/






































































































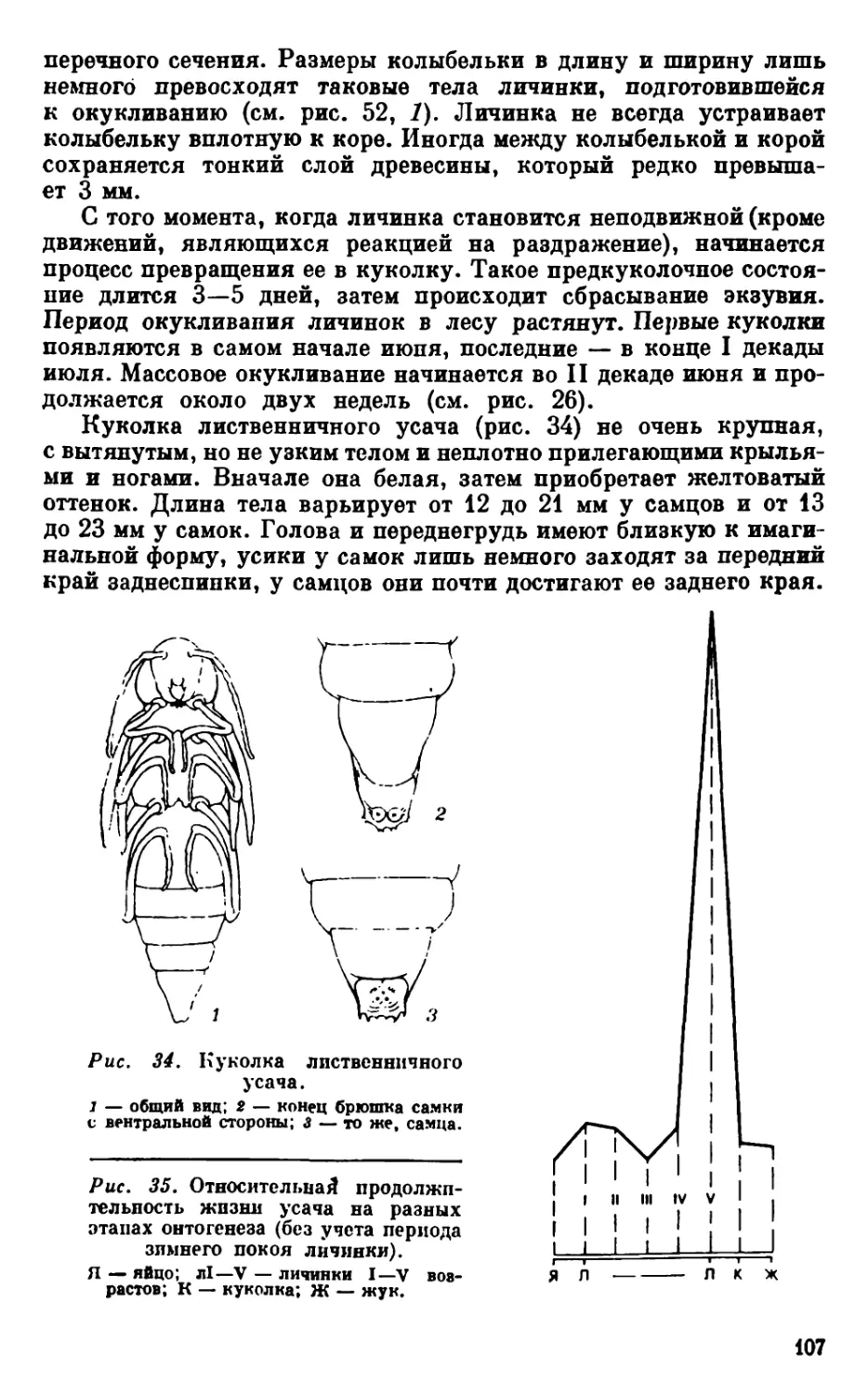



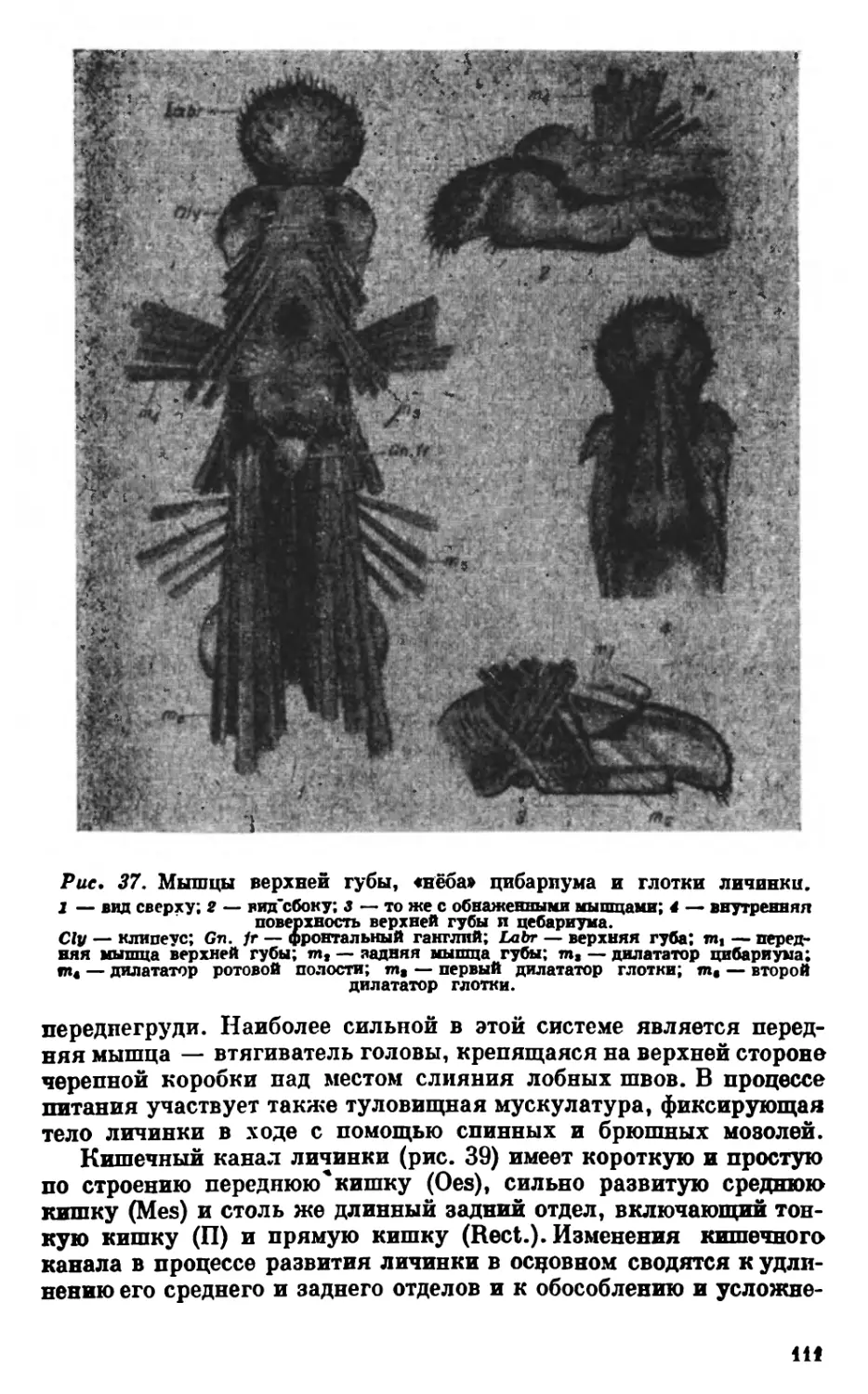

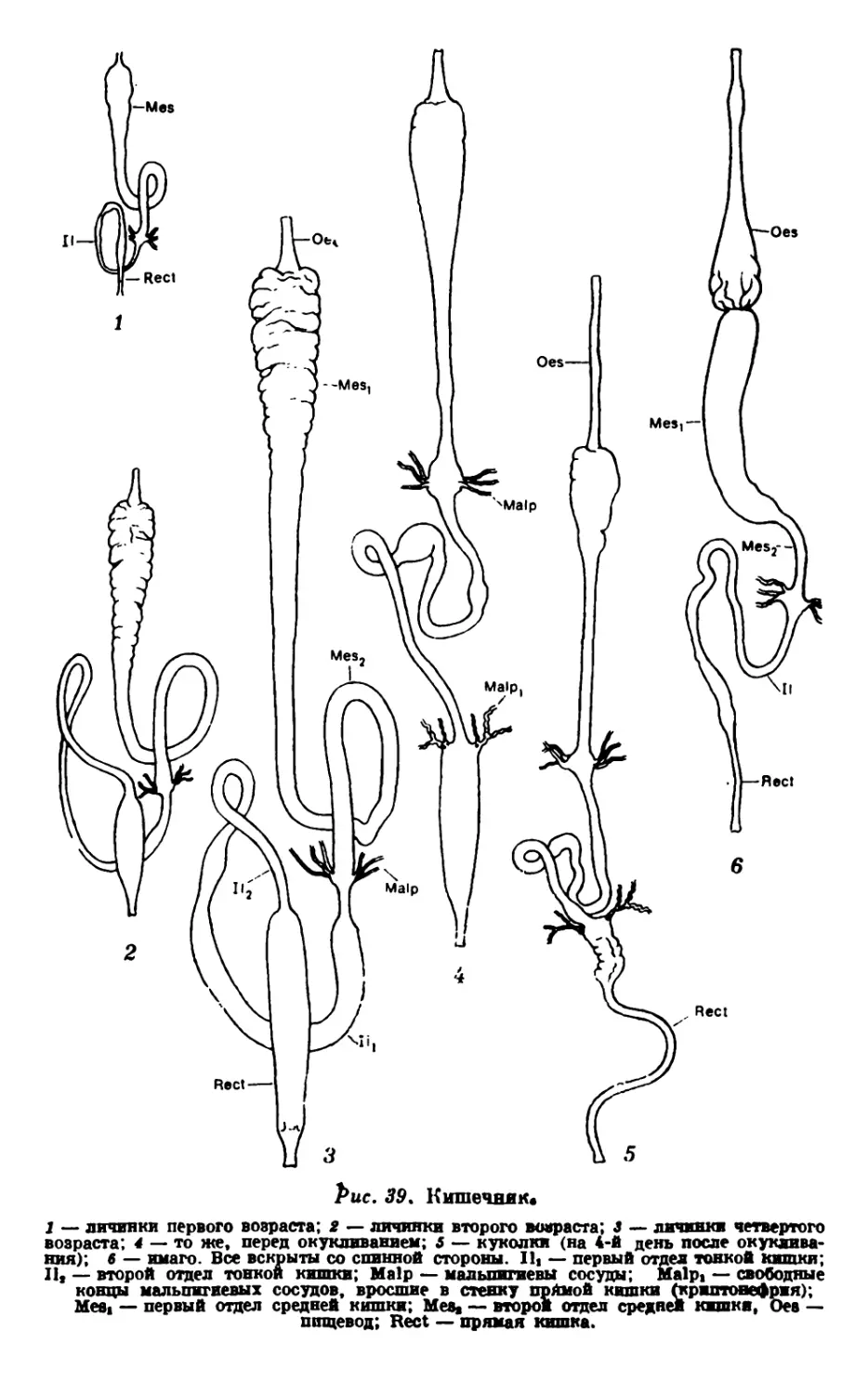



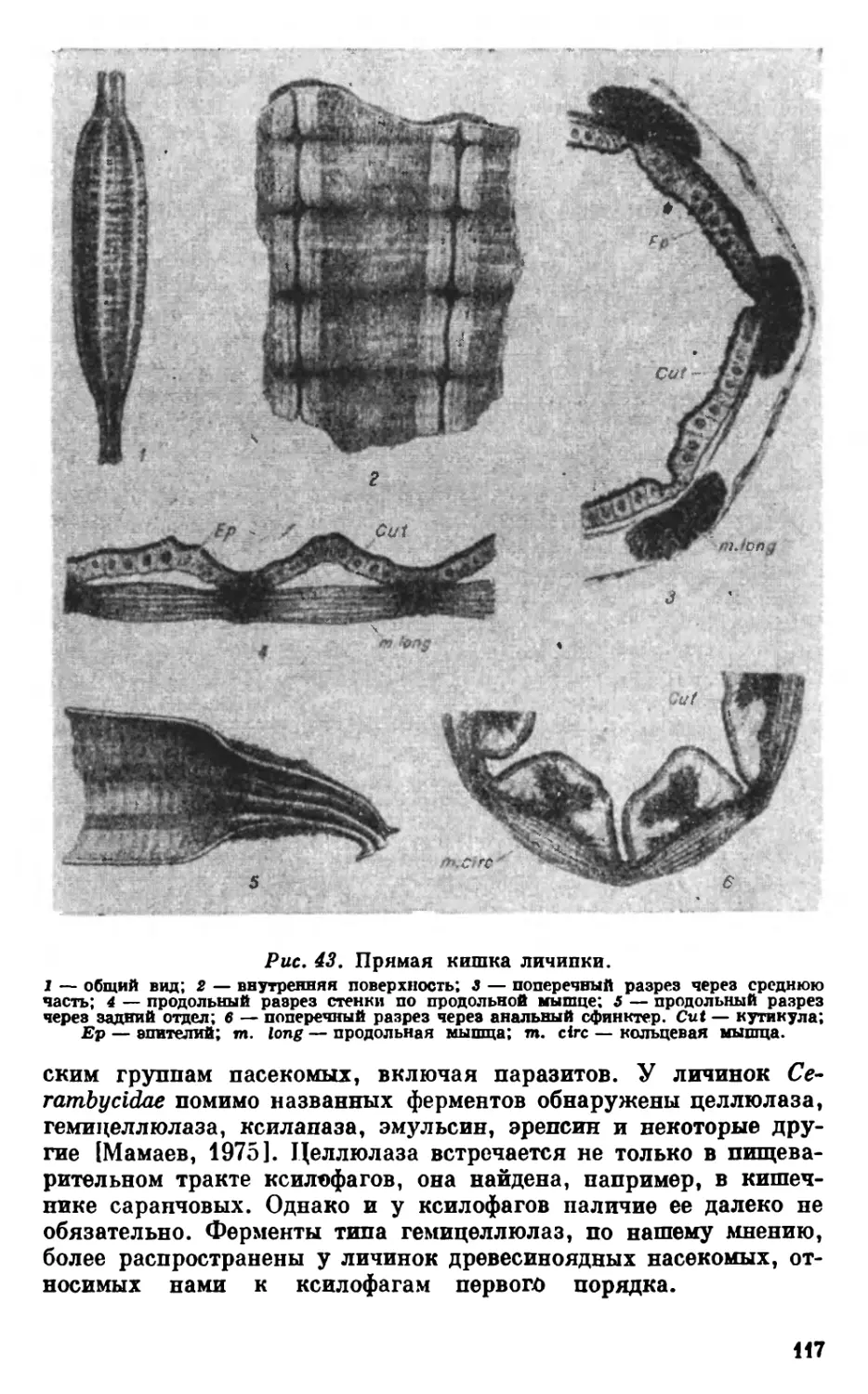












































































Текст
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
сибирский Институт физиологии и биохимии растений
л. С. РОЖКОВ
ДЕРЕВО
И НАСЕКОМОЕ
Ответственный редактор
чл.-кор. АН СССР А. С. Исаев
ИЗДАТЕЛЬСТВО «НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
Новосибирск • 1981
УДК 634.0.453
Рожков А. С. Дерево и насекомое.— Новосибирск:
Наука, 1981.
В книге рассматривается взаимодействие древесных растений
и насекомых-ксилофагов. В порядке введения в ней обсуждаются
также вопросы филогении, морфоэкологической адаптации, фор-
мирования трофических группировок, плотности популяций и мас-
сового размножения насекомых-дендрофагов в целом и ксилофагов
в особенности. В специальной части на примере лиственницы
(Lartx) и лиственничного усача (Xylotrechus altaicus Gobi.) показа-
ны взаимная адаптация дерева и ксилофага, структурные и функ-
циональные защитные преобразования, возникающие у дерева при
повреждении, и механизмы преодоления вредителем его сопротив-
ляемости.
Книга представляет интерес для широкого круга специалистов
в области энтомологии, биоценологии, патологии и защиты леса,
а также физиологии растений.
Табл. 12. Ил. 66. Библиогр. 196.
А натолий Сергеевич Рожков
ДЕРЕВО И НАСЕКОМОЕ
Ответственный редактор
Александр Сергеевич Исаев
Утверждено к печати Сибирским институтом
физиологии и биохимии растений СО АН СССР
Редактор издательства Д. М. Резяпова. Художественный редактор В, И, Желнин
Художник А. Г. Иерх. Технический редактор Г. Я. Герасимчук
Корректоры IH. Я. Михайлова, С. М. Поеу&ина
ИБ 10598
Сдано в набор 17-06.80. Подписано к печати 13.02.81. МН-00503. Формат 60 X 907м*
Бумага офсетная. Обыкновенная гарнитура. Высокая печать. Уел. печ. л. 114-1 п. л.
на мел. Усл. кр. отт. 12,1. Уч.-изд. л. 13,5. Тираж 1700 екз. Закав № 198. Цена 2 р. 30 к.
Издательство «Наука», Сибирское отделение. 630099, Новосибирск, 99, Советская, 18.
4-я типография издательства «Наука». 630077, Новосибирск, 77, Станиславского, 25.
Р 2>0<^____— 527.80.2001050000.
055(02) — 81 © Издательство «Наука», 1981.
ПРЕДИСЛОВИЕ
Мне хотелось раскрыть перед читателями картину глубокой?
взаимной адаптации дерева и насекомого-фитофага, проявляющую
юся во время их взаимодействия. Мне хотелось также показать
удивительную способность древесного растения к самозащите,
к быстрой структурной и функциональной перестройке, к. мобили-
зации защитных средств в критический момент его жизни. Не вся-
кий животный организм способен к подобной реактивности; И мож-
но ли не удивляться, открывая здесь все новые и новые явления,
в том числе относящиеся и к такой «опорожненной» дисциплине,
какой представляется многим анатомия растений? Наконец, глав-
ной задачей, которую я ставил перед собой, было увлечь ботаников»
и зоологов идеей совместных поисков новых путей в защите леса.
Базой для таких поисков должны служить комплексные исследо-
вания патологии и защитных свойств дерева, его взаимодействия»
с насекомыми-фитофагами, экологии резистентности древостоев,,
а также факторов, определяющих градации численности и вспыш-
ки массового размножения вредителей.
В общей части книги я позволил себе высказать некоторые сло-
жившиеся у меня представления о филогении и пищевой специали-
зации насекомых-дендрофагов, сущности вспышек их массового
размножения, фазах развития очагов, о ксилофагии и других воп-
росах лесной энтомологии в целом. Ознакомление с ними помимо
собственного интереса должно помочь пониманию излагаемого
далее представления о взаимодействии дерева и насекомого-
ксилофага.
Исследования, положенные в основу настоящей; книги, прове-
дены в лесах юга Восточной Сибири (Иркутская и Читинская об-
ласти, Бурятская АССР). Начаты они в 1965 г. и продолжались
с перерывами до 1978 г. Помимо деревьев, повреждаемых насеко-
мыми, для выяснения последовательности анатомических и физио-
лого-биохимических защитных преобразований использовались,
также искусственно травмированные лиственницы — аналоги де-
ревьев, поврежденных лиственничным усачом на разных этапах
развития личинок.
&
Выбор в качестве основных объектов лиственницы и листвен-
ничного усача не был случайным. Среди хвойных деревьев листвен-
ница является наиболее устойчивой к повреждению как филлофа-
гами, так и ксилофагами, а лиственничный усач — самым агрес-
сивным из числа последних.
Выражаю глубокую признательность моим постоянным по-
мощникам в экспедиционных и лабораторных исследованиях
Е. С. Степанчук, Г. И. Массель и М. К. Сокову, искренне благо-
дарю члена-корреспондента АН СССР А. С. Исаева за редактирова-
ние книги, профессоров А. И. Воронцова и П. М. Рафеса за про-
чтение разделов рукописи и полезные замечания, докторов
Ф. Кике и Б. Клаузницера за помощь при работе с коллекциями
Берлинского и Дрезденского зоологических музеев, а также весь
коллектив лаборатории энтомопатологии древесных растений
СИФИБР СО АН СССР.
Помещенные в книге рисунки и фотографии оригинальны, вы-
полнены автором.
ОБЩАЯ ЧАСТЬ
НАСЕКОМЫЕ-ФИТОФАГИ
И КСИЛОФАГИЯ У НАСЕКОМЫХ
РАСТИТЕЛЬНОЯДНЫЕ НАСЕКОМЫЕ
Число выявленных в настоящее время видов насекомых при-
ближается к 1 млн. Среди них растительноядные виды, или фито-
фаги, занимают главенствующее положение. По весьма грубым
подсчетам па их долю приходится примерно 68%, хищные и пара-
зитические насекомые (зоофаги) составляют около 15%, а осталь-
ные (17%) питаются неживой органикой (сапрофаги, детритофа-
ги и др.).
На представленной диаграмме (рис. 1) показано относительное
количество фитофагов, сапрофагов в целом и зоофагов в классе
насекомых (без учета подкласса первичнобескрылых) и в отдельных
его надотрядах. Наибольшее число фитофагов содержат отряды
жесткокрылых, чешуекрылых, полужесткокрылых, двукрылых и
прямокрылых.
Пищевая специализация насекомых в процессе исторического
развития шла от потребления разлагающихся растительных орга-
низмов (сапрофитофагии) к
питанию живыми растениями
и от питания продуктами
распада тканей животных
(сапрозоофагии)к хищничест-
ву и паразитизму. Паразити-
ческие виды и потребители
детрита являются крайними
формами трофической адап-
тации насекомых.
Рис. 1. Относительное количест-
во растительноядных видов (а),
видов, живущих за счет неживой
органики (б), хищников и пара-
зитов (в) в классе насекомых4и в
отдельных его надотрядах.
/ — колеоптероиды; 2 — мекоптерои
ды; 3 — гемиптероиды; 4 — ортопте-
роиды; 5 — нейроптероиды; 6 — одо-
натовды; 7 — эфемероиды.
в
5
Растительноядные насекомые занимают первое место по чис-
ленности особей в семейных группах отдельных, видов и по способ-
ности к массовому размножению. Плотность их популяций (чис-
ленность особей на единицу площади) достигает чрезвычайно боль-
ших величин. Так, например, в очагах массового размножения
сибирского шелкопряда количество гусениц, каждая из которых
потребляет до 430 мг хвои в сутки, может превышать 3 млн. на
1 га кедрового леса. И это далеко не предел. Среди тех же чешуе-
крылых есть виды, плотность популяций которых при массовом
размножении исчисляется десятками миллионов особей на гектар
{луговой мотылек, капустная моль, лиственничная листовертка
и др.).
Насекомые-фитофаги являются основными потребителями зеле-
ных растений на Земле.
Пищевая специализация насекомых-дендрофагов
Среди насекомых нашей фауны, для которых источником пищи
.служат семенные растения, депдрофаги и тамнофаги составляют
в целом менее 20% видов. Остальные относятся к числу хортофа-
гов, развивающихся за счет травянистых и мелких кустарничковых
растений. Естественно, что в лесной зоне удельный вес дендро-
тамиофагов много больший. В некоторых типах хвойных лесов,
например в сухих лишайниковых сосняках (беломошниках),
дендрофаги являются господствующими представителями расти-
тельноядных насекомых.
В лесной энтомологии до сих пор не возникало разногласий в
разделении насекомых на основные группы по характеру связей
с кормовыми растениями. К монофагам всегда относили виды,
питающиеся на одной породе, т. е. на представителях одного рода
древесных растений, к олигофагам — виды, которые в выборе
кормовых растений в основном ограничиваются одним семейством,
к полифагам — многоядные насекомые, питающиеся пе только
древесными, но и кустарниковыми и даже травянистыми растени-
ями из отдаленных семейств. В сельскохозяйственной энтомологии
к группе монофагов чаще относят только тех насекомых, которые
развиваются на одном виде растений, а промежуточную группу
с ограниченным выбором кормовых растений разделяют на две
подгруппы — узких и широких олигофагов. Выделение лесных
насекомых, развивающихся на одном виде растений, вероятно,
целесообразно только в порядке особой подгруппы мопофагов.
Анализ литературных данных показывает, что в состав этой под-
группы в основном входят виды, имеющие очень ограниченные
ареалы. К ней условно относят также редких и малоизвестных
дендрофагов.
На территории СССР зарегистрировано около 900 видов насе-
комых, развивающихся на хвойных растениях. Среди них, пи-
тающихся хвоей, почками, молодыми побегами и топкими корня-
6
Рис. 2. Соотношение ко-
личества монофагов (м),
олигофагов (о) и поли-
фагов (п), зарегистриро-
ванных на хвойных в
СССР.
Л — виды, питающиеся хво-
ей, почками, побегами, шиш-
ками и тонкими корнями;
Б — виды, развивающиеся
в тканях ствола, ветвей и
корневых лап (ксилофаги).
м& — развиваются на одном
виде растений; м — разви-
ваются на представителях
одного рода растений; о —
развиваются на представи-
телях разных родов одного
семейства; п — развиваются
на представителях разных
семейств древесных и кус-
тарниковых растений; п, —
помимо деревьев и кустар-
ников повреждают кустар-
нички и травянистые расте-
ния.
190
ми, заметно преобладают монофаги (рис. 2). Количество видов в
обеих подгруппах монофагов составляет около 48%. Среди видов,
питающихся тканями ствола, ветвей и корневых лап, преобладают
олигофаги (46 %).
Наиболее широкие полифаги встречаются среди чешуекрылых.
Особенной популярностью в этом отношении пользуется непарный
шелкопряд (L. dispar L.), питание которого прослежено более чем
на 150 видах древесных, кустарниковых и травянистых растений
[Кожанчиков, 1950]. Очень многоядны подгрызающие совки.
Наряду с травянистыми растениями .они повреждают молодые
деревья, в том числе хвойные, в питомниках и на вырубках. Среди
насекомых, питающихся древесиной и корой, многоядных видов
значительно меньше. Но и здесь есть виды, которые помимо хвой-
ных и лиственных деревьев развиваются на кустарниках. На-
пример, восточный непарный короед (Xyleborus aequalis Reitt.)
повреждает около 50 видов растений, весьма отдаленных система-
тически — от аянской ели до амурского винограда и таволги
(Старк, 1952).
Как у олигофагов, так и у полифагов есть предпочитаемые кор-
мовые растения, причем у первых такие растения обычно постоянны
по всему ареалу, а у вторых в большинстве случаев различаются.
Сосновый шелкопряд (D. pint L.), например, всюду предпочитает
сосну обыкновенную, но может развиваться на ели, лиственнице
и сибирском кедре. В то время как для непарного шелкопряда
излюбленными растениями в различных частях ареала могут быть
тополь, дуб, лиственница и некоторые другие.
Насекомые используют в пищу все органы и части растения,
включая внешнюю мертвую кору (ритидом). Дендрофильных насе-
7
КАРПОФАГИ
комых в этом отношении разделяют на пять основных групп:
вредителей ствола (кормофаги), вредителей корней (р и з о-
ф а г я), вредителей развивающихся побегов (бластофаги),
вредителей листьев (ф и л л о ф а г и), вредителей плодов (к а р-
п о ф а г и). В некоторых случаях в качестве самостоятельных
групп выделяют вредителей ветвей (к л а д о ф а г и), почек
(н е ф р о ф а г и), цветков (а н т о ф а г и), семян (спермо-
ф а г и), а также шишек хвойных (к о и о ф а г и). Границы меж-
ду этими группами нечеткие, почти в каждой из них встречаются
гетеротрофные виды, способные питаться также другими частями
и органами растения.
По характеру потребляемых тканей среди насекомых-дендро-
биоптов достаточно четко выделяются две большие группы: х л о-
р о ф а г и, питающиеся ассимиляционными тканями листьев и
молодых зеленых побегов, и ксилофаги, развивающиеся за
счет тканей ствола и ветвей (рис. 3). Последних иногда разделяют
на флеофагов (питаются корой) и собственно ксилофагов
(питаются древесиной).
Первичные и вторичные вредители леса
С конца прошлого столетия пасекомых-дендрофагов стали раз-
делять на первичных и вторичных вредителей. К первой группе
относили вредителей листьев, ко второй вредителей ствола. Вреди-
телей плодов и семян, почек и побегов, а также корней выделяли
в самостоятельные экологические группы, либо (кроме вредителей
плодов) рассматривали как вредителей молодняка. Разделение
наиболее многочисленной группы вредителей древесных растений
на первичных и вторичных было в известной степени условным.
И все же современное категорическое отрицание даже относитель-
ной правомерности и тем более целесообразности такого разделе-
ния вряд ли оправдано.
Некоторое время назад сделана попытка сохранить термин
«первичные» и «вторичные» вредители, вложив в них новый смысл,
а именно к группе вторичных вредителей относить только те виды
дендрофагов, которые нападают на физиологически ослабленные
и неспособные к сопротивлению деревья, а к
первичным — л исто- и хвоегрызущих вредителей, которые и ранее
рассматривались как один из факторов первичного ослабления
деревьев. При этом считалось, что на развитии филлофагов не ска-
зывается физиологическое состояние и степень жизнестойкости
деревьев. Однако, как отмечает П. М. Рафес (1968), многие иссле-
дования убедительно показывают, что первичные вредители леса
также встречают сопротивление со стороны кормового растения.
Следовательно, термин «первичные» не верен по существу, а за
отсутствием первичных термин «вторичные» лишается смысла.
Нам представляется такой довод не очень убедительным. Расте-
ние не может не оказывать сопротивление любому повреждающе-
му фактору, в том числе (и особенно!) массовым листогрызущим
вредителям. Это результат их многовекового взаимодействия.
Однако в данном случае обсуждению подлежит другой вопрос.
Необходимо решить, развиваются ли хвое- и листогрызущие
насекомые в свойственных им стациях на совершенно здоровых
деревьях и могут ли они в годы массового размножения приводить
эти деревья к первоначальному физиологическому ослаблению,
т. е. делать их доступными для нападения вторичных вредителей.
В лабораторных и естественных условиях установлено, что гу-
сеницы ряда видов чешуекрылых и личинки некоторых пилильщи-
ков лучше развиваются, а имаго оказываются более плодовитыми
при питании листьями и хвоей ослабленных деревьев. При массо-
вом размножении этих вредителей первоначальное накопление их
происходит в древостоях, ослабленных теми или иными фактора-
ми [Руднев, 1958, 196% Thalenhorst, 1953; Рожков, 1958, 1965;
Гримальский, 1959; Ханисламов, 1963; Галкин, 1960, 1963; Ивли-
ев, 1960, 1966; Zwolfer, 1963; Васильева, 1972; и др. 1. Д. Ф. Руднев
считает изменения качества пищи под влиянием погодных, почвен-
ных и антропогенных факторов одной из главных причин в колеба-
9
ниях численности вредителей; В. И. Гримальский эксперимен-
тально установил токсичность эфирных масел хвои здоровых сосен
.для хвоегрызущих насекомых и показал, что при засухе и недоста-
точном азотном питании интенсивность выделения смолы из хвои-
нок уменьшается, а это влечет за собой повышение выживаемости
вредителей. В настоящее время достоверно известно, что первич-
ные очаги размножения целого ряда хвое- и листогрызущих вреди-
телей возникают вблизи старых очагов, гарей, в насаждениях,
расстроенных рубками или ослабленных в результате каких-то
иных причин.
Все это так. Но, во-первых, первоначальное накопление чис-
ленности этих вредителей и последующее развитие их очагов от-
нюдь не одно и то же; во-вторых, вопрос о благотворном влиянии
на хвое- и листогрызущих насекомых физиологического ослабле-
ния деревьев не решен однозначно. Многолетние наблюдения за
динамикой численности серой лиственничной листовертки
(Z. griseana Hbn.) в Швейцарии и Австрии, пихтовой листовертки
(Ch. fumiferana Clem.) в Канаде, сосновой пяденицы (В. piniari-
us L.) и непарного шелкопряда (L. dispar L.) в СССР показывают,
что ослабление насаждения хотя и служит предпосылкой вспышек
массового размножения, однако далеко не всегда достаточно [Во-
ронцов, Семеновский, 1963]. Добавим к этому, что повышение
плотности популяций хвоегрызущих чешуекрылых в резервациях
и собственно в очагах обусловливается различными факторами.
Более того, в период вспышек массового размножения насекомые
приобретают значительно большие физиологические возможности
а преодолении защитных свойств и реакций кормовых растений.
Специальные исследования развития, выживаемости и плодо-
витости массовых хвоегрызущих чешуекрылых в зависимости от
биохимического состава хвои здоровых и ослабленных деревьев,
проведенные Т. Г. Васильевой (1973), со всей очевидностью пока-
зали, что пища влияет определяюще на плодовитость насекомых и
соответственно отражается на дипамике их численности. Но даже
при самом оптимальном сочетании компонентов кормового суб-
страта повышение плодовитости не может само по себе увеличить
плотность популяции в течение двух-трех поколений в сотни раз,
что мы наблюдаехМ в очагах размножения хвое- и листогрызущих
насекомых. Вспышки размножения фитофагов есть результат
повышения выживаемости, сопровождающегося повышением аг-
рессивности насекомых во взаимодействии с используемыми для
питания растительными компонентами биоценоза.
Сказанное особенно наглядно проявляется у ксилофагов, спо-
собных к вспышкам размножения. Лиственничный усач (X. atta-
ints Gebl.), экология которого подробно описана в специальной
части книги, вне очагов размножения, а также во время предвспы-
шечного повышения плотности популяции проявляет себя как ти-
пичный вторичный вредитель. Его очаги чаще всего возникают в
древостоях, обесхвоенных сибирским шелкопрядом. Во время
10
вспышек размножения этот ксилофаг приобретает практически все
особенности, свойственные первичным вредителям.
Все листо- и хвоегрызущие насекомые, включая массовые виды,
обитающие вне резерваций и территорий первичных очагов, раз-
виваются на здоровых деревьях. При этом численность их не под-
нимается до величин, угрожающих деревьям физиологическим ос-
лаблением и заселением вторичными вредителями. Деревья сохра-
няют жизнестойкость при условии ежегодного изъятия части
листовой массы, соответствующей их компенсационным возмож-
ностям. Иные последствия наблюдаются при вспышках размно-
жения филлофагов и. распространении очагов в эти жизнестойкие
древостои.
Среди массовых хвоегрызущих видов чешуекрылых одним из
наиболее изученных на сегодняшний день является сибирский
шелкопряд (Dendrolimus superans sibiricus Tschetv.). Этому вреди-
телю за 70 лет со времени его описания посвящено свыше 300 ра-
бот, в том числе пять монографий [Флоров, 1938; Жохов, 1961;
Коломиец, 1962; Рожков, 1963, 1965; Болдаруев, 1969]. Из этих
работ известно, что первичные очаги сибирского шелкопряда обыч-
но возникают на узкой полосе, граничащей с участками расстро-
енного древостоя, вдоль опушек или в виде небольших куртин в
низкоплотных участках насаждений. Древостои, в которых проис-
ходит первоначальное накопление численности шелкопряда, по-
мимо прочих необходимых для этого условий несомненно отлича-
ются низкой степенью защитных свойств деревьев и благоприятны
в кормовом отношении. Однако уже в процессе развития первого
поколения вредителя площади первичных очагов увеличиваются
во много раз, а в дальнейшем, за счет образования миграционных
очагов, могут охватывать десятки и даже сотни тысяч гектаров ле-
са. Естественно, на такой громадной площади вредитель обесхво-
ивает не только ослабленные деревья, но и вполне здоровые, обла-
дающие высокими защитными свойствами.
В периоды массового размножения хвое- и листогрызущие на-
секомые проявляют себя именно как первичные вредители, вредо-
носность их не просто предшествует заселению деревьев короеда-
ми, усачами, златками, рогохвостами, а является фактором пер-
вичного ослабления деревьев, обеспечивающего благоприятные
условия для развития вторичных вредителей.
Первичными вредителями леса следует
считать лишь тех листо- и хвоегрызущих
насекомых (также немногих ксилофагов),
которые способны давать вспышки массо-
вого размножения. Вторичные вредители
проявляют себя только в ослабленных
древостоях. В 'здоровых лесах и вне очагов размножения
питание всех насекомых-дендрофагов не отражается па состоянии
лесных биоценозов.
11
О сущности вспышек размножения вредителей леса
Причины и сущность вспышек массового размножения насе-
комых, составляющих 80% современного животного мира,—
многосложный вопрос. Он требует всестороннего обсуждения.
Поэтому, имея в виду посвятить этому вопросу специальную рабо-
ту, мы приведем здесь лишь основные представления, сложивши-
еся у нас в итоге многолетних исследований.
Прежде всего мы исходим из того, что не все насекомые-фито-
фаги способны размножаться в массовых количествах и что п е
всякое увеличение плотности популяции
вида есть вспышка размножения. Часть видов
имеет относительно стабильную численность, соответствующую
сравнительному постоянству условий в занимаемой экологической
нише. Они отличаются ограниченными ареалами, обычно не выхо-
дящими за пределы первоначальной территории формирования ро-
да, а их популяции характеризуются отсутствием обособленных
семейных групп и очень слабо выраженным полиморфизмом. Ко
второй группе относится подавляющее большинство растительно-
ядных насекомых, плотность популяций которых может колебать-
ся в значительных пределах. Причем эти колебания целиком или
почти целиком определяются изменениями условий жизни (корм,
конкуренция в семейных группах, энтомофаги, погодные условия).
Наконец, третью относительно небольшую в видовом отношении,
но имеющую наибольшее хозяйственное значение группу составляют
насекомые, которые на определенном уровне плотности популяции
выходят из-под контроля ограничивающих факторов среды и дают
«безусловные* вспышки массового размножения.
Если популяции второй группы насекомых претерпевают толь-
ко два осцилляторных состояния — депрессии и экспрессии, то
третьей группе свойственно еще особое «вспышечное» состояние.
Переход популяций в это состояние, по-видимому, определяется
теми же условиями, которые вызывают повышение их плотности
у всех растительноядных насекомых. Однако в данном случае
вызываемые ими изменения в обмене веществ, достигнув опреде-
ленного уровня, преобразуют не только физиологию, но и этоло-
гию особей. У насекомых появляется стремление к миграциям в
наиболее благоприятные для развития участки биотопа (стации
первичных очагов), в результате чего образуются смешанные се-
мейные группы и, как следствие этого, проявляется «эффект гете-
розиса* — вспышка массового размножения. Условия среды, вы-
зывающие вспышку размножения, неадекватны масштабам пос-
ледней.
Сказанное не отрицает возможности концентрации насекомых
под воздействием стихийных факторов, например в результате
оседания мигрирующих бабочек совки-гаммы под встречную об-
лачность циклонических образований [Пятницкий, 1951|. Глав-
ным здесь остается тенденция к миграциям.
12
Границы между названными выше группами насекомых не мо-
гут быть четкими. Несомненно существуют переходные формы.
При этом вряд ли есть роды, которые бы целиком относились к
третьей группе. Например, среди усачей рода Xylotrechus вспышки
массового размножения известны только у X. altaicus Gebl. Боль-
шинство остальных видов относится ко второй группе, хотя плот-
ность популяций некоторых из них может повышаться до значи-
тельных размеров. А такие эндемичные виды, как X. iteratus Pan.,
X. salvazi Pic., X. basalts Schwzr. и некоторые другие, имеют очень
ограниченные ареалы, относятся к числу редких и, несомненно,
представляют собой тупиковые ветви в филогении рода. В богатом
видами роде волнянок (Limantria) вспышки размножения дают
лишь непарный шелкопряд (L. dispar L.), монашенка (L. monacha
L.) и еще 6—7 видов. Из 11 известных видов хвойных шелкопря-
дов (коконопрядов) рода Dendrolimus только 4 образуют очаги
вспышек размножения, это — сосновый (D. pint L.), сосновый
восточный (D. spectabilis Butl.), большой хвойный (D. superans
Butl.) и пятнистый (D. punctatus Walk.) шелкопряды. Наиболее
грандиозные вспышки размножения среди хвоегрызущих вреди-
телей свойственны сибирскому подвиду большого хвойного шелко-
пряда (D. superans sibiricus Tschetv.).
Вспышки массового размножения присущи только прогрессирую-
щим видам насекомых. для которых характерны большой или уве-
личивающийся ареал, большое количество подвидов, племен и абер-
раций. несоответствие генетической природы условиям среды у гра-
ниц ареала, высокая чувствительность к изменениям условий среды
и значительные колебания плотности популяций в периоды между
вспышками размножения {Рожков. 1965]. Вспышки размножения
свойственны насекомым, развивающимся на живых растениях.—
эволюционно наиболее молодой группе* фитофагов. Это явление
более прогрессивное по сравнению с обычным увеличением численно-
сти (адекватным условиям среды), происходящим при Переходе
популяции в экспрессивное (выраженное в биоценозе) состояние.
Древесиноядные насекомые (ксилофаги) за исключением очень
немногих видов, способных развиваться на вполне жизнеспособ-
ных деревьях, не дают таких вспышек размножения, которые свя-
заны с переходом популяции в стадное состояние и с повышением
физиологической выживаемости особей. Наблюдаемые увеличе-
ния их численности в ослабленных или усыхающих древостоях,
иногда значительные,— результат улучшения условий развития,
в первую очередь трофических.
Что же представляет собой вспышка массового размножения
насекомых-фитофагов? II. М. Рафес (1968) дал. например, следую-
щую формулировку, отражающую незаурядность этого природ-
ного явления: массовым размножением следует называть такое
повышение плотности популяции, которое преодолевает мощность
регулирующих факторов и тем самым нарушает действие стаби-
лизирующих механизмов. Иными словами, массовое размножение
13
ПойуЛяЦии какого-либо вида — это случай нарушения нормаль-
ных биогеоценотических процессов: мощность Механизмов обрат-
ной связи в биогеоценозе оказывается недостаточной.
С точки зрения адаптивного развития прогрессирующих видов <
вспышка массового размножения есть
крайняя форма разрешения противоре-
чий между стремлением вида к воспроиз-
ведению и сохранению потомства и проти-
водействующими условиями среды. Во времй
вспышек размножения в результате гетерозиса в наибольшей сте-
пени проявляется генетический полиморфизм, что способствует
отбору в популяциях.
Вспышки массового размножения правильнее было бы называть
вспышками выживания. Многолетние наблюдения показывают,
что быстрое многократное увеличение плотности популяций про-
исходит здесь не в результате повышения плодовитости, а вслед-
ствие резкого снижения коэффициента смертности. Мы уже писа-
ли 1Рожков, 19651 о том, что по существу до сих пор нет доказа-
тельств того, что вспышки размножения насекомых-фитофагов
являются прямым следствием нарушения нормального (среднего)
соотношения между способностью вида к размножению (макси-
мальной плодовитостью) и сопротивляемостью среды, тормозящей
эту способность. В то же время хорошо известно, что выжива-
емость до имаго многих массовых видов, например, таких, как
непарный шелкопряд, монашенка, сосновый и сибирский шелко-
пряды, во время депрессии численности составляет десятые доли
одного процента, а на первых этапах вспышки размножения до-
стигает 20 и даже 40%.
Абсолютная плодовитость растительноядных насекомых не мо-
жет не изменяться в зависимости от условий среды и в первую
очередь от степени соответствия химического состава потребля-
емых в пищу тканей растений потребностям организма как в отно-
шении питательных, так и защитных веществ (токсинов, репеллен-
тов). Однако эти изменения невелики и не могут сами по себе
вызвать вспышку размножения. Переход популяции в экспрессив-
ное состояние не означает обязательное начало вспышки. Повы-
шение плодовитости является, лишь предпосылкой, одним из усло-
вий возможного перехода популяции в стадное (очаговое) состояние.
В названной выше работе мы недостаточно оценили высказыва-
ния ряда авторов о качественной неравноценности популяций вида
и одной и той же популяции при разной плотности особей. Наши
последующие исследования лиственничного усача, в динамике
численности которого паразиты играют несравнимо меньшую роль,
чем у чешуекрылых, показали, что массовое размножение этого
вредителя является прежде всего результатом повышения его
физиологической стойкости.
Кому принадлежит первое высказывание о возможных генети-
ческих различиях популяции в обычном и очаговом состояниях,
14
нам не известно. В схеме влияния факторов внешней среды и на-
следственных свойств на плотность популяции, составленной 30
лет назад В. Олли с соавторами [Allee е. а., 1949] и используемой
в наших учебных пособиях по лесной энтомологии и экологии на-
секомых [Воронцов, 1963; Яхонтов, 1964], генетические факторы
стоят на одном уровне с экологическими факторами, определяю-
щими повышение выживаемости. В настоящее время многие ис-
следователи считают, что при высокой и низкой плотности попу-
ляции проявляются разные генотипы. Некоторые связывают про-
явление разных генотипов с возникновением уравновешенного*
полиморфизма, после того как популяционные осцилляции ста-
новятся постоянной особенностью данного вида [Chitty, 1957,
1960; Andrewartha, 1959].
По нашему мнению, широкое проявление генетического поли-
морфизма свойственно прогрессирующим видам и осуществляется
оно во время массовых размножений в результате агрегации се-
мейных групп.
Виды, дающие вспышки размножения, обладают обширными
ареалами. Естественно, в пределах последних по могут быть оди-
наково благоприятные условия для их жизни. Поэтому каждый
такой вид бывает представлен большим или меньшим количеством
популяций, обособленных друг от друга и пространственно и ге-
нетически. Более того, популяции этих видов характеризуются
относительной разобщенностью семейных групп. Под последними
мы понимаем элементарные внутрипопуляционные группировки
(демы) особей, постоянно скрещивающихся между собой и потому
генетически более однообразных по сравнению с особями популя-
ции в целом (при одном генофонде последней). Образование семей-
ных групп определяется ландшафтно-экологической неоднород-
ностью территории, занимаемой популяцией. У массовых видов
наибольшая обособленность семейных групп проявляется в пери-
оды между вспышками размножения.
Пока мы можем судить о наличии генетических различий меж-
ду семейными группами в популяции только по проявлению гете-
розиса при концентрации насекомых в первичных очагах вспышек
размножения и по появлению большого количества морфологиче-
ских форм. Последнее особенно примечательно у чешуекрылых.
У сибирского шелкопряда, например, нами описано пять аберра-
ций только одной лиственничной расы [Рожков, 1963], среди кото-
рых две мало отличаются по размерам и окраске от большого япон-
ского шелкопряда (Dendrolimus superans superans Butl.) и от сосно-
вого шелкопряда, а одна крупная форма (ab. olivaceocana) вообще
очень отличается от представителей этого рода.
Таким образом, с наТпей точки зрения, решающим условием
возникновения вспышки размножения является агрегация семейных
групп. Именно в результате концентрации мигрировавших из
изолированных семейных групп особей на территории, где местная
семейная группа имеет повышенную численность, вступают в дей-
15
ФАКТОРЫ
ДИНАМИКИ
ЧИСЛЕННОСТИ
экологиче
абиотические и биотические,
независимые и зависимые от плотности
с к и е
преимущественно
этологические
(миграцил,концентрацил)
генетические
ВЫЖИВАЕМОСТЬ
низкам
повышенная
высокая
плотность
низкая
повышенная
высокая
(в местах концентрации)
очень высокая
ОПРЕДЕЛЯЮЩИЕ
МОМЕНТЫ
эффект
группы
эффект
гетерозиса
ЦИКЛЫ
ДИНАМИКИ
ЧИСЛЕННОСТИ
Рис. 4. Причинная связь в динамике численности фитофагов.
А — в депрессивно-экспрессивном цикле; Б — при переходе в состояние вспышки размножения.
ствие генетические факторы, обусловливающие усиление жизне-
стойкости потомства в одном или нескольких поколениях.
Вспышка размножения есть раскрытие
генетического потенциала вида.
Миграции могут осуществляться отдельными особями, неболь-
шими группами или быть массовыми. В них могут принимать уча-
стие оба пола или только самки, реже одни самцы. Так, в много-
километровых перелетах непарного шелкопряда на юге Восточной
Сибири участвуют, как правило, только неоплодотворенные сам-
ки; в концентрации сибирского шелкопряда, черно-желтой ванес-
сы (Vanessa xanthomelas Esp.) и серой лиственничной листовертки
(Zeiraphera griseana Hbn.) самки являются пионерами. Миграции
самцов, возможно, присущи лишь тем массовым видам, у которых
самки не летают.
Стремление к миграциям у многих видов сохраняется в течение
всего периода вспышки размножения. У фитофагов, развивающих-
ся на темпохвойпых породах и сосне, это явление бывает менее
выраженным, чем у тех из них, которые образуют очаги вспышек
в лиственных и лиственничных лесах.
Вспышка размножения — закономерное явление в становлении
вида. Она отражает объективную связь случайности, заключаю-
щейся в стечении обстоятельств, определяющих явление, и необ-
ходимости — внутренней сущности последнего.
В целом развитие вспышки размножения следует рассматривать
как звенья цепи взаимообусловленных причин, среди которых
определяющими являются «эффект группы» и «эффект гетерозиса»
(рис. 4).
Конкретным проявлением вспышки размножения являются
очаги. Вспышки нередко охватывают огромные территории и мо-
гут включать десятки и сотни очагов, фаз личных по продолжи-
тельности развития. Например, вспышка размножения серой
лиственничной листовертки в 1966—1969 гг. практически охвати-
ла все лиственничные леса Северной Палеарктики, и очаги ее в
каждом регионе различались по характеру образования и дли-
тельности действия. Вспышки размножения сибирского шелко-
пряда обычно также охватывают большие территории. Очаги
этого вредителя в кедровых лесах действуют 3—5 лет (без учета
начального повышения численности), а в лиственничных могут
существовать более двух десятилетий. О закономерностях разви-
тия вспышки размножения на всей охватываемой ею территории
сведений фактически нет ни по одному виду фитофагов.
Собственно очаг проходит две фазы развития — агрега-
ции и образования смешанных семейных групп популяции и
дизъюнкции семейных групп в процессе угасания. Началь-
ное накопление численности (экспрессивное состояние популяции),
как уже говорилось, необязательно завершается возникновением
вспышки размножения.
2 А. С. Рожков
17
КСИЛОФАГИЯ
Вторичные вредители леса практически целиком представлены
насекомыми-ксилофагами. Однако эти термины нельзя считать
синонимами. Среди большой и систематически неоднородной груп-
пы ксилофагов лишь относительно немногие виды с полным пра-
вом могут быть отнесены к вредителям леса. В большинстве своем
это потребители древесины на разных этапах ее распада.
Ксилофаги играют чрезвычайно важную роль в лесу, являясь са-
нитарами и обогатителями почвы органическими веществами.
Под ксилофагией следует понимать пи-
тание древесиной и корой (ксилемой и
флоэмой) живых, отмирающих и мертвых
деревьев до разрушения структуры их
тканей. По резонному заключению Н. П. Кривошеиной и
Б. М. Мамаева (1967), было бы искусственным ограничивать
группу ксилофагов только обитателями собственно древесины,
выделяя особую группу флеофагов [Schedl, 1956], поскольку меж-
ду корой и древесиной как источниками питания различия не столь
значительные. Важным для определения насекомых-ксилофагов
является, кроме того, то, что их личинки развиваются непосред-
ственно в тканях ствола, ветвей и корней. Обгрызание, например,
молодых побегов открыто живущими насекомыми, корней —
почвообитающими видами и тем более высасывание клеточного сока
луба и заболони равнокрылыми и полужесткокрылыми насекомы-
ми не являются ксилофагией.
Состав и филогения ксилофагов
Ксилофагия — одна из древнейших пищевых специализаций
насекомых. Эволюция ксилофагов в целом шла от детрито- и сапро-
фагии к потреблению свежеотмерших, а затем и живых тканей
ксилемы и флоэмы. Часть современных ксилофагов в своем станов-
лении, бесспорно, прошла этап филлофагии (чешуекрылые и рого-
хвосты сем. Siricidae). Питание живыми растениями является вы-
соким достижением насекомых. Оно требует преодоления активно-
го противодействия растения.
Среди насекомых первыми ксилофагами, несомненно, были жу-
ки, а первые жуки, появившиеся па земле, возможно, были кси-
лобионтами. Наиболее древние остатки вымерших жесткокрылых-
архостемат (Archosternata) найдены в отложениях раннепермского
возраста. Представители двух других подотрядов жуков Adephaga
и Polyphaga появились в начале триасового периода. В юре фауна
жуков была уже близкой к современной и представлена в основном
подотрядом разноядных.
По мнению А. Г. Пономаренко (1969), с самого начала своей ис-
тории жуки оказались связанными с ксероморфной хвойной фло-
рой перми; питались они скорее всего гниющей древесиной. Наи-
18
большего развития пермские архостематы получили на территории
Ангариды, откуда активно расселялись на запад к Европе и на
юг в Гондвану. В триасе в подотряде Archostemata появилось
самое древнее из доживших до нашего времени семейство жуков —
Cupedidae. Первичные купедиды имели вид типичных подкорных
жуков. У них было широкое уплощенное тело, окаймленное по бо-
кам параноталиями и расширенной эпиплеврой; маленькая,
спрятанная под переднеспинку голова несла короткие, слабо чет-
ковидпые антенны; ноги были короткими, с широко расставленны-
ми передними и средними тазиками; стерниты довольно выпуклого
брюшка сохранили подвижность. Пищей триасовым купедидам —
дендробионтам служили хвойные, птеридоспермы, цикад офиты и
кордаиты.
Рассматривая основные морфогенетические процессы в эволю-
ции архостемат, А. Г. Пономаренко отмечает, что у жуков, образ
жизни которых связан с древесиной, па голове возникли различ-
ные выросты и борозды, служившие для защиты глаз и антенн.
У некоторых купедид в связи с приспособлением к обитанию в от-
носительно прочной древесине сильно изменились мандибулы.
Концы мандибул у них вывернулись так, что их зубцы оказались
расположенными в вертикальной плоскости, и жук мог использо-
вать все три зубца вместо одного. Появление совершенных манди-
бул сопровождалось у жуков усилением челюстных мышц и вытя-
гиванием в длину и в сторону висков, к которым прикрепляются
эти мышцы.
Совершенствование ксилобионтвости еще в большей степени
отразилось на морфологии личинок, вся жизнь которых проходит
в древесине. У личинок наиболее специализировавшихся совре-
менных ксилофагов — усачей и златок —; размеры головы сокра-
тились, она стала прогнатической и в значительной степени погру-
зилась в переднегрудь. У некоторых видов из переднегруди нару-
жу выдаются лишь часть наличника, верхняя губа и челюсти,
глазки исчезли. Редуцировались ноги, их заменили спинные и
брюшные двигательные мозоли. Покровы тела утратили пигмент.
Значительным преобразованиям подверглись также внутренние
органы личинок, прежде всего — пищеварительный тракт.
Среди жуков имеются семейства, целиком относящиеся к кси-
лофагам. Таковы златки (Buprestidae), короеды (Ipidae), древесин-
ники (Platypodidae), точильщики (Anobiidae), древогрызы (Lyc-
tidae), сверлильщики (Lymexylonidae), ложнокороеды (Bostry-
chidae). Большинство дровосеков, или усачей (Cerambycidae)
являются типичными ксилофагами. Древесиноядпые виды встре-
чаются также в семействах щелкунов (Elateridae), долгоносиков
(Curculionidae), ложносЛоников (A nthribidae), толстощупиков
(Serropalpidae) и ряде других.
Ни в каком другом отряде нет такого количества «ферментатив-
ных» ксилофагов, способных переваривать и усваивать клетчатку.
Тем не менее подавляющее большинство жуков-ксилофагов не имеет
2*
19
целлюлазы и питается за счет легкорастворимых веществ, содер-
жащихся в древесине. Немало среди жуков сапроксилофагов и
ксиломицетофагов. Последних подразделяют па деструктивных
ксиломицетофагов, питающихся древесиной, разрушенной гриба-
ми, и амброзийных ксиломицетофагов, использующих в основном
мицелий грибов. Есть в числе жуков-ксилобиоптов и типичные
мицетофаги.
Некоторые виды жуков могут поселяться на вполне жизне-
способных деревьях («физиологические» вредители) и вызывать их
усыхание. Однако совершенно здоровые деревья подвергаются на-
падению относительно немногих из них, в частности, тех, которые
способны, подобно массовым первичным вредителям, давать
вспышки размножения.
Ксилофагия среди чешуекрылых — явление вторичное. Из их
числа истинно дровесиноядными могут считаться лишь некоторые
представители серии листоверткообразных (Tortricodea ), а именно:
листовертки (Tortricidae), древоточцы (Cossidae), стеклянницы
(Aederiidae) и ряд видов серии огневкообразных (Pyralidodea).
Формирование чешуекрылых-ксилофагов шло от потребления
листьев к питанию почками, молодыми побегами, живой частью
коры (лубом), нежными наружными клетками древесины и, нако-
нец, собственно древесиной (заболонью и ядром). О том, что пере-
ход этих чешуекрылых к ксилофагии начался относительно не-
давно и что осуществляется он именно таким путем свидетельству-
ет следующее: во-первых, подавляющее число их развивается на
живых растениях; во-вторых, гусеницы многих видов живут
внутри почек и молодых побегов либо являются лубоядными;
в-третьих, существует немало видов, питающихся листьями и поч-
ками или лубом и древесиной, а такой широко распространенный
вредитель хвойных деревьев, как еловая шишковая огневка
(Diorictria abietella Schiff.), повреждает, например, у сосен, кору,
луб и древесину ствола, почки, побеги, хвою и шишки [Рожков,
1957].
Рогохвосты являются много более специализированными кси-
лофагами, чем древесиноядные чешуекрылые. Они делятся на две
группы — семейство Xiphidriidae и Siricidae, различающиеся как
в имагинальной, так и в личиночной стадии. Первые приспосо-
бились к развитию в мертвой древесине, часто уже достаточно
мягкой, вторые — в свежих стволах, и нередко нападают на жиз-
неспособные деревья.
В современном представлении, предки низших перепончато-
крылых в личиночной стадии вели наземный образ жизни, пита-
лись растительными остатками, спорами папоротникообразных и
пыльцой голосемянных, в изобилии накапливавшихся в лесной
подстилке.Поедание взрослыми насекомыми пыльцы, по всей веро-
ятности, было тем стимулом, который побудил самок ксиелид от-
кладывать яйца на стробилы хвойных (органы, производящие
пыльцу), а затем и на хвою. Личинки некоторых примитивных
20
пилильщиков из семейства памфилиид (Pamphiliidae) продолжают
еще питаться хвоей, тогда как другие представители их стали
развиваться на листьях деревьев и кустарников покрытосемянных
растений (первые характеризуются более примитивными морфо-
логическими чертами). Гораздо дальше от исходного момента ушли
рогохвосты, откладывающие яйца уже не в зеленые части расте-
ний, а непосредственно в древесину [Малышев, 19661.
Такой путь становления ксилофагииу растительноядных пере-
пончатокрылых, по-видимому, возможен только для Siricidae.
Еще не так давно отделение Xiphydriidae от Siricidae считалось
«явным преувеличением». Основанием тому были не морфологиче-
ские признаки имаго, а сходство образа жизни и внешних призна-
ков личинок [Гуссаковский, 1935]. В настоящее время самостоя-
тельность этих семейств пе вызывает сомнения. А. П. Расницып
(1969), рассматривая эволюцию скрытоживущих Symphyta, счита-
ет, что в надсемействе Siricidea первичной, исходной для остальных
семейств группой могут быть Gigasiriciidae, обладающие множест-
вом примитивных признаков. Xiphydriidae и Siricidae, имеющие
поперечный шов на среднеспинке — должны были отделиться
от Gigasiricidae общим стволом. По нашему мнению, основыва-
ющемуся на различиях в экологии личиночной стадии, разделение
рогохвостообразных на две ветви началось еще со времени на-
земной жизни их общих предков. Развитие Xiphydriidae шло от
Рис, 5. Схема исторического формирования насекомых-ксилофагов.
1 — жесткокрылые, 2 — двукрылые, 3 — чешуекрылые, 4 — ксифидрии, 5 — настоя-
щие рогохвосты.
21
детрито- и сапрофагии к сапроксилофагии и ксиломицетофагии
(далеко не все современные представители семейства являются
настоящими ксилофагами), a Siricidae претерпели в своем развитии
этап питания зелеными частями растений и лишь затем освоили
древесину в качестве кормового субстрата.
Среди двукрылых отсутствуют ферментативные ксилофаги и
деструктивные ксиломицетофаги. Большинство древесиноядных
представителей отряда относится к сапроксилофагам, и немно-
гие — к амброзийным ксиломицетофагам [Кривошеина, Мама-
ев, 19671.
На схеме исторического формирования группы насекомых-
ксилофагов (рис. 5) мы разделили последних на два порядка.
К первому отнесены виды, развивающиеся на вполне жизнеспо-
собных деревьях, не утративших способности к сопротивлению,
ко второй — виды, развивающиеся на отмирающих и мертвых
деревьях.
Некоторые морфоанатомические особенности
личинок ксилофагов
Освоение твердых тканей ствола в качестве субстрата для пита-
ния и развития вызвало изменения внешнего и внутреннего строе-
ния личинок ксилофагов. Эти изменения коснулись многих частей
тела и органов. Тем не менее определить общие для всех ксилофа-
гов признаки, кроме утраты пигмента покровами и усиления верх-
них челюстей, практически невозможно. Различия внешнего вида,
формы головы, сегментов груди и брюшка, степени редукции ног,
строения кишечника и некоторых других органов значительны
даже в пределах одного отряда. Менее всего морфологическая
ксилобионтная адаптация прослеживается в отряде чешуекрылых.
Наибольший интерес представляют особенности строения ро-
товых придатков и пищеварительного тракта личинок ксилофагов.
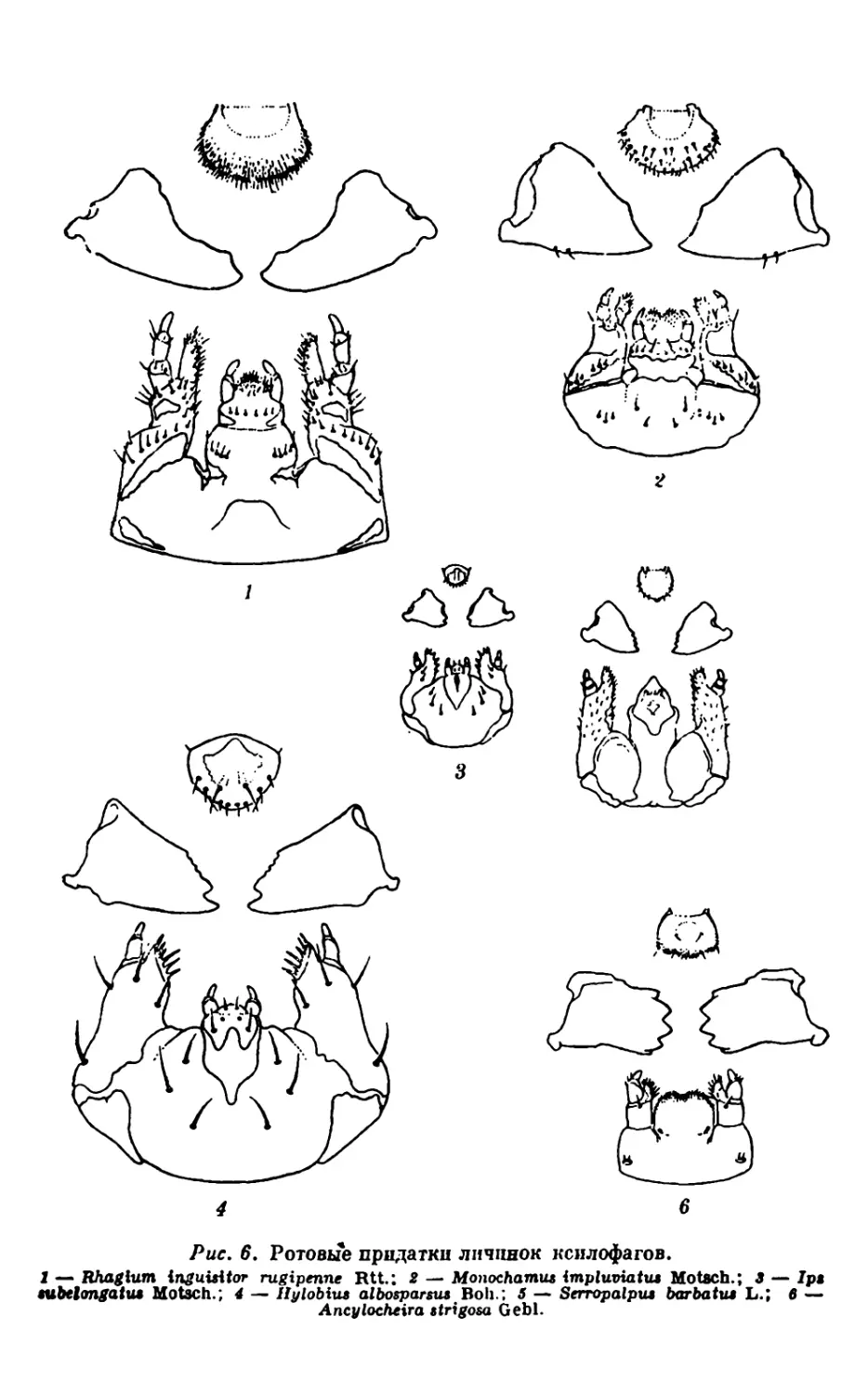
Мы рассмотрим их на примере жуков.
Ротовой аппарат личинок ксилофагов (включая двукрылых)
грызущего типа. Поэтому самой ответственной его частью явля-
ются верхние челюсти (мандибулы). У личинок жуков они отно-
сительно короткие массивные с широким основанием и округлым,
выемчатым или зубчатым режущим краем. Более длинные и слегка
изогнутые верхние челюсти встречаются у некоторых усачей,
питающихся лубом и наружной мягкой частью заболони (рис. 6,
I). Резцовый (вентральный угол режущей части мандибул чаще
всего образует большой зубец, тогда как ое внутренний (дорсаль-
ный) угол тупой, либо несет небольшой зубчик (см. рис. 6, 2, 4,
5). У личинок златок и многих короедов средние зубцы режущей
части выступают сильнее краевых (см. рис. 6, 5, б).
Мандибулы снабжены очень мощными приводящими мышцами,
которые заполняют значительную часть головной капсулы. От-
грызание частиц древесины носит характер либо соскабливания
22
Рис, 6, Ротовые придатки личинок ксилофагов.
1 — Rhaglum inguisitor rugipenne Rtt.; 2 — Monochamus implumatus Motsch.; 3 — /pi
tubelongatu» Motsch.; 4 — Hylobius albosparsus Boh.; 5 — Serropalput barbatu» L.; 6 —
Ancylocheira strigosa Gebl.
(вдоль или поперек древесных волокон), либо подгрызания и пос-
ледующего отщепления групп волокон. Разжевывание уже от-
деленных частиц не является обязательным. У некоторых видов*
питающихся сухой древесиной, дополнительное размельчение
пищи происходит в мышечном желудке.
Нижние челюсти (максиллы) личинок ксилофагов не претерпе-
ли больших изменений. Они слабо склеротизованы и выполняют
при питании ряд функций. Во-первых, максиллы, снабженные
осязательными волосками (трихоидными сенсиллами), ориенти-
руют положение работающих мандибул; во-вторых, поддержива-
ют отщепляемые частицы; в-третьих, подгибаясь внутрь, они
вместе с гипофаринксом проталкивают пищу в ротовое отвер-
стие.
По характеру питания личинок всех ксилофагов можно разде-
лить на три основные группы. Первая из них обладает ферментами,
обеспечивающими усвоение клетчатки стенок трахеид. Эпителий
кишечного канала второй группы не выделяет таких ферментов,
но личинки все жо усваивают большее или меньшее количество
клетчатки за счет симбиотических организмов, содержащихся в
кишечнике (главным образом в специальных криптах) и выраба-
тывающих целлюлазу. Третью группу ксилофагов составляют
виды, усваивающие только растворимые «обычными» ферментами
вещества древесины. Личинки их поглощают большое количество
пищи и соответственно имеют очепь длинный средний отдел ки-
шечника.
На рис. 7 изображены пищеварительные тракты личинок кси-
лофагов с разным типом питания. Наибольшего совершенства в
питании сухой, отмершей многие годы назад древесиной достигли
точильщики. Все они относятся к числу ферментативных ксило-
фагов, имеют мышечный желудок (провентрикулус) для дополни-
тельного размельчения пищи и весьма своеобразную, расширен-
ную переднюю часть средней кишки, которую можно пазвать
«криптовым желудком». Последний в большинстве случаев разде-
лен на отдельные сегменты и имеет многочисленные складки, уве-
личивающие секретирующую поверхность. Ферменты, разлагаю-
щие клетчатку, вырабатываются этим отделом средней кишки.
У пестрого точильщика (Xestobtum rufovillosum De Geer.),
развивающегося в Европе в дубовой древесине, а в Сибири —
в лиственничной, хитиновая выстилка провентрикулуса снабжена
прочными зубцами сложной формы. Стенки его имеют мощные
кольцевые мышцы. Последние не полностью окружают преджелу-
док, а располагаются с двух сторон. Криптовый желудок разде-
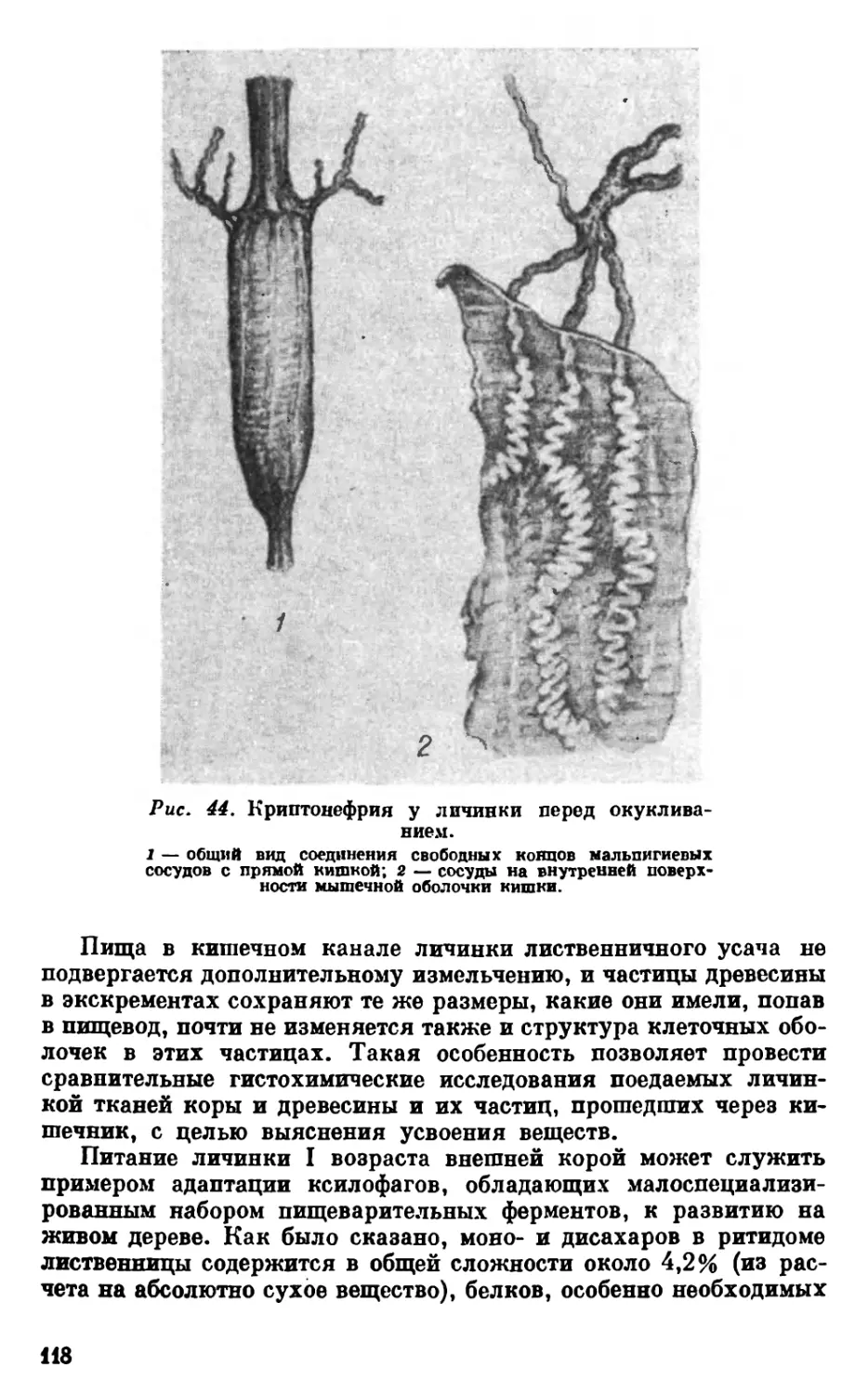
лен на продольные сегменты, средняя кишка относительно ко-
роткая, криптонефрия (врастание концевых частей мальпигие-
вых сосудов в стенки задней кишки) имеет место уже у питающихся
личинок (рис. 7, 1). Возникновение «ранней» криптонефрии, не-
сомненно, является адаптацией точильщика к низкому содержа-
нию воды в пищевом субстрате.
24
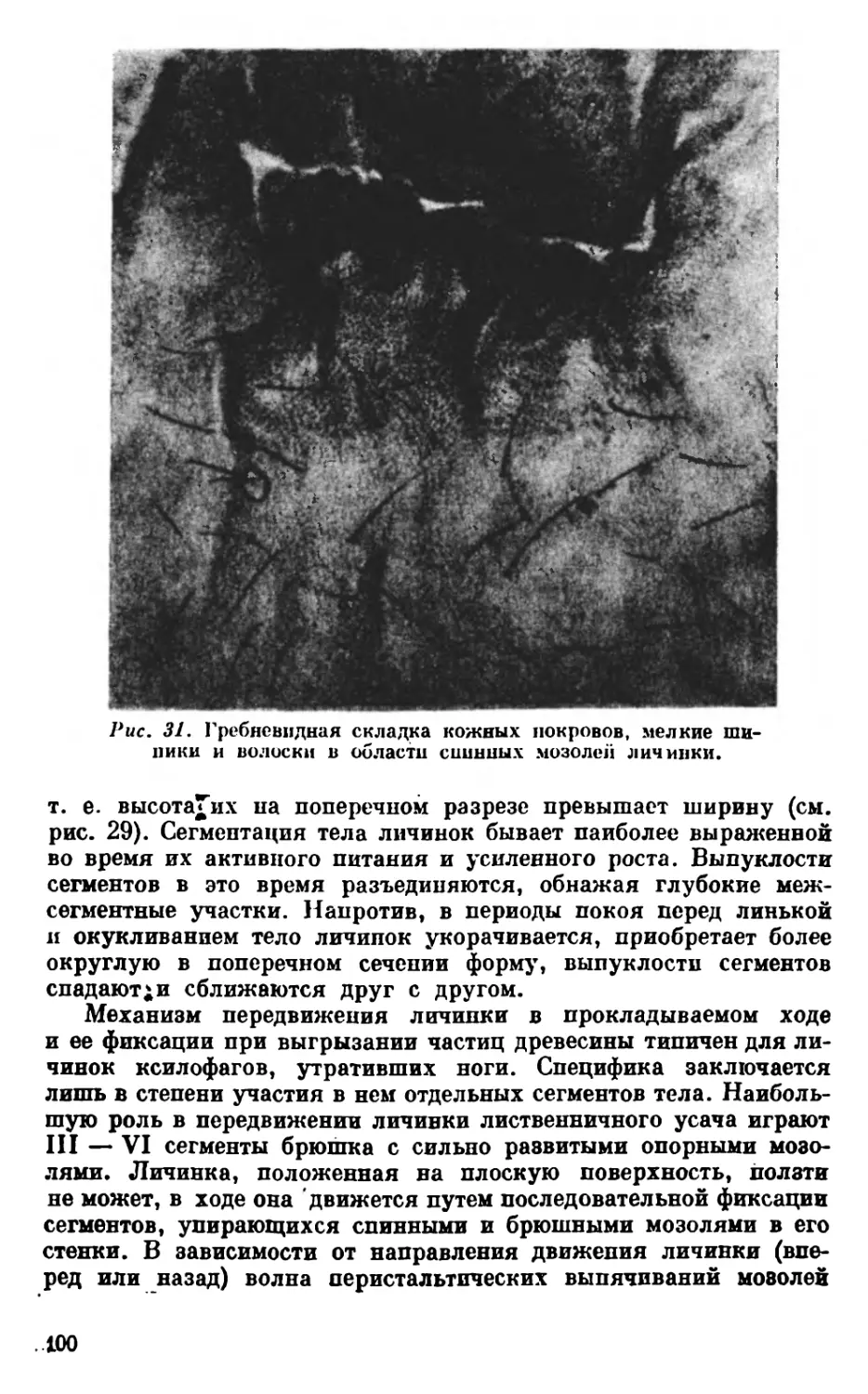
I—Oes
1— Oes
Puc. 7. Кишечник личинок жуков-ксилофагов.
1 — Xatobium futovillosum De Goer.; 2 — Hylobius albosparms Boh.; 3 — Ips. subelongatu*
Motsch.; 4 — Serropalpus barba tvs L.; 5 — Ancylocheira strigosa Gebl.; 6 — Rhagium in-
quisitor rugipenne Rtt.; 7 — Monochamut tutor L. Coes — слепые придатки средней кишки-
Crypt — типичные крипты средней кишки; Cut — шипы на внутренней поверхности про-
вентрикулуса; II — тонкая кишка; Мез — средняя кишка; IffeSiMest — первый и второй
отделы средней кишки; Malp — мальпигиевы сосуды; Malpi — дополнительное соедине-
ние мальпигиевых сосудов с кишечником; Оев — пищевод; Prov — провентрикулус;
Rect — прямая кишка.
Личинки белоточечного долгоносика (Hylobius albosparsus Boh.)
не имеют мышечного желудка и не обладают собственными фер-
ментами, расщепляющими клетчатку. В их кишечнике содержатся
симбионты, вырабатывающие небольшое количество целлюлазы.
Передняя треть средней кишки долгоносика имеет вид объемистого
желудка, снабженного глубокими поперечными складками; зад-
ний отдел средней кишки песет с двух сторон многочисленные
пальцевидные слепые отростки (крипты). Стенки крипт состоят
из одного слоя крупных железистых клеток (см. рис. 7, 2).
Сходное строение кишечного тракта имеют личинки короедов.
У личинок короеда типографа (Ips typographies L.), например, ки-
шечник внешне отличается от такового белоточечного долгоносика
лишь меньшим количеством и иной формой крипт заднего отдела
средней кишки (см. рис. 7, 3).
Своеобразно строение кишечника личинок златок, способных
развиваться в мертвой сухой древесине. Помимо мышечного же-
лудка они обладают парой слепых отростков, каждый из которых
равен по длине средней кишке или даже превышает ее размеры.
Эти слепые отростки являются бродильными вместилищами, слу-
жащими для расщепления клетчатки с участием симбионтов (см.
рис. 7, 5). Вполне возможно, что некоторые виды этой группы зла-
ток вырабатывают собственную целлюлазу.
Значительную группу среди ксилофагов составляют виды,
личинки которых не используют для питания труднорастворимые
полимеры. Как было сказано, их пищеварительный тракт не имеет
мышечного желудка и бродильных камер, но обладает сильно раз-
витой средней кишкой (см. рис. 7, 6, 7). К этой группе относятся
многие усачи, в том числе лиственничный.
Стации и группировки ксилофагов
Заселение жизнестойких деревьев — привилегия очень огра-
ниченного числа насекомых-ксилофагов. Однако и они в основном
используют для питания и развития личинок ослабленные в той
или иной степени деревья. Большинство ксилофагов, личинки
которых в течение всей жизни или на первых этапах развития пи-
таются лубом, заселяют деревья до его полного отмирания.
К числу ксилофагов, заселяющих предпочтительно мертвые
деревья, обычно относят многих короедов, усачей и златок. При
этом мертвыми деревьями помимо сухостоя считаются оставленные
па лесосеках или лесных складах стволы деревьев весенней рубки,
л также свежие ветровальные и буреломные деревья. Между тем
у таких деревьев к моменту лёта большинства видов ксило-
фагов и откладки ими яиц (май — июнь) клетки лубяной и дре-
весной паренхимы сохраняют живой протопласт. В них наблю-
даются все признаки активного метаболизма с заметным увеличе-
нием образования защитных веществ. Вообще паренхимные клетки
древесины срубленного дерева остаются живыми довольно дли-
26
тельное время, большей частью до полного высыхания древесины,
что наступает далеко не скоро, особенно если сохраняется кора
[Яценко-Хмелевский, 1961]. Как показали наши наблюдения,
у срубленных в начале августа деревьев лиственницы даурской
камбии продолжает функционировать до июля следующего года;
в развивающемся годичном кольце формируются травматические
смоляные каналы и сопровождающая их раневая паренхима.
Таким образом, не все срубленные деревья можно относить к
категории мертвых. Свежесрубленные деревья могут в течение
какого-то срока обладать даже большей сопротивляемостью к за-
селению ксилофагами, чем сильно ослабленные деревья на кор-
ню. Именно продолжительным сохранением живого луба и за-
болони объясняется тот факт, что некоторые насекомые не посе-
ляются на усохших деревьях, редко поселяются на усыхающих
п в то же время охотно заселяют наряду с ослабленными, но жизне-
способными деревьями свежезаготовленные бревна, свежий вет-
ровал и бурелом. Примером таких вредителей в лиственничниках
Прибайкалья могут служить большой лиственничный короед
(Ips subelongatus Motsch.) и лиственничная златка (Phaenops gut-
tulata Gebl.).
На сухостойных и срубленных деревьях с отмершим лубом в
период до начала активного разрушения древесины грибами, а так-
же в деревянных конструкциях построек, развивается относитель-
но небольшое число видов.
Снижение устойчивости деревьев к повреждению насекомыми-
ксилофагами может быть вызвано разными причинами. Среди них
особое место занимает старение дерева. Эволюция ксилофагов,
развивающихся на усохших, усыхающих и Ослабленных в разной
степени деревьях, в том числе вполне жизнеспособных, была в
основном связана с естественной утратой резистентности пере-
стойными деревьями и их отмиранием. Стихийные факторы,
вызывающие ослабление или гибель деревьев (пожары, ураганы,
наводнения, вспышки размножения первичных вредителей и’др.),
имели второстепенное значение.
Старение дерева, как и любого организма, заключается в
постепенной утрате резистентности к отрицательному воз-
действию среды (естественный отбор благоприятствует наследст-
венным факторам, противодействующим ослабляющему влиянию
па организм, лишь в том случае, если это влияние возникает в ре-
продуктивном возрасте). Извесно, что устойчивость древесных
растений к повреждению насекомыми-фитофагами в целом повы-
шается с их возрастом, достигает максимума в период интенсивно-
го плодоношения, а затем начинает снижаться по мере утраты
репродуктивной способности. Например, старые и наиболее моло-
дые хвойные деревья под действием вредителей отмирают быст-
рее, чем средневозрастные [Ильинский, 1931 ]. В то же время дав-
но установлено, что те виды короедов, усачей и златок, которые
способны развиваться на деревьях разного возраста — от жердня-
27
на до перестойных, при прочих равных условиях нападают прежде
всего на старые деревья (Прозоров, 1929; Старк, 1929, 1931; Иль-
инский, 1931, 1958; Положевцев, 1953; и др.]. '
Перестойные леса с отмирающими деревьями являются исход-
ной стацией насекомых-ксилофагов. Именно здесь опи постоянно
находили и осваивали разнообразные «жизненные ниши», скла-
дывающиеся на разных стадиях физиологического ослабления
деревьев, утраты ими защитных свойств и реакций, усыхания и
последующего распада тканей ствола.
Среди ксилофагов нет видов, свойственных только гарям, вы-
рубкам или древостоям, ослабленным под влиянием токсичных
выбросов промышленных предприятий. Нет видов, развивающихся
только на ветровальных и буреломных деревьях. Освоение ксило-
фагами древостоев, подвергшихся воздействию тех или иных фак-
торов, включая антропогенные, определяется соответствием ха-
рактера и темпа происходящих здесь у деревьев физиолого-био-
химических изменений определенным этапам их ослабления и усы-
хания в перестойных древостоях — постоянных резервациях
подавляющего большинства видов.
Сказанное в определенной мере относится и к тем ксилофагам,
которых обычно включают в состав вредителей молодняка. Прак-
тически все они развиваются па стволах и ветвях естественно усы-
хающих перестойных деревьев, во но размножаются па них в мас-
совых количествах и не являются вредителями. Для развития
короеда пожарищ (Orthotomicus suturalis Gyll.), например, при
условии быстрого усыхапия деревьев необходимо наличие сочно-
го луба (в течение большей части периода развития личинок)
и относительно тонкой коры. На гарях в сосновых и лиственнич-
ных лесах это один из обычных массовых видов, пионер заселения
топкомерных деревьев. В то же время в естественно усыхающих
древостоях он встречается в виде небольших семейных групп, не
относится к числу видов-первопоселспцев п заселяет толстые су-
чья и обнаженные корневые лапы, сохранившие свежий луб.
В целом же ксилофаги, освоившие молодпяки хвойных пород,
отличаются постоянством видов (особенно долгоносики Pissodes
и Magdalis), что несомненно объясняется наличием естественного
отпада молодых деревьев в процессе формирования древостоев.
В каждом типе нарушенного леса большее или меньшее число
видов ксилофагов находит благоприятные условия для развития.
При этом, вследствие одновременной концентрации значитель-
ного количества особей этих видов и временного выхода их из-
под контроля ограничивающих численность факторов (обилие
субстрата для питания и развития, слабая межвидовая конкурен-
ция, низкая активность энтомофагов), плотность семейных групп
популяций отдельных видов может достигать здесь очень высоких
пределов.
Существует обратная зависимость между численностью видов,
освоивших тот или иной тип нарушенного древостоя, и плотностью
28
особей массовых видов на первых этапах его заселения. Наиболь-
шее количество видов ксилофагов при самой низкой численности
особей в семейных группах популяций встречается на ослаблен-
ных и усыхающих в результате физиологического старения де-
ревьях. Меньше всего видов заселяет неокоренные бревна на лес-
ных складах, но численность особей некоторых из них бывает
здесь столь высокой, что они в течение двух сезонов могут привес-
ти любое количество заготовленного леса в состояние полной не-
пригодности для какого-либо использования.
При обследование нарушенных и девственных хвойных лесов
в зоне строительства Байкало-Амурской магистрали наибольшее
количество видов ксилофагов зарегистрировано на естественно
отмирающих, а также на одиночных ветровальных и буреломных
деревьях (66% общего состава). В то же время плотность поселе-
ния их значительно уступала таковой видов, заселявших вырубки
и гари [Бережных, 1978]. В процессе сбора материала для мо-
нографии «Вредители лиственницы сибирской» [Рожков, Белая,
1966] и в результате последующих исследований, в Прибайкалье
выявлено 92 вида ксилофагов (включая лубо- и древесиноядных
чешуекрылых), развивающихся на этой породе. Из них 83 посто-
янно встречаются на ослабленных, естественно усыхающих де-
ревьях. На яеокоренных лиственничных бревнах, хранившихся
на лесных складах в течение одного сезона, обнаружено только
7 видов
На основании этих исследований, а также других работ, вы-
полненных нашей лабораторией [Рожков и др., 1975; Анисимова,
Соков, 1975], построен график соотношения количества видов
ксилофагов, развивающихся па лиственнице (абсолютные величи-
ны), и плотности популяции
массовых видов (относительные
величины) в разных стациях
Прибайкалья (рис. 8). При по-
строении графика учитывались
все виды ксилофагов, зарегист-
рированные в рассматриваемых
стациях, в том числе те, которые
Рис. 8. Видовое разнообразие и от-
носительная плотность популяций
массовых видов ксилофагов, разви-
вающихся на лиственнице в При-
байкалье.
А — разные стации в целом; В — естест-
венно усыхающие перестойные деревья;
В — гари; Г — древостои, 4ослабленные
и усыхающие под воздействием фтора,
выбрасываемого в атмосферу алюминиевы-
ми заводами; Д —древостои, ослабленные
и усыхающие в результате временного
затопления о заилением почвы; Е — не-
окоренные лесоматериалы на лесных
29
являются для них факультативными, или заселение ими деревь-
ев связано с проявлением каких-то других повреждающих
факторов.
Во всех стациях нарушенного леса, где происходит быстрое
усыхание необратимо ослабленных деревьев, количество харак-
терных видов невелико. Например, в лиственничниках Прибай-
калья, ослабленных и усыхающих под воздействием фтора, выб-
расываемого в атмосферу алюминиевыми заводами, найдено 18 ви-
дов ксилофагов. Массовыми из них можно считать лишь трех:
большого лиственничного короеда (Ips subelongatus Motsch..)
малого лиственничного короеда (Orthotomicus laricis F) и блестя-
щегрудого елового усача (Tetropium castaneum L.). Еще 4 вида
встречаются часто, а остальные относятся к редким и очень редким
Обитание здесь некоторых видов связано не с загазованностью,
а с наличием большого количества перестойных деревьев. В част-
ности, таким видом является лиственничный усач.
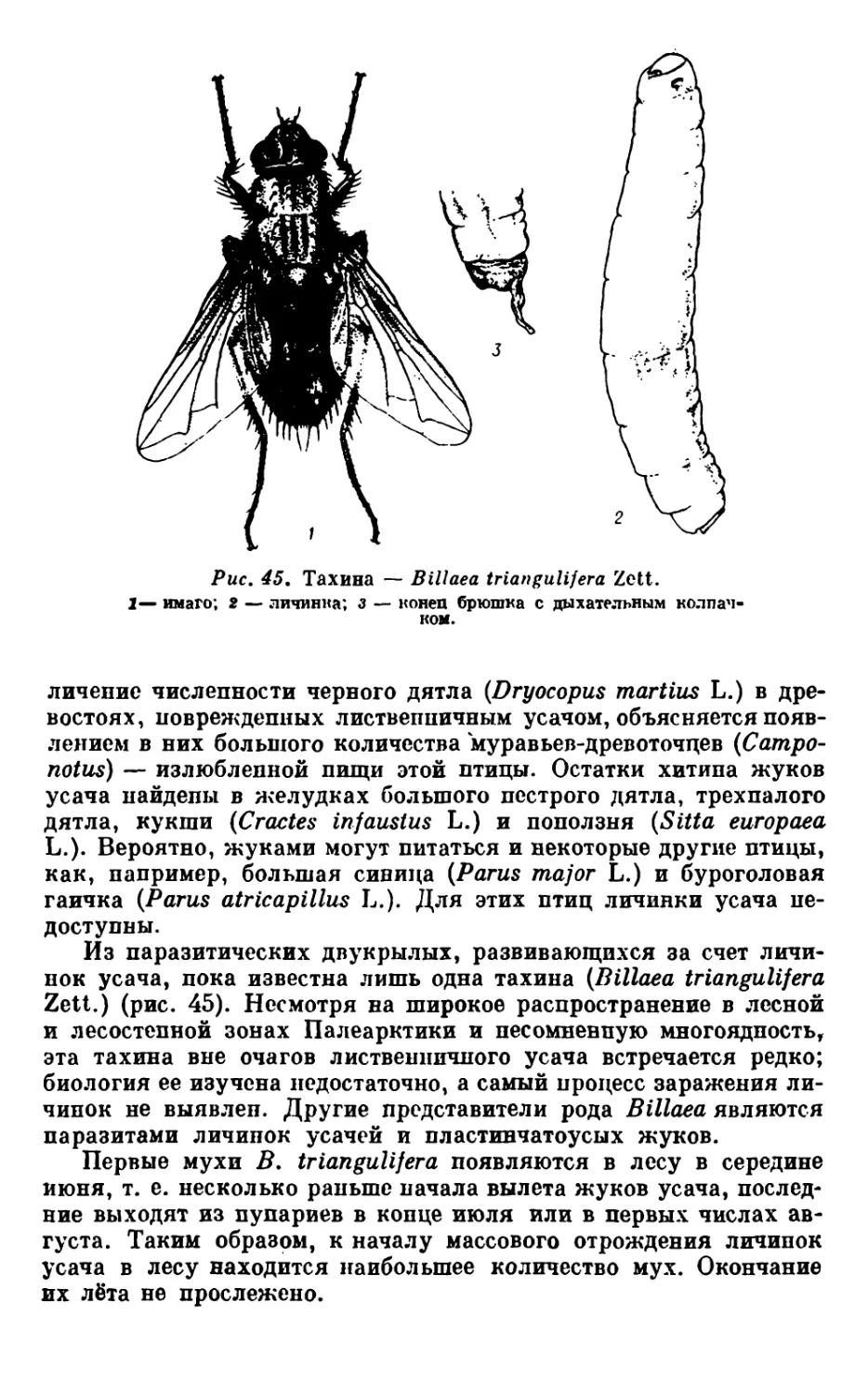
Сведения о ксилофагах гарей любезно представлены нам
Т. А. Агафоновой, много лет занимавшейся их изучением в При-
байкалье. Всего на гарях разной давности возникновения в лист-
венничных лесах этого региона ею зарегистрировано 38 видов .Из них
более или менее постоянно встречается 13, а массовыми во всех
типах пройденных огнем лиственничников являются следующие
7 видов: большой лиственничный короед (Ips subelongatus Motsch.),
короед пожарищ (Orthotomicus suturalis Gyll.), лиственничная злат-
ка (Phaenops guttulata Gebl.), златка пожарищ (Melan ophila acu-
minata Deg.), большой и малый черные хвойные усачи (Monochamu?
urussoii Fisch., М. sutor L.) и сибирский серый длинноусый усач
(Acanthocinus carinulatus Gebl.). Из 25 «необязательных» видов
многие появляются на гарях лишь в послепожарный период диф-
ференциации деревьев па усохшие, усыхающие и восстанавливаю-
щиеся. Усыхание части деревьев протекает медленно и по своему
характеру мало отличается от естественного отмирания старых
деревьев. Возможно, этим объясняется относительно большое ви-
довое разнообразие ксилофагов (при низкой численности особей)
на третий, четвертый и последующие годы после пожара.
Экологическую дифференциацию ксилофагов на виды и группы
видов, последовательно заселяющие деревья в процессе их ос-
лабления и отмирания, в ее полном исходном виде можно наблю-
дать, вероятно, только в естественно усыхающих древостоях.
Различные быстродействующие природные и антропогенные фак-
торы вызывают неодинаковый характер начального ослабления
деревьев и соответственно, определяют разный состав исходных,
производных и завершающих группировок ксилофагов. Даже в
тех случаях, когда во время массового размножения наиболее
агрессивные виды нападают на вполне жизнеспособные деревья,
последующее заселение их другими ксилофагами носит специфи-
ческий характер (см. разд. «Насекомые, сопутствующие листвен-
ничному усачу»).
30
Тем не менее существует определенная группа видов, которые
более Других проявляют себя в качестве первопоселенцев на нез-
начительно ослабленных деревьях. Они не во всех случаях доми-
нируют, но всегда отражают степень и тип ослабления дерева
(комлевый, вершинный, общий).
Л. С. Исаев (1975) сформулировал представление о «скользя-
щей» (ступенчатой) устойчивости дерева к насекомым-ксилофагам
как динамическом процессе его взаимодействия с группировками
последних на разных этапах ослабления и отмирания. Это пред-
ставление очень важно для понимания самих механизмов взаимо-
действия в системе «дерево — ксилофаг».
Имеющиеся у нас данные о динамике резистентности хвойных
деревьевм подвергшихся нападению ксилофагов па разном уровне
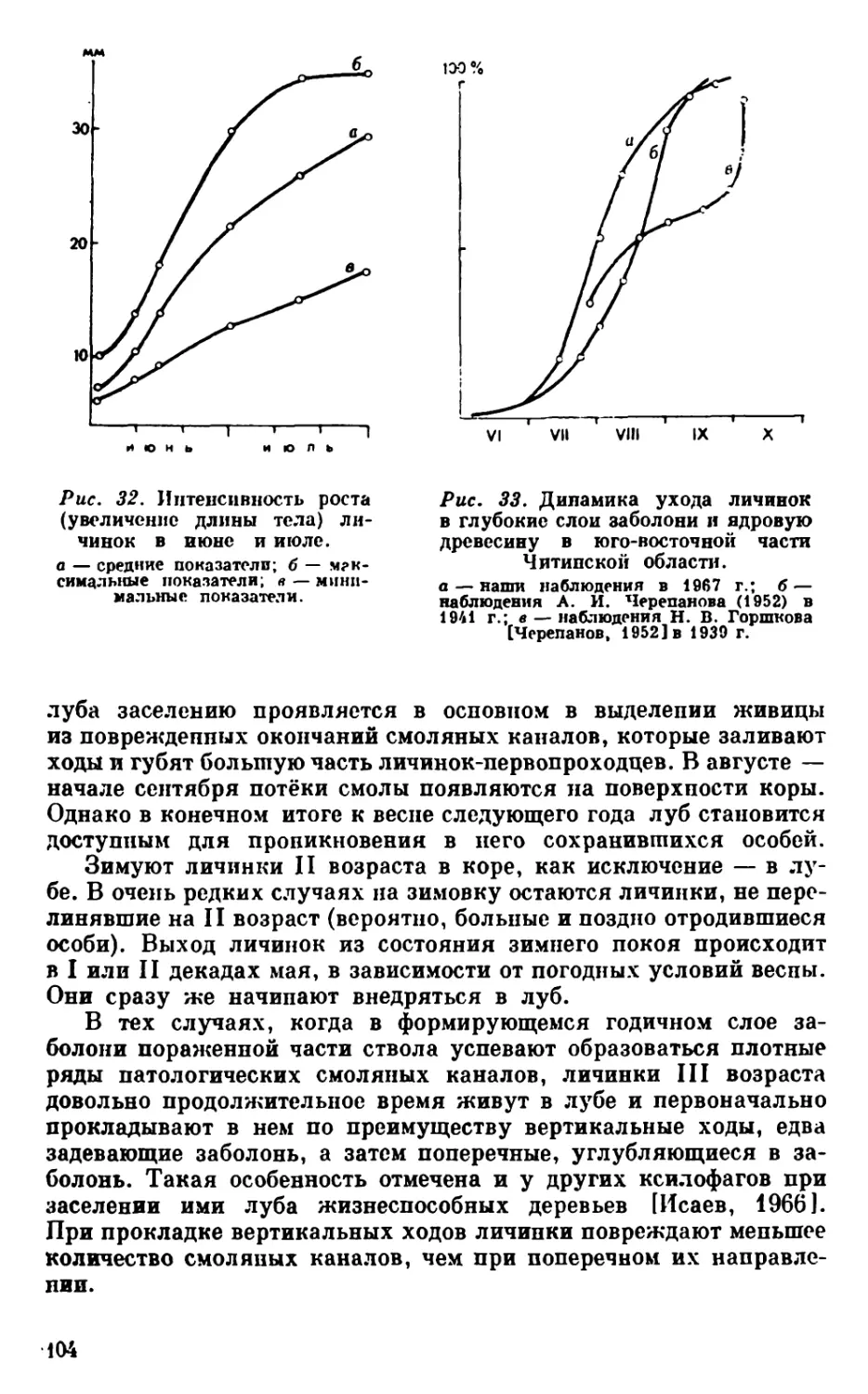
Рис, 9. Сопротивляемость и заселение дерева ксилофагами
на разных этапах его ослабления и отмирания.
П. С.— повышенная сопротивляемость дерева (защитная реакция);
Н. С. — нормальное физиологическое состояние; О. Ж.— ослаблен-
ное, но вполне жизнеспособное; К. С.— критическое состояние; Н. О.—
необратимо ослабленное; П. М. — прекращение метаболических про-
цессов (мертвое дерево); Д. П.— начало деструктивных процессов.
1а — попытки заселения дерева; I — заселение ксилофагами, разви-
вающимися на ослабленных, но вполне жизнеспособных деревьях;
II — заселение ксилофагами, развивающимися на отмирающих де-
ревьях; III — заселение ксилофагами, развивающимися иа свежеот-
мерших деревьях; IV — заселение ксилосапрофагами и ксиломицето-
фагами.
31
физиологического состояния, позволяют графически изобразить
следующие типы изменения сопротивляемости дерева, вплоть до
полной ее утраты, и соответствующую смену экологических груп-
пировок насекомых (рис. 9).
Представленная схема является принципиальной. Тем не менее
характер хода кривых и их соотношение не должны вызывать сом-
нения. Для определения степени устойчивости деревьев к по-
вреждению использованы биохимические показатели^ главным об-
разом изменение состава и соотношения компонентов смолистых
веществ. Приняты во внимание также величины электропровод-
ности луба па переменном токе низкой частоты.
Анализ полученных данных показывает^ что у здоровых де-
ревьев (с нормальными физиолого-биохимическими показателя-
ми) попытки поселения ксилофагов вызывают сильную защитную
реакцию, резко повышающую их резистентность. Преодоление
сопротивляемости таких деревьев доступно очень немногим видам
в периоды массового размножения. Ослабленные, но вполне жиз-
неспособные деревья также сильно реагируют на попытки пре-
одолеть смоляной барьер луба. При этом устойчивость их может
достигнуть состояния здоровых деревьев, затем обычно спадает
ниже исходного уровня (послестрессовая депрессия) и лишь
после этого возвращается к последнему. Деревья той же катего-
рии ослабления, но не оказавшие достаточного сопротивления
ксилофагам, довольно быстро утрачивают защитные свойства.
Массовое заселение их начинается со времени перехода в кри-
тическое состояние, т. е. при снижении общего содержания смол в
лубе (у лиственницы примерно вдвое), сокращении доли участия в
них нейтральных веществ, а в последних — токсичных монотер-
пенов.
Хвойные породы сохраняют некоторую способность реагиро-
вать на повреждение, даже находясь на грани перехода в состоя-
ние необратимого ослабления. Такая реакция уже не препятству-
ет заселению деревьев ксилофагами, в том числе и теми из них,
которые в других условиях входят в состав производных группи-
ровок (см. рис. 9).
СПЕЦИАЛЬНАЯ ЧАСТЬ
В системе «дерево — фитофаг» («хозяин — паразит») осново-
полагающим является их взаимная адаптация. Если бы нам удалось
познать защитные механизмы дерева и механизмы преодоления
фитофагом сопротивляемости последнего, мы бы наполовину ре-
шили проблему создания жизнестойких древостоев.
ЛИСТВЕННИЦА И ЛИСТВЕННИЧНЫЙ УСАЧ
ЛИСТВЕННИЦА (LARIX MILL.)
Хвойные представляют собой наиболее древнюю группу се-
менных растений и, вместе с тем, являются самым обширным клас-
сом среди современных голосеменных. Первые древнейшие хвой-
ные появились свыше 350 млн. лет назад в карбоне, в юре они
достигли наибольшего расцвета. Еще до появления цветковых
растений (в отложениях поздней юры) хвойные были представле-
ны некоторыми из современных родов. Лиственница получила
широкое распространение лишь в неогене, т. е. позднее темно-
хвойных пород.
Лиственница — вершина эволюции хвойных. Ни одно древес-
ное растение не обладает столь высокой приспособленностью к
экстремальным условиям среды. Именно лиственница образует
северную границу распространения древесных растений и в боль-
шинстве случаев ограничивает вертикальное распространение
лесов в горах умеренной зоны северного полушария. В. Ф. Овсян-
ников (1930) писал: «... мы являемся современниками наиболь-
шего расцвета травянистой растительности. За ушедшими в бес-
конечную даль папоротникообразными, оставившими нам в нас-
ледство каменный уголь, нефть и жалких потомков своих, также
выродившихся в травянистые формы, в первую очередь после-
дуют хвойные породы, ...уже приговором истории развития при-
сужденные к удалению с лица Земли...» (с. 4). Это справедливое в
целом заключение нельзя отнести к лиственнице. Фактические
данные о распространении лиственничных * лесов во все периоды
3 А. С. Рожков
33
кайнозойской эры свидетельствуют о том, что к началу широкого
использования этой хвойной породы человеком занимаемая ею тер-
ритория в умеренных областях Европы, Азии и Северной Америки
нисколько не сократилась. Только интенсивная вырубка листвен-
ницы стала причиной сокращения ее исконных территорий и да-
же привела к грани исчезновения некоторых видов в условиях
естественного произрастания (L. principle — rupprechtii Mayr,
L. leptolepis Gord.). Лиственница лучше других хвойных выдер-
живает атмосферу современных городов и вообще более устойчива
к токсичным выбросам промышленности.
О современном расцвете рода Larix свидетельствует помимо
широкого распространения большое количество представленных
в нем гибридных форм и в разной степени выраженных экотипов.
До настоящего времени пет единого мнения о количестве видов
этого рода. Их признается от 10 до 18 [Сукачев, 1924; Ostenfeld,
Larsen, 1930; Gaussen, 1966; Бобров, 1972]. Кроме того, описано
около десятка видов [Сукачев, 1931; Колесников, 1946; Дылис,
1947], большинство из которых теперь считают гибридами.
Е. Г. Бобров (1972) на основе специального изучения коллек-
ционного материала в гербариях и критического анализа наког
пленных в литературе данных по истории и систематике рода
Larix подтвердил видовую самостоятельность 15 лиственниц. Из
них пять распространены в СССР: L. sibtrtca Ledeb., L. gmelinii
Rupr., £. cajanderi Mayr., L. olgensis A. Henry, L. kamtschatica
Rupr. Одновременно он выделил пять групп гибридных ли-
ственниц, сложившихся в процессе интрогрессивной гибридиза-
ции в связи с событиями плейстоцена и новейшего времени. Очер-
ченные автором виды, по-видимому, в наибольшей степени соот-
ветствуют их действительному числу.
Распространение лиственниц,
повреждаемых лиственничным усачом.
Характеристика лесов
На территории Северной Азии наибольшие площади занимают
два вида лиственниц — сибирская (L. sibirica Ledeb.) и даурская,
или Гмелина (L. gmelinii Rupr.). Оба вида являются господст-
вующими в пределах распространения лиственничного усача.
На востоке своего ареала этот вредитель развивается, кроме того,
на камчатской (L. kamtschatica Rupr.) и ольгинской (L. olgensis
A. Henry) лиственницах.
Общая площадь лиственничных лесов СССР определяется в
274,3 млн. га, что составляет 24,6% от лесопокрытой площади,
участие лиственницы в составе лесов закономерно увеличивается
по мере продвижения с запада на восток. Так, в Архангельской
области на долю древостоев с преобладанием лиственницы, при-
ходится 0,4% лесопокрытой площади, в Тюменской области —
9,6, в Красноярском крае — 48,2, а в Якутской АССР — 88,1 %.
34
Общий запас древесины в лиственничных лесах составляет
28,4 млрд, м3, причем большая часть его приходится на спелые и
перестойные древостои [Тихомиров, Фалалеев, 1962].
Сибирская лиственница занимает 14% всей площади листвен-
ничных лесов, имеющих государственное значение. По данным
Е. Г. Боброва (1972), этот вид в европейской части распространен
только па северо-востоке. На Кольском полуострове известно лишь
остаточное местонахождение лиственницы, но уже в Двинско-
Печерском районе она расселена по всей его территории, а в Верх-
не-Волжском районе отсутствует только в крайней юго-западной
части. По Уральскому хребту сибирская лиственница встречается
от Полярного Урала (68° с., ш.) до крайнего юга, включая вер-
ховья рек Белой и Урала. В Западной Сибири вид расселен во
всех флористических районах. Южпая граница его распростра-
нения проходит западнее Тобола, пересекает Обь-Иртышский во-
дораздел (53° с. ш.) и, очерчивая Алтай, спускается к Нарымским
горам, а затем уходит па северо-восток за государственную гра-
ницу. Северная граница ареала сибирской лиственницы проходит
от Полярного Урала и низовьев Оби к низовьям Енисея.. Отсюда
начинается восточная граница вида, пересекающая низовья Ниж-
ней Тунгуски и по долине Подкаменной Тунгуски спускающаяся
к верховьям Лены и центральной части Байкала. На юго-западе
Забайкалья граница распространения этой лиственницы охваты-
вает правобережную часть Селенгинской лесостепи и уходит в
Монголию. Здесь сибирская лиственница поселяется на северных
склонах гор, включая Гобийский Алтай.
Даурская лиственница вместе с лиственницей Каяндера и даль-
невосточными видами занимают около 86% всей площади листвен-
ничных лесов на территории СССР. Из них на долю даурской
лиственницы (если принять за основу схейу распространения восточ-
носибирских и дальневосточных видов, составленную Е. Г. Боб-
ровым) приходится примерно 54%. В эту цифру входит терри-
тория, занятая гибридными формами даурской и камчатской
лиственниц.
Леса из даурской лиственницы распространены па большей
части Восточной Сибири — от описанной границы сибирской ли-
ственницы до низовий Лены (72° с. ш.) и побережья Охотского
моря. Между границами ареалов сибирской п даурской листвен-
ниц проходит относительно широкая полоса их гибридных форм
(L. х czekanovskii Szfer.). Южная граница даурской лиственницы
расположена вне СССР.
Камчатская лиственница распространена на южных Куриль-
ских островах и па Сахалине (кроме его севера). Значительно
большую территорию практически всю южную часть Хаба-
ровского края от р. Уды — занимают гибридные формы камчат-
ской и даурской лиственниц.
Распространение ольгинской лиственницы в советском При-
морье ограничивается небольшой прибрежной территорией и вос-
3*
35
точными предгорьями Сихотэ-Алиня к югу от 45 с. ш. [Бобров,
1972]. Здесь популяция лиственничного усача^ по-видимому, не
выходит из депрессивно-экспрессивного состояния.
Большой удельный вес в лесном фонде имеют лиственничники
горных районов юга Сибири (зона вспышек размножения лист-
венничного усача). В Алтайском крае, например, лиственничные
древостои занимают 18% лесопокрытой площади, в Тувинской
АССР — 54,6, а в Аргуньско-Шилкинских горных лесах — 91 %
[Тихомиров, Фалалеев, 1962; Поздняков, 1975].
На Среднем Урале (западная граница распространения ли-
ственничного усача) чистые лиственничники встречаются отно-
сительно редко. Преобладают смешанные сосново-лиственничные
древостои. Н. А. Коновалов (1968) дает описание 16 типов и групп
типов древостоев, где участие лиственницы в составе от 0,2 и вы-
ше. Наиболее распространенными среди них являются листвен-
ничники и лиственнично-сосняки разнотравные. Они приурочены
к богатым, слабо влажповатым или несколько повышенно увлаж-
ненным почвам па пологих склонах с хорошим дренажом. К числу
обычных типов автор относит также сосняки-брусничники с ли-
ственницей (0,2—0,3 состава), лиственничники бруснично-тра-
вяные, аконитовые и по ручейные.
Для среднетаежной зоны в пределах распространения лист-
венничного усача характерны лиственничники брусничные, раз-
нотравные, лишайниково-толокнянковые, багульниково- и го-
лубично-моховые и сфагновые. Встречаемость усача здесь всегда
большая в древостоях, произрастающих на почвах с повышенным
увлажнением. В высокогорные (подгольцовые) редколесья этой
части ареала он не поднимается. Для района наибольшей вредо-
носности усача, охватывающего горные системы Восточного За-
байкалья, характерно широкое распространение багульниковых,
багульниково-брусничных и сфагновых лиственничников.
В зоне горной лесостепи Тувы, Южного Забайкалья и Монго-
лии господствуют разнотравные типы лиственничников, нередки
парковые леса, разделенные полянами и остепненными участ-
ками.
В Приморском крае наиболее типичными являются заболочен-
ные лиственничники, или мари. В северных частях края (реки
Бикин и Иман) и в бассейне р. Хора в Хабаровском крае они встре-
чаются по широким долинам рек. Аналогичными им на юге
являются так называемые «горные мари» из ольгинской лиственни-
цы, занимающие высокие базальтовые плато на водоразделах рек.
Лиственницы на марях низкорослые, угнетенные, часто с усы-
хающими вершинами. Только по периферии марей деревья при-
обретают нормальную высоту и имеют сравнительно здоровый вид.
Пологие склоны с избыточно увлажненными почвами занимают
парковые лиственничные леса, а сухие склоны — вторичные ли-
ственничники, возникшие после пожара на месте елово-пихтовой
тайги [Куренцов, 19501.
36
Восточным пределом распространения лиственничного усача
является о. Сахалин. Северную часть острова, примерно до широ-
ты 51°30', занимают почти чистые лиственничные леса, местами
представленные редколесьем с подлеском из кедрового стланика,
березы Миддендорфа или багульника. Среднюю часть Сахалина
относят к подзоне темнохвойных лесов-зеленомошпиков с преоб-
ладанием ели. Лиственничники встречаются здесь в низинах на
заболоченных почвах. В бассейне р. Тымь распространены ли-
ственнично-еловые леса, произрастающие и по склонам. Они
занимают довольно большие площади [Криволуцкая, 1958].
В этой части острова наблюдаются вспышки массового размнож-
ния лиственничного усача. На юге Сахалина лиственница не обра-
зует обособленных древостоев.
По всему ареалу лиственничного усача преобладают спелые и
перестойные древостои. Последние являются основной стацией
вредителя в периоды низкой плотности его популяций. По данным
Л. К. Позднякова (1975), перестойные лиственничные древостои
составляют в Иркутской области 53%, в Бурятской АССР — 40,
в Читинской области — 30, в Амурской области — 39, в Хаба-
ровском крае — 34, в Сахалинской области — 54% от общего
запаса древесины в имеющихся на их территории листвен-
ничниках.
Морфология и анатомия сибирской и даурской лиственниц
Сибирская и даурская лиственницы — крупные долговечные
деревья. В благоприятных условиях произрастания достигают в
высоту 40 м при диаметре ствола около 1 м. В Восточной Сибири
лиственницы в возрасте 600—700 лет, не являются редкостью
(рис. 10). Вообще же продолжительность жизни отдельных эк-
земпляров этих видов лиственниц может превышать 900 лет,
а диаметр ствола некоторых из них достигает 1,3 м на высоте
1,5 м от поверхности почвы.
Громадный ареал естественного распространения лиственницы,
простирающийся от Белого моря до побережья Тихого океана и
от сухих степей и полупустынь на юге Сибири до арктической тунд-
ры на севере, обусловливает большое разнообразие лиственнич-
ных лесов. В пределах очерченной территории климатические и
почвенно-грунтовые условия, а также устройство поверхности
чрезвычайно неоднородны, что в свою очередь накладывает отпе-
чаток па формирующиеся здесь лиственничные древостои [Тихо-
миров, Фалалеев, 1962]. Высота и форма деревьев в разных ус-
ловиях произрастания столь различны, что они внешне с трудом
укладываются в рамки вида (рис. И).
Кроны молодых лиственниц в нормальных условиях имеют
коническую или пирамидально-яйцевидную форму, а позже ста-
новятся продолговато-яйцевидными или цилиндрическими. В не-
благоприятных почвенно-климатических условиях у границы
37
Рис. 10. Гигантские лиственницы на о. Ольхоне (оз. Байкал).
распространения древесной растительности лиственницы приобре-
тают вид кустов либо небольших деревец с флаговидными или
плоско-округлыми кронами. Последняя форма крон может возни-
кать также под воздействием других повреждающих факторов.
Например, на остепненных участках юго-западного побережья
оз. Байкал встречаются группы низкорослых лиственниц с иск-
ривленными стволами и широкими плоскими кронами. Такая
форма деревьев является следствием, с одной стороны, частых
ветров, с другой — сильным из года в год повторяющимся пов-
реждением почек лиственничной почковой галлицей (Dasyneura
laricis F. Lw.).
Для большинства видов лиственниц характерно значительное
утолщение комлевой части ствола. Стволы деревьев в сомкнутых
насаждениях ровные, относительно малосбежистые, тогда как в
изреженных древостоях и тем более, у одиночно произрастающих
деревьев утолщение комля бывает большим. По данным Л. К. По-
зднякова и Л. А. Марцинковского, средний коэффициент сбежи-
стости (отношение диаметра пневой части ствола к диаметру его
на высоте 1,3 м) при ступенях толщины ствола от 12 до 44 см
составляет для даурской лиственницы в Якутии 1,22—1,47, для
сибирской лиственницы в Хакасии — 1,26—1,36 [Поздняков,
1975]. У отдельных деревьев сбежистость ствола может быть
много большей. На Камчатке встречаются лиственницы с бутыл-
кообразной формой ствола. Биологическое значение столь высо-
кой сбежистости не совсем ясно. Л. К. Поздняков предполагает,
что утолщение комлевой части способствует повышению устойчи-
вости деревьев, имеющих поверхностную корневую систему,
особенно тех, которые выросли в свободном стоянии и имеют мощ-
ную развитую крону.
Сибирская и даурская лиственницы относятся к деревьям
с поверхностной или почти поверхностной корневой системой.
Особенно это проявляется в зоне распространения многолетней
мерзлоты. Однако и в южных частях ареала глубина залегания
толстых корней у них редко превышает 0,4—0,5 м. Обычно эти
корни стелются у самой поверхности почвы, а ветви третичного
порядка и более тонкие корпи, в том числе придаточные, прони-
кают на глубину до 1 м.
Наиболее характерными признаками лиственницы является
мягкая, опадающая на зиму хвоя, толстая растрескивающаяся
кора с розовой или сиренево-розовой окраской участков свежего
отслоения и твердая тяжелая древесина. Мягкость хвои обуслов-
ливается особенностями строения ее эпидермиса и гиподермы,
цвет коры — присутствием флавоноидных пигментов цианидина
и кверцетина в оболочках клеток, а твердость древесины — силь-
ным развитием поздних трахеид в годичных кольцах и большим
содержанием уплотняющих компонентов — углеводов и смол.
Лиственница цветет рано, когда хвоя только начинает отрас-
тать. Кроющие чешуи женских шишек в это время нежно-розовые
39
Рис. 11. Форма деревьев в разных условиях произрастания.
I — опушка лиственничного леса, 2 — одиночно растущая лиственница с шаровидной
кроной, з — низкорослые лиственницы с плоскоокруглой кроной, 4 — низкорослые
лиственницы с флаговидной кроной.
или зеленовато-желтые с выступающим срединным килем. Они
располагаются спирально на центральной оси и несут в пазухах
семенные чешуи (чешуевидные мегастробилы) с двумя семяпочка-
ми — мегаспорангиями. Молодые шишки с развитыми семенными
чешуями имеют зеленоватую, розово-охристую или красную
окраску, зрелые становятся коричневыми.
Шишки относительно мелкие. Размер их у разных видов и
внутри каждого вида колеблется в значительных пределах. Даже
па одном дереве в урожае одного года максимальная и минималь-
ная длина шишек может различаться в полтора раза и больше
1Дылис, 1961]. Длина шишек сибирской и даурской лиственниц
варьирует от 10 до 40 мм. Семена мелкие (3—4,5 мм), светло-ох-
ристые или коричневатые с длинным крылом.
Микростробилы (мужские «соцветия») цилиндрические, дли-
ной 5—10 мм, состоят из стержня, па котором спирально располо-
жены сильно редуцированные микроспорофиллы с парой микро-
спорангиев (пыльпиков). Микроспорангии до раскрытия желтые,
в вершинной части оранжевые, а после раскрытия становятся
розовыми. Пыльца по имеет летательных воздушных пузырьков.
Хвоя ярко-зеленая с сизоватыми рядами устьиц, заметными
снизу, тупо заострена па конце, в сечении округлая, ромбическая
или треугольная. У сибирской лиственницы хвоя в сечении про-
долговато-округлая, срединный валик с нижней стороны выдается
слабо либо отсутствует. Длина ее 30—50 мм. Хвоя даурской лист-
венницы несколько короче. На удлиненных побегах хвоя распола-
гается поодиночке спирально, на укороченных (брахибластах)—
пучками по 20—40 в каждом.
Наличие брахибластов, способных многократно (у некоторых
видов до 40 лет) возобновлять хвою, является общеродовой чер-
той лиственниц. В первые годы брахибласты имеют полушаровид-
ную форму, затем удлиняются и иногда достигают 20 мм, сохра-
няя диаметр трехлетнего возраста. Брахибласты могут образовы-
вать вторичную хвою в год повреждения. Гипертрофированная
(компенсационная) вторичная хвоя, образующаяся на брахиблас-
тах при дефолиации деревьев насекомыми, а также при сильном
ожоге кроны, достигает в длину НО мм. Спящие почки в основа-
нии толстых ветвей при повреждении дерева формируют в боль-
шинстве случаев брахибласты с пучками гипертрофированной
хвои.
Клетки эпидермиса хвои лиственниц более тонкостенные по*
сравнению с другими представителями Р inaceае, кутикула очень
тонкая; гиподерма представлена одним слоем клеток в виде не-
больших прерывистых участков (кроме ольинской лиственницы,
у которой слой гиподзрмальных клеток сплошной). Ассимиля-
ционная паренхима (мезофилл) состоит из округлых, угловатых и
звездчатых клеток с хорошо развитыми межклетниками. Смоля-
ных каналов всегда два, их эпителиальные клетки толстостенные,
сопровождающие клетки имеются только, со стороны мезофилла.
41
Центральная проводящая система хвои содержит два проводящих
пучка. Эти пучки очень сближены между собой, и разделяющий
их слой склеренхимных волокон представлен всего одним рядом
клеток (у гипертрофированной хвои может быть три ряда таких
клеток). Л. И. Малышев (1958) называет межпучковую ткань
«межпучковой паренхимой» и считает, что она является гомологом
многорядного сильно разросшегося сердцевинного луча, кото-
рый разделил собою на две ветви первоначально единый проводя-
щий пучок, имеющий место в верхней части укороченного побега.
Околопучковая склеренхима состоит из небольшого числа клеток
с простыми порами. Трансфузионная паренхима включает мерт-
вые клетки с утолщенными оболочками и чередующиеся с ними
живые клетки крахмалоносной паренхимы.
Кора листвепницы составляет значительную часть объема ство-
ла. Наибольшей толщины она достигает в комлевой части. По дан-
ным Л. К. Позднякова (1975), у сибирской лиственницы на долю
коры в среднем приходится от 23 до 27% всего объема ствола,
при этом толщина ее на высоте 1,3 м возрастает с 3 см в ступени
диаметра ствола 24 см до 5 см в ступени 40 см. На Камчатке встре-
чаются лиственницы с исключительно толстой корой, достигаю-
щей в нижней части ствола 25 см [Чавчавадзе, Яценко-Хмелев-
ский, 1978].
В молодом возрасте лиственницы имеют гладкую серовато- или
зеяеновато-бурую кору. В дальнейшем она начинает трескаться
и отслаиваться, а в спелом и перестойном возрастах покрывается
продольными трещинами, особенно глубокими в нижней части
ствола. Зона переходной коры начинается относительно высоко,
поэтому некоторые из тех стволовых вредителей, которые у дру-
гих хвойных заселяют только комлевую часть ствола, здесь могут
подниматься до 1/3 его высоты.
Структура внешней (мертвой) коры лиственницы отличается
прежде всего наиболее сильным среди хвойных развитием слоев
тонкостенной пробки и менее выраженными, чем, например, у ели
и кедра, слоями феллоидов. Замечено, что степень развития опроб-
ковевшей (суберинизированпой) ткани зависит от условий произра-
стания дерева, кроме того, образование ее стимулируют повреж-
дения ствола, в том числе «попытки» поселения насекомых-ксило-
фагов. Относительная твердость внешней коры лиственницы
определяется значительным содержанием склеренхимы.
Для луба (флоэмы) лиственницы характерно более или менее
равномерное распределение механических клеток в виде оди-
ночных веретеновидных лубяных волокон. У сосны и кедра в лубе
здоровых деревьев такие волокна отсутствуют, а в старых частях
флоэмы пихты и ели механические элементы представлены укоро-
ченными, асимметричными, иногда ветвящимися склереидами.
Последние собраны группами и неравномерно распределены в
толще луба. У лиственницы механические клетки типа «астро-
склереидов» могут образовываться лишь при повреждении ствола.
42
Паренхимные и ситовидные клетки луба хвойных сходны по
строению. У лиственницы лубяная паренхима начинает раньше
накапливать дубильные вещества и в целом содержит их больше,
чем у сосны, пихты и даже ели. Смол же в крахмалоносных клет-
ках лиственницы меньше, чем у представителей трибы сосновых.
Очень характерным для луба лиственницы помимо лубяных воло-
кон является обилие кристаллоносных клеток. Их тип и особен-
ности распределения могут быть использованы в сравнительных
исследованиях.
Древесина лиственницы обладает рядом отличительных осо-
бенностей строения. По некоторым элементам анатомии она
сходна с древесиной ели (общее меньшее количество продольных
смоляных каналов и увеличение их числа в широких слоях го-
дичного прироста; большая величина многорядных радиальных
лучей и лигнификация их клеток; толстостенпость клеток эпите-
лия смоляных каналов; размер и характер окаймленных пор
между лучевыми и осевыми трахеидами). Тем не менее эти породы
хорошо различаются по степени развития поздней древесины,
по утолщению стенок трахеид, отсутствию зубчатости на стенках
лучевых трахеид и по другим признакам.
Наиболее характерным для лиственницы является узкая
(обычно меньше 20 мм) заболонь, резко отграниченная по окраске
от ядровой древесины. Годичные слои хорошо выделяются благо-
даря различиям в толщине и окраске стенок трахеид поздней и
ранней древесины. Поздняя древесина, как правило, занимает
более четверти ширины годичного слоя.
Трахеиды достигают в длину 1,3—6 мм. Сечение их в ранней
древесине округло-четырехугольное, полость широкая. Поры на
радиальных стенках крупные округлые с широким окаймлением.
В поздней древесине сечение трахеид округлое, что обусловлено
сильным развитием межклетников. У границы годичного слоя они
заметно сдавлены тангентально и имеют щелевидную полость.
Радиальные лучи относительно высокие, однорядные и много-
рядные со смоляным каналом. Основную массу луча составляют
паренхимные клетки, содержащие крахмал п смолы. Фенолы
встречаются в них в небольшом количестве, по быстро накапли-
ваются при поранении ствола. Поры между лучевыми и осевыми
трахеидами мелкие окаймленные с овальными отверстиями. Отвер-
стия окаймленных пор между трахеидами п клетками лучевой
паренхимы щелевидиые.
Продольные смоляные каналы в норме формируются в проме-
жуточной и поздней древесине, патологические каналы образуются
также в ранней древесине. Клетки их эпителия мел кие, относитель-
но быстро склерифицирующиеся, особенно в поздней древесине.
43
Особенности биохимического состава хвои,
коры и древесины лиственницы
Лиственница по содержанию веществ в хвое и их сезонной
динамике отличается от других хвойных и приближается к лист-
венным деревьям умеренных широт. Это обстоятельство вместе с
относительной мягкостью покровных тканей хвои, по-видимому,
и определило переход на лиственницу ряда насекомых с листвен-
ных пород, а также предпочтение ее многими многоядными хло-
рофагами из числа хвое-листогрызущих видов.
Количество воды в хвое сибирской лиственницы в течение веге-
тации (у деревьев разного возраста и в разных условиях их произ-
растания) колеблется от 65 до 80%. Содержание клетчатки неве-
лико, в зависимости от возраста хвои оно составляет 6—10% от
общего количества сухого вещества. Смолистых веществ содер-
жится на 25—45% меньше, чем у сосны, ели и пихты. В то же вре-
мя количество крахмала в хвое лиственницы во второй половине
лета, по данным Б. Н. Вержуцкого с соавторами (1978), в 2—5 раз
превышает уровень его содержания в хвое остальных пород
Сибири.
В табл. 1 приведены сведения о содержании основных резерв-
ных и защитных веществ в хвое лиственницы. Составлена она на
основе данных, взятых из работ Г. Г. Васильевой и А. С. Плеша-
нова (1972, 1975), Б. Н. Вержуцкого с соавторами (1978)j а также
наших исследований [Рожков и др., 1962].
В течение последних двух десятков лет появилось большое
количество публикаций, посвященных изучению химического
состава древесины и коры лиственницы сибирской и даурскцй.
Таблица 1
Биохимический состав хвои лиственницы сибирской
в Прибайкалье, % на сухое вещество
Вещества Содержание в течение вегетации Среднее со- держание по завершении роста хвои
Углеводы:
моносахара 8,4-18,7 9,4
дисахара 1,2—11,1 4,6
крахмал 3,1—9,2 6,3
гемицеллюлоза Азот: 5,8—6,4 6,1
общий 1,5-3,1 2,3
белковый 0,7—2,2 1,3
небелковый 0,2—0,9 1,0
Жиры Фенолы: — 11,6
полифенолы — 1,0
таннины ——* 6,5
Смолы 8,2-14,0 11,9
44
Это объясняется возникшей проблемой комплексного использова-
ния лиственницы в целлюлозно-бумажной, гидролизной и лесохи-
мической промышленности в связи с вовлечением в эксплуатацию
лиственных лесов Сибири и Дальнего Востока.
Кора лиственницы содержит в среднем 81% органических
веществ, немногим более 2% минеральных и 17% влаги. Эти по-
казатели относятся к внутренней части корки, смежной с лубом.
В поверхностных слоях влажность корки колеблется в значитель-
ных пределах в зависимости от условий погоды.
Исследованиями К. И. Анисимовой [Анисимова, 1968; Аниси-
мова и др., 1965] установлено, что в коре лиственницы даурской
из Хабаровского края содержится (от веса сухой коры): целлю-
лозы Кюршнера 19,40%, пентозанов (общих) 6,28, легкогидроли-
зуемых сахаров 8,28, редуцирующих веществ 9,60, лигнина
36,12, метоксильных групп 1,42, полиуроновых кислот 2,12,
веществ, экстрагируемых спирто-бензолом 11,40, экстрагируемых
водой 18,73, золы 2,80%. В составе легкогидролизуемых сахаров
обнаружены: ксилоза 0,76%, арабиноза 5,40, манноза 1,10 и га-
лактоза 1,02. Содержание таннинов варьирует от 9,9 до 15,1%,
в среднем 11,8%. В дубильных экстрактах из коры обнаружены
свободные и связанные фенолокарбоновые кислоты — протока-
техиновая, П-оксибензойная, ванилиновая и сиреневая, а также
катехины — (4-)-катехин, (—)-эпикатехин, три неидентифици-
рованных катехина, кверцетин и цианидин.
Кора разных видов лиственницы довольна близка по составу
п количественному соотношению основных компонентов. Так,
например, в сухой коре лиственницы сибирской из Красноярского
края содержится 20,0—21,0% целлюлозы, 6,2—6,8% пентозанов,
42,1—42,6% лигнина, 11,7—12,1 % веществ, экстрагируемых эфи-
ром, 21,6—22,0 водно-эстрактивных вещеЬтв, в том числе 14,0—
15,0% таннинов [Миронова и др., 1962]; в коре лиственницы
Каяндера из Камчатской области целлюлоза составляет 18,8%,
лигнин — 37,4, таннины — 10,4—19,9% от сухой навески [Ани-
симова, 1968]; в коре лиственницы европейской, по данным
Т. Суэйна (1968), 12% таннинов. В’целом для коры лиственницы
характерно значительное содержание лигнина и водно-экстрак-
тивных веществ.
Следует заметить, что приведенные данные характеризуют
химический состав коры в целом, т. е. внешней коры (ритидома)
п луба вместе взятых. Между тем живой луб биохимически очень
отличается как от внешней коры, так и от древесины. Именно он
представляет собой первую активную линию обороны дерева при
нападении насекомых-ксилофагов.
Во внешней коре лиственницы даурской содержание моносаха-
ров не превышает 2,2%, дисахаров — 2,0; гемицеллюлозы в це-
лом составляют 13,0—14,1% от веса сухих веществ (у поврежден-
ных'насекомыми деревьев содержание их может увеличиваться);
арабиногалактан во внешней коре здоровых деревьев отсутствует.
45
Дубильные вещества внешней коры относятся к классу конденси-
рованных таннинов (флаволаны) и являются продуктами полиме-
ризации катехинов и лейкоантоцианидинов. Содержание смол во
внешней коре колеблется от 2,8 до 7,6% (от веса сухих веществ)
и зависит от физиологического состояния и возраста дерева;
соотношение в смолах внешней коры нейтральных веществ п
кислот 1 : 1,5, в лубе же 1 : 0,6.
Луб представляет собой высокоспециализировапную ткань,
основным назначением которой является проведение органических
веществ из кроны в ствол и корни. Вместе с тем он выполняет
также защитную функцию, особенно ответственную при повреж-
дении дерева.
Влажность луба у здоровых деревьев лиственницы на разной
высоте ствола в течение вегетации относительно постоянна. Обыч-
но опа редко выходит за пределы 55—70% и в среднем составляет
65%. Коллоидно и осмотически связанная (условно свободная)
вода в лубе соответственно составляет 14 и 51%, концентрация
клеточного сока — 14%, его осмотическое давление выражается
такой же величиной бар [Рожков, Габиб-заде, 1978].
Представленные в табл. 2 данные о содержании основных ве-
ществ в лубе лиственницы представляют собой обобщенные ре-
зультаты исследований, выполненных в нашей лаборатории [Ва-
сильева, Плешанов, 1972, 1975; Рожков и др., 1972; Вержуцкий
и др., 1972].
Таблица 2
Биохимический состав луба лиственницы сибирской
в период вегетации в Прибайкалье, % на сухое ве-
щество
Вещества Содержание
пределы колебаний среднее
Углеводы: моносахара 1,2-9,4 5,7
дисахара 1,2-7,3 4,1
крахмал 2,3-19,7 п,з
гемицеллюлоза 8.1-18,5 13,3
Азот: общий 0,3-0,9 0,4
белковый 0,1-0,3 0,2
небелковый 0,2-0,3 0,2
Жиры 1,8-11,1 7,3
Фенолы: полифенолы 1,7-3,2 1,9
тяттикныг 6,0-6,8 6,4
Смолы: всего 5,2-7,4 6,4 2,9
нейтральные 2,7-3,3
Кислоты 1,9-4,5 3,5
46
В сезонной динамике углеводов луба наблюдается следующая
закономерность: содержание моносахаров несколько понижается
во второй половине лета, затем восстанавливается по всей высоте
ствола; дисахара содержатся в наибольшем количестве в комле-
вой части ствола в начале вегетации, а в вершинной и средней его
частях их больше в сентябре; крахмала в лубе по всему стволу
больше во второй половине вегетационного периода в два—четыре
раза, чем в начале лета.
Содержание всех форм азотистых соединений луба в течение
вегетации сокращается и в сентябре составляет лишь 30—50%
от их количества в июне [Васильева, Плешанов, 1975]. Максимум
содержания жиров в лубе (до 10,5%) приходится на зимнее время,
минимум — на апрель. У лиственницы отмечено меньшее колеба-
ние уровня жиров в течение года по сравнению с другими хвой-
ными [Судачкова, 1977].
Эфирные масла хвойных проявляют себя по отношению к
насекомым—дендрофагам как токсины, репелленты и аттрактан-
ты. У лиственницы они изучены лучше других компонентов смол.
Особенно это относится к сибирской лиственнице [Пентегова,
Лисина, 1959; Исаев, Рыжкова, 1968; Рожков и др., 1975; Мас-
сель и др., 1975; Рожков, Степанчук, Массель, 1978] и европей-
ской лиственнице [Sindelar, Skalska, 1965].
В составе эфирных масел лубяных тканей лиственницы преоб-
ладают а-пинеп и Д3-карен; Общее содержание этих соединений в
монотерпеновой фракции здоровых деревьев в июне — августе
составляет 63—74%. Из других монотерпенов большая доля
участия приходится на р-пинен, p-фелландрен и лимонен. Кам-
фен, терпинолен, а-фелландрен, у-терпинен и п-цимол относятся
к минорным компонентам. В сезонном цикле вегетации количество
Д3-карена заметно уменьшается к осени, а наиболее высокое со-
держание а-пинена приходится на июль. Некоторое увеличение
Р-пипена, а-фелландрена и p-феллапдрена наблюдается в конце
вегетации. В целом же количественный состав монотерпенов луба
лиственницы сибирской относительно постоянен (табл. 3) [Рож-
ков и др., 19781.
По высоте ствола лиственницы наблюдается устойчивое уве-
личение общего содержания эфирных масел. Количество Д3-каре-
на увеличивается к вершине дерева, а содержание а-пинепа
уменьшается [Массель и др., 1974].
Исследования углеводородсодержащих фракций экстрактив-
ных смол показали, что основной частью их при фиксации мате-
риала паром являются предельные углеводороды С1о—С14, тогда
как терпеновые соединения дают на хроматограммах минорные
пики. «
Фенольные соединения луба, как и древесины, представлены
в виде моно-, олиго- и полимеров, а именно: оксибензойных и
оксикоричных кислот, кумаринов, флавоноидов, лигнинов и ду-
бильных веществ. У здоровых деревьев содержание флавоноидов
47
Таблица 3
Состав и сезонная динамика монотерпенов в лубе сибирской
лиственницы в Прибайкалье, % от фракции
Состав Июнь Июль Август
а-пинен 24,5±1,2 29,2±1,0 25,3±0,8
камфен 0,4±0,1 0,6±0,1 0,5±0,1
£-пинен 13,9±1,0 14,0±0,8 15,7±0,9
Д3-карен 46,0±2,0 39,0±1,7 37,6±1,7
а-феллацдрен 2,5±0,4 3,8±1,0 4,3±0,8
лимонен 2,7±0,5 2,5±0,4 4,1±0,8
Р-фелландрен 6,5±0,2 7,0±0,6 8,0±0,3
у-терпинен 1,0±0,3 0,8±0,2 0,6±0,2
терпинолен 2,5±0,1 2,0±1,0 2,9±0,2
п-цимол 1,0±0,2 0,9±0,3 0,8±0,2
в лубе в среднем составляет около 2,5% от веса сухого вещества.
Дубильный комплекс луба представлен преимущественно кате-
хинами и конденсированными танпинами. Количество последних
в течение вегетации изменяется: в июле отмечается наибольшее
их содержание, в сентябре — наименьшее. Содержание флобафе-
нов колеблется в пределах 6,0—6,8%. Водно-растворимые поли-
фенолы составляют 6,5% в середине вегетационного периода
и 4,9% в конце вегетации (от веса сухого вещества).
Химии древесины лиственницы посвящено значительное коли-
чество специальных работ и тематических сборников. Многие из
них выполнены с целью определения перспективности использо-
вания отдельных веществ в лесохимической промышленности
(целлюлоза, смолы, полисахариды и др.). Но есть и обстоятельные
исследования биологического направления.
У лиственницы, как уже отмечалось, преобладает ядровая
древесина (до 90%), отличающаяся большим объемным весом
(до 0,833 у свежесрубленных деревьев) и высоким содержанием
уплотняющих веществ. Состав основных химических компонентов
древесины ядра и заболони одинаковый, но содержание их раз-
лично (табл. 4).
Относительно невысокое содержание целлюлозы и лигнина в
ядровой древесине лиственницы компенсируется высоким содер-
жанием арабиногалактана — водно-растворимого полисахарида.
Количество его возрастает по направлению от центра к периферии,
достигая максимума в годичных слоях ядра, граничащих с забо-
лонью [Чочиева, Цветаева, Юрьева и др., 1958].
Функции арабиногалактана до сих пор не ясны и вряд ли
сводятся к защитному действию при повреждении дерева [Антоно-
ва, 1977]. По нашим наблюдениям, эта камедь усваивается ли-
чинками ксилофагов и является для некоторых из них основным
48
Таблица 4
Основной химический состав древесины ядра и заболони даурской лист-
венницы, % на сухое вещество [Цветаева и др., 1958]
Вещества Среднее содержание Вещества Среднее содержание
в ядре | |в заболони в ядре |в заболони
Целлюлоза 35,2 49,2 Манная 7,3 9,4
Лигнин 21,6 25,6 Уроновые кисло- 2,7 2,8
Пентозаны 8,0 6,8 ты
Арабиногалак- 19,5 0,8 Смолы 5,6 3,0
тан Зола 0,5 0,2
питательным веществом в период накопления резервов в жировом
теле.
Общая влажность заболони — кормового субстрата личинок
многих видов насекомых-ксилофагов — варьирует у лиственницы
от 38 до 55%, осмотически связанная (условно свободная) вода
составляет в среднем 34%. Общее содержание азота колеблется в
пределах 0,4—0,8% от сухой древесины. Крахмала в заболони
значительно меньше, чем в лубе,— 0,9—2,4%, в среднем 1,5*
а содержание моносахаридов (галактоза, арабиноза, ксилоза и
глюкоза) варьирует от 5 до 12%.
Из древесины заболони и ядра даурской лиственницы выделе-
ны и идентифицированы четыре флавоноида — дигидрокверцетии*
кверцетин, дигидрокемпферол и кемпферол. Содержание первого
составляет 69%, второго — 11, дигидрокемпферола и кемпферола
вместе — 20% от суммы флавоноидов [Тюкавкипа и др., 1968].
В заболони здоровых деревьев сибирской лиственницы содер-
жание смол, по нашим данным, варьирует от 1,6 до 2,9%, нейт-
ральные вещества составляют в среднем 1,0%, кислоты (жирные
и смоляные)— 1,2%. Смолы образуются в основном в клетках
эпителия смоляных каналов, а при повреждении ствола накапли-
ваются в большом количестве также в преобразованных тра-
хеидах и раневой паренхиме.
Вредители лиственницы и ее устойчивость к повреждению
Как и все древесные породы, лиственницу повреждают насеко-
мые, позвоночные животные, паразитические грибы и бактерии.
Наибольший вред причиняют насекомые и возбудители грибных
заболеваний. Из позвоночных животных большое значение имеют
мышевидные грызуны. Уничтожая опавшие и высеянные семена*
они сильно препятствукйг возобновлению лиственничников.
В Восточной Сибири не обнаруживается различий в видовом
составе и степени вредоносности насекомых, обитающих на сибир-
ской и даурской лиственнице. В Прибайкалье на лиственнице
сибирской нами зарегистрировано 284 вида насекомых. Все они,
4 а. С. Рожков
49
несомненно, встречаются и на даурской лиственнице, по крайней
мере, в западной части Забайкалья.
В зоогеографическом отношении восточносибирская фауна
насекомых-фитофагов лиственницы характеризуется многообра-
зием слагающих ее группировок. Наибольшее место в ней — 133
вида (40,8% общего состава) — занимают транспалеаркты. Евро-
пейско-сибирская группа представлена 71 видом (21,8%), транс-
сибирская — 34 (10,4%), восточносибирская — 57 (17,5%) и го-
ларктическая — 31 видом (9,5%).
По характеру питания фауна насекомых-вредителей листвен-
ницы в основном слагается из олигофагов хвойных (121 вид, или
42,6% состава) и полифагов, развивающихся как за счет хвойных,
так и лиственных древесных пород, кустарников, а в некоторых
случаях й травянистых растений (118 видов, или 41,6% состава).
Монофагов лиственницы зарегистрировано 45 видов (15,8%
состава).
Наиболее вредоносными являются олигофаги хвойных и моно-
фаги лиственницы. Среди первых постоянному надзору подлежат
сибирский шелкопряд (Dendrolimus superans sibtricus Tschetv.),
лунчатый шелкопряд (Selenephera lunigera Esp.), еловая шишко-
вая огневка (Dioryctria abietella F.), большой лиственничный
короед (Ips subelongatus Motsch.), байкальский лесовик, (Dryocoetes
baicalicus Reitt.), короед пожарищ (Orthotomicus suturalis Gyll.)f
лиственничная златка (Phaenops guttulata Gebl.), усачи рода
Monochamus, тонкоусый еловый усач (Tetropium gracilicorne Reitt.),
блестящегрудый усач (Т. castaneum L.), черно-синий рогохвост
(Sirex ermak Sem.) и некоторые другие.
Из числа монофагов лиственницы большое хозяйственное значе-
ние имеют: пяденица Якобсона (Erannis jacobsoni Djak.), листвен-
ничная листовертка (Zeiraphera griseana Hbn), лиственничная
пяденица (Semiothisa continuaria Ev.), алтайский лиственничный
усач (Xylotrechus altaicus Gebl.), заболонник Моравица (Scolytus
morawitzi Sem.), лиственничная муха (Hylemyia laricicola Karl.),
лиственничные галлицы родов Dasyneura и Camptomyia.
Имеются серьезные вредители лиственницы и среди многояд-
ных насекомых, способных размножаться в массе. К ним отно-
сятся, например, непарный шелкопряд (Lymantria dispar L.),
античная волнянка (Orgyia antiqua L.), восточный майский хрущ
(Melolontha hippocastani F.), июньский хрущ (Amphimallon solstir
tialis L.), щелкун блестящий (Selatosomus aeneus L.).
Полное однократное объедание хвои у лиственницы в боль-
шинстве случаев не является пагубным для дерева и даже не сни-
жает его сопротивляемость к заселению стволовыми вредителями.
Обесхвоенные в течение 2—3 лет подряд деревья обычно заселяют-
ся короедами, усачами, златками или рогохвостами, либо гибнут
без вмешательства этих вредителей.
Среди ксилофагов, способных нападать на ослабленные,
но вполне жизнеспособные деревья лиственницы даурской, наи-
50
большее значение имеют лиственничный усач, тонкоусый еловый
усач, большой лиственничный короед, лиственничная златка и
черно-синий рогохвост [Рожков, Бялая, 1966].
Заражение паразитическими грибами ослабленных, усыхаю-
щих и мертвых деревьев лиственницы зависит в основном от тех
же условий, что и заселение их стволовыми вредителями. У лист-
венницы преобладают комлевые (корневые) гнили. Лишь в неко-
торых районах широко распространены стволовые гнили, вызы-
ваемые еловой губкой или стереумом еловым. Распространению
комлевых гнилей лиственницы способствуют низовые пожары,
повреждающие корни дерева, что благоприятствует проникнове-
нию в них грибных инфекций. Существенное значение в жизни
дереворазрушающих грибов имеют также насекомые. Во-первых,
насекомые, повреждая кору, открывают свободный доступ про-
росткам спор к древесине, во-вторых, многие из них являются
пассивными переносчиками спор и даже кусочков мицелия грибов
[Любахский, 1959]. Среди насекомых-ксилофагов есть виды, для
которых развитие грибов в личинковых ходах — необходимое
условие для развития личинок.
Лиственница — наиболее устойчивая порода среди хвойных
как поротношению к дендрофильным насекомым (хвоегрызущим
и стволовым вредителям), так и по отношению к дереворазрушаю-
щим грибам.
Жизнестойкость хвойных деревьев при утрате ассимиляцион-
ного аппарата зависит от их регенерационной способности. Пос-
ледняя определяется частотой естественной смены хвои и соот-
ветственно темпом накопления резервных веществ, необходимых
для ее восстановления. В Восточной Сибири хвоя живет у пихты
6-9 лет, у ели 5—7, у кедровой сосны 4—6, у сосны обыкновен-
ной 3—6 лет, а лиственница сменяет хвою ежегодно.
Все хвойные породы, кроме лиственницы и отчасти сосны*
не выдерживают полной утраты хвои. При анализе литературных
данных о стойкости хвойных пород к засухам, а также к задым-
ленности воздуха — фактору, отражающемуся прежде всего на
деятельности хвои, обнаруживается следующий порядок в распо-
ложении пород: наименее устойчивые пихта и ель, за ними сле-
дует сосна, затем лиственница. В такой же последовательности эти
породы заселяются в ослабленных древостоях стволовыми вредит
телями и дереворазрушающими грибами.
Лиственница занимает особое положение среди хвойных.
Брахибласты ее живут от 2 до 40 лет, ежегодно возобновляя
хвою. Зачатки хвои будущего года развиваются на конусе нара-
стания брахибласта одновременно с хвоей текущего года, и в слу-
чае утраты последней на большей части кроны вторичная хвоя
отрастает в то же лето.
Способность лиственницы ежегодно восстанавливать хвою
обеспечивается накоплением соответствующего запаса ассимиля-
тов в течение лета. По мнению Н. В. Дылиса (1958), способность
быстро развивать ассимиляционный аппарат с минимальной (по
сравнению с другими хвойными породами) 'затратой запасных
веществ сыграла большую роль в эволюции рода Larix и явилась
важным фактором приспособления к суровым условиям среды.
Второй важный фактор устойчивости лиственницы к хвоегры-
зущим насекомым — изменение химического состава хвои, сохра-
нившейся на дереве после объедания насекомыми, а также
вторичной хвои, выросшей в то же лето. Эти изменения приводят
к ухудшению качества хвои как кормового субстрата филлофагов.
Развитие вторичной хвои зависит от времени утраты первич-
ной хвои и от степени повреждения конуса нарастания укорочен-
ных побегов. При гибели на дереве в мае и начале июня 50% и бо-
лее укороченных побегов остальные побеги в дальнейшем произ-
водят вторичную хвою, часто в два раза превышающую длину и
более чем в четыре раза вес нормальной хвои. Вторичная хвоя
содержит на 34% меньше сухого вещества и на 62% меньше
хлорофилла. Особенно велики различия в содержании резервных
веществ в нормальной и вторичной хвое, закончившей рост
(в конце августа). При расчете на сырую массу во вторичной хвое
оказывается углеводов меньше па 44%, в том числе моносахаров
на 61, жиров меньше на 25, белка — па 47% [Рожков, Сендаро-
вич, Данович, 1962; Рожков, 1965].
Увеличение размеров хвои при гибели значительного количе-
ства почек нарастания укороченных побегов является компенса-
ционной реакцией лиственницы, средством восполнения синтези-
рующего аппарата. Наряду с тем недостаток питательных веществ
во вторичной хвое в большей степени отражается на росте и разви-
тии личинок насекомых, особенно в младших возрастах. Именно
низкими питательными качествами вторичной хвои можно объяс-
нить не только предпочтение насекомыми нормальной первичной
хвои, но и сокращение их численности. Например, гусеницы си-
бирского шелкопряда {Dendrolimus superans sibiricus Tschetv.)
I и II возрастов, питаясь только вторичной хвоей лиственницы,
не могут накопить в организме необходимый запас резервных ве-
ществ к началу заморозков и вследствие этого гибнут либо осенью,
либо во время зимовки. Изменением химического состава хвои на
поврежденных деревьях в известной степени можно объяснить
также явление перехода части гусениц сибирского шелкопряда от
2-х к 3-годовому циклу развития [Рожков, 1965].
При однократной значительной (до 80%) дефолиации листвен-
ницы хвоегрызущими вредителями наряду с сохранением, а иног-
да и повышением защитных свойств луба в сохранившейся на
деревьях хвое понижается содержание общего азота, моносахаров
и крахмала. По своему химическому составу такая хвоя прибли-
жается ко вторичной хвое и также представляет собой малопри-
годный пищевой субстрат для фитофагов [Вержуцкий и др., 1970].
Защитные свойства лиственницы по отношению к вредителям
ствола рассматриваются в специальной главе. Пока отметим, что
52
наряду с высокой реактивностью* т. е. способностью тканей ствола
к быстрому структурному и физиологическому преобразованию
в ответ на повреждение насекомыми, для лиственницы характерно
высокое содержание в коре и древесине веществ, обладающих
токсическими и репеллентными свойствами.
В защиту от дереворазрушающих грибов лиственница имеет
мощно развитую кору, содержащую большое количество таннинов,
а ее ядровая древесина богата смолами, таннинами и камедью.
Использование лиственницы
Древесина лиственницы тяжелая, прочная, красивая по тек-
стуре («сильнотекстурная») и естественной окраске. Относительно
узкая заболонь светло-охристая, а широкое ядро красноватое или
коричнево-красноватое с темными контрастными полосами позд-
ней древесины годичных слоев. По механическим свойствам дре-
весина лиственницы много превосходит другие хвойные породы,
по плотности (660 кг/м3) близка к дубовой древесине, несколько
уступает ей по твердости (торцовая — 435 кгс/см2), но превосхо-
дит по прочности на сжатие вдоль волокон (645 кгс/см2).
Как строительный материал лиственница отличается исклю-
чительной долговечностью. О замечательном свойстве ее — сла-
бой подверженности гниению — известно с давних времен.
В Древнем Риме лиственницу использовали для строительства
амфитеатров; в царствование Тиберия (14—37 гг. нашей эры)
сооружали мосты из лиственничных бревен, перевозимых для
этого на большие расстояния. Постройки из лиственницы стоят
столетиями. Так, в Польше костел, построенный в Шидловце,
простоял 500 лет, Обретений костел — свыше 600 лет. Более
трех столетий выстояли лиственничные башни Братского, Илим-
ского и Якутского острогов.
Особое значение имеет стойкость лиственничной древесины в
условиях избыточной и периодически изменяющейся влажности.
Именно это свойство определило широкое использование ее для
постройки кораблей времен парусного флота, которые служили
в 2—2,5 раза дольше построенных из дуба, для строительства
морских плотин (особенно в Голландии), причальных и огради-
тельных сооружений морских и речных портов и свайных фунда-
ментов зданий на зыбких, насыщенных водой грунтах. Известно,;
что большинство зданий, построенных в Венеции в средние века,
стоят на лиственничных сваях; забитые при Петре I лиственничные
сваи Архангельского морского порта за время пребывания в воде
не только не разрушились, но стали еще прочнее. Более того,
захороненные в земле лиственничные стволы могут сохраниться
тысячелетиями. Например, в могильных кургацах скифских пле-
мен Алтая (V—III вв. до н. э.) обнаружены деревянные саркофаги,
выдолбленные из цельных стволов сибирской лиственницы.
53
И в настоящее время указанная особенность лиственничной
древесины не потеряла своего значения в гидростроении, строи-
тельстве мостов, мелком кораблестроении, производстве железно-
дорожных шпал, столбов линии связи и электропередач.
В районах, богатых лесом, особенно в таежной зоне азиатской
части СССР, деревянное малоэтажное домостроение еще долгое
время будет оставаться более экономичным и эффективным по
сравнению со строительством из других материалов. Это относит-
ся не только к сельской местности, но и к поселкам городского
типа. В Сибири лиственницу издавна используют для кладки ниж-
них венцов домов, а нередко возводят стены и целиком из лист-
венничных бревен. В сельской местности ее применяют для
изготовления ответственных деталей при строительстве крупных
производственных и общественных зданий: школ, клубов, живот-
новодческих ферм, овощехранилищ, теплиц и т. д.
В настоящее время лиственницу все шире используют в сто-
лярном производстве, для внутренней отделки помещений и изго-
товления мебели, в том числе высокого качества. Древесину
лиственницы применяют в виде «массива», древесностружечных
и древесноволокнистых плит, строганой фанеры, реже — лущено-
го шпона. Установлены режимы сушки древесины, при которых
не происходит ее растрескивания и коробления.
Для изготовления паркетной клепки у пас пока в основном
используют твердые лиственные породы: дуб, бук, ясень, nnbMf
граб и др. Между тем в США и ряде стран Европы стал популяр-
ным паркет из лиственницы, отличающийся не только высокой
прочностью, но и красивым цветом. Там же нередко делают
полы из неокрашенных лиственничных досок, натираемых спе-
циальным составом [Чавчавадзе, Яценко-Хмелевский, 1978].
Прессованная при определенных режимах древесина листвен-
ницы обладает исключительно устойчивой механической прочно-
стью и служит дешевым заменителем цветных металлов и других
дефицитных материалов в машиностроении.
Широкое вовлечение в промышленную эксплуатацию листвен-
ничных лесов Сибири требует организации полного их использо-
вания — от подсочки выделяемых под вырубку древостоев до пере-
работки лесосечных отходов — сучьев, тонких веток, коры и хвои.
Отходы на лесосеках нередко составляют четвертую часть от
общего объема заготовленной древесины.
При подсочке лиственницы получают ценную прозрачную
живицу, (венецианский терпентин), используемую в лакокрасочной
промышленности, медицине, живописи. Методы подсочки листвен-
ницы, к сожалению, еще недостаточно разработаны, и масштабы
ее относительно невелики. Из древесины и коры лиственницы
путем химической переработки можно получать практически те
же вещества (свыше 100 наименований), для производства которых
пока в основном используют другие хвойные породы. Некоторые
из этих веществ уступают по качеству и трудоемкости извлечения
54
продуктам переработки сосновой и еловой древесины, другие же
превосходят их по содержанию и техническим свойствам. Напри-
мер, кора лиственницы, составляющая 20—29% от объема древе-
сины, содержит до 17 % легкоэкстрагируемых дубильных веществ
[Никифоров, 1968]; по содержанию камеди-арабиногалактана
(до 20% от веса древесины) лиственница не имеет себе равных
среди хвойных [Цветаева и др., 1958]; как мягчитель резиновых
смесей и вулканизаторов лиственничная канифоль не уступает
сосновой, а в некоторых случаях превосходит ее [Часовщиков
и др., 1968]. Рядом преимуществ обладают также некоторые про-
дукты сухой перегонки древесины и коры лиственницы.
Сибирскую и даурскую лиственницы используют в лесопосад-
ках, особенно на излишне увлажненных почвах, а также за их
особенную декоративность широко культивируют в садах и пар-
ках. Под Ленинградом есть знаменитая Линдуловская роща из
сибирской лиственницы, высаженной более 200 лет назад. Деревья
в этой роще достигают 50 м при диаметре ствола 0,6—0,9 м. Хоро-
рошо прижилась сибирская лиственница в чистых и смешанных
насаждениях Шлезвиг-Гольштейна (ФРГ).
В народе лиственницу называют сибирским дубом, лесоведы
же считают ее породой будущего. По мнению В. Ф. Овсянникова
(1930), даурская лиственница заслуживает самого внимательного
к ней отношения как объект, и, пожалуй, единственный, для бу-
дущего лесного хозяйства всего северо-востока Сибири.
ЛИСТВЕННИЧНЫЙ УСАЧ
(XYLOTRECHUS ALTAICUS GERL.)
Лиственничный усач — монофаг лиственницы, единственный
представитель рода Xylotrechus, развивающийся на хвойной по-
роде. Он нападает только на живые, незначительно ослабленные,
а иногда и на совершенно здоровые деревья. В обычных условиях
живет небольшими, плотными малообщающимися между собой
семейными группами; во время вспышек массового размножения
образует очаги, в том числе миграционные, охватывающие в не-
которых частях ареала огромные территории.
Систематическое положение и филогения
Лиственничный усач описан Ф. Геблером — крупным зооло-
гом первой половины прошлого века. Более других этого иссле-
дователя интересовали жесткокрылые, особенно семейство Ceram-
bycidae. В результате участия в ряде экспедиций 1819—1835 гг.
на Алтай, в Прибайкалье и Забайкалье им описано 14 новых ви-
дов усачей, в том числе пять представителей рода Xylotrechus.
Во время поездок в 1833—1835 гг. па Алтай Ф. Геблер нашел,
а затем описал [Gebler, 1836] лиственничного усача как Clytus
altaicus Gebl. Тринадцать лет спустя К. Маннергейм [Маппег-
55
heim, 1849] вторично описал этого усача по экземпляру, пойман-
ному на р. Селенге Н. Поповым, назвав его Clytus popovi Mannh.
Лишь в 1881 г. Л. Гейден в каталоге жесткокрылых Сибири [Неу-
den, 1880—1881] отнес лиственничного усача к роду Hylotrechus
Chevr.
В 1912 г. Е. Рейттер [Reitter, 1908—1916] выделил из рода
Xylotrechus и описал в качестве самостоятельного род Xyloclytus
Reitt., включив в него помимо индокитайско-малайских видов
лиственничного усача (Xyloclytus altaicus Gebl.). За основной от-
личительный признак рода Xyloclytus Е. Рейттер принял форму
члеников усиков и сближенное расположение оснований по-
СЛОДНИХа
Н. Н. Плавильщиков — лауреат Рейттеровской премии по
энтомологии — первоначально (1932) подтвердил наличие родо-
вых различий Xylotrechus л Xyloclytus и предложил следующие
ключи для их распознавания:
род Xyloclytus. Reitt.— членики усиков очень короткие узловато-
треугольные; промежуток между ямками усиков уже про-
межутка между внутренними краями глаз; тело массивное
и коренастое;
род Xylotrechus Chevr.— членики усиков более длинные неузло-
вато-треугольные; промежуток между ямками усиков и внут-
ренними краями глаз равной или почти равной ширины.
Однако в ходе обработки трибы Clytini и подготовки второй
части монографии о жуках-дровосеках фауны СССР (1940}
Н. Н. Плавильщиков пришел к заключению о том, что род Xylo-
trechus Cherv. распадается на два, в значительной степени парал-
лельных цикла форм, характеризующихся величиной глаз, фор-
мой лба, отчасти развитием окологлазных килей:
подрод Xylotrechus s. str.— глаза большие, лоб сужен или сдав-
лен посередине; промежуток между усиковыми ямками
не уже такового между глазами; окологлазные кили
длинные;
подрод Xyloclytus (Reitt.) Plav.— глаза менее развиты, лоб с па-
раллельными сторонами, во всяком случае не сдавлен
и не сужен посередине; промежуток между усиковыми
ямками обычно несколько уже промежутка между глазами;
окологлазные кили короткие, хотя нередко и очень высокие.
По мнению Н. Н. Плавильщикова, в характеристике рейттс-
ровского рода упущен наиболее существенный признак, свойст-
венный видам как Xylotrechus. так и Xyloclytus. а именно форма
лба (имеются в виду лобные кили), строение же усиков никакого
значения в данном случае не имеет. Подрод Xyloclytus характе-
ризуется и географически: в Евразии он представлен почти исклю-
чительно в Индо-Малайской области и в Палеархеарктике, только
один вид распространен в южной части Ангарской и юго-восточной
части Европейско-Обской подобластей Палеарктики. Этим сибир-
ским видом является лиственничный усач (Xylotrechus altaicus
56
Gebl.). Наличие ряда параллельных форм и видов (в обоих под-
родах) с весьма примитивной окраской показывает, что распаде-
ние рода Xylotrechus на два цикла видов произошло чрезвычайно
давно, причем виды подрода Xylotrechus s. str. оказались более
подвижными эволюционно, что доказывается и богатством этого
подрода видами, и его значительно более широким распростра-
нением.
При выборе лиственничного усача в качестве объекта исследо-
ваний у нас не было сомнений в принадлежности его к роду Xylo-
trechus (после разбора этого вопроса Н. Н. Плавилыциковым
только П. II. Тальман 1 некоторое время сохранял за ним рейт-
теровское название), однако вскоре такие сомнения появились.
Явная биологическая и морфологическая специфичность лист-
венничного усача не могла не побудить пас вновь вернуться к воп-
росу о его положении в системе трибы Clytini.
Приводимые ниже данные явились результатом просмотра
жуков-клитов в ряде крупных коллекций, в том числе, хранящих-
ся в Берлинском и Дрезденском зоологических музеях.
Триба Clytini относится к числу наиболее богатых видами и
распространенных во всех зоогеографических областях. По мне-
нию Н. Н. Плавилыцикова (1940), она насчитывает в своем соста-
ве до 50 родов и до 1000 видов. Наиболее богато триба представ-
лена в Индо-Малайской области, где распространено около 430 ви-
дов, причем число известных видов с каждым годом значительно
возрастает. В Палеарктике в узком смысле слова распространено
около 80 видов (8 родов), причем в основном они встречаются в ее
южных частях, тогда как зоне тайги свойственны несколько видов.
По форме эпистери заднегруди триба Clytini разделяется на
две родовые группы: у первой эпистерны относительно короткие
и только в 2—3 раза длиннее своей видимой ширины, у второй—
не менее чем в 4 раза длиннее своей видимой ширины. Централь-
ным родом первой группы является Xylotrechus Chevr., второй —
Chlorophorus Chevr.; оба эти рода космополиты, насчитывают до
150 видов каждый. Лиственничный усач относится к первой родо-
вой группе трибы: длина эпистерн его заднегруди не более чем
в 3 раза превышает их ширину без учета прикрытой надкрыльями
части (рис. 12).
Из семи родов нашей фауны, входящих в родовую группу
усачей с широкими эпистернами заднегруди, три рода — Xylo-
trechus Chevr., Pseudosphegestes Plav. и Clytus Laich.— объединя-
ются сходством ряда признаков. У них короткие усики, вершин-
ные углы члеников которых не вытянуты в заметный зубчик, зад-
1 П. Н. Тальман (1940, 1940а, 1947). Обе статьи 1940 г. опубликованы
лишь на несколько месяцев позднее выхода в свет второй части монографии
по жукам-дровосекам. Тем нс менее автор их был знаком с заключением
II. Н. Плавилыцикова о родовой принадлежности лиственничного усача
(см. примечание в сноске на стр. 1, 1940). В работе 1957 г. П. Н. Тальман
именует усача уже как X ylotrechus altalcus Gebl'.
57
Рис. 12.
1—3 — эпистерна заднегруди: 1 — Xylotrechus rusticus; 2 — X. altaicus} 3 —Chlo-
rophorus figuratus; 4—9—бедро задней ноги: 4 — Brachyclytus. 5— Cyrtaclytus.
6 — Clytus. 7 — Xylotrechus chinensis. 8 — X. rusticus. 9 — X. ibex; 10 — задняя нога
X. altaicus; 11 — усик X. rusticus; 12 — то же X. altaicus; 13 — голова X. altaicus;
14— то же X. chinensis; 15—20— схема головы спереди: is —- X. altaicus. 18 —
X. incurvatus. 17 — X. rusticus. 18— X. hircus. 19— X. grayi. 20 — X. quadripes.
ние бедра слабо утолщенные (см. рис. 12). Первые два рода, не-
сомненно, близки филогенетически.
Род Xylotrechus Chevr., как уже говорилось, характеризуется
наличием нескольких продольных килей на лбу, причем всегда
58
имеется хотя бы один (обычно два) киль посередине лба. Осталь-
ные признаки рода в принятом в настоящее время его объеме
[Плавильщиков, 1940] следующие: щеки короткие; лоб с парал-
лельными сторонами или заметно суженный, как бы сдавленный
посередине; ямки оснований усиков ограничены изнутри про-
дольным килем, который продолжается вперед до вершины лба
и обычно несколько прикрывает изнутри усиковую ямку; проме-
жуток между внутренними краями усиковых ямок и глаз одина-
ково или почти одинаково широкий; усики тонкие или умеренно
толстые, по большей части короткие, у самцов только изредка
достигают середины надкрылий, у самок обычно едва заходят за
их основание; переднеспинка с округленными боками, без пере-
тяжки сзади вершины, посередине диска часто в грубой зернистой
пли шероховатой скульптуре; ноги разнообразны по длине, бедра
более или менее заметно утолщены, задние бедра самцов нередко
заходят за вершину надкрылий.
Как видно, все перечисленные признаки рода Xylotrechus,
исключая форму и скульптуру лба, являются лишь дополнитель-
ными, но отнюдь не характерными. Они свойственны и другим
родам трибы, особенно Pseudosphegestes Plav. и Clytus Laich. Род
Pseudosphegestes*, например, характеризуется отсутствием килей
на лбу, широкими эпистернами заднегруди, поперечным щитком,
сближенными основаниями усиков, очень длинным первым чле-
ником задних лапок и двоякого рода скульптурой (частично гру-
бой) переднеспинки. Все эти признаки, кроме отсутствия лобных
килей, свойственны роду Xylotrechus. Так, широкие эпистерны
заднегруди и поперечный щиток мы находим у большинства видов
подрода Xylotrechus; очень длинный первый членик задних
лапок — признак, свойственный ряду видов того же подрода.
Например, у X. cuneipennis Кг. первый членик задних лапок
в 3 раза длиннее второго и третьего вместе взятых и гораздо длин-
нее всех последующих члеников вместе взятых; двойная скуль-
птура переднеспинки встречается в обоих подродах Xylotrechus,
а сближенные основания усиков — основной признак подрода
Xyloclytus. Более того, у некоторых представителей подрода
Xyloclytus лобные кили (или киль) едва намечены, у большинства
особей X. altaicus Gebl., например, лобный киль имеет вид корот-
кого продольного бугорка, расположенного над клипеусом или
отчасти даже на последнем; окологлазные кили развиты слабо,
иногда едва прослеживаются (X. chinensis Chevr.), а мощные над-
усиковые кили не выходят за пределы усиковой ямки.
• Е. Рейттер [Reitter, 4908—1916] выделил Clytus cinereus Cast, в осо-
бый подрод Pseudosphegestes, который отнес к роду Chlorophorus. При этом
он не учел того, что по форме эпистерн заднегруди этот усач относится к дру-
гой родовой группе трибы Clytini. Н. Н. Плавильщиков (1940) возвел рейт-
теровский подрод в ранг рода Pseudosphegestes Plav., включив в него кроме
Р. cinereus Cast, еще два вида нашей фауны.
59
Выделив и охарактеризовав род Xyloclytus, Е. Рейттер перво-
начально включил в него собственно только два вида — X. chi-
nensis Chevr. и X. popoviMannh.(=altaicus Gebl.).Оба они действи-
тельно обладают отмеченными им признаками, а именно: проме-
жуток между основаниями усиков у них уже промежутка между
внутренними краями глаз, особенно сильно сближены короткие
надусиковые гребни, членики усиков короткие, узловато-тре-
угольные. Однако уже вскоре А. Винклер (Winkler, 1924—1932)
отнес к Xyloclytus 21 вид, а Н. Н. Плавильщиков (1940) назвал
в качестве представителей подрода Xyloclytus еще 10 видов, не
вошедших в каталог А. Винклера. Из этих усачей мы имели воз-
можность исследовать 16 видов и, к нашему удивлению, среди них
кроме X. chinensls Chevr. и X. altalcus Gebl. обнаружили упомя-
нутые признаки рода (Reitt.) и подрода (Plav.) Xyloclytus только
у X. incurvatus Cevr. У такого, например, признанного обоими
энтомологами представителя Xyloclytus, как X. grayi White
culicollis Dalm.), основания усиков оказались расположенными
на таком же расстоянии друг от друга, как и внутренние края
глаз, лоб — суженным посередине, окологлазные кили — хоро-
шо развитыми до нижнего края лба, глаза — большими
(см. рис. 12), членики усиков — не узловато-треугольными.
Столь же очевидные признаки подрода Xylotrechus свойственны
X. magnicollls Fairm., X. dominula White, X. formosanus Schwzr.
Таким образом, с основным признаком, разделяющим рассмат-
риваемые подроды,— параллельностью и суженностью сторон
лба — не всегда находятся в соотношении остальные их признаки,
а именно: размер глаз, степень развития окологлазных килей,
соотношение промежутка между усиковыми ямками и между гла-
зами. Более того, среди видов, относимых в настоящее время
к подроду Xyloclytus, есть такие, у которых лоб сужен, и по этому
признаку они занимают промежуточное положение между типами
подродов — X. (Xyloclytus) altalcus Gebl. и X. (Xylotrechus) sar-
torii Chevr.
H. H. Плавильщиков пришел к совершенно справедливому
заключению о том, что род Xylotrechus является наиболее древним
в трибе, сохранившем и в наши дни ряд архаических видов, что
он весьма гомогенен как в отношении ряда морфометрических
признаков, так и чисто габитуально, но чрезвычайно гетерогенен
в отношении рисунка и окраски надкрылий, обнаруживая в ряду
своих видов прообразы рисунка чуть ли не всех родов трибы.
К этому следует добавить, что в Индо-Малайской области и в Па-
леархеарктике сохранилось много очень обособленных друг от
друга по внутриродовым признакам видов, находящихся к тому
же в состоянии эволюционной депрессии. Объединение их в один
подрод Xyloclytus было искусственным. Необходима ревизия рода
в целом, которая, по нашему мнению, приведет к разделению его
не на два, а на три или даже четыре подрода только в пределах
Южной п Юго-Восточной Азии.
(0
Подрод Xyloclytus нам представляется весьма компактным,
обладающим комплексом четких признаков. В характеристику
его, данную II. Н. Плавилыциковым (1940), необходимо ввести
следующие дополнения: высота нижней доли глаза пе более чем
в два раза превышает высоту щеки от глаза до внутреннего угла
основания челюсти; скульптура переднеспинки двоякая, ее вы-
пуклые элементы зернисто пунктированы, морщинистые, а ос-
тальная поверхность в глубоких точках; бедра задних ног не утол-
щены в дистальной части (см. рис. 12, 10). Форма гениталий —
малонадежный признак для внутриродовых подразделений
(рис. 13, 2—7).
Наиболее близкими видами к лиственничному усачу являются
X. rujobasalis Pic., найденный в Лаосе, и X. basifuliginosus Hell.,
обитающий на Филиппинах. Пять экземпляров первого имеется
в коллекции Берлинского зоологического музея, второй — в Дрез-
денском музее. У обоих видов лобные кили развиты слабо, около-
глазные кили отсутствуют, отношение высоты пижней доли глаза
к высоте щеки от глаза до угла основания челюсти при-
мерно 2—1,5:1, лоб в середине не сужен, промежуток между уси-
ковыми ямками уже промежутка между глазами; скульптура
переднеспинки двоякая, часть ее (особенно выпуклые участки)
зернисто морщинистая, другая часть в глубоких точках с неров-
ными промежутками; бедра задних ног не утолщены.
У X. rufobasalis Pic. надкрылья имеют ту же форму, что и
у лиственничного усача. Рисунок на них также сходен, но более
яркий. Основание и вершина элитр красноватые, светлее средней
части, которая окрашена в темно-бурый цвет; контрастные попе-
речные и угловидная перевязи образованы желтоватыми волос-
ками (см. рис. 13, 9).
X. chinensis Chevr. (см. рис. 13, 70), изначально отнесенный
Е. Рейттером к роду, а Н. Н. Плавилыциковым к подроду Xyloc-
lytus, в действительности относится к близкому, параллельно раз-
вивавшемуся циклу видов. При большом сходстве формы и скульп-
туры лба и размера глаз, он пе вписывается в этот подвид как
габитуально, так и по скульптуре переднеспинки и форме бедер зад-
них ног. Не относится к подроду Xyloclytus и обитающий в Юго-
Восточной Азии X. incurvatus Chevr. (рис. 13, 11). По форме эпи-
стерн заднегруди этот вид занимает одну из наиболее древних
ветвей в филогении рода Xylotrechus. Также следует исключить
из подрода Xyloclytus X. quadripes Chevr. (Индия, Ява), X. grayi
White (Формоза), X. magnicollis Fairm. (Формоза), X. ema-
ciatus H. W. (Япония).
В Палеархеарктике близкородственные лиственничному уса-
чу виды пока не обнаружены, но, возможно, в этой подобласти он
обособился как вид, а уже затем проник в Ангарскую подобласть.
Как считает Е. Г. Бобров (1972), в начале плейстоцена сплошной
ареал лиственницы даурской простирался на юг до гор Северного
Китая, а растительность горных областей Сибири и Северной Мон-
61
Рис. 13.
1 — мужской копулятивный аппарат (продольный разрез через конец брюшка) Cly-
tM\ i—7 — фаллус (эдеагус н фаллобаэа с парамерамн): 2 — СНЫ» arietoidca, 3 —
Cyrtoclytiu capra, 4 — Xylotrechu» aHaictuL 5 — JC. ehinensw, 6 — X. nuiicus, 7 —
X. ibex\ 3—11 — представители подрода Xyloclytus 3 — X. altaieu»* 9 — X. rufoba-
salit, 10 — X. cnincntis, 11 — X. ineuroatue.
голии, по мнению И. M. Крашенинникова (1939), была в это время
очень похожа на современную. Но более вероятно, что формирова-
ние усача протекало в пребореальных мезотермных лесах Север-
ной Азии.
62
Предки лиственничного усача, возможно, были распростране-
ны уже в арктотретичных субтропических лесах Азии и разви-
вались там на каких-то широколиственных породах. В процессе
отступления к югу теплолюбивых деревьев и образования проме-
жуточной зоны мезотермных лесов между крайне северной поло-
сой микротермных (бореальных) и зоной макротермных лесов,
вероятно, и осуществился переход усача на лиственницу вместе
с заболонником Моравица (Scolytus morawitzi Sem.) и другими
монофагами этой породы.
Давнее широкое распространение лиственничного усача в Се-
верной Азии подтверждается в какой-то степени наличием изо-
лированной и, по всей вероятности, автохтонной популяции его
на Урале, а также окраской жуков. Блеклый рисунок на над-
крыльях из-за слабого развития волосяного покрова не является
признаком примитивности, он свидетельствует о длительном влия-
нии Севера. Тип рисунка сохранился, исчезла яркость. Листвен-
ничный усач — один из наиболее молодых и эволюционно про-
грессирующих видов в подроде Xyloclytus.
Географическое распространение
Лиственничный усач отмечался в лиственничных лесах на об-
ширной территории от Урала до берегов Тихого океана. В после-
военные годы был завезен с лесоматериалами в некоторые районы
европейской части СССР, но, видимо, там не прижился. Границы
распространения усача еще недостаточно изучены. В целом его
ареал, по-видимому, укладывается в пределы распространения
сибирского шелкопряда, однако если южная граница ареалов
этих двух вредителей примерно совпадает, то на севере усач вряд
ли проникает за 60-ю параллель. Пока самым северным пунктрм
обнаружения его является р. Вихорева (приток Ангары в Брат-
ском районе Иркутской области), а южным — верховья р. Уссури
в Приморском крае.
На Урале лиственничный усач в единственном экземпляре
найден 17 июля 1896 г. В. В. Редикорцевым (1908) в окрестностях
Екатеринбурга. Лишь в 1960 г. Е. Г. Мозолевская (1964) обна-
ружила этого вредителя, а также пораженные им деревья на Юж-
ном Урале в перестойных лиственничниках Башкирского запо-
ведника. Заповедник основан в 1930 г., и за время существования
неоднократно обследовался энтомологами. Тот факт, что листвен-
ничный усач не обнаружен ими, свидетельствует о чрезвычайной
разреженности уральской популяции и о длительном пребывании
ее в депрессивном состоянии.
Горные лиственничники Южного Урала по своему близки
к южносибирским лиственничникам. Здесь, а также в смежной
части Среднего Урала сформировалась обособленная популяция
усача. В Западной Сибири, кроме Алтая, этот вид не встречается.
На всем пространстве юга Западно-Сибирской равнины и Казах-
63
ского мелкосопочпика отсутствуют горные лиственничники, не-
обходимые для его обитания. Возможность завоза вредителя
вместе с лесоматериалами из Восточной Сибири допустима только
с сооружением Сибирской железной дороги. Однако ко времени
первой находки усача В.В. Редикорцевым Транссибирская маги-
страль была доведена лишь до Красноярска. Повышение числен-
ности усача на территории Башкирского заповедника к 1960 г.
явилось следствием нарушения лесорастительных условий, вы-
званного интенсивной вырубкой леса в 1951—1958 гг. в период
временной передачи заповедной территории Южно-Узянскому
лесхозу.
В Восточной Сибири северная граница ареала лиственничного
усача охватывает Западный и Восточный Саяны, затем поднимает-
ся па север вдоль Ангарского кряжа до 56° с. ш. и примерно
по этой ширине идет к северной оконечности Байкала. Здесь усач
обнаружен в лиственничниках отрогов Байкальского и Баргу-
зипского хребтов. Далее северная граница ареала идет по 55-й
параллели или немного южнее (по южным склонам Станового
хребта) к побережью Охотского моря. Сведения о распространении
усача на Дальнем Востоке недостаточны. Он встречается на Са-
халине (кроме южной части) и в отдельных пунктах Хабаровского
и Приморского краев. Южная граница распространения вида —
от юга советского Приморья до Алтая — находится вне пределов
СССР. В Монголии она охватывает Западный и Восточный Хэн-
тэй, Хангайский хребет и северную часть Монгольского Алтая
[Сергеев, 1948; Гречкин, 1960; Криволуцкая, 1961; Исаев, 1966;
Исаев, Гире, 1975; Любарский, Идрисов, 1970; Ивлиев и др.,
1973; Анисимова, Соков, 1975; Яновский, 1977; Плешанов,
1974; Бережных, 19781.
Ареал усача, таким образом, расположен на территории, за-
нятой четырьмя видами лиственницы — даурской (L. gmelinii
Rupr.), сибирской (L. sibirica Ledb.), ольгинской (L. olgensis
Рис. 14. Ареал лиственничного усача.
64
A.Henry) и камчатской {L.kamtschatica Rupr.), а также несколь-
кими гибридными формами с участием этих видов. Лиственница
каяндера (L. cajanderi Мауг) распространена вне ареала усача
в так называемой центральной камчатской депрессии и спускает-
ся на юг по побережью Охотского моря, по-видимому, не ниже
58° с. ш. [Бобров, 1972]. На ольгинской лиственнице усач найден
лишь один раз в верховьях р. Уссури Л. В. Любарским
и В. В. Шаблиовским в 1936 г. [Сергеев, 1948].
Очерченный в таком виде ареал лиственничного усача (рис. 14)
за недостатком данных носит до некоторой степени условный ха-
рактер. И все же, как нам представляется, нельзя ожидать от по-
следующих исследовании больших исправлений намеченных
границ.
Стация
Мозаичное распространение лиственничного усача в общих
границах ареала определяется орографическими, климатическими
и лесорастительными условиями, значение которых для этого
вида много большее, чем для других представителей Xylotrechus,
встречающихся в таежной зоне Палеарктики. Характерной осо-
бенностью усача является его строгая приуроченность к горным
лесам. Наши исследования показали, например, что в листвен-
ничных лесах Иркутско-Черемховской равнины, несмотря на от-
носительно высокое ее положение (500—600 м над уровнем моря),
усач не появляется даже в годы массового повреждения древо-
стоев сибирским шелкопрядом. В то же время он постоянно встре-
чается в лиственничниках Восточного Саяна и Лено-Ангарского
плато, окружающих эту долину. На территории СССР помимо
горных лесов Урала, Алтая, Восточного и Западного Саяна лист-
венничный усач найден в разном состоянии популяций в Восточно-
Тувинском, Сангиленском и Саяно-Джидинсиом нагорьях, в лист-
венничниках хребта Танну-Ола, Ангарского кряжа, Байкальского
и Баргузинского хребтов, Олекминского становика, Борщовоч-
ного, Станового и Буреинского хребтов.
Типичной стацией усача в обычном состоянии популяций (вне
очагов размножения) по всему ареалу являются низко- и средне-
полнотные перестойные лиственничники, произрастаю-
щие на пологих склонах в нижней части лесного пояса, а также
в поймах небольших рек и ручьев, реже на водораздельных хреб-
тах. Наблюдается явное предпочтение склонов южной и юго-за-
падной экспозиции. Наиболее характерными типами леса явля-
ются багульниковые, багульниково-брусничные и осоково-сфаг-
новые лиственничники с подлеском из березы и ольхи (рис. 15).
Усач обитает также в перестойных древостоях, относящихся
к группе брусничных лиственничников и занимающих среднеув-
лажпенные участки шлейфов горных склонов. В зоне горной лесо-
5а. С. Рожков
65
Рис. 15. Лиственничник ольхогни новый (в подлеске ольха, береза, в покро-
ве брусника, грутанка, багульник, местами мхи) с большим количеством
перестойных и зараженных гнилями деревьев — одна из типичных ста-
ций (резерваций) лиственничного усача.
степи он встречается в разнотравных лиственничниках, причем
в период депрессии локализуется по логам в островах леса, неред-
ко разобщенных значительными открытыми пространствами.
При массовом размножении стацией усача могут служить вы-
сокой олнотные лиственничники и не только перестойные, однако
типы леса в основном остаются те же. В Юго-Восточном Забай-
калье очаги его иногда распространяются в ерниковые листвен-
ничники верхнего лесного пояса гор (рис. 16). Вообще в периоды
вспышек размножения вредитель способен подниматься в горы
на большую высоту, например, в Туве — до 1400 м [Черепанов.
1952], в Монголии — до 1800 м [Гречкин, 1960] пад уровнем
моря.
Генерация и лётные годы
Лиственничный усач имеет двухгодовую генерацию по всему
ареалу 8. Об этом свидетельствуют наши наблюдения на юге и се-
вере Прибайкалья, на юго-востоке Забайкалья и литературные
данные о цикле его развития на юге Приморья, в Амурской об-
ласти, в Туве и на Урале [Сергеев, 1948; Черепанов, 1950; Мозо-
левская, 1964; Исаев, 1966]. Лишь в очень редких случаях при
неблагоприятных условиях развитие усача может затягиваться
па три года.
Исследования А. И. Черепанова (1952) в очаге размножения
усача и в экспериментальных условиях показали, что сумма сред-
недневных (с 7 до 19 часов) температур, при которых происходит
его развитие от яйца до взрослого насекомого (исключая периоды
зимнего покоя), составляет примерно 3724° и что двухгодовая ге-
нерация усача возможна в тех районах, где сумма температур дре-
весины в период развития преимагинальных стадий и воздуха
во время жизни имаго составляет 1870—1900°. Данные о суммах
температур воздуха за вегетационный период (от 5° весной до 10°
осенью) по Прибайкалью [Алисов, 1962] показывают, что коле-
бание сумм температур от 1400° (северная часть Байкальского
хребта) до 1600—1800° (окрестности г. Братска) не отражается
на продолжительности цикла развития усача. Вероятно, по всему
ареалу этот вид развивается при суммах температур воздуха,
8 Двухгодовую генерацию иногда называют двухлетней или даже трех-
летней, имея в последнем случае в виду календарные годы (три лета). Это
вносит определенную путаницу. Двухгодовая генерация есть не что иное,
как 24-месячный цикл воспроизводства имаго нового поколения от времени
появления имаго предыдущего поколения, включая два периода зимнего
покоя.
67
Рис. 16. В ерииковых лиственничниках высокогорий лиственничный усач распространяется
обычно лишь в периоды вспышек массового размножения.
превышающих 1200°, и продолжительности вегетационного пе-
риода от 110 дней и больше.
Лиственничный усач очень чувствителен к качеству кормового
субстрата, поэтому состояние деревьев в большей степени отража-
ется па продолжительности развития его личинок, чем темпера-
турные условия в пределах очерченного ареала. В тех случаях,
когда вредитель поселяется на сильно ослабленных деревьях,
особенно топкомерных, и последние гибнут уже в начале следую-
щего года и быстро высыхают, развитие его может затянуться
на год. В Тункинской долине мы наблюдали затянувшееся раз-
витие лиственничного усача вне очага на перестойной лиственни-
це, срубленной вскоре после ее заселения. Трехгодовой цикл
развития усача — редкое явление. Тем не менее это, по-видимому,
основной источник образования так называемого промежуточного
колена в популяциях.
По одпогодовому циклу усач не развивается, по крайней мере,
в пределах СССР и на севере Монголии. Экспериментально можно
вызвать окукливание личинок за год до их нормального превра-
щения в куколку и выход жуков в соответствующее их лёту вре-
мя. Для этого личинок V возраста перед началом ухода в глубо-
кие слои заболони и ядровую древесину (в начале июля второго
лета развития) необходимо перевести па питание чистой клетчат-
кой. Через несколько дней такие личинки впадают в предку по-
лочное состояние, затем окукливаются. В наших опытах все вы-
шедшие жуки были самцами. Они имели не вполне развитое жи-
ровое тело и недоразвитые семенники. Это свидетельствует о не-
обходимости прохождения личинкой третьего этапа развития,
протекающего во второй половине лета и весной после второй зи-
мовки в глубоких слоях древесины.
На Урале, в Восточной Сибири и в Монголии лиственничный
усач, подобно лиственничной расе сибирского шелкопряда, лета-
ет в четные годы. Промежуточное колено (генерация нечетных
лет) составляет очень небольшой процент в популяциях как в оча-
гах размножения, так и вне их. В Букукинском очаге в 1965 г.
деревья с личинками и куколками промежуточного колена лист-
венничного усача составляли не более 2% от общего числа засе-
ленных деревьев, а количество вышедших пз них имаго до сере-
дины августа (по числу лётных отверстий) не превышало 0,5%
от численности жуков основного колена, появившихся в следую-
щем году. Вполне возможно, что семейные группы усача промежу-
точного колена в очагах размножения утрачивают признаки,
свойственные очаговому состоянию популяции в целом. Они на-
падают преимущественно на перестойные деревья, обладающие от-
носительно слабой сопротивляемостью, и заселяют па них, как
и вне очагов, локальные участки ствола.
Чередование лётных и межлётных годов свойственно в основ-
ном чешуекрылым, в том числе некоторым бластофагам и пефро-
фагам. Так, напрпмер, в Прибайкалье массовый лет побеговьюна
69
Petrova resinella L. происходит только в четные годы [Рожков,
1957]. Среди ксилофагов в Восточной Сибири гряд видов усачей
(Monochamus, Asemum striatum 01.), некоторые златки (Ancylo-
cheira,Chrysobothris, Anthaxia), а также рогохвосты и долгоносики
(Hylobius) имеют двухгодовую генерацию, по ни у одного из них
не наблюдается закономерного преобладания лёта в четные или
нечетные годы. И в этом отношении лиственничный усач имеет сход-
ство с первичными вредителями.
Обычное и очаговое состояние популяции
(агрессивность и характер заселения ствола)
Среди ксилофагов очень немногим видам свойственны вспышки
массового размножения. Речь в даннохм случае идет по о повышении
плотности популяций или их семейных групп вследствие появле-
ния избытка кормового субстрата, а о переходе в стадное (оча-
говое) состояние, характеризующееся рядом физиолого-экологи-
ческих особенностей. Градации плотности особей в семейных груп-
пах, вызываемые изменением трофических условий, далеко не
всегда приводят к качественному изменению состояния популя-
ций и у видов, способных давать вспышки размножения. Для пе-
рехода их в очаговое состояние необходима такая экологическая
ситуация, которая вызывает проявление высшей формы эффекта
группы — стадного состояния. В этом состоянии трофический фак-
тор уже не является столь жестким, ибо диапазон пригодности
корма неизмеримо расширяется.
Лиственничный усач по характеру питания занимает переход-
ную ступень от филлофагов к ксилофагам необратимо ослаблен-
ных, отмирающих деревьев. Для развития его личинок необхо-
димы живые ткани коры и заболони. Физиологическое состояние
деревьев, заселяемых усачом, является поэтому одним из основных
показателей состояния его популяций. Этот фактор, в свою оче-
редь, определяет некоторые экологические, в том числе поведен-
ческие особенности насекомого. Полученные к настоящему вре-
мени сведения о жизни лиственничного усача в очагах размножения
и вне их позволяют дать достаточно четкую характеристику обыч-
ному (депрессивному и экспрессивному) и очаговому состояниям
его популяций.
Депрессивное состояние популяций лиственничного усача ха-
рактеризуется разобщенными и очень небольшими по числу осо-
бей семейными группами, развивающимися на толстомерных де-
ревьях пострепродуктивного возраста (рис. 17). Заселение ствола
происходит по «местному» типу, т. е. не распространяется по всей
высоте зоны толстой и переходной коры, а занимает относитель-
но ограниченный участок в средней, реже в комлевой части ствола
(рис. 18). Эти деревья обладают сравнительно низкими защитными
свойствами, могут заселяться усачом повторно и обычно сразу
после его нападения становятся доступными для поселения дру-
70
Рис. 17. Ствол iiepecTOffHoii лист-
венницы с локальным поселением
усача.
гих ксилофагов, составляющих
в данном случае производную
экологическую группировку, а
в остальных — вне связи с
лиственничным усачом — вхо-
дящих в состав исходных груп-
пировок. Обнаружить присут-
ствие усача в древостое в ус-
ловиях депрессивного состоя-
ния популяции трудно, пос-
кольку оп развивается па едини-
чных деревьях, внешне не от-
личающихся от других пере-
стойных деревьев. Активная
жизнь жуков, покинувших ку-
колочные колыбельки, ограни-
чивается несколькими днями
откладки яиц в теплую сол-
нечную погоду. Остальное вре-
мя они проводят в укрытиях.
Экспрессивное состояние популяций усача отличается от де-
прессивного более выраженным представительством в биоценозе.
Оп нападает уже не только па ослабевшие в результате естествен-
ного старения лиственницы, по и на деревья, преждевременно ут-
ратившие резистентность под воздействием тех или иных факто-
ров. Однако и здесь вредитель предпочитает толстомерные стволы
п заселяет их по тому же «местному» типу. Повышение количе-
ства особей в семейных группах позволяет им одновременно засе-
лять большее или меньшее число угнетенных соседних листвен-
ниц, иногда даже небольшие куртины леса, но они не обретают
способности нападать на деревья, обладающие высокой сопротив-
ляемостью, для преодоления которой необходим иной характер
заселения ствола.
Переход популяции или части ее в экспрессивное состояние не
всегда завершается вспышкой массового размножения. На боль-
шей части ареала повышение плотности популяций усача ограни-
чивается только этим состоянием.
Очаговое (стадное) состояние популяций лиственничного уса-
ча качественно иное. Выше уже говорилось о том, что при массовом
размножении очаги его распространяются в лиственничники, ко-
торые по лесорастительным условиям и характеру древостоев от-
личаются от свойственной ему стации вне очагов. Главной же осо-
бенностью усача в очаговом состоянии является необычная для
71
Гис* 18. Типы заселения ствола лиственничным усачом,
ксилофагов агрессивность, способность заселять устойчивые к пов-
реждению деревья и преодолевать их сопротивляемость.
В очагах размножения образуются «смешанные» семейные груп-
пы вредителя за счет объединения отдельных ранее изолирован-
ных семейных групп. Об этом свидетельствуют: признаки гетеро-
зиса, проявляющиеся в ряде поколений, активная миграция жу-
ков в очаге, обеспечивающая необходимую плотность заселения
деревьев, их массовые перелеты п образование миграционных
очагов, а также возможность развития усача на жизнестойких де-
ревьях в условиях понижения его численности к концу второй
фазы очага, когда заражению подвергаются уже’отдельные деревья
или небольшое их группы.
В очагах размножения усач заселяет лиственницы всех возра-
стов, начиная с тонкомерпых стволов деревьев III класса возраста
(рис. 19). Только в очаговом состоянии он распространяется из
низко- и среднеполнотных лиственничников в древостои с полно-
той 0,7—0,8. Это явление ранее отмечено А. И. Черепановым (1952)
в Читинской области, А. С. Исаевым и Д. А. Тарасовой (1964)
в Туве.
В норме стволы деревьев в очагах размножения заселяются
усачом полосой по одной стороне в пределах толстой, а иногда и
переходной коры (на толстомерпых деревьях обычно — от комля
до первых живых сучьев). При таком характере повреждения со-
храняется возможность обмена органическими и минеральными
веществами между корнями и кроной, а также передвижения воды
и растворов вокруг поранения [Крамер, Козловский, 1963J, бла-
годаря которому клетки луба даже после рассечения его на отдель-
ные участки ходами личинок II и III возрастов остаются некоторое
время живыми. С другой стороны, только благодаря большой про-
тяженности полосы заселения по стволу возможно преодоление
личинками сопротивляемости дерева. Наибольшая относительная
гибель личинок усача от действия выделяющейся смолы бывает в
концевых частях этой полосы. Последняя может быть узкой
(рис. 20), либо охватывает всю южную половину окружности ство-
Рис. 19. Тонкомерный древостой, заселенный усачом в оч.1ге массовою
размножения.
Рис. 20. Лпственпица 70—75-летнего возраста с относительно узкой
полосой поселения усача. Ствол издолблен большим пестрым дятлом
при добывании личинок, углубившихся в ядровую древесину.
ла (рис. 21). Иногда живая часть ствола на некотором протяжении
«сохраняется в виде узкой ленты луба и нескольких наружных
слоев заболони.
Круговое разовое заселение ствола встречается в основном
и первой фазе очага размножения и несомненно является отклоне-
74
Рис. 21. Лиственница 70-летнего возраста с полосой по-
селения усача, охватывающей около половины окруж-
ности ствола. Кора по всей полосе отодрана большим пест-
рым дятлом.
нием от нормы. В этом случае яйца откладываются жуками также
на одной стороне ствола» но в процессе развития личинок их ходы
в одном или нескольких местах охватывают ствол по всей окруж-
ности (см. рис. 18). Такие деревья усыхают ко времени появления
куколок, и иногда луб неповрежденной части ствола остается жи-
вым до осени лётного года усача.
75
Факторы динамики численности
История изучения лиственничного усача со времени его опи-
сания как вида и до последних работ свидетельствует о том, что
всюду для популяций его в периоды между вспышками массового
размножения характерна разобщенность семейных групп и низ-
кая численность особей в них. Иначе говоря, это насекомое в обыч-
ных условиях является редким. Такое явление вполне объяснимо.
Для развития усача необходимы не просто ослабленные до опре-
деленной степени деревья, а такие, которые в течение всего вре-
мени развития личинок могут сохранять жизнеспособность. Вне
очагов размножения усач развивается на деревьях, находящихся
на грани перехода в состояние необратимого ослабления, по такого
медленного перехода, который позволяет ему иногда пов-
торно заселять их один или даже два раза (5—7 календарных лет).
Наиболее соответствуют этому требованию естественно ослабева-
ющие старые деревья. Во время вспышек размножения в лесах,
ослабленных первичными вредителями, пожарами и другими фак-
торами, усач также не заселяет лиственницы, которые быстро
утрачивают свою жизнеспособность.
Таким образом, наличие и количество деревьев, соответству-
ющих по физиологическому состоянию требованиям усача, опре-
деляет присутствие и численность его в древостоях. Вне очагов
размножения трофический фактор является как модифицирующим,
так и регулирующим плотность популяций. В очагах размножения,
когда для вредителя становятся доступными жизнестойкие деревья,
трофический фактор утрачивает на какое-то время ведущее зна-
чение в регулировании его численности. Подобное явление более
выражено при вспышках размножения насекомых-фи л л офагов.
Погодные условия оказывают прямое и косвенное влияние на
численность усача. Для этого насекомого очень важен одновре-
менный массовый вылет жуков, обеспечивающий необходимую плот-
ность откладки яиц по всей заселяемой части ствола в сжатые
сроки. Прохладная дождливая погода задерживает вылет жуков
из кукол очных колыбелек, препятствует концентрации жуков
на заселяемых деревьях, которая происходит только в теплые
солнечные дни. Косвенное влияние погодных условий выражается
в изменении состояния кормового растения. А. И. Черепанов
(1952) приводит ряд наблюдений, показывающих несомненную
связь повышения численности усача с засушливыми годами. В не-
которых же случаях, при низкой резистентности деревьев, жаркая
засушливая погода столь сильно влияет на состояние луба на юж-
ной стороне стволов, что личинки III и IV возрастов бывают
вынуждены покидать зону первоначального заселения и перехо-
дить на северную их часть.
Лиственничный усач, по крайней мере, в условиях массового
размножения не может развиваться изолированными семьями.
Для заселения жизнеспособных деревьев необходима концентра-
76
ция многих жуков, и начальное перенаселение личинок первых
двух возрастов является здесь нормальным явлением. Плотность
поселения на стволе регулируется самками при откладке яиц.
Она редко бывает ниже величины, позволяющей отродившимся
личинкам преодолеть сопротивление смоляного барьера луба.
Вместе с тем у выживших и внедрившихся в заболонь личинок не
бывает и напряженной конкуренции из-за пищи. Вся группа личи-
нок на одном дереве по характеру взаимоотношения между ними
представляет как бы одну семью (гнездо), где конкуренция из-за
пищи отсутствует.
Плотность ходов личинок III и IV возрастов под корой в разных
частях зоны заселения ствола варьирует в больших пределах.
Ходы могут занимать от 2 до 70% общей площади наружных сло-
ев заболони (см. рис. 50, 51). При этом отставание личинок в росте
наблюдается как раз по периферии пораженной зоны, где их плот-
ность, как правило, снижается. Это явление связано с изменением
биохимического состава пограничной с повреждением части забо-
лони.
Жизнь личинок большинства фитофагов, не питающихся во
взрослом состоянии, слагается из двух этапов. Первый из них —
период роста и развития, второй — накопления резервных ве-
ществ, обеспечивающих созревание половых клеток и жизнь има-
го. Лиственничный усач является наиболее ярким представителем
фитофагов, в развитии личинок которых имеется кроме назван-
ных этап «выживания». В этот период, определяющийся необходи-
мостью подавления защитных реакций дерева, гибнет (заливается
в ходах смолой) очень большое количество личинок. В гибели
личинок, освоивших луб и осо-
бенно, проникших в заболонь,
сопротивляемость дерева игра-
ет уже второстепенную роль.
По нашим наблюдениям, вне
очагов размножения выживае-
мость личинок во время прео-
доления ими смоляного барьера
Рис. 22. Выживаемость листвен-
ничного усача на разных этапах
развития.
а — в условиях депрессии численности
(Тункинская долина, 1968, 1972—
1973 гг.); б —в первой фазе очага
размножения (очаги в Читинской об-
ласти, наблюдения П. Н. Тальм ала,
1940); 9 — во второй фазе раввития
очага (Букукинский очаг, 1965—
1967 гг.).
1 — яйца; 2 — личинки, отродившиеся
из яиц; 3 — личинки, проникшие
в луб; 4 — личинки, проникшие в
глубокие слои заболони и ядровую
древесину; 5 — имаго, вышедшие из
куколочных колыбелек.
77
в среднем составляет немногим больше 1/3 особей, отродившихся из
яиц по всей зоне заражения дерева. При этом отрождаемость личи-
нок из яиц более низкая, чем в очагах. Выживаемость усача в про-
цесс© развития от яйца и до имаго (в куколочных колыбельках) в
депрессивном состоянии популяции составляет, по наблюдениям в
Тункинской долине, около 4>7% (рис. 22). Такой показатель с уче-
том возможной гибели жуков во время откладки яиц является пре-
дельно минимальным для обеспечения существования семейной
группы вредителя. В очагах размножения, где усач заселяет жизне-
стойкие деревья, во второй фазе смолой заливается от 55 до 72%
личинок I и II возрастов, в среднем 68,5%. О гибели личинок,
преодолевающих сопротивление дерева в первой фазе развития оча-
га, сведений нет. Исходя из данных П. Н. Тальмана (1940) о вы-
сокой выживаемости усача за все время развития в первом поколе-
нии вспышки размножения (13,2—30,6%), можно предположить,
что гибель молодых личинок выражается здесь значительно мень-
шими цифрами. Выживаемость усача во второй фазе очага в сред-
нем составляет 7,1% от количества отложенных яиц.
Заключение А. И. Черепанова (1950) о том, что в Забайкалье
и в Туве паразитические насекомые пе имеют существенного зна-
чения в ограничении размножения лиственничного усача, вполне
справедливо. Еще меньше гибнет личинок и куколок от патоген-
ных микроорганизмов. И все же роль паразитов, хищников и бо-
лезней в целом в сокращении численности личинок, особенно во
второй фазе вспышки размножения, относительно велика. Если
же рассматривать эффективность врагов усача в гибели той части
личинок, которая избежала участи быть залитой смолой и получи-
ла возможность беспрепятственно развиваться в лубе и заболони
отторгнутой части ствола, то опа выражается уже большими пока-
зателями.
Определить роль хищников в уничтожении яиц лиственнич-
ного усача трудно. Даже при самых тщательных учетах резуль-
таты не могут считаться вполне достоверными по той простой при-
чине, что хищники либо съедают яйца полностью, либо высасы-
вают их, и в этом случае они практически не отличаются от ес-
тественно высохших неоплодотворенпых яиц. В целом, по нашему
мнению, хищники уничтожают относительно небольшое количе-
ство яиц. Во всяком случае их деятельность мало отражается на
плотности отрождающихся личинок и на успехе преодоления
ими сопротивляемости заселяемого дерева.
Гибель личинок усача, напротив, можно учесть достаточно
полно на всех этапах их развития. Наибольшего труда здесь
требует вскрытие ходов во внешней коре, поскольку они очень
мелкие, многочисленные, проходят в разных плоскостях и к тому
же плотно забиты буровой мукой. В 1966 и 1967 гг. мы произвели
учет личинок, залитых смолой в ходах, уничтоженных птицами,
насекомыми, клещами, а также погибших от патогенных микро-
организмов на 26 деревьях, взятых из разных участков Букукин-
78
ского очага (табл. 5). Детальный анализ коры, заболони и ядро-
вой древесины производился на метровых отрезках из нижней,
средней и вершинной частей зараженной зоны ствола. Значитель-
ная доля этой трудоемкой работы выполнена сотрудником нашей
лаборатории М. К. Соковым.
Роль птиц в уничтожении вредных лесных насекомых, в том
числе ксилофагов, при высокой их численности в очагах размно-
жения, считается небольшой или даже почти неощутимой по срав-
нению с полезной деятельностью паразитических и хищных беспоз-
ночочных и патогенных микроорганизмов. Вне условий массового
размножения вредителей участие птиц в регулировании их чис-
ленности расценивается несколько выше. Конечно это правило
не без исключения. По нашим наблюдениям, главными потреби-
телями личинок и куколок лиственничного усача в забайкальских
очагах его массового размножения являются дятлы, а среди них
первое место занимает большой пестрый дятел. В результате
учетов, произведенных в Букукинском очаге, установлено, что
всеми врагами усача там уничтожалось 24,1% личинок и куко-
лок от общего числа отродившихся личинок. Из них на долю дят-
лов приходилось 18,4%. В действительности же полезная роль
дятлов была много большей, поскольку в процессе преодоления
сопротивляемости дерева при внедрении молодых личинок в луб
«естественная» гибель их без участия энтомофагов составляла
в этом очаге 68,5%. Из числа выживших личинок дятлами было
уничтожено 54,0%, паразитическими насекомыми и клещами —
11,8% и патогенными микроорганизмами — 2,2% (см. табл. 5).
Таблица 5
Гибель лиственничного усача в Букукинском очаге в 1966—1967 гг.
Стадия развития, возраст личинок, местонах ождение Пока- затель гибели Залиты смо- лой в ходах Уничтожены дятлами Уничто- жены насекомы- ми и кле- щами Погибли от пато- генных микроор- ганизмов Всего погиб- ло
Личинки младше- А 68,5 1,4 1,3 0,0 71,2
го возраста по Б — — — — —
внешней коре В 55,2-71,9 0,0—1,4 0,9-5,3 —
Личинки средних А Едияичло 12,9 3,3 0,0 16,2
возрастов в лу- Б — 41,0 10,5 0,0 51,5
бе и заболони В — 24,3-79,0 0,0-2,0
Личинки старше- А — 4,1 0,4 0,7 5,2
го возраста, ку- Б — 13,0 1.3 2,2 16,5
колки и моло- дые жуки в яд- ровой древесине В — 1,3-72,5 3,4-57,9 0,0-20,0 —
Личинки всех воз- А *68,5 18,4 5,0 0,7 92,6
растов, куколки и молодые жуки Б — 54,0 11,8 2,2 68,0
Примечание. А — средние показатели гибели, %гот общего числа личинок,
отродившихся из яиц; Б — средние показатели гибели, % (от числа личинок, преодолев-
ших защитное действие смолистых веществ; В— показатели варьирования гибели в раз-
ных частях коры и древесины,от числа проникших в них личинок.
79
Вспышки размножения и характер очагов
Переход популяций лпствеппичпого усача из депрессивного
в экспрессивное состояние происходит в лесах, ослабленных пер-
вичными вредителями, пожарами, рубками, резким нарушением
режима грунтовых и поверхностных стоков и другими фактора-
ми, снижающими резистентность деревьев. Тем не менее действи-
тельные вспышки размножения этого насекомого пока зарегист-
рированы только в лиственничниках, поврежденных сибирским
шелкопрядом и пожарами. В лесах, ослабленных фтористыми со-
единениями, выбрасываемыми в атмосферу алюминиевыми заво-
дами, численность усача вообще не повышается. Здесь он про-
должает заселять отдельные перестойные деревья, тогда как осталь-
ные подвергаются нападению других вредителей, в основном —
короедов.
Массовое размножение усача в лесах, пройденных пожарами,
отмечали П. Н. Тальман (1940), А. И. Черепанов (1952) и А. С. Иса-
ев (1966). Установлено, что на свежих гарях вредитель не разви-
вается. Очаги его возникают в ослабленных древостоях спустя
несколько лет после пожара, а именно — ко времени завершения
дифференциации деревьев па усохшие, усыхающие (с участием
насекомых-ксилофагов) и восстанавливающиеся. Последние и ста-
новятся объектом нападения усача. Сказанное относится в основ-
ном к Читинской области и Тувинской АССР. В ряде других час-
тей ареала пожары не создают благоприятных условий для вспы-
шек размножения лиственничного усача. В лесах Хамар-Дабана,
например, за многие годы наблюдений он не проявлялся в замёт-
ном количестве на гарях и в то же время заселял и губил вполне
жизнеспособные деревья в пойменных лиственничниках, подверг-
шихся временному затоплению с заиливанием почвы, хотя и не
образовывал здесь очагов размножения [Рожков и др., 1975].
Вспышки массового размножения лиственничного усача в юж-
ной части ареала, по-видимому, издавна сопутствуют ослаблению
лесов сибирским шелкопрядом. Очаги его в «шелкопрядниках» бы-
вают поистине грандиозными как по занимаемой площади, так
и по времени действия. Они всегда составляют систему взаимосвя-
занных очагов и могут занимать в целом территорию, исчисляю-
щуюся сотнями тысяч гектар. Надо полагать, что полная многократ-
ная дефолиация лиственниц гусеницами шелкопряда в сочетании
с определенными лесорастительными условиями создает самые
оптимальные условия для развития усача и особенно для выжива-
ния молодых личинок в процессе преодоления ими сопротивля-
емости дерева.
Продолжительное существование очагов размножения усача
(до нескольких десятилетий) обусловливается длительным пребы-
ванием популяции в стадном состоянии (следствие неоднократной
агрегации семейных групп), малой возможностью проявления отри-
цательного эффекта массы (численность особей, развивающихся
30
в стволе, в значительной степени регулируется противодействи-
ем самого дерева), относительно низкой эффективностью парази-
тов п хищников, а также тем, что, как показали исследования,
большие очаги размножения усача возникают в древостоях с из-
начальной пониженной резистентностью деревьев, которые после
ослабления их хвоегрызущими вредителями или другими факто-
рами восстанавливаются медленно. Многие из них заражаются
паразитическими грибами. Например, в Букукппском очаге через
35—36 лет после обесхвоивапия леса сибирским шелкопрядом
большая часть деревьев, в том числе III класса возраста, имела
признаки поражения комлевой части ствола лиственничной фор-
мой еловой губки.Однако такие деревья сохраняли устойчивость
к нападению других видов ксилофагов.
Фазы развития очагов размножения лиственничного усача не-
достаточно изучены. Пока можно с уверенностью считать лишь то,
что период наибольшей плотности популяции вредителя, когда
он заселяет деревья или делает попытки заселения их более или
менее равномерно по всей зоне первичной агрегации семейных
групп, продолжается относительно недолго, а затем сменяется
длительным периодом куртинного повреждения деревьев при много
меньшей, но все же достаточно высокой численности особей в оча-
ге в целом. Эта вторая фаза развития очагов усача (фаза дизъюнк-
ции семейных групп популяции) по продолжительности дейст-
вия и постоянному перемещению зон концентрации вредителя не-
сколько напоминает затяжные очаги размножения лиственнич-
ной расы сибирского шелкопряда в Прибайкалье [Рожков, 1965].
Основные закономерности возникновения и развития очагов
размножения лиственничного усача мы рассмотрим па примере
Букукинского очага, где деятельность вредителя была прослежена
путем анализа поврежденных, но выживших деревьев на протя-
женпп почти 40 лет. Этот очаг сложный, состоящий из ряда пер-
вичных и миграционных очагов, мозаичный по интенсивности
поражения древостоев и пульсирующий по характеру развития.
Площадь очага составляет несколько десятков тысяч гектаров.
Границу его установить невозможно, поскольку он входит в со-
став системы очагов вспышки размножения усача, возникшей в
лесах, повреждавшихся сибирским шелкопрядом в 1930—1942 гг.
на территории Нерчинского, Шилкинского, Балейского, Шело-
пугинского, Оловяпнинского, Борзинского, Приаргунского, Кал-
ганского и Сретенского районов Читинской области.
Появление малоизвестного вредителя сразу на огромной тер-
ритории вызвало большое опасение за судьбу леса. Для изучения
очагов сибирского шелкопряда и лиственничного усача Централь-
ным институтом лесного < хозяйства и Трестом лесной авиации
в 1937 г. была организована Забайкальская комплексная лесоэнто-
мологическая экспедиция. В работах экспедиции приняли участие
также сотрудники кафедры лесной энтомологии Ленинградской
лесотехнической академии (1937—1938 гг.), которые под руковод-
ил, С. Рожков
81
ством IL Н. Тальмана впервые проследили особенности экологии
усача в начальной фазе развития очагов. В 1939—1941 гг. в За-
байкалье действовал постоянный лесопатологический пункт, воз-
главлявшийся А. И. Черепановым. Уже к 1941 г. изучен цикл
развития усача и выяснено состояние его очагов. Стала очевидной
необходимость разработки мероприятий по предотвращениию даль-
нейшего увеличения численности этого опасного вредителя. Од-
нако Великая Отечественная война надолго задержала научные
и практические работы в забайкальских очагах усача (опыта
борьбы с ним до сих пор пет). Общая площадь этих очагов к 1950 г.
превышала 1,5 млн. га. В последующие годы в некоторых из них
имели место рецидивы вспышки размножения вредителя.
На территории Букукипского очага лиственничный усач оби-
тает постоянно. Повреждение им отдельных деревьев, сохранив-
ших жизнеспособность, прослеживается с начала текущего сто-
летия. До вспышки размножения некоторое повышение численно-
сти усача имело место в 1916—1918 гг. Возможно, оно связано с
пожаром 1914 г., охватившим большую площадь лиственничников.
В годы массового размножения сибирского шелкопряда первое
наиболее сильное угнетение (полное обесхвоивание в течение двух
лет подряд) деревья испытали в 1932—1934 гг., а первое массо-
вое (очаговое) заселение деревьев усачом приходилось на его поко-
ление 1934—1936 гг. В некоторых районах зоны вспышки очаги
усача появились в 1936—1938 гг. По-видимому, они были уже
миграционными.
Представленная диаграмма относительной плотности популя-
ции лиственничного усача в Букукинском очаге (рис. 23) постро-
ена по степени зараженности леса в разные годы. Она весьма при-
близительна, особенно для первой половины охватываемого пери-
ода, не отражает абсолютной зараженности леса, но все же дает
представление о характере развития очага. Букукинский очаг
возник в 1934 г. в результате концентрации вредителя с большой
территории. Па участках первоначальной концентрации, как и
па остальной площади очага, количество повреждавшихся уса-
чом деревьев, их возраст и тип заселения ствола до 1934 г. соот-
ветствовали экспрессивному состоянию популяции. Численность
вредителя была ниже, чем в период ее спада в 1948—1958 гг. В пер-
вом очаговом поколении усача часть деревьев повреждена по всей
окружности ствола и в дальнейшем усохла. Особенно много сухо-
стоя появилось в результате вредоносности второго очагового
поколения усача (1936—1938 гг.). Вырубка усохших, а также вы-
живших поврежденных деревьев для местных нужд началась в
1937 г., широкая заготовка дровяного леса проводилась в воен-
ные годы. Это очень затруднило восстановление картины развития
очага.
Пик численности вредителя в Букукинском очаге приходился
на поколение 1936—1938 гг. Усач заселял деревья почти подряд
на больших участках леса. Значительная часть этих деревьев в
82
Рис. 23. Динамика относительной плотности популя-
ции лиственничного усача в Букукинском очаге
в 1930—1968 гг. (возник в древостоях, ослабленных
сибирским шелкопрядом).
а — наличие большого количества деревьев, заселенных уса-
чом по всей окружности ствола; б — годы массового размноже-
ния сибирского шелкопряда; в — первые годы полного объеда-
ния хвои шелкопрядом.
дальнейшем усохла. Очень высокая платность популяции усача
сохранялась и в следующем поколении. О количестве жуков, за-
канчивавших развитие на деревьях в те годы, мы можем судить
по результатам учетов, проведенных А. И. Черепановым (1952)
в другом очаге той же вспышки размножения (Шахтаминская
лесная дача) в 1940 г. На деревьях с диаметром ствола па высоте
груди около 25 см им обнаружено в среднем ио 179 лётных отвер-
стий.
Большая часть жуков, вышедших из стволов в 1940 г., мигри-
ровала из Букукипского очага. Однако спад его численности не
был продолжительным. Уже в конце лета 1946 г. вредоносность
усача вновь возросла, но только на время развития одного поколе-
ния. А. И. Черепанов объясняет этот временный подъем числен-
ности засухой 1944—1946 гг., приведшей к снижению сопротивля-
емости деревьев (то же явление, по его данным, наблюдалось в Ту-
ве в 1946 г.). Последовавший затем глубокий спад плотности по-
пуляции усача охватил большинство очагов вспышки.
В течение 1948—1958 гг. численность усача в Букукинском оча-
ге находилась почти на одном уровне. Деревья заселялись в ос-
новном небольшими куртинами, иногда участками в несколько
гектаров, уже наблюдались случаи нападения жуков на отдельные
деревья. Выживаемость вредителя снизилась (в основном в резуль-
тате истребления личинок и куколок дятлами), на стволах насчи-
тывалось в среднем около 20 лётных отверстий жуков. Затем, на-
чиная с 1958 г., начался новый подъем активности усача по всему
очагу.
Резкое повышение вредоносности усача в поколении 1962—
1964 гг. явилось следствием возникновения в пределах Букукин-
ского очага трех «внутренних» миграционных очагов. Все они воз-
никли в древостоях с большой примесью березы и полнотой 0,5—
0,6, где, судя по поврежденным стволам, не было постепенного
нарастания численности вредителя. Характерным было наличие
кругового типа заселения стволов и, что особенно показательно,
массовая миграция из очага жуков, вылетевших в 1964 г. (до 80
лётных отверстий на одно дерево). Количество повреждавшихся
деревьев на этих участках, как и по всему очагу, сократилось
до уровня 1948—1958 гг.
В поколениях 1964—1968 гг. популяция усача еще сохраняла
очаговое состояние, но находилась уже на грани полной дизъюнк-
ции очень небольших семейных групп. В 1968 г. Букукинский
очаг, по-видимому, угас. Вполне возможно, что в некоторых смеж-
ных очагах семейные группы вступили в фазу депрессии числен-
ности позднее.
Характер заселения деревьев усачом в депрессивном, экспрес-
сивном и очаговом состояниях популяции описан выше. Однако
для выяснения условий возникновения и развития вспышки раз-
множения необходимо рассмотреть еще некоторые экологические
особенности, присущие вредителю в очагах и вне их.
Продолжительная депрессия численности особей в семейных
группах усача, их обособленность сопровождается инбредной
депрессией как в отношении числа оставляемых жизнеспособных
потомков, так и возможности преодоления ими сопротивляемости
дерева. Депрессивное состояние популяций характеризуется: растя-
нутым вылетом жуков из куколочпых колыбелек, что вызывает
большую продолжительность их лёта и слабую выраженность пе-
риода массовой откладки яиц; преобладанием самок в первые дни
лёта п соответственно относительно высоким процентом неоплодо-
творенных яиц, откладываемых жуками (повторное спаривание
у этого насекомого происходит в процессе откладки яиц), о чем
можно судить по отрождению личинок (см. рис. 22); неравномер-
ным развитием личинок до первой зимовки и в период между пер-
вой п второй зимовками; гибелью части молодых личинок, уже
внедрившихся в заболонь, без признаков заражения их патоген-
ными организмами.
Более всего прослежена экология усача во второй фазе разви-
тия очага. В этой фазе наблюдается четко выраженный период
массовой откладки яиц, в течение которого каждое дерево в ос-
новном заселяется за 2—4 дня, а древостой в целом (в пределах
84
семейной группы)— за 6—10 дней. Откладка яиц единичными
особями, в основном на уже заселенные деревья, может продол-
жаться еще до двух и более недель. Соотношение полов в период
массовой откладки яиц близко к 1 : 1, или даже имеет место преоб-
ладание самцов, что обеспечивает относительно полную оплодо-
творенность яиц.
Наиболее существенным отличием очагового состояния попу-
ляции в обеих фазах развития очага является повышенная вы-
живаемость личинок, проникающих в луб и заболонь. Выживае-
мость усача за весь период онтогенеза в начале первой фазы очага
может в 3 раза превышать таковую во второй фазе и в 4,5 раза —
в депрессивном состоянии популяции (см. рис. 22).
Повышение выживаемости личинок усача в первой фазе оча-
гов размножения обусловливается, по-первых, преобладанием
среди них гетерозисных особей, во-вторых, оптимальным сниже-
нием резистентности деревьев и, в-третьих, высокой концентра-
цией жуков при откладке яиц. Расчеты показывают, что для за-
селения дерева полосой по всей высоте ствола до первых живых
сучьев (8—10 м) в течение 3—4 дней при непрерывной откладке
яиц с И до 19 ч достаточно присутствия 20—25 пар жуков. В дей-
ствительности каждая самка откладывает в день не больше 40 яиц,
чаще значительно меньше, и при этом прерывает откладку па про-
должительное время. На заселяемых стволах в часы интенсивной
откладки яиц бывает большее количество особей обоих полов.
Однако во второй фазе развития очага нам не приходилось наблю-
дать концентрацию нескольких сотен жуков на одном дереве, что,
судя по данным П. Н. Тальмана (1940), является характерным
для начального этапа вспышки размножения этого вредителя.
Плодовитость лиственничного усача изучена довольно хорошо
как в депрессивном, так и в очаговом' состоянии популяций. На-
ши исследования и данные П. Н. Тальмана (1940) свидетельству-
ют о том, что переход популяции в очаговое состояние не сопровож-
дается повышением средней плодовитости самок. В очагах наблю-
дается лишь увеличение амплитуды колебания числа откладыва-
емых яиц отдельными особями. В условиях депрессии популя-
ции в яичниках самок развивается в среднем 78 (59—98) яиц,
в первой фазе очага — 64 (51—120), к концу второй фазы — 96
(51-102) яиц.
Существенное значение в жизни усача в период массового раз-
множения имеют групповые миграции. Миграции — результат
повышенной активности, проявляющейся у насекомых только
при переходе в стадное состояние. Вне очагов семейные группы
усача перекочевывают в поисках ослабленных перестойных де-
ревьев на относительно небольшое расстояние. Максимальный пе-
релет зарегистрирован нами в пойме р. Хорглохой (Тункинская
долина), где вредитель появился в угнетенном в результате вре-
менного затопления древостое, расположенном на расстоянии
1—1,5 км от ближайшего заселенного и.М лиственничника. В оча-
85
гах размножения помимо обычного перелета жуков на соседние
деревья или в ближайшие участки леса происходят миграции се-
мейных групп на значительные расстояния, в том числе с преодо-
лением безлесных пространств. Например, в 1942 г. мигрирующие
жуки лиственничного усача обнаружены в степи близ станции
Борзя на расстоянии 20—30 км от ближайшего леса. Наблюдался
массовый перелет жуков через с. Большой Зерентуй (Читинская
область) из леса, находящегося от него в 6 км, через степь в сто-
рону Нерчинского завода (Черепанов, 1952]. Вследствие направ-
ленных перелетов семейных групп и концентрации их в отдельных
участках леса образуются действенные миграционные очаги вре-
дителя.
Вспышки массового размножения лиственничного усача могут
служить примером временного разрешения противоречий между
вынужденной низкой численностью особей в семейных группах,
их разобщенностью в условиях ограниченной кормовой базы и
необходимостью концентрации большого количества особей для
преодоления сопротивляемости заселяемых деревьев. Наши наблю-
дения за поведением, развитием, выживаемостью и плодовитостью
усача в действующих очагах и вне их, литературные данные о его
экологии, а также анализ динамики очагов по сохранившимся
поврежденным деревьям позволяют представить следующую кар-
тину возникновения и развития вспышки (рис. 24).
Вспышка размножения лиственничного усача, как и насеко-
мых, развивающихся за счет зеленых частей растений, в том числе
первичных вредителей леса, есть не что иное, как переход попу-
ляции или части ее в стадное состояние. При этом усач, будучи
ксилофагом, сохраняет большую зависимость от кормовых усло-
вий по сравнению с филлофагами. Значение трофического фактора
в регулировании плотности его популяций в момент вспышки
лишь снижается, но не утрачивается. Состояние луба и древесины
определяет не только их кормовые качества. Для лиственничного
усача степень ослабления защитных возможностей луба и наруж-
ных слоев заболони (изменение количественного соотношения
терпенов и снижение смоляного давления) является решающим
моментом при освоении дерева. Поэтому трофический фактор здесь
следует рассматривать как комплекс условий питания и разви-
тия вредителя.
Конкретным выражением перехода популяции в стадное со-
стояние являются очаги вспышек. Каждый очаг представляет со-
бой комплекс взаимосвязанных семейных групп. Участки леса,
где происходит начальная концентрация вредителя, представляют
собой первичные очаги. Они, как и миграционные, проходят
две фазы — агрегации и дизъюнкции. Вторая фаза может быть
очень продолжительной и протекает на фоне пространственной
дифференцпации кормовых условий. Переход популяции из оча-
гового в депрессивное состояние в системе очагов установить труд-
но. Этот переход пе всегда приурочен к предельному сокращению
кормовой базы.
86
Рис. 24. Цикл вспышки размножения и фазы оча-
гов лиственничного усача.
Характерным для лиственничного усача является возможность
образования новых очагов за счет миграции и концентрации осо-
бей из разных семейных групп в течение всей фазы дизъюнкции.
Именно это обстоятельство определяет пульсирующий характер
и большую продолжительность действия очагов вредителя.
Многие из описанных исследователями «куртинных» и «диф-
фузионных» очагов и очажков усача в разных частях ареала на
самом дело не являются очагами вспышек размножения. Это не
более как переход отдельных семейных групп популяции из де-
прессивного в экспрессивное состояние, и поэтому их можно счи-
тать лишь очагами экспрессии популяции. Вообще на большей
части ареала усача в пределах СССР популяции его постоянно на-
ходятся в депрессивно-экспрессивном состоянии. Основная зона
вспышек размножения вредителя охватывает горные лиственнич-
ники юго-восточной части Забайкалья и юга Средней Сибири.
Вспышка размножения уЬача, приведшая к массовому усыханию
камчатской (курильской) лиственницы, имела место в 50—60-х гг.
на Сахалине (Любарский, Идрисов, 1970]. На материковой части
Дальнего Востока, по наблюдениям Л. А. Ивлиева, Д. Г. Коно-
87
нова и В. П. Масловой (1973), массовых размножений лиственнич-
ного усача на значительных площадях никогда не было. Даже
после колоссальной вспышки размножения сибирского шелко-
пряда в 1943—1944 гг. в лиственничниках Амурско-Зейского
водораздела этот ксилофаг не играл ведущей роли в отмирании
ослабленных древостоев.
Внешнее строение и образ жизни взрослого насекомого
Лиственничного усача в широтах его распространения можно
отнести к средним по размеру представителям семейства. Габиту-
ально он отличается от близких видов Xylotrechus вытянутым,
узким, окрашенным в бурый цвет телом, очень длинными ногами
и бледным рисунком на надкрыльях (см. рис. 13,8). Длина тела
самцов в среднем составляет 17,5 (11 до 20) мм, самок — 19 (11—
23) мм без яйцеклада. Голова и переднегрудь свежих экземпляров
черно-бурые, низ тела также темно-бурый, усики и ноги красно-
бурые или красновато-охристые. Надкрылья серовато-бурые,
в основании и по краям красновато-бурые или охристые; на гра-
нице их передней 1/3 по бокам шва находятся два светлых штриха,
составляющие вместе стреловидное пятно, которое иногда при-
нимает форму суживающейся к концам короткой поперечной по-
лосы; ниже, на границе задней 1/3 надкрылий и близ их верши-
ны, имеются две узкие светлые перевязи. У некоторых экземпля-
ров, кроме того, бывает по 1—2 неясных косых штриха в пле-
чевой (базальной) части надкрылий. Эти штрихи, как и концевые
перевязи, нередко отсутствуют. Светлый рисунок надкрылий
образован белесыми волосками и более светлой кутикулой под
ними.
Голова жука умеренно большая, сильно втянута в передне-
грудь. Лоб с параллельными сторонами, лобные кили очень слабо
выражены, иногда представлены едва выпуклой полосой с редки-
ми волосками (см. рис. 12, 13). Остальная поверхность лба густо
покрыта короткими сероватыми волосками. Щеки короткие, темя
в густой мелкой зернистой пунктировке, разделено слабо наме-
ченной продольной линией. Глаза относительно большие, глу-
боковыемчатые в верхней части (см. рис. 12, 13). Усики 11-членп-
ковые, у самца ненамного длиннее, чем у самки. Членики их не-
сут большое количество щетипковидных обонятельных сенсилл
(свыше 3500 па каждом членике в средней части антенны). Имеют-
ся, кроме того, и плакоидпые сенсиллы, по в меньшем количестве
(рис. 25). Мандибулы, как у типичного древогрыза, имеют вид
треугольных пирамид со слегка загнутой долотовидной верши-
ной и вогнутой задней плоскостью. По внутреннему краю они
несут гребни мелких зубчиков. Галея и лаципия максилл слабо
склеротизировапы, снабжены густыми щеточками волосков. Их
функция у непитающихся жуков, вероятно, ограничивается
выбрасыванием опилок из приоральной полости при выгрызании
88
Рис. 25.
1 — членики антенны жука; 2 — щетинковидные и плаковдные
сенсиллы антенн; з — мандибула; 4 — максилла; 5 — нижняя
губа; 6 — яйцеклад самки в выдвинутом положении и его кон-
цевая часть.
лётного отверстия в коре. Нижняя губа развита слабо, ее внутрен-
ние лопастинки редуцированы, наружные тонкие, ночти пленча-
тые (рис. 25). Максиллярные щупики 4-члепиковые. Вершина
их концевого членика уплощена, слабо хитинлзирована и снаб-
жена погруженными (целоконическими) сенсиллами. Последние
являются не вкусовыми, а обонятельными. О роли этих сенсилл
при откладке яиц сказано ниже. Вероятно, обонятельными яв-
ляются также редкие щетинковидные сенсиллы апикального чле-
ника щупиков. Лабиальные щупики 3-члениковые.
Переднеспинка жука массивная, почти шаровидная, несколько
резче суживающаяся кзади, с передним и задним окаймлениями.
Диск ее выпуклый, округло сужен и слабо килевидно приподнят
у основания. У самцов (у самок не всегда) на диске нечетко обо-
значены средняя и две боковые выпуклости, имеющие в целом
вид трилистника, направленного вперед (см. рис. 13, 8). Поверх-
ность выпуклых участков переднеспинки мелко-и густозернисто
пунктирована, в поперечных морщинках, остальная ее часть
в глубоких точках, местами скрытых прилегающими волосками.
Щиток округло треугольный. Стернальный отросток переднегруди
узкий, заднегруди — широкий; эпнстерны последней не более
чем в три раза длиннее своей видимой ширины (см. рис. 12; 2).
Для усачей трибы Clytini характерны более или менее длин-
ные ноги. У лиственничного усача и близких к нему видов.ноги
очень длинные. У самцов задние ноги превышают общую длину
тела, а их бедра заметно заходят за вершину надкрылий (см.
89
рис. 13,S). Как самцы, так п самки быстро бегают по стволу де-
рева. Их лапки снабжены топкими серповидными коготками,
а «подошвы» члеников несут густые щеточки коротких волосков.
Надкрылья сильно вытянуты, несколько уплощены к вершине,
где срез их образует небольшой апикальный зубец. Вся поверх-
ность надкрылий равномерно и очень густо покрыта короткими
прилегающими волосками. Собственно крылья хорошо развиты.
Их парусность, коэффициент полезного веса и силовая нагрузка
свидетельствуют о более высоких летальных способностях на-
секомого по сравнению с другими усачами, развивающимися на
хвойных в Северной Палеарктике.
Брюшко жука узкое у обоих полов, состоит из семи сегментов.
При этом видимых стернитов только пять (3—7). Первые два
стернита укорочены и скрыты в грудобрюшной складке, а три
последпих у самцов редуцированы, у самок обнажаются только
при откладке яиц. Вся нижняя сторона брюшка покрыта мелкой
неглубокой пунктировкой и густыми полуприлегающими воло-
сками, которые у заднего края каждого сегмента становятся бе-
лыми или светло-золотистыми и образуют поперечные каемки.
Самцы отличаются от самок более коротким концевым (VII)
сегментом брюшка.
Самка лиственничного усача обладает длинным телескопиче-
ским яйцекладом (см. рис. 25). В выдвинутом положении оп
почти удваивает длину брюшка насекомого. В состав яйцеклада
входят три сегмента (VIII — X), из которых X составляет почти
3/4 его длины. Базальный сегмент (VIII) не превышает размера
открытых сегментов брюшка, оп более склеротизирован, чем
остальные сегменты яйцекладки; IX сегмент лишь частично
выдвигается из VIII в момепт откладки яиц. Десятый имеет тон-
кие покровы, его опорная конструкция состоит из четырех тон-
ких продольных стержней, два из которых проходят по всей дли-
не, а два другие расположены в дистальной половине сегмента.
На конце яйцеклада имеется пара церков (см. рис. 26). Они снабже-
ны несколькими длинными «стоячими» щетинками — осязательными
сенсиллами. Считается, что у жуков, как и у большинства других
пасекомых с полным превращением, церки отсутствуют. Тем не
менее описываемые придатки, вероятно, соответствуют церкам.
Гениталии самца (см. рис. 13, 4) типичны для жуков, не обла-
дающих сперматофором и у самок которых половое отверстие рас-
положено в глубокой складке между сегментами, образующими
яйцеклад. Эдеагус уплощен, глубоко разделен в дистальной части
на верхнюю и нижнюю пластины; фаллобаза в виде кольца, пере-
ходящего в широкое общее основание парамер, расположенных
над эдеагусом. Многократное спаривание жуков во время от-
кладки яиц происходит при полувыдвинутом яйцекладе. Эдеагус
вводится в складку последнего между VIII и погруженным в него
основанием X сегмента (IX сегмент в это время целиком погру-
жается внутрь VIII).
90
На большей части ареала усача основной лёт жуков проис-
ходит в июле. В Приморском крае, по данным В. К. Сергеева
(1948), массовое окукливание личинок усача начинается в конце
июля и лёт жуков соответственно смещается на август. В забай-
кальских очагах размножения II. Н. Тальман (1940) находил
первых жуков уже в конце июня, а по окончании массового лёта
единичные особи отмечались им до конца августа. Нам такой про-
должительный лёт усача наблюдать не приходилось. Период
массовой откладки яиц жуками по всему ареалу обычно продол-
жается меньше двух недель, что и обеспечивает возможность
одновременной большой концентрации их па заселяемых деревьях.
Более того, у лиственничного усача имеются какие-то регулирую-
щие механизмы, с помощью которых различия во времени разви-
тия личинок постепенно сокращаются. Особенно это заметно по
продолжительности окукливания личинок, появления жуков
и вылета их из куколочпых колыбелек. Массовый вылет жуков
в пределах одной семейной группы иногда ограничивается 2—3
днями, как это имело место в Букукинском очаге в 1966 г. (рис. 26).
Жуки лиственничного усача живут недолго. Продолжитель-
ность их жизни зависит в основном от погодных условий и соот-
ветственно от темпа откладки яиц. В природе и близких к есте-
ственным лабораторных условиях большая часть особей живет
9—14 дней, а в целом — от 6 до 19 дней. Искусственно срок жиз-
ни имаго может быть продлен более чем в 3 раза.
Короткая жизнь взрослого насекомого ограничивается функ-
цией расселения и размножения. Жуки усача в естественных ус-
ловиях не питаются, и запас питательных веществ, накопленных
в личиночной стадии, быстро расходуется при высокой активности
их во время откладки яиц. В дождливую прохладную погоду
они могут оставаться в куколочных колыбельках до недели, воз-
Рис, 26. Динамика окукливания личинок (а), появления
жуков в куколочных колыбельках (б) и вылета их из ство-
лов заселенных деревьев (в) в юго-восточной части Чи-
тинской области.
а, б, в — по нашим наблюдениям в 1966 г.; alt бь «t — по наблюде-
ниям А. И. Черепанова (1952) в 1940 г.
91
можно, и больше. В теплые солнечные дни жуки, покинувшие
деревья, на которых они развивались, уже на второй день присту-
пают к откладке яиц и нередко через 5—6 дней погибают. После
откладки последних яиц самки обычно погибают на 2—3-й день.
Самцы в среднем живут на 1—2 дня меньше, чем самки.
Афагия — нередкое явление среди насекомых, в том числе
и жесткокрылых. В ряде случаев она свойственна только самкам
(некоторые Scarabaeidae, Buprestidae, Elateridae). У листвен-
ничного усача и самцы и самки пьют только воду. Нектар и пыльца
цветов их не привлекает. Потребление воды (в основном росы)
не питающимися во взрослом состоянии несекомыми известно
у чешуекрылых и ручейников. В экспериментальных условиях
при постоянной температуре около 9° и относительной влажности
воздуха пе менее 90% изолированные жуки лиственничного усача
становятся малоподвижными и живут до 22—30 дней. В опытах
П. Н. Тальмапа (1940), когда жукам вместо воды давался раствор
сахара, продолжительность жизни самцов удлинялась до 58 дней,
а самок — до 70 дней, в среднем для обоих полов — до 40—45
дней. Эти опыты свидетельствуют о том, что в природе жуки гиб-
нут в результате быстрого истощения.
Лиственничный усач заселяет деревья при незначительном
снижении ими резистентности. В лубе таких деревьев общее
количество смолистых веществ и содержание в последних нейт-
ральной фракции относительно близко к норме. Они способны
к быстрой перестройке смолоносной системы и преобразованию
состава монотерпенов в сторону повышения их защитных свойств.
В очагах размножения усач нередко нападает и па вполне
здоровые деревья (по физиолого-биохимическим показателям),
однако в этих случаях жукам далеко пе всегда удается преодо-
леть их сопротивляемость. Следы попыток отродившихся личинок
проникнуть в луб здоровых деревьев мы находили в большом ко-
личестве в Букукинском очаге размножения усача. Как пра-
вило, личинки заливались смолой во внешней коре, п лишь не-
многие особи гибли в лубе. Надо полагать, что нападение жуков
на деревья с исходной высокой резистентностью является откло-
нением от нормы, по механизмы первичной привлекательности
остаются здесь в основном теми же, что и при выборе пригодных
для заселения деревьев. Возможно, при высокой плотности по-
пуляции жуки начинают откладывать яйца па устойчивые деревья,
среди которых имеются ослабленные экземпляры, создающие
привлекательность данного участка древостоя. Именно такое объ-
яснение дают заселению здоровых деревьев короедами в очагах
размножения Р. Таре и Ю. Костер [Gara, Coster, 1968; Coster,
Gara, 1968]. По данным А. С. Исаева (1975), большой концентра-
ции жуков на таких деревьях обычно не происходит из-за быстро-
го прекращения выделения феромонов особями-первопоселен-
цами в условиях высокой активности защитных реакций заселя-
емых деревьев. Поскольку же жуки лиственничного усача при
921
откладке яиц не повреждают деревья, защитные реакции послед-
них не проявляются и соответственно пе тормозится эффект феро-
монов.
/кттрактапты, используемые жуками при выборе пригодных
для заселения деревьев (стимул первичной привлекательности),
п тем более феромоны, обеспечивающие необходимую концентра-
цию жуков для подавления их сопротивляемости (стимул вторич-
ной привлекательности), у представителей семейства Cerambycidae
практпчески не исследовались. Почти все наши представления о
механизмах химической коммуникации усачей в семейных груп-
пах пока строятся по аналогии с короедами, у которых аттракта-
ция и образование феромонов исследованы лучше, чем у других
насекомых-дендрофагов.
Участие а-пипена, лимонена и камфена в первичном привле-
чении лиственничного усача несомненно. Исследования, прове-
денные в нашей лаборатории [Массель и др., 1974; Рожков и др.,
1975; Массель и др., 1975; Плешанов, Массель, 1978], показы-
вают. что у лиственницы в процессе физиологического ослабле-
ния содержание в лубе а-пинена и Д3-карена и их количественное
соотношение изменяются по-разпому, в зависимости от характера,
степени и времени повреждения деревьев или нарушения ус-
ловий пх жизни. В то же время содержание лимонена и камфена
почти во всех случаях ослабления деревьев, приводящего в ко-
нечном итоге к нападению ксилофагов, повышается в большей
пли меньшей степени. Оно может сократиться лишь на какое-то
время, когда в ответ на повреждение увеличивается концентрация
Д3-карена и резистентность дерева временно повышается. Воз-
можно, аттрактивными веществами для лиственничного усача
служат также р-пипен (во второй половице лета содержание его
у ослабленных деревьев повышается, а у здоровых снижается),
мирцен и терпинолен.
В концентрации жуков лиственничного усача, как и других
ксилофагов, поселяющихся на жизнеспособных деревьях, пер-
вичные аттрактанты играют вспомогательную роль. Здесь всту-
пают в действие половые и агрегационные феромоны. Лишь виды,
не образующие больших гнездовых колоний, как, например,
Dendroctonus mtcans Kug., находят пригодные для заселения де-
ревья, используя в основном первичные аттрактанты. Агрегация
жуков в ответ на привлекательность кормовых деревьев просле-
жена у D. frontalis Zimm. [Coster е. а., 1977], хотя есть мнение,
что этот вид реагирует на запах аттрактанта, образующегося в ре-
зультате начального этапа разложения тканей ствола [Heikkenen,
1977]. В очагах размножения лиственничного усача можно на-
блюдать быструю концентрацию жуков на деревьях сразу же за
появлением на них первых особей, тогда как до этого они не при-
влекали жуков, откладывавших яйца на соседних деревьях.
Известно, что у короедов отдельные компоненты агрегацион-
ных феромонов образуются в кишечнике в результате окисления
93
монотерпенов кормового растения. В более сложных случаях жуки
получают с пищей вещества гормональной природы, которые лишь
включают механизм синтеза феромонов [Vite, 1978]. Установлено
также, что для образования веществ вторичной аттрактации
необходимо наличие в кишечнике не только терпенов, но и сахаров
кормового растения, оказывающих синергетическое действие
в процессе синтеза феромонов [Chararas, 1977]. Многими исследо-
ваниями показано, что большинство короедов хвойных способно
выделять половые и агрегационные феромоны лишь в процессе
питания [Vite, Gara, 1962; Jantz, Rudinsky, 1965; Исаев, Гире,
1975]. Наряду с тем у недавно вышедших из-под коры и непитав-
шихся самцов Dendroctonus adjunctus в задней кишке обнаружены
в большом количестве экзобревикомин (феромон, привлекающий
самок этого вида), транс-вербенол, вербенол, миртенол, борнеол
и изоборнеол [Hughes, Renwick, Vite, 1976]. Цис-вербенол най-
ден в кишечнике непитавшихся самок и самцов Pityokletnes curvi-
dens Germ, и P. vorontzovi Jac. [liarring Candace, 1978]. Наконец,
Ю. Рудинским с сотрудниками [Rudinsky е. а., 1977) прослежено
выделение непитавшимися и неспаривавшимися жуками псевдот-
сугового дендроктона наряду с феромонами лимонена. Лимонен
является основным компонентом моноциклических терпенов кор-
мового растения лубоеда — дугласовой пихты и аттрактантом пер-
вичной привлекательности. Присутствие его в кишечнике авторы
объяснить не смогли. Здесь в равной степени можно предположить
как возможность длительного сохранения пищевых терпенов
в неизменной форме, так и восстановление их в организме на-
секомого. Например, в экстракте из жуков Blastophaguspiniperda L.,
у которых был удален кишечник, содержался тот же типичный
набор терпенов, в том числе монотерпеновых углеводородов,
который свойствен лубу сосны, причем окисленные монотерпены
составляли небольшую часть [Francke, Heemann, 1976].
Имаго лиственничного усача не питаются. Личинки последнего
возраста питаются заболонью и, как нами установлено, частично
утилизируют смолистые вещества клеток радиальных лучей. Фе-
ромон агрегации образуется у жуков уже к моменту выхода их
из куколочных колыбелек.
Целенаправленный полет жуков лиственничного усача ви-
зуально зарегистрирован примерно с 200 м от заселяемого дерева.
Вероятно, это не предел. Полагают, что половые и агрегационные
феромоны у ксилофагов служат для привлечения па большом рас-
стоянии, а запах дерева — для ориентации на коротком расстоя-
нии [Vite, Francke, 1976]. При этом феромоны, по-видимому,
улавливаются трихоидными обонятельными сенсиллами, а ат-
трактанты дерева — базикопическими (у лиственничного усача
плакоидными) сенсиллами. У большого соснового долгоносика
(Hylobius abietis L.), например, методом антеннографии установ-
лено, что транс-вербенол возбуждает трихоидпые рецепторные
клетки, а а-пинен—базиконические [Mustaparta, 1975]. У чешуе-
крылых целенаправленный полет самцов к самкам и их химиче-
ской модели зарегистрирован с больших расстояний, например,
у дубового шелкопряда (Antheraea pernyl Guer.)— с 600 м [Пле-
ханов и др., 19771.
В ясную теплую погоду жуки лиственничного усача появля-
ются на стволах деревьев иногда с 9 ч, массовая откладка ими
яиц происходит в период с И до 18 ч, единичные особи пе покидают
деревья до 20 ч (рис. 27). Быстрое нарастание численности жуков
на заселяемом дереве наблюдается только в первый день откладки
Рис. 27. Жуки на стволе лиственницы во время откладки яиц.
яиц в его кору, в следующие 1—2 дня количество жуков если
и увеличивается, то ненамного, а затем начинает сокращаться.
Характер заселения ствола в очагах размножения и вне их рас-
смотрен выше.
Жуки па стволе держатся парами. Самцы лишь на непродол-
жительное время отпускают самок. Даже в момент откладки яйца
они обычно держат их, обхватив передними ногами в верхней
трети надкрылий. Спаривание повторяется после каждого или
после 2—3 отложенных яиц. Если количество самцов па дереве
больше, чем самок, то они затевают драки, и нередко побежден-
ные особи сбрасываются со ствола.
Яйца откладываются поодиночке под чешуйки и в трещины
коры, иногда довольно глубоко. Равномерность размещения яиц
в полосе заражения ствола при высокой концентрации жуков на
дереве регулируется с помощью феромонных меток. Одновременно
с выделением вещества для приклеивания яйца самка выделяет
пахучий секрет, служащий предупреждающей меткой для других
самок. Эти метки «читаются» жуками посредством обонятельных
сенсилл, расположенных не на антеннах, а на щупиках. Наблю-
дая за самками, ползающими по стволу, можно видеть, как они
вначале проверяют щупиками каждую щель в коре, а уже затем
поворачиваются и начинают зондировать ее яйцекладом. Такая
манипуляция продолжается иногда до двух и более минут, тогда
как самый процесс откладки яйца обычно занимает меньше ми-
нуты.
Количество яиц, развивающихся в яичниках самок, варьирует
в значительных пределах. По нашим наблюдениям, они отклады-
вают от 51 до 102 яиц. П. Н. Тальман (1940) зарегистрировал слу-
чаи откладки до 120 яиц. Во второй фазе развития очага у нормаль-
ных крупных самок в яичниках в среднем развивается 96 яиц.
К моменту выхода самки из куколочной колыбельки в ее яйцевых
трубочках вполне сформировавшиеся яйца составляют около
40% от исходного числа ооцитов; в течение первых трех дней
созревает еще примерно 33% яиц, а остальные — в последующие
2—3 дня. Все яйца за время жизни самки созревают, но обычно
после ее смерти в яичпиках остается несколько готовых к от-
кладке яиц. У некоторых особей таких яиц насчитывается до 20
и более, чаще же — от одного до семи.
Морфология и развитие преимагипальных стадий
Яйца лиственничного усача молочно-белые сильно вытянутые
и слегка семечковидные, т. е. передний их конец немного шире
заднего. Длина яиц 2,7—3 мм при ширине в передней трети 0,6—
0,8 мм.. Хорион, как у большинства яиц ксилофагов, тонкий,
участие «приклеивающего» вещества придаточных желез в уплот-
нении наружной оболочки незначительно, однако, по-виднмомуя
96
именно это вещество снижает смачиваемость яиц смолами. Яйца
усача обладают большой способностью всасывать воду, при этом
хорион их растягивается,и они приобретают грушевидную форму.
Способность к растяжению хорион не утрачивает даже в спирто-
формалиновом фиксаторе, где бластема, окружающая желток,
сохраняет первоначальные размеры, а вода заполняет образовав-
шееся пространство между желточной и наружной оболочками
(рис. 28).
Эмбриональное развитие усача нами не прослежено. В целом
оно продолжается около двух педель. По наблюдениям А. И. Че-
репанова (1952), в лабораторных условиях при среднесуточной
температуре 19,2° яйца развиваются 12—16 дней, в лесу — от
14 до 15 дней.
Личинка (рис. 29,30) безногая, до 34 мм длиной, белая с лег-
ким охристо-рыжеватым оттенком. На покровах имеются редкие
рыжеватые волоски. Лицевая часть головы красновато- или черно-
бурая с более темными верхними челюстями. Вся теменная часть
белая или слегка буроватая с маленьким угловидным пятном
над местом слияния лобпых швов.
Голова личинки относитель-
но небольшая, составляет]* *.3/5
ширины переднегруди и наполо-
вину или больше втянута в пос-
леднюю (см. рис. 30). Головная
капсула почти квадратная свер-
ху, достигает в ширину 4 мм у
самок и 3,6 мм у самцов, замет-
но суживается к переднему
краю. Её срединный и лоб-
ные швы очень слабо выраже-
ны. Усики короткие, трехчле-
никовые, их сочленовная мемб-
рана имеет вид широкого бес-
цветного членика, первый чле-
ник короткий красновато-бу-
рый, второй длиннее его/а кон-
цевой членик очень маленький
и тонкий. У личинок первого
возраста усики обнаруживают-
ся с трудом, они почти целиком
погружены в обонятельные ям-
ки. Глазков одна пара. Располо-
жены они кнаружи и несколько
Рис. 28. Разбухшее яйцо листвен-
ничного усача ири большом увели-
чении, рядом нормальные яйца в на-
туральную величину.
97
* А. С. Рожков
Рис. 29. Личинка лиственничного усача.
з — общий вид сверху и сбоку; 2 — поперечный, разрез третьего брюш-
ного сегмента; 4 — двигательные диски (мозоли) брюшных сегментов.
вниз от основания антенн. Верхняя губа прозрачная поперечно-
овальная, снабжена многочисленными щетинками, наличник
(клииеус) также бесцветный прямоугольный. Верхние челюсти
мощные, широко соприкасающиеся, слабо находящие одна на дру-
гую (особенно в первых двух возрастах личинок), с тонким режу-
98
Рис. 30.
1 — голова личинки IV воз-
раста; 2 — усик; з — рото-
вые придатки-
1
2
щим краем, имеющим спереди округлую форму (см. рис. 30). Ниж-
нечелюстные щупики состоят из трех коротких члеников (у личинок
с ортоптеройдным ротовым аппаратом максиллярный щупиконосец,
сливающийся с основанием галеа, нередко принимают за основной
членик щупиков). Внутренняя лопасть нижних челюстей хорошо
развита, ложечковидная, загнута внутрь и вооружена в концевой
части щетинками. Нижнегубпые щупики двухчлениковые. Лабио-
максиллярный комплекс личинки снабжен хеморецепторами, при
этом, по-видимому, но только вкусовыми, но и обонятельными.
Грудь — наиболее широкая часть тела личинки. Ширина ее
на 1/3 превышает общую длину всех трех сегментов, но лишь
немного больше ширины I сегмента брюшка. Боковые стороны
переднегруди округло суживаются к переднему краю. Диск
переднегруди (пронотум) не очень сильно хитинизирован, мелко-
зернистый, с продольной бороздкой л пятью более или менее за-
темненными рыжевато-буроватыми пятнами. Два из них распо-
ложены в передней половине диска, разделены узким промежут-
ком и имеют форму неровных прямоугольников с выемкой по пе-
реднему краю; большое поперечно вытянутое заднее пятно за-
нимает около трети диска, его передний край в большинстве
случаев образует срединный и по два боковых выступа; боковые
пятна небольшие продольные, расположены на аларных долях
тергита. Нередко рисунок бывает выражен очень слабо или даже
отсутствует.
Брюшко личинки состоит из девяти вполне развитых сегментов,
X сегмент имеет вид полного кольца, заключающего в себе три
анальные лопасти, а последние представляют собой рудименты
XI сегмента. Стигмы расположены на I — VIII сегментах (также
на II грудном). Тергнты и стерниты первых семи сегментов брюш-
ка несут двигательные диски, или мозоли, имеющие вид слившихся
парных вздутий. Эти ^оэоли отличаются структурой покровов
и обеспечивают последовательную фиксацию сегментов тела ли-
чинки при ее передвижении в ходе. На рис. 31 представлен участок
спинной мозоли личинки при большом увеличении.
У здоровых, нормально развивающихся личинок сегменты
брюшка в отличие от грудных несколько сужены латерально,
Рис. 31. Гребневидная складка кожных покровов, мелкие ши-
пики и волоски в области сииниых мозолей личинки.
т. е. высота^их на поперечном разрезе превышает ширину (см.
рис. 29). Сегментация тела личинок бывает паиболее выраженной
во время их активного питания и усиленного роста. Выпуклости
сегментов в это время разъединяются, обнажая глубокие меж-
сегментные участки. Напротив, в периоды покоя перед линькой
и окукливанием тело личинок укорачивается, приобретает более
округлую в поперечном сечении форму, выпуклости сегментов
спадают*и сближаются друг с другом.
Механизм передвижения личинки в прокладываемом ходе
и ее фиксации при выгрызании частиц древесины типичен для ли-
чинок ксилофагов, утративших ноги. Специфика заключается
лишь в степени участия в нем отдельных сегментов тела. Наиболь-
шую роль в передвижении личинки лиственничного усача играют
III — VI сегменты брюшка с сильно развитыми опорными мозо-
лями. Личинка, положенная на плоскую поверхность, ползти
не может, в ходе она движется путем последовательной фиксации
сегментов, упирающихся спинными и брюшными мозолями в его
стенки. В зависимости от направления движения личинки (впе-
ред или назад) волна перистальтических выпячиваний мозолей
.100
перемещается либо вперед, либо назад. При перемещении вперед
мозоли VI или VII сегментов брюшка вздуваются и упираются
в верхнюю и нижнюю стенки хода, одновременно подтягиваются
концевые сегменты и выдвигается вперед передняя часть тела.
Затем тот же процесс повторяется при последовательной фиксации
V, IV и III сегментов брюшка.
Ход, прокладываемый личинкой, в некоторых участках может
значительно превышать ширину ее тела, но в высоту он на всем
протяжении обеспечивает возможность одновременной опоры для
спинных и брюшных мозолей. Выпячивание мозолей обусловли-
вается сокращением мышц и нагнетанием гемолимфы в посегмент-
пые отделы перикардиальной и перинейральной полостей. О том,
насколько сильной должна быть фиксация мягкого в целом тела
личинки, можно судить по спрессованности экскрементов, которы-
ми она забивает ход позади себя.
У личинок многих ксилофагов основную роль в закреплении
тела при отгрызании частиц древесины играет переднегрудь,
несущая па спинной и брюшной сторонах различные по форме
опорные структуры. У личинок лиственничного усача, как от-
мечалось, переднегрудной сегмент по высоте уступает брюшным
сегментам и пе способен столь сильно увеличиваться дорсо-вен-
трально. Поэтому при питании личинка фиксируется в ходе в ос-
новном также за счет средних сегментов брюшка.
О количестве липек, претерпеваемых в процессе роста и раз-
вития личинками усачей в целом, как, впрочем, и других ксило-
фагов; известно очень мало. В работах, содержащих морфологи-
ческое описание личинок усачей и определительные таблицы
по ним (Ромадина, 1954; Шаблиовский, 1956; Ильинский, 1962;
Алексеева, Лурье, 1970; Черепанов, Черепанова, 1971, 1973,
1975, 1977; Мамаев, Данилевский, 1975;' Duffy, 1953, 1968; Тер-
рпег, 1968—1969], сведения о возрастных отличиях отсутствуют.
Есть данные о продолжительности генерации некоторых видов
усачей, об изменении сроков развития преимагипальных стадий
в разных географических зонах, о возможности длительной за-
держки развития, например, с 3 до 12 лет, с 2 до 10 лет и с 3 до
20 лет у представителей рода Er gates, Trtchojerus, Chlorophorus
[Плавильщиков, 1932], о том, что первые две линьки личинок
наступают сравнительно быстро и осуществляются у разных ви-
дов за период от 9 дней до полутора месяцев, что в эксперименталь-
ных условиях личинки усачей могут линять многократно, не пре-
вращаясь в куколку [Мамаев, Данилевский, 1975]. Однако ко-
личество возрастов у личинок даже самых массовых видов остается
невыясненным. Нам известна лишь одна работа Е. С. Петренко
(1965), в которой приводятся возрастные отличия личинок Мопо-
chamus sutor L.
Составление определительной шкалы возрастных изменений
личинок лиственничного усача помимо научного интереса важно
в практическом отношении. Каждый возраст личинок соответ-
101
ствует определенному этапу их экологии и характеризуется сте-
пенью причиняемого дереву физиологического г и технического
вреда. Поэтому определение возраста .личинок необходимо, во-
первых, при планировании сроков вырубки зараженных деревьев
как истребительного мероприятия в очагах вредителя, во-вто-
рых, при оценке степени вероятного понижения качества древе-
сины и ее стоимости в течение планируемых сроков рубки. Кроме
того, по соответствию возраста личинок с пребыванием их во внеш-
ней коре и лубе можно судить о состоянии популяции вредителя
п ее агрессивности в следующем поколении.
При двухгодовой генерации лиственничного усача продол-
жительность его личиночной стадии равна почти 23 месяцам.
Фактически личинка развивается не более 7 мес, а остальное время
находится в состоянии холодового оцепенения.-
Личинки лиственничного усача за время развития линяют
4 раза и имеют, таким образом, пять возрастов, достаточно четко
различающихся по ширине головной капсулы. Длина тела явля-
ется весьма относительным показателем возраста личинок, по-
скольку в пределах одного возраста может различаться более
чем в 2 раза и даже быть меньшей, чем у личинок предыдущего
возраста. Переходных величин между максимальной шириной
головной капсулы предыдущего и минимальной последующего
возрастов не обнаружено (табл. 6).
Вызывает некоторое сомнение коэффициент увеличения ши-
рины головной капсулы при линьке с IV на V возраст по макси-
муму и моде. Возможно, у личинок будущих самок существует
пятая линька. Дополнительным основанием для такого предполо-
жения могут служить опыты с вынужденным окукливанием ли-
чинок V возраста, взятых из заболони перед уходом их в ядро-
вую древесину, т. е. за год до нормального окукливания. Если
таких личинок в лабораторных условиях заставить питаться
чистой клетчаткой (фильтровальная бумага), то они вскоре окук-
ливаются и дают только самцов. Иных доказательств существо-
вания у личинок усача пятой линьки пока пет.
Таблица 6
Возрастные показатели личинок лиственничного усача (по из*
мерениям в Букукинском очаге), мм
Воз- раст Местонахождение Длина тела Ширина головной капсулы
мин. макс. мода мин. макс.
I Кора 2,4 4,0 0,7 0,6 0,8
II Кора, луб 4,5 8,5 1,1 1,0 1,2
III Луб 7,0 16,0 1,6 1,5 1,7
IV Заболонь, луб 10,0 25,0 2,2 2,0 2,3
V Заболонь, ядро 21,0 34,0 з,с 2,8 4,0
102
До первой зимовки личинки линяют один раз во второй по-
ловине августа, а после зимовки за 2 мес (с середины мая до конца
II декады июля) претерпевают три линьки. В это время личинки
растут наиболее быстро. В целом же, начиная со II возраста,
развитие их идет неравномерно, и иногда даже на одном дереве
в июне можно встретить личинок III, IV и V возрастов одновре-
менно. Выравнивание развития личинок начинается со времени
углубления их в ядровую древесину и особенно весной после вто-
рой зимовки.
Вылупление личинок усача из яиц на одном дереве обычно
занимает 2—3 дня, иногда затягивается па более продолжительное
время, а в древостое в целом может продолжаться от двух до трех
недель. Причем в зависимости от погодных условий, определяющих
время лёта жуков и откладки ими яиц, начало отрождения личинок
может смешаться с конца II декады июля до конца I декады ав-
густа. Так, например, по наблюдениям А. И. Черепанова (1952),
в 1940 г. в Шахтамипской лесной даче (Читинская область) ли-
чинки начали выходить из яиц 20 июля, тогда как в Букукинском
очаге, по нашим наблюдениям, в 1966 г. первые личинки появились
лишь 6 августа.
Только что отродившиеся личинки не превышают в длину
2,8 мм. Они не используют для питания скорлупу яиц, а сразу же
внедряются в кору и прокладывают в ней извилистые ходы с ко-
роткими ответвлениями. Эти ходы занимают небольшие участки
коры в тангентальной плоскости, но пронизывают всю ее толщу
(см. рис. 49, 7, 3).
Экология личинок I возраста представляет особый интерес.
Именно благодаря их жизнедеятельности, закономерно приводя-
щей к гибели значительную часть особей семейной группы, про-
исходит ослабление защитных свойств луба и обеспечивается воз-
можность оставшимся в живых особям дальнейшее развитие в за-
болони и ядре ствола.
В течение августа и большей части сентября до наступления
ночных заморозков и понижения дневных температур личинки
I возраста, как и перелинявшие на II возраст, периодически про-
кладывают ходы из внешней коры к лубу, выгрызают в нем не-
большие углубления и вновь возвращаются в кору. Луб вокруг
поранения отмирает, образуя бурые пятна (раневые линзочки).
На рис. 49 представлен участок луба с раневыми линзочками в на-
туральную величину (4) и в увеличенном виде (5). Плотность по-
ранений луба личинками варьирует в больших пределах — от
2—3 до 60—70 на одном квадратном дециметре. На одну личинку
приходится в среднем около четырех поранений из расчета на
общее количество личинок, отродившихся в коре, и около 12— из
расчета на количество выживших и внедрившихся в луб личинок.
Фактически одна личинка наносит до девяти поранений.
Механизмы преодоления сопротивляемости дерева личинками
усача рассматриваются в специальном разделе. Противодействие
103
Рис. 32. Интенсивность роста
(увеличение длины тела) ли-
чинок в июне и июле.
о — средние показатели; б — мак-
симальные показатели; в — мини-
мальные показатели.
Рис. 33. Динамика ухода личинок
в глубокие слои заболони и ядровую
древесину в юго-восточной части
Читинской области.
а — наши наблюдения в 1967 г.; б —
наблюдения А. И. Черепанова (1952) в
1941 г.; в— наблюдения Н. В. Горшкова
[Черепанов, 1952] в 1939 г.
луба заселению проявляется в основном в выделении живицы
из поврежденных окончаний смоляных каналов, которые заливают
ходы и губят большую часть личинок-первопроходцев. В августе —
начале сентября потёки смолы появляются на поверхности коры.
Однако в конечном итоге к весне следующего года луб становится
доступным для проникновения в него сохранившихся особей.
Зимуют личинки II возраста в коре, как исключение — в лу-
бе. В очень редких случаях на зимовку остаются личинки, не пере-
линявшие на II возраст (вероятно, больные и поздно отродившиеся
особи). Выход личинок из состояния зимнего покоя происходит
в I или II декадах мая, в зависимости от погодных условий весны.
Они сразу же начинают внедряться в луб.
В тех случаях, когда в формирующемся годичном слое за-
болони пораженной части ствола успевают образоваться плотные
ряды патологических смоляных каналов, личинки III возраста
довольно продолжительное время живут в лубе и первоначально
прокладывают в нем по преимуществу вертикальные ходы, едва
задевающие заболонь, а затем поперечные, углубляющиеся в за-
болонь. Такая особенность отмечена и у других кси.тофагов при
заселении ими луба жизнеспособных деревьев [Исаев, 1966].
При прокладке вертикальных ходов личинки повреждают меньшее
количество смоляных каналов, чем при поперечном их направле-
нии.
104
Заболонь в основном повреждают личинки V возраста в июне
и июле до ухода в ядровую древесину. Часть личинок питается
заболонью в течение всего августа, а некоторые особи — до конца
сентября. Наряду с заболонью они используют для питания и луб.
Этот период, особенно вторая половина июня и первая половина
июля, характеризуется наиболее интенсивным питанием и неве-
роятно быстрым ростом личинок (рис. 32). Суточный прирост их
тела в длину составляет (по расчетным данным) в среднем 0,4 мм,
при максимуме —- 0,8 и минимуме 0,16 мм. При таком темпе раз-
вития общий прирост за месяц (с середины июня по середину
июля) в среднем равен 1/3 длины тела личинок, завершивших
рост.
У личинок в период их наибольшего роста сочленовная мембрана
между сегментами задней части брюшка растягивается, а сами
сегменты становятся выпуклыми, конец брюшка приобретает не-
которое подобие «тачки» короедов вследствие уплощения и даже
впячивания кольцевого X сегмента с анальными лопастями.
«Тачка» служит для отталкивания и уплотнения экскрементов
в ходе.
Ходы, прокладываемые личинками в наружных слоях заболо-
ни, весьма характерны. Они имеют в целом поперечное направле-
ние, пе пересекаются даже при очень плотном расположении,
а изгибаются параллельно друг другу. «Поисковые» ответвления
занимают свободные участки заболони между ходами (см. рис. 50).
Каннибализм у личинок лиственничного усача отсутствует, но
в случае гибели личинки в ходе, соседняя личинка, не получая
предупреждающих сигналов, может проделать ход сквозь ее
тело.
Ширина поражаемой личинками части ствола всегда несколько
превышает таковую полосы, где были отложены яйца. На тонких
стволах в обеих фазах развития очага ходы личинок нередко
охватывают всю их окружность, а в первые 2—3 года вспышки
размножения, круговое заселение стволов, в том числе тол-
стомерных, может даже преобладать над односторонним за-
селением.
У края полосы заражения ходы обычно бывают не столь плот-
ными и часто утрачивают поперечное направление (см. рис. 51).
Противодействие защитного слоя смоляных каналов вынуждает
здесь личинку временами уходить в луб и даже кору, в результа-
те чего ходы в заболони прерываются и имеют вид коротких
отрезков.
Общая протяженность хода личинок двух последних возрастов
в заболони иногда превышает 300 мм при ширине до 13 мм и глу-
бине до 5—6 мм. На 'всем протяжении они плотно забиты
опилками.
Уход отдельных личинок в глубокие слои заболони и в ядро-
вую древесину иногда можно наблюдать уже в I декаде июня,
однако большая их часть покидает наружные слои заболони во
105
второй половине июля и в августе (рис. 33). В это время они до-
стигают предельного размера и в их жизни начинается этап
накопления резервных веществ.
Типичные ходы личинок в ядровой древесине имеют форму
дуги или скобы, расположенной в горизонтальной (поперечной)
плоскости и часто имеющей небольшой прогиб в средней части,
параллельный годичным слоям ядра (см. рис. 52). Встречаются
слабо изогнутые и почти прямые ходы через ядро (обычно в тон-
ких стволах), а также 5-образные. Последние следует считать
разновидностью дугообразного хода, образующегося в результате
того, что личинка при выходе из ядровой древесины направляет
ход в противоположную сторону. Некоторые ходы не проникают
в ядро, а проходят по границе между ним и заболонью. В целом
дугообразные и близкие к ним ходы составляют около 76%. Осталь-
ные ходы нетипичные. К пим относятся прежде всего вертикальные
ходы, которые иногда имеют также дугообразную форму, но
располагаются в вертикальной или сильно наклонной плоскости.
Чаще же на тонких стволах они пронизывают отвесно толщу ядра,
в том числе сердцевину, и затем резко изгибаются в сторону.
Длина ходов в ядровой древесине варьирует от 80 до 230 мм.
Личинки последнего возраста активно питаются глубокими
слоями заболони и ядровой древесиной примерно до середины
сентября. У личинок лиственничного усача, как и у большинства
других ксилофагов, зимующих в личиночной стадии, не наблюда-
ется четкого периода физиологической подготовки к зиме. Листо-
грызущие насекомые, например, прекращают питаться и очищают
кишечник задолго до впадения в состояние зимнего покоя. Ли-
чинки усача могут временно впадать в состояние холодового оце-
пенения-уже в начале сентября, а затем пробуждаться в теплые
дни и питаться вновь. Обычно во второй половине сентября они
уже не пробуждаются и остаются на зиму в описанном выше
дугообразном ходе па разных этапах его прокладки. Рано внед-
рившиеся в ядро личинки успевают проложить к этому времени
большую часть хода и даже выйти в заболонь с другой стороны
-ствола, другие остаются зимовать в самом начале хода, а очень
незначительная часть личинок зимует под корой. Судьба этих
личинок остается невыясненной.
Вторая зимовка личинок, как и их весеннее пробуждение,
по срокам не отличается от первой зимовки. Весной личинки на-
чинают питаться в I декаде мая, иногда только с середины этого
месяца. Они продолжают прокладывать дугообразный ход, в ре-
зультате чего значительно увеличивают содержание резервных
веществ в жировом теле.
Достигнув внутренней поверхности коры, личипки очищают
концевую часть хода от опилок, утрамбовывая их в задней его
части, и приступают к окукливанию. Таким образом, колыбель-
кой (камерой) для куколки служит окончание хода личипки. Эта
часть хода отличается от остальной более округлой формой по-
106
перечного сечения. Размеры колыбельки в длину и ширину лишь
немного превосходят таковые тела личинки, подготовившейся
к окукливанию (см. рис. 52, 7). Личинка не всегда устраивает
колыбельку вплотную к коре. Иногда между колыбелькой и корой
сохраняется тонкий слой древесины, который редко превыша-
ет 3 мм.
С того момента, когда личинка становится неподвижной (кроме
движений, являющихся реакцией на раздражение), начинается
процесс превращения ее в куколку. Такое предкуколочное состоя-
ние длится 3—5 дней, затем происходит сбрасывание экзувия.
Период окукливания личинок в лесу растянут. Первые куколки
появляются в самом начале июня, последние — в конце I декады
июля. Массовое окукливание начинается во II декаде июня и про-
должается около двух недель (см. рис. 26).
Куколка лиственничного усача (рис. 34) не очень крупная,
с вытянутым, но не узким телом и неплотно прилегающими крылья-
ми и ногами. Вначале она белая, затем приобретает желтоватый
оттенок. Длина тела варьирует от 12 до 21 мм у самцов и от 13
до 23 мм у самок. Голова и переднегрудь имеют близкую к имаги-
нальпой форму, усики у самок лишь немного заходят за передний
край заднеспинки, у самцов они почти достигают ее заднего края.
Рис. 34. Куколка лиственничного
усача.
1 — общий вид; 2 — конец брюшка самки
с вентральной стороны; з — то же, самца.
Рис. 35. Относительна^ продолжи-
тельность жизни усача на разных
этапах онтогенеза (без учета периода
зимнего покоя личинки).
Я — яйцо; л1— V — личинки I—V воз-
растов; К — куколка; Ж — жук.
107
У самок стернит IX сегмента, представленного в виде кольца,
окружающего анальный сосочек, образует две -округлые выпук-
лости (см. рис. 34). Тергиты брюшка покрыты редкими коричне-
выми шипиками. На первых шести сегментах шипики располага-
ются в виде двух овалов, обрисовывающих участки бывших мо-
золей личинки; на тергите VII сегмента они располагаются тремя
более или менее ровными рядами, в которых наибольший размер
имеют две (у самцов) или три (у самок) парышипиков, обрамляю-
щих задний край тергита. Выше этого ряда в большинстве случаев
имеется одна пара шипиков. Симметричное расположение шипиков
в рядах, в том числе и в заднем, часто нарушается вследствие их
смещения и появления дополнительных шипиков. Выпуклый
край тергита VIII сегмента несет шесть крупных загнутых внутрь
шипиков, видимых у самцов с вентральной стороны (см. рис. 34).
Развитие куколки длится 2—3 недели в среднем — 16—17
дней. Первые куколки, а именно те, которые формируются в пер-
вой половине июня, развиваются несколько дольше, чем те,
которые появляются во второй половине июня и в июле. Разница
составляет 3—4 дня. Это явление, видимо, нельзя объяснить только
повышением температуры воздуха в течение означенного периода,
поскольку в лабораторных условиях при одинаково повышенной
температуре «ранние» куколки развиваются дольше июльских
на 1—2 дня.
Куколки встречаются до середины июля. Период преоблада-
ния куколочной стадии приходится на вторую половину июня.
В это время появляются уже и первые молодые жуки в куколочных
колыбельках.
Нормальный двухгодовой цикл развития лиственничного уса-
ча захватывает три календарных года (табл. 7). В первый год ак-
Таблица 7
Фенология развития лиственничного усача в юго-восточном части Читин-
108
тивное развитие продолжается с июля по сентябрь, во второй —
с мая по сентябрь и в третий — с мая по июль.
На рис. 35 показана продолжительность развития усача на
разных этапах двухгодового цикла развития. Средняя про-
должительность развития личинок в каждом из первых четырех
возраётов, а также продолжительность развития яйца, куколки
и жизни имаго ограничивается пределами от двух до трех недель,
тогда как жизнь личинки последнего возраста без учета периода
зимнего покоя длится свыше 3 мес. Такое явление свойственно не-
которым другим усачам, например роду Monochamus, при пере-*
ходе от одногодовой генерации к двухгодовой. У них также в этом
случае наиболее долго живет личинка последнего возраста,
перезимовывающая преимущественно у периферии ствола, вблизи
будущего лётного отверстия жука (Костин, 1964].
Строение пищеварительного тракта
и особенности питания личинок
В настоящем разделе рассматриваются некоторые анатоми-
ческие и этологические особенности личинок лиственничного
усача, позволяющие им питаться древесиной живого дерева. Эти
особенности определяются, с одной стороны, наличием относи-
тельно большого количества легкодоступных для усвоения ве-
ществ в живых тканях ствола, с другой — твердостью древесины
лиственницы.
Лиственничный усач относится к той группе ксилофагов, ли-
чинки которых пе вырабатывают фермента, расщепляющего цел-
люлозу, кишечник их не содержит также и симбиотических микро-
организмов, секретирующих этот фермен*г. Питание личинок про-
исходит за счет поглощения большого количества древесины и
извлечения из нее легкоперевариваемых веществ. За время разви-
тия с I по III возраст личинка съедает в среднем 0,8 см3 коры и
луба, в IV и V возрастах только за период жизни в заболони
она съедает около 7,5 см8 древесины и примерно столько же погло-
щает пищи в V возрасте во время развития в ядровой древесине.
За всю жизнь объем съеденной личинкой коры, луба и древесины
составляет в среднем 16 см3, при максимуме 21 см3, что в 18—23 ра-
за превышает ее собственный объем в последнем возрасте.
Как уже говорилось, ходы личинок лиственничного усача на всем
протяжении заполнены плотно спрессованными опилками. Эти
опилки в отличие, например, от таковых личинок Monochamus
почти целиком состоят из экскрементов: частиц древесины, не про-
шедших через кишечный канал, среди них очень мало.
Ротовые придатки личинки представляют высокоспециализи-
рованный аппарат для питания древесиной. Основным его орга-
ном являются мандибулы. Жующий край мандибул не несет зуб*
цов, он широкоокруглый и острый. Для таких мандибул более
109
Рис. 36. Мандибулярные мышцы личинки усача.
подходит название режущий или скоблящий. Личинка не отщепля-
ет частицы древесины, захватывая их поперек волокон, а как бы
надрезает и отрывает тонкие слои, в том числе в продольном на-
правлении трахеид. Можно представить, каким сильным должен
быть нажим челюстей, если учесть относительную остроту их ре-
жущего края и твердость древесины лиственницы. Мандибулы
снабжены чрезвычайно мощными приводящими мышцами (mJ,
заполняющими большую часть головной капсулы. Они крепятся
ко внутреннему краю челюстей, но в основном — к очень сильно-
му сухожилию, отходящему от этого края. Отводящие мышцы
(т2) развиты много слабее (рис. 36).
Верхняя губа (рис. 37) подвижна, снабжена передней и задней
мышцами (mx, m2), позволяющими ей подгибаться внутрь. Дви-
жение губы обеспечивается также мышцами — дилаторами циба-
риума и ротовой полости (m3, т4). При питании личинка произво-
дит «подгребающие» движения верхней губой, иногда довольно
быстрые. Большого развития в ротовом аппарате достигают ди-
латоры глотки (m5, me).
В тот момент, когда мандибулы внедряются в древесину, ниж-
ние челюсти вместе с нижней губой несколько выдвигаются вперед
и отгибаются вниз, обнажая гипофаринкс. Когда же личинка от-
рывает частицу древесины, нижние челюсти подгибаются внутрь,
и гипофаринкс проталкивает ее в ротовое отверстие. Максиллы,
нижняя губа и гипофаринкс снабжены сложной системой мышц,
обеспечивающей их выдвижение, отгибание и втягивание (рис. 38).
Помимо мускулатуры ротового аппарата в отрывании захва-
ченных челюстями частиц древесины большую роль играют про-
дольные спинные и вентральные мышцы тела, втягивающие голо-
ву. Они начинаются от края затылочного отверстия и идут к шей-
ным склеритам, а также к фрагме, спинальным складкам и фурке
но
Рис. 37. Мышцы верхней губы, «нёба» цибариума и глотки личинки,
1 — вид сверху; 2 — видсбоку; з — то же с обнаженными мышцами; 4 — внутренняя
поверхность верхней губы п цебариума.
С1у — клипеус; Gn. jr — фронтальный ганглий; Labr — верхняя губа; mt — перед-
няя мышца верхней губы; ти, — задняя мышца губы; тя — дилататор цибариума;
тп4 — дилататор ротовой полости; т, — первый дилататор глотки; тл — второй
дилататор глотки.
переднегруди. Наиболее сильной в этой системе является перед-
няя мышца — втягиватель головы, крепящаяся на верхней стороне
черепной коробки над местом слияния лобных швов. В процессе
питания участвует также туловищная мускулатура, фиксирующая
тело личинки в ходе с помощью спинных и брюшных мозолей.
Кишечный канал личинки (рис. 39) имеет короткую и простую
по строению переднюю*кишку (Oes), сильно развитую среднюю
кишку (Mes) и столь же длинный задний отдел, включающий тон-
кую кишку (П) и прямую кишку (Rect.). Изменения кишечного
канала в процессе развития личинки в основном сводятся к удли-
нению его среднего и заднего отделов и к обособлению и усложне-
на
Рис. 38. Мышцы максплл и нижней губы личинки. *
Crd — кардо; D. sal — слюнной проток; Нур — гипофаринкс; т. sero. v — вентраль-
ные шейные мышцы; т. Нур — мышцы гипофаринкса; т. lb — лабиальные мышцы;
т. тх — максиллярные мышцы; Р. lb — лабильный щупик; Р. тх — максилляр-
кий щупик; Stp — стипес; Tnt — тенториум.
нию прямой кишки. Наибольшей дифференциации кишечный ка-
нал достигает в IV возрасте личинки. Передний отдел средней киш-
ки к этому времени становится наиболее широким, образуя вме-
стительный желудок с кольцевыми складками, увеличивающими
секретирующую и всасывающую поверхность. Тонкая кишка
подразделяется на два отдела, неодинаковые по толщине, а прямая
кишка обособляется от тонкой резким сужением над ректальным
клапаном (см. рис. 39, 3). В V возрасте по мере снижения интен-
сивности питания у личинки начинается процесс постепенного
укорачивания средней кишки и врастания свободных концов
мальпигиевых сосудов в стенку прямой кишки (криптонефрия).
У куколки сокращение длины средней кишки идет параллельно с
удлинением пищевода, прямая кишка сохраняет некоторое время
расширенное вместилище для экскрементов лишь в области доба-
вочного соединения с пей мальпигиевых сосудов (см. рис. 39, 5).
Кишечник имаго еще более модифицируется. Его передний отдел
образует объемистый тонкостенный зоб, утрачивается крипто-
нефрия, в задней кишке исчезает видимая граница между тонкой
и прямой кишками (рис. 39, б).
Переходя к более детальному рассмотрению анатомического
строения пищеварительного тракта личинки с целью выяснения
особенностей, свойственных ксилофагам, напоминаем, что многие
древесиноядные личинки жуков обладают мышечным желудком,
а также разного рода криптами и «бродильными камерами». По-
112
1 — личинки первого возраста; 2 — лячяякл второго возраста; 3 — личинки четвертого
возраста; 4 — то же, перед окукливанием; 5 — куколки (на 4-й день после окуклива-
ния); 6 — имаго. Все вскрыты со спинной стороны. I!t — первый отдел тонкой кишки;
II, — второй отдел тонкой кишки; Ма!р — мальпигиевы сосуды; Ма1р> — свободные
концы мальпигиевых сосудов, вросшие в стенку прймой кишки (криптонефрия);
Мев1 — первый отдел средней кишки; Мез* — второй отдел средней кишки, Оез —
пищевод; Rect — прямая кишка.
Рис. 40. Кардиальный клапан и средняя кишка личинки.
1 — продольный разрез через кардиальный клапан; 2 — то же, через его кольцевую
складку; з — поперечный разрез через первый отдел средней кишки; 4 — то же, через
ее второй отдел. Gut — хитиновая выстилка пищевода; Ер — эпителий; m. circ — коль-
цевые мышцы; т. long — продольные мышцы; Mes — средняя кишка; Оев — пищевод;
Peritr — перитрофическая мембрана.
добных образований у личинки лиственничного усача нет.
Пищевод личинки (рис. 40, Oes) несколько расширен перед
кардиальным клапаном, но такое расширение не может считаться
зобом, поскольку пища в нем не задерживается. Кардиальный
клапан образован за счет кольцевого утолщения стенки передней
кишки, его складка, покрытая хитиновой кутикулой, не свисает
в полость средней кишки, а отогнута в стороны и прижата к эпи-
телию последней. Средняя кишка по характеру мышечной оболоч-
ки, эпителия и перитрофической мембраны не имеет каких-либо
специфических особенностей (см. рис. 40, 3, 4).
Пилорический отдел задней кишки (рис. 41) является весьма
характерным. Его клапан состоит из трех массивных складок-
створок, расположенных непосредственно в зоне впадения маль-
пигиевых сосудов. При открывании пилорического отдела и пос-
туплении в него пищевой массы из средней кишки створки пило-
рического клапана одновременно прекращают доступ в кишечник
экскретов из мальпигиевых сосудов. Сфинктер в принятом поня-
тии отсутствует. Его роль выполняют мощные кольцевые мышцы
(m. shp.), расположенные по всей длине пилорического отдела,
и глубокие продольные складки эпителия. Xтипизированная
кутикула этих складок покрыта слегка уплощенными зубовидны-
ми шипами, направленными назад (см. рис. 41, 5). По-видимому,
такое строение мускулатуры и внутренней поверхности пилори-
ческого отдела наилучшим образом обеспечивает проталкивание
грубой пищи в тонкую кишку.
Рис. 41. Пилорический отдел задней кишки личинки.
1 — общий вид; 2 — поперечный разрез через пилорический клапан; 3 — продольный
разрез; 4 — поперечный разрез через среднюю часть пилорического отдела; 5 — внут-
ренняя поверхность продольных складок. Cut — кутикула; Ер — эпителий; Malp —
мальпигиевы сосуды; т. circ — кольцевые мышцы; т. long — продольные мышцы;
ти. spfi — мышца сфинктера; S. pyl — продольные складки; V. pyl — пилорический
клапан.
Топкая кишка (рис. 42), в отличие от таковой у личинок листо-
грызущих насекомых, образует пе менее глубокие продольные
складки эпителия, чем пилорический отдел. Они являются их
продолжением. Наибольшего развития складки, увеличивающие
всасывающую поверхность топкой кишки, достигают в среднем и
заднем ее отделах. При переходе в прямую кишку продольные
складки несколько расширяются и образуют очень слабо выра-
женный ректальный клапан. Но существу функцию клапана вы-
полняет весь задний отдел тонкой кишкй, что также связано с
питанием древесиной.
Рис. 42. Топкая кишка и ректальный клапан личинки.
1 — поперечный разреэ через переднюю (а), среднюю (в) и заднюю (с) части тонкой киш-
ки; 2 — то же, при большом увеличении; з — продольный разрез через ректальный
клапан. Cut—кутикула; Ер — эпителий; т. circ — кольцевые мышцы; т. long —
продольные мышцы.
В расширенной прямой кишке происходит дополнительное
обезвоживание и формирование комочков экскрементов. Ее эпи-
телий, кроме зоны сфинктера, не образует продольных складок.
Мускулатура достаточно своеобразная, представлена шестью
мощными тяжами продольных мышц и сильными кольцевыми
мышцами анального сфинктера (рис. 43). У личинок V возраста
возникает криптонефрия — дополнительное соединение мальпи-
гиевых сосудов с прямой кишкой. Свободные концы сосудов вра-
стают в стенку кишки между мышечной оболочкой и эпителием
(рис. 44). Они всасывают воду из прямой кишки и выводят ее
вместе с экскретами в пилорический отдел задней кишки. Обра-
зуется замкнутая система водообеспечения работы мальпигиевых
сосудов, что очень важно при недостатке влаги, получаемой ли-
чинками во время питания ядровой древесиной и, тем более, в
предкуколочный и куколочный периоды онтогенеза. По данным
А. И. Черепанова и Н. Е. Черепановой (1973), за время метамор-
фоза вес организма лиственничного усача снижается на 30%. Это
снижение в значительной мере обусловливается утратой воды.
Основные катализаторы гидролиза углеводов, белков и жи-
ров — амилаза, а-глюкозидаза (мальтаза), Р-фруктофуранозида-
за (инвертаза), пепсин, трипсин и липаза — присутствуют у всех
растительноядных насекомых независимо от биохимических осо-
бенностей их пищевого субстрата. Вероятно, присущими всем
фитофагам являются и многие другие гидролитические ферменты.
Всего, по данным С. С. Ижевского (1974), у насекомых-фитофагов
идентифицировано около 30 пищеварительных фермептов-гидролаз
(«Номенклатура ферментов», 1966) и еще 10—20 ферментов выяв-
лено и названо по гидролизуемому субстрату, но точно не иден-
тифицировано. Большинство из них свойственно всем трофиче-
Рис, 43, Прямая кишка личипки.
1 — общий вид; 2 — внутренняя поверхность; 3 — поперечный разрез через среднюю
часть; 4 — продольный разрез стенки по продольной мышце; 5 — продольный разрез
через задний отдел; 6 — поперечный разрез через анальный сфинктер. Cut — кутикула;
Ер — эпителий; m. long — продольная мышца; т. circ — кольцевая мышца.
ским группам насекомых, включая паразитов. У личинок Се-
rambycidae помимо названных ферментов обнаружены целлюлаза,
гемицеллюлаза, ксилапаза, эмульсин, эрепсин и некоторые дру-
гие [Мамаев, 1975]. Целлюлаза встречается не только в пищева-
рительном тракте ксилофагов, она найдена, например, в кишеч-
нике саранчовых. Однако и у ксилофагов наличие ее далеко не
обязательно. Ферменты типа гемицеллюлаз, по нашему мнению,
более распространены у личинок древесиноядных насекомых, от-
носимых нами к ксилофагам первого порядка.
117
Рис. 44. Криптонефр и я у личинки перед окуклива-
нием.
1 — общий вид соединения свободных концов мальпигиевых
сосудов с прямой кишкой; 2 — сосуды на внутренней поверх-
ности мышечной оболочки кишки.
Пища в кишечном канале личинки лиственничного усача не
подвергается дополнительному измельчению, и частицы древесины
в экскрементах сохраняют те же размеры, какие они имели, попав
в пищевод, почти не изменяется также и структура клеточных обо-
лочек в этих частицах. Такая особенность позволяет провести
сравнительные гистохимические исследования поедаемых личин-
кой тканей коры и древесины и их частиц, прошедших через ки-
шечник, с целью выяснения усвоения веществ.
Питание личинки I возраста внешней корой может служить
примером адаптации ксилофагов, обладающих малоспециализи-
рованным набором пищеварительных ферментов, к развитию на
живом дереве. Как было сказано, моно- и дисахаров в ритидоме
лиственницы содержится в общей сложности около 4,2% (из рас-
чета на абсолютно сухое вещество), белков, особенно необходимых
118
личинкам младших возрастов, во внешней коре ничтожно мало.
В этих условиях дополнительным источником получения пита-
тельных веществ служит, во-первых, живой луб (повреждается
личинкой I возраста периодически и нечасто) и, во-вторых, отми-
рающий луб в процессе его изолирования перидермой. Деятель-
ность личинки стимулирует формирование феллогепа и интенсив-
ное преобразование внешних слоев луба в ритидом. Вследствие
этого к моменту окончательной утраты своих защитных свойств
луб значительно утончается.
Лигнин ритидома, содержащийся главным образом в лубяных
волокнах, суберин оболочек тонкостенной пробки и танпины не
усваиваются личинкой. Конденсированные таннины коры быстро
полимеризуются и уже в пищеводе личинки превращаются в не-
растворимые флобафены. Внешняя кора в норме содержит мало
смолистых веществ, а участки, насыщающиеся живицей в резуль-
тате повреждения луба, личинка обходит. Не затрагивает она так-
же и мелкие вместилища смолы — ритидомные смоляные капсулы.
Среди опилок, не прошедших через кишечник личинки, чаще всего
встречаются частицы коры, содержащие толстостенную пробку.
Личинки, ходы которых в течение осени не проникают к лубу
отстают в росте почти наполовину, по все же линяют на II возраст.
Некоторая часть из пих гибнет. Тем не менее высокий процент
смертности личинок двух первых возрастов определяется гибелью
не отставших в росте, а нормально развивавшихся особей в про-
цессе первоначального повреждения ими луба.
Для питания личинки II возраста луб становится более до-
ступным, а в III возрасте она полностью переходит на питание жи-
выми клетками луба и заболони. Здесь помимо белков, липидов и
углеводов протопласта гидролизу в кишечнике личинки подвер-
гаются пектины первичных клеточных Стенок и межклетного ве-
щества заболони, что свидетельствует о присутствии пектолити-
ческих ферментов. Остается непонятным сокращение или пол-
ное исчезновение смолистых веществ в лубяной паренхиме и в
клетках радиальных лучей заболони, прошедших через пищева-
рительный тракт. Было предположено, что смолы вымываются из
клеток в кишечнике и входят вместе с продуктами выделения маль-
пигиевых сосудов в состав вещества, связующего отдельные час-
тицы в комочках экскрементов. Однако и там смолы гистохимиче-
ски не обнаруживаются, по крайней мере в исходных количе-
ствах.
В ядровой древесине содержится меньше 1% сахаров и крах-
мала. Между тем именно с переходом личинки в ядровую часть
ствола начинается завершающий этап накопления ею резервных
веществ. Источником получения углеводов служит здесь араби-
погалактан. Содержание этого гетерополисахарида в древесине
лиственницы, как уже говорилось, исключительно высокое. У от-
дельных деревьев лиственницы даурской он составляет почти
30% от всех сухих веществ древесины й в основном концентри-
119
руется в ядре [Чочиева и др., 1958]. Исследования А. Ф. Зайце-
вой (1958) показали, что абсолютное содержание'арабиногалакта-
на в ядровой древесине увеличивается от центра к периферии и
становится максимальным на границе с заболонью. По-видимому,
этим объясняется наличие характерного прогиба хода личинки
усача вдоль годичных слоев периферической части ядра. Прове-
денный нами гистохимический анализ частиц ядровой древесины в
экскрементах личинки свидетельствует о полном гидролизе ара-
биногалактана межклетников ранних и поздних трахеид.
Жировое тело личинок, питавшихся во второй половине июля
и в августе ядровой древесиной, уже к началу зимовки достигает
полного развития. Его париетальный и висцеральный слои запол-
няют все полости между органами (см. рис. 29, 2). Опыты показа-
ли, что такие личинки могут оставаться живыми без пищи при
температуре 18—19° до 80 дней. За это время они полностью рас-
ходуют резервные вещества жирового тела и почти наполовину
сокращаются в длину.
Механизмы преодоления усачом сопротивляемости дерева
Активное ослабление деревьев, снижение их, сопротивляемости
заселению достигается насекомыми-ксилофагами несколькими пу-
тями. Основные из них следующие.
1. Общее ослабление сопротивляемости дерева, вызываемое
взрослыми насекомыми во время дополнительного питания. Из
числа усачей такой путь присущ прежде всего представителям
рода Monochamus (в первую очередь М. urussovi Fisch.), из коро-
едов — лесным садовникам {Blastophagus). из долгоносиков —
Hylobius abietis L. и некоторым другим.
2. Ослабление только заселяемого участка ствола за счет гибе-
ли части личинок-первопоселенцев. Наиболее типичным предста-
вителем этой группы ксилофагов является лиственничный усач.
Гибель части личинок в процессе преодоления «смоляного барье-
ра» луба и наружной части заболони имеет место также у некото-
рых других усачей (например, Tetropium). у златок (Phaenops
guttulata Gebl.) и даже у короедов (Scolytus morawitzi Sem.).
3. Снижение или предотвращение токсического действия смол
путем вентиляции и очистки маточных ходов (у некоторых видов
«семейных» ходов) взрослыми насекомыми. Это явление свойст-
венно короедам, развивающимся на жизнеспособных деревьях.
Большое значение имеет здесь также расположение ходов молодых
личинок, которые прокладывают их в прикамбиальной зоне луба
и заболони, где содержится меньше смолистых веществ.
Взаимодействие лиственничного усача и заселяемых им де-
ревьев характеризуется исключительно высокой напряженностью,
и исход этого взаимодействия далеко не всегда бывает однознач-
ным. Анализ неповрежденных деревьев в очагах массового размно-
жения вредителя показал, что многие из них имеют следы «попы-
ток» поселения. Эти деревья оказались достаточно устойчивыми,
и в результате отродившиеся из яиц личинки были залиты смолой
в ходах до проникновения их из внешней мертвой коры в луб.
Приведенные в предыдущих разделах данные о жизни жуков и
личинок лиственничного усача позволяют выделить основные
механизмы преодоления этим насекомым сопротивляемости дере-
ва. Они включают целый ряд адаптаций, проявляющихся, с од-
ной стороны, в концентрированном (групповом) воздействии на
защитную систему дерева, с другой — во избежании токсического
действия смол на личинок. В основе механизмов преодоления
лиственничным усачом защитных свойств и реакций дерева лежат
поведенческие инстинкты, значительно меньшую роль играет фи-
зиологическая адаптация личинок.
Одним из очень важных условий преодоления сопротивляе-
мости дерева лиственничным усачом являются одновременный
массовый вылет жуков из куколочных колыбелек и сжатые сроки
их лвта. При обсуждении этого явления отмечалось, что основной
период откладки яиц жуками повсюду продолжается меньше двух
недель, а массовый вылет их из заселенных стволов в пределах
одной семейной группы иногда ограничивается 2—3 днями. У это-
го насекомого имеются не совсем ясные механизмы, сокращающие
продолжительность развития личинок старших возрастов и куко-
лок, первоначально отставших в развитии от основной массы осо-
бей семейной группы. Это хорошо прослеживается по динамике
окукливания личинок, появления жуков в куколочных колыбель-
ка! и их вылета (см. рис. 26).
Столь же важным условием преодоления усачом сопротивляе-
мости дерева является высокая степень взаимной координации
поведения жуков, обеспечивающей их концентрацию на заселяе-
мых деревьях и одновременную массовую откладку яиц. В кон-
центрации жуков основную роль играют вещества вторичной
аттрактации — агрегационные и половые феромоны дальнего и
ближнего действия. Равномерность откладки яиц по всей засе-
ляемой части ствола достигается за счет феромонных меток, ос-
тавляемых самками.
Период жизни молодых личинок в коре (особенности прокла-
дываемых ими ходов, характер начального повреждения луба)
является наиболее ответственным в преодолении сопротивляемо-
сти дерева. Короеды, например, при прокладке ходов в лубе раз-
рушают окончания большого количества смоляных каналов. Если
дерево обладает достаточной способностью к сопротивлению, то
обычно уже маточные ходы жуков заполняются живицей. Реже
поселения короедов гибнут в начальный период прокладки ходов
личинками (запоздалая реакция дерева). Иная картина наблюда-
ется при заселении дерева лиственничным усачом.
Личинки усача отрождаются из яиц во внешней коре (ритидо-
ме) и в течение довольно длительного времени (до первой зимовки)
наносят лубу небольшие, но массовые повреждения. Судьба дере-
121
ва и личинок решается именно в это время. Живица заливает и
губит значительную часть (в очагах в среднем около 70%) личинок
и нередко по их ходам вытекает на поверхность' ствола. Содержа-
ние активных смол в лубяных каналах сердцевинных лучей начи-
нает сокращаться (общее содержание смолистых веществ в лубе
в это время увеличивается за счет неактивных смол, заполняющих
решетчатые трубки), давление внутри смоляных каналов пони-
жается, вязкость живицы повышается. Все это приводит к тому,
что живица перестает заполнять ходы, проделываемые личинками;
и те из них, которые избежали гибели в начальный период пов-
реждения луба, проникают в последний, а затем и в заболонь.
Гибель большей или мепыпей части личинок является непре-
менным условием заселения лиственничным усачом жизнеспособ-
ных деревьев.
Личипки, преодолевшие сопротивление луба, затем не испыты-
вают или мало испытывают действие смолистых веществ. В тех
случаях, когда в лубе и наружной части заболони сохраняется
значительное количество активных смол, личинки первоначально
могут прокладывать ходы, направленные вдоль ствола. Однако
вскоре ходы принимают типичное для лиственничного усача попе-
речное направление.
Как показали наблюдения, личинки усача используют для
проникновения в луб и заболонь некоторые защитные реакции де-
рева, предотвращающие отрицательное влияние атмосферы па об-
наженные ткани. В частности, они способствуют образованию
тиллоидов, закупоривающих просветы смоляных каналов, а также
других изолирующих структур, снижающих поступление живицы
в прокладываемые ими ходы.
Насекомые, сопутствующие лиственничному усачу
Имеющиеся в литературе сведения о сопутствующих листвен-
ничному усачу ксилофагах в очагах его размножения и вне их
ограниченны и в некоторой степени разноречивы. Это объясняет-
ся, с одной стороны, эколого-географическими различиями в сос-
таве исходных группировок вторичных вредителей лиственницы в
разных частях ареала усача, с другой — отсутствием в публика-
циях данных о состоянии деревьев во время его нападения и о ха-
рактере наносимого им повреждения (местное, одностороннее,
круговое), что имеет первостепенное значение.
По данным А. И. Черепанова (1952), в юго-восточной части
Читинской области деревья, заселенные лиственничным усачом,
часто подвергаются нападению других насекомых. На второе лето
на них нападают короеды: байкальский лесовик (Dryocoetes bai-
calicus Reitt), гравер (Pityogenes chalcographus L.), большой лист-
венничный короед (Ips subelongatus Motsch.) и полосатый древе-
синник (Trypodendron Uneatum Oliv.). На второе же и третье лето
эти деревья заселяются серым сибирским усачом (Acanthocinus
122
carinulatus Gebl.), тонкоусым еловым усачом (Tetropium gracili-
cornae Reitt.), ребристым рагием (Rhagium inquisitor L.), златка-
ми — Phaenops guttulata Gebl., Anthaxia quadripunctata L., Ancylo-
cheira sibirica Fleisch., рогохвостом — Urocerus gigas taiganus
Bern, и другими насекомыми. Надо полагать, что все эти вредите-
ли заселяют деревья, значительно ослабленные до нападения
лиственничного усача, или те, которым усач нанес круговое пов-
реждение в какой-то части ствола. В. Д. Наумов (1960) также отме-
чает широкое участие усачей, златок и короедов в последующем
заселении деревьев, поврежденпых лиственничным усачом; но
не приводит списка видов. В Шимановском районе Амурской об-
ласти А. С. Исаев (1966) обнаружил па усыхающих в результате
повреждения лиственничным усачом деревьях поселения заболон-
ника Моравица Scolytus morawitzi Sem. и усачей рода Monochamus.
Деревья, ослабленные из-за неблагоприятных лесорастительных
условий (высокий уровень вечной мерзлоты и избыточное увлаж-
нение), заселялись здесь лиственничным усачом по комлевому ти-
пу по всей окружности ствола на высоту до 3—4 м и уже на сле-
дующий год становились доступными для развития ксилофагов
производной группировки. Массовое заселение вершинной части
ствола и ветвей заболонником Моравица происходило на третий
год, т. е. в период вылета молодых жуков лиственничного усача.
На севере Прибайкалья в производную группировку ксилофагов
на лиственницах, ослабленных в результате изменения лесора-
стительных условий при вырубке просек и подвергшихся затем
нападению лиственничного усача, входят, по данным Е. Д. Бе-
режных (1978), лиственничная златка, тонкоусый еловый усач и
серый сибирский усач. В работе, посвященной вредителям лесов
Тувы, А. И. Черепанов (1948) называет 11 видов ксилофагов, по-
селяющихся на лиственнице после 2—3-кратпого объедания хвои
гусеницами сибирского шелкопряда. В их число он не включает
лиственничного усача, хотя приводит данные о наличии заражен-
ных им деревьев вне очагов сибирского шелкопряда в тех же райо-
нах. Исследования А. С. Исаева и Д. А. Тарасовой (1964) очагов
сибирского шелкопряда в Туве показали, что наряду с листвен-
ничным усачом в экологических группировках ксилофагов па ос-
лабленных деревьях участвуют сибирская хвойная златка (Ап-
cylocheira sibirica Fleisch.) и серый сибирский усач, а по наблюде-
ниям Т. И. Нальняевой (1960), деревья, зараженные лиственнич-
ным усачом, в процессе усыхания заселяются короедами. В очагах
размножения сибирского шелкопряда на соседней территории
МНР В. П. Гречкин (1960) установил три типа заселения ослаб-
ленных деревьев лиственницы вторичными вредителями-первопо-
селенцами. При одном из^них ствол заселяется синим монгольским
рогохвостом (Sirex mongolorum Sem. et Guss.), полосатым древе-
синником, большим лиственничным короедом и тонкоусым ело-
вым усачом; при втором типе — большим лиственничным короедом
и серым сибирским усачом (70—90% орлабленных деревьев);
123
отдельные группы деревьев заселяются только лиственничным
усачом. Иной характер начального заселения ксилофагами лист-
венниц наблюдался Е. Г. Мозолевской (1964) на Южном Урале.
По ее данным, деревья, не имеющие внешних признаков ослаб-
ления, а также ослабленные и усыхающие, заселяются лист-
венничным усачом совместно с тонкоусым еловым усачом.
В результате наших исследований участия ксилофагов в фор-
мировании группировок, сопутствующих лиственничному усачу
в очагах его массового размножения и вне очагов, установлены
следующие закономерности.
1. В очагах массового размножения лиственничный усач всегда
нападает на деревья первым из ксилофагов и не входит в состав
какой-либо даже самой агрессивной группировки этих вреди-
телей.
2. Одностороннее (полосой) повреждение усачом ствола впол-
не жизнеспособных деревьев в очагах размножения, как правило,
усиливает защитные свойства сохранившейся его части. Такие
деревья становятся на большее или меньшее время недоступными
для повторного заселения их лиственничным усачом и тем более
другими ксилофагами.
3. Поврежденная усачом часть ствола уже до вылета имаго
может заселяться ксилофагами, для развития которых необяза-
тельно наличие живого или усыхающего луба (полосатый древе-
синник, некоторые златки, точильщики). Однако они не прони-
кают в обособившуюся смоляным барьером живую часть ствола.
4. В случае кругового поражения части ствола деревья засе-
ляются на следующий год ксилофагами производной экологиче-
ской группировки по всей его высоте. Эта группировка обычно
включает виды, характерные для исходных группировок ксило-
фагов, формирующихся на естественно усыхающих перестойных
лиственницах, а также на деревьях, необратимо ослабленных в
результате многократной дефолиации первичными вредителями.
5. Вне очагов размножения лиственничный усач нападает на
деревья, находящиеся в состоянии медленного перехода в необра-
тимое физиологическое ослабление, при котором они уже станут
доступными для поселения лиственничной златки, тонкоусого ело-
вого усача, большого лиственничного короеда, большого хвойно-
го и черно-синего рогохвостов и некоторых других. В этих усло-
виях ксилофаги нападают па деревья сразу вслед за лиственнич-
ным усачом, а первые два вида иногда и одновременно с ним. При
локальном повреждении усачом ствола (обычное явление вне оча-
гов размножения) сопутствующие виды заселяют смежную его
часть. В дальнейшем они расширяют общую зону поражения,
и усач может участвовать в повторном заселении дерева, если оно
остается живым.
Приведенные данные позволяют считать, что, по крайней мере
в периоды депрессивного состояния популяций лиственничного
усача, основными сопутствующими ему видами ксилофагов по
124
всему ареалу являются тонкоусый еловый усач и лиственничная
златка.
В последующую группировку ксилофагов — ее, видимо, сле-
дует считать завершающей — входят помимо названных видов
полосатый древесинник, ребристый рагий, серый сибирский усач,
плоский фиолетовый усач (Callidium violaceum L.). В разрушении
древесины ствола погибших лиственниц, как и отмершей его части
у выживших деревьев, в большинстве случаев участвуют мура-
вьи — красногрудый древоточец (Camponotus herculeanus L.) и
золотистоволосый древоточец (С. saxatilis Ruzs.).
Учеты хвоегрызущих насекомых, проведенные в 1966—1967 гг.
в Букукинском очаге, показали, что повреждение усачом деревьев
стимулирует повышение численности гусениц непарного шелко-
пряда (Lymantria dispar L.). Однако популяция его не выходит из
экспрессивного состояния. Другие насекомые — потребители
хвои — здесь либо встречаются в меньших количествах по срав-
нению с близлежащими здоровыми участками леса, либо вовсе от-
сутствуют. Вполне возможно, что это объясняется не только из-
менениями биохимического состава хвои поврежденных деревьев,
но и различиями экологических условий в сравниваемых дре-
востоях.
Враги усача
В число врагов лиственничного усача входят птицы, насеко-
мые (перепончатокрылые, двукрылые, верблюдки), паукообразные
(клещи, лжескорпиопы), круглые черви — нематоды и патоген-
ные микроорганизмы. При столь широком представительстве
общее число потребителей этого усача девелико по сравнению,
например, с усачами, развивающимися под корой мертвых де-
ревьев. Среди энтомофагов лиственничного усача нет специальных
паразитов, и вообще для большинства из них круг жертв и хозяев
не ограничивается только представителями Cerambicidae. Тем не
менее элементы специализации обнаруживаются, например, у та-
хины Billaea triangulifera Zett. и даже у такого многоядного хищ-
ника, как пузатый клещ (Pediculoides ventricosus Newp.). Только
этот хищник способен проникать в ходы усача и даже на какое-то
время,в ожидании выхода жуков, изменять своим хшцным наклон-
ностям.
Из птиц первостепенное значение в истреблении личинок и ку-
колок лиственничного усача имеет большой пестрый дятел (Dend-
rocopos major L.) Два других дятла — трехпалый (Picoides trida-
dylus L.) и малый пестрый {Dryobates minor L.), являющихся обыч-
ными обитателями лиственничников таежного типа, также добы-
вают пищу почти исключительно долблением зараженных стволо-
выми вредителями деревьев. Однако личинок лиственничного
усача они уничтожают в меньшем количестве и притом почти толь-
ко в начальный период их развития в коре и лубе. Заметное уве-
125
Рис, 45. Тахина — Billaea triangulijera Zett.
1— имаго; 2 — личинка; a — конец брюшка с дыхательным колпач-
ком.
личепис числепности черного дятла (Dryocopus martius L.) в дре-
востоях, поврежденных лиственничным усачом, объясняется появ-
лением в них большого количества муравьев-древоточцев (Campo-
notus) — излюбленной пищи этой птицы. Остатки хитина жуков
усача найдены в желудках большого пестрого дятла, трехпалого
дятла, кукши (Cractes infaustus L.) и поползня (Sitta europaea
L.). Вероятно, жуками могут питаться и некоторые другие птицы,
как, например, большая синица (Parus major L.) и буроголовая
гаичка (Parus atricapillus L.). Для этих птиц личинки усача не-
доступны.
Из паразитических двукрылых, развивающихся за счет личи-
нок усача, пока известна лишь одна тахина (Billaea triangulijera
Zett.) (рис. 45). Несмотря на широкое распространение в лесной
и лесостепной зонах Палеарктики и несомненную многоядпостьг
эта тахина вне очагов лиственничного усача встречается редко;
биология ее изучена недостаточно, а самый процесс заражения ли-
чинок не выявлен. Другие представители рода Billaea являются
паразитами личинок усачей и пластинчатоусых жуков.
Первые мухи В. triangulijera появляются в лесу в середине
июня, т. е. несколько раньше начала вылета жуков усача, послед-
ние выходят из пупариев в конце июля или в первых числах ав-
густа. Таким образом, к началу массового отрождения личинок
усача в лесу находится наибольшее количество мух. Окончание
их лёта не прослежено.
По данным А. И. Черепанова (1952), за время своего развития
личинки усача подвергаются заражению тахиной в два срока.
Молодые личинки заражаются ею вскоре после выхода из яиц и
живут до июня — июля следующего года. Они наравне со здоровы-
ми личинками разрушают луб и, проникая под кору, прокладыва-
ют в заболони поперечные ходы. В июне или в начале июля зара-
женные личинки усача погибают, предварительно проделав в коре
выходное отверстие, а покинувшие их личинки тахины окукли-
ваются в буровых опилках ходов. Вторично личинки усача зара-
жаются тахиной в августе следующего года незадолго до их ухода
из заболони в ядровую древесину.
В том, что мухи вылетают из зараженных усачом деревьев
дважды, можно убедиться по остаткам их пупариев в ходах сред-
невозрастных личинок в наружной части заболони и в концевой
части ходов личинок последнего возраста. И все же, видимо, все
опи относятся к одному поколению, все начинают свое развитие
в молодых, только что отродившихся личинках усача. Разные сро-
ки вылета мух объясняются разной продолжительностью разви-
тия их личинок.
Как известно, личинки паразитических и хищных мух-ксило-
бионтов утратили способность к прокладыванию собственных
ходов в древесине. Опи проникают к своим жертвам, используя их
же ходы. При этом хищники способны переносить длительные го-
лодовки, сохраняя жизнеспособность в течение года и более
[Кривошеина, Мамаев, 1967]. У тахин — практически единствен-
ных паразитов личинок усачей из двукрылых — такие возмож-
ности сравнительно ограниченны, зато их молодые личинки
могут продолжительное время оставаться в теле хозяина в состоя-
нии покоя. Для В. triangultfera одпогодовой цикл развития яв-
ляется исходным, и переход ее к двухгодовой генерации есть при-
способление к циклу развития хозяина, у которого годы массово-
го лёта чередуются с годами низкой численности. Продолжи-
тельность развития В. triangulifera зависит от времени линьки
зараженных ею личинок усача: в тех личинках, которые линяют
и начинают питаться лубом до первой зимовки, развитие тахины
завершается в течение одного года, а в личинках, линяющих на
II возраст после зимовки, развитие паразита затягивается еще
на год.
В течение большей части жизни личинка тахины лишена воз-
можности свободно перемещаться в полости тела хозяина. Она
находится в подвешенном состоянии и дышит атмосферным возду-
хом посредством воронковидного колпачка, раструб которого
плотно облегает конец ец брюшка, а трубковидпый стебель откры-
вается через стенку тела хозяина наружу. Дыхательный колпачок
образуется в результате разрастания покровов личинки усача от
краев отверстия, прободаемого паразитом. Только с наступлением
интенсивного гистолиза тканей хозяина личинка тахины, перели-
няв на III возраст, приобретает свободу движения. В это время опа
очень активно питается, буквально в несколько дней заканчивает
развитие и покидает погибшую личинку усача.
Завершение личиночной стадии паразита при одногодовом цик-
ле развития именно в момент линьки личинок усача перед уходом
из наружной части заболони в глубокие слои древесины, вероятно,
связано с описанной ранее предрасположенностью последних в
это время к окукливанию. Паразит в этом случае лишь стимули-
рует развитие гистолиза тканей хозяина.
В одной личинке усача развивается одна, реже две личинки
тахины. Покинув останки хозяина, они окукливаются либо сра-
зу, либо в течение суток. Стадия куколки длится от двух до трех
недель в зависимости от условий погоды. Молодые мухи с нера-
справившимися еще крыльями выходят из ствола через отверс-
тие в коре, проделываемое личинкой усача незадолго до своей
гибели.
Паразитические перепончатокрылые лиственничного усача
представлены ихневмонидами подсемейства Pimplinae*. Xylonomus
sepuleralis Holmgr., X. irrigator F., Xorides collaris Grav., X. ni-
tens Grav., Ephialtes mantfestator L. Все эти наездники, исключая,
может быть, последнего, заражают средневозрастных личинок
лиственничного усача, находящихся под корой (в июне — июле).
Развитие их типично для представителей подсемейства Pimplinae.
Примерно в течение полутора месяцев личинки наездников за-
канчивают питание и, оставив высосанный труп хозяина, кокони-
руются здесь же в его ходе. Зимуют личинки в коконе, окуклива-
ются весной следующего года. Е. mantfestator заражает куколок,
а иногда даже молодых, только что сформировавшихся жуков в ку-
колочных колыбельках. На рис. 46 запечатлен именно момент
заражения эфиальтом молодого жука в колыбельке.
Пузатый клещ (Pediculoides ventricosus Newport) — исключи-
тельно многоядный хищник, уничтожающий яйца, личинок, ку-
колок, а иногда и имаго очень многих насекомых. В качестве его
жертв зарегистрированы представители жесткокрылых, перепон-
чатокрылых, двукрылых и чешуекрылых, обитающих преимуще-
ственно в закрытых местах — среди растительных и животных
остатков, например, в гнездах птиц — дуплогнездников, в дре-
весине, в запасах зерна и т. п. Есть основание полагать, что он
способен нападать и на других животных, в том числе позвоноч-
ных, выступая здесь уже в качестве паразита. Известны случаи
нападения пузатого клеща на человека, у которого он вызывает
образование на коже язвочек [П. И. Шорохов, С. И. Шорохов>
1938]. Наконец, установлена возможность питания пузатого кле-
ща на сапрофитных грибках, развивающихся в ходах насекомых-
ксилофагов [Черепанов, 1948].
, Развитие пузатого клеща как хищника лиственничного усача
подробно прослежено А. И. Черепановым (1948, 1952). По опубли-
кованным им сведениям и отчасти нашим наблюдениям, самки кле-
ща нападают на молодых, только что отродившихся личинок уса-
128
Рис. 46• Наездник Ephialtes manifestator L. в момент откладки яиц и под-
корный лжескорпион из рода Cheltfer.
ча, в меньшей степени питаются его яйцами. Личинок они пред-
варительно умерщвляют, вводя через рану ядовитый секрет, об-
ладающий, возможно, протеолитическим действием (через 20 мин
после первого укола личинка желтеет), а затем уже начинают вы-
сасывать. Брюшко насосавшихся самок сильно раздувается,
приобретает вытянутую, булавовидную, а затем шаровидную фор-
му (рис. 47). У закончивших питание особей объем брюшка уве-
личивается больше чем в 1,5 тыс. раз. На одной личинке усача
обычно встречается несколько самок клеща.
Пузатый клещ является живородящим. В естественных усло-
виях не более чем через две недели после начала питания самка
родит 100—120 вполне сформировавшихся клещей. Среди них
самцов бывает очень немного, иногда не больше 4%. Этого коли-
чества тем не менее достаточно для оплодотворения всех или почти
всех самок. Неоплодотворенные самки способны производить
партеногенетическим путем только самцов. Молодые клещи вско-
ре после появления на свет (до начала питания) приступают к спа-
риванию. Самцы затем погибают либо живут непродолжительное
время, паразитируя на питающихся самках.
Оплодотворенные самки живут в ходах личинок усача, прони-
кая по ним до куколочных колыбелек. Они питаются в это время
слизью, выделяемой перитецием спрофитного грибка Ceratosto-
mella pint Munch., и производят столь же многочисленное потом-
ство, как и при питании животной пищей. Оплодотворенные сам-
ки, скопившиеся в куколочных колыбельках усача, цепляются
за анальные щетинки вышедших из куколок жуков и таким путем
переносятся ими на вновь заселяемые деревья.
По мнению А. И. Черепанова, пузатый клещ не нападает на
взрослых личинок, углубившихся в древесину, и на куколок
9 А. С. РОЖКОВ
129
Рис. 47. Пузатый клещ Pediculoides ventricosus Newport.
1 — молодая самка, 2 — самка с раздувшимся брюшком, 3 — самец,
4 — коготки и подушечка второй — четвертой пар ног.
усача, исключая случаи питания мертвыми, начавшими раз-
лагаться насекомыми. Нам, однако, приходилось находить ку-
колок самок, погибших в результате массового нападения клещей.
Последние буквально сплошной массой покрывали тергиты брюш-
ка куколок и, разрушив покровы, питались сформировавшимися
в нем яйцами.
Отложенные усачом яйца уничтожают кроме пузатого клеща
также личинки верблюдок (Inocellia, Aguila) и подкорный лже-
скорпион из рода Chelifer (см. рис. 46). Заключение П. Н. Таль-
мана (1940) о том, что одним из главных истребителей яиц лист-
венничного усача является муравьежук (Thanasimus formi-
carius L.), по-видимому, чисто умозрительное. Этот муравьежук,
как и два других вида, встречающиеся в пределах ареала листвен-
ничного усача, хищничает главным образом на деревьях, засе-
ленных короедами. Нередок он и в ходах других ксилофагов,
развивающихся под отставшей корой мертвых деревьев. На за-
селяемых лиственничным усачом деревьях личинки муравьежука,
как правило, не встречаются, а для взрослого насекомого яйца
этого вредителя просто недоступны.
В полости тела и кишечнике жуков лиственничного усача об-
наружены нематоды из семейства Tetradonematidae. Встречаются
они редко и вряд ли влияют на плотность его популяции.
Хозяйственное значение лиственничного усача.
Современное состояние, задачи и перспективы борьбы с ним
Лиственничный усач заселяет только живые деревья. По сте-
пени причиняемого деревьям физиологического вреда в периоды
массового размножения он превосходит самых серьезных хвое-
грызущих вредителей лиственницы. Будучи к тому же одним из
опаснейших технических вредителей древесины, усач по хозяй-
ственной значимости не имеет себе равных среди дендрофагов этой
породы.
О вреде лиственничного усача можно судить по известной лесо-
патологам вспышке его массового размножения 1934—1967 гг.,
очаги которой к 1950 г. только в пределах Читинской области за-
нимали свыше 1,5 млн. га. По данным П. Н. Тальмана (1940),
проводившего учет поврежденности леса в одном из очагов этой
вспышки, за три года (1936—1938) вредителем было заселено
в 4,7 раза больше деревьев, чем за предыдущие 18 лет на той же
площади. На многих участках он повредил до 75% всех деревьев,
значительная часть из которых вскоре усохла (вследствие круго-
вого заселения ствола). А. И. Черепанов (1952) отмечает, что в не-
которых очагах той же вспышки погиб почти весь спелый лес.
Например, в Шахтаминской лесной даче запас лиственницы со-
ставлял свыше 3,6 млн. м*. Из этого количества к 1941 г. повреж-
дено 1,98 млн. м*, или 54%, а во втором квартале этой да-
чи — 86%, причем 74% деревьев от всего запаса по кварталу
усохло. Вся Шахтаминская дача вскоре превратилась в кладби-
ще леса.
На Алтае, в горных лиственничниках Тувы, в Саяно-Джидин-
ском нагорье, а также в лесах МНР пока не отмечалось таких ко-
лоссальных по продолжительности и охватываемой территории
вспышек размножения усача. Однако и здесь он занимает первое
место среди ксилофагов и среди видов, наносящих лиственнице
физиологический вред. В ряде районов Тувы, например, листвен-
ничники, поврежденные усачом, являются типичным элементом
пейзажа.
Вспышки массового размножения лиственничного усача имеют
место также на Сахалине. Их возникновение связано в основном
с активностью белополосого шелкопряда (Dendrolimus superans
albolineatus Mats.) — ближайшего родственника сибирского шел-
копряда. По данным Л. JB. Любарского и Р. И. Идрисова (1970),
в равных типах леса Тымовского лесхоза (центральная часть
острова) за период примерно с 1941 г. усач заселил около 47%
лиственничных деревьев. Ив них к 1962 г. сохраняли жизнеспо-
собность около 2%, усыхающие составляли 24%, усохшие (вклю-
9*
131
чая валежник с ходами личинок вредителя) — 74%. Наибольшее
количество деревьев повреждено в 1958—1960 гг. Авторы работы
объясняют массовое усыхание древостоев снижением их сопро-
тивляемости к заселению лиственничным усачом. С увеличением
возраста деревьев (спелые и перестойные лиственничники на тер-
ритории лесхоза составляют 73,9%) постепенно увеличивалась
их зараженность дереворазрушающими грибами. Вследствие это-
го складывались благоприятные условия для развития усача.
Непосредственным толчком к вспышке его размножения, по-ви-
димому, явилась дефолиация леса гусеницами шелкопряда.
Большой урон лесному хозяйству наносит лиственничный усач
и в тех частях ареала, где не бывает вспышек его массового раз-
множения. В периоды перехода популяции из депрессивного
в экспрессивное состояние вредоносность его становится весьма
ощутимой. Особенно это проявляется при ухудшении лесорасти-
тельных условий под влиянием антропогенных факторов (Южный
Урал, западная часть зоны строительства Байкало-Амурской ма-
гистрали, районы интенсивной эксплуатации лесов в Прибайкалье
и в Амурской области).
Завершающий этап развития личинок усача протекает в глу-
боких слоях древесины. В зависимости от толщины ствола ходы
его проникают либо только в периферийную часть ядра, либо
достигают сердцевины (см. рис. 52, 53, 6). Разрушение древесины
поврежденной части ствола гнилями в одних случаях происходит
в течение 2—3 лет, в других она сохраняется почти в неизменном
виде многие годы. Значение деструктивных ксилофагов и ксило-
мицетофагов в поврежденных усачом древостоях в большинстве
случаев невелико.
Древесина из очагов усача по действующим ГОСТам относится
к дровам. По «Временным требованиям» к круглым лесоматериа-
лам, разработанным Л. В. Любарским и Р. И. Идрисовым (1970)
специально для лиственницы, поврежденной лиственничным уса-
чом, она может быть использована на 50—65% как низкокачест-
венный пиловочник (5-й сорт) и технологическое сырье. С таким
заключением можно согласиться только в отношении толстомер-
ного леса, у которого в процессе обработки периферийные пороки
устраняются. При этом выход деловой древесины, по крайней
мере в обследованных нами очагах вредителя в Читинской области,
вряд ли может быть столь высоким. Следует отметить, что в рай-
онах большой вредоносности лиственничного усача поврежден-
ный им лес нередко используется в качестве строительного мате-
риала. В пос. Букука, например, целиком выстроенном из леса,
взятого в очаге размножения усача, трудно найти бревно, брус
или доску, включая феленки дверей и оконные переплеты, без
отверстий, проточенных личинками этого вредителя.
Заслуживают внимания рекомендации названных выше ис-
следователей по способам рубок в усыхающих лиственничниках
Тымовского лесхоза. В основе своей они, несомненно, приемлемы
132
для всех древостоев, поврежденных лиственничным усачом. Ис-
ходя из интересов лесоводства и лесоэксплуатации, с учетом групп
леса, степени повреждения и усыхания деревьев, крутизны и
экспозиции склонов, а также характера почв рекомендуется три
способа рубок: сплошные концентрированные рубки без ограни-
чения размеров лесосек (с непосредственным ежегодным примы-
канием), группово-постепенные двухприемные рубки и санитар-
ные рубки.
Специальные меры борьбы с лиственничным усачом не раз-
рабатывались. Пока они ограничиваются надзором за состоянием
популяций вредителя в потенциальных очагах размножения и
лесохозяйственными мероприятиями. Последние направлены
на профилактику распространения усача в древостоях и снижение
его численности в действующих очагах.
В вонах повышенной и большой вредоносности лиственнич-
ного усача надзор за ним должен осуществляться лесопатологами,
прошедшими специальную подготовку. В задачу надзора входит:
1) наблюдения за состоянием популяции вредителя, 2) выяв-
ление изменений резистентности древостоев, 3) учет числен-
ности хвоегрызущих насекомых, в особенности сибирского шел-
копряда.
Отличительные особенности депрессивного, экспрессивного
и очагового состояний популяций усача рассмотрены в специаль-
ном разделе и отчасти при характеристике вспышек размножения.
Помимо сказанного там важным является также определение жиз-
нестойкости его личинок. Так, например, нахождение личинок II
и тем более III возрастов во второй половине мая и начале июня
межлётного года во внешней коре свидетельствует о снижении их
способности преодолевать сопротивляемость деревьев. Заселение
деревьев, обладающих более высокой резистентностью по сравне-
нию с теми, на которые усач нападает в состоянии депрессии
численности, говорит о повышении его агрессивности, характер-
ном для очагового состояния популяций.
В качестве показателя физиологического состояния (степени
ослабления) деревьев можно использовать электропроводность
луба. Для этих целей сотрудником нашей лаборатории В. А. Марь-
енко разработана и опробована схема портативного полевого при-
бора, работающего на переменном токе низкой частоты [Марьен-
ко, 1976; Агафонова и др., 1978]. Измерение электропроводности
луба следует производить на стационарных пробных площадках
с перестойными, спелыми и приспевающими деревьями. Особое
внимание необходимо уделять состоянию деревьев в засушливые
годы, а также в древостбях, поврежденных хвоегрызущими вре-
дителями и пожарами.
В тех лесхозах, где вспышки размножения усача ранее не на-
блюдались, но возможность возникновения их не исключена (рас-
положены в пределах общей зоны вспышек, имеют перестойные
древостои, где зарегистрированы случаи перехода семейных групп
вредителя в экспрессивное состояние), необходимо также вести
систематический лесопатологический надзор.
Очаги размножения усача возникают так же неожиданно, как и
очаги сибирского шелкопряда, но обнаружить их путем аэро-
визуальных наблюдении значительно труднее. Кроны поврежден-
ных деревьев сохраняют зеленый цвет, по меньшей мере, в течение
двух лет, а часто деревья остаются живыми с наполовину источен-
ным стволом многие годы. Поэтому до появления надежной ап-
паратуры для дистанционной регистрации физиологического ос-
лабления древостоев заменять полностью наземное обследование
аэровизуальным нельзя.
Параллельно с надзором проводятся профилактические меро-
приятия на всей территории потенциальных очагов размножения
усача. Эти мероприятия направлены на предотвращение перехода
его популяции в экспрессивное состояние и на предупреждение
и устранение причин, снижающих устойчивость деревьев. Они
предполагают первоочередное вовлечение в эксплуатацию пере-
стойных древостоеЬ, вырубку отдельных деревьев пострепродук-
тивного возраста, утративших способность противостоять напа-
дению усача, и повышенную охрану лесов от пожаров и насеко-
мых-филлофагов.
При обнаружении даже небольших по площади очагов усача
с признаками начала вспышки размножения (особенно в зоне его
большой вредоносности на юго-востоке Забайкалья) необходимо
срочное проведение крупного лесопатологического обследования
территории всех смежных лесхозов. Такое обследование должна
проводить специализированная экспедиция.
Пока основным способом борьбы с лиственничным усачом
в действующих очагах остается вырубка заселенных им деревьев
и целых участков леса (при куртинно-сплошном заселении). Сро-
ки проведения рубки должны быть приурочены ко времени на-
хождения личинок в лубе и наружных слоях заболони. Сваленные
деревья подлежат немедленной окорке с целью уничтожения
личинок.
Выборка свежезаселенных усачом деревьев очень трудное дело.
Первым признаком активного повреждения личинками луба и
заболони являются тонкие потёки смолы на поверхности ствола.
Обычно такие потёки появляются в начале сентября, а иногда
только в мае следующего (межлётного) года. Между тем клейме-
ние, рубку и окорку деревьев необходимо закончить до первых
чисел июля, до начала массового ухода личинок в глубокие слои
заболони и ядро ствола (см. рис. 33).
В случае вынужденной вырубки заселенных деревьев во вто-
рой половине лета и осенью межлётнего года, а также^в зимнее
время и весной лётного года, их следует удалять из леса до появ-
ления куколок вредителя и подвергать переработке, обеспечи-
вающей гибель личинок.
Одной из первоочередных задач совершенствования лесоза-
щитных мероприятии в зонах повышенной и большой вредонос-
ности лиственничного усача является организация единого цент-
ра по сбору всесторонней информации об этом вредителе и согла-
сованию научных и производственных работ. Все сведения долж-
ны собираться по единой программе, позволяющей их дальнейшую
машинную обработку. В Институте леса и древесины им. В. Н. Су-
качева СО АН СССР выполняется подобная программа по над-
зору за сибирским шелкопрядом.
В задачу центра входит: 1) организация и надзор за деятель-
ностью карантинной службы в районах массового размножения
лиственничного усача с целью предотвращения возможного за-
воза его с лесом в другие районы страны; 2) назначение сроков
и мер борьбы в очагах размножения, а также контроль за прове-
дением лесхозами профилактических мероприятий в лесах с боль-
шим запасом перестойных деревьев и ослабленных хвоегрызущи-
ми насекомыми, пожарами и другими факторами; 3) координиро-
вание исследований, направленных на усовершенствование ме-
тодов надзора за состоянием популяций усача и борьбы с ним.
Центр должен объединить усилия лесоводов, энтомологов, био-
химиков и биофизиков в этом важном деле.
По нашему мнению, при поисках новых путей и способов пре-
дупреждения массового повреждения леса лиственничным усачом
необходимо ориентироваться прежде всего на средства его аттрак-
тации. Использование химических средств борьбы, паразитов
и болезнетворных организмов вряд ли будет иметь успех.
Прошло всего лишь два десятилетия с того времени, когда не-
мецкий биохимик А. Бутенандт впервые выяснил химическую
структуру, а затем и синтезировал половой аттрактант насеко-
мого [Butenandt, 1959, 19621. Этим первым насекомым был туто-
вый шелкопряд (Bombyx mori). Вслед за тем американский иссле-
дователь М. Джекобсои [Jacobson, 1963] выделил в чистом виде
феромон самок непарного шелкопряда (Lymantria dispar L.),
использовав для этого 500 тыс. бабочек. Он назвал его гиптолом.
Количество гиптола, вырабатываемое железами одной самки (око-
ло 0,1 мкг), может привлечь до миллиона самцов. Этот половой
аттрактант также был синтезирован. В 60-е годы изучением при-
влекающих веществ у насекомых увлеклись многие ученые. Те-
перь ежегодно выходит несколько десятков статей с результатами
исследования и испытания природных и синтетических аттрак-
тантов, причем около трети публикаций посвящается практиче-
скому использованию феромонов и веществ первичной привлека-
тельности в защите лесных и сельскохозяйственных растений.
Синтез феромонов4 открывает исключительные возможности
в избирательном снижении плотности популяций только вредящих
видов. Это очень важно в свете современных представлений о вре-
доносности насекомых, их взаимодействии с растениями и уязви-
мости биоценозов.
«5
Методика синтеза феромонов в последние годы значительно
усовершенствована, внесены изменения и в стратегию синтеза,
в использование реакции и реагентов [Katzenellenboden, 1976];
появились обзоры текущего положения и перспектив в области
применения феромонов в борьбе с насекомыми [Джекобсон, 1976;
Campion, 1976; Shorey, 1977; Бероза, 1977]. Целый ряд работ по-
священ аттрактивности половых и агрегационных феромонов кси-
лофагов (в основном короедов) и использованию их в ловушках
и на ловчих деревьях [Dyer, 1975; Vite, 1978; Bakke, Austara,
1978; и др. I.
В конечном итоге определились три главные направления в об-
ласти использования феромонов для борьбы с вредителями:
1) применение половых аттрактантов в качестве приманок в ло-
вушках; 2) нарушение нормальной химической коммуникации
путем насыщения атмосферы феромоном (дезориентация полов
в период спаривания); 3) применение антиагрегационных феро-
монов (репеллентов). В лесозащите наиболее перспективно первое
направление.
Использование полового аттрактанта самок в борьбе с лист-
венничным усачом, как и с другими ксилофагами, у которых для
оплодотворения яиц требуется многократное спаривание, должно
быть эффективным. Откладка какой-то части неоплодотворенных
яиц (самки усача, отложив яйцо, оставляют феромонную метку,
предупреждающую о том, что данный участок коры занят) при-
ведет к гибели личинок, отродившихся из оплодотворенной части
яиц, которые из-за недостаточной численности не смогут преодо-
леть сопротивляемость дерева. Возможно, однако, что более дей-
ственным окажется половой аттрактант самцов. Пока не извест-
но, какие феромоны обусловливают массовую концентрацию жу-
ков обоих полов на заселяемых деревьях. Существует^мнение, что
агрегационные феромоны короедов служат сигналом о наличии
подходящего кормового субстрата и о месте встречи полов [Vite,
1978], т. е. они заменяют как аттрактанты первичной привлека-
тельности, так и половые аттрактанты дальнего действия. Во вся-
ком случае, выяснение химической структуры феромонов лист-
венничного усача и их синтез представляются насущной
задачей.
И все же генеральным направлением в защите леса от вреди-
телей и болезней является создание здоровых, устойчивых древо-
стоев. Эта всеобъемлющая проблема не может быть решена в целом
виде в ближайшее время. Она будет решаться поэтапно, в процессе
изучения отдельных узловых вопросов. К числу таких вопросов
относится повышение устойчивости древесных растений к повреж-
дению насекомыми-ксилофагами.
Изучение защитных свойств и реакций у лесных древесных
растений начато в недавнее время. Но то, что уже известно, дает
основание считать иммунологический подход в защите леса наи-
более перспективным. Выяснение природы иммунитета деревьев
136
открывает возможность целенаправленных поисков путей повы-
шения их устойчивости к вредителям.
Без знания защитных свойств и реакций дерева и его взаимо-
действия с насекомыми-фитофагами нельзя иметь полное пред-
ставление о здоровом дереве, нельзя разработать надежные ме-
тоды диагностики его физиологического состояния, а значит, нель-
зя решить проблему создания жизнестойких лесонасаждений.
ЗАЩИТНЫЕ МЕХАНИЗМЫ У ЛИСТВЕННИЦЫ
ОТ ПОВРЕЖДЕНИЯ НАСЕКОМЫМИ-КСИЛОФАГАМИ
Во взаимодействии дерева и насекомых-фитофагов наибольший
интерес представляют защитные механизмы первого от поврежде-
ния видами, развивающимися в тканях ствола. Только познав их,
можно вести целенаправленные поиски путей повышения рези-
стентности как искусственных лесонасаждений, так и естествен-
ных древостоев. Это тем более важно в наше время, когда хозяй-
ственная деятельность человека, как правило, способствует ак-
тивности именно вторичных вредителей леса.
Средства защиты у древесных растений от повреждения насе-
комыми можно разделить на две группы. Первая из них включает
защитные свойства, присущие деревьям вообще, т. е. вне связи
с повреждением. Таковыми являются: репеллентность выделяе-
мых наружу эфирных масел, защитные свойства (в основном ме-
ханические) покровных тканей — эпидермы и перидермы и ток-
сичность клеток флоэмы и мезофилла. Вторую группу средств
защиты составляют реактивные структурные и функциональные
преобразования, возникающие в ответ на повреждение. Эти пре-
образования проявляются в изменении строения и функций от-
дельных клеток, в повышенном синтезе защитных веществ, в об-
разовании раневой ткани и восстановлении утраченных клеток.
Регуляторные механизмы жизнедеятельности дерева в целом
перестраиваются на поддержание реактивной направленности
метаболизма и 'локальную1 активизацию меристем и росто-
вых процессов.
При написании настоящего раздела в основном использованы
ранее опубликованные нами материалы, переработанные и до-
полненные в соответствии с поставленной задачей. Предлагаемый
обзор является, таким образом, итогом обобщения данных много-
летних исследований анатомических и биохимических защитных
преобразований, возникающих у лиственницы при повреждении
лиственничным усачом, а* также некоторыми другими ксилофага-
ми, повреждающими жизнеспособные деревья.
Кроме деревьев, поврежденных насекомыми, для изучения
защитных преобразований были использованы деревья, получив-
шие искусственные поранения ствола, имитирующие повреждения
<37
луба и заболони личинками лиственничного усача на разных ста-
диях развития.
Большинство защитных реакций у деревьев носит неспецифи-
ческий характер [Рожков, 1971J. Практически все формы струк-
турных преобразований в лубе и заболони, возникающие в резуль-
тате повреждения дерева ксилофагами, можно вызвать путем ме-
ханического повреждения или путем локального ожога ствола.
Но, например, при дефолиации деревьев и очагах размножения
хвоегрызущих насекомых или при общем ослаблении их под влия-
нием вредных выбросов промышленности анатомические измене-
ния в тканях ствола ограничиваются лишь трансформацией не-
которых клеток луба и заболони. Конечно, при этом сокращается
прирост древесины и изменяется соотношение рядов ранних и
поздних трахеид.
Интенсивность защитных преобразований в тканях ствола
зависит не только от характера и степени повреждения, но и от
физиологического состояния дерева. У здоровых и незначительно
ослабленных деревьев, сохраняющих достаточно высокую рези-
стентность, реакции па повреждение проявляются в значительно
большей степени, чем у сильно ослабленных, хотя и жизнеспособ-
ных деревьев. У необратимо ослабленных деревьев эти реакции
либо вовсе не обнаруживаются, либо бывают очень слабыми.
Небезынтересно отметить, что у срубленных деревьев луб и на-
ружная часть заболони остают-
ся живыми в течение продолжи-
тельного времени. Стволы све-
жесрубленных лиственниц пер-
воначально даже сохраняют спо-
собность активно реагировать
на попытки поселения ксилофа-
гов, заливая ходы смолой. В
прикамбиальной зоне древесины
у них могут происходить и не-
которые анатомические преоб-
разования. Особенно долго про-
тивостоят заселению ксилофа-
гами отрезки стволов в том слу-
чае, если их торцы сразу же тща-
тельно залить варом или гуд-
роном. В наших опытах закоп-
Рис. 48. Отрезок ствола листвен-
ницы с залитыми гудроном торцами.
В опытах такие отрезки не заселя-
лись вредителями в течение лета,
а к осени на них развивались бра!
хибласты с пучками гипертрофиро.
ванной хвои.
4 ЗЯ
сервированные таким способом части ствола лиственницы не за-
селялись вредителями в течение всего лета, а к осени на них сфор-
мировались брахибласты с пучками гипертрофированной хвои
(рис. 48).
Преобразование элементов флоэмы и ксилемы.
Раневые новообразование
Анатомические преобразования в лубе и заболони, возникаю-
щие при повреждении дерева ксилофагами, в целом направлены,
во-первых, на образование раневых покровных и замещающих
тканей, а также других изолирующих структур, предотвращаю-
щих деструктирующее воздействие атмосферы на обнаженные жи-
вые клетки, во-вторых — на повышение синтеза защитных ве-
ществ. Последнее у хвойных осуществляется за счет вовлечения
в смоло- и фенолообразующие системы клеток лучевой паренхимы
и трахеид, в норме не содержащих эти соединения, а также кле-
ток раневой (каллюсной) ткани.
По характеру образования раневые элементы, таким образом,
могут быть либо репродуцированными, либо трансформирован-
ными. Первые развиваются из околораневых участков меристем
или их производных — клеток, сохранивших способность к де-
лению, вторые’— результат преобразования клеток, не завер-
шивших дифференциацию. В некоторых случаях преобразованию
подвергаются вполне сформировавшиеся клетки, в том числе
с одревесневшими стенками.
Структурные изменения в ксилеме Ги флоэме деревьев,
подвергшихся повреждению насекомыми-ксилофагами (рис. 49—
64), сопровождаются не менее глубокими изменениями функции
клеток этих тканей. Трахеальные клетки ксилемы, в норме свя-
занные главным образом с функцией проведения воды, превра-
щаются в клетки, продуцирующие и накапливающие защитные
вещества; клетки лучей, проводящиетвглубь древесиныгпластиче-
ские веществами отчасти запасающие их/также могут накапливать
в себе продукты вторичного метаболизма.
Образование изолирующих и замещающих тканей. Небольшие
повреждения луба, например, при попытках внедрения в него
молодых личинок лиственничного усача, быстро изолируются
в результате образования слоя пробки. Клетки изолированного
участка луба (раневые «линзочки») отмирают и пропитываются
смолой (рис. 55, Z). При более значительных повреждениях в око-
лораиевой части луба слои пробки образуются по всей его толще
эа счет деления клеток тяжевой паренхимы и лубяных лучей.
Эти слои'захватывают лубяные волокна, благодаря чему на по-
перечных срезах имеют вид волнистых линий. Содержание смо-
листых веществ в клетках лубяной паренхимы и лучей повышает-
ся, а последние, кроме того, приобретают головчатые окончания
с . расширенным смоляным каналом.
При выгрызании личинками луба до заболони изоляция по-
следней от воздействия внешних факторов осуществляется путем
образования каллюсной ткани. Причем в тех случаях, когда об-
наженный участок заболони невелик, раневая паренхима может
образовываться за счет сохранившихся частиц камбия и клеток
лучевой паренхимы, еще не завершивших дифференциацию. Если
же камбиальный слой на всем участке уничтожается вместе с лу-
бом, а это свойственно большинству лубоядных насекомых, то за-
растание его происходит от краев раны (см. рис. 55, 2). Клетки
раневой паренхимы образуются камбием по всей границе повреж-
дения, и формирующийся из них каллюс «наплывает» па обнажен-
ный участок заболони.
Одновременно с накоплением массы паренхимных клеток в на-
ружной части каллюса формируется раневая меристема, которая
начинает образовывать вторичную ксилему и флоэму. Слой пер-
вичной каллюсной паренхимы оказывается, таким образом, за-
ключенным между поверхностью поврежденной насекомыми кси-
лемы и вновь сформировавшейся древесиной со всеми присущими
ей элементами.
В некоторых случаях каллюсная паренхима оказывается глу-
боко погруженной в древесину. Интересны случаи многократной
активности меристематических клеток, связанной с уничтожением
сформировавшейся каллюсной ткани личинками насекомых, имею-
щих двухгодовой цикл развития, или повреждением ее другими
видами ксилофагов, обычно называемыми «нахлебниками». При
повторном повреждении одного и того же участка луба и заболони
помимо раневой меристемы каллюса, принявшей на себя роль
камбия, в образовании паренхимы активно участвуют клетки
околораневой части годичного слоя заболони. На рис. 55, 3 изо-
бражен поперечный срез краевой части стволика молодой листвен-
ницы, повторно поврежденной лиственничной листоверткой уже
после образования слоя заболони с патологическими смоляными
каналами, в момент развития нового годичного слоя. Здесь от-
четливо видно участие околораневой зоны последнего в формиро-
вании каллюса.
Вообще роль клеток околораневой ксилемы развивающегося
годичного слоя заболони в восполнении утраченных тканей ве-
лика. Благодаря активному делению клеток лучевой паренхимы
и сопровождающих клеток смоляных каналов происходит раз-
растание годичного слоя заболони. При этом направление при-
роста древесины смещается в сторону поранения, так что иногда
узкие хо^ы, протачиваемые насекомыми, заполняются древесной
тканью при незначительном участии каллюсной паренхимы.
Интенсивность одревеснения оболочек паренхимных клеток
каллюса у разных хвойных различна. У лиственницы они пол-
ностью одревесневают в год возникновения. Поры во вторичных
оболочках образуются простые и в зависимости от степени утол-
щения оболочек просматриваются в плане округлыми или щеле-
видными. Щелевидные поры обычно появляются на второй или
третий год.
Поверхностные паренхимные клетки каллюса остаются тонко-
стенными. Они содержат суберин и какое-то время замещают фел-
лему коры. Рбразующийся затем каллюсный луб отличается
обилием склеренхимных элементов. Последние имеют ветвистую
форму (подобно астросклереидам), собраны в пучки и очень часто
бывают окружены слоем пробки.
Гипертрофия клеток радиальных лучей, их активное деление,
образование агрегатных лучей (соединение двух смежных лучей)—
характерная особенность деятельности каллюсного камбия. Эти
преобразования направлены на повышение концентрации хими-
ческих средств защиты в наружном слое заболони поврежденной
части ствола. Аналогично каллюсной паренхиме преобразованные
клетки радиальных лучей накапливают в большом количестве
смолы и фенолы. У сосны радиальная паренхима не одревеснева-
ет, у лиственницы утолщение вторичных оболочек протекает так
же, как в клетках каллюсной паренхимы.
Установить границу между первичной паренхимой каллюса
и трахеидами — производными его меристемы — в большинстве
случаев бывает трудно, поскольку, во-первых, первые трахеи-
дальные элементы появляются среди паренхимный клеток и, во-
вторых, потому, что здесь встречается ряд промежуточных форм
клеток, а также необычные формы трахеид. Для переходной зоны
каллюса характерно, кроме того, большое количество перегород-
чатых трахеид.
В заключение обзора развития и структуры каллюса необхо-
димо рассмотреть вопрос о возможных размерах каллюсообразо-
вания при повреждении ствола ксилофагами.
Хвойные деревья обладают большими возможностями каллю-
сообразования. Но эти возможности реализуются в полной мере
только у деревьев, обладающих высокой резистентностью к по-
вреждению, необходимым запасом резервных веществ в лубе и
активным фотосинтезом. Наибольшего развития каллюсные об-
разования достигают у лиственниц, поврежденных лиственнич-
ным усачом.
Относительно большие обнаженные участки заболони изоли-
руются в результате некроза наружных слоев трахеид, сопровож-
дающегося пропитыванием стенок клеток фенолами (продуктами
их необратимого окисления) и смолами. В тех случаях, когда та-
кая травма изолирована от атмосферного влияния толщей коры,
над ней может развиться сплошной пласт раневой паренхимы,
а затем и дифференцированных элементов древесины. Поврежден-
ная заболонь в ходах лиственничного усача, достигающих в ши-
рину до 1,5 см, нередко в том же году полностью покрывается
каллюсом даже при одностороннем его развитии (см. рис. 55, 4).
Лиственничный усач в очагах размножения, как уже говори-
лось, заселяет ствол полосой от основания до нижней части кроны.
141
Такие деревья не гибнут. В неповрежденной части ствола проис-
ходят преобразования смолоносной системы, предотвращающие
возможность заселения ее ксилофагами, в том числе и самим уса-
чом (исключение представляют перестойные лиственницы, а так-
же деревья, подвергшиеся воздействию вредных выбросов про-
мышленных предприятий, например фтористых соединений). Де-
ревья, поврежденные этим вредителем, служат примером мощной
защитной реакции и больших регенерационных возможностей
у лиственницы. Поврежденная часть ствола зарастает каллюсом
в течение многих лет (см. рис. 54). Относительно редко края кал-
люса, наплывающего от границ повреждения, смыкаются, но даже
в тех случаях, когда неповрежденная часть ствола представляет
собой узкую полосу живого луба и заболони, дерево сохраняет
жизнеспособность. Со временем эта живая «лента» ствола разрас-
тается, а его поврежденная часть разрушается насекомыми и гни-
лями (см. рис. 54). У дерева как бы образуется вторичный ствол,
сохраняющий устойчивость к повреждению ксилофагами в тече-
ние многих десятилетий.
Повышение химических средств защиты обусловливается преж-
де всего образованием патологических смоляных каналов и со-
провождающих их клеток древесной паренхимы, в том числе
трахеид альной.
Патологические смоляные каналы часто называют травмати-
ческими. Мы считаем правильнее все же называть их патологи-
ческими. Дело в том, что до сих пор образование этих каналов
связывалось непосредственно с поранением (trauma — рана),
тогда как они могут формироваться и без травмы ствола, напри-
мер при дефолиации деревьев. Как показали наши исследования,
у лиственницы количество продольных смоляных каналов очень
варьирует даже в тех случаях, когда деревья не имеют следов
повреждения и обладают всеми показателями высокой жизнеспо-
собности. Часть из этих каналов может располагаться не только
в поздней древесине, но и в промежуточной, а иногда и в ранней
древесине годичного кольца. Такие каналы только условно можно
считать нормальными. В некоторых же случаях у деревьев, не
имеющих повреждений ствола, встречаются и явно патологические
смоляные каналы, располагающиеся группами (на поперечном
срезе — короткими цепочками) в ранней древесине [Рожков,
Степанчук, 1970, 1972].
У ряда хвойных — пихты, кедра (Cedrus), тсуги и таксодие-
вых — каналы в древесине ствола в норме отсутствуют и возни-
кают лишь в ответ на поранение. У других хвойных смоляные
каналы в древесине имеются всегда, но количество их увеличива-
ется при повреждении дерева. Очевидная связь смоляных каналов
с повреждениями послужила основанием некоторым исследова-
телям рассматривать все смоляные каналы, встречающиеся в дре-
весине хвойных, как патологические структуры [Thomson, Sif-
ton, 1925; Bannan, 1936]. Это, конечно, крайняя точка зрения.
142
Естественно предположить, что в процессе эволюции некоторые
хвойные обрели способность^ образовывать нормальные, непато-
логические смоляные каналы, принявшие на себя, помимо защит-
ных, какие-то физиологические функции. Трудно представить,
например, что смоляные каналы стебля сосны в стадии его пер-
вичного роста тоже являются патологическими.
На рис. 56 представлены нормальные смоляные каналы лист-
венницы в древесине и лубе на поперечных, тангеита л ьных и ра-
диальных срезах. Продольные смоляные каналы, как нормальные
так и патологические, закладываются в камбиальной зоне путем
преобразования производных веретеновидных инициален в па-
ренхимные клетки. Эти клетки образуют первичную (эпителиаль-
ную) и вторичную обкладки каналов (рис. 57). От нормальных
смоляных каналов патологические каналы отличаются, во-пер-
вых, их большим количеством и образованием в любое время ве-
гетации, а не только в поздней и промежуточной древесине годич-
ного слоя; во-вторых, более широким диаметром просвета и боль-
шим числом эпителиальных и сопровождающих паренхимных
клеток, содержащих не только смолы, но и фенолы, в-третьих,
преобразованием смежных с каналами трахеид в раневые формы
клеток (перегородчатые трахеиды, трахеидальная паренхима),
которые также накапливают смолы и фенолы. В тех случаях, ког-
да в просвет патологического смоляного канала попадает сердце-
винный луч, паренхимные клетки последнего либо участвуют
в формировании эпителия (см. рис. 57, 2), либо сохраняются в ви-
де тяжа, но также выполняют смолообразующую функцию
(см. рис. 57, 6).
При повреждении ствола количество продольных смоляных
каналов увеличивается в десятки и сотнц раз за счет образования
патологических смоляных каналов. У лиственницы на 10 см длины
поперечного сечения годичного слоя заболони приходится обычно
8—15 нормальных каналов^ иногда меньше, а число патологиче-
ских каналов может превышать 3 тыс. При этом они располагают-
ся плотным рядом (часто захватывая весь промежуток между ра-
диальными лучами) или двумя, тремя и даже четырьмя рядами
один над другим (рис. 58).
Количество патологических смоляных каналов зависит от сте-
пени повреждения ствола и исходного физиологического состояния
дерева. При локальном повреждении по мере удаления от пора-
нения количество каналов уменьшается, запаздывают сроки их
появления и нарушается стройность цепочек. У лиственницы, ели
и пихты ровные, сплошные цепочки каналов образуются на боль-
шем околораневом участке ствола по сравнению с сосной и кедром.
В табл. 8 приведены данные о количестве патологических смоля-
ных каналов у деревьев IV класса возраста при искусственном
повреждении ствола высечкой на глубину 1—2-годичных сло-
ев заболони площадью 10 см2 па высоте 1,5 м от основания
ствола.
4 АЗ
Таблица 8
Количество патологических смоляных каналов на 1 мм
длины поперечного среза годичного слоя на разном рас-
стоянии от края поранения (Тункинский район, Бурятская
АССР, 1973—1975 гг.)
Расстояние от края по- ранения. мм Листвен- ница Ель Пихта Сибирский кедр Сосна
Сверху от раны
20 1 8~к 1 1 8-10 1 8-9 1 1 4-5 1 I 4-5
80 1 8-9 ) 1 8-9 | 1 8-9 | 2-3 1 1 2-3
Сбоку от раны
2 16—20 16-20 16-20 4-7 4-7
5 7-11 7—11 7-8 3-5 3-5
10 7—8 7-8 6—7 2-4 2-3
15 6—7 6-7 4 1-2 1-2
20 3-6 с© 1 со 3 1—2 1-2
У заселяемых лиственничным усачом деревьев патологические
смоляные каналы формируются также вне зоны поражения по все-
му стволу (табл. 9). При этом обнаруживается увеличение коли-
чества каналов от основания к вершине ствола и сокращение их
в цепочках от ранней к поздней древесине годичного слоя в ниж-
ней и средней частях ствола.
Таблица 9
Соотношение количества патологических смоляных каналов в цепочках при
четырехрядном расположении их в годичном кольце (все три дерева в массе
заселены личинками лиственничного усача)
Часть ствола Модельные деревья Количество смоляных каналов па 10 см длины поперечного среза годичного слоя, шт.
в пределах полосы пора- жения вне полосы поражения
всего в цепочках* все- го в цепочках*
11 1 2 | 1 3 4 1 2 1 1 3 4
Верхняя (под первы- 1 2392 638 760 265 684 2272 621 625 403 623
ми живыми сучья- 2 2862 660 682 680 840 3237 750 717 910 860
ми) 3 1823 722 646 0 455 1738 600 445 409 284
1 1025 301 345 ИЗ 266 994 585 379 30 0
Средняя 2 1748 608 607 0 533 1283 803 480 0 0
3 753 304 227 70 152 405 360 45 0 0
Нижпяя (на высоте 1 1299 756 483 60 0 470 470 0 0 0
0.3 м пад почвой) 2 637 397 240 0 0 90 90 0 0 0
3 738 323 303 112 0 318 260 58 0 0
* Порядковые померь цепочек от ранней к поздней древесине.
144
Преобразованию подвергаются отчасти и смоляные каналы
радиальных лучей в лубе. Нормальные смоляные каналы луба
представляют собой продолжения радиальных каналов древесины.
Они пронизывают всю толщу активного луба и заканчиваются
слепо в периферической его части. В отличие от смоляных кана-
лов древесных лучей каналы луба в большинстве случаев имеют
более широкий просвет,* особенно в концевой части. У физиологи-
чески ослабленных или получивших травмы деревьев отдельные
смоляные каналы могут значительно расширяться и образовывать
цистоподобные (головчатые) окончания (рис. 59). Такие каналы,;
как патологические структуры, описаны у канадской ели [Thom-
son, Sifton, 1925]. В случаях образования большого количества
продольных патологических смоляных каналов в развивающемся
годичном кольце заболони однорядные радиальные лучи в этом
кольце могут преобразовываться в многорядные (веретеновидные)
и давать начало смоляным каналам луба.
В отмершем лубе внешней коры смоляные каналы облитери-
руются, содержащиеся в эпителиальных клетках смолы исчезают
(сохраняются в оболочках клеток), и в целом следы каналов об-
наруживаются с трудом.
Кроме патологических смоляных каналов у лиственницы при
повреждении насекомыми могут образовываться камедные кана-
лы. Это также продольные ходы, но более короткие, формирую-
щиеся лизигенно вблизи радиальных лучей. Камедные каналы
возникают в годичных кольцах прошлых лет, в которых трахеиды
давно утратили протопласт. Камедь образуется в результате рас-
творения оболочек трахеид. В сформировавшихся каналах на по-
перечном срезе иногда можно видеть сохранившиеся остатки пер-
вичной оболочки клеток (см. рис. 59). Камедные каналы в зоне
раневых преобразований заболони являются по существу камед-
но-смоляными, ибо вскоре после образования они начинают от-
части функционировать как смоляные каналы. Экскреторным
эпителием в них служат тиллоподобные отростки паренхимы ра-
диальных лучей.
У лиственницы в отличие от сосны клетки эпителия нормаль-
ных смоляных каналов, как правило, отмирают полностью или
частично уже в год образования. Установлена определенная за-
висимость между продолжительностью жизни смолоносного эпи-
телия и местом образования каналов в годичном слое ксилемы.
В переходной зоне древесины клетки эпителия функционируют
дольше, чем в поздней древесине, а в ранней древесине, где обра-
зуются только патологические смоляные каналы, они у листвен-
ницы живут до трех лет. Однако в каналах, находящихся непо-
средственно в зоне порайения ствола, вне зависимости от распо-
ложения их в годичном слое, клетки смолоносного эпителия быст-
ро утрачивают жизнеспособность. При этом наблюдается облите-
рирование эпителия каналов, просвет которых в результате пов-
реждения оказался открытым, и закупорка каналов тиллоидами.
10 а. С. Рожков
145
Образование тиллоидов. Мнение о том, что тиллоидообразова-
ние свойственно тем хвойным, которые производят небольшое
количество смолы и у которых оболочки эпителия смоляных ка-
налов быстро одревесневают (Abies, Tsuga), не совсем верно,
и вовсе неверно представление о тиллоидах как нормальном этапе
развития смолоносного эпителия [Эсау, 1969]. Образование тил-
лоидов — защитная реакция дерева па повреждение. Закупори-
вание ими каналов предотвращает проникновение в них воздуха,
под воздействием которого активные молодые клетки эпителия
погибают и облитерируются. Тиллоиды встречаются у всех хвой-
ных, и вторичные оболочки их клеток бывают уплотнены в разной
степени — от незначительного склерифицирования до сильного
одревеснения. Обнаружены также суберинизированные клетки
тиллоидов.
Тиллоиды в большем количестве образуются в патологических
смоляных каналах. В нормальных каналах поздней древесины
они развиваются в случаях поранения до начала склерифициро-
вания смолоносного эпителия. В смоляных каналах радиальных
лучей тиллоиды образуются реже и также только при поврежде-
нии ствола. Процесс формирования тиллоидов включает два эта-
па — разрастание клеток, заполняющих просвет канала, и уплот-
нение их вторичных оболочек. В закупоривании канала обычно
участвует несколько клеток, причем некоторые из них вытягива-
ются даже вдоль канала, но иногда тиллоиды на поперечных
срезах бывают представлены одной или двумя разросши-
мися клетками.
По интенсивности образования тиллоидов лиственница значи-
тельно превосходит сосну. Оболочки клеток эпителия у нее при
этом быстро и сильно утолщаются и одревесневают (см. рис. 59).
У сосны патологические каналы, находящиеся в зоне, прилежа-
щей к месту образования раневой паренхимы, как правило, име-
ют крупные эпителиальные клетки, которые почти полностью
закрывают просвет. Именно в этой зоне наблюдается утолщение
оболочек эпителия и образование тиллоидов. В отличие от лист-
венницы клетки тиллоидов сосны одревесневают слабо и не со-
держат фенолы.
Как уже говорилось, облитерирование эпителия смоляных
каналов происходит при повреждении заболони, открывающем
доступ атмосферному воздействию. Облитерируются клетки, еще
не затронутые процессом одревеснения. В них между сплющенны-
ми оболочками сохраняются смолы, а у лиственницы также и фе-
нолы. При повторном повреждении луба, например лиственнич-
ной листоверткой, после образования патологических смоляных
каналов облитерирование в них эпителия происходит по всему
травмированному участку (см. рис. 59).
Преобразование трахеид и древесной паренхимы. Вокруг па-
тологических смоляных каналов на значительном протяжении
происходит трансформирование трахеид в клетки, способные об-
146
разовывать защитные вещества. Эти клетки вместе со смоляными
каналами образуют насыщенный смолами и фенолами слой древе-
сины, усиливающий действенность химической защиты дерева.
В тех случаях, когда насекомые повреждают луб, не затраги-
вая камбия, трахеиды претерпевают морфологические преобразо-
вания по всей площади травмы; при повреждении заболони ране-
вые трахеидальные элементы возникают в пограничной с травмой
зоне развивающегося годичного слоя.
Раневая ксилема отличается большой продолжительностью
жизни протопласта и активным метаболизмом трахеид. В трахе-
идах часто происходит клеточное деление и образование перего-
родок. В зависимости от степени завершенности первоначального
формирования трахеид образовавшиеся из них перегородчатые
трахеиды могут быть двух типов — тяжевые и септированные.
В простейшем случае раневые трахеиды (при трансформации
трахеид, уже завершивших в основном дифференциацию) отли-
чаются от нормальных продолжительностью сохранения физио-
логической активности. Нередко в заболони пораженной усачом
части ствола можно встретить ранние (весенние) трахеиды, со-
держащие протоплазму и ядро до конца вегетации (рис. 60, 2, 2).
Такие трахеиды могут не иметь каких-либо отличий в строении
стенок. Чаще же при повреждении дерева преобразованию под-
вергаются трахеиды, еще не вышедшие из камбиальной зоны или
находящиеся на ранней стадии дифференциации. Эти трахеиды
по завершении формирования отличаются количеством, распо-
ложением и формой пор. Количество окаймленных пор у раневых
трахеид обычно увеличивается, и они распространяются на тан-
гентальные стенки. Сами поры сокращаются в размере (умень-
шается диаметр камеры), изменяется форма их канала (рис. 61).
Крайней степенью преобразования пор является полная утрата
окаймления и превращение их в простые поры, свойственные па-
ренхимным клеткам.
У раневых трахеид крупные окаймленные поры могут чередо-
ваться с мелкими окаймленными порами, имеющими щелевидные
или пересекающиеся отверстия (см. рис. 61, 5, 4). Весьма инте-
ресны случаи преобразования уже сформировавшихся пор: они
либо разделяются на 2—4 части путем поперечного или радиаль-
ного слияния нависающей части вторичной оболочки с замыкаю-
щей пленкой поры, либо слияние названных элементов поры но-
сит мелкоячеистый характер и в результате образуется структура,
напоминающая ситовидный участок проводящих элементов фло-
эмы (см. рис. 61, 5—8).
Глубина трансформирования трахеид определяется степенью
дифференциации их к моменту повреждения ствола. Они могут
быть преобразованы в тяжевые трахеиды с окаймленными порами
или (в поздней древесине) с редуцированным окаймлением пор,
в септированные трахеиды с простыми порами и в трахеидальную
паренхиму.
10*
147
Самыми обычными среди раневых трахеид являются тяжевые
трахеиды (см. рис. 60, Зх 4). Формироваться они начинают в кам-
биальной зоне, и потому торцевые стенки их члеников имеют
вполне развитую первичную оболочку и содержат 2—4 окаймлен-
ные поры. Поры боковых стенок члеников многочисленные, мел-
кие, слабо окаймленные.
В древесине хвойных септированные трахеиды до сих пор не
обнаружены. Более того, отрицалась даже возможность их обра-
зования. Как нами установлено, септированные трахеиды обра-
зуются у хвойных, по крайней мере у лиственницы, при поврежде-
нии ствола насекомыми. Встречаются они значительно реже дру-
гих форм раневых трахеид. Перегородки в таких трахеидах воз-
никают уже после завершения образования первичной оболочки,
поэтому последняя в них отсутствует (см. рис. 60, 5, 6).
В зоне образования патологических смоляных каналов трахе-
иды замещаются (иногда почти полностью) клетками раневой па-
ренхимы, играющими ведущую роль в образовании химического
защитного пояса в развивающемся годичном слое заболони. У де-
ревьев, поврежденных лиственничным усачом, можно проследить
все переходные формы от тяжевых трахеид к трахеидальной па-
ренхиме (рис. 62, 1—6). Быстрое накопление массы раневой дре-
весной паренхимы происходит также за счет увеличения количест-
ва клеток вторичной обкладки смоляных каналов и роста (иногда
продольного деления) паренхимных клеток радиальных лучей.
Все эти клетки активно используют резервные вещества, содер-
жащиеся в лубе, а также поступающие из кроны ассимиляты на
образование защитных веществ, в первую очередь смол.
В зоне патологических смоляных каналов встречаются и иск-
лючительные случаи, например, преобразование меристематиче-
ских производных трахеид в смешанные тяжи клеток, часть из ко-
торых сохраняет признаки трахеид, а остальные становятся
паренхимными клетками (ем. рис. 62, 4). Столь же, на первый
взгляд, невероятно образование паренхимных клеток внутри сфор-
мировавшейся трахеиды. Окаймленные поры в стенках таких
трахеид не развиваются, а паренхимные клетки имеют простые
поры, в том числе и на торцевых стенках (перегородках), и всег-
да бывают целиком заполнены конденсированными таннинами
(см. рис. 62, 5, 6).
В раневой ксилеме могут трансформироваться также трахеиды,
уже лишившиеся живого протопласта. Вследствие образования
перетяжек, расширения просвета, частичного лизиса и слияния
клеток ровные ряды трахеид превращаются в беспорядочное скоп-
ление клеток разной формы и величины (см. рис. 62, 7, 8). Такие
трахеиды вместе с другими клетками, у которых происходит силь-
ное утолщение вторичных оболочек (см. рис. 60, S), выполняют
роль арматуры в раневых клетках развивающегося годичного
слоя заболони.
Таким образом, в ксилеме и флоэме ствола жизнеспособных
деревьев, поврежденных лиственничным усачом, как и другими
ксилофагами-первопоселенцами, происходит образование слож-
ных изолирующих структур, возмещение утраченных частей спе-
цифическими раневыми тканями, возникновение замещающих
опорных элементов, многократное увеличение смолообразующей
и смолопроводящей систем, а в целом — формирование сплош-
ного защитного пояса из долгоживущих смоло- и фенолопроду-
цирующих клеток.
Физиолого-биохимические преобразования
Физиолого-биохимические защитные реакции у деревьев, под-
вергшихся нападению ксилофагов, выражаются в изменении вод-
ного режима, углеводного и фосфорного обмена, дыхания луба,
осмотического давления в клетках и давления газа в межклетных
полостях и каналах (смоляного давления), в повышении биосин-
теза биологически активных соединений и в накоплении токсинов
и репеллентов [Рожков и др., 1972].
Изучение патологических и защитных преобразований у по-
врежденных лиственничным усачом деревьев проводилось в Бу-
кукинском очаге размножения этого вредителя в Читинской об-
ласти и в Тункинском районе Бурятской АССР в окрестностях
сел Зактуй и Зун-Мурино. Для дополнительных исследований
использованы искусственно поврежденные деревья в возрасте
45—50 лет. В одних случаях поранение заболони производилось
с помощью высечки на глубину до 20 годичных колец (имитация
повреждения деревьев личинками лиственничного усача на вто-
рое лето их развития). Стволы повреждались в конце мая на вы-
соте 1 м от корневой шейки. Площадь поранения (первоначальная
50—70 см2) увеличивалась ежемесячно и к концу вегетации охва-
тывала 1/3 окружности ствола. Анализу подвергались околоране-
вые участки ствола. Эти деревья отнесены нами к первой группе.
В других случаях луб и наружные годичные кольца заболони по-
вреждались с помощью сверла диаметром 1,5—2,0 мм по трафа-
рету. Такие точечные поранения наносились деревьям в III де-
каде мая. На 1 дм2 приходилось 27 отверстий. Поврежденная пло-
щадь также охватывала 1/3 окружности ствола, но при этом у од-
них деревьев она составляла 10,4 дм2 (вторая группа деревьев),
у других — 5,2 дм2 (третья группа деревьев). Такие искусствен-
ные повреждения сходны с повреждением луба и заболони личин-
ками лиственничного усача в первый год их развития. Кроме того,
также в III декаде мая у части модельных деревьев ствол повреж-
дался поперечными запилами с интервалами в 10 см на глубину
луба 0,5 см. Площадь повреждения составляла 10 дм2 и охватыва-
ла 1/3 окружности^ствола (четвертая группа деревьев).
В течение вегетации у лиственниц пораженных и здоровых влажность
тканей определялась весовым методом; содержание свободной и связанной
4А0
воды — рефрактометрическим методом [Гусев, I960]; редуцирующих саха-
ров — антроновым методом [Вечер, Проказов, 1967]; моно- и дисахаров —
методом Бертрана в фиксированном горячим спиртом материале; фосфора —
по Г. Шмидту и С. Тангаузеру [Белозерский, Проскуряков, 1951]; дыха-
ние — манометрическим методом на аппарате Варбурга. Смолистые вещества
экстрагировались хлороформом и петролейным эфиром и делились на кислую
и нейтральную части; кислоты разделялись на жирные и смоляные методом
С. С. Малевской и В. Д. Карнауховой (1956). Выделенные фракции нейт-
ральных веществ из петролейного экстракта луба и заболони наносились
на колонки А12О3 (активностью 1—2), а затем вымывались петролейным эфи-
ром (40—70°) углеводородсодержащие, а этанолом — кислородсодержащие
фракции. Фенольпые соединения, экстрагированные метиловым спиртом
(1968 г.) и ацетоном (1969—1970 гг.) из материала, предварительно обессмо-
ленного хлороформом, определялись по методике Т. Суэйна и В. Хиллиса
[Swain, Hills, 1959), а флавоноиды — методом А. Г. Гусевой и М. Н. Нестюк
(1953). Для построения калибровочной кривой использовался кверцетин.
Качественный состав флавоноидов исследовался хроматографически в сис-
темах бутанол — уксусная кислота — вода (4 : 1 : 5) и уксусная кислота —
муравьиная кислота — вода (10 : 2 : 3). Просмотренные в УФ-свете до и
после проявления парами аммиака и хлористым алюминием пятна флавоно-
идов сравнивались с чистыми флавоноидами по окраске и значению Rf.
Дубильные вещества определялись по методике К. А. Анисимовой (1967).
Срубленные деревья быстро утрачивают сопротивляемость
к заселению ксилофагами. Для выяснения роли водного обмена
в поддержании давления газа в смоляных каналах и способности
заливать живицей раны, наносимые стволовыми вредителями,
в 1970 г. поставлен опыт, показавший, что сваленные весной до
распускания хвои лиственницы (с кроной), комлевая часть кото-
рых тотчас же погружается в воду (горный ключ), не подвергают-
ся в течение лета нападению ксилофагов. У таких деревьев наблю-
даются признаки физиологического ослабления — развивается
укороченная хвоя, слабо развиваются ауксибласты. Но даже
при искусственном заселении их большим лиственничным коро-
едом, жуки в лучшем случае проделывают в коре и лубе короткие
ходы и гибнут в них или покидают дерево из-за выделения живи-
цы. Деревья, срубленные и оставленные на берегу (также в верти-
кальном положении), утрачивают способность сопротивляться
заселению короедами. Хвоя на таких деревьях не распускается,
хотя луб остается живым в течение всего лета.
У искусственно поврежденных деревьев второй группы влаж-
ность луба околораневых участков в I декаде июня несколько по-
вышалась, а затем снижалась [Рожков и др., 1972]. Уже во II де-
каде этого месяца содержание воды в нем снизилось в целом на
9%, а связанной — на 31% при повышении количества свободной
воды на 8%. В это же время — в первый месяц после поранения —
содержание связанной воды в наружной части заболони повыша-
лось на 25%, а обводненность клеток камбиальной зоны остава-
лась меньшей по сравнению с контрольными деревьями (табл. 10).
Подобные изменения влажности тканей наблюдаются также
у деревьев в очагах размножения лиственничного усача в~период
массового отрождения личинок и начала проникновения их из
коры в луб и заболонь. У лиственниц на второй год после заселе-
150
Таблица 10
Изменение содержания воды в тканях ствола при поврежде-
нии (в числителе — здоровые деревья, в знаменателе — по-
врежденные второй группы)
Общая влажность, % Содержание воды, %
свободной связанной
июнь, II июль, III август, нюнь, II июль, III август, III нюнь, II июль, III август.
61,6 1 56,4 58,8 33,7 1 Луб 41,5 I 141’9 1 27,9 14,9 I 1 16,9
- -
56,1 J 5^9 61,0 36,6 1 38,2 | 36,5 | 19,5 19,7 | 24,5
Камбий
85,8 1 62,2 | 1 7|’5 I 70,5 1 59,4 I 58,1 I 1 42,4 1 41,4 I 52,4 | 49,0 | 27»7 I 3°Л I _
19,8 | 18,1 1
37,7 I I 54,9 I Заболонь 1 25,2 I 31,5 I — 1 1 12,5 I | 23,4 I -
1 | 23,5 |
39,4 48,3 1 41,0 30,3 28,6 15,9 18,0 12,4
ния их вредителем, как и у искусственно поврежденных (деревья
первой группы), происходят большие нарушения физиологических
процессов, сопровождающихся резким снижением влажности
луба и заболони в течение июня — августа.
Специальные исследования [Рожков, Габиб-заде, 1978] пока-
зали, что осмотическое давление клеточного сока луба само по
себе не препятствует развитию ксилофагов, наносящих деревьям
физиологический вред, и может рассматриваться лишь как один
из показателей жизнеспособности последних. Нормальное или
повышенное осмотическое давление — признак недоступности по-
селения только для тех видов, которые развиваются на необратимо
ослабленных деревьях.
Для ксилофагов, развивающихся на жизнеспособных деревь-
ях, влажность луба и соотношение связанной и свободной воды
в нем не является показателем доступности поселения.
Поранение луба сопровождается значительным повышением
его дыхательной активности. Сотрудниками нашей лаборатории
[Животова и др., 1971] установлено, что у поврежденных деревьев
второй группы интенсивность дыхания увеличивается более чем
на 70%, у деревьев четвертой группы — на 150% по сравнению
с контролем. В августе интенсивности дыхания пораженного луба
(деревья второй и четвертой групп) значительно ослабляется,
хотя и превышает таковую луба здоровых деревьев. Средние по-
казатели активности окислительной системы пораженного и здо-
рового луба лиственницы сибирской (мкл О2 на 1 г сырого веса
151
в час) были следующими: у здоровых деревьев в июле — 161,51,
в августе — 177,09; у поврежденных деревьев’ второй группы
в июле — 278,08, в августе — 239,77; у деревьев четвертой груп-
пы в июле — 404,30, в августе — 316,99.
Повышение активности окислительной системы пораженного
луба вызывает усиленный расход пластических веществ (угле-
водов) и накопление фосфорных соединений. Так, в лубе исследо-
ванных нами здоровых деревьев *в июне — августе содержание
фосфора составляло 0,1%, а у поврежденных деревьев первой
группы оно возросло к этому времени до 0,13—0,17% (от сухой
навески).
На корреляцию между содержанием углеводов, интенсив-
ностью дыхания и общим содержанием фосфорных соединений
указывают Б. А. Рубин и Б. В. Арциховская (1968). Нарастание
содержания фосфорных соединений приводит к активированию
фосфорилирующих ферментов промежуточного углеводного об-
мена, при этом возрастает потребление сахаров и усиливается
дыхание.
Результаты анализов показали, что накопление фосфорных
соединений и повышенное дыхание пораженного луба вызывают
быстрое его обеднение гидролизуемыми углеводами и гемицел-
люлозой. У заселенных лиственничным усачом деревьев содержа-
ние гемицеллюлоз сокращается в лубе и увеличивается во внутрен-
ней части мертвой флоэмы. По-видимому, это происходит в резуль-
тате образования и оттока в кору камеди, играющей у лиственниц
наряду со смолами большую защитную роль. В коре заселенной
личинками части ствола содержание гемицеллюлоз составляло
18,05%, в лубе — 16,9; вне зоны заселения ствола в коре —
15,6%, в лубе — 18,9; в коре контрольных деревьев — 14,1%,
в лубе — 19,25% от веса сухого вещества.
В ослабленном дереве к моменту заселения его усачом содер-
жание сахаров в прилегающей к лубу части коры значительно
понижено. В лубе зараженной части ствола количество их также
понижается, причем моносахара сокращаются примерно на 40%.
В лубе неэараженной части ствола содержание дисахаров остается
в пределах нормы. В прилегающей к лубу части коры на заражен-
ном участке ствола моносахара составляли в среднем 1,92%,
дисахара — 1,08%; в той же части коры вне зараженного участка
их было соответственно — 2,16 и 1,21%, в коре контрольных де-
ревьев — 2,20 и 1,99%. В лубе зараженной части ствола моноса-
харов содержалось 1,59%,'дисахаров — 7,19; в лубе контрольных
деревьев — соответственно*^,74 и 7,48% от веса сухого вещества.
В местах пораненияствола в лубе и наружной части заболони
деревьев всех групп в начале вегетации наблюдается некоторое
истощение запасов углеводов. Со второй половины лета ткани этих
деревьев отличаются уменьшением содержания крахмала при уве-
личении количества гидролизуемых углеводов. Известно, что при
уменьшении воды в лубе крахмал частично превращается в сахар
4S9.
[Крамер, Козловский, 1963]. Этим, по-видимому, и объясняется
различие в осенних запасах крахмала у здоровых и поврежденных
лиственниц.
Усиление химических средств защиты
В настоящее время к числу защитных химических веществ у
растений относят некоторые алкалоиды, глюкозиды, эфирные
масла, фенолы и продукты их окислительных превращений [Ру-
бин, 1971 ]. Связь между содержанием этих соединений и устой-
чивостью растений выяснялась по поражаемости последних пара-
зитарными грибами и бактериями. О действии глюкозидов и
алкалоидов на насекомых сведений нет. Из числа флавоноидов и со-
единений с их участием некоторые обладают слабыми инсектицид-
ными свойствами. Исследования, проведенные в нашей лаборато-
рии, показали, например, что содержащийся в коре лиственницы
фенольный гликозид антоциан нисколько не препятствует пита-
нию ею личинок насекомых, а флавоноиды из луба лиственницы
(дигидрокемпферол, кемпферол, дигидрокверцетин, кверцетин)
в совокупности оказывают токсическое действие на некоторых
дендрофильных насекомых только в чистом виде и при больших
дозировках. Полимеризованные таннины (флобафены) — нейт-
ральны, по крайней мере, для большинства ксилофагов хвойных.
Тем не менее роль фенольных соединений в предупреждении засе-
ления деревьев ксилофагами очень велика. Они препятствуют раз-
витию в тканях ствола патогенных микроорганизмов и грибовк
являющихся во многих случаях предшественниками древесино-
ядных насекомых.
Локализация смол и фенолов. Луб выполняет главную роль
в защите дерева от насекомых-ксилофагов, а основным средством
этой защиты у хвойных являются смолы. В околораневом участке
луба оболочки всех элементов в большей или меньшей степени
лигнифицируются и к моменту изоляции его пробковой тканью
Клетки заполняются смолами. Смолы появляются также в сито-
видных трубках живого луба^ окружающего раневую линзочку.
Гистохимические исследования [Рожков, Степанчук, 1972].
показали, что быстрое накопление смолистых веществ сопровожда-
ется столь же быстрым расходованием крахмала клетками лубя-
ной паренхимы в зоне образующейся раневой линзочки и за ее
пределами. Впрочем, в последних вскоре после образования ране-
вой линзочки крахмал восстанавливается до нормы, а в разрос-
шихся клетках радиальных лучей и раневой паренхиме, подстила-
ющей линзочку, зерна йрахмала накапливаются в очень большом
количестве.
Для гистохимических исследований мы в основном использовали свежий
материал, но в ряде случаев хорошие результаты получены и при работе
с зафиксированными образцами тканей. При исследовании фенольных сое-
153
динений был испробован ряд реакции. Лучшими из них для тканей листвен-
ницы оказались две — нитрозореакция [Дженсен, 1965] и реакция с хлор-
ным железом в безводном эфире [Прозина, 1960].
Фенольные вещества в разных клетках выявляются по-разному
в зависимости от степени их раневого преобразования. В лубяной
паренхиме здоровых лиственниц они локализуются в виде одного,
а чаще нескольких ярко-оранжевых (при нитрозореакции) и черно-
голубых (при реакции с хлорным железом) сгустков, соединенных
между собой и с пристенным слоем тяжами, образующими сетчатую
структуру. У поврежденных деревьев в клетках лубяной парен-
химы раневой зоны фенольные вещества локализуются чаще всего
в виде одного крупного сгустка в центре клетки, занимающего
нередко значительную ее часть. В ближайших к камбию клетках
лубяной паренхимы поврежденных лиственниц фенольные веще-
ства могут находиться в виде многочисленных мелких капель,
заполняющих всю клетку.
Гомогенно, заполняя всю клетку, фенольные вещества встреча-
ются в лубяной паренхиме формирующейся раневой линзочки
(при точечном поражении), в той части ее, которая затем оттор-
гается от здорового луба, и в клетках толстостенного пробкового
слоя, окружающего уже сформировавшуюся раневую линзочку.
Различия в содержании фенольных веществ в лубе здоровых и
поврежденных лиственниц гистохимически проявляются прежде
всего в радиальных лучах. На срезах луба неповрежденных лис-
твенниц положительная реакция на фенольные вещества иногда
наблюдается лишь в клетках прикорковой части радиальных лучей.
Для луба пораненных лиственниц характерно повышенное со-
держание фенольных веществ во всех клетках лучей.
Пока нет оснований говорить о повышенном содержании фе-
нольных веществ в клетках лубяной паренхимы поврежденных
лиственниц. Лубяная паренхима здоровых лиственниц всегда бо-
гата фенольными веществами. Вместе с тем отмечено, что опроб-
ковению вокруг пораженных участков луба предшествует некото-
рое уменьшение количества фенольных веществ в клетках лубяной
паренхимы.
О качественных различиях фенольных веществ в лубе повреж-
денных и неповрежденных лиственниц пока можно судить лишь по
различной структуре этих веществ, выявляющейся при описанных
гистохимических реакциях. Замечено, что в образцах со здоровых
и поврежденных лиственниц, зафиксированных в 10 %-ном форма-
лине, фенольные вещества бурого цвета выявляются по-разному:
в клетках лубяной паренхимы здоровых лиственниц они распреде-
лены равномерно по всей клетке, в которой хорошо различается
неокрашенная зернисто-сетчатая структура протоплазмы. Лишь
в прикамбиальной и прикорковой частях луба (в клетках лубяной
паренхимы) фенольные вещества от этого фиксатора выявляются
в виде бурых капель разной величины и сетчатая структура прото-
плазмы не просматривается. У пораненных лиственниц фенольные
154
вещества в виде таких бурых капель встречаются при зарастании
раны в лубе (при опробковении), а также в клетках паренхимы
отмирающего луба (прослежено на образцах, взятых с лиственниц,
поврежденных способом изолирования участка коры и луба).
Фенольные вещества в молодых эпителиальных клетках форми-
рующихся патологических смоляных каналов, как правило, не
обнаруживаются. В большем количестве они накапливаются в пер-
вичной внешней лигнизированной обкладке завершивших форми-
рование смоляных каналов, особенно вблизи околораневых участ-
ков заболони. Чем дальше удалены патологические смоляные
каналы от места поранения, тем меньше фенольных веществ обна-
руживается в их первичной обкладке. Раневая (трахеидальная)
паренхима, образующаяся в зоне смоляных каналов, и некоторые
формы преобразованных трахеид накапливают в обилии феноль-
ные вещества, которые при описанных реакциях проявляются
равномерно во всей клетке либо гомогенно (рис. 63, 1, 2, 4, 5)
или в виде гранул (рис. 63, 7), либо концентрируются в виде ка-
пель около стенок (рис. 63, 3, 3). Нередко фенолы образуют круп-
ное тело в центральной части клетки, а их каплевидные образова-
ния располагаются около стенок (рис. 63, 6). Содержание фенолов
повышается также в паренхиме сердцевинных лучей (рис. 63,10).
Образование патологических смоляных каналов сопровожда-
ется появлением зерен крахмала в ранних трахеидах от самой гра-
ницы их с поздними трахеидами предшествующего годичного коль-
ца. В это же время крахмал в виде зерен разной величины — от
очень мелких до относительно крупных — обнаруживается в фор-
мирующихся сопровождающих клетках и эпителиальной обкладке
смоляных каналов прикамбиальиой зоны. Одновременно проис-
ходит заметное уменьшение количества крахмала в клетках бли-
жайшей к камбию лубяной паренхимы.*
При развитии патологических смоляных каналов смолы начи-
нают накапливаться в большом количестве в эпителиальных и
сопровождающих клетках, заменяя крахмал. Чаще в эпителии
крахмала бывает немного, он быстро расходуется на образование
смол (рис. 64* 1—3). При наличии нескольких рядов патологиче-
ских смоляных каналов крахмал в эпителиальных и сопровожда-
ющих клетках первых рядов может отсутствовать» но смолы в боль-
шом количестве всегда заполняют клетки обкладок всех смоляных
ганалов.
В значительном количестве смолы накапливаются также в
клетках радиальных лучей в пределах зоны патологических смоля-
ных каналов (см. рис. 64, 5). Здесь, кроме того, трахеидальная
паренхима, некоторые трансформированные трахеиды, а отчасти и
нормальные поздние трахеиды предыдущего года развития начи-
нают синтезировать крахмал, мобилизуемый затем на образование
защитных веществ.
Крахмал и смолы в заболони здоровых лиственниц встречают-
ся только в клетках радиальных лучей и эпителии смоляных кана-
155
лов, притом в значительно меньшем количестве, чем у поврежден-
ных деревьев.
Помимо зоны формирования патологических смоляных кана-
лов, смолы содержатся в клетках каллюсной ткани, в большом
количестве накапливаются в лубяной паренхиме травмированной
части ствола и особенно в эпителии смоляных каналов сердце-
винных лучей луба.
Изменение содержания и состава защитных веществ в тканях
ствола деревьев, подвергшихся воздействию тех или иных ослаб-
ляющих факторов среды, является одним из важнейших показате-
лей их физиологического состояния и степени резистентности по
отношению к древесиноядным насекомым, паразитарным грибам
и патогенным микроорганизмам. У способных к сопротивлению
деревьев при попытках поселения на них ксилофагов метаболизм
клеток луба и развивающегося годичного слоя заболони, а отчасти
и живых элементов годичных слоев прошлых лет перестраивается
в сторону преимущественного накопления смолистых и фенольных
веществ. В зависимости от исходного физиологического состояния
дерева происходит большее или меньшее усиление системы его
химической защиты. В смолах изменяется соотношение нейтраль-
ных веществ (терпеноидов и алифатических соединений) и кислот
(смоляных и жирных), в составе фенольных соединений повыша-
ется содержание таннинов и увеличивается количество компонен-
тов флавоноидной группы.
Результаты анализа проб, взятых в июне, показали следующее
содержание таннинов в тканях здоровых лиственниц: во внешней
коре их от 9,9 до 13,9%, в лубе — от 6,0 до 6,8%, в камбиальной
зоне— от 0,55 до 1,1% от веса сухого вещества. Со II декады июня
содержание таннинов в камбиальной зоне увеличивается в 2 раза.
В лубе и внешней коре искусственно поврежденных деревьев
второй группы содержание таннинов несколько снижалось ч’з
среднем в лубе — 5,4%, в коре — 9,7%), а в камбиальной зоне
оставалось прежним. В это время в клетках луба вокруг поране-
ний образуются флобафены. В июле количество водорастворимых
таннинов здесь заметно повышается и составляет в среднем 7,5%
(табл. 11).
Содержание таннинов в лубе зависит от степени поранения де-
рева. В лубе деревьев первой группы с весны и до осени отмеча-
ется очень высокое их содержание — от 13,8 до 15,7% от сухого
веса.
Хроматографически (в системе уксусная кислота — муравьиная
кислота — вода, 10 : 2 : 3) в экстракте из луба здоровых деревьев
обнаружено девять веществ, из них одно — дигидрокверцетин
(Rf 0,67). В течение всего вегетационного периода качественный
состав флавоноидов, включающий дигидрокверцетин, не меняется.
Однако содержание сопровождающих их фенокислот уменьшает-
ся. Так, на хроматограммах в июне обнаружено четыре, в июле г
и августе три вещества. Суммарное содержание флавоноидов в лу-
156
Таблица 11
Содержание водорастворимых таннинов и полифенолов в коре, лубе и забо-
лони здоровых и искусственно поврежденных лиственниц в течение
вегетации
Содержание, % от абсолютно сухой навески
Категория модель- Месяц, де- Таннины Полифенолы
ных деревьев када
Кора Луб Камбий Луб | | Заболонь
Здоровые Июнь, I 10,0 6,7 0,55 3,17
» II 13,9 6,8 0,98 1,71 0,18
» III 9,9 6,0 1,10 — —
Июль, II Август, III 12,5 6,3 0,80 1,69 0,23
— — 1,78 0,28
Поврежденные Июнь, II
первой группы — 13,8 — 3,40 0,33
Август, III — 15,7 — 4,80 0,54
второй группы Июнь, I 9,3 5,6 0,55 — —
» II 9,2 5,4 1,37 — —
в III 10,7 5,4 0,83 — —
Июль, II 9,2 7,5 0,82 — —
третьей группы Июнь, I — 5,0 — 1,85 0,11
Июль, II — 6,4 — 1,77 0,36
Август, III — 6,3 — 1,78 0,39
бе здоровой лиственницы сибирской в течение вегетации изменя-
лось от 3,17 до 1,78%, а в заболони — от 0,18 до 0,28 от веса су-
хого вещества (см. табл. И).
У поврежденных деревьев третьей группы к концу вегетации
флавоноидный комплекс усложняется. В июне на хроматограммах
просматривалось четыре пятна (из них одно — дигидрокверцетин),
в июле и августе пять пятен (из них два флавоноида — дигидро-
кверцетин и следы кверцетина с Rf 0,34). Общее содержание фла-
воноидов в лубе деревьев этой группы в течение сезона колебалось
от 1,85 до 1,77%, а в заболони — от 0,11 до 0,39% от веса сухого
вещества. На хроматограммах из экстракта поврежденных лист-
венниц первой группы просматривалось девять пятен, из них три
флавоноида: дигидрокверцетин, кемпферол (Rf 0,51) и следы
кверцетина. Общее содержание флавоноидов в июне в лубе состав-
ляло 3,4%, августе — 4,8; в заболони соответственно — 0,33 и
0,54% от веса сухого вещества.
Хроматографически (в системе бутиловый спирт — уксусная
кислота — вода, 4:1: 5), в метанольном экстракте у лиственницы,
пораженной лиственничным усачом, в лубе обнаружены девять
веществ, из них четыре флавоноида — дигидрокемпферол, кемпфе-
рол, дигидрокверцетин, кверцетин (в сумме 0,075%); в заболони
найден дигидрокверцетин и кемпферол (в сумме 0,06%).
Таким образом, в лубе поврежденной части ствола содержание
таннинов вначале несколько сокращается за счет образования фло-
бафенов, а затем начинает возрастать. Чем сильнее повреждено
Дерево, тем больше накапливается таннинов в его лубе. Степень
повреждения определяет и состав флавоноидов луба: сильно пов-
режденные, но жизнеспособные деревья характеризуются боль-
шим числом их компонентов.
В химическом барьере, препятствующем заселению хвойных
деревьев насекомыми-ксилофагами, смолы и особенно содержащие-
ся в них эфирные масла — терпены играют первостепенную роль.
Именно понижение у деревьев смолообразования делает их до-
ступными для поселения и развития этих насекомых. Жизнеспо-
собные, устойчивые деревья отражают попытки поселения ксило-
фагов либо локализуют зараженную часть ствола путем засмоления
прокладываемых вредителями ходов или изолирования поражен-
ного участка насыщенной смолами раневой тканью.
Даже в тех случаях, когда отродившиеся личинки короедов,
усачей и златок успешно осваивают луб и наружную часть заболо-
ни, в смолах таких деревьев в начальный момент заселения могут
происходить преобразования защитного характера. У жизнеспо-
собных деревьев эти преобразования кроме повышения общего со-
держания смол в клетках лубяной и древесной паренхимы выра-
жаются в изменении соотношения нейтральных веществ и кислот.
Необратимое ослабление деревьев сопряжено с понижением со-
держания в смолах наиболее токсичных компонентов.
Количественные изменения содержания смол в лубе и заболо-
ни соответствуют анатомическим преобразованиям смолоносной
системы и также зависят от степени повреждения и исходного фи-
зиологического состояния дерева. Наши исследования показали.!
что при локальном механическом повреждении ствола, если тако-
вое не приводит к необратимым патологическим процессам, коли-
чество экстрактивных смол из околораневого участка луба и за-
болони повышается либо сразу после поранения (у кедра, напри-
мер, иногда более чем на 70% от контроля), либо нарастает в про-
цессе вегетации (у лиственницы). Соответственно увеличивается
содержание нейтральных веществ (в основном в лубе) и кислот
[Животова и др., 1971]. У деревьев, временно незначительно ос-
лабленных (в эксперименте — точечное повреждение луба и на-
ружного годичного слоя заболони, имитирующее «попытки» по-
селения короедов), также повышается интенсивность смолообра-
зования и осмотический потенциал, усиливающий активность
смоловыделения. Даже у деревьев, получивших невосстановимое
повреждение, например при кольцевом поранении комля, количе-
ство смол первоначально повышается, и они какое-то время сохра-
няют сопротивляемость заселению ксилофагами.
В очагах массового размножения хвоегрызущих вредителей,
как и в древостоях, сильно ослабленных другими факторами*
например при временном затоплении и заилении почвы, заселение
лиственниц, пихт и сосен насекомыми-ксилофагами происходит
при обеднении луба смолами и нейтральными веществами [Мас-
сель и др.4 1974; Рожков и др., 1975].
Повышение общего содержания смол в лубе сосен до 8—10 % ,
а в лубе лиственниц — до 6—7% может быть только временным
явлением, защитной реакцией на повреждение. Происходит оно
за счет интенсивного использования резервных веществ. Содержа-
ние смол у таких деревьев затем либо восстанавливается до нормы,
либо снижается до минимума, угрожающего заселению ксилофага-
ми, или гибелью без участия последних.
Анализы показали, что в экстрактах из луба и камбия искус-
ственно поврежденных лиственниц второй группы и луба деревьев
четвертой группы содержание нейтральных веществ в начале
вегетации сокращается. В июне в лубе и клетках камбиальной
зоны поврежденных деревьев второй группы содержание нейт-
ральных веществ сокращается на 15%, а в лубе деревьев четвер-
той группы количество их сокращается в июне в среднем на 72,3%,
в июле — на 48 % относительно показателей контрольных деревь-
ев. В июле же в клетках камбия деревьев четвертой группы коли-
чество нейтральных веществ превышает контроль в среднем в 3
раза, содержание общего количества кислот — в 4,5 раза; в за-
болони— соответственно в 1,5 и 2 раза.
Изменения в количественном составе нейтральной фракции из
луба поврежденных лиственниц объясняются оттоком живицы че-
рез раневые отверстия в кору и частичным испарением летучей
фракции терпенов, а также их частичным окислением и превраще-
нием в твердые вещества кислотного характера.
В июне в клетках камбиальной зоны поврежденных деревьев
четвертой группы содержание кислот достигает 11,2% от веса су-
хого вещества при 2,5% в контроле. Исследование кислой части
смолистых веществ из луба поврежденных деревьев третьей груп-
пы показало, что в них преобладают смоляные кислоты (3,18% от
веса сухого вещества при 2,04 в контроле).
В лубе и заболони здоровых деревьев в течение вегетации со-
держание жирных и смоляных кислот изменяется незначительно
(табл. 12), тогда как у поврежденных лиственниц в околораневых
участках количество их либо резко увеличивается (деревья первой
группы), либо сокращается (деревья второй группы).
У жизнеспособных деревьев при поранении заболони смоля-
ная блокада предшествует зарастанию раны каллюсом. У деревьев
первой группы в наружных слоях заболони, особенно в ее разви-
вающемся годичном кольце вокруг пораженного участка, содер-
жание смолистых веществ начинает повышаться. В наших опытах
на второй год после поранения ствола общее количество смолистых
веществ в заболони в I декаде июня увеличилось на 87%, нейт-
ральной фракции — на <91 % и кислот — на 86 % относительно
контроля. Луб пораженных деревьев первой группы к июню сле-
дующего года по общему содержанию смолистых веществ почти не
отличался от нормы, но состав последних изменился. Количество
терпенов сократилось на 26%, что объясняется их испарением из
обнаженных усыхающих участков.
Т аблиц Содержание терпеноидов и жирных кислот на второй год после повреждения, % от абсолютно сухой навески а 12 [
Состояние модельных Смола Нейтральные (сумма) Кислоты (сумма ) \ * смоляные /
деревьев Месяц
VI I VII VIII | VI VII | VIII | VI VII | VIII
Луб
Поврежденные первой группы | 5,00 | — 8,82 1,60 1 - 1 4,02 3,4 1,50 | 1,90 j - 1 4,8 - 2,10 775“
Заболонь
1 4,52 | - 6,81 1,72 | -1 3,80 2,8 0,71 | 2,09 - 1 3,01 - 0,10 2,90
Луб
Поврежденные третьей 1 7,40 1 6,62 | 6,40 I 2,9 2,70 | 3,00 4,5 1,32 I 3,92 1,08 | 3,4 - 1,05
группы 3,18 2,84 1 2,35
Заболонь
| 3,04 | 2,92 | 2,88 I 0,86 ! 1 0,85 | 0,98 2,18 0,68 2,07 0,60 | 1,9 0,50
1,50 1,47 1 1,40
Луб
Здоровые 1 5,42 1 6,08 | 6,49 | 2,19 1 2,5 1 2,87 3,23 1,19 2,04 | 3,58 1,24 | 2,34 | 3,62 • 1,39 2,23
Заболонь
1 2,41 1 2,9 | 3,16 0,90 | 1,09 | 1,40 | 1.51 0,41 1,10 |.,8 0,5 I 1,3 1 1,76 • 0,46 1,30
В августе следующего после повреждения года одолорапевые
участки как заболони, так и луба отличаются наибольшим содер-
жанием смолистых веществ и сопутствующих им жирных кислот.
Подобное накопление в конце вегетации смолистых веществ осо-
бенно характерно для пораженнык тканей деревьев четвертой
группы.
В условиях массового размножения лиственничный усач мо-
жет нападать на вполне здоровые деревья, либо на незначительно
ослабленные, когда содержание смолистых веществ и их каче-
ственный состав в лубе и заболони близки к норме. Однако уже в
самом начале активной жизни личинок в концентрации смол про-
исходят существенные изменения. Эти изменения охватывают и
внешнюю кору. В прилегающем к лубу слое коры содержание смол
удваивается. В исследованных пробах заселенной части ствола ко-
личество смол в коре составляло в среднем 12,5%, в лубе — 10,
в заболони — 3,4%. В лубе незаселенной части ствола содержание
смол первоначально сокращается (по-видимому, из-за оттока в
поврежденные ткани), но вскоре начинает повышаться. Ко време-
ни преодоления личинками сопротивляемости луба концентрация
смол в развивающемся годичном слое заболони достигает, предель-
ной величины (14,5—16% от веса сухого вещества), а затем по мере
разрушения заболони содержание смол в ней сокращается. В это
время смолистые вещества в пробах из луба заселенной части ство-
ла составляли в среднем около 18%; в незаселенной части ствола
содержание смол достигло в лубе 8,1%, в заболони — 6,3% от
веса сухого вещества при 5,9 в лубе и 2,7 % в заболони контроль-
ных деревьев (рис. 65).
Рис. 65. Динамика смо-
листых веществ в лубе
и заболони деревьев,
заселенных лиственнич-
ным усачом.
А — луб здорового дерева
(средний уровень содержа-
ния смол), Ai — луб засе-
ленной части ствола, А, —
луб незаселенной части ство-
ла, Б — заболонь здорового
дерева (средний уровень со-
держания смол), Б| — забо-
лонь заселенной части ство-
ла, Б, — заболонь незасе-
ленной части ствола. 1 —
период преодоления сопро-
тивляемости луба отродив-
шимися личинками; 2 —
полное освоение личинками <
луба, 3— освоение личин-
ками заболони, 4 — углуб-
ление личинок в ядровую
древесину.
Ha. С, Рожков
Этапы освоения личинками
луба и заболони
161
Вне очагов массового размножения лиственничный усач засе-
ляет более ослабленные, хотя и вполне жизнеспособные деревья.
В некоторых случаях почти одновременно с ним такие деревья под-
вергаются нападению других ксилофагов-первопоселенцев. Ис-
следование хлороформеппого экстракта из тканей листвеппиц,
заселенных лиственничным усачом и затем лиственничной злат-
кой, показало, что они сохраняют относительно высокое содер-
жание смолистых веществ в лубе и заболони и еще обладают доста-
точной реактивностью. Тем не менее соотношение нейтральных ве-
ществ и кислот в смолах незаселенной части ствола у этих деревьев
свидетельствует о начале процессов перехода их в состояние необ-
ратимого ослабления (табл. 13).
Повреждение деревьев ксилофагами вызывает изменения ка-
чественного и количественного состава углеводородов нейтраль-
ной фракции. В живом лубе заселяющегося участка ствола лист-
венниц (начальная стадия повреждения) преобладающими компо-
нентами нейтральной фракции являются углеводороды СцЩ*,
и С13Н 2ъ. Содержание их у исследованных деревьев колеба-
лось от 22 до 27%, в то время как соединения С10Н22 и СиН30 не
превышали 12% от суммы веществ фракции. В мертвом лубе де-
ревьев, заселенных лиственничным усачом, преобладающим компо-
Таблица 13
Содержание смолистых веществ в лубе и заболони здоровых деревьев и за*
селенных лиственничным усачом и лиственничной златкой, % от абсолютно
сухой навески
Состояние Смолистые вещества Нейтральные (сум- ма) Кислоты (сумма)
VIII. 1969 VI. 1970 VIII. 1969 VI. 1970 VIII 1969. VII. 1970
Луб пораженной части ствола
Заселены в 1969 г. уса- | 17,4 | 4,2 | 4,6 | 1,1 | 12,8 |
чом, в 1970 г. златкой
Заболонь пораженной части ствола
| 5,0 | 1,5 | 1,7 | 0,9 | 3,3 |
Луб незаселенной части ствола
| 7,6 | 2,7 | 2,8 | 2,0 | 4,8 |
Заболонь незаселенной части ствола
| 5,7 | 2,2 | 1,2 | 1,6 | 4,5 |
3,1
0,6
0,7
0,5
Луб
Здоровые
| 6,7 | 5,0 | 2,5 | 3,3
3,9 | 1,9
Заболонь
| 2,9 1,6 | 1,3 | 0,8
1,7 | 0,7
Рис. 66. Процентное со-
держание и состав али-
фатических соединений
в лубе и заболони де-
ревьев, заселенных лист-
венничным усачом.
1 — здоровое дерево; 2 —
заселено вредителем. А —
луб заселенной части ство-
ла; Б — заболонь той же
части ствола.
пентом углеводородной фракции является соединение С14Н30,
отсутствующее в пораженной заболони и в лубе здоровых деревьев.
Анализ показал, что с уменьшением числа атомов углерода в али-
фатических соединениях (от С13— до С10—), содержащихся в мерт-
вом лубе, уменьшается и их количество (рис. 66).
Углеводородсодержащая фракция из луба здоровой лиственни-
цы содержит наибольшее количество алифатических соединений
с молекулярным весом Си— (38%) и С14— (44%).
В камбиальной зоне здоровых деревьев содержатся два непре-
дельных углеводорода С13_ и С14_, причем последний в максималь-
ном количестве — 83%. В результате повреждения в клетках
камбия появляются углеводороды С10__, Си_, С12_, отсутствующие
у здоровых деревьев. Таким образом, под влиянием повреждений
в лубе и камбиальной зоне происходят существенные количествен-
ные и качественные превращения непредельных углеводородов
[Рожков и др., 1972; Рожков и др., 1978].
В заболони лиственниц с локальным повреждением ствола
происходит накопление веществ С1вНм и С11Н14 и уменьшение
алифатических соединений с более высоким молекулярным весом
(14). Наряду с тем отмечено обогащение заболони таких деревьев
Р-ситостерином (25,2% от веса нейтральных веществ при 9,8%
в контроле) и одновременное сокращение содержания его в лубе.
Очевидно, увеличение 0-ситостерина в заболони поврежденных
деревьев является защитной реакцией. Возможно, он выполняет
в какой-то степени роль раневого гормона в преобразовании тра-
хеид. Проведенные в полевых условиях испытания показали, что
0-ситостерин токсичен для личинок и имаго ксилофагов.
В результате спектрофотометрического определения содержа-
ния 0-ситостерина при1тах 250 установлено, что в пораженной за-
болони (лиственницы, поврежденные лиственничным усачом) ко-
личество его достигает 66 % при 18 в контроле (от веса нейтральных
веществ). В мертвом лубе количество ситостерина сокращается до
1,8% при 29,2% в лубе здоровых деревьев. Исследования кисло-
11*
163
родсодержащей фракции из луба искусственно поврежденных
лиственниц (деревья второй группы) показали, что в августе он
значительно богаче Р-ситостерипом, чем луб здоровых деревьев
(32,2% при 23,1% в контроле).
Монотерпепы занимают особое место среди химических средств
защиты хвойпых деревьев от повреждения насекомыми. Прежде
чем перейти к рассмотрению защитных преобразований этих ве-
ществ, приведем основные результаты испытания токсичности ком-
понентов смол.
Токсичность смолистых веществ и механизм их действия на
насекомых-ксилофагов в течение ряда лет исследовала Г. И. Мас-
сель [Рожков и др., 1978; Масссль, 1979]. В опытах ею использо-
ваны имаго и личинки следующих видов ксилофагов: большого
черного елового усача (Monochamus urussovi Fisch.), малого чер-
ного елового усача (Monochamus sutor II.), бархатпо-пятнистого
черного усача (Monochamus saltuarius Gebl.), сибирского длинноусо-
го усача (Acantkocinus carinulatus Gebl.), рагия ребристого (Rha-
giuni inquisitor Rtt.), черного ребристого дровосека (Asemum
striatum IL), таежной хвойной златки (Ancylocheira strigosa Gebl.),
белоточечного долгоносика (Hylobius albosparsus Motsch.), боль-
шого лиственничного короеда (Ips subelongatus Motsch.), щелкуна
блестящего (Selatosomus aeneus IL). Из веществ испытаны а-пи-
нен, 0-пинен, Д3-карен, углеводород- и кислородсодержащие фрак-
ции п-цимол, Dl-борнеол, изоборнеол, борпилацетат, камфара,
моно- и сесквитерпены, нейтральная фракция живицы, живичный
скипидар, смеси некоторых из этих веществ, а также спиртовые
растворы 0-ситостерина, смоляных и жирных кислот.
Результаты опытов показали, что смолистые вещества обладают
весьма неодинаковой токсичностью по отношению к вредителям
в целом и к каждому виду в отдельности. Токсичными для личи-
нок и имаго всех ксилофагов являются Д3-карен, а-пинен, п-
цимол, 0-пинен, борпилацетат, фракции углеводородов, хлори-
рованные монотерпепы и живичный скипидар лиственницы. При
этом как для видов, заселяющих ослабленные, но жизнеспособные
деревья, так и для тех из них, которые развиваются в основном
на необратимо ослабленных и мертвых деревьях, наибольшей ток-
сичностью обладают Д3-карен и а-пинен, наименьшей — жирные
и смоляные кислоты, камфара и борнеол.
По мнению Г. И. Массель, повышенная токисичность Д3-каре-
па може!1 быть обусловлена его легкой окисляемостыо кислородом
воздуха с образованием перекисных форм, обладающих высокой
окислительной способностью. Учитывая значение перекисных со-
единений в окислении сульфгидрильных групп, можно считать, что
более выраженная биологическая активность терпенов в опре-
деленной степени связана с блокированием сульфгидрильных
групп белков. Экспериментально показано, что смесь Р-пинен Н-
Ч- Д3-карен, а-пипен и фракция монотерпенов оказывают тормо-
зящее действие на фермент сукцинатдегидрогеназа, в активном
центре которого имеются сульфгидрильные группы. Очевидно, тор-
можение активности сукцинатдегидрогеназы происходит вслед-
ствие блокирования ее серных групп образующимися перекисны-
ми формами терпенов, что приводит к нарушению структуры фер-
мента. Падение активности фермента сукцинатдегидрогеназы
вызывает нарушение газового обмена в нервных клетках. По-
скольку нервные клетки очень чувствительны к кислородному го-
лоданию, то вследствие развивающегося нарушения дыхания* ней-
ронов (сопровождающегося повреждением протоплазмы) проис-
ходит повышение сорбционных свойств по отношению к виталь-
ным красителям. Нарушение функций нервной системы приводит
к расстройству работы других органов и в конечном итоге к дегид-
ратации организма насекомых в целом.
Патологические и защитные преобразования монотерпенов.
Показатели суммарного содержания смолистых веществ^ которые
всегда высоки в период интенсивных защитных преобразований,
как и соотношение нейтральных веществ, еще не дают полного
представления о степени ослабления дерева и его регенерацион-
ных возможностях. Устойчивость деревьев к нападению стволовых
вредителей в основном определяется содержанием в смолах моно-
терпеновых углеводородов, обладающих, как было сказано, наи-
большей токсичностью для личинок ксилофагов.
Состав и сезонная динамика монотерпенов в лубе здоровых
деревьев лиственницы сибирской рассмотрены ранее (см. табл. 3).
Количественное соотношение монотерпеповых соединений у раз-
ных пород неодинаково. Особенно существенны эти различия Для
основных монотерпенов — Д3-карена и а-пинена. У лиственницы
соотношение Д3-карена и а-пинена в течение летних месяцев выра-
жается в среднем как 4 : 3, у сосны — 3:5, у кедра — 1:7 и у
ели — 1 : 20.
Анализ монотерпеновых углеводородов из луба искусственно
поврежденных сосен и елей показал, что локальное заселение их
жуками сопряжено с увеличением концентрации в монотерпеновой
фракции а-пинена (у сосен — до 78,4%) и резким снижением уча-
стия Д3-карена, у кедров в момент заселения их вредителями кон-
центрация Д3-карена в монотерпеновой фракции находится на
среднем уровне, а у лиственницы — даже на высоком (более 50%
от содержания всех соединений фракции).
Такое, на первый взгляд, несоответствие общепринятому пред-
ставлению о ведущей роли Д3-карена в резистентности дерева по
отношению к стволовым вредителям вполне объяснимо. Высокая
концентрация Д3-карена в монотерпеновой фракции не отражает
действительное содержание его в лубе и заболони. Необходимо
учитывать долю участия в смолах нейтральных веществ в целом,
определяющих содержание монотерпенов. В наших опытах именно
у лиственниц во время нападения ксилофагов количество нейт-
ральных веществ было очень низким и срставляло лишь 26—52%
от контроля.
165
Таблица 14
Содержание Д’-карева и а-пинена в монотерпеновой фракции в соотноше-
нии с общим количеством смол и нейтральных веществ в лубе здоровых,
ослабленных и заселяемых ксилофагами лиственниц
Состояние деревьев Содержание, %
в лубе в монотерпеновой фракции
смол нейтраль* ных веществ Д*-карена а-пинена
Здоровые 4,9 3,1 54,8 18,1
Подверглись временному затоплению, устойчивы к заселению ксилофага- ми 7,1 5,4 55,8 20,0
Ослаблены в результате временного затопления, начали заселяться ксилофагами 2,6 1,0 35,8 40,6
Обесхвоены пяденицей Якобсона, на- чали заселяться ксилофагами 3,0 1,3 16,8 34,3
Данные табл. 14 свидетельствуют о значительном сокращении
общего содержания смол и нейтральных веществ в лубе ослаблен-
ных в результате дефолиации и временного затопления деревьев
(с заиливанием почвы) в момент заселения их стволовыми вреди-
телями. Лиственницы, подвергшиеся затоплению, но сохранившие
устойчивость к заселению ксилофагами, характеризуются высоким
содержанием в лубе смол в целом, нейтральных веществ и Д’-
карена. Поскольку Д3-карен является наиболее эффективным ток-
сическим компонентом защитной системы лиственницы, перестрой-
ка в сторону его увеличения свидетельствует о проявлении защит-
ной реакции у деревьев, о восстановлении и даже усилении на
какое-то время их резистентности к повреждению ксилофагами.
Приведенные показатели устойчивости лиственниц к заселению
ксилофагами получены в июле (в период лёта большинства ксило-
фагов). В августе происходит снижение содержания Д’-карена и
лимонена, повышается концентрация а- и р-пиненов и относи-
тельно нормализуется количество других компонентов монотерпе-
новой фракции. При таком составе монотерпенов степень устой-
чивости деревьев не исключает возможность заселения их вреди-
телями, особенно лиственничным усачом, однако в это время у них
в основном заканчивается период откладки яиц.
Снижение содержания Д’-карена в лубе разных хвойных пород
к моменту утраты деревьями способности противостоять нападению
ксилофагов, в том числе и тех из них, которые обычно входят в со-
став вторичных (производных) экологических группировок, вы-
ражается разными показателями. Так, если у лиственницы в это
время концентрация его в монотерпеновой фракции из луба сокра-
щается в 1,5—3 раза, то у сосны — в 50—180 pas. Одновременное
повышение концентрации а-пинена не компенсирует в данном слу-
чае утраты деревьями защитных свойств.
Механизмы преодоления сопротивляемости дерева лиственнич-
ным усачом рассмотрены ранее. Исключая этого опасного физио-
логического вредителя, а также большого елового лубоеда и,
возможно, немногих других, остальные виды ксилофагов, по дан-
ным Г. И. Массель_(1979), заселяют деревья при следующем сни-
жении в их лубе‘содержания смол: у лиственницы — до 3%
(1,5% нейтральных веществ), у сосны — до 6% (2,5 %'нейтральных
веществ), у пихты — до 8% (4,5% нейтральных веществ).
Из приведенного явствует, что структурные и функциональные
защитные преобразования, возникающие у деревьев в ответ на
повреждение пасекомыми-ксилофагами, могут затрагивать все
живые клетки ствола, а иногда и лишившиеся протопласта. Пре-
образования эти многообразны и сложны, но в конечном итоге
сводятся к регенерации изолирующих структур, увеличению сме-
ло- и фенолообразующих элементов ксилемы и флоэмы и повыше-
нию токсичности и репеллентности смолистых веществ.
Таковы в основных чертах проявления защитных реакций у
лиственницы. Они, как видно, близки у всех хвойных. Конечно,
в идеале следовало бы рассмотреть реакции дерева на фоне транс-
формирования метаболизма в целом, но, к сожалению, об этом мы
мало знаем. В данном случае мы имеем дело не с обычной патоло-
гической физиологией, а со стрессовым явлением, во многих слу-
чаях связанным с использованием всех резервов в борьбе за жизнь.
ЛИТЕРАТУРА
Агафонова Т. А., Анисимова О. А., Марьенко В. А. Большой еловый лу-
боед в хвойных лесах Южного Прибайкалья.— В кл.: Хвойные деревья
и насекомЫе-'дендрофаги. Иркутск, 1978, с. 67—73.
Алексеев А. В., Лурье М. А. Практический определитель личинок усачей
(Coleoptera, СегашЪушаае), обитающих на ели обыкновенной в европейской
часта СССР.— Энтомол. обозр., 1970, т. 49, выл. 3, с. 650—655.
Алисов Б. П. Климат.— В кн.: Атлас Иркутской области. М.— Иркутск,
1962, с. 53-64.
Анисимова К. И. Определение колориметрическим методом дубильных
веществ в корнях Polygonum coriarium Grig.— В кн.: Растительные ресурсы.
Вып. 1. Л., 1967, с. 128—131.
Анисимова К. И. Некоторые виды лиственницы как источник получения
таннидов и камеди и перспективы их использования в народном хозяйстве.—
Автореф. канд. дне. Л., 1968.
Анисимова К. И., Антоновский С. Д., Белозерова Л. А. н др. Лиственни-
ца как источник получения ряда полезных веществ.— В кн.: Растительные
ресурсы. Т. 1, вып. 1, М,—Л., 1965, с. 74—83.
Анисимова О. А., Соков М. К. Роль насекомых в древостоях, ослаблен-
ных токсичными выбросами алюминиевых заводов.— В кн.: Влияние антро-
погенных и природных факторов на хвойные деревья. Иркутск, 1975,
с. 61—84.
Антонова Г. Ф. К вопросу образования арабиногалактана в ксилеме
лиственницы сибирской.— В кн.: Обмен веществ п продуктивность хвойных.
Новосибирск, 1977, с. 53—59.
Белозерским А. Н.9 Проскуряков Н. И. Практическое руководство по
биохимии растений. М., 1951.
Бережных Е. Д. Насекомые-ксилофаги нарушенных хвойных лесов Бай-
кало-Амурской среднетаежной зоны.— В кн.: Хвойные деревья и насекомые
дендрофаги. Иркутск, 1978, с. 87—51.
Бероза М. Аттрактанты и репелленты для борьбы с вредными насекомы-
ми.— В кн.: Стратегия борьбы с вредителями, болезнями растений и сорняка-
ми в будущем. М., 1977, с. 228—256.
Бобров Е. Г. История и систематика лиственниц. Т. 25. Л., 1972.
Болдаруев В. О. Динамика численности сибирского шелкопряда и его
паразитов. Улап-Удэ, 1969.
Васильева Т. Г. Зависимость плодовитости хвоегрызущих чешуекрылых
от биохимического состава хвои поврежденных деревьев.— В кн.: Анатоми-
ческие, гистохимические и биохимические преобразования у лиственницы при
повреждении насекомыми. Иркутск, 1972, с. 57—66.
Васильева Т. Г. Плодовитость массовых хвоегрызущих вредителей в за-
висимости от биохимического состава корма. Автореф. канд. дне. Красно-
ярск, 1973.
Васильева Т. Г., Плешанов А. С. Биохимические особенности деревьев,
поврежденных серой лиственничной листоверткой.— В кн.: Анатомические,
16R
гистохимические и биохимические преобразования у лиственницы при по-
вреждении насекомыми. Иркутск, 1972, с. 26—35.
Васильева Т. Г., Плешанов А. С. Физиология ослабления и отмирания
лиственницы в очагах пяденицы Якобсона.— В кн.: Влияние антропогенных
и природных факторов на хвойные деревья. Иркутск, 1975, с. 179—214.
Вержуцкии Б. Н., Агафонова Т. А., Волкова Л. М. Биохимические изме-
нения в тканях деревьев, поврежденных лиственничной почковой галлицей,—
В кн.: Анатомические, гистохимические и биохимические преобразования у
лиственницы при повреждении насекомыми. Иркутск, 1972, с. 76—81.
Вержуцким Б. Н., Агафонова Т. А., Швец М. М. Место лиственницы
среди голосеменных как кормового объекта пилильщиков (Hymenoptera,
Symphyta) в связи с биохимическим составом хвои подроста.— В кн.: Хвой-
ные деревья и насекомые-дендрофаги. Иркутск, 1978, с. 114—120.
Вержуцким Б. Н., Докнйчук Т. А., Животова М. М. Биохимические изме-
нения в хвое лиственничного подроста, поврежденного побеговым пилиль-
щиком.— В кн.: Информационный бюллетень координационного совета по
физиологии и биохимии растений. Вып. 7. Иркутск, 1970, с. 76—78.
Вечер А. С., Проказов Г. Ф. Определение общего содержания углево-
дов.— В кн.: Физиолого-биохимические исследования растений. Минск,
1967, с. 76—82.
Воронцов А. И. Биологические основы защиты леса. М., 1963.
Воронцов А. И., Семевский Ф. Н. Некоторые вопросы колебания числен-
ности лесных насекомых.— В кн.: Вопросы лесозащиты. Т. 2. М., 1963,
с. 18—22.
Галкин Г. И. Некоторые вопросы формирования резерваций и первичных
очагов сибирского шелкопряда в лесах Красноярского края.— В кн.: Мате-
риалы по проблеме сибирского шелкопряда. Новосибирск, 1960, с. 21—34,
Галкин Г. И. Сибирский шелкопряд в лесах Красноярского края.— Тр.
Вост.-Сиб. НИИ лесной и деревообраб. пром., Красноярск, 1963, вып. 7,
с. 74
Гречкин В. П. Сибирский шелкопряд (Dendrolimus sibiricus Tschelw) —
вредитель лесов Монголии.— Зоол. журн., 1960, т. 39, вып. 1, с. 84—96.
Гримальский В. И. До питания про стшюсть сосновых насаждень против
соснового шовкопряда.— В кн.: Проблеми ентомологн на Украшь Ки>в,
1959, с. 236.
Гусев Н. А. Рефрактометрический метод определения свободной и связан-
ной воды.— В кн.: Некоторые методы исследования водного режима расте-
ний. Л., 1960, с. 60.
Гусева А. Г., Нестюк М. Н. К методике определения флавоновых ве-
ществ в растениях.— Биохимия, 1953, т. 18, вып. 4, с. 480—483.
Гуссаковский В. В. Рогохвосты и пилильщики.— В кн.: Фауна СССР.
Т. 2. Вып. 1. Новая серия, № 1. Перепончатокрылые. М.—Л., 1935, с. 452.
Джекобсон М. Половые феромоны насекомых. М., 1976.
Дженсен У. Ботаническая гистохимия. М., 1965.
Дылис Н. В. Сибирская лиственница. В. 2 (10). М., 1947.
Дылис Н. В. О происхождении брахибласт у Larix.— Докл. АН СССР,
1958, т. 122, № 1.
Дылис Н. В. Лиственница Восточной Сибири и Дальнего Востока .М., 1961.
Животова М. М., Гуменюк М. А., Костромина 3. И. Физиолого-биохими-
ческие изменения в тканях поврежденных деревьев лиственницы сибир-
ской.— В кн.: Информационный бюллетень координационного совета по
физиологии и биохимии растений. Вып. 8. Иркутск, 1971, с. 108—110.
Животова М. М., Рожков А. С. К вопросу моделирования повреждения
насекомыми древесных растений при изучении их патологии и защитных ре-
акции.— В кн.: Информационный бюллетень координационного совета по
физиологии и биохимии. Вып. 5; Иркутск, 1969, с. 101—195.
Животова М. М., РожковА. С., Массель Г. И. Изучение состава нейтраль-
ной фракции экстрактивных смол у лиственниц, пораженных алтайским лист-
венничным усачом.— В кн.: Информационный бюллетень координационного
совета по физиологии и биохимии.— Вып. 8. Иркутск, 1971, с. 110—111.
169
Зайцева А. Ф. Изучение локализации арабогалактана в клеточных стоп-
ках древесины лиственницы на микросрезах.— В кн.: Изучение химического
состава древесины даурской лиственницы. Т. 45. М., 1958, с. 50—60.
Ивлиев Л. А. Сибирский шелкопряд в лесах Дальнего Востока.— В кн.:
Материалы по проблемам сибирского шелкопряда. Новосибирск. I960,
с. 47-59.
Ивлиев Л. А. Сибирский шелкопряд в лесах Дальнего Востока. Автореф.
канд. дис. Владивосток, 1966.
Ивлиев Л. А., Кононов Д. Г., Малахова В. П. Насекомые-фитофаги ре-
ликтовой породы Приморского края лиственницы Любарского (Larix In-
barskii Suk.).— В кн.: Энтомофаупа советского Дальнего Востока. Т. 9 (12).
Владивосток, 1973, с. 20—34.
Ижевский С. С. Функциональные особенности ферментных систем ки-
шечника насекомых-фитофагов.— В кн.: Вопросы экологической физиологии
беспозвоночных. М., 1974, с. 156—175.
Ильинский А. И. К вопросу о закономерностях в размножении короеда
типографа, о методике изучения его н борьбе с ним в лесу. Вып. 2. Харьков,
1931, с. 5-31.
Ильинский А. И. Вторичные вредители сосны и ели и меры борьбы с ни-
ми.— В кн.: Сборник работ по лесному хозяйству ВНИИЛМ. Вып. 36.
М.-Л., 1958, с. 178-228.
Ильинский А. И. Определитель вредителей леса. М., 1962.
Исаев А. С. Стволовые вредители лиственницы даурской. М., 1966.
Исаев А. С., Гире Г. И. Взаимодействие дерева и насекомых-ксилофагов.
Новосибирск, 1975.
Исаев А. С., Рыжкова Т. С. Состав эфирных соединений в лубе лиственни-
цы сибирской и их роль в защитной реакции дерева.— В кн.: Лиственница.
Т. 3. Красноярск, 1968, с. 273—286.
Исаев А. С., Тарасова Д. А. Формирование очагов стволовых вредителей
в лиственничных лесах Тувинской АССР.— Тр. Сиб. техпол. ин-та. Сб. 39.
Лиственница, Новосибирск, 1964, с. 244—253.
Кожанчнков И. В. Волнянки (Opgyidae).— В кн.: Фауна СССР. Т. 12.
Новая серия, № 42. Насекомые чешуекрылые. М.—Л., 1950.
Колесников Б. П. К систематике и истории развития лиственниц секции
Pauciserialis Patschke.— В кн.: Материалы по истории флоры и раститель-
ности СССР. Т. 2. М., 1946, с. 321—364.
Коломиец Е. Г. Паразиты и хищники сибирского шелкопряда. Ново-
сибирск, 1962.
Коновалов Н. А. Типы лесов с участием лиственницы Сукачева на Сред-
нем Урале.— В кн.: Лиственница. Т. 3. Красноярск, 1968, с. 18—27.
Костин И. А. Стволовые вредители хвойных лесов Казахстана. Алма-
Ата, 1964.
Крамер П., Козловский Т. Физиология древесных растении. М., 1963.
Крашенников И. М. Основные пути развития растительности Южного
Урала в связи с палеогеографией Северном Евразии в плейстоцене и голоце-
не.— Сов. ботаника, 1939, № 6—7.
Криволуцкая Г. А. Короеды острова Сахалина. М.—Л., 1958.
Криволуцкая Г. А. Фауна усачей (Coleoptera, Cerambucidae) острова
Сахалина.— Энтомол. обозр., 1961, т. 40, вып. 2, с. 310—314.
Кривошеина К. П., Мамаев Б. М. Определитель личинок двукрылых на-
секомых — обитателей древесины. М., 1967.
Куренцов А. И. Вредные насекомые хвойных пород Приморского края.—
Тр. ДВФ АН СССР. Сер. зоол., Владивосток, 1950. Т. 1 (4), с. 256.
Любарский Л. В. Эколого-географическая характеристика дереворазру-
шающих грибов советского Дальнего Востока.— В кн.: Биологические ре-
сурсы Дальнего Востока. М., 1959, с. 97—108.
Любарский Л. В., Идрисов Р. И. Усыхание лиственницы курильской на
Сахалине.— Сб. трудов ДальНИИЛХ, 1970, вьш. 10, с. 264—368.
Макфедьен Э. Экология животных (цели и методы). М., 1965.
Малсвская С. С., Карнаухова В. Д. Смола в древесине ели.— Ж. при-
кладной химии, 1956,-Вып. 29, 3, с. 401—410.
170
Малышев Л. И. Применение анатомического метода для целей определе-
ния сосен и выявления их филогении.— В кн.: Ботаника. Благовещенск,
1958, с. 107—127.
Малышев С. И. Становление перепончатокрылых и фазы их эволюции.
М.—Л., 1966.
Мамаев Б. М., Данилевский М. Л. Личинки жуков-дровосеков. М.,
1975.
Марьенко В. А. Электропроводность по переменному току как показа-
тель физиологического состояния деревьев.— В кн.: Оперативные информа-
ционные материалы. Иркутск, 1976, с. 34—35.
Массель Г. И. Защитные свойства смолистых веществ у лиственницы.—
В кн.: Анатомические, гистохимические и биохимические изменения у лист-
венницы при повреждении насекомыми. Иркутск, 1972, с. 109—115.
Массель Г. И. Смолистые вещества хвойных и их влияние на насекомых-
ксилофагов. Автореф. канд. дис. Красноярск, 1979.
Массель Г. И., Плешанов А. С., Дубовенко Ж. В. Изменения содержания
монотерпеповых углеводородов у лиственницы и пихты, заселяемых ксило-
фагами в очагах хвоегрызущих вредителей.— В кн.: Проблемы физиологии и
и биохимии древесных растений. Вып. 4. Красноярск, 1974 с. 28—30.
Массель Г. И., Рожков А. С., Пентегова В. А., Вольский Л. Н. Защитные
преобразования состава монотерпсновых углеводородов при повреждении
хвойных деревьев.— Изв. СО АН СССР. Сер. биол., 1975, выи. 2, № 10,
с. 90—95.
Миронова Т. И., Зархина Е. М., Волкова В. И. Изучение и использование
лесосечных отходов и коры лиственницы сибирской.— В кн.: Лиственница.
Т. 29. Красноярск, 1962, с. 217—225.
Мозолевская Е. Г. Стволовые вредители лесов Башкирского заповедни-
ка.— В кн.: Вопросы защиты леса. Вып. II. М., .1964, с. 57—78.
Нальняева Т. И. Материалы о вредителях лиственницы сибирской в
Туве.— Тр. Биол. ин-та СО АН СССР, Новосибирск, 1960, вып. 5, с. 149—156.
Наумов В. Д. Характеристика очагов сибирского шелкопряда и меры
борьбы с ним в условиях Читинской области.— В кн.: Материалы по проб-
леме сибирского шелкопряда. Новосибирск, 1960, с. 93—102.
Никифоров Г. Ф. К экономическому обоснованию нового способа экстра-
гирования тапнидов не лиственничной коры.— В кн.: Лиственница. Т. 3.
Красноярск, 1968, с. 472—478.
Овсянников В. Ф. Хвойные породы. Хабаровск, 1930.
Пентегова В. А., Лисина А. И. Состав скипидара сибирской лиственни-
цы.— Тр. Химико-мед. ин-та Зап.-Сиб. филиала АН СССР, Новосибирск,
1959 т. 6, вып. 13, с. 23—24.
Петренко £. С. Насекомые — вредители лесов Якутии. М., 1965.
Плавилыциков Н. Н. Жуки-дровосеки — вредители древесины. М.—Л.,
1932.
Плавильщиков Н. Н. Жуки-дровосеки. Ч. 2.— В кн.: Фауна СССР. Т. 22.
Новая серия. Насекомые жесткокрылые. Л., 1940.
Плеханов Г. Ф., Елизаров Ю. А., Калашников А. В. н др. О природе сиг-
нала, привлекающего самцов дубового шелкопряда Anlheraea pernyi Guer.—
В кн.: Общие и прикладные вопросы хеморецепции (обоняние). М., 1977,
с. 133-142.
Плешанов А. С. О лесохозяйственном значении вредных насекомых в
зоне Байкало-Амурской магистрали.— В кн.: Тезисы Научной сессии по воп-
росам сельскохозяйственного освоения западного участка БАМ, изучения
растительных ресурсов, их использования и охраны. Иркутск, 1974, с. 24—26.
Плешанов А. С., Массель Г. И. Экологические аспекты заселения очагов
пяденицы Якобсона стволовыми вредителями.— В кн.: Хвойные деревья и
насекомые-дендрофаги. Иркутск, 1978, с. 26—36.
Поздняков Л. К.гДаурская лиственница. М., 1975.
Положенцев П. А. Энтомопнвазия ветровальной сосны.— Научные
записки Воронежского лесохозяйственного ин-та, 1953, вып. 12, с. 126—194.
Пономаренко А. Г. Историческое развитие жеСткокрылых-архостемат.—
171
В кн.: Труды Палеонтологического института АН СССР. Т. 125. М.,1969,
с. 239.
Прозина М. Н. Ботаническая микротехника. М., 19Й0.
Прозоров С. С. Гари в сосновых лесах как очаги заражения.— Тр.
СибНИИСХ, 1929, т. 12, вып. 3, с. 54.
Пятницкий Г. К. К вопросу массовых размножений насекомых.— В кн.:
Вторая экологическая конференция по проблеме «Массовые размножения
животных и их прогнозы. Ч. 3. Киев, 1951.
Расницын А. П. Происхождение и эволюция низших перепончатокры-
лых.— В кн.: Труды Палеонтологического института АН СССР. М., 1969,
с. 196.
РефесП. М. Роль п значение растительноядных насекомых в лесу. М.,1968.
Редикорцев В. В. Материалы к этномофауне Урала.— В кн.: Записки
Уральского общества любителей естествознания. Т. 47, 1908, с. 95—122.
Рожков А. С. Чешуекрылые — вредители почек п побегов соснового мо-
лодняка в Прибайкалье.— Тр. Вост.-Сиб. филиала АН СССР, сер. биол.,
1957, вып. 5, с. 122—137.
Рожков А. С. К динамике очагов массового размножения сибирского шел-
копряда в горных лесах юга Прибайкалья.— Изв. СО АН СССР, Новоси-
бирск, 1958, № 4, с. 125—133. .
Рожков А. С. Сибирский шелкопряд (Систематическое положение, фило-
гения, распространение, экономическое значение, строение и образ жизни).
М., 1963.
Рожков А. С. Массовое размножение сибирского шелкопряда и меры борь-
бы с ним. М., 1965.
Рожков А. С. Обоснование иммунологического направления в защите
леса.— В кн.: Проблемы защиты таежных лесов. Красноярск, 1971,
с. 116—117.
Рожков А. С. Изучение защитных реакций у древесных растений в экс-
перименте.— В кн.: Проблемы защиты таежных лесов. Красноярск, 1971а,
с. 117—118.
Рожков А. С. Преодоление сопротивляемости дерева стволовыми вреди-
телями-первопоселенцами на примере алтайского лиственничного усача.—
В кн.: Анатомические, гистохимические и биохимические преобразования у
лиственницы при повреждении насекомыми. Иркутск, 1972, с. 83—87.
Рожков А. С., Вялая И. В. Вредители ствола.— В кн.: Вредители лист-
венницы сибирской. М., 1966, с. 19—22.
Рожков А. С., Габиб-заде Л. А. Осмотическое давление клеточного сока
луба как показатель возможности заселения деревьев ксилофагамп.— В кн.:
Хвойные деревья и насекомые-дендрофаги. Иркутск, 1978, с. 121—123.
Рожков А. С., Сендарович Б. П., Данович К. Н. О некоторых взаимоот-
ношениях гусениц сибирского шелкопряда (Dendrolimus sibiricus Tschetv.)
и повреждаемой ими лиственницы (Larix sibirica I.edb.).— Тр. Вост.-Сиб.
филиала СО АН СССР, Иркутск, 1962, вып. 35, с. 90—95.
Рожков А. С., Степанчук Б. С. К вопросу о нормальных и патологиче-
ских смоляных каналах у лиственницы.— В кн.: Информационный бюлле-
тень координационного совета по физиологии п биохимии растений. Вып. 7.
Иркутск, 1970, с. 110—111.
Рожков А. С., Степанчук Е. С. Анатомические п гистохимические защит-
ные преобразования у деревьев, подвергшихся нападению стволовых вреди-
телей.— В кн.: Анатомические, гистохимические и биохимические преобра-
зования у лиственницы при повреждении насекомыми. Иркутск, 1972,
с. 87-96.
Рожков А. С.9 Степанчук Е. С. Образование и особенности строения пато-
логических смоляных каналов в древесине хвойных.— В кн.: Влияние антро-
погенных и природных факторов на хвойные деревья. Иркутск, 1975,
с. 241—250.
Рожков А. С., Степанчук Е. С., Массель Г. И. Защитные механизмы от
повреждения пасекомыми-ксилофагами у хвойных деревьев.— В кн.: Хвой-
ные деревья и насекомые-дендрофаги. Иркутск, 1978, с. 5—25.
172
Рожков А. С., Холопова Р. В., Массель Г. И. и др. Патология к резистент-
ность сосны и лиственницы во временно затопляемых пойменных лесах юга
Восточной Сибири.— В кн.: Влияние антропогенных и природных факторов
на хвойные деревья. Иркутск, 1975, с. 95-—140.
Рожков А. С. 9 Швец М. М., Массель Г. И. Биохимические изменения у по-
раженных стволовыми вредителями деревьев.— В кн.: Анатомические, гисто-
химические и биохимические преобразования у лиственницы при повреж-
дении насекомыми. Иркутск, 1972, с. 96—109.
Ромадина К. Г. Древогрызущие личинки жуков-усачей (Coleoptera,
Cerambycidae) долины реки Урала.— Тр. ЗИН АН СССР, 1954, т. 16,
с. 211-228.
Рубин Б. А. Курс физиологии растений. М., 1971.
Рубин Б. А., Арциховская Е. В. Биохимия и физиология иммунитета
растений. М., 1968.
Руднев Д. Ф. Причины снижения устойчивости насаждений и условия
образования очагов вредителей в лесах Украины.— Бюлл. научно-техн, ин-
форм. Укр. ИЗР, 1958, вып. 4, с. 39—41.
Руднев Д. Ф. Влияние физиологического состояния растений на массовое
размножение вредителей леса.— Зоол. журн., 1962, т. 41, вып. 3,
с. 313—329.
Сергеев В. К. О лиственничном усаче (Xylotrechus altaicus Gebl.) на
Дальнем Востоке.— В кн.: Сборник работ Дальневосточного научно-иссле-
довательского института лесного хозяйства и лесоэксплуатации. Вып. 1.
Хабаровск, 1948, с. 171—174.
Сибирский шелкопряд и меры борьбы с ним/Жохов И. И., Гречкин В. П.,
Коломиец Н. Г. и др. М.—Л., 1961.
Старк В. Н. Образование короедных группировок в зависимости от пол-
ноты насаждений.— Защита растений. 1929, т. 5, № 3—4.
Старк В. II. Вредные лесные насекомые. М.—Л., 1931. 428 с.
Старк В. Н. Короеды.— В кн.: Фауна СССР. Т. 31. Новая серия, № 49.
Жесткокрылые. М.—Л., 1952, с. 461.
Судачкова Н. Е. Метаболизм хвойных и формирование древесины. Ново-
сибирск, 1977.
Сукачев В. Н. К истории развития лиственниц.— В кн.: Лесное дело.—
Л.-М., 1924, с. 12-44.
Сукачев В. Н. О двух новых ценных для лесного хозяйства древесных по-
родах.— Тр. и исслед. по леей. $оз-ву и леей, «промышл., 1931, вып. 10,
с. 1—20.
Суэйн Т. Тапнины.— В кн.: Биохимия растений. М., 1968, с. 329—348.
Тальман П. Н. Биология забайкальского усача Xyloclytus altaicus
Gebl.— Тр. Лесотехн, акад., 1940, вып. 57, с. 60—83.
Тальман П. Н. Лесохозяйственное значение забайкальского усача.—
Лесное хозяйство, 1940а, № 7, с. 61—64.
Тальман П. Н. К биологии усачей Monochamus saltuarius Gebl. и М. im-
pluviatus Motsch.— Тр. Лесотехн, акад., 1947, вып. 61, с. 123—134.
Тальман П. Н. Влияние среды и преобразующей ее роли человека в свя-
зи с размножением сибирского шелкопряда.— Тр. Лесотехн, акад., 1957,
выЦ. 81, с. 107.
Тихомиров Б. Н., Фалалеев Э. Н. Характеристика лиственничных лесов
СССР.— В кн.: Лиственница. Т. 29. Красноярск, 1962, с. 5—22.
Тюкавкина Н. А., Лаптева К. И., Модонова Л. Д. и др. Исследование
экстрактивных веществ древесины даурской лиственницы.— В кн.: Феноль-
ные соединения и их биологические функции. М., 1968, с. 72—78.
Фролов Д. II. Сибирский шелкопряд в Восточной Сибири. Иркутск, 1938.
Ханнсламов М. Г. О ведущих условиях начала вспышек хвое- и листо-
грызущих вредителей.— В кн.: Вопросы лесозащиты. Т. 2. М., 1963,
с. 150—154.
Цветаева И. П., Юрьева М. К., Никитин И. И. Особенности химического
состава древесины даурской лиственницы.— В кн.: Изучение химического
состава древесины даурской лиственницы. Т. 45. М.,' 1958, с. 22—30.
173
Чавчавадзе Е. С., Яценко-Хмелевский А. А. Семейство сосновые (Pina-
сеае).-— В кн.: Жизнь растений. М., 1978, с. 350—374.
Часовщиков Г. Л., Ильина М. В., Крашенников И. А., Жовнер Н. А.
Сравнение лиственничной и сосновой канифоли в резинах.— В кн.: Листвен-
ница. Т. 3. Красноярск, 1968, с. 447—451.
Черепанов А. И. Пузатый клещ,— Изв. Зап.-Сиб. филиала АН СССР,
Новосибирск, 1948 т. 2. Серия биол., вып. 2 (зоол.), с. 63-—68.
Черепанов А. И. Главнейшие энтомовредители Тувы и возможные меры
борьбы с ними.— Изв. Зап.-Сиб. филиала АН СССР, Новосибирск, 1950,
т. 3. Серия биол., вып. 2 (зоол.), с. 35—51.
Черепанов А. И. Лиственничный дровосек и борьба с ним. Новосибирск,
1952.
Черепанов А. И., Черепанова Н. Е. Морфология личинок и биология
усачей рода Fvodinus Lee. (Coleoptera, Cerambycidae).— В кн.: Новые и
малоизвестные виды фауны Сибири. Вып. 4. Новосибирск, 1971, с. 17—27.
Черепанов А. И., Черепанова Н. Е. Морфология личинок и биология уса-
чей группы Xylotrechus Ghevr. (Coleoptera, Cerambycidae).— В кн.: Новые и
малоизвестные виды фауны Сибири. Вып. 6. Новосибирск, 1973, с. 21—72.
Черепанов А. И., Черепанова Н. £. Морфология личинок и биология уса-
чей группы Monochamus (Coleoptera, Cerambacidae), населяющих леса Си-
бири.— В кн. Новые и малоизвестные виды фауны Сибири. Вып. 7. Ново-
сибирск, 1973а, с. 38—72.
Черепанов А. И., Черепанова Н. Е. Жуки-дровосеки ивовых лесов Си-
бири. М., 1975.
Черепанов А. И., Черепанова Н. Е. Новое о морфологии и биологии жу-
ков-дровосеков Callichromini — Hosaliini (Coleoptera, Cerambycidae).—
В кн.: Таксоны фауны Сибири. Новосибирск, 1977, с. 137—155.
Чочиева М. М., Цветаева И. П., Юрьева М. К. и др. Распределение ара-
бога лактана в древесине даурской лиственницы.— В кн.: Изучение химиче-
ского состава древесины даурской лиственницы. Т. 45. М., 1958, с. 31—49.
Шаблиовский В. В. Определитель личинок главнейших видов жуков-
дровосеков (Coleoptera, Cerambycidae) — вредителей лесов Дальнего Восто-
ка.— Тр. ДВФ АН СССР. Сер. зоол., Владивосток, 1956, т. 3(6), с. 111—136.
Шорохов II. И., Шорохов С. И. Вредители зерна и зерно продуктов. М.,
1938.
Эсау К. Анатомия растений. М., 1969.
Яновский В. М. Насекомые-дендрофаги Монголии.— В кн.: Насекомые
Монголии. Вып. 5. JL, 1977, с. 30—59.
Яхонтов В. В. Экология насекомых. М., 1964.
Allee W. С., Emerson А. Е., Park О., е. a. Principles of Animan Ecology.—
X. Y., 1949..
Andrewatha II. G. Self — regulating mechanisms in animal populations.—
Aust T. Sci., 1959, v. 22, p. 200-205.
Bakke Alf. Austara Oystein. Granbarkbileens feromoner som vapen i bil-
lekrigen.— Skogen, 1978, Bd 43, X 1, S. 5—7.
Bannan M. W. Vertival resin ductus in the secondary wood of the Abieti-
neae.— New Phytol., 1936, v. 35, p. 11—48.
Butenandt A. Wirkstoffe des Jnsektenreiches.— Naturwissenschaften,
1959, Bd 46, N 15, S. 461-471.
Butenandt A. Fettaikohole als Sexual — Lockstoffe der Schmetterline.—
Fette, Seifen. Anstrichmittel, 1962, Bd 64, X 3, S. 187—192.
Campion D. C. The potential use pheromones in insect control.— Proc.
Sth Brit. Insectic. and Fungic. Conf., Brighton, 1975. v. 3, Nottingham, 1976,
j). 939—946.
Chararas C. Laction synergique des constituants glucidiques et des consti-
tuants terpeniques dans le processus d'attraction secondaire et le mfcanisme
de relaboration des pheromones chez les Scolytidae parasites des coniferes.—
C. r. Acad. Sci., 1977, D284, X 16, p. 1545—1548.
Chitty D. Self-regulation of numbers through changes in viability.— In:
Cold Spring Harbor Symposia on Quantitative Biology, v. 22, 1957,
p. 277—280.
174
Chitty D. Population processes in the vole and their relevance to general
theory.— Canad. I. Zool., 1960, v. 38, p. 99—113.
Coster J. E., Gara Б. I. Studies on the attack behaviour of the southern
pine beetle. II. Response to attractive host material.— Con tri bs. B. Thompson
Inst. 1968, v. 24, N 4, p. 69—75.
Coster J. E., Payne T. L., Hart E. R., Edson L. J. Aggregation of the so-
uthern pine beetle in rcsponce to attractive host trees.— Environ. Entomol.,
1977, v. 6, N 5, p. 725-731.
Duffy E. A. J. A monograph of the immature stages of British and imported
timber beetles (Cerambycidae). London, 1953.
Duffy E. A. J. A monograph of the immature stages of oriental timber
beetles (Cerambycidae). London, 1968.
Dyer £. D. A. Frontalin attractant in stands infested by the spruce beetle
Dendroctonus rufipennis (Coleoptera: Scolytidae).— Canad. Entomol., 1975,
v. 107, N 9, p. 979—988.
Francke W., Heemann V. Das Duftstoff — Bouquet des Groben Waldgart-
ners Blastophagus piniperda L. (Coleoptera: Scolytidae).— Z. angew. Entomol.,
1976, Bd 82, N 2, S. 117—119.
Gara R. I., Coster J. E. Studies on the attack behaviour of the southern
pine beetle. III. Sequence of tree infestation within stands.— Contribs B.Thom-
pson Inst., 1968, v. 24, N 4, p. 77—85.
Gaussen H. Les Gymnospermes actue)les et fossiles. Toulouse, 1966.
Gebler F. Clytus altaicus.— Bull, de la Soc. des Natur de Moscou, 1836,
v. 10, p. 342—343.
Harring Candace M. Aggregation pheromones of the European fir engraver
beetles Pityokteines curvidens, P. spindens and P. vorontzovi and the role of
juvenile hormone in pheromone biosynthesis.— Z. angew. Entomol., 1878,
Bd. 85, N. 3, p. 281—317.
Heikkenen H. J. Southern pine Beetle: hypothesis regarding its primary
attractant.— J. Forest., 1977, v. 75, N 7, p. 408—413.
Heyden L. Catalog der Coleopteren von Sibiria. Berlin, 1880—1881.
Hughes P. R., Renwick J. A.,. Vite J. P. The identification and field bio-
assay of chemical attractants in the roundheaded pine beetle.— Environ. Ento-
mol., 1976, v. 5, N 6, p. 1165—1168.
Jacobson M. Recent progress in the chemistry of the sex attractants.—
Advances in Chemistry Series, 1963, N 41, p. 1—10.
Jantz О. K., Rudinsky J. A. laboratory and field methods for assayiong
olfactory responses of the Douglas — fir beetle, Dendroctonus pseudotsugae
Hopk.— Canad. Entomol., 1965, v. 97, N 9, p. 935—941.
Katzenellenbogen John A. Insect pheromone synthesis: new methodology.—
Science, 1976, v. 194, N 4161, p. 139—148.
Mannerheim K. Insectes coleop teres de la Siberie Orientale nonveaux et
oeu connus.— Bull, de la Soc. des Natur. de Moscou, 1849, v. 22, N 1, 241 p.
Must apart a H. Behavioural responses of the pine weevil Hylobius abietis
L. (Col.: Curculionidae) to odours activating different groups of receptor cells.—
J. Comp. Physiol., 1975, v. A102, N 1, p. 57—63.
Ostenfeld С. IL, Larsen C. Syrach. The species of the genus Larix and their
geographical distribution. Kobenhavn, 1936.
Rao G. S. TSe physiology of digestion in the midgut of Poecilocerus pictus
Fabr. (Orthoptera, Acrididae). I. pH and digestive enzymes.— J. Zool. Soc.
India, 1969, v. 21, p. 67—73.
Reitter E. Fauna Germanica.— Die Kafer des Deutschen Reiches, v. 1—5,
Stuttgart, 1908—1916. 1655 s.
Rudinsky J. A., Morgan M. E.. Libbey L. M., Putman T. B. Limonene re-
leased by the Scolytid beetle Dendroctonus pseudotsugae.— Z. angew. Ento-
mol. 1977, v. 82, N 4, p. 376—380.
Schedi К. E. Breeding habits of arboricole insects in Central Africa.—
Proc. X Internal. Congr., Entomol., 1956 (1958), v. 1, p. 183—197.
Shorey H. H. Current state of the field use of pheromones in insect cont-
rol.— Sem. etude theme Prod, natur. et prot. plant*., 1976, Citta del Vaticano,
1977, v. 385, p. 401—408.
175
Si nd el a r J., Skalska A. Variabilita obsahu siilis vorginech modtinu ev-
ropskeho (Larix decidua Mill.).— Lesn. casop. Ustav v6deck. unform. MZLVH,
1965, Bd 11, IN 1, S. 54—60.
Swain T., Hills W. E. The quantitative analysis of phenolic constituents.—
J. Sci. Food, and Agric., 1959, v. 10, N 1, p. 63r-68.
Teppner 11. Bestimmugstabelle mittel europ aischer Lamiinae — Lar ven
(Co)eoptera, Cerembycidae) mit Bemerkungen zu deren Biologic.— Verhandl.
zooL-bot. Gws. Wien., 1968—1969, v. 108—109, S. 19—58.
Thalenhorst W. Vorzei tiger Zusammenbruch einer Massenvermehrung von
Gilpinia frutetorum F. (Hym., Diprionidae).— Anz. Schadlingskunde, 1953,
Bd 20, N 4, S. 91—93.
Thomson R. G., Sifton H. В. В esin canals in the Canadian spruce (Picea
canadensis (Mill.) B. S. P.) — an anatomical study, espicially in relation to
traumatic effect and their bearing on phylogeny.— Roy. Soc. London, Phil.
Tryns., Ser. B, 1925, Bd 214, S. 63—111.
Vite J. P. Einsatz von Lockstoffen bei der Borkenkaferbekampfung.—>
Allg. Forstzeitschrift, 1978, N 15, S. 428—431.
Vite J. P. I nsek ten hormone im Waldschutz: Erreichtes und Erreichbares.—
Biol, unserer Zeit, 1978a, Bd 8, N 4, S. 112—119.
Vite J. P., Francke W. The aggregation pheromones of bark beetles:
progress and problems.— Naturwissenschaften, 1976, v. 63, N 12, p. 550—555.
Vite J. P., Gara R. I. Volatile attractants from ponderosa pine attacked by
bark-beetles (Coleoptera: Scolytidae).— Contribs. B. Thompson Inst., 1962,
v. 21, p. 251—273.
Winkler A. Catalogue Coleopterorun regionis palaearcticae.— Lief. Wien,
1924—1932, Bd 1 — 13, S. 1698.
Z we If er W. Uber Abxvehreinrichtungen anserer Waldbaume gegen Insekten-
schaden.— Z. angew. Entomol., 1963, Bd 51, N 4, S. 364—370.
Рис. 49.
] — молодая личинка по внешней коре; 2 — личинки, залитые смолой п ходах; 3 —
ходы молодых личинок во внешней коре; 4—5 — раневые линзочки в лубе, образую-
щиеся вокруг повреждения.
Рис. 50. Типичные ходы личинок лиственничного усача в заболони.
Рис. ~/. Ходы личинок усача у 'рая полосы повреждения.
J>uc. 52. Личипка лиственничного усача в куколочной колыбельке. Ти-
пичные ходы личинок в ядровой древесине.
Рис. 53. Нетипичные ходы личинок в ядровой древесине (7 — 7). Отрезок
ствола дерева, повторно заселенного усачом через 20 лет (8).
Рис. L4. 13 ссстаног.пение ствола у деревьев, поврежденных листвен-
ничным усачом.
] — прирост сохранившейся части заболони отсутствует, дерево обречено; 2—з —
широкая и узкая полоса нарастающей заболони: 4 — зарастание калл юсом повреж-
денной части ствола; 5 — дерево, поврежденное 20 лет назад (отмершая часть ствола
источена насекомыми).
Рис. 55. Раненые структуры, изолирующие обнаженные участки луба
ч и заболони.
1 — изолированный перидермой участок луба (раневая линзочка), поврежденного
молодой личинкой усача; 2 - зарастание каллюссм поврежденной части луба и за-
болони; 3 — зарастание каллюссм повторно поврежденного участка луба (каллюсная
ткань формируется за счет деления клеток околораневой зоны развивающегося го-
дичного слоя заболони); 4 — каллюс, заполняющий ход, проделанный личинкой.
Рис. 56. Нормальные смоляные каналы лиственницы в заболони и лубе
1—2 — продольные каналы на поперечном срезе; «3 — 4 -- каналы сердцевинных лу
чей заболони на тангентальных срезах; 5 — 6 -- пересечение продольных и радиаль
ных каналов; 7—9 — каналы сердцевинных лучей луба на тангентальном и попе
речном срезах.
Рис. 57.^Формирование продольных смоляных каналов.
1—4 — образование эпителия и просвета канала; 5 — эпителий формирующегося
канала на продольном срезе; 6 — канал, пересеченны! сердцевинным лучом, b —
луб; nf — трахеиды; с — камбий; гс — смоляной канал; е — клетки эпителия; еш —
клетки эпителия, образующиеся из лучевой паренхимы.
Рис. 58. Патологические смоляные каналы.
1—4 — одно-, двух- и четырехрядные цепочки каналов на поперечных срезах; 5—6 —
смоляной канал на радиальном и тангентальном срезах.
Рис. 59.
1—2 — цистоподобные окончания смоляных каналов в лубе; :>—4 — формирование
лизигенных камедно-смоляных каналов; 5-6 — образование тиллоидов; 7 — обли-
терированный смоляной канал.
Рис. tO. b'] u lj листание трахеид.
1—2 — трахеиды, сохраняющие протоплазму и ядро до конца вегетации; :i — тяже-
вая трахеида на продольном срезе; •/ — то же, на поперечном срезе (видны поры
в перегородках); 5—6— септированные трахеиды; 7 — перегородчатые трахеиды
С измененными порами; 8 — пропзв< диые трахеид с утолщенными вторичными обо-
лочками.
Рис. 61. Преобразование окаймленных нор трахеид в зоне патологических
смоляных каналов (пояснение в тексте).
Рис. 62. Типы раненой трахеи да л иной паренхимы (1—6) и раневое преоб-
разование трахеид, лишившихся протопласта (7, 8).
Ml’.
Рис. 63. Накопление фенолов в нормальных и трансформированных
клетках заболони (пояснение в тексте).
Рис. 64.
1—3 — последовательное накопление крахмала и замена его смолами в эпителиаль-
ных клетках патологического смоляного канала и сопровождающих клетках; 4 —
заполнение смолами просвета канала; 5 — смолы в паренхимных клетках сердцевин-
ных лучей, с — камбий; Ь — луб; h — ранние трахеиды; hf — поздние трахеиды,
1пг — сердцевинные лучи; с! — просвет смоляного канала; егс — эпителий смоляного
канала; сс — сопровождающие клетки, (s) — зерна крахмала, (г) — смолы.
СОДЕРЖАНИЕ
Предисловие..................................................... 3
Общая часть
mupnrn фи 1 и фат и ксилофагия у насекомых.................
пвФительноядные насекомые..................................
Пищевая специализация насекомых-дендрофагов.............
Первичные и вторичные вредители леса....................
О сущности вспышек размножения вредителем леса..........
Кснлофагия ................................................
Состав и филогения ксилофагов...........................
Некоторые морфологические особенности личинок ксилофагов
Стации и группировки ксилофагов...........................
5
S
Специальная часть
Лиственница и лиственничный усач................................
Лиственница (Larix Mill.)...................................
Распространение лиственниц, повреждаемых лиственничным
усачом. Характеристика лесов.............................
Морфология и анатомия сибирской и даурской лиственниц
Особенности биохимического состава хвои, коры и древесины
лиственницы...............................................
Вредители лиственницы и ее устойчивость к повреждению
Использование лиственницы.................................
Лиственничным усач (Xylotrechus altaicus Gebl.)..............
Систематическое положение и филогения.....................
Географическое распространение............................
Стация................................... ................
Генерация и лйтные годы..............................
Обычлое и очаговое состояние популяций................ .
Факторы динамики численности . .Л >
Вспышки размножения и характер очагов................
Внешнее строение и образ жизни взрослого насено|шв
Морфология и развитие преимагинальных стадий . . .
Строение пищеварительного тракта и особенности имгокй
чинок............................................- ♦ ►
Механизмы преодоления усачом сопротивляемости дерШ^.£^ьЙ
Насекомые, сопутствующие лиственничному усачу л >
Враги усача.....................................
Хозяйственное значение лиственничного усача. Со
состояние, задачи и перспективы борьбы с ням
Защитные механизмы у лиственницы от повреждения
ксилофагами....................................
Преобразование элементов флоэмы и ксилемы,
образования................................
Физиолого-биохимические преобразования . .
Усиление химических средств защиты ....
Литература.....................................
33
м
эт