного дыхания — основной меха-
низм терморегуляции у копытных и хищников, способный
обеспечить быстрое охлаждение организма. Этот механизм,
однако, эффективно действует лишь при кратковременных
вспышках активности, вслед за которыми и происходит быстрое
остывание животного. Для того чтобы этот механизм мог эф-
фективно работать на протяжении длительного времени, требу-
ются специальные процессы, противодействующие гипервенти-
ляции (Newman, 1970).
Таким образом, альтернативные потоотделению механизмы
терморегуляции, в большинстве случаев способные действовать
эффективно, оказываются неэффективными при продолжитель-
ной и интенсивной активности животного в условиях высокой
солнечной радиации. Тот факт, что в качестве основного термо-
регуляторного механизма у гоминид эволюционировало потоот-
деление, говорит о том, что ранние гоминиды в течение какого-
то времени вынуждены были существовать именно в таких
условиях. Решение проблемы терморегуляции за счет потоот-
деления, однако, обходится животному дорого. Организм вы-
нужден расплачиваться за высокий уровень потоотделения
повышенной зависимостью от воды. Современный человек мо-
жет выдержать лишь сравнительно небольшую потерю организ-
мом воды (до 10% от массы тела), а выпивать сразу много
воды он не способен (1 л за 10 мин по сравнению со 100 л,
которые за 10 мин может выпить верблюд). Интенсивное пото-
отделение у гоминид породило необходимость держаться побли-
зости от водных источников.
5.3.3. Зависимость от воды
Как уже подчеркивалось, вода играет в тропической эколо-
гии исключительную роль: уровень осадков определяет возоб-
новление источников пищи и их доступность для животных,
140
Глава 5
поверхностные воды определяют существование тех видов, кото-
рые нуждаются в регулярном питье.
Очевидно, это обстоятельство могло иметь целый ряд важ-
ных адаптивных последствий. Прежде всего, если гоминиды
зависели от воды столь же сильно, как современный человек
и большинство приматов, их ареалы должны были ограничи-
ваться зонами с непересыхающими источниками поверхностных
вод. В саваннах же такие источники встречаются очень редко.
Из сказанного следует, что гоминиды должны были либо
как можно полнее и эффективнее (учитывая конкуренцию со
стороны других животных, столь же сильно зависящих от воды)
утилизировать расположенные поблизости от воды источники
пищи, либо вести длительный пищевой поиск на более обшир-
ной территории и возвращаться на центральные участки своего
•ареала для утоления жажды. Пищевой поиск на территории,
заселенной многочисленными видами других животных, и к
Тому же не ориентированный на центральные участки ареала,
был бы малоэффективным. С другой стороны, длительный и
дальний пищевой поиск мог иметь для гоминид важные адап-
тивные последствия, связанные с увеличением уровня дневной
активности и модификацией локомоторной системы (см. гл. 7).
Другими словами, по всей вероятности, должна была устано-
виться положительная обратная связь между тепловым стрес-
сом, пищевым поиском и зависимостью от воды. По мере того
как увеличивался уровень дневной активности гоминид, возра-
стали и их потребности в воде. В результате увеличивались
расстояния, которые они вынуждены были ежедневно покры-
вать, что в свою очередь вызывало дальнейшее усиление как
теплового стресса, так и потребности в воде (рис. 5.7).
Рассматриваемый вопрос еще больше усложняется возмож-
ной связью между потоотделением и отсутствием волосяного
покрова. Нередко высказывается предположение, что потери
воды с потом возрастают при отсутствии на теле волос. По мне-
нию Ньюмена (Newman, 1970), однако, для эффективного
потоотделения отсутствие волос совсем необязательно, но если
волосяного покрова нет, потоотделение становится необходи-
мым. В этой связи Ньюмен полагает, что исчезновение волося-
ного покрова сопровождалось усилением теплового стресса, а
это в свою очередь стимулировало отбор в сторону высокой
эффективности потоотделения. Здесь уместно спросить, почему
вначале должно было произойти исчезновение волосяного покро-
ва? Уилер (Wheeler, 1985) недавно предположил, что необходи-
мой преадаптацией к исчезновению функционального волосяно-
го покрова была бипедия, которая привела к уменьшению
площади поверхности тела, подвергающейся солнечной радиа-
ции, и, следовательно, ослаблению теплового стресса. При
Гоминиды как тропические животные
141
Рис. 5.7. Связь между потребностью в воде и пройденным расстоянием у со-
временного человека; потребность в воде варьирует в зависимости от размеров
тела (Mount, 1979, табл. 7.1). Предполагается, что интенсивность потоотделе-
ния при ходьбе со скоростью 3 км/ч и температуре 35 °C составляет 965 г/ч
вертикальном положении тела возможно рассеивание теплоты
за счет отделения пота и испарения влаги с кожи (за исклю-
чением кожи головы и плеч).
В общем проблема перегрева и проблема воды для гоминид
в тропиках были, по всей видимости, тесно связаны. Новизна
природы терморегуляторных механизмов у человека заставля-
ет думать, что тепловой стресс был одним из основных факторов
отбора, которому подвергались гоминиды экваториальной обла-
сти; что тепловой стресс был тесно связан с высокой активно-
стью гоминид во время длительных и дальних перемещений в
поисках пищи и что ослабление теплового стресса могло быть
«куплено» гоминидами только ценой большей зависимости от
воды с вытекавшими отсюда серьезными последствиями для их
территориального поведения и пищевого поиска. Следует,
однако, отметить, что эта модель не учитывает характер источ-
ников пищи, которые использовались гоминидами. Основное
внимание уделяется в ней «режиму» суточной активности гоми-
нид, который, хотя и не поддается точной оценке по прошествии
столь длительного времени, все же, вероятно, был важным
фактором эволюции этих приматов.
5.4. Палеоэкологические изменения
Обсуждавшиеся выше проблемы выживания в значительной
мере не связаны с каким-либо определенным историческим кон-
текстом, а касаются скорее непреходящих законов термодина-
142
Глава 5
мики. Однако ряд проблем, вставших перед тропическими жи-
вотными, прямо был связан с тем обстоятельством, что условия'
жизни в тропиках не оставались чем-то неизменным, а подвер-
гались в доисторические времена значительным изменениям.
Изменения эти оказали на эволюцию гоминид сильнейшее влия-
ние.
5.4.1. Глобальные изменения климата
Классические представления о климатических и экологичес-
ких изменениях во время эволюции гоминид были сформулиро-
ваны Пенком и Брюкнером (Penck, Bruckner, 1909). На основа-
нии данных, полученных при изучении морен Швейцарских
Альп, эти авторы описали процесс постепенного глобального-
похолодания в конце плиоцена (виллафранкский ярус), за ко-
торым последовали 4 продолжительных периода интенсивного
оледенения, чередовавшиеся с более короткими и теплыми
межледниковыми эпохами. Хотя в текущем столетии эта модель
подвергалась различным модификациям и даже накапливались,
противоречащие ей данные, ее жизнеспособность оказалась
поистине изумительной. Хотя палеоклиматологи давно уже при-
знали гораздо более сложный характер климатических измене-
ний в позднем кайнозое, выяснение природы других процессов,
протекавших в четвертичном периоде, заняло гораздо больше-
времени (Roberts, 1984, р. 25).
Современные данные указывают на то, что постепенное сни-
жение температуры началось в раннем кайнозое (рис. 2.13).
Средняя температура воды у поверхности Северного моря в
палеоцене составляла 16 °C, в эоцене увеличилась приблизи-
тельно до 30 °C, а в раннем олигоцене упала ниже 10 °C (Ви-
chardt, 1978). В миоцене температура вновь поднялась до 15 °C,
но затем опять последовало ее быстрое снижение в плиоцене и
плейстоцене. Такая же картина обрисована Кеннетом (Kennett,
1977) для субантарктического пояса.
С периодом понижения температуры связаны заметные па-
леогеографические изменения, вызванные дрейфом материков.
Эти изменения включали смещения Австралии и Антарктики
в южном направлении и, что имело еще более важное значение,
обособление восточной оконечности моря Тетис, разделявшего-
до тех пор Африканскую и Евразийскую платформы (рис. 5.8).
Это обстоятельство сильно сказалось на циркуляции океанских
течений, а следовательно, и на характере климата, что, вероят-
но, повлекло за собой превращение устойчивых, широко распро-
страненных географически, поросших густым лесом тропических
и субтропических биотопов в гораздо менее устойчивые биотопы
с выраженной сезонностью.
Рис. 5.8. Биографические изменения в Старом Свете в миоцене. В начале этой эпохи (25—18 млн. лет назад) про-
исходило постепенное разделение Африки и Евразии морем. С этого времени между двумя материками суще-
ствовал сохраняющийся н поныне перешеек, более широкое распространение получили лесные биотопы
144
Глава 5
Оледенение в разных географических зонах началось в раз-
ное время. В Антарктике лед образовался около 10 млн. лет
назад, а оледенение, по мнению многих исследований, происхо-
дило 3—4 млн. лет назад (Shackleton, Opdyke, 1976; Mercer,
1978; Roberts, 1984). Мерсер (Mercer, 1978) полагает, что зна-
чительный ледниковый покров в Южном полушарии образовал-
ся около 3,5 млн., а в Северном — примерно 3,2 млн. лет назад.
На протяжении последних 3 млн. лет сформировался зрелый
ледниковый климат. Вначале, вероятно, оледенение было не
столь интенсивным, однако за последний миллион лет его
проявления участились и приняли более выраженный характер.
По-видимому, 800 тыс. лет назад каждые 100 тыс. лет начали
происходить ледниково-межледниковые циклы, а это означает,
что в середине и конце плейстоцена было 8 циклических изме-
нений климата (рис. 2.14) (Roberts, 1984). Признание цикличес-
кого характера климатических изменений в плейстоцене оживи-
ло интерес к астрономическим теориям климатических сдвигов,
впервые сформулированным Миланковичем и Кроллом.
Вообще говоря, в плейстоцене климат был холоднее, чем
сейчас, но и экстремальные ледниковые условия складывались
редко. Ледниково-межледниковые циклы, по всей видимости,
сопровождались постепенным снижением температуры и нача-
лом оледенения, за которым следовала быстрая дегляциация
(Broeker, van Donk, 1970; Roberts, 1984). Это хорошо иллюст-
рируется последним ледниковым циклом, который происходил
в позднем плейстоцене. Он начался около 100 тыс. лет назад
с постепенного падения температуры, а 40 тыс. лет назад
прервался более мягким межледниковым периодом. После этого
сложилась гораздо более суровая ледниковая ситуация, темпе-
ратурный и ледниковый максимумы которой отмечались при-
мерно 18 тыс. лет назад. Затем температура начала быстро,
расти, и 6—8 тыс. лет назад произошла дегляциация (рис.
5.9). Последняя, не закончившаяся еще межледниковая эпоха
началась около 10 тыс. лет назад.
5.4.2. Тропическая палеоэкология
Каким образом отразились рассмотренные климатические и
тектонические изменения на экологии тропиков, в частности
тропиков африканских, где обитали гоминиды? Спад температу-
ры в миоцене привел к возникновению более выраженного ши-
ротного климатического градиента, что сопровождалось частич-
ным «расщеплением» умеренных и тропических сред (Pearson,
1978, р. 170). В высоких широтах падение температуры было
выражено гораздо сильнее, чем в низких.
Гоминиды как тропические животные
14&
Стадии
Предположительная изотопов кислорода
летняя (из глубоководных
Пыльца деревьев. % температура, °C морских отложений)’
Рис. 5.9. Климатические кривые для позднего плейстоцена и голоцена, постро-
енные на основании анализа континентальных н морских отложений (Roberts,
1984). Цифрами со стрелками обозначен возраст отложений в тыс. лет, цифры
в скобках означают, что возраст отложений определен приблизительно. А. Се-
веро-атлантическая океаническая земная кора V 28—14; Б. Озеро Клир-Лейк,
Калифорния; В. Гранд-Пиль, Франция; Г. Северо-атлантическая океаническая
земная кора V 23—82
Согласно классическому представлению, тропики и субтро-
пики в миоцене были сплошь заняты лесом. В последнее время
стало ясно, что именно так обстояло дело в раннем миоцене
(23,5—16,0 млн. лет назад) (Van Couvering, Van Couvering,
1976) и что позднее картина сильно изменилась. Сведений о
том, каковы были экосистемы в Восточной Африке в позднем
миоцене, мало, но, судя по находкам из Форта Тернана
(14 млн. лет) и бассейна озера Баринго в Кении, можно думать,
что леса здесь в это время перестают быть сплошными и их
начинают вытеснять мозаичные саванные биотопы (Van Couve-
ring, 1980; Shipman и др., 1981; Hill, Drake и др„ 1984). Фор-
мирование в Африке саванн с выраженной сезонностью, кото-
рые окончательно утвердились здесь в плейстоцене, по всей
вероятности, заняло более продолжительное время и началось
по меньшей мере 70 млн. лет назад.
Результаты изучения отложений из Сивалика в Северном
Пакистане (около 8 млн. лет) подтверждают это предположе-
ние. По мнению Бэджели и Беренсмейера (Badgeley, Behrens-
meyer, 1980), условия сложившейся тогда гидрологической и
146
Глава 5
геоморфологической ситуации не обеспечивали из-за сезонных
изменений климата возможности существования сплошных лес-
ных или лесистых биотопов, так что в то время наблюдалась
мозаичность растительных сообществ — от лесов до открытых
травянистых пространств.
Несмотря на отсутствие сведений из других частей света,
складывается впечатление, что в миоцене — под влиянием как
тектонических, так и климатических сдвигов — происходили
длительные экологические изменения, приведшие к формирова-
нию более открытых, сухих и разнообразных тропических сред
с выраженной сезонностью.
Такие тропические среды окончательно утвердились в пли-
оцене и плейстоцене. Недостаток сведений не позволяет соста-
вить сколь-либо достоверную картину тропических экосистем в
плейстоцене 25 тыс. лет назад и ранее, однако, судя по отдель-
ным находкам, относящимся к тому периоду, можно предпола-
гать, что в Восточной и Южной Африке, как и в других частях
тропического пояса, к этому времени сложились условия,
характерные для настоящих саванн (Bonnefille, 1972, 1976;
Hendey, 1973; Klein, 1974; Urba, 1974, 1975; Isaac, 1984). Эти
данные указывают и на то, что за последние 5 млн. лет в тро-
пиках плавно сменялись самые разнообразные экологические
среды (подробнее об этом см. гл. 8), однако сведений о связи
между климатическими сдвигами и типом среды очень мало.
После основополагающей работы Соломона (Solomon, 1931)
долго считали, что периоды оледенения в северных широтах
'коррелировали с «плювиальными эпохами» (периодами усиле-
ния осадков) в тропиках. В результате большую популярность
получила точка зрения, согласно которой влажный тропический
лес в плейстоцене был распространен так же или даже более
широко, чем в настоящее время (Roberts, 1984). Проверка дан-
ных, касающихся уровня озерных отложений, на основании
которых и была сформулирована эта гипотеза (Butzer, Isaac
и др., 1972; Street, Grove, 1975), показали, что как для Восточ-
ной Африки, так и для других тропических зон верно как раз об-
ратное. Глобальный спад температуры привел к уменьшению ко-
личества осадков в тропиках и увеличению их сухости, а геогра-
‘фическое расширение зоны тропических лесов, по всей вероят-
ности, связано с ранними стадиями межледниковых эпох.
Сегодня бытует мнение (Flenley, 1979; Hamilton, 1982), что на
высоте, соответствующей уровню оледенения, площадь влажных
тропических лесов в Африке и Америке быстро сокращалась
(рис. 5.10). Этот новый взгляд иа тропическую палеоэкологию
помог объяснить многие биогеографические особенности тропи-
ков, так как стало ясно, что на протяжении длительных перио-
дов времени ^растительные и животные виды оказывались изо-
Гоминиды как тропические животные 14 7‘
лированными на «экологических островках». Подобное чередо-
вание возникновения и ломки географических барьеров сыграло-
важную роль в образовании новых видов и в других эволюцион-
ных изменениях.
Современные представления о тропических экосистемах мож-
но, пожалуй, свести к двум основным положениям: во-первых,
что характерные для современных тропиков сухие саванные
мозаики не являются результатом пожаров, вызванных челове-
ком, или его скотоводческой деятельностью, но образовались
вследствие длительного развития тропических зон, и, во-вторых,
что на протяжении длительных геологических эпох протяжен-
ность и географическое расположение тропических областей
изменялись, а эти изменения в свою очередь оказывали замет-
ное влияние на эволюционные и экологические процессы.
5.4.3. Адаптивные последствия
Как описанные выше изменения тропиков могли отражаться
на стратегии выживания ранних гоминид? Для ответа на этот
вопрос следует особо учитывать два обстоятельства. Во-первых,
на протяжении последних 10 млн. лет происходило непрерывное
(хотя и не всегда равномерное) расширение саванных биотопов..
Говоря экологическим языком, тропические экосистемы прини-
мали все более выраженный сезонный характер, причем доми-
нирующее положение в растительности постепенно занимала'
трава, а фауна приспосабливалась к этим условиям. Выжива-
ние в тропиках в конце третичного и в четвертичном периодах
должно было обеспечиваться развитием в условиях непрерывно
сокращающихся массивов влажного тропического леса либо
специализаций, либо адаптаций к новым средам. В гл. 8 такая
адаптация рассматривается специально, здесь же следует под-
черкнуть, что длительное превращение тропических экосистем
из лесов в открытые степи ставило перед животными серьезные
адаптивные проблемы. Сегодня из палеоэкологических данных
ясно одно: выраженные климатические и экологические сдвиги
не ограничивались только плейстоценом и происходили не
только в высокоширотных зонах, непосредственно подвергшихся
оледенению. Эволюционные последствия долгосрочного эколо-
гического сдвига имели гораздо более широкое эволюционное
значение.
Второе обстоятельство связано скорее не с самим экологиче-
ским сдвигом, а с его последствиями. В результате разрыва не-
прерывного пояса влажного тропического леса и его вытеснения
саваннами формировались изолированные экологические остро-
ва, мозаичные экосистемы и пограничные сообщества или эко-
тоны. Независимо от особенностей этих экологических структур,,..
!0*
148
Глава 5
А
----1 Лишенная растительности
t I каменистая пуст-ыня
Пустынные травы и кустарники
gg'—1 Полупустынная степь с ксерофитными
• j -кустарниками
р‘ -/I Травяные среды
МИ Саванна и (или) кустарниковый лес
Г- - | Открытое редколесье
ffjfeg Средиземноморские кустарники
Недифференцированная растительность
речных долину
Широколиственный влажныйтропический
Недифференцированные лес
высокогорные сообщества
Саванна
Гоминиды как тропические животные
149
Рис. 5.10. Африканские среды обитания: А — настоящее время; Б — конец мио-
цена— плиоцен; В—ледниковый период; Г — межледниковый период. (Воп-
nefille, 1984; Roberts, 1984). Цифрами на рис. Б обозначены места пребывания
ранних гоминид: 1 — Эфиопия; 2 — бассейн озера Туркана; 3 — бассейн озера
Баринго; 4 — Северная Танзания/Южная Кения; 5 — Трансвааль, Южная Аф-
рика
150
Глава 5
их формирование имело серьезные эволюционные последствия.
Изолированные биоценозы (такие, например, как острова
влажного тропического леса в «море» саванны или наоборот)
создают подходящие условия для образования небольших
изолированных популяций, естественный отбор в которых спосо-
бен принимать высокоспецифический характер и протекать
весьма интенсивно, а генетические изменения — происходить
быстро и непредсказуемо. Таким образом, важным следствием
экологического сдвига явилось формирование условий, способ-
ствовавших быстрым эволюционным процессам и появлению и
сохранению новых форм.
Характер распространения разных животных (в особенно-
сти лесных видов) в зависимости от палеоэкологических изме-
нений интенсивно изучался в Африке и Южной Америке.
Постепенное разрушение сплошных массивов влажных тропи-
ческих лесов в Африке, зачастую сохранявшихся только в гор-
ных убежищах, привело к аллопатрическому образованию боль-
шого числа видов (Moreau, 1969; Laurent, 1973, Hamilton, 1982).
То же самое было обнаружено и для многих видов рыб, обита-
ющих в африканских тропических озерах, то сообщавшихся
между собой, то разъединявшихся в результате колебания
уровня дождей в плейстоцене (Beadle, 1974; Greenwood, 1974).
Здесь, как вероятно, и в случае многих других организмов и
экосистем, видообразование обусловлено тем, что любая фраг-
ментация среды может привести к эволюционному новшеству
вследствие как дрейфа генов, так и дивергентного отбора (Мауг,
1963).
Важное эволюционное значение могли иметь и мозаичные
экосистемы, н границы между биотопами, так как экотоны не-
редко дают животным возможность осваивать местность за
пределами тех биотопов, границей которых они, собственно
говоря, и являются. Это — экологические ниши, к которым осо-
бенно хорошо подходят животные, толерантные к широкому
разнообразию биотопов и условий существования (эвритопные
организмы, к числу которых относятся и гоминиды). И наконец,
палеоэкологический сдвиг мог действовать и как дестабилизи-
рующий отбор, открывая путь к эволюционным изменениям.
Можно утверждать, что непостоянство экосистем в простран-
стве и во времени способствует эволюционным изменениям.
Сегодня ясно, что именно такие биогеоценозы и доминировали
в тропической Африке в более поздних эпохах кайнозойской
эры и что гоминиды и их предшественники в ответ на измене-
ние этих экосистем должны были подвергаться естественному
отбору. В общем окружающая среда изменялась в сторону
формирования более открытых биотопов с более выраженным
сезонным характером. Учитывая тропическое происхождение и
Таблица 5.2. Эволюция гоминоидов в Зависимости ot экологических изменений в тропиках
Геологическая Динамика экологических
эпоха сдвигов
Характеристики
популяций
Характер эволюции
Эволюция гоминоидов
Средний миоцен Устойчивый влажный Крупные разнородные Стабилизирующий отбор: Ранние по большей
лес: сплошная однород- популяции с низким, низкая скорость видооб- части древесные аф-
ная среда но постоянным пото- разования и медленные риканские гоминоиды
ком генов эволюционные изменения
Поздний миоцен Увеличение аридиости, Изоляция небольших Дизруптивиый отбор: Дивергенция африкан-
и плиоцен сопровождающееся обра- популяций, сопровож-дрейф генов и локальный ских гоминоидов; воз-
зованием сезонных и са- дающаяся прерывани- отбор, способные вызвать никновение гоминид
ванных сред; мозаичные ем потока генов быстрые эволюционные
среды; фрагментация био- изменения н быстрое вн-
топов н образование ре- дообразование
фугий и экологических
островов
Плиоцеи и плей- Усиление экологической Небольшие локальные Дизруптивиый отбор и Радиация гоминоидов
стоцен неустойчивости н ускоре- популяции, обнаружи- вымирание локальных по- и гоминид
кие экологических сдви- вающие изменчивость пуляцнй, а также пере-
гов и подвергающиеся мещение популяций, при-
влиянию экологиче- водящие к образованию
ских сдвигов разрывов между пере-
крывающимися и смеж-
ными популяциями
Средний мио- Вулканическая и текто- Дальнейший изоляция Усиление прерывистого Возникновение пред-
цен — плейсто- ннческая активность в популяций под влия- потока генов и отбора посылок для эволю-
цен Восточной Африке в ре- нием географических ции восточноафрикан-
зультате образования и факторов ских гоминоидов
растяжения рифта
152
Глава 5
особенности отряда приматов, можно думать, что возникавшие
перед ними проблемы, связанные с выживанием в пространст-
венно разнородной и динамической среде, разрешалась путем
быстрых и радикальных эволюционных изменений.
В табл. 5.2 отражено соответствие между описанными об-
щими тенденциями в экологическом изменении тропиков, дина-
микой эволюционных сдвигов и возникновением гоминид.
Африканские гоминоиды в середине миоцена обитали в слабо
дифференцированных средах в больших гетерогенных популя-
циях с низким, но постоянным уровнем потока генов. Увеличе-
ние аридности, по всей вероятности, сопровождалось расшире-
нием саванных биотопов и вызвало общую тенденцию к адап-
тации к саванным условиям (см. гл. 7). Поскольку расширение
саванны означало также и формирование более дифференциро-
ванной и более изменчивой экосистемы, сочетание изоляции и
дизруптивного отбора должно было вызвать в популяциях
гоминоидов снижение потока генов и все усиливавшуюся дивер-
генцию. Ускорение экологического сдвига в конце третичного и
начале четвертичного периода1, по-видимому, способствовало
дальнейшему углублению этих процессов в результате локаль-
ного вымирания организмов и, следовательно, усиления генети-
ческой и адаптивной разобщенности популяций. Вулканическая
и тектоническая активность в Восточной Африке и в особенно-
сти образование значительных географических барьеров (на-
пример, лавовые потоки) еще больше усугубили общую тенден-
цию к адаптивной и филогенетической дивергенции гоминоидов.
Другими словами, экологическая динамика в Африке на протя-
жении последних 10 млн. лет создала подходящие условия —
как в плане формирования адекватных экосистем, так и в плане
возникновения соответствующих биогеографических предпосы-
лок— для радикальных эволюционных изменений гоминид за
это время. Помимо общих экологических процессов, однако, в
эволюционной экологии гоминид важную роль сыграли и сугубо
специфические характеристики этих млекопитающих. Одна из
таких особенностей заключалась в крупных размерах тела ран-
них гоминид.
1 В настоящее время третичный период разделяют на палеогеновый и нео-
геновый, а четвертичный период обычно называют антропогеновым. — Прим,
перев.
Глава 6
ГОМИНИДЫ КАК КРУПНЫЕ
МЛЕКОПИТАЮЩИЕ
В науке, как и в повседневной жизни, часто не замечают
самого очевидного. Сказанное, например, относится к размерам
животных. Никого не удивляет, что слоны большие, а мыши
маленькие. Поскольку все животные одного и того же вида
имеют приблизительно одинаковые размеры, редко кто задумы-
вается над тем, почему собственно животные достигают именно
этих размеров и каким образом размеры животного связаны с
его поведением, физиологией и адаптацией. Современные дан-
ные, однако, все чаще указывают на то, что размеры тела
животного — важнейший фактор и следствие адаптации (Klei-
ber, 1961; Huxley, 1972; Jarman, 1974; Clutton-Brock, Harvey,
1977b; Eisenberg, 1981; Hervey, Clutton-Brock, 1981; R. D. Mar-
tin, 1981; MacMahon, Bonner, 1983; Peters, 1983). Размеры орга-
низма— не просто конечный результат отбора и адаптации, но
и существенный направляющий фактор эволюции. Еще более
важное значение, по мнению автора этой книги, имеет то обстоя-
тельство, что зависимость между размерами тела и адаптацией
животного можно предсказать (а в отдельных случаях и оце-
нить количественно) на основании некоторых общих геометри-
ческих и биоэнергетических законов. Эти законы позволили
биологам разработать ряд действенных моделей, с помощью
которых можно объяснить некоторые эволюционные процессы.
Для палеоантропологов эти модели ценны тем, что позволяют
провести количественное сопоставление эволюционных и адап-
тивных возможностей гоминид.
До самого последнего времени при изучении эволюции
гоминид на их размеры обращали сравнительно мало вни-
мания. Итненсивно исследовался и широко дискутировался
лишь вопрос о размерах головного мозга, причем давно уже
утвердилось мнение, согласно которому энцефализация (увели-
чение головного мозга) сыграла в эволюции человека исклю-
чительную роль. Не менее важно, однако, отметить, что эволю-
ция человека сопровождалась увеличением не только мозга, но
и размеров тела. Если самые первые гоминиды были очень
небольшими и весили зачастую менее 30 кг, то уже в среднем
плейстоцене некоторые гоминиды имели крупные размеры и
154
Глава 6
массивное телосложение. Изменение размеров тела гоминид —
важный ключ к пониманию эволюции человека.
В этой главе делается попытка показать, что многие уни-
кальные характеристики гоминид и особенности их эволюции
на самом деле вполне правомочно рассматривать как следствие
их крупных размеров и теплокровности. Сделать это можно с
помощью основных законов аллометрии, описания изменений
размеров гоминид в процессе их эволюции, сравнительного изу-
чения коррелятов крупных размеров тела и, наконец, выяснения
вопроса, в какой степени гоминиды способны приспосабливаться)
к более разнообразным экологическим условиям.
6.1. Размеры тела и эволюция
6.1.1. Аллометрия
Аллометрия — область знаний, описывающая изменение про-
порций тела при изменении его размеров (Reeve, Huxley, 1945;
р. 123). Аллометрический рост предполагает изменение формы
тела в результате разной скорости роста отдельных его частей.
Классические представления в этой области были разработаны
Д. Томпсоном в книге «Вопросы роста и формы тела» (D’Arcy
Thompson, 1917), в которой было показано, что вследствие раз-
ной скорости роста отдельных частей тела в организме могут
происходить значительные сдвиги. Таким образом, морфологи-
ческие особенности организма могут прямо определяться его'
размерами, и изменение размеров организма легко может при-
вести к изменению его морфологии.
В последовавших затем работах большое внимание уделя-
лось количественной оценке и объяснению природы этой зави-
симости. Дж. Хаксли (J. Huxley, 1924) первым показал, что
общий закон постоянного соотношения между разными скоро-
стями роста отдельных частей организма может быть выражен
следующей формулой:
где у — некая неизвестная биологическая величина, которую»
можно определить на основании В (масса тела или какой-ни-
будь показатель его размеров) и х и k (константы, определяе-
мые для данной популяции, вида или какого-либо другого так-
сона эмпирически) (Peters, 1983; р. 1). Другими словами, раз-
меры тела организма соответствуют многим другим его
биологическим характеристикам, которые поддаются определе-
нию, если известен коэффициент этого соответствия (основан-
ный на х и k). С тех пор как появилась работа Хаксли, приве-
денная выше формула многократно использовалась для описа*
Гоминиды как крупные млекопитающие
155
Рнс. 6.1. Схематическое изображение того, каким образом изменение линейных
размеров тела может привести к аллометрическим изменениям его размеров
в двух- и трехмерном пространстве (площади поверхности и объема)
ния различных частных случаев, в которых фигурировали
такие факторы, как размер головного мозга, скорость метабо-
лизма, толщина костей, а также разнообразные экологические
взаимодействия (см. ниже).
Зависимость от размеров тела, например, его формы можно
проиллюстрировать некоторыми геометрическими построениями.
Как известно из общей геометрии, если форма не изменяется,
удвоение линейных размеров сопровождается четырехкратным
увеличением площади поверхности (рис. 6.1). Объем тела при
этом увеличивается в 8 раз. Для животного это означает, что,
если форма тела не изменяется, увеличение его высоты в два
раза будет сопровождаться восьмикратным увеличением массы
тканей. Этому геометрическому закону подчиняются не все
аллометрические зависимости, зато все они подчиняются тому
общему принципу, что увеличение размеров организма связано
с изменением других его характеристик неодинаково. При уве-
личении высоты животного в два раза должна изменяться и его
форма, ибо в противном случае произойдет чрезмерное воз-
растание массы тела.
6.1.2. Эволюция млекопитающих и размеры тела
Описанный подход плодотворно используется для объяснения
некоторых основных тенденций эволюции и адаптации. У жи-
вотных вообще, а у млекопитающих в особенности, эволюцион-
ные изменения сопровождались изменением размеров тела; это
прекрасно иллюстрируется эволюцией лошади, чьи предки в
эоцене были величиной с небольшую собаку (Romer, 1959).
Тенденцию к эволюционному увеличению размеров тела нередко
называют законом Копа, и хотя далеко не все организмы в
процессе эволюции укрупняются (альтернативное направление
обсуждается в работе Маршалла и Корручини; Marshall, Сог-
ruccini, 1978), эта тенденция, безусловно, отмечается очень ча-
156
Глава 6
сто (Bonner, 1968). Ее объясняли теми преимуществами, кото-
рые крупные размеры тела дают животному в плане выживания
в опасном и проникнутом конкуренцией мире: увеличение эф-
фективности охоты — для хищников, повышение шансов убе-
жать от хищника — для жертв, а также развитие интеллекта,
совершенствование терморегуляции и метаболизма, увеличение
продолжительности жизни и т. д. (Newell, 1949; Kurten, 1953;
Rensch, 1959; Gould, 1966; Bonner, 1968; Stanley, 1973). Прове-
денные недавно исследования (Eisenberg, 1981; Peters, 1983)
подтвердили, что некоторые из перечисленных факторов дейст-
вительно имеют важное адаптивное значение, однако Стэнли
(Stanley, 1973) занял в этом вопросе более эволюционистскую
позицию. По мнению Стэнли, закон Копа на самом деле пред-
ставляет собой лишь статистическое и вероятностное обобщение.
Стэнли отмечает, что в любом случае в результате естествен-
ного отбора достигаются оптимальные размеры тела — будь то
их увеличение или уменьшение. Преимущественное направление
отбора в сторону укрупнения животных объясняется тем, что
именно крупные размеры тела обычно являются оптимальными
(и необходимыми) для специализации организма, причем такая
специализация редко приводит к дальнейшему образова-
нию видов и появлению новых таксонов. Новые виды, особенно
дающие начало таксонам более высокого порядка, обычно
образуются из адаптивно и морфологически неспециализирован-
ных предков, а это — животные небольших размеров. Пред-
ставители нового таксона также будут небольшими, но обнару-
жат тенденцию к увеличению размеров тела.
Эволюция млекопитающих началась с небольших животных
и включала укрупнение размеров тела. Эта тенденция, однако,
не имела всеобщего характера. Действительно, хотя в любом
ряду поколений обнаруживается тенденция к увеличению мак-
симальных и средних размеров, минимальные размеры и даже,
быть может, мода размеров тела, несмотря на весь разброс
этого показателя, остаются на прежнем уровне. Со временем
появляются виды таксона с более крупными размерами тела, но
это не означает обязательного исчезновения мелких видов. Ги-
потеза Стэнли (1973) разъясняет реальное направление эво-
люции и предсказывает преимущественную, но не абсолютную
тенденцию к эволюционному укрупнению животных. Многочис-
ленные примеры этого явления наблюдались и в эволюции
млекопитающих, когда средние размеры животных увеличива-
лись, а мода этого показателя оставалась в общем невысокой
(рис. 6.2).
Проведенный анализ эволюционных механизмов изменения
размеров позволяет заключить, что новые эволюционные направ-
ления должны зарождаться в популяциях животных с неболь-
Гоминиды как крупные млекопитающие
157
Рис. 6.2. Частотное распределение млекопитающих по размерам тела. (Eisen-
berg, 1981.)
шими размерами тела и что по мере эволюционного развития
в некоторых таксонах должно происходить укрупнение живот-
ных. Остается выяснить, соответствовала ли описанной динами-
ке эволюция размеров тела у гоминид и как с точки зрения
адаптации и отбора можно объяснить то обстоятельство, что
оптимальными для гоминид стали крупные размеры тела.
6.2. Размеры тела у гоминид
Эйзенберг (Eisenberg, 1981) установил, что длина тела
большинства млекопитающих не превышает 32 см (рис. 6.2).
Таким образом, при росте около 172 см современный человек
приблизительно в 5—6 раз крупнее «среднего млекопитающего»
по линейным размерам (и в гораздо большей степени по объе-
му и массе). Таким образом, человек — достаточно крупное1
млекопитающее. Более того, и среди приматов он относится к*
S58
Глава 6
Фис. 6.3. Частотное распределение ви-
дов приматов по весу самок. (Clutton-
Brock, Harvey, 1977а.)
одному из самых крупных ви-
дов (рис. 6.3). Резонно заклю-
чить, что общая эволюционная
тенденция к укрупнению охва-
тывала и гоминид. Поэтому,
вероятно, в эволюции гоминид
участвовали те факторы отбо-
ра, которые «приводят в дей-
ствие» закон Копа и которые
могут способствовать понима-
нию некоторых аспектов эво-
люции этих млекопитающих.
6.2.1. Определение
размеров тела
Проследить эволюцию мас-
сы тела гоминид не так про-
сто. Имеются многочислен
ные данные, касающиеся
массы современного человека
в разных популяциях и (в
меньшей степени) массы ныне живущих приматов (см. Clutton-
Brock, Harvey, 1977а; Jungers, 1985), однако определение
размеров тела ископаемых животных сопряжено с известными
трудностями. Судить об этих трудностях можно по тому раз-
бросу показателей массы тела, которые были рассчитаны для
австралопитековых (табл. 6.1)—от 10 до 43 кг для Australo-
pithecus africanus и от 36 до 80 кг для Australopithecus robus-
tus (Steudel, 1980; Grine, 1981; McHenry, 1982).
Принципы определения массы тела ископаемых животных
.достаточно просты. С помощью корреляционного и регрессион-
ного статистических методов рассчитывается связь между жи-
вой массой и анатомическими характеристиками, которые мож-
но определить и у ныне живущих, и у ископаемых животных.
Для гоминид и приматов рассчитываются следующие основные
зависимости:
1. Позвонки. Площадь поперечного сечения последнего
поясничного позвонка у современного человека коррелирует с
массой тела r=0,72 (McHenry, 1975, р. 686—687). Это значит,
что, зная площадь поперечного сечения позвонка, массу тела
ископаемого гоминида можно определить по следующей фор-
муле:
lg №=0,6 lg V+1,1,
Гоминиды как крупные млекопитающие 15&-
где W—масса тела (в кг), а V — площадь поперечного сечения
последнего поясничного позвонка (в кв. мм).
2. Бедренные кости. Подобным же образом коррелирует у
современного человека с массой тела и размер головки бедрен-
ной кости (r=0,68) (McHenry, 1976, р. 79—80):
W = 0,028 (площадь поперечного сечения головки) +
+ 0,018 (площадь поперечного сечения шейки) +
+ 0,013 (площадь поперечного сечения диафиза) —
— 12,07.
3. Р. Мартин (R. A. Martin, 1980, 1981) выявил у млекопита-
ющих корреляцию (г=0,98) между массой тела и максималь-
ным диаметром головки бедренной кости, а также между мас-
сой тела и максимальным диаметром затылочных мыщелков.
Последнюю зависимость этот автор использовал для оценки’
размеров тела ранних гоминид (г=0,98):
W = 4.0Х10С,
где W—масса тела (в граммах), а С — диаметр затылочного»
мыщелка (в миллиметрах).
4. Лавджой и Хейпл (Lovejoy, Heiple, 1970) используют раз-
меры бедренной кости для оценки роста Australopithecus africa-
nus, а затем на основании роста рассчитывают массу его тела.
Подобным же образом определяет массу тела Australopithecus-
Уолпофф (Wolpoff, 1973), пользующийся для пересчета роста*
(в сантиметрах) в массе (в килограммах) коэффициентом 3,7.
Мак-Генри (McHenry, 1974) также определял рост гоминид и
обсуждал возможность использования этого показателя для
расчета их массы, но сам такие расчеты не делал. Учитывая
проблемы, связанные с эволюционным изменением пропорций
конечностей, можно думать, что определение массы на основа-
нии роста, по всей видимости, дает менее точные результаты,
нежели прямые методы оценки массы.
5. Как подчеркивает Стьюдел (Steudel, 1980), регрессии,
связывающие массу тела с анатомическими характеристиками
отдельных его частей, позволяют определять массу тела в мо-
мент смерти. Люди редко умирают в расцвете сил, а потому
этот показатель может и не отражать живую массу организма.
В качестве альтернативного подхода эта исследовательница
пользуется для расчета размеров тела неполной массой скелета
(табл. 6.2). В неполную массу скелета входит масса основных
частей скелета (черепа, нижней челюсти, обеих бедренных ко-
стей, обеих плечевых костей, таза, крестца и обеих лопаток).
Хотя этот показатель и не соответствует истинной массе тела,
общую массу перечисленных костей можно использовать для
расчета относительных размеров тела. Стьюдел показала, что»
160
Глава 6
Таблица 6.1. Определение
Образец
Место находки
Таксон
Sts 5 Стеркфонтейн A. africaaus
Sts 14 > »
Sts 14
Sts 73 >
— > »
Sk 3981 Сварткранс A, robustus
Sk 82
Sk97 > »
KNM-ER 738 Кооби-Фора Australopithecus Sp. (boi- sei)
KNM-ER 1503 »
OH 5 Олдувай A. boisei
KNM-ER 1472 Кооби-Фора Homo sp.
KNM-ER 1481
неполная масса скелета у разных видов наиболее тесно корре-
лирует с шириной таза (г = 0,96), шириной обеих скуловых ко-
стей (г=0,98), шириной глазных впадин (г=0,97) и окружно-
стью бедренной кости непосредственно под малым вертелом
(г=0,98).
Жене-Варжен (Genet-Vargin, 1966) и Робинсон (Robinson,
1972) также определяли массу австралопитековых, однако под-
робностей об использовавшихся при этом методах не сообща-
ют. Пилбим и Гоулд (Pilbeam, Gould, 1974) пользовались в
своих расчетах разными методами (поскольку эти авторы не
соотносят полученные результаты с конкретными образцами, их
данные в табл. 6.2 не включены). Стрингер (Stringer, 1984) для
определения размеров тела поздних гоминид использовал метод
Стьюдел.
6.2.2. Размеры тела ископаемых гоминид'
Есть веские основания полагать, что метод Стьюдел позволя-
ет оценивать размеры гоминид более адекватно, нежели опреде-
ление с помощью нескольких разных методов некоего «средне-
го» показателя, как это делают некоторые авторы. Во-первых,
метод Стьюдел основан на межвидовых регрессиях, а не толь-
ко на регрессиях, описывающих современного человека. Поэто-
му для описания гоминид он подходит больше. Во-вторых,
коэффициенты регрессии у Стьюдел гораздо выше, чем, напри-
Гоминиды как крупные млекопитающие
161
массы тела ранних гоминид
Установлен- ная масса тела, кг Объект реконструкции т. массь| тела Иеточник
25,4 10,5—27,6 18—22 43,0 37,3 10,7—36,1 49,8 52,7 43,1 Затылочный мыщелок R. A. Martin, 1981 Позвонки McHenry, 1975 Бедро->рост->-вес Lovejoy, Heiple, 1970 Позвонки McHenry, 1976 Бедро->рост->-вес Wolpoff, 1973 Позвонки McHenry, 1975 Бедро McHenry, 1976 » » » »
49,1 52,4 51,3 54,3 » Затылочный мыщелок R. A. Martin, 1981 Бедро McHenry, 1976 > »
мер, у Мак-Генри. И в-третьих, определение размеров по не-
полной массе скелета, по всей видимости, дает возможность
избежать искажений показателей, связанных со смертью орга-
низма. Главным недостатком метода Стьюдел является то,
что он позволяет получать данные в виде неполной массы
скелета, а не истинной массы тела, что исключает их непосред-
ственное использование для биоэнергетических расчетов. Как
бы то ни было, на основании этих данных можно получить
приблизительное представление о том, как изменялись размеры
тела гоминид в процессе эволюции (рис. 6.4).
Из рисунка видно, что самые ранние гоминиды, представлен-
ные грацильными австралопитеками (Australopithecus afaren-
sis и A. africanus), имели небольшие размеры, хотя
около двух млн. лет назад отмечалось их незначитель-
ное и постепенное укрупнение. С появлением Ното
erectus размеры тела гоминид в интервале между 2
и 1 млн. лет назад начали увеличиваться быстрее. Эта
тенденция продолжалась и достигла максимума к концу средне-
го плейстоцена, представленного образцами, которые нередко
рассматриваются как переходные формы между Homo erectus
и Homo sapiens. В промежутке между 100 и 35 млн. лет назад
с появлением анатомически современного человека размеры те-
ла гоминид уменьшались, хотя у современных людей отмечается
значительный разброс этого показателя.
Чтобы объяснить такую динамику размеров гоминид с точ-
ки зрения общих принципов эволюционной экологии, вначале
необходимо изучить экологические корреляты размеров тела.
11—367
Таблица 6.2. Определение неполной массы скелета ранних гоминид
(Steudal, 1981) н их сравнение с ныне живущими гоминоидами1'
Образец Место находки Тайрон Современный примат, с которым данный таксон сопоставим Логарифм неполной массы скелета, определенный на основании:
шири- ны неба ширины обеих скуловых костей ширины глазных впадин окруж- нести бедренной кости
Sts 17 Стеркфонтейн A. africanus Средний орангутан 2 2,39 2,5
Sts 5 » Небольшой орангутан 2 2,39 2,34
SK48 Сварткранс A. robustus Средний или крупный 2,44 2,45
орангутан 2
SKH Небольшой орангутан 2 2,33
SK79 » » Средний орангутан 2 2,4
SK52 > » 2,46
SK46 » Средний орангутан 2,46
TM 1517a Кромдраай » 2,59 2,74
KNM-ER 406 Кообн-Фора A. boisei Крупный орангутан 2,54 2,94 2,77
Sts 19 Стеркфонтейн A. africanus Средний орангутан $ 2,5
KNM-ER 736 Кообн-Фора ? 2,81 2,47
KNM-ER 738 ? 2,47
OH 20 Олдувай A. boisei Небольшой орангутан $ 2,64
SK82 Сварткранс A. robustus » 2,59
SK97 » 2,64
*) Логарифм частичной массы скелета ныне живущих гоминоидов составляет: у Ран 3 —2,50; Pan
Gorilla <3 —2,66; Gorilla 9 —2,47; Pongo 3 —2,66; Pongo 9 —2,47; Homo 3 —2,64: Homo $ —2,56.
9 -2,45;
Гоминиды как крупные млекопитающие
163
6.3. Экологические корреляты размеров тела
6.3.1. Метаболизм
Увеличение размеров тела изменяет характер связей между
различными отделами и функциями организма. Пожалуй, наи-
более сильное изменение в процессе эволюции претерпела
зависимость между размерами тела и энергетическим обменом,
что наглядно отражает классическая кривая Клайбера. Хотя,
очевидно, крупным животным для жизнедеятельности требуется
больше энергии, чем мелким, характер этой зависимости изме-
няется, если вместо абсолютных энергетических показателей
рассматривать относительные. Как видно из рис. 6.5, с увеличе-
нием размеров тела количество энергии, необходимое для под-
держания основного обмена, на единицу массы тела (измерен-
ное по потреблению кислорода за единицу времени), снижает-
ся. На основании этой зависимости можно рассчитать
регрессию, показывающую, что скорость основного обмена —
количество энергии, необходимое для поддержания основных
биологических функций,— прямо зависит от размеров тела (рис.
3,0
•и
9» 12—13
• 10
•14
•15
• —5------ .8
•7
• 6
.3
•2 4*
2,5 2,0 1,5 1,0 0,5 0
Млн. лет назад
Рис. 6.4. Связь между логарифмом неполного веса скелета ископаемых гоми-
нид и геологическим возрастом находки: 1—Australopithecus afarensis; 2 —
A. africanus; 3 — Homo habilis (образец KNM-ER 1470); 4—ранний Homo
(образец KNM-ER 1813); 5 — среднее значение и разброс показателей массив-
ных австралопитековых; 6—8 — Homo erectus (6 — образец KNM-ER 3733; 7 —
образец KNM-ER 3883; 8 — Ява-Сангиран); 9—11 —ранний Homo sapiens (9 —
Петралона; 10 —Кабве; И—Бодо); 12 — средневосточный неандерталец
(Ирхуд); 13—14 — западноевропейский неандерталец (13 — Ла-Феррасси;
14 — Ла-Шапель-о-Сэн); 15 — анатомически современный человек (средний
показатель)
11*
8
Рис. 6.5. Кривая Клайбера — потребление кислорода на единицу массы в за»
висимости от размеров тела. (Schmidt-Nielsen, 1975.)
Интенсивность метаболизма, ккал’Ч
Рис. 6.6. Связь между размерами тела и интенсивностью основного обмена
у животных разных групп. (Schmidt-Nielsen, 1975.)
Гоминиды как крупные млекопитающие 165
6.6) и что в логарифмическом масштабе увеличение массы тела
на единицу требует возрастания скорости обмена всего на 0,75
единицы. Эта зависимость может быть описана следующим
уравнением:
Л1 = /СГ°>75,
где М — скорость основного обмена (ккал/ч), W—масса тела
(г) и К—постоянная.
В появившемся недавно обзоре Питерс (Peters, 1983, р. 45—
46), проведя обстоятельный анализ данных и моделей, касаю-
щихся этого вопроса, показал, что, хотя степенной показатель
этой зависимости, по мнению всех авторов, составляет 0,75, в
действительности зависимость может быть иной. Питерс (Pe-
ters, 1983, рис. 4.1) установил, что в среднем экспонента состав-
ляет 0,738, ее мода — 0,74, а стандартное отклонение — 0,11.
Причины столь постоянной зависимости между рассматривае-
мыми показателями до конца еще не выяснены. Сделанное ра-
нее предположение о том, что она отражает теплопотери,
соответствующие отношению поверхности тела к его объему
(0,67), не подтверждается фактической величиной экспоненты
(0,75) (Kleiber, 1961; Peters, 1983).
6.3.2. Последствия увеличения размеров тела
Каковы бы ни были причины рассмотренной зависимости, из
нее вытекают важные экологические и эволюционные послед-
ствия: увеличение размеров тела сопровождается возрастанием
абсолютных, но снижением относительных потребностей в пище.
Это означает, что с укрупнением животных их зависимость от
источников пищи изменяется, а действие на них естественного
отбора усиливается. Характер этих изменений можно опреде-
лить теоретически, так как известна их энергетическая основа.
Между изменением размеров тела животного и его экологичес-
кими и физиологическими характеристиками существуют кор-
реляционные отношения. Чтобы лучше понять экологические и
эволюционные аспекты изменений размеров тела гоминид, про-
ведем анализ этих корреляций. Все приведенные ниже рассуж-
дения касаются только млекопитающих.
1. Увеличение размеров тела может расширить пищевую ни-
шу животного. Количество энергии, которое ежедневно требу-
ется для поддержания обмена мелкому млекопитающему, соот-
ветствует значительной части его массы (табл. 6.3 и 6.4).
Поскольку увеличение размеров животных сопровождается бо-
лее медленным ростом их энергетических потребностей, более
крупному животному на единицу массы ежедневно требуется
меньше энергии. В экологическом плане это означает, что
мелкие млекопитающие либо должны потреблять богатую
166
Глава 6
Таблица 6.3. Примеры идеальной зависимости между массой тела
и метаболическими потребностями у млекопитающих
Масса тела (М), г Интенсивность основного обмена (R). ккал/день . ллмйА M/R (Х100)
10 2,04 20,4
100 11,7 11,7
1000 67,6 6,67
10 000 389,0 3,89
100 000 2 238,7 2,24
1 000 000 12882,5 1,28
Таблица 6.4. Масса тела, интенсивность осиовиого обмена и метаболическая
эффективность у ныне живущих и ископаемых* приматов
Вид Масса тела (M), г Интенсив- ность основ- ного обмена (R), ккал/день M/R' (Х100)
Callithrix jacchus 241 22,9 9,5
Theropithecus gelada 9 830 384,1 3,9
Pan troglodytes 39 000 1 094 2,8
Australopithecus africanus* 36 000 1 029 2,86
Australopithecus robustus* 59 000 1 499,2 2,54
Homo sapiens sapiens 60 000 1 518 2,53
энергией пищу, либо кормиться беспрерывно, в то время как
более крупные млекопитающие могут обойтись и менее каче-
ственными пищевыми продуктами. Можно предположить также,
что если одни крупные млекопитающие занимают достаточно
широкую пищевую нишу, изобилующую низкокачественными
пищевыми продуктами, то другие могут благополучно сущест-
вовать, питаясь только высококачественной пищей. Это пред-
положение подтверждается тем обстоятельством, что среди
травоядных (то есть млекопитающих, потребляющих низкокаче-
ственную пищу) преобладают крупные животные (Eisenberg,
1981, р. 271—274), а также проведенным Клаттон-Брокком и
Харви (Clutton-Brock, Harvey, 1977) анализом связи между
типом пищи и размером тела у приматов.
2. Увеличение размеров животных сопровождается расшире-
нием территории, по которой они перемещаются в поисках пищи
на протяжении года (индивидуальный участок) или суток. Хотя
повышение массы тела и приводит к снижению относительных
пищевых потребностей животного, его общие или абсолютные
потребности в пище увеличиваются. В результате поиск пищи
животное вынуждено вести на более обширной территории. Это
положение было эмпирически подтверждено для многих млеко-
Гоминиды как крупные млекопитающие
167
Рис. 6.7. Связь между размерами тела и площадью ареалов у млекопитающих
разных групп. (А, Б, В-. Harested, Bunnel, 1979; Г: Glutton-Brock, Harvey,
1977b.)
питающих (McNab, 1963; Harested, Bunnell, 1979), включая и
приматов (Milton, May, 1975; Clutton-Brock, Harvey, 1975b)
(рис. 6.7). Соответствующие расчеты, однако, сильно осложня-
ются разным образом жизни у млекопитающих (одни из иих
живут и ведут поиск пищи группами, другие придерживаются
одиночного образа жизни) и различиями в продуктивности био-
топов. Таким образом, хотя, вообще говоря, укрупнение живот-
ных и сопровождается расширением территории, на которой
они ведут поиск пищи, зависимость эта имеет далеко не линей-
ный характер.
3. Увеличение размеров тела сопровождается возрастанием
мобильности животных. Небольшие млекопитающие менее мо-
бильны, чем крупные, так как сильнее ограничены в расходе
энергии (и подвергаются большему риску стать жертвой хищ-
ника). Это обстоятельство сильно влияет на адаптацию живот-
ных. Чтобы выжить, мелкое млекопитающее должно, например,
существовать на таком клочке земли, который либо предостав-
ляет животному неистощимый источник пищи, либо дает ему
возможность запасать пищевые продукты впрок (поведенчески
или физиологически).
Более высокая мобильность крупных животных делает их
менее зависимыми от подобных ограничений. Они могут пере-
1r8
Глава 6
Размеры жертвы
Рис. 6.8. Модель оптимальных размерон животных, поедаемых хищниками
ходить из одной местности в другую и, следовательно, исполь-
зовать более разнообразные источники пищи, а также источни-
ки, доступность которых в пространстве и времени ограниченна.
Так, одной из стратегий поиска пищи у крупных животных яв-
ляется «кормежка по случаю» пищей, доступной лишь очень
короткое время.
4. Увеличение размеров тела сопровождается изменением
отношений хищник — жертва. Возможности хищника (если
речь идет не о групповой охоте) ограничиваются размерами
жертвы, которую он может поймать (Bourliere, 1963). Опти-
мальные размеры жертвы определяются максимальной энерге-
тической выгодой от ее поимки и минимальным риском полу-
чить при этом травму (рис. 6.8). Таким образом, естественный
отбор способствовал увеличению размеров как животных-жертв
с целью оградить их от хищников, так и хищников —
с целью разнообразить их «меню». Учитывая принцип «жизнь
или обед» (например, кролик убегает от охотящейся на него
лисицы, чтобы спасти свою жизнь, а лисица преследует кроли-
ка, чтобы один раз поесть!), можно думать, что размеры живот-
ных-жертв увеличивались быстрее, чем размеры хищников.
5. Увеличение размеров тела сопровождается повышением
эффективности терморегуляции (уменьшением эффективности
охлаждения организма или увеличением его теплоизоляции и
способности к удержанию теплоты). Этот аспект укрупнения
животных подробно обсуждался в предыдущей главе (см. разд.
5.3). Как уже отмечалось, увеличение размеров тела улучшает
адаптацию к холодным средам и затрудняет адаптацию к жар-
Гоминиды как крупные млекопитающие 169
ким, поскольку задержание теплоты в организме, обусловленное
совершенством его теплоизоляции, в жаркой среде может ока-
заться серьезным недостатком.
6. Увеличение размеров тела, как правило, сопровождается
увеличением продолжительности жизни. У крупных млекопита-
ющих значительный вклад в репродуктивное и энергетическое
состояние всей популяции вносит каждая особь. Это обстоятель-
ство имеет два важных следствия. Первое состоит в том, что,
поскольку развитие организма ограничивается как внутренними,
так и внешними факторами, время, которое ему требуется для
достижения окончательных размеров, у крупных животных
больше, чем у мелких. Во-вторых, увеличение продолжитель-
ности жизни как способ максимально повышать репродуктивный
выход особую важность приобретает для тех животных, которые
затрачивают много времени и энергии на воспроизведение од-
ной особи. Вместе эти два фактора и способствуют более высо-
кой продолжительности жизни крупных животных; есть указа-
ния на то, что у некоторых крупных животных продолжитель-
ность жизни может выступать в качестве одного из факторов
полового отбора. У млекопитающих размеры тела положитель-
но коррелируют с продолжительностью жизни (Eisenberg, 1981),
однако у них обнаруживаются две четко различающиеся тен-
денции в отношении размножения. Первая состоит в воспроиз-
ведении небольшого числа хорошо сформировавшихся (то есть
хорошо развитых и в значительной мере независимых от роди-
телей) детенышей, вторая — в воспроизведении многочисленно-
го беспомощного (то есть очень незрелого и сильно зависящего
от родителей) потомства в каждом помете. Эти дивергентные
стратегии размножения в значительной мере определяют долго-
вечность и выживаемость животных, причем первая из них
связана с более высокой продолжительностью жизни. Вообще
говоря, приматы производят на свет довольно развитых детены-
шей; связь между средними размерами тела взрослых приматов
и регистрировавшейся продолжительностью их жизни представ-
лена на рис. 6.9.
7. Увеличение размеров тела обычно связано с замедлением
темпов размножения. Увеличение продолжительности жизни,
которым сопровождается укрупнение размеров тела у млекопи-
тающих, предполагает и удлинение всех жизненных стадий,
включая беременность и период постнатального развития. Сле-
довательно, увеличивается и интервал между событиями, свя-
занными с процессом размножения, а, стало быть, у крупных
животных общий темп размножения замедляется (рис. 6.10).
Это обстоятельство в некоторой степени может компенсиро-
ваться растяжением репродуктивного периода, но реальная пло-
довитость все равно не может быть высокой. Это обстоятельст-
170
Глава 6
3,0
3,0 4,0
log массы тела, г
5,0
Рис. 6.9. Связь между средними размерами взрослых животных и достоверной
продолжительностью жизни у приматов. (Eisenberg, 1981.)
во в свою очередь имеет важные эволюционные и адаптивные
последствия. У крупных млекопитающих каждый детеныш
представляет собой более значительную «часть» общего репро-
дуктивного выхода данной особи, чем у мелких. А следователь-
но, можно полагать, что каждый детеныш крупных млекопита-
ющих получит больше родительской заботы и что гибель одного
детеныша отразится на общем росте популяции крупных мле-
копитающих более сильно, чем у мелких. Это означает, что
эволюционную уязвимость крупных млекопитающих может
повысить всякий процесс, ухудшающий выживание их детены-
шей и, наоборот, что естественный отбор способствует закрепле-
нию всех адаптивных стратегий, которые повышают выживае-
мость их потомства.
8. Увеличение размеров тела может сопровождаться усиле-
нием общественных отношений у животных. Общественное
поведение у крупных млекопитающих, по-видимому, может
усиливаться за счет двух факторов. Первый из них прямо свя-
зан с обсуждавшимся выше замедлением темпов размножения.
Поскольку каждая особь у крупных млекопитающих вносит
значительный вклад в воспроизведение популяции, повышается
вероятность родственного отбора. Поведение родителей направ-
лено на обеспечение выживания каждого детеныша, а живот-
ные, находящиеся в горизонтальном родстве, обнаруживают по
отношению друг к другу альтруистическое поведение. Отбор в
популяциях крупных млекопитающих идет по способности
узнавать родственников и по способности вести себя по отно-
шению к родственникам иначе, чем к неродственникам. Это и
обеспечивается главным образом за счет социального поведе-
ния. Известно, что социальность, ориентированная на родствен-
ников, среди крупных млекопитающих распространена очень
Гоминиды как крупные млекопитающие
171
Масса тела, кг
Рис. 6.10. Связь между массой тела и темпами размножения у различных
африканских млекопитающих. 1 и 2— прыгунчиковые землеройки; 3— мангуст;
4 — антилопа-левретка; 5 — газель Томпсона; 6—газель Гранта; 7 — импала;
8 — бородавочник; 9 — болотный козел; 10 — гиена; 11 — водяной козел личи;
12 — лев; 13— обыкновенный бубал; 14— обыкновенный водяной козел; 15 —
антилопа-гну; 16 — зебра; 17—буйвол; 18—жираф; 19— носорог; 20 — беге-
мот; 21 — слон. (Western, 1979.)
широко (см. Wilson, 1975). Второй фактор связан с увеличенит
ем продолжительности жизни. Вероятность повторных встреч
одних и тех же животных друг с другом на протяжении жизни
выше у крупных животных. В результате отбор будет способ-
ствовать выживанию тех особей, которые на основании своего
прошлого опыта научатся по-разному реагировать на разных
животных. Таким образом удовлетворяются некоторые условия,
необходимые, по мнению Трайверса (Trivers, 1972), для эволю-
ции реципрокного альтруизма. Можно думать поэтому, что
групповой отбор, кооперация и альтруизм среди крупных
млекопитающих распространены более широко, чем среди мел-
ких.
9. Увеличение размеров млекопитающих может сопровож-
даться укрупнением головного мозга. О тесной связи между
размерами тела и величиной головного мозга было известно
давно (Brody, 1945; Jerison, 1961; Stahl, 1965). Эйзенберг
(Eisenberg, 1981) использовал эту связь для определения так
называемого «коэффициента энцефализации», на основании
которого можно судить, насколько мозг того или иного живот-
ного больше или меньше теоретически рассчитанной величины.
Р. Мартин (R. D. Martin, 1981) показал, что увеличение разме-
ров головного мозга связано с укрупнением тела экспоненци-
альной зависимостью, причем показатель степени составляет
0,75. Это не согласуется с величиной экспоненты 0,67, ранее
172
Глава 6
полученной Джерисоном (Jerison, 1973), и указывает на то,
что размеры мозга связаны с интенсивностью обмена, в особен-
ности у самок во время беременности. Таким образом, чем круп-
нее млекопитающее, тем меньше у него относительные размеры
головного мозга. Учитывая, однако, природу связи между
энергетическим обменом и размерами, можно думать, что высо-
кий коэффициент энцефализации «позволить себе» вправе лишь
немногие мелкие млекопитающие. Пожалуй, еще важнее, что
крупные размеры мозга вряд ли могут дать мелким млекопи-
тающим какие-либо преимущества, так как продолжительность
их жизни невелика. Наоборот, крупные млекопитающие в этом
отношении оказываются в большом выигрыше. Крупный мозг
обходится животному очень дорого (R. D. Martin, 1981), но
зато позволяет ему более гибко реагировать на изменения
окружающей среды. Как уже говорилось, в жизни крупного
млекопитающего случаются ситуации, когда гибкость поведения
может оказаться для него особенно полезной: использование
источников пищи, доступность которых ограничена в простран-
стве и времени, повторные встречи с другими особями и возни-
кающая при этом необходимость вести себя в соответствии с
прошлым опытом, необходимость различать родственников.
Важно и то, что долго живущие организмы должны уметь
противостоять изменениям окружающей среды. Все перечислен-
ные факторы можно причислить к факторам отбора, которые,
вероятно, способствуют энцефализации крупных млекопитаю-
щих.
Делались попытки связать многие из рассмотренных выше
характеристик с широко принятой моделью эволюционных стра-
тегий— моделью r-К-отбора. Еще в ранних работах по теоре-
тической экологии было сделано предположение, что рост
популяций характеризуется некоторой общей закономерностью,
которую можно описать сигмовидной кривой (рис. 6.11) и сле-
дующим уравнением:
R^rNll-4),
\ А /
где R — скорость увеличения популяции в данный момент вре-
мени, г — максимальная скорость экспоненциального роста
популяции или вида, N — число организмов и К — число осо-
бей, существование которых может обеспечить данная среда
обитания (вообще говоря, К соответствует так называемому
переносимому объему среды). На практике это означает, что
рост популяции определяется двумя факторами. Во-первых,
темпом воспроизведения популяции, т. е. свойственным ей уров-
нем рождаемости, и, во-вторых, способностью среды поддержи-
вать эту популяцию. Какой из этих двух факторов играет ре-
Гоминиды как крупные млекопитающие
173
Поддерживающая
емкость среды
Рис. 6.11. Связь между ростом популяции и г- и К-отбором. Если численность
популяции невелика по сравнению с переносимым объемом, скорость роста
популяции определяется ее размером и темпами размножения вида. Когда
численность популяции приближается к переносимому объему, определяющую
роль начинают играть факторы конкуренции
шающую роль, зависит от конкретных условий, в частности от
размера популяции (N). Если популяция невелика, скорость ее
роста главным образом определяется максимальной скоростью
экспоненциального роста (то есть основное значение имеет
показатель г). Если численность популяции значительна, более
важное значение приобретает N и все сильнее на ее росте
сказывается величина К (переносимый объем), которая оказы-
вает на рост популяции сдерживающее влияние. Таким обра-
зом, можно сказать, что скорость роста популяции в разное
время «контролируется» либо г, либо К.
Разработка этой очень простой модели популяции привела
к признанию двух основных стратегий в процессе эволюции.
Если плотность популяций того или иного вида гораздо меньше
переносимого объема, отбор будет направлен на увеличение
способности вида к быстрому размножению. Поскольку пищи
в таких условиях, хотя бы временно, имеется в достатке, темпы
размножения приобретают для вида более важное значение, не-
жели способность к конкуренции за источники пищи. Можно
сказать, что эволюция этого вида идет в направлении г-отбора.
Когда же плотность популяций близка к переносимому объему,
высокие темпы размножения теряют для вида свое значение.
Все больше особей «не могут найти себе места» в популяции,
174
Глава 6
и преимущество в процессе отбора получают те животные,
которые способны эффективно конкурировать за существующие
источники пищи. В этом случае можно говорить, что эволюция
вида идет в направлении /(-отбора.
Хотя, по мнению большинства авторов, эти две эволюцион-
ные стратегии не являются взаимоисключающими, можно пред-
полагать, что в общем эволюция видов происходит преимущест-
венно либо в г-, либо в /(-направлении. Выгоды, которые может
дать виду та или иная эволюционная стратегия, зависят от
конкретных условий окружающей среды, определяющих, кроме
Таблица 6.5. Корреляты г- н К-отбора (Pianka, 1978)
г-отбор Л-отбор
Климат Изменчивый и (или) непредска- Достаточно устойчивый »
зуемый; неопределенный (или) предсказуемый; бо- лее определенный
Смертность Часто катастрофическая, не имеющая направленного харак- тера и не зависящая от плотно- сти популяции Более направленного ха- рактера и зависящая от плотности популяции
Выживаемость Высокая смертность детенышей Более постоянный уро* вень смертности
Размер популяций Изменяющийся во времени, не- уравновешенный; обычно гораз- до меньше переносимого объема среды; ненасыщенные сообще- ства или ненасыщенные части сообщества; экологические пу- стоты; ежегодная реколони- зация Довольно постоянный, уравновешенный, более или менее соответствую- щий переносимому объе- му среды; насыщенные сообщества; в реколони- зации нет необходимости
Внутри- н межви- довая конкуренция Изменчивая, часто — слабая Обычно острая
Отбор способст- вует: Быстрому развитию Высокой максимальной скоро- сти увеличения численности, Г max Снижению репродуктивного возраста Сохранению мелких размеров Одноразовому воспроизведению потомства Воспроизведению многочислен- ного мелкого потомства Более медленному разви- тию Повышению конкуренто- способности Увеличению репродуктив- ного возраста Укрупнению размеров Повторному воспроизве- дению потомства Рождению меньшего чис- ла более крупных дете- нышей
Продолжитель- Короткая, обычно менее одного Более значительная,
ность жизни года обычно больше одного
Отбор приводит к: Увеличению продуктивности года Повышению эффектив- ности
Гоминиды как крупные млекопитающие 175
того, и особенности самой эволюции вида в том или ином на-
правлении (см. табл. 6.5). Замечено, что увеличение размеров
тела часто связано с /(-отбором. Многие виды, становясь круп-
нее, жертвуют темпами размножения (как уже говорилось, оии
понижены у крупных млекопитающих) ради повышения кон-
курентоспособности. Таким образом, можно предполагать, что
укрупнение гоминид в процессе эволюции было обусловлено
более выраженным /(-отбором.
6.4. Адаптивные последствия увеличения размеров гоминид
в процессе эволюции
Как было показано, изменение размеров тела можно рас-
сматривать и как причину, и как следствие адаптации. Отмеча-
лось и то обстоятельство, что значительное изменение размеров
тела у гоминид произошло в течение двух млн. лет. Попытаемся
теперь установить, насколько тесно могло быть связано увели-
чение размеров гоминид с экологическими и эволюционными
характеристиками, которые обсуждались в предыдущем разде-
ле. Все эти характеристики перечислены в табл. 6.6. Для этого
необходимо изучить данные, касающиеся адаптации ранних
гоминид, выяснить, соответствуют ли они теоретическим пред-
положениям, и определить, могли ли способствовать экологичес-
кие факторы развитию у ранних гоминид адаптивных изменений.
С этой целью рассмотрим вначале поиск пищи, а затем —
социальное и репродуктивное поведение ранних гоминид.
6.4.1. Поведение при поисках пищи
Относительно пищевого поведения гоминид (при условии
постоянной доступности пищи) можно сделать три основных
предположения. Во-первых, должно было происходить расши-
рение территорий гоминид. Во-вторых, должно было увеличи-
ваться разнообразие потреблявшейся ими пищи. И, в-третьих,
либо в пищевой рацион должна была включаться менее каче-
ственная пища, либо должно было сокращаться время, которое
они затрачивали на поиск пищи. Некоторые из этих предполо-
жений кратко уже обсуждались ранее (Foley, 1984b).
Совершенно очевидно, что современные люди — охотники
и собиратели — занимают более обширные территории, чем
приматы. Разница может составлять несколько порядков в поль-
зу охотников и собирателей (рис. 6.12). Отчасти это обстоя-
тельство можно объяснить тем, что очень большие территории
занимают охотники и собиратели, живущие в высоких широтах,
.где они являются единственными представителями приматов;
176
Глава 6
Таблица 6.6. Экологические последствия увеличения размеров
тела (Foley, 1984b)
Результаты увеличения
размеров тела
Экологические последствия
Увеличение абсолютного расхо-
да энергии
Уменьшение относительного
расхода энергии
Увеличение мобильности
Повышение задержки теплоты
в организме
Увеличение мышечной силы и
скорости передвижения
Увеличение размеров головного
мозга
Увеличение продолжительности
жизни
Удлинение пренатального пе-
риода
Возрастание пищевых потребностей; расши-
рение ареалов; снижение плотности популя-
ций
Повышение способности выжить при исполь-
зовании низкокачественной пищи
Повышение способности адаптироваться к
холодному климату и использовать более
разнообразные источники пищи
Повышение способности адаптироваться к
холодному климату и увеличение активно-
сти; усиление зависимости от воды; тепло-
вой стресс в тропических средах
Повышение способности сопротивляться
хищнику или убегать от него
Использование разнородных и разобщенных
в пространстве источников пищи; гибкость
поведения; усиление социальности; увеличе-
ние расхода энергии
Удлинение репродуктивного периода, удли-
нение периода развития
Увеличение репродуктивных затрат; сниже-
ние рождаемости; удлинение интервала меж-
ду родами; появление на свет развитого по-
томства
однако и в тропических широтах различия в размерах участков
выражены очень сильно.
И все же, в какой степени расширение ареалов могло быть
связано с изменением размеров тела? Милтон и Мей (Milton,
Мау, 1975) предложили уравнение, описывающее связь между
размерами тела приматов и площадью их территорий с учетом
того обстоятельства, что приматы живут группами:
lgH^=l,231gBW —2,86,
где HRI—общая площадь территории (га) и BW—средняя
масса тела взрослого организма (г). Рассчитанная по этому
уравнению площадь территории современных охотников и соби-
рателей должна составлять 8—10 км2, что гораздо меньше, чем
в действительности (рис. 6.13). Таким образом, обширные тер-
ритории современных охотников и собирателей хотя и соответ-
ствуют предположению об увеличении размеров этих территорий
Гоминиды как крупные млекопитающие
177
Л Эскимосы острова Баффинова Земля
А Современные тасманийцы
А Австралийские аборигены племени Кариера
<0
CJ
s
СО
«с
А Австралийские аборигены племени Гиджингала
А Сан-Кунг
♦ Мартышка гусар
♦ Желтый павиан
♦ Оливковый бабуин
♦ Медвежий павиан
+ Обыкновенный шимпанзе
♦ Горилла
♦ Орангутан
♦ Макак
♦ Серощекий мангобей
♦ Талапойн
♦ Чубастый мангобей
Ф Зеленая мартышка
♦ Гиббон
* Сиаманг
ф Голубая мартышка
Рис. 6.12. Сравнение площади территорий приматов (темные ромбики) и совре^
мениых охотников и собирателей (светлые треугольники)
12—367
478
Глава 6
"Теорети- 2 км
ческая" .(«ооби-
- Территория ф0 ;
4 км
(Кооби-
Фора)
Рис. 6.13. Определение площади территорий пищевого поиска ранних гоминид
иа основании расстояний между местом, где были оставлены орудия, и источ-
ником сырья для их изготовления; представлена также площадь территории,
теоретически рассчитанная по уравнению Милтона и Мея (Milton, Мау, 1975).
Подробные объяснения см. в тексте
с укрупнением тела, однако по своим истинным размерам нам-
ного превосходят теоретически ожидаемые.
Возвращаясь к ранним гоминидам, можно предположить,
что площадь их территории отличалась от площади участков
других приматов не столь разительно. С помощью уравнения
Милтона и Мея можно рассчитать, что площадь территорий ран-
них гоминид колебалась от 3,71 км2 у Australopithecus af are ri-
sis до 7,3 км2 у Homo erectus. Соответствуют ли эти цифры
истинной площади участков этих видов?
Понятно, что определить площадь территорий вида, вымер-
шего более миллиона лет назад,— не простая задача. Для боль-
шинства организмов она вообще невыполнима; однако способ-
ность гоминид к изготовлению орудий позволяет все-таки
произвести для них некоторые расчеты. Для определения мини-
мальных размеров участка можно воспользоваться расстоянием
между источником исходного материала (сырья) для изготов-
ления орудий и тем местом, где готовые орудия были впослед-
ствии оставлены; следует, однако, подчеркнуть, что все подоб-
ные расчеты основаны на том допущении, что территории имели
форму круга. По данным Айзека (Isaac, 1976, р. 561), гомини-
ды, жившие в плиоцене и плейстоцене в бассейне озера Турка-
на, переносили камни на расстояние не менее 4 км, что соот-
ветствует величине территории от 12,56 до 50,2 км2 в зависимо-
сти от того, принимается ли это расстояние за радиус или
диаметр участка. Подобным образом, судя по находкам в
ущелье Олдувай, площадь .критерий обитавших здесь гоминид
Гомнинды как крупные млекопитающие'79'
составляла от 113,1 до 452 км2 (Gowlett, 1984). Находки, отно-
сящиеся к более позднему времени, указывают на то, что впос-
ледствии в поисках сырья для изготовления орудий гоминиды
стали проходить более значительные расстояния (Gowlett,
1984, р. 174). Размеры территорий гоминид, рассчитанные на
основании всех этих данных, однако, меньше, чем теоретически
рассчитанные для приматов с такими же размерами тела (Fo-
ley, 1978; 1984b).
Чем можно объяснить столь обширные участки пищевого по-
иска гоминид? Понятно, что их формирование не связано толь-
ко с увеличением размеров тела, хотя этот показатель, возмож-
но, и определяет отчасти их вариабельность. Не исключено так-
же, что при описании территорий охотников и собирателей
некоторые авторы учитывают и иные, помимо поиска пищи,
формы активности, что, естественно, затрудняет их сравнение
с территориями приматов. Сказанное может относиться и к
приведенным выше оценкам участков ранних гоминид, основан-
ным на расстоянии между местом находки орудия и источником
сырья для его изготовления. Кроме того, многочисленные дан-
ные, касающиеся приматов, получены при изучении популяций,,
занимающих сравнительно продуктивные места обитания (на-
пример, лесные биотопы). Продуктивность открытых биотопов
меньше, и, следовательно, чтобы получить доступ к источникам
пищи, животные должны освоить более обширную территорию.
Размеры участков ранних гоминид, таким образом, могут быть
связаны с малой доступностью источников пищи в саваннах.
Большие размеры территорий могут указывать и на то, что го-
миниды (как предполагается) потребляли низкокачественную
пищу и, стало быть, для поддержания популяции были вынуж-
дены осваивать обширные площади. Не исключена, однако,
возможность, что расширение территорий отражает и изменение
характера питания на этой или более ранней стадии эволюции
человека. Как известно, всеядные и хищные животные занимают
более обширные участки, чем травоядные (Harested, Bunnell,
1979; см. также рис. 6.7), поскольку плотность источников жи-
вотной пищи меньше, чем растительной. Сравнительно большие
участки ранних гоминид, таким образом, могут быть связаны
с увеличением доли мяса в их пищевом рационе. Наконец, сле-
дует отметить и то обстоятельство, что было бы ошибкой при-
писывать всем без исключения гоминидам поиск пищи на обшир-
ных территориях. Вполне возможно, что гоминиды в этом
отношении значительно различались, и при тех методах, кото-
рыми мы располагаем, обнаруживаются лишь самые обширные
из их участков.
Второе предположение, связанное с поиском пищи, состоит
в том, что в пищевом рационе крупных животных должна быть
12*
180
Глава 6
представлена более разнообразная пища. Такое предположение
отчасти оправдано способностью крупных животных переме-
щаться из одной местности в другую и использовать доступные
в течение непродолжительного времени источники пищи, отча-
сти— их способностью потреблять пищу низкого качества.
Некоторый свет на этот вопрос может пролить изучение
того, как используют в настоящее время пищевые ресурсы охот-
ники и собиратели — люди, а также иные приматы. Питерс и
О’Брайен (Peters, O’Brien, 1981) составили список растений,
потребляемых в различных регионах континентальной Африки
человеком, шимпанзе и павианами. Оказалось, что по Африке в
'целом шимпанзе используют в пищу 168 видов растений, павиа-
ны— 163, а человек — 298. Хотя не следует упускать из виду,
что эти данные отчасти отражают общий характер распростра-
нения названных трех родов приматов в Африке, а также дей-
ствие других экологических факторов, они тем не менее указы-
вают на то, что современный человек использует в пищу очень
разнообразные продукты растительного происхождения. Корт-
лэнд (Kortland, 1984) обосновывает противоположную точку
зрения, что из приматов наиболее разнообразная пища требу-
ется шимпанзе. Другим подходом к изучению рассматриваемой
проблемы является определение видового состава растений,
использующихся в пищу в различных популяциях. В табл. 6.7
приводится число видов растений, потребляемых в Африке
разными группами охотников и собирателей — людей и других
приматов. Как видно из таблицы, в целом гоминиды исполь-
зуют в пищу больше видов растений, чем приматы из других
семейств. Кроме того, если в качестве компонента пищевого
рациона учитывать и мясо, человек по плотоядности будет
значительно превосходить прочих приматов. Шимпанзе и павиа-
ны также отчасти плотоядны, но по частоте, с которой они
охотятся на других животных, и по видовому составу своих
жертв они далеко отстают от современных гоминид (Teleki,
1975; Harding, Teleki, 1981) (табл. 6.8).
Нечего и говорить, что определить состав пищевого рациона
гоминид, живших в плиоцене и плейстоцене, гораздо труднее.
Источниками такой информации могут служить остатки иско-
паемых гоминид и археологические находки. Хотя на первый
взгляд кажется, что надежнее всего о пищевом рационе гоми-
нид можно судить по функциональной анатомии зубного
аппарата, на самом деле и морфология, и стертость зубов, к
сожалению, отражают скорее преобладание в питании гоминид
тех или иных продуктов, нежели разнообразие пищевого рацио-
на. Таким образом, несмотря на утверждение Грайна (Grine,
1981), что массивные австралопитеки использовали в пищу
более твердые, но более мелкие объекты, чем грацильные, а
Гоминиды как крупные млекопитающие
181
Таблица 6.7. Число видов растений, используемых |в пищу приматами
и современными охотниками и собирателями
Виды/группы Число видов растений Источник данных
Приматы Желтый павиан (Papio cynocephalus) 14 Post, 1981
Гривистый ревун (Alouatta palliata) 15 Hladik, 1981
Паукообразная обезьяна (Ateles geof- 22 Hladik, 1981
froyi) Белогорлый капуцин (Cebus capicinus) 40 Hladik, 1981
Красномордый лангур (Presbytis se- 12 Hladik, 1981
Пех) Гульман (Presbytis entellus) 23 Hladik, 1981
Обыкновенный шимпанзе (Гомбе) 207 Wrangham, 1977
(Pan troglodytes) Обыкновенный шимпанзе (Pan troglo- 330 Nishida, Llehara, 1983
dytes) (цит. no Teieki, 1981)
Обыкновенный шимпанзе (Pan troglo- 141 Te-leki, 1981
dytes) Современные охотники и собиратели Сан (Г/ви) 45 Silberbauer, 1981
Сан (!Кунг) 85 Lee, 1968
Сан (*Када) 79 Tanaka, 1976
Аборигены западно-австралийских 40 Gould, 1969
пустынь Аборигены Северного Квинсленда 240 Roth, 1901 (цит. no Hay- den, 1981)
также мнение Уокера (Walker, 1981), что австралопитековые
потребляли в основном такой же «фруктовый» (то есть состояв-
ший главным образом из плодов) рацион, что и шимпанзе и
орангутаны, науке до сих пор мало что известно об истинном
разнообразии питания у этих гоминид. По мнению других ав-
торов, специализация зубов австралопитековых, отличающая их
от других и более поздних гоминид, заставляет предполагать,
что пищевые рационы австралопитековых были менее разно-
образными, чем у представителей рода Homo (Jolly, 1970;
Dunbar, 1976).
Немногим достовернее и информация, полученная из археоло-
гических источников. Сохранность ископаемых остатков очень
различна. Кости животных сохраняются лучше растений; одни
растения сохраняются лучше, другие — хуже. Находки, в ко-
торых вместе с остатками ранних гоминид обнаруживались и
остатки макроскопических растений, очень малочисленны
(Isaac, Crader, 1981; Isaac, 1984), и судить о разнообразии пище-
Таблица 6.8. Охотничье поведение приматов и современных охотников и собирателен (НШ, 1982)
Добытчик мяса
Суточное потре- Процент суточ-
п л Добыча мяса на Средняя мае- бление мяса, кг, ного потребления
добыча мяса одного потребите- са взрослой в отношении калорий, покры-
за час, кг ля в день, кг особи, Кг к массе тела ваемый за счет
(кгХЮ-’) мяса
Приматы
Желтый павиан 0,006 0,036 35 0,1 3
Павиан анубис 0,002—0,004 0,016-0,32 25 0,06—0,13 2—5
Шимпанзе — 0,027 35 0,08 3
Охотники и собиратели из племени:
Аче (вооруженный луком) 0,53 1,78 55 3,2 80
Аче (невооруженный) 0,27 — — — —
Янамаме (вооруженный луком) 0,98 0,44 44 0,92 29
1Кунг (вооруженный луком) 0,60—0,66 0,325 44,7 0,73 20,5
Г/ви (вооруженный луком) 0,34 0,32 48,1 0,675 21,5
Мбути (охота с сетью) 0,38 0,8 40 2,025 65
Мбути (лучник) 0,33 40 0,83 27
Бари 0,16—0,25 0,39 Т“ — —
Аибарра — 0,55 40 2,7 26
Гоминиды как крупные млекопитающие 183
вого рациона этих приматов по ним трудно. Находки, когда
вместе с останками гоминид обнаруживались кости других
животных, и наличие на этих костях резанных отметин указы-
вают на то, что ранние гоминиды подвергали животные продук-
ты обработке (Bunn, 1981; Potts, Shipman, 1981; Isaac, 1983;
Potts, 1984a), хотя мнения исследователей о характере и степе-
ни такой обработки очень противоречивы (Binford, 1981). Тем
не менее обилие в этих находках костей с резанными отметина-
ми и разнообразие представленных костями видов животных,
свидетельствуют о том, что гоминиды использовали в пищу
более широкий по видовому составу круг жертв, нежели павианы
« шимпанзе.
Таким образом, вообще, говоря, о разнообразии пищевого
рациона ранних гоминид практически ничего не известно. Круп-
ные размеры гоминид могут свидетельствовать о высоком раз-
нообразии рациона, однако единственным достоверным под-
тверждением этого предположения является, вероятно, исполь-
зование гоминидами в пищу мяса. Третье предположение, свя-
занное с пищевым поиском и во многом дополняющее первые
два, касается возможного перехода гоминид к утилизации
источников пищи низкого качества. Размеры крупных животных
позволяют им потреблять грубую растительную пищу относи-
тельно низкой питательной ценности (Eisenberg, 1981). Лучшим
примером таких животных среди приматов является горилла.
Зто — самый крупный представитель ныне живущих приматов,
питающийся главным образом листьями. Несмотря на отсутст-
вие специализированного пищеварительного аппарата, гориллы
вполне благополучно существуют на такой диете, поскольку
листья всегда имеются в изобилии, а гориллы способны потреб-
лять их в огромных количествах. Кей и Хайлендер (Кау,
Hylander, 1978) сделали еще более общее наблюдение, устано-
вив, что млекопитающие, питающиеся листьями, как правило,
крупнее родственных им плотоядных и насекомоядных живот-
ных. Подобную тенденцию, связанную с увеличением размеров
тела, могли обнаруживать и ранние гоминиды (рис. 6.14). Про-
тивоположные стратегии питания, заключающиеся в увеличении
потребления мяса, с одной стороны, и использовании менее
качественной растительной пищи — с другой, подробно рассмат-
риваются в гл. 8, в которой также делается попытка связать
экологические аспекты укрупнения ранних гоминид со специ-
фическими условиями среды их обитания. Размеры тела — не
просто главный фактор, определяющий поведение при поисках
пищи. Увеличение размеров животных приводит к потере ими
одних адаптивных возможностей и приобретению новых. Какими
из этих новых возможностей животные воспользуются, зависит
от других факторов, в частности от прочих факторов естествен-
184
Глава 6
Дальнейшее увеличение размеров
тела, сопровождающееся более
эффективной утилизацией низко*
качественных пищевых продуктов
Дальнейшее увеличение размеров тела,
сопровождающееся увеличением мышеч.
ной силы, повышением мобильности и
возрастанием потребления мясной пищи
Увеличение размеров теле,
трабующее большого разно-
образия пищевого рациона
и более обширного ареала
t
Мелкие плодоядные
’'родоначальные"
гоминиды
Рис. 6.14. Изменение пищевого поиска гоминид в результате увеличения раз-
меров тела
ного отбора, действующих на данную популяцию, или, пользуясь
принятой в этой книге терминологией, от того, с какими эколо-
гическими проблемами придется столкнуться гоминидам.
6.4.2. Поведение в связи с размножением
Другой формой поведения, через которую размеры тела мо-
гут влиять на адаптацию животных, являются репродуктивное
поведение и связанные с ним социальные взаимодействия. Ясно,
что такое разделение адаптивных эффектов поведения носит
условный характер, поскольку поиск пищи влияет на размноже-
ние, и наоборот. Так, те преимущества в пищевом поиске, кото-
рые животные получают в результате увеличения размеров
тела, нередко сочетаются с менее благоприятными изменениями
их социального и репродуктивного поведения. В качестве край-
него аргумента можно привести то соображение, что необыкно-
венная сложность социального поведения человека — та цена,
которой он вынужден расплачиваться за приобретенные им
преимущества при поисках пищи.
Предположения, которые можно сделать относительно пове-
дения при размножении на основании экологических корреля-
тов, приводятся в табл. 6.6 (разд. 6.4.1). Особенно важное
значение имеют два взаимосвязанных последствия увеличения
размеров животного. Первое состоит в том, что в результате
совместного действия ряда факторов плодовитость животных
должна уменьшаться, а вероятность выживания каждого дете-
ныша увеличивается. Второе заключается в том, что отбор
должен способствовать эволюции более сложного социального
поведения — отчасти вследствие усиления родительской заботы
Гоминиды как крупные млекопитающие
185
Увеличение
размеров тела
Увеличение
периода роста
организации связей, основанных
сообщества на родстве
Рис. 6.15. Предполагаемые связи между увеличением размеров тела и эволю-
цией сложного социального поведения
о детенышах, отчасти — в результате увеличения продолжитель-
ности жизни. Связь между всеми этими переменными изобра-
жена на рис. 6.15.
Изложенной точки зрения придерживается Лавджой (Love-
joy, 1981). По мнению этого автора, эволюция гоминид как груп-
пы животных происходила в сторону укрупнения размеров (в
отличие от эволюции низших узконосых), что привело к сниже-
нию скорости роста популяций. Чтобы выжить, эти приматы
должны были либо обитать в безопасных средах, где было
много пищи и мало хищников, либо усилить заботу о потомстве.
Лавджой считает, что африканские высшие обезьяны последо-
вали по первому пути, а гоминиды — по второму. Шимпанзе,
орангутаны и гориллы обитают в лесистых местностях со слабо
186
Глава 6
выраженной сезонностью. Напротив, гоминиды жили в средах
с сильно выраженной сезонностью и изобиловавших хищника-
ми, однако преодолели это неудобство усилением родительской
заботы о потомстве. Большая родительская забота у людей и,
как можно предполагать, у гоминид — решающий фактор эво-
люции человека. Чтобы материнская особь могла уделять за-
боте о детенышах больше времени, необходимо было ее осво-
бождение по крайней мере от некоторых форм активности,
связанных с пищевым поиском. А это в свою очередь требовало
выполнения двух условий. Во-первых, самец должен был обес-
печить самку и детеныша пищей. Во-вторых, чтобы самец мог
пользоваться выгодами более эффективного поиска пищи,
необходимо было образование моногамной пары. Такие взаи-
модействия наблюдаются у многих птиц. По мнению Лавджоя,
однако, чтобы такая стратегия могла возникнуть в эволюции
человека, необходимо было развитие у него двух неотъемлемых
человеческих признаков — прямохождения (бипедии), которое
позволило ему брать пищу руками и относить ее детенышам,,
и дележа пищи между сородичами независимо от того, участво-
вали они в ее добывании или нет. В результате как следствие
демографических сдвигов, связанных с укрупнением тела, воз-
ник основанный на родственных отношениях дележ пищи в
некоем «наземном гнезде», социальная структура которого осно-
вывалась на приоритете отца. Таким образом, можно думать,
что эволюция социального поведения человека определялась
как увеличением его размеров, так и действием факторов окру-
жающей среды.
Модель Лавджоя хорошо согласуется с предполагаемыми
экологическими и эволюционными последствиями укрупнения
тела (см. разд. 6.3.2). Предсказываемые ею элементы коопера-
тивного поведения можно подкрепить следующими рассужде-
ниями. С увеличением размеров тела, вероятно, увеличивается
продолжительность жизни (см. разд. 6.3.2). В этой связи можно»
думать, что в процессе естественного отбора преимущества
получают те особи, которые способны различать сородичей, ибо»
только в этом случае они в состоянии адекватно вести себя во
время встреч с другими особями, основываясь на собственном!
прошлом опыте конкурентных или кооперативных взаимодей-
ствий с ними. Таким образом, отбор должен был способство-
вать эволюции более сложного и гибкого социального поведе-
ния, которое позволяет животным извлекать пользу из послед-
ствий своей предшествующей активности или, наоборот, нести
от них убыток. Преимущества должны были получать те особиц
которые могли оптимизировать отдачу от вложенных ими &
кооперативное поведение усилий (Houston, Davies, 1985).
В адрес модели Лавджоя, несмотря на все ее достоинства^.
Гоминиды как крупные млекопитающие 187
можно высказать и ряд критических замечаний. Согласно этой
модели, бипедия должна была возникнуть на ранних этапах
эволюции гоминид. Как говорилось в гл. 2 (разд. 2.2.2), обна-
руженная в Летоли цепочка следов свидетельствует о том, что
характерная для человека двуногая походка сформировалась
у гоминид еще приблизительно 3,6 млн. лет назад (Leakey,
1978). Гораздо сомнительнее аргументы, касающиеся возникно-
вения дележа пищи и жилища (Binford, 1981; 1984; Isaac, Сга-
der, 1981; Isaac, 1983; 1984; Potts, 1984b). Кроме того, предска-
зываемое моделью Лавджоя высокое развитие у гоминид
кооперативного и моногамного поведения заставляет думать,
что половой диморфизм у них был выражен слабо; как, однако,
было установлено, у многих из ранних гоминид обнаруживались
четкие половые различия. С другой стороны, не столь резко,
по-видимому, как может показаться на первый взгляд и как
полагает Лавджой, выражен и контраст между гоминидами и
прочими гоминоидами. Если орангутаны и гориллы действитель-
но обнаруживают очень низкие темпы размножения и живут в
«безопасных средах», то шимпанзе, как известно, населяют
саванные биотопы, не слишком сильно отличающиеся от той
•среды, где обитали ранние гоминиды (Suzuki, 1975; McGrew,
Baldwin, Tutin, 1981). Кроме того, и в лесных биотопах перед
животными могут возникать разнообразные проблемы, связан-
ные с пространственной разобщенностью пищевых источников
(Wrangham, 1977).
Тем не менее главное положение гипотезы Лавджоя, что
увеличение размеров тела налагает на животных определенные
ограничения в связи с размножением и что эти ограничения
могут вызвать значительные сдвиги в их социальном поведе-
нии, звучит достаточно убедительно и лишний раз заставляет
вспомнить, что стратегия животных при размножении эволю-
ционировала так же сильно, как и поведение при поисках пищи.
Это положение перекликается с недавно появившимися работа-
ми Тринкауса (Trinkaus, 1981; 1983), который обратил внима-
ние на посткраниальные различия между неандертальцами и
морфологически современным человеком. По мнению Тринкау-
са, эти различия обусловлены противоборствовавшими тенден-
циями отбора, направленными, с одной стороны, на укрупнение
головного мозга, а с другой — на развитие эффективной бипе-
дии. По мере укрупнения мозга возникала проблема появления
на свет детенышей с очень крупной головой, что обусловило
снижение эффективности локомоторной системы у самок и, воз-
можно, значительное увеличение детской смертности. У анато-
мически современного человека, по всей видимости, происходит
сдвиг в сторону более грацильного телосложения, а быть может,
и в сторону укорочения сроков беременности и преждевремен-
188
Глава 6
ного появления на свет новорожденных. Еще раз следует под-
черкнуть, что все описанные сдвиги должны были сопровож-
даться значительным усилением родительской заботы о потом-
стве, что, по мнению Тринкауса, могло играть важную роль в
изменении поведения гоминид в процессе эволюционного фор-
мирования анатомически современного человека.
6.5. Размеры тела, размеры головного мозга
и адаптация гоминид
Таким образом, укрупнение гоминид сопровождалось как
изменением их пищевого поиска (что дало им значительные
адаптивные преимущества), так и изменением их репродуктив-
ного и социального поведения (что столкнуло их с целым рядом
адаптивных проблем). Обе эти формы поведения, однако, тесно
друг с другом связаны одним фактором, сыгравшим в эволюции
человека критическую роль — укрупнением головного мозга.
Связь между размерами тела и величиной головного мозга рас-
сматривалась выше (см. разд. 6.3.2). Было сделано предполо-
жение, что за энцефализацию животные (в особенности самки)
вынуждены расплачиваться очень дорогой ценой (R. D. Martin,
1981) и что энцефализация развивается только в том случае,
если эта цена окупается приносимыми ею выгодами. В случае
мелких млекопитающих это условие редко когда выполнимо
вследствие относительно высокой интенсивности их метаболиз-
ма. В случае крупных млекопитающих энцефализация оказыва-
ется выгодной там, где требуется определенная гибкость пове-
дения. Как уже отмечалось, естественному отбору у гоминид
подвергался как поиск пищи (увеличение разнообразия пище-
вого рациона и повышение способности к утилизации разобщен-
ных источников пищи), так и социальное поведение (усложнение
социальных взаимосвязей). Таким образом, укрупнение головно-
го мозга, по всей вероятности, должно было начаться уже на
самых ранних стадиях эволюции человека. Мартин (Martin,
1983) полагает, что именно так обстояло дело в действительно-
сти и что в плиоцене и плейстоцене гоминиды превосходили по
величине головного мозга живших в то время обезьян (рис.
6.16), а Стрингер (Stringer, 1984) показал к тому же, что укруп-
нение головного мозга у гоминид происходило непрерывно (рис.
6.17).
Следует, однако, подчеркнуть, что изучение всех описанных
взаимосвязей до сих пор не позволило выявить какой-либо
единой причинно-следственной последовательности событий.
В обзоре существующих представлений о характере взаимосвя-
зи между размерами тела и размерами головного мозга у при-
матов Мартин (Martin, 1984) отмечает, что, хотя между разме-
Рис. 6.16. Связь между размерами головного мозга ископаемых гоминид (с уче-
том размеров тела) и геологическим возрастом находки. А. Сопоставление ко-
эффициента энцефализации (отношение размеров головного мозга к размерам
тела) гоминид и млекопитающих вообще; Б. Сопоставление коэффициента
энцефализации гоминид и узконосых обезьян (Martin, 1983). Сокращения:
Аа — Australopithecus africanus-, Hh — Homo habiiis; He — Homo erectus; Hs—
Homo sapiens
Рис. 6.17. Связь между размерами головного мозга ископаемых гоминид
и геологическим возрастом находки. (Stringer, 1984.)
190
Глава 6
рами головного мозга, размерами тела и особенностями пище-
вого поведения у этих животных и прослеживается четкая
зависимость, интерпретировать ее можно по-разному. Мартин
(Martin, 1984, р. 92) рассматривает «гипотезу стратегии пище-
вого поиска» и «гипотезу стратегии энергообмена» в качестве
двух альтернативных подходов к пониманию эволюционных и
экологических аспектов размеров. Согласно первой из этих
гипотез, необходимость укрупнения головного мозга была выз-
вана потребностями при поисках пищи: непредсказуемость и
разобщенность источников пищи вынуждала животных «обза-
водиться» более крупным мозгом. Напротив, вторая гипотеза
предполагает, что в первую очередь отбору подвергался метабо-
лизм материнских особей, причем размеры головного мозга
изменялись при этом как «пассивное следствие стратегии энер-
гообмена материнских особей» (Martin, 1984, р. 93): следствием
стратегии низкоинтенсивного энергообмена явился небольшой
головной мозг, высокоинтенсивного — крупный. Как полагает
Мартин, в известной мере эти две гипотезы можно увязать та-
ким образом, что признание важности отбора в сторону укруп-
нения головного мозга не будет противоречить предположениям
ни о важной роли репродуктивных факторов, ни о значении
пищевого поиска: «...разные стратегии пищевого поиска непос-
редственно сказываются на метаболизме, который в свою оче-
редь оказывает опосредованное влияние на относительные
размеры головного мозга; независимо от того, однако, имеют ли
эти эффекты положительный или отрицательный характер, они
могут сглаживаться модификацией репродуктивных характери-
стик» (Martin, 1984, р. 93).
Таким образом, понимание причин и следствий увеличения
размеров тела позволяет объяснить многие аспекты эволюции
человека. Все предположения, касающиеся расширения террито-
рий, увеличения разнообразия пищевого рациона и усиления
кооперации и родительской заботы у гоминид так или иначе
подтверждаются методами сравнительной биологии или непос-
редственно ископаемыми находками. Однако, как было нагляд-
но показано на примере расширения территорий, в поведении
и адаптации гоминид произошли гораздо более значительные
сдвиги, чем можно предполагать, исходя только из увеличения
размеров тела. Таким образом, для объяснения этих сдвигов
необходимо привлечь и некоторые другие факторы.
Прежде, однако, уместно сказать несколько слов о тех фак-
торах внешней среды, которые могут способствовать укрупне-
нию размеров. На основании проведенного выше анализа, по
всей видимости, можно утверждать, что особенности гоминид,
связанные с их размножением и общественным поведением,
являются результатом укрупнения их размеров, а стратегия
Гоминиды как крупные млекопитающие 191
энергетического обмена в связи с утилизацией источников пи-
щи— его причиной. А следовательно, можно предполагать, что-
какие-то факторы, связанные с источниками пищи и средой оби-
тания гоминид, должны были способствовать увеличению их.
размеров.
Чаще всего укрупнение размеров организмов происходит
вследствие конкуренции (Pianka, 1978), ибо конкуренция — ос-
нова /(-отбора (рис. 6.11). Эволюционную экологию ранних
гоминид можно, таким образом, рассматривать как результат
/(-отбора со всеми присущими ему признаками: способностью»
организмов к острой конкуренции, рождением небольшого числа
детенышей и низкой их смертностью. Неправильно было бы.
думать, что /(-отбор могут вызвать сами по себе специфичес-
кие факторы окружающей среды, ибо зачастую он определя-
ется спецификой организмов и их адаптивными характеристика-
ми. Критическими факторами при этом являются высокий уро-
вень конкуренции и стабильность условий, в которых она
протекает. Если оба этих фактора действуют, отбор, вероятнее
всего, будет проходить по /(-типу. Уровень и характер конкурен-
ции в средах обитания ранних гоминид обсуждаются в гл. 8 и 9:
прежде, однако, следует подробнее рассмотреть одну особен-
ность гоминид, которая в их эволюции сыграла важную роль —
их наземный образ жизни.
Глава 7
ГОМИНИДЫ КАК НАЗЕМНЫЕ ПРИМАТЫ
Пожалуй, наиболее сжато сущность человеческой эволюции
можно выразить словами «человек спустился с дерева на зем-
лю». Человек стал тем, что он есть потому, что это некогда
древесное животное приняло вызов наземного образа жизни.
При этой мысли нередко возникает смутное ощущение, что на
земле условия жизни суровее, чем на деревьях. Зато именно
на земле эволюция развернула свою победную поступь, а те
животные, которые продолжали довольствоваться дарами при-
роды в чистом виде, остались на деревьях. Эволюционный
успех гоминид объясняется тем, что они не побоялись спустить-
ся с трибуны высоколесья на земную арену эволюционных кон-
фликтов и вступить здесь в бой за выживание.
Утверждение, что переход к наземному образу жизни сыг-
рал в эволюции гоминид исключительную роль, представляется
самоочевидным и не требующим доказательств. Вытекает оно
из двух простых наблюдений общего характера: во-первых,
приматы в отличие от людей в большинстве своем ведут древес-
ный образ жизни, и, во-вторых, из всех тех приматов, которые
все-таки перешли к наземному существованию, у одних только
гоминид сформировался в высшей степени специализированный
способ передвижения — бипедия. Столь сильное отклонение
человека от эволюционного и адаптивного развития его ближай-
ших биологических родственников уже само по себе можно
расценивать как достаточное свидетельство его уникальности.
Человеку, быть может, горько будет осознавать, что его
эволюция протекала не слишком гладко. Хотя приматы с само-
го начала вели древесный образ жизни, многие их виды — как
современные, так и вымершие — сумели хотя бы отчасти при-
способиться к наземному существованию. Последние экологи-
ческие и палеоэкологические данные свидетельствуют о важной
роли наземного образа жизни в эволюции некоторых групп
приматов. Переход гоминид к жизни на земле скорее вписыва-
ется в общую тенденцию эволюции приматов, нежели идет с
ней вразрез. Хотя это обстоятельство и может поколебать
представление о том, что эволюция гоминид коренным образом
отличается от эволюции остальных приматов, оно позволяет
Гоминиды как наземные приматы
193
провести более полный анализ эволюционного развития живот-
ных, в частности оценить последствия перехода к наземному
существованию и сравнить альтернативные решения тех проб-
лем, которые наземный образ жизни ставил перед животными.
В настоящей главе делается попытка выяснить, что же озна-
чал для приматов переход к наземному существованию. Почему
и каким образом приматы стали наземными животными, и как
это можно объяснить в свете ископаемых находок и с точки
зрения эволюционной и экологической теорий? Что можно ска-
зать об адаптивных последствиях наземного образа жизни?
В каких условиях протекала эволюция наземного существова-
ния гоминид?
7.1. Эволюция приматов и наземный образ жизни
Приматов обычно считают отрядом млекопитающих, кото-
рый характеризуется древесным образом жизни. Отличительные
признаки этого отряда, рассмотренные в гл. 1, — хватательные
руки и ноги, сильная зависимость от зрительных ощущений,
способность передвигатся путем подвешивания к веткам и
перепрыгивания с одной ветки на другую и т. д. (см. рис.
1.1), — по всей видимости, связаны с выживанием в сложной,
трехмерной древесной среде. Поскольку эти же признаки были
присущи и ранним ископаемым приматам эоцена, можно ду-
мать, что примитивные млекопитающие — предки приматов —
также предпочитали жизнь на деревьях. Когда же в начале
третичного1 периода млекопитающие подверглись адаптивной
радиации, эти места обитания превратились в специализирован-
ную экологическую нишу приматов.
Далеко не все из ныне существующих приматов, однако,
ведут древесный образ жизни. Согласно Клаттон-Броку и Харви
(Clutton-Brock, Harvey, 1977а), из 101 вида приматов, включая
и современного человека, 23,8% являются наземными животны-
ми. Внутри отряда эти виды распределены неравномерно: из
полуобезьян наземный образ жизни ведут лишь два вида, из
широконосых обезьян — ни одного, а из узконосых — 42,9%.
Поскольку узконосые обезьяны — одна из новейших эволюцион-
ных групп приматов, переход к наземному существованию сле-
дует рассматривать как важнейший элемент более поздней
эволюции приматов. С учетом того, что почти половина всех
человекоподобных приматов Старого Света значительную часть
времени проводит на земле, представление об уникальности
наземного образа жизни гоминид и попытки объяснить на его
1 В настоящее время первый период кайнозойской эры обычно называют
палеогеновым. — Прим, перев.
13—367
194
Глава 7
основе все и вся сильно сдают свои позиции. А потому в объяс-
нении нуждаются характерные признаки не именно гоминид, а
узконосых в целом.
7.1.1. Причины перехода к наземному образу жизни
Зачем представителям древесного отряда млекопитающих
нужно было спускаться на землю? Здесь можно выдвинуть две
главные причины (Foley, 1964b). Первая связана с экологичес-
кими законами, вторая — с историей изменения среды обита-
ния. Основное ограничение, связанное с древесным образом
жизни, относится к размерам животных. Свобода и быстрота
движений и доступ к разнообразным источникам пищи лучше
всего обеспечиваются при небольших размерах тела. Как гово-
рилось в предыдущей главе, небольшие размеры тела имеют
и свои недостатки. Приматы «расплачиваются» наземным суще-
ствованием за освобождение от высокого расхода энергии,
свойственного мелким животным. Следовательно, если у при-
матов обнаруживается тенденция к увеличению размеров тела,
должна, по-видимому, обнаруживаться и тенденция ко все
более выраженному наземному существованию. Зависимость
между размерами тела и наземным образом жизни у приматов
изображена на рис. 7.1, который — хотя и включает несколько
Рис. 7.1. Распределение особей по массе тела у древесных (светлые столбцы)
и наземных (темные столбцы) приматов. (Clutton-Brock, Harvey, 1977а.)
Гоминиды как наземные приматы 195
крупных древесных приматов (например, орангутана; см. также
разд. 7.2.2) —дает возможность проследить общую тенденцию
к более выраженному наземному образу жизни с укрупнением
размеров.
Однако сами по себе ограничения размеров тела, связанные
с древесным образом жизни, недостаточны для перехода к на-
земному существованию. Наземный образ существования дол-
жен быть сам по себе необходимым и обеспечивать организму
жизнеспособность. Наземные животные должны располагать
соответствующей средой обитания. Как уже говорилось в гл. 5,
с середины миоцена вследствие климатических изменений пло-
щадь лесных тропических биотопов начала сокращаться. Это
означало, что в остававшихся тропических лесах конкуренция
между видами за места обитания обострялась, и, что еще важ-
нее, появлялись новые разнообразные среды, пригодные для
освоения. Эту палеоэкологическую ситуацию и отражает тенден-
ция в эволюции приматов Старого Света на протяжении пос-
ледних 15 млн. лет.
7.1.2. Эволюция приматов: свидетельства
Ископаемые находки прямо свидетельствуют о переходе
приматов к наземному образу жизни. Хотя этот переход мог
независимо происходить и у стрепсириновых — например, ко-
шачьего лемура {Lemur catta) и некоторых из субфоссильных
лемуров Мадагаскара (Walker, 1967)—ниже он рассматрива-
ется только в группе узконосых обезьян.
Узконосые обезьяны представляют собой подотряд ныне жи-
вущих высших приматов Старого Света. Первые узконосые
обезьяны со всеми присущими им признаками появились в оли-
гоцене около 40 млн. лет назад (Szalay, Delson, 1979). Несмотря
на некоторые сомнительные находки из Пондаунга (Бирма)
(Szalay, Delson, 1979), данные, полученные в Фаюме (Египет),
позволяют составить четкую картину эволюции разных линий
антропоидов Старого Света (рис. 7.2).
Согласно Флиглу и Кею (Fleagle, Kay, 1983), среди ископае-
мых фаюмских приматов можно выделить две основные группы.
Первая представлена примитивными видами Parapithecidae,
вторая — ранними Hominoidea. Первая группа в это время чис-
ленно преобладает, что свидетельствует о ранней радиации
узконосых, до того, как появились ныне живущие формы. Эти
примитивные узконосые вымерли к началу миоцена.
Вторая группа, Hominoidea, ныне живущие потомки которой
представлены человекообразными обезьянами и человеком,
подверглась ранней адаптивной радиации и положила начало
не только ряду таких примитивных форм, как, например, егип-
13»
196
Глава 7
,4^
Л/
Млн. лет назад
Рис. 7.2. Упрощенная кладограмма связей между таксонами высших узконо-
сых обезьян и ископаемыми остатками ранних узконосых обезьян. Pondaun-
ga — антропоидный примат из азиатского эоцена. Олигопитек — древний ант-
ропоид из олигоценовых отложений в Фаюме (Египет) (Szalay, Harvey, 1979;
Fleagle, Kay, 1983.)
топитек, плиопитек и проплиопитек, но и таксонам, продолжав-
шим существовать в миоцене в виде форм, составивших основу
дальнейшей эволюции гоминидов.
Миоценовые гоминоиды представлены многочисленными и
сложными формами, определять таксономическое положение
которых еще труднее, чем классифицировать палеоантрополо-
гический материал. Недавние находки, особенно в Сивалике
(Пакистан), а также использование новых методов и аналити-
ческих приемов произвели в этой области настоящую револю-
цию. Десять лет назад в литературе признавалось существова-
ние трех основных групп миоценовых гоминоидов — рамапитека
как первого гоминида, дриопитековых, от которых происходят
современные человекообразные обезьяны, и предков гиббонов.
Новейшая точка зрения на этот предмет гораздо сложнее (рис.
7.3). Вслед за Пилбимом (Ciochon, Corruccini, 1983; Ward,
Pilbeam, 1983; Pilbeam, 1984), по всей видимости, нужно приз-
нать существование в миоцене нескольких групп крупных гоми-
ноидов.
Первая группа представлена обнаруженными в Африке древ-
ними формами, образовавшимися, возможно, в результате ран-
них олигоценовых радиаций — проконсул и лимнопитек. Это —
примитивные неспециализированные животные, которые пита-
лись фруктами и, по-видимому, вели древесное существование.
Они не обнаруживают специализированных локомоторных адап-
таций, характерных для более поздних гоминоидов, однако
Гоминиды как наземные приматы
197
Рис. 7.3. Упрощенная гипотетическая кладограмма верхнеолигоценовых и мио-
ценовых узконосых. (Szalay, Harvey, 1979; Fleagle, Kay, 1983.)
вполне могут рассматриваться как формы, которые в дальней-
шем отделились от низших обезьян Старого Света (Cercopit-
hecoidea).
Другая группа представлена формами, более разнообразны-
ми по происхождению и более широко распространенными гео-
графически. Для этих крупных приматов с толстым слоем зуб-
ной эмали, иногда обнаруживавших признаки адаптации к ме-
нее лесистым биотопам с более выраженной сезонностью, Пил-
бим (Pilbeam, 1979) впервые предложил считающийся ныне
неадекватным термин «рамаморфы». Географическое распро-
странение и характер эволюции этих приматов, возможно, от-
ражают выход гоминоидов (как и других животных) за пределы
Африки после образования перешейка между Африкой и Евра-
зией. Эти виды нередко обнаруживаются в отложениях среднего
и верхнего миоцена в Юго-Восточной Европе и Азии. Самыми
обычными из них являются сивапитеки. К этой группе относят
и рамапитека, однако Эндрюс (Andrews, 1983) и Пилбим (в пе-
чати) считают, что название рамапитек следует упразднить и
включавшиеся ранее в этот таксон формы относить к сивапите-
кам. Другие филетические линии этой радиации включают
гигантопитека и уранопитека, а также африканского кениапите-
ка. Все большее признание получает мнение, согласно которому
эта радиация знаменует собой разделение предков ныне жи-
вущих африканских и азиатских человекообразных обезьян, и
что орангутан является сохранившимся потомком одной из этих
миоценовых форм.
В своем современном виде «эволюционное древо» гоминои-
дов значительно отличается от прежних вариантов. Во-первых,
198
Глава 7
признается, что в действительности ранние гоминоиды пред-
ставлены гораздо более сложными и разнообразными формами,
чем считалось прежде. Как оказалось, Саймонс и Пилбим
(Simons, Pilbeam, 1965; Pilbeam, 1969) зашли слишком далеко
в своем стремлении упростить таксономическую структуру мио-
ценовых Hominoidea. Как морфологические, так и биогеографи-
ческие данные заставили признать заметные различия между
европейскими, азиатскими и африканскими гоминоидами, обус-
ловленные биогеографическими факторами. Увеличение разно-
образия гоминоидов и колебания «уровня разнообразия» этих
приматов во времени затрагивают ряд интересных экологичес-
ких вопросов, которые рассматриваются ниже в связи с назем-
ным существованием этих животных.
Вторая особенность современного подхода к интерпретации
эволюционных взаимосвязей гоминоидов заключается в том,
что если раньше исследователи, как правило, пытались найти
связь между ископаемыми и ныне существующими формами
(обычно вывести вторые из первых), то сегодня прослеживает-
ся тенденция рассматривать ископаемых гоминоидов как неза-
висимые адаптивные формы и лишь «между делом» попытаться
втиснуть их в рамках взаимосвязей с ныне живущими видами.
Такое изменение подхода представляется очень важным. Как
подчеркивает Пилбим (Pilbeam, 1979), миоценовые гоминоиды
должны рассматриваться в контексте современных им сред
обитания, а не просто как предшественники более поздних ви-
дов. Главное внимание в исследованиях нужно уделять тому,
чем в действительности является тот или иной вид, а не чем
он может стать со временем. Как уже предполагалось в гл. 2
(см. разд. 2.1.3), важную роль в этом изменении подхода сыг-
рали биохимические исследования, главным образом благодаря
тому, что выявленные с помощью молекулярных и иммунологи-
ческих методов более поздние сроки дивергенций прямо
свидетельствовали о невозможности для многих ископаемых
форм быть непосредственными предками ныне живущих видов.
Следующая особенность современного отношения исследова-
телей к миоценовым гоминоидам касается их взаимосвязей с
другой большой ветвью узконосых — мартышкообразными
обезьянами. Согласно классическим представлениям о взаимо-
отношениях между этими двумя надсемействами, низшие узко-
носые (мартышкообразные) стоят на более примитивном и про-
стом, а высшие узконосые (включая, конечно, и людей)—на
более высоком и сложном уровне организации. В результате
родилось предположение, что ранние узконосые должны были
походить скорее на низших, нежели на высших узконосых обезь-
ян, и что ископаемых низших узконосых следует датировать
более ранними сроками.
Гоминиды как ваземные приматы
199
Сегодня эта точка зрения значительно поколебалась. Флигл
и Кей (Fleagle, Kay, 1983) считают, что многие из своих призна-
ков мартышкообразные заимствовали от гоминоидов и что пос-
ледние по своим признакам стоят ближе к предкам узконосых
обезьян. Таким образом, как показано на рис. 7.2, предполагает-
ся, что именно низшие обезьяны отделились от линии гоминои-
дов. Следует также отметить, что ископаемые находки ранних
мартышкообразных чрезвычайно малочисленны (Szalay, Del-
son, 1979, рр. 320—327) и нет бесспорных указаний на то, что
низшие узконосые появились ранее 20 млн. лет назад, т. е. того
времени, когда произошла, по мнению биохимиков, дивергенция
гоминоидов и низших узконосых обезьян. И уж конечно первые
известные нам мартышкообразные жили гораздо позже разде-
ления единого ствола узконосых, если датировать его по фаюм-
ским находкам. Это согласуется и с ископаемыми данными,
свидетельствующими в пользу поздней (конец миоцена — плей-
стоцен) радиации низших узконосых и более ранней (начало —
середина миоцена) радиации гоминоидов.
Все три рассмотренные особенности современного подхода
к эволюции гоминоидов — большое разнообразие форм, лишь
частичная их связь с существующими ныне видами и поздние
сроки эволюции низших узконосых — имеют решающее значение
для понимания той роли, которую играло наземное существова-
ние в эволюционной экологии последовательных адаптивных
радиаций, составляющих историю антропоидов Старого Света
(рис. 7.4).
7.2. Эволюционная экология узконосых обезьян
Характер эволюции узконосых обезьян порождает вопрос,
который в плане эволюционной экологии можно сформулиро-
вать следующим образом: что служило экологической основой
тех многочисленных адаптивных радиаций, которые выявля-
ются по ископаемым находкам? Ответ на данный вопрос по-
зволил бы подвергнуть гоминид, составляющих центральный
предмет обсуждения в этой книге, более глубокому сравнитель-
но-эволюционному анализу.
7.2.1. Адаптивные радиации узконосых обезьян
Старого Света
На рис. 7.4 изображены 6 адаптивных радиаций, хотя в
принципе их число может быть выбрано более или менее про-
извольно — в зависимости от того, на каком таксономическом
уровне их рассматривать. Первой была радиация ранних узко-
носых обезьян в олигоцене, и участвовали в ней примитивные
200
Глава 7
0
40 L
Рис. 7.4. Адаптивные радиации узконосых. (Cronin, 1983; рис. 4.)
группы приматов и ранние предки ныне существующих над-
семейств. Затем происходила радиация ранних гоминоидов,
например, проконсула. В третьей радиации участвовали сива-
питековые, в четвертой — гиббоны, а в результате пятой про-
изошло разделение низших узконосых обезьян (последняя ра-
диация фактически представляет собой две радиации, в кото-
рых участвовали тонкотелые обезьяны, питавшиеся листьями,
и мартышковые с отчасти наземным образом жизни). Произо-
шла, наконец, и радиация крупных африканских гоминоидов,
в том числе и гоминид.
Появление первых антропоидов Старого Света, по всей
вероятности, связано с изменением их питания в сторону по-
едания фруктов. У ныне живущих стрептосериновых приматов
Гоминиды как наземные приматы
201
наблюдается склонность к насекомоядности или всеядности
(Harding, 1981), и, возможно, такого же пищевого рациона
придерживались и некоторые ископаемые виды. С другой сто-
роны, узконосые обезьяны — в значительной мере растительно-
ядные (предпочитающие плоды) животные (Harding, 1981).
Судя по самым древним фаюмским ископаемым, растительно-
ядность была присуща и самым ранним формам и всем при-
митивным семействам этой группы (Fleagle, 1983). Возможно,
эволюционный успех узконосых обезьян объясняется расшире-
нием диапазона адаптации этих приматов к дневному питанию
древесными плодами в широко раскинувшихся олигоценовых
тропических лесах. Такое предположение подтверждается как
анатомическими данными, свидетельствующими о существова-
нии в то время небольших, медленно передвигавшихся древес-
ных четвероногих (Fleagle, 1983), так и данными палеоэколо-
гическими, которые, по мнению Бауна, Крауса и др. (Bown,
Kraus et al., 1982), с убедительностью говорят о широком
распространении в то время истинных лесных биотопов, хотя
Кортлэндт (Kortlandt, 1983) придерживается на этот счет про-
тивоположной точки зрения.
В начале миоцена эта общая тенденция в адаптации узко-
носых достигает апогея. Вначале происходила радиация ранних
гоминоидов в Африке, затем — в других частях Старого Света.
Можно сказать, что нижний и средний миоцен был временем
наиболее широкого распространения и процветания гоминои-
дов. По-видимому, в начале миоцена гоминоиды продолжали
придерживаться тех же адаптивных стратегий, что и в олиго-
цене. Изучение посткраниологического материала (Flegle, Kay,
1983; Morbeck, 1983; Rose, 1983; Walker, Pickford, 1983) сви-
детельствует о том, что миоценовые гоминоиды не обладали
ни одной из тех локомоторных специализаций, которые свойст-
венны современным высшим обезьянам, и скорее всего пред-
ставляли собой неспециализированных древесных четвероно-
гих. Лишь самый крупный из них вид, Proconsul major, обна-
руживает некоторые признаки, характерные для наземного
образа жизни, а остальные, по-видимому, могли в известной
мере подвешиваться к веткам, хотя истинными брахиаторами
не были. Если, как предполагает Грэнд (Grand, 1972), подве-
шивание к веткам и в самом деле дает животным возможность
более эффективно перемещаться между островками пищи,
можно думать, что это обстоятельство имело важные послед-
ствия в плане пищевого поиска. Как показала Айелло в своем
очень детальном исследовании (Aiello, 1981), всех этих древес-
ных четвероногих эпохи миоцена можно разделить на две
большие группы в соответствии с тем, кормились ли они сидя
на ветке или находясь под веткой (рис. 7.5). Включение вида
202
Глава 7
Рис. 7.5. Кормежка на ветке и под веткой (объяснения см. в тексте)
в ту или иную группу в значительной мере зависит от разме-
ров тела — фактора, который, как установила Айелло, играл
в адаптации миоценовых Hominoidea особенно важную роль.
Флигл (Fleagle, 1984), однако, подчеркивает, что, хотя страте-
гия пищевого поиска и характер локомоции могут быть, вооб-
ще говоря, взаимосвязаны, до сих пор удалось выявить лишь
очень немногочисленные примеры тесной корреляции между
типом пищи и способом передвижения.
Изменение размеров тела, способов передвижения и пище-
вого поведения — таковы факторы, которые определяли раз-
деление экологической ниши гоминоидов между многочислен-
ными видами в начале миоцена. Несмотря, однако, на доступ-
ность нескольких экологических ниш, все эти виды занимали
один-единственный биом — лес. Современные данные (Van
Couvering, Van Couvering, 1976; Andrews, Lovd, Evans, 1979;
Pickford, 1983) указывают на то, что тропики в то время были
покрыты огромными, непрерывными и однородными массивами
влажного тропического леса. Этот продуктивный тип среды
обитания и сыграл решающую роль в адаптивной радиации
гоминоидов, происходившей в интервале между 20 и 12 млн.
лет назад.
Как уже говорилось в гл. 5, климатические и географичес-
кие сдвиги в середине миоцена привели к возникновению бо-
лее дифференцированных сред обитания, сужению зоны
влажных тропических лесов и их замещению во многих частях
света более фрагментарными мозаичными биомами, в которых
все более обычными становились редколесье, кустарники и
трава. Эти экологические перемены имели для эволюции мле-
копитающих вообще и узконосых обезьян в частности далеко
идущие последствия. Экологическая проблема, с которой столк-
Гоминиды как наземные приматы 203
нулись среднемиоценовые узконосые, заключалась в необхо-
димости приспособиться к сократившимся по площади, зачас-
тую изолированным лесам и к обширным лесистым и кустар-
никовым биотопам с выраженной сезонной изменчивостью. Ис-
ходя из принципов экологии и учитывая по-прежнему высокое
разнообразие форм узконосых обезьян на протяжении всего
верхнего кайнозоя, можно думать, что в зависимости от тех
специфических условий, в которых оказались в то время при-
маты, эту проблему они решали по-разному. В одних случаях
в результате уменьшения доступности пищевых источников и
сокращения привычных биотопов, конкуренция, по всей веро-
ятности, усиливалась. В других, возможно, вымирание и изо-
ляция приводили к обеднению сообществ видами, конкуренция
между которыми становилась слабой (MacArthur, Wilson,
1967). Отбор адаптивных стратегий, появившихся у антропои-
дов Старого Света в ответ на описанные экологические сдви-
ги, отразился в характере их эволюции.
7.2.2. Альтернативные стратегии
Каковы были ответные реакции приматов на изменение ус-
ловий существования? У неспециализированных древесных
обезьян, передвигавшихся на четырех ногах и представленных
такими ранними гоминоидами, как проконсул, началось сокра-
щение числа видов, распространения и численности. Их место
заняли более специализированные обезьяны, которые могли
либо более эффективно использовать окружающую среду, либо
выживать в широком диапазоне новых биотопов. Одним из
вариантов такого ответа является адаптация сивапитековых.
Рамапитек, сивапитек и гигантопитек — обезьяны, обнаружи-
вавшие типичные признаки, связанные с потреблением грубой
и низкокачественной пищи, доступность которой имела выра-
женный сезонный характер. Это подтверждается главным об-
разом значительной толщиной у них слоя зубной эмали (Pil-
beam, Meyer et al., 1977; Ward, Pilbeam, 1983). Сопутствовав-
ший находкам этих ископаемых попалеоэкологический матери-
ал указывает также на то, что эти обезьяны жили в более су-
хой среде. И наконец, один из рассматриваемых видов, а имен-
но гигантопитек, просуществовавший в Китае, а возможно, и
в других частях Азии до середины плейстоцена, вел, судя по
его размерам, наземный образ жизни. Единственный сохранив-
шийся поныне представитель сивапитековых, орангутан, оби-
тает в настоящее время лишь в нескольких лесных областях
Борнео и Суматры. Хотя этот вид является ныне древесным
животным, не исключено, что у его предков и боковых родст-
венников были более разнообразные среды обитания и спосо-
204
Глава 7
бы передвижения, и что они, вероятно, в большей мере тяго-
тели к наземному образу жизни (Andrews, 1983). Крайне ус-
тоявшийся древесный образ жизни современного орангутана,
возможно, возник в результате прихода его предков в Юго-Вос-
точную Азию в эпоху плейстоцена (McKinnon, 1974). Пост-
краниальные адаптации сивапитековых до сих пор изучены
плохо. Роуз с сотрудниками (Raza, Barry et al., 1983; Rose;
1983; 1984) предположили, что средне- и нижнемиоцено-
вые гоминоиды не обнаруживали локомоторного поведения,
свойственного более поздним и ныне живущим гоминоидам;
они способны были выполнять самые различные движения
и принимать разнообразные позы, но все это служило главным
образом для того, чтобы энергично взбираться на деревья, хотя
у некоторых форм могли осуществляться также подвешивание
руками к веткам, четвероногость, четверорукость и бипедия.
Второй тип ответной реакции, предполагающий более спе-
циализированное использование древесной среды обитания,
обнаружили гиббоны. Предки этой линии изучены особенно
плохо (Fleagle, 1983). Хотя на протяжении долгих лет к гиб-
боновым пытались относить самые различные ископаемые
формы, не исключая и столь древних, как олигоценовые, это
обстоятельство, вероятно, отражает лишь сходство (крани-
альное) гиббонов с очень ранними узконосыми обезьянами.
По единодушному мнению специалистов, подкрепленному био-
химическими даными, эта ветвь гоминоидов, по-видимому, обо-
собилась в интервале между 12 и 16,5 млн. лет назад (см.
рис. 2.4 и 7.3). Эти животные, в настоящее время представлен-
ные семью видами в Юго-Восточной Азии, живут в высоких
ярусах влажного тропического леса, и, будучи чрезвычайно
ловкими брахиаторами, являются среди гоминоидов настоящи-
ми чемпионами по акробатике. Они придерживаются главным
образом плодового пищевого рациона, и, стало быть, своим
пищевым поведением не отличаются от большинства приматов;
при этом локомоторное поведение гиббонов дает им возмож-
ность не нарушать свой пищевой рацион, оставаясь в высоко-
специализированной среде с сильно выраженной конкуренцией
(Chivers, 1977).
Ответная реакция на изменение экологических условий у
африканских человекообразных обезьян была чем-то промежу-
точным между реакциями сивапитековых и гиббонов. Подобно
гиббону, шимпанзе и горилла остались жить в лесных биото-
пах. Однако, они, как, возможно, и некоторые сивапитековые,
начали вести более выраженный наземный образ жизни. Уве-
личение у африканских обезьян размеров тела и развитие у
иих походки с опорой на фаланги пальцев верхних конечностей
наряду с другими адаптациями способствовали более эффек-
Гоминиды как наземные приматы
205
тивному использованию сохранившихся лесных биотопов. Так,
специализация питающейся листьями гориллы, связанная с за-
висимостью от низкокачественной, но легкодоступной пищи,
выражена столь же сильно, как и специализация гиббона. На-
против, пищевая специализация шимпанзе носит более широ-
кий характер. Это, в частности, выражается в способности
шимпанзе использовать пищевые источники, расположенные на
краю леса. В этой связи уместно вспомнить предположение,
что прежде шимпанзе обитали в более открытых средах, вели
более выраженный наземный образ жизни и, подобно орангу-
тану, адаптировались к лесу уже вторично (Kortlandt, 1972).
Позднее другая группа африканских гоминоидов — гомини-
ды — прореагировали на изменение экологии так, как до них
не реагировал никто. Возможно, переход к более выраженному
наземному образу жизни у гоминид (и в известной мере у
шимпанзе) в редколесных биотопах Африки соответствовал
аналогичному изменению образа жизни у некоторых сивапи-
тековых (например, гигантопитека) в конце миоцена в Азии.
Хотя о поведении и экологии гигантопитека известно мало,
этот гоминоид заявляет о себе как о представителе узконосых,
который в позднем кайнозое обнаружил наиболее выраженные
адаптации к наземному существованию, конвергирующие с
аналогичными адаптациями гоминид.
Хотя многие гоминоиды и смогли приспособиться к измене-
ниям экологических условий, начиная с середины миоцена их
разнообразие и географическое распространение идут на спад.
Это разительно контрастирует с эволюционными судьбами
другой группы узконосых обезьян — мартышкообразных
(рис. 7.6). Низшие обезьяны начинают сильно преобладать над
высшими в ископаемых находках, с периода со времени верх-
него миоцена, — обстоятельство, возможно, указывающее на то,
что низшие обезьяны смогли адаптироваться к более сухим и
бедным условиям существования лучше, чем гоминоиды. Так,
многочисленные виды низших узконосых обезьян, в том числе
мартышки (Cercopithecus), павианы и макаки, приспособились
к полуназемному существованию и саванновым биотопам, где
довольствовались сухими листьями и плодами, а в отдельных
случаях даже переходили к абсолютной всеядности.
7.2.3. Экологическая модель дивергенции
Таким образом, 12 млн. лет назад экологическая ниша уз-
коносых в ответ на изменение условий окружающей среды на-
чала расширяться. Отраженные в ископаемых находках дивер-
генция и параллелизм касаются скорее эволюционной эколо-
гии этих животных, нежели их филогении (рис. 7.7). В пред-
206
Глава 7
Миллионы лет назад
Рис. 7.6. Относительное видовое разнообразие низших и высших узконосых
в неогеновом и актропогеновом периодах. (Andrews, 1981.)
ставленной на рисунке схеме сделана попытка проследить пу-
ти развития различных групп приматов в ответ на изменение
характера отбора в то или иное время их существования.
В развитии узконосых уже на самых ранних этапах присут-
ствуют элементы дивергентной и параллельной эволюции,
а также взаимодействие факторов, связанных со средой оби-
тания (лес, редколесье, саванна), пищевым рационом (пита-
ние плодами, листьями, всеядность), древесным и наземным
образом жизни. Общей тенденцией был переход к гораздо бо-
лее выраженному наземному образу жизни, что нашло отра-
жение как в эволюции гоминоидов, так и в их «эволюционном
соревновании» с мартышкообразными.
Зная эту общую тенденцию, можно понять, с какими пробле-
мами сталкивались приматы в своем развитии и за счет каких
процессов происходила их адаптация. Так, представляется оче-
видным, что одна и та же ситуация — увеличение открытости
сред и усиление их сезонности — могла через естественный от-
бор вызывать у разных групп приматов разные ответные ре-
акции. На одну и ту же ситуацию разные виды реагируют по-
разному. Характер ответов зависит от целого ряда факторов.
Кроме того, в известной мере варьировать могут и локальные
условия: так, экологические сдвиги в Евразии, возможно, про-
изошли раньше и были выражены сильнее, чем в Африке, что
повлекло более широкую адаптивную радиацию. Впоследствии
различия могли определяться не столько текущим давлением
отбора, сколько особенностями эволюционной предыстории жи-
вотных. Эволюционное прошлое животного в значительной ме-
5
Листоядные
специализированные
древесные
обезьяны
(тонкотелые)
Плодоядные/
всеядные наземные и
древесные обезьяны
(мартышковые)
Всеядные
наземные
саванновые
гоминоиды
(гоминиды)
Плодоядные
10
Миллионы лет назад
15
20 -
25
30
Специализированные
азиатские наземные
травоядные
Полудревесные
неспециализированные
лесные гоминоиды
(шимпанзе)
Листоядные лесные
наземные гоминоиды
(горилла)
лесные
плодоядные
(орангутан)
Полуназемные обитатели
леса (верхнемиоценсвые
Мелкие древесные
олигоценовые
узконосые обезьяны
Рис. 7.7. Суммарная модель экологической дивергенции узконосых обезьян
208
Глава 7
ре определяет его эволюционное будущее. Так, к примеру, бо-
лее крупные ранние гоминоиды, возможно, были предадапти-
рованы к более выраженному наземному существованию,
а мелкие — к брахиации (Aiello, 1981).
Проведенный краткий анализ эволюционной истории узко-
носых обезьян позволяет сделать два вывода. Во-первых, один
и тот же экологический сдвиг предполагает разнообразие адап-
тивных ответов животных, и, следовательно, разнообразие са-
мого процесса адаптации. Характер же эволюционного сдвига
определяется не только изменением среды обитания живот-
ного, но и другими факторами, включая его эволюционную
предысторию. Во-вторых, гоминиды, перейдя в более открытые
среды и начав вести более выраженный наземный образ жиз-
ни, в своем эволюционном развитии двигались отнюдь не
вразрез с другими приматами, а были вовлечены в широкий
поток эволюции.
7.3. Адаптивные характеристики наземных приматов
Хотя единственным источником прямых сведений об эволю-
ционном прошлом приматов являются ископаемые находки, к
ныне живущим приматам можно относиться в известном смыс-
ле как к подопытным животным в лаборатории эволюции.
В признаках существующих в настоящее время представите-
лей этого отряда отражаются многие из путей и проблем эво-
люции. Сопоставив адаптивные характеристики разных групп
приматов, можно связать их с особенностями сред их обита-
ния и условиями жизни. С помощью такого подхода можно
выяснить, как наземный образ жизни отражается на адапта-
ции приматов. Сравнение наземных приматов с их древесны-
ми «кузенами» позволяет выявить некоторые общие законо-
мерности адаптации и взять их за основу модели возникнове-
ния той группы приматов, которая находится в центре внима-
ния в данной книге — гоминид.
7.3.1. Социоэкология: сравнительный подход
Сравнительный подход в эволюционной экологии предпо-
лагает изучение особенностей адаптации ныне существующих
видов животных (Krebs, Davies, 1981). Сравнение близко род-
ственных видов с целью выявления различий, обусловленных
экологическими переменными, впервые было проделано Круком
(Crook, 1964) у ткачиков и Джарменом (Jarman, 1974) у ан-
тилоп. Для приматов соответствующие исследования начали
Круг и Гартлан (Crook, Gartlan, 1966). Как показали эти ав-
торы, различия адаптивного поведения видов, относящихся к
одной группе, можно объяснить особенностями сред их обита-
Гоминиды как наземные приматы
209
ния, которые в случае антилоп, например, опосредуют свое
влияние через такой фактор, как размеры тела животных.
Предложенный подход оказался эффективным средством про-
верки экологических и эволюционных гипотез, выявления кри-
тических для адаптации особенностей окружающей среды и
оценки поведения животных.
Этот подход, к сожалению, не лишен недостатков и огра-
ничений. Главным из них является соблазн объяснять любое
различие между видами ad hoc1 (Altman, 1974). Если между
двумя видами обнаруживаются различия, для их объяснения
почти всегда можно найти какую-нибудь разницу в условиях
существования, так как идентичных сред обитания не быва-
ет. Пользуясь сравнительным подходом, недостаточно только
выявлять экологические различия; необходимо показать, ка-
ким образом эти различия могут быть связаны с адаптив-
ными особенностями животных.
Еще большая трудность практического порядка заключа-
ется в выборе таксономической категории, внутри которой бу-
дет проводиться сравнение животных (Clutton-Brock, Har-
vey, 1979; Krebs; Davies, 1981). На самом деле этот вопрос
можно рассматривать как один из вариантов общей проблемы
Гальтона (см. Thomas, 1978), касающейся независимости дан-
ных. Если два вида обнаруживают одинаковую адаптацию,
необходимо решить, связано ли это с тем, что они обитают
в одинаковых экологических условиях, или же с тем, что та-
кую же адаптацию обнаруживал их общий предок. Решение
этого вопроса имеет принципиальное значение, когда требуется
провести количественную оценку или статистический анализ
данных. По мнению Клаттон-Брока и Харви (Clutton-Brock,
Harvey, 1977а), адекватным для сравнения является такой
таксономический уровень, на котором выявляется максималь-
ная дисперсия (в случае приматов —это род). Однако при
анализе в более узком таксономическом диапазоне, когда,
например, между собой сравниваются только узконосые обезь-
яны, тенденции адаптивных стратегий выявляются на уровне
видов.
7.3.2. Социоэкология наземных приматов
Клаттон-Брок и Харви (Clutton-Brock, Harvey, 1977 а, b;
Harvey, Clutton-Brock, 1981, 1985) собрали и проанализирова-
ли данные, касающиеся поведенческих и экологических осо-
бенностей разных приматов. Эти данные позволяют изучить
различия между древесными и наземными узконосыми обезья-
нами (рис. 7.8; табл. 7.1). Результаты анализа показывают,
1 Для данного случая (лат.). — Прим, перев.
14—267
210
Глава 7
А. Пищевой рацион
Наземные Древесные
Плодоядные 19 19
Листоядные 3 15
В. Половой диморфизм
н *
Наземные • • •• • • • • • •••
Древесные ••••• * • •
1---1--1---1----l|_—I—j------1-1-----1---1
1,0 1,1 1,2 1,3 1,4 1,5 1,6 1,7 1,8 1,9 2,0
Масса (5/9)
Г, Размер кормящейся группы
Наземные •. . «мни •
Древесные /йшти . •
। 8 'I_।_____]______1_____|_
0 * 204060 80
Число особей
__________I
100 310 330
£. Размер репродуктивной группы
Наземные •
Древесные
•М *••• •
0 20 40 60 80 100
Число особей
Е. Размер популяции
Наземные • ••••• •• • * •• •
Древесные • •• «••• • • •
.И , ________>_______!—
0 20 40 60 80
Число особей
—J______I
100 120
Численное соотношение полов в репродуктивной группе
Наземные
Древесные
1 ,, „ т
13 14
Число самок, приходящихся на одного самца
Гоминиды как наземные приматы
211
3. Социальная организация Наземные Древесные
Группы с большим числом самцов 13 6
Группы с одним самцом 4 10
//.Масса кормящейся группы
Наземные «• ••••mL* • • •
Древесные • •••• ••••••• •• «* *
. " I ._________________________!_________________.
12 3 4
log массы кормящейся группы, кг
К. Площадь территории сообщества
Наземные • • J • •**••• •
Древесные • • • •
lirtiiiiirTiiiiiiiiiliii.il.. u,i । I I i I I I । । 1 । i.J
-2,0 -1,0 0 1,0 2,0
log площади территории, км2
Л. Перемещения за день
Наземные ф , , ффф «••••• •
Древесные • » •• • •
,_______________J,
-1,0 0 1,0 2,0
log расстояния, покрываемого за день, км
М. Плотность популяции
Наземные ф # •<• • •
Древесные • • • • • •••• • а
।____________।____________।_________-.л___________ ।
-1,0 о 1,0 2,0 3,0
log плотности популяции. Л'-км"2
Н. Биомасса
н
Наземные • •••• • . ••••
Древесные • •• .• • •• • ♦ •••• • •
,—:__________।____________। 1__________1_____________1
о 1,0 2,0 3,0 4,0
log биомассы, кг-км-2
Рис. 7.8. Сравнение экологических характеристик наземных и древесных узко-
носых. Сравнение проводится на межвидовом уровне (звездочкой отмечены
средние значения). Хотя, как считают Клаттон-Брок и Харви (Clutton-Brock,
Harvey, 1979), сравнение да этом уровне и сопряжено со статистическими труд-
ностями, оно позволяет выявить у животных некоторые адаптивные тенденции.
(Clutton-Brock, Harvey, 1977а.)
14*
212
Глава 7
Таблица 7.1. Сравнение (соцноэкологических характеристик наземных
и древесных узконосых (см. рис. 7.8)
Пищевой рацион
Масса тела
Половой диморфизм
Размер кормящихся
групп
Размер репродуктивных
групп
Размер популяций
Численное соотношение
полов в репродуктивных
группах
Социальная организация
Масса кормящихся групп
Площадь ареалов сооб-
щества
Дневные перемещения
Плотность популяции
Биомасса
Число листоядных видов среди НУ несравненно
меньше
НУ обычно крупнее ДУ (см. также рис. 7.1)
У НУ выражен несколько сильнее, чем у ДУ, од-
нако разница, возможно, несущественна
Кормящиеся группы НУ гораздо крупнее, чем
У ДУ
Репродуктивные группы НУ крупнее, чем у ДУ
Популяции НУ крупнее, чем у ДУ
У НУ число самок, приходящееся на одного сам-
ца, несколько меньше, чем у ДУ, однако разница
очень незначительна
Группы с большим числом самцов чаще встреча-
ются у НУ, однако четких различий нет
У НУ выше, чем у ДУ
НУ имеют более обширные ареалы, чем ДУ
За день НУ покрывают более значительные рас-
стояния, чем ДУ
У НУ ниже, чем у ДУ
У НУ меньше, чем у ДУ
НУ — наземные узконосые, ДУ — древесные узконосые
что, хотя характеристики этих двух групп животных частично
совпадают, можно выявить некоторые экологические тенденции,
связанные с переходом к наземному существованию. Чтобы по-
ложить эти тенденции в основу модели, которая дала бы воз-
можность объяснить адаптацию разных гоминид и строить соот-
ветствующие гипотезы, необходимо изучить факторы отбора,
ответственные за возникновение упомянутых выше различий.
Одной из выявленных с помощью полученных данных (см.
рис. 7.1) тенденций является уже обсуждавшееся (см.
разд. 7.1.1) увеличение размеров тела с переходом к наземному
образу жизни. Было сделано предположение, что древесный
образ жизни накладывает на размеры тела ограничения,
а наземный образ жизни эти ограничения снимает. В резуль-
тате многие из характеристик наземных приматов могут сов-
падать с характеристиками, связанными с укрупнением разме-
ров вообще (см. гл. 5 — например, большая безопасность от
хищников, более широкий индивидуальный участок, возмож-
ность использовать в пищу более разнообразные и менее
качественные продукты. Действительно, у более крупных при-
матов в пищевом рационе, как правило, повышается относи-
тельное содержание листьев и понижается общее качество
Гоминиды как наземные приматы
213
потребляемых ими продуктов (Clutton-Brock, Harvey, 1977 b;
Sailer, Gaulin et al., 1985). Кроме того, у более массивных
приматов обширнее территории, что можно видеть и на при-
мере наземных узконосых. Как видно из рис. 7.8, в течение дня
наземные приматы совершают и более значительные перемеще-
ния. Поскольку для поддержания жизнеспособности особи или
группы наземных приматов требуются более обширные участ-
ки, у этих приматов прослеживается сопутствующая тенденция
к снижению плотности (числа особей на единицу площади) и
биомассы (массы тела на единицу площади) популяции.
Одной из причин расширения территорий у наземных при-
матов могут быть большие размеры их стад. Это обстоятельст-
во традиционно связывают с защитой от хищников (см. van
Schaik, van Hoof, 1983), однако не исключено и влияние других
факторов. Во-первых, многие наземные приматы нередко жи-
вут в таких средах, где пищевые источники распределены не-
равномерно; Хорн (Horn, 1968) в этой связи предполагает,
что там, где это наблюдается, размер стада должен возрастать
для повышения эффективности пищевого поиска. Согласно
другой гипотезе, в средах с неравномерно распределенными
источниками пищи отбору должны подвергаться более крупные
стада вследствие острой межстадной конкуренции. Если на
одну и ту же территорию с источниками пищи претендуют
два стада животных, победу в конкурентной борьбе одержит
более многочисленное (Wrangham, 1980).
В результате увеличения размеров стад у наземных прима-
тов могут изменяться и их социальная структура, и стратегии
размножения и спаривания. Половой диморфизм у приматов
связан с конкуренцией среди самцов. Особенно острый харак-
тер эта конкуренция носит в стадах с большим числом самцов
и у полигамных видов; у моногамных видов она слабее. В ста-
дах, где осуществляется полигиния1 или, наоборот, стратегия
спаривания со многими самцами, последние борются между со-
бой за обладание самками и, таким образом, отбору подверга-
ются более крупные самцы. Поскольку наземные приматы жи-
вут, как правило, большими стадами, а уровень моногамии со-
ответственно низок (само определение моногамии предполага-
ет небольшое число особей в группе), можно думать, что поло-
вой диморфизм у наземных приматов выражен сильней, чем у
преимущественно древесных видов. Группы с большим числом
самцов более типичны для наземных, нежели древесных, при-
матов.
1 Форма половых отношений, при которой за одни сезон размножения
самец в норме спаривается с несколькими самками. —Прим, перев.
214
Глава 7
6 7
Социоэкономическое соотношение полов, число
самок, приходящихся на одного самца
Рис. 7.9. Связь между выраженностью полового диморфизма и численным соот-
ношением полов у узконосых. (Clutton-Brock, Harvey, 1977b.)
Половой диморфизм тесно связан с другим фактором —i
численным соотношением полов (см. рис. 7.9). Если половой
диморфизм — результат соперничества между самцами, то
следует ожидать, что это соперничество будет особенно острым
в тех стадах, где самцы вынуждены конкурировать за большое
число самок. Дело может обстоять именно так, однако связи
между социоэкономическим соотношением полов и социальной
структурой у узконосых обезьян выявить не удалось (Clutton-
Brock, Harvey, 1977b).
Таким образом, проведенный краткий анализ показывает,
что наземный образ жизни ставит перед всеми узконосыми
обезьянами одинаковые проблемы и дает им одинаковые воз-
можности для их решения, причем разные виды этих живот-
ных справляются с этими проблемами аналогичным образом.
Хотя связать прямыми причинно-следственными отношениями
такие характеристики узконосых, как, например, наземный об-
раз жизни, размеры тела и половой диморфизм не удается,
ясно, что эти и другие их адаптивные признаки тесно и слож-
но переплетены друг с другом и с наземным существованием.
Наземный образ жизни, так или иначе, накладывает отпечаток
на морфологию, поведение и экологию тех приматов, которые
«спустились с деревьев». Грубо говоря, «смоделированные»
наземные узконосые должны обнаруживать крупные размеры
тела, сильно выраженный половой диморфизм, большие раз-
меры стад и обширные территории при низкой плотности попу-
ляции. Такая модель позволяет более плодотворно изучать и
наземный образ жизни гоминид.
Гоминиды как наземные првматы
215
7.4. Наземный образ жизни гоминид
Возникает вопрос, насколько ранние гоминиды соответству-
ют или не соответствуют этим теоретическим требованиям?
Ясно, что из-за скудости ископаемых остатков гоминид и ар-
хеологических находок дать точный ответ на этот вопрос труд-
но, хотя некоторые замечания по этому поводу можно все-та-
ки сделать.
7.4.1. Половой диморфизм, и спаривание
Как уже говорилось в гл. 6, в процессе эволюции гоминиды
становились крупнее. Несомненно, хотя бы частично эта тен-
денция была обусловлена наземным образом жизни. Судить
о выраженности полового диморфизма у гоминид труднее.
Изменчивость ископаемых форм, обусловленная половым ди-
морфизмом, составляет лишь часть непрерывного ряда пере-
ходных форм, различия между которыми определяются много-
численными факторами, в том числе временем существования,
географическим распространением и таксономической принад-
лежностью. Джохансон и Уайт (Johanson, White, 1979) обна-
ружили выраженный половой диморфизм у самого раннего из
известных представителей гоминид — Australopithecus africa-
nus. Если образцы, приписываемые этому таксону, действитель-
но принадлежат одному виду, можно думать, что половой ди-
морфизм в семействе гоминид выражен наиболее сильно имен-
но у его древнейших представителей. В известной мере присущ
половой диморфизм и другим представителям семейства Но-
minidae (табл. 7.2). Как грацильные, так и массивные австра-
лопитековые обнаруживают достаточно большую вариабель-
ность, чтобы можно было предполагать половой диморфизм и
у них. Что же касается ранних представителей рода Ното, то
царящая здесь таксономическая путаница не позволяет с оп-
ределенностью относить обнаруженные ископаемые остатки к
тому или иному полу (хотя образцу KNM-ER 1470 из Кооби-
Фора приписывают мужской пол, а образцу KNM-ER 1813—J
женский). Предполагается существование полового диморфиз-
ма и у Homo erectus—вида, к которому принадлежат некото-
рые из самых крупных и массивно сложенных представителей
известных гоминид (рис. 6.4). Развитие современного человека
с присущими ему анатомическими признаками, по-видимому,
сопровождалось стиранием половых различий.
То обстоятельство, что ранние гоминиды обнаруживали в
той или иной степени половой диморфизм, свидетельствует о
несостоятельности классических представлений о возникновении
устойчивых пар и моногамии в самом начале эволюции чело-
216
Глава 7
Таблица 7.2. Оценка полового диморфизма (в процентном отношении)
у современных гоминоидов и ископаемых гоминид на основании различных
анатомических показателей (Frayer, Wolpoff, 1985)
й „ I * 6 га Й н s Ь
2 OF SS OF га OF = 2 р £ OF
Is 'о сд а х а 'о X X 'о 'о
Зё ‘е' 3§ "е’ Ц в? е
Pan paniscus 118,3 23/25 100,0 24/33 101,2 10/10 100,0 12/17
Pan troglodytes 123,3 84/90 103,7 124/124 104,9 38/58 108,7 22/20
Gorilla gorilla 138,0 153/95 107,0 170/170 120,9 84/54 123,8 47/22
Pongo ptjgmaeus 126,6 48/59 112,0 69/73 115,0 32/29 117,0 12/16
Australopithecus afarensis 127,5 11/5 111,2 6/10 118,4 5/5
A ustralo pithecus africanus 117,3 9/6 109,1 7/3 119,6 126,9 4/4
Homo habilis 125,4 4/7 107,9 4/8 5/10
Homo erectus 113,6 6/15 106,0 10/22 108,1 119,6 10/17
Европейские неан- дертальцы 115,8 13/9; 104,8 12/10 4/1 115,5 115,3 7/8
108,4
Первые люди верх- него европейского 109,8 7/6 105,5 7/8 5/3 8/8
палеолита
века; более правдоподобно звучит предположение, что социаль-
ная организация ранних гоминид не отличалась от таковой
других наземных приматов с такими присущими ей характерис-
тиками, как высокая численность групп и конкуренция между
самцами за обладание самками. Возможно, эта социальная ор-
ганизация изменилась лишь с появлением поздних гоминид,
когда преобладать стала моногамия или полигамия, структури-
рованная в возрастном отношении и носившая менее выражен-
ный характер прямой конкуренции между самцами, исход ко-
торой определялся уже иными, нежели размеры тела, факто-
рами.
Понятно, что попытки пойти в изучении гоминид дальше их
полового диморфизма и получить какую-либо информацию об
их социальном и репродуктивном поведении сопряжены с ог-
ромными трудностями. В ископаемых остатках эти аспекты
жизни ранних гоминид прямо никак не отражены, а достаточ-
но надежных методов их изучения по косвенным данным еще
не разработано. Некоторые заключения, однако, можно сделать
на основании сравнительной анатомии. Вслед за Шортом
(Short, 1979, 1980, 1981) Харкерт и др. (Harcourt, Harvey et al.,
1981) показали, что размер наружных половых органов у прима-
тов варьирует в зависимости от социальной структуры. В про-
стейшем случае эта закономерность проявляется в том, что раз-
Гоминиды как наземные приматы
217
Человек
Шимпанзе
Рис. 7.10. Сравнительные размеры наружных половых органов и тела у совре-
менных крупных гоминоидов. В левой половине рисунка изображены размеры
тела самцов четырех приматов относительно размеров тела самки, а также
сравнительные размеры яичек и полового члена в состоянии эрекции. В правой
половине рисунка изображены размеры тела самок этих же приматов относи-
тельно размеров тела самца, а также сравнительные размеры промежности
и молочных желез. (Martin, Мау, 1981.)
мер яичек возрастает у самцов тех животных, которые живут ста-
дами с большим числом самцов, нежели у животных с единст-
венным самцом в стаде. Это обстоятельство можно объяснить
тем, что в стадах с большим числом самцов конкуренция меж-
ду ними за обладание самками острее, чем в тех популяциях,
где один-единственный самец удерживает монополию на одну
или нескольких самок. Если спаривание непродолжительно и
может быть прервано вмешательством других самцов, опреде-
ленные репродуктивные преимущества дает самцу высокая
продукция спермы. Мартин и Мей (Martin, Мау, 1981) исполь-
зовали эти соображения для анализа возможного характера
спаривания у ранних гоминид (рис. 7.10). Авторы установили,
что размеры яичек у современного человека соответствуют
показателям, характерным для животных, которые живут ста-
дами с одним самцом. Согласуется этот факт и с обсуждав-
шейся выше выраженностью полового диморфизма у гоминид.
218
Глава 7
По мнению Мартина и Мея, которое подтверждается ископае-
мыми находками, «исходная» система спаривания у ранних
гоминид должна была основываться на полигинных группах с
единственным самцом. Полигинной системе спаривания отве-
чают также выраженность половых различий размеров и фор-
мы тела у современного человека (см. рассматривавшийся вы-
ше вопрос о соперничестве самцов из-за самок) н более зна-
чительный репродуктивный вклад в мужское потомство по
сравнению с женским (в полигинных системах «разброс» ре-
продуктивного успеха самцов больше, чем у самок, и, следова-
тельно, с большей выгодой окупаются энергетические затраты
на воспроизведение мужского по сравнению с женским, по-
томства) (Gibson, McKeown, 1952). Таким образом, можно
выдвинуть предварительную гипотезу о том, что репродуктив-
ное поведение ранних гоминид было основано на полигинных
группах, включающих только одного самца. Может показаться,
что эта гипотеза противоречит выводам проведенного выше
анализа, в котором значительное место отводилось кооперации
среди самцов, и, следовательно, обосновывалось возможное
преобладание у ранних гоминид репродуктивного поведения,
связанного с наличием больших стад с многочисленными сам-
цами в каждом из них. В настоящее время репродуктивное по-
ведение у животных сильно варьирует. В сообществах совре-
менных людей преобладает полигиния (Ford, Мау, 1952); ясно,
однако, что социальная организация может изменяться под
влиянием локальных факторов.
Яички, к сожалению, никогда не подвергаются фоссилиза-
ции, и поэтому непосредственно проверить все эти предполо-
жения нельзя. Можно, однако, попытаться разработать модель
социальной организации ранних гоминид, учитывая давление
на них окружающей среды и общие характеристики африканс-
ких гоминоидов.
Гориллы живут полигинными группами, в которых, хотя
и допускается присутствие нескольких самцов, всегда четко
прослеживается доминирование какого-нибудь одного из них,
устанавливающего, таким образом, своего рода репродуктив-
ную монополию. В отличие от горилл обыкновенные шимпанзе
живут более многочисленными группами, ядро которых состав-
ляют самцы, а самки ведут полунезависимый образ жизни,
перемещаясь по частично перекрывающимся территориям со-
седних сообществ. Таким образом, у африканских гоминоидов
ядро социальной группы образуют самцы. Наоборот, у низших
узконосых ядро социальной группы составляют родственные
самки, а самцы переходят из одной группы в другую.
Можно предположить, что «родоначальные» африканские
гоминоиды были в значительной степени полигинными живот-
Гоминиды как наземные приматы 219
ними, жившими небольшими группами в лесных средах. Пе-
реход к существованию в более открытых средах должен был
вызвать в такой системе огромное напряжение, поскольку сам-
цы, вероятно, оказались не в состоянии защищать слишком
большую территорию, и, кроме того, должно было сказываться
значительное давление отбора в сторону увеличения численно-
сти групп, что наблюдается и у других наземных приматов.
Так, у шимпанзе, существование в лесных и редколесных сре-
рах привело к образованию самками более независимых соци-
альных групп, частично вышедших из-под контроля самцов.
В более экстремальных условиях, однако, такая социальная
структура может поставить животных в невыгодное положение
как в плане уязвимости для хищников, так и в плане эффек-
тивности поиска пищи в средах с неоднородным распределени-
ем ее источников. Низшие узконосые решают эту проблему за
счет образования групп, основанных на материнском родствен
включающих большое число самцов; маловероятно, однако,
чтобы система, основанная на родстве по матери и предпола-
гающая перемещения самцов из одной группы в другую, могла
эволюционировать из сообщества гоминоидов, ядро которого
составляют самцы. Противоречия между особенностями соци-
альной структуры у гоминоидов и требованиями наземного об-
раза жизни (или существования в открытых средах) могли
разрешиться лишь единственным способом — за счет усиления
основанной на родственных отношениях кооперации среди
самцов, или, другими словами, за счет отхода от простой по-
лигинии в сторону полигинии, регулируемой родственными от-
ношениями и предполагающей свободное объединение групп
родственных самцов в более крупные группы, внутри которых
прослеживаются более специфические полигинные семейные
структуры. Такая система хорошо соответствует как (предпо-
лагаемым) тенденциям к усилению у гоминид родственных свя-
зей и наблюдающемуся у них стиранию (но не исчезновению)
полового диморфизма, резко выраженного на ранних этапах
эволюции, так и характеристикам наземных приматов, живу-
щих в открытых средах, которые можно предсказать исходя
из принципов эволюционной экологии. Модель эволюции стра-
тегий спаривания у гоминоидов изображена на рис. 7.11.
7.4.2. Плотность популяции
Некоторые из предположений, связанных с пищевым поис-
ком у гоминид, — снижение плотности популяции и расширение
территорий — уже обсуждались в гл. 6. Определять плотность
популяции по ископаемым остаткам трудно: проведенные рас-
четы (Boaz, 1979; R. A. Martin, 1981) смогли дать лишь очень
220
Глава 7
Горилла
Шимпанзе
Сохраниашаяся в сходных
(лесных) средах социальная
структура ''родоначальных"
африканских гоминоидов,
модифицированная увеличением
размеров тела и усилением
полового диморфизма
Более многочисленные группы;
социальная структура по-прежнему
основана на приоритете самцов,
однако самки пользуются большей
независимостью и контроль их
поведения одним самцом
невозможен
Ранние гоминиды
"Родоначальные" африканские
гоминоиды
Небольшие полигинные группы
с одним самцом, обитавшие в
лесных средах; значительный
поповой диморфизм
Отбор более многочис -
ленных групп в сочетании
со все более широким-------
освоением более откры-
тых сред
Более многочисленные группы;
отбор более многочисленных групп
обусловлен освоением саванны, в
результате чего независимость самок
ограничена. Образование "павианопо-
добных** групп с большим числом сам*
цов невозможно из-за отсутствия ядра
группы, состоящего из самок, и сущест-
вования тесных связей между самцами;
полигиния обеспечивается высокой
родственной кооперацией среди самцов,
приводящей к развитию полигинии,
регулируемой родственными взаимо-
связями
Рис. 7.11. Гипотетическая модель эволюции социальных систем у африканских
гоминоидов. В основу модели положено предположение, что под давлением
условий окружающей среды в исходной полигинной системе с «гаремом»
происходит увеличение численности групп и упрочение социальных связей
между самцами.
приблизительную оценку. Однако, в этом отношении представ-
ляет интерес тот факт, что плотность популяций современных
охотников и собирателей чрезвычайно низка; если при расчетах
учитывать размеры тела и доступность источников пищи,
оказывается, что эти популяции характеризуются меньшей био-
массой, чем популяции других наземных приматов (Foley,
1984b) (рис. 7.12).
Плотность популяции и половой диморфизм — переменные,
позволяющие поставить гоминид в один непрерывный ряд с
другими наземными приматами. Есть, однако, и другие пе-
ременные, которые, хотя и являются прямым следствием назем-
ного образа жизни гоминид, заставляют говорить о сущест-
венных отличиях их адаптивной стратегии от адаптивных
стратегий прочих наземных приматов. Переходим к их рассмот-
рению.
7.4.3. Бипедия
Поскольку предки наземных приматов вели древесный об-
раз жизни, можно думать, что главной проблемой для них
было передвижение по земле. Должны были возникнуть но-
вые способы локомоции. Необходима была локомоторная сис-
Гоминиды как наземные приматы
221
103
Наземные приматы (за
• исключением человека)
е Современные тропические
охотники и собиратели
А
___I________________I_______________I
1000 2000 3000
Среднегодовой уровень осадков, мм
Рис. 7.12. Связь между средним годовым уровнем дождей и биомассой в попу-
ляциях тропических охотников и собирателей и наземных приматов. (Foley,
1984b.)
тема, которая удовлетворяла бы двум требованиям: эффектив-
ному и безопасному передвижению и возможности пользовать-
ся передними конечностями для кормежки и других целей.
В отличие от большинства других млекопитающих приматы не
хватают пищу сразу ртом и поэтому должны обладать некото-
рой «ловкостью рук».
У наземных приматов эволюционировало несколько систем
передвижения. Обычно их принято разделять на три главных
типа: хождение на четырех ногах, хождение с опорой на
фаланги пальцев верхних конечностей и бипедия (прямохож-
дение) (рис. 7.13). Наиболее специализированным из этих трех
способов локомоции, требующим значительных анатомических
модификаций, является присущая гоминидам бипедия. Пропор-
циями конечностей (соотношением длины рук и ног) гомини-
ды сильно отличаются от большинства высших и низших обезь-
ян, у которых руки имеют или такую же длину, как и ноги,
или даже длиннее их (см. Napier, Napier, 1967). Изменилась и
стопа; большой палец, выровнявшийся с другими пальцами,
потерял хватательную способность, но его опорные возможно-
сти увеличились. Таз у двуногих должен выдерживать боль-
222
Глава 7
Рис. 7.13. Локомоторные системы наземных узконосых: А. Передвижение на
четырех ногах у низших узконосых обезьян; Б. Походка с опорой на фаланги
пальцев верхних конечностей у африканских человекообразных обезьян; В. Би-
педия гоминид. (Weiss, Мапп, 1975.)
шую нагрузку, чем у четвероногих. Соответственно у гоминид
таз, превратившись в опору для позвоночника, укоротился и
приобрел более шарообразную форму. Стал более массивным,
особенно в нижней своей части, и сам позвоночник, приняв
для оптимального распределения нагрузки S-образный изгиб.
Череп тоже модифицировался, причем большое затылочное
отверстие переместилось вниз. Прямохождение наложило от-
печаток на всю анатомию гоминид. В эволюционном плане
переход к прямохождению означал коренное отличие гоминид
от современных приматов (однако, совсем не означал такого
же радикального отхода от их предков) и, следовательно, рез-
кое усиление давления отбора.
Как кратко отмечалось в гл. 1, впервые признаки бипедии
обнаруживаются у древнейшего из известных ныне гоминид —
Australopithecus aferensis, что указывает на развитие локомо-
торных изменений где-то в самом начале эволюции гоминид.
Тому есть двоякое подтверждение. Это, во-первых, морфология
черепов и посткраниальных остатков, найденных в Хадаре
(Эфиопия) Джохансоном, которые датируются на уровне
3,0 млн. лет (Johanson, Taieb, Coppens, 1982). Частично сохра-
нившийся скелет AL-288, известный в обиходе как «Люси»,
своим общим строением и пропорциями конечностей особенно
Гоминиды как наземные приматы
223
отчетливо указывает на то, что он принадлежал гоминиду, спо-
собному к вертикальной бипедии. Другим подтверждением яв-
ляются отпечатки следов, обнаруженные в вулканическом пеп-
ле в Летоли (Танзания). Эти отпечатки, датируемые возрастом
свыше 3,6 млн. лет (Leakey, Нау и др., 1976; White, Harris,
1977), представляют собой длинную цепочку следов, оставлен-
ных передвигавшимися на двух ногах гоминидами. Отпечатки
эти можно рассматривать как почти прямое доказательство то-
го, что ранние гоминиды были прямоходящими или, во всяком
случае, могли проходить на двух ногах значительные расстоя-
ния.
В связи с вышесказанным возникают два вопроса. Во-пер-
вых, когда появилась бипедия, и, во-вторых, отличалась ли она
у ранних гоминид от бипедии современного человека? Отвечая
на первый вопрос, следует сказать, что бипедия, очевидно, воз-
никла достаточно рано, чтобы ее можно было рассматривать
как один из компонентов исходного комплекса адативных сдви-
гов у гоминид. Если подтвердятся недавние сообщения (Hill,
1985) о челюсти A. afarensis из Табарина (района озера Барин-
го, Кения), датируемой на уровне 5 млн. лет, и, если будет
найден дополнительный посткраниологический материал, впол-
не может оказаться, что бипедия появилась даже еще раньше.
Ответить на вопрос о природе ранней бипедии сложнее.
Согласно сложившемуся за минувшее десятилетие мнению,
бипедия австралопитековых в энергетическом и биохимическом
плане сильно напоминала бипедию современного человека, а вы-
явленные у австралопитековых анатомические особенности
обусловливали лишь очень незначительные функциональные
отличия. Находки из Хадара опровергли это представление.
Джангерс с сотрудниками (Jungers, 1982; Jungers, Stern,
1983; Stern, Susman, 1983) предложили, что прямохождение
A. afarensis отличалось от прямохождения современного че-
ловека. Главным аргументом этих авторов является бедренная
кость образца AL-200 (Люси), которая имеет такую же дли-
ну относительно размеров тела, как и бедренная кость у шим-
панзе. Относительная длина верхних конечностей также ве-
лика. Исследователи считают, что локомоция этого вида в
значительной мере включала и перемещение в древесных сре-
дах. Последнее обстоятельство, по мнению авторов, подтверж-
дается тем фактом, что фаланги пальцев рук у A. afarensis
изогнуты — признак, характерный для древесных приматов.
Таким образом, исследователи заключают, что на ранних эта-
пах эволюции (в интервале между 5,0 и 2,5 млн. лет назад)
полностью двуногая походка у гоминид еще не возникла.
Эту точку зрения оспаривают Уолпофф (Wolpoff, 1983),
Лавджой (Lovejoy, 1979,1980) и Лэтимер (Latimer, 1984). Уол-
24
Глава 7
пофф считает, что Джангерс (Jungers, 1982) не учитывал
всех аллометрических факторов и что, поскольку длина ноги
уменьшается со снижением массы тела непропорционально бы-
стро, AL-200 имеет бедро именно такой длины, которая долж-
на быть у особи данной массы. По мнению Уолпоффа (Wol-
poff, 1983, р. 449), «пропорции конечностей Люси нельзя ис-
пользовать как свидетельство против способности гоминид к
абсолютно современному способу прямохождения». Лэтимер
(Latimer, 1984) также относится с сомнением к выводам Джан-
герса и Стерна, утверждая, что надежнее всего способ локомо-
ции отражает морфология суставов, а в этом отношении сход-
ство A. afarensis с современным человеком очень велико.
Все эти спорные вопросы ждут своего разрешения, однако
уже сейчас с определенностью можно сказать следующее. Об-
ладали ли древнейшие из известных ныне гоминид в точности
таким же прямохождением, что и современный человек, или
нет — главное заключается в том, что их анатомия подверглась
коренной реорганизации, отвечающей требованиям двуногой
локомоции. Australopithecus afarensis был двуногим в большей
степени, чем какой-либо из ныне живущих гоминоидов и, по
всей вероятности, прибегал к прямохождению чаще, чем к ка-
кому-либо другому способу передвижения. Кроме того, прими-
тивные признаки анатомии передней конечности, связанные с
древесным образом жизни, необязательно должны прямо ука-
зывать на время, проводимое на деревьях. Как подчеркивает
Мак-Генри (McHenry, 1982), передние конечности при переходе
к бипедии, вероятно, подвергались менее интенсивному отбору,
чем задние, поэтому задние конечности у ранних бипедов были
более специализированными, чем передние. Именно это и об-
наруживается у A. afarensis.
Приняв таким образом к сведению существование у ранних
гоминид бипедии, можно вернуться к вопросу, почему у них
эволюционировало именно прямохождение, а не какой-нибудь
другой способ локомоции, свойственный другим наземным при-
матам? Как отмечает Мак-Генри (McHenry, 1978), возникнове-
ние бипедии разные авторы объясняли самыми разными причи-
нами: переходом к жизни в саваннах (Darwin, 1871), изготов-
лением орудий (Washburn, 1950, 1960, 1967), пищевой адапта-
цией (DeBrul, 1962; Jolly, 1970), ношением в руках предметов
(Hewes, 1961, 1964; Kortlandt, 1967; Lovejoy, 1981), хищничест-
вом (Ravey, 1978), демонстрациями (Livingstone, 1962), термо-
регуляцией (Wheeler, 1984, 1985) и биоэнергетикой (Taylor,
Rowntree, 1973). В принципе все эти гипотезы можно разде-
лить на две группы: гипотезы, объясняющие возникновение би-
педии теми прямыми преимуществами, которые она дает в
плане локомоции, и гипотезы, рассматривающие ее как побоч-
Гоминиды как наземные приматы 225
ный продукт действия других факторов отбора, направленно-
го, как правило, на хватательную специализацию верхних ко-
нечностей и увеличение связанных с этим преимуществ.
Экспериментальное и теоретическое изучение эффективности
различных локомоторных систем имеет предварительный ха-
рактер. Энергия, которую капуцины и шимпанзе затрачивают
при ходьбе с обычной скоростью, одинакова при передвижении
на четырех и двух конечностях (Taylor, Rowntree, 1973). При
беге с максимальной скоростью, однако, бипедия оборачивает-
ся почти в два раза более высоким расходом энергии по срав-
нению с типичным для млекопитающих передвижением на че-
тырех ногах (Taylor, Schmidt-Nielson, Raab, 1970). Ни одна
локомоторная система приматов по своей энергетической эф-
фективности не может сравниться со свойственным, например,
собаке или корове, передвижением на четырех конечностях,
причем, как предполагают Родмэн и Мак-Генри (Rodman,
McHenry, 1980), критическими в этом отношении являются не
максимальные, а обычные скорости передвижения. По данным
этих авторов, человеческая походка при обычной скорости
ходьбы дает больший энергетический эффект, чем передвиже-
ние шимпанзе на четырех или на двух ногах и, следовательно,
можно думать, что при прочих равных условиях именно эта
походка и должна была эволюционировать.
При оценке биоэнергетики локомоторных систем учитывать
следует не только скорость передвижения. Решающее значение
имеет также время, в течение которого может протекать ло-
комоторная активность. Было установлено (Campbell, 1974;
Nabakov, McClean, 1980; Bortz, 1985), что характерной особен-
ностью современного человека является способность пробегать
за длительное время очень большие расстояния. Этой своей
выносливостью человек резко отличается от других гоминоидов
и вообще от других животных, которые к продолжительному
бегу неспособны (Gericho, Hoffmeyer, 1976). Хоффмейер, Лув
и дю Приз (Hoffmeyer, Louw, du Preeze, 1973) обнаружили,
что у зебры, подвергающейся преследованию, коллапс наступа-
ет после пробегания всего 800 ярдов (780 м), а этнографы со-
общают о способности охотников и собирателей загонять
жертву до полного изнеможения (Dorman, 1925; Silberbauer,
1981; Schell, 1981, цит. по Bortz, 1985). Эти данные позволяют
предполагать, что преимущества, которые дает бипедия орга-
низму в энергетическом плане, связаны прежде всего с увели-
чением его выносливости при продолжительном беге. Это
предположение согласуется с приведенными в гл. 5 теоретиче-
скими соображениями об увеличении расстояний, покрываемых
во время пищевого поиска, при укрупнении размеров тела,
а также с моделью экстремального теплового стресса, пред-
15—367
226
Глава 7
ставленной в гл. 4. Там говорилось, что механизмы терморегу-
ляции у гоминид обнаруживают сильно выраженную специа-
лизацию. Интенсивная дневная активность, включавшая про-
должительный бег и ходьбу от одного источника пищи к дру-
гому, по всей видимости, и была главной движущей силой
отбора, обусловившей у гоминид эволюцию бипедии. Как пола-
гает Бортц (Bortz, 1985), столь высокий уровень физической
активности должен был стимулировать укрупнение головного
мозга гоминид и, следовательно, определял давление отбора
в сторону энцефализации.
Хотя продолжительный и дальний поиск пищи и мог опре-
делять направление отбора в сторону бипедии, ход эволюции
зависит не только от непосредственного давления отбора, но и
от эволюционной предыстории животных (см. разд. 3.3.2). Эво-
люционная предыстория определяет наличие или отсутствие
соответствующего биологического субстрата, на который может
воздействовать отбор, а также влияет на уровень преадаптации
животных. Таким образом, различия локомоторного поведения
у наземных приматов, вероятно, зависят не только от непо-
средственного действия отбора, но и от эволюционного прош-
лого животных.
Необходимые для возникновения бипедии преадаптации
рассматривались разными авторами (Washburn, 1960; Stern,
1971; Tuttle, 1975). Вслед за Роузом (Rose, 1973) прекрасный
анализ этого вопроса провела Айелло (Aiello, 1983), которая,
опираясь на соображения перечисленных выше авторов, пока-
зала, что и у ныне живущих и у ископаемых приматов суще-
ствует определенная связь между размерами тела и способом
передвижения (рис. 7.14). Особого внимания заслуживает раз-
личие, которое Айелло проводит между древесными примата-
ми, кормящимися на ветке или под веткой. Предполагается,
что последний способ кормежки включает необходимые пре-
адаптации у всех современных гоминоидов. По-видимому, су-
ществует определенная связь между пищевым поведением, спо-
собом передвижения и размерами тела, причем «кормежка
под веткой», как правило, в большей степени свойственна бо-
лее крупным приматам. Из ныне живущих приматов этот спо-
соб кормежки характерен для гиббона, орангутана и полубра-
хиаторов Нового Света. Как показала Айелло, представители
миоценовых гоминоидов со средними размерами тела (Procon-
sul sp. и Paidopithex rheanus) сильно напоминают современных
приматов, кормящихся под веткой. Это обстоятельство, а так-
же тот факт, что среди мартышкообразных нет видов, обнару-
живающих этот способ кормежки, заслуживает самого при-
стального внимания.
Разграничение способов кормежки у приматов порождает
"" 1— 1 1 j Мелкие | приматы 1 Приматы, 1 кормящие 1 ся на вет- 1 ках и под 1 ветками 1 II 1 III Средние 1 Крупные приматы 1 приматы Приматы, кормящиеся под ветками ! Специализации крупных высших । обезьян + СХВ/СХЗ — СХЗ 1 1 1 L ... ... ..
_ Древесная четвероногость мелких приматов
Древесная четвероногость приматов средних размеров
Древесная четвероногость крупных приматов (СХВ)
— Древесная четвероногость крупных приматов (полубрахиация приматов Старого Света)
— Древесная четвероногость крупных приматов (полубрахиация приматов Нового Света)
1 — — Смешанная древесная (СХВ) и наземная (СХЗ) четвероногость крупных приматов
— Брахиация
Бипедия • • • • 1
Хождение с опорой на фаланги пальцев верхних конечностей -- - - . 1 1 1 1
Локомоторные
группы миоце-
новых гоминои-
дов (Aiello)
Локомоторные
группы совре-
менных прима-
тов (Rose)
О
10
20 30
40 50
60+
Месса тела, кг
Рис. 7.14. Размеры тела и локомоторные системы приматов (СХВ—сидение на ветках и лазание по ветвям,
СХЗ — состояние на земле и хождение по ней). (Rose, 1973; Aiello, 1981.)
228
Глава 7
Наземный образ жизни
Бипедия
(гоминиды)
Древесные четвероногие
(тонкотелые обезьяны)
Наземная четвероногость
(низшие узконосые обезьяны)
Полуназемное хождение с
опорой на фаланги пальцев
Полуназемная частичная
бипедия (карликовый
шимпанзе)
Полностью наземное хождение
с опорой на фаланги пальцев
(горилла)
Самые ранние
австралолитековые
Полубрвхиация +четеерорукость
(крупные миоценовые
гоминоиды и орангутан)
Древесные четвероногие,
кормившиеся на ветках
(ранние низшие узконо-
сые обезьяны)
Мелкие приматы
Приматы средних и мелких
размеров, кормившиеся___ Полностью древесная
на ветках и под ветками-^брахиация (гиббоны)
(ранние гоминоиды)
Крупные приматы
Рис. 7.15. Модель эволюции локомоторной дифференциации у узконосых
обезьян.
некоторые мысли о процессе дифференциации локомоторной
системы у наземных приматов (рис. 7.15). Ранние узконосые
кормились как сидя на ветке, так и подвесившись к ветке.
Предки низших узконосых, однако, кормились исключительно
сидя на ветках. Когда же некоторые из них перешли к более
выраженному наземному образу жизни, необходимых преадап-
таций к бипедии у них не было. Эволюционная предыстория
низших узконосых, возможно, в сочетании с отбором быстрых
движений, необходимых им по причине мелких размеров для
спасения от хищников, подтолкнула их в сторону четвероно-
гого хождения, а не бипедии.
Иной была эволюция Hominoidea. Большее видовое разно-
образие ранних гоминоидов в нижнем и среднем миоцене
предполагало и большую вариабельность их размеров и спосо-
бов питания. В зависимости от своих размеров ранние виды
гоминоидов прибегали как к первому, так и ко второму спо-
собу кормежки. У ранних гоминоидов, питавшихся сидя на вет-
ке, развивалось передвижение на четырех ногах, характерное
для низших обезьян (и возможно, наземный образ жизни). На-
оборот, для гоминоидов, кормившихся под веткой, эволюция в
сторону четвероногого хождения была закрыта. Для потенци-
ально наземных приматов бипедия была наиболее вероятным
Гоминиды как наземные приматы
229
направлением развития, если, конечно, отсутствовали факторы,
препятствовавшие ее эволюции. В случае гоминид такие фак-
торы не имели сколь-либо существенного значения. Они, одна-
ко, отразились на эволюции африканских понгид. В частности,
большую роль сыграло сохранение способности к древесному
образу жизни в лесу. В этом случае у животных эволюциони-
ровали полубрахиация и наземное хождение с опорой на фа-
ланги пальцев верхних конечностей, что позволяет им и пере-
двигаться по земле, и кормиться на деревьях. Так, небольшие
африканские понгиды — обыкновенный и карликовый шимпан-
зе (Pan. troglodytes и Р. paniscus) ведут полуназемный образ
жизни и передвигаются по земле, опираясь на пальцы. По всей
видимости, именно этим обстоятельством можно отчасти объ-
яснить тесное сходство между ранними австралопитековыми и
карликовым шимпанзе (Zhilman, Cronin et al., 1978).
Более крупные гориллы ведут гораздо более выраженный
наземный образ жизни, однако их тесное филогенетическое
родство с другими африканскими понгидами, которые в свое
время уже приспособились к хождению с опорой на фаланги
пальцев, по всей вероятности, препятствовало развитию у них
бипедии. Не исключено, что размеры тела также налагают ог-
раничения на эволюцию в сторону бипедии.
Таким образом, для ответа на вопрос, почему в определен-
ных условиях у приматов развивалась бипедия, можно сослать-
ся как на общую закономерность, состоящую в том, что одну
и ту же экологическую проблему животные в зависимости от
своей эволюционной предыстории могут решать по-разному,
так и на более специфические процессы. Условиями, необходи-
мыми для развития бипедии, были ограниченные размеры тела
и специфический образ жизни в древесных средах. Низшие
узконосые не удовлетворяли этим условиям, а потому, столк-
нувшись с отбором на наземное существование, остались четве-
роногими. Напротив, этим требованиям исходно удовлетворяли
африканские гоминоиды, но сначала «стремление» сохранить
способность к древесному образу жизни, а затем и увеличение
размеров тела направило одну их группу по альтернативному
пути эволюции — развитию частичной брахиации и хождению
по земле с опорой на фаланги пальцев, т. е. развитию системы
локомоции, характерной для современных африканских понгид.
Другая группа африканских гоминоидов подверглась отбору в
направлении полностью наземного образа жизни. В результате
возникла бипедия. Преадаптация к бипедии означала возмож-
ность ( а может быть, и необходимость) свободного пользова-
ния верхними конечностями, что в данном случае привело к
развитию способностей, зачастую неотделимых от прямохожде-
ния — изготовление орудий, ношение предметов, обработка
'230
Глава 7
пищи и т. д. В связи с предложенной моделью возникает ин-
тереснейший, на наш взгляд, и ждущий своего разрешения во-
прос о том, в какой степени подверглись подобному естествен-
ному отбору другие гоминоиды и соответственно как далеко
эволюционировали они в направлении бипедии?
7.4.4. Убежище
Наземный образ жизни ставит перед животными и иные,
помимо способа передвижения, проблемы. Для животных, ве-
дущих наземное существование, деревья — не просто источник
пищи, но и место, где можно найти приют и убежище. А куда,
в самом деле, прячутся на ночь и укрываются от опасности
дневные наземные приматы?
Большинство из ныне живущих наземных приматов при
всяком удобном случае также пользуются для этого деревьями
(Altmann, Altmann, 1970). Шимпанзе обоих видов устраивают
на деревьях гнезда для ночлега и часто спасаются на деревьях
от опасности. Деревьями пользуются для сна практически все
виды павианов и макак. Как показали С. Олтмэн и Дж. Олт-
мэн (Altmann, Altmann, 1970), такие «спальные рощи» явля-
ются важными очагами социальной активности у павианов
Национального парка Амбосели в Кении. Самый наземный из
видов рода Cercopithecus (мартышки) верветка (Cercopithecus-
aethiops) также спасается на деревьях от наземных хищников
и устраивается на них на ночь (Struhsaker, 1967). Мартышки
гусары (Erythrocebus patas),живущие в местности,бедной дре-
весной растительностью, на ночь во что бы то ни стало стремят-
ся устроиться на деревьях (Hall, 1966). Часто это приводит к
распаду социальной группы, так как ее члены рассредоточива-
ются по редко стоящим деревьям. Лишь самый крупный при-
мат— горилла — строит наземные гнезда (Fossey, Harcourt,
1977). Размеры этого вида не позволяют ему лазать по деревь-
ям, зато надежно защищают его от хищников, за исключением
человека.
Что же, однако, происходит, если деревьев поблизости нет,
как, например, могло обстоять дело на открытых просторах
саванны? Павианы, лишенные доступа к деревьям, ищут для
себя другие убежища. В Эфиопии гамадрилы (Papio hamadry-
as) часто спят на крутых склонах сбросовых уступов Афри-
канской Рифтовой долины (Kummer, 1968), и такое же поведе-
ние обнаруживают оливковые павианы (Papio anubis) в Гил-
гиле (Центральная Кения); в Южной Африке приматы иногда
устраиваются на ночь в пещерах. В общем при отсутствии де-
ревьев, животные, вероятно, ищут другие укрытия, которые
могли бы предоставить им столь же надежную защиту от на-
земных хищников.
Гоминиды как наземные приматы
231
Напрашивается из этих наблюдений и более широкое обоб-
щение: на ночь и в случае опасности наземные приматы стре-
мятся отыскать деревья. Если доступных деревьев нет, они
пытаются найти альтернативные варианты укрытий, которые
могли бы обеспечить им такую же надежную безопасность—t
скалы, пещеры и т. п. Если, как иногда бывает в некоторых
местностях, деревьев или других убежищ мало, острая конку-
ренция за укрытия может привести к отбору приматов в аль-
тернативном направлении.
Развитие такой конкуренции может быть обусловлено це-
лым рядом факторов. Первым и наиболее важным из них яв-
ляется уменьшение доступности деревьев в связи с исчезнове-
нием лесов и освоением приматами все более открытых биото-
пов. Как будет показано в гл. 9, в плиоцене и раннем плейсто-
цене отмечалось большое видовое разнообразие приматов, что,
по-видимому, еще более обостряло конкуренцию. Такие альтер-
нативные формы поведения, как, например, использование на-
земных гнезд, вполне могли выступать в качестве адаптаций,
позволявших избегать конкуренции. Вторым фактором являет-
ся увеличение территории пищевого поиска, связанное с укруп-
нением размеров тела и наземным образом жизни (см.
разд. 7.3.2). С увеличением продолжительности пищевого по-
иска в течение дня и расширением его территории все более
важное значение должны приобретать когнитивные способно-
сти животных: способность к обучению, пространственная па-
мять и способность к принятию решения (см. также Cant, Те-
merin, 1984). Примером таких когнитивных способностей могут
служить «мысленные карты местности» у гамадрилов, позво-
ляющие животным вести эффективный пищевой поиск в средах
с чрезвычайно низкой доступностью воды и мест для ночлега
(Sigg, Stolba, 1981). В процессе эволюции когнитивные спо-
собности могли получить дальнейшее развитие и позволить
животным решать встающие перед ними все новые и новые
проблемы — например, где и как лучше всего устраивать на-
земные гнезда и создавать другие наземные очаги активности.
Отбор в этом направлении еще более усилился в связи с повы-
шением у гоминид специализации локомоторной системы и
увеличением их размеров; таким образом, деревья переставали
быть для ранних гоминид идеальным местом укрытия. Могли
усиливать конкуренцию и такие факторы, как, например, уве-
личение риска заразиться паразитами на постоянно использу-
емых для ночлега деревьях (Hausfater, Meade, 1982); в общем
же отбор среди специализированных наземных приматов в сто-
рону устройства гнезд на земле усиливался именно недостат-
ком «спальных» деревьев.
У современных людей «наземные гнезда» (т. е. жилища и
232
Глава 7
поселения) везде одинаковы и во многих случаях являются
центрами социальной и экономической активности человека.
Поскольку же развитие гоминид неизбежно должно было со-
провождаться и модификацией их жилищ, возникает вопрос,
когда и как произошел этот адаптивный сдвиг?
В большинстве случаев эволюционное развитие основыва-
ется на модификациях предсуществующих адаптаций или на
изменениях функций существующих адаптаций. Так, эволюцию
птичьего полета обычно связывают с предшествовавшей моди-
фикацией перьев, которые вместо первоначальной функции теп-
лоизоляции стали выполнять аэродинамические функции. Ана-
логично и наземные убежища и жилища, по всей вероятности,
возникли в результате усложнения функций ранее существо-
вавших «очагов» дневной активности. По-видимому, существо-
вало несколько типов таких «очагов». Примером одного из них
могут служить тенистые места, где приматы любят отдыхать
днем в промежутках между кормежкой (см. обсуждение во-
проса о формах дневной активности в гл. 5). В качестве других
примеров можно назвать ежедневно посещаемые источники
воды, а также места с богатыми источниками пищи, например
участки земли с определенным типом растительности или (если
в пищу используется мясо) тушей мертвого животного. Про-
должительность пребывания у туши или на водопое, однако, мо-
жет снижаться из-за присутствия хищников. Как бы то ни
было, именно в таких местах могли устраиваться первые на-
земные гнезда.
Развитие наземных очагов как дневной, так и ночной ак-
тивности и возможность сочетания разных форм активности в
одном и том же подобном очаге для гоминид вполне могло
означать решение проблемы убежища в средах, лишенных дре-
весной растительности, а также стать основой дальнейшего
развития жилища. Таким образом, можно сделать предполо-
жение, что с возникновением жилища решилась для гоминид
проблема дневного и ночного убежища и что развитие некото-
рых форм деятельности, связанных с существованием назем-
ных очагов активности (дележ пищи, повышенная социальная
активность и т. д.), явилось побочным результатом развития
жилища.
Это предположение противоречит некоторым недавно вы-
двинутым объяснениям процесса развития жилища. По мнению
Айзека (Isaac, 1978а, b, 1981, 1983), у гоминид жилище — не-
отъемлемый компонент стратегии пищевого поиска. Как утвер-
ждает этот автор, включение в пищевой рацион мяса возмож-
но лишь при развитом разделении труда. С разделением форм
пищевого поиска должно было возрастать значение дележа
пищи, что в свою очередь определялось наличием центра, ку-
Гоминиды как наземные приматы
233
да должна была переноситься пища для последующего дележа.
Таким образом, жилище должно было выполнять центральную
(в прямом и переносном смысле) роль в развитии у ранних
гоминид стратегии пищевого поиска.
Эту модель, ставшую в свое время предметом широкого об-
суждения, резкой критике подверг Бинфорд (Binford, 1981,
1984). Сомнению подверглось как предположение о том, что
включение в пищевой рацион мяса предполагает дележ пищи
и разделение труда, так и интерпретация автором ископаемых
остатков гоминид и археологических находок. По утверждению
Поттса (Potts, 1984b), обнаруженные до сих пор древнейшие
археологические находки имеют мало общего с признаками
жилища современного человека, если не считать скопления
костей животных и обломочного материала. Кроме того, по
мнению Поттса, ввиду угрозы со стороны крупных хищников
гоминиды скорее должны были сокращать, нежели растягивать
свое пребывание у туш мертвых животных.
Эти соображения заставляют сомневаться в том, что бога-
тые источниками пищи участки могли превращаться в жилища.
Поттс (Potts, 1984 b) по-иному объясняет возникновение жили-
ща у гоминид. С помощью компьютерного моделирования ему
удалось показать, что эффективная утилизация туш животных
(павших или убитых на охоте) предполагала создание «скла-
дов» каменных орудий (см. рис. 7.16), т. е. появление мест,
где подвергшиеся или не подвергшиеся обработке каменные
орудия оставлялись гоминидами для того, чтобы использовать
их по мере надобности. Такие склады каменных орудий, к ко-
торым перетаскивались найденные туши животных, должны
были быть разбросаны по всей территории сообщества и скорее
напоминать плиоценовые и плейстоценовые археологические
находки со скоплением камней и костей. Склады каменных
орудий должны были давать гоминидам выгоды в энергетичес-
ком плане. Создание складов орудий и повторные к ним воз-
вращения — с добычей или с пустыми руками — недавно наблю-
далось и у западно-африканских шимпанзе. Беш и Беш (Boesch,
Boesch, 1983) обнаружили такое поведение в связи с исполь-
зованием обезьянами в пищу орехов масличной пальмы:
чтобы добраться до богатого белками ядра ореха, в некото-
рых популяциях шимпанзе пользуются «каменными молот-
ками».
Поттс всячески старается подчеркнуть, что такие склады
орудий еще не являются жилищами. Однако эти скопления;
каменных приспособлений, которые, по-видимому, соответству-
ют еще одному типу очагов активности ранних гоминид, уже
могут рассматриваться как предшественники жилища. И все
же остается непонятным, в какой степени эти скопления были
234
Глава 7
Рис. 7.16. Компьютерное моделирование пищевого поиска ранних гоми-
нид. Оцениваются относительные затраты энергии иа перетаскивание каменных
орудий к тушам мертвых животных и перетаскивание туш по частям к «скла-
дам» каменных орудий. По мнению Поттса, гоминидам было более выгодно
создавать «склады» орудий. Объяснения см. в тексте. (Potts, 1984b.)
«истинными» складами орудий, а не побочным следствием
повторного использования некоторых мест территории. Модель
Поттса, однако, выдвигает на первый план то обстоятельство,
что по мере расширения территории пищевого поиска у гоми-
нид усиливается тенденция к выбрасыванию и повторному ис-
пользованию орудий на всей площади участка (это обстоятель-
ство все время подчеркивается и в данной книге). Если учесть
еще и усиливающуюся тенденцию к устройству наземных гнезд,
можно думать, что такие места повторного использования ору-
дий вполне могли превратиться в «центры» самых разных ти-
пов активности гоминид.
Таким образом, отчетливо прослеживается конвергенция
нескольких процессов. С одной стороны, это — нарастающая
потребность в наземном укрытии, с другой — изменение пище-
вого поиска, вызванное совершенствованием изготовления ору-
дий в связи с использованием в пищу мяса. Объединение этих
двух процессов и обусловило появление предшественников жи-
лища—центров разнообразных форм активности и поведения
гоминид. Главным следствием этого, по всей вероятности, явил-
ся постепенный отход гоминид от типичного для приматов пи-
Гоминиды как наземные приматы
235
Рис. 7.17. Образцы путей, проходимых при пищевом поиске в зависимости от
социальной организации сообщества и типа очагов пространственной активно-
сти. А. Пищевой поиск, при котором единственными очагами активности
являются места ночлега (черные кружочки); Б. Пищевой поиск, предполагаю-
щий наличие «складов» каменных орудий и (или) мест для обработки пищевых
продуктов (черные треугольники) и для ночлега (черные кружочки); В. Пище-
вой поиск современных охотников и собирателей, ориентированный на един-
ственный центр активности; в этом центре протекает вся активность охотников
и собирателей, за исключением непосредственного поиска и перетаскивания
пищи.
щевого поиска, предполагающего повторное использование
многочисленных «складов каменных орудий» в сторону пище-
вого поиска, характерного для современных охотников и соби-
рателей, начинавшегося в одном-единственном центре террито-
рии сообщества и заканчивавшегося возвращением в него же
(рис. 7.17).
Уместно, однако, спросить, на каком этапе эволюции гоми-
нид произошло соединение описанных процессов, т. е., когда
возник пищевой поиск, ориентированный на один центр участ-
ка? Как свидетельствуют ранние археологические находки в
Олдувае, Кооби-Фора и других местах, гоминиды переносили
туши (кости) животных и (или) каменные орудия в некие точ-
ки своей территории. Это вполне могло стать основой страте-
гии пищевого поиска с центральной ориентацией, хотя совсем
необязательно, чтобы центр пищевого поиска становился и
центром других форм поведения, связанных с поиском пищи.
Пока остается неясным, спали ли австралопитековые и (или)
Homo habiiis на деревьях или в каких-либо других укрытиях
и спали ли они в тех же местах, где протекала их активность^
связанная с пищевым поведением.
Плотность приносного материала значительно увеличивает-
ся в местах раскопок, связанных с пребыванием Homo erectus-,
с ними, возможно, связано и первое использование огня челсн
236
Глава 7
веком (Gowlett, Harris et al., 1981). Часто в этих местонахож-
дениях обнаруживается и значительное количество костей. Как,
однако, недавно показал Бинфорд (Binford, 1984), нет никаких
оснований говорить о том, что до появления анатомически со-
временного человека в пищу регулярно использовалось мясо
(т. е. осуществлялся забой животных) или существовало насто-
ящее жилище. Хотя некоторые положения этого автора и мож-
но оспорить (Scott, 1986), сегодня, по-видимому, еще нельзя
установить сколько-нибудь точную дату возникновения жили-
ща в его современном виде. Безусловно, места пребывания
Homo erectus сильно отличаются от жилищ современных охот-
ников и собирателей, но постепенное увеличение плотности
приносного и остеологического материала свидетельствует о
тенденции ко все большей пространственной фокусировке по-
ведения ранних гоминид в плейстоцене. В настоящее время
можно лишь констатировать, что тенденция к централизации
активности зародилась уже на самых ранних этапах эволюции
гоминид.
В этой главе было показано, что переход к наземному су-
ществованию явился важным направлением в эволюции при-
матов Старого Света — направлением, охватившим наряду с
другими приматами и гоминид. С помощью более широкого и
детального анализа наземного образа жизни гоминид удалось
выявить ряд общих адаптивных проблем, с которыми сталки-
вались приматы в своем развитии и определить общие для
всех них адаптивные характеристики. В этой связи на основа-
нии принципов эволюционной экологии были сделаны некото-
рые предположения, позволившие установить, в какой степени
ранние гоминиды вели себя как «типичные» наземные приматы.
Сравнение адаптивных стратегий ранних гоминид и «идеаль-
ных наземных приматов» позволило сделать ряд выводов.
Во-первых, что адаптация к наземному образу жизни явилась
следствием увеличения размеров тела и возникновения специ-
фических палеоэкологических условий. Во-вторых, что адапта-
ция приматов, включая и гоминид, к наземному образу жизни
зависела не только от непосредственного давления отбора в
новой среде обитания, но и от их эволюционной предыстории.
И, в-третьих, что физиологические, экологические и поведенче-
ские характеристики гоминид во многом тождественны соответ-
ствующим характеристикам наземных приматов вообще, одна-
ко, у них обнаруживается и целый ряд специфических адапта-
ций. Так, способы решения гоминидами двух основных проблем
наземного существования — передвижения и убежища за счет
соответственно бипедии и устройства наземных гнезд — яви-
лись принципиальной основой для эволюции многих характе-
ристик, присущих лишь гоминидам и человеку.
Гоминиды как наземные приматы 237
Хотя адаптации эти никоим образом не противоречат прин-
ципам эволюционной экологии, они тем не менее отражают
собой некий крайний случай и не обнаруживаются больше ни
у каких других животных. Для объяснения специфики многих
характеристик гоминид в качестве основного фактора привле-
калось изменение их пищевого поиска. В этом отношении ран-
ние гоминиды в силу ряда причин, обсуждавшихся в предыду-
щих главах, сильно отошли от адаптаций других узконосых
приматов. Стало быть, следует глубже изучить характер пи-
щевого поиска гоминид в непосредственном контексте их среды
обитания — полусухой африканской саванны с выраженной се-
зонной изменчивостью. В такой среде ранние гоминиды подвер-
гались особенно сильному давлению естественного отбора.
Глава 8
ГОМИНИДЫ В СРЕДАХ
С ВЫРАЖЕННОЙ СЕЗОННОЙ ИЗМЕНЧИВОСТЬЮ
При рассмотрении бипедии было сделано предположение,
что адаптация гоминид была обусловлена как непосредствен-
ным давлением естественного отбора, так и их эволюционной
предысторией. Действительно, тропическое происхождение при-
матов, размеры их тела и переход к бипедии были в значитель-
ной мере обусловлены структурными ограничениями, наложен-
ными на их предков. Однако в конечном итоге направление
эволюционных сдвигов определяется требованиями той среды,
в которой те или иные животные обитают в текущий момент
времени. А потому рассмотрим более подробно те специфичес-
кие экологические проблемы, с которыми в процессе развития
сталкивались гоминиды, в частности те препятствия, которые
они вынуждены были преодолевать при использовании необхо-
димых для выживания ресурсов.
Хотя общие адаптации гоминид в тропиках уже обсужда-
лись, те специфические возможности и ограничения, с которы-
ми было связано их существование в этой среде, следует рас-
смотреть подробнее. Экология тропической среды ставила пе-
ред гоминидами проблемы выживания, решение которых
зависело от их адаптации. По всей вероятности, конкретной
средой обитания ранних гоминид была полусухая африканская
саванна. Одной из главных экологических характеристик этого
биома является выраженная сезонная изменчивость—фактор,
оказавший сильное влияние на всю эволюцию гоминид.
В настоящей главе рассматриваются особенности среды
обитания ранних гоминид, сезонные изменения доступности ее
ресурсов, предположительные способы использования этих ре-
сурсов гоминидами и возможные эволюционные последствия.
Вначале, однако, следует выяснить, каким образом внутри по-
пуляции и при взаимодействии между ними вырабатываются
те или иные стратегии пищевого поиска. Основным механизмом
этого процесса является конкуренция.
Гоминиды и сезонная изменчивость сред
239
8.1. Конкуренция
8.1.1. Природа конкуренции
«Конкуренция возникает в том случае, когда два или боль-
шее число организмов мешают друг другу или подавляют
друг друга» (Pianka, 1978, р. 114). Как правило, такое подав-
ление или вмешательство развивается в связи с тем или иным
пищевым ресурсом, в котором нуждаются оба организма, но
запасы которого ограниченны. Конкуренция между организма-
ми может проявляться в той эффективности, с которой каждый
из них использует данный ресурс, причем прямые взаимодейст-
вия между организмами при этом могут почти (или совсем)
отсутствовать. И наоборот, конкуренция может протекать в ви-
де активного вмешательства, когда один из организмов, так или
иначе, пресекает активность конкурента или это делает каждый
из них (Pianka, 1978).
Конкуренция может возникать в сравнительно простых ус-
ловиях. Независимо от характера пищевого ресурса нужно,
чтобы запасы его были ограниченны, т. е., чтобы «спрос» на
него превышал его доступность. Из этого простого условия вы-
текает целый ряд важных экологических последствий.
Главное из них состоит в том, что конкуренция почти всегда
ставит все участвующие в ней организмы в невыгодное поло-
жение. В результате возрастания затрат времени (энергии) на
использование ресурса или вследствие уменьшения его доступ-
ности конкуренция может привести к ухудшению приспособлен-
ности животных и (или) снижению численности популяции.
Многочисленными наблюдениями и опытами было показано, что
в отсутствие конкурентов животные «чувствуют себя» лучше
(см., например, Crowell, 1962; Grant, 1972). А это значит, что
организму выгодно по возможности избегать конкуренции.
В этой связи можно предположить, что ресурсы должны ис-
пользоваться таким образом, чтобы конкуренция либо вообще
не возникала, либо была минимальной. Стремление избежать
конкуренции — основной догмат современной экологии (Colin-
vaux, 1973). Расплата за конкуренцию столь велика, что жи-
вотным, как правило, выгоднее безраздельно пользоваться не-
большим участком «экологического пространства», нежели де-
лить более обширный участок с другими популяциями. Если
организму удастся таким образом избежать конкурентов, он
сможет занять уникальную экологическую нишу. Это — основ-
ной механизм разделения экологического мира на обособлен-
ные ниши, занимаемые разными видами животных с разными
стратегиями пищевого поиска; в конечном итоге это также и
основа эволюционного видообразования. Распределение видов
240
Глава 8
по разным нишам под влиянием конкуренции характеризуется
«принципом исключения Гаузе» (Colinvaux, 1973). Согласно
этому принципу, никакие два вида не могут занимать одну и
ту же экологическую нишу неопределенно долгое время; один
из них будет использовать ресурсы этой ниши более эффектив-
но и развиваться «за счет» другого до тех пор, пока этот дру-
гой вид не исчезнет совсем.
В сущности принцип исключения Гаузе не лишен некоторой
тавтологии и потому не имеет существенной аналитической
ценности, однако он невольно заостряет внимание на том, на-
сколько может быть допустимым в разных условиях «перекры-
вание» адаптивного поведения. Обращает он внимание и на
важное экологическое последствие конкуренции, которое за-
ключается в том, что в любой ситуации поражение в конку-
рентных взаимодействиях означает гибель отдельных особей и
вымирание вида в целом, а победа — монополизацию ресурсов
вместо того, чтобы разделить их с животными других видов.
Окончательным экологическим, а точнее говоря, поведен-
ческим, разрешением ситуации с ограниченными ресурсами яв-
ляется развитие активного вмешательства одного или больше-
го числа организмов в жизнедеятельность другого. Благодаря
такому активному вмешательству особь или популяция может
обеспечить доступность пищевого ресурса в гораздо большей
степени, нежели просто за счет более эффективного поиска.
Примерами такого активного вмешательства может служить
поведение некоторых крупных наземных хищников (Kruuk,
1972; Schaller, 1972) и широко распространенное в мире рас-
тений воздействие на другие организмы токсическими вещест-
вами.
Хотя влиянием конкуренции и можно объяснить многие
структурные и функциональные характеристики экосистемы,
некоторые важные моменты остаются неясными. Все сказанное
выше отражает классические представления о конкуренции, но
и эти представления были приняты не безоговорочно. Особенно
остро встают три вопроса. Во-первых, на каком уровне возни-
кает конкуренция? Во-вторых, насколько важную роль играет
конкуренция в экологии и эволюции. И, в-третьих, каким обра-
зом конкуренция определяет стратегию пищевого поиска?
Первый из этих вопросов связан с необходимостью разли-
чать уровни, на которых могут протекать те или иные экологи-
ческие процессы. Конкуренция может возникать между отдель-
ными особями внутри одной популяции, между отдельными
популяциями одного вида и между видами. По всей видимости,
наиболее острой является внутривидовая конкуренция, так как
в этом случае потребности организмов «перекрываются» в наи-
большей степени. Кроме того, организмы одного вида конкури-
Гоминиды и сезонная изменчивость сред 241
руют между собой не только за пищу и жизненное пространст-
во, но и за возможности, связанные с размножением, а это еще
больше обостряет конкуренцию. Острова конкуренции на этом
уровне, по-видимому, смягчается тесным генетическим родст-
вом и высокой ценой, которую платят все особи за конкурент-
ное вмешательство. Возникает конкуренция и между организ-
мами разных видов, когда они вынуждены делить между со-
бой одно и то же жизненное пространство с ограниченными
ресурсами веществ и энергии. Однако в этом случае острота
конкуренции смягчается видовыми различиями потребностей
организмов и низкой частотой их взаимодействий.
8.1.2. Биологическая значимость конкуренции
Таким образом, характер конкуренции зависит от того уров-
ня, на котором она протекает, причем наиболее остро конкури-
руют между собой особи одного вида. Следовательно, второй
вопрос — вопрос о роли конкуренции в адаптации — встает
особенно проблематично на межвидовом уровне: мнения иссле-
дователей о том, как часто на этом уровне происходят конку-
рентные взаимодействия между животными, очень сильно рас-
ходятся (Wiens, 1977; Diamond, 1978; Pianka, 1978; Schoener,
1983).
Представление о значительной остроте межвидовой конку-
ренции, вероятно, впервые сформулировал Дарвин при разра-
ботке теории естественного отбора (см. гл. 3). Ряд авторов
выступают против этого предположения, утверждая, что кон-
куренция между разными видами либо маловероятна, либо
вообще невозможна. Для обоснования первой из этих точек
зрения некоторые исследователи прибегли к моделированию и
показали, что структуры сообщества, которые обычно рассмат-
риваются как результат конкуренции, могут возникать вследст-
вие случайных процессов. А если так, то конкуренция вообще
не нужна, поскольку разделение биологического мира на дис-
кретные экологические образования может происходить и без
ее участия. Если эти авторы все-таки не исключают возмож-
ности межвидовой конкуренции до конца, то другие исследова-
тели утверждают, что сколь-либо значительная межвидовая
конкуренция в биологических системах может возникать до-
вольно редко. Главный довод в пользу этого утверждения авто-
ры видят в изменчивости сред обитания. Как полагает Уайенс
(Wiens, 1977), среды изменяются столь сильно (под влиянием
как биотических, так и абиотических факторов), что числен-
ность популяций редко соответствует поддерживающей емкости
среды и, что, следовательно, развитие конкуренции за ресурсы
среды обитания маловероятно. Острая нехватка пищи или
16-367
'242
Глава 8
жизненного пространства может возникать лишь эпизодически,
в «переломные периоды», и, хотя в это время развитие конку-
ренции не исключено, последующее «смягчение^ условий из-
менчивой среды скорее всего позволит популяции вернуться в
свое исходное состояние. Против адаптивной роли конкуренции
выступает и то обстоятельство, что благодаря хищникам чис-
ленность популяции остается ниже поддерживающей емкости
среды и тем самым предотвращается развитие конкуренции за
ресурсы местообитания (Connell, 1975).
Таким образом, утверждению о высокой биологической зна-
чимости конкуренции противоречит тот факт, что хищники или
экологические сдвиги не дают популяции превзойти некий
уровень численности, когда начинает ощущаться нехватка ре-
сурсов. Хотя при определенных обстоятельствах это условие и
в самом деле выполняется, было бы ошибочным думать, что
сказанное в полной мере относится ко всем биологическим си-
стемам. Изменчивость среды не исключает конкуренции вооб-
ще, но вызывает лишь преходящие колебания ее выраженнос-
ти. Выраженность межвидовых конкурентных взаимодействий
.варьирует в зависимости от быстроты изменений среды обита-
ния и от того, насколько «перекрываются» потребности видов.
Если «переломные периоды» случаются на протяжении жизни
одного поколения вида достаточно часто, межвидовая конку-
ренция скорее всего будет играть важную роль в его адапта-
ции. Поскольку изменчивость сред связана главным образом
с сезонностью, это условие, как правило, выполняется. Степень
же «перекрывания» экологической ниши определяется отчасти
особенностями среды, отчасти — степенью эволюционной и эко-
логической дивергенции видов. Следовательно, можно думать,
что виды переживают периоды как острой, так и слабой меж-
видовой конкуренции.
8.1.3. Модель конкуренции
Выяснив важное экологическое значение и изменчивый ха-
рактер межвидовой конкуренции, можно задаться вопросом,
каким образом она влияет на стратегию пищевого поиска?
Этот вопрос изучил Шенер (Schoener, 1983), руководствовав-
шийся в своих рассуждениях изменчивостью среды. Шенер
предположил, что существование каждой особи можно рас-
сматривать как «колебание» между двумя крайними, хотя и не
всегда четко разграниченными, фазами или состояниями. Это —
периоды «скудости» и периоды «изобилия» ресурсов (рис. 8.1).
Первые характеризуются высоким уровнем конкуренции, по-
следние — низким. При общем увеличении запасов пищи в сре-
де продуктивность разных пищевых источников меняется не-
Гоминиды и сезонная изменчивость сред
243;
Периоды
скудости
ПИЩИ
Периоды
изобилия
пищи
Расширение пищевых
рационов
с включением в них
низкокачественной,
но имеющейся
в изобилии пищи
Перемещение видов
не источники пищи,
обеспечивающей
наивысшую "отдачу"
Острая конкуренция
Разделение ниш
Монополизация
источников пищи
Неодновременное увеличение
доступности разных источников
пищи, приводящее к перекрыванию
пищевого поиска равных видов
Конвергенция
на "сверхобильных"
источниках
Истощение
пищевых
источников/
/изменение среды
Конвергенция
на источниках пищи
с высокой "отдачей"'
Слабая конкуренция
Перекрывание ниш
Совместная
утилизация
источников пищи
Рис. 8.1. Модель конкуренции в изменяющейся во времени (т. е. сезонной);
среде (по Schoener, 1983)
одинаково. На вновь появляющихся богатых источниках пищи
конвергируют разные виды животных, что приводит к значи-
тельному «перекрыванию» экологических ниш. Особенно часто,
сосредоточиваются животные либо на сверхизобильных пище-
вых ресурсах независимо от их питательной ценности, либо на
ресурсах, позволяющих получать высокую «отдачу» на мини-
мум затраченных усилий при их утилизации. В биологических
сообществах в это время можно наблюдать очень значительное-
«перекрывание» ниш и совместное использование разными ви-
дами одних и тех же источников пищи. С возвращением «го-
лодных времен» (вследствие роста популяции, истощения ис-
точников пищи или изменения окружающей среды) доступность,
этих источников вновь уменьшается. Тогда виды переходят на
те источники пищи, использование которых сулит им высокую
«энергетическую отдачу», а поскольку конкуренция — весьма
дорогостоящая форма активности, зачастую тот или иной вид.
добивается неограниченного доступа к этим ресурсам. Таким
образом, острая конкуренция может приводить к стремлению
избежать конкуренции или вмешательству, монополизированию
источников и вымиранию видов.
Рассмотренная модель обладает рядом неоспоримых до-
стоинств. Она способна объяснить разделение среды обитания-
(только, правда, при условии ее экологической изменчивости).
Модель удовлетворительно работает в нескольких «временных
режимах» и способна объяснять как кратковременные сезонные
16*
246
Глава 8
Таблица 8.1. Палеоэкологические реконструкции африканских
местонахождений ранних гомииид
Местонахожде- ние Палеоэкологическая характеристика
Табарин Берег озера с локально варьирующими элементами саванны (Hill, Drake et al., 1985).
Мидл Аваш Речная долина с интенсивной тектонической активностью, связанной с образованием Восточно-Африканской Рифтовой долины (Kabb et al., 1982; Clark, Kurashina, 1979; Clark et al., 1984).
Летолн Травяная саванна с четко разграниченными влажным и су- хим сезонами (Нау, 1981).
Хадар Озеро и впадавшие в него реки и ручьи с пойменными доли- нами (Aronson, Taieb, 1981; Gray, 1980).
Омо (Шунгара) 2,1 млн. лет назад и позднее; пологие речные берега, заня- тые сухой колючекустарниковой саванной с галерейным (пойменным) лесом и болотами; до этого, возможно, мест- ность была более лесистой (Bonnefille, 1976, 1979; Brown, 1981).
Кооби-Фора 1,6 млн. лет назад и ранее; пресное озеро с поймой, заня- той галерейным лесом и сухой колючей саванной; позднее- вода в озере иногда становилась солононатой (Isaac, 1984).
Олдувай Соленое озеро с поймой, занятой сухой травянистой саван- ной, и впадавшие в него пересыхавшие ручьи и реки; текто- нические изменения, начавшие происходить 1,5 млн. лет на- зад, привели к высыханию озера (Нау, 1976, 1981; Isaac, 1984).
Пенинж Открытая травяная саванна, окружающая соленое озеро и впадавшие в него пресноводные реки (Isaac, 1984).
Чесованья Небольшое озеро с поймой, занятой травяной саванной (Bi- shop, Hill, Pickford, 1978; Isaac, 1984),
Трансвааль Макапансгат, Стеркфонтейн, Сварткранс, Кромдраай и Та- унг — мозаичные саванновые биомы; пачка 3 Макапансгата и пачка 4 Стеркфонтейна соответствуют менее открытым (более сильно поросшим кустарниками или лесом) средам, чем пачка Mb 1 Сварткранса и пачка 5 Стеркфонтейна, что указывает на увеличение сухости климата в этих местах со временем (Vrba, 1975, 1976, 1985).
пологом, в которых преобладающим типом растительности бы-
ла трава. Однако, прежде чем окончательно принять это пред-
положение, следует рассмотреть две альтернативные возможно-
сти. Первая заключается в том, что при воссоздании палеэко-
логической картины реконструируются «макросреды», в то вре-
мя как гоминиды занимали, в них лишь специфические «микро-
биотопы». Так, их ареалы могли ограничиваться более или ме-
Гоминиды и сезонная изменчивость сред
247
Пениыж А
и g _ Ом о (Иунгура) ду ,
к Сварткранс
Кообн-Фора ду
I 7-
X
о
Лотагам
♦ Летали
♦ Хадар
Омо (Жунгура)
Макапансгат f
I--------1
Чесованья А
Илерег ДТ
П паат 1 Олдувея АТ
у Стеркфонтейн
3
Млн. лет назад
Рис. 8.2. Связь африканских местонахождений ранних гоминид с сезонной из-
менчивостью (обсуждение см. в тексте). По вертикальной оси отложена про-
должительность сухого сезона, определенная иа основании палеоэкологиче-
ских реконструкций: черные ромбики — грацильные австралопитековые, свет-
лые треугольники — массивные австралопитековые, темные треугольники —
Ното
нее замкнутыми лесными массивами в поймах непересыхавших
рек. Безусловно, такую возможность исключать нельзя, однако
в современной Восточной Африке леса, произрастающие по
берегам рек и озер, занимают, как правило, столь небольшую
площадь, что едва ли могут составлять достаточно обширный
ареал для вида, представители которого совершают значитель-
ные перемещения (хотя, конечно, они вполне могут составлять
важную часть этого ареала).
Вторая возможность связана с тем, что по тафономическим
причинам в ископаемых находках представлен лишь «сухой
экстремум» экологического диапазона гоминид. Иногда гоми-
ниды рассматриваются как пограничные популяции, которые
в результате конкуренции были вытеснены из более благопри-
ятных влажных биотопов. Хотя ископаемых находок из более
влажных регионов Африки чрезвычайно мало и, следовательно,
непосредственно проверить эту гипотезу трудно, вряд ли она
соответствует действительности. Как свидетельствуют археоло-
гические находки, относящиеся к более позднему плейстоцену,
осваивая другие части Старого Света, гоминиды предпочитали
занимать открытые пространства, и каких-либо указаний на то,
248
Глава 8
что они жили во влажных тропических лесах, ранее конца
плейстоцена не встречается. Если учесть, что археологические
находки (в особенности каменные орудия), относящиеся к ниж-
нему и среднему плейстоцену, очень часто встречаются как раз
в более сухих тропических и субтропических средах, можно
думать, что влажный тропический лес едва ли мог быть излюб-
ленным местом обитания ранних гоминид.
Таким образом, палеоэкологические данные с наибольшей
убедительностью указывают на то, что ранние гоминиды оби-
тали в лесистых и более открытых саванновых средах. Хотя не
исключено, что гоминиды заселяли и другие биотопы, с полной
определенностью можно утверждать, что они обитали в саван-
нах; в этой связи целесообразно выяснить, с какими экологи-
ческими проблемами они могли здесь сталкиваться.
8.3. Экология саванн
Саваннами называют такие зоны тропиков, в которых
главной растительной формой является трава, а иногда име-
ется и открытый древесный или кустарниковый полог (Harris,
1980). Долгое время трактовка термина «саванна» и обознача-
емого им типа среды была очень путанной и неопределенной;
споры велись по поводу того, отражает ли этот термин расти-
тельные или климатические особенности среды, является ли
саванна антропогенной или естественной климаксовой системой
и можно ли использовать этот термин для описания сообществ,
существующих вне тропиков. Большинство этих сомнений к
настоящему времени более или менее удовлетворительно раз-
решены. По мнению большинства авторов, саванна представля-
ет собой устойчивый тропический биом с определенным типом
растительности, встречающейся в зонах с выраженным засуш-
ливым сезоном (Harris, 1980, р. 7). Открытые травяные саван-
ны, занимающие 11% всей территории суши, образуются в ре-
зультате низкой растительной продуктивности, связанной с
длительными периодами засухи.
Саванны встречаются в тропических зонах, в которых годо-
вой уровень дождей составляет 250—2000 мм, а сухой сезон
длится от 2,5 до 10 месяцев. Эти зоны характеризуются также
сильной изменчивостью общего годичного уровня осадков и их
сезонного распределения. Особенности саванн связаны и с ти-
пом почвы и геологических структур, которые могут обусловли-
вать нерегулярный сток вод. Хотя в горах саванновая расти-
тельность может встречаться на высоте до 2000 м над уровнем
моря, в типичном случае саванна — плоская или слегка вол-
нистая топографическая поверхность. Вследствие сухости здесь
нередко возникают пожары, которые играют важную роль в
Гоминиды и сезонная изменчивость сред
249
Саванновый лес
Саванновое редколесье
Травяная саванна
Кустарниковая саванна
Рис. 8.3. Типы саванновых сред (по Harris, 1980)
поддержании структуры растительных сообществ (Harris,
1980).
Саванновые среды сильно различаются по типу раститель-
ности. Хотя все они характеризуются наличием сплошного тра-
вяного покрова, другие компоненты растительности сильно
варьируют. Древесный покров может быть почти непрерывным,
но постепенно разрежается, пока от него не остаются лишь
разбросанные по обширной территории одиночные деревья;
в других случаях нетравяная растительность может быть пред-
ставлена кустарниками и полукустарниками. Встречаются и
саванны, сплошь покрытые одной только травой (рис. 8.3). Тип
саванновой растительности определяется уровнем осадков, ис-
пользованием саванн в качестве пастбища и степными пожа-
рами. Биологическая продукция имеет главным образом на-
земный характер; продуктивность саванн варьирует, однако
среднегодовой уровень первичной продукции составляет здесь
1197 г-м-2-год~1 (Harris, 1980, р. 17). Для сравнения можно
указать, что первичная продукция во влажном тропическом ле-
су и пустыне соответственно 2400 и 200 г-м-2-год-1 (Murphy,
1974).
Хотя уровень первичной продукции невысок, многие саван-
новые среды характеризуются изобилием и многообразием
крупных животных. Характерной и весьма впечатляющей чер-
той современной африканской саванны, а в прошлом и других
.регионов Африки, являются пасущиеся травоядные копытные,
которые нередко собираются здесь в огромные стада. Их био-
масса прямо зависит от среднегодового уровня осадков (Сое,
Cummings, Phillipson, 1976) (рис. 8.4.). Разнообразен и видо-
250
Глава 8
20
Среднегодовое количество осадков^ мм
Рис. 8.4. Зависимость между среднегодовым количеством дождей и биомассой
крупных травоядных животных в полусухих регионах Африки, расположенных
южнее Сахары (по Сое, Cummings, Philltpson, 1976)
1500
вой состав животного мира; во многих регионах сосуществуют
20 и более симпатрических видов животных. Использование
растительных ресурсов четко дифференцируется в соответствии
с видами растений и порядком пастьбы. Процветание крупных
травоядных в саваннах ограничивается сравнительно высокой
численностью хищников.
Таким образом, основные характеристики саванны опреде-
ляются уровнем осадков. Из-за недостатка влаги растительная
пища в ней отличается низкими питательными качествами, за-
то в изобилии имеется животная пища. Однако, пожалуй, глав-
ной отличительной чертой этого биома является ярко выражен-
ная сезонная изменчивость. Количество, распределение и до-
ступность источников как растительной, так и животной пищи
на протяжении года очень сильно варьируют, что можно рас-
ценивать как один из важнейших факторов в связи с адапта-
цией.
8.4. Сезонная изменчивость и доступность пищевых истоков
8.4.1. Сезонный цикл
Различия в доступности источников пищи (высокая доступ-
ность животной пищи и низкая — растительной) в средах оби-
тания ранних гоминид еще резче проявлялись на фоне сезон-
ной изменчивости климата. На рис. 8.5 отражены общие изме-
Гоминиды и сезонная изменчивость сред
251
Влажный сезон
Сухой сезон
Начало
Конец
Начало
Конец
Керни
Листья
Цветки
Плоды
Травы
Крупные
травоядные
Мелкие
млекопитающие
Птицы
Другие
источники лищи
—---------------"1------
Утилизация
энергетических
запасов
Рост новых листьев—
максимальное содержание
питательных веществ
Период наиболее
обильного цветения
Рост корней Утилизация
до максимальных энергетических
размеров запасов
Опадение листьев;
уменьшение содержания
питательных веществ
Увядание цветков
Период наиболее обильного созревания
и наибольшей доступности плодов
Остаются только
ксерофитные
суккуленты
Начало роста — образование листьев Снижение содержания белков
и стеблей
Образование семян - Корневища
Рассредоточение Резидентные виды в местностях
пасущихся видов с травяным покрытием
Молодняк прячется
в растительном покрове
Пасущиеся видь!
сосредоточи ваются
около воды
Высокая смертность
(обилие падали); -
"Взрыв" некоторых
популяций
Изобилие яиц и птенцов
истощение
Снижение активности
и спячка.
Оперившиеся птенцы
обретают независимость
Брачный полет Доступность
термитов меда
______________1
В пересыхающих водоемах
остается рыба
___________1_____________
Начало
Конец
Начало
Конец
Влажный сезон
Сухой сезон
Рис. 8.5. Доступность сезонных источников пищн в тропических африканских
саваннах
нения доступности различных пищевых ресурсов саванновых
сред в зависимости от циклической смены дождливых и засуш-
ливых сезонов. Такие циклы могут повторяться один или два
раза в год, причем длительность каждой фазы цикла подвер-
жена значительным колебаниям. Основные же закономерности
циклов неизменны и, следовательно, позволяют судить о тех
проблемах, с которыми могли сталкиваться ранние гоминиды.
Доступность растительной пищи в Африке недавно изучили
Питерс и др. (Peters, O'Brien, Box, 1984). Растительная про-
дукция носит выраженный сезонный характер. Новые корни
и побеги у трав образуются после их повреждения и в начале
дождливого сезона, когда отмечается быстрый общий рост рас-
тения. С наступлением засушливого времени года рост трав
252
Глава 8
250
Сезон
Рис. 8.6. Число видов съедобных рас-
тений в разное время года в «сухих»
регионах Африки, расположенных
южнее Сахары (по Peters, O’Brien,
Box, 1984)
замедляется, они буреют, а со-
держание белков в них пада-
ет (Harper, 1977). У много-
летних трав образование но-
вых зеленых побегов в сухой
сезон может усиливаться в
результате их ощипывания
животными во время пастьбы
или после степных пожаров.
У однолетних трав образова-
ние листьев сразу прекраща-
ется с созреванием семян. У
растений с расположенными
на поверхности земли или не-
посредственно под нею запа-
сающими органами (полу-
криптофиты) продукция био-
массы сконцентрирована в-
корневищах и клубнелукови-
цах (Harper, 1977). Таким об-
разом, в течение влажного се-
зона животным доступны зна-
чительные количества зеленой
растительной пищи, но потом
эти ресурсы уменьшаются. В
начале сухого сезона доступ-
ны семена, клубнелуковицы и
корневища, причем их интен-
сивное потребление животны-
ми дает вегетативно размножающимся многолетникам опреде-
ленные преимущества в плане естественного отбора. Если су-
хой сезон продолжается долго, продукция зеленой раститель-
ной массы прекращается (если только растения не произраста-
ют в непосредственной близости от непересыхающих источни-
ков воды). Что касается других растительных форм, целесооб-
разно проследить динамику развития у них листьев, корней,
плодов и цветков по отдельности. У большинства видов тро-
пических растений развитие цветков и плодов приходится на
влажные сезоны (Hladik, 1977), что и определяет возможность
их использования в это время года, а также в начале сухих
сезонов (рис. 8.6, табл. 8.2). Некоторые виды растений реаги-
руют на длительную засуху образованием семян и плодов (Har-
per, 1977), что может привести к локальному изобилию пищи в
особенно засушливое время года (Lee, 1968). Большинство ви-
цов тропических растений вечнозеленые (что позволяет им осо-
бенно бурно расти в короткий период вегетации), а следователь-
Гоминиды и сезонная изменчивость сред
253
Таблица 8.2. Число типов растительных продуктов, используемых в пищу
павианами, шимпанзе и людьми в разное время года в Африке (по Peters,
O’Brien, Box, 1984); в скобках —число типов потребляемой растительной
пищи за вычетом продуктов, доступных круглый год (а также тех видов
растений, информация о потреблении которых в то или иное время года
отсутствует)
Сезон
конец сухо- конец влаж- го/начало влажный ного/начало сухой влажного сухого
Цветки Плоды и др. Листья Подземные растений 5(5) 1(1) 0(0) 3(3) 48(46) 118(116) 73(71) 51(49) 22(7) 34(19) 18(3) 20(5) части 55(0) 55(0) 55(0) 55(0)
Всего 130(58) 208(136) 146(74) 129(57)
но, их листья (которые в полусухих средах имеют, как правило,
небольшие размеры) могут использоваться в пищу круглый год,
хотя их химический состав на протяжении года иногда подвер-
жен колебаниям (Hladik, 1977). Корни некоторых травянистых
растений в сухие сезоны функционируют как запасающие орга-
ны и, стало быть, могут служить на протяжении всего засушли-
вого периода ценным источником пищи (Tanaka, 1976; Peters,
et al., 1984; Vincent, 1985).
На сезонные изменения растительности реагируют крупные
травоядные млекопитающие. Эти животные питаются травой и
низко расположенными молодыми древесными побегами. Соот-
ветственно в дождливые сезоны они широко рассредоточивают-
ся по саванне, питаясь обильной, но недолговечной зеленой
растительной массой, а в сухое время года сосредоточиваются
на пастбищах вокруг непересыхающих водных источников
(Western, 1975) (рис. 8.7). Исключением, пожалуй, являются
некоторые виды, способные ощипывать листья и побеги, кото-
рые в сухие периоды иногда рассеиваются по саванне, питаясь
вечнозеленой листвой (Kingdom, 1981).
Совместное действие таких факторов, как недостаток пищи
у травоядных млекопитающих, и, следовательно, увеличение их
смертности вследствие сокращения запасов растительных про-
дуктов, а также пространственной концентрации вторичной
биомассы обеспечивает в сухое время года изобилие пищи для
хищников и животных, питающихся падалью (Houston, 1979).
Хотя крупные травоядные в дождливые сезоны более широко
254
Глава 8
Рис. 8.7. Сезонные изменения биомассы крупных животных в Национальном
парке Амбосели (Кения) (по Western, 1973)
рассредоточены по саванне, многие виды, постоянно обитающие
в данной местности, обзаводятся в это время потомством
(Kingdom, 1981). Детеныши обычно прячутся в траве (Dela-
ney, Happold, 1979) и могут стать добычей хищников, придер-
живающихся определенных стратегий пищевого поиска
(Schaller, Lowther, 1969). Более мелкие животные в сухие
сезоны способны впадать в спячку и размножаются в дождли-
вое время года.
Пищей могут служить также насекомые и их яйца, яйца
птиц и оперившиеся птенцы. Термиты, кружащиеся роями в
брачном полете в начале сезона дождей (Thoillay, 1970), в те-
чение этого короткого периода используются в пищу как шим-
панзе, так и некоторыми современными популяциями людей.
Большинство птиц откладывают яйца либо ближе к концу су-
хого сезона, либо в самом начале дождливого (Owen, 1974).
Хотя они и стараются обезопасить свои гнезда от хищников,
яйца и птенцы могут стать добычей приматов (Teleki, 1981).
Как только птенцы обретают самостоятельность и начинают ле-
тать, они, как правило, становятся недосягаемыми для назем-
ных хищников.
Таким образом, можно видеть, что доступность пищевых
ресурсов на протяжении сезонного цикла значительно изменя-
ется. Следовательно, выживание в саванновой среде зависит от
способности животных получить доступ к источникам пищи в
сильно меняющихся в разное время года условиях.
Гоминиды и сезонная изменчивость сред
255
8.5. Реакция на сезонную изменчивость:
сравнительный анализ
Если доступность пищи в саванне подчиняется сезонным
изменениям, каких адаптивных стратегий могли придерживать-
ся ранние гоминиды? Ответ на этот вопрос можно получить с
помощью общей модели конкуренции, рассмотренной в начале
этой главы, учитывая то известное обстоятельство, что в целом
приматы являются оппортунистическими всеядными животны-
ми, т. е. способны использовать энергию самых разнообразных
пищевых продуктов.
8.5.1. Модель адаптации к сезонному стрессу
Можно предложить очень простую модель альтернативных
стратегий выживания в неблагоприятное время года (рис. 8.8).
При истощении пищевых ресурсов животные, как правило, со-
кращают активность, не имеющую решающего значения для
выживания. Энергия расходуется на пищевой поиск в ущерб
другим формам активности (например, социальной). Так жи-
вотные могут «перебиться» до лучших времен. В крайне не-
благоприятных условиях они способны даже впадать в полную
зимнюю или летнюю спячку, но это явление, как правило, на-
блюдается редко.
Помимо этой общей адаптивной тенденции животные обна-
руживают две альтернативные стратегии выживания. В первом
случае животные «справляются» с неблагоприятным временем
года, включая в свой рацион низкокачественную пищу (то
есть такую, которая имеется в изобилии, но характеризуется
низкой питательной ценностью и переваривается очень медлен-
но). За эту стратегию животные расплачиваются большими за-
тратами времени на пищевой поиск. Вторая стратегия заклю-
чается в освоении ресурсов высококачественной пищи с сезон-
но изменяющейся доступностью, которые в нормальных усло-
виях животными не используются, так как для этого могут по-
Снижение активности
и энергетических
потребностей
Использование
источников
обильной,
но низко-
качественной
пищи
Использование
сезонно
доступных
источников
высоко-
качественной
пищи
Рис. 8.8. Альтернативные типы поведения в «голодное время года»
256
Глава 8
требоваться слишком большие энергетические затраты. Эта
стратегия предполагает значительное изменение пищевого ра-
циона и, таким образом, связана с увеличением разнообразия
пищи, которая потребляется животными на протяжении года.
Понятно, это эта стратегия зависит и от доступности источни-
ков пищи, а это означает, что в тех саванновых средах, в ко-
торых обитали ранние гоминиды, первостепенное значение име-
ла взаимосвязанная доступность источников растительной и
животной пищи. Однако, прежде чем более подробно остано-
виться на этом обстоятельстве, полезно будет рассмотреть во-
прос о существовании описанных стратегий выживания у при-
матов.
8.5.2. Сезонная изменчивость поведения приматов
Снижение активности во время стресса свойственно самым
разным приматам. Особенно выделяется в этом отношении тол-
стохвостый лемур (Cheriogaleus medias'), способный, как из-
вестно, впадать в летнюю спячку. Этот примат, в хвосте у ко-
торого образуются запасы жира, может переживать в малоак-
тивном состоянии длительные неблагоприятные периоды
(Hladik, Charles-Dominique, Peters, 1980). Чаще, однако, при-
маты снижают уровень социальной активности, требующей зна-
чительных расходов энергии. Так, Ли (Lee, 1984) показал, что
зеленые мартышки (Cercopithecus aethiops) в сухие сезоны
меньше времени посвящают играм (рис. 8.9).
Обе альтернативные стратегии выживания обнаруживаются
в поведении ныне живущих приматов как на протяжении срав-
нительно коротких отрезков времени, так и в течение более
длительных периодов. Например, изучавшиеся Сузуки (Suzu-
Рис. 8.9. Сезонные изменения активности зеленых мартышек (Cercopithecus
aethiops) в Национальном парке Амбосели (Кения) (по Lee, 1984). Активность,
требующая значительных затрат энергии, в сухое время года снижается
Гоминиды и сезонная изменчивость сред
257
Сухой сезон Влажный сезон Сухой сезон
Рис. 8.10. Число типов растительных продуктов, используемых в пищу обык-
новенным шимпанзе (Pan troglodytes) в саванновом редколесьи (Касакати,
Танзания) (по Suzuki, 1969)
ki, 1969) шимпанзе Касакати в засушливые сезоны включают
в свой рацион более разнообразную растительную пищу, чем
в благоприятное время года (рис. 8.10). В сухие сезоны эти
шимпанзе относятся к пище менее разборчиво и потребляют
менее качественные пищевые продукты. Аналогично ведет себя
и обыкновенный шимпанзе (Pan troglodytes) в Гомбе (Танза-
ния) и в Габоне (Hladik, 1977), увеличивая в сухие сезоны
долю листьев в своем рационе (рис. 8.11). В более крупном
временном масштабе первая из рассматриваемых стратегий
обнаруживается, например, в поведении гелад (Theropithecus
gelada), которые ныне живут только на эфиопских нагорьях и
являются травоядными животными. Трава — типичный пример
низкокачественной пищи, которая всегда имеется в саваннах в
изобилии. Благодаря адаптивной специализации зубного и ло-
комоторного аппарата гелады способны потреблять значитель-
ные количества травы и могут длительное время выдержи-
вать нехватку другой пищи (табл. 8.3) (Jolly, 1972; Dunbar,
1977, 1983).
Все приведенные выше примеры относятся к приматам,
придерживающимся стратегии потребления в сухое время года
низкокачественной пищи.
Вторая стратегия, нередко требующая от животных гораздо
более гибкого поведения, встречается, по-видимому, реже. Не-
которое оживление охотничьего поведения в сухие сезоны на*
17—367
258
Глава 8
Рис. 8.11. Потребление растительных продуктов обыкновенным шимпанзе (Pan
troglodytes) иа протяжении годового сезонного цикла: А. В Габоне; Б. В На-
циональном парке Гомбе-Стрим (Таизаиия) (по Hladik, 1977)
блюдается у странствующих по саваннам шимпанзе (Susuki,
1975). Кроме того, как показал Данбар (Dunbar, 1983), в бо-
лее сухих и, следовательно, обладающих более выраженной се-
зонной изменчивостью средах чаще прибегают к хищничеству
и павианы (Papio), которые отличаются от гелад большей все-
ядностью (рис. 8.12).
8.6. Стратегии пищевого поиска у гоминид в средах
с сезонной изменчивостью
Наличие у приматов описанных выше стратегий выживания
заставляет задуматься над тем, какую роль сезонная изменчи-
вость могла играть в адаптации ранних гоминид и какое влия-
ние могли оказать эти две альтернативные стратегии на эво-
люционную экологию гоминид. Начнем анализ этих вопросов
с простого описания общей стратегии использования всеядным,
организмом различных источников пищи по мере увеличения
их доступности (рис. 8.13).
Гоминиды и сезонная изменчивость сред_________________259
Таблица 8.3. Потребление растительных продуктов геладами (Theropithecus gelada) в разное время года (по Dunbar, 1977)
Растения Подземные части растения Количество наблюдений (в процентах)
влажный сезон начало сухого конец сухого
сезона сезона
Злаковые травы Листья 93,0 17,3 24,7
Корневища — 6,5 66,9
Семена 69,7 —
Травянистые расте- Листья, стебли 0,2 0,4 2,7
НИЯ Корни, клубни 3,3 0,9 —
Цветки 0,2 3,0 0,1
Семена — 1,3 5,2
Чертополох Листья 0,1 0,8 0,1
Кустарники Листья 0,2 — 0,2
Цветки — —— 0,1
Плоды 2,9 — —.
Насекомые — 0,1 — 0,1
Количество наблюдений 3051 231 3291
8.6.1. Стратегии пищевого поиска в дождливые сезоны
Как видно из рис. 8.5, влажное время года — период, когда
доступны наиболее разнообразные источники пищи. При таких
обстоятельствах гоминиды должны были выбирать пищевые
ресурсы, использование которых давало им максимальную «от-
дачу» (Pyke, Pulliam, Charnov, 1974), т. е., чтобы время, затра-
8
а
Гилгил •
(засушливый год)
0,4 -
Г илгил •
(дождливый год)
0,2
х
с
Амбосели (засушливый год) •
Амбосели (дождливый год) •
Метахара
^Боуп___________________!
Сухость 1 2 3 4 Сухой
климата Влажный экстремум экстремум
Рис 8 12. Хищническое поведение павианов Papio в Восточной Африке (по Dun-
bar, 1984)
171
260
Глава 8
Рис. 8.13. Модель оптимизации и приспособления к условиям для пищевого
поиска гоминид на протяжении сезонного цикла, основанная на сезонных из-
менениях доступности пищевых ресурсов
ченное на поиск и(или) преследование добычи многократно
окупалось ее ценностью. Это соответствует поведению организ-
ма в «период изобилия» в модели конкуренции, изображенной
на рис. 8.1. Какие источники пищи удовлетворяют этим крите-
риям, зависит, помимо прочего, от промежутка времени между
началом дождей и появлением этих источников. Так, термиты
реагируют на наступление дождливого сезона мгновенно
(Thoillay, 1970) и тем самым обеспечивают животных на ко-
роткое время сверхизбыточной пищей в конце «голодного» пе-
риода. Из растений травы первыми реагируют на дожди обра-
зованием новых побегов, и свежая поросль может служить хо-
Гоминиды и сезонная изменчивость сред 261'
рошей пищей, например, геладам (Post, Hausfater, McCuskey,.
1980).
Позднее во влажное время года начинают зацветать и пло-
доносить растения, которые, вероятно, и служили гоминидам*
основным источником пищи в дождливые сезоны, обеспечивая
их легко доступными высококачественными пищевыми продук-
тами, которые и поныне являются излюбленной пищей боль-
шинства приматов (Harding, 1981). Доступной, хотя и требо-
вавшей на поиски ее гораздо большего времени, пищей, могли
служить телята крупных травоядных млекопитающих, появляв-
шиеся на свет во влажное время года и прятавшиеся в траве
(Delaney, Happold, 1979), а также яйца и оперившиеся птенцы
различных птиц (Owen, 1974). В общем с дождями наступает
настоящее изобилие растительной пищи, на что гоминиды, ве-
роятно, реагировали интенсивным использованием соответству-
ющих пищевых ресурсов, прежде чем их доступность начинала
уменьшаться. Другие источники пищи, по-видимому, использо-
вались гоминидами во влажное время года лишь эпизодичес-
ки. Таким образом, в дождливые сезоны гоминиды, по всей
вероятности, придерживались довольно разнообразной, «эклек-
тической», хотя во многом и «травоядной», стратегии пищево-
го поиска, которая в значительной степени «перекрывалась»
со стратегиями пищевого поиска других видов, но не была
связана с острой межвидовой конкуренцией.
8.6.2. Стратегия пищевого поиска в сухое время года
С началом сухого сезона стратегия пищевого поиска гоми-
нид должна была изменяться, так как все обитавшие в саван-
не животные вступали в это время в «голодную фазу» цикла
конкуренции (рис. 8.1), что, по всей вероятности, сопровожда-
лось обострением последней. Конкурентных взаимодействий
животным в это время было выгодно избегать. Цветущие рас-
тения в засушливые сезоны встречаются исключительно редко;
постепенно истощается и урожай плодов. Основным типом рас-
тительной пищи гоминид в начале сухого времени года, по-ви-
димому, были семена трав, а позднее — их корневища (Dun-
bar, 1977). В разгар сезона засухи единственным значительным
источником растительной пищи служили расположенные на
разной глубине корни (Dunbar, 1977). Их доступность отчасти
зависела, вероятно, от умения раскапывать землю палками
(Tanner, 1981). Именно в таких условиях, когда уменьшается
доступность растительной пищи, приобретает решающее значе-
ние использование в пищу мяса. С усилением засухи многие
виды травоядных животных должны были сосредоточиваться
вокруг водных источников, рядом с которыми дольше всего
262
Глава 8
сохранялись пастбища (Western, 1975). В такой ситуации хищ-
ничество могло становиться высокоэффективной стратегией,
позволявшей разнообразить растительный рацион. Гоминиды
могли добывать мясо либо путем прямого хищничества, либо
питаясь животными, убитыми другими хищниками или павши-
ми в результате болезни или истощения.
Таким образом, усиление зависимости от мясной пищи по
мере усиления засухи могло позволить гоминидам решить про-
блему нехватки растительных продуктов и в то же самое время
привести к обострению конкуренции.
8.6.3. Сезонная изменчивость и непредсказуемость
Была ли описанная стратегия питания достаточно эффектив-
ной для того, чтобы гоминиды могли выдерживать разного ро-
да отклонения сезонного цикла? Хорошо известно, что усиле-
ние сезонности в тропиках коррелирует с непредсказуемостью
дождей (Harris, 1980): они могут долго не выпадать, и засуха
иногда сильно затягивается. В столь неопределенных условиях
должна существовать хоть какая-то адаптация, способствую-
щая выживанию. Возникает вопрос: можно ли рассматривать
зависимость гоминид от мясной пищи в качестве такой адапта-
ции к условиям засухи? Отчасти ответ на этот вопрос зависит
от характера пространственных перемещений, но, полагая, что
ареалы гоминид были приурочены к непересыхающим вод-
ным источникам, можно думать, что действительно, по мере
того как увеличивалась смертность животных в связи с исто-
щением источников растительной пищи, питание мясом приоб-
ретало для гоминид в плане выживания все более важное зна-
чение. Не исключено также, что именно в таких стрессовых ус-
ловиях ослабевала альтернативная стратегия питания
растительной пищей.
Адаптивные последствия, по-видимому, имела и противопо-
ложная ситуация — интенсивные и длительные дожди и слиш-
ком короткие сухие сезоны. Некоторые растения в таких ус-
ловиях обнаруживают парадоксальное поведение, формируя
главным образом новые листья и корни, а не репродуктивные
органы, обеспечивающие половое размножение (Harper, 1977);
в результате образуется меньше плодов, а доступность листьев
и травы увеличивается. Это приводит к значительному обога-
щению пищевой базы для определенной части сообщества тра-
воядных животных.
Гоминиды н сезонная изменчивость сред
26»
8.7. Гоминоиды на все сезоны
8.7.1. Разнообразие гоминид и сезонные стратегии
В основе описанной выше стратегии пищевого поиска лежит
то обстоятельство, что ранние гоминиды были полностью оп-
портунистическими организмами, которые использовали ресурсы
среды по мере увеличения их доступности. Кроме того, при
описании этой стратегии предполагалось, что потребности го-
минид и их стратегия выживания всегда и везде были одина-
ковыми. Как, однако, отмечалось, гоминид, обитавших в конце
плиоцена и начале плейстоцена, уже нельзя рассматривать
как однородную группу. Уже в это время они обнаруживают
значительные различия, особенно сильно выраженные в мор-
фологии зубной и жевательной системы, что, по-видимому, сле-
дует учитывать при анализе их пищевого поведения и сезонных
стратегий выживания.
Долгое время существовало мнение, что мегадонтия (нали-
чие крупных зубов) у массивных австралопитековых (рис. 8.14)
явилась результатом потребления низкокачественных рационов
с грубыми и твердыми пищевыми частицами, разжевывание ко-
торых требовало значительных усилий. Джолли (Jolly, 1981)
объяснял это обстоятельство специфической зависимостью мас-
Рис. 8.14. Относительные размеры зубов ранних африканских гоминид: А. Ното
habilis (Олдувай); Б. Australopithecus boisei (Пенинж). Образцы изображены
с одинаковым расстоянием между клыками, что позволяет лучше видеть раз-
личия между размерами зубов заднего ряда ,
264
Глава 8
'Сивных австралопитековых от семян злаковых трав. Уолкер
(Walker, 1981) на основании микроскопического анализа стер-
тости зубов предположил, что существенной частью пищевого
рациона массивных австралопитековых были грубые, твердые
и сухие плоды саванновых растений. О том, что твердые пи-
щевые продукты были важным компонентом рациона этих
гоминид говорят и результаты анализа Грайна (Grine, 1981).
В своем недавнем обзоре этого вопроса Кей (Кау, 1985) сде-
лал предположение, что вся совокупность известных фактов —
размеры тела, толщина зубной эмали, размеры зубов, высота
коронок моляров и микростертость зубов — свидетельствует о
том, что рационы массивных австралопитековых включали
очень твердые компоненты с низким содержанием раститель-
ных волокон, лишь в незначительной степени подвергавшиеся
предварительной обработке резцами (например, плоды или се-
мена с твердыми оболочками). Равным образом можно предпо-
лагать, что, если этим гоминидам в течение длительных перио-
ров приходилось питаться подобными растительными продукта-
ми, массивные зубы могли давать им значительные адаптив-
ные преимущества.
Иную эволюционную тенденцию обнаруживала другая груп-
па ранних гоминид — представители рода Ното. Хотя у ранних
представителей этого рода зубы были крупнее, чем у современ-
ных людей, зубных специализаций, характерных для австрало-
питековых, у них не выявляется. Гораздо менее специализиро-
вана и их жевательная система, что, вероятно, указывает на
большее разнообразие их пищевых рационов (всеядность).
Различия зубной морфологии представляют интерес в связи
с сезонной изменчивостью сред обитания ранних гоминид.
Если предположить, что функции и эффективность зубного ап-
парата подвергались наиболее интенсивному отбору в самые
«голодные времена», можно думать, что различия между гоми-
нидами в этом плане отражают особенности их стратегий пи-
тания в сухие сезоны. Особенности зубной морфологии массив-
ных австралопитековых более всего соответствуют стратегии
использования источников низкокачественной пищи, а зубной
аппарат представителей рода Ното отражает расширение их
пищевой базы и освоение нового пищевого продукта — мяса
позвоночных животных, что повысило выживаемость этих го-
минид в условиях нехватки растительной пищи (рис. 8.15).
8.8. Условия экологического разделения
Предположение, что дивергенция ранних гоминид отражает
различия их стратегий выживания в неблагоприятное время
года, порождает целый ряд важных проблем. Основные из них
Гоминиды и сезонная изменчивость сред
265
Питание низкокачественной
растительной пищей
Массивный
Australopithecus
Оппортунистическая утилизация
сезонно доступных источников
высококачественной пищи
Мясо позвоночных
— Ното-
Подземные запасающие
органы растений
Рис. 8.15. Альтернативные реакции раниих гоминид иа сезонный стресс
касаются вопроса, могли ли условия окружающей среды спо-
собствовать эволюции этих стратегий и почему, собственно, эво-
люционировали эти две стратегии? Другими словами, в каких
условиях могла эволюционировать каждая из этих стратегий
выживания?
Первостепенным и наиболее важным фактором, определяю-
щим, какая из двух стратегий выживания окажется оптималь-
ной в локальных условиях существования, являются сами эти
условия. Там, где в сухие сезоны происходило значительное
увеличение биомассы крупных млекопитающих, в особенности
если разнообразие и продуктивность растений в это время
сильно снижались, идеальным вариантом для гоминид, по всей
вероятности, был бы переход к питанию мясом. Адекватной
моделью такой ситуации может служить Национальный парк
Амбосели (рис. 8.16), где в сухое время года животные сосре-
доточиваются у источников воды, образуя очень плотные по-
пуляции (что ослабляет конкуренцию между хищниками), и
где из-за высокой засоленности почв растительной пищи чрез-
вычайно мало. Подобная среда> по всей видимости, должна
способствовать отбору всеядных организмов.
Такими в Восточной Африке, однако, являются не все са-
ванновые среды. Если позволяют локальные условия, многие
млекопитающие вместо того, чтобы сосредоточиваться в каких-
либо определенных местах, совершают дальние сезонные ми-
грации. Самой известной из подобных экологических систем
является Национальный парк Серенгети в Танзании (рис.
8.17). Плотность популяций, собирающихся в засушливое вре-
мя года на его равнинах, гораздо меньше, а в саванновом лесу
она зачастую вообще не изменяется на протяжении года. В та-
ких условиях переход к мясной пище не имел бы адаптивного
значения, и поэтому можно думать, что в подобных средах го-
миниды питались не мясом, а низкокачественными продуктами.
По всей вероятности, в экосистемах описанного типа австрало-
266
Глава 8
Рис. 8.16. Национальный парк Амбосели (Кения): пример системы крупных
млекопитающих, рассредотачивающейся и концентрирующейся на протяжении
сезонного цикла. Животные рассеиваются по окрестностям Амбосели во влаж-
ное время года, и возвращаются в него с наступлением засушливого сезона.
Соответственно биомасса, и, следовательно, доступность жертв в сухие сезоны
повышаются (по Western, 1973)
питековые должны были легче всего решать проблемы, связан-
ные с выживанием в неблагоприятное время года.
Если сказанное верно, можно предполагать, что на ранних
этапах дифференциации гоминид их популяции, рассеянные по
полусухим регионам африканского континента, придерживались
в зависимости от локальных условий существования альтерна-
тивных способов выживания в засушливые сезоны.
В дифференциации гоминид могли участвовать и другие
факторы. Если экологические условия длительное время оста-
вались неизменными, в разных частях Африки могли образо-
вываться популяции гоминид, придерживавшихся двух альтер-
нативных стратегий выживания. Популяции эти могли обнару-
живать перекрывание ареалов и симпатрию. Различия страте-
гий, раз установившись, вполне были способны ослаблять кон-
куренцию в трудное время года. Хотя «стратегия Ното-» вряд
rm чжгхг'ттп гч оттттталтп тт otl ст и гч агтгпи о v г'ттст тго.пл
Гоминиды н сезонная изменчивость сред
267
Рис. 8.17. Национальный парк Серенгети (Танзания): пример мигрирующей си-
стемы. В засушливое время года крупные млекопитающие мигрируют из са-
занн Серенгети в охотничий заповедник Маасаи-Мара. Соответственно в сухие
:езоны в Серенгети отмечается уменьшение биомассы и доступности жертв (по
Maddock, 1979)
вотной пищи происходил отбор в направлении «австралопите-
ковой стратегии», австралопитековые, по-видимому, вполне
успешно способны были выживать в средах обитания Ното.
Следовательно, можно думать, что географическое распростра-
нение массивных австралопитековых было более широким, чем
распространение Ното (см. также разд. 9.3.1). Отсутствие
прямой конкуренции и отбор, зависящий от частоты генотипа,
способны были обеспечить сосуществование разных форм го-
минид. На такой основе у симпатрических видов часто проис-
ходит в конце концов разделение экологических ниш.
8.8.1. Использование в пищу мяса ранними гоминидами
Как говорилось выше, включение мяса в пищевые рационы
ранних африканских гоминид, по всей вероятности, происходи-
ло под влиянием сезонного стресса. Подобная стратегия пита-
ния должна была сводить к минимуму сезонные колебания до-
268
Глава 8
ступности пищи, а следовательно, и конкуренцию с травоядны-
ми животными. Равным образом она должна была ослаблять и
межвидовую конкуренцию с крупными хищниками, так как для
них засушливые сезоны — время изобилия пищи, а, согласно
модели конкуренции (см. рис. 8.1), «перекрывание» пищевых
потребностей животных допустимо. Для гоминид переход к пи-
танию мясом означал превращение среды обитания с резкой
сезонной изменчивостью и, следовательно, создающей для них
массу проблем в среду, у которой сезонная изменчивость сгла-
живалась благодаря возможности «переключаться» на другую
пищу. Как указывалось в предыдущих главах, за это гоминиды
«расплачивались» повышением уровня общей активности и пи-
щевого поиска, укрупнением своих размеров и коренной пере-
стройкой механизмов терморегуляции.
В настоящее время вопрос о мясном рационе ранних гоми-
нид является предметом острых дискуссий и противоречивых
мнений. Сомнения, высказанные Бинфордом (Binford, 1982,
1983) по поводу предположений о питании ранних гоминид
мясом и о тождественности пищевого поведения ранних гоми-
нид и современных охотников и собирателей, сильно стимулиро-
вали критическую проверку палеонтологических и археологиче-
ских данных. В появившемся недавно обзоре этой проблемы
Потс (Potts, 1984а) показал, что гоминиды действительно под-
вергали животные продукты, включая мясо, обработке. Основы-
ваясь на результатах изучения животных остатков в слое I
Олдувая, против критических высказываний Бинфорда высту-
пают также Банн и Кролл (Bunn, Kroll, 1986). Хотя вопрос о
том, в какой степени мясо было представлено в пищевом раци-
оне ранних гоминид и каким образом они его добывали (охо-
той или поиском павших животных), до сих пор не решен,
значительные скопления костей вместе с каменными орудиями
и характер следов, оставленных рубящим инструментом на
костях, от которых отделяли мясо, по всей видимости, указы-
вают на то, что несколько менее двух млн. лет назад некото-
рые африканские гоминиды уже использовали в пищу разно-
образных млекопитающих всевозможных размеров гораздо ши-
ре, чем это свойственно современным приматам (не считая че-
ловека). Это обстоятельство совсем не означает, что ранние
гоминиды были организованы в сообщества охотников и соби-
рателей, а говорит лишь о том, что они были в значительной
мере всеядными организмами.
Пожалуй, наибольший интерес представляет вопрос, в ка-
кой степени использование в пищу мяса ранними гоминидами
было вызвано сезонным стрессом, или, иначе говоря, насколько
сильнее оно было выражено в засушливые сезоны. К сожале-
нию, пока не существует надежных методов прямого определе-
Гоминиды и сезонная изменчивость сред 269
ния времени года, когда произошла гибель животного, которое,
вероятно, использовалось в пищу гоминидами. Один из возмож-
ных подходов к решению этой проблемы состоит в изучении
видов животных, остатки которых обнаруживаются вместе со
следами деятельности гоминид. По характерным признакам
остатков жертв можно судить о хищническом поведении гоми-
нид (Foley, 1983). Характер пищевого поиска гоминид (хищни-
чество или поедание павших животных) можно, по-видимому,
определять по сезонной изменчивости поведения и распростра-
нения разных видов копытных. Ныне живущие в Восточной
Африке животные обнаруживают значительную вариабельность
поведения (рис. 8.17). Можно думать, что для таких зависев-
ших от воды хищников, какими, по всей вероятности, являлись
ранние гоминиды, быстро бегающие млекопитающие, которые
собирались большими стадами лишь в определенное время го-
да, были доступны только в засушливые сезоны. Остатки этих
животных, обнаруженные вместе с древними орудиями гоми-
нид, могли бы указывать на сезонный характер их хищничес-
кого поведения, однако предварительные данные, полученные
при изучении некоторых из таких находок (рис. 8.18), не по-
зволяют сделать каких-либо определенных выводов. В самых
древних находках не обнаруживается повышенного содержания
«остатков млекопитающих с сезонной изменчивостью поведения,
хотя именно так обстоит дело с находками, относящимися к
ранним местообитаниям Homo erectus. Для решения этого во-
проса требуются дополнительные, более точные данные;
описанный подход, однако, поможет не только решить вопрос,
потребляли ли гоминиды мясо, но и позволит определить, при
каких обстоятельствах мясо могло войти в их рацион. В связи
с нынешними дискуссиями по поводу костных остатков, отно-
сящихся к нижнему африканскому плейстоцену, важно пом-
нить, что, если станет понятно, почему гоминиды начали по-
треблять мясо, наполовину будет решен и сам вопрос о плото-
ядности этих приматов.
8.9. Эволюционные и поведенческие последствия
питания мясом
Существование в средах с сезонной изменчивостью приво-
дит к развитию у животных способности использовать разно-
образные источники пищи, доступность которых изменяется во
времени, а в отдельных случаях — и к возникновению более
специфической, хотя и менее безопасной, способности включать
в пищевой рацион мясо. Чем бы ни были обусловлены подоб-
ные изменения пищевого поведения, у гоминид они оказали
значительное влияние на весь процесс адаптации. Эволюцион-
270
Глава 8
Сезонность перемещений
Сильно выраженная Слабо выраженная Резидентные виды
Зависимые от воды Слон Тростниковый козел Буйвол Бегемот Кистеухая свинья Бушбок Водяной козел
< Частично зависимые со от воды о Зебра Лошадиная антилопа Черная лошадиная антилопа Гну Болотный козел Большой куду
О 2 X ш Слабо зависимые со от воды Бубал Бубал топи Двурогий носорог Однорогий носорог Жираф Газель Томпсона Бородавочник Канна Малый куду Импала Дукер Сасса
Независимые Г азель Г ранта Сернобык Антилопа штейнбок Г еренук Дикдик
Рис. 8.18. Кросс-табулирование современного африканского сообщества круп-
ных млекопитающих в соответствии с сезонной изменчивостью поведения
животных и их зависимостью от воды. Так, например, слоны отличаются чрез-
вычайной изменчивостью сезонного распределения по саваннам н в то же
время нуждаются в регулярном питье. С помощью подобного кросс-табулиро-
вания можно устанавливать виды животных, являющихся потенциальными.
жертвами хищников. Подробнее об этом см. Foley, 1983
ные изменения редко когда не сопровождаются «потрясающи-
ми» эффектами, которые в свою очередь сами могут превра-
титься в факторы отбора и вызвать последующие эволюцион-
ные сдвиги.
Кратко рассмотрим некоторые эволюционные последствия
описанного выше пищевого поиска гоминид.
1. Коэффициент энцефализации. Как уже говорилось, раз-
меры головного мозга у млекопитающих соответствуют разме-
рам тела (см. гл. 6). Хотя эта связь и является статистически1
достоверной, существует целый ряд исключений. Последние,,
как показал Эйзенберг (Eisenberg, 1981), касаются определен-
ных групп животных. У видов, характеризующихся сложной:
стратегией пищевого поиска, т. е. у видов, использующих са-
мые разнообразные источники пищи или пользующихся неожи-
данными и изменчивыми пищевыми ресурсами, относительные-
размеры головного мозга больше, чем у видов, использующих.
Гоминиды и сезонная изменчивость сред
271
повсеместно встречающиеся источники пищи (Eisenberg, 1981,
р. 283). Таким образом, как и предполагалось в гл. 6 при об-
суждении вопроса о разнообразии пищевого рациона, описан-
ные выше изменения стратегии пищевого поиска у гоминид
должны были привести к увеличению размеров головного моз-
га относительно размеров тела. В настоящее время установле-
но, что такое увеличение началось по крайней мере у австрало-
питековых (Martin, 1983) и продолжалось на протяжении всего
плейстоцена. Итак, обусловленное сезонной изменчивостью
среды давление отбора, оказавшее влияние на пищевое поведе-
ние гоминид, в конечном итоге сказалось и на величине их
головного мозга.
2. Дележ пищи. Айзек (Isaac, 1978 а, b) разработал модель
эволюции поведения гоминид, в которой центральная роль от-
водится дележу пищи. Модель эта подверглась резкой критике
(Binford, 1981, 1983, 1984; Gowlett, 1984; см. также гл. 7).
Вполне вероятно, однако, что включение мяса в пищевой ра-
цион повлекло за собой определенные социальные и поведен-
ческие последствия. От растительной пищи мясо, помимо пита-
тельных качеств, отличается «компактностью». Туша или даже
только часть туши типичного саваннового млекопитающего
заключает в себе, как правило, больше пищи, чем может
•съесть одна особь. Таким образом, туша животного является
богатым источником питательных веществ, которым могут од-
новременно воспользоваться несколько особей; в случае расти-
тельных продуктов это бывает редко. В результате появляется
возможность дележа пищи — не в социально-активном смысле,
а просто как следствие изобилия пищи. В такой ситуации от-
бор будет способствовать развитию реципрокного альтруизма
в той форме, которую Айзек называет «допускаемым выпраши-
ванием» пищи. Хотя дележ пищи у примитивных гоминид, ве-
роятно, сильно отличался от активного дележа пищи, свойст-
венного современным охотникам и собирателям, туши мертвых
животных вполне могли становиться объектами интенсивной
социальной активности (подобно тому, как это происходит у
некоторых из ныне живущих приматов). Весьма значительные
скопления костей (например, в местонахождении ELKNN в
Олдувае), образование которых так или иначе связано с дея-
тельностью гоминид, опять-таки подтверждает мнение, что су-
ществование «наземных точек приложения активности» должно
было иметь важные последствия для поведения гоминид. Как
уже отмечалось в гл. 7, образование одних таких точек было
порождено потребностью в убежище; потребление мяса, добы-
того на охоте или предоставленного природой в виде туши
мертвого животного, по всей вероятности, способствовало обра-
зованию других.
272
Глава 8
Сезонность перемещений
Сильно выраженная Слабо выраженная Резидентные виды
Зависимые от воды 5 м Частично зависимые о ОТ ВОДЬ! О f Слабо зависимые от воды ш со П Независимые от воды Strepsiceros Kobus sp.
Equus oldowayensis
Parmelarius altidens Alcelaphini indet. Phacochoerus sp.
Oryx sp. Antidorcas bondi ?
Stylohipparion sp. Kolpochoerus limnetes.
Notochoerus sp.
Сезонность перемещений
Сильно выраженная Слабо выраженная Резидентные виды
Зависимые от воды 5 ш Частично зависимые 5 от воды о о 5 Слабо зависимые и от воды S со та со Независимые от воды Болотные козлы Kobus sigmoidalis Бегемотовые
Damaliscus cf. niro D. aegalaius Giraffa juma Aepyceros Хохлатые дукеры
Antidorcas recki?
Б
Mesochoerus limnetes
Metridiochoerus andrewsr
3. Пространственная организация. Как уже говорилось, и
увеличение размеров тела, и переход к наземному образу жизни
могут привести к расширению участка обитания вида. Такой
же эффект вызывает и использование животной пищи; у хищ-
ников и всеядных животных территории больше, чем у траво-
ядных. Площадь участка определяется, однако, и еще одним
фактором — территориальностью животных. Браун (Brown,
1964) разработал получившую широкое признание модель тер-
риториальности, в основу которой положил сопоставление
«затрат» животных на защиту своей территории или источника
пищи в «выгод» от этой защиты (рис. 8.20). Согласно модели
Брауна, территориальность (определяемая как защита терри-
тории с расположенными на ней источниками пищи, которая,
Гоминиды н сезонная изменчивость сред
273
Сезонность перемещений
Сильно выраженная Слабо выраженная Резидентные виды
Зависимые от воды Hippopotamus gorgops
Частично зависимые от воды Equus oldowayensis Hippotragus gigas
Слабо зависимые от воды Ceratotherium simum
Независимые от воды
Stylohipparion sp.
Kolpochoerus
Сезонность перемещений
Сильно выраженная Слабо выраженная Резидентные виды
Зависимые от воды Elephas recki Redunca Homiocerus Hippopotamus gorgops Tragelaphus Kobus
Частично зависимые от воды Equus aff. grevyi E. oldowayensis Strepsiceros
Слабо зависимые от воды Ceratotherium simum Giraffa camelopardis G. gracilis Taurotragus Aepyceros
Независимые от воды
Stylohipparion Sivatherium olduvai Metridiochoerus
albertensis - meadowsi
Рис. 8.19. Кросс-табулирование предположительной сезонной изменчивости
и зависимости от воды крупных млекопитающих, остатки которых обнаружены
в плиоценовых и плейстоценовых отложениях Восточной Африки: А. Местона-
хождение FLK Zinj Олдувая; Б. Местонахождение KBS Кооби-Фора; В. Верх-
ний слой местонахождения ТК Олдувая; Г. Олоргесаили
как правило, предполагает их безраздельное использование
данной особью или группой животных) развивается только в
том случае, если выгода от монопольного обладания источни-
ком пищи превышает затраты на его защиту. Поэтому терри-
ториальность, по-видимому, может возникнуть только там, где
плотность пищевых ресурсов высока, а их доступность предска-
зуема. Выше, однако, было показано, что, хотя плотность источ-
18-367
274
Глава 8
Высокая
Высокая
мобильность;
передача
информации;
пространственно-
временные
территории
Высокое
рассредоточение
и мобильность
Географически
устойчивая
территориальная
система
Перекрывание
участков обитания
Низкая ------
Низкое
Высокое
Изобилие источников пищи
Рис. 8.20. Модель оптимизации территориального поведения. Монопольное ис-
пользование ареала имеет место в том случае, если польза от этого для живот-
ных превосходит усилия, которые приходится затрачивать на его защиту. Со-
отношение между «пользой» и «затратами» определяется условиями окружаю-
щей среды. Так, например, наибольшей выраженности территориальность
может достигать там, где имеются богатые источники пищи с предсказуемой
доступностью (по Brown, 1970; Dyson, Hudson, Smith, 1978)
ников пищи в саванновых средах с сезонной изменчивостью
зачастую и высока, их доступность редко бывает предсказуе-
мой. Таким образом, вопреки предположениям таких извест-
ных авторов, как, например, Ардри (Ardrey, 1961), территори-
альность в строгом смысле слова в таких условиях вряд ли
могла возникнуть. Условия, в которых доступность пищи пред-
сказать нельзя, скорее должны способствовать развитию высо-
кой мобильности животных и сложных закономерностей их кон-
центрирования и рассредоточения на территории, что действи-
тельно свойственно многим видам.
4. Изготовление орудий. Возраст древнейших находок, сви-
детельствующих о способности гоминид к изготовлению камен-
ных орудий, составляет около 2,3 млн. лет (Isaac, 1984; Har-
ris, 1986), т. е. приблизительно на 3 млн. лет меньше, чем
возраст самых древних из известных гоминид; находки эти бы-
ли обнаружены в тех же самых регионах и средах, что и древ-
нейшие ископаемые остатки гоминид. Долгое время существо-
вало мнение, что изготовление орудий развилось в связи с охо-
той и забоем животных. Одинаково правдоподобно, однако,
звучит и предположение, что изготовление орудий развивалось
в рамках стратегии использования растительной пищи; кроме
Гоминиды и сезонная изменчивость сред
275»
того, известно, что приматы способны охотиться на животных,
и поедать мясо своих жертв без помощи каких бы то ни было-
орудий. Орудия могли использоваться гоминидами и давать им
определенные адаптивные преимущества при разделке туш
очень крупных толстокожих млекопитающих. Такие звери,
обычно неуязвимые для хищников, часто лежат нетронутыми
некоторое время после смерти, так как другие животные не
могут разорвать их кожу и добраться до мяса. Вполне веро-
ятно, что с помощью каменных орудий гоминиды могли пер-
выми начать разделку таких туш и, таким образом, по крайней
мере на первых порах одерживали верх в конкуренции с дру-
гими животными, питавшимися падалью.
По всей вероятности, в начале эволюции плотоядности мясо-
потреблялось гоминидами лишь изредка, «по случаю». Потреб-
ление ими мяса вряд ли было настолько высоким, чтобы в
действие вступали соответствующие механизмы блокады мета-
болизма, описанные Спетом и Спилмэном у современных охот-
ников и собирателей (Speth, Spielmann, 1983). Тем не менее
они, вероятно, значительно превосходили в этом отношении
других приматов, также питавшихся мясом, что, по-видимому„
и привело к существенному изменению их пищевого поиска.
Адаптивные преимущества, полученные гоминидами в резуль-
тате освоения нового богатого источника пищи в среде, которая
в противном случае (особенно в засушливые сезоны) могла бы
предоставить им только низкокачественную пищу, были поисти-
не замечательными. Как только мясо превратилось в потенци-
альный источник пищи, под влиянием отбора зависимость от
него гоминид начала увеличиваться, что привело к еще боль-
шему усилению первоначальной адаптации («автокатализ»).
Истинный ход этих событий разъяснится, когда станет понят-
ным развитие различных аспектов плотоядного поведения ран-
них гоминид. Но главным все-таки остается не то, каким обра-
зом происходил переход к питанию мясом, а тот факт, что
мясоедение позволило гоминидам с пользой для себя разре-
шить важную экологическую проблему — проблему пропитания
в засушливое время года.
Как уже говорилось, развитие новых адаптивных стратегий
и форм поведения влечет за собой последствия, которые заклю-
чаются в модификации других аспектов поведения и адапта-
ции организма. Эти новые адаптации в свою очередь могут
влиять и на развитие других видов, входящих в биологическое
сообщество. Организмы должны постоянно приспосабливаться
к присутствию и жизнедеятельности симпатрических видов и
конкурировать с ними за ресурсы, от которых все они находят-
ся в одинаковой зависимости. Это и составляет основу коэво-
люции.
18*
Глава 9
ГОМИНИДЫ И ЭВОЛЮЦИЯ СООБЩЕСТВА
Рассуждения о человеческой эволюции нередко основаны на
интуитивном допущении, что она происходила «в самом конце»,
т. е. после того, как все остальные животные эволюционировать
уже «закончили». В книгах и учебниках по теории эволюции
возникновение гоминид рассматривается в заключительных
разделах, а гоминид зачастую представляют как верхний слой
глазури на филогенетическом торте. Такое допущение, однако,
ошибочно. В тот период, когда появились и эволюционировали
гоминиды, значительным эволюционным изменениям подверга-
лись и многие другие виды животных, которые приобрели свой
современный вид в то время, когда появились гоминиды, или да-
же позднее. Как мы уже видели, гоминиды адаптировались в
соответствии с общими принципами адаптации, присущими все-
му биологическому миру, а их эволюция происходила не в отры-
ве от эволюции других видов. Адаптивные стратегии наподобие
тех, которые, как описывалось в предыдущей главе, развива-
лись в ответ на сезонную изменчивость окружающей среды,
должны были изменять среду обитания других видов, и, следо-
вательно, вызывать их эволюционные изменения, которые в
свою очередь должны были сказываться на эволюции ранних
гоминид.
В основе концепции коэволюции и эволюции сообщества ле-
жит представление о том, что факторы естественного отбора,
действующие в разных популяциях, реципрокно связаны. В на-
стоящей главе будут прослежены эволюционные взаимодейст-
вия между гоминидами и другими видами животных, с которы-
ми они вели симпатрическое сосуществование. Подобные взаи-
модействия имеют последствия двоякого рода. Во-первых, они
определяют общую структуру биологического сообщества, кото-
рая в свою очередь сама превращается в важный фактор отбо-
ра. Во-вторых, они определяют конкретные эволюционные со-
бытия и направления, такие, например, как дивергенция, видо-
образование и вымирание видов. С помощью этого подхода мы
покажем, что адаптивные сдвиги у гоминид, а в конечном итоге
и развитие современного человека, представляют собой не не-
кие «вневременные» эволюционные процессы, но вполне опре-
Гоминиды и эволюция сообщества
277
деленные реакции на проблемы выживания в конкретном эво-
люционирующем сообществе.
Как, однако, отмечалось в предыдущей главе, изучение ко-
эволюции осложняется одним обстоятельством, которым нельзя
пренебречь. Важно помнить не только тот факт, что в плиоцене
и плейстоцене с гоминидами сосуществовали и коэволюциони-
ровали многие другие организмы, но и то, что сами гоминиды
представляли собой разнообразную и изменчивую группу млеко-
питающих. Считается, что по меньшей мере два вида гоминид
жили в Хадаре (Olson, 1981), по меньшей мере два вида гоми-
нид представлены в ископаемых сообществах Восточной Афри-
ки, датируемых в интервале от 3 до 1,3 млн. лет назад, и два
вида обитали в Южной Африке в интервале от 2 до 1 млн. лет
назад. Таким образом, на протяжении по крайней мере двух
млн. лет сосуществовали (возможно, симпатрически) не менее
двух видов гоминид. Коэволюция должна была происходить не
только между гоминидами и другими животными, но и между
разными популяциями гоминид. Поэтому в настоящей главе по-
мимо анализа взаимосвязей между гоминидами и другими мле-
копитающими африканских саванн будут обсуждаться и эколо-
гические основы адаптивной радиации гоминид и последующее
вымирание некоторых их видов.
9.1. Сообщество гоминид
9.1.1. Определение
Прежде всего необходимо определить, к какому экологичес-
кому и эволюционному сообществу принадлежали ранние гоми-
ниды. Отчасти это было уже сделано в предыдущей главе, где
было показано, что ранние гоминиды населяли саванны (а, мо-
жет быть, и другие местности) Восточной и Южной Африки.
Однако, сообщество — понятие более широкое, нежели просто
географическое местообитание, и скорее охватывает собой эко-
логически взаимодействующие организмы (Pianka, 1978, р. 4).
Представление об экологических взаимодействиях как основе
сообщества было развито Ван Куверинг. Термином «сообщест-
во» эта исследовательница обозначает «совокупность растений
и животных, связанных между собой определенными динамиче-
скими взаимодействиями (Van Couvering, 1980, р. 272). Такое
определение сообщества предполагает его вычлененность из
остального сложного и непрерывного биологического мира, что
в противном случае давало бы возможность произвольного тол-
кования сообщества в зависимости от конкретных научных ин-
тересов автора. Соответственно определение сообщества гоми-
нид требует выявления значимых экологических связей, которы-
278
Глава 9
Масаса (2)
Parapapio (5)*
Papio (9)
Dinopithecus (1)*
Gorgopithecus (1)F
Theropithecus (4)
Cercopithecus (1)
Australopithecus (2)*
Pan (2)
Lycaeon (1)
Leecyaena (1)*
Euryboas(2)*
Hyaena (4)
Crocuta (2)
Machairodus (3)*
Homotherium (2)*
Panthera(4)
Anancus (2)*
Stegodon (1 )*
Stegotetrabelodon (1)*‘
Primelephas (1)*
Mammuthus (1)*
Loxodonta (2)
Deinotherium (1)*
Chalicotherium (i)*
Oiceros (1)
Dicerorhinus (1)
Brachypotherium (1
Hipparion (2)*
Hexaprotodon (2)*
Hipopotamus (3)
Giraffa (3)
Sivatheriufn (1) *
Equus(d)
Taurotragus
Tragelaohus
Eotragus
Ugandax*
Pelorovis*
Simatherfum*
Hemibos*
Sylvicapra
Cephalophus
Redunca
Kobus
Menelikia*
Thalerocerus*
Nyanzachoerus (61
Metridiochoerus (3)
Kolpochoerus (6)‘
Stylochoerus (1)*
Notochoerus (3)*
Potamochoeroides (11
Phacochoerus(4)
Sus (1)
Patamocheerus (4)
H у I о choerus (1)
Makapania*
Nimidocapra*
Tossunnoria*
Gazella
Antidorcas
Madoqua
Ourebia
Oreotraous
Raphicerus
A₽pycerus,
Beatragus *
Parmularius
Damaliscus
Oreonagor
Coonochaetes
Megalotragus
Oryx
Hippotragus
**>Ренгы и потеН^*"
Рис. 9.1. Экологическое сообщество ранних гоминид: основные роды крупных
млекопитающих в Африке в плиоцене и раннем плейстоцене. Вымершие роды
отмечены звездочкой (по Maglio, 1978)
ми гоминиды были связаны с другими симпатрическими с ними
организмами.
Динамические взаимодействия гоминиды имели с теми вида-
ми, которым они служили пищей, которые им самим служили
пищей, и видами, с которыми они конкурировали. Подобные
взаимоотношения могли складываться у гоминид по преимуще-
ству с другими крупными млекопитающими и растениями. Сле-
довательно, предметом нашего анализа должно стать сообщест-
во крупных млекопитающих восточноафриканских саванн. Хотя
динамические взаимодействия должны были связывать гоминид,
и с другими организмами — к примеру, с паразитами или насе-
комыми,— следов в ископаемых остатках они оставить не могли..
На рис. 9.1 представлены современные ранним гоминидам виды,,
классифицированные в соответствии с типом экологических
взаимоотношений, которыми они были связаны с гоминидами.
9.1.2. Видовое разнообразие сообщества
Основным вопросом, возникающим в связи с сообществом
гоминид, является вопрос о том влиянии, которое оно могло
оказать на эволюцию гоминид и их адаптацию. Это значит, что
Гоминиды и эволюция сообщества
279
необходимо изучить и описать структуру этого сообщества.
Пожалуй, лучше всего воспользоваться для этого концепцией
разнообразия или видового богатства сообщества, т.е. числом
видов в сообществе — общим или соответствующим различным
экологическим критериям, например трофическому уровню или
размерам тела животных. Таким образом, основным предметом
нашего анализа будут виды.
По богатству видов сообщества сильно варьируют. В таких
сообществах, как, например, влажный тропический лес, число
видов, приходящееся на единицу площади, велико. В арктичес-
ких сообществах или тундре этот показатель сравнительно не-
высок. Подобная вариабельность, однако, подчиняется опреде-
ленной закономерности, которую для млекопитающих недавно
продемонстрировал Эйзенберг (Eisenberg, 1981, р. 219). Тропи-
ческие сообщества более разнообразны, чем сообщества уме-
ренных широт, а континентальные сообщества богаче видами,
чем островные. Число видов на континентальных островах
больше, чем на океанических (рис. 9.2). Кроме того, районы
с высокой продуктивностью и устойчивым климатом богаче
видами, чем области с противоположными характеристиками.
Вероятно, для каждого типа сообщества имеется оптимальное
или равновесное число видов, определяемое различными эколо-
Рис. 9.2. Линии регрессии для зависимости между числом видов нелетающих
млекопитающих и общей площадью географической территории (no Eisenberg
1981, рис. 38)
280
Глава 9
Таблица 9.1. Гипотетические факторы, ответственные за видовое разнообразие
сообщества (по Foley, 1984b; Pianka, 1978 с изменениями)
Фактор
Эволюционное время
Экологическое время
Временная неоднород-
ность
Пространственная неод-
нородность
Продуктивность
Хищничество
Разрежение
Конкуренция
Объяснение
Видовое разнообразие увеличивается со временем
в результате видообразования до тех пор, пока не
наступит насыщения сообщества видами
Видовое разнообразие увеличивается со временем
в результате рассредоточения и освоения новых
биотопов
Изменчивость климата в разных временных мас-
штабах — от сезонных до геологических; ее не-
предсказуемость снижает видовое разнообразие
Наоборот, пространственная неоднородность от-
крывает большие возможности для специализации
и, таким образом, приводит к увеличению разно-
образия
Высокая продуктивность увеличивает как разме-
ры основания, так и высоту трофической пирами-
ды и, таким образом, способствует увеличению ви-
дового разнообразия
Хищничество часто зависит от встречаемости жертв
и, следовательно, поддерживает видовое разнооб-
разие сообщества, предотвращая доминирование
какого-либо одного вида
Снижение численности видов в сообществе способ-
ствует повышению его видового разнообразия
Увеличение видового разнообразия сообщества
стимулирует развитие конкурентоспособности;
в результате получает преимущества большое чис-
ло видов с высокоспециализированной конкурен-
тоспособностью
гическими характеристиками. Это—первый вывод, который
напрашивается из работ Мак-Артура и его сотрудников и кото-
рый лежит в основе одной из нескольких теорий видового раз-
нообразия (см. табл. 9.1). Отклонения от теоретического равно-
весия, примером которых могут служить обедненные в видовом
отношении тропические области южной Индии, можно объяс-
нить историческими процессами, включая недавнее исчезнове-
ние видов вследствие человеческой деятельности, изменения
окружающей среды или действия специфических экологических
факторов. В случае южной Индии таким важным фактором
является выраженный муссонный климат.
Видовое равновесие экосистемы достигается тогда, когда эта
экосистема насыщается видами. По существу это означает, что
Гоминиды иэволюция сообщества
281
Низкое
Высокое
---разнообразие разнообразие —
Насыщенность
видами
Ненасыщенность
видами
Климатическая неустойчивость/ /низкая продуктивность Климатическая устойчивость/ /высокая продуктивность
Исторически изменившиеся среды ?
Рис, 9.3. Факторы изменения видового богатства сообщества
все потенциальные экологические ниши этой экосистемы заня-
ты и что новый вид может внедриться в нее только за счет дру-
гого вида. Границы биомассы экосистемы зависят от ее ресур-
сов. Популяции приближаются к этим границам, увеличи-
ваясь в размерах. В результате экосистема (быстро?) насыща-
ется особями, хотя видами на этой стадии своего существова-
ния она может и не быть насыщена. Со временем, однако, особи
тех видов, которые способны более эффективно использовать
имеющиеся ресурсы и которым, следовательно, требуется мень-
ше экологического пространства, вытесняют менее специализи-
рованных особей. Таким образом, с течением времени экологи-
ческие ниши сужаются, а видовое разнообразие экосистемы уве-
личивается до тех пор, пока не будет достигнуто ее видовое
насыщение или равновесие. Колебания условий окружающей
среды замедляют этот процесс. Соответственно видовое богат-
ство экосистемы можно связать с ее видовой насыщенностью
и разработать общую модель (рис. 9.3), учитывающую как дей-
ствие непосредственных, неоэкологических факторов, так и влия-
ние исторических событий, таких, например, как изменение
уровня моря или климата.
Какое значение имеет видовое богатство экосистемы для
отдельных видов, составляющих сообщество? Насыщенность
и большое видовое разнообразие экосистемы, вероятнее всего,
указывают на существование в ней острой конкуренции — в пер-
вую очередь среди организмов, способных эффективно исполь-
зовать ресурсы узкой экологической ниши. В ненасыщенных
282
Глава 9
экосистемах шансы оппортунистических видов на выживание
повышаются, а их экологические ниши в среднем становятся
шире.
9.1.3. Современные и ископаемые африканские сообщества
Проведенный анализ вопроса о видовом богатстве экосисте-
мы заставляет предполагать, что африканские сообщества,
в состав которых входили ранние гоминиды, по всей вероятно-
сти, были сравнительно богаты видами млекопитающих. Ими,
безусловно, богаты и современные африканские саванновые
экосистемы. По данным Эйзенберга (Eisenberg, 1981, р. 219),
видовая плотность млекопитающих в Восточной Африке дости-
гает 1,98х 10~4/км2. Столь же высокое видовое разнообразие,
сравнимое с соответствующим средним показателем для тропи-
ков (0,76Х10-4/км2), отмечается и на уровне семейств и родов
(Maglio, 1978).
Необычайно высокое видовое богатство африканских саванн
можно объяснить общими принципами Эйзенберга. Восточная
Африка является частью континентального массива, это — тро-
пическая область, ее первичная продуктивность относительно^
биомассы высока, и в плейстоцене по сравнению с другими
регионами здесь вымерло не так уж много видов. Возникает,
однако, вопрос, имела ли плиоценовая и плейстоценовая фауна
Восточной Африки такую же экологическую структуру и видо-
вое разнообразие, как и современная фауна.
Ответ на этот вопрос дает сравнение видового богатства
ископаемых сообществ с видовым разнообразием современных
экосистем. Такое сравнение, очевидно, заставляет задуматься
о том, в какой степени ископаемые отражают реальный состав
живых организмов той эпохи, к которой они относятся. Это свя-
зано с объемом выборки, образованием палимпсеста и разной
сохранностью остатков; все эти факторы необходимо учитывать
при интерпретации экологического сходства и различия между
экосистемами. В общем современные экосистемы богаче вида-
ми, чем ископаемые (рис. 9.2). Среднее число видов в восточно-
африканских заповедниках составляет 70,2, что выше соответ-
ствующего показателя для большинства ископаемых сообществ.
Отчасти, однако, такое положение вещей может быть связано
с размерами животных. Известно, что мелкие животные реже
подвергаются фоссилизации, чем крупные (Behrensmeyer, Dec-
hant Boaz, 1980), и, следовательно, в ископаемых скоплениях
представлены слабее. Если современные и ископаемые экосисте-
мы рассматривать без мелких животных, различия между пока-
зателями их видового разнообразия в значительной мере сгла-
живаются— в среднем 46,8 и 35,9 видов для современных
Гоминиды и эволюция сообщества
283
(табл. 9.2) и плио-плейстоценовых местонахождений Восточной
Африки соответственно. Кроме того, если из анализа исключить
также небольшие ископаемые, которые нельзя считать достаточ-
но представительными, то средний показатель видового разно-
образия плиоценовых и плейстоценовых сообществ повышается
до 43,1 вида. Если учесть к тому же возможность неполной
представленности ископаемой фауны в находках, можно думать,
что видовое богатство обеих экосистем было примерно оди-
наковым.
Что, однако, можно сказать о структуре ископаемых и сов-
ременных сообществ? Представлено ли их видовое богатство
одними и теми же группами животных? Эндрюс и др. (Andrews,
Lorg, Evans, 1979) и Ван Куверинг (Van Couvering, 1980) раз-
работали методы определения среды обитания форм, представ-
ленных в ископаемых скоплениях, по профилю их экологиче-
ской структуры. Сообщества характеризуются различным соот-
ношением разных групп животных; относительное число траво-
ядных видов выше в сообществах травяных местообитаний, а от-
носительное число фруктоядных — в лесных сообществах. Опре-
делив подобные соотношения для современных сообществ, их
можно сравнить с ископаемыми сообществами. Если экологи-
ческий профиль последних окажется близким к экологическому
профилю современных сообществ, можно говорить и о сходстве
структуры ископаемых и современных сообществ.
Для представленного ниже анализа экологических профи-
лей были выбраны пять групп млекопитающих: приматы, круп-
ные хищники, представители мегафауны (носороги, хоботные,
бегемоты и жирафы), свиньи и крупные, быстро бегающие тра-
воядные (лошади и полорогие). Эти группы соответствуют как
таксономическим категориям, так и способу питания животных.
Чтобы сделать результаты сравнения современных и ископае-
мых сообществ как можно более надежными, мелкие виды мле-
копитающих в анализ не включались. Данные по составу вос-
точноафриканских сообществ были заимствованы из работы
Уильямса (Williams, 1971). О видовом составе ископаемых сооб-
ществ судили по описаниям фауны главных плиоценовых и
плейстоценовых местонахождений Восточной Африки. Все под-
вергнутые анализу сообщества — как современные, так и иско-
паемые— относятся или относились к одному и тому же широ-
кому типу экосистем — травяным или редколесным саваннам.
Сообщества тропических лесов в анализ не включались.
При сравнении фауны современных и ископаемых сообществ
выявляется ряд интересных закономерностей (табл. 9.3 и 9.4;
рис. 9.4—9.6). В современной фауне преобладают быстро бегаю-
щие травоядные — пасущиеся полорогие и лошади. Именно они
и образуют те огромные стада, которые придают неповторимый
Таблица 9.2. Сравнение структуры современных н ископаемых
африканских сообществ (по числу видов млекопитающих)
Группа животных
Средний про- Средний про- Среднее число Средний про-
_ Средний про- цент от обще- Среднее число цент от числа видов в иско- цент от числа
среднее число цент от числа го числа ви- видов в иско- видов круп- паемых сооб- видов круп-
видов в совре- видов круп- дов млекопи- паемых сооб- ных млекопи- ществах, со- ных млскопи-
менных эко- ных млекопи- тающих в со- ществах тающих в ис- держащих не тающих в ис-
системах тающих временных (N=24) копаемых со- менее 30 ви- копаемых со-
экосистемах обществах дов (N=17) обществах
Мелкие приматы 1,4 3,0 2,0 111 0,21 0,6 0,3 0,7
Крупные приматы 3,8 8,1 ’ 5,4 4,5 12,6 5,9 13,7
Мелкие хищные 15,0 32,1' 21,0 . 44 7 3,2 8,9 4,4 10,2
Крупные хищные 5,9 12,6 ’ 8,4 3,7 10,3 4,9 11,4
Хоботные 0,8 1,7 1,1 2,2 6,1 2,3 5,3
Носороги 0,9 1,9 1,3 1,5 4,2 1,7 3,9
Лошадиные 1,1 2,4 1,6 1,7 4,7 1,6 3,7
Бегемотовые 0,8 1,7 1,1 1,7 4,7 1,9 4,4
Свиные 1,5 3,2 2,1 4,0 11,2 4,5 10,4
Жирафовые 0,9 1,9 1,3 2,8 7,8 3,2 7,4
Мелкие полорогие 6,4 13,71 9,1
Средние полорогие 4,1 14,4 8,8 ,30,8 5,8 .20,5 . 10,3 . 28,8 . 12,4 28,7
Крупные полорогие 3,9 8,3 5,6
Крупные млеко- 46,8 (46,5) 99,4 66,7 35,9 99,9 43,1 99,8
питающие
Все млекопитаю- 70,2 — — — — — —
щие
Таблица 9.3. Состав фауны современных восточноафриканских сообществ (по Williams, 1971)
Сообщество Прниаты Крупные хищники Мегафауиа Свиньи Крупные, быстро бегающие травояд- ные
N11 1-%- N 1 % N 1 % N 1 % N 1 %
Абердарес 4 18,2 3 13,6 2 9,1 2 9,1 11 50,0
Амбосели 4 11,8 5 14,7 4 11,8 2 5,9 19 55,9
Марсабит 5 20,8 6 25,0 3 12,5 1 4,2 9 37,5
Масаи-Мара 7 17,1 6 14,6 4 9,8 3 7,3 21 51,2
Меру 6 14,3 6 14,3 5 11,9 3 7,1 22 52,4
Найроби 6 18,2 6 18,2 3 9,1 1 3,0 17 51,5
Накуру 4 16,0 5 20,0 2 8,0 0 0,0 14 56,0
Самбуру/Исиоло 5 15,2 6 18,2 4 12,1 1 3,0 17 51,5
Тсаво 5 13,5 6 16,2 3 8,1 1 2,7 22 59,5
Озеро Маннара 5 20,8 3 12,5 4 16,7 1 4,2 11 45,8
Микумн 3 12,5 5 20,8 4 16,7 1 4,2 11 45,8
Нгоронгоро 2 7,7 5 19,2 4 15,4 2 7,7 13 50,0
Руаха 5 16,1 5 16,1 4 12,9 1 3,2 16 51,6
Серенгети 7 16,7 6 14,3 4 9,5 2 4,8 23 54,8
Кидепо 4 13,3 5 16,7 4 13,3 1 3,3 16 53,3
Мерчисон 6 22,2 4 14,8 4 14,8 2 7,4 11 40,7
Парк Королевы Елизаветы 10 34,4 14 13,8 3 10,3 3 10,3 9 31,0
X 5,2 17,0 5,4 16,6 3,6 11,9 1,6 5,1 15,4 49,3
Таблица 9.4. Состав фауны ископаемых сообществ в Восточной
Африке (по Leakey, 1972; Coppens, Howell,
1978; Johanson, Taieb, Coppens, 1982; Harris, 1983)
Сообщество Приматы Крупные хищники Мегафауна Свиньи Крупные, быстро бегающие травояд- ные
N 1 % N 1 % N 1 % N 1 % N 1 %
Восточный берег озера Туркана, CU2 3 9,4 3 9,4 11 34,4 3 9,4 12 37,5
Восточный берег озера Туркана, CU3 8 16,3 7 14,3 11 22,4 3 6,1 20 40,8
Восточный берег озера Туркана, CU4 9 16,4 7 12,7 12 21,8 4 7,3 23 41,8
Восточный берег озера Туркана, CU5 7 17,1 4 9,8 9 22,0 4 9,8 17 41,5
Омо-Усно 8 22,2 9 25,0 10 27,8 5 13,9 4 П,1
Омо, пачка В 12 28,5 3 7,1 10 23,8 6 14,3 11 26,2
Омо, пачка С 10 18,5 9 16,7 11 20,4 7 1,0 17 36,5
Омо, пачка D 6 21,4 3 10,7 10 35,7 3 10,7 6 21,4
Омо, пачка Е 7 23,3 3 10,0 7 23,3 3 10,0 10 33,3
Омо, пачка F 8 19,5 7 17,1 10 24,3 4 9,8 12 29,3
Омо, пачка G 9 16,1 8 14,3 13 23,2 6 10,7 20 35,7
Омо-Калам 6 16,7 2 5,6 7 19,4 6 16,7 15 41,6
Хадар 5 10,6 9 19,1 12 25,5 3 6,4 18 38,3
Олдувай, пласт I (L) 3 11,1 3 11,1 8 29,6 3 11,1 10 37,0
Олдувай, пласт I (М) 3 10,0 3 10,0 6 20,0 5 16,7 13 43,5
Олдувай, пласт I (U) 0 0,0 2 8,7 7 30,4 3 13,0 11 47,8
Олдувай, пласт II (М) 1 3,1 1 3,1 7 21,9 7 21,9 16 50,0
X 6,2 15,3 4,9 12,0 9,5 25,0 4,4 11,8 13,8 35,2
Гоминиды н эволюция сообщества
287'
Самбу ру/Исиоло
П X М СТ
Озеро Маниара
Нгоронгоро
Рис. 9.4. Структура сообществ современных восточноафриканских экосистем-,
(национальные парки и охотничьи заказники). На графиках изображены про-
фили относительного разнообразия разных групп крупных млекопитающих.
Сокращения: П — приматы, X — хищники, М — мегафауиа, С — свиньи, Т —
крупные травоядные (полорогие и лошадиные)
облик современным саваннам. Эти животные иногда составля-
ют в сообществе до 59,5% от общего числа видов крупных мле-
копитающих; в среднем этот показатель равен 49,3%- В иско-
паемых же сообществах средняя относительная частота встре-
чаемости остатков крупных, быстро бегающих травоядных
составляет 35,8%. Иначе говоря, в современной фауне явно’
преобладает одна экологическая и таксономическая категория:
288
Глава 9
Восточный берег
озера Туркана
Омо-Усно
Восточный берег
озера Туркана
Восточный берег
отера Туркана
01 । 1 .
П X м С т
П X м с т
Омо, пачка !
Хадар FT
' Олдувай, слой I
La/
__J____I___I__I
Олдувай,
_слой 11 (М)
П X М С т
Рис. 9.5. Воссозданная по ископаемым находкам структура сообществ плиоце-
новых и плейстоценовых экосистем. Сокращения такие же, как и на рис. 9.4
животных, чего нельзя сказать об ископаемых сообществах.
Различия между этими сообществами выявляются и в отноше-
нии других экологических категорий животных. Ископаемые
экосистемы характеризуются большим абсолютным и относи-
тельным видовым разнообразием свиней и представителей мега-
фауны и большим абсолютным видовым богатством приматов
(в особенности крупных). С другой стороны, число видов круп-
ных хищников в среднем больше в современных сообществах.
Следующий момент, заслуживающий внимания, заключается
в том, что если структура современных сообществ варьирует
сравнительно слабо, то различия между ископаемыми сообще-
ствами выражены сильно (рис. 9.4 и 9.5). В особенности выде-
ляются ископаемые находки из Омо, характеризующиеся отно-
Гоминиды и эволюция сообщества
289
О______।____।____।____।
40 Г Среднее (все ископа-
емые сообщества)
О ----1___|____1
П X М С Т
Рис. 9.6. Профили общего экологического разнообразия современных и иско-
паемых (плиоцен—плейстоцен) сообществ Восточной Африки. Сокращения
такие же, как и на рис. 9.4
сительно высокой представленностью приматов и низкой — по-
лорогих и лошадей, что дает основание относить их к совершен-
но иному типу сообщества. В этой связи возникает вопрос, не
обусловлены ли, по крайней мере частично, те различия, кото-
рые обнаруживаются между ископаемыми и современными со-
обществами, фоссилизацией, а не экологическими процессами.
Можно думать, что именно по этой причине в ископаемых на-
ходках недостаточно представлены мелкие млекопитающие, и
точно так же объяснять отсутствие в них остатков мелких по-
лорогих. Но правдоподобно будет звучать и утверждение, что
определенные группы животных — в особенности свиньи и пред-
ставители мегафауны — в плиоцене и плейстоцене были гораздо
разнообразнее, чем в настоящее время и превалировали тогда
как в абсолютном, так и в относительном плане.
Ископаемые остатки могут недостаточно отражать видовой
состав представленных ими сообществ в связи с различной со-
хранностью погребенных в осадочных породах животных, но
вследствие образования палимпсеста картина может искажать-
ся и в другую сторону. Не исключено, что состав ископаемых
сообществ отражает не какой-то кратковременный этап жизни
сообщества (подобно приводимым в литературе перечням совре-
19—367
290
Глава 9
Рис. 9.7. Моделирование влияния, которое оказывает образование палимпсеста!
на экологическое разнообразие. Метод заключается в сведении воедино переч-
ней видов, характерных для различных восточноафриканских экосистем, и ана-
лизе общего видового разнообразия «результирующего» сообщества. Каждая,
кривая соответствует одному такому сведению воедино состава фауны 10 раз-
личных экосистем. Предполагается, что эти операции с экосистемами имити-
руют влияние долгосрочных экологических изменений на процесс фоссилизации.
Использовались перечни современного фаунистического состава национальных
парков и заповедников Восточной Африки. Сообщества животных для каждо-
го сведения воедино выбирались наугад (по Williams, 1971)
менных животных), а постепенное накопление остатков живот-
ных за длительные периоды времени, когда вполне могли про-
исходить изменения окружающей среды. В результате в одной
и той же палеонтологической находке будут обнаруживаться
животные, которые жили в разное время. Некоторое представ-
ление о такой возможности дает «сведение воедино» фаунисти-
ческого состава нескольких современных сообществ и анализ,
видового разнообразия образующегося в результате такого све-
дения «результирующего сообщества». Сведя воедино фауни-
стический состав выбранных наугад десяти разных сообществ
и проследив изменение при этом видового разнообразия «ре-
зультирующих сообществ», можно видеть (рис. 9.7), что если
общая величина этого показателя и в самом деле увеличивает-
ся, то изменения составляющих сообщество экологических групп
животных выражены не столь значительно, чтобы можно было
Гоминиды и эволюция сообщества
291
говорить о какой-либо аналогии с разнообразием в плиоцене
и плейстоцене. Хотя этот вывод не может считаться оконча-
тельным, так как сегодня невозможно определить, насколько
географическая изменчивость современных животных в Восточ-
ной Африке сравнима с таковой в плиоцене и плейстоцене,
в известной мере он подкрепляет предположение о существова-
нии структурных различий между плио-плейстоценовыми и сов-
ременными саванновыми сообществами крупных млекопитаю-
щих.
Рассмотренные данные позволяют провести лишь сравни-
тельно грубую оценку структуры сообществ и, по всей вероят-
ности, искажены различными тафономическими факторами и
смешением ископаемого материала, относящегося к различным
эпохам; так, материал, обнаруженный в Омо, относится к пе-
риоду в более чем 2 млн. лет, а за это время, как известно,
происходили значительные экологические сдвиги (Bonnefille,
1976, 1985).
9.1.4. Контекст эволюционного развития ранних гоминид
Большое видовое разнообразие и структура сообщества ран-
них гоминид позволяют сделать некоторые предположения об
экологическом, «конкурентном» и эволюционном характере той
среды, в которой у них развивались первые адаптации. Соглас-
но принципам, обсуждавшимся в предыдущей и начале этой
глав, большое видовое разнообразие сообщества предполагает
высокий уровень конкуренции, т. е. либо прямые конкурентные
взаимодействия между видами, либо устойчивую структуру эко-
логической ниши, обусловленную стремлением избежать кон-
куренции, которое само возникает в результате конкуренции.
Вследствие насыщения экологического пространства особями
и видами интенсивность отбора в обоих случаях должна быть
высокой. Можно думать, что отбор этот будет оказывать в ос-
новном стабилизирующий эффект на общую структуру сооб-
щества. Каждый вид строго «контролируется» его взаимодей-
ствиями с видами, занимающими смежные с ним ниши. Однако
любой вид, способный вырваться из-под такого контроля, может
оказать огромное влияние на все сообщество в целом, вызвав
быстрые эволюционные изменения широкого ряда конкурирую-
щих и взаимозависимых видов.
Исходя из общего видового богатства африканских плиоце-
новых и плейстоценовых сред, в которых жили ранние гомини-
ды, можно сделать предположение, что существовавшая в них
острая конкуренция, по всей вероятности, способствовала усиле-
нию естественного отбора. В пользу этого предположения гово-
рят и различия между современной и древней фауной. Если
19*
292
Глава 9
современные сообщества крупных млекопитающих представле-
ны главным образом травоядными, потребляющими низкокаче-
ственные в пищевом отношении травы, то плиоценовые и плей-
стоценовые сообщества были богаче видами, занимавшими ни-
ши с более высококачественной растительной пищей — напри-
мер, включали больше видов крупных приматов и свиней. Сле-
довательно, можно думать, что самые острые эволюционные
взаимодействия складывались именно между «разборчивыми
в пище» животными — всеядными и плодоядными, к которым
относились и ранние гоминиды (следует помнить, что именно
.в этот период отмечались наибольшее разнообразие и наиболее
быстрое эволюционное изменение гоминид — от первых австра-
лопитековых до позднего Ното erectus). Таким образом, харак-
тер эволюции гоминид соответствует характеру эволюционных
изменений и экологической структуре сообщества в целом.
Чтобы подкрепить фактами предположение об острых взаи-
модействиях гоминид с их симпатрическими конкурентами, не-
обходимо рассмотреть конкретные взаимоотношения между раз-
ными видами животных. Для этого нужно более подробно изу-
чить изменчивую природу самих гоминид в этот период вре-
мени.
9.2. Факты, свидетельствующие о разнообразии гоминид
Хотя в гл. 2 при рассмотрении общих направлений эволюции
приматов и отмечалось существование нескольких симпатричес-
ких видов ранних африканских гоминид, все остальное изложе-
ние основывалось главным образом на том специально не ого-
варивавшемся допущении, что гоминиды являлись однородной
группой, сталкивавшейся с одними и теми же экологическими
проблемами и прибегавшей к их одинаковому адаптивному раз-
решению. Это, однако, — явное упрощение ситуации. Безуслов-
но, более широкий и общий взгляд на ранних гоминид, позво-
ляющий сравнить их с другими группами животных, имеет свои
преимущества; необходимо, однако, учитывать и тот факт, что
на протяжении плиоцена и плейстоцена гоминиды представля-
ли собой неоднородную группу африканских приматов и обна-
руживали значительную вариабельность. Разнообразие гоминид
заслуживает самого пристального эволюционного и экологиче-
ского изучения.
Сосуществование двух или большего числа видов предпола-
гает наличие двух или более адаптивных стратегий и соответст-
венно двух или более экологических ниш. Это в свою очередь
заставляет думать, что не все гоминиды сталкивались с одними
и теми же экологическими проблемами, а если все же экологи-
1,ческие проблемы и были для всех гоминид одинаковыми, то
Гоминиды и эволюция сообщества
293’,
адаптивное их решение различалось. Чтобы подкрепить это-
предположение и включить конкуренцию между гоминидами:
в модель их эволюционного развития, разнообразие гоминид,
и их сосуществование следует изучить по ископаемым остаткам.
9.2.1. Уровни изменчивости
Можно выделить несколько уровней изменчивости и, следо-
вательно, несколько уровней разнообразия гоминид. Изменчи-
вость на уровне подвидов проявляется в существовании гетеро-
генных популяций, характеризующихся, однако, непрерывным
потоком генов. С другой стороны, изменчивость может стать
настолько высокой, что поток генов прервется и образуются
новые виды. Если это будет происходить в течение достаточно
длительного времени, различия станут настолько существенны-
ми, что систематики вынуждены будут признать образование
раздельных таксонов более высокого уровня, например родов.
Все эти уровни изменчивости и разнообразия прослеживаются
и в африканских ископаемых находках.
Как говорилось в разд. 2.2.2, обычно предполагается, что на
протяжении плиоцена и плейстоцена в Африке существовали
два рода ранних гоминид — Australopithecus и Ното. Первые
австралопитеки, по всей видимости, являются самыми первыми
формами гоминид вообще; судя по анатомическим признакам,
два названных рода гоминид являются представителями двух
дивергентных эволюционных направлений. При этом австрало-
питеки характеризуются небольшим шарообразным черепом
с тонкими черепными костями, но зачастую хорошо выраженны-
ми костными гребнями и выступами, а также с мощным зубо-
челюстным аппаратом; представителям второго рода, Ното,
присущ более объемистый и удлиненный череп и менее массив-
ный зубочелюстной аппарат.
Представители каждого из этих родов обнаруживали значи-
тельную изменчивость, заставляющую говорить о существова-
нии нескольких видов (рис. 2.8 и 2.10). Изменчивость носит как
хронологический, так и пространственный характер. Из древ-
нейших африканских местонахождений — Табарина, Летоли
и Хадара — известен примитивный вид — Australopithecus afaren-
sis. Высказываются, однако, мнения, что многочисленные об-
разцы из Хадара относятся к разным таксонам. Как считает
Олсон (Olson, 1981), еще 3 млн. лет назад в Восточной Африке
жили по крайней мере два разных вида гоминид. Немного стар-
ше находки грациального Australopithecus africanus из Южной
Африки. От него сильно отличаются массивные австралопите-
ковые. Эти гоминиды, основные признаки которых совпадали
с общими родовыми характеристиками, отличались более круп-
294
Глава 9
ними размерами, более крепким телосложением и чрезвычайно
сильно развитой черепной мускулатурой. Наиболее «яркие» об-
разцы этого типа были найдены в Восточной Африке (Australo-
pithecus boisei) и Хауэлом (Howell, 1978) в Южной Африке
(Australopithecus robustus и Australopithecus crassidens).
Род Ното обнаруживает меньшее разнообразие. Для плио-
цена н нижнего плейстоцена описаны лишь два вида — Ното
habiiis и Homo erectus, по мнению некоторых исследователей в
ископаемом материале из Кооби-Фора представлен и третий вид
(Homo ergaster?) (Howell, 1978; Walker, Leakey, 1978). Homo
habiiis превосходит грациальных австралопитековых общими
размерами и размерами черепной коробки. Еще более вмести-
тельным черепом, крупными размерами тела и массивным те-
лосложением характеризуется Homo erectus.
Следует, однако, учитывать, что изменчивость отмечается
и внутри этих видов и что исследователи не пришли еще к еди-
ному мнению, связана ли изменчивость гоминид на этом уровне
с половым диморфизмом, как это предполагалось для ископае-
мых образцов из Хадара и Летоли (Johanson, White, 1979; Whi-
te, Johanson, Kimbel, 1983), или отражает географическое и
хронологическое разнообразие представителей единого поли-
морфного вида (Tobias, 1980).
9.2.2. Сосуществование
Таким образом, в интервале времени от 4 до 1 млн. лет
назад существовало по крайней мере семь видов гоминид, четы-
ре или пять из которых жили в Восточной Африке и несколь-
ко — в Южной. Кроме того, хотя в ряде случаев можно просле-
дить постепенные эволюционные переходы (например, переход
от австралопитековых легкого телосложения к австралопитеко-
вым более массивного склада или замещение Homo habiiis ha-
biiis на Н. erectus), существование этих видов в значительной
мере перекрывалось во времени (рис. 9.8). Можно полагать, та-
ким образом, что на протяжении плиоцена и плейстоцена в со-
обществах африканских саванн сосуществовало не менее двух
видов гоминид. Остается лишь определить, насколько тесно со-
существовали разные виды гоминид и в какой степени перекры-
вались их экологические ннши. В частности, являлись ли ранние
гоминиды полностью симпатрическими видами или они просто
соседствовали в пределах какой-то области или всего африкан-
ского континента в целом.
Симпатрию гоминид более или менее детально изучил
Хауэлл (Howell, 1978, рр. 222—223). Он установил, что в Сварт-
крансе, Олдувае (пласты I и II), Кооби-Фора (отложения над
и под слоем туфа (KBS) и Омо (слои G-H) остатки Homo sp.
Гоминиды н эволюция сообщества
295
1,0 г
A. afarensis A. africanus A. boisei A. robustus A. crassidens Н. habilis Я erectus
Сварткранс?
Хадар
Пенинж
Кромдрай 1
Чесованьп Сварткранс
Олдувай
Кооби-Фора
Омо
Омо?
Стеркфонтейн
Макапансгат
I Хадар?
| (неопр. вид)
Летоли
Табарин
Сварткранс А
Олдувай
Нариокотоме
. Кооби-Фора
Стеркфонтейн?
Омо
Олдувай
Кооби-Фора
5,0
'°ис. 9.8. Предположительное время существования разных видов гоминид
(Australopithecus и Ното) в Африке в плиоцене и плейстоцене
(предположительно Н. habilis) представлены в тех же самых
геологических отложениях, что и остатки массивных австрало-
питековых. В Омо, кроме того, Australopithecus boisei встреча-
ется в тех же пластах, что и Australopithecus africanus, а в Коо-
би-Флора и Олдувае симпатрическими видами были Ното
erectus и Australopithecus boisei. Таким образом, можно заклю-
чить, что ранние гоминиды не только в течение длительного
времени сосуществовали в одних и тех же биогеографических
.районах, но и обнаруживали симпатрию в том смысле, какой
этому термину придают экологи, изучающие прямые взаимодей-
ствия живых организмов.
Судя по ископаемым свидетельствам, в Африке гоминиды
лретерпели адаптивную радиацию в плиоцене и плейстоцене.
Кроме того, они обнаруживали широкую географическую и вре-
менную симпатрию, и, следовательно, должны были взаимодей-
ствовать экологически. Разнообразие форм гоминид можно ис-
пользовать для анализа специфических коэволюционных взаи-
моотношений— взаимоотношений между разными видами гоми-
296
Глава 9
нид и между гоминидами и другими видами — и выяснения той
роли, которую эти взаимоотношения играли в эволюции
гоминид.
9.3. Экология сообщества и коэволюция
Итак, в той информации об экологическом сообществе ран-
них гоминид, которой мы пока располагаем, можно выделить
три главных момента. Во-первых, что этому сообществу было
присуще в целом высокое разнообразие видов крупных млеко-
питающих; во-вторых, что экологическая группа, к которой при-
надлежали ранние гоминиды, характеризовалась гораздо боль-
шим видовым разнообразием, чем теперь. И в-третьих, что го-
миниды подвергались адаптивной радиации. Эти факты можно
взять за основу анализа той роли, которую в эволюции гоми-
нид, очевидно, играли межвидовые взаимоотношения. Такой
анализ предполагает как выяснение вопроса, соответствовала
ли эволюция гоминид общему характеру эволюции сообщества,
так и изучение конкретных конкурентных и коэволюционных
взаимоотношений животных.
Предположение о параллельном адаптивном развитии го-
минид и эволюции сообщества тропических африканских саванн
высказывалось и ранее (Foley, 1984b). Повторяющийся харак-
тер развития сообществ — от установления новых условий ок-
ружающей среды до достижения некоторого уровня устойчи-
вости— был подмечен многими исследователями. Важным мо-
ментом при этом, как подчеркивал Хоффмэн (Hoffman, 1979),
является не эволюция сообществ в целом (сообщество в целом
не может быть единицей отбора), но те общие свойства индиви-
дуальных и межвидовых взаимодействий, которые присущи
всем экосистемам и которые и обусловливают установившуюся
схему развития сообщества.
Один из образцов такой схемы изображен на рис. 9.9. Когда
возникает новая окружающая среда, ее осваивают виды, харак-
теризующиеся высокими темпами размножения и расселения.
Эти виды редко когда могут использовать ресурсы среды доста-
точно эффективно, однако благодаря высокой скорости размно-
жения, устойчивости к широкому диапазону условий и отсутст-
вию конкурентов на какое-то время получают определенные-
экологические преимущества. На ранних стадиях образования
новой среды доминировать в ней будут такие виды; по этой при-
чине, а также из-за того, что для освоения среды другими вида-
ми требуется время, ее видовое разнообразие (богатство) будет
низким. Однако по мере того, как новая среда насыщается осо-
бями и новыми видами, преимущества начинают переходить от
нейтральных, менее специализированных видов к более спе-
I оминиды и эволюция сообщества
297
Стадия 1 Стадия 2 Стадия 3
Новые условия окружающей среды Колонизация оппорту- нистическими видами Низкое видовое разнообразие Увеличение специализации и адаптивная радиация Разделение ниш Максимальное видовое разнообразие Стабилизация ниш Усиление конкурентного исключения и вымирание видов Упрочение положения новых менее специализированных видов Снижение видового разнообразия
--------------------------- Время-------------------------*-
Рис. 9.9. Модель эволюции сообщества, основанная на характере изменения
конкуренции в новых условиях окружающей среды (по Foley, 1984b)
циализированным, способным использовать ресурсы среды бо-
лее эффективно. В зависимости от географических и временных
факторов либо эти биотопы будут осваиваться новыми видами,,
пришедшими извне, либо естественный отбор будет способст-
вовать эволюции тех видов, которые могут эффективнее всего!
выживать в условиях все более обостряющейся конкуренции.
На этой стадии увеличивается видовое разнообразие сообщест-
ва, в особенности за счет видов, использующих высококачествен-
ные продукты питания. Эта тенденция сопровождается дальней-
шим разделением экологических ниш, что приводит к развитию
двоякого рода процессов. Во-первых, выживание в более узкой
экологической нише предполагает усиление зависимости живот-
ных от источников низкокачественной пищи, а также и от источ-
ников высококачественной, и, во-вторых, численность видов
с широкими адаптациями сокращается, и они в конце концов,
вымирают. Увеличение скорости вымирания видов может
повлечь за собой снижение видового разнообразия сообщества.
Кроме того, на поздних этапах развития сообщества по мере
увеличения вторичной биомассы возрастают пищевые преимуще-
ства животных, питающихся падалью, и хищников.
Можно ли сопоставить изменения видового разнообразия и
структуры африканских саванновых сообществ с описанной
схемой? Как видно из рис. 9.10, на котором в графическом
виде изображен характер изменения видового богатства афри-
канских сообществ в позднем кайнозое, разнообразие видов,
увеличивалось с плиоцена до нижнего плейстоцена, а затем
снижалось. Такая динамика была свойственна как общему
видовому разнообразию африканских сообществ, так и разно-
образию отдельных экологических и систематических групп
животных. Особый интерес вызывает сначала возрастание, а в-
•298
Глава 9
Рис. 9.10. Изменения видового разнообразия крупных африканских млекопи-
тающих в верхнем кайнозое (по Foley, 1984b; данные Maglio, 1978)
дальнейшем резкое снижение разнообразия видов, питавшихся
высококачественной пищей, и лишь незначительное уменьшение
на протяжении всего верхнего плейстоцена разнообразия тра-
воядных видов, потреблявших низкокачественную пищу. Разно-
образие хищных также быстро увеличивалось, а затем сильно
снижалось (этот факт также заслуживает внимания; см.
разд. 9.5.2).
Таким образом, можно утверждать, что африканские иско-
паемые находки свидетельствуют об увеличении и последую-
щем снижении видового разнообразия животных на протяже-
нии последнего миллиона лет, что соответствует теоретическим
предположениям, сделанным на основании разработанной мо-
дели эволюции сообщества в целом. Кроме того, адаптивная
радиация гоминид, при которой их видовое разнообразие дости-
гало максимума в интервале от 2 до 1 млн. лет назад, указы-
вает на то, что с теоретически ожидаемой картиной совпадает
и эволюция гоминид как всеядных организмов, потреблявших
высококачественную пищу и живших популяциями с небольшой
плотностью. Сказанное не означает, что эволюцию гоминид
можно было бы предсказать; это, очевидно, — гораздо более
сложная задача, связанная с другими факторами, рассмотрен-
ными в предыдущих главах настоящей книги. Тем не менее
можно предполагать, что гоминиды заняли именно те экологи-
ческие ниши, которые им «следовало» занять в процессе разви-
тия нового сообщества, и, заняв эти ниши, они стали составной
частью целостного сообщества африканской саванны.
Место, которое гоминиды занимают в этом сообществе, од-
нако зависит и от других специфических факторов, в частности
ют характера доступных источников пищи и присутствия или
Гоминиды и эволюция сообщества
299
отсутствия конкурентов. В этом можно убедиться, изучив эво-
люционные и экологические характеристики гоминид и их непо-
средственных конкурентов.
9.3.1. Коэволюция
Под эволюцией обычно подразумевают реципрокное дейст-
вие естественного отбора (Ehrlich, Raven, 1964). По сути дела
это означает, что, если какой-нибудь вид под влиянием давле-
ния отбора, обусловленного симпатрическим видом, подвергается
эволюционному изменению, последнее в свою очередь приводит
к ухудшению среды обитания второго вида и, естественно, уси-
лению на него давления отбора и его последующему эволюци-
онному изменению. В качестве классического примера можно
привести эволюцию у животных-жертв способности более эф-
фективно избегать хищников, что в свою очередь вызывает
усиление действия естественного отбора на хищников в смысле
повышения их способностей ловить жертв и т. д. Более общим
примером коэволюции может служить сосуществование парази-
тов и их хозяев и конкурирующих между собою животных. Ян-
зен (Janzen, 1980) предложил более узкое определение коэво-
люции, ограничив это понятие только теми эволюционными
изменениями, которые предполагают прямую и неизбежную
межвидовую зависимость; несомненно, однако, что с точки зре-
ния эволюционно-экологических перспектив более плодотворно
рассматривать механизмы эволюции животных в терминах дав-
ления отбора. Такой подход позволяет уподобить эволюцию
игре, целью которой является репродуктивный успех, а игроки
(особи и популяции) придерживаются разных стратегий. Как
и во всех «серьезных» сложных играх, стратегия данного игрока
определяется в первую очередь тем, что делают другие игроки.
Именно в таком смысле и используется в настоящей книге тер-
мин «коэволюция».
Рассмотрение эволюции гоминид в связи с другими, симпа-
трцческими с ними, видами животных следует начинать с ана-
лиза динамики численности наземных приматов. Как уже гово-
рилось, ранние гоминиды представляли собой лишь одну из
нескольких групп приматов, перешедших к наземному образу
жизни, и, поскольку экологические потребности всех этих жи-
вотных во многом совпадали, их симпатрическое сосуществова-
ние должно было, по всей вероятности, сопровождаться значи-
тельными взаимодействиями. Африканские саванны населяли
две основные группы наземных приматов — павианы и гомини-
ды. Другие группы приматов, например, мартышки из рода Сег-
copithecus, в прежние времена, возможно, вели более выражен-
ный наземный образ жизни.
330 Глава 9
Динамика численности и географического распространения
павианов и гоминид обнаруживает ряд интересных закономер-
ностей. Под численностью здесь и далее подразумевается числом
особей, принадлежащих к данному виду. Среди наземных при-
матов самым распространенным видом в начале плейстоцена
был Theropithecus oswaldi — гигантская гелада. В Восточной
Африке этот вид численно превосходил другую главную группу
павианов — род Papio (Jolly, 1972). В настоящее время карти-
на совершенно изменилась. Ныне представители рода Papio
встречаются в Восточной и Южной Африке повсеместно, а рас-
пространение Theropithecus, представленного лишь одним ви-
дом (Т. gelada), ограничивается только некоторыми районами
в эфиопских нагорьях.
Резко менялась и численность ископаемых гоминид (Howel,
1978). Если считать, что ископаемые материалы дают нам объ-
ективную и надежную информацию, то можно заключить, что
численность австралопитековых, по-видимому, почти в два раза
превосходила численность представителей рода Ното. Остатки
Australopithecus boisei в отложениях над слоем туфа KBS
в Кооби-Фора встречаются в два раза чаще, чем остатки Ното,
и такая же картина наблюдается в Формации Шунгура в Омо>
(Howell, 1978), Независимо от тафономических условий остат-
ки Australopithecus встречаются чаще, чем остатки Ното и в
пещерных местонахождениях в Трансваале (Brain, 1981). Са-
мым разительным примером может служить пачка I Сварткран-
са, где на три ископаемые особи Ното приходится 87 ископае-
мых особей Australopithecus robustus.
Столь высокую абсолютную и относительную численность
австралопитековых по крайней мере отчасти можно объяснить
их более массивным телосложением и, следовательно, лучшей
сохранностью после захоронения; вместе с тем это обстоятель-
ство удерживает нас от слишком поспешного заключения, что
представители Ното — предки современного человека — неиз-
бежно должны были превосходить австралопитековых конку-
рентоспособностью. Для объяснения изменчивой судьбы адап-
тивного успеха гоминид целесообразно было бы проследить их
адаптации к различным конкретным условиям. Особенно важны
здесь факты, связанные с питанием и морфологией зубов этих
организмов. Похоже, что как Australopithecus, так и Theropithe-
cus были более специализированными формами, чем родствен-
ные им таксоны, Ното и Papio. Представители ныне живущего
вида, Theropithecus gelada, питаются главным образом травой
и корневищами (Dunbar, 1977). Эта адаптация оказала значи-
тельное влияние на зубную морфологию гелады. Как показал
Джолли (Jolly, 1972) (см. гл. 4 и особенно рис. 4.3), передние
зубы этих животных сильно редуцированы, а задние — увеличе-
Гоминиды и эволюция сообщества
301
яы и уплощены. Кроме того, в соответствии со специализиро-
ванной наземной кормежкой у гелад претерпели некоторые
изменения адаптации, связанные с передвижением и принятием
позы (Jolly, 1972; Dunbar, 1977). Своей зубной морфологией
эти животные разительно отличаются от других павианов, по-
требляющих более разнообразные рационы с преобладанием
•фруктов. Такие же различия отмечаются и между Ното и Aust-
ralopithecus, в особенности, массивными австралопитековыми.
У последних задние зубы также были сильно увеличены, а пе-
редние редуцированы (см. рис. 8.14).
Все эти данные указывают на то, что Theropithecus и Aust-
ralopithecus являлись более специализированными родами, при-
чем особенно резко их специализация выражалась в отношении
потребления грубых мелких и низкокачественных пищевых
продуктов, широко встречающихся в саваннах. Как уже говори-
лось в гл. 8, такая специализация развивалась в ответ на дав-
ление отбора, связанного с сезонностью саванновых сред. На-
против, Ното и Papio — менее специализированные таксоны.
В плиоцене и самом начале плейстоцена более специализиро-
ванные таксоны превосходили численностью родственные им ме-
нее специализированные таксоны и, по всей видимости, дости-
гали наибольшего адаптивного успеха. Это обстоятельство
удовлетворительно вписывается в рассмотренную выше общую
модель эволюции сообщества (рис. 9.9).
Господство Australopithecus и Theropithecus совпадает с пе-
риодом максимального разнообразия видов. Их специализации
и, вероятно, сужение пищевых ниш эволюционировали как часть
-общей тенденции саваннового сообщества к более сильному
разделению экологических ниш. Эти виды подвергались такому
же давлению отбора, как и другие травоядные виды, и эволю-
ционировали в ответ на увеличение конкуренто- и жизнеспособ-
ности непрерывно расширявшейся фауны крупных млекопитаю-
щих. Благодаря своей специализации к потреблению такой
пищи, которая встречалась повсеместно и в изобилии, эти виды
превратились в обычный компонент фауны саванн. Напротив,
Ното и Papio не пошли по пути специализации, а заняли более
широкие и менее специализированные экологические ниши. Та-
кая iадаптация должна была значительно ограничивать числен-
ность этих видов, которые, по всей вероятности, жили популя-
циями с низкой плотностью и обширными территориями.
Связь двух линий развития, обнаруженных у ранних гоми-
нид— австралопитековых и рода Ното — с альтернативными
стратегиями выживания уже рассматривалась на предыдущих
страницах с подчеркиванием особой важности стратегии пище-
вого поиска для эволюционных процессов. Не менее важным
фактором, однако, является и степень специализации животных
302
Глава 9
независимо от характера пищи, которую они поедают. При том:
подходе, который занимает центральное место в этой главе
и основан на рассмотрении сообщества в целом, решающее зна-
чение имеет то обстоятельство, что тенденция к специализации
и сужению экологических ниш у гоминид находится в соответ-
ствии с такой же тенденцией у всех остальных крупных млеко-
питающих. Таким образом, у ранних гоминид вновь обнаружи-
ваются явления, совпадающие с теоретическими предсказания-
ми, сделанными на основании общих эволюционных и экологи-
ческих принципов. Хотя адаптивная стратегия гоминид специфи-
ческим образом определялась потребностями их предков и кон-
кретными особенностями их среды обитания, сравнение их
эволюции с характером эволюции сообщества в целом пред-
ставляется весьма плодотворным. Эволюция гоминид имеет
коэволюционную природу. Ранние гоминиды изменялись в ответ
па происходившие вокруг них сдвиги, и их изменения, безуслов-
но, в свою очередь влияли на эволюцию других, симпатриче-
ских им видов. Сказанное удалось продемонстрировать только’
для одной группы организмов — наземных приматов; возможно,
однако, что такой же характер эволюции свойствен и другим:
группам животных. Как бы то ни было, важно помнить, что-
эволюцию ранних гоминид до конца можно понять только на
фоне эволюции экологического сообщества в целом.
9.4. Образование и вымирание видов
Сказанное в этой и предыдущей главах позволяет заклю-
чить, что разнообразие ранних африканских гоминид могло оп-
ределяться особенностями их пищевого поведения и степенью-
их специализации. И то и другое следует рассматривать как
в контексте локальных условий обитания гоминид, так и в кон-
тексте эволюции всего саваннового сообщества в целом. А это-
вплотную подводит нас к другой области эволюционной эколо-
гии— области видообразования, изучающей, каким образом
происходит дивергенция популяций и образование новых, обо-
собленных, видов. По мнению Данбара (Dunbar, 1983), на при-
мере гоминид можно понять, почему перед разными популяция-
ми животных могли вставать разные экологические проблемы
и каким образом животные могли по-разному решать одни и те
же экологические проблемы. В свете представленных выше дан-
ных ждут своего решения следующие вопросы: как протекает
процесс видообразования, как часто видообразование имело»
место в ходе эволюции гоминид? И какое влияние этот основ-
ной процесс эволюции оказывает на взаимодействия между по-
пуляциями, т. е. на их сосуществование или вымирание?
Гоминиды н эволюция сообщества 303»
9.4.1. Типы видообразования
Отмечающееся ныне широкое разнообразие жизненных форм.'
является результатом дивергенции популяций. Если этот про-
цесс приводит к разделению генофонда и прекращению скрещи-
вания между популяциями (интербридинга), он означает видо-
образование. Касаясь разных видов гоминид, можно заклю-
чить, что они были генетически изолированы и преследовали
разные адаптивные стратегии. Прежде чем перейти к анализу-
дивергенции гоминид, следует подробнее рассмотреть современ-
ные представления о видообразовании.
Вопрос о том, каким образом образуются новые виды, в по-
следнее время широко обсуждался (см. Eldredge, Gould, 1972;;
Stanley, 1975; White, 1978; S. J. Gould, 1980; Maynard Smith,.
1982; Mayr, 1982). Видообразование разные авторы обыкновен-
но разделяют на анагенез и кладогенез. Анагенез предполагает
образование нового вида в результате изменения существующе-
го вида во времени. При этом отношения между старым и но-
вым видами имеют, как правило, характер связи между пред-
ком и потомком. Так, например, обычно считают, что Homo sa-
piens эволюционировал из Homo erectus в результате анагене-
за. Образовавшиеся таким путем виды в некотором смысле
являются «псевдовидами», так как образуются за счет постепен-
ной трансформации старых видов, и четкой границы между
ними провести нельзя. Напротив, кладогенез предполагает обра-
зование новых видов в результате разделения исходного вида
на несколько (чаще всего на две) отдельных линий. В качестве-
примера можно привести понгид — общих предков шимпанзе-
и горилл. Предполагается, что таким образом произошло и раз-
деление Australopithecus и Ното, которому ниже будет уде-
ляться основное внимание. По мнению некоторых авторов, вся-
кий процесс видообразования можно считать кладогенезом, ибо-
в тех случаях, когда увеличения числа эволюционных линий не
происходит, новые виды образуются только из какой-то части-
исходной предковой популяции (Eldredge, Cracraft, 1980).
Видообразование предполагает развитие репродуктивной изо-
ляции и развитие новых способов существования. Пожалуй, са-
мым простым и наиболее широко распространенным способом
возникновения репродуктивной изоляции является географиче-
ское разделение (аллопатрическое видообразование). Если меж-
ду популяциями длительное время существуют физические
барьеры, вероятнее всего, произойдет разделение этих популя-
ций— либо в результате дрейфа генов, либо вследствие адапта-
ции к различным локальным условиям. Когда барьеры исчеза-
ют и популяции вновь перекрываются (т. е. становятся симпат-
рическими), они уже настолько различаются, что успешный
304
Глава 9
Рис. 9.11. Аллопатрическое видообразование: образование новых видов как
результат географической изоляции. А. Длительное сосуществование дочер-
них видов после исчезновения географических барьеров. Б. Один вид вытесняет
другой
интербридинг становится невозможным. Следовательно, можно
говорить об образовании новых видов, которых в будущем ожи-
дает либо симпатрическое сосуществование, либо конкурентное
исключение (рис. 9.11).
Видообразование может произойти и в симпатрических по-
пуляциях. Предложено несколько механизмов такого процесса
(табл. 9.5). Все это — биологические механизмы, предотвращаю-
щие возможность появления фертильного потомства у опреде-
ленных особей популяции. Хотя теоретически эти меха-
низмы представляются- достаточно обоснованными, прове-
рить или наблюдать их действие в природе затруднительно.
Независимо от симпатрического или аллопатрического харак-
тера видообразования самым важным его аспектом, пожалуй,
является связь между двумя основными его компонентами —
новыми адаптациями и репродуктивной изоляцией. Согласно
классическим дарвинистским представлениям, репродуктивная
изоляция развивается в результате возникновения новых адап-
таций. Популяции начинают приспосабливаться к новым усло-
виям среды, изменяясь в тех или иных направлениях. Это при-
водит к образованию репродуктивных барьеров. Другими сло-
вами, адаптация предшествует репродуктивной изоляции.
И новые виды, стало быть, образуются постольку, поскольку
Гоминиды и эволюция сообщества 305
Таблица 9.5. Механизмы видообразования (факторы, приводящие
к репродуктивной изоляции) (по Ayala, Valentine, 1979)
Аллопатрическое видообразование
Происходит, когда популяции изолированы друг от друга географическими
барьерами (например, горами, реками и т. п.).
Симпатрическое видообразование
Репродуктивная изоляция может иметь место даже в том случае, когда попу-
ляции занимают одну и ту же территорию:
1. Презиготическая изоляция (предотвращающая образование гибридных зи-
гот):
экологическая изоляция (животные предпочитают разные места обита-
ния, что предотвращает их спаривание);
временная изоляция (спаривание происходит в разное время дня или
года);
этологическая изоляция (спаривание невозможно из-за поведенческих
различий между особями);
механическая изоляция (спаривание невозможно из-за анатомических
различий);
гаметическая изоляция (несмотря на спаривание, образование зиготы
невозможно).
2. Постзиготическая изоляция (гибриды на разных стадиях развития менее
жизнеспособны, чем остальное потомство).
появляются новые экологические ниши. В последнее время, од-
нако, различные авторы выдвинули предположение об обратной
последовательности событий в процессе видообразования. Спер-
ва происходит географическая или генетическая изоляция по-
пуляций, в результате чего они начинают эволюционировать
в разных направлениях, переходя к новым адаптивным страте-
гиям. Таким образом, репродуктивная изоляция предшествует
адаптации. В пользу этого предположения свидетельствует тот
факт, что в больших популяциях с непрерывным потоком генов
естественный отбор вряд ли может достигать настолько значи-
тельной силы, чтобы «толкнуть» виды на новые адаптации.
Говоря о репродуктивной изоляции небольших популяций, мож-
но предполагать, что сила естественного отбора в них окажется
значительной и новые адаптации будут формироваться доста-
точно быстро, чтобы избежать «растворения» в большом гено-
фонде.
Таким образом, образование новых видов, по-видимому,
включает как отбор в направлении новых адаптивных страте-
гий, так и возникновение адекватных демографических усло-
вий. Пожалуй, самый важный вывод, который можно сделать
из рассмотрения видообразования у гоминид, состоит в том,
что видообразование — не просто процесс изменений популяций
во времени и приобретения ими соответствующих анатомиче-
ских и поведенческих характеристик и что оно определяется не
20—367
306
Глава 9
только действующими силами естественного отбора, но что в-
значительной мере зависит и от пространственного распределе-
ния популяций. Другими словами, видообразование имеет
и пространственное измерение.
9.4.2. Видообразование у гоминид
Видообразование зависит от того или иного типа изоляции
и дивергентного локального давления естественного отбора.
В гл. 8 было высказано предположение, что дивергентные ло-
кальные условия существования гоминид обеспечивались раз-
личными типами экосистем; одни из них включали «рассредо-
точивающиеся и концентрирующиеся» системы крупных млеко-
питающих, а другие — «мигрирующие» системы тех же
животных. Важную роль могли играть и другие факторы—на-
пример, локальные особенности почвы, оказывающие огромное
влияние на характер растительности и, следовательно, доступ-
ность растительной пищи. Подобные локальные различия име-
ют эволюционное значение только в том случае, если они свя-
заны с более или менее выраженной изоляцией животных. Как
отмечалось выше, чаще всего это условие выполняется за счет
географической изоляции.
В этой связи интересно отметить, что на важнейших этапах
эволюции ранних гоминид восточноафриканские среды, по всей
вероятности, полностью удовлетворяли всем перечисленным
выше условиям. Во-первых, как уже говорилось, все большая
неустойчивость глобального климата в начале ледникового
периода должна была сопровождаться значительными экологи-
ческими колебаниями и «борьбой» между лесными и открыты-
ми биомами. В гл. 5 было показано, что изолированные остро-
ва саванны вполне могли обеспечивать все необходимые усло-
вия как для возникновения гоминид в целом, так и для их
последующего дробления на ряд родственных таксонов. Во-вто-
рых, в самой Восточной Африке образовывалась система Боль-
шой Рифтовой Долины и происходило расширение ее дна, что
сопровождалось высокой тектонической и вулканической актив-
ностью. Такие геологические процессы, как формирование реч-
ной сети, образование огромных сбросовых уступов и выброс
лавы и пепла, способствовали установлению более изолирован-
ных условий существования, чем в предшествующие времена,,
когда существовала геологическая и экологическая ста-
бильность.
Сказанное позволяет предположить, что популяции ранних
гоминид зачастую оказывались в изоляции и в соответствии
с обсуждавшимися выше экологическими механизмами по-раз-
Гоминиды и эволюция сообщества
307
Australopithecus Ното
Предковые
гоминиды
Рис. 9.12. «Классическая» модель видообразования у гоминид — простая ди-
вергенция на австралопитековых и представителей рода Ното — предков более
поздних гоминид
ному реагировали на локальные условия существования. Такой
подход позволяет понять видообразование у гоминид лучше
всего.
Дивергенция гоминид обыкновенно мыслится как «разделе-
ние на Ното и Australopithecus-». Главное событие, приведшее
к разделению этих двух основных линий, произошло на каком-
то отрезке времени в далеком прошлом, и последовавшее затем
развитие соответствовало довольно простой эволюционной схе-
ме (рис. 9.12). Более вероятным, однако, представляется, что
в результате совместного действия дивергентного отбора и гео-
графической изоляции на протяжении длительного времени по
всему ареалу обитания ранних гоминид в Африке образовыва-
лись популяции, адаптация которых к окружающим условиям
сильно различалась. Поскольку в разных частях материка, ве-
роятно, существовали среды с одинаковыми условиями, можно
думать, что и адаптивные тенденции, свойственные обитавшим
в них гоминидам, также были во многом похожими. Происхо-
дившие затем экологические сдвиги, возможно, нередко приво-
дили либо к локальному вымиранию популяций, либо к их слия-
нию, что означало стирание резких различий. Общим резуль-
татом этого процесса, по всей вероятности, было не простое эво-
люционное раздвоение, а радиация на многочисленные формы,
зачастую носившая параллельный характер в связи со сход-
ными экологическими условиями (рис. 9.13). Как отмечалось
выше, эволюция ранних гоминид представляет собой сложное
переплетение разных решений разных экологических проблем,
полярно противоположных решений одной и той же проблемы
и одинаковых решений сходных проблем существования. В до-
20*
308
Глава 9
Рис. 9.13. Модель радиации ранних гоминид в зависимости от условий окру-
жающей среды. Разделение гоминид произошло в результате усиления сезон-
ного характера среды и под влиянием ее локальных экологических условий,
что привело к образованию локальных популяций гоминид с параллельной
адаптацией
вольно скудных ископаемых находках отражены, по-видимому,
лишь отдельные эпизоды радиации гоминид.
Такой подход позволяет решить некоторые проблемы, свя-
занные с ископаемыми находками ранних гоминид. Новые на-
ходки, похоже, не только не упрощают, а наоборот, усложняют
существующие представления об эволюции гоминид, так как раз-
ные ископаемые образцы и скопления обнаруживают подчас
совершенно независимые характеристики. Этого как раз и сле-
дует ожидать в свете изложенных выше эволюционных и эколо-
гических соображений. Кроме того, возможно, не всегда «сра-
батывает» в случае ископаемых остатков и беспощадная «брит-
ва Оккама»1: «чем проще филогения, тем она лучше». Так, на-
пример, образцы из Хадара, быть может, относятся к несколь-
ким видам гоминид, и необязательно считать, что все эти виды
, 'Уильям Оккам (ок. 1285—1349)—английский средневековый философ.
Согласно принципу «бритвы Оккама», понятия, которые нельзя свести к ин-
туитивному и опытному знанию, необходимо удалять из науки. — Прим, пере в.
Гоминиды и эволюциясообщества
309
1,®
I
Н. erectus
A. boisei
Восточно-
африканские
A. robustus
1,5
5
« 2,0
а>
С
3,0 -
3,5-
Н. habilis
грацильные
гоминиды
A. africanus
Ветвь южноафриканских
австралопитековых
A. afarensis
Второй вид из Хадара
|(? ранние массивные
_ формы)
Рис. 9.14. Теоретическая модель адаптивной радиации ранних гоминид в Аф-
рике, предполагающая высокий уровень видообразования. Спорными аспекта-
ми модели являются существование второго вида гоминид 3 млн. лет назад
и ранее и разделение восточноафриканских и южноафриканских форм на раз-
ные ветви
находятся в родстве с более поздними Ното или Australopithe-
cus. В отдельных случаях, возможно, массивные формы эволю-
ционировали независимо. Аналогичным образом восточно- и юж-
ноафриканские формы гоминид, возможно, не представляют со-
бой единой; под давлением одинаковых факторов естественного
отбора они могли эволюционировать параллельно. В заключение
хочется отметить, что разнообразие ранних форм Ното в Вос-
точной Африке может указывать на то, что процесс, аналогич-
ный описанному выше, происходил и внутри этого рода.
Предположение, что ранние гоминиды подвергались парал-
лельной эволюции, можно привлечь для объяснения их филоге-
нии. Массивных австралопитековых обычно рассматривают как
единую ветвь гоминид (см. гл. 2, и особенно рис. 2.11), хотя
признается существование двух аллопатрических видов этих
приматов. Такая картина, однако, представляется слишком
упрощенной. Если массивное телосложение возникло как реак-
ция на определенные условия окружающей среды, то Australo-
pithecus boisei может иметь более тесные родственные связи
с другими восточноафриканскими гоминидами, в том числе и с
родом Ното, нежели с южноафриканским Australopithecus ro-
bustus (рис. 9.14). Кроме того, недавняя находка в местности
к западу от озера Туркана (Кения) гоминида массивного тело-
310
Глава 9
сложения, жившего, по-видимому, 2,5 млн. лет назад, что почти
на 0,5 млн. лет превышает возраст других известных восточно-
африканских массивных австралопитековых (Walker, Leakey et
al., 1986), а также предположение, высказанное Олсоном (Ol-
son, 1981), что ископаемые находки из Хадара возрастом
3 млн. лет включают остатки массивного австралопитека, гово-
рят о возможности более ранней и независимой эволюции адап-
таций, связанных с массивным телосложением. Это означает,
что филогения гоминид представляла собой гораздо более
сложную картину, чем обыкновенно считают (рис. 9.14). Фило-
гения, признающая возможность дивергенции в прошлом и ро-
да Ното, потребует пересмотра систематики и номенклатуры
гоминид, не исключающего и восстановление рода Zinjanthropus.
Сказанное проливает определенный свет на происхождение
разнообразия гоминид. За счет чего, однако, поддерживалось
это разнообразие и не только поддерживалось, но и усилива-
лось, что заключалось, например, в усугублении различий меж-
ду представителями Ното и австралопитековыми? Как отмеча-
лось в гл. 8, одним из основных механизмов поддержания раз-
личий между популяциями, а в конечном счете и между видами
является стремление избежать конкуренции. Если в результате
расширения популяций, обусловленного адаптивным успехом,
устанавливается контакт между двумя популяциями с разной
степенью специализации, обе они могут получить дополнитель-
ные преимущества за счет избежания конкуренции и перекры-
вания экологических ниш. Это приводит к дальнейшему измене-
нию признаков, причем в обеих популяциях отбор будет способ-
ствовать выживанию тех особей, которые наиболее сильно
отличаются от особей другой популяции, что еще больше уве-
личит морфологическую и адаптивную дивергенцию. Если на
ранних этапах дифференциацию популяций на новые виды
может облегчать их географическое разделение, то в дальней-
шем дивергенция может усиливаться за счет симпатрии.
Еще одним механизмом, усиливающим различия между по-
пуляциями, является отбор, связанный с частотой адаптации, ко-
торый происходит в том случае, когда преимущество той или
иной адаптивной стратегии зависит от частоты возникновения
данной адаптации. Чем реже возникает адаптация, тем больше
преимуществ имеет адаптивная стратегия. В результате отбора
преимущественной стратегии частота адаптации увеличивается,
что неизбежно приводит к уменьшению преимуществ данной
адаптивной стратегии и в конечном итоге — к переходу преиму-
ществ к исходной адаптации, которая становится все более ред-
кой. Иногда между частотой адаптаций устанавливается равно-
весие; в других случаях частота адаптаций колеблется. В ре-
зультате в одной и той же экосистеме могут сосуществовать
Гоминиды и эволюция сообщества
311
альтернативные адаптации. Так упрочение доминирующего по-
ложения австралопитековых в экосистеме сопровождалось обо-
стрением их конкурентных взаимоотношений друг с другом.
В такой ситуации преимущества должны были получать попу-
ляции Ното, что соответственно сопровождалось и увеличением
их частоты. В некоторый момент времени, однако, когда пита-
тельные ресурсы, использовавшиеся Ното, начали истощаться,
равновесие в экосистеме вновь нарушилось в пользу австрало-
питековых. Таков возможный способ поддержания видового раз-
нообразия в экосистемах.
Итак, образование новых видов гоминид явилось результа-
том двух альтернативных путей развития, основанных на раз-
ных формах пищевого поиска. Выбор того или иного альтерна-
тивного пути зависел от локальных условий, но коль скоро путь
этот определялся, конкуренция, изменение признаков, отбор,
связанный с частотой адаптации, и филогения надежно обеспе-
чивали его сохранение и совершенствование. Таким образом,
можно утверждать, что эволюция гоминид, хотя она и не была,
как первоначально предполагалось, полностью поступательным
процессом с одной линией развития, во всяком случае носила
характер истинной адаптивной радиации.
9.4.3. Пространственное распределение гоминид
Сказанное выше свидетельствует о важной роли в эволюци-
онных процессах пространственного распределения популяций.
К сожалению, в ископаемых находках очень неполно отражены
повторяющиеся особенности локальной изменчивости гоминид,
составляющие суть их адаптивной радиации и представляющие
значительный интерес для понимания всей их эволюции. С дру-
гой стороны, большой интерес представляет и пространственная
изменчивость гоминид, позволяющая проследить характер их
эволюции в масштабе всего африканского континента.
Допустим, что видообразование является по преимуществу
локальным событием. Виды эволюционируют не в масштабе все-
го континента в целом; новые адаптации возникают локально,
а широкое расселение их носителей происходит уже после этого.
В процессе такого расселения особи могут подвергаться воздей-
ствию новых факторов естественного отбора, и, таким образом,
их географическое распространение будет сопровождаться эво-
люционными изменениями. Этот процесс и находит отражение
в «пространственных» особенностях ископаемых остатков го-
мцнид.
На рис. 9.15 изображено изменение географического распро-
странения гоминид на протяжении четырех млн. лет. Следует
сразу оговориться, что ископаемый материал, использованный
312
Глава 9
при составлении рисунка, фактически получен из двух афри-
канских регионов — Трансвааля (Южная Африка) и Большой
Рифтовой долины (Восточная Африка), поэтому предлагаемая
реконструкция во многом основана на экстраполяции.
Предполагается, что гоминиды появились где-то в Восточной
Африке не позднее 6 млн. лет назад. Однако, учитывая общее
географическое распространение африканских гоминоидов, мож-
человекообразные
обезьяны
Популяции раннего
примитивного
Australopithecus
afarensis
A. afarensis
. Африканские
I человекообразные
обезьяны
Африканские
3—2 млн. лет
A. africanus
из юге
Рассеянные популяции
ранних массивных
ауст ралопитековых
Расселение
A afarensis
у в Южную Африку и
трансформация в
Australopithecus
africanus
Г рацильные
на востоке
Ранние
массивные
формы
на востоке
Гоминиды и эволюция сообщества 313
но думать, что центр их расселения располагался ближе к запа-
ду. В промежутке между 5 и 3 млн. лет назад гоминиды, по
всей вероятности, были относительно однородной группой, а их
распространение ограничивалось восточной частью африканско-
го континента. В интервале от 3 до 2 млн. лет назад в жизни
гоминид произошла одна из значительных перемен. Они распро-
странились на юг. Можно предположить, что во время этого
расселения «родоначальная» форма гоминид, грацильный Aust-
ralopithecus afarensis, претерпела эволюционные изменения, ко-
торые привели к возникновению Australopithecus africanus
в южной части материка, а возможно, и похожих на него гра-
цильных форм на востоке (представленных ископаемым мате-
риалом из Омо и образцами KNM-ER 1805 и 1813 из Кооби-
Фора).
Около 2 млн. лет назад произошла еще одна крупная пере-
мена. Восточноафриканские гоминиды разделились на две вет-
ви, которым удалось не только выжить, но и преуспеть в эволю-
ционном отношении: одна из них продолжала развитие австра-
лопитековых, вторая положила начало эволюции Ното habilis.
Этот последний вид расселился затем на юг, где он изредка
встречается в виде ископаемых остатков. Грацильные австра-
лопитековые в Южной Африке также претерпели эволюцион-
ные изменения, на сей раз в сторону большей массивности.
Около 2 млн. лет назад доминирующей формой гоминид был
Australopithecus robustus. Менее ясно, однако, произошло ли за-
тем расселение массивных форм австралопитековых на север
или же имела место параллельная эволюция грацильных авст-
ралопитековых в Восточной Африке.
Около 1,5 млн. лет назад в Африке произошло еще одно эво-
люционное событие — появление Homo erectus. Этот вид быстро
расселился не только по всему африканскому континенту, но и
по всему тропическому поясу Старого Света, положив тем са-
мым конец периоду разнообразия гоминидных форм, ибо ко вре-
Рис. 9.15. Крупномасштабные изменения географического распространения
и ход эволюции ранних гоминид в Африке. Как видно из рисунка, эволюцион-
ный процесс зависит и от пространственных факторов, и его нельзя сводить
к простым филогенетическим схемам. А. Дифференциация африканских гоми-
иоидов и возникновение гоминид. Б. Образование популяций гоминид (Austra-
lopithecus afarensis) в Восточной Африке. В. Дифференциация древнейших
гоминид — появление первых популяций со специализациями, свойственными
массивному телосложению. Г. Расселение гоминид в Южную Африку и эволю-
ция Australopithecus africanus. Д. Появление Ното в Восточной Африке и его
частичное расселение в Южную Африку. Появление в Восточной и Южной
Африке популяций массивных австралопнтековых. Е. Эволюция и расселение
Homo erectus и выживание популяций поздних массивных австралопитековых.
#14
Глава 9
мени расцвета Homo erectus (1 млн. лет назад) все австралопи-
тековые вымерли.
Следует отметить, что приведенное описание рассеяния гоми-
нид в Африке в ходе их эволюции имеет предположительный
характер и никоим образом не может считаться окончатель-
ным. Эта схема отличается и более консервативным характером
по сравнению с рассмотренным в предыдущем разделе широ-
ким, многовариантным видообразованием у гоминид. Для объ-
яснения полученных к настоящему времени данных можно
предположить и другие модели, однако необходимо подчерк-
нуть, что данные, полученные при изучении географического
распространения ископаемых остатков гоминид, слишком отры-
вочны для того, чтобы можно было провести основательный
анализ этого вопроса. Тем не менее, хотя частные его аспекты
наверняка еще будут уточняться, незыблемым остается одно:
эволюционный процесс и эволюционные события имеют не толь-
ко временное, но и пространственное измерение, и это обстоя-
тельство должно учитываться при разработке эволюционных
моделей, которые обычно ограничиваются вырванными из про-
странства «генеалогическими схемами».
9.4.4. Вымирание
Другим, помимо видообразования, основным эволюционным
процессом является вымирание видов. Какими факторами бы-
ло обусловлено вымирание австралопитековых или какого-либо
другого вида гоминид? Согласно классическим представлениям,
австралопитековые вымерли, не выдержав конкуренции с более
жизнеспособным (более смышленым) родом Ното. Это, однако,
неполный ответ. Сам факт вымирания массивных австралопи-
тековых из-за неспособности выдержать конкуренцию не вызы-
вает сомнений. В конце концов, это — закономерность, прису-
щая вымиранию видов вообще. Совершенно необязательно, что-
бы виновниками гибели австралопитековых были другие гоми-
ниды. Это могли быть совсем иные животные. Учитывая только
что рассмотренную модель дивергенции, можно думать, что на
поздних стадиях развития два типа гоминид экологически пе-
рекрывались довольно незначительно. В самом деле, стремление
избежать конкуренции должно было «отталкивать» их друг от
друга. В результате такого отталкивания, однако, каждый из
обоих типов гоминид с необходимостью должен был включаться
во все обострявшуюся конкуренцию с другими видами, не от-
носившимися ни к гоминидам, ни вообще к приматам. В слу-
чае Homo erectus такими видами могли быть плодоядные, все-
ядные и крупные хищники. Уолкер (Walker, 1984) предположил,
что к тому времени Н. erectus присоединился к экологической
Гоминиды и эволюция сообщества
315
группе крупных хищников, что вызвало в этой группе значи-
тельные эволюционные изменения. Безусловно, в то время уже
существовали и эволюционировали различные крупные виды
отряда хищных млекопитающих.
Напротив, Australopithecus robustus и A. boisei, по всей ве-
роятности, конкурировали с травоядными. Травоядные в то вре-
мя сами подвергались быстрым эволюционным изменениям,
сопровождавшимся увеличением их видового разнообразия.
Не исключено, что именно из-за постоянно возраставших жизне-
способности, численности и видового разнообразия травоядных
как австралопитековые, так и теропитековые подверглись зна-
чительным экологическим ограничениям и в конце концов вы-
мерли. Проблемы, с которыми столкнулись и которые так и не
разрешили австралопитековые, сводились к тому, что эти гоми-
ниды оказались зажатыми между более жизнеспособными и бо-
лее специализированными травоядными, с одной стороны, и бо-
лее жизнеопосо1бными всеядными — с другой. Их эволюционный
выбор был крайне ограниченным.
Обращаясь к общей модели эволюции сообщества, представ-
ленной на рис. 9.9, можно утверждать, что около 1 млн. лет
назад наступила третья стадия развития саваннового сообще-
ства-стадия увеличения скорости вымирания видов. Развитие
структуры сообщества за счет разделения экологических ниш
привела к такому их сильному сужению, что специализация
к питанию травой в саванновых средах стала для приматов
невозможной. Соответственно, как видно из рис. 9.10, специали-
зированные наземные приматы терпят крах, и выживают из них
лишь самые всеядные виды.
Если Homo erectus и впрямь как-то способствовал вымира-
нию других наземных приматов, скорее всего он выступал при
этом в роли хищника, <а не прямого их конкурента. Так, в сред-
неплейстоценовом местонахождении Олоргесаилли .(Кения) ос-
татки пятидесяти особей Theropithecus найдены вместе с
ашелыскими1 ручными топорами; поэтому Шипмэн (Shipman
1982) и ряд других авторов высказали предположение, что в их
гибели прямо повинны гоминиды. Однако, по мнению Бинфорда
и Тодда (Binford, Todd, 1982), нет никаких оснований думать,
что деятельность гоминид не ограничивалась простым перетас-
киванием в это место туш уже погибших гелад.
И возникновение австралопитековых как новых видов, и их
эволюционная история, и вымирание должны рассматриваться
на фоне экологического сообщества в целом. Изучение гоминид
в отрыве от других, симпатрически сосуществовавших с ними
1 Ашель — одна из эпох раннего палеолита, названная по месту эо Фран-
ции, где были найдены типичные для нее орудия. — Прим, перев.
316
Глава 9
видов неизбежно приводит к переоценке их экологических
взаимодействий и более узкой и антропоцентрической интерпре-
тации их эволюции.
9.5. Темпы эволюции
В этой книге была сделана попытка проследить связь между
характеристиками гоминид и эволюционной и экологической
средой, в которой они вынуждены были бороться за существова-
ние. Эта связь может быть документирована фактическим мате-
риалом. До сих пор, однако, представления о ходе эволюции
гоминид во времени довольно неопределенны. Основное внима-
ние исследователи уделяли соответствию отдельных эпизодов
и характеристик общим принципам эволюции, а не последова-
тельности эволюционных событий во времени. И совсем не изу-
чался вопрос о том, носили ли описанные выше эволюционные
изменения постепенный и непрерывный характер или же то был
более скачкообразный и прерывистый процесс. Попытки бо-
лее точно определить временную последовательность событий
в эволюции человека позволяют понять, насколько задержива-
лось решение возникавших перед гоминидами адаптивных
проблем, и можно ли говорить о том, что гоминиды всегда на-
ходили оптимальные решения этих проблем. Вот почему так
важно рассмотреть темпы эволюции гоминид.
9.5.1. Проблема скорости эволюционного процесса
Последние несколько лет вопрос о темпах эволюции обсуж-
дался особенно остро (Eldredge, Gould, 1972; Stanley, 1979;
S. J. Gould, 1980). Ископаемые остатки ранних гоминид исполь-
зовались как для подтверждения (Gould, 1977), так и для опро-
вержения (Cronin, Boaz et al., 1981) представлений об эволю-
ции как прерывистом равновесии. Эти споры, подогреваемые
главным образом палеонтологами, сосредоточились на вопросе
о том, происходила ли эволюция постепенно (а может быть, и с
постоянной скоростью) или же она отмечена периодами быстрых
изменений, сменяющимися продолжительными периодами за-
стоя. Свидетельства в пользу прерывисто-равновесной модели
эволюционных изменений использовались как доказательства
того, что характер эволюции определяется не естественным от-
бором, а другими механизмами (Eldredge, Gould, 1972; Stanley,
1979; S. J. Gould, 1980; Vrba, 1980).
Мейнард Смит (Maynard Smith, 1982) предположил, что раз-
вернувшиеся споры фактически можно свести к основным и про-
изводным вопросам. Вопрос о формах эволюции является основ-
ным, а вопрос о ее темпах—производным. Согласно классичес-
Гоминиды и эволюция сообщества
317
ким дарвинистским представлениям, эволюция действует на
уровне отдельной особи. Такой формой эволюции можно объяс-
нить любые эволюционные изменения, ъключая и образование
новых видов. Соответственно, поскольку адаптация и отбор тес-
но взаимосвязаны, видообразование является одной из сторон
адаптивных тенденций данной ветви.
В настоящее время, однако, эта точка зрения оспаривается.
Ряд авторов (Stanley, 1975, 1979; Eldredge, Cracraft, 1980;
С. J. Gould, 1980; Vrba, 1980) считают, что механизмы видооб-
разования отличны от механизмов, действующих на уровне от-
дельных особей. Новые виды возникают (как правило, быстро)
•в небольших изолированных популяциях в результате изоляции,,
а появляющиеся при этом новые адаптации отличаются по на-
правлению от адаптивных тенденций родительских популяций.
Таким образом, образование новых видов не связано с какой-
либо направленностью эволюционных изменений в предковых
популяциях. Поскольку этот феномен предполагает разный ха-
рактер внутривидовых изменений (за счет естественного отбора)
и межвидовых изменений (отбор видов), его нередко называют
«расщеплением» механизмов эволюции.
Выше уже было высказано предположение, что небольшие изо-
лированные популяции, по всей видимости, играют важную роль
в видообразовании (см. разд. 9.4.1), но это не означает, что
данная функция популяций обязательно независима от общих
адаптивных тенденций. Дрейф генов — не единственный про-
цесс, действующий на небольшие популяции. Высокого уровня
•способен достигать и естественный отбор, причем факторы его
могут быть такими же, как и факторы, действующие на круп-
ные популяции, а небольшие размеры генофонда позволяют мел-
ким популяциям модифицироваться достаточно быстро. Высо-
кая скорость эволюции небольших популяций, в которых обра-
зуются новые виды, совсем не означает, что естественный отбор
не является главным механизмом эволюционных изменений.
Предположение о «расщеплении» механизмов видообразова-
ния, связанного с микроэволюционными процессами, находит
главное подтверждение в скорости эволюции. Анализ ископае-
мых моллюсков из донных отложений озера Туркана позволил
Уильямсону (Williamson, 1981) предположить, что длитель-
ные периоды застоя (стабилизации) видов нельзя объяснить
•естественным отбором и что, следовательно, движущей силой
эволюции являются какие-то другие, нежели естественный от-
бор, механизмы. Таковы лишь некоторые из общих соображений
о факторах, определяющих скорость (темпы) эволюционного
процесса.
Считается, что скорость эволюции определяют три фактора;
споры между исследователями возникают по поводу значения
318
Глава 9
+
того или иного фактора для темпов эволюционного процесса.
Одним из этих факторов является интенсивность и направление
отбора; если интенсивность отбора невысока или если он имеет
сильно выраженный стабилизирующий характер, скорость эво-
люционных изменений будет низкой. Если новые признаки бла-
гоприятны или возникают в период экологических сдвигов, ско-
рость эволюции будет высокой. Согласно классическому дарви-
низму, скорость эволюции определяется главным образом ин-
тенсивностью отбора, чем, однако, нельзя объяснить постоянст-
во или низкую величину скорости эволюционного процесса.
Другими факторами скорости эволюции являются частота и ха-
рактер мутаций, а также размер популяции и характер ее вос-
произведения. По мнению Гулда и др., если популяции, удов-
летворяющие всем этим условиям, подвергаются быстрым эво-
люционным изменениям, то значение естественного отбора
уменьшается.
9.5.2. Темпы эволюции гоминид
Какими же были темпы эволюции гоминид? Этот вопрос изу-
чался разными авторами. По мнению Гулда (Gould, 1977), ни
у одного из ныне известных видов ранних гоминид никаких эво-
люционных изменений не обнаруживается, и, следовательно,,
эволюция человека целиком отвечает модели прерывистого рав-
новесия. Эти представления развивали также Элдредж и Тэт-
терсолл (Eldredge, Tattersall, 1982), которые по ископаемым
остаткам гоминид выявили в эволюции человека периоды быст-
рых изменений и стабилизации: между Australopithecus afaren-
sis, A. boisei, A. robustus и Homo erectus (все они на протяже-
нии приблизительно 1 млн. лет практически не изменялись)
существуют резкие морфологические разрывы. Райтмайер
(Rightmire, 1981) произвел количественную оценку периода ста-
билизации Homo erectus, однако Уолпофф (Wolpoff, 1984) под-
верг его данные сомнению.
Подобный критический анализ модели прерывистого равно-
весия для эволюции гоминид провели Кронин, Боаз и др. (Cro-
nin, Boaz et al., 1981). В отличие от Гулда эти авторы полага-
ют, что ископаемые остатки очень неполны и что морфологиче-
ские разрывы между видами связаны с отсутствием соответст-
вующих геологических отложений. Эти исследователи считают
также, что им удалось выявить в эволюции гоминид некоторые
устойчивые тенденции. Примером одной из таких тенденций
является плавное изменение некоторых анатомических призна-
ков при переходе от Australopithecus afarensis к A. africanus,
примером другой—промежуточное положение образца
KNM-ER 1470 между родами Ното и Australopithecus (Walker,.
Гоминиды и эволюция сообщества 319
1976); еще один случай иллюстрирует другой образец из
Кооби-Фора, KNM.-ER 1813, который рассматривается как пере-
ходная форма. Но, пожалуй, наиболее изученный образец зна-
чительных и постепенных эволюционных сдвигов у гоминид
представляет собой переход от Homo erectus к Homo sapiens.
Хотя по поводу терминологии в этом случае и возникали неко-
торые споры (что, возможно, уже само по себе свидетельствует
о постепенности этого перехода!), европейские и африканские
находки, относящиеся к среднему плейстоцену, обнаруживают
непрерывный ряд переходных форм между этими двумя хроно-
видами. Кронин, Боаз и др. (Cronin, Boaz et al., 1981) допуска-
ют возможность отдельных прерывистых событий в эволюции,
в частности появление массивных австралопитековых, хотя, как
показал Алан Билсборо (Bilsborough, личное сообщение),
в Южной Африке между грацильными и массивными австрало-
питековыми обнаруживается ряд переходных форм.
Отсутствие четких морфологических критериев и стратигра-
фическая путаница не позволяют сделать каких-либо определен-
ных выводов относительно темпов изменений в пределах эволю-
ционной ветви гоминид. По всей видимости, можно все-таки
заключить, что скорость эволюции гоминид не была постоянной.
Со временем она менялась, причем на протяжении одного и то-
го же периода разные характеристики гоминид эволюциониро-
вали с разной скоростью. Сам по себе факт изменяющейся ско-
рости эволюционного процесса не отрицает дарвиновской эво-
люции и скорее порождает вопрос о возможной связи между
непостоянными темпами эволюции гоминид, с одной стороны,
и характером естественного отбора и конкретными экологиче-
скими условиями, в которых жили ранние гоминиды, с другой.
Можно выделить три периода сравнительно быстрых эволю-
ционных изменений у гоминид: появление массивных австрало-
питековых, появление Homo erectus и появление анатомически
современного человека. Эти события не были «мгновенными»,
но они развивались быстрее, чем предшествовавшие стадии эво-
люции. Всем этим событиям могла предшествовать еще одна
стадия, отмеченная появлением первых прямоходящих гоминид,
однако из-за скудности ископаемого материала проверить это
предположение нельзя.
Первое из названных событий произошло 2 млн. лет назад,
второе — около 1,6 млн. лет назад и третье — в интервале меж-
ду 0,1 и 0,05 млн. лет назад. В соответствии с представленны-
ми в настоящей главе моделями можно думать, что эти события
связаны с периодами острой конкуренции и эволюционной ак-
тивности других животных. К сожалению, в связи с недостат-
ком хронологически достоверного ископаемого материала про-
верить это предположение трудно. Как видно из рис. 9.16, плио-
320
Глава 9
100 %
Период быстрого
эволюционного
изменения гоминид
Рис. 9.16. Изменение фауны крупных млекопитающих в Африке в верхнем кай-
новое (по Maglio, 1978)
цен — период появления массивных австралопитековых — харак-
теризовался очень высокой скоростью обновления фауны (при-
близительно 75% на уровне родов), в то время как уровень
эндемизма оставался постоянным, а скорость вымирания видов
снижалась. Все это указывает на то, что плиоцен в целом был
временем эволюционных изменений. В пользу этого свидетельст-
вует и анализ характера эволюции наиболее изученных групп
ископаемых животных. Хорошо изученные свиньи и слоны
(White, Harris, 1977; Harris, 1983) подвергались в эту эпоху
сильным эволюционным изменениям. Следовательно, появление
массивных австралопитековых совпало со становлением многих
других видов. По мнению Врба (Vrba, 1985), утверждающего,
что развитие эволюционных изменений гоминид и полорогих в
Южной Африке было тесно связано, этот период быстрых эво-
люционных изменений определялся происходившими в то время
сильными климатическими сдвигами и изменением африканских
сред. Как свидетельствуют глубоководные морские отложения,
Гоминиды и эволюция сообщества
321
в интервале между 2,5 и 2,0 млн. лет назад отмечалось одно из
глобальных понижений температуры (Thunell, Williams, 1983)..
Очевидно, прерывистые эволюционные изменения фактически
можно рассматривать как реакцию организмов на новые и ин-
тенсивно воздействовавшие факторы отбора, обусловленные
экологическими изменениями и для гоминид опосредованные
эволюционными изменениями сообщества африканской саванны.
Труднее проследить изменение фауны в плейстоцене, так как
соответствующие отрезки времени здесь гораздо короче. Тем не
менее установлено, что около 1,6 млн. лет назад в Олдувайском
ущелье произошло сильное изменение фауны, а именно основа-
тельное обновление ее видового состава. Это событие совпало-
с появлением Homo erectus. По мнениюУолкера (Walker, 1984),
з этот период произошли значительные сдвиги в системах «хищ-
ник— жертва» крупных млекопитающих, сопровождавшиеся'
«упадком» древних крупных травоядных и охотившихся на них
хищников. Можно думать, что под влиянием экологического
давления, которое рассматривалось в гл. 7, гоминиды «примкну-
ли» к группе крупных хищников во время эволюционной и эко-
логической неустойчивости и что быстрые эволюционные изме-
нения, происходившие внутри рода Ното, были лишь одной из
сторон этого процесса.
Период, последовавший за появлением Homo erectus, был
временем относительной морфологической стабильности. В это
время началось, однако, и расселение гоминид за пределы Аф-
рики. Homo erectus был конечным продуктом эволюционной
и экологической динамики африканских сред обитания в плио-
цене и нижнем плейстоцене. Решения адаптивных проблем,
с которыми сталкивались ранние гоминиды и которые в конеч-
ном итоге обусловили появление Homo erectus, создали основу
для быстрого изменения географического распространения го-
минид, что поставило их перед лицом новых экологических
проблем. Решение этих новых проблем в свою очередь явилось
основой адаптивных, поведенческих и эволюционных изменений,
которые произошли в плейстоцене позднее. Выяснив, насколько
тесно связана эволюция ранних гоминид с эволюционной дина-
микой африканской саванны, остается кратко рассмотреть неко-
торые последствия перехода гоминид в новые среды обитания.
21—376
Глава 10
ГОМИНИДЫ КАК РАССЕЛЯЮЩИЕСЯ
ЖИВОТНЫЕ
Настоящая книга посвящена главным образом ранним го-
минидам. Согласно современным представлениям, эволюция
гоминид, во всяком случае на самом раннем и, пожалуй, наи-
более «продуктивном» этапе, протекала в Африке. Как отмеча-
лось в предыдущих главах, гоминиды ближе всего стоят к аф-
риканским человекообразным обезьянам, а, кроме того, по мно-
гим характеристикам могут быть связаны с экологической дина-
микой африканских сред. Следует, однако, помнить, что на этом
этапе эволюции гоминид сложилась основа того, что составляет
не менее важный аспект их эволюционной экологии — освоение
гоминидами всех основных типов сред на земном шаре и их
расселение по всем континентам. В этой главе в свете некоторых
общих принципов эволюционной экологии кратко рассматрива-
ются различные аспекты географического распространения го-
минид.
Теоретической предпосылкой представленных ниже рассуж-
дений является положение, что распространение всех животных
в большей или меньшей степени ограничивается определенными
местами обитания. В этих местах обитания животные находят
себе пищу и убежища, в то время как другие среды — вследст-
вие физических барьеров или конкуренции — они эффективно
использовать не могут. Существуют, однако, некоторые общие
принципы, определяющие, в какой степени тот или иной вид
может быть стенотопным (т. е. привязанным к определенному
местообитанию) или эвритопным (т. е. способным существовать
в разнообразных условиях среды). Похоже, что на ранних эта-
пах эволюции гоминиды были привязаны к определенным афри-
канским биотопам, однако на протяжении плейстоцена эта при-
вязанность ослабевала. Возникают важные вопросы: что сдела-
ло возможным расселение гоминид и каковы были экологичес-
кие последствия этого расселения?
10.1. Исход из Африки
Нужно признать, что из-за обрывочности исследований, та-
фономических факторов и отсутствия надежных методов дати-
Гоминиды как расселяющиеся животные 32Э
ровкй критических периодов среднего и верхнего плейстоцена
ход расселения гоминид из Африки изучен плохо, хотя некото-
рые обобщения по этой проблеме сделать все-таки можно.
10.1.1. Последовательность расселения
1. Нет каких-либо надежных свидетельств появления гоми-
нид за пределами африканских регионов, расположенных к югу
и юго-востоку от Сахары, ранее чем 1,5 млн. лет назад.
2. Есть свидетельства присутствия гоминид в африканских
регионах к югу от Сахары и в Юго-Восточной Азии в промежут-
ке между 1,5 и 1,0 млн. лет назад. На основании этого можно
предположить также, что к этому времени гоминиды колонизи-
ровали и промежуточные тропические регионы Старого Света,
включая части Среднего Востока и полуостров Индостан. Это
предположение, однако, нуждается в дополнительных доказа-
тельствах, хотя и есть указания на пребывание гоминид в место-
нахождении Убейдия в Израиле с очень древней датировкой
(около 2 млн. лет) (Repenning, Fejfar, 1982). Равным образом
подверглись сомнениям (Matrsu’ura, 1982; Pope, 1983; Sartono,
1982) и сделанные ранее предположения о присутствии остатков
гоминид в отложениях Джетис и ископаемой фауне Явы, да-
тируемых 1,9 млн. лет назад или меньше (Curtis, 1981). В на-
стоящее время представляется наиболее вероятным, что древ-
нейшие следы пребывания гоминид за пределами Африки не
старше 1,5 млн. лет.
3. Считается, что освоение гоминидами теплых умеренных
зон, включая и средиземноморские области Европы, началось
в промежутке между 1,0 и 0,7 млн. лет. Хотя и высказывались
многочисленные предположения о более раннем освоении Евро-
пы (см., например, Turner, 1984, р. 198), древнейшие из извест-
ных находок, обнаруженные в Изерния-ла-Пирета (Италия),
датируются возрастом 1,0 млн. лет (Coltorti, Cremacshi et al.,
1982). В общем можно сказать, что колонизация гоминидами
теплых умеренных зон Старого Света произошла на границе
нижнего и среднего плейстоцена.
4. В интервале между 700 тыс. и 300 тыс. лет назад Ното
erectus колонизировал более северные умеренные среды.
5. В промежутке между 300 тыс. и 40 тыс. лет назад гомини-
ды включили в свой ареал обитания северные широты (хотя, по
всей вероятности, области с резко выраженным арктическим
климатом остались ими не охваченными); этот период совпал
с эволюцией Homo erectus в Н. sapiens.
6. В интервале между 40 тыс. и 10 тыс. лет назад гомини-
дами были колонизированы практически все остальные области
земного шара. Ко времени резкого похолодания были освоены
21
324
Глава 10
и зоны с резким арктическим климатом. Благодаря этому го-
миниды проникли в Новый Свет и очень быстро расселились по
всей его территории; в это время или, быть может, чуть'раньше
гоминиды достигли Австралии и Океании.
7. 10 тыс. лет назад незанятыми гоминидами оставались
лишь полярные зоны и некоторые из отдаленных островов. Эти
территории постепенно осваивались в более поздние доистори-
ческие и исторические времена.
10.1.2. Экологические аспекты расселения гоминид
На описанный ход расселения гоминид, безусловно, влиял
ряд факторов. Главными из них были географические и эколо-
гические барьеры (влияние этих факторов зависело от потребно-
стей видов). Географические барьеры, в особенности океан, го-
ры и ледники, очевидно, сыграли важную роль в задержке ос-
воения гоминидами Австралии. Помимо этих географических
барьеров на характере расселения гоминид сказались и эколо-
гические факторы.
На рис. 10.1 изображена последовательность колонизации
ранними гоминидами разных местообитаний. Можно видеть, что
этот процесс определялся не только температурой воздуха, ибо,
как свидетельствуют полученные недавно данные, гоминиды
.начали переселяться в умеренные широты еще до того, как
приобрели способность существовать в дождевом тропическом
лесу. По всей вероятности, более важное значение имели струк-
тура биотопов и типы источников пищи. Следовательно, колони-
зация биотопов происходила в порядке, совпадающем с умень-
шением их сходства с саванной — исходной средой гоминид.
Степи умеренных широт, хотя и отличались от саванны характе-
ром сезонности и температурой воздуха, оказались для гоминид
более подходящей средой, чем непроницаемые для солнечных
лучей густые тропические леса. Кроме того, гоминиды предпочи-
тают те среды, в которых обнаруживается ряд определенных
экологических характеристик. Как видно из рис. 10.2, в таких
средах наблюдается большая величина отношения первичной
продуктивности к биомассе, а не просто высокая доступность
энергии. Это заставляет предполагать, что источники пищи, ко-
торые использовали представители Homo erectus, представляли
собой либо организмы с высокой скоростью роста и размноже-
ния, либо животных, которые питались такими видами. Эти ор-
ганизмы, как правило, являются высокоценными продуктами
питания, но доступны лишь в течение коротких отрезков време-
ни, что еще раз указывает на тесную связь эволюции с харак-
тером сезонности сред (Foley, 1984b, см. также гл. 8).
Гоминиды как расселяющиеся животные 325
Прежде чем перейти к рассмотрению других факторов, сле-
дует отметить, что радиация человека происходила не в отрыве
от других видов животных. Ранние гоминиды покидали Африку
вместе с другими животными, одновременно ведя с ними острую
конкуренцию. Не так давно Тернер (Turner, 1984) провел пло-
дотворное сравнение колонизации высоких широт гоминидами
и другими животными. Любопытно, что Homo erectus и ряд
других видов (лев, леопард, гиена и волк) достигли умеренных
широт Евразии примерно в одно время (Turner, 1984, табл. 8.2).
В этой связи можно думать, что гоминиды были лишь частью
некоего общего биогеографического процесса, в котором участ-
вовали разнообразные виды и который замедлялся и ускорялся
одними и теми же факторами. Таким образом, представление
о «неповторимости» гоминид вновь оказывается иллюзорным.
Вероятнее всего, факторы окружающей среды в наибольшей сте-
пени благоприятствовали организмам с определенными призна-
ками; к числу этих признаков, по-видимому, относятся крупные
размеры, хищническое поведение и социальность. Виды с подоб-
ными характеристиками, несмотря на свое тропическое проис-
хождение, обладали высокой адаптивной способностью к освое-
нию новых сред обитания.
Что лежало в основе такой способности животных? Ответ
на этот вопрос позволяют дать некоторые общие экологические
принципы. Во-первых, известно, что хищники обладают более
широкой приспособляемостью (эвритопностью), нежели траво-
ядные. Мясоедение требует меньшей специализации и меньше
локальных адаптаций, чем питание растительной пищей. Для
хищника нет особой разницы между оленем и антилопой, равно
как и между передними и задними ногами этих копытных. Иное
дело — растения, которые сильно варьируют структурно и ло-
кально и использование которых в пищу предполагает существо-
вание более специализированных и узких экологических ниш.
Таким образом, можно предполагать, что географическая ра-
диация хищников в отличие от радиации травоядных протекала
без сколько-нибудь выраженного видообразования или морфо-
логического изменения. Лучше всего это предположение иллю-
стрируется повсеместным незначительным разнообразием круп-
ных хищных млекопитающих, все виды которых занимают весь-
ма обширные географические ареалы, и очень высоким локаль-
ным разнообразием крупных травоядных. Это обстоятельство,
а также сходство между радиацией гоминид и крупных хищни-
ков заставляют предполагать, что эти организмы сильно зависе-
ли от мясной пищи и что независимо от того, какое место зани-
мало мясоедение в жизни гоминид в тропической саванне, оно
в значительной степени обусловило их успех в освоении новых
сред.
Глава 10
Рис. 10.1. А. Расселение ранних гоминид из Африки; кривыми линиями обозначены предположительные маршру-
ты расселения (с соответствующими датами в млн. лет назад). Б. Расселение анатомически современных людей
из Африки (указаны приблизительные даты — в тыс. лет назад) и образование древнейших популяций Homo sa-
piens, известных по ископаемым находкам: белые кружочки — предки анатомически современных людей в афри-
канских региоиах, расположенных южнее Сахары; черные кружочки — европейские и западноазиатские популя-
ции неандертальцев; горизонтальные и вертикальные прямые — восточноазиатские популяции
328
Глава 10
Рис. 10.2. Экологические характеристики местообитаний, освоенг
гоминидами
Гоминиды как расселяющиеся животные 329
Согласно второму из рассматриваемых общих экологических
принципов, экзогения часто дает организму значительные адап-
тивные преимущества. Нередко считают, что экологический ус-
пех организма определяется тесной и гибкой адаптацией к ло-
кальным условиям. Правильность этого утверждения, однако, не
исключает того, что экологический успех требует от организма
расплаты. Цена расплаты — ширина занимаемой им экологиче-
ской ниши. Как правило, локальная адаптация достигается
за счет специализации и избегания и недопустимости конку-
ренции. Этот процесс — обычная и успешная стратегия вида,
существующего в устойчивых условиях неизменяющегося со-
общества. Нередко, однако, адаптивные преимущества получа-
ют вновь появляющиеся в сообществе (экзогенные) виды, обла-
дающие способностью утилизировать самые разнообразные эко-
логические ниши. Хотя эффективность использования ими каж-
дой из этих ниш может быть ниже, чем ее утилизация каким-
либо аборигенным видом, использование ресурсов разных ниш
дает этим видам значительную компенсацию. Примеры такой
стратегии, обнаруживающейся как у животных, так и у челове-
ка, хорошо известны. Это — расселение благородного оленя
в Новой Зеландии, крупного рогатого скота — в Африке, к югу
от Сахары, и кроликов — в Австралии. Можно поэтому предпо-
лагать, что как экзогенные организмы с мало специализирован-
ным пищевым поиском гоминиды пользовались в умеренной
Евразии (а затем и в Австралии, и Новом Свете) определенны-
ми экологическими преимуществами. Аборигенные виды умерен-
ных широт, по всей вероятности, не смогли конкурировать с эти-
ми пришельцами, и в этом состоит возможная причина выми-
рания многих крупных млекопитающих в плейстоцене.
В качестве третьего общего принципа для анализа расселе-
ния гоминид можно использовать некоторые положения эколо-
гической физиологии и, в частности, правило Аллена. Давно из-
вестно, что вследствие существования связи между теплоотда-
чей, объемом тела и площадью его поверхности отбор в холод-
ных средах способствует выживанию организмов с крупными
размерами тела и относительно короткими конечностями.
В этой связи любопытно отметить, что, как показали Тринкаус
(Trinkaus, 1981) и Стрингер (Stringer, 1984), из всех известных
популяций человека самыми короткими (относительно размеров
тела) дистальными отделами конечностей обладали неандер-
тальцы. В этом отношении они сближаются с живущими ныне
в высоких широтах популяциями людей (рис. 10.3). К сожале-
нию, соответствующий посткраниальный материал для других
представителей раннего Homo sapiens и Homo erectus отсутст-
вует, однако совершенно ясно, что по крайней мере некоторые
популяции гоминид приспособились к новым холодным средам.
Эво
Глава 10
Рнс. 10.3. Связь между средней годовой температурой воздуха и средним зна-
чением берцово-бедренного указателя (длина большеберцовой кости/длина
бедренной кости Х100) в популяциях современных людей. Как видно нз гра-
фика, у людей, живущих в более теплом климате, длина дистальных отделов
конечностей, как правило, больше. Судя по ископаемым остаткам гоминид,
пропорции конечностей у неандертальцев приближались к соответствующим
показателям современных популяций, приспособившихся к холодному климату,
а пропорции конечностей древнейших представителей анатомически современ-
ных людей — к соответствующим показателям тропических популяций. Это —
еще одно указание на тропическое происхождение анатомически современного
человека (данные Trinkaus, 1981) (по Stringer, 1984)
за счет «классических» физиологических механизмов. Не исклю-
чена возможность, что успешному географическому распростра-
нению человека хоть в какой-то мере способствовало увеличе-
ние размеров тела у Homo erectus.
Хотя расселение гоминид как часть общего биогеографиче-
ского процесса во многом объясняется общими биологическими
принципами, ряд вопросов остается все еще нерешенным. Вы-
живание в высоких широтах поставило перед гоминидами весь-
ма специфические проблемы; решением этих проблем, по всей
видимости, и обусловлены некоторые характеристики, присущие
только гоминидам.
Как показал Торренс (Torrence, 1983) в своей прежней ра-
боте, живущие в высоких широтах охотники и собиратели по-
стоянно подвергаются «временному стрессу»: существование
в высоких широтах связано с вполне определенной проблемой
нехватки времени («окна») на утилизацию ресурсов. Заслужи-
вают внимания два предположения. Первое связано с высокой
сезонной изменчивостью источников пищи, которые в одно вре-
мя года более чем обильны, а в другое — крайне скудны. Вто-
Гоминиды как расселяющиеся животные
331
80
0|____I____!____I____I----1---Г
0 10 20 30 40 50 60
О 50 100 150 200 250 300
Общее количество техноединиц
Б
Рис. 10.4. Связь между географической широтой местности, разнообразием
орудий (Д) и сложностью орудий (Б) у современных охотников и собирате-
лей (по Torrence, 1983)
Общее число типов орудий
рое предположение учитывает значительную изменчивость про-
должительности светового дня на протяжении года и недостаток
времени на поиск пищи и другие формы активности зимой. Об-
щие экологические принципы позволяют привести некоторые
теоретические соображения о том, каким образом может быть
разрешена проблема «временного стресса».
Если источники доступны, но время на их утилизацию огра-
ниченно, отбор, очевидно, будет благоприятствовать стратегиям,
которые предполагают: 1) использование источников пищи
с высокой «энергетической отдачей» и (или) 2) высокую эф-
фективность утилизации источников. Первая стратегия часто
сопровождается увеличением хищничества, которое почти навер-
няка стало главным способом питания гоминид в высоких ши-
ротах. Вторая стратегия связана с повышением эффективности
хищничества. «Охотничьи успехи» гоминид в целом зависели
от эффективности соответствующих технологий, а следователь-
но, отбор по эффективности пищевого поведения должен был
протекать в форме технического развития. Здесь уместно отме-
тить два обстоятельства. Во-первых, Торренсу удалось ясно
показать, что у современных охотников и собирателей просле-
живается тесная связь между сложностью технических средств
и широтой, на которой живет данная популяция (рис. 10.4).
Популяции, живущие в высоких широтах, располагают более
разнообразными орудиями, и орудия эти сложнее по устройству
и состоят из большего числа компонентов. Во-вторых, хотя раз-
витие техники изготовления орудий в плейстоцене имело общий
и глобальный характер, наиболее сильно оно было выражено
332 Глава 10
именно в высоких широтах; орудия, найденные в разных частях
тропиков, оставались простыми. Так, индустрия оббитых галек
в Юго-Восточной Азии продержалась на протяжении всего-
плейстоцена, и лишь в голоцене на смену ей пришла технология
микролитов. Таким образом, освоение высоких широт подверга-
ло гоминид воздействию новых факторов отбора, успешное про-
тивостояние которым зависело от технического прогресса. Это,,
вероятно, в свою очередь способствовало развитию общих по-
знавательных способностей, ибо, как показал Гоулетт (Gowlett,.
1984), изготовление каменных орудий требует сложных логиче-
ских операций и большого воображения. Таким образом, если,,
согласно современным представлениям, эволюция анатомически-
современных людей протекала в Африке, то их расселение в вы-
соких широтах можно рассматривать и как важное проявление-
давления естественного отбора, и как следствие этого эволюци-
онного сдвига.
ЗАКЛЮЧЕНИЕ
Попытка подвести итог продолжавшейся более 5 млн. лег
эволюции нескольких сильно различавшихся между собою ли-
ний— непростая задача. Не исключено, что истинная природа
ранних гоминид навсегда останется скрытой от нас, и мы даже
не сможем оценить, насколько приближаются к ней разрабаты-
ваемые нами модели. Эти модели и их проверка дают, однако,
единственную возможность подойти к пониманию причин, обус-
ловивших эволюцию того неповторимого вида, к которому отно-
симся мы сами. Если экология, поведение и эволюция ранних
гоминид не получили на страницах этой книги должного осве-
щения, остается по крайней мере надеяться, что в ней достаточ-
но убедительно показана роль биологической теории и модели-
рования в палеоантропологии.
Главный вывод, который можно извлечь из представленных
в настоящей книге данных и рассуждений, заключается в том,
что возникновение вида с признаками, расценивающимися нами
как сугубо «человеческие», не было неизбежным и предопреде-
ленным заранее результатом телеологического эволюционного
процесса, но явилось теоретически объяснимым следствием
существования предков человека в «подходящих» экологических
и эволюционных условиях. Гоминиды и люди утвердились на
Земле потому, что, подвергаясь действию основных биологиче-
ских и эволюционных факторов и процессов, благодаря прису-
щим им адаптивным характеристикам они могли «решать»
проблемы, возникавшие перед ними в определенных филогенети-
ческих и экологических обстоятельствах. В этой книге была
сделана попытка показать в свете основных принципов эволю-
ционной экологии, что в соответствии с этими обстоятельствами
и протекала эволюция гоминид.
С этой точки зрения ранние гоминиды предстают как суще-
ства, сильно отличающиеся от тех в высшей степени честолю-
бивых, но недалеких полуобезьян, которых так любят изобра-
жать в научно-фантастической литературе. Ни один из аспектов
эволюции не носит промежуточного, переходного или зачаточ-
ного характера, приобретающего смысл лишь в контексте более
поздней человеческой истории. Скорее наоборот, всякое эволю-
.-334
Заключение
ционное изменение поддается объяснению с точки зрения его
непосредственной, краткосрочной эффективности или той степе-
ни, в которой оно способствует решению текущих проблем вы-
живания и воспроизведения. Прошлое человека, как и всякого
.другого вида, нужно анализировать в контексте тех событий
и процессов, которые развивались тогда же, в прошлом, а не
.в свете последующих эволюционных явлений. Именно эти собы-
тия и процессы давно прошедших времен, составлявшие движу-
щую силу человеческой эволюции, обсуждались и моделирова-
лись в настоящей книге. К числу главных из этих факторов
относятся характеристики, унаследованные гоминидами и их
предками в результате предшествовавшего эволюционного раз-
вития,— размеры тела и признаки млекопитающих и приматов.
Как было показано, это филогенетическое наследие в значитель-
ной мере определило ход последующей эволюции гоминид. Оно
явилось тем субстратом, на который должен был воздействовать
«естественный отбор. Реакции ранних гоминид на встававшие
перед ними проблемы были обусловлены их предысторией.
Урок, который эволюция на примере гоминид преподносит дру-
гим животным, можно, пожалуй, свести к следующему: если
хотите стать гоминидами, учтите, что то, с чего вы начинаете,
имеет такое же важное значение, как и то, к чему вы стреми-
тесь!
Если филогенетическое наследие можно назвать внутренней
движущей силой эволюции, то окружающая среда составляет
«е внешнюю движущую силу, а следовательно, эволюцию чело-
века необходимо рассматривать и в этом аспекте. Как было
показано, человеческая эволюция и необходимые для нее пред-
посылки определялись разнообразными условиями окружающей
среды. На эволюцию гоминид наложили свой отпечаток и тро-
пические среды Старого Света, и африканские саванны с сезон-
ной изменчивостью, и плейстоценовые среды с умеренным, но
все более холодным климатом; здесь играли роль и характер
изменений среды обитания, и неустойчивость экологии. Стало
совершенно ясно, однако, что окружающая среда — не просто
пассивный фон эволюции, как и не самостоятельный фактор мо-
дификации видов, но сама составлена из эволюционирующих
видов, конкурирующих организмов и постоянно изменяющихся
экологических взаимоотношений. Это — сложная паутина взаи-
мосвязей между организмами, не просто развивающаяся в од-
ном направлении в силу «экологического детерминизма», но са-
ма подвергающаяся сложным эволюционным изменениям.
Что можно сказать об адаптивных стратегиях ранних гоми-
нид? По современным представлениям, эти стратегии возникли
главным образом в Африке — отчасти в результате радиации
африканских гоминоидов, отчасти в результате развития сооб-
Заключение
335
щества коэволюционировавших африканских млекопитающих.
О таком более локальном развитии ранних гоминид заставляют
говорить сегодня многочисленные данные, которые должны за-
острить взимание исследователей на изучении роли африкан-
ских сред в формировании первичных этапов гоминидной эво-
люции. Эти данные должны способствовать и скорейшему при-
знанию огромной эволюционной значимости исхода гоминид из
Африки. От экзогении в значительной мере может зависеть как
равновесие экологических систем, так и характер адаптивных
стратегий. К сожалению, за рамки рассматриваемого вопроса
выходит то любопытное обстоятельство, что на всем протяже-
нии человеческой предыстории основным способом сохранения
и увеличения популяций гоминид было рассредоточение, а не
интенсификация. Рассредоточение, по всей видимости, является
самой обычной стратегией выживания у большинства животных
видов, и в этом отношении ранние гоминиды исключения не со-
ставляли. Интенсификация существующих адаптаций, вероятно,
становится составной частью предыстории и истории человечест-
ва в более позднее время.
Важнейшее значение для эволюции гоминид помимо их «аф-
риканской природы» имело прямохождение. Прямохождение —
рано появившийся, в той или иной степени присущий всем гоми-
нидам признак, который наложил отпечаток на всю их анато-
мию. Если раньше считалось, что толчок к человеческой эволю-
ции дало укрупнение головного мозга с соответствующим повы-
шением умственных способностей, то теперь полагают, что ре-
шающую роль в этом сыграл тип передвижения. Ископаемые
свидетельства этого обстоятельства заострили внимание палео-
антропологов на вопросе, что делали ранние гоминиды на двух
ногах?
Прежде чем ответить на этот вопрос, следует вспомнить, что
гоминидных линий было несколько. Можно спорить по пово-
ду точного числа видов гоминид, но тот факт, что в интервале
между 3 и 1 млн. лет назад одновременно существовало не ме-
нее двух видов этих приматов, никаких сомнений в настоящее
время не вызывает. Пожалуй, за два последних десятилетия
это — самое выдающееся заключение палеоантропологии, ибо
оно доказывает, что эволюция человека происходила так же,
как и эволюция других видов, и что гоминиды могли иметь раз-
ный облик. Адаптации гоминид явились теоретически объясни-
мым результатом эволюционных и экологических процессов,
а потому не исключено, что адаптации эти менялись в ходе
эволюции и были неоднозначными. Особенное впечатление про-
изводит мысль о том, что подобные эксперименты эволюция
могла проводить и в более раннее время, создавая гоминидопо-
добные формы, с более поздними видами непосредственно не
•336
Заключение
связанные. Быть может, когда-нибудь будут получены ископае-
мые свидетельства и этого предположения.
Что же касается количества существовавших видов гоминид,
то, возможно, еще не все они открыты. Поскольку видовое раз-
нообразие лишь оживляет палеоантропологическую картину, то
чем больше их будет открыто — тем лучше! Учитывая все ска-
занное в этой книге, можно, однако, думать, что любой вновь
обнаруженный вид в той или иной мере будет повторять уже
известные адаптивные признаки гоминид. Как было показано,
Australopithecus и Ното отражают собой соответственно тенден-
ции к специализации и генерализации. Это — альтернативные
пути эволюции гоминид, результат межвидовых взаимодействий
и локальной конкуренции. Возможно, что подобные тенденции
возникали и в других частях Африканского материка; это долж-
но найти отражение в ископаемом материале.
Соответственно Australopithecus и Ното обнаружили и раз-
ные адаптивные характеристики. Australopithecus проявлял тен-
денцию к увеличению размеров и более массивному телосложе-
нию, а Ното — к большей грацильности. Подобные две дивер-
гентные тенденции прослеживаются, однако, и внутри одной
эволюционной линии, приведшей к возникновению современного
-человека. Такое повторение тенденций показывает, каким обра-
зом экологические проблемы и адаптивные изменения «прелом-
ляются» через одни и те же эволюционные процессы. Похоже,
что больше всего о ходе человеческой эволюции можно узнать
из общего характера развития, данных по изменчивости разме-
ров тела, географическому распространению и разнообразию.
Как показывает вся эта книга, история человеческой эволю-
ции— не просто вопрос поисков и находок захватывающих во-
ображение окаменелостей и построения генеалогических схем,
но в первую очередь — выяснение того, как жили животные
в прошлом: чем они питались, каким было их социальное пове-
дение и какими физиологическими характеристиками они обла-
дали. Именно эти вопросы составляют суть эволюции как чело-
века, так и любого другого вида. А это означает, что находка
и таксономическое описание ископаемых видов гоминид являют-
ся лишь отправной точкой для выяснения происхождения чело-
века. Такое же большое значение имеет и воссоздание образа
жизни гоминид, ибо это — субстрат, на который воздействует
естественный отбор, и одновременно результат этого воздейст-
вия. Таким образом, для изучения человеческой эволюции необ-
ходимо привлекать все области эволюционной биологии. Круг
вопросов, касающихся эволюции человека, никоим образом не
должен ограничиваться «чисто человеческими характеристика-
ми» или искусственно сужаться по соображениям «неполноты
.ископаемого материала».
Заключение
337
Опубликовав «Происхождение видов» и «Происхождение че-
ловека», Чарлз Дарвин нанес первый удар по представлениям
об уникальности человека среди всех остальных обитающих на
нашей планете видов. Значительный ущерб нанесли этим пред-
ставлениям и обнаруженные в ископаемом материале виды, от-
части заполнившие собой разрыв между человеком и другими
животными. Пьедестал, на который мы хотели вознести самих
себя, еще более пошатнулся в результате дальнейшего изуче-
ния животных: их поведения — в природе, их анатомии — в сек-
ционном зале, их биохимии — в пробирке. Эти исследования
внесли огромный вклад в антропологию и вообще в биологию,
в известной мере изменив наши представления и об учении
Дарвина. Суть эволюционной теории составляет не утвержде-
ние, что все виды одинаковы, но что все они образуются в ре-
зультате одних и тех же биологических процессов под влиянием
естественного отбора. Теперь, когда биологическая и эволюци-
онная природа гоминид установлена, главная задача палеоант-
ропологов заключается, пожалуй, не в том, чтобы опровергнуть
миф о неповторимости человеческого вида, ибо все виды по
своему неповторимы, но в том, чтобы показать, каким образом
эта неповторимость могла возникнуть под влиянием процессов,
одинаково воздействующих на всю живую материю. Сознавая
свою «неповторимость», человек должен помнить, что он — всего
лишь еще один неповторимый вид.
22—367
ЛИТЕРАТУРА
Aronson J. L., Taieb M. 1981. Geology and paleogeography of the Hadar hominid
site, Ethiopia. In: Rapp C, and Vondra C. F. (eds.), Hominid Sites: their
Geologic Settings, AAAS Selected Symposium No. 68, Boulder, Co: West-
view Press, pp. 165—196.
Aiello L. C. 1981. Locomotion in the Miocene Hominoidea. In: Stringer С. B.
fed.), Aspects of Human Evolution, Symposia for the Study of Human
Biology No. 12, London: Taylor and Francis, pp. 63—98.
Allen E. et al. 1976. Against sociobiology, New York Review of Books (13 No-
vember), 182.
Altmann S. 1974. Baboons, space, time and energy, American Zoologist, 14,
221-248.
Altmann S., Altmann J. 1970. Barboon Ecology, Chicago: University of Chicago
Press.
American Journal of Physical Anthropology. 1982. Vol. 57(4).
Andrews P. J. 1981. Species diversity and diet in apes and monkeys during the
Miocene. In: Stringer С. B. (ed.), Aspects of Human Evolution, Symposia
for the Study of Human Biology No. 12, London: Taylor and Francis,
pp. 63—98.
Andrews P. J. 1983. The natural history of Sivapithecus. In: Ciochon R. and
Corruccini R. (eds.), New Interpretations of Ape and Human Ancestry, New
York: Plenum Press, pp. 441—464.
Andrews P. J. 1984a. The descent of man, New Scientist (3 May), p. 24—25.
Andrews P. J. 1984b. An alternative interpretation of the characters used to
define Homo erectus. Courier Forschungsinstitut Seckenberg, 69, 167—175.
Andrews P. J., Tobien H. 1977. New Miocene locality in Turkey with evidence on
the origins of Ramapithecus and Sivapithecus, Nature, 268, 699—701.
Andrews P. J., Lord J. M., Evans E. N. M. 1979, Patterns of ecological diversity
in fossil and mammalian faunas, Biological Journal of the Linnean Society,
11, 177—205.
Anon. 1983. Oncogenes, reductionism and all that, Nature, 301, 369.
Ardrey R. 1961. African Genesis, London: Atheneum.
Ayala F. J., Valentine J. IF. 1979. Evolving: the Theory and Processes of Orga-
nic Evolution, Menlo Park, Ca.: Bejamin Cummings.
Badgeley C., Behrensmeyer A. K. 1980. Palaeoecology of middle Siwalik sedi-
ments and faunas, North Pakistan, Palaeogeography, Palaeoclimatology and
Palaeoecology, 30, 133—155.
Beadle L. C. 1974. The Inland Waters of Tropical Africa: an Introduction to Tro-
pical Limnology, London: Longman,
Behrensmeyer A. K. 1978. Taphonomic and ecologic information from bone
weathering, Paleobiology, 4, 150—162.
Behrensmeyer A. K. 1983. Patterns of natural bone distribution on recent land
surfaces: implications for archaeological site formation. In: Clutton-Brock J.
and Grigson C. (eds.), Animals and Archaeology: 1. Hunters and their Prey,
B. A. R. International Series No. 163. Oxford: B. A. R., pp. 93—106.
Литература
339
Behrensmeyer А. К., Dechant Boaz D. E. 1980. The recent bones of Amboseli
Park, Kenya, in relation to East African paleoecology. In: Behrens-
meyer A. K. and Hill A. P. (eds.), Fossils in the Making, Chicago: University
of Chicago Press, pp. 72—93.
Behrensmeyer A. K-, Hill A. P. (eds.) 1980. Fossils in the Making, Chicago: Uni-
versity of Chicago Press.
Behrensmeyer A. K-, Gordon K. D., Yanagi G. T. 1986. Trampling as a cause of
bone surface damage and pseudo-cutmarks, Nature, 319, 768—771.
Bernor R. L. 1983. Geochronology and zoogeographic relationships of Miocene
Hominoidea. In: Ciochon R. L. and Corruccini R. S. (eds.), New Interpreta-
tions of Ape and Human Ancestry, New York: Plenum Press, pp. 149—164.
Binford L. R, 1967. Smudge pits and hide smoking: the use of analogy in
archaeological reasoning American Antiquity, 32, 1—12.
Binford L. R. 1977. For Theory Building in Archaeology, London and New York:
Academic Press.
Binford L. R. 1978a. Dimensional analysis of behavior and site structure: learning
from an eskimo hunting stand, American Antiquity, 43, 330—361.
Binford L. R. 1978b. Nunamuit Ethnoarchaeology, New York and London:
Academic Press.
Binford L. R. 1979. Organization and formation processes: looking at curated
technologies, Journal of Anthropological Research, 35, 255—273.
Binford L. R. 1980. Willow smoke and dogs’ tails: hunter—gatherer settlement
systems and archaeological formation systems, American Antiquity, 45,
4—20.
Binford L. R. 1981. Bones: Ancient Men and Modern Myths, New York and Lon-
don: Academic Press.
Binford L. R. 1983. In Pursuit of the Past, London: Thames and Hudson.
Binford L. R. 1984. Faunal Remains from Klasies River Mouth, New York and
London: Academic Press.
Binford L. R., Binford S. R. 1966. A preliminary analysis of functional variabi-
lity in the Mousterian of Levallois facies, American Anthropologist, 68,
238—295.
Binford L. R., Sabloff J. 1982. Paradigms, systematics and archaeology, Journal
of Anthropological Research, 38, 137—153.
Binford L. R., Todd L. C. 1982. On arguments for the «butcherin» of giant gela-
das, Current Anthropology, 23, 108—110.
Birch F. 1951. Recent work on the radioactivity of potassium and some related
geophysical problems, Journal of Geophysical Research, 56, 107—126.
Bishop W. W., Miller J. (eds.) 1972. Calibration of Hominoid Evolution, Edin-
burgh: Scottish Academic Press.
Bishop W. W., Hill A., Pickford M. 1978. Chesowanja: a revised geological
interpretation. In: Bishop W. W. (ed.), Geological Background to Fossil Man,
Edinburgh: Scottish Academic Press, pp. 309—327.
Boaz N. 1979. Early hominid population densities: new estimates, Science, 206,
592—595.
Boesch C., Boesch H. 1983. Optimization of nut-cracking with natural hammers
by wild chimpanzees, Behaviour, 83, 265—285.
Bonnefille R. 1972. Associations polliniques actuelles et quaternaires en Ethiopie
(vallees de 1'Awash et de 1’Omo), Thesis, University of Paris VI.
Bonnefille R. 1976. Palynological evidence for an important change in the
vegetation of the Omo Basin between 2,5 and 2 million years ago. In:
Coppens Y. et al. (eds.), Earliest Man and Environments in the Lake Rudolf
Basin: Stratigraphy, Paleoecology, and Evolution, Chicago: University of
Chicago Press, pp. 421—431.
Bonnefille R. 1979. Methode palynologique et reconstitutions paleoclimatiques au
Cenozoique dans le Rift Est Africain, Bulletin de la Societe Gfeologique,
France, 21, 331—342.
22*
340
Литература
Bonnefille R. 1984. The Evolution of East Asian Environment, ed, R. Dr White,
Centre of Asian Studies, University of Hong Kong.
Bonnefille R. 1985. Evolution of the continental vegetation: the palaeobotanical
record from East Africa, South African Journal of Science, 81, 267—270.
Bonner J. B. 1968. Size change in development and evolution, Journal of Palae-
ontology, 42, 1—15.
Bordes F. 1961. Typologie du paleolithique ancien et modern, Publications de
1’Institut de Prehistoire de 1’Universite de Bordeaux Memoir No. 1, Bordeaux:
Imprimeries Delmas.
Bortz IF. M, 1985. Physical exercise as an evolutionary force, Journal of Human
Evolution, 14, 145—155.
Bourlitre F. 1963. Specific feeding habits of African carnivores, African Wildlife,
17, 21—27.
Bown T. M., Kraus M, J., Wing S. L., Fleagle J. G., Tiffney В. H., Simons E. L.,
Vondra C. F. 1982, The Fayum primate forest revisited, Journal of Human
Evolution, 11, 603—632.
Boyde A., Martin L. 1984. The microstructure of primate dental enamel. In: Chi-
vers D., Wood B. A. and Bilsborough A. (eds.), Food Acquisition and Pro-
cessing in Primates, New York: Plenum Press, pp. 341—367.
Brace C. L. 1964. The fate of the classic Neanderthals: a consideration of hominid
catastrophism, Current Anthropology, 5, 3—43.
Brain С. K. 1981. The Hunters or the Hunted: an Introduction to African Cave
Taphonomy, Chicago: University of Chicago Press.
Brauer G. 1984. A craniological approach to the origin of anatomically modern
Homo sapiens in Africa and implications for the appearance of modern
Europeans. In: Smith F. and Spencer F. (eds.), The Origins of Modern
Humans: a World Survey of the Fossil Hominids, New York: Alan Liss,
pp. 327—410.
Brody S. 1945. Bioenergetics and Growth, Baltimore: Reinhold.
Broeker W. S., van Donk J. 1970. Insolation changes ice volumes and 18» record
in deep-sea cores, Reviews of Geophysics and Space Physics, 8, 168—
198.
Brose D. S., Wolpoff M. H. 1971. Early Upper Palaeolithic man and late Middle
Palaeolithic tools, American Anthropologist, 73, 1156—1194.
Brown F. H. 1981. Environments in the Lower Omo Basin from one to four
million years ago. In: Rapp C. and Vondra C. F. (eds.), Hominid Sites:
their Geologic Settings, AAAS Selected Symposium No. 63, Boulder Co.:
Westview Press, pp. 149—164.
Brown J. L. 1964. The evolution of diversity in avian territorial systems, Wilson
Bulletin, 76, 160—169.
Buchardt B. 1978. Oxygen isotope palaeotemperatures from the Tertiary period
in the North Sea area, Nature, 275, 121—123.
Bunn H. T. 1981. Archaeological evidence for meat-eating by Plio-Pleistocene
hominids from Koobi Fora and Olduvai Gorge, Nature, 291, 575—577.
Bunn H. T., Kroll E. M. 1986. Systematic butchery by Plio/Pleistocene hominids
at Olduvai Gorge, Tanzania, Current Anthropology 27(5) (in press).
Butzer K. W. 1972. Environment and Archaeology (2nd edition), London:
Methuen.
Butzer K. W. 1982. Archaeology as Human Ecology, Cambridge: Cambridge
University Press.
Butzer K. W., Isaac G. L., Richardson I. L., Washbourn-Kamau C. 1972. Radio-
carbon dating of East African lakes, Science, 175, 1069—1076.
Cain A. J. 1964. Animal Species and Evolution, London: Hutchinson.
Campbell B. 1974. Human Evolution: an Introduction to Man’s Adaptations,
Chicago: Aldine.
Cant I. G. H., Temerin L. A. 1984. A conceptual approach to foraging adaptations
in primates. In Rodman P. S. and Cant J. G. H. (eds.), Adaptations for
Литература
341
Foraging in Nonhuman Primates, New York: Columbia University Press,
pp. 304—342.
Cavalli-Sforza L., Feldman M. 1981. Cultural Transmission and Evolution, Prin-
ceton N. J.: Princeton University Press.
Chivers D. 1977. The feeding behaviour of siamang (Symphalangus syndactylus).
In: Clutton-Brock T. (ed.), Primate Ecology, New York and London: Acade-
mic Press, pp. 355—383.
Ciochon R. L. 1983. Hominid cladistics and the ancestry of modern apes and
humans. In: Ciochon R. and Corruccini R. (eds.), New Interpretations of Ape
and Human Ancestry, New York: Plenum Press, pp. 781—843.
Ciochon R. L., Chiarelli A. B. (eds.) 1980. Evolutionary Biology of the New World
Monkeys and Continental Drift, Plenum Press: New York.
Ciochon R. L., Corruccini R. S. (eds.) 1983. New Interpretations of Ape and
Human Ancestry, New York: Plenum Press,
Clark I. D., Kurashina H. 1979. Hominid occupation of the east-central highlands
in the Plio-Pleistocene, Nature, 282, 33—39.
Clark J. D., Asfaw B., Assefa G., Harris J. W. K-, Kurashina H., Walter R. C„
White T. D., Williams M.A. J. 1984. Palaeoanthropological discoveries in the
Middle Awash Valley, Ethiopia, Nature, 307, 421—428.
Clark J. G. D. 1968. World Prehistory: a New Outline (2nd edition), Cambridge:
Cambridge University Press.
Clarke D. L. 1968. Analytical Archaeology, London: Methuen.
Cloudsley-Thomson J. L. 1975. Terrestrial Environments, London: Croom-Helm.
Clutton-Brock T. H., Harvey P. 1977a. Primate ecology and social organization,
Journal of the Zoological Society of London, 183, 1—39.
Clutton-Brock T. H., Harvey P. 1977b. Species differences in feeding and ranging
behaviour in primates. In: Clutton-Brock T. H. (ed.), Primate Ecology, New
York and London: Academic Press, pp. 557—584.
Clutton-Brock T. H., Harvey P. 1979. Comparison and adaptation, Proceedings of
the Royal Society Series, B205, 547—565.
Clutton-Brock T. H., Harvey P. 1981. Primate home range size and metabolic
needs, Behavioural Ecology and Sociobiology, 8, 151—155.
Coe M. D., Cummings D. H., Phillipson J. 1976, Biomass and production of large
herbivores in relation to rainfall and primary production, Oecologia, 22,
341—354.
Colinvaux P. 1973. Introduction to Ecology, New York: Wiley.
Coltorti M., Cremaschi M., Delitala M. C., Esu D., Fornaseri M., McPherron A.
et al. 1982. Reversed magnetic polarity at an early Lower Palaeolithic site
in Central Italy, Nature, 300, 173—176.
Connell J. H. 1975. Producing structure in natural communities. In: Cody M. L.
and Diamond J. M., Ecology and Evolution of Communities, Cambridge
Mass: Harvard University Press, pp. 460—490.
Conroy G. C., Pilbeam D. 1975. Ramapithecus: a review of its hominid status. In:
Tuttle R. (ed.), Palaeoanthropology: Morphology and Palaeoecology, The
Hague: Mouton, pp. 59—86.
Coppens Howell F. C. 1976. Mamalian faunas of the Omo group: distribu-
tional and biostratigraphical aspects. In: Coppens Y., Howell F. C., Isaac G.
and Leakey R. E. F. (eds.), Earliest man and environments in the Lake
Rudolf Basin, Chicago: University of Chicago Press, pp. 177—192.
Cronin J. E. 1983. Apes, humans and molecular clocks: a reappraisal. In: Cio-
chon R. and Corruccini R. S. (eds.), New Interpretations of Ape and Human
Ancestry, New York: Plenum Press, pp. 115—138.
Cronin J. E., Boaz N. T-, Stringer С. B., Rak Y. 1981. Tempo and mode in hominid
evolution, Nature, 292, 113—122.
Crook J. H., Gartlan J. S. 1966. Evolution of primate societies, Nature munica-
tion in weaver birds (Ploceinae), Behaviour Supplement, 10, 1—178.
342
Литература
Crook J. H., Gartlan J. S. 1966. Evolution of primate societies, Nature, 210,
1200—1203.
Crowell K. L. 1962. Reduced inter-specific competition among the birds of Ber-
muda, Ecology, 43, 75—88.
Curio E. 1973. Towards a methodology of teleonomy, Experieniia, 29, 1045—
1058.
Curtis G. H. 1981. Establishing a relevant time-scale in anthropological and
archaeological research, Philosophical Transactions of the Royal Society,
B292, 7—20.
Darwin C. 1859. Origin of Species, Reprinted Penguin Books, 1968. [Имеется
перевод: Ч, Дарвин. Происхождение видов. — М.: Сельхозиздат, 1952.]
Darwin С. 1871. Descent of Man and Selection in Relation to Sex, London:
Murray. [Имеется перевод: Ч. Дарвин. Происхождение человека и половой
отбор. В издании: Ч. Дарвин. Сочинения. Под общей редакцией Л. С. Бер-
га, Н. И. Вавилова и др. Том 5, 1953.]
Dawkins R. 1976. The Selfish Gene, London: Oxford University Press.
Dawkins R. 1982a. The Extended Phenotype, San Francisco: Freeman.
Dawkins R. 1982b. Replicators and vehicles. In: King’s College Sociobiology
Group (eds), Current Problems in Sociobiology, Cambridge: Cambridge
University Press, pp. 45—64.
Day M. H. 1971. Postcranial remains of Homo erectus from Bed IV, Olduvai
Gorge, Tanzania, Nature, 232, 383—387.
Day M. H., Molleson T. 1973. The Trinil femora. In: Day M. H. (ed.), Human
Evolution, Symposium for the Study of Human Biology No. 11, London:
Taylor and Francis, pp. 127—154.
DeBrul E. L. 1962, The general phenomenon of bipedalism, American Zoologist,
2, 205—208.
Delaney M. J., Happold D. C. D. 1979. Ecology of African Mammals, London:
Longman.
DeVore I., Washburn S. L. 1963. Baboon ecology and human evolution. In:
Howell F. C. and Bourliere F. (eds.), African Ecology and Human Evolution,
Chicago: Aldine.
Diamond J. M. 1977. Distributional strategies. In: Allen H., Jones R. and Gol-
son J. (eds.), Sunda and Sahul, New York and London: Academic Press,
pp. 295—315.
Diamond J. M. 1978. Niche shifts and the rediscovery of inter-specific competi-
tion, American Scientist, 68, 322—331.
Dobzhansky T. 1974. Chance and creativity in evolution. In: Ayala F. J. and
Dobzhansky T. (eds.), The Philosophy of Biology, Berkeley: University of
California Press, pp. 309—339.
Dorman S. 1925. Pygmies and Bushmen of the Kalahari, London: Seeley Ser-
vice and Co.
Dunbar R. I. M. 1976. Australopithecine diet based on a baboon analogy, Journal
of Human Evolution, 5, 161—167.
Dunbar R. I. M. 1977. Feeding ecology of gelada baboons: a preliminary report.
In: Clutton-Brock T. (ed.), Primate Ecology, New York and London: Acade-
mic Press, pp. 251—273.
Dunbar R. I. M. 1982. Adaptation, fitness and the evolutionary tautology. In:
King’s College Sociobiology Group (eds.), Current Problems in Sociobiology,
Cambridge: Cambridge University Press, pp. 9—28.
Dunbar R. I. M. 1983. Theropithecines and hominids: contrasting solutions to
the same ecological problem, Journal of Human Evolution, 12, 647—658.
Dyson-Hudson R., Smith E. 1978. Human territoriality: an ecological reasses-
sment, American Anthropologist, 80, 21—41.
Efremov I. A. 1940. Taphonomy: a new branch of paleontology, Pan-American
Geologist, 74, 81—93.
Литература
343
Ehrlich Р. R., Raven P. H. 1964. Butterflies and plants: a study in coevolution,
Evolution, 18, 586—608.
Eichna L. W., Bean W. F., Bean W. B., Shelley IF. B. 1945. The upper limits of
heat and humidity tolerated by acclimatized men working in hot environ-
ments, Journal of Industrial Hygiene and Toxicology, 27, 59—84.
Eisenberg J. 1981. The Mammalian Radiations: an Analysis of Trends in Evolu-
tion, Adaptation and Behaviour, London: Athlone Press.
Eldredge N-, Cracraft J. 1980. Phylogenetic Patterns and the Evolutionary Pro-
cess, New York: Columbia University Press.
Eldredge N., Gould S. J. 1972. Punctuated equilibrium: an alternative to phyletic
gradualism. In: Schopf T. J. M. (ed.), Models in Paleobiology, San Fran-
cisco: Freeman, pp. 82—115.
Eldredge N., Tattersall R. 1982. The Myths of Human Evolution, New York:
Columbia University Press.
Ellen R. 1982, Environment, Subsistence and System: the Ecology of Smallscale
Social Formation, Cambridge: Cambridge University Press.
Eyre S. R. 1968. Vegetation and Soils: a World Picture, London: Arnold.
Fagan B. 1978. People of the Earth (2nd edition), Boston: Little, Brown and Co.
Fleagle J. G. 1983. New interpretations of the phyletic position of Oligocene
hominoids. In: Ciochon R. and Corruccini R. S. (eds ), New Interpretations
of Ape and Human Ancestry, New York: Plenum Press, pp. 181—210.
Fleagle J. G. 1984. Primate locomotion and diet. In: Chivers D., Wood B. and
Bilsborough A. (eds.), Food Acquisition and Processing in Primates, New
York: Plenum Press, pp. 105—118.
Fleagle I. G., Kay R. F. 1983. Locomotor adaptations of Oligocene and Miocene
hominoids and their phyletic implications. In: Ciochon R. and Corruccini R. S.
(eds.), New Interpretations of Ape and Human Ancestry, New York: Plenum
Press, pp. 301—324.
Fleischer R. L., Hart H. R., Jr. 1972. Fision-track dating: techniques and prob-
lems. In: Bishop W. W. and Miller J. (eds.), Calibration of Hominoid
Evolution, Edinburgh: Scottish Academic Press, pp. 135—170.
Flenley J. R. 1979. The Equatorial Rainforest: a Geological History, London:
Butterworth.
Foley R. 1978. Incorporating sampling into initial research design: some aspects
of spatial archaeology. In: Cherry J., Gamble C. and Shennan S. (eds.),
Sampling in Contemporary British Archaeology, B. A. R. British Senes
No. 50, Oxford: B. A. R., pp. 49—66.
Foley R. 1981. Aspects of variability in palaeoecological studies. In: Sheridan A.
and Bailey G. (eds.), Economic Archaeology, B. A. R. International Series
No. 96, Oxford: B. A. R„ pp. 67—76.
Foley R. 1982. A reconsideration of the role of predation on large mammals in
tropical hunter-gatherer adaptation, Man (N. S.), 17, 383—402.
Foley R. 1983. Modelling hunting strategies and inferring predator behaviour
from prey attributes. In: Clutton-Brock J. and Grigson C. (eds.), Animals
and Archaeology: 1. Hunters and their Prey, B. A. R. International Series
No. 163, Oxford: B. A. R„ pp. 63—76.
Foley R. 1984a. Putting people into perspective. In: Foley R. (ed.), Hominid
Evolution and Community Ecology: Prehistoric Human Adaptation in Bio-
logical Perspective, New York and London: Academic Press, pp. 1—24.
Foley R. 1984b. Early man and the Red Queen: tropical African community
evolution and hominid adaptation. In: Foley R. (ed.), Hominid Evolution
and .Community Ecology: Prehistoric Human Adaptation in Biological
Perspective, New York and London: Academic Press, pp. 85—110.
Foley R. 1985. Optimality theory in anthropology, Man (N. S.), 20, 222—242.
Ford C. S., Beach F. A. 1952. Patterns of Sexual Behaviour, London: Eyre and
Spottiswood.
Forde C. D. 1934. Habitat, Economy and Society, London: Methuen.
344
Литература
Fossey D., Harcourt A. 1977. Feeding ecology of free-ranging mountain gorillas
(Gorilla gorilla beringei). In: Clutton-Brock T. (ed.), Primate Ecology, Lon-
don: Academic Press, pp. 415—449.
Frayer D. W„ Wolpoff M. H. 1985. Sexual dimorphism, Annual Review of Anthro-
pology, 14, 429—473.
Friday A. E. 1981. Hominoid evolution: the nature of the biochemical evidence.
In: Stringer C. (ed.), Aspects of Human Evolution, London: Taylor and
Francis, pp. 1—24.
Futuyma D. I. 1979. Evolutionary Biology, Sunderland Mass: Sinauer Asso-
ciates.
Gamble C. 1981. Scratches on the palaeolithic record, Nature, 291, 533—534.
Gannt D. G. 1983. The enamel of Neogene hominoids: structural and phyletic
implications. In: Ciochon R. and Corruccini R. S. (eds.), New Interpreta-
tions of Ape and Human Ancestry, New York: Plenum Press, pp. 249—
298.
Genet-Vargin E. 1966. Conjures sur 1’allure generale des Australopitheques, Bul-
letin de la Societe Prehistorique Francaise, 63, cvi—cvii.
Gericho M., Hoffmeyr J. 1976. Aetiology and treatment of capture stress and
myopathy in springbok (A. marsupialis), South African Journal of Science,
72, 28.
Gibson J. R., McKeown T. 1952. Observations in all births (23970) in Birmin-
gham, 1974, 6, Birthweight, duration of gestation and survival related to
sex, British Journal of Social Medicine. 6, 152—158.
Gifford-Gonzalez D. P., Damrosch D. B., Datnrosch D. R., Pryor ThunenR.L.
1985. The third dimension in site structure: an experiment in trampling and
vertical dispersal, American Antiquity, 50, 803—818.
Gillespie R., Gowlett J. A. I., Hall E. T., Hodges R. E. M. 1984. Radiocarbon
measurement by accelerator mass spectrometry: an early selection of dates.
Archaemetry, 26, 15—20.
Goodall J. 1963. Feeding behaviour of wild chimpanzees: a preliminary report,
Symposia of the Zoological Society of London, 10, 39—47.
Goodall J. van Lawick. 1970. Tool-using in primates and other vertebrates. In:
Lehrman P., Hinde R. and Shaw E. (eds.), Advances in the Study of Beha-
viour, London: Academic Press, pp. 195—249.
Goodall J. van Lawick. 1971. In the Shadow of Man, London: Collins. [Имеется
перевод: Джейн ван Лавик-Гудолл. В тени человека.—М.: Мир, 1974.]
Goodman М. 1976. Towards a genealigical description of the primates. In: Good-
man M. and Tashien R. E. (ed.), Molecular Anthropology, New York: Plenum
Press, pp. 321—353.
Goodman M., Baba M. L., Darga L. L. 1983. The bearing of molecular data on
the cladogenesis and times of divergence of hominoid lineages. In: Ciochon R.
and Corruccini R. S. (eds.), New Interpretations of Ape and Human Ancestry.
New York: Plenum Press, pp. 67—86.
Gould R. 1969. Subsistence behaviour among the western desert aborigines,
Oceania, 39, 253—274.
Gould R. 1980. Living Archaeology, Cambridge: Cambridge University Press.
Gould S. J. 1966. Allometry and size in ontogeny and phylogeny, Biological
Reviews, 41, 587—640.
Gould S. J. 1977. Ever Since Darwin: Reflections in Natural History, London:
Penguin.
Gould S. J. 1980. Is a new and general theory of evolution emerging? Paleobio-
logy, 6, 119—130.
Gould S. J., Lewontin R. 1979. The spandrels of San Marco and the panglossian
paradigm: a critique of the adaptationist programme, Proceedings of the
Royal Society, London, B205, 581—598.
Gould S. J., Vrba E. S. 1983. Exaptation — a missing term in the science of
form, Paleobiology, 8, 4—15.
Литература
345
Gowlett J. A. J. 1984. Mental abilities of early man: a look at some hard evidence.
In: Foley R. (ed.), Hominid Evolution and Community Ecology: Prehistoric
Human Adaptation in Biological Perspective, New York and London: Aca-
demic Press, pp. 167—192.
Gowlett J. A. J., Harris J. W. K-, Walton D., Wood B. A. 1981. Early archaeolo-
gical sites, hominid remains, and traces of fire from Chesowanja, Kenya,
Nature, 294, 125—129.
Grand T. I. 1972. A mechanical interpretation of terminal feeding, Journal of
Mammalogy, 53, 198—203.
Grant P. R. 1972. Convergent and divergent character displacement, Biological
Journal of the Linnean Society, 4, 39—68.
Gray В. T. 1980. Environmental reconstruction of the Hadar Formation (Afar,
Ethiopia), PhD thesis, Cape Western Reserve University.
Greenwood P. N. 1974. The cichlid fishes of Lake Victoria, East Africa: the
biology and evolution of a species flock, Bulletin of the British Museum
(Natural History) Zoological Supplement No. 6.
Grine F. 1981. Trophic differences between «gracile» and «robust» australopithe-
cines: a scanning electron microscope analysis of occlusal events, South
African Journal of Science, 77, 203—230.
Grove A. T. 1967. Africa South of the Sahara, London: Oxford University
Press.
Haeckel E. 1876. The History of Creation, London: Murray. [Имеется перевод:
Эрнст Геккель. Естественная история миротворения. Полный перевод со
II немецкого издания. Т. 1—2. Спб. 1914.]
Hall К. R. L. 1962. Numerical data, maintenance activities and locomotion in
the wild chacma baboon (Papio ursinus), Proceedings of the Zoological
Society of London, 139, 181—220.
Hall K. R. L. 1966. Behaviour and ecology of the wild patas monkey, Erythro-
cebus patas, in Uganda, Journal of the Zoological Society, London, 148,
15—87.
Hamilton A. 1982. Environmental History of East Africa, New York and London:
Academic Press. ‘
Hamilton W. D. 1964. The genetical theory of social behaviour, Journal of
Theoretical Biology, 7, 1—52.
Hanson С. B. 1980. Fluvial taphonomic processes: models and experiments. In:
Behrensmeyer A. K. and Hill A. P. (eds.), Fossils in the Making, Chicago:
University of Chicago Press, pp. 156—181.
Harcourt A., Harvey P. H., Larson S. G-, Short R. V. 1981. Testis weight body
weight and breeding system in primates, Nature, 293, 55—57.
Harding R. S. O. 1981. An order of omnivores: non-human primate diets in the
wild. In: Harding R. S. O. and Teleki G. (eds.), Omnivorous Primates, New
York: Columbia University Press, pp. 191—214.
Harding R. S. O., Teleki G. (eds.). 1981. Omnivorous Primates, New York:
Columbia University Press.
Harested A. S., Bunnell F. L. 1979. Home range and body weight: a re-evaluation,
Ecology, 60, 389—402.
Harper I. 1977. Population Biology of Plants, New York and London: Academic
Press.
Harris D. R. 1980. Tropical savanna environments: definition, distribution, diver-
sity and development. In: Harris D. R. (ed.), Human Ecology in Savanna
Environments, New York and London: Academic Press, pp. 3—30.
Harris J. M. (ed.). 1983. Koobi Fora Research Project Vol. 2, The Fossil Ungu-
lates: Proboscidea, Perissodactyla, and Suidae, Oxford: Clarendon Press.
Harris !. W. K. 1983. Cultural beginnings: Plio-Pleistocene archaeological occu-
rences from the Afar, Ethiopia, The African Archaeological Review, 1,
3—31.
Harris J. W. R. 1986. Archaeological evidence bearing вн an understanding of
346
Литература
adaptive behaviours of Late Pliocene hominids, The Longest Record: The
Human Career in Africa, A Conference in Honour of J. Desmond Clark
Berkeley, California, April 1986 (abstract).
Harvey P. H. 1985. Intra-demic group selection and the sex ratio. In: R. M. Sib-
ley and R. H. Smith (ed.),Behavioural Ecology: Ecological Consequences
of Adaptive Behaviour, Oxford: Blackwell, pp. 59—74.
Harvey P. H., Clutton-Brock T. H. 1981. Primate home range size and metabolic
needs, Behavioural Ecology and Sociobiology, 8, 151—155.
Harvey P. H., Clutton-Brock T. H. 1985. Life history variation in primates,
Evolution, 39, 559—581.
Hausfater G., Meade В. J. 1982. Alternation of sleeping groves by yellow baboons
(Papio cynocephalus) as a strategy for parasite avoidance, Primates, 23,
287—297.
Hay R. L. 1976. Geology of Olduvai Gorge, Berkeley: University of California
Press.
Hay R. L. 1981. Lithofacies and environments of Bed I, Olduvai Gorge, Tanza-
nia. In: Rapp G. and Vondra C. F. (eds.), Hominid Sites: their Geologic
Settings, AAAS Selected Symposium No. 63, Boulder Co: Westview Press,
pp. 25—56.
Hayden B. 1981. Subsistence and ecological adaptations of modern hunter/
gatheres. In: Harding R. S. O. and Teleki G. (eds.), Omnivorous Primates,
New York: Columbia University Press, pp. 344—421.
Hendey Q. B. 1973. Fossil occurrences at Langebaanweg, Cape Province, Nature,
244, 13—14.
Hewes G. W. 1961. Food transportation and the origins of bipedalism, American
Anthropologist, 63, 687—710.
Hewes G. W. 1964. Hominid bipedalism: independent evidence for food-carrying
theory, Science, 146, 416—418.
Hill A. P. 1975. Taphonomy of contemporary and Late Cenozoic East African
vertebrates, PhD Dissertation, University of London.
Hill A. P. 1978. Hyaenas, bones, and fossil man, Kenya Past and Present, 9,
8—12.
Hill A. P. 1979. Disarticulation and scattering of mammal skeletons, Paleobio-
logy. 5. 261—274.
Hill A. P. 1984. Hominids and hyaenas: taphonomy and hypothesis testing. In:
Foley R. (ed.), Hominid Evolution and Community Ecology: Prehistoric
Human Adaptation in Biological Perspective,' New York and London: Acade-
mic Press, pp. Ill—128.
Hill A. P. 1985. Early hominid from Baringo, Kenya, Nature, 315, 222—224.
Hill A. P., Drake R., Tauxe L., Monaghan M., Barry J. C., Behrensmeyer A. K.,
Curtis G., Fine Lacobs B., Johanson N., Pilbeam D. 1985. Neogene palaeon-
tology and geochronology of the Baringo basin, Kenya, Journal of Human
Evolution, 14, 759—774.
Hill K. 1982. Hunting and human evolution, Journal of Human Evolution, 11,
521—544.
Hladik С. M. 1977. Chimpanzees of Gabon and chimpanzees of Gombe: some
comparative data on diet. In: Clutton-Brock T. H. (ed.), Primate Ecology,
New York and London: Academic Press, pp. 481—501.
Hladik С. M. 1981. Diet and the evolution of feeding strategies among forest
primates. In: Harding R. S. O. and Teleki G. (eds.), Omnivorous Primates,
New York: Columbia University Press, pp. 215—254.
Hladik С. M., Charles-Dominique P., Peters J. J. 1980. Feeding strategies of five
nocturnal prosimians in the dry forest of the west coast of Madagascar. In:
P. Charles-Dominique (ed.), Nocturnal Malagasy Primates, New York: Aca-
demic Press, pp. 433—444.
Hodder I. 1982. The Present Past: an Introduction to Anthropology for Archaeolo-
gists, London: Batsford.
Литература
347
Hoffman А. 1979. Community palaeocology as an epiphenomenal science, Paleo-
biology, 5, 357—379.
Hoffmeyr J., Louw G., du Preeze S. 1973. Incipient capture myopathy as revealed
by blood chemistry of chased zebras, Madoqua Series I (7), 45—50.
Horn H. S. 1968. The adaptive significance of colonial nesting in the Brewer’s
blackbird (Euphagus cyanocephalus), Ecology, 49, 682—694.
Houston A. F., Davies N. B. 1985. The evolution of co-operation and life history
in the dunnock, Prunella modularis. In: R. M. Sibley and R. H. Smith (eds.),
Behavioural Ecology: Ecological Consequences of Adaptive Behaviour, Ox-
ford: Biackwell, pp. 471'—487.
Houston D. C. 1979. Scavenging adaptation in the Serengeti. In: Sinclair A. R. E.
and Norton-Griffiths M. (eds.), Serengeti: Dynamics of an Ecosystem, Lon-
don: Academic Press, pp. 263—286.
Howell F. C. 1978. The Hominidae. In: Maglio V. and Cooke H. B. S. (eds.),
Evolution of African Mammals, Cambridge Mass: Harvard University Press,
pp. 154—248.
Hutchinson G. E. 1957. Concluding remarks, Cold Spring Harbour Symposium
on Quantitative Biology, 22, 415—427.
Huxley J. 1924. Constant differential growth ratios and their significance, Natu-
re, 114, 895—896.
Huxley J, 1972. Problems of Relative Growth (2nd edition), New York: Dover.
Huxley T. H. 1863. Evidence as to Man’s Place in Nature, London: Williams and
Nor gate.
Imbrie J., Imbrie К. P. 1979. Ice Ages: Solving the Mystery, London: Mac-
millan.
Ingram Y., Mount Y. 1975. Man and Animals in Hot Environments, New York:
Springer.
Isaac G. 1965. The stratigraphy of the Peninj Beds and the provenance of the
Natron Australopithecus mandible, Quaternaria, 7, 101—130.
Isaac G. 1967. Towards an interpretation of occupation debris: some experiments
and observations, Kroeber Anthropological Society Papers, 37, 31—57.
Isaac G. 1971a. Whither archaeology? Antiquity, 45, 123—129.
Isaac G. 1971b. The diet of early man: aspects of archaeological evidence from
Lower and Middle Pleistocene sites in Africa, World Archaeology, 2, 278—
298.
Isaac G. 1972, Chronology and tempo of culture change during the Pleistocene:
In: Bishop W. W. and Miller J, (eds.), Calibration of Hominoid Evolution,
Edinburgh: Scottish Academic Press, pp. 381—430.
Isaac G. 1976. Olorgesailie, Chicago: University of Chicago Press.
Isaac G. 1978a, Food sharing and human evolution: archaeological evidence
from the Plio-Pleistocene of East Africa, Journal of Anthropological Re-
search, 34, 311—325.
Isaac G. 1978b. The food-sharing behavior of protohuman hominids, Scientific
American, 238(4), 90—108.
Isaac G. 1981. Archaeological tests of alternative models of early hominid beha-
viour: excavation and experiments, Philosophical Transactions of the Royal
Society, London, B292, 177—188.
Isaac G. 1983. Bones in contention: competing explanation for the juxtaposition
of artefacts and faunal remains, In: Clutton-Brock J. and Grigson C. (eds.),
Animals and Archaeology: 1, Hunters and their Prey, B. A. R. International
Series No. 163, Oxford: B. A. R., pp. 3—19.
Isaac G. 1984. The archaeology of human origins: studies of the Lower Pleisto-
cene in East Africa 1971—1981, Advances in World Archaeology, 3, 1—79.
Isaac G., Crader D. 1981. To what extent were early hominids carnivorous?
An archaeological perspective, In: Harding R. S. O. and Teleki G. (eds.),
Omnivorous Primates, New York: Columbia University Press, pp. 37—
103.
348
Литература
Janzen D. H. 1980. When is it coevolution? Evolution, 34, 611—612.
Jarman M. V., Jarman P. J. 1973. Daily activity of impala, East African Wildlife
Journal, 11, 75—93.
Jarman P. J. 1974. The cocial organization of antelope in relation to their eco-
logy, Behaviour, 48, 215—267.
Jerison H. J. 1961. Quantitative analysis of evolution of the brain in mammals,
Science, 133, 1012—1014.
Jerison H. J. 1973. Evolution of the Brain and Intelligence, New York: Academic
Press.
Jochirn M. A. 1981. Strategies of Survival, New York: Academic Press.
Johanson D. C., Edey M. 1981. Lucy: the Eeginings of Mankind, New York:
Simon and Schuster. [Имеется перевод: Д. Джохансон, М. Иди. Люси.
Истоки развития человеческого. — М.: Мир, 1984.]
Johanson D. С., White Т. D. 1979. A systematic assessment of early African
hominids, Science, 203, 321—330.
Johanson D. C., Taieb M., Coppens Y. 1982. Pliocene hominids from the Hadar
Formation, Ethiopia (1973—1977): stratigraphic, chronologic, and palaeoen-
vironmental contexts, American Journal of Physical Anthropology, 57, 373—
402.
Johnson S. C. 1981. Bonobo: generalized hominid prototype or specialized insular
dwarfs? Current Anthropology, 22, 363—375.
Jolly C. 1970. The seed-eaters: a new model of hominid differentiation based on
a baboon analogy, Man, 5, 5—26.
Jolly C. 1972. The classification and natural history of Theropithecus (Simopi-
thecus) (Andrews 1916) baboons of the African Plio-Pleistocene, Bulletin of
the British Museum (Natural History) Geology, 22, 1—123.
Jungers W. L. 1982. Lucy’s limbs: skeletal allometry and locomotion in Austra-
lopithecus afarensis, Nature, 297, 676—678.
Jungers W. L. 1985. Body size and scaling og limb proportions in primates, In:
Jungers W. L. (ed.), Size and Scaling in Primate Biology, New York:
Plenum Press, pp. 345—381.
Jungers W. L., Stern J. T. 1983. Body proportions, skeletal allometry and loco-
motion in the Hadar hominids: a reply to Wolpoff, Journal of Human Evo-
lution, 12, 673—684.
Kalb J. E., Oswald E. B., Tebedge S., Mebrate A., Tola E., Peak D. 1982. Geology
and stratigraphy of Neogene deposits, Middle Awash Valley, Ethiopia, Natu-
re, 298, 17—25.
Kaplan D. 1978. The Sociobiology Debate, New York: Harper and Row.
Kavanagh M. 1984. A Complete Guide to Monkeys, Apes and Other Primates.
London: Jonathan Cape.
Kawai M. 1965. Newly aquired pre-cultural behaviour of the natural troop of
Japenese monkeys on Koshima Islet, Primates, 6, 1—30.
Kay R. F. 1981. The nut-crackers — a new theory of the adaptations of the
Ramapithecinae, American Journal Anthropology, 55, 141—151.
Kay R. 1985. Dental evidence for the diet of Australopithecus, Annual Review of
Anthropology, 14, 315—343.
Kay R. F., Hylander W. L. 1978. The dental structure of mammalian folivores
with special reference to primates and Phalangeroids (Marsupala), In: Mont-
gomery G. G. (ed.), The Ecology of Arboreal Folivores, Washington D. C.:
Smithsonian Institution Press.
Keeley L. H. 1978. Experimental Determination of Stone Tool Uses, Chicago:
University of Chicago Press.
Keeley L., Toth N. 1981. Microwear polishes on early tools from Kenya, Nature,
293, 464—465.
Kennedy G. E. 1984. The emergence of Homo sapiens: the post-cranial evidence,
Man, 19, 94—110.
Kennett J. P. 1977. Cenozoic evolution of antarctic glaciation, the circum-
Литература
349
anatarctic ocean, and their implications on global palaeooceanography, Jour-
nal of Geophysical Research, 82, 3843—3860.
Kingdon J. 1981. East African Mammals Volume III, London: Academic Press.
King’s College Sociobiology Group (eds.). 1982. Current Problems in Sociobio-
logy, Cambridge: Cambridge University Press.
Kleiber M. 1961. The Fire of Life: an Introduction to Animal Energetics, New
York: Wiley.
Klein R. G. 1974. Environment and subsistence of prehistoric man in the
Southern Cape Province, South Africa, World Archaeology, 5, 249—289.
Klein R. G. 1983. The stone age prehistory of southern Africa, Annual Review of
Anthropology, 12, 25—48.
Kortlandt A. 1967. Experimentation with chimpanzees in the wild, In: Starck D.,
Scheinder R. and Kuhn H. J. (eds.), Neue Ergebnisse der Primatologie,
Stuttgart: Fischer, pp. 208—224.
Kortlandt A. 1972. New perspectives on ape and human evolution, Amsterdam:
Stichting voor Psychobiologie.
Kortlandt A. 1983. Facts and fallacies concerning Miocene ape habitats, In:
Ciochon R. and Corruccini R. S. (eds.), New Interpretations of Ape and
Human Ancestry, New York: Plenum Press, pp. 465—514.
Kortlandt A. 1984. Habitat richness, foraging range and diet in chimpanzees and
some other primates, In: Chivers D., Wood B. A. and Bilsborough A. (eds.),
Food Acquisition and Processing in Primates, New York: Plenum Press,
pp. 119—160.
Krebs J., Davies N. B. 1981. Introduction to Behavioural Ecology, Oxford:
Blackwell.
Krebs J., Davies N. B. (eds.). 1984. Behavioural Ecology (2nd edition); Oxford:
Blackwell.
Kretzoi M. 1975. New ramapithecines and Pliopithecus from the Lower Pliocene
of Rudabanya in northedstern Hungary, Nature, 257, 578—581.
Kruuk H. 1972. The Spotted Hyaena, Chicago: University of Chicago Press.
Kukla G. J. 1975. Loess stratigraphy in Central Europe, In: Butzer K. W. and
Isaac G. (eds.), After the Australopithecines, The Hague: Mouton, pp. 99—
188.
Kummer H. 1968. Social Organization of the Hamadryas Baboon, Chicago:
University of Chicago Press.
Kurten B. 1953. On the variation and population dynamics of fossil and recent
mammal populations, Acta Zoologica Fennica, 76, 1—122.
Ladell W. S. S. 1964. Terrestrial animals in humid heat: man, In: Dill D. B.
(ed.), Handbook of Physiology 4. Adaptation to the Environment, Baltimore:
Williams and Wilkins, pp. 625—659.
Latimer B. 1984. The pedal skeleton of Australopithecus afarensis, American
Journal of Physical Anthropology, 63, 182 (abstract).
Laurent R. F. 1973. A parallel survey of equatorial amphibians and reptiles in
Africa and South America, In: Meggars B. J., Ayensu E. S. and Duck-
worth W. D. (eds.), Tropical Forest Ecosystems in Africa and South Ame-
rica: a Comparative Review, Washington: Smithsonian Institution Press,
pp. 259—266.
Layton R., Foley R. in press. Human origins, In: Medawar P. (ed.), New Fron-
tiers of Science, London: Rainbow Press.
Le Gros Clark IF. E. 1949. History of the Primates, London: British Museum
(Natural History).
Le Gros Clark W. E. 1964. The Fossil Evidence for Human Evolution (2nd edi-
tion), Chicago: University of Chicago Press.
Leakey L. S. B. 1967. An early Miocene member of the Hominidae, Nature, 213,
155.
Leakey M. D. 1972. Olduvai Gorge, Volume 3, Cambridge: Cambridge Univer-
sity Press.
350
Литература
Leakey М. D. 1978. Pliocene footprints at Laetoli, Northern Tanzania, Antiquity,
52, 133.
Leakey M. D., Hay R. L., Curtis G. H., Drake R. E., Jackes M. K-, White T. D.
1976, Fossil hominids from the Laetolil beds, Nature, 262, 460—466.
Lee P. C. 1984. Ecological constraints on the social development of vervet mon-
keys,'Behaviour, 91, 254—262.
Lee R. B. 1968. What hunters do for a living, or, how to make out on scarce
resources. In: Lee R. B. and DeVore I. (eds.), Man the Hunter, Chicago:
Aldine, pp. 30—48.
Lee R. B., DeVore I. 1968, Problems in the study of hunters and gatherers. In:
Lee R. B. and DeVore I. (eds.), Man the Hunter, Chicago: Aldine, pp. 3—12.
Lewis G. 1934. Preliminary notice of new man-like apes from India, American
Journal of Science, 27, 161.
Lewontin R. 1978. Adaptation, Scientific American, 239(3), 157—169.
Livingstone F. B. 1962. Reconstructing man’s Pliocene pongid ancestor, Ameri-
can Anthropologist, 64, 301—395.
Lovejoy С. O. 1979. A reconstruction of the pelvis of AL—288 (Hadar Forma-
tion), American Journal of Physical Anthropology, 50, 460 (abstract).
Lovejoy С. O. 1980. Hominid origins: the role of bipedalism, American Journal
of Physical Anthropology, 52, 250 (abstract).
Lovejoy С. O. 1981. The origin of man, Science, 211, 341—350.
Lovejoy С. O., Heiple K. G. 1970. A reconstruction of the femur of A. africanus,
American Journal of Physical Anthropology, 32, 33—40.
Lumsden C. J., Wilson E. O. 1980. Translation of epigenetic rules of individual
behaviour into ethnographic patterns, Proceedings of the National Academy
of Sciences, U.S.A., 77, 4382—4386.
MacArthur R. H., Pianka E. R. 1966, On optimal use of a patchy environment,
American Naturalist, 100, 603—609.
MacArthur R. H., Wilson E. O. 1967. The Theory of Island Biogeography, Prin-
ceton: Princeton University Press.
McGrew IF., Baldwin P. L, Tulin С. E. G. 1981. Chimpanzees in a hot, dry,
open habitat: Mount Assirik, Senegal, West Africa, Journal of Human Evolu-
tion, 10, 227—244.
McHenry H. M. 1974. How large were the australopithecines? American Journal
of Physical Anthropology, 40, 329—340.
McHenry H. M. 1975. Fossil hominid body weight and brain size, Nature, 254,
686—688.
McHenry H. M. 1976. Early hominid body weight and encephalization, American-
Journal of Physical Anthropology, 45. 77—84.
McHenry H, M. 1978. Fore- and hindlimb proportions of Plio-Pleistocene homi-
nids, American Journal of Physical Anthropology, 49, 15—22.
McHenry H. M. 1982. The pattern of human evolution: studies on bipedalism,
mastication and encephalization, Annual Review of Anthropology, 11, 151 —
173.
McHenry H. M., Corruccini R. S. 1981. Pan paniscus and human evolution,
American Journal of Physical Anthropology, 54, 355—367.
McKinnon J. R. 1974. The behaviour and ecology of wild orang-utan (Pmgo
pygmaeus). Animal Behaviour, 22, 3—74.
MacMahon T. A., Bonner J. T. 1983. On Size and Life, San Francisco: Freeman.
McNab В. K. 1963. Bioenergetics and the determination of home range size,
American Naturalist, 97, 130—140.
McNab В. K- 1974. On the energetics of endotherms, Ohio Journal of Science, 74,
370—380.
Maddock L. 1979. The «migration» and grazing succession, In: Sinclair A. R, E.
and Norton-Griffiths M. (eds.), Serengeti: Dynamics of an Ecosystem, Lon-
don: Academic Press, pp. 104—209.
Maglio V. J. 1978. Patterns of faunal evolution, In: Maglio V. J. and!
Литература
351
Cooke Н. В. S. (eds.), Evolution of African Mammals, Cambridge, Mass.:
Harvard University Press, pp. 603—620.
Marshall L. G., Corruccini R. S. 1978. Variability evolutionary rates and allometry
in dwarfing lineages, Paleobiology, 4, 101—119.
Martin R. A. 1980. Body mass and basal metabolism of extinct mammals, Compa-
rative Biochemistry and Physiology, 66a, 307—314.
Martin R. A. 1981. On extinct hominid population densities, Journal of Human
Evolution, 10, 427—428.
Martin R. D. 1981. Relative brain size and basal metabolic rates in terrestrial
vertebrates, Nature, 293, 57—60.
Martin R. D. 1983. Human Brain Evolution in an Ecological Context (52nd
James Arthur Lecture on the Evolution of the Brain), New York: American
Museum of Natural History.
Martin R. D. 1984. Body size, brain size and feeding strategies. In: Chivers D.,
Wood B. and Bilsborough A. (eds.), Food Acquisition and Processing in
Primates, New York: Plenum Press, pp. 73—104.
Martin R. D., May R. 1981. Outward signs of breeding, Nature, 293, 7—9.
Matsu’ura S. 1982. A chronological framework for the Sangiran hominids, Bulle-
tin of the Natural Science Museum of Tokyo, D8, 1—53.
Maynard Smith J. 1976a. Evolution and the theory of games, American Scientist,
64, 41—45.
Maynard Smith J. 1976b. Game theory and the evolution of fighting. In: May-
nard Smith J., John Maynard Smith on Evolution, Edinburgh: Edinburgh
University Press, pp. 8—28.
Maynard Smith J. 1978. Optimization theory in evolution, Annual Review of
Ecology and Systematics, 9, 31—56.
Maynard Smith I. 1982. Evolution Now: a Century after Darwin, London:
Macmillan.
Maynard Smith J. 1983. Evolution and the Theory of Games, Cambridge: Cam-
bridge University Press.
Mayr E. 1963. Animal Species and Evolution, Cambridge, Mass.: Harvard Uni-
versity Press.
Mayr E. 1982. Questions concerning speciation, Nature, 296, 609.
Mellor D. 1974. New archaeology for old, Cambridge Review, 95, 71—72.
Mercer J. H. 1978. West antarctic ice sheet and COj greenhouse effect: a threat
of disaster, Nature, 259, 321—325.
Milton K.. May M. L. 1975. Body weight, diet and home range area in primates,
Nature, 259, 459—462.
Montagu A. 1964. Comments on the human revolution by C. F. Hockett and
R. Ascher, Current Anthropology, 5, 160—161.
Morbeck M. E. 1983. Miocene hominoid from Rudabanya: implications from the
postcranial skeleton, In: Ciochon R. and Corruccini R. S. (eds.), New Inter-
pretations of Ape and Human Ancestry, New York: Plenum Press, pp. 369—
404.
Moreau R. E. 1969. Climatic changes and the distribution of forest vertebrates
in West Africa, Journal of Zoology, 158, 39—61.
Mount L. N. 1979. Adaptations to a Thermal Environment, London: Arnold.
Murphy P. 1974. Primary productivity in tropical ecosystems, In: Leith H. and
Whittaker R. H. (eds.), Primary Productivity of the Biosphere, Berlin: Sprin-
ger, pp. 217—234.
Nabakov P., McClean M. 1980. Ways of native running, Coevolution Quarterly,
4, 21.
Napier J. 1971. Roots of Mankind, London: George Allen and Unwin.
Napier J., Napier P. 1967. Handbook of Living Primates, London: Academic
Press.
Newell N. D. 1949. Phyletic size increases — an important trend illustrated by
fossil invertebrates, Evolution, 3, 103—124.
352
Литература
Newman R. W. 1970. Why man is such a sweaty and thirsty naked animal:
a speculative review, Human Biology, 42, 12—27.
Nishida T., Uehara S. 1983. Natural diet of chimpanzees (Pan troglodytes
schweinfurthi): long-term records from the Mahale Mountains, Tanzania,
African Studies Monographs, 3, 109—130.
Oakley К. P. 1966. Frameworks for Dating Fossil Man (2nd edition), London:
Weidenfield and Nicholson.
Oakley K. P-, Campbell B., Mollison T. H. 1977. Catalogue of fossil hominids:
Africa (2nd edition), London: British Museum (Natural History).
Oakley К. P., Montagu M. F. A. 1949. A reconsideration of the Galley Hill
skeleton, Bulletin of the British Museum (Natural History), Geology, 1(2),
25—48.
Olson T. 1981. Basiocranial morphology of the extant hominoids and Pliocene
hominids: the new material from the Hadar Formation, Ethiopia, and its
significance in early hominid evolution and taxonomy. In: С. B. Stringer
(ed.), Aspects of Human Evolution, Symposia for the Study of Human
Biology No. 12, London: Taylor and Francis, pp. 99—128.
Oster G. F., Wilson E. 0. 1978. Caste and Ecology in the Social Insects, Prince
ton: Princeton University Press.
Owen D. F. 1974. What is Ecology? London: Oxford University Press.
Passingham R. 1982. The Human Primate, San Francisco: Freeman.
Patterson B., Behrensmeyer A. K., Sill W. D. 1970. Geology and fauna from a
new Pliocene locality in Northwestern Kenya, Nature, 226, 918—921.
Pearson R. 1978. Climate and Evolution, London: Academic Press.
Penck A., Bruckner E. 1909. Die Alpen im Eiszeitalter, Leipzig.
Peters R. H. 1983. The Ecological Implications of Body Size, Cambridge: Cam-
bridge University Press.
Peters C. R-, O'Brien E. M. 1981. The early hominid plant food niche: insights
from an analysis of plant exploitation by Homo, Pan and Papio in eastern
and southern Africa, Current Anthropology, 22, 127—140.
Peters C. R., O’Brien E. M., Box E. O. 1984. Plant types and seasonality of
wild plant foods, Tanzania to southwestern Africa: resources for models of
the natural environment, Journal of Human Evolution, 13, 397—414.
Pianka E. 1978. Evolutionary Ecology (2nd edition), New York: Harper and
Row. [Имеется перевод: Пианка Э. Эволюционная экология. — М.: Мир,
1981.].
Pickford М. 1983. Sequence and environment of the Lower and Middle Miocene
hominoids of western Kenya. In: Ciochon R. L. and Corruccini R. S. (eds.),
New Interpretations of Ape and Human Ancestry, New York: Plenum Press,
pp. 421—440.
Pilbeam D. 1968. The earliest hominids, Nature, 219, 1335.
Pilbeam D. 1969. Tertiary Pongidae of East Africa: evolutionary relationships,
and taxonomy, Peabody Museum of Natural History Bulletin, 31, 1—185.
Pilbeam D. 1972. The Evolution of Man, London: Macmillan.
Pilbeam D. 1975. Middle Pleistocene hominids. In: Butzer K. W. and Isaac G.
(eds.), After the Australopithecines, The Hague: Mouton, pp. 809—856.
Pilbeam D. 1979. Recent finds and interpretations of Miocene hominoids, Annual
Review of Anthropology, 8, 333—352.
Pilbeam D, 1984. The descent of hominoids and hominids, Scientific American,
250(3), 60—69.
Pilbeam D. 1986. Distinguished lecture: hominoid evolution and hominoid origins,
American Anthropologist, 88, 295—312.
Pilbeam D., Gould S. J. 1974. Size and scaling in human evolution, Science, 186,
892—901.
Pilbeam D., Vaisnys J. R. 1975. Hypothesis testing in palaeoanthropology. In:
Tuttle R. H. (ed.), Palaeoanthropology: Morphology and Palaeoecology, The
Hague: Mouton, pp. 3—14.
Литература
353
Pilbeam D., Mayer G. E., Badgeley C., Rose M. D., Pickford M., Behren-
smeyer A. K.., Shah S. M. I. 1977. New hominoid primates from the Siwaliks
of Pakistan and their bearing on hominoid evolution, Nature, 270, 689—
695.
Pope G. G. 1983. Evidence for the age of the Asian Hominidae, Proceedings of
the National Academy of Sciences, U.S.A., 80, 4988—4992.
Post D. 1981. Activity patterns of yellow baboons (Papio cynocephalus) in the
Amboseli National Park, Animal Behaviour, 29, 357—374.
Post D., Hausfater G., McCuskey S. 1980. Feeding behaviour of yellow baboons
(Papio cynocephalus): relationship to age, gender and dominance rank, Folia
Primatologica, 34, 170—195.
Potts R. 1982. Lower Pleistocene Site Formation and Hominid Activity at Oldu-
vai Gorge, Tanzania, PhD Dissertation, Harvard University.
Potts R, 1984a. Homnid hunters? Problems of identifying the earliest hunter/
gatherers. In: Foley R. (ed.), Hominid Evolution and Community Ecology:
Prehistoric Human Adaptation in Biological Perspective, London: Academic
Press, pp. 129—166.
Potts R. 1984b. Home bases and early hominids, American Scientist, 72, 338—
347.
Potts R., Shipman P. 1981. Cutmarks made by stone tools on bones from Oldu-
vai Gorge, Tanzania, Nature, 291, 577—580.
Pyke G. H., Pulliam H. R., Charnov E. L. 1977. Optimal foraging: a selective
review of theory and tests, Quarterly Review of Biology, 52, 137—154.
Rak Y. 1983. The Australopithecine Face, New York: Academic Press.
Raup D. M., Stanley S. 1978. Principles of Paleontology (2nd edition), San
Francisco: Freeman.
Ravey M. 1978. Bipedalism: an early warning system for Miocene hominoids,
Scioxico 109 372
Raza S. M., Barry J. C., Pilbeam D., Rose M. D., Shah S. M. I., Ward S. 1983.
New hominoid primates from the Middle Miocene Chinji Formation, Pottvar
Plateau, Pakistan, Nature, 305, 52—54.
Reader J. 1981. Missing Links, London: Collins.
Reeve E. С. B., Huxley J. S. 1945. Some problems in the study of allometric
growth. In: Le Gros Clark W. E. and Medawar P. (eds.), Essays on Growth
and Form, Oxford: Clarendon.
Rensch B. 1959. Evolution Above the Species Level, New York: Columbia Uni-
versity Press.
Repenning C. A., Fejfar O. 1982. Evidence for earlier date of ’Ubeidiya, Israel,
hominid site, Nature, 299, 344—347.
Rightmire G. P. 1981. Patterns in the evolution of Homo erectus, Paleobiologv,
7, 241—246.
Rightmire G. P. 1984. Homo sapiens in sub-Saharan Africa. In: Smith F. and
Spencer F. (eds.), The Origins of Modern Humans, New York: Alan Liss,
pp. 295—326.
Roberts N. 1984. Pleistocene environments in time and space. In: Foley R. (ed.),
Hominid Evolution and Community Ecology: Prehistoric Human Adaptation
in Biological Perspective, London: Academic Press, pp. 25—54.
Robinson J. T. 1972. Early Hominid Posture and Locomotion, Chicago: Univer-
sity of Chicago Press.
Roche H. 1980. Premiers outils tailies d’Afrique, Societe d’Ethnographie, France.
Roche H., Tiercelin J. I. 1977. Decouverte d’une Industrie lithique ancienne in
situ dans la formation d’Hadar, Afar, central Ethiopie, Comptes rendus de
I’Academie des Sciences, 284—D, 1871—1874.
Rodman P. S., McHenry H. M. 1980. Bioenergetics and the origin of hominid
bipedalism, American Journal of Physical Anthropology, 52, 103—106.
Romer A. S. 1959. The Vertebrate Story, Chicago: University of Chicago Press.
Rose M. D. 1973. Quadrupedalism in primates, Primates, 14, 337—357.
23—367
364
Литература
Rose М. D. 1983. Miocene hominoid postcranial morphology: monkey-like, ap
like, neither or both? In: Ciochon R. and Corruccini R. S. (eds.), New Int<
pretations of Ape and Human Ancestry, New York: Plenum Press, pp. 405
420.
Rose M. D. 1984. Food acquisition and the evolution of positional behaviour: t
case of bipedalism. In: Chivers D., Wood B. and Bilsborough A. (eds
Food Acquisition and Processing in Primates, New York: Plenum Pre:
pp. 509—524.
Rosenzweig M. L. 1968. Net primary productivity of terrestrial community
prediction from climatological data, American Naturalist, 102, 74—76.
Roih W. 1901. Food: its search, capture and preparation, North Queensland Ethr
graphy Bulletin No. 3.
Rothenbuhler W. C. 1964. Behaviour genetics of nest cleaning in honey be
American Zoologist, 4, 111—123.
Sailer D. L., Gaulin S. J. C., Booster J. S., Kurland. J. A. 1985. Measuring i
relationship between dietary quality and body size in primates, Primat
26, 14—27.
Sarich V. M. 1971. A molecular approach to the question of human origins. 1
Dolinhow P. and Sarich V. M. (eds.), Background for Man, Boston: Lit
Brown, pp. 60—81.
Sarich V. M. 1983. Appendix: retrospective of hominoid macromolecular systen
tics. In: Ciochon R. and Corruccini R. S. (eds.), New Interpretations of A
and Human Ancestry, New York: Plenum Press, pp. 137—150.
Sarich V. M., Wilson A. C. 1967. Immunological time scale for hominoid evo
tion, Science, 158, 1200—1203.
Sarich V. M., Wilson A. C. 1973. Generation time and genomic evolution in f
mates, Science, 179, 1144—1147.
Sartono S. 1982. Characteristics and chronology of early men in Java, Cong
International de Paleontologie Humaine, Nice Pretirage, pp. 491—533.
Schaller G. B. 1972. The Serengeti Lion, Chicago: University of Chicago Pre
Schaller G., Lowther G. R. 1969. The relevance of carnivore behaviour to
study of early hominids, Southwestern Journal of Anthropology, 25, 30’
341.
Schell T. 1981. An American finds out that running with Pygmies is no sn
challenge, Sports Illustrated, 11, 6—7.
Schmidt-Neilson K. 1975. Animal Physiology: Adaptation and Environrm
Cambridge: Cambridge University Press.
Schoener T. W. 1983. The controversy over interspecific competition, Ameri
Scientist, 70, 586—595.
Scott K- 1986. Review of Faunal Remains from Klasies River Mouth by L. R. I
ford, Journal of Archaeological Science, 18, 89—91.
Semenov S. A. 1964. Prehistoric Technology, Bath: Adams and Dart. [Имее
перевод: С. А. Семенов. Первобытная техника (Опыт изучения древней!
орудий н изделий по следам работы).—М. — Л.: Изд-во АН СС
1957.]
Seyfarth R. М., Cheney D. L., Marler Р. 1980. Monkey responses to three di
rent alarm calls: evidence of predator classification and a semantic com:
nication, Science, 210, 801—803.
Shackleton N-, Opdyke N. D. 1976. Oxygen-isotope and palaeomagnetic sti
graphy of equatorial Pacific core V28—238, Late Pliocene to Latest I
stocene. In: Cline R. M. and Hays J. D. (eds.), Investigation of Late Qur
nary Paleoceanography and Paleoclimatology, Geological Society of An
ca Memoir No. 145, 449—464.
Shipman P. 1982. Taphonomy of Ramapithecus wickeri at Fort Ternan Ke:
University of Missouri Museum Brief No. 26.
Литература
355
Shipman Р. 1983. Life History of a Fossil, Cambridge, Mass: Harvard Univer-
sity Press.
Shipman P., Walker A. C., Van Couvering J., Hooker P., Miller J. 1981. The Fort
Ternan hominoid site, Kenya: geology, age, taphonomy and palaeoecology,
Journal of Human Evolution, 10, 49—72.
Short P. V. 1979. Sexual selection and its component parts: somatic and genital
selection, as illustrated by man and the great apes, Advances in the Study
of Behaviour, 9, 131.
Short R. V. 1980. The great apes, Journal of Reproduction and Fertility Supple-
ment, 28, 3—11.
Short R. V. 1981. Sexual selection in man and the great apes. In: Graham С. E.
(ed.), Reproductive Biology of the Great Apes, London: Academic Press,
pp. 319—341.
Sibley C., Ahlquist J. 1984. The phylogeny of hominoid primates as indicated by
DNA-DNA hybridization, Journal of Molecular Evolution, 20, 2—15.
Sigg H., Stolba A. 1981. Home range and daily march in a hamadryas baboon
troop, Folia Primatologica, 36, 40—75.
Silberbauer G. 1981. Central Kalahari Hunter—Gatherers, Cambridge: Cambridge
University Press.
Simons E. 1961. The phyletic position of Ramapithecus, Postilla, 57, 1—9.
Simons E., Pilbeam D. 1965. Preliminary revision of the Dryopithecinae (Pon-
gidae, Anthropoidea), Folia Primatologica, 3, 81—152.
Skelton R. R., McHenry H. M., Drawhorn G. M. 1986. Phylogenetic analysis of
early hominids, Current Anthropology, 27, 21—43.
Solomon J. D. 1931. The geology of the implementiforous deposits in the Nakuru
and Naivasha basins and the surrounding area in the Kenya Colony. In:
Leakey L. S. B., The Stone Age Cultures of the Kenya Colony, Cambridge:
Cambridge University Press, pp. 245—266.
Speth J. D., Spielmann K. A. 1983. Energy source, protein metabolism, and
hunter—gatherer subsistence strategies, Journal of Anthropological Archaeo-
logy, 2, 1—31.
Stacey F. D. 1969, Physics of the Earth, New York: Wiley.
Stahl W. R. 1965. Organ weights in primates and other mammals, Science, 150,
1039—1042.
Stanley S. M. 1973. An explanation of Cope’s rule. Evolution, 27, 1—26.
Stanley S. M. 1975. A theory of evolution above the species level, Proceedings
of the National Academy of Sciences, U.S.A., 72, 646—650.
Stanley S. M. 1979. Macroevolution: Pattern and Process, San Francisco:
Freeman.
Stern !. T. 1971. Functional Myology of the Hip and Thigh of Cebid Monkeys
and its Implications for the Evolution of Erect Posture, Biblioteca Primato-
logia No. 14, Basel: Karger.
Stern J. T. 1976. Before bipedality, Yearbook of Physical Anthropology, 19,
59—68.
Stern J. T., Susman R. L. 1983. The locomotor anatomy of Australopithecus
afarensis, American Journal of Physical Anthropology, 60, 279—317.
Steudel K. 1980. New estimates of early hominid body size, American Journal
of Physical Anthropology, 52, 63—70.
Street F. A., Grove A. T. 1975. Environmental and climatic implications of Late
Quaternary lake-level fluctuations in Africa, Nature, 261, 385—390.
Stringer C. 1984. Human evolution and biological adaptation in the Pleistocene.
In: Foley R. (ed.), Hominid Evolution and Community Ecology: Prehistoric
Human Adaptation in Biological Perspective, London: Academic Press,
pp. 55—84.
Strugnell R. G., Pigot C. D. 1978. Biomass, shoot production and grazing of
23*
356
Литература
two grasslands in the Ruwenzori National Park, Uganda, Journal of Ecology,
66, 73—96.
Struhsaker T. T. 1967. Ecology of vervet monkeys (Cercopithecus aethiops) in
Masai-Amboseli Game Reserve, Kenya, Ecology, 48, 891—904.
Suzuki A. 1969. An ecological study of chimpanzees in savanna woodland, Pri-
mates, 19, 103—148.
Suzuki A. 1975. The origin of hominid hunting: a primatological perspective. In:
Tuttle R. H. (ed.), Socioecology and Psychology of Primates, The Hague:
Mouton, pp. 259—278.
Szalay F. S., Delson E. 1979. Evolutionary History of the Primates, London:
Academic Press.
Tanaka J. 1976. Subsistence ecology of Central Kalahari San, In: Lee R. B. and
DeVore I. (eds.), Kalahari Hunter—Gatherers, Cambridge. Mass.: Harvard
University Press, pp. 98—119.
Tanner N. M. 1981. On Becoming Human, Cambridge: Cambridge University
Press.
Taylor C. R., Rowntree V. J. 1973. Running on two legs or four: which consumes
more energy? Science, 179, 186—187.
Taylor C. R., Schmidt-Nielson K., Raab J. L. 1970. Scaling of energetic cost of
running to body size in mammals, American Journal of Physiology, 219.
1104.
Teleki G. 1975. Primate Subsistence pattern: collector—predators and gatherer—
hunters, Journal of Human Evolution, 4, 125—184.
Teleki G. 1981. The omnivorous diet and eclectic feedings habits of chimpanzees
in Gombe National Park, Tanzania. In: Harding R. S. O. and Teleki G.
(eds.), Omnivorous Primates, New York: Columbia University Press,
pp. 303—343.
Tkoillay J. M. 1970. L’exploitation par les oiseaux de essaimages de fourmis et
termites dans une zone du contact savane-forSt en Cote d’Ivoire, Alauda, 38,
255—273.
Thorne A., Macumber P. 1972. Discoveries of Late Pleistocene man at Kow
Swamp, Nature, 238, 316—319.
Thomas D. H. 1978. Figuring Anthropology, New York: Holt, Rhinehart and
Winston.
Thompson D’Arcy. 1917. On Growth and Form, Cambridge: Cambridge Univer-
sity Press.
Thunnell R. C., Williams D. F. 1983. The stepwise development of Pliocene—
Pleistocene paleoclimate and paleooceanographic conditions in the Mediterra-
nean oxygen isotope studies of DSDP Sites 125 and 132. In: Meulen-
kamp J. E. (ed.), Reconstruction of Marine Paleoenvironments, Utrecht Micro-
palaeontological Bulletin No. 30, pp. Ill—127.
Tinbergen N. 1963. On aims and methods of ethology, Zietschrift fur Tierpsycho-
logie, 20, 410—433.
Tite M. S. 1972. Methods of Physical Examination in Archaeology, London:
Academic Press.
Tobias P. 1980. «Australopithecus afarensis» and «А. africanus»: critique and
alternative hypothesis, Palaeontologica Africans, 23, 1—17.
Torrence R. 1983. Time budgeting and hunter—gatherer technology. In: Bailey G.
(ed.), Hunter—Gatherer Economy in Prehistory: a European Perspective,
Cambridge: Cambridge University Press, pp. 11—22.
Trinkaus E. 1981. Neanderthal limb proportions and cold adaptation. In: Strin-
ger C. (ed.), Aspects of Human Evolution, London; Taylor and Francis.
Trinkaus E. 1982. The Shanidar Neanderthals, London: Academic Press.
Trinkaus E. 1983. Neanderthal postcrania and the shift to modern humans. In:
Trinkaus E. (ed.), The Mousterian Legacy, B. A. R. International Series
No. 164, Oxford: B. A. R„ pp. 165—200.
Литература
357
Trinkaus Е., Howells IF. W. 1979. The neanderthals, Scientific American, 241 (6),
118—133.
Trivers R. 1972. Parental investment and sexual selection. In: Campbell B. G.
(ed.), Sexual Selection and the Descent of Man, Chicago: Aldine, pp. 136—
179.
Turner A. 1984. Hominids and fellow-travellers: human migration into high
latitudes as part of a large mammal community. In: Foley R. (ed.), Homi-
nid Evolution and Community Ecology: Prehistoric Human Adaptation in
Biological Perspective, London: Academic Press, pp. 183--218.
Tuttle R. H. 1975. Knuckle-walking and knuckle-walkers: a commentary on some
recent perspective on hominoid evolution. In: Tutlle R. 11. (ed.), Primate
Functional Morphology and Evolution, The Hague: Mouton, pp. 203—212.
Uzell T., Pilbeam D. 1971. Phyletic divergence dates of hominoid primates:
a comparison of fossil and molecular data, Evolution, 25, 615—635.
Van Couvering Z. A. H. 1980. Community evolution in luist Africa during the
Late Cenozoic. In: Behrensmeyer A. K. and Hill A P. (eds.), Fossils in the
Making, Chicago: University of Chicago Press, pi». 272- 297.
Van Couvering J. A. H., Van Couvering J. A, 197(1. I'.nrly Miocene mammal
fossils from East Africa: aspects of geology, fnunistics, and palaeoecology.
In: Isaac G. and McGown E. (eds.), Human Origins: Louis Leakey and the
East African Evidence, Menlo Park, Ca.: W. A. Benjamin, pp. 155—207.
Van Schaik С. P., Van Hoof J. 1983. On the ultimate causes of primate social
systems, Behaviour, 85, 91—117.
Vincent A. 1985. Underground Plant Foods and Subsistence in Human Evolution,
PhD Dissertation, University of California, Berkeley.
Vita-Finzi C., Higgs E. S. 1972. Prehistoric economy in the Mount Carmel area
of Palestine, Proceeding of the Prehistoric Society, 36, I—37.
Vrba E. 1974. Chronological and ecological implications of the fossil Bovidae
at Sterkfontein archaeological site, Nature, 250, 19--23.
Vrba E. 1975. Some evidence on chronology and palneoecoiogy at Sterkfontein,
Swartkrans and Kromdraai from the fossil Bovidae, Nature, 254, 301 —
304.
Vrba E. 1976. The fossil Bovidae of Sterkfontein, Swartkrans, and Kromdraai,
Memoirs of the Transvaal Museum, 21, 1 —166.
Vrba E, 1980. Evolution, species and fossils: how does life evolve? South Afri-
can Journal of Science, 76, 61—84.
Vrba E. 1985. Ecological and adaptive changes associated with early hominid
evolution. In: Delson E. (ed.), Ancestors: the Hard Evidence, New York:
Alan Liss, pp. 63—71.
Walker A. C. 1967. Patterns of extinction among the subfossil Madagascan
lemuroids. In: Wright H. E. and Martin P. S. (eds.), Pleistocene Extinctions:
the Search for a Cause, New Haven: Yale University Press, pp. 425—432.
Walker A. C. 1976. Remains attributed to Australopithecus in the East Rudolf
succession. In: Coppens Y. et al. (eds.), Earliest Man and Environment in
the Lake Rudolf Basin, Chicago: University of Chicago Press, pp. 484—
489.
Walker A. C. 1981. Dietary hypotheses an human evolution, Philosophical Trans-
actions of the Royal Society, London, B292, 47—64.
Walker A. C. 1984. Extinction in hominid evolution. In: Nitecki M. H. (ed„),
Extinctions, Chicago: University of Chicago Press, pp. 119—152.
Walker A. C., Leakey R. E. 1978. The East Turkana hominids, Scientific Ameri-
can, 239(2), 54—66.
Walker A., Leakey R. E., Harris J. M„ Brown F. H. 1986. 2.5 Myr Australopithe-
cus boisei from west of Lake Turkana, Kenya, Nature, 322, 517—522.
Walker A. C., Pickford M. 1983. New postcranial fossils of Proconsul africanus
and Proconsul nyanzae. In: Ciochon R. and Corruccini R. S. (eds.), New
358
Литература
Interpretations of Ape and Human Ancestry, New York: Plenum Press,
pp. 325—354.
Ward S. C., Pilbeam D. 1983. Maxillofacial morphology of Miocene hominoids
from Africa and Indo-Pakistan. In: Ciochon R. and Corruccini R. S. (eds.),
New Interpretations of Ape and Human Ancestry., New York: Plenum Press,
pp. 211—238.
Washburn S. L. 1950. The analysis of primate evolution with particular reference
to the origin of man, Cold Spring Harbour Symposium on Quantitative Bio-
logy, 15, 67—78.
Washburn S. L. 1960. Tools and human evolution, Scientific American, 203(3),.
3—15.
Washburn S. L. 1967. Behaviour and the origin of man, Proceedings of the Royal
Anthropological Institute, 3, 21—27.
Washburn S. L., Lancaster C. S. 1968. The evolution of hunting. In: Lee R. B.
and DeVore I. (eds.), Man the Hunter, Chicago: Aldine, pp. 293—303.
Weiner J. S., Hellman K. 1960. The sweat glands, Biological Reviews, 25, 141 —
186.
Weiss M. L., Mann A. E. 1975. Human Biology and Behaviour: an Anthropolo-
gical Perspective, Boston: Little, Brown and Co., p. 89.
Western D. 1973. The Structure, Dynamics and Changes of the Amboseii Ecosys-
tem, PhD dissertation, University of Nairobi.
Western D. 1975. Water availability and its influence on the structure and dyna-
mics of a large mammal ecosystem, East African Wildlife Journal, 13,
365—380.
Western D. 1979. Size, life history and ecology in mammals, African Journal of
Ecology, 17, 185—205.
Wheeler P. 1984. The evolution of bipedality and loss of functional body hair in
hominids, Journal of Human Evolution, 13, 91—98.
Wheeler P. 1985. The loss of functional body hair in man: the influence oi ther-
mal environment, body form and bipedality, Journal of Human Evolution,
14, 23—28.
White M. 1978. Modes of Speciation, San Francisco: Freeman.
White T. D., Harris J. 1977. Suid evolution and correlation of African hominid
localities, Science, 198, 13—21.
White T. D., Johanson D. C., Kimbel W. H. 1983. Australopithecus africanus:
its phyletic position reconsidered. In: Ciochon R. and Corruccini R. S. (eds.),
New Interpretations of Ape and Human Ancestry, New York: Plenum Press,
pp. 721—780.
Wiens J A 1977. On competition and variable environments, American Scientist,
65, 590—597.
Williams G. C. 1966. Adaptation and Natural Selection, Princeton: Princeton
University Press.
Williams J. G. 1971. Guide to the Parks of East Africa, London: Collins.
Williamson P. G. 1981. Palaeoecological documentation of speciation in Cenozoic
molluscs from Turkana basin, Nature, 293, 437—443.
Wilson D. S. 1980, The Natural Selection of Populations and Communities,
Menlo Park, Ca.: Benjamin Cummings.
Wilson E. O. 1975. Sociobiology: the New Synthesis, Cambridge, Mass.: Harvard
University Press.
Wolpoff M. H. 1973. Posterior tooth size, body size and diet in South African
gracile australopithecines, American Journal of Physical Anthropology, 39,
375—394.
Wolpoff M. H. 1983, Lucy’s little legs, Journal of Human Evolution, 12 , 443—
453.
Wolpoff M. H. 1984. Evolution in Homo erectus: the question of stasis, Paleobio-
logy, 10, 389—406.
,Литература 359
Wood В. 1984. The origins of Homo erectus, Courier Forschungsinstitut Senken-
berg, 69, 99—111.
Wrangham R. W. 1977. Feeding behaviour of chimpanzees of Gombe National
Park, Tanzania. In: Clutton-Brock T. (ed.), Primate Ecology, London: Acade-
mic Press, pp. 504—537.
Wrangham R. W. 1980. An ecological model of female bonded primate groups,
Behaviour, 75, 262—300.
Wynne-Edwards V. C. 1962. Animal Dispersion in Relation to Social Behaviour,
Edinburgh: Oliver and Boyd.
Yellen J. E. 1977. Archaeological Approaches to the Present: Models for Reconst-
ructing the Past, London: Academic Press.
Zihlman A. L., Cronin J. E., Cramer D. L., Sarich V. M. 1978. Pygmy chimpanzee
as a possible prototype for the common ancestor of humans, chimpanzees and
gorillas, Nature, 275, 744—746.
Zihlman A. L., Lowenstein J. M. 1983. Ramapithecus and Pan paniscus: signifi-
cance for human origins. In: Ciochon R. and Corruccini R. S. (eds.), New
Interpretations of Ape and Human Ancestry, New York: Plenum Press,
pp. 677—694.
БЛАГОДАРНОСТИ
Приношу благодарность Jared М. Diamond, автору статьи,
напечатанной в сборнике «Sunda and Sahul» (New York — Lon-
don, 1977) (Diamond, 1977) и издательству Academic Press за
разрешение перепечатать отрывок из этой работы (стр. 296—
297 сборника). Выражаю также глубокую признательность
следующим организациям и авторам за разрешение воспроиз-
вести иллюстрации и таблицы из опубликованных ими работ:
Academic Press for figs 7.12, 9.9 and 9.10 from figs 4.2, 4.5 and
4.6, and Tables 6.5 and 9.10 from Tables 4.1 and 1.3 (Foley, 1984
a and b); Academic Press and the authors for fig 6.7D from fig 6
(Clutton-Brock, Harvey, 1977), fig 8.3 from fig 4 (Harris, 1980),
fig 8.11 from fig 4 (Hladik, 1977), figs 2.14, 2.15, 5.9 and 5.10 from
figs 2.1, 2.2, 2.3 and 2.4 (Roberts, 1984), figs 6.17 and 10.3 from
figs 3.3 and 3.2 (Stringer, 1984) and Table 8.3 from Table 2 (Dun-
bar, 1977); the American Anthropological Association and the
authors for fig 8.21 from fig 1 p. 26 (Dyson-Hudson, Smith, 1978)
and for fig 2.4 from fig 3 p. 302 (Pilbeam, 1986); the American
Geophysical Union for fig 2.13 from fig 6 (Kennett, 1977) copy-
right by the American Geographical Union; the American Museum
of Natural History’s James Arthur Committee for fig 6.16 from
fig 16 (R. D. Martin, 1983); American Scientist for fig 7.16 from
the graph in fig 8 (Potts, 1984b); Annual Reviews Inc. for Tab-
le 6.4 from Tables 6 and 7 (Frayer, Wolpoff, 1985) (c) 1985 by An-
nual Reviews Inc., The Athlone Press for figs 1.3, 6.2 and 9.2 from
figs 4, 43 and 38 (Eisenberg, 1981); Blackwell Scientific Publica-
tions Ltd for fig 6.10 from fig 3 (Western, 1979); Dr. Bonnefille
for fig 5.9 from fig 1 (Bonnefille, 1984); Cambridge University
Press for figs 6.5 and 6.6 from figs 6.9 and 6.11 (Schmidt-Nielson,
1975); The Ecological Society of America for fig 6.7a, b (c) from
fig 1 (Harested, Bunnell, 1979); Harvard University Press for
fig 9.16 from fig 30.6 (Maglio, Cooke, 1978); Little, Brown and
Co for fig 7.13 from p. 89 (Weiss, Mann, 1975) Macmillan Jour-
nals Ltd and the authors for fig 7.10 from fig 1 (Martin, May,
1981) copyright (c) 1981 Macmillan Journals Ltd; Plenum Publi-
shing Corporation and the authors for fig 2.8 from figs 8 and 10
(White, Johanson, Kimbel, 1983); Princeton University Press for
Table 6.4. from a table in figure 1, p. 278 (Smith, 1954); the Royal
Anthropological Institute of Great Britain and Ireland for fig 5.4
from fig 7 p. 401 (Foley, 1982); and Taylor and Francis Ltd and
the authors for fig 7.14 from fig 1 (Aiello, 1981) and fig 7.6 from
fig 1 (Andrew, 1981).
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Абердарес 285, 287
Австралия, освоение гоминидами 324
Адаптация
-— гоминид 64—67
— единица 80—81
— пределы 81—84
— природа 79—80
— как процесс решения задач 84—85
— и совершенство 81—84
— уровни 79—80
Адаптивная радиация
— гомииид 296, 307—311
— узконосых обезьян 199—203
Азия, освоение гоминидами 323
Активность млекопитающих, характер из-
менения 135—139
Аллеиа правило 329
Аллометрия 154—155
Альтруизм 77. 172
Амбосели 230, 256, 259, 265—266, 285, 287
Анагенез 303
Антропоморфизм, нежелательность 85—88
Археология 105—106
Африка
— видовое разнообразие 282
— выход гоминид за ее пределы 322—324,
335
•— полеоэкология 144—147
— и происхождение человека 52
Африканская природа гоминид, значение
для их эволюции 334—335
Берцово-бедренный указатель 330
Биотопы
— приматов 125—126
— тропические 131—133
— этапы освоения 328
Бипедия 47—48
— развитие 220—230
— и социальное поведение 186
— и терморегуляция 138—139
Бодо 51, 163
Брокен Хилл, см. Кабве
Вертешеллеш 51
Вид, определение 16
Видовое разнообразие 278—282
— африканских млекопитающих в позднем
кайнозое 297—298, 320
— в восточной Африке 282
— гомииид 309—311
— млекопитающих 279
— в плиоцене и плейстоцене 282—291, 297—
298
— теории 279—282
Видообразование
— у гоминид 306—311
— типы 303—306, 317—318
— н эволюция сообщества 296—298
Временной стресс 303—33!
Вымирание
— гоминид 314—316
и эволюция сообщества 297
Габон 257—258
Гадеб 46
Геологического времени шкала 32—34
Гиббоны 26, 29, 196
Глубоководные морсьяс отложения 320
Гомбе 80, 257—258
Гоминиды
— бипедия 220—230
— видовое разнообразие 292—293
— видообразование 306—31!
— время существования видов 294—295
— дальний пищевой поиск 139, 225
— дележ пищи 270—271
— жилища 230—236
— зависимость от воды 139—141
— изготовление орудий 274—275
— и изменения окружающей среды 148—
149
— изменчивость 293—295
— интенсивность обмена веществ 165
— ископаемые остатки, места находок 46
— контекст эволюционного развития 290—
292
— как крупные млекопитающие 153—191
— и люди 30—3!
— как наземные приматы 192—237
— определение 30—31
— оптимальная стратегия пищевого поиска
259—260
— освоение новых территорий 322—332
— отпечатки стоп 45. 187, 223
— первоначальные 38—41
— питание мясом 184, 262, 264—270
— плейстоценовые 48—52
— плиоценовые 45—48
— плотность популяции 219—220
— поведение 56—67
— поведение в связи с размножением 184—
188
— половой диморфизм 215—219
— и постоянство окружающей среды 147,
150—152
— предпочитаемые биотопы 245—248
— проблемы адаптации 120—121
— происхождение 4!—45
— пространственная организация 272—274
— пространственное распределение 311—314
— размеров тела оценка 157—163
— размеры зубов 263
— размеры мозга 188—191
— и сезонная изменчивость 258—275
— симпатрия 296
— состав рациона 165, 175—177, 178—181
— сосуществование 294—296
— социальная эволюция 219—221
— социальное поведение 184—188
— среда обитания 244—248
— стратегия пищевого поиска в дождливые
сезоны 259—261
— стратегия пищевого поиска в сухое вре-
мя года 26!—262
— стратегия спаривания 216—221
— таксономия 30
362
Предметный указатель
— темпы эволюции 318—321
— и тепловой стресс 135—139
— как тропические животные 123—152
— убежища 230—237
— участок обитания, величина 176—179
— филогения 52—56
— характер изменения активности 135—139
— численность групп 216—219
— эволюционные последствия размеров те-
ла 175—190
— экологические предпосылки видового
разнообразия 264—267
— экологическое сообщество 277—292
Гоминоиды
— локомоторное поведение 223
— миоценовые 45—46
— олигоценовые /96
— половой диморфизм 215 ' j
— социальная ^милюция z20
— темпы размножения и социальное по-
ведение 184—188
— эволюция 196—199
Группой отбор 78—79
Дарвиновская революция в естествознании
337
Датировка следов расщепления 37
Датировки способы 34—38
Джетнс отложения 323
Дожди
— и биомасса 221
— в тропической экологии 128—131
Дриопитековые 196
Европа, освоение гоминидами 323, 325—327
Единообразия принцип 108—109
Естественный отбор 72—79
— как дифференциальное воспроизведение
72—73
— единица 76—79, 84
— и конкуренция 242
— ограничении 73—76
— приводящий к адаптации 79—81
<Жизнь или обед», принцип 168
Жилище 188—189, 230—236
Зависимость от воды 139—141
Зубная эмаль 42
Изерния-ла-Пирета 323
Изменчивость 293—294
Илерет 46, 247
Импала, см. Aepyceros
Интенсивность основного обмена и размер
тела 163—105
Ископаемые гоминиды
— места находок 46—51
— распространение 46—51, 126—127
Ископаемые остатки
— двойственность происхождения 112, 290—
291
— в палеоантропологии 96—98
Ископаемые приматы, распространение
126—127
Кабве 51, 53, 163
Када-Гона 59
Калий-аргоновый метод датировки 36—37
Капапои 46
Касакати 257
Кидепо 285, 287
Кислорода изотопы стабильные, использо-
вание для датировки 37—38
Кладогенез 303
Клайбера кривая 163—164
Конкурентное исключение 304
Конкуренция 239—244
Кооби-Фора 46—49, 59, 65. 162, 178, 215, 235,
246—247, 273, 294—295, 313, 319
Копа закон 155—156
Коэволюция
— у гоминид и других млекопитающих 299
— у гомииид и приматов 299—302
— определение 299
— в пределах семейства Hominidae 302—
316
Кромдраай 46, [62, 246
Крупных хищников группа 321
Культура
— неадекватность термина 18—20
— определение 18—19
Ланиамок 46
Лев, см. Panther а
Летоли 45—46, 51, 187, 223, 246—247, 293—
295
Локомоторное поведение
---и бипедия 222—230
— — и пищевое поведение 202
---и эволюция узконосых обезьян 200—
202
Лотагам 40, 45—46, 247
Лукейно 46
Люди
— как животные 21—23
— место в природе 21
— определение 30—31
— как приматы 23—24
— происхождение современно1 л человека
52
— сходство с другими видами 16—18
— таксономия 31
Люси 45, 222—224
Макапансгат 46—47, 49, 246—247
.Макроэволюция 317—318
Маниара (озеро) 285, 287
Марсабит 2Ь5, 287
Масаи Мара 285, 287
Массивные австралопитековые. см. Austra-
lopithecus
Мауер 51
Мегафауна 284—289
Мелка-Контуре 46
Меру 285. 287
Мерчисон 285, 287
Мидл Аваш 46. 48, 246
Микуми 285, 287
Миоценовые гоминоиды 42—45, 196—199
Млекопитающие, эволюция и размеры тела
155—157
Мобильность животных 167—168
Модели (при изучении прошлого) 113—116
Мозаичные экосистемы 147, 150
Мозга размеры
— у гоминнд 188—190
— и обмен веществ у материнских особей
190
— и размеры тела 171—172, 188—[90
Молекулярные часы 39, 101—1С2
Моногамия 185, 216—217
Мутации 74
Мяса использование в пищу
— у гоминид 64—66, 103—104, 118—119
— у приматов 65—66, 101
— и сезонная изменчивость 261—262, 267—
269
Наблюдения при изучении прошлого, пря-
мые и косвенные 107—108
Наземный образ жизни 192—193
Предметный указатель
363
— у гоминид 215—236
Найроби 285, 287
Накуру 285, 287
Нариокотоме 46, 49
Нгоронгоро 285, 287
Неандертальцы 51—52, 104
— размеры тела 163
Недостающее звено 38
Неодарвинизм 72
Низшие узконосые обезьяны, эволюция
195—197, 202—203
Новый Свет, освоение гоминидами 324, 327
Огонь 235—236
Однолинейная модель эволюции 96—98
Олдувай 46—49, 51, 59, 65, И2, 162, 178,
235, 245, 263, 268, 271. 273, 286, 288, 294—
295, 321
Оледенение 142, 144, 147, 149
Олоргесаили 273, 315
Омо 46—47, 246—247, 286, 288, 295, 300, 313
Оптимальности принципы 88—90
Организменный уровень 70
Орудий изготовление
— древнейшее 59
— в ходе эволюции человека 105—106, 274
— у шимпанзе 59, 233
Освоение новой среды 296—297
— гоминидами 322—332
— крупными млекопитающими 325, 329
— и экология 324—332
/(-Отбор 172—175, 191
г-Отбор 172—174
Отпечатки стоп 45, 187, 223
Охота 64—66
-Охота-собирательство 102—104
Охотники-собиратели
— как модель эволюции челоевка 103—105
— охотничье поведение 183
— пищевой поиск, ориентированный иа
единственный центр активности 230—236
— состав рациона 178—181
— технология 330—331
Павианы
— адаптация к сезонной изменчивости 257—
259
— гнездовое поведение 230
— как модель в палеоантропологии 100
— охотничье поведение 179—183
— состав рациона 180
— см. также Papio, Theropithecus
Палеоантропология
— аналогии 98—10!
— и ископаемые остатки 96—98
— методы ] 16—120
— «модельные виды» 99—100
— и приматы 98—99
— сравиеине с другими науками 107—108
— и этнография 103—105
Палеоклимат
— глобальные изменения 142—144
— в плейстоцене 146
— и тектонические изменения 142
— циклы 144
Палеомагнетизм, использование для дати-
ровки 37
Палеоэкологические изменения
— влияние на эволюцию гоминид 141—152,
320—321
— и наземный образ жизни 194—195
Пенинж 46—47, 49, 134, 245—247, 263
Петралона 51, 163
Пильтдаунская находка 114
Питание мясом, см. мяса употребление в
пищу
Питание семенами у ранних гоминид, ги-
потеза 100
Пищевой поиск 175—184
Пнщи дележ 271
Пищи качество и размеры тела 165—166
Плейстоценовые среды обитания 57—59,
144—146
Поведение и генетика 92—94
Поведенческая эволюция 20
— у гоминид 94
Поведенческая экология 90—94
Полигиния 213, 218—219
Половой диморфизм у гоминид 215—219
Половые органы, размеп. и социальная си-
стема 216—217
Поидауиг 195
Потоотделение 133
Прерывистого равновесия теория 316—318
Приматов эволюция 25—26, 39—41
— и локомоторное поведение 201—202
— н наземный образ жизни 194—199
— и распространение 126—127
— и тектонические изменения 25
Приматы
— видовое разнообразие в плиоцене и плей-
стоцене 284—289
— классификация 27—29
— наземный образ жизни 193—195
— в палеоантропологии 98—102
— и питание мясом 65—66
— и предпочитаемые биотопы 125—126
— происхождение 39
— размеры тела 158
— распространение 24—26, 123—127
— сезонная изменчивость поведения 256—
258
— социоэкология 208—215
— участок обитания величина 166—167, 177
— характеристика 23—30
Продолжительность жизни 169
Продуктивность/биомасса, отношение 324,
328
Пятнистая гиена, см. Crocuta
Радиоуглеродный метод датировки 36—37
Размножения темпы 170—172
Расселение гоминид 261—265. 322—327
Растителная пища 179, 25!—252
Редукционизм 20—21
Резанные отметины 184
Ретроспективный взгляд иа эволюцию че-
ловека 68—69
Решение задач, в теории эволюции 84—94
Родительская забота о потомстве 64, 188—
189
Родственный отбор 76—77
Роста скорость 64
Руаха 287, 295
Саванна
— определение 132
— развитие 144—147
— ресурсы 25!—255
— сезонная изменчивость 250—255
— экология 248—250
Самбуру/Исиоло 285, 287
Сванскомб 51
Сварткранс 47, 49, 162, 246—247
Свиньи 283—284, 287—289. 320
Сезонная изменчивость
— и адаптивные реакции 255—258
— и видовое разнообразие гоминид 264—267
— и доступность пищевых ресурсов 251—
254
— и конкуренция 242—244
— и крупные млекопитающие 269—270
364
Предметный указатель
— и освоение новых территорий 324
— и питание мясом 261—262
— плиоценовой и плейстоценовой фауны
272—273
— сред обитания ранних гоминид 244—248
— в тропиках 128—132
Семлики слои 59
Серенгети 267, 285, 287
Сивалик 145, 196
«Склады» каменных орудий 233—234
Совершенство
— и адаптация 81—82
— пределы 81
Совокупная приспособленность 77
Сообществ структура
— в восточной Африке 282—289
— в плиоцене и плейстоцене 282—289
Сообществ эволюция 296—298, 316
Сообществ экология 296—299
Социальные отношения 171—172
Социобиология 91
Социоэкология
— наземных приматов 209—215
— сравнительный метод 208—209
Сравнителньый подход 10!
Среды обитания 70—71
— ранних гоминид 244—248
Стеиотопный виц 32?
Стеркфонтейн 47, 49, 160, 162, 246—247
Стратегия 85—88
Суточные перемещения 166
Табарин 40, 45, 223, 246, 293
Таксономия 38
Таунг 46
Тафономия 111—113
Тела размеры
— у гоминид 157—162
— и качество пищи 165—166
— и наземный образ жизни 194—195
— и г- и К-отбор 173—175, 190—191
— и отношения «хищник—жертва» 167—
168
— и продолжительность жизни 169
— и размеры мозга 171—172, 188—190
— и социальные отношения 170—171
— и темпы размножения 169—170
— и терморегуляция 168—169
— в эволюции гомннид 175—190
— и эволюция 154—157
— экологические корреляты 163—175
Темпы эволюции
— гоминид 318—321
— и изменения окружающей среды 151—
152, 320
— полорогих 320
— постановка проблемы 316—318
Теория (при изучении прошлого) ИЗ—116
Теория средних значений 109—111
Теория эволюции
— и г- и Л-отбоо 173
— прерывистое равновесие 316—318
— и теория игл 93—94
— см. также Естественный отбор
Тепловой стресс 135
Термолюминесцентный метод датировки
37
Терморегуляция
— И бипедия 137
— у гоминид 133—135
— н зависимость от воды 139—140
— у млекопитающих 133
— и охота 135
— и утрата шерстяного покрова 139
— и пропорции тела 138
— и размеры тела 168—169
— и характер активности 135—139
Территориальное поведение 272—274
Тетис (море) 142—143
Технология 54—59
Травоядные 284—289
Трансвааль, места находок ранних гоми-
иид 246
Тропики
— биомасса и продуктивность 130
— биотопы 131—133
— и распространение приматов 123—127
Тропическая палеоэкология 144—147
Тропическая экология 127—133
Трофическая структура 71—72
Тсаво 285, 287
Туркана (озеро) 45, 149, 286, 288, 309, 317
Убежище 230—237
Убейдия 323
Узконосые обезьяны
— адаптивная радиация 199—203
— видовое разнообразие 205—206
— локомоторное поведение, эвглючця А'ч ••
230
— и наземный образ жизни 208—215
— социоэкология 209—215
— эволюционная экология 199—208
— эволюция 196—199
— экологическая дивергенция 207
Уникальность
— всех видов 16
— исторические факторы 18
— уровни 15
— природа человека 15—18, 121
Уранового ряда изотопы, использование
для датирования 37
Участок обитания 166—167, 177—179
Фаюм 195—196
Фенотипический набор 89
Филогенетическое наследие 334
Форт Тернан 112, 145
Фтор, датирование по его содержанию 35—
36
Хадар 45—46, 48, 222—223, 246—247, 277,
293—295, 310
«Хищник — жертва» отношения
— и размеры тела 167—168
Хищные (отряд) 283—284, 287—288. 298
Человек 31
Чесованья 47, 49, 246—247
Шимпанзе
— адаптация к сезонному стрессу 256—258
— гнездовое поведение 230
— изготовление орудий 233
— локомоторное поведение 228, 230
— как модель в палеоантропологии 99
— охотничье поведение 182
— состав рациона [80—183
— суточная активность 136
— сужение» термитов 59
— эволюция 204—205
Шкала
— геологическая временная 32—34
— эволюции человека 32—41
— эволюционных процессов 32—34
Эволюционная экология 70—94
Эволюция
— и ископаемые остатки 96—98
— н филогения 96—97
Эволюция человека
— однолинейная модель 96—98
Указатель латинских названий
365
— резюме 67—68
— хронология 38—41
— шкала 32—41
Эвритопиый вид 322
Эгоистичный ген 77
Экзогения, адаптивные преимущества 329,
335
Экологическое сообщество
— определение 277
— сообщество гомииид 277—278
Экология 70—72
Электронный парамагнитный резонанс
(ЭПР), использование для датировки
37
Энцефализации коэффициент 171—172, 270—
271
Ява 53, 323
Язык
— гомннид 66—67
— приматов 19, 100—10!
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Aegyptopithecus 43, 195—197
Aepyceros, характер изменения активности
136
Apis mellifera 92
Australopithecus
— вымирание 314—316
— питание 263—264
— происхождение 46—48
— пространственное распределение 313—314
— размеры тела 158—162
— состав рациона 180—183
— филогения 53—55, 307, 309—31!
— afarensis 46—47, 50,53—54, 161, 163, 178,
216, 223—224, 293, 295, 309, 313, 318
— africanus 47, 50, 53—54, 158—163, 189, 216,
293. 295. 309, 313, 3U
— boisei 47, 54, 160, 162, 294—2S5, 300, 309,
3l5, 318
— crassidens 47, 294 , 309
— robustus 47, 50. 54, 158, 160, 162, 294, 300,
309, 313, 315, 318
Australopithecus — Homo, дивергенция 307—
311
Cercopithecus
— адаптация к сезонным изменениям 256
— убежища 230
Crocuta crocuta, характер изменения актив-
ности 136
Dryopithecus 43—44
Hrythrocebus patas 125, 230
Gigantopithecus 42—44, 197, 203
Gorilla
— локомоторное поведение 228—229
— суточная активность 136
— эволюция 204—205
Ното
— питание 264
— половой диморфизм 215—216
— происхождение 46—52
— пространственное распределение 311—
314
— состав рациона 180—184
— филогения 52—56
— характеристики 48—52
— Australopithecus — Ното, дивергенция
307-31!
— erectus 16—17, 49—50, 54—56. 67, 97. 161,
163, 189, 215—216, 235—236, 269, 292, 294—
295, 303, 309, 313—315, 318—319, 321, 323—
325, 329—330
— ergaster 294
— habiiis 48—50, 54—56, 67, 97, 163, 189, 216»
294—295, 309
— sapiens 30—31, 51—52, 56, 67, 97. 104, 161.
163, 189, 303, 319, 323, 329
— sapiens sapiens 15—16, 30—31. 52, 55, 97,
138
Kenyapithecus 43—44, 112, 197
Limnopithecus 45, 196
Macaco fuscata
— отмывание клубней 19, 93
— распространение 123—125
Масаса mulatto 123
Масаса sylvanus 125
Oligopithecus 43, 196
Ouranopithecus 43—44, 197
Paidopithex rheanus 226
Panthera leo, характер изменения активно-
сти 136
Papio
— и гоминиды 300—301
— хищническое поведение 65, 258—259
Pliopithecus 38, 44, 196—197
Pongo 29. 197, 203—204. 216—217
Proconsul 45. 197, 201, 203, 226
Propliopithecus 196
Ramapithecus 42—44, 97, 112, 196—197, 203
Siuaplthecus 42—44, 197, 203
Theropithecus
— адаптация к сезонной изменчивости 257
— и гоминиды 300—301, 315
— как модель для изучения эволюции че-
ловека 100
— суточная активность 136
Zinianthropus 310
ОГЛАВЛЕНИЕ
Предисловие редактора перевода .................................... 5
Предисловие........................................................ 7
Введение...........................................................11
Глава 1. Проблема человеческой уникальности........................15
1.1. Отличаются ли люди от других существ?.....................15
1.2. Наше место в природе......................................21
Глава 2. Рассказ об эволюции человека..............................32
2.1. Шкала эволюции человека...................................32
2.2. Ископаемые гоминиды.......................................41
2.3. Поведение гоминид.........................................56
2.4. За рамкамн повествования..................................67
Глава 3. Эволюционная экология.....................................70
3.1. Эволюция и экология.......................................70
3.2. Естественный отбор........................................72
3.3. Отбор и адаптация.........................................79
3.4. Адаптация как процесс решения задач.......................85
Глава 4. Пути в прошлое............................................95
4.1. Палеоантропология.........................................95
4.2. Природа прошлого.........................................107
4.3. Установление связей между прошлым и настоящим . 109
4.4. Методы изучения прошлого.................................113
4.5. Аналитическая палеоэкология..............................120
Глава 5. Гоминиды как тропические животные........................123
5.1. Эволюция гоминид и тропики ..............................123
5.2. Особенности тропической экологии.........................127
5.3. Ранние гоминиды в условиях тропиков......................133
5.4. Палеоэкологические изменения.............................141
Глава 6. Гоминиды как крупные млекопитающие.......................153
6.1. Размеры тела и эволюция..................................154
6.2. Размеры тела у гоминид...................................157
6.3. Экологические корреляты размеров тела....................163
6.4. Адаптивные последствия увеличения размеров гоминид в процес-
се эволюции....................................................175
6.5. Размеры тела, размеры головного мозга и адаптация гоминид 188
Глава 7. Гоминиды как наземные приматы............................192
7.1. Эволюция приматов и наземный образ жизни.................193
7.2. Эволюционная экология узконосых обезьян..................199
Оглавление 367
7.3. Адаптивные характеристики наземных приматов .... 208
7.4. Наземный образ жизни гоминид.........................215
Глава 8. Гоминиды в средах с выраженной сезонностью изменчивостью 238
8.1. Конкуренция..........................................239
8.2. Среда обитания ранних гоминид........................244
8.3. Экология саванн......................................248
8.4. Сезонная изменчивость и доступность пищевых истоков . 250
8.5. Реакция на сезонную изменчивость: сравнительный анализ 255
8,6. Стратегии пищевого поиска у гоминид в средах с сезонной
изменчивостью.................................................258
8.7. Гоминоиды на все сезоны...................................263
8.8. Условия экологического разделения.........................264
8.9. Эволюционные н поведенческие последствия питания мясом 269
Глава 9. Гоминиды и эволюция сообщества............................276
9.1. Сообщество гоминид........................................277
9.2. Факты, свидетельствующие о разнообразии гоминид . . 292
9.3. Экология сообщества и коэволюция..........................296
9.4. Образование и вымирание видов.............................302
9.5. Темпы эволюции............................................316
Глава 10. Гоминиды как расселяющиеся животные......................322
10.1. Исход из Африки..........................................322
Заключение.........................................................333
Литература....................................................... 338
Благодарности......................................................360
Предметный указатель...............................................361
Указатель латинских названий.......................................364