/






















































































































































































































































































































































































































































































































Текст
РУКОВОДЯЩИХ ФОРМ
ИСКОПАЕМЫХ
ФАУНЫ И ФЛОРЫ
ЗАПАДНОЙ
СИБИРИ
ГО СГЕОЛТЕХИЗДАТ
ЗАПАДНО-СИБИРСКОЕ ГЕОЛОГИЧЕСКОЕ УПРАВЛЕНИЕ
ТОМСКИЙ ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ПОЛИТЕХНИЧЕСКИЙ ИНСТИТУТ имени С. М. КИРОВА
АТЛАС
РУКОВОДЯЩИХ ФОРМ
ИСКОПАЕМЫХ ФАУНЫ И ФЛОРЫ
ЗАПАДНОЙ СИБИРИ
ТОМ I
Под редакцией Л. Л. ХАЛФИНА
ГОСУДАРСТВЕННОЕ НАУЧНО-ТЕХНИЧЕСКОЕ ИЗДАТЕЛЬСТВО
ЛИТЕРАТУРЫ ПО ГЕОЛОГИИ И ОХРАНЕ НЕДР
МОСКВА 1955
АВТОРЫ:
А. В. Аксарин, А. Р. Ананьев, Р. Н. Бенедиктова,
М. Г. Горбунов, Р. Т. Грацианова, Л. И. Егорова,
В. А. Ивания, Л. Н. Краевская, П. С. Краснопеева,
И. В. Лебедев, М. П. Ломовицкая, О. К. Полетаева,
Л. А. Рагозин, Г. П. Радченко, М. А. Ржонсницкая,
А. Г. Сивов, В. Д. Фомичев, В. К. Хал-
фина, Л. Л. Халфин и С. В. Чернышева
ПРЕДИСЛОВИЕ
Значительные потребности промышленности в минеральном сырье
и крутой подъем сельского хозяйства требуют быстрейшего и полного
освоения минерально-сырьевых ресурсов, что, в свою очередь, обуслов-
ливает огромный разворот геологических работ, проводимых в Совет-
ском Союзе. Успешное выполнение этих работ в значительной степени
зависит от правильной и научно обоснованной методики проведения их.
Современная геологическая служба применяет многочисленные методы
при своих исследованиях. Среди них, как известно, большое значение
имеет палеонтологический метод, позволяющий наиболее правильно
и обоснованно проводить стратиграфическое расчленение геологических
толщ.
Тем не менее в последние годы было отмечено значительное отстава-
ние палеонтолого-стратиграфических работ, недооценка палеонтологиче-
ского метода и ограниченное использование его при геологических иссле-
дованиях.
Учитывая необходимость усиления использования палеонтологии
в практике полевых работ и составления справочников-определителей
в виде атласов руководящих форм, Западно-Сибирское геологическое
управление по инициативе бывшего главного геолога Управления
И. Н. Звонарева в соответствии с директивами Министерства геологии
предусмотрело в плане своих работ 1950 г. выполнение специальной те-
мы по составлению «Атласа руководящих форм ископаемой фауны
и флоры Западной Сибири». Эту большую работу намечено было выпол-
нить путем привлечения широких кадров специалистов-палеонтологов
томских высших учебных заведений, ВСЕГЕИ и палеонтологического ка-
бинета Западно-Сибирского геологического управления.
В составлении атласа приняли участие специалисты-палеонтологи
различных учреждений: Западно-Сибирского геологического управления,
Томского политехнического института, Томского государственного уни-
верситета, ВСЕГЕИ, Западно-Сибирского филиала Академии наук.
Общее руководство работой по составлению атласа и редактирование
его было возложено на Л. Л. Халфина.
При составлении атласа были использованы как фактические мате-
риалы, хранящиеся в ЗСГУ, ВСЕГЕИ и томских вузах, так и литера-
турные данные. Иллюстрационный материал был подготовлен палеонто-
логическим кабинетом Западно-Сибирского геологического управления.
Характер имеющегося материала в значительной мере определил со-
держание атласа, территорию, объем статей и степень детальности
отдельных описаний. В частности, совершенно не освещены вопросы
палеонтологии четвертичных отложений Западно-Сибирской низменности.
Охарактеризована в основном Саяно-Алтайская горная область.
По замыслу атлас является руководством, с помощью которого гео-
логи, работающие на территории Западной Сибири, смогут выполнять
предварительные палеонтологические определения в полевых условиях.
,ч
С другой стороны, он отражает современное состояние изученности иско-
паемой фауны и флоры Западной Сибири по состоянию на 1952 г. Кри-
тическое обсуждение материалов, помещенных в атласе, будет способ-
ствовать дальнейшему развитию работ в области палеонтологии и стра-
тиграфии Западной Сибири.
Работа была выполнена в весьма короткий срок следующими авто-
рами: А. В. Аксариным, А. Р. Ананьевым, Р. П. Бенедиктовой,
М. Г. Горбуновым, Р. Т. Грациановой, Л. И. Егоровой, В. А. Иванин,
Л. Н. Краевской, П. С. Краснопеевой, И. В. Лебедевым, М. П. Ломо-
вицкой, О. К. Полетаевой, Л. А. Рагозиным, Г. П. Радченко, М. А. Ржон-
сницкой, А. П. Ротаем, А. Г. Сивовым, В. Д. Фомичевым, В. К. Халфи-
ной, Л. Л. Халфиным и С. В. Чернышевой и отредактирована Л. Л. Хал-
финым.
Большая работа по подготовке атласа к печати и по переработке
иллюстрационного материала была выполнена под руководством
ст. теолога И. М. Мягкова сотрудниками палеонтологического кабинета
ЗСГУ Г. А. Сибирцевой, Н. П. Мельниковой и К. Н. Шмидт.
Всем указанным лицам руководство Западно-Сибирского геологиче-
ского управления выражает благодарность.
Так как атлас, несомненно, имеет свои недостатки, просьба ко всем
лицам и организациям прислать свои замечания для использования их
в дальнейшей практической работе.
Западно-Сибирское геологическое управление
ВВЕДЕНИЕ
Л. Л. ХАЛФИН
При подготовке атласа мы не могли по ряду причин ставить задачу
полного охвата всех групп ископаемых животных и растений, известных
в Западной Сибири. Так, в атласе совершенно не представлены разделы
микропалеонтологии, палеокарпологии и палеонтологии позвоночных.
Сводки по этим разделам, подобные настоящему атласу, желательны,
и их составление — задача ближайшего будущего.
Для палеонтологической характеристики той или иной системы мы
приводим не все группы ископаемых, известные в отложениях этой
системы, а лишь важнейшие, наиболее распространенные. Так, для кем-
брия приводятся материалы, главным образом по трилобитам и архео-
циатам, отчасти по водорослям, тогда как брахиоподы совсем опущены;
в Западной Сибири они известны, содержат в своем составе весьма ори-
гинальные формы, но распространены слабо и изучены плохо. Для
палеонтологической характеристики девона использованы брахиоподы,
мшанки, различные целентераты, растения и некоторые другие группы,
но совершенно исключены гониатиты, встречающиеся в западносибир-
ском девоне очень редко.
Не включены в атлас также и некоторые распространенные, но недо-
статочно изученные группы, например остракоды угленосных отложе-
ний ’, моллюски третичного и четвертичного периодов.
В атласе отсутствует четвертичная система: палеонтологически она
характеризуется главным образом пыльцой и семенами растений и ко-
стями млекопитающих, а четвертичные моллюски Западной Сибири
почти не изучены.
Атлас содержит материалы, которые публикуются впервые или пред-
ставляют собой в корне пересмотренные и подвергнутые ревизии опубли-
кованные данные. Это относится, например, к кембрийским трилобитам,
среди которых преобладают вновь описываемые роды и виды, разделяю-
щиеся на ряд новых стратиграфических комплексов; основная работа по
этой группе ископаемых выполнена А. Г. Сивовым. Впервые публи-
куются интересные материалы М. А. Ржонсницкой по руководящим фор-
,мам из числа брахиопод нижнего девона и Эйфеля Салаира. Новым
по своему характеру является описание верхнепалеозойской флоры,
сделанное Г. П. Радченко: придерживаясь узкого понимания объема
вида, Г. П. Радченко дает более детальную, чем в опубликован-
ных материалах, классификацию палеозойских растений, что значительно
расширяет список руководящих форм в составе этой флоры. Боль-
шое количество новых форм описано и среди других групп ископаемых,
в частности среди кишечнополостных, по неопубликованным материалам
А. П. Жмаева и Э. 3. Бульванкер.
1 Во время составления этого атласа не была еще закончена работа М. И. Ман-
дельштама, посвященная изучению ископаемых остракод Кузбасса.
5
Наряду с этим в атласе широко использованы и опубликованные
материалы, из которых заимствованы диагнозы ряда руководящих форм.
Среди них имеются достаточно полные и недавно опубликованные ра-
боты, доступные для заинтересованных лиц, например работа В. П. Не-
хорошева по девонским мшанкам Алтая, работа Л. Л. Халфина по
нижнедевонской фауне Горного Алтая и некоторые другие. Наличие
таких работ позволило дать описания ряда руководящих форм в виде
очень кратких диагнозов, причем имелось в виду, что в случае надоб-
ности читатель имеет возможность обратиться к первоисточникам.
Наоборот, формы, описанные в труднодоступных литературных материа-
лах, а тем более формы новые даются в атласе с подробными описа-
ниями. Так, с большими подробностями, приближаясь к монографи-
ческому, дается описание верхнепалеозойской флоры (Г. П. Радченко),
поскольку в этом описании, по существу, впервые и в противополож-
ность литературным данным к этой флоре применяется принцип дроб-
ных видов, что требует в ряде случаев разъяснения различных деталей.
В связи с этим нужно отметить, что достигнуть полного единообразия
в описаниях родов и видов не удалось ввиду большого количества уча-
стников работы. Это несущественное неудобство, однако, вполне компен-
сируется тем, что большой круг участников обеспечивает достаточно
компетентный отбор и описания руководящих форм из очень разнообраз-
ных групп ископаемых животных и растений
Особые трудности встретились при подготовке раздела, посвященного
силурийским брахиоподам: из Западной Сибири описано очень немного
форм из состава этой группы ископаемых, но в ряде литературных
источников имеются списки, в которых приведены многие важные и об-
щеизвестные руководящие формы. Не включить их в атлас было невоз-
можно, но описания их сибирских представителей никем не давались,
а коллекции, по которым эти формы установлены, в настоящее время
недоступны. Поэтому характеристики этих форм составлены по описа-
ниям их представителей из других областей и стран; равным образом
и их изображения заимствованы из литературы, не относящейся к геоло-
гии Западной Сибири.
В отношении иллюстраций, приведенных в справочнике, нужно сде-
лать общее разъяснение. Разумеется, мы во всех случаях стремились
приводить изображения (оригинальные или заимствованные из литера-
туры) местных экземпляров, но порой приходилось отступать от этого
правила: во-первых, в тех случаях, когда вообще не представлялось воз-
можности получить такие изображения; во-вторых, в том случае, когда
сибирские представители того или иного вида описаны по очень непол-
ным экземплярам, не дающим надежного представления о диагностиче-
ских признаках, или приведенные в литературе изображения неудовлет-
ворительны. В этом случае мы считали более полезным дать изображе-
ния того или иного -вида по экземплярам, хотя и не происходящим из
Западной Сибири, но достаточно полно представляющим его характер-
ные особенности.
Как известно, Саяно-Алтайская область на юго-западе и на востоке
выходит за административные границы Западной Сибири. Тем не менее
так как эта область представляет собой естественную, хотя и внутренне
сложную, геотектоническую единицу, мы считали невозможным исклю-
чить из атласа матералы, относящиеся к Рудному Алтаю, Минусинской
котловине, Западному Саяну и другим частям Саяно-Алтайской области,
лежащим за пределами административных границ Западной Сибири. Но
мы полностью исключили Полярный Урал, который лежит внутри этих
границ: в геологическом и палеонтологическом отношении он представ-
ляет собой единое целое с Уралом.
Описанию руководящих форм предпослан особый раздев, посвящен-
ный морфологии групп, описанных в атласе. Содержание этого раздела
6
также подчинено основной задаче атласа: в нем даются объяснения тех
элементов морфологии растений и животных и специальных терминов,
которые фигурируют в описаниях. Таким образом, этот раздел представ-
ляет собой лишь краткий справочник по указанным вопросам.
Описания руководящих форм даются по системам, за исключением
верхнего палеозоя, в котором выделены в особую главу угленосные отло-
жения. Описаны лишь роды и виды, описания более крупных системати-
ческих единиц (семейства, отряды и т. д.) не приводятся: эти сведения
(так же, как и дополнительные сведения по морфологии) могут быть
почерпнуты из общего руководства по палеонтологии, например из
«Основ палеонтологии» К. Циттеля и «Палеоботаники» А. Н. Кришто-
фовича.
В атласе применяется дробная родовая классификация, принятая
ныне в палеонтологии. Эта классификация мало известна широким кру-
гам геологов, а в геологической литературе, особенно старой, обычно
употребляются родовые названия в широком объеме. Однако отказаться
от новой классификации невозможно даже в издании, подобном настоя-
щему атласу. Это, конечно, создает некоторые неизбежные трудности:
геологи не найдут в атласе таких привычных названий, как Productus
punctatus, Splrlfer verneulll, Splrlfer martlanovl, Amplexus altalcus,
Pachypora cervlcornls и др. Все эти формы, разумеется, помещены
в атласе, но описаны под названиями, соответственно: Echlnoconchus
punctatus, Cyrtosplrifer verneulll, Platyrachella martlanovl, Triplasma
altalca, Thamnopora cervlcornls и т. д.
В конце книги, наряду с родовым и видовым указателями, помещен
указатель, разъясняющий старое (широкое) и новое (узкое) понимание
объема отдельных родов.
Синонимика описанных форм в атласе, разумеется, не приводится, но
когда, по правилам синонимики, старое, широко известное название ока-
зывается замененным новым и еще мало популярным, первое указы-
вается как синоним. Например, после названия рода Ajaclcyathus ука-
зывается, что его представители до недавнего времени описывались под
родовым названием Archaeocyathus. Это применяется и по отношению
к некоторым, наиболее широко известным названиям видов, когда эти
названия -изменены на основании новейших исследований.
Наименование форм в атласе дается несколько упрощенно по сравне-
нию со специальными палеонтологическими изданиями. Так, родовые
и видовые названия приводятся без указания года установления соот-
ветствующего рода или вида; типичные виды в родовых диагнозах ука-
зываются в сопровождении ныне принятого родового названия, а не
того, под которым они были первоначально опубликованы, и т. д.
Описанию руководящих форм предпосылаются краткие стратиграфи-
ческие очерки соответствующих отложений. В них приводится краткая
палеонтологическая характеристика систем, указываются основные лите-
ратурные источники и дается разъяснение тех местных стратиграфиче-
ских единиц, которые упоминаются в описаниях руководящих форм.
Подробнее других дается лишь очерк стратиграфии кембрийской системы,
написанный А. Г. Сивовым по новым материалам.
Издание, подобное настоящему атласу, охватывающее чрезвычайно
большой и с различной степенью детальности изученный материал, к тому
же подготовленное в короткий срок, конечно, не свободно от ряда недо-
статков. Этот атлас является лишь первым шагом, в дальнейшем пред-
стоит подготовка дополнительных выпусков к нему, а через несколько
лет, в течение которых, несомненно, будут получены в большом коли-
честве новые и важные материалы по палеонтологии и стратиграфии
Западной Сибири, — расширенное издание атласа. Кроме того, на оче-
реди стоит подготовка полных определителей по тем группам ископае-
мых, которые изучены уже достаточно полно, например по девонским
брахиоподам 1 и верхнепалеозойской флоре, подготовка справочника по
микрофауне и микрофлоре Западной Сибири, по палеокарполопии и дру-
гие справочные издания.
Сейчас, подводя итоги работы по подготовке настоящего атласа,
я не могу не упомянуть с признательностью еще раз о той готовности
содействовать этому начинанию, которую я встретил со стороны всех
участников работы и особенно со стороны коллектива кафедры истори-
ческой геологии Томского политехнического института. Работа в целом
могла быть осуществлена лишь благодаря поддержке, которую она
встретила со стороны главного геолога Западно-Сибирского геологиче-
ского управления И. Н. Звонарева и директора Томского политехниче-
ского института им. С. М. Кирова проф. А. А. Воробьева.
В заключение остановимся на некоторых указаниях по части сбора
органических остатков и их предварительных определений; некоторые ив
этих замечаний и указаний являются элементарными, но, как показы-
вает опыт, порой при сборах фауны и флоры геологи не соблюдают
и самых элементарных правил.
Определения остатков ископаемой фауны и флоры и достоверность
или, во всяком случае, надежность стратиграфических выводов из этих
определений в большой степени зависят от тщательности сборов. Нередко
в распоряжение палеонтолога, который должен дать стратиграфическое
заключение, поступает совершенно случайно и беспорядочно взятый ма-
териал, иногда лишь несколько обломков окаменелостей, исключающие
возможность сколько-либо точного их определения. В сибирских усло-
виях, когда мы имеем дело с большим количеством ископаемых фаун,
изученных очень слабо или совсем не изученных, такие «сборь!» часто не
позволяют сделать вывод о возрасте данной фауны или флоры. Совер-
шенно исключив из рассмотрения сборы органических остатков для
каких-либо специальных палеонтологических исследований, а также (в со-
ответствии с содержанием настоящего справочника) не останавливаясь
на отборе образцов для микропалеонтологических и микроботанических
анализов, мы приведем ниже ряд указаний, которыми следует руковод-
ствоваться при сборах фауны в обычных условиях работы геологической
партии.
Прежде всего, несколько замечаний в отношении поисков органиче-
ских остатков. В настоящее время большинство геологов вполне ясно
осознало все значение и всю важность использования палеонтологиче-
ского материала при геологическом картировании и проведении поисково-
разведочных работ. Известно, что в ряде случаев те или иные толщи
считались «немыми», лишенными органических остатков, только потому,
что поисками этих остатков не занимались. Сейчас, когда столь сущест-
венно возросли требования к палеонтолого-стратиграфической обоснован-
ности составляемых геологических карт, организация поисков и сборов
органических остатков должна входить в прямые обязанности каждого
геолога, ведущего съемку или поисково-разведочные работы.
Не отрицая возможности существования осадочных толщ, действи-
тельно лишенных органических остатков, в общем случае нужно считать
обязательными поиски фауны в любых карбонатных и обломочных отло-
жениях. Отметим некоторые особые случаи. Прежде всего, остановимся
на поисках фауны и флоры в сильно измененных, метаморфизованных
(мраморизованных) и перемятых известняках; это—древние, кембрийские
и докембрийские толщи или более молодые, попавшие в зоны смятия.
1 Из числа девонских брахиопод, которые изучены полнее, чем какая-либо дру-
гая группа ископаемых, и по которым имеется достаточно большая литература, в на-
стоящем атласе приведены лишь очень немногие, наиболее важные формы; это сде-
лано с целью предоставить больше места материалам, впервые публикуемым или
подвергшимся существенной ревизии.
8
Поиски фауны в таких известняках обычно связаны с значительными
трудностями, но Имеют большое значение.
Археоциаты, одна из наиболее распространенных групп кембрийской
фауны, часто с большим трудом различимы или совсем не различимы
в свежих изломах, но более или менее отчетливо видны на выветрелой
поверхности. Следует, однако, иметь в виду, что археоциаты, незамет-
ные на свежих изломах, часто становятся хорошо различимы и опреде-
лимы в прозрачных шлифах; поэтому, если установлено (путем осмотра
выветрелых поверхностей обнажения), что данный известняк является
археоциатовым, необходимо взять достаточное количество образцов этого
известняка для расшлифовки.
Трилобиты, составляющие важнейшую группу кембрийской фауны,
нередко обладают очень мелкими размерами; поэтому надо внимательно
осматривать разломьт известняка, иначе остатки мелких трилобитов могут
ускользнуть от взгляда наблюдателя.
Особое внимание, следует обращать на поиски граптолитов в глини-
стых сланцах силурийского возраста. Граптолиты составляют важнейшую
в стратиграфическом отношении группу силурийской фауны. Но встре-
чаются они лишь в виде тонких отпечатков, далеко не всегда бросаю-
щихся в глаза. Возможно, этим объясняется тот факт, что до настоящего
времени мы знаем мало местонахождений граптолитов в Западной Си-
бири (Рудный Алтай, Салаир).
Массивные, желвакообразные или неправильной формы, колонии
широко распространенной в известняках силура и девона группы стро-
матопор геологи нередко принимают за куски слоистого известняка, в то
время как эта группа кишечнополостных сейчас достаточно хорошо из-
учена и может быть полезной для решения стратиграфических вопросов.
Не изжито у нас также предубежденное и скептическое отношение
к мшанкам, которые, действительно, в течение длительного времени не
играли какой-либо роли в стратиграфии, но, благодаря трудам советских
палеонтологов, разработавших новые методы их изучения, мшанки сейчас
могут служить надежными стратиграфическими индикаторами. Поэтому
следует уделять должное внимание поискам и сборам как строматопор,.
так и мшанок.
Часто совершенно недостаточное внимание уделяется геологами поис-
кам органических остатков в красноцветных континентальных и лагунно-
континентальных отложениях девонской системы. Не говоря уже о воз-
можности находок в этих отложениях прослойков с морской фауной,
в них нередко содержатся остатки рыб и рыбообразных бесчелюстных
(в виде чешуй, обломков панцырей), а также остатки растений — как
поилофитов, так и более высоко организованных групп. Вполне возможны
находки пластинчатожаберных моллюсков, ракообразных и близких
к ракообразным групп и др. Подобные находки очень важны и могут
помочь надежному стратиграфическому расчленению этих порой очень
мощных и однообразных толщ.
Фауна угленосных верхнепалеозойских отложений Западной Сибири
бедна и однообразна; она представлена главным образом пластинчато-
жаберными моллюсками и остракодами. Встречаются они обычно в по-
родах тонкозернистых (аргиллиты и алевролиты), чаще всего в кровле
угольных пластов, прослоев углистых пород или слоев, обогащенных
растительными остатками.
В отношении количества отбираемых образцов можно сказать сле-
дующее: чем больше их взято, тем быстрее и точнее будут сделаны
определения, тем надежнее будут полученные стратиграфические выводы.
Если фауна обнаружена в толще, возраст которой является неизвестным
или спорным, нужно эту фауну собрать в большом количестве, не огра-
ничиваясь несколькими, подвернувшимися под руку, экземплярами. При
9
плохой сохранности фауны для ее определения также потребуются боль-
шие сборы.
В собранной коллекции состав фауны данного пункта должен быть
представлен возможно более полно. Наряду с крупными, бросающимися
в глаза формами, должны собираться и мелкие, наряду с формами,
встречающимися в изобилии, и более редкие формы (и вообще органи-
ческие остатки).
Нужно также иметь в виду, что нередко фауна различных (часто
соседних) слоев одной и той же толщи имеет совершенно различный
состав, причем состав фауны каждого отдельного слоя пород очень одно-
образен (один-два вида); это характерно, например, для многих нижне-
девонских толщ Алтая. Если геолог собирает даже большое количество
образцов из одного слоя и все они принадлежат одному-двум видам,
и если эти виды оказываются новыми, то вывод о возрасте этой фауны
может быть сделан лишь очень приближенный. В то же время сбор'ы
фауны в соседних слоях или соседних обнажениях могут значительно
обогатить коллекцию.
Во всех случаях, если имеется возможность, сборы должны произво-
диться послойно: из каждого слоя и из каждого пункта фауна или флора
должна собираться отдельно и иметь особую этикетку *. Соблюдение
этого правила абсолютно необходимо; из истории изучения ископаемой
фауны и флоры можно привести примеры, когда смешение образцов,
происходящих из различных пунктов, приводило к большим трудностям
и ошибочным стратиграфическим построениям.
Наибольшее значение имеют образцы, взятые из коренных обнаже-
ний, так как они точно привязываются к определенным слоям разреза.
Но большого внимания заслуживают также сборы образцов, котооые
в результате выветривания содержащих фауну известняков и мергелей
подвергаются естественной, природной препарировке. Такие образцы
нередко обладают прекрасной сохранностью и в изобилии встречаются по
пологим склонам, у подножья обнажений или в русле речек. Даже в тех
случаях, когда образцы не могут быть точно привязаны к определенным
слоям, они должны собираться в достаточном количестве, так как, бла-
годаря хорошей сохранности, облегчают определение фауны, заключен-
ной в породе.
Если порода содержит органические остатки в большом количестве,
кроме образцов, которые удалось извлечь на месте или собрать в есте-
ственно отпрепарированном виде, следует брать эту породу большими
кусками, чтобы извлечь из нее фауну в лаборатории.
При сборах фауны в конгломератах образцы, взятые из цемента,
должны иметь особую этикетку; особыми этикетками должны снабжаться
также экземпляры, взятые из каждой гальки или глыбы (в базальных
конгломератах вполне возможно присутствие в виде галек пород различ-
ного возраста).
Встречающаяся в ископаемом состоянии фауна беспозвоночных
в большинстве своем обладает известковым скелетом (археоциаты, ко-
раллы, мшанки, брахиоподы, моллюски, трилобиты и некоторые другие).
Такой скелет (раковины, панцыри и т. п.) хорошо сохраняется в извест-
няках и мергелях; но в песчаниках и глинистых породах он часто бывает
полностью выщелочен. В этом случае на месте, скажем, створки рако-
вины брахиоподы остается в породе пустота и два отпечатка—наружной
поверхности створки и ее внутренней поверхности (так называемое
внутреннее ядро створки). Расколов кусок породы, можно увидеть оба
эти отпечатка; необходимо' обязательно брать оба отпечатка, так как
они дополняют друг друга; один позволяет установить характер скульп-
1 Самое тщательное и достаточно полное этикетирование образцов совершенно не-
обходимо во всех случаях.
10
туры и иные детали внешней поверхности, а другой — те или иные эле-
менты внутреннего строения. Равным образом при сборе растительных
остатков рекомендуется брать отпечатки и противоотпечатки листьев.
В этикетках должны быть сделаны соответствующие пометки, чтобы
при разборке коллекции и последующем определении парные отпечатки,
относящиеся к одному объекту, могли быть сразу отделены от осталь-
ных образцов.
С большой тщательностью должен производиться отбор органических
остатков из керна буровых скважин; необходимо отобрать все органиче-
ские остатки, которые оказались в керне, разумеется документируя
стратиграфическое положение (глубину взятия) каждого из них. Вообще
при производстве геологоразведочных работ, в тех случаях, когда полез-
ное ископаемое залегает среди фаунистически охарактеризованных пород
и, следовательно, палеонтологический метод должен являться повседнев-
ным рабочим методом работы партии, сборы фауны и флоры должны
быть особо тщательными, систематическими, обязательно послойными
и обильными.
Настоящий атлас может придать сборам фауны и флоры целеустрем-
ленный характер: проводить поиски тех окаменелостей, которые позво-
лили бы сразу же, хотя и в первом приближении, решить вопрос о воз-
расте содержащих данную фауну отложений. В первую очередь такую
задачу может поставить перед собой каждый геолог в том случае, когда
он имеет дело с отложениями, возраст которых предположительно или
приближенно известен, и когда, следовательно, стоит задача подтвердить
или уточнить этот возраст. В этом случае количество руководящих форм
окажется сравнительно ограниченным, и, следовательно, облегчается
задача непосредственного сравнения встречающихся в данном пункте
ферм с описанными и изображенными в атласе руководящими ископае-
мыми. Нужно, однако, при этом иметь в виду, что в условиях геологиче-
ского маршрута, как правило, можно сравнивать найденные формы
с описанными лишь по внешним признакам, чего далеко не всегда доста-
точно для надежного определения их, поэтому дальнейшее уточнение
определений необходимо во избежание возможных ошибок.
В процессе определения органических остатков могут встретиться раз-
личного рода трудности, зависящие от факторов как частного характера
(например, степень сохранности фауны или флоры, количество собран-
ного материала и т. п.), так и от обстоятельств характера более общего,
таких, например, как степень изученности данной фауны или флоры.
Последнее обстоятельство в условиях Сибири имеет большое значе-
ние, так как наряду с фаунами и флорами, которые к настоящему мо-
менту достаточно хорошо изучены и состав которых более или менее
известен, мы нередко встречаемся с фаунами еще не изученными, не опи-
санными и порой очень своеобразными и самобытными, не имеющими
достаточно близких аналогов среди известных фаун других областей
и стран.
Во втором случае определение фауны и ее возраста требует длитель-
ного времени, использования обширной литературы и должно выпол-
няться специалистом. Наоборот, в случае когда в процессе работы
партии встречены в том или ином пункте органические остатки, принад-
лежащие фауне (или флоре), уже изученной по сборам из других пунк-
тов или районов, каждый геолог вполне может справиться с задачей
предварительного определения этих остатков. Решить эту задачу помо-
жет и настоящий атлас.
Многие группы ископаемых для определения их представителей тре-
буют изготовления и использования ориентированных шлифов; таковы
археоциаты, строматопоры, тетракораллы, табуляты, мшанки, а также
многие водоросли. Правда, и среди этих групп имеются очень типичные
представители, легко определяющиеся макроскопически с точностью до
11
рода (а иногда и до вида), причем порой уже родовое определение может
быть полезным для стратиграфических выводов, хотя обычно и доста-
точно широких. Но в общем случае все эти группы требуют изучения их
представителей в прозрачных шлифах, которые без труда могут быть
изготовлены, при наличии в распоряжении экспедиции шлифовальной
мастерской. В главе, посвященной характеристике морфологии различ-
ных групп ископаемых, для каждой из них указывается, каким образом
должны быть ориентированы изготовляемые шлифы, а также перечис-
ляются и характеризуются те элементы строения представителей данной
группы, которые должны в первую очередь приниматься во внимание
при родовых и видовых определениях.
Из числа тех групп ископаемой фауны, предварительные определе-
ния которых, в общем случае, не требуют изготовления шлифов, важней-
шими (наиболее распространенными) являются трилобиты, брахиоподы
и пластинчатожаберные моллюски. В отношении трилобитов необходимо
иметь в виду следующее: полные панцыри этих животных встречаются
сравнительно редко; обычно мы находим в изолированном вйде их части
(хвостовые отделы, части головного отдела). И хотя для многих трило-
битов в строении хвостового отдела имеются различные характерные
диагностические признаки, но наиболее дифференцированным строением
обладает их головной отдел, в особенности глабель. Следовательно,
наибольшее количество признаков, по которым определяются трилобиты,
относится к строению их головного отдела. Поэтому при сборах три-
лобитовой фауны следует обращать особое внимание на поиски именно
этой части их панцыря или (что, конечно, еще лучше) полных пан-
цырей.
Среди брахиопод также имеется значительное количество форм, обла-
дающих характерными внешними признаками (очертания и форма рако-
вины, характер арей и скульптуры и т. д.), позволяющими по ним делать
видовые определения. Однако во многих случаях, если не в большинстве,
оказывается необходимым выяснить те или иные детали внутренного
строения раковины. Многие из этих элементов внутреннего строения при-
урочены к макушечной части створок, другие располагаются на различ-
ных частях их внутренней поверхности.
Методика изучения внутреннего строения брахиопод зависит от того,
в каком состоянии сохранились они, т. е. от того, имеем' ли мы дело
с сохранившимися створками или только с их отпечатками.
В первом случае достаточно легко устанавливается наличие или
отсутствие, а также характер зубных пластин, перегородок, септалиума
и т. п. (см. стр. 41). Для этого достаточно слегка срезать (пришлифо-
вать), хотя бы с помощью подпилка, макушку той или иной створки
(или обеих створок) и рассмотреть полученный срез (пришлифованную
поверхность), на котором при достаточной сохранности фауны будут
видны в разрезе детали строения (см. рис. 81, 83, 87 на стр. 41—42).
Может быть, придется подвергнуть такой обработке не один, а несколько
экземпляров данного вида. Пришлифованную поверхность полезно слегка
протравить соляной кислотой, в результате чего картина разреза стано-
вится более отчетливой.
Отпечатки внутренней поверхности (внутренние ядра) изолированных
створок весьма благоприятны для наблюдений многих деталей внутрен-
него строения брахиопод (а также пластинчатожаберных моллюсков);
но нужно помнить, что на отпечатках как внутренних, так и внешних все
элементы рельефа той и другой поверхности створки представляются
в обратном, негативном виде (см. рис. 90 на стр; 43). С отпечатков
и внутренних ядер рекомендуется делать слепки, которые будут пред-
ставлять собой реставрацию внешней и внутренней поверхностей
створки и, следовательно, позволят более полно уяснить признаки и осо-
бенности данной формы. Такие слепки могут быть сделаны с помощью
12
любого пластичного материала, например пластилина, размягченного
воска, парафина, сургуча и т. п.
Сказанное выше в отношении брахиопод в общих чертах может
относиться и к определению пелеципод (пластинчатожаберных мол-
люсков) .
Граптолиты часто приходится изучать при значительных увеличениях
(с помощью бинокулярной лупы).
Если в процессе картирования или при выполнении других геологи-
ческих работ будет встречена фауна или флора, в составе которой пол-
ностью отсутствуют формы, включенные в настоящий справочник, это
будет служить указанием, что открыта какая-то новая, не изученная или
ранее в Западной Сибири не обнаруженная фауна. В этом случае она
должна быть собрана в достаточном количестве и передана для опреде-
ления специалистам. В том случае, если будут обнаружены какие-либо
из слабо распространенных в Западной Сибири групп ископаемых
(иглокожие, головоногие моллюски, ракоскорпионы и т. д.), следует так-
же произвести возможно более обширные и тщательные сборы.
В заключение нужно сделать предупреждение, что предварительные
определения являются делом очень серьезным, требующим большого вни-
мания и тщательного сравнения определяемых образцов с описанными
формами. Нередко те или иные дигностичесюие особенности форм
являются далеко не бросающимися в глаза; существует большое коли-
чество форм, внешне сходных, но в действительности различных и имею-
щих различное стратиграфическое распространение.
При недостаточной опытности и недостаточной внимательности воз-
можны ошибочные определения и, следовательно, ошибочные стратигра-
фические выводы. Поэтому предварительные определения, особенно вы-
полняемые в процессе полевых работ, впоследствии необходимо прове-
рить с помощью палеонтолога. В случаях особо ответственных, когда
палеонтологическими определениями будут руководствоваться для ориен-
тировки дальнейших поисково-разведочных работ, консультация специа-
листа должна быть обязательной.
ЭЛЕМЕНТЫ МОРФОЛОГИИ
ВАЖНЕЙШИХ ГРУПП ИСКОПАЕМЫХ
ЖИВОТНЫХ И РАСТЕНИЙ
П. С. КРАСНОПЕЕВА
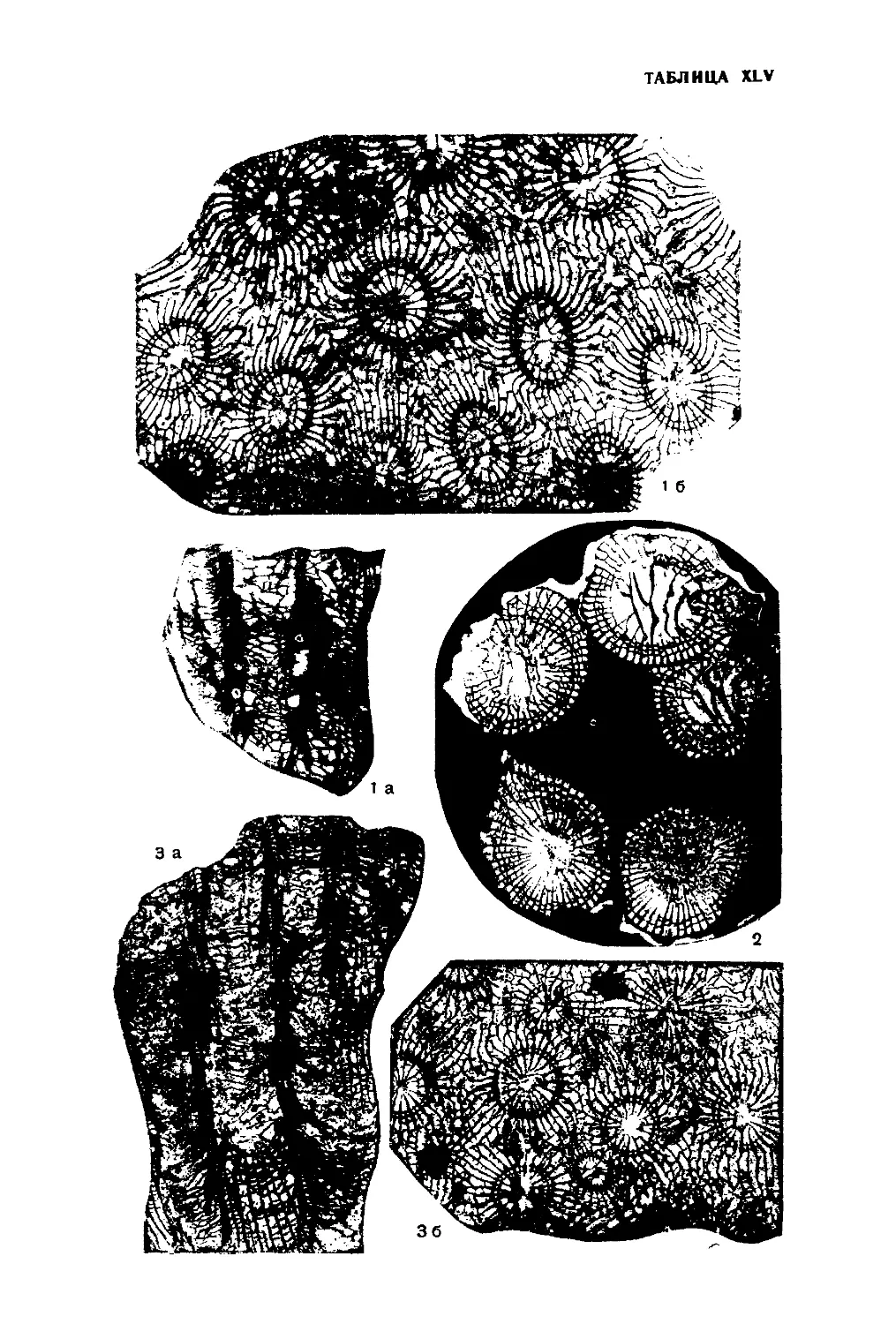
Тип Archaeocyathi. Археоциаты
Археоциаты являются древней, полностью вымершей группой орга-
низмов. Их геологическое распространение ограничивается нижним
и средним кембрием. Некоторые исследователи сближают археоциат
с губками и рассматривают их как подтип типа пористых. Основанием
для этого, в первую очередь, является пористый характер скелета архео-
циат.
Археоциаты делятся на два класса — Regularia (правильные) и 1гге-
gularia (неправильные). К первому классу относятся одиночные, редко
колониальные организмы, имеющие кубкообразную или производную от
нее форму тела.
Кубок правильных археоциат образован двумя пористыми стенками,
пространство между которыми (интерваллум) бывает заполнено септами
и другими скелетными образованиями. Мягкое тело животного поме-
щалось между стенками, а в некоторых случаях частично заполняло
внутреннюю полость кубка и выходило за его пределы. На это указывает
наличие у некоторых форм известковых пленок поверх наружной стенки
(роды Tegerocyathus, Erbocyathus), а также скелетных элементов в цен-
тральной полости кубка (род ? Retecyathus). Кроме того, наблюдаемая
в ряде случаев кайма из радиальнолучистого кальцита на наружной
и внутренней стенках кубков, возможно, возникает в результате заме-
щения органической ткани археоциат (см. рис. 125).
Ко второму классу относятся также двухстенные кубки, пространство
между стенками у которых заполнено серией радиально расположенных
трубочек многоугольного или округлого сечения. Настоящих стенок
у многих представителей этого класса нет. Каждая ячейка или трубка,
входящая в состав кубка, имеет пористые стенки. Трубки имеют наклон
от наружной поверхности кубка к внутренней полости. Типичными пред-
ставителями этого класса являются роды Syrlngocnetna и Archaeocyathus
(рис. 1—6).
Рис. 1. Род Syringocnema
Та у I.
Поперечное сечение кубка.
Увеличено
Рис. 2. Род Syringocnema
Т а у 1.
Продольное сечение куб-
ка. Увеличено
Р ис. 3. Род
Syringoc-
nema
Тау I.
Наружная
стенка
кубка.
Увеличе-
но
2 Зак. 2024
17
Наиболее разнообразным по составу и важным в геологическом отно-
шении является первый класс археоциат. Скелет представителей этого
класса состоит из двух пористых стенок — наружной и внутренней, в про-
странстве между которыми расположены опорные элементы скелета —
Рис. 4. Syrin-
gocnema
Т а у 1.
Внутренняя
стенка куб-
ка. Увеличе-
но
Рис. 5. Archaeocyathus
atlanticus Bill.
Поперечное сечение куб-
ка. Увеличено
Рис. 6. Archaeocyathus atlanti-
cus В II 1.
Продольное сечение кубка.
Увеличено
септы, днища, диссепименты и синаптикулы; элементы скелета у разных
родов присутствуют в различных сочетаниях. Имеющиеся данные
говорят за то, что археоциаты имели внутренний скелет. Об этом свиде-
тельствует наличие у некоторых родов тонких пленок поверх известко-
вых стенок кубка (рис. 7), а также в центральной полости (см. рис. 14
и 1556).
Рис. 7. Род Tegerocyathus
К г a s п., gen. п.
Наружная стенка с поровыми
каналами и пленкой и сложная
внутренняя стенка. X 4,5
Рис. 8. Род AJacicyathus
а—поперечное сечение
кубка, видны пронизан-
ные каналами (ситовид-
ные) внешняя и внут-
ренняя стенки и септы;
б—внешняя стенка;
в—участок внутренней
стенки между двумя
септами. X 3
Рис. 9. Thalamocyathus
G о г d.
а—щелевидные поровые
отверстия на внутренней
стенке кубка. X 5,5
В настоящей работе предложена новая схема классификации архео-
циат с септами (стр. 75, табл. 2). В этой классификации особое
значение придается характеру поровой системы стенок кубков. Важней-
шая роль этой системы, отражающей способ питания организмов, при-
18
знается многими исследователями. На это указывает, например, факт
установления семейств Ajacicyathidae, Ethmophyllidae, Thalamocyathi-
dae. '
У археоциат с септами по характеру пор установлено три основных
типа стенок кубков: а) тонкие, ситовидные, б) более толстые, пронизан-
ные поровыми каналами, в) несущие в себе щелевидные горизонтальные
отверстия.
Характер строения наружной и внутренней стенок кубка археоциат
принимается за руководящий признак при выделении семейств
(рис. 8—10). Встречающиеся иногда одностенные кубки во многих слу-
чаях рассматриваются как стеблевые части обычных двухстенных кубков
(рис. 11).
Рис. 10. Ethmophyllum Meek
а—поперечное сечение кубка;
б—продольное сечение кубка;
поровые каналы внутренней
стенки коленчато изогнуты. X 4,5
Рис. 12. Род Thala-
mocyathus G о г d
А—поперечное сече-
ние кубка; Б—про-
дольное сечение куб-
ка. В основании куб-
ка тонкая ситовидная
внутренняя стенка,
выше—сечение гори-
зонтальных колец;
а—участки септ в
продольном сече-
нии. X 4
Рис. 11 Род Aja-
cicyathus В е d f.
Схема развития
кубка. В нижней
части кубка одна
стенка, х 12
Некоторые наблюдения показывают, что на ранних стадиях индиви-
дуального развития большинство археоциат имеет ситовидное строение
стенок; более сложная внутренняя стенка с шелевидныМ|И отверстиями,
открытыми е сторону центральной полости кубка, развивается позднее
(рис. 12).
Септы представляют собой вертикальные пористые перегородки,
разделяющие междустенное пространство (интерваллум) на более или
менее равные камеры (см. рис. 7). Количество септ и характер пор у раз-
личных видов бывают различны. Относительное количество септ в кубке
является одним из наиболее важных признаков вида; отношение числа
септ к диаметру кубка в миллиметрах представляет собой так называе-
2*
19
мый септальный коэффициент — существенный элемент видового диаг-
ноза. Однако величина септального коэффициента меняется в процессе
роста кубка; более или менее постоянной она становится лишь на взрос-
лых стадиях развития цилиндрического кубка; поэтому сопоставления по
септальному коэффициенту рекомендуется делать для сечений примерно
одинакового диаметра.
Днищами называются горизонтальные пористые перегородки,
имеющиеся в интерваллуме кубков многих археоциат обоих классов
(рис. 13).
Рис. 13. Род Co cinocyathus
Born.
Поперечное скошенное сечение
неполного кубка:
а—септы; б—днища. X 5
Рис. 14. Род Loculocyathus
V о 1 о g d.
а—продольное сечение кубка;
б—поперечное сечение кубка;
в—пузырчатая ткань. X 3,8
Диссепименты или пузыр-
чатая ткань представляет собой не-
пористые тонкие известковые пленки
(рис. 14).
Синаптикулы — стержневид-
ные перемычки между септами
(рис. 15).
Наличие или отсутствие того или
иного элемента скелета является ха-
рактерным признаком рода; эти при-
знаки могут быть не выражены лишь
Рис. 15. Род Annulofungia Krasn., на самых ранних стадиях онтогенеза,
gen. п. Характер строения вышеуказанных
Поперечное сечение кубка; скелетных элементов и их пор, а так-
а-синаптикулы. х 4,7 же относительное количество их
в кубке являются руководящими признаками вида.
Кроме вышеуказанных признаков, при определении вида принимается
во внимание так называемый коэффициент массивности кубка, т. е. от-
ношение радиуса кубка к ширине скелетной части кубка. Ширину ске-
летной части кубка составляют толщина его стенок плюс ширина про-
странства между ними (интерваллума). В процессе роста поперечник
кубка увеличивается быстрее, чем ширина интерваллума, благодаря
20
чему коэффициент массивности тоже увеличивается, хотя и в незначи-
тельной степени.
Например, для вида Ethmophyllum tuvaense К г a s п. вычисление
коэффициента массивности производится следующим образом:
диаметр кубка 6 мм
число септ....................60
ширина скелетной части кубка . 1,7 мм
6°
септальный коэффициент =-^-=10,0
коэффициент массивности кубка=-ру-«2.
В. К. ХАЛФИНА
Тип Coelenterata. Кишечнополостные
КЛАСС HYDROZOA
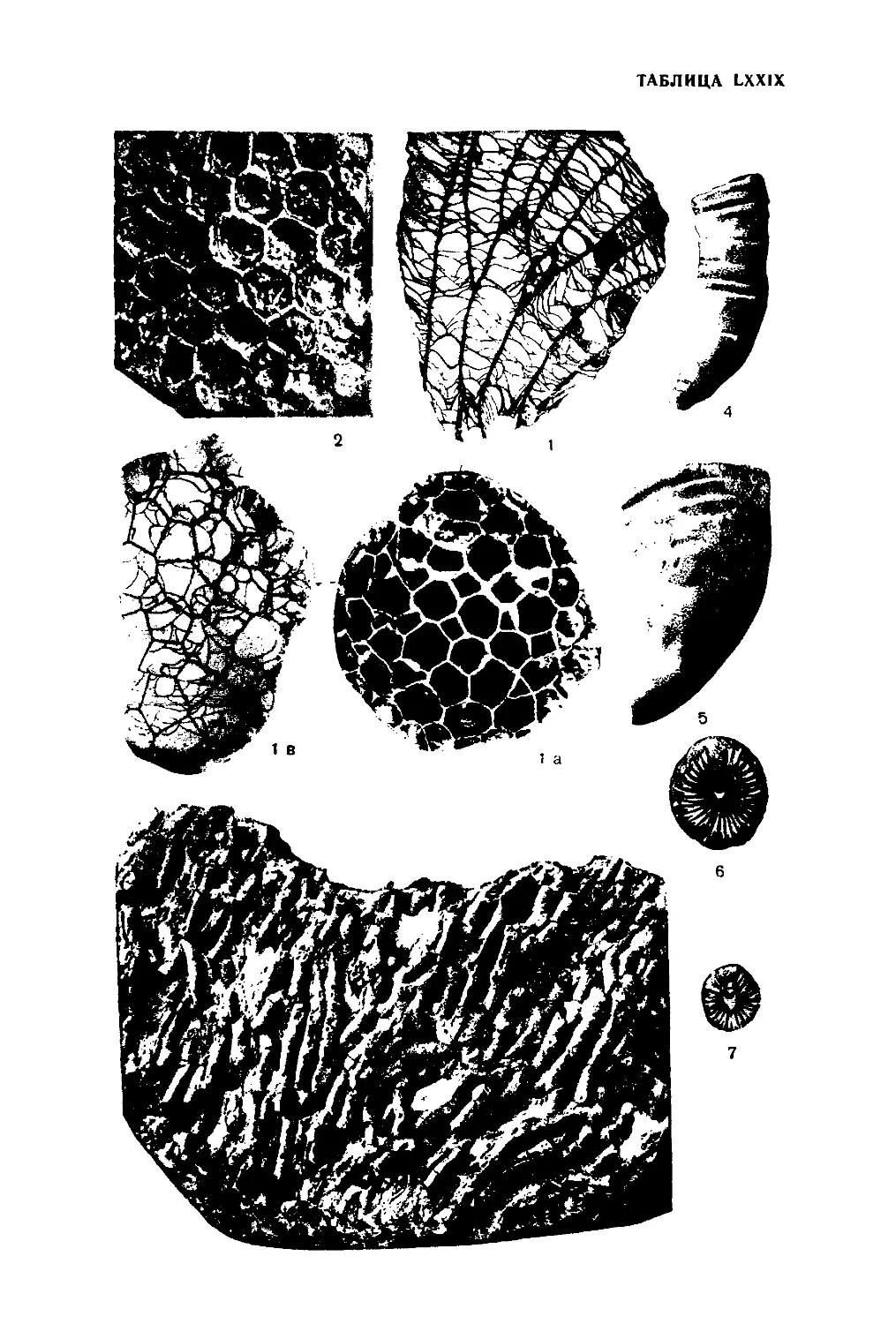
STROMATOPOROIDEA. СТРОМАТОПОРОИДЕЙ
Систематическое положение Stromatoporoldea — вымершей группы
морских организмов, сближаемых с современными гидрактимиями
и миллепорами, до сих пор твердо не установлено. Stromatoporoldea
были развиты главным образом в палеозое, но имели значительное рас-
пространение также и в мезозое (В. И. Яворский, 1947).
Нередко строматопороидеи слагают целые пласты (например, амфи-
поровые известняки), но чаще они сопутствуют другим целентератам,
как это наблюдается в Кузбассе, где «обычно их было не больше, чем
и других форм, а именно: Favosites, Heliolites, Alveolites, Chaetetes,
колониальных кораллов из группы Rugosa и др.» (В. И. Яворский, 1931).
Иллюстрации в приведенном ниже описании строматопороидей заим-
ствованы из монографии Никольсона (Nicholson, 1886—1892).
Строение скелета строматопороидей нужно изучать под микроско-
пом *, причем необходимо иметь не менее двух шлифов — вертикальный
или радиальный и перпендикулярный ему — тангенциальный (шлифы
просматриваются в проходящем свете с небольшими увеличениями; удоб-
нее всего 10—20-кратное увеличение).
Скелет полипняка (ценостеум) образован концентрическими пластин-
ками или ляминами и радиальными столбиками (рис. 16). Эти элементы
скелета находятся в различных соотношениях друг с другом. Так, у рода
Actlnostroma концентрические пластинки образуются в результате сра-
стания отходящих от столбиков в процессе их роста горизонтальных от-
ростков (связки, arms — рис. 17), у других родов лямины развиваются
независимо от столбиков (род Stromatoporella и др.) и т. д.
У некоторых родов концентрические пластинки тонкие, однородные,
у других — толстые и в продольном сечении обнаруживают вдоль сред-
ней линии светлую щель или темную полоску.
Расстояние между двумя соседними концентрическими пластинками
называется межпластинчатым промежутком или интерляминарным про-
странством (см. рис. 16г).
Радиальные столбики бывают различной длины: у одних родов они
соединяют только две концентрические пластинки {Clathrodlctyori),y дру-
гих — проходят через ряд пластинок и межпластинчатых промежутков
{Actlnostroma). В осевой части столбиков иногда имеется зона, окра-
Макроскопические признаки второстепенны.
21
шейная или в более темный, или в более светлый тон. Многие авторы
называют это образование осевым каналом (рис. 18), с чем не согласен
Леконт (Lecompte, 1952), полагающий, что появление этих зон связано
с неравномерным распределением в столбиках органического вещества;
этому образованию он не придает диагностического значения.
Рис. 16. Actinostroma clathratum
Nich.
Продольный разрез:
а—радиальный столбик; б—концент-
рическая пластинка (лямина); в—ячея;
г—межпластинчатый промежуток
(интерляминарное пространство), у 17
Рис. 18. Actinostroma clathratum
Nich.
Поперечное сеченне:
а—радиальный столбик; б—осевой
канал, х 60
Рис. 17. Actinostroma clathratum
Nich.
Тангенциальный разрез:
а—радиальные столбики с осевыми
каналами; б—связки. X 17.
Рис, 19. Parallelopora capitata G о 1 d t.
a—ампула с перегородками; б—зооидные
трубки и днища в них (продольный раз-
рез). X 14
Пространство, заключенное между двумя концентрическими пластин-
ками и двумя радиальными столбиками, называется ячеей, ячейкой или
петлей (см. рис. 16в).
В ценостеуме у некоторых строматопороидей, наряду с ячеями,
имеются более крупные овальной или удлиненноовальной формы пусто-
ты, снабженные тонкими перегородками. Эти пустоты иногда называют
ампулами (рис. 19). Леконт (1952) связывает их с астроризами (см.
ниже).
У ряда родов (Stromatopora, Parallelopora) имеются более или менее
вертикально ориентированные зооидные трубки, снабженные тонкими
22
перегородками, которые называются днищами или табулями (рис. 20).
Днища могут быть горизонтальными, прямыми или слегка выпуклыми,
или косо направленными. Леконт называет зооидные трубки камерами
и также связывает их с астроризами (см. ниже).
В росте скелета Stromatoporoldea часто наблюдаются периодические
перерывы. В литературе применяется термин лятилямина для «тех тол-
-стых слоев, которые являются результатом перемежаемости роста»
(Н. Лебедев, 1896). Таким образом, границы лятилямины отвечают оста-
новкам роста колоний (рис. 21). «Обычно по этой плоскости* стромато-
поры легко раскалываются, а в выветрелом состоянии сечения лятиля-
мин легко заметны простым глазом в виде ряда концентрических или
волнистых полос» (Рябинин, 1934).
Микроскопическое строение скелетной ткани строматопорид может
быть плотным (Actinostroma и др.), пронизанным мельчайшими кана-
лами (рис. 22), зернистым или пористым (Stromatoporella, Stromato-
рога и др.).
На поверхности концентрических пластинок у многих представителей
этой группы ископаемых имеются неглубокие бороздки или желобки, от-
ходящие от одного центра и расположенные в виде звездчатых фигур —
астроризы. Каждая отдельная бороздка или желобок называется астро-
ризальным каналом (рис. 23 и 24). Астроризальные каналы самостоя-
тельных стенок не имеют; роль их играют концентрические пластинки
и радиальные столбики, ограничивающие канал (рис. 24 и 25).
Астроризы часто располагаются (на каждой последующей пластинке)
беспорядочно по отношению друг к другу, но в некоторых случаях они
образуют вертикальные ряды, располагаясь строго одна над другой.
В последнем случае для всей вертикальной системы имеется общий
центр — осевой канал, который обычно открывается наружу в виде
круглого отверстия на вершине конического бугорка — сосочка (рис. 26);
эта вертикальная система астрориз называется астроризальным цилин-
дром (см. рис. 25). В некоторых случаях в астроризальных каналах на-
блюдаются прямые, косо расположенные или несколько искривленные
перегородки. Астроризы развиты не у всех родов (например, у рода
Labechia их нет).
У форм древовидных — Amphipora, Idlost roman др.—имеется неред-
ко очень широкий осевой канал2, снабженный наклонными, порой
искривленными, иногда воронкообразными днищами (рис. 27).
Формы полипняков разнообразны: пластинчатые, желвакообразные,
шарообразные, грушевидные. Встречаются также полипняки в виде ко-
рок, обрастающие другие предметы (инкрустирующие формы) и т. д.
На нижней поверхности у некоторых из них имеется морщинистая ба-
зальная эпитека — известковый чехол, одевающий колонию снизу. При-
крепление к субстрату происходило или всей нижней поверхностью или
при помощи небольшой ножки. Часто бывает, что один и тот же вид
характеризуется различными способами роста ценостеума. По данным
В. Н. Рябинина (1951), способ роста ценостеума строматопор зависит
от среды их обитания.
На поверхности полипняка могут наблюдаться многочисленные поры,
многочисленные мелкие круглые бугорочки — концы радиальных столби-
ков. Часто поверхность бывает украшена довольно крупными кониче-
скими возвышениями— сосочками (см. рис. 26), могут быть развиты
бородавчатообразные бугорки — туберкулы (рис. 28—29). На наружной
поверхности, кроме того, часто наблюдаются астроризы (см. выше).
Среди строматопороидей выделяют две группы — гидрактиноидную
и миллепороидную.
1 Пограничная между лятиляминами поверхность.
" М. Леконт (1952) считает эти каналы астрорнзальными.
23
Рис. 21. Stromatopora concentrica
G о 1 d f.
a—лятилямина. Нат. вел.
Рис. 20. Idiostroma romeeri .Nich.
Продольное сечение:
а—зооидные трубки и днища в них.х!6
Рис. 23. Parallelopora dartingtonensis
Cart.
Астроризы с длинными и ветвящимися
каналами. хЗ
Рис. 22. Stromatoporella eifellensis
Nich.
а—каналы в радиальных столбиках
и концентрических пластинках;
б—ячеи, х 33
Рис. 24. Stromatopora antiqua Nich.
et Mur.
Поперечное сечение:
а - астрорнзальные каналы, х 17.
«Гидрактиноидная группа характеризуется ясно различимыми гори-
зонтальными и вертикальными скелетными элементами (лямины и ради-
альные столбики); микроскопическое строение ткани плотное. Зооидные
трубки не найдены» (Рябинин, 1934).
«Миллепороидная группа характеризуется отсутствием правильной
сетки в вертикальном сечении. Лямины толстые, неправильные; радиаль-
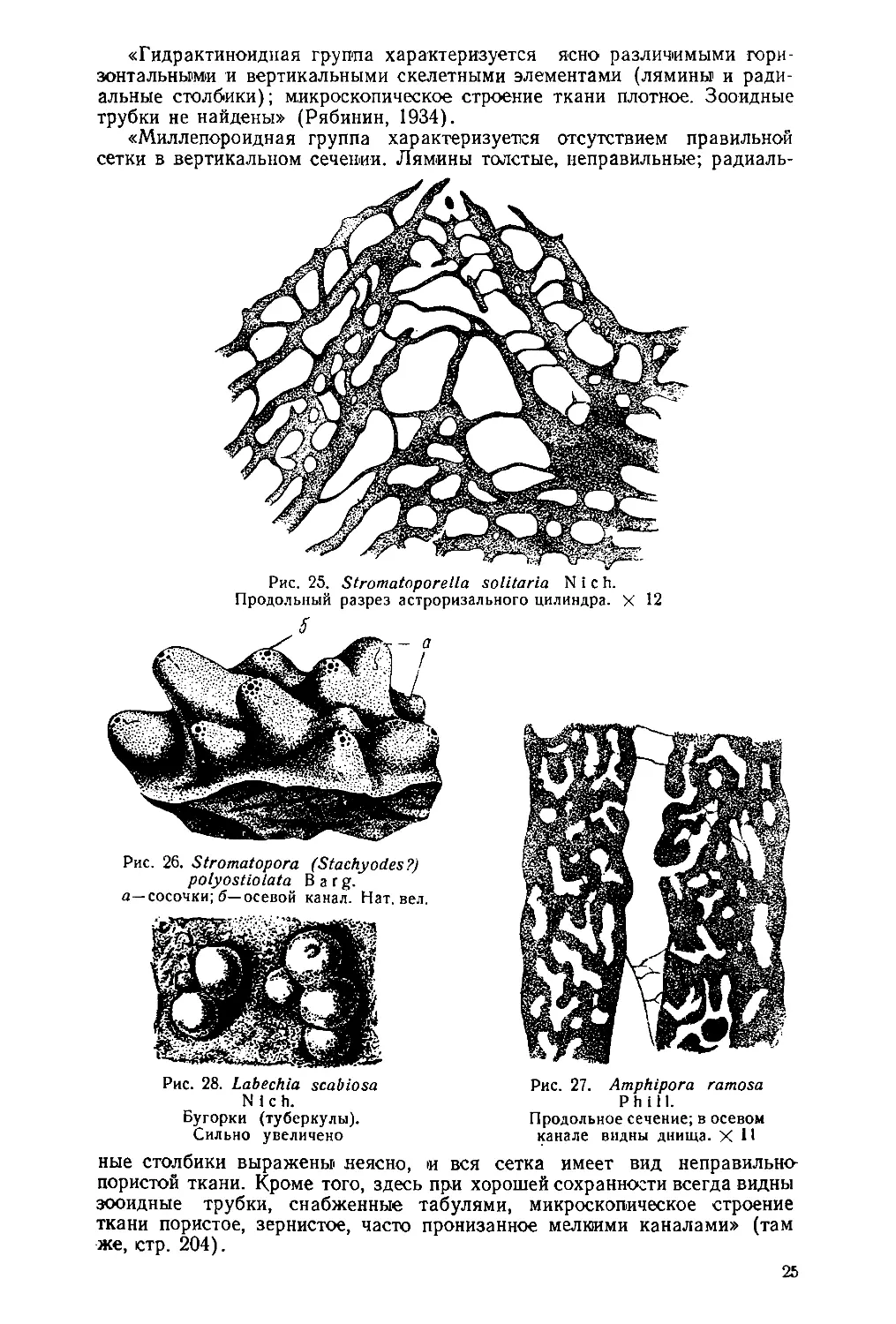
Рис. 25. Stromatoporella solitaria N i с h.
Продольный разрез астроризального цилиндра. X 12
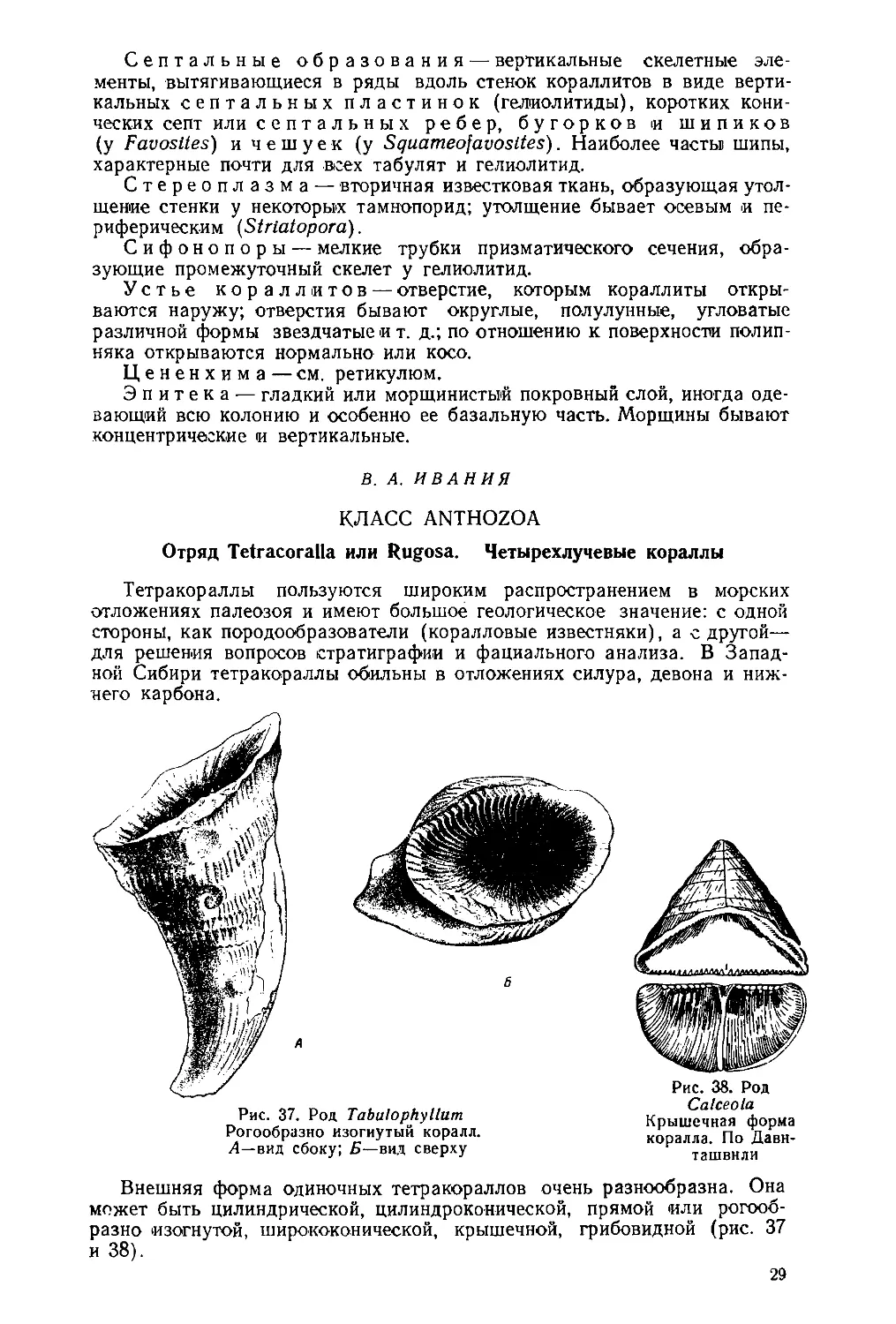
Рис. 26. Stromatopora (Stachyodes?)
polyostiolata Barg.
а— сосочки; б— осевой канал. Нат. вел.
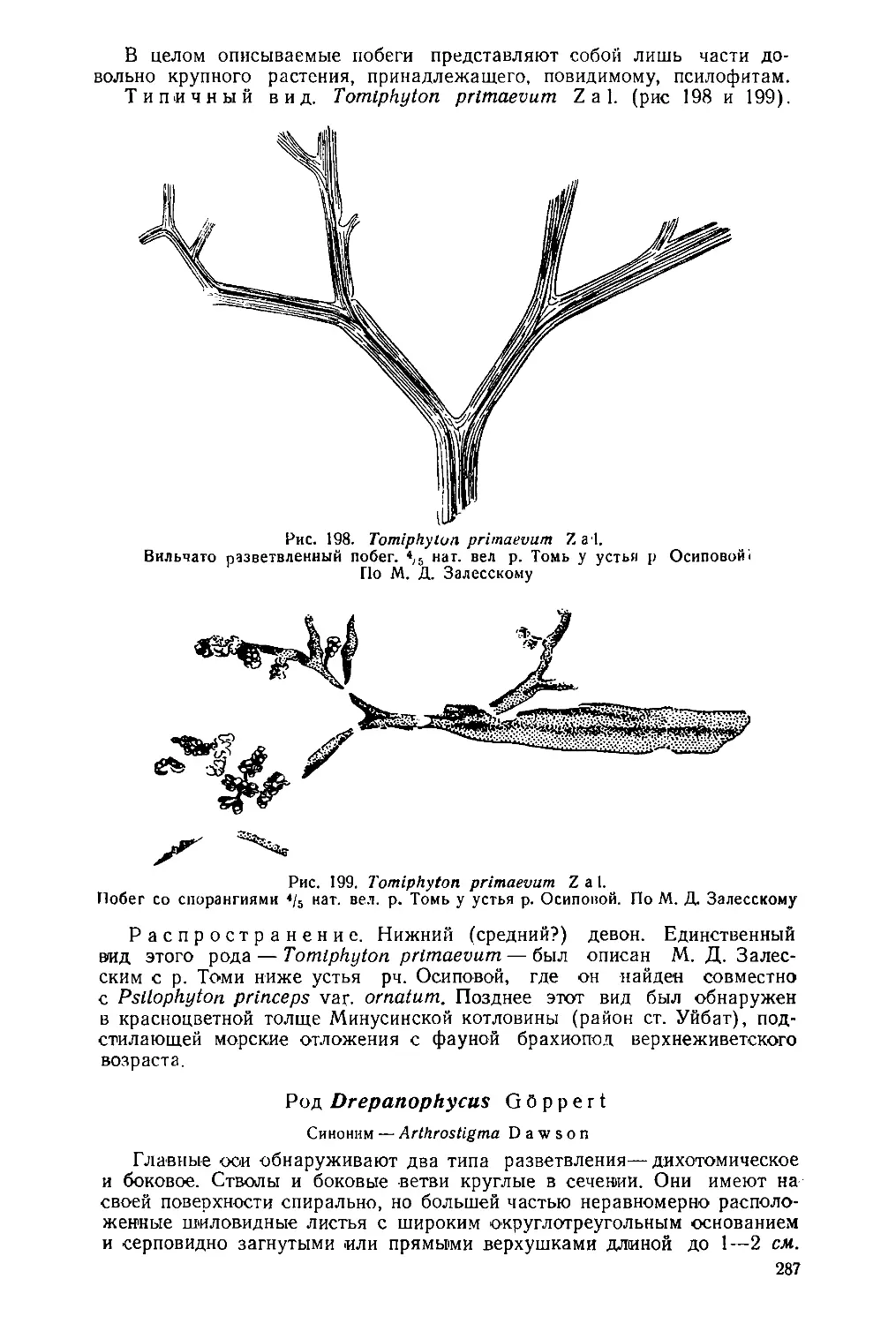
Рис. 28. Labechia scabiosa
N i с h.
Бугорки (туберкулы).
Сильно увеличено
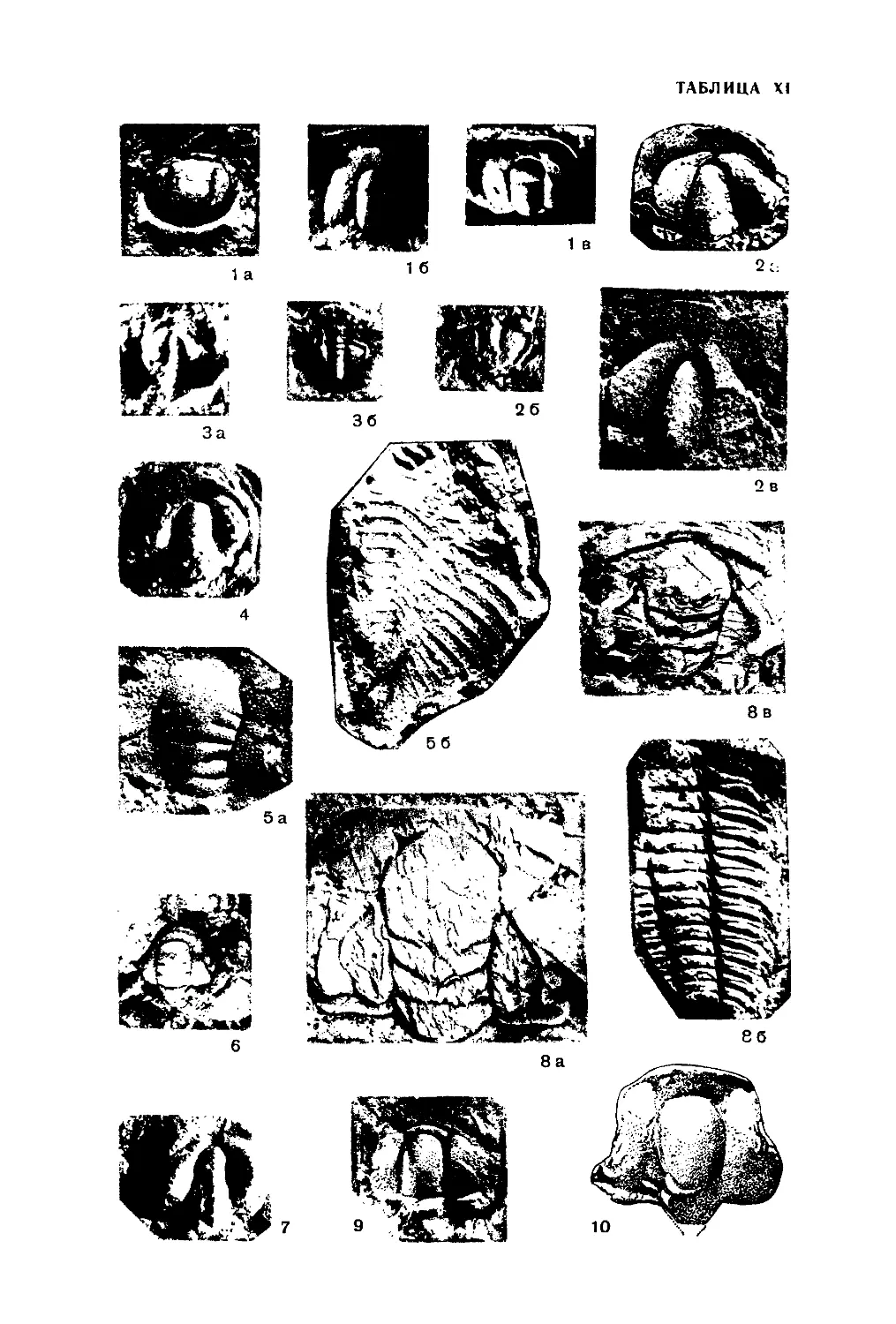
Рис. 27. Amphipora ramosa
Ph ill.
Продольное сечение; в осевом
канале видны днища. X П
ные столбики выражены неясно, и вся сетка имеет вид неправильно-
пористой ткани. Кроме того, здесь при хорошей сохранности всегда видны
зооидные трубки, снабженные табулями, микроскопическое строение
ткани пористое, зернистое, часто пронизанное мелкими каналами» (там
же, стр. 204).
25
Строматопороидеи в Западной Сибири широко распространены в си-
луре и девоне и дают ряд руководящих форм. Но их сборы и поиски
требуют от геолога большого внимания, так как строматопороидеи часто
не обладают бросающимися в глаза внешними признаками и легко могут
быть приняты за обломки слоистого известняка.
Рис. 29. Labechia scabiosa Nich.
Наружная поверхность, туберкулы. X 2,2
Л. И. КРАЕВСКАЯ
КЛАСС ANTHOZOA
ПОДКЛАСС TABULATA И ГРУППА HELIOLITIDA
Среди палеозойских Coeleneterata большое развитие в Западной Си-
бири имеют табуляты и гелиолитиды. Эти группы, долгое время не имели
стратиграфической ценности вследствие недостаточной изученности, запу-
танности их систематики и потому, что объем вида у большинства родов
этих групп понимался чрезмерно широко.
Работы советских исследователей (Л. Б. Рухина, Б. Б. Чернышева,
Б. С. Соколова) явились большим вкладом в дело изучения этих групп.
Особенно большое значение имеют работы Б. С. Соколова «Систематика
и история развития палеозойских кораллов Anthozoa Tabulata» и «Табу-
ляты палеозоя Европейской части СССР», в которых дается критический
пересмотр, новая классификация и новые формулировки диагнозов этих
групп.
Для определения видов Tabulata и Heliolitida недостаточно внешних
признаков, необходимо знать внутреннее строение, для чего изготов-
ляются шлифы — поперечный и продольный. Детали строения скелета
по шлифам изучаются с помощью лупы и микроскопа. Все части скелета
кораллов имеют значение для систематики; при определении необходимо
обращать внимание на особенности строения и размеры днищ, характер
и размеры кораллитов, характер септальных образований и т. д. (см.
ниже объяснения этих терминов).
Б. С. Соколов (1950) дает следующий диагноз для группы табулят
в целом:
«Колониальные кораллы, сложенные округлыми или полигональными
мономорфными замкнутыми кораллитами, всегда отличающимися незна-
чительными размерами и образующими массивные, ветвистые, кустистые
или стелющиеся дерновидные полипняки; кораллиты находятся или в со-
общении друг с другом посредством пор, соединительных трубок и пла-
стин — СТОЛОНОВ, ИЛИ ЯВЛЯЮТСЯ свободными ОТ ЭТИХ связей И ТОЛЬКО СО-
прикасаются стенками, которые во всех случаях являются самостоятель-
ными у каждого кораллита; цененхима отсутствует; внутренние скелет-
26
ные образования отличаются большой простотой; главнейшими являются:
септальный аппарат, представленный различным количеством почти не
дифференцированных септальных пластинок, ребрышек, шипиков, бугор-
ков или чешуек, и днища, представленные горизонтальными, изгибаю-
щимися и расщепленными пластинками; пузырчатая ткань и столбик
наблюдаются редко».
Приводим также диагноз группы Heliolitida по Б. С. Соколову:
«Колониальные полипняки пластинчатой, вздутой или ветвистой фор-
мы, сложенные цилиндрическими кораллитами, разделяющимися проме-
жуточным скелетом (ретикулюм), имеющим трубчатое или пузырчатое
строение; септальные образования в виде шипов и шиповатых пластин,
простирающихся за пределы стенок у форм с трубчатым ретикулюмом
и ограниченных стенкой у форм с пузырчатым ретикулюмом; количество
септ строго фиксировано и только у одного рода превышает 12; сообще-
ние между полостями кораллитов и трубок ретикулюма отсутствует;
кораллиты большею частью подразделены днищами, в промежуточном
скелете развиваются диафрагмы и пузыри; вертикальные скелетные эле-
менты всегда окрашены в более светлые тона, чем горизонтальные».
Ниже дается объяснение специальных терминов, наиболее часто
употребляемых при описании табулят и гелиолитид (рис. 30—36).
Аутопоры — см. кораллиты.
Рис. 30. Род Favosites
а—массивная колония; 6—призматические ячейки с порами
на стенках и днищами. Из Циттеля
Диафрагмы — поперечные перегородки в трубках ретикулюма
(см. след, стр.) у гелиолитид.
Диссепименты — см. пузырчатая ткань.
Днища — поперечные перегородки в кораллитах. Типы днищ: гори-
зонтальные, аркообразные, цепочные, воронкообразные, неполные и т. д.
Некоторые авторы называют днища потолочками или табулами.
Колония или полипняк — соединение отдельных индивидов
в связную постройку. Колонии бывают массивные, когда кораллиты
плотно прилегают друг к другу; ветвистые, когда изгибается и ветвится
тело полипняка при плотном соединении кораллитов (Thamnopora,
Striatopora и др.); кустистые, когда кораллиты свободно расходятся
в стороны или связываются посредством особых соединительных образо-
ваний. По форме колонии бывают: сферические, полусферические, пла-
стинчатые, древовидные, стелющиеся и т. п.
Кораллиты (у гелиолитид аутопоры)—отдельные индивиды
колонии. По форме кораллиты бывают: цилиндрические с эллиптическим
или круглым сечением, призматические, прямые или изгибающиеся. Рас-
положение кораллитов бывает параллельным, радиальным, стелю-
щимся и т. п.
27
Мезопоры — см. промежуточные трубки.
По липняк — см. колония.
Поры — соединительные отверстия овальной или округлой формы,,
посредством которых сообщаются друг с другом полости смежных ко-
раллитов у многих табулят. Поры бывают: угловые (Palaeofavosltes}
и стенные (Favosltes) или и те и другие (Mesofavosites); на стенках они
могут образовывать вертикальные ряды или располагаться беспорядочно.
Промежуточные трубки или мезопор ы — мелкие приз-
матические трубки между более крупными цилиндрическими коралли-
тами (аутопорами), например у рода Halysttes.
Рис. 31. Кустистая ко-
лония рода Syringopora
Цилиндрические ко-
раллиты с соедини-
тельными трубками.
По Циттелю
Рис. 32. Колония рода
Halysites с цепным
расположением цилинд-
рических кораллитов.
По Циттелю
Рис. 33. Продольное се-
чение колонии рода
Thamnopora:
к—кораллиты со стенка-
ми, утолщенными стерео-
плазмой; р—поры. Уве-
личено. По Никольсону
4 б*
Рис. 34. Род
Halysites.
Поперечное
сечение:
а—аутопоры
с шипами;
б—мезопо-
ры. Х4. По
Шимеру и
Шроку
Пузырчатая
Рис. 35. Род Halysites
Продольное сечение:
а—аутопоры; б—мезо-
поры; в—днища.
Увеличено
Рис. 36. Род
Alveolites
А—поперечное
и Б—продоль-
ное сечения:
а—шипы;
б—днища;
в—стенные по-
ры. X б. Из
Циттеля
ткань или диссепим
е н т ы — скелетная ткань
в виде мелких, пересекающихся, часто вздутых пластинок.
Ретикулюм — промежуточный скелет у гелиолитид, имеющий
трубчатый или пузырчатый характер; в старых работах назывался «це-
ненхима».
Соединительные трубки — горизонтальные соединительные
выросты в виде тонких цилиндрических трубочек, связывающих корал-
литы друг с другом (Syringopora)-, соединительные трубки образуют пра-
вильные ряды или располагаются беспорядочно.
Солении — форма соединительных образований, промежуточная
между порами и соединительными трубками, например у рода Multiso-
lenia.
28
Септальные образования — вертикальные скелетные эле-
менты, вытягивающиеся в ряды вдоль стенок кораллитов в виде верти-
кальных септальных пластинок (гелиолитиды), коротких кони-
ческих септ или септальных ребер, бугорков и шипиков
(у FavosU.es) и чешуек (у Squameofavosltes). Наиболее часты шипы,
характерные почти для всех табулят и гелиолитид.
Стереоплазма — вторичная известковая ткань, образующая утол-
щение стенки у некоторых тамнопорид; утолщение бывает осевым и пе-
риферическим (Striatopora).
С иф оно поры — мелкие трубки призматического сечения, обра-
зующие промежуточный скелет у гелиолитид.
Устье кораллитов — отверстие, которым кораллиты откры-
ваются наружу; отверстия бывают округлые, полулунные, угловатые
различной формы звездчатые и т. д.; по отношению к поверхности полип-
няка открываются нормально или косо.
Цененхима — см. ретикулюм.
Эпитека — гладкий или морщинистый покровный слой, иногда оде-
вающий всю колонию и особенно ее базальную часть. Морщины бывают
концентрические и вертикальные.
В. А. ИВАНИЯ
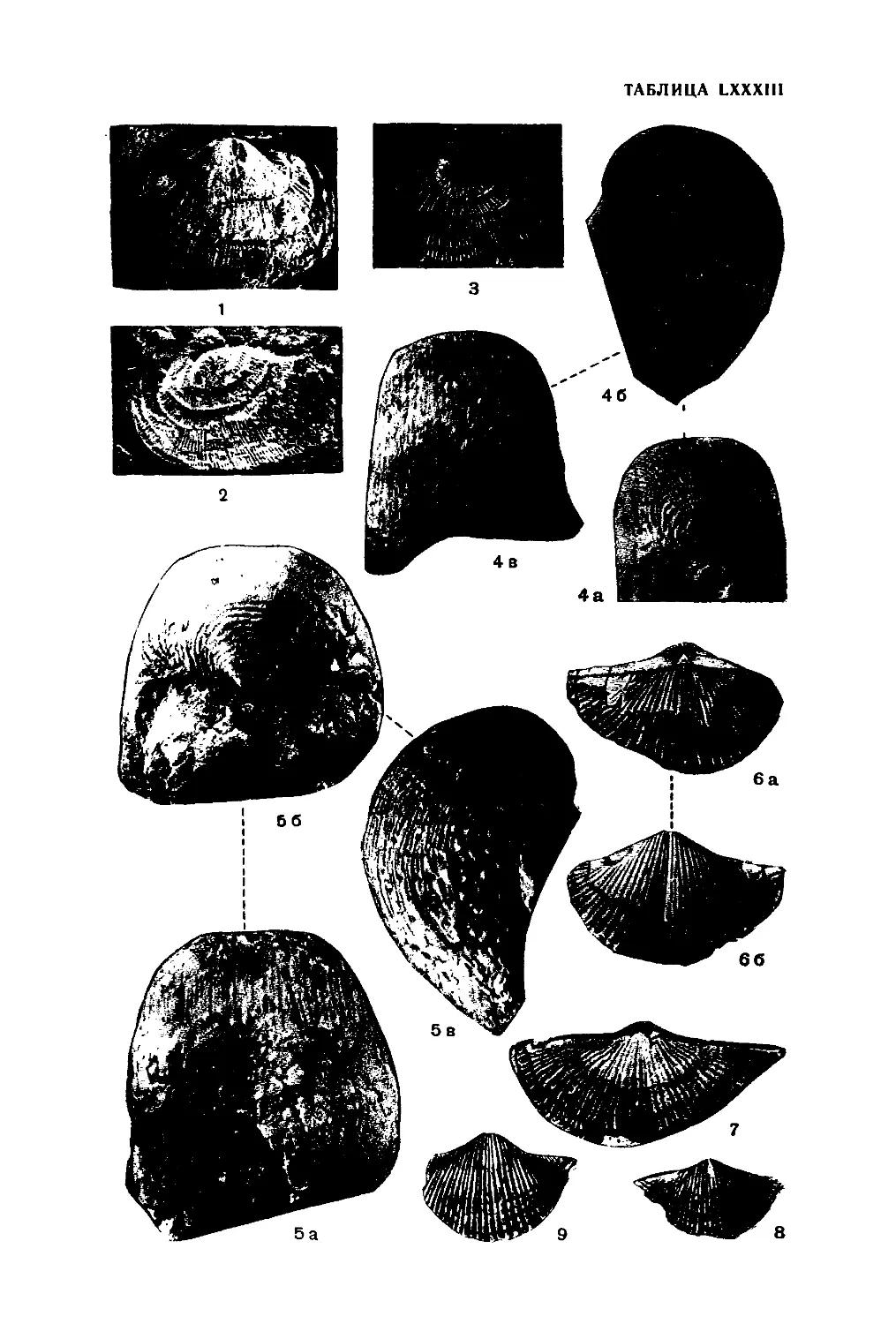
КЛАСС ANTHOZOA
Отряд Tetracoralla или Rugosa. Четырехлучевые кораллы
Тетракораллы пользуются широким распространением в морских
отложениях палеозоя и имеют большое геологическое значение: с одной
стороны, как породообразователи (коралловые известняки), а с другой—
для решения вопросов стратиграфии и фациального анализа. В Запад-
ной Сибири тетракораллы обильны в отложениях силура, девона и ниж-
него карбона.
Рис. 37. Род Tabulophyllum
Рогообразно изогнутый коралл.
А—вид сбоку; Б—вид сверху
Рис. 38. Род
Calceola
Крышечная форма
коралла. По Дави-
ташвили
Внешняя форма одиночных тетракораллов очень разнообразна. Она
может быть цилиндрической, цилиндроконической, прямой или рогооб-
разно изогнутой, ширококонической, крышечной, грибовидной (рис. 37
и 38).
29
В результате бесполого размножения (почкования) возникают коло-
нии, особи которых называются кораллитами. Наряду с нормальным
почкованием, когда новая особь возникает из некоторой группы клеток
материнского организма, у тетракораллов наблюдается так называе-
мое парисидальное почкование, которое представляет собой продольное
деление материнского организма на две дочерние особи. Разные спосо-
бы почкования имеют важное систематическое значение. Форма колоний
разнообразна: массивные колонии, когда призматические кораллиты
тесно прижаты друг к другу (рис. 39); астреевидные, когда соприкасаю-
щиеся кораллиты утрачивали внешнюю стенку так, что септы соседних
кораллитов срастались между собой или терялись в массе пузырчатой
ткани (рис. 40); кустистые или ветвистые, состоящие из цилиндрических
кораллитов, находящихся на некотором расстоянии друг от друга
(рис. 41).
Рис. 39. Род Phillipsastraea
Массивная колония
Рис. 40. Род Ра-
chyphyllum
Астреевидная ко-
лония
Рис. 41. Ветвистая
колония
Колонии, образовавшиеся путем парисидального почкования, назы-
ваются псевдоколониями, так как кораллиты их сохраняли все особен-
ности внешней формы одиночных кораллов, из которых они развились.
Внешняя стенка одевает мягкое тело животного и скрывает внутрен-
ние части скелета. Порой она может быть тонкой; в других случаях по-
верх стенки отлагается более или менее толстый поперечноморщинистый
чехол или эпитека. Иногда внешняя стенка образуется не внешним кле-
точным слоем (эктодермой), а сросшимися внешними краями септ и имеет
вид так называемого ободка (см. рис. 56).
На верхнем конце кораллов и кораллитов обычно имеется углубле-
ние, называемое чашкой. Форма чашки может быть бокаловидной, с от-
весными стенками, острыми краями, плоским, выпуклым или вогнутым
дном. При наклонных стенках она становится воронковидной. Иногда
чашка может иметь широкий или узкий отворот на краях и т. п. (рис. 42
и 43).
Внутренний скелет тетракораллов имеет достаточно сложное строение.
Важнейшими его элементами являются вертикальные перегородки
(септы), днища и пузырчатая ткань. В целом усложнение внутреннего
скелета происходило в процессе развития отряда тетракораллов. Давита-
швили в «Курсе палеонтологии» (1949 г.) указывает, что «у древнейших
нижнесилурийских форм интерсептальный или межперегородочный
30
внутренний скелет состоит только из днищ. Это так называемые одно-
зонные кораллы. Позже, в верхнем силуре, появляются двухзонные ко-
раллы, у которых, в дополнение к септам и днищам, есть еще перифери-
Рис. 42. Вид чашки Chonophyllum
canadeuse сбоку. По Сошкиной
Рис. 43. Род Pseudomphyma.
Разрез чашки. По Сошкиной
ческая зона пузырчатой ткани. Еще позже, в карбоне, развиваются
трехзонные формы, обладающие, кроме только что упомянутых скелет-
ных элементов, еще столбиком и другими более или менее сложными
центральными скелетными образованиями» (см. рис. 44, 51, 52 и 55).
Рис. 44. Схема однозонных (а), двухзонных (б)
и трехзонных (в) тетракораллов. По Давиташвили
Септы, вертикальные перегородки, имеют важное систематическое
значение (их строение, форма, длина и т. д.). Септы состоят из септаль-
ных трабекул, которые, в свою очередь, состоят из волокон. У древних
Rugosa трабекулы септ крупные и хорошо видны простым глазом. Ряды
Рис. 45. Строение
септ из параллель-
ных трабекул.
По Сошкиной
Рис. 46.
Строение
септ из вееро-
образных
трабекул.
По Сошкиной
Рис. 47. Закручивание септ
в центре и каринированные
септы. По Сошкиной
их обычно направлены от периферии к оси кораллита, в большей или
меньшей степени поднимаясь вверх. От расположения трабекул зависит
рисунок септ внутри тела на поперечных разрезах, например — строение
септ из параллельных трабекул (рис. 45) или строение септ из веерооб-
разных трабекул (рис. 46).
Иногда септы на своей боковой поверхности имеют так называемые
карины, лейсты или поперечные балочки — в виде бугорков, горизон-
31
тальных и вертикальных ребер. Это местные разрастания отдельных тра-
бекул, придающие большую прочность септам (рис. 47). Иногда осевые
концы септ сильно удлинялись, и септы, превышая радиус кораллита,
закручивались в центре (см. рис. 47). Септы могут быть плойчато изо-
гнуты (рис. 48 и 49). В некоторых случаях они редуцированы в перифе-
рической зоне (рис. 49). Длина септ у различных родов тетракораллов
колеблется в широких пределах; часто наблюдается чередование септ
двух порядков — длинных и коротких.
Днища — важнейшие горизонтальные скелетные элементы. Они раз-
личны у разных семейств и родов: горизонтальные, колоколовидно выпук-
лые, вогнутые (рис. 50).
Рис. 48. Плойчатое
изгибание септ.
По Сошкиной
Рис. 49. Четковид-
ные септы, реду-
цированные в пе-
риферической
зоне. По Сошкиной
Рис. 60. Непол-
ные выпуклые
днища. По
Сошкиной
Днища, доходящие до стенки коралла, называются полными, в про-
тивоположном случае — неполными (см. рис. 50). Они могут быть про-
стыми или расщепленными (рис. 51 и 52). При сильном усложнении дни-
ща становились войлокообразно расщепленными (рис. 53).
Рис. 52. Двухзонный коралл
Phillipsastraea с расщеплен
ными днищами и развитой
пузырчатой тканью.
Рис. 51. Однозонный
коралл с полными
днищами
Рис. 53. Войлоко-
образные днища.
По Сошкиной
Пузырчатая ткань обычно сосредоточена в периферической зоне. Со-
стоит она из наклонных, выпуклых к оси и вверх пластинок. Последние,
нарастая друг на друга, образовывали более или менее крупнопузырча-
тую ткань в периферической зоне (зона пузырей) пли выполняли всю
полость кораллита.
32
У некоторых девонских Rugosa наблюдаются еще и так называемые
подковообразные пузырьки. Они образуют цилиндрической формы зону
внутри коралла. Эта зона состоит из выпуклых вверх, насаженных друг
на друга вздутых пузырьков, к которым прикрепляется веер септальных
трабекул (рис. 54).
Внутренняя стенка развивалась у многих кораллов обычно на границе
зоны днищ и зоны пузырей. Образуется она либо путем слияния утол-
щенных частей септ, либо развитием кольца подковообразных пузырьков,
либо путем вторичного отложения известкового вещества в этой зоне
(рис. 55).
Рис. 54. Схема соотношения
подковообразных пузырьков и
веерообразного расположения
септальных трабекул. По Сош-
киной
Рис. 55. Трехзонный ко-
ралл Lithostrotion.
Вверх у—поперечное
сечение кораллита,
вниз у—его продольный
разрез: а—зона пузырей;
б—зона днищ; в—столбик.
По Давиташвили
Рис. 56. Стадии
онтогенеза че-
тырехлучевого
коралла
В осевой части у многих тетракораллов развивается столбик. Разли-
чают ложный столбик, образованный закрученными концами септ или
выпуклостями днищ, и истинный столбик, представляющий собой само-
стоятельный скелетный элемент (рис. 55).
Для наблюдения различных элементов скелета, характер и строение
которых лежат в основе определений тетракораллов, необходимо изготов-
ление, по крайней мере, двух шлифов — продольного и поперечного.
Однако надо иметь в виду, что скелет тетракораллов претерпевает зна-
чительные изменения в процессе роста особей, поэтому для более точного
определения необходимо проследить эти изменения (выяснить онтогенез
коралла), что достигается путем изготовления ряда поперечных шлифов
на различных расстояниях от его основания (рис. 56).
3 Зак. 2024 33
Л. Л. ХАЛФИН
Тип Bryozoa. Мшанки
Мелкие колониальные животные, мшанки, по внешнему облику напо-
минают некоторых кишечнополостных, но по внутренней организации
наиболее близки к брахиоподам, с которыми их нередко объединяют
в один тип. Колонии мшанок (зоарии) имеют различное строение и со-
стоят из ячеек (зооеций) неодинаковой формы. В палеозойских (S—CJ
отложениях Зап. Сибири широко распространены мшанки отрядов Cryp-
tostomata и Trepostomata и одного семейства из отряда Cyclostomata.
Представители первого отряда характеризуются тем, что у них внеш-
нее отверстие (устье) каждой ячейки открывается в особое преддверие
(вестибюль), которое уже далее сообщается с внешней средой (рис. 57).
Для представителей отряда Trepostomata и палеозойских Cyclostomata
характерны длинные трубчатые ячейки с поперечными перегородками
(диафрагмами) внутри, благодаря чему колонии этих мшанок напоми-
нают колонии некоторых табулят (см. стр. 26), от которых отличаются,
прежде всего, очень малыми размерами ячеек (рис. 58). Изучение и оп-
ределение мшанок требует применения микроскопа.
в
Рис. 57. Схема строения
ячеек мшанок из отряда
Cryptostomata
а—полость ячейки; ,
б— вестибюль; в—гемн-
септа. По Давиташвили
HiHiiliiiiiilliiiuiiilhiiiimiliiiiil
Рис. 58. Продоль-
ное сечение мшан-
ки из отряда
Trepostomata. Вид-
ны трубчатые
ячейки с диафраг-
мами. X 14. По
Басслеру
Рис. 60. Соеди-
нение прутьев
посредством
анастомоз
По А. И. Ники-
форовой
Рис. 59. Колония
мшанки рода Fenestella
а—устья ячеек,
разделенные килем;
б-петля; в—прутики,
соединенные перекла-
динами. Сильно
увеличено. Схема
6
Из отряда Cryptostomata наиболее важным является семейство Fene-
stellidae. Колония мшанок этого семейства (рис. 59) состоит из полых
прутьев, которые внутри разделены на ячейки; устья последних распо-
лагаются с одной стороны — лицевой или ячеистой, противоположная
(тыльная) сторона прутьев лишена устьев. Соседние прутья обычно сое-
диняются друг с другом при помощи перекладин (см. рис. 59) или ана-
стомоз; в последнем случае это соединение достигается непосредственно
соприкосновением волнисто изогнутых прутьев (рис. 60). В том и дру-
гом случае получается сетчатая колония, прутья и перекладины которой
образуют петли.
Размеры различных элементов скелета колонии (прутьев, перекладин,
петель и др.) имеют диагностическое значение (рис. 61). У многих родов
ячейки в прутьях расположены двурядно; в этом случае на лицевой сто-
роне прутьев, между двумя рядами отверстий, ведущих в вестибюли
ячеек1, обычно проходит продольное ребро — киль (см. рис. 59), кото-
рый может быть гладким или бугорчатым. У представителей некоторых
1 Эти отверстия обычно называются устьями, хотя у Cryptostomata собственно
устье лежит в глубине вестибюля (см. рнс. 57).
34
родов (Hemitrypa, Isotrypa и др.) с этими бугорками и килями связано
развитие особой защитной сетки (рис. 62), располагающейся над основ-
ной сеткой, образованной прутьями. У некоторых родов ячейки в прутьях
Рис. 61. Род Fenestella. Тангенциальное сечение
сетчатой формы. X 16
a-а—количество прутьев на 10 мм лирины
зоарии (4 прута); b-b — количество --петель на
10 мм длины зоарии (3 петли); с-с-количество
ячеек на 5 мм длины прута (4 ячейки); d-d—ши-
рина прута; е-е—длина петли; /-/—ширина
петли; k-k—ширина перекладины; т-т—диаметр
устья ячейки; п-п—расстояние между устьями
ячеек; о-о—диаметр бугорка; г-г— расстояние
между центрами бугорков; s-s—ширина киля.
По А. 14. Никифоровой
Рис. 62. Тангенциальное сече-
ние колонии рода Hemitrypa
с защитной сеткой (слева), х 10.
По В. П. Нехорошеву
Рис. 63. Часть колонии
мшанки рода Poly рога. X 12.
Из Циттеля
66
а
Рис. 64. Поперечное сечение
колоний мшанок семейства
Fenestellidae.
а—полость ячейки; б—киль. По
В. П. Нехорошеву
Рис. 65. Мезопоры (а).
По А. И. Никифоровой
располагаются более чем в два ряда, порой без определенного порядка
(рис. 63).
Определение представителей фенестеллид и близких семейств мша-
3* 35
нок требует наблюдения при значительных увеличениях лицевой и тыль-
ной поверхности колонии и продольного и поперечного сечения прутьев;
практически это возможно с помощью двух прозрачных шлифов—попе-
речного (рис. 64) и косо-тангенциального. Косо-тангенциальный шлиф
изготовляется (по предложению В. П. Нехорошева) таким образом, что
его плоскость частью совпадает с лицевой поверхностью прутьев, частью
сечет их на различной глубине, частью совпадет с тыльной поверхностью
прутьев (см. рис. 61 и 62). Дополнительно может быть полезен косо-
продольный шлиф — сечение, идущее вдоль прута нормально к лицевой
поверхности последнего.
Колонии мшанок отряда Trepostomata очень различны по форме —
ветвистые, массивные, листовидные, коркообразные. Ячейки, как уже
отмечалось, длинные, трубчатые, с поперечными перегородками (диа-
фрагмами). Во внутренних частях (во внутренней зоне) колоний ячейки
непосредственно соприкасаются друг с другом, благодаря чему имеют
призматическую форму и тонкие стенки. Во внешней или перифериче-
ской зоне ячейки резко отгибаются (рис. 58) наружу, часто становятся
цилиндрическими, их стенки утолщаются, и ячейки как бы раздвигаются.
Пространство между ними заполнено или плотной тканью, или особыми
промежуточными (интерзооэциальными) структурами; среди последних
наиболее обычны мелкие трубочки — мезопоры (рис. 65). Для изучения
мшанок этого отряда также необходимы три шлифа — тангенциальный
(касательный к поверхности колонии), поперечный (секущий ячейки по-
перек) и продольный (секущий ячейки вдоль).
Из отряда Cyclostomata в палеозое Западной Сибири распространено
семейство Fistuliporidae; мшанки этого семейства имеют такие же, как
у Trepostomata, трубчатые ячейки, но последние остаются цилиндриче-
скими на всей их длине и обладают устьями с лунариями (см. ниже).
Приведенные ниже объяснения некоторых терминов, употребляемых
при описании мшанок, заимствованы у В. П. Нехорошева (1948).
«Акантопоры — трубчатые иглы, расположенные между устьями
ячеек. Встречаются у многих представителей отрядов Trepostomata
и Cryptostomata (рис. 66).
Рис. 66. Акантопо-
ры(а). По А. И.
Никифоровой
Рис. 67. Устья яче-
ек с лунариями. X 20
По В. П. Нехорошеву
Гемисепты верхняя и нижняя — пластинчатые выступы
в ячейках у представителей отряда Cryptostomata, расположенные на
верхней стенке у сочленения ячейки с вестибюлем, а на нижней — против
этого сочленения. При жизни колонии гемисепты, вероятно, служили
для прикрепления мускулов для высовывания венчика щупальцев нару-
жу и втягивания обратно в полость ячейки — вестибюля (см. рис. 57).
Диафрагмы (перегородки) — тонкие пластины, пересекающие
ячейки в мшанках отрядов Cyclostomata и Trepostomata и мезопоры
у представителей последнего отряда. У некоторых форм диафрагмы бы-
вают не только сплошные, но и продырявленные и при этом иногда утол-
щенные.
Капилляры — тонкие трубочки, пронизывающие скелетную ткань
зоарии. Особенно ясно бывают выражены на неячеистой поверхности
сетчатых форм, но встречаются и в других формах.
36
Рис. 68. Пузырчатая ткань,
выполняющая промежутки
между ячейками. X И-
По В. П. Нехорошеву
Лунарии — складки скелетной ткани в стенке ячейки у устья, вы-
раженные в виде полулунной выемки или серповидного выступа; часто
встречаются у представителей отряда Cyclostomata (рис. 67).
Мезопорй — угловатые неправильной формы полости, располо-
женные между ячейками в периферической части зоарии отряда Тгерр-
stomata и обычно имеющие многочисленные диафрагмы (см. рис. 65).
Пятна (макулы)—плоские, вогнутые или слегка выпуклые об-
разования на поверхности зоарий некоторых форм мшанок, покрытые
плотной известковой тканью, окруженной более крупными устьями ячеек,
или же скопления мезопор, акантопор или более крупных устьев ячеек.
Иногда такие пятна становятся сильно выпуклыми, имея характер хол-
миков (бугорков); в таких случаях они носят наименование бугорков
(монтикул). Строение таких бугрков-монтикул такое же, как и пятен;
и пятна и бугорки обычно расставлены
через более или менее правильные проме-
жутки на расстоянии 3—6 мм одно от
другого.
Сечение ячеек наблюдается в тан-
генциальных сечениях у сетчатых форм;
бывает весьма разнообразно и имеет важ-
ное систематическое значение, представ-
ляя существенный видовой признак в се-
мействах Fenestellidae и Acanthocladiidae.
Выделяются следующие основные типы
сечения ячеек: четырехугольное, пяти-
угольное, треугольное, полулунное и эл-
липтическое. Между ними имеются мно-
гочисленные промежуточные типы сечений.
Струйки (струйчатость) —
тончайшие гребешки, покрывающие не-
ячеистую поверхность прутьев и иногда
промежутки между устьями ячеек на
ячеистой поверхности Fenestellidae. В основном струйки образованы спе-
циальным рядовым распределением капилляров, пронизывающих извест-
ковую ткань основания ячеек. При большей толщине основания рядовое
распределение капилляров утрачивается, и они распределяются равно-
мерно по всей известковой ткани, в результате чего неячеистая поверх-
ность бывает равномерно покрыта мельчайшими бугорками (около ка-
пилляров) — грануляцией.
Ткань пузырчатая (везикулярная) — скелетная ткань, со-
стоящая из мелких неправильной формы пузырьков, окруженных тонкой
известковой оболочкой. Выполняет промежутки между ячейками у неко-
торых семейств отрядов Cyclostomata и Cryptostomata и иногда служит
для укрепления оснований колоний» (рис. 68).
В последнее время М. И. Шульга-Нестеренко разработала наиболее
совершенный метод изучения мшанок, важным элементом которого яв-
ляется исследование микроструктуры скелетных тканей. Этот метод дает
наиболее точные определения и очень повышает стратиграфическую
ценность мшанок.
Л. Л. ХАЛФИН
Тип Brachiopoda. Плеченогие
Брахиоподы — морские одиночные двусторонне симметричные жи-
вотные, ведущие донный образ жизни. Древнейшие брахиоподы известны
из верхнепротерозойскмх отложений. Наибольшего развития они до-
стигли в палеозое; в современной фауне играют второстепенную роль.
37
отложении нижнего и среднего палеозоя.
п'кр
Ш
Рис. 69. Раковина брахиопод. '/s нат. вел.
бр. с.—брюшная створка; сп. с—спинная
створка; м.—макушка; з. кр.—задний или замоч-
ный край; п. кр.—передний край; д,—длина,
ш.— ширина, т.—толщина раковины; з. р.—знаки
роста; а-а— плоскость симметрии. По Циттелю
В Западной Сибири брахиоподы сравнительно немногочисленны в кемб-
рийских отложениях, но для силура, девона и нижнего карбона являются
важнейшей группой ископаемых: пользуясь очень широким распростра-
нением, они дают большое количество руководящих форм и поэтому
играют выдающуюся роль в стратиграфической параллелизации морских
Тело животного заключе-
но в двустворчатую ракови-
ну (рис. 69). Каждая створка
и раковина в целом являются
двусторонне симметричными.
Каждая створка снабжена
макушкой — несколько обо-
собленной, более или менее
приостренной, нередко клю-
вообразно загнутой частью,
которая является начальной,
наиболее ранней частью
створки: от макушечной ча-
сти створка в дальнейшем
нарастала, увеличиваясь в
размерах. Рост раковины
имел прерывистый характер,
и на поверхности створок
обычно ясно различимы кон-
центрически располагающие-
ся линии нарастания, отве-
чающие остановкам роста.
Раковина у брахиопод не-
равностворчатая: брюшная
(вентральная) створка обла-
дает более развитой макуш-
кой, обычно снабженной фораменом — отверстием для выхода ножки,
с помощью которой животное прикрепляется к какому-либо подводному
предмету; противоположная створка называется спинной (дорзальной).
Тот край раковины или створки, на котором располагается макушка, на-
зывается задним или замочным, так как у более развитой группы бра-
Х'иопод (класс замковые *) этот край снабжен особым шарнирным
устройством — замком (см. ниже). На рис. 69 показаны обозначения
краев раковины и положение основных ее измерений.
Многие руководящие формы из числа брахиопод могут быть опреде-
лены на основании внешних признаков раковины, другие требуют обяза-
тельного изучения внутреннего строения.
Остановимся сначала па внешних признаках, имеющих диагностиче-
ское значение. Такое значение имеют общие очертания раковины, отно-
шение ширины к длине и относительная выпуклость створок.
Наряду с двояковыпуклыми, часто встречаются раковины, у которых
одна из створок (брюшная или спинная) плоская или вогнутая. В опи-
саниях и диагнозах таких раковин сначала характеризуется спинная
створка, затем — брюшная, т. е. выражение «раковина выпукло-вогну-
тая» означает, что спинная створка выпуклая, а брюшная вогнутая; вы-
ражение «раковина плоско-выпуклая» показывает, что спинная створка
плоская, брюшная выпуклая, и т. д. •
Характер выпуклости (или вогнутости) также бывает различен:
в одних случаях выпуклость (или вогнутость) более или менее равно-
1 Класс беззамковых брахиопод очень слабо распространен в Западной Сибири;
по этой причине его представители, отличающиеся от замковых рядом особенностей,
здесь не рассматриваются.
38
мерная, плавная, в других поверхность створки образует (обычно близ
переднего и боковых краев) резкий, иногда под прямым углом, перегиб;
в этом случае говорят о коленообразном характере изгиба створки.
Рис. 70. Характер выпуклости раковины (сечения2.по плос-
кости симметрии)
а—двояковыпуклая; 6—плоско-выпуклая; в—выпукло-плос-
кая; г—вогнуто-выпуклая; д—выпукло-вогнутая; е—вогнуто-
выпуклая с коленообразным изгибом створок; ж—выпукло-
вогнутая с коленообразным изгибом створок
На рис. 70 даны продольные (по плоскости симметрии) сечения ра-
ковин с различной относительной выпуклостью створок. У беззамковых
брахиопод часто (особенно среди кембрийских форм) встречаются ра-
ковины, у которых створки имеют характер конуса или колпачка, благо-
даря тому, что у них макушки лежат несколько впереди замочного края
(рис. 71).
Рис. 72. Характер призамочных (кардинальных)
углов
а—округленные; б—прямые; в—острые
Рис. 71. Род Discina Dall.
Неправильно коническая
форма створок; беззамковые
брахиоподы
Характер замочного края также играет немаловажную роль при опре-
делении брахиопод. Замочный край может быть прямой или изогнутый,
короткий или длинный. Часто длина замочного края определяется по от-
ношению к наибольшей ширине раковины. В связи с длиной замочного
края находится характер призамочных или кардинальных углов рако-
вины: они могут быть округленные, прямые или оттянутые в остроконе-
чия (рис. 72).
Часть поверхности створки, лежащая у кардинальных углов, нередко
обособляется или благодаря очертаниям, или (чаще) благодаря тому,
что она является плоской или значительно1 менее выпуклой, чем осталь-
ная поверхность створки, в этом случае говорят о более или менее раз-
витых ушках; между ушками располагается обычно наиболее вздутая
часть створки, называемая умбональной частью.
Вдоль замочного края на обеих створках или только на одной (в этом
случае, как правило, на брюшной) у многих групп брахиопод разви-
ваются особые площадки, более или менее резко отграниченные от
остальной поверхности створки — так называемой арен. Обычно (за
очень редким исключением) арея брюшной створки бывает развита зна-
чительно больше, чем арея спинной створки *. Наличие или отсутствие
арей, их размеры и характер имеют большое диагностическое значение
(рис. 73). Помимо тонкой штриховки, которая часто не сохраняется,
ареи у некоторых брахиопод бывают снабжены мелкими зубчиками или
1 Когда в описаниях и диагнозах характеризуется арея без указания створки,
имеется в виду арея створки брюшной.
39
насечками (кренуляция) по всей длине ареи или лишь в средней части
(рис. 74).
Посередине ареи брюшной створки располагается треугольное отвер-
стие для выхода ножки—дельтирий. Дельтирий может быть открытым
Рис. 73. Арея
брюшной
створки с от-
крытым дель-
тирием и зуба-
ми.‘По Уолкотту
Рис. 74. Brachyprion S h а 1.
Зазубренность (кренуляция)
нижиего края ареи
Рис. 75. Непрямой
замочный край.
По Голлу
ф— форамен;
д—дельтидиаль-
ные пластинки;
з—зубы
Рис. 76. Род Spirifer Sow.
Синус и седло. По Голлу
(см. рис. 73) или в различной мере
Чаще всего он прикрывается сверху
Рис. 77. Различные формы язычка
синуса у представителей семейства
Camarotoechiidae. По Дэвидсону
и различным способом прикрыт,
(со стороны макушки) одной сво-
дообразной (выпуклой) пластин-
кой, которая называется псевдо-
дельтидием (см. рис. 85), или за-
растает с боков и снизу (со сто-
роны замочной линии), прикры-
ваясь парными дельтидиальными
пластинками, между которыми
образуется заметный шов; в по-
следнем случае в верхней части
остается круглое или овальное
отверстие (рис. 75). Аналогичные
дельтирию и псевдодельтидию, но
иначе называемые образования
могут присутствовать и на арее
спинной створки.
Из других внешних признаков
брахиопод большое значение
имеют синус и седло, т. е. продольные вдавленность (углубление) и воз-
вышение, развивающиеся на створках вдоль плоскости симметрии
(рис. 76). Обычно синус присутствуем на брюшной, а седло на спинной
створках, но встречается (например, у многих пентамерид) и обратное
расположение седла и синуса; в некоторых случаях на обеих створках
развиваются синусообразные вдавленности. Наличие или отсутствие си-
нуса и седла, их положение, степень развития и характер являются при-
знаками, учитываемыми видовыми и родовыми диагнозами. В случае
наличия синуса очертания отвечающей ему части переднего края обра-
зуют так называемый язычок синуса, размеры и форма которого часто
имеют диагностическое значение (рис. 77).
Скульптура наружной поверхности створок у брахиопод очень разно-
образна. Наряду с гладкими раковинами, несущими только знаки роста,
40
имеются раковины с различными радиальными и концентрическими
элементами скульптуры — складками, ребрами, струйками, валиками,
морщинами. В некоторых случаях дополнительно развиваются бугорки,
чешуйки, шипы. Наряду со скульптурой первого порядка, состоящей из
достаточно грубых (во всяком случае, легко различимых невооружен-
ным глазом) элементов, нередко наблюдается очень тонкая и далеко не
всегда сохраняющаяся скульптура в виде нитевидных струек, мельчай-
ших иголочек, шипиков, сосочков и т. д. Различные типы скульптурных
элементов и различные их комбинации нередко дают важные диагности-
ческие признаки, позволяющие распознавать различные виды.
Не останавливаясь на микроструктуре вещества створок, отметим, что
у некоторых групп брахиопод это вещество бывает пористым, причем
пористость обычно легко различима в простую лупу. Пористость имеет
различную природу и иногда свойственна только тому или иному слою
вещества створки, но для предварительных определений часто бывает
достаточно установить лишь ее наличие. В описаниях брахиопод такая
пористость нередко отмечается как «точечное строение» вещества рако-
вины.
Переходя к характеристике тех элементов внутреннего строения рако-
вины брахиопод, которые составляют минимум, необходимый для пред-
варительных определений, напомним, прежде всего (см. «Введение»,
стр. 13), что методика их изучения определяется состоянием сохранения
материала, именно: внутренние ядра створок позволяют наблюдать непо-
средственно (хотя и в негативном виде) многие детали внутреннего
строения, тогда как целые раковины требуют применения специальных
методов, в первую очередь метода параллельных сечений (рис. 78).
"888880(3
Рис. 78. Серия поперечных разрезов примакушечной
части рода Camarotoechia. Брюшная створка (вверху)
с зубными пластинами. В спинной створке (внизу)—раз-
деленная замочная пластина, срединная септа и септа-
лиум. По Уэллеру
Рис. 79. Внутреннее строе-
ние брюшной створки
з.—зубы; з. м.—закры-
вающие мускулы (аддук-
торы); о. м— открываю-
щие мускулы (дидук-
торы). По Голлу и Клерку
Рис. 80. Внутреннее
строение брюшной
створки
з.—зубы; з. п.—
зубные пластинки
По Л. Ш. Давиташ-
вили
Рис. 81. Попе-
речное сечение
макушечной
части брюшной
створки
з. п.—зубные
пластинки;
д. п.—дельтири-
альная пластин-
ка
Замок раковины брахиопод состоит из двух зубовидных выростов,
расположенных на замочном крае по бокам дельтирия (рис. 79 и 80);
в спинной створке им отвечают углубления, называемые зубными лун-
ками. Во многих случаях (но не всегда) в макушечной полости брюшной
створки имеются две пластинки, связанные с зубами и потому называе-
мые зубными пластинками (рис. 80 и 81). Иногда зубные пластинки
соединяются поперечной дельтириальной пластинкой, располагающейся
параллельно арее (см. рис. 81).
41
У некоторых брахиопод в брюшной створке имеется располагающаяся
по плоскости симметрии срединная перегородка или септа; она может
иметь различный характер и различную степень развития — от высокой
и тонкой пластинки до низкого гребня или валика. На внутренних ядрах
септа и зубные пластинки наблюдаются непосредственно в виде соот-
Рис. 82. Внут-
реннее ядро
брюшной створки
з. п.—зубные
пластинки;
з. м.—откры-
вающие муску-
лы; о. м.—за-
крывающие
мускулы.
По Шухерту и
Куперу
С
Н.П-
Рис. 83. Внут-
реннее ядро
брюшной створки
з. и.—зубные
пластины;
с,—срединная
перегородка
(септа).
По Голлу
Рис. 84. Попереч-
ное сечение (при-
шлифовка) маку-
шечной части
брюшной створки
з. п.—зубные плас-
тины; с.—септа
(срединная перего-
родка)
Рис. 85. Строение
спондилиума у ро-
да Cyrtina Dav.
с.-септа; з. п_зуб-
ные пластины
а—арея;
п. д.—псевдодель-
тидий.
По Дэвидсону
Рис. 85. Внутреннее строение створок
у рода Conchidium L.
с—септа и з. п. —зубные пластины,
образующие спондилиум; с. п,—сеп-
тальная пластина; к. п.— крураль-
ная пластина.
ветствующих углублений (рис. 82 и 83),
но они легко могут быть обнаружены
путем пришлифовки макушки и у це-
лых раковин (рис. 84). При одновре-
менном присутствии септа и зубные
пластины могут быть или несвязанны-
ми друг с другом (см. рис. 84) или сое-
диняться, образуя так называемый
спондилий; у разных групп брахиопод
спондилий может иметь в деталях раз-
личное устройство (рис. 85—87).
В спинной створке непосредственно
под макушкой у многих брахиопод
располагается замочная пластинка
(рис. 90), цельная или двураздельная.
Иногда эта пластинка снабжена отвер-
Рис. 87. Строение спондилиума у
рода Cyrtina Dav. (а) и у рода
Conchidium L. (б) в поперечных
сечениях (пришлифовки макушечной
части брюшных створок); см.
рнс. 85 и 86
стием, которое называется висцераль-
ным фораменом.
К замочной пластинке прикрепля-
ются (когда они имеются) ручные под-
держки; они составляют известковый
скелет рук — двух мясистых бахромча-
тых спирально свернутых выростов,
располагающихся по бокам от ротового
отверстия и служащих органами дыха-
ния и собирания пищи. Ручные под-
держки имеют различные устройства и
степень развития (рис. 88): простые и
сравнительно короткие, часто крючковидные отростки (сем. Rhynchonel-
lidae и др.), длинные известковые ленты, поддерживающие руки на всем
их протяжении и образующие спиральные конусы с различным количе-
ством витков (сем. Spiriferidae, Atrypidae, Athyridae и др.), различным
образом устроенные петли (сем. Terebratulidae и др.).
Если определение (с помощью пришлифовки) общего типа ручных
42
поддержек обычно не представляет больших трудностей (рис. 89), то
выяснение деталей строения ручного аппарата требует длительной обра-
ботки большого количества образцов, обладающих хорошей сохранностью.
В спинной створке (так же,
как и в брюшной) может при-
сутствовать срединная септа;
при наличии двураздельной за-
мочной пластинки септа может
соединяться с обеими частями
последней, причем возникает
треугольное ложкообразное уг-
лубление, называемое септа-
лиумом. На рис. 90 и 91 изо-
бражены септалиумы на внут-
реннем ядре спинной створки и
в виде пластилинового слепка с
такого внутреннего ядра и в ви-
де поперечного сечения септа-
лиума на поверхности пришли-
фовки макушечной части
Рис. 88. Типы ручных поддержек
а—спиральный; б—петлевидный; в—крючко*
видный (круральиый). По Н. Н. Яковлеву
створки.
У некоторых брахиопод
(например, из семейства Реп-
tameridae) в спинной створке
наблюдается устройство, ана-
логичное спондилиуму. Это так называемый круралиум, образуемый кру-
ральными пластинками (к которым, так же как и замочной пластинке
других групп брахиопод, прикрепляются ручные поддержки) и двумя
септальными пластинками в случае слияния последних (рис. 92). В дру-
гих случаях септальные пластинки остаются разобщенными (рис. 93).
Рис. 90. а—септалиум и замочная пластинка иа
внутреннем ядре спинной створки; б—пластилиновый
слепок с внутреииего ядра, подобного изображен-
ному на рис. а; с.—септа, образующая септалиум;
з. л.—замочная пластинка; з. л.—зубная лунка. По Голлу
Рис. 89. Ракови-
на рода Atry-
ра L. с пришли-
фованной спин-
ной створкой;
виден разрез
спирального ко-
нуса ручных
поддержек
В спинной створке многих брахиопод, в центре замочной линии, рас-
полагается замочный отросток (рис. 94 и 96), имеющий различное строе-
ние и часто поддерживаемый срединной септой или валиком. К дисталь-
ному концу замочного отростка прикрепляются открывающие мускулы
(рис. 95).
Система открывающих (дидукторы) и закрывающих (аддукторы)
мускулов схематично показана на рис. 95. Места прикрепления мускулов
на внутренней стороне створок представляют собой некоторые углубле-
ния (на ядрах — выпуклости) весьма различных размеров и очертаний
(см. рис. 96). В спинной створке имеются только отпечатки аддукторов,
43
в брюшной — тех и других, причем отпечатки открывающих мускулов
охватывают располагающиеся в средней части мускульной площадки от-
печатки аддукторов. В спинной створке некоторых групп брахиопод, на-
сс. 'сп
Рис. 91. Септалиум
в поперечном се-
чении
Рис. 92. Круралиум
в поперечном се-
чеиин
с. с.-спинная створ-
ка с круралиумом;
к. п.—круральиые
пластинки;
б. с,—брюшная
створка со спондн-
лиумом; з. п.—зуб-
ные пластинки
с. с.—спинная ст-
ворка с септалиу-
мом з. п.—замоч-
' ная пластинка;
б. с.—брюшная ст-
ворка с зубными
пластинками
Рес.93. Септальные
(с. п.) и крураль-
ные (к. п.) плас-
тинки на попереч-
ном сечении спин-
ной створки (с. с.);
б. с,—брюшиая
створка со
спондилиумом
Рис. 94. Внутренняя поверх-
ность спинной створкн рода
Productus Sow.
з. о.—замочный отросток;
б. о.—брахиальные отпечатки;
с. в.—срединный валик.
По Циттелю
Рис. 95. Схема расположенья
и прикрепления мускулов
о. м.—открывающие мускулы
(дидукторы); з. м.—закрываю-
щие мускулы (аддукторы);
вверху —форамен.
По Л. Ш. Давиташвили
Рис. 96. Спинная створка с внутренней стороны (а)
з. о.—замочный отросток; з. м— закрывающие
мускулы; с. в,—срединный валик; б—внутрен-
нее ядро с теми же элементами строения.
По Дэвидсону
ряду с отпечатками мускулов, присутствуют отпечатки рук, называемые
брахиальными отпечатками и имеющие обычно почковидные очертания
(см. рис. 94).
44
Таковы элементы морфологии раковины брахиопод, знакомство с ко-
торыми необходимо для того, чтобы приведенные далее диагнозы руко-
водящих форм могли быть понятны и использованы для предваритель-
ных определений.
Л. Л. ХАЛФИН
Тип Arthropoda. Членистоногие
КЛАСС TRILOBITA. ТРИЛОБИТЫ
Распространение класса трилобитов ограничено палеозоем; трилобиты
нижнего палеозоя представляют собой одну из важнейших в стратигра-
фическом отношении группу животных. В Западной Сибири трилобиты
широко распространены в кембрийских отложениях, для стратиграфии
которых они наиболее важны.
Двусторонне симметричное тело животного со спинной стороны было
покрыто прочным панцырем, который обычно достаточно хорошо сохра-
няется в ископаемом состоянии. Общая схема строения панцыря, отра-
жающая строение тела трилобита в целом, изображена на рис. 97.
Рис. 97. Панцирь трило-
бита
а—головной отдел;
б—туловищный отдел;
в—хвостовой отдел
(пигидий); г—спинные
бороздки. По Л. М. Да-
виташвили
Рис. 98. Головной щит трилобита (объяснения
в тексте)
Характерным для всех трилобитов является трехдольное строение их
тела: двумя продольными бороздками (спинные бороздки) оно делится
на среднюю или осевую часть (рахис) и две боковые — плевральные
части. В передне-заднем направлении тело животного делится на голов-
ной отдел (цефалон), туловищный отдел (торакс) и хвостовой отдел
(пигидий). Головной и хвостовой отделы состоят из слившихся члеников
(сегментов); членики туловища соединены подвижно.
Осевая часть головного щита носит название глабели, плевральные
части называются щеками (рис. 98). У большинства трилобитов каждая
щека подразделяется лицевым швом на подвижную и неподвижную.
Нередко подвижные щеки и средняя часть головного щита, состоящая
из глабели и неподвижных щек и называемая кранидием, встречаются
изолированно. На рис. 98 показано строение головного щита; для обо-
значения отдельных его частей и элементов приняты следующие наи-
менования:
а — кранидий; б — подвижная щека; в — неподвижная щека; г —
глабель; д — предглабелярное поле; е — базальные лопасти глабели;
ж—з — глабелярные лопасти; и — глабелярные бороздки; к — спинные
45
бороздки; л — передне-боковые ямки; м — затылочная борозда;
н — затылочное кольцо; о — задне-боковая борозда; п — задне-боко-
вая кайма; р— фронтальная борозда; с — фронтальная кайма (лимб,
валик); т — лицевой шов; у — глаз; ф — глазная крышка (пальпе-
бральная лопасть); х — глазной валик; ц — щечное остроконечие.
Членики туловища могут иметь различное строение; в частности, сег-
менты плевр бывают гладкими или каждый из них несет косую бороздку
(плевральные бороздки). Количество члеников туловищного отдела у три-
лобитов различно (от 2 до 42).
Хвостовой щит в редких случаях полностью утрачивает следы сегмен-
тации (род Illaenus), а иногда и не делится на осевую и плевральные
части (род Bumastus). Обычно же слившиеся членики этого щита хо-
рошо различимы; разделяющие их бороздки на плевральных частях пи-
гидия называются межплевральными или интерплевральными борозд-
ками; в некоторых случаях на плевральных сегментах пигидия имеются
плевральные бороздки.
Поверхность панцыря или отдельных его частей может нести различ-
ного рода скульптуру (бугорки, ямки, бороздки, шипы).
Спинной панцырь, одевая с верхней стороны тело трилобита, по
краям подвертывается на брюшную сторону, образуя так называемый за-
ворот (рис. 99). У некоторых трилобитов вдоль заворота в пределах
Рис. 99. Разрез панцыря трилобита по плоскости симметрии.
а—головной отдел; б—туловищный отдел; в—хвостовой от-
дел, г—заворот; д—гипостома. По Л. Ш. Давиташвили
головного щита и параллельно краю последнего проходит борозда (суб-
кранидиальный желоб). Под головным щитом, причленяясь к внутрен-
нему краю заворота, располагается щитообразная пластинка — гипо-
стома.
Класс трилобитов в зависимости от положения лицевых швов
(рис. 100) делится на два отряда: Opisthoparia (заднещечные) и Рго-
Рис. 100. Схема строения головного отдела трилобитов,
а—без лицевых швов; б—заднещечных и в—переднещеч-
ных. По Л. Ш. Давиташвили
paria (переднещечные). Кроме того, часто обособляется под названием
Hypoparia группа трилобитов, у которых лицевой шов отсутствует или
невидим сверху.
Л. Л. ХАЛФИН
Тип Mollusca. Мягкотелые
КЛАСС PELECYPODA ИЛИ LAMELLIBRANCHIATA
ПЛАСТИНЧАТОЖАБЕРНЫЕ МОЛЛЮСКИ
Обширный тип мягкотелых распадается на пять классов, из которых
в ископаемом состоянии в Западной Сибири наиболее распространен-
ным является класс пластинчатожаберных моллюсков. Представители
46
этого класса являются водными донными организмами, ведущими сво-
бодный или прикрепленный образ жизни. Они населяют как морские,
так и в различной степени опресненные и пресные водоемы. В большин-
стве своем пелециподы — обитатели мелководных бассейнов.
В горной части Западной Сибири остатки пелеципод многочисленны
в морских отложениях девона и нижнего карбона, а также в угленосных
отложениях верхнего палеозоя и, отчасти, мезозоя. В пределах Западно-
Сибирской низменности местами пелеципод достаточно много в пресно-
водных третичных и четвертичных
в морских фациях, главным обра-
зом на Крайнем Севере.
Тело животного заключено
в двустворчатую известковую ра-
ковину, покрытую роговым поверх- а
ностным слоем (периостракум).
В отличие от брахиопод, у пла-
стинчатожаберных каждая створ-
ка, взятая в отдельности, асим-
метрична; раковина в целом не-
редко обладает двусторонней сим-
метрией, причем плоскость сим-
метрии лежит между створками.
Обе створки могут быть развиты
одинаково (раковина равноствор-
чатая), или одна из них более вы-
пуклая, чем другая (раковина не-
равностворчатая) .
Измерения раковины показаны
на рис. 101. Верхний, или замоч-
ный, край может быть прямым
или непрямым; у неравномускуль-
ных пелеципод (см. ниже) при
наличии прямого замочного края
по бокам выпуклой макушечной
части (умбональная часть створ-
ки) обычно развиваются плоские
или слабо выпуклые ушки
(рис. 102). Нижний край иначе
называется брюшным.
Макушки в большинстве слу-
чаев бывают смещены к перед-
нему краю (см. рис. 101 и др.),
реже — к заднему (см. рис. 107),
благодаря этому передний конец
раковины обычно короче заднего. В
отложениях, реже они встречаются
Рис. 101. А—левая створка с внеш-
ней стороны
а-а—длина раковины; d-б—высота
раковины; Б—левая створка с внут-
ренней стороны: 'гетеродонтный за-
мок, отпечатки переднего и заднего
замыкающих мускулов; простая ман-
тийная линия; В-вид раковины со
стороны верхнего (замочного) края;
в-в—толщина раковины. По Л. Ш. Да-
виташвили
зависимости главным образом от бо-
лее или менее значительного смещения макушек к переднему (или к зад-
нему) краю раковина пелеципод является в различной мере неравносто-
ронней. В некоторых случаях макушки бывают смещены на передний край;
тогда говорят о краевом или терминальном их положении (рис. 103).
Степень и характер выпуклости створок у пелеципод различны.
У многих пелеципод на каждой створке более или менее резко обособ-
ляется область наибольшей выпуклости, чаще всего проходящая от ма-
кушки к нижнезаднему краю; в описаниях она часто называется диаго-
нальным возвышением (рис. 104). Вдоль диагонального возвышения
часто проходит отчетливый, округленный или острый, киль. Во многих
случаях впереди диагонального возвышения располагается заметное уг-
лубление (умбовентральная вдавленность). При наличии такого углубле-
ния (вдавленности) очертания брюшного края образуют вогнутость (синус).
47
Створки называются правой и левой. Их определение производится
так: раковина помещается макушками кверху и передним краем от наблю-
дателя; при такой ориентировке справа будет располагаться правая
створка, слева — левая.
Раскрывается раковина при помощи пассивного органа, называемого
связкой или лигаментом (рис. 105). Связка лежит вблизи макушек или
Рис. 102. Род Pterinea Gold!. Слева —левая створка с
прямым замочным краем и хорошо развитыми ушками; спра-
ва—та же створка с внутренней стороны; видна связковая
арея с параллельными замочному краю связковыми бо-
роздками и отпечаток заднего замыкающего мускула.
По Циттелю
Рис. 103. Род МуШагса
Hall. Макушечная
часть правой створкн
с внутренней стороны.
Краевое положение
макушки; связковая
арея, макушечные и
задние зубы. По Голлу
Рис. 104. Род Modiomor-
pha Hall. Левая створ-
ка с диагональным воз-
вышением, у мбовентраль-
ной вдавленностью
и синусом брюшного
края. По Голлу
Рис. 105. Вер-
тикальное сече-
ние раковины
(схема); поло-
жение связки
(а) и замыкаю-
щих муску-
лов (б).
вдоль всего замочного края и может быть внешней или внутренней или
отчасти внутренней и отчасти внешней одновременно. Внешняя связка
располагается на особой связковой площадке, внутренняя — в углубле-
ниях (см. рис. 107) или бороздах на утолщенном замочном крае—на
так называемой замочной площадке (см. рис. 102 и 103).
Закрывается раковина при посредстве замыкающих или сводящих
мускулов (аддукторов), места прикрепления которых, имеющие вид
углублений, обычно хорошо заметны на внутренней поверхности створок.
Класс пластинчатожаберных разделяется на два подкласса по харак-
теру и степени развития замыкающих мускулов. У представителей под-
класса равномускульных (Isomyaria) примерно в одинаковой мере раз-
виты передний и задний мускулы (см. рис. 101, 106, 107), а у предста-
вителей подкласса неравномускульных (Anisomyaria) передний мускул
редуцируется нередко до полного исчезновения (см. рис. 1026).
Помимо отпечатков мускулов, на внутренней стороне створок, обычно
хорошо различима так называемая мантийная линия, представляющая
собой след прикрепления мантии к створке. Мантийная линия бывает
простая (см. рис. 1016 и 107) или образует в задней ее части различной
глубины бухту или синус (см. рис. 106).
Большинство пелеципод обладает замком, способствующим более
прочному сочленению створок, хотя имеются и беззубые раковины. Замок
образован чередующимися зубами и зубными лунками, расположенными
на замочной площадке. Типы строения замка у пелеципод разнообразны.
Мы отметим лишь те из них, которые свойственны помещенным в данном
справочнике родовым группам
48
У равномускульных пелеципод важнейшими типами замка являются:
а) таксодонтный, представляющий собой серию мелких, более или
менее однотипных зубчиков (рис. 107);
б) гетеродонтный, представленный небольшим числом зубов различ-
ной длины, толщины и формы (см рис. 101, 106); в нем различают ле-
жащие под макушкой кардинальные зубы и расположенные более или
б
Рис. Юб.Род Avimactra А п d г.
Левая створка с внутренней
стороны: гетеродонтный замок;
кардинальные (а) и боковые
(б) зубы; связковая лунка (в);
отпечатки переднего и заднего
замыкающих мускулов (г); ман-
тийная линия (б) с небольшим
синусом. По В. П. Колесникову
Рис. 107. Род Nucula Lam.
Правая створка с внутренней
стороны: таксодонтный замок;
отпечатки переднего (а) и зад-
него (в) замыкающих мускулов;
связковая лунка (б); простая
мантийная линия. По Лагузену
менее параллельно замочному краю, впереди и позади макушек, боко-
вые зубы; количество тех и других неодинаково у различных групп пеле-
ципод.
У неравномуокульных пелеципод зубы или совсем отсутствуют, или
представлены небольшими и немногочисленными простыми зубчиками
или ребрышками (см. рис. 103), реже имеют иной характер.
Скульптура наружной поверхности створок у морских пелеципод
разнообразна и нередко очень типична для представителей того или
иного вида; у пресноводных форм она более однообразна, и поверхность
створок чаще всего снабжена лишь концентрическими линиями и зна-
ками роста.
При определении пелеципод большую роль играют детали внутрен-
него строения (замок, отпечатки мускулов и т. д.). Лучше всего эти де-
тали наблюдаются на изолированных створках, извлекаемых из рыхлых
или слабо диагенетизированных отложений. В палеозое Западной Сибири
такие породы обычно отсутствуют, и наиболее благоприятным для опре-
делений состоянием сохранения пелеципод являются отпечатки их изоли-
рованных створок, обычно встречаемые в песчано-глинистых породах.
Исключение представляют угленосные отложения, в которых, как пра-
вило, встречаются лишь скульптурные ядра створок, не позволяющие
наблюдать детали внутреннего строения.
Г. П. РАДЧЕНКО
РАСТЕНИЯ
В большинстве случаев при определении ископаемых растений при-
ходится пользоваться отпечатками их листьев, затем стеблей или ство-
лов, особенно тех, которые сохранили детали внешней скульптуры,
и, наконец, различных органов размножения. Приводимая ниже характе-
ристика внешних признаков растений касается именно этих их элемен-
тов — применительно к тем классам и группам вымерших растений, ко-
торые наиболее распространены в верхнепалеозойских и мезозойских* 1
отложениях Саяно-Алтайской области.
1 Некоторые специальные термины, использованные в описаниях псилофитов
(девон) и мезозойской флоры, объяснены в этих описаниях. Прим. ред.
I 3.1К. 2024 19
Наиболее широко представлены в этих отложениях следующие классы
и группы ископаемых растений:
- 1. Папоротниковые.
2. Членистостебельные (хвощевые).
3. Плауновые.
4. Папоротникообразные семенные (птеридоспермы).
5. Кордаитовые.
6. Саговообразные (цикадофиты).
7. Гинкговые.
8. Хвойные.
9. Растения неопределенного систематического положения.
10. Органы размножения.
Тип Pteridophyta. Папоротникообразные
КЛАСС FILICALES. ПАПОРОТНИКИ
От этих растений в ископаемом виде обычно сохраняются на породе
отпечатки листьев, а чаще всего — отпечатки их незначительных обрыв-
ков.
У настоящих папоротников листья в большинстве случаев сложные,
т. е. сильно расчленены на доли или перья. Значительно реже настоящие
папоротники имеют простые, т. е. нерасчлененные листья (роды Chlro-
pteris, Hausmannld). Целый лист, простой или сложный, называется у па-
поротников вайей.
Сложные вайи имеют, как и простые листья, более или менее тол-
стый черешок, которым они прикреплялись к корневищу или стеблю.
Черешок этот, продолжаясь в лист, у одних форм сразу же разделяется
на многочисленные стержни перьев, радиально расходящиеся из оконча-
ния черешка (роды Laccopteris, Matonldium), у других вильчато разде-
ляется на два равнозначащих развилка стержня вайи (роды Campto-
pterls, Dictyophyllum) и, наконец, у третьих, наиболее часто встречаю-
щихся папоротников, продолжение черешка в листе прямое или слабо
извилистое и носит название главного стержня вайи (рахис).
Вайи, характеризующиеся первым или радиальным типом располо-
жения стержней, несущих листовые пластинки, называются дланевид-
ными или веерообразными. Части вайи, относящиеся к радиальным
стержням, можно называть перьями, так как они имеют обычно перистое
строение.
Вайи, обладающие одним центральным стержнем, называются пери-
стыми; этот тип листьев является наиболее распространенным у папорот-
ников. Перистые вайи бывают двух типов: простоперистые, у которых по
обе стороны главного стержня вайи расположены листовые образования
или перышки, обычно уменьшающиеся к основанию и верхушке вайи
(рис. 108), и сложноперистые, у которых на главном стержне располо-
жены перья, т. е. перистые образования со своими стержнями, напоми-
нающие целые вайи (рис. 109).
Если перышки расположены на стержнях, которые отходят прямо от
главного стержня вайи, то такие стержни называются стержнями первого
порядка, а вся такая вайя-—дважды перистой. Если перышки сидят на
стерженьках, которые отходят от стержней первого порядка, то вайя
называется трижды перистой, а названные стерженьки — стержнями вто-
рого порядка (рис. 109). Могут быть и четырежды перистые вайи, обла-
дающие стержнями третьего порядка. Часто стержни, на которых распо-
ложены перышки, называются стержнями последнего порядка. Такими же
названиями обозначаются перья различных порядков.
Перышки могут быть цельными, с волнистыми краями, с городча-
тыми краями, надрезанными на лопасти или, наконец, рассеченными на
50
доли (рис. НО). По способу их прикрепления к несущему стержню раз-
личают перышки трех основных типов: сфеноптероидные, пекоптероид-
ные и невроптероидные.
Невроптероидные перышки прикрепляются к стержню только не-
большой, средней частью своего основания, часто только одной точкой
его; поэтому основание у таких перышек обычно является сердцевидным
с закругленными нижними углами. Невроптероидные перышки всегда
бывают цельными, очень редко с волнистыми краями. У настоящих папо-
ротников невроптероидные перышки встречаются крайне редко.
Рис. 108. Образец
простоперистой
вайи папоротника
Рис. 109. Участок трижды перистой вайи
<1—перо первого порядка; б—перышко; в—перо
второго порядка; г—стержень пера первого по-
рядка; д—стержень пера второго порядка; е—
стержень вайи
Перышки пекоптероидной формы прикреплены к стержню всем своим
основанием. Обычно они цельные или с городчатыми краями, но нередко
бывают и надрезанными на лопасти.
Перышки сфеноптероидной формы, изменяясь в очертании от почти
круглых до почти линейных, к основанию сужены или даже слегка пере-
тянуты. Сфеноптероидные перышки редко бывают цельными; в большин-
стве случае они рассечены на доли.
Внешние очертания или общий абрис перышек, включая все их ло-
пасти и доли, могут быть весьма различны; этот признак считается весьма
важным морфологическим и систематическим признаком ископаемых
папоротников. Следует обращать внимание также на то, как располо-
жены перышки на стержне: слиты ли они своими основаниями друг
с другом в общую пластинку или нет, соприкасаются ли они своими осно-
ваниями или нет, расставлены они на стержне или нет. Кроме того, от-
мечают, под каким углом наклонены перышки к стержню.
Жилкование перышек папоротников отличается значительным разно-
образием (рис. ПО). Различают следующие основные типы жилкования:
веерное, без выделения главной жилки, когда тонкие одинаковые жилки
лучисто расходятся от основания перышка и по пути ветвятся; перистое,
когда выделяется более или менее толстая главная или, как ее называют
у папоротников, средняя жилка, в обе стороны от которой отходят боко-
вые жилки, в свою очередь ветвящиеся; ложноперистое жилкование ха-
рактеризуется тем, что одна, центральная, сильно извилистая жилка
вильчато делится поочередно то в одну, то в другую сторону и ее корот-
кие ветви как бы соответствуют боковым жилкам. Наконец, некоторые
типы пекоптероидных перышек характеризуются сетчатым жилкованием.
4 51
Зеленые неплодущие вайи папоротников называются стериль-
ными. Плодущие экземпляры называются спороносными или
фертильными. Вместилища спор, или спорангии, располагаются
всегда на нижней стороне листовой пластинки перышек, причем у разных
форм папоротников их форма и взаимное расположение строго законо-
мерны. Спорангии могут располагаться поодиночке или же в виде кучек.
Рис. ПО. а, б— сфеноптероидные перышки; в, г, д, к,
л—пекоптероидные перышки; е, и—сложно-пекоптеро-
идные перышки; ж—невроптероидное перышко; з—цик-
лоптероидное перышко; а, б, в, г, д, е, ж—перистое жил-
кование; и—ложноперистое жилкование; з—веерное
жилкование; к—параллельное жилкование; л—сетчатое
жилкование; г—волнистые края перышек; д—городчатые
края перышек; а, е, и—лопастные края; , б—перышко
рассечено на доли
так называемых сорусов. Детали строения спорангиев, имеющие большое
значение в систематике современных папоротников, очень редко сохра-
няются в ископаемом состоянии.
КЛАСС PTERIDOSPERMAE. ПАПОРОТНИКООБРАЗНЫЕ
СЕМЕННЫЕ
От этих растений в ископаемом состоянии обычно сохраняются обуг-
ленные остатки или чаще отпечатки на породе их сложных листьев,
близко напоминающих вайи папоротников и также называемых вайями.
Подавляющее большинство папоротникообразных семенных (птеридо-
спермов) имеют либо простоперистые, либо дважды перистые вайи, мор-
фология и элементы которых таковы же, как у вайи настоящих папорот-
ников (см. выше).
Особенно характерны для птеридоспермов сфеноптероидный, невро-
птероидный и циклоптероидный типы перышек (см. рис. 110 б, ж, з).
Первый тип был уже описан выше. Перышки циклоптероидного типа
отличаются от перышек невроптероидных только тем, что они не продол-
говатые, как невроптероидные, а круглые или почти круглые. Перышки
у птеридоспермов обычно имеют значительные размеры.
Своеобразной особенностью некоторых птеридоспермов являются так
называемые промежуточные перышки, располагающиеся непосредственно
на главном стержне вайи между стержнями боковых перьев.
Жилкование перышек птеридоспермов, при сохранении основных его
типов, часто очень густое, сложное. Нередко наблюдаются дополнитель-
ные жилки, вступающие в перышко по бокам средней жилки непосред-
ственно из стержня пера или вайи. У некоторых форм известно еще более
52
своеобразное жилкование, характеризующееся тем, что вместо средне!!
жилки в перышко вступает пучок тонких сближенных параллельных
жилок, как бы заменяющих среднюю жилку; пучок этот прослеживается
на некотором расстоянии, а затем веерообразно расходится. Это одна из
разновидностей веерного жилкования. Иногда жилкование невроптероид-
ных перышек может быть сетчатым.
КЛАСС ARTICULATAE. ЧЛЕНИСТОСТЕБЕЛЬНЫЕ (ХВОЩЕВЫЕ)
От хвощевых в ископаемом виде сохраняются обугленные остатки или
отпечатки наружной стороны стеблей, часто вместе с листьями, сидев-
шими в мутовках на узлах, отпечатки целых или неполных мутовок
листьев, отпечатки колосьев спороношения, каменные ядра или слепки
внутренней полости трубчатых полых стеблей хвощевых, сохраняющие
скульптуру внутренних стенок этой поло-
сти, и, наконец, отпечатки диафрагм, пред-
ставляющих собой горизонтальные пере-
городки внутри стебля, разделяющие вну-
треннюю его полость на членики.
У всех членистостебельных стебли раз-
делены на правильные участки или чле-
ники, которые называются междоузлиями
(рис. 111). Последние отделяются один
от другого узкими пережимами или утол-
щенными кольцами, называемыми узла-
ми. Последним внутри стебля соответ-
ствуют поперечные горизонтальные пере-
городки— диафрагмы. У одних форм
диафрагмы были рыхлые, при гибели ра-
Рис. 111. Типы листвы хвощевых.
А—облиствение типа AsterophyHi-
tes', Б—облиствение типа Annularia;
а—междоузлия; б—узлы; в—мутов-
ки (или кольца) листьев; г—средин-
ная жилка
стения сгнивали и в ископаемом состоя-
нии не сохранялись; у других форм, на-
против, они были жесткими, даже частич-
но минерализованными, и хорошо1 сохра-
нились в ископаемом виде, выжимаясь из
стебля при его расплющивании в виде
изолированных кружков.
Листья у членистостебельных собраны
в мутовки, т. е. розетки, располагающиеся сразу над узлами и окружаю-
щие стебель. По расположению листьев в мутовках все членистосте-
бельные разделяются на две большие группы. У одних все листья
в каждой мутовке направлены в сторону верхушки стебля, окружая его
со всех сторон и образуя подобие метелки (ботанический тип Astero-
phyllites). У других все листья в каждой мутовке распростертые при-
мерно в одной плоскости и расходятся лучисто или в бока от стержня
(ботанический тип Annularia).
Основными систематическими признаками, принятыми в палеобота-
нике при разделении членистостебельных на роды и виды при отсутствии
спороношений, считаются характер облиствения побегов и форма
листьев. Листья могут быть совершенно свободны до самого основания,
сращены друг с другом на различном протяжении их длины или, на-
конец, путем срастания образовывать солистья, имеющие вид опахал
или сложных листьев.
Важным признаком считается также расположение листьев в каждой
мутовке, независимо от того, являются ли они свободными или сращен-
ными: они могут тесно прилегать к стеблю или окружать его свободно,
наподобие колокола. Сращенные нижние части листьев, если только
они не составляют солистьев, образуют влагалище, окружающее стебель;
в зависимости от длины сращенной части листьев влагалища могут иметь
форму воротника, чашки, колокола, бокала или узкого цилиндра.
53
Листья могут срастаться друг с другом непосредственно краями или
при помощи так называемых комиссуральных складок, которые, наподо-
бие узких перепонок, связывают основания смежных листьев.
У большинства членистостебельных листья узкие, ланцетные, ленто-
видные или шиловидные. Посередине каждого листа пробегает одна
срединная жилка, иногда очень плохо выраженная (см. рис. 111 г).
Исключение представляют хвощевые из палеозойской группы клинолист-
ников или сфенофиллов, обладающих клиновидными листьями, харак-
теризующимися веерным жилкованием.
Каменные ядра или слепки внутренней полости стволов или стеблей
представляют отпечатки внутренних стенок стебля; эти отпечатки обычно
несут на поверхности многочисленные продольные ребра, представляю-
щие собой отпечатки сердцевинных лучей стебля. Желобки или бороздки
между ребрами соответствуют проводящим пучкам стебля.
Органы спороношения членистостебельных встречаются либо в виде
спороносных колосьев или шишек, располагавшихся на верхушках стеб-
лей и ветвей, либо в виде мутовок спороносцев, расположенных на стебле
между мутовками зеленых листьев. И в том и в другом случае спорангии
располагаются на концах тонких столбиков или спороносцев.
КЛАСС LYCOPODIALES. ПЛАУНОВЫЕ
а б
Рис. 112. а—листовые рубцы, располо-
женные в вертикальных рядах—орто-
стихах; б—листовые подушки с листовы-
ми рубцами, расположенные в спираль-
ных рядах—парастихах
систематическими в палеоботанике,
тикальных рядах — ортостихах, или
В ископаемом виде сравнительно редко встречаются остатки облист-
венных стволов или ветвей плауновых, поскольку колоннообразные
стволы этих растений еще при жизни утрачивали листья и облиственной
v них оставалась только самая крона дерева, образованная тонкими,
многократно вильчато делящими-
ся ветвями. Обычны отпечатки
коры стволов или ветвей плауно-
вых, лишенных листьев, но сохра-
нивших скульптурные или рельеф-
ные образования, всегда имею-
щиеся на поверхности стволов
этих растений.
Скульптурные образования бы-
вают двух типов. Первый тип —
это рубцы от опавших листьев на
коре, прикреплявшихся своими
основаниями непосредственно к
поверхности ствола. Рубцы эти
представляются в виде углубле-
ний на отпечатках или в виде
слабых бугорков на самой щэре.
У разных родов плауновых они
по-различному располагаются,
имеют разные размеры и очерта-
ния; все эти признаки являются
/бцы могут располагаться в вер-
спиральных рядах — парастихах,
или одновременно в тех и других (рис. 112).
Внутри рубца от прикрепления листа обычно наблюдаются один или
два небольших рубчика округлой или треугольной формы, соответствую-
щие проводящим пучкам (сосудам), проходящим из ствола в каждый
лист и протягивающимся вдоль последних в виде одной или двух жилок
листа. Поэтому такие рубчики называют рубчиками листового следа. По
бокам от этих рубчиков иногда заметны еще рубчики, так называемые
парихны.
Второй тип скульптурных образований — листовые рубцы — распола-
гаются на так называемых листовых подушках, представляющих собой
54
небольшие выпуклости или бугорки различной формы на коре дерева.
Обычно листовые подушки имеют удлиненную форму. Они могут распо-
лагаться также либо в ортостихах, либо в парастихах.
У некоторых плауновых над листовым рубцом наблюдается неболь-
шой круглый точечный рубчик, соответствующий так называемой языч-
ковой камере, в которой у живых растений помещался небольшой язы-
чок или лигула. По наличию или отсутствию лигулы все плауновые раз-
деляются на две большие естественные группы — язычковые плауновые
(ligulatae) и безъязычковые плауновые (eligulatae).
Следует иметь в виду, что при сгнивании или вообще утере стволом
самых верхних слоев коры до погребения его в осадок скульптурные
образования теряются, и на отпечатках можно наблюдать только грубые
и продолговатые следы листовых рубцов или стигмы.
Тип Gymnospermae. Голосеменные
КЛАСС CORDAITALES. КОРДАИТОВЫЕ
Растения, относящиеся к вымершему классу кордаитовых, все без
исключения являлись крупными деревьями, ветви которых несли бес-
черешковые, т. е. сидячие, цельные, обычно крупных размеров удлинен-
ные листья с правильным жилкованием, состоящим из слегка веерно
расходящихся, почти параллельных жилок. Отпечатки таких листьев и
встречаются чаще всего в ископаемом состоянии. Древесины их нередко
сохраняются достаточно хорошо, особенно если они целиком минерали-
зованы известью или кремнеземом. В этих случаях может быть хорошо
изучено анатомическое строение.
У листьев кордаитовых отмечают следующие основные элементы: об-
щие очертания их листовой пластинки, форму верхушки листа, форму
его основания (широкое, узкое, прямое, вогнутое, выемчатое), местопо-
ложение линии наибольшей ширины листа, толщину жилок, степень их
веерообразного расхождения, прямизну или изогнутость, жилок, нали-
чие ложных жилок, представляющих собой узкие желобки на нижней
стороне листа между настоящими жилками, густоту жилкования, угол
подхода жилок к краям листа и т. д.
КЛАСС CYCADOPHYTA. САГОВООБРАЗНЫЕ (ЦИКАДОФИТЫ)
Листья или облиственные побеги саговообразных принято называть
вайями. Основное отличие их от ваий папоротников заключается в пре-
имущественном развитии простых долей листа, которые достигают у них
довольно крупных размеров и по форме часто сильно отличаются от
перьев папоротников. Нередко их называют поэтому сегментами. Все
вайи саговообразных имеют прямой стержень вайи, переходящий в ос-
новании в черешок.
Вайи бывают простые, цельные, имеющие вид крупного линейного
или ланцетного листа (роды Taeniopieris, Nllssonia) или, чаще, просто-
перистые, несущие два ряда сегментов или перьев, прикрепляющихся
непосредственно к стержню вайи. Способ прикрепления сегментов или
перьев к стержню вайи у саговообразных часто бывает своеобразным,
присущим только растениям этого класса. Так, у одних форм сегменты
прикрепляются к верхней стороне стержня, а не к его бокам, так что
с верхней стороны стержень совершенно или почти совершенно закрыт
листовыми пластинками (роды Nllssonia, Dioonites, Ptilophyllum).N дру-
гих форм сегменты прикреплены ближе к верхней стороне стержня
при помощи особых мозолистых утолщений (роды Zamites, Otozamltes)
Наконец, у третьей группы форм сегменты у основания сужены и имеют
более или менее короткий черешок, с помощью которого они прикреп-
ляются к стержню вайи или к оси побега (Yavorskia).
55
Жилкование сегментов у саговообразных сводится к двум основным
типам: параллельному, с очень редким ветвлением отдельных жилок, и
сетчатому. У некоторых форм сегменты столь узки, что каждый из них
имеет только одну простую или две параллельные грубые жилки (роды
Cycadites. Pseudocycas и др.).
КЛАСС GINKGOALES. ГИНКГОВЫЕ
В ископаемом состоянии встречаются обычно в виде отпечатков изо-
лированных черешковых, почти круглых, вееровидных или клиновидных
листьев, характеризующихся, правильным расходящимся или веерным
жилкованием.
Черешок у листьев гинкговых обычно длинный и тонкий, но у неко-
торых форм (Phoenicopsis и Czekanovskla) отсутствует. В подавляющей
массе листья гинкговых разделены на доли, веерообразно расходящиеся
из конца черешка. У многих форм доли, в свою очередь, дихотомически
или вильчато разделены на лопасти различной формы. В каждую ло-
пасть проходит несколько тонких, часто почти параллельных жилок.
Основными морфологическими признаками, на которых основывается
систематика ископаемых гинкговых, являются: прикрепление листа, сте-
пень рассеченности листа на доли, форма, количество и размеры послед-
них, рассеченность долей листа на лопасти, характер жилкования, осо-
бенно в конечных лопастях.
КЛАСС CONIFERALES. ХВОЙНЫЕ
В ископаемом состоянии обычно находятся отпечатки облиственных
тонких (верхушечных) побегов или ветвей, реже изолированных листьев,
либо, еще реже, отпечатки шишек или отдельных чешуй.
Листья у хвойных называются хвоей; они обычно мелкие, шиловид-
ные, ромбовидные или чешуевидные, реже ланцетные, линейные или
кинжаловидные, с острой верхушкой, массивные, круглые, граненые или
же сплющенные. На оси побега хвои располагаются густо, двурядно
или по спирали и часто бывают изогнуты.
Жилки у листьев настоящих хвойных часто незаметны, вследствие ко-
жистой их структуры. Но у ряда форм, характеризующихся более круп-
ными и плоскими хвоями (роды Elatocladus, Cephalotaxopsis), бывает
заметна одна тонкая срединная жилка.
СЕМЕНА
Семена ископаемых голосеменных у нас обычно сохраняются в виде
отпечатков на породе, которые очень часто сопровождаются обугленными
остатками самих семян. Часто встречающиеся в ископаемых флорах
Западной Сибири ископаемые семена отличаются большим разнообра-
зием.
Они могут быть разделены на четыре большие группы соответственно
их основным морфологическим признакам:
1. Плоские семена, снабженные специальными летательными приспо-
соблениями, так называемой крылаткой или крыльями, в связи с чем
такие семена называют также летучками. Крылатки бывают самого раз-
личного устройства, размеров и формы. У одних семян крылатка обра-
зует одно крупное асимметричное крылышко, в одном конце которого
расположено само семя. У других форм семена снабжены с обеих своих
сторон перепончатыми крыловидными выростами, выемчатыми наверху
и внизу; подобные семена называют билатеральными, т. е. дву-
сторонними. В большинстве случаев крыловидные выросты разра-
стаются либо в стороны, либо кверху, т. е. в передней части семени.
56
2. Плоские или слабо выпуклые семена, окруженные густыми воло-
сками, крючечками или нитевидными выростами, которые помогали пе-
реносу семян потоками, ветром и животными.
3. Семена наподобие плода с косточкой, обладающие мясистой внеш-
ней оболочкой, называемой саркотестой, и более плотной внутренней
оболочкой или оклеростестой, в которую было заключено само «ядро»
семени. При погребении в осадке подобные семена могли сильно расплю-
щиваться, причем мясистая саркотеста, раздаваясь в стороны, образо-
вывала концентрические ядру семени выросты, окружавшие семя почти
по всей его периферии и тем самым имитировавшие крылатку семян
первой группы.
4. Семена, обладающие очень тонкой саркотестой и достаточно твер-
дым двояковыпуклым или даже эллипсоидальным ядром. Они вовсе не
имели крыльев или других летательных при-
способлений. Иногда эти семена могли
иметь продольный киль или грани и были
снабжены шиловидным выростом.
У семян различают следующие части:
тело семени или плод; более или менее удли-
ненную, напоминающую горлышко фляги
или бутылки верхушку семени, иногда очень
короткую, в которой располагался узкий
цилиндрический, открывающийся наружу
Рис. 113.4—семя с крылаткой;
Б—семя с мясистой саркоте-
стой; а—крылатка; б— семявход
или микропиле; в—верхушка
семени или нуцеллус; г—плод;
д—остаток или рубчик от се-
мяножки; «—расплющенная
саркотеста
проход — семявход или микропиле; место
прикрепления или основание семени, кото-
рое на отпечатках представляется либо в ви-
де рубчика (углубления) от стерженька или
семяножки, к которой прикреплялось семя, либо в виде небольшого со-
сочковидного выступа, представляющего собой остаток части семяножки,
(рис. 113).
ГЛАВА I
КЕМБРИЙСКАЯ СИСТЕМА
А. Г. С И ВО В
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Общая характеристика отложений и палеонтологических остатков
кембрия Саяно-Алтайского региона
Кембрийские отложения, занимающие обширные пространства
в Саяно-Алтайской области, в подавляющей своей массе выражены гео-
сипклинальными образованиями, достигающими мощности не менее
20 км.
Эти образования, относящиеся ко всем трем отделам системы, харак-
теризуются многообразием фаций, среди которых фация рифовых из-
вестняков, богатых остатками археоциат, водорослей, реже трилобитов,
строматопор и брахиопод является наиболее типичной и пользуется
почти повсеместным распространением. Не меньшим развитием поль-
зуются эффузивно-обломочные фации, в осадочных компонентах кото-
рых тоже, но много реже, обнаруживаются окаменелости, главным
образом трилобиты, иногда брахиоподы, губки и радиолярии.
Эти две группы фаций как в горизонтальном, так и вертикальном
сечениях характеризуются сложными взаимосвязями и географически
распространены неравномерно. Так, фации археоциатово-водорослевых
известняков преобладают в составе кембрийских отложений Кузнецкого
Алатау, Салаира, а эффузивно-обломочные свойственны преимущест-
венно Горному Алтаю, Восточному Саяну и, особенно, Западному Саяну,
а также его отрогам, спускающимся в Тувинскую котловину.
Правда, в составе первой из этих фаций появляются элементы вто-
рой и наоборот, но при всем этом в продольном и поперечном профилях
главная фация всегда является преобладающей и с большим постоян-
ством сохраняет свои индивидуальные особенности.
За последние годы в важнейших регионах Саяно-Алтайской области
были проведены относительно детальные геологические исследования,
в результате которых кембрийские отложения подверглись дробному
расчленению с выделением в их составе многих местных стратиграфиче-
ских единиц; в большей части последних были обнаружены органические
остатки. Среди них особенно обильными и многообразными оказались
археоциаты, трилобиты и водоросли, которые явились предметом отно-
сительно детального изучения, правда далеко не равномерного и не за-
вершенного.
Важнейшие группы кембрийской фауны Саяно-Алтайской области —
археоциаты и трилобиты, — будучи весьма разнообразными по составу
видов и родов, характеризуются резко выраженными чертами самобыт-
ности. В этом можно легко убедиться на основе анализа хотя бы мате-
риалов сотрудников палеонтологических кабинетов Томского политехни-
ческого института имени С. М. Кирова и Западно-Сибирского геологи-
ческого управления, где длительное время проводилось изучение кемб-
58
рийской фауны Западной Сибири. Согласно этим материалам, только
среди трилобитовой фауны установлено и частично описано большое ко-
личество новых семейств, родов и видов.
Таблица I
Семейства Роды Виды
всего • в том числе всего в том числе всего в том числе
новых ранее извест- ных новых ранее извест- ных новых ранее извест- ных
21 3 18 56 45 и 154 150 4
Из табл. 1 видно, что не только видовой, но и родовой состав трило-
битовой фауны Саяно-Алтайской области почти целиком местный. Что же
касается археоциатовой фауны, то она в смысле обилия и многообразия
форм оказалась единственной в своем роде не только в Советском
Союзе, но и во всем мире, поэтому нет ничего удивительного в том, что
в ее составе новыми являются в большинстве случаев и крупные таксо-
номические единицы.
Однако несмотря на столь ярко выраженный эндемичный состав па-
леонтологических остатков кембрия Саяно-Алтайской области, значение
их для стратиграфии очень велико. Среди археоциатовой и особенно три-
лобитовой фаун обособляются строго индивидуальные комплексы; гори-
зонтальное распространение и последовательный восходящий порядок
многих из них проверены достаточно надежно детальным картированием
отложений, содержащих эти комплексы.
В составе нижнего кембрия отчетливо обособляются следующие ком-
плексы в восходящем порядке: аккольский — водорослевый, чингин-
ский — трилобитовый, гавриловский и камешковский — археоциатовые,
бабаковский — трилобитовый и обручевский — археоциатовый. В сред-
нем кембрии в том же порядке устанавливаются санаштыкгольский три-
лобитовый и археоциатовый комплексы, большеербинский и орлиногор-
ский трилобитовые комплексы. В верхнем кембрии выделяются салаир-
ский и порношорский трилобитовые комплексы, причем первый из них —
ранний, а последний — поздний.
Стратиграфическое положение трилобитовых и археоциатовых
комплексов и их общий состав
Аккольский комплекс, охарактеризованный пока лишь остатками во-
дорослей и среди них родом Osagia, типичен для одноименной серии ',
развитой по южным и северным склонам Западного Саяна. Нижнекем-
брийский возраст этого комплекса определяется условно, а его горизон-
тальное распространение пока остается не выясненным. Можно лишь
полагать, что в Горной Шории, в бассейне рч. Матур, левого притока
Абакана, установленные в развитой там карбонатной толще, выделен-
ной в свое время в матурскую формацию (А. Г Сивов, 1940), остатки
водорослей Osagia, Collenia, Conophyton и редкие примитивные архео-
циаты (?) окажутся эквивалентными аккольскому комплексу.
Чингинский комплекс трилобитов и гавриловский комплекс архео-
циат в совместном проявлении установлены в чингинской серии, распро-
страненной по юго-западной окраине Куртушибинского хребта Западного
Термин «серия» в настоящем издании применяется для обозначения единиц
региональной стратиграфии, ограниченных снизу и сверху поверхностями тектоно-
денудационных перерывов. На рис. 114—116, иллюстрирующих настоящую работу
А. Г. Сивова, они называются формациями. Свиты и горизонты являются единицами,
подчиненными сериям. Прим. ред.
59
Саяна (рч. Элэхем и Чинга, правые притоки Улухема). Эта серия, сло-
женная эффузивно-осадочной зеленокаменной толщей с микрокварци-
тами, располагается с резким несогласием на отложениях аккольскоп
серии, представленной празинитовыми сланцами и кварцитами при ред-
ком участии мраморов.
В известняках чингинской серии обнаружены водоросли и архео-
циаты, в том числе характерные для гавриловского комплекса Orbi-
cyathus mongollcus и Ajaclcyathus immanls\ в сланцевых толщах упомя-
нутой серии найдены трилобиты -Schagonaria1 Pol., Redllchlna
Lerm., Pagetina Lerm., Bergeroniellus L e г m., входящие в чингин-
ский комплекс. Элементы этого комплекса установлены также в Восточ-
ном Саяне, в окрестностях дер. Камешки, в глинистых сланцах с Redli-
china Lerm., Pagetina Lerm., Pagetla Wale., Bergeroniellus Lerm.,
которые, между прочим, здесь перекрываются известняками с характер-
ным камешковскйм комплексом археоциат.
Гавриловский комплекс археоциат наиболее типичен для одноимен-
ной серии известняков, развитых по восточному склону Салаира. Из
этих известняков известны представители родов Orblcyathus, Astero-
cyathus, Urcyathus, Bronchocyathus и сопутствующий им Ajaclcyathus
immanls. К сожалению, остатки трилобитов в гавриловских известняках
пока не обнаружены.
В других районах Саяно-Алтайской области аналоги гавриловского
комплекса археоциат обнаружены в известняках чингинской серии юго-
западной окраины Западного Саяна (бассейн среднего течения рч. Б. Кы-
заСа), а также в Кузнецком Алатау по р. Мрассу (верхнемрасская сви-
та), в Восточном Саяне (бирюсинские известняки), на восточном склоне
Кузнецкого Алатау (нижнечесноковская толща).
Как было сказано выше, камешковский комплекс археоциат типично
представлен в известняках одноименной толщи, распространенной в окре-
стностях д. Камешки Восточного Саяна. В упомянутых известняках
давно были установлены такие характерные представители археоциато-
вой фауны, как Ajaclcyathus tomlcus, A. patulus, A. acutus, A. tjlzkli,
Coscinocyathus dlanthus, C. calathus, Thalamocyathus G о г d. и др. От-
сюда известны также трилобиты из семейства Protolenidae, оставшиеся
не изученными.
Типичные представители камешковского комплекса археоциат уста-
новлены еще Толлем в известняках с. Торгашино (северо-западная
окраина Восточного Саяна) без точной привязки их к разрезу и даже
местонахождению, о чем постоянно забывали последующие исследова-
тели, ссылаясь на определение Толля. Эту оговорку необходимо сделать
потому, что в эталонном разрезе с. Торгашино (Торгашинский хребет)
археоциаты, переполняющие там так называемый торгашинский извест-
няк, сопутствуемые брахиоподами (Kutorglna) и трилобитами (Koote
niella), являются близкими по составу не к камешковскому, а к обру-
чевскому и санаштыкгольскому комплексам, что будет показано не-
сколько позже. В отношении же толлевского или нижнеторгашинского
горизонта с остатками археоциат камешковского комплекса (Ajcicyathus
acutus Born., A. proskurjakovl Toll., A. patulus Born., Coscinocya-
thus dlanthus Born., C. corblcula Born., C. elongatus Born., Thalatno-
cyathus Gord.) следует заметить, что он в эталонном разрезе с. Торга-
шино со времени исследований Толля никем не был обнаружен.
Вместе с тем камешковский комплекс археоциат теперь уверенно
устанавливается в верхнечесноковской толще восточного склона Кузнец-
кого Алатау (рч. Б. Ерба), в карчитской свите Горной Шории и многих
других пунктах.
1 В списках фауны, приводимых А. Г. Сивовым, фигурируют новые роды; все они
описаны ниже. Прим. ред.
60
Можно без преувеличения сказать, что рифовые известняки с камеш-
ковскими археоциатами являются наиболее распространенными отложе:
ниями нижнего кембрия Саяно-Алтайской области. Однако их выделе-
ние сопряжено с некоторыми трудностями, обусловленными тем, что эле-
менты камешковского комплекса археоциат нередко играют очень боль-
шую роль и в составе гавриловского комплекса. В силу этих обстоя-
тельств отложения с двумя указанными комплексами не всегда легко
расчленяются, особенно в условиях мелкомасштабных геологических
съемок. Несмотря на это, нижнекембрийский возраст гавриловского
археоциатового, чингинского трилобитового и камешковского археоциа-
тового комплексов не вызывает никаких сомнений. Достаточно сказать,
что с камешковскими археоциатами, более поздними в сравнении с дву-
мя первыми комплексами, ассоциируют Protolenidae, в том числе род
Protolenus.
Бабаковский трилобитовый комплекс в типичном выражении уста-
новлен в бассейне рч. Табат, по ключу Бабак, на северном склоне За-
падного Саяна, в известняках бабаковской серии, отношения которой
с подстилающими ее отложениями чингинской серии остались не выяс-
ненными. Но в то же время известняки бабаковской серии с ясным не-
согласием и базальной брекчией в основании перекрываются эффузив-
ными накоплениями нижнемонокской серии.
В известняках бабаковской серии (кл. Бабак) установлены следующие
родовые группы трилобитов: Granularia Р о 1., Babakovia S i v., Tabato-
pygellina S i v., Arbataspis S i v., Proerbia (?) L e г m., Chondragraulos
L e г m., Ixooteniella L e г m., Erbia Lerm., представленные большей
частью новыми видами; из новых видов отметим Erbia gigantea J е g о г.,
а из ранее описанных — Granularia obrutchevl Pol.
В отдельных пунктах распространения (кл. Известковый системы
р. Табат, нижнее течение кл. Герасимова — притока рч. Монока) извест-
няки бабаковской серии при сравнительно редком присутствии трилоби-
тов переполняются остатками археоциат, в том числе: Ajacicyathus
dwigthl, A. proscurjakovi, A. disparvallum, A. otychtaschensis, Ethmo-
phyllum cl. gracile, E. ratum, E. ulmense, Thalamocyathus (Cyclocyathus)
kusnetzovi и др. По общему составу эта археоциатовая фауна прибли-
жается к фауне обручевского комплекса.
Несколько в ином проявлении трилобиты бабаковского комплекса
выражены в известняках каянчинской серии, распространенной в бас-
сейне р. Катуни Горного Алтая, покоящейся там, с несогласием и ба-
зальным конгломератом в основании, на известняках баротальской и
древнепорфиритовой серии докембрия (М. К. Винкман, 1949).
В каянчинской серии из трилобитов установлены Bergeroniellus
L е г m., Pagetlna L е г т., Pagetla Wale., Cobboldla Lerm., Granu-
laria Pol., Erbia gigantea Jegor., Chondragraulos Lerm., Arbataspis
S i v. и, возможно, Bonnia (?) Wale. Перечисленным трилобитам со-
путствуют археоциаты, близкие по составу к археоциатам бабаковской
серии Западного Саяна, но в сопровождении Archaeocyathus subradiatus
и Erbocyathus heterovallum, т. е. форм, типичных для обручевского ком-
плекса ’.
Однако наиболее типично отложения с обручевским комплексом ар-
хеоциат, изредка сопутствуемых трилобитами бабаковского комплекса,
представлены на восточном склоне Кузнецкого Алатау в бассейнах ре-
чек Б. Ербы и Биджи. В первом из названных пунктов на размытой по-
верхности чесноковской толщи, содержащей остатки археоциат камеш-
ковского комплекса, с базальной брекчией в основании располагаются
П. С. Краснопеева, изучавшая археоциат нз этого пункта, не исключает воз-
можности присутствия в каянчинской серии двух не вполне одновозрастных комплек-
сов— трилобитов и археоциат. Прим. ред.
61
известняки ербинской «формации» (свиты) или, по номенклатуре
П. С. Краснопеевой (1937), полициатового горизонта. В этих известня-
ках давно были известны археоциаты и среди них: Erbocyathus obru-
tchevi, Ethmophyllum edelsteinl, ассоциирующие, как показывают недав-
ние исследования, с Archaeocyathus subradtatics, а также с трилобитами,
в частности, с Kooteniella slatkovskii (Schm.).
Последний совместно с Erbia sibirica (Schm.) и брахиоподами
Kutorgina cingulataeformis L е г m., по данным Е. В. Лермонтовой
(1940), присутствует в известняках Долгий Мыс, откуда в свое время
были определены Erbocyathus obrutchevi и Е. heterovallum.
К юго-западу от долины рч. Б. Ербы карбонатная фация ербинской
свиты постепенно замещается терригеновыми накоплениями, иногда с по-
кровами эффузивов. В этой фации известняки появляются в виде раз-
личной мощности линз, располагающихся на разных стратиграфических
уровнях, причем в районе рч. Биджи некоторые линзы богаты остатками
археоциат, оставшихся не изученными, и, значительно реже, трилоби-
тов, например: Granularia obrutchevt Pol., Proerbla (?) bidzhenla Siv.,
Babakovia S i v., Arbataspis S i v.
He менее типично обручевский комплекс археоциат выражен в из-
вестняках березовской «формации» (свиты), распространенной в Горной
Шории по рч. Амзас (Эмзас). Основание этой свиты не выходит на
дневную поверхность, и доступные для наблюдения ее нижние члены
слагаются кислыми эффузнвами, их туфами, постепенно сменяемыми
конгломератами, на которых и лежат интересующие нас известняки
с археоциатами. Здесь установлены: Ajaclcyathus amplus, A. clarus,
Erbocyathus obrutchevt, Ethmophyllum ratum, E. katunicum, Retecyathus
kuzminl, Spirocyathus subradiatics, Tegerocyathus beybuluki и др. С ними
ассоциируют трилобиты и, гораздо реже, брахиоподы (в их числе —
Kutorgina cingulataeformls Lerrn.). Трилобиты оказались настолько
плохой сохранности, что не поддаются определению даже до рода.
Как указывалось выше, в известняках разреза с. Торгашино, объеди-
ненных ныне К- В- Радугиным в торгашинскую «формацию» (серию).
лежащую с несогласием на подстилающих ее породах, трилобиты по со-
ставу весьма близки к бабаковскому комплексу. Здесь найдены: Koote-
niella slatkovskii (Schm.), Erbia sibirica (Schm.), которым сопут-
ствуют брахиоподы Kutorgina ctngulataeformis L e г m. и Kutorgina cf.
irbensis L e r m.
В известняках торгашинской серии установлено около 5—6 литологи-
ческих горизонтов с археоциатами, предварительное изучение которых
показывает, что среди них преобладают представители родов Ethmo-
phyllum и Tegerocyathus, при заметном участии видов, относящихся
к родам Ajaclcyathus и Coscinocyalhus. Не останавливаясь на описании
других пунктов распространения трилобитов бабаковского и археоциат
обручевского комплексов, отметим, что они, встречаясь то совместно, то
порознь, характеризуют разнофациальные отложения, возраст которых
вряд ли моложе верхов нижнего кембрия.
Для основания среднего кембрия Саяно-Алтайского региона харак-
терны санаштыкгольский комплекс трилобитов и одноименный комплекс
археоциат. В типичном проявлении эти фаунистические комплексы об-
наружены в известняках кл. Санаштыкгол на юго-западной окраине За-
падного Саяна, где они входят в состав терригенно-эффузивной верхне-
монокской серии с базальным конгломератом в основании, с угловым
несогласием налегающей на пирогенную нижнемонокскую серию, оха-
рактеризованную остатками спикуль губок, относящихся к роду Proto-
spongia. В санаштыкгольских известняках из трилобитов установлены:
Poliellina Р о 1., Poliellaspts L е г m., Kootenia W а 1 с., Chondragraulos
anomalls S i v„ Chakasskia P о 1., Klotziella Raymond и, возможно,
Erbia L e r m.
62
Фауна археоциат с Ethmophyllum, Ajaclcyathus (в частности, коло-
ниальных), а также Annulocyathus, Carlnocyathus, Erbocyathus, Tegero-
cyathus несет черты резкого обновления, правда, при заметном сохране-
нии элементов обручевского комплекса, но при почти полном исчезнове-
нии представителей камешковского комплекса археоциат. Кроме того,
именно в санаштыкгольских известняках были впервые обнаружены
строматопоры и те оригинальные с елочной скульптурой брахиоподы,
о которых сообщил К. В. Радугин (1937).
Санаштыкгольский комплекс трилобитов теперь точно установлен на
восточном склоне Кузнецкого Алатау, в окрестностях д. Б. Ербы. Здесь
в южном крыле антиклинали, на размытой поверхности чесноковской
толщи, располагаются известняки Лощенкова лога, составляющие глав-
ную часть потехинской «формации» (свиты). В известняках обнаружены
трилобиты Poliellina Р .о 1., Dinesus Etheridge, Chakasskia Pol.,
ассоциирующий с брахиоподами, в частности с Jamesella grandis Lerm.
Известные отсюда археоциаты изучены недостаточно, но все же присут-
ствие рода Ethmophyllum и особенно родов Annulocyathus, Tegerocyathus
и Cosclnocyathellus указывает на принадлежность их к санаштыкголь-
скому археоциатовому комплексу.
Вполне возможно, что близкими к санаштыкгольскому трилобито-
вому комплексу окажутся трилобиты известняков рч. Ирбы (Восточный
Саян), располагающихся как будто .стратиграфически выше пирогенной
толщи, близкой по стратиграфическому положению и литологическому
составу к нижнемонокской серии Западного Саяна. В ирбинских извест-
няках трилобиты представлены родами Kootenlelllna Siv., Llostracus
А п g., Chondragraulos Lerm. (в том числе Chondragraulos anomalls
Siv.); с трилобитами там ассоциируют брахиоподы Jamesella grandis
Lerm.
Присутствие в санаштыкгольском трилобитовом комплексе Chondra-
graulos anomalts позволяет объединить в этот комплекс и трилобитов
мондыбашской серии, развитой в Горной Шории по рч. Амзас (Эмзас).
Упомянутая серия, сложенная тонкообломочными породами, при замет-
ном участии эффузивов, но слабом развитии известняков, лежит через
базальную брекчию на эффузивах бачатской серии, которые, в свою оче-
редь, располагаются, повидимому, несогласно на известняках вышеопи-
санной березовской свиты, охарактеризованной археоциатами обручев-
ского комплекса.
Отложения мондыбашской серии относительно богаты остатками три-
лобитов, брахиопод и, реже, археоциат, причем последние оказались
плохой сохранности и определениям не подвергались. Но зато среди бра-
хиопод быстро были выявлены те с характерной елочной скульптурой
формы, о которых говорилось выше и которые, как и по кл. Санаштык-
гол, в мондыбашской серии ассоциируют с трилобитами санаштыкголь-
ского комплекса. Среди них, кроме Chondragraulos anomalls S i v., уста-
новлено еще несколько новых видов, принадлежащих к этому же роду,
а также роды Paradoxides В г о п g п., Llostracus A n g., Babakovla S i v.,
Acrocephalops P о u 1 s., Pagetla Wale, и др.
Таким образом, синхронность отложений мондыбашской и верхне-
монокской серий отчетливо выявляется на основе палеонтологических
данных, литологического состава и особенно стратиграфического поло-
жения. '
Из вышеизложенного видно, что санаштыкгольский комплекс трило-
битов, будучи в видовом отношении резко самобытным, характеризуется
присутствием типично среднекембрийских родовых форм, из которых
в первую очередь могут быть названы следующие: Paradoxides
В г о п g n., Dinesus Ether., Acrocephalops Poulsen, и Klotziella
R a у m.
Правда, сегодня мы еще не можем твердо оказать, какая из палеон-
63
тологических зон среднекембрийского стандарта Западной Европы или
Северной Америки будет соответствовать санаштыкгольским комплексам
трилобитов и археоциат, но зато уверенно можем утверждать, что
в Саяно-Алтайском регионе заключающие их отложения лежат страти-
графически ниже осадков с большеербинским трилобитовым комплексом.
Действительно, в бассейне рч. Б. Ербы (восточный склон Кузнецкого
Алатау) в верхах упомятутой выше потехинской свиты Лощенкова лога
отчетливо обособляется толща большеербинских темных известняков
с остатками трилобитов. Они исключительно богато представлены в пра-
вобережье рч. Б. Ербы, где потехинская свита, лишенная, к сожалению,
в нижних и средних своих частях органических остатков, налегает не-
посредственно на известняки ербинской свиты с обручевским комплек-
сом археоциат.
Среди трилобитов верхов потехинской свиты наиболее характерными
родовыми формами являются следующие: Olenoides Meek., Kootenia
W а 1 с., Dorypyge Dames, Erbia Lerm., Corynexochus Angelin,
Pagetia W a 1 c., Chondragraulos Lerm., Prohedinella S i v., а также
агностиды и из них — Peronopsls С о г d а. С этими формами крайне
редко встречаются археоциаты, остающиеся и поныне неизученными.
Большеербинский комплекс трилобитов достаточно отчетливо выяв-
ляется и на Салаире. Там, в районе горы Орлиной, на размытой поверх-
ности эффузивов бачатской серии, синхронной, вероятно, нижнемонок-
ской серии Западного Саяна, лежит песчано-конгломератовая свита
(формация по М. А. Усову), которая детальным картированием про-
слежена вплоть до нижнего течения рч. Юрман. Здесь в составе этой
свиты, кроме терригенных накоплений, участвуют светлосерые известняки
с остатками трилобитов, далеко несовершенной сохранности. Но все же
среди них были определены роды Olenoides и Dorypyge (?), т. е. формы
большеербинского комплекса.
В районе горы Орлиной Саламра отчетливо выявляется орлиногор-
ский трилобитовый комплекс. Он маркирует отложения так называемой
орлиногорской «формации» (свиты), лежащей трансгрессивно на подсти-
лающих породах, в том числе на песчано-конгломератовой свите с остат-
ками Olenoides.
В основании орлиногорской свиты располагается слой хорошо про-
мытого базального конгломерата, сменяемого выше песчаниками, вклю-
чающими мелкие линзы известняка и переслаивающимися с алевроли-
тами и синхронными амигдолофирами с ясно выраженной подушечной
текстурой. Стратиграфически выше с ясным несогласием располагаются
светлосерые известняки, которые К. В. Радугин (1945) в свое время вы-
делял в «березовскую формацию» и сопоставлял с известняками одно-
именной серии рч. Амзас Горной Шорни.
Отложения собственно орлиногорской свиты прорваны, силлами
и штоками лабрадоровых порфиритов, как будто не затрагивающими
известняков «березовской формации», но участвующими в строении так
называемой гурьевской «формации» (свиты), развитой к северу от
Гурьевского завода и сложенной там, преимущественно, сланцами при
небольшом участии средне- и грубообломочных накоплений (М. А. Усов,
1936).
То, что отложения орлиногорской и гурьевской «формаций» (свит)
представляют собой лишь фациальное выражение одной стратиграфиче-
ской единицы, доказывается одинаковым составом встречающихся в них
органических остатков, в частности трилобитов, о которых будет сказано
ниже. Относительно же «березовской формации» необходимо сделать
следующие замечания: она переименовывается здесь в верхнеорлино-
горский известняк, который не является синхронным известнякам бере-
зовской «формации» Горной Шорни, и на Салаире не повсеместно отде-
ляется несогласием от нижележащих пород.
64
Во многих пачках орлиногорской свиты присутствуют трилобиты, из
которых приведем наиболее часто встречающиеся формы: Belovta Р о 1.,
Pesaina Pol., Orlovleila Lerm., Anomocare A ng., Acrocephalites (?)
W a 11 e r., Peronopsls С о r d a, Phoidagnostus Whitehouse, Lejopyge
Co rd a, Grandagnostus Howell. Последний род в ассоциации с Ano-
mocare A n g. присутствует и в отложениях гурьевской свиты. В верхне-
орлиногорских же известняках, вообще скудных органическими остат-
ками, пока установлены Anomocare gigantea Lerm. (nomen nudum),
Orlovleila conlca S i v., Lejopyge sp.
Совершенно очевидно, что состав трилобитов болыпеербинского и, тем
паче, более молодого орлиногорского комплексов является типично сред-
некембрийским. Достаточно указать на присутствие в составе первого
комплекса таких характерных для этой эпохи родов, как Olenoides и,
особенно, Dorypyge, а в составе второго — упомянутых агностид.
Указания В. Окулича Okulitch (1948) на то, что якобы в Северной
Америке род Dorypyge ассоциирует с типично нижнекембрийскими три-
лобитами, нуждаются в проверке.
Аналоги болыпеербинского трилобитового комплекса (Anomocare,
Olenoides, Solenopleura) и орлиногорского (Peronopsls, Orlovleila, Acro-
cephalites (?) установлены и в Горном Алтае, но там они обнаружены
в гальках и валунах базального конгломерата, перекрытого терриген-
ными осадками неизвестного возраста.
В Горном Алтае с большей уверенностью к орлиногорскому комп-
лексу могут быть отнесены трилобиты ишимской серии, сложенной пес-
чаниками и аргиллитами и распространенной-в бассейне р. Сары-Кокша,
где она с резким несогласием и базальным конгломератом в основании
лежит на карбонатных породах с обручевским комплексом археоциат.
Среди трилобитов ишимской серии установлены Prohedlnla Lerm., Ну-
pagnostus J а е k е 1.
Салаирский комплекс трилобитов характерен для одноименной серии,
распространенной на восточном крыле Орлиногорской антиклинали Са-
лэира. В основании серии лежит мелкогалечниковый базальный конгло-
мерат, состоящий из галек подлежащих пород орлиногорской свиты
и прорывающих ее лабрадоровых порфиритов. Стратиграфически выше
базального конгломерата располагается толща переслаивающихся алев-
ролитов, тонкозернистых песчаников, аргиллитов, почти по всему раз-
резу переполненная остатками трилобитов, реже брахиопод, мшанок (?)
и птеропод.
В настоящее время в составе трилобитов выделено и описано около
42 новых видов, из которых лишь семь форм принадлежат к ранее
известным родам, а остальные группируются вокруг новых родов. Назо-
вем лишь важнейшие родовые группы: Orlovleila Lerm. (nom nud.),
Koldlniella Lerm. (nom. nud.), Pesala Wale, et R ess er, Anomoca-
riopsls S i v., Acrocephalites Waller., Kanlnlella S i v., Anomocare A n g.,
Proampyx S i v., Metisasplna S i v., Coosia W a 1 c., Abakanpleura S i v.,
Couselllna S i v., Cephalocoella R a у m., Homagnostus Howell, Dlplag-
nostus J a e k e 1.
Приведенный список родовых форм салаирского комплекса в сравне-
нии с ранее предложенным (А. Г. Сивэв, 1940) заметно уточнен и рас-
ширен. В связи с этим, а также в связи с исправлением ошибки, допу-
щенной в свое время в отношении определения стратиграфического поло-
жения нижеописываемой толсточихинскэй «формации» (свиты), охарак-
теризованной типично верхнекембрийским комплексом трилобитов и ле-
жащей выше салаирской серии, возраст содержащихся в последней три-
лобитов уточнен.
Не вдаваясь в детальный анализ, укажем лишь, что присутствующие
в их составе родовые формы Coosia Wale, и Cephalocoella R а у m.
характеризуют самые нижние части дрюсбачского (Dresbachian) яруса
5 Зак. 2024 65
верхнего кембрия Северной Америки (В. Howell и др., 1944). С этими
формами там ассоциируют роды Coosella (В. Howell и др., 1944) и
Metisaspis Rasetti (Т. Rasetti, 1946), которые в салаирском комплексе
выражены их производными, представленными соответственно родами
Coosellina S i v. и Metlsasplna S i v. Вместе с тем нельзя было не при-
нять во внимание материалов В. Хоуэлла и Лочман (В. Howell and Ch.
Lochman, 1939), утверждавших, что новоземельская фауна так называе-
мого озарка, в том числе Pesaia W а 1 с. et R е s s е г, близка по возрасту
фауне первой снизу зоны (зона Elvinia) второго снизу — франконского
(Franconian) яруса верхнего кембрия Северной Америки.
По мнению палеонтологов Северной Америки, в отрезок времени,
отвечающий зоне Elvinia, там впервые появляются родовые группы
Homagnostus Howell и Acrocepha'.ites Waller. (В. Howell и др.,
1944), которые, как известно, в других стандартах кембрия отмечаются
и в позднем среднем кембрии (Е. В. Лермонтова, 1940).
Таким образом, ориентируясь на американский верхнекембрийский
стандарт, возраст трилобитов салаирского комплекса сейчас устанавли-
вается в рамках низов верхнего кембрия, а не его верхов, как это ранее
принималось (А. Г. Сивов, 1940). Этому не противоречат и ассоциирую-
щие с трилобитами брахиоподы, принадлежащие, в частности, к родам
Eoorthis Wale., Blllingsella Hall et Clarke, Orusia Wale.
Аналоги салаирского комплекса трилобитов отчетливо устанавли-
ваются в арбатской серии, типично представленной в разрезе по рч. Руд-
ной Кене в Западном Саяне (А. Г. Сивов, 1950). Там эта серия распо-
лагается трансгрессивно на породах среднего кембрия и слагается грубо-
и среднеобломочными накоплениями с маломощными линзами известня-
ков. В известняковых линзах, лежащих в основании арбатской серии,
присутствуют трилобиты, в том числе Abakanopleura S i v. и Coosellina
S i v., а в выше лежащих линзах — брахиоподы и среди них — Llnnar-
ssonella Wale, и Billingsella Hall et Clarke.
Салаирский комплекс столь же точно установлен и в составе горно-
шорской серии, распространенной в Горной Шории по рч. Амзас (Эмзас).
Там названная серия лежит через базальную брекчию на эффузивах
верхов нижнего кембрия и слагается в нижней своей части средне- и тон-
кокластическими породами, включающими в себя линзы известняков
с остатками Coosellina S i v., Homagnostus Howell, Orlovlella L e r m.
в ассоциации с брахиоподами рода Eoorthis Wale.
Горношорский комплекс трилобитов типично представлен в верхней
части горношорской .серии. Крайне важно подчеркнуть, что нижние
части последней, сложенные терригенными накоплениями, постепенными
взаимопереходами связаны с ее верхними горизонтами, выраженными
тонкослоистыми («слоеный пирог» по К- В. Радугину) известняками,
богатыми остатками трилобитов, реже брахиопод, и члениками Pelmato-
zoa.
Из трилобитов здесь установлены Chuanglopsis S i v., Kazelllna S i v.,
Emsurlna S i v., Talbotina L о c h m a n, Maladiopsls S i v., Dolgatella
S i v., Ktngstonta Wale., Neoacrocephalltes S i v., Schorlella S i v.,
Pseudagnostus cyclopyge (Tullberg), Agnostus pisiformis Linn. s. 1.
Заметим, что ранее род Chuanglopsis S i v. отождествлялся с родом
Chuangia Wale., известным в верхнем кембрии Китая, а род Kazelllna
S i v.—с родом Kazella Wale, et R е s s е г, распространенным в «озар-
ке» Новой Земли (А. Г. Сивов, 1940).
Уточнение состава трилобитов горношорского комплекса отнюдь не
меняет сложившихся представлений о его верхнекембрийском возрасте;
наоборот, этот возраст подтверждается рядом вновь определенных
форм — Pseudagnostus cyclopyge, Agnostus pisiformis, — а также родом
Emsurlna, относящимся к сем. Cheiruridae Salter, и родом Schorlella,
принадлежащим к сем. Dikelocephalidae Miller.
66
Сопоставление
с разрезом Сред-
ней Сибири
биостратиграфаческие
подразделения
Корреляция нижнекембрийских отлоЖений Саяно
И1
4
по £ В
Лермонтовой
1
Палеонтологические
комплексы
Горный Алтай (верхнее
течение р Катуни)
Салаир
(Гурьевский район)
восточный склон
,л • • А/л„лл , Кузнецкого Алатау За
(дерхн.течениер.Мрассы)\ £рба-Асуь13) у
Горная Шория
Санаштык-
гольский ?
горизонт
Е
Е
(кзлициатовыйр)
____ и верхнекутор
гитбый(^т\)
горизонты
Е
5J
НиЖнекутор-
гиновый
горизонт
Синьская
толща
5J
а”-
5
Е
Санаштыкгольский
археоциатобый
и трилобитовыи
баба кодекий комплекс три
лобитов Granulana, trbia.
Proerbia, Beroerwmttus. Pagetint.
Cobboldia, Babakov из
Обручедский комплекс
археоциат - Brbocyathus
Камешковский комплекс
археоциат
Ajaclcyathus acutus
A Ijizfai, Coscinocyatnus
dlanthus и dp. ; редко три-
лобиты и водоросли
Читинский комплекс
трилобитов fedlichina,
Pagetla, Pagetina, Schaaonaria
dergeronlellus
Пестроцвгт-
ная толща
Слои с
Olenelhdae
Габрилавский комплекс
археоциат
Ajaclcyathus tmmanis,
Asterocyathus salalrtcus,
Orbicyathus mongolicus,
Tersia nodosa; иногда
трилобиты; бодорасли-
Osagio, Epiphytan
толща
'Vmsstoj Уэасская„ формация'
Известняки
Сары-кок-ша
Каянчинскоя
формация
бландинская
/, формация n
верхнемрасская
„ формация "
Адиакская. формация ‘
Матурская
формация
-Ц Гавриловская
формация
Устьсеминскоя
формация
новопросветовская
.формация’
Печеркинская
,, формация’
березовская
„ формация*
Карчитская
и узасская
формации
Карчитская. формация"
верхнемрассКоя-
мрасская фор-
мации и
адиакская свита •
V
водоросли Osagta
Quosimilis, Collenia
и археоциаты (?)
Ербинская
формация
Верхняя
чесноковская
свита
НцЖняя
чесноковская-(Т)
и енисейская
О' E5.J ЕЗ* □« (33* ЕЛ)// ЕЕ
Рис. 114. Схема соотношений местных и бностратиграфическнх единиц нижнего кембрия Саяно-Ал
/-метаморфизованные песчаники; 2—алевролиты и глинистые сланцы; 3-известняки: /—конгломераты; 5-углнсто-кремяистые сланцы: 6 — микрокварпиты; 7— преимущественное разе
известняки; 9-преимущественное ра «витке зеленокаменнык эффузИвив основного ряда; Ю—графитизированные сланцы. Н—эеленокаменные пироксеновые порфириты. /2-тонкослоис
рованной фазы состояния; /5-кварцнты; /б—микродиориты и габбро каирского типа; /7— гипербаэиты; /^-положение в разрезе остатков археоциат, водоросл
Зак. 2<124
Корреляция нижнекембрийских отлоЖений Саяно-Алтайской области
ерхнее
'HU)
Салаир \Горная Шория
(Гурьевский район) \рер*н. течение р. Мрассы) ‘'PpPpPsa^ckCoj
восточный склон ' 7 i восточный Саян
кузнецкого Алатау Западный Саян [ район селений Камеш-
ки и н. Кур я ты
бирюсинска-торгашин-
ский район
толща
Формация'
Мазасская-
березобская
„ формация"
чнекая
ця
новопросветовская
„формация'
'.кая
ця
Аккальская
формация
аерхнемрасск.ая-
мрасская фор-
мации и
адиакская свит
Гаврилобская
формация
Ч Матурская
г формация
ки
-Шо
оормация'
^формация1
кая
ция"
хкая
Анчешедская. формации
Ербинская
формация
верхнетореошин
ский известняк
-&ЧЧ карчитская
' 1 и узасская
формации
Верхняя
чесноковская
свита /
НиЖнеторгошин-
ский известняк
Кызас-
екая
свита
Свита кремнисто-
, глинисты*
сланцев
нижняя
чеснокодская-(?)
и енисейская
свиты
Мало- | *
шумен- (
ская 1
свита
I*
©и f J биртсинский
-Р-т^т известняк
3 Известнякобо-
граувакковая
свита
тюбильская формация
Эффузивная
формация
(ИЗ* E3J I Is Ок ESH E3S ЕЗ® EI3* ЕЗ® ЕЗ® (ЕЗл-ЕЗ® EZJ® S®
?ма соотношений местных и бностратиграфических единиц нижнего кембрия Саяно-Алтайского региона
известняки; 4 -конгломераты; 5-углнсто-кремнистые сланцы,- б-микрокваринты; 7- преимущественное развитие туфов зеленокаменных эффузивов основного ряха; S—доломнтизнровакные
«иищго ряда; !0 графитизированные сланцы; 11— эеленикаменные пироксеновые порфириты; /2-тонк<>слчи4гые алевролиты; /5-празиниты; /^-альбитофиры i ..„.римизи.
<оодиориты и габбро каирского типа; /7— гнпербазнты; /^-положение а разрезе остатков археоциат, водорослей и трилобитов; /S-поверхностн несогласий и перер
Корреляция среднекембрийских отлоЖений Саяно-Алтайской области
Палеонтологические
комплексы
Сапаштыкеолыкиц комплекс
тешюбигов-Лоееие, PoUellma
AaiKiuides, Скопдгодпшйвкгя
йй1а,АспсмМорз thakaiskia,
eS&WtymteiieUiHa/'agetui
ТамшпАгалск. комплекс шао
UparAnmilDCgathus. ТедегЪ-
Stbrnophyltum. £rbo
Protospongia
Anomocare gigantea,
Orloirieua
II
Каянчинский ярус Cm,
Палеонтологичес-
кие комплексы
Западный Саян
{северный склон)
Горная шория
(район рч Амзас)
восточный
Саян
(районрч. М.Нрба)
Санаштыкгольские
археоциатовые
и трилобитовые
Комплексы
бальшеербинскии комп-
лекстрилобигоб Olenoi-
dec frbiq Dory руде Aero-
nopsis. koo tenia, erohe -
dineaa, Pagetig_____
Canaup
(гора Орлиная и
прилегающие к
ней участки)
восточный склс ч
Кузнецкого Алая. $
(район рч. б.Ерйа,
Belovia,l/ssoiria,Orloiri- ,
ellpPeronopsipAnomo- §
care, Phoiaagnostus, -ч
Grandagnostus.lejopygi
Рона
Puradojtides
forthhammen
Бабаков-
ская фор-
мация
Зона
Lejopyge
laevigata
Рис. 115. Схема соотношений местных и бностратнграфнческнх единиц среднего кембрия Саяно-Алтайского региона
/—силлы лабрадоровых порфиритов; 9-силлы (?) авгитовых порфиритов; J—светлые известняки; 4—темные известняки, частью *лоистые;5- хонгломера > явные песчаники я грапелиты;
4—песчаники; 7—песчаники с .альбитовым* гравелитом в основании; S— валунчатые конгломераты. 9- алевролиты к глинистое слаьиы. 10-гранитиид^ миинского типа; //-сиениты
и макгериты кирсинского типа; 12— авгнювые порфириты м их туфы; 13—спилиты с редкими туфами; /4-лиднты; /5-альбитофиры иер«>ифиры. их туфы, мессами с мелкыалечни-
новым конгломератом в основании; 16—диориты я габбро каирского типа; 17—местоположение в раареэе органических остатков; 13-несогласия регионального типа; 19—местные
несогласия
tpounckai
.Форматы^
Сопоставле-
ние с Севера-
Аглонгичес-
кой провин-
цией Зап
бвралы
Верхние части горношорской серии, охарактеризованные горношор-
ским комплексом трилобитов, вполне сопоставимы с так называемой
толсточихинской свитой, типично представленной на участке горы Орли-
ной Салаира и выраженной там премущественно известняками
(М. А. Усов, 1936). На западном крыле Орлиногорской антиклинали эти
известняки лежат трансгрессивно, с базальным песчаником в основании,
на верхнеорлиногорских известняках и на подстилающих последние по-
родах орлиногорской свиты. На восточном же крыле описываемой анти-
клинали толсточихинские известняки непосредственно налегают на пес-
чано-сланцевые образования салаирской серии, причем характер соотно-
шений между сериями остался невыясненным.
Повсеместно в толсточихинских известняках, которые иногда оши-
бочно именуются орлиногорскими, обнаруживается богатая фауна три-
лобитов и среди них: Acrocephalltes Waller., Mataninella S i v., Hani-
vella S i v., Apatokephallna S i v., Artokephalus S i v., Portentosus J e-
g о r., Kingstonla Wale., Emsurlna S i v., Kazelllna S i v., Pseudagnostus
cyclopyge (Tullb.), Agnostus plslformls Linn. s. 1. Последние три рода
и два вида указывают на несомненный верхнекембрийский возраст фауны
и принадлежность ее к горношорскому комплексу.
О биостратиграфических подразделениях кембрия Саяно-Алтайского
региона и их возрасте
Рассмотренные комплексы археоциат и трилобитов в известной мере
помогают не только сопоставлять заключающие их отложения в преде-
лах Саяно-Алтайского региона, но и объединить местные стратиграфиче-
ские единицы в последовательные разномасштабные единицы биостра-
тиграфического характера. Соотношения между последними и местными
стратиграфическими единицами видны на стратиграфических колонках,
в графической форме отражающих типовые разрезы системы отдельных
участков Саяно-Алтайского региона (рис. 114—116). Эти биостратигра-
фические единицы, подчиненные отделам, мы считает возможным наиме-
новать ярусами.
В составе нижнего кембрия Саяно-Алтайского региона представ-
ляется возможным выделить в восходящем порядке следующие ярусы:
аккольский—Ст1'(?), чингинский—Стр и каянчинский—Спи3; в сред-
нем кембрии соответственно: верхнемонокский— Спи1, большеербин-
ский — Cm22 и орлиногорский — Cm23; наконец, в верхнем кембрии —
салаирский — Ст3* и горношорский — Ст32.
Название каждого из перечисленных ярусов совпадает с наименова-
нием той местной стратиграфической единицы, отложения и палеонтоло-
гически^ остатки которой, по сравнению с другими объединяемыми в со-
ответственный ярус местными стратиграфическими единицами, изучены
лучше. Вместе с тем известно, что до сего времени в Западной Европе,
где впервые был выработан биостратиграфический и геохронологический
стандарт кембрия, его отделы принято подразделять на зоны, которых
в каждом отделе устанавливается от четырех до пяти и которые не мо-
гут быть уверенно выделены в Саяно-Алтайском регионе, ибо они, как
и ярусы указанного региона, являются местными биостратиграфиче-
скими единицами.
В то же время, ориентируясь главным образом на последовательную
смену палеонтологических ассоциаций в зонах западноевропейского раз-
реза кембрия, мы пытаемся, хотя бы приблизительно, сопоставить с этим
разрезом ярусы кембрия Саяно-Алтайского региона. Поскольку видовой
состав наших трилобитов в подавляющей массе новый, поставленная
задача решается на основе сопоставления главным образом родового
состава трилобитовых комплексов, среди которого, как это было видно
из предыдущего изложения, выявляется немало руководящих форм.
5* 67
Литологический состав отложений интересующих нас ярусов, а также
характер соотношений между ними рассматривается здесь без особых
деталей.
Аккольский ярус—Cmi’(?)—выделяется условно, ибо встре-
чающиеся в его отложениях водоросли, в том числе Osagla, Collenia
и Conophyton, остаются слабо изученными. Вместе с тем отложения
этого яруса очень характерны по своему литологическому составу, бу-
дучи почти всюду представлены глубоко метаморфизованными породами
Корреляция верхнекембрийских отлоткений
Саяно-Алтайской области
Ьиосграгиграфические подразделения
важнейшие комплексы 1яшгл/
трилобитов ру
-о
too
ХЮ
300
400
S00
BOO
ТОО
too
ООО
кч!,0
й
12
1.3
I _
Западный склон
(северный склон)
И
Салаир
(гора Орлиная)
Ш
Горная Шароя
(рч Амзас)
Горно •
шорская
форма-
ция
..формам*
Горношорскцй комплекс fmsu-
rina, Chuangioosis Acrocephalites,
Pseudagnostus. nemopleundae,
Hingstonia, Kazelluta, Schoriella
Салаирский комплексOrlouiella
Kohtima, Acrocephalites льасапо-
pleura,Coosia, Homognostus, Hetisa-
sputa,kcniniella Anomocare Anomo-
'^curiopsis Cooselltna, Pesaia
iMl L" .13 ЕдЗЗ I— 13 ,,y
Eap КЗ* Г~5~1 Д "
Рис. 116. Схема соотношений местных и биостратиграфических единиц верхнего
кембрия Саяно-Алтайского региона
/—основные разности эффузнвов и их туфов; 2-изнестияки: 3—песчаники; 4 -мелкогалечннковые конгло-
мераты; 5—валунчатый конгломерат и осадочные брекчии; 6—лабрадоровые порфириты силловой фацни,
7—кератофиры я нк туфы; ^-местоположение органических остатков в разрезе; 9 алевролиты и гли-
нистые сланцы; /^-несогласия регионального типа; // — местные несогласия
типа праэинитовых сланцев и мраморизованных известняков. С теми
и другими породами то в больших, то в меньших количествах тесно
связаны кварциты, вероятно осадочного происхождения. Отложения
аккольского яруса поверхностями несогласия отделяются от более древ-
них пород, относимых к докембрию, и более молодых отложений несом-
ненно нижнекембрийского возраста.
Ч инг и некий ярус—Спи2,—выраженный двумя резко различ-
ными фациями—эффузивно-обломочной (чингинская серия) и карбо-
натно-эффузивной (чесноковская толща), повсеместно располагается
с несогласием на подлежащих породах, причем его интенсивно дислоци-
рованные и тонко рассланцованные отложения обнаруживают отно-
сительно слабый метаморфизм, отвечающий по своему характеру эпи-
зоне Грубенмана. Однако эффузивы яруса всюду находятся в зеленока-
менном состоянии, а микроструктура обломочных пород большей частью
является бластокластической; карбонатные породы всегда заметно мра-
моризованы.
Из разнообразных пород чингинского яруса наиболее характерны
пироксеновые порфириты и микрокварциты, пространственно приурочен-
ные к любым фациям яруса. Для выделения этого яруса решающую
роль играют трилобиты чингинского комплекса и в их числе—Bergero-
nlellus, Pagetlna, Schagonarla, Redllchlna, Pagetia ornatlformis S i v.,
а также археоциаты камешковского и гавриловского комплексов.
68
Археоциаты и трилобиты позволяют не только уверенно установить
нижнекембрийский возраст чингинского яруса, но сопоставить его с типо-
выми разрезами Сгщ некоторых зарубежных стран и, особенно, Средней
Сибири. На территории последней интересующий нас отдел кембрия
недавно расчленен на нижний — кэрфейский (алданский) ярус и верх-
ний—ленский (протоленусовый) ярус, причем внутри первого (слои
с Olenellidae по Е. В. Лермонтовой) лежат археоциатовые известняки,
а верхи второго (зона Protolenus по Е. В. Лермонтовой), завершаю-
щиеся куторгиновой толщей, перекрываются переходными слоями (ката-
линская толща по О. В. Флеровой), относимыми условно к низам сред-
него кембрия (Е. В. Лермонтова, 1951). Е. В. Лермонтова идентифици-
ровала среднесибирский разрез Спи с разрезом Cm2 Южной Австралии,
в основании которого, лежащем с резким несогласием на докембрийских
известняках, выступают тоже археоциатовые известняки, сменяемые
в вертикальном сечении отложениями зоны Protolenus в низах и слоями
с Redllchina в верхах, отсутствующими в Средней Сибири (Е. В. Лер-
монтова, 1951).
В Западном Саяне представители сем. Protolenidae (род Bergero-
niellus) в ассоциации с родом Redllchina маркируют один и тот же гори-
зонт, который, возможно, лежит выше горизонта с гавриловским комп-
лексом археоциат, проявляющих близкое родство с южноавстралийскими
формами. В Восточном Саяне горизонт с Redllchina перекрывается гори-
зонтом с камешковским комплексом археоциат, многие виды которых
тождественны южноевропейским формам, с одной стороны (Сардиния),
и близко родственны южноавстралийским — с другой.
В отличие от Южной Австралии, горизонт с камешковскими архео-
циатами содержит представителей сем. Protolenidae.
Из этих сравнений видно, что восходящая последовательность палеон-
тологических горизонтов чингинского яруса явно не совпадает с последо-
вательностью подобных горизонтов и зон заметно укороченного разреза
нижнего кембрия Южной Австралии.
В свою очередь, элементы кэрфейского яруса и почти весь ленский
ярус, за исключением верхнекуторгиновых слоев (Е. В. Лермонтова,
1951) Средней Сибири, вероятно, полностью будут соответствовать чин-
гинскому ярусу, если принять во внимание, что в его нижнем архео-
циатовом (гавриловском) горизонте, возможно синхронном рифовым
известнякам основания нижнего кембрия Средней Сибири, были обнару-
жены примитивные представители сем. Protolenidae (адиакская свита
Горной Шории). Таким образом, зона Protolenus международной биост-
ратиграфической шкалы по своему объему далеко не отвечает чингин-
скому ярусу, являющемуся в Саяно-Алтайской области вторым ярусом
нижнего кембрия, укладывающимся в возрастном отношении, вероятно,
во вторую треть нижнекембрийской эпохи.
Каянчинский яру с—Спи3. Отложения этого яруса преимуще-
ственно карбонатные при слабом развитии кластических накоплений, но
при заметном содержании эффузивов, находящихся в диагенетизирован-
ной фазе, располагаются резко трансгрессивно на подлежащем фунда-
менте, покрывая то докембрийские, то различные горизонты нижнекемб-
рийских отложений. В связи с этим становится возможным объяснить
отсутствие верхнекуторгинового, а может быть и верхов нижнекуторги-
нового горизонтов ленского яруса Средней Сибири на территории
Саяно-Алтайской области, уничтоженных денудацией перед отложением
осадков каянчинского яруса.
Известный на территории Средней Сибири верхнекуторгиновый гори-
зонт не является синхронным одноименному горизонту Саяно-Алтайской
области, выделенному там Е. В. Лермонтовой (Лермонтова, 1951), ибо
последний литологически, стратиграфически и палеонтологически пред-
ставляет то же самое, что и полициатидовый горизонт (Лермонтова,
69
1951), он же, по другим авторам, полициатовый (Краснопеева, 1937),
принятый за основу для обособления каянчинского яруса, характерного
присутствием обручевского археоциатового и бабаковского трилобито-
вого комплексов.
Состав первого из них пока не может быть строго индивидуализиро-
ван, ибо даже такая характерная для него форма, как Erbocyathus
(ранее Polycyathus) obrutchevl, ныне известна в более молодых отложе-
ниях. Но все же в совокупности представители родов Erbocyathus,
Ethmophyllum, Archaeocyathus, реже Tegerocyathus, при заметном уча-
стии форм камешковского комплекса, определяют известную самостоя-
тельность обручевского комплекса '.
В составе бабаковского трилобитового комплекса важнейшее биостра-
тиграфическое значение имеют виды родов Babakovia, Granularia, Taba-
topygelllna, Arbataspls, Chondragraulos, Erbia, Kooteniella, Bonnia (?),
Cobboldia, Proerbla, Bergeroniellus и др.
Представители рода Erbocyathus, Kooteniella slatkovskll и Erbia
sibirica в сопровождении Jamesella grandis обнаруживаются не только
в одном слое, но и в одном образце (район рч. Б. Ербы, восточный
склон Кузнецкого Алатау). Присутствие рода Bergeroniellus в составе
трилобитов каянчинского яруса отнюдь не свидетельствует о соответст-
вии его возраста зоне Protolenus международной биостратиграфической
шкалы нижнего кембрия. Появление Erbia, Chondragraulos и Babakovia,
представители которых получили наибольшее развитие в среднем кемб-
рии, дает больше оснований считать каянчинский ярус (он же верхнеку-
торгиновый горизонт Е. В. Лермонтовой для Западной Сибири, он же
полициатидовый или полициатовый горизонт) среднекембрийским или,
точнее, переходным от нижнего кембрия к среднему.
С другой стороны, нельзя не считаться и с тем, что нижнекембрий-
ские археоциаты камешковского комплекса и того же возраста трилобиты
из родов Proerbla. Cobboldia, Bonnia (?) в составе фауны каянчинского
яруса все же занимают большое место. Правда, Proerbla и Cobboldia
представлены новыми видами, отличающимися многими деталями от
видов названных родов, известных хотя бы в куторгиновой толще средне-
сибирского разреза нижнего кембрия (Е. В. Лермонтова, 1951), но это
естественно, ибо она несколько древнее каянчинского яруса.
В Северо-Атлантической провинции Западной Европы отложения
с палеонтологическими ассоциациями, подобными обручевскому архео-
циатовому и бабаковскому трилобитовому комплексам, либо вовсе не
отлагались, либо оказались размытыми перед отложением среднего
кембрия, в основании которого, как известно, местами лежат базальные
конгломераты (С. Бубнов. 1934).
В общем, в Саяно-Алтайском регионе за счет каянчинского яруса,
отвечающего концу нижнего кембрия, разрез последнего значительно
удлиняется, причем граница отдела, возможно, совпадает с его грани-
цей в Средней Сибири, если упомянутые выше «переходные слои», завер-
шающие ленский ярус (Е. В. Лермонтова, 1951), считать эквивалент-
ными каянчинскому ярусу.
Верхнем о нокский ярус — Cm21, — составляя основание сред-
него кембрия Саяно-Алтайского региона, характеризуется сложным
фациальным профилем. В одних районах нижняя часть яруса слагается
эффузивами громадной мощности (до 4 км), а верхняя — обломочно-
карбонатными породами с маломощными покровами эффузивов, которые
в этом ярусе находятся в диагенетизированной фазе состояния. В других
1 П. С. Краснопеева отмечает, что на рч. Б. Ерба (гора Долгий Мыс) археоциаты
обручевского комплекса были обнаружены совместно с брахиоподами верхнекуторгино-
вого горизонта Е. В. Лермонтовой Прим. ред.
70
же районах отложения яруса представлены только карбонатными поро-
дами мощностью не более нескольких сот метров.
Нижняя и верхняя части яруса во многих участках его распростране-
ния отделяются друг от друга резким несогласием параллельного типа;
подобное несогласие в весьма ослабленной форме и далеко, вероятно, не
повсеместно проявляется и в лежачем боку яруса. При этом его верхние
слои, охарактеризованные остатками археоциат и трилобитов санаштык-
гольского комплекса, в ряде пунктов, например, в бассейне рч. Б. Ербы
(восточный склон Кузнецкого Алатау), лежат бесспорно стратиграфи-
чески выше осадков каянчинского яруса с остатками археоциат обручев-
ского комплекса и трилобитов — бабаковского.
В связи с этим следует отметить, что обручевский комплекс археоциат
является древнее санаштыкгольского, а не наоборот, как полагает
И. Т. Журавлева (Журавлева, 1951). Больше того, археоциаты сана-
штыкгольского комплекса и сопутствующие им трилобиты одноимен-
ного комплекса (кл. Санаштыкгол), будучи много моложе фауны зоны
Protolenus, а не наоборот, как считала Н. В. Покровская (Н. С. Зайцев
и Н. В. Покровская, 1950), в совокупности указывают на среднекембрий-
ский возраст вмещающих эту фауну отложений, стратиграфически отве-
чающих верхней части верхнемонокского яруса.
Из его археоциат представители родов Annulocyathus, Carlnocyathus,
Ethmophyllum, Tegerocyathus и колониальные Ajaclcyathus имеют самое
важное биостратиграфическое значение для Саяно-Алтайского региона.
Из трилобитов для него характерны Poliellina, Polielldspis, Chakas-
skia, Kooieniellina, Chondragraulos anomalls, Kootenla abakanica и осо-
бенно Acrocephalops, Klotziella, Dinesus и Paradoxldes suboelandlcus.
Казалось, что опираясь на вид Paradoxldes suboelandlcus и род
Dinesus, в известной мере можно было бы рассматривать заключающие
их отложения верхнемонокского яруса эквивалентными соответственно
зоне Paradoxldes oelandicus и зоне Paradoxldes tessini стандартного раз-
реза среднего кембрия Скандинавского полуострова, при учете указаний
Уайтхауза, что последняя из названных зон отвечает какой-то части зоны
Dinesus типового разреза среднего кембрия Австралии (Е. W. White-
house, 1936, 1939).
Однако, не исключена возможность и более древнего, чем зона Рага-
doxides oelandicus, возраста верхнемонокского яруса. В самом деле,
в его отложениях среди трилобитов присутствует род Acrocephalops,
впервые появляющийся в Северной Америке в четвертой снизу зоне
Glossopleura—Kootenia среднего кембрия (В. Е. Howell и др., 1944).
С ней сопоставляется трилобитовая фауна каратауского горизонта Ка-
захстана (А. К- Ившин, 1949), в составе которой присутствует Kootenla
abakanica, характерная и для трилобитовых ассоциаций верхнемонок-
ского яруса.
Кроме того, известно, что в типовых кембрийских разрезах Австралии,
Северной Америки и Западной Европы археоциаты характерны только
для нижнего кембрия и верхнюю его границу не переходят. Но в Саяно-
Алтайском регионе археоциаты пышного расцвета достигают как раз
в верхнемонокском ярусе и, следовательно, могут быть признаны
реликтовой для него фауной, понижающей его возраст, по крайней мере,
до низов среднего кембрия. Это тем более вероятно, что археоциаты,
верхнемонокского яруса в большей степени, чем трилобиты, связаны
с предшествующими ассоциациями каянчинского яруса самых верхов
нижнего кембрия.
В литологическом отношении каянчинский и верхнемонокский ярусы
также тесно связаны: отнесенные к низам верхнемонокского яруса эффу-
зивно-осадочные накопления с характерными для них лидитами с остат-
ками Protospongta (нижнемонокская и бачатская серии) могут в той же
мере принадлежать к верхам каячинского яруса. В будущем граница
71
между рассматриваемыми ярусами несомненно будет исправлена на
основе более детального изучения свойственных им палеонтологических
остатков.
Большеербинский яру с—Cm22,—поводимому, целиком сла-
гается осадочными породами, среди которых карбонатные породы пред-
ставляют наиболее характерную фацию. С ней чаще всего связываются
относительно разнообразные органические остатки и в их числе — трило-
биты, в составе которых важнейшее значение имеют следующие: О1е-
noides, Dorypyge, Prochedlnella, Pagetla, Erbia granulosa и Peronopsis
из группы Peronopsis fallax var. n. Опираясь только на трилобитов, воз-
раст заключающих их отложений большеербинского яруса мог бы быть
определен в промежутке между концом нижнего кембрия и почти концом
среднего.
Действительно, по мнению Е. В. Лермонтовой, Erbia granulosa
является характерной формой для низов среднего кембрия, точнее —
прлициатидового горизонта (Е. В. Лермонтова, 1940), ныне отнесенного
мною к верхам нижнего кембрия, a Peronopsis из группы Peronopsis
fallax, как известно, типичен для самых высоких зон среднего кембрия
Скандинавии.
Исправляя ошибку Е. В. Лермонтовой в отношении вертикального
распространения Erbia granulosa и учитывая, что Peronopsis fallax
в большеербинском комплексе трилобитовой фауны представлен новым
варьететом, возрастной диапазон Erbia granulosa правильнее определять
в рамках конца первой половины и начала второй половины среднего
кембрия.
Это как раз будет соответствовать стратиграфическому положению
большеербинского яруса, лежащего местами непосредственно на отложе-
ниях верхнемонокского яруса (рч. Б. Ерба, верхи потехинской свиты).
Вместе с тем не утрачивают связи и фауны этих ярусов; эта связь под-
держивается, в частности, археоциатами, изредка встречающимися
в отложениях большеербинского яруса. Последний, будучи трудносопо-
ставимым с типовыми разрезами среднего кембрия зррубежных и оте-
чественных стандартов, пока остается характерным только для Саяно-
Алтайского региона или даже для отдельных его участков.
Орлиногорский ярус—Ст23. В этом ярусе, помимо осадоч-
ных накоплений, разделенных внутренними несогласиями и лежащих,
вероятно, повсеместно с крупным перерывом на более древних породах
(в том числе на породах большеербинского яруса), некоторую роль
играют эффузивы, в частности лабрадоровые порфириты.
Весьма характерны для отложений этого яруса трилобиты, в составе
которых, наряду с чисто местными родами — Orlovlella, Belovia, —
немало руководящих форм, например: Peronopsis fallax, Enetagnostus
kjerulfl, Photdagnostus bltuberculatus, Grandagnostus glandlformis, Lejo-
pyge armata var. n.
Орлиногорский ярус, содержащий только что перечисленных трило-
битов, в самой нижней части вполне может быть сопоставлен с зоной
Paradoxides forchammeri и даже с зоной Lejopyge laevigata Западной
Европы. Однако верхние подразделения яруса, выраженные верхнеор-
тиногорскими известняками с остатками Anomocare glgantea Lerm.
и новыми родами Belovia и Orlovlella, удлиняют разрез среднего кемб-
рия Саяно-Алтайского региона. Вот почему здесь и выделяется третий
(орлиногорский) ярус с возрастом, отвечающим самым верхам среднего
кембрия.
Салаврский яру с—Cm31—всюду представлен только осадоч-
ными и, притом, преимущественно обломочными породами, налегающими
местами с несогласием на подстилающие их отложения.
Отложения салаирского яруса богаты органическими остатками,
в частности трилобитами, среди которых Kanlniella, Pesala, Cephalocoe-
72
Ila, Coosellina, Coosia, Homagnostus, Acrocephalites, Abakanopleura и ряд
видов родов Orloviella и Koldlnlella имеют для него руководящее значе-
ние. В составе трилобитовой фауны этого яруса присутствуют также
роды Апотосаге и Diplagnostus, представители которых во всей Северо-
Атлантической провинции характерны только для второй половины
среднего кембрия и границу самой верхней его зоны (Lejopyge laevigata)
не переходят.
Однако как род Апотосаге, так и род Diplagnostus, будучи представ-
лены в салаирском ярусе новыми видами, являются типичным примером
реликтовых форм на общем резко обновленном фоне фауны этого яруса,
несомненно более молодой, чем фауна зоны Lejopyge laevigata. Доста-
точно вспомнить, что на Салаире отложения салаирского яруса лежат
бесспорно стратиграфически выше осадков, эквивалентных зоне Lejopyge
laevigata (орлиногорской «формации»). Вместе с тем, поскольку фауны
дробных подразделений лучше всего изученных верхнекембрийских раз-
резов Западной Европы (нижние зоны) и Северной Америки (нижний
дрюсбачский ярус) остаются трудносопоставимыми с фаунами салаир-
ского яруса, его возраст приближенно определяется здесь в рамках пер-
вой половины верхнекембрийской эпохи.
Горношорский ярус-— Cm32 — слагается преимущественно
известняками, которые в одних районах (рч. Амзас в Горной Шории)
через тонкие взаимопереходы связаны с песчано-глинистыми накопле-
ниями салаирского яруса, в других же районах (гора Орлиная на
Салаире) с несогласием налегают на отложения орлиногорского яруса.
Отложения горношорского яруса наиболее богаты органическими остат-
ками, в том числе трилобитами, остающимися и поныне слабо изучен-
ными. Но все же из них роды Kingstonia, Chuanglopsis, Emsurlna,
Apatokephalina, Artokephalus, Kazelllna, Pseudagnostus cyclopyge, Agnos-
tus pisiformis (s. 1.) являются наиболее часто встречающимися и наибо-
лее характерными руководящими формами для этого яруса.
Обращает на себя внимание сочетание не только в одном слое, но
даже в одном образце Agnostus pisiformis или Pseudagnostus cyclopyge,
характерных для нижних зон верхнего кембрия Западной Европы,
с родом Emsurlna (из сем. Cheiruridae) или родом Artokephalus (из
сем. Remopleuridae), близко родственные представители которых марки-
руют главным образом самые верхние части верхнего кембрия и, из-
редка, появляются даже в низах ордовика. Это обстоятельство создает
известные трудности при сопоставлении фауны верхнего кембрия Запад-
ной Европы с фауной горношорского яруса, крайне специфичной и лишь
приблизительно позволяющей определить возраст вмещающих ее отло-
жений в рамках второй половины верхнекембрийской эпохи.
Тесная связь осадков горношорского яруса с отложениями салаир-
ского яруса подкрепляет верхнекембрийский возраст первого.
Горношорским ярусом кембрийская система Саяно-Алтайского реги-
она, видимо, не завершается, ибо на его территории все отчетливее
выявляются отложения с фаунами явно древнее трилобитовых ассоциа-
ций самых низов ордовика (Euloma—Niobe); однако фауна этих,
повидимому, поздпекембрийских отложений пока совершенно не изу-
чалась.
Изложенная выше биостратиграфическая схема кембрия Саяно-
Алтайского региона лишь в крупных звеньях, отвечающих по стратигра-
фическому диапазону отделам, претендует на относительную точность.
Что же касается ярусов, то они далеко не столь уверенно выявляются на
всей площади региона. Тем более последовательность объединяемых
в ярусы местных стратиграфических единиц во многих случаях опреде-
лена в качестве первого приближения к действительности.
Вполне возможно, что позже будут уточнены стратиграфические
соотношения, например между верхнемонокской и мондыбашской
73
сериями, ныне объединяемыми в верхнемонокский ярус. Такого рода
уточнения могут быть осуществлены главным образом на основе более
детального изучения палеонтологических остатков, характерных для
упомянутых серий, так как они разобщены значительными простран-
ствами. То же самое можно сказать и о внутренней стратиграфии горно-
шорского, чинганского и других ярусов кембрия интересующего нас
региона.
Все это, конечно, является крупным недостатком предлагаемой схемы,
но все же она, во-первых, с достаточной полнотой и четкостью отражает
ход развития Саяно-Алтайского региона в целом за кембрийский период
и, во-вторых, позволяет уточнить вертикальное распространение ряда
руководящих форм хотя бы из числа трилобитовой фауны. Так, в лите-
ратуре прочно укоренилось представление, что представители сем. Remo-
pleuridae Corda являются характерными только для нижнего силура. Но
теперь можно уверенно сказать, что архаичные формы этого семейства,
например род Artokephalus, появляются в конце верхнего кембрия
(горношорский век). Вертикальное распространение Pseudagnostus
cyclopyge завершается где-то в верхах верхнего кембрия (горношорский
ярус), а не его низах, как считали до сего времени на основе изучения
кембрия Западной Европы.
Род Redltchlna, ранее считавшийся характерной формой то для низов
среднего кембрия, то для верхов зоны Protolenus нижнего кембрия
(Е. В. Лермонтова, 1951), в действительности типичен для слоев, лежа-
щих во всяком случае ниже горизонта с камешковским комплексом
археоциат второй трети нижнего кембрия (чингинский ярус).
Наконец, появились высказывания и о том, что археоциаты Саяно-
Алтайского региона границы нижнего кембрия не переходят (Н. С. Зай-
цев и Н. В. Покровская, 1950); в действительности же они, как это
видно из вышеизложенного, существуют до второй половины среднего
кембрия (большеербинский ярус).
Характеристика важнейших родовых и видовых представителей
археоциат, трилобитов и водорослей приводится ниже.
П. С. КРАСНОПЕЕВА
Тип Archaeocyathi. Археоциаты
Археоциаты широко распространены в известняковых фациях ниж-
него и низов среднего кембрия Западной Сибири, характеризуясь исклю-
чительным богатством и разнообразием форм. В настоящее время в со-
ставе археоциатовой фауны Западной Сибири выделяются четыре ком-
плекса, характеристика которых дана частично в приведенной выше об-
зорной статье А. Г. Сивова и в опубликованной работе П. С. Красно-
пеевой (1947):
гавриловский комплекс 1 - г
камешковский комплекс / 1
обручевский комплекс Ст, или CmJ
саиаштыкгольский комплекс CmJ или Ст,
Ниже приводятся описания важнейших представителей сибирских
археоциат; эти описания содержат лишь важнейшие диагностические
признаки; остальные признаки видов (например, количество рядов пор
между смежными септами, их размеры и т. п.) показаны на рисунках
и фотоснимках.
74
Таблица 2
Семейство Признаки семейства—характер пористоои стенок кубка Признаки родов—скелетные образования в ннтерваллуме
наружная стенка внутренняя стенка септы септы и днища септы и синап- тнкулы септы и пузыр- чатая ткань
Archaeocyathidae Taylor Поры Поры Род Ajicicyathus (подроды Urcyathus и Orbicyathus) Род Coscinocyathus (подрод: Asterocyathus^ Род Nochoroicyathus Род Archaeo- fungia Род Loculo- cyathus
Ethmophyllidae Okul. Поры Паровые каналы Роды Ethmophyllum и Clathrocyathus Род Coscin> cyathellus Род Formosocyathus Род Pycnoido- cyathus
Cai inocyathidae fam. n. Поровые каналы Поровые каналы Род Tegerocyathus Krasn. gen. n. Род Carinocyathus
Tlialamocyathidae Zhuravl. Поры Желобообразные кольца Род Thalamocyathus Род Bronchocyathus Рид Sygmojim- gia
Annulocyathidae fam. n. Поровые каналы Желобообразные кольца Род Annulocyathus Рол Annulofun- gia gen. n.
Sygmocyathidae fam. n. Желобообраз- ные кольца Желобообразные кольца Род Sygmocyathus Род Sygmocoscinus
Tuniulocyathidae fam. n. Поровые каналы Поры Род Tumulocy thus Род Erbocyathus
Описание представителей рода Cosclnocyathus выполнено С. В. Чер-
нышевой, которая также принимала участие в подборе иллюстраций.
На предыдущей странице приведена схема классификации архео-
циат1 класса Regularia (табл. 2).
КЛАСС REGULARIA. ПРАВИЛЬНЫЕ АРХЕОЦИАТЫ
СЕМЕЙСТВО AJACICYATHIDAE TAYLOR
Семейство характеризуется ситовидными наружной и внутренней
стенками кубков и такими же септами. Кроме септ, в промежутке ме-
жду стенками кубков могут присутствовать днища, синаптикулы и пу-
зырчатая ткань.
Род Ajacicyathus Bedford
Синоним—Archaeocyathus у многих авторов; колониальные формы описывались
под родовым названием Densocyathus
Обычно одиночные, редко колониальные формы. Наружная стенка
ситовидная, гладкая или слабо ребристая. Внутренняя стенка ситовид-
ная, гладкая или с килевидными междупоровыми промежутками. Поры
внутренней! стенки крупнее пор наружной стенки. В пространстве между
стенками присутствуют только септы. Виды, входящие в данный род,
ранее (начиная с Борнемана, 1886 г.) относились к роду Archaeocyathus.
Однако типичный вид последнего рода — Arch, profundus Bill, обла-
дает в интерваллуме наряду с септами пузырчатой тканью, что и яв-
ляется отличием рода Archaeocyathus от рода Ajacicyathus.
Широко распространенный род главным образом в камешковском и
гавриловском комплексах.
Типичный вид. Ajacicyathus ajax (Taylor).
Ajacicyathus acutus (Borneman n)
Табл. I, фиг. 3; рис. 117
Рис. 117. Ajacicyathus acu-
tus (Bor n.).
Поперечное сечение кубка,
X10. По Бедфорду
Кубок конической формы. На наружной
стенке между септами иногда выражены слабо
продольные ребра. Внутренняя стенка несколь-
ко толще наружной и имеет более крупные
поры. Септы прямые, равномерной толщины.
При диаметре кубка 2,7 и 5,0 мм септальный
коэффициент 5,2 и 5,0. Коэффициент массив-
ности кубка в среднем 2,5.
От близкого по септальному коэффициенту
A. patulus var. aspera отличается меньшей мас-
сивностью внутренней стенки.
Распространение. Камешковский
комплекс. Река Нижняя Терсь и рч. Б. Ерба
(гора Чеснокова) в Кузнецком Алатау; р. Ка-
рагач в системе р. Ульмень Северного Алтая;
с. Торгашино в Восточном Саяне; р. Кеняда
в Анабарском массиве.
1 Предлагаемая автором классификация является рабочей. Автор занят ее даль-
нейшей разработкой. В табл. 2 включены не все известные роды правильных архео-
циат. Прим. ред.
76
Ajaclcyathus facilis (Vologdin)
Рис. 118
Толщина наружной и внутренней стенок конического кубка почти
одинакова. При диаметре кубков 3,5; 10,0 и 13,5 мм септальный коэффи-
циент соответственно 1,0; 0,9 и 1,3. Коэффи-
циент массивности скелетной части кубка ко-
леблется в пределах 1,7—3,0.
От похожего по наименьшему септальному
коэффициенту A. subtills (V о 1.) описанный
вид отличается отсутствием пленок на стенках
кубка. От похожего по септальному коэффи-
циенту A. grandis (V о 1.) отличается близкой
толщиной стенок кубка.
Распространение. Г авриловский ком-
плекс. Река Черная Уса против пос. Верхняя
Ивановка и бассейн р. Нижняя Терсь в Куз-
нецком Алатау; с. Ариничево на Салаире;
р. Шивелик-Хем в Туве; Южный Урал, район
е. Рысаево.
Рис. 118. Ajaclcyathus fa-
cilis (V о 1.)
Поперечное сечение кубка,
Хб,5. р. Шивелик-Хем, Тува
Ajaclcyathus itnmanis (Vologdin)
Табл. I, фиг. 1
Кубок очень крупных размеров, достигающий в поперечнике 200 лш.
Внутренняя стенка несколько толще наружной. Ширина скелетной части
кубка незначительна и колеблется в пределах 1,0—-2,5 мм. Коэффициент
массивности кубка 8,0—10,0. Наблюдаемые обломки кубка в большин-
стве случаев имеют почти прямые, параллельные между собой стенки.
От похожего по крупным размерам кубка и незначительной ширине
скелетной части кубка A. clarus (V о 1.) описанный вид отличается мень-
шей массивностью внутренней стенки кубка и отсутствием ребер на на-
ружной стенке.
Распространение. Гавриловский комплекс. Белая горка
в окрестностях сел Горскино, Ариничево, Гавриловское на Салаире;
р. Кии и перевал между кл. Банным и
р. Ч. Усой в Кузнецком Алатау; р. Коура
в Северном Алтае; бассейн р. Кийзас в си-
стеме р. Абакана в Западном Саяне; р. Ши-
велик-Хем в Тувинской области.
Ajaclcyathus tomicus (Vologdin)
Рис. 119
Кубок узкоконический. Наружная стенка
тонкая, с продольными ребрами в проме-
жутках между соседними септами. Вутрен-
р. Белокаменка в системе
to-
Рис. 119. Ajaclcyathus
micus (Vol.)
Поперечное сечение кубка,
Х15. Гора Чеснокова, рч.
Б. Ерба, Кузнецкий Алатау
Распространение,
бассейн р. Нижняя Тсрсь в
няя стенка толще наружной и с более круп-
ными порами. Септы тонкие, равномерной
толщины. Септальный коэффициент при
диаметре кубка 2,0; 2,4; 3,9 и 8,0 лш соот-
ветственно 13,5; 11,2—13,5; 12,0 и 12,0.
Камешковский комплекс. Рч. Б. Ерба и
_ Кузнецком Алатау; р. Малый Таз в Горной
Шории; с. Торгашино в Восточном Саяне; Алтай, северное побережье
Телецкого озера.
77
Ajacicyathus clarus (Vologdin)
Табл. I, фиг. 2; рис. 120
Кубок близкий к цилиндрическому. Наружная стенка очень тонкая.
с продольными ребрами.
v Г Г Т Г Гтттггу
Рис. 120. Ajacicyathus cla-
rus (Vol.)
Поперечное сечение части
к)бка, Хб, р. Н. Терсь,
Кузнецкий Алатау
Внутренняя стенка приблизительно в три раза
толще наружной. Септы тонкие у наружной
стенки и резко утолщены у внутренней. Интер-
валлум узкий, не более 1 мм ширины. При
диаметре кубка 8,0 и 22,0 мм септальный коэф-
фициент 11,0 и 10,0.
Распространение. Камешковский
или гавриловский комплекс. Рч. Шивелик-Хем
в Туве; р. Мазас в системе р. Мрассу и бассейн р. Нижняя Терсь в Куз-
нецком Алатау.
Ajacicyathus suchlensls К г a s п о р е е v a, sp. п.
Рис. 121
Кубок характеризуется тонким строением всех скелетных элементов.
Толщина обеих стенок и септ почти одинакова. Септы прямые, тонкие,
без утолщений. Коэффициент септальности при диаметре кубка 1,5; 4,5;
6,7 и 11 —12 лии соответственно 20; 18; 17 и 15—17. Коэффициент мас-
сивности кубка 3,5.
От A. tomicus отличается большим септальным коэффициентом и
меньшей толщиной внутренней стенки.
Распространение. Предположи-
тельно гавриловский комплекс. Р. Черная
Уса против пос. Верхняя Ивановка и кл. Су-
хой в Кузнецком Алатау.
Рис. 121. Ajacicyathus
suchiensis К г a s п. sp. п.
Поперечное сечение куб-
ка, У 5. кл. Сухой, при-
ток р. Усы. Кузнецкий
Алатау
Рис. 122. Ajacicyathus Sa-
na stykgolensis (у о 1.)
Поперечное сечение раз-
ветвляющегося кубка, У5,5.
кл. Санаштыкгол, Западный
Саян
Ajacicyathus sanaschtykgolensls (Vologdin)
Синоним — Densocyathus sanaschtykgolensis
Рис. 122
Колониальная форма. Отдельные особи колонии имеют круглый по-
перечник. Септы слабо утолщены у обеих стенок.
У обособленных индивидов при поперечнике 3,5 мм септальный коэф-
фициент 3,5. Ширина скелетной части кубка 0,6 мм. Коэффициент мас-
сивности кубка 1,5.
Распространение. Санаштыкгольский комплекс, р. Карагач
(система р. Ульмень, Северный Алтай); ключ Санаштыкгол в системе
р. Абакана (Западный Саян).
78
Ajacicyathus patulus (Bornemann)
Табл. I, фиг. 4
Внутренняя стенка конического кубка массивнее наружной, участки
стенки между порами на ней килевидные. При диаметре кубков 5,8 и
5,0—6,0 мм септальный коэффициент соответственно 3,1 и 4,0. Коэффи-
циент массивности кубка около 2,0.
От близкого по септальному коэффициенту A. proscurlakovl отли-
чается меньшей массивностью внутренней стенки и более равномерной
толщиной септ.
Распространение. Камешковский комплекс, с. Торгашино и
д. Камешки в Восточном Саяне и Усинский район Кузнецкого Алатау.
Ajacicyathus patulus var. aspera Krasno-
p e e v a
Рис. 123
От типичный формы отличается большей мас-
сивностью внутренней стенки и большим коэффи-
циентом септальности, который при диаметре
кубка 3,0 мм равен 5,0. Концы септ замет-
но утолщены у внутренней стенки. Ширина ске-
летной части кубка равна около одной трети его
радиуса.
Распространение. Камешковский и об-
ручевский комплексы. Улус Бей-Булук и Лощен-
ков лог близ рч. Б. Ербы в Кузнецком Алатау;
приток р. Б. Сумульты, Алтай.
Рис. 123. Ajacicyathus
р dulus var.aspera Krasn.
Поперечное сечение куб-
ка, х5. Лощенков лог,
рч. Б. Ерба, Кузнецкий
Алатау
рч. Аксааскан, правый
Ajaclcyahtus proscurlakovl (Toll)
Табл. I, фиг. 5; табл. Ill, фиг. 6
Кубок близкий к цилиндрическому. Внутренняя стенка массивная,
усложнена системой крючковидных отростков, ввиду чего возникает пред-
положение о наличии в ней упрощенных поровых каналов. Септы утол-
щены у внутренней стенки. При диаметре кубка 7,0; 10,0 и 10,3 мм сеп-
тальный коэффициент соответственно равен 3,3; 2,8 и 2,7. Ширина ске-
летной части кубка от 2,2 до 2,7 мм. Коэффициент массивности кубка
примерно 2,0.
От похожего по массивности внутренней стенки A. patulus var. aspera
отличается меньшим септальным коэффициентом.
Распространение. Камешковский и гавриловский комплексы.
Дер. Камешки, улус Бей-Булук; р. Малый Таз и р. Кийзас в Горной Шо-
рни; д. Ариничева на Салаире; Алтай, бассейн р. Катунь. Торгашинская
свита в Восточном Саяне.
Ajaclcyathus salebrosus (Vologdin)
Табл. I, фиг. 6
Кубок конической формы. Внутренняя стенка массивная, с бугорча-
тыми междупоровыми промежутками. Септы равномерной толщины.
При диаметре кубка 5,4; 6,2; 7,0 и 10,0 мм септальный коэффициент со-
ответственно равен 7,2; 8,0; 7,5 и в пределах 7,3—8,3. Коэффициент мас-
сивности кубка в среднем 2,0—3,0.
Распространение. Бассейн рр. Нижняя Терсь и Большая
Ерба в Кузнецком Алатау; р. Малый Таз в Горной Шории; кл. Ипполи-
тов в системе р. Абакана Западного Саяна; р. Аномас, пос. Ивановский
в Северном Алтае; юго-восточный берег оз. Хара-Усу в Западной Мон-
голии. Комплекс неясен.
79
Подрод Urcyathus Vologdin
Внутренняя стенка пористая, образует резко выраженные острые или
округленные ребра, приуроченные к промежуткам между септами, что и
является особенностью этого подрода.
Типичный вид. Ajaclcyathus (Urcyathus) asteroldes Vol.
Ajaclcyathus (Urcyathus) asteroldes Vologdin
Табл. I, фиг. 7
Кубок правильный, с тонкими стенками и такими же септами. На
внутренней стенке выражены заостренные ребра. Септы равномерной
толщины. Септальный коэффициент при диаметре кубка 3,0—4,0 мм ра-
вен около 5,1. Коэффициент массивности кубка 2,0.
Распространение. Гавриловский комплекс. Салаир, с. Гор-
скино; близкие виды встречены в окрестностях с. Гав-
риловского и д. Ариничевой.
Ajaclcyathus (Urcyathus) intracostatus Vologdin
Рис. 124
Рис. 124. Ajacicy-
athus (Urcyathus)
intracostatus Vol.
Поперечное сече-
ние неполного куб-
ка. Х2. р. Н.Терсь
в системе р. Кии
Кубок узкоконической формы, с тонким скелетом.
Наружная и внутренняя стенки ситовидные, одинако-
вой толщины. Внутренняя стенка несет на себе за-
остренные, резко выраженные продольные ребра. Сеп-
ты равномерной толщины. Септальный коэффициент
при диаметре кубка 14,0 мм равен приблизительно 7,0.
Коэффициент массивности кубка примерно 3,5.
Распространение. Гавриловский комплекс, р. Нижняя Терсь
в Кузнецком Алатау.
Подрод Orblcyathus Vologdin
Кубок характеризуется резкими поперечными складками, захваты-
вающими одновременно наружную и внутреннюю стенки, что является
особенностью данного подрода.
Типичный в и д. Ajaclcyathus (Orblcyathus) mongollcus V ol.
Ajaclcyathus (Orbicyahtus) cf. mongollcus Vologdin
Табл. I, фиг. 8
Поперечные складки кубка резкие, расположены через 1,5—2,0 мм.
Расстояние между крайними выступами складок в среднем 4,0—5,0 мм.
Септальный коэффициент при диаметре кубка в 16,0 мм равен 5,0.
Распространение. Гавриловский комплекс, с. Горскино (Бе-
лая Горка), с. Гавриловское и д. Ариничева на Салаире; рр. Кия и Бело-
каменка в Кузнецком Алатау. Близкие виды найдены в Усинском районе
Горной Шории.
Ajaclcyathus (Orblcyathus) talovkensts Кг a s пор ее v a, sp. n.
Рис. 125
Кубок с круглым поперечным сечением. Наибольший наблюдаемый
диаметр кубка 13 мм. Наружная стенка ситовидная, толщиной 0,03 мм,
с 3—4 рядами пор между смежными септами. Внутренняя стенка мае-
сивная, в два раза толще наружной стенки. Септы
концы их утолщены у внутренней стенки.
Септальный коэффициент при поперечниках
кубков в 9,0; 10,0 и 17,0 мм соответственно ра-
вен 8,4; 7,6 и 6,0. Коэффициент массивности куб-
ка колеблется в пределах 2—5, увеличиваясь по
мере увеличения поперечника кубка.
прямые, ситовидные;
Рис. 125. Ajacicyathus (Orbicyathus) talovkensis К г a s n., sp. n.
a—поперечное сечение части кубка; б—продольное сечение кубка.
Х4. р. Ч. Уса, вблизи устья р. Таловки
Распространение, р. Черная Уса выше устья рч. Сухая Та-
ловка.
Род Loculocyathus Vologdin
Наружная и внутренняя стенки кубка массивные, ситовидные. Про-
странство между стенками заполнено септами и пленками пузырчатой
ткани.
Пленки пузырчатой ткани различной длины. В некоторых случаях от-
дельные пленки занимают промежутки только между двумя септами,
в других они пересекают несколько септ. Пленки очень часто бывают
развиты и в центральной полости кубка.
Типичный вид. Loculocyathus profundus (В i 11.)
Loculocyathus irregularis (Toll)
Синоним — Loculocyathus tolli Vol.
Табл. II, фиг. 1
Кубок близкий к цилиндрическому. Наружная и внутренняя стенки
массивные, с порами. Поры внутренней стенки крупнее пор наружной
стенки. Септы также массивные, утолщенные у обеих стенок. Проме-
жутки между септами заполнены пленками пузырчатой ткани. Реже
пленки заполняют собой более широкие промежутки. Пузырчатая ткань
иногда наблюдается также и в центральной части кубка и приурочи-
вается преимущественно к его основанию. Септальный коэффициент
при диаметре кубков до 10 мм равен 3,0—4,0.
Распространение. Камешковский и гавриловский комплексы,
р. Нижняя Терсь и другие районы Кузнецкого Алатау; *д. Камешки и
с. Торгашино в Восточном Саяне; д. Ариничева на Салаире.
Род Archaeofungia Taylor
Синоним — Sibirecyathus Vol.
Наружная стенка ситовидная, внутренняя ситовидная или несколько
усложненная. В интерваллуме выражены септы, соединенные между со-
бой синаптикулами.
Типичный вид. Archaeofungia sibirica (Т о 11).
6 Зак. 2024 81
Archaeofungia sibirica (Toll)
Синоним — Archaeocyathus sibiricus Toll
Рис. 126
Форма кубка близка к цилиндрической. Внутренняя стенка имеет
слабо выраженные шиповидные образования, выступающие вверх во
Рис. 126. Archaeofungia sibirica (Toll). Поперечное
сечение части кубка, х9. Лощенков лог, рч. Б. Ерба,
Кузнецкий Алатау
внутреннюю полость кубка. Септы многочисленные, слабо волнистые
в поперечном сечении. При диаметре кубка 11,0 мм септальный коэффи-
циент равен 8,2.
Распространение. Камешковский и санаштыкгольский (?)
комплексы, сс. Торгашино и Камешки в Восточном Саяне и Лощенков
лог в районе рч. Б. Ербы Кузнецкого Алатау.
Archaeofungia dlsseplmentalls (Vologdin)
Рис. 127
Внутренняя стенка с крючкообразными отростками, равными числу
септ. Септы слабо .волнистые, в местах соединения их с синаптикулами
имеются утолщения. При диаметре кубка 10,0 мм септальный коэффи-
циент равен около 6,0.
Рис. 127. Archaeofungia dissepimentalis (V о 1.) Поперечное сечение части
кубка, х8. р. Шивелик-Хем, Тува
От близкого вида A. sibirica отличается более грубым строением
внутренней стенки и более редкими септами.
Распространение. Камешковский и санаштыкгольский комп-
лексы. рч. Шивелик-Хем в Туве; Лощенков лог в районе рч. Б. Ербы
Кузнецкого Алатау; ключ Санаштыкгол в Западном Саяне.
Род Coscinocyathus Во г п ешапп
Наружная стенка кубка тонкая, ситовидная. Внутренняя стенка тоже
ситовидная, несколько массивнее наружной. Нередко на ее внутренней
82
поверхности наблюдаются шипики. Поры внутренней стенки крупнее
пор наружной стенки или одинаковых размеров.
Септы правильные, ситовидные, равномерной толщины или утолщены
близ стенок. Кроме септ, в промежутках между стенками имеются
днища; они в большинстве случаев выпуклые. Реже наблюдаются пло-
ские днища; они часто имеют утолщения в месте пересечения с септами
(гребенчатые днища у рода Nochorolcyathus). Расстояния между дни-
щами различны у разных видов.
Типичный вид. Coscinocyathus dlanthus Born.
Coscinocyathus dlanthus Bornemann
Табл. II, фиг. 2
Кубок конический или кеглеобразный. Днища выпуклые. Септы рав-
номерной толщины. Септальный коэффициент при диаметре кубка
12,0 мм равен 5,8. Коэффициент массивности кубка 2,6.
Распространение. Камешковский и гавриловский комплексы,
д. Камешки и с. Торгашино в Восточном Саяне; р. Полтавка и другие
пункты в Кузнецком Алатау; д. Ариничева на Салаире.
Рис. 129. Coscinocyathus cancel-
lathus Born, я—поперечное
сечение части кубка, х5; б—
продольное сечение части куб-
ка, х8. р. Шивелик-Хем, Тува
Рис. 128. Coscinocyathus dlanthus''var. stabila
К r a s n., var. n. a—поперечное сечение части куб-
ка, \5; б—продольное сечение части кубка, х5;
в—продольное скошенное сечение кубка, Х4.
р. Шивелик-Хем, Тува
Cosctnocyahtus dlanthus var. stabila Krasnopeeva, var. nov.
Рис. 128
Кубок обычно узкоконический, близкий к цилиндрическому, реже
ширококонический. Днища выпуклые. Септы правильные, имеют утол-
с# 83
щения у наружной стенки. Септальный коэффициент при диаметре куб-
ков 20,0 и 17,0 мм равен соответственно 7,8 и 5,8. Коэффициент массив-
ности в среднем около 4,0. Характерно большое количество зачаточных
септ.
Распространение. Камешковский комплекс, р. Шивелик-Хем
в Тувинской области; кл. Ипполитов и Герасимов в системе р. Аба-
кана— Западный Саян; бассейн р. Малый Таз в Горной Шории.
Cosclnocyathus cancellatus Bornemann
Рис. 129
Кубок бочковидный или цилиндрический. Днища выпуклые. Харак-
терной является узкая скелетная часть кубка. Септы равномерной тол-
щины. Септальный коэффициент при диаметре кубка 12,0 мм равен 10,2.
Коэффициент массивности кубка приблизительно 5,0.
Распространение. Камешковский комплекс, с. Торгашино и
д. Камешки в Восточном Саяне; р. Полтавка в центральной части Куз-
нецкого Алатау; р. Шивелик-Хем в Тувинской области.
Cosclnocyathus sptnosus Vologdin
Табл. II, фиг. 3; табл. III, фиг. 1
Кубок конической формы. Днища выпуклые. Септы правильные, рав-
номерной толщины. Септальный коэффициент при диаметре кубка 5,0 мм
равен 8,5. Коэффициент массивности скелетной части кубка приблизи-
тельно 2,5.
Распространение. Камешковский комплекс, с. Торгашино
в Восточном Саяне; р. Нижняя Терсь в Кузнецком Алатау.
Подрод Asterocyathus Vologdin
Кубок одиночный и колониальный с ситовидными наружной и внут-
ренней стенками. Внутренняя стенка несет на себе резкие продольные
ребра, приуроченные к промежуткам между септами, чем этот подрод и
отличается от рода Cosclnocyathus. В интерваллуме, кроме септ, присут-
ствуют днища. Этим признаком данный подрод отличается от подрода
U г cyathus.
Типичный вид. Cosclnocyathus (Asterocyathus) salalrlcus Vol.
Cosclnocyathus (Asterocyathus) salalrlcus Vologdin
Табл. Ill, фиг. 4
Кубок узкоконический, диаметром до 5 мм. Перегородки размещены
несколько неравномерно. Септальный коэффициент 2,5—3,0. Днища тон-
кие и тонкопористые, размещены неравномерно и часто непараллельны
друг другу. Внутренняя стенка с заостренными продольными ребрами.
Распространение. Гавриловский комплекс, с. Горскино и
д. Ариничева, Салаир.
СЕМЕЙСТВО ETHMOPHYLLIDAE OKULITCH
Наружная стенка простая, ситовидная. Внутренняя стенка массив-
ная, пронизана поровыми каналами, располагающимися в один ряд ме-
жду смежными септами. Септы обычного типа.
Род Ethmophyllum Meek
Колониальные формы относились к роду Sajanocyathus
Род Ethmophyllum включает в себя значительное количество видов,
характеризующихся наличием поровых каналов во внутренней стенке.
84
Стенки поровых каналов без пор. Типичные поровые каналы рода
Ethmophyllum в вертикальном сечении внутренней стенки имеют V-об-
разную форму. Но имеются группы с поровыми каналами, направлен-
ными косо вверх в центральную полость или почти
горизонтальными. Такие упрощенные поровые каналы
наблюдаются в большинстве случаев у форм с менее
массивной внутренней стенкой.
Ethmophyllum disparvallum (Vologdin)
Рис. 130
Рис. 130. Ethmo-
phyllum disparval-
lum Vol. Попе-
речное. несколько
скошенное сечение
кубка, У 5. р. Ши-
велик-Хем, запад-
ная часть Тувы
Форма кубка коническая. Поровые каналы внутрен-
ней стенки упрощенные. Септы прямые, концы их утол-
щены у обеих стенок. При диаметрах кубков 4—5; 6,0;
8,8 и 12,0 мм септальный коэффициент равен 4,2; 4,1;
3,6 и 5,0. Коэффициент массивности кубка от
2,2 до 4,0.
Распространение. Камешковский и санаш-
тыкгольский комплексы, р. Шивелик-Хем в Туве; р. Аксааскан, правый
приток р. Большой Сумульты в Горном Алтае; кл. Санаштыкгол в За-
падном Саяне.
Ethmophyllum kourense Krasnopeeva, sp. n.
Табл. V, фиг. 4; рис. 131
Кубок узкоконической формы. Основной его особенностью является
узкая скелетная часть. При диаметре кубка 13,0 и 25,0 мм септальный
коэффициент соответственно равен 9,0 и 6,5. Коэффициент массивности
кубка 6,1 и 6,8.
Рис. 132. Ethmophyllum tuvaense Kias.i., sp. п.
a—поперечное скошенное сечение кубка; б—про-
дольное сечение кубка, х4,5., р. Шивелик-Хем.
Тува
Рис. 131. Ethmophyllum
kourense Krasn., sp. п.
а—поперечное сечение
части кубка; б—про-
дольное сечение части
кубка, Хб,5. р. Коура,
Горная Шорня
Преимущественно обручевский комплекс.
Распространение.
Северный Алтай, бассейн р. Ульмень.
Ethmophyllum tuvaense Krasnopeeva, sp. n.
Табл. V, фиг. 5; рис. 132
Кубок узкоконической формы. Внутренняя стенка с шипами; поро-
вые каналы V-образные. Септы слабо утолщены у обеих стенок. При
85
диаметре кубка 5,2; 5,6; 6,0 и 6,5 мм септальный коэффициент соответ-
ственно равен 9,7; 9,0; 10,0 и 9,3. Коэффициент массивности соответ-
ственно равен 2,9; 2,8; 3,0 и 2,3. От близкого Ethm. karaganlcum отли-
чается наличием шипов.
Распространение. Предположительно обручевский комплекс,
р. Коура ниже устья р. Акшна, Северный Алтай; р. Шивелик-Хем в Ту-
винской области.
Ethmophyllum ratum Vologdin
Табл. VI, фиг. 3; рис. 133
Узкоконический кубок. Поровые каналы внутренней стенки близки
к горизонтальным. Септы равномерной толщины. При диаметре кубка
3,6 и 10,0 мм септальный коэффициент равен 7,5 и 8,0. Коэффициент
массивности кубка неодинаков и колеблется в пределах 1,5—3,5.
Рис. 133. Ethmophyllum ratum Vol.
а—поперечное сечение кубка; б—продольное сечение кубка;
Хб. р. Амзас, Горная Шория
Распространение. Преимущественно обручевский комплекс.
Ряд пунктов в Горном Алтае; р. Шивелик-Хем в Туве; р. Амзас в Гор-
ной Шорин.
Ethmophyllum katuntcum К г a s п о р е е v a, sp. п.
Табл. V, фиг. 2
Кубок близкий к цилиндрическому. Поровые каналы внутренней
стенки близки к горизонтальным. Септы слабо утолщены у обеих стенок.
86
При диаметре кубков 2,6; 3,6 и 6,0 мм септальный коэффициент равен
6,5; 5,5 и 6,3. Коэффициент массивности кубка около 2,0.
От близкого Ethmophyllum raturn отличается более разреженными
септами.
Распространение. Обручевский и санаштыкгольский ком-
плексы. кл. Санаштыкгол в системе р. Абакана в Западном Саяне; из-
вестен также из Горного Алтая.
Ethmophyllum amzassense К г a s п о р е е v a, sp. п.
Табл. IX, фиг. 3; табл. X, фиг. 3
Кубок близкий к цилиндрическому, иногда неправильный.
Наружная стенка тонкая, ситовидная. В промежутке между двумя
смежными септами расположено обычно три ряда пор. Септы прямые,
утолщенные у обеих стенок, поры мелкие.
Внутренняя стенка пронизана поровыми каналами, ориентирован-
ными от интерваллума наклонно к центральной полости на значитель-
ном расстоянии и на коротком расстоянии — косо вверх. Поровые ка-
налы прикрыты защитной пленкой, образовавшейся за счет разветвле-
ния концов стенок поровых каналов. Септальный коэффициент при диа-
метре кубка 18,0 мм равен 7,0. Коэффициент массивности кубка 3—4.
От представителей рода Clathrocyathus описанный вид отличается
более крупными порами наружной стенки и более массивными стенками
поровых каналов внутренней стенки.
Распространение, р. Амзас в Горной Шории.
Ehmophyllum aff. ussovi (Vologdin)
Синоним — Sajanocyathus ussovi
Табл. VI, фиг. 4; рис. 134
Колониальная форма. Детали строения поровых каналов внутренней
стенки не установлены. Септы тонкие, местами неправильные. В некото-
Рис. 134. Ethmophyllum aff. ussovi Vol. Попереч-
ное сечение кубка в начальной стадии ветвле-
ния, хб. р. Ульмень, Северный Алтай
рых экземплярах между септами наблюдаются редкие палочковидные
перемычки. Септальный коэффициент для отдельных индивидов в сред-
нем равен 8,0. Коэффициент массивности кубка приблизительно 2—3.
Распространение. Санаштыкгольский комплекс, р. Карагач
в системе р. Ульмень, Северный Алтай; кл. Санаштыкгол в системе
р. Абакана, Западный Саян.
Pop, Clathrocyathus Vologdin
Синоним — Tercyathus Vol.
Род характеризуется мелкопористой наружной стенкой кубка и
V-образными резко несимметричными поровыми каналами во внутренней
стенке. Часть порового канала, отходящая непосредственно от интервал-
87
лума вниз, значительно длиннее, чем часть канала, ориентированная
косо вверх, в центральную полость, причем в укороченной части поровые
каналы у большинства видов разветвляются.
В отличие от рода Ethmophyllum стенки поровых каналов внутренней
стенки кубка рода Clathrocyathus имеют крупные поры, посредством
которых сообщаются между собой.
Представители рода Clathrocyathus, имеющие на наружной стенке
поровые каналы, должны быть выделены в самостоятельный род.
Род Clathrocyathus объединяется с родом Tercyathus, так как они
имеют близкое строение наружной и внутренней стенок кубка.
Типичный вид. Clathrocyathus fossaengulatus Vо 1
Clathrocyathus fossaengulatus Vologdin
Табл. V, фиг. 3; рис. 135
Рис. 135. Clathrocyathus
fossaengulatus V ol. По-
перечное сечение части
кубка, Х5. 5. р. Шиве-
лик-Хем,’’ Тува
Кубок близкий к цилиндрическому. На внут-
ренней стенке кубка со стороны, обращенной
в центральную полость, находится тонкопористая
пленка. Септы сравнительно тонкие, утолщены
у обеих стенок. При диаметре кубков 16,0; 15,0;
6,3 и 6,0 мм септальный коэффициент соответ -
ственно равен 7,2; 6,5—7,0; 7,8 и 6,6. Коэффи-
циент массивности кубка колеблется в пределах
1,8—2,5.
Распространение. Санаштыкгольский
комплекс, рр. Лебедь и Катунь, выше д. Усть-Семы,
Алтай; кл. Санаштыкгол в Западных Саянах и р. Шивелик-Хем в Туве.
Clathrocyathus kenlensis Krasnopeeva, sp. n.
Табл. VII, фиг. 1; рис. 136
Форма кубка коническая. Внутренняя стенка на стороне, обращенной
в центральную полость, имеет тонкопористую пленку. Септы многочислен-
ные, слабо утолщены у стенок. Септаль-
ный коэффициент кубков при диаметре
5,8 и 4,2 мм равен 10,2 и 14,2. Коэффи-
циент массивности кубка
2.
Рис. 136. Clathrocyathus kenlensis Krasn., sp. п.
a—поперечно-косое сечение кубка, ХЗ; б—продольное сечение кубка, v7.
Устье рч. Верхняя Кеня (система р. Абакан). Кузнецкий Алатау
От близкого вида Clathrocyathus fossaengulatus Vol. отличается
более сгущенными септами.
88
Распространение. Санаштыкгольский комплекс, р. Верхняя
Кеня, у ее устья в Западном Саяне.
Clathrocyathus altaicus Vologdin
Рис. 137
Форма кубка неизвестна. Наружная стенка тонкопористая, с продоль-
ными ребрами у наружных краев септ. Внутренняя стенка массивная;
толщина ее почти достигает ширины септ. На стороне, обращенной
Рис. 137. Clathrocyathus altaicus Vol.
а—поперечное сечение кубка; б—поперечное сечеиие того же кубка;
в—вид внутренней стенки в продольном сечении кубка; г—вид наружной
стенки в продольном сечении кубка, Хб. р. М. Таз в Горной Шории
(а, в); р. Лебедь в Северном Алтае (б, г)
в центральную полость, внутренняя стенка имеет тонкую пористую
пленку. Септы тонкие; концы их у наружной стенки утолщены.
Септальный коэффициент при диаметре кубков 3,5; 7,0; 8,2 и 12,0 мм
равен 18,0; 12,7; 14,0 и 13,0. Коэффициент массивности кубка приблизи-
тельно 1,5.
Распространение. Преимущественно санаштыкгольский ком-
плекс. р. Малый Таз в Горной Шории; кл. Санаштыкгол в Западном
Саяне; р. Лебедь в. Северном Алтае.
Род Coscinocyathellus Vologdin
Синоним — Ethmocosclnus Simon
Кубок имеет простую ситовидную наружную стенку и пористые пере-
городки. Внутренняя стенка построена по типу рода Ethmophyllum.
Форма поровых каналов в деталях не установлена. В интерваллуме,
кроме септ, присутствуют днища.
Типичный вид. Cosclnocyathelli^s parvus Vol.
Coscinocyathellus utukensis К г a s n о p e e v a, sp. n.
Табл. VI, фиг. 2; рис. 138
Форма кубка не установлена. Характерно строение внутренней стенки
и днищ: внутренняя стенка с упрощенными поровыми каналами и ши-
пами; днища плоские, с резкими утолщениями в месте пересечения их
89
с септами. Септы утолщены у обеих стенок. Септальный коэффициент при
диаметре кубка 7,0 мм равен 9,1. Коэффициент массивности кубка
приблизительно 3,5.
Распространение. Гора
Утук-Кая в Тувинской области. Комп-
лекс неясен.
Coscinocyathellus loscenkovi
2мм
a » —*
Рис. 138. Coscinocyathellus utukensis
К г a s п., sp. п.
а—продольное скошенное сечение
части кубка, X10; б—поперечное сече-
ние кубка, х5. Гора Утук-Кая в си-
стеме р. Хемчик, Тува
Krasnopeeva, sp. п.
Табл. IV, фиг. 3
Форма кубка не установлена. Дни-
ща плоские, утолщены в месте пересе-
чения их септами; септы утолщены у
наружной стенки. Септальный коэффи-
циент при диаметре кубков 4,5 и 6,0 мм
равен 7,1 и 6,5. Коэффициент массив-
ности кубка не более 2,0. От Coscino-
cyathellus utukensis отличается отсут-
ствием шипов на внутренней стенке
кубка и несколько более редкими
септами.
Распространение. Предпо-
ложительно санаштыкгольский ком-
плекс. Лощенков лог близ рч. Б. Ербы,
Кузнецкий Алатау.
СЕМЕЙСТВО CARINOCYATHIDAE KRASNOPEEVA, FAM. N.
Наружная стенка кубка сложного строения; она содержит в себе си-
стему поровых каналов различной формы. Каналы в вертикальном сече-
нии V-образной формы или воронкообразные, открыты широким концом
в наружное пространство. Воронкообразные поровые каналы обычно бы-
вают прикрыты тонкой пленкой с чрезвычайно мелкими порами, попереч-
ник которых измеряется сотыми долями миллиметров.
Септы правильные, ситовидные. Внутренняя стенка построена по типу
стенки рода Ethmophyllum, т. е. включает в себя систему поровых кана-
лов V-образной формы или близких к горизонтальным. В интерваллуме,
кроме септ, присутствуют днища.
Род Те ger осу at hus Krasnopeeva, gen. n.
Наружная стенка кубка включает в себя систему поровых каналов.
Поровые каналы коленчато изогнутые, чаще воронкообразные, открыты
широким концом кнаружи. В последнем случае наружная стенка бывает
прикрыта тонкой пористой пленкой; поперечник пор на ней измеряется
сотыми долями милиметра. Септы правильные, пронизаны порами,
расположенными в чередующемся порядке. Внутренняя стенка и ее по-
ровые каналы построены по типу рода Ethmophyllum. Таким образом,
род Tegerocyathus отличается от рода Ethmophyllum характером внешней
стенки.
Типичный вид. Tegerocyathus abakanensls (Vol.).
Распространение. Преимущественно обручевский и санаштык-
гольский комплексы.
Tegerocyathus erblensls Krasnopeeva, sp. n.
Табл. VIII, фиг. 5; рис. 139
Кубок ширококонический. Наружная стенка с воронкообразными по-
ровыми каналами, прикрыта мелкопористой пленкой. Поровые каналы
90
внутренней стенки упрощенные. Септы правильные, несколько утолщен-
ные у обеих стенок и с очень мелкими порами. При поперечниках кубка
20,0 мм септальный коэффициент равен 3,5. Ширина скелетной части
кубка 3,4 мм, коэффициент массивности кубка приблизительно 30.
Распространение. Камешковский комплекс, рч. Б. Ерба в Куз-
нецком Алатау; р. Шивелик-Хем в Туве.
Рис. 139. Tegerocyathus erbiensis
К г a s п„ sp. п.
б—продольное сечение кубка,х7; а—по-
перечное сечение части кубка, Х4. Го-
ра Чеснокова, рч. Б. Ерба
Рис. 140. Tegerocyathus altaicus
S. Т с h е г п., sp. п. Поперечное слабо
скошенное сечение части колонии, Х5.
р. Карагач, правый ’приток р. Ульмень,
Горная Шория
Tegerocyathus altaicus S. Т с h е г и у с h е v a, sp. п.
Табл. VI, фиг. 5; рис. 140
Колониальная форма. Строение стенок такое же, как у Tegerocyathus
erbiensis. Септы утолщены у обеих стенок. Септальный коэффициент при
диаметре кубков 2,8; 3,0; 4,0 и 4,2 мм соответственно равен 5,0; 5,3; 5,5
и 5,9. Поровые каналы внутренней стенки с массивными стенками, ха-
рактер их строения не установлен. Коэффициент массивности скелетной
части кубка колеблется в пределах 1,2—1,4.
Распространение. Обручевский комплекс, р. Карагач, правый
приток р. Ульмень в Северном Алтае; р. Малый Таз в Горной Шории.
Tegerocyathus dlploperforatum (Vologdin)
Рис. 141
Кубок близкий к цилиндрическому. Наружная стенка с воронкообраз-
ными поровыми каналами прикрыта мелкопористой пленкой. Поровые
каналы внутренней стенки V-образные. Септы равномерной толщины.
Септальный коэффициент при диаметре кубка 4,2; 5,7; 6,0 и 8,0 мм равен
соответственно 6,2; 4,7; 5,0 и 5,5. Скелетная часть кубков массивная;
коэффициент массивности кубков около 1,5.
Распространение. Обручевский комплекс. Ряд пунктов в Гор-
ном Алтае; кл. Санаштыкгол в Западном Саяне; рч. Ерба в Кузнецком
Алатау.
Tegerocyathus tannu-ola Krasnopeeva, sp. n.
Табл. VII, фиг. 2; рис. 142
Кубск узкоконической формы. Поровые каналы наружной и внутрен-
ней стенок V-образные. Септы утолщены у наружной стенки. Септальный
91
коэффициент при диаметре кубка 4,5 мм равен 6,7. Скелетная часть
кубка довольно массивна, коэффициент массивности равен около 1,9.
Рис. 141. Tegerocyathus diplo-
perforatum {V о 1.)
«-поперечное сечение части
кубка, Х5, 5; б—касательное
сечение наружной стенки,
х20. р. Ерба, Кузнецкий Алатау
Рис. 142. Tegerocyathus tannu—ola
К г a s n., sp. п.
а—поперечное сеченне кубка, X 2,5;
б—продольное сечение кубка, Х10.
р. Шивелик-Хем, западная часть Тувы
Распространение. Обручевский комплекс, р. Шивелик-Хем
в Туве.
Род Carinocyathus Vologdin
Наружная и внутренняя стенки имеют сложное строение с поровыми
каналами. В интерваллуме, кроме септ, присутствуют прямые или слабо
выпуклые мелкопористые днища. Последний признак отличает род
Carinocyathus от рода Tegerocyathus.
Поровые каналы внутренней стенки у всех видов рода Carinocyathus
V-образные. Наружная стенка указанного рода имеет V-образные и во-
ронкообразные поровые каналы, открытые широким концом кнаружи.
В последнем случае, как и у рода Tegerocyathus, стенка бывает при-
крыта пористой пленкой.
Типичный вид. Carinocyathus loculatus Vol.
Carinocyathus loculatus Vologdin
Рис. 143
Кубок узкоконический. Стенки с V-образными поровыми каналами.
Септы слабо утолщены у наружной стенки. Септальный коэффициент при
Рис. 143. Carinocyathus loculatus Vol.
а—поперечное сечение части кубка; б—продоль-
ное сечение части кубка, Х12. Устье р. В. Кеня,
приток р. Абакан
диаметре кубка 8,0 мм равен 7,5. Днища тонкие, редкие, плоские. Коэф-
фициент массивности кубка около 2,5.
Распространение. Санаштыкгольский комплекс, р. Верхняя
Кеня в Западном Саяне; р. Лебедь в Северном Алтае.
92
Cartnocyathus loculattformls Krasnopeeva, sp. n.
Табл. VII, фиг. 4; рис. 144
Кубок конической формы. Обе стенки построены по типу стенок Саг.
loculatus. Септы равномерной толщины. При диаметре кубков 2,5 и
5,6 мм число септ равно 20 и 47, септальный коэффициент равен соот-
б
Рис. 144. Cartnocyathus loculattformls К г a s n., sp. ш
а—продольное сечение части ’ кубка, показывающее
детали строения стенок, Х20; б—продольное скошенное
сечение кубка, уб. кл. Ипполитов в системе р. Абакан
ветственно 8,0 и 8,4. Днища плоские, утолщены в
с септами. Коэффициент массивности кубка — в
новное отличие от Car. loculatus — наличие утол-
щений на днищах.
Р аспространение. р. малый Таз в Гор-
ной Шории; кл. Ипполитов в системе р. Абакана.
Cartnocyathus bericullcus Krasnopeeva, sp. n.
Рис. 145
Наружная стенка в продольном и косом сече-
ниях состоит из S-образных элементов, образую-
щих на ее внешней поверхности небольшие вы-
ступы. Внутренняя стенка включает в себя
V-образные поровые каналы. Септы утолщены
у внутренней стенки. Септальный коэффициент
при диаметре кубков 2,5; 5,2; 6,0 и 10,0 мм соот-
ветственно равен 8,0; 10,0; 8,6 и 11,0. Ширина
скелетной части кубка 1,0 мм. Коэффициент мас-
сивности колеблется от 2,0 до 3,5.
Распространение. Камешковский и
местах пересечения их
пределах 2,0—3,0. Ос-
Рис. 145. Cartnocyathus
bericullcus К г a s п-> sp. п.
Поперечное скошенное
сеченне кубка; нат. вел.
рч. Белокаменка в си-
стеме р. Кин
санаштыкгольский комплексы. Бассейн рр. Нижняя Терсь и Белокаменка
в системе р. Кии, Кузнецкий Алатау; р. Малый Таз в Горной Шории.
Cartnocyathus slblrtcus Krasnopeeva, sp. n.
Табл. VIII, фиг. 1; рис. 146
Узкоконический кубок. Наружная стенка с воронкообразными поро-
выми каналами прикрыта пористой пленкой. Поровые каналы внутренней
93
стенки V-образные. Септы слабо утолщены у обеих стенок. Септальный
коэффициент при диаметре кубка 30,0 мм равен 3,0—3,5. Днища тонкие,
плоские. Коэффициент массивности кубка около 3,5.
Рис. 146. Carinocyathus sibiricus Krasn., sp. n.
a—поперечное сечение части кубка; б—продольное сечение части
кубка, X 4,5. р. Шивелик-Хем, Тува
Распространение. Камешковский комплекс, р. Шивелик-Хем
в Туве.
Род Erbocyathus ZJi u г a v 1 е v а
Синоним — Polycyathus
Колониальные формы с наружной стенкой, свойственной роду Tege-
rocyathus, и с простым строением внутренней стенки кубка.
Типичный вид: Erbocyathus obrutchevt (Vol.).
Erbocyathus obrutchevt (Vologdin)
Синоним — Polycyathus obrutchevt
Рис. 147
Колониальная форма. В поперечном сечении у большинства экземпля-
ров колония имеет вид цепочки с неправильными звеньями. Септы пря-
Рис. 147. Erbocyathus obrutchevt Vol.
а—поперечное сечение части кубка. Х3,5; б—продоль-
ное сечение части кубка, X 5. р. Ульмень—приток
р. Бии, Северный Алтай
мые, равномерной толщины; в местах почкования кубков они искрив-
ляются и раздваиваются. Септальный коэффициент меняется в зависимо-
сти от стадии почкования. Коэффициент массивности кубка непостоянен.
94
Распространение. Обручевский комплекс, д. Сухая Ерба
и рч. Б. Ерба в Кузнецком Алатау; р. Ульмень в Северном Алтае.
СЕМЕЙСТВО THALAMOCYATHIDAE ZHURAVLEVA
Наружная стенка кубка простая, ситовидная. Внутренняя стенка сло-
жена серией лежащих одно на другом горизонтальных колец V- и S-об-
разного сечения. В месте скрепления колец с септами у некоторых видов
выражены нормальные поры. В интерваллуме присутствуют септы,
иногда в сочетании с днищами или синаптикулами.
Род Thalamocyathus G'ordon
Синоним — Cyctocyathus Vol.
Наружная стенка кубка ситовидная, а внутренняя представляет собой
серию желобообразных колец. В интерваллуме присутствуют правильные,
ситовидные септы. У внутренних концов септ на стенке имеются округ-
ленные поры (см. рис. 149).
Типичный вид. Thalamocyathus trachealls G о г d.
Этот род может быть разделен на три группы видов: I группа —
кольца внутренней стенки имеют S-образное сечение; II группа — кольца
имеют V-образное сечение; III группа — V-образные кольца открыты не
вверх, а вниз.
I ГРУППА
Thalamocyathus terslensts (Vologdin)
Табл. IX, фиг. 4
Кубок узкоконический. Наружная стенка ситовидная. Септы правиль-
ные, ситовидные, одинаковой толщины. Внутренняя стенка кольцевидная;
сечение колец S-образное. Не исключена возможность, что такая форма
сечения является результатом неполного и скошенного сечения желобо-
образных колец. Характерным признаком данного вида является самый
меньший септальный коэффициент для всех представителей рода Thala-
mocyathus G о г d. При диаметре кубков 5,6; 5,6—6,0 и 20,0 мм септаль-
ный коэффициент равен 3,0; 3,0—3,5 и 3,0. Коэффициент массивности
кубка в среднем около 2,0.
Распространение, кл. Ипполитов и Герасимов в Западном
Саяне; с. Гавриловское и д. Ариничева на Салаире; пос. Ивановский на
Алтае; бассейн р. Нижняя Терсь в Кузнецком Алатау.
II ГРУППА
Thalamocyathus kuznetsovl (Vologdin)
Рис. 148
Узкоконический кубок. Септы тонкие, равномерной толщины. Сеп-
тальный коэффициент при диаметре кубков 4,5; 6,5; 9,0 и 15,0 мм соот-
ветственно равен 6,6; 5,0; 6,0 и 6,2. Коэффициент массивности кубка от
2,4 до 4,1.
Распространение. Обручевский и камешковский комплексы,
рч. Б. Ерба в Кузнецком Алатау; с. Торгашино в Восточном Саяне;
кл. Герасимов в Западном Саяне; р. Малый Таз в Горной Шорин;
с. Горскино на Салаире; р. Шивелик-Хем в Туве.
Thalamocyathus gerassimovensis Krasnopeeva, sp. n.
Табл. VIII, фиг. 4; рис. 149
Кубок узкоконический. Септы тонкие, равномерной толщины. Септаль-
ный коэффициент при диаметре кубков 5,0; 6,5—7,0 и 8,0 мм равен
95
Соответственно 7,4; 7,4—8,0 и 7,7.
равен 2,0.
Коэффициент массивности кубка
Рис. 148. Thalamocyathus kuz-
net soot (V о 1.) .'
Поперечное сильно скошенное
сечение кубка, X 5. р. Шиве-
лик-Хем, Тува
Рис. 149. Thalamocyathus
gerassimovensis К г a s п., sp. п.
Поперечное скошенное сече-
ние кубка, X 5. кл. Ипполи-
тов в среднем течении
Распространение. Камешковский и гавриловский комплексы,
с. Горскино на Салаире; р. Белокаменка в системе р. Кии, Кузнецкий
Алатау; кл. Герасимов, вблизи устья, в Западном Саяне.
Thalamocyathus undatus S. Tschernycheva, sp. и.
Табл. VIII, фиг. 6; табл. IX, фиг. 5
Диаметр кубка 7,3 мм. Наружная стенка ситовидная. Септы пра-
вильные, равномерной толщины, местами изогнутые, с зачаточными
синаптикулами. Септальный коэффициент равен 7,5. Внутренняя стенка
кольцевидная с шипами. Ширина скелетной части 1,6 мм. Коэффициент
массивности кубка 2,5.
От Thalamocyathus splnosus описанный вид отличается строением
септ.
Распространение, р. Катунь, между рр. Чепош и Сема.
Thalamocyathus loculatus (Vologdin)
Табл. X, фиг. 1
Форма кубка неизвестна. Кольца внутренней стенки несут на себе
шипы, обращенные вверх в центральную полость. Септы утолщены
у внутренней стенки. Септальный коэффициент при диаметре кубков
9,4 и 10,0 мм равен 11,0. Коэффициент массивности кубка равен 3,0.
Распространение, с. Торгашино в Восточном Саяне. Комплекс
неясен.
Thalamocyathus splnosus Krasnopeeva, sp. n.
Табл. VII, фиг. 5; рис. 150
Кубок конической формы с поперечноморщинистой ситообразной на-
ружной стенкой. Кольца внутренней стенки имеют неясно выраженное
96
V-образное сечение. На поверхности колец, выходящих в центральную
полость, расположены шипы. В продольном сечении можно видеть, что
внутренняя стенка вблизи основания кубка тонкая и не имеет кольцеоб-
Рис. 150. Thalamocyathus spinosus Krasn., sp. n.
<a—продольное сечение кубка, б—поперечное сечение кубка, X 5,5. рч. Шивелик-Хем,
Тува
Thalamocyathus
Рис. 151.
jakovlevi (V о L)
а—продольное сечение ча-
сти кубка, х 4; б—попе-
речное сечение кубка. хЗ,5.
р. Шивелик-Хем. Тува
разного строения. Септы утолщены на концах у обеих стенок. Септаль-
ный коэффициент при диаметре кубка 9,5 мм равен 3,8. Коэффициент
массивности кубка 2,0.
Распространение. Камешковский комплекс, р. Шивелик-Хем
ч Туве.
HI ГРУППА
Thalamocyathus jakovlevt (Vologdin)
Табл. VIII, фиг. 2; рис. 151
Кубок воронкообразный, конический. Жело-
бообразные кольца, слагающие внутреннюю
стенку, открыты вниз, а не вверх, как у всех
других видов этого рода. Септы слабо утол-
щены у наружной стенки. Септальный коэффи-
циент при диаметре кубков 6,5 и 16,0 мм соот-
ветственно равен 5,2 и 3,6. Коэффициент мас-
сивности кубка по мере увеличения попереч-
ника увеличивается от 1,8 до 2,5.
Распространение. Камешковский комплекс, д. Камешки в Во-
сточном Саяне; р. Шивелик-Хем в Туве.
Род Bronchocyathus Bedford
Синоним — Salairocyathus Vol.
Наружная стенка кубка ситовидная; внутренняя стенка сложена
желобообразными горизонтальными кольцами с V-образным сечением их.
В интерваллуме, кроме септ, присутствуют днища.
Типичный вид. Bronchocyathus sygmoldeus В е d f.
Зак. 2024
97
Bronchocyathus erbiensis Krasnopeeva, sp. n.
Рис. 152
Конический кубок. Днища тонкие, плоские, утолщенные у наружной
стенки и в месте пересечения их септами. Септальный коэффициент при:
диаметре кубка 2,6 мм равен 3,8. Коэффициент массивности кубка:
равен 2;1.
Рис. 152. Bronchocyathus erbiensis К г a s n., sp. п.
а—поперечное сечение части кубка, х 9,5; р. М. Таз, Горная Шория;
б—скошенное сечение кубка, нат. вел.; р. Ерба, Кузнецкий Алатау
Распространение. Комплекс неясен. Кузнецкий Алатау, Лощен-
ков лог в районе рч. Б. Ербы.
Bronchocyathus suchiensis Krasnopeeva, sp. n.
Табл. IX, фиг. 1; табл. X, фиг. 4
Кубок близкий к цилиндрическому. Диаметр его 17,0 мм. Септ около1
125. Септальный коэффициент приблизительно 7,4—8,0. Коэффициент
массивности примерно 3,5. Наружная стенка тонкая, ситовидная.
Толщина стенки 0,07 мм, диаметр пор 0,07 мм, ширина промежутков не
превышает 0,05 мм. Внутренняя стенка сложена желобообразными коль-
цами. У внутренних концов колец на стенке выражены поры. Расстояние
между кольцами приблизительно 0,10 мм. Наибольшая толщина колец
у их основания равна 0,1 мм. Днища плоские; с мелкими порами. Диа-
метр пор 0,06—0,09 мм, ширина промежутков 0,02—0,03 мм, в месте-
пересечения днищ с септами видны утолщения; форма их близка к ромбо-
видной. Местами наблюдается гребневидное сечение днищ в месте пере-
сечения их с септами, обусловленное наличием наклонных пластинок
вблизи утолщений.
Распространение, кл. Сухой, вблизи впадения его в р. Чер-
ную Усу.
СЕМЕЙСТВО ANNULOCYATH1DAE KRASNOPEEVA, FAM. N.
Наружная стенка кубка с поровыми каналами; внутренняя стенка
с желобообразными кольцами, образующими щелевидные горизонталь-
98
ные отверстия. У известных в настоящее время родов сем. Annulocyathi-
dae в интерваллуме присутствуют септы (род Annulocyathus) и септы
с синаптикулами (род Annulofungia).
Род Annulocyathus Vologdin
Наружная стенка с V-образными поровыми каналами, внутренняя
с горизонтальными щелевидными отверстиями. Горизонтальные кольца,
образующие внутреннюю стенку, имеют V- и S-образное сечение. В ин-
терваллуме присутствуют только септы; они ситовидные, правильные.
Типичный вид. Annulocyathus pulcher Vol.
Распространение. Предположительно санаштыкгольский комп-
лекс. рр. Белокаменка и Нижняя Терсь в Кузнецком Алатау; кл. Санаш-
тыкгол в Западном Саяне.
Annulocyathus lavrenovae Krasnopeeva
Табл. Ill, фиг. 2; рис. 153
Узкоконический кубок. Наружная стенка его содержит в себе систему
поровых каналов с промежутками S-образного сечения. Внутренняя
стенка сложена горизонтальными кольцами S-образного сечения. Септы
равномерной толщины с мелкими порами и широкими промежутками
а 6
Рис. 153. Annulocyathus lavrenovae К г a s n.
а—продольное сечение части кубка, X 5; б
поперечное сечение кубка, X 15. Лощенков
лог, р. Ерба
между ними. Септальный коэффициент при диаметре кубка 3,8 мм равен
9,0. Коэффициент массивности кубка равен 3,0. От Annulocyathus pulcher
отличается S-образным сечением колец внутренней стенки и деталями
строения наружной стенки (рис. 153а).
Распространение. Гавриловский и санаштыкгольский (?) ком-
плексы. с. Гавриловское на Салаире; Лощенков лог близ рч. Б. Ерба
в Кузнецком Алатау.
Род Annulofungia Krasnopeeva, gen. n.
Наружная стенка с V-образными поровыми каналами, внутренняя -
с щелевидными горизонтальными порами. Септы соединены между собой
синаптикулами, что является отличием данного рода от рода Annulo-
cyathus.
Типичный вид. Annulofungia taylorl К г a s n.
7*
99
Annulofungia taylorl Krasnopeeva
Табл. VIII, фиг. 3; рис. 154
Кубок узкоконический. Наружная стенка включает в себя серию мел-
ких V-образных поровых каналов. Внутренняя стенка сложена кольцами
V-образного сечения. Септы тонкие, равномерной толщины, соединены
Рис. 154. Annulofungia taylori К г a s n.
а—поперечное сечение кубка, X 3,5; б—продоль-
ное сечение кубка, X 10. Лощенков лог, р. Ерба
между собой синаптикулами. Септальный коэффициент при диаметре
кубка 9,6 мм равен 7,8. Коэффициент массивности кубка равен 3.
Распространение. Камешковский комплекс. Лощенков лог
близ рч. Б. Ерба в Кузнецком Алатау; с. Торгашино в Восточном
Саяне (?).
КЛАСС IRREGULARIA. НЕПРАВИЛЬНЫЕ АРХЕОЦИАТЫ
СЕМЕЙСТВО ARCHAEOCYATHIDAE OKULITCH
Состав этого семейства требует пересмотра, так как строение скелет-
ных элементов его представителей не изучено с достаточной деталь-
ностью. Ниже приведены описания видов, у которых кубок в перифери-
ческой части сложен тесно соприкасающимися трубочками, наружные
концы которых у многих форм образуют наружную стенку, а внутренние
концы — внутреннюю стенку. В этом случае наружная и внутренняя
стенки кубка являются несамостоятельными. Стенки ячеек пористые.
Ячейки ориентированы радиально и косо вниз от наружного пространства
к центральной полости кубка. Иногда внутренние концы их, загибаясь на
коротком расстоянии вверх, образуют самостоятельную внутреннюю стенку.
Наружная стенка у представителей рода Archaeocyathus принимается
за самостоятельную в том случае, когда поперечник пор и их количество
не соответствует поперечнику концов ячеек, выходящих на наружную по-
верхность кубка, например у Archaeocyathus rete (Т а у 1.). Некоторые
представители сем. Archaeocyathidae имеют днища (род Claruscyathus
Vol.) или пузырчатую ткань (род ? Retecyathus Vol.).
Ячейки иногда бывают заполнены пузырчатой тканью. Стенки ячеек
массивные и имеют слоистое строение.
1 См. сноску на стр. 76.
юо
Род Archaeocyathus Billings
Синоним — Spirocyathus Hind
Характерным признаком рода является массивное, слоистое строение
стенок трубочек. Сечение трубочек многоугольное или неправильно
округленное.
Типичный вид. Archaeocyathus atlantlcus Bill.
Archaeocyathus aff. subradiatus (Vologdin)
Рис. 155
Кубок имеет узкоконическую форму; наблюдавшийся диаметр его
20,0 мм. Диаметр ячеек у внутренней стенки 0,35 мм, а у наружной
0,49 мм. Ячейки в нижней части кубка заполнены пленками пузырчатой
Рис. 155. Archaeocyathus aff. subradiatus (Vol)
а—поперечное сечение кубка, v 4; б—продольное
сечение ближе к основанию кубка. 5. р. Кара-
гач—правый приток р. Ульмень
ткани. Ширина скелетной части кубка 3,5 мм. Поперечник центральной
полости равен 2,3 мм. Внутренняя стенка самостоятельная.
Распространение. Камешковский и обручевский комплексы.
Улус Бей-Булук и д. Камешки в Восточном Саяне: р Карагач, правый
приток р. Ульмень, Алтай.
Род? Retecyathus Vologdin
Кубок правильный, узкоконический. Наружная и внутренняя стенки
самостоятельные. Промежуточное пространство заполнено ячейками и
пузырчатой тканью. Скелетные элементы иногда присутствуют и в цен-
тральной полости кубка.
Типичный вид. Retecyathus kuzminl Vol.
101
Retecyathus kuzmlnl Vologdin
Табл. IX, фиг. 2
Кубок конический. Наблюдавшийся диаметр кубка 5,0 и 7,0 мм.
Ячейки, слагающие интерваллум, пересечены многочисленными пленками
пузырчатой ткани. Во внутренней полости тоже развита скелетная ткань,
но более массивная. Ширина скелетной части кубка 2,0 мм. Поперечник
центральной полости 1,3 мм.
Распространение. Обручевский, гавриловский и др. комплексы.
Ряд пунктов в Горном Алтае.
СЕМЕЙСТВО SYRINGOCNEMIDAE TAYLOR
Кубок двухстенный, одиночный или колониальный, построен системой
призматических трубчатых ячеек, плотно прилегающих друг к другу.
Ячейки ориентированы косо вниз от наружного пространства к централь-
ной полости. Концы их, подобно роду Archaeocyathus, в большинстве слу-
чаев открываются в виде крупных отверстий в наружное пространство
и в центральную полость. Стенки ячеек тонкие, плотные и сложены од-
нородной тканью.
Сем. Syringocnemidae включает в себя два рода — Syrlngocnema
и Protopharetra. От представителей сем. Spirocyathidae они отличаются
тонкими стенками ячеек, имеющими однородное сложение.
Род Syrlngocnema Taylor
Кубок двухстенный, одиночный или колониальный. Промежуточное
пространство целиком заполнено трубочками, имеющими многоугольное
сечение. Стенки трубочек тонкие, однородные, пористые.
Типичный вид. Syrlngocnema favus Т а у 1.
Рис. 156. Syrlngocnema
eleganta Т а у 1. Попереч-
ное сечение, X 4. р. Уль-
мень—приток р. Бии
Syrlngocnema eleganta Taylor
Рис. 156
Диаметр кубка 8,0 мм. Ширина промежу-
точного пространства между стенками 2,8 мм.
Диаметр ячеек у внутренней стенки 0,17 мм, у
наружной 0,35 мм. Стенки трубочек тонкие,
плотные. Ширина скелетной части кубка 3,0 мм.
Поперечник центральной полости 2,2 мм.
Распространение. Обручевский и
санаштыкгольский комплексы. Бассейн рр. Уль-
мень и Карагач в Горном Алтае; кл. Сана-
штыкгол в Западном Саяне.
Л И. ЕГОРОВА, М. П ЛОМОВИЦКАЯ, О. К. ПОЛЕТАЕВА,
А. Г. С И ВО В
Тип Arthropoda. Членистоногие
КЛАСС TRILOBITA. ТРИЛОБИТЫ
Трилобиты составляют важнейшую группу кембрийской фауны За-
падной Сибири. Монографического описания этой группы ископаемых
нет; опубликованные материалы представляют собой несколько заметок.
Изучение кембрийских трилобитов Западной Сибири далеко не закон-
чено. Тем не менее уже сейчас установлен ряд последовательных
102
комплексов, характеризующих различные стратиграфические подразделе-
ния кембрия:
чинпинский Спи2;
бабаковский СгП|3,
санаштыкгольский Cm21,
большеербинский Cm22,
орлиногорский Cm23,
салаирский Cm31,
горношорский Cm32.
Характеристика состава и распространение этих комплексов даны в об-
щем обзоре кембрийской системы Западной Сибири. Приведенные ниже
описания руководящих форм охватывают лишь незначительную часть
богатой и своеобразной фауны. Из них Л. И. Егоровой принадлежит
описание нового рода Portentosus с соответствующими видами и неко-
торые новые виды и разновидности других родов, М. П. Ломовицкой,
О. К. Полетаевой описаны из горношорского комплекса Pagetla schorlen-
sis, Paradoxldes suboelandlcus, род Parasolenopleura с двумя видами
и Chondragraulos schorlensls. Все остальные формы описаны А. Г Си-
зовым .
А Г Н О С Т И Д Ы
Эта обширная группа мелких трилобитов, у которых головной и хво-
стовой щиты примерно одинаковых размеров и очертаний, парные
глаза и лицевые швы отсутствуют, а туловище состоит
из двух члеников (рис. 157), нередко встречается в кем-
брийских отложениях Западной Сибири, но изучена еще
недостаточно. В настоящее время эта группа имеет слож-
ную и дифференцированную систематику, состоит из
ряда семейств, подсемейств и родов. Для иллюстрации
ниже приводится описание западносибирского предста-
вителя рода Peronopsls С о г d а, краткий диагноз кото-
рого, по Е. В. Лермонтовой, таков: «Передняя лопасть
глабели простая, но шире остальных лопастей глабели.
У основания глабели — маленькие дополнительные лопа-
сти» (Циттель, стр. 938).
Peronopsls 'Из группы Peronopsls fallax Linnarsson
Табл. XI, фиг. 1
Рис. 157. Общее
строение паи-
цыря трилобита
из группы аг-
ностид, X 3.
По Берри
Головной щит небольших размеров (длина 3 мм), прямоугольной
формы. Глабель длинная, выпуклая; сзади она выпукла сильнее, вперед
понижается. На глабели имеются три поперечных бороздки, которые
делят ее на лопасти. Передняя бороздка хорошо выражена, идет поперек
глабели, несколько оттянута назад в средней части и отделяет переднюю
часть глабели. Две другие бороздки выражены очень слабо. Посредине
глабели имеется маленький бугорок, у основания — маленькие треуголь-
ные дополнительные лопасти. Краевая кайма узкая, огибает весь голов-
ной щит, отделяясь от него отчетливой бороздкой. Спинные борозды
широкие, резко выражены. Щеки выпуклые.
Хвостовой щит широкий (длина 3 мм, ширина 4 мм), окружен
широкой выпуклой каймой. Краевая борозда широкая и глубокая. На
задних боковых углах пигидия имеются небольшие шипы. Осевая часть
пигидия большая, шире боковых (плевральных) частей, параболических
очертаний, выпуклая. В передней части на ней находится продольный
бугорок. Спинные бороздки ясно выражены, широкие и глубокие.
Распространение. Большеербинский и орлиногорский комп-
лексы (Ст22-3). Алтай, верховья рч. Верхняя Еланда; восточный склон
Кузнецкого Алатау, рч. Б. Ерба; Салаир, гора Орлиная.
103
СЕМЕЙСТВО PAGETIDAE KOBAYASHI
Ниже описываемые представители этого семейства, согласно класси-
фикации Е. В. Лермонтовой, составляют особую II группу, у которой
отчетливо развиты спинные и затылочная борозды и ясно сегментирован
пигидий.
Род Pagetia Walcott
Мелкие пропарные трилобиты с головными и хвостовыми щитами
одинаковой величины, окруженными узкой, плоской каймой. Туловище
состоит из двух сегментов. Глабель спереди заострена. Затылочное
кольцо и предпоследний членик рахиса пигидия вытянуты в длинные
шипы. Бока пигидия нерасчлененные, гладкие.
Типичный вид. Pagetia bootes Walcott.
Распространение. Спи—Cm2 Северной Америки, Средней
и Западной Сибири.
Pagetia ferox Lermontova
Табл. XI, фиг. 3
Очень мелкие формы с узкоконической глабелью, заметно приподня-
той вблизи затылочного кольца. Треугольного очертания, сильно вздутые
щеки поднимаются от заднебоковых бороздок к маленьким глазам и да-
лее своими узкосерповидными передними концами слегка охватывают
передний конец глабели. Щеки впереди глабели разделены узкой верти-
кальной бороздкой. Спинные борозды отчетливые, глубокие. Затылочное
кольцо, будучи отделено короткой и очень мелкой затылочной бороздкой,
является выпуклым, широким и оттянуто в недлинный, быстро заостряю-
щийся шип. Равномерно изогнутая валикообразная фронтальная кайма
с характерными вертикальными насечками отделена от передней части
глабели ясной краевой бороздкой.
Полукруглый хвостовой щит с гладкими боками обладает выпуклым
рахисом, ясно сегментированным в передней части. Рахис, несколько на-
висая над задним краем щита, снабжен шипом, от которого сохранился
лишь след его прикрепления.
Распространение. Большеербинокий комплекс (Cm22),
рч. Б. Ерба, восточный склон Кузнецкого Алатау; Ст2 Анабарского
массива.
Pagetia schoriensis Poletaeva, sp. n.
Табл. XI, фиг. 2
Глабель неширокая, длинная, впереди постепенно заостряющаяся.
Затылочное кольцо оттянуто в длинный шип, от которого обычно сохра-
няются только основание или передняя половина. Шип идет вначале косо
вверх, а затем поворачивается вдоль туловища. Затылочная борозда
более ясно выражена против боков глабели. Глубокие спинные бороздки
сливаются впереди глабели и образуют широкую и глубокую борозду,
разделяющую неподвижные щеки. Краевой лимб широкий и плоский,
сохраняющий свою ширину и на боках головного щита. Неподвижные
щеки имеют вид двух конических вздутий. Глазные валики тонкие,
хорошо выраженные. Пигидий имеет узкую, выпуклую, с отчетливыми
сегментами осевую часть, гладкие плевры и неширокий лимб. Шипик на
последнем сегменте пигидия у наших образцов не обнаружен: невиди-
мому, он не сохранился.
Распространение. Санаштыкгольский комплекс (Ст2‘)
рч. Мондыбаш, Горная Шория.
104
Pagetia ornatlformis S i v о v, sp. n.
Табл. XII, фиг. 13
Этот вид, устанавливаемый только на основании достаточно хорошо
сохранившегося пигидия, проявляет исключительно характерные при-
знаки. Очень маленький пигидий по своим очертаниям сходен с пигидием
Pagetla ferox, но в отличие от последнего у Pagetla ornatlformis рахис
хвостового щита более выпуклый и резче отделен глубокими спинными
бороздками с боков. Рахис особенно резко вздут и вздернут в своей зад-
ней части, вблизи последнего сегмента, от которого отходит короткий
и толстый шип. Гладкая поверхность пигидия украшена крупными изящ-
ными бугорками, которые располагаются вдоль внутренних краев пиги-
дия, непосредственно примыкающих к спинным бороздкам.
Распространение. Чингинский комплекс (Ст!2). Восточный
Саян, д. Камешки.
Род Pagetina Lermontova
Головной щит имеет то же строение, что и у рода Pagetla. Хвостовой
щит у рода Pagetina обладает сильно расчлененным коническим рахисом,
отделенным ясными дорзальными бороздками от расчлененных боков, на
которых постоянно присутствуют как плевральные, так и межплевраль-
ные бороздки.
Типичный вид. Pagetina rjonsnitzkll Lerm.
Распространение. Зона Protolenus Средней Сибири; Ст.-’
и СгП|3 Западной Сибири.
Pagetina katunica Poletaeva, sp. и.
Табл. XI, фиг. 7
Глабель узкая, коническая, округленная спереди, далеко не доходит
до краевой каймы, имеет две пары бороздок; передняя из них широкая,
но слабо заметная. Спинные бороздки четкие, становятся более мелкими
близ переднего края глабели.
Фронтальный край широкий, чуть выпуклый, слегка расширяется
впереди глабели. Неподвижные щеки вздутые, сильно поднимаются от
заднебоковых бороздок к глазам и затем широким треугольным вздутием
идут к краевой борозде, часто окружая с боков суженный передний конец
глабели. Впрочем, у других экземпляров передние концы вздутых щек
срезаются краевой бороздой на одном уровне с глабелыо. Затылочное
кольцо широкое, вытянуто в недлинный, быстро заостряющийся шип.
Хвостовой щит со слабо выделяющимся, но ясно сегментированным ра-
хисом, выпуклый. Осевая часть рахиса переходит в задний шип, на об-
разце сохраняется след его прикрепления. Плевры хвоста слабо сегмен-
тированы, причем у краев иногда видны межплевральные бороздки.
Кайма толстая.
Pagetina katunica отличается от близкой к ней формы Pagetina
pritnaeva из д. Камешки Восточного Саяна и Анабарского района глав-
ным образом строением хвостового отдела и некоторыми деталями
в строении глабели. У Pagetina katunica на глабели присутствуют четко
выраженные глабелярные бороздки, чего не наблюдается у Pagetina
pritnaeva. Пигидий у Pagetina katunica сегментирован менее отчетливо,
чем у Pagetina pritnaeva.
Распространение: Бабаковский комплекс (СгП|3) Горный
Алтай, р. Катунь, в 2,5 км выше устья р. Семы.
Род Cobboldia Lermontova
Мелкие трилобиты, близкие к роду Hebediscus (Whitehouse, 1936)
с головным щитом, несколько большим, чем пигидий, и тремя туловищ-
ными сегментами. Глабель субцилиндрическая, выпуклая сзади и упло-
105
щенная впереди. Три пары глабелярных бороздок слабо заметны, спин-
ные борозды очень глубокие. Неподвижные щеки выпуклые в передне-
заднем направлении, оканчиваются спереди на одном уровне
с заостренным концом глабели, оставляя впереди широкое вогнутое пред-
глабелярное поле. Фронтальная кайма шнуровидная. Глаза длинные.
Затылочное кольцо со срединным бугорком. Пигидий округлотреуголь-
ный, с выпуклым, неясно сегментированным рахисом. Бока пигидия рас-
членены, причем на них различаются как плевральные, так и межплев-
ральные сильно сглаженные бороздки.
Типичный вид. Cobboldia dentata Lerm.
Распространение. Cm,. Восточная Сибирь, Алтай.
Cobboldia altalca Poletaeva, sp. n.
Табл. XI, фиг. 4
Кранидий очень маленький, с равномерно округленным передним
краем. Глабель почти цилиндрическая, очень слабо расширяется позади
и широко закругляется спереди. Гладкая предглабелярная площадка
плоская и лишь в передней половине очень слабо вогнута; против гла-
бели на ней имеется неясная и неглубокая вдавленность. Ни каймы, ни
валика не имеется, лишь фронтальный край кранидия чуть приподнят.
Спинные бороздки нечеткие, широкие, различной глубины в зависимости
от выпуклости кранидия. У переднего конца глабели они быстро выкли-
ниваются, так что передний конец глабели постепенно сливается с пред-
глабелярной площадкой и кажется суживающимся спереди. Затылочное
кольцо и борозда хорошо выражены; кольцо несет бугорок. Непо-
движные щеки широкие, слабо выпуклые. Выпуклость наиболее сильно
выражена в задней части.
По общему облику описанная форма сильно напоминает Cobboldia
dentata Lerm. из Сгщ Восточной Сибири. У Cobboldia altalca глабель
менее выпукла; слабее выражены спинные бороздки; отсутствуют крае-
вая борозда и валик; на хвосте отсутствуют зубчики.
Распространение. Бабаковский комплекс (Ст(3). Алтай, пра-
вый борт долины р. Катуни, в 3,3 км выше устья р. Семы.
СЕМЕЙСТВО DINESIDAE LERMONTOVA
Род Proerbia Lermontova
По Е. В. Лермонтовой, данный род характеризуется субквадратной
формой кранидия, с выгнутым вперед, несколько угловатым посередине
передним краем и почти цилиндрической, очень выпуклой глабелью. Ее
передний конец притуплен; на ней заметны четыре пары коротких
борозд, из которых две задние, направленные назад, более резкие, чем
передние. Задняя пара доходит до глубокой затылочной борозды и отре-
зает по маленькой треугольной дольке на заднебоковых углах глабели.
Затылочное кольцо плоское, но сильно приподнятое от затылочных бо-
розд и оттянутое в мощный шип.
Неподвижные щеки сравнительно узкие, особенно в участках, лежа-
щих впереди глазных крышек. Последние сравнительно длинные, изогну-
тые и примыкают к глабели несколько выше верхней пары глабелярных
бор >здок. Передние ветви лицевых швов длиннее задних, параллельны
продольной оси кранидия. Наиболее характерной частью кранидия яв-
ляется широкая обратнотрапецоидальная предглабелярная площадка,
расчлененная на три шаровидных дольки; она обрамлена спереди выпук-
лой, равномерной ширины фронтальной каймой. Поверхность кранидия
покрыта относительно крупными бугорками. Туловищный и хвостовой
щиты неизвестны.
Типичный вид. Proerbia prisca Lerm.
Распространение. Нижний кембрий. Сибирь.
1 06
Proerbia (?) bidzhenia Sivov, sp. n.
Табл. XI, фиг. 9
Средней величины форма, сходна с типичным видом этого рода. Од-
нако у Proerbla bidzhenia глабель впереди округлена, причем на ней
заметны лишь две пары глабелярных бороздок, из них задняя пара,
сильно наклонная, отрезает пару треугольных долек у основания гла-
бели более мелких, чем дольки у Proerbla prlsca. Но самое главное раз-
личие между сравниваемыми видами заключается в том, что у Proer-
bla (?) bidzhenia на относительно широкой и выпуклой предглабелярной
площадке отчетливо выявляется лишь серединная шаровидная долька,
тогда как у Proerbla prlsca таких долек три и две из них располагаются
по бокам предглабелярной площадки. Наконец, у Proerbia (?) bidzhenia
вместо оттянутого шипа на затылочном кольце присутствует бугорок,
а поверхность покрыта более многочисленной и резче выраженной гра-
нуляцией.
Распространение. Бабаковский комплекс (Cmi3). рч. Биджа,
восточный склон Кузнецкого Алатау; близкий вид встречен в известняках
кл. Бабак, на северном склоне Западного Саяна.
Род Erbia Lermontova
Согласно исследованиям Е. В. Лермонтовой, род Erbia близко сходен
с австралийским родом Dinesus Е t h е г i d.
Мелкие формы рода Erbia обладают полукруглым головным щитом,
рассеченным глубокими и широкими спинными бороздками на три резко
выдающиеся части. Глабель цилиндрическая, выпуклая и спереди округ-
лая или слабо притупленная.
Из глабелярных борозд развиты лишь бороздки задней пары; корот-
кие, глубокие, наклоненные к затылочной борозде, они отрезают пару
маленьких треугольных долек у основания глабели.
Затылочно кольцо во многих случаях с крупным, торчащим вверх,
шипом. Предглабелярная площадка, ограниченная с боков то мелкими,
то глубокими расходящимися ответвлениями спинных бороздок, имеет
трапецоидальную форму; она либо вдавленная, либо выпуклая. Глаза
маленькие, торчащие. Неподвижные и подвижные щеки выпуклые. Фрон-
тальная кайма шнуровидная, вздутая. Щечные углы закруглены, иногда
с маленьким шипиком.
Туловище с выпуклой осью состоит из 11—13 сегментов, с прямыми
бороздчатыми плеврами, образующими резкий коленчатый перегиб,
и оканчивающиеся небольшими коготками. Пигидий очень маленький,
поперечноовальный, с коротким рахисом (1—2 сегмента) и слабо расчле-
ненными боками. Поверхность гранулированная.
Типичный вид. Erbia sibirica (Schmidt).
Распространение. Cmi3—Cm2. Западная Сибирь.
Erbia sibirica (Schmidt)
Табл. XI, фиг. 10
У этого вида наиболее полно выражены характерные признаки рода,
но предглабелярная площадка у него плоская, а глабель спереди притуп-
лена. Глаза располагаются ближе к заднему краю кранидия,.чем к пе-
реднему.
Распространение (поЕ. В. Лермонтовой). Низы Ст2; харак-
терная форма верхнего горизонта торгашинского известняка близ г. Крас-
ноярска и горы Долгий Мыс восточного склона Кузнецкого Алатау;
бабаковский комплекс Ст!3 (по А. Г Сивову).
107
Erbia granulosa Lermontova
Табл. XV, фиг. 10
Вид близок к предыдущему. Очень выпуклый небольшой кранидий,.
рассеченный спинными бороздками на три почти равных части. Глабель
спереди округлена и почти упирается в краевую кайму. Вследствие этого
предглабелярная площадка впереди глабели почти исчезает, а сохра-
няется на ее боках в виде двух выпуклых треугольных долек, соединен-
ных узкой перемычкой посередине. Спинные борозды умеренной глу-
бины, в передней части расходящиеся. Неподвижные щеки выпуклые,
с торчащими на них бобовидными маленькими глазными крышками,
расположенными несколько впереди середины глабели. Поверхность
покрыта грубыми бугорками. Затылочное кольцо с небольшим бугорком.
Отличается от Erbia sibirica характером переднего конца глабели, более
узким предглабелярным полем и наличием резко выраженной глануля-
ции.
Распространение. Большеербинский комплекс (Ст22). Восточ-
ный склон Кузнецкого Алатау, рч. Б. Ерба.
Erbia gigantea Jegorova, sp. п.
Табл. XII, фиг. 15; табл. XV, фиг. 5
Кранидий средней величины, квадратный, с прямым передним краем,
сильно выпуклый. Глабель округлоугловатая спереди, сильно выпуклая.
Задние углы глабели срезаны в небольшие продолговатотреугольные
лопасти. Предглабелярное поле состоит из двух треугольных площадок,
расположенных по обе стороны глабели. Фронтальный валик четкий,
почти прямой. Спинные борозды глубокие, у передних боков глабели они
разветвляются и своими косыми расходящимися передними продолже-
ниями ограничивают предглабелярную площадку. Затылочное кольцо
вздутое, сильно развитое. Затылочная борозда глубокая, посередине
имеет небольшой коленообразный изгиб.
Неподвижные щеки широкие, сильно выпуклые. Глазные крышки
бобовидной формы, небольшие, сильно приподнятые, расположенные
несколько впереди середины глабели. Лицевые швы впереди глаз идут
параллельно друг другу, а позади глаз довольно сильно расходятся.
Поверхность кранидия покрыта крупной и мелкой грануляцией. Erbia
gigantea имеет большое сходство с Erbia granulosa L е г m., но отли-
чается от последней большой величиной, более округлой формой глабели
и отсутствием бугорка на затылочном кольце.
Распространение. Бабаковский комплекс (Стр3). Горный
Алтай, Катунь, в 2,5 км выше р. Семы; Западный Саян, кл. Бабак,
система рч. Табат.
Род Chakasskia Poletaeva
Небольших размеров формы с достаточно выпуклым кранидием, резко
рассеченным глубокими и широкими спинными бороздками на три части.
Глабель узкая, длинная, заметно расширяющаяся в основании и посте-
пенно сужающаяся по направлению к ее переднему краю, который,
будучи равномерно округлен, упирается в фронтальную кайму.
Три пары глабелярных бороздок, расположенные в задней и средней
части глабели, короткие, но глубокие, причем задняя пара, косо спу-
скаясь к затылочному кольцу, отрезает треугольные лопасти на глабели,
подобные лопастям рода Erbia. Глубокие спинные бороздки, почта па-
раллельные в передней и средней части кранидия, слабо расходятся
вблизи боков затылочного кольца. Последнее ровное, широкое, без шипа
и отделено от глабели глубокой затылочной бороздой. Неподвижные
щеки широкие, треугольной формы, со срезанной передней вершинкой.
108
Глазные крышки небольшие, расположены косо по отношению к гла-
бели и продолжаются в глазные валики, которые, достигая переднего
края неподвижных щек, оканчиваются вблизи фронтальной каймы, под
острым углом к ней. Фронтальная кайма имеет форму узкого валика,
который на середине почти сливается с глабелью, а на боках становится
более четким и более широким. На поверхности кранидия располагаются
мелкие и редкие бугорки до четырех штук на 1 мм.
К семейству Dinesidae этот род отнесен условно.
Типичный вид. Chakasskla minussensis Pol.
Распространение. Санаштыкгольский комплекс (Ст2‘). Запад-
ный Саян, Кузнецкий Алатау.
Chakasskla minussensls Poletaeva
Табл. XIII, фиг. 12
Диагноз рода соответствует описанию вида.
Распространение. Санаштыкгольский комплекс (Cm21). Кл.
Санаштыкгол, приток р. Абакана в юго-западной части Западного Саяна.
Близкий вид известен также из известняков восточного склона Кузнец-
кого Алатау, в бассейне рч. Б. Ерба.
СЕМЕЙСТВО GRANULARIDAE LERMONTOVA
Род Granularia Poletaeva
Различной, но преимущественно крупной величины трилобиты, обла-
дающие широкоовальным спинным щитом, с равновеликими головным
и хвостовым щитами, с выгнутыми краевыми лимбами на них. Туловище
с слабо сужающейся к заднему концу осевой частью, состоит из 10
и более одинаковых сегментов. Плевры узкие, плавно изгибаются вниз
и в передней своей части рассечены очень глубокой, но узкой бороздкой.
Глабель обратногрушевидная или булавовидная, с тремя, реже одной,
парами глубоко врезанных глабелярных бороздок, имеющих вид попе-
речных ямок; иногда присутствует и четвертая, передняя пара, почти
всегда слабо выраженная. Глазные крышки довольно большие, лежат
в задней половине длины, головного щита и, вероятно, соединяются
с глабелью глазными валиками; лицевые швы повидимому пропарного
строения.
Хвостовой щит широкополукруглого очертания, почти с прямым пе-
редним краем, выпуклой средней частью и колоколообразно вогнутым
краевым лимбом. Ось сравнительно узкая, слабо сужается к заднему
концу и состоит из большого числа сегментов (до 15), на середине каж-
дого из которых располагается ясно выраженный бугорок. Бока пиги-
дия разделены плоско-выпуклыми ребрами, разделенными бороздками,
которые по положению их в отношении борозд рахиса можно принять
за плевральные. От переднего края пигидия к заднему сегменты плевр
постепенно отгибаются назад, так что там они лежат уже параллельно
оси пигидия.
Скульптура панцыря этих трилобитов состоит из многочисленных
различной величины бугорков, обычно округлых и составляющих не-
сколько систем.
Типичный вид. Granularia obrutchevt Pol.
Распространение. Сшь Западная и Средняя Сибирь.
Granularia obrutchevi Poletaeva
Табл. XI, фиг. 5
Характеристика вида соответствует описанию рода. Изображенный
Здесь образец происходит из известняков кл. Бабак Западного Саяна.
У этого образца повреждена фронтальная кайма кранидия, остальные
109
же его элементы проявляют полное сходство с типичным видом
с рч. Биджи. Ассоциирующий совместно с нашим образцом хвостовой
щит более крупных размеров, чем у типичного вида.
Распространение. Характерная форма для бабаковского комп-
лекса трилобитов (Cmi3). Северный склон Западного Саяна, кл. Бабак;:
рч. Биджа (полициатовый горизонт) — восточный склон Кузнецкого.
Алатау.
Granularia babakovia Sivov, sp. n.
Табл. XIII, фиг. 10
Крупные формы с характерной булавовидной сильно выпуклой и длин-
ной глабелью, заметно расширяющейся на середине и затем слегка:
сужающейся в передней части; здесь глабель становится чуть плоской,
а ее передний край слегка притупленным. В задней половине глабель
расчленена двумя парами ясных глабелярных бороздок; бороздки задней
пары, будучи более глубокими и широкими на боках глабели, сливаются
на середине последней и становятся более мелкими и узкими. Задние
лопасти глабели на боках несколько сужаются, а к середине слегка
расширяются.
Передняя пара борозд глабели, начинаясь в некотором удалении от
ее боков, проявляется в форме резких и широких полулунных ямок, еле
заметно сливающихся на середине глабели.
Широкий лимб состоит из слегка вогнутого предглабелярного поля,,
по середине которого, проникая непосредственно к переднему концу гла-
бели, выступает чуть заметный бугорок; несколько вогнутая вперед, угло-
ватая фронтальная кайма почти не сохранилась. Сходящимися от зад-
них углов глабели, узкими, но глубокими спинными бороздками она
резко отделяется от плоских неподвижных щек, видимо расширяющихся
впереди и сливающихся здесь с предглабелярным полем. Затылочное
кольцо широкое, равномерное и отделено от глабели ровной и неглубокой
затылочной бороздкой.
Вся поверхность кранидия покрыта бугорками, которые более тонкие
и равномерные на глабели и более грубые и неодинаковой величины на
щеках и предглабелярной поле.
Отличается от типичного вида булавовидной формой глабели, несу-
щей только две пары глабелярных бороздок, а также присутствием не-
резко выраженного бугорка, расположенного на середине предглабеляр-
ного поля.
Распространение. Характерная форма для бабаковского комп-
лекса (СгП)3) кл. Бабак, северный склон Западного Саяна. Близкая
форма недавно обнаружена в полициатовом горизонте известняков в бас-
сейне рч. Б. Ерба — восточный склон Кузнецкого Алатау.
СЕМЕЙСТВО PARADOXIDAE EMMRICH
Род Paradoxldes В г о п g n i а г t
Крупные трилобиты с панцырем заметной выпуклости. Головной щит
большой, с небольшими подвижными щеками, с длинными щечными
остроконечиями и неширокой фронтальной каймой. Глабель к переднему
концу расширяется, доходит до фронтальной каймы, в задней части
имеет 2—3 поперечные бороздки. Глаза большие, более или менее узкие,
дугообразные, до глабели не доходят.
Лицевые швы расходящиеся: позади глаз более короткие, впереди
более длинные. Туловище длинное, чаще узкое, с многочисленными (от
семи до 23) сегментами, с узкой косой бороздкой на каждом из них.
ПО
Острые концы плоских плевральных сегментов загнуты назад. Хвостовой
щит очень маленький, не расчлененный на сегменты.
Типичный вид. Paradoxides tesslni В г о n g n.
Распространение. Средний кембрий.
Paradoxides suboelandlcus Poletaeva, sp. n.
Табл. XI, фиг. 8
Глабель имеет две бороздки. Предглабелярная площадка по бокам
довольно широкая. Против середины глабели она суживается и на очень
короткий промежуток прерывается совсем, так что глабель в середине
отделена от краевого валика только широкой бороздкой, ограничивающей
валик, но не упирается в него. У очень юных экземпляров предглабеляр-
ная площадка имеет еще большую ширину и не прерывается в середине.
Глазные крышки широкие, и длинные. Они незаметно сливаются с корот-
кими глазными валиками, а сзади почти достигают туловища и отде-
лены от него лишь узкими заднебоковыми лопастями. На туловище
сохранилось только 15 сегментов, но можно думать, что их было 16—17
В наружной половине плевр сегменты постепенно спускаются вниз,
а также постепенно разделяются и заостряются, образуя серповидные
концы. Плевральные бороздки кончаются на изгибе плевр.
От близкого вида — Paradoxides stblrlcus Lerm. описываемый вид
отличается соотношением глабели с лимбом и краевым валиком,
а именно: у Р. suboelandlcus глабель на большом протяжении сопри-
касается с лимбом, почти сливаясь с ним, а у Р. stblrlcus, кроме того,
передние ветви лицевых швов, вследствие сильного нависания лимба над
боками кранидия, образуют большие внутренние утлы.
Распространение. Санаштыкгольский комплекс (Ст2’)
рч. Мондыбаш, Горная Шорня.
Род Schagonaria Р о 1 е t а е v a, gen. и.
Мелкие и средней величины формы, близко родственные представите-
лям рода Paradoxides (см. стр. НО), с одной стороны, и роду Redllchlna
(см. стр. 112) —с другой. Описываемый род характеризуется длинной,
цилиндрической, умеренно вздутой и чуть расширяющейся вперед гла-
белью, равномерно закругленный конец которой не доходит до фронталь-
ного валика. Последний отделен от глабели узким треугольным высту-
пом. В задней половине глабель расчленена двумя парами глубоких
глабелярных бороздок. Бороздки задней пары несколько наклонены
назад и не доходят до середины глабели. Бороздки передней пары, бу-
дучи параллельны бороздкам задней пары, сливаясь на середине гла-
бели, отделяют на ней лопасти равной ширины как на боках, так и на
середине. Спинные борозды неглубокие, отчетливые, но вблизи передней
части глабели они остановятся несколько мельче.
Предглабелярная площадка небольшая, на боках широкая, а впереди
глабели сильно суживается. Краевой валик равномерно широкий, пло-
ский, широкоокруглый и несколько выпуклый на середине против гла-
бели. С внутренней стороны он ограничен очень тонкой и мелкой борозд-
кой. Между валиком и передним концом глабели располагается не на
всех образцах различимое узкое и небольшое вздутие, отходящее от
валика и иногда доходящее до переднего конца глабели. Неподвижные
щеки узкие, плоские.
Глазные крышки большие, широкие и, будучи ограничены с внутрен-
ней стороны мелкой бороздкой, почти вплотную подходят к глабели
в месте ее наибольшей ширины, затем косо опускаются назад и подхо-
дят вплотную к затылочному кольцу, в некотором удалении от глабели.
Лицевые швы впереди глаз расходятся, затем на уровне краевого валика
11!
поворачивают друг к другу и срезают на боках краевой валик. Позади
глаз направление лицевых швов неизвестно, но они, вероятно, здесь очень
короткие.
Затылочное кольцо широкое, невысокое и несколько сужается на
боках. Шипа или бугорка не имеется; затылочная борозда неширокая,
отчетливая. Подвижные щеки широкие, с широким плоским валиком,
переходящим в длинный щечный шип. Шип отходит от щеки немного
выше щечного угла. Туловище, состоящее более чем из восьми сегментов,
быстро сужается назад, второй и третий сегменты утолщены. Пигидий
неизвестен.
По строению глабели и, особенно, глазных крышек описанный род
несколько напоминает род Redllchlna (см. ниже), но у последнего глаз-
ные крышки примыкают к глабели, и она в переднем своем конце сильно
сужается. От рода Paradoxld.es род Schagonarla отличается строением
глабели, но имеет с ним сходство в строении неподвижных щек и отчасти
плевр туловищного отдела.
Типичный вид. Schagonarla tannuola Р о 1 е t. sp. n.
Распространение. Читинский комплекс (Cmi2). Тувинская
область.
Schagonarla tannuola Poletaeva, sp. n.
Табл. XII, фиг. 1
Родовой диагноз соответствует описанию вида.
Распространение: Чингинская формация (Cmi2). Правобе-
режье р. Улухема, в 4—5 км к северо-западу от Шагонара, Тувинская
область.
СЕМЕЙСТВО REDLICHIDAE POULSEN
Род Redllchlna Lermontova
Различных, но преимущественно крупных размеров формы, очень
близкие к роду Redlichla Cossmann. Почти прямоугольный кранидий
рода Redllchlna характеризуется выпуклой и ясно конической глабелью,
постепенно сужающейся к переднему краю, где она, будучи заметно при-
остренной, окаймляется широким предглабелярным полем. Последнее,
заметно расширяющееся на боках и сужающееся посередине, является
выпуклым и отделяется от фронтальной каймы глубокой и широкой крае-
вой бороздкой, покрытой ясно выраженной поперечной штриховкой.
Фронтальная кайма широкая, полукруглая, слегка сужается к бокам
и слабо наклонена в сторону предглабелярного поля.
На глабели заметны две или три пары глабелярных бороздок. Непод-
вижные щеки очень узкие. Большие дугообразные глаза спереди подхо-
дят почти вплотную к глабели, а сзади отделены от нее участком
неподвижных щек. Затылочное кольцо широкое и снабжено длинным
шипом. Совместно встреченная гипостома продолговатая, выпуклая,
с параллельными краями и парой обращенных назад зубчиков. Туло-
вищный и хвостовой щиты неизвестны.
Типичный вид. Redllchlna vologdlnl Lerm.
Распространение. Спи2. Восточный и Западный Саяны.
Redllchlna vologdlnl Lermontova
Табл. XII, фиг. 5
Характеристика вида отвечает приведенному выше диагнозу рода.
Распространение. Чингинский комплекс (Cmi2). д. Камешки
(Восточный Саян) и правый берег р. Улухема близ г. Шагонар (Тувин-
ская область).
112
Redllchina sayanica Sivov, sp. n.
Табл. XII, фиг. 2
Мелкие формы, обладающие выпуклой субконической глабелью со
слабо сохранившимися на ней весьма короткими глабелярными борозд-
ками, из которых только две задние пары более или менее удается раз-
личить при небольших увеличениях.
Узкая валикообразная фронтальная кайма, почти одинаковой ши-
рины на всем протяжении, отделена от предглабелярного поля глубокой
бороздкой. Предглабелярное поле, более выпуклое на боках, в середине
становится плоским и принимает треугольное очертание, с основанием,
направленным к фронтальной кайме.
Выпуклое затылочное кольцо, отделенное от глабели широкой и глу-
бокой затылочной бороздкой, вытянуто в длинный шип. Неподвижные
щеки и глазные крышки не сохранились. Этот вид обнаруживает замет-
ное сходство с типичным видом рода, отличаясь от него небольшими
размерами, присутствием двух пар глабелярных бороздок и особенно-
стями строения предглабелярного поля.
Распространение. Характерная форма чингинского комплекса
(Cmi2). Западный Саян, правобережье р. Улухема, в 4—5 км к северо-
западу от г. Шагонара; Восточный Саян, д. Камешки.
СЕМЕЙСТВО SOLENOPLEURIDAE ANGELIN
Род Solenopieura Angelin
В этот род, требующий пересмотра, здесь включаются преимуще-
ственно мелкие формы с эллиптическим спинным щитом, с большим
головным щитом и маленьким пигидием.
Округленноконическая, короткая, сильно выпуклая глабель ограни-
чена глубокими спинными бороздками. На глабели присутствуют 2—3
пары то резких, то слабо заметных бороздок. Неподвижные щеки выпук-
лые, умеренной ширины. Глаза маленькие, располагаются в средней
части кранидия и нередко глазными валиками соединяются, с глабелью.
Лицевые швы впереди глабели сходящиеся. Фронтальный край состоит
из относительно узкого предглабелярного поля и выпуклой валикообраз-
ной ^фронтальной каймы, у некоторых видов расширяющейся посередине
и несколько вздернутой кверху. Подвижные щеки небольшие. Поверх-
ность часто гранулирована.
Туловище с выпуклой осью и коленчато изогнутыми сегментами.
Плевры на внешней части оканчиваются тупо и рассечены узкой борозд-
кой.
Типичный вид. Solenopieura holometopa An g.
Распространение. Cm2, почти повсеместно; в Cmi, Cm3 и Si1
имеются очень близкие группы, отношение которых к роду Solenopieura
требует выяснения.
Solenopieura holometopa Angelin
Табл. XII, фиг. 4
Вид характеризуется яйцевидной глабелью с слабо развитыми глабе-
лярными бороздками. Предглабелярное поле посередине весьма узкое.
Бугорок на затылочном кольце отсутствует. Поверхность покрыта гру-
быми бугорками, особенно многочисленными и крупными на неподвиж-
ных щеках.
Распространение. Средний кембрий. Скандинавия.
Примечание. Описание и изображение типичного вида дается для удобства
сравнений с западносибирскими видами.
8 Зак. 20’4 113
Solenopleura djalnensis Lermontova var. altaica
Jegorova, var. n.
Табл. XII, фиг. 7
Кранидий умеренно выпуклый, с округленноконической выпуклой гла-
белью, с очень слабыми глабелярными бороздками. Неподвижные щеки
выпуклые, умеренно широкие, к переднему краю равномерно сужаются
и, сливаясь впереди глабели, образуют слегка выпуклое, весьма узкое
предглабелярное поле. Глазные крышки маленькие, тянутся почти па-
раллельно переднему краю кранидия.
Фронтальная кайма, слегка расширяющаяся посередине, выпуклая,
валикообразная, отделена резкой краевой бороздкой от предглабеляр-
ного поля. Затылочное кольцо выпуклое, с небольшим срединным бугор-
ком.
Часть сохранившегося туловища с прямыми плеврами, рассеченными
прямой, глубокой и широкой бороздкой. Описанная разновидность имеет
очень близкое сходство с видом Solenopleura djalnensis, известным
с Джайна (притока р. Лены). Но у ленской формы неподвижные щеки
и предглабелярное поле более широкие, глазные валики выражены
слабо, а поверхность кранидия с слабой грануляцией.
Распространение. Болыпеербинский (?) комплекс (Ст22).
Горный Алтай, в 2,5 км к юго-востоку от Шиловки.
Род Parasolenopleura Poletaeva, gen. n.
Трилобиты среднего и малого размеров. Характерной для рода
является глабель — длинная, узкая, слабо коническая, более или менее
усеченная спереди и округленная на переднебоковых углах. Глабель по
высоте несколько превышает широкие и значительно вздутые щеки
и слегка выпуклый лимб. На глабели имеются то более резкие, то слабо
выраженные короткие бороздки, иногда они могут отсутствовать вовсе.
Затылочное кольцо сохраняет на всем своем протяжении одинаковую
ширину и имеет маленький бугорок. Затылочная бороздка хорошо выра-
жена на всем протяжении. Фронтальный валик округлый, выпуклый,
иногда чуть расширяющийся на середине головного щита. Глазные
валики тонкие резкие.
Лицевые швы впереди глазных крышек параллельны друг другу или
слегка расходятся, а позади глазных крышек они расходятся довольно
сильно. Хвостовой щит поперечно вытянут, насчитывает четыре резко
выраженных сегмента на оси и плеврах. Исключение представляет по-
следний, задний, сегмент плевр, который очень мал и плохо заметен. На
первых трех сегментах плевр резко выражены широкие плевральные
бороздки, кроме того, присутствуют менее широкие межплевральные
бороздки. В наружной части хвостового щита плевры изгибаются назад
и сливаются с узкой ровной краевой каймой.
Типичный вид. Parasolenopleura subconsoclalis Pol.
Распространение. Cm2. Западная Сибирь (Горная Шория).
Parasolenopleura subconsoclalis Poletaeva, sp. n.
Табл. XII, фнг. 11
Диагнозом для данного вида является приведенный выше диагноз
рода. Видовым отличием Р. subconsoclalis служит весьма слабое
проявление глабелярных бороздок или полное отсутствие их.
Распространение. Санаштыкгольский комплекс (Ст/)',
рч. Мондыбаш, Горная Шория.
114
Parasolenopleura schortensls Poletaeva, sp. n.
Табл. XII, фиг. 10
Головные щиты среднего и малого размеров, трапецеидального
очертания. Глабель неширокая, усеченноконическая, с слегка округлен-
ными передними углами. На ней насчитываются две пары коротких,
довольно отчетливых косых борозд. Третья (передняя) пара борозд если
и имеется, то проявляется весьма слабо. Затылочное кольцо едва за-
метно расширяется в середине и снабжено маленьким бугорком. Заты-
лочная бороздка хорошо выражена. Спинные бороздки узкие, глубокие,
окружают глабель с боков и спереди, где они становятся менее глубо-
кими.
Неподвижные щеки широкие, равные по своей ширине глабели, но не
достигающие ее высоты. Перед глабелью они сливаются, образуя слабо
выпуклое неширокое предглабелярное поле. Фронтальный валик прямой,
выпуклый, довольно широкий, отделенный от предглабелярной площадки
глубокой, но узкой краевой бороздкой. Глазные крышки маленькие, рас-
положены немного позади середины длины глабели. Глазные валики от-
четливые, тонкие, идущие от глаз слегка косо к глабели и подходящие
к ней сзади переднебоковых углов.
Этот вид отличается от других представителей данного рода своим
прямым фронтальным краем, вследствие чего включается в род Paraso-
lenopleura пока условно.
Распространение. Санаштыкгольский комплекс (Сгщ1)
рч. Мондыбаш, Горная Шория.
Род Acrocephalites* W а 11 е г i u s
Чрезвычайно характерные и широко распространенные в кембрии За-
падной Сибири трилобиты, обладающие выпуклым кранидием, имеющим
форму шестиконечной звезды, концы которой лежат в разных плоско-
стях. По срединной продольной оси весь кранидий килеобразно припод-
нят. Глабель большая, сужающаяся кпереди с слабо притупленным
фронтальным краем. Две, реже три пары резких глабелярных бороздок
сильно наклонены назад. Затылочное кольцо резкое, заметно расширяю-
щееся посередине, где оно оттянуто в длинный острый шип.
Неподвижные щеки широкие, сильно приподнятые от глубоких спин-
ных бороздок к глазным крышкам и затем постепенно сливающиеся
с предглабелярным полем. Посередине последнего имеется шарообраз-
ное вздутие, не распространяющееся на выпуклую фронтальную кайму.
Она посередине утолщена и оттянута в длинный, направленный вперед
шип.
Глазные крышки резко изогнуты, часто стебельчаты, с глазными вали-
ками, расположенными почти посередине кранидия. Передние ветви лице-
вых швов, при взгляде сверху, в виде двух прямых линий сходятся
к фронтальному концу кранидия. Туловище состоит из 17—25 сегментов;
сегменты плевр с закругленными концами; интерплевральные бороздки
глубокие. Пигидий маленький. Поверхность панцыря гранулирована.
Типичный вид. Acrocephalites stenometopa A n g.
Распространение. Cm2 и Cm3. Западная Европа, Сибирь, Се-
верная Америка.
Acrocephalites regularls Lermontova
Табл. XIII, фиг. 13
Средней величины формы с несколько выпуклой на боках глабелью,
передний конец которой округлотреугольный. Глабель рассечена тремя
1 Некоторые авторы относят этот род к сем. Dokimocephalidae.
8* 115
парами косых глабелярных бороздок, из которых две задние пары наи-
более резкие и длинные. Спинные борозды глубоко вдавлены; от них
круто вздымаются неподвижные щеки, на которых ближе к заднему
краю кранидия располагаются глазные крышки.
Предглабелярное поле узкое, выпуклое, с небольшим вздутием по-
середине. Фронтальная борозда глубокая, широкая на боках и преры-
вающаяся на очень коротком расстоянии против глабели. Фронтальная
кайма, заметно расширяющаяся посередине, слегка здесь вздута и сзади
непосредственно соединяется с вздутием предглабелярного поля. Заты-
лочное кольцо ровное, узкое, отделено широкой и глубокой затылочной
бороздкой, краевые концы которой, плавно закругляясь вперед, пересе-
кают борозды заднебоковых лопастей неподвижных щек. У изображен-
ного на табл. XIII, фиг. 13 экземпляра фронтальный и затылочный шипы
не сохранились.
Поверхность кранидия покрыта правильными и выпуклыми бугор-
ками, более грубыми на лимбе и неподвижных щеках и более тонкими
на глабели.
Распространение. Характерная форма для салаирского ком-
плекса (Cm31). Салаир, гора Орлиная.
Acrocephalites elandiensis Jegorova, sp. n.
Табл. XII, фиг. 3
Вид близкий к Acrocephalites regularls; отличается от последнего неко-
торыми существенными деталями.
У Acr. elandiensis общая выпуклость кранидия выражена сильнее,
чем у Acr. regularls; на глабели заметны только две пары слабых глабе-
лярных бороздок. Неподвижные щеки относительно широкие и плавно
вздымаются от узких, но глубоких спинных бороздок к глазным крышкам.
Предглабелярное поле более широкое, почти без вздутия посередине;
фронтальная бороздка, более глубокая и широкая, прерывается посе-
редине на более значительном расстоянии. Фронтальная кайма, чуть
расширяющаяся посередине, продолжается в длинную и прямую иглу.
Затылочное кольцо более широкое, отделено более глубокой и широ-
кой затылочной бороздкой, внешние концы которой, под тупым углом
направленные вверх к спинным бороздкам, резко срезают заднебоко-
вые лопасти. Затылочная игла не сохранилась. Поверхность покрыта
редкими, но высокими бугорками, особенно обильными в фронтальной
части кранидия.
Распространение. Орлиногорский (?) комплекс (Сгп23). Район
д. Верхней Бланды (Горный Алтай).
Acrocephalites mlnimalis Sivov, sp. n.
Табл. XIII, фиг. 9
Очень небольшой экземпляр с недостаточно полно сохранившимся
кранидием. Усеченноконическая, с притупленным передним’краем гла-
бель рассечена тремя парами резких глабелярных бороздок, из которых
задняя пара очень длинная, косо направлена к глубокой затылочной
бороздке. Затылочное кольцо без шипа, но сильно расширено и утол-
щено посередине. Предглабелярная площадка выпуклая, субтреуголь-
ного очертания, без срединного бугровидного возвышения, причем ее
передний, суженный, край продолжается в толстую прямую иглу.
Валикообразная фронтальная кайма у основания иглы прерывается
и слегка отгибается вперед. Фронтальная краевая бороздка мелкая
и также прерывается у основания иглы. Неподвижные щеки плохо
сохранились, но заднебоковые лопасти хорошо выражены, длинные,
валикообразные и отделены глубокой заднебоковой бороздкой, расши-
ряющейся кнаружи.
116
Поверхность покрыта редкими, беспорядочно расположенными бу-
горками.
По отсутствию бугровидного возвышения на середине предглабе-
лярной площадки, треугольной ее форме и форме фронтальной иглы
описанный вид резко отличается от всех известных видов рода Асго-
cephalites.
Распространение. Характерная форма горношорского ком-
плекса (Cm32), Салаир, толсточихинская свита горы Орлиной.
Род Neoacrocephatites Sivov, gen. n.
Известен только кранидий, обнаруживающий некоторое сходство
с кранидием рода Acrocephalltes. Но у рода Neoacrocephalites глабель
почти трапецеидальная, килеватая, слабо сужающаяся впереди, с сильно
притупленным, почти прямолинейным передним краем. Три пары гла-
белярных бороздок хорошо заметны, причем две задние пары, будучи
оттянуты назад, вырезают на глабели грушевидной формы лопасти.
Неподвижные щеки широкие, сильно приподняты от узких и мелких
спинных бороздок к глазным крышкам и затем заметно ниспадают
к глабелярному полю. Последнее посередине широкое, вздутое и отде-
лено от глабели прямой, резко вдавленной бороздкой, которая на боках
сливается со спинными бороздками. Фронтальная кайма дугообразно изо
гнутая, шнуровидная, посередине несколько расширяется и отгибается
назад. Фронтальная бороздка узкая и глубокая; по бокам вздутия пред-
глабелярного поля, как бы на переднем продолжении спинных бороздок,
но отделяясь от них, образует глубокие, небольшие ямки.
Затылочное кольцо узкое, валикообразное, с небольшим шипом
посередине; оно отделено широкой затылочной бороздкой, мелкой и почти
прямой посередине и резко изгибающейся вперед в направлении к спин-
ным бороздкам; не сливаясь с последними, она на боках оканчивается
глубокими удлиненными ямками. Глазные крышки большие, но плохо
сохранившиеся; передние их концы продолжаются в почти прямые глаз-
ные валики, пересекающие неподвижные щеки около переднего края
глабели. Передние ветви лицевых швов короткие, сходящиеся; положе-
ние задних ветвей лицевых швов неизвестно; не сохранились и задне-
боковые лопасти кранидия.
Т и п и ч н Ы1 й вид. Neoacrocephalites iogensis S i v.
Распространение. Характерная форма горношорского ком-
плекса (Сш32). Горная Шория.
Neoacrocephalites togensis Sivov, sp. n.
Табл. XII, фиг. 8
Видовой диагноз соответствует описанию рода. Вид характерен для
горношорского комплекса (Ст32), обнаруженного в одноименной форма-
ции бассейна рч. Амзас (Эмзас) Горной Шории.
Род Abakanopleura Sivov, gen. n.
Мелкие трилобиты с выпуклой ширококонической глабелью, ограни-
ченной глубокими спинными бороздками. На глабели две пары вдавлен-
ных глабелярных бороздок, особенно резко' выраженных на ядрах, при-
чем задняя пара на боках на коротком расстоянии прямая, а затем резко
отгибается назад и тянется почти до затылочной бороздки; передняя
пара очень короткая и прямая.
Неподвижные щеки узкие (менее ’/4 ширины глабели в средней ее
части), выпуклые, круто вздымаются от спинных бороздок. Затылочное
кольцо более широкое посередине и отделяется глубокой, полого изогну-
той затылочной бороздкой.
117
Глазные крышки не сохранились, но они, вероятно, большие и про-
должались в глазные валики, остатки которых, пересекая неподвижные
щеки, примыкают почти к переднему широко округленному концу гла-
бели.
Фронтальная кайма выпуклая, относительно широкая, чуть наклонена
назад, отделена от неподвижных щек и узкой предглабелярной площад-
ки полого изогнутой фронтальной бороздкой, с выпуклостью, обращен-
ной назад. Поверхность кранидия, исключая щиты, с очень тонкой
грануляцией. Туловищный и хвостовой щиты неизвестны.
Описанный род очень близок к верхнекембрийскому американскому
роду Aposolenopleura (Raymond, 1937). Однако полного сравнения
этого американского рода с саяноалтайски?/ родом провести не удается
вследствие плохой сохранности у последнего заднего края кранидия.
Поэтому род Abakanopleura пока выделяется условно.
Род Abakanopleura своими узкими неподвижными щеками, узким
и плоским предглабелярным полем, относительно широкой, слегка на-
клоненной назад фронтальной каймой, а также широко округленным
передним концом глабели резко отличается от Solenopleura.
Типичный вид: Abakanopleura kenensls S i v.
Распространение. Cm.?. Западная Сибирь.
Abakanopleura kenensls S i v о v, sp. n.
Табл. XIII, фиг. 8
Диагноз вида полностью соответствует описанию рода. Данный вид
очень близок к американскому верхнекембрийскому виду Aposoleno-
pleura? brevlfrons Rasetti (Journal of Paleont., 1944, № 3, стр. 239,
табл. 36, фиг. 46), но у последнего неподвижные щеки субтреугольные
и более широкие, фронтальная кайма узкая, а поверхность кранидия
почти гладкая.
Распространение. Салаирский комплекс (Cm31). Салаир, гора
Орлиная. Очень сходный образец, но без сохранившейся фронтальной
каймы известен из арбатской «формации» (нижнее течение рч. Рудной
Кени, впадающей в Абакан слева, Западный Саян)
Род Mataninella Sivov, gen. n.
Средней величины трилобит; известен только по нескольким крани-
диям несовершенной сохранности.
Глабель, занимающая большую часть кранидия, сильно выпуклая,
шарообразная, чуть сужающаяся к переднему краю и ниспадающая
в этом направлении. При косом освещении заметна одна пара очень
слабых, но весьма характерных глабелярных бороздок. Они на коротком
расстоянии вначале протягиваются параллельно заднему краю, а затем
почти под прямым углом резко отгибаются назад и отсекают у основания
глабели почти прямоугольные лопасти. Затылочная бороздка глубокая,
резкая; затылочное кольцо не сохранилось. Спинные бороздки узкие, на
задней половине кранидия глубокие, а близ переднего края глабели
постепенно затухают и сливаются с резкой фронтальной бороздкой.
Неподвижные щеки субтреугольные, широкие в своем основании, но
затем около середины резко сужаются и полностью исчезают вблизи
переднего края глабели. Заднебоковые лопасти узкие, валикообразные
и отделены глубокими бороздками. Фронтальный проволокообразный
валик дугообразно изогнут, ровный, слегка отогнут вперед. Строение
передних ветвей лицевых швов неясно; задние ветви лицевых швов под
очень тупым углом тянутся к щечным углам. Сохранились лишь следы
глазных крышек, которые располагаются против середины глабели. По-
верхность покрыта редкими, но резкими бугорками.
118
Выпуклая шарообразная глабель и форма неподвижных щек сбли-
жают представителей описанного рода с верхнекембрийским американ-
ским родом Onchonotus (Raymond, 1924), но у последнего присутствует
узкое предглабелярное поле, и фронтальная кайма посередине в форме
треугольника оттянута вперед. Гораздо большее сходство род Mata.nl-
nella имеет с верхнекембрийским американским родом Matanlnla (Ra-
setti, 1944), однако у последнего глабель овальная, без глабелярных
бороздок, гладкая, неподвижные щеки гораздо уже в основании, а фрон-
тальная бороздка полого дугообразная. Впрочем, полное сравнение аме-
риканского рода Matanlnla с нашим родом, вследствие неполной сохран-
ности кранидия у последнего, сделать не представляется возможным.
Типичный вид. Matantnella escharoida Sivov.
Распространение. Горношорский комплекс (Ст32). Салаир.
Matanlnella escharoida Sivov, sp. n.
Табл. XII. фиг. 12
Описание рода полностью соответствует характеристике вида.
Распространение. Толсточихинская свита горы Орлиной,
Салаир (горношорский комплекс- -Ст32).
СЕМЕЙСТВО DORYPYGIDAE KOBAYASHI
Род Tabatopygellina Sivov, gen. n.
Средней величины трилобиты с весьма характерным субтреугольного
очертания кранидием. Выпуклая, грушевидная, без каких-либо следов
глабелярных бороздок глабель равномерно расширяется к переднему
краю, где она упирается в мелкую фронтальную бороздку. Спинные бо-
розды глубокие и относительно широкие в задней и средней части кра-
нидия, в передней его части становятся более мелкими и заканчиваются
здесь небольшими, но ясно заметными ямками.
На затылочном кольце, заметно расширяющемся посередине и посте-
пенно сужающемся к бокам, располагается отчетливо выраженный бу-
горок. Неподвижные щеки узкие и длинные. Глаза небольшие и, будучи
почти параллельны спинным бороздкам, располагаются в передней части
неподвижных щек. От глаз протягиваются косо направленные глазные
валики, которые примыкают к глабели чуть позади спинных ямок.
Неполно сохранившаяся фронтальная кайма представляет собой
узкий, острый валик, нависающий в виде острой кромки над глабелью.
Описанный род по общему строению кранидия заметно сходен с ро-
дами Bonnia Walcott, Bonniella R е s s е г и, особенно, Kootenla
Walcott. Но у последнего глабель удлиненноцилиндрическая, с слабо
заметными 2—3 парами глабелярных бороздок, а глаза относительно
длинные. От рода Bonnia род Tabatopygellina отличается величиной,
а также строением глазных крышек, которые у него являются весьма
небольшими и продолжаются в глазные валики. Кроме того, у рода
Tabatopygellina спинные бороздки в передней части заканчиваются ям-
ками, чего не наблюдается у рода Bonnia.
Что касается рода Bonniella, то он своими небольшими размерами,
присутствием грануляции и глабелярных бороздок на глабели, строе-
нием фронтальной каймы и другими признаками заметно отличается от
рода Tabatopygellina.
Типичный вид. Tabatopygellina babakovlensls Sivov.
Распространение. Бабаковский комплекс (Ст!3). Западный
Саян.
119
Tabatopygellina babakoviensis S i v о v, sp. n.
Табл. Х1Г, фиг. 9
Родовой диагноз соответствует описанию вида.
Распространение. Характерная для бабаковского комплекса1
форма (Cmi3). Северный склон Западного Саяна, кл. Бабак (система
рч. Рабата); верховья рч. Биджи восточного склона Кузнецкого Алатау.
Род Kootenia Walcott
Е. В. Лермонтова указывает следующие важнейшие признаки этого
рода. Продолговатоовальный спинной щит характеризуется большим
полукруглым головным щитом и почти равным ему по величине пиги-
дием. Глабель удлиненноцилиндрическая, нередко слегка пережатая на
боках, иногда незаметно расширяющаяся к притупленному переднему
концу, упирающемуся в глубокую фронтальную бороздку. Две-три пары
глабелярных бороздок развиты слабо. Глубокие спинные бороздки за-
канчиваются у передних концов едва заметными ямками. Неподвижные
щеки узкие и длинные. Глазные крышки относительно длинные, прямые,
параллельные спинным бороздкам. Фронтальная кайма шнуровидная,
иногда представляет собой приподнятую кромку,' которая перегибается
над серединой глабели. Поверхность головного щита нередко гранули-
рована.
Туловище состоит из семи сегментов с бороздчатыми плеврами. Пи-
гидий с большим выпуклым рахисом и более или менее слитыми сегмен-
тами плевр, полукруглого очертания. Интерплевральные бороздки на
пигидии слабые или отсутствуют, но плевральные бороздки отчетливые
и делят его бока на ряд плоских или округлых ребер. Пигидий окружен
плоской каймой, которая продолжается в ряд зубцов или шипов различ-
ной длины и формы. Ребра и борозды плевр на кайму не переходят.
Типичный вид. Kootenia dawsoni Walcott.
Распространение. Cm,—Cm2 Северной Америки; Cm21 Запад-
ного Саяна и Кузнецкого Алатау.
Kootenia abakanlca Poletaeva
Табл. ХШ, фиг. 3
Кранидий умеренно выпуклый, с цилиндрической глабелью, слегка
суженной ямками спинных бороздок у самого переднего ее конца. На
глабели три пары глабелярных бороздок, из которых передняя пара
очень слабо выражена. Глазные крышки длинные, слабо изогнутые, почти
параллельны спинным бороздкам. От переднего конца глазных крышек,
косо пересекая небольшой ширины неподвижные щеки, протягиваются
глазные валики, упирающиеся в глабель чуть позади спинных ямок.
Фронтальная кайма в виде узкого валика перегибается против сере-
дины переднего края глабели и отделяется от нее глубокой бороздкой.
Встречающийся совместно пигидий характеризуется отчетливо выражен-
ным рахисом и выпуклыми ребрами на боках. На ребрах слабый след
интерплевральных борозд. Плевральные бороздки глубокие. Кайма пло-
ская, с небольшими шипиками. Поверхность кранидия украшена тонкой
сеточкой.
Распространение. Вид характерен для санаштыкгольского
комплекса (Спь1). кл. Санаштыкгол, приток Абакана на юго-западной
окраине Западного Саяна.
Род Kooteniella Lermontova
По Е. Б. Лермонтовой, род Kooteniella, будучи близким к роду
Kootenia, отличается от последнего рядом характерных признаков.
120
У рода Kooteniella выпуклая яйцевидная глабель резко пережата
ямками спинных бороздок у переднего конца. Она то гладкая, то с очень
слабыми глабелярными бороздками. Неподвижные щеки узкие, выпук-
лые. Глазные крышки маленькие, слегка наклонные и располагаются
большей частью посередине. Затылочное кольцо нередко с шипом. Фрон-
тальная кайма нитевидная. Подвижные щеки плоские, иногда с недлин-
ными, отогнутыми назад щечными шипами.
Пигидий с выпуклым рахисом и сегментами плевр, образующими
округлые ребра, которые пересекают краевую кайму и непосредственно
переходят в краевые шипы. Кайма намечается углубленными в виде
ямок концами плевральных бороздок, не переходящих через кайму;
у ряда видов Kooteniella краевая кайма пигидия выражена очень слабо.
Типичный вид. Kootenlella slatkovskll (Schmidt).
Распространение. Cm21, с. Торгашино, северо-западная окраи-
на Восточного Саяна; восточный склон Кузнецкого Алатау; Западный
Саян.
Kootenlella slatkovskll (Schmidt)
Табл. XII, фиг. 6, табл. XIII, фиг. 14
Диагноз вида в точности соответствует описанию рода. Пигидий со-
стоит из пяти сегментов, кроме конечного, а краевых шипов — по пять
с каждой стороны. Туловищные сегменты прямые, с резким коленчатым
перегибом у наружных острых концов и глубокой продольной бороздкой.
Поверхность гладкая.
Распространение. Бабаковский комплекс (Ст!3). Характерная
форма для торгащинских известняков Восточного Саяна и горы, Долгий
Мыс восточного склона Кузнецкого Алатау и кл. Бабак в Западном
Саяне.
Род Kooteniellina Sivov, gen. n.
Различной величины, но преимущественно крупные трилобиты, зани-
мающие промежуточное положение между родами Kootenia Walcott
и Doryруge Dames.
Гладкая, сильно выпуклая, овально удлиненная и широкая глабель
рода Kooteniellina слегка пережата в передней части глубокими и яв-
ственно заметными ямками спинных борозд. Передний край глабели
округлен и упирается в шнуровидную фронтальную кайму, равномерно
обрамляющую кранидий. Неподвижные щеки узкие, слабо выпуклые.
Прямые неподвижные глазные крышки продолжаются в глазные валики,
которые косо пересекают неподвижные щеки и примыкают к глабели чуть
сзади ямок спинных бороздок. Затылочное кольцо, слабо расширяющееся
посередине и сужающееся на боках, имеет толстый, вздернутый кверху шип.
Полукруглый пигидий с большим выпуклым рахисом состоит из
4—5 сегментов, кроме конечного. Сегменты плевр более или менее сли-
тые, выражены в виде плоских ребер, пересеченных косыми бороздками,
подобными таковым у вида Kootenia abakanica (см. стр. 120). Пигидий
у описываемого рода окружен слегка вогнутой каймой, на которую
ребра и бороздки не переходят. Кайма продолжается в тонкие наружные
зубчики, часто неполностью сохраняющиеся.
Строением неподвижных щек, отсутствием глабелярных бороздок,
маленькими глазами, особенной формой глабели и характером фронталь-
ной каймы род Kooteniellina заметно отличается от рода Kootenia. С дру-
гой стороны, род Kooteniellina, будучи близким к роду Dorypyge, по
форме глабели, по строению других элементов кранидия, особенно пиги-
дия, резко от последнего рода отличается.
Типичный вид. Kooteniellina tubaenla S i v.
Распространение. Санаштыкгольский комплекс (Ст2'). Вос-
точный Саян.
121
Kootenlelllna tubaenla Sivov. sp. n.
Табл. XIII, фиг. 2
Характеристика рода соответствует описанию вида.
Распространение. Санаштыкгольский комплекс (Cm21). Вос-
точный Саян, окрестности д. Ирбы и Торгашино.
Род Olenoides Meek
Вопрос о наименовании рода, его систематике и объеме остается дис-
куссионным. Мы относим этот род к семейству Dorypygidae, а его объем
ограничиваем нижеследующим диагнозом.
Преимущественно крупные трилобиты с почти одинаковыми по вели-
чине головным и хвостовым щитами.
Глабель выпуклая, цилиндрическая, несколько расширяющаяся к пе-
реднему концу, где она, будучи округленной или угловатой, упирается
в фронтальную кайму. Глабель рассечена тремя парами глабелярных
бороздок, причем бороздки задней пары всегда заметно отгибаются на-
зад. Неподвижные щеки умеренной ширины, слабо выпуклые. Глазные
крышки небольшие, слегка изогнутые, узкие. Глазные валики круто
изогнутые, оканчиваются у ямок в передних частях спинных бороздок.
Передние ветви лицевых швов почти параллельны, задние расходятся
и направлены к щечным углам. Фронтальная кайма узкая, шнуровидная
почти на всем протяжении, ровная. Затылочное кольцо умеренно выпук-
лое, чуть расширяющееся посередине, где располагается небольшой шип.
Туловище состоит из семи сегментов, с плеврами, рассеченными ко-
сыми бороздками; внешние концы плевр имеют острые шипы, оттянутые
назад.
Пигидий с выпуклым, почти цилиндрическим рахисом, на котором от-
четливо выделяются 4—5 сегментов с бугорком на каждом из них. Сег-
менты плевр неслитые, причем рассечены ясными интерплевральными и
плевральными бороздками, из которых последние глубокие и косые (от-
клоняются назад). Краевая кайма пигидия слабо выражена, и по ее
краю располагаются различной длины и формы шипы.
Типичный вид. Olenoides nevadensls (Meek).
Распространение. Cmt и Cm2 Северной Америки и Сибири.
Olenoides convexus 1 Lermontova
Табл. XIII, фиг. 4
Крупные формы, почти во всех деталях тождественные типичному
виду. Полукруглый кранидий с выпуклой цилиндрической глабелью,
расчлененной тремя парами бороздок, из которых передняя и средняя
пары почти прямые, а задняя —• слабо наклонная. Глабель, постепенно
расширяясь в направлении к переднему краю, спереди округлена и упи-
рается в фронтальную кайму. Глазные крышки небольшие, с глазными
валиками, косо пересекающими неподвижные щеки и заканчивающи-
мися около ямок спинных бороздок.
Пигидий с широким и выпуклым рахисом, с четырьмя сохранивши-
мися сегментами, на которых видны слабо выраженные бугорки. Бока
пигидия резко расчленены, причем на сегментах отчетливо проявляются
интерплевральные и косые плевральные бороздки. По краю пигидия
располагаются четыре пары относительно коротких шипов.
Распространение. Большеербинский комплекс (Cm22). Вос-
точный склон Кузнецкого Алатау, бассейн рч. Б. Ерба.
1 Под этим же видовым названием в 1948 г. Разетти описал другой вид рода
Olenoides; согласно правилу приоритета, вид Разетти должен быть переименован.
А. Г Сивов.
122
Olenoides convexus Lermontova var. altalca
Jegorova, var. n.
Табл. XIII, фиг. 6
Эта разновидность отличается сужающейся в направлении к перед-
нему краю глабелью, рассеченной тремя парами глабелярных бороздок,
причем бороздки всех трех пар косо направлены назад; неподвижные
щеки более узкие. Кроме того, у Olenoides convexus vai. altalca по
внешнему краю пигидия, состоящего из четырех сегментов, распола-
гаются три пары коротких шипов.
Выделение этой разновидности является первым шагом к выяснению
изменчивости представителей вида Olenoides convexus-, лишь после изу-
чения изменчивости может быть окончательно решен вопрос о значении
и самостоятельности данной разновидности.
Распространение. Большеербинский (?) комплекс (Cm22).
Горный Алтай, близ д. Шиловки.
Род Dorypyge Dames
Род, близкий к Kootenia (см. стр. 120) и Olenoides (см. стр. 122).
Характерным признаком рода Dorypyge является боченкообразная или
яйцевидная, сильно выпуклая глабель, которая слегка сужается впереди
ямок в спинных бороздах близ ее переднего конца. На глабели слабо
выражены три пары коротких, прямых бороздок. Неподвижные щеки не-
широкие, слабо вздутые. Глазные крышки небольшие. Затылочное
кольцо, расширяющееся посередине и сужающееся на боках, нередко
оттянуто в шип. Фронтальная кайма очень узкая. Подвижные щеки с не-
длинными щечными шипами. Туловище состоит из восьми сегментов,
причем плевры рассечены глубокой и широкой прямой бороздкой.
Пигидий с выпуклым рахисом, состоящим из 4—5 сегментов. Сег-
менты на боках пигидия слитые, рассечены глубокими и широкими пле-
вральными бороздками, продолжаются в шипы, из которых два задних
сильно выделяются по своей длине и толщине. Интерплевральные бо-
розды очень слабые или совершенно отсутствуют.
Типичный вид. Dorypyge rlchthojeni Dam е s.
Распространение. Cm2 Европы и Азии.
Dorypyge rlchthofenl Dames
Табл. XIII, фиг. 11
Средней и небольшой величины формы с боченкообразной глабелью
и тремя парами коротких и очень слабых бороздок на ней. Пигидий
с длинными и толстыми заднебоковыми шипами. Скульптура в виде гру-
бых и редких бугорков.
Распространение. Форма характерна для среднего кембрия
(нижняя часть второй половины) Китая, Кореи и Манчжурии. Близкие
формы встречены в большеербинском комплексе (Cm22) восточного скло-
на Кузнецкого Алатау (рч. Б. Ерба).
Род Babakovia Sivov, gen. n.
Этот род характерен крупным, полукруглого очертания кранидием со
слабо изогнутым фронтальным краем и узкой шнуровидной фронтальной
каймой; последняя на боках несколько расширяется и отгибается назад.
Глабель выпуклая, гладкая, с наибольшей шириной посередине и не-
сколько сужающаяся к округленному переднему концу, упирающемуся
в мелкую, но явственно заметную фронтальную борозду. Спинные бо-
розды узкие, глубокие, с отчетливо заметными округленными ямками
у передних концов. Неподвижные щеки полутреугольные, резко сужаю-
123
щиеся впереди и отделенные от фронтальной каймы неглубокой борозд-
кой. Глазные крышки полулунные, небольшие, почти параллельные
спинным бороздам. Затылочное кольцо почти треугольного очертания,
с бугорком или шипом посередине. Оно отделяется от заметно сужен-
ного заднего конца глабели мелкой и узкой затылочной бороздкой. Пе-
редние ветви лицевых швов короткие, почти параллельные; задние ветви
длинные и направлены в щечные углы.
Ассоциирующие с кранидием хвостовые щиты обладают большим
выпуклым рахисом, состоящим из 5—7 сегментов. На сегментах плевр
видны как интерплевральные, так и плевральные бороздки; сегменты
продолжаются в длинные толстые шипы, постепенно отклоняющиеся на-
зад, причем из них два задних значительно длиннее и тоньше остальных.
Описанный род занимает промежуточное положение между родами
Kooteniellina Siv. (стр. 121) и Dorypyge Dames (см. выше). Однако
у рода Dorypyge на глабели присутствуют глабелярные бороздки, хотя
они и слабо заметны. Кроме того, род Dorypyge отличается от рода
Babakovia строением фронтального края кранидия и, особенно, харак-
тером пигидия. По этим же признакам род Babakovia отличается от
рода Kooteniellina.
Типичный вид. Babakovia dorypygaejormis S i v.
Распространение. Бабаковский и санаштыкгольский ком-
плексы (СгП|3—Cm21)- Западный Саян, восточный склон Кузнецкого Ала-
тау, Горная Шория, Салаир.
Babakovia dorypygaeformis Sivov, sp. n.
Табл. XVI, фиг. 4
Характеристика рода соответствует описанию вида.
Распространение. Бабаковский комплекс (Ст,3). рч. Биджа -
восточный склон Кузнецкого Алатау; рч. Табат — северный склон За-
падного Саяна.
СЕМЕЙСТВО ELL1PSOCEPHALIDAE MATTHEW
Род Chondragraulos Lermontova
Диагноз по Е. Б. Лермонтовой: «Очертания кранидия (без подвиж-
ных щек) округлые. Удлиненноконическая, умеренно выпуклая, спереди
притупленная глабель ограничена узкими, но глубоко врезанными дор-
зальными бороздками и обрамлена выпуклым дугообразным валиком,
образованным широкими неподвижными щеками, непосредственно пере-
ходящими во фронтальный лимб одинаковой с ним выпуклости, но с не-
большим вздутием посередине.
В скользящем свете иногда удается разглядеть границу фронтальной
каймы, а на щеках — изогнутые глазные валики, идущие от передних
углов глабели к маленьким глазам, лежащим близко к заднему краю
кранидия. Затылочное кольцо валикообразное, неширокое. На глабели
три пары слабых бороздок, редко заметных». К семейству Ellipsocepha-
lidae отнесен условно.
Типичный вид. Chondragraulos minussensis Lerm.
Распространение. Cm,—Cm2 Западной и Восточной Сибири.
Chondragraulos babakovensts Sivov, sp. n.
Табл. XIII, фиг. Г, табл. XIV, фиг. 15
Довольно крупная форма, по строению кранидия очень близкая к ти-
пичному виду рода. Однако у Chondragraulos babakovensis спинные бо-
роздки вблизи передних углов глабели и в ее передней части более мел-
кие. Вместе с тем, у описываемого вида поверхность кранидия вовсе ли-
124
шена грануляции, но зато у него гораздо отчетливее проявляются глаз-
ные валики, косо идущие от передних углов глабели к маленьким глаз-
ным крышкам, расположенным почти у заднего края кранидия; затылоч-
ная бороздка является также более глубокой.
Chondragraulos babakovensls проявляет значительное сходство с не-
которыми представителями американского рода Syspacephalus (Resser,
1936) из верхов нижнего кембрия, но отличается от них строением пред-
глабелярного поля и большими размерами.
Распространение. Характерная форма бабаковского ком-
плекса (Cmi3). Северный склон Западного Саяна, кл. Бабак, система
рч. Табат.
Chondragraulos (?) schorlensis Poletaeva, sp. n.
Табл. XIII, фиг. 7
Головной щит субквадратный. Глабель узкая, длинная, равномерно
выпуклая, цилиндрической формы, лишь очень слабо суживающаяся
спереди, где она усечена. Глабелярные борозды отсутствуют.
Распространение. Санаштыкгольский комплекс (Ст2').
рч. Мондыбаш, Горная. Шория.
Chondragraulos (?) anomalis Sivov, sp. п.
Табл. XIII, фиг. 5
Крупная форма, по строению переднего края кранидия близкая к пре-
дыдущему виду. Однако у Chondragraulos anomalis на длинной субкони-
ческой килеватой глабели заметны три пары глабелярных бороздок,
проявляющихся в виде коротких вдавленностей и сливающихся с глубо-
кими и широкими спинными бороздками. Они у переднего края глабели
заканчиваются ямками, соединенными одна с другой узкой, неглубокой
и угловатой бороздкой. Кроме того, у описываемого вида неподвижные
щеки более широкие и более выпуклые, а передний край глабели углова-
тый. Наконец, затылочное кольцо, несколько расширяющееся посередине,
отделяется узкой, расплывчатой затылочной бороздкой.
Распространение. Санаштыкгольский комплекс (cmg1). кл. Са-
наштыкгол в Западном Саяне. рч. М. Ирба в восточном Саяне, рч. Амзас
в Горной Шории.
Род Kingstonia Walcott
Типично верхнекембрийские мелкие трилобиты с выпуклым гладким
цефалоном полукруглого в плане очертания и без щечных углов. Глабель
почти квадратная, спереди округленная и отделена слабо выраженными
спинными бороздками, развитыми лишь у самого заднего края. Непо-
движные щеки умеренно широкие, выпуклые. Глазные крышки маленькие,
выдвинутые либо вперед, либо расположенные посередине длины крани-
дия, или позади его срединной линии. Передние ветви лицевых швов
параллельные, задние диагональные. Фронтальная кайма узкая и часто
как бы наложенная на панцырь.
Пигидий небольшой, почти треугольного или полукруглого очертания,
гладкий, без краевой каймы. Рахис выпуклый и короткий, узкий, очерчен
отчетливыми, неглубокими спинными бороздками.
Типичный вид. Kingstonia apton Wale.
Распространение. Верхний кембрий (низы) Северной Америки;
горношорский комплекс (Ст32) Западной Сибири.
Kingstonia globosa Sivov
Табл. XIV, фиг. 2
Относительно крупных размеров кранидии этого вида характеризуется
почти шаровидной формой, несколько вытянутой в продольном направле-
нии. Вдоль переднего края кранидия тянется сравнительно широкая, но
125
плоская фронтальная кайма, внутренний край которой обрамляется
проволокообразным возвышением; такое же возвышение, но более плос-
кое, располагается и по переднему краю каймы. Спинные бороздки почти
не развиты и проявляются в виде чуть заметных ямок в задней части
кранидия. Вследствие этого граница между глабелью и неподвижными
щеками почти на всем протяжении кранидия остается неясной. Затылоч-
ное кольцо очень узкое, отделенное едва заметной затылочной бороздкой.
Неподвижные щеки продолжаются в длинные, оттянутые назад заднебо-
ковые лопасти; заднебоковые бороздки почти не выражены.
Глазные крышки маленькие, сдвинутые почти к заднему краю и рас-
положены косо к спинным бороздкам. Передние ветви лицевых швов
длинные, прямые и параллельные; задние несколько короче, расходятся
и секут задний край под острым углом. Туловищный щит и пигидий
неизвестны.
Описанный вид по шаровидной форме кранидия, почти полному
отсутствию спинных бороздок, расположению глазных крышек вблизи
заднего края кранидия резко отличается от типичного вида (Klngstonla
aplon). Среди других представителей рода Klngstonla характеризуемый
сибирский вид обнаруживает ближайшее сходство с Klngstonla tumlda
(Resser, 1938, табл. 12, фиг. 11, стр. 83). Но последний вид обладает
более мелкими размерами кранидия, имеет более узкую фронтальную
кайму и короткие заднебоковые лопасти, но дорзальные бороздки у него
сильно развиты и относительно длинные.
Распространение. Одна из наиболее характерных форм горно-
шорского комплекса (Ст32). Горная Шория, бассейн рч. Эмзас.
Klngstonla salatrlca Sivov, sp. n.
Табл. XIV, фиг. 1
Относительно крупный кранидий полукруглого очертания, слегка
вытянутый в ширину. Глабель, заметно вздутая посередине, отделена
отчетливыми спинными бороздками, которые относительно широкие и глу-
бокие у заднего края кранидия и постепенно затухают только вблизи
переднего конца глабели. Фронтальная кайма узкая; она слегка расши-
ряется посередине кранидия и резко сужается в направлении к его бо-
кам. Глазные крышки очень маленькие и расположены около середины
кранидия.
Хвостовой щит, встреченный совместно с кранидием, выпуклый, глад-
кий, вытянут в ширину, с прямым передним краем. Рахис широкий, очень
маленький и заметно отграничивается с боков широкими, расплывчатыми
спинными бороздками.
Этот вид отличается от других известных представителей рода
Klngstonla большими размерами кранидия, длинными и хорошо выражен-
ными спинными бороздками и фронтальной каймой, резко сужающейся
на боках.
Распространение. Является одним из важных элементов горно-
шорского комплекса (Сгпз2). Толсточихинская свита горы Орлиной,
Салаир.
Klngstonla patchinenstformls Sivov, sp. n.
Klngstonla cf. paichinensis, С и в о в, 1940
Табл. XIV, фиг. 3
Рассматриваемый вид обнаруживает некоторое сходство с Klngstonla
salatrlca, но отличается от последней вытянутым в ширину кранидием,
окаймленным более узкой фронтальной каймой. Кроме того, у Klngstonla
salatrlca спинные бороздки в виде ямок намечаются лишь у заднего
края и на уровне глаз полностью исчезают.
126
Хвостовой щит с более длинным рахисом, отграниченный с боков
и сзади ясно выраженными спинными бороздками.
От Kingstonia paichinensis Kabayashi сибирский вид отличается
более задним положением глаз и коническим, менее широким рахисом
пигидия.
Распространение. Горношорский комплекс (Сш32). Рч. Эмзас
в Горной Шории.
СЕМЕЙСТВО OLEN1DAE BURMEISTER
Род Prohedinia Lermontova
Почти цилиндрическая или заметно сужающаяся вперед глабель
несет три-четыре пары бороздок, из которых особенно характерны бо-
роздки задней пары. Каждая из бороздок задней пары, начиная от
спинной, быстро разделяется на две ветви, из которых задняя направ-
лена косо назад, а передняя — горизонтально, поперек глабели. Перед-
ние ветви обеих бороздок иногда соединяются между собой, благодаря
чему на глабели ими и задними ветвями бороздок обособляется треуголь-
ное поле, обращенное вершиной назад (см. табл. XIV, фиг. 5).
Другие диагностические признаки этого рода: косые вдавленности на
неподвижных щеках, идущие от внутренних концов глазных валиков
к задним концам глазных крышек, и широкие вдавленности на предгла-
белярном поле у передних углов глабели. Как эти вдавленности, так
и обособляемые ими вздутия на неподвижных щеках и предглабеляр-
ном поле выражены в различной мере у разных видов: у одних они очень
отчетливы, у других обнаруживается лишь тенденция к образованию этих
вдавленностей и вздутий. Остальные признаки рода менее характерны
и приведены ниже в видовых описаниях.
Типичный вид. Prohedinia attenuata Lerm.
Распространение. Cm2 Западной и Восточной Сибири.
Prohedinia solida Poletaeva, sp. n.
Табл. XIV, фиг. 5
Глабель широкая, почти цилиндрическая, слегка сужающаяся вперед.
На переднем конце она прямо срезана, вследствие чего имеет спереди
угловатое очертание. Глабель умеренно и равномерно вздута, с тремя
парами глубоких бороздок. Бороздки задней пары соединяются со спин-
ными бороздками и обычным для рода Prohedinia образом разветвля-
ются. Задние ветви этих бороздок ограничивают на боках глабели пер-
вую пару резких, выпуклых глабелярных лопастей. Передние ветви
первой пары глабелярных бороздок очень мелкие, тонкие, горизонталь-
ные, соединенные друг с другом и не всегда с задними ветвями своих
борозд; обычно они имеют вид тонкой поперечной черточки, пересекаю-
щей среднюю часть глабели.
Глабелярные бороздки передней пары расположены против глазных
валиков и не соединяются со спинными бороздками, что является одной
из характерных особенностей данного вида.
Спинные борозды равномерно глубокие, узкие, слегка извилистые
(в связи с огибанием глабелярных лопастей). Затылочное кольцо высо-
кое, неширокое, чуть расширяется посередине. Неподвижные щеки ши-
рокие и неравномерно вздутые. От внутреннего конца глазного валика
проходит широкая косая вдавленность, которая делит щеку на две
неравные вздутые части: более значительную около глазных крышек
и меньшую в заднем внутреннем углу щеки, близ глабели. Глазные ва-
лики прямые, тонкие, высокие, шнуровидные, расположенные в некото-
ром углублении.
Лицевые швы впереди глаз сначала почти параллельные, затем
округло поворачивают друг к другу; задние ветви лицевых швов сильно
127
расходятся. Предглабелярное поле широкое; против передних углов
глабели на нем имеются широкие нечеткие впадины, которые делят поле
на три выпуклые части: две на боках и одну против глабели. Краевая
бороздка резко выражена, глубокая. Краевая кайма узкая, высокая,
четкая.
Распространение. Орлиногорский комплекс (Cm23). Алтай,
рч. Инырга (Чойский аймак Горно-Алтайской области).
Род Prohedinella Sivov, gen. n.
Мелкие трилобиты, внешне в известной мере напоминающие род
Prohedinia L е г т.
Кранидий выпуклый, почти квадратный, с слабо выгнутым вперед
передним краем. Цилиндрическая, широкая и сильно выпуклая глабель
имеет две явственно выраженные пары глабелярных бороздок, из ко-
торых задняя пара более длинная и косо направлена назад. Концы бо-
роздок, дугообразно преломляясь у заднего края глабели, отрезают от
нее грушевидной формы дольки. Передняя пара глабелярных бороздок
короткая и также отгибается назад.'Глабель окаймляется глубокими, но
узкими спинными бороздками, сливающимися у переднего края глабели,
которая там несколько притупленная. Затылочное кольцо треугольного
очертания, расширяющееся посередине и отсюда постепенно переходящее
в шип.
Неподвижные щеки умеренной ширины, постепенно сужающиеся
к фронтальному краю, где они незаметно спиваются с выпуклой, равно-
мерной ширины предглабелярной площадкой, отделенной от фронталь-
ного валика мелкой, но широкой фронтальной бороздкой. Передние
ветви лицевых швов длиннее задних, сходящиеся; задние ветви лицевых
швов короткие и, вероятно, направлены в щечные углы. Глазные
крышки, расположенные против передней пары глабелярных бороздок,
мелкие, бобовидные.
Типичный вид. Prohedinella erbiensis Sivov
Распространение. Cm2. Западная Сибирь.
Prohedinella erbiensis Sivov, sp. n.
Табл. XI, фиг. 6
Родовой диагноз полностью соответствует описанию этого вида.
Распространение. Большеербинский комплекс (Cm22).
Рч. Б. Ерба, восточный склон Кузнецкого Алатау.
СЕМЕЙСТВО CORYNEXOCHIDAE ANGELIN
Род Poliellina Poletaeva
Спинной щит эллиптически вытянутый, головной щит значительно
больше, чем пигидий. Выпуклая, слабо расширяющаяся в направлении
к переднему краю глабель упирается в валикообразную фронтальную
кайму и, будучи здесь ограничена почти прямой линией, имеет округло-
угловатую форму. Глабель рассечена тремя парами резких глабелярньих
бороздок; бороздки задней и передней пары соединяются посередине
глабели. Глазные крышки небольшие, угловато изломанные, распола-
гаются приблизительно параллельно глабели и вместе с соответственным
участком неподвижных щек являются сильно приподнятыми. От перед-
него конца глазных крышек, косо пересекая неподвижные щеки, отходят
глазные валики, примыкающие к глабели несколько впереди передней
пары глабелярных бороздок.
128
На спинных бороздках, достаточно глубоких в задней части крани-
дия и более мелких в передней, несколько впереди передних концов глаз-
ных валиков присутствуют небольшие, но глубокие ямки. Неподвижные
щеки умеренно широкие.
Туловище состоит из 12 коленчато изогнутых сегментов, плевральные
части которых несут глубокую плевральную бороздку. Она отходит от
переднего конца сегмента, примыкающего непосредственно к осевой
части туловища. На месте слияния с осевой частью туловища сегмент
несколько расширяется и приобретает характер треугольного утолщения.
Пигидий очень маленький, состоит из трех сегментов со слабыми
плевральными и интерплевральными бороздками.
Близок американский среднекембрийский род Poliella Walcott,
1916, и особенно Corynexodldes Rasetti, 1948.
Типичный вид. Pollelllna lermontovl Pol.
Распространение. Спь1. Западный Саян и Кузнецкий Алатау.
Pollelllna lermontovl Poletaeva
Табл. XIV, фиг. 4
Родовой диагноз соответствует описанию вида.
По общему характеру спинного щита и кранидия, по глубине глабе-
лярных бороздок Pollelllna lermontovl очень сходна с американским
видом Poliella balus. Главные различия между ними следующие.
У Р. lermontovl глабель спереди имеет угловатую форму, рассечена
тремя парами глабелярных бороздок, окаймлена сильно суженной на ее
середине фронтальной каймой. У Poliella balus глабель спереди широко
округлена, имеет четыре пары глабелярных бороздок и более равномерно
окаймлена широкой краевой каймой. Глаза у первой формы имеют
маленькую величину, туловище состоит из большого количества сегмен-
тов и более сложного строения в сравнении с Poliella balus.
Распространение. Характерная форма санаштыкгольского
комплекса (Cm21). Западный Саян, кл. Санаштыкгол, левый приток
р. Абакана; восточный склон Кузнецкого Алатау, рч. Б. Ерба, по Ло-
щенкову логу.
Род Poliellaspis Lermontova
Этот род, по указаниям Е. В. Лермонтовой, занимает промежуточное
положение между родами Poliella и Pollelllna.
Диагноз рода, по Е. В. Лермонтовой, таков: глабель у Poliellaspis
массивная, цилиндрически прямоугольная, с слегка вогнутыми боками
и угловатая в передней части. На глабели присутствует лишь одна пара
глабелярных бороздок, глубоко вдавленная, изогнутая назад и соеди-
няющаяся посередине глабели. Затылочное кольцо узкое, выпуклое, со
срединным возвышением. Глазные крышки почти прямые, продолжаю-
щиеся в глазные валики, косо пересекающие неподвижные щеки. Послед-
ние широкие и выпуклые. Фронтальная кайма валикообразная, узкая
посередине и утолщенная на боках.
Туловищные сегменты с резкими треугольными возвышениями
у внутреннего конца плевральных бороздок. Пигидий маленький (два
сегмента, кроме конечного), с толстым рахисом и сравнительно широкой,
ясно отграниченной каймой.
Типичный вид. Poliellaspis sayanicus (Pol.)
Распространение. Cm21. Западный Саян.
Poliellaspis sayanicus (Poletaeva)
Табл. XIV, фиг. 9
Диагноз рода соответствует описанию вида.
Вид Poliellaspis sayanicus близок к виду Pollelllna lermontovl, но
9 Зак. 2024 1 29
отличается от последней меньшей величиной, цилиндрически прямоуголь-
ной формой глабели, одной парой глубоких, слившихся глабелярных
бороздок и другими признаками.
Распространение. Характерный элемент санаштыкгольского
комплекса (Cm21). Кл. Санаштыкгол, приток р. Абакана в юго-запад-
ной части Западного Саяна.
СЕМЕЙСТВО ANOMOCARIDAE POULSEN
Род Anomocare Angelin
Объем этой сложной родовой группы, распространенной в Атланти-
ческой провинции, а также в Западной Сибири, остается далеко не
выясненным.
К роду Anomocare мы относим сравнительно крупные формы, обла-
дающие равными или почти равными по размерам головным и хвосто-
вым щитами.
Кранидий с большой и выпуклой, почти всегда килеватой глабелью,
рассеченной тремя парами бороздок. Фронтальный край состоит из
плоского, очень широкого предглабелярного поля и сравнительно узкой
фронтальной каймы, отогнутой кверху. Неподвижные щели умеренной
ширины, сильно выпуклые на боках. Подвижные щеки с короткими
щечными остроконечиями. Передние ветви лицевых швов прямые, расхо-
дящиеся. Глазные крышки длинные, дугообразные, начинаются на неко-
тором расстоянии от затылочного кольца и близко подходят к глабели
в передней ее части; здесь они продолжаются в глазные валики, косо
срезающие сильно суженные неподвижные щеки и примыкающие почти
к переднему концу глабели. Затылочное кольцо, более широкое посере-
дине и сужающееся к бокам, с небольшим шипом или бугорком. Туло-
вище состоит из 10 сегментов с плеврами, рассеченными продольной
бороздкой.
Пигидий с выпуклым рахисом, имеющим до семи сегментов, плевры
которых слитые, но более или менее с резкими плевральными бороздками,
нередко пересекающими краевой лимб. Последний неширокий, вогнутый.
Типичный вид. Anomocare laeve A n g.
Распространение. Верхи Cm2, почти повсеместное; в Западной
Сибири известен и в верхнем кембрии.
Anomocare laeve Angelin
Табл. XIV, фиг. 6; табл. XV, фиг. 2
Неподвижные щеки относительно широкие; затылочное кольцо с ши-
пом. Пигидий большой, с нешироким вогнутым лимбом, на который
ребра и плевральные бороздки не переходят.
Распространение. Характерная для Cm2 (зона Paradoxides
forchhammeri ). Скандинавия.
Близкие западносибирские формы описаны ниже.
Anomocare salalrensls 1 Lermontova
Табл. XV, фиг. 4
Характеристика вида по Е. В. Лермонтовой: кранидий очень близок
к генотипу, но на затылочном кольце бугорок, а не шип. Фронтальная
кайма посередине расширяется в виде обращенного назад треугольника.
Поверхность мелко гранулирована, на предглабелярном поле струйчата.
1 Anomocare salairensis Lermontova, 1940=Anomocare batchatiensis Sivov,
1940.
130
к этому диагнозу можно добавить и другие существенные особен-
ности, отличающие данный вид от Апотосаге laeve: у Апотосаге salal-
rensis глабель в верхней части заметно притуплена, а неподвижные
щеки гораздо уже, чем у скандинавской формы.
Распространение. Салаирский комплекс (Cm31). Салаир, гора
Орлиная.
Апотосаге (?) gravis Jegorova, sp. n.
Табл. XIV, фиг. 7
Средний величины кранидий с заметно выпуклой и килеватой гла-
белью, плавно округленной в передней части, в направлении к которой
она постепенно сужается. На глабели отчетливо выражены три пары
косо направленных бороздок, из которых задняя пара, будучи более глу-
бокой и резче наклоненной назад, отрезает треугольные выпуклые доль-
ки у основания глабели. На затылочном кольце, сильно расширяющемся
посередине и сужающемся на боках, имеется небольшой бугорок. Не-
подвижные щеки умеренной ширины.
Фронтальный край состоит из плоского, сильно суженного перед
глабелью предглабелярного поля и плоской ровной фронтальной каймы,
несколько отогнутой назад.
Глазные крышки длинные, узкие, дугообразно изогнутые. Передние
концы их близко подходят к глабели и соединяются с ней короткими
глазными валиками. Задние концы крышек также близко подходят к гла-
бели. Поверхность тонко гранулирована.
Данная форма по строению глазных крышек, неподвижных щек и гра-
нуляции поверхности кранидия близка к Апотосаге salalrensls, отли-
чаясь от этого вида формой глабели, небольшой шириной предглабеляр-
ного поля и иным строением затылочного кольца.
Распространение. Большеербинский (?) комплекс (Cm22).
Горный Алтай, 2,5 км к юго-востоку от д. Шиловки.
Род Anomocariopsis Sivov, gen. n.
Род, очень близкий к Апотосаге (см. выше), но отличается от него
рядом существенных признаков.
Кранидий у рода Anomocariopsis почти прямоугольный, вытянутый
в длину, с умеренно выпуклой длинной глабелью, широкоокругленной
впереди. На глабели две пары глубоких, коротких глабелярных бороз-
док, из которых задняя пара заметно оттянута назад. Неподвижные
щеки узкие, выпуклые и, заметно сужаясь впереди, сливаются с фрон-
тальным лимбом. Последний выпуклый и состоит из широкого предгла-
белярного поля субпрямоугольного очертания, вытянутого в ширину
и слегка вздутого посередине и по бокам.
Фронтальная кайма субтрапецоидальная, узкая, отделена от предгла-
белярного поля глубокой краевой бороздкой, прерывающейся против
середины глабели. Здесь развивается небольшое треугольного очертания
вздутие, основанием соединенное со вздутием середины предглабелярной
площадки. Глазные крышки скобообразные, умеренной длины, передними
и задними концами близко подходят к глабели; глазные валики отсутст-
вуют. Передние ветви лицевых швов длинные, сходящиеся; рассекая
фронтальный лимб, образуют выпуклость, обращенную наружу. Заты-
лочное кольцо шнуровидное, более широкое посередине; оно отделено
глубокой затылочной бороздкой. Заднебоковые лопасти не сохранились.
Туловищный и хвостовой щиты неизвестны.
Типичный вид. Anomocariopsis salalrensls (S i v.) (1940).
Распространение. Сгпз1. Салаирский комплекс. Западная Си-
бирь, гора Орлиная, Салаир.
9* 131
Anomocariopsis salairensis Sivov
Табл. XIV, фиг. 12
Характеристика вида и распространение полностью соответствуют
описанию рода.
Род Coosia Walcott
Удлиненноэллиптический спинной щит с средней ширины осевой
лопастью и почти одинаковыми по величине головным и хвостовым
щитами.
Головной щит полукруглый, с широким вогнуто-выпуклым фронталь-
ным лимбом, без краевой каймы. Глабель выпуклая, ширококониче-
ская, удлиненная, со слабыми или отсутствующими глабелярными бо-
роздками. Затылочное кольцо почти плоское и одинаковой ширины как
на боках, так и посередине; оно отделено мелкой и узкой затылочной
бороздкой. Неподвижные щеки узкие, около ширины глабели; сзади
они отделяются глубокими заднебоковыми бороздами от субтреуголь-
ного строения заднебоковых лопастей. Глазные крышки обычно неболь-
шие, но у некоторых форм длинные, дугообразно изогнутые, с глазными
валиками. Передние ветви лицевых швов длинные, сходящиеся; задние
более короткие и протягиваются почти параллельно заднему краю, круто
обрезая последний около щечных углов.
Туловище состоит из многочисленных сегментов (12) с плевральными
лопастями, слегка отогнутыми назад. Пигидий широкий, с коротким
выпуклым рахисом, на котором выделяются 2—3 слабых сегмента, кроме
конечного; бока пигидия гладкие.
Типичный вид. Coosia superba Wale.
Распространение. Cm3 Северной Америки; салаирский комп-
лекс (Cm31) Салаира.
Coosia radugini Sivov
Табл. XIV, фиг. 8
Вид, близкий к типичной форме, но отличается от нее более круп-
ными размерами кранидия, слабее выраженными спинными бороздками,
отделяющими глабель от более широких и плоских неподвижных щек.
Кроме того, у Coosia radugini полулунные глазные крышки своими пе-
редними концами близко подходят к переднему краю глабели. И, нако-
нец, почти у основания широкого полукруглого лимба протягивается
слабо изогнутая краевая бороздка, отделяющая как бы предглабеляр-
ное поле валикообразной формы.
Распространение. Салаирский комплекс (Сшз1). Гора Орли-
ная, Салаир.
Род Hanivella Sivov, gen. n.
Маленький кранидий с выпуклой боченкообразной, почти прямоуголь-
ной глабелью, несколько сужающейся к переднему краю, без глабеляр-
ных бороздок. Спинные бороздки неглубокие, но резкие. Неподвижные
щеки узкие, плоские и на уровне притупленного переднего края глабели
срезаются мелкой краевой бороздкой. Затылочное кольцо широкое,
отделено слабо выраженной затылочной бороздкой которая на боках
отгибается в переднем направлении.
Предглабелярное поле выпуклое, прямоугольное, вытянуто в ширину
и слегка наклонено назад, вследствие чего оно как бы нависает над гла-
белью. Глазные крышки длинные, дугообразные, достигают переднего
края глабели; задние концы глазных крышек оканчиваются на уровне
затылочной бороздки. Передние ветви лицевых швов короткие, субпарал-
132
дельные, прямые. Заднебоковые лопасти сохранились плохо; они выпук-
лые, постепенно сужаются наружу и отделены чуть заметными задне-
боковыми бороздками.
Описанный род своей почти прямоугольной глабелью и длинными
глазными крышками проявляет некоторое сходство с китайским верхне-
кембрийским родом Hanlvla (Kobayashi, 1933). Но у последнего на гла-
бели заметны две пары глабелярных бороздок, неподвижные щеки очень
широкие, предглабелярное поле отогнуто вперед, затылочное кольцо
с шипиком посередине.
Типичный вид. Hanivella primaeva Sivov.
Распространение. Горношорский комплекс (Сш32). Салаир.
Принадлежность этого рода к сем. Anomocaridae требует подтвер-
ждения.
Hanivella primaeva Sivov, sp. n.
Табл. XV, фиг. 1
Описание рода вполне соответствует характеристике вида.
Распространение. Горношорский комплекс (Ст32). Толсточи-
хинская свита горы Орлиной, Салаир.
СЕМЕЙСТВО DIKELOCEPHALLIDAE MILLER
Род Schoriella Sivov, gen. n.
Вытянутый в длину кранидий с заметно выпуклой, длинной, усечен-
ноконической глабелью, отделенной с боков узкими, но глубокими спин-
ными бороздками. На глабели три пары вдавленных глабелярных бороз-
док, из которых две передние очень короткие и чуть отгибаются назад.
Задняя пара более длинная, с ямками на боках, резко отгибается в нап-
равлении к глубокой и ровной затылочной бороздке. Затылочное кольцо
узкое, шнуровидное. Глазные крышки большие, дугообразные, передними
своими концами подходят очень близко к спинным бороздкам около
передней пары глабелярных бороздок глабели; задние концы крышек,
будучи несколько утолщенными, достигают затылочной бороздки. Паль-
пебральные бороздки глубокие, впереди и сзади сливаются со спинными
бороздками, вследствие чего плоские, умеренной ширины полулунные
неподвижные щеки исчезают несколько впереди середины глабели.
Фронтальный лимб большой и состоит из широкой плоско-выпуклой,
слегка ниспадающей вперед предглабелярной площадки и плоской, ров-
ной, полого дугообразной, более узкой фронтальной каймы, чуть накло-
ненной назад. Фронтальная краевая бороздка узкая, полого изогнутая,
с выпуклостью, направленной в сторону переднего края кранидия.
Передние ветви лицевых швов длинные, под углом 70э расходятся от
передних концов глазных крышек. Строение заднебоковых лопастей
и задних ветвей лицевых швов неизвестно.
Описанный род своим широким и большим лимбом, большими дуго-
образными глазными крышками, вдавленными глабелярными бороздками
на выпуклой глабели обнаруживает сходство с верхнекембрийсиим аме-
риканским родом Rlchardsonella в новом понимании объема этого рода
(Rasetti, 1944). Однако у Rlchardsonella предглабелярная площадка
и бороздка с вертикальной штриховкой, передние концы глазных крышек
примыкают непосредственно к глабели, несколько сзади притупленного
ее переднего конца, а передние ветви лицевых швов начинаются от спин-
ных бороздок, а не от глазных крышек, как у Schoriella.
Род Schoriella весьма близок также к роду Conokephallna Brogger,
известному из среднего и верхнего кембрия Зап. Европы. Но у рода
Conokephallna предглабелярная площадка неширокая, а равная ей по
ширине фронтальная кайма валикообразная. Неподвижные щеки впереди
133
постепенно сужаются и сливаются с предглабелярной площадкой, а не
срезаются пальпебральной бороздкой, как у рода Schoriella.
Типичный вид. Schoriella schorlca S iv., sp. n.
Распространение. Горношорский комплекс (Cm32). Горная
Шор и я.
Schortella schorlca Sivov, sp. n.
Табл. XTV, фиг. 10
Диагноз рода полностью соответствует описанию вида. Кранидий
вида очень близок к Conokephallna olenorus West. var. aspinosa Ler-
montova из верхнекембсийских отложений хребта Караулах северо-
восточной части Сибири. Однако хараулахская форма очень маленькая,
поичем предглабелярная площадка кранидия очень узкая, а пальпе-
бральные бороздки, отделяющие большие дугообразные глазные крышки,
не срезают передние узкие части неподвижных щек и не сливаются со
спинными бороздками.
Распространение. Характерный элемент горношорского комп-
лекса (Ст32). Горная Шория, бассейн рч. Амзас (Эмзас).
СЕМЕЙСТВО ASAPHISC1DAE RAYMOND
Род Pesaia Walcott et Ress er
Авторы, установившие этот род1, коротко останавливаются на его
диагностических признаках, подчеркивая, что по строению кранидия он
сходен с родом Euloma, но отличается большей шириной кранидия и ха-
рактером фронтального лимба. Более пезкие особенности имеются
в строении пигидия, отнесенного к роду Pesaia. Другие авторы подчер-
кивают сходство рода Pesaia с родом Elvinia Walcott2.
Типичный в ид. Pesaia exsculpta Wale, et R e s s.
Распространение. Новая Земля, пеоеходные горизонты от
верхнего кембрия к нижнему силуру; Салаир (Сгпз1, салаирский комп-
лекс).
Pesaia avia Sivov, sp. n.
Табл. XIV, фиг. 11
Описание этого вида дается подробно с целью до некоторой степени
уточнить характеристику рода.
Кранидий крупный, субпрямоугольный, умеренно выпуклый, вытяну-
тый в длину. Глабель ширококоническая, короткая, слегка притупленная
впереди, с одной задней парой глабелярных бороздок, сильно оттянутых
назад и отделяющих на заднебоковых углах глабели почковидные
лопасти. Спинные бороздки широкие, глубокие, причем передние концы
их оканчиваются небольшими округленными ямками. От них, отграничи-
вая передний конец глабели от лимба, проходит неглубокая, но широкая
бороздка.
Неподвижные щеки умеренно широкие (’/з ширины глабели в сред-
ней ее части), выпуклые, впереди плавно сливаются с предглабелярной
площадкой, а сзади, несколько расширяясь, отделяются широкой и глу-
бокой бороздкой от заднебоковых лопастей, внутренние концы которых
срезаются боковыми концами затылочного кольца. Последнее, широкое
и плоское посередине, резко сужается на боках; затылочная бороздка
широкая, полого изогнутая, более глубокая и вдавленная в заднебоко-
вых углах и более мелкая посередине. 1 2
1 Walcott and Resser, 1924.
2 Howell and Cr. Lochman, 1939.
134
Фронтальный лимб широкий, выпуклый и состоит из равных по ши-
рине предглабелярного поля и фронтальной каймы, отделенных широкой
полого изогнутой бороздкой. Фронтальная кайма полого дугообразная,
более широкая посередине, слегка оттянута назад и чуть наклонена
вперед.
Передние ветви лицевых швов длинные, субпараллельные, сигмооб-
разно изгибаются вокруг предглабелярной площадки и фронтальной
каймы: рассекая переднебоковые концы последней, становятся сходящи-
мися. Глазные крышки умеренной ширины, полулунные, располагаются
почти против середины глабели и продолжаются в косо направленные
глазные валики.
Пигидий, ассоциирующий с кранидием, полукруглого очертания,
с ясно выраженным широким, выпуклым и коническим рахисом, с тремя
заметными сегментами. Плевры с прямыми слитными ребрышками, не
переходящими на выпуклую и узкую краевую кайму.
Pesala avia заметно отличается от типичного вида, у которого непо-
движные щеки более широкие, предглабелярная площадка ужей углова-
тая, а пигидий с рахисом, достигающим заднего края каймы.
Распространение. Весьма характерная форма для салаирского
комплекса (Сгпз1). Салаир, гора Орлиная.
Род Kaniniella Sivov, gen. n.
Этот род близок к родам Pesala (см. выше) и Orlovlella (см.
стр. 136).
Кранидий субквадратный, умеренно выпуклый. Глабель с почти
параллельными боками, слабо килеватая, слегка сужающаяся вперед, но
с широкоокругленяым передним концом.
Глабелярные бороздки отсутствуют. Спинные бороздки неглубокие, но
четкие. Затылочное кольцо широкое, ровное и отделено слабо выражен-
ной затылочной бороздкой. Неподвижные щеки широкие, умеренно вы-
пуклые и впереди сливаются с фронтальным лимбом. Последний состоит
из широкого выпукло-вогнутого предглабелярного поля, отделенного
тонкой, но четкой краевой бороздкой от узкой, почти прямолинейной
краевой каймы, отогнутой кверху.
Глазные крышки умеренной величины, полулунного очертания, рас-
положены параллельно спинным бороздкам почти посередине кранидия.
Передние ветви лицевых швов субпапаллельные, длинные, слабо расхо-
дящиеся; они, окаймляя фронтальный лимб, сигмообразно изгибаются
вокруг его отдельных элементов. Задние ветви лицевых швов короткие
и от задних концов глазных крышек почти под прямым углом направ-
ляются наружу, разрезая узкие заднебоковые лопасти.
Туловищный и хвостовой шиты неизвестны.
Типичный вид. Kaniniella alata (Siv.).
Распространение. Сгпз1. Западная Сибирь.
Kaniniella alata (Sivov)
Табл. XIV, фиг. 14
Характеристика вида соответствует описанию рода. Kaniniella alata
по общему очертанию кранидия, широким неподвижным щекам, отсут-
ствию глабелярных бороздок на глабели имеет ближайшее сходство
с Kanlnla lata (Walcott and Resser, 1924), известной из озаокских слоев
Новой Земли. Однако у новоземельского вида глабель субконическая,
более выпуклая, отделена более резкими спинными бороздками и обла-
дает менее широким предглабелярным полем и ясно расходящимися,
относительно короткими передними ветвями лицевых швов. Глазные
крышки расположены косо к спинным бороздкам и соединены с гла-
белью глазными валиками.
135
Вместе с тем Kanlnlella alata по форме лимба п глабели и общему
очертанию кранидия обнаруживает некоторые черты сходства с Anomo-
care salairensis (см, стр. 130), отличаясь от последнего вида отсутствием
глабелярных бороздок, более широкими неподвижными щеками, отно-
сительно мелкими глазными крышками и формой передних ветвей лице-
вых швов.
Распространение. Характерная форма салаирского комплекса
(Сгпз1). Салаир, гора Орлиная.
Род Orlovleila Lermontova (nom. nud.)
В свое время Е. В. Лермонтова среди кембрийских трилобитов Сала-
ира установила род Orlovleila, но не привела описания и иллюстраций
его представителей. А. Г Сивов (1940), установив в том же пункте не-
сколько видов, принадлежащих к упомянутому роду, ограничивается
очень краткими характеристиками некоторых неполных экземпляров.
Позднее были найдены образцы хорошей сохранности, принадлежность
которых к роду Orlovleila не вызывает сомнений и которые позволяют
дать описание этого рода.
Различных размеров эллиптического очертания спинной щит рода
Orlovleila характеризуется очень большим головным щитом и крайне
небольшим пигидием.
Головной щит полукруглый, вытянутый в ширину, обрамленный вдоль
наружного края валикообразной выпуклой фронтальной каймой, не-
сколько расширенной и вздернутой в ее средней части. Глабель субко-
ническая. сильно выпуклая, либо с притупленным, либо приостренно-
окпугленным передним концом. Глабелярные бороздки отсутствуют.
Спинные бороздки, ограничивающие глабель с боков, резкие и глубокие,
но близ переднего конца глабели они становятся очень мелкими, слабо
выраженными.
Неподвижные щеки очень узкие (’Л ширины глабели), ио выпуклые
и впереди постепенно вливаются в предглабелярную площадку. Она
плоско-выпуклая, заметно ниспадает к слегка нависающему назад фрон-
тальному валику, от которого отграничивается ясной, слабо изогнутой
краевой бороздкой. Затылочное кольцо более широкое посередине и не-
сколько отогнутое вперед, к бокам заметно сужается, где и срезает
заднебоковые лопасти. Затылочная борозда посередине мелкая, но
широкая, на боках становится глубокой и при слиянии со спинными
бороздками иногда заканчивается небольшими ямками.
Подвижные щеки умеренной ширины, выпуклые, полукруглые, без щеч-
ных остпокочечий. Передние ветви лицевых швов вначале под тупым углом
расходятся наружу, но при пересечении фронтального валика принимают
сходящееся направление. Задние ветви лицевых швов диагональные,
более копоткие и режут щечные углы. Заднебоковые лопасти узкие,
проволоковидные и отграничены краевыми бороздками, расширяющимися
к бокам. Глазные крышки небольшие, полулунные, располагаются посе-
редине кранидия и иногда сопровождаются глазными валиками.
Туловищный шит состоит не менее, чем из 12 сегментов, с очень
выпуклой осью, горизонтальными плеврами и округленными концами.
Сегменты туловища и плевральных частей рассечены мелкими плевраль-
ными и более глубокими интерплевральными бороздками.
Пигидий очень маленький, с выпуклой короткой осью (1—2 сегмента)
и неслитыми наружными концами плевральных сегментов.
Головной шит Orlovleila по общим очертаниям кранидия и, особенно,
субконической глабели с отсутствующими на ней глабелявными борозд-
ками, обнаруживает весьма близкое сходство с подом Orlovla (Walcott
et Resser, 1924), известным из озаркских слоев Новой Земли. Но у пос-
леднего рода неподвижные щеки и предглабелярная площадка более
136
широкие, а фронтальная кайма более узкая и не отгибается назад, как
у рода Orlovlella. Кроме того, у Orlovta затылочная бороздка к бокам
затухает, отсутствуют глазные валики, а пигидий большой, обрамленный
плоской краевой каймой.
Типичны йвид. Orlovlella typlca S i v.
Распространение. Салаирский комплекс (Спь1), орлиногор-
ский комплекс (Сш23). Сибирь.
Orlovlella typlca Sivov, sp. n.
Табл. XV, фиг. 3
Небольшой трилобит с конической, округленной впереди глабелью,
без глазных валиков и относительно выпуклой, слегка ниспадающей
вперед предглабелярной площадкой.
Распространение. Салаирский комплекс (Cm31). Салаир, гора
Орлиная.
Orlovlella conlca Sivov
Табл. XIV, фиг. 13
Форма, близкая к типичному виду, но с более крупным кранидием,
с заметно килеватой, конической, приостренной впереди глабелью. Пред-
глабелярная площадка ясно выпуклая и слегка ниспадает вперед.
Распространение. Салаирский комплекс (Сгпз1). Салаир, гора
Орлиная.
Orlovlella truncata Sivov
Табл. XVI, фиг. 3
Вид, близкий к предыдущему, нос субконической, сильно притупленной
впереди глабелью. Предглабелярная площадка резче ниспадает к перед
нему краю. Присутствуют выпуклые, короткие глазные валики.
’ Вид встречается совместно с предыдущей формой.
Orlovlella elandiensis J е g о г о v a, sp. п.
Табл. XII, фиг. 14
Кранидий субтреугольных очертаний, с плавно закругленным перед-
ним краем. Общая выпуклость глабели небольшая и несколько меняется
у различных экземпляров. Глабель конической формы с округленным
передним концом. Поперечных борозд на глабели не видно. Фронталь-
ный лимб неширокий. Спинные борозды глубокие и резкие по бокам
глабели. Затылочное кольцо и затылочная борозда хорошо выражены.
Неподвижные щеки в передней части узкие, позади глаз сильно
расширяются. Глазные крышки небольшие, полулунной формы. Лицевые
швы впереди глаз изгибаются дугообразно, сначала слегка наружу, за-
тем внутрь; позади глаз они сильно расходятся.
Хвостовой щит, предположительно отнесенный к этому виду, неболь-
шого размера, вытянутый в поперечном направлении (длина почти вдвое
меньше ширины), заметно выпуклый. Ось пигидия короткая, почти ци-
линдрическая; число колец на оси небольшое. Вдоль края хвостового
щита тянется неширокая, плоская или слабо выпуклая кайма, сужаю-
щаяся слегка посередине, против заднего конца оси.
Распространение. Орлиногорский (?) комплекс (Cm23). Алтай,
верховья рч. Еланды.
Род Metisaspina Sivov, gen. n.
Неполный крупный, сильно выпуклый кранидий этого рода характе-
ризуется длинной, конической, килеватой глабелью, отграниченной
С боков широкими и глубокими спинными бороздками. Последние на
137
переднебоковых углах заканчиваются сильно вдавленными и крупными
ямками, вследствие чего передний край глабели сильно пережат и ее
передний конец приобретает угловатоконическую форму. Имеется одна
пара глабелярных бороздок; бороздки косо протягиваются назад и внутрь
глабели, а затем под тупым и резким углом поворачивают в направлении
к ямкам затылочной борозды. В связи с этим отграниченные глабеляр-
ными бороздками лопасти глабели приобретают характерную треуголь-
ную форму с вершинами, обращенными к фронтальному краю кранидия.
Неподвижные щеки субтреугольной формы, выпуклые, с шириной
у основания, почти равной ширине заднего края глабели. Кпереди они
резко сужаются и, достигая половины ширины глабели у ее переднего
края, постепенно вливаются в широкий фронтальный лимб. Последний
состоит из выпуклой, узкой, вытянутой в ширину угловатой предглабе-
лярной площадки и несколько более широкой фронтальной каймы. Она
выпуклая, широкодугообразная, заметно расширяется посередине, слегка
отгибается вперед и отделяется от предглабелярной площадки широкой,
глубокой, но полого изогнутой бороздкой, которая против середины гла-
бели становится очень мелкой.
Затылочное кольцо сохранилось неполностью; его боковые узкие
валикообразные концы оттянуты вперед и срезают узкие валикообразяые
заднебоковые лопасти, отделенные широкими и глубокими заднебоко-
выми бороздками.
Затылочная бороздка широкая и мелкая посередине, а на боках более
узкая, сильно вдавленная. Передние ветви лицевых швов длинные, сиг-
мообразно изгибаются вокруг предглабелярной площадки и фронтальной
каймы, а рассекая последнюю у ее переднего края, становятся слабо
сходящимися. Глазные крышки не сохранились.
Описанный род глубокими спинными бороздками, заканчивающимися
в переднебоковых углах глубокими ямками, а также формой глабели
с пережатым передним концом и валикообразными заднебоковыми ло-
пастями, отделенными широкими и глубокими спинными бороздками,
близко напоминает американский верхнекембрийский род Metisaspls
(Rasetti, 1946). Но у последнего неподвижные щеки одинаковой ширины
по всей их длине, предглабелярная площадка и равная ей фронтальная
кайма с одинаковой выпуклостью и разграничены почти прямой и глу-
бокой бороздкой. Поверхность кранидия рода Metisaspls, за исключением
бороздок, покрыта туберкулами и грануляцией.
Mettsasplna по общим очертаниям кранидия и особенно по форме
фронтального лимба обнаруживает некоторые черты сходства с родом
Pesala (см. стр. 134). Но у последнего глабель ширококоническая с мин-
далевидными лопастями на ее задних боках, неподвижные щеки почти
одинаковой ширины по всей их длине, а ямки в переднебоковых углах
спинных бороздок очень мелкие. Кроме того, у Pesala имеются глазные
валики, а предглабелярная площадка менее выпуклая и не угловатая,
как у Mettsasplna.
Типичный вид. Mettsasplna anomala Sivov.
Распространение. Салаирский комплекс (Ст3’). Салаир.
Mettsasplna anomala Sivov, sp. n.
Табл. XV, фиг. 6
Характеристика рода полностью соответствует описанию вида.
Распространение. Характерный компонент салаирского комп-
лекса (Сгпз1)- Салаир, гора Орлиная.
СЕМЕЙСТВО ILLAENIDAE HAWLE ЕТ CORDA
К этому широко известному семейству А. Г. Сивовым условно отно-
сится род Koldlnlella, характеризующийся сходством в строении крани-
дия с родом Illaenus (см. стр. 174).
138
Род Koldiniella Lermontova (nom. nud.)
Среди кембрийских трилобитов Саяно-Алтайской области Е. В. Лер-
монтова установила новый род — Koldiniella, близкий к роду Koldlnla
Walcott and Resser, 1924), известному из озаркских слоев Новой Земли.
Однако диагностические признаки и изображения представителей рода
Koldiniella Е. В. Лермонтовой приведены не были. Позднее А. Г. Сивов
(1940), приведя изображения некоторых представителей рассматривае-
мого рода, коротко останавливался на его характеристике. Поэтому не-
обходимо подробно охарактеризовать этот важный и широко распрост-
раненный род.
Род Koldiniella объединяет различные по величине формы, у которых
кранидий сильно выпуклый, эллиптического или шаровидного очертания,
гладкий, рассеченный слабо развитыми спинными бороздками. Фронталь-
ный край кранидия слегка изогнут и окаймлен относительно широкой,
слабо выпуклой фронтальной каймой, которая образует с поверхностью
кранидия отчетливый тупой угол и отделяется от глабели тонкой, но чет-
кой краевой бороздкой. Глабель гладкая, занимает почти всю ширину
кранидия и отделена в задней своей части слабо выраженными спин-
ными бороздками, исчезающими в направлении к переднему краю.
Неподвижные щеки выпуклые и в той части, где они отделены от
глабели спинными бороздками, на уровне глазных крышек, очень узкие.
Позади глазных крышек неподвижные щеки несколько расширяются, обра-
зуя небольшие, узенькие заднебоковые лопасти. Затылочное кольцо уз-
кое и отделено едва заметной затылочной бороздкой. Глазные крышки
маленькие, узкие и располагаются ближе к заднему краю кранидия. Пе-
редние ветви лицевых швов почти прямые, в передней части кранидия
немного расходятся и слегка изгибаются внутрь в пределах краевой
каймы. Задние ветви лицевых швов очень короткие, сильно расходятся
и с задним краем образуют острый угол.
Пигидий полукруглого очертания, выпуклый, вытянутый в поперечном
направлении, с почти прямым передним краем и с отчетливо выражен-
ным, длинным и не сегментированным рахисом. Рахис немного не дости-
гает заднего края пигидия, обрамленного слабо выпуклой краевой кай-
мой. Поверхность последней не выдается над поверхностью боков пиги-
дия, представляя как бы их продолжение.
Типичный вид. Koldiniella mltella Sivov.
Распространение. Низы Cm3 (салаирский комплекс).
Koldiniella mltella S i v о v
Табл. XV, фиг. 7
Крупный, выпуклый и вытянутый в ширину кранидий, со спинными
бороздками, выраженными только в задней его части. Небольшие и уз-
кие глазные крышки располагаются ближе к середине кранидия. Пиги-
дий полуклуглый с широким рахисом, отделенным вполне заметными
дорзальными бороздками.
По общей форме кранидия и хвостового щита Koldiniella mltella
очень близка к типичному виду рода Koldlnla *, отличаясь от последнего
более широкой и уплощенной фронтальной каймой, короткими и чуть
заметными дорзальными бороздками, быстро затухающими к переднему
краю, а также большей шириной глабели у ее основания.
Форма, характерная для салаирского комплекса (Cm31, Салаир, гора
Орлиная).
Koldiniella bulba Sivov
Табл. XV, фиг. 8
Вид очень близкий к вышеописанному, но отличается от него меньши-
ми размерами кранидия, обладающего удлиненноокруглой формой, а также
1 Koldlnla typa Wale, et R e s s. из озаркских отложений Новой Земли.
бет
более отчетливыми спинными бороздками, протягивающимися почти до
середины кранидия. Затылочная бороздка отсутствует. Хвостовой щит
с более узким рахисом, ограниченным менее выраженными спинными
бороздками.
Вид является характерной формой салаирского комплекса (Cm31, Са-
лаир, гора Орлиная).
СЕМЕЙСТВО LEIOSTEG1DAE BRADLEY
В это семейство мы» пока условно включаем новый род Chuanglopsis,
обнаруживающий заметное сходство с верхнекембрийскими родами
Chuangla w а 1с. и Bercinella Fred.
Род Chuangiopsis Sivov, gen. n.
Представители этого рода известны лишь по кранидиям и совместно
встречающимся с ними пигидиям.
Кранидий очень небольшой, субтрапецоидальный, реже почти прямо-
угольный, слегка вытянутый в ширину. Глабель с очень слабыми сле-
дами глабелярных бороздок, длинная, впереди округленная или слабо
притупленная, субконическая, заметно расширяющаяся в основании.
Спинные борозды резкие, узкие, заметно затухающие у переднего конца
глабели и вдавленные у ее заднего края. Затылочное кольцо выпуклое,
заметно сужающееся на боках, с маленьким бугорком посередине; заты-
лочная бороздка мелкая, слегка изгибающаяся на боках.
Неподвижные щеки широкие, выпуклые, довольно круто вздымаются
от спинных бороздок, чуть ниспадают к переднему краю, а у заднего
края, несколько расширяясь, отделяются глубокой заднебоковой борозд-
кой, от узких шнуровидных заднебоковых лопастей.
Фронтальная кайма в виде выпуклого, почти прямолинейного валика
обрамляет спереди кранидий; против глабели она несколько расширяется,
под очень тупым углом отгибается вперед, вследствие чего между нею
и глабелью образуется очень узкое предглабелярное поле, отделенное
резкой краевой бороздкой. Последняя на боках срезает неподвижные
щеки. Глазные крышки маленькие, расположены несколько сзади сре-
дины глабели; впереди они продолжаются в глазные валики, которые,
косо рассекая неподвижные щеки, примыкают к глабели почти у перед-
него ее конца. Передние ветви лицевых швов короткие, почти прямые
и субпараллельные; задние ветви лицевых швов длинные, диагональные
и тянутся к щечным углам.
Пигидий очень маленький, полукруглый, вытянутый в ширину, с почти
прямым передним краем и относительно узкой краевой каймой, отделен-
ной изнутри бороздкой. Рахис выпуклый, конический, с двумя ясно вы-
раженными сегментами (кроме конечного). Бока пигидия слабо сегмен-
тированы, причем ребрышки переходят и на кайму.
Этот род по общему очертанию кранидия, его размерам, широким
щекам и строению фронтальной каймы обнаруживает близкое сходство
с родом Bercinella F г е d., но у последнего глабель почти прямоугольная,
глабелярные бороздки и глазные валики отсутствуют, пигидий неиз-
вестен.
Большое сходство Chuanglopsis имеет также с родом Chuangla
W а 1 с., но представители этого китайского рода—крупные формы с тра-
пецеидальной глабелью, упирающейся в узкую, рельефную, прямую
кайму. Пигидий без каймы, причем рахис менее выпуклый, чем у
Chuanglopsis.
Типичный вид. Chuanglopsis sibirica (Siv.).
Распространение. Горношорский комплекс (Ст32) Западной
Сибири.
140
Chuanglopsls slblrica (Sivov)
Табл. XV фиг. 9
Характеристика вида полностью соответствует описанию рода.
Распространение. Характерный вид горношорского комплекса
(Ст32). Горная Шория, бассейн рч. Эмзас.
Chuanglopsls batiaeformis (Sivov)
Табл. XV, фиг. 15; табл. XVI, фиг. 8
Вид близок к предыдущему, но отличается притупленным передним
концом глабели, более широкой, брускообразной, почти прямой фрон-
тальной каймой, более глубокой затылочной бороздкой и отсутствием
на затылочном кольце бугорка.
Р а с пространен и е. Встречается совместно с предыдущим видом.
Chuanglopsls speclosa (Sivov)
Табл. XVI, фиг. 9
Вид близок к Chuanglopsls slblrica, но глабель, с отсутствующими
глабелярными бороздками, упирается в проволоковидную, широко изо-
гнутую фронтальную кайму, отделенную позади резкой краевой борозд-
кой. Глазные крышки располагаются посередине кранидия и продол-
жаются в глазные валики, которые, будучи почти параллельными перед-
нему краю кранидия, примыкают к переднему округленному концу гла-
бели.
Распространение. Совместно с двумя предыдущими формами.
СЕМЕЙСТВО PLETHOPELTIDAE RAYMOND
Род Kazellina Sivov, gen. n.
Трилобиты, относимые к этому роду, характеризуются вытянутым
в длину кранидием почти прямоугольного очертания, исключая задне-
боковые лопасти. Глабель выпуклая, длинная, субконическая или суб-
цилиндрическая, с широкоокругленным передним краем, без глабеляр-
ных бороздок; она отделена вполне заметными спинными бороздками,
которые являются относительно резкими в задней половине кранидия, но
затем становятся более слабыми, особенно у переднего конца глабели.
Неподвижные щеки очень узкие, примерно одинаковой выпуклости
с глабелью и вблизи переднего конца последней постепенно сливаются
с умеренно широким, субпрямоугольным, вытянутым в ширину, выпук-
лым лимбом. Фронтальная кайма отсутствует.
Затылочное кольцо широкое, выпуклое, отделено почти прямой заты-
лочной бороздкой, постепенно1 исчезающей при движении к спинным
бороздкам. Заднебоковые лопасти узкие, большие, валикообразные
и снаружи слегка оттянуты назад. Заднебоковые борозды! мелкие, но
широкие, затухают около дорзальных бороздок и сливаются с ними.
Глазные крышки умеренной величины своими передними концами
примыкают к месту слияния неподвижных щек и лимба. Передние ветви
лицевых швов прямые, параллельные; задние — диагональные, длинные,
оканчивающиеся в щечных углах.
По важнейшим признакам строения кранидия род Kazellina чрезвьр-
чайно близок к роду Kazelia (Walcott and Resser, 1924) из так назы-
ваемых озаркских слоев Новой Земли. Крайне схематичная характеристика
рода Kazelia не позволяет сделать детальных сравнений его с родом
Kazellina. Тем не менее у последнего спинные, затылочная и заднебоко-
вые бороздки выражены гораздо резче, глазные крышки более длинные,
расположены посередине кранидия.
141
Формой либма, узкими неподвижными щеками и положением глаз-
ных крышек род Kazelltna весьма близок к американскому верхнекемб-
рийскому роду Monocheilus R е s s е г, но у последнего заднебоковые
лопасти гораздо уже, на глабели присутствуют следы глабелярных бороз-
док и лимб менее широкий.
Типичный вид. Kazelltna amsasstensts (S i v.).
Распространение. Горношорский комплекс (Cm32). Горная
Шория, Салаир.
Kazelltna amsasstensts (Sivov.)
Табл. XVI, фиг. 1
Диагноз вида полностью соответствует характеристике рода.
Распространение. Один из важных видов горношорского
комплекса (Ст32). рч. Амзас (Эмзас), Горная Шория.
Kazelltna gigantea Sivov, sp. n.
Табл. XVI, фиг. 7
Вид очень близок к вышеописанному, но отличается от него весьма
крупными размерами, почти цилиндрической глабелью, передний край
которой резче отделен сходящимися вокруг него спинными бороздками.
Последние имеют почти одинаковую глубину на всем протяжении. Заты-
лочная бороздка широкая, расплывчатая.
Распространение. Горношорский комплекс (Ст32). Гора Орли-
ная, Салаир.
Kazelltna tarta Sivov, sp. n.
Табл. XV, фиг. 16
Вид весьма близок к предыдущему, но глабель усеченноконическая,
более короткая, ограниченная глубокими и широкими спинными борозд-
ками. Затылочная бороздка широкая и глубокая, выражена лишь посе-
редине; затылочное кольцо является более узким, особенно на боках.
Передние ветви лицевых швов субпараллельные. Глазные крышки не
сохранились.
Распространение. Вид очень характерен для горношорского
комплекса (Ст32). Горная Шория, бассейн рч. Амзас.
СЕМЕЙСТВО REMOPLEUR1DAE CORDA
Род Apatokephalina Sivov, gen. п.
Крупный, довольно выпуклый и вытянутый в длину кранидий имеет
субпрямоугольное очертание. Глабель длинная, выпуклая, с угловато
приостренным передним краем. Она рассечена парой длинных глабеляр-
ных бороздок, круто изгибающихся назад и отрезающих на глабели
длинные и широкие субтреугольной формы лопасти, с вершинами, обра-
щенными в сторону переднего края. Спинные бороздки мелкие, узкие,
затухают около переднего края глабели. Затылочное кольцо широкое,
плоское и отделено ровной, но тонкой и узкой затылочной бороздкой.
Неподвижные щеки узкие, плоские, миндалевидной формы и целиком
располагаются в задней части кранидия. Они окаймляются длинными,
выпуклыми, дугообразными глазными крышками, передние концы кото-
рых примыкают почти к переднему краю глабели, а задние достигают
затылочной бороздки. Предглабелярное поле узкое, почти плоское, слабо
выпуклое в продольном направлении, оттянутое вперед и отделено от
глабели глубокой, но узкой, угловатой бороздкой. Фронтальная кайма,
видимо, отсутствует. Передние ветви лицевых швов короткие, начинаются
от спинных бороздок и затем под тупым углом расходятся наружу. Зад-
ние ветви лицевых швов и заднебоковые лопасти не сохранились.
142
Представители описанного рода занимают, повидимому, промежуточ-
ное положение между известными верхнекембрийскими родами Rlchard-
sonella (Rasetti, 1944) и Apatokephaloldes (Raymond, 1924). Однако на-
дежного сравнения описанного рода с двумя названными выше провести
невозможно вследствие того, что род Apatokephallna установлен только
по кранидиям, и его самостоятельное систематическое значение, а также
принадлежность к семейству Remopleuridae требуют дополнительных
доказательств.
Типичный вид. Apatokephallna bruta Sivov.
Распространение. Горношорский комплекс (Ст32). Салаир.
Apatokephallna bruta Sivov, sp. n.
Табл. XV, фиг. 13
Характеристика вида полностью соответствует описанию рода.
Распространение. Горношорский комплекс (Ст32). Толсточи-
хинская свита горы Орлиной, Салаир.
Род Artokephalus Sivov et Jegorova, gen. n.
Мелкие и средней величины трилобиты с субовальным очертанием
кранидия и относительно длинной глабелью, упирающейся в плоскую или
валикообразную фронтальную кайму. Глабель выпуклая, равномерно спа-
дающая к бокам и круто обрывающаяся книзу, на переднем конце она
рассечена одной, реже двумя-тремя парами глабелярных бороздок, из
которых задняя пара более длинная и резко отгибается к затылочному
кольцу.
Длинные, дугообразные глазные крышки начинаются от мелкой, ров-
ной затылочной бороздки и спереди примыкают к глабели несколько
позади переднего округленного ее конца. Между средней частью глабели
и глазными крышками располагаются почти плоские полулунной формы
неподвижные щеки, отграниченные как от первой, так и вторых мелкими
и узкими бороздками, передние концы которых вливаются в глубокую
и широкую краевую бороздку впереди глабели.
Затылочное кольцо широкое, валикообразное. Передние ветви лице-
вых швов очень короткие, прямые и параллельные. Заднебоковые лопасти
узкие и короткие.
По строению переднего края кранидия, направлению передних ветвей
лицевых швов, небольших неподвижных щек представители рода Arto-
kephalus проявляют сходство с родом Apatokephaloldes Raymond,
однако у последнего глазные крышки небольшие, фронтальный лимб
гораздо шире, глабель менее растянута между глазными крышками и ме-
нее выпуклая посередине, заднебоковые лопасти очень длинные, субтре-
угольной формы.
Типичный вид. Artokephalus minimus Sivov.
Распространение. Горношорский комплекс (Ст32). Салаир.
Artokephalus minimus Sivov, sp. n.
Табл. XVI, фиг. 6
Очень небольших размеров кранидий с выпуклой и широкой посере-
дине глабелью, заметно сужающейся к округленному переднему краю;
на глабели одна пара резких косых глабелярных бороздок. Фронталь-
ная кайма узкая, выпуклая, иногда валикообразная.
Этот вид по форме глабели, длинным дугообразным крышкам и узким
неподвижным щекам обнаруживает сходство с американским верхне-
кембрийским видом Richardsonella elongata (Rasetti, 1948), но у послед-
него имеется предглабелярное поле, окаймленное фронтальной каймой;
последняя отделена краевой бороздкой, вдоль которой располагается тон-
кая штриховка. Передние ветви лицевых швов широко и диагонально
расходящиеся.
143
Распространение. Важная форма горношорского комплекса
(Cm32). Толсточихинская свита горы Орлиной, Салаир.
Artokephalus squalarls Sivov, sp. п.
Табл. XV, фиг. 11
Вид близок к предыдущему и отличается от него сильно вздутой,
почти шаровидной, резко расширяющейся в передней части глабелью,
рассеченной тремя парами глабелярных бороздок. Неподвижные щеки
более широкие. Фронтальная кайма не сохранилась. Затылочное кольцо
широкое, плоское и отделено мелкой, но широкой, расплывчатой заты-
лочной бороздкой.
Распространение. Совместно с предыдущим видом.
Artokephalus suburceus J е g о г о v a, sp. п.
Табл. XV, фиг. 12 и 14
Вид очень близок к Artokephalus minimus, но отличается от него
более крупными размерами кранидия и глабелью, округленноугловатой
на переднем конце и рассеченной тремя парами глабелярных бороздок,
направленных назад.
Распространение. Совместно с предыдущими формами.
Род Portentosus Je gorova, gen. n.
Кранидий среднего размера, почти плоский. Глабель длинная, округ-
локоническая, копнообразная; иногда у основания она имеет параллель-
ные бока.
На глабели наблюдаются три пары слабо выраженных поперечных
борозд. Неподвижные щеки широкие, полукруглой формы. Глазные
крышки дугообразные, длинные, широкие, почти плоские. Задние концы
их отходят от затылочного кольца и подходят близко к переднему концу
глабели. Лицевые швы дугообразно вогнуты наружу и в передних ветвях
расходятся. Имеется широкая, чуть наклонная кайма и неширокое пред-
глабелярное поле.
Типичный вид. Portentosus brevis Jegorova.
Распространение. Cm3. Западная Сибирь (Салаир).
Portentosus brevis Jegorova, sp. n.
Табл. XVI, фиг. 5
Ширина кранидия несколько превышает длину за счет больших не-
подвижных щек. Глабель копнообразная, умеренно выпуклая. Передний
узкий и удлиненный конец глабели незаметно сливается с предглабеляр-
ным полем. Три пары поперечных борозд слабо выражены и имеют вид
удлиненных ямочек, направленных косо назад.
Фронтальный лимб состоит из небольшого, почти плоского предгла-
белярного поля и возвышающейся над ним плоской каймы, ограничен-
ной неглубокой бороздкой. Спинные борозды неглубокие, нитевидные.
Затылочное кольцо широкое, ниже уровня глабели. Затылочная борозда
неглубокая, ясно выражена. Неподвижные щеки широкие, по ширине
равные половине ширины глабели.
Глазные крышки узкие, полулунные, длинные, сильно изогнутые. Они
простираются от заднего края до переднего конца глабели, близко под-
ходя к ней. Передние ветви лицевых швов не соприкасаются с передней
частью глабели и проходят к предглабелярному полю, дугообразно
изгибаются внутрь, а затем расходятся к переднему краю.
Распространение. Горношорский комплекс (Сгп32). Салаир,
гора Орлиная.
144
СЕМЕЙСТВО CHEIRURIDAE SALTAR
ПОДСЕМЕЙСТВО CHEIRURINAE RAYMOND
Род Emsurina Sivov, gen. n.
Крупный, полукруглого очертания, вытянутый в ширину кранидий
•с выпуклой, почти прямоугольной, широкой глабелью и ясными пропар-
ными лицевыми швами.
Глабель с параллельными боками и широкоокругленным передним
краем упирается в проволоковидную дугообразную фронтальную кайму.
Глабель рассечена тремя парами коротких глабелярных бороздок, из
которых задняя пара несколько оттянута назад. Спинные бороздки узкие,
глубокие и в передней части окаймляют глабель. Затылочное кольцо
узкое, ровное, с очень небольшим срединным бугорком; затылочная
борозда глубокая и почти прямая.
Неподвижные щеки умеренно выпуклые, треугольной формы, заметно
сужающиеся впереди, исчезающие вблизи переднего края глабели. Зад-
небоковые лопасти небольшие, выпуклые, валикообразные и отделены
глубокими бороздками. Глазные крышки маленькие, почковидные, распо-
лагаются около переднего края глабели. Передние ветви лицевых швов
очень короткие, сходящиеся; задние ветви лицевых швов диагональные
и режут неподвижные щеки выше округленных щечных углов.
Описанный род близок к роду Ptlekla Barton, но у последнего
глабель полукруглая или даже субконическая, с более резкими оттяну-
тыми назад глабелярными бороздками. Неподвижные щеки более широ-
кие, с резкой грануляцией и оттянутыми в шипы щечными углами. Заты-
лочное кольцо без срединного бугорка; затылочная бороздка слегка
прерывается посередине. Фронтальная кайма более широкая, а глазные
крышки удлиненные.
Типичный вид. Emsurina sibirica Sivov.
Распространение. Горношорский комплекс (Cm32). Салаир.
Emsurina sibirica Sivov, sp. п.
Табл. XVI, фиг. 2
Описание рода полностью соответствует видовой характеристике.
Распространение. Горношорский комплекс (Ст32). Толсточи-
хинская свита горы Орлиной, Салаир.
77. С. КРАСНОПЕЕВА
ALGAE. ВОДОРОСЛИ
В известняках кембрия значительным развитием пользуются извест-
ковые водоросли неопределенного систематического положения.
Род Epiphyton Chapman
Слоевище кустообразное, в целом эллипсоидальной или шаровидной
формы, состоит из дихотомически ветвящихся известковых нитей, круг-
лых в сечении. Толщина нитей у различных видов колеблется в преде-
лах 0,02—0,07 мм.
Типичный вид. Epiphyton fasclculatum С h а р m.
Распространение. Спи—Ст2.
30 Зак. 2024 1 45
Epiphyton fasciculatum Chapman
Рис. 158. Epiphyton fa-
sciculatum Cha pm. X30.
рч. Б. Ерба, Кузнецкий
Алатау
Рис. 158
Слоевище пучкообразное, состоит из дихото-
мически ветвящихся нитей. Толщина нитей 0,05
0,07 мм. Ветвление нитей происходит через про-
межутки, равные в среднем 0,35 мм и, реже, —
через 0,15—0,20 м. Наибольшая толщина нитей
перед их ветвлением, а наименьшая — непосред-
ственно после ветвления.
Вид пользуется повсеместным распростране-
нием в известняковых фациях кембрия.
Epiphyton tenue Vologdin
Рис. 158
Слоевище кустообразное. Толщина нитей, со-
ставляющих слоевище, 0,02—0,03 мм. Дихотомия
нитей частая, через промежутки, равные 0,09--
0,15 мм 1И, реже, 0,25 мм.
Распространение. Кембрийские известия
ки Горной Шории. Алтая, Салаира, а также Урала
и Монголии.
Род Tubophyllum Krasn opeeva, gen. n.
Слоевище стелющееся, образовано длинными
трубочками, переплетающимися между собой.
Типичный вид. Tubophyllum victori Krasn.,
sp. n.
Распространение. Нижний кембрий За-
падной Сибири.
Рис. 159. Epiphyton
tenue V о 1. X 30. Об-
ручевский горизонт,
рч. Б. Ерба, Кузнец-
кий Алатау
0 /
Рис. 160. Tybophyllum vic-
tor! Krasn., sp. П.Х28. Гор;
Мартюхина, рч. Б. Ерба,
Кузнецкий Алатау
Tubophyllum victori Krasnopeeva, sp. п.
Рис. 160
Слоевище стелющееся; трубочки, слагаю-
щие слоевище, имеют поперечник 0,15—0,05 мм.
Распространение. Гора Мартюхина
в районе с. Б. Ерба и рч. Белокаменка в си-
стеме р. Кии (Кузнецкий Алатау).
Род Osagia Т w е п h о f е I
Концентрически слоистые, шарообразные,
удлиненные или лепешкообразные водоросле-
вые структуры, образованные чередованием
светлых и более темных (обогащенных органи-
ческим материалом) слоев. Нарастание слоев
происходит от центра во все стороны; центром
роста является какое-либо постороннее тело
или целые группы их. Род Osagia относят
к группе онколитов, которые обычны в волноприбойной зоне современ-
ных морей. Истинная природа озагий до настоящего времени не выяс-
нена; некоторые ученые сопоставляют их с современными водорослевыми
конкрециями, образованными главным образом синезелеными водо-
рослями.
146
Род Osagia широко распространен в палеозойских известняках, в За-
падной Сибири, главным образом в кембрии и, возможно, в протерозое.
Osagia ovoslmills Vologdin
Табл. XVI, фиг. И
Слоевище шарообразной или эллипсоидальной формы с поперечни-
ком, в среднем, 2—3 мм; центральная часть его заполнена кальцитом.
Толщина кальцитовых светлых и более темных слоев колеблется в пре-
делах сотых и десятых долей миллиметра.
Распространение. Кембрийские отложения Западного Саяна
и Кузнецкого Алатау.
Osagia lamellosa Krasnopeeva
Табл. XVI, фиг. 10
Слоевище шарообразной формы, поперечником до 10 мм. Характер-
ной особенностью данного вида является волнистый характер слоев,
расположенных пачками (или зонами): в каждую пачку, при толщине
ее около 0,5 мм и диаметре слоевища 10,0 мм, входит в среднем 7—9 тон-
ких слоев; толщина каждого слоя не превышает 0,07 мм; толщина белых
кальцитовых слоев меньше, чем темных. Своеобразной особенностью
этого вида является также наличие в слоевище радиально расположен-
ных светлых полос, напоминающих собой радиальные каналы.
Распространение. Нижний кембрий, район рч. Б. Ерба (Куз-
нецкий Алатау).
Род Collenia Walcott
Род Collenia относится к группе строматолитов. Это водорослевые
слоистые структуры различной формы — от полого слоистой до столбча-
той. Виды рода Collenia выделяются по форме строматолита, степени
изогнутости слоев в центре и у периферии экземпляра, а также и их мик-
роструктуре
Распространение. Протерозой — нижний палеозой.
Collenia kundatensis Krasnopeeva, sp. n.
Табл. XVII, фиг. 1
Характер залегания водорослевых известняков не установлен. Наблю-
даемый образец имеет размеры 40 X 20 см и высоту 20 см. Колония об-
разована столбчатыми слоистыми структурами, имеющими слабо вы-
пуклые слои. Диаметр каждой столбчатой структуры 2,0—12,0 см. Гра-
ницы отдельных столбчатых структур выражены слабо. Слои образованы
тонкокристаллическмм кальцитом различной окраски. Слоистость непра-
вильная; толщина темных слоев колеблется от 1,0 до 3,0 мм; толщина
светлых слоев от 0,7 до 1,2 мм. Отчетливо выражена микрослоистость
и местами сгустковая структура: в темных слоях выделяются светлые
микрослои, а в светлых—темные. Толщина микрослоев измеряется со-
тыми долями миллиметра.
В состав темных слоев и микрослоев входит тонкокристаллический
кальцит со значительной примесью глинистого и, вероятно, железистого
вещества. В светлых слоях примесей значительно меньше.
1 «Роды» и «виды», на которые делятся онколиты и строматолиты, являются искус-
ственными объединениями, так как и те и другие представляют собой водорослевые
конкреции, образуемые не отдельными видами, а целыми колониями различных
водорослей (см. В. П. Маслов. Принципы номенклатуры и систематики строматоли-
тов. Изв. АН СССР, сер. геол., 1953, №'4, стр. 105—112). Прим. ред.
10 147
Отличительными особенностями описанного вида являются: 1) мас-
сивность каждого столбчатого образования и пологий загиб слоев,
2) тонкая слоистая микроструктура слоев, местами сгусткового харак-
тера, и 3) неясность границ каждого столбика.
Сходство с Collenia burlatica Masi, выражается во внешней форме
строматолита и общем характере его слоистости, отличие — в наличии
микрослоистости и в отсутствии облекания нижних слоев верхними.
Coll, burlatica Masi, установлена в среднем кембрии р. Бирюсы и вер-
ховьев р. Лены.
Распространение. Cm2, р. Кундат у устья кл. Соболинки
в Мариинской тайге (Кузнецкий Алатау).
ГЛАВА //
СИЛУРИЙСКАЯ СИСТЕМА
Л.Л. ХАЛФИН
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Силурийские отложения распространены в пределах Саяно-Алтайской
области не менее широко, чем отложения других палеозойских систем,
но стратиграфия их выяснена далеко не полно. Основной причиной яв-
ляется -слабая изученность содержащейся в этих отложениях фауны.
Литература, посвященная этой фауне, исчерпывается несколькими рабо-
тами, частью имеющими характер заметок, по граптолитам (Аверьянов,
1932), трилобитам (Вебер, 1923 и 1928; Ломовмцкая, 1936; Чернышева,
1937 и 1951), брахиоподам (Бубличенко, 1927; Лазуткин, 1936; Б. Б. Чер-
нышев, 1937) и табулятам (Б. Б. Чернышев, 1937а; Радугин, 1936); два
вида строматопор описаны В. И. Яворским (1929) и один вид К. В. Ра-
дугиным (1931).
Кроме того, в ряде геологических работ приводятся более или менее
значительные списки силурийской фауны из отложений различных райо-
нов, представляющие собой результаты предварительных определений.
Неопубликованные палеонтологические работы, посвященные силурий-
ской фауне Западной Сибири, также немногочисленны.
Несмотря на столь слабую изученность этой фауны, выяснено, что
силурийская система в Саяно-Алтайской области представлена доста-
точно полно обоими ее отделами, внутри которых выделяются подчинен-
ные им стратиграфические единицы, хотя и не во всех случаях датируе-
мые с точностью до яруса. Задачи настоящего очерка, имеющего харак-
тер краткого введения к описанию руководящих форм, позволяют огра-
ничиться указаниями на отложения, возраст которых установлен по до-
статочно надежно определенной фауне (см. табл. 3); это, прежде всего,
отложения, из которых фауна описана, хотя бы частично, а затем, отло-
жения, возраст которых установлен по наиболее типичным и легко
определяемым формам, вроде Bilobites bilobus или Conchldlum knlghti.
В качестве нижнего члена силурийских отложений Саяно-Алтайской
области обычно указывается широко распространенная толща значи-
тельно метаморфизованных песчано-глинистых пород преимущественно
зеленого, с фиолетовыми прослоями, цвета, известная в литературе под
названием зелено-фиолетовой или западноалтайской толщи («формации»).
Возраст этой толщи определяется по стратиграфическим соотношениям
ее с фаунистически охарактеризованными отложениями верхов кембрия
и среднего ордовика, так как отложения зелено-фиолетовой формации,
как правило, лишены органических остатков или, очень редко, содержат
скудную, не подвергавшуюся сколько-либо серьезному изучению фауну.
149
Лишь в самое последнее время в пределах юго-западной окраины
Куртушибинского хребта Западного Саяна в отложениях этой толщи
собрана более значительная фауна, состоящая, однако, по данным
А. Г. Сивова, почти исключительно из весьма своеобразных пелеципод,
не имеющих близких аналогов среди описанных форм. Охарактеризован-
ные фауной трилобитов отложения низов нижнего силура известны лишь
в Кузнецком Алатау. Это, прежде всего, песчано-глинистые отложения
северо-западной оконечности Кузнецкого Алатау (в системе рр. Золотой
Китат, Кожух и Тайдон), содержащие смешанную кембрийско-силурий-
скую фауну, повидимому, с преобладанием кембрийских элементов в виде
представителей рода Solenopieura A n g. (см. стр. 113), семейства Dikelo-
cephalidae (см. стр. 133).
Таблица 3
Отде- лы 1 Ярусы j i Опорные стратиграфические горизонты
Даунтонский и лудловский Остракодовый горизонт Салаира с брахиоподами Wilso- nella tarda, Gypidula procerula, Howellella crispa, Prot- athyris didyma и лр., различными целентератами и ооракодами (Leperditia и др.)
S2—готландий Уинлокский Частью брахиоподовые и коралловые известняки Салаира, Алтая, Западного Саяна с Clathrodlctyon vesiculosum. Heliolites interstinctus, Favosltes gotlandicus, Bilobites bilobus и лр.; брахиоподовые и коралловые известняки Тувинской области с Tuvaella rackowskii, Camarotoechia nahvkini. Howellella crispa, Favusites gotlandicus н другие; конхидиумовые банки Горного Алтая с Conchi- dium knighti.
Лландоверский Граптолитовые сланцы Салаира и Рудного Алтая с Diplo- graptus bellulus, Diplograptus sinuatus, Monograptus distans, Rastrites longispinus и др.
Карадокский Веберовский горизонт Салаира с Illaenus salairlcus, Amphilichas, sniatkowi, Cheirurus weberi и другими трилобитами
S аз Ландейльский Известковистые песчаники и сланцы Салаира с Dalmanella cf. testudinaria, Paurorthis cf. part'а и другими брахио- подами
О =Х о. о Ц Аренигскф! Граптолитовые сланцы Салаира с Didymograptus и Phyllograptus 1
Тремадокский ! i ! Отложения бассейна р. Амзас (Горная Шория) с трило- | битами: Parabolinella argentiensis и различные местные представители родов Promegalaspides, Maj а и др. Отложения северо-западной окраины Кузнецкого Алатау со смешанной кембро-силурийской фауной трилобитов (представители родов Solenopieura, Euloma и др.)
Однако в составе этой фауны появляются такие преимущественно
силурийские группы, как семейство Cheiruridae (см. стр. 178) и особен-
но характерный для этих отложений род Euloma An g. (рис. 161 и 162),
у представителей которого «глабель выпуклая, короткая, с резко очер-
150
пенными боковыми лопастями, напоминающими таковые у Calymene.
Впереди глабели широкий лимб. Глаза большие. Пигидий короткий
и широкий» (К- Циттель. Основы палеонтологии, 1934 г., стр. 942).
На юге Кузнецкого Алатау, в Горной Шории (бассейн р. Амзас),
обнаружены отложения, в которых нижнесилурийские трилобиты господ-
Рис. 161. Кранидий, подвижная щека и пигидий рода
Euloma A n g. Нат. вел. По Питтелю
ствуют, но еще встречаются кембрийские элементы; трилобиты бассейна
р. Амзас — Parabolinella argentlensls Kob., Protnegalaspld.es robustus
Lomov., Maja subsulcata Lomov, и др.—указывают на верхнетрема-
докский возраст содержащих эту фауну отложений.
На Салаире, в бассейне р. Сухой, известны отложения аренигского
Рис. 162. Кранидий пред-
ставителей рода Euloma
из отложений нижнего
тремадока Кузнецкого
Алатау. Нат. вел.
По О. К. Полетаевой
Рис. 163. Didymograptus
bifldus Hall (по М. Э.
Янишевскому) Арениг-
ский ярус Салаира.
Увеличено
яруса с граптолитами из родов Phyllograptus и Didymograptus-, на
рис. 163 приводится изображение салаирского экземпляра Didymograp-
tus, заимствованное у М. Э. Янишевского.
Одним из наиболее точно определенных стратиграфических горизон-
тов силура Западной Сибири является так называемый веберовский го-
ризонт с богатой и разнообразной фауной, из состава которой изучены
и описаны трилобиты (В. Н. Вебер, 1928 и 1948; М. П. Ломовицкая,
1936); этот горизонт на северо-восточном склоне Салаира (г. Орлиная
и другие пункты) представлен известняками с Illaenus salalricus Web.,
151
Bumastus maccallumi Salt., Amphillchas sniatkowi W e b., Cheirurus
weberl Lomov, и др., указывающими на принадлежность этого гори-
зонта к карадокскому ярусу. Стратиграфические аналоги этого горизонта
известны'в Горной Шории (бассейн р. Амзас), а также в Горном Алтае,
где они содержат требующую дополнительного изучения фауну брахио-
под, целентерат, трилобитов.
Наиболее древний горизонт верхнего силура в Саяно-Алтайской об-
ласти представлен черными граптолитовыми сланцами с Dlplograptus
bellulus Torn q., Dlplograptus slnuatus N i c h., Climacograptus scalarls,
Monograptus distans, Pastrites longlnsplnus и др. Эти граптолитовые
сланцы известны в Рудном Алтае и на Салаире (г. Орлиная); они отно-
сятся к лландоверскому ярусу. На рис. 164 воспроизводится схематичное
изображение верхнесилурийских граптолитов Рудного Алтая по
В. Н. Аверьянову.
Рис. 164. Граптолиты лландоверского яруса
Рудного Алтая (по В. Н. Аверьянову):
a-Clim acograptus rectangularls; б—Diplo-
graptus tamariscus; в—Monograptus dis-
tans; г- Rastrites peregrinus; d—Rastrites
longinspinus. Сильно увелич.
Широким распространением
пользуются верхнесилурийские
отложения с обильной и разно-
образной фауной брахиопод,
целентерат и трилобитов,
частью, вероятно, одновозраст-
ные с лландоверскими грапто-
литовыми сланцами, но глав-
ным образом относящиеся к бо-
лее высоким горизонтам гот-
ландия. На Салаире и по юго-
западной окраине Кузбасса эти.
отложения содержат гастропод,
брахиопод (Bllobites bllobus L.
и др.), но особенно обильны в
них различные целентераты
(Clathrodlctyon vesiculosum N i с h. et Mur., Heliolites interstlnctus
Linn e, Favosites gotlandicus L a m., Favosites forbesi E. et H., Halysltes
и др.).
На Алтае и в Западном Саяне брахиоподовые и коралловые фации
низов и средины верхнего силура широко распространены, но их фауна
детальному изучению не подвергалась. В Тувинской области сюда отно-
сятся отложения бассейна р. Элегест, р. Улу-Хем и ряда других пунктов.
Они содержат богатую фауну брахиопод, целентерат и трилобитов, опи-
санную Б. Б. Чернышевым (1937, 1937а) и Н. Е. Чернышевой (1937);
из браихопод наиболее важны Tuvaella radkowskll Тс hern., Stegerhyn-
chus decimplicatus Sow. var. angadlensis Tch e r n., Camarotoechla
nalivkint T c h e r n. и некоторые другие; из табулят присутствуют Favo-
sites hlsingerl Е. et Н., Favosites gotlandicus L a m. и др.
Наиболее высокие горизонты верхнего силура представлены на Са-
лаире (юго-западная окраина Кузбасса) так называемым остракодовым
горизонтом; это главным образом темносерые известняки с остракодами
(преимущественно, род Leperdltla), строматопорами (Stromatopora
typica Ros. и др.) и разнообразными брахиоподами, описанными
П. С. Лазуткиным (1936): Wilsonella tarda (Barr.), Gypidula procerula
(Barr.), Protathyris didyma (D a 1 m.) и др. По заключению П. С. Ла-
зуткина, эти известняки относятся к лудловскому и даунтонскому яру-
сам, т. е. частью принадлежат к нижнему девону.
Обычно к верхам S2 относят также известняки с. Сары-Чумыш (юж-
ная окраина Кузбасса), из которых Н. Л. Бубличенко описал брахиопод
Nallvklnia griinewaldtlaejormis (Peetz), Camarotoechla vesta (Barr.),
Delthyrts elevata D a 1 m. и некоторые другие. Нужно, однако, заметить,
что это определение возраста известняков с. Сары-Чумыш (гора Гля-
день) является слабо обоснованным, и в настоящее время имеются,
152
правда требующие подтверждения, данные, говорящие о более молодом
(девонском) возрасте этих известняков.
В Горном Алтае (бассейн р. Черный Ануй) распространены верхне-
силурийские известняки, переполненные раковинами Conchldlum knlghti
S о w.; невидимому, аналогичные известняки имеются и в верхнем силуре
Салаира.
Детальное изучение силурийской фауны Западной Сибири должно
рассматриваться как задача ближайшего будущего; оно позволит уточ-
нить и детализировать стратиграфию силурийской системы Саяно-Алтай-
ской области.
Тип Coelenterata. Кишечнополостные
Представители этого типа широко распространены в силурийских
отложениях Западной Сибири, но до настоящего времени не изучались.
Они представлены строматопорами, табулятами, гелиолитидами и тетра-
кораллами; некоторые важнейшие представители трех первых групп опи-
саны ниже.
В. К. ХАЛФИНА
КЛАСС HYDROZOA
STROMATOPOROIDEA. СТРОМАТОПОРОИДЕИ
Силурийские строматопороидеи Западной Сибири известны почти ис-
ключительно по далеко неполным спискам, приводимым в некоторых гео-
логических работах. Лишь два вида описаны В. И. Яворским (1929) из
верхнесилурийских отложений Салаира и один — К. В. Радугиным (1931)
из нижнего силура Горного Алтая.
Ниже дается характеристика двух видов, фигурирующих в списках
в качестве наиболее распространенных и типичных.
СЕМЕЙСТВО CLATHRODICTYONIDAE KOHN
Род Clathrodictyon Nicholson et Murie
Форма ценостеума изменчивая, чаще встречаются пластинчатые
и желвакообразные формы. С нижней стороны ценостеум часто имеет
морщинистую базальную эпитеку с небольшой площадкой для прикреп-
ления, а на верхней стороне — обычно хорошо развитые астроризы
и небольшие бугорки — свободные концы радиальных столбиков. Скелет
состоит из радиальных столбиков и концентрических пластин (лямин).
Радиальные столбики соединяют только две соседние пластинки, иногда
часть столбиков бывает недоразвита. Строение ткани плотное.
Типичный вид. Clathrodictyon vesiculosum Nich. et Mur.
Распространение. Редко кембрий, часто силур, реже девон.
Clathrodictyon vesiculosum Nicholson et Murie
Табл. XVIII, фиг. 1
Ценостеум полушарообразный или шарообразный, иногда пластинча-
тый. Диаметр его обычно равен 50—55 мм. Астроризы маленькие, рас-
стояние между их центрами 3—5 мм, обыкновенно они располагаются
одна над другой и имеют общий осевой канал, но на поверхности возвы-
шений не образуют (Nicholson, 1886—1892, стр. 148).
В радиальном шлифе видны близко расположенные концентрические
пластинки, на 1 мм их насчитывается 9—10; каждая пластинка на всем
протяжении мелко волнисто изогнутая (гофрирована). Радиальные стол-
бики берут начало в местах изгибов лямин. Развитие каждого из них
153
ограничено одним межпластинчатым промежутком, но некоторые из них
не достигают соседней пластинки.
Благодаря такому характеру радиальных и горизонтальных элемен-
тов скелета, радиальное сечение под микроскопом в целом выглядит, как
своеобразная пузырчатая (кружевная) ткань с длиной ячей до 0,12 мм.
В тангенциальном шлифе видны астроризы и овального очертания раз-
резы, радиальных столбиков.
Распространение. Широко распространенная верхнесилурий-
ская форма, в частности встреченная в верхнеоилурийских отложениях
бассейна р. Черневой Бачат — юго-западная окраина Кузбасса. Имеется
ряд близких видов с подобным пузырчатым (кружевным) характером
радиального сечения; все эти формы тоже верхнесилурийские; Clathro-
dictyon. vesiculosum отличается от них более тесно сближенными пластин-
ками и более мелкими ячейками.
СЕМЕЙСТВО STROMATOPORIDAE NICHOLSON
Род Stromatopora G о 1 d f u s s
Ценостеум массивный или пластинчатый. Обычно имеется эпитека.
Почти всегда имеются лятилямины.
Радиальные и горизонтальные элементы скелета обычно плохо выра-
жены, благодаря чему в радиальном сечении правильная сетка отсут-
ствует. Радиальные столбики все же развиты более отчетливо, чем кон-
центрические пластины. Зооидные трубки в большинстве случаев отчет-
ливы и снабжены днищами. Большей частью имеются астроризы. Строе-
ние скелетной ткани мелкопористое или зернистое, поры, располагаются
почти вплотную друг к другу.
Типичный вид. Stromatopora concentrlca Gо 1 d f.
Распространение. Силур—девон, в последнее время описаны
из юры и верхнего мела.
Stromatopora typlca Rosen
Табл. XVIII, фиг. 2
Ценостеум дискоидальный или полусферический, с хорошо развитой
морщинистой эпитекой. На поверхности видны несложно ветвящиеся
астроризы, центры которых удалены друг от друга на 6—9 мм. В. Н. Ря-
бининым из Монголии описаны крупные образцы, имеющие длину
150 мм, ширину 140 мм и высоту 80 мм.
Из Кузбасса В. И. Яворским описаны более мелкие экземпляры
(диаметр наибольшего из них 53 мм).
У этого вида всегда присутствуют лятилямины, их толщина равна
1,5—2 мм.
Вертикальные и горизонтальные элементы скелета почти одинаково
хорошо выделяются. Зооидные трубки снабжены днищами. На 2 мм
насчитывается 6—8 зооидных трубок, диаметр их около 0,1 мм. Ткань
скелета мелкопористая.
Распространение. Stromatopora typlca пользуется широким
распространением и известна из силурийских отложений Эстонии, По-
долии, Тувы, Ферганы и др. Это существенно уинлокская форма.'^По
В. И. Яворскому, в Кузбассе она пользуется массовым распространением
в отложениях остракодового горизонта.
Л. И. КРАЕ ВСКАЯ
КЛАСС ANTHOZOA
ПОДКЛАСС TABULATA И ГРУППА HELIOLITIDA
СЕМЕЙСТВО HALYSITIDAE EDWARDS ET HAIME
Колония состоит из цилиндрических кораллитов, имеющих цепочное
петлеобразное срастание друг с другом. До недавнего времени в составе
154
этого семейства указывался лишь один широко известный род — Halysi-
tes Fisch. Исследования Б. Б. Чернышева и особенно Б. С. Соколова
показали, что этот род может быть разделен, по меньшей мере, на четыре
самостоятельных рода. Весьма популярные в геологической литературе
виды Halysites escharoides и Halysites catenularia потеряли свое значение
в стратиграфии, так как под этими названиями описывались различные
формы. Эти виды нуждаются в ревизии, поэтому они не приводятся ни-
же, хотя в литературе по геологии Западной Сибири неоднократно упо-
минались.
Следует отметить, что сибирские хализитиды почти совсем не изу-
чены.
Род Palaeohalysites Tchernychev
Полипняк состоит из длинные кораллитов, срастающихся двумя сто-
ронами. Цепочные ряды кораллитов в поперечном разрезе образуют
петли, весьма различающиеся у отдельных видов. Кораллиты в попереч-
ном сечении круглые или эллиптические. Стенки кораллитов плотные,
покрыты эпитекой. Днища горизонтальные, правильные. Между коралли-
тами отсутствуют промежуточные трубки — мезопоры; этим данный род
отличается от рода Halysites. Септальные образования развиты в виде
шипиков, которые иногда образуют в центре кораллита ложный столбик.
Типичный вид. Palaeohalysites gothlandlcus (Y a b е).
Распространение. Ордовик — венлок повсеместно.
Palaeohalysites gothlandlcus (Y a b е)
Табл. XVIII, фиг. 4
В поперечном сечении колония характеризуется неправильной сеткой,
образованной цепочками кораллитов. Длина петель колеблется от 4 до
13 мм. В сторонах петель насчитывается от одного до 12 кораллитов.
Поперечное сечение кораллитов слегка эллиптического, почти круглого
очертания. Шипы короткие, наблюдаются редко. Днища многочисленные,
правильные, горизонтальные, редко слабо изогнутые; расстояние между
днищами около 0,3—0,6 мм.
Распространение. Один из наиболее распространенных силу-
рийских видов, особенно в лландовери—венлоке; в старой литературе
описывался нередко под другими названиями. В настоящее время изве-
стен в Западной Европе и в Азии, в том числе на Алтае и па севере Си-
бири.
Palaeohalysites parallelus Schmidt
Рис. 165
Полипняк имеет неправильную форму и состоит из однообразных
кораллитов, которые располагаются в длинные, местами параллельные,
цепочки; в каждую из таких цепочек входит
от двух до 20 ячеек. Петли, образованные
цепочками кораллитов, различных размеров;
ширина их от 3 до 5 мм, длина от 5 до
32 мм. Количество кораллитов в каждой
стороне петли колеблется от одного до 12.
Кораллиты в поперечном сечении прямо-
угольные, удлиненные, с почти параллель-
ными сторонами, что является характерным признаком вида. Днища пол-
ные, либо горизонтальные, либо слабо выпуклые, довольно частые; на
5 мм длины 7—8 днищ. Шипы сохраняются редко.
Близкий вид Halysites gracilis (Hall) отличается размерами.
Распространение. Низы S2 Прибалтики и северных районов
СССР; в Западной Сибири встречен в Северном Алтае.
Рис. 165. Palaeohalysites para-
llelus Schm. Поперечное
155
СЕМЕЙСТВО FAVOS1T1DAE DANA
Род Favosites Lamarck
Полипняки массивные или древовидные, уплощенной или неправиль-
ной формы, реже ветвистые. Кораллиты призматические, плотно срастаю-
щиеся своими стенками. На стенках кораллитов расположены соедини-
тельные поры, имеющие овальное очертание. Эти поры образуют 1—2"
или более вертикальных рядов. Толщина стенок у одного вида постоянна,
что является важным признаком для разделения видов. Днища тонкие,
полные, горизонтальные, распределены весьма неравномерно. На внут-
ренней поверхности стенок развиваются септальные образования в виде
рядов шипиков или бугорков.
В верхнем ордовике и готландии встречается близкий род — Palaeo-
favosites Т w е n h., отличающийся тем, что соединительные поры у него
располагаются не на гранях, а на ребрах призматических кораллитов.
Род Squameofavosltes В. В. Тс hern, отличается наличием чешуек
вместо шипов (см. стр. 197).
Типичный вид. Favosites gothlandlcus Lam.
Распространение. Верхний силур-—пермь, повсеместно.
Favosites htsingeri М.-Е dwards et Haime
Табл. XVIII, фиг. 3
Полипняк дисковидной формы, иногда значительных размеров; более
редки округлые или цилиндрические колонии, которые, повидимому, ха-
рактеризуют молодые стадии. Кораллиты правильно призматические,
обычно шестигранные и чаще всего равны между собой. Диаметр корал-
литов 1,0—1,3 мм, иногда до 1,5 мм. Стенки сравнительно тонкие, тол-
щина их 0,04—0,06 мм. Днища полные, большей частью прямые, перпен-
дикулярные к стенкам; на 5 мм длины от 10 до 25 днищ. Наиболее рас-
пространенное расстояние между соседними днищами 0,25—0,30 мм. По-
ры круглые, мелкие, поперечником 0,10—0,15 мм, на каждой стенке
кораллита располагаются обычно в два, реже в один или три ряда. Ши-
пы тонкие, длинные, загнуты вверх; число их обычно равно 12.
Наряду с типичными Favosites htsingeri существует целый ряд нети-
пичных представителей, являющихся промежуточными формами между
этим видом и многими другими; однако нетипичные формы в литературе
описаны под названием Favosites htsingeri, что делает границы вида
F. hlsingeri несколько расплывчатыми и затрудняет его определение.
Распространение. Favosites hlsingeri является характерным
представителем фауны верхнего силура почти всех районов Евразии
и Северной Америки (чаще всего лландовери — венлок). В Кузнецком
бассейне Б. Б. Чернышев указал на присутствие этого вида в острако-
довом горизонте.
Favosites gothlandlcus Lamarck
Табл. XIX, фиг. 1
Полипняк крупный, массивный, обычно полусферической или плоской
формы. Кораллитьи в поперечном сечении правильно многоугольные, бо-
лее или менее однородные по величине. В одной колонии диаметр корал-
литов изменяется от 2,0 до 3,0 мм. Стенки тонкие, прямые, всегда раз-
делены четко выраженным швом. Днища полные, прямые, обычно гори-
зонтальные; расстояние между ними равно половине диаметра кораллита,
на 5 мм длины приходится от трех до пяти днищ. Шипы редкие, корот-
кие, и обычно перпендикулярны к стенкам кораллитов. Поры круглые,
диаметром 0.2—0,3 мм, расположенные в 1—2 ряда на каждой стенке
155
кораллита. В случае двух рядов поры несколько смещены друг относи-
тельно друга. На 5 мм длины в каждом ряду насчитывается 7—8 пор.
Каждая пора окружена приподнятым краем.
Наибольшее сходство Favosites gothlandlcus имеет с среднедевонским
видом Favosites goldfussl d’O г b. и его вариететами. Различия между
этими близкими видами следующие.
Favosites gothlandlcus
Полипняк крупный, массивный
Диаметр кораллитов от 2,0 до 3,0 мм
Однородность и правильная многогран-
ность формы кораллитов
Тонкость стенок, всегда разделенных
хорошо выраженным швом
Favosites goldfussl
Полипняк мелких размеров, округлый
Диаметр кораллитов от 2,0 до 2,5 мм
Неоднородность кораллитов по разме-
рам
Более толстые стеики со слабо выра-
женным или отсутствующим швом
Различие в окраске стенок и днищ
Более частые диища
Более густые поры
Указания на нахождение Favosites gothlandlcus в девоне, особенно
в среднем его отделе, вызывают большие сомнения.
Распространение. Повсеместно распространенный верхнесилу-
рийский вид. Указания на присутствие его в нижнем девоне Западной
Сибири требуют проверки.
Favosites forbesl М.-Е dwards et Haime
Табл. XVIII, фиг.
Полипняк округленный, сферической, сильно вздутой полушаровидной
или несколько уплощенной формы; размеры небольшие. Неоднородные
по величине кораллиты располагаются радиально, причем крупные, попе-
речником до 2,0 мм, окружены более мелкими кораллитами, поперечни-
ком 0,6—1,2 мм\ встречаются кораллиты и переходных размеров (между
1,2 и 2,0 мм). Стенки тонкие, прямые, изредка в продольном сечении изо-
гнутые; толщина стенок около 0,05 мм, редко достигает 0,10 мм. Днища
полные, тонкие, прямые или слабо изогнутые. Толщина днищ обычно
в пределах 0,3—0,5 мм, расстояние между днищами 0,3—0,9 мм. Шипы
хорошо развиты, тонкие, короткие, частые. Поры круглые, средних раз-
меров, располагаются в два ряда.
В литературе, особенно у старых исследователей, под названием
Favorites forbesl описаны нередко близкие, но различные виды.
Распространение. Верхний силур, повсеместно. В Западной
Сибири — на Алтае и в Салаире.
СЕМЕЙСТВО HELIOLITIDAE LINDSTROM
Род Heliolites Dana
Диагноз рода по Б. Б. Чернышеву (1951, стр. 87): «Полипняки раз-
личной формы: пластинчатые, полусферические, грибообразные, почти
сферические и клубневидные на небольшой ножке, реже ветвистые. Ко-
раллиты цилиндрические, в поперечном сечении круглые или овальные,
со складчатыми и зазубренными на внутренней стороне стенками или со
стенками цельноокругленньгми, не складчатыми. У одних видов септы
хорошо развиты и достигают центра аутопор (кораллитов), у других ви-
дов— рудиментарны и часто отсутствуют. Количество септ обычно 12.
Иногда септы и днища несут шипы. Днища многочисленные.
Кораллиты окружены цененхимой, состоящей из призматических тру-
бок (сифонопор) с многочисленными днищами, обычно более частыми,
157
чем в кораллитах. Новые кораллиты возникают в толще цененхимьи из
нескольких трубок или путем деления».
Типичный вид. Heliolites porosus G о I d f u s s.
Распространение. Встречается повсеместно в оилурийских
и девонских отложениях.
Heliolites decipiens (М’С о у)
Табл. XIX, фиг. 2; рис. 166
Полипняк массивный, неправильно сферической или полусферической
формы, с отдельными округлыми наростами. Кораллиты круглые или
широкоовальные, равные между собой в каждом отдельном полипняке;
диаметр кораллитов 1,0—1,2 мм, ра-
стояние между ними 1—2 мм. Септы
отсутствуют или рудиментарны. Дни-
ща очень тонкие, умеренно частые,
более редкие, чем в трубках ценен-
химы; они редко прямые, обычно
слабо вогнутые, выпуклые или изви-
листые. Расстояние между днищами
в пределах одного кораллита ме-
няется от 0,45 до 0,90 мм.
Сифонопоры неоднородные по-
вел ичине и призматические по фор-
Рис. 166. Heliolites decipiens М’Соу
а—продольное сечение; б—поперечное
сечение, хЗ,5
ме, имеют пяти- или шестиугольные, реже четырехугольные очертания.
Диаметр их колеблется от 0,2 до 0,4 мм и, реже, до 0,7 мм. Число рядов
их между кораллитами меняется от одного до трех.
Стенки кораллитов в поперечном разрезе правильно закруглены, тон-
кие, постоянны по толщине и не утолщены по сравнению со стенками
сифонопор. Расстояние между днищами в сифонопорах от 0,25 до
0,35 мм.
Распространение. Верхний силур, преимущественно венлок—
лудлоу. В Сибири—на Алтае, в Салаире, в Западном Саяне.
Heliolites interstinctus (L i п п ё)
Табл. XIX, фиг. 3; рис. 167
Полипняк округлой, шаровидной формы, с диаметром 7 -8 см.
Круглые в поперечном сечении кораллиты сближены между собой.
иногда до соприкосновения. Стенки
кораллитов зазубренные; расстояние
между кораллитами меньше их диа-
метра, а диаметр в пределах 1,6—
1,8 мм. Внутри каждого кораллита
зачаточный столбик, 12 длинных септ
и горизонтальные, редко изогнутые
днища; на 5 мм длины 20 днищ.
Мелкие правильно полигональные
сифонопоры, с диаметром от 0,3 до
0,5 мм, характеризуются толстыми
стенками. Днища в сифонопорах го-
Рис. 167. Heliolites interstinctus L.
а—продольное сечение; б—поперечное
сечение, Х3,5
ризонтальны или слабо выпуклы и более часты, чем в кораллитах.
От близкого среднедевонского Heliolites porosus Goldf. описы-
ваемый вид отличается более сближенными кораллитами, меньшим диа-
метром сифонопор, наличием у кораллитов зачаточного столбика и более
коротких шипов.
Распространение. Верхний силур, повсеместно. В Западной
Сибири — Алтай, Салаир, Западный Саян. Указания на нахождение
Н. interstinctus в нижнем девоне требуют проверки.
158
Тип Brachiopoda. Плеченогие
В силурийских отложениях Саяно-Алтайской области брахиоподы
распространены широко и известны из многих пунктов. В большинстве
случаев их изучение ограничивалось предварительными определениями,
на основании которых в различных геологических работах приведены
списки форм. Описания имеются в работах П. С. Лазуткина
(1936)—брахиоподы остракодового горизонта Кузбасса, Б. Б. Черны-
шева (1937)—брахиоподы верхнего силура Тувинской области
и Н. Л. Бубличенко (1927) — брахиоподы верхнего силура с. Сары-
Чумыш (юго-западная окраина Кузбасса).
Из упомянутых работ и списков форм, приведенных в других работах,
выбраны наиболее типичные руководящие формы, характеристики кото-
рых по литературным данным составлены В. К. X а л ф и н о й. Изобра-
жения форм, сибирские представители которых не описаны, а лишь упо-
минаются в списках, заимствованы из различных литературных источ-
ников.
Распространение охарактеризованных ниже видов дается лишь для
Саяно-Алтайской области; большинство из этих видов распространено
в отложениях соответствующего возраста других областей СССР
и Западной Европы.
СЕМЕЙСТВО BILOBITIDAE SCHUCHERT ET COOPER
Род Bilobites Linne
Раковина маленькая, с коротким замочным краем, с двухлопастным
передним краем и с глубоким синусом на обеих створках. Треугольная
арея с открытым дельтирием. Зубы и зубные лунки слабо развитые;
замочный отросток маленький и простой.
Поверхность раковины покрыта радиальными ребрами. Особенно
характерен двухлопастной характер переднего края.
Типичный вид. Bilobites bilobus Linn.
Распространение. Нижний силур — нижний девон.
Bilobites bilobus L i n п ё
Табл. XIX, фиг. 4
Раковина маленькая, с сильно выемчатым передним краем. Замочная
линия прямая, она короче ширины раковины. Ушки прямоугольные.
Наибольшая ширина находится у переднего края раковины. На обеих
створках имеется хорошо выраженный глубокий синус. Брюшная створка
более выпуклая, чем спинная. Поверхность створок покрыта радиаль-
ными ребрышками двух порядков: в промежутках между более длин-
ными ребрами вставляются одно или два более коротких ребрышка.
В брюшной створке — зубы и зубные пластинки.
Распространение. Средний готландий горы Орлиной,
Салаир.
СЕМЕЙСТВО ORTHIDAE WOODWARD
Род Tuvaella В. В. Tchernychev
«Раковина полукруглого очертания, с прямым замочным краем, кото-
рый соответствует наибольшей ширине ее. Замочные углы оттянуты
в виде длинных остроконечий. Боковой профиль плоско-выпуклый или
неравно двояковыпуклый. Брюшная створка всегда более выпукла, чем
спинная, и несет ясно выраженное седло, начинающееся от самой
макушки. В спинной створке имеется довольно широкий и глубокий
синус. Арея брюшной створки длинная и низкая, а в спинной створке
она отсутствует...
159
Поверхность раковины покрыта простыми резкими складками, пере-
сеченными концентрическими следами нарастания. Вещество раковины
тонковолокнистое, не точечное. Внутри брюшной створки—резкие зубы,
едва заметные слабые круральные ямки и эллиптическое мускульное
поле. Внутри спинной створки — короткий толстый замочный отросток
в виде простого гребня»... (Б. Б. Чернышев, 1937, стр. 11).
Характерной особенностью рода является закрытый псевдодельти-
диумом дельтирий и отсутствие ареи в спинной створке.
Типичный вид. Tuvaella raCkovskll Т с h е г n.
Распространение. Верхний силур Тувинской области.
Tuvaella raikovskll Tchernychev
Табл. XX, фиг. 1
Диагноз вида отвечает приведенному выше диагнозу рода; количество
ребер от 32 до 46 на каждой створке.
Размеры представителей этого вида: длина 1 —1,5 см и ширина
1,5—2 см. Совместно встречающийся вид Tuvaella gigantea Tchern.
•отличается более крупными размерами и большим количеством ребер.
Распространение. Верхний силур Тувинской области.
СЕМЕЙСТВО CAMAROTOECH1IDAE SCHUCHERTET LE VENE
Род Camarotoechia Hall et Clarke
Ринхонеллоидная радиальноребристая раковина с синусом и возвыше-
нием; ребра начинаются от макушек; в спинной створке септалиум,
в брюшной — зубные пластины.
Распространение. Силур—карбон, повсеместно.
Camarotoechia vesta Barrande
Табл. XIX, фиг. 5
«Раковина округленноромбоидальная. Спинная створка более выпук-
лая. Макушка брюшной створки невысокая, загнутая... Синус широкий,
неглубокий, начинается на половине длины раковины. Седло едва наме-
чается. У взрослых экземпляров седло и синус более резко выра-
жены...» (Бубличенко, 1927, стр. 991).
Поверхность покрыта отчетливыми, начинающимися от макушки
ребрышками, разделенными промежутками такой же ширины, как
и ребра. Количество ребрышек на каждой створке 22—26, в синусе их
насчитывается 6—8 штук.
Распространение. Верхний силур, с. Сары-Чумыш; остракодо-
вый горизонт Салаира.
Camarotoechia nalivkinl Tchernychev
Табл. XIX, фиг. 6
«Раковина овального очертания, с наибольшей шириной на середине
длины. Длина раковины равна ширине или немного меньше. Брюшная
створка слабо вздутая, с небольшой острой умеренно загнутой макуш-
кой... Синус неглубокий... и становится заметным на расстоянии
5—6 мм от макушки. У молодых экземпляров он отсутствует или слегка
заметен лишь у лобного края. Язычок синуса невысокий, широкий, дуго-
образный.
Спинная створка более выпуклая, чем брюшная, с тупой макушкой
и пологим седлом, ясно выраженным только у лобного края. Скульптура
состоит из тонких, простых, округленных радиальных складочек, разде-
ленных бороздками несколько более широкими, чем они сами. Каждая
створка несет от 26 до 32 складок, причем в синусе располагаются
5—10 складок, которым соответствуют 6—11 на седле. Наиболее часто
160
в синусе насчитываются 6—8 складок». (Чернышев, 1937, стр. 35). Раз-
мер раковины 1—1,5 см.
Распространение. Верхний силур Тувы и Монголии.
Род Wilsonella Nikiforova
«Полукубичеокая многоребристая вздутая форма. Часто толщина
превышает ширину и длину раковины. Синус и возвышение слабо раз-
виты. Синус переходит на лобном крае в высокий трапецеидальный язы-
чок. Соединение створок на лобном крае и по бокам зубчатое.
Поверхность створок покрыта радиальными ребрами, которые, начи-
наясь немного ниже макушки, становятся продольнобороздчатыми на
лобном крае.
В брюшной створке—тонкие зубные пластины, поддерживающие тол-
стые зубы. В спинной створке—разобщенная замочная пластина и септа,
образующая септалиум. Кардинальный 1 отросток отсутствует» (О. И. Ни-
кифорова, 1937, стр. 33).
Внешне близкими родами являются Hypothyrldtna Buck., отличаю-
щаяся отсутствием септы в спинной створке, и Unclnulus В а у 1 е, у кото-
рого имеется замочный отросток, септа не образует септалиума и замоч-
ная пластина неразделенная.
Типичный вид: Wilsonella wllsoni (Sow.).
Распространение. Верхний силур—нижний девон.
Wilsonella tarda (В а г г а и d е)
Табл. XX, фиг. 3 и 4
«Раковина двояковыпуклая, овального или округленнопятиугольного
очертания. Ширина почти равна длине. Спинная створка более выпукла,
чем брюшная, но встречаются и равновыпуклые формы Макушки
створок маленькие. Синус и возвышение отчетливо выражены только
у лобного края.
Поверхность складчатая. Складки начинаются на некотором расстоя-
нии от макушки и хорошо заметны у переднего края. Они покрывают
синус, седло и бока раковины. Примакушечная часть гладкая
Синус углубляясь и расширяясь по направлению к переднему краю,
образует там трапецеидальный язычок тем больший, чем толще рако-
вина Поверхность покрыта округленными радиальными складками,
которые становятся хорошо заметными только в передней части рако-
вины .. Примакушечная часть раковины гладкая ...» (П. С. Лазуткин,
1936, стр. 28).
Ребра в синусе и на седле более крупные, чем на боках створки.
В синусе 2—3 складки, а на седле 3—4. На боках створки распола-
гается от двух до пяти складок.
Распространение. Остракодовый горизонт Кузбасса.
Р >д Stegerhynchus F о е г s t е
Внешний вид, как у Camarotoechla Н. et Cl. (см. выше), но в спин-
ной створке имеется замочный отросток.
Типичный вид. Stegerhynchus whltll (Hall).
Распространение. Силур.
Stegerhynchus decemplicatus (So we г by) var. angaiiensis
Tchernychev
Табл XIX, фиг. 8
«Раковина небольшая, поперечнопятиугольного очертания Брюш-
ная створка умеренно выпуклая, с небольшой острой загнутой макушкой.
1 Замочный.
11 Зак. 2024
161
Синус хорошо выражен, довольно широкий, глубокий и начинается почти
у самой макушки. Язычок высокий, трапецеидальный, с острозубчатой
вершиной Спинная створка вздутая, с высоко поднятым двусклад-
чатым седлом. Складки начинаются от самой макушки, угловатые, про-
стые, разделенные промежутками, равными или несколько более широ-
кими, чем они сами. Седло образовано двумя складками, которым соот-
ветствует одна, лежащая посередине синуса.
Очень редко удается встретить образцы, у которых в синусе две
складки, а на седле три. С каждой стороны седла и синуса распола-
гается от пяти до девяти складок (обычно 7—8). Поверхность раковины
у хорошо сохранившихся экземпляров покрыта концентрическими зиг-
загообразными следами нарастания» (Б. Б. Чернышев, 1937, стр. 29).
Размеры раковины: длина 0,5—1,5 см, ширина 1 —1,5 см.
От типичного Stegerhynchus decemplicatus (Sow.), распространенного
в Европе, отличается большим числом складок и более широкой рако-
виной, имеющей ясное поперечнопятиугольное очертание.
Распространение. Верхний силур Тувы и Западной Монголии.
Stegerhynchus ulukhemensls Tchernychev
Табл. XIX, фиг 7
«Раковина небольшая, округленнопятиугольного очертания, с наи-
большей шириной, лежащей между серединой ее и лобным краем
Брюшная створка умеренно выпуклая, слегка уплощенная близ центра.
Синус неширокий, хорошо выражен и начинается на расстоянии 4—6 мм
от макушки. Язычок синуса высокий, трапецеидальный. Макушка
маленькая, острая, торчащая Спинная створка более выпуклая, чем
брюшная, часто вздутая. Седло довольно высокое, резко приподнятое,
уплощенное и начинается так же, как и синус, вблизи макушки.
Складки, покрывающие поверхность створок, начинаются от самых
макушек. У одних экземпляров они резко угловатые, у других — округ-
ленноугловатые. В синусе располагается от одной до пяти складок
(более часто 2—3), которым соответствуют от двух до шести складок,
образующих седло. С каждой стороны седла и синуса насчитывается
5—7 складок...» (Б. Б. Чернышев, 1937, стр. 33). Размеры в тех же
пределах, что и у предыдущего вида.
Распространение. Верхний силур Тувинской области.
СЕМЕЙСТВО PENTAMERIDAE M’COY
Род Conchidium L1 п п ё
Достигающие крупных размеров раковины с непрямым замочным
краем. Макушка брюшной створки длинная, клювообразная. Спинная
створка значительно меньше брюшной. В брюшной створке — спонди-
лиум и длинная срединная септа, в спинной — разобщенные септальные
пластинки. Поверхность створок покрыта радиальными ребрами. Синус
и седло обычно не выражены.
Типичный вид. Conchidium bilocular е L i n п ё.
Распространение. Верхний силур—средний девон.
Conchidium knightl S о w е г b у
Табл. XX, фиг. 2
«Большая, вздутая, неравностворчатая, более или менее удлиненная,
остроскладчатая форма. Синус и возвышение отсутствуют. Брюшная
створка сильно вздутая, изогнутая, равномерно округленная. Примаку-
шечная часть сильно вздутая, загнутая, сверху приплюснутая. Макушка
широкая, загнутая. Дельтириум узкий. Спинная створка слабо вздутая,
в центре приплюснутая, округленная.
Радиальные складки многочисленные, резкие, правильные, изредка
дихотомирующие, широкие, крышеобразные. У лобного края они очень
высокие и резкие, к макушке постепенно сглаживаются и округляются.
Число их достигает 40, но часто меньше. Следы нарастания редкие, рез-
кие. иногда ступенчатые.
В брюшной створке высокая и длинная септа, почти доходящая до
лобного края обычно она равна высоте зубных пластин, но иногда
несколько больше. Толщина септы сравнительно невелика. Поверхность
раскола ее почти гладкая, покрытая очень тонкими, более или менее
изогнутыми линиями нарастания» (Д. В. Наливкин, 1930, стр. 44).
У Conchldtum vogulicum V е г п. высота септы значительно меньше
высоты зубных пластин, линии нарастания на септе грубые и прямые,
и септа значительно толще, чем у С. knlghti Sow. Среднедевонские
формы (группа С. baschklricum Vern.), близкие по внешнему виду,
отличаются значительно более короткой и более высокой септой.
Распространение. Верхний силур Горного Алтая: повидлмому,
распространен также в верхнем силуре Салаира.
Род Gypidula Hall
Описание см. на стр. 262.
Gypidula procerula (Barrande)
Табл. XXI, фиг. 1
«Раковина развита по ширине, овального или округленноромбоидаль
ного очертания, неравностворчатая, двояковыпуклая. Замочный край
прямой, с округленными углами, короче наибольшей ширины рако-
вины.
Брюшная створка равномерно и значительно более выпуклая, чем
противоположная, правильно дугообразноизогнутая. Наибольшая выпук-
лость в средней части. Макушка небольшая, округленная, изогнутая.
Арея на одних экземплярах узкая, едва заметная, на других же отсут-
ствует или неясная. Дельтириум треугольный, открытый. Возвышение,
проходящее вдоль середины брюшной створки, становится заметным от
самой макушки или на некотором расстоянии от нее. Посередине
возвышения проходит неглубокая пологая вдавленность. На боковых
сторонах заметно по одной округленной складке. Остальная, крайняя,
часть боковой поверхности не складчатая.
Спинная створка. менее выпуклая, наибольшая выпуклость прихо-
дится на промежуточную часть.
Макушка маленькая, притупленная, слабо загнутая. Синус, начинаю-
щийся от макушки или на некотором расстоянии от нее, пологий, широ-
кий, отчетливо ограниченный с боков. У переднего края он образует
небольшой трапецоидальный язычок.
Посередине синуса проходит округленная невысокая складка, соответ-
ствующая срединной бороздке возвышения .. На боковых сторонах
створки имеется по одной широкой, округленной, слабо заметной складке,
которые у наиболее крупных экземпляров иногда дихотомируют»
(П. С. Лазуткин, 1936, стр. 26).
Кроме радиальных складок, на поверхности раковины имеются много-
численные, очень тонкие, концентрические знаки роста. Размеры: длина
от 7 до 11 мм, ширина 7—10 мм, толщина 5—9 мм.
Распространение. Остракодовый горизонт Кузбасса.
1Г 1ЬЗ
СЕМЕЙСТВО SPIRIFERIDAE KING
Род Eospirifer Schuchert
Описание см. на стр. 241.
Eospirifer tuvaensts Tchernychev
Табл. XXI, фиг. 2
«Раковина овального очертания с наибольшей шириной, лежащей
примерно на середине длины. Длина раковины обычно значительно пре-
восходит ширину; реже попадаются экземпляры, у которых она почти
равна ширине или даже немного менее ее.
Брюшная створка выпуклая, сильно изогнутая, с небольшой острой
загнутой макушкой. Синус плоский, мелкий, начинается от самой
макушки, слабо расширяется к лобному краю и на всем протяжении
остается одинаково глубоким. С боков он ограничен двумя небольшими,
слабо расходящимися или почти параллельными, слегка приподнятыми
валиками. Язычок небольшой, трапецеидальный или трапецеидально-
округленный.
Арея небольшая, треугольная, несколько короче наибольшей ширины
раковины и сильно изогнута.
Спинная створка умеренно выпуклая. . с резко приподнятым, упло-
щенным или слабо округленным седлом. Макушка очень маленькая.
Арея узкая, еле заметная.
Раковина гладкая, складки совершенно отсутствуют. Поверхность
покрыта очень тонкими нитевидными радиальными струйками, в числе
около 10—12 на 1 мм. Новые струйки возникают путем резкого дихото-
мирования. Хорошо видны концентрические следы нарастания, которые
местами становятся очень резкими, ступенчатыми» (Б. Б. Чернышев,
1937, стр. 46).
Размеры: длина 14—27 мм, ширина 18—21 мм. Внутри брюшной
створки — две толстые зубные пластины. Септы нет.
Распространение. Верхний силур Тувы и Монголии.
Род Howellella Kozlowsky
Синоним — Crispella К о z 1.
Во всех отношениях подобен роду Delthyrls D а 1 m. (см. стр. 241), но
без срединной перегородки в брюшной створке.
Типичный вид. Howellella crispa (Н i s.).
Распространение. Верхний силур.
Howellella crispa (Н i s i n g е г)
Табл. XX, фиг. 5
«Раковина маленькая, поперечноовального очертания, с округлен-
ными замочными углами. Наибольшая ширина ее лежит около середины
длины.
Брюшная створка более выпуклая, чем спинная, с небольшой уме-
ренно загнутой макушкой. Арея небольшая, вогнутая. Синус хорошо
выражен, раза в два шире первых межреберных пространств.
Спинная створка. с тупой маленькой макушкой и чрезвычайно
маленькой ареей. Седло отчетливо выражено, округлое, но очень широ-
кое. С каждой стороны седла располагаются 2—3 округлые простые
толстые складки, а по обе стороны синуса 3—4» (Б. Б. Чернышев, 1937,
стр. 49).
Кроме радиальной скульптуры, поверхность створок покрыта черепит-
чатыми тонкими концентрическими пластинками, по краю которых, пер-
164
пендикулярно им, располагаются тонкие шипики. При сближенных
пластинах создается впечатление прерывистой радиальной струйчатости.
Распространение. Верхний силур Монголии и Тувы, острако-
довый горизонт Кузбасса.
Howellella angustiplicata (К о z 1 о w s к у)
Табл. XX, фиг. 6
Двояковыпуклые, неравностворчатые раковины округленноромбои-
дального или поперечноовального очертания, с прямым замочным краем,
равным наибольшей ширине раковины или чуть меньше.
Макушка брюшной створки маленькая. Арея треугольная, вогнутая.
Дельтириум треугольный. Синус начинается от макушки; ограничиваю-
щие его складки более широкие и отчетливые, чем остальные.
Арея спинной створки едва заметна. Высокое и округленное или
трапецоидальное седло начинается от макушки и заметно расширяется
у переднего края. Органичивающие седло бороздки более глубокие
и широкие, чем остальные. Седло и синус гладкие. На боковых частях
створок количество складок варьирует: чаще на брюшной створке насчи-
тывается четыре ребра, реже три и пять (не считая складок, ограничи-
вающих синус), а на спинной—4—5, реже три.
Кроме радиальных складок, поверхность створок покрыта тонкими
черепитчато расположенными концентрическими пластинами, ширина
которых уменьшается у переднего края. В средней части раковины их
насчитывается 4—5 на 1 мм длины, а у переднего края 7—8. По краю
каждой пластины имеются вертикально расположенные шипики. Когда
концентрические пластины сближены, создается впечатление, что рако-
вина орнаментирована прерывистыми радиальными струйками.
Размеры: длина не больше 14 мм, ширина 19 мм.
От Howellella crlspa Hi s. отличается большим количеством складок
на боках раковины, более широкой ареей, менее правильными концентри-
ческими пластинами и более плоским седлом спинной створки.
Распространение. Остракодовый горизонт Кузбасса.
СЕМЕЙСТВО ATRYPIDAE GILL
Род Nativkinia Bublitchenko
«Форма раковины яйцевидная или яйцевидно приплющенная. Обе
створки приблизительно равновыпуклые. Замочный край короткий
й изогнутый. Макушка брюшной створки умеренной величины, снаб-
жена на вершине отверстием. В передней части створки наблюдается
большей частью слабо выраженное срединное углубление. На спин-
ной ему соответствует слабо намечающееся срединное возвышение.
Поверхность радиальноребристая» (Н. Л. Бубличонко, 1927, стр. 982)
Характерной особенностью рода является отсутствие концентри
ческой скульптуры. В брюшной створке — зубы и хорошо развитые зуб-
ные пластины; в спинной — слабо развитая срединная септа. Спираль-
ные конусы ручных поддержек обращены вершинами к центру спин-
кой створки.
Род Grilnewaldtia Т s с h е г п. отличается формой раковины (брюшная
створка более выпуклая), наличием наряду с радиальной скульптурой
скульптуры концентрической и внутренним строением (вершины конусов
ручного аппарата обращены в сторону брюшной створки). От рода
Atrypa D а 1 m. отличается присутствием зубных пластин, равновыпук-
лыми створками и отсутствием концентрической скульптуры.
Типичный вид. Nallvklnia grunewaldttaeformls (Peetz)
Распространение. Верхний силур. Кузбасс.
165
Nallvklnta griinewaldtiaeiormis (Peetz)
Табл. XX, фиг. 7
Видовые признаки отвечают приведенному выше родовому диагнозу
Размеры: длина 14—26 мм, ширина 13—22 мм, толщина 8—16 мм.
Совместно встречающаяся Nallvklnta slblrica В u b 1. отличается более
вздутыми створками и более развитыми седлом и синусом.
Распространение. Верхи S2. с. Сары-Чумыш.
СЕМЕЙСТВО ATHYRIDAE PHILLIPS
Род Protathyris Kozlowsky
«Двояковыпуклая, сильно варьирующая в контурах раковина. Брюш-
ная створка без ареи, макушка с треугольным отверстием для ножки.
В типичных видах синус развит на обеих створках, но может отсутство-
вать как на одной из них, так и на обеих. Поверхность створок покрыта
простыми концентрическими морщинами, сгущающимися у лобного края.
В брюшной створке зубы поддерживаются вогнутыми зубными пла-
стинами. .. В спинной — большая треугольная замочная пластина, про-
дырявленная висцеральным форамоном» (О. И. Никифорова, 1937
стр. 61).
От рода Whltfieldla Hall et Clarke отличается отсутствием ясно
развитой срединной септы и септалиума.
Типичный вид. Protathyris praecursor К о z 1.
Распространение. Верхний силур.
Protathyris didyma (D а 1 m a n)
Табл. XXI, фиг. 3 и 4
«Внешне очертания раковины различны, но чаще всего они бывают
округленнопятиугольной или округленноромбоидальной формы. Обычно
они несколько больше вытянуты в длину, но часто длина и ширина их
почти одинаковы или даже, в некоторых случаях, последняя бывает не-
сколько больше. Наибольшая ширина соответствует середине раковины.
Обе створки выпуклые, причем спинная более выпукла, чем брюшная,
и выпуклость больше у удлиненных форм, чем у широких. .. Синус
брюшной створки выражен отчетливо и начинается обычно от середины
раковины. .. Макушка брюшной створки небольшая, загнутая. На
спинной створке имеется слабо выраженное седло, более заметное у пе-
реднего края. Иногда на середине седла наблюдается продольная вдав-
ленность, более замепная у переднего края» (П. С. Лазуткин, 1936,
стр. 48). Поверхность покрыта концентрическими пластинками.
Размеры: длина раковины не превышает 13,5 мм, ширина 13 мм.
Распространение. Остракодовый горизонт Кузбасса.
41. П. Л ОМОВ И ИКАЯ
Тип Arthropoda. Членистоногие
КЛАСС TRILOBITA. ТРИЛОБИТЫ
К настоящему времени изучены трилобиты верхнего тремадока Гор-
ной Шории и веберовского горизонта (карадок) Салаира. Последние
описаны в опубликованных работах В. Н. Вебера (1923, 1928) и М. П. Ло-
мовицкой (1936).
Ниже приводятся описания важнейших трилобитов обоих горизонтов.
Иногда описания основываются на определении очень неполных экзем-
пляров; в этих случаях даются не диагнозы соответствующих видов,
а описания фактических находок.
166
СЕМЕЙСТВО SOLENOPLEURIDAE ANGELIN
Род Solenopieura Angelin
Описание см. на стр. 113.
Solenopieura amsassensis Poletaeva, sp. n.
Табл. XXI, фиг. 6
Головной щит небольшого размера, с субтреугольным очертанием,
г усеченным и сильно округленным передним краем. Он сильно выпук-
лый, поэтому легко подвергается деформации. Глабель очень сильно
вздута и значительно возвышается над щеками. Спереди глабель равно-
мерно округлена. Глабелярные борозды на наших образцах не наблюда-
лись. Затылочное кольцо выражено резко, узкое. Против неподвижных
щек на заднебоковой кайме наблюдаются небольшие вздутия, опущенные
назад и соответствующие перегибу книзу плевральных частей туловища.
Ни шипа, ни бугорка на затылочном кольце не имеется. Затылочная бо-
роздка глубокая. Краевой валик выпуклый, неширокий.
Неподвижные щеки неширокие, сильно выпуклые, с крутыми боками,
соединяющиеся перед глабелью. Глазные крышки маленькие, располо-
жены почти посередине длины головного щита, немного ближе к его
заднему краю. Глазные валики тонкие, но ясные, расположены слегка
косо. Передние ветви лицевого шва косо режут с боков краевой валик.
Скульптура мелкоточечная; бывает видна очень редко. Длина головного
щита 4,0 мм; ширина 3,8—4,7 мм.
От близкого вида Solenopieura ctncta Lerm., распространенного
в верхнем кембрии Северо-Восточного Казахстана и в нижнетремадок-
ских отложениях северо-западной оконечности Кузнецкого Алатау, отли-
чается большей величиной.
Распространение. Верхний тремадок. Горная Шория.
р. Амзас.
СЕМЕЙСТВО OLENIDAE BURMEISTER
Род Parabolinella В г б g g е г
Глабель прямоугольного очертания, иногда слегка суживающаяся
спереди, с 2—3 парами бороздок, из которых две последние (задние)
пары очень наклонные. Предглабелярная площадка очень большая. Глаза
находятся почти на середине расстояния между передним и задним краем
головного щита; с передним углом глабели их соединяют глазные валики.
Лицевые швы идут внутрь и назад от переднего края к глазам и от-
сюда — наружу и назад к заднему краю. Число члеников туловища и хво-
стового щита не менее 14. Хвостовой щит широкий, округлый, окружен-
ный каймой, с плеврами, не имеющими остроконечий.
Распространение. Верхний кембрий — тремадок.
Parabolinella argentinensis Kobayashi
Табл. XXI, фиг. 7
Головной щит полукруглого очертания, с длинными щечными остро-
конечиями. На глабели насчитывается три пары наклонных борозд. За-
тылочное кольцо узкое, с срединным бугорком. Предглабелярная пло-
щадка выпуклая, с узким краевым валиком. Глаза расположены немного
ближе к переднему краю головного щита (по отношению к его сере-
дине), очень близко к глабели и соединяются с ее передними углами вы-
пуклыми наперед валиками. Передние ветви лицевого шва идут от перед-
него края внутрь к глазам. На некоторых участках головного щита
имеется струйчатость. Длина головного щита 4,5 мм, ширина 8,5 мм.
167
Распространение. Тремадок. В Западной Сибири — Горная'
Шория, р. Амзас.
СЕМЕЙСТВО DIKELOCEPHALIDAE MILLER
Род Dikelocephalopsis Poletaeva
Крупные трилобиты с широким головным щитом, с длинными щеч-
ными остроконечиями. Глабель угловатая, усеченноконической формы,
с тремя парами коротких борозд. Предглабелярная площадка более или
менее вогнутая. Краевая кайма очень широкая и плоская. Неподвижные
щеки довольно широкие, сливаются с предглабелярной площадкой. Глаза
большие. Имеются глазные валики. Лицевые швы впереди глаз вначале
несколько расходятся, а затем круто поворачивают друг к другу и сли-
ваются на краевой кайме. Позади глаз они значительно отходят один
от другого и косо идут назад.
Хвостовой щит такой же величины, как и головной, округленнопяти-
угольной формы. Его плевры сильно бороздчаты; плевральные бороздки
широкие, интерплевральные — узкие, более слабые. Осевая часть хвосто-
вого щита спереди широкая, но довольно быстро суживается и закруг-
ляется, далеко не доходя до заднего края. На ней насчитывается
5—6 сегментов. Краевая кайма хвостового щита широкая, слегка вогну-
тая, вытянутая сзади в два остроконечия, промежуток между которыми
имеет округленнотреугольную форму.
Типичный вид. Dikelocephalopsis amzassensls Pol.
Распространение. Верхний тремадок.
Dikelocephalopsis amzassensls Poletaeva, sp. п.
Табл. XXI, фиг. 5
Головной щит значительной величины, широкий у основания и на
одну треть суживающийся спереди. Глабель типичной для рода формы,
с тремя парами коротких борозд, с большим между ними промежутком.
Задняя пара, самая длинная, имеет вид двух глубоких, косо назад на-
правленных продолговатых ямочек, не доходящих до спинных борозд.
Средняя пара менее глубокая и более короткая, в виде слегка вытяну-
тых ямочек, также не соединенных со спинными бороздками и направ-
ленных несколько косо назад и к середине глабели. Третья пара борозд
также представляет собой ямочки, еще менее глубокие и совсем корот-
кие. Затылочная бороздка довольно глубокая. Краевая кайма очень ши-
рокая и плоская.
Неподвижные щеки довольно широкие, сливаются с предглабелярной
площадкой. Глазные крышки не сохранились, но, судя по расположению
глазных валиков и форме лицевых швов, они широко расставлены, рас-
положены посередине длины головного щита и имеют среднюю вели-
чину. Лицевые швы впереди глаз вначале несколько расходятся, а затем
круто поворачивают друг к другу, сливаются и образуют спереди на
краевой кайме тупой угол. Позади глаз они значительно отходят один от
другого, косо идут назад. На предглабелярной площадке сохранились
следы поперечной струйчатости.
Хвостовой щит одинаковой величины с головным, округленнопяти-
угольной формы, с двумя широкими остроконечиями. Плевры сильно
бороздчаты. В передней половине хвостового щита плевральная часть
является наибольшей частью сегмента; в задней его половине ширина
плевр сокращается. В передней части хвостового щита плевры имеют ко-
ленчатый изгиб; задние сегменты отгибаются назад без коленчатого
перегиба. Осевая часть спереди широкая, суживающаяся в задней части,
с 5—6 сегментами. Лимб широкий, слегка вогнутый. Сзади он становится
еще шире и вытянут в два короткие острые прямые шипа, имеющие очень
168
широкие основания. Лимб покрыт тонкой и слабой поперечной струй-
чатостью. Длина головного щита 16,1 мм, ширина 31,5 мм.
Распространение. Верхний тремадок, Горная Шория, р. Амзас
СЕМЕЙСТВО ASAPHIDAE BURMEISTER
Род Promegalaspid.es Westergard
Щит трилобита плоский или умеренно выпуклый. Головной щит полу-
овальный. Глабель цилиндрическая, часто слабо расширяющаяся вперед,
с округленными передними углами. У основания она несколько расши-
рена за счет выдающихся на неподвижные щеки базальных лопастей.
Иногда на глабели замечается слабый продольный киль. Глабелярные
борозды в большинстве случаев имеются, но часто плохо заметны. Зад-
ние глабелярные (базальные) лопасти, по сравнению с остальными, раз-
виты и ограничены хорошо, выступают за пределы глабели и имеют суб-
овальную или треугольную форму.
Затылочное кольцо и затылочная борозда выражены слабо. Спинные
борозды неглубокие, но четкие. Краевая кайма узкая, плоская как на
головном, так и на хвостовом щитах и отделяется бороздкой. Подвижные
щеки заканчиваются длинными остроконечиями. Глаза большие полу-
лунные. Передние ветви лицевого шва краевые или почти краевые;
встречаются у переднего края, образуя тупой угол.
Краевая кайма на ядрах покрыта струйчатостью. Плевры туловища
с диагональными бороздками на сегментах, выклинивающимися или
к концу сегментов или к их перегибу. Длинные шипы последнего туло-
вищного сегмента — характерный признак рода. Спинные бороздки вы-
ражены хорошо.
Осевая туловищная лопасть равняется примерно одной трети ширины
всего туловища, хорошо отграничена от плевр. Хвост довольно большой.
На осевой лопасти наблюдаются 3—4 сегмента, разделенные хорошо вы-
раженными бороздами; остальные сегменты слились. Сегменты плевр
с отчетливыми бороздками. На краях плевр имеются валики.
От типичных представителей рода Promegalaspldes West., распро-
страненных в верхнем кембрии Швеции, представители рода Promegala-
spldes из Горной Шории отличаются меньшим количеством сегментов на
осевой лопасти хвоста и отсутствием бугорка на глабели.
Распространение. Верхний кембрий, тремадок.
Promegalaspldes robustus Lomovitskaja, sp. n.
Табл. ХХИ, фиг. 9
Глабель взрослых экземпляров продолговатая, очень слабо расширяю-
щаяся вперед и более заметно — назад, что происходит за счет сравни-
тельно сильно выдающихся задних глабелярных (базальных) лопастей.
Глабель более молодых экземпляров менее вытянута в длину и еще
менее расширяется в передней части. У большинства образцов на глабели
намечается продольный киль и с каждой стороны насчитывается по пять
поперечных борозд. Задние глабелярные лопасти овальные и сравни-
тельно сильно развиты; кроме них имеются еще четыре пары глабеляр-
ных лопастей, тоже овального очертания.
Передние ветви лицевого шва типичны для данного рода. Задние
ветви, направляясь назад и в то же время слабо кнаружи от задних кон-
цов глаз, очень скоро затем резко поворачивают кнаружи и некоторое
время идут параллельно или почти параллельно заднебоковой борозде;
наконец, они довольно круто поворачивают назад и секут заднебоковую
борозду под острым углом.
Отличительными признаками вида являются: сравнительно большие
размеры, пять пар овальных боковых лопастей глабели, из которых зад-
169
ние вздутые и большие. Длина головных щитов от 11,4 до 22,0 мм;
ширина до 32,0 мм.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Promegalasptd.es angeltnl Lomovitskaja, sp. n.
Табл. XXII, фиг. 11
Глабель удлиненная, немного расширена и слабо вздута в передней
части, занимающей несколько более одной трети всей глабели. У основа-
ния глабель также значительно расширена. Борозды глабели выражены
слабо и на наших образцах, представленных ядрами, различаются с тру-
дом. Они узкие и неглубокие, по четыре с каждой стороны и образуют
каждая слабо выпуклую вперед дугу. Первая и вторая (считая от перед-
него края глабели) борозды глабели короткие, достигают примерно одной
трети ширины глабели, слабо опущены назад. Третья пара очень длин-
ная, доходящая или почти доходящая до затылочного кольца, косо на-
правлена назад. Четвертая пара борозд, ограничивающая базальные ло-
пасти, косо опущена назад.
Базальная лопасть овальная, выражена для данного рода хорошо.
У Р. angeltnl наблюдается своеобразное развитие базальной лопасти:
с каждой стороны глабели обнаруживается еще маленькая дополнитель-
ная лопасть грушевидной формы, с направленной назад более широкой
частью. Она находится в углу между задним краем глазной крышки
и глабелью и ограничивается с одной стороны спинной бороздой и с дру-
гой — задней ветвью лицевого шва. Затылочное кольцо в середине за-
метно расширяется и образует выступ назад, суживаясь на концах.
Направление передних ветвей лицевого шва типично для данного
рода. Задние ветви идут от нижнего угла глаз косо назад, по направле-
нию к заднему углу головного щита, на очень близком расстоянии от
заднебоковой борозды, затем под косым углом секут последнюю. Хво-
стовые щиты имеют полуовальную форму. Плевральные части хвоста,
как и у остальных горношорских видов данного рода, совершенно отчет-
ливые, несут посередине мелкую широкую борозду.
Осевая часть, быстро суживаясь, доходит до краевой каймы. Ее
первые четыре сегмента отчетливо разделены бороздами, остальные
слиты. Краевая кайма довольно широкая, слегка вогнутая. Наружная
поверхность ее гладкая; на внутренней замечается отчетливая струйча-
тость.
Отличительными признаками вида являются: довольно большие раз-
меры, неширокая глабель и характер ее борозд, дополнительные базаль-
ные лопасти глабели и направление задней ветви лицевого шва.
Длина головного щита 20,2 мм, ширина 30,0 мм.
Отличия от Promegalaspldes majusculus Lomov, приводятся при
описании последнего.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Promegalaspldes majusculus Lomovitskaja, sp. n.
Табл. XXII, фиг. 12
Крупный трилобит, по величине превышающий все другие известные
виды сибирских Promegalaspldes. Глабель в фронтальной части несколько
расширяется и слабо вздувается. Основание глабели также в незначи-
тельной степени расширяется за счет выступающих, вздутых, округлых
базальных лопастей. Выше базальной наблюдается еше одна лопасть —
овальная, менее выпуклая, чем базальная, и меньших размеров. Имеется
по четыре боковых борозды глабели. Затылочное кольцо суживается
к краям глабели и в середине сильно выдается назад.
Впереди глабели, очень близко к ней, наблюдается плоская краевая
кайма. Передняя ветвь лицевого шва краевая. Задняя ветвь идет от ниж-
.1.70
него угла глаз по кривой непосредственно по направлению к заднебоко-
вому углу кранидия.
Длина глабели 15—19 мм, ширина 10—12 мм.
Отличительными признаками вида являются сравнительно крупные
размеры и направление задней ветви лицевого шва от нижнего угла глаз
непосредственно к внутреннему, а не к внешнему краю головного шита.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Promegalasptdes elongatus L о m о v i t s k a j a, sp. n.
Табл. XXII, фиг. 10
Глабель длинная, узкая, с почти параллельными боковыми краями,
не расширяющаяся вперед. Следов глабелярных борозд и боковых лопа-
стей глабели не обнаруживается: повидимому, они были развиты слабо
или совершенно отсутствовали. Базальные лопасти глабели имеют суб-
треугольную форму, с вершинкой, направленной вперед, вытянуты
в длину, слабо вздуты и не сильно выдаются вбок. Вследствие этого
глабель у основания расширяется только в незначительной степени.
Передние ветви лицевого шва, встречаясь перед глабелью под тупым
углом, образуют небольшое остроконечие. Задние ветви от нижнего угла
глаз направляются к наружной боковой стороне головного щита, затем
поворачивают и секут под острым углом заднебоковую борозду.
Длина головных щитов 15,5—16,0 мм, ширина 10,5 мм.
Отличительными признаками вида являются: удлиненная глабель, от-
сутствие боковых лопастей глабели, слабо вздутые, вытянутые в длину
базальные лопасти, направление задней ветви лицевого шва.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Род Maja Lomovitskaja, gen. п.
Данный род очень близок к описанному выше роду Promegalasptdes
West.
Для рода Maja характерна совокупность следующих признаков: длин-
ные щечные остроконечия; субцилиндрическая форма глабели; наличие
2—3 пар четких глабелярных борозд, причем каждая бороздка последней
пары имеет дугообразную форму с выпуклостью, обращенной к спинным
бороздкам, и идет почти параллельно последним; наличие на более или
менее близком расстоянии от глабели краевого валика, узкого и выпук-
лого, отделенного четкой бороздкой; узкая ось туловища; отчетливые
борозды на плевральных сегментах туловища; узкая ось хвостового щита
с восьмью четкими сегментами; отчетливые борозды на плеврах хвосто-
вого щита; узкая краевая кайма хвостового щита.
Род Maja отличается от рода Promegalasptdes меньшими размерами,
характером последней пары глабелярных борозд, наличием вблизи гла-
бели выпуклого краевого валика, меньшим расширением глабели у осно-
вания, более узким рахисом туловища, вдвое большим количеством сег-
ментов на рахисе хвостового щита и некоторыми менее существенными
деталями.
Типичный вид. Maja subsulcala Lomov.
Распространение. Верхний тремадок.
Maja subsulcala Lomovitskaja, sp. n.
Табл. XXI, фиг. 9
Глабель близкая к цилиндрической, удлиненная, с округленными пе-
редними углами, слабо расширяющаяся к передней своей части и еще
более слабо — у основания. На глабели наблюдается слабый продоль-
ный киль. Имеется три пары глабелярных борозд. Третья пара борозд,
ближайшая к затылочной борозде, вертикальная и характерна для рода
п \
в целом. Вторая — резкая, четкая, хотя и неглубокая, доходит до спин-
ных борозд; каждая борозда слабо направлена назад. Бороздки перед-
ней пары не доходят до спинных борозд, очень короткие и имеют вид
маленьких прямых черточек.
Боковые лопасти очень неотчетливы и неопределенны. Затылочное
кольцо в средней части несколько опущено назад. Затылочная и спинные
бороздки узкие, неглубокие, но четкие. На очень небольшое расстоянии
от глабели, отделяясь от нее четкой бороздкой, проходит неширокий,
слабо выпуклый краевой валик. Лицевые швы неизвестны.
Длина головного щита 9 мм.
Отличительными признаками вида являются удлиненная глабель со
слабым расширением в передней ее части и у основания, чуть намечен-
ный на глабели киль, слабо выпуклый краевой валик, расположенный
близко к глабели, и затылочное кольцо, направленное в своей средней
части назад.
Очень близкими к данному виду являются Maja sulcata Lomov,
и Maja venusta Lomov. Сравнения приводятся при описании этих видов.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Maja sulcata Lomovitskaja, sp. n.
Табл. XXI, фиг. 8
Данный вид тесно связан с Maja subsulcata Lomov.
Отличительными признаками Maja sulcata являются: широкая глабель
без киля, три пары глабелярных лопастей, краевой валик, расположен-
ный почти вплотную к глабели. Длина головного щита 12 мм.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Maja venusta Lomovitskaja, sp. n.
Табл. XXI, фиг 11
Глабель несколько расширяется вперед. Борозды и лопасти глабели
аналогичны таковым у М. subsulcata Lomov., за исключением того, что
борозд у описываемого вида на одну пару меньше. Этот вид очень бли-
зок к виду Maja subsulcata Lomov.
Отличительными признаками М. venusta от последнего вида являются:
большие размеры, большая ширина глабели, отсутствие сужения средней
части глабели и меньшее, на одну пару, количество борозд глабели.
Кроме того, вторая пара глабелярных борозд здесь не доходит до спин-
ных бороздок. В целом отличительными признаками вида являются:
сравнительно широкая глабель, не суживающаяся в средней части, и ха-
рактер затылочного кольца: на его концах имеются вздутия, а средняя
часть уплощена.
Распространение. Верхний тремадок. Горная Шория, р. Амзас
Maja cyllndrtca Lomovitskaja, sp. n.
Табл. XXI, фиг. 10
Вид очень близок к М. subsulcata L omov., но отличается цилиндри-
ческой формой глабели, не суживающейся в средней части. Кроме того,
у М. cyllndrica вторая пара глабелярных борозд не доходит до спинных,
как это имеет место у М. subsulcata. Полностью сохранившийся хвосто-
вой щит с последним туловищным сегментом вполне соответствует родо-
вой характеристике. Длина головного щита 10,5 мм.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
СЕМЕЙСТВО TRINUCLEIDAE EMMRICH
Род Clavatellus Poletaeva, gen. n.
Трилобит небольших размеров. Глабель сильно расширяется в сере-
дине или около середины в передней ее половине и суживается спереди
172
и сзади, причем сзади значительно сильнее. Таким образом, глабель при-
обретает грушевидную или булавовидную форму. Борозды глабели
ясные, не сливающиеся посередине. Лимб перед глабелью достигает зна-
чительной ширины и имеет продольное вздутие на середине головного
шита.
Неподвижные щеки широкие. Глаза маленькие, широко расставлен-
ные. Глазные валики отчетливые, прямые, горизонтальные, примыкают
к наиболее широкой части глабели. Осевая лопасть узкая. Плевры туло-
вища широкие, с прямыми продольными бороздками на каждом сегменте,
без шипов. Пигидий маленький, широкий.
Описываемый род очень близок к роду Orometopus В г о g., отли-
чаясь от него следующими признаками: наличием ясных глабелярных
борозд, более сильным расширением глабели спереди, наличием значи-
тельной ширины лимба перед глабелью и наличием продольного вздутия
на лимбе.
Типичный вид. Clavatellus globosus Pol.
Распространение. Верхний тремадок.
Clavatellus globosus Poletaeva, sp. n.
Табл. XXII, фиг. 1
Головной щит у основания очень широкий и наполовину суживаю-
щийся на уровне глаз. Глабель сильно сужена в задней части и более
чем в полтора раза расширяется в передней половине, но к переднему
концу она опять несколько суживается и сильно закругляется, приоб-
ретая грушевидную форму. Спереди она усечена, а у некоторых образцов
даже слабо вогнута. На боках глабели имеются три пары отчетливых,
очень коротких, но довольно глубоких бороздок. Затылочное кольцо узкое,
ясно выраженное. Заднебоковые бороздки на всем протяжении четкие,
тонкие и довольно глубокие, затылочная борозда несколько мельче.
Спинные бороздки узкие, но глубокие и везде одинаково четкие.
Неподвижные щеки широкие, плоские. Ниже глазных валиков на них
заметна слабая тонкая скульптура в виде струек, направленных от гла-
бели к наружным нижним углам головного щита. Неподвижные щеки
сливаются спереди в широкий лимб, окружающий глабель.
Против середины глабели на лимбе наблюдается продольный киль.
Глазные крышки маленькие, расположенные приблизительно посередине
длины головного щита. Глазные валики тонкие, очень четкие, прямые
и горизонтальные, примыкают к наиболее широкой части глабели. Лице-
вые швы впереди глаз идут вперед, параллельно друг другу, а позади
глаз направлены наклонно к наружным углам головного щита.
Длина головного щита 2,8 мм, ширина у основания 8,5 мм.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
Clavatellus amsassensts Р о 1 с t а е v a, sp. п.
Табл. XXII, фиг. 2
Спинной щит плоский, широкий, имеет малые размеры и яйцевидную
форму.
Головной щит очень близок к таковому Clavatellus globosus Pol.,
отличаясь от него следующими признаками: глабель еще сильнее сужена
сзади, имеет только две пары борозд—таких же коротких и глубоких, как
у С. globosus, но более заметно наклоненных назад; на лимбе
впереди глабели киль проявляется более ясно; осевая часть туловища
очень узкая, невысокая; плевральные части широкие, плоские, без шипов,
с продольными, тонкими прямыми бороздками на сегментах; пигидий не-
большой, треугольный.
Ширина головного щита у основания 6,4 мм, длина 2,8 мм.
Распространение. Верхний тремадок. Горная Шория, р. Амзас.
173
Род Orometopus В г б g g е г
Мелкие трилобиты с большим головным щитом. Глабель суживается
у основания; глабелярные бороздки, не доходящие до ее середины. Не-
подвижные щеки широкие. Глаза небольшие. Глазные валики выражены
хорошо. Пигидий широкий, маленький.
Распространение. Кембрий — нижний силур.
Orometopus subpraenuntlus Poletaeva, sp. n.
Табл. XXII, фиг. 3
Головной шит значительно шире туловища, широкий у основания
и значительно более узкий спереди. Глабель удлиненная, суживающаяся
у основания и угловатоокруглая спереди. По бокам ее у большинства
образцов имеются по три коротких широких бороздки, далеко не доходя-
щих до середины глабели. Задняя пара борозд заметно наклонена назад.
Затылочное кольцо резко выраженное, с высоким бугорком на середине.
Затылочная и заднебоковые борозды сплошные. Глабель окружена ши-
рокими, довольно глубокими спинными бороздками, которые впереди
глабели много мельче, но остаются отчетливыми. Перед глабелью
наблюдается широкий лимб, перегибающийся вниз под прямым или
тупым углом.
Неподвижные щеки широкие, сливающиеся с лимбом впереди гла-
бели. У основания глабели на неподвижных щеках иногда бывают видны
небольшие вздутия. Глазные крышки небольшие, имеют форму запятой.
Глазные валики расположены поперек неподвижных щек, слегка вы-
пуклы вперед. Лицевые швы впереди глаз постепенно поворачивают друг
к другу, но, повидимому, не сливаются. Позади глаз лицевые швы рас-
ходятся наружу, а затем резко поворачивают назад.
Туловище и хвостовой щит малого размера и вместе взятые имеют
яйцевидную форму с несколько суживающимся задним концом. Туло-
вище состоит из десяти сегментов, тупо усеченных у своих наружных
концов. Каждый сегмент плевр имеет широкую и не очень глубокую бо-
роздку. Кончики первых двух сегментов немного опущены назад,
а остальные постепенно выпрямляются до горизонтального положения.
Хвостовой щит очень широкий, состоит из четырех сегментов.
Ширина головных щитов у основания от 4,3 до 7,8 мм; длина от 1,5
до 2,5 мм.
Распространение. Верхний тремадок. Горная
Шория, р. Амзас.
Рис. 168. Род lllae-
nus D a I m а п.
Общий вид; схема.
По В. Н. Веберу
Типичный
СЕМЕЙСТВО 1LLAENIDAE HAWLE ET CORDA
Род lllaenus D а 1 m а п
Рис. 168
Головной и хвостовой щиты широкие и короткие.,
без лимба и краевой каймы. Глабель ограничена не-
ясно, глабелярные борозды отсутствуют, спинные бо-
розды очень короткие. Глаза небольшие, гладкие. Ту-
ловище состоит из 8—10 сегментов. Хвостовой щит
без ребер, с короткой, слабо намеченной осевой ло-
пастью, не ограниченной сзади.
вид: lllaenus crasslcaudatus Wahl.
Распространение. Силур, повсеместно.
lllaenus salairicus Weber
Табл. XXII, фиг. 5
Головной щит сильно выпуклый, причем в продольном направлении
выпуклость больше, чем в поперечном. Глабель имеет самостоятельную,
174
выпуклость только в задней части, где она ограничена короткими спин-
ными бороздками, не доходящими до линии, соединяющей глаза, и слабо
расходящимися сзади. Глаза удлиненные, слабо выпуклые. Для вида ха-
рактерно, что глаза сильно смещены вперед, ближе к переднему, чем
к заднему краю головного щита.
Туловище состоит из десяти сегментов. Осевая его лопасть (рахис)
почти вдвое шире боковых (плевральных), равномерно и сильно вы-
пукла. Боковые части резко перегнуты почти под прямым углом. Длина
туловища равна ширине осевой лопасти. Хвостовой щит слабо выпуклый,
широкий; осевая лопасть намечена только широко расставленными вдав-
ленностями на переднем крае щита.
Длина головного щита 5,3 мм, ширина 8,1 мм.
Распространение. Карадок. Салаир, р. Черневой Бачат.
Illaenus ovtfortnis Warburg
Табл. XXII, фиг. 6
Головной щит широкий, сильно продольно перегнутый. Резкий пере-
.иб профиля характерен для данного вида: передняя часть головного
щита, а также задняя и боковые части хвостового круто загибаются вниз.
Глабель в задней части сильно выпуклая. Спинные борозды головного
щита очень отчетливые. Глаза расположены вблизи заднего края голов-
ного щита. У хвостового щита имеется широкий заворот.
Длина головного щита 8,2 мм, ширина 11,0 мм.
Распространение. Карадок. В Западной Сибири—Салаир,
р. Черневой Бачат.
Род Bumastus Murchison
Рис. 169
Очень близок к роду Illaenus (см. выше), отличаясь от него в основ-
ном очень широкой осевой лопастью, почти не отделенной от бо-
ковых частей. Некоторые виды имеют вогнутую кай-
му на хвостовом- щите, изредка встречается узенькая
кайма на головном щите. Осевая лопасть хвостового
щита не очерчена.
Типичный вид. Bumastus barrlensls М u г с h.
Распространение. Силур, повсеместно.
Bumastus maccalumi Salter
Табл. XXII, фиг. 4
Хвостовой щит параболической формы, слабо вы-
пуклый в продольном направлении и сильно — в по-
перечном. Передний край сильно изогнут наперед. На
поверхности наблюдаются правильно расположенные
многочисленные ямки.
Рис. 169. Род Би-
тах tus М и г с h.
Общий вид. Нат.
вел. По В. Н. Уэл-
леру
Длина хвостовых щитов 18,0 и 11,5 мм, ширина 19,5 и 11,0 мм.
Распространение. Карадок. В Западной Сибири—Салаир,
Черневой Бачат.
СЕМЕЙСТВО HOLOTRACHELIDAE WARBURG
Род Holotrachelus Linnarsson
Головной щит выпуклый, с краевой каймой. Глабель выпуклая, без
борозд. Глаза расположены в передней половине головного щита. Зад-
няя ветвь лицевого шва пересекает задний край головного щита около
щечных углов. Плевры туловища с бороздками. Хвостовой щит малень-
кий. с плеврами, оканчивающимися свободно.
175
Типичный вид. Holotrachelus punctlllosus Т 6 г п q.
Распространение. Верхи нижнего силура.
Holotrachelus punctlllosus Т б г n q u i s t
Рис. 170
Глабель яйцевидная, сильно выпуклая в обоих направлениях, без бо-
розд, со слабым продольным килем. Спереди глабель круто понижается.
Наибольшая ширина затылочного кольца находится на его середине. За-
тылочное кольцо очень слабо
отделено от глабели, так как
затылочная борозда выражена
очень плохо и более углублена
лишь у боков глабели. Задне-
боковая борозда широкая и
глубокая. Впереди глабели
Рис. 170. Holotrachelus punctlllosus имеется выпуклая неширокая
Tfirnqu. Глабель, х 1.5-,По В. Н. Веберу вертикальная предглабелярная
площадка, ограниченная спе-
реди выпуклым круглым валиком, покрытым продольными струйками.
Неподвижные щеки выпуклые. На них заднебоковой бороздой отде-
ляется выпуклая кайма, расширяющаяся кнаружи; последняя при
взгляде сбоку имеет вид площадки, наклоненной вперед к затылочной
борозде щеки. Глаза большие, расположены в передней части головного
щита. Лицевые швы пересекают задний край головного щита у щечных
закругленных углов. Скульптура точечная.
Длина головных щитов от 3,5 до 30,0 мм.
Распространение. Карадок. В Западной Сибири—Салаир.
р. Черневой Бачат.
СЕМЕЙСТВО ISOCOLIDAE ANGELIN
Род Isocolus Angelin
Головной щит выпуклый, глабель с двумя парами глубоких борозд.
Маленькие глаза расположены близко к переднему краю глабели. Туло-
вище имеет шесть сегментов. Хвостовой щит полукруглый, с немногими
сегментами.
Типичный вид: Isocolus sjogreni A n g.
Распространение. Верхи нижнего силура.
Isocolus aff. sjogreni Angelin
Табл. XXII, фиг. 7
Головной щит полуцилиндрический, выпуклый, с отвесными боками
и передней частью. Поперечных борозд глабели имеются две пары;
третья (передняя) пара едва различима. Борозды продолжаются на одну
треть ширины глабели, перпендикулярно оси головы, резко выражены
и оканчиваются ямками. Иногда задняя пара под прямым углом повора-
чивает назад. Затылочное кольцо равно ширине задней лопасти глабели.
Затылочная борозда глубокая. Спинные борозды выражены отчетливо.
Неподвижные щеки падают к бокам отвесно, выпуклые, треугольного
очертания. Глаза сильно выдвинуты наперед, находясь против передних
борозд. Длина глаз равна ширине лопастей глабели. Глазные крышки
расположены у спинных борозд. Хвостовой щит полукруглый, с выпук-
лой, притупленной на конце осевой лопастью, слабо сегментированный.
Длина головных щитов 4,5—5,5 мм, ширина 4.0—6,2 мм.
Распространение. Карадок. Салаир, р. Черневой Бачат.
176
семейство lichadidae corda
Род Amphillchas Raymond
Широкая глабель, имеется пара боковых лопастей. Затылочное кольцо
без боковых лопастей, образует сплошную полосу. Осевая лопасть хво-
стового щита сзади не очерчена.
Типичный вид. Amphillchas lineatus (A n g.)
Распространение. Нижний силур.
Amphillchas sniatkovl Weber
Табл. XXII, фиг. 8
Головной щит перегнут спереди в продольном направлении под углом
45°. Борозды глабели тонкие. Боковые лопасти почти не поднимаются
над остальной частью глабели. Скульптура состоит из бугорков различ-
ной величины, удлиненных и местами круглых. На осевой лопасти хвоста
имеются два кольца. Второе из них ограничено сзади бороздой, не до-
ходящей до спинных борозд. После второй пары плевр осевая лопасть
сильно суживается. На конце лопасти спинные борозды, ее ограничиваю-
щие, пропадают. Плевры с свободными концами; на первых двух парах
имеются продольные борозды. Длина глабели по окружности 40,0—
47,5 мм.
Распространение. Карадок. Салаир, р. Черневой Бачат.
Amphillchas batchatlcus Weber
Рис. 171
Головной щит перегнут спереди в продольном на-
правлении под углом около 65°. Глубина и резкость
борозд глабели уменьшаются от переднего края к за-
тылочной борозде. Средняя лопасть глабели ограни-
чена сзади параллельными бороздами, не продолжаю-
щимися до затылочной борозды. Не доходя до пере-
гиба, центральная лопасть глабели расширяется слабо
и вблизи переднего края — очень резко. В попереч-
ном направлении она слабо выпукла сзади и силь-
но — спереди. Боковые лопасти по всей длине сохра-
няют свою ширину, и их самостоятельная выпуклость
более всего выражена на перегибе. Затылочное коль-
цо узкое, очень длинное.
Скульптура состоит из округлых бугорков, редко
встречаются вытянутые. От A. sniatkovl отличается
меньшим продольным перегибом головного щита и
меньшей заостренностью кранидия при взгляде сверху.
Борозды глабели у A. batchatlcus глубже, затылочное
скульптура состоит в основном из округлых бугорков, тогда как
у A. sniatkovl они удлиненные. Длина глабели по окружности 16—26 лиг.
Распространение. Карадок. Салаир, р. Черневой Бачат.
Рис. 171. Amphili-
chus batchatlcus
Web. Глабель.
Нат. вел. По В. Н.
Веберу
кольцо
Род Acrolichas Foerste
Весьма близок к предыдущему роду, отличаясь от него некоторыми
признаками. У Acrolichas хвостовой щит расщепляется на остром конце
осевой лопасти, а продольные борозды глабели резкие и доходят до за-
тылочной борозды. У Amphillchas осевая лопасть хвостового щита сзади
не очерчена, продольные борозды глабели менее глубокие и менее рез-
кие, неполные и до затылочной борозды не доходят.
Типичный вид. Acrolichas cucullus (Meek et Worth.).
Распространение. Нижний силур.
12 Зак. 2024 1 77
Acroltchas cucullus Meek et Worthen
Рис. 172
В продольном
глабели подходят
направлении головной щит сильно перегнут. Лопасти
к затылочной борозде под прямым углом. Трехсостав-
ные боковые лопасти глабели имеют самостоятельную
выпуклость, слабую в задней части глабели и боль-
шую — в передней. На перегибе центральная лопасть
глабели имеет вздутие. Затылочная борозда глубокая
и резкая. Скульптура представлена круглыми бугор-
ками различной величины. Три бугорка, расположен
ные в передней части центральной лопасти, значитель-
но крупнее остальных. Длина глабели по окружно-
сти 8,7 мм.
Распространение. Карадок. В Западной
Сибири — Салаир, р. Черневой Бачат.
СЕМЕЙСТВО CHEIRURIDAE SALTER
Род Cheirurus В е у г i с h
Рис. 172. Acroli-
chas cucullus
Meek et Worth.
Глабель, x 2.
По В. H. Веберу
Головной щит полукруглый. Глабель выпуклая,
выдающаяся наперед, большая, занимающая больше
одной трети ширины щита. Имеются три пары глабе-
лярных борозд, задняя отшнуровывает треугольные
базальные лопасти; передние борозды прерываются
в средней части глабели. Глаза маленькие, расположены довольно да-
леко от глабели, неподвижные щеки заканчиваются остроконечиями. Ту-
ловище с одиннадцатью сегментами, хвостовой щит с шестью или семью
шипами, почти одинаковой величины.
В девоне (в частности, в нижнем девоне Алтая) распространен близ-
кий род—Crotalocephalus Salt. (см. стр. 273), отличающийся непре-
рывными, пересекающими глабель, передними бороздами.
Типичный вид. Cheirurus exsul В е у г.
Распространение. Силур — низы нижнего девона.
Cheirurus weberl Lomovitskaja
Рис. 173
Глабель округленночетырехугольной формы. Борозды глабели глубо-
кие. Передние пары борозд продолжаются более, чем на одну треть ши-
рины глабели. Задняя пара борозд соединяется, образуя язычок, спу-
Рис. 173. Cheirurus weberl Lomov. Головной щит, х 1.5.
Салаир. По В. Н. Веберу
скающийся до затылочной борозды. Передняя лопасть глабели узкая,
задние — треугольные. Подвижные щеки образуют остроконечия. Глаза
выпуклые, расположены против средних лопастей глабели.
178
На осевой лопасти хвостового щита насчитываются три выпуклых
кольца; спереди наблюдается сочленовое кольцо. Передние плевры с про-
дольными короткими, но глубокими бороздами. Имеются еще две задних
пары плевр небольших размеров. Первая пара отделена от второй резкой
и глубокой бороздой, более мелкой кнаружи. Поверхность первой плевры
покрыта мелкими, густо расположенными бугорками. Длина глабелн
14,5—5,5 мм, ширина 12,0—13,0 мм.
Распространение. Карадок. Салаир, р. Черневой Бачат.
12*
ГЛАВА ///
ДЕВОНСКАЯ СИСТЕМА
Л. Л. ХАЛФИН
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Девонские отложения широко развиты в пределах Саяно-Алтайской
области и представлены всеми отделами и ярусами системы. Они содер-
жат обильную и разнообразную фауну, а в лагунно-континентальных
фациях — и флору. Наибольшим распространением пользуются брахио-
поды и кишечнополостные, местами многочисленны мшанки, пластин-
чатожаберные моллюски и трилобиты. Брахиоподы являются наиболее
изученной группой, которой в основном пользуются при стратиграфиче-
ском расчленении девонских отложений Западной Сибири.
В составе девонской фауны Западной Сибири обособляются два
основных фациальных типа; один из них приурочен к известняковым от-
ложениям, другой — к песчаникам и глинистым сланцам. Одновозраст-
ные фауны этих двух типов нередко совершенно различны по составу;
это особенно характерно для нижнего девона: в нижнедевонских извест-
няках распространена фауна богемско-уральского типа (Петц, 1901),
а в песчано-глинистых отложениях — своеобразная лептодонтелловая
фауна, состоящая почти целиком из местных групп и форм (Халфин,
1948).
Местные группы и формы преобладают в составе девонской фауны
Западной Сибири, и среди них имеется ряд важнейших руководящих
ископаемых. Наряду с ними в некоторых фаунах широко распространены
европейские формы; американские и южноазиатские формы играют вто-
ростепенную роль.
Древнейшая девонская фауна, в составе которой содержатся еще
многочисленные верхнесилурийские элементы и которая по этой причине
может рассматриваться как переходная между силуром и девоном, из-
вестна в пределах северной части Горного Алтая и северо-восточного
склона Салаира; это фауна известняков д. Соловьихи на Алтае (Хал-
фин, 1948, стр. 144) и нижней части крековских известняков Салаира.
Указанная фауна состоит преимущественно из брахиопод, среди которых
могут быть отмечены Atrypa umbra Barr., Spirlfer jaschel Roem.,
Rhynchosplrlna formosa (Hall) и переходящие стратиграфически выше
Cymostrophla stephanl (Barr.) и Atrypa supramarglnalls Kha 1 f.
Несколько более молодая фауна в известняковой фацин представлена
формами из крековских известняков Салаира: Gypidula kayserl (Peetz),
Atrypa semiorbicides Laz., Tryplasma altalca (Dyb.) и др. Известняки
с аналогичной фауной имеются также в Рудном Алтае, близ с. Курьи.
Того же возраста, но иного состава фауна характеризует ганинскую
серию в Северном Алтае. (Халфин, 1948, стр. 17). В нижней части этой
180
серии залегает известняк с многочисленными представителями рода
Rensselaerla Hall et Cl. (Rensselaerla navlcelllformts К h a 1 f. и др.).
Atrypa supramrglnalls Kha If. и др.; выше этот известняк постепенно
переходит в мергелистые песчаники и сланцы, содержащие так называе-
мую первую лептодонтелловую фауну с брахиоподами (Levenea taento-
lata К h а 1 f Leptaena bouel Barr.. Leptodontella rotundata К h a 1 f.,
Schuchertella altalca К h a 1 f., Eosplrlfer pseudotogatus К h a 1 f.,
Delthyrls prelamellosa Hall) и другими группами ископаемых. Вен-
чается ганинская серия отложениями с фауной преимущественно пластин-
чатожаберных моллюсков — Paramnlgenla blcarlnata К h а 1 f., Laurskla
attenuata К h a 1 f. и др.
При двучленном делении нижнего девона ганинская серия (так же,
как и крековские известняки в целом) может быть отнесена к жедин-
скому ярусу; более точно, однако, будет отнести к этому ярусу лишь от-
ложения со смешанной силуро-девонской фауной (известняки д. Со-
ловьихи и нижняя часть крековских известняков), тогда как ганинскую
серию и в основном крековские известняки следует отнести к зигенскому
ярусу.
Отложения кобленцского яруса также представлены двумя основными
фациями. Известняковая фация развита на Салаире; это пестеревские
слои 1 с брахиоподами и кораллами; Camarotoechla nympha (Ваг г.)
var. pseudollvonica, Karplnskla conjugula T s c h e r n., Eosplrlfer secans
(Bar г.) и др. Известняки с кобленцской фауной известны, также в Ле-
киногорском районе Рудного Алтая (Ф. Н. Чернышев, 1893).
В Горном и Рудном Алтае кобленцские отложения представлены глав-
ным образом песчано-глинистыми толщами с разнообразной фауной
(Халфин, 1948). В Горном Алтае они разделяются на ннжнекобленцские
(кондратьевская толща со второй лептодонтелловой фауной) и верхне-
кобленцские (медведевская толща с третьей лептодонтелловой фауной).
В кондратьевской толще массовым развитием пользуются: из брахио-
под — Chonetes rectlsplna К h а 1 f., Leptaena bouel Barr., Leptodontella
rotundata К h a 1 f., Ussovia prlma (K h a 1 f.); из пластинчатожаберных
моллюсков — Mytllarca elongata К h a 1 f., Toechomya subclrcularls
К h a 1 f., Praemyophorla acutlrostrls К h a 1 f., Tolmala squamosa Khali.,
Lelopecten rectangularis К h a 1 f., Archaeocardium scroblculare К h a 1 f.
и др.; из трилобитов — Probolium altalcum (К h a 1 f.), Phacops subcrlstata
Kha If.; из табулят— Pleurodlctyum cf. problematlcum Goldf.
В медведевской толще верхнекобленцского возраста преобладают бра-
хиоподы; Chonetes pseudobohemica Kha If., Leptaena bouel Barr.,
Brachyprlon nallvklnt Kha If., Leptodontella planuscula (Kha If.), Con-
chidium celstseptosum Kha If., Acrosplrlfer korovlnt Kha If., Acrosptrlfer
pseudofallax К h a 1 f. и др.
Присутствие отложений, эквивалентных кондратьевской и ганинской
толщам, доказано в последнее время и в Рудном Алтае (Левенко, 1950).
Главнейшими областями развития палеонтологически охарактеризо-
ванных среднедевонских отложений являются окраины Кузбасса, Салаир,
Минусинская котловина и Алтай.
Отложения эйфельского яруса наиболее полно представлены по юго-
западной окраине Кузбасса; здесь различают несколько фаций эйфель-
ских отложений с различной фауной.
На основании изучения брахиопод М. А. Ржонсницкая выделяет
(снизу вверх) два горизонта эйфельских отложений этого района:
1. Салаиркинские слои с Paraspirifer (?) gurjevskensi's Rzon.
1 Согласно новейшим данным (М. А. Ржонсницкой, Г. Н. Карцевой, Г. С. Харина
и др.), под названием «пестеревские слои» обычно объединяются отложения различ-
ного возраста. Необходима тщательная ревизия имеющихся материалов по фауне
этих известняков.
181
2. Шандинокие слои с массовым развитием крупных раковин Conchidium
ex gr. pseudobaschklricum Tschern. и Stropheodonta uralensls Vern.
По простиранию эти известняковые горизонты замещаются обломоч-
ной чумыщской толщей с Eoretlcularla deretmsi (Oehl.), Elythlna salal-
rlca Rzonsn. и др.
Эйфельские отложения Минусинской котловины В. С. Мелещенко раз-
деляет на толчаковскую красноцветную свиту (внизу) и таштыпскую
свиту сложного состава (вверху) В последней имеются известняковые
горизонты с массовым развитием Acrosplrlfer subgregarlus Rzonsn.
Эйфельские отложения распространены также в Горном Алтае (Гра-
цианова, 1950). Возможно, к самым низам Эйфеля относятся известняки
Кукушкиной сопки близ с. Лосиха в Рудном Алтае с лептодонтелловой
фауной (Бубличенко, 1927).
Эйфельские отложения Западной Сибири охарактеризованы богатой
фауной строматопор, табулят и тетракораллов.
Живетские отложения наиболее полно развиты также по юго-запад-
ной окраине Кузбасса, где М. А. Ржонсницкая выделяет в их составе
четыре горизонта (снизу вверх):
1. Мамонтовские слои с характерным спирифером Lazutklnla татоп-
tovensls (L a z.).
2. Акарачкинские слои с гониатитами и брахиоподами.
3. Керлегешские (хаскотирисовые) слои с Denckmanella datnest
(Н о 1 z.) и Chascotharls salalrlca Rzonsn.
4. Сафоновские (индоспириферовые) слои с Indosplrlfer pseudowtl-
llamsl Rzonsn., Atrypa bodlnl Mans., Unclnulus trlcostatus (Peetz),
Unctnulus angularls (Ph ill.) и др.
Наиболее высокий горизонт живетского яруса — чиэлевый (с Асго-
splrlfer cheehiel) — по юго-западной окраине Кузбасса представлен, по-
видимому, лишенными фауны отложениями. Наиболее типично он раз-
вит по северо-западной окраине Кузбасса (Халфин, 1937), в Горном
Алтае (Наливкин, 1938) и в Минусинской котловине (Штукенберг,
1886). Для него наиболее характерны спириферы группы Acrosplrlfer
cheehiel Коп.), Platyrochella martlanovl (Stuck.), Unclnulus korovlnl
Kha If., Schellwlenella umbraculum (Sc hl.) и др.
В Горном Алтае Л. Л. Халфин делит чиэлевый горизонт на две части:
слои с Acrosplrlfer cheehiel внизу и слои с Lamelllsplrtfer mucronatus
вверху.
В Минусинской котловине ниже чиэлевого горизонта залегает мощ-
ная толща преимущественно красноцветных отложений, разделяющихся
на ряд свит. Таков же характер среднедевонских отложений в Горной
Шории.
Нижнефранокие отложения в Западной Сибири представлены не-
сколькими фациями, охарактеризованными различной фауной. Наиболее
распространенной и наилучше изученной из числа нижнефраноких фаун
является так называемая фаленовая фауна (по имени руководящей
формы — Anathyrls phalaena (Phi 11.). Отложения с этой фауной раз-
виты по южной, восточной и северной окраинам Кузбасса (Халфин,
1932; Халфина, 1940) и на Алтае.
Фауна состоит из брахиопод, среди которых руководящее значение
имеют: Cyrtosplrlfer achmet N а 1., Cyrtosplrifer tentlculum (Vern.),
Gilrlchella zlckzack (Roem.), Anathyrls phalaena (Phi 11.), атрипы
группы A. reticularis и др. Местами (например, у Черепанова брода на
р. Яя, северная окраина Кузбасса) эта фауна ассоциирует с различными
кораллами.
Примеры других фаций нижнефранских отложений:
коралловые известняки с. Зарубина (северо-западная окраина Куз-
басса) с Schliiterla emstl Wed., Prlsmatophyllum vogultcum В u 1 v.,
Alveolites suborblcularis L., Thamnopora cervlcornls (В 1 a i n v.) и др.;
182
песчаники д. Асколад (там же), переполненные раковинами Eoretlcu-
larla mlcrosplnosa К h а 1 f.;
черные сланцы с бухиолами (Buchlola retrostrlata Buch, и др.) Руд-
ного Алтая (близ ст. Локоть).
Несколько более высокое стратиграфическое положение занимают
гониатитовые известняки района Николаевского рудника (Рудный Алтай),
залегающие в виде линз среди эффузивов, а также гониатитовые песча-
ники северо-западной окраины Кузбасса (фауна их не описана).
Отложения с фаленовой фауной широко распространены в Горном
Алтае (Халфин, 1948).
Переходными к верхнефранским отложениям в пределах северо-запад-
ной окраины Кузбасса являются так называемый атриповый горизонт
с брахиоподовой фауной, в составе которой наиболее важная руководя-
щая форма — Anathyrls tyzhnovl К h а 1 f., и залегающий выше глубокин-
ский известняк с богатой фауной брахиопод и кораллов. Аналоги послед-
него распространены в Рудном Алтае близ Николаевского рудника
и в районе г. Камень на р. Оби (Краевская, 1936). Среди брахиопод
этого горизонта наиболее распространенными являются: Gypidula comls
(Ow.), Hypothyrldina cubotdes (Sow.), Pugnax acuminata (Mart.),
Cranaena romtngerl Hall и др. Очень многочисленны также кораллы,
например: Peneckiella darvlnl (Freeh), Phllllpsastraea pentagona
(G о 1 d f.), Solominellcb soshklnae I v a n i а и др.
Верхнефранские отложения распространены по северо-восточной
и северо-западной окраинам Кузбасса (Халфин, 1933) и содержат харак-
терную монстровую фауну с Anathyrls monstrum К h а 1 f., Anathyrls
ussovi К h a 1 f., Cyrtosplrlfer ussovi К h a 1 f., Cyrtosplrlfer khalfinl I v„
Indosplrtfer sibtrlcus К h a 1 f.
На границе между верхнефранским и фаменским ярусами происхо-
дит отчетливое обновление фауны — исчезают представители родов
Gilrlchella Р а е с к. и Indosplrtfer Grab., циртоспириферы группы
Cyrtosplrlfer ussovi, анатирисы, атрипы.
Фаменская фауна (Халфин, 1933) представлена главным образом
брахиоподами, среди которых важнейшее руководящее значение имеют
продуктусы группы Pllcatlfera praelonga (Sow.), а также Cyrtosplrlfer
verneulll (Murch.), Cyrtosplrlfer archiacl (Mu г ch.) и некоторые дру-
гие. Отложения с этой фауной известны по северо-восточной и северо-
западной окраинам Кузбасса, в Рудном Алтае близ с. Тарханокого
и в северной части Горного Алтая в бассейне ключа Филаретова (черем-
шанская толща).
По окраинам Кузбасса и близ с. Тарханского отложения фаменского
яруса перекрываются отложениями с переходной девонско-карбоновой
фауной, содержащей наряду с верхнедевонскими формами, спириферов
группы Splrlfer tornacensis Коп. (тарханская свита Рудного Алтая,
абышевская зона Кузбасса).
Средне- и верхнедевонские отложения Западной Сибири в ряде райо-
нов в большом количестве содержат мшанок, среди которых преобладают
представители семейства Fenestellidae (Нехорошев, 1948).
Наряду с морскими девонскими отложениями, фауна которых охарак-
теризована выше, в пределах Саяно-Алтайской области широко распро-
странены лагунно-континентальные, преимущественно красноцветные от-
ложения; местами они содержат органические остатки. Девонские отло-
жения этого фациального типа по окраинам Кузбасса, по р. Томи, в Ми-
нусинской котловине и на Алтае нередко охарактеризованы разнообраз-
ной флорой, в составе которой А. Р. Ананьев выделяет комплексы, харак-
теризующие каждый из трех отделов системы (см. стр. 279).
Для стратиграфического расчленения красноцветных девонских отло-
жений Западной Сибири большое значение будут иметь остатки рыб,
которые до настоящего времени не собирались сколько-нибудь система-
183
Таблица 4
От- делы Ярусы Опорные стратиграфические горизонты
Фаменский Верхняя красноцветная толща Кузбасса. Прелонгусовый горизонт Кузбасса с брахиоподами Plicatifera praelonga, Cyrtospirifer ver- neuili, Cyrtospirifer archiaci и др. Черемшанская формация Гор- ного Алтая, тарханская (частью) — Рудного Алтая
D3 Франский Монстровый горизонт Кузбасса с брахиоподами: Anathyris monst- rum, Cyrtospirifer ussovi, Cyrtospirifer khalfini и промежуточная толща c Anathyris ussovi, Indospirifer sibiricus, Lamellispirifer mucronatiformis Атриповый горизонт c Anathyris tyzhnovi Кузбасса н глубокинский горизонт Кузбасса, Рудного Алтая и района г. Камня с брахиопо- дами (Gypidula comis, Hypothyridina cuboides, Pugnax acuminata и др.) и кораллами (Peneckiella darwini, Phillipsastraea penta- gona, Solominella soshkinae и др.) Зарубинский коралловый известняк Кузбасса с Schluteria emsti, Prismatophyllum vogulicum и др. и фаленовый горизонт Кузбасса и Горного Алтая с Anathyris phalaena, Cyrtospirifer achmet, Cyrtospirifer tenticulum и др.
d2 Живетский ! Чиэлевый горизонт Кузбасса, Горного Алтая и Минусинской котло- вины с Acrospirifer cheehiel, Schellwienella umbraculum, Platy- rochella martianovi, Schizophoria striatula и др. Индоспириферовый горизонт (сафоновские слои) Кузбасса и Салаира с Indospirifer pseudowilliamsi, Atrypa bodini, Uncinulus tricosta- tus и др. Хаскотирисовый горизонт (керлегешские слои) Кузбасса (Салаира) с Chascothyris salairica, Denkmanella damesi и др. Акарачкинские слои Кузбасса (Салаира) Мамонтовские слои Кузбасса (Салаира) с Lazutkinia mamontoviensis и др.
Эйфельский Шандинские конхидиумовые известняки (Conchidium ex gr. pseudo- baschkiricum) и салаиркинские брахиоподовые известняки (Uncinu- lus parallelepipedus, Atrypa subspinosa и др.) и их аналог — чумышская террагенная толща с Eoreticularia dereimsi и Elythina salairica Кузбасса и Салаира; таштыпская свита Минусинской котловины с Acrospirifer subgregarius
Di Кобленцский Пестеревские известняки Кузбасса1 (Салаира) с Karpinskia conju- gula, Eospirifer secans, Camarotoechla nympha и др.; крюковские известняки Лениногорского района Рудного Алтая с Phacops altaicus; песчано-слаицевые отложения Горного Алтая, разделяю- щиеся на верхнекобленцскне (медведевская толща) с Leptaena bouei, Leptodontella planuscula, Brachyprion nalivkini н др. и нижнекобленцские (кондратьевская толща) с Leptaena bouei, Leptodontella rotundata, Pterinea (Tolmaia) squamosa, Archaeo- cardium scrobiculare, Probolium altaicum и др.
Жединский Крековские известняки Кузбасса (Салаира) с Gypidula kayseri, Atrypa semiorbicides, Tryplasma altalca и др. Ганинская серия Горного Алтая с Leptaena bouei, Eospirifer pseudotogatus, Rens- selaeria navicelliforniis, Paramnigenia bicarinata и др. Соловьихинский известняк Горного Алтая со смешанной силурийско- девонской фауной: Atrypa umbra, Rhynchospirina formosa, Spirlfer jaschei и др.
1 См. сноску на стр. 352.
184
тически. В Минусинской котловине (Д. Обручев, 1941) установлено на-
личие панцирных рыб (Asterolepls slblrica D. О b г u t с h. и др.), а также
хрящевых и различных костных рыб, что позволило надежным образом
выделить среднедевонские, франские и фаменские отложения среди
красноцветного минусинского девона.
По стратиграфии девонских отложений Западной Сибири накопился
достаточно большой материал, позволивший перейти к различного рода
обобщениям и выяснению закономерностей их образования. Так, исследо-
вания Западно-Сибирского геологического управления (М. К- Винкман,
А. Б. Гинцингер, А. С. Калугин и др.) в Горном Алтае показывают, что
выделенные различными методами местные стратиграфические подраз-
деления могут быть объединены в три последовательные комплекса, раз-
деленные перерывами с явным угловым несогласием, именно: караку-
дюрокая серия (D^, березовская серия (эйфель) и куротинская серия
(верхи D2 — низы Г)з).
Для девона Минусинской котловины В. Д. Мелещенко устанавли-
вает определенную ритмичность, выделяя шесть «циклов», каждый из
которых сложен лагунно-континентальными осадками и эффузивами
и венчается карбонатными свитами. Еще ранее определенную ритмич-
ность в строении девонских отложений северных окраин Кузбасса уста-
новил А. В. Тыжнов, описав характер и последовательность ритмов для
геосинклинального (северо-западная окраина Кузбасса) и платформен-
ного (северо-восточная окраина) типов этих отложений и приведя их
корреляцию.
Из числа стоящих на очереди задач по дальнейшему уточнению стра-
тиграфии девона Саяно-Алтайской области отметим следующие, как
имеющие наибольшее практическое значение:
1. Салаир и юго-западная окраина Кузбасса — согласование и увязка
накопившихся материалов, ревизия фауны ряда горизонтов, в первую
очередь остракодового и пестеревских известняков.
2. Рудный Алтай — детальное изучение фауны девонских отложений;
необоснованное, но упорно отстаиваемое отрицание наличия в Рудном
Алтае нижнего девона (Н. Л. Бубличенко и др.) опровергается недавно
опубликованной работой А. И. Левенко (1950), но это лишь первый шаг
в направлении разработки детальной стратиграфии девона этого района.
3. Северо-западная окраина Кузбасса и Минусинская котловина —
дополнительное исследование ритмичности девонских отложений северо-
западной окраины Кузбасса и Минусинской котловины и, в особенности,
выяснение характера контактов последовательных ритмов (циклов).
В табл. 4 приведены некоторые примеры типичных палеонтологически
охарактеризованных девонских отложений Западной Сибири.
Тип Coelenterata. Кишечнополостные
В. К. ХАЛФИНА
КЛАСС HYDROZOA
STROMATOPOROIDEA. СТРОМАТОПОРОИДЕИ
СЕМЕЙСТВО ACTINOSTROMIDAE NICHOLSON
Род A ctinostroma Nicholson
Ценостеум пластинчатый или в виде желваков неправильной формы.
Радиальные столбики и концентрические пластинки образуют в радиаль-
ном сечении правильную сетку, причем радиальные столбики пересекают
большое число пластинок. В столбиках часто наблюдается осевой канал.
Ячейки прямоугольные или четырехугольные. Многочисленные связки
185
в тангенциальном сечении имеют четырех-, шести- и даже восьмилуче-
вое расположение. Строение ткани плотное. Астроризы узкие.
Типичный вид. Acllnostroma clathratum N i с h.
Распространение. Силур—девон, реже — кембрий. Повсе-
местно.
Actlnostroma clathratum Nicholson
Табл. XXIII, фиг. 1
«Форма обычно сфероидальная или полусфероидальная, часто непра-
вильная и различных размеров.. Ценостеум сложен плотными ради-
альными столбиками и отходящими от них концентрическими пласти-
нами, своими пересечениями образующими прямоугольную сетку. Ради-
альные столбики проходят непрерывно через много концентрических пла-
стинок (до 30 и более).. Радиальные столбики чаще расположены
в шахматном порядке. Отростки этих столбиков, в целом образующие
концентрические пластинки, не плоские, а несколько выпуклые с верх-
ней и нижней стороны и утолщающиеся у столбика.
Лучистые ветви или arms. ., дающие в тангенциальном сечении
столь характерное шестилучевое соединение между столбиками, есть
не что иное, как сечение этих отростков. При ином расположении стол-
биков (не шахматном.—В. X.) сечение получается четырех, пяти- и
восьмилучевым..
При хорошей сохранности экземпляров поверхность их украшена ма-
ленькими бугорками, представляющими выходы концов радиальных
столбиков. Возвышений (mamelons) на поверхности не имеется. На 1 мм
приходится 3 —4 радиальных столбика и 4—6 концентрических пласти-
нок. Диаметр столбиков «/гв—5/га мм (0,14—0,18 мм), реже 6/28 мм
(0,21 мм), в двух случаях 2/28 мм (0,07 мм) и в одном — 3/28 лои
(0,11 мм). Толщина пластинок около 3/28 мм (0,11 мм). Радиальные
столбики в поперечном сечении круглые. В некоторых из них ясно ви-
димы осевые каналы. Пластинки прямые или слегка изогнутые. Ткань
плотная» (Яворский, 1930, стр. 476).
На некоторых экземплярах наблюдались астроризы, которые у этого
вида часто отсутствуют. В. И. Яворский указывает, что расстояние ме-
жду центрами астрориз 11—13 мм, в одном случае 8 мм.
Широко распространенный в Кузбассе вид Actlnostroma mamontovl
Y a v о г. (табл. XXIII, фиг. 2), характеризующий самые верхние гори-
зонты Эйфеля, отличается от A. clathratum меньшими размерами элемен-
тов скелета (диаметр радиальных столбиков 0,06—0,08 мм, на 1 мм при-
ходится пять радиальных столбиков и столько же пластин) и наличием
сдвоенных концентрических пластинок.
Распространение. A. clathratum пользуется весьма широким
географическим распространением; в Кузбассе она встречена в ряде
мест в среднедевонских отложениях.
Actlnostroma piriformis V. К h а 1 f i n a, sp. n.
Табл. XXIII, фиг. 3; табл. XXV, фиг. 3
Ценостеум грушеобразной формы. Высота его 31 мм, наибольший
поперечник 32 мм, а ширина у основания колонии 15 мм. На нижней
стороне ценостеума имеется маленькая площадка, при помощи которой
он прикреплялся к посторонним предметам. Поверхность усеяна мелкими
круглыми бугорками (концы крупных радиальных столбиков).
Характерными особенностями данного вида являются: 1) наличие
двух типов радиальных столбиков — очень толстых, достаточно длинных
и тоненьких, коротких; 2) наличие таких же тонких, как столбики второго
порядка, концентрических пластинок; 3) шестилучевое строение связок.
Толстые' столбики многочисленны и пересекают максимум 20 пласти-
нок, чаще же 7—8—10. Расстояние между ними в большинстве случаев
186
не превышает толщины самого столбика; на 1 мм приходится три стол-
бика или семь столбиков на 2 мм.
Столбики второго порядка тоненькие и короткие. Они пересекают
2—3 пластинки, но часто ограничены в своем развитии одним межпла-
стинчатым промежутком. Тоненькие столбики встречаются очень редко
и нельзя установить закономерность в их распределении.
Концентрические пластинки тонкие, толщиной 0,07—0,11 мм, реже
0,13 мм; на 1 мм их насчитывается 5—6. Ячейки сетки в целом прямо-
угольные, но часто встречаются и круглые.
В тангенциальном шлифе видны округлой формы сечения крупных
радиальных столбиков диаметром от 0,18 до 0,26 мм, чаще же 0,23—
0,24 мм. Осевого канала в столбиках нет. Поперечные сечения столбиков
второго порядка встречаются крайне редко. Многочисленны обрывки
астроризальных каналов шириной 0,23—0,26 мм.
Наличие радиальных столбиков двух порядков свойственно Acttno-
stroma blfarium Nich. из D2 Западной Европы и из низов D3 Тимана,
но у вида Никольсона столбики двух порядков совершенно отчетливы,
причем толстые столбики отстоят друг от друга на. таком расстоянии,
что между ними умещается до шести рядов столбиков второго порядка.
Такое расположение столбиков отличает A. blfarium от описываемого
вида. От A. contextual Barr, из девона Чехии, очень близкой на первый
взгляд форме, у которой имеются толстые, близко расположенные ради-
альные столбики и тонкие концентрические пластинки, Actinostroma
piriformis отличается наличием столбиков второго порядка и более гру-
быми скелетными элементами вообще.
Распространение. Средний девон (эйфель). Салаир.
Род Gerronostroma Yavorsky
Подобен Actinostroma Nich., но отличается отсутствием связок, при-
сутствием невысоких астроризальных цилиндров и иным способом роста
колонии (обрастает концентрическими слоями другие организмы).
Строение скелетной ткани плотное.
Типичный вид. Gerronostroma elegans Y a v о г.
Распространение. Девон.
Gerronostroma elegans Yavorsky
Табл. XXIII, фиг. 4
Радиальные столбики и концентрические пластинки образуют отчет-
ливую сетку. Радиальные столбики длинные, они пересекают ряд пласти-
нок. Последние раза в 2—3 тоньше радиальных столбиков. Ячейки чаще
всего прямоугольные, но встречаются и овальной формы. Пластинки
в местах отхода от столбиков утолщены. В межпластинчатых промежут-
ках местами наблюдаются косо расположенные тонкие известковые пе-
регородки, видимые и в тангенциальном сечении. Имеются невысокие
астроризальные цилиндры, расстояние между их центрами 10—15 мм.
В тангенциальном сечении видны округлые поперечные разрезы ради-
альных столбиков и такие же разрезы астроризальных цилиндров.
Связки отсутствуют. На 1 мм насчитывается 3—4 радиальных столбика
и 6—7 пластинок, местами число их уменьшается до трех. Лятилямины
не наблюдались.
Распространение. Средний девон. Кузнецкий бассейн, окрест-
ности с. Бачат.
Gerronostroma concentrlca Yavorsky
Табл. XXIV, фиг. 1
Колония обрастает концентрическими слоями кораллы (форма обра
стаемого предмета обусловливает форму колонии). Сетка отчетливая.
187
Радиальные столбики толстые и пересекают ряд концентрических пла-
стинок, имеющих примерно такую же толщину, что и столбики. Харак-
терной особенностью вида является местами наблюдаемое косое пере-
сечение столбиков с. пластинками. В межпластинчатых промежутках из-
редка встречаются выпуклые тонкие известковые перегородки. В попе
речном разрезе столбики имеют круглую форму. На 1 мм насчитывается
2—3 (последнее чаще) радиальных столбика и 3 концентрических пла-
стинки.
От Actlnostroma clathratum N i с h. отличается способом роста и от
сутствием связок.
Распространение. Нижний девон. Кузбасс, окрестности с. Том-
ский Завод (р. Томь-Чумыш).
СЕМЕЙСТВО CLATRODICTYONIDAE KOHN
Род Clathrodictyon Nicholson et Murie
Описание см. на стр. 153.
Clathrodictyon praetenerum Yavorsky
Табл. XXIV, фнг. 2
«Ценостеум сложен весьма тонкими (0,04—0,07 мм) радиальными
столбиками с отходящими от них столь же тонкими концентрическими
пластинками. При начальном росте колонии она прикреплялась нижней
своей поверхностью к субстрату. Волокна ткани плотные. Радиальные
столбики вполне развиты. Распространение их ограничено одним меж-
пластинчатым промежутком. На 1 мм их насчитывается до пяти. У не-
которых столбиков имеется двойное основание. Концентрические пла-
стинки слабо волнисто изогнутые, кроме того, они мелкоморщинисты,
причем в местах стыка двух морщин от них отходят радиальные стол-
бики. ..
На промежуток в 1 мм помещаются четыре пластинки; там, где пла-
стинки сближены, число их доходит до пяти, и, напротив, там, где они
более удалены друг от друга, их помещается три на 1 мм. В межпла-
стинчатых промежутках проходят выпуклые известковые перегородки.
Местами, при большом увеличении, ясно наблюдается более темная ли-
ния, проходящая посередине концентрических пластинок и от этих по-
следних переходящая в радиальные столбики. Лятиляминарность отсут-
ствует.
В тангенциальном разрезе видны радиальные столбики в поперечном
сечении. Они округлой или овальной формы. Местами между ними про-
ходят выше отмеченные известковые перегородки. Там, где разрез про-
ходит близко к основанию столбиков, видны, но весьма редко, очень
мелкие полые кольца, отвечающие двойному основанию столбиков.
Астроризы отсутствуют» (Яворский, 1931).
Распространение. Кузбасс, левый берег р. Чумыша, в районе
устья р. Таловки — средний девон. Среднедевонские отложения Урала и
бассейна р. Печоры.
Род Clathrocoilona Yavorsky
Подобен роду Clathrodictyon, от которого отличается присутствием
пустот круглой или овальноудлиненной формы. В пустотах имеются тон-
кие перегородки. Чаще всего форма ячей круглая. Имеются астрори-
зальные цилиндры. Скелетная ткань плотная.
Типичный вид. Clathrocoilona abeona Y a v о г.
Распространение. Средний девон в Кузбассе; нижний и сред-
ний девон в бассейне р. Колымы.
188
Clathrocollona abeona Yavorsky
Табл. XXIV, фиг. 4
Ценостеум пластинчатый или желвакообразный. Лятилямины отсут-
ствуют. Радиальные столбики соединяют только две соседние пластинки;
последние толще столбиков. Посередине каждой пластинки проходит от-
четливая белая линия, нигде не переходящая в столбики. Пластинки
иногда раздваиваются (дихотомируют); срединные белые полоски дихо-
томируют с ними вместе. Ячеи круглой или овальной формы. Кроме
того, имеются более крупные, чем ячеи, пустоты. В некоторых круглых
пустотах известковые перегородки располагаются так, что создается
впечатление, будто пустоты заполнены пузырчатой тканью. Астроризы
хорошо развиты, ветвящиеся; расположены вертикальными рядами.
Астроризальные каналы снабжены днищами.
На 1 мм насчитываются 3—4 столбика (местами меньше) и 2—3 пла-
стинки. Диаметр ячей 0,17 мм, длина овальных ячей до 0,5 мм. Диаметр
астроризальных каналов до 0,5 мм. Расстояние между центрами астро-
риз 10—12 мм.
Распространение. Средний девон. Салаир, в ряде пунктов по
р. Степной Бачат и на Тыргане — между вершинами рр. Абы и Сур-
таихи.
СЕМЕЙСТВО STROMATOPORIDAE NICHOLSON
Род Stromatopora G о 1 d f u s s
Описание см. на стр. 154.
Stromatopora concent г lea G о 1 d f u s s
Табл. XXVI, фиг. 2
Ценостеум разнообразной формы (массивный, желвакообразный
и т. д.), достигающий крупных размеров. Лятилямины всегда присут-
ствуют, их толщина от 1,5 до 3,0 мм. Зооидные трубки вертикальны,
диаметр их 0,14—0,17 мм. Днища горизонтальны и многочисленны.
Астроризы редки, небольших размеров; расстояния между их центрами
от 7 до 10 мм. Скелетные волокна мелкопорнстые.
Распространение. Географически широко развитая форма;
в Кузбассе известна в ряде пунктов в отложениях среднего девона.
Stromatopora hiipschll Bargatzky
Табл. XXIV, фиг. 3
Ценостеум пластинчатый или сфероидальный. Лятиляминарность от-
сутствует. В. И. Яворский (1951 г., стр. 10) так характеризует эту форму:
«Вертикальный разрез показывает очень хорошо развитые зооидные
трубки различной формы и размеров. В некоторых из них сохранились
днища, расположенные перпендикулярно к стенкам зооидных трубок.
Радиальные столбики существуют как отчетливая структура. только
в тех участках скелета, где зооидные трубки образуют вертикальные
ряды.
В тангенциальном сечении видна грубая петельность червеобразной
структуры, в которой нет следов радиальных столбиков, как отчетливо
выделяющихся элементов. Там же видны зооидные трубки в поперечном
их сечении. В большинстве они круглые. Продольные и косые сечения
придают им разные очертания. Диаметр зооидных трубок 0,033—0,16 лш.
Внутренняя структура скелетных волокон ткани грубопористая. Астро-
ризы, очень маленькие для этого вида, в наших экземплярах распозна-
ются в тангенциальном сечении с трудом.
Как в вертикальном, так и в тангенциальном сечении хорошо выде-
ляются Syrlngopora sp.».
189
От европейских экземпляров этого вида формы из Кузбасса отли-
чаются несколько более мелкими скелетными элементами.
Распространение. Slromatopora hiipschll— форма, широко
известная в отложениях среднего девона. В Кузбассе, по данным
В. И. Яворского, она пользуется достаточно широким распространением
как в эйфеле, так и в живете.
СЕМЕЙСТВО STROMATOPORELL1DAE LECOMPTE
Род Stromatoporella Nicholson
Ценостеум чаще всего пластинчатый. Радиальные столбики и кон-
центрические пластинки достаточно отчетливы. Обычно столбики не вы-
ходят за пределы одного межпластинчатого промежутка. Зооидные
трубки неправильные, короткие, сравнительно немногочисленные, с уме-
ренным количеством днищ. Астроризы хорошо развиты и часто распо-
лагаются вертикальными рядами, имея общий осевой канал.
Тангенциальный шлиф скорее напоминает таковой у представителей
рода Stromatopora, но имеются разрезы радиальных столбиков, часто
с осевым каналом в центре. Поверхность концентрических пластинок
часто бывает усеяна мелкими бугорками, имеющими отверстие на вер-
шинке; в поперечном разрезе эти туберкулы выглядят, как полые коль-
ца, и присутствие их является характерным признаком рода. Скелетные
волокна тонкопористые и пронизаны тоненькими каналами.
Типичный вид. Stromatoporella granulata N i с h.
Распространение. Силур—девон; в среднем девоне Кузбасса
этот род пользуется широким распространением.
Stromatoporella sniatkovl Yavorsky
Табл. XXV, фиг. 1
Отчетливо развиты радиальные столбики и слабо волнистые морщи-
нистые концентрические пластинки. Радиальные столбики, имеющие та-
кую же толщину, как и пластинки, в своем развитии ограничены одним
межпластинчатым промежутком. «...Морщинки пластинок приходятся
в местах отхода их от радиальных столбиков» (Яворский, 1931, стр. 13).
В межпластинчатых промежутках изредка наблюдаются очень тонкие
выпуклые известковые перегородки. Толщина радиальных столбиков и
пластинок до 0,10 мм. На 1 мм насчитываются три, чаще четыре концен-
трических пластинки.
Астроризы отсутствуют. Зооидные трубки не наблюдались. Волокна
ткани мелкопористые и пронизаны каналами.
В тангенциальном шлифе обильны разрезы бугорков в виде полых
колец и сечения радиальных столбиков.
Распространение. Средний девон. Кузбасс, правый берег
р. Чумыша, в 3 кии выше устья р. Таловки.
Stromatoporella loutouguini Yavorsky
Табл. XXVI, фиг. 1
Ценостеум пластинчатый, крупных размеров. Скелетные волокна
тонкопористые и пронизаны сетью гонких каналов.
«Концентрические пластинки, как и радиальные столбики, хорошо
развиты. На промежуток в 1 мм помещается 4—5, редко шесть столби-
ков. Число пластинок на таком же промежутке 4—5. Столбики, прохо-
дящие между пластинками, дают ячеистую структуру скелета, причем
ячейки квадратной, чаще же овальной формы. В ячейках часто наблю-
даются нитевидной формы известковые перегородки.
В вертикальном сечении можно видеть, что толщина пластинок и ра-
диальных столбиков не постоянна в одном и том же экземпляре. Наблю-
дается как бы чередование участков с тонкими и толстыми пластинками
190
и столбиками. В первом случае они достигают толщины 0,05 мм, во вто
ром —0,10 мм и даже до 0,15 мм. В этом же сечении видны трубки
Syrlngopora yavorskyt Т с h е г п. var. dellcata Т с h е г n.
В тангенциальном сечении видны круглые точки, отвечающие попе-
речному сечению радиальных столбиков. Тут же видны трубки Syringo
рога в поперечном разрезе. Между столбиками местами заметны соеди-
няющие их, отмеченные выше, нитевидные перегородки. Характерными
для этого рода служат наблюдаемые в этом сечении полые кольца, внут-
ренний диаметр которых достигает 0,05 мм.
Астроризы, судя по тому, что наблюдались они не во всех образцах,
видимо редкие. Каналы их прямые, ветвящиеся. Диаметр каналов
0,25 мм, но некоторые доходят до 0,50 мм. Они снабжены днищами. Рас-
стояние между центрами астрориз 8—10 и до 12 мм» (Яворский, 1951,
стр. 243).
Распространение. Юго-западная окраина Кузбасса; ряд пунк-
тов. Единичные экземпляры относятся к хаскотирисовому горизонту,
большинство — к индоспириферовому горизонту живетского яруса.
Stromatoporella duplex V. Kh а 1 f i n a, sp. n.
Табл. XXV, фиг. 2
Ценостеум пластинчатый. Толщина концентрических пластинок и ра-
диальных столбиков одинакова. Последние занимают только один мсж-
пластинчатый промежуток. Чаще всего столбики располагаются парами
(сдвоенные): два столбика почти рядом (расстояние между ними 0,Io-
О. 13 мм), затем через значительный (0,18—0,25 мм), а иногда и совсем
большой (0,52—0,70 мм) промежуток — опять пара столбиков и т. д.
Но наряду со сдвоенными столбиками часто встречаются и одинарные.
Концентрические пластинки сильно волнообразно изогнуты; толщина
их 0,10—0,17 мм. Ширина межпластинчатых*промежутков различная (от
0,11 до 0,43 мм)-, в местах сильных изгибов концентрических пластинок
она очень мала, а порой и совсем сходит на нет — межпластинчатые
промежутки выклиниваются. Концентрические пластинки не сплошные,
часто они прерываются между двумя сближенными столбиками, и тогда
два соседних межпластинчатых промежутка соединяются между собой.
В ячейках округлой формы иногда наблюдаются тоненькие, чаще гори
зонтальные, перегородки. В тангенциальных шлифах астроризы не на
блюдались, а в вертикальных — весьма отчетливы. На 1 мм насчиты-
вается обычно четыре столбика (реже 5—6) и от трех до пяти концен-
трических пластинок.
В тангенциальном сечении видны поперечные разрезы радиальных
столбиков и астроризальных цилиндров. Расстояние между центрами
последних 11 —15 мм. Многочисленны полые кольца; их диаметр 0,20—
0,30 мм, а диаметр внутренней полости кольца —0,10—0,13 мм. Строе-
ние скелетной ткани обычное для строматопорелл.
У Stromatoporella tchernyshevl Y a v о г., близкой по ряду признаков
формы, более грубые скелетные элементы, отчетливо выраженных астро-
ризальных цилиндров нет и астроризы с густой сетью длинных каналов
(до 15 мм длины). От других строматопорелл этот вид отличается еще
более существенно.
Распространение. Средний девон. Салаир, д. Мамонтово.
КЛАСС ANTHOZOA
Л. Н. КРАЕВСКАЯ
ПОДКЛАСС TABULATA И ГРУППА HELIOLITIDA
В девонских отложениях Западной Сибири большое развитие имеют
группы табулят и гелиолитид. Среди этих групп имеется ряд руководя-
щих форм; важнейшие из них описаны ниже. Диагнозы видов девонских
191
табулят и гелиолитид взяты из работы Б. Б. Чернышева «Силурийские
и девонские Tabulata и Heliolitidae окраин Кузнецкого бассейна» (1951),
а также из материалов палеонтолога Западно-Сибирского геологиче-
ского управления А. П. Жмаева, которому принадлежат и изображения
новых форм.
В отношении геологического распространения ряда видов, описанных
Б. Б. Чернышевым, нужно сделать следующее замечание. Изучая кол-
лекции, собранные рядом геологов, Б. Б. Чернышев указывал страти-
графическое положение для вновь устанавливаемых им видов в соответ-
ствии с данными этих геологов. В частности, ряд форм им описан из от-
ложений, относимых к верхнему силуру (остракодовый горизонт). Од-
нако последующие исследования, выполненные палеонтологами Западно-
Сибирского геологического управления, показали, что во многих случаях
эти виды встречаются не в силурийских, а в нижне- и среднедевонских
отложениях. Дальнейшие наблюдения покажут, имеем ли мы дело с ви-
дами широкого стратиграфического распространения, или к остракодо-
вому горизонту верхнего силура были в ряде случаев ошибочно отне-
сены отложения, имеющие более молодой возраст. На основании имею-
щихся данных мы в настоящее время склоняемся ко второму предполо-
жению.
СЕМЕЙСТВО FAVOSITIDAE DANA, ЕМ. SOKOLOV
Род Favosites Lamarck
Описание см. на стр. 156.
Favosites placentas R о m i n g е г
Табл. XXVI, фиг. 3
Полипняк дискоидальный или в виде неправильных волнистых пла-
стин полусферической формы, сравнительно небольших размеров. Ко-
раллиты неодинаковые по величине: диаметр их обычно 0,5—0,8 мм,
иногда менее 0,5 мм, еще более редко около 1 мм. Стенки умеренно тол-
стые, толщиной 0,05 мм. Днища полные, прямые или слабо вогнутые,
неравно удаленные друг от друга. Расстояние между соседними дни-
щами колеблется от 0,2 до 1,6 мм. На 5 мм длины насчитывается 4—7
днищ. Шипы немногочисленные, короткие. Поры круглые редкие, до-
вольно большие, диаметром 0,2 мм, располагаются в один ряд на каж-
дой стенке кораллита.
Наибольшее сходство имеют с видом Favosites nltellus Winch., на
присутствие которого в Di Кузбасса указал Г. Г Петц (1901, стр. 179),
но отличается внешней формой полипняка и большим диаметром корал-
литов.
Распространение. Имеет массовое развитие в живетском
ярусе среднего девона Америки и Азии; в Западной Сибири — Алтай,
Салаир, Кузнецкий бассейн.
Favosites aculeatus Tchernychev
Табл. XXVII, фиг. 1
«Полипняк массивный. Поперечник кораллитов 1,5—2 мм. Стенки
тонкие, не более 0,1 мм толщины, на внешней поверхности с тонкими,
слабо вогнутыми поперечными морщинками. На 5 мм насчитывается
5—7 днищ. Поры круглые, диаметром 0,22 мм, расположенные в 1—2
или три ряда. Шипы многочисленные, длиной 0,4—0,5 мм». (Б. Б. Чер-
нышев, 1951, стр. 28).
Среднедевонский Favosites radiatus Rom. отличается ямками на
днищах и линейными бороздками между рядами шипов. Сходный Favo-
sites turkmenensis var. blrica Ruch, из верхнего силура Туркестана-отли-
чается наличием только одного ряда пор на стенках кораллитов.
Распространение. Средний девон юго-западной окраины- Куз-
басса и Центрального Салаира.
192
Favosltes festivus Tchernychev
Табл. XXVI, фиг. 5
«Полипняк массивный, кораллиты 5—6-гранные, однородные по вели-
чине, поперечником 0,4—0,6 мм. Толщина стенок 0,06 мм (каждой).
Днища в числе 20—25 на 5 мм длины. Поры круглые или широкооваль-
ные, диаметром 0,15—0,24 мм, расположенные в один ряд в количестве
8—14 на 5 мм. Шипы многочисленные, мелкие, треугольной формы»
(Б. Б. Чернышев, 1951, стр. 26).
Обнаруживает большое сходство с F. inosculans N i с h. из нижнего
девона Западной Европы и F. flstulosus Т с h е г п. из верхнего силура
Новой Земли.
«Favosltes inosculans N i с h. отличается отсутствием шипов, более
мелкими и частыми порами, более редкими и неправильными днищами.
Favosltes flstulosus Т с h е г п. обладает более крупными кораллитами
(0,6—0,8 мм), менее частыми днищами и более постоянным диаметром
пор, которые пропорционально меньше» (Чернышев, там же).
Распространение. Встречается в нижнем девоне Салаира;
Б. Б. Чернышев указывает этот вид из отложений с. Сары-Чумыш
(см. стр. 152), относимых к остракодовому горизонту S2. Б. С. Соколов
считает, что F. festivus является по своему облику видом девонским,
а не силурийским.
Favosltes obllquespinus Tchernychev
Табл. XXVII, фиг. 2
Массивный полусферической или неправильной формы полипняк.
Кораллиты неодинаковые по величине, крупные преобладают. Мелкие
кораллиты, вероятно, являются более молодыми. Поперечник кораллитов
обычно около 2,5 мм и несколько больше. Стенки прямые и довольно
толстые, толщина их до 0,12 мм. Днища обычно полные, прямые или
изогнутые, по 5—15 на 5 мм длины.
«Шипы хорошо развиты, довольно многочисленные, толстые, заостряю-
щиеся к концу и, как правило, все направлены кверху, образуя со
стенкой угол в 40°. В продольных сечениях между каждой парой днищ
наблюдается только один шип. Длина шипов не более 1,0 лии, обычно
около 0,5 мм. На 5 мм длины от семи до 14 шипов Поры круглые...
диаметром 0,25—0,30 мм. На стенках кораллитов в зависимости от их
ширины располагаются один или два ряда пор... они несколько смещены
(когда их два) относительно друг друга; на 5 мм длины 6—7 пор»
(Б. Б. Чернышев, 1951, стр. 25).
Близкий среднедевонский вид Favosltes argus Hall отличается более
мелкими, редкими, неправильно расположенными порами и иначе
ориентированными шипами. Среднедевонский Favosltes hamlltonlae
Hall отличается реже расположенными порами и днищами и слабо
развитыми шипами.
Распространение. Руководящий вид в эйфельском ярусе
Салаира.
Favosltes obllquespinus Tchernychev var. zolotainskll
Z h m a e v, var. n.
Табл. XXVII, фиг. 3; рис. 174
Эта разновидность отличается большим диаметром кораллитов, боль-
шим количеством рядов пор и их диаметром и тем, что у разновидности
не наблюдается закономерного расположения шипов по одному между
соседними днищами.
Типичная форма F. obllquespinus имеет поперечник кораллитов
2,5 мм, а разновидность—от 3,3 до 3,5 лои; типичная форма имеет один
или два ряда пор, а вариетет—2—3 ряда.
13 Зак. 2024 1 93
Распространение. Эта разновидность установлена А. П. Жмае-
вым в Эйфеле Центрального Салаира.
Рис. 174. Favosites obliquespintis var. zolotainskli
Z h m a e v, var. n.
а—продольный разрез, X 8; б—поперечный разрез, X 8
Favosites polyniorphus (G о 1 d f u s s)
Табл. XXVIII, фиг. 1
Полипняк клубневидный, в виде сравнительно небольших неправиль-
ных масс или в виде наростов на посторонних телах. По величине
кораллиты не равны, в поперечнике 1,5—1,8 мм; бывают и более мелкие
кораллиты, относящиеся к молодым индивидам. Толщина стенок
колеблется в пределах от 0,05 до 0,25 мм1 в разных частях одной и той
же колонии.
Днища различны в одном полипняке: прямые, перпендикулярные
к стенкам, и косые, вогнутые, выпуклые и неправильно изгибающиеся.
На 5 мм длины от семи до 14 днищ. Расстояние между соседними дни-
щами от 0,1 до 1,2 мм. Шипы редкие, тонкие, направлены вверх. Поры
круглые, диаметром 0,25 и, редко, 0,20 мм. Закономерно расположены
в один ряд. Расстояние между порами в пределах 0,2—0,7 мм.
Распространение. Средний девон Европы и Азии. В Салаире
и на Алтае характеризует Эйфель.
Favosites polytnorphus (Gold fuss) var. crontgera (d’Orb igny)
Табл. XXIX, фиг. 1
От типичной формы отличается более мелкими и равномерными
в поперечном сечении кораллитами, более тонкими стенками (толщина
в пределах 0,05—0,15 мм) и более многочисленными шипами, которые,
как и у типичной формы, наблюдаются в виде тонких чешуек, загнутых
вверх. Следы шипов в поперечном сечении имеют вид тонких коротких
горизонтальных черточек и точек; к тому же по размерам шипы у этой
разновидности немного крупнее, чем у типичного вида.
Распространение. В тех же горизонтах, что и типичная форма.
Favosites rzonsnlckyae Tchernychev
Табл. XXVIII, фиг. 2
Полипняк массивный, большой, неправильной формы. Кораллиты*
одинаковые по величине, 5—7-сторонние, в поперечнике 1,5 мм. Стенки
тонкие и прямые. Днища полные, слабо вогнутые; на 5 мм длины
1 По Б. С. Соколову толщина стенок меняется от 0,2—0,25 мм до 0,35—0,4 мм
и может достигать 0,5 леи.
194
6—9 днищ. Шипы не наблюдались. Поры круглые, диаметром
0,2—0,3 мм, располагаются в один ряд и, как исключение, в два ряда,
быстро переходящие в один; на 5 мм длины около семи пор.
Близкий среднедевонский Squameofavosltes alpen.en.sls Winch, отли-
чается более редкими прямыми днищами и родовым признаком—наличием
редких чешуек (squamae), а сходен формой полипняка, однородностью
и размерами кораллитов, расположением пор в один ряд. От средне-
девонского Favosites intermedlus S t. описываемый вид отличается
меньшим поперечником кораллитов, более редкими и правильными дни-
щами; в остальном эти два вида вполне сходны.
Распространение. Б. Б. Чернышевым установлен в отложе-
ниях, относимых к остракодовому горизонту верхнего силура; по иссле-
дованиям палеонтологов Западно-Сибирского геологического управления,
вид является характерной формой нижнего девона в Салаире.
Favosites brusnitzinl Peetz
Рис. 175
Рис. 175. Favosites brusnitzinl Peetz,
а—продольный разрез; б —попереч-
ный разрез. X 6. Салаир, ключ Фо-
кин—левый приток р. Хвощевки в
ее среднем течении
Диагноз вида по Б. Б. Чернышеву: «Полипняк массивный, уплощен-
ный, неправильной формы, нередко достигает больших размеров: 4,5 см
высоты и 15 X 13 см в поперечнике. Нижняя поверхность полипняка по-
крыта морщинистой эпитекой. Корал-
литы неоднородны, 4—8-угольные,
обычно неправильных очертаний,
причем крупные и мелкие кораллиты
перемешаны в беспорядке; попереч-
ник кораллитов колеблется от 0,5
до 1,0 мм (крупные преобладают).
Стенки прямые в поперечном и
продольном направлениях, толщина
их 0,05—0,06 мм. Днища полные,
прямые, реже косые, изогнутые (чаще
вогнутые). На 5 мм длины от девяти
до 16 днищ. Шипы редкие и корот-
кие. Поры круглые, диаметром
0,2 мм, располагаются в один про-
дольный ряд на каждой стенке ко-
раллита; на 5 мм длины их 12.
Favosites nitlda Charm, отличается несколько меньшим попереч-
ником кораллитов (от 0,50 до 0,75 мм), более многочисленными шипами,
несколько меньшим диаметром пор (0,17 мм) и несколько более частыми
днищами (15—20 на 5 мм длины)» (1951, стр. 28).
Распространение. Руководящий вид для нижнего девона
Западной Сибири (Салаир, Алтай).
Favosites porfirievi Tchernychev
Табл. XXIX, фиг. 3
Диагноз вида по Б. Б. Чернышеву: «Полипняк массивный, образован-
ный радиально расходящимися от небольшого основания кораллитами...
Кораллиты неоднородные, причем более крупные, 7—8-сторонние, с по-
перечником 2,5—3,0 мм, окружены более мелкими, 4—6-сторонними,
имеющими в поперечнике 1,5—2,0 мм. Стенки тонкие, с грубыми, сбли-
женными морщинками на внешней поверхности», поэтому ребра, обра-
зованные гранями кораллитов, представляют волнистую кривую.
Днища полные, прямые; на 5 мм приходится три дниша. Шипы
отсутствуют. Поры круглые, диаметром 0,25—0,30 мм, расположенные
в 1—2—3 ряда на каждой стенке. Чаще всего бывает два ряда пор;
13* 195
в таких случаях поры приближены к ребрам; на 5 мм длины приходится
4—5 пор.
Большое сходство имеет Favosltes forbesl Е d w. et H a i m e, от ко-
торого отличается характером ребер кораллитов, украшением стенок,
расположением пор и более редкими днищами.
Распространение. Руководящий вид нижнего девона юго-за-
падной окраины Кузнецкого бассейна и Центрального Салаира.
Favosltes endygaschensls Tchernychev
Табл. XXX, фиг. 1
«Древовидный полипняк с короткими толстыми ветвями, поперечни-
ком до 5 см. Кораллиты в поперечных сечениях неоднородные, округ-
ленно полигональные, поперечником 1,0—1,6 мм. Днища полные, прямые
или изогнутые, в количестве 7—-12 на 5 мм длины. Стенки толстые;
толщина их от 0,1 до 0,2 мм. Шипы хорошо развиты, но короткие,
толстые и тупые. В шлифах шипы видны в виде прямоугольников
с округленными углами. Поры круглые или широкоовальные, попереч-
ником около 0,3 мм. На каждой стенке кораллитов располагается один
ряд пор в количестве около пяти на 5 мм длины» (Б. Б. Чернышев, 1951,
стр. 36).
Близкий описываемому вид Squameofavosltes radlclformts R о m i п-
g e г из девона Северной Америки отличается более неоднородными и
правильными кораллитами, а также шипами в виде чешуек.
Имеет сходство с Favosltes polymorphus G о 1 d f. и его вариететами,
от которых отличается формой колоний, очертанием кораллитов в по-
перечном сечении, формой пор и рядом менее значительных признаков.
Распространение. Руководящий вид для• эйфеля Салаира,
Алтая и окраин Кузнецкого бассейна.
Favosltes goldfusst d’O г b i g п у
Табл. XXVI, фиг. 4
Полипняк небольших размеров, полусферической, сферической или
несколько вытянутой формы. Кораллиты неоднородные по размерам,
правильно полигональные, радиально расходящиеся; крупные кораллиты
преобладают. Диаметр кораллитов обычно 2,0—2,5 мм. Стенки прямые,
плотные, утолщенные, со слабо выраженным или отсутствующим швом.
Днища полные, прямые, частые. Шипы развиты. Поры круглые, диамет-
ром около 0,20—0,25 мм, располагаются в два ряда.
Favosltes goldfussl d’O г b. имеет значительное сходство с верхие-
силурийским F. gothlandlcus (см. стр. 156), от которого отличается
меньшими размерами полипняка, меньшим диаметром и неоднородностью
кораллитов, более толстыми стенками со слабо выраженным или от-
сутствующим швом, более густыми порами и более частыми днищами.
Распространение. Типичный среднедевонский вид. В Запад-
ной Сибири характерен для эйфельского яруса (так же, как и его много-
численные разновидности).
Favosltes petrovttcht Z h m а е v, sp. n.
Табл. XXX, фиг. 3; рис. 176
Полипняк массивный. Кораллиты 3—7-угольные, в основной массе
одинаковых размеров; преобладает диаметр 1,5—2,2 мм, толщина сте-
нок от 0,20 до 0,25 мм. Днища полные, косые и вогнутые, редко прямые;
на 5 мм 8-—И днищ. Поры мелкие, многочисленные, расположены
в 1- 2 ряда; диаметр пор в среднем 0,20—0,15 мм. Шипы сплошь
усеивают стенки кораллитов; в продольном и поперечном сечениях они
имеют вид небольших бугорков; на 5 мм приходится 10—12 шипов.
196
Отличается от всех известных видов очень толстыми стенками, мно-
гочисленными мелкими порами, массой шипов, покрывающих почти
сплошь стенки кораллитов.
Распространение. Нижний девон в Центральном Салаире.
Подрод Dictyofavosites Tchernychev
«От типичных Favosites представители подрода Dictyofavosites отли-
чаются только характером днищ. Днища у этого подрода в соседних
кораллитах располагаются на одних уровнях, являясь как бы продолже-
нием одно другого в виде общей пластины для всего полипняка. В про-
дольных сечениях, таким образом, получается довольно правильная
сетка» (Б. Б. Чернышев, 1951, стр. 36).
Рис. 176. Favosites petrovitchl Z hmaev.,
sp. n.
а—продольный разрез; б—поперечный раз-
рез. X 9. Салаир. кл. Петрович — левый
приток р. Павловки.
Рис. 177. Favosites (Dyc-
tyofavosites) salairicus
'1 c h e r n. Продольный
разрез, X 8.4. Салаир,
кл. Мишуков — левый
приток р. Северный
Мочег . в его верхнем
течении
Типичный вид. Favosites (Dictyofavosites) salairicus Т с h е г n.
Распространение. По исследованиям палеонтологов Западно-
Сибирского геологического управления, типичный вид этого подрода
характеризует нижний девон Центрального Салаира и юго-западной
окраины Кузбасса. Б. Б. Чернышев описал этот вид из отложений, отно-
симых к верхнему оилуру (остракодовый горизонт), что требует проверки.
Favosites (Dictyofavosites) salairicus Tchernychev
Табл. XXX, фиг. 4; рис. 177
«Полипняк массивный, достигающий значительных размеров. Корал-
литы однородные по величине, поперечником в разных частях полипняка
1,0—1,5 мм. Стенки довольно тонкие. Днища полные, прямые или слабо
вогнутые, расположенные в кораллитах на одной высоте. На 5 мм насчи-
тывается 4—9 днищ. Шипы очень редкие. Поры овальные, поперечником
0,24X0,42 мм, расположенные в один ряд, на расстоянии 0,24—0,90 jmjh.
Характер расположения днищ и очертание пор служат отличительным
признаком этого вида» (Б. Б. Чернышев, 1951, стр. 37).
Распространение. См. выше, в описании подрода.
Род Squameofavosites Tchernyschev
Полипняк полусферической, неправильной фермы. Кораллиты раз-
личны: то однородные, то неоднородные. От типичных представителей
Favosites этот род отличается септальными образованиями: вместо кони-
197
ческих шипов у него имеются чешуи, располагающиеся продольными
рядами на стенках кораллитов под порами. Чешуи имеют округленно-
прямоугольную или треугольную форму.
Типичная форма. Squameofavosltes bohemlcus (Р о с t а).
Р ас простр аие н ие. Нижний и средний девон.
Squameofavosltes bohemlcus (Р о с t а)
Рис. 178
Диагноз по Б. Б. Чернышеву: «Полипняк грибообразный. Кораллиты
поперечником 2,0—2,6 мм. Стенки довольно толстые (около 0,1 мм).
Рис. 178. Squameofavosltes bohemlcus Р о с t а
а—продольное сечение кораллита; б и в—стен-
ки кораллитов с порами; г—поперечные сече-
ния кораллитов, видны чешуи; д — видны про-
дольные и поперечные сечения чешуи в раз-
резах кораллитов, х 8. По Б. Б. Чернышеву
Днища полные, прямые и изо-
гнутые или неполные, изгибаю-
щиеся и соединяющиеся с со-
седними; на 5 мм их от 12 до
20. Поры круглые, диаметром
0,2 — 0,3 мм, расположенные
в 1—2 и три, редко в четыре
ряда, в количестве 11 —12 на
5 мм длины. Септальные обра-
зования представлены чешуй-
ками треугольной или округ-
ленно-прямоугольной формы,
длиной до 0,5 мм. Ширина че-
шуй у основания 0,12—0,55 мм,
толщина 0,04—0,12 мм. Чешуи
расположены под порами и
вогнуты по ширине кверху; на
5 мм длины их от 10 до 16».
Распространение. Ха-
рактеризует нижний девон Се-
вера Сибири, а для Западной
Сибири Б. Б. Чернышевым определен из отложений, относимых к остра-
кодовому горизонту (близ с. Томский Завод). По исследованиям палеон-
тологов Западно-Сибирского геологического управления, характеризует
нижний девон Салаира.
СЕМЕЙСТВО THAMNOPOR1DAE SOKOLOV
Род Thamnopora Steininger
В литературе обычно фигурирует под названием Pachypora L i п d s t г.
Полипняк ветвистый, образован свободными цилиндрическими или
срастающимися друг с другом ветвями. Кораллиты фавозитоидного
облика, веерообразно отходят от оси ветвей и открываются перпендику-
лярно к их поверхности. Сечение полигональное. Стереоплазм этическое
утолщение нарастает постепенно от оси к периферии. Днища горизон-
тальные, сравнительно редкие. Шипы развиты слабо. Поры крупные, рас-
положенные большей частью в один ряд.
Род Pachypora L i n d s t г. отличается от рода Thamnopora своеобраз-
ным слоистым, почти сплошным стереоплазматическим утолщением ске-
летных элементов, а также утолщенным ветвистым полипняком и нередко
косо открывающимися устьями кораллитов (как у рода Strlatopora).
Типичный вид. Thamnopora cervlcornls (В 1.).
Распространение. В девоне род Thamnopora пользуется исклю-
чительным распространением, в силуре он редок, тогда как род Pachy-
pora стратиграфически не выходит за пределы силура.
198
Thamnopora cervicornls (В 1 a i n v i 11 e)
Табл. XXIX, фиг. 2
Синонимы: Favosltes cervicornls и Pachypora cervicornls
Полипняк ветвистый, с цилиндрическими или слегка уплощенными
ветвями. Кораллиты полигональные, иногда несколько неоднородные,
с поперечником около 1,5 мм. Стенки на внутренней стороне утолщены
стереоплазмой, вследствие чего отверстия кораллитов становятся округ-
ленными и несколько суженными. Шипы видны очень редко в виде не-
больших заостренных бугорков. Поры круглые, диаметром около 0,20—
0,25 мм, располагаются неправильно в один ряд на расстоянии от 0,6 до
0,9 мм. Днища тонкие, прямые, перпендикулярные к стенкам, или косые,
изогнутые, иногда соединяющиеся с соседними; между днищами расстоя-
ние 0,5—1,0 мм.
У различных авторов имеется различное понимание данного вида,
в связи с чем под этим названием описаны разные виды; по этой при-
чине трудно установить истинное распространение его. В Западной Си-
бири часто встречается в среднем и низах верхнего девона (Алтай,
Салаир, Кузбасс).
Thamnopora alta (Tchernychev)
Табл. XXXI, фиг. 1
Полипняк древовидный, с диаметром ветвей до 3 см. Кораллиты
вблизи поверхности полипняка (на расстоянии одной трети диаметра
ветви) резко поворачивают к поверхности и образуют с ней прямой угол;
с этого места стенки сильно утолщаются; толщина стенок увеличивается
до 0,5—0,7 мм. Поперечник кораллитов около 1,5 мм, иногда достигает
2 мм. Шипы редкие, толстые и длинные (до 0,4 мм). Поры круглые,
диаметром до 0,3 мм, расположены в один ряд на расстоянии чаще всего
около 1 мм.
От близкого вида Thamnopora cervicornls (Bl.) этот вид отличается
большей толщиной стенок, величиной кораллитов, наличием резкого
изгиба кораллитов и сильным утолщением стенок после этого изгиба,
а также характером пор.
Распространение. Распространенный вид в среднем девоне
Сибири и Казахстана.
Род Striatopora Hall
«Полипняк древовидный, ветвящийся, с цилиндрическими ветвями.
Кораллиты открываются косо к поверхности полипняка, полигональные,
изгибающиеся от оси полипняка к поверхности, с которой образуют ост-
рый или почти прямой угол. Чашечки довольно глубокие, несколько ко-
сые, с острыми краями и ясными продольными струйками внутри. Стенки
внутри полипняка тонкие, но утолщающиеся к поверхности, причем
у одних видов стенки утолщаются постепенно и остаются умеренно тол-
стыми, тогда как у других утолщение настолько сильно, что висцераль-
ная полость кораллита становится очень небольшой.
В последнем случае в шлифах в середине полипняка видны сечения
кораллитов с тонкими стенками, окруженные более или менее широкой
каймой сечений с очень сильно утолщенными стенками (обычно висце-
ральные полости в этой зоне почти или совсем не видны). Днища пол-
ные и редкие. Поры хорошо развиты, но у большинства видов непра-
вильно расположены. Шипы редкие» (Б. Б. Чернышев, 1951, стр. 52).
Типичный вид. Striatopora flexuosa Hall.
Распространение. Верхний силур — девон, повсеместно.
199
Striatopora kuznetskiana Tchernychev
Табл. XXX, фиг. 2
Диагноз вида по Б. Б. Чернышеву: «Полипняк представляет сетку,
образованную дихотомирующими и срастающимися ветвями, поперечни-
ком до 2,5 мм. -Поперечник кораллитов на поверхности 0,8—1,0 мм, се-
чения кораллитов 5—6-угольного очертания. Стенки быстро утолщаются
к поверхности. Днища полные, косые, редкие; часто днища не заметны
на значительных расстояниях; на 1 мм длины до трех днищ. Поры круг-
лые, редкие, расположенные в один ряд на каждой стенке; чашечки
умеренно глубокие, со слабыми струйками».
Отличается от всех близких форм малым числом отверстий коралли-
тов на поверхности колоний, формой полипняка и способом роста.
Распространение. Нижнедевонские отложения Кузнецкого
бассейна и Салаира.
СЕМЕЙСТВО ALVEOLITIDAE DUNCAN, ЕМ. SOKOLOV'
Род Alveolites Lamarck
«Полипняк массивный, реже пластинчатый или ветвистый, достигаю-
щий иногда значительных размеров. Кораллиты плотно прилегают друг
к другу, обычно изогнуты и направлены косо к поверхности. На поверх-
ности полипняка и в поперечных разрезах кораллиты разной формы:
удлиненные, округленнотреугольные, овальные, полулунные, четырех-
угольные (с верхней выпуклой стороной, нижней вогнутой и сходящи-
мися, слегка вогнутыми, боковыми сторонами), неправильно полигональ-
ные, удлиненнопрямоугольные. Стенки кораллитов тонкие, иногда непра-
вильно утолщенные, несущие довольно крупные, обычно немногочислен-
ные поры.
Днища тонкие, полные, горизонтальные или изогнутые, многочислен-
ные; шипы очень разнообразные: наиболее часто в каждом кораллите
видны один или три продольных ряда сильно развитых шипов, причем
остальные шипы рудиментарны или совсем отсутствуют. У других видов
шипы мелкие, одинаковой длины или совсем отсутствуют».
Типичный вид. Alveolites suborblcularls Lamarck.
Распространение. Представители рода Alveolites часто встре-
чаются в верхнем силуре и девоне всех стран, в том числе и Западной
Сибири.
Alveolites malllleuxl S а 1 ё е
Табл. XXVIII, фиг. 3
Полипняк массивный, неправильной формы или в виде пластин. Корал-
литы почти равной длины и ширины, полигональные, иногда овальные,
полулунные; по величине неоднородные в одном полипняке, в попереч-
нике до 1 мм. Стенки тонкие, местами утолщенные. Днища полные,
многочисленные, прямые или косые, реже вогнутые или выпуклые; рас-
стояние между днищами варьирует таким образом, что на 5 мм длины
насчитывается 10—20 днищ. Поры круглые, диаметром 0,20—0,25 мм,
расположены в один ряд на расстоянии 0,2—0,5 мм друг от друга (дру-
гие расстояния случайны). Шипы короткие и немногочисленные.
Этот вид формой чашечек весьма напоминает представителей рода
Favosites и занимает промежуточное место между Favosites и Alveolites.
Распространение. Живетский ярус Западной Европы. В За-
падной Сибири обычно характеризует живетский ярус, редок в эйфель-
ском и франском ярусах окраин Кузбасса.
1 Диагнозы рода и видов взяты из работы Б. Б. Чернышева «Силурийские и де-
вонские Tabulata и Heliolitida окраин Кузнецкого угленосного бассейна», 1951.
200
СЕМЕЙСТВО SYRINGOPORIDAE EDWARDS ET HAIME, EM. SOKOLOV
Род Syringopora Gold fuss
«Полипняк кустовидный или образованный сближенными, но редко
соприкасающимися кораллитами. Кораллиты... длинные, цилиндрические,
обычно более или менее изгибающиеся, но в общем идущие параллельно
или почти параллельно друг другу. Расстояние между кораллитами
у разных видов колеблется в широких пределах... Отдельные кораллиты
соединены между собой при помощи полых поперечных трубок.
Днища воронкообразные, с осевой трубкой, которая иногда запол-
нена вторичными горизонтальными, косыми или воронкообразными дни-
щами. У некоторых видов днища имеют очень неправильную форму
и сильно изогнуты. Внутри кораллитов на стенках располагаются про-
дольными рядами или в беспорядке хорошо развитые или рудиментар-
ные шипы.
Внешняя поверхность кораллитов несет концентрические складочки,
пересеченные у некоторых видов продольными бороздками... В ранней
стадии развития полипняк имеет вид стелющихся трубок, напоминаю-
щих представителей рода Aulopora Goldf.» (Чернышев, 1951, стр. 71).
Типичный вид. Syringopora reticulata Goldf.
Распространение. Силур—пермь, повсеместно.
Syringopora elfeliensls Schluter
Табл. XXXII, фиг. 2
Кораллиты длинные, обычно прямые или несколько изогнуты, диамет-
ром 4—5 мм. Расстояние между кораллитами в одном полипняке варьи-
рует в пределах до двух диаметров. Толщина стенок 0,2—1,0 мм. Соеди-
нительные трубки толстые и очень редкие. Днища воронкообразные,
с осевой трубкой, в которой изредка наблюдаются вторичные днища.
Шипы небольшие, немногочисленные,
Близкий вид Syringopora abdita Vern. отличается более глубокими
и частыми воронкообразными днищами и более резкими концентриче-
скими складочками на внешней поверхности стенок.
Распространение. Руководящий вид для среднего девона.
В Западной Сибири встречается в эйфельском и живетском ярусах
(в последнем редко).
Syringopora yavorskyl Tchernychev
Табл. XXXII, фиг. 5
«Кораллиты прямые, диаметром 0,6—0,7 мм, на расстоянии 0,2—
2,0 мм (чаще 0,5—1,0 мм). Толщина стенок 0,1 мм. Соединительные
трубки редкие, диаметром 0,4 мм. Днища воронкообразные, глубокие,
с осевой трубкой. Шипы не обнаружены. Этот вид встречается только
в симбиозе со строматопорами» (Б. Б. Чернышев, 1951, стр. 76).
От близкого Syringopora compacta Bill, отличается сильной сбли-
женностью кораллитов и более частыми соединительными трубками.
Распространение. Характеризует эйфельский ярус юго-запад-
ной окраины Кузбасса.
СЕМЕЙСТВО AULOPOR1DAE FROMENTEL, ЕМ. NICHOLSON
Рол Aulopora Goldfuss
«Полипняк стелющийся, ползучий, составленный трубчатыми корал-
литами, образующими сетчатые или ветвистые колонии... Кораллиты
цилиндрические или в форме рожков, прикрепляются к посторонним
предметам... Днища очень редкие или отсутствуют... Септы в виде едва
201
заметных продольных гребней. Стенки кораллитов всегда сплошные, без
пор» (Б. Б. Чернышев, 1951, стр. 81).
Типичный вид. Aulopora serpens G о 1 d f.
Распространение. Силур—пермь, повсеместно.
Aulopora slblrica Tchernychev
Табл. XXVII, фиг. 4
Стелющийся полипняк. Кораллиты цилиндрические, удлиненные, резко
изогнутые в передней части у чашечки. Отверстия кораллитов круглые,
несколько суженные, направленные вертикально, подняты на высоту
около 1 мм. Диаметр их 0,5—0,6 мм. Диаметр кораллитов 0,6—0,8 мм.
длина 1,8—3,0 мм. Стенки умеренно толстые, с тонкими морщинками.
Каждый кораллит дает начало обычно двум (редко одному) новым, ко-
торые отходят в самой верхней части материнского индивида, из-под
чашечки.
От сходного вида Aulopora heckerl Тс hern, сибирский вид отли-
чается меньшей поднятостью отверстий кораллитов, менее грубыми скла-
дочками на внешней поверхности стенок кораллитов, а также характе-
ром почкования. У Aulopora heckerl новые кораллиты отходят от сред-
ней части материнской особи, а у Aulopora slblrica—от верхней.
Распространение. Установлена в среднедевонских отложе-
ниях Кузнецкого бассейна. В Центральном Салаире образует целые
прослои в отложениях среднего девона.
ГРУППА HELIOLITIDA
СЕМЕЙСТВО HELIOLITIDAE LINDSTROM
Род Heliolites Dana
Описание см. на стр. 157.
Heliolites vulgaris Tchernychev
Табл. XXXI, фиг. 2
Полипняк массивный, полусферический, в поперечнике чаше всего до
8 см. Кораллиты круглые, со слегка ограниченными, незазубренными
стенками, диаметром 1,5—2,0 мм. Расстояние между ними варьирует от
1,3 до 4,0 мм. Стенки тонкие и лишь немного толще стенок сифонопор.
Септы отсутствуют. Днища прямые или слегка изогнутые; на 5 мм от
четырех до восьми днищ. Сифонопоры 5—6-гранные, в поперечнике
0,3—0,6 мм. Между двумя кораллитами от трех до 12 рядов сифонопор.
Днища в сифонопорах умеренно частые; на 5 мм их 8—-9 штук.
Имеет сходство с Heliolites decipiens М’Соу и Heliolites bohemlcus
Wentz, Heliolites decipiens отличается более мелкими в поперечном
сечении кораллитами, более многочисленными днищами в кораллитах
и сифонопорах и меньшим количеством рядов сифонопор между корал-
литами. Heliolites bohemlcus W. имеет более крупные в поперечном сече-
нии и сближенные кораллиты, более многочисленные днища в коралли-
тах и сифонопорах.
Распространение. Часто встречается в среднем девоне Запад-
ной Сибири (Салаир, Алтай).
Heliolites yavorskyt Tchernychev
Табл. XXXII, фиг. 4
Полипняк грибообразной формы, с короткой ножкой. Кораллиты круг-
лые, без следов огранения или зазубренности, диаметром 0,7—0,8 мм;
расстояние между кораллитами 0,2—1,0 мм. Септальные образования
неизвестны. Днища прямые или слабо изогнутые; на 1 мм длины 3—4
днища. Стенки кораллитов умеренно толстые и толще стенок сифонопор.
Сифонопоры 5—6-гранные, неоднородные по величине, поперечником
202
0,15—0,30 мм (обычно около 0,2 мм). Между соседними кораллитами
2—3 ряда сифонопор. Днища в сифонопорах прямые; на 1 мм длины их
5—6.
Очень близок формой полипняка Н. barrandei var. spongo.tdes
L i n d s t., который отличается наличием септальных образований и бо-
лее мелкими кораллитами с продольноскладчатыми стенками. Heliolites
declpiens М’Соу отличается более крупными кораллитами, сильно уда-
ленными друг от друга, присутствием (хотя и очень редко) септальных
образований.
Распространение. Остракодовый горизонт окраин Кузбасса по
Б. Б. Чернышеву; по исследованиям палеонтологов Западно-Сибирского
геологического управления — в нижнем девоне Центрального Салаира.
Heliolites insole ns Tchernychev
Табл. XXXII, фиг. 1
Полипняк массивный. Кораллиты неправильно округленных очерта-
ний. Зазубренность их стенок носит разнообразный характер у разных
индивидов; диаметр 0,9—1,2 мм (чаще 1 мм); расстояние между корал-
литами 0,2—0,7 мм. Днища прямые, расположены на более или менее
равных расстояних друг от друга; на 5 мм длины семь днищ. Септы уме-
ренно развиты, тонкие и короткие. Сифонопоры неправильные и неодно-
родные по величине, в поперечнике 0,15—0,50 мм. Местами сифонопоры
очень длинные и изогнутые; на 5 мм длины в сифонопорах 14 днищ.
Вокруг каждого кораллита расположено И-—16 сифонопор; между двумя
кораллитами от двух до четырех рядов сифонопор.
Наиболее близок верхнесилурийский Heliolites interstlnctus var. Irre-
gularis Wenzel, который отличается более крупными и более сближен-
ными кораллитами и более многочисленными, неправильно расположен-
ными и изогнутыми днищами; иногда днища кораллитов сифонопор
у последнего вида составляют параллельные линии.
Распространение. Широко распространен в Эйфеле Салаира,
но встречается и в более высоких горизонтах среднего девона.
Heliolites tomensls Tchernychev
Табл. XXXI, фиг. 3
Массивный полипняк неправильной формы с выпуклой верхней поверх-
ностью и широкой поверхностью прикрепления. Кораллиты круглые,
с пескладчатыми краями; диаметр 1,9—2,1 мм; между кораллитами рас-
стояние 0,3—1,2 мм. Толщина стенок 0,1—0,2 мм. Септы короткие, дли-
ной до 0,3 мм и в поперечных сечениях наблюдаются в виде шипов. На
5 мм приходится 6—7 днищ. Днища прямые, слегка вогнутые или вы-
пуклые; на 5 мм их от шести до семи. Сифонопоры правильно полиго-
нальны, в поперечнике 0,3—0,5 мм, обычно 0,4 мм; на 5 мм в сифонопо-
рах 11—14 днищ. Между кораллитами 1—4 ряда сифонопор (диагноз
вида по Б. Б. Чернышеву).
Сходные виды: Heliolites barrandei Р е n., Н. dalnterll Е t h. et N i с h.,
H. interstlnctus L.
«Heliolites barrandei отличается менее правильно круглым очертанием
кораллитов поперечником 1,4—1,7 мм, сильно зазубренными стенками
кораллитов и более длинными септами. Кроме того, вокруг каждого ко-
раллита намечается венец сифонопор, несколько вытянутых по его
радиусам.
Heliolites Interstlnctus отличается несколько более мелкими в попе-
речном сечении и менее сближенными кораллиФами, более длинными
септами и зазубренными стенками кораллитов; последнее отличие
является наиболее важным» (Б. Б. Чернышев, 1951, стр. 91).
Распространение. Характеризует средний девон Салаира; по
Б. Б. Чернышеву — живетский ярус.
203
Heliolites porosus (G о 1 d f u s s)
Табл. XXXII, фиг. 6; табл. XXXIII, фиг. 1
Полипняк грибообразный, почти сферический, с короткой ножкой
или в виде сильно выпуклых неправильных бугров. Кораллиты круглые,
правильно зазубренные; диаметр 1,0—1,4 мм (мелкие встречаются редко);
расстояние между кораллитами 1—3 мм... Стенки кораллитов умеренно
толстые, толще стенок сифонопор. Септы умеренно длинные, длина их
неодинакова; количество септ в каждом кораллите 12. Днища прямые,
редко изогнутые; на 5 мм длины 6—8 днищ. Сифонопоры неоднородные,
поперечник от 0,25 до 0,50 мм. Между двумя кораллитами от двух до
девяти рядов сифонопор. Днища в сифонопорах более многочисленны,
чем в кораллитах (диагноз вида по Б. Б. Чернышеву).
В литературе под названием Н. porosus Goldf. описаны разные
виды.
Наиболее близок Heliolites vulgaris В. В. Тс hern. (см. стр. 202)
с разновидностями; отличается отсутствием септ и другими деталями
строения.
Распространение. Heliolites porosus характерен для живет -
ского яруса среднего девона в Кузбассе (по Б. Б. Чернышеву); воз-
можно его присутствие в верхах эйфельского яруса.
Heliolites nagorskyi Z h m а е v, sp. n.
Табл. XXXIII, фиг. 2; рис. 179
Полипняк массивный, почти сферический. Кораллиты круглые или
несколько овальные, неправильно зазубренные, диаметром от 1,5 до
2,1 мм, иногда до 2,5 мм (последние встречаются редко). Стенки корал-
литов толще стенок сифоно-
пор. Расстояние между со-
седними кораллитами колеб-
лется от 0,5 до 1,5 мм. Сеп-
ты умеренно длинные, длина
их менее половины радиуса
кораллита. Количество септ
в каждом кораллите 12.
Днища слабо вогнутые, ино-
гда не доходят до противо-
положной стенки и соединя-
ются с соседними днищами;
на 5 мм длины приходится
7—10 днищ. Сифонопоры в
поперечном сечении из-за
Рис. 179. Heliolites nagorskyi Zhtnaev, sp. п. сильно утолщенных стенок
а—продольный разрез, х 9; б—поперечный (0,1 мм) кажутся округлыми,
разрез, X 8. Салаир, р. Хайрюзовка, среднее Поперечник 0,3--0,5 ММ. Me-
течение ЖДу дВумя кораллитами от
одного до шести рядов сифо-
нопор. Днища в сифонопорах прямые или косые, реже изогнутые;
на 5 мм — от 11 до 16 днищ. От близких Heliolites interstinctus L.,
Н. tomensis Т с h е г n., Н. porosus Goldf. и от других видов Heliolites
nagorskyi отличает сочетание признаков: количество днищ на 5 мм
в сифонопорах и кораллитах, толстые стенки сифонопор и кораллитов,
неправильная зазубренность стенок последних.
Распространение. Характерный вид среднего девона в Цент-
ральном Салаире.
204
Heliolites gluchovl Z h m a e v, sp. n.
Табл. ХХХШ, фиг. 3; рис. 180
Полипняк неправильно сферической формы. Кораллиты круглые,
с правильно зазубренными стенками; диаметр от 1,25 до 1,40 мм в одном
и том же полипняке. Стенки кораллитов умеренно толстые и равны' тол-
щине стенок сифонопор. Расстояние
между кораллитами от 0,5 до 1,2 мм,
чаще всего 0,7—0,8 мм. Септы име-
ют вид треугольных выростов. Коли-
чество септ в каждом кораллите 12.
Днища полные, прямые, перпендику-
лярные к стенкам кораллитов, до 11
на 5 мм длины. Сифонопоры круг-
лые, правильно зазубренные,неодно-
родны по величине и очертаниям,
в поперечнике 0,2—0,4 мм. Между
двумя кораллитами от двух до четы-
рех рядов сифонопор. Днища прямые,
редко слабо вогнутые или выпуклые;
на 5 мм длины— 18—19 днищ.
От сходного Heliolites porosus
G о 1 d f. отличается короткими сеп-
тами, большим количеством днищ
Рис. 180. Heliolites gluchovi
Z h ni а е v, sp. п.
а—продольный разрез, X 9,5; б—по-
перечный разрез, X 9,3. Салаир,
р. Глухая—левый приток р. Амфимов
Мочег
в кораллитах и сифонопорах. От
Heliolites decipiens М’С о у отличается развитыми септами, меньшим
расстоянием между соседними кораллитами, большим количеством днищ
в сифонопорах.
Распространение. Установлен в нижнем девоне Центрального
Салаира.
Heliolites salalrlcus Tchernychev
Табл. XXXII, фиг. 3
Массивный крупный полипняк поперечником до 18 см. «Кораллиты
неправильно широкоовальные, без следов зазубренности стенок. Попе-
речник их равен 1,0—1,4 мм. Расстояние между соседними кораллитами
варьирует от 0,1 до 1,1 мм, но наиболее часто равно 0,4—0,7 мм. Стенки
тонкие, толщиной максимум 0,1 мм (обычно меньше этой величины).
Септы полностью отсутствуют... Днища прямые или чуть вогнутые; на
5 мм 7—8 днищ. Сифонопоры неправильно полигональны и неодно-
родны по величине; поперечник от 0,2 до 0,6 мм. Между двумя корал-
литами 1—2 ряда сифонопор; днища в последних прямые, полные,
в количестве 10—12 на 5 мм длины (диагноз вида по Б. Б. Черны-
шеву) .
Распространение. Описан его автором из верхнего силура
Центрального Салаира, палеонтологами Западно-Сибирского геологи-
ческого управления — из нижнего девона Центрального Салаира. Возра-
стное положение требует проверки.
Heliolites khvosievtensis Z h m а е v, sp. n.
Табл. ХХХШ, фиг. 4; рис. 181
Полипняк массивный, достигающий значительных размеров. Корал-
литы круглые, в поперечнике 1,0—1,2 мм, расстояние между ними от 0,9
до 2,3 мм, редко больше или меньше этих размеров. Стенки кораллитов
и сифонопор по толщине совершенно одинаковы. Септальных образова-
ний не обнаружено. Днища полные, обычно вогнутые, редко прямые;
иногда днища до противоположной стенки не доходят и соединяются
с соседними; на 5 мм длины — от 11 до 14 днищ.
205
Сифонопоры полигональные, неоднородны по величине, в попереч-
нике 0,3—0,5 мм. Между двумя кораллитами от трех до пяти сифоно-
пор; днища в них полные, косые или вогнутые, реже прямые в количе-
стве 14—17 на 5 мм.
От наиболее близкого верхнесилурийского Heliolites deciplens М'С о у
отличается меньшим количеством сифонопор между кораллитами, боль-
Рис. 181. Heliolites khvoscevlensis Zhmaev,
sp. n.
а—продольный разрез; б— поперечный разрез.
X 8. Салаир, ключ Смородиновый — левый
приток р. Хвощевки
шим количеством днищ на 5 мм в сифонопорах и кораллитах и отсут-
ствием даже слабой огранки кораллитов.
Распространение. Установлен в эйфельском ярусе Централь-
ного Салаира.
Л. Н. КРАЕВСКАЯ
ПОДКЛАСС ZOANTHARIA
Отряд tetracoralla или rugosa. тетракораллы
(нижний и средний девон)
СЕМЕЙСТВО PHOLIDOPHVLLIDAE WDKD
Объединяет кораллы, септы которых построены из трабекул, разъеди-
няющихся в осевой части. В полости коралла, кроме септ, развиты толь-
ко более или менее усложненные днища.
Род Tryplasma Lonsdale
Табл. XXXV, фиг. 182
«Одиночные кораллы цилиндрической формы, иногда с корневыми
выростами и резкой двойной ребристостью на эпитеке... Характерны
многочисленные пережимы, и вздутия. Септы состоят из вертикальных
рядов трабекул, периферические части которых слиты не только в верти-
кальном направлении, но и по всей внутренней поверхности коралла. От
слияния трабекул на периферии образуется широкая стереозона».
Расщепление трабекул создает на днищах и внутренней стенке корал-
ла шиповидные образования. В продольном разрезе через септу эти
образования имеют вид. шипов, а в поперечном разрезе выглядят точ-
ками. Днища более или менее правильно горизонтальные. Пузырчатой
ткани не имеется.
«Девонские представители рода отличаются от силурийских отсут-
ствием резких пережимов, продольной ребристостью эпитеки и значитель-
но более узкой стереозоной». (Диагноз рода по Е. Д. Сошкиной.)
206
Ниже приводятся диагнозы крупных представителей видов рода
Tryplasma — Tr. devoniana и Tr. altaica,—присутствие которых в Сиби-
ри зафиксировано разными авторами. Первым автором, установившим
присутствие крупных Tryplasma в Сибири, был Дыбовский; он предложил
для них название Amplexus altaicus. Под этим названием они широко
известны в сибирской геологической литературе. Взаимоотношения двух,
названных выше близких видов и даже возможное их тождество могут
быть установлены лишь специальными исследованиями.
Э. 3. Бульванкер оставляет за этими видами название рода Pholldo-
phyllum, как более распространенное в геологической литературе.
Типичный вид. Tryplasma aequalls (L о n s d.).
Распространение. Силур—средний девон. Европа, Азия.
Tryplasma devoniana (S о s h k i n а)
Табл. XXXV, фиг. 3
Одиночные кораллы цилиндрической формы, часто значительной ве-
личины (от 1 до 10 см). Внешняя стенка покрыта слабо выраженной
продольной бороздчатостью. Небольшие пережимы и вздутия отчетливо
видны. Стереозона узкая. Сеп-
тальные трабекулы неправильно
изогнуты и не всегда заметны.
Днища тонкие, широкие, го-
ризонтальные или слегка изо-
гнутые, простые или слабо рас-
щепленные, иногда у краев не-
много выпуклые; на 10 мм при-
ходится их 5—7. Короткие сеп-
ты почти одинаковы, но все же
местами можно разделить их
на два порядка; на периферии
они утолщены стереоплазмой,
образующей ободок. Количе-
а
Рис. 182. Tryplasma hedstroml Wdk.
а—продольный разрез; б—поперечный разрез.
X 4. Восточный склон Урала. По Е. Д. Сош-
ство септ зависит от диаметра киной
кораллитов; при диаметре 5 см число их достигает 80—85. Tryplasma
devoniana обладает сильной изменчивостью строения днищ и септаль-
ных трабекул; наиболее постоянным признаком является слабое разви-
тие стереозоны. (Диагноз вида по Е. Д. Сошкиной.)
Распространение. Встречается на Урале в D2 и Di, в Салаире
и Кузбассе — руководящая форма для Dj; особенно часто встречается
в кобленцском ярусе.
Рис. 183. Tryplasma altaica
Dybowski.
Часть'поперечного разреза,
2/з нат. вел.
Tryplasma altaica (Dybowski)
Табл. XXXIV, фиг. 1; табл.
XXXV, фиг. 2; рис. 183
В литературе обычно указывается
под названием Amplexus altaicus
«Коралл одиночный, очень крупный,
цилиндрической, слегка изогнутой формы.
В поперечном сечении описываемого вида,
кроме септ и перерезанных днищ, других
скелетных образований нет. Септы вто-
рого порядка по величине почти равны
септам первого порядка и только местами
едва отличаются от них. Довольно длинные, зубовидные, сильно утол-
щенные стереоплазмой септы тесно соприкасаются наружными кон-
цами. .. В поперечном сечении большую плоскость коралла заполняют
тонкие, горизонтальные, слегка расщепленные днища. На 10 мм прихо-
дится около 7—8 днищ» (Бульванкер, 1938).
207
Отличается от других представителей рода главным образом боль-
шим диаметром и длиной коралла, сильно утолщенными септами и их
большим количеством.
Распространение. Нижний девон Салаира и Кузбасса.
Tryplasma hercyntca (Roemer)
Табл. XXXV, фиг. 4
Одиночные мелкие кораллы иногда образуют негустые псевдоколонии
путем развития «парисидальных» почек. В таких псевдоколониях корал-
литы прочно удерживали особенности внешней формы одиночных корал-
лов. Чашка глубокая, бокаловидная, с отвесными стенками, плоским
дном и толстыми краями. Кораллиты цилиндрические, с мелкими взду-
тиями и нерезкой продольной ребристостью. Септальные трабекулы длин-
ные, ободок широкий (четверть-треть радиуса). Септы двух порядков, но
септы второго порядка развиты очень слабо, иногда почти незаметны.
Количество септ первого порядка 30—33. Днища тонкие, горизонтальные,
слабо расщепленные.
От Tryplasma devonlana (см. выше) отличается меньшими размерами
кораллитов, более удлиненной цилиндрической формой их, большей дли-
ной септ, большей шириной ободка и более косым расположением сеп-
тальных трабекул, отчего на поперечном разрезе всегда заметно четко-
видное строение септ, нехарактерное для Tryplasma devonlana.
Распространение. На Урале и Кузбассе характеризует нижний
девон.
СЕМЕЙСТВО KODONOPHYLL1DAE WEDEKIND
Род Calceola Lamarck
«Этот род выделяется обычно по внешней туфлеобразной форме,
с одной крышечкой. Важной особенностью его является строение септ из
слоев трабекул, образующихся на всей поверхности чашки, т. е. в пери-
ферической и осевой частях. Кроме септ, в полости коралла развиваются
днища, простые горизонтальные или слабо вогнутые, иногда более или
менее сильно расщепленные. Кроме крышечной формы, отличается
полным слиянием септ в ободок, так что границы между ними почти
исчезают». (Диагноз рода по Е. Д. Сошкиной.)
Типичный вид. Calceola sandallna Lam.
Распространение. Средний девон, повсеместно.
Calceola sandallna Lam. var. acuminata M a n s u у
Табл. XXXVI, фиг. 1 и 2
Общая форма узкая и удлиненная. Угол при основании не превышает
45—50°, большей частью равен 30°. Крышечка близкая к полукруглой по
очертаниям.
Р а с п р ос т р а н ен и е. В Западной Европе этот вариетет распро-
странен в средней части стрингоцефаловых слоев; на Урале — в нижне-
живетских слоях; в Китае—в эйфеле (Е. Д. Сошкина, 1949). В Запад-
ной Сибири, р. Чумыш и Верхнебердский район — в Эйфеле.
СЕМЕЙСТВО SPONGOPHYLL1DAE HILL
Род Grypophyllum Wedekind
К этому роду относятся преимущественно одиночные цилиндрической
формы, кораллы, но изредка и колониальные, ветвящиеся. Септальный
аппарат состоит из очень тонких или слабо и неправильно утолщенных
септ. Септы обычно полные, двух порядков. Септы первого порядка дохо-
дят до центра, а септы второго порядка ограничены краевой зоной. Там,
208
где септы первого порядка исчезают, у внешней стенки образуются крае-
вые пузыри. Межсептальный аппарат резко делится на краевую и осе-
вую зоны. Краевая зона построена из одних пузырей, а осевая зона —
из горизонтальных днищ со срединным вдавлением, которое может от-
сутствовать у крайних членов рода.
Типичный вид Crypophyllum gracile W е d е k.
Распространение. Средний девон и верхи нижнего девона.
Grypophyllum gracile Wedekind
Табл. XXXVI, фиг. 4; рис. 184
Одиночные цилиндрические кораллы, слабо рогообразно изогнутые.
Чашка бокаловидная, с острыми краями, с диаметром 9—13 мм. Изредка
встречаются в форме несильно разветвленных колоний. Тонкие септы до-
ходят почти до центра; количество
кораллитов; при диаметре 12—
14 мм септ первого порядка на-
считывается 25—26. Перифериче-
ские концы септ утолщают стенку
коралла. Септы второго порядка
зачаточны, имеют вид зубчиков на
внешней стенке, выходящих за
границу стереозоны. Центральное
вдавление днищ у этого вида вы-
ражено неясно.
Пузырчатая зона построена из
5—6 вертикальных рядов пузы-
рей средней величины, которые
круто наклонены к оси или почти
септ зависит от величины' диаметра
Рис. 184. Crypophyllum gracile W е d е k.
а—продольный разрез; б—поперечный
разрез. X 4. Северный Урал, р. М. Паток.
По Е. Д. Сошкниой
вертикальны (вздутием обращены к оси). Днища горизонтальные или
слабо изогнутые; на 10 мм приходится 20—22 днища, при поперечнике
кораллита 12—14 мм они резко отграничены от зоны пузырей, изредка
расщепляются и группируются в пучки, не более 2—3 в каждом.
Близкий Grypophyllum regressum W е d е к. отличается сильной редук-
цией септ на периферии и значительно более мелкими пузырями. Сход-
ный Grypophyllum cynacanthlnum Soshk. отличается закручиванием
осевых концов септ, ясным срединным вдавлением и значительной сбли-
женностью днищ.
Распространение. Живетский ярус среднего девона Западной
Европы и СССР. Указания на нахождение в Di2 и D2* требуют проверки.
Grypophyllum pestereviensis Z h m а е v, sp. n.
Табл. XXXVI, фиг. 5
Ветвистые колонии, достигающие значительных размеров. Корал-
литы цилиндрические, слабо изогнутые. Внешняя стенка тонкая. При
диаметре 9—10 мм насчитывается от 42 до 48 септ. Септы первого
порядка доходят или почти доходят до центра; периферические части
их слабо утолщены, местами они слабо изогнуты. Закручивания внут-
ренних концов септ не наблюдается. Септы второго порядка обычно едва
выступают в виде мелких зубчиков за границу стереозоны, редко дости-
гают '/2—2/з Длины септ первого порядка.
Пузырчатая зона состоит из 2—5 рядов пузырей. Днища очень слабо
вогнутые или почти прямые. Они резко отграничены от зоны пузырей
и почти не расщепляются. Срединное вдавление днищ у этого вида не
выражено.
На ранних стадиях роста наблюдаются только вогнутые днища, при-
мыкающие непосредственно к внешней стенке, при отсутствии пузырей.
Позднее появляются пузырчатая ткань, зачатки септ первого порядка;
14 Зак. 2024 209
Далее днища теряют свою вогнутость, становясь более плоскими. Септы
второго порядка появляются на самых зрелых стадиях роста. Пузыри
тоже изменяются, приобретая с возрастом вытянутость параллельно оси,
причем выпуклость пузырей обращается к ней. Отличия этого вида от
Gr. fasclculum указаны в описании последнего (см. ниже).
Р а с п р ос т р а н ен и е.'Нижний девон Салаира и Кузбасса.
Grypophyllum fasclculum Zh m а е v, sp. n.
Табл. XXXVI, фиг. 3
Ветвистые колонии, состоящие из цилиндрических, слабо изогнутых
кораллитов с резкой продольной ребристостью эпитеки на зрелых стадиях
роста. Пережимы и вздутия намечены слабо. Септы слабо волнисто
изогнуты, значительно утолщены стереоплазмой на периферии и посте-
пенно утоняются к оси. Септы первого порядка доходят до центра
и в некоторых кораллитах имеют слабую тенденцию к закручиванию.
Септы второго порядка хорошо развиты и доспигают !/2—2/з длины септ
первого порядка.
Зона днищ резко отграничена от зоны пузырей. Днища простые, ред-
кие, прямые или слабо вогнутые. Пузыри несколько вытянуты парал-
лельно оси и выпуклостью обращены вверх или к оси.
От сходного Crypophyllum tenue W е d е к. этот вид отличается мень-
шим диаметром кораллитов, соответственно меныпим количеством септ
и хорошо развитой ребристостью эпитеки. От Gr. pesterevlensis отли-
чается хорошо развитыми септами второго порядка, большим количест-
вом септ и рядов пузырей.
Распространение. Эйфельский ярус Западной Сибири (Са-
лаир).
Род Stenophyllum'Y^ma.v.sh&\is^x, em. Wedekind
Одиночные кораллы цилиндрической формы или массивные колонии
с субцилиндрической и неправильно призматической формой кораллитов.
Чашка плоско воронковидная, глубина ее при диаметре 30 мм — около
10 мм. Тонкие септы первого порядка достигают центральной зоны, где
иногда бывают клубкообразно закручены. Около внешней стенки септы
часто становятся толще. ААежсептальный аппарат в центральной зоне со-
стоит из почти горизонтальных днищ, расположенных близко друг
к другу; в центре каждого днища имеется узкое углубление. Перифери-
ческая зона из пузырей.
Характерные черты рода по Е. Д. Сошкиной (1949, стр. 127): «Пол-
ное развитие тонких септ с едва намеченным утолщением в той или дру-
гой части их; сильное развитие септ второго порядка; резкая граница
между зонами днищ и пузырей; слабая вогнутость, сильная расщеплен-
ность и срединное вдавливание днищ».
Типичный вид. Stenophyllum diluvianum A m a n s h.
Распространение. Эйфель и нижняя половина живетского
яруса. СССР и Западная Европа.
Stenophyllum vogullcum Bulvanker
Табл. XXXVII, фиг. 1
Массивные колонии из тесно соприкасающихся призматических ко-
раллитов; чашка коническая, глубина ее равна величине диаметра.
Септы двух порядков, в количестве от 40 до 44. Тонкие и прямые септы
первого порядка на расстоянии примерно 2/з своей длины начинают
веретенообразно утолщаться, достигая максимума утолщения у внешней
стенки; до центра они немного не доходят, осевые концы их могут за-
кручиваться или загибаться. Главная септа длиннее других; остальные
210
септы расположены преимущественно гребневидно по отношению к ней.
Длина септ второго порядка более половины длины септ первого по-
рядка.
Зона днищ занимает примерно '/з диаметра кораллита. Днища тон-
кие, сильно сближенные, почти горизонтальные, иногда изогнутые и пу-
зырчато расщепленные. В широкой периферической зоне мелкие одно-
образные пузыри, выпуклостью обращенные вверх. (Диагноз вида по
Э. 3. Бульванкер.)
От близкого Stenophyllum altum S о s h к. отличается более корот-
кими и менее утолщенными септами, имеющими гребневидное располо-
жение, чего нет у Stenophyllum vogullcum.
Распространение. Средний девон. Урал, Кузбасс, Салаир.
Stenophyllum gorskti В u 1 v а n к е г, sp. n.1
Табл. XXXVII, фиг. 2
Колония массивная, состоящая из призматических кораллитов, диа-
метр которых варьирует от 4 до 7 мм. Септы двух порядков, тонкие,
хорошо развитые, в количестве 30—32; у наружной стенки септы сильно
утолщаются. Септы первого порядка доходят до центра, но в центре не
закручиваются. Септы второго порядка местами достигают ’/г длины, септ
первого порядка. Узкая периферическая зона заполнена 1—2 рядами
пузырей, вытянутых параллельно оси и выпуклостью обращенных вверх.
Широкая центральная зона имеет вогнутые, слегка расщепленные
днища с центральным углублением; на 10 мм насчитывается 18 днищ.
(Диагноз вида по Э. 3. Бульванкер.)
От близкого вида Stenophyllum slbiricum (см. ниже) отличается
меньшим диаметром кораллитов, соответственно этому меньшим коли-
чеством септ, более тонкими септами и меньшим количеством рядов пу-
зырей периферической зоны.
Распространение. Кобленцский ярус юго-западной окраины
Кузбасса и Центрального Салаира.
Stenophyllum slbiricum В u 1 v а n к е г, sp. n.
Табл. XXXVIII, фиг. 1
Характеристика вида по Э. 3. Бульванкер (1938): «Массивная коло-
ния, состоящая из шестигранных кораллитов диаметром 8—15 мм. Септы
обоих порядков, в количестве 32—40, тонкие, ровные. Септы первого по-
рядка сходятся в центпе, образуя несильное центральное завивание.
Септы второго порядка достигают 3/4 длины первых. Периферическая
зона имеет 5'—7 рядов крупных, почти одинаковых по величине пузырей.
Центральная зона состоит из пузыреобразных расщепленных, сильно
вогнутых днищ со срединным углублением».
От близкого Stenophyllum devonicum Bulv. (см. ниже) отличается
сильной вогнутостью днищ, наличием срединного углубления в днищах.
Распространение. Эйфельский ярус Кузбасса.
Stenophyllum devonlcum В u 1 v а n к е г, sp. n.
Табл. XXXVIII, фиг. 2
Массивные колонии небольших размеров, неправильной формы. Ко-
раллиты призматические (4—6-гранные), отделены друг от друга ясной
стенкой; диаметр их 12—14 мм. Септы в количестве 36—40 обоих поряд-
ков хорошо развиты, тонкие у периферии и утолщенные к центру. Септы
первого порядка доходят почти до центра, но центрального закручивания
1 Э. 3. Бульванкер, относя эту форму к роду Grypophyllum, предлагает для нее
название Grypophyllum slbiricum, так как видовое название gorskii в применении
к роду Grypophyllum использовано ею для одной уральской формы.
14* 211
не образуют. Септы второго порядка имеют длину, равную 2/з и более
длины септ первого порядка. Днища слабо вогнутые, почти горизонталь-
ные и слабо расщепленные; очень редко имеют неясно выраженное
срединное углубление, но, как правило, его нет. Широкая перифериче-
ская зона заполнена пузырями, образующими 4—7 рядов; пузыри не-
больших размеров, выпуклостью обращены вверх и к оси. (Диагноз
вида по Э. 3. Бульванкер).
От близкого Stenophyllum sibiricum В u 1 v. (см. выше) отличается
утолщением внутренних концов септ, отсутствием центрального закручи-
вания септ, характером днищ — отсутствием срединного углубления
у них; последнее наблюдается крайне редко и выражено неотчетливо.
Распространение. Эйфельский ярус Западной Сибири.
Stenophyllum salalricum Z h m а е v, sp. n.
Табл. XXXIX, фиг. 1
Массивные колонии с призматическими кораллитами, диаметр кото-
рых 10—11 мм. Септьр двух порядков, количество их 45—48. Септы пер-
вого порядка постепенно утоняются от периферии к оси, иногда имеют
веретенообразную форму; в центре концы их не изгибаются и располо-
жены' радиально. Септы второго порядка длинные, достигают 3/< длины
септ первого порядка. Днища занимают небольшую часть полости ко-
раллита, слабо вогнутые и расщепленные. Широкая периферическая зона
заполнена пузырями. У внешней стенки пузыри обращены выпуклостью
вверх и к оси, к центру они уплощаются, становясь вытянутыми парал-
лельно оси и выпуклостью обращаясь к ней. Количество рядов пузырей
от 7 до 10.
От наиболее сходного Stenophyllum altum S о s h к. отличается мень-
шим диаметром кораллитов, большим количеством септ при равном диа-
метре кораллитов, и большей толщиной септ. От Stenophyllum sibiricum
(см. выше) отличается меньшим диаметром кораллитов, большим коли-
чеством септ при равном диаметре кораллитов, большим количеством ря-
дов пузырей, более толстыми септами. Кроме того, у St. salalricum не
наблюдается тенденции к центральному закручиванию септ первого по-
рядка, что имеет место у St. sibiricum.
От Stenophyllum devonicum (см. выше) отличается меньшим диамет-
ром кораллитов, большим числом септ при равных диаметрах, большим
количеством рядов пузырей.
От Stenophyllum urallcum S о s h к. отличается меньшим диаметром
кораллитов, меньшим количеством септ, радиальным их расположением.
Распространение. Эйфельский ярус юго-западной окраины
Кузбасса.
Stenophyllum soshkinae Z h m а е v, sp. n.
Табл. XXXIX, фиг. 2
Массивные колонии значительных размеров. Внешняя стенка покрыта
резкой продольной ребристостью. При диаметре 7—8 мм кораллиты
имеют 36—40 септ обоих порядков; иногда они причленяются к внешней
стенке большими треугольными утолщениями. Септы первого порядка
доходят до центра. Септы второго порядка равны 3/4 длины первых.
Днища занимают !/з полости коралла, вогнутость их различна; на 10 леи
длины приходится 34—35 днищ. Широкая периферическая зона заполнена
пузырями в количестве 8—9 вертикальных рядов.
Наиболее близкий вид Stenophyllum salalricum Zhmaev отличается
большим диаметром кораллитов и большим количеством септ; сходство
выражается в большом количестве рядов пузырей и в отсутствии тен-
денции к закручиванию септ.
Распространение. Совместно с предыдущим видом.
212
Род Pseudospongophyllum Zhmaev, gen. n.
Виды этого рода образуют массивные колонии. Кооаллиты на взрос -
лых стадиях имеют диаметр в пределах 2,5—6,0 мм. Стенка кораллитов
выражена хорошо и имеет ясную продольную ребристость; имеются пере-
жимы и вздутия. Септа двух порядков. Септы первого порядка почти
доходят до центра и расположены радиально или гребневидно. Септы
второго порядка различной длины; они могут быть зачаточными, но
иногда достигают ’/г—2/з длины септ первого порядка. Редукции септ не
наблюдается. В осевой части — простые, слабо вогнутые, нерасщеплен-
ные днища. Зона пузырей узкая, с 1—3, редко четырьмя рядами пузырей;
пузыри средних размеров.
Близкие роды — Spongophyllum Е d w. et Н a i m е и Fasclphyllum
Schluter. От последнего отличается менее толстым ободком, более
тонкими септами и характером пузырчатой ткани. От Spongophyllum
отличается отсутствием редукции септ.
Типичный вид. Pseudospongophyllum mdsslvum Zhmaev.
Распространение. Средний девон Западной Сибири.
Pseudospongophyllum tnasslvum Zhmaev, sp. и.
Табл. XL, фиг. 1
Небольших размеров массивные колонии, состоящие из призматиче-
ских кораллитов. Септы тонкие, причленяются к внешней стенке при
помощи треугольных утолщений. Септы первого порядка доходят до
центра или почти до центра, но там не закручиваются. Септы второго
порядка достигают ’/а и более длины септ первого порядка. Ободок
иногда очень тонок, иногда достигает значительной толщины. Числосепт
около 30, при диаметре кораллитов 4—6 мм.
Днища редкие, слабо вогнутые; на 10 мм длины их насчитывается от
14 до 18 в соответствии с диаметром кораллитов. Пузыри вытянуты па-
раллельно оси и выпуклостью обращены вверх и к оси; количество' рядов
пузырей от одного до трех, редко четыре. Граница между зонами пузы-
рей и днищ резкая.
Распространение. Эйфельский ярус юго-западной окраины
Кузбасса.
Pseudospongophyllum tomense Zhmaev, sp. n.
Табл. XL, фиг. 2
Массивные, достигающие иногда значительных размеров колонии, со-
стоящие из призматических кораллитов небольшого диаметра. Септы
первого порядка до центра не доходят и расположены радиально, иногда
гребневидно. Септы второго порядка зачаточные на всех стадиях разви-
тия коралла; оци имеют вид зубчиков на внешней стенке. Очень редко
некоторые из них становятся длинными, достигая ’/.з длины септ первого
порядка. Общее количество септ от 25 до 38.
Днища редкие, слабо вогнутые, почти плоские: срединное углубление
на днищах не выражено. КотичествО' днищ на 10 мм длины до 20, при
диаметре кораллитов 3—5 мм. Пузыри уплощенные, вытянутые парал-
лельно оси, выпуклостью обращенные вверх и к оси, расположены в че-
тыре вертикальных ряда. Граница зоны пузырей и днищ резкая.
Близкие виды; Spongophyllum haluslto’des Ether, var. minor
S о s h k., Stenophyllum gorskti В u 1 v., Pseudospongophyllum masslvum
Zhmaev, но от всех их данный вид отличается сочетанием своих харак-
терных признаков.
Распространение. Характерный вид в нижнем девоне юго-
западной окраины Кузбасса и Центрального Салаира.
213
Род Spongophyllum М. Edwards et Haime
Колонии массивные или ветвистые, с диаметром кораллитов от 3 до
20 мм. Чашка воронковидная, неглубокая, с крутыми стенками и до-
вольно широким дном. «Внешняя стенка кораллитов всегда хорошо раз-
вита и имеет ясную продольную ребристость. Разветвление колонии про-
исходит путем отделения почек от края материнской чашки.
Септы первого порядка всегда доходят до оси, осевые концы их тон-
кие. Периферические концы большинства септ на взрослых стадиях не
доходят до внешней стенки. У некоторых видов, однако, таких редуци-
рованных на периферии септ немного.
Септы второго порядка развиты неодинаково. На внешней стенке на
продолжении каждой септы изнутри всегда есть утолщение, имеющее на
поперечном разрезе вид треугольника или зубца.
В онтогенезе большинства видов на самых ранних стадиях септы
имеют полное развитие и не редуцированы на периферии. Однако у неко-
торых видов... редукция септ на периферии начинается так рано, что
первую стадию с полными септами уловить трудно... Внутрен-
ней стенки нет. Осевая часть кораллитов содержит днища вогну-
той формы. Вогнутость их может быть значительной или небольшой,
тогда они приближаются к горизонтальным... Зона пузырей узкая (1—3
ряда), резко отделена от зоны днищ»... Пузыри чаще крупные, приплюс-
нутые, очень круто наклоненные к оси, иногда вертикальные. (Диагноз
вида по Е. Д. Сошкиной).
Наиболее близок по редукции септ на периферии и типу колонии род
Endophyllum Е d w. et Haime из верхнего девона Европы и Западной
Сибири. Но Spongophyllum отличается от него вогнутыми, без дополни-
тельных пластинок днищами и отсутствием внутренней стенки.
Распростран.ение. Нижний и средний девон. Западная Европа,
Урал, Западная Сибирь.
Spongophyllum origlnalis Z h m а е v, sp. n.
Табл. XLI, фиг. 3
Массивные колонии значительных размеров. Кораллиты 4—6-гранные,
диаметром 2,5—3,0 мм, имеют 20—24 септы обоих порядков. Все септы
редуцированы и наблюдаются только на внешней стенке в виде неболь-
Рис. 185. Spongophyllum minimum
Z h m a e v, sp. n.
a—продольный разрез, X 5,4; д'—попе-
речный разрез, X 6. Центральный Са-
лаир, р. Правая Березовка
ших зубчиков. В поперечном раз-
резе иногда наблюдается подобие
внутренней стенки, образованной
стенками пузырей; там, где внут-
ренняя стенка хорошо выражена,
можно видеть на ней остатки
септ. Весьма редко немногие сеп-
ты идут от периферии к оси. Дни-
ща слабо вогнутые, почти пло-
ские. Пузыри несколько вытянуты
параллельно оси и довольно круп-
ные, расположены в один, редко
в два ряда.
Характером днищ описывае-
мый вид напоминает Spongophyl-
lum halysltold.es Ether, forma
minor S о s h k., но отличается
меньшим диаметром кораллитов,
меньшим количеством рядов пузырей, меньшим количеством септ.
Распространение. Характерный вид для нижнего девона юго-
западной окраины Кузбасса.
214
Spongophyllum minimum Z h m a ev, sp. n.
Табл. XXXIX, фиг. 3; рис. 185
Колония массивная, с призматическими кораллитами диаметром 3—
4 мм; септ обоих порядков 22—26; центрального завивания септ нет.
Периферическая зона состоит из 1—3 рядов пузырей. Днища слегка рас-
щепленные, вогнутые, с ярко выраженным центральным углублением.
От близкого вида Stenophyllum gorskli В u 1 v. отличается меньшим
диаметром кораллитов, меньшим количеством септ и большим количест-
вом рядов пузырей (до трех рядов).
Распространение. Нижний девон Салаира.
Род Fasciphyllum Schluter
«Кораллы колониальные. Кораллиты цилиндрические или призматиче-
ские... без резких пережимов и вздутий. Чашка бокаловидная, довольно
глубокая. Септы первого порядка почти доходят до оси. Септы
второго порядка хорошо развиты. Септальные трабекулы парал-
лельные, поднятые вверх и внутрь. Толстые периферические
части септ плотно срастаются, образуя ободок, в котором на некоторых
стадиях онтогенеза отчетливо наблюдается разделение на внешнюю зону'
и вставленные в нее утолщенные части септ с темными точками от раз-
резов столбиков.
Днища более или менее широкие, вогнутые... Пузырчатая ткань резко
отделяется от зоны днищ и состоит из 1—5 рядов пузырей, выпуклостью
обращенных к оси» (Сошкина, 1949).
От сходного по наличию ободка и пузырчатой ткани, а также по
ходу онтогенеза рода Neocolumnaria Soshkina из среднего девона
Урала отличается более простыми днищами. Отличия от близкого по ходу
онтогенеза и наличию ободка рода Solominella I v а n i а даны при его
описании (см. стр. 225).
Распространение. Нижний и средний девон. Западная Европа,
Урал, Западная Сибирь.
Fasciphyllum petshorense Soshkina
Табл. XL, фиг. 3
Ветвистые, реже массивные колонии. Кораллиты ветвистых колоний
с редкими пережимами и вздутиями, цилиндрические, прямые или изо-
гнутые. «Чашечки очень глубокие, с почти отвесными стенками, тонкими
краями и воронковидным дном». Септы первого порядка почти доходят
до центра; «они расположены перисто или гребневидно по отношению
к одной септе, доходящей до оси и утолщенной на осевом конце». Септы
второго порядка немного короче. На периферии имеется ободок от слия-
ния утолщенных септ с внешней стенкой; «ширина ободка около поло-
вины радиуса. Внутрь от ободка развиты 2—3 ряда пузырей... Граница
зоны днищ и пузырей резкая...» Днища редкие, слабо расщепленные,
вогнутые. Зона днищ занимает менее половины' коралла. На 10 мм при-
ходится от 17 до 22 днищ, при диаметре кораллитов от 4,5 до 10,0 мм.
(Диагноз вида по Е. Д. Сошкиной).
От сходного Fasciphyllum halliaforme S о s h к. отличается колониаль-
ной формой роста, более широким ободком и более простыми, слабо во-
гнутыми днишами.
Распространение. Руководящий вид кобленцского яруса Урала
(вместе с Karplnskla conjugula); в Западной Сибири встречается в осг-
ракодовой толще1 по р. Томь-Чумышу и в Центральном Салаире — в
нижнедевонских отложениях.
1 По данным Палеонтологического кабинета Западно-Сибирского геологического
управления, эти отложения имеют нижнедевоиский возраст.
215
СЕМЕЙСТВО COLUMNAR1DAE OKULITCH
Род Columnaria G о 1 d f u s s
Кораллы колониальные. Колонии массивные и ветвистые с мелкими
кораллитами. Длинные или короткие тонкие септы с расширенными
основаниями образуют двуслойный ободок. Днища полные, простые,
колоколовидно выпуклые у силурийских видов, горизонтальные или не-
правильно изогнутые — у девонских. Пузырчатая ткань отсутствует.
(Диагноз рода по Е. Д. Сошкиной).
Силурийские виды рода Columnaria имеют длинные или короткие
септы и обычно узкий ободок. Девонские виды рода характеризуются
более или менее укороченными тонкими септами на ранних стадиях онто-
генеза. С возрастом на них развивается периферическое утолщение, ко-
торое разрастается в сплошной периферический ободок.
По данным Е. Д. Сошкиной (1949), для нижне- и среднедевонских
видов характерна двусторонняя симметрия в расположении септ и ясное
выступание одной противоположной септы.
От сходных по наличию ободка из расширенных оснований септ и по
ранним стадиям онтогенеза среднедевонских родов Fasclphyllum Schlu-
ter, Neocolumnarla Soshkina и от верхнедевонското рода Solominella
I v a n i а род Columnaria легко отличается по полному отсутствию пу-
зырчатой ткани.
Типичный вид. Columnaria alveolata Gо 1 d f.
Распространение. Силур—-девон. Повсеместно.
Columnaria vulgaris Soshkina
Табл. XLI, фиг. 1; рис. 186
Колония слабо ветвящаяся, состоящая из очень длинных тонких
цилиндрических кораллитов со слабыми пережимами и вздутиями. На
внешней стенке — слабая " ~
о °
б
продольная бороздчатость. Диаметр попереч-
ника кораллитов изменяется от 5,5 до
8,5 мм\ в поперечном разрезе форма
кораллитов округленная, округленно-
многоугольная. Септы двух порядков
не доходят до центра, кроме одной
сильно выступающей; длина септ не-
одинакова. Они слабо утолщены на
осевых концах и в большинстве случа-
ев имеют радиальное расположение.
На периферии утолщение септ стерео-
плазмой образует ободок шириной бо-
лее 1 мм, а в некоторых кораллитах —
до 2 мм. Септы второго порядка едва
выступают за границу ободка.
Днища тонкие, горизонтальные, из-
изгибами и расщеплением; на 10 мм
или меньше. Пузырчатой
Рис. 186. Columnaria vulgaris
Soshkina
а—продольный разрез; б—попереч-
ный разрез, х 4. Северный Урал.
По Е. Д. Сошкиной
редка с небольшими местными
длины насчитывается 10 днищ, редко больше
ткани нет.
Распространение. Верхние горизонты живетского яруса на
Урале и в Западной Сибири.
Columnaria rhenana Freeh
Табл. XLI, фиг. 2
Кораллы образуют ветвистую колонию, с диаметром кораллитов от 6
до 10 мм. Септы двух порядков, общее количество их от 36 до 40. Септы
первого порядка почти достигают центра, причем главная и противопо-
ложная септы всегда длиннее остальных и нередко соединяются друг
216
с другом. Септы второго порядка местами равны '/г длины первых.
Внутренние окончания септ тонкие; по направлению к периферии септы
утолщаются и у наружной стенки кораллита образуют стереоплазматп-
ческий ободок.
Днища правильно расположенные, вогнутые; на 10 мм насчитывается
12 днищ. Пузырчатая ткань отсутствует.
От близкой Colummarla vulgaris S о s h к. отличается большей дли-
ной септ первого порядка, почти доходящих до центра, и ясным двусто-
ронне симметричным расположением септ с удлиненными главной и
противоположной септами.
Распространение. Верхние стрингоцефаловые слои в Западной
Европе и в Кузбассе.
Род Loyolophyllum Chapman
Массивные кораллы с небольшими кораллитами, тонкими септами,
полными, горизонтальными или блюдцевидными днищами и с неполной
серией вертикально удлиненных пузырей, лежащих вдоль стенки в меж-
септальных промежутках. Характерным признаком рода является наличие
весьма непостоянной зоны пузырей.
Типичный вид. Loyolophyllutn croswelll Chapman.
Распространение. Девон.
Loyolophyllum urense Z h m a e v, sp. n.
Табл. XXXVI, фиг. 6
Небольших размеров массивные колонии. Кораллиты призматические,
с довольно резкими очертаниями. Наибольший диаметр их достигает
4,5 мм. Разграничить септы на септы первого и второго порядков почти
не удается. Наибольшее количество септ 14—16; они иногда тонкие,
иногда утолщены стереоплазмой, почти доходят до центра. Ободок до-
вольно толстый. Пузыри вытянуты параллельно оси и выпуклостью обра-
щены к оси, а реже — вверх и к оси. Во взрослых кораллитах всегда
имеется один часто прерывающийся ряд пузырей. Днища плоские или
слабо вогнутые; на 10 мм длины их от 10 до 16.
От сходного Loyolophyllum croswelll Chapman отличается большим
диаметром кораллитов, большим количеством септ и большим коли-
чеством днищ на 10 мм длины.
Распространение. Характерная форма для эйфельского яруса
юго-западной окраины Кузбасса. Имеются указания на находки этого
вида в эйфеле Салаира.
Род Pseudochonophyllum Soshkina
Кораллы образуют небольшие колонии, иногда одиночные формы.
Внешняя форма кораллитов субцилиндричеокая, реже бокаловидная.
Чашечки представляют собой маленькую бокаловидную ямку с выпуклым
дном и широким, более или менее горизонтальным отворотом. В средней
части, а иногда у периферии септы сильно утолщены, к оси становятся
более тонкими и на осевых концах снова утолщаются и часто закручи-
ваются. Часто септы каринированы. Септы обоих порядков длинные.
Пузыри мелкие, многочисленные, тонкие, расположены горизонтально.
Днища тонкие, сближенные, расщепленные войлокообразно, вогнутые.
Типичный вид. Pseudochonophyllum pseudohellanthold.es
Scherzer.
Распространение. Нижний и средний девон Чехии, Урала и
Западной Сибири.
217
Pseudochonophyllum pseudohellanthold.es Scherzer
Табл. XLII, фиг. 4
Небольшие колонии или одиночные кораллы, широко конической или
цилиндрической формы; только на взрослых стадиях септы каринированы
и спирально закручены в осевой части. Расположение септ радиальное.
Днища разнообразно изгибаются, но большей частью слабо вогнуты.
Пузырчатая зона изменчива, в зависимости от возраста; на взрослых
стадиях развития пузырчатая зона расширяется. Число рядов пузырей
от 10 до 16; число днищ на 10 мм длины от 25 до 60. Эти величины
изменяются в зависимости от диаметра кораллитов, а диаметр колеблется
в пределах ог 18 до 30—40 мм. Число септ обоих порядков до 72. (Диаг-
ноз вида по Е. Д. Сошкиной).
Распространение. Кобленцский и эйфельский ярусы Урала,
Центрального Салаира, юго-западной окраины Кузбасса и Алтая, а также
Западной Европы.
СЕМЕЙСТВО THAMNOPHYLLIDAE SOSHKINA
Род Thamnophyllum Р е n е с к е
К этому роду относятся одиночные или колониальные кораллы неболь-
ших размеров, нередко рогообразно изогнутые, с глубокой чашечкой.
Септы двух порядков хорошо развиты. Осевые концы септ тонкие, иногда
разрываются и никогда не доходят до центра. Периферическая зона со-
стоит из подковообразных пузырей, неравных по размерам и по располо-
жению: у наружной стенки более мелкие пузыри, в центральной зоне —
более крупные. Днища горизонтальные, простые или слабо расщепленные.
Типичный вид. Thamnophyllum stachei Hornes.
Распространение. Род Thamnophyllum существовал в течение
всего девона.
Thamnophyllum trlgemme (Quenstedt)
Табл. XXXV, фиг. 5; табл. XXXVIII, фиг. 3
Небольшие ветвистые колонии, состоящие из тонких цилиндрических
кораллитов, на наружной поверхности которых хорошо заметна продоль-
ная ребристость. Ветвление идет путем образования 1—5 почек в чашке.
Чашка бокаловидная, с плоским дном. Эпитека тонкая, легко разрушаю-
щаяся. Септы первого порядка по длине тянутся на две трети радиуса
кораллита, септы второго порядка вдвое короче. В зоне пузырей септы
утолщены, а в зоне днищ они утоняются и изгибаются. Септальные
трабекулы веерообразны в зоне пузырей. Днища горизонтальные, до-
вольно редкие, иногда расщепляются. Зона пузырей сложного строения.
Е. Д. Сошкина (1949) характеризует эту зону:
«...На периферии виден один ряд горизонтальных плоских диссепи-
ментов, связывающих внешнюю и внутреннюю стенки. Они часто разру-
шаются вместе с внешней стенкой или, может быть, совсем не разви-
ваются. Дальше внутрь идет один ряд правильных подковообразных
пузырьков, тесно насаженных друг на друга, вздутием обращенных вверх
и очень редко расщепляющихся на два пузырька. По обе стороны вдоль
кольца подковообразных пузырьков — два концентрических кольца внут-
ренней стенки. Внутрь от внутреннего кольца идет ряд слабо выпуклых,
наклонных к оси пластинок, образующих зону, переходную к днищам».
Некоторое сходство по кустистой форме колонии Thamnophyllum
trlgemme (Quenstedt) имеет с Thamnophyllum monozonatum
(Soshk.), но у последнего утолщение септ значительнее.
Распространение. Характерная форма для среднего девона,
в особенности его живетского яруса. Урал и Кузбасс.
218
В. А. ИВАН ИЯ
Отряд Tetracoralla или Rugosa. Тетракораллы
(верхний девон)
СЕМЕЙСТВО PENECKIELLIDAE SOSHKINA
Род Peneckiella S о s h k i п a
Ветьнстые и массивные колониальные кораллы с небольшим попереч-
ником кораллитов и хорошо развитой эпитекой. В поперечных сечениях
септы веретеновидно утолщены, слабо расщеплены на поперечные балочки
и всегда оставляют в осевой части коралла большую или меньшую сво-
бодную зону.
Днища полные, простые, плоские или слабо вогнутые с немногочис-
ленными дополнительными пластинками на краях и в середине, которые
опираются на днища. Пузырчатая зона построена из одного, редко двух
рядов правильных, приплюснутых сверху толстостенных пузырей. Развита
внутренняя стенка. Ранние стадии онтогенеза характеризуются короткими,
далеко не доходящими до осевой части септами и толстыми внешними
стенками кораллитов.
Типичный вид. Cyathophyllum minus R о е m.
Распространение. Средний — верхний девон. Западная Европа,
СССР, Малая Азия.
Peneckiella darwlni (Freeh)
Табл. XLII, фиг. 1
Массивная колония призматических мелких кораллитов с резкой
наружной и не всегда ясной внутренней стенками. При диаметре корал-
литов 2,0—3,5 мм имеется 18—20 септ двух порядков, сильно различаю-
щихся по длине, местами распадающихся на поперечные балочки.
Септы первого порядка длинные, утолщенные к периферии, с нитевидно
тонкими, изогнутыми осевыми концами. Септы второго порядка короткие,
доходящие лишь до внутренней стенки.
Днища тонкие, изменяющиеся от слабо вогнутых до слабо выпуклых,
полные, простые, редко расщепленные (на 5 мм длины насчитывается
20—22 днища). Пузырчатая зона состоит из одного ряда довольно
правильных, слегка приплюснутых сверху пузырей, иногда осложненных
1—2 неполными рядами мелких пузырьков.
Характерны короткие септы на ранних стадиях онтогенеза, удлиняю-
щиеся по мере роста кораллитов.
Наблюдается большая индивидуальная изменчивость представителей
вида: меняется толщина септ в периферической части от умеренных до
сильно утолщенных; днища изменяются от выпуклых через горизонтальные
до вогнутых и иногда расщепляются.
Близкий по форме колонии, малым размерам и очертаниям коралли-
тов и слабому развитию пузырчатой ткани вид Peneckiella tabulata
(В u 1 v.) из среднефранских отложений Кузбасса отличается более ред-
кими днищами.
Распространение. Среднефранские отложения северо-западной
окраины Кузбасса (глубокинский известняк, окрестности с. Соломине);
верхнеживетские и нижнефранские отложения Урала; D31 Малой Азии.
Peneckiella tabulata Bulvanker
Табл. XLII, фиг. 2
Массивная колония из мелких призматических кораллитов с резкими,
довольно толстыми внешними и тонкими внутренними стенками. При
диаметре кораллита в 2—4 мм имеется 20 почти не распадающихся на
219
поперечные балочки септ двух порядков, сильно различающихся по длине.
Септы первого порядка длинные, нитевидно тонкие в центре, утолщаются
к периферии. Септы второго порядка равны ’/з—2/г, длины септ пер-
вого порядка и доходят до внутренней стенки.
Зона днищ широкая. Днища полные, тонкие, выпуклые, простые, редко
расщеплены. Пузыри мелкие, вздутые, расположены в один, реже в два
неполных ряда. Характерно удлинение септ в процессе онтогенеза.
Отличия от близкого вида Penecklella darwlnl указаны выше.
Распространение. Среднефранские отложения северо-западной
окраины Кузбасса (глубокинский известняк, окрестности с. Соломине)
Род Megaphyllum Soshkina
Массивные и ветвистые колониальные ругозы. На поверхности корал-
литов отчетливы продольные ребра, тонкие линии нарастания и неболь-
шие пережимы и вздутия. Чашка бокаловидная, без отворота на краях.
Периферические концы септ двух порядков более или менее утолщены.
Осевые концы тонкие, изогнутые, значительно не достигающие оси.
Зона днищ широкая. Днища неполные, слабо выпуклые, плоские или
слабо вогнутые в осевой части, расщепленные в пучки; дополнительные
пластинки на краях крупные и хорошо развиты. К ним и присоединяются
днища, не имеющие дополнительных пластинок в середине. Пузырчатая
зона состоит из довольно мелких, вздутых, круто наклонных к оси пузырь-
ков. На поперечных разрезах линии пересечения пузырей взаимно
пересекаются. На ранних стадиях онтогенеза характерны короткие септы
и толстые внешние стенки.
Сходный по ходу онтогенеза род Penecklella Soshkina отличается
простыми, полными, нерасщепленными днищами и слабым развитием
дополнительных пластинок на их краях.
От близкого рода Campophyllum Е d w. et Н a i m е отличается бокало-
видной формой чашки, несколько большей длиной септ и однообразными
по величине пузырями.
Сходный с Megaphyllum род Macgeea W е b s t. отличается присутст-
вием подковообразных пузырей.
Род Schluterla Wed. из среднего девона Германии и верхнего девона
Урала и Кузбасса, сходный с родом Megaphyllum характером днищ и
пузырей, отличается более длинными, доходящими до оси, септами и тем,
что дополнительные пластинки развиваются и в середине, а также длин-
ными септами на ранних стадиях онтогенеза.
Типичный вид. Megaphyllum pashiense S о s h k.
Распространение. Верхи живетского и низы франского ярусов
Урала; нижнефранские отложения Кузбасса.
Megaphyllum pashiense Soshkina
Табл. XLII, фиг. 3
Ветвистая колония состоит из довольно длинных, слабо ветвящихся
кораллитов. Чашка неглубокая, бокаловидная, с острыми отвесными
краями и широким плоским дном. При диаметре кораллита 10—13 мм
имеется 50—56 септ двух порядков, различающихся по длине. Септы
первого порядка тонкие и изогнутые, утолщенные на периферии, равны
7з длины радиуса кораллита. Септы второго порядка тоже утолщены на
периферии и достигают ’/4—’/з длины септ первого порядка.
Зона днищ широкая, расщепляется на широкие, плоские, слабо во-
гнутые или слабо выпуклые в середине и узкие, наклонные к оси по пе-
риферии днища (число днищ на 5 мм длины равно 8—10). Пузырчатая
зона состоит из 3—4 рядов мелких, вздутых, круто наклоненных к оси
пузырей.
220
Близкие виды Megaphyllum cyllndricum S о s h к. и Megaphyllum
catavense S о s h k. из франских отложений Урала отличаются: пер-
вый — более короткими септами первого порядка и резкой внутренней
стенкой, второй—более широкой зоной пузырей *. От всех других видов
рода описываемый вид отличается широкими пучковидными днищами.
Распространение. Нижнефранские отложения северных окраин
Кузбасса (зарубинский известняк близ с. Зарубино и фаленовый гори-
зонт у Черепанова брода на р. Яя). Верхнеживетские и франские отло-
жения Урала.
СЕМЕЙСТВО NEOCOLUMNARIIDAE SOSHKINA
Род Phillipsastraea d’ Orbigny
Кораллы с непарасидальным почкованием. Внешняя стенка коралли-
тов развита полностью или местами редуцирована, и тогда септы сосед-
них кораллитов соединяются. Чашка глубокая, с высоким кашечным ва-
ликом и с широким отворотом краев. Септы тонкие или утолщенные к пе-
риферии, часто веретеновидно распадающиеся на поперечные балочки.
Септальные трабекулы веерообразные. На ранних стадиях онтогенеза
септы тонкие и длинные. Днища полные или неполные, с более или менее
ясной осевой надстройкой из дополнительных пластинок. Пузырчатая
зона широкая, из однообразных рядов пузырей, из которых наруж-
ные почти горизонтальны или слабо наклонны.
Близкий род Schluterla Wed. отличается более глубокой бокаловид-
ной чашкой без отворота краев, параллельными септальными трабеку-
лами, отсутствием коленчатого изгибания септ, более узкой пузырчатой
зоной.
Типичный вид. Phillipsastraea radlata Mart.
Распространение. Франские отложения. СССР, Западная
Европа, Малая Азия.
Phillipsastraea sedgwickl (Edwards et Haime)
Табл. XLIII, фиг. 4
Колония массивная. Кораллиты призматические с ясными внешними
стенками. Чашка с неглубокой узкой центральной ямкой, с отвесными
стенками, выпуклым дном и с горизонтальным отворотом, окаймляю-
щим ее снаружи. При диаметре кораллита от 9 до 16 мм в среднем
имеется 36—42 тонких или утолщенных по периферии, распадающихся
на поперечные балочки септ двух порядков, различающихся по длине.
Септы первого порядка длинные, почти доходящие до центра, с тон-
кими, изогнутыми внутренними концами, иногда соединяющимися между
собой, с тенденцией к завиванию, утолщенные веретеновидно или на
периферии за счет распадения на поперечные балочки.
Септы второго порядка равны 3/4—2/з длины септ первого порядка и
почти не отличимы от них по толщине. Днища, занимающие от */з ДО ’/4
поперечника кораллита, довольно тонкие, в общем слабо выпуклые,
расщепленные, осложненные дополнительными пластинками на перифе-
рии и в осевой зоне. На 5 мм длины приходится 12—16 днищ. Пузырча-
тая зона широкая и состоит из 2—7 рядов некрупных, вздутых, тонких
пузырей, почти горизонтальных, более крупных на периферии и более
мелких, круто наклоненных у границы с зоной днищ.
Для ранних стадий онтогенеза характерны едва заметная внешняя
стенка и тонкие длинные септы, утолщающиеся с возрастом.
Близкий по очертаниям кораллитов, длинным распадающимся на по-
перечные балочки септам и типу дпищ вид Phillipsastraea lazutkini
1 Отличия эти не существенны, и Е. Д. Сошкина оба эти вида считает синони-
мами Meg. pashiense.
221
(В u 1 v.) из нижнефранских отложений Кузбасса отличается сильным
расщеплением септ на периферии.
Phllllpsastraea schafferl Penecke из верхнего девона Малой Азии,
сходный по строению септ, днищ и пузырей, отличается сильно зигзаго-
образной внешней стенкой, короткими септами второго порядка, расщеп-
лением септ и наличием более резкой внутренней стенки.
Распространение. Верхний девон Англии, Германии, Малой
Азии; франские отложения Урала; нижнефранские отложения Кузбасса
(зарубинский известняк; фаленовый горизонт).
Phillipsastraea lazutklnl (Bulvanker)
Табл. XLIV, фиг. 1
Колония массивная. Кораллиты призматические с ясными внешними
стенками. При диаметре кораллитов 14—20 мм имеется 36—48
(иногда 54) распадающихся на поперечные балочки, мало различаю-
щихся по длине, веретеновидно утолщенных и расщепленных на перифе
рии септ двух порядков.
Днища, занимающие */4 поперечника кораллита, плосковыпуклые,
довольно тесно сближенные, расщепленные. Пузырчатая зона состоит
из вздутых, почти горизонтальных у периферии и наклонных у оси пузы-
рей, расположенных в 6—8 рядов. Расщепление септ появляется на позд-
них стадиях онтогенеза.
Близкий по расщеплению септ на периферии вид Phllllpsastraea
schafferl из D3 Малой Азии отличается более тонкими, менее много-
численными септами, более короткими септами второго порядка, сильно
зигзагообразно изогнутой внешней стенкой и наличием резкой внутрен-
ней стенки.
Отличия от Phllllpsastraea sedgwicki приведены при описании этого
вида.
Распространение. Нижнефранские отложения Кузбасса (зару-
бинский известняк близ с. Зарубино).
Phtlllpsastraea lateseptata I v a n i а
Табл. XLIV, фиг. 2
Массивная колония состоит из довольно крупных призматических
кораллитов с довольно толстыми внешними стенками и длинными,
распадающимися на поперечные балочки, довольно толстыми септами
двух порядков, мало различающимися по длине.
Септы первого порядка длинные, доходящие или слегка не доходя-
щие до центра. Их внутренние кончики тонкие, изогнутые, иногда соеди-
няющиеся друг с другом или с септами второго порядка. Они резко,
неравномерно и значительно утолщаются к периферии. Септы второго
порядка равны 4/5, реже 3/4 длины первых.
Днища довольно тесно сближенные, выпуклые и расщепленные.
Пузырчатая зона состоит из нескольких рядов мелких вздутых пузырей,
почти горизонтальных на периферии и наклонных у зоны днищ.
В онтогенезе характерно раннее появление внешней стенки, ее силь-
ное развитие и раннее утолщение септ с последующим их распадением
на поперечные балочки.
Диаметр кораллитов 12—13 мм. Число септ 26—34.
Близкий по типу колонии, очертаниям кораллитов, распадающимся
на поперечные балочки септам, строению днищ и пузырей вид Phlllipsa-
straea sedgwicki отличается более тонкими септами, которые на взрос-
лых стадиях начинают разъедаться, менее утолщенными стенками и
несколько иным ходом онтогенеза. На ранних стадиях его, в противопо-
ложность Phllllpsastraea lateseptata, характерно позднее появление
внешней стенки, ее сравнительно слабое развитие и появление тонких
удлиненных септ.
222
Распространение. Нижнефранские отложения Кузбасса (верх-
няя пачка зарубинского известняка, близ с. Зарубино).
Phllllpsastraea pentago па (G о 1 d f u s s)
Табл. XLIII, фиг. 2
Массивная колония мелких призматических кораллитов с резкой на-
ружной и внутренней стенками. При диаметре 3—5 мм имеются 20—22
веретеновидно утолщенных септы, распадающиеся на поперечные ба-
лочки (трабекулы). Септы первого порядка длинные, доходящие или
слегка не доходящие до центра, с тонкими заостренными или более утол-
щенными внутренними кончиками; они веретеновидно или более равно-
мерно утолщенные в наружных 2/3 длины. Септы второго порядка равны
'/г или немного больше длины первых, имея их толщину между стен-
ками. Они доходят до внутренней стенки, образованной утолщениями
септ и пузырей.
Зона днищ шире зоны пузырей. Днища тонкие, расщепленные на
взрослых стадиях развития, плоские, слабо выпуклые или слабо вогну-
тые, иногда с развитием дополнительных пластинок в осевой зоне. Пу-
зырчатая зона узкая и состоит из 2—3, реже четырех рядов мелких,
вздутых пузырей.
Для ранних стадий онтогенеза характерны довольно длинные септы,
внутренняя стенка расположена близко к настоящей (узкая зона пузы-
рей), простые днища. С возрастом удлиняются и утолщаются септы,
внутренняя стенка отодвигается ближе к центру, увеличивая ширину
зоны пузырей, но последняя узка и на взрослых стадиях; днища рас-
щепляются.
Велика индивидуальная изменчивость вида; особенно изменчивы
септы — их толщина, длина, степень распадания на трабекулы; меняется
и густота расположения днищ.
От близкого вида Phllllpsastraea mlcrommata (A. Roe m.) отли-
чается более резкой наружной стенкой, более широкой зоной днищ и
узкой зоной пузырей.
Phllllpsastraea mlcrastraea Penecke из D3 Малой Азии, сходный
мелкими кораллитами, строением септ и днищ, отличается меньшим ко-
личеством рядов пузырей.
Распространение. Франские отложения Западной Европы и
Мугоджарских гор; верхний девон Англии. В Кузбассе этот вид изве-
стен лишь из среднефранских отложений (глубокинский известняк близ
с. Соломино).
Phllllpsastraea mlcrommata (A. Roemer)
Табл. XLIII, фиг. 1
Массивная колония мелких призматических кораллитов с неправиль-
ной, иногда неясной наружной и ясной внутренней стенками. При диа-
метре 3—5 мм имеется 20—22 септы двух порядков, веретеновидно утол-
щенных и распадающихся на поперечные балочки. Септы первого по-
рядка длинные, своими нитевиднотонкими внутренними кончиками до-
стигающие центра кораллитов. Септы второго порядка, равные 3/4—4/s
длины первых, доходят до внутренней стенки.
Зона днищ узкая. Днища чаще полные, в осевой зоне крышеобразно
выпуклые или плоские, с выпуклыми дополнительными пластинками:
дополнительные пластинки на краях днищ развиты редко. Пузырчатая
зона из 3—5 рядов мелких вздутых пузырьков.
В онтогенезе увеличивается ширина зоны пузырчатой ткани и внут-
ренняя стенка сдвигается к центру так, что в своем индивидуальном раз-
витии этот вид проходит стадию Phllllpsastraea pentagona G о 1 d f. с уз-
кой пузырчатой зоной.
223
Наиболее изменчивы септы; по толщине и степени распадения на по-
перечные балочки.
От близкого по типу колонии строению септ, днищ и пузырей вида
Phillipsastraea pentagons, отличается малым поперечником зоны днищ,
более широкой зоной пузырей и более сильным распаданием септ на тра-
бекулы.
Распространение. Среднефранские отложения Кузбасса
(окрестности с. Соломине; высота Голый Мыс у д. Абышевой); франский
ярус Германии.
Род Schluteria Wedekind
Колонии ветвистые или массивные. Размножение происходит путем
непарисидального почкования. Чашка глубокая, бокаловидная, с ост-
рыми без отворота краями. Эпитека развита. Длинные септы двух по-
рядков утолщены на периферии и не распадаются на поперечные ба-
лочки. Септальные трабекулы параллельны. На ранних стадиях онтоге-
неза септы тонкие и длинные.
Днища полные или чаще неполные, усложненные дополнительными
пластинками у краев, часто и в середине. На продольном разрезе они
имеют вид одного вздутия у оси или двух вздутий на перегибах днищ.
Пузырчатая зона из 1—3, реже 4—5 вертикальных рядов пузырей, из
которых наружные ряды расположены почти горизонтально или слабо
наклонно.
От близкого рода Phillipsastraea d’Orb. отличается глубокой бока-
ловидной чашкой с широкой центральной ямкой, без отворота краев,
параллельными септальными трабекулами, строением днищ и более уз-
кой зоной пузырей. Отличия от рода Megaphyllum даны при его опи-
сании.
Типичный вид. Schluteria emstl Wed.
Распространение. Живетские отложения Германии; франский
ярус Европейской части СССР и Урала; нижнефранские отложения Куз-
басса.
Schliiterla emstl Wedekind
Табл. XLIII, фиг. 3
Колония ветвистая. Кораллиты цилиндрические, соприкасающиеся
или не соприкасающиеся. Их наружная поверхность с нежной продоль-
ной штриховкой. Чашка довольно глубокая, без отворота. При диаметре
кораллита 6—11 мм имеется 36—48 септ двух порядков, не распадаю-
щихся на поперечные балочки. Септы первого порядка длинные, септы
второго порядка короткие и равны не более ‘/г длины первых.
Днища редкие, типичные для рода, с дополнительными валикообраз-
ными пластинками над осевой частью и по краям. Пузырчатая зона из
2—3 рядов пузырей, из которых внешние почти горизонтальные.
Распространение. Живетские отложения Германии; живетские
и франские отложения Печорского края; франские отложения западного
склона Урала; нижнефранские отложения Кузбасса (зарубинский из-
вестняк; фаленовый горизонт).
Schliiterla striata I v a n i а
Табл. XLV, фиг. 2; табл. XLVI, фиг. 2
Колония ветвистая. Слабо изогнутые цилиндрические кораллиты
иногда срастаются своими боковыми стенками попарно, образуя на по-
перечном разрезе цепочку. Эпитека тонкая. Наружная поверхность ко-
224
раЛлйтов покрыта тонкой продольной бороздчатостью, отчего на попе-
речном разрезе внешняя стенка фестончата. Чашка глубокая, бокало-
видная, с острыми отвесными краями и слабо вогнутым дном.
При диаметре кораллита 11 —15 мм имеется 46—58 септ двух поряд-
ков, не расщепленных поперечными балочками, различающихся по
длине. У внешней стенки септы сливаются своими основаниями, утол-
щая ее. Септы первого порядка обычно не доходят до оси, оставляя там
довольно широкое, свободное пространство, иногда достигают оси и
даже слегка закручиваются в центре. Они всегда утолщены в перифери-
ческой части, постепенно утончаясь к осевой, где становятся нитевидно-
тонкими. Септы второго порядка равны '/4-—'/з длины первых.
Днища, занимающие 2/з—3/4 поперечника кораллита, тонкие, плоско-
выпуклые, обычно широкие и полные, с дополнительными надстройками
на перегибах. В ряде случаев дополнительные пластинки наблюдаются
в осевой зоне и имеют вид кольцевых валиков (на 5 лии длины насчи-
тывается 6—10 днищ).
Пузырчатая зона состоит из 1—6 рядов некрупных вздутых пузырей,
которые на, границе с зоной днищ на поперечных разрезах дают тонкую
линию внутренней стенки. Пузыри внешнего ряда более крупные, почти
горизонтальные; остальные — более мелкие, круто наклонные.
Характерны длинные тонкие септы на ранних стадиях онтогенеза.
Индивидуальная изменчивость: признаком, выдерживающимся у всех
кораллитов, является постепенное утолщение септ к периферии. Измен-
чивы длина септ, строение дополнительных пластинок и число рядов
пузырей.
Наиболее близок по внутреннему строению вид Schlilterla kostetskae
S о s h к. из франских отложений Урала, от которого Schliiterla striata
отличается ребристой и фестончатой внешней стенкой, всегда постепен-
ным утолщением септ к периферии, их большим количеством и обычно
более полными и более выпуклыми днищами.
Распространение. Нижнефранские отложения Кузбасса (фа-
леновый горизонт у Черепанова брода на р. Яя).
Род Solominella I v а п 1 а
Кораллы колониальные; колония ветвистая. Периферические части
септ сильно расширены и слиты в довольно широкий ободок, местами
прерывающийся пузырями. Днища выпуклые, расщепленные. Присут-
ствует пузырчатая ткань. В процессе индивидуального развития ободок
начинает рассасываться снаружи, от периферии, образованием у наруж-
ной стенки пузырей.
Этот род близок к родам Columnaria Goldf. и Neocolumnarla
S о s h к., верхнедевонским потомком которых он, вероятно, и является.
От первого он отличается развитием пузырчатой ткани и более слож-
ными днищами; от второго — способом рассасывания ободка и появле-
ния пузырчатой ткани. Ободок у рода Solominella рассасывается сна-
ружи во внутрь, а не наоборот, как это характерно для рода Neocolum-
naria, и пузырчатая ткань наслаивается на ободок снаружи, от наруж-
ной стенки.
От рода Fasclphyllum S с h 1 й t е г, характеризующегося развитием
пузырчатой ткани, наслаивающейся на ободок изнутри, род Solominella,
как и Neocolumnarla, отличается выпуклыми и усложненными днищами
и наслоением пузырчатой ткани на ободок снаружи.
Конвергентно сходный род Thamnophyllum Р е n е с к е отличается от-
сутствием подковообразных пузырей и присутствием эпитеки.
Типичный вид. Solominella soshklnae Ivan Га.
Распространение. Франский ярус Кузбасса (глубокинский
известняк).
15 Зак. 2024 225
Solomlnella soshklnae I v a n i a
Табл. XLVI, фиг. 3
Ветвистая колония состоит из некрупных цилиндрических, довольно
тесно сближенных, прямых или несильно изогнутых кораллитов (длина
их до 45 мм). Чашка бокаловидная, глубокая, с отвесными стенками и
плоским дном. При диаметре кораллита 9 мм имеется 30—40 простых
или слабо расщепленных на поперечные балочки септ, сильно различаю-
щихся по длине. Септы первого порядка длинные, слегка изогнутые, до-
стигающие центра у юных и не достигающие его у взрослых кораллитов.
Их внутренние кончики шиповидно или клиновидно заостренные, тонкие,
иногда изогнутые и соединяющиеся, сильно утолщаются по направле-
нию к наружной стенке и вместе с утолщенными септами второго по-
рядка образуют ободок. Септы второго порядка равны */2—2/з длины
первых и имеют одинаковую с ними толщину у наружной стенки.
Толщина ободка, образованного расширенными наружными концами
септ, равна */г радиуса кораллита у юных и в ‘А у взрослых форм.
С возрастом ободок начинает рассасываться посередине, с образованием
там одного, реже двух неполных рядов мелких пузырей. Часто он сохра-
няется у взрослых форм в виде двух узких каемок, разделенных пузы-
рями.
Днища тонкие, выпуклые, чаще полные, слабо расщепленные (на
5 мм длины насчитывается 8—12 днищ). Пузырчатая зона очень узкая,
из 1—2 неполных рядов мелких пузырей, выпуклостями обращенных
вверх и расположенных внутри ободка.
В онтогенезе важны: большое сходство ранних стадий с родом Co-
lumnaria; септы молодых стадий доходят до оси; постепенное исчезно-
вение ободка с возрастом и превращение его во внутреннюю стенку.
Возможно, что возрастным признаком является образование крышевид-
ного поднятия в осевой части днищ.
Изменчивы длина и характер утолщения септ; степень выпуклости
днищ.
Распространение. Среднефрапские отложения (глубокинский
известняк) северо-западной окраины Кузбасса (с. Соломине).
СЕМЕЙСТВО THAMNOPHYLLIDAE SOSHKINA
Род Pachyphyllum Edwards et Haime
Колонии астреев1идные. Почкование парисидальное. Чашки коралли-
тов с глубокой срединной ямкой, окруженной высоким валиком, на ко-
тором выступают гребши септ. Эпитека редуцирована и поверхность
кораллита ребристая от выступающих септ. Последние явственно распа-
даются на поперечные балочки, веретеновидно утолщенные. Септальные
трабекулы веерообразные.
Днища плоские, чаще полные, слабо усложненные расщеплением и
дополнительными пластинками. Всегда развиты подковообразные пу-
зырьки, на которые с эпитеки перенесено прикрепление септальных тра-
бекул, образующих валик вокруг чашечной ямки. Пузырчатая ткань за-
нимает широкую зону между подковообразными пузырьками соседних
кораллитов и состоит из плоских, разных по величине пузырей.
Астреевидная колония, обусловленная отсутствием эпитеки у отдель-
ных кораллитов, и наличие подковообразных пузырьков, к которым при-
крепляются веерообразные септальные трабекулы, образующие чашеч-
ный валик — признаки, отличающие этот род от всех сходных с ним
родов. Наиболее близки роды Macgeea W е b s t. и Thamnophyllum Р е-
песке. Первый отличается от рода Pachyphyllum одиночной формой
роста, а второй — ветвистыми колониями.
Типичный вид. Pachyphyllum devonlense Е dw. et Haime.
226
Распространение. Верхний девон, повсеместно. На Алтае он
впервые встречен и в верхнеживетских отложениях.
Pachyphyllum devontense Edwards et Haime
Табл, XLV, фиг. 1
Колония астреевидная, с ясным парисидальным почкованием. Чашка
с высоким валиком, наружная стенка отсутствует. Внутренняя стенка
резкая, круглая. Хорошо развиты септы двух порядков, сильно распа-
дающиеся на поперечные балочки и различающиеся по длине. Септы
первого порядка длинные, почти доходящие до центра, изогнутые, не-
сильно веретеновидно утолщенные у зоны днищ и подковообразных пу-
зырей. Септы второго порядка тоже изогнутые, утолщенные в осевой
части, доходящие до зоны днищ, равные 2/3 длины первых. В кораллите
насчитывается 38—42 септы.
Днища слабо выпуклые, расщепленные, редкие. На границе зоны
днищ и пузырей всегда развит ряд подковообразных пузырей. Пузырча-
тая зона из некрупных, слабо наклоненных пузырей.
Близкий по астреевидной колонии и по строению септ Pachyphyllum
ibergense Roem. из D3 Германии отличается меньшим числом септ и
более крупными пузырями.
Распространение. Франские отложения Западной Европы и
Тимана; нижнефранские отложения Кузбасса (фаленовый горизонт).
Pachyphyllum Ibergense A. Roemer
Табл. XLV, фиг. 3; рис. 187
Колония астреевидная. Чашка с высоким валиком. Наружная стенка
отсутствует. Внутренняя стенка резкая, круглая. В кораллите насчиты-
вается 26—34 септы двух порядков, сильно
различающиеся по длине, расщепленные на
поперечные балочки. Септы первого порядка
длинные, почти доходящие до центра, тон-
кие у оси и на периферии, веретеновидно
утолщенные близ внутренней стенки. Более
короткие септы второго порядка не выхо-
дят за пределы последней. Септы соседних
кораллитов чаще соединяются, реже чере-
дуются.
Днища выпуклые, расщепленные, редкие.
Подковообразные пузыри резкие, располо-
жены в один ряд. Пузырчатая ткань из до-
вольно крупных слабо наклонных пузырей.
Отличия от близкого вида Pachyphyllum
devontense указаны при его описании.
Распространение. Франский ярус
Германии; нижнефранские отложения Куз-
басса (фаленовый горизонт); на Тимане из-
вестен из верхней части франского яруса.
Рис. 187. Pachyphyllum iber-
gense A. Roem. Поперечное
сечение. Сильно увеличено
СЕМЕЙСТВО ENDOPHYLLIDAE HILL
Род Endophyllum Edwards et Haime
Одиночные и колониальные формы с редуцированной или сильной
эпитекой. Характерна редукция септ на периферии. Днища полные или
неполные, с опущенными книзу краями, осложненные дополнитель-
ными пластинками. Пузырчатая ткань сильно развита, состоит из круп-
ных, круто наклоненных пузырей.
Типичный вид. Endophyllum abdltum Е d w. et Haime.
Распространение. D2—D3, повсеместно.
15*
227
Endophyllum abditum Edwards et Haime
Табл XLIV, фиг. 3
Колония массивная. Кораллиты крупные, призматические. Наружная
стенка толстая, изогнутая в продольном разрезе и мелкозазубренная
в поперечном вследствие развития редуцированных септ. Внутренняя
стенка тонкая. Септы редуцированы на периферии. Септы первого по-
рядка тонкие или слегка утолщенные к периферии, изогнутые и длинные.
Септы второго порядка имеют вид коротких шиповидных отростков.
Иногда обрывки септ лежат на пузырях или у наружной стенки, зубчато
утолщая ее. При диаметре кораллита 12—24 мм имеются 48—76 септ
двух порядков.
Днища тонкие, редкие, выпуклые, со срединной вдавленностью; концы
их загнуты вниз, упираясь в пузыри или на нижележащие днища. Пу-
зырчатая зона из круто наклоненных, почти вертикально стоящих удли-
ненных пузырей, местами с дополнительными мелкими пузырьками.
Endophyllum weberl Lebedew из франских отложений р. Ухты,
почкующиеся колонии которого по утолщенной внешней стенке и реду-
цированным септам сходны с Endophyllum abditum, отличается формой
роста, более короткими септами, более простыми днищами и узкой зо-
ной пузырей.
Распространение. D3 Англии; среднефранские отложения
Кузбасса (высота Голый Мыс у д. Абышево, окрестности с. Соломино).
Endophyllum macconnelll Whiteaves
Табл. XLVI, фиг. 1
Крупный одиночный коралл, неправильно волчковидной или широко-
конической формы с ясными стадиями роста, выражающимися в резких
вздутиях и пережимах. Чашка более или менее глубокая, с плоским или
слабо приподнятым дном и почти отвесными краями. Многочисленные
септы (их 88 при диаметре коралла 30лл<) двух порядков сильно варьи-
руют по своему устройству, всегда разорваны или редуцированы на пе-
риферии, равномерно тонкие или слабо утолщенные к периферии. На-
ряду с радиальным типом расположения септ наблюдаются случаи,
когда расположение септ подчиняется двусторонней симметрии. Длина
септ тоже изменчива: порой они достигают центра и здесь закручи-
ваются или переплетаются с днищами, в других случаях они более ко-
роткие и оставляют свободное осевое пространство в. центре коралла.
Днища полные или неполные, более или менее плоские, расположены
.пучками и усложнены дополнительными пластинками. Пузырчатая зона
состоит из нескольких рядов вздутых, удлиненных, крупных и мелких
пузырей, круто наклонных к оси.
Близкий по строению септ Endophyllum ehlersl Fe n t. et Fe n t. из
верхнего девона Северной Америки отличается более неправильными
днищами, более многочисленными и более мелкими пузырями.
Распространение. Верхний девон Полярного Урала, Франции,
Канады; в Кузбассе — нижнефранские отложения (фаленовый гори-
зонт) Черепанова брода на р. Яя.
П. С. КРАСНОПЕЕВА
Тип Bryozoa. Мшанки
Мшанки широко распространены в средне- и верхнедевонских отло-
жениях Кузбасса, Алтая, Минусинской котловины. Они описаны в ряде
работ В. П. Нехорошева (1926, 1926а, 1948) и П. С. Краснопеевой (1935).
В недавно опубликованной работе В. П. Нехорошева (1948) приведены
обширные материалы по богатой фауне девонских мшанок Алтая; из
этой работы, в основном, и заимствованы приведенные ниже диагнозы
или сокращенные описания некоторых руководящих форм.
228
Отряд trepostomata
СЕМЕЙСТВО BATOSTOMELLIDAE ULRICH
Род Lioclema Ulrich
«Зоария ветвистая, пластинчатая, шаровидная или нарастающая; на
поверхности часто имеются бугорки или пятна. Устья ячеек округленные
или неправильно петалоидные, разделенные многочисленными углова-
тыми мезопорами, которые в одних видах на поверхности открытые,
в других закрытые. Диафрагмы в ячейках редкие, в мезопорах много-
численные. Акантопоры, многочисленные и резко выраженные в одних
видах, в других, наоборот, редкие и мелкие» (Нехорошее, 1948, стр. 58).
Стенки соседних ячеек слившиеся.
Типичный вид. Lioclema punctata Hall.
Распространение. Силур — нижняя пермь. Алтай, Минусин-
ская котловина. Северная Америка.
Lioclema yakovlevl Schoenmann
Табл. XLVII, фиг. 1
«Зоария массивная, весьма разнообразной формы, натечная, гроздье
видная, ветвистая, однослойная или многослойная. На поверхности
имеются мелкие бугорки. Устья округленные, окружены одним, реже
двумя рядами угловатых мезопор. Акантопоры распределены неравно-
мерно, местами обильны, в других случаях редки. Диафрагмы в ячей-
ках редкие, в мезопорах весьма обильные. Расстояние между цент-
рами бугорков около 3,0—3,5 мм.
Никакой закономерности в распределении устьев ячеек не наблю-
дается, и при наличии крупных мезопор иногда бывает довольно1 затруд-
нительно отличить их от устьев ячеек. На 2 мм насчитывается 6—8 усть-
ев и столько же мезопор. Диаметр устьев в среднем 0,15—0,18 мм. Ме-
зопоры зачастую равного с устьями диаметра; наиболее мелкие имеют
в диаметре всего лишь 0,06 мм. Обычно мезопоры многоугольные, но
есть и округленные. Вокруг устьев имеется по 1—2 ряда мезопор.
Акантопоры расположены без определенного порядка; наиболее ча-
сто около каждого устья наблюдается по одной акантопоре или по две,
расположенных на противоположных сторонах перистом устьев. В от-
дельных участках акантопор значительно больше и около каждого устья
располагаются по три или даже по четыре акантопоры. Акантопоры
обычно располагаются в местах скрещений стенок ячеек и мезопор и
внутрь ячеек не вдаются. Диаметр акантопор 0,04—0,05 мм. Стенки
ячеек у поверхности утолщаются до 0,02 мм, глубже они значительно
тоньше» (Нехорошее, 1948, стр. 62—63).
Распространение. Верхи среднего девона Минусинской котло-
вины и Барзасского района Кузбасса; верхи среднего девона и основа-
ние верхнего девона Горного Алтая (Чуйский тракт, урочище Бельге-
баш; ущелье Кизыл-Шин на р. Бугусун).
Lioclema bugusunlca Nekhorochev
Табл. XLVII, фиг. 2
«Зоария ветвистая, диаметром от 5 до 30 мм. Устья округленные,
окруженные 1—2 рядами мелких мезопор. Акантопоры весьма редкие.
Периферическая часть зоарии обычно значительно шире осевой.
Устья округленноовальные, диаметром от 0,17 до 0,22 мм. В отдель-
ных участках как будто устья крупнее, в других мельче, но выяснить
какую-либо закономерность в их распределении не удалось; На 2 мм
насчитывается 7—7,5 устьев. Промежутки между устьями заполнены
1—2 рядами мелких угловатых и округленных мезопор диаметром от
0,03 до 0,10 мм.. Акантопоры исключительно редки...
229
Мезопоры в продольных и поперечных сечениях пронизывают всю
периферическую часть зоарии, имея вид тонких, длинных цилиндриче-
ских трубочек, в которых в более тонких ветвях зоарии на 0,5 мм насчи-
тывается до семи диафрагм. В ячейках диафрагмы в периферической
части довольно частые и в более тонких ветвях на 1 мм насчитывается
6—8 диафрагм. В более толстых зоариях диафрагмы более редкие» (Не-
хорошее, 1948, стр. 66).
Встречающаяся совместно Lloclema polenovl N е k h. отличается бо-
лее крупными устьями и более короткими мезопорами.
Распространение. Верхи среднего девона, Горный Алтай
(ущелье Кизыл-Шин на р. Бугусун в Юго-Восточном Алтае).
СЕМЕЙСТВО HETEROTRYPIDAE ULRICH
Род Leptotrypa U1 rich
«Зоария обрастающая, часто принимающая форму трубки, диско-
идальную и другие формы. Трубки зооэций тонкие, многоугольные, с ред-
кими диафрагмами; акантопоры редкие и мелкие. Мезопор не имеется»
(Нехорошее, 1934, стр. 430). Стенки соседних ячеек сливаются (границы
между ними не видны).
Распространение. Силур — девон.
Leptotrypa tubullformls Schoenmann
Рис. 188
Тонкие, прямые, пустые внутри веточки, образованные слабо изги-
бающимися ячейками. Устья ячеек слегка округленно многоугольные.
Акантопоры незначительных размеров, расположены в углах ячеек. По-
Рис. 188. Leptotrypa tubuliformis S с h.
а—скошенное поперечное сечение колонии,
X 20; б—скошенное поперечное сечение
колонии, X 15. Верхи среднего девона. Бар-
засский район Кузбасса
перечник зоарии до 1,2 мм. Высота ячеек в среднем 0,2 мм. Поперечник
устьев ячеек 0,1.2—0,17 мм. На 5 мм длины зоарии около 30 устьев. По-
перечник акантопор 0,03—0,04 мм, число их вокруг каждой ячейки равно
2—3. Ширина центральной полости 0,65—0,80 мм.
230
Распространение. Верхи D2 Минусинской котловины (д. Са-
рагаш на левом берегу Енисея) и Кузбасса (Барзасский район)
Отряд Cryptostomata
СЕМЕЙСТВО FENESTELL1DAE KING
Род Fenestella Цо n s d а 1 е
«Зоария веерообразная или воронковидная, ячеистая на одной сто-
роне. Прутья соединяются перекладинами, образуя сетчатую поверх-
ность. Устья ячеек распределены в два ряда, разделенные гладким или,
чаще, бугорчатым килем. Неячеистая поверхность прутьев струйчатая»
(Нехорошее, 1948, стр. 81).
Для предварительных определений представителей рода Fenestella
и других фенестеллид большое значение имеют следующие измерения:
количество прутьев на 10 мм\ количество петель на 10 мм (вдоль пру-
тьев); количество ячеек на 5 мм длины прута. В приведенных ниже ви-
довых диагнозах эти три показателя даются в последовательной цифро-
вой записи как основные размеры.
Типичный вид. Fenestella plebeja М’С о у.
Распространение. Силур — пермь, повсеместно.
Fenestella vera Ulrich
Табл. XLVIII, фиг. 1
«Размеры: 22—24/13—14//22—24. Сетка пра-
вильная, сравнительно мелкая, прочная. Килевая
линия1 прямая. Киль покрытый бугорками»
(Нехорошей, 1948, стр. 86).
Близкая по основным размерам сетки нижне-
карбоновая Fenestella multtsplnosa U1 г. отли-
чается зигзаговидной килевой линией.
Распространение. Верхи D2 Горного
Алтая (рч. Коргончик в Западном Алтае; ущелье
Кизыл-Шин на р. Бугусун в Юго-Восточном Ал-
тае; урочище Бельгебыш на Чуйском тракте
в Центральном Алтае). Верхи среднего девона
Северной Америки.
Fenestella buratlnensls Krasnopeeva
Рис. 189
«Размеры: 15—16/4—6//21—23. Сетка тонкая,
неправильная, крупная. Килевая линия прямая, ячейки четырехугольного
сечения» (Нехорошее, 1948, стр. 90).
В верхнем девоне и нижнем карбоне Рудного Алтая распространена
Fenestella quadrulla N е k h., близкая по общему строению, но отличаю-
щаяся более мелкой сеткой (основные размеры сетки Fenestella qua-
dra Ila: 20—24/8— 12//22—24).
Распространение. Низы верхнего девона. Горный Алтай (Чуй-
ская степь, падь Буралы).
Род Hemitrypa Phillips
«Зоария состоит из двух сеток: основной, имеющей такое же строе-
ние, как сетки Fenestella, и расположенной над нею защитной сетки
с мелкими однородными отверстиями, число которых обычно соответ-
ствует числу устьев ячеек основной сетки. Сильно развитые бугорки на
1 Килевой линией называется линия, которая в тангенциальном сечении прутьев
делит два ряда ячеек. Прим. ред.
Рис. 189. Fenestella bura-
tinensis К г a s n.
Тангенциальное и попе-
речное сечения, X 19.
Горный Алтай, Чуйская
степь, падь Бураты
231
киле основной сетки вверху расщепляются, давая отростки, которые, сли-
ваясь с отростками соседних бугорков того же прута и соседних прутьев,
образуют мелкосетчатую защитную поверхность в виде правильных гре-
бешков, расположенных вдоль прутьев и над промежутками между
прутьями, соединенных поперечными чешуйками» (Нехорошее, 1948,
стр. 92—93).
Подрод Hemltrypella N е k h. отличается трубчатой формой колоний.
Типичный вид. Hemltrypa oculata Р h i 11.
Распространение. Силур — нижняя пермь.
Hemltrypa cornea Nekhorochev
Табл. XLVIII, фиг. 2
«Размеры: 17—20/13—15//16—18. Зоария роговидная; неячеистая
поверхность прутьев покрыта многочисленными мелкими и крупными
бугорками. Перекладины на неячеистой поверхности развиты сильнее
прутьев. Сечение ячеек округленнопятиугольное. Килевая линия зигзаго-
видная» (Нехорошее, 1948, стр. 95).
Распространение. Нижний девон и низы эйфельского яруса
Рудного Алтая (окрестности Змеиногорска, окрестности с. Лосиха и вер-
ховья рч. Золотухи).
Hemltrypa devonlca Neckorochev
Табл. XLVIII, фиг. 3
«Размеры: 23—25/18—22//23—25. Зоария мелкосетчатая, воронкооб-
разная, по удалении от основания сильно расширяющаяся. Неячеистая
поверхность бугорчатая. Среднее сечение ячеек пятиугольное» (Нехоро-
шее, 1948, стр. 96).
Распространение. Средний девон Кузбасса; низы верхнего
девона Горного Алтая (ущелье Кизыл-Шин на р. Бугусун).
Род Semicoscinium Prout
«Зоария обычно воронкообразная, с наружной ячеистой поверхно-
стью. Прутья несут по два ряда ячеек и соединены анастомозами или
перекладинами. На неячеистой поверхности прутья обычно резко изогну-
тые и их истинное направление неразличимо. Вся неячеистая поверх-
ность нередко килеватая и покрытая многочисленными мелкими бугор-
ками. На ячеистой поверхности на прутьях имеются весьма высокие
кили, кверху расширяющиеся. Сечения ячеек в тангенциальных шлифах
четырехугольные или овальные, и ряды ячеек разделены прямой или
волнистой, но не ломаной килевой линией» (Нехорошее, 1948, стр. 100).
Типичный вид. Semicoscinium rhomboideum Prout.
Распространение. Силур — нижний кембрий.
Semicoscinium altalcum Nekhorochev
Табл. IL, фиг. 1
«Размеры: 11 —13/4—5//17—18. Зоария воронкообразная, сетка проч-
ная, прутья сильно изогнутые, в связи с чем кили почти до самой вер-
шины волнисто изогнутые. Сечение вершин килей ромбическое. Неячеи-
стая поверхность покрыта многочисленными крупными бугорками» (Не-
хорошее, 1948, стр. 102). Для этого вида особенно характерны крупные
размеры сетки.
Распространение. Данный вид широко распространен в низах
D3 Горного Алтая (рр. Бугусун и Чаган-Узун).
232
Serntcosclnlum rhombtcum Ulrich var. altaica Krasnopeeva
Табл. X-LVIII, фиг. 4
«Размеры: 18/9—10//20—21. Сетка сравнительно мелкая, прутья
резко изгибающиеся, кили имеют ромбическое сечение с острой верши-
ной» (Нехорошее, 1948, стр. 103).
Распространение. Верхи среднего девона Горного Алтая
(р. Коргончик в Кортонском хребте, Западный Алтай).
Semicosclnlum altchedaticum Nekhorochev
Табл. IL, фиг. 2
«Размеры: 12—14/8—10//20. Сетка сравнительно мелкая; прутья
слабо волнистые, ясно различимые на неячеистой поверхности. Кили
умеренной высоты, кверху сильно расширенные и закругленные, несущие
по краям внутри с каждой стороны по ряду бугорков такого же строе-
ния, как и бугорки на неячеистой поверхности» (Нехорошее, 1948,
стр. 107).
Распространение. Верхи среднего девона Горного Алтая (ур.
Бельгебаш и р. Бугусун), количественно господствует среди других
мшанок. В низах D3 Горного Алтая известны близкие формы, отличаю-
щиеся теми или иными показателями основных размеров сетки.
Род Reteporlna d’O г b i g п у
«Зоария сетчатая; прутья, волнисто изгибаясь, соединяются через
правильные промежутки анастомозами. На прутьях по два ряда устьев
ячеек, разделенных гладким килем. Неячеистая поверхность гладкая
или покрытая крупными бугорками, иногда килеватая» (Нехорошев,
1948, стр. 116).
Типичный вид. Reteporlna striata Hall.
Распространение. Нижний девон — нижний карбон. Азиат-
ская часть СССР, Западная Европа, Северная Америка.
Reteporlna grandlformls Krasnopeeva
Табл. IL, фиг. 3
«Размеры: 11 —12/4—5//17—19. Сетка крупная, прутья резко волни-
сто изогнутые даже не ячеистой поверхности, анастомозы широкие. Не-
ячеистая поверхность килеватая, пронизанная многочисленными мел-
кими капиллярами» (Нехорошев, 1948, стр. 120).
По основным размерам близка к Reteporlna ubensls Nekh. из низов
эйфельского яруса Рудного Алтая, но отличается прямыми прутьями на
ячеистой поверхности.
Распространение. Низы верхнего девона Горного Алтая
(ущелье Кизыл-Шин на р. Бугусун).
Тип Brachiopoda. Плеченогие (брахиоподы)
Стратиграфия морских девонских отложений Западной Сибири раз-
работана главным образом на основании изучения брахиопод, которые
составляют наиболее важную и наиболее распространенную группу в со-
ставе фауны этих отложений. Их изучением и описанием занимался ряд
авторов (см. список литературы), из числа которых пять человек при-
няли участие в составлении характеристик руководящих форм для на-
стоящего атласа.
Очень своеобразна лептодонтелловая фауна брахиопод нижнего де-
вона Алтая, недавно подробно описанная Л. Л. Халфиным (1948), по-
этому из ее состава ниже приводятся лишь немногие, наиболее важные
руководящие формы. Более обычного (богемско-уральского) типа фауна
233
нижнего девона Салаира и юго-западной окраины Кузбасса. В свое
время она была описана Г. Г Петцем (1901), но это описание устарело,
и в настоящее время эта фауна монографически изучается М. А. Ржон-
сницкой (1952). Нижнедевонская фауна указанного типа известна ме-
стами и на Алтае, в частности в Рудном Алтае (Ф. Н. Чернышев, 1893).
Такой же характер носит и эйфельская фауна, которая наиболее
обильно представлена в Салаире, но распространена и на Алтае (Гра-
цианова, 1950). Преимущественно на Салаире (и по юго-западной окра-
ине Кузбасса) развиты также нижнеживетские отложения; брахиоподы
этих отложений частично описаны М. А. Ржонсницкой (1937).
Значительно более широко распространена верхнеживетская чиэле-
вая фауна (Кузбасс, Алтай, Минусинская котловина), которая является
одной из наиболее полно изученных девонских фаун Западной Сибири.
Достаточно хорошо изучены брахиоподы верхнего девона, в составе
которых выделяется ряд стратиграфических комплексов, важнейшими
из которых являются:
1. Фаленовая (или супрафаленовая) фауна — Кузбасс, Горный Ал-
тай, район г. Камня на р. Обь; нижнефранские отложения.
2. Фауна глубокинского известняка — Кузбасс, Рудный Алтай, рай-
он г. Камня; среднефранские отложения.
3. Монстровая фауна — Кузбасс; верхнефранские отложения.
4. Прелонгусовая фауна — Кузбасс, Горный Алтай, Рудный Алтай;
фаменские отложения.
Л.Л. ХАЛФИН
БРАХИОПОДЫ НИЖНЕГО ДЕВОНА АЛТАЯ
Древнейшая из фаун, которую уже можно отнести к девону, хотя
она и характеризуется смешением силурийских и девонских форм, из
вестна в северо-западной части Горного Алтая у с. Соловьиха. Она
имеет и в географическом отношении смешанный характер: наряду
с большим количеством местных форм в ней большую роль играют бо-
гемско-уральские элементы.
Более молодые нижнедевонские фауны Алтая имеют весьма само-
бытный характер. Формы европейского типа в них хотя и достаточно
многочисленны, но представлены они в большинстве случаев не тожде-
ственными, а лишь очевидно близкими видами, причем формы богемско-
уральского фациального типа редки, и наряду с ними присутствуют
формы рейнского фациального типа.
По массовому развитию представителей рода Leptodontella К h а 1 f.
эти фауны получили название лептодонтелловых. Выделено три после-
довательные лептодонтелловые фауны, характеризующие в восходящем
стратиграфическом порядке различные подразделения разреза нижне-
девонских отложений Алтая, именно: ганинскую серию и кондратьев-
скую и медведевскую толщи. Возможно, этот же тип фауны местами
(например, в Рудном Алтае) характеризует и самые низы эйфеля.
Богемско-уральского типа фауна распространена на Алтае очень
слабо и известна пока лишь в двух пунктах (в Рудном Алтае близ
с. Курьи и близ бывшего Крюковского рудника).
СЕМЕЙСТВО DALMANELLIDAE SCHUCHERT
Род Levetiea Schuchert et Cooper
Двояковыпуклая раковина округлого или субквадратного очертания.
Замочный край прямой, меньше наибольшей ширины раковины. Брюш-
ная створка более выпукла, чем спинная, с хорошо развитой треуголь-
ной ареей и седлообразным возвышением вдоль плоскости симметрии.
Спинная створка несет ясно выраженный, узкий синус. Скульптура со-
234
стоит из многочисленных тонких радиальных пебрышек. Мускульное
поле в брюшной створке сердцевидное, в спинной — четырехдольное. Ве-
щество створок пористое.
Большое сходство имеет с силурийским родом Dalmanella Hall.,
который отличается меньшей выпуклостью спинной створки, пучкообраз-
ным характером ребер и некоторыми деталями внутреннего строения.
Внешне очень близок также род Isorthls Kozl., отличающийся оди-
наковой выпуклостью створок.
Типичный вид. Levenea subcarlnata (Hall)
Распространение. Верхний силур — средний девон, повсе-
местно.
Levenea taenlolata К h а 1 f i п
Табл. L, фиг. 1
Средних размеров раковина, изометричная или слегка развитая по
ширине. Наибольшую ширину имеет вблизи замочного края; к перед-
нему краю слегка сужается. Передний край в средней части прямой.
Брюшная створка имеет невысокую, треугольную, вогнутую арею
с открытым дельтирием; срединное возвышение ясное, округленное.
Спинная створка слабо выпуклая, с очень низкой ареей и неглубо-
ким, но ясно выраженным синусом. Радиальные ребрышки очень тонкие,
округленные, многочисленные (до 200 штук на каждой створке).
Очень близка нижнедевонская Levenea subcarlnata (Hall), которая
указывается и для кобленцских (пестеревских) известняков Салаира;
она отличается округлыми очертаниями раковины: линия наибольшей
ширины лежит на равном расстоянии от замочного и переднего краев.
Распространение. Ганинская серия Горного Алтая, окрестно-
сти с. Соловьиха.
СЕМЕЙСТВО STROPHOMENIDAE KING
Род Leptaena D а 1 m а п
Вогнуто-выпуклая раковина с гладкими ареями и характерным, рез-
ким и угловатым, коленообразным перегибом створок близ переднего и
боковых краев; позади этого перегиба поверхность створок слабо выпук-
лая, почти плоская и покрыта грубыми концентрическими морщинами.
Через всю поверхность створок проходят очень многочисленные и очень
тонкие радиальные ребрышки (струйки).
Род Cymostrophla отличается прерывистым характером концентриче-
ских морщин (образованы межреберными вздутиями), округлым (а не
угловатым) перегибом поверхности створок и немногочисленными, ши-
роко расставленными радиальными ребрами.
Типичный вид. Leptaena rhotnboldalls W i 1 с k.
Распространение. Нижний силур — нижний карбон, повсе-
местно.
Leptaena bouel (В а г г a n d е)
Табл. L, фиг. 2—9
Крупная, трапецеидального очертания раковина, обладающая всеми
отмеченными выше признаками рода и особенно характеризующаяся
развитием на брюшной створке, близ переднего края, широкого синуса,
по бокам которого поверхность створки резко приподнята и образует
характерный бордюр, наиболее высокий у переднебоковых углов и по-
степенно понижающийся к призамочным углам. Поверхность спинной
створки следует поверхности брюшной.
Наличие синуса и седла и вообще более сложный рельеф створок
235
хорошо отличает описываемый вид от Leptaena rhomboidalls W i I с к.
(см. стр. 257); труднее отличить его от Leptaena rhomboidalls var. si-
nuata (см. стр. 244).
Распространение. Одна из важнейших руководящих форм
нижнего девона, хотя и поднимается до самых низов Эйфеля в Рудном
и Горном Алтае. Широко распространена в нижнем девоне Западной
Европы.
Род Cymostrophia A. Williams
Вогнуто-выпуклая раковина с длинным прямым замочным краем и
невысокой ареей, несущей зазубренность. Характерна скульптура: тон-
кие, но резкие радиальные ребра, разделенные широкими, плоскими про-
межутками. В каждом межреберном промежутке располагаются попе-
речные вздутия, имеющие полулунную форму и обращенные выпукло-
стью к макушке; в целом эти вздутия создают картину неправильных
концентрических морщин.
Силурийский род Rafinesquina Hall отличается отсутствием описан-
ной выше скульптуры.
Типичный вид. Cymostrophia stephanl (В а г г a n d е).
Распространение. Нижний силур — нижний девон, СССР и
Западная Европа.
Corrugatella stephanl (В а г г a n d е)
Табл. L, фиг. 10; табл. LI, фиг. 7
Во взрослом состоянии раковина обладает резким, но округленным
коленообразным изгибом створок близ переднего и боковых краев. Кон-
центрические морщины, образованные межреберными вздутиями, распо-
лагаются только в примакушечной и средней частях створок, не распро-
страняясь за пределы коленообразного изгиба.
Распространение. Одна из характерных форм для богемско-
уральского типа фауны нижнего девона СССР и Западной Европы;
в Западной Сибири встречается на Салаире и Алтае.
Род Brachyprion S h а 1 е г
Раковина имеет такое же строение, как и у представителей рода
Stropheodonta Hall (см. стр. 244), но зазубренность имеется только на
прилегающей к дельтирию средней части ареи, причем зубчики располо-
жены по нижнему краю последней.
Типичный вид. Brachyprion leda (Bill).
Распространение. Верхний силур — нижний девон. СССР, За-
падная Европа, Северная Америка.
Brachyprion nallvklnl К h а 1 f i n
Табл. L, фиг. 12
Крупная, вогнуто-выпуклая раковина с замочным краем, составляю-
щим наибольшую ее ширину. Ширина равна длине. Боковые края суб-
параллельны, передний край округленный. Арея брюшной створки невы-
сокая (ее высота около 1,5 мм), треугольная. В брюшной створке име-
ются короткие, расходящиеся зубные пластины; мускульная площадка
сердцевидного очертания. В спинной створке двураздельный замочный
отросток. Наружная поверхность с тонкими и многочисленными ради-
альными ребрышками, в промежутках между которыми располагаются
нитевидные струйки (в количестве 2—5 штук в каждом межреберном
промежутке).
236
Распространение. Нижний девон Горного и Рудного Алтая;
в медведевской толще (верхнекобленцские отложения окрестностей
д. Александровки) встречается массами вместе с вадом Brachyprion
latus Kha If. (табл. L, фиг. 11; табл. LI, фиг. 8), который отличается
сильным развитием раковины по ширине.
Род Leptodontella К h а 1 f i п
Выпукло-вогнутая раковина полукруглого и полуэллиптического
очертания с длинным прямым замочным краем. Призамочные углы
обычно образуют небольшие ушки, иногда превращающиеся в короткие
остроконечия. Створки, почти плоские в примакушечной и средней части,
близ переднего и боковых краев образуют резкий и острый (под прямым
углом) перегиб. Обе створки снабжены невысокими зазубренными
ареями. Мускульная площадка в брюшной створке большая, треуголь-
ная. В спинной створке — четырехлопастной замочный отросток, четы-
рехдольное мускульное поле и обычно хорошо выраженные брахиальные
отпечатки. Скульптура состоит из тонких радиальных ребрышек и струек.
Строение вещества раковины точечное.
Род Strophonetla Hall отличается отсутствием брахиальных отпе-
чатков, двулопастным замочным отростком и обычно отсутствием рез-
кого коленообразного изгиба поверхности створок. Род Leptodonta nom.
nov. (=Ochlertla Kha If., 1948) обладает обратной выпуклостью створок.
Типичный вид. Leptodontella caudata (Schnur).
Распространение. Нижний — средний девон. Западная
Европа, Южная Азия. Пользуется массовым развитием в нижнем девоне
Алтая; указывается из нижнего девона Дальнего Востока.
Leptodontella rotundata К h а 1 f i n
Табл. LI, фиг. 1 и 2; рис. 190
Небольшая раковина с замочным краем, составляющим наибольшую
ширину и образующим маленькие острые ушки; боковые края выпуклые;
_______передний край в средней части вогнутый. Наружная по-
Оверхность створок покрыта тонкими радиальными реб-
рышками в количестве 50—60 штук на каждой створ-
ке; в промежутках между ними располагается по 2—4
нитевидных струйки. Замочный отросток массивный.
Рис. 190. Leptodon- Внешне к описываемой форме очень близка евро-
К h^a ИГ Нат^вел пейская среднедевонская Strophonella anaglypha
Kays., отличающаяся правильно округленным перед-
ним краем, менее развитыми ушками и обычно относительно более ко-
ротким замочным краем (равен или меньше наибольшей ширины).
Отличия от Leptodontella planuscula указаны ниже.
Распространение. В массовом количестве встречается в га-
нинской серии и кондратьевской толще Горного Алтая.
Leptodontella planuscula (Kh а 1 f i n)
Табл. LI, фиг. 3; рис. 191
Этот вид отличается от Leptodontella rotundata
трапецеидальными очертаниями раковины: боковые
края почти прямые, образующие с замочным краем
углы примерно в 60°; замочный отросток не массив-
ный, он состоит из двух ветвей, из которых каждая
в свою очередь образует две доли; брахиальные от-
печатки выражены отчетливо.
Распространение. Руководящая форма
ских отложений Алтая (медведевская толща).
Рис. 191. Leptodontella
planuscula (К h а 1 f.)
Внутреннее строение
брюшной створки.
Нат. вел.
для верхнекобленц-
237
Род Schuchertella Girt у
Достигающая крупных размеров раковина с длинным замочным
краем, составляющим наибольшую ширину. Брюшная створка от выпук-
лой до вогнутой, обладает прямой треугольной ареей; дельтирий закрыт
выпуклым псевдодельтидием. Зубные пластины отсутствуют. Спинная
створка равномерно выпуклая. Наружная поверхность покрыта тонкими,
но резкими прямыми радиальными ребрами, увеличивающимися в коли-
честве путем вставления.
Внешне наиболее близок род Schellwlenella Thom. (см. стр. 257),
отличающийся наличием хорошо развитых зубных пластин. Род Strepto-
rhynchus King обладает коротким замочным краем и сильно выпуклой
брюшной створкой.
Типичный вид. Schuchertella lens (White).
Распространение.• Девон—пермь, повсеместно.
Schuchertella altatca К h а 1 f i n
Табл. LI, фиг. 4 и 5
Раковина выпукло-вогнутая или выпукло-плоская, крупных размеров,
слегка развитая по ширине. Замочный край равен максимальной ширине,
призамочные углы прямые. Боковые и передний края очерчены плавной
кривой. Арея брюшной створки сравнительно невысокая, арея на спин-
ной створке не развита. Скульптура обычного для данного рода характера.
Внешне к описываемому виду близка SchnellwleneUa umbraculum
Schl. (см. стр. 257), отличающаяся более высокой ареей брюшной
створки и наличием хорошо развитых зубных пластин.
Распространение. Нижний девон и, возможно, низы эйфель-
ского яруса Горного и Рудного Алтая.
СЕМЕЙСТВО CHONET1DAE HALL ET CLARKE
Род Chonetes Fischer
Раковина вогнуто-выпуклая, развитая по ширине, обычно полукруг-
лого, полуэллиптического или округленнопрямоугольного очертания. Пря-
мой замочный край составляет наибольшую ширину раковины. Макушки
слабо развиты. Арея брюшной створки невысокая, с закрытым дельти-
рием. Вдоль ареи располагается один ряд шипов. Наружная поверхность
радиальноребристая или радиальноструйчатая.
Ряд близких родов отличается отсутствием радиальной скульптуры
или развитием седла и синуса.
Типичный вид. Chonetes variolata d’O г b.
Распространение. Силур—пермь, повсеместно.
Chonetes pseudobohetnlca К h а 1 f i n
Табл. LI, фиг. 6; табл. LII, фиг. 1
Раковина средних размеров, изометричная или развитая по ширине
(отношение ширины к длине достигает 2); боковые и передний края пра-
вильно округленные. Брюшная створка слабо и равномерно выпуклая,
с очень маленькой макушкой. Арея очень низкая, шипы тонкие, сохра-
няются редко. Поверхность створок покрыта очень тонкими дихотоми-
рующими радиальными струйками в количестве до 100 штук на каждой
створке.
Распространение. Верхнекобленцские отложения (медведев-
ская толща) Горного Алтая. Встречается совместно с другими видами,
отличающимися лишь значительным развитием раковины по ширине.
238
Chonetes rectlspina К h a if i n
Табл. LII, фиг. 2
Раковина небольшая, округленнотреугольного очертания, которое
обусловлено оттянутым и суженным передним краем. Вздутая брюшная
створка седлообразно приподнята вдоль плоскости симметрии; спинная
створка равномерно вогнутая. Шипы немногочисленные, различной длины
(их длина возрастает по мере движения от макушки к кардинальным
углам), направлены перпендикулярно к замочному краю. Скульптура
состоит из тонких дихотомирующих радиальных ребрышек в количестве
50—60 штук на каждой створке.
Близок американский вид Chonetes hemisphaerica Hall, отличаю-
щийся отсутствием седлообразного возвышения и широкоокругленным
передним краем раковины.
Распространение. Нижнекобленцские отложения Горного
Алтая (кондратьевская толща).
СЕМЕЙСТВО PENTAMERIDAE М’СОУ
Род Conchidium LI п и ё
Описание см. на стр. 162.
Conchidium celslseptosum К h а 1 f i n
Табл. LII, фиг. 3; табл. LIII, фиг. 6; рис. 192
Раковина небольших для рода Conchidium разме-
ров, развитая по длине, сильно неравностворчатая.
Брюшная створка значительно выпуклая, с длинной,
клювообразно загнутой макушкой, под которой рас-
полагается явственно ограниченная арея с узким
дельтирием посередине. Спинная створка слабо вы-
пуклая, округленного очертания, с слабо развитой ма-
кушкой. Седло и синус отсутствуют. Наружная по-
верхность створок покрыта резкими, грубыми, исчеза-
Рис. 192. Con-
chidium cel sisep-
tcsum К h a 1 f.
Характер септы
в брюшной створ-
ке. Нат. вел.
ющими вблизи макушек радиальными ребрами в количестве 16—20 штук
на каждой створке. Септа в брюшной створке очень высокая.
Распространение. Верхнекобленцские отложения Горного
Алтая (медведевская толща).
Рис. 193. Ussovia
prima К h a I f.
С|роение замоч-
ной пластинки.
Увеличено
СЕМЕЙСТВО CAMAROTOECHI1DAE SCHUCHERT ET LE VENE
Род Ussovia К h а 1 f i и
Раковина субкубической формы, с сильно вздутой спинной и почти
плоской брюшной створками. Седло и синус развиты лишь близ перед-
него края. Наружная поверхность створок покрыта про-
стыми, округленными радиальными ребрами, начинаю-
щимися от макушек. В брюшной створке массивные
зубы; зубные пластины рудиментарны. В спинной
створке — септалиум, прикрытый парной пластинкой
с небольшим висцеральным фораменом (ркс. 193).
Среди палеозойских ринхонеллоидных раковин
имеется ряд родов, внешне подобных данному, но не
обладающих закрытым септалиумом, являющимся
основным отличительным признаком рода Ussovia.
Типичный вид. Ussovia prima (К h а 1 f.).
Распространение. Нижнекобленцские отложения Горного
Алтая.
239
Ussovia prlina (К h a If i n)
Табл. LII, фиг. 4—6
Диагноз этого вида совпадает с приведенным выше родовым описа-
нием. Количество ребер — около 30 на каждой створке; из них семь
ребер проходят по плоскому дну синуса, образующего язычок прямо-
угольного очертания, и восемь ребер лежат на плосковерхом седле спин-
ной створки.
Распространение. Кондратьевская толща Горного Алтая.
СЕМЕЙСТВО ATRyPIDAE GILL
Род Atrypa D а 1 m а п
Очень обширный род, из состава которого в последнее время выде-
лено значительное количество групп в качестве самостоятельных родов
или подродов.
Раковина неравностворчатая: брюшная створка от выпуклой до
почти плоской или слабо вогнутой в средней и передней части, спинная—
всегда выпуклая. Скульптура состоит из радиальных ребер или складок,
пересеченных знаками роста. Очень часто знаки роста имеют пластин-
чатый характер и в точках пересечения с радиальными ребрами и склад-
ками образуют чешуйки, бугорки, а иногда и шипы.
Типичный вид. Atrypa reticularis L i n п ё.
Распространение. Силур—девон, единичные находки из ниж-
него карбона. В Западной Сибири не поднимается выше границы между
франским и фаменским ярусами.
Atrypa umbra Barrande
Табл. LII, фиг. 7 и 8
Раковина маленькая, плоско-выпуклая, почти круглого очертания.
Брюшная створка слабо выпуклая, крышеобразная; спинная створка
почти плоская, снабженная отчетливым синусом. Поверхность створок
покрыта округленными радиальными ребрами в количестве 10—12 на
каждой створке.
Очень близка нижнедевонская Atrypa subleplda Vern., отличаю-
щаяся главным образом многочисленными и ясными знаками роста.
Распространение. Верхние горизонты S2 и переходные между
силуром и девоном отложения СССР и Западной Европы. В Западной
Сибири найдена в известняках с. Соловьиха (Горный Алтай).
Atrypa supramarglnalls К h а 1 f i n
Табл. LII, фиг. 9
Раковина изометричная, округленнопятистороннего очертания. Брюш-
ная створка слабо выпуклая. Синус начинается на некотором расстоянии
от макушки, к переднему краю сильно расширяется и углубляется,
образуя хорошо выраженный язычок; по дну синуса проходит одно
ребро. Спинная створка сильно и равномерно выпуклая. В пределах
седла располагаются 2—4 ребра. Боковые части створок покрыты не-
многочисленными крупными, округленными дихотомирующими ребрами,
в промежутках между которыми часто вставляются ребра второго по-
рядка.
От наиболее близкой верхнесилурийской Atrypa marginalls D а 1 m.,
обладающей более многочисленными ребрами, описываемый вид отли-
чается наличием лишь одного ребра в синусе.
Распространение. Переходные между силуром и девоном от-
ложения и низы нижнего девона. Горный Алтай, окрестности с. Со-
ловьиха.
240
Семейство spiriferidae kino
Род Eospirifer Schuchert
Замочный край короче наибольшей ширины, кардинальные углы
округленные. Арея брюшной створки треугольная, -вогнутая. Поверхность
створок нескладчатая или с радиальными складками, не распространяю-
щимися на седло и синус. Тончайшие радиальные струйки покрывают
всю наружную поверхность створок. В брюшной створке — хорошо раз-
витые зубные пластины.
Типичный вид. Eospirifer radiatus (Sow.).
Распространение. Силур—нижний девон, реже эйфельский
ярус, повсеместно.
Eospirifer pseudotogatus К h а 1 f i n
Табл. LIII, фиг. 1 и 2
Раковина крупная, поперечноэллиптического очертания, с умеренно
и одинаково выпуклыми створками. Длина замочного края составляет
3/5 ширины раковины. Арея умеренно высокая, треугольная, вогнутая.
Синус и седло начинаются от самых макушек. Радиальные складки
отсутствуют; радиальная струйчатость отчетливая.
Очень близкие виды, отличающиеся главным образом длинным замоч-
ным краем, распространены в верхнем силуре и нижнем девоне Запад-
ной Европы.
Распространение. Ганинская серия Горного Алтая.
Род Delthyris D а 1 m а и
Замочный край различной длины; призамочные углы от округленных
до оттянутых в остроконечия. Арея брюшной створки треугольная,
обычно высокая, вогнутая, редко почти прямая. Раковины обычно ра-
диально складчатые, складки немногочисленные, широкие и округленные,
незаходящие на седло и в синус. Знаки роста многочисленные, имеют
черепитчатый характер. Пластинки нарастания несут тончайшие радиаль-
ные струйки. В брюшной створке хорошо развиты септа и зубные пла-
стины.
Близкий к описываемому силурийский род Howellella К о z 1.
(см. стр. 164) отличается отсутствием свиты в брюшной створке. Род
Cyrtinopsls Scup. (см. стр. 251) отличается наличием в брюшной створке
спондилиума.
Типичный вид. Delthyris elevata D а 1 m.
Распространение. Силур—нижний девон, редко эйфельский
ярус, повсеместно.
Delthyris perlatnellosa Hall
Табл. LIII, фиг. 3—5
Раковина сильно развита по ширине, нередко с оттянутыми в остро-
конечия кардинальными углами. Арея умеренно высокая, вогнутая, резко
ограниченная. Седло и синус начинаются от макушек. Радиальные
складки широкие, округленные, присутствуют в количестве 4—6 по
каждую сторону от седла и синуса. Септа в брюшной створке высокая
и длинная.
Распространение. Нижняя половина нижнего девона Северной
Америки и Горного Алтая (ганинская серия и кондратьевская толща).
Spirtfer (Delthyris?) jaschei Roemer
Табл. LII, фиг. 10 и 11
Раковина развита по ширине, полуэллиптического очертания. Замоч-
ный край длинный, но призамочные углы закругленные. Брюшная створка
16 Зак. 2024 241
сильно выпуклая. Арея высокая, треугольная, слабо вогнутая, отчетливо
ограниченная. Синус начинается от макушки, неширокий, но достаточно
глубокий, с округленным дном. Спинная створка слабо выпуклая, с не-
широким округленным седлом начинающимся от макушки и ограничен-
ным глубокими вдавленностями. По каждую сторону седла и синуса рас-
полагается по две округленных радиальных складки.
Распространение. Нижнедевонская форма. Переходные между
силуром и девоном отложения Горного Алтая; нижний девон Салаира.
Имеются требующие проверки указания (Бубличенко, 1927) на присут-
ствие этого вида в верхнесилурийских отложениях горы Глядень близ
с. Сары-Чумыш (южная окраина Кузбасса).
Род Acrospirifer Wedekind
Раковина обычно крупных размеров, с длинным замочным краем,
полукруглого или поперечно полуэллиптического очертания. Арея брюш-
ной створки невысокая или умеренно высокая. Седло и синус сильно раз-
виты, гладкие. На боковых частях створок — широкие радиальные
^кладки. В брюшной створке сильно развиты зубные пластины.
Типичный вид. Acrosplrlfer prlmaevus (Stein.).
Распространение. Di—D2. Повсеместно.
Acrosplrlfer pseudofallax (К h a 1 f i n)
Табл. LIII, фиг. 7; табл. LIV, фиг. 2
Раковина округленного очертания, достигает крупных размеров, не-
сколько развита по ширине. Замочный край длинный, но несколько
меньше наибольшей ширины раковины. Боковые и передний края очер-
чены очень плавной кривой. Брюшная створка с умеренно высокой ареей
треугольного очертания. Макушка слегка загнута. Синус начинается от
макушки, глубокий и узкий, дно его слегка закруглено. Слабо выпуклая
спинная створка обладает высоким крышеобразным седлом. Радиальные
складки на боковых частях створок широкие, округленные, вполне от-
четливо выражены на спинной створке и часто еле заметны на брюшной
(внутренние ядра брюшных створок гладкие); по каждую сторону седла
лежат 3—5 радиальных складок.
Распространение. Широко распространен в верхнекобленцских
отложениях Горного и Рудного Алтая совместно с видом Acrosplrlfer
korovtnl (Kh а 1 f.) (табл. LIII, фиг. 9; табл. LIV, фиг. 1), который отли-
чается сильным развитием раковины по ширине.
СЕМЕЙСТВО RHYNCHOSPIRINIDAE SCHUCHERT
Род Rhynchospirina Schuchert et Le Vene
Раковины небольшие, удлиненноовальные, двояковыпуклые, с непря-
мым замочным краем и круглым фораменом на загнутой макушке
брюшной створки. Поверхность створок покрыта'радиальными ребрами.
Вещество раковины тонкопористое.
Типичный вид. Rhynchosplrtna fortnosa (Hall).
Распространение. Верхний силур—нижний карбон, повсе-
местно.
Имеется ряд внешне очень сходных родов , отличающихся деталями
внутреннего строения; из них в девоне Западной Сибири широко распро-
странен род Retzla King, представители которого обладают расщеплен-
ной трубкой в макушечной полости брюшной створки.
242
Rhynchosptrtna formosa (Hall)
Табл. LII, фиг. 12
Раковина небольшая, слегка развитая по длине, округленнопятиуголь-
ного очертания, с почти одинаково выпуклыми створками. Макушка
брюшной створки невысокая, загнутая, прободена большим фораменом.
О6р створки значительно выпуклы и снабжены неглубокими и узкими
синусами (на брюшной створке синус развит более отчетливо). Поверх-
ность створок покрыта округленными радиальными ребрами в количе-
стве от 16 до 23 на каждой створке.
От многочисленных и разнообразных представителей рода Retzia
King, встречающихся в девоне Западной Сибири, этот вид внешне от-
личается наличием продольных бороздок — синусов на обеих створках.
По этому признаку он приближается к некоторым представителям рода
Retzia из S2 и D! Западной Европы, но отличается от них или меньшим
количеством, или округленным (а не угловатым) характером радиаль-
ных ребер.
Распространение. Верхние горизонты S2 и низы Di Северной
Америки, Центральной Азии, Алтая. Переходные силуро-девонские отло-
жения Горного Алтая (известняк д. Соловьихи).
СЕМЕЙСТВО CENTRONELL1DAE WAAGEN
Род Rensselaeria Hall et Clarke
Раковина удлиненноовальная до изометричной, двояковыпуклая,
с непрямым замочным краем. Брюшная створка более выпукла, чем
спинная. Острая загнутая макушка прободена небольшим фораменом.
Седло и синус не развиты. Наружная поверхность створок радиально
ребристая. Вещество раковины тонкопористое.
Имеется ряд внешне близких родов, различающихся деталями вну-
треннего строения. В нижнедевонских отложениях Горного Алтая встре-
чены многочисленные мелкие раковины, условно отнесенные к этому
роду; их внешним отличием от типичных Rensselaeria является очень
слабое развитие радиальных ребер — до полной их редукции.
Типичный вид. Rensselaeria ovoldes (Eaton).
Распространение. Нижний девон, повсеместно.
Rensselaeria navicelllformis К h а 1 f i n
Табл. LIII, фиг. 8
Небольшая, почти изометричная раковина; наибольшей ширины до-
стигает ближе к замочному краю. Обе створки равномерно выпуклые,
брюшная более сильно, чем спинная, без всяких признаков седла и си-
нуса. Макушка брюшной створки длинная, острая, загнутая и нависаю-
щая над макушечной частью спинной створки. Скульптура состоит из
многочисленных (до 24 на каждой створке) простых, тонких радиальных
ребрышек, отчетливо выраженных близ краев створок и исчезающих уже
в их средней части. Вещество раковины тонкопористое.
Очень близка североамериканская Renss. mutabllls, отличающаяся
равновыпуклыми створками и более тонкой ребристостью.
Распространение. Низы ганинской серии Горного Алтая.
16*
243
М. А. РЖОНСНИ ИКАЯ
БРАХИОПОДЫ НИЖНЕГО И СРЕДНЕГО ДЕВОНА КУЗБАССА 1
СЕМЕЙСТВО STROPHOMENIDAE KING
Род Stropheodonta Hall1 2
Stropheodonta uralensis (V e г n e u i 1)
Табл. LIV, фиг. 3
Большая вогнуто-выпуклая, округленная или поперечновытянутая
форма. Брюшная створка сильно выпуклая, особенно в средней части.
Арея длинная, мелкозубчатая. Макушка большая, широкая, сильно
загнутая. Спинная створка вогнутая. Поверхность брюшной створки
в передней половине неправильно продольноволнистая. На хорошо
сохранившемся поверхностном слое видны тонкие слабо изгибающиеся
струйки и концентрические линии нарастания. На внутреннем слое ра-
ковины видны более крупные, округленные радиальные ребра.
Распространение. Средний девон Урала, Средней Азии, Новой
Земли и Вайгача. В Кузнецком бассейне встречается вместе с Concht-
diutn ex gr. pseudoba'schklricum Tschern. в эйфельских отложениях
юго-западной окраины.
Род Leptaena D а 1 m а п
Описание см. на стр. 235.
Leptaena rhomboldalls W i 1 с k е n s var. sinuata Rzonsnickaj a
Табл. LIV, фиг. 4
Крупная, почти равностворчатая трапецеидальная форма, с резким
коленообразным перегибом у переднего края. Замочный край прямой,
длинный, с оттянутыми ушками. Толщина раковины увеличивается по
направлению к переднему краю. Макушка маленькая, незагнутая за за-
мочный край. Арея длинная, узкая. У переднего края брюшной створки
развит отчетливо выраженный синусообразный изгиб, которому в спин-
ной створке соответствует широкое возвышение. Поверхность раковины
покрыта многочисленными радиальными струйками, пересеченными кон-
центрическими морщинами.
Отличается от Leptaena rhomboldalls W i 1 с к. (см. стр. 257) нали-
чием синусообразного изгиба у переднего края. Leptaena bouel Barr,
(см. стр. 236) от описываемого вида отличается наличием килеобразного
возвышения на брюшной створке и вдавленности посередине спинной
створки и иным профилем раковины.
Распространение. Распространенная форма в живетском
ярусе (слои с Indospirifer pseudowilltamst R z о п.) юго-западной окраины
Кузнецкого бассейна.
1 В этом разделе описаны брахиоподы (руководящие формы) из девонских отло-
жений, развитых по северо-восточному склону Салаира, т. е. слагающих юго-запад-
ную окраину Кузбасса. Такого же характера отложения с той же фауной развиты
местами и во внутреииих частях Салаира. Прим. ред.
2 Характеристика рода Stropheodonta: раковина вогнуто-выпуклая с длинным пря-
мым замочным краем и зазубренными ареями. Скульптура состоит из тонких радиаль-
ных ребрышек и струек. Прим. ред.
244
СЕМЕЙСТВО PENTAMERIDAE M'COY
Род Gypldula Hall
Описание см. на стр. 262.
Gypldula kayserl (Peetz)
Табл. LIII, фиг. 10
Средних размеров грубоскладчатая округленнопятиугольная форма
с сильно вздутой, изогнутой брюшной створкой и слабо выпуклой
округленной спинной. Макушка брюшной створки довольно большая,
загнутая. Посередине брюшной створки проходит возвышение, отчетливо
выраженное в передней половине створки. По срединному возвышению
проходят две крупные, округленные складки, а по середине синуса
спинной створки — одна резко выраженная складка; по бокам створок
расположено по две округленных складки, из которых на спинной
створке одна пара крупных складок ограничивает синус.
Отличается от Gypldula acutolobata (Sand b.) простыми, нерасщеп-
ляющимися складками.
Распространение. Встречена в крековских слоях жединского
яруса юго-западной окраиньи Кузнецкого бассейна; довольно распро-
страненная форма. Кроме того, известна из нижнего девона Германии.
Род Conchidium L i n п ё
Описание см. на стр. 162.
Conchidium ex gr. pseudobaschkiricum Tschernyschev
Табл. LV, фиг. 4 и 5
Крупная, вздутая, удлиненная, сильно неравностворчатая форма.
Брюшная створка сильно выпуклая, с длинной загнутой макушкой. Спин-
ная створка округленная, умеренно выпуклая. Синус и возвышение не
развиты. Поверхность раковины покрыта многочисленными, резкими,
плоскоокругленными, иногда раздваивающимися ребрами. Септа сред-
ней величины, достигает половины длины раковины, заканчивается
дугообразным изгибом1.
Распространение. Близкие формы пользуются широким рас-
пространением в среднем девоне Урала и Средней Азии (эйфельский
и живетский ярусы). В Кузнецком бассейне является руководящей фор-
мой для шандинских слоев (верхи эйфельского яруса), встречается также
в линзах известняка в эйфельских отложениях по р. Чумыш. Известна
из синхроничных отложений Центрального Салаира.
СЕМЕЙСТВО CAMAROTOECHIIDAE SCHUCHERT ET LE VENE
Род Camarotoechia Hall et Clarke
Описание см. на стр. 160.
Camarotoechia nympha pseudolivonica asiatica subsp. n.
Табл. LIV, фиг. 6
Средних и довольно больших размеров, вздутая, вытянутая в длину,
груборебристая форма. Брюшная створка почти плоская, с широким
и пологим синусом в передней части. Макушка маленькая, загнутая.
Спинная створка сильно вздутая, с круто падающими боками; возвыше-
ние широкое, округленное. Передний край изогнут в виде дугообраз-
ного язычка с резко зубчатым верхом. По бокам раковины в примаку-
1 Близкий по внешним признакам верхнесилурийский Conhidium knighti Sow.
(см. стр. 162) отличается более низкой и более длинной септой в брюшиой створке.
Прим. ред.
245
шечной части развиты довольно глубокие, сглаженные луночки. Поверх-
ность раковины покрыта 20 простыми ребрами, очень тонкими и пло-
скими в примакушечной части и грубыми, угловатыми у переднего края.
На боках спинной створки ребра дугообразно изогнуты.
Распространение. Встречается в эйфельском ярусе юго-запад-
ной окраины Кузнецкого бассейна. Типичная форма, пользуется широким
распространением в нижнем девоне Урала, Средней Азии, Богемии и Гер-
мании.
Camarotoechia lazutklnt Rzonsnickaja, sp. n.
Табл. LIII, фиг. 11; табл. LIV, фиг. 9
Небольшая округленнотреугольная, умеренно вздутая тонкоребристая
форма. Синус и возвышение развиты только в передней половине ство-
рок. Язычок средней высоты, дугообразный. Поверхность раковины по-
крыта очень тонкими округленнотреугольными, не дихотомирующими реб-
рами, по 30—40 на каждую створку. Ребра слегка расширяются по на-
правлению к переднему краю.
Camarotoechia bifera Р h i 11. отличается крылатой формой раковины
и дихотомированием ребер. Camarotoechia kayseri (Р а е с к.) обладает
еще более тонкими и многочисленными ребрами.
Распространение. Живетский ярус юго-западной окраины
Кузнецкого Бассейна — акарачкинские и сафоновские слои.
Род Uncinultts Bayle
Округленнопятиугольные, часто полукубические, вздутые формы
с прямым передним краем и приплюснутыми боками. Синус и возвыше-
ние более или менее хорошо развиты. Ребра многочисленные, плоско-
округленные, у переднего края разделенные короткими продольными бо-
роздками.
Внутри брюшной створки присутствуют хорошо развитые зубные пла-
стины; в спинной створке — септа с массивным замочным отростком
и неразделенная замочная пластина '.
Типичный вид. Unctnulus subwilsoni (Orbigny).
Распространение. Девон, преимущественно нижний и средний.
Широко распространенный род в СССР, Западной Европе, Азии и Аме-
рике.
Unctnulus dubroviensis Rzonsnickaja, sp. n.
Табл. LV, фиг. 1
Небольшая, округленнопятиугольная, сильно вздутая форма. Синус
ограничен крупными, округленными складками, раздваивающимися
у переднего края; посередине синуса почти от самой макушки проходит
крупное округленное ребро, а по бокам его еще по одному, более тон-
кому и короткому. Возвышение резко ограниченное, с продольной борозд-
кой посередине, разделяющей его на две складки, которые у переднего
края еще раз раздваиваются. Боковых ребер около пяти на каждую сто-
рону створки. У шва ребра вилкообразно расщепляются. Примакушечная
часть раковины гладкая.
Отличается от Unctnulus goldfussl (S с h п и г) более грубой ребри-
стостью и раздвоенным бороздкой возвышением. Unctnulus goldfussl
var. slgnata S c h n u г, которая имеет аналогичное раздвоенное возвы-
шение, отличается значительно более тонкой ребристостью.
Распространение. Сафоновские слои (известняки с Indospirifer
pseudowilliamsi R z о п.) жйветского яруса Кузнецкого бассейна.
1 Наиболее близкий род Hypothyridina Buckm. (см. стр. 248) отличается отсут-
ствием септы в спииной створке. Прим. ред.
246
Uncinulus tricostatus (Peetz)
Табл. LIV, фиг. 5
Небольшая, сильно вздутая, почти равностворчатая, округленно-
пятиугольная форма. Синус пологий, отчетливо выраженный только
в передней части створки. Язычок высокий, прямоугольный. Возвышение
невысокое, плоское, ясно ограниченное. Поверхность раковины покрыта
низкими, округленными ребрами, начинающимися от самой макушки.
Срединных ребер от трех до пяти, боковых 12 на каждую сторону створки.
От Uncinulus paralleleplpedus Bronn отличается более вздутой рако-
виной, меньшим числом ребер в синусе и отсутствием складок, ограни-
чивающих синус.
Распространение. Встречается совместно с предыдущим видом.
Uncinulus angularls (Phillips)
Табл. LIV, фиг. 8; табл. LV, фиг. 2
Небольшая, широкая, пятиугольная, умеренно вздутая форма с при-
тупленным передним краем. Синус широкий, ограниченный по бокам до-
вольно резкими складками. Спинная створка почти плоская; возвышение
невысокое плоскоокругленное, ограниченное от боковых частей глубо-
кими бороздками. Поверхность раковины покрыта резкими дихотомирую-
щими ребрами, начинающимися от самой макушки. Число срединных
ребер от четырех до шести, боковых насчитывается по 5—7 на каждую
сторону створки, причем все боковые складки отходят от складок, огра-
ничивающих синус.
Весьма близкий Uncinulus pentagonus Kays, отличается слабо раз-
витыми синусом и возвышением, большим количеством более тонких
ребер и совершенно ровным, прямым швом на переднем крае.
Распространение. Руководящая форма для живетокого яруса.
Встречается на Урале, в Средней Азии и в Западной Европе. В Кузнец-
ком бассейне пользуется широким распространением в сафоновских слоях
живетского яруса (известняки с Indospirifer pseudowilllamsi Rzon.).
Uncinulus subcordiformls (S c h n u r)
Табл. LV, фиг. 3
Небольшая, округленнопятиугольная, умеренно вздутая форма с при-
тупленными боками и передним краем. Синус широкий, глубокий, не-
резко ограниченный, с довольно высоким, но не вздернутым язычком.
Возвышение низкое, плоскоокругленное. Поверхность раковины покрыта
низкими сглаженными ребрами, которых в синусе и на возвышении от
восьми до 14 и по бокам каждой створки 12—14.
Распространение. Характерная руководящая форма для жи-
ветского яруса Урала и Западной Европы. В Кузбассе встречается
в сафоновских (индоспириферовых) слоях живетского яруса.
Uncinulus paralleleplpedus (Bronn)
Табл. LIV, фиг. 7
Небольшая, умеренно вздутая, округленнопятиугольная форма с при-
тупленными боками и передним краем. Синус резко отграниченный от
боков округленными складками. Язычок высокий, с праллельными бо-
ками и дугообразным верхом. Возвышение плоскоокругленное, отграни-
ченное от боковых сторон отчетливо выраженными бороздками. По-
верхность раковины покрыта тонкими плоскими ребрами: по 8—10
в синусе и на возвышении и по 12—15 на каждую сторону створки.
Отличается от близкого Uncinllus subcordiformls (Schnur) более
резко ограниченными синусом и возвышением и более резкими ребрами.
Распространение. Встречается в среднем девоне юго-западной
247
окраины Кузбасса, особенно многочисленна в отложениях эйфельского
яруса. Известна из эйфельского яруса Урала и Средней Азии, а также
из эйфельского и живетского ярусов среднего девона Западной Европы.
Род Hypothyridina Bu с k m а’п1
Hypothyrldlna procuboldes (Kayser)
Табл. LV, фиг. 6
Небольшая шарообразная форма с неодинаково выпуклыми створ-
ками. Брюшная створка почти плоская, с широким, пологим синусом,
изгибающим передний край в виде прямоугольного язычка. Спинная
створка сильно вздутая, с невысоким, слабо заметным возвышением.
Примакушечная часть гладкая, ребра довольно тонкие, развитые лишь
в передней половине раковины. Срединных ребер 7—8, боковых 7—10 на
каждой стороне створки.
Hypothyrldlna coronula D rev erm. отличается своеобразными зуб-
чатыми краями раковины.
Распространение. Живетский ярус юго-западной окраины
Кузнецкого бассейна, Урала, Западной Европы и средний девон Индо-
Китая.
СЕМЕЙСТВО ATRYPIDAE GILL
Род Atrypa D а 1 m а и
Описание см. на стр. 340.
Atrypa semiorbiotdes Lazutkin, sp. п.
Табл. LV, фиг. 11—13
Удлиненноовальная или округленная, почти равностворчатая, двояко-
выпуклая форма. Замочный край изогнутый, меньше наибольшей шири-
ны раковины. Брюшная створка более выпуклая, чем спинная, с наиболь-
шим вздутием в средней части. Макушка небольшая, загнутая за замоч-
ный край. Синус и возвышение почти не выражены. Радиальные ребра
округленные, более резкие в средней части и у переднего края, увеличи-
ваются в числе на различных расстояниях от макушки раздвоением
и появлением новых в промежутках между старыми. Концентрические
знаки нарастания многочисленные, пластинчатые. Внутри брюшной
створки имеются массивные зубы, отходящие от внутренней поверхности
боковых сторон и не поддерживающиеся зубными пластинами.
Распространение. Встречается в крековских известняках же-
динского яруса юго-западной окраины Кузнецкого бассейна.
Atrypa bodlnt М a n s и у
Табл. LV, фиг. 9 и 10
Небольшая, округленная, равностворчатая форма. Замочный край
короткий, изогнутый. Брюшная створка одинаково или немного менее
выпукла, чем спинная створка. Макушка небольшая, широкая, незагну-
тая. Передний край ровный или очень слабо изогнут в сторону спинной
створки. Поверхность раковины покрыта довольно грубыми, округлен-
ными, радиальными ребрами, которых насчитывается у переднего края
до 23—25 на каждой створке. Концентрические знаки нарастания много-
численные, чешуйчатые. Внутри брюшной створки присутствуют хорошо
развитые зубные пластины.
1 Диагноз рода Hypothyrldlna: раковина субкубическая или почти шарообразная;
широкий плоский сииус образует высокий язычок; простые радиальные ребра часто
иесут близ переднего края продольную бороздку; в брюшной створке — зубные пла-
стины, в спинной — разделенная замочная пластинка; ни в брюшной, ни в спинной
створках иет септы. Прим. ред.
248
Atrypa tubaecostata Paeck. (см. стр. 259) отличается от описывае-
мого вида прямым замочным краем, более плоской раковиной и отчет-
ливо выраженным синусом. Atrypa aspera Schloth. характеризуется
меньшим числом более грубых складок.
Распространение. Распространенная форма в живетских отло-
жениях Кузнецкого бассейна (слои с Indospirifer pseudowilliamsi
Rzon.) и Китая.
Atrypa subspinosa Lazutkin, sp. п.
Табл. LVI, фиг. 1
Средних размеров, почти равностворчатая, умеренно вздутая округ-
ленная раковина с почти прямым замочным краем. Брюшная створка
равномерно выпуклая. Макушка небольшая, выступающая над замоч-
ным краем, загнутая. Арея маленькая, треугольная; дельтириум прикрыт
с боков дельтидиальными пластинами, с круглым отверстием для выхода
ножки посередине. Синус пологий, широкий, заметный лишь вблизи пе-
реднего края. Спинная створка несколько более вздута, чем брюшная.
Возвышение неотчетливо выраженное.
Поверхность раковины покрыта довольно грубыми, округленными,
радиальными ребрами, увеличивающимися в числе по направлению к пе-
реднему краю дихотомированием и вставлением. Концентрические знаки
нарастания имеют вид правильных, широко расставленных округленных
складок, которые в местах пересечения с радиальными ребрами обра-
зуют бугорки. Кроме того, вся поверхность раковины покрыта тончай-
шими концентрическими струйками.
Отличается от Atrypa splnosa Hall менее многочисленными, более
грубыми ребрами и не черепитчатой, а складчатой концентрической
скульптурой.
Распространение. Распространенная форма в отложениях
эйфельского яруса юго-западной окраины Кузнецкого бассейна.
Род Karpinskia Tscherny'ch e’v’
Karplnskia conjugula Tschernyschev
Табл. LVII, фиг. 11
Небольшая, вздутая, сильно удлиненная, сжатая с боков радиально-
ребристая форма с трапецеидальным поперечным сечением. Брюшная
створка слабо выпуклая, плоская или слегка вогнутая. Макушка боль-
шая, острая, торчащая. Спинная створка сильно вздутая, с крутопадаю-
щими боками. Поверхность раковины покрыта довольно многочислен-
ными плоскоокругленными дихотомирующими ребрами, изгибающимися
на боках. Внутри брюшной створки присутствуют довольно толстые зуб-
ные пластины, в спинной створке — разобщенные замочные пластины
и низкий срединный валик.
Karplnskia fedorovl Tschern. отличается более тонкими и много-
численными ребрами и резко выраженной концентрической скульптурой.
Karpinskia consuelo Gort. характеризуется более вздутой раковиной
и более резкими и грубыми ребрами.
Распространение. Весьма характерная руководящая форма для
нижнего девона. Встречается в кобленцском ярусе юго-западной окраины
Кузнецкого бассейна. Известна из нижнего девона Урала, Средней Азии
и Карнийских Альп.
1 Диагноз рода: «Ринхонелловидные, двояковыпуклые, сильно вытянутые в длину
раковины с трапецеидальным поперечным сечением. Поверхность радиальиоребристая.
В спинной створке имеется срединная септа» (Циттель, Основы палеонтологии, 1934,
стр. 524). Прим. ред.
249
Род Carinatina N а 1 i v к i п
Широкая, плоская, почти равностворчатая раковина, округленного
или удлиненноовального очертания. Замочный край длинный, прямой,
с довольно высокой, резко ограниченной ареей. Брюшная створка с киле-
образным срединным возвышением; спинная створка более выпуклая,
с продольным синусом. У переднего края взрослых экземпляров имеется
шлейфообразное продолжение краев раковины, обычно изогнутое
в складку. Внутри брюшной створки присутствуют зубные пластины,
в спинной—разобщенные замочные пластины. Характер спиральных под-
держек такой же, как у Atrypa.
Типичный вид. Carinatina aritnaspa (Е i с h w.).
Распространение. Нижний и средний девон Европы и Азии.
В СССР — Урал, Средняя Азия и Кузбасс.
Carinatina eudokia Lazutkin, sp. п.
Табл. LVI, фиг. 2
Большая, плоская, полукруглая или поперечновытянутая форма,
с длинным прямым замочным краем и с широким шлейфом в передней
части. Макушка маленькая, широкая, острая. Арея длинная, высокая, тре-
угольная, с широким дельтириумом, закрытым двумя дельтидиальными
пластинами. Вдоль середины брюшной створки проходит слабое килеоб-
разное возвышение. Спинная створка со слабой вогнутостью посередине.
Поверхность створок до шлейфа покрыта 30 довольно грубыми, округ-
ленными дихотомирующими ребрами. Шлейф покрыт лишь концентриче-
скими линиями нарастания. У переднего края на-шлейфе имеется широ-
кая концентрическая складка.
Carinatina aritnaspa Е i с h w. отличается от описываемой формы
меньшим количеством более грубых ребер, более короткой ареей, ра-
диальноребристым шлейфом и более резко выраженным синусом и воз-
вышением.
Распространение. Встречается в верхнеэйфельокихизвестняках
с Conchidium ex gr. pseudobaschklrlcum T s c h e r n. юго-западной
окраины Кузнецкого бассейна.
СЕМЕЙСТВО SPIRIFERIDAE KING
Род Eospirifer Schuchert
Описание см. на стр. 241.
Eospirifer secans В а г г а п de
Табл. LV, фиг. 8; табл. LVI, фиг. 3—4
Средних размеров поперечно эллиптическая, умеренно вздутая форма.
Замочный край короче наибольшей ширины раковины, с округленными
углами. Боковые края острые, язычок умеренной высоты, округленно-
трапецоидальный. Брюшная створка умеренно вздутая, с слабо загну-
той макушкой и с довольно высокой резко ограниченной ареей. Синус
узкий и глубокий в примакушечной части, значительно расширяющийся
и выполаживающийся по направлению к переднему краю. В примаку-
шечной части синус ограничен двумя округленными складками, совер-
шенно не выраженными в передней половине раковины.
Посередине спинной створки проходит плоскоокругленное возвыше-
ние, ограниченное по бокам глубокими бороздками. Боковые складки
развиты только вблизи макушки, по 2—3 на каждую сторону створки.
Вся поверхность раковины покрыта тонкими радиальными струйками.
Внутри брюшной створки массивные зубные пластины.
Eospirifer secans (Ваг г.) отличается от близкого Eospirifer togatus
(Barr.) наличием складок в примакушечной части раковины, меньшей
250
вздутостью створок и острыми боковыми краями. Весьма близок Eospi-
rifer pseudotogatus Kha If. (см. стр. 241), который отличается от Eospi-
rifer secans (Barr.) отсутствием складок.
Распространение. Eospirifer secans (Barr.) встречен в отло-
жениях кобленцского и эйфельского ярусов юго-западной окраины Куз-
нецкого бассейна; кроме того, известен в кобленцском ярусе Урала.
Центрального Богемского массива и Германии и в нижнем девоне Кир-
гизии и Италии.
Род Cyrtinopsis Scupin
Крупных и средних размеров грубоскладчатые раковины, с более или
менее высоко!', вогнутой ареей. Синус и возвышение гладкие. Боковые
складки крупные, резкие, угловатые. Поверхность раковины покрыта
многочисленными, пластинчатыми концентрическими знаками нараста-
ния. Строени. вещества раковины неточечное.
Внутри брюшной створки имеется двойной спондилиум, образованный
соединением зубных пласт?»' срединной септой.
Типичный вид. Cytllnopsis undosus (Schnur).
Распространение. Нижний и средний девон (эйфельский ярус)
Кузбасса и Германии.
Cyrtinopsis nalivkini Rzonsnickaja
Табл. LVI, фиг. 7
Средних и довольно крупных размеров сильно неравностворчатая
форма с гладким синусом и возвышением. Замочный край длинный, со-
ответствующий наибольшей ширине раковины, с острыми замочными
углами. Язычок умеренной высоты, округленнотрапецоидальный. Брюш-
ная створка значительно больше спинной, округленнотреугольного очер-
тания. Макушка небольшая, слабо загнутая над высокой, треугольной,
вогнутой ареей. Дельтириальное отверстие сравнительно узкое, треуголь-
ное, открытое. Синус широкий, глубокий, округленно угловатый, по бокам
ограниченный двумя более крупными, чем боковые, округленными склад-
ками.
Спинная створка поперечновытянутая, трапецоидальная, слабо выпук-
лая. Возвышение резко ограниченное, широкое, низкое, плоское. Боковые
складки крупные, резкие, по 4—5 на каждую сторону створки.
Поверхность раковины покрыта многочисленными пластинчатыми
концентрическими знаками нарастания. Строение вещества раковины во-
локнистое. Внутри брюшной створки имеется спондилиум, образованный
слиянием зубных пластин и тонкой септальной пластины; у основания
спондилиума расположен трубкообразный отросток — тихоринум.
Описываемая форма отличается от Cyrtinopsis undosus, описанного
Шнуром из эйфельского яруса Германии, большим числом боковых скла-
док и более низкой ареей. Весьма близка форма из нижнего девона
Алтая, описанная Л. Л. Халфиным как Delthyrls perlatnellosa Con г. как
по внешним признакам, так и по внутреннему строению !.
Распространение. Cyrtinopsis nalivkini встречается в отложе-
ниях эйфельского яруса юго-западной окраины Кузнецкого бассейна.
Род Eoreticularia N а 1 i v k i п
Небольших и средних размеров, округленные или поперечноовальные
вздутые гладкие формы. Замочный край короткий, с округленными
углами. Арея вогнутая, часто неясно ограниченная. Синус и возвышение
развиты в различной степени. Поверхность створок покрыта многочислен-
1 Алтайские представители вида Delthyris perlamellosa Hall не имеют споиди-
лиума в брюшиой створке (см. табл. LIII, фиг. 4). Прим., ред.
251
ними тонкими, тесно расположенными концентрическими линиями без
игл, иногда пересеченными тонкой радиальной струйчатостью.
Зубные пластины, находящиеся в брюшной створке, иногда расхо-
дятся, иногда почти параллельные. Септа отсутствует.
Типичный вид. Eoreticularla indifferens (Barrand е).
Распространение. Силур — девон Европы и Азии.
Eoreticularla dereitnsl (О е h 1 е г t)
Табл. LV, фиг. 7
Небольших и средних размеров, умеренно вздутая гладкая форма,
поперечноовального, округленного или округленноромбоидального очер-
тания. Замочный край немного короче наибольшей ширины раковины,
с округленными углами. Макушка небольшая, загнутая. Арея невысокая,
треугольная, слегка вогнутая. Синус неширокий, ограниченный по бокам
слабо выраженными округленными складками. Возвышение узкое, невы-
сокое, ограниченное бороздками. Язычок невысокий, узкий, дугообраз-
ный. Поверхность раковины гладкая. Микроскульптура состоит из много-
численных, тонких концентрических линий. Иглы отсутствуют. Внутри
брюшной створки имеются хорошо выраженные, слегка расходящиеся
зубные пластины.
От Eoreticularla curvata (S ch loth.) отличается менее вздутой у пе-
реднего края раковиной, невысоким язычком и более резко ограничен-
ными синусом и возвышением.
Распространение. Эйфельский ярус Кузнецкого бассейна
(чумышская толща) и Испании.
Род Elythina Rzonsnickaja
Поперечноовальные или округленные раковины крупных и средних
размеров. Замочный край немного меньше наибольшей ширины, с округ-
ленными углами. Синус и возвышение гладкие. Боковые складки низкие,
широкие, иногда очень слабо выраженные.
Микроокульптура состоит из тонких удлиненных игл, разбросанных
в шахматном порядке по всей поверхности раковины. Концентрические
линии нарастания частые, особенно у переднего края. Внутри брюшной
створки развиты длинные и довольно массивные зубные пластины и низ-
кий срединный валик. Мускульный выступ имеет своеобразный рисунок
в виде елки, главный ствол которой соответствует впечатлениям замы-
кающих мускулов.
От рода Elytha Fred, отличается микроскульптурой и характером
мускульных отпечатков.
Типичный вид. Elythina salairica Rzonsnickaj а.
Распространение. Средний девон Западной Сибири: эйфель-
ский ярус Алтая, эйфельский и живетский ярусы юго-западной окраины
Кузнецкого бассейна.
Elythina salatrlca Rzonsnickaj а.
Табл. LVI, фиг. 5 и 6; табл. LVII, фиг. 3 и 4
Крупных и средних размеров, полукруглого или поперечноовального
очертания форма с гладкими синусом и возвышением. Замочный край
длинный, но немного короче наибольшей ширины раковины, с округлен-
ными замочными углами. Синус неглубокий, с округленным дном, не-
сколько расширяющийся к переднему краю. Возвышение округленное,
отчетливо ограниченное, довольно высокое. Складки низкие, широкие,
более резко выраженные на спинной створке; на брюшной створке они
очень слабо развиты и часто отсутствуют. По каждую сторону от синуса
и возвышения имеется по 5—6 складок.
252
Микроскульптура состоит из тонких удлиненных игл, расположенных
в шахматном порядке. Внутри брюшной створки присутствуют массив-
ные, довольно длинные расходящиеся зубные пластины и низкий средин-
ный валик. Мускульный бугор, на котором расположены отпечатки от-
мыкающих и замыкающих мускулов, имеет своеобразный рисунок в виде
елки, главный ствол которой соответствует отпечаткам замыкающих
мускулов, а ответвления — отмыкающих.
Сильно изменчивая форма, которая варьирует в очертаниях раковины
от удлиненной, округленной до сильно поперечновытянутой; кроме того,
варьирует характер возвышения: острое, округленное, с срединной
бороздкой посередине.
Весьма близкая Elyihina grlgorievi (Bub 1.) отличается более сильно
загнутой макушкой, наличием широких, округленных складок, ограничи-
вающих синус, и слабо выраженными складками на спинной створке.
Ранее данная форма определялась как Splrlfer undifer Roe m., от
которого она, однако, отличается более длинным замочным краем, более
узким синусом, иным характером мускульного ядра и микроскульптурой.
Распространение. Эйфельский ярус Кузнецкого бассейна
(песчано-сланцевая чумышская толща).
Род Indospirifer Grabau
Описание см. на стр. 269.
Indospirifer pseudowilliamsl Rzonsnickaja
Табл. LVII, фиг. 6—8
Средних размеров, округленнотреугольная, умеренно вздутая форма.
Замочный край короче наибольшей ширины раковины, с округленными
замочными углами. Макушка небольшая, узкая, загнутая. Арея средней
высоты, треугольная. Синус резко ограниченный, довольно широкий и
глубокий, с дугообразным язычком. Возвышение резко ограниченное, вы-
сокое, округленное. Ребра резкие, иногда у переднего края раздваиваются.
Число срединных ребер 3—5, боковых 7—8.
Вся поверхность раковины покрыта тонкими прерывистыми струйками,
веерообразно расходящимися из промежутков на ребра и пересекающи-
мися концентрическими линиями. Внутри брюшной створки присутст-
вуют хорошо выраженные расходящиеся зубные пластины.
Indospirifer padaukpinensls var. dagon (Reed) имеет простые боко-
вые ребра и в большем количестве (9—10). Indospirifer padaukpinensls
var. maoerhchuanensls Grabau отличается лишь простыми, не раздваи-
вающимися боковыми ребрами.
Распространение. Живетский ярус юго-западной окраины Куз-
нецкого бассейна (сафоновские слои).
Род Lazutkinia Rzonsnickaja
Небольших и средних размеров, сильно неравностворчатые, ребри-
стые или гладкие формы. Брюшная створка более длинная, значительно
выпуклая. Арея высокая, вогнутая, с большим открытым дельтириумом.
Арея спинной створки низкая, треугольная, с открытым нототириумом1.
Синус и возвышение отсутствуют или имеют вид узкой продольной
бороздки. Ребра довольно многочисленные, плоскоокругленные.
Внешний слой раковины покрыт тонкими многочисленными концент-
рическими струйками. Там, где внешний слой раковин несколько потерт,
видны тонкие радиальные струйки, заходящие и на ребра и в проме-
жутки. Внутри брюшной створки имеются слабо развитые, короткие
1 Нототириум — треугольное отверстие на арее спинной створки, аналогичное
дельтирнуму брюшной створки. Прим. ред.
253
зубные пластины, типа дельтириальных килей. В спинной створке при-
сутствуют массивный замочный отросток и круральные пластины, сходя-
щиеся ко дну створки. Септа отсутствует или слабо развита.
От рода Emanuella Grab а и отличается наличием зубных пластин
и отсутствием игл на внешней поверхности раковины.
Типичный вид. Lazutkinla mamontovensis (Lazutkin).
Распространение. Средний девон юго-западной окраины Куз-
нецкого бассейна.
Lazutkinla mamontovensis (Lazutkin)
Табл. LVII, фиг. 1 и 2
Небольших и средних размеров сильно неравностворчатая форма.
Брюшная створка значительно вздутая, удлиненная, с сильно пентамеро-
образно изогнутой примакушечной частью. Арея высокая, вогнутая,
с большим открытым дельтириальным отверстием. Синус в виде узкой
тонкой бороздки. Спинная створка полукруглого очертания без явно вы-
раженного возвышения, но ребра в средней части створки более крупные,
чем остальные.
Поверхность раковины покрыта тонкими плоскоокругленными реб-
рами в количестве 23—27 на каждую створку. Кроме того, на цельной
поверхности раковины имеются хорошо выраженные тонкие концентри-
ческие струйки; на нескольких потертых экземплярах заметны тончай-
шие радиальные струйки. Внутри брюшной створки присутствуют слабо
развитые, очень короткие зубные пластины типа дельтириальных килей.
В спинной створке имеется замочный отросток и круральные пластины.
Весьма своеобразная форма.
Распространение. Мамонтовские слои среднего девона юго-за-
падной окраины Кузнецкого бассейна.
СЕМЕЙСТВО CENTRONELLIDAE WAAGEN
Род Denckmanella Schuchert et Le Vene
Крупных и средних размеров удлиненноовальные или округленные,
вздутые, почти равностворчатые гладкие формы. Макушка маленькая,
загнутая, с круглым отверстием. Арея не развита. Поверхность створок
гладкая. Строение вещества раковины волокнистое. Внутри брюшной
створки имеются зубные пластины. Ручные поддержки длинные, петле-
видные.
Типичный вид. Denckmanella circularls (Holzapfel).
Распространение. Живетский ярус Кузбасса, Чукотского
полуострова, Урала, Средней Азии и Западной Европы.
Denckmanella damesl (Holzapfel)
Табл. LVII, фиг. 5
Гладкая форма средних и довольно крупных размеров, сильно или
умеренно вздутая, удлиненноовальная. Обе створки почти одинаково
выпуклы. Макушка небольшая, загнутая, иногда почти соприкасающаяся
с макушкой спинной створки. Передний край совершенно ровный; синус
и возвышение отсутствуют. Поверхность раковины гладкая. Строение
раковины волокнистое.
Отличается от Denckmanella circularls (Holz.) более вздутой и удли-
ненной раковиной.
Распространение. Руководящая живетская форма, распростра-
ненная в Кузбассе (керлегешские слои), на Урале, Чукотском полуострове
и в Западной Европе.
254
Род Chascothyris Holzapfel
Формы крупных и средних размеров, овальные или округленные,
двояковыпуклые, гладкие. Макушка небольшая, острая, широкая, торча-
щая. Арея неясно ограниченная, с большим открытым дельтириумом.
Синус и возвышение слабо развитые или отсутствуют. Строение вещества
раковины точечное. Внутри брюшной створки короткие, расходящиеся
зубные пластины. В спинной створке — петлевидные ручные под-
держки с длинным тонким отростком (pilum). Септа отсутствует в обеих
створках.
Т и п и ч н ы й в и д. Chascothyris barroisi Holzapfel.
Распространение. Живетский ярус Кузбасса, Средней Азии,
Урала и Западной Европы.
Chascothyris salairica Rzonsnickaja, sp. n.
Табл. LVII, фиг. 9 и 10; табл. LVIII, фиг. 1 и 2
Гладкая форма средних и довольно крупных размеров, умеренно
вздутая, удлиненноовальная или почти округленная. Брюшная створка
более выпукла, чем спинная, с небольшой широкой, острой или слабо
загнутой макушкой. Арея с большим треугольным дельтириумом. Спин-
ная створка округленная, слабо выпуклая, почти плоская. Синус и воз-
вышение отсутствуют. Поверхность раковины гладкая. Строение раковины
точечное. Внутри брюшной створки присутствуют тонкие, расходящиеся
зубные пластины.
От Chascothyris tschernyschewi Holzapfel отличается менее широ-
кой раковиной и более плоской спинной створкой.
Распространение. Характерная руководящая форма для кер-
легешских слоев живетского яруса юго-западной окраины Кузнецкого
бассейна.
В эйфельских отложениях Минусинской котловины (таштыпская
свита) массовым развитием пользуется вид Acrospirifer (?) subgregarlus
Rzonsnickaj а.
Acrospirifer (?) subgregarlus Rzonsnickaj a
Табл. LXIII, фиг. 11
«Небольших и средних размеров умеренно и значительно вздутые
раковины, преимущественно поперечноовального и реже удлиненного
очертания. Замочный край немного короче наибольшей ширины рако-
вины, с округленными замочными углами. Язычок средней высоты,
округленно-дугообразной формы.
Брюшная створка более сильно вздута, чем спинная, с наибольшей
выпуклостью в примакушечной части. Макушка значительно загнутая,
клювообразная. Арея треугольная, средней высоты, резко отграниченная
от боковых частей, с продольной и вертикальной штриховкой. Дельти-
риальное отверстие треугольное, открытое. Синус хорошо развитый,
с плоскоокругленным дном, ширина и глубина его варьирует, по бокам
он ограничен двумя, несколько более резкими, чем остальные, складками.
Спинная створка поперечноовального очертания, умеренно вздутая.
Макушка маленькая. Арея низкая, но отчетливо развита. Возвышение
довольно высокое, округленное, не широкое, отчетливо отграниченное от
боковых частей бороздками.
Поверхность раковины покрыта отчетливо выраженными округлен-
ными складками, разделенными друг от друга узкими бороздками. Число
складок у отдельных экземпляров варьирует от пяти до семи, большин-
ство форм имеет 5—6 складок, некоторые более крупные экземпляры
имеют семь и редко восемь складок с каждой стороны створок. Кроме
того, вся поверхность раковины покрыта многочисленными тонкими кон-
255
центрическими линиями нарастания, на которых заметны тончайшие
штрихи. Эти линии нарастания особенно многочисленны близ переднего
края, где они часто имеют пластинчатый вид.
Внутри брюшной створки присутствуют хорошо развитые довольно
тонкие зубные пластины». (М. А. Ржонсницкая. Сборник «Палеон-
тология и стратиграфия», 1952, стр. 147—148).
Р. Т. Г РА ЦИ АН ОБА
БРАХИОПОДЫ ВЕРХИ ЕЖИВЕТСКИХ ОТЛОЖЕНИИ
Верхнеживетская фауна Западной Сибири очень богата и свое-
образна. В ней совершенно отсутствуют руководящие формы широкого
географического распространения (Strlngocephalus burtlnl и др.), а роль
важнейшей характерной группы играет группа Splrlfer (Acrosplrlfer)
cheehlel Коп. (Халфин, 1937а), по которой вся эта фауна получила на-
звание чиэлевой фауны. Она распространена в различных районах Гор-
ного Алтая, в Минусинской котловине, по северо-западной окраине Куз-
басса. Из Минусинской котловины она описана А. Штукенбергом (Stuc-
kenberg, 1886), из Кузбасса — Л. Л. Халфиным (1937), из юго-восточ-
ного Алтая — Д. В. Наливкиным (1938), отнесшим ее к верхнему девону.
СЕМЕЙСТВО SCHIZOPHORHDAE SCHUCHERT
Род Schizophoria King
Раковина обычно крупная, округленных или округленноугловатых
очертаний, более или менее вздутая, обычно неравностворчатая. Замоч-
ный край короче наибольшей ширины раковины. Арея низкая, треуголь-
ная. Макушка брюшной створки небольшая, треугольная, загнутая или
прямая. Спинная створка вздута обычно более, чем брюшная. Поверх-
ность створок покрыта тонкими трубчатыми ребрами, иногда с полыми
шипами. Строение вещества створок точечное. В брюшной створке —
зубы и зубные пластины, в спинной — простой замочный отросток.
Типичный вид. Schizophoria resuplnata Martin.
Распространение. Силур—пермь, повсеместно.
Schizophoria strlatula (S с h 1 о t h e i m)
Табл. LVIII, фиг. 3
Раковина средних размеров, поперечноовальных или округленных
очертаний, вздутая. Брюшная створка менее вздутая, с короткой, не-
высокой, но хорошо развитой ареей. Синус и седло умеренно развитые,
не ограниченные. Поверхность створок покрыта тонкими, одинаковыми
ребрами, в области кардинальных углов изгибающимися параллельно
замочному краю.
Распространение. Широко распространенная как в вертикаль-
ном, так и в горизонтальном отношении форма. В Кузбассе встречается
в среднем и верхнем девоне.
Род Isorthis Kozlowski
Раковина двояковыпуклая, округленных очертаний. Спинная створка
выпукла несколько более брюшной и несет на себе слабый синус, кото-
рому на брюшной створке соответствует седло. В брюшной створке при-
сутствуют хорошо развитые зубы и зубные пластины. В спинной створке
имеется замочный отросток. Скульптура в виде тонких радиальных ребер.
Близкий род Dalmanella Hall et Clarke отличается почти пло-
ской спинной створкой и пучкообразным расположением ребер. Наиболее
близкий род Levenea S с h u с h. et Соор. см. стр. 234) отличается
более выпуклой брюшной створкой.
256
Типичный вид. Isorthls szajnochal Kozlowski.
Распространение. Верхний силур — нижний девон Европы и
Северной Америки. В Западной Сибири широко распространен в сред-
нем девоне.
Isorthls sibirica (К h а 1 f i n)
Табл. LVIII, фиг. 4
Раковина довольно крупная, округленночетырехугольных или попереч-
ноэллиптических очертаний; наибольшую ширину имеет посередине
длины. Створки выпуклы почти одинаково. Брюшная створка имеет
умеренно высокую, вогнутую арею, открытый дельтирий и плоское седло,
ограниченное слабыми вдавленностями. Спинная створка имеет очень
низкую прямую арею и синус. Поверхность створок покрыта тонкими,
округленными ребрами и редкими, грубыми концентрическими знаками
роста.
Распространение. Вид широко распространен в верхнеживет-
ских отложениях Кузбасса и Горного Алтая.
СЕМЕЙСТВО STROPHOMENIDAE KING
Род Schellwienella Thomas
Раковина выпукло-вогнутая, по форме, очертаниям и скульптуре
вполне аналогичная раковинам рода Schuchertella G i г t у (см. стр. 238),
но обладающая хорошо развитыми зубными пластинами.
Внешне к описываемому близок род Orthotetes Fisch., отличаю-
щийся наличием септы в брюшной створке.
Типичный вид. Schellwienella inflata (White et Whitfield).
Распространение. Верхний силур — нижний карбон. Повсе-
местно.
Schellwienella umbraculum (S с h 1 о t h е i m)
Табл. LVIII, фиг. 5 и 6; табл. LIX, фиг. 6
Раковина обычно крупная, полукруглых или округленно прямоуголь-
ных очертаний. Спинная створка выпуклая, брюшная вогнутая или
плоская. Замочный край равен наибольшей ширине раковины. Арея
брюшной створки довольно высокая, прямая, откинутая. Синус и седло
слабо развиты или отсутствуют. Иногда на спинной створке присут-
ствует продольная вдавленность. Поверхность створок покрыта тонкими,
высокими радиальными ребрами и резкими концентрическими знаками
роста. Радиальные ребра увеличиваются в количестве путем вставления.
Нижнедевонская Schuchertella altaica Kha If. (см. стр. 238) внешне
близка, но отличается более низкой ареей брюшной створки и отсут-
ствием зубных пластин.
Распространение. Девон, повсеместно (кроме Америки). Ука-
зания на присутствие этого вида в силуре сомнительны. В Западной
Сибири •— руководящая форма верхнеживетских отложений.
Род Leptaena D а 1 m а п
Описание см. на стр. 235.
Leptaena rhomboidalls W i I с k е n s
Табл. LVIII, фиг. 8
Раковина средних размеров, субквадратных очертаний, слегка вы-
тянутая в ширину, коленообразно изогнутая. Прямой замочный край
равен наибольшей ширине раковины. Брюшная створка слабо выпуклая,
с низкой ареей. Спинная створка плоская или слегка вогнутая. Поверх-
ность раковины покрыта очень тонкими радиальными ребрами. При-
17 Зак. 2024 257
макушечная часть до перегиба покрыта грубыми концентрическими мор-
щинами.
Отличия от нижнедевонской Leptaena bouel Barr, и нижнеживетской
Leptaena rhomdoldalis var. slnuata указаны в описаниях этих видов
(см. стр. 235 и 244).
Распространение. Девон, повсеместно. В Западной Сибири
этот вид является руководящим для верхнеживетских отложений.
СЕМЕЙСТВО CAMAROTOECHI1DAE SCHUCHERT ET LE VENE
Род Uncinulus В а у 1 е
Описание см. на стр. 246.
Uncinulus korovinl К h а 1 f i n
Табл. LVIII, фиг. 7
Раковина средних размеров, вздутая, сильно неравностворчатая,
округленнопятиугольных очертаний, слегка вытянутая в ширину. Седло
и синус развиты близ переднего края. Язычок высокий, плоский, с парал-
лельными боковыми краями. Поверхность створок покрыта плоскоокруг-
ленными ребрами, разделенными узкими промежутками. Ребра сглажи-
ваются в примакушечной части створок. Близ краев раковины на каждом
ребре появляется одна продольная бороздка. Число ребер в синусе и на
седле обычно равно четырем, часто трем и редко пяти. По каждую сто-
рону седла и синуса находится 6—9 ребер.
Близкий вид Uncinulus subslgnatus Reed отличается тем, что ребра
прослеживаются до самой макушки, синус и седло развиты уже в при-
макушечной части.
Распространение. Верхнеживетские отложения Кузбасса и Гор-
ного Алтая.
СЕМЕЙСТВО ATRYP1IDAE GILL
Род Atrypa D а 1 m а п
Описание см. на стр. 240.
Atrypa ex gr. reticularis L i n n ё
Табл. LIX, фиг. 1
Широкий вид Atrypa reticularis L i n n ё в последнее время разделен-
ия большое количество новых видов, причем название Atrypa reticularis
L i n п ё сейчас употребляется только для силурийских форм.
Девонские представители группы Atrypa reticularis Linn, отличаются
крупными размерами раковины, грубой сетчатой скульптурой, слабо вы-
пуклой брюшной и вздутой спинной створками и толстой, загнутой
макушкой.
Atrypa desquamata Sow. отличается более тонкой скульптурой и тор-
чащей макушкой.
Распространение. В Западной Сибири атрипы этой группы
широко распространены в среднем девоне и франском ярусе.
A trypa desquamata S о w е г b у
Табл. LIX, фиг. 2
Раковина крупная, удлиненноовальных или округленных, иногда суб-
квадратных очертаний. У молодых экземпляров обе створки слабо и оди-
наково выпуклы. Взрослые формы имеют более вздутую спинную створку.
Синус и седло отсутствуют или слабо заметны только возле переднего
края. Псевдоарея неясно ограниченная, низкая. Макушка брюшной
створки острая, высокая, торчащая. Ребра многочисленные, тонкие,
округленные, правильные, дихотомирующие. Концентрические знаки
роста редкие, линейные, слабо заметные.
258
Атрипы группы Atrypa reticularis L i n п ё отличаются более грубой
скульптурой и толстой, загнутой макушкой.
Распространение. Средний девон Западной Европы, Урала и
Средней Азии. Распространенная руководящая форма.
Atrypa tubaecostata Paeckelmann
Табл. LIX, фиг. 3 и 4
Раковина средней величины, с одинаково выпуклыми створками.
Макушка брюшной створки высокая, острая. Форамен и псевдоарея
хорошо видны. Замочный край прямой, короткий. Для вида характерна
скульптура, состоящая из трубкообразных ребер, разделенных широкими,
округленными промежутками, и многочисленных густых знаков роста,
представляющих собой концентрические пластинки, черепитчато налегаю-
щих одна на другую.
Atrypa desquamata Sow. отличается более тонкими ребрами и линей-
ным характером концентрических знаков роста.
Распространение. Средний и верхний девон Европы. В Запад-
ной Сибири очень широко распространена в верхнеживетских отложениях.
СЕМЕЙСТВО SPIRIFERIDAE KING
Род Acrosplrlfer W е d е k i п g
Описание см. на стр. 242.
Acrosplrlfer cheehiel К о п i п с к
Табл. LIX, фиг. 5; табл. LX, фиг. 1, 2, 5
Раковина большая, вытянутая в ширину, округленнотреугольных,
трапецеидальных или полуэллиптических очертаний. Створки сильно и
почти одинаково выпуклы. Замочный край равен наибольшей ширине
раковины. Арея низкая, с почти параллельными краями. Синус глубокий,
резко ограниченный, гладкий, с дугообразным язычком. Седло высокое,
узкое, округленнотреугольное, гладкое. По каждую сторону седла и
синуса находится 6—9 грубых, плоскоокругленных, тесно расположенных
ребер. Поверхность раковины покрыта резкими, пластинчатыми знаками
роста. При хорошей сохранности на краях пластинок роста видны мел-
кие шипики.
Распространение. Верхние горизонты среднего девона Запад-
ной Сибири и Юго-Восточного Китая. Распространенная руководящая
форма.
Род Theodossia Nalivkin
Раковина средних размеров, умеренно вздутая, округленных очерта-
ний. Замочный край короче наибольшей ширины раковины. Брюшная
створка более выпуклая. Арея треугольная, невысокая. Синус мелкий,
седло плоское, слабо ограниченное. Вся поверхность раковины покрыта
узкими, плоскими многочисленными ребрами. В брюшной створке
имеются зубные пластины.
От Cyriosplrifer N а 1. отличается округленными призамочными угла-
ми, короткой ареей, плоскими ребрами и отсутствием дельтириальной
пластины.
Типичный вид. Theodossia anossoffl (V е г п е u i 1).
Распространение. Средний девон и франский ярус Европы,
Азии и Америки.
Theodossia (?) schmtdti (Stuckenberg)
Табл. LIX, фиг. 7
Раковина слегка вытянутая в ширину, овальных очертаний. Замочный
край короткий, призамочные углы округленные. Створки почти одина-
17* 259
ково выпуклы. Макушка брюшной створки тонкая, загнутая. Арея не-
высокая, треугольная. Синус слабо ограниченный, с дугообразным языч-
ком. Седло довольно высокое. Ребра не дихотомирующие, высокие. Коли-
чество ребер в синусе и на седле равно семи, по каждую сторону седла
и синуса — 15, причем срединные и боковые ребра одинаковы по вели-
чине.
Theodossia anossovl V е г п. отличается толстой, сильно загнутой
макушкой брюшной створки и большим количеством плоскоокругленных
ребер.
Распространение. Верхнеживетские отложения Минусинской
котловины.
Род LamelUspirifer N а 1 i v k i п
Раковина средних размеров, треугольных или трапецеидальных очер-
таний, сильно вытянутая в ширину. Арея желобообразная, заканчиваю-
щаяся шипами. Синус и седло иногда гладкие, часто со срединными реб-
ром и бороздкой, по бокам которых порой появляются дополнительные
ребра и бороздки. Ребра резкие, грубые, округленные, простые. Харак-
терны резкие, частые, пластинчатые знаки роста. Иногда заметна радиаль
ная струйчатость. В брюшной створке присутствуют расходящиеся зуб-
ные пластины.
Типичный вид. LamelUspirifer mucronatus (Hall).
Распространение. Средний и верхний девон. Географически
распространен широко.
LamelUspirifer mucronatus (Conrad)
Табл. LX, фиг. 3 и 4
Раковина средних размеров, умеренно вздутая, треугольных очерта-
ний, сильно вытянутая в ширину. Арея низкая, желобообразная, заканчи-
вающаяся шипами. Синус и седло узкие, резко ограниченные, гладкие.
По каждую сторону седла и синуса находится 8—15 простых, тесно рас-
положенных, округленноугловатых ребер. Концентрические знаки роста
густые, пластинчатые, черепитчато налегающие друг на друга.
Верхнедевонские LamelUspirifer ales К h а 1 f. и LamelUspirifer mucro-
natlformls Kha If. (см. стр. 266) отличаются, прежде всего, расчленен-
ным седлом и наличием ребер в синусе.
Распространение. В Западной Сибири (Алтай) — руководящая
форма для самых верхов D2.
Род Platyrochella Fenton et Fenton
Раковина довольно крупная, обычно вытянутая в ширину, субтра-
пецоидальных очертаний. Замочный край равен наибольшей ширине
раковины, арея обычно высокая, треугольная, прямая или вогнутая.
Синус и седло хорошо развитые, гладкие, с присутствующей иногда на
склонах зачаточной ребристостью. Вся поверхность покрыта тончайшими
радиальными струйками или рядами очень мелких бугорков. Радиальные
ребра простые, округленные. В брюшной створке присутствуют зубные
пластины и дельтириальная пластина, закрывающая верхнюю часть
дельтирия.
Близкие по форме представители рода Brachyspirifer W е d е к. отли-
чаются совершенно гладкими седлом и синусом и отсутствием дельти-
риальной пластины.
Типичный вид. Platyrochella maebridei (Calvin).
Распространение. Средний и верхний девон Сев. Америки,
средний девон Западной Сибири.
260
Platy rochella tnartianovi (Stuckenberg)
Табл. IL, фиг. 5; табл. LVIII, фиг. 9; табл. LX, фиг. 6
Раковина довольно большая, вытянутая в ширину, полуэллиптических
очертаний. Створки выпуклы значительно и почти одинаково. Замочный
край равен наибольшей ширине раковины. Брюшная створка имеет
прямую или слегка загнутую макушку и высокую, треугольную, хорошо
ограниченную, прямую или несколько вогнутую арею. Синус и седло
хорошо развитые, гладкие. По каждую сторону седла и синуса находятся
от 12 до 20 простых, округленных ребер.
Похожие по внешней форме верхнедевонские Cyrtospirifer tentlcu-
lutn (Ver n.) и Cyrtospirifer verneuill (Mure h.) отличаются полной
ребристостью седла и синуса.
Распространение. Вид широко распространен в Западной Си-
бири и является руководящим для верхнеживетских отложений.
Platyrochella klzylschlnlca G г a t i a nov a, sp. n.
Раковина средних размеров, вытянутая в ширину, полуэллиптических
или субтреугольных очертаний. Замочный край равен наибольшей ширине
раковины. Створки выпуклы довольно значительно. Макушка брюшной
створки слегка загнута над высокой, хорошо ограниченной, треугольной,
несколько вогнутой ареей. Синус глубокий, широкий, с невысоким, дуго-
образным язычком. Седло широкое, высокое, хорошо ограниченное. На
склонах седла и синуса находятся по два или три радиальных ребра;
верх седла и дно синуса гладкие. По каждую сторону седла и синуса на-
ходятся 18—22 тонких, простых, округленных ребра.
Наиболее близкий вид — Platyrachella martjanovl (Stuck.) отли-
чается отсутствием ребер на седле и в синусе.
Распространение. Встречается в верхнеживетских отложениях
Юго-Восточного и Центрального Алтая.
В. А. И В АН И Я И Л. Н. КРАЕВСКАЯ
БРАХИОПОДЫ ВЕРХНЕГО ДЕВОНА
Фауна брахиопод Западной Сибири претерпела на протяжении верх-
него девона ряд достаточно резких изменений, позволяющих выделить
в ее составе несколько ассоциаций, характеризующих различные страти-
графические горизонты. Основание франского яруса охарактеризовано
фаленовой (или супрафаленовой) фауной, названной так по важнейшей
руководящей форме — Anathyris phalaena (Phil 1.). В этой фауне на-
ряду с большим количеством местных форм имеется достаточно много
европейских видов. Характерно для нее также наличие ряда форм, кото-
рые, или ближайшие аналоги которых, в других странах встречаются
в более древних отложениях. Эта фауна описана в работах И. П. Толма-
чева (1915), Л. Л. Халфина (1931, 1932) и В. К. Халфиной (1940).
Среднефранские отложения характеризуются фауной, наиболее типич-
но представленной в глубокинском известняке Кузбасса, аналоги которого
развиты также в Рудном Алтае (близ с. Николаевского) и в районе
г. Камня на р. Обь. Эта фауна характеризуется такими видами, как
Gypidula comis (Owen), Gypidula obi Kr., Hypothyrldtna cuboldes
(Sow.), Pugnax acuminata (Mart.), Cranaena romlngerl Hall и др.
Она описана в работе Л. Н. Краевской (1936).
Для верхнефранских отложений Кузбасса характерна так называемая
монстровая фауна; она состоит почти целиком из местных форм, из
которых важнейшими являются группа Anathyris monstrum К h а 1 f.
и вид Cyrtospirifer ussovi К h а 1 f. Описана в работе Л. Л. Халфина
(1933).
‘261
Фауна фаменских отложений Западной Сибири известна под назва-
нием прелонгусовой по имени наиболее важной группы из ее состава —
Productus praelongus Sow. Она распространена в Кузбассе и Рудном
Алтае, в последнее время обнаружена в Горном Алтае (Грацианова,
1950). Описана в работе Л. Л. Халфина (1933).
Для детальной стратиграфии верхнедевонских отложений Западной
Сибири особенно важны представители родов Anathyrls Peetz, и Суг-
tosplrlfer N al.; их описанию посвящены работы Л. Л. Халфина (1933а)
и В. А. Ивании (1948).
Нижеследующие описания руководящих форм сделаны Л. Н. Краев-
ской, за исключением рода Cyrtosplrlfer, представители которого описаны
В. А. Ивания.
СЕМЕЙСТВО PRODUCT1DAE GRAY
Род Plicatifera S о w е г b у
В девонских отложениях Азии, в частности Западной Сибири, распро-
странена группа продуктусов, известная под названием Productus
praelongus So w., в состав которой входят Pr. vlangalll, Pr. melsterl и др.
В последнее время эта группа относится к роду Plicatifera.
Внешне это небольших и средних размеров формы с хорошо разви-
тыми ушками, с тупой, широкой, загнутой макушкой. Примакушечная
часть покрыта неправильными концентрическими морщинами. Поверх-
ность раковины радиальноскладчатая или бугорчатая.
Типичный вид. Plicatifera pllcatllls (Sow.).
Распространение. Верхний девон - карбон, повсеместно.
Plicatifera praelonga (S о w е г b у)
Табл. LX, фиг. 7—10
Раковина почти прямоугольной формы. Замочный край равен или не-
сколько длиннее наибольшей ширины раковины. Брюшная створка
сильно вздутая, с узенькой ареей, не всегда заметной под круто загнутой
макушкой. Посередине синуса, имеющего характер широкой и плоской
вдавленности, лежит округленное ребро (валик), не всегда отчетливо
выраженное. Скульптура состоит из тонких, коротких, прерывистых
радиальных ребрышек, которые представляют собой сильно удлиненные
основания тонких шипов, обычно не сохраняющихся (обломанных). Про-
дольный ряд значительно более толстых шипов располагается вдоль
линии симметрии, на срединном валике. Концентрические морщины по-
крывают ушки и примакушечную часть раковины.
Наиболее важным отличительным признаком вида является наличие
ряда крупных бугорков (основания шипов) на срединном валике.
Распространение. Фаменский ярус и переходные от девона
к карбону отложения Кузбасса, Горного и Рудного Алтая, Казахстана,
Западной Европы.
СЕМЕЙСТВО PENTAMERIDAE M’COY
Род Gypldula Hall
Небольших размеров, вздутые, округленные, неравностворчатые рако-
вины шлемообразной формы. Более выпукла брюшная створка с четко
заметным возвышением. Спинная створка менее выпукла, с синусом, об-
разующим иногда у переднего края язычок. На брюшной створке нахо-
дится чуть заметная арея, которая иногда отсутствует. Поверхность
створок гладкая или радиальноскладчатая. Для внутреннего строения
характерны расходящиеся септальные пластины в спинной створке
и короткий спондилиум в брюшной створке.
262
Близкий род Sleberella О е h 1 е г t отличается наличием в спинной
створке короткой септы, образованной сходящимися септальными пла-
стинами.
Типичный вид. Gypidula occldentalls (Hall).
Распространение. Верхний силур — верхний девон, повсе
местно.
Gypidula comls (Owen)
Табл. LX, фиг. 14
Раковина средних размеров, округленная, неравновыпуклая, с корот-
ким и непрямым замочным краем. Наибольшая вздутость в примакушеч-
ной части. На спинной створке имеется синус, образующий у переднего
края вздернутость в виде язычка. Поверхность покрыта грубыми корот-
кими (заметны лишь близ переднего края) складочками в количестве от
трех до шести на каждой створке.
Близкий по общей форме раковины вид Gypidula globosa (S с h n и г)
отличается отсутствием складочек.
Распространение. Верхний девон Северной Америки, Европы
и Азии. В Западной Сибири — характерный вид для глубокинского гори-
зонта верхнего девона (Кузбасс, Рудный Алтай, район города Камня на
р. Обь).
Gypidula obi Krajevskaia
Табл. LXI, фиг. 1
Гладкие, небольших размеров, умеренно вздутые, почти равновыпук-
лые, поперечновытянутые раковины. Лобный край прямой или слабо
изогнут в сторону брюшной створки. Брюшная створка вздута не-
сколько больше спинной, возвышение четко не отграничено, арея вы-
ражена слабо. Спинная створка слабо вздута, у взрослых экземпляров
почти плоская. Синус ясно не отграничен, слабо намечаясь у лобного
края. В брюшной створке имеется короткая срединная септа.
Наиболее близкие виды «Pentamerus» Integer Barr, и «Pentame-
rus» glaber T s c h.; от них Gypidula obi отличается очень небольшими
размерами. По общей форме данная форма напоминает вышеописанную
G. comls, от которой отличается отсутствием складок, несколько мень-
шими размерами и меньшей вздутостью раковины.
Распространение. Западная Сибирь, совместно с предыдущим
видом.
СЕМЕЙСТВО CAMAROTOECH1IDAE SCHUCHERT ET LE VENE
Род Hypothyridina В u с k m а п
Округленнокубическая форма раковины. Возвышение и синус в при-
макушечной части слабо заметны, но у лобного края выражены отчет-
ливо. Бока обеих створок крутые, почти вертикальные. Шов острозубча-
тый. Поверхность обеих створок покрыта плоскими, широкими склад-
ками, разделенными у лобного края продольной бороздкой.
От близких по внешним признакам родов Unctnulus King. (см.
стр. 246) и Wilsonella Niki ph. (см. стр. 161) отличается отсутствием
септы в спинной створке.
Типичный вид. Hypothyridina cuboldes (S о w.).
Распространение. Средний и верхний девон, повсеместно.
Hypothyridina cuboldes (S о w е г b у)
Табл. LXI, фиг. 2; табл. LXII, фиг. И
Вид обладает вышеуказанными признаками рода. Количество средин-
ных складок у него более пяти, чем он отличается от сходной по внеш-
нему виду Hypothyridina venustula (Hall).
263
Распространение. Франский ярус, почти повсеместно. В За-
падной Сибири главным образом глубокинский горизонт.
Hypothyrldlna tschingisica Krajevskaia
Табл. LX, фиг. 11 и 12
Раковина небольшая, вздутая, округленнопятиугольная, неравноствор-
чатая, с почти прямым замочным краем. На умеренно вздутой брюшной
створке синус средней ширины, не очень глубокий, начинающийся близ
макушки. Высоко вздернутый язычок перпендикулярно отогнут, с парал-
лельными боками и плоским верхом. Спинная створка более вздутая. Не-
высокое возвышение, начинаясь от самой макушки, постепенно рас-
ширяется, у лобного края вздернуто. Вся поверхность раковины покрыта
многочисленными, средней толщины, округлыми ребрами, доходящими до
макушки.
По общей форме близка к Н. cubold.es и Н. venustula. Отличается от
них размерами, количеством складок, угловатостью язычка. Очень близка
к И. cuboldes var. semlluklana N а 1. по размерам и очертаниям, отли-
чаясь более высоким и узким язычком, характером и количеством ребер.
Распространение. Один из характерных видов глубокинского
горизонта Западной Сибири.
Род Pugnax Martin
Раковины средних размеров, вздутые или уплощенные, неравноствор-
чатые, различных очертаний. Поверхность створок гладкая или складча-
тая. В брюшной створке две короткие зубные пластины. В спинной
створке септы нет.
Типичный вид. Pugnax acuminata (Martin).
Распространение. Девон—пермь, повсеместно.
Pugnax acuminata (Martin)
Табл. LX, фиг. 13
Раковина сильно вздутая, имеет форму трехгранной пирамиды. Резко
изогнутая брюшная створка отличается широким синусом. Язычок под-
нимается вверх почти перпендикулярно и оканчивается более или менее
острой дугой. Спинная створка вздутая, двускатная, с наибольшим воз-
вышением посередине створки. Поверхность раковины гладкая или
имеются зачаточные складки в синусе у лобного края.
Распространение. Средний девон — верхний карбон в Запад-
ной Европе. В Сибири — франский ярус (главным образом глубокинский
горизонт).
Pugnax voroni N а 1 i v k i n
Табл. LXI, фиг. 3
Раковина маленькая, вздутая, по облику напоминающая трехгранную
пирамидку. Брюшная створка плоская, сильно изогнутая, с широким и
довольно глубоким синусом. Язычок резко вздернут вверх и заканчи-
вается острым углом. Спинная створка сильно вздутая, двускатная,
с острым возвышением посередине. Поверхность раковины гладкая,
только в синусе имеются 2—3 коротких складочки.
Распространение. Франский ярус Центрального девонского
поля Европейской части Советского Союза. В Западной Сибири — один
из распространенных видов глубокинского горизонта.
СЕМЕЙСТВО TEREBRATULIDAE GRAY
Род Cranaena Hall et Clarke
Раковина теребратулоидного облика, округленнопятиугольного очерта-
ния, небольших и маленьких размеров. Синус или синусовидная вдавлен-
264
ность имеется или отсутствует. Поверхность створок гладкая, имеются
концентрические знаки роста. Строение вещества створок тонкопористое.
Имеется ряд внешне близких родов, различающихся деталями внутрен-
него строения.
Типичный вид. Cranaena rotningeri Hall.
Распространение. Девон — карбон.
Cranaena rotningeri Hall
Табл. LXI, фиг. 9
Раковина пятиугольная, большей частью умеренно и равномерно вы-
пуклая, удлиненная; длина иногда равна ширине. Размеры средние, чаще
маленькие. Передний край прямой или с выемкой, изгибающийся в сто-
рону спинной створки. Створки выпуклы одинаково, иногда брюшная
створка более выпукла, чем спинная. Макушка брюшной створки слабо
удлиненная, загнутая, несет форамен. От лобного края идет небольшая
вдавленность, которая доходит или почти доходит до средины створки.
Спинная створка мало отличается от брюшной, являясь, естественно, не-
сколько меньшей по длине. Имеется также синусовидная вдавленность,
не доходящая до макушки. Поверхность обеих створок гладкая, часто по-
крыта концентрическими знаками роста.
Распространение. Верхи среднего девона Северной Америки;
глубокинский известняк Западной Сибири.
СЕМЕЙСТВО SPIRIFERIDAE KING.
Род Eoretlcularia N а 1 i v k i п1
Описание см. на стр. 251.
Eoretlcularia (?) microsplnosa К h а 1 f i n
Табл. LXI, фиг. 5
«Крупная, почти изометричная, слегка развитая по ширине, округлых
очертаний раковина; замочный край составляет */з—2/з максимальной
ширины. Брюшная створка более выпукла, чем спинная. Вентральная
арея невысокая, треугольная, вогнутая; зубные пластины хорошо развиты.
Синус начинается вблизи макушки, плоский, с округленным дном; седло
невысокое. Очень плоские и глубокие, исчезающие у кардинальных углов
радиальные складки лежат на боковых частях створок (4—5 по каждую
сторону седла и синуса). Поверхность створок с очень тонкими и густыми
пластинками роста, по свободным краям которых сидят тончайшие
шипики» (Халфин, 19506, стр. 14).
Наиболее близок Splrifer sublaevlgatus V К h а 1 f. из нижнефранских
отложений Кузбасса; отличается большим количеством более отчетливых
складок и плоскодонным синусом. По форме раковины и характеру
складчатости близок среднедевонский Splrifer undlfer R о е m.
Распространение. В массовом количестве присутствует
в аскольдовском песчанике (основание франского яруса) близ с. Зару-
бино (северо-западная окраина Кузбасса).
’ Автор этого рода (Д. В. Наливкин, 1930, стр. 133—135) отнес к нему весьма
различные формы (в частности, Splrifer undlfer Roem., Splrifer simplex Ph ill).
Если объем рода ограничить спириферами типа Splrifer indlfferens Barr, и счи-
тать характерным для него отсутствие шипов на пластинках роста, то формы, подоб-
ные Eoretlcularia microsplnosa, должны быть исключены из его состава и, повиди-
мому, выделены в особый род, который отличается от рода Elytha Fred, простыми
(р не двойными) иглами и отсутствием септы в брюшной створке, а от рода
Elythina Rzonsn. (см. стр. 252)—характером микроскульптуры и отсутствием
в брюшной ствопке срединного валика. Прим. ред.
265
Род LamelUspirifer N a I i v к i n
Описание см. на стр. 260.
LamelUspirifer mucronatlformis К h a 1 f i в
Табл. LXI, фиг. 7
Раковина прямоугольного или трапецеидального очертания, с сильно
и почти одинаково выпуклыми створками. Призамочные углы оттянуты
в остроконечия. Макушка брюшной створки невысокая, острая, сильно
загнутая; низкая арея ограничена вполне параллельными краями. Синус
начинается от самой макушки, ограничен рельефными ребрышками, глу-
бокий; в синусе лежат обычно три, реже одно или пять ребер. Седло
спинной створки невысокое, но ограничено резкими и глубокими борозд-
ками, расчленено соответственно ребристости синуса. По каждую сторону
седла и синуса располагаются 8—12 округленных, всегда простых
радиальных ребрышек. Знаки роста густые, пластинчатые.
Среднедевонский LamelUspirifer mucronatus (Con г.) (см. стр. 260)
отличается нерасчлененным седлом и гладким (без ребер) синусом.
Нижнефранский LamelUspirifer ales К h а 1 f. обладает более развитой по
ширине раковиной, большим количеством ребер, слабо выпуклыми
створками и очень высоким седлом.
Распространение. Одна из наиболее распространенных форм
промежуточной толщи (верхи франского яруса) северо-западной окраины
Кузбасса.
Род Cyrtospirifer N а 1 i v k 1 п
Это очень важная группа спириферов, в составе которой имеется ряд
руководящих форм.
Постоянными диагностическими признаками рода, выдерживаю-
щимися у всех его видов, являются: присутствие в примакушечной части
брюшной створки дельтириальной пластины, соединяющей зубные пла-
стинки, и скульптура из радиальных плоскоокругленных ребер, простых
на боковых частях раковины и дихотомирующих в синусе и на седле.
Признаками непостоянными, сильно изменяющимися у различных видов
являются: характер ареи (ее ширина, высота, степень вогнутости); глу-
бина синуса и высота седла; очертания раковины.
Типичный вид. Cyrtosplrlfer verneulll (М u г с h.).
Распространение. Главным образом верхний девон, редко
верхи среднего девона и переходные от девона к карбону отложения. По-
всеместно.
Cyrtosplrlfer achmet N а 1 i v k i n
В литературе по геологии Западной Сибири часто фигурирует под названием
Spirifer disjunctus Sow.
Табл. LXI. фиг. 6; табл. 1L, фиг. 4
Раковина крупная, субтрсугольная, сильно поперечновытянутая (из-
вестны экземпляры с длиной раковины 2 см и шириной 10 см). Арея
низкая, желобообразная, с параллельными краями. Углы ее оттянуты
в острые ушки. Синус узкий и резкий. Седло узкое, резкое и слабо вы-
пуклое.
От сходного по низкой параллельнокрайной арее вида Cyrtosplrlfer
disjunctus Sow. из франских отложений Европейской части Советского
Союза и Западной Европы отличается субтреугольными очертаниями
раковины, более сильно развитой по ширине.
Распространение. Нижнефранские отложения Западной Си-
бири и Казахстана.
266
Cyrtospirifer curvirostris I van i a
Табл. LXI, фиг. 4
Раковина крупная, трапецоидальная, поперечновытянутая, сильно
вздутая в примакушечной части. Макушки довольно крупные, сильно
клювообразно загнутые, почти соприкасающиеся. Замочный край прямой
и длинный, равен наибольшей ширине раковины. Арея очень длинная,
низкая, с параллельными краями, резко ограниченная; отношение ее
высоты к длине 1 : 8. Синус ясный, имеющий у макушки вид узкой и до-
вольно глубокой бороздки, сильно расширяется и уплощается у лобного
края, где ширина его почти равна ширине боковой части раковины.
Седло невысокое, резко ограниченное, дугообразное в поперечном сече-
нии.
Очень вздутая лишь в умбональной части раковина, довольно круп-
ные, сильно клювообразно загнутые, почти соприкасающиеся макушки,
низкая параллельнокрайная арея — признаки, отличающие этот вид от
всех близких видов.
Распространение. Фаменские отложения Кузбасса.
Cyrtospirifer verneuili (Murchison)
Табл. LXI, фиг. 11
Раковина средних размеров или довольно крупная, трапецоидальная
или полукруглая, умеренно выпуклая. Замочный край прямой и длинный,
с острыми углами, равный наибольшей ширине раковины. Арея треуголь-
ная, умеренно высокая (отношение ее высоты к длине от 1 5 до 1 :3).
Синус и седло резкие. Скульптура обычная для рода.
Сибирские представители этого вида отличаются большой изменчи-
востью в отношении глубины и ширины синуса, высоты седла и харак-
тера ребристости; на основании этих вариаций выделяется ряд разновид-
ностей данного вида (Ивания, 1948).
По очертаниям раковины, характеру синуса и скульптуре сходен
с Cyrtospirifer disjunctus (Sow.) из франских отложений Европейской
части СССР и Западной Европы, от которого легко отличается треуголь-
ной ареей. Отличия от Cyrtospirifer archiaci (Mu г ch.) указаны в описа-
нии последнего.
Распространение. Широко распространен в фаменских отло-
жениях, изредка встречаясь и в франских. Кузбасс.
Cyrtospirifer barzassensis Т у z h п о v, sp. n.
Табл. LXI, фиг. 10
Раковина довольно крупная, широкотреугольная, поперечновытяну-
тая. Длинный прямой замочный край с острыми углами равен наиболь-
шей ширине раковины. Арея треугольная, довольно высокая. Синус
уплощерный, посередине его проходит резкая острая складка или киле-
видное ребро, бока которого покрыты радиальной скульптурой. Седло
высокое, остро килевидное, соответствующее срединной складке синуса.
Скульптура обычная.
Оригинальное строение синуса и седла легко отличают этот вид от
всех других.
Распространение. Нижнефранские отложения Кузбасса.
Cyrtospirifer archiaci (Murchison)
Табл. LXI1, фиг 1
Раковина средних размеров или довольно крупная, вздутая, неравмо-
створчатая, полукруглая, развитая по ширине. Длинный и прямой замоч-
ный край равен наибольшей ширине раковины. Призамочные углы пря-
мые. Высокая вогнутая арея имеет вид треугольника несколько большей
ширины, чем ширина раковины: боковые концы ее продолжаются за пре-
267
делы раковины в виде тонких пластинок. Синус довольно глубокий,,
широкий и резко ограниченный. Седло невысокое, округленное в попереч-
ном сечении.
От Cyrtosplrlfer verneulll (Mure h.) отличается менее крылатой
и более вздутой раковиной, прямыми призамочными углами, более вогну-
той, резко ограниченной ареей, боковые части которой не заканчиваются
ясно в пределах раковины, а продолжаются в виде двух тонких пласти-
нок и вне ее.
От близкого по очертаниям раковины и строению синуса и седла
Cyrtosplrlfer tschernyschewl К h а 1 f i п из фаменских отложений Запад-
ной Сибири отличается большей вздутостью раковины, длинным замоч-
ным краем, прямыми призамочными углами и резко ограниченной ареей.
Распространение. Фаменские отложения Европы и Азии.
Cyrtosplrlfer tschernyschewl К h а 1 f i n
Табл. LXII, фиг. 2
Раковина среднего размера или небольшая, почти изометричная,
с сильно выпуклой брюшной створкой, слегка развита по ширине. Очер-
тания ее меняются от округленнотреугольных до округленночетырех-
угольных и полукруглых. Замочный край короткий и прямой, с закруг-
ленными углами. Арея треугольная, нерезкая, слегка вогнутая. Синус
умеренно глубокий, резкий и широкий. Седло невысокое, резкое и
широкое.
От сходного по сильно выпуклой брюшной створке, очертаниям рако-
вины и строению синуса и седла Cyrtosplrlfer archlacl (Mure h.) этот
вид отличается менее загнутой, неясно отграниченной ареей, коротким
замочным краем, округленными призамочными углами.
Cyrtosplrlfer tschernyschewl var. slblrica I v a n i а (табл. LXI, фиг. 8)
отличается от типичной формы более резко выраженными седлом и сину-
сом.
Распространение. Широко распространен в фаменских отло-
жениях Кузбасса.
Cyrtosplrlfer tenticulum (V е г n е u i I)
Табл. LXI, фиг. 12; табл. LXII, фиг. 3
Раковина крупная или средних размеров, неравностворчатая, развита
по ширине. Брюшная створка высокая, пирамидальная, спинная слабо
выпукла. Длинный прямой замочный край с острыми углами равен наи-
большей ширине раковины. Арея очень высокая, треугольная, прямая
или лишь слегка вогнутая. Синус широкий, умеренно глубокий. Седло
плоское, неясное.
Пирамидальная брюшная створка, высокая, почти прямая треуголь-
ная арея, умеренно глубокий синус и плоское седло—признаки, отли-
чающие Cyrtosplrlfer tenticulum (V er и.) от всех близких видов.
Распространение. В Западной Сибири Cyrtosplrlfer tenticulum
(V ern.) распространен преимущественно в нижнефранских отложениях;
единичные экземпляры встречаются в верхнефранских и даже в фамен-
ских отложениях. Франские отложения Европы.
Cyrtosplrlfer khalflnl I v a n i а
Табл. LXII. фиг. 4—6; табл. LXIII, фиг. 1 и 2
Раковина небольшая или довольно крупная, неравностворчатая, раз-
витая по ширине. Брюшная створка высокая, пирамидальная. Боковые ее
части имеют вид почти плоских треугольников. Спинная створка трапе-
цеидальная, почти плоская. Длинный прямой замочный край равен наи-
большей ширине раковины. Арея очень высокая (высота составляет
*/2—’/з длины ее основания), треугольная, прямая. Синус угловатый,
268
узкий, резкий и глубокий, треугольный в поперечном сечении. Седло
высокое, узкое, резкое, вздернутое над плоской поверхностью спинной
створки, округленное в поперечном сечении.
По очертаниям раковины, прямой высокой арее и почти плоской спин-
ной створке очень сходен с Cyrtosplrifer tenticulum (Ver п.), от которого
отличается: более крылатой и более плоской брюшной створкой (боко-
вые части которой имеют вид плоских треугольников); узким, углозатым
и резким синусом; более крылатой и более плоской спинной створкой:
высоким, узким и резким седлом.
Руководящий вид для верхнефранских отложений Кузбасса (мон-
стровый горизонт).
Cyrtosplrifer ussovi К h а 1 f i п
Табл. LXIII, фиг. 3 и 4
Раковина крупная или средних размеров, по очертаниям приближаю
щаяся к Cyrtosplrifer tenticulum. Наибольшая ширина у замочного края,
концы которого заостренные или слегка закругленные. Субпирамидаль-
ная брюшная створка с высокой прямой треугольной ареей. Синус,
идущий от макушки, неглубокий, обычно плоский. Спинная створка слабо
выпуклая, седло плоское. Характерной является скульптура, состоящая
из более крупных, дихотомирующих ребер в синусе и на седле и более
тонких простых на боках раковины. Часто наблюдается искривленность,
асимметричность раковины: макушка брюшной створки повернута
в сторону, искривлена, арея имеет вид неправильно криволинейной
плоскости. Эта деформация раковины объясняется прикрепленным об-
разом жизни.
Распространение. Является руководящим видом для верхне-
франских отложений Кузбасса (монстровый горизонт).
Род Indospirifer G г a b а и
Раковина поперечноовальная, реже округленная, более или менее
вздутая. Замочный край равен или чуть короче наибольшей ширины
раковины. Арея треугольная, вогнутая. Макушка брюшной створки
острая, загнутая. Синус и седло хорошо развиты. Вся поверхность
створок покрыта немногочисленными, довольно грубыми, округленными
ребрами. Имеет характерную микроскульптуру, состоящую из тонких,
коротких струек, располагающихся веерообразно на боках ребер.
В местах пересечения с концентрическими знаками роста эти струйки
делаются тонкобугорчатыми. В брюшной створке находятся большие,
расходящиеся зубные пластины.
Типичный вид. Indospirifer padaukptnensls (Reed).
Распространение. Девон Азии: средний девон Индо-Китая
и Китая; средний и верхний девон Кузбасса. Очень близок верхнедевон-
ский род Gurichella Paeck. (см. ниже), отличающийся только микро-
скульптурой.
Indospirifer stblrlcus К h а 1 f i n
Табл. LXII, фиг. 7—10; табл. LXIII, фиг. 9
Раковина умеренно развитая по ширине, округленнопятиугольного
или трапецеидального очертания. Призамочные углы слегка закруглены,
прямые или несколько оттянутые. Арея брюшной створки в виде низкого
треугольника, вогнутая. Синус начинается от самой макушки, достаточно
широкий и глубокий, резко ограничен двумя широкими ребрами; по дну
синуса проходит ребро. Спинная створка менее выпукла, чем брюшная;
седло невысокое, с крутыми боками, плосковерхое, несет продольную
борозду. По каждую сторону седла и синуса лежат 5—8 простых, округ-
ленных радиальных ребер. На седле, в синусе и в межреберных про-
269
межутках — обычная для рода Indospirifer тонкая веерообразная скульп-
тура.
Эта специфическая скульптура отличает данный вид от представи-
телей группы Giirichella ziczac (Roem.) (см. ниже), которые во всех
остальных отношениях совершенно сходны с ним.
Распространение. Очень обилен в отложениях промежуточной
толщи (верхи франского яруса) ряда районов по северо-западной
окраине Кузбасса.
Род Giirichella Paeckelmann
Раковины небольшие, умеренно вздутые, округленнопятиугольные,
с прямым замочным краем, который короче или равен наибольшей
ширине раковины. Замочные углы округленные, реже острые. Арея
брюшной створки треугольная, низкая, всегда вогнутая. Поверхность
створок покрыта округленными резкими складками. Седло и синус редко
совершенно гладкие, обычно в синусе лежит одно ребро, которому на
седле соответствует продольная бороздка. В брюшной створке зубные
пластины.
Типичный вид. Giirichella deflexa (Roem.).
Распространение. Средний (?) и верхний девон Европы
и Азии. В Западной Сибири в отложениях франского яруса широко рас-
пространены спириферы, относившиеся к группе Spirifer ziczac (Roe m.)
(табл. LXIII, фиг. 5—6; табл. LXIII, фиг. 12), принадлежащей к роду
Giirichella *; для них особенно характерно наличие в синусе одного или
трех ребер и соответствующего количества бороздок на седле.
Giirichella (?) vratschinsktae Kraevskaja
Табл. LXIII, фиг. 13
Раковина крупная, округленнопятиугольная, почти равностворчатая,
умеренно вздутая, с округленными призамочными углами. Замочный
край прямой, короче наибольшей ширины раковины. На брюшной створке
от самой макушки идет узкий неглубокий синус, постепенно расширяю-
щийся к лобному краю и переходящий в длинный язычок. Арея неболь-
шая, низкая, слабо вогнутая. На спинной створке невысокое четкое воз-
вышение, вздернутое у лобного края. Поверхность створок покрыта не-
многими плоскоокругленными складочками, разделенными узкими
бороздками; имеются узкие и густо расположенные пластинки роста 1 2.
В синусе и на седле складки слабо заметны.
От близкого G. deflexa данный вид отличается более крупными раз-
мерами и характерным длинным язычком синуса.
Распространение. Характерный вид в комплексе фауны глубо-
кипского горизонта верхнего девона Западной Сибири и Рудного Алтая
СЕМЕЙСТВО ATHYRIDAE M’COY
Род Anathyris Peetz
Раковина двояковыпуклая, развитая по ширине, с длинным прямым
замочным краем, вдоль которого на брюшной створке обычно бывает
развита невысокая арея с параллельными краями. Макушка брюшной
створки снабжена круглым фораменом. На брюшной створке всегда
имеется синус, а на спинной — седло различного строения: простое или
несущее продольную вдавленность, у некоторых видов достигающую
1 Точная родовая принадлежность этих спириферов, а также их взаимоотношение
с родом Indospirifer (см. стр. 269) требуют выяснения, но их руководящее значение
для франских отложений Кузбасса не вызывает сомнений. Прим. ред.
2 Принадлежность этого вида к роду Giirichella сомнительна; судя по характеру
знаков роста, он скорее относится к роду Lamellispirifer (см. стр. 260). Прим. ред.
'27P,
ширины и глубины синуса брюшной створки. Поверхность створок по-
крыта густыми пластинчатыми знаками роста. В брюшной створке на-
ходятся хорошо развитые зубы и зубные пластины, в спинной — замочная
площадка, прободенная круглым отверстием (так называемым висце-
ральным фораменом).
На ранних стадиях роста раковина имеет форму, свойственную рако-
винам рода Athyrls М’Соу.
Типичный вид. Anathyrls phalaena (Phil 1.)
Распространение. В Европе род Anathyrls известен во всех
трех отделах девона. В Западной Сибири появляется в самых верхах
живета и особенно характерен для франского яруса.
Anathyrls phalaena (Phillips)
Табл. LXIII, фиг. 7 и 8
Во взрослом состоянии раковина сильно вытянута по ширине, крыла-
тая, с длинным прямым замочным краем и хорошо выраженной ареей
брюшной створки. Широкий и глубокий синус ограничен отчетливыми
складками. Вдоль седла спинной створки проходит широкая и глубокая
вдавленность, подобная синусу брюшной створки; обе створки благодаря
этому выглядят почти одинаково. Юные экземпляры имеют раковину
круглого очертания, с неглубокими продольными бороздками вдоль пло-
скости симметрии на обеих створках. Совместно встречающийся
Anathyrls peetzl Kha If. отличается нерасчлененным седлом.
Распространение. В Западной Европе вид Anathyrls phalaena
распространен в нижнем девоне и в низах среднего девона. В Запад-
ной Сибири он является руководящей формой для низов франского
яруса (окраины Кузбасса, Алтай). Л. Л. Халфин считает, что верхне-
девонские сибирские представители не тождественны западноевропей-
скому кобленцско-эйфельскому виду Anathyrls phalaena и обособляет их
под названием Anathyrls supraphalaena.
Anathyrls tyzhnovl К h a 1 f i п
Табл. LXIV, фиг. 3
Раковина достаточно крупная, поперечновытянутая, с длинным
прямым замочным краем. Призамочные углы округленные, реже прямые.
Невысокая арея брюшной створки отчетливо выражена. Брюшная
створка значительно менее выпукла, чем спинная. Синус начинается не
от самой макушки, а по направлению к переднему краю очень резко
расширяется и углубляется; близ переднего края ширина синуса дости-
гает половины ширины раковины. Седло спинной створки очень высокое,
простое (без продольной вдавленности); общая форма спинной створки
крышеобразная. Молодые раковины имеют круглые очертания и слабо
развитые синус и седло.
Наиболее близкий вид — нижнефранский Anathyrls flmbrlata К h а 1 f.,
отличающийся одинаково выпуклыми створками и умеренно развитыми
седлом и синусом.
Распространение. Среднефранские отложения окраин Куз-
басса (атриповый горизонт северо-западной окраины и, возможно,
средняя красноцветная толща Барзасского района).
Anathyrls monstrum К h а 1 f i n
Табл. LXIII, фиг. 10; табл. LXIV, фиг. 2
Раковина очень крупная, с гипертрофированно развитым синусом
брюшной створки, благодаря чему при взгляде со стороны этой створки
она представляется двулопастной. Призамочные углы закруглены, арея
не развита. Синус начинается на некотором довольно значительном рас-
стоянии от макушки и сразу же приобретает характер широкой плавной
271
вдавленности с округленным широким дном и крутыми боками. По дну
синуса проходит узкая острая бороздка, прослеживающаяся до самой
макушки. Спинная створка очень выпуклая, седло несет довольно
широкую, но неглубокую вдавленность. Поверхность створок с обычными
для рода Anathyrls пластинчатыми знаками роста. Молодые экземпляры
имеют облик представителей рода Athyrls М’С о у.
Распространение. Руководящая форма верхнефранских отло-
жений Кузбасса, главным образом прибрежных фаций (северо-восточная
окраина бассейна). По северо-западной окраине Кузбасса в отложениях
того же возраста (промежуточная толща) распространен близкий вид —
Anathyrls ussovi Kh а 1 f. (табл. LXIV, фиг. 1), отличающийся менее глу-
боким и более угловатым синусом, отсутствием бороздок в синусе и на
седле, остроконечными призамочными углами.
Л,. Л. ХАЛФИН
Тип Arthropoda. Членистоногие
КЛАСС ТРИЛОБИТЫ
В нижнедевонских отложениях Саяно-Алтайской области трилобиты
достаточно многочисленны и разнообразны; они описаны в работах
Ф. Н. Чернышева (1893), Г. Г. Петца (1901), Н. Е. Чернышевой (1951),
Л. Л. Халфина (1948). Преобладают среди них представители семейств
Phacopidae и Dalmanitidae, местами (например, в нижнекобленцских
отложениях Горного Алтая) встречающиеся в массовом количестве. Зна-
чительно реже трилобиты встречаются в среднем и верхнем девоне. Ниже
охарактеризованы некоторые наиболее типичные нижнедевонские виды
трилобитов.
СЕМЕЙСТВО PROETIDAE CORDA
Род Proetus S t е i п i n g е г
Эллиптического очертания тело трилобита состоит из полукруглых и
почти равных головного и хвостового отделов (последний несколько
меньше) и туловища, образованного 10 члениками. Глабель обычно
слегка сужается вперед и почти достигает краевого валика, слабо расчле-
нена; обычно бывает выражена задняя (базальная) пара лопастей. Глаза
полулунные, сильно приближены к глабели. Лицевые швы начинаются на
половинном расстоянии между щечными углами и глабелью.
Типичный вид. Proetus conclnnus D а 1 m.
Распространение. Силур—пермь, повсеместно.
Proetus carinatus К. h а 1 f i n
Табл. LXIV, фиг. 4
Характеризуется узкой и очень высокой осевой частью, ограниченной
глубокими спинными бороздками, и значительно сужающейся вперед
глабелью, имеющей до некоторой степени грушевидные очертания. Зад-
ние лопасти глабели представляют собой округленные вздутия; средняя
и передняя пара глабелярных борозд отсутствуют полностью. Глабель
сильно вздута, в задней ее части килеватая. Затылочная борозда вы-
пукла вперед, затылочное кольцо с двумя боковыми лопастями.
Щеки умеренно выпуклые, с щечными остроконечиями, прижатыми
к плеврам. Глаза расположены в непосредственной близости к глабели.
Плевральные сегменты с бороздками. Полукруглый хвостовой щит харак-
теризуется чрезвычайно узким рахисом, состоящим из 8—9 члеников.
Распространение. Нижнекобленцские отложения (кондратьев-
ская толща) Горного Алтая (окрестности с. Куятан) и Рудного Алтая
(Левенко, 1951, стр. 72).
.272
СЕМЕЙСТВО CHEIRURIDAE SALTER
Род Cheirurus Bey rich
Описание см. на стр. 178.
Cheirurus strabo Weber
Табл. LXIV, фиг. 5
Расширяющаяся вперед глабель несет три пары борозд, из которых
задние косо направлены назад и вместе с затылочной бороздой отсекают
характерные треугольные базальные лопасти глабели. Бороздки передней
и средней пары параллельны, идут поперек глабели; они глубокие и от-
четливые, но затухают к средней части глабели; каждая из них представ-
ляет собой небольшую дугу, обращенную выпуклостью вперед.
Неподвижные щеки умеренно выпуклые, с небольшими щечными
остроконечиями, направленными параллельно оси тела животного.
Глаза располагаются против передней лопасти глабели и скошены под
углом 45° к оси последней, что является одним из характерных призна-
ков вида.
Распространение. Нижний девон Ферганы; нижнекобленцские
отложения Горного Алтая (кондратьевская толща, окрестности
с. Куяган).
Род Crotalocephalus S а 11 е z
Этот девонский род отличается от силурийского Cheirurus (см.
стр. 178) тем, что передняя и средняя борозды глабели пересекают по-
следнюю не прерываясь.
Crotalocephalus sternbergi Boeck
Табл. LXV, фиг. 6
Глабель слабо расширяющаяся вперед и слабо выпуклая, пересечена
двумя дугообразно отогнутыми назад поперечными бороздами. Третья
(задняя) пара глабелярных борозд отсекает треугольные базальные
лопасти. Умеренно выпуклые неподвижные щеки вертикально обрываются
в глубокие спинные бороздки и полого опускаются назад и вбок. Задняя
ветвь лицевого шва идет почти параллельно задней стороне головного
отдела.
Распространение. Нижнекобленцские отложения (кондратьев-
ская толща) Горного Алтая. Широко распространен в нижнем и сред-
нем девоне Западной Европы.
СЕМЕЙСТВО PHACOPIDAE CORDA
Род Phacops Е m m г i с h
Спинной панцырь вздутый. Головной щит полукруглый, с округлен-
ными щечными углами и узким лимбом. Глабель большая, сильно рас-
ширяется вперед, нерасчлененная или с одной (задней) парой бороздок;
поверхность глабели гранулирована. Глаза большие, с очень крупными
фасетками. Хвостовой щит меньше головного, округлого очертания.
Типичный вид. Phacops latifrons Bronn.
Распространение. Силур—девон, редко нижний карбон, по-
всеместно.
Phacops subcrlstata К. h а 1 f i n
Табл. LXV, фиг. 4 и 5
Головной щит со слегка оттянутыми назад округленными щечными
углами. Глабель вздутая, выдается вперед и нависает над лимбом; она
снабжена одной парой бороздок, покрыта грубыми бугорками. Глаза
18 Зак. 2024 273
значительно ниже глабели. Боковые части субкранидиального желоба
с обеих сторон снабжены мелкими зазубринами. Хвостовой щит парабо-
лического очертания; его осевая часть сильно выпуклая, состоит из
10—11 сегментов.
В кобленцских отложениях Рудного Алтая (район бывшего Крюков-
ского рудника) распространен Phacops altaicus Т s с h е г п. (Ф. Н. Чер-
нышев, 1893, стр. 202), отличающийся более выступающей вперед гла-
белью с более грубой грануляцией ее и ровными (не зазубренными)
краями субкранидиального желоба (табл. LXIV, фиг. 6).
Распространение. Нижне- и верхнекобленцские отложения
Горного Алтая (кондратьевская и медведевская толща). Окрестности
с. Куяган и д. Александровки.
СЕМЕЙСТВО DALMANIT1DAE DELO
Род Probolium О о h 1 е г t
Головной щит снабжен щелочными остроконечиями и фронтальным
выростом, трехлопастным или бифуркирующим. Расширяющаяся вперед
глабель несет три пары глабелярных бороздок; дистальные концы гла-
белярных лопастей передней и средней пары сливаются. Глаза большие,,
полулунные в плане. Хвостовой щит треугольный или параболический,
с ровными краями; количество члеников рахиса свыше 16. Поверхность
панцыря усеяна мелкими бугорками.
Отличием от рода Dalmanttes Barr, является наличие фронтального
выроста.
Типичный вид. Probolium nasutum (Con г.)
Распространение. Нижний девон северного полушария; руко-
водящая группа.
Probolium altalcum (Khalfin)
Табл. LXV, фиг. 1—3; рие. 194
Крупных размеров трилобит, достигающий 15 см в длину. Длина
головного отдела немного превышает 2/5 его ширины. Фронтальный
* вырост трехлопастной; средняя лопасть раз-
вита значительно сильнее, чем боковые.
Края головного щита снабжены двумя-тремя
широкими зубчиками по каждую сторону
основания фронтального выроста. Длина
щечных остроконечий достигает длины го-
ловного отдела. Хвостовой щит с коротким
осевым остроконечием; количество колец ра-
хиса достигает 20—22 штук.
Рис. 194. Probolium altaicum
<К h a I f.). Нат. вел.
Наиболее близкие виды известны из Di*
Северной Америки; они отличаются строе-
нием фронтального выроста и другими при-
знаками.
Распространение. В нижнекоб-
ленцских отложениях (кондратьевская тол-
ща) Горного Алтая пользуется массовым распространением (окрестно-
сти с. Куяган).
Л. Л. ХАЛФИН
Тип Mollusca. Мягкотелые
КЛАСС LAMELLIBRANCHIATA. ПЛАСТИНЧАТОЖАБЕРНЫЕ
МОЛЛЮСКИ ИЛИ ПЕЛЕЦИПОДЫ
Наиболее богатой из всех известных в Западной Сибири являете®
фауна пелеципод нижнего девона Алтая; она содержит ряд важных руко-
274
водящих форм, частично описанных ниже (см. также Л. Л. Халфин.
1948). Пелециподы распространены также в девоне Кузбасса и Минусин-
ской котловины, но изучены пока очень неполно.
СЕМЕЙСТВО PTERINOPECTENIDAE NEWELL
Род Leiopecten К h а 1 f i n
Раковина неравностворчатая, сильно неравносторонняя, с прямым
замочным краем и нормально развитыми ушками. Левая створка более
выпукла, чем правая, с более развитой макушкой. Связковая площадка
несет несколько продольных борозд, образующих под макушкой тупой
стропилообразный перегиб. Поверхность створок гладкая.
Типичный вид. Leiopecten rectangularis К h а 1 f.
Распространение. Нижний девон; массовым развитием поль-
зуется в нижнем девоне Алтая.
Наиболее близкий род — Pseudavlculopecten New., имеющий своих
представителей в девоне Западной Сибири, отличается наличием радиаль-
ной скульптуры.
Leiopecten rectangularis К h а 1 f i n
Табл. LXVI, фиг. 5
Дополнительно к родовым признакам характеризуется небольшими
прямоугольными ушками и сравнительно коротким замочным краем, рав-
ным примерно */2 длины раковины.
Близким видом является североамериканский нижнедевонский Leio-
pecten umbonatus, отличающийся более длинным замочным краем и боль
шими размерами ушек.
Распространение. Нижний девон Горного Алтая (ганинская
серия и кондратьевская толща). Окрестности сс. Куяган и Соловьиха.
СЕМЕЙСТВО PTERINEIDAE DALL
Род Pterinea G о 1 d f u s s
Раковина косая, неравносторонняя, неравностворчатая: правая створ-
ка плоская, или вогнутая, левая выпуклая. Замочный край прямой. Ушки
нормально развиты, переднее значительно меньше заднего. Вдоль замоч-
ного края располагается связковая площадка с рядом продольных бо-
розд. Зубы в виде коротких поперечных ребрышек под макушками и бо-
лее длинных, косо направленных назад, позади макушек. По характеру
скульптуры выделяется несколько подродов. В нижнем девоне Алтая
широко распространен подрод Tolmaia с тонкими радиальными ребрами.
Типичный вид. Pterinea laevls Goldf.
Распространение. Силур—карбон, повсеместно.
Pterinea (Tolmaia') squamosa Kh a I f i n
Табл. LXVI, фиг. 1 и 2
Раковина крупных размеров, почти одинаково развитая по высоте
и длине. Замочный край длинный. Переднее ушко маленькое, треуголь-
ное, заднее большое, крылообразное. Связковая арея несет до 15 про-
дольных борозд. Поверхность левой створки покрыта тонкими, но рез-
кими радиальными ребрами, в широких промежутках между которыми
располагается по одному более короткому и тонкому ребрышку второго
порядка. Знаки роста образуют на радиальных ребрах резкие чешуйки;
промежутки между знаками роста покрыты тончайшей концентрической
струйчатостью. Скульптура правой створки аналогична, но более слабая.
Очень близка западноевропейская нижнедевонская Pterinea lorana
Fuchs, отличающаяся лишь несколько более высокой раковиной, боль-
шими размерами переднего ушка и отсутствием глубокой бухтьи, ограни-
чивающей заднее ушко у нашего вида.
18* 275
Распространение. Нижний девон Горного Алтая (кондратьев
ская толща). Окрестности с. Куяган.
СЕМЕЙСТВО AMBONYCHI1DAE MILLER
Род Mytilarca Hall
Раковина равностворчатая, высокая, клиновидно суживающаяся
к макушкам (митилоидная), с небольшим задним ушком, которое сверху
ограничено косо направленным, коротким, прямым замочным краем.
Макушки острые, направлены вперед, лежат на переднем конце замоч-
ного края. Вдоль замочного края располагается связковая площадка
с продольными бороздками. Под макушками находятся мелкие карди-
нальные зубы; задние зубы короткие, косо направлены назад (табл. LXV
фиг. 7). Поверхность створок с концентрическими знаками роста.
Типичный вид. Mytilarca chetnungensis (Con г.)
Распространение. Силур — нижний карбон. СССР, Западная
Европа, Северная Америка. В массовом количестве встречается в ниж-
нем девоне Алтая совместно с родом Plethomytllus, отличающимся от-
сутствием кардинальных зубов.
Mytilarca elongata К h а 1 f i n
Табл. LXV, фиг. 8
Раковина значительных размеров, развитая по высоте, почти прямая,
удлиненнояйцевидных очертаний. Заднее ушко небольшое, но выражено
отчетливо. Верхняя половина переднего края в плане ‘Слегка вогнутая,
в остальном раковина очерчена выпуклыми краями. Макушки длинные,
острые, значительно подняты над замочным краем. Створки выпуклы
умеренно и плавно. Кардинальные зубы таксодонтного типа распола-
гаются под макушками; на заднем ушке лежат боковые зубы в виде
2—3 коротких ребрышек.
Распространение. Обычна для кондратьевской толщи Горного
Алтая совместно с другими представителями этого рода, отличающимися
очертаниями и формой раковины.
СЕМЕЙСТВО LAURSKI1DAE KHALFIN
Род Laurskia К. h а 1 f i п
Раковина, развитая по высоте, митилоидного типа:
клинообразно суживающаяся к макушке и расширяю-
щаяся книзу. Острые макушки направлены вперед.
Передний край ниже макушки вогнут, остальные края
выпуклые. В макушечной области каждой створки
имеется внутренняя перегородка для прикрепления пе-
реднего мускула. Наружная поверхность с концентри
ческими линиями роста.
Морфологически вполне подобен описываемому
мезо-кайнозойский род Drelssensia Ben. В морских
отложениях девона (в частности, Западной Сибири)
распростра1нен внешне сходный род Mytilarca Hall,
отличающийся совершенно иным устройством замоч-
ного края (см. выше).
Типичный вид. Laurskia attenuata К h а 1 f.
Распространение. Лагунные отложения же-
динского яруса Горного Алтая.
Lciurskla attenuata К h а 1 f i n
Рис. 195
Раковина небольшая, высокая и узкая: высота вдвое превышает
длину. В остальном полностью отвечает приведенному выше родовому
276
Рис. 195. Laurskia
attenuata К h а 1 f.
Левая створка с
внутренней сто-
роны; s—перего-
родка для при-
крепления перед-
него мускула.
Увеличено
диагнозу. Встречающиеся совместно виды отличаются более широкой
раковиной.
Распространение. Г анинская серия Горного Алтая, окрестности
с Соловьиха.
СЕМЕЙСТВО TRIGON1IDAE LAMARCK
Род Praemyophoria К h a I f i n
Раковина косочетырехугольного или субтрапецоидальнсмо очертания,
равностворчатая, неравносторонняя. Макушки направлены вперед, лежат
на переднем конце прямого замочного края. От макушки к нижнезаднему
углу направляется более или менее резко выраженный киль, отделяю-
щий треугольное задневерхнее поле от остальной поверхности створки.
В левой створке три зуба, в правой два, реже три. Наружная поверх-
ность створок покрыта густыми и тонкими концентрическими линиями.
Как и повсюду, этот распространенный девонский род в Западной
Сибири встречается совместно с генетически близким к нему родом
Toechomya, который отличается более округленными очертаниями рако-
вины и отсутствием килей на створках.
Т и п и ч н ы й в и д. Praemyophoria acutirostris К h a I f.
Распространение. Нижний девон, повсеместно
Praemyophoria acutirostris К h a I f i n
Табл. LXV, фиг. 11
Раковина достигает значительных размеров, субквадратного очерта-
ния с высокими острыми макушками, удаленными от переднего края на
расстояние ’А—‘А длины раковины. Высота раковины равна ее длине.
Передний край широкоокругленный, брюшной край слабо выпуклый,
задний прямой. Створки умеренно выпуклы; кили округленные, но очень
отчетливые. На наружной поверхности имеются обычные концентриче-
ские линии.
Распространение. Этот вид широко распространен в нижне-
кобленцских отложениях Горного Алтая (кондратьевская толща); там же,
в верхнекобленцских отложениях, встречается Praemyophoria elongata
Bench., отличающаяся скошенной раковиной, почти параллельным на-
правлением переднего и заднего краев и очень узким задневерхним
полем. В отложениях кобленцского яруса Горного Алтая широко рас-
пространены также представители рода Toechomya Clarke (например,
Toechomya subclrcularls Kh а 1 f. — табл. LXV. фиг. 9, 10), отличающиеся
прежде всего, отсутствием килей на створках.
СЕМЕЙСТВО ARCHAEOCARDIIDAE KHALF1.X
Род Archaeocardium К h а 1 f I п
Раковина крупная и толстостенная, равностворчатая, неравносторон-
няя, округлого очертания. Макушки высокие, длинные повернуты вперед.
Замочный край непрямой; массивная замочная площадка снабжена кар-
динальными и боковыми зубами. Скульптура наружной поверхности со-
стоит из радиальных рядов очень мелких точечных ямок, как бы из пре-
рывистых радиальных бороздок, промежутки между которыми имеют
характер плоских радиальных ребер (рис. 196).
Близких родов нет; весьма характерная скульптура составляет отли-
чительную особенность данного рода.
Типичный вид. Archaeocardium scrobiculare К h а 1 f.
Распространение. Верхний силур-- нижний девон Чехия
и Горный Алтай.
277
Archaeocardlum scrobiculare Khalfin
Табл. LXVI, фиг. 3 и 4
Раковина крупная, почти изометричная (слегка развита по ширине).
Замочный край слабо изогнутый, позади макушек почти прямой. Ма-
кушки слегка смещены к переднему краю. Задняя сторона раковины
развита несколько больше, чем передняя. Выпуклость створок умерен-
Рис. 196. Род Archaeocardlum.
Характер скульптуры. s/4 нат. вел. По Барранду
ная, передняя половина их более выпукла, чем задняя. Скульптура на-
ружной поверхности обычная для рода Archaeocardlum.
Распространение. Archaeocardlum scrobiculare обычен в нижне-
кобленцских отложениях (кондратьевская толща) Горного Алтая. Очень
сходные, родственные виды известны из верхнего силура и нижнего
цевона Чехословакии.
СЕМЕЙСТВО AMNIGENI1DAE KHALFIN
Род Paramnigenia Khalfin
Раковина крупная, развитая по длине, с макушками, смещенными
к переднему краю. Замочный край длинный, прямой, беззубый. Брюш-
ной край субпараллельный замочному, снабжен широким синусом. Диаго-
нальное возвышение выражено отчетливо, впереди него лежит широкая
и глубокая вдавленность. Наружная поверхность с концентрическими
линиями роста.
Лагунно-пресноводный девонский род Amnlgenla Hall (встреченный,
в частности, совместно с характеризуемым) отличается, прежде всего,
более удаленными от переднего края макушками.
Типичный вид. Paramnigenia blcarlnata К h а 1 f.
Распространение. Жединский ярус Горного Алтая, отложения
лагунной фации.
Paramnigenia blcarlnata Khalfin
Табл. LXV1, фиг. 6
Очертания раковины приближаются к округленному прямоугольнику.
Макушки смещенны к самому переднему краю, который от них
опускается почти вертикально, обладая очень слабой выпуклостью. Синус
278
брюшного края очень широкий (охватывает почти всю длину брюшного
края). Диагональное возвышение имеет характер киля, впереди которого
лежит очень широкая вдавленность, отделенная от переднего края
вторым, широкоокругленным возвышением. Высота раковины составляет
3/5 ее длины.
Встречающаяся совместно с этим видом Paramnigenla ussovi
Khalf. отличается большим развитием раковины по длине и расшире-
нием ее назад.
Распространение. Ганинская серия Горного Алтая, окре-
стности с. Соловьиха.
А. Р. АНАНЬЕВ
РАСТЕНИЯ
Растительные остатки нередко встречаются в континентальных
и лагунно-континентальных, часто красноцветных, девонских отложе-
ниях Саяно-Алтайской области (окраины Кузбасса, Горная Шория,
Алтай, Минусинская котловина и др.). Они имеют большое значение для
стратиграфии этих отложений, тем более что какие-либо другие органи-
ческие остатки в осадках данного фациального типа очень редки. Следует
обратить внимание, что остатки растений в девонских отложениях часто
представлены небольшими, не бросающимися в глаза отпечатками, по-
этому нередко игнорируются геологами.
Девонская флора Западной Сибири почти не изучена. Однако работы
.последних лет показали, что девонские отложения Западной Сибири
богаты ископаемыми растениями, которые вполне пригодны для страти-
графического расчленения этих отложений, а отдельные местонахожде-
ния (на р. Енисее у Торгашино, на р. Кара-Чумыш в Салаире) по богат-
ству растительных остатков и их сохранности становятся в ряд с класси-
ческими местонахождениями, описанными в литературе.
Особенно богаты остатками растений девонские отложения Минусин-
ской котловины и южной окраины Чулымо-Енисейской впадины. Часто
встречаются, но обладают менее полной сохранностью девонские расти-
тельные остатки в Кузнецком Алатау, Салаирском кряже, по окраинам
Кузбасса и на Алтае.
В настоящее время можно выделить следующие руководящие ком-
плексы, установленные в различных районах горной части Западной
Сибири.
1. Нижний девон
Zosterophyllum myretonianum Р е п h а 11 о w
Taenlocrada dechenlana (G о е р р.) Кг. et Wе у 1.
Psllophyton Goldschmldtll Halle
Psllophyton prlnceps Dawson
Hostlmella sp.
Aphyllopterls robusta (Daw s.)
Barlnophyton Obrutschevli Ananiev
Drepanophycus spinaeformis G о e p p.
P. Goldschmldtll и T. dechenlana найдены, в частности, на Алтае
в системе рек Песчаной и Куваш вместе с морской кобленцской фауной.
2. Средний девон
Psllophyton prlnceps Dawson
Psllophyton sibiricum Zalessky
Asteroxylon sibiricum К г у s h t
Hostlmella sp.
279
Tomlphyton prlmaevum Z a 1.
Protolepldodendron Scharyanum KrejGi
Aneurophyton germanlcum Kraus, et W e у 1.
Отложения с этим комплексом повсеместно залегают непосредственно
под верхнеживетскими морскими отложениями, содержащими фауну
брахиопод, мшанок и др.
3. Верхний девон
Растения верхнего девона подразделяются на два комплекса.
Фаменский комплекс
Archaeopterls Halliana (G о е р р.) Dawson
Archaeopterls obtusa Lesquereux
Archaeopterls sp.
Archaeopterls sibirica Zalessky
Комплекс переходный от девона к карбону
Helenlella Theodorl Zalessky
Sphenophyllum subtenerrlmum N a t h о г s t
Asterocalamltes scroblculatus (S ch loth.) Ze i Iler
Rhacophyton incertum (Daws.) Kraus, et W e у 1.
Anelmltes acadlca Dawson
Фаменский комплекс растений был обнаружен главным образом
в туранской красноцветной свите Минусинской котловины.
Второй комплекс установлен в игрышинских слоях, залегающих на
границе туранской и минусинской свит как в Минусинской котловине,
так и по южной окраине Чулымо-Енисейской впадины.
Класс Psilophytales. Псилофиты
Род Zosterophytlum Р е п h а 11 о w
Считается наиболее полно изученным нижнедевонским растением
с проводящей системой из кольцевидных трахеид1. Растение маленькое,
состоящее из обильно разветвленного корневища и скудно разветвлен-
ных, скученных, прямо стоящих, безлистных наземных побегов. Послед-
ние близ верхушек несут рыхлые спороносные колоски. Почковидные,
округлые или округлоклиновидные спорангии располагаются вокруг оси
колоска спирально и имеют недлинную ножку.
Типичный вид. Zosterophytlum myretonlanum Р е n h а 11 о w.
Распространение. Верхний силур — нижний девон. В литера-
туре имеются не вполне достоверные указания о распространении рода до
верхнего девона включительно.
Zosterophytlum myretonlanum Р е п h а 11 о w
Табл. LXVII, фиг. 1
Растение образует кустистое корневище из лентовидных частей, от
которого плотным пучком отходят вверх слабо разветвленные безлистные
побеги. Ветвление имеет вид дихотомического разветвления в сочетании
с системой разветвления, напоминающей букву «Н». Спороносные органы
представляют собой рыхлые колоски, состоящие из спирально сидящих
на коротких ножках, овальных или почковидных спорангиев, причем со-
седние спорангии почти не соприкасаются.
1 Трахеиды — удлиненные клетки, слагающие древесину у псилофитов, хвойных
и некоторых других групп растений; стенки трахенд имеют различного характера
утолщения — кольцевые, лестничные и др. Прим. ред.
280
Близкие видыс Zosterophyllum australlanum Lang et Cookson
и Z. rhenanum Krausel et Weyland отличаются: первый, из-
вестный только по спороносным колоскам, более плотным их строением,
второй — более плотным строением спороносных колосков и более про-
стым строением растения, не образующего пучков.
Распространение. Нижний девон Шотландии, северо-восточ-
ной окраины Орского района Южного Урала; река Енисей, правый берег,
в 1 км от с. Торгашино (район города Красноярска).
Род Taeniocrada D. White
Принадлежащие этому роду остатки прежде считались лентообразной
водорослью и относились к роду Hallserltes Sternberg. После от-
крытия проводящей системы из лестничных трахеид и концевых споран-
гиев это растение было отнесено к псилофитам.
Остатки рода Taeniocrada всегда встречаются в виде массовых ско-
плений обильно дихотомически разветвленных лентовидных осей шири-
ной до 1,5 см, имеющих посредине узкую полоску — проводящий пучок
из трахеид.
Побеги или совершенно голые, или покрыты многочисленными, очень
мелкими, прямыми, штрихоподобными волосками или шипиками, при-
легающими к осям в продольном направлении.
Спорангии имеют концевое или боковое расположение. Если конеч-
ные части побегов с спороносными органами встречаются изолированно
от самого растения, их невозможно отличить от других родов псилофитов
{Psllophyton, Asteroxylon и др.). Виды с штрихоподобными волосками
приближаются к родам Psllophyton, Asteroxylon и др. и отличаются от
них почти микроскопическими размерами волосков или шипиков.
Типичный вид. Taeniocrada dechenlana (G б р р.) Kraus, et
Weyl.
Распространение. Нижний девон.
Taeniocrada dechenlana (Go р pert) Krausel et Weyland
Табл. LXVII, фиг. 6; табл. LXXIII, фиг. 2
Растение образует заросли из длинных, плоских, более или менее
лентовидных и гладких побегов, многократно дихотомически разветвляю-
щихся и достигающих ширины 1,5 см. Верхушечные части побегов
иногда свернуты в спираль. Проводящий пучок из трахеид хорошо вы-
ступает посредине «ленты» побега в виде полоски шириной 0,5— 1,0 мм.
Спорангии овальной формы имеют в длину 3—7 мм и располагаются
на концах побегов по 1—2, но часто до 10 в рыхлой гроздевидной кисти.
Конечные веточки со спорангиями с трудом отличаются от таковых
Dawsonites arcuatus Halle. Taeniocrada dubla Kraus, et Weyl.
отличается от описываемого вида круглым сечением побегов и штрихо-
подобными волосками на них. Taeniocrada Langl Stockmans отли-
чается боковым расположением спорангиев на побегах.
Распространение. В Рейнской области Германии и в Бельгии
встречается в верхнежединских и нижнекобленцских отложениях. В За-
падной Сибири обильные скопления частей лентовидных дихотомически
ветвящихся побегов этого вида встречены в Северо-Минусинской котло-
вине— в 2,5 км западнее озера Салбат и Горном Алтае — в бассейне
рр. Песчаная и Куваш. В районе г. Красноярска найдены обрывки подоб-
ных побегов, которые, вероятно принадлежат этому же виду.
Род Psilophyton Dawson
Одно из ранних наземных сосудистых растений небольших размеров,
обитавшее, повидимому, на сырых местах. От ползучего корневища
поднимаются тонкие прямые наземные побеги диаметром в основании не
281
более 0,5—0,75 см; постепенно утоняясь кверху, они разветвляются
дихотомически (вильчато) или симподиально на более тонкие веточки.
Верхушки этих побегов в юных стадиях роста свернуты в спиральные
завитки. Настоящие листья отсутствуют, и вместо них на поверхности
побегов развиваются колючки (боковые выросты или эмергенцы). Ко-
жица побегов несет устьица. В центре побега развит простой тонкий
проводящий пучок из кольчатых трахеид. Спорангии в виде простых
овальных мешочков располагаются на концах побегов.
От рода Asteroxylon (см. ниже) отличается негустыми и торчащими
в стороны колючками и отсутствием листообразных выростов, от рода
Drepanophycus (см. стр. 297) — значительно более тонкими побегами и
более тонкими и короткими колючками.
Типичный вид. Psllophyton prlnceps Daws.
Распространение. Нижний и средний девон (возможно верх-
ний силур — средний девон).
Psllophyton prlnceps Dawson
Табл. LXVII, фиг. 2; табл. LXVIII, фиг. 3;
табл. LXIX, фиг. 12; табл. LXX, фиг. 3;
табл. LXXII, фиг. 3
Сравнительно тонкие, толщиной не более 0,75 см, наземные побеги,
поднимаясь от горизонтального ползучего корневища, вскоре повторно
узкодихотомически ветвятся и заканчиваются тонкими веточками, часто
свернутыми в молодом возрасте в спираль. Конечные веточки несут на
концах одиночные или попарно сидящие, поникавшие книзу, овальные
многослойные гладкие мелкие спорангии. На поверхности побегов, кроме
прерывистой морщинистости или бороздчатости, имеются сравнительно
негусто расположенные колючки, покрывающие как старые части стволов,
так и более молодые, хотя на последних сохраняются значительно реже.
Наиболее часто в ископаемом состоянии встречаются лишь обрывки
различных частей наземных побегов.
Близкие виды отличаются от данного вида: Psllophyton Goldschmld-
tli — боковым разветвлением побегов и более тонкими колючками;
Drepanophycus splnaejormts (тонкие ветви) — более грубыми и более
широкими у основания колючками. При плохой сохранности материала
все три вида становятся неразличимыми, особенно когда побеги имеют
одинаковую толщину.
Распространение. Нижний девон Восточной Канады и Норве-
гии; нижний и средний девон ряда других стран. В Западной Сибири
(Минусинская котловина, Кузнецкий Алатау, Алтай) характеризует
нижнюю половину девона.
Psllophyton Goldschmldtll Halle
Табл. LXVII, фиг. 5; табл. LXIX, фиг. 4—9; табл. LXX, фиг. 4; табл. LXXII, фиг. 4
Тонкие фрагменты круглых в поперечном сечении побегов до
0,5—0,7 см толщины в основании; в длину достигают более 50 см и
обнаруживают характерную систему разветвления. Прямо стоящая
главная ось растения обнаруживает боковое (симподиальное) ветвление.
Боковые ветви дихотомически ветвятся на сравнительно коротком рас-
стоянии от главной оси и далее снова многократно дихотомически и
симподиально разветвляются под довольно широкими углами. В целом
вся система разветвления растения является широко раскинутой, на от-
печатках отдаленно напоминая оголенную вайю папоротника. Главная
ось, а также нижние части боковых ветвей покрыты длинными (до 4 мм),
тонкими и нежными колючками (эмергенцами), направленными к осям
ветвей перпендикулярно. Проводящая система из кольчатых трахеид
образует в центре осей круглый пучок толщиной до 0,5 мм, который на
282
Рис. 197. Psilophyton
siblricum Z а 1.
Участок стволика с
почти боковым вет-
влением. 4/5 нат. вел.
р. Томь у устья
р. Осиповой
некоторых отпечатках выделяется в виде тонкой средней полоски и за-
метен невооруженным глазом.
Галле (Halle, 1916), Крейзель и Вейланд (Krausel und Weyland, 1930)
считают, что длинные и тонкие побеги Р. Goldschmldtll вряд ли могут
считаться прямо стоящими, воздушными; скорее это растение обитало
в водной среде.
Отличие от сходных видов см. выше, в описании вида Р. princeps.
Распространение. Нижний девон Норвегии, нижний кобленц
Германии, верхний кобленц Бельгии, кобленц (слои
стретмор) Великобритании. В Западной Сибири
встречается в кобленцском ярусе Алтая (р. Песча-
ная), на р. Кара-Чумыш в юго-восточной части Са-
лаира, на восточном склоне Кузнецкого Алатау
(система р. Терси), в юго-восточном углу Чулымо-
Енисейской впадины (на правом берегу р. Енисея
у с. Торгашино в основании качинской свиты).
Psilophyton siblricum Zalessky
Рис. 197
Оси имеют толщину до 0,4.....0,5 см, ветвятся
симподиально. По обеим сторонам осей под острым
углом отходят более тонкие чередующиеся веточки,
носящие характер настоящих боковых разветвле-
ний моноподия. На поверхности осей и их боковых
разветвлений колючки замечены не были, хотя их
присутствие предполагается на основании кое-где
наблюдавшихся бугорков или шипиков. Органы
спороношения неизвестны.
В связи с недостаточной изученностью всех при-
знаков данной формы, приходится ее рассматривать
как установленную условно. Но такого характера остатки часто встре-
чаются в отложениях нижнего и среднего девона Западной Сибири,
иногда совместно с Psilophyton prtnceps.
Род Asteroxylon Kidston et Lang
Наземные побеги, отходящие от ползучего гладкого, местами виль-
чато ветвящегося корневища, достигают в диаметре 1 см. Главные оси
побегов разветвляются вильчато только в верхушечной части, но имеют
довольно многочисленные ветвящиеся, отходящие со всех сторон, а не
только в одной плоскости, боковые веточки моноподиального типа. По-
верхность как главных осей, так и боковых ветвей, покрыта со всех
сторон густо расположенными щетиновидными и черепитчато налегаю-
щими листьями, как у современного плауна. Конечные тонкие веточки
воздушных побегов, а также подземные разветвления были гладкими,
без листьев. На концах первых, поводимому, располагаются мелкие,
грушевидной формы, многослойные спорангии.
Центральный цилиндр побегов состоит из пучка древесины, имеющей
в разрезе звездочкообразную форму. Растение достигает высоты 40 см.
Типичный вид. Asteroxylon Macklei К i d s t. et Lang.
Распространение. Средний девон.
Согласно известной реконструкции Asteroxylon elberfeldense, без-
листные дихотомически разветвленные побеги, известные ранее под
названием Hostimella hostimensls, должны рассматриваться как конеч-
ные молодые веточки этого вида. В эту же реконструкцию в качестве
различных частей растения вошли фрагментарные остатки, ранее опреде
283
лившиеся из отдельных местонахождений Западной Европы как Psilo-
phyton prlnceps, Thursophyton Mlllerl, Aphyllopteris sp.
В связи с этим, находимые за пределами Западной Европы!, разроз-
ненные, морфологически различные части побегов рода Asteroxylon
стало невозможно относить к известным видам (A. Mackiel, A. elberfel-
dense). А. Н. Криштофович предложил, впредь до обнаружения доста-
точно полных экземпляров, все фрагментарные остатки из Западной
Сибири, принадлежащие данному роду, именовать — Asteroxylon
slbiricum.
Asteroxylon slbiricum Kryshtofovich
Табл. LXVIII, фиг. 5; табл. LXIX, фиг. 3, 10, 11
Фрагменты наземных побегов толщиной около 1 см или меньше,
густо покрытые щетиновидными листьями. Если листья на побегах не
сохранились, то вместо них видны многочисленные, так же густораспо-
ложенные, точечные выступы, создающие вид шагреневой кожи.
Вполне возможно, что эти остатки, часто встречаемые в девонских
отложениях Западной Сибири, при открытии более полно сохранившихся
экземпляров могут оказаться, в связи с географической обособленностью,
действительно относящимися к местному виду и не принадлежать ни
A. Mackiel, ни A. elberfeldense.
Распространение. Средний девон. Остатки данного вида
были встречены в Кузнецком Алатау к западу от Уленьской котловины,
в Минусинской котловине (водораздел рек Узунжул и Неня, район
ст. Уйбат) в Казахстане (Киргизская степь)
Род Hostimella Potonie et Bernard
Выше (стр. 283) отмечено, что Крейзель и Вейланд доказали принад-
лежность части остатков, относившихся ранее к роду Hostimella, к виду
Asteroxylon elberfeldense Kraus, et W e у 1. Но в связи с тем, что от
верхнего силура до среднего девона изолированные растительные
остатки типа Hostimella встречаются часто и должны как-то описываться
и систематизироваться, оказалось необходимым сохранить это наимено-
вание хотя бы для обозначения чисто морфологической группы остатков.
В род Hostimella обычно объединяются тонкие, совершенно голые оои
с преобладающим дихотомическим (вильчатым) и подчиненным симпо-
диальным (боковым) разветвлением, причем дочерние ветви образуют
углы от узких до широких. Часто в местах разветвлений на оси наблю-
даются почковидные образования, представляющие собой, вероятно,
зачаточные ответвления, расположенные в другой плоскости.
Близкий, также искусственный, род Aphyllopteris N a t h о г s t отли-
чается от рассматриваемого преобладающим симподиальным, реже
моноподиальным разветвлением побегов, но на отдельных участках
иногда присутствует и дихотомическое разветвление. Встречаются
остатки ветвей, обнаруживающих смешанное разветвление (дихотомиче
ское и симподиальное), и в этом случае решать вопрос об их родовой
принадлежности трудно.
Изолированные конечные голые участки побегов родов Psllophyton,
Asteroxylon и Thursophyton от рода Hostimella отличить невозможно.
Типичный вид. Hostimella hostlmensis Pot. et Bern
Распространение. Верхний силур — средний девон.
Hostimella sp.
Табл. LXV11I. фнг. 16, 6. 7; табл. LXX, фиг. 16; табл. LXX1, фиг. 2; табл. LXXII, фиг. 2
В Западной Сибири растительные остатки, отвечающие приведенному
выше диагнозу рода Hostimella, часто встречаются в отложениях ниж-
него и среднего девона Минусинской котловины. Кузнецкого Алатау,
281
южной окраины Чулымо-Енисейской впадины, Салаира, окраин Куз-
басса и Алтая.
Распространение. Верхний силур — средний девон. В Запад-
ной Европе, Европейской части СССР, на Урале и в Западной Сибири
характеризуют нижний и средний девон; в Австралии -- с верхнего
силура.
Род Aphyllopterls N a t h о г s t
Под родовым названием Aphyllopterls условно объединяются гетеро-
генные остатки растений из различных отделов девонской системы, сход-
ные между собой в морфологическом отношении.
Все они характеризуются одним общим признаком — голыми, оимпо-
диально или моноподиально ветвящимися осями, напоминающими мор-
фологически оголенные части вайи папоротников или их крепкие глав-
ные оси без листовых пластинок.
Сходство с родами Loganlella S t о 11 е у, Dlcranophyton Zalessky,
Hostlmella Pot. et Bern., Asteroxylon К i d s t. et Lang, (конечные
ответвления) большое, и нередко они различаются с трудом, особенно
при несовершенной сохранности образцов и их фрагментарности. Роды
Loganlella и Dlcranophyton отличаются короткими боковыми придатками
на оси; кроме того, первый отличается пучкообразным характером боко-
вых ветвей, а у второго на поверхности осей видны точечные микровы-
росты. Род Hostlmella отличается преобладающим дихотомическим раз-
ветвлением побегов.
Типичный вид автором рода выделен не был.
Распространение. Встречается во всех отделах девона.
Aphyllopterls robusta (Dawson)
Табл. LXXI, фиг. 1; табл. LXXV, фиг. 5; табл. L.XXVI, фиг. 7
Крепкая и гладкая главная ось растения имеет чередующиеся боко-
вые разветвления, а вблизи верхушки разветвляется дихотомически
(вильчато). Боковые ветви, в свою очередь, разветвляются симподиально
или дихотомически. Органы спороношения в виде голых заостренных
спорангий на ножке сидят на верхушках растений гроздьями. В центре
главных стволов установлено наличие древесины из кольцевидных тра-
хеид в виде цилиндрического пучка.
Доусон отнес этот вид, встреченный в нижнем девоне Канады, к роду
Psllophyton на основании наличия на поверхности осей микроскопических
выростов, обычно плохо сохраняющихся и поэтому часто не различимых.
Эти микровыросты в сочетании с симподиальной системой разветвления
послужили М. Д. Залесскому основанием для отнесения данного вида
к роду Dlcranophyton Z а 1., установленному им в низах верхнего девона
восточного склона Урала. Однако, ввиду невыясненности природы упо-
мянутых микровыростов, целесообразно присоединиться к мнению тех
палеоботаников, которые считают возможным временно отнести данный
вид к сборному роду Aphyllopterls Nathorst.
Распространение. Нижний девон Восточной Канады, Бельгии,
Западной Сибири (правый берег р. Енисея, окрестности с. Торгашино
в районе г. Красноярска).
Род Protobarinophyton Ananiev, gen. n.
Водорослеподобное растение с круглыми дихотомически ветвящимися
осями, достигающими в нижней части 7 мм толщины. Поверхность осей
гладкая, в центре их имеется тонкий (диаметром до 0,5 мм) проводя-
щий пучок из кольцевидных трахеид. Спороносные органы в виде дву-
рядных шишкообразных колосков располагаются на концах дихотомиче-
ски разветвленных осей растения. Колоски достигают в длину 4 см. и
285
в ширину 1,5 см. Спорангии имеют дисковидную форму, крупные, распо-
лагаются на оси колоска плотно двумя рядами, направленными в одну
сторону. В отпечатке боковой стороной спороносный колосок напоминает
односторонний гребешок.
Растение обитало в водной среде, а спороносные органы поднимались
над водой.
Похожий род Pectlnophyton Н о е g отличается однорядным располо-
жением спорангиев на спороносном колоске. Род Barinostrobus Kraus,
et Weyl. отличается спиральным расположением спорангиев вокруг
оси спороносного колоска. Род Barlnophyton White отличается пери-
стым расположением спороносных колосков на оси.
Типичный вид. Protobarinophyton Obrutschevli (Ananiev)
п. comb.
Распространение. Нижний девон.
Protobarinophyton Obrutscheuii (Ananiev) n. comb.
Табл. LXVII, фиг. 3 и 4; табл. LXVIII, фиг. 2; табл. LXX, фиг. 6 и 7;
табл. LXXI, фиг. 3 и 4; табл. LXXII, фиг. 1; табл. LXXIII, фиг. 1
Водорослеподобное растение с круглыми в поперечном сечении, глад-
кими, редко дихотомически разветвленными осями, заканчивающимися
двурядными спороносными колосками. Оси, имеющиеся в коллекции, до-
стигают в длину 30 см, их ширина (3—7 мм) мало изменяется снизу
вверх. Внутри они содержат цилиндрический проводящий пучок из коль-
цевидных трахеид диаметром 0,5 мм. Спороносные колоски располага-
ются на концах дихотомически разветвленных осей. Длина спороносных
колосков 1,5—3,7 см, ширина 0,7—1,2 см.
Двурядно расположенные спорангии дисковидной формы диаметром
до 0,5 см- С брюшной стороны они несут продольный шов для раскры-
вания. Стенки спорангиев упругие, с косо-поперечной волокнистой струк-
турой. Ось спороносных колосков в центре имеет проводящий пучок.
С брюшной стороны она прикрыта смыкающимися спорангиями, со
спинной стороны голая.
Сходный вид Protobarinophyton dawsonl (Kraus, et Weyl.>
n. comb, из нижнего девона Восточной Канады отличается значительно
более узкими спороноаными колосками. Barlnophyton rlchardsont
(Daws.) White и В. cltrulltforme Arnold из верхнего девона Се-
верной Америки отличаются перистым расположением спороносных ко-
лосков на оси растения.
Распространение. Нижний девон Западной Сибири; Рыбин-
ская впадина у Торгашино, Северо-Минусинская впадина у Каскыра,.
Тувинская впадина.
Род Tomiphyton Zalessky
Круглые в поперечном сечении побеги, обычно сплющенные на кон-
цах, неколько раз последовательно широковильчато ветвятся. У основа-
ния побеги имеют толщину 8—12 мм, но при последующих разветвле-
ниях сужаются до 2 мм. Первая дихотомия проходит под углом 75°,
последующие разветвления под более острыми углами —50—70°. Другие
части этого растения имеют прямые стволы с симподиальным ветвлением.
В центре побегов имеется сосудистый пучок, иногда хорошо заметный
даже на отпечатках в виде более плотной морщинистости. В случае хо-
рошей сохранности на поверхности побегов наблюдаются овальные
бугорки или шипики.
Спорангии, перемежаясь, располагаются на концах изогнутых ветвей
с одной или с обеих сторон. Они имеют овальную форму; длина их около-
4 мм, ширина до 2,5 мм.
286
В целом описываемые побеги представляют собой лишь части до-
вольно крупного растения, принадлежащего, повидимому, псилофитам.
Типичный вид. Tomlphyton primaevum Z а 1. (рис 198 и 199).
Рис. 198. Tomlphyton primaevum Zal.
Вильчато разветвленный побег. 4;5 нат. вел р. Томь у устья р Осиповой!
По М. Д. Залесскому
Рис. 199. Tomlphyton primaevum Zal.
Побег со спорангиями */5 нат. вел. р. Томь у устья р. Осиповой. По М. Д. Залесскому
Распространение. Нижний (средний?) девон. Единственный
вид этого рода — Tomlphyton primaevum — был описан М. Д. Залес-
ским с р. Томи ниже устья рч. Осиповой, где он найден совместно
с Psllophyton prlnceps var. ornatum. Позднее этот вид был обнаружен
в красноцветной толще Минусинской котловины (район ст. Уйбат), под-
стилающей морские отложения с фауной брахиопод верхнеживетского
возраста.
Род Drepanophycus G б р р е г t
Синоним — Arthrostigma Dawson
Главные оси обнаруживают два типа разветвления— дихотомическое
и боковое. Стволы и боковые ветви круглые в сечении. Они имеют на
своей поверхности спирально, но большей частью неравномерно располо-
женные шиловидные листья с широким округлотреугольным основанием
и серповидно загнутыми или прямыми верхушками длиной до 1—2 см.
287
При плохой сохранности отпечатков часто вместо листьев видны их осно-
вания в виде округлых бугорков.
На хорошо сохранившихся образцах иногда виден тонкий проводя-
щий пучок из кольцевидных трахеид, причем сам пучок (срединная
полоска) можно видеть невооруженным глазом, тогда как трахеиды
различимы только на микроскопических препаратах. Наряду со стериль-
ными (неплодоносными) осями встречаются плодоносные оси, которые от
первых отличаются тем, что имеют на листьях овальные спорангии
диаметром от 2—3 до 5—6 мм.
От сходных родов Psllophyton Dawson и Thursophyton Nathorst
описываемый отличается более толстыми и более грубыми осями и соот-
ветственно более крупными шиловидными листьями. В тонких осях все
три рассматриваемых рода отличаются друг от друга с трудом.
Типичный вид. Drepanophycus sptnaeformis G о р р.
Распространение. Нижний и средний девон (главным обра-
зом, в нижнем девоне). В Западной Сибири встречен в основании качин-
ской красноцветной свиты на правом берегу р. Енисея, в окрестностях
с. Торгашино (район г. Красноярска).
Drepanophycus sptnaeformis G б р р е г t1
Табл. LXVIII, фиг. 1а; табл. LXX, фиг 1а и 8
Типичный вид рода. Характеризуется теми же признаками, что и сам
род (см. выше). В качестве видовых признаков следует добавить нерав-
номерность распределения шипов (или листьев) на поверхности осей,
сравнительно небольшую их густоту и треугольную форму.
От Psllophyton prlnceps Dawson отличается более толстыми побе-
гами и соответственно более крупными листьями. От представителей
родов Asteroxylon и Thursophyton отличается также большей толщиной
побегов и иным расположением листьев: у описываемого вида листья
торчащие, а не прилегают к стволу. От Drepanophycus gaspianus Daws,
отличается нерегулярным расположением листьев и меньшей их плот-
ностью, от Drepanophycus splnosus (Krejti) Кг. et W. более жест-
кими листьями.
Распространение. Кобленцский ярус в Германии, Бельгии
и Шотландии, нижний девон в Норвегии, Восточной Канаде и Китае.
В Западной Сибири встречен в основании качинской красноцветной
свиты на правом берегу р. Енисея в окрестностях с. Торгашино (район
г Красноярска).
КЛАСС LYCOPODIALES. ПЛАУНОВЫЕ
Род Protolepidodendron Krejci
В этот род объединяются небольшие девонские растения, состоящие
из тонких одиночных или дихотомически разветвленных побегов, густо
покрытых линейными листьями, прикрепляющимися к осям раздутым
основанием и в большинстве случаев заканчивающимися на концах виль-
чатым раздвоением (бифуркацией).
При плохой сохранности образцов или в особых случаях захороне-
ния вильчатость листьев бывает не видна: в первом случае концы
листьев могут быть обломанными, во втором — листья располагаются
перпендикулярно к плоскости отпечатка и, будучи погруженными в по-
роду «ребром», не обнаруживают вильчатости. На поверхности стволов
обычно наблюдаются овальные или закругленноромбовидные бугорки,
1 Образцы, изображенные на табл. LXVIII, фиг. 1а и табл. LXX, фиг. 1а, неотли-
чимы от Psllophyton. rectissimum Н б е g. Прим, автора.
288
расположенные густо и спирально и являющиеся местами прикрепления
листьев. Эти бугорки отдаленно напоминают листовые подушки рода
Lepldodendron, хотя и не несут каких-либо признаков листовых рубцов
или других знаков. Центральный проводящий пучок из лестничных тра-
хеид имеет треугольную форму.
Продолговатые или овальные, повидимому без ножки, спорангии рас-
полагаются на верхней поверхности листьев.
Типичный вид. Protolepldodendron scharyanum К г е j t i.
Распространение. Нижний и средний девон.
Protolepldodendron scharyanum Krejci
Табл. LXX, фиг. 2; табл. LXXV, фиг. 3 и 4; табл. LXXVI, фиг. 1 и 2
Является наиболее полно изученным видом рода, происходящим из
среднего девона Чехии и Германии.
Растение имеет горизонтальное корневище, от которого поднимаются
до 0,5—1,0 см толщиной дихотомически разветвляющиеся облиственные
стволы. Чаще всего на одном и том же образце можно наблюдать по
краям узкие листья, бифуркирующие только у самого кончика, а на
самом стволе — спирально расположенные овальноромбовидные рубцы.
Овальные спорангии не были собраны в спороносные колоски, а рас-
полагались на верхней поверхности некоторых листьев.
В центре стволов имеется солидный проводящий пучок треугольной
формы из лестничных трахеид.
Нижнедевонский вид Protolepldodendron wahnbachense Kraus, et
W e у 1. отличается наличием на стволах, вместо овальноромбовидных
рубцов, коротких гребней от низбегания листьев на ствол и листьями,
бифуркация которых начинается на половине длины листа или даже
раньше.
Распространение. Средний девон Чехии, Германии, Китая
(Юннань), Алтая (район Змеиногорска и окраина Чуйской степи), вос-
точного склона Кузнецкого Алатау, Минусинской котловины. В Чуйской
степи растение найдено под морскими слоями с фауной брахиопод
•верхнеживетского возраста.
Род Heleniella Zalessky
Кора стволов покрыта продольными ребрами, между которыми рас-
полагаются волнистые бороздки. Листовые рубцы приурочены к расши-
ренным частям ребер и чередуются между собой на смежных ребрах.
М. Д. Залесский выделил эти растительные остатки в особый род из
группы сигиллярий с волнистыми ребрами, на основании того, что ори-
гинальная волнистость их ребер приближает эти растения к некоторым
лепидодендронам. М. Д. Залесский высказал предположение, что род
Heleniella можно рассматривать как предковую группу сигиллярий
и лепидодендронов.
Типичный вид. Heleniella Tschlrkovaeana Zal.
Распространение. Верхний девон — нижний карбон.
Heleniella Theodorl Zalessky
Табл. LXXVIII, фиг. 7
Характеристика данного вида совпадает с родовой характеристикой.
Тонкие стволы, иногда дихотомически разветвленные, покрыты корой,
на поверхности которой образовались волнистые продольные ребра
и разделяющие их волнистые бороздки. Волнистость обусловлена законо-
мерным расширением и сужением ребер. В самых широких местах, не-
сколько ниже неясных листовых рубцов, ширина ребер достигает 1,5 мм,
а в пережимах она становится не более 0,75 мм. Листовые рубцы
19 Зак. 2024 289=
не установлены и нигде не изображены, хотя М. Д. Залесский в описа-
нии типичных образцов вада, происходящих из верхнего девона Донбас-
са, о них упоминает. Неясность вопроса с листовыми рубцами послу-
жила некоторым палеоботаникам основанием для присоединения Hele-
nlella Theodorl к роду Lepldodendropsis Lutz или в качестве особого
вида L. Theodorl, или в качестве синонима к виду L. Hlrmerl.
До проверки оригиналов М. Д. Залесского и изучения новых сборов
из местности, откуда происходят эти оригиналы, лучше пока воздержать-
ся от переименования вида, тем более что название Helenlella Theodorl
стало широко популярным в нашей геологической литературе.
От близких нижнекаменноугольных сигиллярий (Slglllarla younglana
К i d s t о n, S. canobiana Kids ton и H. Tchirkovaeana Zalessky)
отличается малой величиной стволов, мелкими ребрами и неясной формой
листовых рубцов.
Распространение. В Донбассе характеризует верхний девон.
Многочисленные остатки этого вида, встреченные на правом берегу
р. Чулым у совхозного поселка Игрыш в Минусинской котловине, стра-
тиграфически залегают между туранской и минусинской свитами и ха-
рактеризуют самые верхние горизонты девона или самые низы нижнего
карбона.
КЛАСС ARTICULATAE. ЧЛЕНИСТОСТЕБЕЛЬНЫЕ
ПОДКЛАСС PSEUDOBORNIALES. ПСЕВДОБОРНИЕВЫЕ
Род Pseudobornia N a t h о г s t
Довольно крупное растение с массивным горизонтальным главным
побегом, от которого вверх поднимались прямые, более тонкие побеги,
несущие в мутовках по четыре дланевидно разделенных листа, причем
отдельные доли расщеплены на узкие, бахромчато изрезанные по краям
лопасти. Плодоносящие шишки достигали 30 см длины.
Поверхность стеблей неясно членистая и неясно ребристая.
От сходных родов Asterocalamltes S с h i m р., Equlsetltes Sternberg
и др. отличается сложностью листьев и неясной членистостью и ребри-
стостью побегов.
Типичный вид. Pseudobornia urslna Nathorst.
Распространение. Верхний девон Медвежьего острова.
Pseudobornia cf. urslna Nathorst
Табл. LXXHI, фиг. 3; табл. LXXV, фиг. 1
Является единственным видом рода и поэтому его видовая характе-
ристика полностью совпадает с родовой характеристикой.
В Минусинской котловине в песчаниках красноцветной туранской
свиты во многих местах встречаются в изобилии отпечатки крупных ство-
лов и тонких ветвей с почти гладкой, неясно ребристой и неясно члени-
стой поверхностью. Так как встреченные до сих пор остатки залегают
в грубозернистых породах аллохтонно, то на них не сохранились ни
листья, ни органы плодоношения, которые могли бы вполне надежно
подтвердить наличие в Западной Сибири Pseudobornia urslna Nath.
Однако большое количество отпечатков стволов из Минусинской котло-
вины обнаруживает полное морфологическое сходство со стволами
Pseudobornia urslna с Медвежьего острова.
Распространение. Верхний девон Медвежьего острова и Мину-
синской котловины; в Минусинской котловине вид присутствует повсе-
местно, где имеются отложения туранской красноцветной свиты.
290
ПОДКЛАСС SPHENOPHYLLALES. КЛИНОЛИСТНИКОВЫЕ
Род Sphenophyllum Brongniart
Тонкие стебли и ветви не превышают в диаметре 1 см и обычно из-
меряются единицами миллиметров.
Стебель членистый, ребристый, обильно ветвящийся. Клиновидные,
суживающиеся к основанию листья располагаются в узлах мутовками
в числе трех или кратном трем. Располагаются они в соседних узлах
супротивно, не чередуясь (лист против листа). Листья веерообразно
пронизаны дихотомически ветвящимися жилками, цельные, надрезанные
или глубоко рассеченные, на узкие дольки. В побегах расположен цен-
тральный проводящий пучок, утолщающийся путем вторичного роста.
Органы спороношения в виде колосков, но встречаются виды, у которых
спорангии сидели просто в пазухах листьев обычных мутовок.
От родов Catamites и Asterocalamltes отличается тонким стеблем
и клиновидной формой листьев.
Типичный вид. Sphenophyllum cunetfolium (Sternberg)
Zeil.
Распространение. Верхний девон — пермь.
Sphenophyllum subtenerrlmum N athorst
Табл. LXXVII, фиг. 5 и б
Стебли имеют диаметр обычно 1—4 мм и в редких случаях достигают
12 мм. На отпечатках стеблей почти всегда выступают три продольных
ребра. Длина междоузлий изменчива и достигает 34 мм. В узлах стебли
всегда утолщены, что является характерным для рода.
Листья очень глубоко рассечены на очень узкие, почти волосовидные
дольки, имеющие срединную жилку, различимую лишь в лупу. Число
листьев в мутовках точно не установлено, но, повидимому, было боль-
ше, чем у сходных карбоновых видов.
Близкий вид Sphenophyllum tenerrimum (Е 11 i n g s h.) S t u г из
нижнего карбона отличается более длинными междоузлиями и слабее
выраженными утолщениями в узлах.
Распространение. Верхний девон Медвежьего острова. В За-
падной Сибири встречен в Северо-Минусинской котловине на правом
берегу р. Чулым у нижнего конца совхоза Игрышинского в переходных
слоях от туранской свиты к минусинской свите.
ПОДКЛАСС CALAMARIALES. КАЛАМИТЫ
Род Asterocalamltes S с h i m р е г
Древовидное растение имело полые, разветвленные, членистые ство-
лы, достигающие нередко значительных размеров. .Многочисленные
сосудистые пучки, расположенные вокруг центральной полости, образуют
на слепках (отливах) с этой полости округлые ребра и бороздки, кото-
рые при переходе через узлы не чередуются. Листья расположены мутов-
чато, не срастаются у основания; встречаются на более тонких конеч-
ных побегах, имеют длину до 10 см и повторно вильчато ветвятся.
Органы спороношения состоят из многочисленных спорофилловых
мутовок, собранных в шишковидный колос.
В ископаемом состоянии чаще всего встречаются внутренние ядра
(отливы внутренней полости) стволов.
От наиболее близкого нижнекаменноугольного рода Catamites отли-
чается тем, что у последнего ребра и бороздки чередуются в соседних
междоузлиях.
Типичный вид. Asterocalamltes scroblculatus (S ch loth.'
Ze ill.
Распространение. Верхний девон—нижний карбон.
19*
291
Asterocalamltes scroblculatus (Schlotheim) Zeiller
Табл. LXXVI, фиг. 3—5
Наиболее часто в ископаемом состоянии встречаются ядра внутрен-
них полостей стволов. Толщина таких отливов достигает 16,5 см. Ответ-
вления от главного ствола частые, но положение ветвей не особенно
правильное; боковые ветви могут давать свои ответвления. Листья рас-
полагаются мутовками в узлах, повторно вильчато разветвляются на
узкие дольки, достигая 10 см в длину. Спорофиллы сидят мутовчато,
в количестве шести или несколько более в каждой мутовке.
Распространение. Впервые появляется в верхнем девоне, ши-
рокое распространение получает в нижнем карбоне Европы и Малой
Азии. В Западной Сибири в игрышинскмх слоях, пограничных между
туранской и минусинской свитами. Широко распространен в минусинской
свите нижнекарбонового возраста.
КЛАСС ANEUROPHYTALES
Род Aneurophyton Krausel et We у land
Довольно крупное растение, состоящее из прямого, постепенно сужи-
вающегося кверху ствола, который лишь в самой верхней части дает
обильные боковые ответвления, напоминающие внешне довольно круп-
ные вайи папоротника трижды перистого типа. Перья последнего порядка
носят характер листовых органов; они снабжены мелкими двух- или трех-
кратно дихотомически разветвленными перышками со слегка загнутыми
кончиками.
Близкий род Rhacophyton С г е р i п отличается признаками, перечис-
ленными при его описании (см. ниже).
Типичный вид. Aneurophyton germanicum Kraus, et Weу 1.
Распространение. Средний и верхний девон Евразии и Север-
ной Америки.
Aneurophyton germanicum Krausel et Weyland
Табл. LXVIII, фиг. 4; табл. LXIX, фиг. 1 и 2; табл. LXX, фиг. 5
От ствола растения отходят крупные трижды перистые листовидные
вайи, напоминающие вайи папоротника. От главного стержня вайи (ра-
хиса?) отходят в стороны чередующиеся перья первого порядка, от ко-
торых, в свою очередь, отходят простые, узкие перья второго порядка.
Последние не имеют проводящих пучков и низбегают на ось разветвле-
ния первого порядка в форме влагалища. Перья второго порядка отсы-
лают еще более мелкие многочисленные перышки, обнаруживающие
одно-, двух- и трехкратное дихотомическое деление на узкие сегменты
со слегка загнутыми кончиками. Длина сегментов измеряется всего
несколькими миллиметрами.
Плодоносные вайи имеют такое же строение, как и стерильные, но
менее расчленены. Небольшие овальные спорангии располагаются пуч-
ками на мелких дихотомических разветвлениях.
Распространение. Средний девон в Эльберфельде в Германии.
В Западной Сибири фрагментарные остатки вида обнаружены в среднем
девоне Южно-Минусинской котловины (лог Тустужул на водораздела
рек Узунжул и Неня). А. Н. Криштофович отметил присутствие этого
вида в составе древнедевонской флоры Казахстана.
Род Rhacophyton С г е р i п
Дважды перистая вайя состоит из крепкой главной оси (рахиса)
« чередующихся боковых разветвлений (перьев первого порядка), несу-
щих на себе двурядно расположенные, так же чередующиеся листочки
292
или перышки, разделенные более или менее глубоко дихотомически на
линейные сегменты, слегка изогнутые на концах. Нежные листочки часто
не сохраняются на своих осях, особенно если растение захоронено аллох-
тонно.
Спороносная вайя отличается от стерильной супротивно расположен-
ными и сросшимися у основания на главной оси перьями первого поряд-
ка, несущими два споранпиевых, симметрично расположенных органа,
состоящие из несколько раз дихотомически разветвленной ножки, при-
чем последние разветвления заканчиваются многочисленными мелкими,
не превышающими 2 мм длины веретенонидными спорангиями.
Близкий нижнекарбоновый род Rhodea Goeppert и некоторые кар-
боновые виды рода Sphenopteris Brongniart по характеру дихото-
мически рассеченных листочков практически неотличимы от рассматри-
ваемого рода, но резко отличаются от него характером органов спороно-
шения. Наиболее близкий девонский род Aneurophyton Kraus, et
Weyl. отличается трижды перистой вайей, значительно меньшей рас-
члененностью листочков, более простым разветвлением спороносных
перьев.
Типичный вид. Rhacophyton incertum (Dawson) Kraus,
et W e у 1.
Распространение. Верхний девон.
Rhacophyton incertum (Dawson) Krausel et Weyland
Табл. LXXV, фиг. 2; табл. LXXVI, фиг. 6; табл. LXXVII, фиг. 1—4
Как типичный вид рода, это растение характеризуется всеми призна-
ками последнего, указанными выше.
Распространение. Верхний девон Донецкого бассейна, Бель-
гии и Северной Америки. Верхние горизонты верхнего девона Западной
Сибири — на правом берегу р. Чулыма у совхоза Игрышинского (Севе-
ро-Минусинская котловина), у с. Мариновки, Назаровского района,
Красноярского края (южная окраина Чулымо-Енисейской впадины), что
стратиграфически отвечает самым верхним горизонтам верхнего девона
или нижним горизонтам нижнего карбона.
КЛАСС FILICALES. ПАПОРОТНИКИ
Род Aneimites Dawson
Достаточно крупные продольнобороздчатые оси, повторно дихотоми-
чески разветвляясь, заканчиваются тонкими осями, переходящими в ко-
роткие перья, состоящие из яйцевидных или округлоклиновидных перы-
шек; жилкование веерное, жилки дихотомирующие.
Сходный по форме перышек род Adlantites Goeppert emend.
К i d s t о п отличается трижды и даже четырежды перистой вайей и чере-
дующимся расположением перышек на перьях последнего порядка;
перышки лишь в отдельных случаях могут быть сгруппированы в пучки,
тогда как у рода Aneimites это является правилом.
Типичный вид. Aneimites acadlca Dawson.
Распространение. Верхний девон — нижний карбон.
Aneimites acadlca Dawson
Табл. LXXVII, фиг. 7; табл. LXXVIII, фиг. 1-6
Толстая, повторно дихотомически ветвящаяся ось с тонкой продоль-
ной бороздчатостью заканчивается тонкими разветвлениями, на концах
которых размещаются перья с несколькими широкояйцевидными или
клиновиднояйцевидными перышками. Перышки цельные или слегка
неравномерно надрезанные, часто асимметричные, пронизаны дихотоми-
рующими, веерно распростертыми жилками.
293
Сходные виды рода Adiantites G о е р р е г t emend. К i d s t о п отли-
чаются сложностью строения вайи и преобладающим перистым располо-
жением перышек на перьях последнего порядка.
Распространение. Верхний девон в Западной Сибири (р. Чу-
лым, правый берег у совхоза Игрышингкого в Новоселовском районе)
и у с. Мариновки, Назаровокого района, Красноярского края (в погра-
ничных слоях между туранской и минусинской свитами); нижний карбон
Северной Америки.
Род Arckaeopteris Dawson
Сложный лист (вайя), нередко достигающий длины 60—90 см, всегда
имеет дважды перистое строение. Сильная ось или рахис вайи отсылает
двурядно в стороны длинные линейные перья первого порядка, несущие
также двурядно расположенные перышки (сегменты) овально-клиновид-
ной формы с густым веерным жилкованием. Эти перышки у одних видов
цельные, у других— в различной степени рассеченные.
Ось вайи гладкая или продольнополосчатая, причем в некоторых
случаях можно видеть, как из-под наружного слоя коры обнажается
чрезвычайно характерная скульптура в виде тесно сближенных парал-
лельных ребер, соединенных поперечными перемычками или морщинками,
придающими поверхности рахиса точечную скульптуру.
Спороносные перья размещаются в нижней (проксимальной) части
вайи. Они напоминают стерильные перья, но отличаются полной редук-
цией листовой пластинки, вместо которой на оси пера сидят непосред-
ственно или на коротких черешках одиночно, или в группах по 2—3,
гладкие веретеновидные или овальные спорангии. В одном случае твердо
установлена гетероспория (разноспоровость), что Крейзель и Вейланд
склонны считать типичным для рода в целом.
Типичный вид. Archaeopterls hibernlca (Forbes) Dawson.
Распространение. Верхний девон северного полушария; в За-
падной Сибири — не менее трех видов.
Archaeopterls Halliana (Goeppert) Dawson
Табл. LXX1V, фиг. 1 и 2
Стерильные '(неплодущие) перышки цельнокрайние, иногда слабо рас-
сечены, округлоклиновидной формы, часто с волнистыми краями. Они
располагаются на боковых перьях вайи очень густо, так что каждое
последующее перышко наполовину перекрывает предыдущее. Длина
перышек, как правило, меньше 2 см. Плодущие (спороносные) перышки
простые или вильчатые, несущие сидячие спорангии (по 5—10 на
перышке).
Близкий вид A. hibernlca (Forbes) Dawson отличается менее
плотным расположением перышек (они лишь слегка перекрывают друг
друга), более крупными их размерами, обычно превышающими 2 см,
и спорангиями, располагающимися на черешках.
Распространение. Верхний девон северного полушария.
В Западной Сибири — в туранской свите Северо-Минусинской котло-
вины (правый берег р. Енисея в Новосельском районе).
Archaeopterls obtusa Lesquereux
Табл. LXXIV, фиг. 3
Стерильные перышки цельнокрайные, клиновидной формы или вееро-
образные, с более или менее прямыми боковыми краями. Они распола-
гаются на боковых перьях вайи более или менее отдаленно друг от друга.
Длина перышек обычно больше 2 см. Плодущие перья большей частью
простые, несут сидячие спорангии от 8 до 20 штук на каждом пе-
рышке.
294
Сходная по размерам перышек форма A. hlbernlca отличается более
тесным их расположением (соседние перышки слегка перекрывают друг
друга) и спорангиями на черешках. A. Halltana отличается меньшим
размером перышек и значительно более густым их расположением.
Распространение. Верхний девон северного полушария. В За-
падной Сибири — туранская свита Минусинской котловины.
Archaeopterls sibirica Zalessky
Рис. 200 и 201
Дважды перистые вайи значительных размеров имеют продольно-
ребристую главную ось или рахис, достигающий толщины 6 мм в нижней
части. Боковые перья почти супротивные, достигающие иногда 11 см
длины. На боковых ветвях двурядно сидят листочки или перышки
Рис. 200. Archaeopterissibirica Zal.
'Отпечаток части вайн —очер-
тания и жилкование перышек. °/10
нат. вел. р. Красная—приток р. Яя,
Кузбасс (по М. Д. Залесскому)
Рис. 201. Archaeopteris sibirica Zal.
Часть вайи со стерильными и плодоносными
перьями. 9/,о нат. вел. р. Красная—приток р. Яя,
Кузбасс. Верхний девон (по М. Д. Залесскому)
последнего порядка, клиновидные по форме. Постепенно сужаясь к осно-
ванию, перышки принимают характер черешка, низбегаюшего на ось
пера. Жилкование перышек веерообразное. Перышки сильно изменчивы—
цельные или рассечены на доли, до бахромчатого состояния. Плодущие
перышки сильно изменены, несут мелкие спорангии овальной формы.
Близкие A. macilenta L е s q. и A. flssllis S с h m а 1 h. отличаются тем,
что имеют более постоянную форму перышек, всегда так или иначе рас-
сеченных; цельные перышки у них, как правило, отсутствуют.
Распространение. Верхний девон северо-восточной окраины
Кузбасса (р. Красная, приток р. Яя).
Archaeopteris sp.
Табл. LXXIV, фиг. 4—7
Очень часто в ископаемом состоянии встречаются утратившие кору
стержни вайи папоротника, обнаруживающие характерную для рода
Archaeopteris скульптуру в виде тесно сближенных продольных рядов
295
точек или ямок. Такие стержни часто встречаются среди аллохтонных
скоплений растительных остатков, претерпевших длительный перенос
водой и лишившихся не только перьев и перышек, но и наружного слоя<
коры. Хотя подобные остатки не допускают видовых определений, уже-
установление их принадлежности к роду Archaeopterls позволяет с точ-
ностью до отдела определить их геологический возраст.
В Западной Сибири они встречаются в аллохтонном залегании
в туранской свите Минусинской котловины.
Г Л AHA 1 V'
НИЖНИЙ КАРБОН
Л. Л. ХАЛФИН
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Главнейшими областями развития морского нижнего карбона в За-
падной Сибири являются Кузбасс и связанный с ним Горловский бас-
сейн и Колывань-Томская складчатая дуга. Нижнекаменноугольные отло-
жения окаймляют угленосную площадь Кузбасса. Они содержат богатую
и разнообразную фауну, из состава котовой наиболее изучены кишечно-
полостные (Габуния, 1919; Толмачев, 1924 и 1931; Фомичев, 1931),
мшанки (Нехорошев, 1926) и брахиоподы (Толмачев, цит. соч.; Ротай,
1938).
А. П. Ротай делит эти отложения на пять биостратиграфических зон
(снизу вверх):
1. Абышевская — состоящая из известняков и песчаников, часто не-
мых в палеонтологическом отношении, и характеризующаяся значитель-
ными изменениями состава в различных районах ее развития. Фауна
носит смешанный характер: наряду с верхнедевонскими спириферами
группы Cyrtosplrifer verneuili (Murch.) в ней присутствуют нижнекар-
боновые виды из группы Splrifer tornacensls Коп., а также Productus
(Dictyoclostus) burlingtonensls Н а 11 и др.
2. Тайдонская — сложена серыми и темносерыми известняками и лишь
в Барзасском и Анжеро-Судженском районах замещается (частично или
полностью) песчаниками и аргиллитами. Содержит богатую и разно-
образную фауну турнейского яруса: Splrifer tornacensls Коп., Splrifer
usslensis То 1m., представителей рода Syrlngothyris Winch., Dictyo-
clostus burlingtonensls (Hall), Caninla cyllndrlca Seoul, и др.
3. Фоминская — почти повсеместно представлена темносерыми изве-
стняками с включениями черных кремней. Чрезвычайно богатая фауна
характеризуется смешением турнейских и визейских форм и очень круп-
ными размерами многих ее представителей. Особенно характерны: спи-
риферы группы Splrifer cinctus Коп., некоторые представители рода
Syringothyrls, Lltostrotion grande Т о 1 m.; появившиеся здесь продуктиды
Dictyoclostus deruptus (Rom.), Dictyoclostus robustus (To Im.), а также
Caninla ussovi G a b. и другие переходят в вышележащую зону.
4. Подъяковская—сложена темносерыми известняками, сильно загряз-
ненными примесью глинистого и песчаного материала. Многие формы
брахиопод переходят в эту зону из фоминской, но фауна уже имеет
типично выраженный визейский характер: Dictyoclostus robustus
(ТоIm.), Dictyoclostus deruptus (Rom.), Hemltrypa proutana U 1 r.,
Llthostrotion slbiricum G a b., Llthostrotion Irregulars P h i 11. var. bala-
khonkaensis Fom., Caninla ussovi G a b. и др.
297
5. Верхотомская — повсеместно образована мелкозернистыми зеле-
ными или зеленовато-серыми, нередко известковистыми песчаниками
с прослоями известняков. В составе фауны преобладают формы, перехо-
дящие из подъяковской зоны; наиболее обычны Dlctyoclostus robustus
(Т о 1 m.), Echlnoconchus punctatus (М art.) и некоторые другие.
А. П. Ротай три первые из перечисленных выше зон относит к тур-
нейскому ярусу, а две последние — к визейскому. Р. Н. Бенедиктова
установила, что все пять зон выделяются также в нижнем карбоне Гор-
ловского бассейна и характеризуются в целом теми же группами бра-
хиопод; фауна здесь более бедная, чем в Кузбассе, но не обнаруживает
существенных отличий.
Фауна нижнего карбона Колывань-Томской складчатой области изу-
чена в районе г. Томска, где по р. Томи имеется хороший разрез этих
отложений. Они представлены здесь преимущественно глинистыми слан-
цами, которым подчинены прослои алевролитов и песчаников. В некото-
рых слоях содержится обильная фауна мшанок, брахиопод и пластин-
чатожаберных моллюсков, описанная в 1915 г. М. Э. Янишевоким, но
требующая сейчас более детального изучения.
К числу наиболее распространенных форм в этой фауне относятся
Splrlfer stblrlcus Janich., Sptrlferlna octopllcata Sow. var. concentrtca
J a n i c h., Schuchertella tomsklensls Janich., Fenestella plebeja M’C о у,
Polypora slblrica J a n. и некоторые другие.
He все отложения указанного разреза являются морскими; местами
в них появляются слои с наземной флорой: Asterocalamltes S с h i m р.,
Archaeopterls grandlfolla Tchirk., Angaropteridlum cardlopteroldes
S c h m a 1 h.
Нижнекаменноугольные отложения из района г. Томска протяги-
ваются на юго-запад и обнажаются на р. Оби в районе горы Камня, где,
по данным Л. Н. Краевской и М. П. Ломовицкой (1935), наряду
с турнейскими, имеются более молодые (средне- или даже верхнекарбо-
новые) морские осадки.
В морской фации нижний карбон имеется также в Рудном Алтае, где
разрез его детально изучен В. П. Нехорошевым. В отложениях С, Руд-
ного Алтая преобладают глинистые и песчанистые сланцы большой
мощности с подчиненными горизонтами эффузивов и известняков.
К нижнему карбону здесь относится верхняя часть тарханской свиты,
в частности ретепориновая толща с многочисленными мшанками, среди
которых особенно характерна Reteporina altalca Nekh. (Нехорошей,
1926а). Выше залегает бухтарминская свита с Splrlfer tornacensls Коп.,
Syringothyrls sp. и другими брахиоподами и мшанками, которая пере-
крывается ульбинской свитой с многочисленными мшанками (Polypora
slblrica Janich., Hemttrypa proutana U 1 г. и др.), брахиоподами и мол-
люсками. Возраст всех этих отложений от переходного девоно-карбоно-
вого до нижневизейского включительно.
В Минусинской котловине нижний карбон представлен континенталь-
ными отложениями с наземной флорой.
В последнее время все больше появляется материалов в пользу отне-
сения к нижнему карбону острогской свиты, залегающей в основании
угленосной толщи Кузбасса ’.
В. Д. ФОМИЧЕВ
Тип Coelenterata. Кишечнополостные
Кишечнополостные широко распространены в отложениях нижнего
карбона Кузбасса; их описание содержится в работах К. Е. Габуния
(1919), И. П. Толмачева (1924, 1931) и В. Д. Фомичева (1931).
1 Широкое совещание, состоявшееся в июле 1954 г., после всестороннего обсужде-
ния этого вопроса вполне единодушно признало нижнекарбоновый возраст острогской
свиты доказанным (см. т. II).
298
КЛАСС ANTHOZOA
ПОДКЛАСС TABULATA
СЕМЕЙСТВО FAVOSITIDAE EDWARDS ET HAIME
Род Michelinia Koninck
Дискоидальной или выпуклой формы полипняки, часто значительной
величиньи; на нижней стороне имеют эпитеку с корнеобразными отрост-
ками. Кораллиты многоугольные; перегородки заменены вертикальными
струйками на стенках. Поры в стенках размещены беспорядочно. Днища
многочисленные, расщепленные, выпуклые кверху. Крупноячеистые коло-
нии представителей этого рода легко распознаются среди прочих табу-
лят. Реже встречаются несколько разветвленные полипняки с коралли-
тами округлого сечения.
Типичный вид. Michelinia favosa Коп.
Распространение. Карбон — пермь; особенно часто встречается
в карбоне, редко — в перми. В Западной Сибири этот род известен пока
только из нижнекаменноугольных отложений.
Michelinia ienulsepta Phillips
Табл. LXXIX, фиг. 1
Полипняки имеют форму более или менее высокого, обычно несколько
изогнутого конуса, прикрепляющегося к субстрату узким концом. Тол-
стая эпитека их покрыта продольными складками и поперечной морщини-
стостью. Кораллиты неправильно многогранной формы, с тремя или
четырьмя гранями у молодых, семью и восемью — у более взрослых.
Наибольший поперечник их равняется 9 мм, но рядом с крупными корал-
литами встречаются и другие, значительно меньших размеров (молодые)
Соответственно общей форме полипняка отдельные кораллиты очень
высоки. Стенки их пронизаны порами, расположенными не совсем пра-
вильно. Днища многочисленны, расщеплены, различно сближены, гори-
зонтальны или наклонны, обычно выпуклы кверху.
Вместе с колониями небольших размеров в Кузбассе имеются и более
крупные (наиболее крупный экземпляр без отломанной вершины конуса
имеет в длину 10 см, а верхний диаметр до 6 см). Угол конусности у куз-
нецких М. ienulsepta сильно варьирует: наряду с полипняками с острым
углом при вершине имеются формы с широкой верхней поверхностью,
т. е. с гораздо большим числом кораллитов при той же высоте конуса,
что и в первом случае. Форма полипняка, по мере его роста, из кони-
ческой нередко превращается в неправильно цилиндрическую.
Распространение. Кузбасс — тайдонская и фоминская зоны.
Широко распространена в нижнем карбоне других областей СССР,
а также в Англии и Бельгии.
Michelinia megastoma Phillips
Табл. LXXIX, фиг. 2
В пределах этого вида намечаются две нерезко разграниченные
группы полипняков:
1. Форма А. Более выпуклые колонии, имеющие форму шарового
сегмента. Диагональ кораллитов, равная 7—9 мм у молодых колоний
С поперечником полипняка в 35—45 мм, значительно больше у крупных
с поперечником 10—12 см. Толстая стенка веерообразно расходящихся
кораллитов становится более тонкой в периферических частях больших
колоний. Днища, в соответствии с выпуклой формой колоний, довольно
сильно выпуклы кверху; ширина их (видно в продольном сечении)
обычно не превышает половины поперечника кораллитов.
299
2. Форма В. Плоские, лепешкообразные колонии округлой или не-
правильно эллипсоидальной формы. Диагональ кораллитов достигает
в среднем 11 —12 мм, т. е. того же порядка, что и у представителей пре-
дыдущей группы. Молодые экземпляры обеих групп не отличимы друг
от друга. В соответствии с плоской невысокой формой колонии и днища
внутри кораллитов здесь более плоские; ширина их больше половины по-
перечника кораллитов (видно в продольном сечении).
Кораллы группы А развиты преимущественно на востоке и юго-во-
стоке Кузнецкого бассейна, в то время как кораллы группы В — пре-
имущественно на западе и северо-западе его и в Горловском бассейне.
Распространение. В Кузбассе встречается совместно с преды-
дущим видом, а также в подъяковской зоне. Нижний карбон Подмосков-
ной котловины, Казахстана, Англии и Бельгии.
СЕМЕЙСТВО SYRINGOPORIDAE EDWARDS ET HAIME
Род Syringopora G о 1 d f u s s
Описание см. на стр. 201.
Представители этого рода широко распространены в нижнем карбоне
Кузбасса и Горловского бассейна и относятся к нескольким видам.
Syringopora ramulosa G о 1 d f u s s
Табл. LXXIX, фиг. 3
Крупные кустистые полипняки неправильной формы. Отдельные
кораллиты их длинные, в разной мере изогнутые (сильнее в местах при-
крепления соединительных трубок), расположены местами почти парал-
лельно, местами слегка радиально, на расстоянии в среднем около 2 мм
друг от друга. Диаметр отдельных кораллитов около 2,5 мм. Соедини-
тельные трубки отстоят друг от друга на 5—10 мм, редко более. Поверх-
ность кораллитов покрыта слабыми поперечными складочками. Днища
многочисленны, расщеплены, имеют форму глубоких воронок.
Вследствие изгибания кораллитов расстояние между ними не посто-
янно. Наряду с сближенными кораллитами попадаются такие, расстояние
между которыми достигает 6—7 мм. Соединительные трубки толстые.
Распространение. Syringopora ramulosa пользуется широким
географическим распространением, встречаясь в отложениях турнейского
и визейского ярусов; в Кузбассе главным образом в тайдонской и фо-
минской зонах.
ПОДКЛАСС ZOANTHARIA
Отряд Tetracoralla или Rugofea
СЕМЕЙСТВО ZAPHRENTIDAE EDWARDS ET HAiME
Род Stereophrentis F о m i t s h e v
Небольшие конические кораллы. Главная септа лежит в углублении
(фоссуле). Септы первого порядка доходят до оси коралла и, срастаясь
внутренними концами, обрамляют фоссулу. Септы второго порядка
короткие или отсутствуют. Днища многочисленные, приподнятые по оси
коралла, доходят до наружной стенки. Столбик и пузырчатая ткань
отсутствуют.
Этот род объединяет нижнекарбоновые виды, ранее относившиеся
к роду Zaphrentis R a f. et Cliff., установленному для среднедевонских
форм, но видимо, не имеющие генетической связи с карбоновыми видами.
Типичный вид. Stereophrentis delanouei Е d w. et H.
Распространение. Нижний карбон (турнейский и визейский
ярусы).
ЗОН
Stereophrentls konincki Edwards et Haime
Табл. LXXIX, фиг. 4—7
Кораллы конические, с почти круглым поперечным сечением. Боковая
поверхность их со слабо заметной продольной бороздчатостью и с попе-
речными утолщениями — знаками нарастания. Длина кораллов достигает
40(45) мм. Угол конусности сильно изменчив. Максимальный диаметр
поперечного сечения 25 мм, но встречаются также и узкие кораллы, у ко-
торых, при той же длине, диаметр поперечного сечения не превышает
13 мм (имеются и промежуточные экземпляры). Вместе с крупными
кораллами в изобилии встречаются более мелкие.
Фоссула расположена на выпуклой стороне коралла. Чашечки неглу-
бокие. Септы первого порядка почти прямые, иногда несколько волнооб-
разно (S-образно) изогнуты. Они слабо утолщены у наружного края;
в главных квадрантах более толстые, чем в противоположных. Число их
различно в разных сечениях и зависит от диаметра кораллов: при диа-
метре 22—26 мм максимальное число их достигает 46—47. Септы второго
порядка хорошо развиты, имеются и в молодых сечениях.
Фоссула расширена к внутреннему концу и почти целиком выполнена
стереоплазмой, густо отлагающейся по ее стенке. Это обильное отложе-
ние стереоплазмы не позволяет наблюдать главную септу; последняя во
всяком случае не длиннее септ второго порядка, так как во внутренней
части фоссулы, обычно свободной от стереоплазмы, ее не видно. Фоссула
более длинная в молодых сечениях, когда она захватывает и осевую
часть коралла, чем во взрослых, где она часто только доходит до оси.
В продольном сечении днища неправильные, часто крупнопузырчатые,
очень приподняты к оси коралла и круто падают к фоссуле и в противо-
положную сторону.
Распространение. Западная Сибирь (тайдонская и фоминская
зоны Кузбасса), Урал, Англия и Бельгия.
В нижнекаменноугольных отложениях Кузбасса род Stereophrentls
представлен рядом видов, внешне близких к описанному и отличающихся
главным образом различными деталями внутреннего строения (в первую
очередь отсутствием септ второго порядка).
Род Caninia Michelin
Одиночные конические и цилиндро-конические кораллы. Септы пер-
вого порядка до оси коралла не доходят, в главных квадрантах утол-
щены. Главная перегородка сильно укорочена, расположена в фоссуле.
Часто имеются септы второго порядка.
Днища хорошо развиты, широкие, слегка выпуклые кверху. Пузырча-
тая ткань всегда присутствует во взрослых стадиях развития кораллов.
Кроме мелких диссепиментов пузырчатой ткани, расположенных между
септами, иногда присутствует кольцо диссепиментов, отделяющее перего-
родки от стенок. В поперечном и продольном сечениях достаточно резко
обособляются зона пузырчатой ткани и зона днищ (табл. LXXX,
фиг. 1—3).
Типичный вид. Caninia cornucoplae Mich, из нижнего карбона.
Распространение. Карбон, преимущественно нижний отдел.
Для нижнего карбона Кузбасса чрезвычайно характерным многочислен-
ные, порой очень крупные представители этого рода (табл. LXXX,
фиг. 4).
Caninia cylindrica S с о u 1 е г
Табл. LXXX, фиг. 1—3
Форма кораллов удлиненноконическая, слегка искривленная и почти
цилиндрическая в верхней части. На наружной поверхности их имеются
лродольная штриховатость и поперечные кольцевые борозды и вздутия.
В поперечном сечении ясно различимы три зоны:
301
1. Периферическая, сложенная из очень больших, частично сильно
удлиненных пузырьков; участками наблюдаются более мелкие пузырьки.
Септы эту зону не пересекают, но во внутренней ее части на продолже-
нии септ обоих порядков имеется ряд зубчиков, отходящих во внутрь от
выпуклой стороны пузырьков.
2. Промежуточная, занятая септами. В главных квадрантах септы
покрыты стереоплазмой. Фоссула намечается более короткими главной
и 2—4 соседними с ней перегородками первого порядка. Септы в общем
имеют радиальное направление и только соседние с фоссулой обращены
внутренними концами в ее сторону. Септы второго порядка слабо выра-
жены.
3. Центральная, свободная от септ, шириной около 0,15 диаметра
сечения.
В продольном сечении периферическая часть занята крупными
пузырьками, вытянутыми параллельно стенке коралла. Среди крупных
пузырьков в беспорядке расположены более мелкие. Ширина зоны
пузырчатой ткани достигает 0,25 сечения коралла. Осевая часть сечения
выполнена днищами, выпуклыми кверху; края их отогнуты книзу,
особенно круто в сторону фоссулы; стереоплазма густо покрывает зону,
пограничную между пузырьками и днищами, со стороны фоссулы. Длина
кораллов достигает 150 мм и более, диаметр поперечного сечения до
43 мм, число перегородок 60.
Вышеприведенное описание характеризует коралл во взрослой стадии
развития.
В молодых стадиях все септы толстые, доходят до стенки коралла
и не доходят до оси его. Затем появляются крупные длинные диссепи-
менты, отделяющие перегородки от стенки коралла, а утолщение (стерео-
плазма) сохраняется лишь на септах главных квадрантов. В еще более
взрослых стадиях зона пузырчатой ткани расширяется и строение ее
усложняется, а стереоплазма утолщает лишь главную и несколько бли-
жайших к ней септ. Септы второго порядка все время слабо развиты.
Распространение. Турнейские и нижневизейские отложения
СССР (в частности, Кузбасса) и Западной Европы.
Canlnla ussovi G a b u n i а
Табл. LXXX, фиг. 9
Крупные, удлиненноконические или почти цилиндрические кораллы,
слабо изогнутые. Боковая поверхность их с продольной ребристостью
и с резкой поперечной штриховатостью.
В поперечном сечении (во взрослой стадии развития) выделяются
три зоны:
1. Периферическая, занятая наружными концами септ первого и вто-
рого порядка и пузырчатой тканью. Ширина ее может достигать
0,25—0,35 радиуса. Стереоплазма сюда не проникает. Пузырчатая ткань
мелкоячеиста. Пузырьки расположены интерсептально: они мелкие, до-
вольно правильной четырехугольной формы и иногда совершенно не
прерывают септ, которые в таких случаях доходят до стенки коралла.
Обычно же вдоль самой стенки появляются длинные (часто очень длин-
ные) узкие пузыри, прерывающие перегородки. От выпуклой стороны их
отходят короткие зубчики на продолжении септ обоих порядков (такого
же вида, как и у С. cyllndrlca).
2. Промежуточная, занятая внутренними концами септ первого
и второго порядка (общая длина последних достигает 0,5 длины септ
первого порядка). Септы второго порядка свободны на всем своем про-
тяжении (в этой зоне) или же соединены с септами первого порядка
более или менее частыми перемычками; в одном и том же шлифе встре-
чаются и свободные и соединенные септы второго порядка. Укорочены
302
главная и противоположная септы, так что иногда их можно смешать
друг с другом.
3. Центральная, свободная от септ и занятая днищами. Как правило,
С. ussovi встречается в Кузбассе в более или менее деформированном
виде; особенно сдавлена периферическая часть сечений, к которой при-
урочена область развития длинных пузырей: обычно тут наблюдаются
лишь мелкие обломки диссепиментов и концов септ. Стенка коралла
сохраняется еще реже.
В продольном сечении осевая часть занята днищами, не очень сбли-
женными: среднее расстояние между ними 2 мм. Они выпуклы кверху
(в средней части их иногда наблюдается некоторая вдавленность), по
краям более круто отогнуты книзу, а в месте соединения с пузырчатой
тканью снова заметно отгибаются кверху. Днища простого типа, не
пузырчаты, продолжаются через большую часть сечения коралла. Пери-
ферическая часть сечения занята пузырчатой тканью: пузырьки длинные
и узкие, вытянуты кверху и выпуклы во внутрь. Стереоплазматическая
стенка довольно хорошо выражена на границе между днищами и пузыр-
чатой тканью.
Распространение. Нижневизейские отложения Кузбасса.
СЕМЕЙСТВО L1THOSTROTIONTIDAE GRABAU
Род Llthostrotion L h и у d
Колониальные кораллы из цилиндрических или призматических корал-
литов с полностью развитыми стенками. Септы двух порядков. Почти
всегда имеется пузырчатая ткань; диссепименты ее не прерывают септ.
По оси кораллитов наблюдается грифелевидный пластинчатый столбик.
Хорошо развитые днища приподняты к столбику.
Типичный вид. Llthostrotion striatum F1 еm.
В Кузнецком бассейне, наряду с массивными полипняками, встре-
чаются и кустистые колонии этого рода, образованные цилиндрическими
кораллитами.
Распространение. Карбон, главным образом нижний; повсе-
местно. Из турнейских отложений не известен. Появление Llthostrotion
совпадает с началом визейского века.
Llthostrotion (Eolithostrotlonella) slbiricum G a b u n i а
Табл. LXXX, фиг. 5 и 6
Массивные полипняки сложены тесно соприкасающимися коралли-
тами; поперечник кораллитов достигает 8—10 мм, редко больше. Наряду
со взрослыми имеются более молодые кораллиты, меньших размеров.
Полипняки достигали значительных размеров, с поперечником до 250 мм
и высотой до ПО мм, но есть и более мелкие колонии. Тонкая стенка
полипняка с поперечной бороздчатостью волнообразно переходит с од-
ного индивида на другой. Чашечка обычно неглубокая, с широким,
полого опускающимся во внутрь бортом. Тонкие септы обоих порядков
спускаются по нему невысокими ребрышками, начинаясь у стенки или
несколько отступая от нее, и снова поднимаются кверху в осевой части
кораллитов, примыкая к тонкому пластинчатому столбику. Фоссула неза-
метна.
В поперечном сечении кораллиты обычно 5—6-угольного очертания.
В очень крупных кораллитах число сторон может доходить до восьми.
Во взрослых кораллитах имеется периферическая зона крупных пузырей;
ширина ее варьирует и может достигать 0,35 поперечника кораллитов.
Хорошо развитая внутренняя стенка отделяет периферическую зону от
внутрилежащей. Внутри последней развиты септы обоих порядков: септы
первого порядка в количестве 12—16 (до 18) обычно доходят до центра
и вес или только часть их — то столбика.
303
Столбик имеется не во всех сечениях кораллитов. Он тонкий, пластин-
чатый или чечевицеобразный, продолжается своими вытянутыми концами
в две подходящие к нему с противоположных сторон септы первого
порядка. В ряде кораллитов эти две септы просто срастаются, продол-
жаясь одна в другую, и не образуют утолщенного столбика. Есть, нако-
нец, сечения, в которых ни столбика ни срастания септ не наблюдается.
Наружные концы септ первого порядка доходят до внутренней стенки
или же продолжаются дальше, во внешнюю пузырчатую зону, и могут
достигать стенки кораллита.
Септы второго порядка имеют форму коротких зубчиков, развитых от
внутренней стенки кораллита по направлению к центру; они могут про-
должаться на некоторое расстояние в противоположном направлении,
в зону пузырчатой ткани. Имеются сечения кораллитов без септ второго
порядка. Стенка кораллитов тонкая. Ряд следов пересечения днищ, обра-
щенных выпуклостью к центру, связывают септы; они располагаются
рядами (дугами). Имеются и диссепименты, обращенные выпуклостью
к периферии; они расположены у самой внутренней стенки, вне зоны,
занятой следами пересечения днищ.
В продольном сечении периферические части кораллитов заняты
крупноячеистой пузырчатой тканью. Пузырьки располагаются 1—2 ря-
дами и вытянуты по направлению от периферии к оси кораллитов, пер-
пендикулярно к стенке. Средняя часть сечения занята днищами. Послед-
ние приподняты в осевой части и опущены по краям. Приближаясь
к пузырчатой ткани, они еще круче начинают отгибаться книзу, довольно
часто при этом расщепляются, а затем, в месте соприкосновения
с пузырьками, несколько отгибаются кверху.
По оси кораллитов проходит столбик. Последний развит неодинаково
з разных кораллитах и колониях: он то продолжается непрерывной
линией через все сечения кораллита, то несколько прерывается, не вполне
доходя от одного днища до другого, что сразу же сопровождается выпо-
лаживанием днищ в центральной их части. Могут быть, наконец, слу-
чаи. когда столбик прерывается и на некоторых участках совсем отсут-
ств ует.
Наличие крупноячеистой пузырчатой ткани позволяет отнести этот
вид к роду Lithostrotlon лишь с известными оговорками. Точнее будет
отнесение его к близкому роду Eollthostrotionella Z h i z h i n а, отличаю-
щемуся от Lithostrotlon, по наличию крупных, прерывающих септы пу-
зырьков пузырчатой ткани.
Распространение. Характерен для нижневизейской подъяков-
ской зоны Кузбасса; в верхнетурнейской фоминской зоне присутствует
Lithostrotlon grande Т о 1 m., отличающийся прежде всего более круп-
ными (до 2 см в поперечнике) кораллитами (табл. LXXXI, фиг. 1, 2).
Lithostrotlon irregulare Phillips var. balachonkaensls Fomitchev
Табл. LXXX, фиг. 7 и 8
Полипняки состоят из длинных, цилиндрических, слабо изгибающихся
кораллитов, находящихся на расстоянии 0,5 (и меньше) диаметра корал-
лита друг от друга. Стенка кораллитов с тонкой поперечной морщини-
стостью и со слабо заметной продольной ребристостью. Столбик тонкий,
пластинчатый. В поперечном сечении диаметр отдельных кораллитов
достигает 5—6 мм. Число септ первого порядка 20 (редко 22): две из
них подходят с противоположных сторон к столбику, срастаясь с ним,
остальные несколько не доходят до него (за немногими исключениями).
Длинные септы второго порядка лишь немного короче септ первого по-
рядка; они далеко вдаются во внутрь за периферическое кольцо пузырча-
той ткани, состоящей из одного ряда пузырьков. Внутренняя стенка,
304
соответствующая внутренней границе пузырчатой зоны, отчетливо наблю-
дается в ряде кораллитов.
В продольном сечении наблюдается по одному ряду пузырьков вдоль
стенок ячей, столбик, занимающий осевое положение, и тонкие, припод-
нятые к столбику днища (около двух на 1 мм).
От типичных L. Irregulare Р h i 11. эта кузбасская разновидность отли-
чается более длинными септами второго порядка, которые почти равны
по длине септам первого порядка, и несколько меньшим числом септ, при
том же диаметре кораллитов.
Распространение. Кузбасс, подъяковская зона.
П. С. КРАСНОПЕЕВА
Тип Bryozoa. Мшанки
Отряд Trepostomata
СЕМЕЙСТВО BATOSTOMELLIDAE ULRICH
Род Tabulipora Young
Колонии различной формы (массивные, ветвистые и т. д.). Мезопоры
различных размеров, распределены неравномерно. Диафрагмы продыряв-
ленные, чем это род отличается от рода Stenopora L о n s d.
Распространение. Карбон—пермь, повсеместно.
Tabulipora elbaschensls Krasnopeeva, sp. n.
Табл. LXXXI, фиг. 3; рис. 202
Колония двуслойносимметричная, ланцетовидная, с одного конца
ее — два двуслойносимметричных пласта. Ячейки резко изгибающиеся
вблизи основания. Диафрагмы прободенные; на длину одной ячейки
Рис. 202. Tabulipora. elbaschensls Krasn., sp. n.
a—сечение, касательное к поверхности колонии; б—поперечное сечение колонии.
Турнейский ярус Салаира
насчитываются 3—5 диафрагм. Акантопоры крупные, по 2—4 вокруг
ячейки. Мезопоры различных размеров, по 3—5 вокруг каждой ячейки.
Толщина зоарии с одного конца 2,5 мм, с другого — около 5,0 мм. На
поверхности окаменелости наблюдаются пятна более крупных ячеек.
Диаметр устьев ячеек 0,5 мм, на пятнах — 0,7 мм. Ближе к основанию
в стенках ячеек наблюдается темная разделяющая линия. Толщина сте-
нок ячеек 0,03—0,10 мм. В продольном сечении ячейки трубковидные;
своими основаниями они отходят от общего слоя мезотеки, толщина ко-
торой около 0,4 мм. Ячейки резко изгибаются вблизи основания и выхо-
дят на поверхность почти под прямым углом.
Некоторое сходство описанная форма обнаруживаете Stenopora ovata
L о n s d., но последняя имеет цилиндрическую колонию, состоящую из
двух концентрических слоев; кроме того, у нее нет диафрагм в ячейках.
20 Зак. 2024 305
St. intercalarta U1 г. имеет ветвистую колонию; ячейки у нее отходят
радиально от ее воображаемой оси; диафрагмы приурочены к осевой
части колонии.
Распространение. Турнейский ярус Салаира (р. Каменный
Елбаш, выше дер. Хариной в системе р. Бердь).
Отряд Cryptostomata
СЕМЕЙСТВО FENESTELLIDAE KING
Род Fenestella Lonsdale
Описание см. на стр. 231.
Fenestella rudls Ulrich
Табл. LXXXI, фиг. 4; табл. LXXXII, фиг. 5
Основные размеры: 14—15/9—11//18—20. Ячейки пятиугольного сече-
ния в основании. Киль и шипы выражены отчетливо. Зоария веерообраз-
ная. На ячеистой поверхности прутьев наблюдается резко выраженный
киль с шипами. Прутья прямые, толщиной примерно 0,35 мм. Неячеистая
поверхность прутьев и перекладин покрыта бугорками различной вели-
чины. Кроме того, на неячеистой поверхности прутьев имеются массив-
ные шипы, сходные с шипами ячеистой поверхности. Поперечник шипов
около 0,05 мм; число их на длину одной петли равно трем. Поперечные
перекладины килевидные на неячеистой поверхности; ширина их около
0,35 мм. Длина петель 0,7—0,8 мм, ширина 0,35 мм. Устья ячеек круг-
лые, с поперечником 0,15 мм. На длину одной петли приходится четыре
ячейки.
Распространение. Нижний карбон Кузбасса — фоминская,,
подъяковская и верхотомская зоны (р. Выдриха, ниже дер. Беловой;
правый берег р. Томи у с. Верхотомского); ульбинская свита Рудного
Алтая; турнейский ярус Туркестана; слои кеокук Северной Америки.
Fenestella regalis Ulrich
Табл. LXXXI, фиг. 5; табл. LXXXII, фиг. 3
Основные размеры: 9—10/4—5//12—14. Колония с крупными петлями.
Неячеистая поверхность колонии покрыта тонкой струйчатостью и прони-
зана тонкими капиллярами. Ячейки в основании пятиугольного сечения.
Прутья прямые, несколько утолщенные в месте слияния их с переклади-
нами. Перекладины массивные, пониженные по отношению к прутьям.
Поперечник устьев ячеек 0,2 мм.
В размерах составных частей зоарии наблюдается сходство
с F. virgosa Е i с h w. Основное отличие от F. virgosa — более короткие
и в то же время более широкие петли и меньшее количество ячеек на
длину одной петли.
Распространение. Турнейский ярус Салаира (р. Каменный
Елбаш, выше дер. Хариной, система р. Бердь); слои кеокук Северной
Америки.
Род Polypora М’ С о у
Отличается от рода Fenestella (см. стр. 231) отсутствием срединного
киля и наличием на прутьях нескольких рядов ячеек. Между устьями-
иногда имеются ряды бугорков.
Типичный вид. Polypora dendroIdes М’С о у.
Распространение. Силур—пермь, повсеместно.
Polypora sibirica Janischewsky
Табл. LXXXII, фиг. la. 16
Основные размеры: 4—6/11//11 —12. Число рядов ячеек на пруте 4—5..
Сетка весьма крупная, неправильная. Форма ячеек ромбическая, перехо-
306
дяшая в шестиугольную. Перед дихотомией число рядов ячеек на пруте
увеличивается до 7—8.
Распространение. Встречается в визейском ярусе окрестностей
г. Томска, в ульбинской свите Рудного Алтая и в визейских отложениях
Средней Азии.
Род Reteporina d’Orbigny
Описание см. на стр. 233.
Reteporina altalca Nekhorochev
Табл. LXXXII, фиг. 4
Этот вид образует ряд разновидностей, основные размеры которых
таковы:
Прутьев на 10 мм ширины колонии Петель на 10 мм длины прута Ячеек на 5 мм
20 13—14 22—23
18—19 10—11 22 23
17-18 9 22-24
15—16 6-7 22—23
Все разновидности данного рода имеют близкое число ячеек, в то
время как количество прутьев и петель подвергается сильным колеба-
ниям.
Распространение. Верхи тарханской свиты Рудного Алтая.
Род Hemitrypa Phillips
Описание см. на стр. 231.
Hemitrypa proutana Ulrich
Табл. LXXXH, фиг. 2
Основные размеры: 24—28/22//22—25. Сетка мелкая, правильная.
Форма ячеек в основании пятиугольная.
Распространение. Нижнекаменноугольные отложения Кузнец-
кого бассейна (фоминская, подъяковская и верхотомская зоны), ульбин-
ская свита Рудного Алтая, турнейокие отложения Средней Азии.
Р. Н. БЕНЕДИКТОВА
Тип Brachiopoda. Плеченогие
Для стратиграфии морского нижнего карбона Западной Сибири наи-
большее значение имеют брахиоподы. Их описанию посвящены работы
М. Э. Янишевского (1915) и И. П. Толмачева (1924 и 1931). Некоторые
данные о нижнекаменноугольных брахиоподах Западной Сибири имеются
в работах В. Н. Лебедева (1916), а также Л. Н. Краевокой и М. П. Ло-
мовипкой (1935). На основании изучения брахиопод А. П. Ротай (1938)
расчленил нижний карбон Кузбасса на пять биостратиграфических зон
(см. стр. 297).
СЕМЕЙСТВО STROPHOMENIDAE KING
Род Schuchertella G i г t у
Описание см. на стр. 238.
Schuchertella tomskiensis Janischewsky
Табл. LXXX1II, фиг. 1—3
Очертания раковины усеченнояйцевидные. Замочный край короче наи-
большей ширины раковины; у призамочных углов имеются ясные ушки.
20* 307
Брюшная створка снабжена достаточно высокой треугольной ареей с за-
крытым дельтирием; зубные пластины редуцированы. Слепка выпуклая
спинная створка имеет двураздельный замочный отросток. Поверхность
створок покрыта тонкими, но резкими радиальными ребрами, которые
увеличиваются в числе путем вставления новых ребрышек между ранее
появившимися.
Распространение. В изобилии встречается в нижнекарбоновых
сланцах г. Томска.
СЕМЕЙСТВО PRODUCTIDAE GRAY
Род Dictyoclostus М u i r-W о о d
Под этим названием из рода Productus Sow. выделены формы, ранее
объединявшиеся в виде группы Productus semiretlculatus Mart. Для
рода Dictyoclostus характерна скульптура, состоящая из дихотомирую-
ших (изредка вставляющихся) радиальных ребер, которые в висцераль-
ной части (задняя часть створок) пересечены концентрическими вали-
ками, благодаря чему создается сетчатый рисунок. Шипы беспорядочно
разбросаны по всей поверхности брюшной створки, сгущаясь на ушках.
В спинной створке—массивный трехлопастной замочный отросток.
Типичный вид. Dictyoclostus semiretlculatus (Martin).
Распространение. Карбон—пермь, повсеместно.
Dictyoclostus burllngtonensts (Hall)
Табл. LXXXIII, фиг. 4
Раковина средних размеров, округленноквадратных очертаний. Замоч-
ный край по длине близок наибольшей ширине раковины, имеются от-
четливые, достаточно большие ушки. Брюшная створка вздутая и сильно
продольно изогнута, с довольно острой макушкой. Синус начинается
в пределах задней трети раковины, глубина и ширина его различна
у различных экземпляров; обычно он широкий и плоский, нередко почти
исчезает. Спинная створка в задней части почти плоская, затем изги-
бается почти под прямым углом, вместе с брюшной створкой образует
шлейф. Скульптура представлена правильными тонкими ребрами; зад-
няя часть створок, кроме того, снабжена концентрическими складками;
шипы разбросаны беспорядочно.
От сходного вида Dictyoclostus deruptus (Rom.) отличается мень-
шими размерами, меньшим развитием раковины в длину, более тонкой
ребристостью.
Распространение. Является характерной формой для турней-
ских осадков нижнего карбона Кузбасса; наиболее типичные представи-
тели вида — в тайдонской и фоминской зонах, несколько уклоняющиеся
формы — в абышевской и подъяковской зонах. Встречается в Казахстане
и на Урале, в верхах турнейского и низах визейского ярусов. Слои бур-
лингтон Северной Америки.
Dictyoclostus deruptus (Romanowsky)
Табл. LXXXIII, фиг. 5
Раковина крупных размеров округленнопрямоугольных очертаний,
обычно сильно развитая в длину, вздутая; замочный край составляет наи-
большую ширину раковины. Брюшная створка в профиль круто изогну-
тая, с вертикально падающими боками, несет неглубокий синус. Спин-
ная створка с резким коленообразным изгибом. Скульптура представлена
тонкими радиальными ребрами, пересеченными в примакушечной части
раковины. концентрическими складочками. Шипы разбросаны беспоря-
дочно. Отличия от Dictyoclostus burllngtonensts указаны выше.
308
Распространение. Руководящая широко распространенная
форма верхов турнейского и низов визейского ярусов Казахстана. В Куз-
бассе и Горловском бассейне — фоминская и подъяковская зоны.
Dictyoclostus robustus (Tolmatchoff)
Табл. LXXXV, фиг. 6
Раковина обычно крупная, вздутая, расширяющаяся к переднему
краю. Брюшная створка правильно и сильно изогнута, со слабо заметным
синусом или без него. Макушка большая, высокая, с загнутым клювом.
Раковина покрыта округлыми, изредка дихотомирующими ребрами, кон-
центрическая складчатость в примакушечной части выражена неясно.
Следы игол разбросаны по всей поверхности створки.
Распространение. Распространен в осадках турнейского (фо-
минская зона) и визейского ярусов (подъяковская и верхотомская зоны)
Кузбасса.
Род Echinoconchus Weller
Объединяет продуктусов группы Productus punctatus Mart. Харак-
терна скульптура: широкие и плоские, лентообразные концентрические
валики, усаженные многочисленными мелкими шипами, которые располо-
жены неправильными концентрическими рядами; на каждой ленте шипы
заднего ряда крупнее остальных.
Типичный вид. Echinoconchus punctatus (Mart.).
Распространение. Карбон—пермь повсеместно.
Echinoconchus punctatus (Martin)
Табл. LXXXV, фиг. 7
Раковина обычно крупная, почти изометричная, округлых очертаний.
Замочный край короче наибольшей ширины раковины. Ушки небольшие,
плоские. Макушка круто загнутая. Синус брюшной створки широкий, не-
глубокий, часто очень слабо выраженный. Спинная створка очень слабо
и правильно вогнутая. Концентрические складки многочисленные, почти
плоские; на них располагаются многочисленные мелкие бугорки (основа-
ния шипов), располагающиеся на валиках неправильными рядами
(см. выше—диагноз рода).
Распространение. Встречается в глинистых сланцах нижнего
карбона г. Томска. Является характерной формой для подъяковской
и верхотомской зон визейского яруса Кузбасса. Широко распространен
в визейских и намюрских отложениях других областей СССР и в Запад-
ной Европе. Рядом авторов указывается и из более высоких горизонтов
карбона.
СЕМЕЙСТВО SPIRIFERIDAE KING
Р од Splrifer S о w е г b у
Двояковыпуклая раковина с прямым длинным замочным краем.
Брюшная створка снабжена хорошо развитой, но невысокой ареей
с параллельными краями или в виде треугольника, сильно вытянутого по
основанию. Седло и синус нормально развиты. В брюшной створке нахо-
дятся хорошо развитые зубные пластины. Вся поверхность створок
покрыта радиальными ребрами.
Типичный вид. Splrifer striatus Mart.
Прежде очень широко понимаемый род Splrifer на протяжении по-
следнего времени разделен на целый ряд самостоятельных родов, раз-
личающихся как внутренними, так и внешними признаками (например,
Cyrtosplrifer N а 1. — см. стр. 266; Lamelllsplrifer N а 1. — см. стр. 260;
Platyrachella F е n t. et F е n t. — см. стр. 260 и ряд других).
ЗП9
Splrlfer tornacensis Kon in с к
Табл. LXXXIH, фиг. 6
Раковина средних размеров, развитая по ширине. Замочный край
равен наибольшей ширине раковины; призамочные углы обычно оття-
нуты в остроконечия. Брюшная створка умеренно выпуклая, с ясным
синусом. Последний, начинаясь от макушки, постепенно расширяется
к переднему краю. Арея невысокая, с почти параллельными крами. Седло
спинной створки невысокое, но ясно ограниченное. Скульптура представ-
лена многочисленными простыми, изредка дихотомирующими ребрами
и тонкими концентрическими струйками. В брюшной створке — хорошо
развитые зубные пластины.
Отличия от сходного Splrlfer usslensls То 1m. указаны ниже, в описа-
нии этого вида.
Распространение. Splrlfer tornacensis Kon. — широко распро-
страненная турнейская форма. В изобилии встречается во всех зонах
турнейского яруса Кузбасса и Горловского бассейна, особенно характе-
рен для тайдонской зоны. Присутствует также в бухтарминокой свите
Рудного Алтая.
Splrlfer usslensls Tolmatchoff
Табл. LXXXIH, фиг. 8 и 9
Раковина небольшая, развитая по ширине, имеет полукруглые очер-
тания. Призамочные углы нередко оттянуты, в остроконечия. Обе створки
сильно выпуклые. Брюшная створка с массивной макушкой, с узким
неглубоким синусом, несущим вблизи переднего края до трех ребер.
Арея низкотреугольная. Седло спинной створки неясно выраженное, несет
до четырех ребер. Скульптура представлена многочисленными простыми,
округлыми, довольно грубыми ребрами, пересекающимися знаками на-
растания.
От сходного Splrlfer tornacensis Коп. описываемый вид отличается
более треугольной формой ареи и характером седла и синуса.
Распространение. Встречается в турнейских отложениях Куз-
басса, особенно характерен для тайдонской зоны; реже отмечается
в осадках абышевской зоны.
Splrlfer из группы Splrlfer clnctus Ко n i nс k
Табл. LXXXIV, фиг. 1. и 2; табл. LXXXV, фиг. 2
Раковина крупная или очень крупная, округленнопятиугольного или
округлого очертания, чаще несколько развитая по ширине. Брюшная
створка с маленькой макушкой, равномерно и умеренно выпуклая. Арея
широкая, с субпараллельными краями. Синус вблизи макушки очень
неясный, сильно расширяется к переднему краю; обычно неглубокий;
нередко исчезает вовсе. Спинная створка так же выпукла, как и брюш-
ная; седло неясно отграниченное от остальной поверхности створки.
Поверхность раковины покрыта многочисленными невысокими, дихотоми-
рующими ребрами и тончайшими радиальными и концентрическими струй-
ками.
Распространение. Splrlfer clnctus и близкие к нему формы
широко распространены в верхнетурнейоких отложениях ряда стран
и областей СССР. В списках А. П. Ротая в качестве руководящей формы
для фоминской зоны Кузбасса приводится новый вид из этой группы —
Splrlfer grandis Ro tai; у И. П. Толмачева (1924) из нижнего карбона
Кузбасса описан Splrlfer subcinctus Коп. Присутствуют спириферы этой
группы и в фоминской зоне Горловского бассейна.
310
Splrlfer sibiricus Janichewsky
Табл. LXXXIII, фиг. 7
Раковина развита по ширине, с острыми призамочными углами.
Брюшная створка с невысокой ареей и узким, ясно выражении синусом,
начинающимся от самой макушки. Скульптура состоит из угловатых
радиальных ребер, образующих пучки (по 2—3 ребра в каждом). На
седле и в синусе располагаются по 6—8 ребер, по каждую сторону седла
и синуса — по 18 ребер. Ребра пересечены густыми концентрическими
пластинчатыми знаками роста. Внутреннее строение не описано, благо-
даря чему точная родовая принадлежность этого вида неизвестна.
Распространение. Часто встречается в нижнекарбоновых слан-
цах г. Томска.
Splrlfer slbericus N. Lebedev
Табл. LXXXIV, фиг. 3—6
Раковина довольно крупных размеров, сильно неравностворчатая.
Брюшная створка значительно выпуклая, пирамидальной формы, с высо-
кой, прямой или слегка вогнутой ареей. Синус ясно обособленный, несет
10—12 простых ребер. Спинная створка менее выпукла, чем брюшная,
трапецоидального очертания, с высоким, резко обособленным седлом,
несущим 10—12 ребер. Скульптура представлена многочисленными про-
стыми ребрами, покрывающими всю поверхность раковины; близ перед-
него края на 5 мм приходится 4—5 ребер. В брюшной створке хорошо
развитые зубные пластины.
От других видов рода Splrlfer отличается очень высокой ареей.
Внешне очень напоминает некоторые верхнедевонские циртоспириферы,
например, Cyrtospirifer archiaci М u г с h. и Cyrtospirifer tentlculum
Vern. (см. стр. 268), от которых отличается отсутствием дельтириаль-
ной пластинки.
Распространение. Splrlfer slbericus N. Lеb. является руково-
дящей формой турнейских отложений (кассинские слои) северо-восточ-
ного Казахстана; в изобилии встречается в абышевской зоне Горловского
бассейна.
Примечание. Под одним и тем же видовым названием описаны двумя
авторами два различных вида (см. предыдущий вид — Spirifer sibiricus Jani ch.).
Если будет доказана принадлежность вида М. Э. Янишевского к роду Spirifer
(в узком смысле), то вид, описанный Н. Лебедевым, — Spirifer sibiricus—должен
быть переименован.
Род Syringothyris Winchell
Раковина обычно крупная, пирамидальной формы. Последняя обу-
словлена присутствием очень высокой треугольной, прямой или почти
прямой ареи на брюшной створке. Синус и седло четкие. Скульптура
представлена довольно грубыми округленными радиальными ребрами
и тонкими концентрическими знаками нарастания. Синус и седло глад-
кие. Внутри открытого дельтириума имеется развитая дельтириальная
пластинка с трубкой на внутренней стороне (табл. LXXXIV, фиг. 8); зуб-
ные пластины хорошо развиты.
Внешне близкий род Pseudosyrinx Well, отличается отсутствием
трубки на дельтириальной пластине.
Распространение. Очень характерная группа для отложений
нижнего карбона. Распространена повсеместно. В нижнем карбоне Куз-
басса в осадках тайдонской и фоминской зон турнейского яруса
и в подъяковской зоне визейского яруса встречается ряд близких между
собой видов: Syringothyris cuspldata Mart. (табл. LXXXIV, фиг. 7),
Syringothyris typa Winch., Syringothyris altalca T о 1 m.
311
СЕМЕЙСТВО SPIRIFERINIDAE DAVIDSON
Род Spiriferina d ’ О r b i g n у
Спирифероидная раковина с немногочисленными широкими складками
и точечным строением вещества створок, что отличает этот род от сход-
ного рода Delthyrls D а 1 ш. (см. стр. 241); в брюшной створке находятся
зубные пластины и срединная септа.
Распространение. Карбон—юра.
Spiriferina octopllcata (S о w е г b у) var. concentrtca Janischewsky
Табл. LXXXV, фнг. 3—5
«Раковина полукруглого очертания с закругленными ушками, благо-
даря чему наибольшая ширина ее приходится не на самый замочный
край. Арея брюшной створки довольно высокая, немного вогнутая
Поверхность брюшной створки покрыта довольно резкими складками,
располагающимися по сторонам угловатого синуса, ширина которого
превышает ширину борозд между складками, по крайней мере, вдвое.
Число складок по каждую сторону седла от четырех до шести, реже до
семи, а еше реже—до восьми.
На спинной створке, менее выпуклой, чем брюшная, в середине нахо-
дится более значительная угловатая складка, соответствующая синусу
брюшной створки; по сторонам ее — угловатые складки, аналогичные
складкам брюшной створки. Поверхность раковины покрыта пра-
вильно располагающимися резкими пластинами нарастания, придаю-
щими ей черепитчатый вид» (Янишевский, 1915, стр. 47).
Распространение. Часто встречается в нижнекарбоновых гли-
нистых сланцах г. Томска.
РОДОВОЙ УКАЗАТЕЛЬ
Жирным шрифтом отмечены страницы, на которых даются описания родов.
Abakanopleura— 65, 66, 73, 117, 118
Acrocephalites —65, 66, 67, 73, 115, 117
Acrocephalops—63, 71
Acrolichas— 177
Acrospirifer — 242, 259
Actinostroma — 21, 23, 185, 187
Adiantites — 293
Ajacicyathus— 18, 19, 62, 63, 71, 75, 76
Alveolites — 21, 28, 200
Amnigenia — 278
Amphilichas —177
Amphipora — 23
Anathyris — 262, 270, 271, 272
Aneimites — 293, 294
Aneurophyton — 292, 293
Annularia — 53
Annulocyathus — 63, 71, 75, 99
Annulofungia — 20, 75, 99
Апотосаге — 65, 73, 130, 131
Anomocariopsis — 65, 131
Apatokephallna — 67, 73, 142, 143
Apatokephaloldes — 143
Aphyllopteris — 284, 285
Apnsolenopleura — 118
Arbataspis — 61, 62, 70
Archaeocardium — 278
Archaeocyathus — 17, 70, 76, 100, 101, 102
Archaeofungia — 75, 81
Archaeopterls — 294, 295, 296
Arthrostigma — 287
Artokephalus — 67, 73, 74, 143
Asterocalamltes — 290. 291, 298
Asterocyathus — 60, 75, 84
Asterophyllites — 53
Asteroxylon — 281, 282, 283, 284
Athyris —271, 272
Atrypa —43, 165, 240, 248, 250, 258
Aulopora — 201
Avimactra — 49
Babakovia —61, 62, 63, 70, 123, 124
Barinophyton — 286
Barinostrobus — 286
Belovia—65, 72
Bercinella — 140
Bergeroniellus — 60, 61, 68, 69, 70
Billingsella — 66
Bilobites — 159
Bonnia —61, 70, 119
Bonniella — 119
Brachyprion — 40. 236
Brachyspirifer — 260
Bronchocyathus — 60, 75, 97
Bumastus — 46, 175
Catamites — 291
Calceola — 29, 208
Calymene — 151
Camarotoechia — 41, 160, 161, 245
Campophyllum — 220
Camptopteris — 50
Caninia — 301
Carinatina — 250
Carinocyathus — 63, 71, 75, 92
Cenbalocoelia — 65. 72
Cephalotaxopsis — 56
Chaetetes — 21
Chakasskia — 62, 63, 71, 108
Chascothyris — 255
Cheirurus— 178, 273
Chiropteris — 50
Chondragraulos — 61, 63, 64, 70, 71, 124-
Chonetes — 238
Chuangia — 66, 140
Chuangiopsis — 66, 73, 140
Claruscyathus — 75, 100
Clathrocoilona — 188
Clathrocyathus — 87, 88
Clathrodictyon — 21, 153, 188
Clavatellus — 172
Cobboldia — 61, 70, 105
Collenia — 59, 68, 147
Columnaria — 216, 225, 226
Conchidium — 42, 162, 239, 245
Conokephalina — 133
I Conophyton — 59, 68
| Coosella — 66
Coosellina — 65, 66, 73
Coosia — 65. 73, 132
Corynexochus — 64
Corynexodides— 129
Coscinocyathellus — 63, 75, 89
Cosclnocyathus 20, 62, 75, 76, 82, 84
Cranaena — 264
Cr'sneila — 164
Crotalocephalus— 178, 273
Cycadites — 56
Cyclocyathus — 95
Cymostrophi a — 235, 236
Cyrtina — 42
Cyrtinopsis — 241, 251
Cyrtospirifer — 259, 262, 266, 309
Czekanowskia — 56
Dalmanella — 235, 256
Polm^nites — 274
Delthyris— 164, 241, 312
Denckmanella — 254
Densocyathus — 76
Dicranophyton — 285
Dictyoclostus — 308
Dictyofavosites— 197
313
Dictyophyllum — 50
Dictyostrophia — 236
Didymograptus — 150, 151
Dikelocephalopsis— 168
Dinesus —63, 71, 107
Dioonites — 55
Diplagnostus — 65, 73
Discina —39
Dolgaiella — 66
Dorypyge — 64, 65, 72, 121, 123, 124
Dreissensia — 276
Drepanophycus — 282, 287
Echinoconchus — 309
Elatocladus — 56
Elvinia — 134
Elytha —252, 265
Elythina — 252, 265
Emanuella — 254
Emsurina — 66, 67, 73, 145
Endophyllum —214, 227
Eolithostrotionella — 304
Eoorthis — 66
Eoreticularia — 251, 265
Eospirifer — 164, 241, 250
Epiphyton — 145
Equisetites— 290
Erbia —61, 62. 64, 70, 107, 108
Erbocyathus — 17, 63, 70, 94
Ethmocoscinus — 89
Ethmophyllum— 19, 62, 63, 70, 71 75, 84,
88, 90
Euloma — 134, 150
Fasciphyllum — 213, 215, 216, 225
Favosites —21, 27, 29, 156, 192, 197, 200
Fenestella —34, 35, 231, 306
Formosocyathus — 75
Gerronostroma — 187
Grandagnostus — 65
Granularia — 61, 70, 109
Griinewaldtia — 165
Grypophyllum — 208, 211
Gurichella—183, 269, 270
Gypidula — 163, 245, 262
Haliserites — 281
Halysites — 28, 152, 155
Haniva — 133
Hanivella — 67, 132
Hausmannia — 50
Hebediscus — 105
Heleniella — 289
Heliolites —21, 157, 202
Hemitrypa — 35. 231, 307
Hemitrypella — 232
Holotrachelus — 175
Homagnostus — 65, 66, 73
Hostimella — 284, 285
Howellella — 164, 241
Hypagnostus — 65
Hypothyridina—161, 246, 248, 263
Idiostroma — 23
Illaenus—46, 138, 174
Indospirifer—183, 253, 269, 270
Isocolus — 176
Isorthis — 235, 256
Isotrypa — 35
Kaniniella —65, 72, 135
Karpinskia — 249
314
Kazelia—66, 141
Kazellina— 66, 67, 73, 141, 142
Kingstonia — 66, 67, 73, 125
Klotziella — 62, 71
Koldinia — 139
Koldiniella —65, 73, 138, 139
Kootenia —62, 64, 119, 120, 121, 123
Kooteniella —60, 61, 70, 120, 121
Kooteniellina —63, 71, 121, 124
Kutorgina — 60
Labechia — 23
Lascopteris — 50
Lamellispirifer — 260, 266, 270, 309
Laurskia — 276
Lazutkinia — 253
Leiopecten — 275
Lejopyge — 65
Leperditia — 150
Lepidodendron — 289
Leptaena — 235, 244, 257
Leptodonta — 237
Leptotfontella — 234, 237
Leptotrypa — 230
Levenea — 234, 256
Linnarssonella — 66
Lioclema — 229
Liostracus — 63
Lithostrotion — 33, 303, 304
Loculocyathus — 20, 75, 81
Loganiella — 285
Loyolophyllum — 217
Macgeea — 220, 226
Maja—150, 171
Mataninella — 67, 118, 119
Mataninia — 119
Matonidium — 50
Megaphyllum — 220, 224
Meladiopsis — 66
Mesofavosites — 28
Metisaspina—65, 66, 137, 138
Metisaspis — 66, 138
Michelinia — 299
Modiomorpha — 48
Monocheilus— 142
Multisolenia — 28
Mytilarca — 48, 276
!
Nalivkinia — 165
Neoacrocephalites — 66, 117
Neocolumnaria — 215, 216, 225
Nilssonia —55
Nochoroicyathus — 75, 83
Nucula — 49
Olenoides — 64, 65, 72, 122, 123
Onchonotus — 119
Orbicyathus — 60, 75, 80
Orlovia—136, 137
Orloviella — 65, 66, 72, 73, 135, 136. 137
Orometopus—173, 174
• Orthotetes — 257
I Orusia — 66
Osagia —59, 68, 146, 147
Otozamites — 55
Pachyphyllum — 30, 226
Pachypora — 198
Pagetia — 60, 61, 63, 64, 72, 104. 105
Pagetina — 60, 61, 68, 105
Palaeofavosites — 28
Palaeohalysites — 155
Parabolinella — 167
Paradoxides —63, 110, 111, 112
Parallelopora — 22
Paramnigenia — 278
Parasolenopleura — 103, 114, 115
Pectinophyton — 286
Peneckiella— 219, 220
Peronopsis — 64, 65, 72, 103
Pesaia — 65, 66, 72, 134, 135
Pesaina — 65
Phacops — 273
Phillipsastraea — 30, 32, 221, 224
Phoenicopsis — 56
Phoidagnostus — 65
Pholidophyllum — 207
Phyllograptus — 150, 151
Platyrochella—260, 309
Plethomytilus — 276
Plicatifera — 262
Poliella — 129
Poliellaspis—62, 71, 129
Poliellina —62, 63, 71, 128, 129
Polycyathus — 94
Polypora — 35, 306
Portentosus — 67, 103, 144
Praemyophoria — 277
Proampyx — 65
Probolium — 274
Productus — 44, 308
Proerbia —61, 70, 106
Proetus — 272
Prohedinia — 64, 65, 127, 128
Prohedinella — 72, 128
Promegalaspides—150, 169, 170, 171
Protathyris — 166
Protobarinophyton — 285
Protolenus — 69
Protolepldodendron — 288
Protopharetra— 102
Protospongia — 62, 71
Pseudaviculopecten — 275
Pseudobornia.— 290
Pseudochonophyllum — 217
Pseudocycas — 56
Pseudomphyma — 31
Pseudospongophyllum — 213
Pseudosyrinx — 311
Psllophyton— 281, 284, 285, 288
Pterinea — 48, 275
Ptilophyllum — 55
Pugnax — 264
Pycnoidocyathus — 75
Rafinesquina — 236
Redlichia — 69
Redlichina —60, 68, 69, 74, 111, 112
Rensselaeria—181, 243
Retecyathus—17, 100, 101
Reteporina — 233, 307
Retzia — 242, 243
Rhacophyton — 292, 293
Rhodea — 293
Rhynchospirina — 242
Rlchardsonella — 133, 143
Sajanocyathus — 84
Salairocyathus — 97
Schagonarla — 60, 68, 111, 112
Schellwienella — 238, 257
Schizophoria — 256
; Schluteria — 220, 221, 224
Schoriella — 66, 67, 133, 134
Schuchertella — 238, 257, 307
Semicoscinium — 232
Sibirecyathus — 81
Sieberella — 263
Solenopleura — 65, 113, 118, 150, 167
Solominella — 215, 216, 225
Sphenophyllum — 291
Sphenopteris — 293
Spirifer — 40, 309, 311
Spiriferina — 312
Spirocyathus — 101
Spongophyllum — 213, 214
Squameofavosites — 29, 197
Stegerhynchus— 161
Stenophyllum — 210
Stenopora — 305
Stereophrentis — 300, 301
Streptorhynchus — 238
Striatopora — 29, 198, 199
Stromatopora — 22, 23, 24, 154, 189, 190
Stromatoporella — 21,23, 190
. Stropheodonta — 236, 244
I Strophonella — 237
Sygmocoscinus — 75
Sygmocyathus — 75
Sygmofungia — 75
i Syringocnema—17, 18, 102
Syringopora — 28, 189, 201, 300
! Syringothyris — 297, 298, 311
i Syspacephalus — 125
| Tabatopygellina — 61, 70, 119
Tabulophyllum — 29
Tabulipora — 305
Taeniocrada — 281
Taeniopteris — 55
Talbotina — 66
Tegerocyathus— 17, 18, 62, 63, 70, 71, 75,
। SO, 92
' Tercyathus — 87, 88
Thalamocyathus— 18, 19, 60, 75, 95
Thamnophyllum — 218, 225, 226
Thamnopora — 28, 1S8
Theodossia — 259
Thursophyton — 284, 288
Toechomya — 277
Tolmaia — 275
Tomiphyton — 286
Try plasma — 206, 207
Tubophyllum — 146
Tuvaella — 159
I Tumulocyathus — 75
I Uncinulus—161, 246, 258,263
I Urcyathus — 60, 75, 80, 84
Ussovia — 239
Whitfieldia— 166
Wilsonella — 161, 263
Yavorskia — 55
Zamites — 55
Zaphrentis — 300
Zosterophyllum — 280
ВИДОВОЙ УКАЗАТЕЛЬ
Жирным шрифтом отмечены страницы,
abakanensis Tegerocyathus — 90
abakanica Kootenia — 71, 120, 121
abdita Syringopora — 201
abditum Endophyllum — 227, 228
abeona Clathrocoilona—188, 189
acadica Aneimites — 280, 293
achmet Cyrtospirifer — 182, 184, 266
aculeatus Favosites—192
acuminata Pugnax—183, 184, 261, 264
acutilobata Gypidula — 245
acutirostris Praemyophoria—181, 277
acutus Ajacicyathus — 60, 76
aequalis Tryplasma — 207
alata Kaniniella—135, 136
ales Lamellispirifer — 260, 266
alpenensis Squameofavosites—195
alta Thamnopora — 199
altaica Cobboldia—106
altaica Reteporina — 298, 307
altaica Schuchertella—181, 238, 257
altaica Syringothyris — 311
altaica TrypLsma—180, 184, 207
altaicum Probolium—181, 184, 274
altaicum Semicoscinium — 232
altaicus Amplexus—207
altaicus Clathrocyathus — 89
altaicus Phacops—184, 274
altaicus Tegerocyathus — 91
altchedaticum Semicoscinium — 233
altum Stenophyllum — 211, 212
alveolata Columnaria — 216
amplus Ajacicyathus — 62
amsassense Ethmophyllum — 87
amsassensis Clavatellus—173
amsassensis Dikelocephalopsis — 168
amsassensis Solenopieura—167
amsassiensis Kazellina — 142
anaglypha Strophonella—237
angelini Promegalaspides — 170
angularis Uncinulus— 182, 247
angustiplicata Howellella — 165
anomala Metisaspina — 138
anomalis Chondragraulos — 62, 63, 71, 125
anossoffi Spirifer — 259, 260
antiqua Stromatopora—24
apion Kingstonia—125, 126
archiaci Cyrtospirifer—183, 184, 267, 311
arcuatus Dawsonites — 281
argentinensis Parabolinella—150, 151, 167
argus Favosites—193
arimaspa Carinatina — 250
armata Lejopyge — 72
aspera Atrypa — 249
asteroides Ajacicyathus (Urcyathus) — 80
316
на которых даются описания видов.
atlanticus Archaeocyathus— 18, 101
attenuate Laurskia—181, 276
attenuata Prohedinia — 127
australianum Zosterophyllum — 281
avia Pesaia — 134, 135
babakovensis Chondragraulos—124, 125
babakovensis Tabatopygellina—119, 120
babakovia Granularia — 110
balus Poliella—129
barrandei Heliolites — 203
barrandei var. spongoides Heliolites — 203
barriensis Bumastus—175
barroisi Chascothyris — 255
barzassensis Cyrtospirifer — 267
baschkiricum Conchidium—163
batchaticus Amphilichas—177
batiaeformis Chuangiopsis—141
batschatensis Апотосаге—130
bellulus Diplograptus—150, 152
bericulicus Carinocyathus — 93
beybuluki Tegerocyathus —62
bicarinata Paramnigenia — 181, 184, 278
bidzhenia Proerbia — 62, 107
bifarium Actinostroma — 187
bifera Camarotoechia — 246
bilobus Bilobites—144, 150, 152, 159
bifidus Didymograptus—151
biloculare Conchidium—162
bituberculatus Phoidagnostus — 72
bodini Atrypa—182, 184, 248
bohemicus Heliolites — 202
bohemicus Squameofavosites — 198
bootes Pagetia—-104
bouei Leptaena— 181, 184, 235, 244, 258
brevifrons Aposolenopleura—118
brevis Portentosus—144
brusnitzini Favosites—195
bruta Apatokephalina — 143
bugusunica Lioclema — 229
bulba Koldiniella—139
buratiensis Fenestella — 231
buriatica Collenia — 148
burlingtonensls Dictyoclostus — 297, 308
burtini Stringocephalus — 256
calathus Coscinocyathus — 60
canadense Chonophyllum — 31
cancellatus Coscinocyathus — 83, 84
canobiana Sigillaria — 290
capitata Parallelopora — 22
cardiopteroides Angaropteridium — 298
carinatus Proetus — 272
catavense Megaphyllum — 221
catenularia Halysites — 155
caudata Leptodontella—237
celsiseptosum Conchidium—181, 239
cervlcornls Favosites—199
cervicornis Pachypora—199
cervicornis Thamnopora—183, 198, 199
cheehiei Acrospirifer—182, 184, 256, 259
chemungensis Mytilarca — 276
cincta Solenopleura—167
cinctus Spirifer — 297, 310
cingulatiaeformis Kutorgina — 62
circularis Denckmannia — 254
citrulliforme Barinophyton — 286
clarus Ajacicyathus — 62, 77, 78
clathratum Actinostroma — 22, 186, 188
comis Gypidula—183, 184, 261, 263
compacta Syringopora—201
concentrica Gerronostroma—187
concentrica Stromatopora — 189
concinnus Proetus — 272
conica Orloviella—65, 137
conjugula Karpinskia — 181, 184, 215, 249
Consuelo Karpinskia — 249
contextum Actinostroma—187
convexus Olenoides—122, 123
convexus var. altaica Olenoides—123
corbicula Coscinocyathus — 60
cornea Hemitrypa—232
cornucopiae Caninia — 301
coronula Hypothyridina — 248
crassicaudatus Illaenus—174
crispa Howellella—150, 152, 164, 165
croswelli Loyolophyllum — 217
cuboides Hypothyridina — 183, 184,261,263,
264
cuboides var. semilukiana Hypothyridina —
264
cucullus Acrolichas— 177, 178
cuneifolium Sphenophyllum — 291
curvata Eoreticularia — 252
curvirostris Cyrtospirifer — 267
cuspidata Syringothyris— 311
cyclopyge Pseudagnostus — 66, 67, 73, 74
cylindrica Caninia — 297, 301
cylindrica Maja — 172
cylindricum Megaphyllum — 221
cynacanthium Grypophyllum — 209
daintreli Heliolites — 203
damesi Denckmanella — 182, 184, 254
dartingtonensis Parallelopora — 24
darwini Peneckiella—183, 184, 219, 220
dawsoni’ Protobarinophyton — 286
dawsoni Kootenia—120
decemplicatus Stegerhynchus—162
decemplicatus var. angaciensis Stegerhyn-
chus— 152, 161
decheniana 1'aeniocrada— 279, 281
decipiens Heliolites — 202, 203, 205, 206
deflexa Giirichella— 270
delanouei Stereophrentis — 300
dendroides Polypora — 306
dentata Cobboldia — 106
dereimsi Eoreticularia—182, 184, 252
deruptus Dictyoclostus — 297, 308
desquamata Atrypa — 258, 259
devoniana Tryplasma — 207, 208
devonica Hemitrypa — 232
devonicum Stenophyllum — 211, 212
devoriense Pachyphyllum — 227
dianthus Coscinocyathus — 60, 83
dianthus var. stabila Coscinocyathus — 83
didyma Protathyris — 150, 152, 166
diluvianum Stenophyllum—210
dipioperloratum i egerocyathus — 91, 92
disjunctus Cyrtospirifer — 266, 267
disjunctus Spirifer — 266
disparvallum Ajacicyathus — 61
disparvallum Ethmophyllum — 85
dissepimentalis Archaeofungia — 82
distans Monograptus—150, 152
djalnensis Solenopleura—114
djainens.s var. altaica Solenopleura — 114
dorypygeaeformis Babakovia — 124
dubia Taeniocrada—281
dubroviensis Uncinulus — 246
duplex Stromatoporella — 191
dwighti Ajacicyathus — 61
edelsteini Ethmophyllum — 62
ehlersi Endophyllum — 228
eifelensis Syringopora — 201
eifeliensis Stromatoporella — 24
elandiensis Acrocephalites—116
I elandiensis Orloviella—137
I elbaschensis Tabulipora — 305
elberfeldense Asteroxylon — 283, 284
elegans Gerronostroma — 187
eleganta Syringocnema — 102
; elevata Delthyris— 152, 241
; elongata Mytilarca—181, 276
; elongata Praemyophoria — 277
elongata Richardsonella—143
elongatus Coscinocyathus — 60
elongatus Promegalaspldes—171
emsti Schlflteria — 182, 184,224
endygashensis Favosites—196
erbiensis Bronchocyathus — 98
erbiensis Prohedinella —128
erbiensis Tegerocyathus — 90, 91
escharoida Mataninella—119
escharoides Halysites—155
eudokia Carinatina — 250
exsculpta Pesiia—134
exsul Cheirurus—178
facilis Ajacicyathus — 77
fallax Peronopsis — 72, 103
fasciculatum Epiphyton—145, 146
fasclculum Grypophyllum — 210
favosa Michelinia—299
favus Syringocnema—102
fedorovi Karpinskia — 249
ferox Pagetia — 104
festivus Favosites — 193
fimbriata Anathyris — 271
fissilis Archaeopteris — 295
fistulosus Favosites—193
flexuosa Striatopora—199
forbesi Favosites — 157, 196
formosa Rhynchospirina—180, 184, 242, 243
fossaengulatus Clathrocyathus — 88
gaspianus Drepanophycus — 288
gerassimovensis Thalamocyathus — 95, 96
german com Anevrophyton — 280, 292
gigantea Anomocare — 65
gigantea Erbia — 61, 108
gigantea Kazellina—142
gigantea Tuvaella — 160
glaber Pentamerus — 263
glandiformis Grandagnostus — 72
glcbosa Gypidula — 263
globosa Kingstonia — 125
globosus Clavatellus—173
317
gluchovi Heliolites — 205
goldfussi Favosites—157, 196
goldfussi Uncinulus — 246
goldfussi var. signata Uncinulus — 246
goldschmidtii Psilophyton— 282, 283
gorskii Stenophyllum — 211, 213, 215
gothlandicus Favosites—150, 152, 155, 156,
196
gothlandicus Palaeohalysites —155
gracile Ethmophyllum — 61
gracile Grypophyllum — 209
gracilis Halysites—155
grande Lithostrotion — 297
grandifolia Archaeopteris — 298
grandiformis Reteporina — 233
grandis Ajacicyathus — 77
grandis Jamesella — 63, 70
grandis Spirifer — 310
granulata Stromatoporella—190
granulosa Erbia — 72, 108
gravis Anomocare—131
grigorievi Elythina — 253
grOnewaldtiaeformis Nalivkinia— 152, 165,
166
halliaforme Fasciphyllum — 215
Halliana Archaeopteris — 280, 294, 295
halysitoides f. minor Spongophyllum— 213,
214
hamiltoniae Favosites—193
heckeri Aulopora — 202
hemisphaerica Chonetes — 239
hercynica Tryplasma — 208
heterovallum Erbocyathus — 61, 62
hibernica Acrhaeopteris — 294, 295
hisingeri Favosites—152, 156
holometopa Solenopleura—113
hostimensis Hostimella — 283, 284
hupschii Stromatopora — 189, 190
ibergense Pachyphyllum — 227
ijizkii Ajacicyathus — 60
immanis Ajacicyathus — 60, 77
incertum Rhacophyton — 280, 293
indifferens Eoreticularia — 252
indifferens Spirifer — 265
inflata Schellwienella — 257
inosculans Favosites—193
insolens Heliolites — 203
integer Pentamerus — 263
intercalaria Stenopora — 306
intermedius Favosites—195
interstinctus Heliolites— 150,152, 158, 203,
204
interstinctus var. irregularis Heliolites — 203
intracostatus Ajacicyathus (Urcyathus) — 80
irbensis Kutorgina — 62
irregulare Lithostrotion — 305
irregulare var. balakhonkaensis Lithostro-
tion — 297, 304
irregularis Loculocyathus — 81
jakovlevi Thalamocyathus — 97
jaschei Spirifer (Delthyris) — 180, 184, 241
karaganicum Ethmophyllum—86
katunica Pagetina — 105
katunicum Ethmophyllum — 62, 86
kayseri Camarotoechia — 246
kayseri Gypidula—180, 184, 245
kenensis Abakanopleura—118
keniensis Clathrocyathus — 88
318
khalfini Cyrtospirifer — 183, 184, 268
khvosieviensis Heliolites — 205, 206
kizylschinica Platyrochella — 261
kjerulfi Enetagnostus — 72
knighti Conchidium—149, 150, 162, 163
konincki Stereophrentis — 301
korovini Acrospirifer—181, 242
korovini Uncinulus— 182, 258
kostetskae Schluteria — 225
kourense Ethmophyllum — 85
kundatensis Collenia—147
kuzmini Retecyathus — 62, 101, 102
kuznetskiana Striatopora — 200
kuznetzovi Thalamocyathus — 61, 95, 96
laeve Anomocare—130
laevis Pterinea—275
lamellosa Osagia—147
langi Taeniocrada—281
lata Kaninia—135
lateseptata Phillipsastraea — 222
latifrons Phacops—273
latus Brachyption — 237
lavrenovae Annulocyathus — 99
lazutkini Camarotoechia — 246
lazutkini Phillipsastraea — 221, 222
leda Brachyprion — 236
lens Schuchertella — 238
lermontovi Poliellina — 129
lineatus Amphilichas — 177
loculatiformis Carinocyathus — 93
loculatus Carinocyathus — 92, 93
loculatus Thalamocyathus — 96
longispinus Rastrites — 150, 152
lorana Pterinea — 275
loscenkovi Coscinocyathellus—90
loutouguini Stromatoporella — 190
macbridei Spirifer — 260
maccalumi Bumastus—152, 175
macconnelli Endophyllum — 228
macilenta Archaeopteris — 295
mackiei Asteroxylon — 283, 284
maillieuxi Alveolites — 200
majusculus Promegalaspides—170
mamontovensis Lazutkinia—182, 184, 254
mamontovi Actinostroma — 186
marginalis Atrypa — 240
martianovi Platyrochella—182, 184, 261
massivum Pseudospongophyllum — 213
megastoma Michelinia—299
meisteri Productus—262
micrastraea Phillipsastraea — 223
micrommata Phillipsastraea—223
microspinosa Eoreticularia—265
milleri Thursophyton—284
minimalis Acrocephalites—116
minimum Spongophyllum — 215
minimus Artokephalus—143, 144
minus Cyathophyllum—219
minussensis Chakasskla — 109
minussensis Chondragraulos—124
mitella Koldiniella — 139
mongolicus Ajacicyathus (Orbicyathus) —60.
80
monozonatum Thamnophyllum — 218
monstrum Anathyris — 183, 184, 261, 271
mucronatus Lamellispirifer— 182, 260, 266
mucronatiformis Lamellispirifer—184, 260,
266
multispinosa Fenestella — 231
mutabilis Rensselaeria — 243
myretonianum Zosterophyllum — 279, 280
nagorskii Heliolites — 204
nalivkini Brachyprion— 181, 184, 236
nalivkini Camarotoechia — 150, 152, 160
nalivkini Cyrtinopsis—181, 251
nasutum Probolium — 274
navicelliformis Rensselaeria—181, 184, 243
nevadensis Olenoides— 122
nitellus Favosites—192
nitida Favosites—195
nympha Camarotoechia — 184
nympha var. pseudolivonica Camarotoe-
chia — 181, 245
obi Gypidula — 261, 263
obliquespinus Favosites—193
obliquespinus var. zolotainskii Favosites —
193, 194
obrutchevi Erbocyathus — 62, 70, 94
obrutchevi Granularia — 61, 62, 109
obrutchevi Polycyathus — 94
obrutschevii Protobarinonhyton — 279, 286
obtusa Archaeopteris — 280, 294
occidentalis Gypidula — 263
octoplicata var. concentrica Spiriferina —
298, 312
oculata Hemitrypa — 232
olenorus var. aspinosa Conokephalina — 134
originale Spongophyllum — 214
ornatiformis Pagetia — 68, 105
otychtaschensis Ajacicyathus — 61
ovata Stenopora — 305
oviformis Illaenus — 175
ovoides Rensselaeria — 243
ovosimilis Osagia —147
padaukpinensis Spirifer — 269
padaukpinensis var. dagon Indospirifer — 253
padaukpinensis var. maoerhchuanensis Indo-
spirifer — 253
paichinensiformis Kingstonia —126
paichinensis Kingstonia — 126, 127
parallelepipedus Uncinulus— 181, 184. 247
parallelus Palaeohalysites — 155
parvus Coscinocyathellus—89
pashiense Megaphyllum — 220
patulus Ajacicyathus — 60, 79
patulus var. aspera Ajacicyathus — 79
peetzi Anathyris — 271
pentagona Phillipsastraea — 183, 184, 223
pentagonus Uncinulus — 247
perlamellosa Delthyris—181, 241, 251
perlamellosa Delthyris—181, 241, 251
pestereviensis Grypophyllum — 209, 210
petrovitchi Favosites—196, 197
petshorense Fasciphyllum — 215
phalaena Anathyris—182, 181, 261, 271
piriformis Actinostroma — 186, 187
pisiformis Agnostus — 66, 67, 73
placentus Favosites—192
planuscula Leptodontella— 181, 184, 237
plebeja Fenestella — 298
plicatilis Placitifera-—262
polenovi Lioclema — 229
polymorphus Favosites—194, 196
polymorphus var. cronigera Favosites—194
polyostiolata Stromatopora — 25
porfirievi Favosites—195
porosus Heliolites—158, 204, 205
praecursor Protathyris—166
praelonga Plicatifera— 183. 184, 262
praelongus Productus — 262
•praetenerum Clathrodictyon — 188
prima Ussovia— 181, 239, 240
primaeva Hanivella — 133
primaeva Pagetina — 105
primaevum Tomiophyton — 280, 287
primaevus Acrospirifer — 242
princeps Psilophyton — 279, 282, 283, 288
princeps var. ornatum Psilophyton — 287
prisca Proerbia — 106, 107
problematicum Pleurodictyon—181
procerula Gypidula — 163
procuboides Hypothyridina— 248
profundus Archaeocyathus — 76
profundus Loculocyathus — 81
proscuriakovi Ajacicyathus — 60, 61. 79
proutana Hemitrypa — 297, 298, 307
pseudobaschkiricum ex gr. Conchidium —
182, 184, 244, 245
pseudobohemica Chonetes— 181, 238
pseudofallax AcroSpirifer—181, 242
pseudohelianthoides Pseudochonophyllum —
217, 218
pseudotogatus Eospirifer—181, 184, 241,
250. 251
pseudowilliamsi Indospirifer—182,184, 244,
246, 247, 249, 253
pulcher Annulocyathus — 99
punctata Lioclema — 229
punctatus Echinoconchus — 298, 309
punctatus Productus — 309
punctillosus Holotrachelus—176
quadrulla Fenestella — 231
raikowskii Tuvaella — 150, 152, 160
radiata Phillipsastraea — 221
radiatus Eospirifer — 241
radiatus Favosites—192
radiciformis Squameofavosites— 196
radugini Coosia — 132
ramosa Amphipora — 25
ramulosa Syringopora — 300
ratum Ethmophyllum — 61, 62, 86, 87
rectangularis Leiopecten—181, 275
rectispina Chonetes—181, 239
regalis Fenestella — 306
regressum Grypophyllum — 209
regularis Acrocephalites—115, 116
resupinata Schizophoria — 256
rete Archaeocyathus—100
reticularis Atrypa— 182, 240, 258, 259
reticulata Syringopora — 201
retrostriata Buchiola—183
rhenana Columnaria — 216
rbenanum Zosterophyllum — 281
rhombicum var. altaica Semicoscinium — 233
I rhomboidalis Leptaena — 235, 236, 244, 257
I rhomboidalis var. sinuata Leptaena — 236,
244, 258
। rhomboideum Semicoscinium — 232
I richardsoni Barinophyton — 286
richthofeni Dorypvge — 123
rjonsnitzkii Pagetina—105
robusta Aohyllopteris — 279, 285
robustus Dictyoclostus — 297, 298, 309
robustus Promegalaspides — 151, 169
roemeri Idiostroma—24
romingeri Cranaena — 183, 261, 265
rotundata Leptodontella — 181, 184, 237
n’dis Fenestella — 306
rzonsnickyae Favosites — 194
I salairensis Апотосаге —130, 131, 136
319
salairensis Anomocariopsis—131, 132
salairica Chascothyris—182, 184, 255
salairica Elythina—182, 184, 252
salairica Kingstonia — 126
salairicum Stenophyllum — 212
salairicus Coscinocyathus (Asterocyathus) —
84
salairicus Favosites (Dictyofavosites) — 197
salairicus Heliolites — 205
salairicus lllaenus—150, 151, 174
salcbrosus Ajacicyathus — 79
sanaschtykgolensis Ajacicyathus — 77
sanaschtykgolensis Densocyathus — 77
sandalina Calceola— 208
sandalina var. acuminata Calceola — 208
sayanica Redlichina — 113
sayanicus Poliellaspis—129
scabiosa Labechia — 25, 26
scalaris Climacograptus—152
schafferi Phillipsastraea — 222
scharyanum Protolepldodendron — 280, 289
schmidti Theodossia — 259
schorica Schoriella — 134
schoriensis Chondragraulos — 103, 125
schoriensis Pagetia—103, 104
schoriensis Parasolenopleura—115
scrobiculare Archaeocardium—181, 184, 278
scrobiculatus Asterocalamites — 280, 291,292
secans Eospirifer—181, 184, 250 . 251
sedgwicki Phillipsastraea — 221. 222
semiorbicides Atrypa—180, 184, 248
semiretlculatus Dictyoclostus — 308
semiretlculatus Productus — 308
serpens Aulopora — 202
sibericus Spirifer (Cyrtospirifer?)—311
sibirica Archaeofungia — 81, 82
sibirica Archaeopteris — 280, 295
sibirica Asterolepis—185
sibirica Aulopora — 202
sibirica Chuangiopsis—140, 141
sib:rica Emsurina — 145
sibirica Erbia — 62. 70, 107, 108
sibirica Isorthis — 257
sibirica Nalivkinia—166
sibirica Polypora — 298, 306
sibiricum Asteroxylon — 279, 284
sibiricum Grypophyllum — 211
sibiricum Lithostrotion (Eolithostrotio-
nella) — 297, 303
sibiricum Psilophyton — 279, 283
sibiricum Stenophyllum — 211, 212
sibiricus Archaeocyathus — 82
sibiricus Carinocvathus — 93. 94
sibiricus Indospirifer— 183, 184, 269
sibiricus Paradoxides — 111
sibiricus Spirifer — 2P8, 311
simplex Spirifer — 265
sinuatus Diplograptus — 150, 152
sjorgeni Isocolus — 176
slatkowskii Kooten:ella — 62, 70. 121
sniatkovi Ampbilichas — 150, 152, 177
sniatkovi Stromatoporella—190
solida Prohedinia — 127
solitaria Stromatoporella — 25
sosbkinae Solominella—183, 184, 225, 226
soshkini Stenophyllum — 212
speciosa Chuang:opsis—141
spinaeformis Drepanophycus — 279, 282, 288
spinosa Atrvpa — 249
spinosus Coscinocvathus — 84
spinosiis Thalamocvathus — 96, 97
squalaris Artokephalus—144
320
squamosa Pterinea (Tolmaia) — 181, 184,
275
stachei Thamnophyllum — 218
stenometopa Acrocephalites — 115
stephani Corrugatella — 180, 236
sternbergi Crotalocephalus — 273
strabo Cheirurus — 273
striata Reteporina — 233
striata Schliiteria — 224, 225
striatula Schizophoria— 184, 256
striatum Lithosrotion — 303
striatus Spirifer — 309
subcarinata Levenea — 235
subcinctus Spirifer — 310
subcircularis Toechomya — 181, 277
subconsocialis Parasolenopleura — 114
subcordiformis Uncinulus — 247
subcristata Phacops—181, 273
subgregarlus Acrospirifer—182, 184, 255
sublaevigatus Spirifer — 265
sublepida Atrypa — 240
suboelandicus Paradoxides — 71, 103, 111
suborbicularis Alveolites—183, 200
subpraenuntius Orometopus—174
subradiatus Archaeocyathus — 61, 62, 101
subsignatus Uncinulus — 258
subspinosa Atrypa—181, 184, 249
subsulcata Maja—151, 171, 172
subtenerrimum Sphenophyllum — 280, 291
subtil's Ajacicyathus — 77
suburceus Artokephalus — 144
subwilsoni Uncinulus — 246
suchiensis Ajacicyathus—78
suchiensis Bronchocyathus — 98
sulcata Maja — 172
superba Coosia— 132
supramarginalis Atrypa—180, 181, 240
supraphalaena Anathyris — 271
sygmoideus Bronchocyathus — 97
szajnochai Isorthis — 257
tabulate Peneckiella — 219
taeniolata Levenea—181, 235
talovkensis Ajacicyathus (Orbicyathus)—80,
81
tannuola Schagonaria — 112
tannu-ola Tegerocyathus — 91, 92
tarda Wilsoniella— 150, 152, 161
tarta Kazellina — 142
taylori Annulofungia — 99, 100
tchemyshevi Stromatoporella — 191
tenerrimum Sphenophyllum — 291
tenticulum Cyrtospirifer— 182, 184, 261, 268,
269, 311
tenue Epiphyton — 146
tenue Grypophyllum — 210
tenuisepta Michelinia — 299
tersiensis Thalamocyathus — 95
tessini Paradoxides—111
testudinaria Dalmanella — 150
theodori Heleniella — 280, 289
togatus Eospirifer — 250
togens's Neoacrocephalites — 117
tolli Loculocyathus — 81
tomense Pseudospongophyllum — 213
tomensis Heliolites — 203, 204
tomicus Ajacicyathus — 60, 77, 78
tomskiens:s Schuchertella — 298, 307
tomacensis Spirifer— 183, 297, 298, 310
trachealis Thalamocyathus — 95
tricostatus Uncinulus—182, 184,247
trigemme Thamnophyllum — 218
iruncata Orloviella —137
tschernyschewi Chascothyris — 255
tschernyschewl Cyrtospirifer — 268
tschernyschewi var. sibirica Cyrtospirifer —
268
tschingisica Hypothyridina — 264
tschirkovaena Heleniella — 289
tubaecostata Atrypa — 249, 259
tubaenia Kooteniellina—121, 122
tubuliformis Leptotrypa — 230
tumida Kingstonia—126
turkmensis var. birica Favosites—192
tuvaense Ethmophyllum — 21, 85
tuvaensis Eospirifer—164
typa Koldinia — 139
typa Syringothyris — 311
typica Orloviella — 137
typica Stromatopora— 152, 154
tyzhnovi Anathyris— 183, 271
ubensis Reteporina — 233
ulmense Ethmophyllum — 61
ulukhemensis Stegerhynchus—162
umbonatus Leiopecten — 275
umbra Atrypa— 180, 184, 240
umbraculum Schellwienella — 182, 184, 238,
257
undatus Thalamocyathus — 96
undifer Spirifer — 253, 265
undosus Cyrtinopsis — 251
uralensis Stropheodonta— 182, 244
uralicum Stenophyllum — 212
urense Loyolophyllum — 217
ursina Pseudobornia — 290
ussiensis Spirifer — 297, 310
ussovi Anathyris—183, 272
ussovi Caninia — 297, 302
ussovi Cyrtospirifer— 183, 184, 261, 269
ussovi Ethmophyllum — 87
ussovi Paramnigenia — 279
ussovi Sajanocyathus — 87
utukensis Coscinocyathellus — 89, 90
variolata Chonetes — 238
venusta Maja — 172
venustula Hypothyridina — 263, 264
vera Fenestella — 231
verneuili Cyrtospirifer — 183, 184, 261, 266,
267, 268, 297
vesiculosum Clathrodictyon — 150, 152, 153
vesta Camarotoechia—152, 160
victori Tubophyllum — 146
virgosa Fenestella-—306
vlangalii Productus — 262
vogulicum Conchidium—163
vogulicum Prismatophyllum—182, 184
vogulicum Stenophyllum — 210, 211
vologdini Redlichina —112
voroni Pugnax — 264
vratschinskiae Giirichella (?)—270
vulgaris Columnaria — 216, 217
vulgaris Heliolites — 202, 204
wahnbachense Protolepidodendron — 289
weberi Cheirurus—150, 152, 178
weberi Endophyllum — 228
whitii Stegerhynchus—161
wilsoni Wilsonella — 161
yakovlevi Lioclema — 229
yavorskii Heliolites — 202
yavorskii Syringopora — 201
yavorskii var. delicata Syringopora — 191
youngiana Sigillaria — 290
ziczac Giirichella— 182, 270
ziczac Spirifer — 270
21 Зак. 2024
дополнительный родовой указатель
В результате уточнения и дифференциации палеонтологической систематики многие
известные широким геологическим кругам руководящие формы стали в палеоитоло и-
ческой литературе фигурировать под новыми родовыми натваниями; например, Rhyncho-
nella cuboides Sow. теперь называется Hypothyridina cuboides S о w.; Productus puncta-
tus Mart. —Echinoconchus punctatus Mart, и т. д. Из широко известного рода Spirifer
Sow. выделились многочисленные самостоятельные роды—Acrospirifer, Brachythyris,
Elythina, Theodossia и многие другие.
Пересмотр систематики и уточнение синонимики многих форм повлекли за собой
аналогичные последствия; так, родовое название Ajicicyathus употребляется для форм,
которые обычно относились к роду Archaeocyathus, а название Arch.aeocyath.us— для форм,
относимых ранее к роду Spirocyathus и т. д.
Приведение сколько-либо полной синонимики родов и видов в издании, подобном
настоящему „Атласу', невозможно. Но для облегчения пользования, наряду с видовым
и родовым указателями, дается небольшой дополнительный указатель, в котором для
некоторых новых или измененных родовых названий указываются названия, ранее упот-
реблявшиеся в литературе по геологии и палеонтологии Западной Сибири. Например,
читатель захочет познакомиться по «Атласу* с описанием формы, которая в использо-
ванной им литературе фигурирует под названием Favosites cervicornis. По указателю
видов на стр. 316 он находит форму .cervicornis 1 hamnopora' с указанием соответ-
ствующей страницы, где эта форма описана. По дополнительному указат лю он может
убедиться, что это именно та форма, которая ему нужна: на стр. 323 указано, что
в «Атласе* под родовым названием' Thamnopora приводится форма, которая ранее опи-
сывалась под' родовыми названиями Favosites и Pachypora.
Дополнительным родовым указателем, в котором учтена лишь часть синонимики
родов,’можно пользоваться только в сочетании с видовым указателем. Например, если
в дополнительном указателе на стр. 323 приведена синонимика .Theodossia—Spirifer',
то это не значит, что более позднее родовое название .Th:odossia‘ полностью отвечает
старому названию .Spirifer'-. в данном случае имеется в виду лишь определенный вид—
Theodossia sch.mid.ti Stuck, из среднего девона Минусинской котловины, который
в литературе прежних лет обычно фигурирует под названием Spirifer schmidti.
Abiella — Posidonomya
Acrolichas — Lichas
Acrospirifer — Spirifer
A jacicyathus — Archaeocyathus, Densocya-
thus
Amphilichas — Lichas
Angaridium — Psygmophyllum
Angarocarpus — Samaropsis
Angaropteridium — Neurogangamopteris
Anomocariopsis — Anomocare
Anthraconaia — Anthracomya
Anthraconauta — Anthracomya
Archaeocyathus — Spirocyathus
Avonia — Productus
Brachyprion — Stropheodonta
Brachyspirifer — Spirifer
Brachythyris — Spirifer
Bronchocyathus — Salairocyathus
Camarotoechia — Rhynchonella
Conchidium — Pentamerus
Crotalocephalus — Cheirurus
Cymostrophia — Stropheodonta, Strophomena
Cyrtinopsis — Spirifer
322
Cyrtospirifer — Spirifer
Dictyoclostus — Productus
Dictyofavosites — Favosites
Echinoconchus — Productus, Pustula
Elythina — Spirifer
Eoreticularia — Spirifer, Reticularia
Eospirifer — Spirifer
Erbia — Solenopleura
Erbocyathus — Polycyathus
Ethmophyllum — Sajanocyathus
Grypophyllum — Cyathophyllum
Giirichella — Spirifer, Adolfia
Gypidula — Pentamerus
Hypothyridina — Rhynchonella, Hypothyris
Indospirifer — Spirifer
Isorthis — Orthis, Dalmanella
Kaniniella — Kaninia
Kootenia — Neolenus
Kooteniella — Dorypyge
Kazellina — Kazelia
Lamellispirifer — Spirifer
Lazutkinla — Spirifer
Leptodontella — Strophonella
Levenea — Orthis, Dalmanella
Microdonta — Palaeomutela
Microdontella — Posidonomya, Abiella
Mrassiella — Posidonomya
Nalivkinia — Atrypa, Karpinskia
Pachyphyllum — Phillipsastraea
Palaeohalysites — Halysites
Paraschizoneura — Schizoneura
Peronopsls — Agnostus
Phillipsastraea — Acervularia, Cyathophyllum
Platyrochella — Spirifer
Plicatifera — Productus
Poliellaspis — Poliellina
Probolium — Dalmanites
Prokopievskia — Anthracomya, Anthraconauta
Protathyris — Athyris
Prynadaeopteris — Sphenopteris, Pecopteris
Pseudospongophyllum — Cyathophyllum
Pugnax — Rhynchonella
Schellwienella — Orthotetes, Schuchertella
Sphenobaiera — Baiera
SchlUteria — Cyathophyllum
Squameofavosites — Favosites
Stenophyllum — Cyathophyllum
Thalamocyathus — Cyclocyathus
Theodossia — Spirifer
Thamnopora — Favosites, Pachypora
Tryplasma — Amplexus
Uncinulus — Rhynchonella
Ussovia — Wilsonella
Wilsonella — Rhynchonella, Wilsonia
Yaworskiella — Yaworskia
ЛИТЕРАТУРА
В приведенном ниже списке литературных источников помещены работы, упоми-
наемые в тексте «Атласа»; в основном это работы по палеонтологии и стратиграфии
Западной Сибири. Этот список не претендует иа полноту: авторы, участвовавшие
в составлении «Атласа», в различном количестве приводят ссылки на литературу.
В виде исключения в списке помещены работы (в том числе иностранные) о фауне
других областей и стран: ряд форм в настоящем «Атласе» описывается впервые, по-
этому авторы соответствующих разделов считают необходимым дать сопоставление
западносибирских форм с их аналогами или сходными формами.
В списке приведены работы, опубликованные по 1951 г. включительно; в виде
исключения этот список дополнен некоторыми, опубликованными в 1952 и 1953 гг.
Аверьянов Б. А. Граптолиты с Рудного Алтая. Изв. Всес. геол-разв. объедин.,
т. I, вып. 8. 1932.
Бубличенко Н. Л. О некоторых среднедевоиских брахиоподах Алтая. Изв.
Геол, ком., т. XLV1, № 10. 1927.
Бубличенко Н Л. Фауна брахиопод нижнего палеозоя окрестностей с. Сара-
Чумышского (Кузнецкий бассейн). Изв. Геол, ком., т. XLVI, № 8. 1927.
БульванкерЭ. 3. Среднедевоиские кораллы группы Rugosa западного склона
Кизеловского района. Труды ЦНИГРИ, вып. 10. 1939.
Вебер В. Н. Силурийские трилобиты из Кузнецкого бассейна. Изв. Геол, ком.,
т. XII, № 5—9. 1923.
Вебер В. Н. Новые находки нижнесилурийских трилобитов в Азиатской части
СССР. Ежегодн. Русск. палеонтолог, об-ва, т. VI. 1926.
Вебер В. Н. Верхиеордовикские трилобиты из Киргизской степи и Кузнецкого
бассейна. Изв. Геол, ком., т. XVII, № 3. 1928-
Вебер В. Н. Нижнесилурийские трилобиты. Монографии по палеонтологии СССР,
т. XIX, вып. 1. 1948.
ВинкманМ. К- Новые данные о возрасте гипербазитов Алтая. Доклады АН
СССР, № 1, т. XVIII. 1949.
Габуния К. Е. Материалы к изучению фауны кораллов из нижнекаменноуголь-
ных отложений около деревни Ройки по р. Томи. Изв. Сиб. отд. Геол, ком., т. I,
вып. 3. 1919.
Грацианова Р. Т. Новые данные о фауне и стратиграфии девона Горного
Алтая. Труды Горно-геол, ин-та Зап.-Сиб. филиала АН СССР, вып. 10. 1950.
Журавлева И. Т. О возрасте археоциатовых горизонтов Сибири. Доклады АН
СССР, т. 30, № 2. 1951.
Зайцев Н. С. и Покровская Н. В. О строении смежных частей Западного
Саяна и Тувы. Изв АН СССР, сер. геол., № 6. 1950.
И ван ия В. А. Некоторые представители Cyrtospirifer N а 1. из девона Голого
мыса (Кузбасс). Труды Томского гос. университета им. В. В. Куйбышева, сер. геол.,
т. 99. 1948.
Ива ни я В. А. Материалы к изучению кораллов Черепанова брода на р. Яя.
Ученые записки Томского гос. университета им. В. В. Куйбышева, вып. 13. 1950.
И в ш и и Н. К- Среднекембрийские трилобиты Бощекуля (Северо-Восточный
Казахстан). Акад, наук Казахской ССР, Ин-т геол. наук. 1949.
КраевскаяЛ. Н. и Ломовицкая М. П. О каменноугольных отложениях
района г. Камня. Вестник Зап.-Сиб. геол, треста, -вып. 4. 1935.
Краев с кая Л. Н. Материалы к изучению девонских отложений Чингисского
района. Матер, по геологии Зап.-Сиб. края, № 35. 1936.
Краснопеева П. С. Мшаики среднего и верхнего девона Алтая. Матер, по
геологии Зап.-Сиб. края, № 20. 1935.
Краснопеева П. С. Водоросли и археоциаты древнейших толщ Потехинского
планшета Хакассии. Матер, по геол. Красноярского края, вып. 3. 1937.
Краснопеева П. С. Основные комплексы археоциат Западной Сибири. Вестник
ЗСГУ, № 6. 1947.
324
Криштофович А. Н. Палеоботаника. Госгеолиздат. 1941.
Лазуткин П. С. Верхнесилурийские брахиоподы остракодового горизонта юго-
западной окраины Кузнецкого бассейна. Труды ЦНИГРИ, вып. 80. 1936.
Лебедев Н. Stromatoporoldea по Nicholson’y в связи с русской литературой
по этому предмету. Изв. Геол, ком., т. XV, № 8—9. 1896.
Лебедев Н. Spiriferidae из каменноугольных отложений Донецкого бас-
сейна и некоторых других районов России (продолжение). Изв. Екатеринославского
горного ин-та, вып. 1. 1916.
Левен ко А. И. К стратиграфии девонских отложений Рудного Алтая. Бюлл.
Московского об-ва испытателей природы. Отдел геол., т. XXVI, вып. 1. 1951.
Лермонтова Е. В. Arthropoda. Атлас руководящих форм ископаемых фаун
СССР. Кембрий. Госгеолиздат. 1940.
Лермонтова Е. В. Нижнекембрийские трилобиты и брахиоподы Восточной
Сибири. Госгеолиздат. 1951.
Ломовицкая М. П. Некоторые новые данные о географическом распростране-
нии веберовского горизонта в северо-восточной части Салаира. Матер, по геологии
Зап.-Сиб. края, вып. 35. 1936.
Мелещенко В. С. О некоторых вопросах стратиграфии девонских отложений
Минусинской котловины. Сборник «Палеонтология и стратиграфия». ВСЕГЕИ. 1953.
Наливки н Д. В. Брахиоподы среднего и верхнего девона Туркестана. Труды
Геол, ком., нов. сер., вып. 180. 1930.
Наливкин Д. В. Верхний девон Горного Алтая. Матер. ЦНИГРИ, общая сер.,
сборник 3. 1938.
Нехорошев В. П. Нижнекамениоугольные мшанки Кузнецкого бассейна. Изв.
Геол, ком., т. XLIII, № 10. 1926.
Нехорошев В. П. Алтайские Reteporinae тарханской свиты. Изв. Геол, ком.,
т. XLVI, № 8. 1926а.
Нехорошев В. П. Некоторые девонские мшанки Кузнецкого бассейна. Изв.
Геол, ком., т. XLVI, № 10. 19266.
Нехорошев В. П. Класс Bryozoa (Циттель. Основы палеонтологии). ОНТИ.
1934.
Нехорошев В. П. Девонские мшанки Алтая. Палеонтология СССР, т. III,
ч. 2, вып. 1. АН СССР. 1948.
Никифорова О. И. Брахиоподы верхнего силура Среднеазиатской части СССР.
Монографии по палеонтологии СССР, т. XXX, вып. 1. 1937.
Обручев Д. В. Материалы по девонским рыбам СССР. Труды Палеонтология,
ин-та АН СССР, т. VIII, вып. 4. 1941.
П е т ц Г. Г. Материалы к познанию фауны девонских отложений окраин Кузнец-
кого угленосного бассейна. Труды Геол, части каб., т. IV. 1901.
Полетаева О. К- Фауна кембрийских трилобитов санаштыкгольского изве-
стняка Западного Саяна. Матер, по геологии Зап.-Сиб. края. Вып. 35. 1936.
Радугии К. В. Новости сибирской палеонтологии. О Beatricea sp. из Сибири.
Изв. Зап.-Сиб. геол.-разв. упр., т. XI, вып. 1. 1931.
Радугин К- В. Некоторые целентераты из нижнего силура Горной Шории.
Матер, по геологии Зап.-Сиб. края, вып. 35. 1936.
Радугин К. В. О соотношении кембрия и докембрия в Горной Шории. Про-
блемы советской геологии, № 4. 1937.
Радугин К. В. Coelenterata среднего девона окрестностей с. Лебедянского.
Изв. Томского индустриальн. ин-та, т. 56, вып. VI. 1938.
Радугин К. В. О фазах тектогенеза среднего и верхнего кембрия. Вопросы
геологии Сибири. АН СССР, т. 1. 1945.
Ржонсницкая М. А. Брахиоподы ипдоспириферового горизонта (живетский
ярус) юго-западной окраины Кузнецкого бассейна. Труды ЦНИГРИ, вып. 97. 1937.
Ржонсницкая М А. и др. Материалы к изучению фауны таштыпской свиты
Минусинской котловины. Сборник «Палеонтология и стратиграфия». ВСЕГЕИ. 1952.
Ржонсницкая М. А. Спирифериды девонских отложений окраии Кузнецкого
бассейна. Труды ВСЕГЕИ. 1952.
Ротай А. П. Стратиграфия нижнекаменноугольных отложений Кузнецкого бас-
сейна. Труды ЦНИГРИ, вып. 102. 1938.
Рябинин В. Н. Stromatoporoldea (Циттель. Основы палеонтологии). ОНТИ.
1934.
Рябинин В. Н. Строматопороидей Эстонской ССР (силур и верхи ордовика).
Труды Всесоюзн. нефт. научио-исслед. геол.-разв. ин-та, новая серия, вып. 43. 1951.
Сивов А. Г. Верхний кембрий Саяно-Алтайской области. Труды Научн. конфе-
ренции по нзуч. и освоен, произв. сил Сибири, т. II, г. Томск. 1940.
Сивов А. Г. Верхнекембрийская арбатская формация. Изв. Томского политех-
нического ин-та, т. 65, вып. 2. 1950.
Соколов Б. С. Систематика и история развития палеозойских кораллов Antho-
zoa Tabulata. Вопросы палеонтологии, т. I. 1950.
Соколов Б. С. Табуляты палеозоя Европейской части СССР. Ч. I—IV. Гос-
топтехиздат. 1951, 1952.
Сошкина Е. Д. Девонские кораллы Rugosa Урала. Труды Палеонтологич.
ин-та АН СССР, т. XV, вып. 4. 1949.
325
Толмачев И. П. Верхнедевонская фауна с р. Усы в Алтайском Горном округе.
Труды Геол части каб., т. VIII. 1915.
Толмачев И. П. Нижнекамениоугольная фауна Кузнецкого угленосного бас-
сейна. Ч. I. Матер, по общ. и прикл. геол., вып. 25. 1924.
Толмачев И. П. Нижнекаменноугольная фауна Кузнецкого угленосного бас-
сейна. Ч. II. Издание Главн. геол.-разв. упр. 1931.
Фомичев В. Д. Новые данные о нижнекаменноугольных кораллах Кузнецкого
бассейна. Труды Главн. геол.-разв. упр., вып. 49. 1931.
Халфин Л. Л. Верхнедевонские брахиоподы Черепанова брода на р. Яе. Труды
Научио-исслед. угольн. ин-та Востокугля, сер. Г, вып. 2. 1931.
Халфин Л. Л. Нижнефранские брахиоподы окраин Кузнецкого каменноуголь-
ного бассейна и Горловского угленосного района. Изв. Зап.-Сиб. геол.-разв. треста,
т. XII, вып. 3. 1932.
Халфин Л. Л. Верхний девон села Жарковского на реке Яе. Труды Научно-
исслед. угольн. ин-та Кузбассугля. 1933.
Халфин Л. Л. Род Anathyris Peetz в сибирском девоне. Сборник по геол.
Сибири. Г. Томск. 1933а.
Халфин Л. Л. Сибирский девон и группа Spirifer cheehiel Коп. Изв. Томского
индустриальн. ин-та, т. 55. 1937.
Халфин Л. Л. Среднедевонские брахиоподы с. Лебедянского Аижеро-Суджеп-
ского района. Изв. Томского иидустриальи. ин-та, т. 57. 1937а.
X а л ф и и Л. Л. Пелециподы иижнего девона Алтая. Труды Научной конферен-
ции по изуч. и освоен, произв. сил Сибири, т. II. г. Томск. 1940.
Халфин Л. Л. Нижнедевонские спирифериды Алтая. Изв. Томского политехни-
ческого ин-та им. С. М. Кирова, т. 62. 1944.
Халфин Л. Л. Фауна и стратиграфия девонских отложений Горного Алтая.
Известия Томского политехнического ин-та им. С. М. Кирова, т. 65, вып. I. 1948.
Халфин Л. Л. Индоспириферы верхнего девона Кузнецкого бассейна. Труды
Горио-геол. ии-та Зап.-Сиб. филиала АН СССР, вып. 10. 1950а.
Халфин Л. Л. Новые спириферы девонских отложений северно-западной окраи-
ны Кузнецкого бассейна. Труды Горно-геол, ии-та Зап.-Сиб. филиала АН СССР,
вып. 10. 19506.
Халфина В. К. О верхнедевонской фауне с. Яя — Петропавловского (Кузбасс).
Труды Научи, конференции по изуч. и освоен, произв. сил Сибири, т. II. г. Томск.
1940.
Халфина В. К. Некоторые средиедевонские Stromatoporoidea юго-западной
окраины Кузбасса. Труды Горно-геол, ин-та Зап.-Сиб. филиала АН СССР, вып. 13.
1953.
Циттель К. Основы палеонтологии; ч. I, Беспозвоночные. ОНТИ. 1934.
Чернышев Б. Б. Силурийские и девонские Tabulate Монголии и Тувы. Труды
монгольской комиссии АН СССР, № 30. 1937.
Чернышев Б. Б. Силурийские брахиоподы Монголии и Тувы. Труды монголь-
ской комиссии АН СССР, № 29. 1937а.
Чернышев Б. Б. Силурийские и девонские Tabulate и Heliolitidae окраины
Кузнецкого бассейна. Госгеолиздат. 1951.
Чернышев Ф. Н. Материалы к изучению алтайской девонской фауны. Изв.
Геол, ком., т. XI. 1893.
Чернышева Н. Е. Силурийские и девонские трилобиты Монголии и Тувы.
Труды монгольской комиссии АН СССР, № 28. 1937.
Чернышева Н. Е. Верхпесилурийские и девонские трилобиты Кузнецкого бас-
сейна. Госгеолиздат. 1951.
Шейнман Ю. М. Trepostomata из среднего девоиа Минусинского уезда. Ени-
сейской губ. Изв. Геол, ком., т. XL1V, № 10. 1926.
Штукеиберг A. Mater, z. Kenntn. d. Fauna d. devon. Ablager. Sibiriens'. Труды
СПБ АН, VII серия, т. XXXIV, № 1, 1886.
Яворский В. И. Силурийские Stromatoporoidea. Изв. Геол, ком., т. 48, вып. I.
1929.
Яворский В. И. Actinostromidae из девонских отложений окраин Кузнецкого
бассейна и Урала. Изв. Главн. геол.-разв. упр., т. XLIX, № 4. 1930.
Яворский В. И. Некоторые девонские Stromatoporoidea из окраин Кузнецкого
бассейна, Урала и других мест. Изв. Всес. геол.-разв. объедии., т. I, вып. 94. 1931.
Яворский В. И. Девонские Stromatoporellidae и их роль в деле изучения стра-
тиграфии Кузбасса. Доклады АН СССР, т. 39. № 9. 1943.
Яворский В. И. Девонские Stromatoporella и их значение для стратиграфии.
Вопросы палеонтологии, т. I. 1950.
Яворский В. И. Некоторые палеозойские строматопороидеи. Госгеолиздат.
1951.
Янишевский М. Э. Глинистые сланцы, выступающие около г. Томска. Труды
Геол, ком., новая серия, вып. 107j 1915.
Яковлев Н. Н. Класс Anthozoa — коралловые полипы (Циттель. Основы пале-
онтологии). 1934.
Borneman J. G. Die Versteinerungen des cambrischen Schichtensyst. der Insel
Sardinien. Nova Acta Ac. Leop.-Carol. Bd. LI und LVI. 1886, 1891.
326
В г 6 g g е г. Огл paradoxidesskifrene ved Krenling Nyt. Magazin for Naturviden-
saaberne, Bd. 24, Ht. I. 1877.
Cherny schev B. Calceola from the Devonian Beds of the Salair Range.
Ежегодн. Рус. Палеонтол. Об-ва, т. VIII. 1930.
Halle Т. G. Lover devonian Plants from Roragen in Norway. Kungl. Svenska
Vetenskapsakademiens Handlingar, Bd. 57, N 1, Stockholm. 1916.
Howell В. E. and Ch. Loch man. Succession of late Cambrian faunas in the
Northern Hemisphere. Journal Paleontology, Vol. 13, N 1. 1939.
Howell В. E. и др. Correlation of the Cambrian formations of North America.
Bulletin of the Geological Society of America, vol. 55, N 8. 1944.
Kobayashi T. Japanese Journal ox Geology and Geography, vol. XI, N 1—2.
1933.
Krausel R. und Weyland H. Die Flora des deutschen Unterdevon. Abhand-
lungen der Preussischen Geologischen Landesanstalt. Neue Folge, Heft 131, Berlin.
1930.
Krausel R. und Weyland H. Neue Plancenfunde im Reinischen Unterdevon.
Paleontographica. Bd. 80, Abt. B. 1935.
Krausel R. und Weyland H. Pflancenreste aus dem Devon von Nord America
Paleontographica, Bd. LXXXVI, Abt. B. Stutgart. 1941.
Lecompte M. Les Stromatoporoides du Devonien moyen en superieur du Bassin
de Dinant. I—II part. 1951—1952.
Nicholson H. A. Monograph of the British Stromatoporoides. Pal. Soc. 1886—
1892.
Oku 1 itch V. J. Lower Cambrian Pleospongia from the Purcell Range of Bri-
tish Columbia, Canada. Journal of Paleontology, vol. 22, N 3. 1948.
Rasetti F. Upper Cambrian trilobites from the Levis conglomerate. Journal of
Paleontology, vol. 18, N 3. 1944.
Rasetti F. Early Upper Cambrian trilobites from Western Gaspe. Journal of
Paleontology, vol. 20, N 5. 1946.
Rasetti F. Middle Cambrian trilobites from the conglomerates of Quebek. Journal
of Paleontology, vol. 22, N 1. 1948.
Raymond E. New Upper Cambrian and Lower Ordovician trilobites from Ver-
mont. Proceeding Boston, vol. 37, N 4. 1924.
Raymond E. Upper Cambrian and Lower Ordovician Trilobita and Ostracoda
from Vermont. Bull, of the Geological Society of America, vol. 48, N 8. 1937.
R e s s e г Ch. Second contribution to nomenclature of Cambrian trilobites. Smithso-
nian Misc. Coll., N 9, vol. 95. 1936.
R ess er Ch. Cambrian system (Restricted) of the Soothern Appalachians. GeoL Soc.
of America' sp. papiers, N 15. 1938.
Walcott Ch. Cambrian Trilobites. Smiths. Misc. Collec., vol. 64, N 5. 1916.
Walcott Ch. and Resser Ch. Trilobites from the Ozarkian sandstones of the
Island of Novaya Zemlya. Report of the scient results of the Norvegian expedition to
Novaya Zemlya, 1921, N 24. 1924.
Whitehouse F. W. The Cambrian Faunes of North-Eastern Australia. Mem.
Queensland Mus., vol. XI, t. I. 1936.
Whitehouse F. W. The Cambrian Faunas of North-Eastern Australia. Ibid.,
Part III. 1939.
ТАБЛИЦЫ
ИЗОБРАЖЕНИЙ И ОБЪЯСНЕНИЯ К НИМ
ТАБЛИЦА I
Фиг. I. Ajaclcyathus immanls (Vologdin) Стр. 77
a—поперечное сечение кубка; б—продольное скошенное сече-
ние кубка. X 4. Тува, рч. Шивелик-Хем. Гавриловский комплекс.
Фиг. 2. Ajaclcyathus clarus (Vologdin) . . Стр. 7 3
Часть крупного кубка, X 3,8. Тува. рч. Шивелик-Хем, Камеш-
ковский или гавриловский комплекс.
Фиг. 3. Ajaclcyathus acutus (Bornemann) Стр. 76
Поперечные сечения кубков: а— X 4, между гг. Салаиром
и Гурьевском; б— X 4,5, Горная Шория, кл. Сухой, приток
р. Черной Усы. Камешковский комплекс.
Фиг. 4. Ajaclcyathus patulus (Bornemann) . Стр. 79
а—продольное скошенное сечение кубка; б—поперечное сече-
ние кубка. X 3. Восточный Саян, с. Торгашино. Камешковский
комплекс.
Фиг. 5. Ajaclcyathus proscurlakovl (Toll) . . Стр. 79
Поперечное сечение кубка, X 2. Восточный Саян, с. Торга-
шино. Камешковский комплекс.
Фиг. 6. Ajaclcyathus salebrosus (Vologdin) Стр. 79
Поперечное сечение кубка, X 5. Р. Катунь между рр. Чепош
и Сема. Комплекс неясен.
Фиг. 7. Ajaclcyathus {Urcyathus} a>steroldes Vologdin Стр. 80
Поперечное слабо скошенное сечение части кубка, X 6. Са-
лаир (Белая Горка), с. Горскино. Гавриловский комплекс.
Фиг. 8. Ajacicyathus (Orbtcyalhus) cf. mongollcus Vologdin Стр. 80
Продольное сечение части кубка, X 5. Кузнецкий Алатау,
устье рч. Белокаменки, притока р. Кии. Гавриловский комплекс.
ззэ
ТАБЛИЦА I
ТАБЛИЦА II
Фиг. 1. Loculocyathus irregularis (Toll) Стр. 81
a—-продольное сечение кубка; б—поперечное сечение кубка.
X 3,8. Кл. Сухой, приток р. Черной Усы в Горной Шории.
в—продольное сечение кубка, X 6. Северо-Восточный Салаир,
район с. Ариничево.
Камешковский и гавриловский комплексы!.
Фиг. 2. Coscinocyathus dianthus Bornemann . Стр. 83
а—поперечное сечение кубка,X 7; б—касательное сечение
кубка X 7. Центральная часть Кузнецкого Алатау, р. Полтавка.
в—продольное сечение части кубка, X Ю. Восточный Саян,
дер. Камешки.
Камешковский и гавриловский комплексы.
Фиг. 3. Coscinocyathus splnosus Vologdin . Стр. 84
Продольное сечение кубка, X 3,8. Кузнецкий Алатау, р. Ниж-
няя Терсь. Предположительно камешковский комплекс.
332
ТАБЛИЦА II
3
ТАБЛИЦА HI
Фиг. 1. Coscinocyathus spinosus Vologdin . Стр. 84-
a — поперечное окошенное сечение кубка, X 6. Р. Нижняя
Терсь, приток р. Кии.
б—продольное скошенное сечение кубка, X 3,8. Между гг. Са-
лаиром и Гурьевском.
Камешковский и гавриловский комплексы.
Фиг. 2. Annulocyathus lavrenovae Krasnopeeva Стр. 99
Поперечное сечение кубка, X 5. Кузнецкий Алатау, к югу от
Лощенкова лога, рч. Большая Ерба.
Фиг. 3. Archaeofungia sp. . Стр. 99
Скошенное поперечное сечение кубка, X 4. Тува, рч. Шивелик-
Хем.
Фиг. 4. Coscinocyathus (Asterocyathus) salairicus Vologdin Стр. 84
Поперечное сечение кубка, X 3,8. Между гг. Салаиром и
Гурьевском. Гавриловский комплекс.
Фиг. 5. Asterocyathus sp. . . Стр. 84
Разветвляющийся кубок, поперечное сечение, X 3. Салаир,
Белая Горка в окрестностях г. Горскино. Гавриловский комплекс.
Фиг. 6. Ajacicyathus proscurlakovl (Toll) . . Стр. 79
Поперечное сечение части кубка, X 3,8. Горная Шория, кл. Су-
хой, приток р. Черной Усы.
334
ТАБЛИЦА 111
ТАБЛИЦА IV
Фиг. 1. Annulocyathus sp.
Стр. 99
Скошенное продольное сечение кубка, X 4. Кузнецкий Алатау,
р. Нижняя Терсь в системе р. Кии.
Фиг. 2. Ethomophyllum sp.
Стр. 84
Поперечные скошенные сечения кубков, X 3,8. Горная Шория,
кл. Сухой, приток р. Черной Усы.
Фиг. 3. Cosclnocyathellus losienkovl Krasnopeeva, sp. n. Стр. 90
a—поперечное и б — поперечное скошенное сечения кубка.
ХП. Кузнецкий Алатау, к югу от Лощенкова лога, рч. Боль-
шая Ерба.
336
ТАБЛИЦА IV
22 Зак. 2024
ТАБЛИЦА V
Фиг. 1. Coscinocyathellus aff. losienkovl Krasnopeeva, sp. n. Стр. 90
Поперечное скошенное сечение кубка, X 3,8. Горная Шория,
кл. Сухой, приток р. Черной Усы.
Фиг. 2. Ethmophyllum katunlcum Krasnopeeva, sp. n. Стр. 86
a—поперечное сечение; б—продольное скошенное сечение части
кубка. X 3,5. Р. Катунь между устьями рр. Чепош и Сема.
Фиг. 3. Clathrocyathus fossaengulatus Vologdin Стр. 88
Поперечное сечение кубка, X 4,2. Тува, рч. Шивелик-Хем. Са-
наштыкгольский комплекс.
Фиг. 4. Ethmophyllum kourense Krasnopeeva, sp. n. Стр. 85
a—продольное сечение кубка; б—поперечное сечение кубка.
X 3. Горная Шория, р. Ульмень.
Фиг. 5. Ethmophyllum tuvaense Krasnopeeva, sp. n. Стр. 85
a—продольное сечение кубка; б—поперечное сечение кубка.
X 4. Тува, рч. Шивелик-Хем. Камешков.?кий—санаштыкгольский
комплексы.
338
ТАБЛИЦА V
22:
ТАБЛИЦА VI
Фиг. 1. Ethmophyllum s.p. . . . . Стр. 84
Поперечное сечение кубка, X 3,5. Р Катунь между устьями
р,р. Чепош и Сема. Обручевский комплекс (?).
Фиг. 2. Coscinocyathellus utukensis Krasnopeeva, sp. n. Стр. 89
Поперечное сечение кубка, X 4. Гора Утук-Кая в системе
р. Хемчик. Комплекс неясен.
Фиг. 3. Ethmophyllum ratum Vologdin Стр. 86
а—поперечное сечение, X 6,5; б—продольное скошенное сече-
ние кубка, X 5. Алтай, р. Ульмень. Преимущественно обручев-
ский комплекс.
Фиг. 4. Ethmophyllum aff. ussovi Vologdin Стр. 87
Поперечное сечение разветвляющегося кубка, X 4. Горная
Шория, р. Ульмень. Санаштыкгольский комплекс.
Фиг. 5. Tegerocyathus altaicus S. Т с h е г п у с h е v a, sp. п. Стр. 191
Поперечное сечение кубка, X 3. Горная Шория, р. Карагач.
340
ТАБЛИЦА VI
5
ТАБЛИЦА VII
Фиг. 1. Clathrocyathus keniensis Krasnopeeva Стр. 88
Поперечное скошенное сечение кубка, X 7. Устье р. Верхняя
Кеня у Абаканского завода.
Фиг. 2. Tegerocyathus tannu-ola Krasnopeeva, sp. п. Стр. 191
a — продольное сечение кубка, X 4; б — поперечное сечение
кубка, X 8,5. Тува, рч. Шивелик-Хем.
Фиг. 3. Coscinocyathus sp. п. (с пленками пузырчатой ткани) Стр. 82
а—поперечное сечение кубка; б— продольное сечение кубка.
X 3,8. Кл. Сухой, приток р. Черной Усы.
Фиг. 4. Carinocyathus loculatiformis Krasnopeeva, sp. n. Стр. 93
Скошенное сечение кубка, X 3,5. Среднее течение кл. Ипполи-
това в системе р. Абакан. Камешковский — санаштыкгольский
комплексы.
Фиг. 4. Ethmophyllum atnsassense Krasnopeeva, sp. n. Стр. 9 6
a—продольное сечение кубка, X 4,5; б—поперечное сечение
кубка, X 4,2. Тува. рч. Шивелик-Хем. Камешковский—санаштык-
гольский комплексы.
342
ТАБЛИЦА VII
ТАБЛИЦА VIII
Фиг. 1. Carinocyathus siblricus Krasnopeeva, sp. n. Стр. 93
Часть поперечного сечения кубка; видно днище и пленка на
наружной стенке, X 3. Тува, рч. Шивелик-Хем. Камешковский
комплекс.
Фиг. 2. Thalamocyathus jakovlevi (Vologdin) Стр. 97
Продольное сечение кубка, X 3,8. Тува, рч. Шивелик-Хем. Ка-
мешковский комплекс.
Фиг. 3. Annulofungta taylorl Krasnopeeva Стр. 100
Поперечное сечение, X 5.’Кузнецкий Алатау, к югу от Лощен-
кова лога, рч. Большая Ерба. Камешковский комплекс.
Фиг. 4. Thalamocyathus gerassimovensis Krasnopeeva, sp. n. Стр. 95
Скошенное сечение кубка, Х3,5. Кл. Герасимов, среднее тече-
ние, приток р. Монок.
Фиг. 5. Tegerocyathus erbiensis Krasnopeeva, sp. n. Стр. 90
Поперечное сечение кубка, X 3,5. Тува, рч. Шивелик-Хем.
Фиг. 6. Thalamocyathus undatus S. Тс h е г п ус h е v a, sp. п. Стр. 96
Поперечное сечение кубка, X 5. Р. Катуиь, между рр. Чепош
и Сема.
344
ТАБЛИЦА VIIT
ТАБЛИЦА IX
Фиг. 1. Bronchocyathus suchiensis Krasnopeeva, sp. n. Стр. 98
a—продольное скошенное сечение кубка; б—поперечное сече-
ние кубка. X 3. Горная Шория, кл. Сухой в системе р. Черной
Усы. Гавриловский комплекс.
Фиг. 2. Retecyathus kuzminl Vologdin
Стр. 102
а—продольное сечение части кубка; б—поперечное сечение
кубка. X 5,5. Р Ульмень, левый приток р. Бии. Обручевский
комплекс.
Фиг 3. Ethmophyllum amsassense Krasnopeeva, sp. n. Стр. 187
a—поперечное сечение части кубка, внутренняя стенка с плен-
кой; б—продольное сечение части кубка; в—касательное сечение
к внутренней стенке. X 12. Горная Шория, р. Амзас вблизи устья
кл. Алгаин. Обручевский комплекс.
Фиг. 4. Thalamocyathus tersiensls (Vologdin)
Стр. 95
Поперечное сечение кубка, X 16. Горная Шория, кл. Сухой,
приток р. Черной Усы. Камешковский и гавриловский ком-
плексы.
Фиг. 5. Thalamocyathus undatus S. Т с h е г п у с h е v a, sp. п. Стр. 96
Поперечное сечение кубка, X 6. Р. Катунь между устьями
рр. Чепош и Сема. Обручевский комплекс.
-346
ТАБЛИЦА IX
ТАБЛИЦА X
Фиг. 1. Thalamocyathus loculatus (Vologdin) . Стр. 9&
Поперечное сечение части кубка, X 12. В интерваллуме тре-
щина. Кузнецкий Алатау, р. Нижняя Терсь, приток р. Кии.
Фиг. 2. Ethmophyllum. sp. .... Стр. 84
Поперечное сечение кубка, X 5,5. Р. Катунь между устьями
рр. Чепош и Сема.
Фиг. 3. Ethmophyllum amsassense Krasnopeeva, sp. п. Стр. 87
Скошенное сечение кубка, X 12. Горная Шория, р. Амзас
вблизи устья кл. Алгаин. Обручевский комплекс.
Фиг. 4. Bronchocyathus suchiensis К г a s п о р е е v a, sp. n. . Стр. 98
а—поперечное скошенное сечение части кубка, X 5; б--про-
дольное сечение части кубка, видны гребенчатые днища, поры
и кольца на внутренней стенке; в—поперечное сечение части
кубка, видны детали строения днищ; г — продольное скошенное
сечение части кубка, видны мелкие поры на днищах между сеп-
тами. Горная Шория, кл. Сухой, левый приток р. Черной Усы_
Гавриловский комплекс.
348
ТАБЛИЦА XI
Фиг. 1. Peronopsls ex gr. P. fallax (Linnarsson). Стр. 103
a—хвостовой щит, X 5,5; б—головной Щит, X 7. Алтай, рч.
Верхняя Еланда. Большеербинский и орлиногорский комплексы.
в — головной щит. Восточный склон Кузнецкого Алатау,
рч. Большая Ерба.
Фиг. 2. Pagetia schoriensis Р о 1 е t а е v a, sp. п. Стр. 104
а—головной щит, X 7; б—хвостовой щит, X 4; в—головной
щит, X 7. Горная Шория, р. Мондыбаш. Санаштыкгольский
комплекс.
Фиг. 3. Pagetia ferox Lermontova Стр. 104
а—головной щит; б—хвостовой щит. X 5. Восточный склон
Кузнецкого Алатау, рч. Большая Ерба. Большеербинский ком-
плекс.
Фиг. 4. Cobboldia altaica Poletaeva, sp. n. Стр. 10&
Головной щит, X 4. Горный Алтай, р. Катунь. Бабаковский
комплекс.
Фиг. 5. Granularia obrutchevi Poletaeva . Стр. 10t>
а—кранидий, X 1,2; б—хвостовой щит, X 1,1- Западный Саян,
кл. Бабак. Бабаковский комплекс.
Фиг. 6. Prohedinella erbiensis Sivov, sp. n. Стр. 128
Кранидий, ХЗ. Восточный склон Кузнецкого Алатау, рч. Боль-
шая Ерба. Большеербинский комплекс.
Фиг. 7. Pagetina katunica Poletaeva, sp. n. Стр. 105
Головной щит, X 5- Горный Алтай, р. Катунь. Бабаковский
комплекс.
Фиг. 8. Paradoxides suboe'andlcus Poletaeva, sp. n. Стр. Ill
a—кранидий, X 2,5; 6—туловищный щит с хорошо сохранив-
шейся плевральной частью, X 1,5; в—кранидий с хорошо сохра-
нившимися глазными крышками, X 3,5. Горная Шория, р. Мон-
дыбаш. Санаштыкгольский комплекс.
Фиг. 9. Proerbla (?) bidzhenia Sivov, sp. n. . Стр. 107
Кранидий, X 3. Восточный склон Кузнецкого Алатау, верхнее
течение р. Биджи. Бабаковский комплекс.
Фиг. 10. Erbia sibirica (Schmidt) Стр. 107
Оригинал Лермонтовой из верхнеторгашинского известняка
близ г. Красноярска. X 1,5.
350
ТАБЛИЦА XI
ТАБЛИЦА XII
'Фиг. 1. Schagonaria tannuola Poletaeva, sp. n. Стр. 112
a—почти полный кранидий, X 3; б—часть кранидия, X 2,5.
Тувинская область, правобережье р. Улухема, к северо-западу
от города Шагонара. Чинпинский комплекс.
Фиг. 2. Redllchlna sayanlca S i v о v, sp. n. . Стр. 113
Неполный кранидий, несколько деформированный, X 1,2.
Тувинская область, правобережье р. Улухема, к северо-западу
от города Шагонара. Чингинский комплекс.
Фиг. 3. Acrocephalltes elandlensis J ego го v a, sp. n. Стр. 116
Почти полный кранидий, X 2. Алтай, район дер. Верхней Елан-
ды. Орлиногорский (?) комплекс.
Фиг. 4. Solenopleura holometopa Angelin Стр. ИЗ
Швеция. Средний кембрий. По Ангелину.
Фиг. 5. Redllchlna vologdlnl Lermontova . . Стр. ИЗ
а—кранидий с сохранившейся глазной крышкой; б—деформи-
рованный кранидий. Нат вел. Восточный Саян, дер. Камешки.
Чингинский комплекс.
Фиг. 6. Kooteniella slatkovskll (Schmidt) Стр. 121
а—.почти полный кранидий и г—ассоциирующий с ним пиги-
дий, X 2,5. Из известняков ербинской «формации» восточного
склона Кузнецкого Алатау.
б — кранидий, нат. вел.; в — небольшой пилидий с хорошо со-
хранившимися шипами, нат. вел. Восточный Саян, дер. Торга-
шино. Верхнеторгашинский известняк. Оригиналы Лермонтовой.
•Фиг. 7. Solenopleura djainensis Lermontova var. altalca J e go-
rova, var. n. . . Стр. 114
Полный хорошо сохранившийся кранидий, XI,5- Горный
Алтай, дер. Шиловка. Большеербинский (?) комплекс.
Фиг. 8. Neoacrocephalltes togensls Sivov, sp. n. Стр. 117
Неполный кранидий с хорошо сохранившейся заметной глаз-
ной крышкой и валиком на левой неподвижной щеке, X 3. Гор-
ная Шория, р. Амзас. Горношорский комплекс.
Фиг. 9. Tabatopygelllna babakoviensls Sivov, sp. n. . Стр. 120
Кранидий, X 1.2. Западный Саян, кл. Бабак. Бабаковский
комплекс.
Фиг. 10. Parasolenopleura schoriensls Р о 1 е t а е v a, 'sp. п. Стр. 115
Кранидий,Х2,5. Горная Шория. Санаштыкгольский комплекс.
• Фиг. 11. Parasolenopleura subconsocialis Poletaeva Стр. 114
Кранидий, X 4. Горная Шория. Санаштыкгольский комплекс.
• Фиг. 12. Matanlnella escharoida Sivov, sp. n. . Стр. 119
Кранидий, X 22. Салаир, гора Орлиная. Горношорский ком-
плекс.
Фиг. 13. Pagetia ornatiformis Sivov, sp. n. Стр. 105
Хвостовой щит с хорошо заметным бугорком на рахисе, X 6.
Восточный Саян, дер. Камешки. Чингинский комплекс.
• Фиг. 14. Orloviella elandlensis J е go го v a, sp. п. Стр. 137
Кранидий,XU5. Алтай, верховья рч. Бланды. Орлиногорский
(?) комплекс.
Фиг. 15. Erbia glgaritea J е go г ov a, sp. n. . . . Стр. 108
Хорошо сохранившийся кранидий, X 1,5. Горный Алтай,
р. Катунь. Бабаковский комплекс.
;352
ТАБЛИЦА XII
23 Зак. 2Г24
ТАБЛИЦА XIII
Фиг. 1. Chondragraulos babakovlensls Sivov, sp. n. Стр. 124
Кранидий, X 3. Западный Саян, кл. Бабак. Бабаковский ком-
плекс.
Фиг. 2. Kooteniellina tubaenla Sivov, sp. n. Стр. 122
a—кранидий с обломанным затылочным шипом; б—неполно
сохранившийся хвостовой щит. X 1,2. Восточный Саян, рч. Ирба.
Санаштыкгольский (?) комплекс.
Фиг. 3. Kootenia abakanica (Poletaeva) Стр. 120
а—почти полный кранидий; б—неполный хвостовой щит.
X 3,3. Западный Саян, кл. Санаштыкгол. Санаштыкгольский
комплекс.
Фиг. 4. Olenoides convexus Lermontova Стр. 122
а—кранидий; б—пигидий. X 2. Восточный склон Кузнецкого
Алатау, рч. Большая Ерба. Большеербинский комплекс.
Фиг. 5. Chondragraulos (?) anomalis Sivov, sp. n. Стр. 125
Кранидий, нат. вел. Восточный Саян, рч. Малая Ирба. Са-
наштыкгольский комплекс.
Фиг. 6. Olenoides convexus Lermontova var. altaica Jegorova,
var. n. . Стр. 123
a—хорошо сохранившийся кранидий, X 1,2; б—пигидий,
X 1,8. Горный Алтай, близ дер. Шиловки. Большеербинский
комплекс.
Фиг. 7. Chondragraulos (?) schoriensls Poletaeva Стр. 125
Несколько поврежденный кранидий. Горная Шория, р. Мон-
дыбаш. Санаштыкгольский комплекс.
Фиг. 8. Abakanopleura kenensls Sivov, sp. n. Стр. 118
a—кранидий с плохо сохранившимся затылочным кольцом;
б—кранидий с заметными глабелярными бороздками. X 2. За-
падный Саян, рч. Рудная Кеня и Салаир, гора Орлиная. Сала-
ирский комплекс.
Фиг. 9. Acrocephalltes mlnimalis Sivov, sp. n. Стр. 116
Неполный кранидий, X 4,5. Салаир, гора Орлиная. Горно-
шорский комплекс.
Фиг. 10. Granularia babakovia Sivov, sp. п. Стр. 110
Кранидий, X 1,1- Западный Саян, кл. Бабак. Бабаковский
комплекс.
Фиг. 11. Dorypyge rlchthofenl Dames . . Стр. 123
а—кранидий; б—хвостовой щит. Нат. вел. Оригиналы Да-
меса. Китай. Средний кембрий.
Фиг. 12. Chakasskla mlnussensls Poletaeva Стр. 109
Кранидий, X 3,7. Западный Саян, кл. Санаштыкгол. Санаш-
тыкгольский комплекс.
Фиг. 13. Acrocephalltes regularts Lermontova Стр. 115
Неполный кранидий, X 1,3- Салаир, гора Орлиная. Салаир-
ский комплекс.
Фиг. 14. Kooteniella slatkovskll (Schmidt) . Стр. 121
Хорошо сохранившийся кранидий, -нат. вел. Западный Саян,
кл. Бабак. Бабаковский и большеербинский комплексы.
354
ТАБЛИЦА XIII
23’
ТАБЛИЦА XIV
Фиг. 1. Kingstonia salalrlca Sivov, sp. n. Стр. ’"(>
a—кранидий; 6—пигидий.X 1,2. Салаир, гора Орлиная. Гор-
ношорский комплекс.
Фиг. 2. Kingstonia globosa Sivov . Стр. ,Q5
а—кранидий, X 1,8; б—образец в полупрофиль, X 2; в—кра-
нидий, Х2,1. Горная Шория, р. Амзас. Горношорский комплекс.
Фиг. 3. Kingstonia palchlnenstfortnls Sivov, sp. n. Стр. Г26
(^Kingstonia cf. paichinensls — Сивов, 1940).
a — кранидий; 6 — пигидий. X 1,2. Горная Шория, рч. Амзас.
Горношорский комплекс.
Фиг. 4. Pollelllna lermontovl Poletaeva . Стр. 129'
а—почти полный, слегка свернувшийся экземпляр, без по-
движных щек; б—хвостовой щит; в—кранидий. X 3,3. Западный
Саян, кл. Санаштыкгол. Санаштыкгольский комплекс.
Фиг. 5. Prohedinia solida Poletaeva, sp. n. Стр. 127
a—почти полный кранидий; б—кранидий с сохранившимися
заднебоковыми лопастями. Нат. вел. Алтай, рч. Ынырга. Орли-
ногорский комплекс.
Фиг. 6. Anomocare laeve Angelin . Стр. 130
Неполный кранидий. Скандинавия. Средний кембрий—верхи.
По Ангелину.
Фиг. 7. Anomocare gravis Jegorova, sp. n. Стр. 131
Кранидий с ассоциирующим с ним хвостовым щитом, X 2,7.
Горный Алтай, в 2,5 км к юго-востоку от дер. Шиловки. Боль-
шеербинский (?) и орлиногорский (?) комплексы.
Фиг. 8. Coosia radugini Sivov, sp. n. Стр. >Q
Коанидий, нат. вел. Салаир, гора Орлиная. Салаирский
комплекс.
Фиг. 9. Poliellaspis sayanicus (Poletaeva) Стр. 129'
Кранидий, X 3,3. Западный Саян, кл. Санаштыкгол. Санаш-
тыкгольский комплекс.
Фиг. 10. Schoriella schorlca Sivov, sp. n. . Стр. 134
Кранидий, X 2,5 и неполный экземпляр Klngstonla globosa.
Горная Шория, рч. Амзас. Горношорский комплекс.
Фиг. 11. Pesala avia Sivov, sp. n. . Стр. 134
Коанидий, нат. вел. Салаир, гора Орлиная. Салаирский ком-
плекс.
Фиг. 12. Anomocariopsis salairensls Sivov Стр. 132
Коанидий, X 1,8. Салаир, гора Орлиная. Салаирский ком-
плекс.
Фиг. 13. Orlovlella conlca Sivov . Стр. 137
Коанидий, нат. вел. Салаир, гора Орлиная. Салаирский ком-
плекс.
Фиг. 14. Kaniniella alata (Sivov) Стр. 135
Коанидий, нат. вел. Салаир, гора Орлиная. .Салаирский ком-
плекс.
Фиг. 15. Chondragraulos babakovlensls Si'rov, so. n. . Стр. 124
Почти полный кранидий, X 2. Восточный склон Кузнецкого-
Алатау, рч. Большая Ерба. Большеербинский комплекс.
356
ТАБЛИЦА XIV
ТАБЛИЦА XV
Фиг. 1. Hanivella primaeva Sivov, sp. n. . Стр. 133
Кранидий, X2. Салаир, гора Орлиная. Горношорский ком-
плекс.
Фиг. 2. Anomocare laeve Angelin . Стр. 130
а—пигидий; б—кранидий с затылочным шипом. Скандинавия.
Средний кембрий —верхи. По Ангелину.
Фиг. 3. Orlovleila typlca Sivov, sp. n. Стр. 137
a—почти полный спинной щит, нат. вел.; б—тот же экзем-
пляр, X 1,5- Салаир, гора Орлиная. Салаирский комплекс.
Фиг. 4. Anomocare salairensls Lermontova Стр. 130
а—почти полный кранидий; б—кранидий с поперечной штри-
ховкой вдоль фронтальной каймы. Нат. вел. Салаир, гора Орли-
ная. Салаирский комплекс.
Фиг. 5. Erbia gigantea J е go го v a, sp. п. Стр. 108
a — почти полный кранидий, X 2,5. Алтай, р. Катунь.
б — нат. вел. Западный Саян, кл. Бабак.
Бабаковский комплекс.
Фиг. 6. Metisasplna anomala Sivov, sp. n. . Стр. 138
Кранидий, нат. вел. Салаир, гора Орлиная. Салаирский ком-
плекс.
Фиг. 7. Koldiniella mltella Sivov Стр. 139
а—неполный кранидий, нат. вел.; б—хвостовой щит, ассоции-
рующий с кранидием, нат. вел.; в—почти полный кранидий
другого экземпляра, но того же самого вида, X 1.5. Салаир.
гора Орлиная. Салаирский комплекс.
Фиг. 8. Koldiniella bulba Sivov . Стр. 139
Кранидий с заметными спинными бороздками, X 2,2. Салаир,
гора Орлиная. Салаирский комплекс.
Фиг. 9. Chuangiopsts sibirica (Sivov) Стр. 141
а—кранидий; б—пигидий. X 2. Горная Шория, рч. Амзас.
Горношорский комплекс.
Фиг. 10. Erbia granulosa Lermontova . Стр. 108
Кранидий с хорошо заметной глазной крышкой на правой
неподвижной щеке, X 1,5. Восточный склон Кузнецкого Алатау,
рч Большая Ерба. Большеербинский комплекс.
Фиг. 11. Artokephalus squalarls Sivov, sp. n. Стр. 144
Неполный кранидий, X 1,5. Салаир, гора Орлиная. Горно-
шорский комплекс.
Фиг. 12 и 14. Artokephalus suburceus Je go г ov a, sp. n. . Стр. 144
J2—неполный кранидий, X 1,5; И—более полный кранидий,
X 2,5. Салаир, гора Орлиная. Горношорекий комплекс.
Фиг. 13. Apatokephalina bruta Sivov, sp. n. . Стр. 143
Кранидий, X 1,5. Салаир, гора Орлиная. Горношорский ком-
плекс.
Фиг. 15. Chuanglopsis batiaeformls (Sivov) Стр. 141
Кранидий, X 2,2. Горная Шория, рч. Амзас. Горношорский
комплекс.
Фиг. 16. Kazelllna tarta Sivov, sp. n. . Стр. 142
Кранидий, нат. вел. Горная Шория, рч. Амзас. Горношорский
комплекс.
358
ТАБЛИЦА XV
ТАБЛИЦА XVI
Фиг. 1.
Фиг. 2.
Фиг. 3.
Фиг. 4.
Фиг. 5.
Фиг. 6.
Фиг. 7.
Фиг. 8.
Фиг. 9.
Фиг. 10.
Фиг. 11
Kazellina amsasslensis (Sivov) Стр. 142
Кранидий, нат. вел. Горная Шория, рч. Амзас. Горношорский
комплекс.
Emsurina slblrica S i vo v, sp. n. . . Стр. 145
Почти полно сохранившийся кранидий, X 2. Салаир, гора
Орлиная. Горношорский комплекс.
Orlovlella truncata Sivov . . Стр. 137
Кранидий, X 2,5. Салаир, гора Орлиная. Салаирский ком-
плекс.
Babakovia dorypygeformis Sivov, sp. n. . Стр. 124
a—кранидий, б—хвостовой щит. Нат. вел. Восточный склон
Кузнецкого Алатау, рч. Биджа. Бабаковский комплекс.
Portentosus brevis Jegorova, sp. n. Стр. 144
a—хорошо сохранившийся кранидий; б—неполный кранидий.
Х5. Салаир, гора Орлиная. Горношорский комплекс.
Artokephalus minimus Sivov, sp. n. Стр. 143
a—почти полный кранидий; б—кранидий другого экземпляра
с тонкой грануляцией. X 3,5. Салаир, гора Орлиная. Горно-
шорский комплекс.
Kazellina gigantea Sivov, sp. n. Стр. 142
Кранидий, нат. вел. Салаир, гора Орлиная. Горношорский
комплекс.
Chuanglopsls batiaeformls (Sivov) . Стр. 141
Кранидий, X 2. Горная Шория, рч. Амзас. Горношорский
комплекс.
Chuanglopsls speclosa (Sivov) Стр. 141
Кранидий, X 1,2. Горная Шория, рч. Амзас. Горношорский
комплекс.
Osagia lamellosa Krasnopeeva, X 6. Стр. 147
Кузнецкий Алатау, рч. Большая Ерба.
Osagia ovosimllls Vologdin, X 5. Стр. 147
Кембрийские отложения Кузнецкого Алатау и Западного
Саяна.
360
ТАБЛИЦА XVI
ТАБЛИЦА XVII
Фиг. 1. Collenia kundatensls Krasnopeeva, sp. n. Стр. 147
a — общий вид колонии в поперечном сечении, уменьшено
в 1,5 раза; б—продольное сечение части столбика, уменьшено
в 1,5 раза; в—микроструктура слоев, X 7. Мариинская Тайга»
р. Кундат.
362
ТАБЛИЦА XVIf
'б 1 R
ТАБЛИЦА XVIII
-Фиг. 1. Clathrodlctyon vesiculosum Nicholson et Murie Стр. 153
a—тангенциальный разрез; б—вертикальный разрез. X Ю—12.
Верхний силур. По Никольсону.
Фиг. 2. Stromatopora typlca Rosen Стр. 154
а--тангенциальный разрез; б—вертикальный разрез. X Ю-
Верхний силур. По Яворскому.
Фиг. 3. Favosltes hlslngerl М. Edwards etHaime . Стр. 156
а—поперечный разрез; б—продольный разрез. X 3. Бассейн
р. Уксуная. Верхний силур.
Фиг. 4. Palaeohalysltes gothlandtcus Ya be . . Стр. 155
Поперечный разрез, X 2,5. Салаир, р. Уксунай.
Фиг. 5. Favosites forbesi М. Edwards etHaime . . Стр. 157
а—поперечный разрез; б—продольный разрез. X 2,5. Салаир,
левый берег р. Бердь, в 1,5 км выше заимки Похолкина.
364
ТАБЛИЦА XVIir
26
ТАБЛИЦА XIX
Фиг. 1. Favosltes gothlandicus Lamarck . Стр. 156
a—поперечный разрез; б—продольный разрез. Х2,5. Горный
Алтай, верховье правого притока р. Каинды-Одру (нижний).
Верхний силур.
Фиг. 2. Heliolites declplens (М’Соу) . . Стр. 158
а—поперечный разрез; б—продольный разрез. X 3. Горный
Алтай, р. Ануй.
Фиг. 3. Heliolites tnterstinctus (L i n п ё) . . Стр. 158
Поперечный разрез, X 3. Горный Алтай, верховье правого при-
тока р. Каинды-Одру (нижний).
Фиг. 4. Btlobites bilobus L i n п ё Стр. 159
a—брюшная створка; б—спинная створка; в—вид со стороны
замочного края. Увеличено. Верхний силур. По Дэвидсону.
Фиг. 5. Camarotoechia vesta Barrande Стр. 160
а—брюшная створка; б—спинная створка; в—вид со стороны
замочного края. Нат. вел. Р Сара-Чумыш. Верхний силур. По
Н. Л. Бубличенко.
Фиг. 6. Camarotoechia nallvkint Tchernychev Стр. 160
а—'Передний край; б—профиль; в—брюшная створка. Нат. вел.
Тува. Верхний силур. По Б. Б. Чернышеву.
Фиг. 7. Stegerhynchus ulukhemensts Tchernychev Стр. 162
а — брюшная створка; б — спинная створка; в — передний
край. Тува. Верхний силур. По Б. Б. Чернышеву.
-Фиг. 8. Stegerhynchus decempllcatus (S о w е г b у) var. angeclensls
Tchernychev Стр. 161
a — брюшная створка; б — спинная створка; в — передний
край. Нат. вел. Тува. Верхний силур. По Б. Б. Чернышеву.
.366
ТАБЛИЦА XIX
ТАБЛИЦА XX
Фиг. 1. Tuvaella raikovskii Tchernychev Стр. 160
a — брюшная створка; б — спинная створка; в — профиль;
г — передний край. Нат. вел. Тува, р. Элегест. Верхний силур.
По Б. Б. Чернышеву.
Фиг. 2. Conchidiutn knighti Sow er by . Стр. 162
a — брюшная створка в двух проекциях. Нат. вел. По Д. В. На-
ЛИВК'ИНу.
б — вид раковины сбоку. Нат. вел.
Верхний силур.
Фиг. 3. Wilsonela tarda (Barra nde) . Стр. 16!
Брюшная створка, X 1,5. Кузбасс, р. Черневой Бачат, окрест-
ности горы Орлиной. Остракодовый горизонт. По П. С. Лазут-
кину.
Фиг. 4. WllsoneUa tarda (Barrande) Стр. 161
Вид со стороны спинной створки и переднего края. По Дэвид-
сону.
Фиг. 5. Howellella crispa (Hi sin ger) . Стр. 164
a — вид co стороны спинной створии. По Дэвидсону.
б — брюшная створка. Тува. По Б. Б. Чернышеву.
Верхний силур.
Фиг. 6. Howsllella aagustlplicata (Ко z low sky). . . Стр. 165
a — брюшная створка; б — спинная створка. X 1,5. Кузбасс,
р. Черневой Бачат, окрестности горы Орлиной. Остракодовый
горизонт. По П. С. Лазуткину.
Фиг. 7. Nallvklnla grilnewaldtiaejormis (Peetz) Стр. 166
а — брюшная створка; б — с,тинная створка; в — передний
край. Нат. вел. Южная окраина Кузбасса, р. Сары-Чумыш. Верх-
ний силур. По Н. Н. Бубличенко.
,368
ТАБЛИЦА XX
24 Зак. 2024
ТАБЛИЦА XXI
Фиг. 1. Gypidula procerula (Barrande) Стр. 163
a — брюшная створка; б — спинная створка; в—-профиль.
X 1,5. Кузбасс, р. Черневой Бачат, окрестности горы Орлиной.
Верхний силур. По П. С. Лазуткину.
Фиг. 2. Eospirifer tuvaensis Tchernychev .Стр. 164
а — брюшная створка; б — спинная створка. Нат. вел. Тува,
р. Элегест. Верхний силур. По Б. Б. Чернышеву.
Фиг. 3. Protathyrls didyma (D а 1 m а п) Ст,р. 166
Верхний силур. По Дэвидсону.
Фиг. 4. Protathyrls didyma (D а 1 m a n) . . Стр. 166
Раковина в трех проекциях. Кузбасс. Остракодовый гори-
зонт. По П. С. Лазуткину.
Фиг. 5. Dikelocephalopsis amsassensis Poletaeva, sp. n. Сто. 168
a — головной щит; б — хвостовой щит. X 1 >5. Горная Шория,
р. Амзас. Верхний тремадок.
Фиг. 6. Solenopleura amsassensis Р о let a ev a, sp. n. . Стр. 167
Головной щит, Х4. Горная Шория, р. Амзас. Верхний тре-
мадок.
Фиг. 7. Parabollnella argentinensls Kobayashi . . Стр. 167
Головной щит, X 4. Горная Шория, р. Амзас. Верхний тре-
мадок.
Фиг. 8. Maja sulcata Lomovitskaja, sp. n. . Стр. 172
Головной щит, X 1,5. Горная Шория, р. Амзас. Верхний тре-
мадок.
Фиг. 9. Maja subsulcata L от о v i t s k a j a, 'sp. n. . Стр. 171
Неполный головной щит, ХГЗ. Горная Шория, р. Амзас.
Верхний тремадок.
Фиг. 10. Maja cyltndrica L о m о v i t s k a j a,'sp. n. . . Стр. 172
a — головной щит; б — хвостовой щит. ХГ5. Горная Шория,
р. Амзас. Верхний тремадок.
Фиг. 11. Maja venusta Lo m о v i t s k a j a, sp. n. . Стр. 172
Почти полный экземпляр, X 2. Горная Шория, р. Амзас. Верх-
ний тремадок.
370
ТАБЛИЦА XXI
24
ТАБЛИЦА XXII
Фиг. 1. Clavatellus globosus Poletaeva, sp. n. Стр. 173
Головной щит и часть сегментов осевой лопасти, Х$- Горная
Шория, р. Амзас. Верхний тремадок.
Фиг. 2. Clavatellus amsassensts Р о 1 е t а е v a, sp. n. . Стр. 173
Почти полный экземпляр, X 5. Горная Шория, р. Амзас.
Верхний тремадок.
Фиг. 3. Orometopus subpraenunllus Poletaeva, sp. n. Стр. 174
a — головной щит и туловище; б — головной щит. Х4. Гор-
ная Шория, р. Амзас. Верхний тремадок.
Фиг. 4. Bumastus maccaluml Salter . Стр. 175
Хвостовой щит сверху (а) и сбоку (б). Нат. вел. Салаир,
р. Черневой Бачат. Карадок. По В. Н. Веберу.
Фиг. 5. Illaenus salalrlcus Weber Стр. 174
а — хвостовой щит; б—часть головного щита и туловища,
X 2. Салаир, р. Черневой Бачат. Карадок. По В. Н. Веберу.
Фиг. 6. Illaenus oviformls Warburg Стр. 175
а — головной щит; б — хвостовой щит. Нат. вел. Салаир,
р. Черневой Бачат. Карадок. По В. Н. Веберу.
Фиг. 7. Isocolus aff. sjogrenl A n ge 1 i n . . . Стр. 176
a — глабель; 6 — головной щит сбоку. X 2. Салаир, р. Черне-
вой Бачат. Карадок. По В. Н. Веберу.
Фиг. 8. Amphilichas sniatkovi Weber . . Стр. 177
а — головной щит сверху; б — головной щит сбоку; в — перед-
няя часть глабели. Нат. вел. Салаир, р. Черневой Бачат. По
В. Н. Веберу.
Фиг. 9. Promegalaspid.es robustus L о m о v i t s k a j a, sp. n. Стр. 169
Неполный головной щит, X 1,5. Горная Шория, р. Амзас.
Верхний тремадок.
Фиг. 10. Promegalaspides elongatus L о m о v i t s k a j a, sp. n. Стр. 171
Головной щит, нат. вел. Горная Шория, р. Амзас. Верхний
тремадок.
Фиг. 11. Promegalaspides angellnl Lom о v i t s k a j a, sp. n. Стр. 170
Неполный головной щит, X Горная Шория, р. Амзас.
Верхний тремадок.
Фиг. 12. Promegalaspides majusculus L о m о v i t s ka j a, sp. n. Стр. 170
Головной щит, нат. вел. Горная Шория, р. Амзас. Верхний
тремадок.
372
nxx vUHiravi
ТАБЛИЦА XXIII
Фиг. 1. Actinostroma clathratum Nicholson . Стр. 186
a — вертикальный разрез, ХЮ; б — тангенциальный разрез,
X 5. Кузбасс, окрестности с. Бачат. По В. И. Яворскому.
Фиг 2. Actinostroma mamontovl Yavorsky . . Стр. 186
а — тангенциальный разрез; б — вертикальный разрез. X Ю.
Кузбасс, район дер. Мамонтово. По В. И. Яворскому.
Фиг. 3. Actinostroma piriformis V. К h а И i n a, sp. n. . Стр. 186
а — поперечный разрез, X Ю; б—продольный разрез, X5.
Кузбасс, дер. Мамонтово. Средний девон.
Фиг. 4. Gerronostroma elegans Yavorsky Стр. 187
а — тангенциальный разрез; б — вертикальный разрез (рисунок
на таблице повернут на 90°). Х5. Кузбасс, окрестности с. Ба-
чаты. Средний девон. По В. И. Яворскому.
374
ТАБЛИЦА XX11I
1 а
2а 26
ТАБЛИЦА XXIV
Фиг. 1. Gerronostroma concentrlca Yavorsky Стр. 187
a — тангенциальный разрез; б — вертикальный разрез. ХЮ.
Кузбасс, р. Томь-Чумыш. Средний девон. По В. И. Яворскому.
Фиг. 2. Clathrodictyon praetenerum Yavorsky . Стр. 188
а — тангенциальный разрез; б — вертикальный разрез. ХЮ.
Кузбасс, р. Томь-Чумыш. Средний девон. По В. И. Яворскому.
Фиг. 3. Stromatopora hupschil Bargatzki . . Стр. 189
а — тангенциальный разрез; б — вертикальный разрез. ХЮ.
Кузбасс, р. Бачат, р. Томь-Чумыш. Средний девон. По
В. И. Яворскому.
Фиг. 4. Clathrocollona abeona Yavorsky . Стр. 189
а — тангенциальный разрез; б — вертикальный разрез. ХЮ.
Кузбасс, р. Салаирка. Средний девон. По В. И. Яворскому.
376
ТАБЛИЦА XXIV
1 б
2 а
2 б
ТАБЛИЦА XXV
Фиг. 1. Stromatoporella sniatkovl Yavorsky Стр. 190
a — тангенциальный разрез; б — вертикальный разрез; ХЮ.
Кузбасс, р. Чумыш. Средний девон. По В. И. Яворскому.
Фиг. 2. Sromatoporella duplex V. Khalfina, sp. n. . Стр. 191
a — поперечный разрез, X Ю; б — продольный разрез, X Ю;
в — продольный разрез, X 5. Кузбасс (юго-западная окраина),
дер. Мамонтово. Средний девон.
Фиг. 3. Actlnostroma piriformis V. Khalfina, sp. n. Стр. 186
Продольный и поперечный разрезы, ХЮ. Кузбасс, дер. Ма-
монтово. Средний девон.
378
ТАБЛИЦА XXV
1 б
2а 26
ТАБЛИЦА XXVI
Фиг. 1. Stromatoporella loutougulnl Y av or sky Стр. 190'
a — тангенциальный разрез; б — вертикальный разрез. X Ю.
Юго-западная окраина Кузбасса. Живетский ярус среднего-
девона. По В. И. Яворскому.
Фиг. 2. Stromatopora concentrica G о 1 d f u s s . Стр. 189
a — тангенциальный разрез; б — вертикальный разрез. ХЮ-
Кузбасс. По В. И. Яворскому.
Фиг. 3. Favosites placentas Rom in ger . Стр. 192
a — поперечный разрез, X 2; б — продольный разрез, X 2,2.
Центральный Салаир. Живетский ярус среднего девона.
Фиг. 4. Favosites goldfussl d’O г b i g п у . . Стр. 196
a — поперечный разрез; б — продольный разрез. X 2. Цент-
ральный Салаир. Эйфельский ярус среднего девона.
Фиг. 5. Favosites festlvus Tchernychev . . Стр. 193-
а — поперечный разрез; б — продольный разрез. X 4. Кузбасс,,
левый берег р. Чумыш, гора Глядень. Кобленцский ярус ниж-
него девона.
380
ТАБЛИЦА XXVI
1а 1б
2а 26
ТАБЛИЦА XXVII
Фиг. 1. Favosites aculeatus Tchernychev . Стр. 192
a — продольный разрез, X 2,4; б — поперечный разрез, X 2,3.
Центральный Салаир. Средний девон.
Фиг. 2. Favosites obliquesplnus Tchernychev Стр. 193
а — продольный разрез; б — поперечный разрез. X 3. Куз-
басс, правый берег р. Чумыш, гора Кызых. Эйфельский ярус
среднего девона. По Б. Б. Чернышеву.
Фиг. 3. Favosites obliquesplnus Tchernychev var. zolotalnsklt
Zhmaev, var. n. . . Стр. 193
a — продольный разрез; б — поперечный разрез. X 2. Салаир,
ключ Золотаинский, правый приток р. Павловой. Эйфельский
ярус среднего девона.
Фиг. 4. Au'opora sibirica Tchernychev, ХЗ. . Стр. 202
Юго-западная окраина Кузбасса, р. Артышта. Живетский ярус.
382
ТАБЛИЦА XXVII
1 а 16
ТАБЛИЦА XXVIII
Фиг. 1. Favosites polymorphus (Goldfus's) Стр. 194
a—-внешний вид колонии, нат. вел. По Лекомту. б — продоль-
ный разрез, Х2,4; в — поперечный разрез, X 2,3. Томская
область, Туганский район, левый берег р. Щербак.
Средний девон.
Фиг. 2. Favosites rzonsnicktyae Tchernychev . Стр. 194
а — продольный разрез, X 2; б — поперечный разрез, X 2,3.
Центральный Салаир, бассейн р. Тогул. Нижний девон.
Фиг. 3. Alveolites maillleuxi S а 1 ё е . . Стр. 200
а — продольный разрез, X 2,3; б — поперечный разрез, X 2,2.
Центральный Салаир, Октябрьское месторождение. Средний
девон.
Д81
ТАБЛИЦА XXVIlf
25 Зак. 2(24
ТАБЛИЦА XXIX
Фиг. 1. Favosites polymorphus (G о 1 d f u s s) var. cronlgera (d’Or-
b i g n у) Стр. 194
a — внешний вид колонии, нат. вел. По Лекомту.
б — продольный разрез, X 2; в — поперечный разрез, X 2.
Кузбасс, правый берег р. Кондомы, в 1 км выше ул. Абрамов-
ского. По Б. Б. Чернышеву.
Средний девон.
Фиг. 2. Thamnopora cervlcornls (В 1 a i n v i 1 1 е) . Стр. 199
а — внешний вид колонии, нат. вел.; б — поперечный разрез,
X 5,5; в — продольный разрез, X 5,5. Девон. По Б. Б. Чер-
нышеву.
Фиг. 3. Favosites porflrievl Tchernychev Стр. 195
а — продольный разрез; б — поперечный разрез. X 3. Салаир,
рч. Тихобаевка. Нижний девон.
386
ТАБЛИЦА XXIX
2'i*
ТАБЛИЦА XXX
Фиг. 1. Favosites endygaschensls Tchernychev . Стр. 196
a — продольный разрез; б — поперечный разрез. X 3. Кузбасс,
левый берег р. Чумыш, выше устья р. Эндыгаш. Эйфельский
ярус. По Б. Б. Чернышеву.
Фиг. 2. Strlatopora kuznetskiana Tchernychev . Стр. 200
Внешний вид, нат. вел. Кузбасс. Нижний девон. По Б. Б. Чер-
нышеву.
Фиг. 3. Favosites petrovltchl Z h m а е v, sp. n. . . Стр. 196
a — продольный разрез; б — поперечный разрез. X 2. Салаир,
кл. Петрович, левый приток р. Павловки. Нижний девон.
Фиг. 4. Favosites (Dlctyofavosltes) salalricus Tchernychev Стр. 197
a — продольный разрез; б — поперечный разрез. X 2. Цент-
ральный Салаир, верховья р. Берди. Нижний девон.
388
ТАБЛИЦА XXX
ТАБЛИЦА XXXI
Фиг. 1. Thamnopora alta (Tchernychev) . Стр. 199
a — внешний вид, нат. вел.; б — продольный разрез, X 3;
в — поперечный разрез, X 3. Кузбасс, р. Бачат. Живетский ярус
среднего девона.
Фиг. 2. Heliolites vulgaris Tchernychev Стр. 202
а — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса. Средний девон. По Б. Б. Чернышеву.
Фиг. 3. Heliolites tomensls Tchernychev Стр. 203
а — продольный разрез, Х3,2; б — поперечный разрез, Х3,3.
Юго-западная окраина Кузбасса, правый берег р. Томь-Чумыш.
Средний девон. По Б. Б. Чернышеву.
390
ТАБЛИЦА XXXI
ТАБЛИЦА ХХХП
Фиг. 1. Heliolites tnsolens Tchernychev . Стр. 203
а — продольный разрез; б — поперечный разрез.X 2. Централь-
ный Салаир. Живетский ярус среднего девона.
Фиг. 2. Syrlngopora elfeliensis S chi liter . Стр. 201
а — продольный разрез; б — поперечный разрез. Юго-западная
окраина Кузбасса, р. Чумыш. Средний девон. По Г Г. Петцу.
Фиг. 3. Heliolites salairicus Tchernychev Стр. 205
а — продольный разрез; б — поперечный разрез. X 3. Салаир,
р. М. Березовая. По Б. Б. Чернышеву.
Фиг. 4. Heliolites yavorskyl Tchernychev Стр. 202
а — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса. Нижний девон. По Б. Б. Чернышеву.
Фиг. 5. Syrlngopora yavorskyl Tchernychev. Стр. 201
а — продольный разрез; б — поперечный разрез. X 3. Кузбасс,
дер. Михайловка, правый берег, р. Кара-Чумыш. Эйфельский
ярус среднего девона. По Б. Б. Чернышеву.
Фиг. 6. Heliolites porosus (G о 1 d f u s s) . Стр. 204
Внешний вид колонии, нат. вел. Юго-западная окраина Куз-
басса, левый берег р. Артышты. Живетский ярус среднего
девона. По Б. Б. Чернышеву.
392
ТАБЛИЦА XXXI!
ТАБЛИЦА XXXIII
Фиг. 1. Heliolites porosus (Gold fuss) . Стр. 204
a — продольный разрез; б — поперечный разрез. Х2. Салаир-
ский кряж, бассейн р. Верди в среднем течении. Живетский ярус
среднего девона.
Фиг. 2. Heliolites nagorskll Z h m a e v, sp. n. . Стр. 204
a — продольный разрез; б — поперечный разрез. X 2. Салаир,
р. Хайрюзовка, среднее течение. Эйфельский ярус среднего
девона.
Фиг. 3. Heliolites gluchovl Z h m a e v, sp. n. . Стр. 205
a — продольный разрез; б — поперечный разрез. X 3. Салаир,
р. Глухая, левый приток р. Анфимов Мочег. Нижний девон.
Фиг. 4. Heliolites khvo'scevlensls Z h m а е v, sp. n. Стр. 205-
a — продольный разрез, X2; 6 — поперечный разрез Салаир,.
кл. Смородиновый, левый приток р. Хвощевки. Средний девон.
394
ТАБЛИЦА ХХХПГ
ТАБЛИЦА XXXIV
Фиг. 1. Tryplasma altaica (Dy bow ski) Стр. 207
a — продольный разрез; б — поперечный разрез. Нат. вел.
Центральный Салаир, рч. Глухая. Нижний девон.
39 6
ТАБЛИЦА XXXIV
ТАБЛИЦА XXXV
Фиг. 1. Род Tryplastna Lonsd. ... . Стр. 206
Общий вид коралла, нат. вел. По Э. 3. Бульванкер.
•Фиг. 2. Tryplastna altalca (Dy bow ski) Стр. 207
a — продольный разрез; б — поперечный разрез. Нат. вел.
Салаир, рч. Глухая, левый приток р. Анфимов Мочег. Нижний
девон.
Фиг. 3. Tryplastna devontana (Soshkina) Стр. 207
а — продольный разрез; б — поперечный разрез. X 2. Юго-
Западное Присалаирье. Нижний девон.
Фиг. 4. Tryplastna hercyntca (Roemer) . Стр. 208
а — продольный разрез; б — поперечный разрез. X 4. Левый
берег р. Черневого Бачата у крековской мельницы. Нижний
девон. По Э. 3. Бульванкер.
Фиг. 5. Thamnophyllum trlgemme (Quenstedt) Стр. 218
Поперечный разрез. Центральный Салаир.
Г398
ТАБЛИЦА XXXV
ТАБЛИЦА XXXVI
Фиг. 1. Calceola sandalina Lamarck var. acuminata M a n su у Стр. 208
a — крышечка; б — внешний вид коралла. Нат. вел. Живетский
ярус среднего девона. По Е. Д. Сошкиной.
Фиг. 2. Calceola sandalina Lamarck var. acuminata M a n suy Стр. 208
а и 6 — внешний вид в двух проекциях; в — крышечка.
Нат. вел. Центральный Салаир. Живетский ярус среднего
девона.
Фиг. 3. Grypophyllum [asclculum Z h m а е v, sp. n. . Стр. 210
a — продольный разрез; б — поперечный разрез. X 3. Кузбасс,
р. Томь, выше устья р. Кара-Чумыш. Средний девон.
Фиг. 4. Grypophyllum gracile Wedekind Стр. 209'
а — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса. Живетский ярус среднего девона.
Фиг. 5. Grypophyllum pesterevtensis Z h m а е v, sp. n. . Стр. 209
a — продольный разрез; б — поперечный разрез. X 3,5. Куз-
басс, в 3 км северо-западнее дер. Ново-Пестерево. Средний
девон.
Фиг. 6. Loyolophyllum urense Z h m а е v, sp. n. . . Стр. 217
a — продольный разрез; б — поперечный разрез. X 3. Юго-за-
падная окраина Кузбасса, южнее дер. Мамонтово. Эйфельский
ярус среднего девона.
4 СО
ТАБЛИЦА XXXVI
26 Зак. 2024
ТАБЛИЦА XXXVII
Фиг. 1. Stenophyllum vogulicum Bulvanker . Стр. 210
a — внешний вид колонии, нат. вел.; б — продольный разрез,
X 1,5. По Э. 3. Бульванкер.
в — поперечный разрез, Х2,4; г — продольный разрез, X 2,4.
Салаир, среднее течение р. Верди.
Средний девон.
Фиг. 2. Stenophyllum gorsku В u 1 v a n к е г, sp. п. Стр. 211
a — продольный разрез; б — поперечный разрез. X 4. Салаир,
р. Черневой Бачат. Нижний девон. По Э. 3. Бульванкер.
ТАБЛИЦА XXXVII
ТАБЛИЦА XXXVHI
Фиг. 1. Stenophyllum siblricum В u 1 v a n к e r, sp. n. . Стр. 211
a — продольный разрез; б •- поперечный разрез. Центральный
Салаир. Эйфельский ярус среднего девона.
Фиг. 2. Stenophyllum devonlcum В u I v а п к е г, sp. n. . Стр. 211
а — продольный разрез; б — поперечный разрез. X 3,5. Куз-
басс. Правый берег р. Томь-Чумыша. Эйфельский ярус среднего
девона. По Э. 3. Бульванкер.
Фиг. 3. Thamnophyllum trigemme (Quenstedt) Стр. 218
а — продольный разрез; б — поперечный разрез. X Цент-
ральный Салаир. Живетский ярус среднего девона.
404
ТАБЛИЦА XXXVIII
ТАБЛИЦА XXXIX
Фиг. 1. Stenophyllum, salalrlcum Zhmaev, sp. n. Стр. 212
a — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса, в 4 км южнее дер. Мамонтово.
Эйфельский ярус среднего девона.
Фиг. 2. Stenophyllum soshkinae Zhmaev, sp. n. Стр. 212
a — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса, в 2 км южнее дер. Ново-Пестерево.
Эйфельский ярус среднего девона.
Фиг 3. Spongophyllum minimum (Zhmaev), sp. п. . Стр. 215
а — продольный разрез; б — поперечный разрез. Центральный
Салаир. Нижний девон.
406
ТАБЛИЦА XXXIX
ТАБЛИЦА XL
Фиг. 1. Pseudospongophyllum masslvum Zhmaev, sp. n. . Стр. 213
a — продольный разрез; б — поперечный разрез. X 3. Юго-за-
падная окраина Кузбасса, р. Бачат, в 2,5 км выше дер. Шынды.
Эйфельский ярус среднего девона.
Фиг. 2. Pseudospongophyllum tomense Zhmaev, sp. n. Стр. 213
a — продольный разрез; б — поперечный разрез. X 3. Юго-
западная окраина Кузбасса, р. Томь-Чумыш в районе с. Томский
завод. Нижний девон.
Фиг. 3. Fasciphyllum petshorense Soshkina. Стр. 215
а — продольный разрез; б — поперечный разрез. X 4. Коб-
ленцский ярус нижнего девона. По Е. Д. Сошкиной.
408
ТАБЛИЦА XL
ТАБЛИЦА XLI
Фиг. 1. Columnaria vulgaris Soshkina Стр. 216
a — продольный разрез; б — поперечный разрез. X 3. Левый
берег р. Кара-Чумыш, у дер. Белый Камень (Юго-западный
Кузбасс). Живетский ярус среднего девона.
Фиг. 2. Columnaria rhenana F г е ch. . Стр. 216
а — продольный разрез; б — поперечный разрез. X 4- В 4,5 км
от дер. Сафоновой, Кузбасс. Живетский ярус среднего девона.
•Фиг. 3. Spongophyllum origlnalis Zhmaev, sp. n. Стр. 214
a — продольный разрез; б — поперечный разрез. X 3. Юго-за-
падная окраина Кузбасса, левый берег р. Кара-Чумыша
у дер. Красная поляна. Нижний девон.
410
ТАБЛИЦА XU
1 б
2а
ТАБЛИЦА XLH
Фиг. 1. Peneckiella darwinl (Freeh) . Стр. 219
a — продольный разрез; б — поперечный разрез. X 2. Северо-
западная окраина Кузбасса, рч. Большая Стрельна у с. Соло-
мине. Горизонт глубокинского известняка верхнего девона.
Фиг. 2. Peneckiella tabulata Bulvanker, sp. n. Стр. 219
a — продольный разрез; б — поперечный разрез. X 2. Северо-
западная окраина Кузбасса, рч. Большая Стрельна у с. Соло-
мине. Горизонт глубокинского известняка.
Ф'иг. 3. Megaphyllum pashiense Soshkina . Стр. 229
а — продольный разрез; б — поперечный разрез. X 2. Север-
ная окраина Кузбасса, р. Яя у Черепанова брода. Нижнефран-
ские отложения верхнего девона.
Фиг. 4. Pseudohonophyllum pseudohellanthoides Scherzer Стр. 218
а — продольный разрез; б — поперечный разрез. X 4. Девон.
По Е. Д. Сошкиной.
412
ТАБЛИЦА XLII
ТАБЛИЦА XLIII
Фиг. 1. Phillipsastraea mlcrommata (A. Roemer) Стр. 223
a — продольный разрез; б — поперечный разрез. X 2. Кузбасс,
рч. Большая Стрельна у с. Соломине. Горизонт глубокинского
известняка верхнего девона.
Фиг. 2. Phillipsastraea pentagona (G о 1 d f u s s) . Стр. 223
a — продольный разрез; б — поперечный разрез. X 2. Кузбасс,
рч. Большая Стрельна у с. Соломино. Горизонт глубокинского
известняка верхнего девона.
Фиг. 3. Schlilterla emstl Wedekind Стр. 224
а — продольный разрез; б — поперечный разрез. X 2. Северо-
западная окраина Кузбасса, рч. Большая Стрельна у с. Зару-
бине. Нижнефранские отложения верхнего девона.
Фиг. 4. Phillipsastraea sedgwlckl (Edwards et Haime) Стр. 221
a — продольный разрез; б и в — поперечный разрез. X 2. Се-
веро-западная окраина Кузбасса, рч. Большая Стрельна у с. За-
рубино. Нижнефранские отложения верхнего девона.
414
ТАБЛИЦА XLIII
ТАБЛИЦА XL1V
Фиг. 1. Phillipsastraea lazutkini (В u 1 v а п к e г) Стр. 222
a — продольный разрез; б и в — поперечный разрез. X 2. Се-
веро-западная окраина Кузбасса, рч. Большая Стрельна у с. За-
рубино. Нижнефранские отложения верхнего девона.
Фиг. 2. Phillipsastraea lateseptata I v a n i а . . Стр. 222
a — продольный разрез; б — поперечный разрез. X 2. Северо-
западная окраина Кузбасса, рч. Большая Стрельна у с. Зару-
бино.
Фиг. 3. Endophyllum abdltum Edwards et Haime . Стр. 228
a — продольный разрез; б — поперечный разрез. X 2. Северо-
западная окраина Кузбасса, правый берег рч. Большая
Стрельна у с. Соломино.
-416
ТАБЛИЦА XLIV
27 Зак. 2024
ТАБЛИЦА XLV
Фиг. 1. Pachyphyllum devontense Edwards et Haime. Стр. 227
a — продольный разрез; б — поперечный разрез. X 2. Север-
ная окраина Кузбасса, правый берег р. Яи у Черепанова брода.
Нижнефранские отложения верхнего девона.
Фиг. 2. Schluteria striata I v a n i а Стр. 224
Поперечный разрез, X 2. Северная окраина Кузбасса, правый
берег р. Яи у Черепанова брода. Нижнефранские отложения
верхнего девона.
Фиг. 3. Pachyphyllum ibergense A. Roem е г . Стр. 227
а — продольный разрез; б — поперечный разрез. X 2. Север-
ная окраина Кузбасса, правый берег р. Яи у Черепанова брода.
Нижнефранские отложения верхнего девона.
418
ТАБЛИЦА XLV
ТАБЛИЦА XLVI
Фиг. 1. Endophyllum macconnelll Whiteaves Стр. 228
а и б — продольные разрезы; в, г и д — поперечные разрезы.
X 2. Северная окраина Кузбасса, правый берег р. Яи у Черепа-
нова брода. Нижнефранские отложения верхнего девона.
Фиг. 2. Schluteria striata I v a n i а . . Стр. 224
a — продольный разрез; б — поперечный разрез. X 2. Северная
окраина Кузбасса, правый берег р. Яи у Черепанова брода.
Нижнефранские отложения верхнего девона
Фиг. 3. Solomlnella soshklnae I v a n i а Стр. 226
a — поперечный разрез; бив — продольные разрезы. X 2. Се-
веро-западная окраина Кузбасса, с. Соломине. Среднефранские
отложения верхнего девона.
420
ТАБЛИЦА XLVl
ТАБЛИЦА XLVII
Фиг. 1. Liocletna yaKOvlevi Schoenmann . . Стр. 229
a — внешний вид колонии, нат. вел. Горный Алтай, Чуйская
степь при выходе р. Бугусун из ущелья Кизыл-Шин. Основание
верхнего девона.
б — шлиф, X 5. Северо-западная окраина Кузбасса, Барзас-
ский район.
в — тангенциальное сечение, X 80; г — тангенциальное сечение,
X 20; д — поперечное сечение, X 20. Горный Алтай. По
В. П. Нехорошеву.
Фиг. 2. Liocletna bugusunlca Nekhoroschev Стр. 229
а — обломок толстой ветвистой колонии, нат. вел.; б — про-
дольное сечение, ХЮ; в — тангенциальное сечение, X 20. Гор-
ный Алтай, р. Бугусун, ущелье Кизыл-Шин. Верхи среднего
девона. По В. П. Нехорошеву.
422
ТАБЛИЦА XLVII
26
ТАБЛИЦА XLVIII
Фиг 1. Fenestella vera Ulrich . . Стр. 231
a — тангенциальное и поперечное сечения, X Ю- Северо-
Западная Монголия, р. Чигерты. Средний девон.
б — тангенциальное сечение, Х22. Центральный Алтай,
р. Бельгебаш. По В. П. Нехорошеву и П. С. Краснопеевой.
Фиг. 2. Hemitrypa cornea Nekhoroschev . Стр. 232
Тангенциальное сечение, X Ю. Рудный Алтай. Средний девон.
По В. П. Нехорошеву.
Фиг. 3. Hemitrypa devonica Nekhoroschev . Стр. 232
а — тангенциальное сечение неячеистой поверхности, видны
бугорки различной величины, X 46; б — то же, X 20; в — танген-
циальное сечение ячеистой поверхности, ХИ- Горный Алтай.
Чуйская степь, при выходе р. Бугусун из ущелья Кизыл-Шин.
Верхи верхнего девона. По В. П. Нехорошеву.
Фиг. 4. Semtcoscinlum rhomblcum Ulrich var. altaica Krasno-
peeva . Стр. 233
a — тангенциальное сечение, X 15; б — поперечное сечение,
X 20. Горный Алтай, Кортонский хребет, р. Коргончик. Верхи
среднего девона.
424
ТАБЛИЦА XIA III
ТАБЛИЦА XLIX
Фиг. 1. Semicoscinitim altalcum Nekhoroschev Стр. 232
a — тангенциальное сечение неячеистой поверхности, видны
бугорки, а в более глубоком сечении — пузырчатая ткань;
б — тангенциальное сечение, видны кили, ячейки и неячеистан
поверхность; в — поперечное сечение. XII. По В. П. Нехоро-
шеву.
Фиг. 2. Setnlcosclnium altchedaticum Nekhoroschev Стр. 233
а — тангенциальное сечение; б — поперечное сечение. X Ю-
Горный Алтай, р. Бугусун, ущелье Кизыл-Шин. Верхи среднего
девона.
Фиг. 3. Reteporina grandiformis Krasnopeeva Стр. 233
а — тангенциальное сечение, прутья более сильно изогнуты на
неячеистой поверхности, X 15; б—поперечное сечение одного
прутика, X 16. Горный Алтай, р. Бугускун, ущелье Кизыл-Шин.
Низы верхнего девона.
Фиг. 4. Cyrtospirifer achrnet N а 1 i v k i п. Нат. вел. Стр. 266
Кузбасс. Верхний девон, фаленовый горизонт.
Фиг. 5. Platyrochella martlanovl (Stuckenberg). Нат. вел. Стр. 261
Горный Алтай. Средний девон, чиэлевый горизонт.
426
ТАБЛИЦА XLIX-
1 а
2 б
2а
ТАБЛИЦА L
Все изображения даны в натуральную величину
Фиг. 1. Levenea taeniolata Khalfin . . Стр. 235
Отпечаток брюшной створки. Горный Алтай. Нижний де-
вон, ганинская свита.
Фиг. 2 и 3. Leptaena bouel (Barrande) Стр. 235
Отпечаток и внутреннее ядро брюшной створки. Горный
Алтай. Нижний девон, кондратьевская свита.
Фиг. 4. Leptaena bouel (Barrande) . . Стр. 235
Отпечаток брюшной створки. Горный Алтай. Нижний де-
вон, медведевская свита.
Фиг. 5 и 6. Leptaena bouel (Barrande) . Стр. 235
Внутренние ядра брюшной и спинной створок. Горный
Алтай. Нижний девон, ганинская свита.
Фиг. 7—9. Leptaena bouel (Barrande) Стр. 235
Брюшная створка, вид раковины со стороны переднего
края и профиль раковины. По Барранду.
Фиг. 10. Cymostrophla stephanl (Barrande). . . Стр. 236
Брюшная створка в двух проекциях. По Барранду.
Фиг. 11. Brachyprlon latus К h а 1 f i n . . Стр. 237
Внутреннее ядро брюшной створки. Горный Алтай. Ниж-
ний девон, медведевская свита.
Фиг. 12. Brachyprlon nalivklnl Khalfin. . Стр. 236
Внутреннее ядро брюшной створки. Горный Алтай. Ниж-
ний девон, медведевская свита.
428
ТАБЛИЦА L
10а
ТАБЛИЦА LI
Все изображения, кроме фиг. 7, даны в натуральную величину
Фиг. 1 и 2. Leptodontella rotundata К h а 1 f i п Стр. 237 Горный Алтай. Нижний девон, ганинская свита.
Фиг. 3. Leptodontella planuscula (К h а 1 f i n). Стр. 237 Горный Алтай. Нижний девон, медведевская свита.
Фиг. 4 и 5. Schuchertella altalca К h а 1 f i n . . Стр. 238 Отпечаток и внутреннее ядро брюшной створки. Горный Алтай. Нижний девон, ганинская свита.
Фиг. 6. Chonetes pseudobohemlca К h а 1 f i п Стр. 238 Брюшная створка. Горный Алтай. Нижний девон, медве- девская свита.
Фиг. 7 Cymostrophla stephanl Barrand е. Стр. 236 Характер скульптуры, увеличено. По Барранду.
•Фиг. 8. Brachyprlon latus К h а 1 f i n . Стр. 237 Внутреннее ядро спииной створки. Горный Алтай. Нижний девон, медведевская свита.
-430
ТАБЛИЦА LI
ТАБЛИЦА LI I
Все изображения даны в натуральную величину
Фиг. 1. Плитка песчаника с многочисленными отпечатками раковин
Chonetes pseudobohetnica К h а 1 f i п и Chonetes subextensa
К h a 1 f i n. . Стр. 238
Горный Алтай. Нижний девон, медведевская свита.
Фиг. 2. Chonetes rectisplna К h а 1 f i п Стр. 239
Многочисленные отпечатки изолированных створок. Гор-
ный Алтай. Нижний девон, кондратьевская свита.
Фиг. 3. Conchidium celsiseptosum К h а 1 f i n . Стр. 239
Ядро брюшной створки. Горный Алтай. Нижний девон,
медведевская свита.
Фиг. 4—6. Ussovia prima (Kha 1 fin) . Стр. 240
Ядро брюшной створки, характер седла и профиль дефор-
мированного образца. Горный Алтай. Нижний девон, кон-
дратьевская свита.
Фиг. 7 и 8. Atrypa umbra Barrande . Стр. 240
Горный Алтай. Нижний девон, соловьихинский известняк.
Фиг. 9. Atrypa supramarginalis К h а 1 f i n . Стр. 240
Горный Алтай. Нижний девон, соловьихинский известняк.
Фиг. 10 и 11. Spirifer (Delthyris?) jaschel Roemer. . Стр. 241
Горный Алтай. Нижний девон, соловьихинский известняк.
Фиг. 12. Rhynchosplrina formosa (Hall) . Стр. 243
Горный Алтай. Нижний девон, соловьихинский известняк.
432
ТАБЛИЦА LII
1
28 Зак. 2024
ТАБЛИЦА LIII
Все изображения, кроме фиг. 5 и 8, даны в натуральную величину
Фиг. 1 и 2. Eospirifer pseudotogatus Khalfin Стр. 241
Отпечаток брюшной створки и внутреннее ядро спинной
створки. Горный Алтай. Нижний девон, ганинская свита.
Фиг. 3—5. Delthyrls perlamellosa Hall. . Стр. 241
3 — ядро брюшной створки; 4 — ядро брюшной створки
со срезанной макушкой, видны септа и зубные пластины;
5 — неполный отпечаток спинной створки — характер скульп-
туры, увеличено. Горный Алтай. Нижний девон, ганинская
свита.
Фиг. 6. Conchidium celsiseptosum Khalfin . Стр. 239
Ядро спинной створки. Горный Алтай. Нижний девон,
медведевская свита.
Фиг. 7. Acrospirifer pseudofallax (Khalfin) Стр. 242
Внутреннее ядро раковины со стороны брюшной (а)
и спинной (б) створок. Горный Алтай. Нижний девон, мед-
ведевская свита.
Фиг. 8. Rensselaeria navicelllformis Khalfin, Х2 • Стр. 243
Горный Алтай. Нижний девон, низы ганинской свиты.
Фиг. 9. Acrospirifer korovini (Khalfin) . Стр. 242
Внутреннее ядро спинной створки. Горный Алтай. Ниж-
ний девон, медведевская свита.
Фиг. 10. Gypidula kayseri (Peetz). Стр. 245
Салаир. Нижний девон, крековские слои.
Фиг. 11. Camarotoechia lazutkini Rzonsnickaja, sp. n. Стр. 246
Салаир. Средний девон, акарачкинские и сафоновские слои.
431
ТАБЛИЦА LIII
ТАБЛИЦА LIV
Фиг. 1.
Фиг. 2.
Фиг. 3.
Acrospirifer korovtni (Khalfin). Стр. 242
Внутреннее ядро брюшной створки, нат. вел. Горный Алтай.
Нижний девон, медведевская свита.
Acrospirifer pseudofallax (Khalfin), нат. вел. Стр. 242
Горный Алтай. Нижний девон, медведевская свита.
Stropheodonta uralensls (Verneuil). Стр. 244
Ядро брюшной створки в двух проекциях, нат. вел. Салаир.
Средний девон, шандинские слои.
Leptaena rhomboldalls W i 1 с kе n s var. slnuata Rzonsn ic-
Фиг. 4.
k a j а . Отпечаток брюшной створки, нат. вел. Салаир. ( сафоновские слои. Стр. 244 Средний девон,
Фиг. 5. Unctnulus tricostatus (Peetz), X 2. Салаир. Средний девон, сафоновские слои. Стр. 247
Фиг. 6. Camarotoechia nympha (В а г г a n d е) var. pseudolivonica (В а г- г a n d е), X 2 Стр. 245 Салаир. Нижний девон, пестеревские слои.
Фиг. 7. Unctnulus paralleleplpedus (Bronn), Х2 Салаир. Средний девон, салаиркинские слои. Стр. 247
Фиг. 8. Unctnulus angularls (Phillips), X 2 Салаир. Средний девон, сафоновские слои. Стр. 247
Фиг. 9. Camarotoechia lazutkinl Rzonsnickaja, sp. n., X 2 Стр. 246 Салаир. Средний девон, акарачкинские и сафоновские слои.
436
ТАБЛИЦА LIV
ТАБЛИЦА LV
Фиг. 1 . Unclnulus dubroviensis Rzonsickaja, sp. п. Нат.
вел. Салаир. Средний девон, сафоновские слои. Стр. 246
Фиг. 2. Unclnulus angularls (Phillips). X 2. Салаир. Средний девон, сафоновские слои. Стр. 247
Фиг. 3. Unclnulus subcordlformis Schnur. Нат. вел. Салаир. Средний девон, сафоновские слои. Стр. 247
Фиг. 4 и 5. Conchidium ex gr. pseudobaschkirlcum Tscherny- s c h e v . Стр. 245 4a и 46 — внешний вид раковины в двух проекциях; 5 — раковина, расколотая по плоскости симметрии — харак- тер септы. Нат. вел. Салаир. Средний девон, шандинские слои.
Фиг. 6. Hypothyridlna procuboldes (Kayser), X 2 Салаир. Средний девон. Стр. 243
Фиг. 7 Eoretlcularla dereimsl (О е h 1 е г t). Нат. вел. Салаир. Средний девон, чумышская толща. Стр. 252
Фиг. 8. Eospirifer secans Barrande. Нат. вел. Салаир. Нижний девон, пестеревские слои. Стр. 250
Фиг. 9. At гура bodlnl Mansuy, Х2 Салаир. Средний девон, сафоновские слои. Стр. 248
ф|иг. 20. Atrypa bodlnl Mansuy, X 2 Салаир. Средний девон, сафоновские слои. Стр. 248
Фиг. 1 1 -- -13. Atrypa semlorbicides Lazutkin, sp. п. Нат. вел. Салаир. Нижний девон, крековские слои. Стр. 248
438
ТАБЛИЦА LV
ТАБЛИЦА LVI
Фиг. 1. Atrypa subspinosa Lazutkin, sp. n., XL5. Стр. 249
Салаир. Средний девон, салаирские слои.
Фиг. 2. Carinatina eudokia Lazutkin, sp. n. Нат. вел . Стр. 250
Салаир. Шандинские слои.
Фиг. 3 и 4. Eosplrlfer secans Barrande Стр. 250
3 — нат. вел.; 4 — характер микроскульптуры, сильно уве-
личено. Салаир. Нижний девон, пестеревские слои.
Фиг. 5 'И 6. Elythina salairica Rzonsnickaj а. Стр. 252
5 — нат. вел.; 6 — характер микроскульптуры, сильно уве-
личено. Салаир. Нижний девон, пестеревские слои.
Фиг. 7. Cyrtlnopsis nallvkinl Rzonsnickaja. Нат. вел. Стр. 251
Салаир. Нижний девон — низы среднего девона.
ТАБЛИЦА LVb
ТАБЛИЦА LVII
Фиг. 1 и 2.
Фиг. 3 и 4.
Фиг. 5.
Фиг. 6—8.
Фиг. 9 и 10.
Фиг. 11.
Lazutklnia mamantovensis (Lazutkin). Нат. вел. Стр. 254
Салаир. Средний девон, Мамонтовские слои.
Elythina salalrica Rzonsnickaja Стр. 252
4 — ядро брюшной створки; 3 — характер мускульного
бугра. Нат. вел.
Denkmanella damesi (Holzapfel). Нат. вел. Стр. 254
Салаир. Средний девон, керлегешские слои.
Indospirifer pseudowllliamsi Rzonsnickaja Стр. 253
б, 7 — нат. вел.; 8—характер микроскульптуры, сильно
увеличено. Салаир. Средний девон, сафоновские слои.
Chascothyrls salalrica Rzonsnickaja, sp. n. Нат.
вел. Стр. 255
Молодые экземпляры. Салаир. Средний девон, сафонов-
ские слои.
Karpinskla conjugula Tschernyschev. Нат.
вел. Стр. 249
Салаир. Нижний девон, пестеревские слои.
442
таблица lvh
,7 а
ТАБЛИЦА LVIII
Все изображения даны в натуральную величину
Фиг. I и 2. Chascothyris salairica Rzonsnickaja, sp. n. Стр. 255
Взрослые раковины. Салаир. Средний девон, керлегеш-
ские слои.
Фиг. 3. Schizophoria striatula (Schlotheim) Стр. 255
Вид раковины со стороны брюшной створки (а) и перед-
него края (б). Северо-западная окраина Кузбасса, с. Лебе-
дянское. Средний девон, чиэлевый горизонт.
Фиг. 4. Isorthis slblrica (Khalfin). Стр. 257
а — спинная створка; б — брюшная створка; в—вид со
стороны замочного края. Северо-западная окраина Кузбасса,
с. Лебедянское. Средний девон, чиэлевый горизонт.
Фиг. 5. Schellwienella umbraculum (Schlotheim) Стр. 257
Внутреннее ядро брюшной створки. Северо-западная
окраина Кузбасса, с. Лебедянское. Средний девон, чиэлевый
горизонт.
Фиг. 6. Schellwienella umbraculum (Schlotheim) Стр. 257
Спинная створка. Северо-западная окраина Кузбасса,
с. Лебедянское. Средний девон, чиэлевый горизонт.
Фиг. 7. Unctnulus korovini Khalfin . . Стр. 258
Северо-западная окраина Кузбасса, с. Лебедянское. Сред-
ний девон, чиэлевый горизонт.
Фиг. 8. Leptaena rhombotdalls W i 1 с k е n s . . Стр. 257
Северо-западная окраина Кузбасса, с. Лебедянское. Сред-
ний девон, чиэлевый горизонт.
фиг. 9. Platyrochella martlanovl (Stuckenberg) Стр. 261
Брюшная створка. Минусинская котловина, с. Бейское.
Средний девон, чиэлевый горизонт.
ТАБЛИЦА LVIII
ТАБЛИЦА LIX
Все изображения, кроме фиг. 4, даны в натуральную величину
Фиг. 1. Atrypa ex gr. reticularis Linne Северо-западная окраина Кузбасса, девон, чиэлевый горизонт. Стр. 258
с. Лебедянское. Средний
Фиг. 2. Atrypa desquamata Sower by Стр. 258
Северо-западная окраина Кузбасса, девон, чиэлевый горизонт. с. Лебедянское. Средний
Фиг. 3. Atrypa tubaecostata Paeckelmann Стр. 259
Северо-западная окраина Кузбасса, девон, чиэлевый горизонт. с. Лебедянское. Средний
Фиг. 4. Atrypa tubaecostata Paeckelmann Стр. 259
Характер скульптуры, увеличено. Северо-западная окраина
Кузбасса, с. Лебедянское. Средний девон, чиэлевый горизонт.
Фиг. 5. Acrospirifer cheehlel (Ко n i nc k) Стр. 259
Северо-западная окраина Кузбасса, девон, чиэлевый горизонт. с. Лебедянское. Средний
Фиг. 6. Schellwlenella umbraculum (Schlotheim) Стр. 257
Вид раковины сбоку. Северо-западная окраина Кузбасса, с. Ле-
бедянское. Средний девон, чиэлевый горизонт.
Фиг. 7 Theodossla (?) schmidtl (Stuckenberg) Стр. 259
Минусинская котловина, с. Бейское. Средний девон, чиэлевый
горизонт.
446
ТАБЛИЦА LIX
ТАБЛИЦА LX
Все изображения даны в натуральную величину
Фиг. 1. Acrospirifer cheehiel (Коп i пск) Стр. 259
Вид со стороны переднего края. Северо-западная окраина
Кузбасса, с. Лебедянское. Средний девон, чиэлевый горизонт.
Фиг. 2. Acrospirifer cheehiel (Коп i nek) var. altalca Khal-
f i n . . Стр. 259
Внутреннее ядро брюшной створки. Горный Алтай, окре-
стности с. Куяган. Средний девон, чиэлевый горизонт.
Фиг. 3. Lamellispirifer mucronatus (Conrad) . Стр. 260
Внутреннее ядро брюшной створки. Горный Алтай, окре-
стности с. Куяган. Средний девон, чиэлевый горизонт.
Фиг. 4. Lamellispirifer mucronatus (Conrad) . . Стр. 260
Внутреннее ядро спинной створки. Горный Алтай, окре-
стности с. Куяган. Средний девон, чиэлевый горизонт.
Фиг. 5. Acrospirifer cheehiel (Koni nek) Стр. 259
Вид раковины сбоку. Северо-западная окраина Кузбасса,
с. Лебедянское. Средний девон, чиэлевый горизонт.
Фиг. 6. Platyrochella martlanovl (Stuckenberg). Стр. 261
Минусинская котловина, с. Бейское. Средний девон, чиэле-
вый горизонт.
Фиг. 7—10. Pllcatlfera praelonga (Sow er by) . Стр. 262
7 — вид брюшной створки сбоку; 8 — брюшная створка;
9—вид брюшной створки с переднего края; 10—характер
синуса брюшной створки с серединным валиком. Севере
западная окраина Кузбасса, с. Жаркове кое. Фаменский ярус
верхнего девона.
Фиг. 11. Hypothyrldlna tchingislca Kraewskaja Стр. 264
а—вид со стороны переднего края; б—брюшная створка.
Правобережье р. Оби в районе г. Камня. Глубокинский
горизонт верхнего девона.
Фиг. 12. Hypothyrldlna tchingislca Kraewskaja . Стр. 264
Спинная створка. Правобережье р. Оби в районе
г. Камня. Глубокинский горизонт верхнего девона.
Фиг. 13. Pugnax acuminata Martin Стр. 264
а — спинная створка; б—вид сбоку; в — вид со стороны
переднего края. Франский ярус верхнего девона. По
Ф. Н. Чернышеву.
Фиг. 14. Gypidula comis (Owen) Стр. 263
а — брюшная створка; б — вид со стороны переднего края;
в—вид сбоку; г—вид со стороны заднего края. Право-
бережье р. Оби в районе г. Камня. Глубокинский горизонт
верхнего девона.
-448
ТАБЛИЦА LX
29 Зак. 2024
ТАБЛИЦА LXI
Все изображения, кроме фиг. 10, даны в натуральную величину
Фиг. 1. Gypidula obi Kraewskaja Стр. 265
а — брюшная створка; б — спинная створка; в — профиль;
г — вид с переднего края. Правобережье р. Оби в районе
г. Камия. Глубокииокий горизонт франского яруса.
Фиг. 2. Hypothyridina cuboides (Sow er by) Стр. 263
a — вид co стороны переднего края; б — спинная створка;
в — вид сбоку. Франский ярус верхнего девона. По Ф. Н. Чер-
нышеву.
Фиг. 3. Pugnax voronl N а 1 i v k i п Стр. 264
a — профиль; 6 — вид co стороны переднего края. Право-
бережье р. Оби в районе г. Камня. Глубокинский горизонт
франского яруса верхнего девона.
Фиг. 4. Cyrtospirifer curvlrostrls I v a n i а . Стр. 267
a — вид сбоку; б — вид со стороны макушки. Кузбасс, правый
берег р. Иня у дер. Абышево. Фаменский ярус верхнего девона.
Фиг. 5. Eoreticularia mlcrospinosa Khalfin Стр. 265
а — ядро брюшной створки; б — спинная створка. Северо-
западная окраина Кузбасса, с. Зарубино. Основание франского
яруса верхнего девона.
Фиг. 6. Cyrtosplrlfer achmet N а 1 i v k i п Стр. 266
Кузбасс, правый берег р. Иня у дер. Абышево. Фаленовый
горизонт верхнего девона.
Фиг. 7 LamelUspirifer mucronatiformis Khalfin Стр. 266
а — спинная створка; б - вид с заднего края; в — вид
с переднего края. Кузбасс. Верхи франского яруса верхнего
девона (промежуточная толща).
Фиг. 8. Cyrtospirifer tchernychevl Khalfin var. slblrica Стр. 266
a — брюшная створка; б — вид со стороны ареи брюшной
створки; в--спинная створка; г — вид сбоку. Кузбасс, правый
берег р. Иня у дер. Абышево. Фаменский ярус верхнего девона.
Фиг. 9. Cranaena romlngerl Hall Стр. 265
а — брюшная створка; б — спинная створка; в — боковой
профиль; г — передний край. Правобережье р. Оби в районе
г. Камня. Глубокинский горизонт франского яруса верхнего
девона.
Фиг. 10. Cyrtospirifer barzassensis Т у zh п о v, sp. n. . Стр. 267
Брюшная створка. Слегка увеличено. Кузбасс, Барзасский
район. Нпжнефрапские отложения верхнего девона.
Фиг. 11. Cyrtosplrlfer verneulll Murchison Стр. 267
а — брюшная створка; б — вид со стороны ареи; в — вид
сбоку. Кузбасс, правый берег р. Ини у дер. Абышево. Фамеп-
ский ярус верхнего девона.
Фиг. 12. Cyrtosplrlfer tenticulum (V е г n е u i 1) . . Стр. 268
а - - вид со стороны ареи; б вид сбоку. Кузбасс, правый
берег р. Яя ниже с. Ишим. Франский ярус верхнего девона.
450
ТАБЛИЦА LXI
ТАБЛИЦА LXII
Все изображения, кроме особо отмеченных, даны в натуральную величину
Фиг. 1. Cyrtospirifer archiaci Murchison Стр. 237
а — брюшная створка; б—вид со стороны ареи; в — вид
сбоку. Кузбасс, правый берег р. Ини у дер. Абышево. Фамен-
ский ярус верхнего девона.
Фиг. 2. Cyrtospirifer tchernychevl К h а 1 f i п Стр. 268
а — брюшная створка; б — вид со стороны ареи; в — вид
сбоку. Кузбасс, правый берег р. Ини у дер. Абышево. Фамен-
ский ярус верхнего девона.
Фиг. 3. Cyrtospirifer tenticulum (V е г n е u i 1) Стр. 268
а -арея брюшной створки; б—синус брюшной створки.
Кузбасс, правый берег р. Яя ниже с. Ишим. Фаленовый
горизонт франского яруса верхнего девона.
Фиг. 4—6. Cyrtospirifer khalfint I v a n i а . Стр. 268
4a — арея брюшной створки; 46 — вид сбоку на брюшную-
створку; 5 — арея брюшной створки, слегка увеличено;
6 — синус брюшной створки. Кузбасс, правый берег р. Ини
у дер. Абышево. Монстровый горизонт верхнего девона.
Фиг. 7—10. Indospirifer sibiricus К h а 1 f i n . Стр. 269
7 — внутреннее ядро брюшной створки; 8 — разновид-
ность alata Kha Hin: а — брюшная створка, б — спинная
створка; 9 — брюшная створка молодого экземпляра;
10 — спинная створка другого экземпляра. X 2. Северо-запад-
ная окраина Кузбасса. Верхи франского яруса верхнего девона.
Фиг. 11. Hypothyrldlna cuboldes (Sow er by) . Стр. 263
a — вид co стороны переднего края; б — вид сбоку. Право-
бережье р. Оби в районе г. Камня. Глубокинский горизонт-
франского яруса верхнего девона.
452
ТАБЛИЦА LXII
ТАБЛИЦА LXI 11
Все изображения, кроме особо отмеченных, даны в натуральную величину
Фиг. 1 и 2. Cyrtosplrlfer khalfini I v а п i а . Стр. 268
Спинная створка. / — слегка увеличено. Кузбасс, правый
берег р. Пни у дер. Абышево. Монстровый горизонт верхнего
девона.
Фиг. 3 и 4. Cyrtosplrlfer ussovi Khalfin Стр. 269
<3 — арея брюшной створки; 4 — брюшная створка со
стороны синуса, слегка увеличено. Кузбасс, правый берег
р. Ини у дер. Абышево. Монстровый горизонт верхнего
девона.
Фиг. 5 и 6. Giirichella zlczac (Roemer) Стр. 270
Франский ярус. По Ф. Н. Чернышеву.
Фиг. 7 и 8. Anathyris phalaena (Phillips). Стр. 271
7 — брюшная створка; 8а— брюшная створка; 86—спин-
ная створка; 8в — вид с заднего края. Кузбасс, устье р. Тай-
дон. Фаленовый горизонт франского яруса.
Фиг. 9. Indospirifer slbirlcus Khalfin 269
Спииная створка, X 2 Кузбасс. Верхи, франского яруса
верхнего девона (промежуточная толща).
Фиг. 10. Anathyris monstrum Khalfin Стр. 271
Спинная створка. Северо-восточная окраина Кузбасса,
с. Жарковское. Монстровый горизонт верхнего девона.
Фиг. 11. Acrospirifer (?) subgregarius Rzonsnickaja Стр. 255
а — брюшная створка; б — спинная створка. Минусинская
котловина. Эйфельский ярус среднего девона, таштыпская
свита. Копия по М. А. Ржонсницкой.
Фиг. 12. Giirichella ziczac Roemer Стр. 270
Брюшная створка. Кузбасс. Франский ярус верхнего
девона.
Фиг. 13. Giirichella (?) vratschinskiae Kraewskaja . Стр. 270
а — вид со стороны брюшной створки; б — профиль.
Правобережье р. Оби в районе г. Камня. Глубокинский
горизонт франского яруса верхнего девона.
454
ТАБЛИЦА LXIII
ТАБЛИЦА LXIV
Все изображения даны в натуральную величину
Фиг. 1. Anathyris ussovi Khalfin . Стр. 272
Раковина в трех положениях. Северо-восточная окраина Куз-
басса, с. Марковское. Верхи франского яруса верхнего девона
(промежуточная толща).
Фиг. 2. Anathyrls monstrum Khalfin Стр. 971
а — брюшная створка; б — вид раковины сбоку. Северо-восточ-
ная окраина Кузбасса, с. Марковское. Монстровый горизонт
верхнего девона.
Фиг. 3. Anathyrls tyzhnovi Khalfin Стр. 271
а — спинная створка; б — брюшная створка; в — вид со сто-
роны заднего края. Северо-восточная окраина Кузбасса. Средина
франского яруса верхнего девона (атриповый горизонт).
Фиг. 4. Proetus carlnatus Khalfin . Стр. 272
Горный Алтай, окрестности с. Куяган. Нижний девон, кобленц-
ский ярус (кондратьевская свита).
Фиг. 5. Cheirurus strabo Weber Стр. 273
Горный Алтай, окрестности с. Куяган. Нижний девон, кобленц-
ский ярус (кондратьевская свита).
Фиг. 6. Phacops altalcus Tschernyschew . Стр. 273
Рудный Алтай, бывший Крюковский рудник. Нижний девон,
кобленцский ярус.
456
ТАБЛИЦА LXiV
ТАБЛИЦА LXV
Все изображения даны в натуральную величину
Фиг. 1—3.
Фиг. 4 и 5.
Фиг. 6.
Фиг. 7.
Фиг. 8.
Фиг. 9 и 10.
Фиг. 11.
Probolium altaicum (Khalfin) Стр. 274
1 -отпечаток головного щита; 2—внутреннее ядро
пигидия с отпечатком заворота; 3—внутреннее ядро пиги
дия. Горный Алтай, окрестности с. Куяган. Нижний девон,
кобленцский ярус (кондратьевская свита).
Phacops subcristata Khalfin . Стр. 273
Горный Алтай, окрестности с. Куяган. Нижний девон,
кобленцский ярус (кондратьевская свита).
Crotalocephalus sternbergi Boeck . Стр. 273
Горный Алтай, окрестности с. Куяган. Нижний девон,
кобленцский ярус (кондратьевская свита).
МуШагса plethomytiloldes Khalfin Стр. 276
Внутреннее ядро правой створки, отпечаток замка. Гор-
ный Алтай, окрестности с. Куяган. Нижний девон, кобленц-
ский ярус (кондратьевская свита).
МуШагса elongata Khalfin Стр. 276
Ядро правой створки. Горный Алтай, окрестности с. Куя-
ган. Нижний девон, кобленцский ярус (кондратьевская
свита).
Toechomya subcircularis Khalfin . Стр. 277
Горный Алтай, окрестности с. Куяган. Нижний девон,
кобленцский ярус (кондратьевская свита).
Praemyophoria acutirostris Khalfin Стр. 277
Ядро правой створки. Горный Алтай, окрестности с. Куя-
ган. Нижний девон, кобленцский ярус (кондратьевская
свита).
458
ТАБЛИЦА LXV
11
9
8
ТАБЛИЦА LXVI
Все изображения даны в натуралыную величину
-Фиг. 1 и 2. Pterinea (Tolmaia) squamosa Kha If in Стр. 275
1 — отпечаток правой створки; 2 — отпечаток левой створ-
ки. Горный Алтай, окрестности с. Куяган. Нижний девон,
кобленцский ярус (кондратьевская свита).
Фиг. 3 и 4. Archaeocardtum scrobiculare К h а 1 f i п Стр. 278
Внутреннее ядро левой створки в двух проекциях. Горный
Алтай, окрестности с. Куяган. Нижний девон, кобленцский
ярус (кондратьевская свита).
Фиг. 5. Lelopecten rectangularls Kha 1 fin . Стр. 275
Ядро левой створки. Горный Алтай, окрестности с. Куя-
ган. Нижний девон, кобленцский ярус (кондратьевская
свита).
-Фиг. 6. Paramnigenla blcarlnata Kha If in . Стр. 278
Ядро левой створки. Горный Алтай, окрестности
с. Соловьихи. Нижний девон, ганинская свита.
460
ТАБЛИЦА LXVt
ТАБЛИЦА LXVII
Фиг. 1. Zosterophyllum myretonlanum Penhallow Стр. 280
Отпечатки двух кустистоподобных растений, дающих вверх
и вниз от сплетенного корневища узкие, дихотомически раз-
ветвленные оси, X 0,75. Правый берег р. Енисея в окрестностях
г. Красноярска (Торгашино). Верхи нижнего девона.
•Фиг. 2. Psllophyton prlnceps Dawson Стр. 282
Отпечаток дважды дихотомически разветвленной оси, покры-
той шипами, X 2,25. Западный склон Кузнецкого Алатау, система
р. Средней Терси, левый приток р. Левой Магонаковой. Нижний
девон.
Фиг. 3. Protobarlnophyton Obrutschevii (Ananiev) Стр. 296
Отпечаток спороносного колоска, вид сбоку, нат. вел. Правый
берег р. Енисея в окрестностях г. Красноярска (Торгашино).
Верхи нижнего девона.
Фиг. 4. Protobarlnophyton Obrutschevii (Ananiev) Стр. 296
Отпечаток двух спороносных колосков со спорангиями, вид
с брюшной стороны, нат. вел. Правый берег р. Енисея в окре-
стностях г. Красноярска (Торгашино).
Фиг. 5. Psllophyton Goldschmidtit Halle Стр. 282
Отпечаток оси с боковым разветвлением на метаморфическом
сланце, пат. вел. Северный Алтай, левый берег р. Песча-
ной, в 1,5 км выше дер. Барагаш в Шебалинском районе. Верхи
нижнего девона.
Фиг. 6. Taenlocrada dechenlana (Goeppert) Krausel et Wey-
I a n d Стр. 281
Дихотомически или симподиально разветвленные лентовидные
оси. Немного уменьшено. Правый берег р. Енисея в окрестностях
г. Красноярска (Торгашино). Нижний девон.
-462
ТАБЛИЦА LXVH
ТАБЛИЦА LXVIII
Фиг. la. Drepanophycus spinaeformis Goeppert Стр. 288
Противоотпечаток оси с боковым разветвлением. Подобные
оси О. Хег выделил в особый вид — Psllophyton rectissimum
Н о е g.
Фиг. 16. Hostlmella sp. Стр. 284
Два остатка дихотомически разветвленных осей, несущие
в местах разветвления «овальные тела», нат. вел. Р. Енисей,
правый берег в окрестностях г. Красноярска (Торгашино).
Нижний девон.
Фиг. 2. Protobarlnophyton Obrutschevii (Ananiev). Стр. 286
Два спороносных колоска в виде отпечатка с брюшной
стороны; вдоль спорангиев видны линии растрескивания,
нат. вел. Р. Енисей, правый берег в окрестностях г. Красно-
ярска (Торгашино). Нижний девон.
Фиг. 3. Psilophyton prlnceps Dawson Стр. 282
Дихотомически разветвленная ось, не сохранившая шипов,
нат. вел. Северный Алтай, правый берег р. Песчаной,
в 1,5 км выше дер. Барагаш, в Шебалинском районе. Ниж-
ний девон.
Фиг. 4. ? Aneurophyton germanicum Krausel et Weyland
Стр. 292
Отпечаток оси первого порядка с редуцированными осями
второго порядка, низбегающими влагалищеобразно, нат. вел.
Лог Тустужул на водоразделе рек Узунжул—Пеня в Южно-
Минусинской котловине. Средний девон.
Фиг. 5. Asteroxylon (?) sibiricum Kryshtofovich Стр. 284
Отпечаток голой тонкой оси, свернутой на конце в спираль,
нат. вел. Лог Тустужул на водоразделе рек Узунжул—Неня
в Минусинской котловине. Средний девон.
Фиг. 6 и 7. Hostlmella sp. Стр. 284
Отпечатки двух голых осей, разветвленных дихотомически
симподиалыно. Нат. вел. Лог Тустужул на водоразделе рек
Узунжул—Неня в Минусинской котловине. Средний девон.
464
ТАБЛИЦА LXVIII
30 Зак. 2)24
ТАБЛИЦА LXIX
Фиг. 1 и 2. Aneurophyton germanicum Krausel et Wey-
1 a n d Стр. 292
Рисунки двух отпечатков осей третьего порядка с плохо
сохранившимися дихотомически рассеченными перышками,
нат. вел. Лог Тустужул на водоразделе рек Узунжул—
Неня в Минусинской котловине. Средний девон.
Фиг. 3. Asteroxylon (?) sibiricum Kryshtofovich Стр. 284
Рисунок с отпечатка оси, густо покрытой игловидными
чешуйчато сплетающимися листоподобными органами,
нат. вел. Лог Тустужул на водоразделе рек Узунжул—Неня
в Минусинской котловине. Средний девон.
Фиг. 4—9. Ps<iophyton Goldschmidtii Halle Стр. 282
4 й 5 — отпечатки осей с боковыми ответвлениями и тон-
кими длинными шипами, нат. вел. Торгашино. Нижний
девон.
6—9 — рисунки с отпечатков, показывающие части рас-
тения с широкими разветвлениями боковых ветвей, нат.
вел. Юго-Восточный Салаир, р. Кара-Чумыш у пасеки
колхоза «Кузбасс». Нижний девон.
Фиг. 10 и 11. Улиткообразно завитые голые концевые части растения,
вероятно, принадлежащие Asteroxylon (?) sibiricum (см.
фиг. 3). Нат. вел. . . Стр. 284
Лог Тустужул в Южно-Минусинской котловине. Сред-
ний девон.
Фиг. 12. Psilophyton prlnceps Dawson Стр. 282
Отпечаток дихотомически разветвленной оси с проводя-
щим пучком и характерными для этого вида шипами, нат.
вел. Лог Тустужул в Южно-Минусинской котловине.
Средний девон.
466
ТАБЛИЦА LXIX
6
12
ТАБЛИЦА LXX
Фиг. la. Drepanophycus spinaeformls Goeppert Стр. 288
Комбинированный рисунок с отпечатка и противоотпечатка
(последний изображен на таблице LXVIII, фиг. 1а). Нат. вел.
Торгашино. Нижний девон.
Фиг. 16. Hostlmella sp. . .Стр. 284
Рисунок с противоотпечатка, изображенного на таб-
лице LXVIII, фиг. 16. Голые дихотомически симподиалыные оси
в местах разветвлений несут «овальные тела». Нат. вел. Торга-
шино. Нижний девон.
Фиг. 2. Protolepldodendron scharyanutn Krejti Стр. 289
Плохо сохранившийся отпечаток дихотомически симподиаль-
но разветвленной оси с бертериоидными листовыми подушками
и узкими листьями. Бифуркирующие кончики последних не
сохранились. Нат. вел. Лог Тустужул в Южно-Минусинской
котловине. Средний девон.
Фиг. 3. Psllophyton princeps Dawson Стр. 282
Рисунок с отпечатка, несущего характерные для данного вида
шипы. Остаток, возможно, окажется псилофитоновой стадией
роста Asteroxylon slblrlcum. Нат. вел. Лог Тустужул в Южно-
Минусинской котловине. Средний девон.
Фиг. 4. Psllophyton Goldschmidtll Halle Стр. 282
Отпечатки обрывков главных осей, несущих боковые раз-
ветвления, редко сохранившиеся шипы и бугорки от шипов. Нат.
вел. Торгашино. Нижний девон.
Фиг. 5. ?Aneurophyton germanlcum Krausel et Weyland Стр. 292
Рисунок с отпечатка, воспроизведенного на таблице LXVIII,
фиг. 4. Нат. вел. Лог Тустужул в Южно-Минусинской котло-
вине. Средний девон.
Фиг. 6. Protobarinophyton Obrutschevli (Ananiev) Стр. 286
Рисунок с отпечатка неполно сохранившегося спороносного
колоска со спинной стороны. Нат. вел. Торгашино. Нижний
девон.
Фиг. 7. Protobarinophyton Obrutschevli (Ananiev) Стр. 286
Рисунок с отпечатка поникшего спороносного колоска, вид
сбоку. Нат. вел. Торгашино.. Нижний девон.
Фиг. 8. Drepanophycus spinaeformls Goeppert Стр. 288
Рисунок с отпечатка обрывка дихотомически разветвленной
оси, снабженной длинными толстыми шипами. Нат. вел. Торга-
шино. Нижний девон.
468
ТАБЛИЦА LXX
ТАБЛИЦА LXX1
Фиг. 1. Aphyllopterls robusta (Dawson) Стр. 285
a — рисунок с отпечатка голой оси с симподиально-боковым
разветвлением; б — верхушка образца фиг. 1а. Нат. вел. Торга-
шино. Нижний девон.
Фиг. 2. Hostlmella sp. Стр. 284
Голая дихотомически разветвленная ось, возможно составляю-
щая верхушечную часть Asteroxylon sibiricum (стадия роста
Hostlmella). Нат. вел. Лог Тустужул в Южно-Минусинокой кот-
ловине. Средний девон.
Фиг. 3. Protobarlnophyton Obrutschevii (Ananiev) Стр. 286
Рисунок с образца, изображенного на табл. LXVII, фиг. 3.
Торгашино. Нижний девон.
Фиг. 4. Protobarlnophyton Obrutschevii (Ananiev) Стр. 286
Рисунок части образца, изображенного в полном виде на
табл. LXXIII, фиг. 1. Верхушечная часть голых осей с рас-
пускающимся спороносным колоском. Нат. вел. Торгашино.
Нижний девон.
470
ТАБЛИЦА LXXl
ТАБЛИЦА LXXII
Фиг. 1. Protobarinophyton Obrutschevli (Ananiev) Стр. 286
Отпечатки параллельно ориентированных голых, местами дихо-
томически разветвленных осей растения, расположенных на об-
ратной стороне плиты мергелистого песчаника, с так же ориенти-
рованными осями этого же растения, изображенного на
табл. LXXIII, фиг. 1. Х0,8. Торгашино. Нижний девон.
Фиг. 2. Hostimella sp. Стр. 284
Отпечаток голой дихотомически разветвленной верхушки не-
известного растения. XI,7. Р. Енисей, левый берег между
деревнями Усть-Ерба и Каскыр. Нижний девон.
Фиг. 3. Psilophyton prtnceps Dawson Стр. 282
Отпечаток обрывка оси с шипами и остатками шипов в виде
бугорков с образца, рисунок которого дан на табл. LXX,
фиг. 3. XI,9. Лог Тустужул в Южно-Минусинской котловине.
Средний девон.
Фиг. 4. Psilophyton Goldschmidtii Halle Стр. 282.
Плохо сохранившийся на метаморфическом сланце отпечаток
оси с боковым разветвлением. От тонких шипов сохранились
лишь бугорки. Нат. вел. Северный Алтай, левый берег р. Песча-
ной в 1,5 км выше дер. Барагаш, Шебалинский район. Нижний
девон.
472
ТАБЛИЦА LXXIl
4
ТАБЛИЦА LXXIII
Фиг. 1. Protobarinophyton Obrutschevli (Ananiev) Стр. 286
Отпечатки голых, редко дихотомически разветвленных осей
с распускающимися на концах спороносными колосками (вверху
снимка). Х0,7. Торгашино. Нижний девон.
Фиг. 2. Taeniocrada dechenlana (Goeppert) Krausel et Wey-
land Стр. 281
Отпечаток лентовидных, дихотомически разветвленных осей.
Нат. вел. Северо-Минусинская котловина, в 2,5 км западнее
оз. Салбат. Нижний девон.
Фиг. 3. Pseudobornia cf. urslna Nathorst Стр. 290
Отпечаток ствола с неясной членистостью и ребристостью. Нат.
вел. Северо-Минусинская котловина, левый берег р. Убей у верх-
него конца дер. Медведеве. Верхний девон.
-474
ТАБЛИЦА LXXI1I
1
ТАБЛИЦА LXXIV
Фиг. 1 и 2. Archaeopteris Halltana (Goeppert) Dawson Стр. 294
Фиг. 3.
Плохо сохранившийся на красном песчанике отпечаток,
показывающий рахис вайи с двоякоперистым расположением
перьев первого порядка, на которых смежные перышки
сильно перекрывают друг друга (фиг. 1). Участок пера
первого порядка лучшей сохранности (фиг. 2). Нат. вел.
Северо-Минусинская котловина, на водоразделе рек Енисея
и Убея, юго-восточнее села Новоселове. Верхний девон.
Archaeopteris obtusa Lesquereux
Стр. 294
Рисунок отпечатка пера на ложнослоистом красном песча-
нике. Нат. вел. Левый берег Енисея, южнее дер. Кокоревой
в Северо-Минусинской котловине. Верхний девон.
Стр. 295
Фиг. 4—7. Archaeopteris sp.
Обескоренные скульптурные оси, показывающие вертикаль-
ные ряды точечных углублений, образованных поперечными
морщинками. На фиг. 7 внизу виден слой наружной коры.
Нат. вел. Северо-Минусинская котловина, р. Убей у дер.
Медведеве. Верхний девон.
476
ТАБЛИЦА LXXIV
ТАБЛИЦА LXXV
Фиг. 1. Pseudobornia cf. urslna Nathorst . Стр. 290
Отпечатки крупных и мелких осей растения со следами
окатаиности, обнаруживающих неясную членистость и ребри-
стость. Нат. вел. Северо-Минусинская котловина, левый берег
р. Убея у верхнего конца дер. Медведево. Верхний девон.
Фиг. 2. Два спорангия, вероятно, принадлежащих Rhacophyton
incertum. X 9 ... . Стр. 293
Увеличение сделано с табл. LXXVII, фиг. 26.
Фиг. 3 и 4. Protolepidodendron scharyanum Krejci Стр. 289
Отпечаток тонкого ствола с бергериоидными подушками и
узкими, листьями: фиг. 3 — нат. вел.; фиг. 4— Х2- Северный
Алтай, район г. Змеиногорска (между дер. Гусинкой I и II).
Средний девон.
Фиг. 5. Aphyllopteris robusta (Dawson) Стр. 285
Анатомическое строение проводящего пучка древесины
с образца на табл. LXXI, фиг. 1; видны кольчатые тра-
хеиды. X 50- Торгашино. Нижний девон.
478
ТАБЛИЦА LXXV
ТАБЛИЦА LXXVI
-Фиг. 1
Фиг. 3—
Фиг. 6.
-Фиг. 7.
1 2. Protolepldodendron scharyanum К г е j t i Стр. 289
Отпечаток тонкого ствола с бергериоидной скульптурой и
узкими листьями по бокам: фиг. 1 — нат. вел.; фиг.. 2— Х2.
Северный Алтай, район г. Змеиногорска (между дер. Гусин-
кой I и II). Средний девон.
5. Asierocalamltes scroblculatus (Schlotheim) Z е i 1-
1 e г . . Стр. 292
Отпечатки ядер стволов. Нат. вел. Р. Чулым, правый берег
у совхоза Игрышинского в Северо-Минусинской котловине.
Верхний девон — нижний карбон.
Rhacophyton incertum (Dawson) Kraiisel et Wey-
land Стр. 293
Отпечаток рахиса вайи с боковыми перьями последнего
порядка, на которых мелкие перышки не сохранились. Нат.
вел. Р. Чулым, правый берег у совхоза Игрышинского
в Северо-Минусинской котловине. Верхний карбон.
Aphyllopterts robusta (Dawson) Стр. 285
Анатомическое строение проводящего пучка древесины
с образца на табл. LXXI, фиг. 1; видны кольчатые трахеиды.
X 37,5. Торгашино. Нижний девон.
-480
ТАБЛИЦА LXXVI
31 Зак. 2и24
ТАБЛИЦА LXXVII
Фиг. 1
Фиг. 4.
Фиг. 5
Фиг. 7.
3. Rhacophyton Incerlutn (Dawson) Кг ausei et Wey-
1 a n d Стр. 293
Отпечатки рахисов ваий с боковыми перьями последнего
порядка. На фиг. 2а и 26 очерчены спорангии, лежащие
совместно с обрывками осей, принадлежащих данному виду.
Нат. вел. Р. Чулым, правый берег у совхоза Игрышинского
в Северо-Минусинской котловине. Верхний девон—нижний
карбон.
Группа спорангий, изображенная на табл. LXXVII, фиг. 2а,
совместно с остатком Rh. Incertum. ХЗ Стр. 293
[ 6. Sphenophyllutn subtenerrlmum Nathorst . Стр. 291
Отпечатки двух ребристых мутовчатых стеблей с характер-
ными утолщениями в узлах. На фиг. 5 у верхней мутовки
видны листья, глубоко рассеченные на нитевидные дольки.
Х2,5. Р. Чулым, правый берег у совхоза Игрышинского
в Северо-Минусинской котловине. Верхний девон — нижний
карбон.
Aneimltes acadlca Dawson Стр. 293
Отпечаток плодоносной веточки с двумя парами семян
в оболочках (плюсках). Х1>6. Р. Чулым, правый берег у сов-
хоза Игрышинского в Северо-Минусинской котловине. Верх-
ний девон — нижний карбон.
482
ТАБЛИЦА LXXVII
ТАБЛИЦА LXXVIII
Фиг. 1—6. Anelmites acadlca Dawson Стр. 293
1 — отпечаток почти полного растения с толстым корне-
вищем (?) и кустисто расходящимися, вильчато разветвлен-
ными вайями, концы которых обломаны; в левой верхней
части, рядом с осью, справа лежит отпечаток слабо рассечен-
ного перышка с веерным жилкованием; 2 — конечная часть
вайи с гроздевидно расположенными перышками, с веерным
жилкованием; 3 и 4 — отпечатки отдельных опавших асим-
метричных перышек, слабо рассеченных по краю и с густым
веерным жилкованием; 5 — отпечаток оси более крупной
вайи, чем на фиг. 1; 6 — попарно сидящие семена, обернутые
в оболочку (увеличено с образца, изображенного на
табл. LXXVII, фиг. 7). 1—5 — нат. вел.; 6—ХЗ. Р. Чулым,
правый берег у совхоза Игрышинского в Северо-Минусин-
ской котловине. Верхний девон — нижний карбон.
Фиг. 7. Heleniella Theodorl Zalessky . Стр. 289
Отпечаток внутренней поверхности коры с обратным изо-
бражением волнистых ребер и бороздок. Видны листовые
рубцы округлоромбовидной формы, сливающиеся своими
узкими концами в правильные дорожки. Нат. вел. Р. Чилым,
правый берег у совхоза Игрышинского в Северо-Минусинской
котловине. Верхний девон — нижний карбон.
481
ТАБЛИЦА LXXVIII
6
7
2
ТАБЛИЦА LXXIX
Фиг. 1.
Фиг. 2.
Фиг. 3.
Фиг. 4—7.
Michelinia ienulsepta Phillips . . Стр. 298
а — вид колонии сверху, нат. вел. Кузбасс, р. Нижняя
Терсь. По И. П. Толмачеву.
б — продольный разрез, нат. вел.; в — поперечный разрез,
нат. вел. Кузбасс, дер. Ройка на р. Томи. Нижний карбон.
По В. Д. Фомичеву.
Michelinia megastoma Phillips . Стр. 298
Вид колонии сверху. Нат. вел. Кузбасс, р. Кара-Чумыш.
Нижний карбон. По И. П. Толмачеву.
Syrlngopora ramulosa G о 1 d f u s s . Стр. 309
Общий вид колонии. Нат. вел. Нижний карбон. По
И. П. Толмачеву.
Stereophrentls konlncki Edwards et Haime. Стр. 30!
4 — общий вид коралла. Нат. вел. Кузбасс, р. Тайдон.
5 — то же, широкая форма. Нат. вел. Кузбасс, р. Тайдон.
6 — поперечный разрез (под чашечкой), Х5; 7 — попереч-
ное сечение (у вершины конуса), Х5- Нижний карбон. По
В. Д. Фомичеву.
486
ТАБЛИЦА LXXIX
7
ТАБЛИЦА LXXX
Фиг. 1—3. Caninia cylindrica Scouler Стр. 301
1 — продольное сечение; 2 — поперечное сечение; 3 — по-
перечное сечение. Нат. вел. Кузбасс, р. Иня выше дер. Кат-
ковой. Нижний карбон. По В. Д. Фомичеву.
Фиг. 4. Caninia sp. Стр. 301
Общий вид коралла. Нат. вел.
Фиг. 5 и 6. Lithostrotion (Eolithostrotionella) sibiricum G a b u n i а
Стр. 303
5 — вид колонии сверху; 6 — поперечное сечение. Нат.
вел. Кузбасс, р. Артышта. Нижний карбон. По И. П. Тол-
мачеву.
Фиг. 7 и 8. Lithostrotlon Irregulare Phillips var. balachonkaensis
Fomichew Стр. 304
7 — продольное сечение, X5; 8 — поперечное сечение,
X 2,5. Кузбасс, р. Томь ниже дер. Балахонки. Нижний кар-
бон. По В. Д. Фомичеву.
Фиг. 9. Cainlnia ussovi G a b u n i а Стр. 302
Поперечное сечение (молодая стадия развития). Кузбасс,
р. Иня. Нижний карбон. По В. Д. Фомичеву.
488
ТАБЛИЦА LXXX
ТАБЛИЦА LXXXI
Фиг. 1 и 2. Lithostrotlon grande Tolmachew
Стр. 304
1 — вид колонии сверху; 2 — продольное сечение. Нат. вел.
Кузбасс, р. Артышта. Нижний карбон. По И. П. Толмачеву.
Фиг. 3. Tabultpora elbaschensls Krasnopeeva, sp. n. Стр. 305
Фиг. 4.
Фиг. 5.
a — поперечное сечение колонии, показывающее две срос-
шиеся двуслойные плоские веточки; б — тангенциальное се-
чение к поверхности колонии, показывающее поперечное
сечение ячеек. X Ю. Салаир, р. Каменный Елбаш, выше
дер. Хариной. Турнейский ярус нижнего карбона.
Fenestella rudls Ulrich
Стр. 306
Тангенциальное сечение. ХЮ- Салаир. Горловский район.
Нижний карбон.
Fenestella regalls Ulrich
Стр. 306
Тангенциальное сечение, Х&- Салаир, р. Каменный Елбаш„
выше дер. Хариной. Турнейский ярус нижнего карбона.
490
ТАБЛИЦА LXXXF
4
ТАБЛИЦА LXXXII
Фиг. 1. Polypora sibirica Yanischevsky Стр. 306
a — рисунок, показывающий характер сетки и размещение
устьев ячеек. Х2,7; б — отпечаток 1колонии, по М. Э. Янишев-
скому. Окрестности г. Томска. Глинистые сланцы карбона.
Фиг. 2. Hemltrypa proutana Ulrich. Стр. 306
Тангенциальное сечение. ХЮ. Кузбасс. Верхи турнейского
яруса нижнего карбона. По В. П. Нехорошеву.
Фиг. 3. Fenestella regalls Ulrich . Стр. 307
Тангенциальные сечения: а — ХЮ; б — X 6,5. Салаир, р. Ка-
менный Елбаш, выше дер. Хариной. Турнейский ярус нижнего
карбона.
Фиг. 4. Reteporlna altalca Nekhoroschev . . Стр. 307
Тангенциальное и поперечное сечения. ХЮ. Рудный Алтай.
Верхи тарханской свиты. По В. П. Нехорошеву.
Фиг. 5. Fenestella rudls Ulrich Стр. 306
Тангенциальное сечение. Х25. Нижний карбон.
492
ТАБЛИЦА LXXXIf
4
ТАБЛИЦА LXXXIH
Все изображения даны в натуральную величину
Фиг. 1—3.
Фиг. 4.
Фиг. 5.
Фиг. 6.
Фиг. 7.
Фиг. 8 и 9.
Schuchertella tomsklensls Yanischevsky Стр. 307
1 — внутреннее ядро брюшной створки; 2 — внутреннее
ядро спинной створки; 3 — отпечаток брюшной створки, по-
казывающий характер ареи. Окрестности г. Томска. Глини-
стые сланцы нижнего карбона.
Dlctyoclostus burllngtonensls (Hall) Стр. 308
Брюшная створка в трех положениях.
Dlctyoclostus de/uptus (Romanovsky) Стр. 308
Брюшная створка в трех положениях.
Splrlfer tornacensis Koninck Стр. 310
а — спинная створка; б — брюшная створка. Кузбасс. Тур-
нейский ярус нижнего карбона. По И. П. Толмачеву.
Splrlfer stblrlcus Yanischevsky Стр. 311
Внутреннее ядро брюшной створки. Окрестности г. Томска.
Глинистые сланцы нижнего карбона.
Splrlfer usslensls Tolmatchoff Стр. 310
Брюшные створки. Кузбасс. Турнейский ярус нижнего
карбона. По И. П. Толмачеву.
494
ТАБЛИЦА LXXXIII
ТАБЛИЦА LXXXIV
Все изображения, кроме фиг. 5, даны в натуральную величину
Фиг. 1 и 2. Splrifer из группы Spirifer cinctus Koninck Стр. 310
Кузбасс. Верхнетурнейские отложения нижнего карбона
(фоминская зона). По И. П. Толмачеву.
Фиг. 3—6. Splrifer Siberians N. Lebedev Стр. 311
3 — вид раковины со стороны ареи; 4 — брюшная створка;
5 — вид раковины сбоку, X 2; 6— вид раковины со стороны
замочного края. Горловский бассейн. Низы турнейского
яруса нижнего карбона (абышевская зона).
Фиг. 7. Syrlngothyrls cuspldata Mar tin Стр. 311
a — вид сбоку; б — вид со стороны переднего края. Куз-
басс, р. Тайдон. Нижний карбон. По И. П. Толмачеву.
Фиг. 8. Syrlngothyrls sp. Стр. 311
Брюшная створка с пришлифованной макушечной частью;
видны зубные пластины и дельтириальная пластинка с труб-
кой на внутренней стороне. Нижний карбон.
496
ТАБЛИЦА LXXXIV
32 Зак. 2024
ТАБЛИЦА LXXXV
Все изображения даны в натуральную величину
Фиг. 1 Syringothyris cf. cuspldata М а.г tin . Стр. 311
а — вид со стороны замочного края; б — вид со стороны
спинной створки. Кузбасс, дер. Ройка. Нижний карбон.
Фиг. 2. Splrlfer ex gr. Splrlfer clnctus Kon i nek Стр. 310
Нижний карбон. Кузбасс. По И. П. Толмачеву.
Фиг. 3—5. Spiriferlna octopllcata (S о w е г b у) var. concentrica Y a n i-
schewsky Стр. 312
3 — внутреннее ядро брюшной створки; 4 — отпечаток не-
полной спинной створки; 5 — отпечаток брюшной створки.
Окрестности г. Томска. Глинистые сланцы нижнего карбона.
Фиг. 6. Dlctyoclostus robustus Tolmatchoff Стр. 309
а — брюшная створка со стороны макушки; б — вид со
стороны спинной створки. Кузбасс. Нижний карбон. По
И. П. Толмачеву.
Фиг. 7. Echinoconchus punctatus (Martin) Стр. 309
а — вид со стороны брюшной створки; б — вид со стороны
спинной створки; в — брюшная створка сбоку. Нижний кар-
бон. По Т. Г. Сарычевой и А. Н. Сокольской.
лхххт vhHirayj,
ОГЛАВЛЕНИЕ
Предисловие . 3
Введение. Л. Л. X а л ф и н 5
Элементы морфологии важнейших групп ископаемых животных и растений
Археоциаты. П. С. Краснопеева 17
Строматопороидеи. В. К. X а л ф и н а 21
Табуляты и гелеолитиды. Л. Н. Краевская 26
Тетракораллы. В. А. И в а н и я 29
Мшанки. Л. Л. X а л ф и н 34
Брахиоподы. Л. Л. X а л ф и н 37
Трилобиты. Л. Л. Халфин 45
Пластинчатожаберные моллюски. Л. Л. Халфин 46
Растения. Г П. Радченко 49
Глава I
КЕМБРИЙСКАЯ СИСТЕМА
Стратиграфический очерк. А. Г Си во в 58
Археоциаты. П. С. Краснопеева 74
Трилобиты. Л. И. Егорова, М. П. Л о м о в и ц к а я, О. К. Полетаева,
А. Г. С и в о в 102
Водоросли. П. С. Краснопеева 145
Глава II
СИЛУРИЙСКАЯ СИСТЕМА
Стратиграфический очерк. Л. Л. Халфин 149
Строматопороидеи. В. К. Халфина 153
Табуляты и гелиолитиды. Л. Н. Краевская 154
Брахиоподы. 159
Трилобиты. М. П. Ломовицкая 166
Глава Ill
ДЕВОНСКАЯ СИСТЕМА
Стратиграфический очерк. Л. Л. X а л ф и и 180
Строматопороидеи. В. К. Халфииа 185
Табуляты и гелиолитиды. Л. Н. Краевская 191
Тетракораллы нижнего и среднего девона. Л. Н. Краевская 206
Тетракораллы верхнего девона. В. А. И в а н и я 219
Мшанки. П. С. Краснопеева 228
Брахиоподы нижнего девона Алтая. Л. Л. Халфии 234
Брахиоподы нижнего и среднего девона Кузбасса. М. А. Ржонсницкая 244
Брахиоподы верхнего живета. Р. Т. Грац Панова 256
Брахиоподы верхнего девона. В. А. И в а н и я и Л. Н. К р а е в с к а я 261
Трилобиты. Л. Л. Халфин 272
Пластинчатожаберные моллюски. Л. Л. Халфии 274
Растения. А. Р. Ананьев 279
501
Глава IV
НИЖНИЙ КАРБОН
Стратиграфический очерк. Л. Л. Халфин 297
Кишечнополостные. В. Д. Фомичев 298
Мшанки. П. С. Краснопеева 305
Брахиоподы. Р. Н. Бенедиктова 307
Родовой указатель 313
Видовой указатель 316
Дополнительный родовой указатель 322
Литература 324
Таблицы изображений и объяснения к ним 329
ОПЕЧАТКИ
Стр. Строка Напечатано Следует читать
22 Рис. 16 повернуть на 180*
24 Рис. 20 повернуть иа 180°
29 Рис. 38 повернуть на 180°
38 Рис. 69 1/2 нат. вел. Нат. вел.
57 6 сверху оклеростестой склеротестой
68 Рис. 116, колонка I Западный склон Западный Саян
118 8 сверху ЩИТЫ борозды
145 1 сверху Saltar Salter
175 Рис. 169 По В. Н. Уэллеру По Ульриху
176 Рис. 170 Глабель Кранидий
184 1 снизу на стр. 352 на стр. 181
206 15 снизу фиг. 182 фиг. 1. Рис. 182
216 Рис. 186 повернуть на 180°
236 8 сверху A. Williams Caster
236 17 сверху скульптуры. скульптуры и зазубрен-
ности ареи.
236 19 сверху Нижний силур — Девон
нижний девон
236 21 сверху Corrugatella Cymostrophia
237 21 сверху Ochlertia Oehlertia
241 20 снизу СВИТЫ септы
278 Рис. 196 3/4 нат. вел. Справа сильно
увеличено.
279 9 снизу Barlnophyton Protobarinophyton
291 16 сверху cuneifolium verticillatum S с h I о t h.
(Sternberg)
314 12 сверху Lascopteris Laccopteris
314 22 снизу Haniva Hanivia
320 5 сверху Corrugatella Cymostrophia
322 20 сверху стр. 316 стр. 317
334 11 сверху Стр. 99 Стр. 81
340 2 снизу Стр. 191 Стр. 91
342 5 сверху Стр. 191 Стр. 91
342 4 снизу Фиг. 4. Ethmophyllum Фиг. 5. Thalamocyathus
amsassense splnosus
346 10 сверху Стр. 187 Стр. 87
Зак. 2024
По тех нческому недосмотру в сноске на стр. 181 ошибочно пометена фамилия М. А. Ржои-
сницкой
АТЛАС РУКОВОДЯЩИХ ФОРМ ИСКОПАЕМЫХ
ФАУНЫ И ФЛОРЫ ЗАПАДНОЙ СИБИРИ
Той I
Редактор издательства В. Н. Никитина
Техн, редактор О. А. Гурова
Корректор А. Н. Шеманова
Сдано в набор 27/VII 1955 г.
Подписано в печать 30/XI 1955 г.
Формат бумаги 70><108,/ie-
Печ. л. 43,1 + вкл. 0,68. Уч.-изд. л. 37,8.
Бум. л. 16. Т 08672. Тираж 3000 экз.
Зак. № 2024. Цена 28 р. 45 к.
Картфабрика Госгеолтехиздата