/



























































































































Похожие




Текст
INSTITUT ZOOLOGIQUE DE L’ACADEMIE DES SCIENCES DE L’URSS
NOUVELLE SERIE n* S
FAUNE DE L’URSS
Sous la direction de S. A. SERNOV, Membre de Г Academie Redige par A. A. STACKELBERG
INSECTES
DERMAPTERES
par G. /. BEY-BIENKO
EDITION DE 1’ACADEMIE DES SCIENCES DE I’URSS
MOSCOU • 1936 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ №
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ Редактор А. А. ШТАКЕЛЬБЕРГ
НАСЕКОМЫЕ
КОЖИСТОКРЫЛЫЕ
Составил
Г. Я. БЕЙ-БИЕНКО
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА . 1936 • ЛЕНИНГРАД
Напечатано по распоряжению Академии Наук СССР Июнь 1936 г.
Непременный секретарь академик Н. Горбунов
Редактор издания А. А. Штакельберг
Технический редактор и ученый корректор Н. Г. Редько
Сдано в набор 19 января 1936 г. — Подписано к печати 26 нюня 1936 г.
Х-+-240 стр.
Формат бум. 72Х1Ю см- — 151/g печ. л. — Уч.-авт. л. 19.48. — 49870 зн. в печ. л.
Тираж 1500.— Ленгорлит № 16475. — АНИ № 1121. — Заказ № 228
Типография Академии Наук СССР. Ленинград, В. О., 9 линия, 12
ПРЕДИСЛОВИЕ
Предлагаемая работа представляет собой обзор фауны Dermaptera восточной части Палеарктической области; в нее вошли все известные до сих пор виды, зарегистрированные в пределах данной территории. Виды, ограниченные в своем распространении только западной частью Западной Европы и Средиземноморьем, мы полностью исключили из данной работы, руководствуясь при этом с одной стороны тем, что по ним имеются, если не полные, то, во всяком случае, обстоятельные сводки, касающиеся отдельных стран (см. напр. Chopard, 1922), и с другой — тем, что довольно хорошая изученность распространения видов в западной Палеарктике позволяет не рассматривать такие виды, возможность нахождения которых в СССР исключена. Совершенно иное положение имеет место в отношении фауны Dermaptera восточной Палеарктики; по ней нет ни одной более или менее обстоятельной сводки, не говоря уже о том, что сопредельные с СССР территории Китая, характеризующиеся богатейшей фауной, стали изучаться только в самое последнее время. Вследствие этого у нас нет никаких оснований считать, что ряд видов, известных из собственно Китая (Ганьсу, Сычуань и северо-восточные провинции) не будет найден и у нас, в частности, на территории Уссурийского края. Кроме того, мы считали необходимым включение в данную работу ряда видов из палеарктического Китая и потому, что там лежит центр современного видового разнообразия, сохранивший с древних времен ряд форм, родичи которых были прежде широко распространены по Евразии и сыграли весьма существенную роль в образовании современной фауны Западной Европы и Средиземноморья. Анализ современной фауны как СССР, так и Палеарктической области в целом, был бы невозможен без учета фауны Палеарктического Китая, а это потребовало критического изучения последней и включения ее в данную работу. Для более быстрой ориентировки виды, уже зарегистрированные для СССР, отмечены в определительных таблицах и описаниях звездочкой.
Наконец, мы считали необходимым дать более широкий очерк истории фауны Dermaptera Палеарктической области, так как данная группа животных резко отличается в этом отношении от многих других групп существенными особенностями экологии, что накладывает на ее
VI
ФАУНА СССР — КОЖИСТОКРЫЛЫЕ
эволюцию и географическое распространение печать значительного своеобразия.
Материалом для данной работы, помимо литературных источников, явились богатые коллекции Зоологического Института Академии Наук СССР, включающие в себе также коллекцию А. П. Семенова-Тян-Шанского; кроме того, нами были использованы небольшая личная коллекция и коллекция б. Кавказского Музея; ссылки на эти коллекции в тексте сопровождаются сокращенными их наименованиями по следую
щей схеме:
Коллекции Зоологич. Института....... колл. ЗИН
„ А. П. Семенова-Тян-Шанского . . колл. С.-Т.-Ш.
„ б. Кавказского Музея ...... колл. Кавк. М.
„ автора..................... колл. Г. Б.-Б.
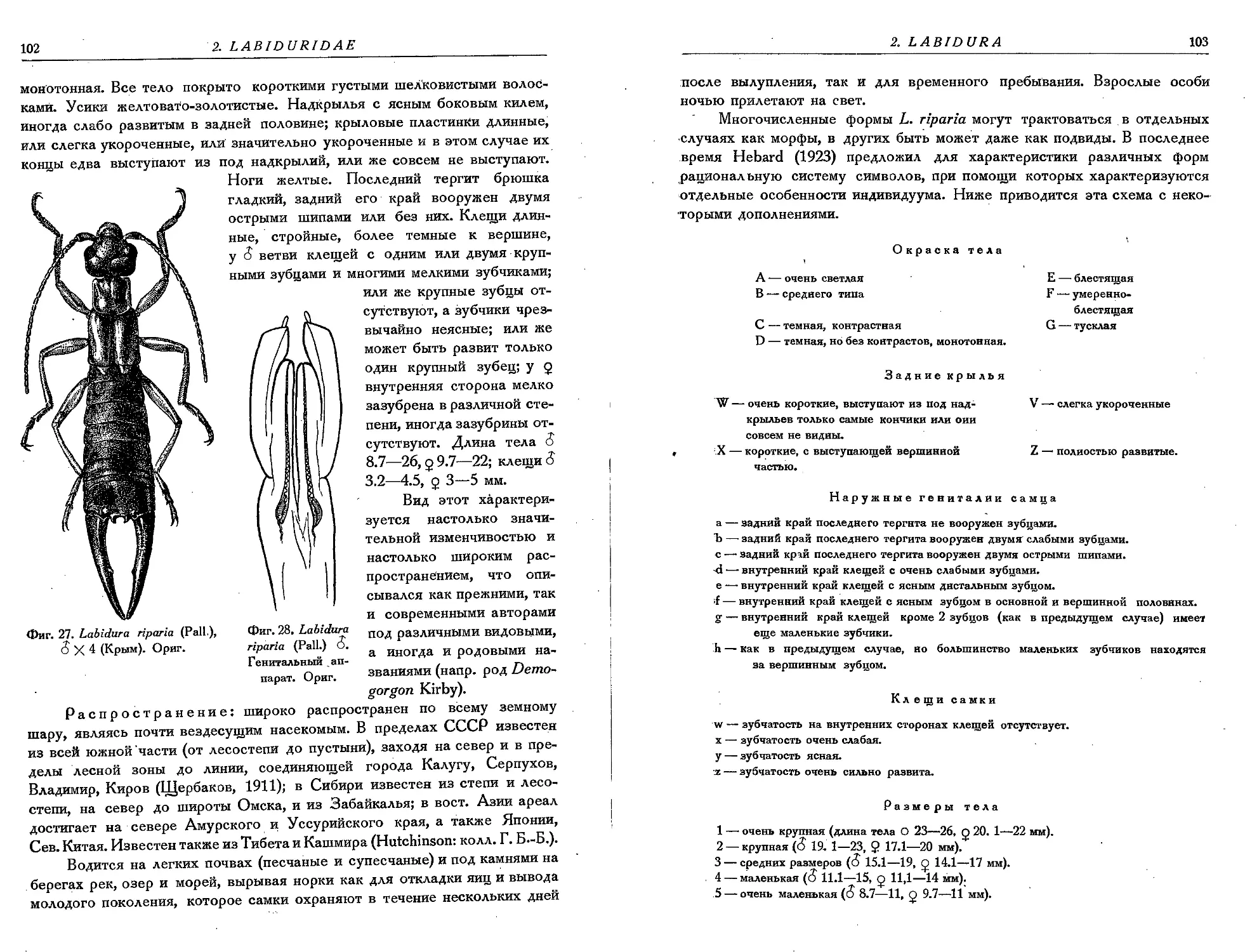
Измерения даны только для длины тела (без клещей) и отдельно для клещей.
В заключение считаем своим долгом выразить благодарность Дирекции Зоологического Института в лице академика С. А. Зернова, предоставившего нам для обработки богатый коллекционный материал Института, а также А. П. Семенову-Тян-Шанскому, как за любезное предоставление весьма ценной, специально подобранной им коллекции, так и за проявление особого внимания и интереса к нашей работе и ряд ценных указаний. Кроме того, автор очень обязан А. В. Мартынову, предоставившему для изучения типы описанных им ископаемых Dermaptera и за некоторые ценные указания, а также зав. Ортоптерологическим отделением Института — Э. Ф. Мирам за неизменно предупредительное отношение к нашей работе.
Рисунки для данной работы были выполнены главным образом художником Ф. И. Гуняевым; часть рисунков (детали строения и пр.) выполнены автором.
Г. Бей-Биенко
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
[1. Подотряд Archiderm apter а]
[II. Подотряд Arixenina]
Ш. Подотряд Dermaptera s. str.
1. Надсем. PROTODERMAPTERA
1. Сем. Pygidicranidae
1. Подсем. Anataeliinae
1. Род Challia Burr
1. Ch. fletcheri Burr............ ............... 82
2. Сем. Labiduridae
1. Подсем. Psalinae
1. Род Anisolabis Fieb.
1. A. (in sp.) maritima (Gene)............................... 89
2. A. (Apolabis) marginalis (Dohrn).......................... 91
3. A. (Mongolabis) setshuana B. - Bienko ..................... 93
4. A. (M.) cavaleriei Borelli................................. 94
5. A. (Eubo -ellia) annulipes (Lucas)......................... 96
6. A. (E.) plebeja (Dohrn)................................... 97
7. A. (E.) pallipes Shiraki ........................... . 98
8. A. (E.) moesta (Serv.) . ................................. 98
9. A. (?) pectoralis (Esch.) ............................. 100
2. Подсем. Labi dur inae
2. Род Labidura Leach.
1. L. riparia (Pall.) . .................................. 101
3. Род Nala Zacher
1. N. lividipes (Duf.)...................................... 104
3. Подсем. Parisolabiпае
4. Род Pseudisolabis Burr.
1. P. burri Borelli . ..................................... 106
VIII ФАУНА СССР-КОЖИС ТО КРЫЛИЕ
2. Надсем. EUDERMAPTERA
3. Сем. Labiidae
1. Подсем. Labiinae
1. Род Labia Leach.
1. L. minor (L.)...................................................... 110
2. L. curvicauda.(Motsch.)............................................. 110
2. Род Prolabia Burr
1. Pr. arachidis (Yers.) .................................; . . . . 112
3. Род Nesogaster Verb.
1. N. nigritus (Shiraki)............................................ 114
2. N. lewisi (Burr) . ............................................ . 115
2. Подсем. Chelisochinae
4. Род Chelisoches Scudd.
1. Ch. morio (F.) .................................................. 116
5. Род Proreus Burr
1. Pr. simulans (Stal)...................... ...................... 118
6. Род Hamaxas Burr
1. H. kozlovi Sem. et B. - Bienko .’................................... 119
4. Сем. Forficulidae
1. Подсем. Forficulinae
1. Род Forficula L.
1. F. schlagintweiti (Burr)....................................... . 130
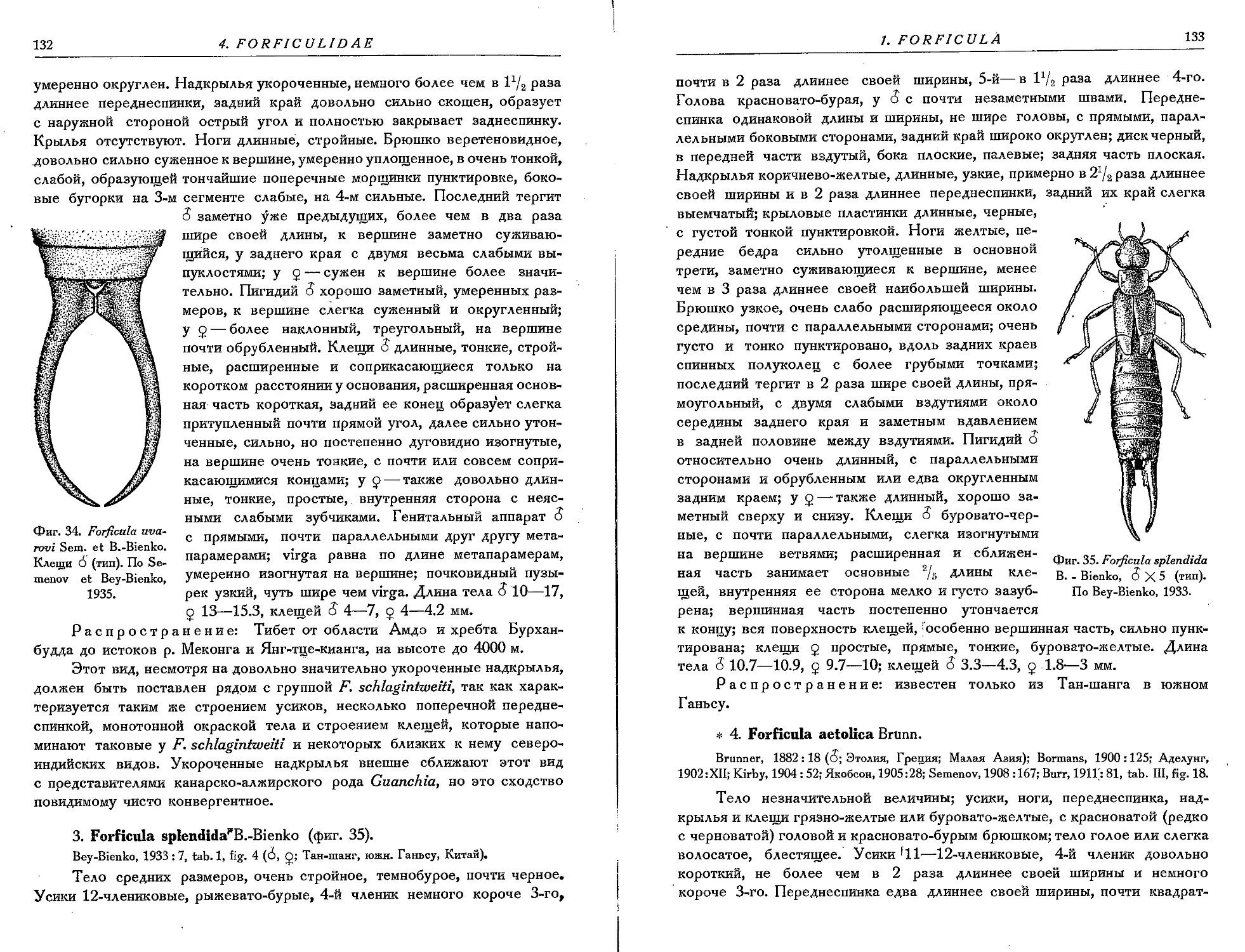
2. F. uvarovi Sem. et B. -Bienko.................................. . 131
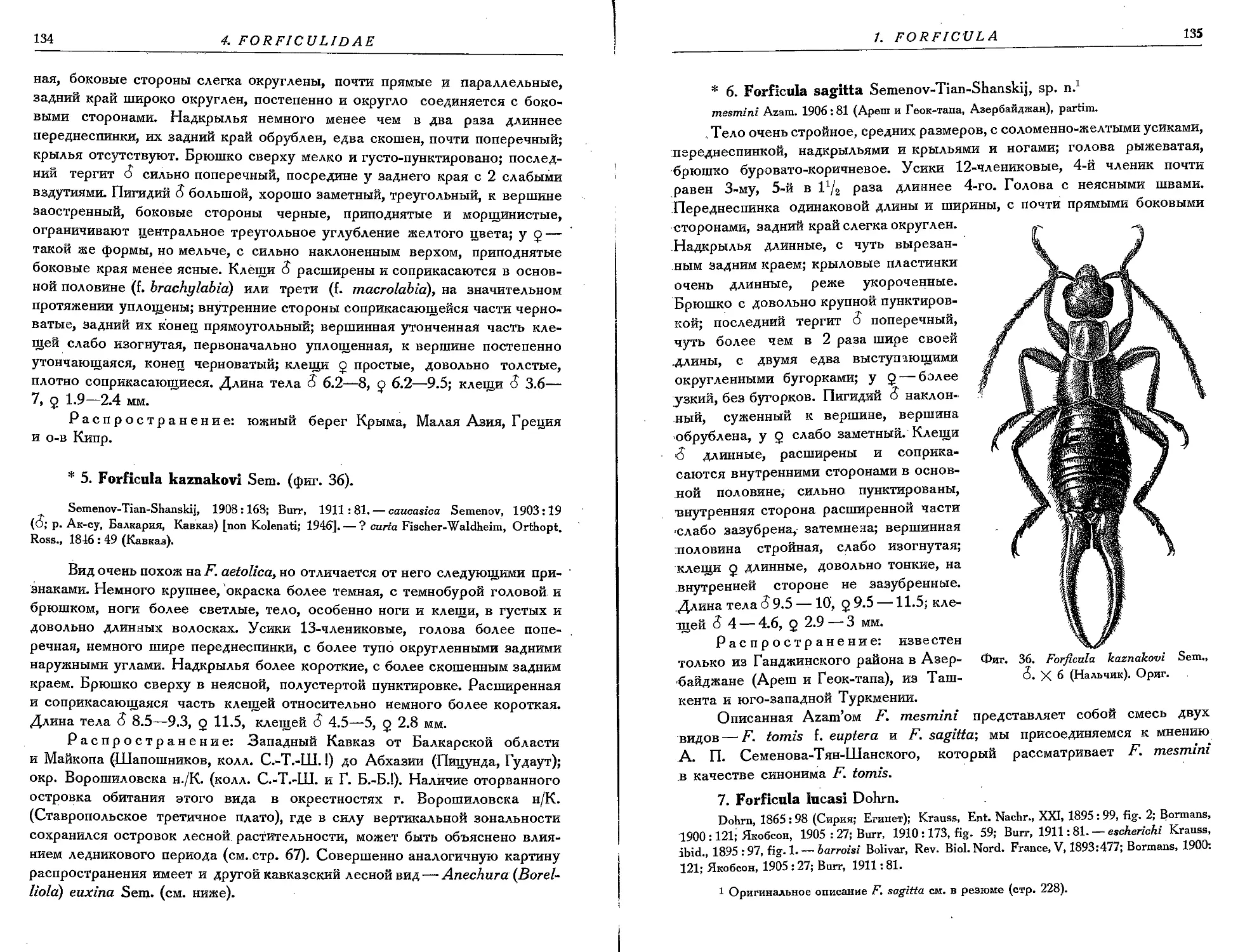
3. F. splendida B. - Bienko........................................... 132
4. F. aetolica Br. - Watt............................................ 133
5. F. kaznakovi Sem....................................... •............ 134
6. F. sagitta Sem. . . ....................... . . . ............• 135
7. F. lucasi Dohrn . . . . . . . . . ................................... 135
8. F. mandarina Borelli.............................................. 136
9. F. tomis Koi......................................................... 137
10. F. robusta Sem................................................... 139
11. F. macrobasis B. - Bienko . .................................. .... 141
12. F. ambigua Burr..................................................... 142
13. F. sinica B. - Bienko................................................ 142
14. F. mikado Burr ............................. . . ................. 143
15. F. berezovskii B. - Bienko .'........................................ 144
16. F. smyrnensis Serv................................................... 145
17. F. davidi Burr .................................................. . .' 146
18. F. decipiens Gene................................................... 147
СИСТЕМАТИЧЕСКИМ УКАЗАТЕЛЬ
IX
19. F. hirida Fisch. ..................................................... 148
20. F. vicaria Sem......................................................... 149
21. F. burriana Sem........ ............................................ 150
22. F. subauricularia B. - Bienko . . ................................... 150
23. F. auricularia L. ..................................................... 152
24. F. pubescens Serv.................................................... 154
2. Род MesasioMa Sem.
1. M. hemixanthocara Sem................................................. 156
3. Род Oreasiobia Sem.
1. O. stoliczkai (Burr)................................................... 159
2. O. fedtschenkoi (Sauss.).............................................
4. Род Anechura Scudd.
1. A. (Odontopsalis) potanini B. - Bienko................................. 167
2. A. (O.) forficuliformis Sem. et B. - Bienko............................ 168
3. A. (O.) japonica (Borm.).............................................. 169
4. A. (O.) harmandi (Burr)................................................ 171
5. A. (in sp.) sven-hedini B. - Bienko.................................... 172
6. A. (in sp.) quelparta Okamoto.......................................... 173
7. A. (in sp.) bipunctata (F.)........................................ 174
8. A. (in sp.) asiatica Sem............................................... 176
9. A. (in sp.) filchneri (Burr.).......................................... 178
10. A. (Burriola) semenovi (B.-Bienko)................................... . 179
11. A. (B.) tibetana (Sem. et B.-Bienko)................................... 181
12.. A. (B.) przevalskii (Sem.)............................................. 182
13. A. (B.) redux (Sem.)................................................ • 183
14. A. (Borelliola) euxina (Sem.)........................................• 184
5. Род Chelidura Latr.
1. Ch. aptera Charp. ..................................................... 187
6. Род Eumegalura B. - Bienko
1. E. mandarina B. - Bienko............................................... 188
7. Род Apterygida Westw.
1. A. media (Hagenb.)..................................................... 190
8. Род Chelidarella Verb.
1. Ch. acanthopygia (Gene)............................................... 192
2. Ch. mutica (Krauss) .................................................. 193
3. Ch. transsilvanica Ebner............................................. 193
9. Род Allodahlia Verb.
1. A. scabriuscula (Serv.)............................................... 195
2. Подсем. Opisthocosmiinae
10. Род Timomenus Burr
1. T. inermis Borelli..................................................... 1^
2 T. unidentatus Borelli................-................................. 198
X ФАУНА СССР —КОЖИСТОКРЫЛЫЕ
3. Т, komarovi (Sem.)............. ............................. 199
4. Т. aesculapius (Burr)....................................... 200
11. Род Eparchus Burr \
1. E. insignis (Haan)........................................... 202
2. E. oberthuri Borelli.................................... . 203
3. E. yezoensis (Mats, et Shir.)................................. 204
12. Род Narberia Burr
1. N. tuberculata Borelli...................................... 205
13. Род Sondax Barr
1. S. pitanini B. - Bienko.................................... 207
ВВЕДЕНИЕ
ХАРАКТЕРИСТИКА ОТРЯДА
Отряд уховерток или кожистокрылых (Dermaptera — Euplexoptera) представляет собой сравнительно небогатую видами группу насекомых •со следующими признаками.
Насекомые с неполным превращением, с более или менее уплощенным и удлиненным телом, голова с направленными вперед жующими ротовыми органами, лапки трехчлениковые (современные формы), или 4—5-члениковые (мезозойский подотряд Archidermaptera), передняя пара крыльев, если имеется, то превращена в твердые, лишенные жилкования и . сильно укороченные надкрылья, задние крылья, если развиты, в покое •складываются вдоль и два' раза поперек, в большей своей части скрыты под надкрыльями, лишь выступая из под них в виде короткой крыловой пластинки, представляющей собою твердое хитиновое основание передней часуги крыльев; или же насекомые совершенно лишены крыльев; брюшко снабжено на конце парой крупных нечленистых твердых придатков — клещей или же имеет короткие членистые церки (подотряд Archiderma-ptera), стернальные и тергальные полукольца брюшка, соединяясь между собой, образуют зигзагообразную линию.
1. СТРОЕНИЕ ТЕЛА
Тело маленькое или умеренных размеров, заметно, в некоторых случаях сильно (Apachyus, Platy labia) уплощено, удлиненное и более или менее стройное, с довольно твердыми наружными покровами.
Голова (фиг. 1,3) с Направленным вперёд ртом, уплощенная, более или менее сердцевидная, вариируя от почти треугольной до пятиугольной; задний край обычно прямой или слегка вырезан около середины. Верхняя сторона обычно имеет ясно выраженные швы (sutura), из которых один поперечный шов располагается между внутренними сторонами глаз и отделяет лоб, расположенный кпереди от этого шва, от затылка, лежащего кзади от него; кроме поперечного шва имеется и затылочный или продольный шов, идущий от средины поперечного шва до средины заднего края головы, деля затылок на правую и левую часть. Степень развития
Фауна СССР. Кожиетокрылые 1
2
КОЖИСТОКРЫЛЫЕ—DERMAPTERA
швов имеет значение для распознавания видов и даже родов. Глаза умеренной величины или (у подотряда Arixenina) значительно редуцированы, слабо выступают, располагаясь около середины боковых сторон головы. Простые глазки отсутствуют у многих видов или же пред-
голова
переднеспикка
анальный угол надкрылий
надкрылья
крыловые пластинки
боковые бугорки брюшка
1-й членик лапки
2-й членик лапки
3-й членик лапки
коготки
Фиг. ,1. Тело уховертки сверху. (По Burr, 1910).
усики лоб
сложи, глаза!
затылок
затыл. шов
передн. часть переднеспиика средний. борозда, задняя часть переднеспинки плечи надкрылий; щиток
шовный край
боковой край.
задний край*
заднее бедро*
задняя голень.
задняя лапка
последний тергит-
пигидий
клещи
ставлены незначительными, слабо заметными, обычно более или менее полулунными или округлыми вдавлениями, располагающимися близ внутреннего края глаз у боковых концов поперечнго шва; однако у многих представителей наиболее примитивно организованного сем. Pygidicranidae глазки лучше выражены и хорошо заметны. Наличник маленький,, отделен от лба прямым швом и обычно ясно подразделен поперечной бороздкой на переднюю часть (anteclypeus) и заднюю часть (postclypeus). Ротовые органы (фиг. 3) обычного грызущего типа, не представляют каких-либо особенностей. Верхняя губа в виде небольшой, несколько закругленной спереди пластинки; верхние челюсти умеренной вели
ВВЕДЕНИЕ
3
чины, более или менее треугольные, несколько загнутые, на конце с двумя зубцами, а у представителей подотряда Arixenina с тремя зубцами и с покрытой густыми щетинками внутренней стороной. По бокам передней части головы, между основанием усиков и основанием верхней челюсти
грудные пластинки
последний етеринт
переднегрудная
среднегрудная
заднегрудная
предпоследний етеринт
пигидий
Фиг. 2. Тело уховертки снизу. (По Впгг> 1910).
имеется, подобно тому, как это имеет место у Orthoptera и' Blattodea, маленький треугольный склерит, так называемый трохантин верх-н их челюстей (trochantinus mandibular is, ,,Basimandibulare“ немецких авторов). Нижниечелюсти также обычного типа, состоя как и у других насекомых из наружной жевательной лопасти или г а л е и, внутренней жевательной лопасти или л я цини и и челюстных щупалец, укрепленных на общем основном членике; наружные лопасти стройные, длинные, без ясных зубцов, внутренние лопасти примерно такой же формы, как и наружные, но на вершине вооружены двумя зубцами; челюстные щупальцы пятичлениковые, с маленьким дополнительным 1*
К О ЖИ С Т О К Р Ы ЛЫ E—D E R М АР T E R A
Фиг. 3. Голова уховертки снизу. А — усик; G — наружная жевательная лопасть нижних челюстей; Gen — щек ; Lac — внутренняя жевательная лопасть нижних челюстей; Lig— язычек; Lp — губные щупальцы; Md — верхняя челюсть; Mnt — подбородок; Мр — челюстные щупальцы; Ос — сложные глаза; Pgl — наружные жевательные лопасти нижней губы; Ptl — добавочны-i вершинный сегмент на щупальцах — подотельсон. Ориг.
сегментом на5-мчленике(podotelson VerhoefРа). Нижняя губа состоит из маленького непарного язычка (ligula), получившегося в результате срастания внутренних лопастей, далее из пары наружных жевательных лопастей (paraglossa) и трехчлениковых губных щупалец, укрепленных на довольно маленьком подбородке и, также как и челюстные щупальцы, несущих на конце podotelson. Строение ротовых органов почти не имело до сих пор приложения к систематике отряда, вследствие того, что детали строения ротовых органов мало отличаются у различных групп и родов уховерток.
Усики с весьма различным числом члеников, колеблющимся от 8 примерно до 50; 1-й их членик наиболее длинный, обычно несколько расширенный на вершине, иногда почти булавовидный; 2-й. членик у всех современных форм всегда очень маленький, иногда плохо заметный; 3-й — чрезвычайно изменчивый как по форме, так и по длине — цилиндрический, конический или почти булавовидный; 4-й членик обычно короче 3-го, а 5-й всегда немного длиннее 4-го; остальные членики одинаковой длины или постепенно удлиняющиеся к вершине усика. Форма члеников может быть различной — цилиндрическая, овальная, веретеновидная, коническая (в последнем случае основание членика более узкое чем вершина) и полубулавовидная. Относительная длина 3—5-го, а иногда и 1-го, а также форма как указанных, так и остальных члеников имеют большое значение в систематике уховерток.
Грудь (фиг. 1, 2, 4) весьма явственно отделена от головы; пере дн грудь сверху прикрыта большой цельной плоской передне-
ВВЕДЕНИЕ
спинкой обычно более или менее четырехугольной формы; у крылатых ф,орм задний край переднеспинки округлен, у бескрылых — прямой или почти прямой; боковые стороны прямые или выгнутые, образуя с перед?-ней и задней сторонами углы, очень часто широко округленные и неясные;:
иногда переднеспинка овальная или почти круглая; передняя часть (prozona) обычно более или менее выпуклая, а задняя (metazona) плоская^
вдоль середины часто имеется продольная борозда (sulcus medianus). Средне-спинка (фиг. 4) у крылатых форм скрыта под надкрыльями, у бескрылых хорошо заметна, хотя несколько прикрыта задней частью переднеспинки, имеет форму обычно слегка поперечной простой пластинки, с прямым или немного вырезанным задним краем; бока среднеспинки у некоторых бескрылых родов имеют продольный, более или менее косо расположенный киль. Среднеспинка у крылатых форм, в отличие от переднеспинки, подразделена на несколько склеритов; наиболее крупный передний склерит имеет вид поперечной пластинки и представляет собой собственно спинку (scutum). К средней части задней стороны scutum примыкает маленький сильно хитинизированный щиток, имеющий треугольную форму и сильно суживающийся и заостряющийся кзади в виде шипа. По бокам заднего конца среднеспинки имеются выступы—место прикрепления передней пары крыльев. Заднеспинка (фиг. 4) у бескрылых форм представляет собой простую, несколько поперечную пластинку, прямую спереди и более или менее дуговидно вырезанную сзади. У крылатых форм она скрыта под над-
Фиг. 4. Грудь уховертки (Dorn lineare Eschs.) сверху. Pro, Mez и Met — переднеспинка, среднеспинка и заднеспинка; Sc — спинка; Sol — щиток; р. Scl — постскутелюм. (По Packard, 1883, с изменениями).
крыльями и подобно среднеспинке дифференцирована на несколько склеритов, именно крупный scutum и маленький scutellum, который имеет также треугольную форму, но в отличие от среднеспинки обычно заострен спереди; между задним краем собственно заднеспинки и передней частью первого сегмента брюшка располагается короткое, поперечное нежноперепончатое образование. Если надкрылья имеются, но сильно укорочены, заднеспинка лишается описанной структуры, становясь почти такой же, как у совершенно бескрылых форм; в этом случае задний край ее не прикрыт
надкрыльями.
Плевры расположены почти горизонтально. Два наиболее крупные склерита располагаются один над другим, причем верхний склерит —
6 КОЖИСТОКРЫЛЫЕ —DERM APTER A
эпимер удлиненный, чешуевидный, несколько более крупный, чем расположённый под ним эпистерн; последний более или менее треугольный, слегка наклонный и суженный к тазику.
Г р у д к а (фиг. 2) состоит из трех грудных пластинок или стернитов, располагающихся таким образом, что основная (задняя) часть их расположена между тазиками ног. Переднегрудь или переднегрудная пластинка более или менее удлинена, расширена кпереди, а перед задним краем сужена вследствие помещения здесь основания передних ног; иногда переднегрудь имеет почти параллельные боковые стороны. Средне-грудь более или менее округленная, в зависимости от стройности тела или поперечная, т. е. ее ширина превосходит длину, или удлиненная; задний край ее округлен или прямой. Заднегрудь достигает наибольшей величины; наибольшая ее ширина находится в передней части, так что задняя ее часть суживается и выступает в виде лопасти, .кзади от мысленной линии, соединяющей между собой вертлуги, причем степень выступания может быть различной у разных родов; задний край весьма изменчив — прямой, округленный или вырезанный. Форма грудных пластинок и особенно характер заднего края заднегруди имеют большое значение в систематике отряда.
Передняя пара крыльев (фиг. 1) лишена жилкования, кожистая и превращена в укороченные надкрылья, Закрывающие собой средне- и заднегрудь; при нормальной степени их развития они более или менее четырехугольные, вытянуты вдоль груди Сверху, тесно соприкасаясь между собой вдоль шовного (внутреннего) края, образуя таким образом шов; боковые или костальные их стороны более или менее параллельные друг другу; верхняя их сторона называется диском, а свешивающаяся по бокам и охватывающая здесь бока груди часть образует боковую или костальную часть; обе эти части у многих родов резко отделены друг от друга острым продольным килем. Наружный передний угол-— п л е ч и надкрылий более или менее выступают у видов с вполне развитыми надкрыльями и крыльями. У полнокрылых форм анальный угол надкрылий, располагающийся у основания шовного края, прямоугольный, так что если надкрылья сложены, то среднеспинка полностью бывает закрыта, однако у- видов с сильно укороченными надкрыльями, а также и у тех полнокрылых форм, у которых анальный угол округлен (Pyg'.dicranidae и др.), между основаниями надкрылий, у средины заднего края передне-спинки остается незакрытой часть среднеспинки, именно щиток. Вершина надкрылий обрублена, т. е. с совершенно прямым (виды с недоразвитыми крыльями), или со слегка вырезанным задним краем (полнокрылые формы); у видов с сильно укороченными надкрыльями задний край может быть более или менее скошен, так что между задними краями надкрылий образуется треугольная выемка. Степень укорочения надкрылий может быть весьма различной; крайний случай — полное их отсутствие, что свойственно довольно многим видам; сильно укороченные надкрылья
ВВЕДЕНИЕ
7
имеют вид двух широко расставленных маленьких пластинок по сторонам среднеспинки (некоторые виды Anisolabis), или же эти надкрылья соприкасаются внутренними сторонами, срастаются со среднеспинкой, а спереди открывают большой щиток (Chelidura)', при большей степени развития надкрылий последние могут вполне соприкасаться по всей длине внутреннего края, не открывая щиток и лишь только оставляя незакрытой “большую или меньшую часть, а иногда и всю заднеспинку.
Фиг. 5. Заднее крыло обыкновенной уховертки (Forficula auricularia) а. р. — краевое поле; с. ар. — конечное поле; п — узелок; sq — роговая пластинка. Жилки: R — радиус, М — медиана, Си — кубитус, А3 — первая анальная, А2 — вторая анальная, ch. pr. — пред краевая дуга или делящая жилка. Ориг.
Задние крылья или просто крылья (фиг. 5) часто полностью отсутствуют или сильно укорочены; у полнокрылых форм они в спокойном состоянии сложены продольно в виде веера вдоль жилок, а затем два раза поперек, будучи в этом случае скрыты под надкрыльями и выступая из под них только в виде т. н. крыловых пластинок, которые представляют собой только конец твердой роговой пластинки (squama), располагающейся вдоль передней стороны крыла в его основной половине. В расправленном состоянии крылья очень широкие и представляют собой тонкую полукруглую перепонку. Передний край крыльев у самого основания имеет тонкое, кожистое узкое поле, т. н. краевое или предкостальное поле (area .praecostalis). За ним следует твердая роговая пластинка (squama), кончающаяся у средины переднего края, а отсюда к концу крыла идет подвижно причлененное к роговой пластинке так наз. конечное поле (campus apicalis); в месте их подвижного сочленения передний край крыла несколько вырезан, образуя узелок (nodus); вдоль роговой пластинки проходят несколько продольных жилок: обычно, субкостальная (subcosta — Sc), р а д и а л ь-
8
КОЖИСТОКРЫЛЫЕ —DERM APT ERA
ная (radius —R), иногда имеющая ветвь — сектор радиуса (RS), медиана (М) и кубитальная (Си). Несколько отступя назад от узелка,, также на конце роговой пластинки, располагается т. н. по'дпорка (fulcrum), где подвижно соединены с концом роговой пластинки основания десяти ветвей 2-й анальной жилки (А2). Что касается нескольких жилок, расположенных в перепончатой передней части крыла (кпереди от ветвей А2), то гомологизация их затруднительна; очень возможно, что они представляют собой продолжение М и R. Таким образом,, перепончатая часть крыла в значительной мере занята радиальными ветвями А 2* Кроме продольных жилок имеется еще одна жилка, идущая дугообразно от вершинной части переднего края вдоль всего наружного края до основания крыла — это предкраевая дуга (chorda praemar-ginalis). Складывание крыла поперек происходит сначала через узелок, и подпорку, а затем вдоль предкраевой дуги, несколько внутрь от нее. Характер жилкования крыльев, хотя и может иметь значение в систематике высших групп отряда (семейств и подсемейств), но в силу трудности препаровки и исследования совершенно не используется при определении.
Ноги обычного бегательного типа, относительно короткие или у некоторых Chelisochinae и Opisthocosmiinae длинные и стройные. Бедра обычно довольно заметно сжаты с боков, у некоторых групп (большинство Pygidicranidae) с коротким продольным килем; впрочем у многих видов передние бедра довольно сильно утолщены и даже вздуты. Г олени обычно чуть изогнуты, сжаты с боков; верхний их край близ вершины у некоторых Chelisochinae характеризуется наличием ясной продольной борозды. Лапки у всех современных форм 3-члениковые (у ископаемых мезозойских Archidermaptera 4—5-члениковые); первый их членик (metatarsus) обычно длиннее остальных; второй членик наиболее короткий, различной формы, цилиндрический, сердцевидно расширен или с узкой длинной лопастью снизу, выступающей под 3-й членик и не видной сверху; третий членик обычно равен примерно половине первого членика, в большинстве случаев цилиндрический; на конце третьего членика имеется пара изогнутых коготков, присоски между которыми почти у всех уховерток отсутствуют; только у Diplatys и немногих других, родов между коготками имеется маленькая присоска (arolia), которая, однако, у коллекционных экземпляров так сильно ссыхается, что не может быть широко использована для определения. Лапки обычно покрыты густыми волосками или щетинками. Задняя пара ног наиболее крупная по размерам, передняя — наименьшей величины, хотя разница в общем относительно небольшая. Строение ног и особенно лапок имеет весьма, большое значение в систематике отряда.
Брюшко (фиг. 1 и 2) представляет собой наиболее удлиненную часть тела, с довольно плотными покровами, обычно более или менее уплощенное или же почти цилиндрическое, часто веретеновидное, т. е.
ВВЕДЕНИЕ
9
расширенное посредине и суженое к основанию и к вершине; у $ 2 форма брюшка менее изменчива, почти всегда с несколько суженным к вершине задним концом. По числу члеников брюшко устроено различно у 3<5 и 2 £; у <5 5 оно состоит из 10 видимых тергитов и 9 стернитов, у £ £ из. 8 тергитов и 7 стернитов. Меньшее по сравнению с тергитами число стернитов объясняется тем, что 1-й членик брюшка плотно срастается с заднегрудью, причем его стернит недоразвит; что касается ££, то у последних 8-й и 9-й членики брюшка представлены в виде маленьких, скрытых кожистых пластинок, как бы вдавленных внутрь 7-го кольца и прикрытых снизу 7-м стернитом, а сверху 7-м тергитом и поэтому незаметных снаружи. Весьма характерно, что тергиты брюшка по бокам опускаются вниз и прикрывают с боков стерниты, совершенно закрывая таким образом плевры, отчего брюшные сегменты при рассматривании сбоку представляют собой систему треугольных выступов, вдающихся друг в друга на подобие двух пил, сложенных зубцами внутрь, так что последние внедряются в промежутки между противоположными зубцами. Последний тергит, морфологически у обоих полов являющийся 10-м тергитом, наибольший по величине, прямоугольный или трапециевидный, сверху, особенно у $ S, часто с бугорковидными или сосковидными выступами перед задним краем, а иногда по бокам с выступающими хитиновыми складками (напр. у Forficula. auricularia). Предпоследний видимый стернит, морфологически являющийся у $ 9-м, а у £ — 7-м стернитом, также очень большой, несколько суженный кзади, часто называется субгенитальной пластинкой; он почти полностью, а иногда совсем закрывает последний стернит. Если предпоследний стернит приподнять, то под ним будет обнаружен последний стернит, морфологически являющийся у обоих полов 10-м стернитом; он разделен посредине на две части — правую и левую. Бока 3-го и 4-го тергитов сверху у обоих полов многих видов имеют особые бугорковидные, несколько удлиненные выступы, так. наз. боковые бугорки. Кроме этих бугорков брюшко может иметь по бокам некоторых сегментов различные выступы, бугорки, кили, длинные шипы и пр.; у многих Psalinae бока сегментов выступают назад, в виде острого угла и часто снабжены продольными бороздками и килями; в отличие от боковых бугорков эти признаки свойственны только 33. По характеру скульптуры хитина брюшко вариирует от почти гладкого до зернистого или сильно пунктированного.
От последнего, т. е. 10-го сегмента, отходят два длинных сильно-хитинизированные одночленистые отростка — так. наз. клещи (forcipes — фиг. 1—2), морфологически являющиеся видоизмененными церками других насекомых. Как показали исследования Heymons’a (1895), эти церки эмбриологически закладываются на 11-м сегменте и поэтому собственно являются придатками 11-го, а не 10-го сегмента; однако, в дальнейшем развитии они смещаются и уже у личинки 1-го возраста они расположены так же, как у взрослого насекомого. Форма клещей чрезвычайно измен-
10
КОЖИ С ТОКРЫЛЫЕ — DERMAPTERA
А
Фиг. 6. Опистомеры О О. А— Labidura riparia (сем. Labidwidae)-, В — Kalocrania sp. (сем. Pygidicranidae)-, р — пигидий, ш — метапигидий, t — тельсон. (По Burr, 1911).
нива у разных видов; клещи могут соприкасаться у основания или же быть широко расставлеными здесь, будучи стройными, почти цилиндрическими, коническими, дуговидно- или волнистоизогнутыми, вооруженными различными зубцами и выступами, сильно расширенными и более или менее уплощенными у основания, асимметричными и пр. Однако, это морфологическое разнообразие свойственно только <5 <5, тогда как £ £ в этом отношении чрезвычайно однообразны; только у некоторых Cheli-sochinae клещи £ £ также могут быть сильно специализованными, отличаясь при этом от <5 $ соответствующего вида настолько, что £ £ принимались за <5 <5 и описывались как особые виды.
Позади средины последнего тергита брюшка выступает сильно хити-низированный орган, располагающийся между основаниями клещей — это пигидий (pygidium). Форма его чрезвычайно изменчива у разных родов и видов, причем у £ £ он устроен более однообразно, чем у <5 <5: он или выступает назад, или же простирается вниз почти не выступая, или же срастается с опущенным вниз задним концом последнего тергита. Иногда последний тергит сильно выступает назад, срастаясь своей вентральной частью с пигидием и образуя так. наз. сквамопигидий (squa-
mopygidium) (Apachyidae, Esphalmeninae). Если пигидий отпрепарировать, то на его нижнем (заднем) конце можно заметить другую хитиновую пластинку — метапигидий (metapygidium), за которой следует слабо хити-низированный, сильно редуцированный тельсон (telson). Все эти три пластинки называются вместе также опистомерами (opisthomeres) (фиг. 6). Является ли этот тельсон действительным гомологом настоящего тельсона, представлящего собой следующее за 11-м сегментом тергаль-ное образование, не имеющее в отличие от других члеников мезодермальных зачатков, вопрос еще не решенный, хотя некоторые авторы допускают данную гомологию.
Наличие на брюшке тех или иных структур, к также его форма, характер последнего тергита, предпоследнего стернита и пигидия, степень развития всех опистомер и форма клещей имеют громадное значение в систематике Dermaptera.
Половые органы $ развиваются на 9-м стерните, в виде парного или непарного полового аппарата (фиг. 8). Что касается £ £, то у последних нет наружных половых придатков и только у наиболее низко организованного сем. Pygidicranidae (исключая подсем. Pyrarginae) ££ имеют на 8-м и 9-м стернитах прикрытые 7-м стернитом две пары отростков, из которых первая пара больших размеров отходит от 8-го, а вторая —
ВВЕДЕНИЕ
И
Фиг. 7. Гонапофизы из рода Kalocrania (сем. Pygidicranidae) Gp8 — гонапофизы 8-го стернита брюшка, Gp9—гонапофизы 9-го стернита. (По Zacher, 1911, с изменениями).
меньших размеров — от9-го стернита; это гонапофизы (gonapophyses) являющиеся гомологами яйцеклада у ближайших отрядов (фиг. 7).
Если изобразить брюшко при помощи применяемых для этой цели формул, то различные полы уховерток будут иметь такую формулу:
т I. II. III. IV. V. VI. VII. VIII. IX. X. С. XI. т
II. III. I V. V. VI. VII. VIII. IX. X. 1
I. II. III. IV. V. VI. VII. (8-9) X. С. XI. т
2 II. Ill IV. V. VI. VII. (8 -9) X. 1
Цифрами отмечены номера тергитов (сверху) и стер-нитов (снизу), арабскими — скрытые части; в скобках отмечены рудименты, буква С — церки, буква Т — тельсон, жирными цифрами — склериты, принимающие участие в образовании полового аппарата. Эта формула несколько условна, так как церки собственно являются придатками 11-го, а не 10-го сегмента и приводится здесь только‘ для лучшей ориентировки в строении брюшка.
Переходя к внутреннему строению необходимо отметить, что оно, за исключением полового аппарата и
некоторых других деталей, в общем сходно со строением-других насекомых, имеющих неполное превращение.
Пищеварительный канал извитой, состоит из тонкостенного пищевода, длинного зоба, простирающегося через весь грудной отдел до 2-го брюшного сегмента и заканчивающегося слабым жевательным желудком; внутренняя поверхность последнего снабжена, в отличие от многих насекомых, только шестью хитиновыми щетинками, но лишена хитиновых зубов; средняя кишка не имеет, в отличие от Orthoptera, дивертикул, прямая, отделена от передней шестью короткими клапанообразными выступами; задняя кишка сначала более тонкая, оканчивающаяся более широкой прямой кишкой, открывающейся на последнем стерните перед пигидием. Органы выделения состоят из 8—10 мальпигиевых сосудов, группирующихся в два пучка. Описанные выше боковые бугорки на 3-м и 4-м брюшных тергитах представляют собой особые пузырьки, стенки которых снабжены железами; из этих желез пахучая жидкость
12
КОЖИСТОКРЫЛЫЕ —DERM APT ER A
накопляется в пузырьках, причем когда мышцы брюшка сокращаются и когда одновременно при помощи особой мышцы открывается выводное отверстие пузырька, пахучая жидкость поступает наружу. Биологическое значение их неясно — многие авторы высказывали соображения, что пахучие железы, выделяя пахучие вещества, способствуют отталкиванию животных, делающих попытки питаться уховертками (птицы и др.),‘ однако эти соображения не подтверждены фактами.
Кровеносная система как и у прямокрылых, характеризуется наличием дополнительных пульсирующих органов — головных ампул, помещающихся во лбу, у основания усиков и посылающих в усик трубковидный, сосуд, снабжающий кровью усики.
Дыхательная система характеризуется наличием двух пар главных трахейных стволов, проходящих вдоль тела; расширенные участки трахей —• воздушные пузыри, свойственные Orthoptera, совершенно отсутствуют у уховерток, анастомозы между трахеями довольно сильно развиты. Дыхалец имеется 10 пар, из которых 2 пары расположены на груди и 8 пар на брюшке; на первом грудном сегменте дыхальца достигают наибольшей величины, на втором они также довольно хорошо заметны, а на брюшке они очень маленькие и едва заметные. Так как 3-я пара располагается На первом сегменте брюшка, слившемся с заднегрудью, то некоторые старые исследователи (Westwood), считая этот сегмент за заднегрудь, определяли число стигм на груди равным 3-м парам, что резко отличало-бы уховерток от других насекомых.
Нервная система очень напоминает таковую у Orthoptera и состоит из трех грудных и шести брюшных ганглиев, а также надглоточного и подглоточного узлов в голове, от которых отходят нервы к ротовым органам и органам чувств (усикам и глазам).
Половая система уховерток весьма своеобразна, так как у самцов (фиг. 8) надсем. Protodermaptera она состоит из двух совокупительных органов (penis), что наблюдается также у поденок (Agnatha); в этом отношении уховертки сильно отличаются от настоящих Orthoptera. У надсем. Eudermaptera один совокупительный орган атрофирован. Так как строение полового аппарата имеет весьма большое значение в систематике отряда, мы даем его более подробное описание.
Располагается половой аппарат самца на 9-м стерпите брюшка; он состоит (фиг. 8) из пары симметричных хитиновых удлиненных образований — парамер, разделенных на две половины — основная половина, прикрепляющаяся к 9-му сегменту, называется п р о п а р а-мерами, а вершинная — метапарамерами. Пропарамеры сильно хитинизированы только снаружи и имеют вид длинных, узких пластинок, отделенных от метапарамер перетяжкой с наружной стороны, иногда очень слабо выраженной. Метапарамеры чрезвычайно изменчивы по форме и размерам и представляют собой сильно хитинизированную пластинку; иногда на конце ее имеется маленькая дополнительная лопасть, так. наз. э п и м е
ВВЕДЕНИЕ
13
р и т. Собственно penis представлен так называемым препуциальным мешечком (saccus praeputialis); их два у Proiodermaptera и один у Еи-dermaptera', они прикреплены к парамерам (собственно к пропарамерам) и имеют вид мягкого мешечка, часто снабженного .хитиновыми пластинками, или иногда серией маленьких зубчиков. Внутри penis имеется тон-
Фиг. 8. Схема полового аппарата О. А.— надсем. Protodermaptera; В — надсем. Eudermaptera, Ch. j — хитиновые утолщения; Ch. 2 — хитиновые пластинки; D — зубчики; D. ej — семяизвергательный канал; М — метапарамеры, Р — пр жарамеры, S.pj. — собственно penis в выправленном состоянии, S.p2— он же в покое; V] — вирга со спиральной структурой; V2 — простая форма вирги; Vg — выступающий конец вирги; V. г. — почковидный пузырек (По Burr, 1915, с изменениями).
кая трубка, называемая виргой (virga), представляющая собой часть семе-извергательного канала (ductus ejaculatorius); вирга очень изменчива по структуре и размерам, может иметь вид простой трубки или же при основании иметь тонкую спиральную выкладку. Основание вирги соединяется с почковидным пузырьком (vesiculus reniformis), представляющим как бы расширение вирги; почковидный пузырек значительно более короткий, чем вирга, часто имеет спиральную выкладку и кроме того иногда снабжен снаружи хитиновыми придатками в виде пластинок и выступов. Характер соединения вирги с почковидным пузырьком довольно изменчив — в виде простого постепенного перехода первой в почковидный пузырек, в виде дуги или даже угла. Задний (или нижний)
14
КОЖИСТОКРЫЛЫЕ-DERMAPTERA
конец почковидного пузырька продолжается назад в виде тонкого, как вирга, канала, представляющего собой семеизвергательный канал.
Наибольшее значение в систематике отряда имеет наличие или отсутствие парного penis, форма метапарамер, длина и характер вирги и почковидного пузырька, а также характер их соединения друг с другом, и, наконец, наличие тех или иных хитиновых включений на собственно penis и на почковидном пузырьке.1
Кроме сказанного выше в отношении наружного и внутреннего строения уховерток, необходимо упомянуть про так. наз. манубрий (manubrium), представляющий собой утолщение хитина у основания предпоследнего стернита <5 сверху (т. е. изнутри), состоящего из прозрачной мембраны, поддерживаемой на хитиновом остове. Строение манубрия имеет значение в систематике подсем. Protodermaptera.
Что касается полового аппарата самки, то последний состоит из пары яичников, каждый с несколькими политрофическими трубочками; вполне созревшие яичники занимают почти всю полость брюшка от 2-го до 7-го сегмента. От каждого яичника сзади отходит вниз по короткому яй ц ев о ду мезодермального происхождения (так наз. парные яйцеводы), соединяющиеся в один общий или непарный яйцевод (эктодермального происхождения), открывающийся наружу особым отверстием — гонопорой у заднего края 7-го стернита. Выше уже говорилось, что 8-й и 9-й сегменты брюшка самки скрыты внутри 7-го сег-
1 Для выделения и отпрепарирования полового аппарата самца с целью изучения, без чего нельзя обойтись при определении некоторых родов, особенно в подсем. Psalinaet исследуемое насекомое, если оно сухое, предварительно размачивается во влажной камере (на сыром песке); если оно недавно поймано и еще не высохло или сохранялось в спирту, размачивание не производится. После того, как насекомое станет мягким и гибким, его укладывают на спинную сторону и прижимая пинцетом брюшко и клещи, осторожно приподнимают тонкой иглой предпоследний стернит; эту манипуляцию более удобно осуще_ ствлять на торфяной пластинке или в ванночке с восковым дном, где насекомое предварительно прочно укрепляется путем обкалывания булавками. Приподняв вверх предпоследний стернит, легко можно обнаружить торчащий генитальный аппарат, помещающийся в маленьком углублении, откуда он может быть легко вынут при помощи тонкого пинцета. Для того, чтобы сделать постоянный препарат парамеры вывариваются в кипящем 10°/о растворе едкого кали или натра до того момента, когда они освободятся от прилегающих к ним мышечных волокон и пр. После этого парамеры помещаются для обезвоживания в крепкий спирт (не ниже 96°), налитый в часовое стекло, где остаются не менее 10—15 минут; так как крепкий спирт легко поглощает влагу, часовое стекло необходимо прикрывать плоским куском стекла. Затем для просветления парамеры помещаются в гвоздичное или бергамотное масло, или же в карбол-ксилол; просветление продолжается минуты 2—3 и считается.законченным, если, поместив часовое стекло на черный фон, препарат будет выглядеть совершенно темным (если он будет казаться беловатым от покрывающей его эмульсии, то это значит, что просветление еще не закончилось). После этого препарат, с помощью шпателя, переносится на предметное Стекло, а затем его сверху покрывают каплей канадского бальзама и накрывают покровным стеклом. К предметному стеклу приклеивается этикетка с датой поимки данного экземпляра и с названием вида. Приготовленный таким образом препарат хорошо сохраняется и может быть легко изучен с помощью микроскопа, бинокуляра или, если че требуется выяснение деталей, сильной лупы.
ВВЕДЕНИЕ
15
мента; редуцированные 8-й и 9-й стерниты, располагающиеся над
7-м стернитом ограничивают собою сверху генитальную полость, нижняя часть которой ограничена 7-м стернитом; таким образом генитальная полость представляет собой пространство, заключенное между 7-м стернитом снизу и 8—9-м стернитами сверху. Как раз выводное от
Фиг. 10. Клещи Forficula tomis S; а — f. brachylabia, b — f. macrolabia. Ориг.
Фиг. 9. Клещи Forficula auricularia S', а — f. brachylabia, b — f. macrolabia.
Ориг.
верстие яйцевода и открывается в самом переднем конце этой половой полости, на границе, где внутренняя сторона заднего края 7-го стернита соединяется с задним краем 8-го стернита. Имеющийся семеприемник, расположенный над 8-м стернитом, также открывается в половую полость отверстием, но уже независимо от гонопоры. Половая полость открывается наружу у заднего конца 7-го стернита в виде щели, расположенной между задним концом 7-го стернита и 10-м стернитом.
Весьма важной и интересной с биологической точки зрения является чрезвычайно сильная морфологическая изменчивость многих видов уховерток. Наиболее обычные случаи ненаследуемой изменчивости касаются строения клещей самца и степени развития надкрылий. Диморфная изменчивость клещей <5 выражается как в отношении их длины, так и формы; здесь разли
чают две формы — f. brachylabia, когда клещи <5 сильно укорочены,
причем вершинная утонченная половина в этом случае оказывается часто более изогнутой, и f. macrolabia, когда клещи сильно удлинены, в соответствии с чем изменяется пропорция отдельных частей, а вершинная часть у многих видов станов?.тся прямой (фиг. 9—10). Соот
ветственно этим формам у некоторых видов коррелятивно изменяется общий габитус тела и даже строение конца брюшка и пигидия. Особенно в сильной степени это выражается у Anechura (Borelliola) euxina Sem. (фиг. 11—12), у которого f. brachylabia характеризуется более коренастым телом, и, что особенно важно, — сильнейшим изменением строения последнего тергита брюшка; последний характеризуется появлением на нем весьма сильных пластинкообразных выступов, отсутствующих у f. macrolab'a-, кроме того и клещи приобретают совершенно другой тип строения, так что обе эти формы внешне настолько сильно отличаются друг от друга, что могут быть приняты за два разных вида и даже рода (что фактиче
16
КО ЖИСТОКРЫЛЫЕ —DERM APTER А
ски и имело место). При этом, невидимому, у разных видов преобладает одна какая-либо форма — macrolabia vlicr brachylabia, хотя первая в общем является более редкой, чем вторая; в отношении Forficula auricularia •есть указания, что f. macrolabia попадается главным образом на островах и в горах. В некоторых случаях одна из указанных форм может напоминать ближайшие роды — напр. Oreasiobia stoliczkae f. brachylabia напоминает виды очень близкого рода Anechura.
Биометрические исследования Bateson’a (1892) над обыкновенной уховерткой, а позднее обстоятельное изучение этого вопроса Д. Дьяконовым (1925) показали, что сообразно наличию двух типов клещей получается и двувершинная кривая распределения. Это обстоятельство могло бы послужить доводом к тому, что f. macrolabia и f. brachylabia представляют собой различные генотипы, т. е. формы, имеющие различные наследственные задатки. Однако, Дьяконов и Huxley (1927) показали, что в менее благоприятных условиях преобладают самцы f. brachylabia и кроме влия-лия этих условий нельзя было обнаружить какой-либо зависимости между преобладанием той или иной формы и другими факторами.
Математический анализ вопроса, проделанный Дьяконовым в специальной статье,1 дал возможность доказать, что в случае, если норма реакции органа изменяется не прямо пропорционально изменению внешних условий, а так, что в наименее благоприятных условиях возрастание величины органа происходит очень медленно, затем в условиях средней благоприятности это возрастание происходит с наибольшей скоростью (возрастая в отдельных случаях при этих условиях так, что эти переходные классы условий практически могут быть неуловимыми), и, наконец, в условиях наибольшей благоприятности — снова очень медленно, то в этом случае генотипически однородный материал при статистическом исследовании размеров того или иного органа, имеющего сложную норму реакций, .дает двувершинную кривую.
Таким образом, диморфизм самцов уховерток в отношении клещей ле связан с наличием двух наследственных форм — ff. macrolabia и bra-chylabia, а представляет собой такой тип изменчивости, когда наибольшая скорость возрастания размеров клещей в течение постэмбрионального развития происходит в условиях средней благоприятности существования индивидуумов; раз наибольшая изменчивость клещей происходит в таких классах средних условий, которые по отношению к остальным .классам имеющимся в природе составляют наименьшее число случаев, особи, имеющие средние размеры клещей встречаются наименее часто.
Необходимо, однако, отметить, что в некоторых случаях ясной двувершинной кривой может и не получиться; так, в работе КиЫ’я (1928) приведено много таких примеров, причем автор объясняет это весьма неблагоприятными условиями существования вида, внешним выражением
1 Дьяконов, Д. М. Диморфная изменчивость как результат сложной реакционной нормы. Изв. Бюро Генет., Евген., 4, 1926: 97—104.
ВВЕДЕНИЕ
17
чего является преобладание среди собранных особей f. brachylabia. Несмотря на это все же кривые имеют явно асимметричный характер, резкй обрываясь в своей левой части (характеризующей f. brachylabia) и весьма плавно сходя на нет в правой части.
В отношении степени развития надкрыльев и крыльев различают £ brachyptera и f. eupiera (Семенов-Тян-Шанский, 1935); к первой отно-
Фиг. 11. Anechura (Borelliold) euxina Sem., f. brachylabia, О (окр. Майкопа). X 5. Ориг.
Фиг. 12. Anechura (BoreUiola) euxina Sem., f. macrolabia, О (Bo-рошиловск. н/К.). X 5. Ориг.
сятся особи с укороченными в разной степени крыльями и коррелятивно слегка укороченными надкрыльями, а ко второй — особи с вполне развитыми крыльями, сильно выступающими из под надкрылий в виде .длинных крыловых пластинок, причем надкрылья в этом случае имеют резко выступающие плечи и слегка вырезанный задний край. Известны также и такие факты, когда совершенно бескрылый вид, без каких-либо признаков не только крыльев, но и надкрылий, изредка дает вполне кры-.латую форму; достоверный случай подобного рода описан Рап1е1’ем для Anisolabis (Euborellia) annulipes. Наконец, необходимо отметить также чрезвычайно сильную морфологическую изменчивость у такого вида, как Labidura riparia, имеющего большое количество различных форм, для рационального обозначения которых предложена целая система символов {см. описание этого вида).
Фауна СССР, Кожистокрылые 2
18 КОЖИСТОКРЫ ЛЫЕ—DERM APTER А
В большинстве своем эти формы, видимо, являются не наследственными модификациями, причем крайние из них часто связаны между собой: постепенными переходами; поэтому они должны трактоваться ка,к морфы, зависящие в своем появлении от экологических условий. Однако, этот вопрос серьезно никем еще не изучался.
Чрезвычайная изменчивость уховерток сильно затрудняет определение и вообще выяснение систематического положения и значения отдельны^ форм; невнимательное и неосторожное отношение к этой изменчивости может повлечь за собой ряд грубейших ошибок в определении. Достаточно указать на то, что единственный известный вид рода Labidura, именно L. riparia, был описан прежде под четырьмя родовыми и более чем: под тридцатью видовыми названиями.
2. ПРЕВРАЩЕНИЕ
Уховертки относятся к насекомым с неполным превращением, характеризующимся тем, что из яйца выходит молодое насекомое, внешне очень похожее на взрослых особей и отличающееся от Них только меньшими размерами тела, отсутствием крыльев и некоторыми деталями внешней морфологии; путем последовательных линек молодое насекомое' постепенно превращается во взрослое. Благодаря тому, что молодые особи отличаются от взрослых не так сильно, как у насекомых с полным превращением, термин „личинка", употребляемый для обозначения фазы, предшествующей взрослому состоянию, у уховерток не может считаться правильным; однако, он широко вошел в практику и поэтому, в дальнейшем, молодые особи нами будут обозначаться как личинки.
Уховертки, видимо, в большинстве случаев имеют четыре возраста, личинок, хотя можно предполагать, что у низших групп, как напр. Proto-dermaptera число их может быть большим; четыре возраста личинок отмечены у Forficula tomis (Песоцкая, 1927), Chelisoches morio (Burr, 1910) и Forficula auricularia (Chapman, 1917 b).
Впрочем, необходимо отметить, что в предыдущей своей работе Chapman (1917 а) отмечает наличие у обыкновенной уховертки 6-ти возрастов личинок; точно так же и Nel (1929), хотя и не касается специально вопроса о числе возрастов у личинок обыкновенной уховертки, но определенно отмечает для нее 6 возрастов. Таким образом, вопрос о числе возрастов у личинки остается нерешенным; очень возможно, что переход, в следующий возраст не обязательно связан с увеличением числа члеников усиков и внешние отличия между возрастами могут носить другой характер, тем более, что Nel для самок личинок старших возрастов отмечает изменения в числе видимых сегментов брюшка (см. ниже). Поэтому допуская вместе с большинством исследователей наличие у личинки, по крайней мере у представителей надсем. Eudermaptera, 4-х возрастов, мы, однако, должны подчеркнуть некоторую условность этого допущения.
ВВЕДЕНИЕ
19
Личинки 1-го возраста у всех указанных выше видов имеют 8-чле-никовые усики, причем интересно, что это число члеников свойственно личинкам столь различных родов как Chelisoches и Forficula-, первый во взрослом состоянии имеет 20—21, а виды рода Forficula—12— 14 члеников. Увеличение числа члеников происходит во время линьки за счет разделения только одного 3-го членика на несколько новых члеников, что резко отличает уховерток от настоящих Orthoptera, у которых во время линьки происходит разделение не одного, а нескольких члеников на два новых.
В зависимости от того, какое число члеников в усиках имеет взрослое насекомое, третий членик с каждой линькой у разных видов делится на разное число члеников. В качестве иллюстрации приведу следующую сравнительную таблицу для указанных выше видов.
Виды Число члеников в усиках по возрастам
I II III IV Imago
1. Chelisoches morio . . 8 12 16 19 20—21
2. Forficula tomis .... 8 10 10 11 12
3. Forficula auricularia . 8 10 11 12 13—14
Так как увеличение числа члеников в усиках происходит за счет только 3-го членика, то из этой таблицы вытекает, что у Ch. morio этот членик разделяется при линьке во II возраст на 5 члеников, в III возраст также на 5, в IV — на 4, а при линьке во взрослую стадию — на 2—3 членика; у F. tomis и F. auricularia соответственно происходит разделение на 3 членика, далее при линьке в III возраст число члеников остается прежним, но зато 3,4 и 5 членики увеличиваются в длине или же у F. auricularia членик делится пополам, при линьке в IV возраст и в стадию imago 3-й членик у F. tomis делится только на две части, а у F. auricularia при последней линьке этот членик делится на две или три части.
Чрезвычайно интересным является то обстоятельство, что у многих представителей наиболее примитивного сем. Pygidicranidae личинки имеют многочленистые церки; так, у личинок рода Diplatys число члеников в зависимости от возраста достигает 14—45, увеличиваясь с каждым последующим возрастом, причем первый, т. е. основной членик имеет наибольшую длину; при линьке в последний личиночный возраст все остальные членики отпадают, так что личинка последнего возраста имеет одночлениковые церки; через оболочку оставшегося членика можно заметить будущие клещи. При линьке во взрослое насекомое оставшийся основной членик превращается в клещи. Таким образом, клещи уховерток представляют собой одночленистые церки. Представители всех остальных семейств всегда имеют даже в I возрасте, а согласно Heymons’y (1895) — 2»
20
КО ЖИСТО КРЫЛЫЕ—DЕRМАРТЕRА
даже в эмбриональном состоянии—одночлениковые церки, в виде почти на всем протяжении одинаково тонких, прямых или в старших возрастах чуть изогнутых клещей, причем отличия между самцом и самкой едва уловимы только в последнем возрасте; лишь при достижении взрослой стадии клещи приобретают резкий половой диморфизм, столь характерный для Dermaptera.
Характерные для многих видов боковые бугорки на 3-м и 4-м терги-тах брюшка отсутствуют у личинок младших возрастов, появляясь только в старших возрастах; впрочем Rehn (1903) показал, что у неотропической Psalis americana Palis, бугорки, будучи развитыми у личинок, исчезают при переходе в imago.
Что касается развития надкрылий и крыльев, то хорошо заметные зачатки их появляются, начиная с IV возраста, в виде оттянутых назад боковых сторон средне- и заднеспинки, причем Chapman (1917 а) отмечает, что в предшествующем возрасте какие-либо признаки зачатков надкрылий и крыльев у обыкновенной уховертки отсутствуют. Зачатки крыльев на заднеспинке имеют характерное жилкование в виде прямых, радиально расходящихся от основания, не ветвящихся, приподнятых полосок.
Выше говорилось, что брюшко самок взрослых уховерток характеризуется по сравнению с самцами меньшим числом видимых сегментов, так как 8-й и 9-й сегменты недоразвиты и скрыты внутри 7-го сегмента. Однако, у личинок первых трех возрастов Forficula auricularia, по исследованиям Nel (1929), брюшко имеет нормальную сегментацию, как у самца, и только при достижении последнего личиночного возраста 8-й стернит (и тергит?) почти полностью закрывается 7-м; после линьки во взрослое состояние кроме 8-го сегмента оказывается скрытым и 9-й сегмент.
Во время линьки оболочка тела трескается сверху вдоль средины груди и насекомое постепенно освобождается от нее. Продолжительность личиночной фазы может быть весьма различной; в условиях средней полосы европ. части СССР личинки первых возрастов Forficula tomis живут около двух недель, тогда как III и IV возраста, идущие на зимовку, живут не менее полгода. В условиях тропиков продолжительность каждого личиночного возраста для таких видов как Diplatys greeni Burr и D. ger-staeckeri Dohrn равна примерно 10 дням.
3. ОБРАЗ ЖИЗНИ И ЭКОЛОГИЯ
Уховертки являются по преимуществу ночными насекомыми, скрывающимися днем в различных укромных местах (под камнями, под опавшей листвой, в различных щелях и пр.), собираясь здесь иногда довольно значительными по числу особей группами. Ночью же они становятся деятельными, оживленно ползая в поисках пищи и пр. Однако, немногие виды могут быть деятельными и днем; так, наша малая уховертка (Labia minor) летает при солнечном освещении, особенно часто около навозных
ВВЕДЕНИЕ
21
куч, а тропическая черная уховертка {Chelisoches morio), по наблюдениям Terry на Гавайских островах (1905), является весьма подвижным насекомым, обитающим влажные места на плантациях сахарного тростника, где она наблюдалась ползающей на листьях в поисках пищи в самые жаркие часы дня.
Длинное, тонкое и гибкое тело уховерток позволяет им проникать во всевозможные укромные места, что при ночном образе жизни большинства видов делает этих насекомых днем мало заметными. Виды, имеющие вполне развитые крылья, могут хорошо летать, несмотря на то, что отдельные виды прибегают к этому способу передвижения редко {Forficula auricularia). Наиболее часто у нас можно наблюдать летающими два вида — малую уховертку {Labia minor) и прибрежную уховертку {Labidura riparid), причем оба эти вида ночью, особенно при облачной теплой погоде, прилетают на свет. Вообще нужно сказать, что хорошо летающие виды ночью очень часто прилетают на свет при наличии соответствующих условий погоды. Весьма оригинально то, что и длиннокрылая форма (f. euptera) у видов с нормально атрофированными крыльями также хорошо может летать, прилетая ночью на свет (напр. F. tomis f. euptera).
Наличие у уховерток весьма своеобразного органа — клещей вызывало у многих исследователей вопрос о том, какую биологическую функцию имеет этот орган. В этом отношении существуют различные взгляды, причем очень часто те или иные предположения основаны больше на остроумных предположениях, чем на прямых наблюдениях. Так, существует мнение, что клещи служат для развертывания и складывания задних крыльев; однако, наблюдения (Morgan, 1924) говорят против этого; многие авторы говорят о клещах как об органе, служащем для удержания самцом самки во время спаривания, но и в этом случае прямые наблюдения говорят скорее против этого допущения. Однако, несомненно, что клещи представляют собой орган, используемый для защиты и нападения (Cadeau de Kerville, 1905; Morgan, 1924), причем очень характерно, что в том и другом случае уховертки принимают позу скорпиона, поднимая заднюю часть брюшка с клещами кверху и загибая последние вперед, так, что концы клещей располагаются над головой и грудью. Мускулатура, управляющая клещами, настолько сильная, что насекомое может проколоть клещами кожу человека до крови: такие случаи описаны для Forficula auricularia и Titanolabis colossea (Burr, 1910). Кроме того, Baer (1904) сообщает, что относительно слабая южно-американская Doru lineare Esch, может причинить местное воспаление кожи при помощи своих клещей, чему вероятно также способствует наличие на них септического начала. Клещи используются также для удержания добычи во время ее поедания. Необходимо, однако, отметить, что некоторые виды уховерток, напр. Forficula macrobasis (см. специальную часть), имеют такого типа клещи, что концы их не могут даже соприкасаться друг с другом, так что ни о какой защите и нападении с помощью их не может быть и речи.
22
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
В отношении кормового режима, уховертки являются вполне многоядными насекомыми, питаясь как животной, так и растительной пищей, употребляя живые и гниющие растения, животные остатки, а также нападая на мелких личинок или взрослых насекомых, червей, пауков и т. п., причем для ряда видов отмечается явное предпочтение животной пище. Liistner (1914), изучавший пищевой режим обыкновенной уховертки (F. auricularia) по содержимому зоба и желудка пойманных на воле особей, обнаружил, что характер пищи меняется от места, где была поймана та или иная особь. Обычно пища состоит из мертвых частей растений и, кроме того из живущих на листьях грибков, а если особи были пойманы под корой деревьев, то в значительной степени из особой водоросли — Cysti-coccus humicola. Однако, если тем или иным видам уховерток представляется возможность питаться живыми растениями, то они становятся подчас заметными вредителями (F. auricularia, F. tomis, Anechura asiatica и др.). Виды, относящиеся к особому подотряду Arixenina одно время считались паразитами млекопитающих; однако, повидимому, они являются свободноживущими пещерными обитателями.
По крайней мере виды, живущие в умеренных широтах, имеют одногодовую генерацию. Обыкновенная уховертка {Forficula auricularia) начинает спариваться в конце лета — начале осени, причем в этом случае самец и самка пятятся друг к другу задними концами тела до соприкосновения, после чего самец изгибает брюшко вокруг своей продольной оси, так что нижняя сторона конца последнего соприкасается с нижней стороной конца брюшка самки; в результате этого, по наблюдениям некоторых исследователей, копулирующая пара образует одну прямую линию с головами, направленными в противоположную сторону; в таком положении пара остается несколько часов. Спустя несколько дней происходит откладка яиц, в одну общую кучу, причем яйца эти у ряда видов (F. auricularia, F. tomis, Labidura riparia и др.) помещаются в гнезде в виде норки или ямки в земле (фиг. 13), которую самка (F. tomis) начинает рыть еще до откладки. У F. tomis самка откладывает яйца один раз в количестве 50—80 штук; у других видов исследователи отмечают многократную откладку. Кроме откладки яиц осенью, которые таким образом зимуют, самка обыкновенной уховертки, перезимовав, продолжает яйцекладку и весной. У других видов, напр. F. tomis яйцекладка производится только в конце весны—начале лета. Самцы обыкновенной уховертки в отличие от самок, видимо, плохо переживают зиму, так как некоторые исследователи отмечают наличие весной в соответствующих местах многочисленных мертвых самцов совместно с живыми самками. Весьма интересно наличие у ряда видов уховерток (F. tomis, F. auricularia и др.) заботы о потомстве, которая выражается в том, что самка охраняет отложенные ею яйца, находясь при них в течение всего периода развития, повидимому, совсем не питаясь в это время и живя вместе с отродившимися личинками еще 4—б дней (F. tomis). Некоторые исследователи (Verhoeff,
ВВЕДЕНИЕ
23
1913) даже говорят о том, что самка обыкновенной уховертки остается с вылупившимися личинками до момента превращения последних в iftiago; однако^ другие авторы (Песоцкай, 1927) высказывают предположение, что здесь, видимо, имело место более раннее окрыление в колоний личинок одной сймки, которая й была смешана поэтому с матерью остальных еще не окрылившихся личинок. Так как самцы и неотлоЖившйе еще яиц самки охотно питаются яйцами себе подобных, указанная выше охрана самкой отложенных ею яиц имеет несомненно большое значение для сохранения потомства. Кроме того, Песоцкая отмечает, что самка F tomis, не оставляя
ни на минуту отложенные ею яйца,
с Места на место и „облизывает" последние. Поставленный указанным автором опыт (к сожалению, только однократный) показал, что если взять самку из гнезда, оста-вйв яйца без попечений матери, то в этом случае большинство яиц тускнеют, покрываются плесенью и до 90% их погибает.
Вылупление у наших видов происходит примерно через 2—4 недели после яйцекладки (по Песоцкой у F. tomis в средней полосе СССР
часто ощупывает, перекладывает
Фиг. 13. Самка обыкновенной уховертки в гнезде с яйцами. (По Fulton из Beier, 1933).
через 12—16 Дней, по Riihl
у обыкновенной уховертки через 26 дней). В зависимости от особенностей годового цикла развития зимующая фаза может быть у разных видов различной; обыкновенная уховертка, как уже отмечалось выше, зимует в фазе яйца й imago, a.F. tomis в фазе личинки последних двух возрастов.
В бтношении стаций обитания необходимо отметить, что характер их у большинства видов более или менее сходен. Многие виды встречаются группами или по одиночке в сырых местах под камнями, под опавшей листвой, в гниющих пнях деревьев (Anechura bipunctata), под корбй, по берегам морей в гниющих растительных остатках (Anisolabis maritima) и забираясь даже в плоды, помещения человека, а также встречаясь на цветах и пр. Некоторые виды, напр. Labidura riparia, обитая в умеренных широтах, песчаные почвы по берегам рек или с высокими грунтовыми водами, устраивают в земле наклонные норки до 1/i м длиной, живя в них по одиночке. Однако, в условиях юга (в Индии), этот же вид обитает различные укромные места вроде щелей в коре деревьев; такая разница в экологии, повидимому, объясняется тем, что в условиях песчаных почв на севере создается более теплый, чем в окружающей обета-
24
К ОЖИ СТОКРЫЛЫE — DЕRМАРТ Е R А
новке, микроклимат, позволяющий существовать здесь данному виду,, широкое распространение которого в тропиках говорит о его термофиль--ности. Подобная смена стаций обитания по зонам отмечалась другими исследователями (Бей-Биенко ’) и для других групп насекомых. Если в условиях влажной тропической зоны L. riparia не устраивает норок, в земле, в отличие от того, что она делает в умеренных широтах, то этот-факт был бы крайне интересен в качестве показателя значительной изменчивости биологии вида на протяжении ареала его распространения. В отношении одного индийского вида {Forficula decolyt) отмечается, что он живет под камнями на берегах ручьев, почти в воде. Некоторые тропические виды, имеющие очень плоское тело {Apachyus и вероятно Plafylabia) живут, видимо, постоянно под корой деревьев, так что и особенности их. морфологии приспособлены к условиям жизни под корой.
Громадное большинство видов связано теснейшим образом с лесной, растительностью, что впрочем ясно из характеристики указанных выше условий обитания уховерток. Кроме того, характер стаций, населяемых, уховертками, говорит также и о том, что они являются влаголюбивыми: насекомыми. Выше уже отмечалось, что такой казалось ксерофильный вид, как L. riparia, обитая песчаные почвы, живет по берегам рек; точно» так же такой „ксерофил" как Anechura asiatica, обитая сухие каменистые склоны (напр. в горах Каратау в Казахстане) может быть обнаружена там: только в том случае, если под камнями сохранилась сырая почва. Так как настоящих ксерофилов среди уховерток, видимо, нет и оба упомянутые вида по сравнению со всеми известными остальными являются, наименее влаголюбивыми, необходимо притти к выводу, что вообще весь отряд представляет собой влаголюбивую группу насекомых.
Точно так же и рассмотрение географического распространения отряда Dermaptera в сопоставлении с климатическими особенностями дает некоторое представление об экологии этих насекомых. Наибольшего» богатства в отношении видового состава уховертки ^достигают в тропической зоне, тогда как в умеренных широтах число видов сокращается, буквально до единиц; напр., во всей европейской части СССР (без Крыма и Кавказа) известно всего только 9 видов, в лежащем к югу Средиземноморье 45 видов, а на территории значительно менее обширного Бельгийского Конго в Африке — 75 видов;2 более того, северная граница распространения Dermaptera в Евразии как отряда проходит между 54—бб° с. ш. (фиг. 17). Указанная картина дает основание рассматривать уховерток как очень теплолюбивую группу насекомых. Однако, такое толкование останется весьма односторонним, если не попытаться взглянуть на богатство тех или иных фаун с учетом других основных экологических факторов,.
1 Б е й-Б и е н к о, Г. К вопросу о зонально-экологическом распределении саранчевых в Западно-сибирской и Зайсанской низменностях. Тр. Защ. Раст., 1, № 1,1930 : 51—90.
2 М е п о z z i, С. Contribute alia conoscenza dei Dermatteri del Congo Belga. Rev. ZooL Bot. Afr., XXVII, 1935:15—32.
ВВЕДЕНИЕ
25
особенно осадков и растительного покрова. Для этой цели мы воспользуемся таблицей количественного распределения видов в Палеарктической области приведенной ниже (стр. 44). При сопоставлении таких территорий, как Туранская низменность (5 видов) и Иран (8 видов), характеризующихся очень сухим и жарким, в южных частях субтропическим климатом, с Зап. Европой (без Средиземноморья —16 видов), лежащей в более холодном климате, бросается в глаза наличие здесь в 2—3 раза большего числа . видов; примерно аналогичная картина получается при сопоставлении фауны » Монголии (5 видов) и соседней во много раз меньшей территории Уссурийского края (9 видов). В выбранных здесь примерах значительным богатством фауны уховерток отличаются области, имеющие более холодный климат, но зато характеризующиеся значительно большим количеством осадков, позволяющим развиваться здесь лесной растительности.
Таких примеров можно было бы привести очень много; здесь мы ограничимся указанием на то, что и в условиях тропической зоны наибольшего богатства фауна уховерток достигает в областях, имеющих влажный климат и достаточно богатую лесную растительность (экваториальная Африка, южная Азия, Малайский архипелаг и др.); в качестве иллюстрации к сказанному сопоставим число видов, известных из восточной, более влажной части северной Индии (Ассам, Сикким, Бутан, Бенгалия) с годовым количеством осадков от 1300 до 2000 и более мм, с числом видов населяющих западную часть Сев. Индии, лежащую примерно на той же широте (Пенджаб, Синд, Раджпутан), но имеющей только от 200 до 600 мм осадков. В первом случае число видов достигает 70, во втором — только 9; если же из восточной части оставить только одну низменную Бенгальскую провинцию, то и здесь число видов достигает почти 30, несмотря на то, что взятая территория во много раз меньше чем сравни" ваемые западные провинции.
Весьма характерно, что обыкновенная уховертка по опытам W. Morgan’а (1926) хорошо плавает, причем взрослые особи, находившиеся на поверхности воды 24 часа, были также активны, как и в момент, когда они были опущены в воду; кроме того, и погружение их в воду на короткое время переносится легко. В связи со сказанным нельзя не сопоставить приведенный выше пример с индийским представителем того же рода, как и обыкновенная уховертка — F. decolyi, живущим по берегам ручьев, почти непосредственно в воде.
Резюмируя все сказанное, необходимо сделать вывод, что решающим экологическим фактором для этой группы насекомых является степень увлажнения как обитаемых ими территорий, так и населяемых ими стаций, причем уховертки легко переносят непосредственное пребывание в воде.
В связи с высказанными здесь соображениями следует отметить, что недавно Вульф1 показал аналогичную зависимость между видовым
1 В у л ь ф, Е. В. Опыт деления земного шара на растительные области на основе количественного распределения видов. Тр. Прикл. Ботан., Генетики и Селекции, сер. 1, № 2,1934.
26 КОЖИСТОКРЫЛЫЕ —DERM APTERA
разнообразием мировых флор и степенью увлажнения занимаемых ими областей; эта параллель между уховертками и растениями идет и дальйгё если иметь в виду то, что наиболее увлажненные территорий занимаются Лесной растительностью, где одновременно имеет место и наиболее богатая фауна Dermaptera.
Было бы, однако, неправильно отсюда сделать вывод, что температурные условия для уховерток имеют небольшое значение; отсутствие этих насекомых в арктической зоне и наличие сравнительно небольшого количества в лесных областях умеренных широт говорит против этого; оптимум условий существования для них создается во влажном и жарком климате. Однако, основным экологическим фактором, имеющим особенно большое значение для данной группы насекомых, несомненно, является влажность климата или, по крайней мере, обитаемых ими стаций (Labidura riparia, Anechura asiafica).
В заключение остается сказать несколько слов о роли человека в распространении уховерток. Уже беглое знакомство с распространением последних показывает, что человек сыграл громадную роль в расселений многих видов по всему земному шару, причем в новых условиях эти виды очень часто находили для себя благоприятные условия, иногда, видимо, даже лучшие, чем это имело место на их родине, так как на вновь заселенных территориях они иногда размножались в таких массах, что становились бичом местного сельского хозяйства. Насколько значительна роль человека в расселении этих насекомых говорит то, что в СССР имеется не менее б видов завезенных человеком (Anisolabis maritima, A. pectoralis, A. annulipes, Labidura riparia, Labia minor, Prolabia arachidis), невидимому, из тропических стран. В Западной Европе к этим видам необходимо прибавить еще Nala lividipes и некоторые другие виды. Кроме того, обыкновенная уховертка (Forficula auricularia) была завезена из Европы в Сев. Америку, Новую Зеландию и другие страны, а в расселении F. tomis на восток до зап. Сибири, несомненно, также сыграл значительную роль человек (Бей-Биенко, 1927). Наконец, все перечисленные из СССР шесть видов широко распространены по всему земному шару, не говоря уже о многих других видах, расселенных с помощью человека в ряде тропических и субтропических стран.
Столь значительная роль человека в расселении многих видов не совсем понятна, хотя такие виды, как Anisolabis maritima и Labidura riparia, живущие по берегам морей или рек, т. е. в непосредственной близости от водных торговых путей, весьма легко могут быть занесены на корабли и другие суда вместе с товарами, хранившимися на берегу. Вероятно биологические особенности уховерток, заключающиеся в том, что они на день забираются в различные укромные места, обеспечивают им попадание вместе с товарами, особенно с такими, как плоды и фрукты, лесом, живыми растениями и пр. на транспортные суда, при помощи которых они и расселяются по торговым путям. Указанная биологическая особенность
ВВЕДЕНИЕ
27
является, видимо, причиной того, что, попав в новые места, уховертки быстро прививаются здесь; ведя скрытый образ жизни, они не приобретают на вновь заселенной территории серьезных врагов и подчас находят здесь даже более благоприятную экологическую обстановку (напр. F. auricularia, являющаяся серьезным вредителем в Новой Зеландии и Сев. Америке).
Здесь нельзя не упомянуть об интересном соображении W. Morgan’а (1926), отметившего недавнее появление большого количества особей обыкновенной уховертки в районе г. Портлэнда в штате Орегон, о роли дождей в расселении этого вида. Основываясь на своих опытах, показавших, что уховертка легко переносит пребывание в воде, а также на том, что этот вид живет во всяких укромных местах под различными предметами, этот автор приходит к выводу, что вовремя дождя уховертки смываются совместно с различными предметами, попадают с потоками воды в ручьи и канавы и благодаря этому переносятся в новые еще ненаселенные ими места.
Несомненно, некоторую роль в расселении уховерток человеком имеет и то обстоятельство, что человек в своей хозяйственной деятельности иногда создает для отдельных видов подходящие местообитания. Так, F. tomis в СССР наблюдалась в погребах, омшаниках, в пожнивных остатках (стебли подсолнечника), в сырых домах и пр., причем, видимо, в некоторых из этих мест (омшанники, пожнивные остатки) они проводят и зиму; примерно в аналогичных условиях встречается и обыкновенная уховертка. Живя в непосредственной близости от человека, эти виды и были развезены последним, причем так как первый из них является жителем внутренних частей континента, не доходя своим естественным ареалом до берегов морей, он естественно не мог быть так широко развезен, как обыкновенная уховертка; наоборот, соприкосновение естественного ареала последней с приморскими странами обеспечило ей возможность расселения по морским путям во многие отдаленные страны.
Вместе с тем необходимо отметить, что даже широкораспространенные человеком виды все же отсутствуют в некоторых страна^ несмотря на то, что они могли быть туда завезены; напр., обыкновенная уховертка отсутствует во всей восточной и юго-восточной Азии; точно так же Rehn отмечает отсутствие Labidura riparia на западном побережье Сев. Америки. Все это говорит о том, что здесь имеется еще много неясных сторон, которые могут быть выяснены только при тщательном исследовании.
4. КЛАССИФИКАЦИЯ И КОЛИЧЕСТВЕННАЯ ХАРАКТЕРИСТИКА ОТРЯДА
Классификация и систематика отряда долгое время находились в крайне неразработанном состоянии; достаточно сказать, что весь разнородный комплекс видов отряда относился к одному семейству Forficulidae, причем ряд видов, ныне часто относящихся к различным семействам и даже надсемействам, объединялись в один род (напр. Apterygida,
28
КОЖИ С ТОКРЫЛЫЕ —DERMAPTERA
Opisthocosmia и др.). Только с начала XX столетия исследования Verhoeff а, Zacher’a и Burr’a дали возможность вывести классификацию отряда из того хаотического состояния, в котором она находилась прежде. Однако, если современная группировка на семейства, основанная главным образом на обобщающей работе Burr’a (1915—16), не вызывает особых возражений и может рассматриваться как достаточно естественная, то этого нельзя сказать про более мелкие систематические категории, особенно подсемейства и роды, которые требуют еще значительных исследований для того, чтобы достичь большей естественности и совершенства; особенно это касается родовых подразделений в подсем. Psalinae и подразделений на подсемейства в сем. Forficulidae.
В настоящее время классификация отряда, учитывая и ископаемое мезозойское семейство Protodiplatyidae, представляется в таком виде.
Отряд Dermaptera
I. Подотряд Archidermaptera (j-) nov.
1. Сем. Protodiplaiyidae (1 род и 1 вид).
II. Подотряд Arixenina
1. Сем. Arixeniidae (1 род и 2 вида).
III. Подотряд Dermaptera s. str., s. Forficulina
А. Надаем. Protodermaptera
1. Сем. Pygidicranidae (24 рода и около 140 видов).
2. Сем. Labiduridae (40 родов и подродов и около 200 видов).
3. Сем. Apachyidae (2 рода и 13 видов).
В. Надсем. Eudermaptera
4. Сем. Labiidae (38 родов и около 270 видов).
5. Сем. Forficulidae (68 родов и около 260 видов).
Подотряд Archidermaptera характеризуется членистыми церками и 4—5-члениковыми лапками; подотряд Arixenina представляет собой группу, невидимому, пещерных насекомых, несущих следы дегенерации и характеризующихся сильно редуцированными глазами, сильно уплощенными и вооруженными тремя зубцами мандибулами, слабо хитинизированными, едва клещеобразными церками и полной бескрылостью. Что касается подотряда Dermaptera s, str., то наиболее примитивная его группа — надсем. Protodermaptera характеризуется главным образом двойным половым аппаратом самца, тогда как в надсем. Eudermaptera половой аппарат за счет атрофии его половины превратился в непарный орган.
В настоящее время во всем отряде известно примерно 174 рода и около 900 рецентных видов; таким образом, весь отряд представлен относительно весьма небольшим числом видов при одновременно значительной родовой дифференциации, так как в среднем на один род прихо
ВВЕДЕНИЕ
29.
дится 5.2 вида с колебаниями у разных родов от одного до 45—50 видов (Diplatys, Forficula, Labia). Наибольшим богатством отличается Индомалайская и Неотропическая' области, а наименьшим — Неоарктическая.
5. СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ЭВОЛЮЦИЯ ОТРЯДА DERMAPTERA В СВЯЗИ С ПАЛЕОНТОЛОГИЧЕСКИМИ ДАННЫМИ И ГЕОГРАФИЧЕСКИМ РАСПРОСТРАНЕНИЕМ
Отряд уховерток долгое время рассматривался многими авторами только в качестве семейства отряда прямокрылых (Orthoptera), несмотря на то, что некоторые исследователи, основываясь на особенностях морфологии, выделяли их уже давно в особый отряд (Burmeister, Westwood, Kirby и др.). Накопление к началу XX столетия сведений по морфологии, анатомии и развитию этой группы, при одновременном открытии близких к ним групп насекомых, послужили причиной окончательного выделения их в особый отряд; так как Dermaptera и близкий к нему отряд Hemime-roidea (представители которого известны в качестве паразитов грызунов в Африке) оказались весьма отличными от группы прямокрылых s. lato, некоторые авторы (Мартынов1) выделяют Dermaptera ts. Hemimeroidea в особый надотряд Dermapteroidea — уховерткообразные. Основанием к этому является своеобразный характер надкрылий (сильно укороченных по сравнению с задними крыльями), своеобразное жилкование и способ складывания задних крыльев, особенности внутренней анатомии, в частности строение полового аппарата, и некоторые морфологические признаки, особенно строение конца брюшка. Однако, наличие у уховерток неполного превращения (эпиморфоз), типично-грызущего ротового аппарата и складывающихся веерообразно вдоль продольных жилок задних крыльев (последние впрочем кроме этого складываются еще поперечно) сближает Dermapteroidea с двумя другими надотрядами— таракано-образными (Blattopteroidea s. Blattaeiformes), и прямокрылообразными (Orthopteroidea), благодаря чему эти три надотряда выделяются современными систематиками в особый подотдел — Polyneoptera. Таким образом и теперь уховертки сближаются с теми отрядами, которые в большинстве своем относились прежде к отряду прямокрылых. Однако, это сближение указывает только на очень отдаленное родство, восходящее вероятно только к первичным представителям отдела Polyneoptera, три указанные ветви которых, в том числе Dermapteroidea, начали дифференцироваться еще в Палеозое; за это говорит открытие в последнее время в Пермских отложениях Канзаса в Соед. Штатах особой группы насекомых— Protelytroptera, проливающей некоторый свет на эволюцию интересующего нас отряда, не говоря уже о том, что в Палеозое уже имелись настоящие Blattodea, представленные несколькими вымершими семействами.
1 Martynov, А. То the knowledge of fossil insects from Jurassic beds in Turkestan. Изв. Росс. Акад. Наук, 1925: 577.
30
КОЖИСТ О К Р Ы Л Ы Е — D Е RM АР Т Е R А
Поскольку отряд Protelytroptera, впервые описанный Tillyard’oM,1 из нижне-пермских отложений Канзаса, обнаруживает, по сравнению со всеми известными палеозойскими насекомыми, наибольшее сходство с Derma-ptera, необходимо несколько остановиться на особенностях, говорящих о филогенетической близости этих отрядов.
Представители Protelytroptera характеризуются некоторым внешним сходством с отрядом жуков, но имеют членистые церкви и поэтому не могут быть отнесены к Holometabola. Передние крылья длинные, полностью закрывающие брюшко, но значительно короче задних крыльев, сильно специализованы и превращены в надкрылья, выпуклые сверху, с зернистой или шероховатой поверхностью; жилкование у отдельных семейств ясное, у других слабо развитое, и тогда с немногими продольными жилками. Крылья кроме обычного веерообразного типа складывания вдоль многочисленных продольных жилок в анальной области, характеризуются наличием недалеко от середины переднего края у конца R3 так наз. узелка (nodus); здесь крыло может сгибаться, складываясь поперечно, так что вершинная часть подгибается вниз и внутрь, благодаря чему они полностью закрываются надкрыльями; кроме этого узелка, имеющегося и у настоящих Dermaptera, в месте разветвления анальных жилок (А3 и А2), сзади и недалеко от узелка имеется утолщение, располагающееся как раз на линии, по которой происходит поперечное складывание крыла, благодаря чему характер крыла также приобретает большое сходство с Dermaptera. Ноги довольно короткие, лапки пятичленистые. Брюшко оканчивается парой коротких церков, состоящих из немногих члеников.
Своеобразное строение заднего крыла, могущего складываться вдвое и подгибаться под низ, и характер специализации жилкования, напоминающего современных Dermaptera, при одновременном учете остальных особенностей (особенно членистых церков) позволило Tillyard’y сблизить этот отряд с Dermaptera', кроме того, строение церков и пятичленистые лапки весьма напоминают мезозойского представителя Dermaptera — Protodi-platys, не говоря уже о том, что личинки некоторых современных Pygidi-cranidae имеют также членистые церки (клещи). Указанный автор поэтому рассматривает отряд Protelytroptera как родоначальную группу, из которой развились настоящие Dermaptera. Судя по тому, что этот отряд оказался сильно дифференцированным на ряд семейств (в настоящее время известно 16 видов, относящихся к 10 родам и 5 семействам), можно думать что он возник еще в верхнем Карбоне.
Точка зрения Tillyard’а, однако, подверглась критике со стороны F. М. Carpenter’a,2 который усматривает в данном отряде производное от верхне-карбоновых тараканообразных насекомых, развивавшихся совер-
1 Tillyard, R. J. Kansas Permian Insects. Part 13. The new order Protelytroptera. Amer. Journ. Sei., XXI, 1931: 232—266.
2 C arp en ter, F. M. The lower Permian Insects of Kansas. Part. 6. Proc. Amer. Arts a. Sci., 68, n° 11, 1933: 461—487.
ВВЕДЕНИЕ
31
щенно самостоятельно в направлении превращения передних крыльев в элитры, подобно Dermaptera и Coleoptera', в качестве основного довода выдвигается отсутствие более значительного сходства между Protelytroptera и Dermaptera, чем оно фактически выражено, а также некоторое сходство жилкования надкрылий Protelytroptera с палеозойскими Blattodea. Эта точка зрения вряд ли более обоснована, чем таковая Tillyard’a, тем более, что характер заднего крыла у Protelytroptera имеет все же больше отличий от Blattodea, чем от Dermaptera. С другой стороны, если наличие сходства между Protelytroptera и Blattodea в отношении характера надкрылий в действительности имеет место (достаточных оснований у Саг-penter’a для этого не приведено, кроме сравнения надкрылья гипотетической родоначальной формы Protelytroptera с Blattodea), то и в этом случае это не может полностью отклонить точку зрения первого автора, так как сходство между данными тремя группами насекомых в палеозое может быть не случайным, а иметь глубокие филогенетические корни.
Точно так же мы не склонны разделять и мнения Zeuner’a,1 который сближает Protelytroptera с палеозойскими Protocoleopter а и тех и других выводит из Protoblaitoidea. Protocoleopter а представляют собой по Мартынову2 3 древнюю фазу эволюции разных групп современных жуков и сколько нибудь резкое разграничение Coleoptera от Protocoleoptera совершенно невозможно. Если стать на точку зрения Zeuner’a, то в таком случае мы должны будем сблизить Protelyroptera с представителями Coleoptera s. lato, что находится в резком противоречии с особенностями морфологии Protelytroptera, в частности с наличием у них членистых церков и совершенно отличного способа складывания крыльев.
Совершенно неприемлемым является и заключение Zeuner’a о том, что как Protelytroptera, так и Coleoptera, Dermaptera, Blattodea и Orthoptera, т. е. часть представителей отдела Neoptera, ведут свое начало через Protoblaitoidea от Palaeodictyoptera; после блестящих обобщений Мартынова по морфологии крыла,® рассматривающего Palaeodictyoptera как весьма специализированную группу, имеющую совместно с Odonata и Agnatha совершенно особый тип крыла (отдел Palaeoptera), заключение Zenner’a свидетельствует лишь о неуяснений им основных особенностей крыла и его эволюции.
Нельзя забывать также и того обстоятельства, что уже в юре водились настоящие Dermaptera, имеющие подобно Protelytroptera членистые церки, 5-члениковые лапки и намеки на жилкование на надкрыльях. Так недавно описанный мезозойский представитель отряда Dermaptera —
1 Z eu пег, Fr. Die Stammesgeschichte der Kafer. Ueber die Beziehungen der Form der Organe zu ihrer Funktion. I. Palaeont. Zeitschr., 15, 1933: 280—311.
2 Мартынов, А. О новых ископаемых насекомых Тихих Гор. Отдел Neoptera. Труды Геолог. Муз. Акад. Наук, VHI, 1930: 190—196.
3 Martynov, A. Ober zwei Grundtypen der Flugel bei den Insekten und ihre Evolution. Zeitschr. Morph. Oekol. d. Tiere, 4, Heft 3, 1925.
32
КОЖИСТОКРЫЛЫЕ —DERMAPT ERA
Protodiplatys fortis Mart, (юрские отложения Туркестана) и выделенный его автором в особое семейство Protodiplatyidae,1 характеризуется некоторыми признаками, общими с представителями отряда Protelytroptercr, сюда относятся короткие членистые церки, намеки на жилкование на надкрыльях и пятичленистые лапки задних ног (передние и средние лапки, повидимому, четырехчлениковые). Эти признаки таким образом более напоминают Protelytroptera и Blattodea, чем современных Dermaptera-, однако, наиболее характерная особенность последнего отряда — сильно укороченные надкрылья и скрытые под ними крылья — выражена и у Protodiplatys fortis, что не вызывает сомнений в принадлежности этой формы к отряду Dermaptera', что касается пятичлениковых лапок, то этот признак не может служить доводом к выделению особого отряда (ср. современных представителей отряда Coleoptera, отдельные семейства которого имеют от трех до пяти члеников в лапках), а членистые церки, как уже говорилось, свойственны личинкам некоторых современных Dermaptera. Правильнее все же рассматривать этот вид в качестве представителя особого вымершего подотряда, являющегося связующим звеном между современными Dermaptera и их палеозойскими предками, для которого мы предлагаем название Archidermaptera subord. nov.
Таким образом, имеется некоторое основание рассматривать отряд Protelytroptera в качестве если не настоящих предков, то, во всяком случае, близкой к ним группы насекомых, давших позднее настоящих Dermaptera-, исходной же группой для Protelytroptera несомненно являлись какие-то таракано-образные формы, близкие к верхнекарбоновым Proto-blaitoidea.
Характер распространения некоторых современных Protoder-maptera (см. ниже) дает основание предполагать наличие уже в мезозое, начиная по крайней мере с юры, настоящих Dermaptera s. str., именно относящихся даже к некоторым, существующим еще в настоящее время, родам. К сожалению, из мезозоя пока известен только один представитель, повидимому, настоящих Dermaptera, именно Semenoviola obliquotruncata Mart., описанный из юрских сланцев ю, Казахстана.2 3 * Эта форма была сближена с некоторыми современными Coleoptera-, однако С. Bolivar8 весьма убедительно показал, что эту форму необходимо отнести к уховерткам, что совпадает и с нашим мнением, основанном на тщательном изучении типа, любезно предоставленного в наше распоряжение А. В. Мартыновым; при этом весьма характерно, что по форме суженной кзади головы и очень толстым усикам этот вид напоминает представителей наиболее примитивного семейства
1 Martynov, Изв. Росс. Акад. Наук, 1925: 572—575, fig. 3.
2 Мартынов, А. В, Об одном интересном ископаемом жуке из юрских сланцев северного Туркестана. Русск. Энт. Обозр., XIX, 1925: 73, табл. I.
3 Bolivar, С. Sobre una nuova familia de Coleopteros. Eos, II, 1926: 191 (202—204,
fig. 1—3).
ВВЕДЕНИЕ
35
Pygidicranidae.1 Почти несомненно, что в лице Semenoviola мы имеем юрского представителя настоящих уховерток (подотр. Dermaptera s. str). Наличие уже в юре настоящих Dermaptera, имеющих вместо членистых церков одно-члениковые клещи, факт чрезвычайно важный и интересный, проливающий некоторый свет на время появления настоящих Dermaptera и косвенно указывающий на возможность появления в то время даже некоторых существующих еще до сего времени родов, современное распространение которых без этого допущения невозможно объяснить. Окончательное решение этого вопроса может быть дано только в результате соответствующих палеонтологических находок; однако, в высказанном предположении нет ничего невероятного, так как известны многие случаи существования многих родов современных растений еще в начале мезозоя (Araucaria, Ginkgo и др.).
Помимо указанных выше мезозойских представителей, в настоящее время известен ряд ископаемых форм и из третичных отложений (около -30-ти видов). При этом особенно существенно то, что уже из нижнего олигоцена, именно из Балтийского янтаря, был описан вид (Forficula praecursor Burr), относящийся не только к современному и к тому же высокоорганизованному роду, но являющийся одновременно близким к трем современным видам— F. senegalensis Serv. из Африки, F. beelzebub Burr из Сев. Индии (Burr2), и F. sagitta из Средней Азии и Закавказья. Далее, нижне-олигоценовая Forficula (?) pristina Burr, без сомнения, весьма близка к современному восточно-азиатскому подроду Odontopsalis Burr из весьма специализированного рода Anechura, который в системе стоит еще выше, чем род Forficula. Очень характерно, что подрод Odontopsalis является наиболее примитивным в роде Anechura, причем такие его виды, как Anechura (Odontopsalis) potanini и А. (О.) forficuli-jormis, являясь наименее специализованными видами в данном цодроде, имеющими даже черты переходные к роду Forficula, одновременно обнаруживают значительную близость к вышеуказанной F. pristina, что заставляет трактовать этот вид в качестве представителя данного подрода. Нельзя не отметить при этом, что F. pristina была отнесена к роду Forficula с вопросом при одновременном, однако, указании на наличие у этого вида некоторых особенностей, свойственных данному роду (как раз таких которые свойственны двум указанным выше современным видам); так как во время описания F. pristina 1911) наши знания о роде Anechura были чрезвычайно незначительны, причем указанные выше виды — А. (О.)
1 В пользу того, что данный вид является представителем Dermaptera s. str. говорят следующие особенности строения: форма головы, наличие на переднеспинке продольной срединной борозды (sulcus medianus), сближенные ноги с сильно утолщенными бедрами, присутствие на крыловых органах хотя и слабой, но заметной поперечной борозды, разграничивающей надкрылья от треугольных коротких крыловых пластинок, и наличие вершинных концов клещей (остальная их часть деформирована).
2 В u г г, М. Dermaptera preserwed in Amber, from Prussia. Trans. Linn. Soc. Lond., 2 ser., XI, 1911: 145—150, pl. I.
Фаун* СССР, Козвистокрылые 5
34
КОЖИСТОКРЫЛЫЕ—DERMAPTERA
pofanini и A. (О), forficuliformi's были описаны только в самые последние годы, естественно, что правильное определение положения в системе данной ископаемой формы было невозможно. Изложенные факты говорят о том, что весь отряд в целом представляет собой весьма консервативную группу насекомых.
Очень важным обстоятельством является нахождение многих ископаемых видов там, где современная фауна Dermaptera крайне обеднена. На такой незначительной территории как северо-восточная часть современной Пруссии было найдено 12 различных видов только в одном янтаре;, точно так же в миоцене Флориссана в Сев. Америке было обнаружено 11 видов, относящихся к уже вымершему роду Labiduromma, повидимому,. близкому к Forficula. Правда, указанными двумя примерами почти исчерпываются наши сведения об ископаемых уховертках, так как для всех других мест пока известно всего меньше десятка ископаемых видов; однако,, учитывая то, что в ископаемом состоянии смогла случайно сохраниться только ничтожная часть существовавших прежде видов нельзя не говорить. о действительном богатстве третичной фауны Dermaptera в умеренных широтах северного полушария.
Также весьма замечательно, что наиболее специализированный современный вид — обыкновенная уховертка (Forficula auricularia) была, недавно обнаружена в ископаемом состоянии в плиоцене Дурхэма в Англии (Lesne1).
Таким образом, приведенный фактический материал, подчеркивая. большую древность отряда, говорит одновременно о том, что даже самое высокоорганизованное семейство—Forficulidae — появилось не позднее начала третичного времени, что хорошо увязывается и с характером. распространения данного семейства (см. ниже). Но помимо того нахождение в нижнем олигоцене представителей современных высокоорганизованных родов и даже близких к современным формам видов говорит о медленном темпе , эволюции группы, т. е. об уже указанной нами ее консервативности. Такой вывод подтверждается также и анализом современных картин распространения некоторых более примитивных семейств, отряда; эти картины можно понять только при одном допущении, что некоторые современные роды Dermaptera возникли еще в мезозое, именно в юре. Для того, чтобы это было ясным, рассмотрим более подробно географическое распространение отдельных групп, и родов отряда.
Прежде всего необходимо отметить широкое, почти космополитное распространение семейств Pygidicranidae) Labiduridae, Labiidae, свойственных главным образом тропикам Старого и Нового Света, включая Австралию. Несколько аберрантное семейство Apachyidae наседает влажные тропики Африки, южной Азии, включая Малайский архипелаг, и Сев. Австралии с Новой Гвинеей, а сильно уклонившиеся сем. Arixeniidae
1 Lesne, Р. Quelques insectes du pliocene,superior du comte de Durham. Bull. Mus. Hist. Nat. Paris, 1920: 388.
ВВЕДЕНИЕ.
35
известно только с Малайского архипелага. Совершенно иную картину дает наиболее высокоорганизованное сем. Forficulidae — оно довольно значительно представлено в умеренных широтах Евразии и почти отсутствует в Австралии, если не считать наличия там только одного эндемичного вида, относящегося к неотропическому роду Dora Burr.
Очень характерной особенностью всего надсем. Protodermaptera является наличие многочисленных примеров широких разъединений ареалов как среди отдельных родов, так и подсемейств (фиг. 14). Так, род Diplatys (сем. Pygidicranidae) свойствен влажным тропикам Азии, Африки и Америки, род Echinosoma из того же семейства — также влажным тропикам южн. Азии, Африки и сев. Австралии, род Forcipula (сем. Labiduridae) распространен так же, как и Diplatys, род Anisolabis из того же семейства, имеющий даже один эндемичный вид в Средиземноморье, широко распро-транен по всему земному шару и пр. Перечисленные роды связаны в своем распространении с наиболее влажными тропическими областями. Однако, помимо этих примеров, имеющих многочисленных аналогов среди других животных и среди растений, необходимо указать на чрезвычайно своеобразное разъединение в ареале рода Pseudisolabis (подсем. Parisola-binae), несколько видов которого известны из Сев. Индии, включая Кашмир, один вид с остр. Явы и один вид из Новой Зеландии; при этом оказывается, что на Новой Зеландии водятся очень близкие роды Pariso-labis и Parisopsalis, тогда как в Индии и вообще в Азии у них отсутствуют ближайшие родичи. Все же подсемейство Parisolabinae, состоящее всего из четырех родов, свойственно Нов. Зеландии с Австралией, Сев. Индии, остр. Яве и Южн. Америке. Очень своеобразный характер распространения имеет подсем. Anataelinae, состоящее всего только из двух родов, из которых один свойственен Канарским островам, а другой сев.-вост. Китаю.
Указанные особенности распространения различных таксономических групп надсем. Protodermaptera (семейств, родов), говорят о весьма большой древности всего надсемейства, что хорошо увязывается и с морфологическими его особенностями, носящими более примитивные черты по сравнению с надсем. Eudermaptera. Сюда относится, во-первых, парный половой аппарат самца; кроме того сем. Pygidicranidae, несомненно являющееся наиболее древним среди современных Dermaptera, характеризуется присутствием хорошо развитых глазков, наличием у личинок многих родов членистых церков, а также и наличием у самок большинства родов гона-пофизов. Одновременно надсемейство характеризуется чрезвычайной бедностью многих групп родами, что особенно относится к сем. Pygidicrani-dae; из восьми подсемейств в последнем только два — Pygldicraninae (свойственное всей тропической зоне) и Pyrarginae (свойственное только Центр, и Южн. Америке) имеют больше двух родов — в первом случае их 9, а во втором 4; остальные подсемейства состоят только из одного — трех родов. Сказанное в значительной мере относится и к сем. Labiduridae, 3*
36
КОЖИС ТО К Р Ы ЛЫ E—D Е RM АР ТЕ RA
из шести подсемейств которого только одно — Psalinae характеризуется родовым и особенно подродовым многообразием, тогда как все остальные состоят из одного — четырех родов и только в подсем. Brachylabinae их насчитывается до десятка; также и семейство Apachyidae состоит всего из двух родов.
Чрезвычайно широкое распространение как семейств, так подсемейств и родов, при одновременном наличии весьма обширных разъединений в их распространении, охватывающих такие територии как Австралия—Азия, Сев. Австралия — Азия — Африка и т. д., а также морфологическая примитивность, бедность родами и, во многих случаях, видами и, наконец, наличие резких морфологических границ между различными таксономическими группами — все это говорит о том, что в лице надсем. Protodermaptera мы имеем остаток когда-то весьма богатой представителями и древней группы, возникшей в тот период истории земли, когда основные материковые массы не были так разобщены, как это имеет место, начиная с юры и мела, и кончая современным периодом.
Таким образом, географическое распространение Protodermaptera показывает, что это очень древняя группа, вероятно, ведущая свое происхождение с того периода жизни земли, когда материковые пространства были объединены или в виде одной общей суши, или же при помощи „материковых мостов". Если стать на точку зрения Вегенера,1 согласующуюся с огромным количеством биогеографических фактов, то в этом случае происхождение современных семейств Protodermaptera необходимо выводить по крайней мере с юрского времени, т. е. когда Австралия еще была соединена с Индией; это дает возможность понять такие разъединения ареалов, как в случае с родами Pseudisolabis, Echinosoma, Apachyus и др. Сюда же нужно отнести и такой род, как Antisolabis (сем. Labidu-ridae), виды которого известны из Южн. Африки и Австралии—здесь имеет место также юрское разъединение, так как Австралия была связана с Южн. Африкой только до юры.
В отношении таких родов как Diplatys, Forcipula и др., свойственных влажным тропикам Африки, южн. Азии, Австралии и Америки, то основываясь на соображениях Вегенера или Арльдта,2 необходимо сделать вывод, что разъединение ареалов произошло не позднее или немного позднее эоцена, когда окончательно утратилась связь между Южн. Америкой и Африкой, а последней через Мадагаскар с Индией.
Переходя к рассмотрению распространения в надсем. Eudermaptera, необходимо отметить, что менее специализированное семейство Labiidae хотя и является космополитным обладает только двумя вполне космополитными родами — Labia и Spongovostox’, остальные роды имеют более узкое распространение, причем из них только род Chaetospania распро-
1 Wegener, A. Die Entstehung der Kontinente und Ozeane. 4 Aufl., 1929.
2Arldt, Th. Die Entwicklung: der Kontinente und ihrer Lebenwelt. Leipzig, 1907; он же Handbuch der Palaeogeographie. 1.1919,
Фиг. 14. Ареалы распространения некоторых родов надсем. Protodermaptera. Distribution of'some’genera of the superfam. Protodermaptera. Diplatys ( ), Esphalmenus (-), Antisolabis (— „ — „ —), Pseudisolabis (—. — . —). Ориг.
38 КОЖИСТОКРЫЛЫЕ —DERM APTERA
странен от Африки через южную Азию до Австралии. Наиболее специализированное подсем. Chelisochinae распространено наиболее узко, достигая наибольшего разнообразия на островах Индо-Малайской области, с немногими видами в Африканской и Австралийской области.
Наличие на Малайском архипелаге многих общих или близких с юго-восточной частью Азиатского материка родов и видов, если считать вместе с Molengraff’oM,1 что здесь постоянный архипелаг существует с начала третичного времени, причем соединение с материком постоянно прерывалось глубоким морем, говорит о том, что эта группа была еще в палеогене широко распространена в юго-восточной оконечности Азии и образование системы островов способствовало здесь, через изоляцию, массовому образованию эндемичных видов и даже немногих родов (напр. Kinesis и Lamprophorus). Так как отделение островов от материка закончилось согласно представлениям геологов только в конце третичного времени, а может быть и в начале четвертичного, и так как в мезозое архипелаг, повидимому, покрывался в разных частях морем, необходимо притти к заключению, что эта группа, по крайней мере, на Малайском архипелаге представлена сравнительно молодыми видами и даже родами.
’Что касается другого подсемейства — Labiinae, то для него характерно космополитное распространение, как будто говорящее об его относительно большой древности, что подтверждается также их меньшей, по сравнению с no^e.Chelisochinae специализованностью подсем. Labiinae и наличием космополитных или вообще широко распространенных родов (см. вышеприведенные примеры с родами Labia, Spongovostox и Chaeto-spania).
Переходя к сем. Forficulidae, необходимо отметить, что наличие у последнего своеобразной специализации лапок и меньшего, не превышающего 15, количества члеников в усиках, при одновременной весьма слабой морфологической дифференциации подсемейств и даже отдельных родов, говорит о том, что оно должно рассматриваться как наиболее специализованное и филогенетически наиболее юное. Распространение семейства вполне подтверждает эту точку зрения. Особенно интересно отсутствие эндемичных представителей этого семейства в Австралии; исключение составляет только один вид — Dora spiculiferum Kirby, относящийся к американскому роду; остальные виды как Forficula auricularia L. и Eulanon bipartitus Kirby несомненно были сюда завезены. Точно так же во всем семействе нет ни одного примера вполне космополитного рода; более того, все известные случаи наиболее широко распространенных родов охватывают территории или бывшие только недавно геологически связанными между собой, или же по каким-либо другим причинам имевшие видовой обмен друг с другом. Мы не будем здесь касаться распространения отдельных подсемейств, так как такие из них, как Opisthocosmiinae
1 Molengraff. Modern deep sea research in the East Indian Archipelago. Geogr. . Journ., XLVII, 1921.
ВВЕДЕНИЕ
39
л Forficulinae нам представляются мало естественными, тем более, что некоторые американские роды, относимые к последнему, вероятно должны быть выделенными в особое подсемейство, тогда как Anechurinae должно быть слито с Forficulinae', однако, все же необходимо отметить, что ряд подсемейств (Eudohrninae, Ancistrogastrinae и Diaper asticinae) ограничены в своем распространении только одной зоогеографической областью (первое — Индо-Малайской, второе Неотропической, третье Африканской). В отношении же распространения отдельных родов необходимо указать на несколько примеров разъединений ареалов, проливающих свет на происхождение и возраст этих родов.
Так, род Dora, известный из Америки и наиболее богато представленный в южном материке имеет, как уже упоминалось, один вид в южн. .Австралии (фиг. 15); такое разъединение говорит о том, что уже в начале третичного времени (эоцен), когда южная часть Южн. Америки была соединена через Антарктику с Австралией, этот род был широко распространен в южном полушарии от Америки через Антарктику до Австралии. Высказанное соображение подтверждается нахождением в Антарктике ископаемых представителей современных растений, распространенных теперь в Южн. Америке и Австралии, не говоря о других фактах аналогичных разъединений.1 Отсутствие данного рода в Южн. Африке, которая, по воззрениям геологов, потеряла связь с Антарктическим материком в юрском периоде, говорит о том, что этот род возник между мелом и началом палеогена.
Другие примеры разъединений показывают на существование связей в более поздние времена. Так, широко распространенный в Евразии род Anechura, центр многообразия которого находится в Китае, представлен также одним видом в Мексике;2 такое разъединение указывает на то, что .виды рода Anechura существовали в более северных частях Сев. Америки, .но благодаря каким-то причинам сохранился только один вид в Мексике (фиг. 16).
Весьма интересную картину распространения обнаруживают роды Mesochelidura, Chelidurella и Apterygida, 'вмды которых известны с одной стороны из Европы, а с другой — из Южн. Африки, в частности (для первого рода) с Пиренейского полуострова и Южн. Африки (фиг. 16).
Довольно сходную с этими родами картину распространения имеет и род Forficula (фиг. 15), который, помимо нескольких десятков видов в Евразии и Сев. Африке, представлен также немногими видами в тропической Африке. Учитывая генетическую близость всех указанных родов необходимо притти к выводу, что все они являются выходцами из север
1 Ср. Вульф, Е. В. Введение в историческую географию растений. Труды Прикл. .Ботан., Генет. и Селекции, прил. 52, Ленинград, 1932: 275.
2 Мексиканский вид (Л. vara Scudd.), изученный нами по 2 55, и 2 ^2’ представляет -собою самостоятельный подрод, близкий к подроду Anechura in sp.
40
КОЖИСТОКРЫЛЫЕ—DERMAPTERA
ного полушария (см. ниже) и смогли проникнуть в Африканскую область,, только после исчезновения моря Тетис, т. е. не раньше миоцена.
Разобранные нами примеры говорят, что и сем. Forficulidae является относительно древним, возникновение которого относится к концу мезозоя (мел) или к началу палеогена, что подтверждается также и тем, что уже для средины палеогена (нижний олигоцен) доказано существование некоторых современных родов этого семейства. За отсутствие более глубоких корней у данного семейства достаточно убедительно говорит его современное распространение, показывающее отсутствие необходимых для этого картин распространения отдельных родов и подсемейств.
Таким образом, резюмируя все сказанное выше о распространении отряда и о палеонтологических находках его представителей, необходимо притти к выводу, что весь отряд в целом является очень древним, причем корни его восходят еще к палеозою (верхний карбон, нижняя пермь), где в лице вымершего отряда Protelytroptera можно усматривать исходную или близкую к ней группу насекомых. В мезозое уже появляются Derma~ ptera, относящиеся к особому ныне вымершему и примитивному подотряду Archidermaptera, отличавшемуся от современных представителей (подотр.. Dermaptera s. str.) пятичлениковыми лапками и членистыми церками. В это же время, видимо не позднее юры, появляются первые представители уже подотряда Dermaptera s. str., относящиеся к надсем. Protoder-maptera и вероятно к подсем. Labiinae из надсем. Eudermaptera, причем тогда же возникли и некоторые существующие по настоящее время роды.. Последовавшее вслед за этим раздробление Гондванского материка, выразившееся в отделении Австралии от Индии, и Африки от Австралии способствовало разъединению ареалов существовавших тогда родов; но так как влажный тропический климат и лесная растительность сохранились в экваториальной части отделившихся материков до настоящего времени, некоторые из возникших здесь еще в мезозое родов сохранились до современного периода. Вероятно в мезозое же происходит и окончательное-вымирание подотр. Archidermaptera, вытесненного более совершенными представителями подотр. Dermaptera s. str.
В начале третичного времени (эоцен) или быть может к концу мезозоя появляется наиболее высокоорганизованное семейство Forficulidae, некоторые современные роды которого возникли не позднее эоцена (Dora) или раннего олигоцена (Forficula). Это семейство в течение третичного времени достигло особенного расцвета в умеренных широтах северного-полушария, будучи представлено большим количеством видов, чему способствовал влажный, теплый и ровный климат и наличие обширных пространств лесной растительности.
К ближайшему рассмотрению фауны Dermaptera умеренных широт мы и перейдем в следующей главе.
В заключение нашего обзора эволюции отряда естественно задать вопрос—чем можно объяснить столь значительную консервативность этой.
фиг. 15. Ареалы распространения некоторых родов из сем. Forflctilidae. ТЬе area of distribution of some g-enera^of the fam. Forficulidae, 1. Dora (--------------------------); 2. Forficala (—„—). Ориг,
42
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
группы насекомых, отдельные роды которых сохранились до настоящего времени еще с юры и, во всяком случае, с начала третичного периода? Ответ на это надо искать в экологии уховерток. В соответствующей главе мы подчеркнули, что наиболее характерной экологической особенностью отряда в целом является с одной стороны его тесная зависимость от условий увлажнения среды, с другой—исключительная приуроченность к укромным местам и скрытый образ жизни. Наиболее обычной комбинацией этих условий является поверхностный слой почвы, где под камнями, под гниющими остатками деревьев, а иногда и непосредственно в вырытых в земле норках данные насекомые находят для себя оптимум условий. Как раз такие условия представляют собой весьма монотонную в экологическом смысле среду, одновременно укрытую от непосредственного воздействия ряда внешних экологических факторов. Это наложило на организацию отряда сильнейший отпечаток, выразившийся в образовании специальных приспособлений для такого скрытого образа жизни, что в конечном итоге способствовало односторонней специализации и явилось тормозом в эволюции отряда. Быть может этими же обстоятельствами необходимо объяснить и относительно чрезвычайную бедность отряда видами.
6. СОСТАВ И ПРОИСХОЖДЕНИЕ ФАУНЫ DERMAPTERA ПАЛЕАРКТИЧЕСКОЙ ОБЛАСТИ
Фауна Dermaptera Палеарктической области значительно беднее сопредельных с ней областей — Восточной и Африканской, но во много раз богаче Неоарктической. Количественное распределение' видов и их абсолютное число на территории Палеарктической области наглядно выявляется из нижеследующей таблицы на стр. 44—45 (цифры указывают число видов).
При рассмотрении этой таблицы бросается в глаза наличие двух центров многообразия—Средиземноморского на западе, почти совпадающего с границами Средиземноморской подобласти многих авторов, и восточноазиатского, совпадающего в значительной степени с Палеархе-арктической1 подобластью Палеарктики (фиг. 17). При этом оказывается, что в обоих случаях максимумы видового разнообразия отряда совпадают с максимумами таких характерных родов как. Anechura и Forficula. Однако, Средиземье и Зап. Европа населены видами по преимуществу эндемичными (28 эндемиков из 45 видов) с узким ареалом распространения и с наличием всего только 9—10-ти широкораспространенных по Средиземью и Зап. Европе форм, тогда как восточно-азиатский центр помимо сравнительно большого количества эндемичных форм характеризуется наличием и чисто-тропических родов, которые находят здесь северную границу распространения (напр. Timomenus, Eparchus, Narberia, Sondax,
1 Семене в-Т я н-Ш а н с к и й, А. П. Границы и подразделения Палеарктической •области для наземных животных, на основании изучения географического распределения жесткокрылых (Coleoptera). Труды Зоол. Инет. Акад. Наук, 1935, с картой.
г S 4 § 1 ° 8 Л 9 1
S О i * c 0 0 ₽0 1 I о о 0 i о 1 1 1 о - ——— со о
g V 1 I о ( " ' S > о
1 о ir Y 4/ ' \\\ 1 k /-! ? 1 1 1 ] о
§ XV .» V^X"- SY’xh V>4y 5^ \ 1 S ! 1 14 |
2" ' rv" 4 u fefc= 7 1 1 1 1 4J О
00 o°4n °СГ7 1 ' J/>vT 1 1 1 1 00 - о i
10 ~)f 2j) 7x 1 \ 0 • **— i i « I , - , О 1
i л / |VJ И ( i । ‘ Г«Л> - _.. - о
О 0 g^gVi/3 lY 1 о 4 1 । /у / > i 4 1 1 1 см
О **? - * 1 0 ib Jk 1 1 1 1 1 о
Q Xj। r i 1 1 1 1 . 1 С4 О
о 1 k? 1 . ( i « i i , о
*0 С ( i ‘-'vj ГК e <o ? I s |. 4 О
° ? <Г ^3 Й гЙ\. 5 ll 1 . । i । । - СО о л
° ^r! ’’Jkf &Г । । < । । о
- V) C5> о fife I i i । । г 1 о
g n. X_j \ i I i । I । о , ф
| 1 । ।
Mi 70 60 40 i 20 0 20 40
Фиг. 16, Ареалы распространения некоторых родов из сем. Forficulidae» The area of distribution of „some genera of the fam. Forficulidae. 1. Anechura (——), 2. Apterygida (—.—3. Timo menus („—Ориг.
44
Арктическая зона Зап. Европа Средиземье Восточная Европа
Всего Атлантические о-ва Крым Кавказ Всего В том числе! СССР
I. Pygidicranidae
1. Anataelia Bol — — 1 1 — — — —
2. Challia Burr. — — — —• — — — —
II. Labiduridae
3. Anisolabis Fieb 2 5 3 3 2
4. Labidura Leach __ 1 1 1 1 1 1 1
5. Nala Zacher — — 1 .— — — — —
6. Pseadisolabis Burr. — — — — — — — —
III. Labiidae
7. Labia Leach 1 2 2 1 1 1 1
8. Prolabia Burr — 1 1 — — —. — —
9. Nesogaster Verb — — — — — — —
10. Chelisoches Scudd. . — — — — — — ——
11. Proreus Burr — — — — • — — — —
12. Hamaxas Burr — — — — — — — —
IV. Forficulidae 13. Isolabella Verb 1 -
14. Forficula L 1 3 14 1 4 5 2 2
15. Guanchia Burr — — 5 4 — — —
16. Mesasiobia Sem — — — — —
17. Oreasiobia Sem — — — — — — —
18. Apterygida Westw — 1 1 — — — 1 1
19. Chelidurella Verb — 3 1 — — 2 2
20. Anechura Scudd — 1 6 ?1 2 2 2
21. Pseudochelidura Verb — 1 1 —. — — —
22. Lithinus Burr — 1 — — —
23. Perirrhytus Burr — — 2 2 — — — —
24. Chelidura Latr — 2 1 — — — — —
25. Mesochelidura Verh —. 1 — — —— —
26. Eumegalura B.-Bienko — — — — — — —
27. Allodahlia Verh — —_ .— —— —
28. Timomenus Burr — — — — — —
29. Eparchus Burr. — — — — — — — —
30. Narberia Burr — —— — — .— —-• —
31. Sondax Burr. — — — — — •— — —
Всего видов . . 1 16 45 15 9 11 9 9
В том числе эндемичных . . . — 3 28 9 — 2 —
Allodahlia, Nesogaster и др.). При этом и эндемичные виды оказываются иногда представителями тропических родов (напр. Sondax potanini, Ерат-chus yezoensis, Anisoldbis sp. sp. и др.).
Мы уже указывали на то, что как средиземноморский, так и восточный центр характеризуются преобладанием в своем составе видов рода Anechura
45
1 Иран 1 Туранская низменность Монголия и сопред. страны Сибирь с Сахалином Горы Средней Азии Кашмир ‘ Палеархеарктическая подобласть Вся Палеарктика СССР
Всего Тибет Япония Уссурийский край Кореи, Манчжурия и с.-в. Китай Зап. Китай
1
— — — — — — 1 — — — 1 — 1 —
2 1 - 1 7 5 2 5 2 12 4
1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 — — — — — 1 —— 1 —— 1 —— 1 —-
—- — — — — 1 — — — — — — 1 —
1 1 1 1 1 2 1
— — — 1 — — 1 — 1 1 — — 1 1
— — — — — 2 — 2 —— — 2 —
— — — — — — 1 — 1 — — 1 —
— — ' — — — 1 — 1 —- — — 1 —
— — 1 — — — — — — — — — 1 —
1
3 2 1 3 2 3 14 3 2 2 5 8 31 8
5 —
—— — — — 1 — —• — — — — • 1 1
— — — — 1 1 — — — — — 2 1
1 1
—• — - — - ——. — п . — — —. 3 2
1 1 2 2 1 1 12 7 2 2 2 б 18 5
—— — — — — 1 — —— . — 1 —
1 —
— — • 2
2 —
—- — — — • — — — — — — — 1 —
— — — — 1 — 1 1
— — — — — — 2 1 — —. 1 1 2 —
— — — — — — 4 — — 1 4 — 4 1
— — — — — 3 — 1 — 2 — 3 —
— — — — <— — 1 — — — 1 — 1 —
— — — — •— — 1 — — — — 1 1 —
8 5 5 9 б 8 54 12 18 9 24 20 106 26
— — 1 — 2 3 30 4 4 5 11 84 5
и Forficula. Однако, это сходство простирается и дальше; дело в том, что ряд средиземноморских и европейских видов и подродов генетически весьма близок к китайским, и вообще восточно-азиатским, видам и подродам. Так, например, северо-итальянская Anechura orsinii чрезвычайно близка к китайско-тибетской A. sven-hedini; европейская Forficula auri-
46
КОЖИСТОКРЫЛЫЕ—DERMAPTERA
cularia, несомненно являющаяся древним представителем европейской фауны, так как она была найдена в ископаемом состоянии в плиоцене Дурхэма в Англии, имеет в восточной Азии целую группу близких, но несколько более примитивных видов (F. subauricularia, F. vicaria,. F. burriana). Средиземноморская F. smyrnensis сходна с китайско-тибетской F. davidi, а восточно-европейская F. tomis имеет чрезвычайно близкий вид на востоке — F. robusta, к которому в свою очередь примыкает F. macrobasis; точно также балкано-кавказский подрод Borelliola рода. Anechura представлен в Китае и Тибете весьма близким подродом — Burriola.
Таким образом, мы видим, что генетическое родство Средиземноморской и вообще европейской фауны Dermaptera с восточно-азиатской фауной чрезвычайно велико.
Такое явление не представляет собой чего-либо нового, так как известны многочисленные факты подобного амфиконтинентального распространения для других групп животных и для растений; большое количество таких фактов собрано Бергом.1 Приведем некоторые из них. Голубая сорока (Cyanopolius cyanus), свойственная восточной Азии, представлена на Пиренейском полуострове весьма близким видом — С. cooki; дубы (Quercus-sp. sp.), встречаясь в Европе и не переходя на востоке Уральский хребет, появляются на востоке Азии вновь в виде близких видов, начиная с восточного Забайкалья; очень характерно при этом, что в ископаемом состоянии дубы были найдены и в Сибири (напр., плиоцен Бухтармы на Алтае). Есть случаи наличия совершенно идентичных видов — напр. саранчевое — СаШр~ tamus ictericus, свойственное Пиренейскому полуострову и Сицилии и отсутствующее во всей остальной части Европы, в Зап. Сибири и Ср. Азии, появляется в виде той же самой формы на Алтае и распространено по всей южной части Сибири до Тихого океана и Сев. Китая.
Однако, эти факты все же представляют собой в других группах, животных и растений, кроме уховерток, единичные случаи, многочисленные только в их сумме, так как в образовании современной фауны и флоры восточной Азии и Средиземья сыграли большую роль совершенно различные элементы (см. ниже); поэтому обе эти территории и относятся к различным зоо- (и фито-)географическим подобластям — Средиземноморской на западе в Палеархеарктической на востоке. Совершенно иная картина имеет место в отношении Dermaptera — восточный и западные центры современного многообразия этих насекомых имеют свыше 60%. видов, относящихся к общим родам, не говоря уже о том, что число генетически близких видов также довольно велико; кроме того и некоторые роды и породы из обоих центров весьма близки друг другу.
Все эти факты объясняются двумя основными причинами—тесной экологической зависимостью Dermaptera от влажности климата и лесной
1 Б е р г, Л. С. Рыбы бассейна Амура. Зап. Акад. Наук, VIII сер., XXIV, № 9,. 1909: 253.
ВВЕДЕНИЕ
47
растительности с одной стороны и наличием в третичное время на территории современных ^центральных и умеренных частей Евразии благоприятных экологических условий для существования этих насекомых — с другой.
Палеоботанические данные (Engler,1 Кузнецов,2 3 4 Криштофович® и др.) говорят о существовании в третичное время на территории современных умеренных широт Евразии в начале более теплого, а затем умеренного влажного климата с богатой, в северных широтах по преимуществу широколиственной, а в южных тропической и субтропической лесной растительностью, в состав которой входили такие растения, которые, отсутствуя ныне на этих же местах, сохранились или в Средиземье, или особенно в Восточной Азии. В качестве примера можно указать на представителей рода Quercus (см. выше), рода Liquidambor виды которого (напр. L. euro-раеа) известны из олигоцена Западного Казахстана, Европы и пр., а сейчас сохранились только в количестве четырех видов (один в Малой Азии, один в Восточной Азии и два в сев. и центр. Америке); различные виды чинара (Platanus), сохранившегося теперь в Средиземье, горах Средней Азии, Зап. Гималаев и Америке и пр. Для характеристики климатических условий в третичное время приведем данные для юго-западного Казахстана и Зап. Европы, полученные Коровиным и Неег’ом на основании сравнения вымершей растительности с ныне живущими родичами.
Юго-западный Казахстан (по Коровину 4)
Осадки за год 'Средняя годовая температура Температура наиболее холодного месяца Общая характеристика
Олигоцен-миоцен . . 1000 12° 4° Умеренно-теплый влажный
Соврем, эпоха 100 7.2 -11.2 Умеренно-холодный сухой
Что касается Зап. Европы, то по Неег’у (1868)5 средние годовые температуры в различные эпохи третичного периода были таковы.
1 Е n g 1 е г, A. Versuch einer Entwicklungsgeschichte der Pflanzenwelt seit der Tertiar-periode. I, 1879.
2 Кувнецов, H. И. Принципы деления Кавказа на ботанико-географические провинции. Зап. Акад. Наук, VIII сер., XXIV, № 1, 1909.
3 Криштофович, А. Н, Основные черты развития третичной флоры Азии. Ивв. Гл. Бот. Сада, XXIX, № 3-4, 1930.
4 Кор овин, Е. К вопросу о палеоэкологнческнх сменах в Средней Авии. Вопр. Эколог, и Биоценолог., 1934 : 40.
5 Цитир. по Кузнецову, Н. И. Loc. cit.: 67.
48
КОТКИС ТОКРЫЛЫЕ—DERMAPTERA
Олигоцен Верхний миоцен
Северная Италия .... 22° 20
Швейцария 20.5 18.5
Данциг 16 —
Силезия — 15
Таким образом, средние температуры здесь были еще выше; что касается условий увлажнения, то судя по характеру растительности, здесь .должны были выпадать обильные осадки.1
Из рассмотрения этих данных становится совершенно очевидным, что условия середины третичного периода в районе нынешнего засушливого юго-западного Казахстана, где современная фауна уховерток ограничивается всего 2—3 видами, были весьма благоприятны для данных насекомых и мы вправе ожидать наличия здесь богатой фауны последних. За это говорит с одной стороны нахождение 12-ти различных ископаемых видов в олигощене Вост. Пруссии и 1б-ти рецептных видов в Зап. Европе (без Средиземья), а с другой — наличие в современный период богатой фауны Derma-ptera в тех местах Европы, которые характеризуются аналогичными условиями, т.е. влажным умеренным климатом и наличием лесной растительности.
Указанные выше факты, а также находки ископаемых остатков широко? лиственного третичного леса во многих других ныне засушливых частях Евразии говорят о том, что в третичное время умеренная северная часть Евразии, ныне почти лишенная фауны уховерток, была как в отношении климата, так и характера растительности, весьма благоприятной для этих влаголюбивых насекомых и многие виды их имели широкое распространение по всей Евразии.
Постепенное похолодание климата к концу третичного времени и создание большей сухости в центральных частях Евразии способствовало отступанию многих видов в двух направлениях — к Средиземью на западе и в сторону восточной Азии на востоке (фиг. 17). Объясняется это тем, что климатические условия на указанных территориях все время оставались более или менее благоприятными для существования уховерток. Так, Зап. Китай и Япония располагаются в области, где годовое количество осадков выражается цифрою в 600—2000 мм. Точно так же наш Уссурийский край, Корея и Манчжурия, хотя и имеют 450—600 мм осадков,
1 Мы исключили из рассмотрения данные Коровина для миоцен-плиоцена, так как согласно Ярмоленко они основаны на верхне-меловой флоре (Бот. Журн. СССР, 20, 1935: 94); что касается критики методов Коровина, Неег’а и др., то необходимо отметить, что их данные ценны как попытки уяснить климатические особенности прошлого даже несмотря на неизбежную при этом схематизованность; метод же рекомендуемый Ярмоленко (сравнение данных фациального анализа и растительных остатков) требует еще своих исследователей.
Фауна СССР, Кожнстокрылые
4
50
КОЖИСТОКРЫЛЫЕ — DERMAPTERA
но вследствие того, что они носят муссонный характер и 75% их приходится на лето, последнее характеризуется чрезвычайной сыростью, что способствует развитию здесь богатейшей растительности. Что касается Средиземноморского центра, то и здесь осадки выпадают периодически; почти отсутствуя летом, они настолько обильны осенью, зимой или весной,, что это дает возможность существования своеобразной вечно-зеленой лесной растительности. Среднее количество осадков для всего Средиземноморья (т. е. включая азиатские и африканские части) Th. Fischer1 исчисляет в 760 мм; но кроме того имеется ряд областей, где количество' осадков повышается до 1000—2000 мм и более (Понтическая провинция: Кавказа, Далмация и вообще Адриатическое побережье Балканского полуострова, центральная Италия и пр.) и где поэтому сохранилась богатая лесная растительность третичного типа.
Немудрено поэтому, что похолодание климата и особенно уменьшение количества осадков в центральной части Евразии, т. е. на территории нынешней вост. Европы, Казахстана и Сибири, способствовало постепенному стягиванию фауны уховерток в указанных двух направлениях; в результате этого широко распространенные роды и виды постепенно исчезли в центральных частях их прежнего евразийского ареала, что способствовало дифференциации видов на две самотоятельные формы—г Средиземноморскую и Восточно-азиатскую, давшие в конце концов самостоятельные виды.
Таким образом, современные два центра многообразия уховерток в Евразии являются производными одной общей третичной фауны, начавшей дифференцироваться на восточную и западную части во вторую-половину третичного времени (повидимому начиная с миоцена) в связи: с похолоданием климата и уменьшением количества осадков в центральных частях их ареалов. Только таким образом и можно объяснить столь большое сходство средиземноморской и восточно-азиатской фауны.
Можно было бы, впрочем, приписать основную роль в такой дифференциации видов ледниковому периоду, что несомненно и имело место,, как это показал Берг2 в отношении многих других животных и растений.. Однако, в данном случае мы совместно с Семеновым-Тян-Шанским (1935) не можем придавать этому фактору решающего значения для уховерток,, так как тогда необходимо было бы допустить, что дифференциация на самостоятельные виды й даже подроды (Burriola и Borelliola) произошла только за послеледниковое время, с чем, конечно, никак нельзя согласиться, учитывая значительную консервативность группы. С другой стороны несмотря на наличие в восточном и западном центрах близких видов, среди Dermaptera в отличие от многих других групп животных,, нет ни одного вида, который являлся бы амфипалеарктом. Кроме того, необходимо отметить, что виды, идущие более глубоко на север, как
1 Fischer Th. Das Klima der Mittelmeerlander. Peterm. Mitt., 58, 1879.
2 Б e p г, op. cit. (1909).
ВВЕДЕНИЕ
51
в восточном, так и западном центре, являются и наиболее генетически близкими друг другу; сюда относится Forficula tomis на западе и F. robusta на востоке. Все это говорит о том, что дифференциация началась значительно раньше ледникового периода, захватив первоначально наиболее теплолюбивые формы, которые в последующем своем движении по направлению к двум современным центрам многообразия ушли наиболее далеко от области своего сплошного распространения; наименее же теплолюбивые формы, к которым, повидимому, необходимо отнести родоначальную форму для F. robusta и F. tomis, подверглись дифференциации только во вторую половину неогена, вероятно в конце его, в результате чего они, дифференцировавшись на два вида, сохранили до сих пор черты наибольшей генетической близости. Данным примером исчерпываются все факты наибольшей генетической близости между восточными и западными формами, так как остальные виды, несмотря на их явную близость, успели дифференцироваться значительно резче, что только и можно связывать с более ранним началом их дивергенции. Поэтому есть все основания утверждать, что уже начавшееся в миоцене ухудшение климатических условий должно было в значительной степени коснуться многих наиболее теплолюбивых и влаголюбивых форм.
Дальнейшее похолодание климата, закончившееся наступлением ледникового периода, способствовало еще большему стягиванию фауны Der-maptera в указанные два центра, в результате чего в северной части Евразии эта группа совершенно исчезла, тогда как Средиземье на западе и восточная Азия на востоке превратились в два центра — реликтовой третичной фауны, подвергнувшейся в зависимости от местных условий, большей или меньшей трансформации, но сохранившей, несмотря на громадную территорию, отделяющую их, признаки весьма значительного и глубокого родства.
Выяснив причины весьма большого генетического сходства в фауне уховерток Средиземноморского и Восточно-азиатского центра, коснемся вопроса о происхождении фауны уховерток Палеарктической области в целом и попытаемся выяснить первичный центр ее происхождения, а также влияние соседних с Палеарктикой фаун — Африканской и Индийской.
Прежде всего необходимо подчеркнуть, что наиболее характерные виды в фауне уховерток Палеарктической области, именно представители родов Forficula и Anechura, несомненно являются автохтонными элементами данной фауны. Это доказывается^тем, что род Anechura совсем отсутствует’ в Африканской и Индийской области. Что касается рода Forficula, то примерно 80% его видов свойственны только Палеарктической области; примерно 7 видов известны из Африканской области и 2—3 вида заходят в низменные части Индии, с одним эндемичным видом на о-ве Цейлоне; такое распространение говорит в пользу того, что колыбелью этого рода является также Палеарктическая область, тем более, что достоверные виды данного рода известны еще из олигоцена Вост. Пруссии. С другой
4*
52
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
стороны в эоцене и, по крайней мере, на протяжении большей части олигоцена Европа была отделена от Африки древним, сохранившимся еще с мезозоя, Средиземным морем — Тетис, простиравшимся от западных областей Европы до Индостана (Неймайр1); только в миоцене или в верхнем олигоцене это море отделилось от бассейна Индийского океана, превратившись во внутренний водный бассейн, сохранившийся до настоящего времени в виде современного Средиземного моря. Наличие водной поверхности, отделявшей Европу и Азию от Африки до конца олигоцена, несомненно было большим препятствием для обмена фауны между указанными двумя сушами; а раз представители рода Forficula существовали в Европе уже в янтаре (нижний олигоцен или даже верхний эоцен), ясно, что они являлись аборигенами северного полушария. Поэтому нельзя допускать, что род Forficula зародился в Африканской области; наоборот, несомненно, что виды этого рода в Африканской области являются потомками тех форм, которые смогли проникнуть туда из Европы только начиная с миоцена (или верхнего олигоцена). Как раз, начиная, главным образом, с миоцена и имел место весьма значительный обмен фауны и флоры между Европой и Африкой, чему способствовало образование в восточной части Древнего Средиземья (Попов, 1927) суши, соединившей эти два материка.2
Весьма важным обстоятельством, позволяющим уяснить более детально центр происхождения указанных родов, является факт существования в восточной Азии таких видов из родов Forficula, Anechura и Oreasiobia, которые являются как бы промежуточными между этими родами. В качестве примера можно указать на гималайскую О. stoliczkae и F. schla-gintweiti, которые, несмотря на то что .они относятся к двум самостоятельным и хорошо отличимым рбдам, обнаруживают явную генетическую близость друг к другу. Аналогичным примером является тибетская Anechura forficuliformis, имеющая, как показывает само название, сходство с видами рода Forficula и, кроме того, с родом Apterygida.
Существование на востоке Азии промежуточных форм между столь хорошо отличающимися в остальных частях своих ареалов родами при одновременном наличии здесь большего количества представителей этих родов, чем в Средиземноморье и Зап. Европе говорит о том, что центром их происхождения является вост. Азия. Действительно, общее количество видов рода Forficula в Палеарктической части восточной Азии и Гималаях достигает цифры свыше 20, а во всем Средиземноморье с Европой —14; что касается рода Anechura, то последний представлен в Европе и Средиземноморье всего лишь б-ю, а в восточной Азии 12-ю видами. Раз это так, то совершенно очевидно, что первоначальной колыбелью этих двух родов является древнейшая азиатская суша, расположенная в нынешней
1Неймаяр,М. История земли. II. СПб. 1903: 422.
2 П о п о в , М. Г. Основные черты истории развития флоры Средней Азии. Бголл. Ср. Аз. Гос. Унив., XV, 1927: 239—289. Мензбир, М. А. Очерк истории фауны Европейской части СССР. М.-Л., 1934: 82,86.
ВВЕДЕНИЕ
53
восточной Азии и названная S,uess’oM Ангарским материком, где в настоящее время усматривается центр зарождения фауны и флоры умеренных широт северного полушария (Дильс;1 Сушкин;2 Uvarov;8 Попов, op. cit. и др.), причем некоторые .фитогеографы (Попов) обозначают всю территорию занятую ангарской флорой в виде особой ботанической области (Область флоры Гинкго), противопоставляя ей область Древнего Средиземья.
Разобрав происхождение основного ядра фауны Dermaptera Палеарктической области, коснемся тех элементов ее, которые нами еще не рассматривались.4 Прежде всего неоходимо отметить, что как в восточном (включая и сопредельные территории), так и в западном центре имеется ряд родов и видов, генетически группирующихся вокруг разобранных нами родов Anechura и Forficula. Сюда относятся:
Восточныйцентр Западный центр
1. Род Mesasiobia .1 вид 1. Род Guanchia . . 5 видов
2. „ Oreasiobia . ... . 2 вида! и подвид 2. „ Pseudochelidura 1 „
3. „ Eumegalura . . 1 вид 3. „ Chelidara 2 „
4. „ Anechura 4. „ Mesochelidura 1 „
а) подрод Odontopsalis 4 вида 5. „ Perirrhytus . . 2 „
б) „ Burriola . . . 4 „ 6. „ Anechura
а) подрод Borelliola 3 „
7. „ Apterygida . . 1 „
8. „ Chelidurella . . 3 „
Роды Guanchia и Apterygida чрезвычайно близки к Forficula, а роды Eumegalura, Pseudochelidura, Chelidura, Mesochelidura, Chelidurella и Pe-rirrhy.us являются явными родственниками рода Anechura-, что касается Mesasiobia и Oreasiobia, то они имеют черты сходства как с Anechura, так и Forficula, являясь как бы морфологически специализированными производными промежуточного между указанными родами типа, что относится особенно к Oreasiobia. Большинство этих родов свойственно только Палеарктике, не встречаясь в соседних областях; лишь только роды Mesochelidura, Chelidurella и Apterygida представлены немногими видами в Африке, преимущественно в ее южной части.
Помимо перечисленных выше десяти родов, образующих совместно с Anechura и Forficula ядро ангарской фауны, остается еще ряд видов, которые являются несомненными выходцами из смежных тропических
1 Д и ль с , Л. География растений, перев. с нем., 1908.
2 С у ш к и н , П. П. Зоологические области средней Сибири и ближайших частей нагорной Азии, и опыт истории современной фауны Палеарктической Азии. Бюлл. Моск. О-ва Исп. Прир., 1929: 53—55.
3 U v аг о v, В. Р. Composition and origin of the Palaearctic fauna of Orthoptera. X-e Cong. Intern. Zool., 1930: 1519.
4 Род Isolabella с одним видом нз Греции нами исключен из последующего рассмотрения в связи с тем, что он весьма плохо изучен и систематическое положение его совершенно неясно.
54
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
фаун. Часть из этих видов находят в пределах Палеарктики только северную границу распространения; сюда относятся, за одним исключением, виды свойственные только восточному центру, а именно:
Восточный центр Западный центр
Anisolabis............. 3 вида Anisolabis........... 1 вид
Proreus.................1 „
Allodahlia ....... 2 „
Eparchus . ............ 2 „
Narberia .......... 1 „
Timomenus ............. 1 „
Другая часть, хотя и относится к тропическим родам, но представлена в Палеарктике эндемичными видами и даже родами; громадное большинство последних свойственно также восточному центру.
Восточный центр' Западный центр
Challia ....... Anataelia . ... . . . . . 1 вид
Anisolabis . . 2 „ Anisolabis .... ... 2 „
Pseudisolabis . . . . - • 1 ,, ? Lithinus . . . . . . 11
Hamaxas . . 1 ,,
Nesogaster . . . . • • 2 „
Timomenus . ... • • • 1
Son dax • • 1 „
Eparchus • • 1
Всего . . . . 4 вида.
Кроме этих двух групп имеется и третья группа тропических видов — именно видов завезенных человеком и акклиматизировавшихся в новых условиях. Однако, эту группу мы рассмотрим позднее, обратив первоначально наше внимание на две первые группы.
Прежде всего необходимо отметить, что широкораспространенные в тропиках роды заходят в Палеарктику только в пределы восточной ее части, характеризующейся влажным климатом и наличием богатой лесной растительности; единственным исключением является род Anisolabis.
Такая картина объясняется тем, что восточная часть палеарктической Азии непосредственно примыкает к влажным тропическим областям южной Азии, благодаря чему создаются благоприятные условия для проникновения на север наиболее экологически пластичных тропических форм, которые не имея экологических преград, проникают даже до северо-восточного Китая, Уссурийского края и Манчжурии (Timomenus komarowi, Eparchus и др.). Очень вероятно, что большинство этих форм населяло Восточную Азию вплоть до Уссурийского края еще в третичное время; однако, не исключена возможность, что некоторые виды проникли так далеко на север и в послетретичную эпоху, тем более, что особи одного и того же вида
1 Генетические взаимоотношения данного рода нам неясны и поэтому мы его исключаем из последующего рассмотрения.
ВВЕДЕНИЕ
55
из северных частей своих ареалов не дают даже подвидовых отличий от особей, распространенных в Индо-Малайской области.
Что касается Средиземноморского центра, то в силу изоляции последнего от влажных тропических областей пустынями Сев. Африки и Передней Азии, являющимися для уховерток, особенно для тропических видов, непреодолимой экологической преградой, сюда, конечно, не могут проникнуть представители тропической фауны, что и накладывает на фауну Dermaptera Средиземноморья печать значительной изоляции от сопредельных фаун. Совершенно иная картина получилась бы, если бы среди уховерток имелись ксероф ильные формы, для которых пустыни и полупустыни не являются непреодолимой экологической преградой; как раз в отличие от Dermaptera ксерофильные африканские группы животных и растений и сыграли громадную роль в образовании современной фауны и флоры Средиземья (см. ниже).
Единственный тропический вид в Средиземьи—Anisolabis (Euborellia) moesta, распространенный кроме того и в восточной Африке, повидимому, проник из последней в Средиземье в тот период, когда обе эти .области не были полностью изолированы друг от друга пустынями, как это имеет место в настоящее время. Такой благоприятный для миграции уховерток период существовал в неогене, о чем будет сказано ниже.
Наибольший интерес представляет рассмотрение группы эндемичных видов, относящихся к тропическим родам. Наличие, особенно в восточном центре, довольно большого количества эндемичных видов, являющихся представителями тропических родов, говорит о том, что третичная фауна Dermaptera умеренных широт северного полушария характеризовалась присутствием, помимо основных ангарских форм, также и тропическими элементами. При этом похолодание климата и последовавшее за ним стягивание фауны в два центра естественно должно было коснуться и тропических элементов, которые, в процессе отступания в более теплые южные зоны, оставили в наиболее подходящих в климатическом отношении местах, именно в Средиземье и особенно в восточной Азии, отдельных своих представителей. В процессе дальнейшей эволюции эти представители под влиянием ряда благоприятных для видообразования обстоятельств дали ряд местных форм, являющихся как бы свидетелями прежнего характера фауны. Нахождение в балтийском янтаре (нижний олигоцен) представителей надсемейства Protodermaptera/ (гем. Pygidicranidae и Labiduridae}, являющегося чисто тропической группой и не имеющего корней на Ангарском материке, вполне подтверждает данное допущение, тем более, что наличие в третичной, особенно палеогеновой, фауне и флоре Евразии чисто тропических групп животных и растений является общепризнанным фактом. Таким образом, группа видов, относящихся к тропическим родам, должна рассматриваться в качестве реликтовых эндемиков.
В отличие от ангарского ядра Палеарктической фауны, реликтовые эндемики восточного и западного центра не обнаруживают сколько-нибудь
56
КОЖИСТОКРЫЛЫЕ—DERMAPTERA
значительного родства друг с другом, относясь генетически к удаленнымг друг от друга родам или видам. Так средиземноморские Anisolabis (Gelo~ tolabis) maxima Brulle и Anisolabis (in sp.) mauritanica Luc. имеют родственные виды в тропической Африке, тогда как восточно-азиатские эндемичные Anisolabis, относясь к другим подродам (Apolabis, Mongolabis и Euborellld), генетически связаны с представителями соседней Индийской фауны. В этом обстоятельстве нет ничего удивительного, так как восточная Азия находилась под влиянием соседней тропической индийской фауны,, а Западная Европа под чрезвычайно ослабленным влиянием соседней африканской. Что касается родов, свойственных восточному центру, то большинство их распространено в Индо-Малайской области, причем род Pseu-disolabis кроме' того известен и из Австралийской области.
Большой интерес представляют роды Challia и Anataelia, которые: являются единственными во всей мировой фауне представителями особого, подсемейства Anataeliinae из сем. Pygidicranidae-, первый род известен: из Ю. Кореи и Сев. Китая, второй — с Канарских островов, т. е. в данном случае мы имеем по представителю из восточного и западного центра-Все сем. Pygidicranidae представляет собой исключительно тропическую группу, наиболее примитивные и наиболее многочисленные формы которой сохранились по преимуществу в Африке и тропической Азии; поэтому центр зарождения данной группы надо усматривать в области древнейшей суши, располагавшейся к югу от Ангарского материка — Гондванском материке. Повидимому, в лице подсем. Anataeliinae мы-имеем остаток весьма, древних представителей сем. Pygidicranidae, получивших возможность проникновения на Ангарский материк еще в мезозое и трансформировавшихся в особую специфическую для северного полушария группу, которая: уже в третичное время под влиянием климатических изменений почти целиком вымерла, сохранившись лишь только в расколотом на два близкие: рода состоянии. Поскольку все сем. Pygidicranidae является, за исключением данных двух родов, чисто тропическим по своему распространению,, требования к жаркому климату у него несомненно более повышены и поэтому надо полагать, что начало выпадения в третичное время в центральной части Евразии представителей подсем. Anataeliinae имело место раньше,, чем у ангарской группы видов, в силу чего один из указанных родов сохранился в самой южной части Палеарктики, притом только в виде: островной формы. Здесь уместно напомнить, что как раз фауна Канарских, островов сохранила следы миоценовой и даже олигоценовой фауны.. Что касается путей и границ проникновения рассмотренных нами тропических элементов в умеренные широты и ангарских форм — в тропические области (фиг. 18), то прежде чем коснуться разбора этого вопроса, необходимо подчеркнуть следующее важное обстоятельством довольно значительное представительство ангарской фауны Dermaptera. в Африканской области (роды Forficula, Apterygida, Mesochelidura и Che-lidiirelld) при почти полном отсутствии влияния тропической африканской
58
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
фауны на состав современной фауны западного центра; и, наоборот, значительное участие в образовании фауны восточного центра тропической (Индо-Малайской) фауны при одновременном чрезвычайно ослабленном влиянии ангарской фауны на состав соседней к восточному центру тропической фауны, сказавшемся только в проникновении сюда двух-трех видов из рода Forficula.
Наиболее ясным является вопрос с особенностями обмена фаун на востоке. Мы уже указывали, что восточная часть Палеарктики непосредственно прилегает к влажным тропическим областям Индии и Южного Китая, что обеспечивает беспрепятственную миграцию наиболее подвижных тропических форм, в пределы подходящих территорий восточной Палеарктики; этим объясняется наличие здесь большого количества не только широко распространенных, но и эндемичных видов, относящихся к тропическим родам. Казалось бы, что указанные обстоятельства могли быть благоприятными и для обратного процесса — заселения ангарскими формами соседних тропических областей; однако, в действительности мы этого не наблюдаем — только единичные виды и притом единственного рода Forficula оказались способными достичь южных частей Индийской области. Причины этого, повидимому, заключаются в том, что род Forficula является одним из наиболее древних элементов ангарской фауны, существовавшей еще в то время, когда в более южных частях Евразии имелся тропический климат; поэтому некоторые наиболее теплолюбивые виды данного рода оказались способными ужиться и в современных тропических областях.
Что касается причин диаметрально противоположного положения на западе, то прежде чем найти их, дадим краткую характеристику распространения интересующих нас родов западного центра. Род Anisolabis широко распространен в тропиках, причем ближайшие родичи двух западнопалеарктических эндемиков — A. mauritanica и A. maxima водятся в тропических областях Африки. В отношении родов Forficula, Apterygida, Mesochelidura и Chelidurella уже указывалось, что три последние из них распространены, за исключением одной восточно-африканской Apterygida cavallii Вог., только в южной субтропической части Африки. Род же Forficula, помимо нескольких видов свойственных африканской части Средиземья, представлен рядом видов на территории всей тропической Африки, отсутствуя, однако, в пустынных северных областях. Таким образом, все четыре рода изолированы от внеевропейских частей своих ареалов широкими пространствами пустынь Сев. Африки и Передней Азии, являющихся непреодолимой для них экологической преградой. Совершенно естественно поэтому, что все эти роды смогли проникнуть в Африканскую область только тогда, когда она не была изолирована от Европы при помощи- морских бассейнов или пустыни. Выше мы имели случай указать, что уже в миоцене происходили значительные миграции животного и растительного мира,1 чему способствовало исчезновение древнего Средиземного моря
1 Ср., Попов, op. cit. (1927), Мензбир, op. cit. (1934).
ВВЕДЕНИЕ
59
и установление непосредственной связи между Европейско-Азиатским и Африканским материками. В частности, нельзя не отметить крайне интересные соображения М. Попова (1927), который с большой убедительностью доказывает, что ксерофильная флора Средиземья (в широком смысле этого слова) есть преимущественно послепалеогеновое производное ксерофильной флоры южной Африки, получившей возможность проникновения в Средиземье после исчезновения Тетиса, т. е. начиная примерно с миоцена. При этом одновременно с ксерофильной флорой по столовым возвышенностям Восточной Африки шел обмен и мезофильной флоры между Южной Африкой, с одной стороны, и Средиземьем, с другой, так что в южной и возвышенной восточной Африке акклиматизировались и некоторые мезофильные средиземноморские элементы.
Несомненно в этот период и имело место проникновение, с одной стороны, рода Anisolabis в Средиземноморье, а с другой стороны — представителей родов Forficula, Apterygida, Chelidurella и Mesochelidura из Европы в Африку. Большинство проникших форм под влиянием местных условий трансформировалось в самостоятельные виды, отличающиеся от современных родственных им видов. Но кроме них имеется один вид — Anisolabis (Euborellid) moesta, который, повидимому, проник в Средиземье из вост. Африки, сохранив при этом на новой родине все свои видовые особенности; с другой стороны, африканская Forficula senegalensis чрезвычайно близка к южно-туранской и закавказской F. sagitta и, быть может, последняя представляет собою только подвид первой (Semenov-Tian-Shanskij, 1936). Такйм образом, современное распространение этих родов является показателем существования в прежние эпохи благоприятных условий для обмена фаунами между тропической и южной Африкой с одной стороны, и Европой — с другой.
Однако, наличие в Африке всего только пяти общих с Средиземьем и Зап. Европой родов заставляет предполагать, что условия для миграций здесь были значительно менее благоприятными, чем в восточной Азии. Причины этого, повидимому, заключаются в том, что после исчезновения древнего Средиземного моря, открывшего возможность для обмена фаунами между Европой и Азией, с одной стороны, и Африкой, с другой, уже в конце третичного времени вплоть до современной эпохи, гипсоносные пласты палеогеновых отложений Тетиса подверглись развеиванию и размыванию, превратившись в современные песчаные пустыни. Благодаря этому к югу и к востоку от Средиземья создались непреодолимые для многих групп животных, особенно таких влаголюбивых как Dermaptera, экологические преграды, что и послужило причиной полной изоляции последнего от соседней Африканской области. Кроме того, и в начале неогена, климатические условия северной Африки несомненно были более благоприятными для миграций ксерофилов, чем мезофилов, о чем говорят более многочисленные факты участия как раз ксерофильной африканской флоры и фауны в образовании современного био-географического лика Средиземья.
60
КО ЖИСТОКРЫЛЫЕ —DERMAPTERA
Несмотря на наличие указанных выше палеогеографических факторов, препятствовавших обмену фаун между Европой и Африкой, все же остается непонятным, почему из Европы в Африку смогли прорваться представители четырех родов, а из Африканской области в Средиземье только один: род — Anisolabis и то всего в количестве трех видов. Вероятные причины этого явления заключаются, повидимому, в том, что немногие тропические виды, если и достигали при своих миграциях до Средиземья, то, в силу своеобразного климатического режима последнего (периодичность в распределении осадков) и, быть может, благодаря мегатермным своим качествам: не смогли здесь надолго ужиться. Представители же ангарских родов Apterygida, Mesochelidura и Chelidurella смогли обосноваться в Африке, повидимому, только потому, что южная ее часть располагается в такой климатической зоне, в которой только и может иметь место наиболее близкий к европейским условиям климат. Что касается рода Forficula, то наличие довольно значительного числа его видов в тропических областях Африки несомненно объясняется теми же причинами, как и заселение им тропических частей Азии, т. е. его повышенной требовательностью к теплу, как одного из наиболее ранних третичных представителей подсем. Forficulinae,
В заключение необходимо отметить, что в отношении таких родов как Apterygida, Mesochelidura и Chelidurella, можно было бы допустить, основываясь на их распространении, африканское происхождение и рассматривать их европейских представителей в качестве потомков тех мигрантов, которые проникли в Европу из Африки. Однако, мы категорически отрицаем возможность такого допущения, исходя из того, что эти роды генетически близки к родам Anechura и Forficula и не могут рассматриваться от них отдельно; поскольку в лице родов Anechura и Forficula мы имеем несомненных и наиболее характерных представителей ангарского ядра фауны Палеарктики, постольку мы сюда же должны будем отнести и упомянутые выше роды. Более того, мы склонны рассматривать все подсем. Forficulinae в качестве выходцев с Ангарского материка, отнеся роды не имеющие связи с этой колыбелью фауны Евразии к другим подсемействам. Однако от окончательного суждения мы пока отказываемся, так как необходимо прежде всего произвести монографическое изучение всего сем. Forficulidae.
Коснемся теперь третьей группы тропических видов, завезенных человеком. Таковых довольно много; большинство их крайне широко распространено в тропиках Старого и Нового Света.
Восточный центр Западный центр
Anisolabis .... 3 вида Anisolabis .... 2 вида
Mala.............1 „ Nala ...... 1 „
Labidura.........1 „ Labidura..............1 „
Labia............1 „ Labia.................2 „
Prolabia.........1 „ Prolabia..............1 „
Chelisoches .... 1 „
Итого............ 8 видов 7 видов
ВВЕДЕНИЕ
61
В отличие от двух предыдущих тропических групп видов, данная группа характеризуется как в восточном, так и западном центре одинаковым числом представителей, кроме того относящихся в большинстве случаев к одним и тем же видам. Отличия заключаются только в том, что в западном центре имеется один вид — Labia minor, отсутствующий на востоке, а в восточном центре имеются только два вида—Chelisoches morio и Anisolabis plebeja, отсутствующие на западе. При этом, за исключением только двух далеко идущих к северу форм {Labia minor и Labidura riparid) все остальные виды строго ограничены в своем распространении границами Средиземноморской подобласти на западе и Палеархеарктической — на востоке, будучи, однако, широкораспространенными в тропиках.
Эта группа видов представляет значительный интерес благодаря тому, что центры ее многообразия в Палеарктике совпадают с современными центрами многообразия аборигенной фауны Палеарктики. Такое совпадение не случайно и объясняется теми же причинами, которые позволили сохраниться в Средиземье и вост. Азии представителям древней третичной фауны Dermaptera, именно наличием в данных двух центрах благоприятных для уховерток климатических особенностей. Благодаря торговым сношениям человека, ряд видов смог с помощью последнего перешагнуть через зоогеографические границы и экологические преграды и заселить такие территории, с благоприятными для этих видов экологическими условиями, куда многие из них не смогли бы проникнуть естественным путем; по крайней мере это можно сказать про западный центр.
Нам осталось рассмотреть еще один фактор, именно горообразовательный процессы, охватившие Евразию во вторую половину третичного периода, особенно в миоцене. Этот фактор способствовал значительному видообразованию как среди основного автохтонного ядра фауны, так и среди выходцев из соседних тропических областей — Африканской и Индийской.
Значительная роль горообразовательных процессов в качестве фактора, стимулирующего видообразование, отмечалась уже давно. Однако, помимо этого А. П. Семеновым-Тян-Шанским (in litt.) было высказано чрезвычайно интересное соображение, касающееся роли этих процессов в качестве могущественного фактора, способствовавшего вертикальному перемещению фауны, населявшей древние пенеплены в области третичной складчатости в совершенно новые для нее условия; эта мысль была подхвачена и применена рядом исследователей (Кузнецов, Уваров, и др.)1 для объяснения некоторых особенностей современной фауны в горных областях.
Анализ современного распространения уховерток в Палеарктической области, как мы видели, показал, что фауна последних в третичное время
'Кузнецов, Н. Н. К вопросу о вертикальном распределении животных форм :в Туркестане. Бюлл. Ср.-Аз. Гос. Унив., 2, 1923:33—34; Uvarov, В. Р. Orthoptera of the mountains of Palaearctic region. Contr. а Г etude du Peupl. des Hautes Mount., Paris, 1928 : 5—6.
62
КОЖИСТОКРЫЛЫЕ —DERM APTER A
была, по крайней мере, в более умеренных широтах более однородной, чем это имеет место в настоящее время; генетическая близость ряда современных западных видов и родов к восточно-азиатским является отголоском этой особенности. Точно так же Криштофович1 отмечает большое однообразие для умеренных широт и палеогеновой лесной флоры.
Горообразовательные процессы в тех областях, куда во вторую половину третичного времени, повидимому, уже начали стягиваться представители данной фауны (юг Зап. Европы, вост. Азия) явились также могущественным фактором, способствовавшим дифференциации фауны. Благодаря этому фауна, с одной стороны, была раздроблена возникшими горными системами наряд территорий, взаимная связь между которыми была затруднена или полностью нарушена, а с другой — виды, населявшие области,, охваченные орогеническими процессами, были перемещены с сушей вверх. То и другое способствовало значительному формообразованию, выразившемуся в конечном счете в появлении ряда узко-распространенных видов. К числу видов, обязанных своим происхождением орогеническим процессам второй половины третичного времени, необходимо отнести на западе виды рода Chelidurella, один вид которого свойствен только Альпам, а другой Карпатам. Но помимо того имело место даже образование специальных родов — таковым несомненно является монотипичный род 'Pseudochelidura и род Chelidara с двумя видами. Оба эти рода чрезвычайно близки к роду Anechura и имеют крайне ограниченные ареалы распространения, обитая только высокогорные области юго-западной Европы, не ниже 1000 м над уровнем моря; при этом они лишены крыльев и имеют в различной степени атрофированные надкрылья. Некоторые из образовавшихся видов, впоследствии под влиянием похолодания климата в ледниковое время принуждены были спуститься вниз, что и способствовало их расселению на значительной территории, после чего наступившее потепление заставило часть их вернуться в горные области, оставив, однако, завоеванные низменные места за собой; к таким видам относится Chelidurella acanthopygia и вероятно Forflcula tomis, Anechura asiaiica, A. bipunctata.
Что касается восточного центра, то здесь результаты воздействия орогенических процессов, в силу большего богатства фауны и меньшего значения ледниковых влияний, сохранились до настоящего времени в большом числе случаев. Такие роды, как Oreasiobia и Mesasiobia, свойственные горам Центральной Азии, как Eumegalura, свойственная горам Сычуани, как подрод Burriola рода Anechura с высокого плато Тибета, несомненно являются трансформированными потомками тех видов из родов Forficula, Anechura и близких к ним, которые населяли прежде низменности или выравненные и превращенные в пенеплены области древнейшей, дотретич-ной складчатости. Примеры с видами еще более многочисленны. Сюда нужно отнести многие виды рода Forficula, максимум которых локализуется
1Криштофович, op. cit., 1930: 395—398.
ВВЕДЕНИЕ
6>
в области третичной складчатости, особенно на Гималаях и сопредельных частях Китая, причем большинство видов имеет крайне узкое распространение. Достаточно указать, что среди восьми западно-китайских видов этого рода и десяти гималайских нет ни одной общей формы, причем только’ один вид из каждой группы достигает Тибета; точно так же из пяти северо-китайских -видов имеется только единственный общий вид с провинцией .Сычуань и Тибетом. Одновременно и виды из подродов Odonto-psalis и Anechura s. str. рода Anechura дают в горных областях Китая ряд узкораспространенных форм, характеризующихся ясной тенденцией к атрофии надкрылий и крыльев.
Что касается тропических элементов, то к ним принадлежат такие виды, относящиеся к чисто тропическим родам, как Sondax potanini из Сычуани, единственный родич которого (5. repens) известен из Мадраса в южн. Индии, кашмирская Pseudisoldbis burri, ближайшие родичи которой известны из предгорий сев. Индии и даже из Австралийской области и, повидимому, некоторые виды из рода Anisolabis. Предки этих видов до начала горообразовательных процессов, вероятно, населяли территорию вост. Азии и, будучи поднятыми вместе с сушей вверх, трансформировались в самостоятельные виды.
В результате такого воздействия горообразовательных процессов третичного времени на фауну Dermaptera наиболее богатыми по числу видов оказались страны, где эти процессы протекали особенно интенсивно. В противоположность этому, всюду на территории умеренных частей Палеарктической области, где процессы горообразования не имели места, число видов Dermaptera выражается буквально единицами. Однако в большинстве случаев это объясняется уже другими причинами, разобранными нами выше, именно тем, что горные области являются с одной стороны аккумуляторами атмосферной влаги, а с другой — климатическими границами и поэтому здесь смогли создаться такие экологические условия, которые способствовали переживанию неблагоприятных изменений климата, имевших место во вторую половину третичного времени и в ледниковую эпоху. Как раз восточный и западный центры современного разнообразия фауны Dermaptera в Палеарктической области и являются по преимущесту горными странами.
Таким образом в результате нашего разбора состава и происхождения фауны Dermaptera Палеарктической области -мы пришли к следующим выводам.
1. Современная фауна Dermaptera Палеарктической области, состоящая примерно из 106 видов, имеет два центра наибольшего видового разнообразия — Восточный, охватывающий Палеархеарктическую подобласть и сопредельные страны, и — 3 а п ад н ы й, охватывающий Средиземье и южную часть Зап. Европы; центральные части области, характеризующиеся суровым континентальным климатом, имеют весьма обедненную фауну не более, чем с 8—10 видами.
«4
КО ШИС ТОКРЫЛЫЕ — DERMAPTERA
2. В образовании современной фауны Dermaptera Палеарктической области приняли участие следующие элементы:
а) Основное древнее ядро ангарской фауны во главе с родами Forficula и Anechura, зародившееся на древнейшей азиатской суше — Ангарском материке, где филогенетическая нить жизни ни разу не прерывалась; к ангарским элементам относится свыше 65°/0 всего видового состава фауны в Палеарктике.
в) Представители соседних тропических фаун, причем восточная часть Палеарктики оказалась под сильным влиянием Индийской фауны, а западная— в чрезвычайно ослабленной степени — Африканской.
с) Группа видов, завезенных человеком по торговым путям вместе с товарами и акклиматизировавшихся преимущественно в области Средиземья на западе и восточной Азии на востоке.
3. Аборигенная фауна восточного и западного центров, несмотря на значительную удаленность их друг от друга, характеризуется весьма существенной генетической близостью, что выражается:
а) Преобладанием в восточном и западном центрах представителей родов Anechura и Forficula, составляющих около 5О°/о всего видового состава фауны Dermaptera в Палеарктике.
в) Наличием многих генетически близких друг к другу видов, относящихся к указанным родам и ограниченных в своем распространении или восточным, или западным центром, причем виды восточного центра обнаруживают черты более примитивной организации, чем родственные им ви^ы в западном центре. Однако, общие для указанных двух центров виды совершенно отсутствуют.
с) Наличием двух генетически близких друг к другу подродов из рода Anechura — Burriola и Borelliola, а также нескольких, хотя и различных родов, но близких к роду Forficula и Anechura.
4. Значительная генетическая близость между фауной Dermaptera восточного и западного центров видового многообразия, относящихся к совершенно различным зоогеографическим подобластям Палеарктики, резко отличают данную группу животных от многих других групп. Это своеобразие современной фауны Dermaptera Палеарктической области может быть понято только при учете следующих факторов.
а) Экологии данных насекомых, характеризирующихся с одной стороны •скрытым образом жизни, а с другой — теснейшей зависимостью их от таких климатических условий, которые одновременно обеспечивали бы и наличие достаточного теплого вегетационного периода, и лесной растительности.
Ь) Геоисторических явлений, выразившихся в изменении, в предшествовавшие современному состоянию эпохи, климатического режима с одной стороны и очертания и рельефа всех территорий, занятых в настоящее время Палеарктической областью — с другой.
с) Развитием в историческую эпоху широкого товарообмена между различными странами, что обеспечило завоз в наиболее благоприятные
ВВЕДЕНИЕ
65
.для Dermaptera в климатическом отношении области с умеренным климатом ряда преимущественно тропических видов.
5. В первую половину третичного времени территория нынешней Палеарктической области характеризовалась, по крайней мере, в умеренных широтах, более или менее однородной на значительном протяжении фауной, состоявшей как из ангарских, так и из тропических элементов, причем среди последних представлены были вероятно только восточные ^(индийские) типы.
6. Начиная с миоцена фауна уховерток претерпевает весьма значит тельные изменения под влиянием следующих трех основных факторов:
а) Общего похолодания климата и сокращения количества осадков, особенно в центральных умеренных частях Евразии, выбившего из состава фауны ряд наиболее теплолюбивых и влаголюбивых форм и способствовавшего стягиванию последних в два центра — восточно-азиатский и средит земноморско-европейский, сохранивших до настоящего времени благоприятные климатические условия для данных насекомых; в результате этого более или менее однородная до-миоценовая фауна была расколота на две территориально крайне удаленные, но генетически близкие фауны,
Ь) Исчезновения древнего Средиземного моря — Тетис и установлвт ния непосредственной связи между южной Евразией и Сев. Африкой, открывшего путь для миграции и обмена фаунами, в силу чего некоторые ангарские формы (роды Forficula, Apterygida, Mesochelidura и Chelidurella) перешагнули в сев. Африку, а отсюда проникли в числе немногих видов даже в тропическую ее часть, откуда в свою очередь по тем же путям проникли в Средиземье весьма немногие африканские элементы {Anisolabis). Так как эта связь была более благоприятна для миграции ксерофиль-ных элементов, то как в Африканскую область, так и в Средиземье смогли проникнуть только весьма немногие виды уховерток, чему в значительной степени также способствовали процессы эрозии и выветривания гипсоносных отложений Тетиса, на месте которых образовались песчаные пустыни; благодаря этому фауна Dermaptera современного Средиземья оказалась совершенно изолированной от соседней африканской области.
с) Орогенических процессов, способствовавших поднятию низменной фауны и раздроблению ее на ряд частей, в которых дальнейшая эволюция протекала более или менее автономно, что способствовало образованию ряда узко-распространенных видов и родов.
7. В четвертичный период фауна Dermaptera претерпела дальнейшие изменения под влиянием следующих факторов:
а) Ледникового периода, выбившего из состава умеренных широт Евразии последние остатки третичной фауны Dermaptera, в результате чего умеренные широты были полностью лишены представителей данной группы насекомых; одновременно это способствовало с одной стороны еще большему стягиванию фауны в указанные выше два центра, а с другой-спусканию вниз и широкому расселению тех высокогорных видов,
Фауна СССР, Кожистокрылые 5
66
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
которые возникли в третичное время вследствие орогенических процессов-в более северных частях, где в последующее время влияние ледникового периода было более заметным.
Ь) Потепления климата в послеледниковый период, что послужило причиной обратного заселения освободившихся из под ледникового влияния территорий немногими видами.
с) Деятельности человека и развития широких торговых сообщений, между самыми отдаленными странами, что, благодаря своеобразным экологическим особенностям уховерток, способствовало завозу в современные центры европейско-азиатской культуры ряда совершенно чуждых данным странам форм.
В заключение считаем необходимым отметим, что мы рассматриваем наш опыт истории современной фауны Dermaptera Палеарктической области: лишь как первое приближение; совершенно очевидно, что данная нами схема, как всякая схема, не может объяснить всей суммы фактов — нами были разобраны только основные, наиболее важные и наиболее достоверные явления. Дальнейшее накопление фактов позволит уточнить и развить ряд тех сторон, которые нами или освещены только в самом приближенном, схематичном виде, или же совершенно не подвергались обсуждению.
7. ХАРАКТЕРИСТИКА ФАУНЫ DERMAPTERA СССР
Анализ состава и происхождения фауны Dermaptera Палеарктики целиком приложим к территории СССР, занимающей весьма значительную часть данной области; однако, центром нашего внимания была восточноазиатская и средиземноморская фауна и некоторые специфические особенности фауны СССР, в том числе указанные А. П. Семеновым-Тян-Шан-ским [(1935), не были отмечены. Прежде всего необходимо указать, что в состав фауны Dermaptera СССР входит только 26 видов, т. е. часть от всего наличного числа видов в Палеарктике, хотя территория СССР’ по отношению ко всей области составляет несколько больше половины. Такая бедность объясняется, как ясно из изложенных выше фактов, тем, что СССР как на востоке, так и на западе почти не соприкасается с двумя современными центрами разнообразия уховерток; лишь только Крым и Кавказ с одной стороны и Уссурийский край с другой являются обедненными форпостами данных центров в Союзе, где число видов, несмотря на незначительность их территории, достигает в каждом случае 9—11 видов, из которых некоторые являются даже эндемиками.
Характерно при этом то, что территориально близкие фауны Кавказа и Крыма обнаруживают значительное несходство; на это обращает особое внимание А. П. Семенов-Тян-Шанский, связывая данный факт с особенностями генезиса Крымской и Кавказской фаун; в частности необходимо отметить, что на Кавказе, именно в его западной части, представляющей
ВВЕДЕНИЕ
67
собою область реликтового третичного леса,1 и сопредельных территориях водятся два характерные Кавказские эндемика — Anechura (Borelliola) euxina и Forficula kaznakovi, являющиеся эпигонами некогда несомненно более богатой Кавказской фауны. В отличие от Кавказа, Крым эндемиков не имеет совершенно, хотя крымская F. aetholica, распространенная также в Малой Азии и на Балканах, генетически близка к F. kaznakovi. Кроме того, весьма характерно отсутствие в Крыму такого широко распространенного преимущественно горного вида как Anechura (in sp.) bipunctata-, последнее обстоятельство говорит о том, что данный вид проник на Кавказ по горам Малой Азии; изоляция же Крыма от ближайших горных систем пространствами южных степей не позволила ему достичь Крыма в соответствующий благоприятный момент (ледниковый период). Весьма замечательно, что изолированный островок обитания двух характерных эндемиков Зап. Кавказа — А. (В.) euxina и F. kaznakovi имеется также несколько к северу от хребта, именно в области Ставропольского поднятия, где в силу вертикальной зональности основной формацией в центральной, наиболее высокой, части являются не степи, а лесостепь. Существование этого островка обязано несомненно ледниковому периоду, когда, в силу вертикального смещения зон, пониженное пространство современных степей, отделяющих главный хребет от Ставропольского плато, было занято лесной растительностью, благодаря чему данные два вида получили возможность широкого распространения на север; впоследствии, с исчезновением лесной растительности в наиболее пониженных частях, эти виды, смогли удержаться к северу от Главного хребта только в области Ставропольского поднятия.
Наличие на Кавказе двух эндемичных видов, отсутствующих в Крыму, является еще одним подтверждением давно высказанного тем же автором2 соображения о том, что горные части этих областей в течение Кайнозойской эры ни разу не были друг с другом в связи, в силу чего и фауна их развивалась автономно.
В остальном фауна Кавказа представляет собой довольно разнородную группу видов; так F. sagitta, распространенная в вост. Закавказье, известна также и из Туранской низменности; наоборот Anechura Ырип-ctaia, распространенная в восточных частях Кавказа, отсутствует в Средней Азии, a F. tomis свойственна как Туранской низменности, так и югу Европейской части СССР. Остальные виды, кроме F. smyrnensis, свойственной Зап. Кавказу и распространенной в восточном Средиземьи до Корсики, не представляют особого интереса, будучи широко распространенными.
Что касается Крыма, то несмотря на отсутствие здесь эндемичных форм, он представляет собою известный интерес в связи с присутствием
'Кузнецов, Н. И., op. cit. (1909).
2 Семенов, А. Несколько соображений о прошлом фауны и флоры Крыма. Зап. Акад. Наук, VIII сер., № 6, 1899.
5
68 КОЖИСТОКРЫЛЫЕ—DERM APTERA
трех типично-средиземноморских видов, крымский ареал которых оторван от их общего ареала; сюда относятся F. aetholica, F. smyrnensis и Aniso-labis (Euborellia) moesta. Эти три вида являются свидетелями существования древней, но ныне уже утраченной связи с Балканским полуостровом через Добруджу, восходящей к нижнему плиоцену и миоцену1 или, по воззрениям некоторых авторов, с Малой Азией.2 Остальные шесть видов являются широко распространенными и наиболее обычными видами, при чем два из них—представители рода Anisolabis населяют берега морей И несомненно были сюда завезены.
Если обратиться к фауне европейской части, то из девяти свойственных ей видов, три вида (F. auricularia, Labia minor и Labidura riparid) представляют собою группу наиболее широко распространенных видов, интересных лишь в том отношении, что они идут на север дальше, чем все остальные виды, причем один вид заходит даже в Арктическую зону. Другие три вида {Apterygida media, Chelidurella acanthopygia и Ch. transsyl-vanica), являясь лесными западно-европейскими элементами, заходят только в пределы лесостепной и лесной Украины, отсутствуя во всей остальной европейской части СССР. Из оставшихся трех видов—Forficula tomis, Anechura bipunctata и A. asiatica — наибольший интерес представляет последний, заходящий в пределы европейской части только др южного Заволожья, тогда как остальная часть ареала этого вида охватывает всю Среднюю Азию и сопредельные страны.
В отношении Средней Азии необходимо отметить, что низменная ее часть, несмотря на южное положение, имеет всего только пять видов, из которых наиболее характерными и обычными являются Anechura asiatica и Labidura riparia', такой широкораспространенный вид, как обыкновенная уховертка (F. auricularia) здесь совершенно отсутствует. Остальные виды представлены весьма редкими Forficula sagitta и Labia minor, а также F. tomis, которая, видимо, переносит здесь чрезвычайно сухой климат только потому, что приурочена к побережьям озер и берегам рек. Горная часть Средней Азии, хотя имеет всего шесть видов, т. е. на один вид больше, чем Туранская низменность, однако, из их числа два вида, относящиеся к тому же к особым родам, являются эндемичными (Oreasiobia fedtschenkoi и Mesasiobia hemixanthocara). Кроме того, весьма интересно, что в горах Копет-дага встречается и обыкновенная уховертка (F. auricularia), отсутствующая во всей остальной части Средней Азии и, таким образом, имеющая в Копет-даге крайнюю восточную границу своего распространения в Ср. Азии (Семенов, 1901); присутствие этой уховертки в Копет-даге повидимому является отголоском влияния соседей Гирканской провинции, сохранившей черты неогеновой флоры.® В осталь
1 Архангельский, А. Д. Геологическое строение СССР. Европейская и Среднеазиатские части. Л.—М., 1932:261, 265, 272, карты 58—59.
а В у л ь ф, Е. Происхождение флоры Крыма. Зап. Крымск. Общ. Естеств., IX, 1926.
®Гроссгейм, А. А. Флора Талыша. Тифлир, 1926.
ВВЕДЕНИЕ ............................ 69
ном горная часть отличается от низменной только отсутствием таких видов, как Labia minor и F. sagitta^ впрочем, нахождение в горах Первого из них весьма вероятно.
Присутствие двух эндемичных видов, относящихся даже к эндемичным родам, при одновременной чрезвычайной бедности фауны уховерток Горной части Средней Азии, говорит о том, что эта фауна,' повйдимому, прежде была Значительно более богатой и ее бедность есть явление вторичное. Наличие в горах лесной растительности, благоприятной для существования видов, и громадная протяженность Средней Азии, где можно было бы ожидать богатую фауну Dermaptera, заставляет предполагать, что это обеднение имело характер катастрофы; тем более, что в расположённых более к северу территориях, как напр. Зап. Европа, число видов уховерток превышает в три раза число видов, свойственных горам Средней Азии. Единственное объяснение этому явлению Можно дать, лишь основываясь на весьма своеобразных условиях Средней Азии В целом, выражающихся в том, что горные системы, благоприятные для жизни уховёрток, здёсь опоясаны пустынями, непригодными для существования огромного большинства видов; при этой йустынйые территории Средней Азии представляют собой достаточно древнее явление, существовавшее еще в неогене.1 Учитывая это, можно высказать предположение, что вертикальным перемещением существующих и пб сиё время обширных высокогорных ледников, всё Зоны не только были смещены вниз, но некоторые из них в результате столкновения с низменными пустынями могли выпасть или весьма сократиться, вследствие чего ряд видов уховерток оказался стиснутым с однбй стороны МежДу неблагоприятными для них пуётынйМи, а с другой — Областями, подВержённЫми прямому воздействию ледников; это обстоятельство йе Могло не способ-’ ствоватэ йсчёзйовенйю ряда видов.
Вышесказанное соображение Хорошо увязывается с Известными фактами выпадения лесной Зойы в некоторых горных областях, непосредственно примыкающих к пустыням или полупустыням. Так, Сапожников2 3 * показал для Монгольского и юго-ВосточногО Алтая отсутствие переход-нЫХ звёйьёв от растительности степных и поЛупустыйных долин к альпийскому поясу; точйо так же наши исследования сРВМестно с проф. В. И. Барановым8 показали йепрерывное распространение от полупустыни до высокогорной тундры И ледйиков в юго-востойном Алтае некоторых кСерофильных форм Orthoptera. Общеизвестным примером В’ыпадеййя лёсной ЗОйы в горах, соприкасающихся с полупустыйей, является Тарба-Гатайский хребет. Подтверждением данного предположений является И тот
1 См. Попов, dp; cit. (1927); Су ш кии, др’, ей. (193й);
2 Сапожников, В. В. Монгольский Алтай в истоках Иртыша и Кобдо. Томск, 1911:351.
3 Баранов, В. И. и Бе й-Б и е н к о, Г. Я. Опыт фито-экологической харак-
теристики местообитаний Orthoptera saltatotia на Алтае. Изв! Зап. Сиб. Геогр. о-ва, V, 1926.
70
КОЖИСТОКРЫЛЫЕ —DERMAPTERA
факт, что такой свойственный Туранской низменности вид, как Anechura asiatica, обитает также и всю горную часть Средней Азии, причем центр происхождения его, видимо, находится здесь же, так как он распространен, кроме Средней Азии, только в сопредельных частях Монголии, Ирана и Тибета, не говоря уже о том, что весь род Anechura тесно связан с горными странами. Раз это так, то в лице A. asiatica можно усматривать единственный горный вид, преодолевший неблагоприятные условия низменных полупустынь во время смещения ледников вниз и получивший возможность после исчезновения ледников не только сохраниться здесь, но даже расселиться по пустыням и полупыстыням на значительное расстояние от своей первоначальной родины, являясь таким образом свидетелем условий, некогда заставлявших переселяться эти горные виды в область засушливой низменности.
Приспособление A. asiatica к низменным полупустыням в значительной мере должно было зависить от весьма редкой среди уховерток способности уживаться в сухом климате; эта способность является специфическим свойством A. asiatica, выработавшемся, несомненно, не в последнее время, а в течение всего процесса эволюции вида, так как трудно допустить, чтобы приспособление к столь необычному для Dermaptera климатическому режиму появилось у данного вида только в сравнительно недавнее время; с этим не вяжется медленный темп эволюции группы и ее консервативность. Anechura asiatica и теперь населяет области высокогорных инверсионных степей и полупустынь, каковыми являются Тян-Шанские сырты, высокие нагорья Памира и Каракорума, т. е. как раз те территории, где необходимо усматривать центр возникновения данного вида. По всей вероятности, перемещения ледников только способствовали перемещению данного вида вниз — в область низменных и подгорных полупустынь, где он нашел весьма сходные с высокогорной полупустыней экологические условия, благодаря чему после сокращения площади оледенения, он не только сохранился, но и широко распространился в Туранской низменности.
Переходя к рассмотрению фауны Сибири, отметим полное отсутствие в ней эндемичных форм. Западная ее часть имеет всего только пять видов, распространяющихся сюда из восточной части Европы, центральная— всего только один вид — Anechura bipunctata (впрочем здесь возможно нахождение Labia minor и Labidura riparia), а самая восточная, включая Сахалин и Камчатку, шесть видов. Среди последних замечательно нахождение на Камчатке двух тропических видов — Anisolabis pectoralis и Prolabia arachidis, несомненно завезенных сюда человеком; кроме того, необходимо отметить наличие на Сахалине как Prolabia arachidis, так и Anechura (Odbntopsalis) harmandi и Forficula robusta’, последний известен также из Уссурийского края, Манчжурии и сев.-вост. Китая, а предыдущий— из Японии и Уссурийского края. Что касается Забайкалья, то отсюда пока известен всего только один вид — Labidura riparia.
ВВЕДЕНИЕ
71
Таким образом, мы видим, что в наиболее суровой центральной и сопредельной с ней восточной части Сибири фауна уховерток почти сходит на нет, тогда как самый запад и самый восток характеризуются наличием несколько большего числа видов.
Что касается Уссурийского края, то, несмотря на его относительно весьма незначительные размеры, мы встречаем здесь девять видов. Оставляя в стороне четыре вида, в распространении которых сыграл значительную роль человек, чему в данном случае благоприятствовало приморское положение края, обратим наше внимание на остальные пять видов. Из них четыре вида — Forficula robusta, F. vicaria, Anechura (OdontOpsalis) japonica и A. (O.) harmandi здес^ находят свою северную границу распространения,1 а остальная часть их ареала охватывает в трех случаях Японию и также в трех случаях Сев. Китай. Последний вид (Timo-.menus komarovt), в отличие от всех предыдущих,- является представителем тропического рода, распространенного кроме восточной Азии также и в Индо-Малайской области; при этом весьма характерно, что Т. komarovi, минуя Японию, распространен в сев. Китае и на Формозе. Наличие здесь сравнительно богатой фауны и даже захождение одного тропического вида объясняется тем, что Уссурийский край расположен на периферии восточно-азиатского центра многообразия видов в Палеарктике, что более детально было разобрано выше.
Такова краткая характеристика особенностей фауны Dermaptera СССР.
В заключение необходимо отметить, что вышеперечисленным списком в 26 видов фауна Dermaptera СССР вряд ли исчерпывается. Надо •ожидать нахождение некоторых дополнительных видов как в Уссурийском крае, так вероятно и в Закавказье. 8
8. ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ УХОВЕРТОК
Уховертки, будучи, как это было показано выше, немногочисленной ио количеству видов группой насекомых, имеют по сравнению с остальным комплексом отрядов относительно небольшое хозяйственное значение. Однако, эта группа все же включает в себя некоторое количество видов, которые имеют определенное и подчас весьма существенное экономическое значение для отдельных культур и хозяйственно-ценных растений.
Среди 26 видов этого отряда, известных из пределов СССР, имеют хозяйственное значение 4 вида; из них наиболее существенную роль играют два вида — азиатская уховертка (Anechura asiatica) и огородная уховертка (Forficula tomis). Что касается первого вида, то он является вредителем семян дикорастущего каучуконоса — тау-сагыза (Scorzonera tau-saghyz) в горах Каратау в ю. Казахстане, являющегося в настоящее
1 Один из этих 4-х видов — А. (О.) japonica проникает немного севернее, достигая .Николаевска на Амуре.
72 КОЖИС ТОКРЫЛЫЕ —DERMAPTERA
время существенным источником семенной продукции, необходимой для: закладки плантаций этого весьма ценного каучуконоса; вред выражается в выгрызании рылец и пестиков в цветах или иногда в полном выедании завязей в молодых бутонах. Поскольку проблема производства отечественного каучука имеет чрезвычайно большое народно-хозяйственное значение, нельзя не признать; что азиатская уховертка имеет хотя и специфическое, но весьма большое экономическое значение, тем более, что урожай семян тау-сагыза снижается более чем в три раза на зараженных, уховерткой участках (Правдин, 1934).
Огородная уховертка является наоборот чрезвычайно многоядным вредителем, имеющим особенно существенное значение для огородных культур (картофель, свекла, соя, мак, всходы подсолнечника, капуста,, томаты), а также Для бахчевых (огурцы, тыквы, дыни), табака и цветоводства, нанося повреждения как в личиночном, так и во взрослом состоянии путем обгрызания листьев, подгрызания стеблей или даже полного съеданий всходов и молодых высадок. Кроме того, этот вид отмечен как вредитель семенников сахарной свеклы на Украине, выгрызающий полузрелые семена; забираясь также в слабые плоды яблони он может и здесь причинять некоторый^ но незначительный вред.
Что касается других известных из СССР вредных видов, то один из них — обыкновенная уховертка (F. auricularia) вредит на Украине й в Зап. Европе огородным растениям и цветоводству, объедая живые листья, цветы и полузрелые семена; кроме того, этот вид имеет значение для кукурузы и других злаков (выедаются полузрелые семена) й плодовых деревьев. Будучй завезен в Сев. Америку, он стал там даже карантинным объектом в связи с сильными повреждениямй, наносимыми им питомникам декоративных и других деревьев; в Новой Зеландии, куда, этот вид был также завезен, он является бичом местного плодоводства. Кроме того, отмечены случаи выгрызания этим видом ходов в печеном хлебе.
Последний из известных в СССР вредных видов — уховертка Федченко (Oreasiobia fedtschenkoi) является вредителем табака в горных частях Средней Азйй (Чймган).
Таким образом, даже не бстанавлйваясь на некоторых извееТнЫх за пределами СССР вредителях, необходимо констатировать, Что уховертки не являются Индифферентной для Человека группой насекомых; хозяйственное значение их иногда может быть настолько существенным,, что отсутствие тех или иных контролирующих мероприятий может поставить под угрозу получение урожая с повреждаемых культур.
9. СПИСОК ЛИТЕРАТУРЫ ПО DERMAPTERA СССР И СОПРЕДЕЛЬНЫХ СТРАН
Предлагаемый список не представляет собой полной библиографии по Dermaptera СССР и сопредельных стран; в этот список вошли лишь
ВВЁДЁНИЕ
73
наиболее важные работы; поэтому различные мелкие заметки, за немногими исключениями, не включены сюда, а ссылки на них в случае необходимости даются при описании того или иного вида.
Все работы расположены по латинскому алфавиту, так как на русском языке опубликовано лишь весьма небольшое число работ. Русские работы, начинающиеся С буквы Ш и Щ приводятся под буквой S, а с буквы Я — под буквой J.
[A de lung, N.] А д е л у н г, Н. 1902. Новый для фауны Российской Империи вид уховерток — Forficula aetolica. Ежегод. Зоол. Муз., VII: XII—XIII.
A z a m, J. 1906. Description d’une espece et d’une variete de Forficules de Russie. Bull. Soc. Ent. France, 1906:80—82.
Bateson, W. and Brindley, H. H. 1892. On some cases of variation in secondary sexual characters statistically examined. Proc. Zool. Soc. London: 586—589.
Beier, M. Dermaptera in: Biologie der Tiere Deutschlands. Berlin, Lief. 36, Teil 26: 169—231, 51 fig.
. [B e y-B i e n k o, G. J.] Б e й-Б и e н к о, Г. Я. 1926. Заметки по некоторым прямокрылым Палеарктической Азин. Тр. Сиб. C.-Хоз. Акад., VI, № 8:1—-13.
---- 1930. Очерк фауны Dermaptera и Orthoptera Омского округа. Тр. 3anj. Раст., Сер. Энтомол., 1, № 1 :161—177.
В е y-B i е n к о, G. J. 1933. Forficulidae in: Schwedisch-chinesische wissenschaftliche Expedition nach den nordwestlichen Provinzen Chinas, unter Leitung von Dr. Sven-Hedin und Prof; Sin Ping-chang; Arkiv Zool., 25 A, № 20: 1—9, tab. I.
---- 1934. Studies on the Dermaptera of the province of Sechuan, China. Ann. Mag. Nat.
Hist., 10, XIII: 401—425, 12 figs.
---- 1935. Dermaptera in: Wissenschaftliche Ergebnisse Niederlandischen Expeditionen in den Karakorum, Zoologie, Leipzig: 208—212.
В e y-B i e n к o, G. J. et S e m e n о v-T i a n-S h a n s к у, A. P. — vide Semenov.
Borelli, A. 1909. Nuove forficule del Kashmir. Bull. Mus. Zool. Anat. Comp. Torino, XXIV, № 603:1—4.
---- 1912. Dermapteres nouveaux ou peu connus du Museum de Paris. Bull. Mus. Hist. Nat. Paris, 1912, № 4:1—20, fig.
---- 1915. Di alcuni Dermapteri della Cina. Boll. Mus. Zool. Anat. Comp. Torino, XXX, № 698:1—6.
—— 1921. Dermapteres nouveaux du Museum de Paris. Bull. Mus. Hist. Nat. Paris, 1921 :77— 83, 153—161.
---- 1927. Dermatteri raccolti nell’Estremo-Oriente dal Prof. L. Silvestri. Boll. Lab. Zool.
Portici, XX: 60—78, fig.
Bormans, A. 1900. Forficulidae in: Das Tierreich, Berlin, 11 :1—142, 47 fig.
Brindley, H. 1918. Notes on certain parasites, food and capture by birds of the common earwig (Forficula auricularia L;). Proc. Cambridge Phil. Soc., XIX: 167—177.
Brunner von Wattenwyl K. 1882. Prodromus der europaischen Orthopteren, Leipzig (Forficiilaria: 1—26, tab. 1).
Burr, M. 1903. Notes on the Forficularia. —• VII. Some hitherno unpublished descriptions of new species by the late M. A. de Bormans. Ann. Mag. Nat. Hist., 7, XI: 231—241.
---- 1904. Observation oa the Dermatoptera, including revision of several genera and descriptions of new genera and species. Trans. Ent. Soc. Lond., 1904:277.
---- 1907 a. A preliminary revision of the Forficulidae (s. str;) and of the Chelisochidae* families of Dermatoptera. Ibid., 1907 : 91—134, pl. IV.
—— 1907 b. Dermaptera in: Filchner, Expedition China-Tibet, Zool.-bot. Ergebn., Hexapoda, Berlin: 57—58, tab. III.
74
КОЖИСТОКРЫЛЫЕ
Burr, М. 1908. Notes on Forficularia. — XIII. A revision of the Brachglabidae. Ann. Mag. Nat. Hist., 8, II: 246.
---- 1909. Notes on the classification of the Dermaptera. Deutsch. Ent. Zeitschr., 1909:320, pl. IV.
---- 1910a. The fauna of British India, including Ceylon and Burma. Dermaptera. London: 1—217, I—XVIII, 10 plates.
---- 1910 b. A preliminary revision of the Labiduridae, a family of the Dermaptera. Trans. Ent. Soc. London, 1910 :161—203, pls. XLVI—XLVII.
---- 1911. Genera Insectorum, fasc. 122, Dermaptera. Bruxelles:!—112, 9 plates.
---- 1912 a. Contribution to our knowledge of Indian earwigs. Journ. Asiat. Soc. Bengal, VII: 771—800.
---- 1912b. Die Dermapteren des k. k. naturhistorischen Hofmuseum in Wien. Ann. Hofmus. Wien, XXVI: 63—108.
---- 1915 a. The opisthomeres and the gonapophyses in the Dermaptera. Trans. Ent. Soc. Lond. 1915 : 257—268.
---- 1915b. Notes on the manubrium of the ninth sternite in the male earwig. Ibid., 1915 : 268— 272, tab.
---- 1916. On the male genital armature of the Dermaptera. Journ. R. Mier. Soc., 1915:413— 447, pl. V—IX; 521—546, pl. X—XII; 1916:1—18, pl. I—IV.
В u x t о n, P. A. 1921. Earwigs from Mesopotamia and N. W. Persia. Journ. Bombay Nat. Hist. Soc., XXVII: 963—964.
Chapman, T. A. 1917a. Notes on early stages and life history of the earwig (Forficula auricularia L.). Ent. Rec. a. Journ. Variat., XXIX: 25—29, 3 pls.
---- 1917b. Further notes on the earwigs. Ibid., XXIX: 177—180.
Chopard, L. 1922. Faune de France. 3. Orthopteres et Dermapteres, Paris: 1—122, 466 fig.
[Djakonov, D. M.] Дьяконов, Д.М. 1923. Заметки о кастрации, паразитах и регенерации клещей у уховерток. Изв. Биолог. Инет. Пермск. Унив., 1, 9—10:159—164.
Djakonov, D. М. 1925. Experimental and biometrical investigations on dimorphic variability of Forficula. Journ. Genet., XV: 201—232.
D о h r n, H. 1863. Versuch einer Monographic der Dermapteren. Stett. Ent. Zeit., XXIV: 35— 66, 309—322.
---- 1864. Id., op. cit., XXV: 285—296, 417—429.
---- 1865. Id., op. cit. XXVI: 68—99.
---- 1867. Id., op. cit. XXVIII: 341—343.
Dufour, L. 1828. Recherches anatomiques sur les Labidoures. Ann. Sc. Nat., XIII: 337—366, tab. XIX-XXII.
F i e b e r, Fr. 1853. Synopsis der europaischen Orthopteren. Lotos, III: 92, 252—258. Также отдельно, Prag, 1854 :1—78, 1—4.
F i s c h e r, L. H. 1854. Orthoptera europaea. Lipsiae : 1—454, I—XX, 18 tab.
Fischer von Waldheim, C. 1846. Orthoptera Imperii Rossici. Mosquae: 1—443, 37 tab.
Gadeau de Kerville, H. 1905. Notes sur les fonctions de la pince des insectes Orthop-teres de la famille des Forficulides. Bull. Soc. Zool. France, XXX: 53—6.
Gene, G. 1832. Saggio di una Monographia del Forficula indigene. Ann. Sc. Nat. Regno Lombardo-Veneto, Padova, II: 215—228.
H ebard, M. 1923. Studies in Indian Dermaptera. Mem. Dep. Agric. India, VII: 195—242, pls. XIX—XXL
H e у m о n s, R. 1895. Die Embryonalentwickelung von Dermapteren und Orthopteren unter besonderer Beriicksichtigung der Keimblatterbildung, monographisch bearbeitet. Jena, VI -+-136 pp., 12 tab.
Huxley, J. S. 1927. Discontinuous variation and heterogeny in Forficula. Journ. Genetics, XVII: 309—327.
ВВЕДЕНИЕ
75
[Jacobson, G. G.]. Я к о б с о н, Г. Г. 1905 in: Якобсон, Г. Г. и Бианки, В. Л. Прямокрылые и ложносетчатокрылые Российской Империи и сопредельных стран. СПб. 1903— 1905; Dermatoptera : 6—28.
Kirby, W. Е. 1891. A revision of the Forficulidae, with description of new species in the British Museum. Journ. Linn. Soc., Zool., XXIII: 502—531, pl. 12.
---- 1904. A synonymic catalogue of Orthoptera, I. London : 1—501 (Forficulidae: 1—58).
К i 11 a г у, M. 1849. Orthopteres observes dans les steppes des Kirgises. Bull. Soc. Nat. Moscou, XXII: 437—442.
[Kolosov, J. M;]. Колосов, Ю. M. 1928. Каталог прямокрылых Среднего Урала. Свердловск, изд. Энтом. Каб. Уральск. Политехи. Института: 1—8.
Kuhl, W. 1928. Die Variabilitat der abdominalen Korperanhange von Forficula auricularia. Unter Beriicksichtigung ihrer normalen and abnormen Entwicklung, nebst einem Anhang uber die Geschlechtsbiologie. Zeitschr. Morph. Okol. Tiere, 12:299—532, 69 figs.
L ii s t n e r, G. 1914. Die Nahrung des Ohrwurmes (Forficula auricularia L.) nach der Inhalt seines Kropfes. Zentralbl. Bakt., 40 : 482—514.
Matsumura, S. undShiraki, T. 1905. Monographic der Forficuliden Japans. Journ. Sapporo Agr. Coll., II: 75—86, 3 figs.
Morgan, W. D. 1924. Notes on the function of the forceps of earwigs. Proc. Indiana Acad. Sc., 33, 1923:303—307, 7 figs.
---- 1926. A note on the mode of distribution of earwigs. Ibid., 36:332—333.
N e 1, R. J. 1929. Studies in the development of the genital and the genital ducts in Insects. I. Female of Orthoptera and Dermaptera. Quart. Journ. Mier. Soc. 73: 20—85, 4 pls., 5 figs. [Pesotzkaja, E.]. П e с о ц к а я, E. 1927. К биологии уховерток Forficula tomis Koi. и Labidura riparia Pall. Изв. Научи. Инстит. Лесгафта, XII, 2:51—61.
[Pravdin, Th.] Правдин, Ф. "1934. Азиатская уховертка н борьба с ней. Советск. Каучук, 5 : 31—34.
[R о d z j а n к о, V. N.] Родзянко, В. Н. 1897. О паразитизме личинок мухи Roeselia antiqua Meig. внутри личинок уховертки Forficula tomis Koi. Труд. Русск. Энт. Общ., XXXI: 72-86.
Saufesure, Н. 1874. Путешествие в Туркестан А. П. Федченко. Прямокрылые. Изв. И. Общ. Люб. Ест., Антр. и Этногр., XI, 4:1—-52,1 tab.
Scudder, S. Н. 1876. Critical and historical notes on the Forficulariae. Proc. Bost. Soc. Nat. Hist., XVIII: 287—332.
[S h u g u г о v, A. M.] Ш у г у p о в, А. М. 1909. Материалы к познанию ортоптерофауны России. Зап. Новоросс. Общ. Естествоисп., XXXIV: 109—124-
---- 1912. Материалы к изучению географического распределения прямокрылых в Таврической губ. Ibid. XXXVII: 145—167 (Forficulodea : 146—148).
Schtscherbakov, Т. 1903. Neue Daten zur geographischen Verbreitung von Forficula tomis Koi. und Labidara riparia pallipes Fabr. Zool. Anzeig., XXXIII: 97—101.
Щербаков, Ф. C. 1911. Несколько новых данных о географическом распространении уховерток Российской Империи (Orthoptera, Forficulodea). Русск. Энт. Обозр., XI: 271-276.
---- 1912. Dermatoptera коллекции В. П. Мочульского. Ibid., XII: 349—355.
---- 1913. Список уховерток (Dermatoptera) Крыма. Зап. Крымск. Общ. Исп. Прир. П., 1912:1—6 (отд. отт.)
Семенов [Тя н-Ш а н с к и й], А. П. 1901 а. Русские виды родов Anechura Sc. и Forficula L. и их географическое распределение. Труд. Русск. Энт. Общ., XXXV: 185—201. ---- 1901b. Первый палеанарктический представитель рода Opisthocosmia Dohrn. Русск. Энт. Обозр., 1 : 98—100.
---- 1901с. К географическому распространению Forficula auricularia L., Ibid., 1:103—104. ---- 1903. Заметки об уховертках (Dermatoptera) русской фауны. Ibid., Ill: 196—199.
Семенов-Тян-Шанский, А. П. 1935. Общий обзор фауны кожистокрылых (Dermatoptera) СССР. Известия Акад. Наук, 1935: 825—830.
76
КОЖЙСТОКРЫЛЫЕ
Semenov [Т i a n-S h a n s к i j], A. 1902. Dermatoptera nova aut minus cognita, I. Rev. Russe d’Ent., II: 99—102, 2 fig.
---- 1903. Id. II, Ibid., Ill: 19—21.
---- 1908. Id., Ill, Ibid. VIII: 159—173.
—— 1936. Notes sur quelques Dermatopteres. Eds, X
S e m e n о v-T i a n-S h a n s к y, A. et В e y-B i ё n к o, G. 1934. Dermapteres de la Mongolie. Bull. Soc. Ent. France, XXXIX: 55—58, 1 fig.
------ 1935. Les Dermapteres du Tibet. Eos. X : 221—232, 6 figs.
Serville, Aud. 1839. Histoire naturelie des irisectes Orthopteres. Paris, 1—766, 14 tab. (Forficulariae : 9—-55, tab. I).
S h i г a к i, T. 1905. Neue Forfictilideri Japans. Trans'. Sappofo Nat. Hist. Soc., I, part. 1 : 91— 96, tab. III.
---- 1906. Neue Forficuliden und Blattiden Japans. Ibid. I, part. 2 :1—14.
---- 1928. Dermapteren aus dem Kaiserreich Japan. Insecta Matsumur., Ill: 1—25, 9 fig.
Verhoeff, K. W. 1902a. Ueber Dermapteren. I. Versuch eines netien natiirlichen Systems Zool. Anzeig., XXV: 181-208.
---- 1902b. Id. 2. Neue ungeflugelte Eudermapteren-Gattungen. Sitz.-Ber. GeSelsch. Naturf. Fr. Berlin, 1901, № 1:7—18.
---- 1903. Ueber die Endsegmente des Korpers der Chilopoden, Dermapteren und Japygi-den. Nova Acta Acad. Caes.-Leop. Carol., Halle, XXXI.
---- 1913. Ueber Dermapteren. Zeitschrift fur wissensch. Insektenbiologie, IX.
Werner, F. 1901. Die Dermapteren- und OrthopterenfaunajKleinaSieas. Sitz.- Bet. K; Akad. Wiss. Wien, Math.-nat. Classe, GX: 259—306.
Wnukowsky, W. 1926. Zur Fauna der Orthopteren tad Dermatopteren des Bezirks Kamenj. Mitt. Miinchn. Ent. Ges., 16 : 87—92.
Zach er, F. 1911. Studien uber das System der Protodermapteren. Zool. Jahrb., Abt. Syst., XXX: 304—400, 80 figs.
---- 1912. Das mannliche Copulationsorgan urid das System der Eudermaptereft. Zeitschr. Wiss. Insectenb., VIII: 276—284.
---- 1917. Die Geradfliigler Deutschlands und ihre Verbreitung, Jena: 1—287 (Dermaptera'. 46—59), 1 Karte.
[Zubovskij, N. N.]. Зубовский, H. H. 1897. Прямокрылые С.-Петербургской губ. Ежегод. Зоол. Муз. И. Ак. Наук, II: 162—214 (Dermatoptera: 165, 176—179).
’ СПЕЦИАЛЬНАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ
1 (2). Дапки ног 4—5-члениковые. Надкрылья с немногими продольными приподнятыми полосками (жилками?). Церки тонкие, короткие, членистые. . ». . . . . . .1. Подотряд Archidermaptera nov. ( i) 2 (1). Лапки ног 3-члениковые. Надкрылия, если развиты, всегда без продольных приподнятых полосок, лишь у немногих форм с продольным боковым килем. Церки одночлениковые, большие, превращены в клещи.
3 (4). Глаза сильно редуцированные. Верхние челюсти с тремя зубцами в вершинной части; внутренний край густо покрыт щетинками. Церки слабо хитинизированные, образующие несовершенные клещи. Совершенно бескрылые формы ......... 2. Подотряд Arixenina.
4 (3). Глаза вполне развитые. Челюсти с двумя зубцами на вершине, без щетинок на внутреннем крае. Церки сильно хитинизированные, образующие сильные клещи. Крылатые или бескрылые формы ....
............................ . 3. Подотряд Dermaptera s. str.
I. Подотряд ARCHIDERMAPT ERA nov. (-*-)
Усики с довольно длинным 2-м члеником, равным по длине 3-му. Надкрылья довольно длинные, достигающие до 2-го членика брюшка, с немногими продольными приподнятыми полосками (жилками?).1 Ноги короткие, бедра толстые, по крайней мере последние из них с двумя килями; лапки ног более чем 3-члениковые, задние 5-ти, передние и средние 4-х или 5-члениковые. Церки короткие, стройные, с немногими (не более 10-ти) члениками, из которых основной наиболее длинный.
Этот подотряд устанавливается нами для ископаемого мезозойского семейства Protodiplatyidae, известного пока всего только по одному виду —
1 Благодаря любезности А. В. Мартынова, я имел возможность изучить тип Proto-diplatys fort is Mart., хранящийся в Палеозоологическом Институте Академии Наук СССР; немногочисленные продольны^ ребрышки на надкрыльях, повидимрму, представляют собой жилки; однако, сохранность этой части типа не позволяет решить вопрос более определенно. Во всяком случае, наличие продольных жилок на надкрыльях вполне вероятно, так как у палеозойских Protelytroptera эти жилки, как правило, хорошо выражены.
78
КОЖИС ТОКРЫЛЫЕ
Protodiplatys fortis Mart, из юрских отложений южного Казахстана (с. Галкино, горы Каратау).
Вид этот представляет большой научный интерес, так как является промежуточной формой между современными подотрядами и тараканообразными предками типа палеозойских Protelytroptera, что и послужило причиной выделения ее в особый подотряд; сильно укороченные надкрылья, короткие, с толстыми бедрами ноги и удлиненное, стройное брюшко не вызывают сомнения в принадлежности этой формы к отряду Dermaptera. Присутствие у Protodiplatys килей на бедрах сближает его с современным сем. Pygidicranidae из подотр. Dermaptera s. str., которое как раз характеризуется тем, что личинки многих видов имеют сегментированные церки.
Нахождение этой формы указало путь, по которому шла эволюция отряда Dermaptera’, поэтому совершенно несомненно, что в дальнейшем в мезозойских отложениях будут найдены другие представители, родственные данной ископаемой форме.
IL Подотряд ARIXENINA
Burr, 1911:6—7.
По внешнему виду напоминает бескрылых уховерток. Тело покрыто густыми волосками. Голова сердцевидная, не плотно соединяющаяся с грудью; глаза сильно рудиментированы; верхние челюсти с тремя зубцами в вершинной части, внутренняя сторона густо покрыта жесткими щетинками. Усики с очень коротким 2-м члеником. Переднеспинка почти полукруглая, поперечная. Надкрылья и крылья отсутствуют. Лапки ног трех-члениковые. Церки одночлениковые, густо волосистые, слабо хитинизован-ные, образующие несовершенные слабые клещи. Половой аппарат отличается от настоящих уховерток тем, что имеет форму цилиндра или трубки, а не уплощен в спинно-брюшном направлении; пропарамеры очень слабо, метапарамеры едва сильнее хитинизированные; собственно penis парный, но одна половина его сильно рудиментирована, не вооружена, тогда как функционирующая половина вооружена хитиновыми включениями.
Этот подотряд включает всего только одно семейство Arixeniidae с одним родом Arixenia Jordan и всего только двумя видами известными с Малайского архипелага; один вид этого рода (A. esau Jord.) описан как паразит рукокрылых (Chiroptera), найденный на теле последних. Однако, другой вид, открытый несколько позднее (Л. jacobsoni Burr), оказался обитателем пещер, живущим на покрывающем пол пещеры помете (гуано) и ведущим хищнический образ жизни, подобно настоящим уховерткам (Burr a. Jordan, 1913). Очень возможно, что нахождение A. esau на теле Chiroptera представляет собой случайное явление и оба вида являются пещерными обитателями, „паразитический" облик которых (редукция глаз и пр.) явился в результате жизни в пещерах.
I. AR CH I DERMAPTERA, 11. ARIXENINA
79
Представление об Arixenina как о насекомых ведущих паразитиче-кий (точнее — полупаразитический) образ жизни настолько укоренилось в литературе, что в ряде новейших руководств (Chopard, 1922; Handlirsch, 1926; Beier, 1933) эта группа до сих пор фигурирует как пример полу-паразитов в отряде Dermaptera и совершенно забывается работа Burr a. Jordan, из которой следует, на основании наблюдений Е. Jacobson’a, что образ жизни A. jacobsoni не имеет ничего общего с паразитизмом.
III. Подотряд DERMAPTERA s. str. (= FORFICULINA)
Dermaptera Kirby, Trans. Linn. Soc. London, XI, 1815 : 87. — Dermatoptera Burmeister, Handb. Ent., II, 1838 : 743. — Euplexoptera Westwood, Introd. Class. Ins., 1,1839 : 398. — For-ficularia Brunner, 1882:1.—Foificulina Burr, 1911:9.
Голова более или менее сердцевидная или округленно-пятиугольная, глаза вполне развиты; верхние челюсти с двумя зубцами на вершине, внутренняя сторона без щетинок. Усики с очень коротким 2-м члеником. Переднеспинка разнообразная, обычно более или менее прямоугольная, реже округленная, от несколько удлиненной до поперечной. Надкрылья и крылья вполне развиты, или в различной степени укороченные, или совсем отсутствуют. Лапки ног 3-члениковые. 3-й и 4-й тергиты брюшка часто по бокам несут по бугорку. Церки сильно хитинизированные, крупные, одночлениковые, превращенные в сильные клещи. Половой аппарат 3 с парным (надсем. Protodermaptera) или непарным (надсем.£пс?егтар/ега) penis.
Этот подотряд включает подавляющее большинство видов, группирующихся в два надсемейства, пять семейств и множество подсемейств, относящихся как к современным, так и ископаемым формам. Среди последних из мезозоя известен пока только один род и вид (Semenoviola obliquotruncata Mart, из юрских отложений ю. Казахстана), а из третичных отложений около 30 видов (начиная с нижнего олигоцена), относящихся как к современным, так и к вымершим родам.
В пределы Палеарктической области заходят четыре семейства; только одно семейство Apachyidae здесь отсутствует.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ НАДСЕМЕЙСТВ И СЕМЕЙСТВ
1 (4). Генитальный аппарат 3 двойной, с парным penis, основание которого укреплено у вершины пропарамер. Пигидий 3 простой. (Надсем. Protodermaptera).
2 (3), Опистомеры состоят из трех хорошо развитых частей, из которых пигидий чуть больше двух остальных члеников взятых вместе. Бедра у большинства подсемейств сжаты с боков, с продольными килями.1
................................1. Сем. Pygidicranidae (стр. 81).
1 Этот признак отсутствует у южно-американского подсем. Pyrarginae и палеотропического подсем. Echinosomatinae, которые, однако, характеризуются сильным развитием на теле коротких щетинкоподобных волосков.
80 HI. DERMAPTERA
3 (2). Опистомеры состоят только из двух ясных члеников, из которых верхний — пигидий — в 2—6 раз длиннее другого. Бедра цилиндрические, без килей . . ................2. Сем. Labiduridae (стр. 83).
4 (1). Генитальный аппарат $ простой, с непарным penis, основание которого укреплено у основания пропарамер. Пигидий 3 часто вооружен шипами и отростками. 2-й членик лапок простой или сердцевидно расширен, или же на нижней стороне е узкой лопастью, продолжающейся под 3-й членик. (Надсем. Eudermaptera).
5 (6). 2-й членик лапок простой, не расширен сердцевидно или же с нижней стороны несет узкую лопасть, продолжающуюся под 3-й Членик. Генитальный аппарат 3 с заостренными на вершине, часто оттянутыми метапарамерами......................3. Сем. Labiidae (стр. 108).
6 (5). 2-й членик лапок сердцевидно расширен. Генитальный аппарат 3 с более или менее округленными на вершине метапарамерами. . . .
..............................4. Сем. Forficulidae (стр. 120).
Помимо этой таблицы, пользование которой затруднено, вследствие необходимости изучения полового аппарата 3, мы прилагаем другой, значительно более упрощенный, синопсис, который хотя и не даёт полной и наиболее надежной характеристики семейств, но при его помощи можно будет определять без особого труда всех представителей отряда, вошедших в данную работу.
1 (6). 2-й членик лапок не расширен сердцевидно, простой или снизу с узкой лопастью, продолжающейся под 3-й членик. Усики обычно более чем 15-члениковые.
2 (3). Бедра сжатые с боков, е продольными килями. Самки имеют гона-пофизы......................... 1. Сем. Pygidicranidae.
3 (2). Бедра цилиндрические, без килей. Самки без гонапофизов.
4 (5). Задний конец последнего тергита брюшка подогнут вниз и сращен с пигидием, который образует наклонную и направленную своим нижним концом вперед пластинку, заключенную между основаниями клещей и не видную при рассматривании сверху, Надкрылья вполне развитые, или укороченное, или совсем отсутствуют..................
.........................................2. Сем. Labiduridae.
5 (4). Последний спинной сегмент брюшка не срастается с пигидием. Пигидий 3 часто вооружен шипами и отростками, свободный, заметный сверху, если-же вертикальный, то его нижний конец не направлен вперед. Надкрылья вполне развитые.......... ,3. Сем. Labiidae.
6 (1). 2-й членик лапок расширен сердцевидно, значительно шире остальных, Усики 10—15-члениковые. ........ 4. Сем. Forficulidae.
I. Надсемейство PROTODERMAPTERA
Генитальный аппарат самца двойной, с парным penis, основание которого укреплено у вершины пропарамер (фиг. 7) (только у африканского
ТАБЛИЦА СЕМЕЙСТВ
81
подсем. Karschiellinae один penis рудиментарный); пррпарамеры сильно хитинизированные, большие, отделены от метапарамер ясной перетяжкой или бороздой (только у палеотропического подсем. Echinosomaiinae граница между пропарамерами и метапарамерами слабо выражена). Опистомеры состоят из трех члеников: метапигидия, тельсона и пигидия, из которых наиболее сильно развит пигидий, а предыдущие два опистомера более коротки или (Labiduridae) сильно редуцированы, образуя один общий членик? Пигидий простой, без выростов. Глазки часто имеются; ветви А2 частот с внутренней дополнительной ветвью. Клещи у личинок некоторых родов, многочленистые.
Данное, надсемейство является, по сравнению с следующим надсем. Eudermaptera, более примитивным, что особенно подчеркивается присут-: ствием глазков, которые у высших Dermaptera совершенно отсутствуют или едва заметны, двойным половым аппаратом самца мало дифференцированными опистомерами (Pygidicranidae) и многочлениковыми клещами у личинок некоторых родов. ‘ '
Представители надсем. Protodermxiptera являются в огромном своем большинстве жителями тропиков как Старого, так и Нового Света; вполне достоверных аборигенов в умеренщх широтах известно всего несколько видов — все остальные виды, населяющие умеренные широты, являются видами, завезенными из тропиков.
Надсем. Protodermaptera состоит из трех семейств—Pygidicranidae И Labiduridae, заходящих в пределы Палеарктической области, и Apachyi-dae, свойственного только тропикам Старого Света.
1. Семейство PYGIDICRANIDAE
, • . , Голова уплощенная, , не выемчатая по заднему краю. Усики 15—50-ти-; члениковые; первый членик усиков обычно с килем на верхней стороне. Глазки имеются. Надкрылья обычно развиты, реже укороченные или совсем отсутствуют вместе с крыльями (только у корейского рода Challia Burr, Канарского Anataelia Bolivar и тропического Acnodes Burr). Бедра у большинства родов сжаты с боков, с продольными килями; только у не-рассматриваемых здесь неотропического подсем. Pyrarginae и палеотропического подсем. Echinosomatinae кили отсутствуют. Лапки простые; их 2-й членик не расширен сердцевидно и не продолжен в виде лопасти под 3-й (только у рода Tagalina Dohrn из Австралийской области и Филиппин 2-й членик лапок сердцевидно расширен). Брюшко цилиндрическое, часто сужено перед серединой и расширено к вершине. Все три опистомера-хорошо развиты; метапигидий и тельсон, взятые вместе, почти равны по длине пигидию (фиг. 6 В). Самки имеют гонапофизы (фиг. 7), исключая подсем. Pyrarginae. Личинки многих родов характеризуются многочлениковыми клещами (церками).
Фауна СССР, Кожистокрылые 6
82
1. PYGIDICRANIDAE
Семейство это делится на много подсемейств, из которых только подсем. Anataelinae заходит в Палеарктику.1
1. Подсем. ANATAELINAE
Тело стройное, совершенно бескрылое. Глаза маленькие; усики с 16-— 25 члениками, в коротких волосках. Заднегрудная пластинка сзади обрублена. Бедра сжаты с боков, с продольными килями. Личинки с несегменти-рованными церками.
Известно всего два вида, относящихся к двум родам, из которых один, рассматриваемый ниже, водится в Корее и Сев. Китае, а другой на Канарских островах.
1. Род CHALLIA BURR.
Burr, 1904:286; Burr, 1911:11.
Тип рода: Challia fletcheri Burr.
Средних размеров. Усики по крайней мере с 16 члениками, в коротких волосках; 1-й членик длинный; 2-й короткий, 3-й длинный, немного длиннее 4-го и 5-го, остальные членики удлиненные. Переднеспинка почти квадратная; среднеспинка почти квадратная, по бокам с ясным продольным килем,, задний край ее слегка дуговидно вырезан; заднеспинка широкая, с сильной дуговидной вырезкой по заднему краю. Переднегрудная пластинка, удлиненная, слегка суживается перед задним краем; среднегрудь и заднегрудь широкие, последняя с прямым задним краем. Ноги стройные,, довольно длинные. Брюшко в волосках, без бугорков по бокам 3-го и 4-го члеников, суженное у 3 по средине, цилиндрическое у g; последний тергит вздут у 3, квадратный у g, с шаровидными бугорками у обоих полов. . Клещи <3 с соприкасающимися ветвями, толстые, уплощенные, несколько расширены и дуговидно-изогнуты перед вершиной; у g простые, прямые.
Известен только, один вид.
1. Challia fletcheri Burr
Burr, 1904:286,3, g (Вей-хай-вей, с.-вост. Китай; Да-лин-суэн, Тунг-юн и остр. Тамгой, южн. Корея); Burr, 1911:11, tab. 1, fig. 10,
Тело довольно крупное, рыжевато-темное, сверху грязно-желтое с темными отметинами или весь верх темный. Ноги грязновато-желтые,, с темными полосками. Последний тергит брюшка сильно пунктирован, посредине с бороздкой, более глубокой у 3, задний край с 5 шаровидными бугорками, из которых 3 маленькие, а 2 более крупные. Клещи при осно
1 Описанная Burr’ом из „Закаспийской области" Kalocrania semenoffi (Sitzungsber. Ges. Nat. Freunde, Berlin, 5,1912: 312, fig. 1) несомненно тропического происхождения;; указание на Закаспийскую область очевидно основано на неверной этикетке.
/. CHALLIA
83
вании грязновато-желтые, в остальной части темные, у $ более толстые, внутренний край перед серединой приподнят кверху в виде тупого угла, далее сильно уплощенные, слегка изогнутые, перед вершиной параллельные, внутренний край зазубрен, на самой вершине сильно загнутые, перекрещивающиеся; у g — прямые, параллельные, тонкие, цилиндрические, внутренний край у основания мелко зазубрен, перед вершиной с несколькими заостренными зубчиками, у самой вершины загнутые, перекрещивающиеся. Длина тела 3 15, § 14.5—16; клещей 3 5, 2 4.5—5 мм.
Распространение: известен из сев.-вост. Китая и ю. Кореи у побережья Желтого моря.
Этот вид и род являются единственными представителями сем. Pygidicranidae в Палеарктической Азии, причем распространение всего подсемейства Anataelinae говорит о большой древности данного рода и всего подсемейства в целом, что подтверждается также И чрезвычайной морфологической замкнутостью двух известных родов при одновременной бедности видами. Несомненно в лице корейской Challia fletcheri и Канарской Anataelia canariensis Во!, мы имеем уцелевших до настоящего времени представителей некогда, видимо, богатой и широкораспространенной группы уховерток.
2. Сем. LABIDURIDAE
Г олова часто выемчата по заднему краю; глазки имеются или отсутствует. Усики с различным числом члеников, от 10—15 (поде. Parisolabinae vi.Brachylabinae} до 32—35 (Labidurinae), 3—4-й членики короткие, обычно короче 1-го и 3-го, 5-й членик часто также короткий. Надкрылья и крылья у большинства видов совсем отсутствуют или надкрылья зачаточные, а крылья отсутствуют; реже надкрылья вполне развиты, в этом случае часто с продольными боковыми килями. Бедра цилиндрические, без килей; 2-й членик лапок простой, не расширен сердцевидно и не продолжен под 3-й в виде лопасти. Брюшко у большинства родов постепенно расширяется кзади или по средине, особенно у 3, редко с параллельными сторонами; бугорки на боках 3-го и 4-го члеников обычно отсутствуют; задний конец последнего спинного полукольца подогнут вниз и сращен с пигидием, заключенным между основанием клещей и образующим вертикальную треугольную пластинку; при рассматривании сверху пигидий не виден, так как он не только расположен вертикально, но своим нижним концом направлен, иногда очень сильно, вперед, так что линия сращения пигидия с задним концом последнего тергита брюшка выступает в виде более или менее острого клина; место сращения последнего тергита и пигидия часто не имеет даже заметного шва. Опистомеры состоят только из двух члеников — очень крупного пигидия и в 2—6 раз меньшего метапигидия; последний собственно представляет собой рудимент не только метапигидия, но и 6*
84
2. LABID URIDAE
тельсона (фиг. 6 a). Самки не имеют гонапофизов.1 Клещи различной формы, часто асимметричные, у личинок не сегментированы.
Это семейство включает в себя ряд подсемейств, из которых только 3 заходят в пределы Палеарктики, а остальные распространены по преимуществу в тропиках Старого и Нового света. Многие роды являются космополитами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ И РОДОВ
1 (2). Среднегрудная пластинка с округленным задним краем.2 Клещи на поперечном разрезе треугольные, у £ обычно асимметричные. Ману-брий очень длинный, при основании узкий, на вершине расширенный. Генитальный аппарат S без почковидного пузырька и часто без virga (фиг. 20—22, 25). [1. Подсем. Psa.lin.ae]. — Заднегрудная пластинка обрублена сзади. Брюшко не очень сильно расширено за срединой, далее слегка суживающееся к вершине. (Совершенно бескрылые или с зачаточными надкрыльями; редко с вполне развитыми надкрыльями и крыльями) ...........................1. Anisoiabis Fieb.
2 (1). Среднегрудная пластинка с обрубленным задним краем. Манубриум короткий, не более чем в 2 раза длиннее своей ширины, не расширен на вершине. Генитальный аппарат S с почковидным пузырьком и virga (фиг. 28).
3 (6). Усики 25—35-члениковые. Переднегрудная пластинка сужена кзади, средне- и заднегрудные пластинки такой же длины, как их ширина. Надкрылья вполне развитые (2. Подсем. Labidurinae).
4 (5). Крупнее (длина тела 9—26 мм). Ноги длинные, задние бедра в 1 х/2 раза длиннее переднеспинки; 1-й членик задних лапок длиннее 3-го. Метапарамеры на вершине слегка расширены и округлены, с маленькой дополнительной лопастью................2. Labidura Leach.
5 (4). Мельче (длина тела 7.5—13 мм). Ноги относительно короткие, задние бедра не длиннее переднеспинки; 1-й и 3-й членики задних лапок одинаковой длины. Метапарамеры очень сильно заострены на вершине, без дополнительной лопасти.................... . 3. Nala Zach.
6 (3). Усики 10—15-члениковые. Переднегрудная пластинка с параллельными боковыми сторонами; средне- и заднегрудная пластинки шире своей длины. Маленькие, совершенно бескрылые формы. (3. Подсем. Parisolabinae). Последний тергит брюшка очень короткий, у S чуть сужен к вершине. — Переднеспинка поперечная, более чем в полтора раза длиннее своей ширины, с прямым задним краем....................
......................................... 4. Pseudisolabis Burr.
1 Исключая представителей палеотропического подсем. Allostethinae.
2 Только у Ново-Каледонского рода Spondox Burr среднегрудная пластинка обрублена сзади.
ТАБЛИЦА РОДОВ
85
1. Подсем. PSALINAE
Тело различной величины, от маленького и стройного до очень крупного. Усики со многими (15—30) Мелкими члениками, 4, 5 и 6-й членики короткие, короче 3-го. Переднегрудь не сужена кзади, с почти параллельными боковыми сторонами; среднегрудь округлена по заднему краю (за исключением только одного рода — Spondox Burr с Новой Каледонии, у которого среднегрудь сзади обрублена); заднегрудь обрублена сзади или же вытянута в округленную лопасть. Надкрылья и крылья отсутствуют или надкрылья зачаточные, а крылья отсутствуют, или же надкрылья и крылья вполне развиты (гл. обр. у видов Нового Света). Брюшко расширено у средины или к заднему концу. Клещи на поперечном разрезе треугольные, толстые, довольно сближенные, у 5 обычно асимметричные, с редкими, мелкими, иногда неясными зубчиками по внутреннему краю. Манубрий очень длинный, узкий при основании, расширен к вершине, прямоугольный. Генитальный аппарат <3 с различной у разных родов и подродов длиной метапарамер; вирга если имеется, то простая; почковидный пузырек всегда отсутствует.
Очень большое количество видов, внешне чрезвычайно сходных друг с другом; поэтому распознавание видов по наружным морфологическим признакам крайне затруднено и подчас невозможно, так как многие представители крайне плохо изучены. Систематика группы в прежней время основывалась главным образом на отличиях в степени развития надкрылий; однако дальнейшие исследования показали, что виды с сильно укороченными надкрыльями и отсутствующими крыльями могут давать и длиннокрылую форму. В связи с этим ряд форм, в прежнее время считавшихся самостоятельными видами и даже относившихся к различным родам, в действительности оказались принадлежащими к одному и тому же виду (напр. обычно короткокрылая A. stali Dohrn иногда дает длиннокрылую форму, известную прежде под названием Psalis plebeja Dohrn и долго считавшуюся не только самостоятельной, но даже относящейся к, другому роду — Psalis).
Внешнее однообразие группы, однако, не соответствует чрезвычайному разнообразию в строении полового аппарата самца; использование этого признака позволило в значительной мере разобраться во взаимоотношениях многих родов и видов; однако, некоторые авторы (Zacher, Burr) слишком переоценили значение полового аппарата для систематики подсемейства, одновременно игнорируя наружную морфологию, что привело к мало обоснованному дроблению родов. В частности большое значение придавалось в качестве родового критерия присутствию или отсутствию вирги, тогда как несомненно близкие виды часто характеризуются отсутствием virga у одного из них и присутствием у другого. Наибольшее значение в классификации подсемейства имеет строение метапарамер,
86
2. LABIDURIDAE
которые по своей форме изменяются от лопастевидно удлиненных, с отношением длины к ширине как (3—4): 1, до почти квадратных или трапециевидных, длина которых не превосходит их ширину; к промежуточной группе относятся виды с промежуточной формой метапарамер. Этот признак был положен Вшт’ом (1916) как основа для выделения весьма большого количества родов. Однако, формы, относящиеся к таким „ррдам“ настолько сходны между собой, что многие неотличимые друг от друга по внешним признакам виды оказались представителями разных „родов". С другой стороны длина метапарамер у некоторых весьма близких форм может быть различной, а тип их строения оказывается одним и тем же, так что если стать на точку зрения Burr’a, то эти виды пришлось бы отнести к разным родам, что, конечно, было бы неправильно; примерами таких видов являются Anisolabis pacifica Erichs, и A. formosana Bor.
Все это заставляет нас трактовать многие роды, установленные Zacher’oM (1911) и Вшт’ом (1916) только как подроды. В результате этого ряд родов характеризуется почти космополитным распространением; однако в этом нет ничего странного, так как вся группа Protodermaptera несомненно мезозойского происхождения и широкое распространение родов говорит о глубокой их древности; с другой стороны и многие установленные Zacher’oM и Вигг’ом роды также имеют весьма широкое распространение.
Правильная классификация подсемейства может быть дана только после его тщательной ревизии во всесветном масштабе; тогда будут более ясными взаимоотношения отдельных видовых групп, хотя и сейчас совершенно ясно, что родовых подразделений для имеющихся видов сделано больше, чем это необходимо.
1. Род ANISOLABIS FIEB.
Fieber, 1853:257; Bormans 1900:41; Kirby, 1904:17; Якобсон, 1905:23; Burr, 1911:28. — Forcinella Dohrn, 1862: 226. — Brachylabis Dorhn, 1864:292 (partim).—Eubo-rellia Burr, Proc. U. St. Nat. Mus., XXXVIII, 1910:448; Burr, 1911:30. — Apolabis Burr, 1916:538. — Mongolabis Zacher, 1911:385.
Тип рода—A. marltima Gene.
Тело выпуклое, довольно вытянутое, обычно коренастое. Усики с 15—25 члениками, постепенно утончающиеся к вершине; 3-й членик цилиндрический, равен половине длины 1-гб членика, так же, как и 4-й, короткий, оба не длиннее своей ширины, 5-й немного длиннее своей ширины, последующие членики постепенно удлиняются. Голова выпуклая, длиннее своей ширины. Переднеспинка прямоугольная, почти квадратная; среднеспинка поперечная, заднеспинка с дуговидно-выемчатым задним краем. Надкрылья и крылья полностью отсутствуют, или надкрылья зачаточное; редко те и другие вполне развиты. Грудные пластинки довольно узкре, длиннее своей ширины; заднегрудь несколько выступает между
7. ANISOLABIS
87
задними вертлугами, задний край обрублен. Брюшко уплощенное, довольно широкое, постепенно расширяется кзади, достигая наибольшей ширины около конца второй трети его длины, далее к вершине слегка сужено; боковые бугорки отсутствуют или имеются; боковые стороны 6-го и 7-го сегмента у 5 выпуклые, часто вытянуты назад в виде острых углов, с продольной морщинистостью, или же с продольными килями; эти признаки у 2 выражены очень слабо. Последний тергит у $ широкий, почти четырехугольный, шире своей длины, но уже предыдущего тергита; у £ — сходной формы, но несколько уже; предпоследний стернит $ округлен или округленно треугольный, редко слегка вырезанный по заднему краю, у 2 он шире. Клещи у 8 двух форм: сближенные или почти сближенные друг с другом, толстые, почти прямые, суживающиеся к концу; или же довольно широко расставленные у основания, прямые при основании, а затем сильно и резко загнутые внутрь, правая ветвь часто загнута сильнее, чем левая, так что клещи становятся асимметричными; у 2 клещи соприкасаются, прямые, толстые, суживающиеся к вершине.
Весьма большое количество видов, населяющих как Старый, так Новый Свет. Ряд видов изучен слабо, причем для некоторых неизвестно строение полового аппарата <5; поскольку надежное определение вида без выяснения строения его полового аппарата, при современном состоянии наших знаний, невозможно, очень важно при обнаружении этих видов дать описание и . рисунок гениталий <5.
Род Anisolabis понимается нами широко и включает свыше десяти подродов, из которых четыре заходят в пределы Палеарктической области; однако ни одного эндемичного подрода здесь нет, хотя есть несколько эндемичных видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ И ВИДОВ i
1 (2). Метапарамеры длинные, ланцетовидные, в 3—4 раза длиннее своей ширины, с почти параллельными сторонами, едва или совсем не заострены на вершине (фиг. 20). Совершенно бескрылые насекомые. (1. Подрод Anisolabis Fieb. in sp.). —Усики одноцветные, буроватожелтые, 22—25-члениковые. Клещи $ сильно изогнутые, особенно правая ветвь, расставлены при основании. Переднеспинка одноцветная. Тело сверху в редких разбросанных точечных вдавлениях (фиг. 19)
................................* 1. A. (in sp.) maritlma Gen6
2 (1). Метапарамеры короткие, не более чем в 2 раза длиннее своей ширины, на вершине часто заостренные (фиг. 21, 22, 25).
.3 (8). Метапарамеры в 1 У2—2 раза длиннее своей ширины, на вершине заострены; наибольшая их ширина приходится на основную половину (фиг. 21, 22). Совершенно бескрылые формы.
1 В определительную таблицу не включена, как недостаточно описанная, A. pectoralis с Камчатки (см. стр. 100).
88
2. LABIDURIDAE
4:(5). Наружная сторона метапарамер S-образно изогнутая; наибольшая ширина их недалеко от основания (фиг. 21). Брюшко умеренно расши-/ ренное. Клещи 5 немного расставлены, не соприкасаются при осно-вании. (2. Подрод Apolabis Burr). — Каштаново-бурый, усики серобурые, их 13—15-й или 15—16-й членики желтые. Бедра двуцветные,. Клещи <3 умеренно изогнутые, почти симметричные. Метапарамеры довольно короткие, в 1х/2 раза длиннее своей ширины, с очень маленькой короткой, округленной и оттянутой лопастью на вершине (фиг. 21).
...................................2. A. (A.) marginalis Dohrn.
5 (4). Метапарамеры с прямыми или почти прямыми сторонами, трапециевидные, наружная их сторона прямо-или тупоугольная; наибольшая ширина их около середины (фиг. 22). (3. Подрод Mongolabis Zacher)., 6 (7). 3-й членик усиков почти равен 4-му и 5-му, взятым вместе. Боковые стороны 5—8-го тергитов брюшка 5 сильно заостренные, с резким килем и продольной морщинистостью. Предпоследний стернит брюшка <5 четырехугольный, почти с параллельными боковыми сторонами и с ясной дуговидной вырезкой по заднему краю...............
.......................... . . 3. А. (М.) setshuana B.-Bienko..
7 (6). 3-й членик усиков на одну треть длиннее 4-го. Боковые стороны тергитов брюшка 5 тупоугольные, без резкого продольного киля. Предпоследний стернит брюшка 5 округленно-треугольный, с обрубленным задним краем . . . ..................4. А. (М.) cavaleriei Вог-
8 (3). Метапарамеры не длиннее или едва длиннее своей ширины, часто короче ее, неправильно округленные или четырехугольные, на вершине не заострены; penis часто с сильными зубчиками, расположенными в ряд прямых линий (фиг. 25). Совершенно бескрылые, или с зачаточными надкрыльями, редко с вполне развитыми надкрыльями и крыльями формы. (4. Подрод Euborellia Burr).
9 (10). Совершенно бескрылая (очень редко вполне крылатая) форма. Бока 6—9-го сегментов 5 острые, с сильным килем и грубой продольной морщинистостью. Метапарамеры трапециевидные, наружный их край тупоугольный. Тело маленькое, стройное, блестяще-черное; ноги желтоватые, обычно с черными колечками (фиг. 24) . * 5. А. (Е.) annulipes Luc.
10 (9). Рудименты надкрылий имеются в виде пластинок по бокам среднеспинки или же надкрылья и крылья вполне развиты. Метапарамеры с почти дуговидно округленным наружным краем.
И (14). Мельче (длина тела 8—10 мм). Усики темные, со светлыми 1—3 предвершинными члениками. Ноги светложелтые, голени и бедра иногда с темными колечками. Брюшко 5 с продольным килем на боках 7—10-го сегментов.
12 (13). Тело черное или коричневато-черное. Переднегрудная пластинка с округленным задним краем. 3-й членик усиков более чем в 2 раза длиннее 2-го. Ноги желтоватые, голени и бедра с черноватыми колечками .................. ....................6. А. (Е.) plebeja Dohrn.-
1. ANISOLABIS
89
13 (12). Тело сверху каштаново-коричневое. Переднегрудная пластинка обрублена по заднему краю. 3-й членик усиков не более, чем в 1 х/2—2 раза длиннее 2-го. Ноги светложелтые, иногда с 'темным основанием голеней................................7. А. (Е.) pallipes Shir.
14 (11). Крупнее (длина тела 11—15 мм). Усики одноцветные, темнобурые. Ноги с темными, на вершине желтоватыми бедрами. Брюшко 5 с продольным килем на боках б—10-го сегментов (фиг. 26). . . .
..........................................* 8. А. (Е.) moesta Serv.
1. Подрод Anisolabis Fieb.
Fieber, 1853: 257; Zacher, 1911 : 373; Burr, 1916 : 534. — Forcinella Dohrn, 1862: 226..
Совершенно бескрылые виды. Метапарамеры длинные, ланцетовидные, в 3—4 раза длиннее своей ширины, с параллельными сторонами, на вершине не заострены. Виды средних или крупных размеров, довольно коренастые.
Этот подрод включает сравнительно немного видов, распространенных только в Старом Свете. Виды отличаются как по наружным признакам, так и по внутренним, особенно строением метапарамер и присутствием или отсутствием вирга.
* 1. Anisolabis (in sp.) maritima (Gene). Приморская уховертка (фиг. 19, 20).
Gene, 1832: 224 (Генуя, Италия; Ницца, Франция; гора Лебанон, Сирия) [Forficula] Serville, 1839:27 (Forficesilis); Fischer, Orthopt. Europ., 1853:68, tab. 6, fig'. 4 [Forficula (Labidura)]; Fieber, 1853 : 257 (Anisolabis); Brunner, 1882 : 9, tab. 1, fig. 2; Kirby, 1904 :17 Якобсон, 1905 :23; Zacher, 1911:3.
Тело черное, довольно коренастое, средних размеров. Усики 22— 25-члениковые, буровато-желтые, одноцветные. Переднеспинка сверху черная, гладкая, с немногими вдавленными точками, со слабой продольной линией. Ноги одноцветно-желтые. Брюшко довольно широкое, без боковых бугорков на 3-м и 4-м сегментах, боковые стороны спинных полуколец, у $ оттянуты назад, вытянуты в виде углов, которые на первых 3—4 сегментах слабо выражены, почти отсутствуют, а на последующих 5—8 сегментах хорошо выражены, с продольной складчатостью и часто с продольным килем; у g боковые стороны гладкие, не выступают углом; вся поверхность брюшка покрыта редкими разбросанными точечным^ вдавлениями, особенно заметными по задним краям сегментов; наибольшая ширина брюшка находится примерно около его середины. Последнее спинное полукольцо <5 прямоугольное, с почти прямыми боковыми сторонами, примерно в 2 раза шире своей длины, с довольно широким продольным вдавлением посредине; задний край почти прямой; у g в общем такой же формы, но. несколько сужено кзади и более короткое. Предпоследнее брюшное полукольцо с довольно сильно выступающим тупоугольным на вершине задним краем. Низ тела более светлый, чем верх,'.
90
2. LABIDURIDAE
от красновато-коричневого (брюшко) до палево-желтого (грудь), брюшко в грубых густых вдавленных точках. Клещи 5 не соприкасаются при основании, в основной трети толстые, прямые, на поперечном разрезе треугольные, несколько расширенные, асимметричные, так как правая ветвь резко и сильно загнута внутрь около средины, так что вершинная половина образует почти прямой угол с основной половиной и обычно налегает сверху на вершинную часть левой ветви; левая ветвь постепенно заги
Фиг. 19. Anisolabis (in sp.) maritima (Gene), $. X 4 (Батум). Opиг.
Фиг. 20. Anisolabis (in sp.) maritima (Gene), 5. Генитальный аппарат. Ориг.
бается от основания к вершине; внутренняя сторона в мелких бугорковидных зубчиках; клещи g простые, симметричные, сближены друг с другом, почти прямые, постепенно суживающиеся к вершине. Генитальный аппарат $ не имеет virga, метапарамеры прямые, очень постепенно суживающиеся к вершине и слегка расширенные до середины. Длина тела S 14.5—23, g 21—23; клещей $ 3—3.5, g 3.5—4. 25 мм.
Этот вид является представителем той группы видов подрода Anisolabis, которая характеризуется отсутствием virga в генитальном аппарате самца; сюда относится большинство видов. Виды, характеризующиеся присутствием virga, весьма немногочисленны и известны только из Африки и Австралийской области.
/. ANISOLABIS
91
Распространение: повидимому является аборигеном Средиземья, так как известен из многих мест по берегам Средиземного моря, встречаясь также на южном берегу Крыма (Шугуров, 1912) и на Черноморском побережье Кавказа; вид широко развезен по всему земному шару по торговым путям и является почти космополитом, будучи известным из всех частей света. В Европе известен кроме Средиземья также из Венгрии и Англии; в Азии из б. Томского уезда (?), Уссурийского края (Щербаков, 1911), Японии, Кореи и всей тропической зоны.
Обычно населяет побережья морей, живя под камнями и растениями; однако, встречается изредка и вдали от моря, иногда даже на значительной высоте в горах (до 1800 м). Экологическая связь вида с морскими побережьями видимо в значительной степени облегчила широкое расселение вида торговыми и другими кораблями по всему земному шару. Являясь в общем достаточно стабильным в отношении морфологического строения и окраски видом, A, maritima, однако, вариирует в известных пределах, в частности в размерах тела; южные формы обычно крупнее, чем северные (напр. крымские SS не длиннее 14.5—15 мм, тогда как тропические экземпляры достигают 23 мм). Кроме того, известны особи самцов, характеризующиеся простыми, как у самки, клещами; изредка даже попадаются особи, имеющие усики с палевыми колечками.
2. Подрод Apolabis Bun-
Barr, 1915 : 538. — Anisolabis auct. (partim).
Тип подрода — Apolabis hottentota Dohrn (Ю. Африка).
Сходен c Anisolabis. Брюшко умеренно расширенное. Надкрылья и крылья отсутствуют. Клещи 5 не соприкасающиеся при основании. Гениталии <5 с умеренно-удлиненными метапэрамерами, длина которых в 1х/2—2 раза превосходит их наибольшую ширину, сильно сужены к вершине и заострены, но не загнуты. Наибольшая ширина приходится на основную половину, но не само основание; наружная их сторона S-образно изогнутая.
Этот подрод включает около десятка видов, свойственных, за исключением рассматриваемого ниже A. marginalis Dohrn, Эфиопской области. Все виды, за исключением западно-африканской A. isomorpha Вог., характеризуются отсутствием virga.
2. Anisolabis (Apolabis) marginalis (Dohrn) (фиг. 21).
Dohrn, 1864: 288 (S; Япония, Батавия, Ява) [Forcinella]; Bormans, 1900:50, fig. 20; Kirby, 1904:18; Якобсон, 1905:25; Burr, 1911:29; Borelli, 1927:65, fig. II; Shiraki,1928:7; Burr, 1916 : 538, tab. I, fig. 14 (Apolabis). — fallax Shiraki, 1905 : 94, fig. 4.
Тело средних размеров, каштаново-бурое, слабо-блестящее. Усики 19-члениковые, двуцветные, серо-бурые, с светложелтым 13—15-м или 15—16-м члениками; 4—5-й членики конические, короткие, слегка
92
2. LABIDURIDAE
Фиг. 21. Anisolabis (Apolabis) marginalis (Dohrn), 3. Генитальный аппарат. (По Borelli, 1927).
длиннее своей ширины, 6-й членик на одну треть длиннее своей ширины. Голова шире своей длины, с ясным продольным швом, со светлой передней частью наличника. Переднеспинка одноцветная, почти квадратная, чуть шире своей длины. Надкрылья и крылья отсутствуют. Ноги грязно-желтые, основная половина бедер более темная; основание голеней также иногда темное. Брюшко слабо расширяющееся к 7-му членику, далее слегка сужено, очень тонко пунктировано, с немногими более грубыми точками в задней части сегментов; бугорки на 3-м и 4-м члениках слабо выраженные; боковые стороны 5—9-го сегментов 5 вытянуты сзади в виде острых углов, задняя часть 6—7-гО сегментов с продольным килем; последний тергит 3 немного темнее чем предыдущие, в редкой пунктировке, примерно на одну треть шире своей длины, по всей длине с глубокой продольной срединной ^бороздой, но без ясных бугорковидных выступов перед задним краем; у g бороздка более слабая; предпоследний стернит тонко пунктирован и слегка морщинистый, слегка суживающийся к заднему краю, последний очень широко закруглен. Клещи 3 слегка расставлены при основании, слабо изогнутые, зазубренные в основной половине внутренней стороны, пунктированные, умеренно асимметричные; правая ветвь в вершинной трети дуговидно загнута под тупым углом к продольной оси тела, левая ветвь перед вершиной немного загнута внутрь; клещи g простые, внутренняя сторона в основной половине с довольно сильными зубчиками. Метапарамеры с оттянутым закругленным концом, образующим маленькую округленную лопасть; собственно
penis со многими хитиновыми зубчиками; virga отсутствует. Длина тела 3 g 12—19.5; клещей 3 3—3.9. g 3.5 мм.
A. marginalis является единственным представителем подрода Аро-labis в восточной Азии — все остальные виды свойственны Африке; это обстоятельство, кажущееся на первый взгляд странным, однако не находится в противоречии с фактами распространения многих других животных (напр. Gryllotalpa africana Palis.)
Распространение: Восточная Азия от Японии и Кореи (Shiraki) до Формозы и Явы.
3. Подрод MongoJabis Zacher.
Zacher, 1911:385; Burr, 1916: 543. ,
Тип подрода — Mongolabis woodward! Burr (Зап. Австралия). , Как Anisolabis и Apolabis, но отличается строением метапарам еров; Совершенно бескрылые виды. Брюшко 3 расширено у вершины. Метана^
7. ANISOLABIS
93
рамеры немного длиннее своей ширины (около 1*/2 раз), трапециевидные, с прямыми или с почти прямыми сторонами, наружная их сторона прямо-или тупоугольная; наибольшая ширина метапарамер находится около их средины; собственно penis несет мелкие зубчики, не образующие, однако, правильных рядов.
Этот подрод был установлен Zacher’oM в качестве рода для двух видов из Зап. Австралии, причем дифференциальными признаками были взяты, кроме упомянутых выше, еще ряд более мелких особенностей, в частности присутствие зубчиков, а также хитиновых утолщений на собственно penis’e и пр. Однако, как показал Burr, если довести систему Zacher’a до ее логического конца, то необходимо будет установить чрезвычайно большое количество родов (в нашей трактовке подродов), каждый с очень небольшим количеством видов; при этом несомненно близкие морфологически виды окажутся представителями различных подродов. Поэтому трактовку данного подрода Zacher’oM нельзя признать правильной. В настоящее время известно всего 6 видов, относящихся к подроду Mongolabis, из которых 3 водятся в Восточной Азии, охватывая территорию сев. и сев.-зап. Китая и Формозы. Ниже рассматриваются только два вида, заходящие в пределы Палеарктики; не исключена возможность обнаружения и других видов.
3. Anisolabis (Mongolabis) setsbuana B.-Bienko (фиг. 22, 23).
Bey-Bienko, 1934:402, figs 1—2 (5, Тацзинлу, сев. Сычуань).
Тело довольно маленькое, со слабо, но ясно расширенным перед вершиной брюшком, одноцветно красновато-темно-коричневое. Усики более чем 15-члениковые, 3-й членик почти равен 4-му и 5-му, взятым вместе; последние очень короткие, почти равны друг другу. Голова одинаковой длины и ширины, чуть шире переднеспинки, с неясными швами. Передне-спинка чуть шире своей длины, со слабой продольной срединной линией. Ноги грязновато-палевые, основная половина всех бедер затемнена; передние бедра очень толстые и короткие. Переднегрудная пластинка в 2’/3 раза длиннее своей ширины по заднему краю. Брюшко без боковых бугорков, очень тонко и густо пунктировано; максимальная его ширина находится около 6—7 сегментов; боковые стороны 5—8 тергитов <5 с сильными килями, заострены, с ясной продольной морщинистостью, особенно на 7—8 сегментах; у д бока брюшных сегментов простые; последний тергит сужен к вершине, в рассеянной пунктировке, у $ с ясной продольной срединной бороздой, у 2 заметной только в основной половине, почти в 2 раза шире своей длины, с прямым задним краем; предпоследний стернит у <5 поперечный, четырехугольный, с ясной дуговидной вырезкою по заднему краю и с почти параллельными боковыми сторонами, у £ — сужен к вершине, широко-треугольный. Клещи $ короткие, расставленные при основание, асимметричные, сильно расширенные в основной трети, постепенно, сужйвающиеся к вершине; левая ветвь прямая в основных двух третях, слабо
94
2. LABIDURIDAE
загнутая в вершинной трети, правая ветвь изогнута более сильно, в основной трети прямая, в остальной части постепенно дуговидно загнута,.
перекрывая своим концом вершину левой ветви и образуя с продольной осью тела тупой угол (около 110°)} внутренне стороны клещей с очень слабыми зубчиками. Клещи £ простые, несколько уплощенные и сильно суживаются на вершине. Метапарамеры чуть больше чем в 172 раза длиннее своей ширины, с совершенно прямыми сторонами, с заостренной
но не оттянутой вершиной, наружная сторона тупоугольная; собственно penis с мельчайшими зубчиками в вершинной части,.
Фиг. 22. Anisolabis (Mongolabis) setshuana B.-Bienko, $ Генитальный аппарат. Ориг.
без хитиновых пластинок и утолщений. Длина тела $ 9.8, £ 8.1 — 9.4; клещи $ 2.0, g 2.2 — 2.4 мм.
Этот вид был описан в качествее представителя рода Anisolabis в прежнем понимании последнего. По структуре гениталий он является несом-
Фиг. 23. Anisolabis {Mongolabis)' setshuana B.-Bienko, S-. Клещи. (По Bey-Bienko, 1934).
ненным представителем подрода Mongolabis, отличаясь, однако^, от вСех видов строением предпоследнего стернита брюшка самца'.
Распространение: известен пока только из Тац-зынлу в сев. Сычуани.
Изучение генитального аппарата единственного извест-
ного самца этого вида позволило- установить интересное отклонение, заключающееся в том,, что virga оказалась развитой1 только на одном penis’e, тогда
как другой не имеет никаких следов присутствия virga. Это обстоятельство
лишний раз подтверждает взгляд на то; что присутствие и отсутствие virga является признаком изменчивым не только в пределах подрода, но
также и вида и даже отдельных индивидуумов. 4
4. Anisolabis (Mongolabis) cavaleriei Bor.
Borelli, 1921:10 (<5, Киао-Чжоу, сев.-вост. Китай)) Borelli; 1927: 68, fig. 3.
Крупнее, чем предыдущий, со слабо и очень постепенно расширяющимся брюшком. Усики 17-члениковые, три первых членика рыжеватые,, последующие буроватые; 4-й членик на одну треть короче 3-го, 7-й почти такой же длины, как 3-й. Голова немного'длиннее своей ширины, красновато-коричневая, с бурыми щупальцами и наличником; швы слабо заметны. Переднеспинка рыжевато-бурая, прямоугольная; передний ее край чуть шире головы, передние две-трети с продольной бороздкой. Ноги буровато-желтые. Брюшко- с ясными боковыми бугорками на 3-м и 4-м сегмен-
1. ANISOLABIS
95
тах, каштаново-рыжее, очень тонко пунктировано, с немногими более сильными точками, слабо расширяющееся от 1-го к 8-му сегменту; бока спинных полуколец у <5 тупоугольные, морщинистые, но без продольного киля, у g одинакового устройства с 3. Последний тергит брюшка 3 с продольной срединной бороздкой, перед задним краем с 2-мя боковыми округленными тупыми выступами, по бокам снизу с хорошо выраженным продольным килем; у $ — более узкий, без бугорков; предпоследний стернит округленно-треугольный, суженный к заднему краю, морщинисто-пункти-рован, задний край у $ прямой, у» 2 слегка округлен. Клещи 3 довольно стройные, при основании расставленные, умеренно расширенные в основной половине, очень постепенно суживающиеся к вершине, немного асимметричные; левая ветвь прямая, за исключением слабо изогнутой вершинной четверти, первая ветвь, начиная от средины постепенно дугообразно изогнута, образует с продольной осью тела очень тупой (около 135°) угол; внутренний край слегка зубчатый. Клещи £ простые, стройные,, к вершине утончающиеся. Пигидий слабо заметный. Метапарамеры как у М. setshuana, но чуть короче, немного менее чем в 1 1 /а раза длиннее своей ширины, вершина их остро-треугольная, с закругленным концом,, наружная сторона тупоугольная, как у М. setshuana, но более округлена; собственно penis с многочисленными мельчайшими хитиновыми зубчиками, распределенными неравномерно по всей поверхности; virga отсутствует.. Длина тела 3 18.5, 2 15.9; клещи <5 3, 2 2.9 мм.
Распространение: известен пока только из сев.-вост. (Киао-Чжоу) и зап. Китая (Юннань).
4. Подрод Euborellia Burr.
Burr, Proc. и. St. Nat. Mus., XXXVIII, 1910:448; Zacher, 1911:379; Barr, 1915:544. -Boreilia Burr, 1909 : 325 (nec Rehn 1906).
Тип подрода — E. moesta Serv.
Внешне чрезвычайно сходен с предыдущими подродами, отличаясь от них гл. образом строением гениталий самца. Надкрылья отсутствуют или' если имеются то степень развития их вариирует у различных видов от сильно укороченных, представленных только в виде боковых чешуй [напр. А. (Е.) moesta] до вполне развитых, с развитыми крыльями. Изредка у совершенно бескрылых видов попадаются особи с вполне развитыми надкрыльями и крыльями напр. у А. (Е.) annulipes', кроме того у отдельных видов наблюдается довольно значительная изменчивость в степени, развития надкрылий среди отдельных индивидуумов [напр. у американского вида А. (Е.) janeirensis Dorhn]. Брюшко стройное, у 3 не расширенное или. очень слабо расширенное за срединой. Метапарамеры очень короткие, широкие, не длиннее или едва длиннее, иногда короче своей ширины,, на; вершине тупые или прямоугольные — в последнем случае они кажутся;
96
2. LABIDURIDAE
обрубленными; наружная их сторона округлена или совершенно прямая, образует с дистальной стороной прямой угол. Собственно penis часто имеет ряды сильных зубчиков, особенно в дистальной части, однако, крупные изолированные хитиновые утолщения отсутствуют; virga не развита. Строение клещей такого же типа, как у предыдущих, с аналогичной изменчивостью их у разных видов.
Подрод Euborellia наиболее близок к подроду Mongolabis, с которым он очень сходен в строении генитального аппарата самца. В отношении географического распространения он является космополитом, достигая особен-, ного разнообразия в тропиках Старого и Нового Света. В пределы Палеарктиче-, ской области заходят немногие виды, из которых в СССР и пограничных с ней частях водится всего 4 вида.
* 5. Anisolabis (Euborellia) annulipes (Lucas) (фиг. 24).
Lucas, Ann. Soc. Ent. France (2) V, 1847: LXXXIV (3, g; Ботанический сад в Париже) (Forfi-cesila); Brunner, 1882 : 8; Kirby, 1904:18; Якобсон, 1905 : 23;. Burr 1911:29; Burr 1916: 545 (Euborellia).
Размеры средние для рода; тело черное блестящее, стройное. Усики 16-члени-ковые, с красноватым 1-м члеником, остальные серовато-бурые исключая нескольких предвершинных члеников, которые беловато-желтого цвета; изредка светлыми могут быть и немногие другие членики; 3-й членик короткий, немного длиннее 2-го и по крайней мере в два раза короче 1-го. Переднеспинка почти квадратная, такого
- Фиг. 24. Anisolabis (Euborellia) ап- же цвета> как тело или иногда светлая.. nulipes (Luc.), оХ7. (Крым). Ориг. Надкрылья и крылья полностью отсутствуют или исключительно редко вполне, развиты. Ноги грязно-желтоватые, бедра обычно с черным кольцом, голени иногда затемнены при основании. Передне-грудная пластинка с обрубленным задним краем. Брюшко с почти параллельными сторонами, без бугорков на 2-м и 3-м сегментах; бока 6—9 тергитов у 3 угловидно оттянуты, с продольным килем и морщинками, у g простые. Последний тергит брюшка почти в два раза длиннее своей ширины, со слабым продольным вдавлением посредине, боковые стороны почти параллельные. Клещи 3 почти соприкасаются при основании, довольно толстые, по-. - степенно суживающиеся к вершине, умеренно изогнутые, асимметричные;
7. ANISOLABIS
97
правая ветвь загибается почти под прямым углом к продольной оси тела и перекрывает более умеренно изогнутую левую ветвь; у £ клещи прямые, сближенные, удлиненно-конические. Метапарамеры довольно длин
ные, заметно длиннее своей ширины, с тупоугольным наружным краем. Длина тела 5 9—11, £ 12—14, клещей S 2, g 3—3.5 мм.
Этот вид является единственным бескрылым представителем подрода Euborellia в Палеарктической области; ближайшие родичи вида известны из Африки.
Распространение: широко распространен по земному шару, видимо, благодаря развозу с товарами. В СССР известен только с южного
берега Крыма, Черноморского побережья Кавказа (Батум: колл. Г. Б.-Б.).
и из Уссурийского края (колл. ЗИН!). За пределами СССР известен из
многих частей Европы (особенно из Средиземья, но также и из Германии и Англии — в двух последних случаях не являясь, однако, постоянным элементом фауны), Африки» Австралии, Америки и Азии (от Малой Азии через южную часть материка до востока — Японии и Китая).
Встречается под камнями, мертвыми стволами деревьев и пр. Интересной особенностью вида, подтверждающей правильность включения его в подрод Euborellia, характеризующийся развитием у огромного большинства видов надкрылий, является наличие у этого вида, кроме основной бескрылой формы, полнокрылых особей (Pantel, 1912), известных, однако, по очень немногим экземплярам. Этот факт также крайне интересен тем, что данный вид является абсолютно бескрылым в нормальном состоянии, т. е. дошедшим в этом отношении до крайнего предела
Фиг. 25. Anisolabis (Euborellia) plebeja (Dohrn), 3. Генитальный аппарат. (По Ме-
редукции органа и несмотря на это дающий иногда вполне nozzi, 1927).
крылатых особей.
б. Anisolabis (Euborellia) plebeja (Dohrn) (фиг. 25)
Dohrn, 1863:322 ($, остр. Ява) (Labidura); Bormans, 1900 : 38 (Psalis); Burr 1911 : 32; Hebard, Proc. Acad. Nat. Sc. Philad., LXXIX, 1927 : 27 (Euborellia). — stdli Dohrn, 1864:286 (5, остр. Ява) \Forcinella\; Bormans, 1900: 55 (Anisolabis); Burr, 1911 :31 (Euborellia) Shiraki, 1928:6, fig. 3a.— minuta Caudell, journ. N. York Ent. Soc., XV,1907:168 (Anisolabis).
Тело маленькое, черное, блестящее. Усики черные, за исключением красноватых основных двух члеников и беловатых двух-трех вершинных члеников; 3-й членик усиков длинный, в три раза длиннее 2-го и почти равен 1-му членику. Надкрылья имеются в виде маленьких овальных пластинок по бокам среднеспинки; длина их равна длине среднеспинки и значительно превосходит свою ширину, или же надкрылья и крылья вполне развиты. Ноги желтоватые, бедра и голени с черноватыми кольцами. Переднегрудная пластинка с округленным задним краем, заднегрудная пластинка со слегка выемчатым дугообразно задним краем. Брюшко стройное,
Фауна СССР, Кожистокрылые 7
98
2. LABIDURIDAE
слабо расширяющееся. Клещи 3 с несоприкасающимися ветвями, в основной половине треугольные, прямые, толстые, в вершинной половине утонченные, цилиндрические, сильно изогнутые в вершинной трети, правая ветвь изогнута сильнее чем левая и располагается своим концом над вершиной последней; у g. ветви толстые, треугольные, прямые, утончающиеся, на вершине цилиндрические, и здесь слегка изогнутые. Метапарамеры с округленным наружным краем. Длина тела 3 8—9, g 9—10; клещей 3 1.2—2, g 1—2 мм.
Распространение: широко распространен, являясь одним из обычных видов в Индо-Малайской области; также известен с островов Индийского и Тихого океана, Африки, Америки и восточной Палеарктики— Японии и северо-вост. Китая (Киао-Чжоу, Borelli, 1915).
Недавно Buxton (1921) показал для Месопотамии (Амара) Psalis femoralis Dohrn; последний настолько близок к А. (Е.) plebeja, что быть может является синонимом этого вида. Экземпляры Buxton’a были на определении у Dr. A. Borelli, причем последний отметил, что они отличаются отсутствием металлического оттенка на надкрыльях, т. е. как раз тем признаком, которым характеризуется длиннокрылая форма А. (Е.) plebeja-, очень вероятно, что данное указание необходимо будет отнести к А. (Е.) plebeja, что в таком случае значительно расширит ареал этого вида к северо-западу.
7. Anisolabis (Euborellia) pallipes Shir.
Shiraki, 1905:93, fig. 3 (3; Такасадо, Япония); Burr, 1911:31 (Euborellia); Shiraki, 1928 : 7, fig. 36 (Euborellia).
Очень сходен с E. plebeja Dohrn как по размерам, так и по окраске, отличаясь лишь только более коротким 3-м члеником усиков, который примерно только в 11/2 раза длиннее 2-го и почти в два раза короче 1-го, прямым задним краем переднегрудной и среднегрудной пластинки и одноцветными палево-желтоватыми ногами, лишь только иногда с немного затемненным основанием голеней.
Распространение: Япония, Формоза и юго-восточный Китай.
* 8. Anisolabis (Euborellia) moesta (Serv.) (фиг. 26)
Serville, 1839:28(3, g; Италия, южн. Франция, Сардиния) [Forficesila]; Brunner 1882 : 9 (Anisolabis); Bormans, 1900:44; Kirby, 1904 :18; Якобсон, 1905:23; Burr, 1909: 325 (Boreilia); Burr, 1911:31 (Euborellia); 1915 :545, tab. XII, fig. 19—taurica Fischer-Waldheim 1846:47 (Крым) (Foificesila).
Тело довольно крупное для подрода, слабо блестящее, черное, с умеренно-расширенным брюшком. Усики 18-члениковые, бурые, иногда с немногими светлыми члениками, 3-й членик почти в два раза длиннее 2-го и более чем в два раза короче 1-го. Переднеспинка чуть расширенная кзади, совершенно черная или же с грязно-желтыми боковыми сторонами, с ясной продольной бороздой посередине. Надкрылья имеются в виде сильно укороченных пластинок, располагающихся по бокам средне^ спинки. Ноги рыжевато-желтые, за исключением бедер, которые черного
1. ANISOLABIS
99
или темного цвета с рыжевато-желтой вершиной. Переднегрудная пластинка обрублена по заднему краю. Низ тела красновато-бурый. Брюшко постепенно расширяется от основания к средине, а затем едва суживается к вершине, почти с параллельными сторонами в предвершинной части; бугорки на боках 3-го и 4-го тергитов очень маленькие, слабо развитые, иногда отсутствуют; бока 4—9-го тергитов у $ угловатые, на 6—9-м углы более острые, с ясным продольным килем, заходящим и на последний тергит; последний тергит почти прямоугольный, в два раза шире своей длины, с продольной срединной бороздкой по заднему краю в глубоких морщинистых точках. Вся поверхность брюшка покрыта очень густыми, мелкими, но ясными точечными вдавлениями. Предпоследний стернит округленно треугольный в густых, довольно грубых точках. Клещи $ короткие, толстые, асимметричные, слегка расставлены при основании, сильно расширенные в основной половине, постепенно суживающиеся к вершине; левая ветвь в основных двух третях прямая, треугольная, в вершинной трети слегка изогнута, цилиндрическая; правая ветвь изогнута сильнее, так что ее вершинная треть образует с продольной осью тела прямой угол; у 2 клещи постепенно утончающиеся к вершине и сильно загнутые; внутренние стороны со слабыми или неясными зубчиками. Метапарамеры без ясно выраженных углов, с совершенно дугообразной наружной стороной. Длина тела S 11—14, g 11—15; клещей $ 2.2—2;5, g 2—3 мм.
Описанный из Крыма вид — Forficesila taurica Fisch.-W. был сведен Якобсоном (op. cit.) в синонимы к А. (Е.) moesta; однако, некоторые позднейшие авторы (Kirby, Burr и др.) рассматривали оба вида как самостоятельные, против чего, однако, имеются очень серьезные возражения. Именно отличия от Е. moesta, указанные Fischer-Waldheim’ом, настолько незначительны, что не выходят за пределы изменчивости вида; с другой стороны Е. moesta является типичным средиземноморским видом, и нахождение ее в Крыму вполне вероятно. К сожалению позднейшие исследователи не находили этого вида в Крыму.
Распространение: Средиземноморские страны от Пиренейского полуострова и Сев. Африки до Малой Азии и Крыма; вост. Африка...
7
100
2. LABIDURIDAE
Встречается под камнями, на цветах и пр. По сообщению Silvestri (Boll. Lab. Zool. Portici, IV, 1910), этот вид поедает куколок листоеда Galerucella luteola.
Недостаточно описан и неизвестно к какому подроду относится.
9. Anisolabis (subg.?) pectoralis (Esch.).
Eschscholtz, Entomograph., 1, 1822:82 (Камчатка) [Forficuld\\ Bormans 1900:50; Kirby, 1904 :19; Якобсон, 1905 : 23; Burr, 1911:30.
Тело черно-бурое. Усики 15-члениковые; 1-й, 3-й, 13-й и 14-й членики, бока переднеспинки, а иногда ее передний и задний края, переднегрудь и ноги желтые. Клещи расширены при основании, почти прямые, только на концах загнутые, зазубренные по внутреннему краю. Длина (с клещами) 16 мм.
Вид этот с момента его описания никем не исследовался. Поэтому данный вид можно будет включить в тот или иной подрод только после тщательного изучения экземпляров из типичной местности.
Распространение: известен только с Камчатки. Так как род Anisolabis является почти исключительно тропическим, рассматривать данный вид в качестве эндемичного для Камчатки нет никаких оснований, тем более, что отсюда известен другой широко распространившийся при помощи торговли тропический по происхождению вид — Prolabia arachidis Yers, из сем. Labiidae (см. ниже); другие представители отряда отсюда неизвестны. Очень вероятно поэтому, что A. pectoralis завезен на Камчатку из тропических стран и быть может даже описан под другим названием, так как оригинальное описание Eschscholtz’a довольно схематично и может подойти к ряду видов. Уяснение систематического положения вида возможно только на типичном материале с Камчатки, который, к сожалению, отсутствует. Однако, судя по характеру описания, можно полагать, что этот вид относится к подроду Euborellia или к близким подродам, но никак не к Anisolabis in sp.
2. Подсем. LABIDURINAE
Тело не-уплощенное, или слабо уплощенное, почти цилиндрическое, средних и крупных размеров. Усики со многими члениками (25—35), 4, 5 и 6-й членики короткие, почти одинаковой длины. Надкрылья вполне развитые, с ясными продольными килями по бокам. Переднегрудная пластинка немного сужена кзади, средне- и заднегрудная пластинки такой же длины, как их ширина, обрублены по заднему краю. Брюшко с параллельными сторонами или слегка расширяется кзади, почти цилиндрическое, обычно без бугорков по бокам 3-го и 4-го тергитов. Клещи удлиненные, слабоизогнутые, цилиндрические, у 5 расставлены при основании, симметричные. Пигидий во много раз длиннее метапигидия. Метапарамеры более или меиее
2. LABIDURA
101
с параллельными сторонами, узкие, очень заостренные, с длинной virga; почковидный пузырек со спиральной структурбй. Манубрий прямоугольный, короткий, не более чем в два раза длиннее своей ширины, не расширен на вершине.
Это подсемейство включает в себя всего только 4 рода, распространенных в тропиках Старого и Нового Света и заходящих в числе только двух видов в пределы Палеарктики.
2. Род LABIDURA LEACH.
Leach, Edinbourgh Encycl., IX, 1815 : 48; Якобсон, 1905:22; Burr, 1911:26; Burr 1915:443. — Forficesila Serville, Ann. Sc. Nat., XII, 1831:32.—Demogorgon Kirby, Journ. Linn. Soc. Lond., Zool., XXIII, 1891:513.
Тип рода — Labidura riparia Pall.
Тело крупное. Усики 25—35-члениковые, 3-й сегмент длинный, почти равен 1-му сегменту, 4-й и 5-й короткие, такой же длины, как 3-й, последующие членики постепенно удлиняющиеся, но даже вершинный едва достигает длины 3-го членика. Переднеспинка более или менее квадратная. Крылья вполне развиты, сильно выступают из под надкрылий или иногда укороченные. Ноги длинные, стройные, задние бедра в 11/2 раза длиннее переднеспинки; первый членик задних лапок длиннее третьего чЛеника, цилиндрический. Последний тергит брюшка $ большой, почти квадратный, у д более узкий. Предпоследний тергит у <5 суженный, почти обрубленный на вершине; у g этот стернит округленно-треугольный. Пигидий скрытый. Клещи <5 несоприкасающиеся при основании, вооруженные зубчиками и шипами или же без них; у g — сближенные, почти соприкасающиеся, прямые. Метапарамеры на вершине слегка расширены и округлены, с маленькой дополнительной лопастью.
Известен всего только один чрезвычайно изменчивый вид, описанный под. многими видовыми и даже родовыми названиями.
*1. Labidura riparia (Pall.). — Прибрежная уховертка (фиг. 27, 28).
Pallas, Reise, Russ. Reiche, II; 1773 : 727 (Берега p. Иртыша в Зап. Сибири) [Forficula); Dohrn 1863 : 313; Brunner, 1882 : 5 (L. riparia Pall, et var. inermis); Bormans, 1900 : 33—36 (L. riparia Pall, et subsp. pallipes, pluvialis, erythrocephala, japonica, livida); Якобсон, 1905 : 22 (L. riparia Pall, et subsp. pallipes, japonica, livida); Burr, 1910 : 38—39 (L. riparia et L. ben-galensis Dohrn); Burr, 1911:36—37; Hebard, 1923: 207—210; Shiraki, 1928:8—9 (L. riparia et L. japonica). —pallipes Fabricius, Syst. Ent., 1775 : 270 (Forficula). — bidens Olivier, Encycl. Meth. VI, 1791:466 (Forficula).—gigantea Fabricius, Ent. Syst., II, 1793:1 (Forficula). — icterica Serville, 1839:25 (Forficula).—gigantea var. japonica Haan, Verb. Nat. Gesch. Nederl. Qv. Bezitt., Orthopt. 1842 : 240 (Forficula).
Чрезвычайно изменчивый вид как по окраске, так особенно в отношении размеров тела, степени развития крыловых пластинок (задних крыльев), строения последнего тергита брюшка и клещей <5. Окраска от очень светложелтой, одноцветной до буровато-темной, контрастная или
102
2. LABIDURIDAE
монотонная. Все тело покрыто короткими густыми шелковистыми волосками. Усики желтовато-золотистые. Надкрылья с ясным боковым килем, иногда слабо развитым в задней половине; крыловые пластинки длинные,
или слегка укороченные, или значительно укороченные и в этом случае их
концы едва выступают из
Фиг. 27. Labidura riparia (Pall ), <5 X 4 (Крым). Ориг.
под надкрылий, или же совсем не выступают. Ноги желтые. Последний тергит брюшка гладкий, задний его край вооружен двумя острыми шипами или без них. Клещи длинные, стройные, более темные к вершине, у 5 ветви клещей с одним или двумя крупными зубцами и многими мелкими зубчиками;
Фиг. 28. Labidura riparia (Pall.) <?. Генитальный .аппарат. Ориг.
или же крупные зубцы отсутствуют, а зубчики чрезвычайно неясные; или же может быть развит только один крупный зубец; у 9 внутренняя сторона мелко зазубрена в различной степени, иногда зазубрины отсутствуют. Длина тела 5 8.7—26, g 9.7—22; клещи S 3.2—4.5, 2 3—5 мм.
Вид этот характеризуется настолько значительной изменчивостью и настолько широким распространением, что описывался как прежними, так и современными авторами под различными видовыми, а иногда и родовыми названиями (напр. род Demo-gorgon Kirby).
Распространение: широко распространен по всему земному шару, являясь почти вездесущим насекомым. В пределах СССР известен из всей южной части (от лесостепи до пустыни), заходя на север и в пределы лесной зоны до линии, соединяющей города Калугу, Серпухов,
Владимир, Киров (Щербаков, 1911); в Сибири известен из степи и лесостепи, на север до широты Омска, и из Забайкалья; в вост. Азии ареал достигает на севере Амурского и Уссурийского края, а также Японии, Сев. Китая. Известен также из Тибета и Кашмира (Hutchinson: колл. Г. Б—Б.).
Водится на легких почвах (песчаные и супесчаные) и под камнями на
берегах рек, озер и морей, вырывая норки как для откладки яиц и вывода молодого поколения, которое самки охраняют в течение нескольких дней
2. LABIDURA
103
после вылупления, так и для временного пребывания. Взрослые особи ночью прилетают на свет.
Многочисленные формы L. riparia могут трактоваться в отдельных случаях как морфы, в других быть может даже как подвиды. В последнее время Hebard (1923) предложил для характеристики различных форм рациональную систему символов, при помощи которых характеризуются отдельные особенности индивидуума. Ниже приводится эта схема с некоторыми дополнениями.
Окраска тела
А — очень светлая В — среднего типа
Е — блестящая F — умеренно-
С — темная, контрастная
D — темная, но без контрастов, монотонная.
блестящая G — тусклая
Задние крылья
W — очень короткие, выступают из под надкрыльев только самые кончики или оии совсем не видны.
V — слегка укороченные
X — короткие, с выступающей вершинной
Z — полностью развитые.
частью.
Наружные гениталии самца
а — задний край последнего тергнта не вооружен зубцами.
Ъ — задний край последнего тергита вооружен двумя слабыми зубцами.
с — задний край последнего тергита вооружен двумя острыми шипами.
ч! — внутренний край клещей с очень слабыми зубцами.
е — внутренний край клещей с ясным дистальным зубцом.
f — внутренний край клещей с ясным зубцом в основной и вершинной половинах.
g — внутренний край клещей кроме 2 зубцов (как в предыдущем случае) имеет еще маленькие зубчики.
h — как в предыдущем случае, но большинство маленьких зубчиков находятся за вершинным зубцом.
Клещи самки
w — зубчатость на внутренних сторонах клещей отсутствует.
х — зубчатость очень слабая.
у — зубчатость ясная.
х — зубчатость очень сильно развита.
Размеры тела
1 — очень крупная (длина тела О 23—26, g 20. 1—22 мм).
2 — крупная (S 19. 1—23, 9 17.1—20 мм).
3 — средних размеров (<5 15.1—19, g 14.1—17 мм).
4 — маленькая (<5 11.1—15, g 11,1—14 мм).
5 — очень маленькая (<5 8.7—11, g 9.7—11 мм).
104
2. LABIDURIDAE
В соответствии с указанными обозначениями, различные формы вида,, прежде фигурировавшие как отдельные виды, будут иметь следующие, характеристики.
Типичная L. riparia — BF /— )ех/?)
L. riparia morpha ebumea Sem. — AE, X e 5.
L. riparia morpha inermis Br.-W. — DCWa, d (или fg), xy
L. riparia morpha bidens Ol. — DF Z— c\ ex
w
L. riparia subsp. (?) japonica Haan BF — a.
Интересно отметить, что subsp. (?) japonica, по свидетельству Shiraki,, только и встречается в Японии, тогда как остальные формы там отсутствуют. Это говорит за то, что чрезвычайная изменчивость вида, однако, находится в какой-то связи с географическими и экологическими условиями. Если некоторые из этих форм распространены повсюду (напр. japo-nica), то быть может это является следствием завоза, благодаря чему теряется географический критерий для точного уяснения таксономического значения указанных форм. Во всяком случае данный вид является удивительным примером приспособляемости к различным внешним уело-» виям, позволившей ему завоевать весь земной шар; в связи с чрезвычайно широким колебанием этих внешних условий в различных частях ареала находится, повидимому, и исключительная изменчивость вида. Было бы поэтому крайне желательно использовать данный вид для генетических исследований.
3. Род NALA ZACHER.
Zacher, Ent. Rundsch., XXVII, 1910: 29; Burr, 1911:35; 1915 : 443.—Paralabidura Burr,. 1910 b:184.
Тип рода — Nala lividipes Duf.
Как Labidura, но меньших размеров; тело буровато-черное. Передне-спинка длиннее своей ширины. Ноги относительно короткие, задние бедра не длиннее переднеспинки; 1-й и 3-й членики задних лапок одинаковой длины.. Надкрылья всегда с ясно выраженными по всей длине боковыми килями. Метапарамеры очень сильно заострены и оттянуты на вершине, без дополнительной лопасти.
Небольшое количество видов, распространенных от Средиземноморья и Африки через южную Азию до Японии; Австралия (завоз); в нашей фауне — 1 вид.
1. Nala lividipes (Duf.) (фиг. 29)
Dufour, Ann. Sc. Nat, XIII, 1829 : 340 (Forficula)', Bormans, 1900 : 36 (Labidura); Zacher,. Ent. Rundsch., XXVII, 1910:29; Burr, 1911 :36; Burr, 1915 :444, Tab. IX, fig. 10. — pallipes
3. NALA
105-
Dufour (пес Fabricius), Ann. Gen. Sc. Bruxelles, VI, 1820:316, tab. 96, fig. 7 (<?, g; Нижняя Каталония, Испания). — meridionalis Serville, 1839:26 (Forficula). — dufouri Scudder,. 1876:328; Brunner, 1882:7.
Тело буровато-черное, матовое с такого же цвета головой. Усики 25-члениковые, темные. Переднеспинка с округленным задним краем и задними боковыми углами, так что бока дуговидные; цвет буровато-черный с падево-желтыми, иногда голубоватыми боками. Надкрылья желтовато-коричне-
вые, с боковыми килями, развитыми по всей длине надкрылий; крылья того же цвета, вполне развитые или укороченные. Ноги серовато-желтые, иногда местами более темные. Брюшко гладкое, черноватое, слегка расширяющееся к заднему концу; последний тергит $ более чем в два раза шире своей длины; у g более узкий, сужен к вершине; предпоследний стернит широкий, обрублен на вершине. Клещи 8 широко расставлены при основании, цилиндрические, слабо изогнутые, постепенно утончающиеся к вершине, в вершинной половине с зубовидным бугорком, иногда отсутствующим; у g ветви клещей соприкасаются, простые, цилиндрические. Длина тела с? 7—11, g 6.5—10; клещей S 1.5—3.2; g 1.5—2 мм.
Распространение: Средиземно-морскъе страны (Пиренейский п-ов, южная Франция, Италия, Сев. Африка), тропи* ческая Азия до южного Китая и Формозы, Япония и северо-вост. Китай (Киао-чжоу); Гавайские острова; Австралия. Столь широкое распространение обязано завозу по торговым путям; центром происхожде-
Фиг. 29. Nala lividipes (Duf.), 5 X 7' (Sanaa, Yemen). Ориг.
ния вероятно является Индомалайская область, так как только оттуда
и известны немногие остальные виды рода.
3. Подсем. PARISOLABINAE
Полностью бескрылые, мелкие формы. Усики 10—15-члениковые,. 2-й членик очень маленький, 3-й длинный, но короче 1-го, 7-й и последуют щие членики не короче 3-го. Г олова с неясными швами. Переднегрудная, пластинка с параллельными сторонами; средне-и заднегрудная пластинки шире своей длины, с обрубленным задним краем. Брюшко без боковых: бугорков. Клещи S расставлены у основания, Пигидий примерно только-
106
2. LABIDURIDAE
в два раза длиннее метапигидия. Генитальный аппарат $ имеет virga, которая снабжена при основании почковидным пузырьком. Манубрий короткий, не более чем в два раза длиннее своей ширины, не расширенный к вершине.
Известно всего четыре рода, из которых один свойствен Ю. Америке, два других Новой Зеландии и Австралии, а четвертый Новой Зеландии, острову Яве и Северной Индии до Кашмира. Такое распространение говорит о большой древности данных родов и всего подсемейства в целом.
4. Род PSEUDISOLABIS BURR
Burr, Ann. Mag. Nat. Hist., (8) II, 1908 : 254; Burr, 1910a: 102; Burr, 1911: 39.
Тип рода — Pseudisolabis walkeri Burr из Новой Зеландии.
Тело густо и очень тонко пунктировано, покрыто снизу прилегающими короткими волосками. Усики 15-члениковые, 1-й членик утолщающийся к вершине, 3-й цилиндрический, 4-й округлен, очень короткий, последующие конические, за исключением вершинных 4—5-го члеников. Голова в задней половине правильно-четырехугольная. Переднеспинка поперечная, в 1г/2 раза шире своей длины, все четыре стороны прямые; среднеспинка без боковых килей, с параллельными сторонами, задний край почти прямой; среднеспинка сзади слегка дуговидно вырезана. Ноги с толстыми бедрами, особенно первая пара; голени и лапки тонкие, 2-й членик лапок длинный, равен вершинному, в два раза короче 1-го. Брюшко S расширено к средине, суживающееся к вершине. Клещи тонкие, не вооруженные шипами или зубцами, у $ конические в основной половине, а затем загнутые внутрь, расставленные при основании, у £ почти прямые, соприкасающиеся, сильно утончающиеся и заостренные к вершине.
Известно небольшое количество видов, из которых один водится в Новой Зеландии, один на остр. Яве, а другие в Сев. Индии и Кашмире. Столь своеобразное распространение данного рода является несомненным показателем глубокой его древности, восходящей вероятно к тому периоду, когда Австралия еще не была отделена от Азиатского континента.
В пределы Палеарктики заходит 1 плохо изученный вид.
1. Pseudisolabis burn Вог. (фиг. 30).
Borelli, 1909:1 (<5 £; между Сринагаром и Гильгитом, сев.-вост.' Кашмир); Burr, 1910 : 103, fig. 35, Bey-Bienko, 1935 : 209; tenera Burr, 1910a : 104, fig. 36.
Черный или черно-коричневый. Усики темнокоричневые. Голова блестящечерная или чуть коричневатая, лоб выпуклый. ПереднеспинКа прямоугольная, такого же цвета как голова. Бедра черные или темнокоричневые, иногда желтоватые при основании и на вершине; голени буровато-коричневые, иногда с желтоватыми концами; лапки желтоватые. Брюшко <5 более или менее расширяется к серёдине, бока 5—7 сегментов угловидные, иногда с килем; последний спинной сегмент $ немного суживается
4. PSEUDISOLABIS
107
к вершине, слегка выпуклый по средине, с двумя боковыми слабыми бугорковидными вздутиями, расположенными как раз над основанием клещей; у g последний сегмент заметно суживается к вершине, боковые бугорки выражены слабее. Клещи 5 в основной половине прямые, более
толстые при основании, заметно утончающиеся к средине, вершинная поло-.
вина довольно резко загнута внутрь или же клещи, начиная с вершинной
трети, постепенно загибаются внутрь, не образуя резкого угла с прямой основной частью; у g клещи простые, соприкасающиеся, сильно утончающиеся к вершине. Длина тела 5 8.8—11.5, g 8—11.9; клещи $ 1.3—2.2, g 1—1.3 мм.
Весьма изменчивый вид, быть может представляющий собою совокупность двух видов.
Распространение: Кашмир и Пенджаб до высоты 2800 м.
.II. Надсемейство EUDERMA-PTERA
Генитальный аппарат самца простой, с непарным penis, основание которого укреплено в основной части пропарамер (фиг. 8); пропарамеры слабо хитинизированные, укороченные, не отделены от метапарамер бороздой, со слабой перетяжкой или без нее, так что граница между парамерами слабо выражена. Опистомеры состоят только из одного ясного крупного членика — пигидия; метапигидий и тельсон атрофиро-
Фиг. 30. Pseudisolabis burn Borelli, (5X6 (Sonomarg, Kashmir). Ориг.
ваны, сильно укорочены или совсем отсутствуют; пигидий часто с выступами или шиповидными отростками. Манубриум у известных в этом отношении видов короткий, маленький, примерно в 11/2 раза шире своей длины, округленный на вершине. Глазки отсутствуют или представлены в виде плохо заметных вдавленных точек. Продольные жилки крыловой чешуи (R и М) часто соединены друг с другом поперечной жилкой; ветви А2 иногда без дополнительных ветвей. Клещи у личинок не сегментированы.
Это надсемейство включает только два семейства — Labiidae и Forfi-
culidae, причем большинство жителей умеренных широт приходится на долю последнего. По своей организации Eudermaptera стоят выше Protoder-maptera, за что говорят также характер распространения и слабая .дифференцированность некоторых подсемейств и родов надсемейства
108
3. LABIIDAE
и особенно сем. Forficulidae', последнее должно рассматриваться как наиболее высоко организованное и юное, хотя достоверные современные роды его. уже известны из нижнего олигоцена.
3. Сем. LABIIDAE
От очень мелких до умеренно-крупных размеров. Усики 10—23-чле-никовые. Второй членик лапок простой, цилиндрический (подсем. Labiinae)' или же с нижней стороны несет узкую лопасть, продолжающуюся под третий членик и поэтому незаметную при рассматривании сверху (подсем. Chelisochinae)-, у некоторых мелких форм эта лопасть бывает трудно заметна вследствие густых волосков, покрывающих лапки. Генитальный аппарат 5 с сильно заостренными, иногда оттянутыми на вершине метапарамерами. Пигидий 5 часто с длинными шипами или выростами. Переднеспинка часто расширяется кзади. Надкрылья развиты, иногда с килями, крылья развиты или отсутствуют.
Данное семейство включает в себя два подсемейства — Labiinae и Chelisochinae, считавшиеся прежде самостоятельными семействами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ И РОДОВ
1 (6). Второй членик лапок простой, цилиндрический. (1. Поде. Labiinae). 2 (5). Надкрылья без продольных килей по наружному краю.
3 (4). Усики с цилиндрическими, довольно однородными члениками; 4-й и 5-й членики почти такой же длины, как 3-й .... 1. Labia Leach.
4 (3). Усики с довольно разнородными члениками, 4-й членик значительно короче 3-го, почти шаровидный, 5-й членик конический или грушевидный, заметно короче и толще 3-го............. 2. Prolabia Burr.
5 (2). Надкрылья с ясным продольным килем по наружному краю. Членики усиков более или менее конические, с сильно утонченными концами. Передние бедра сильно утолщенные ..... 3. Nesogaster Verh.
6 (1). Второй членик Лапок с нижней стороны несет узкую лопасть, продолжающуюся под 3-й членик. (2 Поде. Chelisochinae).
7 (10). Тело удлиненное, стройное, от средних до крупных размеров (10— 19 мм). 3-й членик усиков в два раза длиннее своей ширины.
8 (9). Крупная. Усики толще, 3-й членик почти в два раза длиннее 4-го и заметно длиннее 5-го, все три членика слегка расширены к вершине или по крайней мере некоторые из них конические..................
• • • ............... 4. Chelisoches Scudd.
9 (9). Средних размеров. Усики очень тонкие, 3-й членик немного длиннее 4-го и едва длиннее 5-го, все членики цилиндрические ....
..............................................5. Proreus Burr.
10 (7). Тело маленькое (7—9.5 мм), широкое. 3-й членик усиков короткий, не более чем в 11/2 раза длиннее своей ширины . 6. Hamaxas Burr.
1. LABIA
109
1. Подсем. LABIINAE
Мелкие или очень мелкие, реже средних размеров виды. Усики с 10— '20 члениками. Второй членик лапок простой, цилиндрический. Генитальный аппарат $ более сложный, с длинной virga, обычно превышающей по длине метапарамеры, часто снабженный на переднем конце хитиновыми пластинками, почковидный пузырек отсутствует или имеется; penis у вершины часто с различными по форме хитиновыми пластинками или мелкими зубчиками.
Большое количество родов, свойственных почти исключительно тропическим странам Старого и Нового Света. В пределы СССР и сопредельных с ним стран заходят только три рода с немногими видами.
1. Род LABIA LEACH
Leach, Edinbourg Encycl., IX, 1813 :118; Burr, 1911 : 55. — Copiscelis Fieber, 1853 : 257.
Тип рода — Labia minor L.
Тело маленькое. Усики с удлиненными, цилиндрическими, довольно однородными по размерам и форме члениками; 4-й и 5-й членики почти или совсем такой же длины, как 3-й. Г олова гладкая, с параллельными сторонами, щеки не вздутые, швы отсутствуют или очень неясные; глаза маленькие, не длиннее 1-го членика усиков. Переднеспинка почти квадратная, задний край почти прямой. Надкрылья без килей, крылья у большинства видов вполне развитые. Брюшко с параллельными сторонами, умеренно уплощенное. Ноги не очень длинные, лапки тонкие, такой же длины как голени, 1-й и 3-й членики почти одинаковой длины, 2-й маленький. Клещи 3 разнообразно устроенные, обычно расставленные при основании. Пигидий 3 обычно выступает, чрезвычайно изменчивый у различных видов формы.
Известно около 50 видов этого рода, распространенных во всех частях света. В пределы Палеарктической области заходят только 2 вида.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Клещи 3 длинные, тонкие, слегка изогнутые, с редкими зубчиками на внутренней стороне. Предпоследний брюшной стернит 3 посредине заднего края с длинным тонким торчащим выступом, хорошо заметным при рассматривании сверху. Тело желтовато-бурое....................
......................................... . .* 1. L. minor L.
1 (1). Клещи 3 короткие, сильно треугольно расширенные при основании, в вершинной части сильно, почти под прямым углом, загнуты внутрь. Предпоследний брюшной стернит 3 простой. Темнокоричневая или рыжая с более темными надкрыльями и крыльями. . ....................
-...........................................2. L. curvicauda Motsch.
ПО 3. LABIIDAE
* 1. Labia minor (L.). Малая уховертка (фиг 31).
Linne, Syst. Naturae, I (ed. 10), 1758:423 (Европа) [Forficula]; Brunner, 1882 :10,. tab. I, fig-. 1; Kirby, 1904 : 25; Якобсон, 1905 : 23; Burr, 1911: 55, tab. 5, fig. 7.
Тело довольно крупное для рода, стройное, желтовато-бурое, матовое, в коротких волосках, иногда очень темное, с черной головой. Усики 12-ти. члениковые, буроватые, на концах светлее, 4-й членик чуть толще 3-го, но равен ему по длине, цолуконический, 5-й членик немного длиннее 3-го. Голова с незаметными швами. Переднеспинка не длиннее своей ширины,, слегка расширенная кзади, передний край заметно уже головы, задний край широко округлен, округло соединяется с боковыми сторонами. Надкрылья большие, широкие, красновато-бурые; крыловые пластинки длинные, широкие, уплощенные, немного длиннее половины длины надкрылий,, такого же цвета как последние или светлее. Ноги желтые. Брюшко умеренно уплощенное, слегка расширено по средине, рыжевато-или желтовато-бурое, при основании по бокам, включая боковые бугорки, черноватое; последний тергит $ с двумя маленькими бугорками у заднего края, у £ простой; предпоследний стернит $ посредине заднего края с длинным, тонким, торчащим шиповидным выступом, хорошо заметным при рассматривании насекомого и сверху. Пигидий <5 вертикальный, при рассматривании сзади почти квадратный, без выступов и бугорков; пигидий g почти вертикальный,, заметно суженный книзу. Клещи <5 при основании расставлены, длинные,, тонкие, слегка изогнутые, с редкими маленькими зубчиками на внутренней стороне, из которых наиболее крупный находится у основания, у £ — простые, соприкасающиеся. Длина тела $ Q 4—6.5, клещей $ 1.3—2.5,. 2 0.8—1.5 мм.
Характерный выступ на заднем крае предпоследнего стернита брюшка $ часто принимается при невнимательном рассмотрении за пигидий.
Распространение: широко распространен, являясь почти космополитом. В пределах СССР известен почти из всей европейской части,, на севере до Ленинграда, юга Карельской области, Горького, Сарапула и Свердловска, а на юге встречаясь также в Крыму и на всем Кавказе,, включая Закавказье; из азиатской части известен для зап. Сибири только из Омска, а из пределов Средней Азии — только из окр. Ташкента (Г. Б.-Б );; данные из восточной Сибири и Дальнего Востока отсутствуют. За пределами СССР водится во всей Европе (на севере до Швеции), Малой Азии, Сирии, Африке, Сев. и Южн. Америке; для восточной Азии сведения отсутствуют, а для тропической ее части указан с острова Цейлона (Burr, 1912).
В противоположность многим другим уховерткам, этот вид летает днем и вечером, часто массами над кучами навоза, удобренными навозом полями и пр.
2. Labia curvicauda (Motsch.)
Motschulsky, Bull. Soc. Nat. Moscou, XXXVI, 1863:2, tab. II, fig. 1 (Гора Нура-эллиа^ Цейлон) [Fo/^cesz'Za]; Dohrn, 1864:428; Kirby, 1904 : 26; Burr, 1910:118, fig. 38; Burr,, 1911:56; Shiraki, 1928:11. ,
Z. LABIA
111’
Тело черновато-коричневое или рыжеватое, очень маленькое. Усики с 14 (?) члениками, черноватые или красновато-бурые, с немногими палевыми или желтоватыми члениками; 1-й членик булавовидный, 4-й почти равен и чуть толще 3-го, 5-й почти равен 3-му. Г олова слегка вырезана по заднему краю, черная или темнобурая. Переднеспинка длиннее своей ширины,,
уже головы, с параллельными сторонами, черная, бурая или светложелтая. Надкрылья черные или темнобурые, в коротких волосках; крылья длинные, такого же. цвета как и надкрылья. Ноги грязновато-желтые, бедра обычно с черноватыми кольцами. Брюшко довольно сильно уплощенное, рыжеватое или черноватое, у J с параллельными сторонами, у расширенное около середины и суживающееся к вершине. Последний тергит брюшка i поперечный, с продольным срединным вдавлением и боковыми бугорками, у g — более узкий, с менее заметным вдавлением. Предпоследний стернит округлен по заднему краю. Клещи 5 короткие, расставленные и сильно треугольно расширенные при основании, в вершинной части загнуты внутрь ПОЧТИ ПОД прямым углом, у 2 простые, соприкасающиеся, толстые, треугольные, суживающиеся к вершине. Пигидий 5 уплощен, короткий, широкий, почти четырехугольный, обрубленный по заднему краю, обычно слабо выступает; у 2 — скрытый. Длина тела 5 2 4—5.5, клещей 0.6—1 мм.
Распространение: широко распространен в тропиках Старого и Нового Света (южная Азия, Африка, Америка и Океа-
Фиг. 31. Labia minor (L). 5x1 (Крым). Ориг.
ния), заходя на востоке до Формозы, Японии и Южного Китая, а на
западе до острова Мадейры; возможно нахождение его в Южно-Уссурий-
ском крае.
Чрезвычайно, изменчивый по окраске и отчасти строению клещей 5 вид; клещи 5 характеризуются различной степенью изогнутости и расширения при основании.
Примечание. Щербаковым (1911) был описан, но не был назван еще один вид из данного рода — Labia sp. п. из Тифлиса, известный только по одному плохо сохранившемуся экземпляру. По описанию данного автора.. этот. вид. характеризуется гораздо более длинным, чем L. minor пигидием,.
уплощенными клещами, менее резко выраженным отростком на последнем, стерните брюшка и наличием двух больших вздутых складок на последнем.
112
3. LABIIDAE
тергите. Что представляет собой эта форма, сказать трудно; очень возможно, что это есть только уклоняющаяся от типа особь весьма обычной в Закавказье L. minor.
2. Род PROLABIA BURR.
Burr, Deutsch. Ent. Nat.-Bibl., II, 1911: 60; Burr 1911: 57.
Тип рода — Prolabia arachidis Yers.
Весьма сходен c Labia, но тело обычно более коренастое, усики с разнородными члениками, 4-й членик конический или грушевидный, заметно короче и толще 3-го, последующие членики конические, с тонким основанием.
Известно более полутора десятка видов, свойственных главным образом тропической Америке; немногие виды известны из Индо-Малайской области, а один — космополит, заходящий и в пределы СССР.
*1. Prolabia arachidis (Yersin) (фиг. 32).
Yersin, Ann. Soc. Ent. France, VIII (3), 1860:509, tab. X, figs. 33—35 (Марсель, Франция) (Forficula); Brunner, 1882:21 (Chelidura); Bormans, 1900:117 (Apterigida); Kirby, 1904:44; Якобсон, 1905:26; Burr, 1910:123, fig. 82 (Labia); Burr, 1911:57. — nigripennis Motschulsky, Bull. Soc. Nat. Moscou, XXXVI, 1863:1 (Forficula).—gravidula Gerstacker, Arch. £. Naturg., XXXV, 1869 : 221 (Apterygida).
Тело темнокоричневое или каштановое, голое, блестящее. Усики 12—13-членикбвые, рыжевато-бурые. Голова рыжевато-коричневая, с тонкими, но ясными швами. Переднеспинка квадратная, или чуть поперечная, с прямым задним краем и с узкими палевыми каемками по бокам. Надкрылья вполне развитые, черные или рыжевато-коричневые, довольно короткие, (не более, чем в 1х/2 раза длиннее переднеспинки, с обрубленным задним краем; крылья отсутствуют. Ноги грязновато-желтые, бедра иногда с черноватым кольцом около основания, 2-й членик лапок с очень маленькой лопастью, заметной лишь при очень сильном увеличении. Брюшко гладкое, задние края всех сегментов с очень короткими волосками; 5—8-й сегменты у S и иногда клещи слегка пунктированы. Последний тергит почти квадратный, без ясных бугорков. Клещи $ расставлены у основания, короткие, стройные, цилиндрические, слабо изогнутые; внутренняя сторона с очень маленьким, плохо заметным зубцом у самого основания и с маленьким зубцом в вершинной трети; клещи g короткие, простые, немного изогнутые к вершине. Пигидий 3 относительно хорошо заметный, короткий, не длиннее своей ширины, сильно наклонный, немного суживающийся к вершине, задний край слегка дуговидно вырезан. Длина тела 3g 6.8—10; клещей 3 1.9—2.8, g 1.7—2 мм.
Присутствие на 2-м членике лапок очень маленькой лопасти, заметной только при сильном увеличении, послужило причиной тому, что этот вид считался многими авторами за представителя рода Apterygida из сем. Forficulidae-, однако, у последнего 2-й членик лапок сильно расширен
2. PRO LABI A, 3. NESOG ASTER
113
в обе стороны и не может быть сравниваем с той маленькой лопастью,’ Которая имеется на 2-м членике лапок у Р. arachidis.
Распространение: почти космополит; обитает тропики Старого и Нового Света (Азия, Африка, Америка, Австралия с Океанией),- развозится кораблями и нередко попадает в Зап. Европу (портовые города^ .Англии и Франции) и восточную Азию до Южно-Уссурийского края (окр. Владивостока, колл. С.-Т.-Ш.!), Сахалина (Супруненко, колл. ЗИН) и чатки (колл. ЗИН).
о-ва
Кам-
Burr,
Burr
3. Род NESOGASTER VERB.
Verhoeff, 1902: 191; Burr, 1908: 42; 1911: 48. — Nesogastrella Verhoeff, ibid.: 193.
Тип рода — Nesogaster dollchus с острова Целебес.
Тело сильно блестящее, гладкое, голое. Усики 12—20-члениковые, 1-й членик толстый, расширяющийся к вершине (булавовидный), 3-й членик длинный, тонкий, несколько утолщающийся к вершину, 4-й примерно, в два раза короче 3-го, довольно толстый, веретенообразный, 5-й — ясно длин? иее, чем 4-й, но короче 3-го, грушевидный или веретенообразный, последующие членики постепенно удлиняющиеся^ грушевидные или веретенообразные, с утонченным, почти стебельчатым основанием. Г олова гладкая, сверху шаровидно выпуклая, швы отсутствуют. Пе- <
реднеспинка правильно четырехугольная, все фиг 32< proiaJ}ia arachidis (Yers.), ее стороны, включая заднюю, совершенно $ X10 (Сахалин).Орйг.
прямые, углы не закруглены; верхняя сторона
выпуклая, за исключением уплощенных боков; уплощенная часть более широка у заднего края переднеспинки, так что выпуклость имеет форму треугольника, направленного вершиной кзади. Надкрылья вполне развитые, удлиненные или укороченные, квадратные, с ясным продольным килем по всему наружному краю, задний край обрублен; крылья отсутствуют, редко вполне развиты. Бедра, особенно передняя пара, сильно утолщены; лапки длинные и тонкие, почти равны по длине голеням, 1-й членик длиннее, чем 2-й и 3-й вместе взятые. Брюшко довольно уплощенное, с ясными бугорками на боках 3-го и 4-го сегментов. Последний тергит 3 прямоугольный, широкий, задний край утолщен; у j—более узкий. Предпоследний стернит почти четырехугольный, задний край широко закруглен. Пигидий S хорошо заметный, у многих видов очень длинный; уд — маленький; Клещи
Фауна СССР, кожистокрылые 8
114
3. LABIIDAE
3 обычно длинные, тонкие, слабо изогнутые, с зубцами или же расширен-’, ные в основной части; у g— короткие толстые, при основании на внутренней стороне обычно с довольно глубокой вырезкой и зубчиками.
Характерной особенностью рода является сильно блестящее тело,, утолщенные передние бедра, форма члеников усиков и присутствие киля на надкрыльях. Известно около десятка видов, свойственных главный образом Индо-Малайской области; один вид известен из Австралии, а два. из Японии.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Крупнее (длина тела 13.5—14 мм). Пигидий 3 треугольный, широкий ........................................... 1. N. nigritus Shir.
2 (1). Мельче (длина тела не более 10 мм). Пигидий 3 ланцетовидный, острый......................................... 2. N. lewisi Borm.
1. Nesogaster nigritus (Shiraki).
Shiraki 1905 : 91, fig-. l.(<5, g; Иозанкеи,- остр. Иезо, сев. Япония) [Labidurodes]; Burr, 1911:32; Shiraki 1928: 14.
Тело крупное, блестяще-черное, Усики 20-члениковые, грязно-черные,. 3-й членик такой же длины, как 1-й, но тоньше; 4-й и 5-й членики очень маленькие, шаровидные. Переднеспинка почти прямоугольная, едва достигающая ширины головы, задний край слабо округлен, почти прямой. Надкрылья длинные, почти в два раза длиннее переднеспинки; крылья вполне развитые. Ноги черные с каштаново-бурыми лапками. Брюшко чуть расширяющееся от 3—4-го членика к основанию последнего тергита,. почти с параллельными сторонами, снизу каштаново-бурое. Последний тергит 3 широкий, почти квадратный, у g — слегка суживается к вершине..-Клещи 3 тонкие, длинные, слабо изогнутые, широко расставлены при основании, на внутреней стороне с одним средним небольшим зубчиком и с немногими более мелкими зубчиками; у g — соприкасающиеся, короткие. Пигидий короткий, но хорошо заметный, широкий, у 3 треугольный, у g почти квадратный. Длина тела 3 g 13.5—14; клещей 3—12, g —3.8 мм.
Этот вид был первоначально отнесен его автором к роду Labidu.ro-des из сем. Lahiduridae-, Burr (1911:32) отнес его с вопросом к тому же самому роду, a Shiraki (1928:14) к роду Nesogaster. От типа рода Nesogaster он несколько отличается более многочленистыми усиками, с шаровидным 5-м члеником и округленным задним краем переднеспинки; однако, 'ПО другим своим признакам он несомненно напоминает Nesogaster. Быть может данный вид совместно со следующим образует самостоятельный род, близкий к роду Nesogaster, к сожалению, оба эти вида нам неизвестны ш натуре.
Распространение: известен только с о-ва Иезо, Сев. Япония^
3. NESOGASTER
115
2. Nesogaster lewisi (Bormans).
Bormans in: Burr, Ann. Mag. Nat. Hist. (7), XI; 1903:234 (3: Хако, Япония) \Spongiphord\-, Burr, 1911:32; Burr, 1911: 52 (Spongovostox); Shiraki 1928: 14. — cingularis Shiraki, 1906:8 (Labidurodes).
Сходен c N. nigritus, но мельче. Буровато-черный, матовый; усики, брюшко, лапки и клещи немного светлее, голые. Усики не менее чем 14-члениковые (20-члениковые?). Переднеспинка такой же длины и ширины как голова, задний край округлен. Брюшко очень нежно пунктировано с почти параллельными сторонами; задние края всех члеников слабо зернистые. Последний тергит 3 трапециевидный, немного шире своей длины, суженный к вершине, рыжевато-бурый, задний край утолщенный.. Клещи <3 широко расставлены при основании, довольно тонкие, почти одинаковой толщины на всем протяжении, почти прямые, сближающиеся примерно в третьей четверти их длины; или же немного изогнутые и перекрещивающиеся; внутренный край со слабыми редкими зубчиками около одного маленького срединного зубца. Длина 3 9—12, j 7.8; клещей 3 6.3— 8.5, $ 5 мм.
Хотя этот вид был отнесен недавно Shiraki (1928) к роду Nesogaster,. есть некоторое сомнение в принадлежности его к данному роду, тем более, что Burr (1911) отнес его к роду Spongovostox Burr, хотя и со знаком вопроса; род Spongovostox характеризуется отсутствием продольного киля на надкрыльях и более близок к роду Labia, отличаясь от последнего поперечной головой с хорошо выраженными швами и выпуклыми большими глазами, превышающими или равными по длине 1-му членику усиков; 4-й их членик почти такой же длинный как 3-й, остальные членики цилиндрические. К сожалению, описания Shiraki и Burr’a крайне кратки;, первый автор ограничивается только указанием на размеры и строение пигидия 3, сближая вид в остальном с N. nigritus.
Распространение: известен только из Японии (о-ва Иезо и Гондо).
2. Подсем. CHELISOCHINAE
Тело от маленького до крупного, довольно коренастое, умеренно уплощенное. Усики с 15—23, реже 12—14, цилиндрическими или почти цилиндрическими члениками, 4-й членик обычно короче 3-го, часто почти конический. Переднеспинка обычно расширяется кзади. Надкрылья и крылья вполне развиты или укороченные. 2-й членик лапок несет с нижней стороны узкую лопасть, продолжающуюся под 3-й членик и незаметную сверху;, иногда лопасть слабо заметна благодаря покрывающим 2-й членик густым волоскам. Генитальный аппарат 3 с простой, обычно короткой virga,. снабженной при основании узкими хитиновыми лопастинками; основание вирги или слегка расширено, или же имеет более или менее ясно выраженный базальный пузырек; собственно penis на вершине без хитиновых, пластинок и без мелких зубчиков.
8
116
3. LABIIDAE
Это подсемейство долгое время рассматривалось в качестве самостоятельного семейства; однако, изучение полового аппарата показало чрезвычайное сходство в этом отношении между Chelisochidae и Labiidae, что наряду с другими чертами, указывающими на их большую близость, заставило Burr’a (1916) рассматривать обе эти группы только как подсемейства одного сем. Labiidae.
Сравнительно небольшое количество родов, известное из данного подсемейства, свойственно главным образом Индомалайской области, с немногими видами и родами, заходящими в тропики Африки и Австралии с Океанией; в Палеарктическую область заходят только три вида, относящиеся к трем весьма плохо отличающимся друг от друга родам.
4. Род CHELISOCHES SCUDD.
Scudder, 1876 : 295; Burr, 1907 : 128; 1911 : 65.—Lobophora Serville, 1839 : 32.
Тип рода — Chelisoches morio F.
Тело довольно крупное, удлиненное. Усики 15—22-члениковые. 2-й членик заметно короче расстояния между внутренними сторонами глаз, 3-й членик в два раза длиннее своей ширины, почти в два раза длиннее 4-го и заметно длиннее 5-го, все три членика слегка расширены к вершине, или по крайней мере некоторые из них конические; 6-й не длиннее 3-го, остальные членики удлиненные. Голова выпуклая, со слабо заметными швами. Переднеспинка такой же ширины как голова, со слабо расходящимися кзади боковыми сторонами; задний край широко округлен. Надкрылья и крылья хорошо развиты; первые без боковых килей. Ноги довольно короткие, голени уплощены, вершинная половина с продольной бороздкой сверху; лапки очень короткие, в густых коротких волосках. Переднегрудная пластинка с параллельными сторонами, слегка суживается у основания; среднегрудная пластинка почти квадратная, сзади обрубленная; заднегрудная пластинка поперечная, обрубленная сзади. Брюшко -с параллельными сторонами, довольно уплощенное, с ясными боковыми бугорками; последний тергит S поперечный, прямоугольный, у g— слегка сужен; предпоследний стернит широко округлен. Клещи расставлены при основании, уплощенные, массивные, удлиненные, без крупных зубцов, но с мелкими, иногда неправильными зубчиками. Пигидий 3 маленький; у g крупнее, умеренно выступающий.
Около десяти видов, известных из Индомалайской, Африканской и Австралийской области с Океанией; один вид развозится кораблями и иногда попадает в портовые города Европы и Вост. Азии.
1. Chelisoches morio (F.)— Черная уховертка
Fabricius, Syst. EntomoL, 1775: 270 (остр. Отахейти, Таити) [forficula]; Bormans, 1900 : 85; Kirby, 1904 : 33; Якобсон, 1905 : 24; Burr, 1910 ; 135, fig. 45; Burr, 1911 : 65. — rufi-tarsis Serville, 1839 : 33 (Lobophora). — comprimens Scudder, 1876 : 252.
4. CHELISOCHES, 5. PROREUS
117
Тело блестяще-черное, вариирующее до буровато-черного или реже коричнево-желтое. Усики 20—21-члениковые, перед концом со светлым кольцом. Лапки желтые. Последний тергит S гладкий, посредине перед задним краем несколько вдавлен, с двумя продольными возвышающимися ребрышками в средней части этого вдавления, перед основанием каждой ветви клещей со слабым вздутием; у о — строение последнего тергита такое же, но продольные ребрышки менее выражены. Клещи $ двух типов: 1) ветви расставлены при основании, довольно короткие, очень широкие и плоские, с сильно выступающими, разнородными зубчиками на внутренней стороне при основании или в основной половине, а затем слабо изогнутые, с немногими слабыми зубчиками или без них; 2) ветви уплощенные, длинные, но менее широкие, с широким, плоским зубчатым расширением на внутреней стороне при основании, а затем постепенно изогнутые, с одним или несколькими маленькими зубчиками; у j клещи менее уплощены, почти соприкасающиеся, сначала почти прямые, а затем слабо изогнутые, внутренняя сторона слегка зазубрена. Пигидий $ маленький, широкий, слабо заметный, у 2 — более выступающий, широкий, задний край выступает в виде лопасти с прямыми сторонами, вершина которой сильно вырезана, а боковые углы оканчиваются маленьким бугорком или тупым выступом. Метапарамеры расширены посредине, наружный их край дуговидно изогнут, к вершине постепенно заостряющиеся; virga короткая, с двумя хитиновыми пластинками при основании. Длина тела 3 12—17.5, 2 14—18.5; клещей 3 3—7.5, 2 4—8 мм.
Р а с п р о с т р а н е н и е: Индомалайская область, Новая Зеландия, вост. Африка, Океания; завозится в портовые города восточной Азии до Японии (Щербаков, 1911) и Зап. Европы (Англия). Первоначальная родина—Индомалайская область, откуда он распространился при помощи кораблей в другие страны.
Чрезвычайно активный вид, вероятно дневного образа жизни, населяющий сырые места.
5. Род PROREUS BURR
Burr, 1907 : 129; 1910 : 136; 1911: 64.
Тип рода — Proreus simulans Stal.
Очень близок к роду Chelisoches, отличаясь более тонкими и длинными усиками, 3-й членик немного длиннее 4-го Jh едва длиннее 5-го, все членики цилиндрические, никогда не бывают коническими или булавовидными. Тело меньших размеров, более стройное; окраска всегда красноватых или коричневатых тонов.
Около десятка видов, свойственных Индомалайской области; один вид заходит в Японию. Род этот настолько близок к роду Chelisoches, что при монографическом изучении всей группы может возникнуть необходимость их слияния. При этом, однако, необходимо иметь в виду, что ряд
118
3. LABI I DAE
Видой, отнесенных к роду Proreas, на самом деле могут оказаться представителями рода Hamaxas (см. ниже), так как отличия, установление Burr’ом для родов Chelisoches и Proreus от Hamaxas, заключающиеся в том, что первые два рода имеют уплощенные голени, снабженные в вершинной половине бороздкой, а последний не имеет такой бороздки на голенях — не могут •считаться достаточно основательными; представители одних и тех же видов из рода Hamaxas характеризуются довольно значительной изменчивостью в строении голеней и попадаются особи, у которых голени характеризуются признаками, приписываемыми роду Proreus (и Chelisoches).
1. Proreus simulans (Stal).
Stal, Kongl. Svenska Freg. Eugen. Resa, 1.1860 : 302 (<5; остр. Ява) (Forficula). Bormans, 1900:37 (Chelisoches); Burr,. 1907:131; 1910: 137, fig. 85; 1911: 64, tab. 5, fig. 21, tab. 6, , fig. 3. — modesta Stal, ibid., 1860 : 302 (Forficula).
Тело средний размеров, стройное, оранжево- или каштаново-красное. Усики буроватые, тонкие, длинные, 20-члениковые, 1-й членик очень длинный, длиннее 2-го, 3-го и 4-го взятых вместе, 3-й цилиндрический, немного, но ясно длиннее 4-го и едва длиннее 5-го, оба также цилиндрические. Голова светло-кирпично-красная, гладкая, с ясными резкими сутурами, боковые стороны за глазами суживаются кзади, задний край заметно выемчатый посредине. Переднеспинка чуть короче головы, заметно длиннее своей ширины, обычно ясно расширяющаяся кзади, задний край широко округлен. Надкрылья длинные, довольно узкие, гладкие, светложелтые, с узкой продольной темной полосой вдоль наружного и внутреннего края; крылья вполне развитые, такого же цвета, как надкрылья, или отсутствуют. Ноги довольно короткие, оранжево-или грязно-желтые, голени уплощенные, с бороздкой в вершинной трети. Брюшко с параллельными сторонами, темно-каштаново-красное, темнее у основания, нежно пунктированое; боковые бугорки ясные; последний тергит с? широкий, прямоугольный, светлокрасный, задний край обрублен и затемнен, с вдавлением посредине, на дне которого имеются два маленьких бугорковидных возвышения; боковые края этого углубления несут также по бугорку; у g — сходного строения, но уже. Клещи 3 расставлены при основании, слегка уплощены, но не расширены, стройные, почти прямые и лишь только в вершинной половине — трети слегка загибаются друг к другу, внутренний край с острым •треугольным зубцом, расположенным на расстоянии одной трети от основания (иногда очень слабо выраженным), или посредине и иногда со вторым маленьким зубчиком перед вершиной; у g—-расставлены при основании, почти прямые, весь внутренний край слегка зазубрен. Пигидий у обоих полов очень короткий, тупой, поперечный, не выступающий. Длина тела 3 10—12, g 8.5—10; клещей 3 3, g 5 мм.
., Распространение: широко распространен в Индомалайской области от Индии, Бирмы, Аннама до Борнео, Явы, Суматры, Филиппин, южного Китая и Формозы, заходя на севере в пределы Палеарктической Азии (Япония). •
5. PROREUS, 6. HAMAXAS
119
Изменчивый вид. Боковые стороны переднеспинки у одних особей параллельные, у других расходящиеся кзади; вооружение клещей также изменчиво. Что касается формы с неразвитыми крыльями (описанной Stal’eM из Гонг-Конга, под названием Forficula modesta) то, быть может, эта форма представляет самостоятельный вид.
6. Род HAMAXAS BURR.
Burr, 1907 : 133; Burr, 1910 : 147; Burr, 1911 : 67.— Chelisoches auct., part.
Тип рода — Hamaxas feae Borm, из Индомалайской и Австралийской области.
Сходен с предыдущими, но мельче, тело более коренастое и широкое. Усики 15—19-члениковые, 3-й членик довольно короткий, не более чем в 1х/2 раза длиннее своей ширины, цилиндрический или полуконический, 4-й членик округленный, короче 3-го. Голова с неясными сутурами. Переднеспинка чуть длиннее своей ширины. Надкрылья и крылья вполне развиты, первые без боковых килей. Ноги короткие, в густых коротких волосках, бедра Толстые. Брюшка короткое, уплощенное, часто в волосках; боковые бугорки имеются; последний тергит S' широкий, у заднего края посредине обычно с продольными короткими приподнятыми ребрышками или с бугорками; у £ — узкий. Пигидий маленький. Клещи S толстые, короткие, расставленные при основании, полуцилиндрические или умеренно уплощенные, на внутренней стороне с зубцами, к вершине загнутые.
Как уже указывалось, род Hamaxas чрезвычайно близок к Chelisoches и особенок Proreus, причем, как показал’НеЬагб (1927), ряд видов, описанных в качестве представителей рода Proreus, на самом деле должен быть отнесен к роду Hamaxas.
Род этот включает более полудесятка видов из Индомалайской области, Новой Гвинеи и Новой Зеландии; один вид заходит в пределы .Центральной Монголии.
1. Hamaxas kozlovi Sem. et B.-Bienko (фиг. 33).
Setnenov-Tian-Shanskij et Bey-Bienko, 1934: 56, fig. 1 (S хребет Дунду-сайхан, центр. ’ Монголия).
Тело крупное для рода, рыжевато-бурое, голова, усики, боковые стороны переднеспинки и задний край последнего тергита брюшка черноватые. Усики 13 (?)-члениковые; 1-й членик чуть длиннее трех последующих взятых вместе, 3-й членик цилиндрический, равен по длине 5-му, 4-й членик короткий, полушаровидный. Голова чуть шире переднего края пе-'реднеспинки. Переднеспинка с полуокругленным передним и задним краем, боковые стороны в передних двух третях прямые, далее слегка расхоДЯ-щиеся, образующие с задним краем прямой широкозакругленный утоЛ.
120
4.FORF ICULIDAE
Надкрылья с широко округленными плечами и прямым обрубленным задним краем. Лапки ног с плохо заметной лопастью на 2-м членике. Брюшко ,с густой тонкой пунктировкой, бока 5—8-го тергитов у заднего края с сериями из 5—9 маленьких килевидных бугорков; последний тергит $ чуть более чем в два раза шире своей длины, прямоугольный, у заднего края посредине с четырьмя продольными приподнятыми параллельными друг другу ребрышками, расположенными на равном расстоянии друг от друга; бока брюшных стернитов в золотисто-желтых волосках, среди которых располагаются длинные редкие щетинки. Клещи 5 толстые, при основании полуцйлиндрические, в вершинной половине умеренно загнутые внутрь; внутренний край с тремя зубцами, из которых один расположен у самого основания, средний перед срединой, а задний — наиболее крупный — немного за срединой клещей. Пигидий 5 хорошо заметный, выступающий,, почти вертикальный, кверху слегка расширеный, посредине с двумя слабо выступающими бугорками. Длина тела <5 9.5, клещей 2 мм. g неизвестна..
Распространение: известен пока только с хребта Дунду-сайхан в центр. Монголии.
4. Сем. FORFICULIDAE
Усики с 10—15 члениками, 4-й членик равен по длине 3-му или короче его; членики обычно цилиндрические или почти цилиндрические^, реже почти конические, довольно длинные, 2-й членик лапок расширен с обеих сторон, более или менее сердцевидный, заметно шире 1-го и 3-гб члеников. Генитальный аппарат с удлиненными, на вершине округленными и притупленными, реже с умеренно заостряющимися, но не оттянутыми и не утонченйЫми, концами метапарамер; почковидный пузырек хорошо развит; virg-a почти прямая или умеренно S-образно изогнутая; собственно-penis без хитиновых зубчиков и пластинок.
Наиболее обширное по числу видов семейство, довольно богато,, в отличие от остальных семейств, представленное также и в умеренных широтах, где имеется много эндемичных родов и видов. В филогенетическом отношении это наиболее молодое семейство, видимо, достигшее наибольшего разнообразия в третичный период; современные роды, обитающие умеренные широты северного полушария, являются вероятно остатками третичной фауны широколиственных лесов, покрывавших в то время умеренные широты.
Систематика семейства находится в сравнительно слабо разработанном состоянии, так как группировки на подсемейства, существовавшие до настоящего времени, крайне поверхностны и не вполне необоснованы.. Достаточно сказать, что в 1907 г. Burr установил 11 подсемейств, сведя их в 1911 г. к 8, а в 1915 г. — к 6—7, кроме того ряд родов относился в разное время к разным подсемействам. При этом отличия между наиболее широко представленными в умеренных широтах подсемействами For--
4: FORFICULIDAE
121.
ficulinae и Anechurinae при дальнейшем изучении группы оказались настолько условными и не; реальными, что еслибы доводить это разделение до его логического конца, то тогда мы принуждены были бы различные подроды и даже виды одного и того же рода относить к различным подсемействам (напр. подрод Borelliola и Anechura в роде Anechura, различные виды подрода Odontopsalis, относящегося к тому же роду и пр.). Объясняется это тем, что Burr придавал большое значение в классификации семейства ширине грудных пластинок; но этот признак в лучшем случае может считаться только родовым, так как он коррелятивно связан со стройностью тела, а последняя в значительной мере зависит от степени развития надкрыльев. Так как у свойственного гл. обр. Палеарктической обл. „подсемейства" Anechurinae большинство родов представлено короткокрылыми или даже совсем бескрылыми коренастыми видами, то вполне понятно, что грудные пластинки у этих видов оказываются широкими; однако из этого правила имеются и исключения — ряд короткокрылых видов имеют умеренно широкие грудные пластинки. Исследование полового аппарата у короткокрылого рода Mesasiobia, с широкими грудными пластинками, относившегося прежде к. „подсемейству" Anechurinae,. показало большую близость данного рода к Forficula, что довольно хорошо увязывается и с характером клещей самца; в данном случае мы имеем пример конвергентного сходства, связанного с общими условиями жизни (род Mesasiobia, подобно многим „Anechurinae" связан с горными местностями). Одновременно необходимо отметить, что в лице такого вида, как Oreasiobia stoliczkai, относившегося раньше к роду Anechura, мы имеем вид несомненно близкий к видам группы F. schlagintweiti, являющимся типичными представителями данного рода; поэтому нельзя не подчеркнуть большой морфологической близости О. stoliczkai к видам рода Forficula. Наконец, половой аппарат самца, как показали исследования Zacher’а и Burr’а, у указанных двух подсемейств одинакового типа.. На основании сказанного мы считаем совершенно необоснованным отделение подсемейства Anechurinae от Forficulinae и объединяем их в одно подсемейство Forficulinae, рассматривая род Anechura и его родственников как дериват подсем. Forficulinae, связанный с горными условиями жизни.
Сказанное выше в известной мере относится и к подсемейству Opi-sthocosmiinae, отличия которого от Forficulinae заключаются в форме брюшка, в более удлиненных члениках усиков и в суженном кзади последнем спинном сегменте самца. Однако, как показал Burr, для ’двух последних признаков имеется ряд исключений; с другой стороны описанный нами из сев. Сычуани вид, относящийся к роду Sondax (S. potanini) и являющийся несомненным сородичем индийского S. repens Burr, на основании формы брюшка и усиков, должен быть отнесен к подсем. Forficulinae (s. lato), причем по характеру стернальных пластинок он весьма напоминает представителей'бывшего подсем. Anechurinae, что совершенно несвойственно-подсем. Opisfhocosmiinae. Таким образом, два вида одного и того же рода.
.122
4. FORFICULIDAE
должны были бы быть отнесены к различным подсемействам. В данном случае мы, однако, отказывается от каких-либо изменений в трактовке подсемейства Opisthocosmiinae на том основании, что это подсемейство является почти исключительно тропическим и вряд ли целесообразно производить перегруппировки, основываясь только на немногих палеарктических видах; однако, отметить искусственность и произвольность деления необходимо.
В пределах Палеарктической области известно около 70 видов, относящихся примерно к двум десяткам родов, из которых большинство свойственно Зап. Европе и особенно восточной Азии, где это семейство достигает значительного разнообразия.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ
1 (24). Надкрылья всегда имеются, вполне развитые или укороченные; крылья имеются или отсутствуют.
2 (19). Брюшко заметно уплощенное, на поперечном разрезе овальное; последний тергит $ поперечный, не суживающийся кзади (только у немногих видов рода Forficula умеренно суживающийся). Усики с относительно короткими члениками; 1-й членик короче половины длины головы, 4-й обычно заметно короче 3-го. Клещи более массивные, при основании у 5 обычно, у £ всегда значительно более толстые, чем на вершине. (1. Подсем. Forficulinae).
3 (18). Надкрылья без продольного киля по наружному краю, гладкие или слегка пунктированные.
4 (7). Клещи $ при основании расширенные, очень сближенные или соприкасающиеся.1
5 (6). Надкрылья вполне развитые или немного укороченные, всегда прикрывают заднеспинку. Голова сверху одноцветная . .1. Forficula L.
6 (5). Надкрылья сильно укороченные, не прикрывающие заднеспинку, задний край которой дугообразно вырезан. Голова сверху двуцветная, с черным лбом и желтым затылком ...... 2. Mesasiobia Sem.
7 (4). Клещи S широко расставлены, никогда не соприкасающиеся при основании; основание клещей цилиндрическое, реже умеренно расширено и уплощено.
8 (15). Пигидий $ короткий, умеренно выступающий, или полускрытый.
9 (10). Клещи у $ f. macrolabia очень длинные, тонкие, при самом основании на внутренней стороне с выступающим, пластинчатым расширением, в остальной части без зубцов и выступов, при рассматривании с боку не загнуты вниз. Надкрылья вполне развитые. Усики с длинными члениками; 4-й членик в 21/2—3 раза, 5-й — более чем в 3 раза длиннее своей ширины....................3. Oreasiobia Sem.
1 Только у части особей Forficula schlagintweiti клещи <5 могут быть умеренно расставленными при основании, но тогда тело совсем черное, а задние голени расширены около •средины.
ТАБЛИЦА РОДОВ
123
10(9). Клещи 3 без пластинчатого расширения на внутренней стороне у основания, иногда сильно волнистые или с зубцами. Надкрылья вполне развитые или укороченные.
11 (17). Брюшко 3 почти с параллельными сторонами или расширяется к вершине.
12 (16). Тело более или менее коренастое. Среднегрудная пластинка поперечная или по крайней мере одинаковой длины и ширины. Пигидий 3 широкий. Брюшко сверху и клещи голые или в редких волосках.
13 (14). Брюшко 3 почти с параллельными сторонами, или едва расширяется к вершине. Надкрылья вполне развитые или в различной степени укороченные, но всегда их длина по наружному краю не меньше их ширины, а щиток не виден или очень маленький . . .
...........................................4. Anechura Scudd.
14 (13). Брюшко 3 сильно расширяется от основания к вершине, так что его наибольшая ширина находится у самой вершины. Надкрылья сильно укороченные, их ширина не менее чем в 1х/г раза превосходит их длину по наружному краю; внутренние^ стороны едва соприкасаются, ограничивая крупный щиток...............5. Chelidura Latr.
15 (8). Пигидий 3 очень длинный, такой Же длины, как средние бедра, сильно расширен у основания и сильно сужен к вершине. Надкрылья укороченные, с косым задним краем ., . . 6. Eumegalura B.-Bienko.
16(12). Тело очень стройное. Среднегрудная пластинка удлиненная. Пигидий 3 удлиненный, сильно выступает. Брюшко сверху и клещи в-густых, довольно длинных' волосках. Клещи 3 тонкие, слабо изогнутые, в профиль прямые . ...................7. Apterygida Westw.
17 (11). Брюшко 3 с округлыми боковыми сторонами, сильно расширено посредине и сильно суживается к вершине. Клещи 3 тонкие, в профиль прямые, без зубцов или только с маленьким бугорком при основании сверху. Надкрылья сильно укороченные ...................
......................................... 8. Chelidurella Verb.
18 (3). Надкрылья с сильным продольным килем по наружному краю, сильно морщинистые, часто в мелких бугорках. Клещи 3 при основании широко расставлень!........................9. Allodahlia Verb.
19 (2). Брюшко цилиндрическое, сверху выпуклое, на поперечном разрезе круглое; последний тергит 3 у многих родов сужен к вершине. Усики с длинными члениками, 1-й членик длиннее половины длины головы, 4-й равен или едва короче 3-го. Клещи тонкие, у 3 сверху часто с зубцами, у д длинные и очень тонкие, почти нитевидные, особенно в вершинной половине. (2. Подсем. Opisthocosmiinae).
20 (21). Последний тергит брюшка 3 поперечный, прямоугольный, не очень сильно наклонный. Клещи при основании умеренно расставлены, не соприкасаются. 1-й членик лапок не длиннее или едва длиннее 3-го.
........................................ 10. Timomenus Burr.
124
4. FORFICULIDAE
21 (20). Последний тергит $ узкий, трапециевидный, сильно сужен кзади,, наклонный. Клещи $ при основании соприкасаются или очень сближены, почти соприкасаются. 1-й членик лапок примерно в два раза, длиннее 3-го.
22 (23). Переднеспинка одинаковой длины и ширины. Бока 6—9-го тергитов брюшка $ не загнутые, без выступающих назад углов. Клещи $ соприкасающиеся при основании ........ 11. Eparcbus Burr.
23 (22). Переднеспинка длиннее своей ширины. Бока 6—9-го тергитов S загнутые, с выступающим назад углом. Клещи $ слегка расставлены при основании................................... 12. Narberia Burr.
24 (1). Совершенно бескрылые виды. Последний тергит брюшка S резко суживается кзади. Клещи $ почти соприкасающиеся при основании., . . ..............................................13. Sondax Burr.
1. Подсем. FORFICULINAE
Усики с цилиндрическими или полуконическими члениками, довольно^ короткие и толстые; 1-й членик короткий, значительно короче головы; 4-й членик короче, иногда значительно, 3-го членика. Брюшко более или менее уплощенное сверху вниз, так что на поперечном разрезе оно имеет вид поперечного овала; последний сегмент <5 поперечный, прямоугольный,, не суживающийся или едва суживающийся кзади. Ноги короткие. Надкрылья и крылья вполне развиты, или крылья отсутствуют, а надкрылья нормальные или в различной степени укороченные. Клещи обычно массивные, различно устроенные, при основании у & обычно, у j всегда, значительно более толстые, чем на вершине. Пигидий S часто заметно выступает, у многих видов поперечный.
Это подсемейство в отличие от всех остальных групп отряда представлено в умеренных широтах северного полушария, именно в Палеарктической области, большим количеством видов, со многими эндемиками; многие роды характеризуются наличием большого количества видов. Фауна уховерток Палеарктической области почти на 2/3 состоит из представителей данного подсемейства, причем только на долю двух самых обширных родов — Forficula и Anechura приходится около половины видов.
1. Род FORFICULA LINNE.
Linne, Syst. Naturae, 1 (ed. 10), 1758:423; Brunner, 1882:12; Bormans, 1900:119;; Семенов, 1901:192; Kirby, 1904:49; Якобсон 1905: 27; Burr, 1907:109; 1910:164; 1911 :80; 1916:14.
Тип рода — Forficula auricularia Linne.
Тело довольно стройное. Голова гладкая, выпуклая сверху, сутуры обычно хорошо заметны. 3-й членик усиков длинный, почти такой же длины
1. FORFICULA
125
как 1-й, слегка утолщенный к вершине или цилиндрический; 4-й короче 3-го, часто значительно, иногда почти шаровидный; 5-й немного длиннее 4-го, остальные постепенно удлиняющиеся. Переднеспинка более или менее прямоугольная, спереди прямая, сзади округлена или прямая. Надкрылья хорошо развиты или слегка укорочены, но всегда прикрывают задне-спинку, гладкие, без килей; задний край обрублен, редко несколько вырезан или скошен; крылья вполне развитые, укорочены или отсутствуют. Ноги умеренно стройные. Брюшко довольно уплощенное, наибольшая его ширина посредине, суженное к вершине, особенно у J, с ясными боковыми бугорками; последний тергит $ поперечный, задний край гладкий или с двумя бугорками или выступами, у § — простой, узкий. Клещи <5 всегда утолщены, расширены и соприкасаются в основной части, а далее более или менее резко утончены и изогнуты; расширенная часть у различных видов различной длины, короткая или длинная; у g клещи простые, прямые, соприкасающиеся. Пигидий S маленький, шаровидный, реже ^сильно выступает, удлиненный; у g— простой, узкий. Метапарамеры почти или -совсем прямые, узкие, одинаковой ширины по всей их длине, в большей части параллельны друг другу; virga различной длины, постепенно утолщается по направлению к основанию, соединяясь с почковидным пузырьком обычно без дуги или угловидного изгиба.
Наиболее богатый по количеству видов род, включающий около полусотни видов, распространенных в Европе, почти всей Африке иАзии,(см. фиг. 15); один вид — F. auricularia развезен по всему свету. Большинство видов свойственно южной части Центральной Азии, от провинций Ганьсу и Сычуань в зап. Китае до Гималаев, и северной, пригималайской части Индии; второй центр многообразия расположен в Средиземьи. Что касается непалеарктической Африки, то оттуда известно до 7 видов; всего только 2—3 представителя данного рода известны из низменных частей Индомалайской области. Таким образом, центр происхождения рода, несомненно, надо искать на Ангариде; современный центр видового разнообразия лежит в южной части Восточной Азии, которая характеризуется, помимо наибольшего видового разнообразия, также наличием ряда видов, являющихся несомненными ближайшими родичами многих широко распространенных в Палеарктике видов как F. auricularia, F. tomis и др. Виды, населяющие Африку и Азию, несомненно являются позднейшими выходцами из восточной Азии и Средиземья. Достоверный представитель этого рода (F. praecursor Burr) известен еще из нижнего олигоцена (Балтийский янтарь) и обнаруживает большое сходство с африканской Ft senegalensis Serv., северо-индийской F. beelzebub Burr и турано-закав-казской F. sagitta Sem.
Систематика видов основывается гл. образом на строении переднеспинки, усиков, степени развития крыльев и особенно на строении клещей <5; хорошие отличия имеются также в половом аппарате <5, именно в длине virga и в характере соединения ее с. почковидным пузырьком.
126
4. FORFICULIDAE
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Тело сплошь глубоко черного цвета. Переднеспинка значительно шире своей длины. Задние голени расширены к средине, а отсюда суживаются к вершине. 4-й членик усиков немного короче 3-го, 5-й равен 3-му. Клещи $ расширены и соприкасаются только у самого: основания, иногда расставлены как у Anechura, но тогда не изогнуты волнисто...................................1. F. schlagintweiti Burr.
2 (1). Тело если с примесью черного цвета, то по крайней мере голова или бока переднеспинки и надкрылья, или крыловые пластинки, или ноги светлые. Переднеспинка только у немногих видов поперечная (ср. F. uvarovi, sinica, mandarina, davidi). Задние голени нормальные, примерно одинаковой ширины на всем протяжении.
3 (12). Тело голое или покрыто редкими волосками.
4 (5). Надкрылья укороченные, менее чем в Р/д раза длиннее переднеспинки, с сильно скошенным задним краем, образующим с наружной стороной острый угол; крылья отсутствуют. Последний тергит брюшка <5 немного суживается кзади. Клещи <5 очень тонкие, соприкасающиеся только у самого основания, а затем дуговидно изогнутые (фиг. 34)............................2. F. uvaroVi Sem. et B.-Bienko.
5 (4). Надкрылья вполне развиты, более чем в 1У!2~2 раза длиннее переднеспинки; задний их край срезан под прямым углом к наружной стороне. Последний тергит брюшка 8 с параллельными боковыми сторонами.
6 (40). Клещи 8 на внутренней стороне без изолированно-расположенного зубца, лишь только соприкасающаяся у основания ветвей часть иногда оттянута на своем заднем конце в виде тупоугольного зубца.
7 (37). Последний тергит брюшка без продольных килевидных складок по бокам у заднего края. 4-й членик усиков удлиненный, цилиндрический или булавовидный, примерно в два раза длиннее своей ширины.
8 (26). Клещи <5 расширены и соприкасаются по крайней мере в основной трети, у f. brachylabia почти в основной половине.
9 (13). Пигидий 8 большой, сильно выступающий, хорошо заметный.
10 (11). Тело темнобурое, почти черное, очень стройное. Крылья вполне развитые. Пигидий 8 с параллельными сторонами, длиннее своей ширины, вершина почти обрублена. Брюшко с очень густой и тонкой пунктировкой, задние края сегментов с линией более грубых точек (фиг. 35)..................................3.F. splendida B.-Bienko.
11 (10). Тело желтоватое, маленькое, умеренно коренастое. Крыловые пластинки отсутствуют. Пигидий 8 треугольный, заостренный на вершине, с приподнятыми и несколько утолщенными краями. Брюшко сверху в мелких густых точках . . . • • . . * 4. F. aetolica Brunn..
1. FORFICULA
127
12 (3). Тело, особенно ноги, брюшко снизу и клещи в густых длинных волосках. Надкрылья слегка укорочены, с немного скошенным задним краем, образующим с наружной стороной умеренно острый угол. Брюшко сверху едва пунктировано. Пигидий и клещи $ как у предыдущего вида (фиг. 36)....................* 5. F. kaznakovi Sem.
13 (9). Пигидий <5 слабо выступающий, маленький, часто едва заметный.
14 (21). Крылья вполне развитые, выступают из-под надкрылий.
15 (20). Переднеспинка не шире или едва шире своей длины, одинаковой ширины с головой. Клещи <5 слабо изогнутые, с удлиненно овальным промежутком между внутренними сторонами ветвей.
16 .(17). Тело очень стройное, соломенно-желтое. Надкрылья одноцветные,. без пятен и полос. Клещи $ сильно пунктированные, очень стройные^ Брюшко каштаново-коричневое, с довольно сильной пунктировкой .
............................................* 6. F. sagitta Sem.
17 (16). Тело умеренно-коренастое, красновато-или смоляно-бурое. Надкрылья с небольшим пятном или полосой. Клещи гладкие или слегка пунктированные в вершинной части.
18 (19). Надкрылья с овальным светлым пятном. Брюшко очень тонко пунктировано, у <5 сильно расширено посредине. Переднеспинка одноцветная, красноватая или желтая. Пигидий очень короткий, тупой, бугорковидный.....................................7. F. lucasi Dohrn.
19 (18). Надкрылья с длинной продольной светлой полоской. Брюшко густо, но довольно грубо пунктировано, особенно у $, с почти параллельными сторонами. Переднеспинка с двумя продольными темными полосками. Пигидий $ почти треугольный, относительно длинный . . .
.......................................* 9. F. tomis f. euptera..
20 (15). Переднеспинка поперечная, на одну треть шире своей длины, заметно более широкая чем голова. Клещи <5 сильнее изогнуты, почти с круглым промежутком между внутренними сторонами ветвей. . .
...........................................8. F. mandarina Вог.
21 (14). Крылья не выступают из под надкрылий, полностью отсутствуют или зачаточные. Брюшко с густой и грубой пунктировкой.
22 (25). Тело коренастое, крупное. Задний внутренний конец расширенной части клещей $ прямо- или чуть тупоугольный, без выступающей назад округленной лопасти; длина расширенной части у f. brachyla-Ыа не превышает половину длины клещей.
23 (24) Тело менее коренастое, умеренно крупное. Внутренний угол глаз широко округлен, очень тупой. Клещи $ с параллельными наружными сторонами, вершинная половина у f. brachylabia слегка дуговидно изогнутая. Крыловые пластинки иногда вполне развитые, в этом случае надкрылья с резкой светлой продольной полосой (фиг. 10, 37) .................................... * 9. F. tomis Koi.
24(23). Тело очень коренастое, более крупное. Внутренний угол глаз,, почти прямоугольно выступающий, менее округленный. Клещи J
128
4. FORFIGULIDAE
посредине расширенной части с вогнутыми наружными сторонами;, далее у f. brachylabia сильно дуговидно изогнутые; наибольшее расстояние между наружными сторонами вершинной части клещей у f. brachylabia значительно шире расстояния между наружными сторо-ронами расширенной части клещей (фиг. 39,40) . * 10. F. robusta Sem.
25 (22). Тело стройное, средних размеров. Задний внутренний конец'расширенной части клещей 8 выступает назад в виде тупой округленной лопасти; длина расширенной части у f. brachylabia занимает больше половины длины клещей (фиг. 41) . . . 11. F. macrobasis B.-Bienko.
26 (8). Клещи 8 расширены и соприкасаются менее чем в их основной трети, часто только ,у самого основания. Крылья вполне развитые;
27 (35). Надкрылья одноцветные, без пятна.
28 (36). Последний тергит брюшка 8 с двумя очень слабыми округлыми бугорковидными выступами, располагающимися у заднего края перед основанием клещей; или выступы совсем отсутствуют.
29 (32). Пигидий 8 маленький, слегка выступающий или полускрытый, значительно короче чем расширенная часть клещей. Клещи 8 с со-. прикасающимися и параллельными друг другу внутренними cTopoi нами расширенной части.
30 (31). Наружная сторона клещей $ без маленького бугорка у самого основания. Голова не шире переднеспинки, швы почти незаметные. Переднеспинка одинаковой длины и ширины, бока слегка округлены! Тело каштановое, брюшко рыжевато-каштановое, очень тонко пунктированное ................................. 12. F. ambigua Burr.
31 (30). Наружная сторона клещей 8 с маленьким бугорком при основании у заднего наружного угла последнего тергита брюшка. Г олова большая, шире переднеспинки, швы хорошо развиты. Переднеспинка почти в 172 раза шире своей длины. Тело буровато-черное, брюшко блестяще-черное, сильно пунктированное . . 12. F. sinica B.-Bienko.
32 (29). Пигидий 8 очень большой, сильно выступающий, не длиннее своей ширины, к вершине суживающийся. Расширенная и сближенная часть основания клещей очень короткая, немного длиннее пигидия, внутренние стороны заднего конца расширенной части выступают, в виде широкого треугольного зубца, раздвинуты выступающим пигидием и не соприкасаются друг с другом.
33 (34). Крупнее. Усики голые; 3-й членик почти цилиндрический, чуть более толстый у вершины чем у основания . . . 14. F. mikado. Burr.
34 (33). Мельче. Усики в густых волосках; 3-й членик цилиндрический в основных двух третях и сферически вздут в вершинной трети . .
................................. 15. F. berezovskii B.-Bienko.
35 (27). Надкрылья с круглым желтым пятном перед серединой. Брюшко сверху и клещи очень густо и сильно пунктированы.. Тело крупное, с рыжевато-красной головой и смоляно-бурым брюшком и клещами.
. . . . . . . . . . . . . . .... . * 16. F. smyrnensis Serv.
7. FORFICULA
129
36 (28). Последний тергит брюшка <5 у заднего края перед основанием клещей с двумя сильными, заостренными, иногда сильно удлинен-. ными, выступами. Переднеспинка квадратная или чуть поперечная; задний край слегка округлен. Клещи $ расширены и соприкасаются только у самого основания. Тело с примесью черного цвета, брюшко густо пунктировано (фиг. 42)..................... . 17. F. davidi Burr.
37 (7). Последний тергит брюшка по бокам с килевидной продольной складкой перед задним краем, более ясной у 8. 4-й членик усиков короткий, овальный, не длиннее или едва длиннее своей ширины. Брюшко $ грубо пунктировано. Клещи 8 расширены и соприкасаются в основной четверти или трети, далее суживаются к средине и сильно загнуты внутрь.
38 (39). Переднеспинка чуть шире своей длины. Крылья отсутствуют. Клещи 8 в вершинных двух третях с сильно дугообразно изогнутыми ветвями, образующими здесь почти правильный круг.....................
........................................18. F. decipiens Gene.
39 (38). Переднеспинка одинаковой длины и ширины. Крылья вполне развитые. Клещи 8 с угловидно загнутыми внутрь ветвями, ограничивающими пространство ромбической или квадратной формы . . .
.............................................19. F. lurida Fisch.
40 (6). Клещи 8 на внутренней стороне с изолированным зубцом, расположенным на некотором расстоянии от сближенного и соприкасающегося основания ветвей.
41 (48). Тело голое. Крыловые пластинки имеются. Клещи 8 расширены и утолщены не более чем в основной трети их длины; внутренний зубец расположен перед серединой клещей. Тело довольно крупное.
42 (47). Усики с удлиненными основными члениками; 4-й членик чуть короче 3-го, цилиндрический, в 2—3 раза длиннее своей ширины. Последний тергит брюшка 8 по бокам без выступающей боковой треугольной складки.
43 (46). Пигидий 8 очень короткий, полускрытый. Клещи 8 при основании соприкасаются, не разделены друг от друга вклинивающимся пигидием; расширенная их часть занимает около одной трети их длины.
44 (45). Клещи 8 сверху и снизу без продольной бороздки, с более тол-• стой вершинной половиной (фиг. 43) .... * 20. F, vicaria Sem.
45 (44). Клещи 8 сверху с сильной, снизу с более слабой продольной бороздкой, вершинная их половина более тонкая . 21. F. burriana Sem.
46 (43). Пигидий 8 длинный, большой, округленно треугольный, сильно выступающий, при рассматривании в профиль сильно выдается над уровнем клещей. Клещи 8 при основании отделены друг от друга вклинивающимся пигидием; расширенное основание клещей очень короткое, занимает не более одной шестой их длины (фиг. 44) . . .
................................22. F. suhauricularia B.-Bienko.
Фауна СССР, Кожистокрылые 9
130 4. FORFICULIDAE
АП (42). Усики с более короткими члениками; 4-й членик очень короткий,, полушаровидный, почти в два раза короче 3-го и едва длиннее своей ширины. Последний тергит <5 по бокам с треугольной выступающей: складкой (фиг. 9, 45).........................* 23. F. auricularia L.
48 (41). Тело покрыто редкими волосками. Крыловые пластинки имеются. Клещи <5 расширены и утолщены, по крайней мере в основной половине внутренний зубец расположен посередине или за серединой клещей. Тело очень маленькое..................24. F. pubescens Serv-
1. Forficula schlagintweiti (Burr).
Burr, 1904:313 (<5, g; Лахол, южн. Тибет; Дарджилинг, Сикким, сев. Индия) [Anechura];.-Burr, 1910:166, fig. 54; 1911: 81; Hebard, 1923: 225,. tab. XX, figs. 21—22. — beebei Burr,. 1912a: 792.
Тело крупное, одноцветно-черное, гладкое и блестящее. Усики 12-чле-никовые, 4-й членик немного короче 3-го, примерно в 2.5 раза длиннее своей ширины, 5-й членик равен по длине 3-му, более чем в 3 раза длиннее своей ширины. Голова совершенно гладкая, черная, с неясными швами.. Переднеспинка поперечная, почти в 1V2 раза шире своей длины, боковые стороны слегка округлены, задняя — также округлена; общее очертание переднеспинки четырехугольное, но все углы округлены. Надкрылья широкие, большие, совершенно гладкие, черные; крыловые пластинки длинные, черные, иногда отсутствуют. Ноги длинные, интенсивно-черные, включая покрытые густыми рыжеватыми волосками лапки; задние голени расширены к средине, а отсюда суживающиеся к вершине; передние и средние голени также расширены к средине, но в меньшей степени-Брюшко черное, совершенно гладкое, наибольшая его ширина около середины; боковые бугорки очень ясные; последний тергит $ поперечный,, чуть суженный к вершине или если клещи расставлены у основания,, то с параллельными боковыми сторонами; бугорки у заднего края слабые;-, у g последний тергит более узкий, без бугорков. Пигидий и клещи <5 двух типов: 1) —: клещи расширены и соприкасаются у самого основания, а далее дуговидно загнуты; пигидий короткий, в виде округлого бугорка; 2) — клещи расставлены у основания, не соприкасаются (как у рода Anechura)^ внутренние стороны в основной половине зазубрены, основная половина более толстая, суживающаяся к средине, вершинная половина загнута внутрь под очень тупым округленным углом к продольной оси основной половины; пигидий прямоугольный, задний край прямой, по бокам с маленьким зубчиком; пигидий g простой, бугорковидный, клещи g простые, прямые, тонкие. Длина тела с? 8:—16.5, g7.8—14.5, клещей $2.6—3.75, g2.2—3.5 мм. .
Распространение: Сев. Индия, от Сиккима на востоке до Пенджаба на западе, Тибет и Кашмир (Bey-Bienko 1935); встречается в горах до 3500 м и выше.
7. FORFICULA
131
Этот вид характеризуется весьма своеобразной изменчивостью в Строении конца брюшка <5; в одних случаях клещи имеют типичное ля рода Forficula строение, а в других — попадаются особи с расставленными при основании клещами, в силу чего изменяется строение последнего тергита и пигидия, причем иногда исчезают и крыловые пластинки; такие особи несколько напоминают виды Anechura- Одновременно особенности строения усиков (длинные, однородные основные членики), совершенно не дифференцированная окраска тела, длинные надкрылья и строение клещей <5 говорят за то, что этот вид должен рассматриваться как один из наиболее примитивных видов Forficula. Наконец, нельзя не отметить явной генетической близости данного вида к видам Oreasiobia, которых он напоминает характером своих усиков, поперечной переднеспинкой и отсутствием пунктировки на теле; особенно велико сходство с О. stoliczkai, которая даже по окраске тела напоминает F. schlagintweiti. Основываясь на этих фактах можно было бы слить все указанные роды в один; однако, это было бы неправильно, так как все виды Forficula отличаются от видов родов Anechura и Oreasiobia некоторыми существенными признаками, полностью никогда не проявляющимися у представителей двух последних родов. Кроме того, если слить все эти роды в один то получится настолько разнородный комплекс видов, что в нем можно будет разобраться только после разделения его на три самостоятельные группы, т. е. фактически роды; наконец, во всем этом комплексе имеются всего только два вида, несколько сглаживающие отличия между данными тремя группами, и нет никаких оснований пренебрегать той далеко зашедшей морфологической дифференциаций на эти самостоятельные и весьма характерные группы, которая и положена в основание разделения на роды. Географическая локализация F. schlagintweiti как раз на той территории, которая расположена близ центра зарождения родов Forficula, Anechura и Oreasiobia, где больше всего сохранилось древних третичных форм, а также указанные выше морфологические особенности данного вида, говорят за то, что в его лице необходимо усматривать одну из наиболее примитивных форм Forficulidae в Палеарктике, близкую к тем родоначальным формам, которые впоследствии дифференцировались в самостоятельный род Oreasiobia и быть может Anechura.
2. Forficula uvarovl Sem. et B.-Bienko (фиг. 34).
Semenov-Tian-Shanskij et Bey-Bienko, 1935:227 (<?, ряд мест из Тибета).
Весьма напоминает по строению усиков и отчасти по характеру переднеспинки предыдущий вид, но отличается многими признаками. Тело довольно стройное, почти одноцветное, темно-рыжевато или черновато-бурое, голое, довольно блестящее, голова, усики и клещи желтоваторыжие, ноги буровато-желтые. Усики 12-члениковые, строение их как у предыдущих видов. Голова с довольно ясными швами. Переднеспинка слегка поперечная, боковые стороны прямые, параллельные, задний край 9*
132
4. FORFICULIDAE
вые оугорки на э-м
Фиг. 34. Forficula uva-rovi Sem. et B.-Bienko. Клещи 3 (тип). По Semenov et Bey-Bienko, 1935.
умеренно округлен. Надкрылья укороченные, немного более чем в 1% раза длиннее переднеспинки, задний край довольно сильно скошен, образует с наружной стороной острый угол и полностью закрывает заднеспинку. Крылья отсутствуют. Ноги длинные, стройные. Брюшко веретеновидное, довольно сильно суженное к вершине, умеренно уплощенное, в очень тонкой, слабой, образующей тончайшие поперечные морщинки пунктировке, боко-сегменте слабые, на 4-м сильные. Последний тергит 3 заметно уже предыдущих, более чем в два раза шире своей длины, к вершине заметно суживающийся, у заднего края с двумя весьма слабыми выпуклостями; у g — сужен к вершине более значительно. Пигидий 3 хорошо заметный, умеренных размеров, к вершине слегка суженный и округленный; у g — более наклонный, треугольный, на вершине почти обрубленный. Клещи 3 длинные, тонкие, стройные, расширенные и соприкасающиеся только на коротком расстоянии у основания, расширенная основная часть короткая, задний ее конец образует слегка притупленный почти прямой угол, далее сильно утонченные, сильно, но постепенно дуговидно изогнутые, на вершине очень тонкие, с почти или совсем соприкасающимися концами; у 2 — также довольно длинные, тонкие, простые, внутренняя сторона с неясными слабыми зубчиками. Генитальный аппарат 3 с прямыми, почти параллельными друг другу метапарамерами; virga равна по длине метапарамерам, умеренно изогнутая на вершине; почковидный пузырек узкий, чуть шире чем virga. Длина тела 310—17, 2 13—15.3, клещей 3 4—7, 2 4—4.2 мм.
Распространение: Тибет от области Амдо и хребта Бурхан-будда до истоков р. Меконга и Янг-тце-кианга, на высоте до 4000 м.
Этот вид, несмотря на довольно значительно укороченные надкрылья, должен быть поставлен рядом с группой F. schlagintweiti, так как характеризуется таким же строением усиков, несколько поперечной передне-спинкой, монотонной окраской тела и строением клещей, которые напоминают таковые у F. schlagintweiti и некоторых близких к нему североиндийских видов. Укороченные надкрылья внешне сближают этот вид с представителями канарско-алжирского рода Guanchia, но это сходство невидимому чисто конвергентное.
3. Forficula splendida^B.-Bienko (фиг. 35).
Bey-Bienko, 1933:7, tab. 1, fig. 4 (3, 2>‘ Тан-шанг, южн. Ганьсу, Китай),
Тело средних размеров, очень стройное, темнобурое, почти черное. Усики 12-члениковые, рыжевато-бурые, 4-й членик немного короче 3-го,
1. FORFICULA
133
почти в 2 раза длиннее своей ширины, 5-й—в l1^ раза длиннее 4-го. Голова красновато-бурая, у S с почти незаметными швами. Переднеспинка одинаковой длины и ширины, не шире головы, с прямыми, парал-
лельными боковыми сторонами, задний край широко округлен; диск черный,
в передней части вздутый, бока плоские, палевые; задняя часть плоская. Надкрылья коричнево-желтые, длинные, узкие, примерно в 2% раза длиннее
своей ширины и в 2 раза длиннее переднеспинки, выемчатый; крыловые пластинки длинные, черные, с густой тонкой пунктировкой. Ноги желтые, передние бедра сильно утолщенные в основной трети, заметно суживающиеся к вершине, менее чем в 3 раза длиннее своей наибольшей ширины. Брюшко узкое, очень слабо расширяющееся около средины, почти с параллельными сторонами; очень густо и тонко пунктировано, вдоль задних краев спинных полуколец с более грубыми точками; последний тергит в 2 раза шире своей длины, прямоугольный, с двумя слабыми вздутиями около середины заднего края и заметным вдавлением в задней половине между вздутиями. Пигидий S относительно очень длинный, с параллельными сторонами и обрубленным или едва округленным задним краем; у g — также длинный, хорошо заметный сверху и снизу. Клещи $ буровато-черные, с почти параллельными, слегка изогнутыми на вершине ветвями; расширенная и сближенная часть занимает основные 2/в Длины клещей, внутренняя ее сторона мелко и густо зазуб-
задний их край слегка
Фиг. 35. Forficula splendida В. - Bienko, <?Х5 (тип). По Bey-Bienko, 1933.
рена; вершинная часть постепенно утончается
к концу; вся поверхность клещей, - особенно вершинная часть, сильно пунктирована; клещи g простые, прямые, тонкие, буровато-желтые. Длина тела 5 10.7—10.9, g 9.7—10; клещей 5 3.3—4.3, g 1.8—3 мм.
Распространение: известен только из Тан-шанга в южном Г аньсу.
* 4. Forficula aetolica Brunn.
Brunner, 1882 : 18 (<5; Этолия, Греция; Малая Азия); Bormans, 1900:125; Аделунг, 1902:ХП; Kirby, 1904 : 52; Якобсон, 1905:28; Semenov, 1908 :167; Burr, 1911: 81, tab. Ill, fig. 18.
Тело незначительной величины; усики, ноги, переднеспинка, надкрылья и клещи грязно-желтые или буровато-желтые, с красноватой (редко с черноватой) головой и красновато-бурым брюшком; тело голое или слегка волосатое, блестящее. Усики fll—12-члениковые, 4-й членик довольно короткий, не более чем в 2 раза длиннее своей ширины и немного короче 3-го. Переднеспинка едва длиннее своей ширины, почти квадрат
134
4. FORFICULIDAE
ная, боковые стороны слегка округлены, почти прямые и параллельные, задний край широко округлен, постепенно и округло соединяется с боковыми сторонами. Надкрылья немного менее чем в два раза длиннее переднеспинки, их задний край обрублен, едва скошен, почти поперечный; крылья отсутствуют. Брюшко сверху мелко и густо-пунктировано; последний тергит $ сильно поперечный, посредине у заднего края с 2 слабыми вздутиями. Пигидий 3 большой, хорошо заметный, треугольный, к вершине заостренный, боковые стороны черные, приподнятые и морщинистые, ограничивают центральное треугольное углубление желтого цвета; у g— такой же формы, но мельче, с сильно наклоненным верхом, приподнятые боковые края менее ясные. Клещи <5 расширены и соприкасаются в основной половине (f. brachylabia) или трети (f. macrolabia), на значительном протяжении уплощены; внутренние стороны соприкасающейся части черноватые, задний их конец прямоугольный; вершинная утонченная часть клещей слабо изогнутая, первоначально уплощенная, к вершине постепенно утончающаяся, конец черноватый; клещи g простые, довольно толстые, плотно соприкасающиеся. Длина тела 5 6.2—8, g 6.2—9.5; клещи $ 3.6— 7, g 1.9—2.4 мм.
Распространение: южный берег Крыма, Малая Азия, Греция и о-в Кипр.
* 5. Forficula kaznakovi Sem. (фиг. 36).
Semenov-Tian-Shanskij, 1908:168; Burr, 1911:81. — caucasica Semenov, 1903:19 (<5; p. Ак-су, Балкария, Кавказ) [non Kolenati; 1946]. — ? carta Fischer-Waldheim, Orthopt. Ross., 1846 : 49 (Кавказ).
Вид очень похож на F. aetolica, но отличается от него следующими признаками. Немного крупнее, окраска более темная, с темнобурой головой и брюшком, ноги более светлые, тело, особенно ноги и клещи, в густых и довольно длинных волосках. Усики 13-члениковые, голова более поперечная, немного шире переднеспинки, с более тупо округленными задними наружными углами. Надкрылья более короткие, с более скошенным задним краем. Брюшко сверху в неясной, полустертой пунктировке. Расширенная и соприкасающаяся часть клещей относительно немного более короткая. Длина тела <5 8.5—9.3, g 11.5, клещей $ 4.5—5, g 2.8 мм.
Распространение: Западный Кавказ от Балкарской области и Майкопа (-Шапошников, колл. С.-Т.-Ш.!) до Абхазии (Пицунда, Гудаут); окр. Ворошиловска н./К. (колл. С.-Т.-Ш. и Г. Б.-Б.1). Наличие оторванного островка обитания этого вида в окрестностях г. Ворошиловска н/К. (Ставропольское третичное плато), где в силу вертикальной зональности сохранился островок лесной растительности, может быть объяснено влиянием ледникового периода (см. стр. 67). Совершенно аналогичную картину распространения имеет и другой кавказский лесной вид — Anechura (Borel-liold) euxina Sem- (см. ниже).
1. FORFICULA
135
* 6. Forficula sagitta Semenov-Tian-Shanskij, sp. n.1 mesmini Azam. 1906 : 81 (Ареш и Геок-тапа, Азербайджан), partim.
, Тело очень стройное, средних размеров, с соломенно-желтыми усиками, переднеспинкой, надкрыльями и крыльями и ногами; голова рыжеватая, брюшко буровато-коричневое. Усики 12-члениковые, 4-й членик почти равен 3-му, 5-й в Р/2 раза длиннее 4-го. Голова с неясными швами. .Переднеспинка одинаковой длины и ширины, с почти прямыми боковыми
сторонами, задний край слегка округлен. Надкрылья длинные, с чуть вырезанным задним краем; крыловые пластинки очень длинные, реже укороченные. Брюшко с довольно крупной пунктировкой; последний тергит 3 поперечный, чуть более чем в 2 раза шире своей .длины, с двумя едва выступающими округленными бугорками; у 2— более узкий, без бугорков. Пигидий <5 наклонный, суженный к вершине, вершина обрублена, у 2 слабо заметный. Клегци 3 длинные, расширены и соприкасаются внутренними сторонами в основной половине, сильно, пунктированы, ’внутренняя сторона расширенной части слабо зазубрена, затемнена; вершинная половина стройная, слабо изогнутая; клещи Q длинные, довольно тонкие, на внутренней стороне не зазубренные. Длина тела 3 9.5 — 10, 2 9.5 —11.5; клещей 3 4 — 4.6, 2 2.9 — 3 мм.
Распространение: известен только из Ганджинского района в Азербайджане (Ареш и Геок-тапа), из Ташкента и юго-западной Туркмении.
Фиг. 36. Forficula kaznakovi Sem., 3. X б (Нальчик). Ориг.
Описанная Azam’oM F. mesmini представляет собой смесь двух видов — F. tomis f. euptera и F. sagitta', мы присоединяемся к мнению
А. П. Семенова-Тян-Шанского, который рассматривает F. mesmini в качестве синонима F. tomis.
7. Forficula lucasi Dohrn.
Dohrn, 1865:98 (Сирия; Египет); Krauss, Ent. Nachr., XXI, 1895:99, fig. 2; Bormans, 1900 :121; Якобсон, 1905 : 27; Burr, 1910:173, fig. 59; Burr, 1911:81. — escherichi Krauss, ibid., 1895 : 97, fig. 1. — barroisi Bolivar, Rev. Biol. Nord. France, V, 1893:477; Bormans, 1900: 121; Якобсон, 1905:27; Burr, 1911:81.
1 Оригинальное описание F. sagitta см. в резюме (стр. 228).
136
4. FORFICULIDAE
Тело средних размеров, каштаново-бурое. Усики 11—12-члениковые, довольно толстые, буровато-желтые. Голова рыжая или желтая, с неясными швами, выпуклая. Переднеспинка немного шире своей длины,, рыжеватая или желтая, боковые стороны прямые, параллельные, задний край широко округлен. Надкрылья рыжевато-красные, с большим желтоватым овальным пятном, расположенным в передней половине или посредине диска, задний край заметно вырезан. Крыловые пластинки длинные, сильно выступающие, или сильно укороченные, слегка выступающие из под надкрылий, грязновато-желтые или рыжеватые. Ноги грязноватожелтые. Брюшко каштаново-бурое, очень мелко пунктированное; последний: тергит 8 поперечный, прямоугольный. Пигидий <5 очень короткий, тупой,, бугорковидный. Клещи рыжеватые, у <5 сильно уплощенные и расширенные в основной половине (f. brachylabia) или основных двух-пятых (f. macrolabia) их длины, с слабо зазубренными внутренними сторонами, задний, конец расширенной части прямоугольный; вершинная часть тонкая, умеренно или (f. macrolabia) слабо изогнутая, постепенно утончающаяся к самому концу; клещи Q простые, соприкасающиеся, на внутренней стороне без зубчиков. Длина тела 8 10—17, 9 11—14; клещей 8 5—11.5,. 2 2.5—3.5 мм.
Распространение: ареал распространения этого вида очень обширен в меридиональном направлении, но довольно узок в широтном, охватывая Сев. Африку от острова Джебра у Тунисского побережья: и оазиса Гурара в сев. Сахаре (Krauss), через Алжир до Египта; Сирия, Синай, Палестина, Зап. Иран (Арабистан, колл. С.-Т.-Ш1), Иранский Белуджистан (Керман) и Бирма; таким образом большая часть ареала, располагается в южной части Палеарктической области, выходя, однако, за пределы последней и охватывая часть Индомалайской области.
Вид этот характеризуется значительной изменчивостью в степени развития крыловых пластинок, которые у некоторых особей могут быть сильно укороченными и едва выступать из под надкрылий. Эта форма (описанная прежде как особый вид — F. barroisi Bol., сведенный затем А. П. Семеновым-Тян-Шанским в синономы к F. lucasi) одновременно характеризуется и несколько более короткими надкрыльями, а также расположением светлой полосы на надкрыльях ближе к их срединной части; оба этих признака находятся, видимо, в коррелятивной зависимости оТ степени развития крыльев.
8. Forficula mandarina Borelli.
Borelli, 1915 :2 (Cj; Сычуань).
Тело умеренно-крупное, красно-бурое. Усики рыжевато-красно-бурые, 4-й членик более короткий чем 3-й, 5-й членик равен 3-му. Голова заметно длиннее своей ширины, рыжевато-красно-бурая, блестящая, с темножелтой верхней губой, швы слабо заметные. Переднеспинка такого же цвета как усики и голова, с желтыми боковыми сторонами, почти прямоугольная,
1. FORFICULA
137
более широкая чем голова и на одну треть шире своей длины, боковые стороны прямые, немного расходящиеся кзади, задний край и задние углы округлены. Надкрылья каштаново-рыжие, тусклые, заметно вырезанные по заднему краю, длинные, в два раза длиннее переднеспинки. Крыловые пластинки сильно выступают, длиннее переднеспинки, такого же цвета как надкрылья. Ноги желтовато-рыжие. Брюшко красно-бурое, с очень тонкой едва заметной пунктировкой и с серией более сильных точек вдоль задних краев сегментов, заметно расширяющееся от основания к 6-му сегменту, суживающееся к вершине; боковые бугорки сильно развитые, особенно на 3-м сегменте. Последний тергит более светлый, у 3 прямоугольный, в три раза шире своей длины, с двумя большими округленными выступами около заднего края, перед основанием клещей. Задний край прямой. Пигидий слабо заметный, сзади слегка округленный. Клещи 3 темножелтые, с буроватыми точками, основная треть уплощена и расширена, со слегка зазубренными внутренними сторонами, задний конец расширенной части образует прямой угол; далее почти цилиндрические, сильно изогнутые к вершине, ограничивая своими внутренними сторонами почти круглое пространство. Длина тела 3 12.5, клещей 4.5 мм. Самка неизвестна.
Распространение: известен пока только из Сычуани.
* 9. Forficula tomis (Koi.) (фиг. 10, 37, 38).
Kolenati, Melet. Entom., V, 1846:74, tab. XVII, fig. 6 (3, g; Грузия Армения). \Chelidura\-, Brunner, 1882 :17 (partim); Bormans, 1900 :124 (partim); Семенов 1901a : 196 (partim); Kirby, 1904:52 (partim); Якобсон, 1905:27, tab. II (partim); Burr, 1911:81.— tomis var. scidens Kolenati, ibid., 1846 : 74, 79 (Chelidura).—hellmanni Kittary, Bull. Soc. Nat. Moscou, II, 1849 : 438, tab. II, fig. 1—2. — pomerantzevi Semenov, 1903 : 20. — mesmini Azam 1906:81 (partim).
Тело довольно крупное, умеренно коренастое, красно- или смолянобурое. Усики буровато-желтые, 12-члениковые, 4-й членик в два раза длиннее своей ширины, на 1/s короче 3-го; 5-й членик почти в 11/2 раза длиннее 4-го и немного длиннее 3-го. Голова буровато-красная; глаза с очень широкоокругленным и тупым внутренним углом, образованным внутренней и задней сторонами. Переднеспинка одинаковой длины и ширины, или чуть поперечная, с прямыми, едва расходящимися боковыми сторонами, задний край широко округлен; боковые, а иногда передний край и срединная продольная полоска, грязно-желтые. Надкрылья почти в два раза, или у f. pomerantzevi Sem. ( = f. eupterd) немного более чем в два раза длиннее своей ширины, с поперечным, обрубленным задним краем, смоляно-или красно-бурые, обычно с большим продольным светлым пятном, часто занимающим почти все надкрылье и особенно резким у f. euptera. Крылья сильно укорочены, скрыты под надкрыльями или реже у f. euptera вполне развитые, сильно выступают из под накрылий, почти сплошь желтые. Ноги грязно-желтые. Брюшко 3 с почти параллельными сторонами, в густой густой пунктировке; последний тергит 3 прямо
138
4. FORFICULIDAE
угольный, поперечный, в 2х/2—3 раза шире своей длины, с двумя слабыми вздутиями посредине у заднего края. Пигидий маленький, треугольный, с наклонной верхней стороной. Клещи 5 расширены и соприкасаются в основной [половине (f. brachylabia), или трети (f. macrolabid), с почти
параллельными на всем протяжении наружными сторонами, внутренняя сторона [соприкасающейся части неправильно зазубрена, сзади оканчи-
Фиг. 37. Forficula tomis Koi., £ X 4,5 Фиг. 38. Forficula (Алтай). Ориг. tomis KoL, <5. По-
ловой аппарат. Ориг.
вается прямым или слегка притупленным, но не закругленным углом; вершинные части клещей умеренно тонкие, слегка загнуты только у самой вершины, параллельные друг другу, цилиндрические; клещи Q — простые, соприкасающиеся, довольно тонкие. Длина тела 5 11.5—17, $ ’11.5,—15;
клещей <5 5—9, $ 3.5— 4 мм.
Распространение: южная и центральная часть вост; Европы на юг до Крыма, на запад до Бессарабии, захватывая всю степную и лесостепную полосу Украины до Могилева-Подольского (Г адд, колл. С.-Т.-Ш!) и Каменец-Подольска (показание Belke: F. decipiens), вся лесостепь РСФСР, южная часть лесной зоны на севере до с. Королево близ Витебска (колл. С.-Т.-Ш.!), Москвы, с. Щекотова близ Ярославля (Яковлев, колл. С.-Т.-Ш.!), Владимира, Горького, Кирова, Перми и района Сверд-
ловска; вся южно-степная часть от границе Украиной (Новочеркасск, колл. С.-Т.-Ш.!) до Кизляра (колл. Г. Б.-Б.!), весь Кавказ от Сухума (колл. Кавк. Муз.!) до Талыша, заходя и в Турецкую Армению (Эрзерум, Карс— колл* ЗИН!), Астраханский край, зап. Казахстан (пос. Чижинский, колл. ЗИН!), зап. Сибирь на севере до лесостепи (самый северный пункт — Омск), Зайсан, южный Алтай (Ульбинское, колл. ЗИН!), Туранская низменность {берег Аральского моря в районе Чимбая, колл. Г. Б.-Б.!), горы центральной Азии от Семиречья (Лепсинск, Токмак, Алма-ата, Чилик — колл.
1. FORFICULA
139
ЗИН!) до долины Зеравшана, Памира и вост. Тян-шаня (Урумчи). Таким образом, в северной части своего ареала этот вид приурочен к низменностям, а на юге поднимается в горы, причем в пределы Туран-ской низменности заходит только в северную ее часть, широко распространяясь, однако, по Тянь-шанской горной системе и доходя даже до Памира и Урумчи. В пределы Зап. Сибири (Омск), видимо, был недавно завезен волной переселенцев, хлынувших из европейской части в Сибирь в период 1917—20 годов; в изобилии встречался в г. Омске в жилых, особенно сырых домах, заползая во влажное белье и одежду, варенье и пр.
Отмечен для ряда мест в качестве вредителя огородных культур (картофель, свекла, соя, мак, всходы подсолнечника, капуста, томаты), бахчевых (огурцы, тыквы, дыни), табака, цветоводства (астры, хризантемы); обгрызает листья, подгрызает стебли, а иногда съедает нацело растения, особенно всходы и молодые высадки. Кроме того, отмечен как вредитель семенников свеклы на Украине, выедающий полузрелые семена; также иногда забирается в слабые плоды, причиняя незначительный вред плодоводству (яблоня). Вредит как во взрослом, так и в личиночном состоянии. В условиях лесостепи РСФСР зимует в фазе личинки 3—4-го возрастов; окрыление начинается в мае и длится до июня. С начала июня производится яйцекладка, причем самка откладывает в вырытую в земле или навозе неглубокую ямку от 50 до 80 яиц, из которых дней через 12—16 вылупляются личинки. Самка охраняет отложенные яйца и живет с личинками после их отрожДения дня 4—6, так что большую часть своей жизни личинки ведут вполне самостоятельный образ жизни. Как большинство уховерток, и F. tomis является ночным насекомым; днем в массах забирается под кору деревьев и в другие укромные места.
* 10. Forficula robusta Sem. (фиг. 39, 40).
Semenov-Tian-Shanskij, 1908 :166 (О, g; о-в Аскольд около Владивостока; Владивосток, ряд мест в Корее; Ашихе и Ван-ли-хо-тун в Манчжурии); Burr, 1911:81. — ? scud-deri Bormans, Ann. Soc. Espan. Hist. Nat., IX, 1088: 514 (g; Япония). — scudderi Shiraki, 1928:20. — Forficula tomis auct., partim.
Очень близок к F. tomis, но отличается от нее следующими признаками. Тело крупнее и более коренастое. Внутренний угол глаз, образованный внутренней и задней сторонами, почти прямой, менее округленный. Переднеспинка относительно длиннее, по крайней мере у <5 немного длиннее своей ширины. Надкрылья обычно без светлого широкого продольного пятна, желтовато-бурые. Пигидий $ значительно меньше выступающий, более наклонный, предпоследний стернит брюшка немного крупнее. Клещи $ при основании немного более расширенные, наружные стороны в основной половине ясно вогнуты; верх внутреннего края расширенной части с ясной приподнятой шероховатой складкой начинающейся почти от основания клещей, затем идущей по внутреннему краю и проходящей над задним углом расширенной части несколько отступя
140
4. FORFICULIDAE
от вершины угла в виде скошенной приподнятости, которая исчезает у начала утонченной вершинной половины клещей; вершинная половина клещей у f. brachylabia ясно дуговидно изогнута на всем протяжении, так что весь наружный край клещей от основания до вершины S-образно изогнут, у f. macrolabia вершинная часть слабо изогнута. Длина тела <5 14.5—18, Q 15—23.5, клещей <5 7.5—14, 9 4.6—5.4 мм.
Распространение: весь Уссурийский край на север до Хабаровска (колл. ЗИН!), о. Сахалин (колл. ЗИН!), Корея, Манчжурия, сев. Китай и Япония.
Фиг. 39. Forftcula robusta Sem. <5, f. macrolabia X 4 (Славянка, Уссур. край);— фиг. 40. Она же, <5, f. brachylabia X 4 (та же местность). Ориг.
Этот вид долго смешивался с F. tomis и впервые был основательно описан А. П. Семеновым-Тян-Шанским. Что касается F. scudderi, птисяя-ной Bormans’oM из Японии, то это описание, если даже оно действительно относится к F. robusta, не может быть принято, так как основано на самке (которые в роде Forficula очень часто неотличимы даже у далекостоящих видов) и собственно должно рассматриваться как nomen nudum.
1. FORFICULA
141
11. Forficula macrobasis B.-Bienko (фиг. 41).
Bey-Bienko, 1934:423, fig. 12 (<5; Сигусян, на границе Тибета и Сычуани).
Тело довольно стройное, средних размеров, желтовато-коричневое. Усики (?). Голова с очень ясными швами, темно-красновато-коричневая. Переднеспинка одинаковой длины и ширины, немного уже головы, с прямыми параллельными боковыми сторонами, задний край сильно округло выступает, образуя с боковыми сторонами очень тупые, округленные углы; диск коричневый, с желтоватыми боковыми сторонами. Надкрылья немного менее чем в ПД раза длиннее переднеспинки, с обрубленным задним краем, грязно-желтовато-коричневые. Крылья отсутствуют. Ноги грязно-желтовато-коричневые, вершина бедер ^неясно затемнена, передние бедра сильно утолщенные. Брюшко желтовато-коричневое, слегка расширяющееся к 6-му сегменту, сильно и густо пунктировано, с ясными боковыми бугорками; последний тергит $ черновато-коричневый, короткий, примерно в 3 раза длиннее своей ширины, с параллельными сторонами, с относительно редкой, но глубокой пунктировкой, задняя половина с двумя неясными вздутыми выпуклостями в срединной части. Пигидий короткий, слабо выступающий, треугольный. Клещи $ очень большие, у f. brachylabia сильно расширенные и уплощенные в основных двух третях, с прямыми соприкасающимися здесь внутренними, неясно зубчатыми, сторонами; наружная сторона расширенной части очень по
степенно и округло вогнута внутрь, а вершинная утонченная часть дугообразно выпуклая, так что весь наружный край от основания до вершины ясно S-образно изогнут; задние концы расширенной части выступают назад в виде округленных коротких лопастей, не соприкасающихся друг с другом вершинами и разделенных угловидным промежутком; вершинная не расширенная часть клещей цилиндро-коническая, округло загнутая, постепенно суживающаяся к вершине; клещи $ у f. macrolabia неизвестны. Длина тела $ 9.8, клещи 4.2. Самка неизвестна.
Распространение: известен только из Сигусяна на границе Тибета и Сычуани.
Этот оригинальный вид является несомненным ближайшим родичем F. tomis и особенно F. robusta, представляя по строению своих клещей дальнейшую модификацию и усложнение того типа, который выражен у этих видов. Удлинение расширенной части клещей, при одновременном образовании S-образной изогнутости наружной их стороны, ясно намечается у F. robusta, но у F. macrobasis оно достигает почти крайних возможных пределов такой модификации.
Фиг. 41. Forficula macrobasis B.-Bienko, 3. Клещи (тип). По Bey-Bienko, 1934.
142
4. FORFICULIDAE
12. Forficula ambigua Burr.
Burr, 1904:321 (<5, g; Дарджилинг, Сикким, Вост. Гималаи); Burr 1910:171, fig. 58; Burr, 1911:81; Shiraki, 1928:19.
Тело маленькое, довольно стройное, каштановое. Усики 12-члениковые, 4-й членик относительно длинный, полуконический. Голова темнокрасная, выпуклая, швы почти отсутствуют. Переднеспинка квадратная,, боковые стороны слегка округлены, задний край не выступает, слегка, округлен, постепенно переходит в боковые стороны, не образуя углов;, диск темнокаштановый, с палевыми боками. Надкрылья и крылья хорошо развитые, длинные, гладкие, грязновато-желтые, задний край надкрылий поперечно обрублен. Ноги короткие, светложелтые. Брюшко с параллельными сторонами, слабо уплощенное, темно-рыжевато-каштановое, каждый сегмент темнее при основании, чем на вершине, очень тонка пунктированный; последний тергит S немного шире своей длины, четырехугольный, несколько наклонный кзади, с продольным вдавленйем по средине, особенно в задней половине и с двумя слабо выпуклыми выступами перед задним краем около средины; задний край прямой; у $ — более, узкий, с плохо выраженными выступами перед задним краем. Пигидий $ очень короткий, конический. Клещи S со слабо уплощенными ветвями, расширенная часть занимает только самое основание клещей, соприкасаясь внутренними, слегка зазубренными сторонами только на очень коротком расстоянии, с тупоугольным задним концом; далее тонкие, постепенно утончающиеся к вершине, очень слабо изогнутые; у Q—клещи простые, короткие, соприкасающиеся. Длина тела $ 9.5. 2 8.2—9; клещей <5 2.5—3, Q 2 мм.
Распространение: Сев. Индия, Тонкин, Формоза, Китайское: побережье от южной части до Киао-чжоу на севере (Borelli, 1916).
13. Forficula sinica B.-Bienko.
Bey-Bienko, 1934:421, fig. 11 ($, g; долина Кусёр, Мунгу-чиути, Сычуань).
Тело маленькое, довольно стройное, буровато-черное. Усики рыжевато-коричневые, 3-й членик немного длиннее 4-го, но чуть короче 5-го, Голова большая, красновато-бурая, с широко-закругленными в задней части боковыми сторонами, заметно шире переднеспинки, с ясными швами, с гладкой выпуклой поверхностью. Переднеспинка темнобурая,, с буровато-палевыми боковыми сторонами, сильно поперечная, почти в 1 т/2 раза шире своей длины, со слегка округленными боковыми и задними краями, задние боковые углы очень широко округлены. Надкрылья широкие,, в два раза длиннее переднеспинки, буровато-желтые, с поперечно-обрубленным задним краем; крыловые пластинки довольно длинные, чуть короче переднеспинки, такого же цвета как надкрылья, но немного более темные или у 2 буровато-черные. Ноги рыжевато-бурые. Брюшко слабо расширенное к середине и отсюда слегка суживающееся к вершине, сверху
1. FORFICULA
143
блестяще-черное, co слабым красноватым оттенком, снизу темно-красно-вато-бурое; с очень ясной и густой пунктировкой, исключая задние концы каждого сегмента, которые имеют более редкую пунктировку; последний тергит $ немного более чем в 2 раза шире своей длины, прямоугольный, с неокругленными прямыми задними боковыми углами, задняя часть с двумя очень ясными срединными сферически выступающими вздутиями и с глубоким вдавлением между ними; у g последний тергит сильно сужен к вершине, с менее приподнятыми вздутиями. Пигидий $ относительно короткий, только частично видимый сверху, .заметно более широкий, чем его длина, задний край округленно вырезан, с остро-высту-пающими наружными углами; уд — более узкий, с прямым задним краем. Клещи рыжевато-бурые; у $ длинные, тонкие, сильно расширенные только у самого основания, внутренние края расширенной части очень слабо зазубрены, задний конец с тупым, не округленным углом, но без зубца; далее цилиндрические, постепенно утончающиеся, слабо изогнутый, с очень редкими и короткими волосками; наружная сторона у самого основания с маленьким округленно-треугольным бугорком, располагающимся у самого заднего края последнего тергита, близ задних наружных его углов; у g клещи простые, с ясным килем по наружной стороне в основной половине. Длина тела $ 8.9, g 10—11.9, клещей $ 3, g 2.4—2.6 мм.
Распространение: Сычуань в сев.-зап. Китае.
Этот вид очень хорошо отличается от всех остальных видов рода Forficula присутствием маленьких бугорков у основания клещей <5 снаружи; большая голова и широкая переднеспинка также весьма характерны для вида.
14. Forficula mikado Burr.
Burr, 1904:319 (<5; центр. Япония); Burr, 1907:112, tab. Ill, fig. 9; 1911:81; Shiraki, 1928:20. — longipyga Matsumura et Shirakii, 1905 : 84. fig. 2 (Apterygida). — diminuta Matsumura et Shiraki, 1905:84, fig. 3 (Chelidura).
Тело коренастое, средних размеров, буровато-каштановое, надкрылья и крылья светлее. Усики 12-члениковые, 4-й членик немного короче 3-го. Голова буровато-красная, с неясными швами. Переднеспинка широкая,, едва уже головы, буровато-красная, со светлыми сторонами, задний край и задние боковые утлы округлены. Надкрылья одноцветные, буроватые или буровато-желтые; крылья вполне развитые, буровато-желтые. Ноги длинные, бурые или буровато-желтые. Брюшко буро-каштановое, блестящее, у $ за срединой слегка расширено, с ясными боковыми бугорками;, последний тергит S поперечный, короткий и широкий, посредине вдавлен, с неясной бороздкой, задний край посредине вогнутый, по бокам, над основанием килей, с тупыми, бугорками; у д — последний тергит более узкий, с неясными бугорками. Пигидий $ крупный, трапециевидный, не длиннее своей ширины, сильно выступающий, хорошо заметный, несколько сужен к вершине, задний край обрублен и со слабыми боковыми малень
144
4. FORFICULIDAE
кими бугорками. Клещи $ расширены и соприкасаются только у самого основания, расширенная часть не длиннее или едва длиннее пигидия, оканчивается сзади тупоугольным выступом; остальная часть прямая, без зубцов, уплощенная, на самой вершине изогнутая, но не перекрещивают щаяся; у g— простые, соприкасающиеся. Длина тела $ g 8—11, клещей 3—6 мм.
Распространение: Япония.
15. Forficula berezovskii B.-Bienko
Bey-Bienko, 1934 : 419, fig. 10 (c5; Люнанфу, Ходзигоу, пров. Сычуань).
Очень близок к предыдущему виду по строению клещей 6. Тело довольно маленькое, относительно стройное. Усики волосатые, желтовато-коричневые, 10 (?)-члениковые, 3-й членик длинный, на вершине расширен и вздут булавовидно, 4-й членик полуконический, немного короче 3-го. Голова большая, ясно выпуклая, с довольно хорошо заметными швами, блестяще-черная, с широко-округленными задними боковыми углами. Переднеспинка немного уже головы, почти одинаковой длины и ширины, с широко закругленным задним краем и задними углами, черноватая, со светлыми боковыми сторонами. Надкрылья одноцветные, желтовато-коричневые, немного менее чем в два раза длиннее переднеспинки, с поперечно обрубленным задним краем; крылья вполне развитые, такого же цвета как надкрылья, умеренно выступающие. Ноги грязновато-желтые, передняя пара более темная. Брюшко коричневато-черное, слабо расширено к 6-му сегменту и сужено к вершине; все членики очень густо пунктированы, особенно в передней их части; бугорки вполне ясные, не пунктированные; последний тергит очень короткий, примерно в 4 раза шире своей длины, с параллельными боковыми сторонами, сверху в срединной части перед задним краем с двумя низкими, вздутыми, округлыми выступами, между которыми имеется ясное вдавление, но отсутствует продольная бороздка. Пигидий и клещи как у F. mikado; вершина пигидия без боковых бугорков, а расширенная основная часть клещей заканчивается на своей вершине прямоугольным выступом, несколько приподнятым кверху и поэтому заметным даже при рассматривании несколько сбоку и сверху; самая вершина этого выступа чуть оттянута и заострена. Длина тела S 8.0, клещей 3.5. Самка неизвестна.
Распространение: известен только из Люнанфу, пров. Сычуань, зап. Китай.
Этот вид очень близок к F. mikado по строению клещей и пигидия, образуя с последним естественную группу и отличаясь от него строением 3-го членика усиков, окраской, некоторыми деталями в строении последнего тергита и клещей, а также наличием на усиках хорошо заметных волосков. Несколько расставленные при основании клещи 3 сближают эти два вида с видами рода Anechura.
/. FORFICULA
145
* 16. Forficula smyrnensis Serv.
Serville, 1839:38 ($; Смирна, Малая Азия); Fischer, 1853 : 71, tab. 6, fig. 8, 8 a. Brunner, 1882:14; Bormans, 1900:123; Семенов, 1901a: 194; Kirby, 1904:51; Якобсон, 1905 : 27; Burr, 1911 : 82.—biguttata Fischer-Waldheim, 1846 : 40, tab. 1 fig. 1 (<^), partim).
Тело крупное, смоляно-или каштаново-бурое. Усики 12-члениковые, рыжевато-грязно-желтые, 4-й членик довольно длинный, в 2 раза длиннее своей ширины и почти равен 3-му. Голова красно-бурая или желтоваторыжая, с резко выраженными швами. Переднеспинка почти квадратная, одинаковой длины и ширины или чуть шире, с прямыми параллельными боковыми сторонами и со слабо округленным, иногда почти прямым, задним краем, рыжевато- или буровато-желтая, с грязно-желтыми боковыми сторонами. Надкрылья длинные, с параллельными сторонами, буровато-каштановые или буровато-черные, с крупным округлым палево-желтым пятном в передней половине; задний край чуть дугообразно вырезан, почти прямой; боковые, подвернутые вниз, стороны надкрылий желтоватые. Крыловые пластинки длинные, палево-желтые, иногда грязноватые, с зачерненным внутренним краем. Ноги рыжевато-желтые. Брюшко смоляно- или каштаново-бурое, блестящее, с очень густыми, хорошо заметными точками; последний тергит 5 сильно поперечный, в 21/2—3 раза шире своей длины, прямоугольный, с чуть округленно-выступающим задним краем, срединные бугорки у заднего края над основанием клещей хорошо выражены, но низкие, тупые; у $ последний тергит более узкий, в основной половине суживающийся по направлению к вершине, а от средины с параллельными сторонами, так что задние наружные углы как у $ прямые, бугорки слабые. Пигидий $ слабо заметный, почти вертикальный, у 2 при рассматривании сверху и сзади кажется треугольным, совершенно вертикальный. Клещи смоляно-бурые, у $ в довольно грубых и не очень густых точках, расширены и соприкасаются только у самого основания, внутренняя соприкасающаяся часть с редкими зубчиками, не длиннее ширины клещей около их средины, далее цилиндрические, довольно толстые, слабо утончающиеся к концу, очень постепенно и слабо загнутые; клещи Q простые, довольно сильно уплощенные, без грубых точек, наружный край утонченный, почти килевидный, внутренний, край мелко-зазубренный. Длина тела $ 14—18, 'Q 13—19; клещей S 5—8, 2 3.5—5 мм.
Распространение: Средиземье от Корсики и южных Балкан до Сирии (Бейрут и Ладакия, Burr, 1912), Малой Азии, включая Турецкую Армению, южного берега Крыма, Черноморского побережья Кавказа от Уч-дюре на севере до Сухума и Батума на юге (колл. С.-Т.-Ш.!) (фактически вероятно и дальше до Малой Азии) и Нухинского района в Азербайджане. Отсутствие указаний для местностей, лежащих между Черноморским побережьем и Азербайджаном, повидимому, объясняется слабой изученностью ареала данного вида в Закавказье.
Фауна СССР, Кожистокрылые Ю
146
4. FORFICULIDAE
17. Forficula davidi Burr (фиг. 42).
Burr, Ent. Mon. Mag., 2 ser., XVI, 1905: 85 (5, J; Мупин, Сычуань, Зап. Китай); Burr, 1907 :112, tab. IV, fig. 8; Burr, 1911 : 81; Bey-Bienko, 1933 : 6, tab. 1, fig. 3.
Тело довольно крупное, каштановое, бурое или черное. Усики 12-чле-никовые, черные, с бурыми вершинными члениками, или сплошь бурые,. 4-й членик немного короче 3-го, почти в два раза длиннее своей ширины,.
5-й членик немного длиннее 3-го. Голова буровато-красная, иногда очень темная,, швы неясные. Переднеспинка квадратная или чуть поперечная, с прямыми, параллельными боковыми сторонами и слабо округленным, почти прямым задним краем; диск черный, вдоль боковых сторон с грязно-желтыми полосками или реже одноцветно рыжевато-бурый, передняя часть вздута, задняя уплощена, с ясной, тонкой,, поперечной морщинистостью. Надкрылья большие, широкие, немного более чем в два раза длиннее переднеспинки, одноцветно-черные, или золотисто-каштановые, или каштаново-коричневые; крыловые пластинки имеются, обычно длинные, такого же цвета как надкрылья. Ноги бурые или черные, реже желтовато-бурые, концы голеней и лапки в густых, очень коротких золотистых волосках. Брюшко довольно толстое, каштановое, рыжее или красно-вато-черное сверху и снизу, в мелкой и гу-
фиг.42. Forficula davidiburr, 3x5 стой пунктировке; последний тергит 3 (Сычуань), по Bey-Bienko 1933. прямоугольный, в 2—3 раза шире своей длины, посредине у заднего края с двумя хорошо выраженными, сильно приподнятыми, сосковидными бугорками или отростками, у более мелких форм достигающими наибольшего размера; у £ бугорки слабо выражены. Клещи 3 черные или бурые, расширенные и соприкасающиеся только у самого основания, реже расширенная и соприкасающаяся часть занимает до одной пятой длины клещей; внутренний край соприкасающейся части мелко зазубрен; остальная часть, тонкая, длинная, ясно уплощенная, слабо и очень постепенно загнутая внутрь; у д — простые, прямые, сближенные на всем протяжении. Пигидий 3 удлиненный, прямоугольный, обычно хорошо заметный, значительно, или только немного короче расширенной части клещей, задний край при рассматривании снизу заметно вырезан, с заостренными боковыми углами. Длина тела 3 9—14.5, 2 13—18; клещей 3 4.3—11.2, £ 3.8—5 мм.
1. FORFICULA
147
Распространение: от северо-вост. Китая (Киао-чжоу, Borelli, 1915) до пров. Ганьсу и Сычуань в зап. Китае и области Амдо в сев.-вост. Тибете.
Этот вид характеризуется довольно значительной изменчивостью как в окраске, так и в размерах тела и строении последнего тергита и клещей 3. Особи с более короткими клещами, характеризующиеся относительно более длинным расширенным основанием, повидимому, представляют собой f. brachylabia-, особенно мелкие особи $$ кроме того характеризуются очень сильным развитием выступов на последнем тергите брюшка. В генетическом отношении этот вид, повидимому, близок к F. smyrnensis, хотя оба вида резко отличаются друг от друга многими признаками.
18. Forficula decipiens Gene
Gene 1832:228 (3, g; Генуя и Лигурийские Альпы, Сев. Италия); Brunner, 1882:17; Bormans, 1900:125; Kirby, 1904:51; Якобсон 1905 : 28; Burr, 1911: 78. —pallidicornis Brulle, Exped. Scient. Moree, III (2), 1832:82, tab. 29, fig. 2. — laminigera Costa, Atti Acad. Sc. Napoli, IX (6), 1881 : 38, fig. 9 (Apterygida)-, Kirby, 1904:52.
Тело довольно коренастое, желтое, с более темным брюшком. Усики желто-бурые, 13-члениковые, с короткими члениками; 1-й членик толстый и очень короткий, не более чем в два раза длиннее своей ширины, 3-й членик цилиндрический, примерно в l1^ раза длиннее своей ширины, 4-й членик почти шаровидный, довольно значительно короче 3-го, последующие членики заметно более толстые чем 3-й членик. Голова ржаво-красная. Переднеспинка заметно шире своей длины, с прямыми, немного расходящимися кзади боковыми сторонами, задний край слабо выступает, умеренно округлен, диск посредине буровато-желтый, по бокам более светлый. Надкрылья довольно короткие, в 11/2 раза длиннее переднеспинки, желтоватые, с прямым поперечным задним краем; крылья сильно укороченные, скрыты под надкрыльями. Ноги желтоватые. Брюшко буровато-рыжее или грязно-желтое, густо и довольно крупно пунктировано; последний тергит 3 сильно поперечный, прямоугольный, по заднему краю обычно зачерненный, со слабыми срединными вздутиями у заднего края и с ясными, умеренно приподнятыми складками по бокам; последний тергит g более узкий, слегка суженный к вершине без ясных боковых складок, но с довольно ясным продольным килевидным утолщением, располагающимся наклонно по бокам тергита в задней его половине и образующим с задним краем прямой, чуть выступающий бугорковидный угол. Пигидий 3 короткий, сверху обычно плохо, снизу хорошо заметный, нижняя сторона на вершине по бокам с двумя острыми короткими выступами; у g — более удлиненный, хорошо заметный. Клещи 3 у более обычной f. brachylabia сильно расширены и уплощены при основании так, что расширенная часть занимает немного менее трети или же четверть длины клещей, с сильно зазубренными внутренними сторонами; далее постепенно утончающиеся к вершине и полукругло изогнутые, образуя своими внутренними сторонами почти правильный круг; у более редкой f. macrolabia расширенная часть отно-10*
148
4. FORFICULIDAE
сительно более короткая, а вершинная часть менее сильно изогнутая, длинная; клещи Q простые, при основании довольно толстые, к вершине постепенно утончающиеся. Длина тела 3 11—13, Q 10—12, клещей 3 3—8, 2 2.2—2.5 мм.
Присутствие на последнем тергите 3 боковых складок (подобно тому как это имеет место и у F. auricularia), при наличии характерной структуры усиков делает этот вид весьма легко распознаваемым.
Распространение: южная Европа от Пиренейского до Балканского полуострова (Испания, южн. Франция, Сардиния, вся Италия, южн. Швейцария, Балканский полуостров на север до южн. Далмации и Семи-градья в Румынии) (Burr, 1912), Сев. Африка, Малая Азия до Трапезунда. Указание для Подолии (Каменец-Подольск) (Belke, 1859), как и указание для Сарепты (Becker, 1864) несомненно относятся к F. tomis. Таким образом, вопрос о нахождении этого вида в СССР остается еще открытым, хотя вполне вероятно его обнаружение в Зап. Закавказье, так как этот вид известен из Трапезунда, т. е. сравнительно недалеко от границы СССР.
Водится под камнями и на кустах, часто совместно с F. auricularia.
19. Forficula lurida Fischer.
Fischer, 1853 :75, tab. 6, fig. 12 ab (3, европ. Турция); Brunner, 1882:16, fig. 4 ABC Bormans, 1900:123; Kirby, 1904:51; Якобсон, 1905:27; Burr, 1911:82. — orientalis Burr, Ann. Mag. Nat. Hist., (7) VI, 1900: 82, tab. 4, fig. 7; Kirby, 1904: 52.
Тело грязно-желтое, иногда почти смоляно-бурое. Усики 13-члени-ковые, 2-й членик довольно длинный, хорошо заметный, чуть более чем в два раза короче 3-го, 3-й членик чуть расширен к вершине, цилиндрический, такой же длины как 5-й, 4-й членик короткий, едва длиннее свРей ширины, примерно в два раза короче 3-го. Голова ржаво бурая, иногда почти или совсем черная, швы очень резкие, глубоко-вдавленные. Переднеспинка почти квадратная, с грязно-желтыми боковыми сторонами; задний край умеренно округлен. Надкрылья грязновато-желтые, иногда буроватые, с обрубленным задним краем; крыловые пластинки вполне развитые, грязно-желтые, с бурой оторочкой. Ноги грязно-желтые, одноцветные. Брюшко умеренно расширенное посредине, слегка суживающееся к вершине, в густой грубой пунктировке; последний тергит с поперечной морщинистостью и с двумя округлыми бугорками перед задним краем, очень слабыми у Q; боковые стороны тергита в задней половине с продольной килевидной едва приподнятой складкой, у 2 менее развитой, чем <5; пигидий слабо выступающий, тупой. Клещи 3 довольно сильно уплощенные, их соприкасающаяся часть занимает не более одной четверти их длины, с зазубренным внутренним краем, постепенно суживающиеся к середине, далее округло изогнутые, ограничивая'внутренними сторонами почти правильный круг; задний конец расширенной части тупоугольный, наружная сторона от основания к вершине слегка S-образно изогнута, или же клещи более
1. FOR FI CULA
149
удлиненные, около середины изогнуты под очень тупым углом, наружные стороны параллельные друг другу, внутренние — ограничивают ромбоидальное пространство; клещи g едва зазубрены изнутри, простые. Длина тела <5 11.5—14, g 11—13; клещей 5 3—3.4, g 2.5—3.2 мм.
Распространение: Южные Балканы, Малая Азия, Сирия, Палестина, юго-зап. Иран (Арабистан, колл. С.-Т.-Ш.!).
* 20. Forficula vicaria Sena. (фиг. 43).
Semenov, 1902:99, fig. 1 (Cj; Косен и Син-часо, Корея); Kirby, 1904:50; Semenov-Tian-Shan-skij, 1908:169; Burr, 1911:82.
Тело довольно маленькое, ржаво, или темно-бурое. Усики 12-члениковые, грязножелтые, с длинными тонкими члениками, 4-й членик длинный, цилиндрический чуть более чем в два раза длиннее своей ширины, почти равен 3-му; 5-й членик в П/г раза длиннее 4-го, почти в четыре раза длиннее своей ширины, последующие членики постепенно удлиняющиеся, в четыре-пять раз длиннее своей ширины. Голова красновато-рыжая или рыжевато-бурая, иногда довольно темная, швы слабо выраженные, лобные стерты, затылочный слабо заметный. Переднеспинка чуть уже головы почти квадратная, боковые стороны пря мые, параллельные, задний край широко округлен, слабо выступающий; диск рыже-вато-желтый, иногда бурый с светложелтыми боковыми сторонами. Надкрылья довольно длинные, буровато-желтые; крыло
вые пластинки развитые, НО слабо высту- ®Hr- 43. Forficula vicaria Sem., пающие ИЗ ПОД надкрылий, В несколько раз <5 б(Славянка,Уссур. край). Ориг. короче переднеспинки, такого же цвета как надкрыльБярюшко мелко и густо пунктировао, ржаво-бурое, иногда темнобурое; последний тергит <5 прямоугольный, в 2—-2У2 раза шире своей длины» задний край чуть выступающий, почти прямой, два срединных бугорак у заднего края слабо вздутые, низкие, боковые стороны тергита простые, без возвышающейся складки; у g — сужен и наклонен кзади, бугорки у заднего края неясные. Пигидий почти вертикальный, при рассматриваний сверху почти невидный. Клещи 5 в основной половине уплощены, в основной четверти сильно расширены и соприкасающиеся, с довольно редко зазубренными внутренними сторонами, далее к средине постепенно суживающиеся и вооруженные на внутренней стороне перед концом расширенной
15Э
4. FORFICULIDAE
части, т. е. почти у средины клещей, маленьким, изолированно расположенным зубцом; вершина соприкасающейся основной части образует очень тупой угол, пространство между этим углом и изолированным зубцом с редкими, маленькими зазубринами; вершинная часть клещей цилиндрическая, слабо утончающаяся и слабо изогнутая к концу; клещи g простые, сближенные, на внутренней стороне почти не зазубренные или очень мелко зазубренные. Длина тела <5 9—11, 2 10—11; клещей 3 4—4.5, £> 3 мм.
Распространение: Южно^Уссурийский и Амурско-Уссурийский край на север до р. Амур (Нижне-Тамбовское; Синдинское озеро, колл. ЗИН!), Корея, Манчжурия (Bey-Bienko, 1930) и сев.-вост. Китай (Borelli 1915).
Этот вид чрезвычайно близок к F. burriana Sem., F. subauricularia B.-Bienko и F. auricularia L. (см. ниже) и может рассматриваться как форма, близкая к прототипу, из которой развились и остальные упомянутые виды (см. об этом F. auricularia).
21. Forficula burriana Sem.
Semenov-Tian-Shanskij, Ent. Monthly Mag-az., (2), XVIII, 1907:232 (;£, 2> Лян-чоу, пров. Ганьсу, северо-зап. Китай). — vicaria Burr, 1907 b: 57 (пес Semenov).
Очень похож на F. vicaria Sem., но отличается следующими признаками. Голова желтоватая, темя с вдавлением в виде двух слабых ямок, переднеспинка меньшей величины, крыловые пластинки выступают из под надкрылий еще слабее чем у F. vicaria, клещи 3 в вершинной половине очень слабо утончающиеся, Сверху с сильной, снизу с немного более слабой продольной бороздкой, у основания соприкасающиеся на коротком расстоянии, внутренние стороны менее остро зазубрены, у своего основания сильно скошены, зубец на внутреннем крае расположенный за расширенной частью близ ее конца, слегка притуплен, прямоугольный. Усики, ноги, переднеспинка по бокам и сзади, надкрылья, крыловые пластинки, основная часть клещей, за исключением внутренней стороны, соломенно-желтые; Q чуть темнее. Длина тела 3 10.9, клещей 2.2 (размеры 2 неизвестны).
Распространение: пров. Ганьсу в сез.-зап. Китае.
Наиболее существенным отличием этого вида от F. vicaria является строение клещей 3, особенно присутствие на них продольных борозд; все остальные признаки, особенно окраска, при изучении большой серии экземпляров (данный вид пока известен только по 3 экземплярам) могут оказаться несущественными.
22. Forficula subauricularia B.-Bienko (фиг. 44).
Bey-Bienko, 1934:417, fig. 9 (3; долина Тункгохо, пров. Сычуань, Китай).
Близок к предыдущим двум видам по строению усиков и к F. duricu-laria по строению клещей. Тело рыжевато-бурое. Усики черновато-бурые
1. FORFICULA
151
Фиг. 44. Forficula subauricularia В.- Bien-ko, $ (тип). Клещи. По Bey-Bienko, 1934.
в основной половине и рыжевато-бурые на вершине (число члеников неизвестно), 4-й членик немного короче 3-го, 5-й членик в три раза длиннее •своей ширины, со слегка расширенной вершинной частью, чуть длиннее 3-го. Голова уплощенная, темно-красновато-бурая, с очень слабыми швами. Переднеспинка чуть уже головы, почти одинаковой длины и ширины, со слабо округленными боковыми сторонами, задний край широко округлен; .диск буровато-черный, с желтоватыми боковыми сторонами. Надкрылья .длинные, в два раза длиннее переднеспинки, рыжевато-бурые, с прямым задним краем; крыловые пластинки такого же цвета, как надкрылья, сильно выступающие. Брюшко буровато-черное, посредине и снизу рыжевато-бурое, сверху очень густо пунктировано; последний тергит <5 почти в три раза шире своей длины, боковые стороны прямые, слегка, но заметно сближающиеся по направлению к вершине, без треугольных выступающих складок, около средины у заднего края с двумя заметно вздутыми выпуклостями и вдавлением между ними. Пигидий $ длинный, почти равен по длине расширенному и сближенному основанию клещей, хорошо заметный, сильно выступающий, при рассматривании сверху суживающийся и округленный на вершине, при рассматривании в профиль сильно расширенный и утолщенный к вершине, сама вершина сильно выступает над уровнем клещей, нижняя поверхность с параллельными сторонами, со слабыми заостренными выступами на задних боковых углах, задняя сторона вертикальная, при рассматривании сзади расширяющаяся книзу. Клещи S длинные, основная часть уплощена и расширена на очень коротком расстоянии, немного превышающем длину пигидия; внутренние стороны
^расширенного основания имеют вид широкого треугольного выступа, плотно соприкасающегося с пигидием, но не соприкасаются друг с другом, будучи отделены пигидием и лишь только вершины треугольных выступов сближены друг с другом; далее от основания к вершине постепенно суживающиеся, слабо загнутые, цилиндрические, на внутренней стороне с маленьким, заостренным, изолированным зубцом, удаленным от расширенной основной части на расстояние заметно превышающее длину самой расширенной основной части. Длина тела $ 12.1, клещей, 7.4 мм. Самка неизвестна.
Распространение: известен пока только из долины Тункгохо (пров. Сычуань в сев.-зап. Китае).
По характеру клещей F. subauricularia больше напоминает обыкно венную уховертку (F. auricularia), чем два предыдущих вида (F. vicaria и F. burriana). Однако отсутствие боковых складок на последнем тергите брюшка и однородные членики при основании усиков говорят о меньшей
152
4. FORFICULIDAE
специализованности и некоторой примитивности этого вида по сравнению с F. auricularia при одновременном сходстве в указанных отношениях с F. vicaria a F. burriana. Исходя из сказанного можно было бы говорить о том, что F. subauricularia представляет собой промежуточное звено между такой относительно более примитивной группой как F. vicaria и F. burriana с одной стороны и F. auricularia — с другой; такая трактовка,, однако, вряд ли была бы правильной — идеально промежуточные формы обычно скоропроходящи в природе и более часто являются достоянием, палеонтологии, а с другой стороны наш вид характеризуется сильной специализованностью в отношении строения пигидия. Поэтому более правильно рассматривать этот вид как дериват какой то вымершей промежуточной между F. vicaria и F. auricularia формы, специализация которого пошла совсем по другому пути, чем у F. auricularia.
Наличие у Вост. Азии нескольких очень близких друг к другу видов,, характеризующихся сохранением примитивных особенностей строения,, еще лишний раз говорит о том, что данная группа Dermaptera достигала в третичное время большого разнообразия в Вост. Азии; ярким свидетельством этого являются дошедшие до нас указанные три вида, являющиеся как бы живыми ископаемыми.
* 23. Forficula auricularia Linne. — Обыкновенная уховертка (фиг. 9,. 45, 46).
Linne, Syst. Naturae, I (ed. 10), 1758:423 (Европа); Fischer-Waldheim, 1846 : 38t Fischer, 1853:74, tab. 6, fig. 11, 11 ab; Brunner, 1882:12; Bormans, 1900:122; Семенов,. 1901a: 195; Семенов, 1901b : 103; Kirby, 1904:49; Якобсон, 1905 : 27, табл. 3; Burr, 1911:82 Burr, 1916 :14, tab. Ill, fig. 13; Бей-Биенко, 1932:34.
Средних размеров или довольно крупная, ржаво- или смоляно-бурая. Усики 14-члениковые, 1-й членик желтый, остальные рыжевато- или темнобурые, реже все членики грязновато-желтые; 4-й членик значительно короче 3-го и чуть длиннее своей ширины, обычно почти шаровидный^ 5-й членик чуть короче 3-го, в два, или немного менее, раза длиннее своей ширины, почти цилиндрический; последующие членики, начиная с 6-го, толщей значительно длиннее 3-го или 5-го члеников. Голова рыжая, иногда красноватая, или же лоб буровато-черный, а затылок красновато-бурый,, швы довольно хорошо заметные. Переднеспинка почти квадратная, реже слегка поперечная, с прямыми параллельными боковыми сторонами и слабо выступающим, немного округленным задним краем; диск бурый со светлыми боковыми сторонами. Надкрылья грязно-желтые или светлобурые, примерно в два раза длиннее переднеспинки, задний край едва вырезан или. прямой, крыловые пластинки сильно выступают, длиннее половины длины переднеспинки, желтоватые, с буроватым внутренним краем, так что иногда, при особой резкости окраски создается впечатление желтого пятна на пластинках. Ноги грязно-желтые. Брюшко мелко и густо пунктировано,, у $ с хорошо развитыми боковыми бугорками, довольно стройное; последний тервит S прямоугольный, более чем в два раза шире своей длины,.
1. FORFICULA
153-
боковые стороны с уплощенной треугольной приподнятой складкой, задний край слегка выступает, два срединных вздутых бугорка у заднего края хорошо заметные, но слабо приподнятые; у g тергит более длинный, немного суживающийся к вершине, боковые стороны без треугольных складок,,
но с ясными косорасположенными, килеобразно приподнятыми линиями в задней половине, срединные бугорки у заднего края слабые. Пигидий
короткий, маленький,почти вертикальный, снизу на вершине по бокам с маленькими заостренными выступами. Клещи <5 у f. brachylabia расширены и соприкасаются в основной четверти, далее на внутренней стороне недалеко от конца соприкасающейся части с сильным изолированным треугольным зубцом, утонченная часть сильно дуговидно изогнутая, ограничивая внутренними с т о р о нами почти правильный круг; у f. macro labia—равны или длиннее половины тела, соприкасаются только у самого основания, далее постепенно суживающиеся к вершине, зубец на внутренней стороне более удален от основания, располагаясь от конца соприкасающейся части на расстоя-
Фиг. 45. Forficula auricularia L., <5X6 (Москва). Ориг.
Фиг. 46. Forficula auricularia L., 3., Генитальный аппарат. Ориг.
нии в 1х/2—2 раза превышающем длину этой части, утонченная часть
умеренно дуговидно изогнута, ограничивая своими внутренними сторонами сильно удлиненный овал; клещи g простые, соприкасающиеся, заметно уплощенные. Генитальный аппарат 3 с очень короткой virga, более короткой чем метапарамеры, соединяющейся с передним концом почковидного пузырька без всякого угла; метапарамеры снаружи дуговидно выпуклые. Длина тела 3 9.5—14, g 10—16; клещей 3 4.9, g 3.5—5 мм.
Распространение: чрезвычайно широко распространенный вид, свойственный почти всей Европе (на севере до Фарерских островов и. юго-зап. Лапландии), Сев. Африке, включая Мадеру и Канарские острова, Сев. Америке, Бразилии, Антильским островам, Новой Зеландии, всей.
154
4. FORFICULIDAE
Малой Азии, Сирии и сев. и зап. Ирану до Тегерана (колл. Г. Б.-Б.!).. В пределах СССР ареал распространения охватывает почти всю европейскую часть от Крыма, Сев. Кавказа и Нижней Волги на юге до сев. части Ленинградской области, заходя даже в Финляндию и южную Карелию, а восточнее — до Москвы, р. Немды близ г. Уржума (Шугуров, 1909) и Перми (Колосов); далее обнимает весь Кавказ, включая Закавказье (поднимаясь на большую высоту в горы) и соединяясь таким образом с Малой Азией и северо-западным Ираном, затем в виде языка идет по северному Ирану, заходя в Копет-Даг до меридиана г. Ашхабада, но полностью отсутствуя в остальной части Средней Азии. В пределах Сибири известен только из западной ее части и при том из немногих мест [с. Па-влоградка бл. Омска (Бей-Биенко, 1930) и г. Камень (Wnukowsky, 1928)], достигая на востоке г. Камня; из Казахстана неизвестен, за исключением района между нижним течением Волги и Урала (Kittary, 1849). Первоначальной родиной является Палеарктическая область, так как этот вид обнаружен в плиоценовых отложениях Англии; из Европы он был завезен по торговым путям в Америку и Новую Зеландию. Что касается указаний на местонахождение этого вида в Индии и Японии, то они, повидимому, сомнительны.
Один из обычнейших видов уховерток, днем встречающийся под камнями и корой деревьев и в прочих укромных местах иногда целыми сообществами, а ночью очень деятельный. Спаривание начинается в конце лета; самка с начала осени и даже весной откладывает яйца кучками, находясь до вылупления личинок около яиц. На зиму большинство самцов обычно отмирает, тогда как самки доживают до весны. Первое время личинки живут с матерью, а затем после смерти последней самостоятельно. Обыкновенная уховертка является иногда довольно серьезным вредителем разнообразных культур, так как неоднократно отмечалась во взрослом и личиночном состоянии как вредитель различных хлебных злаков и кукурузы, огородных растений (особенно цветной капусты и капустных семенников), цветоводства (гвоздики, розы, георгины, хризантемы и пр.), хмеля и бобовых; обгрызает живые листья и особенно цветы, а также выедает полузрелые семена злаков. Кроме того вредит плодам, особенно яблоне и персику, забираясь в плоды. Поселяясь в домах и постройках, забирается во влажное белье и одежду и выедает ходы в печеном хлебе. Вредная деятельность отмечалась для Зап. Европы, Украины, Новой Зеландии (где этот вид является бичом плодоводства) и Сев. Америки.
Burr (1907) отмечает, что f. macrolabia этого вида более часто встречается на островах и в горах, тогда как f. brachylabia более обычна в полудомашних условиях.
24. Forficula pubescens Serv.
Serville, Hist. Nat. Orthopt., 1839 : 46 (<5, <^); остр. Сардиния); Fischer, 1853 : 77, tab. VI, fig. 15 a—f; Brunner, 1882 :14; Kirby, 1904: 51; Якобсон, 1905:28; Burr, 1911:82, tab III. fig. 12.— seiulosa Fieber, 1853:256.—yersini Yersin, Ann. Soc. Ent. France, 3 (IV), 1856:741.
2. MESASIOBIA
155
Тело маленькое, стройное, бурое или грязно-желтое, в разбросанных волосках. Усики 12-члениковые, грязно-желтые, 3-й членик почти в 12/2 раза длиннее 4-го, последний полуконический, немного длиннее своей ширины, 5-й членик едва длиннее, но заметно толще 3-го, примерно в два раза длиннее своей ширины. Голова рыжеватая или буровато-рыжая, с ясными швами. Переднеспинка грязно-желтая или буроватая, с полупросвечивающими боковыми сторонами, задний край слабо выступает, округлен. Надкрылья довольно короткие, в Р/г раза длиннее переднеспинки, желтоватые, с умеренно скошенным задним краем; крыловые пластинки отсутствуют. Ноги одноцветные, желтоватые. Брюшко очень мелко и густо пунктировано, боковые бугорки на 3-м тергите слабые, на 4-м крупные, черные; последний тергит <5 сильно поперечный, короткий, по бокам без приподнятых складок, у заднего края около средины с двумя слегка вздутыми бугорками; у g последний тергит без ясных бугорков у заднего края, более узкий и более длинный. Пигидий $ g очень маленький, незаметный. Клещи $ расширены и уплощены на протяжении немного превышающем половину их длины, далее утонченные и умеренно дуговидно изогнутые, в довольно редких, но удлиненных и хорошо заметных волосках; внутренние стороны расширенной части соприкасаются только в основной ее трети или реже половины, а далее очень постепенно расходящиеся и у конца расширения вооружены сильным, широко треугольным зубцом, хотя и расположенным в пределах расширенной части, но изолированным от соприкасающегося основания; клещи g простые, тонкие, соприкасающиеся. Генитальный аппарат S с длинной virga, которая примерно в 1% раза длиннее метапарамер; метапарамеры с совершенно прямыми параллельными наружными сторонами. Длина тела 3 4—7.5, g 5—7; клещей 8 3—4, g 2 мм.
Распространение: известна только из Средиземья, от Пиренейского полуострова, южной Франции, Италии и островов Сицилии, Сардинии и Маллёрки до Балканского полуострова и Сирии (Ладакия, Burr, 1912), а в Сев. Африке—из Алжира и Марокко.
По строению клещей 8 этот вид должен быть сближен с группой F. auricularia, характеризующейся наличием изолированного зубца на внутренней стороне клещей. Отсутствием приподнятой складки на последнем тергите сближает этот вид с более примитивными формами группы (F. vicaria и др.), но волосатое тело и отсутствие крыловой пластинки говорит о большей специализации вида; поэтому F. pubescens может рассматриваться как специализованный дериват данной группы.
2. Род MESASIOBIA SEM.
Semenov-Tian-Shanckij, 1908 :163; Burr, 1911:71.
Тело длинное, довольно стройное. 4-й членик усиков едва короче 3-го, цилиндрический. Голова резко двуцветная. Надкрылья сильно укороченные, не закрывающие полностью заднеспинку, задний дугообразно-
156
4. FORFICULID AE
вырезанный край которой совершенно свободен; длина надкрылий не превышает длину переднеспинки. Крылья отсутствуют. Среднегрудная пластинка округлена, одинаковой длины и ширины. Брюшко почти с параллельными сторонами, чуть расширенное посредине, с ясными боковыми бугорками на 3-м и 4-м тергитах; последний тергит $ у заднего края посредине с двумя сильными выступами. Клещи <5 при основании сильно расширены, внутренние стороны основания расширенной части сближены,, почти или совсем соприкасаются (f. brachylabia), далее постепенно утончающиеся, без зубцов, в профиль несколько волнистые (f. macrolabia) или прямые (f. brachylabia). Пигидий узкий. Генитальный аппарат $ весьма сходен с таковым рода Forficula, но соединение virga с почковидным пузырьком при помощи сильной дуги напоминает род Anechura, метапарамеры длинные, тонкие, параллельные друг другу, перед вершиной слегка расширенные, а затем к самому концу заметно суженные; собственно penis, достигает только конца второй трети длины метапарамер; вирга длинная, S-образно изогнутая, тонкая.
Этот интересный род был отнесен его автором к „подсемейству" Cheli-durinae, затем Burr (1911) отнес его к A.nechurinae, сблизив таким образом с родом Anechura и его ближайшими родичами. Однако, по характеру клещей и гениталий $ род Mesasiobia должен быть сближен с родом Forficula, от которого он главным образом и отличается сильно-укороченными: надкрыльями, причем даже характерные сильно-развитые на последнем тергите брюшка S выступы не могут служить абсолютным признаком,, отличающим этот род от Forficula, так как у F. davidi (см. выше) эти выступы также хорошо развиты, сильно напоминая таковые у Mesasiobia’, у отдельных особей F. davidi, именно у более мелкой f. brachylabia, эти выступы даже сильнее развиты,' чем у соответствующей формы Mesasiobia,. Это сходство между F. davidi и Mesasiobia настолько велико, что возникает даже мысль о довольно значительной генетической близости между указанным видом и Mesasiobia. Наконец, род Mesasiobia, подобно огромному большинству видов рода Forficula, приурочен к области третичной складчатости в Азии. На основании всего сказанного выше можно предполагать,, что род Mesasiobia представляет собой короткокрылый дериват рода Forficula.
Известен только один вид.
* 1. Mesasiobia hemixanthocara Sem. (фиг. 47, 48).
Semenov-Tian-Shanskij, 1908 :165 (3, (J); ущелье p. Каргалинки, озеро Джасылкуль-и верховья р. Кашка-су в Заилийском Алатау, Семиречье); Burr, 1911 :71, tab. VI, figs. 17 et 17 a, b, c, d, e.
Тело смоляно-черное. Усики 11—-13-члениковые, грязновато-желтые,, членики довольно толстые, 4-й членик немного менее чем в 2 раза длиннее своей ширины, 5-й членик заметно длиннее 3-го, примерно в 2х/г раза длиннее своей ширины. Голова двуцветная, передняя часть до затылочных.
2. MESASIOBIA
157
швов черная, затылок у <5 соломенно-желтый; у £ окраска головы менее контрастная, спереди черная,‘у затылочных швов затемнена, а далее на затылке рыжевато-желтая; швы хорошо заметны. Переднеспинка одинаковой длины и ширины, почти одинаковой ширины с задним концом головы, боковые стороны прямые, параллельные, задний край слабо
округлен, в средней части почти прямой; диск черный, боковые стороны немного светлее, темнобурые. Надкрылья со скошенно обрубленным задним краем, едва длиннее своей ширины, тесно соприкасающиеся внутренними сторонами. Ноги грязновато - желтые, одноцветные или со слегка затемненными вершинами бедер. Брюшко смоляно-черное, в средней части •сверху слегка буровато-смоляное, очень мелко и густо пунктировано; последний тергит S сккьпо поперечный, с параллельными сторонами; у заднего края посредине с двумя выступами, имеющими у f. brachylabia вид
Фиг. 47. Mesasiobia hemixanthocara Sem., <5x4 (Окр. Алма-ата). Ориг.
Фиг. 48. Mesasiobia hemixanthocara Sem., $. Генитальный аппарат.
Ориг.
сильных, но умеренно приподнятых бугорков, а у f. macrolabia эти выступы
очень длинные, уплощенные, направленые вверх и несколько назад, разделенные друг от друга узкой вырезкой, имеющей параллельные стороны; последний тергиг £ простой, с двумя слабыми бугорками. Пигидий 3 сверху почти незаметный, не выступающий, сильно наклонный, на вершине обрубленный, при рассматривании снизу также плохо заметный, на вершине с двумя тупыми и короткими боковыми зубцами; у 2 — выступающий, хорошо заметный, на вершине с тремя зубчиками. Клещи S у f. macrolabia длинные, достигают длины брюшка, расширенная их основная часть несколько наклонена к внутреннему краю, конец расширенной и сближенной
3 58
4. FORFICULIDAE
части очень тупоугольный, далее клещи довольно резко суживающиеся, перед концом основной трети вполне цилиндрические и отсюда довольно резко загнутые вниз и назад, а от середины слегка загибаются кверху, так что вершинная часть почти горизонтальная; при рассматривании сверху утонченная часть слегка изогнута дугообразно, с соприкасающимися или очень сближенными концами; у f. brachylabia клещи не длиннее половины брюшка, расширенная основная часть горизонтальная, заходящая за вершину основной трети, при рассматривании в профиль клещи прямые; клещи g простые, соприкасающиеся, довольно длинные, после основания суженные. Длина 5 12—15.5, g 11.5—12.5, клещей 8 4—8.5, g 3.4—3.5 мм.
Распространение: известен из Заилийского Алатау к югу от г. Алма-ата и окрестностей Кульджи в Китайской Джунгарии (Регель, колл. ЗИН1), встречаясь на высоте 1500—2800 м.
3. Род OREASOBIA SEMENOV-TIAN-SHANSKIJ gen. n. i
Anechura auct., partim.
Тип рода — Forficula fedtschenkoi Sauss.
Усики 12-члениковые, членики длинные; 3-й членик едва достигает длины 5-го членика, в 3 и более раза длиннее своей ширины, 4-й членик немного короче 3-го, в 2х/2—3 раза длиннее своей ширины, 5-й членик в 3J/2—4 раза длиннее своей ширины. Переднеспинка поперечная. Ноги длинные. Надкрылья и крылья вполне развитые, последние иногда почти полностью скрыты под надкрыльями. Брюшко слегка расширено около середины, боковые бугорки на 3—4-м члениках вполне развитые; последний тергит <5 очень короткий, сверху по бокам с двумя бугорками или отростками. Пигидий <5 вертикальный, широкий, несколько выступающий в виде сильно утолщенной пластинки. Клещи у 8 в профиль прямые или слегка волнисто изогнутые, длинные, тонкие, цилиндрические, только у самого основания изнутри умеренно расширенные, образуя здесь уплощенный выступ, внутренние стороны которого не соприкасаются, широко расставлены друг от друга. Генитальный аппарат (известен только у О. fedtschenkoi) S с длинными, чуть суживающимися к вершине, почти параллельными метапарамерами, собственно penis доходит почти до самой вершины метапарамер; вирга соединяется с почковидным пузырьком при помощи очень сильно загнутой дуги.
Этот род более близок к Forficula чем к Anechura, хотя с последним его сближают не соприкасающиеся клещи <5. Особенно большое сходство имеет данный род с той группой видов из рода Forficula, куда относятся F. schlagintweiti, тогда как сходство с Anechura является чисто внешним, тем более, что характер клещей Oreasiobia, несмотря на то, что они расставлены при основании, легко может быть выведен из клещей типа Forficula. В частности расширенное основание клещей образо-
1 Оригинальное описание рода дано в резюме (стр. 228).
3. О REASI0B1A
159-
вано как и в роде Forficula за счет уплощенного выступа на внутренней стороне, внутренне края которого также прямые и даже у отдельных особей имеют очень слабые зубчики; лишь только то обстоятельство, что внутренние края этих выступов не соприкасаются, создает впечатление сходства с представителями рода Anechura. Замечательно также и то, что ареал распространения рода Oreasiobia частично захватывает ареал распространения группы F. schlagintweiti, причем в обоих случаях ареалы располагаются в области третичной складчатости (Гималаи и Тян-Шань). Учитывая все сказанное, а также наличие и некоторых других особенностей, сближающих данный род с Forficula, именно с группой F. schlagintweiti, можно допустить, что род Oreasiobia представляет собой спе-циализованное производное одной из примитивных групп рода Forficula (современными представителями которой являются F. schlagintweiti и близкие виды), выделившееся из данной группы в области Зап. Гималаев и Тян-Шаня, повидимому, еще в период третичных горообразовательных процессов и эволюционировавшее совершенно в другом направлении, чем виды рода Forficula.
Род этот представлен всего только двумя видами.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ПОДВИДОВ
1 (2). Голова одноцветно черная. Клещи голые, у <5 без конического выступа над расширеннной частью при основании. Последний тергит брюшка S с двумя маленькими бугорками или у f. brachylabia без них..............................................1. О. stoliczkai Burr.
2 (1). Голова рыжевато-красная. Клещи волосатые, у de коническим выступом при основании над расширенной частью, направленным своим концом в сторону пигидия. Последний тергит брюшка $ обычно с двумя очень сильными, длинными, направленными назад коническими отростками, очень редко только с двумя бугорками..............
......................................* 2. О. fedtschenkoi Sauss. а (Ь). Надкрылья без светлых пятен, одноцветно черные..........
...............................2а. О. fedtschenkoi calciatii Bor.
b (а) Надкрылья с двумя светлыми пятнами или реже целиком светлые .....................* 2в. О. fedtschenkoi fedtschenkoi Sauss.
1. Oreasiobia stoliczkai (Burr) [emend.].
Burr, 1912 : 792 (3, Башар, сев. Индия) [Anechura]; Hebard, 1923 : 224, tab. XX, figs. 19—20.
Тело голое, довольно стройное, черное, с более или менее бурыми боковыми сторонами переднеспинки, ногами и брюшком, реже особенно у 2 только с рыжевато-бурым оттенком; клещи красновато-рыжие. Голова с тонкими, но хорошо заметными, особенно у 2 швами, черная. Перед-
160
4. FORFICULIDAE
неспинка co слабо округленным задним краем. Надкрылья длинные, черные. Ноги без длинных волосков, но лапки покрыты очень короткими, золотисто-желтыми волосками. Крыловые пластинки сильно выступают, реже скрыты под надкрыльями. Брюшко гладкое, без ясной пунктировки; конец брюшка S включая пигидий и клещи двух типов; 1) f. macrolabia — последний тергит по заднему краю с двумя хорошо выраженными, короткоконическими бугорками, пигидий хорошо заметный, выступающий, умеренно широкий, чуть расширенный к вершине, задний конец при рассматривании сверху прямой, задняя сторона почти вертикальная, в виде почти не расширяющейся кверху четырехугольной пластинки, верхний конец которой при рассматривании сзади слегка дуговидно вырезан; клещи длинные, широко расставлены при основании, тонкие, голые, в профиль слегка волнисто изогнутые, расширенное извнутри основание несколько приподнято кверху, образуя по бокам пигидия треугольные лопасти, конический выступ сверху расширенного основания отсутствует; 2) f. brachylabia — последний тергит без ясных бугорков; пигидий скрыт расширенным основанием клещей; клещи короткие, сильно дуговидно загнуты внутрь, так что вершинная треть образует почти прямой угол с продольной осью основания клещей, расширены в основной части, но соприкасаются только у самого пигидия, расширенное основание очень постепенно суживается к середине, не образуя на внутренней стороне резкой грани с цилиндрической основной частью; наружный край клещей .S'-образно изогнут. Последний тергит g заметно суживающийся к вершине, сверху со слабыми боковыми выпуклостями и с несколько вдавленным пространством между ними; пигидий сильно наклонный; клещи простые, соприкасающиеся, на внутренней стороне слегка зазубренные. Длина тела 3 12.5—13.5, 2 11—13.5; клещей 3 f. macrolabia 6.9—9, f. brachylabia 3.4, g 3.6— 4.9 мм.
Распространение: известен только из сев.-зап. Индии (Сутлей-дистрикт и Пенджаб) и Кашмира (Сынгам и Сономарг, колл. ЗИН!).
Этот вид в своей обычной форме весьма напоминает О. fedtschenkoi calciaiii, тогда как характер клещей 3 у f. brachylabia создает впечатление сходства с видами рода Forficula. Однако, это сходство обманчиво, так как у всех видов рода Forficula клещи 3 никогда не имеют того строения, которое свойственно f. brachylabia данного вида; расширенная и соприкасающаяся основная часть клещей у видов рода Forficula всегда резко отделена от утонченной основной части, не говоря уже о том, что внутренняя сторона расширенной части, если последняя достигает хотя-бы 1/4—длины клещей, всегда прямая, а не дуговидно загнута. Несмотря на это нельзя не подчеркнуть отмеченного и на предыдущих страницах сходства между О. stoliczkai с одной стороны и видовой группой Forficula schlagintweiti с другой.
О. stoliczkai была описана и фигурировала прежде как представитель рода Anechura. Однако, не отрицая вероятной генетической близости ее
4. OREASIOBIA
1|Я
к предкам рода Anechura, необходимо все же отметить, что О. stoliczkai совершенно не укладывается в указанный выше род, тогда как строение усиков, отсутствие скульптуры на теле, характер переднеспинки, длинные .и стройные ноги и особенно характер клещей 3 не вызывают сомнений в том, что он должен быть поставлен рядом с О. fedtschenkoi—типом, рода Oreasiobia.
*2. Oreasiobia fedtschenkoi (Sauss.) (фиг. 49, 50).
Saussure, 1874:6, tab. 2, figs. 1—2 (<5, g; гора Наубид близ Варзаминора, оз. Искандер-куль и ущелье Джиджикрут, Зеравшанский хребет; уроч. Караказук и ущелья Джиптык, Чибурган и Каракол, Алайский хребет) [Forficula]; Bormans, 1910 :103 {Anechura}; Семенов, 1901а :191; Якобсон, 1905:25; Burr, 1911 :73 {Anechura}; Bey-Bienko, 1933 : 5.
Тело бурое или каштановое, коренастое, крупное. Голова рыжеватокрасная. Переднеспинка со слабо округленным задним краем, с грязножелтыми боковыми сторонами. Надкрылья бурые, с овальным, крупным грязно-желтым пятном в передней половице или совсем одноцветно грязножелтые (morpha pallidipennis Sem.); задний край обрублен. Крыловые пластинки, если выступают, то с буроватым внутренним и задним краем, и со светлым пятном в остальной части. Ноги грязно-желтые, в длинных волосках, концы бедер темнобурые. Брюшко гладкое, без ясной пунктировки, сверху голое, снизу в коротких волосках; последний тергит 3 сверху по бокам с двумя очень длинными, остро-коническими, параллельными друг другу и направленными назад отростками; очень редко эти отростки отсутствуют и вместо их имеются два небольших бугорка; у g — простой в задней половине посредине слегка вдавлен. Пигидий <5 очень широкий, несколько выступающий над уровнем последнего тергита в виде сильно уплощенной пластинки; пигидий £ сильно наклонный, удлиненный, но довольно широкий, задний край его при рассматривании снизу с едва заметной выемкой, почти прямой, с заостренными боковыми углами. Клещи в длинных, довольно густых волосках, у <5 при основании сверху над расширенной внутренней стороной с небольшим коническим отростком, направленным вверх и несколько внутрь и приближенным своим концом к наружной стороне пигидия; у /. macrolabia — очень длинные, примерно равны по длине телу без головы, в профиль слегка волнисто изогнутые, основная половина при рассматривании сверху с расходящимися от основания и здесь изогнутыми ветвями; у f. brachylabia значительно более короткие, едва длиннее переднеспинки и надкрылий взятых вместе, в профиль прямые, основание слабо изогнутое, с более слабым коническим выступом сверху на внутренней стороне; клещи Q длинные, уплощенные, с тонким слегка загнутым концом. Генитальный айпарат 3 с длинной, суживающейся и почти исчезающей к концу виргой; длина ее почти в l1/^ раза превосходит длину метапарамер и примерно в 8 раз — длину почко-
Фауна СССР, Кожистокрылые И
162
4. FORFICULIDAE
видного пузырька; почковидный пузырек с крупными хитиновыми пластинками на концах. Длина тела 3 j 9—18, клещей 3 4 —11, 2 4— 6.5 мм
Распространение: горы центр. Азии от Китайской Джунгарии,.. Заалийского Алатау (Алма-ата, колл. Г. Б.-Б.!), Александровского хребта
Фиг. 49. Oreasiobia fedtschenkoi fedtschenkoi (Sauss.), 5x4 (Чим-ган). Ориг.
Фиг. 50. Oreasiobia fedtschenkoi fedtschenkoi (Sauss.). 3. Генитальный аппарат.
Ориг.
(ущелье рр. Каирган и Сулута, колл. ЗИН!) и южн. Каратау (ур. Су-алма,. Правдин!) на севере — до Гиссарского и Зеравшан-ского хребтов на юго-востоке и Памира на юге; встречается на высоте не ниже 1000—1500 м, доходя 40 альпийской зоны.
Этот подвид характеризуется довольно значительной изменчивостью; особенно это относится к окраске надкрылий,, к длине выступающих из под надкрылий крыловых пластинок, которые иногда могут быть совершенно скрытыми, и к строению последнего тергита брюшка 3,, на котором иногда могут совершенно отсутствовать столь характерные для О. fedtschenkoi длинные конические отростки. Особи лишенные таких отростков настолько похожи на О. stoliczkae, что
отличать эти два вида
можно только по характеру окраски и по наличию или отсутствию длинных волосков на теле и особенно на клещах. Очень вероятно, что такие особи 3 3, у которых последний тергит брюшка не имеет конических отростков, представляет собой атавистическую форму; за это говорит то, что они крайне редки и пока известны всего только в двух экземплярах, хотя серия .изученных нами нормальных особей достигает многих сотен.
О. fedtschenkoi была отмечена и как вредитель (повреждала табак, в Чимгане близ Ташкента).
4. ANECHURA
163
2 a. Oreasiobia fedtschenkoi calciatii (Bor.).
.calciatii Borelli, 1909 (g; дорога от Сринагара в Гильгит, Кашмир) [Anechura]; Burr, 1910:159, fig. 95 (Anechura); Bey-Bienko, 1935 : 210.
Очень сходна во всех отношениях с типичной формой (см. выше),, отличаясь лишь только одноцветной черной окраской надкрылий; крыловые пластинки отсутствуют или если слегка выступают, то также черные,, брюшко обычно более темное. Размеры как у типичной формы.
Распространение: известна только из Кашмира, с высоты 1600—2900 м.
Этот подвид был описан первоначально в качестве самостоятельного вида, но морфологически он совершенно сходен со среднеазиатской формой; поэтому правильнее его рассматривать только в качестве подвида. Замечательно, что характер окраски надкрылий у данного подвида имеет более примитивные черты и весьма сходен с таковой у О. stoliczkai и видов, группы Forficula schlagintweiti1 доходящей в своем распространении также до Кашмира. Это говорит за то, что в лице О. fedtschenkoi calciatii мы имеем более примитивную форму, сохранившую больше сходных признаков с родоначальной группой из рода Forficula, чем О. fedtschenkoi fedtschenkoi.
4. Род ANECHURA SCUDD.
Scudder, 1876:289; Bormans, 1900:100 (partim); Семенов, 1901a: 185 (partim); Kirby, 1904:406 (partim); Якобсон, 1905 : 24 (partim); Burr, 1911: 73; Bey-Bienko, 1933: 5.—Burriqla Semenov, 1908 :159. — Bqrelliqla Semenov, 1908:159. — Burriola Burr, 1911 : 703. — Odonio-psalis Burr, 1904:315.
Тип рода Anechura bipunctata F.
Тело голое, или снизу в редких и коротких волосках. Усики с довольно короткими члениками, 4-й членик заметно, часто значительно,, короче 3-го членика, обычно не более чем в два раза длиннее своей ширины, полуконический; 5-й членик не более чем в три раза длиннее своей ширины, Переднеспинка более или менее поперечная, с прямыми, и параллельными, боковыми сторонами. Надкрылья без боковых килей, развиты в разной, степени: у подродов Anechura, Anechurella и Odontopsalis вполне развитые, у Burriola и Borelliola сильно укороченные, но всегда соприкасаются .своими внутренними сторонами и не открывают спереди щитка. Крылья вполне развиты или отсутствуют. Среднегрудная пластинка широкая,, поперечная или одинаковой ширины и длины. Брюшко с ясными боковыми бугорками на 3-м и 4-м сегменте, уплощенное, слегка расширенное около середины, а далее к вершине немного суживающееся, или у $ в вершинной половине почти с параллельными сторонами; последний тергит чуть расширен к вершине или с параллельными боковыми сторонами, сверху у заднего края около задних боковых углов с выступами или выпуклостями, имеющими различную более или менее неправильную форму,, но никогда не в виде правильно-конических, длинных острых и параллельных друт другу отростков. Пигидий 3 широкий, поперечный, не выступающий, с отвесным задним краем или же выступающий в различной степени;.
И*
164
4. FORFICULIDAE
у «J очень маленький. Клещи голые или в редких коротких волосках, у 8 при основании широко расставлены, при рассматривании в профиль волнисто-изогнуты или прямые, но в последнем случае в основной половине с зубцом или с расширением на внутренней стороне у основания; внутренняя сторона при основании обычно не зазубрена; клещи у g простые, соприкасающиеся. Генитальный аппарат 5 обычно со сближающимися к вершине метапарамерами, virga соединяется с почковидным пузырьком под углом, иногда несколько округленным; почковидный пузырек с маленькими хитиновыми придатками.
Род Anechura нами трактуется более широко по сравнению с прежними исследователями, придававшими большое значение степени развития надкрылий и рассматривавшими этот признак в качестве родового критерия. Однако дальнейшие исследования и открытие ряда новых видов в Китае и Тибете — вероятном первичном центре рода — выявили ряд промежуточных форм, напр. Anechura (Burriola) semenovi, А. (В.) tibetana и др., которые по характеру надкрылий, несколько укороченных, но более развитых чем у собственно Burriola не могли быть отнесены ни к Anechura, ни к Burriola. Одновременно первый из указанных видов имеет очень сходные с настоящими Anechura волнисто изогнутые клещи 8. Исследование полового аппарата 8 показало также, что типичные Burriola и Anechura, не говоря уже об указанных выше видах, представляют собой Труппу с одинаковым типом метапарамер и с одинаковым характером соединения virga с почковидным пузырьком; правда, многие виды Anechura s. str. в этом отношении еще не изучены, но указанное выше однообразие в строении гениталий 8 говорит об очень большой генетической близости Anechura я Burriola s. str. Наконец, и у некоторых Anechura s. str. имеется тенденция к укорочению надкрылий. Все это заставляет рассматривать две указанные группы в качестве только подродов одного и того же рода. Однако все многообразие форм, близких к Anechura, не укладывается в указанные два подрода; поэтому необходимо выделение еще нескольких подродов, как правило, уже выделенных прежними исследователями, но впоследствии сведенных в синонимы с родом Anechura или Burriola.
Таким образом в нашей трактовке род Anechura состоит из 5-ти подродов, из которых подрод Anechura свойствен всей Палеарктической Евразии, подрод Anechurella nov.1 — Мексике, подрод Burriola — Тибету и сопредельным частям Китая, подрод Odontopsalis — Восточной Азии и подрод Borelliola — Кавказу и Балканам.
Род Anechura является по преимуществу горным; только немногие виды населяют равнинные части.
1 Anechurella subgen. п. Очень напоминает подрод Anechura in sp., отличаясь более стройными усиками, их 4-й членик в 2—2.5 раза длиннее своей ширины, последний тергит 8 сверху гладкий, без бугорков, но снизу по бокам с коротким тупоконическим зубцом; клещи 8 волнисто-изогнутые, но внутренняя сторона в основной части со многими мелкими зубчиками.
Тип подрода — Anechura vara Scudd. из Мексики.
4. ANECHURA
T65
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ И ВИДОВ
1 (18). Надкрылья вполне развитые или чуть укороченные, по крайней мере в 11/2 раза длиннее переднеспинки. Крылья развиты или отсутствуют.
2 (9). Клещи 8 в профиль совершенно прямые (1. Подрод Odontopsalis Burr).
3 (6). Крыловые пластинки отсутствуют. Пигидий 3 сильно выступает в виде пластинки.
4 (5). Ноги светлые. Пигидий 8 короткий, на вершине не загнут кверху, в профиль не виден. Клещи 8 с маленьким тупоугольным зубцом на внутренней стороне (фиг. 51).............1. А. (О.) potanini B.-Bienko.
5 (4). Ноги буровато-черные. Пигидий 8 длинный, на вершине загибается вверх и здесь совершенно вертикальный, при рассматривании в профиль выступает над уровнем клещей. Клещи 8 без зубца, при основании несколько расширены............................................
......................2. А. (О.) forficuliformis Sem. et B.-Bienko.
6 (3). Крыловые пластинки имеются, сильно выступают из под надкрылий, со светложелтым пятном, занимающим почти всю пластинку. Пигидий 8 не выступает, с отвесным задним краем.
7 (8). Брюшко сверху ясно, довольно сильно, пунктировано. Клещи 8 на внутренней стороне перед срединой с одним крупным широким треугольным зубцом, направленным несколько вниз и внутрь (фиг. 52).
................................... * 3. А. (О.) japonica Borm.
8 (7). Брюшко сверху с очень слабой, почти отсутствующей пунктировкой. КЛещи 8 сверху с двузубым выступом, направленным несколько вверх и внутрь............................* 4. А. (О.) harmandi Burr.
9 (2). Клещи 8 при рассматривании в профиль сильно волнисто-изогнутые (2. Подрод Anechura Scudd. in sp.).
10 (17). Крыловые пластинки вполне развитые, выступают из под надкрылий. Пигидий 8 почти не выступает, с отвесным задним краем, у j — простой, на вершине обрубленный или слегка дуговидно выемчатый. Клещи 8 толстые, не сплюснутые с боков.
11 (14). Надкрылья одноцветные, без светлого пятна в передней части. Клещи 8 при основании цилиндрические, сверху у заднего края последнего тергита без выступающего вверх зубца.
12 (13). Голова с ясными швами. Ноги короткие, передние бедра сильно утолщенные. Клещи 8 короткие (фиг. 53).............................
............................5. A. (in sp.) sven-hedini B.-Bienko.
13 (12). Голова с неясными швами. Ноги длинные, стройные. Клещи <5 длинные................................6. A. (in sp.) quelparta Okam.
14 (11). Надкрылья темные, с резким светложелтым пятном в передней части. Клещи 8 при основании сверху обычно с сильно выступающим вверх треугольным зубцом.
16б
4. FORFICULIDAE
15
16
17
18
19
20
21
22
23
24
25
(16). Голова по крайней мере на затылке рыжевато-красная. Бедра светлые или иногда только на вершине затемнены. Тело более коренастое и крупное (фиг. 54)...........* 7. A. (in sp.) bipunctata F.
(15). Голова сплошь глубоко-черная. Бедра обычно черные или темные, иногда со светлыми концами. Тело меньших размеров, более стройное.
.....................................*8. A. (in sp.) asiatica Sem.
а (в). Светлое пятно надкрылий крупнее, простирается кзади за их средину. Окраска ног и брюшка более темная, обычно буроваточерная или черная (фиг. 55) . *8а. A. (in sp.) asiatica asiatica Sem. в (а). Светлое пятно надкрылий маленькое, не заходит кзади за их средину. Окраска ног и конца брюшка более светлая, обычно красновато-бурая...........8 b. A. (in sp.) asiatica zubovskii Sem.
(10). Крылья отсутствуют. Пигидий 5 сильно вступает в виде уплощенной пластинки, у J на вершине с двумя боковыми и одним срединным зубчиком. Клещи <3 в основной трети сильно сплющены с боков............t....................9. A. (in sp.) filchneri Burr.
(1). Надкрылья сильно укороченные, короче переднеспинки или едва равны ей, но тогда их задние концы сильно косо срезаны. Клещи $ без зубцов или со слабым зубчиком.
(26). Клещи на вершине с короткими волосками. Среднегрудная пластинка поперечная. Передние бедра стройные, не вздутые, длиннее переднеспинки (3. Подрод Burriola Sem.).
(23). Надкрылья примерно равны по длине переднеспинке, с сильно скошенным задним краем. Тело крупное, темное, с темнобурой, почти черной переднеспинкой.
(22). Клещи сильно волнисто изогнутые, у самого основания снаружи с маленьким остро-коническим шипом. Голова черная или со слабым коричневым оттенком (фиг. 56) .... 10. А. (В.) semenovi B.-Bienko.
(21). Клещи 8 почти прямые, не изогнутые волнисто, снаружи у основания без шипа. Голова буровато-коричневая (фиг. 57).............
.............................11. А. (В.) tibetana Sem. et B.-Bienko. (20). Надкрылья короче переднеспинки, с поперечно-срезанным задним краем. Тело маленькое, буровато-рыжее, или по крайней мере переднеспинка светлая, золотисто-желтая.
(25). Пигидий 8 снизу по бокам с сильным горизонтально расположенным длинным отростком. Клещи 8 при рассматривании сверху почти прямые, от основания к вершине постепенно сближающиеся» клещи g более толстые и длинные (3.8—4.1 мм).....................
.....................................12. А. (В.) przewalskii Sem.
(24). Пигидий 8 снизу по . бокам со слегка выступающими углами, без длинного отростка. Клещи более тонкие, у 8 от основания к вершине слегка дуговидно изогнутые, на внутренней стороне снизу с маленьким зубчиком перед срединой; у g более короткие (3.5—3.6 мм).
.........................................13. А. (В.) redux Sem.
4, ANECHURA
167
26 (19). Клещи на вершине голые. Среднегрудная пластинка одинаковой длины и ширины. Передние бедра короткие и толстые, в основной половине несколько вздутые, к основанию резко, к вершине более постепенно суживающиеся, не длиннее переднеспинки (4. Подрод Borel-liola Sem.). — Брюшко сверху гладкое с очень неясной, практически отсутствующей пунктировкой. Пигидий 5 короткий, верхняя его сторона сильно наклонная, образует с задней (нижней) стороной тупой угол, вооруженный по бокам маленьким зубчиком. Клещи 5 без зубца на внутренней стороне, у f. macrolabia заметно-волнисто изогнутые, у f. brachylabia с расширенными в основной половине и приподнятыми кверху внутренними сторонами (фиг. 11—12)......................
........................................* 14. А. (В.) euxina Sem.
1. Подрод Odontopsalis Burr.
®urr, 1904: 315. — Apterygida auct. (partim). — Anechura auct. (partim).
Тип подрода — Anechura (Odontopsalis) harmandi Burr.
Брюшко слабо расширено около средины или почти с параллельными боковыми сторонами. Надкрылья длинные, в l'/g—2 раза длинней переднеспинки, вполне развитые или едва укороченные, внутренняя их сторона не короче переднеспинки. Нрги довольно короткие, передние бедра заметно утолщенные, слегка вздутые. Клещи S в профиль совершенно прямые.
Фиг. 51. Anechura (Odontopsalis) potanini B.-Bienko, s (чип). Клещи. По Bey-Bienko, 1934.
1. Anechura (Odontopsalis) potanini B.-Bienko (фиг. 51).
Bey-Bienko, 1934 : 406, fig. 3 (<5; Тацзинлу, пров. Сычуань, сев.-зап. Китай).
Тело средних размеров, стройное, буровато-черное. Усики рыжевато-желтые, 13-члениковые, 3-й членик в 1]/2 раза длиннее 4-го и равен 5-му, цилиндрический, 4-й и 5-й членики полуконические. Голова красноватобурая, уплощенная сверху, с тонкими, но ясными швами. Переднеспинка грязно-бурая, чуть уже головы, с параллельными и прямыми боковыми •сторонами, немного шире своей длины, задний край очень слабо округлен. Надкрылья относительно короткие, примерно в 11/2 раза длиннее цереднеспинки, грязно-бурые, со слабо скошенным задним краем; крылья отсутствуют. Переднегрудная пластинка сильно расширена спереди, среднегрудная пластинка округленная, одинаковой длины и ширины. Ноги рыжевато-желтые. Брюшко с почти параллельными боковыми сторонами, буровато-черное, с красноватым оттенком, гладкое, без ясной пунктировки, боковые бугорки слабые, особенно на 3-м тергите; последний
168
4. FORFICULIDAE
тергит примерно в два раза шире своей длины, с двумя слабыми сферическими выпуклостями у заднего края, расположенными перед основанием внутренней стороны клещей; боковые стороныклещей параллельные, задний край посредине умеренно дуговидно вырезан; пигидий очень широкий,, сильно выступающий, на вершине слегка широко-дуговидно вырезан; задняя сторона почти вертикальная, суживающаяся книзу, посредине боковых сторон с маленьким острым бугорком и со слабой поперечной приподнятостью между этими бугорками. Клещи 3 короткие, цилиндрически-кониче-ские, слегка расширенные у основания и постепенно суживающиеся к вершине, внутренняя сторона перед серединой с маленьким тупоугольным бугорком, а перед самым основанием очень слабо зазубрена; в основной половине прямые, с параллельными наружными сторонами, а далее к вершине очень постепенно изогнутые, концы не соприкасаются, широко расставлены друг от друга. Самка неизвестна. Длина тела 3 11, клещей. 3 2 мм.
Распространение: известен пока только из окр. Тацзинлу в пров. Сычуань, сев.-зап. Китай.
Хотя этот вид довольно значительно отличается от типа подрода Odontopsalis, однако, эти отличия не выходят за пределы видовых, так как по строению клещей и коротким ногам он должен быть отнесен к указанному подроду; по стройности тела он очень напоминает А. (О.) japonica.:
2. Anechura (Odontopsalis) forficul’formis Sem. et B.-Bienko.
Semenov-Tian-Shanskij et Bey-Bienko, 1935 : 222 (3; речка Дза-чу, бассейн Янг-тце— кианг, Кам, Вост. Тибет; р. Серг-чу, бассейн Хуан-хе, Вост. Тибет).
Тело стройное, смоляно-черное. Внешне очень сходен с предыдущим видом. Усики буровато-желтые, 12—13-члениковые, 4-й членик немного короче 3-го, 5-й членик чуть длиннее 3-го, в 2]/2—3 раза длиннее своей ширины. Голова рыжая, большая, сверху немного выпуклая, с тонкими,., но ясными швами. Переднеспинка заметно уже головы, черная с грязножелтыми боками, одинаковой длины и ширины, боковые стороны прямые, с заметно приподнятыми краями, параллельные, задний край широкоокругленный. Надкрылья такого же цвета как диск переднеспинки немного укороченные, примерно в 11/2 раза длиннее переднеспинки,, густо и сильно пунктированные, задний край слегка скошен, крылья отсутствуют. Ноги желтовато-бурые, передние бедра довольно утолщенные.. Брюшко смоляно-бурое, почти черное, чрезвычайно слабо и мелко пунктированное, почти гладкое, у 3 почти с параллельными боковыми сторонами, последний тергит 3 примерно в два раза шире своей длины,, смоляно-черный, у заднего края с 2 слабыми бугорками, задний край слегка выступающий, с чуть выемчатой, почти прямой средней частью,, расположенной между бугорками. Пигидий 3 длинный, широкий, сильно выступающий в виде пластинки, постепенно загибающийся кверху и на. самой вершине вертикальный, загибаясь к заднему концу последнего.
4. ANECHURA
169'
тергита, вершина слегка расширена, задний, вернее верхний, край,, слегка выемчатый; при рассматривании в профиль пигидий виден в виде торчащей над уровнем основания клещей пластинки. Клещи S короткие, при основании расширенные и уплощенные, но широко расставленные, будучи отделены друг от друга широким пигидием, далее постепенно суживающиеся, цилиндрические, на большем протяжении прямые и лишь
на самой вершине слегка загнутые внутрь концы не соприкасаются. Самка неиз-; вестна. Длина тела S 9.9—12.4, клещей 3.5—3.8 мм.
Распространение: вост. Тибет.
Вид этот внешне очень похож на А. (О.) potanini, с которым он сходен также по слегка укороченным надкрыльям. Клещи S, благодаря их некоторой расширенности и уплощенности при основании, делают этот вид сходным с представителями рода Forficula-, однако, благодаря тому, что клещи широко расставлены, этот вид не может быть отнесен к указанному роду. Весьма интересно, что А. (О.) potanini, и А. (О.) forficuliformis весьма напоминают ископаемую нижне-олигоценовую (Балтийский янтарь) форму — Forficula (?) pristina Burr, которая сходна с ними как в отношении стройности тела, так особенно в отношении строения клещей и пигидия. Сходство это несомненно не случайное и говорит о том, что данная группа видов в морфологическом отношении является наиболее примитивной в роде Anechura (некоторая укороченность надкрылий очевидно представляет собой более позднее явление), причем и здесь эта примитивность связана с морфологической близостью
Фиг. 52. Anechura (Odontopsalis) japonica (Borm.), (?X5. (Славянка, Уссур. край). Ориг.
к роду Forficula. Наличие в нижнем олигоцене вида, который должен быть отнесен к подроду Odontopsalis рода Anechura говорит о том, что уже в то время имела место дифференциация сем. Forficulidae на некоторые'
процветающие и теперь роды.
* 3. Anechura (Odontopsalis) japonica (Borm.) (фиг. 52).
Bormans, Ann. Soc. Espan. Hist. Nat., IX, 1880 : 512 (Япония, на берегу моря), {Forficula)-, Bormans, 1900:114 (Apterygida) Якобсон, 1905 : 26; Burr, 1904: 316 (Odontopsalis)-, Burr, 1911: 73 (Anechura)-, Shiraki, 1928:16; Bey-Bienko, 1933 : 3, tab. I, fig. 1. — eoa Semenov, 1902 :100, fig. 2. t- athymia Rehn, Proc. U. S. Nat. Mus., XX-VII, 1904 :540 (Apterygida)..
170
4. FORFICULIDAE
Тело удлиненное, довольно стройное, буровато-красное, или каштаново-бурое, или иногда смоляно-бурое, почти черное. Усики 12-члениковые, рыжевато-или буровато-желтые, 4-й членик полуконический, в два раза длиннее своей ширины, немного короче 3-го, 5-й членик немного длиннее 3-го, цилиндрический. Голова рыжая или черная, швы заметные. Переднеспинка практически такой же ширины, как голова, слегка поперечная, с прямыми, почти параллельными боковыми сторонами, задний край широко округлен, но слабо выступает; диск каштаново-бурый или почти черный, с более светлыми полосками на боках. Надкрылья примерно в два раза длиннее переднеспинки, с параллельными наружными сторонами, такого же цвета как переднеспинка, без светлых пятен. Крыловые пластинки хорошо выступают, на внутренней стороне буроватые, в остальной части заняты крупным желтоватым пятном, иногда не очень ясным. Ноги грязно-желтые. Брюшко слегка расширено около средины или у 3 с почти параллельными сторонами, сверху густо, довольно крупно и очень ясно, снизу более тонко пунктировано, у основания, особенно по бокам, включая боковые бугорки, черноватое или сплошь смоляно-бурое. Последний тергит <5 прямоугольный, не менее, чем в два раза шире своей длины, у заднего края над основаниями клещей с довольно сильным неправильной формы выступом, разделенным на два очень сближенных бугорка, из которых наружный более острый, чем внутренний; у g последний тергит с двумя очень слабыми, едва выступающими бугорками, средина задней половины тергита несколько вдавлена. Пигидий 3 почти не выступающий, с вертикальной, посредине несколько бугорковидно-вздутой задней стороной, у g сильно наклонный, иногда почти вертикальный, треугольный, суженный книзу, довольно большой. Клещи 3 довольно длинные, при основании немного расширенные и вооружены сверху слабо выступающим бугорком, далее по направлению к вершине очень постепенно суживающиеся, сначала почти параллельные друг другу и прямые, а примерно от средины постепенно дуговидно изогнутые к вершине, с сближенными или соприкасающимися концами; внутренняя сторона снизу перед серединой с сильным, широким, уплощенно-треугольным зубцом, направленным внутрь и несколько вниз; клещи £ длинные, умеренно расширенные, в вершинной части тонкие, с загнутыми концами, внутренние стороны почти не зазубрены. Длина тела 3 12.5—14.5, 2 10—12.5; клещей 3 4.5—6.5, 2 4.5—4.8 мм.
Распространение: Уссурийский край на север до Имана (колл. ЗИН!), и вдоль побережья Японского моря, огибая хребет Сихота-алин, до Николаевска на Амуре (колл. ЗИН!); Япония, Корея, Манчжурия (Гунчасулин, колл. ЗИН!) и провинции Гань-су, Сычуань и Шенси в Китае; отсутствие данных о нахождении на Сахалине и в северо-восточном Китае, видимо, объясняется плохой изученностью этих территорий.
Вид этот представлен в двух цветовых формах/ из которых одна, более светлая (буровато-красная или каштаново-бурая) распространена
4. ANECHURA
171
на севере ареала, тогда как другая, смоляно-бурая, почти черная форма, изредка встречаясь в северной части ареала, населяет преимущественно внутренний Китай. Рассматривать эти формы в качестве подвидов было бы преждевременно, так как по крайней мере на севере ареала они встречаются совместно.
* 4. Anechura (Odontopsalis) harmandi (Burr).
Burr, 1904i 316 (<5, g; Центр. Япония) [Odontopsalis]; Burr, 1911: 73; Burr, 1916 : 8, tab. II, fig. 17; Shiraki, 1928: 16; Furukawa, Proc. Imp. Acad., IX, 4, 1933:194, fig. 1—3. — Jewisi Burr, 1904:317 (Odontopsalis); Burr, 1911: 73; Shiraki, 1928 :16.
Тело более коренастое, чем у предыдущего, также представлено в двух цветовых формах — темнокаштановой и смоляно-черной. Усики как у А. (О.) japonica, но более темные, буровато-желтые. Переднеспинка яснопоперечная, боковые стороны чуть округлены, практически прямые. Надо-крылья и крылья как у А. (О./ japonica. Ноги каштановые или каштановобурые. Брюшко более уплощенное, посредине расширенное, или же у более коренастых особей даже слегка расширяющееся к вершине, сверху очень мелко и слабо пунктированное, почти гладкое, вдоль задних концов сегментов с редкими более крупными точками. Последний тергит $ короткий, сильно поперечный, бугорки ПО' бокам над основанием клещей округлены, не раздвоены, на самом конце с коротким сосочком; задний край тергита посредине слегка дуговидно выемчатый* Пигидий S не выступает, задняя сторона вертикальная^ посредине слегка выпуклая. Клещи сверху у самого основания без бугорка, но несколько отступя от основания с сильным двузубым зубцом, направленным вверх, а затем загнутым внутрь. Генитальный аппарат S со слегка дуговидно-изогнутыми наружными сторонами метапарамер, virga длинная, более чем в два раза длиннее метапарамер, соединена с почковидным пузырьком под очень сильной узкой дугой; почковидный пузырек на переднем конце с довольно крупной хитиновой пластинкой. Длина тела 9.2—14, д 12.5—13.8; клещей <3 2.5— 4, <3 2.5—3.
Распространение: Япония, от южной части острова Нипон до Иезо, остр. Сахалин [с. Маука в южной части (колл. С.-Т.-Ш.!) и с. Тунайчи, Охотское побережье (колл. ЗИН)], Владивосток (Burr, 1912).
Описанная Вигг’ом из Японии Odontopsalis lewisi рассматривается нами как f- macrolabia А. (О.) harmandi', эта форма характеризуется, как это свойственно уховерткам, кроме большей длины клещей S, также и более стройным телом. Сам автор этих видов при их описании высказал соображение о том, что с?. A. lewisi представляет собой только особую форму A. harmandi. Точно такое же предположение высказал Shiraki, a Furukawa рассматривал их как один вид.
Этот вид зимует в Японии во взрослом состоянии и был находим весной в земле на глубине до 13 см. Взрослые самки и личинки были находимы весной в гнездах, устроенных среди гальки и песка в овраге.
172
4. FORFICULIDAE
Личинки в условиях неволи выкармливались пыльцой, взятой с цветов,, и вероятно на воле питаются мелкими организмами. В экологическом отношении А. (О.) harmandi является лесным и горным видом и иногда встречается на деревьях.
2. Подрод Anechura Scudd.
' Scudder, 1876:289; Bormans, 1900:100 (partim); Kirby, 1904 : 40 (partim); Burr,. 1911: 73 (partim). .
Тип подрода — Anechura (in sp.) bipunctataY.
Надкрылья длинные, вполне развитые, в 11/2—2 раза длиннее переднеспинки; крылья имеются или отсутствуют. Передние бедра толстые.. Клещи 3 при рассматривании в профиль волнисто изогнутые, довольно короткие, обычно с сильными зубцами. Последний тергит брюшка 3 с сильными бугорками или выступами сверху, снизу по бокам без зубца. Тело коренастое.
5. Anechura (in sp.) sven-hedini B.-Bienko (фиг. 53).
Bey-Bienko, 1934:4, tab. I, fig. 2 (3; сев.-вост. Сычуань, Китай).
Тело желтовато-бурое, иногда более темное. Усики рыжевато-бурые,. 3-й членик цилиндрический, в: вершинной трети несколько расширен, равен по длине 5-му, 4-й и 5-й членики полуконические. Голова с ясными швами, рыжеватая. Переднеспинка чёрновато-бурая, с грязно-палевыми боковыми сторонами, ясно поперечная, боковые Стороны сначала параллельные, от середины постепенно и дуговидно соединяющиеся с задней стороной, которая также широко округлена. Надкрылья почти в два раза длиннее переднеспинки, одноцветные, грязно-желтые, без светлых пятен,, с очень редкой пунктировкой; крыловые пластинки имеются, но короткие, немного выступающие из под надкрылий, такого же цвета как последние.. Ноги короткие, с сильно утолщенными передними бедрами, рыжевато-бурые. Брюшко с нежно пунктированными передними члениками, у 3 довольно сильно расширено перед вершиной, у д—стройное, от середины постепенно суживающееся к вершине. Последний тергит 3 широкий и короткий, у заднего края перед основанием клещей с двумя боковыми вздутиями,, слегка разделенными на два бугорка, у д простой, с неясными бугорками, суженный кзади. Пигидий короткий, слегка выступающий, задняя сторона, отвесная, около средины с очень тупым, широко-коническим бугорком; у 9 — сильно наклонный, удлиненный, слегка суженный к вершине, задний конец слегка вырезан, и с заостренными боковыми углами. Клещи 3 сильно изогнутые в двух плоскостях, сначала от основания немного приподнимаются вверх, затем сильно загнуты вниз и назад и далее, за срединой, горизонтальные, но загнутые уже в горизонтальной плоскости друг к другу; основная часть цилиндрически-коническая, сверху у заднего края, последнего тергита без треугольного зубца, .внутренняя сторона снизу;
4. ANECHURA
около середины с сильным острым треугольным зубцом; клещи g простые, тонкие, за исключением слегка изогнутой вершинной части прямые,
слегка расширенные у основания и постепенно суживающиеся к вершине. Длина тела 3 12.2, g 7.2—10.2, клещей 3 3.9, g 2—2.6 мм.
Распространение: провинция Сычуань в Китае от северо-восточной ее части до г. Тацзинлу и область Амдо в сев.-вост. Тибете.
Этот вид в морфологическом отношении весьма близок к северноитальянской A. orsinii Gene, причем сходство распространяется и на характер окраски, почти совершенно тождественной у обоих видов. Осо-
бенно замечательно сходство в строении осно-. вания клещей <3, которое у A. orsinii также цилиндрически-коническое, без сильного зубца сверху, нормально хорошо развитого у остальных видов подрода Anechura, а лишь только со слабым бугорком. Такая близость в отношении морфологии и окраски этих широко разобщенных видов распространяется и дальше — у A. sven-hedini крыловые пластинки очень короткие, наиболее укороченные по сравнению со всеми полнокрылыми видами, а у A. orsinii они полностью отсутствуют; если еще принять во внимание некоторую обособленность этих видов, -образующих как бы особую группу в подррде Anechura, то вполне можно будет допустить, что указанные два вида являются и генетически близкими друг другу. Это обстоятельство лишний раз подтверждает наше представление об. умеренной Евразии как о первоначальной родине рода Anechura в целом, откуда и происходило рас-
Фиг. 53. Anechura (in sp.) sven-hedini B.-Bienko, <3 X 3.5, (тип). По Bey-Bienko, 1933.
селение видов в остальные части Евразии; A. orsinii таким образом является
производным какого-то широко распространенного в третичное время вида, близкого одновременно и к Л. sven-hedini.
6. Anechura (in sp.) quelparta Okam.
Okamoto, Bull. Agric. Exper. Stat., Korea, I, n° 2, 1922:53, tab. VIII, fig. 5 (3, g; о-в Квельпарт, Корея).
Тело голое, каштаново-бурое, два основных членика усиков, боковые стороны переднеспинки и надкрылий, крыловые пластинки и ноги грязножелтые. Усики 12-члениковые, 3-й членик длинный, почти цилиндрический, 4-й равен примерно двум-третям длины 3-го, 5-й такой же длины как 3-й, цилиндрический, довольно толстый. Голова выпуклая, швы неясные. Переднеспинка четырехугольная, такой же ширины, как голова, поперечная, боковые стороны прямые, задний край округлен. Надкрылья ^широкие, крыловые пластинки имеются. Ноги длинные, тонкие, 1-й членик
174
4.FORFICULIDAE
лапок длиннее чем 2-й и 3-й взятые вместе. Брюшко уплощенное, расширенное около средины, суживающееся к вершине; 3-й и 4-й сегменты с ясными боковыми бугорками; последний тергит у 8 с крупным бугорком на каждой стороне, у 2 — простой, без бугорков; предпоследний стернит широкоокругленный. Пигидий 8 очень короткий и широкий, у 2 довольно большой, треугольный. Клещи 8 сильно волнисто-изогнутые, с сильным и толстым зубцом около средины, у 8—, простые. Длина тела в 15, g 14.5, клещей 8 6, g 4.5 мм.
Распространение: известен только с острова Квельпарт, к югу от Корейского полуострова.
Систематическое положение этого вида не совсем ясно, так как нам он известен только по довольно схематическому описанию его автора. Последний сближает данный вид с ,,Anechura“ crinitata Shir, с Формозы; однако этот вид выделен в особый род Taipinia и вряд ли близок к A. quelparta. По всей вероятности он примыкает более всего к A. sven-hedini.
* 7. Anechura (in sp.) bipunctata (F.) (фиг. 54, 58).
Fabricius, Species Insect., I, 1781:340 (2; Италия); [Forficula]; Brunner, 1882:19, tab. I, fig, 5. (Anechura); Bormans, 1900 :101; Семенов, 1901a : 190; Kirby,1904 : 40; Якобсон, 1905 : 25, табл. Ill; Burr, 1911: 74, tab. V, fig. 5. — bipunctata orientalis Bormans 1900 :102.-— biguttata Fabricius, Ent. System., 1793 : 2 (8) (Forficula); Fischer, 1853 : 72, tab. VI, figs. 9, 9ab. — anthracina Kolenati, Melet. Ent., V, 1846:73, tab. XVII, fig. 5 (Chelidura).
Тело довольно крупное, коренастое, темнобурое, иногда почти черное. Усики 12-члениковые, толстые, красновато-бурые, 5-й членик едва длиннее или такой же длины как 3-й, последующие членики одинаковой длины. Голова выпуклая, передняя ее часть примерно до лобных швов и у основания затылочного шва черная, задняя часть красновато-рыжая или иногда темнобурая, но всегда с красноватым оттенком и никогда не бывает глубоко черной; швы хорошо заметные. Переднеспинка черновато-бурая, с рыжеватыми боками, или значительная часть переднеспинки рыжая и только по середине темнобурая; заметно, но немного шире своей длины и слегка уже головы, с немного приподнятыми, прямыми или чуть округленными боковыми сторонами, задний край широко округлен, задние боковые углы узко-округлены. Надкрылья без пунктировки, немного более чем в 1г/2 раза- длиннее переднеспинки, с резким желтым, слегка удлиненным пятном в передней половине и желтыми боковыми подогнутыми сторонами. Крыловые пластинки хорошо развитые, на внутренней стороне затемненные, в остальной части с желтым пятном. Ноги одноцветные, грязно-желтые, иногда концы бедер темные. Брюшко широкое, гладкое,, значительно уплощенное, у 8 перед задним концом довольно сильно, у 2 слабее расширено; последний тергит 8 сильно поперечный, очень короткий, в 3—4 раза шире своей длины, у заднего края по бокам перед основанием клещей с неправильной формы поперечными бугорками,, наружная часть которых сильнее приподнята и заострена, несколько отде-
4. ANECHURA
175
Фиг. 54. Anechura (in sp.) bipunctata . (F.), $ X 5. (Алтай). Ориг.
лена от округло вздутой и слабо приподнятой внутренней части, так что с каждой стороны имеется по два разнородных бугорковидных выступа» у g последний тергит более длинный, слегка суженный кзади, с ясной продольной вдавленной линией посредине. Пигидий <5 едва выступает, задняя сторона отвесная, четырехугольная, суживающаяся книзу. Клещи $ при основании сверху с сильным, широко треугольным выступом, на внутренней стороне снизу около средины с зубцом, у 2 на внутренней стороне с многочисленными мелкими зубчиками. Метапарамеры узкие, к вершине несколько утонченные, с прямым наружным краем, вирга очень длинная, почти в три раза длиннее метапарамер. Длина тела $ 10—16, 2 9—14, клещей 8 4.5—7, 2 3.5—5 мм.
Распространение: горные части Зап. Европы от Пиренеев, через горы южной Франции, до Альп, подымаясь отсюда по горным системам восточной Франции (Савойя, Юра, Вогезы) до Эльзаса и по южной Германии (Тюрингия, Дрезден) и северной части Чехо-Словакии до верхней Силезии; южная граница проходит через о-в Сардинию и Сицилию, сев. Италию, сев. Балканы, Малую Азию, захватывая Кавказ от< южного Закавказья до северных предгорий Главного хребта (вид отсутствует, однако, вдоль Черноморского побережья) и проникая к северу до Провалья в Донецком бассейне (колл. ЗИН!); совершенно отсутствует в остальной части СССР к западу от Волги (указание Fischer-Waldheima, повторенное последующими авторами для окр. Москвы, очень сомнительно), но появляется вновь в восточном Приуральи (известна
пока только из б. Сарапульского уезда). Далее на восток ареал охватывает средний Урал до Свердловска, южную часть Зап. Сибири на север до линии Омск (колл. ЗИН!), Татарск, Новосибирск, проходя на восток, через Алтай по северным склонам Саян до р. Ангары (Иркутск, колл. ЗИН!),. а на юге захватывая сев. Казахстан от Кокчетавского района до юго-западной части Алтайского массива, далде идет вдоль Иртыша до пос. Ульбин-ского (колл. ЗИН!), Колбинского хребта и отсюда на Саурский хребет, достигая сев.-зап. Монголии. Таким образом из этого обзора видно, что-ареал вида разорван в европейской части СССР; кроме того, в азиатской
176
4. FORFICULIDAE
части южная граница распространения вероятно проходит от Кокчетав-». ского района через гористую часть вост. Казахстана (Карагандинский и Каркаралинский районы), достигая с одной стороны Алтая, а с другой Тарбагатайского хребта.
Почти всюду этот вид является горным жителем, встречаясь под камнями, под корой деревьев, в гнилых пнях и пр.; однако, вполне достоверные данные по Зап. Сибири (Омск, Новосибирск и пр.) показывают, что он . по крайней мере, в Зап. Сибири живет и на равнине.
Описанная Bormans’oM форма orientalis (Малая Азия), характеризующаяся более светлым цветом, более крупным пятном на надкрыльях и менее изогнутыми клещами, не может считаться самостоятельным подвидом, так как она не имеет самостоятельного ареала распространение, встречаясь от Пиренеев до Алтая, а может быть и во всем ареале вида и является вероятно только морфой.
* 8. Anechura (in sp.) asiatica Sem. (фиг. 55).
Семенов, 1903:197; Semenov 1908:169; Burr 1911: 74. — orientalis Семенов, 1901a: 187 (nee Bormans) [c?, Сарепта и Оренбург, Чули, Копет-Даг; Аму-Дарья близ Чарджуя; долина Зерашвана между Иори и Дашты-казы; между Урмитаном и Мадм; Алайский хребет; Каратегин в Таджикистане; Монита, Каз-Казган, Чагарлы и АнДа-сай, близ сев. берега оз. Балхаш; между Суйдун и Хоргосом, р. Ачал и Гучан, Китайская Джунгария сев.-вост. Иран; данные для Арарата ошибочны]; Якобсон, 1905 : 24.
Тело небольшое, умеренно коренастое, черное. Усики 12-члениковые, черные в основной половине, постепенно буреющие к концу, 3-й и 5-й членики примерно одинаковой длины, 4-й заметно короче. Голова сплошь одноцветно черная, с довольно ясными швами. Переднеспинка чуть шире своей длины, черная, с грязно-желтыми боковыми сторонами, задний край слегка округлен, боковые стороны прямые, параллельные. Надкрылья почти в два раза длиннее переднеспинки, черные, с резким довольно крупным, удлиненным желтым пятном, расположенным в передней части и переходящим несколько кзади от средины надкрылий; боковые подогнутые стороны также желтые; крыловые пластинки хорошо развитые, на внутренней стороне затемнены, остальная часть занята желтым пятном. Ноги обычно с черными бедрами и грязно-желтыми голенями. Брюшко гладкое, блестящее, черное или чуть буроватое, почти с параллельными сторонами, не очень коренастое, последний тергит 5 сверху по бокам перед основанием клещей с неправильной формы поперечным выступом, наружный и внутренний края которого примерно одинаково приподняты, но не резко отделены друг от друга и не образуют самостоятельных бугорков; у последний тергит простой, со слегка вздутыми слабыми выступами перед основанием клещей, с тонкой продольной линией посредине. Пигидий 5 едва выступает, с отвесной задней стороной. Клещи 5 при основании сверху с умеренно-выступающим, иногда слабым зубцом _или бугорком, на внутренней стороне около середины с ясно выраженным
4. ANECHURA
177
зубцом; у j на внутренней стороне без ясных зубчиков. Длина тела 610— 716.4; Q 10—15; клещей $ 3.7—7.1, 2 3.2—4.5 мм.
Распространение: юго-восток европейской части СССР (Сарепта, Оренбург ?, степи между южным течением Волги и Урала), пустынные степи Казахстана до Мугоджар (колл. Г. Б.-Б.1), оз. Чалкар-тениз
(Аделунг, 1906), сев. берега оз. Балхаш и Зайсана (р. Джемини, колл. ЗИН!) на севере и, до южной окраины песков Муюн-кум на юге (Актыр-тюбе, колл. ЗИН!), горы и предгорья Средней Азии и Казахстана, от Каратау (Курпеш и Атабаево, колл. Г. Б.-Б.!) и невысоких частей Александровского хребта (ущелье Терес-булак и ущ. Сулутор, колл. ЗИН!) на севере до Копет-Дага, гор южного Таджикистана и Памира (колл. -ЗИН!) на юге, встречаясь также в степных межгорных понижениях и заходя даже в Туранскую низменность (Чарджуй, берег Аральского моря и -Ашхабад, колл. ЗИН!); восточная часть Ирана от Хорасана на севере до Кермана на юге, северо-зап. Монголия, Китайская Джунгария и далее на восток по Тян-Шаню до Урумчи (Bey-Bienko, 1933). В северных частях ареала встречается в пустынных степях, а на юге преимущественно в горах.
Один из наиболее сухолюби
Фиг. 55. Anechura (in sp.) asiatica asiatica Sem., <5 X 5 (Каратау). Ориг.
вых видов, приспособившийся к жизни и в пустынных степях, где он, однако, приурочен преимущественно к берегам
рек или озер.
Вредит в горах Каратау естественным зарослям|каучуконоса, тау-сагыза, выедая цветы и незрелые семена (Правдин 1935).
8а. Anechura (in sp.) asiatica zubovskii Sem.
Семенов, 1901a: 188 (<5, Ладак, долина p. Драс, Кашмир); Якобсон, 1905:25; Burr, 1911: 74, tab. 9, fig. 6; Bey-Bienko, 1935 : 209.
Очень сходна с типичной формой, отличаясь лишь только наличием несколько меньшего светлого пятна на надкрыльях, не переходящего
Фауна СССР, Кожнстокрылые 12
178
4. FORFICULIDAE
кзади за их средину, более светлой, красновато-бурой окраской вершинной половины брюшка и ног и весьма слабо выраженным, иногда практически отсутствующим срединным зубцом на клещах <5. Длина тела как у типичной формы.
Распространение: Кашмир и повидимому юго-зап. Тибет (указания прежних авторов на нахождение здесь A. bipunctata относятся вероятно к данному подвиду); кроме того промежуточная форма между типичной asiatica и zubovskii встречается иногда в юго-вост. Таджикистане Кемперт-тепэ и Хазрет-баба, колл. ЗИН!) и на Памире (Бадаксан,. колл. ЗИН!).
Этот подвид был описан и долго рассматривался как самостоятельный вид; однако, весьма незначительные отличия и наличие переходных к A. asiatica форм говорят за то, что в данном случае мы имеем лишь-только подвид, имеющий в отличие от типичной формы очень узкий ареал распространения. Очень характерно, что и другой среднеазиатский вид—•• Oreasiobia fedtschenkoi имеет в Кашмире также самостоятельный подвид.. Таким образом, в данном случае имеется пример почти аналогичной географической дифференциации; лишь только захождение Anechura asiatica в степи (что несомненно представляет собой вторичное и, повидимому, только послеледниковое явление) отличает ареал данного вида в целом от ареала Oreasiobia fedtschenkoi.
9. Anechura (in sp.) filchneri (Burr).
Burr, 1908:58, tab. Ill, fig. 8 (5; Лян-чжоу, пров. Ганьсу, сев.-зап. Китай) \Odon~ topsalis\-, Burr, 1911: 73; Bey-Bienko, 1933 : 2.
Тело буровато-черное, с светложелтыми надкрыльями, переднеспинкой и ногами. Усики темнобурые, голова совершенно черная, со слабыми швами. Переднеспинка немного шире своей длины, с прямыми, чуть расходящимися или параллельными боковыми сторонами, без более светлых полосок по бокам, задний край широко округлен, слабо выступающий. Надкрылья в 1^2 раза длиннее переднеспинки, одноцветные, задний край прямо-обрубленный, крылья отсутствуют. Брюшко сверху с мелко и густо пунктированными передними половинами члеников, задняя половина члеников неясно, или у 2 совсем не пунктированная, бока брюшка с торчащими волосками. Последний тергит 5 с сильными торчащими кверху боковыми выступами у заднего края, разделенными в верхней половине на две остро-треугольные лопасти, из которых наружная более широкая чем вну -тренняя, у д обычной формы, без ясных выступов. Пигидий <5 сильно выступающий, в виде несколько суженной и слегка округленной или почти прямой на вершине пластинки, вооруженный по заднему краю одним срединным и двумя боковыми маленькими зубчиками, иногда зубчики неясные; у 2 пигидий на вершине как у $ с тремя маленькими, одним срединным и двумя боковыми зубчиками. Клещи $ при основании сплющены с боков и сильно расширены сверху вниз, расходящиеся; далее, у конца
4. ANECHURA
179
расширенной части, тонкие, загнутые внутрь и вниз, к вершине постепенно и слегка суживающиеся, при рассматривании сверху кажущиеся прямыми; основная часть сверху с сильным широко-треугольным зубцом, а снизу почти у самого основания с маленьким тонким шипиком, срединная часть клещей без зубца; клещи g простые, на внутренней стороне не зазубренные. Длина тела 8 11, g 10.8—12; клещей 8 4.5, g 2.5—3.3 мм.
Распространение: провинции Ганьсу и Сычуань в зап. Китае.
Этот вид может рассматриваться в качестве наиболее специализо-ванного представителя подрода Anechura, характеризующегося, кроме утраты крыльев, также и своеобразной структурой клещей 8, сплющенных у основания с боков и лишенных около их середины внутреннего зубца, что резко отличает данный вид от всех остальных. Burr отнес его к подроду Odontopsalis, но его генетическая близость с типичными Anechura несомненна.
3. Подрод Burriola Sem.
Semenov-Tian-Shanskij, 1908:159; Burr, 1911: 70.
Тип подрода — Burriola przevalskii Sem.
Надкрылья сильно укороченные, менее чем в l1^ раза длиннее переднеспинки, не закрывают заднеспинку, или, если доходят до ее заднего края, то их задний край сильно скошенно обрезан. Передние бедра стройные, не вздутые, ясно длиннее переднеспинки. Среднегрудная пластинка ясно шире своей длины. Брюшко сильно уплощенное. Последний тергит 8 чуть расширен кзади, сверху без сильных бугорков, лишь только со слабыми округленными вздутиями. Клещи 8 на вершине с короткими волосками.
Этот подрод свойствен Тибету и сопредельным частям Китая, не заходя, однако, далеко от границ Тибета. В настоящее время известно всего 4 вида в этом подроде, морфологически чрезвычайно напоминающем следующий подрод — Borelliola.
В лице подрода Burriola мы имеем высокогорный дериват Anechura, возникший вероятно после поднятия и образования высокого плато Тибета в миоцене из одновременно поднятых и переработанных в сторону коротко-крылости видов типа Anechura s. str.
10. Anechura (Burriola) semenovi (B.-Bienko) (фиг. 56, 59).
Bey-Bienko, 1934 : 408, figs. 4—5 (8; перевал Хунчао, пров. Сычуань, Китай; р. Дза-чу, Кам, истоки Янг-тце-кианг, В. Тибет).
Тело одноцветно-коричневато-черное, крупное, коренастое. Усики 12-члениковые, основной членик черный, остальные темно-красноватокоричневые. Г олова с ясными швами, черная или с легким коричневым оттенком. Переднеспинка сильно поперечная, примерно в 11/2 раза шире 12*
180
4. FORFICULIDAE
своей длины, с прямыми параллельными боковыми сторонами и со слабо выступающим широко-округленным задним краем. Надкрылья сильно укороченные, равны или едва длиннее переднеспинки, заходящие сзади до заднего конца метанотума, сильно скошенно-обрубленные по заднему краю, с неясной и редкой пунктировкой. Переднегрудная пластинка сильно расширена спереди, передний край примерно в два раза шире заднего; среднегрудная пластинка сильно поперечная, почти в два раза шире своей длины, с очень широко округленным, почти прямым задним краем. Ноги темно-красновато-коричневые. Брюшко <5 широкое, чуть более расширенное перед концом, с почти параллельными боковыми сторонами, с очень густой, но слабой и неясной, при слабом увеличении незаметной пунктировкой. Последний тергит 8 кзади слегка расширен, с сильными, выступающими наружу треугольными бугорками, расположенными на задних наружных углах, задний край прямой; у g последний тергит простой, суженный кзади, без бугорков. Пигидий $ с вертикальным задним краем, постепенно суживающийся книзу, со сферическим вздутием посредине, у g выступающий, довольно узкий, слегка суживающийся к вершине. Клещи 8 очень сильно волнисто-изогнутые, слегка винтообразные у самого основания, снаружи с большим остроконическим выступом, а сверху перед задним краем последнего тергита с резко срезанным основанием, в остальной части без следов
Фиг. 56. Anechura (Burriola) seme- Зубцов ИЛИ бугорков, Вершинная ПОЛОВИНа novi B.-Bienko $ X 5 (тип). Ориг. с короткими, довольно густыми волосками;
клещи 2 простые в основной половине на внутренней стороне с неправильными, довольно крупными зубчиками. Генитальный аппарат <5 со слегка дуговидными наружными сторонами метапарамер, virga в V/2 раза длиннее последних, соединяется с почковидным пузырьком с помощью широкой дуги. Длина тела 8 11.7—12, 2 Ю.5, клещей 8 5.3—5.9, 2 4.2 мм.
Распространение: провинция Сычуань в северо-зап. Китае и восточный Тибет.
Этот своеобразный вид по характеру клещей 8 может быт отнесен к Anechura s. str., но по своим сильно укороченным надкрыльям значительно отличается от представителей указанного подрода. При этом среди
4. ANECHURA
181
Фиг. 57. Anechura (Burriola) tibetana Sem. et B.-Bienko, <5 x9 (тип). Клещи. По Semenov et Bey-Bienko, 1935.
всех известных представителей подрода Anechura нет таких видов, с которыми можно было бы сблизить данный вид, тогда как среди видов подрода Burriola имеется один вид — А. (В.) tibetana (см. ниже), который несмотря на значительные отличия в строении клещей $ должен рассматриваться как очень близкий к А. (В.) semenovi вид; это сходство настолько велико, что самки данных видов морфологически не отличимы и могут быть распознаны только по окраске — впрочем и это отличие при дальнейшем изучении большого материала может оказаться фиктивным. А так как А. (В.) tibetana с другой стороны характеризуется большим сходством в отношении строения клещей $ с А. (В.) przevalskii, то таким образом отнесение А. (В.) semenovi к подроду Anechura нарушило бы целостность группы генетически близких видов. Поэтому в качестве характерного признака подрода Burriola взято не строение клещей, как в случае с подродами Anechura и Odontopsalis, а степень развития надкрылий, причем, так как А. (В.) semenovi является по совокупности своих признаков промежуточным между Anechura и Burriola видом, хотя и более близким к последнему подроду, пришлось прежний род Burriola рассматривать в качестве не более как подрода Anechura.
11. Anechura (Burriola) tibetana Sem. et B.-Bienko (фиг. 57).
Semenov et Bey-Bienko, 1935: 225, figs. 3—4 (3. P* Серг-чу, бассейн Хуан-хэ, внутр. Тибет) [Burriola)-
Очень похож на А. (В.) semenovi по размерам тела, окраске и степени развития надкрылий и отличается только по буровато-коричневой, не черной, голове и по строению конца брюшка и клещей 3. Последний тергит 3 слегка расширен кзади, как и у А. (В.) semenovi, но без сильных выступающих наружу треугольных бугорков на задних наружных углах, у заднего края перед основанием клещей несколько утолщен и вздут, задний край посредине между этими вздутиями несколько приподнят и утолщен. Клещи 3 толстые, короткие, почти прямые, основная четверть с параллельными наружными сторонами, далее по направлению к вершине постепенно и немного сближающиеся, без зубцов, исключая самое основание сверху, которое вооружено небольшим крючковидным отростком, в нормальном положении клещей скрытым под концом последнего тергита; кончик клещей со слабо отделенным тупоугольным бугорком, направленным внутрь, у основания которого перед самым концом клещей имеется группа густых коротких
182
4. FORFICULIDAE
волосков; вершинная половина клещей как и у предыдущего вида с короткими волосками снаружи. Генитальный аппарат почти не отличим от такового у Л. (В.) semenovi. Длина тела 3 12.3—13.1, Q 11.2—13.5, клещей 3 5.7—6.8,
2 5 мм.
Распространение: известен пока только из внутреннего Тибета (бассейн реки Хаун-хе).
По внешнему виду, особенно по строению клещей 3, А (В.) tibetana очень напоминает А. (В.) przevaiskii, отличаясь от последнего, кроме окраски ясно развитых швов на голове и больших размеров тела, также и строением пигидия 3, который у первого такого же типа, как у А. (В.) semenovi, а у второго несет
снизу по бокам сильные отростки
12. Anechura (Burriola) przevaiskii (Sem.) (фиг. 60).
Semenov, 1908:160 (3, горы Бурхан-будда, северо - восточн. Тибет; р. Нурынджанба, Амдо, сев.-вост. Тибет) [Bur-riola]; Burr 1911:70, tab. VI, fig. 19.
Тело довольно маленькое, смолянорыжее или рыжевато-коричневое, голова часто черная, переднеспинка золотистожелтая, надкрылья та-
кие же, но грязноватые. Голова с очень неясными исчезающими швами. Перед-
Фиг. 58—60. Генитальный аппарат 3 (при одном увеличении). 58 — Anechura (in sp.) bipunciata-, 59 — A. (Burriola) semenovi
(B.-Bienko); 60 — A. (B.) przevaiskii (Sem.). Ориг.
неспинка поперечная, немного менее, чем в 1% раза шире своей длины,
с прямыми параллельными боковыми сторонами и широко-закругленным слабо выступающим задним краем. Надкрылья очень сильно укороченные, наружный край заметно, видимая сверху часть не менее чем в два раза, короче переднеспинки, не прикрывающие заднеспинку, большая часть которой, включая задний дуговидно вырезанный край, совершенно свободная. Брюшко сверху с неясной, исчезающей пунктировкой и с немногими крупными точками вдоль задних краев сегментов, у 3 слегка расширено около средины, а далее чуть сужено к последнему тергиту, который имеет почти параллельные боковые стороны и устроен так же, как у A. (B.)tibe-
4. ANECHURA
183
tana. Пигидий 5 с вертикальным, посредине несколько бугорковидно вздутым задним краем, нижние боковые углы которого вооружены длинным, поперечно направленным тонким отростком; пигидий g довольно узкий, выступающий. Клещи 8 без зубцов, у f. macrolabia перед самым основанием параллельные, далее к вершине постепенно сближающиеся, со слегка S-образно изогнутым наружным краем, вершинная половина при рассматривании в профиль немного приподнимается кверху; у f. brachylabia —- прямые, слегка более короткие; клещи g довольно тонкие, при основании слабо расширенные, в вершинной половине утонченные, с острым концом, внутренние стороны слегка зазубрены. Генитальный аппарат 8 с более длинной virga, которая в два раза длиннее метапарамер и соединяется с почковидным пузырьком под ясным углом. Длина тела 8 9—10.8, g 8.7—9.8; клещей 5 4—5.4, g 3.8—4 мм.
Распространение: восточный Тибет от хребта Бурхан-будда на севере до области Кам на юге.
Этот вид внешне представляет собой уменьшенную копию А. (В.) tibetana, отличаясь признаками, указанными в определительной таблице и при описании систематического положения упомянутого вида,
13. Anechura (Burriola) redux (Sem.).
Semenov-Tian-Shanskij, 1908:160, | 162 (<5, g; сев. склоны гор Сининг, пров. Ганьсу сев.-зап. Китай) [Burriola]; Burr, 1911 : 70, tab. VI, fig. 18.
Очень похож по окраске и размерам тела на А. (В.) przewalskii, отличаясь следующими признаками. Тело чуть меньших размеров, глаза чуть менее выступают, брюшко сверху без заметной пунктировки. Последний тергит 8 с параллельными боковыми сторонами и с менее скошенным по бокам, почти прямым задним краем, без ясного вдавления перед серединой заднего края, но с сильной продольной бороздкой посредине, боковые вздутия у заднего края более широко расставлены, помещаясь почти на боковых углах. Пигидий 8 сзади посредине без вздутия, с ясным килем, снизу по обеим сторонам с очень слабым зубчиком. Клещи более стройные, у 8 слегка дуговидно изогнутые от основания к вершине, перед срединой на внутренней стороне снизу со слабым зубчиком; у g более короткие, чем у А. (В.) przevaiskii: Длина тела 8 g 8.5—9.5; клещей 8 4—4.2, -g 3.5—З.б мм.
Распространение: от северных склонов Сининских гор в пров. Ганьсу до озера Куку-нор (Semenov et Bey-Bienko 1935).
Этот вид по структуре клещей 8 сильно отличается от всех остальных видов и представляет собой особую специализованную ветвь подрода Burriola, обнаруживая в то же время некоторое сходство с А. (В.) przevaiskii; самки обоих видов чрезвычайно похожи друг на друга и с трудом отличаются только по длине клещей и по степени выпуклости глаз.
184 4. FORFICULIDAE
4. Подрод Borelliola Sem.
Semenov-Tian-Shanskij, 1908 :160 — Burriolla Burr, 1911: 70 (partim).
Тип подрода — Borelliola euxina Sem.
Внешне очень сходен с представителями подрода Burriola, отличаясь утолщенными, более короткими и несколько вздутыми в основной, половине передними бедрами, к основанию резко, к вершине постепенно суживающимися и не превышающими длину переднеспинки; среднегрудная пластинка не поперечная, одинаковой длины и ширины; надкрылья также сильно укороченные, у всех известных видов не закрывают заднеспинку;. пигидий $ слегка или довольно значительно выступающий; клещи голые..
Подрод Borelliola был установлен в роде Burriola Семеновым-Тян-Шанским для одного вида, описанного им прежде в качестве представителя рода Chelidurella. В настоящее время этот подрод заключает в себе три. вида, из которых один известен с Кавказа, а два других с Балкан. Морфома-тически он очень сходен с подродом Burriola и может рассматриваться как. ближайший родич последнего; современный ареал данных подродов, охватывающий с одной стороны Тибет с сопредельным Китаем, а с другой — Кавказ и Балканы, есть, повидимому, остаток некогда более обширного ареала, объединявшего указанные территории; современное распространение подродов есть пример дизъюнкции, связанной с вымиранием видов в промежутке ареала. Впрочем можно рассматривать эти два подрода, и как параллельные ветви в роде Anechura, имевшие генетически весьма близких друг другу предков и развивающиеся благодаря поднятию исходных длиннокрылых форм во время третичных горообразовательных процессов в направлении брахиптеризма.
* 14. Anechura (Borelliola) euxina (Sem.) (фиг. 11, 12).
Semenov-Tian-Shanskij in: Adelufig, Ежег. Зоол. Муз. Акад. Наук XII, 1907: 1922 (Sr р. Ацгара, Абхазия) [Chelidurella]; Semenov, 1908:163; Burriola (Borelliola); Burr, 1911 : 70 (Burriola).
Тело средних размеров, коричнево-каштановое или иногда буроватокоричневое. Усики 13-члениковые, рыжевато- или почти соломенно-желтые, с довольно короткими члениками, 5-й членик в два, или чуть больше,, раза длиннее своей ширины. Голова кирпично-красная или рыжеватая, швы тонкие, но хорошо выраженные. Переднеспинка умеренно поперечная, с прямыми и параллельными боковыми сторонами и широко округленным, слабо выступающим задним краем, у более темных форм с желтоватыми боковыми сторонами. Надкрылья очень сильно укороченные, немного короче переднеспинки, далеко не закрывающие заднеспинку, с прямым поперечным задним краем. Ноги рыжевато- или почти соломенно-желтые. Брюшко без ясной густой пунктировки, у 3 от средины до заднего конца почти с параллельными сторонами. Последний тергит 3 сильно вариирует; у f. macrolabia заметно расширенный от основания к вер
4. ANECHURA
185
шине, задние наружные углы сильно выступают наружу в виде тупых бугорков, сверху без выступов; у f. brachylabia боковые стороны тергита параллельные, без выступающих наружу бугорков на задних углах, сверху в задней половине по бокам перед основанием клещей с сильно выступающей вверх треугольной, крупной и косо расположенной пластинкой, задний край которой постепенно опускается к средине заднего края тергита, так что при рассматривании сзади эти пластинки имеют вид расположенных под тупым углом друг к другу выступов; у g последний тергит простой, сверху в средней части плоский, со слабой, короткой продольной бороздкой. Пигидий 8 у f. macrolabia более широкий, умеренно выступающий, у f. brachylabia более узкий, сверху не видный; верхняя его сторона сильно наклонена книзу, у f. brachylabia почти вертикальная, образует с задней (у f. brachylabia нижней) стороной ясный угол, обозначенный поперечным килеобразным выступом, со слабым зубчиком на боках, так что весь пигидий имеет вид очень широкого клина; пигидий £ очень короткий, не выступающий. Клещи 8 у f. macrolabia длинные, тонкие, цилиндрические, к вершине постепенно суживающиеся, в профиль ясно, но умеренно волнисто-изогнутые, при рассматривании сверху в основной трети немного расходящиеся, а далее слегка округло загнутые внутрь и отсюда умеренно сближающиеся по направлению к вершине; основание сверху со слабой бугоркообразной выпуклостью, внутренняя сторона в срединной трети с ясно выступающей килевидной складкой, несколько более выступающей перед началом вершинной горизонтальной части клещей (рудимент внутреннего зубца); у f. brachylabia клещи короткие, очень толстые, едва длиннее задних бедер, в основной половине сильно уплощены, расширены и с параллельными прямыми наружными сторонами, внутренняя сторона немного отступя от основания с сильной приподнятой кверху продольной складкой, постепенно снижающейся к концу расширенной части клещей у средины последних, вершинная часть, составляющая примерно 2/5 длины клещей, умеренно дугообразно загнута внутрь, с уплощенной внутренней стороной, концы клещей не соприкасающиеся, умеренно расставлены. Клещи 2 сильно расширены и уплощены, удлиненно-треугольные, их длина не более чем в 21/2 раза превосходит ширину у самого основания, внутренняя и наружная стороны прямые, исключая загнутой и утонченной вершины, внутренняя сторона почти не зазубрена. Генитальный аппарат 8 со слабо сближающимися, тонкими, длинными, одинаковой ширины на всей их длине метапарамерами (очень напоминающие таковые у Mesasiobia hemixanthocara и являющиеся как бы переходными по типу к роду Forficula); virga очень короткая^ немного короче метапарамер и едва в два раза длиннее почковидного пузырька. Длина тела 8 9—11.7, 2 8.8—9.5; клещей 8 3.2—6, 2 2.8—мм.
Распространение: ареал этого вида охватывает две разъединенные друг от друга территории —одна из них, очень незначительных размеров, находится в центре Ставропольского плато (окр. V. Вороши-
186
4. FORFICULIDAE
ловска на К., колл. ЗИН!, С.-Т.-Ш! и автора!), другая занимает северо-западн. Кавказ от Майкопа, Лабинской и Кисловодска (Щербаков, 1911) на северных склонах Кавказского хребта через всю лесную до субальпийской зоны (долина Загдан в верховьях Б. Лабы, колл. С.-Т.-Ш), и юго-западные склоны до побережья Черного моря; распространение здесь еще слабо изучено, так как вид пока известен только из Гагр (Альпийская гостиница, колл. С.-Т.-Ш!), Нового Афона (колл. С.-Т.-Ш!) и района г. Сухума (речка Ацгара).
Этот вид является типичным обитателем лесной зоны, встречающимся под корой деревьев, под камнями и т. п. местах; он может рассматриваться как Понтический, т. е плиоценовый реликт. Тесная связь данного вида с лесной растительностью дает ключ к пониманию характерного ареала вида, разъединенного на две не однородные по размерам части. Вероятно в условиях Ставропольского плато этот вид является реликтом, занесенным сюда с главного Кавказского хребта подобно Forficula kaznakovi (см. выше) во время ледникового периода, способствовавшего понижению зоны лесной растительности и расширению ареала вида на север; в наступивший ксеротермический период леса отступили в горы и, сохранившись на севере только в условиях Ставропольского поднятия, дали здесь приют этому виду, существующему и до настоящего времени.
Диморфизм самцов в отношении строения клещей (ff. macrolabia и brachylabia), коррелятивно связанный с строением последнего тергита брюшка и отчасти пигидия, выражается у этого вида настолько резко и необычно, что обе формы могут быть при беглом знакомстве отнесены не только к различным видам, но даже к различным родам, что уже вводило некоторых авторов в заблуждение (ср. напр. Щербаков, 1911, который упоминает о новом роде с Сев. Кавказа, намеченном к описанию, но так и оставшимся неописанным).
5. Род CHELIDURA LATR.
Latreille, Fam. Regne Anim. 1825 : 410. — Chelidoura Burmeister, Handb. Entom., II, 1838 : 755; Bormans, 1900:105 (partim); Verhoeff, 1902:187; Kirby, 1904 : 42; Якобсон, 1905 : 25 (partim); Burr, 1911: 69.
Тип рода — Chelidura aptera Charp.
Тело очень коренастое. Усики 10—12-члениковые, с овальным 3-м и 4-м члениками. Переднеспинка поперечная. Надкрылья очень сильно укороченные, значительно короче переднеспинки, поперечные, открывающие спереди крупный щиток и едва соприкасающиеся своими внутренними сторонами. Все грудные пластинки сильно поперечные. Брюшко сильно расширяющееся от основания к вершине, достигая наибольшей ширины на самой вершине; последний тергит очень широкий, во много раз превосходящий свою длину. Пигидий не выступает, у S с вертикальным задним краем. Клещи сильно расставленные при основании, дуговидно изогнутые, но не волнистые, без сильных зубцов.
Два вида из Зап. Европы, из которых один доходит на восток до Альп.
5. CHELIDURA, 6. EUMEGALURA
187
1. Chelidura aptera (Charp.)
Charpantier, Horae Ent., 1825 : 69 (Forficula)-, Dohrn, 1867 : 342; Kirby, 1904: 42; Якобсон, 1905: 26; Burr, 1911:69, tab. VI, figs. 9, 9a.
Тело каштаново-бурое. Усики 13-члениковые. Голова рыжая. Переднеспинка с прямым задним краем, грязновато-бурая, с светлыми боками. Ноги буровато-желтые. Брюшко сильно пунктировано, последний тергит 5 у заднего края над основанием клещей с округленными сферическими выпуклостями. Клещи <5 без бугорковидного выступа на внутренней стороне, с малозаметным бугорком сверху у основания; клещи f. macrolabia длинные, слабо изогнутые, почти прямые, f. brachylabia короткие, изогнутые кольцеобразно; клещи £ простые, соприкасающиеся, почти прямые и лишь на конце слегка изогнутые. Длина тела <5д 10—14, клещей $ 5—10, 2 3—3.5 мм.
Распространение: от Пиренеев через южную Францию до южных Альп. Встречается под камнями и под корой в лиственных лесах.
Недостаточно описан и вероятно относится к другому роду: Chelidura thoracica Fisch.-W. (Orthopt. Imp. Ross., 1846:50), характеризующийся матово-черным цветом тела, с желтыми щупальцами, боковыми краями надкрылий и ногами; переднеспинка желто-бурая, надкрылья отсутствуют. Брюшко вальковатое, очень расширенное кзади, черное. Клещи короткие. Длина тела 9, клещей 2.25 мм (пол ?). — Финляндия. Очень возможно, что этот вид относится к роду Anisolabis или его ближайшим рОдичам, за что говорит окраска тела и характер брюшка. Однако, точное его выяснение абсолютно невозможно, так как автор даже не указывает пол экземпляра, по которому было сделано описание. 6
6. Род EUMEGALURA B.-BIENKO
Bey-Bienko, 1934 : 412.
Тип рода — Eumegalura mandarina B.-Bienko.
Очень близок к Anechura, особенно к подроду Burriola, и характеризуется следующими особенностями. Переднеспинка поперечная, со слабо округленным задним краем. Надкрылья сильно укороченные, с сильно скошенно-обрезанным задним краем, но полностью закрывающие задне-спинку. Среднегрудная пластинка широкая, примерно в 11/2 раза шире своей длины. Ноги относительно длинные. Пигидий S чрезвычайно длинный, клиновидный, сильно выступающий между клещами, равен по длине средним бедрам, очень широкий у основания и сильно суживающийся к вершине, самый конец слегка расширен и притуплен. Клещи S длинные, тонкие, в профиль немного волнисто изогнутые, при основании на внутренней стороне сверху с широким, прямоугольным выступом, не образующим,
188
4. FORFICULIDAE
однако, отдельного зубца, в остальной части без признаков зубцов или бугорков; вершинная часть снаружи с короткими, густыми волосками.
Этот весьма своеобразный род, несмотря на очень необычную структуру пигидия <5 несомненно является ближайшим родичем рода Anechura, особенно подрода Burriola, с которым он сходен в отношении:
ряда существенных признаков; сюда относятся укороченные надкрылья,, характер клещей, наличие на последних густых, коротких волосков в вер-
Фиг. 61. Eumegalura manda-rina B.-Bienko, $ X 6 (тип). По Bey-Bienko, 1934.
шинной половине снаружи и довольно длинные ноги. Известен всего один вид.
1. Eumegalura mandarina B.-Bienko (фиг. 61).
Bey-Bienko, 1934:413, fig. 7(5; перевал Сумпан,. пров. Сычуань, сев.-зап. Китай).
Тело темно-красновато-бурое, почти черное. Усики 12-члениковые. Голова с ясными швами, почти черная. Переднеспинка в 11/2 раза шире своей длины, со слегка округленными боковыми и задней сторонами. Надкрылья с очень редкой пунктировкой, одинаковой длины с пе-реднеспинкой. Крылья отсутствуют. Ноги темнобурые. Брюшко заметно уплощенное, широкое, слегка расширенное к 6-му сегменту, в очень тонкой и довольно густой пунктировке. Последний тергит 5 очень короткий и широкий, примерно в 4 раза шире своей длины, с двумя маленькими тупыми бугорками с каждой стороны над основанием клещей; срединная часть со слабым вдавлением перед задним краем и с короткой продольной, бороздкой на дне этого вда-вления. Клещи S в три раза длиннее пигидия,, постепенно суживающиеся к вершине, при рассматривании сверху дугообразно изогнутые
в основных трех четвертях, вершинная четверть, за исключением слегка, загнутого внутрь конца, прямая. Длина тела 5 (без пигидия) 9.5, клещей 6.9, пигидий 2.4 мм. Самка неизвестна.
Распространение: известен только из пров. Сычуань, где был обнаружен на большой высоте, примерно в 3000 м.
Интересно отметить, что по характеру окраски и некоторым деталям строения (напр. степень развития швов, конфигурация и степень развития надкрылий) этот вид обнаруживает особенное сходство с Anechura (Burriola) semenovi и А. (В.) tibetana; возможно он представляет собой крайне своеобразный дериват той сходной группы, из которой развились с одной стороны подрод Burriola, а с другой род Eumegalura’, однако, так как
7. APTERYGIDA
189
последний характеризуется совершенно необычным пигидием 8, резко отличающимся от такового у всех остальных родов семейства, рассматривать этот род в качестве подрода Anechura было бы нецелесообразно.
7. Род APTERYGIDA WESTW.
Westwood, Introd. Classif. Ins. II, 1840 : 44; Bormans, 1900: 109 (partim); Kirby, 1904: 44 (partim); Якобсон 1905:26 (partim); Burr, 1911:80.
Тип рода — A. media Hagenb.
Тело стройное. Усики с довольно короткими члениками, 4-й членик почти конический, едва в два раза, 5-й примерно в 21/2 раза длиннее своей ширины, цилиндрический. Надкрылья вполне развитые, крыловые пластинки отсутствуют. Среднегрудная пластинка длиннее своей ширины. Брюшко слабо уплощенное, на поперечном разрезе коротко-овальное, боковые стороны 8 почти параллельные; последний тергит 3 простой, у заднего края с исчезающими срединными бугорками. Пигидий S сильно выступающий, не поперечный, довольно узкий. Клещи 3 тонкие, слабо изогнутые, при основании широко расставлены и не уплощенные, в профиль прямые, на внутренней стороне с одним или двумя зубцами. Генитальный аппарат 3 такого же типа, как в роде Forficula.
' Род Apterygida до недавнего времени понимался крайне широко и был сборным, включая в себя представителей не только различных родов, но и семейств — достаточно указать, что Prolabia arachidis (см. выше), из сем. Labiidae относилась Bormans’oM к данному роду. В современном понимании этот род включает только три вида, из которых один свойственен Европе, а другие — южной и центральной Африке; остальные виды, относимые некоторыми современными авторами к этому роду, в действительности не могут быть отнесены сюда (напр. весьма своеобразные виды е широким пигидием, установленные Shiraki с о-ва Формозы), так как при включении этих видов в данный род получается совершенно разнородная группа видов, генетически не связанных друг с другом. Что касается африканских видов, то помимо очень большого их морфологического сходства с типом рода, распространяющегося и на половой аппарат самца, имеются и биогеографические показатели, позволяющие трактовать африканских представителей в качестве действительно генетически близких к типу рода видов, несмотря на кажущуюся неестественность ареала данного рода. В качестве аналога распространения рода Apterygida можно указать на такой вполне естественный род, как Ectobius Steph, (из отр. Blattodea), имеющий в принципе то же самое распространение, как и род Apterygida в нашей его трактовке; действительно, виды рода Ectobius в большом числе известны из Европы (с немногими видами, заходящими в пределы западной Азии), но, отсутствуя подобно роду Apterygida в пустынях Сев. Африки, в обилии появляются в непалеарктической ее части. Разрыв ареала в обоих случаях может быть объяснен тем, что как Apterygida,
190
4. FORFICULIDAE
так и род Ectobius, являются тесно связанными с лесной растительностью мезофилами, в силу чего в условиях пустынь Сев. Африки не встречают подходящей для них экологической обстановки. Единственный палеарктический вид характеризуется следующими особенностями.
* 1. Apterygida media (Hagenb.) (фиг. 62).
Hagenbach, Symb. Faunae. Insect. Helvet., 1822 :16, figs. 6—7 (Швейцария) [Forficalafa Bormans, 1900 :117; Якобсон, 1905 : 26, табл. III.—albipennis Fischer, 1853 : 77, tab. VI, figs. 14,
Фиг. 62. Apterygida media Hagenb. 3 X 9 (Польша). Ориг.
14 ab; Brunner, 1882 : 21 (Chelidurd)-, Kirby, 1904: 44; Burr, 1911:80, tab. VII, fig. 13; Burr, 1916:14, tab. Ill, fig. 1.
Тело грязно-желтое, в довольно длинных волосках. Усики 12-члениковые, грязновато-желтые. Голова рыжеватая, с ясными сутурами. Переднеспинка чуть уже головы, почти квадратная, боковые стороны прямые, параллельные, задний край не выступающий, слабо округлен; диск грязновато-желтый, с просвечивающими, почти бесцветными боковыми сторонами. Надкрылья немного более чем в l1^ раза длиннее переднеспинки, желтоватые, полупросвечивающие, задний край поперечно обрублен. Крылья имеются, но сильно укорочены, скрыты под надкрыльями и просвечивающие через них. Ноги грязновато-желтые. Брюшко рыжевато-желтое, сильно блестящее, в мельчайшей густой пунктировке, бугорки на 3-м и 4-м тергите, а иногда и пространство около них, черные; последний тергит 3 прямоугольный, довольно удлиненный, примерно в два раза шире своей длины, задний край посредине едва выемчатый; у 2 — суживающийся кзади, с прямым задним краем. Клещи волосатые, у 3 на внутренней стороне близ основания и около сре-
дины с маленькими зубцами; клещи о простые, соприкасающиеся. Генитальный аппарат <5 с прямыми метапарамерами, наружные стороны их параллельные; virga почти такой же длины, как метапарамеры, соединяется с передним концом почковидного пузырька без
всякого угла, с маленькой хитиновой пластинкой в передней половине. Длина тела 3 6—10, £ 7—9, клещей 3 3.5—5, 2 2.2—2.5 мм.
Распространение: большая часть Европы от Пиренейского полуострова до южных Балкан (самые южные местонахождения — Корфу и Афины), на север до Англии, Бельгии, сев. Франции, южн. Швеции; вся южная часть Средней Европы от Вюртенбурга, Чехо-Словакии до верх
8. CHELIDURELLA
191
него течения р. Одер в южном Бранденбурге (Ramme, 1920) и Новой Александрии в Польше (Семенов, 1903), спускаясь по восточным Карпатам на юг и захватывая лесостепь Украины от Каменец-Подольска на западе до р. Псел на востоке (Гадяч, колл. ЗИН!) и заходя далее в Бессарабию; указание для Кавказа, основанное на отожествлении Chelidura curia Fisch.-W. с A. media (Якобсон, 1905) несомненно неправильно, так как западная граница вида проходит через Балканский полуостров, Бессарабию и лесостепь Советской Украины; очень вероятно, что Ch. curia относится к F. kaznakovi, так как последняя напоминает A. media отсутствием крыловых пластинок, окраской, размерами и волосатым телом (см. F. kaznakovi). Встречается под корой деревьев и на цветах в лесных местностях.
8. Род CHELIDURELLA VERH.
Verhoeff, 1902 :187; Kirby, 1904 : 43; Burr, 1911: 75. — Chelidura auct., partim.
Тип рода — Chelidurella acanthopygia Gene.
Тело довольно коренастое, значительно расширенное за срединой. 4-й членик усиков почти овальный значительно короче 3-го, 5-й членик едва равен 3-му. Переднеспинка поперечная, почти прямоугольная, задний край прямой. Надкрылья сильно укороченные, открывающие щиток, не закрывают полностью заднеспинку, задний дугообразно вырезанный край которой совершенно свободен, крылья отсутствуют. Среднегрудная пластинка одинаковой длины, и ширины. Брюшко у <5 около средины значительно расширенное, сильно суживающееся кпереди и особенно кзади. Последний тергит $ у заднего края с двумя широко расставленными бугорками, расположенными над основанием клещей, у g бугорки неясные, почти или совсем отсутствуют. Пигидий <5 по крайней мере у основания широкий. Клещи $ тонкие, в профиль прямые, при основании широко расставленные, довольно сильно дуговидно загнутые к вершине и здесь почти или полностью соприкасаются; внутренняя сторона совершенно без зубцов или у южно-африканской Ch. purcelli Burr при основании с довольно сильным зубцом.
Известны 4 вида, из которых три свойственны Средней Европе, а четвертый — южной Африке.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Мельче (длина тела 6.5—11 мм). Пигидий 3 сильно торчит в виде длинного, загнутого вверх и здесь почти вертикального рога; у g при рассматривании снизу немного суживающийся к вершине, длиннее своей ширины. Последний тергит брюшка 3 с двумя сильными бугорками у заднего края.......................* 1. Ch. acanthopygia Gene.
192
4. FORFICULIDAE
2 (1). Крупнее (длина тела 8—15.5 мм). Пигидий $ короткий, едва выступающий, задняя сторона отвесная, с бугорком; у g при рассматривании снизу не суживающийся к вершине. Последний тергит брюшка 5 со слабо приподнятыми, плохо заметными бугорками у заднего края.
3 (4). Тело черновато-бурое. Пигидий <5 снизу с крупным выступающим бугорком, у 2 немного длиннее своей ширины, задний край слегка вырезан, с оттянутыми в виде слабых зубцов задними боковыми углами. Клещи <3 постепенно дуговидно загнутые............................
..............................................2. Ch. mutica Krauss.
4 (3). Тело светлокоричневое. Пигидий <3 посредине со слабым, совсем плоским бугорком, у g почти в два раза длиннее своей ширины, с прямым обрубленным задним краем. Клещи 5 посредине изогнуты под слабым, но ясным углом (фиг. 63)......................................
......................................* 3. Ch. transsilvanica Ebner.
* 1. Chelidurella acanthopygia (Gene).
Gene, 1832:228 (южная Европа) \Forficula\-, Fischer, 1853: 83, tab. VI, figs. 20, 2va-d; Brunner, 1882: 24 (Chelidura)-, Bormans 1900 :107; Якобсон, 1905:26; Verhoeff, 1902:187; Burr, 1911: 76, tab. VIII, fig. 2.
Тело рыжевато-бурое или светлокоричневое. Усики 13-члениковые, . желтоватые или буроватые. Голова рыжеватая или буровато-рыжая. Переднеспинка с бесцветными, полупрозрачными боковыми сторонами. Надкрылья сильно поперечные, наружная сторона их более широкая чем внутренняя, сросшаяся со среднеспинкой, ограничивают спереди крупный, сильно поперечный щиток. Ноги желтоватые. Брюшко заметно блестящее, очень мелко, густо и неглубоко пунктированное, задние концы сегментов с немногими более крупными и грубыми точками рыжевато-бурое, по бокам при основании и боковые бугорки грязновато или смоляно-бурые; последний тергит $ очень короткий, сильно поперечный, более крупно пунктированный, задний край между бугорками прямой, а кнаружи от них скошен назад, соединяясь с боковыми сторонами под очень тупым углом или дугообразно; бугорки довольно сильно приподнятые, в профиль хорошо заметные; последний тергит 2 немного более длинный, с непараллельными, несколько сходящимися кзади боковыми сторонами, сильно наклонен кзади, без ясных бугорков. Пигидий S при основании широкий, по направлению к вершине сильно суживающийся, сама вершинная часть удлинена и сильно выступает в виде торчащего и сильно загнутого вверх рога, заметного даже при рассматривании в профиль; пигидий 2 ПРИ рассматривании сверху не выступает, почти вертикальный, суживающийся книзу, при рассматривании снизу слабо выступающий, длиннее своей ширины, несколько суживающийся к вершине и здесь обрублен. Клещи 3 постепенно дуговидно изогнутые к вершине, у основания сверху с плохо заметной бугоркообразной приподнятостью, у 2 простые, при основании широкие, чуть расставленные,
8. CHELIDURELLA
193
к вершине постепенно суживающиеся, на внутренней стороне не зазубренные.
Длина тела 6.5—10, 2 7—99; клещей 8 3.5—5.5, g 2—23 мм.
Распространение: большая часть Зап. Европы, от Пиренеев, средн, и сев. Франции, Бельгии, Голландии и Англии через всю Германию и Чехо-Словакию до границы вост. Пруссии с Литвой (район Кенигсберга и Олецко), и Польши вплоть до Новой Александрии (колл. ЗИН!), Бело-вежья (Roznowska, 1934), Зап. Украины и Семиградья в Румынии; с юга охватывает сев. Балканы и сев. Италию (Пьемонт), а на севере доходит до Дании, Швеции и Латвии; восточная граница вида плохо изучена; очень возможно, что этот вид будет обнаружен в Белоруссии. Для западной части советской Украины имеется указание В. М. Дирша (in litt.), но без более точного обозначения местонахождения; желательно подтверждение этого указания.
Весной встречается под камнями, опавшей листвой и пр., а летом на деревьях, особенно хвойных; поднимается высоко в горы.
2. Chelidurella mutica (Krauss).
Krauss, Verb, zool.-bot. Ges. Wien, XXXVI, 1886 :138, tab. figs. 2,2 ABC (3, Q; Монте Вальдо и долина Лагарина в Тироле) \Chelidura\‘, Bormans, 1900: 107; Якобсон, 1905:26; Verhoeff, 1902:187; Burr, 1911:76.
Очень сходен с Ch. acanthopygia, но отличается немного большими размерами, более темной и одноцветной (черновато-бурой) окраской тела, за исключением головы, клещей и ног, которые, как и у предыдущего вида, желтоватые или рыжеватые; бугорки на последнем тергите брюшка 8 слабые, слегка приподнятые, незаметные в профиль; пигидий 8 короткий с совершенно вертикальным задним краем, снизу с довольно крупным, выступающим и заметным при рассмотрении сверху или снизу бугорком; пигидий у д при рассматривании снизу более широкий, к вершине не суживающийся, задний край слегка дугообразно вырезан и вооружен по бокам слабыми зубчиками, по одному с каждой стороны. Клещи 8 изогнуты более сильно, ограничивая своими внутренними сторонами слабо удлиненное, широко овальное, а иногда почти круглое пространство; основание клещей сверху без бугорка. Длина тела 8 8—11.5, 2 8.5—11; клещей 8 3—4.5, 2 2.2—2. 5 мм.
Распространение: известен только из Тироля.,
* 3. Chelidurella transsilvanica Ebner (фиг. 63).
Ebner, Wien, Ent. Zeitung, XLIX, 1932:16, figs. (8, Гетценберг близ Германн-штадта и Резинар, Семиградье, Сев. Румыния).
Очень близок к Ch. mutica, отличаясь окраской тела, строением пигидия 8 и 2> формой клещей 8 и немного большими размерами. Тело светлокоричневое, сходное по окраске с Ch. acanthopygia, основание брюшка по бокам и боковые бугорки зачернены. Пигидий 8 как у Ch. та-
Фауна СССР, Кожистокрылые 13
194
4. FORFICULIDAE
tica, но бугорок очень слабый, совсем плоский и расположен посредине задней вертикальной стороны; пигидий у 2 ПРИ рассматривании снизу с параллельными боковыми сторонами, заметно длиннее своей ширины,, с прямым, обрубленным задним краем и без боковых зубцов. Клещи $
Фиг. 63 Chelidurella transsilvanica Ebner.
<5x5.5. (Каменец-Подольск). Ориг.
у основания сверху без бугорка как и у Ch. mutica, но около средины изогнуты под слабым, но ясным углом. В остальном как Ch. mutica. Длина тела 8 10—13, 2 И-5—15.5, клещей <5 4—4.5, 2 4—4.6 мм.
Распростран ение: известен только из Трансильванских Альп в Румынии (близ Германнштадта) и окрестностей Каменец-Подольска (1 S,. В. Якубовский! колл. ЗИН!) в Прикарпатской части Советской Украины. Столь отрывочные данные являются следствием еще плохой изученности распространения вида, описанного к тому же только в 1932 г.; однако* нахождение его в окр. Каменец-Подольска показывает, что ареал распространения данного вида охватывает полосу от Трансильванских Альп на юго-западе, по крайней мере до восточных предгорий средней части собственно Карпатских гор на северо-востоке. Вероятнее всего этот вид является карпатским эндемиком и распространен по всей Карпатской гор-
ной системе от Трансильвании до северной оконченности собственна Карпат.
9. Род ALLODAHLIA VERH.
Verhoeff, 1902:194; Burr, 1911: 71.
Тип рода — Allodahlia scabriuscula Serv.
Усики 12—13-члениковые, 4-й и 5-й членики короче 3-го. Переднеспинка шире своей длины. Надкрылья широкие, с сильно выступающими плечами, с сильным продольным килем по всей длине наружного края; поверхность надкрылий зернистая, морщинистая или сильно пунктирована. Грудные пластинки широкие, короткие. Ноги длинные и стройные. Пигидий 8 короткий, поперечный, у обоих полов выпуклый. Клещи 5 широко
9. ALLODAHLIA
195
расставлены и не соприкасаются при основании, длинные, тонкие, различно вооружены, у j простые, прямые, соприкасающиеся.
Известно полдесятка видов, свойственных главным образом горным частям Британской Индии, но распространенных также на Индо-Малайском архипелаге, Филиппинах, в южном Тибете; один приводимый ниже вид доходит до северовосточного Китая.
1. AllodaUia scabriuscula (Serv.).
Servielle, Hist. Nat. Orthopt., 1839 :38 (<5; „Южная Америка" — несомненно по ошибке) [Lor, icula]; Bormans, 1900 :103 (Anechura); Burr, 1910a : 150, fig. 49; Burr, 1911: 71, tab. VI ,figs. 13, 13a.
Тело крупное, коренастое, тусклое, черновато-каштановое, более или менее шероховатое. Усики 12—13-члениковые, черные. Голова выпуклая, с очень ясными швами. Переднеспинка с заостренными и выступающими вперед боковыми передними углами, боковые и задняя сторона широко округлены, не образуют между собой ясных углов, передняя часть с немногими тупыми бугорками, весь диск густо пунктирован. Надкрылья сильно шероховатые, с дуговидно выступающими боковыми сторонами, несколько суженные к заднему краю,густо покрыты маленькими бугорками и отдельными разбросанными крупными вдавленными точками, задний край слегка выемчато-обрезанный. Крыловые пластинки хорошо выступают, морщинистые, шероховатые. Бедра пунктированы. Брюшко широкое, несколько расширенное около средины, вся верхняя сторона густо и тонко, нижняя еще более мелко, пунктированы. Последний тергит <5 короткий, поперечный, слегка наклонный, без бугорков в срединной части сверху, но с выступающими задними наружными углами; задний край утолщен; у £ — более узкий, задний край выпуклый, боковые углы не выступают. Пигидий о короткий, с обрубленным задним краем, очень тупой, с маленьким тупым бугорком на каждом углу, оканчивающимся коротким острым шипом; у g — пигидий в виде короткого тупого бугорка. Клещи 8 от основания сильно расходящиеся в стороны и почти сразу загнуты вниз и назад, длинные, тонкие, внутренняя сторона снизу в основной половине тонко Зазубрена, далее на внутренней стороне в конце второй трети с сильным, довольно длинным, острым зубцом; вершинная треть сильно заостряющаяся к концу, слабо изогнутая, концы перекрещиваются друг с другом; у £ почти соприкасающиеся, прямые и параллельные в двух третях их длины, а затем внезапно утонченные и очень слабо загнутые, с перекрещивающимися концами. Метапарамеры толстые, короткие,_с дугообразно-выпуклым наружным краем; virga очень длинная-, примерно в 4 раза длиннее метапарамер, соединяющаяся с почковидным пузырьком при помощи дуги, резко загнутой у своего основания под прямым углом; собственно penis с хитиновыми включениями. Длина тела 8 11—14, g И—13, клещей 8 8—15, g б—7 мм.
13*
196
4. FORFICULIDAE
Распространение: Британская Индия, включая Тенассерим и Бирму, Тонкин, Малайский архипелаг (Ява, Суматра^ Борнео), Филиппинские о-ва, о-в Формоза, южный Китай, на севере достигая пров. Сычуань и сев.-вост. Китая (Киао-Чжоу).
2. Подсем. OP1STHOCOSM1INAE
Усики с удлиненными, стройными члениками, 1-й членик длиннее половины длины головы (у рода Sondax он короче, но тогда тело совершенно бескрылое), не короче или чуть короче 2-го, 3-го и 4-го члеников взятых вместе; 4-й членик равен или едва короче 3-го. Брюшко сверху выпуклое, цилиндрическое, на поперечном разрезе круглое или почти круглое. Последний тергит 8 обычно довольно длинный, у многих родов сильно сужен к вершине, обычно только немного шире своей длинны. Клещи у большинства родов тонкие, у <3 сверху часто с вертикальными выступами или зубцами, у g длинные и очень тонкие, одинаковой толщины на всем протяжении или немного утонченные к вершине, почти нитевидные, особенно в вершинной половине. Генитальный аппарат 8 с толстыми, снаружи дуговидно выступающими метапарамерами; virga длинная, часто выступает за передний конец собственно penis’a, последний часто снабжен хитиновыми включениями.
Как уже упоминалось в описании семейства Forficulidae, данное подсемейство не может быть охарактеризовано только одним каким-либо признаком. Так весьма характерные усики с удлиненными члениками свойственны большинству, но не всем родам; точно так же и характер последнего тергита 8 у немногих родов почти такого же типа, как у Forficulinae. Поэтому указанные признаки, помогая в определении подсемейства, не могут, однако, служить абсолютным критерием для его характеристики. В. результате наиболее надежными признаками необходимо признать форму брюшка, не уплощённого сверху и поэтому цилиндрического, на поперечном разрезе круглого, и строение клещей самки, как описано выше. Быть может было бы правильнее оба эти подсемейства слить в одно, но от этого приходится отказаться до ревизии всего сем. Forficulidae, так как подсем. Opisthocosmiinae, будучи довольно богатым, представлено в Палеарктике только очень немногими видами.
Все подсемейство представлено более чем 20-ю родами, включающими большое количество видов, распространенных в тропиках Америки, Африки и особенно Азии и заходящих отчасти в восточную Палеарктику, но отсутствующих в Австралии.
10. Род TIMOMENUS BURR
Burr, 1907а: 96; Burr, 1911 : 93.— Opisthocosmia auct., partim.
Тип рода-— Timorr.enus oannes Burr из Ассама и Тонкина.
Тело довольно крупное и относительно коренастое. Усики 12—13-чле-никовые, 1-й членик усиков длинный, толстый, более расширенный к вершине, почти булавовидный, остальные членики значительно тоньше, цилин
10. TIMOMENUS 197
дрические, 4-й членик почти такой же длины, как 3-й. Надкрылья и крылья вполне развиты, первые довольно широкие, с сильно округленными выступающими плечами. Ноги довольно короткие, передние бедра толстые; голени сверху в вершинной части несколько уплощенные, особенно передняя и средняя пара; лапки довольно короткие, 1-й членик не длиннее или едва длиннее 3-го. Брюшко довольно широкое, слегка расширенное за срединой, а затем суженное; бока сегментов выпуклые. Последний тергит <5 поперечный, прямоугольный, не сужен кзади. Пигидий 8 короткий и выпуклый. Клещи 3 немного расставлены при основании, длинные, стройные, цилиндрические, различно вооруженные или совсем без зубцов и выступов.
Известно около 15 видов этого рода, распространенных от Уссурийского края до Британской Индии (фиг. 16). В пределы Палеарктики заходят 4 вида, из которых 3 пока известны только из сев.-вост. Китая.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ.
1 (2). Клещи 8 длинные, тонкие, без зубцов. Бока 6-го и 7-го тергита брюшка 8 сзади оттянуты в виде тупых углов, из которых первый снабжен бугорком. Тело смоляно-черное.............1. Т. inermis Вог.
2 (1). Клещи 8 вооружены зубцами.
3 (4). Клещи 8 вооружены сильным зубцом только на верхней стороне. Бугорки на боках 5—9-го тергитов брюшка отсутствуют. Тело красновато-бурое, брюшко густо, но слабо пунктировано..................
.........................................2. Т. unidentatus Вог.
4 (3). Клещи 8 кроме сильного зубца сверху вооружены также маленьким зубцом на внутренней стороне.
5 (6). Тело смоляно-черное, с желтовато-бурыми надкрыльями. Брюшко сверху морщинисто-пунктированное. Клещи 8 сверху близ основания вооружены сильным зубцом. Бока 5—6 сегментов брюшка 8 сзади с бугорками, следующие сегменты без бугорков (фиг. 64, 65) . . .
...................................... * 3. Т. komarovi Sem.
6 (5). Тело рыжевато-бурое. Брюшко сверху гладкое. Клещи 8 сверху вооружены сильным зубцом в конце основной трети. Бока 6—9-го сегментов брюшка 8 вооружены сзади загнутыми, крючковидными бугорками .
...................................... 4. Т. aesculaplus Burr.
1. Timomemis inermis Bor.
Borelli, 1915 : 5 (5; Киао-Чжоу, сев.-вост. Китай). — ? simplicis Shiraki, 1928 : 24, fig. 8.
Тело смоляно-черное. Усики с булавовидным 1-м члеником, остальные цилиндрические, 4-й чуть короче 3-го, 5-й равен 3-му; первые три членика черные, остальные смоляно-бурые. Голова со слабо заметными швами, смоляно-бурая, ротовые части смоляно-черные. Переднеспинка с светлобурыми боковыми сторонами, одинаковой ширины с головой, длина чуть превосходит ширину, боковые стороны умеренно округленные, задняя
198
4. FORFICULIDAE
сторона и боковые углы сильно округленные. Надкрылья каштановые, тусклые, в два раза длиннее переднеспинки; крыловые пластинки чуть короче переднеспинки, смоляно-черные. Ноги смоляно-черные, передние бедра короткие, утолщенные, 1-й членик лапок почти равен по длине 3-му. Брюшко смоляно-черное, морщинистое, блестящее, слегка расширяющееся от основания к 6-му сегменту, а далее по направлению к вершине умеренно суженное; 6-й и 7-й тергиты по бокам сзади оттянуты в виде тупых углов, из которых 6-й снабжен маленьким бугорком; последний тергит <3 почти квадратный, чуть шире своей длины, наклонный, по средине у заднего края с треугольным вдавлением, а по бокам вдавления перед основанием клещей с маленьким бугорком; задний край между ветвями клещей прямой, утолщенный, по бокам несколько скошен. Пигидий <3 чуть шире своей длины, слабо выступающий. Клещи слегка пунктированные, смоляно-черные, длинные, тонкие, цилиндрические, прямые в основной половине, далее слабо загнутые внутрь, без зубцов; основание ветвей умеренно расставлено. Самка неизвестна. Длина тела <5 14.5, клещей 7 мм.
Распространение: известен пока только из Киао-Чжоу в сев.-вост. Китае и о-ва Формозы (?).
Описанный с Формозы Т. simplicis Shiraki по всей вероятности представляет собой синоним данного вида, так как описания обоих авторов не отличаются друг от друга; повидимому работа Borelli осталась неизвестной Shiraki.
2. Timomenus umdentatus Bor.
Borelli, 1915 : 4 (3; Киао-Чжоу, сев.-вост. Китай).
Тело красновато-бурое. Усики с булавовидным 1-м члеником, остальные цилиндрические, 4-й членик чуть короче 3-го, 5-й одинаковой длины с 3-м; цвет красновато-бурый, с более светлым 1-м члеником. Голова красновато-бурая с темнобурой верхней губой, швы слабо заметные. Переднеспинка каштановая, боковые стороны светлобурые, чуть уже головы и немного шире своей длины, боковые стороны слегка округлены, задний край и задние углы сильно округлены. Надкрылья красновато-рыжие, матовые, более чем в два раза длиннее переднеспинки; крылья красноватобурые, равны по длине переднеспинке; ноги красновато-бурые, голени и лапки более светлые, покрыты волосками, передние бедра толстые, 1-й членик лапок равен по длине 3-му. Брюшко красноватое, задние края сегментов более темные, умеренно расширенное к 6-му сегменту, далее слегка суживающееся, блестящее и слабо, но густо пунктированное; бока 5—9-го тергитов без бугорков; последний тергит 3 почти квадратный, слегка суженный в задней части, умеренно наклонный, с треугольным вдавлением посредине у заднего края, по бокам вдавления перед основанием клещей с продольной приподнятой складкой, задний край посредине между основаниями клещей прямой, утолщенный, по бокам несколько скошен. Пигидий 8 слабо выступающий, прямоугольный, значительно шире своей
10. TIMOMENUS
199
длины, сзади выпуклый. Клещи $ длинные, тонкие, при основании умеренно расставленные, в основной четверти прямые и параллельные, на поперечном разрезе треугольные, далее сверху вооружены треугольным зубцом, сначала загибаются внутрь, а затем сначала расходясь, дуговидно изогнуты и утончаясь к вершине, перекрещиваются здесь своими концами; внутренние стороны слегка зазубрены на протяжении немного превышающем половину .длины клещей. Самка неизвестна. Длина тела 3 11, клещей 6 мм.
Фиг. 64—65. Timomenus komarovi (Sem.) : 64 —’<5 (тип X 5); 65 — (X 5) (Яковлевка, Уссур. край). Ориг.
Распространение: Киао-Чжоу в сев.-вост. Китае.
Этот вид напоминает F. komarovi, отличаясь более светлой окраской, отсутствием бугорков по бокам сегментов брюшка и отсутствием зубца на внутренней стороне клещей 3.
* 3. Timomenus komarovi (Sem.) (фиг. 64, 65, 66).
Семенов, 1901b : 982 figs. (3, долина Садегю, сев. Корея) [Opisthocosmia]; Семенов, Русск. Энт. Обозр, 1, 1901:259; Burr 1907а: 97; Burr, 1911:93.—femorata Matsumura, Thous. Ins. Japan, Add. 1, 1912:1, tab. 1, fig. 2 (Apterygida); Shiraki, 1928:22, 23.
200
4. FORFICULID АЬ
Тело длинное, стройное, смоляно-черное. Усики 12-члениковые,. к-концу заметно более бледные. Голова бурая или рыжая, слабо блестящая. Переднеспинка бурая, со светлыми, просвечивающимися боками, немного уже головы, боковые стороны слабо, задняя сторона сильно, округлены. Надкрылья широкие, кзади слегка суживающиеся, буровато-желтые или почти золотисто-желтые, гладкие, матовые. Крыловые пластинки сильно выступающие, почти равны половине длины надкрылий, одноцветные,, такого же цвета как надкрылья. Брюшко сверху слабо морщинисто-пунктировано, довольно узкое, у 5 бока 5-го и 6-го сегментов с острым бугорком у заднего края, более крупным на 6-м сегменте; брюшко Q по бокам без маленьких бугорков у заднего края; последний тергит 5 с почти параллельными сторонами, посредине и по бокам у заднего края с широким вдавлением, срединное вдавление имеет с каждой стороны по бугорку;, л задний край между клещами прямой, по
6 бокам, кнаружи от них слегка скошен;
последний тергит g к вершине суживаю-щийся, только посредине у заднего края слегка вдавлен. Клещи бурые, у $ очень Фиг. 66. Timomenus komarovi (Sem), длинные и тонкие, длиннее половины тела, Клещи $ при рассматривании в основной половине слегка расходящиеся, сбоку и сверху. Ориг. почти прямые, а далее постепенно утончающиеся, правильно дуговидно изогнутые,, с перекрещивающимися концами, близ основания сверху с сильным, направленным вверх и назад, длинным зубцом, перед самой срединой на внутренней стороне с небольшим шиповидным зубцом, внутренняя сторона клещей от этого зубца до основания с мелкими зубчиками; клещи Q простые,, тонкие, соприкасающиеся, на внутренней стороне незазубренные. Длина тела $ 12.6, £ 11.9—12.5; клещей $ 7.9, Q 6.6 мм.
Распространение: от Уссурийского края (Яковлевка; остров Петров, колл. ЗИН!) через Корею, Манчжурию (Bey-Bienko, 1930) до северо-вост. Китая (Киао-Чжоу, Borelli, 1915) и о-ва Формозы.
Этот вид обнаруживает большое сходство с Т. oannes Burr из Ассама и Тонкина, отличаясь главным образом характером клещей 3, которые у Т. oannes не имеют мелких зубчиков на внутренней стороне,, а внутренний шиповидный зубец расположен немного за срединой клещей,, ближе к их вершине чем к основанию.
4. Timomenus aesculapius (Burr).
Burr, Bol. Soc. Espan. Hist. Nat., 1905:236 (c5; Мария Басти, Бутан, сев. Индия)> [O/nsfftocosmza]; Burr, 1907а: 121 (Eparchus); Burr, 1910 :196, fig. 79; Burr, 1911: 93.
Довольно сходен с T. komarovi, но чуть стройнее, рыжевато-бурого, цвета. Усики красно-бурые, переднеспинка слегка сужена кзади, задний край округлен, более узкий чем передний. Надкрылья тусклые, менее
10. TIMOMENUS
201’.
широкие, не суженные кзади. Крыловые пластинки длинные. Брюшко гладкое, у 8 бока 6—9-го сегментов вооружены сзади маленькими, загнутыми, крючковидными бугорками. Последний тергит <5 почти квадратный,, гладкий, сильно наклоненный, слегка суживающийся кзади, задний край прямой. Клещи $ значительно длиннее половины тела, у основания несколько сближенные, но не соприкасающиеся, постепенно расходящиеся от основания и постепенно дуговидно-загнутые в вершинной половине,, сверху в конце основной трети с сильным, сжатым, треугольным вертикальным зубцом, внутренняя сторона от этого зубца до основания слегка зазубрена, перед вершиной второй трети с небольшим острым шиповидным зубцом; при рассматривании в профиль слегка волнисто загнутые в основной трети, а далее до вершины горизонтальные. Самка неизвестна. Длина тела 8 11, клещей 7 мм.
Распространение: известна из Бутана в сев. Индии и сев.-вост. Китая до Киао-Чжоу (Borelli, 1915); из промежуточных пунктов не; приводился, но вероятно широко распространен и здесь.
11. Род EPARCHUS BURR
Burr. 1907а: 120; 1910:190а; 1911: 94
Тип рода — Eparchus insignis Haan.
Усики 10—12-члениковые, стройные, 1-й членик относительно очень длинный, тонкий, почти не расширенный к вершине, остальные членики немного более тонкие, 3-й и 4-й членики примерно одинаковой длины, взятые вместе довольно значительно короче 1-го, совершенно цилиндрические. Переднесйинка почти или совсем такой же ширины как ее длина. Надкрылья гладкие, широкие, с сильно округленными выступающими плечами. Крылья вполне развитые; ноги длинные и тонкие, передние бедра чуть толще остальных, лапки длинные, 1-й членик немного длиннее 2-го и 3-го взятых вместе. Брюшко веретеновидное, особенно у 8, суженное при основании, расширенное около средины и сильно суживающееся к вершине; бока 5—9-го сегментов у 8 при рассматривании сверху не загнуты и не заострены у заднего края, но у некоторых видов с бугорками. Последний тергит длинный, суживающийся к вершине, задний край прямой или слегка скошен по бокам. Клещи 8 соприкасающиеся по крайней мере у основания или слегка расставлены, тонкие, длинные, различно воружен-ные, обычно с сильным вертикальным выступом или зубцом с верхней стороны у основания или близ него, реже без выступа (Е. inermis Heb. и Е. oberthuri Вог.), более или менее дуговидно загнутые; клещи у Q очень тонкие, прямые, на самой вершине слегка загнутые.
Этот род включает более полудесятка видов, распространенных от Брит. Индии, Индомалайского архипелага до Филиппин, Японии и сев.-вост. Китая; в пределы вост. Палеарктики заходят 3 вида.
202
4. FORFICULIDAE
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
1 (2). Клещи $ сильно дуговидно изогнуты, снабжены сверху у основания округленным, почти шаровидным, бугорком. Бока б—8-го сегментов брюшка 8 с сильными, толстыми, коническими бугорками, сильно выступающими на 6-м и 7-м сегментах...............1. Е. insignis Haan.
2 (1). Клещи 8 слабо изогнутые, почти прямые, сверху на всем протяжении без бугорков или зубцов. Бока 6—8-го сегментов брюшка без бугорков.
3 (4). Последний тергит брюшка 8 суживающийся к вершине. Клещи 8 без зубчиков на внутренней стороне, простые . 2. Е. oberthuri Вог.
4 (3). Последний тергит брюшка 8 почти прямоугольный, с параллельными боковыми сторонами. Клещи 8 на внутренней стороне с маленьким зубчиком близ основания и посредине.................................
............................ 3. Е. yezoensis Mats, et Shir.
1 . Eparchus insignis (Haan).
Haan, Verb. Nat. Gesch. Nederl. Averz. Bezitt., Orthopt., 1842: 243, tab. XXIII, fig. 14 (8, Q, остр. Ява) (Forficula)', Dohrn, 1865:81 (Opishocosmia)-, Bormans, 1900:96; Burr, 1907a : 121; 1910a: 192, fig. 99; 1911:94, tab. VIII, figs. 11 tab.
Тело красновато-каштановое, глубоко темно-красное или иногда почти черное. Усики 12-члениковые, 9-й и 10-й членики иногда более светлые. Голова с очень тонкими швами и с двумя очень слабыми бугорками у заднего края. Переднеспинка чуть длиннее своей ширины, каштановая или очень темнокрасноватая, задний край широко закруглен. Надкрылья каштановые, обычно светлые или рыжеватые на плечах или почти грязножелтые. Крылья желтые, с бурым пятнышком, иногда пойти отсутствующим или же иногда представленным в виде темной косой полосы, которая может быть очень широкой, почти полностью вытесняющей основной желтый цвет. Ноги черновато-красные. Брюшко черное или очень темно-красновато-бурое, иногда более светлое посредине, у 8 бока 6—8-го, а иногда и 5-го сегментов на заднем крае с сильными толстыми коническими бугорками, расположенными под прямым углом к телу и более сильно выступающими на 6-м и 7-м сегментах, у 2 — без бугорков. Последний тергит 8 сильно суживающийся и очень сильно наклонен к заднему концу, с 2-мя тупыми возвышениями над основанием клещей. Клещи сближены, почти соприкасаются у самого основания и здесь сверху с вертикальным, на конце почти шаровидным, а при основании более тонким, бугорком; отсюда клещи сначала расходятся, а от средины вновь сходятся, скрещиваясь на вершине, образуя сильную дугу, начинающуюся близ основания клещей и кончающуюся на их вершине; внутренняя сторона с очень слабыми зубчиками; клещи g простые, на внутренней стороне без зубчиков. Метапарамеры толстые, почти с прямыми наружными сторонами, virga более чем в два раза длиннее метапарамер, соединяется с основанием
11. EPARCHUS
203
(нижним концом) почковидного пузырька под прямым углом; средина собственно penis со слабо-выраженной, бороздчатой хитиновой областью. Длина тела <5 9.5—11.8, 9 9—11.5; клещей 3 4—5.8, 9 3.7—4.5 мм.
Распространение: Британская Индия, включая Бирму, Индо-Малайский архипелаг, северо-вост. Китай до Киао-Чжоу (Borelli, 1915).
Этот вид характеризуется значительной изменчивостью в окраске тела, имея две крайние цветовые формы; одна из них глубоко темнокрасная, с почти полностью желтыми крыловыми пластинками, без или со слабым темным пятнышком; другая почти черная, с глубоко темнокрасными, почти черными надкрыльями, всегда более светлыми на плечах, а крыловые пластинки с широкой косой полоской.
2 . Eparchus oberthuri Borelli
Borelli, 1912:19, fig. 4 (3, 9; Бутан, сев. Индия).
Тело довольно крупное для рода, коричневое. Усики 11-члениковые, .желтовато-бурые, 9-й и 10-й членики беловатые. Голова красновато-коричневая, ротовая часть бурая, швы глубокие, особенно затылочный. Переднеспинка смоляно-бурая, блестящая, одинаковой длины и ширины, почти полукруглая, боковые стороны грязно-желтые. Надкрылья коричневые, гладкие, чуть более чем в два раза превосходят длину переднеспинки, в плечах округленные, более широкие чем на вершине, которая прямо обрублена. Крылья немного длиннее одной трети длины надкрылий, такого же цвета как последние, с мелким желтым пятном на вершине у внутреннего края. Ноги красновато-желтые, 1-й членик лапок в 11/2 раза длиннее 2-го и 3-го взятых вместе. Брюшко смоляно-бурое, нежно пунктировано, расширенное посредине; бугорки на 3-м и 4-м тергитах хорошо выраженные; бока сегментов у <5 и 9 лишены бугорков или выступов. Последний тергит у 3 суженный к вершине, в два раза шире своей длины, •сильно наклонный, слегка вдавленный посредине перед задним краем, с каждой стороны, со слабым выпуклым бугорком перед основанием клещей. Пигидий незаметный. Клещи рыжевато-красные, у 3 соприкасающиеся при основании, тонкие, слегка расходящиеся от основания к концу 2-й трети, в вершинной трети сначала прямые, параллельные, а на конце загнутые и перекрещивающиеся; совершенно лишены зубцов или бугорко-видных выступов и лишь на внутренней стороне в основной части с очень слабыми мельчайшими зубчиками; внутренняя сторона 2-й трети слегка вырезана так, что ширина клещей в вершинной трети несколько превосходит ширину до 2-й трети; клещи 9 почти соприкасающиеся, без зубчиков на внутренней стороне, прямые, на вершине изогнутые и перекрещивающиеся. Длина тела 3 9 12.5, клещей $ 7, 9 5 мм.
Распространение: известен из Бутана в сев. Индии и сев.-вос-точного Китая (Киао-Чжоу; Borelli, 1915).
204
4. FORFICUL1DAE
Этот вид хорошо отличается от остальных видов рода простыми^ совершенно лишенными зубцов и бугорков, клещами. L %
3 . Ер archus yezoensis (Mats, et Shiraki)
Matsumura et Shiraki, 1905:80, fig. 1 (5, Саппоро, остр. Иезо, сев. Япония^ (Labia); Burr, 1911: 57 (Labia); Shiraki, 1928: 25.
Усики 12-члениковые, грязно-коричневые, 1-й членик довольно длин- ' ный, грушевидный, 3-й почти конусовидный, длиннее 4-го, 10-й и 11-й членики желтые. Голова каштаново-коричневая, выпуклая, с хорошо выраженными швами. Переднеспинка немного длиннее своей ширины,, с параллельными боковыми сторонами, задний край слегка округлен, задние боковые углы сильно округленные, округло соединяют боковые стороны с задней. Надкрылья очень широкие, с сильно выступающими плечами, крыловые пластинки длинные. Ноги грязнокоричневые, с желтоватыми голенями и лапками, 1-й члёник лапок немного длиннее 3-го, 2-й очень маленький. Брюшко без бугорков на боках 5—8-го сегментов, умеренно расширенное к средине; последний тергит с? почти прямоугольный, боковые стороны почти параллельные, задний край между основанием клещей прямой, по бокам кнаружи от клещей слегка скошен. Пигидий $ маленький, но хорошо заметный, выступающий. Клещи $ очень длинные, почти равны по длине телу, тонкие, довольно-широко расставленные при основании, слегка расходящиеся отсюда, а затем к средине слегка загибающиеся внутрь, образуя в основной половине весьма слабую дугу, далее, от средины параллельные, на самом конце загнуты внутрь и перекрещиваются; внутренние стороны близ основания с маленьким зубчиком и с таким же зубчиком близ средины; клещи g простые, длинные, тонкие,, при основании немного расставленные, не соприкасающиеся. Длина тела $ 8—10, g 10—11; клещей $ 8—9, g 5—б мм.
Распространение: известен только с острова Иезо, сев. Япония.
Этот вид отличается от других видов рода строением последнего тергита брюшка $ и относительно коротким 1-м члеником лапок и хотя отнесен Shiraki к данному роду, в действительности может оказаться представителем совсем другого рода; оба указанные признака сближают данный вид с,видами рода Timomenus. Кроме того в оригинальном описании Matsumura и Shiraki 2-й членик лапок обозначен как „cylindrisch,. sehr klein“; это обстоятельство и послужило причиной отнесения данного вида в род Labia, что было затем повторено и Вигг’ом в 1911 г.; однако, невидимому, описание лапок неточно, так как Shiraki впоследствии отнес данный вид к сем. Forficulidae. По всей вероятности 2-й членик лапок, у данного вида слабо расширен и при невнимательном рассмотрении может быть принят за цилиндрический. Отсутствие материала по данному виду, к сожалению, не позволяет выяснить его систематическое положение-более точно.
t
11. EP ARCHUS, 12. NARBE RIA
4
12. Род NARBERIA BURR
Burr, 1911:94—95.
Тип рода — Narberia biroi Burr с Новой Гвинеи.
Очень близок к Eparchas, отличаясь строением переднеспинки, которая довольно значительно длиннее своей ширины, с сильно округленным задним краем; надкрылья и крылья как у Ер archus] брюшко 5 умеренно расширенное посредине, бока 6—9-го сегментов на заднем крае с выступающим назад углом, загнутые или со слабым зернистым килем, последний тергит умеренно суживающийся к вершине; клещи 3 слегка расставлены при основании, без зубцов и выступов сверху, несколько утолщенные, умеренно изогнутые.
Известно 4 вида этого рода, распространенные от Бирмы до Индо-Малайского архипелага и до северо-вост. Китая; в пределах Палеарктики известен пока только один вид.
1. Narberia tuberculata Borelli
Borelli, 1921:160 (3 g; Пин-фа, Киао-Чжоу, Malay States Mus., Singapur, XVII, 1932:200.
сев. вост. Китай);
Borelli, Journ. Feder,
Тело рыжевато- или черновато-бурое. Усики 11-члениковые, сероватожелтые или черновато-бурые, 1-й членик более темный. Голова иногда блестяще-черная, с хорошо выраженными швами. Переднеспинка чуть шире головы, почти прямоугольная, задняя сторона и задние углы сильно округленные. Надкрылья в два раза длиннее переднеспинки, с параллельными боковыми сторонами, задний край прямой; крыловые пластинки хорошо выступают. Ноги грязновато-желтые или иногда бедра черные, 1-й членик лапок в 1г/2 раза больше чем 2-й и 3-й взятые вместе. Брюшко тонко пунктировано; бока б—9-го сегментов 3 тупоугольные сзади, со слабым зернистым килем, более ясным на 8-м и 9-м сегментах, оканчивающимся маленьким бугорком; у g без морщинистого киля. Последний тергит при основании в два раза шире чем на вершине, выпуклый, сильно наклоненный кзади, посредине перед задним краем с вдавлением, задний край слабо . округлен; бока в задней трети с ясным килем, снабженным бугорками. Пигидий 3 слабо заметный, конический. Клещи 3 сильно, изменчивые, у типичной формы горизонтальные, в первой трети слегка изогнутые кнаружи и с приподнятой вверх внутренней стороной, далее утончающиеся и почти цилиндрические, изгибаясь дуговидно почти до вершины; перед вершиной с треугольным расширением на внутренней стороне, а далее вновь утончающиеся, перекрещивающиеся; внутренняя сторона от основания до треугольного расширения мелко зазубрена; у f. parallela Bor. — прямые, параллельные, почти цилиндрические, слегка изогнутые и сближенные в вершинной части, внутренняя сторона слегка зазубрена, и снабжена маленьким зубцом перед вершиной; у f. recurva Bor.—клещи
206
4. FORFICULIDAE
слегка изгибаются в основной половине кверху, затем изгибаются книзу, образуя в их вершинной половине чуть более сильную дугу, внутренняя сторона слабо зазубрена по всей длине и лишена предвершинного зубца; у f. sinuosa Bor. — сближены и сильно загибаются кверху в основной половине, далее загнуты книзу, расходясь и утончаясь до вершинной четвертиу где они слегка расширены и снабжены маленьким зубцом и вновь изгибаются кверху, перекрещиваясь на вершине; внутренняя сторона слабо зазубрена Клещи g простые, тонкие, g 4—8 мм.
из Киао-Чжоу в сев.-вост.
Китае и с о-ва Борнео.
Чрезвычайно изменчивый по окраске и по строению клещей вид, очень напоминающий по характеру клещей Eparchus oberthuri Bor.
от основания до предвершинного расширения. Длина тела 3 11—15, g 12.4; клещей S б—8.7, Распространение: известна только
13. Род SONDAX BURR
Burr, 1910а: 177; 1911:89
Тип рода — S. repens Burr из Мадраса в южн. Индии.
Полностью бескрылый. Голова широкая. Переднеспинка сильно поперечная, с прямыми чуть расходящимися кзади боковыми сторонами, задняя сторона слегка округлена, почти прямая, образующая с боковыми сторонами прямые, почти не округленные углы. Заднеспинка с дуговидно-вырезанным задним краем. Брюшко с ясными боковыми бугорками сверху на 3-м и 4-м члениках, расширенное перед BejpniHHofi и далее суженное к вершинному концу. Последний тергит наклонный, суженный к* вершине. Клещи 3 сближенные у основания, почти или совсем соприкасающиеся, далее расходящиеся, дуговидно изогнутые, с соприкасающимися или перекрещивающимися концами; основание на внутренней стороне с выступающим кверху, почти вертикальным зубцом; клещи g простые, довольно толстые, сильно суживающиеся и заостренные к вершине.
Этот род известен только по двум видам, из которых один распространен в южн. Индии, а другой в пров. Сычуань в сев.-зап. Китае. Несмотря на то, что оба вида несомненно генетически близки друг другу, один из них, именно S, potanini, резко отличается от другого как раз такими признаками, которые являются или промежуточными между подсем. Forficulinae и Оpisthocosmiinae или типичными для одного из этих подсемейств, К числу первых признаков необходимо отнести строение клещей, которые у типа рода — S. repens совершенно сходны по строению со многими родами подсем. Оpisthocosmiinae, напр. с родом Eparchus^ у сычуан-ского вида S, potanini клещи, несмотря на большое сходство с клещами предыдущего вида, несколько напоминают по характеру таковые у рода Forficula и лишь только наличие направленного кверху зубца у основания отличает их от типичных Forficula, С другой стороны грудные пластинки у *5. potanini широкие, напоминают в этом отношении род Anechura и близ-
13. SONDAX
207
кие роды, тогда как у S, repens, судя по значительно большой стройности тела, грудные пластинки должны быть примерно такими же как у типич-
ных Оpisthocosmiinae. По характеру усиков S. не отличается от Forficulinae} то же самое надо
potanini также ничем сказать и про клещи
самки. Таким образом данный род является как бы промежуточным между
Forficulinae »и Opisthocosmiinae и может быть отнесен как к тому, так и другому подсемейству.
1. So nd ах potanini B.-Bienko (фиг. 67).
Bey-Bienko, 1934 : 414, fig. 8 (3, g; Тацзинлу, дров. Сычуань, сев.-зап. Китай).
Тело маленькое, коренастое, не- * сколько уплощенное буроваточерное. Усики грязно-палево-рыжеватые, 12 (?)-чле-никовые, 1-й членик короткий, 4-й членик немного короче 3-го. Голова со слабыми, но ясными швами, рыжеватая, чуть шире переднеспинки, с сильно округленными задними боковыми углами. Переднеспинка в 1ж/2 раза шире своей длины. Среднегрудная пластинка почти в два раза шире своей
Фиг. 67. Sondax potanini B.-Bienko > (J X 6.5 (тип). По Bey-Bienko, 1934.
длины. Ноги короткие, ^грязно желтоваторыжие. Брюшко гладкое, с очень неясными редкими точками. Последний тергит $
суженный кзади, с двумя очень слабыми
выпуклостями над основанием клещей, значительно шире своей длины> у g такой же, но без ясных выпуклостей. Пигидий S удлиненный, суженный к вершине. Клещи $ в основной четверти расширены, утолщены и сильно сближены, но не соприкасаются, внутренняя сторона расширенной части у ее заднего конца приподнята кверху, образуя острый почти вертикальный зубец; остальная часть цилиндрическая, суживающаяся и дугообразно
изогнута от своего основания к вершине; клещи g простые, толстые,, сильно суживающиеся к вершине и слегка изогнутые на концах. Длина тела 6 7.4—8.5, g 9.8—11.5; клещей 6 3—3.4, g 3.3—3.7 мм.
Распространение: известна только из пров. Сычуань в сев.-зап. Китае.
SUMMARY
The present part of the „Fauna" deals with the Dermaptera of USSR and adjacent countries; it has been based on critical studies of the world literature as well as upon a rich material on this order of insects. The fundamental part of this material is presented by the precious collections of the Zoological Institute of the Academy of Sciences of USSR which include most interesting materials obtained by the expeditions of the famous Russian explorers of Central Asia (N. Przewalski, C. Potanin, P. Kozlov). They made th*e Dermapterous fauna of the least studied Asiatic area to be known more precisely. Moreover, a vast private collection was kindly placed at the author’s disposal by Dr. A. Semenov-Tian-Shanskij, containing types of the species described by its proprietor. At last a small author’s collection, as well as that of the Caucasian Museum, were also used.
The monograph includes whole Dermapterous fauna of the Eastern part of the Palaearctic region, i. e. all the USSR, Mongolia, China up to the Szechuan province, Korea, Manchuria, Tibet, Kashmir, West China and Japan. Farthermore, the data on Iran, Asia Minor and West Europe, approximately up to the Alps are subjoined. From this vast aera only 78 species of Dermaptera are known at present, and they all are included in the present „Fauna".
Of this umber 61 species have been studied by the author, while the rest 17 are included in the „Fauna" only on the base of their descriptions in the literature; 26 species are recorded at* present from the USSR, and they are marked with an asterisk.
Besides the systematical part, with keys and descriptions of the species, and a general morphological part, there are given chapters on the biology and ecology of earwigs, on their systematical position and evolution in connection with the palaeontological data and geographical distribution, on the composition and origin of the Dermapterous fauna in the Palaearctic region as well as chapters on the characterictic of the fauna of USSR and on the economic importance of earwigs. A short account of the chief chapters is given herein.
LIFE HISTORY AND ECOLOGY
Earwigs are for the most part nocturnal insects, inhabiting different secluded places. As regard the food earwigs are fully omnivorous. They may
DERMAPTERA
209
be either carnivorous, attacking small insects, worms, spiders and so on, or feed both on the plant and animal remains, and growing plants as well.
In the temperate latitudes they have one generation a year, the hibernating stage being either egg (Forficula auricularia) or larva (F. tomis).
Maternal care of progeny is a remarkable feature for these insecte (Verhoeff 1913; Pesotzkaja, 1927 and some old authors). The most important ecological peculiarity of earwigs is their preference for moisture and that they largely depend on the moistened conditions of stations. Vast majority of species is connected with forest biocenoses, characterized, as known, by increased moisture. There they live under bark of trees, beneath dead leaves, under stones and in the rotten wood (Anechura bipunctata). Several species inhabit the banks of rivers and seashores (Labidura riparia, Anisolabis maritima and some others). Even such a xerophilous species as Anechura asiatica, that lives on dry rocky slopes, is found only under stones where some damp soil is preserved. Morgan’s investigation (1926) showed that Forficula auricularia is a good swimmer and may float for 24 hours or more.
The most diversity of Dermaptera is observed in tropical regions. The Belgian Congo, for instance, is inhabited by 75 species (Menozzi, 1935), while in the foir more extensive Mediterranean subregion there are 45 species and in the European part of USSR (the Crimea and Caucasus excluded) only 9; at last from the Arctic region of Eurasia we know, but at present a single species only. It depends, however, chiefly on the moisture, but not on the temperature conditions so that the most humid woody regions are the richest in number of the species of the order.
Dry and desert regions of the tropical zone include in their fauna a trifle of species, while forest regions situated not only in tropical zone, but in temperate countries also, are characterized by considerably greater number of species. There are about 70 species of earwigs in the Eastern part of North India (Assam, Sikkim, Bhutan, Bengalia) with the annual amount of rainfall from 1300 to 2000 mm and more, while only 9 species belong to the adjoining Western provinces (Punjab, Sind, Radjputan) where the rainfall amounts to 200—600 mm a year. Hot and dry Iran inhabit 8 species, deserts of Central Asia 5 and the vast Mongolia 5 too, while the comparatively small and more northern Ussuri region in Far East Siberia has 9 species and the West Europe (excluding its Mediterranean part) even 16 species; the last mentioned regions characterized by relatively cold climatic conditions but have considerably higher amount of rainfall and covered by large forest areas.
Thus, all Dermaptera are hygrophilous insects and depend in their world distribution on the humidify condition rather than on the temperature.
A number of species has been introduced by man all over the globe; for the most part they are those species the area of which stretches for the maritime countries or the sea-shores alone.
Фауна СССР, Кожистокрылые
14
210
- SUMMARY
SYSTEMATICAL' POSITION AND CLASSIFICATION OF THE ORDER DERMAPTERA
I *.
The order Dermaptera, together with the order Hemimeroidea (the latter is. treated as an indepedent order according to Dr. A. Martynov’s interpretation), forms a natural group Dermapt er oidea, which unites - with two other groups — Blattopteroidea and Orthopteroidea—-into a general section Polyneoptera of the subclass Pterygota (Martynov, 1925).
The classification of the order Dermaptera is here interpreted in a somewhat different manner than that by other investigators (Burr, Zacher, Verhoeff). The whole order is divided into three suborders: (1) Archider-maptera, created for the fossil Mezozoic species with 4- or 5-jointed tarsi and segmented cerci, (2) Arixenina and (3) Dermaptera s. str., or Forficulinae For further division in superfamilies and families see page 215—216.
The classification of the family Forficulidae proved to be different too. The author does not consider it convenient to detain the subfamily Anechurinae, because the genus Anechura and its nearest allies are extremely close to the genus Forficula, according to the author’s opinion. Among the new species recently discovered in the Central Asia and China there are members nearly, intermediate between the two above mentioned genera, for instance, Anechura (Odontopsalis) forficuliformis. The Himalayan Forficula schlagintweiti is unquestionably related to Oreasiobia stoliczkai, although the genera Forficula and Oreasiobia are quite distinct and valid; the brachylabic condition of the last mentioned species is characterized by features of the genus Forficula and, on the other hand, the former species appears sometimes in a condition which may be called Anechuroid.
Some genera of the order are regarded more narrowly, and the author thinks to treat them but as subgenera. Both genera Anisolabis and Anechura are to be included there. The full discussion about this question is given in the Russian text.
About 900 recent and approximately 30 fossil species of Dermaptera are known at present.
EVOLUTION OF THE ORDER DERMAPTERA IN RELATION WITH THE PALAENTOLOGICAL DATA AND GEOGRAPHICAL DISTRIBUTION
S
The authentic members of the order Dermaptera are known from the Mezozoic deposits, viz. from the Jurassic of the S. Kazakstan. One of the species — Protodiplatys fortis Mart. — is the member of a primitive and extinct suborder Archidermaptera m., characterized, like Blattodea, by (4-) 5- jointed tarsi and segmented cerci. The other known species — Semenoviola obliquotruncata Mart. —г is, however, in all probability belonging to the suborder Dermaptera s. str.; this species was primarily described as a member of the order Coleoptera (Martynov, 1925) but the author’s examination of the type specimen showed that it is a member of Dermaptera (see also C. Bolivar, .1926).
DERMAPTERA
211
More than 25 species are known from the Tertiary times, so that in the Baltic amber (upper Eocene or lower Oligocene) there are found species belonging not only to the recent genera of the most specialized and highly developed family Forficulidae, but also those allied to the recent species themselves; thus, Forficula praecursor Burr related to the recent African F. senegalensis, to the Central Asiatic F. sagitta Sem. and to the Indian F. beelzebul (Burr, 1911); Forficula (?) pristina Burr seems to be allied to the recent jEast Asiatic Anechura (Odontopsalis) forficuliformis and A. (O.) potanini.
These facts show that the order Dermaptera is rather ancient, but that in the beginning of the Tertiary there appeared already some forms belonging to the recent genera and allied to the recent species. Evidently all the recent families, and even some of the recent genera, were present at the beginning of the Tertiary and might already found in the middle of the Mezpzoic (the Jurassic). That supposition is confirmed by some pecularities in the geographical distribution of certain recent genera, especially that of the Protodermaptera, A number of genera are characterized by wide disjunctions of their areas of distribution, sometimes extending over an enormous territory. For instance, the genus Diplatys, the most primitive one, is peculiar to the humid tropics of Asia, Africa and America (fig. 14); the members of the genera Echinosoma and Apachyus are also found in humid tropics of South Asia, Africa and North Australia, while the species of the genus Pseudisolabis occur in North India, Java and New Zealand, and the genus Antiso lab is in South Africa and Australia etc. These disjunctions may only become clear, if admitted that the given genera were already present and flourished all over the occupied territory already in the Jurassic or, in some cases in the beginning of Eocene.
There are some examples of disjunctions even in the family Forficulidae.. The genus Doru represents by a number of species in both Americas and a single species is known from South Australia; the widely distributed Palaearctic genus Anechura is represented by a single species in Mexico;, the Palaearctic genera Forficula, Apterygida, Mesochelidura and Chelidurella. are also known from tropical and South Africa.
If the aforesaid facts are confronted to the recent discovery of a particular order Protelytroptera in the Permian of Kansas (Tillyard, 1931), being^ an intermediate link between the Palaeozoic Blattoid insects and recent Dermaptera, then the idea of the great antiquity of Dermaptera will be still more demonstrative. It should be remembered that there is a connective link between those insects in the form of the suborder Archidermaptera.
All the above cited data show that roots of the order Demartera reach the Palaeozoic viz. Lower Permian or even Upper Carboniferous, while the roots of certain recent families and genera lead to the. Jurassic and to the end of the Mezozoic or to the beginning of the: Tertiary.
14*
212
SUMMARY
Dermaptera seem to be characterized by a slow rate of evolution. The causes of such conservatism of the group should be looked for in the ecology of the earwigs. The uniformity and peculiarity of the ecological environment in the earwig’s stations and the high and constant requirements of these insects for a moisture are the probable reason of this conservatism.
COMPOSITION AND ORIGINE OF THE PALAEARCTIC FAUNA OF DERMAPTERA
The whole Dermapterous fauna of the Palearctic region comprises about 106 species belonging to 31 genera (see p. 44—45). It is very peculiar that there are two principal centres with maximum number of species, viz* the Western centre, including the Mediterranean subregion and adjacent part of Western Europe, and the Oriental centre, with the Ussuri region, Japan, Korea, Manchuria, North China and Tibet (see map 17). 45 species are known from the Mediterranean subregion, of which 28 are endemic, and 54, with 30 endemic ones, from the Oriental centre.
In spite of the vast space separating the two centres, their fauna is characterized by a considerable genetic affinity. The most typical elements in both centres are the genera Forficula and Anechura, 14 and 6 species for the Western centre and 14 and 12 species for the Eastern centre respectively. Round these two genera the allies are grouping, of which in the West the genera Guanchia, Pseudochelidura, Chelidura, Chelidurella, Mesochelidura, Perirrhytus and Apterygida are characteristic, while in the East, including the adjoining areas, are Mesasiobia, Oreasiabia, Eumegalura. Moreover, there are some closely allied species in the both centres, namely Anechura (in sp.) sven-hedini from China and A. (in sp.) orsinii from Northern Italy, Forficula robusta and F. macrobasis from East Asia and F. tomis from the European part of USSR and adjacent countries, F. vicaria, F. burriana and F. subauricularia in the East and E auricularia in the West. It is very interesting that the last mentioned species is an ancien member of the European fauna, because it was found to be fossil in Pliocene of Durham in England (Lesne, 1920). In the majority of cases the Eastern species are morphologically more primitive than their Western allies.
The group of genera and species mentioned above represent an ancient autochtonous or Angaran elements of the Palaearctic fauna. Besides this principal group a number of tropical species and, at last, some species introduced by man compose the Dermapterous fauna of the Region.
In accordance with an analysis of the recent and fossil fauna and comparing it with the palaeogeographical data and modern idea on the natural history of Eurasia, the author draws the following general conclusions, concerning the composition and origin of the fauna of Dermaptera of the Palaearctic region.
1. There are two centres of maximum differentiation in the recent Dermapterous fauna of the Palaearctic region, namely the Eastern or Oriental,
DERMAPTERA
213
represented chiefly by the Palaearchearctic subregion of Semenov-Tian-Shansky (1935), and the Western one, embracing the Mediterrannean subregion and adjacent parts of Western Europe.
2. The following elements took part in the formation of the fauna of the order in the Palaearctic region.
a. The principal ancient nucleus of Angaran fauna, with genera Forficula, Anechura and their allies, sprung up on the ancient Asiatic land — the Angaran continent.
b. Members of the adjacent tropical fauna, that is to say, in the East the Indian fauna (genera Timomenus, Sondax, Allodahlia and others) and in the West the African one (genus Anisolabis).
c. The group of species introduced by man and established chiefly either in the East or in the West of Eurasia (Labidura riparia, Nala lividipes, Anisolabis maritima, Prolabia arachidis, Labia curvicauda and others).
3. The autochtonous fauna of the Eastern and Western centres in spite of their extreme remoteness is characterized by a considerable genetic affinity expressed:
a. By the predominant genera Forficula and Anechura in both centres which comprise about 50 per cent of the whole number of species of the order Dermaptera in the Palaearctic region.
b. By the presence of many closely allied species, belonging to the indicated genera (Anechura orsinii in the West and A. sven-hedini in the East, Forficula auricularia and F. vicaria, F. burriana, F. subauricularia etc.); the Eastern species are as a rule morphologically less specialized than their allies from the Western centre.
c. By the presence in both centres of a group of genera, though distinct, but closely related to Anechura and Forficula.
4. A considerable genetic affinity between the faunas of both the Eastern and Western centres belonging, according to the modern idea, to the distinct zoo- and phytogeographical subregions of the Palaearctic region, is peculiar to the order Dermaptera, which makes it essentially different and opposed to many other groups of animals. The outstanding features of the Dermapterous fauna may be understood only when the following principal facts are taken into account.
a. Ecology of insects with hidden life habits and their close relation to the climatic condition, especially moisture requirement.
b. Climatic changes, on the one hand, and changes in the outlines and of the relief of Eurasian continent on the other, brought about in the prior geological periods.
c. Introduction of a number of species, chiefly tropical ones, into the temperate regions with most favorable climatic conditions; associated with the truck increase among different countries during the historical period.
5. During the first half of the Tertiary the territory of the present Palaearctic region, at least in its more temperate latitudes, was characterized by more
214
SUMMARY
or less homogeneous fauna composed both of Angaran and tropical elements, amongst the latter only the Eastern (Indian) types seemed to be represented.
6. From the beginning of the Miocene the homogeneous fauna of the Dermaptera underwent considerable changes influenced by the following main factors.
a. Climatic conditions becoming colder and precipitation reduced, especially in the central temperate parts of the continent, resulted in the extermination of some of the most thermophilous and hygrophylous forms, the latter being concentrated into two centres, East-Asiatic and Mediterranean - European ones, where climatic conditions remained rather favourable for earwigs up to the present time. Thus, the more or less homogenous pre - Miocene fauna was divided into two faunas though immensely removed from each other but closely allied genetically.
b. Disappearance of the ancient Mediterranean sea — the Thetis — resulted in a direct connection between the Eurasian and Ethiopian continents, that gave the wide opportunity for invasions and exchange of the faunas. Owing to it some Angaran forms (the genera Forficula, Apterygida, Che-lidurella and Mesochelidurd) invaded the African continent and penetrated as far as its southern part. By the same routs a few tropical Ethiopian forms came inthe Mediterranean region (the genus Anisolabis). That connection, however was most favourable for invasion of more xerophilous animals than earwigs and, therefore, the exchange was somewhat detained, which was also due to the process of erosion of the Thetis gypsiferous depositions instead of which sand deserts were formed. These causes lead to complete isolation of the Dermapterous fauna of the Mediterranean subregion from the adjacent Ethiopian region.
c. In the middle of Tertiary, especially in Miocene, when in relation with the enormous orogenetic processes the immense ranges of Eurasiatic mountains arose, the plain fauna was lifted to great altitudes and brocken off to a number of places where further evolution accomplished more or less independently. It favoured the formation of some endemic species and genera (Oreasiobia, Mesasiobia, Eumegalura, Pseudochelidura, Chelidura, subgenera Burriola and Borelliola and others).
7. During the Quarternary period fauna of the Dermaptera underwent some futher evolution in relation with the following factors.
a. The Glacial period exterminated the last remnants of tropical elements from the fauna of the Eurasian temperate latitudes and favoured the further isolation- of the Eastern and Western centres. On the other hand, it converted those centres into two recent relic areas. Moreover, the Glacial period influenced the coming down and wide distribution of the mountainous species originated with the orogenetic processes [Anechura asiatica, A. (Borelliola) euxina and others].
b. Climatic conditions became warmer after the Glacial period and a back migration of certain species took place.
DERMAPTERA
215
т
c. Introduction of a number of new species was due to the man, appearance and trade increase among the remotest countries (see § 4, c) including the oldest centres of the European (viz. Mediterranean) and Asiatic civilization.
SYSTEMATICAL PART
1 (2). Tarsi 4 or 5-jointed, Elytra with sparse elevated longitudinal stripes
(veins?). Cerci thin, short, segmented
.......................... 1. Suborder Archider
tera nov.
(+).
2 (1). Tarsi 3-jointed. Elytra, if developed, without elevated stripes. Cerci unsegmented, large, modified into forcipes.
3 (4). Eyes much reduced. Mandibles with three teeth in th er apical part, their inner margin densely clothed with bristles. Cerci feebly chitinized,
forming incipient forcipes. Totally apterous . 2. Suborder Arixenina.
4 (3). Eyes well developed. Mandibles with two apical teeth, without bristles
on inner margin. Cerci strongly chitinized, modified into horny forceps.
Fully winged, subapterous or totally apterous.....................
3. Suborder Dermaptera s. str.
1. Suborder ARCHIDERMAPTERA nov.
Antennae with 2nd joint relatively long, equal in length to the 3d Elytra relatively long, reaching the second abdominal segment, with sparse elevated longitudinal stripes (veins?).. Legs short, femora swollen; at least posterior pair with two longitudinal keels; tarsi more than 3-jointed, hind tarsi 5-jointed, anterior and middle tarsi 4- or 5-jointed. Cerci short, thin, segmented, with few (not more than 10) joints, first joint being the largest.
This suborder is erected by us for a single known fossil species, Protodiplat у s fortis Mart., from the Jurassic beds in Southern Kazakstan (Galkino, Kara-tau mountains).
The discovery of this species is of great scientific importance as it
represents an intermidiate form between the recent Dermaptera and their Blattoid ancestors, like Palaeozoic Protelytroptera (which is characterized by
5-jointed tarsi and segmented cerci) and shows the way of evolution of the
order Dermaptera.
II. Suborder ARIXENINA s
One member of this suborder — Arixenina esau Jord. — was described as a parasite of Chiroptera, but Jacobson’s observations on the second known species — A. jacobsoni Burr—has shown thatjt lives in caves and has carnivorous habits (Burr and Jordan, 1913). Therefore A. esau is also probable to be a cavernicous species and its occurence on Chiroptera may be accidental. The „parasitical" habits of the members of this suborder are probably the result of an adaptation to the life in caves.
Known from the Malay Archipelago only.
216
SUMMARY
Ш. Suborder DERMAPTERA s. str.
1 (4). Genital apparatus of d double, with a paired penis, basal part of the latter situated at apex of proparameres. Pygidium of c? simple. (Superfam. Protodermaptera).
2 (3). Opistomeres contain three well developed joints of which pygidium is a little larger than two other joinst taken together. Femora compressed, usually keeled1 ......................1. Fam. Pygidicranidae.
3 (2). Opistomeres consists of two distinct joints of which pygidium is 2 to 6 times larger than the rest. Femora cylindrical, without keels ....
. . ................................... 2. Fam. Labiduridae.
4 (1). Genital apparatus simple, with a single penis, basal part of the latter placed at base of proparameres. Pygidium of 8 often with complex processes. Superfam. Eudermapterd).
5 (6). Second tarsal joint simple, not dilated, or with a narrow lobe produced beneath the third joint. Genital apparatus of 8 with apex of metaparameres acute, often pointed . . . ..............3. Fam. Labiidae.
6 (5). Second tarsal joint dilated, :pordiform, with a lobe on each side. Genital apparatus of 3 with apex of metaparameres not acute........
.........................................4. Fam. Forficulidae.
Superfamily PROTODERMAPTERA
I. Fam. PYGIDICRANIDAE
* R ►
1 (1). Totally apterous. Metasternum posteriorly truncate. Antennae 16- to 25-jointed, setaceous. Larvae with unsegmented cerci. (Subfam. Anataelinae). Mesonotum with lateral keels. Forceps in 8 with contiguous branches, somewhat dilated and arcuate before apex . 1. Challia Burr.
1. Genus CHALLIA BURR.
1 (1). Reddish-brown. Last tergite strongly punctured, with a median longitudinal furrow, more developed in 8, and five globose tubercles in both sexes. Forceps testaceous at base, darkened in other parts; inner margin in 8 elevated, in shape of an obtuse angle before middle, then strongly depressed. — North-Eastern China and Southern Korea at the coast of the Yellow Sea.......................1. Ch. fletcheri Burr.
2. Fam. LABIDURIDAE
1 (2). Mesosternum rounded posteriorly.2 Forceps with trigonal branches, often asymmetrical in 8. Manubrium very long, narrowed at base, broadened at apex. Genital apparatus of <5 without reniform vesicle arid
л
1 Except in the Palaeotropical subfamily Echinosomatinae and in the Neotropical sub-
II
2 In the New-Caledonian genus Spondox Burr mesosternum truncate posteriorly.
DERMAPTERA
217
often without virga. (1. Subfam. Psalinae}. Metasternum truncate posteriorly. Abdomen feebly dilated beyond the middle and slightly narrowed apically. (Totally apterous or subapterous, rarely with fully developed elytra and wings) .................................1. Anisolabis Fieb.
2 (1). Mesosternum truncate posteriorly. Manubrium short, not more than twice as long as broad, not broadened at apex. Genital apparatus of 5 with well developed reniform vesicle and virga.
3 (6). Antennae with 25 to 35 joints. Prosternum narrowed posteriorly, meso-and metasternum as long as broad. Elytra well developed. Pygidium about 6 times as long as the rest part of opistomeres. (2. Subfam. Labidurinae).
4 (5). Larger. Legs long, hind femora half again as long as pronotum; hind . metatarsus longer than the third tarsal joint. Metaparameres slightly dilated and rounded at apex, with a small apical lobule . .................
..................................................2. Labidura Leach.
5 (4). Smaller. Legs relatively shorter, hind femora not longer than pronotum, hind metatarsus and third tarsal joint subequal in length. Metaparameres strongly acute at apex, without apical lobule....................
.....................................................3. Nala Zacher.
6 (3). Antennae with 10 to 15 joints. Prosternum with parallel lateral matgins, meso- and metanotum broader than long. Totally apterous. Pygidium twice as long as the rest parts of opistomeres. Head with very distinct postfrontal and median sutures. (3. Subfam. Parisolabinae), Last tergite very short, in 5 slightly narrowed apically. Pronotum strongly transverse, hind margin straight......................4. Pseudisolabis Burr.
I. Subfam. PSAUNAE
1. Genus ANISOLABIS FIEB.
1 (2). Metaparameres long, lanceolate, 3 to 4 times as long as broad, with more or less parallel lateral margins, apex not or foebly acute. Totally apterous.. (1. Subgen. Anisolabis in sp.). Antennae unicolorous, dirty yellow, with 22 to 25 joints. Forceps of 5 with remote branches, strongly incurved,.
specially the right one, which has a rectangular projection on its inner margin. Pronotum unicolorous, with a fine median line. Abdomen sparsely covered with impressed points. — Black Sea coast of USSR
and Ussuri region; cosmopolitan (figs. 19 and 20)
2 (1). Metaparameres short, not more than twice as long as broad, often
acute at apex.
3 (8). Metaparameres 11)2 to 2 times as long as broad with maximum width in basal half, apex acute. Totally apterous.
4 (5). Outer margin of metaparameres S-shaped, incurved, their maximum width is practically at base. Abdomen moderately dilated. Forceps of
218
SUMMARY
8 with branches slightly remote at base. (2. Subgen. Apolabis Burr.). Chestnut-brown, antennae greyish-brown with yellow 13 to 15 or 15 to 16 joints; femora bicolorous. Forceps of 8 moderately incurved, almost
symmetrical. Metaparameres relatively short, half as long as broad,
with a small, rounded lobule on apex.—East Asia from Japan and
Korea to Formosa and Java (fig. 21)
. 2. A. (A.)
arginalis (Dohrn).
11
5 (4). Metaparameres trapezoidal, with maximum width in the middle; outer
side angulate, with straight margins. (3. Subgen. Mongo lab is Zacher). 6 (7). 3d antennal joint equal in length to the 4th and 5th taken together.
Sides of 5 to 8th abdominal segments in 8 strongly acute, with a sharp keel and longitudinal striation. Penultimate sternite in 8 tetragonal, practically parallel-sided, hind margin with a broad excision in the middle. — Szechuan prov., western China (fig. 23—24).............. .
.....................................3. A. (M.) setshuana B.-Bienko.
7 (6). 3d antennal joint one and one third as long as the 4th. Sides of abdominal segments in 8 obtusely angulate, without a sharp keel. Penultimate sternite in c? roundly triangular, hind margin truncate.— Kiao-Tchau and Yunnan, China - . . . . 4. A. (M.) cavaleriei Borelli.
8 (3). Metaparameres not or a little longer than broad, often slightly broader than long, irregularly rounded or tetragonal; apex not acute; penis often with strong chitinized denticles. Totally apterous or elytra reduced to lateral flaps on the sides of mesonotum, rarely fully winged. (4. Subgen. Euborellia Burr).
9 (10). Totally apterous (very seldom fully winged). Sides of 6th to 9th abdominal segments in 8 acute and strongly keeled, with coarse longitudinal rugosities. Metaparameres trapezoidal, with external margin obtusely angulate. Small, graceful, shining-black; legs yellowish, usually banded with black. — Southern Crimea, Black-Sea coast of Caucasus
and Ussuri region; cosmopolitan (fig. 26) . 5. (A.) E. annulipes (Luc.)
10 (9). Small rudimentary elytra present on the sides of mesonotum or elytra and wings fully developed. Metaparameres with outer margin practically rounded.
11 (14). Smaller and more graceful (length of body 8 to 10 mm). Antennae blackish, except 1 to 3 subapical joints which are whitish. Femora paleyellow, unicolorous, or banded with black. Abdomen of 8 with sides of 7th to 10th segments keeled.
12 (13). Body black or brownish-black. Prosternum with hind margin rounded. 3d antennal joint more than twice as long as the] 2nd. Femora and tibiae usually banded with black. — Japan and North-Eastern China; tropics of the Old and New World (fig. 25) . 6. A. (E.) plebeja (Dohrn) 13 (12). Body chestnut-brown. Prosternum truncate posteriorly. 3d antennal joint not more than half again or twice as long as the 2nd. Legs unicolorous, tibiae sometimes with darkened basal part.—Japan, Formosa and South-Eastern China ........ 7. A. (E.) pallipes Shiraki. *
DERMAPTERA
219
14 (11). Larger and more robust (length of body 11 to 15 mm). Femora blackish, with yellowish apex. Antennae unicolorous, dark-brown. Abdomen of 5 with sides of 6th to 10th segments keeled. — Eastern Europe from the Iberian Peninsula to the Southern Crimea; Eastern Africa (fig. 26) .......................................... 8. A. (E.) moesta (Gene).
2. Subfamily LABIDURINAE
2. Genus LABIDURA LEACH.
1 (1). A single cosmopolitan and extremely variable species (figs. 27, 28). .....................................................1. L. riparia (Pall.).
3. Genus NALA ZACHER. 4
1 (1). Dark chesthut- or blackish-grey, dull. Abdomen in 3 somewhat dilated towards apex. Forceps of $ cylindrical, feebly incurved, inner margin with a small tooth in apical half. — Southern Europe, Africa, Australia, tropical Asia, Japan and North-Eastern China (fig. 29) . . . . . * .
............................................ 1. N. lividipes (Duf.).
3. Subfam. PARISOLABINAE
4. Genus PSEUDISOLABIS BURR
1 (1). Black or blackish-brown, scarcely pubescent. Antennae dark-brown. Abdomen witha few long bristles at sides. Pygidium of $ with a small tubercle in the middle. — Kashmir and Punjab (fig. 30)...................• .
.....................................................1. P. burri Borelli.
Superfamily EUDERMAPTERA
3. Fam. LABHDAE
I
1 (6). Second tarsal joint simple, cylindrical. (1. Subfam. Labiinae).
2 (5). Elytra not keeled along the external sides.
3 (4). Antennal joints subcylindrical, subequal 'in length; 4th and 5th joints each practically as long as the 3d.....................1. Labia Lech.
4 (3). Antennal joints heterogeneous: cylindrical, clavate and piriform or subconical; 4-th and 5th joints each shorter than the 3rd, often conical ................................................2. Prolabia Burr.
5 (2) Elytra keeled along the external sides. Antennal joints subconical or clavate, with strongly attenuate apices. Anterior femora greatly swollen.
.......................................r. 3. Nesogaster Verb.
6 (1). Second tarsal joint with a narrow lobe produced beneath the 3-d joint. (2. Subfam. Chelisochinae).
V (10). Body elongated, graceful, from medium to large (10 to 19 mm). 3rd antennal joint twice as long as broad.
220
SUMMARY
8 (9). Large. Antennae thicker, 3rd joint practically twice as long as the 4th and distinctly longer than the 5th, all three joints more or less thickened towards apex or, at least, some of them conical....................
................................................ 4. Chelisoches Scudd.
. 9 (8). Size medium. Antennae very thin, all joints cylindrical; 3rd joint a little longer than the 4th and scarcely longer than the 5th. 5. Proreus Burr.
10 (7). Body robust, small (7 to 9.5 mm). 3rd antennal joint short, not more than half as long as broad..................................6. Hamaxas Burr-
I. Subfam. LABIINAE
1. Genus LABIA LEACH
1 (2). Yellowish-brown. Forceps long, thin, in 3 slightly incurved, with, sparse minute teeth on the inner margin, in 9 more than twice as long as broad at base. Penultimate sternite in S with very distinct, elongated process, well visible from above. — European part of USSR, including Crimea and Caucasus, Transcaucasia, Western Siberia and vicinity of Tashkent; almost cosmopolitan, but not known from East Asia (fig. 31).
inor (L.)..
2 (1). Brown or chestnut, with darker elytra and wings. Forceps short and, thick, strongly dilated near the base, in 5 strongly incurved in the apical part, sometimes forming practically a right angle, in 9 not more than twice as long as broad at base. Penultimate sternite in simple. —Widely distributed all over tropics of the Old and New World; known also from Japan in the East and from the island Madeira in the West.
.........................................2. L. curvicauda (Motsch.)..
11
2. Genus PROLABIA BURR
1 (1). Wings undeveloped. Pygidium of $ short, not longer than broad, slightly narrowed apically, hind margin with a feeble emargination. Darkbrown or castaneous, with black or brown elytra.—Ussuri region, Sakhalin and Kamtshatka; widely distributed all over tropics and subtropics of the Old and New World (fig. 32) .... 1. P. arachidis (Yersin).
3. Genus NESOGASTER VERH
1 (2) Larger (length of body 13.5 to 14 mm). Pygidium of S triangular, broad.. — Isle Yezo, Northern Japan . ........ 1. N. nigritus (Shir.)..
2(1). Smaller (length of body not more than 10 mm). Pygidium of о lanceolate, sharpened. — Isles Yezo and Gondo, Japan . 2. N. lewisi (Borm.)..
2*. Subfam. CHELISOCHINAE
4. Genus CHELISOCHES SCUDD. К
1 (1). Antennae with 20 to 21 joints. Body black, varying to dull brown or umber, legs black, with orange or yellowish tarsi. Pronotum subqua
DERMAPTERA
221
drate, lateral margins scarcely divergent posteriorly. — Oriental region, East coast of Africa, New Zealand, Australia and Oceania; Japan, sometimes introduced in Western Europe .... 1. Ch. morio (Fabr.).
5. Genus PROREUS BURR
1 (1). Body orange-red, with unicolorous brown antennae; elytra orange-yel-
low, with a narrow dark band along the inner and outer margins. Forceps long, slender, sub cylindrical, almost straight, in S gently incurved apically — Widely distributed in the Oriental region, including Southern
China and Formosa; Japan
1. P. simulans (Sial).
6. Genus HAMAXAS BURR *
1 (1). Practically naked, reddish-brown. Sides of 5 to 8th abdominal tergites with series of small keeled tubercles, near the hind margin; last tergite with four longitudinal keel-shaped parallel ridges in the middle before the hind margin. Inner margin of forceps in $ armed with three teeth in the basal two-thirds (fig. 33). Central Mongolia............
..................................1. H. koziovi Sem. & B.-Bienko.
4. Family FORFICULIDAE a
1 (24). Elytra always present, well developed or abbreviated in different degrees. Wings developed or completely absent.
2 (19). Abdomen distinctly depressed, so that the cross-section is transver-sely-oval; last abdominal tergite in 3 transverse, usually not narrowed posteriorly. Antennae with relatively short joints; 1-st joint shorter than half the length of the head, 4th often shorter than the 3rd. Forceps more robust and stout, basal part in $ usually, in g always considerably thicker than the apical one. (1. Subfam. Forficulinae).
3 (18). Elytra not keeled along the external margin, smooth or slightly punctured.
4 (7). Forceps of 5 with branches contiguous or subcontiguous at base;1 basal part dilated and depressed.
5 (6). Elytra well developed or slightly abbreviate, always covering the meta-notum. Wings developed’ or absent. Head unicolourous above. Metaparameres parallel-sided or regularly narrowing apically................
................................................... 1. Forficula L.
6 (5). Elytra strongly abbreviate, not covering metanotum, hind margin of the latter is quite free, bow-shaped, excised; wings absent. Head bico-lorous. Metaparameres very long, parallel-sided, slightly broadened before apex....................................( . . . 2. Mesasiobia Sem.
1 In some male specimens of the Himalayan Forficula schlagintweiti Burr branches of forceps are moderately remote at base but in that case the body is entirely black and the hind tibiae are heavy at the middle, whence they are «narrowed strongly toward their apices.
222
SUMMARY
7 (4). Forceps of 8 with branches remote at base;1 basal part cylindrical^ rarely moderately depressed and dilated.
8 (15). Pygidium of 5 short, moderately prominent or practically invisible.
9 (10). Forceps of 5 in f. macrolabia with very long and thin branches, basal part of which is armed with produced lamellate dilatation on the inner * margin; the rest part cylindrical, slightly incurved, seen in profile not bowed downwards in the basal part, without teeth or denticles. Elytra well developed............................... . 3. Oreasiobia Sem., gen. nov.
10 (9). Forceps of 3 without lamellate dilatation on the inner margin of the basal part, often strongly undulating. Elytra well developed or abbreviate.
11 (17). Abdomen in $ practically parallel-sided or regularly broadened toward apex.
12 (16). Body more or less robust. Mesosternum transverse or at least as broad as long. Pygidium of 8 broad, transverse. Upper surface of abdomen and forceps without or with sparse and short hairs.
13 (14). Abdomen practically parallel-sided or slightly broadened to apex. Elytra well developed; if abbreviate, exposing little or no scutellum, their length along the external margin equal or greater than their maximum width..................................................4. Anechura Scudd.
«4 Г '
14 (13). Abdomen strongly dilated from base to apex, so that its maximum-width is at apex. Elytra strongly abbreviate, very short, not less than half as broad as long, exposing a broad scutellum..........................
..................... . .....................5. Chelidura Latr.
15 (8). Pygidium of 8 unusually large, equal in length to middle femora^ strongly broadened at base and narrowing apically. Elytra abbreviate,
6
with oblique hind margin
egalura B.-Bienko.
16 (12). Body very slender. Mesosternum elongated. Pygidium of 8 elongated,
strongly prominent. Abdomen and forceps in dense, relatively long hairs. Forceps of 8 slender, slightly incurved . 7. Apterygida Westw.
17 (11). Abdomen strongly broadened in the middle, then narrowing to apex,, lateral margins roundly-convex. Elytra strongly abbreviate. Forceps of 3 thin, unarmed, or with a feeble tooth at base above, seen in profile
straight............................................
8. Chelidurella Verb.
18 (3). Elytra with a strong keel along the outer margin, sometimes strongly rugose and covered with small tubercles. Forceps of 8 with branches very remote at base............................... 9. AUodahlia Verh.
19 (2). Abdomen not much depressed, so that the surface is convex andL the cross-section circular; last tergite often tapering. Antennae with elongated joints, 1st joint longer than half the length of the head, 4th equal or a little shorter than the 3rd. Forceps slender and long, in 8
1 Except the brachylabic condition of Himalayan Oreasiobia stoliczkai Burr, but in that case body brownish-black and pygidium concealed.
DERMAPTERA
223;
often with teeth on the upper surface, in Q very thin, practically filiform,, especially in the apical halE (2. Subfam. Opisthocosmiinae).
20 (21). Abdomen slightly dilated before apex, practically cylindrical; last tergite of 5 transverse, nearly rectangular, moderately sloping. Forceps with branches moderately remote at base. Anterior femora swollen, metatarsus not longer or a little longer than 3rd tarsal joint. Pronotum as broad as long, rounded posteriorly...............10. Timomenus Burr.
21 (20). Abdomen spindle-shaped; last tergite of $ narrow, strongly sloping, narrowing apically. Branches of forceps contiguous or ^subcontiguous at base. Anterior femora slender; metatarsus about twice as long as the 3rd tarsal joint.
22 (23). Pronotum about as broad as long. Sides of abdomen not recurved but often provided with tubercles. Forceps of 5 contiguous at base . . .
............................................. . 11. Eparchus Burr.
23 (23). Pronotum narrow, longer than broad. Sides of 6th to 9th abdominal segments recurved, acute in 5. Forceps of <5 subcontiguous at base
..................................................12. Narberia Burr.
24 (1). Totally apterous. Last abdominal tergite of $ sloping, narrowed posteriorly. Forceps of S with branches subcontiguous at base and with a sharp, practically vertical tooth in the basal fourth..............
..................................................13. Sondax Burr.
I. Subfam. FORFICULINAE
1. Genus FORFICULA L.
1 (2). Colour dull black. Pronotum considerably broader than long. Hind tibiae swollen at the middle, whence narrowed strongly to their apices. Antennae with 4th joint slightly shorter than the 3rd, 5th joint equal to the 3rd. Forceps of $ dilated just near the base only, strongly bowed, or sometimes with branches slightly remote at base (like to species of the genus Anechura). — North India from Sikkim in the East
to Punjab and Kashmir in the West; Tibet. . ......................
........................................1. F. schlagintweiti (Burr).
2 (1). At least the head or lateral margins of pronotum and elytra, or wings and legs light coloured. Pronotum transverse only in few species (cf. uvarovi,. sinica, mandarina, davidi). Hind tibiae normal, practically of equal breadth on whole length.
3 (12). Body not covered with long and dense hairs, practically naked.
4 (5). Elytra abbreviate, less than half as long as the pronotum, with hind margin strongly oblique, forming a sharpened angle with the external side; wings absent. Last abdominal tergite of <5 moderately narrowed apically. Forceps of 5 very slender, contiguous just near the base only, then regularly arcuate. Body entirely naked. — Tibet (fig. 34). .
2. F. uvarovi Sem. & B.-Bienko.
.224
SUMMARY
5 (4). Elytra well developed, more than half as long as pronotum, their hind margin transverse, not forming a sharpened angle with external sides.
6 (40). Forceps of S without isolated tooth on inner margin.
7 (37). Last abdominal tergite without longitudinal keel-shaped fold on sides before its hind margin. 4th antennal joint elongated, cylindrical or clavate, about twice as long as broad.
8 (26). Forceps of $ dilated and contiguous through at least one third of their length, in f. brachylabia practically in basal half.
9 (13). Pygidium of о large, strongly prominent, quite visible.
10 (11). Dark brown, very slender. Wings fully developed. Pygidium of $ parallel-sided, distinctly longer than broad, apex subtruncate. Abdomen with very dense and fine punctuation, hind margin of dorsal segments with a line of more coarse points. — Gan-su province, North-Western China (fig. 35)..................................3. F. splendida B.-Bienko.
11 (10). Dirty or brownish yellow, small, moderately robust. Wings absent. Pygidium of $ triangular, about as long as broad, lateral margins elevated and rugose. Abdomen with dense and fine punctuation. — Southern Crimea, Asia Minor, Greece and Cyprus . . . . I 4. F. aetolica Br.-W.
12 (3). Body densely pilose, especially legs, lower surface of abdomen and forceps. Elytra slightly abbreviate, hind margin moderately ( oblique, forming a slightly sharpened angle with lateral side. Wings absent. Brownish yellow, small. Abdomen scarcely punctured above. Pygidium and forceps as in the preceding species. — Western Caucasus from Balkaria and Maikop to Abkhasia; on a small isolated area near Stavropol (fig. 36).........................................5. F. kaznakovi Sem.
13 (9). Pygidium of S feebly prominent, small, often invisible.
14 (21). Wings developed, protruded from beneath of elytra.
15 (20). Pronotum not broader or a little broader than long, equal in width to head. Forceps of $ with interspace between inner margins of branches elongate-ovate.
16 (17). Body extremely slender, pale-yellow. Elytra unicolorous, not spotted. Forceps strongly punctured, very slender. Abdomen chestnut-brown, with relatively strong punctuation. — Eastern Transcaucasia, South-Western Turkmenia and Tashkent).....................6. F. sagitta Sem., sp. nov.
17 (16). Body moderately robust, reddish- or jet-brown. Elytra with a large pale spot or stripe on the disc. Forceps smooth, or slightly punctured in apical part only.
18 (19). Elytra with oval spot on the disc. Abdomen exceedingly minutely punctulate, in 3 distinctly dilated near the middle. Pronotum unicolorous, reddish or yellow. Pygidium of 3 very short, blunt, tubercular. — North Africa, including Egypt and Northern Sahara, Syria, Palestina, Arabistan and Kerman in Iran, Burma......................7. F. lucasi Dohrn.
J.9 (18). Elytra with a very long pale stripe on the disc. Abdomen coarsely punctured, especially in 5, practically parallel-sided. Pronotum with two
DERMAPTERA
225
11
longitudinal brownish stripes. Pygidium of 3 relatively long, narrowed apically, almost triangular, strongly sloping..........................
........................ ....... 9a. F. tomis (Koi.) f. euptera.
20 (15). Pronotum transverse, one and one third as broad as long, decidedly broader than the head. Forceps of <5 with inner space between the inner margins of branches practically circular.—-Szechuan province, Western China..............................................8. F. mandarina Borelli.
21 (14). Wings not protruding from beneath the elytra, entirely absent or rudimentary. Abdomen with dense and coarse punctuation.
22 (25). Body large, robust. Forceps of 5 in f. brachylabia dilated not more than in basal half; the dilated part ending in an obtuse or straight angle, without roundly produced lobule.
23 (24). Body less robust, not very large. Inner angle of eyes broadly rounded, very obtuse. Forceps of 5 with subparallel external margins, apical part in f. brachylabia moderately bow-shaped incurved. — European part of USSR from Moscow, Vitebsk, Vladimir, Gorkij, Kirov, Perm and Sverdlovsk in the North to Western Ukraine, the Crimea and Ciscaucasia in the South, Bessarabia, Caucasus, Turkish Armenia, Astrakhan district, Western Siberia, Kazakstan, mountains of Central Asia from Sun-garian Alatau to Zeravshan Valley, Pamir and Eastern Thien-Shan (figs. 10, 37, 38)........................................ 9. F. tomis (KoL).
24 (23). Body strongly robust and very large. Inner angle of eyes less rounded, practically rectangular. Forceps of 3 with concave external margins of dilated part; apical part in f. brachylabia strongly bowed, so that the whole external margin is strongly S-shaped incurved. — Ussuri region from Vladivostok to Khabarovsk, Sakhalin, Korea, Manchuria, North-Eastern China and Japan (figs 39 and 40) . . . . 10. F. robusta Sem.
25 (22). Body relatively small and slender. Forceps of <5 in f. brachylabia dilated in the basal three-fifths; the dilated part ending in a broad, roundly produced lobule. Head a little broader than pronotum. Anterior femora very short and thick. Coloration yellowish-brown.—1 Szechuan province, Western China (fig. 41). . . . 11. F. macrobasis B.-Bienko.
26 (8). Forceps of <5 dilated and contiguous less than in their basal third, often just near their base only. Wings fully developed.
27 (35). Elytra unicolorous, not spotted.
28 (36). Last abdominal tergite of 3 with two very feeble spherical inflations, the roots of forceps; or these inf la-
III
placed near the hind margin over tions entirely absent.
29 (32). Pygidium of 3 small, slightly prominent of practically invisible, considerably shorter than dilated part of forceps. Forceps of 3 with inner margins of dilated part contiguous, parallel to each other,
30 (31). External margin of forceps in 5 without a small tubercle at base. Head not broader than pronotum, with sutures nearly obsolete. Pronotum as broad as long, sides slightly convex. Body castaneous, abdomen Фауна СССР, Кожистокрылые 15
226 SUMMARY
**• * ► 4
reddish-chestnut, very finely punctured. — From Kiao-Tchau in North-Eastern China, Tonkin, Formosa arid Northern India . . '.............
...........................12. F. ambigua Burr.
31 (30). External margin of forceps in 8 with a small tubercle at base. Head large, broader than pronotum, sutures very distinct. Pronotum practically half as broad as long, sides slightly convex. Body brownish-black, abdomen shining black, strongly punctulate. — Szechuan province, Western China.....................................13. F. sinica B.-Bienko.,
32 (29). Pygidium of 8 very large, strongly prominent, practically as long as broad, moderately narrowing posteriorly, hind margin subtruncate. Forceps of 8 with basal dilated part very short, slightly remote at base,, scarcely longer than pygidium and furnished with an obtuse, broad and triangular tooth on the apex of dilated part.
33 (34). Larger. Brownish-chestnut, with brownish-red head. Antennae not setaceous; 3rd joint practically cylindrical. — Japan......................
................................................14. F. mikado Burr.
34 (33). Smaller. Brownish-black, with shining black head. Antennae setaceous, 3rd joint distinctly clavate. — Szechuan province, Western China
. .....................................15. F. bferezdvskii B.-Bienko.
35 (27). Elytra with rounded pale spot in anterior part. Upper surface of abdomen and forceps with very dense and coarse punctuation. Large, head reddish, pronotum and legs pale-yellow, abdomen and forceps jet-brown. — Corsica, Southern Balkans, Syria, Asia Minor including Turkish Armenia, Nukha districh in Eastern Transcaucasia, Black-Sea coast of Caucasus from Batum in the South to Nbvorossijsk in the North, Southern Crimea.................................... 16. F. smyrnensis Serv.
36 (28). Last abdominal tergite of $ with two strong, in smallest specimens very long, in normal specimens obtusely-conical projections placed near the hind margin over the roots of forceps. Pronotum as broad as long or slightly transverse, hind margin faintly rounded, chestnut-brown or black, abdomen densely punctulate. — From Kiao-Tchau in North-Eastern China to Gan-su and Szechuan provinces in Western China; Amdo district in North-Eastern Tibet (fig. 42)................17. F. davidi Burr.
37 (7). Last abdominal tergite with longitudinal keel-shaped fold on sides before the hind margin, more distinct in 8. 4th antennal joint, short, ovate, not or slightly longer than broad. Abdomen in 8 punctured. Forceps of 8 dilated and contiguous in baSal fourth or third, then narrowing towards the middle and there strongly bowed inwards.
38 (39). Pronotum slightly broader than long. Wings absent. Branches of forceps in 8 roundly bowed inwards, so that the interspace between them is ovate or circular. — Southern Europe from Spain in the West to Balkan Peninsula, including Southern Dalmatia, and Transsylvania in the East, Asia Minor to Trebizond, North Africa............................
..............................................18. F. decipiens Gene.
DERMAPTERA
227 ч
39 (38). Pronotum as broad as long. Wings fully developed. Branches of forceps in 5 angularly bowed inwards^ in the middle, so that the interspace between them is rhomboidal or quadrate. — Southern Balkans, Asia Minor, Syria, Palestina, Arabistan in South-Western Iran...................
................................................19. F. lurid a Fischer, *
40 (6). Forceps of 3 with an isolated tooth on the inner margin placed at some distance from the contiguous and dilated basal part; in f. brachylabia the distance being sometimes very short but more or less distinct.
41 (48). Body hairless, relatively large. Wings developed, protruding from beneath the elytra. Forceps of, 3 dilated and depressed not more than in the basal third of their length; an isolated tooth situated before the middle of forceps.
42 (47). Antennae with elongated joints; 4th joint slightly shorter that the 3rd, cylindrical, from two to three times ar long as broad. Last abdominal tergite without any longitudinal keel-shaped triangular fold on sides before the hind margin.
43 (46). Pygidium of 3 very short, practically indistinct. Forceps of 3 contiguous at base, not divided from each other by pygidium; dilated part long, occupying basal third or practically half the length of forceps.
44 (45). Forceps of 3 without longitudinal furrow on upper and lower surface, apical part thicker. — Far East Siberia from lower course of Amur to Vladivostok, Korea, Manchuria, North-Eastern China (fig. 43) ....
. ............................................. 20. F. vicaria Sem.
45 (44). Forceps of 3 with distinct longitudinal furrow on the upper and lower surface, the former is more developed, than the latter; apipal part slender. — Gan-su province, North-Western China............................
................................................21. F. burriana Sem.
46 (43). Pygidium of 3 long, large, roundly triangular, strongly prominent, seen in profile apex markedly produced over the level of forceps. Forceps of 3 with inner margins of the basal part not touching each other, divided by prominent pygidium; dilated part very short. — Szechuan province, Western China (fig. 44) . . 22. F. subauricularia B.-Bienko.
47 (42). Antennae with shorter basal joints; 4th joint scarcely longer than broad, practically spherical, nearly half as long as the 3rd. Last abdominal tergite with longitudinal keel-shaped fold on the sides before the hind margin, triangular and more distinct in 3. — The whole of Western Europe, European part of USSR from the Crimea, Northern Caucasus and Astrakhan in the South to Leningrad, Karelia, Moscow, Urzhum and Sverdlovsk in the North, the whole of Caucasus, Asia Minor, Syria, Northern and Western Iran to Teheran, Kopet-Dagh to Askhabad, Western Kasakstan between Wolga and Ural and Western Siberia to Omsk and Kamen; not distributed in Central Asia, Central and Eastern Siberia (figs. 9, 45 and 46) . . . . 23. F. auricularia L.
15*
о
228
SUMMARY
48 (41). Body small, covered with sparse hairs. Wings absent. Forceps of 3 dilated and depressed at leastjn the basal half; inner tooth disposed in the middle or behind the middle of forceps. — Southern Europe from Iberian Peninsula, Southern France, Corsica, Sardinia, Mallorka and Sicily to Balkan Peninsula and Syria; Algeria and Marocco .....
24. F. pub esc ens Serv.
Forficula sagitta Semenov-Tian-Shanskij, sp. n.
? Forficula mesmini Azam, Bull. Soc. Ent. Fr., 1906: 81 (g, partim).
3. F, senegalensi Serv. similis et proxime affinis, sed minor, capite, pro-noto, elytris, alarum squamis pallidioribus, his stramineis, pronoto margine l?asali minus arcuato, elytris longioribus, alarum squamis in forma euptera longitudine elytrorum sesqui tantum brevioribus; abdomine tergito ultimo medio in parte posteriore plus minusve profunde utrinque leviter impresso; forcipis cruribus parte basali minus elongata. g differt a <3 iisdem signis atque in F. senegalensi. Forma euptera: alis omnino explicatis, aptis ad volandum, squamis elongatis elytrorum longitudine subsesqui brevioribus. Forma bra-chyptera: alarum squamis longitudine elytrorum multo brevioribus, pronoto et imprimis elytris et squamis alarum obscurioribus, brunnscentibus. Long. 3 (sine forcipe) 9.5—-10, elytrorum 2.5—3, forcipis 4—4.5; g (sine forcipe) 9.5—11.5, elytrorum 2.5—3, forcipis 3 mm.
Hab. inde a Transcaucasia orientali ad Turkestan occidentale usque; district Aresh: Geok-tapa (A. Shelkovnikov et R. Schmidt: 28 VII et 20 VIII 1901:1 3, 2 g); Muganj (J. Schreiner: 1913:1 3); ad fl. Sumhar (Turcomania occid.) (V. Michajlov: 27IV—27—V 1917; 1 g formae brachypterae); Tashkent, in horto urbano (N. Zarudnyj: 15 IV 1916: (3). — 6 specimina (3 3, 3 j). (coll. A. Semenov-Tian-Shanskij).
2. Genus MESASIOBIA SEM.
1 (1). Jet-black, posterior part of head, antennae and legs dirty-yellow. Pronotum as broad as long, hind margin practically straight. Abdomen with excedingly minute punctures, last tergite of 3 with two strong rugose projections in the middle, very long in f. macrolabia. — Kuldja in Chinese Sungaria and Transilian Alatau (figs 47, 48)..........................
1. M. h emix ant ho car a Sem.
3. Genus OREASIOBIA SEMENOV, gen. nov.
Genus generi Anechura Scudd. proximiim, a quo discedit antennarum articulis magis elongatis; tergito abdominali ultimo in 3 supra marginem posticum utrinque tuberculo valido acuto conico vel etiam fere spiniformi instructo; pygidio late transversal! et prominulo, forcipe 3 longo et gracili, in forma normali (macrolabia) corpore sine capite plerumque longiore, cruribus tenuibus, cylindricis, fere horizontalibus, inter se sat late distantibus, sed basi duobus dentibus validis uno altero superposito, introrsum directis armatis, ad apicem tantum introrsum curvatis, pilis longis obsitis.
DERMAPTERA
229
Typus generis: Forficula fedtschenkoi Stiuss. 1874, montanis Asiae centralis propria. [Semenov-Tian-Shanskij].1
1 (2). Head unicolorous, black. Forceps hairless, basal dilated part in 8 without a conical projection above. Last abdominal tergite of 3 with two small tubercles or (in f. brachylabia) they are indistinct. Body, including antennae, pronotum, elytra and legs black or brownish-black. — Sutlej district and Punjab in North-Western India; Kashmir......................
..............................................1. 0. stoliczkai (Burr).
2 (1). Head reddish. Forceps with long hairs, basal dilated part in <5 with a conical projection above. Last abdominal tergite of 3 usually armed with two very strong and long conical projections........................
.........................................2. 0. fedtschenkoi (Sauss.).
a (b). Elytra without a light spot, unicolourous black. — Kashmir........
.............................2. a. 0. fedtschenkoi calciatii (Bor.).
b (a). Elytra with a large dirty yellow spot on the disc, or completely dirty-yellow. — Mountains of Central Asia from Chinese Sungaria, Trans-ilian Alatau, Alexandrovsky range and Karatau in the North to Hissar and Zeravshan ranges in the South-West and Pamir in the South (figs. 49 and 50)..............2b. 0. fedtschenkoi fedtschenkoi (Sauss.).
4. Genus ANECHURA SCUDD.
1 (18), Elytra well developed or slightly abbreviated but at least half again as long as the pronotum; wings developed or absent.
2 (9). Forceps of 5 seen in profile quite straight. (1. Subgen. Odontopsalis Burr).
3 (6). Wings undeveloped. Pygidium of <5 strongly prominent, lamellate. Forceps of 8 short, robust.
4 (5). Legs pale-reddish. Pygidium of 3 short, broad, apical part not incurved, nor elevated above the level of forceps. Forceps of 3 not dilated in basal part, with a small tooth on the inner margin before the middle. — Szechuan province, Western China (fig. 51).........................
........................................1. A. (0.) potanini B.-Bienko.
5 (4). Legs brownish-black. Pygidium of 8 longer, apical part strongly elevated, seen in profile produced above the level of forceps. Forceps of 8 moderately dilated at base, without inner tooth. — Eastern Tibet.................
.......................2. A. (0.) forficuliformis Sem. & B.-Bienko.
6 (3). Wings developed, protruded from beneath? the elytra, usually with a yellow spot, occupying practically whole wing. Pygidium of 8 not prominent, vertical. Forceps of 8 long, slender.
7 (8). Dorsal surface of abdomen with distinct, moderately] coarse punctuation. Forceps of 8 with a single broad triangular tooth on the inner
4
1 We give the genus Oreasiobia, established by D-r A. Semenov-Tian-Shanskij another eaning than does the Author; we consider the presence of conical processes on the last tergit
in the 8 and hairy forcipes as features of specifical value. We moreover include in the same genus also Anechura stoliczkai Burr from North India and Kashmir.
II
230
SUMMARY
margin before middle. — Far East Siberia from lower course of Amur in the North to Vladivostok, Japan, Korea, Manchuria and Chensi, Gan-su and Szechuan provinces in China (fig. 52).......................
..........................................3. A. (0.) japonica (Borm.).
8 (7). Dorsal surface of abdomen with very feeble puncturation. Forceps of 5 with a bidentate oblique projection on the upper margin in basal third. — Vladivostok, Sakhalin and Japan from Yezo to Nippon. . . .
. . ......................................4. A. (0-) harmandi (Burr).
9 (2) Forceps of 3 seen in profile undulating, strongly bowed downards in the basal part. (2. Subgenus Anechura in sp.)1.
10 (17). Wings develcped, protruded from beneath the elytra. Pygidium of S practically not prominent, with vertical hind margin, in g simple, truncate or slightly excised at apex. Forceps of 3 thick, not flattened laterally.
11 (14). Elytra unicolorous, not spotted. Forceps of 5 cylindro-conical at base, without basal tooth on their upper surface.
12 (13). Head with distinct sutures. Legs short, anterior femora greatly swollen. Forceps of 3 short. — Szechuan province in Western China and district Amdo in North-Eastern Tibet (fig. 53)...............................
.................................5. A. (in sp.) sven-hedini B.-Bienko.
13 (12). Head with indistinct sutures. Legs long, slender. Forceps of 3 long. — Island Quelpart in Southern Korea . . 6. A. (in sp.) quelparta Okam.
14 (11). Elytra dark, with a big yellow spot in the anterior part. Forceps of 3 usually with a very strong basal triangular tooth on the upper surface.
15 (16). Head reddish, at least on the occiput. Femora not darkened or darkened in the apical portion only. Body more robust and large.— Mountains of Western Europe from Pyrenees, Southern France, Alpes to Savoy, Jura, Vosges and Alsace in Eastern France; Southern Germany. Northern Czechoslovakia and Silesia in the North to Sardinia, Sicily, Northern Italiy, Northern Balkans in the South, Asia Minor, Caucasus, Western Ciscaucasia, NE. European part of USSR, Ural mountains, southern part of Siberia from Omsk, Tatarsk and Novosibirsk in the North and Irkutsk in the East to Northern. Kazakstan, Altai mountains, Kol-binsky range, Saur mountains and North-Eastern. Mongolia in the South; not distributed in Crimea, in the largest part of European USSR and in Central Asia (fig. 54 and 58) . 7. A. (in sp.) bipunctata (Fabr.).
1 The Mexican Anechura vara Scudd. belongs to an independent subgenus wit-h the following characters. Anechurella, subg. n. Closely allied to the subgenus Anechura in sp. and differs as follows: antennae with more slender joints, 5th joint usualy longer than the 3rd; last abdominal tergite of 3 smooth, without lateral projections above, but with an obtusely conical small tooth on lateral sides just at the lower surface. Forceps of 3 undulate but inner margin with distinct srriall denticles in the basal part. Type: Anechura (Anechurella) vara Scudd., 1876, from Puebla, Mexico.
4
DERMAPTERA
231
I
I'
16 (15). Head entirely black. Femora usually black or dark-brown, apex not darkened, rarely femora unicolarly reddish-yellow. Body smaller, more slender ...................................8. A. (in sp.) asiatica Sem.
-a (b) Light spot of elytra large, extending beyond the middle of the disc. Abdomen and legs darker, usually brownish-black or black. —Sarepta, Orenburg and lower course of Volga in South-Eastern. European part of USSR, Kazakstan from lower course of Ural river, Mugodzhary mountains and the lake Tshalkar-Tenis to the lakes Saisan and Balkash in the North, Mujunkum desert, mountains of Southern Kasak-stan and Central Asia from Kara-tau and Alexandrovsky range in the North to Kopet-Dagh, Southern Tadzhikistan and Pamir in the South, Chinese Sungaria to Urumchi in North-Western Mongolia, Eastern Iran from Khorasan in the North to Kirman in the South (fig. 55).
................................8 a. A. (in sp.) asiatica asiatica Sem.
Ъ (a). Light spot of elytra small, not passing the middle of the disc. Abdomen and legs less dark, the former usually reddish brown in apical part, the latter reddish-yellow. — Kashmir and SWestern Tibet. . . .
..............................8 b. A. (in sp.) asiatica zubovskii Sem.
17 (10). Wings undeveloped. Pygidium of 5 strongly prominent in the form of a flattened plate, in g with two lateral and ope median spinules on apex. Forceps of <5 strongly flattened laterally in basal third; lower margin with a small thin tooth at base of forceps. Blackish-brown, with yellowish pronotum, elytra and legs. — Gan-su and Szechuan provinus, China . .
.......................................9. A. (in sp.) filchneri (Burr).
18 (1). Elytra strongly abbreviate, shorter than pronotum; if subequal in length, their hind margin obliquely truncate. Forceps of 3 without or with a very feeble tooth. '
19 (26). Forceps covered with short hairs in apical part. Mesosternal plate transverse. Anterior femora slender, not inflated, longer than pronotum. (3. Subgenus Burriola Sem.)
20 (23). Elytra nearly equal in length to pronotum, hind margin strongly oblique. Body large, dark, with pronotum dark-brown, practically black. Last abdominal tergite slightly widened posteriorly.
21 (22). Forceps of 3 strongly undulating, with a small acute, conical tooth on external surface near lateral side of the last tergite. Head black or with a feeble brownish shade. — Szechuan province, Western China, and E.Tibet (fig. 56,59).................. 10. A. (B.) semenovi (B.-Bienko).
22 (21). Forceps of 3 not undulating, practically straight, without conical tooth at base. Head dark-brown.— Basin of Hoang-ho, Tibet (fig. 57).
..............................11. A. (B.) tibetana Sem. & B.-Bienko.
23 (20). Elytra shorter than pronotum, hind margin transversely truncate. Body small, reddish-brown or at least pronotum not darkened, gol-den-yello^. Abdomen of 5 slightly widened in the mididle, then scarcely narrowed apically.
1
232
SUMMARY
24 (25). Pygidium of $ with two long, transverse, horizontal processes at the sides. Forceps of 5 unarmed, seen from above practically straight,, regularly convergent apically; in g more robust and long (3.8 to 4.1 mm.). — Eastern Tibet from Bur khan-Buddha in the North to district Kham;
on the South (fig. 60).................12. A. (B.) przewalskii (Sem.).
25 (24) Pygidium of S without horizontal processes, slightly angulate on the sides. Forceps of $ with a small tooth on the lower inner side before^
the middle, more slender, slightly arcuate; in g shorter (3.0 to 3.6 mm.). — From Sining mountains, Northern Gan-su, to the region of lake Kuku*
nor, North-Eastern Tibet
26 (19) Forceps hairless. Mesosternum as broad as long. Anterior femora short, thick, somewhat inflated in basal half, not longer than pronotum
(4 Subgenus Borelliola Sem.). Upper surface of abdomen smooth, with indistinct punctuation. Pygidium of S short. Forceps of 5 without tooth on inner margin, in f. macroladia slightly undulating, slender, in f. bra-* chylabia with dilated and elevated inner margins of basal half. — Western Caucasus from Kislovodsk and Maikop to Sukhum, Novy Afon and, gagry in Abkhasia; an isolated area placed on Stavropol plateau, (figs., 11 and 12)..........................................................
..........................................14. A. (B.) euxina (Sem.}
5. Genus CHELIDURA LATR.
1 (1). Forceps of S without tuberculiform projection on inner margin, in f. macrolabia long, slightly incurved, practically straigt, in f. brachylabia short, roundly-incurved. — Southern Europe from Pyrenees and Southern France to Southern Alps........................ 1. Ch. aptera (Charp.).
I
6. Genus EUMEGALURA B.-BIENKO
1 (1). Dark reddish-brown, almost black. Pronotum transverse, with sides and hind margin slightly rounded. Forceps of 3 slender, three times as long as pygidium, slightly undulating, apical part covered with dense short hairs on the outer surface.— Szechuan province, Western China (fig. 61)......................................l.E. mandarina B.-Bienko.,
7. Genus APTERYGIDA WESTW.
1 (1). Dirty-yellow. Wings present but completely concealed and do not protrude from beneath the elytra. Abdomen with very fine and dense punctuation. Forceps of 3 with small tooth on the inner margin at the base and near the middle. — The largest part of Western Europe, including Southern. England, Belgium, Northern France and Southern Sweden in the North, Poland, Bessarabia and Ukraine from Kamenetz-Podolsk
to the river Psel in the East (fig. 62) . . . 1. A.
edia (Hagenb.).
11
DERMAPTERA
233
8. Genus CHELIDURELLA VERH.
1
1 (2). Smaller (6. 5 to 11 mm.). Pygidium of 5 strongly produced, apex bowed upwards in shape of a long, subvertical horn; in g seen from below moderately narrowing apically, longer than broad. La stabdominal tergite of c? with two relatively strong tubercles at hind margin. — The largest part of Western. Europe from Pyrenees, Piemont and Northern Balkans in the South to Northern France, Belgium, Holland, England, Denmark, Southern Sweden, Eastern Prussia and Latvia in the North,
Poland, including Western Ukraine, Soviet Ukraine and Rumania . , ....................................... 1. Ch. acarthopygia (Gene)
2 (1). Larger (8 to 15.5 mm.). Pygidium of 3 short, scarcely prominent, hind margin vertical, with a tubercle; in g seen from below not narrowing apically. Last abdominal tergite of 5 with faintly elevated, almost
indistinct tubercles r
3 (4). Blackish-brown. Pydidium of d* with a large prominent tubercle; in g a little longer than broad, hind margin slightly excised, with sharpened and pointed lateral angles. Forceps of 3 regularly arcuate.— Tyrol............................................. 2. Ch. mutica (Krauss).
4 (3). Light brown. Pygidium of 3 with a feeble, quite flat tubercle; in g about twice as long as broad, hind margin truncate. Forceps of 3 with branches angularly bowed inwards in the middle.—Transsylvanian. Alps in Rumania and Kamenetz-Podolsk, Ukraine (fig. 63)...................
.......................................3. Ch. transsilvanica Ebner.
9. Genus ALLODAHLIA VERH.
1 (1). Blackish-chestnut, dull, scabrous. Elytra covered with a dense mass, of small tubercles and a few larger points. Pygidium of 3 with a spinule at each angle. — Northern part of British India, Tenasserim, Burma, Tonkin, Malay Archipelago, Philippine Islands, Formosa and China from Southern part to Kiao-chau on the North-East............................
..........................................1. A. scabriuscula (Serv.)
2. Subfam. OPISTNOCOSMIINAE
10. Genus TIMOMENUS BURR.
1 (2). Forceps of 3 long, slender, unarmed. Sides of 6th and 7th abdominal tergites produced into an obtuse angle, of which the former is furnished with a small tubercle. Body jet-black. — Kiao-Tchau in North-Eastern, China (and Formosa?) .....................................1. T. inermis Bor.
2 (1). Forceps of 3 armed with teeth.
3 (4). Forceps of 3 armed with a single large tooth on the upper surface. Sides of 5th to 9th abdominal tergites without tubercles. Body reddish
brown, Eastern
abdomen feebly and densely punctulate. — Kiao-Tchau, North-
China ................................ 2. T. unidentatus Bor.
234
SUMMARY
4 (3). Forceps of 3 with a large tooth on the upper and small tooth on the inner margin.
5 (6). Jet-black, with yellowish-chestnut elytra. Abdomen densely punctured above. Forceps in 3 with upper tooth placed at base. Sides of the 4th and 5th tergites in 3 armed with a tubercle. — South Ussuri region, Korea, Manchuria, Kiao-Tchau in North-Eastern China and Formosa (figs. 64 and 65)..................................4 T; komarovi (Sem.).
6 (5). Reddish-brown. Abdomen smooth; sides of 6-th to 9th tergites in 3 produced into small hooked tubercle. Upper tooth of forceps in 3 situated at the end of basal third. — Bhutan in Northern India, and North-Eastern China...........................5. T. aesculapius Burr.
II. Genus EPARCHUS BURR
1 (2). Forceps of 3 with branches subcontiguous at base, then curved outwards and in the apical half inwards, thus enclosing an oval space; upper margin with a rounded tubercle near the base. Sides of 6th to 8th abdominal tergites in.3 with a strong conical tubercles. — British India, including Burma, Malay Archipelago, North-Eastern China...................
........................................... - J. E. insignis (Haan).
2 (1) Forceps of 3 feebly incurved, practically straight, upper margin unarmed. Sides of abdominal tergites simple.
3 (4). Last abdominal tergite of 3 narrowing apically. Forceps of 3 simple, unarmed. Brown abdomen finely punctulate. — Bhutan in Northern India, and North-Eastern. China ... •........................2. E. oberthuri Bor.
*
4 (3) Last abdominal tergite of 3 subquadrate. Forceps of 3 very long, inner margin with a smal tooth at base and in the middle. Pronotum a little broader than long, sides parallel. — Insel Yezo, Northern Japan . . .
...................................3. E. yezoensis (Mats. & Shiraki).
12. Genus NARBERIA BURR
1 (1). Head with very distinct sutures. Sides of 6th to 9th abdominal tergites of 3 with granulous longitudinal keel, more developed and furnished with a small tubercle on the 8th and 9th tergites. Forceps of 3 denticulate on inner margin, strongly variable. — North-Eastern. China and Borneo...........................................1. N. tuberculata Bor.
13. Genus SONDAX BURR
1 (1). Brownish-black, head reddish. Pronotum half again as broad as long; mesosternal plate about twice as broad as long. Abdomen smooth. — Szechuan province, Western China (fig. 65) . . 1. S. potanini B.-Bienko-
I
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Названия подсемейств, семейств и высших групп выделены жирным шрифтом; прочие названия — прямым шрифтом, причем роды и подроды начинаются с заглавной буквы» синонимы и прежние родовые названия для видов — курсивом. Жирными цифрами обозначена страница, на которой дано описание.
INDEX
The names of subfamilies, families and higher groups are printed in fat characters; other names in Roman type; synonymies and the former generic names for species in italics. The page on which the description occurs is indicated by thickened numerals.
Acnodes 81
acanthopygia, Chelidura 192
acanthopygia, Chelidurella 62, 68, 191, 192, 193, 233
acanthopygia, Forficula 192
aesculapius, Eparchus 200
aesculapius, Opisthocosmia 200
aesculapius, Timomenus 197, 200, 234
aetolica, Forficula 67, 126, 133, 224
albipennis, Apterygida 190
albipennis, Chelidura 190
Allodahlia 44, 54, 123, 194, 213, 222, 233
AHostethinae 84
ambigua, Forficula 128, 142, 226
americana, Psalis 20
Anataelia 44, 54, 56, 81
Anataeliinae 35, 56, 82
Ancistrogastrinae 39
Anechura 33, 39, 42, 43, 44, 46, 51,- 52, 53, 60, 63, 64, 121, 122, 163, 172, 211, 212, 213, 222, 229
Anechurella 164, 230
Anechurinae 39, 121, 210
Anisolabis 7, 44, 53, 54, 58, 59, 60, 63, 65, 84, 86, 87, 89, 92, 100, 210, 2P, 217
annulipes, Anisolabis (Euborellia) 17, 88, 95, 96 218
annulipes, Anisolabis 26
annulipes, Euborellia 96
annulipes, Forficesila 96
Antisolabis 36, 37, 211
anthracina, Chelidura 174
Apachyidae 10, 28, 34, 36, 81
Apachyus 1, 24, 36, 211
Apolabis 56, 86, 83, 91, 92
aptera, Chelidura 186, 187, 232
aptera, Forficula 187
Apterygida 27, 39, 43, 44, 52, 53, 56,58, 59, 60, 65,123, 189, 211, 212, 214, 222, 232 ,
arachidis, Apterygida 112
arachidis, Chelidura 112
arachidis, Forficula 112
arachidis, Labia 112
arachidis, Prolabia 26, 70, 100, 112, 189, 213, 220
Archidermaptera 1, 8, 28, 40, 77, 210, 211, 215
Arixenia 78, 79
Arixeniidae 28, 34
Arixenina 2, 3, 22, 28, 77, 78, 210, 215
asiatica, Anechura in sp. 22, 24, 26, 62, 68, 70$ 71, 166, 176,178, 209, 214, 231
aihymia, Apterygida 169
auricularia, Forficula 9, 15, 16, 18, 19, 20, 21, 22, 27, 34, 38, 68, 72,124,130,151,152, 209, 212,213,227 л
barroisif Forficula 135
beebei, Forficula 130
beelzebub, Forficula 33, 125
bengalensis, Labidura 101
berezovskyi, Forficula 128, 144, 226
236
УКАЗА ТЕЛЬ
bidens, Forficula 101
bi dens, Labidura, var. 101
biguttata, Forficula 145
biguttata, Forficula 174
bipartitus, Elaunon 38
bipunctata, Anechura in sp. 23, 62, 67, 68, 70, 163, 166, 172, 174, 178, 209, 230
bipunctata, Forficula 174
biroi, Narberia 205
Boreilia 95
Borelliola 46, 50,53, 64, 121, 163, 164, 167,184, 214
Brachylabinae 36, 83
Brachylabis
Brachylabis, partim 86
burri, Pseudisolabis 63, 106, 219
burriana Forficula 46, 129, 150, 151, 152, 212, 213, 227
Burriola 46, 50, 53, 62, 64, 163, 164, 166, 179, 184, 187, 188, 214
calciatii, Anechura 161
calciatii, Oreasiobia 163, 229
canariensis, Anataelia 83
caucasica, Forficula 134
cavaleriei, Anisolabis (Mongolabis) 88, 94, 218
cavalii, Apterygida 58
Chaetospania 36, 38
Challia 44, 56, 81, 82, 216
Chelidura 7, 44, 53, 62, 123, 186, 191, 212, 214, 222, 232
Chelidurella 39, 53, 56, 58, 59, 60, 62, 65,123, 191, 211, 212, 214, 222, 233
Chelidurmae
Chelissochs 19, 44, 60, 108, 116, 220
Chelisochnae 8, 10, 38, 108, 115, 220
chingalaris, Labidurodes 115
collosea, Titanolabis 21
com.prim.ens, Chelisoches 116
Copiscelis 109
curta (?'Chelidura) 134, 191
curvicauda, L bia 110, 213, 220
curvicauda, Forficesila 110
davidi, Forficula 46, 129, 146, 156, 226
decipiens, Forficula 129, 138, 146, 226
decolyi, Forficula 24, 25
Dermaptera 28, 49, 57, 77, 79, 210, 215, 216
Dermatoptera 79
Demogorgon 101 ,
Diaperasticinae 39
diminuta, Chelidura 143
Diplatys 8, 19, 29, 35, 36, 37, 211
dolichus, Nesogaster 113
Doru 35, 40, 41, 211
dufouri, Labidura 105 eburnea, Labidura, var. Echinosoma 35, 36, 211
Echmosomatinae 81, 216
Elaunon
eoa, Anechnra 169
Eparchus 42, 44, 54, 124, 200, 223, 234
erythocephala, Labidura 101
esau, Arixenia 78, 215
escherichi, Forficula 135
Esphalmeninae 10
Esphalmenus 37
Euborellia 56, 86, 88, 95, 100
Eudermaptera 12, 13, 28, 35, 36, 40, 79, 80
107, 216, 219
Eudohrninae 39
Eumegalura 44, 53, 62, 123, 187, 212, 214, 222
Euplexoptera 1, 79
euxina, Anechura (Borelliola) 15, 17, 67, 167
184, 214, 232
euxina, Burriola 184
euxina Chelidurella 184
fall ax, Anisolabis
feae, Hamaxas 119
fedtschenkoi, Anechura 161
fedtschenkoi, Oreasiobia 68, 72, 158, 159, 161.
178, 229
fedtschenkoi, Forficula 161
femoralis, ,,Psalis" 98
femorata, Apterygida 199
filchneri, Odontopsalis
filchneri, Anechura in sp. 166, 178, 231
fletcheri, Challia 82, .83, 216
Forcinella 86, 89
Forcipula 35, 36
Forficeslla 101
Forficula 29, 39, 40, 41, 42, 44, 51, 52, 53, 56, 57, 59, 60, 62, 64, 65, 122, 124, 210, 211, 214, 221, 223
Forfi ularia 79
Forficulidae 28, 40, 43, 44, 80, 120, 210, 211, 221
Forficulina 28, 79, 210
Forfxculinae 39, 121, 122,124, 196, 221, 223
forficuliformis, Anechura (Odontopsalis) 33, 34,
52, 165, 168, 169, 210, 221, 229
formos na, Anisolabis 86
fortis, Protodiplatys 77, 78, 215
gerstaeckeri, Diplatys 20
gigantea, Forficula 101
gravidula, Apterygida 112
greeni, Diplatys 20
Guanchia 44, 53, 212
УКАЗА ТЕЛЬ
237
Мм
Hamaxas 44, 54,108, 119, 220, 221
iharmandi, Anechura (Odontopsalis) 70, 71, 165,
167, 171, 230
harman di, Odontopsalis 171
Jiellmanni, Forficula 137
Hemxmeroidaea
hemixanthocara, Mesasiobia 68, 156, 228
herculeana, Labidura, var.
hottentotta, Apolabis 91
icterica, Forficesila 101
Inermis, Eparchus 201
inermis, Labidura, var. 101
inermis, Timomenus 197, 233
insignis, Eparchus 200, 202, 234
insignis, Opisthocosmia 202
insignis, Forficula 202
Isolabella 44
isomorpha, Apolabis
jacobsoni, Arixenia 78, 79, 215
janeirensis, Anisolabis (Euborella) 95
japonica, Anechura (Odontopsalis) 70, 165, 168, 169, 230
japonica Labidura, var. 101
japonica, Odontopsalis 169
japonica, Forficula 169
Kalocrania 11
.Karschiellinae 81
kaznakovi, Forficula 67, 127, 134, 191, 224
Kinesis 38
komarovi, Opisthocosmia 199
komarovi, Timomenus 54, 71, 197, 199, 234
kozlovi, Hamaxas 119, 221
Labia 29, 36, 38, 44, 60,108, 109, 218, 220
Labidura 44, 60, 84, 101, 217, 219
Labiduridae 28, 34, 35, 36, 44, 55, 80, 81, 83, 216
Labidurinae 83, 84, 100, 217, 219
Labidurodes, part. 114
Labiduromma 34
Labiidae 28, 34, 36, 44, 80, 100, 108, 218
Labiinae 38, 40, 108, 109
Iaminiger a, Apterygida 147
lamin:gera, Forficula 147
Lamprophorus 38
lewisi, Anechura 171
lewisi, Nesogaster 114, 115, 220
lewisi, Odontopsalis 171
lewisi, Spongiphora 115
lewisi, Spongovostox 115
lineare, Doru 21
Lithinus 44, 54
livida, Labidura, var. 101
lividipes, Forficula 104
lividipes, Labidura 104
lividipes, Nala 26, 104, 213, 219
Lobophora 116
longipyga, Apterygida 143
lucasi, Forficula 127, 135, 224
lurida, Forficula 129, 148, 227
macrobasis, Forficula 21, 46, 128„ 141, 212, 225
mandarina Eumegalura 129, 186, 187,232
mandarina, Forficula 127, 136, 225
marginalis, Anisolabis (Apolabis) 88, 91, 92, 218
marginalis, Apolabis 91
marginalis, Forcinella 91
maritima, Anisolabis in sp. 23, 26, 86, 87, 89,
90, 209, 213, 217
maritima, Forficesila 89
maritima, Forficula 89
mauritanica, Anisolabis 563
maxima, Anisolabis 56, 58
media, Apterygida 68, 189, 190, 191, 232
media, Forficula 190
meridionalis, Forficula 105
Mesasiobia 44, 53, 62, 121, 122, 155, 156, 212,
221, 228
mesmini, Forficula 135, 137
Mesochelidura 39, 44, 53, 56, 58, 59, 65, 211,
212, 214
mikado, Forficula 128, 143, 144, 226
minor, Labia 20, 21, 26, 60, 68, 70, 109, 110,
220
minor, Forficula 110
minuta, Anisolabis 97
modest a, Forficula 118
moesta, Anisolab's 98
moesta, Anisolabis (Euborellia) 59, 68, 89, 95,
98, 99, 219
moesta, Borelia 98
moesta, Euborellia 95, 98
moesta, Forficesila 98
Mongolabis 56, 86, 88, 92
morio, Chelisoches 18, 19, 21, 60, 116, 221
morio, Forficula 116
mutica, Chelidura 193
mutica, Chelidurella 192, 193, 194, 233
Nala 44, 60, 84, 104, 217, 219
Narberia 42, 44, 54, 124, 205, 223, 234
Nesogaster 44, 54, 108, 113, 219, 220
Nesogastrella 113
nigripenn s, Forficula 112
nigritus, Labidarodes 114
nigritus, Nesogaster 114, 220
oannes, Timomenus 195
oberthuri, Eparchus 201, 202, 203, 234
obliquotruncata, Semenoviola 32, 79, 210
'238
УКАЗА ТЕЛЬ
Odontopsalis 33, 53, 121, 163, 164, 165, 167 Opisthocosmia 28
Opisthocosmiinae 8, 38, 121, 196, 233
Oreasiobia 44, 53, 62, 122, 158, 159, 210, 212, 214, 222, 228
orientalis, Forficula 148
orientalis, Anechura, var. 174, 176
orsinii, Anechura 45, 173, 212, 213
pacifica, Anisolabis 86
pallidicornis Forficula 147 pallipes Duf. Forficula pallipes F., Forficula 101 pallipes, Anisolabis 104 pallipes, Anisolabis (Euborellia) 89, 98, 218, pallipes, Euborellia 98
Paralabidura 104
parallela, Narberia, var.
Parisolabinae 35, 83, 84, 105, 217, 219
Parisolabis 35
Parisopsalis 35
pectoralis, Anisolabis 26, 70, 87, 100
pectoralis, Forficula 100
Perirrhytus 44, 53, 212
Platylabia 1, 24,
plebeja, Anisolabis (Euborellia) 88, 97, 98, 218
plebeja, Euborellia 97
plebeja, Labidura 97
plebeja, Psalis 85,97
pluvialis, Labidura 101
pomerantzevi, Forficula 137
potanini, Anechura 167
potanini, Anechura (Odontopsalis) 33, 34, 165, 167, 169, 211, 229
potanini, Sondax 44, 63, 121, 206, 207, 234
praecursor, Forficula 33, 125, 211
pristina? (Forficula) 33, 169, 211
Prolabia 44, 60, 108, 112,219, 220
Proreus 44, 108, 117, 119, 220, 221
Protelytroptera 28, 29, 30, 31, 32, 40, 78, 211, 215
Protodermaptera 12, 13, 14, 28, 34, 35, 36, 37, 40, 55, 79, 80, 211, 216
Protodiplatyidae 28, 32
Protodiplatys 30, 32
przevalskii, Anechura (Burriola) 166, 181, 182, 183, 232
przevalskii, Burriola 179, 182
Psalinae 9, 28, 36, 84, 85, 217
Psalis
Pseudisolabis 35, 36, 37, 44, 54, 84, 106, 211, 217, 219
Pseudochelidura 44, 53, 62, 212, 214
pubescens, Forficula 130, 154, 228 I
, Pygidicranidae 2, 6, 8, 10, 19, 28, 30, 33, 34
35, 44, 55, 56, 79, 80, 81, 216
Pygidicraninae
Pyrarginae 10, 35, 81, 216
quelparta, Anechura in sp. 165, 173, 230
recurva, Narberia, var.
redux, Anechura (Burriola) 166, 183, 232
redux, Burriola 183
repens, Sondax 63, 121, 206
riparia, Labidura 10, 17, 21, 22, 23, 24, 26, 27, 60, 68, 70, 101, 102,209, 213
riparia, Forficula 101
I robusta, Forficula 46, 51, 70, 71, 128, 139, 212,
| 225
rufitarsis, Lobophora 116
sagitta, Forficula 33, 59, 67, 68, 69, 125, 127,
135, 211, 224, 228
scabriuscula, Allodahlia 194, 195, 233
scabriuscula, Anechura 195
scabriuscula, Forficula 195
schlagintweiti, Anechura 130
schlagintweiti, Forficula 52, 121, ~!126, 130, 159, 210, 223
scindens, Chelidura, var. 137
scudderi, Forficula 139, 140
sechuana, An’solabis
semenoffi, Kalocrania
semenovi, Anechura (Burriola) 164, 166, 179»
180, 181, 188, 231
semenovi, Burriola
Semenovida
senegalensis, Forficula 33, 59,125
setshuana, Anisolabis (Mongolabis) 88, 93, 94, 95,218
simplicis, Timomenus 197,198
setulosa, Forficula 154
simulans, Chelosoches 118
simnlans, Proreus 117, 118, 221
simulans, Forficula 118
singularis, Labidurodes
sinica, Forficula 128, 142, 226
sinuosa, Narberia, var.
smirnensis, Forficula 46, 67, 68, 128,145, 226
Sondax 42, 44, 53, 124, 196, 206, 213, 223, 234
spiculiferum, Dora 38
splendida, Forficula 126, 132, 133, 224
Spondox 84, 85, 216,
Spongovostox 36, 38
stall, Anisolabis 85
stdli Euborellia 97
stali, Forcinella 97
stoliczkae, Anechura 159
УКАЗА ТЕЛЬ
239’
stoliczkai, Oreasiobia 16, 52,121, 159, 160, 210, 229
subauricula ria, Forficula 46, 129, 150, 151, 152, 212, 213, 227
sven-hedini, Anechura in sp. 45, 165, 172, 173, 212,213,230
Tagalina 81
taurica, Forficesila 98, 99
tenera, Pseudisolabis 106
thoracica (Chelidura)
tibetana, Anechura (Burriola) 164, 166, 181, 188, 231
tibetanai Burriola. 181
Tifnomenus, 42, 43, 44, 54, 123, 196, 213, 222, 233
Titanolabis
tomis, Chelidura 137
tomis, Forficula 15, 18, 19, 20, 21, 22, 23, 27> 46,51,62,68,127,137,139,140,209,212,225
transsilvanica, Chelidurella 68, 192, 193, 233 tuberculata, Narberia 205, 234
unidentatus, Timomenus 197,198, 233
uvarovi, Forficula 126, 131, 223
vara, Anechura
vara, Anechura (Anechurella)
vicaria, Forficula 16, 71, 129, 149, 151,152,212, 227
walkeri, Pseudisolabis
woodwardi, Mongolabis 92
yersini, Forficula 154
yezoensis, Eparchus 44, 204j 234
yezoensis, Labia 204
zubovskii, Anechura in sp. 166, 177, 178, 231
СОДЕРЖАНИЕ
Стр.
Предисловие ................................................................ V
Систематический указатель видов.........................-.................. VII
Общая часть
1. Характеристика отряда и строение тела.............................
2. Превращение.......................................................
3. Образ жизни и экологии............................................
4. Классификация и количественная характеристика отряда......... . .
5. Систематическое положение и эволюция отряда в связи с палеонтологиче-
скими данными и географическим распространением............. . . .
6. Состав и происхождение фауны Dermaptera Палеарктической области . . . 7. Характеристика фауны Dermaptera СССР.............................
8. Хозяйственное значение уховерток . ...............................
9. Список литературы по Dermaptera СССР и сопредельных стран . . . *
18
30
27
29
42
71
72
Специальная часть
Таблица для определения подотрядов
I. Подотряд Archidermaptera ............................................. 77
IL „ Arixenina............................ . . ......................... 78
III. „ Dermaptera s. str........... .............................. . 79
I. Надсем. Protodermaptera................................... . 80
1. Сем. Pygidicranidae ... ......................................... 81
2. Сем. Labiduridae.................................................. 83
IL Надсем. Eudermaptera ............................................ 107
3. Сем. Labiidae .................................................. 109
4. Сем. Forficulidae ....................•.......................... 120
Резюме (Summary) ......................................................... 208
Алфавитный указатель латинских названий.................................. 235