/



































































































































































































































































































































































































































































































Текст
Е. Н.СИНСЫАЯ К П m НЦЙла
ВСЕСОЮЗНАЯ ОРДЕНА ЛЕНИНА АКАДЕМИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ НАУК ИМЕНИ В. И. ЛЕНИНА ВСЕСОЮЗНЫЙ ОРДЕНА ЛЕНИНА НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ РАСТЕНИЕВОДСТВА ИМ. Н. И. ВАВИЛОВА Е. Н. СИНСКАЯ Историческая география HghbTgPHON флоры (НА ЗАРЕ ЗЕМЛЕДЕЛИЯ) Под редакцией академика ВАСХНИЛ Д. Д. Брежнева ИЗДАТЕЛЬСТВО „KOJIOC" ЛЕНИНГРАД 1969
УДК 631.5/.9 581.9.001 Настоящая работа Е. Н. Синской, издаваемая и серии научных трудов ВИР, представляет собой историко-географический обзор мировой культур- ной флоры. Из книги читатель узнает, какие куль- турные растения дала человечеству каждая гео- графическая область, в какие страны и какими путями эти растения распространялись. В книге много внимания уделено областям древней земледельческой культуры: Закавказью и Среднеазиатским республикам с сопредельными странами Передней и Юго-Западной Азии, Рус- ской равнине и Сибири. Отдельные разделы по- святцены культурной флоре стран Восточной и Южной Азии, Западной Европы, а также Африки и Нового Света. Книга предназначена для работников научно- исследовательских учреждений, преподавателей вузов, агрономов-селекционеров и семеноводов, а также для историков и археологов, интересую- щихся вопросами древнего земледелия. Замечания и пожелания просим направлять по адресу: Ленинград, Цен гр, ул. Герцена, 44, Всесоюзный ордена Ленина научно-исследоватыь- ский институт растениеводства им. Н. И. Вави- .лова. 4-3-168 а-68
Дорогой памяти неутомимого пуаешесаввннина, лучшего эна- аона нульаурных расаений- Нинолая Ивановича Вавилова, развиаию идей его посвящает насаоя~ций труд aemор
ПРЕДИСЛОВИЕ На протяжении многих десятилетий умы человечества привлекали вопросы географии и истории земледелия, уяснение которых крайне важно при ин- тродукции растений. Этим вопросам посвятила свою научную деятельность выдающийся ученый, доктор биологических и сельскохозяйственных наук, грофессор Е. Н. Спнская. Предлагаемый вниманию читателей труд Е. Н. Синской — «Историческая география культурной флоры (на заре земледелия)» — является результатом обобщения полувековых исследований автора, а также монографических ра- бот, археологических данных и экспедиционных изысканий советских и зару- бежных ученых. Изучение географии культурных растений было начато в нашей стране и осуществлялось в широком плане академикоч Н. И. Вавиловым. По его теории видообразовательный процесс имеет место повсюду на земном шаре, человек среди местной <~лоры всегда находит растения, пригодные для вве- дения,в культуру. Вместе с тем Вавилов создал учение о центрах происхожде- ния культурных растений, где протекали наиболее интенсивно видообразова- тельные процессы и откуда растения расселялись в другие области земного шара. Н. И. Вавилов не закончил работу по установлению границ центров про- исхождения, не успел уточнить исторические связи между ними и завершить анализ культурных флор очагов. В своих докладах и беседах с товарищами до работе он делился мыслями о необходимости разобраться в истории куль- турной флоры всего мира. Е. H. Синская была ближа~~шим соратником Н. И. Вавилова в течение всего периода его работы в институте Прикладной ботаники и новых культур, в дальнейшем преобразованном во Всесоюзный институт растениеводства. Н. И. Вавилов высоко ценил Е. Н. Синскую как крупного ученого ботаника- географа. Е. Н. Синская в ВИР возглавляла работу отдела масличных, отдела кор- MQBbIx культур, а в последние годы — отдела систематики, географии и эколо- гии (культурных растен~ий и гербария. Е. Н. Сипская организавала и лично при- нимала участие во многих экспедициях по Советскому Союзу (Кавказ, Ал- .тай, Средняя Азия, Полесье, Поволжье, лесостепная зона европейской части CCCP), уделяя особое, внимание Северному Кавказу и Закавказью — родине многих видов сельскохозяйственных растений. Ею была осуществлена также экспедиция в Японию. Е. Н. Синская продолжала начатые Н. И. Вавиловым исследования по географии культурных растений в связл с нх происхождением. По мысли Е. Н. Синской вместо понятия «очаг» целесоабраано принять более широкое — «область», Она пишет в одной из своих работ, что Н. И. Ва-
вилов не только ничего не имел против этого тврмина, но и высоко ценил концепцшо М. Г. Попова о,7~ревнесредиземноморской флористической области и считал эту концепцию полезной для уточнения исгоричеокой географии куль- турной флоры. Для освещения поставленных вопросов Е. Н. Синская пользуется не только ботаническими методами исследования, но и археологическими и исто- рическими данными. Основной костяк современной культурной флоры создавался на заре зем- леделия, и~историческая география культурных растений неразрывно связана с древнейшими цивилизациями. В последние десятилетия археология, и в особенности советская, дала не- Mало материала, который во многих случаях позволяет по-новому осветить ход развития древних земледельческих культур и на фоне их взаимоотноше- ний определить время, место появления и пути распространения отдельных культурных растений и их ассортиментов. В итоге экспериментального изу- чения и селекционного освоения богатейших фондов растительных ресурсов, собранных Н. И. Вавиловым и его сотрудниками, получены данные, позво- ляющие уточнить пути расселения и распространения некоторых растений по земному шару. До сих пор не было ни одной крупной работы, рассматривающей гео- графию культурных растений в связи с историей древнего земледелия. На- стоящая книга представляет собой первый историко-географический обзор мировой культурной флоры. В ней показано, какие культурные растения дала человечеству каждая географическая область, в какие страны и по каким путям эти растения отсюда распространялись. Особенно много внимания Е. Н. Синская уделяла областям древней земледельческой культуры в Совет- ском Союзе — Закавказью, Среднеазиатским республикам, а также сопредель- ным странам Передней и Юго-Западной Азии. Впервые в самостоятельную область развития культурной флоры ею выделены тропическая и 10жная Аф- рика. Е. Н. Синская подробно рассматривает аборигенные культурные расте- ния Русской равнины, Сибири и внесредиземноморских стран Западной Европы. Естественно, что некоторые теоретические положения, высказанные Е. Н. Синской в этой работе, являются дискуссионными. В целом же книга отражает уровень современных знаний по географии культурных растений. B рукопись, как принято при посмертном издании произведений крупных уче- ных, не было внесено каких-либо существенных дополнений и изменений. Но- менклатура растений дается в соответствии с Index Pewensis, внутривидо- вые таксоны — по «Культурной флоре СССР» и ряду монографических работ. Схематическая карта географических областей исторического развития культурной флоры составлена по материалам Е. Н. Снискай кандидатом биологических наук А. А. Филатенко. Книга представляет несомненно большой интерес для работников научно- исследовательских учреждений, агрономов-селекционеров, преподавателей ву- зов, а также историков и археологов, занимающихся вопросами древнего земледелия. Академик ВАСХНИЛ Д. Д, Б р е ж н е ь
ПРИНЦИПЫ И МЕТОДЫ УСТАНОВЛЕНИЯ ИСТОРИКО-ГЕОГРАФИЧЕСКИХ ОБЛАСТЕЙ ПРОИСХОЖДЕНИЯКУЛЬТУРНОЙ ФЛОРЫ АРХЕОЛОГИЧЕСКИЕ СВЕДЕНИЯ В СВЯЗИ С ИСТОРИЕЙДРЕВНЕГО ЗЕМЛЕДЕЛИЯ Историческое прошлое человечества изучается археологами главным образом по вещественным документам, обнаруживае- мым при раскопках древних поселений людей и их погребений. Археологи проводят исследования в так называемом куль- турном слое, состоящем из органических остатков и изделий человека, сохранившихся на местах древних поселений. Современная наука имеет достаточное представление о раз- витии человека. Первые следы его появления относятся к на- чалу четвертичного периода (плейстоцену). Древнейший чело- век был еще во многом звероподобен, и должен был пройти значительный период, прежде чем он мог приступить к первым опытам возделывания растений. В истории человеческого рода различают следующие основ- ные этапы развития: 1) обезьянолюди (питекантроп, синан- троп); 2) древние люди (неандерталец); 3) новый человек (неоантроп — ископаемый и современный) . В истории человеческой культуры различают три основных века: 1) каменный; 2) бронзовый; 3) железный. Каменный век делится на древний — палеолит — и новый — неолит. Палеолит подразделяется на нижний, средний и верхний периоды; они, в свою очередь, делятся на эпохи, получившие названия по тем местностям Западной Европы, где произво- дились раскопки; эти названия сохраняются в археологической литературе и других стран. Нижний палеолит подразделяется на шелльскую (примерно 300 — 100 тыс. лет до н. э.) и ашельскую эпохи. Слои ашель- ской эпохи обнаружены в СССР (Закавказье). Земледелия в нижнем палеолите не было. Это — эпоха «со- бир ательства», а в конце — и охоты. В шелльскую эпоху человек жил в субтропических лесах— пещеры еще не использовались в качестве жилищ. Огонь, ве- роятно, еще не был известен, так как на стоянках, относящихся к этому времени, нет следов костров. Основным источником питания был сбор диких плодов, орехов, семян, корней, клуб- ней и т. д. Лес в это время играл особенно большую роль в
жизни первобытного человека. Местом возникновения человека считают тропики, но его эволюция происходила более быстрыми темпами, когда ему пришлось жить (вследствие постепенного похолодания) в субтропических лесах. Значение леса для пер- вобытного человека показал кавказский дендролог П. 3. Вино- градов-Никитин (1929). Так называемый собирательский период, начавшийся еще в нижнем палеолите, продолжается вплоть до настоящего вре- мени. Имеются данные, что еще в XIX в. (а может быть и те- перь) в районе озера Чад местное население собирало семена различных дикорастущих злаков, относящихся к родам Pani- сит, Setaria, Digitaria, Echinochloa и др., которые составляли главный источник пропитания. Собирательство как подсобный источник добывания продуктов питания сохранило свое значе- ние и в наше время в тех местностях, где имеется особое оби- лие плодов, ягод, орехов, грибов и т. д. Сбор диких плодов практикуется и сейчас довольно часто в некоторых районах Кавказа, Сибири и на Дальнем Востоке. Морские водоросли, выбрасываемые морем во время отлива,— большое подспорье в хозяйстве беднейшего населения Японии. В пустыне Калахари кочевники собирают дикорастущие арбузы, жители Огненной Земли — моллюсков и т. п. До прихода европейцев большая часть Северной Америки была населена индейскими племенами, которые продолжают заниматься собирательством. И в настоя- щее время в области Великих озер индейцы собирают колоски тускароры — дикорастущего водяного риса (Zizania aquatica L.), а в некоторых более южных местностях (Калифорния) делают из желудей муку и выпекают своеобразные лепешки, которые считаются очень питательными. В ашельскую эпоху значение собирательства постепенно уменьшается и возрастает значение охоты. Средний палеолит подразделяется на позднеашельскую и мустьерскую эпохи. Многие исследователи считают, что в позднеашельскую эпоху первобытный человек (неандерталец) обладал огнем. Он уже одевался в звериные шкуры и начал селиться в пещерах. В мустьерскую эпоху человек от стадного образа жизни пе- реходит к первобытнообщинному. Начинается разделение труда между полами. Мужчины занимаются охотой, женщины— домашними работами, собирательством и, возможно, земледе- лием — около жилищ. На смену палеантропу (неандертальцу) приходит неоантроп. Этот важный перелом падает на поздне- мустьерскую эпоху и следующую за ней ориньякскую, которая относится уже к верхнему палеолиту — на этом рубеже начи- нается эпоха «средней дикости». Верхний палеолит подразде- ляется на три эпохи: ориньякскую, солютрейскую и мадленскую. Ориньякская эпоха — это во многом промежуточная эпоха между средним и верхним палеолитом (поздним мустьером)
В это время зарождается искусство — находят главным образом женские статуэтки. В солютрейскую эпоху каменные и костяные орудия совер- шенствуются. На женских статуэтках — наряды и украшения (шляпки, браслеты, бусы, пояса). В это время начинается пе- реход к родовому строю и культу родоначальников. Типичные жилища — землянки. В мадленскую эпоху преобладают костяные орудия. Резьба по кости достигает в лучших образцах замечательного совер- шенства. Часты изображения животных. Основной объект охоты — олень. Появляются жировые светильники. Мезолит — это переходный период от палеолита к неолиту. Мезолит подразделяется на азильскую и тарденуазскую эпохи. В азильскую эпоху фауна уже послеледниковая. Сходна с фауной мадленской эпохи. Наблюдается дальнейшее усовер- шенствование орудий — появляются лук и стрелы. В тарденуаз- скую эпоху они получают широкое распространение. Харак- терны микролиты — мелкие каменные орудия геометрической формы ". На этом рубеже, переходном от палеолита к неолиту, нахо- дят кости домашней собаки. Приручение первого животного было большим достижением. Собака охраняла жилище и помо- гала на охоте. Стоянки тарденуазской эпохи открыты на обшир- ных территориях (в СССР — в Крыму и Закавказье). Неолит подразделяется на кампиньинский период и период позднего неолита. Яля кампиньинского периода из орудий характерны макро- литы (крупные орудия геометрической формы), главным обра- зом топоры. Примитивная глиняная посуда (керамика) имела заостренное дно. К этому периоду приурочивают начало прими- тивного скотоводства (находки костей лошади и быка, по типу отличавшихся от современных и приближающихся к диким родичам). В это время имеется и примитивное земледелие. В кампиньинских стоянках находят каменные мотыги, песты и зернотерки. В первых из исследованных стоянок в Кампиньи во Франции нашли зерно ячменя. Начался период мотыжного зем- леделия. Эта стадия в земледелии еще и поныне сохраняется в некоторых странах. Так, в 1928 — 1929 гг. в Японии, по нашим личным наблюдениям, не применялось плужной обработки, а весь инвентарь земледельца состоял из ручных железных инст- рументов (мотыга, лопата, вилы). Измельчение, перетирание зерен в каменных зернотерках примитивного типа бытовало очень долгое время — в Европе внлоть до римской эпохи. '" Приведенная выше периодизация предложена на основании археологи- ческих исследований в Западной Европе и, если ею пользуются за пределами Западной Европы, то чисто условно из-за отсутствия до настоящего времени своей периодизации археологических памятников.
Начало земледелия относят к неолиту. Но небольшие опыты посадок или посева растений около жилищ, возможно, имели место и в более ранние периоды. В литературе приводятся факты в пользу предположения, что в ряде случаев сначала человек культивировал растения для ритуальных целей и ме- дицинского использования, а уже потом стал их разводить в бо- лее широком масштабе и использовать как продукт питания. В пользу такого предположения говорит история некоторых растений, введенных в культуру гораздо позднее, что позволило иметь о них точные данные. Например, сельдерей выращи- вался сначала для ритуальных целей, позднее для лечебных целей и уже гораздо позже начал употребляться в пищу. Та- ких примеров можно привести немало, но тем не менее не сле- дует думать, что так дело обстояло со всеми культурными растениями. Для столь широких обобщений нет достаточных оснований. Поздний неолит характеризуется распространением полиро- ванных орудий, сверл. В раскопках этого времени открыты це- лые мастерские кремневых орудий. Для Средней Европы этого периода характерны так называемые свайные постройки, осо- бенно хорошо изученные на озерных отложениях Швейцарии. Век свайных построек в Швейцарии относят ко времени о1. 10000 до 4000 лет до н. э. Их продолжали строить и в брон- зовый век. В свайных постройках неолитического периода нахо- дили пшеницу, просо, ячмень, лен, мелкосемянные горох и че- чевицу и другие растения. В то время уже человек изготовлял ткани, веревки. Из домашних животных найдены останки коровы, овцы, козы, лошади и собаки. Энеолит выделяют как промежуточную эпоху между поздниAI каменным веком — неолитом и бронзовым веком. Медные ору- дия предшествовали бронзовым. Характерна расписная кера- мика (узоры наносились кистью). Каменные орудия в энеолите преобладали над медными. В это время уже появляются боль- шие глинобитные жилища, приспособленные для совместной жизни первобытных общин. Мотыжное земледелие (мотыги каменные) приобретает боль- шое значение в хозяйстве. Занимаются им женщины. При рас- копках часто находят глиняные статуэтки женщин-родоначаль- ниц. Стадия энеолиФа в разных странах проходила неодновре- менно: раньше всего, вероятно, в Месопотамии, Египте и южном Иране (конец IV тысячелетия до н. э.). В Анатолии, в северном Иране и в южной Туркмении энеолит относится к III тысяче- летию до н. э., а в Закавказье — ко второй половине III тысяче- летия. Характер кавказских памятников медного века близок медному веку северо-западного Причерноморья и Балканского полуострова. 10
Медный век развивался в разных областях земного шара независимо, автохтонно. Древнейшие медные рудники были сна- чала открыты в Испании, Франции, Австрии, Венгрии, Англии, а затем в ряде других стран. Переход от меди к бронзе и более широкое ее распростране- ние были обусловлены значительно более низкой температурой плавления бронзы, что облегчало литье, и большей твердостью изделий из бронзы. Для этого времени характерно сооружение особых каменных погребальных памятников (дольмены, мен- гиры, кромлехи) . В Швейцарии, кроме неолитических свайных построек, от- крыты и свайные постройки бронзового века. Скотоводство в период свайных построек бронзового века было основой хо- зяйства. Для Италии были характерны террамары — свайные постройки на болотах, в которых найдены бронзовые и камен- ные орудия. Жители террамар занимались скотоводством и земледелием. В это время появились укрепления (земляные валы); уже есть сравнительно хорошие ткани и крашеная шерсть. Земледелие оставалось мотыжным (мотыги с камен- ными и бронзовыми наконечниками). Характерно появление бронзовых серпов (раньше были только деревянные с кремне- выми зубьями и вкладышами). В конце II и начале I тысячелетий до н. э. скотоводство начало отделяться от земледелия. Пока скота было мало, он кормился вблизи жилищ, а когда количество скота сильно воз- росло и он уже не мог прокормиться около жилищ, его стали отгонять на дальние пастбища. Скотоводство в это время во многих местностях превратилось в основной источник благо- состояния, земледелие часто приобретает подсобный харак- тер. Рубеж бронзового и железного веков относится к началу 1 тысячелетия до н. э., хотя, конечно, этот переход не был впол- не синхронным для разных стран. Железный век характеризуется ростом земледелия и пере- ходом от мотыжного к плужному земледелию. Прежде чем по- явились настоящие сохи и плуги, человек пользовался прими- тивными орудиями, сделанными из заостренных древесных стволов. В истории человека можно наметить три периода использо- вания зерновых растений. 1. Кашный. Первобытное земледелие знало лишь такие ра- стения, зерно которых употреблялось только в вареном виде (каша или похлебка). Древнейшие кашные растения — просо, ячмень. Древнейшие пшеницы — полба и однозернянка — при- годны только для такого способа употребления. Значительная 11
часть человечества (Китай, Япония) еще до сих пор вместо хлеба употребляет вареный рис. 2. Период пресного хлеба. Хлеб выпекается прямо на рас- каленных камнях или в примитивных печах. В ряде стран Ближнего Востока такой способ сохранился и до настоящего времени. 3. Период кислого хлеба. Кислый хлеб стали выпекать в более поздний период человеческой истории и прежде всего на севере, где из-за холодного климата рано научились делать настоящие печи (вроде «русской»), в которых хлеб хорошо пропекался. В истории земледелия всех стран прежде всего удивляют те огромные успехи в создании набора культурных растений, которые сделаны были первобытными земледельцами именно на заре земледелия — в периоды неолита и ранней бронзы, когда земледельческая техника была на самом примитивном уровне (мотыжное земледелие) . Как можно видеть из приведенных ниже обзорных очерков по отдельным странам, именно в этот ранний период вошли в культуру наиболее важные растения, составляющие теперь основу питания людей. Ведь еще в неолите вошли в культуру пшеницы (и пленчатые, и голозерные), ячмень, просо, горох, чечевица, бобы, чина, вика, лен, конопля. В глубочайшей древ- ности была освоена человеком культура риса, хлопчатника, сои, кукурузы, финиковой пальмы, винограда и многих других пло- довых и овощных р астений. Возникает вопрос, почему в последующие эпохи, несмотря на усовершенствование земледельческих орудий, пополнение ассортимента культур (особенно полевых) шло менее интенсив- ными темпами. Ассортимент стал расширяться больше за счет увеличения числа культурных видов овощных, кормовых, орна- ментальных растений, но после мотыжного земледелия в куль- туру уже не вводилось пищевых растений, равных по значению пшенице, ячменю и кукурузе, или таких важнейших технических культур, как хлопчатник и лен. Прежде всего следует отметить, что здесь имела значение огромная длительность периода мотыжного земледелия. При всем примитивизме неолитической земледельческой техники имел большое значение фактор времени. Естественный и искус- ственный отбор, а также агросреда, создаваемая первобытным земледельцем, воздействовали на дикорастущий исходный мате- риал в течение тысячелетий и настолько глубоко изменяли при- роду растения, что во многих случаях образовались разрывы между культурными видами и их дикорастущими родоначаль- никами. Отыскание дикорастущих родоначальников для ряда основ- ных и наиболее древних культурных видов все еще составляет трудно разрешимую задачу. 12
ФАКТОРЫ, ИМЕЮЩИЕ РЕШАЮЩЕЕ ЗНАЧЕНИЕ В ПРОЦЕССЕИЗМЕНЕНИЯ ПРИРОДЫ РАСТЕНИЙ ПРИ ВВЕДЕНИИ ИХВ КУЛЬТУРУ Человек вначале брал для окультуривания растения непо- средственно из окружающей его растительности. Основным фактором, влиявшим на изменение природы ди- каря, способствующим превращению его в культурное растение, была, по-видимому, улучшенная человеком почва, как фон вы- явления полезной изменчивости, и последующий искусственный отбор. Первобытный человек выращивал растения на таком пи- тательном субстрате, который отличался от естественной обста- новки их произрастания. У многих народов, у которых мотыжное земледелие сохра- нилось вплоть до нашей эпохи, имело и все еще имеет большое значение применение местных удобрений (фекалий, рыбных от- бросов, морских водорослей и т. д.). В эпоху примитивного мотыжного земледелия не было на- стоящей полевой культуры, а небольшие участки вблизи жилищ издревле тщательно обрабатывались, интенсивно удобрялись и больше походили на огороды, чем на поля. И теперь еще в странах с примитивной техникой земледелия нет резкой грани между огородничеством и полеводством. Примитивный отбор имел значение уже на первых ступенях окультуривания растений. Издревле имело место стремление к более или менее одновременному сбору продукции (семян, клуб- ней, плодов и т. п.), когда многие растения достигают хозяйст- венной годности. Собранный для размножения растений мате- риал (семена, плоды, клубни и т. п.) снова высевался или вы- саживался, и этим путем шел отбор на выравнивание сроков созревания или достижения хозяйственной годности. В условиях культуры в результате отбора человеком расте- ния постепенно утратили свойственные их дикорастущим родо- начальникам приспособления для более быстрого распростра- нения (летучки, придатки на семенах и плодах) . У хлебных злаков в культуре возникли формы с несложными колосовыми стержнями и прочно прикрепленными колосками, у сорных ра- стений в процессе их окультуривания появились формы со слабо выраженной парусностью семян и плодов. В естественных условиях произрастания выгоднее недружное, постепенное прорастание семян. При этом, если одна порция семян прорастала при неблагоприятных условиях и погибала, то другая могла прорасти при более благоприятных условиях и обеспечить продолжение жизни вида. Благодаря отбору человек постепенно создавал популяции с дружной всхожестью. Также постепенно отбирались и формы, наиболее отзывчивые на повышенное плодородие почвы. Чело- век, естественно, предпочитал такие экземпляры, где съедобная 13
часть была крупнее и отличалась лучшими вкусовыми каче- ствами. Например, в горных лесах Кавказа в изобилии растет дикая черешня с горькими плодами, но встречаются отдельные экземпляры и с вкусной, сладкой мякотью. В культуру вошли только сладкоплодные черешни. Этот отбор был проведен, не- сомненно, еще на ранних ступенях окультуривания дикорасту- щих плодовых растений. Для культурных растений характерны увеличенные размеры и большая изменчивость (полиморфизм) именно той части ра- стения, которая имеет пищевую или иную хозяйственную цен- ность. Специализация в использовании растения во многих случаях развивалась постепенно. В странах с примитивным земледелием часто наблюдается потребление растения целиком — не одного, а многих его ор- ганов. Специализация в ходе истории часто выражается в различ- ных формах: например, исторически наиболее старыми являются те формы сельдерея, у которого употребляются в пищу и листья и не особенно толстые корни, а затем появились формы сельде- pcs; с крупными корнеплодами и листоплодные формы — с утол- щенными мясистыми черешкамп. Уже в условиях одного географического района на огородах вблизи жилищ древнего человека совместно произрастали раз- ные виды, разновидности и формы, которые в естественных усло- виях растут более или менее изолированно. При совместной их культуре не исключена возможность переопыления и возник- новения разнообразных гибридных форм, из которых затем че- ловек отбирал лучшие. Бессознательный отбор постепенно заме- нялся сознательным. Надо отметить, что гибридизация начала применяться очень рано, за несколько тысяч лет до открытия пола у растений. Еще в древней Ассирии при разведении финиковой пальмы приме- няли искусственное опыление. Большое значение имели сроки посева. Высев в разные сроки был часто равносилен по значению переносу в чуждые клима- тические условия. В приспособлении к погодным условиям разных сроков по- сева изменялись требования растений к условиям существова- ния — возникали разнообразные формы, различающиеся особен- ностями стадийного развития. Большинство культурных растений по сравнению с их дико- растущими родичами отличается повышенной влаголюбивостью (их структура в большей степени носит характер мезоморфный или гигроморфный по сравнению с более ксероморфной струк- турой их диких родоначальников). Еще на заре земледелия че- ловек начал применять орошение. Очень рано возникло оно в предгорных и горных местностях, где водный режим можно
было легко регулировать, отводя канавки от многочисленных горных ручьев и ручейков. Позднее возникли сложные оросительные сооружения в реч- ных долинах. Такие сооружения были характерны уже для древ- нейших государств двуречья и Египта — за несколько тысяч лет до нашей эры. Условия орошения создавали особый режим для возделы- ваемых растений и вместе с отбором способствовали «пере- делке» растений, которые становились все менее похожими на своих диких предков. Кочевые племена (им было не совсем чуждо и земледелие) могли переносить семена и плоды растений в более отдаленные местности и этим способствовали их распоостранению и после- дующему изменению под влиянием новых условий существова- ния. Еще в доисторический период человек на плотах и утлых лодчонках начал плавать и по морям, перенося растения на еще более значительные расстояния от очага их первоначального порождения. В Западной Европе и отчасти в нашей стране долгое время «миграционным теориям» приписывали чрезмерное значение. Многие завоевания культуры обычно считались привнесенными извне по отношению к данным народам и местностям. Советская наука отводит миграции и заимствованиям более скромную роль, считая, что все расы и все народы способны развивать и создавать культуру; одни и те же прогрессивные достижения часто возникают независимо в разных местах, как результат внутреннего развития, и совсем не обязательно в ре- зультате «влияний». В то же время роль таких влияний нацело не отрицается. Хорошей иллюстрацией этому служит развитие земледелия на земном шаре; вся совокупность известных исто- рических и археологических фактов свидетельствует о том, что земледелие возникало «политопно», независимо, в разных мест- ностях и у разных народов земного шара, хотя обмен земле- дельческими навыками имел несомненно большое значение. Что касается ассортимента культурных растений, то здесь обмен и заимствование играли также немалую роль, ибо гео- графическая среда в каждой области порождала различные растения. Многие народы сделали вклад в фонд культурных ра- стений, многие растения были введены в культуру независимо во многих местах, но вместе с тем есть много и таких растений, которые вошли в культуру в определенном первичном очаге, в определенной геообласти, а оттуда распространились в разные страны. Каким способом распространялись в древности культурные растения? Новейшие исследования (Е. Ю. Кричевский, 946; М. С. Дунин, 1952) показали, что «миграционная теория» при- ложима отнюдь не ко всем фактам и событиям древнейшей исто- рии человеческого общества.
Нам кажется, культурные растения могли распростра- няться из одного поселения в соседнее, от одного племени к со- седнему без особых «отдаленных миграций». Во время охот- ничьих странствований, перегонов скота и т. п. было достаточно встреч для передачи разных «новшеств» и сведений, чтобы за год они распространились, скажем, на 50 — 100 км. Если же при- нять во внимание огромную длительность эпохи мотыжного земледелия в истории человечества, то становится ясно, почему культурные растения еще в глубокой древности в течение тыся- челетий могли распространиться на огромные расстояния от мест своего первичного зарождения и введения в культуру. Еще быстрее стали распространяться всякие «новшества», когда люди начали передвигаться на лошадях, верблюдах, мулах. Нам кмкется важным с точки зрения исторического про- цесса изменчивости культуры растений, что они вначале обычно распространялись медленно, постепенно. В связи с этим действие климатического фактора также носило последователь- ный характер. Процесс изменчивости при этом несколько похо- дил на изменчивость дикорастущих растений при их медленном естественном распространении в определенном географическом направлении. В более поздние времена, когда передвижение по суше и по морю стало обычным делом, зачатки культурных растений пере- носились на огромные расстояния, и тогда в связи с внезап- ными резкими переменами условий чаще возникали возмож- ности для более быстрых скачкообразных изменений без пере- ходных форм. Настоящее полеводства развилось лишь в начале железного века, когда появилась возможность расчищать леса и распахивать целинные степи, занимать значительные площади под посевы. Главным образом с этих пор начала свое развитие и обшир- ная группа сорнополевых растений, которые стали расселяться по культурным землям помимо желания человека. Под влия- нием новой среды обитания природные свойства новых поселен- цев на культурных землях изменялись в сторону окультурива- ния, многие из этих растений впоследствии были замечены чело- веком и стали обьектами уже сознательного культивирования. Возникли так называемые вторичные культуры, к которым из древнейших представителей относят рожь и овес. Свое назва- ние (вторичные) они получили в противопоставление более древ- ним первичным культурам, к которым относятся две подгруппы: 1) происшедшие в результате непосредственного перенесения в культуру диких растений и 2) рудеральные. Мы думаем, что, помимо переноса растений в новые условия, в агросреду, и сознательный отбор начал применяться рано и, может быть, даже на заре земледелия (когда участки были малы и легче было подметить особенности отдельных растений) играл большую роль, чем позднее, когда период первобытного 16
огородничества закончился и когда при возникновении больших полевых массивов выбирать стало труднее. В ходе дальнейшего исторического развития, после отделе- ния огородничества от полеводства и развития садоводства, с наибольшей эффективностью стал проводиться сознательный отбор прежде всего овощных и плодовых растений, именно на этих растениях начала применяться еще в глубокой древности и гибридизация (искусственное опыление финиковых пальм в Ассирии) . МЕТОДЫ, ПРИМЕНЯЕМЫЕ ПРИ ИЗУЧЕНИИ ПРОИСХОЖДЕНИЯКУЛЬТУРНЫХ РАСТЕНИЙ И ПУТЕЙ ИХПЕРВИЧНОГО РАСПРОСТРАНЕНИЯ -В данном труде автор старался насколько возможно исполь- зовать исторический метод в приложении к вопросам о местах введения растений в культуру и путях их расселения. Историче- ский метод в его широком понимании включает археологический. Роль исторического метода, применяемого в узком смысле (использование письменных документов), для разработки исто- рии культурной флоры начинает в последнее время уменьшаться, а значение собственно археологического метода (использованпе материальных документов из раскопок: остатков растений и Нх отпечатков, земледельческих орудий, веревок, растительных тка- ней и их отпечатков, изображений растений и т. п.) возрастает в связи с новыми крупными успехами археологии, в особенности советской. Исторический метод включает в себя также и лингвистиче- ский. Существование многочисленных местных названий куль- турного растения на древних языках или современных названий, происходящих от древнего корня, свидетельствует о древности культурного растения. Сохранившееся древнее название (или корень слова), чуждое языку той страны, где оно в настоящее время возделывается, помогает часто найти родину этого ра- стения, откуда оно позднее распространилось в другие страны. Лингвистический метод имеет более узкое применение, чем археологический, но тем не менее читатели местами встретят в этой книге и примеры применения лингвистического метода. Лучший знаток культурной флоры и ее истории академик Н. И. Вавилов считал основным методом исторической геогра- фии культурных растений ботанический. Применяя ботанический метод, исследователь может поль- зоваться следующими критериями. 1. Наличие дикорастущих предков или близких дикорасту- щих родичей. Для установления первичного очага вхождения п культуру наиболее достоверным критерием является тожде- ство или чаще — очень большое сходство с эндемичной дикора- стущей формой, которая и может быть принята за непосред- 17
ственного родоначальника данного культурного растения. На- пример, близость эндемичной армянской культурной люцерны к горным эндемичным дикорастущим формам говорит за то, что культурная люцерна здесь аборигенна и произошла из мест- ных дикорастущих горных люцерн. Дикорастущие формы мор- кови из Афганистана и советских Среднеазиатских республик ботанически, по совокупности признаков, чрезвычайно близки к местным сортам культурной моркови, на этом основании можно считать, что здесь находится один из первичных очагов введения моркови в культуру. Наличие в условиях примитив- ного земледелия Афганистана большого разнообразия форм моркови, переходных от дикорастущих к настоящим культур- ным, подтверждает это предположение. Эндемичные для Европы (равнинных ее частей) формы дико- растущей моркови близки по совокупности признаков (листьев H соцветий) к европейским сортам культурной моркови и при- нимаются за родоначальников европейского подвида. Дикорастущие виды люцерны из Средне-Юго-Западной Азии ближе к азиатским культурным ее сортам, а культурные виды люцерны из стран Передней Азии и Европы имеют своих родо- начальников в горах Передней Азии (главным образом Закав- казье и прилегающая к нему Турецкая Армения). Японская редька — Raphanus raphanistroides (Mal.1по) Sinsk. — произошла от местной дикой редьки того же вида, очень сходного с культурными японскими сортами, а европейский вид редиса Raphanus sativus L. ближе к дикорастущим видам, рас- пространенным на берегах Средиземного моря. Следовательно, происхождение культурного растения может быть монотопным (в одном районе) или политопным (в разных районах), монофилетичным (от одного родоначальника) или полифилетичным (от нескольких родоначальников). Вид всегда относительно монофилетичен, т. е. происходит от какого-либо определенного вида, но возникновение вида может идти (и обычно идет) политопно — от различных форм исходного вида (от разных подвидов, разновидностей и, во вся- ком случае, от многих, а не от одной особи). Дикорастущий вид может дать начало не одному, а несколь- ким молодым видам (например, от вида, близкого к современ- ной Medicago hemicycla, по нашему мнению, произошли и М. соеги1еа, М. ~praesativa, и каждый из этих видов самостоя- тельно входит в культуру). Понятие культура следует отличать от понятия вид. куль- тура — это растениеводческий, а не ботанический термин. Куль- тура может быть представлена и одним видом (например, лен— Linum usitatissimum), а также несколькими родственными ви- дами, сходными по своему использованию, характеру агротех- ники. Например, редисы — это одна культура, но видов редисов — несколько. 18
Культурная люцерна синяя считается определенной культу- рой, но ботаники признают, что сборный вид Medi~cago sativa sensu lato объединяет несколько близких видов, которые входили в культуру в различных местах независимо друг от друга. Всеми этими данными и соображениями исследователю не- обходимо руководствоваться при определении первичных очагов происхождения культурных видов, при уточнении мест, где они ьводились в культуру, и путей их распространения. Если же в местной дикорастущей флоре непосредственного родоначальника культурной формы не обнаружено, то наличие диких более отдаленных родичей все же говорит в пользу пред- положения об аборигенном происхождении культурных эндеми- ков. Так, в Закавказье и сопредельных странах Передней Азии не имеется такой дикорастущей формы, которую можно было бы считать родоначальником 42-хромосомных пшениц. Современ- ные дикорастущие пшеницы Передней Азии представляют собой боковое филогенетическое ответвление и могут быть признаны прямыми родоначальниками только культурных однозернянок и некоторых двузернянок. Однако в других географических обла- стях нет никаких дикорастущих пшениц и это дает основание предполагать, что Передняя Азия — родина всего рода пше- ницы, а 42-хромосомные пшеницы либо возникли в культуре, либо их непосредственные дикорастущие предки вымерли. 2. Гибридизация и амфидиплоидия. Если имеются основа- ния предполагать гибридное происхождение культурного расте- ния, то в местной дикорастущей флоре ищут предполагаемых родителей. Веским доказательством гибридного происхождения является наличие естественных гибридов между предполагае- мыми исходными формами. Например, В. А. Рыбин (1936) счи- тает, что культурная слива возникла в странах Передней Азии, главным образом на Кавказе, путем естественной гибридизации двух аборигенных переднеазиатских дикорастущих видов— терна и алычи. На» естественные гиб- риды этих двух видов, по ряду существенных признаков напо- минающие культурную сливу. Как таковая, слива Ргипиз dome- :stllca в диком состоянии не обнаружена. Гибриды двух исходных видов — терна и алычи — дали начало новому виду — сливе и форме амфидиплоида — путем присоединения хромосом или генома одного вида к геному другого вида. дикорастущие предки горчицы сарептской — Brassiica juncea, рапса — В. napus — и брюквы — В. napobrassiea — в настоящее время нигде не встречаются. По-видимому, они никогда и не существовали, так как нескольким ученым в Азии и в Европе удалось эти виды получить синтетическим путем при гибриди- зации. При этом были получены амфидиплоиды: соединение ге- номов горчицы черной — В. nigra — и сурепицы яровой — В. cam- pestris — дало В. juncea, а через соединение геномов различных форм сурепицы и капусты (В. о1егасеа) образовались амфиди-
плоидные формы рапса и брюквы. Эти амфидиплоиды широко распространены в культуре. Для определения географического местонахождения первич- ного образования этих видов надо изучить очаги возникновения и области разнообразия тех видов, которые при гибридизации образовали амфидиплоиды. 3. Полиплоидия. Изучение плоидности культурных видов, а также их дикорастущих родичей, нередко дает очень ценные данные для установления первичных и вторичных очагов проис- хождения культурных растений и путей их расселения. Если в роде имеется полиплоидная серия, то обычно древ- ними, родоначальными членами в серии признаются диплоидные виды. Сосредоточение диплоидных видов в Передней Азии, в част- ности в Закавказье и по берегам Каспия, указывает на прпуро- ченность именно к этим странам первичного очага видообразо- вания подрода Falcago многолетних люцерн, где вошли в куль- туру и откуда распространились виды люцерны переднеазиат- ского происхождения на север и на запад. Наличие в определенной географической области полного полиплоидного ряда указывает также на первичность данного очага формообразования или видообразования и на значительную давность здесь формообразовательного процесса (чтобы 06- разовалась полиплоидная серия, нужно соответствующее время). На родине пшениц в Передней Азии имеется в культуре и в дикорастущем состоянии полиплоидная серия пшениц (2n,=- =14, 28, 42). Подобная же серия имеется на Кавказе для жит- няков. Это говорит о том, что Передняя Азия является также очагом происхождения и распространения для видов этой группы. Аналогичная полиплоидная серия имеется в Передней Азии и для рода Aegilaps. По мнению ряда авторов, виды Aegi- laps принимали участие в происхождении мягкой пшеницы через образование амфидиплоида: геном Aegilops (2n= 14) + геном пшеницы (2n =28) =геном мягкой пшеницы (2n =42). Рас- пространение в Передней Азии и видов пшеницы, и видов Aegi- lops делает возможным развитие гибридизационных процессов между видами обоих родов. Культурный батат (Ipomoea batatas) — вид гексаплоидный и в дикорастущем состоянии неизвестен. Предполагается, что Ipomoea batatas — амфидиплоид, возникший путем гибридиза- ции диплоидного и тетраплоидного видов. В Америке распро- странена вся полиплоидная серия видов батата, что и подтвер- ждает теорию его американского происхождения. Все эти виды, нередко распространенные в одних и тех же районах, в том числе и диплоидные, отличаются нескрещиваемостью между со- бой или дают нежизнеспособные гибриды. Подобная генетиче- ская обособленность близких видов указывает на их древность (на развитие генетической обособленности требуется значитель- 20
ное время) и на первичность американского очага происхожде- ния батата. 4. Метод видов-анализаторов. Этот метод разработан для злаков японским исследователем Кихара (Н. Кйага, 1954), а для крестоцветных много сделал в этом отношении другой японский ученый Моринага (Morinaga). Согласно точке зре- ния Кихара, более древними являются основные диплоидные виды-анализаторы.. Соединения геномов этих видов дают На- чало производным, более молодым видам — амфидиплоидам. Так, кариологическое изучение видов Aegilops выявило суще- ствование 9 диплоидных видов-анализаторов. Парные соедине- ния геномов этих видов в различных комбинациях образуюг кариотипы аллотетраплоидных видов. Первичный очаг видообра- зования, сог,гласно этой теории, представляет собой область рас- пространения основных видов-анализаторов. В роде Aegi ops большинство аллотетраплоидных видов имеют сравнительно ши- рокое распространение, а 9 основных.диплоидных видов-анализа- торов сосредоточены на сравнительно ограниченной территории. Эти основные виды приурочены к Передней Азии, а частью- к Балканам, т. е. к территории, флора которой сформировалась под сильным влиянием переднеазиатской культурной флоры. Кихара предполагает, что возникновение аллотетраплоидных видов относится к глубокой древности — к тому периоду, когда Африка и Европа еще не были разделены морем, но это вопрос спорный. Тем не менее метод видов-анализаторов заслуживает большого внимания при определении основного первичного ареала рода. Для таких видов, которые возникли как амфидиплоиды или амфиполиплоиды путем соединения геномов двух видов при спонтанной гибридизации, нет смысла искать дикорастущего непосредственного родоначальника. Он никогда не существовал. Для установления первичного очага возникновения амфиди- плоидного вида и понимания его полиморфизма нужно знать географию и полиморфизм исходных видов-анализаторов. 5. Ограничение или отсутствие у культурного растения спо- собности к генеративному размножению. Многие культурные растения, например батат, ананас, маниак, ока, улыбка и др., размножаются, в основном, вегетативно — способность размно- жаться семенами ими утрачена полностью или проявляется лишь в особых условиях. Такой процесс перехода к вегетатив- ному размножению может завершиться лишь по истечении дол- гого периода времени, и существование многих подобных видов в стране указывает на древность здесь земледелия и на возмож- ности расселения отсюда культурных растений в течение мно- гих веков в страны с более молодым земледелием. 6. Возникновение культурных растений в условиях возделы- вания. В настоящее время накоплено достаточное количество фактов, которые показывают, что в условиях культуры дико-
растущие формы более или менае быстро изменяются. Часто наряду с постепенными и медленными изменениями появляются и резкие уклонения. У некоторых родов (цитрусовые, разные виды орнаментальных растений и др.) такие скачкообразные изменения (спарты) возникают особенно часто. Поэтому нет ни- чего удивительного в том, что некоторые виды культурных ра- стений возникли в условиях культуры и не имеют непосред- ственных родоначальников в дикорастущей флоре. Во многих случаях культурные растения имеют своих родоначальников среди сорняков, а не среди культивируемых растений (декоративные васильки, рыжик, сурепица, горчица сарептская и белая и др.). Следовательно, новые виды и формы могут воз- никать в условиях культуры в качестве сорняков независимо от желания человека, а впоследствии подобные формы обра- щают на себя внимание земледельца и он начинает их использо- вать и улучшать. Наличие специализированных сорняков говорит о древности культуры растения в данной стране, ибо на специализацию тре- буется тем более длительный период времени, чем совершеннее erp специализация, т. е. форма взаимного приспособления куль- турного растения и его засорителя. Например, наличие высоко- специализированных засорителей в посевах льна-долгунца на северо-западе европейской части СССР указывает на автохтон- ность здесь льна-долгунца. древний реликтовый (теперь уже почти исчезнувший) лен- прыгунец — Lingam сгерйапз — сопровождается своим высоко- специализированным сорняком Camelina crepitans. У культурных растений существуют, таким образом, не только первичные (в очагах введения в культуру от диких родо- начальников или в очагах исходного гибридизационного про- цесса, как у Brassica juncea), но и реликтовые ареалы. Эти ре- ликтовые ареалы бывают всегда разорванными — островными; такие же реликтовые ареалы имеют и соответствующие релик- товые сорняки. В ряде случаев трудно ориентироваться в местонахождении первичного ареала того или иного культурного растения за от- сутствием дикорастущего или сорного родоначальника. Это бывает при следующих обстоятельствах: 1) дикорастущий родо- начальник вымер; 2) культурное растение произошло от сор- ного; в этом случае надо искать в дикой природе родоначаль- ника сорняка, а не родоначальника производного от него куль- турного растения; 3) культурное растение произошло от сорного, а последнее вымерло; 4) культурное растение возникло от куль- турного родоначальника при возделывании; 5) культурное ра- стение в форме амфидиплоида возникло в природе или в усло- виях культуры путем гибридизации двух видов. В четырех послед- них случаях дикорастущего родоначальника никогда не суще- ствовало. .22
Во многих местностях можно воочию наблюдать, как дико- растущие родоначальники культурных растений вымирают вследствие уничтожения естественных местообитаний, к которым они приспособились. Например, дикорастущая форма японской редьки обитает по берегам Тихого океана на площадках, увлаж- няемых стекающими с окружающих скал водами, на мягкой на- носной почве, а таких пригодных для произрастания дикой редьки мест становится все меньше — их занимают огородами и застраивают. Вероятно, скоро дикий родоначальник японской редьки совершенно исчезнет. 7. Наличие примитивных культурных форм с признаками дикарей. Дикорастущие формы определенных культурных расте- ний часто характеризуются рядом. определенных признаков. На- пример, дикорастущие и близкие к ним примитивные формы корнеплодов имеют обычно удлиненный, тонкий, погруженный в почву твердый корнеплод с большим содержанием сухого вещества. Для таких корнеплодов характерна поперечная бо- роздчатость в верхней части и сетчатая кожура. Если дикие родоначальники неизвестны, то за первичные очаги введения в культуру корнеплодов принимаются такие местности, к кото- рым приурочены примитивные эндемичные культурные формы корнеплодов с явными признаками дикарей. На севере Запад- ной Европы (в Норвегии, Швеции, Финляндии), а также в ФРГ, ГДР имеются в культуре еще не так давно более широко рас- пространенные, чем теперь, примитивные формы репы (тельтов- ская, серая, черная) и редьки. В других странах таких форм нет или они носят явно заносный характер. Мы полагаем, что эти виды корнеплодов ведут свое происхождение из упомяну- тых стран Европы. 8. Степень окультуривания. В эволюции растения от дикаря к высококультурному типу можно в ряде случаев наметить сте- пени постепенного окультуривания, например, в эволюции куль- турной люцерны *. Прослеживание подобных степеней или эта- пов эволюции растений облегчает установление очагов их про- исхождения и путей расселения. 9. Значение рядов в изменчивости признаков. Исчерпываю- щее изучение изменчивости каждого признака культурных ра- стений и их родоначальников позволяет выразить эту изменчи- вость в виде ряда вариантов. У близких видов эти ряды состав- лены из сходных вариантов и названы Н. И. Вавиловым гомоло- гическими. Особенно большое значение для выяснения вопросов исто- рической географии растений имеют такие ряды, где можно раз- личить более древние, примитивные признаки, и более молодые, указывающие на высокую степень окультуривания. Например, * См. книги: Е. Н. С и н с к а я. Динамика вида. Селькозгиз, 1948, стр. 171 — 184; Купьтурная флора СССР, т. 13, Сельхозгиз, 1950, стр. 95.
у свеклы, репы, редьки, редиса и отчасти у моркови и брюквы изменчивость формы корнеплода выражается следующим ря- дом: островеретеновидный, туповеретеновидный, длинноцилин- дрический, палочковидноизогнутый, удлиненносвекловидный, короткоцилиндрический, кеглевидный, удлиненноовальный, обратнояйцевидный, грушевидный, волчковидный, округлооваль- ный, плоскоокруглый, округлый, плоскореповидный. Этот ряд начинается с наиболее древних, примитивных форм и заканчи- вается самыми молодыми сортами корнеплодов с плоской формой. Страны, являющиеся очагами первичного введения вида в культуру и местом, где эволюция его шла в течение долгого времени, имеют весь ряд форм вида. Страны, где произрастают плоские формы, заимствовали их из стран с более примитивным земледелием. Эта схема ясно прослеживается в странах прими- тивного земледелия. Нужно всегда учитывать роль отбора, ко- торый может привести к исчезновению примитивных форм. Тем не менее применяемые с осторожностью данные о рядах вариан- тов в каждой стране являются полезным материалом при реше- нии вопросов о местонахождении первичных очагов происхожде- ния и ранних путях распространения культурных растений. 10. Интегральные виды. Родоначальные культурные формы, как и дикорастущие виды, давшие начало нескольким или мно- гим молодым видам, от которых отходят филогенетические ветви в нескольких направлениях, часто имеют так называемую инте- гральную или синтетическую конституцию. Они сочетают в себе признаки как общие родственным древним (исчезнувшим и су- ществующим формам), так и некоторым производным более мо- лодым видам. Такими интегральными видами являются европейская иско- паемая пшеница Triti~cum antiquorum, совмещающая в себе черты нескольких современных видов пшеницы, реликтовые пше- ницы Закавказья — Т. macha и Т. ра1еосо1сЫсит, эфиопская— Т. аейлр1сит — рудименты интегрального вымершего вида Т. ~protomacha. В Восточной Азии, где находится первичный очаг развития и расселения подсемейства РгипоЫеае, распростра- нены интегральные виды последнего, совмещающие в себе при- знаки различных близких видов или даже родов, например Агте- niaca holosericea, А. тите и др. Ископаемый мак из свайных построек Швейцарии сочетает в себе признаки видов Papaver setigerum и Р. somniferum. В Восточной Азии, особенно в Китае, распространены не только многие интегральные виды, но и интегральные роды, со- стоящие из видов с чертами родственных родов. По В. Шаферу (1956), роды Primula (первоцвет), Androsace (проломник) в Европе четко различаются, в Китае их границы как бы расплываются. Род Nom!ocharis имеет переходный ха- рактер между родами Lilium (лилия) и Fritillar!a (рябчик).
Род Saxifraga (камнеломка) и Chrysosplenium (селезеночник) вне Восточной Азии четко различаются между собой, а в Китае эти роды связаны переходами. Род Aquilegia (водосбор) в го- рах 1(итая представлен своим древним исходным видом, кото- рый совсем не имеет шпорцев. Некоторые китайские виды рода Aconitum (аконит) так близки к роду Delphinium (живокость), что, вероятно, являются общими предками обоих родов. Таких примеров можно привести еще не мало. 11. Эндемизм и различные степени его проявления. Древние интегральные типы часто являются реликтами — остатками древ- них исчезнувших флор (хотя вовсе не исключена возможность возникновения подобных форм и в наше время).-Но далеко не всякий реликт представляет собой интегральный тип; реликт мо- жет быть и специализированной формой. Понятие реликт также не совпадает с понятием эндем: на- пример, европейская полба Triticum зре11а является эндемом Средней Европы, но не реликтом, ибо ее происхождение отно- сится к сравнительно недавнему периоду. Рыжик с легко раскрывающимися створками — Camelina crapitans и осыпающийся лен-прыгунец — Linum icrepitans— встречаются теперь очень редко (если не исчезли совсем), по- этому их называют реликтами. Но эти виды нельзя назвать эндемами, ибо они имеют довольно широкий, хотя и прерыви- стый (точечный) ареал. В первичных очагах зарождения и распространения культур- ных видов часто встречаются реликты и эндемы, но интеграль- ные типы для них более характерны и особенно показательны. Понятие эндемизма несколько условно и требует пояснения. Форма- и видообразовательные процессы имеют место везде, где может существовать растительность. Однако некоторые об- ласти отличаются особой интенсивностью этих процессов. Там чаще встречаются интегральные типы и эндемичные виды. В очагах особо интенсивного видообразования часто встре- чаются и эндемичные примитивные (монотипные) роды. Родина пшениц — Восточное древнесредиземноморье (Передняя Азия), и здесь же находится очаг распространения такого родственного им примитивного рода с интегральной конституцией, как Haynal- dia (который до известной степени соединяет в себе признаки пшеницы и ржи). Здесь встречаются и другие роды злаков с по- добного рода конституцией, например монотипный род Hete- ranthelium имеет переходные признаки к другим родам и видам злаков. Вследствие формообразовательных процессов на всей до- ступной для растительности поверхности земли, явления энде- мизма наблюдаются везде, но проявляются в разной степени. Наиболее глубокая степень эндемизма — это эндемизм видов и родов, и такой эндемизм в основном чаще развит в очагах нги- более интенсивного развития дикорастущей и культурной флоры.
Однако эта географическая закономерность не имеет характера абсолютно точного закона. Некоторые роды имеют растекаю- щийся характер видообразования. Например, виды рода Rapha- nus (редисы и редьки) распространены на протяжении обшир- ного, как бы линейного, ареала по берегам Средиземного моря и на восточном конце ареала — по морским берегам Японии, Ко- реи и 1(итая. Восточная и западная части этого ареала на tore как бы связываются приморскими культурными видами R. indi- cus и R. caudatus. Последний приурочен к островам Индонезии. У этого рода нельзя выделить особого очага интенсивного видо- образования. По мере распространения вида из первичного очага в ре- зультате приспособления к новым условиям среды, а у культур- ных растений — агросреды (создаваемой человеком на опреде- ленном природном фоне) могут возникнуть и новые виды, а чаще образуются новые эндемичные подвиды, экотипы, эндемичные разновидности и эндемичные местные сорта (следует подчерк- нуть, что под эндемичными сортами подразумеваются только местные, а не селекционные сорта — последние часто создаются на базе инорайонного исходного материала и понятие эндемизма на них не распространяется). В нашем понимании в той или иной степени эндемизм имеется всюду. Если нет эндемичных ви- дов, TQ имеются эндемичные подвиды и экотипы или только разновидности, или только местные сорта. Изучая различные степени проявления эндемизма, мы получаем возможность срав- нивать между собой отдельные геообласти по интенсивности видо- и формообразовательного процесса. Этот метод одинаково применим и к дикорастущей, и к культурной флоре. Дробная внутривидовая систематика, которую Н. И. Вави- лов назвал дифференциальной и которую он первый начал раз- рабатывать в применении к культурным растениям, имеет боль- шое значение в географии растений, открывая возможности для сравнения флор различных геообластей по интенсивности формо- образовательного процесса (по степеням проявления энде- мизма). Н. И. Вавилов, особенно в начале своей научной деятель- ности, склонен был придавать большое значение эндемизму от- дельных признаков. Он считал, что в первичных очагах видо- образования и массового введения в культуру различных видов преобладают доминирующие признаки, а на краях ареала ра- стения тех же видов обычно имеют рецессивные признаки. Сход- ное положение высказывал И. В. Мичурин, который считал, что признаки древних форм растений имеют склонность доминиро- вать при скрещивании над признаками более молодых произ- водных форм. Как выяснилось в результате накопления боль- шого количества фактов, эта закономерность в общем верна, но для целей определения направления миграции (естественной или при посредстве человека) ею пользоваться следует очень осто- 26
рожна. Доминирующие признаки могут развиваться и на краях ареалов. Наблюдения над географической изменчивостью отдельных признаков имеют гораздо меньшее значение, чем выявление по- рядка смены форм, определяемых всей совокупностью призна- ков — смены конституций. 12. Постепенная изменчивость в определенном географиче- ском направлении, указывающая на пути миграции (для дико- растущих) и распространение форм в культуре из первоначаль- ного очага в различные стороны по ареалу. Не останавливаясь детально на географических закономерностях смены состава растительных популяций (зональных экотипов и переходных форм) у дикорастущих растений отметим, что и в культуре при постепенном распространении растения картина географи- ческой изменчивости состава популяций обнаруживает те же закономерности. Особенно яркие географические правильности в смене форм наблюдаются при распространении растений в ус- ловиях культуры, но помимо воли человека — у сорняков. Еще недавно можно было наблюдать, как постепенно изменялся со- став популяций сорного рыжика на Русской равнине (Восточно- Европейской) по направлению с юга на северо-запад и как постепенно в этом направлении формировалась конституция ти- пичного льняного рыжика — Сате11па linicola (теперь почти исчезнувшего). Если в культуру входят сорные формы, то эко- лого-географическое изучение сорняков помогает определить предков, место происхождения и пути распространения культур- ной формы. Аналогично, сходство с местным дикорастущим эко- типом указывает на происхождение местного сорта, а родство с инорайонным экотипом указывает, откуда этот сорт был за- несен. Не только у дикорастущих растений, но в ряде случаев и у культурных последовательность смены форм и постепенная изменчивость состава популяций и признаков указывает на на- правление их расселения. Ряды переходных форм культурного льна от индийских карликовых льнов к абиссинским карлико- вым формам и от индийских крупноплодных льнов к крупно- плодным средиземноморским льнам указывают, что маслич- ный лен постепенно распространился из Индии в Эфиопию и страны Средиземноморья. Эндемичные льны Сирии и Кипра за- нимают промежуточное положение между льнами Индии и ти- пичными средиземноморскими формами крупносемянного льна. Это свидетельство того, что Сирия и Кипр были промежуточ- ными инстанциями на пути исторического распространения льна. 13. Резкие «вспышки» (или постепенное усиление) полимор- физма на краях ареалов при естественном расселении или раз- ' Подробно об этом см. в ки. Е. Н. Синской «Динамика вида», Сельхоз- гиз, 1948. 27
витие полиморфизма при переносе человеком растений в резко отличные условия. В природе при переходе от одной зоны, от одного вертикального пояса к другим, виды и экотипы как бы разлагаются: появляются новые признаки и новые их сочета- ния '". В культурной флоре нередко очаги интенсивного форма- и видообразования имеются на краях ареалов вида или рода. На- пример, средиземноморский род Brassica образует на восточном конце своего ареала — в Японии, Корее, Китае и в меньшей сте- пени — в Индии крупный вторичный очаг видообразования. По А. И. Филову (1960), зоны развития разнообразия высоко- культурных форм располагаются вне или на краю областей распространения их диких родичей (это Филов показывает на примере бахчевых растений). Выводы, полученные ботаническим методом, должны быть подтверждены историческим методом. Возможна и нужна также проверка ботаническим методом многих положений, установ- ленных исключительно на основе исторических данных. Таким образом, ботаники-географы могут с пользой применять исто- рический метод в географии растений, а историки — материалы, добытые ботаническим методом. 14. Одичание культурных форм. Картину исторического рас- пространения культурных растений могут несколько завуалиро- вать процессы одичания культурных форм. Одичавшие формы в каждом конкретном случае следует отличать от дикорастущих родоначальников местных культурных сортов. Знакомство с гео- графией и историей каждого культурного вида с более или ме- нее исчерпывающей полнотой исключает возможность такого рода ошибок. С географической точки зрения явления одичания интересны тем, что они обычно указывают на близость комплекса экологи- ческих условий района одичания с условиями первичного очага происхождения данного вида. Интересные данные 06 одичании томатов в Африке получены известным французским исследователем мировой культурной флоры Шевалье (А. Chevalier, 1953). Процессы одичания томатов имеют место и на Кавказе, чаще в Закавказье. На огородах разводят там только крупно- плодные сорта томата, а одичавшие его формы всегда имеют мелкие вишневидные плоды. Местные жители (как и Шевалье) утверждают, что из семян, прошедших через кишечник человека или животного, вырастают формы томата наследственно мелко- плодные. Однако соответствующими специальными опытами это положение не проверено. В связи с тем, что одичалые формы часто обладают устойчи- вой наследственностью и их бывает трудно вновь переделать 28 ~ См. Е. Н. С ин ск а я. Динамика вида. Сельхозгиз, 1948.
в культурные, они заслуживают более детального эксперимен- тального изучения. Не исключена возможность их обратного во- влечения в культуру в качестве исходного материала для се- лекции. Процессы одичания должны быть учтены и изучены для того, чтобы не смешивать одичавшие формы с примитивными сор- тами первых этапов земледелия или с их дикими родоначаль- никами. Яля правильного решения вопросов, относящихся к уста- новлению очагов происхождения культурных растений, и для прослеживания путей их распространения в ранние периоды земледельческой культуры — на заре земледелия — нужно поль- зоваться всеми указанными выше методами и критериями в их совокупности. Превалирование в исследовании одного какого- либо метода или критерия неизбежно ведет к одностороннему и узкому пониманию действительности и к неправильным вы- водам. ИСТОРИКО-ГЕОГРАФИЧЕСКИЕ ОБЛАСТИ ПРОИСХОЖДЕНИЯАБОРИГЕННЫХ КУЛЬТУРНЫХ ФЛОР ЗЕМНОГО ШАРА Следует подчеркнуть, что в понятие «географическая область культурной флоры» мы вкладываем историко-географическое содержание. В настоящее время вследствие все усиливающегося обмена культурными растениями и между близкими, и между далекими странами области возделывания многих культурных растений чрезвычайно расширились. Особенно ярко это сказалось на Америке, где своеобразная аборигенная культурная флора теперь пополнилась огромным количеством видов внеамерикан- ского происхождения, которые стали обычной и неотъемлемой частью культурного видового ассортимента Нового Света. Соот- ветственно и в Старом Свете прочно вошли в культуру и в быт многие культурные растения, имеющие родину в Новом Свете. Qo работ Н. И. Вавилова и объединенного им коллектива исследователей географии культурных растений в современном понимании не было. Более ранние работы по распространению и происхождению отдельных возделываемых растений еще не могли создать пред- ставления о культурной флоре. Это понятие начало склады- ваться со времени первой большой экспедиции Н. И. Вавилова в Иран и прилегающие горные районы Средней Азии (1916 г.) и получило окончательное развитие в результате ряда последую- щих экспедиций Н. И. Вавилова и его сотрудников. В настоящее время география культурных растений состав- .ляет особый раздел фитогеографии. Многие идеи и понятия об- щей фитогеографии нашли свое отражение и в географии куль- 29
турной флоры, но в своем специфическом преломлении. Среди возделываемых растений нашлись и эндемики, и реликты, вы- явились яркие и очень интересные закономерности простран- ственного распределения видов и форм и их географической из- менчивости. Одной из важнейших задач исследователей в этой области стало установление первичных ареалов культурных ра- стений, где они были введены в культуру из местной флоры (или занесены из других стран в глубокой древности, еще на заре земледелия) и составили набор видов и форм, использованный пионерами земледелия в начальный период его возникновения на данной территории. Н. И. Вавиловым (1935) было установлено восемь основных очагов происхождения и расселения культурной флоры: 1 — Ки- тайский; II — Индийский; Па — Индо-Малайский; III — Средне- азиатский; IV — Переднеазиатский; V — Средиземноморский; VI — Абиссинский (Эфиопский); VI I — Южномексиканский и Центральноамериканский; VIII — Южноамериканский; VI IIa— Чилоанский; VI 116 — Бразильско-Парагвайский. Согласно исследованиям Н. И. Вавилова (1940), для возник- новения крупного очага исторического развития культурной флоры необходимы два условия: 1) богатство местной флоры такими растениями, которые пригодны для введения в культуру, и 2) наличие древней земледельческой цивилизации. Как выяснилось впоследствии, эти два условия не всегда совпадают. Например, в Древнем Египте земледелие начало развиваться в глубочайшей древности, но та небольшая терри- тория, где жили древнейшие земледельцы, — дельта Нила— могла дать очень мало растений для введения в культуру: это были только некоторые прибрежные растения (папирус, чуфа, эрагростис). Почти все остальные культурные растения еще в доисторические времена попали в Египет из соседних стран Передней Азии и Средиземноморья, в меньшей степени — из более южных областей Африки. В другой стране древнего земледелия — Месопотамии (Междуречье, Двуречье) земледелие с самого своего возникно- вения базировалось не на аборигенных растениях из прибрежий рек Тигра и Евфрата. Древнейшие земледельцы Месопотамии культивировали растения, занесенные из предгорий соседних стран Передней Азии. Таким образом, ни Египет, ни Месопотамия не являются самостоятельными очагами происхождения древней культурной флоры. Египет входит в Средиземноморский, а Месопотамия в Переднеазпатский очаги Вавилова. Отсюда видно, что тер- ритория очага неоднородна и, кроме того, весьма обширна. Границы очагов уточнялись Н. И. Вавиловым по мере на- копления относящихся сюда фактов. И постепенно становилось ясно, что этот термин уже не вполне соответствует содержанию понятия. Термин «очаг» создает у людей, мало знакомых с пред- 30
метом, представление о сравнительно небольших территориях, что не соответствует идее Вавилова. Н. И. Вавилов не успел закончить работу по уста- новлению границ очагов, уточнить исторические связи между очагами и провести сравнительный анализ культурных флор разных очагов. Он работал над этими вопросами, но откладывал их решение до окончания всей намеченной программы экспеди- ций (куда входили и Индия, и Китай, и Южная Африка, и дру- гие страны). Он считал, что видообразовательный процесс имеет место везде на земном шаре и что едва ли не всюду человек среди местной флоры может найти растения, пригодные с той или иной целью для введения в культуру. Очаги отличаются от остальных территорий только особой интенсивностью видообразовательного процесса п особенно боль- шим количеством вошедших там в культуру растений, массо- вым их окультуриванием. Н. И. Вавилов интересовался историей земледелия и введе- нием растений в культуру во всех странах. Научное наследие Н. И. Вавилова позволяет наметить, в каких направлениях сле- дует развивать его историко-географические идеи. Мы имеем основания считать, что вместо понятия «очаг» целесообразно принять термин «область». В пользу последнего термина говорит и то, что он является принятым в общей фито- географии. Н. И. Вавилов ничего не имел против этого термина. Ему очень нравилась концепция М. Г. Попова о Древнесредизем- номорской флористической области, он разделял идеи М. Г. По- пова в этом отношении, рассматривая их как весьма полезные для уточнения исторической географии культурной флоры. Продолжая работу Н. И. Вавилова, мы стремились выявить более широкие географические связи. Уточняя состав абориген- ных культурных растений в каждом очаге, мы старались вы- явить взаимовлияние и взаимопроникновение культурных флор основных очагов между собой. Поэтому вводится более широкое новое понятие — историко-географическая область, или геогра- фическая область исторического развития культурной флоры. Мы говорим именно об областях историко-географических, а не об областях современного распространения культурных ра- стений. Следует помнить, что первичный ассортимент культурных растений связан с развитием на территории данных областей древнейших земледельческих цивилизаций. Культурные расте- ния — продукт природы и в значительной степени — деятель- ности человека. Здесь нет прямой зависимости между богат- ством аборигенной дикорастущей флоры, подходящими для окультуривания видами и уровнем развития земледелия. Дико- растущая флора Алтая очень богата, но в культуру взято еще мало видов и форм. Это произошло, помимо других причин, и 31
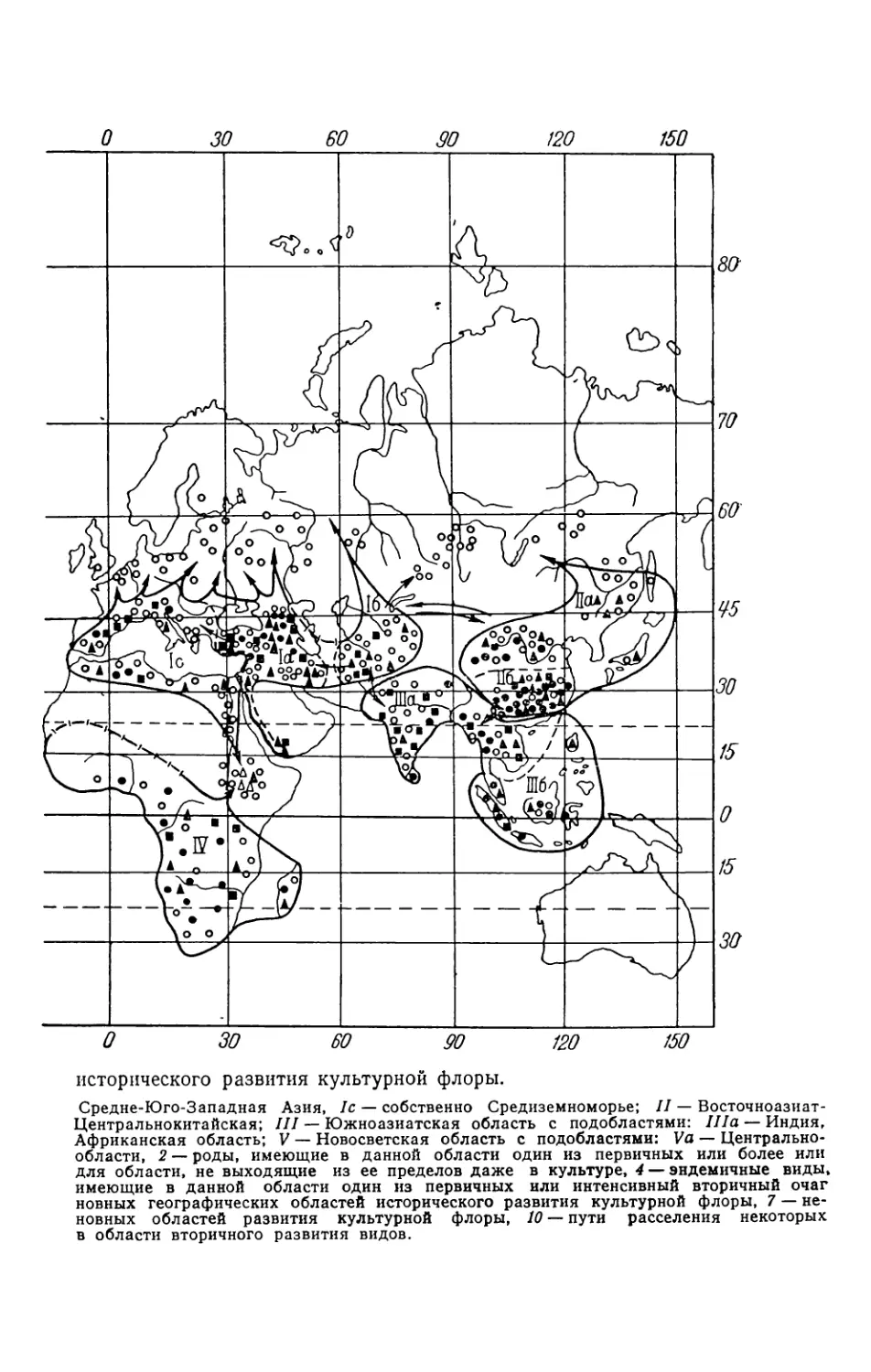
�~ ° Я аЗ д j ~5 о .. у ----,у у уц , ~ ~y е I е/ ф i '>г ф'/ Ф j rl l °: . ° ю ° ) ° ° ° ° в ° ° ° ° ° е ° ° J °° ° ° ° ° ° ° ° ° е ! ° е ° ° ° ! r 'Р ,IELL%?K4~ ЯЯЯ@~~ЯЯ , ИЫЫИЬРаЯ ИИИИЙГМ
70 ЗО f/0 исторического развития культурной флоры. Средне-Юго-Западная Азия, Ic — собственно Средиземноморье; II — Восточноазиат- Центральнокитайская; III — Южноазиатская область с подобластями: IIIa — Индия, Африканская область; V — Новосветская область с подобластями: Va — Центрально- области, 2 — роды, имеющие в данной области один из первичных или более или для области, не выходящие из ее пределов даже в культуре, 4 — эндеыичные виды, имеющие в данной области один из первичных или интенсивный вторичный очаг новных географических областей исторического развития культурной флоры, 7 — не- новных областей развития культурной флоры, 10 — пути расселения некоторых в области вторичного развития видов.
потому, что Алтай не является областью интенсивного развития древнего земледелия. В 10жной Америке флора тропических лесов (гилей) не менее богата видами, чем области сухих наго- рий и прибрежных равнин. Однако по ряду причин тропические гипеи не были очагами развития древних цивилизаций, и соот- ветственно ассортимент важнейших аборигенных культурных ра- стений Америки в основном сложился вне области господства тропических гилей. Мы предлагаем различать пять основных географических областей исторического развития культурной флоры и несколько подо бл астей (см. схему) . 1. Древнесредиземноморская с тремя подобластями: 1) Пе- редняя Азия (Переднеазиатский очаг Н. И. Вавилова); 2) Средне-Юго-Западная Азия (Среднеазиатский очаг Н. И. Ва- вилова); 3) собственно Средиземноморье (Средиземноморский очаг Н. И. Вавилова). II. Восточноазиатская с двумя подобластями: 1) Северо- Восточноазиатская (японо-маньчжурская); 2) Юго-Восточно- Центральнокитайская. I I I. IОжноазиатская с двумя подобластями: 1) южные районы Китая, собственно Индия, Цейлон и Индокитай; 2) Ма- лакка и Малазия. IV. Африканская. V. Новосветская с двумя подобластями: 1) Центральномек- сиканская; 2) Южноамериканская. Области и подобласти — это очаги наиболее интенсивного форма- и видообразования, откуда шло массовое распро- странение культурных растений в соседние и в отдаленные страны. Различия между областью и подобластью особенно легко уясняются при детальном рассмотрении огромной Древнесреди- земноморской области, которая образовалась на месте исчез- нувшего древнего средиземного моря Тетиса и по его прежнему побережью. Территории, не вошедшие в пять основных географических областей исторического развития культурной флоры, мы пред- лагаем назвать областями влияния. Области влияния отличаются от основных областей (и под- областей): 1) менее богатой и часто менее древней аборигенной дикорастущей флорой; 2) исторически менее древним земледе- лием; 3) отсутствием или меньшим количеством эндемичных культурных растений (не вышедших за пределы рассматривае- мых областей); 4) меньшим количеством видов (и форм) расте- ний, вошедших в культуру из местной дикорастущей флоры; 5) отсутствием (или очень малым количеством) таких абориген- ных родов, в которые входят виды культурных растений; 6) або- ригенные культурные растения не составляют основного, самого важного с экономической точки зрения ассортимента; 7) области
влияния представляют собой в большей степени центры вселе- ния, чем расселения культурных растений. Однако и в дикорастущей флоре каждой области влияния действуют видо- и формообразовательные процессы, хотя и с гораздо меньшей степенью интенсивности, чем в основных об- ластях исторического развития культурной флоры. В каждой области влияния человек брал из окружающей его флоры те или иные растения для введения в культуру, и каждая область влияния, помимо заносных растений, имеет хотя бы небольшое количество аборигенных культурных растений. Областью влияния Яревнесредиземноморской области яв- ляются Средняя и Северная Европа (самые южные части Запад- ной Европы входят в собственно Средиземноморье). Н. И. Вавилов не успел уделить достаточного внимания об- ластям влияния, и эта часть исторической географии культурной флоры остается наименее разработанной. Подобласти можно делить дальше на провинции и округа. Такая детализация не является задачей нашей работы, поэтому в ней только иногда даются общие указания на возможность дальнейшего подразделения. Австралия и Полинезия дали очень мало культурных растений и поэтому эти страны не включены в наш краткий обзор. Ниже следует обзор установленных нами областей и под- областей исторического развития культурной флоры. 1.,Яревнесредиземноморская область исторического разви- тия культурной флоры. В нашем понимании древнее Средизем- номорье включает собственно Средиземноморье, Переднюю Азию и Средне-Юго-Западную Азию, т. е. прибрежные страны и острова по берегам Средиземного моря в Европе и Африке, Аравию, Закавказье, Малую Азию, Иран, Узбекистан, Турмени- стан, Таджикистан, Афганистан и Северо-Западную Индию. Это примерно соответствует границам флористической Древнесреди- земноморской области в понимании М. Г. Попова, но может быть несколько шире, ибо мы сюда включаем также Западный Китай и Кашмир. Древнесредиземноморская флористическая область располо- жена на бывшем дне и по прежним берегам тропического Среди- земного моря — Тетиса. Истоки дикорастущей флоры этой об- ласти находятся, с одной стороны, в мезофильной лесной флоре, имеющей основную область развития в Восточной Азии, а с дру- гой стороны — в африканской ксерофильной флоре. В результате контакта элементов этих двух, более древних, флор и дальнейшего их развития в своеобразных условиях тер- ритории, освободившейся после усыхания Тетиса, сложилась вполне оригинальная древнесредиземноморская флора. Что ка- сается культурной флоры Древнесредиземноморской области, то представители ее в основном произошли от аборигенных диких родоначальников. Из других крупных областей формирования
аборигенных культурных флор туда в древности, на заре земле- делця, было занесено сравнительно мало культурных растении. Напротив, многие растения средиземноморского происхожде- ния теперь широко распространены по всему миру: пшеница, овес, тимофеевка, люцерна, клевер, бобы, горох, чечевица, чина, нут, лен, свекла, репа, морковь, капуста, финиковая пальма (ко- торая, вероятно, впервые введена в культуру шумерами), гра- нат, горчица белая, кориандр, кресс-салат, салат-латук и энди- вий, артишок и многие другие. Первичные ареалы некоторых средиземноморских видов культурных растений и их ближайших дикорастущих предков ,доходят до Индии и даже до крайнего востока Азии.,Яревне- средиземноморский род свекла (Beta) имеет примитивную форму с конституцией дикорастущего растения и в Индии. При- :морский род редька и редис — предки современных Raphanus— был распространен по берегам Тетиса до Восточной Азии и имеет теперь дикого представителя и производную от него куль- турную форму на восточном побережье Японии и Восточного Ки- тая и т. д. Такие роды надо считать более древними, чем те, у которых дикие представители распространены в границах, ука занных нами для Древнесредиземноморской области культурной флоры. У ряда культурных растений не сохранилось дикорасту- щих родоначальников, или они занимают теперь сильно реду- цированный ареал. Так, лен представлен в Индии, несомненно, очень древней группой эндемичных сортов, здесь же вошедших в культуру, но дикорастущий родоначальный вид Linum angusti- fo1ium теперь сюда не доходит — очевидно, этот вид раньше имел более широкий ареал. Состав аборигенной культурной флоры в крупных подобла- стях Древнесредиземноморской области — Средне-Юго-Запад- ной Азии, Передней Азии и собственно Средиземноморья рас- смотрен нами в соответствующих разделах. Помещенные в конце каждого раздела обзоры ассортимента культур дают ясное пред- ставление о том, что эти три подобласти являются частями еди- ной крупной области: основной костяк видов, родов и семейств культурных растений здесь общий. Однако каждая подобласть имеет и свои характерные особенности. 1. Подобласть Передняя Азия. Включает Закавказье, Малую Азию, Аравию, северо-западный Иран и, по мнению некоторых исследователей, горный Туркменистан. Земледелие здесь воз- никло в глубокой древности у жителей среднего горного пояса и предгорий, которые затем перенесли земледельческие навыки в Месопотамию, и на пойменных равнинах Тигра и Евфрата развивалась земледельческая культура протошумеров, затем шумеров (или су.меров) и аккадов в первых государствах— Ассирии и Вавилоне — с мощными ирригационными сооруже- ниями, с развитыми отраслями растениеводства (полеводством, садоводством и огородничеством) и с большим ассортиментом
культурных растений. Ассиро-Вавилонская земледельческая культура непосредственно связана с позднейшими земледельче- скими культурами сюбаров и хеттов — предков грузин. Грузин- ских племен было много; особенно большое историческое значе- ние имеет земледельческая культура колхов. С земледельческой культурой сюбаров и хеттов тесно свя- зана земледельческая культура Урарту, продолжением которой является культура Древней Армении, а культуры Урарту и Ар- мении тесно взаимосвязаны с культурами Индии и Ирана. Передняя Азия является родиной или главной ареной раз- вития следующих родов, к которым относятся важные виды культурных растений: пшеницы, ржи, нута, чечевицы, гороха, эспарцета, люцерны, рыжика, ляллеманции, свеклы, граната, монотипных родов мушмулы (Mespilus) и айвы. Культурные растения следующих родов имеют в странах Передней Азии один из первичных очагов или более или менее крупный вторичный очаг своего развития (видообразования): овес, вика, дыня, клевер, груша, слива, вишня и черешня, мин- даль, инжир, боярышник, орешник, рябина и др. В странах Передней Азии насчитывается около 40 эндемич- ных культурных видов, перечисленных в сводном списке в соот- ветствующем разделе. Это очень много по сравнению с соб- ственно Средиземноморьем и особенно по сравнению со Средне- Юго-Западной Азией. По нашему подсчету, более 70 культурных видов имеют один из первичных очагов или вторичный очаг своего развития в стра- нах Передней Азии. Передняя Азия — это крупнейший очаг распространения культурных растений на запад и на север в Европу, на восток— в страны Средне-Юго-Западной Азии, а также один из важней- ших очагов происхождения и одомашнивания сельскохозяй- ственных животных. С. Н. Боголюбский считает Африканский очаг одомашнива- ния животных более важным, чем Переднеазиатский, многие же авторы придерживаются противоположного мнения. Осо- бенно много доводов приводится в пользу доместикации овцы и лошади в Передней и Средней Азии. 2. Подобласть Средне-Юго-Западная Азия. В эту географи- ческую подобласть развития культурной флоры включены гор- ные и предгорные районы Узбекистана, Таджикистана, Афгани- стана, Северо-Западной Индии и Западный Китай. Иран зани- мает промежуточное положение, ибо его территория является ареной развития переднеазиатской и средне-юго-западноазиат- ской культурных флор, причем северо-западный Иран включен нами в Переднюю Азию. Областями влияния Средне-Юго-Запад- ной Азии являются Киргизия, Казахстан и Сибирь (в особен- ности Западная). Влияние на Сибирь было и непосредственное и позднее — через обитателей Русской равнины. И обратно — на
русскую равнину культурные растения могли распространяться через Сибирь. Подразделение этой подобласти культурной флоры на провин- ции довольно затруднительно, поэтому в данной книге оно не дается. Отметим только, что наличие малоокультуренных форм, часто имеющих черты дикорастущих родоначальников, харак- терно в особенности для горных районов Афганистана и приле- гающих горных районов северо-западной Индии и Таджики- стана. Это в полной степени увязывается с примитивным харак- тером земледелия в этих странах в недалеком прошлом (в Афганистане земледелие стоит на низком уровне и по сей день, а в горном Таджикистане было таковым вплоть до Ок- тябрьской революции). Развитое земледелие на равнинах, основанное на ирригации, позднее, чем в предгорьях и в горных долинах, выступило на арену истории, но до недавнего времени сохраняло примитив- ные черты в Хивинском оазисе (где развитие земледелия свя- зано с историей высокой культуры древнего Хорезма) и в За- падном Китае. Здесь развились на орошаемых землях своеоб- разные формы культурных растений, обнаруживающие, однако, тесное родство с более примитивными формами горных районов. Например, сорта люцерны Кашгара и Хивинского оазиса пред- ставляют собой формы, конституция которых формировалась в условиях поливной культуры жаркого равнинного и долинного климатов; но эти формы обнаруживают явное родство с более примитивными культурными горными сортами и дикорастущими горными формами. То же можно сказать и о льне, моркови, репе и др. Таким образом, грубо можно поделить всю подобласть Средне-Юго-Западной Азии на две основные части: 1) террито- рия более древнего горного земледелия с более примитивным характером культурной флоры и большей пестротой популяций, 2) территория менее древнего земледелия — поливные земли с более окультуренными формами. Средне-Юго-Западная Азия не является областью возникно- вения или наиболее интенсивного развития тех родов, к которым относятся культурные растения. Это ясно видно при сравнении списков растений в конце соответствующих разделов книги. 3. Подобласть собственно Средиземноморье. Эта подобласть включает Грецию, юг Италии, Южную Францию, Испанию,. Португалию и Северную Африку до южной границы Сахары. Сирия, Палестина и Иемен являются территориями, переход- ными от собственно Средиземноморья к Передней Азии, но ближе к последней по составу культурной флоры. Начало земледелия в Средиземноморье относится к доисто- рическим временам — развилось еще в эпоху неолита. Бада- рианцы (предшественники древних египтян), пеласги (древней- шие обитатели Греции), этруски и иберы (древние народы
Апеннинского и Пиренейского полуостровов) были земледель- цамии. Крита-микенская земледельческая культура уже стояла на высоком уровне и была предшественницей греческой и римской «ультур. В Средиземноморье (в узком объеме этого понятия) сосредо- точено большое разнообразие форм и сортов культурных расте- ний. Но эта подобласть Древнесредиземноморской области не является ареной особенно интенсивного родо- или видообразо- вания культурных растений и в данном отношении далеко усту- пает Передней Азии. В странах собственно Средиземноморья выработались вы- сококультурные крупноплодные и крупносемянные сорта бобо- бых, плодовых, льна масличного и других растений в отличие от более мелкосемянных и мелкоплодных примитивных сортов Средне-Юго-Западной и Передней Азии. Важнейшие области влияния Древнесредиземноморской 06- ласти: Русская равнина и Средняя и Северная Европа. Абори- генная дикорастущая флора Р у с с к о й р а в н и н ы (Восточно- Европейской равнины) дала пока немного для культуры. Основ- ной ассортимент культурных растений сложился здесь из видов возделываемых растений, распространившихся главным образом из Передней Азии [пшеница мягкая и твердая, полба, сорные овсы вместе с полбой, давшие потом начало культурному овсу (Avena sativa), вероятно, ячмень, частично лен — степные ме- жеумки, все зерновые бобовые, репа, огурец, морковь, сельдерей, кориандр и др.)]. Эти растения могли распространиться как через Крым и Се- верный Кавказ, так и через Балканы, куда доходили границы распространения высокой придунайской — поднепровской земле- дельческой культуры Ш — П тысячелетий до н. э. (Триполья). В Средней и Северной Европе земледелие воз- никло в глубокой древности, в эпоху неолита. Аборигенная дико- растущая флора дала сравнительно мало выходцев в культуру. Основной набор культурных растений сложился из элементов переднеазиатской культурной флоры. Из Передней Азии пришли в Европу полба-эммер, полба-спельта, однозернянка (сначала в качестве сорняка полбы), прототип мягкой пшеницы Т. anti- quorum, ячмень, лен, зерновые бобовые, огурец, многие пло. .довые. Распространение переднеазиатских растений шло через Бал- каны в юго-западную часть СССР. В III — II тысячелетии до и. э. имело место развитие трипольской культуры на Балканах, на территории, прилегающей к Днепру, Бугу, Днестру, Дунаю. Эта земледельческая культура была самой высокой для своего вре- мени в Европе. Она способствовала усилению связей Средней и Северной Европы с Передней Азией через Балканы и со стра- нами, расположенными на восток от трипольских территорий.
Собственно Средиземноморье, конечно, оказало значительное влияние на состав культурных растений в Европе, и северную границу подобласти собственно Средиземноморья не так легко провести. Из западного Средиземноморья распространился по Европе овес песчаный (Avena strigosa), вероятно, мак, из разных мест Средиземноморья — капуста. Однако в гораздо большей степени, чем виды, в более позднее время из Средиземноморья в страны Средней Европы распро- странились высококультурные, крупносемянные и крупноплод- ные сорта, которые выработались в собственно Средиземноморье из ранее широко распространенных мелкосемянных и мелко- плодных форм. Из привнесенных элементов в Средней Европе могли соз- даться в единичных случаях и виды — примером служит европей- ская полба-спельта, которая возникла в приальпийской зоне от пшеницы типа мягкой, ведущей происхождение из Передней Азии. Что касается эндемичных форм и сортов, то в Северной и Средней Европе их имеется большое количество. Земледелие в странах Северной и Средней Европы очень древнее и в раскопках обнаружены примитивные формы пше- ницы, ячменя, мака и других культур, которые представляют ценный материал для освещения вопросов филогении многих культурных растений. Таким образом, Северная и Средняя Европа, хотя в основ- ном и считаются областью влияния Древнесредиземноморской области исторического развития культурной флоры, но вместе с тем являются и ареной первичного видо- и формообразования культурных растений, хотя и не столь интенсивного, как в основ- ных областях. II. Восточноазиатская область исторического развития куль- турной флоры. Исторически известными древними земледельче- скими культурами Восточной Азии являются древняя иньская китайская цивилизация в Северном Китае по течению реки Хуанхэ и культура племени чжоу в центре современной про- винции Шэньси. Однако несомненно, что и в Южном и Юго-Восточном Китае земледелие возникло также в глубокой древности. Восточноазиатская область является обширной ареной видо- и формообразования не только дикорастущих, но и очень боль- шого количества культурных растений. Многие роды плодовых растений всецело сформировались в Восточноазиатской области: мушмула азиатская (Eriobotrya), айва азиатская (Chaenomeles), абрикос, персик, кинкан, актини- дия, лимонник и другие. Многие роды возникли в Восточной Азии, а отсюда распространились и в другие области, где име- лись условия для вторичного видообразования, и из этих родов одни виды дали начало культурным растениям в Восточнои 40
Азии, а другие — во вторичных центрах видообразования в Азии и Европе. Количество эндемичных культурных и используемых видов в Восточной Азии чрезвычайно велико. Многие ценные культур- ные виды, относящиеся к родам, родина которых находится вне Восточной Азии или с точностью не определена, имеют основной очаг происхождения и развития в Восточной Азии. К таковым культурным видам относятся просо, чумиза, пайза, гаолян, соя, фасоль адзуки, ячмень азиатский (Hordeum humile), японская и китайская редьки и др. Культурная флора Восточной Азии недостаточно изучена и поэтому только грубо и предварительно можно провести ее де- ление на две подобласти. 1. Северо-Восточноазиатская подобласть, включающая Япо- нию, Корею, дальний Восток СССР, прилегающие районы Ки- тая, районы по Хуанхэ, Северо-Восточный Китай. Здесь сложи- лись скороспелые сорта сои, кунжута, риса, холодостойких цит- русовых, холодостойкие сорта чая, голозерного овса, азиатской мушмулы, актинидии, многих видов яблони, японской айвы, ви- дов груши, японской сливы, японского, маньчжурского и сибир- ского абрикосов, многих сортов декоративной сливовишни, япон- ской хурмы, некоторых видов лоха, каштана, грецкого ореха, цицании широколиственной, женьшеня, японской редьки, япон- ских групп моркови, репы, огурца, лука татарки, некоторых дру- гих видов лука и др. 2. Юго-Восточно-Е~ентральнокитайская подобласть включает Юго-Восточный Китай, районы к югу от реки Янцзы, провинции Сычуань, Юньнань, Гуандун (последняя — уже переход к Юж- ной Азии), восточную часть Тибетского нагорья. С ней связано происхождение азиатского ячменя, видов чая, кинкана, некото- рых цитрусовых, абрикоса муме, многих других видов плодовых. III. Южноазиатская область исторического развития куль- турной флоры. Включает Индию, Е~ейлон, Индокитай, Малакку и Малезию. Сюда примыкают и тропические районы Южного Китая. 10жная Азия дала мировому земледелию такие важныекуль- туры как рис, чай, цитрусовые, банан, особый масличный лен, распространившийся отсюда в страны Средиземноморья и Эфио- пию, виды фасоли (маш), большое разнообразие форм лобик (долихоса) и коровьего гороха. Здесь предполагается родина голубиного гороха, видов канавалии и других бобовых. В Юж- ной Азии (Индия) сосредоточено огромное разнообразие сортов кунжута, сафлора и масличных крестоцветных из рода Brassica. Здесь родина или один из первичных очагов происхождения огурца, многих клубнекорнеплодов, пачули и других аромати- ческих, родина сахарного тростника, сахарной пальмы, карда- мона, мускатного ореха, черного перца, гвоздичного дерева, ко- ричного дерева, имбиря и многих других пряных растений, юж- 41
ных видов бамбука, коикса, хлебного дерева, видов цитрусовых, мангового дерева, мангустана, дуриана, мига, древовидного' хлопчатника и др. Культурная флора Южной Азии богата эндемичными родами. и видами возделываемых растений. В Южной Азии находится, по С. Н. Боголюбскому (1959), третий по значению очаг доместикации животных. Многие культурные растения, зародившиеся в Южной Азии,. при расселении на север — в Китай — развили там формовое ц сортовое разнообразие. Еще не наступило время для детального районирования Южноазиатской области культурной флоры, но мы предвари- тельно намечаем две крупные подобласти. Первая подобласть включает южные районы Китая, соб- ственно Индию, Индокитай, Цейлон; здесь родина риса, куриного проса, маша и других южноазиатских зерновых бобо- вых, индийских масличных крестоцветных из рода Brassica, огурца, индийского редиса, небольшого количества видов цитру- совых, древовидного хлопчатника, видов чая, сахарного трост- ника и др. Здесь находится первичный очаг вхождения в куль- туру особого масличного льна, отсюда льны распространились. в Эфиопию и Средиземноморье. В Индии — область интенсив- ного формообразования кунжута и сафлора. Вторая подобласть включает Малакку и Малезию; здесь, родина хлебного дерева, коикса, такка, многих цитрусовых, пом- пельмуса, мангустана, дуриана, основной очаг вхождения в куль- туру кокосовой пальмы, мускатного ореха, черного перца, саго- вого дерева и др. Точно распределить видовой набор культурных растений. между основными подобластями и произвести более дробное районирование последних в Южной Азии пока невозможно из-за недостаточной изученности южноазиатской культурной флоры Эта работа, вероятно, будет в основном выполнена учеными, Индии. IV. Африканская область исторического развития культурной флоры. История и география культурных растений этой области слабо изучены. Поэтому еще нет достаточного материала для подразделения ее на подобласти, хотя последние, несомненно, существуют. Африканская область является ареной развития следующих родов, из которых вышли важные культурные расте- ния: кунжут, нуг, колеус, негритянское просо, сорго, коровий го- рох, арбуз, кола, индигофера, масличная пальма. Другие роды имеют здесь одну из областей своего первичного развития: хлоп- чатник, рис, банан. В Юго-Восточной Африке сосредоточен второй по значению очаг первичного вхождения в культуру хлопчатника — отсюда вышел хлопчатник гуза (Gossypium ЙегЬасеигп L.), который из Африки распространился в Азию.
Яля культурной флоры Африки характерны древнейшие связи ее с Азией. В странах Африки, особенно в Южной, имеется много мел- ких эндемичных культур, не получивших распространения за ее пределами. Для растений действительно африканского происхождения характерно то, что они в основном являются растениями корот- кого дня. Те же виды, которые хотя и эндемичны для Африки, но корни происхождения которых надо искать в Древнесредиземно- морье, относятся к растениям длинного дня (таковы Brassica carinata, Анрепа abyssinica, Crambe abyssinica и др.). Обширная и неоднородная область с обобщенным названием Африканская располагается южнее Сахары. Можно назвать ее и Южноафриканской, так как север Африканского континента входит в другую крупнейшую область исторического развития культурной флоры — Яревнесредиземноморскую. Возможно, что следовало бы выделить и Южноафриканскую подобласть с Капской провинцией, но для этого пока нет доста- точного материала. Эфиопия не является самостоятельной областью развития культурной флоры — это область влияния Древнесредиземномор- ской и Африканской областей исторического развития культур- ной флоры. Освоение древним населением Африки ресурсов раститель- ного и животного мира восходит к глубокой древности. С. Н. Боголюбский (~1959) считает, что возникновение живот- новодства в Африке относится к V и возможно даже к VI тыся- челетию до н. э. По его мнению, большинство домашних живот- ных этого континента, а именно: крупный рогатый скот, овца, верблюд и буйвол (но не лошадь) — происходят от местных африканских прародителей. Однако данные о распространении этих животных в мезолите очень скудны, и многие исследователи предполагают, что все они в глубокой древности попали сюда из Азии. Во всяком слу- чае то, что нам известно по данному вопросу, лишний раз сви- детельствует о древнейших культурных связях Азии и Африки. В Эфиопскую (Абиссинскую) обл а сть влияния Древнесредиземноморской и Африканской областей историче- ского развития культурной флоры включены Эфиопия, Эритрея, отчасти Судан и прилегающие с востока, юга и запада районы. Точно границы наметить трудно. Корни восточноафриканской земледельческой культуры вос- ходят к глубочайшей древности, отсюда происходят и истоки .земледельческой культуры Египта. древнейшие поселенцы Египта явились с юга и сначала обосновались в Верхнем Египте, а уже много позднее стала заселяться дельта Нила. В Египте развилась своеобразная культура, достигшая для своего времени очень высокого уровня, в частности земледелие там вступило на
путь интенсивного развития еще в древности. Однако долина Нила дала очень мало видов, пригодных для введения в куль- туру. Культурная флора Древнего Египта не аборигенна: она сло- жилась под влиянием собственно средиземноморской и передне- азиатской культурных флор. Для истории Эфиопии и прилегающих стран, наоборот, ха- рактерно замедленное развитие земледелия. Последнее и ныне остается в Эфиопии на сравнительно примитивном уровне. Соот- ветственно в Эфиопии могли сохраниться формы, распространив- шиеся в далекие времена из других стран, но сохранившие при- митивные черты, например эфиопская пшеница. Вследствие древности земледельческой культуры в этой стране здесь име- лось время для развития из привнесенных извне элементов очень. своеобразных эндемичных видов и форм. Особенно примеча- тельно наличие вторичных очагов развития особого (эфиопского) льна, вида эфиопской пшеницы, вида эфиопского ячменя, под- вида эфиопской (абиссинской) полбы, оригинальных сорных форм кресс-салата, форм пажитника, горчицы черной, чечевицы и других бобовых. Эти растения ведут начало от культурных переднеазиатских или древнесредиземноморских элементов, но прошли в Восточной Африке долгий путь развития. Как и во всякой географической области, называемой нами областью влияния, имеются здесь и такие аборигенные культур- ные виды, которые происходят не от элементов местной дикора- стущей (или сорной) флоры. К аборигенным видам в Эфиопии относятся тефф (Eragrostis abyssinica), нуг (Guizotia abyssi- nica), абиссинская горчичная капуста (Brassica carinata), абис- синский овес (Avena abyssinica), чат (Catha edulis), кофейное дерево, абиссинский ямс, некоторые виды Coleus, строительный молочай. Из этих растений абиссинский овес и абиссинская ка- пуста относятся к древнесредиземноморским родам и, воз- можно, культурные абиссинские эндемичные формы развились из сорных, распространившихся сюда из стран Древнего Среди- земноморья. Напротив, молочай, кофе, чат, нуг, виды колеуса и индигоферы — это элементы африканской флоры. Эфиопия вместе с прилегающими к ней странами Восточной Африки нахо- дится на стыке древнесредиземноморской и южноафриканской культурно-флористических областей, и резких границ при пере- ходе на севере к Средиземноморью и на юге — к южноафрикан- ской культурно-флористической области здесь нет. Ч. Новосветская область исторического развития культурной флоры. Древнее земледелие Америки основано почти исключи- тельно на аборигенных растениях, взятых из местной дикорасту- щей флоры. В огромной Новосветской области намечены две крупные подобласти. 1. В Центральномексиканскую подобласть развития культур- ной флоры входят Мексика, Гватемала, Гондурас, Никарагуа„ 44
Коста-Рика, Панама, северные части Венесуэлы и Колумбии. Земледельческие цивилизации — старотолтекская в северной Мексике, перешедшая к ацтекам, а в Центральной Америке ци- вилизация майя — оставили в наследство много культурных расте- ний. Эта область является родиной кукурузы (впрочем, некото- рые авторы склонны считать, что кукуруза — выходец из Южной Америки), среднеамериканского хлопчатника — упланда (Gossy- ршт hirsutum), колумбийско-панамского хлопчатника (G. tri- cuspidatum), некоторых видов тыквы, многих видов фасоли (многоцветковой, лима и тепари), многих дикорастущих видов картофеля, мексиканского томата (Physalis aequata), обыкно- венного перца, гваюлы, дынного дерева, авокадо, по мнению ряда авторов, махорки, воскового дерева, агавы, юкки, сапаты, многих видов кактуса, чайота, мексиканского амаранта (хуа- утли), лебеды, какао, ачиоте, многих декоративных растений (георгин, цинний, бархатцев, туберозы и др.), чиа — масличного шалфея и др. По С. Н. Боголюбскому (1959), в Центральной Америке находится четвертый по значению очаг доместикации животных: альпака, ламы, морской свинки, мускусной утки. Северная Америка, с одной стороны, представляет собой об- ласть влияния мексиканской и центральноамериканской культур- ной флоры, а с другой стороны, является родиной многих куль- турных растений, и, может быть, ее следовало бы считать само- стоятельным очагом. Здесь много эндемичных элементов. Отсюда вышли культур- ный подсолнечник, земляная груша, многолетний люпин, крупно- плодная клюква, североамериканский кендырь, североамерикан- ские виды ежевики, малины, крыжовника и многие декоративные и плодовые деревья. 2. Южноамериканская подобласть развития культурной флоры включает Боливию, Перу и Чили, Эквадор и Колумбию. Культурные растения создавались здесь на базе исторического развития древних цивилизаций — доинкской и инкской в Перу и Боливии, арауканской культуры в Южном Чили, культуры чибча — в Колумбии. В Южноамериканской подобласти Н. И. Вавиловым наме- чены следующие особо интересные очаги видообразования и формообразования культурных растений. Горные андийские обл асти Перу, Боливии и других андийских районов с прилегающими предгорьями и равнинами. Это родина культурного картофеля Solanum andigenum и близ- ких к нему культурных видов той же группы, многих видов ди- кого картофеля, своеобразных клубнеплодов ока и улыбка, настур- ции, ачира, лебеды квиноа и каньява, амаранта кичунча, люпина изменчивого, хлопчатника (Gossypium barbadense), табака, хин- ного дерева, дынной груши, перуанского перца, дикорастущего опушенного перца, маранхильо, крупнозерной кукурузы куска,
видов томата и, вероятно, культурного томата, тыквы (Cucurbita maxima) и др. Это большей частью растения короткого дня. Ч и л о а н с к и й о ч а г, или провинция Арауко, — тихоокеан- ская прибрежная местность в Южном Чили и остров Чилое с влажным климатом — родина культурного картофеля (Solanum tuberosum), мадии, чилийской земляники, особой вымершей культуры исчезнувшего теперь костра — мунго, араукарии. Ха- рактерны растения длинного дня. Область древней колумбийской культур ы ч и б ч а (Боготское плоскогорье Андийского хребта) — родина аракачи (Arracacia esculenta), колумбийского перца, видов кар- тофеля и др. примыкает к провинции Арауко и нерезко от нее отгр аничена. Провинция лесной тропической Америки (Бра- зилия, Лаплатская низменность). Здесь область происхождения земляного ореха, маниока, ананаса, маланги, фейхоа, маранты, каучукового дерева — гевеи, американского ореха (Bertho! letia), некоторых видов какао, парагвайского чая, видов картофеля группы Commersoniana и др. Границы этих провинций не резкие, за исключением провин- ции Арауко. Деление это грубое и предварительное. Для многих родов Южная Америка — это огромная арена видо- и формообразования. Примером подобных полиморфных и широко распространенных групп являются виды культурного и дикорастущего картофеля. Изучением географического распро- странения этих видов занимается С. М. Букасов. И хотя данная группа видов Solanum еще не изучена с исчерпывающей полно- той, но если и другие группы аборигенных культурных южноамериканских растений будут исследованы хотя бы при- близительно в таком же масштабе и с таким же применением разносторонних методов классификации, то задача точного и детального районирования южноамериканской культурной флоры станет легко разрешимой. В данной книге подробно излагается историческая география культурных флор тех стран умеренной и субтропической Евр- азии, которые входят в Древнесредиземноморскую область исто- рического развития культурной флоры, а также тех стран, куль- турная флора которых в основном сформировалась под влия- нием Древнесредиземноморской области. Таким образом, в центре внимания находится культурная флора Советского Союза и прежде всего очаги древнего земле- делия в Закавказье и Средней Азии с включением некоторых сопредельных стран, ибо флористические границы могут не сов- падать с административными. Русская равнина и Сибирь рассматриваются как области влияния Передней Азии. Земледелие Восточной Сибири сложи- лось под сильным влиянием Восточной Азии.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ ПЕРЕДНЕЙ АЗИИ Передняя Азия в широком понимании (Закавказье, северо- западный Иран, горный Туркменистан, Малая Азия, Аравия, Сирия, Иемен) — это одна из важнейших древних областей раз- вития и распространения человеческой культуры. Исследователи культурной флоры присоединяют к Передней Азии и Пенджаб. Передняя Азия является и поныне областью распространения многих дикорастущих родичей культурных растений. Из Передней Азии вышли многие культурные растения, кото- рые затем получили широкое распространение. ЗЕМЛЕДЕЛИЕ И КУЛЬТУРНЫЕ РАСТЕНИЯВ ДРЕВНЕМ ЗАКАВКАЗЬЕ Неоднократно высказывавшиеся предположения о «недав- нем» заселении Закавказья и превалирующей роли пришлых народов в развитии закавказской культуры опровергнуты новей- шими исследованиями в области археологии. Человек появился в Закавказье в глубочайшей древности, следы его существования обнаружены здесь в слоях, относя- щихся к плейстоцену. Фауна Закавказья в конце третичного периода была похожа на фауну Западной Европы той же эпохи. По данным армянских историков, по-видимому, Закавказье долго было лесистой страной. Конечно, и в то время существо- вали голые скалы и осыпи с ксерофильной растительностью, а на равнине и по южным склонам гор — участки нагорных степей и полупустынь, но и лесов, главным образом на северных скло- нах, было много. Сохранившиеся и доныне в л~:ственных лесах Кавказа обшир- ные заросли плодовых и древесных пород и кустарников обеспе- чивали питание первобытного человека в период собирательства. На то, что земледелие в Закавказье существовало в раннем неолите, указывают найденные здесь А. Н. Каландадзе каменные зернотерки, песты и частично отшлифованные орудия, которые могли служить наконечниками мотыг. У подножья горы Богутлу С. Н. Замятниным и М. 3. Панич- киной в 1946 г. вскрыта целая мастерская неолитических орудий. Неолитические стоянки открыты также недалеко от 1~утапси, около Сочи, в районе Гудауты и в других местах. В неолити- 47
ческом селище Кистрик, близ Гудауты, археологами найдены круговые зернотерки (А. Л. Лукин, 1950). В археологических остатках кургана Диди-Гудзуба (Зугдид- ский район) исследованы различные слои: от неолитических до эпохи греческой колонии. В нижних слоях здесь найдены камен- ные зернотерки и кремневые орудия. В этих слоях обнаружены плоды каштана, бука восточного, дуба имеретинского, лещины обыкновенной. Эти остатки говорят о том, что в тот период дико- растущая флора была близка современной. Из остатков культурных растений здесь (в слое ранней бронзы) были найдены: просо, могар, многорядный ячмень, голо- зерная пшеница типа мягкой, полба, похожая на полбу маха, полба, похожая на древнеколхидскую полбу; ржи здесь обнару- жено не было (В. Л. Менабде, 1940, 1948; Б. А. Куфтин, 1944, 1949, 1950). Эти находки указывают на значительное разнообразие куль- турных растений, возделываемых в Западной Грузии (в Кол- хиде) в эпоху неолита и позднее. Кроме пленчатых пшениц, здесь были обнаружены и голые пшеничные зерна. В. Л. Ме- набде думает, что в эту эпоху, может быть, голозерная пшеница еще не существовала, а просто у части ископаемых зерен пленки истлели, не сохранились, что, конечно, весьма правдоподобно. Особенно интересно нахождение в ископаемом состоянии таких по своему типу примитивных представителей пшениц, которые являются одними из родоначальных форм современных культур- ных пшениц и которые в то же время еще и ныне встречаются на полях Грузии в качестве реликтов древней земледельческой культуры этой страны, — речь идет здесь о пшенице маха и древнеколхидской полбе. Некоторые исследователи рисуют себе процесс образования вида Triticum macha Рекарг. et Menabde несколько иначе, чем это представлено нами в работе о происхождении пшеницы (Е. Н. Синская, 1955). По их мнению (например, М. М. Якуб- цинер, 1959; В. Л. Менабде, 1948 и др.), Т. macha не происходит непосредственно от другого вида пшеницы (от вымершего инте- грального вида Т. protomacha), процесс видообразования, ве- роятно, был осложнен здесь гибридизацией с образованием амфидиплоида с какой-то диплоидной пшеницей — однозернян- кой или с диплоидным видом Aeoilops. Если в данном случае при гибридизации одним из исходных видов была Т. ра1еосо1сЫсит Menabde (как предполагает М. М. Якубцинер), то это очень родственный Т. macha вид и сам может быть назван интегральным, и он также является произ- водным видом от основного интегрального вымершего вида Т. protomacha: Если в формировании гексаплоидных пшениц и принимали участие однозернянки или виды эгилопсов, то все же в резуль- тате возникла пшеница, а не новый вид эгилопса. Интегральный
вид может образоваться по типу не усложненного гибридизацией процесса, но может быть и полиплоидом, в частности и амфи- диплоидом. В области интенсивного видо- и формообразования процессы полиплоидии и амфидиплоидии проходят интенсивно, поли- плоиды и амфидиплоиды встречаются часто. Сравнительно недавно открыт в сообществе с Т. timopheevi Zhuk. новый гексаплоидный (2п =42) вид пшеницы Т. zhulcov- slcyi Menabde et Eritz. Он очень похож по морфологическим признакам на Т. timopheevi. Это обстоятельство подтверждает нашу мысль, что при своем возникновении родоначальные и про- изводнь1е виды в полиплоидной серии сравнительно слабо раз- нятся между собой, сильное расхождение имеет место позднее. Открытие Т. zhukovskyi говорит о том, что в Передней Азии эво- люция этой группы двузернянок продвинулась уже вплоть до образования полного полиплоидного ряда, вероятна, по схеме: древняя закавказская дикорастущая однозернянка (Т. boeoti- <cum Boiss.) — 2n = 14, закавказская двузернянка (Т. at arati- <сит Jakubz.) — 2n = 28, полба Тимофеева (Т. timopheevi)— 2n = 28, полба Яуковского (Т. zhukovskyi, амфидиплоид)— 2п =42. На этой схеме последнее звено филогенетического ряда- полиплоидной серии — не представляет собой прямого продол- жения предыдущего видообразовательного процесса, который на этом этапе осложняется гибридизацией. Полба Тимофеева и полба Жуковского скрещиваются между собой, но в гибридах проявляется стерильность. По В. Л. Менабде (1957), все три родственных вида, входя- щие в состав одного фитоценоза — пшеницы зандури, хорошо обособлены в половом отношении. Межвидовая гибридизация внутри фитоценоза здесь практически отсутствует, что и делает возможным их сосуществование без нарушения видовой целост- ности каждого вида. При возникновении Т. zhukovskyi путем амфидиплоидии барьер между видами создается сразу, без длительного отбора. Возможность сосуществования поддерживается здесь и фе- нотипическим уподоблением Т. timopheevi и Т. zhukovskyi, ко- торое также возникло, вероятно, в данном случае в короткий срок и не было результатом длительной и постепенной работы отбора. Таким образом, весь процесс приспособления к комплексу климатических, почвенных и биотических факторов (СЕВ) мо- жет ускоряться при осложнении основного формообразователь- ного процесса гибридизацией. Последняя и в древнейших очагах происхождения культурных растений, несомненно, оказала влия- ние на филогенетическую историю отдельных ветвей и видов. Несмотря на накопление новых фактов, палеолит и неолит Закавказья изучены еще фрагментарно. Значительно больше 49
сведений о развитии земледельческой культуры можно почерп- нуть из материалов, относящихся к более новым отложениям медного (энеолита), бронзового и железного веков. Особенно хорошо изучено энеолитическое и со слоями ранней бронзы поселение у Шенгавита близ Еревана (Е. А. Байбуртян, 1936 — 1938). Здесь в эпоху энеолита уже были жилища из сыр- цовых кирпичей на каменном фундаменте. К. круглой централь- ной комнате примыкали прямоугольные помещения. В середине центральной круглой комнаты обычно помещался большой ка- мень, который служил основанием столба, поддерживающего крышу. Около этого камня стоял невысокий круглый очаг, укра- шенный рельефным орнаментом. Близ очага находились камен- ные зернотерки, терочки и песты. В крупных сосудах и около очагов были найдены остатки колосьев, а также зерна ячменя и пшеницы, которые, по определению М. Г Туманяна, были более близки к дикорастущим формам, чем современные культурные сорта. Исследования М. Г. Туманяна (1944) показали, что най- денные зерна собраны со смешанного посева ячменя и пшеницы. М. Г. Туманян указывает, что энеолитические хлебные злаки имеют несколько более гигроморфное строение, чем современные широко распространенные сорта этих культур. На этом основа- нии М. Г. Туманян и Б. Б. Пиотровский (1949) приходят к за- ключению, что климат Закавказья в эпоху энеолита был более влажным, чем теперь. Во всех образцах, взятых при раскопках поселения Шенга- вит, обнаружена была древняя круглозерная пшеница, которая„ следовательно, была здесь широко распространена в бронзовом веке. Родственный вид пшеницы возделывался в каменном веке- в свайных постройках Швейцарии и получил название ТгШсит чи1даге antiquorum Heer. В Закавказье, как будет показано ниже, эта пшеница с конца бронзового века стала постепенно исчезать. В Армении она была обнаружена в эпоху Урарту, т. е. спустя 2000 лет после шенга- витской культуры, но только уже как редкая примесь к мягкой пшенице. Закавказье, и, вероятно, именно колхида, является областью происхождения основного филогенетического ствола рода Triti- сшп. В этот основной ствол входят все важнейшие, наиболее распространенные виды пшениц, за исключением однозернянок,. дикорастущих двузернянок и полбы Тимофеева. Прародителем этих пшениц является древний, интегральный, т. е. совмещаю- щий в себе признаки разных других более молодых видов, куль- турный вид, названный В. Л. Менабде Т. protomacha. Этот древ- ний колхидский мезофит распространился широко по Средней и Северной Европе, где дал начало вымершему теперь виду Т. antiquorum Heer, а, по нашему мнению, также и на юго-запад в Эфиопию, где от Т. protomacha произошла древняя эфиопская 50
пшеница Т. protoaethiopicum, а от последней современный вид эфиопской пшеницы Т. aethiopicum Jakubz. Переднеазиатский интегральный вид Т. protomacha распространился и дальше на восток, прежде всего в Месопотамию, где в новых условиях от него пошло большое разнообразие форм пшениц Юго-Западной Азии (особенно в Таджикистане, Афганистане и Северо-Запад- ной Индии) '"" В Закавказье в настоящее время сохранились только дери- ваты древнего полиморфного вида Т. protomacha в виде двух реликтовых, эндемичных для современной Колхиды видов— Т. macha и Т. paleocolchicum. Среди ископаемых пшениц Шенгавита М. Г Туманян во многих образцах обнаружил спельтовидную пшеницу с ломким колосом. Вероятно, это была форма, близкая к Т. paleocolchi- cum, а может быть, и настоящая Т. spelta, посевы которой со- хранились в Иране. В Шенгавите вместе с древними круглозерной и спельтовид- ной пшеницами везде находили и древний круглозерный шести- рядный ячмень. Этот ячмень родствен описанному Геером иско- паемому ячменю из неолитических свайных построек Швейца- рии — Hordeum hexastichum sanctum и Н. hexastichum densum. Ячмень и пшеница, вероятно, всюду или очень во многих странах возделывались в смеси. Они, по-видимому, относились к одному эколого-морфологическому мезофильному виду. Общие признаки для древних пшеницы и ячменя следующие: карлико- вый рост, прочная неполегающая соломина, плотный, короткий, безостый колос, мелкое округлое зерно. древний, теперь вымерший в Европе, как и Т. antiquorum, прототип современных культурных ячменей попал, вероятно, че- рез Балканы в Среднюю и Северную Европу из Передней Азии. Еще более развитая энеолитическая культура представлена в раскопках в Армении близ селения Элар (Е. А. Лалаян и Е. А. Байбуртян, 1927 — 1928). В поселении Элар ниже слоев урартского периода были открыты ямы для хранения зерна. В этих ямах найдены остатки пшеницы. Материалы, собранные в эларских раскопках, свиде- тельствуют также о значительном развитии скотоводства в эпоху энеолита. В составе стада в то время начинает преобладать мелкий рогатый скот, что связано с развитием полукочевых форм скотоводства. При росте поголовья скота около селений уже не хватает пастбищ для прокорма животных, и скот начинают отгонять на удаленные от поселений горные луга в равнинах, предгорьях и нижних горных поясах. Земледелие начинает отделяться от скотоводства. * Детально вопросы эволюции и распространения пшениц изложены а книге Е. Н. Снискай «Происхождение пшеницы» (1955). 51
Энеолитические стоянки эларского типа отнюдь не являются единичным явлением, но, напротив, обнаружены во многих ме- стах Армении. В раскопках П. Ф. Петрова у села Малаклю около Игдыра в энеолитическом слое были найдены каменные зернотерки очень. больших размеров, указывающие, что здесь на муку и крупу перерабатывались большие количества зерна. В раскопках на холме KIoJIb-Тапа около Нахичевани, где, по-видимому, находи- лись и поселение, и могильник, были откопаны человеческие кости и вместе с ними керамика, кости животных и также обуг- ленные зерна ячменя и пшеницы (Б. Б. Пиотровский, 1949). По данным Б. Б. Пиотровского (1944), во многих урартских и средневековых крепостях Армении нижние слои указываюг на преемственность культур, ибо там найдены и слои с энеоли- тической культурой, близкие к эларским (Повинарская кре- пость, Армавирский холм, Двин, Гарни и др.). В раскопках Б. А. Куфтина в Триалети в самом нижнем слое. крепости Ахыллар около села Башташени найдены относящиеся к энеолиту пластинки из кремня, которые служили вкладышами в составном серпе, и ладьевидные зернотерки. В Очамчирском энеолитическом поселении также откопаны каменные вкладыши для серпов и зернотерки. По свидетельству Б. Е. Пиотровского, очамчирские энеолити- ческие материалы сходны с рядом других древних поселений,. обнаруженных в 1(олхиде. В энеолитических слоях Закавказья находили медные кин- жалы, ножи, кремневые и обсидиановые наконечники стрел„ золотые украшения. Земледелие, возникшее уже в неолите (а может быть, и еще раньше), развивалось и в энеолите, но произ- водство орудий земледелия еще долго оставалось на примитив- ной ступени каменного века; и в энеолите наряду с разнообраз- ными предметами из меди все еще продолжали существовать каменные мотыги, хотя и лучше отточенные и отполированные. Большой интерес представляют раскопки энеолитических слоев крепости Гарни в Армении (Б. Н. Аракелян, 1951). Для истории культурной флоры важна найденная здесь косточка абрикоса, что говорит о древности существования этой культуры в Армении. Здесь было обнаружено много каменных зерноте- рок, обсидиановые и кремневые вкладыши для серпов, медный серп. Наряду с земледелием древнейшее население Гарни зани- малось и животноводством. 1( началу бронзового века скотоводство Закавказья достигает особо интенсивного развития. Роль удаленных горных пастбищ возрастает. Экономическое значение скотоводства в этот период было, по-видимому, больше, чем земледелия, отделившегося от скотоводства. Однако состав возделываемых культур был в это время разнообразен: при раскопке кургана бронзового века в Зугдидском районе Западной Грузии были обнаружены зерна
полбы маха, ячменя, проса, могара, а также были найдены каш- таны, желуди, орехи, буковые орешки и др. Интересные находки более поздних эпох (между серединой II и началом I тысячелетия) были сделаны в раскопках в устье реки Моквы, к северо-западу от Очамчиры и также близ Сухуми (А. Н. Соловьев, 1950). В этих древних поселениях была обна- ружена так называемая текстильная керамика. Такое название получили сосуды с отпечатками тканей на глине. По Б. А. Куфтину (1950), большая стоянка с текстильной керамикой у Очамчиры относится приблизительно к VI — Ч вв. до н. э., но там же открыты и значительно более древние слои, где также найдена текстильная керамика. Установлено, что жи- тели этой стоянки занимались соляными промыслами. За последнее время были проведены геологические исследо- вания грунтов побережья и морского дна. Результаты этих ис- следований заставляют считать очамчирскую стоянку более древней, чем предполагали сначала. В соответствии с геологи- ческими данными ее следует отнести к середине или ко второй половине II тысячелетия до н. э., это значит, что ее обитатели жили задолго до античного периода Колхиды. Условно эту стоянку называют энеолитической, хотя никаких медных орудий здесь не было обнаружено. Исследования очамчирских памятни- ков продолжаются, и, возможно, там будет добыт новый архео- логический материал, интересный для освещения истории сель- ского хозяйства в Закавказье. Отпечатки тканей на глиняных сосудах из-под Очамчиры и Сухуми и из других мест говорят о большой древности куль- туры льна в Колхиде. Несомненно, что ткачество достигло весьма высокого уровня в Колхиде задолго до эллинской эпохи и затем приобрело славу в эту эпоху. Текстильная керамика открыта и в культурном слое поздне- неолитической стоянки Кистрик в Абхазии. Сосуды, сформиро- ванные на льняной тканевой основе, заключенной внутри стенки, найдены также в Степанакертском кургане (1-я половина II ты- сячелетия до н. э.). Таким образом, археология позволяет проследить историю культуры льна в Закавказье с середины II тысячелетия до н. э. и до эллинского периода. Льняное ткачество было распростра- нено в Колхиде задолго до эллинской эпохи, там имеется свой аборигенный озимый лен. О колхидском льне писали многие гре- ческие авторы. Геродот сообщает, что колхидское полотно называлось у эл- линов сардинским, т. е. будто бы оно происходило из Сардинии. Б. А. Куфтин (1950) считает, что название «сардинский», приводимое Геродотом, ошибочно, и вместо «сардинский» нужно читать «сардский». По словам Б. А. Куфтина, колхидский лен, по- падая в Грецию через малоазиатские сарды, мог получить от них название «сардский», которое, однако, превратилось в sardon,
-поскольку колхидское полотно могло носить в Греции еще еги- петское название sindon — «тонкая ткань». Он сообщает также о большом разнообразии колхидских ископаемых льняных тканей: от грубой парусины до тонкого полотна. «Это сортовое разнообразие тканей, — пишет Куфтин,— указывает одинаково как на многообразие потребностей у насе- ления в их бытовом применении, так и на специализацию и воз- можности прогрессивного развития этого производства и мест- ного ткацкого станка в доантичной Колхиде». Ботанические данные отнюдь не находятся в противоречии с данными археологии и истории. В Колхиде до сих пор сохра- нился реликтовый озимый лен, который культивируется и в Ма- лой Азии. С этой точки зрения легко допустить, что название льна у Геродота «сардинский» было связано с малоазиатскими сардами, и греки могли получать лен и полотно не только из собственно Колхиды, но и из Малой Азии, где, несомненно, куль- тура льна также возникла в глубокой древности и была осно- вана на одной и той же ботанической форме льна. Какой же именно лен возделывался в Древней Колхиде? jlv- кий родоначальник культурных льнов — Linum angustifolium ныне произрастает в прибрежных средиземноморских странах. Его типичные формы являются озимыми (иногда даже многолет- ними), склонны к простратности (особенно до цветения), имеют большое количество ветвей, отходящих от основания главного стебля. Эти особенности свойственны и культурному передне- азиатскому озимому льну. Последний, как и дикий L. angustifo- lium, цветет и при весеннем посеве, но при посеве осенью в усло- виях Черноморского побережья дает лучшие результаты. При весеннем посеве первое время лен «мучается», ветвится, а затем цветет очень недружно, отчего и созревание растянуто. От дико- растущего родоначальника отличается нерастрескивающимися коробочками (вернее, слабо растрескивающимися). В настоящее время этот совершенно особый лен имеет островное распро- странение. Наиболее значительные площади под его посевами обнаружены П. М. Жуковским в Малой Азии, недавно его можно было встретить в окрестностях Ленкорани, и теперь еще он, хотя и очень редко, высевается в Мегрелии. Небольшие по- севы этого льна имеются кое-где в Баварии. Разорванный ареал уже указывает, что этот лен — древнее, некогда широко рас- пространенное, а теперь реликтовое культурное растение. Под- тверждением широкого распространения этого льна в древности служит открытие его Геером (Heer, 1872) в неолитических свай- ных постройках Швейцарии. Геер сначала определил его как L. angustifolium, но позднее отказался от этого определения и стал считать его озимым льном, сходным с малоазиатским и за- кавказским. Геер дал хороший рисунок и описание этого льна. Коробочки у него не растрескивающиеся, чем он отличается от angustifolium, и очень мелкие — даже мельче, чем у совре-
менного озимого льна из Малой Азии и Мегрелии. И в неолити- ческую эпоху в Швейцарии этот лен был прядильным растением (были найдены не только семена, стебли и коробочки, но и льняные изделия), и в настоящее время в самом большом очаге его культуры — в Малой Азии — он используется именно как прядильное, а не как масличное растение. Интересно, что у местного населения в тех глухих уголках Закавказья, где еще сохранилась его культура, отношение к. этому льну особое. Он считается священным растением, возделы- вается отдельно, большей частью на огородах, и хранится особо. Об этом говорят старики, которые, впрочем, очень неохотно со- общают сведения о нем. Очевидно, он и теперь сохранил скорее ритуальное, чем хозяйственное значение. Вероятно, этот лен, как и другие культурные растения, лишь позднее стал использо- ваться как прядильное растение, но в народных преданиях до сих пор сохранилась память о первоначальной цели его возде- лывания. Мы думаем, что в льняном производстве колхи использовали именно этот лен. Он распространен и теперь в местностях с влажным климатом. Это ярко выраженный мезофит. Влажный. «колхидский» климат был раньше распространен на более об- ширной территории, чем современная область распространения колхидской флоры, и правильнее этот реликтовый лен называть колхидским, а не малоазиатским, как это принято в ботаниче- ской литературе. Родина этого льна — область прежнего рас- пространения колхидской флоры, а не Средняя Европа. В пользу такого предположения говорит то, что именно в Передней Азии (Малая Азия и Закавказье) он дает отхождение филогенетиче- ских ветвей в разных направлениях (см. схему). В Анатолии типичный простратный озимый лен, имеющий сравнительно длинную стадию яровизации, связан постепенными малозаметными переходами с яровыми анатолийскими кудря- шами и межеумками. Это сравнительно гигроморфная ветвь. В Армении и Грузии простратный колхидский лен (также че- рез ряд постепенных переходов) дал начало ранним армяно- грузинским горным кудряшам — сравнительно ксероморфной ветви. От него же имеется постепенный переход к поздним даге- стано-азербайджанским кудряшам, а от них — также с постепен- ными переходами — к дагестано-азербайджанским межеумкам, которые еще недавно имели распространение в предгорьях. В этой среднеспелой ветви идет и постепенное укрупнение коро- бочек. Группа предгорных дагестано-азербайджанских межеумков. родственна украинским так называемым степным межеумкам. Последние отличаются от лесостепных межеумков 1~азахстана, Поволжья и среднерусских областей (Тульская, Орловская) луч- шей облиственностью, большей частью несколько замедленным
Схелт происхождения и распространения льнаиз Колхидского очага дикорастущий лен (Linum angustifolium) Колхидский озимый стелющийся лен Анатолийские кудряши Армяно-грузинскиеи дагестанскиеранние кудряши ,дагестано-азербайджан- ские поздне-среднеспе- лые кудряши Анатолийские межеумки(возможно, частью) Я,агестано-азербайджан- ские межеумки с круп- ными коробочками Западные долгунцы(частично): Голландия,Франция, Бельгия Украинские степныемежеумки ростом и развитием, более крупными коробочками и семенами, что и роднит их со льнами предгорий Азербайджана и Qare- стана. При изучении, например, коллекции местных образцов льна из Ставропольского края "' были обнаружены нечувствительные переходы от типичных степных межеумков к предгорным льнам Азербайджана и Дагестана. Вероятно, в пределах 1~авказа и севернее — на Украине имеет место сравнительно молодой формообразовательный про- цесс, развившийся в результате постепенного расселения древ- него колхидского льна с юго-запада на северо-восток и на северо- запад в условиях весеннего посева. Расселение этого льна в Западную Европу шло, по-видимому, через Балканы с юго-востока на северо-запад, затем он достиг южной Германии (сохранившиеся островки культуры в Баварии) и Швейцарии и, вероятно, Бельгии и Франции. Из последних двух стран имеются образцы культурного льна, приближающе- гося к простратному колхидскому и вместе с тем имеющего пере- ходные черты к L. angustifolium (растрескивающиеся коро- бочки). В Швейцарии озимый лен колхидского типа найден в неолите. В озерной области свайных построек Швейцарии лен был распространен и в более древние времена, на что указывает * В настоящее время культура льна там почти псчезла, и нельзя найти местных форм.
сравнительно высокий уровень ткачества в тех слоях, где он был обнаружен. Следовательно, и в Колхиде, откуда он пришел в Среднюю Европу, начало его культуры имело место не позднее нового каменного века. На примере льна показана эффективность одновременного применения археологического и ботанического методов для ре- шения вопроса о происхождении культурного растения и путях его распространения. К льнам, в настоящее время распространенным в Закавказье, уже нельзя применять ботанический метод. Теперь культура льна в Закавказье сильно сократилась. Яагестано-азербайджанские предгорные межеумки почти исчезли, исчезают и настоящие гру- зино-армянские ранние горные кудряши и другие местные сорта. Добавочные данные о развитии древнего земледелия в За- кавказье получены при раскопках других поселений. Очень интересны, особенно с точки зрения истории садовод- ства, исследования Я. И. Гиммеля (1940) в долине реки Ганджа- чай (Азербайджан) в поселении бронзового века. Здесь открыты две каменные зернотерки, из них одна боль- ших размеров с пестиком. В этих раскопках были найдены хо- рошо сохранившиеся довольно крупные косточки миндаля, по- хожие на плоды современного культурного миндаля. В районе раскопок в ущелье Люлели открыт сад миндаля, который, по мнению Я. И. Гиммеля, является здесь «пережитком» древней культуры. В другом месте обнаружены заросли гранатового кустар- ника и кладбище отмерших в разное время толстых стволов и корней граната. Погребенные стволы расположены последова- тельно снизу вверх в 4 — 5 этажей. Гиммель объясняет это свое- образное расположение длительным периодом образования на- сыпи курганов. Это давало возможность гранату постепенно расти вверх. Стволы в нижних своих частях постепенно отмирали и образовывали по мере их засыпки все новые корневые системы и все ближе к поверхности земли. долговечность граната, неоднократно отмечавшаяся в бота- нической литературе, сделала возможным сохранение этих полу- ископаемых зарослей до нашего времени. Я. И. Гиммель отмечает большое культовое и символическое значение граната в странах Передней Азии и Средиземноморья, возможно, такое значение гранат имел и у древних племен Азер- байджана. В древнем поселении к западу от Ханлара в слое, относя- щемся к самому началу 1 тысячелетия до н. э., обнаружены на- ряду со следами земледелия следы древнего садоводства. Однако орудия все еще примитивны: здесь обнаружены каменные зерно- терки и кремневые вкладыши от серпов. По данным других рас- копок тех же приблизительно лет можно установить, что в это время возделывались аборигенные сорта пшеницы и также
полба-эммер. В сосудах этого периода находились косточки винограда и персика. Наряду с тем найдено было много семян и остатков дикорастущих растений: косточки каркаса кавказ- ского, семена мари и нескольких видов лебеды. Возможно, что марь и лебеду употребляли в это время в пищу, хотя и не воз- делывали: собирательство, конечно, еще сохраняло некоторое зн а чение. В слоях верхней бронзы 1 тысячелетия до н. э. было обнару- жено два типа серпов: 1) более примитивный, составной из кремневых или обсидиановых вкладышей, закрепленных в дере- вянной или костяной основе; 2) кованый металлический серп не- большого размера на деревянной рукоятке. Мотыги в это время употреблялись деревянные с каменными или бронзовыми нако- н ечн ика ми. В кургане около Ханлара найдены были остатки примитивной молотилки, которая представляла собой плоскую дубовую доску с углублениями на нижней стороне. В эти углубления вставля- лись куски кремня. Сходные молотилки примитивного типа были откопаны в несколько более поздних слоях в Алавердском районе Армении. В это время разводился крупный рогатый скот, овцы, козы, свиньи. Все больше используется лошадь. Для обслуживания скотоводства на горных пастбищах сооружают каналы и во- доемы. Чтобы понять историю земледелия 1 тысячелетия до н. э. в Закавказье, остановимся кратко на истории Армении. Армянское нагорье (средняя высота 1500 — 1800 м над ур. м.) расположено между Анатолийским плоскогорьем и Иранским нагорьем. На Армянском нагорье берут начало самые большие реки Передней Азии — Тигр и Евфрат, которые обеспечивали сообщение Армянского нагорья с Месопотамией и облегчили преемственность культуры Месопотамии и более поздней урарт- ской и собственно армянской культуры. Климат здесь довольно суровый, континентальный, особенно на северо-западе, где на .Эрзурумском нагорье зимы отличаются продолжительностью, здесь нередки сильные морозы (до 40'). В юго-восточной части нагорья расположено плато с более мягким климатом и непро- должительными зимами. Разнообразие климатов (земледелие поднимается в горы до 2600 л над ур. м. и в отдельных случаях даже несколько выше) делает возможным возделывание здесь разнообразных культурных растений. Культура собственно Армении является непосредственным продолжением культуры Урарту. По Б. Б. Пиотровскому (1944), первые сведения об Урарту относятся к XIII в. до н. э. и найдены в надписях ассирийского царя Салманасара. В то время урартского государства еще не было, существовал только союз племен, во главе которых были племена Урарту (иначе халды) и Наири. В середине IX в. до 58
н. э. этот союз племен образовал уже довольно сильное государ- ство, которое воевало с Ассирией. Племена были местными— аборигенными — этническими образованиями. Им (племенам) и другим «горным народам» Передней Азии принадлежит огром- ная роль в развитии культуры Передней Азии, в частности— земледельческой. Связи урартской культуры с культурой Асси- рии несомненны, они проявились как в искусстве, так и в других сторонах человеческой деятельности. Сначала ассирийцы разо- ряли урартов и наири, а затем Ассирия начала уступать и силе и влиянии Урарту. Наиболее древняя территория урартов распо- ложена между озерами Ван и Урмия, позднее урарты достигли Евфрата, а на севере утвердились в Закавказье. Администра- тивный центр Урарту был перенесен в Закавказье, где теперь находится селение Армавир. Много памятников VIII в. до н. э. обнаружено в окрестностях озера Севан. Урартское государство пережило Ассирию, которая перестала существовать после па- дения ее столицы Ниневии, осаждавшейся вавилонянами, мидя- нами и скифами в 612 г. до н. э. В начале VI в. до н. э. под натиском мидян перестало суще- ствовать и государство Урарту. В середине VI в. до н. э. на западе Урарту создался новый союз племен, во главе которого стояли уже армены. История арменов начинается еще до паде- ния Урарту, как история одного из племен, входивших в его со- став. Позднее армены (и другие близкие племена, из которых хай даже получили преобладание над арменами) смешались с урартами, восприняли и продолжили культуру Урарту. Отсюда древнее название Армении — Хайастан. Развитие урартской культуры относгтся к железному веку (найдены были железные серпы). Урартская культура достигла высокого уровня. Страна была густо заселена, в ней было много сел и городов. До наших дней сохранились остатки величествен- ных сооружений, крепостей и жилищ, оросительных каналов. По данным Б. Б. Пиотровского (1944), здесь была высоко развита земледельческая культура. Амбары ломились от запасов, остав- шихся от урожаев прежних лет, главным образом пшеницы, ячменя, проса. Было сильно развито скотоводство, в связи с этим даже сеяли кормовые травы для лошадей. Судя по тому, что на Армянском нагорье сохранились эндемичные сорта культур- ной люцерны, приближающиеся по морфологии и биологии к местным дикорастущим предгорным и горным люцернам, можно предположить, что на корм сеяли именно люцерну, хотя на это нет прямых указаний. Наше предположение о культуре аборигенной люцерны на Армянском нагорье еще в эпоху Урарту подтверждается также тем, что до самого последнего времени там в горных районах из кормовых растений имелась только лю- церна. О ней местные старожилы говорят, что «ионджа»— армянское название люцерны — возделывается здесь «с незапа- мятных времен», а эспарцет только недавно стал проникать и 59
з горные районы (не считая Ахалкалакского района, где он, по-видимому, давно возделывался). Природные леса Армянского нагорья были истреблены еще до появления урартов, а часть страны, может быть, была и искони безлесной. Урарты усиленно насаждали леса и рощи (особенно платановые), плодовые сады и виноградники. ~ля характеристики состава возделываемых растений в Урарту много дали продолжающиеся и теперь раскопки в кре- пости Кармир-Блур недалеко от Еревана (Б. Б. Пиотровский, 1944). Материалы Кармир-Блура датируются VIII — VII вв. до н. э. Около очагов здесь находились зернотерки и каменные чаши с пестами. В этих каменных чашах или просто в углубле- ниях, выдолбленных в скале, которая служила полом, произво- дилось рушение зерна, затем зерно перетирали в каменных зер- нотерках. В крепости были обнаружены обширные кладовые с пшеничным зерном и другими запасами. Согласно определению М. Г Туманяна, в Кармир-Блуре найдены мягкие пшеницы- ТгШсит аеШит L., очень похожие на современные армянские сорта. На основании детального изучения материалов из Кармир- Блура и других древних поселений М. Г Туманян установил, что в древности на территории Армении культивировалось несколько видов пшеницы и в том числе такие формы, которые являются очень древними, представляют собой прототип наших современ- ных культурных пшениц. В некоторых образцах пшеницы из Кармир-Блура поражала чистота и однородность материала и отсутствие сорняков. Иско- паемые пшеницы типа мягкой имели зерна средних размеров с хорошо сохранившимся хохолком, с бороздкой и резко выра- женным зародышем. Имеются колоски с опушенными и голыми колосковыми чешуями; было обнаружено большое разнообразие форм колосковой чешуи, колоски в большинстве нежной струк- туры, с двумя, реже с одним зерном, обычно яйцевидной формы, стержень колоса по бокам неопушенный. Преобладали коротко- остистые или безостые колосья. Помимо описанных выше форм мягкой пшеницы в Кармир-Блуре в двух образцах было обнару- жено более 200 зерен древнейшего вида круглозерной пшеницы. Судя по раскопкам в Шенгавите (энеолит — начало бронзового века), эта вымершая форма пшеницы была в более древние времена широко распространена, а в период господства урартов стала уже большой редкостью и встречалась только как при- месь. То же относится и к шестирядному круглозерному ячменю, который был найден в Шенгавите в большом количестве, а в эпоху урартов уже не возделывался. Помимо упомянутых выше форм пшеницы в Кармир-Блуре, спельтовидная пшеница была найдена только в одном образце (несколько колосков), а в Шен- гавите ее было много. Недалеко от Нахичевани, в раскопках холма Кюль-Тапа (О. А. Абибуллаев, 1953) найдены обугленные зерна пшеницы, бО
которые отнесены к концу неолита — началу энеолита. По опре- ,делению М. М. Якубцинера — это зерна карликовой пшеницы, похожей на индийскую круглозерную пшеницу. И здесь древней- шей является круглозерная пшеница, по-видимому, родственная T. antiquorum и Т. protomacha. После тщательного осмотра ископаемых пшениц и ячменей из Кармир-Блура М. Г Туманян пришел к заключению, что кли- мат эпохи Урарту был несколько суше, чем в предшествующем бронзовом веке. Некоторые остатки растений были определены им как рожь. По мнению М. Г. Туманяна это была, вероятно, сорная, а не культурная рожь. Однако Б. Б. Пиотровский (1950) пишет определенно о «запасах» ржи в крепости Кармир-Блур. .Это не могли быть запасы сорной ржи. Значит, рожь культиви- ровалась в Урарту, что представляет большой исторический ин- терес. K со>калению, детально эта ископаемая рожь не изучена. В Кармир-Блуре найдены были значительные запасы пленча- того шестирядного ячменя в сосудах и зернохранилищах. Зерна ячменя из Кармир-Блура, полученные от Б. Б. Пиот- ровского, были исследованы знатоком ячменя Ф. Х. Бахтеевым, который отмечает, что среди них обнару>кены как пленчатые, так и голозерные ячмени. Длина зерен пленчатого ячменя изменялась от 4,5 до 9,0 мм, а ширина — от 2,0 до 3,5 л~л~. Среди пленчатых имелись здесь и своеобразные бутылковидные формы с длинной основной ще- тинкой. Среди современных сортов ячменя форм с бутылковид- ным зерном не встречается. Длина зерен голозерного ячменя из Кармир-Блура варьиро- вала от 4,0 до 7,5 л~м, а ширина — от 2,0 до 4,0 мм. Округло- овальные зерна не были единичными (на рисунке они выгля- .дят менее округлыми, чем швейцарский ископаемый ячмень Геера) . По Ф. Х. Бахтееву, ячмень из Кармир-Блура крупнее, чем ячмень из Боспора, но между ними имеется заметное сходство: и там и здесь отмечено наличие особых бутылковидных зерновок. Большой интерес представляет смешанный запас в большом глиняном сосуде из внутреннего помещения здания. По заклю- чению В. А. Петрова, запас этот состоял из зерен того же шести- рядного ячменя, семян двух видов бобовых — нута, француз- ской чечевицы — и масличного крестоцветного иыдау. Поиски параллелей в этнографической литературе показали, что у MHorHx племен западных отрогов Гималаев и E северо- западной Индии, сохранивших архаические приемы возделыва- ния сельскохозяйственных культур, до настоящего времени су- ществует обычай вести смешанные посевы. Так, смешанные по- севы нута и французской чечевицы распространены в Афгани- стане, а в некоторых провинциях Индии ведут смешанные посевы шестирядного ячменя, нута и индау, т. е. именно той комбинации видов, запас семян которых был обнаружен в Кармир-Блуре.
Некоторые ученые существование смешанных посевов обь- ясняют тем, что основная чистая культура засорялась каким- нибудь сорняком, пригодным для использования. Находка, сделанная в Кармир-Блуре, и этнографические па- раллели В. А. Петрова дают материал для обсуждения этого вопроса. Возможно, что смешанные культуры, представлявшие определенный хозяйственный интерес в суровых (в данном слу- чае — по засушливости) климатических условиях, были распро- странены несравненно шире и поддерживались вполне созна- тельно. Мы присоединяемся к мнению В. А. Петрова о том, что сме- шанные посевы всегда характерны для определенной стадии земледелия. Смеси могут иногда возникать и путем засорения основной культуры, например смесь льна и засоряющих его ин- дау и особой формы сурепицы. Собирая семена такой смеси, че- ловек высевает ее снова. Таким образом, смеси, возникшие по- мимо воли человека, входят в культуру. Еще недавно в горной Армении можно было встретить смешанные посевы масличного льна и рыжика, который, будучи первоначально сорняком льна, «навязался» человеку в использование. Вместе с тем несомненно, что древний земледелец и сознательно высевал смеси, чтобы га- рантировать себе урожай за счет того или иного компонента в. условиях непостоянного климата (не обязательно засушливого, как указывает В. А. Петров). Не исключена, впрочем, возмож- ность и того, что в иных случаях смесь дает более высокий уро- жай благодаря взаимному благоприятному влиянию компонен- тов при совместном произрастании. Однако этот вопрос еще. слабо изучен. В Закавказье и теперь имеются в культуре сме- шанные посевы разных видов пшеницы (однозернянка и полба Тимофеева, пшеница дика и мягкие пшеницы, мягкие и твер- дые пшеницы и др.) . Л. Л. Яекапрелевич (1960) считает, что самые древние по- севы в Закавказье были смешанными и в настоящее время ди- кие однозернянки часто произрастают вместе с двузернянкамк и диким ячменем — первобытный человек вряд ли умел разли- чать эти виды и, несомненно, собирал и затем высевал их вме- сте. И только в эпоху поздней бронзы человек стал переходить к относительно чистой культуре зерновых злаков. Смешанные культуры были характерны (это наблюдалось еще недавно, в 30-х годах нашего столетия) также для Таджи- кистана. В Кармир-Блуре найдено также просо. Был обнаружен со- суд с отверстием в днище, заполненный зернами и соломой ита- льянского проса (Setaria italica Р. В.). У многих колосков чешуи растопырены и неплотно облегают зерно. Вместе с тем в сосуде были и пустые колоски, и выпавшие из них зерна. Это наблю- дается в том случае, если необмолоченное зерно держать в воде, пока оно намокнет, а затем просушить. Зерно подвергаюг 62
намачиванию и высушиванию для двух целей: при подготовке к переработке на крупу (что теперь применяется к овсу) и для приготовления солода. Б. Б. Пиотровский (1944) пришел к за- ключению, что в данном случае имело место изготовление со- .лода из проса гоми для пивоварения. М. Г. Туманян пишет, что в раскопках доурартской эпохи просо в Армении не было обнару>кено. Нахождение его в большом количестве в Кармир-Блуре и в раскопках последующего периода (Звартноц, Двин и др.) на- водит на мысль, что культура проса в Армении не аборигенная, а занесена сюда с востока народами Урарту. Но стоит ли говорить о занесении непосредственно с востока, если просо было обнаружено в неолите соседней Грузии? Б. Б. Пиотровский предполагает, что обитателям Кармир- Блура был известен лен, так как найден рулон обгоревшей ткани мешочного типа, похожей на льняную. Один из сообщающихся сосудов в Кармир-Блуре был напол- нен семенами чистого индау. Как было сказано выше, семена этого же растения найдены также и в смеси с ячменем и нутом. Индау относится к таким культурам, которые раньше имели более широкое распространение, чем в настоящее время. Теперь индау встречается в Закавказье не часто, как сорняк льна. Впрочем М. Г. Туманян находит, что в Кармир-Блуре найден кунжут, а не индау. Нам это не совсем понятно: семена индау легко отличить от семян кунжута. Но нет ничего удивитель- ного, если был найден и кунжут. По другим источникам, эта культура в Закавказье и сопредельных странах считается весьма древней. Кроме того, кунжут, как и индау, является древним растением и обычным компонентом смешанных посевов в Ин- дии и Таджикистане. Позднее Б. Б. Пиотровский (1948) сооб- щил, что кунжутовые жмыхи (уже без ссылок на сомнительность определения) были найдены в большом количестве при дальней- ших раскопках Кармир-Блура. В полевой культуре Урарту видное место занимали зерно- вые бобовые. Выше говорилось о находках нута и французской чечевицы. 06а растения и ныне возделываются в Армении. Фран- цузская чечевица, по словам старожилов, в прежние годы была распространена значительно шире. В Закавказье встречаются массивы дикорастущих видов нута. М. Г. Туманян определил третье бобовое, найденное в Кар- мир-йлуре, как конские бобы. Не случайно здесь была обнару- жена наименее окультуренная, мелкосемянная форма конских бобов, которая теперь сохранилась в культуре лишь в странах примитивного земледелия, а некогда была широко распростра- нена в Евразии. В одном из сосудов найден запас семян рыжика мелкоплод- ного (семейство крестоцветных), который и сейчас произрастает дико на склонах предгорий и нижних поясов гор в Армении и
Грузии. Мелкоплодный рыжик в сосуде из Кармир-Блура был смешан с семенами и стручками двух других видов крестоцвет- ных сорняков. На этом основании В. А. Петров утверждает, что семена мелкоплодного рыжика были здесь собраны не из диких зарослей, а из засоренного посева. Если это так, то данный факт представляет большой интерес для решения вопроса о происхо- ждении местного культурного масличного рыжика, который, не- сомненно, представляет собой эндемичный аборигенный вид Camelina caucasica (Sinsk.) Vass. и нигде, кроме Закавказья, пока не встреченный. Этот вид рыжика вышел в чистую куль- туру из посевов местного масличного льна — армянского горного кудряша. Судя по находке в Кармир-Блуре, исходной формой для С. саисаэ1са был мелкоплодный рыжик — Camelina micro- carpa Andrz., который некогда из дикорастущего состояния пе- решел в сорное, а затем в процессе эволюции при возделывании вместе со льном дал начало сорному кавказскому рыжику С. caucasica. Последний вошел в чистую культуру там, где ус- ловия для него были более благоприятны, чем для его смеси с масличным льном. На развитие садоводства и бахчеводства в Урарту указы- вают находки в Кармир-Блуре остатков плодов алычи, граната, семян арбуза. Особенно широко было развито в Урарту виноградарство. В раскопках обнаружено большое количество обугленных ви- ноградных лоз. Найдены были также и остатки винограда, хотя и в малом количестве; это объясняется тем, что к моменту ги- бели Кармир-Блура в начале августа виноград еще не созрел. Во время раскопок 1945 — 1950 гг. были открыты и исследованы большие кладовые, предназначенные для хранения вина. Вся территория этих кладовых была занята огромными сосудами- карасами, наполовину вкопанными в земляной пол. Все вскры- тые в Кармир-Блуре материалы говорят о высоком развитии виноделия в Урарту. Техника земледелия также была высокой. Основные земле- дельческие орудия изготовлялись из железа. В Кармир-Блуре найдены серпы и вилы, наконечники мотыги или сошники и дру- гие предметы из железа. В конце VII в. мидяне (609 г. до н. э.) положили конец Урарту. В ослаблении Урарту играли роль и скифы (И. М. Дья- конов, 1951). С нашей точки зрения, здесь важно указать на соприкосновение скифской и закавказской культур, что еще раз подтверждает возможность проникновения культурных растений из Закавказья в более северные страны. Господство мидян сменилось господством персов. Персидский царь Кир в 580 г. до н. э. покорил Мидийское царство, затем завоевал Вавилон. Воз- никло обширное персидское государство; культуры переднеази- атских народов при этом скрещивались и взаимно ассимилирова- лись, что способствовало обмену опытом и в области земледе- 64
РАЗВИТИЕ ЗЕМЛЕДЕЛИЯ В БЛИЖАЙШИХ ОБЛАСТЯХ ВЛИЯНИЯП ЕРЕДНЕАЗИАТСКОЙ КУЛЬТУРНОЙ ФЛОРЫ В СССР Развитие земледелия в Дагестане и на Северном 1(авказе. История сельского хозяйства и вообще история культуры Да- гестана изучена еще довольно фрагментарно. Установлено, од- нако, что земледелие и здесь развивалось в глубокой древности. Уже в первой половине II тысячелетия до н. э. основой хозяй- ства современных племен Дагестана (аварцев, даргинцев и др.) являлось земледелие. Палеолит Дагестана изучен слабо. Известны орудия мустьер- ского типа и неолитические памятники, но немало древних па- мятников северо-восточного кавказа в основном относится к бронзовому веку. В раскопках поселения, расположенного около современного селения Каякент, были найдены каменные зернотерки, песты, кремневые вкладыши серпов и остатки зерен пшеницы, которая была определена В. А. Петровым как твердая пшеница. культурные слои Каякента отнесены к первой половине II тысячелетия до н. э. (А. П. круглов, 1946). Во второй поло- вине II тысячелетия до н. э. земледелие продолжает иметь круп- Синская Е. Н ° б5 лия, а также распространению культурных растений. Таким об- разом, растения, имевшие первоначальный очаг происхождения в Грузии, Армении и непосредственно прилегающих территориях, могли широко распространяться на восток и на запад. В частности, люцерна даже получила свое название по имени Мидии (Medicaoo, Herba medica), но неизвестно, где она раньше появилась в культуре: в Урарту (как думаем мы) или в Ин- дии. В Грецию и далее в Европу люцерна проникла, по-види- мому, из Индии. С древнейших времен народы Закавказья имели торговые отношения с народами, населявшими берега Эгейского моря. В VII в. до и. э. эта торговля приняла такие значительные раз- меры, что греки основали здесь колонии: Фазис (на месте Поти), Диоскурию (где теперь Сухуми) и Питиунт (около устья реки Бзыбь). В греческих источниках VI — IV вв. до н. э. имеются указа- ния на возделывание в Грузии пшеницы, полбы, ячменя, ореш- ника, каштанов, груши, яблони, винограда, инжира, граната, вишни, овощных культур и др. Греки вывозили из Грузии лен. Рионский (колхидский) лен считался лучшим материалом для изготовления сетей. Государство колхов существовало раньше VI в. до н. э., но культура льна на Черноморском побережье возникла, несом- ненно, еще с неолита. Через греческие колонии также могло идти распространение местных сортов культурных растений на запад и на север.
ное значение. Согласно данным А. П. Круглова, в раскопках не- далеко от современного селения Джемикент открыты не только зернотерки и вкладыши для серпов, но и хранилища зерна— большие куполообразные ямы. Земледелие характерно и для более поздних периодов истории дагестанской культуры. Это подтверждают многие предметы каякентско-хорочоевской куль- туры, найденные во многих районах Дагестана (горные селения Урма, Хорочой, Маджалис, Дженгутай и др.). Согласно исследованиям 3. А. Никольской и Е. М. Шиллинг (1952), А. Н. Генко (1930) и других авторов, земледельческие термины и обряды в Дагестане носят следы глубокой древности. До самого последнего времени кое-где в Дагестане сохранились аграрные культы. Для племен Дагестана характерна общность названий па- хотных орудий (аварцы — пуруц, табасаранцы — уруц, лезги- ны — цюрец, багулалы — перец и т. д.). Интересно разнообразие типов пахотных орудий в Дагестане (3. А. Никольская и Р. М. Шиллинг, 1952). Орудие состоит из пяти основных частей: дышла, ручки-стойки с особым сучком-ру- кояткой, служащим пахарю для опоры; пяты, или полоза; желез- ного лемеха, надетого на переднюю часть пяты; особых дере- вянных ушек для расширения борозды, рыхления комьев и дру- гих целей. В Дагестане с глубокой древности существует особая само- бытная система земледелия — террасная. У аварцев было три вида террас: 1) террасы, на которые дополнительно нанесена почва, удобренная навозом и золой; 2) террасы, на которые поч- ву не наносят, но»х удобряют золой и навозом; 3) естествен- ные речные террасы, также в той или иной степени удобренные. Орошение производится посредством мелких временных ка- налов, подводящих воду непосредственно к орошаемому уча- стку. При орошении с помощью временных мелких каналов соз- дается рыхлая почва. Этот народный старинный способ ороше- ния входит теперь в практику колхозов. Для Дагестана издревле было характерным трехъярусное ис- пользование каждой террасы: первый верхний ярус составляют плодовые деревья; в их междурядьях выращивают кукурузу (второй ярус); в третьем — овощные культуры, картофель. Дагестан не отличается таким богатством эндемичных куль- турных растений, как Закавказье. Однако здесь довольно велико разнообразие эндемичных сор1ов (например, своеобразные яч- мени, сходные с абиссинскими формами). Культурная флора Дагестана изучалась Н. И. Вавиловым, Т К. Ениным, М. М. Якубцинером и некоторыми другими исследователями. На Северном Кавказе развитие культуры шло, по-видимому, более замедленными темпами по сравнению с Дагестаном и осо- бенно с Закавказьем. Археологически Северный Кавказ изучен недостаточно. Известны находки, относящиеся еще к эпохе ниж- бб
него палеолита (около Майкопа на реке Фортепьянке, у селе- ния Ходжах), и несколько более поздние находки у станиц Иль- ской, Смоленской, в окрестностях Горячего Ключа и др. Пещер- ные стоянки обнаружены близ Сочи. На территории Кабарды, по данным Е. И. Крупнова (1946), найдены следы неолита. Еще больше обнаружено остатков, от- носящихся к последующим эпохам (материалы стоянок и посе- лений на реке Кенже, близ Затишья, в окрестностях Нальчика, по оврагам рек Шамушки, Малки и Баксана). При раскопках Нальчикского кургана и в Агубековском поселении найдены следы примитивного земледелия, относящегося ко второй по- ловине 1 1 тысячелетия до н. э. В Доменcком поселении бронзового века близ Нальчика най- дены каменные мотыги, зернотерки, вкладыши для серпов. В ма- териалах этого поселения много сходства с курганами Кубани (ст. Новосвободная близ Майкопа). Пышный расцвет бронзовой культуры приходится здесь на начало 1 тысячелетия до н. э. Предметы, найденные при раскопках кобанской культуры (Заю- ково, Кызбурун и др.), напоминают предметы скифской куль- туры. Население здесь сначала располагалось в предгорьях. За- тем пришло время скотоводческого освоения высокогорий, а земледелие стало р аспростр аняться в соседние степные р айоны. Примитивное мотыжное земледелие в центральных районах Северного Кавказа изучалось по материалам раскопок энео- литического Лугового поселения, Долинского поселения в Ка- барде (начало II тысячелетия до н. э.) и по другим памятникам. Относящиеся к нижнему слою Алхастинского поселения камен- ные мотыги показали, что первобытный характер земледелия сохранялся здесь до эпохи поздней бронзы. В первой половине 1 тысячелетия до н. э. развитие производительных сил дало толчок дальнейшему развитию земледелия. Вместо древнего деревянного серпа с кремневыми вкладышами (Луговое посе- ление) появился более совершенный бронзовый серп (Галиа~- ский могильник Фаскау в Дигории, Эмейское поселение в Север- ной Осетии, могильник у горы Бык и др.). Зернотерки и первые жернова небольших размеров, вероятно, свидетельствуют о плужном земледелии в поселениях предскифского и скиф- ского времени. В раскопках скифского времени обнару>кены крупные гру- шевидные корчаги, которые, вероятно, слу>кили для хранения муки и зерна; горшки свидетельствуют о приготовлении каш. На поселениях, расположенных почти на границах предго- рий и равнин, откопаны местами крупные хранилища — подзем- ные закрома. Вероятно, главными культурами на Северном Кавказе в эпоху раннего железа были ячмень, пшеница и просо. Ячмень можно считать древнейшим хлебным злаком в Кабарде. Кос- венным доказательством древности возделывания здесь ячменя б?
служит односложное название его «хьэ», это слово сохранилось в кабардинском языке с того древнего периода, когда предшест- венники кабардинцев говорили еще на языке, состоящем из од- носло>кных слов — корней (Н. Ф. Яковлев, 1949). Интересен факт сохранения в черкесском и кабардинском языках таких односложных корней слов, как «дзе» — зерна, злак, «изье» — земля, почва, «мье» — дикое яблоко, кислица (плод), «зе» — кизил, «де» — орех и др. Находки ячменя, мягкой и твердой пшеницы неоднократно отмечались в раскопках на северо-западном Кавказе. Помимо хлебных злаков, в хозяйстве аборигенных кавказ- ских горцев имели большое значение сначала дикорастущие, а позднее и культурные плодовые и ягодные растения: груши, яблони, орех — лещина, терн, алыча, кизил и др. У чеченцев и ингушей дикая груша считалась священным де- ревом и грушевые деревья щадились при сплошной вырубке леса, что способствовало образованию довольно больших чистых грушевых насаждений, из которых некоторые сохранились и до нашего времени. Таким образом, основной ведущей формой хозяйственной деятельности уже безусловно оседлых >кителей горной, предгор- НоН полос и прилегающих степей Северного Кавказа в первой половине 1 тысячелетия до н. э. было пашенное земледелие вме- сте с кочевым (яйлачным) животноводством. Имеются достоверные исторические данные, что в V в. до н. э. на территории современного Краснодарского края, Ростов- ской области и Крыма возделывались мягкая пшеница, рожь, ячмень, просо, бобовые и другие полевые культуры. Рожь была в раскопках в смеси с пшеницей. К. А. Фляксбергер (1934, 1940) указывает, что на террито- рии Краснодарского края, в Ростовской области и в Крыму в V в. до н. э. сеяли мягкую пшеницу, рожь, ячмень, просо, кон- ские бобы. Он полагает, что наши современные местные, гак на- зываемые стародавние, мягкие пшеницы Краснодарского края, Ростовской области и Крыма и вообще районов по побережью Черного моря, по-видимому, ведут свое происхождение от тех мягких пшениц, которые здесь возделывались уже полторы- две тысячи лет назад. То же самое можно сказать относительно ячменя и проса. По Л. И. Лаврову (1952), в период неолита на северо-запад- ном Кавказе земледелие (мотыжное) существовало на побере- жье Черного моря. Здесь мотыжное земледелие (подсечного типа) продолжало иметь значение с конца II до начала 1 тыся- челетия до н. э. В предгорьях в этот период постепенно разви- вается отгонное скотоводство. В период с VII в. до н. э. по III в. н. э. значение охоты и со- бирательства уменьшается, а роль земледелия (одновременно с кочевым скотоводством и рыболовством) возрастает. 68
В степных районах Северного Кавказа, в местностях, наибо- лее удобных для земледелия, по берегам рек, в это время разви- вается пашенное земледелие с зерновым ассортиментом культур: мягкая пшеница, просо, ячмень, чина. В раннем средневековье в степях и предгорьях сеяли просо, а также, по-видимому, овес, пшеницу, ячмень, бобы и горох. В Х в. черкесы выращивали так- же и лен. «После татарского нашествия,— пишет Л. И. Лавров,— па- хотное земледелие (вслед за периодом временного сокращения) существовало в степях и в предгорьях. В горах во многих местами сохранился еще мотыжный способ обработки земли. В степях и предгорьях господствовала переложная, а в горах — подсечная система земледелия. Сеяли главным образом просо. Кроме того, в этот период выращивали ячмень и незадолго до того появив- шийся на Северном 1(авказе овес. В горах посевы проса отсутствовали. В Абхазии в это время был известен лен...». По заключению Л. И. Лаврова, «население северо-западного 1(авказа являлось носителем древнейшей земледельческой куль- туры». Новейшие исследования Н. Анфимова (1953) освещают многие детали в истории культуры и земледелия Кубани. Со- гласно этому автору использование меди населением северо-за- падного Кавказа датируется концом III тысячелетия до н. э. Во второй половине II тысячелетия до н. э. на Кубани возникает мощный очаг металлургии и металлообработки. Население за- нималось в основном скотоводством, мотыжным земледелием и металлургией. В первой половине 1 тысячелетия до н. э. племена, заселяв- шие северо-западный Кавказ, перешли на более высокую сту- пень культуры. ~Келезо здесь начинает распространяться в VIII — VII в. до н. э. и в последующее время полностью выте- сняет бронзу. У части племен развитие производительных сил обусловило переход к кочевому скотоводству. Другие пастушеские племена переходят к оседлому образу жизни, и их культура прогрессирует на основе земледелия— совершается переход от мотыжного к плужному земле- делшо. Скифы не входили в состав постоянного населения степей Прикубанья, но время от времени проникали сюда (через Дер- бентский проход и, возможно,— через Белореченский перевал). Основным населением северо-западного Кавказа были меоты и горские племена Закубанья и Черноморского побережья. Меоты жили не только по восточному побережью Азовского моря, в дельте реки 1(убани, но и по среднему течению 1(убани, лево- бережью, правобережью и в предгорьях Закубанья — прибли- зительно до впадения в 1(убань Лабы.
О сарматских элементах на Кубани во II — 1 вв. до н. э. из- вестно, что сарматы были кочевниками и довольно скоро сме- шались с меотами. Исследования Н. Анфимова (1953) в бассейне реки Кубани показали, что во второй половине 1 тысячелетия до н. э. земле- делие стояло здесь на высоком уровне, не ниже, чем в антич- НЫХ КОЛОНИЯХ. Аборигенное население северного Причерноморья вошло в со- прикосновение с греческой античной культурой при появлении сначала отдельных греческих мореплавателей, а затем коло- нистов. В V в. до н. э. греческие города-колонии объединились в Боспорское государство, расцвет которого падает на IV в. до н. э. Здесь варварская культура племен Северного Кавказа и Восточного KpbIMB вступила в тесное соприкосновение с антич- ной культурой, поэтому историк С. А. ЯКебелев называет Бос- порское государство «греко-скифским». Греки Боспорского государства были главным образом торговцами, а не земледель- цами. Земледелием же в Боспоре и на прилегающей к нему тер- ритории продолжали заниматься аборигенные племена, и зем- ледельческие продукты становятся важным предметом экспорта. По заключению Д. П. Каллистова (1949), появление греков не повлекло за собой существенных перемен в местном сельском хозяйстве и в составе культур. Сорта мягкой пшеницы, испокон веков культивировавшиеся в Северном Причерноморье, продол- жали здесь возделываться на всем протяжении античного пе- риода. Торговые связи боспорских греков были очень широки, и в этих связях виден новый путь распространения сортов куль- турных растений Северного Кавказа в различные страны. При исследовании Кубанских городищ были найдены большие ка- менные жернова и слабо изогнутые серпы. Серпы встречаются при мужских скелетах. Вероятно, жатва была мужским тру- дом. Зерно хранилось в ямах и больших глиняных сосудах. В Боспоре применялся примитивный греческий плуг — -изо- бражение его имеется на моыете II в. до н. э. Основной частью этого плуга была рассоха, на деревянный конец которой наде- вался (вероятно, железный) сошник, от верхнего конца подни- малась стойка с деревянной рукояткой. При помощи рукоятки пахарь направлял плуг. Между сош- ником и рукояткой укреплялся деревянный изогнутый грядиль, соединявший рассоху с дышлом, в которое впрягались волы. Возможно, что такого типа плуг бытовал и у меотско-сармат- ских племен. Состав зерновых культур был довольно разнообразный. Осо- бенно часто встречались пшеница, ячмень и просо — они были найдены в Заурупском, Усть-Лабинском, Краснодарском и дру- гих городищах. Из пшениц представлена только мягкая, твердой пока нигде не найдено. 70
Интересно, что в раскопках встречалиеь и зерна ржи, но всегда в смеси с пшеницей. Очевидно, это была сорная рожь, ко- торая тогда еще не вышла в самостоятельную культуру. Зерна ячменя из Боспора бь|ли рассмотрены Ф. Х. Бахтее- вым. У пленчатых длина была от 4,0 до 5,0 мм, ширина 3,5 мм, толщина 2,5 л~м; у голозерных — длина 5,5 мл~, ширина 3,5 мм, толщина 2,5 мл~. Имелись зерна и более удлиненной формы. Встречались отдельные бутылковидные зерна. Последние отли- чались от зерновок современных культурных форм ячменя тем, что имели своеобразную плодоножку. Позднее Ф. Х. Бахтеев обнаружил подобные бутылковидные формы ископаемого яч- меня и в раскопках Урартского периода (Кармир-Блур) в Ар- мении (Ф. Х. Бахтеев, 1956). Примесь подобных бутылковидных форм была отмечена Ф. Х. Бахтеевым и в раскопках в Азербайджане в Узерлик-Тепе (близ Агдама), и в Кюль-Тапа (близ Нахичевани) — в слоях, относящихся к III — II тысячелетиям до н. э. Ячмень с бутылко- видными зерновками, вероятно, представляет собой древнюю форму, теперь исчезнувшую; может быть, она была некогда эндемична для Передней Азии. Среди ископаемого ячменя свай- ных построек Европы таких форм не обнаружено. Просо составляло один из важнейших пищевых продуктов тогдашнего населения. В раскопках были открыты крупорушки, на Семибратном городище найдена миска с обуглившейся про- сяной кашей. Близ Майкопа в раскопках найдены бобы и горох. Из техни- чееких культур в Боспорскую эпоху, вероятно, возделывались конопля и рами. Было развито и садоводство. В раскопках боспорских поселений находили косточки сливы ц алычи. По данным античных авторов, здесь росли груши, яб- локи, гранаты. В раскопках встречалось много костей домашних животных: коров, свиней, овец, лошадей. Развитие земледелия в Крыму. Многими исследователями предполагается, что на месте впадины, занятой современным Черным морем, во второй половине третичного периода, в нео- гене, существовала суша, получившая название Понтиды. Гор- ный Крым в ту эпоху был северным краем Понтиды, а это зна- чит, что существовала прямая материковая связь Крыма - Ма- лой Азией, Балканами и Кавказом. Вопрос об эндемизме флоры Крыма и значении Понтиды разработан в трудах Н. И. Рубцова (1959, 1960, 1961). Н. И. Рубцов насчитывает около 50 видов, относящихся к крымско-западнокавказскому географическому элементу крым- ской флоры. Особенно большое родство флор установлено для горной ча- сти Крыма и Новороссийского ботаника-географического округа (эспарцет, житняк, люцерна, пажитник и др.).
Крымский полуостров является древней ареной человеческой культуры, и на его территории открыты палеолитические стоянки. По сведениям, приводимым С. Н. Бибиковым (1946), поздне- палеолитические поселения в Крыму открыты в Шанкобе и в гроте Мурзак-Коба. В раскопках раннетаврского поселения Уч-Баш (Х — VIII вв. до н. э.) найдены зернотерки, оправы для серпов, много пше- ницы и ячменя, косточки кизила, терна, вишни, желуди (С. Ф. Стржелецкий, 1952). А. С. Коварский (1929) отмечает, что первобытные черты в крымском земледелии сохранились вплоть до современной нам эпохи. Еще совсем недавно там бытовали самые прими- тивные земледельческие орудия: мотыга и самодельный дере- вянный плуг сабан. Уборку проводили серпом, косой или рас- тения выдергивали руками. Хлеба при уборке не связывали в снопы, а укладывали кругами, которые назывались басиле- ками. Сверху на басилеки накладывали камни, чтобы уберечь хлеб от разбрасывания ветром. Посевы производили большей частью в «чаирах» — участках, окруженных изгородями из кам- ней или хвороста. Состав зерновых культур также указывает на древний ха- рактер крымского земледелия. Кое-где в культуре еще недавно имелась однозернянка, а как сорняк она поныне встречается в посевах мягкой пшеницы, иногда вытесняя последнюю. Встре- чается и дикорастущая однозернянка. На сортовом составе крым- ских пшениц сказывается влияние Малой Азии. В Крыму имеется особая крупноплодная сорная форма озимого рыжика — Сате- lina silvestris ssp. pilosa var. intermedia Sinsk. По Ф. gI,. Лихо- носу (1954), крымские сорта яблони имеют общие черты строе- ния цветков с крымской дикорастущей яблоней. Недавно описан новый вид (возможно, подвид) крымского эндемичного эспарцета Onobrychis jailae N. Tschern., который еще не испытывался в культуре. В KpbIMcKO-Новороссийской 60- танической провинции встречаются некоторые сухолюбы (жит- няк — Agropyron pinifolium Nevski, железистая люцерна— Mediloago glandulosa David., пажитник меловой — Trigonella cretaeea Grossh. и др.), интересные для использования в каче- стве кормовых растений. С этой точки зрения, интересен так- же мало изученный вид эспарцета, распространенный в север- ной части кавказского Причерноморья,— ОпоЬгусЫз miniata Меч. Крым почти не дал эндемичных видов культурных растений, за исключением крым-сагыза. Однако в Крыму имеются свои аборигенные сорта многих растений: пшеницы, винограда, льна масличного и др. Северный Кавказ и Крым представляют собой переходные области. В дикорастущей флоре на Северном Кавказе преобла- дают европейские элементы, а в Крыму (вместе с побережьем 72
Черного моря в окрестностях Новороссийска) к ним примешаны и средиземноморские. В отношении культурной флоры — это в основном область влияния Передней Азии. КУЛЬТУРНАЯ ФЛОРА НЕКОТОРЫХ ЗАРУБЕЖНЫХ СТРАН,ОТНОСЯЩИХСЯ К ПЕРЕДНЕЙ АЗИИ Сирия и Палестина отличаются процессами интенсивного формообразования ксероморфных видов и экотипов. Эти страны всеми исследователями включаются в Переднеазиатскую под- область,1~ревнесредиземноморской области. Ниже дается крат- кий обзор сирийско-палестинской культурной флоры, неболь- шое место отведено также и Йемену. Йемен представляет собой область влияния переднеазиатской и восточноафриканской куль- турных флор и очень интересен как очаг развития предельно скороспелых форм многих культурных растений. Основное ядро переднеазиатской культурной флоры приуро- чено к Закавказью, сопредельным частям Ирана и Малой Азии. Об аборигенных растениях последней можно составить пред- ставление по обзору эндемичных форм всей Передней Азии, по- мещенному ниже после обзоров отдельных стран. Сирия и Палестина. 1~лимат Сирии и Палестины частично пустынный с очень малым количеством осадков и частично средиземноморский с дождливой зимой и сухим летом. Эйг показал, что дикорастущая флора Палестины отно- сится к Переднеазиатской провинции Средиземноморской onëàñòè (А. Eig, 1933). Археологическими изысканиями установлено, что человек жил в этих местах еще в эпоху палеолита. Судя по найденным предметам, в IV тысячелетии до н. э. к северо-востоку от Мерт- вого моря обитал оседлый земледельческий народ. В III и II тысячелетиях до н. э. здесь сложилась под влия- нием Шумера, Вавилона и Египта культура ханаанеев. Ханаан был густо заселен, и основным занятием жителей было земле- делие, а в отсталых горных и степных районах — земледелие и скотоводство. Сеяли ячмень, просо, пшеницу. Уже в III тысяче- летии до н. э. здесь разводились финиковая пальма, виноград, смоковница, гранат, маслина, лен. Маслина разводилась в Па- лестине до прихода туда евреев. ~ое-где находят остатки древних маслобоек, иногда в таких сухих местах, где теперь почти ничего не растет. Это говорит о более влажном климате и о развитии орошения в,1~ревней Палестине, о более благо- приятном комплексе условий земледелия, чем в настоящее время. Земледельческая техника была примитивной. Основным зем- ледельческим орудием служила мотыга, затем уступившая ме- сто примитивному плугу. Уборка зерновых производилась арха- ичными серпами с кремневыми зубьями.
Древнееврейские племена, завоевавшие Палестину в сере- дине II тысячелетия до н. э., использовали культурное наследие Ханаана. В первые века заселения Палестины евреями, кроме перечисленных выше культурных растений, имели значение пол- ба-эммер, чечевица, бобы, фиги, некоторые овощи и даже, по свидетельству историков (В. И. Авдиев, 1948), какие-то «кор- мовые травы». Из бобовых растений в Палестине была популярна чече- вица, о ней имеются многочисленные упоминания в библии, но, поскольку в библии отражен и более древний быт, начало воз- делывания чечевицы, вероятно, относится здесь к более глубокой древности. Яблоня упоминается в библии редко. g. Г Редер (1950) упоминает об известных географических названиях: Бет- Таппуая («Дом яблони»), Таппуая (яблоня) и Ен-Таппуая. По Д. Г. Редеру, «местоположение первого из этих городов может быть в точности установлено. Он находился около Хеброна, что в высшей степени показательно. Хеброн до завоевания евреями Палестины был хеттским городом. А мы знаем, что в Малой Азии, откуда хетты проникли в Палестину, яблоня была самым обычным деревом». И этими данными подтверждается связь Сирии и Палестины с другими странами Передней Азии, где на- ходились очаги древней земледельческой культуры. О земледельческом хозяйстве Сирии имеются интересные данные, относящиеся к римскому периоду (Tchalenko, 1953, 1958). Этот период в горных долинах Сирии и на наго- рьях продолжался с 1 — II вв. н. э. до IV — начала VII в. н. э. Затем жители ушли из прежде населенных мест. Некоторые авторы объясняют уход населения эрозией почвы как след- ствием вырубки лесов и временного потепления климата. Другие исследователи утверждают, что больших лесов здесь и не было, а климат почти не изменялся. Более существенно указание на то, что своей необычайной плодородностью почва обязана почвообразовательным процес- сам, связанным с распадом известковых пород. Теперь такая хо- рошая почва сохранилась только на равнинах, где сеется хло- пок. Раньше все горы были покрыты подобной почвой, что и обусловило некогда распространение селений по всему горному массиву. Судя по распределению селений, наибольшая интен- сивность освоения известкового массива приходится на V— VI вв. н. э. Древнее сельское хозяйство Сирии было основано на планта- циях оливкового дерева (иногда с добавлением виноградной лозы) и производстве оливкового масла. Оливковые плантации занимали самые высокие нагорья, а также южные и западные склоны. Ассортимент культурных растений Сирии и Палестины не отличается особенной самобытностью и обнаруживает родст- венные связи, с одной стороны, с Передней Азией, с другой— 74
с собственно Средиземноморьем. В основном для них характер- ны раннеспелые и ксероморфные формы тех видов культурных растений, которые распространены и в соседних странах Вос- стока и Запада. Из них отметим следующие. Люцерна сирийско-палестинская — Medicago syriaco-palestinica Sinsk.— вид, имеющий родственные черты, с одной стороны, с йеменской люцерной — М. jemenensis Sinsk., с другой стороны — с южными итальянскими формами европей- ской люцерны — М. eusativa Sinsk. П а ж и т н и к а р а 6 с к и й — Trigonella arabica РеШе — кое- где возделывается в Палестине. Сирия и Палестина считаются многими ботаниками родиной александрийского к л е в е р а (берсима) — Trifolium ale- xandrinum L., так как там распространены дико Т. berytheum Boiss. и Т. vavilovii Eig, которые рассматривались как его бли- жайшие родичи. Открытие Е. Г. Бобровым на Кавказе еще более близкого к берсиму вида — Т. apertum Bobr. — заставляет пересмотреть этот вопрос. Нут — Cicer arietinum L. — в Палестине представлен особой палестинской группой среднеспелых сортов — proles ~palestini- cum G. Рор. Кроме того, в Палестине распространен эндемич- ный вид дикорастущего нута — С. judai~cum Boiss.— и пред- ставлен особыми разновидностями другой дикий вид нута— С. ~pinnatifidum Jaub. ct Sp. Конские ба бы — Faba vulgaris Moench — в Сирии пред- ставлены эндемичной формой — f. syriaca Murat. Г о р о х С и р и и образует сирийскую эколого-географическую группу (Л. И. Говоров, 1937), отличающуюся низкорослостью, скороспелостью и ветвистостью Эти формы близки к азиатским пелюшкам и классифицируются Л. И. Говоровым как Pisum sativum ssp. asiaticum proles syriacum Gov. Имеющий более широкий средиземноморский ареал вид л ю- п и н а — Lupinus termis Forsk. произрастает в диком состоянии и в Сирии. Ч и н а н у т о в а я — Lathyrus ciicera L., вероятно, вошла в культуру в Сирии, ибо только там она используется как пи- щевой продукт и только там известна ее культура в сколько- нибудь значительных размерах. В Сирии возделывается подвид зэр. syriacus Zalk. кроме того, здесь возделывается на корм еще особый вид чины — L. gorgoni Раг1. В диком состоянии в Сирии распространен ближневосточ- ный вид д в у 3 е р н я н к и — Triticum йсоосЫез Коегп. Ди- корастущая двузернянка в Палестине была открыта Ааронсоном в 1906 г. Н. И. Вавилов считал (1931), что сирийско-палестин- ская двузернянка имеет общего предка с культурной двузерняп- кой, но не является непосредственным родоначальником послед- ней. Еще позднее М. М. Якубцинер (1932), Е. Н. Макушина- Горощенко (1948) и другие установили, что предком закавказской 75
двузернянки Т. timopheevi является не сирийско-палестинская дикая двузернянка, а закавказская двузернянка — Т. araraticum, очаги которой обнаружены М. М. Якубцинером в Закавказье в нескольких местах. В Сирии распространена также дикора- стущая однозернянка, представленная здесь ssp. thaoudar Flaksb. Т в е р д ы е п ш е н и ц ы представлены здесь особыми фор- мами с коротким густым колосом и округлыми зернами (по- видимому, древнего типа) — proles horanicum Vav. По кругло- зерности они напоминают шарозерную пшеницу, а по плотному колосу — наиболее примитивные формы Эфиопии. В Израиле распространен дикорастущий вид о в с а — Ачепа sterilis L. Возможно, что нменно здесь находится и первичный очаг А. byzantina С. I<och, что еще не доказано. Поэтому пока А. byzantina условно причислена к средиземноморским, а не и переднеазиатским видам. О в е с в и з а н т и й с к и й — А. byzantina — в Палестине обра- зовал особый палестинский очень раннеспелый тип с крупным зерном (А. И. Мордвинкина, 1960). Я ч м е н и представлены в Сирии и Палестине особой группой низкорослых, раннеспелых, плотнокустовых и засухоустойчивых сортов — рго1ез syria~co-palestinicum (Orl.) Vav. et Bacht. А. И. Купцов установил существование следующих экотипов с а ф л о р а: 1) экотип окрестностей Наблуса; 2) экотип районов gaMac~' и Тель-Керама; 3) экотип окрестностей Назарета. По А. И. Купцову, коллекционные образцы сафлора из Сирии слабее других поражались сафлорной мухой. Л е н в Сирии и Палестине представляет собой особую группу довольно низкорослых раннеспелых и средних по вели- чине семян форм, которые являются переходными к настоящим средиземноморским льнам Кипра. Г о р ч и ц а ч е р н а я — Brassica nigra Косй — в Сирии и Палестине образует особую ксероморфную, довольно позднюю, с ясно выраженной розеткой форму — var. rigida Sinsk., не- сколько приближающуюся к малоазиатской черной горчице (В. nigra var. orientalis Sinsk.). Гор ч и ца бел а я — Sinapis alba L. — распространена в Па- лестине среди посевов пшеницы, образуя здесь эндемичную форму S. alba var. orientalis Sinsk. с очень ранним ветвлением и бледно-желтыми лепестками. Горчица на масло в Сирии не использовалась, это было лекарственное растение. Из расти- тельных масел издревле здесь имело значение оливковое масло. Культура оливкового дерева, как уже было сообщено выше, является здесь древней и, возможно, аборигенной. К р е с с - с а л а т — Lepidium sativum L. — представлен в Си- рии и Палестине особой позднеспелой формой с ясно выражен- ной розеткой — var. tardif lorum М. Stichenkova (эта форма встречается изредка и в Иране). 76
К у н ж у т — Sesamum indicum L. — в Палестине, помимо формы, явно заимствованной пз Малой Азии, представлен еще эндемичной 4-плодолистиковой, более или менее фасциирован- ной формой. М о р к о в ь — Паисия sativus Zagor. — в Сирии и Палестине представлена особым подвидом ssp. зуг1асиз Zagor. (без пла- стидной окраски). Эта морковь встречается также в Закавказье. С в е к л а — Beta vulgaris L. — представлена двумя формами. Первая, с красным полосатым черешком и рсзовой мякотью, эндемична для Сирии, Палестины и о. Кипра. Вторая, листвен- ная форма, с зеленой окраской черешка и белым или розовым корнеплодом встречается также и на о. Крите. По мнению В. Т. Красочкина (1960), листовая свекла вошла в культуру на островах Кипр и Крит, оттуда попала в Сирию, а из Сирпш- в Двуречье. Во всяком случае, свекла — древняя сирийскач культура. Древние евреи знали ко р и андр. Древнее название этого растения — gali — упоминается в библии. Ботаник Швейнфурт считает, что кориандр был введен в культуру в Сирии или в Египте. Р е в е н ь с м о р о д и н н ы й — ЯЬеит ribes L. — очень рас- пространен в Сирии и Иране. Вероятно, пз этих стран он был занесен в Европу. Б а кл а ж а н — Solanum melongena L. — представлен в Си- рии особой агроэкологической группой — proles palestinicum Fil. Растения низкорослые, густо опушенные, но позднеспелые и не- засухоустойчивые, что не типично для сирийских форм. Формы этого типа распространены также в Южном Китае. В Сирии распространена особая группа сортов культурной ф и с т а ш к и — Pistacia vera L. — фисташка алеппская, или сп- рийская. При кратком просмотре ассортимента культур Сирии видно, что на него преобладающее влияние оказала Передняя Азия (люцерна, берсим, однозернянка и двузернянка, морковь, кресс- салат, нут, горчица черная, кунжут и др.). Ясны также связи с собственно Средиземноморьем и глав- ным образом с восточной его частью: Кипром и Критом (свекла), Египтом (Luipinus termis) и также Средиземноморьем, взятым более широко, включая и Северную Африку (маслина). Большинство растений представлено в Сирии низкорослыми, более или менее ксероморфными и раннеспелыми формами, но имеются и исключения (среднеспелые: нут, черная горчица). Йемен. Климат знойный, в горной зоне — менее жаркий. Зем- леделие большей частью орошаемое. На культурную флору Йемена оказала большое влияние близость Африки, которая раньше была соединена с Аравией. Многие растения, характерные для земледелия Эфиопии и Эри- треи, возделываются в Йемене. K таким культурам относится 77
кофе, который издревле культивируется в Иемене (но страной его первичной культуры считается Эфиопия), абиссинский овес и др. Иеменский оазис имеет характерный ассортимент тех куль- турных растений, для которых он большей частью отнюдь не яв- ляется первичной родиной, но которые на его полях представ- лены особыми, встречающимися только здесь, крайне раннеспе- лыми tj низкорослыми формами. К их числу относятся сле- дующие. Л юц ер н а Й ем ен с к а я (Medicago jemenensis Sinsk.)— карликовая малолетняя люцерна (1 — 2-летняя), имеющая род- ственные связи с армянской многолетней люцерной (М. praesa- tiva Sinsl<.), самая короткостадийная из всех известных люцерн этой группы. В Иемене имеется 2 подвида — зэр. eujemenensis Sinsk. и ssp. subarabica Sinsk. Последний подвид имеет некото- рые переходные черты к средиземноморскому виду М. polia Vass. П а жити и к й е м е н с к и й [Trigonella jemenensis (Serp.) Sinsk.] — эндемичный карликовый вид, ближе, чем к другим, стоящий к абиссинской культурной тригонелле, но отличаю- щийся от нее по форме бобов и карликовостью. Очень скороспелый а р а в и й с к и й г о р о х, который некото- рые ботаники выделяют в особый вид — Pisum abyssiniicum Bra UrI. Из культурных растений сем. крестоцветных в Иемене от- крыты: особая эндемичная форма очень раннеспелой и карли- ковой с у р е п и ц ы — Hrassica campestris var. jemenensis Sinsk. Эта форма детально не исследована, возможно, представляет собой особый вид. Г о р ч и ц а (Brass',са juncea Czern.) представлена в Иемене особой очень раннеспелой и низкорослой формой. И е м е н с к а я п о л б а (Triticum dic(>coon Schrank), относя- щаяся к абиссинскому подвиду, выделена К. А. Фляксбергером как особая форма f. jemeni Flaksb., отличающаяся большей скороспелостью, чем обычные растения той же разновидности var. arras Idochst. Т в е р д ы е п ш е н и ц ы Иемена представляют собой особую экологическую разность низкорослых эфемеров. По скороспе- лости занимают первое место в культуре на юге. Ботанически йеменские твердые пшеницы родственны пшеницам Эфиопии. Я ч м е н и представлены в Иемене скороспелыми крупнозер- ными эфемерами, составляющими proles jemeniicum (Orl.) Vav. et Baicht. Б а к л а ж а н ы (Solanum melongena L.) представлены в йемене и в соседних районах Аравии особой агроэкологической группой форм — iproles arabicum Fil. Растения низкорослые, сильно опушенные, засухоустойчивые, но среднеспелые и имею- щие черты дикорастущих предков. Иран. Хотя дикорастущая флора Ирана чрезвычайно разно- образна и богата, но среди культурных растений здесь не вы- 78
явлено большого количества ярких эндемиков. Северо-западный Иран примыкает к Передней Азии, а восточный — к средней и Юго-Западной Азии. В Иране однако создались если не виды, то своеобразные формы некоторых культурных растений: карли- ковые формы кукурузы, некоторые эндемичные формы дыни и других растений. В Иране очень разнообразны мягкие пшеницы. Большой инте- рес с точки зрения исторической географии культурной флоры представляет открытие Куккуком (Н. Kuickuicli, Е. Shiemann, 1957, см. также М. М. Якубцинер, 1959) в западном Иране района распространения спельты (Triti~cum spelta L.). В восточ- ном направлении от основного района распространения спельты наиболее удаленное ее местонахождение указано в Исфахане Спельта растет здесь в горной местности с длинной, холодной и снежной зимой и холодными ветрами весной. В области рас- пространения спельты в Иране до сих пор живут древние кочев- ники — бахтиары. С этим племенем связана реликтовая куль- тура спельты. Несмотря на то, что туземцы ее ценят за непри- хотливость к условиям произрастания, площади под спельтой сокращаются. Спельта в основном представлена в Иране озимой формой, но встречаются и яровые сорта. В Иране имеется непрерывная серия промежуточных форм- от голозерной пшеницы до типичной спельты — с очень разно- образными формами чешуй. Куккук и Шиманн не считают, что именно бахтиарское на- горье является родиной спельты. Если принимать новейшую тео- рию происхождения спельты как амфидиплоида, возникшего путем спонтанной гибридизации видов Аер1орз с двузернянкой Triticum dicoccoides (что подтверждается экспериментальным получением спельты при гибридизации Т. diicoocoides и Ае. squar- rosa L.), то они правы, так как виды Aegilops на этом нагорье отсутствуют. В соседнем Луристане, где климат гораздо мягче, встре- чаются Aegilops triuncialis L. и Ае. cylindrica Host, но не был найден Ае. squarrosa. Последний распространен к северу от хребта Эльбурс и в других местах в Передней Азии. Таким образом, родиной спельты приходится признать не Иран, а Переднюю Азию, пока без уточнения основного очага в пределах Переднеазиатской подобласти. В Иране из древних реликтовых культур, по данным тех же авторов, которые писали о новых местонахождениях спельты, до сих пор на бахтиарском нагорье сеют эммер — Triti~cum diicoc- con, представленную здесь остистыми формами. Куккук приво- дит персидское название эммера «Paranj». Он пишет, что очажки культуры эммера в Передней Азии тесно связаны с армянскими поселениями. Армяне приготовляют из полбы суп, называемый ими «Ach». Автор встретил посевы эммера на севере Иранского Азербайджана близ Джульфы, где и теперь живет много армян. 79
Фенотипически эммер из Азербайджана и эммер из страны бахтиаров очень сходны. Внутривидовая изменчивость эммера здесь незначительна. Здешний эммер возделывается как яровая культура. В северо-западном Иране в высокогорной местности в окрест- ностях Исфахана недавно найден оригинальный новый 28-хро- мосомный вид пшеницы — Т. ispahanicum Heslot. Но описаншо М. М. Якубцинера (1959), оригинальные черты этого вида напоминают черты однозернянки (одноостистой), ко- лосковые чешуи прпближаются к таковым Т. ipoloni~cum L., ко- лосья ломкие, как у полбы. Таким образом, эта пшеница совме- щает признаки различных видов подобно Т. ma~cha. Ареал этого вида еще не вполне уточнен. go известной степени этот вид может быть назван интеграль- ным. Значение его в филогенетике пшениц станет яснее, когда удастся установить либо автополиплоидный, либо аллополи- плоидный его характер, а также найти исходные виды, если бу- дет доказана его гибридная природа. Во всяком случае находка этого вида повышает значение Передней Азии как центра разно- образия видов пшенццы. Интенсивные процессы формообразования дикорастущей и сорной ржн протекают и по настоящее время на территории Ирана. Куккук (1956) собрал в большом количестве примитив- ные формы ржи, затем изучавшиеся Кранцем (franz, 1957). Последний дает интересную картину по высотному распростра- нению сорной ржи в Иране: на высоте 1000 — 1320 м над ур. м.— пшеница с малой примесью ржи; 1520 — 1740 м — пшеница с большой примесью ржи; 2000 м — рожь с небольшим количе- ством пшеницы; 2360 м над ур. м. — рожь с единичными пше- ничными растениями. Среди сорной ржи оказались многолетние, озимые, двуручки и яровые формы. В иранских популяциях рожь варьировала по многим при- знакам. В Иране часто встречаются формы с окрашенными уш- ками, что характерно для армянской дикой Secale anatolicum Boiss. Огромные различия были обнаружены по ломкости и по самофертильности. Среди популяций ржи, которую Кранц опре- делял как Secale vavilovii Grossh., им были обнаружены клей- стогамные растения. Из Ирана (Н. А. Базилевская, 1960) были введены в куль- туру некоторые декоративные растения: сирень — Syringa vul- garis L., жасмин — Philadeliphus согопагшз L. и др. ЗЕМЛЕДЕЛЬЧЕСКИЕ КУЛЬТУРЫ МЕСОПОТАМИИ(ДРЕВНЕГО ДВУРЕЧЬЯ) древнее Двуречье не является первичным очагом вхождения в культуру основного ядра аборигенной переднеазиатской флоры, а представляет собой область влияния окружающих гор- 80
ных очагов древней земледельческой культуры. В последнее время археологами получено много нового интересного мате- рпала по культурным растениям Древнего Двуречья. В глубокой древности (6000 лет до н. э. и еще раньше) на- роды хетта-иберийской группы занимали в Передней Азии об- ширные территории и распространялись отсюда далеко на запад в Южную Европу и Северную Африку. Это были так называе- мые яфетические народы (шумеры, или сумеры) — в Месопо- тамии, иберы (баски) — на Пиренейском полуострове, этруски- на Апеннинском, пеласги — предшественники греков — на Бал- канском, хетты — в Малой Азии и т. д. Среди очагов высокой культуры, созданных этими народами, переднеазиатский очаг является важнейшим. Древнейшие поселения в средней части Месопотамии отно- сятся к верхнему неолиту, датируемому около начала 1Ч тыся- челетия до н. э. 5Кители в этот период селились на островах, которые возвышались над болотами. Поселения здесь уже рас- полагались на искусственных земляных насыпях. Вместе с тем известно, что их обитатели занимались осушением окружающих болот и создали осушительно-оросительную систему примитив- ного типа. Жители таких частью естественных, частью искус- ственных островов занимались охотой и рыбной ловлей. Однако уже в эту эпоху, названную эпохой Эль-Обеида (по месту рас- копок недалеко от Ура), или протошумерийской, существовало и скотоводство, и земледелие. Из животных были приручены овца, коза, свинья. Здесь сеяли ячмень и полбу-эммер (Т. dicoc- con). Земледелие было весьма примитивно. Мотыги были камен- ные, серпы глиняные, снабженные кремневыми вкладышами. Однако в общем уровень культуры говорил о ее давней преем- ственности. На это указывают довольно хорошие хижины пз глины, камней и даже кирпича (кирпичные очаги и каменные косяки для дверей); сохранились украшения и женские ста- туэтки. Земледельческая культура Эль-Обеида уже настолько высока, что не соответствует тому уровню цивилизации, когда земледелие только возникает. Есть основание предполагать, что шумерийцы (вернее, сна- чала протошумерийцы) пришли в Месопотамию (Двуречье) из соседних областей. Американский историк Г. Чайльд (G. Childe, 1952) считает, что «неолитическая революция», т. е. переход к земледелию и скотоводству, произошла не в самом Шумере, а за его преде- лами. По мнению Чайльда, первые земледельцы проникли в Ме- сопотамию из Сирии, с востока, а возможно, и с юга. Древней- шим земледельческим поселением в Месопотамии Чайльд считает Хассуну — поселение к западу от Тигра, в Ассирии. Это поселение относится к переходному периоду от неолита к медному веку. Наиболее примечательным орудием, найденным в раскоп- 81
ках этого поселения, является деревянный серп с каменнымхг зубцами, скрепленными асфальтом. Общность типа керамики указывает, что уже существовали широкие связи жителей Мес')- потамии с Сирией, Палестиной и Нижним Египтом. Чайльд ука- зывает на несомненность широких сношений с народами на тер- ритории теперешней Армении, откуда доставлялся в Месо- потамию обсидиан, употреблявшийся для производства орудий Существование этой связи с Арменией (по Чайльду) под- тверждается рядом исследователей, указывающих на большую связь древнейшей культуры Месопотамии с соседними горными областями. Имеется предположение, что именно из горных об- ластей, расположенных на севере и востоке, и пришли первые земледельцы Месопотамии. Начало земледелия в этих горных странах, по-видимому, относится к глубочайшей древности— 10 ООО — 15ООО лет. Большие реки и пойменные равнины было значительно труд- нее осваивать, чем небольшие речки и ручьи предгорий нижних и средних горных поясов. Регуляция паводков на больших ре- ках требует громоздких сооружений, а полив отводом и запру- живанием горных ручьев весьма доступен совсем маленьким группам и даже отдельным земледельцам. Поэтому имеется много данных в пользу того, что первые опыты возделывания ра- стений были приурочены именно к горным местностям, а уже оттуда опыт земледелия распространялся на прилегающие рав- нины. Так было, по-видимому, и в Месопотамии. В настоящее время многие археологи предполагают, что оро- шаемое земледелие Месопотамии отнюдь не носит первичного характера — здесь имело место преемственное развитие, начало которому положили народы окружающих горных областей, в том числе и горцы Закавказья. В Месопотамии в отличие от Древнего Египта ирригацион- ные сооружения предназначались не только для орошения и по- вышения плодородия почвы посредством удобрения речным илом. Здесь особое значение получила система ирригации для предупреждения губительных последствий наводнений во время половодья бурных, стремительно текущих с гор основных круп- ных рек — Тигра и Евфрата. Поэтому с течением времени в Ме- сопотамии раньше, чем во всех соседних областях, земледельче- ская культура достигла высочайшего для того времени уровня на базе широко развитого речного орошения. Вслед за протошумерийской культурой следует период (эпоха Урука) дальнейшей колонизации болотных областей среднего и южного Двуречья и развитие уже гораздо более ин- тенсивного земледелия, что особенно относится к следующей, третьей эпохе (период Джемдет-Наср). В Урукский и Джемдет- Насрский периоды государство еще не сложилось. В это время здесь возделывались пшеница, полба, ячмень. Имеются также указания на культуру сорго. Развивалось и скотоводство (при- 82
ручение быка и осла). Следы настоящей шумерийской культуры отмечаются наряду с протошумерийскими останками еще в эпоху Урука и даже встречены в раскопках Эль-Обеида. Период Джемдет-Наср является переходным к настоящей шумерийской культуре, получившей полный расцвет в III тыся- челетии до н. э., в период возникновения древнейшего рабовла- дельческого общества. Дальнейшее развитие сельского хозяй- ства и ремёсел приводит к возникновению торговли. Уже на- ходки в Джемдет-Наср указывают на связи северной Месопо- тамии с южной и средней, а также Эламом (юго-западный Иран) и Сирией (северной). У шумеров скотоводство и земле- делие достигли высокой степени развития. Разводили тогда короткорогих и длпннорогих быков, коз разных пород, курдюч- ных и мериносовых овец, ослов и даже антилоп, которых иногда впрягали в плуги. Лошадь у них появилась позднее — во II ты- сячелетии до н. э. Ряд условий благоприятствовал здесь развитию интенсив- ного земледелия: плодородные пойменные почвы, обилие воды, благоприятный климат. Однако потребовался огромный проме- жуток времени на освоение этой территории сначала путем осу- шения болот, а затем — устройства оросительной системы. Тех- ника земледелия отставала от темпов развития орошения. В те- чение многих веков здесь землю обрабатывали мотыгой, а за- тем примитивным деревянным плугом. По Г Читая (1941), наиболее раннее неоспоримое доказа- тельство применения пахотного орудия имеется в древнейших шумерийских отпечатках (за 3500 лет до н. э.) из царского могильника в Уре. Это орудие нельзя назвать совершенно при- митивным, и, значит, должен существовать предыдущий период его совершенствования, начало которого, по мнению Г Читая, относится к началу IV тысячелетия до н. э. Известно, что в ур- ский период уже пользовались животными в качестве тягловой силы для колесных экипажей, а может быть, и для земледельче- ских работ. Обнаруженное пахотное орудие состояло (по Г Читая) из подошвы, изогнутого составного дышла и рукоятки. Г Читая считает, что такая конструкция пахотного орудия и есть тот основной прототип, который получил широкое распространение по всей Передней Азии и, в частности, у картвелокавказских народов. Как указывает Грозни (F Нгокпу, 1938), в древнешумерий- ском языке знак, изображающий пахотное орудие, тождествен изображению урского плуга. Таким же образом изображается плуг и в китайском пиктографическом письме (XII — до VI вв. до н. э.). Цитируемый автор указывает, что древние египтяне и китайцы заимствовали пахотное орудие у шумеров и что шу- меры первые, раньше других народов Востока, научились делать мотыгу, а затем — пахотное орудие. Они же были пионерами в металлургии, плотничестве и клинописи. Однако что касается
первой мотыги, то ее изготовили не древние жители Месопота- мии, а скорее горные народы, которые позднее, но еще в древ- нейшие времена спустились с нагорий, прилегающих к Месо- потамии, и перенесли земледелие на пойменные равнины Тигра и Евфрата. В течение тысячелетий шумерийские земледельцы пользова- лись примитивными серпами из глины, а позднее из дерева куда вставляли острые зубья из кремня. Медные орудия появ- ляются в эпоху Джемдет-Наср, но металл очень медленно внед- ряется в сельское хозяйство. Гарлан (J. Harlan, 1955) обнаружил в Ираке между Мосу- лом и Сулейманией смесь колосков однозернянки, двузернянки и ячменя. Эти находки датируются V тысячелетием до н. э. Они свидетельствуют о том, что в древности в Месопотамии суще- ствовали смешанные посевы. Здесь же были найдены вкладыши из обсидиана и кремня для серпов — значит, жать хлеба человек научился на заре земледелия. Найденный материал изучался специалистами копенгаген- ского музея. Интересно обнаружение здесь промежуточных по морфологическим признакам форм пшениц — между Т. boeoti- сит и Т. monococcum, между Т. dicoocoides и Т. йсоссоп- и промежуточных форм ячменя — между дикорастущим ячме- нем (Hordeum зроп1апеип1) и двурядным культурным ячменем (Н. distiichum L.). В 195б г. вышел обстоятельный труд А. И. Тюменева «Госу- дарственное хозяйство Древнего Шумера», где имеется очень много сведений о развитии сельского хозяйства Шумера. Дан- ные о развитии государственного и храмового хозяйства в зна- чительной степени отражают ход развития сельского хозяйства в Шумере вообще. На материалах, собранных А. И. Тюменевым, виден постепенный переход от мотыжного земледелия (дере- вянные мотыги) к плужной обработке почвы при помощи дере- вянного, похожего на соху плуга. В документах эпохи раннего Ура были названы те же глав- ные зерновые культуры, что и в последующие эпохи: на первом месте стоял ячмень, затем шли пшеница и полба-эммер. Зерно этих трех культур шло не только в пищу людям, но и на корм рабочим быкам — в записях храмовых хозяйств того времени встречаются точные указания, сколько именно ячменя, пшеницы и полбы-эммера отпущено для этой цели. Ячменем кормили также овец, предназначенных на убой. В эпоху раннего Ура хлеб выпекали преимущественно из ячменя, из ячменя приготовляли также солод и пиво. В храмовом хозяйстве Шуруппака (XXVII — XXVI вв. до н. э. — эпоха, хронологически непосредственно следующая за ранним Уром) ячмень, как и в Лагаше и позднейшем Уре, яв- ляется главной зерновой культурой. 84
Имеются указания, что в эпоху Лагаша (вторая половина III тысячелетия до H. э.) ячмень иногда высевали на лесных участках между деревьями (A. И. Тюменев, 1956). О ботанических формах шумерийских пшеницы и ячменя известны следующие м атер и алы. Персиваль (J. Percival, 1936) исследовал образец мелко- зерной полбы, который был найден Меллоуэном (Ма11оюеп1 в четырех милях к востоку от Ниневии в Арпахиахе. Эти зерна относились к периоду 4000 лет до н. э. или к еще более ранней эпохе. У Персиваля дан рисунок зерен (удлиненные зерновкп). I(aK видно, это был остистый эммер. В сосуде из раскопок древнего шумерийского дома в Джем- дет-Наср в нескольких милях от I(пша американский исследо- ватель Фильд (Н. Field) в раскопках 1926 г. обнаружил пше- ницу, не похожую на полбу-эммер. Эта ископаемая пшенища была представлена тупоносыми, округло-овальными зернами с ясно заметной спинной выпуклостью, похожими на современ- ную Т. turgidum L. На рисунке, приложенном к статье Перси- валя, однако, видно, что зерна шумерийской пшеницы мелкие— гораздо мельче современной Т. turgidum. Эти зерна неодинаковы по форме, но все они относительно короткие. Одно зерно совсем округлое — очень похожее на зерно ископаемой пшеницы из свайных построек Швейцарии, которой Геер дал название Т. an- tiquorum Heer. Да и вообще все зерна шумерийской пшеницы гораздо больше похожи на Т. antiquorum, чем на современную Т. turgidum. Персиваль отмечает, что эти зерна напоминают абиссинскую полбу-эммер (Т. йсоссоп). О. Филипс (О. Philips) выразил мнение, что зерна шумерийской пшеницы напоминают карликовую пшеницу — Т. compaictum Host. Г. Пик (Н. Peake, 1927) отмечает, что мнения экспертов расходятся по вопросу видового определения шумерийской пше- ницы из Джемдет-Наср. Он приводит мнение Персиваля, что эта пшеница относится к Т. turgidum, в то время как тот же автор находил ее похожей на абиссинский эммер — Т. сотрас- turn. По сообщению Пика, один американский ботаник определил ее, как мягкую пшеницу, а Стапф (D. Stapf) думает, что шуме- рийская ископаемая пшеница представляет собой карликовую пшеницу — Т. сотрас1ит. Если припомнить все литературные указания на характер- ные черты и видовую принадлежность ископаемой швейцарской пшеницы Т. antiquorum, то там имеет место такой же разнобой в описаниях, но каждый автор там был по-своему прав, находя сходство Т. antiquorum с каким-либо определенным из ныне существующих видов пшениц, и вместе с тем каждый из авто- ров все же находил, что полного совпадения признаков ни с одним из известных видов у этой пшеницы нет. В Т. antiquo- гит Геером и другими исследователями открыт древний инте-
гральный вид, отличавшийся огромным полиморфизмом и сов- мещающий в своих многочисленных формах, а также до извест- ной степени в каждой из отдельных форм признаки различных существующих в настоящее время видов пшеницы. Чаще всего формы этой пшеницы имели сходство с современной карликовой пшеницей — Т. сотрас1ит, которая, в свою очередь, очень близка к мягкой. Кроме признаков мягкой пшеницы, у ископае- мой швейцарской пшеницы ряд исследователей отмечает нали- чие признаков английской пшеницы (Т. turgidum), утверждая в то же время, что ее нельзя идентифицировать ни с одной из разновидностей современной Т. turgidum, а также с полбой- эммером. Как выяснено исследованиями грузинского знатока куль- турных растений В. Л. Менабде (1948), ископаемая европейская пшеница Т. antiquorum очень близко стоит к грузинским пшени- цам Т. macha и Т. paleocolchicum, сохранившим древние черты строения. Близкие к последним формы обнаружены в неолите Закавказья. Эти «живые ископаемые» грузинские пшеницы и теперь еще характеризуются полиморфизмом и наличием боль- шого количества общих признаков и форм, которые свидетель- ствуют о сходстве этих пшениц с несколькими современными ви- дами. Согласно теории В. Л. Менабде, древняя полиморфная пшеница, которую он называет Т. protomacha, широко распро- странилась за пределы Закавказья и вообще Передней Азии и дала начало также очень полиморфной ископаемой европейской Т. antiquorum, которая ныне вымерла, но положила начало мно- гим европейским современным формам. древние пшеницы из Закавказья и соседних областей Пе- редней Азии распространились, по нашему мнению, широко не только на северо-запад в Европу, но и на юго-запад в Египет и Эфиопию, а также на восток — в Месопотамию, куда эти пше- ницы были занесены в древности переселенцами из передне- азиатских горных областей. Такой путь распространения мог скорее и легче осуществиться, чем более удаленный путь через Балканы в Среднюю и даже Северную Европу, так как верховья Тигра и Евфрата находятся в пределах древней Армении и из- вестно, что издревле существовали широкие связи между древ- нимц >кителями Месопотамии и соседними горными племенами Закавказья и других соседних стран Передней Азии. Откуда пришел в Месопотамию ячмень — не ясно, но, ве- роятно, из соседних горных стран Передней Азии, может быть из Закавказья. В статье Персиваля (1936) имеются рисунки зерен ископае- мого ячменя из Арпахиаха, найденные Меллоуэном в том же поселении, где была найдена и пшеница, похожая на Т. antiquo- гшп. Эти зерна неодинаковой формы и между ними есть до- вольно округлые.
По-видимому, в Месопотамии еще в период Джемдет-Наср культивировалось сорго. Некоторые ботаники сомневаются в правильности определения этой культуры. Мы, однако, счи- таем вполне естественным здесь существование сорго, ибо те- перь уже достаточно твердо установлено, что связи Передней Азии и, в частности, Месопотамии и Средней Азии с Африкой возникли в чрезвычайно древнее время. Сорго пришло в Месо- потамию, как мы думаем, из Африки через Сирию, а не из Египта. В Древнем Египте существование культуры сорго не установлено с достаточной достоверностью. В ряде документов третьей династии Ура упоминается «про- сяная мука» (A. И. Тюменев, 1956). Неясно, возделывалось лп там настоящее просо или эта мука приготовлялась из сорго. Важное значение в питании жителей Шумера во все эпохи имели бобы. Бобы, как огородное растение, упоминаются в до- кументах государственных хозяйств периода Джемдет-Наср вместе с луком, в документах этого периода говорится о «бобо- вом хлебе». Первое документальное упоминание о вигне в Шумере имеет ту же дату, что и первое упоминание о кунжуте, т. е. около 2350 лет до н. э. Род Vigna Savi — африканского происхождения. И вигна, и кунжут, вероятно, достигли Индии одним и тем же путем- из Африки через Двуречье. Таков же был, вероятно, и путь сорго. Население Шумера издревле потребляло растительное масло. О растительном масле, вероятно кунжутном, упоминается еще в документах храмовых хозяйств периода Джемдет-Наср. В документе Шуруппака (позднее раннего Ура) упоминается о поле под кунжутом (сезамом) и имеются данные о поступле- нии на склады и расходовании сезама наряду с эммером и дру- гими продуктами. В документах царского хозяйства третьей династии Ура указывается, что на корм мелкому скоту наряду с прочими кор- мами отпускалось и растительное масло, по-видимому, кунжут- ное, так как нет сведений о существовании в то время какого- либо другого растительного масла. Вполне достоверно, что кун- жут был известен шумерам примерно в 2350 г. до н. э. (Haudri- icourt et Hedin, 1953). Лен, по-видимому, шел только на волокно. Упоминаний о льняном масле нет. Начало культуры льна в Шумере точно неизвестно. В эпоху третьей династии Ура были распространены шерстяные и льняные одежды, что привело к возникновеншо профессии ткачей. Огородничество стало развиваться в Шумере с древних времен. В хозяйстве храма Бау (точно установить год не уда- лось) разводили лук, огурцы, бобы, репу и пряные растения: укроп, анис, тмин, шафран и др. 87
Лук известен по документам )~жемдет-Насра под названием -'зип1». В значительно более позднюю эпоху (вторая половина !II тысячелетия до н. э.) в хозяйстве храма Бау разводились различные сорта лука, между прочим специальный сорт, упо- треблявшийся на корм быкам. В архиве царских хозяйств третьей династии Ура лук на- равне с другими овощами (репа, бобы, пряные травы, огурцы) постоянно фигурирует среди продуктов, поступающих в цен- тральные склады и выдаваемых оттуда (А. И. Тюменев, 1956). Помимо многочисленных указаний на развитие огородниче- ства в Месопотамии в книге А. И. Тюменева, об этом имеются данные и в других исторических трудах. В тексте Урукагины (царь, возведенный на престол около 2400 г. до и. э.), по И. М. дьяконову (1951), упоминается огород — Ki-sum, что значит «место лука», и бахча — Ki-ukus, что значит «место огурцов». Эти данные очень интересны, так как свидетельствуют о большой древности культуры лука. Что касается огурца, то, по А. И. Филову (1960), здесь воз- делывался не Cucumis sativus, а какое-то другое подобное огурцу растение. О листовой свекле в книге А. И. Тюменева не говорится. Она, по-видимому, появилась в Месопотамии позднее. Листовая свекла не упоминается и в документах Старого Вавилона в VIII в. до н. э. По Мейснеру (Meissner, 1920), листовая свекла указывается в списке тех растений, которые возделывались в огородах вавилонского царя Меродах-Баладана (722 — 711 гг. до н. э.) наряду с салатом, кресс-салатом, чесноком, тмином, кориандром и репой. Листовая свекла отмечалась в списке под названием Silqa. Лёу (J. Еёг, 1881) предполагает, что название Silqa не вавилонского происхождения, а заимствовано из ара- мейского языка. Он считает, что в Вавилон листовая свекла была занесена из Сирии, а в Сирию — со средиземноморских островов. Лёу считает свеклу завозным растением еще и по- тому, что ее название впервые упоминается в списке растений садов царя Меродах-Баладана, а в более древних медицинских и магических вавилонских текстах ее нет. В записках императора Юлиана Отступника есть запись о том, что употреблять свеклу в пищу запрещено жрецом сирий- ской богини кибелы. Эта запись представляет собой косвенное доказательство того, что свекла широко культивировалась в Сирии. Из Сирии она легко могла быть завезена и в Месопо- тамию, согласно предположению В. Т. 1~расочкина (1960). По- явление свеклы в Месопотамии, по-видимому, относится к вави- лонскому периоду. В хозяйстве Шумера огромное значение принадлежало фи- никовой пальме. В некоторых документах раннего Ура упоми- нается о финиковых плантациях «sa-òà-gid». И в эту эпоху и позднее финиковые деревья использовались для получения пло-
дов и как строительный материал. Финики употреблялись све- жие и сушеные, из них получали сахар; ферментированный сок дерева давал вино; из волокон коры вили веревки, из листьев делали корзины, изготовляли одежду, а нежные совсем моло- дые отпрыски использовались в пищу. В более поздние эпохи в Ассирии культура финиковой пальмы была широко распространена, что доказывается мно- жеством изображений дерева, листьев и плодов в виде скульп- турных украшений на стенах и на утвари в древних храмах Ассирии. На скульптурах видно, что ассирийские садоводы ис- кусственно опыляли собранной пыльцой пестичные деревья фи- никовой пальмы, хотя, конечно, не имели ясного представления о поле у растений. По данным, относящимся ко второй половине III тысячеле- тия до н. э., имелись плантации фиговых и гранатовых деревьев. Фиги и гранаты обрабатывались в виде особых хлебцев. После финиковой пальмы гранатовое и фиговое деревья имели наибольшее значение в садоводстве Шумера (А. И. 1'ю- менев, 1956 и другие источники) . Виноградарство было распространено в древнем Уруке мало. Точно не установлено, существовал ли тогда пиктогра- фический знак для винограда. В раннем Уре виноград был, видимо, широко распростра- нен, о чем говорит обилие имен, в состав которых входил слу- живший для обозначения винограда термин. В документах ран- него Ура говорится о виноградарях и виноградниках. Из вино- града приготовляли вино, и, кроме того, лоза употреблялась для плетения. Терновник использовался дикорастущий. Во времена третьей династии Ура для сбора его посылались специальные партии рабочих (А. И. Тюменев, 1956). В документах древнего Урука несколько раз отмечено изо- бражение ветвистого деревца или кустарника, которое, как полагают, является прообразом более поздней идеограммы «Sinig», обозначающей тамарикс. Это дерево имело большое значение в южной Месопотамии и в последующие эпохи в связи с бедностью ее древесной флоры. Роль тамарикса в жизни жи- телей южной Месопотамии характеризуется в басне, где тама- рикс ведет спор с финиковой пальмой. Финиковая пальма гор- дится тем, что из ее ствола делают сельскохозяйственные ору- дия и утварь для царского стола. Тамарикс в ответ ей называет себя «отцом земледельца», он соглашается, что многие пред- меты вырезают из ее ствола, но из его побегов изготовляют мо- тыги для копки каналов и для работ по орошению полей. Стволы тамарикса шли также на изготовление досок. Тамарикс получали со специальных лесных участков, уход за которыми в эпоху Лагаша (вторая половина III тысячеле- тия до н. э.) поручался особому персоналу — людям при тама-
рпксе. Из документов третьей династии Ура видно, что иногда тамарикс культивировался вместе с финиковой пальмой. Известен был и ряд других древесных пород, употребляв- шихся для изготовления плугов и других орудий. Специальные виды деревьев шлп на судостроение. I(poMe тамарикса, в качестве строительных деревьев упо- требляли кедр и кипарис. Имеются сведения о доставке в Шу- мер самшита, а, может быть, он произрастал и в самом Шумере (А. И. Тюменев, 1956) . В документах архаического Урука в самых разнообразных вариантах имеется изображение тростника, впоследствии изо- бражавшегося идеограммой «gi». Из тростника изготовля- лись корзины, служившие для перевозки и хранения докумен- тов и продуктов, маты, циновки, паруса и даже мелкие суда. Тростник, имевший большое хозяйственное значение в южной Месопотамии, упоминается и в документах раннего Ура — из него изготовляли сети и корзины. Тростник занимал видное место и в храмовом хозяйстве Шуруппака (позднее раннего Ура) . Древнешумерийская культура на юге Месопотамии развива- лась все время во взаимодействии с семитской культурой акка- дов в средней части Месопотамии и получила название Шумеро- Аккадской. Эта культура легла затем в основу культур Вави- .лона и Ассирии. В III тысячелетии до н. э. Вавилон входил в состав Шумеро- Лккадского государства. Шумерийская, затем вавилонская куль- тура была основой для ассирийской культуры. Ассирия вначале занимала небольшую территорию по верхнему течению реки Тигра и на севере граничила с Армянским нагорьем. С очень древних времен здесь жили субарейцы, которые были близки к курритам северо-западной часги Месопотамии, урартам Закав- казья и другим горным племенам, жившим к востоку и к северу от Тигра. I( началу II тысячелетия до н. э. субарейцы, смеши- ваясь с семитами, образовали народ Ассирии. 1(лимат Ассирии более суров по сравнению с центральной и южной Месопотамией. Поэтому площадь под пашней здесь была меньше. Однако в Ассирии возделывались те же культуры, как в Вавилоне и раньше — в Шумера-Аккадском царстве. Здесь было развито садоводство (террасные сады). В Ассирии возде- лывали абрикосы, фиги, сливы, айву, виноград, фисташку. I( VII a. до н. э. относится ассирийский травник. Развитие асси- рийской культуры относят к 11 тысячелетию и первой половине 1 тысячелетия до н. э. 1(ак самостоятельное государство Асси- рия перестала существовать в 605 г. до н. э. Культура Ассирии была тесно связана с государством Ми- танни (XVIII в. до н. э.) и малоазиатской хеттской культурой (II — начало 1 тысячелетия до н. э.). В Малой Азии не было больших рек и широких плодородных равнин. Однако и здесь 90
существовало орошение — на малых горных речках. Это обусло- вило развитие в Хеттском царстве культуры зерновых хлебов, садоводства и огородничества. На основе всех использованных источников нами составлен список культурных растений, возделывавшихся в Шумере и Ассирии (табл. 1) . Культурные растения древней Месопотамии не были тузем- ными. Местности, непосредственно прилегавшие к руслам Тигра и Евфрата, дали для использования в шумерском хозяйстве не- многое: тростник, тамарикс, терновник и др. Из приведенного списка видно, что культурная флора древних народов Месопотамии была в основном передне- азиатская — пришла в Месопотамию из соседних горных стран. Связи с Восточной Азией незначительны и сомнительны. Связи с Африкой выражаются в переносе оттуда в Месопотамию кунжута, вигны, сорго. Вероятно, через Месопотамшо эти куль- туры двигались из Африки в Индию. О происхождении шумерийских культур льна и репы трудно сказать. Эти культуры имеют первичные центры и в Индии (северо-западная Индия), и в Передней Азии. Хотя многими историками древнейшие земледельческие культуры Индии и Месопотамии считаются более или менее синхронными, но в отношении своего древнего ассортимента культурных растений оба древних очага земледелия были почти независимы. При этом через Месопотамию шло расселение в Индии (и, возможно, в Китае) некоторых культур (кунжут, сорго, вигна) из Африки, обратного влияния Индии на состав культурных растений Месопотамии почти незаметно. Из культур приведенного списка позднее появились (в асси- рийский период): свекла, салат, кресс-салат, кориандр, олив- ковое дерево, айва, абрикос'"', фисташка. Из нашего очень краткого и поэтому несколько схематиче- ского очерка можно видеть, что культура в целом и земледель- ческая, в частности, развивалась в Двуречье Тигра и Евфрата, в Малой Азии и в соседних областях на основе последо- вательной преемственности и теснейшего взаимодействия и переплетения культур соседствующих и исторически сменяю- щих друг друга народов, населявших страны Древнего Востока. Нас особенно должна интересовать история земледельческой культуры именно народов Двуречья (Месопотамии) и Малой Азии вследствие теснейшей связи ее с древнейшей культурой Закавказья. '" Аор~икос в двуречье (Месопотаышо) пришел, вероятно, не непосред- ственно из Восточной Азии, а из Закавказья (Армении).
Таблица 1 Культурные растения Шумера, Вавилона и Ассирии Про искожденн Распространение 1(ультура Шумер, Вавилон, Асси- рия То же Передняя Азия Шумер Шумер, Вавилон, Асси- рия То же Восточная Азия Африка Бобы ЯЕЕгЕЕа Кунжут Лен Вавилон, Ассирия Передняя Азия Передняя и Средняя Азия Передняя Азия Укроп Кориандр Тмин Анис Шафран Финиковая пальма Передняя Азия и Севе- ро-Восточная Африка Передняя Азия 1 111жЕЕР Гранат Виноград Передняя и СредняяАзияМестный Терновник Тамарикс Кедр Местный и, вероятно, привозной Восточная Азия Передняя Азия Местный Вавилон, Ассирия Средиземноморье Восточная Азия Средиземноморье Передняя Азия Ячмень (основная зерно- вая культура) Пшеница Эммер Просо,(под вопросом) Сорго ~ I)'Ii Чеснок (:) гурец (вероятно, не настоящий) Репа Салат Кресс-салат Кипарис Самшит Тростник (для плетения и построек) Тростник для пряностей сахарный Свекла листовая Абрикос Оливковое дерево Айва Фисташка Шумер, Вавилон, Асси- рия Вавилон, Ассирия Шумер, Вавилон, Асси- рЕ1Я То же Шумер (Вавилон и Ассирия неточно) Lo же Шумер, Вавилон, Асси- рия Шумер Шумер (Вавилон и Ассирия неточно) То же Передняя Азия Африка Передняя Азия (может быть, Индия) Средняя Азия Индия? Передняя Азия?
ОБЗОР ВИДОВ И ФОРМ АБОРИГЕННЫХ КУЛЬТУРНЫХРАСТЕНИЙ ПЕРЕДНЕЙ АЗИИ В ЦЕЛОМ Для крупных областей исторического развития культурной флоры мы даем обзор аборигенных культурных растений. Этп обзоры характеризуют географические области в отношении эндемизма их культурных растений, эндемизма качественного и количественного. Прежде всего в этих обзорах отмечаются такие роды, кото- рые возникли в данной географической области и в состав ко- торых входят культурные растения. Роды, включающие в свой состав культурных представителей, могут иметь основной очаг развития в данной области, но место зарождения рода, его первичный очаг, может находиться в другой области. Это же относится и к видам, которые могли возникнуть в данной об- ласти или имеют здесь только главный очаг современного фор- мообразования. Эндемичными видами культурных растений мы называем виды, возникшие в данной области и нигде больше в культуре не встречающиеся. Меньшей степени эндемизма соответствуют такие культур- ные растения, которые в данной географической области пред- ставлены не эндемичными видами, а только эндемичными фор- мами или сортами. Собирая для наших обзоров материал из многих литератур- ных источников, мы не смогли избежать известной несогласо- ванности в названиях и рангах систематических единиц. Мы приводим формы культурного вида под именем разновидности (varietas), подвида (subspecies), ветви (proles) и т. д. бранная работа не ставит своей задачей разработку вопросов система- тики и введения единообразия в номенклатуру; каждую форму мы приводим так, как она фигурирует у того автора, у кото- рого мы заимствуем материал, и лишь в отдельных случаях сопровождаем этот материал критическими замечаниями в от- ношении классификации. Однако для нашего дела такая «разно- голосица» не особенно вредна. В подавляющем большинстве случаев «подвид» или «пролес» указывает на более крупный ранг, чем «разновидность» или «сорт» и на более интенсивное г.роявление эндемизма. Начиная обзор с зерновых культур, прежде всего отметим Переднюю Азию как родину пшениц — рода ТгЖсит L.— в целом. Затем рассмотрим бобовые и некоторые другие куль- туры (масличные, прядильные, кормовые и т. д.). Пшеница. I. Виды пшеницы, эндемичные для Передней Азии: 1) дикорастущая однозернянка Урарту — Т. urartu Тит.— Ар- мения; 2) закавказская двузернянка — Т. araraticum Jakubz.— Закавказье (Армения, Азербайджан); 3) сирийско-палестинская двузернянка — Т. йсоссоЫез Koern. — Сирия и Палестина;
4) полба Тимофеева — Т. timopheevi Zhuk. — Западная Грузия (Мегрелия); 5) полба Жуковского — Т. zhukovskyi Menabde ct Eritz. — Грузия; 6) дика — Т. persicum Vav. — Грузия (меньше в Дагестане, Северной Осетии и Азербайджане); 7) полба маха — Т. macha Dekapr. et Menabde — Западная Грузия (Леч- хуми); 8) колхидская полба — Т. ра1еосо1сЫсит Menabde — За- падная Грузия (Лечхуми, Имеретия); 9) пшеница ванская— Т. vavilovii Jakubz. — Древняя Армения; 10) пшеница исфаган- ская — Т. ispahanicum Heslot — Иран; 11) пшеница туранская— Т. turanicum Jal~ubz. — Дагестан, Малая Азия, Сирия, Иран, Средняя Азия. Туранская пшеница в основном занимает переднеазиатский ареал и в Среднюю Азию, вероятно, перешла из Передней Азии. II. Пшеница, эндемичная для Передней Азии и Балкан,— дикорастущая однозернянка — Т. boeoticum Boiss. — Закавказье, Малая Азия, Аравия, Балканы. III. Виды пшеницы, имеющие первоначальный ареал и очаг возникновения в Передней Азии, но позднее широко распростра- нившиеся и теперь имеющие многоостровной ареал, или вымер- шие, или имеющие одноостровной ареал: 1) культурная одно- зернянка — Т. monococcum L. — Малая Азия, Закавказье, Крым, Балканы, горы Средней Европь1; в настоящее время встречается редко; 2) полба-спельта — Т. spelta L. — Передняя Азия, Сред- няя Европа (см. выше — o6 Иране); 2) полба-эммер — Т. йаос- соп — Закавказье, Месопотамия, Западный Иран, Индия, Эфио- пия, Балканы, Средняя Европа, Испания (Наварра), Марокко, Волжско-Камские районы; 4) абиссинская пшеница — Т. аей1о- picum Jakubz. — удаленный одноостровной ареал: Эфиопия, Эритрея; 5) ископаемая пшеница Геера — Т. antiquorum Неег— занимала раньше широкий ареал от Северной до Южной Ев- ропы и в Средиземноморье, теперь не встречается. IV. Виды пшеницы, в настоящее время широко распростра- ненные, с полифилетическим происхождением отдельных совре- менных форм, но происходящие от переднеазиатского корня не- посредственно или через производные виды, исчезнувшие ин- тегральные виды, близкие к переднеазиатским (Т. protoaethi~o- pi~cum и Т. antiquorurn): 1) пшеница мягкая — Т. aestivum L.= Т. vulgare Host sensu lato (вместе с Т. compa~ctum) — Передняя Азия, в том числе Кавказ, Юго-Западная Азия, особенно горные районы Афганистана, северо-западная Индия и Таджикистан, Средняя Европа; позднее распространилась по всему земному шару; 2) пшеница твердая — Т. durum Desf. (с включением Т. turgidum) — Средиземноморье, Передняя Азия (Закавказье), степные и лесостепные районы европейской части СССР (не- много в Иране и в Индии) . В этом списке отсутствует так называемая польская пше- ница — Т. polonicum L., не имеющая самостоятельного ареала 94
и распространенная спорадически в областях культуры видов Т. durum и Т. aethiopicum. Из числа самостоятельных видов исключены также Т. turgidum (включен в Т. durum) и Т. сот- pactum (включен в Т. aestivum). Если включить ареал Т. sphaeroooocum Репс. — Пенджаб— в состав стран Передней Азии (что соответствует мнению мно- гих ботаников), то в Передней Азии будем иметь 12 эндемич- ных видов, из них 8 — для Закавказья, 1 — для Пенджаба, 1— для Сирии и Палестины, 1 — для Ирана. Дикорастущая однозернянка, выделенная нами по типу ареала в отдельную группу, по существу также вид передне- азиатский (вне Передней Азии встречается редко и небольшими островками). Остальные виды также ведут свое происхождение от древ- него переднеазиатского корня или непосредственно, или через промежуточные звенья, или с участием гибридизации. Из 8 эндемичных для Закавказья видов пшеницы 5 видов гриходится на Грузию. Если принять во внимание, что сюда входят наиболее древние виды, в числе которых сохранились исходные интегральные формы, что там распространены и не- эндемичные виды, представленные особыми грузинскими фор- мами, то по богатству видами и формами пшениц Грузии при- надлежит первое место на земном шаре. Из широко распространенных видов пшеницы грузинские и вообще закавказские мягкие пшеницы выделены В. Л. Менабде в самостоятельную географическую расу — Т. aestivum transcau- casicum Menabde, характеризующуюся остевидными образова- ниями на зубцах колосковой чешуи. В составе закавказской «расы» (подвида?) мягкой пшеницы В. Л. Менабде различает 5 ветвей (пролесов): 1),proles sieco- camipestre Flal<sb. — засушливая степная; 2) proles medio — топ- 1апо — silvaticum Menabde — среднегорно-лесная; 3) proles meso — cachethi~cum .V1enabde — влаголюбивая кахетинская; 4) ~proles ~col~chicum Menabde — колхидская; 5) proles hyper- montaneum Menabde — высокогорная. Согласно данным В. Л. Менабде, особым полиморфизмом и наибольшей концентрацией видов пшениц отличается область, занятая в основном колхидской ветвью мягких пшениц. М. М. Якубцинер и другие тритикологи считают Закавказье очагом развития пшеницы тургидум. По данным М. М. Якубци- нера (1956), в XIX в. тургидум здесь возделывался в чистых по- севах (в Абхазии, Месхети, Лечхуми). В составе закавказских типичных твердых пшениц, не похо- жих на Т. turgidum, имеются эндемичные озимые сорта. Колхидский тип твердых пшениц — sap. turgidoides Flaksb. представляет собой формы, переходные к Т. turgidum. Эгилопс. Передняя Азия является основным очагом развития рода Aegilops. Виды этого рода широко распространены и
в Закавказье. Экспедицией И. Д. Мустафаева были найдены в советском Азербайджане, в Карабахе и в пограничных с Ира- ном местностях Ае. cylindrica Host, Ae. biuncialis Vis., Ае. triun- cialis L., Ае. squarrosa L., Ае. ovata L., Ae. mutica B~oiss., Ае. speltoides Tausch., Ае. variabilis Eig, Ае. ~comosa Sibth. et Sm., Ае. со1итпаг1з Zhu,<., Ae.uniaristata Vis., Ae. ventricosa Tausch., Ae. caudata L. П. М. Жуковский (1961) считает, что в Закавказье имеется около 2~~ всего количества видов Aegilops. Здесь же на границе Ирана найдены Т. spelta и очень много спельтоидных форм пшеницы. Род Aegilops весьма близок к роду Triti~cum. Известны многочисленные естественные гибриды между видами этих ро- дов и получены амфидиплоиды со всеми основными признаками мягкой пшеницы от соединения геномов тетраплоидной пшеницы и диплоидного вида Aegilops (28+14=42), аналогично полу- чена синтетическим путем спельта (Т. арейа). По нашему мнению, Triticum и Aegilops — роды относительно молодые с синхронным развитием. Aegilops не может быть родоначальным родом для Triticum, и при получении синтетических форм (амфидиплоидов) мы имеем только процесс насыщения конституции пшеницы неко- торыми признаками Aegilops (интрогрессия). Однако значение видов Aegilaps очень велико как исходного материала для селекции, и эти богатейшие ресурсы еще не ис- пользованы селекционерами-практиками. Ячмень. В Передней Азии представлен, по классификации Ф. Х. Бахтеева, только видом евразиатским — Hordeum euro- asiati~cum Vav. et ВасЫ., а в составе последнего вида — передне- азиатским подвидом, который имеет основной ареал на Кавказе и в Малой Азии, но распространен также в Европе и Сибири. В составе этого подвида в Передней Азии имеются следую- щие ветви (пролесы): 1) анатолийский — proles anatolicum (Orl.) Vav. et; 2) западнокавказский — proles occiden- tali-icaucassicum (Orl.) Vav. et ВасЫ.; 3) дагестанский — proles daghestani~cum (Bykovetz.) Vav. et Bacht. 4) апшеронскпй— proles apsheronicum Vav. et ВасЫ.; 5) армянский — iproles arme- niacum (Orl.) Vav. et; 6) армяно-грузинский — proles arme- nia~oo-georgicum Vav. et ВасЫ.; 7) горно-кавказский — proles montano-ñàucàécum Vav. et ВасЫ. Уже один этот перечень указывает на экотипическое разно- образие ячменей Передней Азии. Морфологическое разнообра- зие также значительно. Передняя Азия и особенно Закавказье, несомненно, яв- ляется, если и не первичным, то очень важным очагом развития культурных ячменей. Рожь. Весь род Secale L., несомненно, переднеазиатского про- исхождения. Следующие виды ржи эндемичны для Передней
Азии или имеют там основной ареал: 1) рожь Куприянова— S. kutprijanovii Grossh. — опушки и поляны горных лесов и круп- нотравные луга Северного Кавказа (заповедник), Абхазии, Азербайджана, Армении и Югославии (разорванный ареал в виде редких островов); 2) рожь анатолийская — S. anatolicum Boiss. (по мнению ряда авторов этот вид и предыдущий — под- виды S. montanum Guss.) — Армения, Иран (неточно); 3) рожь Вавилова — S. vavilovii Grossh. — Армения, Азербайджан; 4) рожь предковая — S. ancestrale Zhul'. — Анатолия; 5) рожь дигорская — S. dighori~cum (Vav.) Roshev. — Северная Осетия (Дигория); 6) рожь даралагезская — S. daralagesi Thum. — Ар- мения; 7) рожь реснитчато-чешуйная — S. iciliatoglume (Boiss.) Grossh. — Курдистан. Secale daralagesi и S. ciliatoglume — виды сомнительные, но приведены здесь как показатели большого разнообразия ржи в Закавказье. После Второй мировой войны участники экспедиций уже не встречали этих «видов». Вероятно, под их названиями фигуриро- вали или одичавшие, или очень редкиетеперь исчезнувшие формы. Что касается S. vavilovii, то этот «вид» не значится в по- следнем «Определителе кавказской флоры» А. А. Гроссгейма. Мы видели растения, соответствующие диагнозу А. Л. Гросс- гейма, по обочинам посевов и дорог и предполагаем, что эти довольно редко распространенные формы однолетней узколист- ной ржи следует включить в вид сорной ржи — S. segetale. Рожь культурная — S. cereale L., к которой очень близка сорная рожь — S. segetale (между ними существует вся серия постепенных переходов и по степени ломкости колоса и по дру- гим признакам), имеет наибольшее разнообразие форм в Пе- редней Азии (Закавказье, Иран и Малая Азия). При раскопках в польском городе Гута (данные К. Молден- гауера, 1958) на глубине 250 — 290 м были найдены семена лом- кой ржи, которую определили как S. ancestrale Zhuk. Находка датируется около 400 г. н. э. (эпоха римского владычества). Автор считает, что эта рожь возделывалась, в чем он несомненно ошибается. Она была распространена как сорняк и найдена вместе с семенами пшеницы, проса, вики, конопли и различных сорных трав. Если только определение Молденгауера верно, то аналогичные зерна ржи, по его сообщению, хранятся в ботани- ческом музее в Берлине и были найдены в Восточных Альпах и в Богемском лесу. Зерна датированы 200 — 400 г. н. э. Несом- ненно, что родина этой своеобразной ржи Передняя Азия. Рас- пространение ее на запад автор правильно объясняет существо- вавшими в То время широкими торговыми связями римлян с расположенными к востоку странами. Рожь сорная афганская — S. afghanicum (Vav.),oshev.— c очень ломким колосом, встречается и в Туркменистане, т. е. захватывает и переднеазиатский ареал (в широком понимании). 4 Синская Е. Н. 97
Эта форма ржи слабо изучена, как и дигорийская рожь. Послед- няя, помимо особенной ломкости колоса (колос ломается еще в незрелом состоянии), не проявляющейся так резко ни у одной из известных форм ржи, не является специализированным сор- няком — засорителем посева ячменя и пшеницы, но встречается и по межам на полях картофеля и овощных культур. С нашей точки зрения, дигорийская рожь интересна как тип полурудерального растения, более древнего, чем настоящая сор- ная рожь — Secale segetale. Эта рожь засоряет не только зерно- вые, но также встречается по огородам и вблизи жилищ. Раз- ано~кается главным образом опадающими на землю колосками, а не путем посева вместе с засоряемым растением. Нежным гигроморфным строением резко отличается от других видов сорной ржи. Вероятно, это очень древняя форма, может быть, вначале приуроченная к скалам (первичным местообитанием многих сорных растений были каменистые почвы). Еще более .древней формой является лесной гигрофильный вид ржи, про- 11зводными которого в настоящее время считают рожь Куприя- нова и очень близкие к ней виды S. dalmati~cum Vis. (Далмация) и S. montanum Guss. (западная часть Средиземноморья). Р Ю. Рожевиц (1947) правильно считает S. anatolicum, S. dal- maticum и S. montanum относительно молодыми видами, отде- .лившимися от S. 1<и~рг1]апач|1; их скорее следует считать подви- .дами последней. Рожь африканская — S. afriicanum Stapf — имеет сходство с S. montanum и входит в цикл ее подвидов (или видов). S. sil- ;estre Host, распространенная по пескам от Венгрии до Афга- нистана, более других видов обособлена, но не так уж сильно, чтобы нарушать общее впечатление монолитности всего рода. Предки этого рода были лесными мезофитами, самый древ- ний по конституции вид S. 1'ирг1 anovii сохранился теперь в об- ласти влияния колхидской мезофильной флоры. Овес. Род Анепа L., несомненно, имеет в Передней Азии :область своего прогрессивного развития. Здесь сталкиваются ареалы 28-хромосомных видов (например, дикорастущего Л. harbata Pott.) и 42-хромосомных: культурного византийского овса A. byzantina С. Koch и его дикого родича А. sterilis L., .а также овсюгов A. ludovi~ciana Dur. и A. fatua L. Возникнове- ние 42-хромосомных из 28-хромосомных овсов связано если не с Передней Азией, то во всяком случае с восточной частью Средиземноморья (в широком объеме этого понятия). Большой филогенетический интерес представляют А. ludo- viciana п А. sterilis, имеющие переходные черты и к А. sativa и к А. fatua, и и абиссинским 28-хромосомным видам. Передней Азии свойственны следующие эндемичные формы византийского овса А. Ьууапбпа (по А. И. Мордвинкиной): I) ранний палестинскии, относится к подвиду еп-hyzantina 1ordv. 2) анатолийский — сорный (Малая Азия) относится
к подвиду еи-byzantina Mordv.; 3) марашский — сорный (сеа=р- ная Сирия), относится к подвиду nodiipubescens Malz. Обыкновенный культурный овес А. sativa L. имеет, по A. И. Мордвинкиной, в Гlередней Азии следующие эндемичные типы: 1) тпп армянских горных овсов, засоряющих посевы полбы, пшеницы и ячменя; 2) тип иранских овсов, занесенный из Турецкой Армении. В Малой Азии и Закавказье среди сорных и культурных озимых овсов иногда встречаются формы с переходными чер- тами к А. sativa, А. fatua и А. Iudoviciana, а также переходные формы между A. sativa и A. byzantina (Малая Азия). Просо. Род Pan icum L. имеет древнее происхождение. Р. milia~ceum L. найден в грузинском неолите. В Армении просо появилось позднее. В Передней Азии, по классификации В. Н. Лысова, просо представлено особой переднеазиатской эко- лого-географической группой (см. стр. 150). Чумиза. Вид чумизы — Setaria italica (L.) Р. В. — появился в Грузии позднее, чем в 10жной Европе (по исследованиям В. Л. Менабде — в XV в.). В. Л. Менабде выделил сорную грузинскую чумизу в само- стоятельный вид S. ketzi<hovelii Menabde et Eritz. и высказал предположение, что грузинская культурная чумиза могла про- изойти от этого вида. П. М. Жуковский придерживается проти- воположной точки зрения и считает, что сорный вид здесь про- изошел, подобно фатуоидам, от культурного. Недостаточно ясно, насколько грузинская чумиза отличается от других географических форм чумизы; это прежде всего надо установить и, кроме того, необходимо провести детальное срав- нительное изучение форм сорной чумизы и ее распространения в других странах. Такие сведения пока отсутствуют. В Закавказье, и в особенности в Грузии, имеются ценные местные сорта этой культуры, интересные как исходный мате- риал для селекции (Л. Яекапрелевич, 1960). Кукуруза. Кукуруза — Zea mays L. — начала возделываться в Грузии во второй половине XVI в. или, может быть, позднее— в XVII в. Предполагают, что кукуруза попала в Грузию из страны лазов (Чанети) . Первое достоверное свидетельство о культуре кукурузы в Грузии имеется в «Географии Грузии» Вахушти (который жил до 1724 г.). В то время кукуруза была уже широко распространенным культурным растением в Грузии. Из Грузии кукуруза проникла в Азербайджан в конце XVIII в., но распространялась там медленно, как и в Армении, где в начале XIX в. была еще малоизвестна. Ее тогда сеяли только на огородах как овощную культуру (употреблялись по- чатки в вареном виде) . Армянский низкорослый местный сорт кукурузы близок к персидскому типу и мог быть занесен в Армению из Ирана.
Ежа. Род Dactylis L. представлен на Кавказе двумя видами. j~j3 них древнейший и родоначальный 14-хромосомный вид 13. aschersoniana Graebn. обнаружен нами на Северном Кав- казе (окрестности Майкопа, Бештау) и в Закавказье (bop- жоми). При выселении из-под полога леса образуется произ- водный 28-хромосомный более молодой вид D. glomerata L., который на кавказе представлен огромным многообразием эко- логических разностей и морфологических форм. Важнейшие из них: 1) экотип лесных полян Северного Кавказа (здесь и в на- стоящем совершается переход от D. aschersoniana к D. glome- га1а — пример скачкообразного видообразования, вызванного внезапной сменой среды); 2) экотип пониженных местообитаний лесной зоны; 3) группа экотипов субальпийской зоны; 4) армян- ский экотип степных местообитаний и др. Ареал D. aschersoniana большой, хотя и разорванный, и образование D. glornerata идет во многих областях, где имеются широколиственные леса с полянами. Однако Кавказ является од- ним из крупных очагов развития вида D. glomerata. Костер. Из рода Bromus L. на Кавказе наиболее широко рас- пространен костер береговой — В. riparius Rehm., представлен- ный здесь очень ценными в кормовом отношении экотипами, из которых вводятся в культуру и более других изучены следую- щие: 1) шунтукский предгорный; 2) нагорностепной из восточ- ных районов Краснодарского края (по реке Урупу), по-види- мому, сходен с шунтукским, но отличается большим полимор- физмом; 5) учкуланский — с поливных сенокосов по рекам Кубани, Учкулану и Уллукану. По всему Кавказу в субальпийской и альпийской зонах ши- роко распространен близкий вид — В. variegatus М. В., связан- ный многими переходными формами с В. riparius. Овсяница. Овсяница луговая — Festuca pratensis L. — харак- теризуется огромным богатством форм и представляет большой интерес как исходный материал для окультуривания в селекции. Испытывались в культуре и оказались практически интерес- нымп армянский нагорный экотип (Лори) и предгорностепные экотипы Северного Кавказа. Кавказ — область интенсивного формообразования овсяницы луговой. Тимофеевка. Тимофеевка луговая — Phleum pratense 1 .— также представлена рядом интересных для селекции форм. Из них введены в культуру: 1) нагорно-армянский экотип (лорий- скпй); 2) нагорно-грузинский (бакурианский); 3) предгорные экотипы северо-западного Кавказа. Вопреки установившемуся мнению мы считаем, что область развития, а возможно и происхождения, вида Ph. tpratense на- ходится не в лесной зоне Европы, а в горных странах Восточ- ного Средиземноморья, в особенности в Передней Азии. Райграс. Райграс высокий — Arrhenatherum elatius М. et К.— на Кавказе представлен ценными кормовыми формами, которые 100
вводятся в культуру. Из них отметим: 1) райграс высокий на- торной Армении (лорийский); 2) райграс высокий северо-запад- ного Кавказа (Лагонаки). Особенно обильно распространен райграс многолетний пастбищный — Lolium регеппе L., на Северном Кавказе (во- сточные районы Краснодарского края и Ставропольский край). Он еще мало изучен. В окрестностях р. Шунтук растет его слабо остистая форма. �итняк. Род Agropyron Gaertn. представлен на Кавказе ря- дом эндемичных форм, которые возводятся в ранг видов часто с недостаточным основанием, но представляют значительный теоретический и практический интерес. Житняк ширококолосый — А. cristatum (L.) Gaertn. — o6pa- зует большие массивы в Талинском районе Армении. Яля За- кавказья (Арзни, окрестности Севана) А. А. Гроссгеймом вы- делен эндемичный вид А. dagnae Grossh. В Армении на горных лугах этим же автором найден эндемичный вид житняка А. pu- berulum (Boiss.) Prok., обнаруживающий родство с А. ipinifolium Nevsl<i (значительные массивы в окрестностях Геленджика и Новороссийска) и очень похожий на крымский житняк— А. Iponticum Nevski. В Закавказье обнаружены 14, 28 и 42-хромосомные формы ширококолосых житняков, еще слабо дифференцированные и детально слабо изученные. Молиния. Молиния прибрежная — Мо11п1а litoralis Host— растет дико в Западной Грузии (в Колхиде) и используется как кровельный материал. Раньше существовала даже культура этого растения (А. K. Макашвили, 1951). Горох. Род Pisum L. происходит из Передней Азии. Скальные многолетние и однолетние виды Pisum не имеют прямого отношения к происхождению культурного гороха. Эти виды (Р. formosum Boiss., Р. au~cheri Jaub. et Spaich, Р !>umile B~oiss. et Noe и Р. fulvum Sibth. et Sm.) имеют переднеазиатские ареалы. Наиболее близкий родич культурного гороха — Р. ela- tius (М. В.) Stev., хотя и распространен шире — в области gpea- него Средиземноморья от Индии до Италии и по северному побе- ре>кью Африки, но определенную конституцию дикорастущего растения сохраняет только в странах Передней Азии — в Закав- казье, Малой Азии. Сирии и Палестине. Закавказье, несомненно, представляет собой первичный очаг вхождения гороха в культуру. Здесь встречаются в дикорасту- щем состоянии формы с признаками культурных растений и обратно — в культуре имеются формы, по совокупности призна- ков близкие к дикарям. К таковым относятся ахалкалакский го- рох, в разной пропорции засоряющий вику (Vicia sativa), иногда в таком количестве, что создается впечатление совмест- ной культуры. Эта форма гороха (вар. transicau~casicum Gov.) имитирует вику по величине зерна и форме листьев. Вероятно,
на первык этапах своей истории «ультурный горох существовал совместно с викой, и виколистная его форма является релик- товой. Мы представляем себе развитие рода Pisum согласно прила- гаемой схеме. 1(омплекс признаков Р агъепзе Е. представляет собой из- вестный исторический этап в развитии культурных видов, и рез- кой границы с типичным Pisum sativum здесь нет. В настоящее время комплекс пелюшки (Pisum arvense) г. основном распространен в Передней, Средней, Юго-Западной Азии и Северной Африке (ssp. asiati~cum Gov.) . .Но и среди типичного посевного гороха Pisum sativum sensu stricto имеются мелкосемянные формы с пигментированными семенами, т. е с частью признаков Р arvense. Эти формы более примитивны„ чем самые молодые светлосемянные и крупносемянные формы типичного Р. sa ti vuln. В Передней Азии имеются формы, переходные от азиатских пелюшек (ssp. asiaiicllrn.) к типичным формам посевного го- роха — Р sativuln ssp. oomlnune proles огеоапа1о11сит Gov. (Восточная Анатолия). В Сирии распространен горох, близкшг к азиатским пелюшкам — Р. sativum sap. asiaticum proles syria- сит Gov. Нут. Род Cicer L. имеет основную область развития и проис- хождения в Передней Азии. Из 28 видов нута в странах Перед- ней Азии (Малая Азия, Сирия, Палестина, Месопотамия, За- кавказье и Иран) распространено 12. 1(ультурный нут — С1сег arietinum L. — рассматривается мо- нографом рода М. Г Поповым в подроде Pseudononis секции Nonocicer. I( этой секции, кроме Ci~cer arietinum, относятся два псреднеазиатскпх II очень близких к нему дикорастущих вида: С. pinnatifidum ЛапЬ. et Sipach и С. judaicum Boiss., а также один вид, эндемичный для Эфиопии,— С. cuneatum I-lochsi. По- следний близок и к С, arietinum и, вероятно, происходит также из Передней Азии. I(aKIIe-то формы секции Monoclicer очень давно распространились в горы, которые начинаются в Сирии и Палестине, а затем переходят в Эфиопию. Из этих занесен- ных переднеазиатских форм под влиянием долгого воздействия иных условий выработался эндемичный вид Восточной Аф- рики — С. cuneatum. С. pinnatifidum — ближайший родич культурного нута, очень сходен с последним и распространен в странах Передней Азии: Малой Азии, Месопотамии, Сирии и Палестины. Отсюда же ведет происхождение и культурный нут С. arietinl.'ò. В Передней Азии распространены следующие эндемичные группы (пролесы культурного нута) по классификации Г М. По- H0B0H. 1. Восточный подвид — ssp. orientale G. Рор. включает Иран- скую группу — proles iranicum G. Рор. Эта переднеазиатская 102
Предполагаемая схема развития рода Pi'sum i.. Вымерший мезофильный многолетний дикорастущий предок Промежуточные звенья Pisum elatius (мезофильный) Многолетниескальные виды Однолетниескальные виды Ксерофильный (Pisum elatius) Сорный виколистный (ssp. transcaucasicum G0v.) Горох в чистойкультуре Pisum abyssinicumBraun. Горох с комплексом признаков пелюшки (Pisum arvense ssp. asiaticum Gov.) Горох с комплексомпризнаков Pisum sati-vum ssp. соп1пыпеGov. (семена мелкие,пигментированные) Горох ssp. commune Gov. светлосемянные и крупносемянные формы ветвь распространена в Азербайджане, Нахичеване п Северо-За- падном Иране — примитивная темносеменная форма. 2. Евразиатский подвид — ssp. eurasiaticum G. Рор.; вклю- чает Палестинскую группу — proles palestirticum G. Рор.; ipac- пространена в Сирии, Палестине, Марокко; Турецкую — proles tuvci~cum G. Polyp., распространена в Малой Азии; Закавказ- скую — proles trartscaucasicum G. Рор. — в Армении, Азербай- джане, на Северном Кавказе (на Украине распространилась позднее) . Надо отметить, что подвиды в эгой классификации слишком сборные, а пролесы скорее соответствуют хорошим подвидам. Вика. Род Vicia L. древними корнями уходит в лесную флору Восточной Азии, но в Древнесредиземноморской области, в том числе в странах Передней Азии, имеет центр интенсивного вто- ричного развития. 103
Так называемая французская чечевица — V ervilia Willd.— представляет собой древнюю культуру Закавказья (Ахалка- лаки), которая теперь исчезает и становится реликтовой. Она возделывается местами в Малой Азии, Сирии и Палестине, а также в странах Средиземноморья и Юго-Западной Азии В диком состоянии V ervilia распространена в Закавказье, Ма- лой Азии, Сирии, Иране, Средней Азии, на Балканах и в Испа- нии. Более близкие к дикорастущим культурные формы V ervi- lia (grex mediterraneae Bar.) — карликовые, скороспелые, мелко- цветковые, распространены в Сирии, Палестине, Малой Азии и других странах Восточного Средиземноморья (Греция, 1(рит) „ заходят и в Испанию. В Передней Азии, несомненно, находится первичный очаг вхождения в культуру V. ervilia, но, вероятно„ введение в культуру имело место и в других странах, где рас- пространены дикорастущие формы этого вида. Вика паннонская — V ipannonica Crantz — представляет собой также редкую (реликтовую) культуру, островки которой сохранились еще в Закавказье (Ахалкалаки) и которая в сорном состоянии распространена во многих районах 1(авказа. Гамс считает родиной этой культуры ПаннонЕю (Венгрия), но, судя по ареалам близких видов, богатству форм в Закав- казье, родина самого вида и первичный очаг культуры нахо- дится в Армении, 1(урдистане, Северо-Западном Иране. Вика посевная — V sativa L. — широко распространена в странах Передней Азии в сорном, одичалом и, по-видимому, в диком состоянии и представлена эндемичными культурными формами, детально изученными А. Ю. Тупиковой (результаты исследований опубликованы пока неполностью). дикорастущие формы, наиболее близкие к V. sativa, обнаружены А. Ю. Тупи- ковой в Туркменистане, который включается в число стран Пе- редней Азии. Вика мохнатая — V villosa Roth. — также широко распро- странена в сорном и дикорастущем состоянии на 1(авказе и в дру- гих странах Передней Азии. Самые зимостойкие формы этой вики распространены на Северном 1(авказе. А. Ю. Тупиковой (1960) выявлено богатое разнообразие эко- типов и большой внутриэкотипический полиморфизм у сорных видов вики посевной, паннонско~~, узколистной, мохнатой. Инте- ресно, что здесь обнаружены полуозимые формы посевной вик;1 и формы вики узколистной. Родоначальный вид V. angustifolia L. дает ряд переходных форм к вике посевной (V sativa). По-видимому, здесь вика посевная местами распространена не только в сорном состоянии, но представлена на лугах настоящими дикорастущими фор- мами. Многолетние виды представлены в Передней Азии огром- ным разнообразием еще слабо изученных видов и форм. Из них сейчас вводится в культуру вика изменчивая — V variabilis Freyn et Sint. 104
Конские бобы. Faba vulgaris Moench имеют, по-видимому, по- литопное вхождение в культуру в разных местах Древне- средиземноморской области, на обоих противоположных концах которой имеются реликтовые формы, очевидно близкие к родо- начальникам вида и первым культурным формам: F. vulgaris pliniana Trabut (в Алжире) и F. vulgaris ssp. ~paucijuga Nurat. (в Индии). Во многих странах встречаются примитивные мелко- и темно- семянные формы, которые, судя по раскопкам, некогда были везде распространены. Крупносемянные формы возникли позд- нее, разновременно в отдельных странах. В странах Передней Азии находится один из первичных оча- гов культуры конских бобов. Там имеются эндемичные группы форм: очень примитивные мелкосемянные дагестанская и сва- нетская, раннеспелые палестинская и аравийская (Иемен, Эфио- пия). Анализ этой культуры затрудняется обособленностью эко- логической классификации от таксономической (В. С. Мура- това). Чечевица. В странах Передней Азии распространены все дико- растущие виды рода чечевицы Lens Adans.: L. kotsichyana Alef., эндемичный для Месопотамии; L. 1~епИси1а Л1е1., в странах Пе- редней Азии и собственно Средиземноморья; 1.. nigriicans Gedr., в странах Передней Азии и собственно Средиземноморье, 1. orientalis Hand.-Nazz., в странах Передней Азии, а также в Средней Азии и Афганистане. Последний вид сходен с куль- турной чечевицей. Передняя Азия, таким образом, является областью концентрации видов рода Lens n, вероятно, областью erp происхождения. Наиболее близкий к культурной чечевице вид L. lenticula имеет восточносредиземноморский ареал и в Юго-Западную Азию не заходит — далеко на восток в Азии заходит только более далекий вид L. nigrilcans. Уже по одному этому можно предполагать, что область происхождения и пер- вичный очаг культуры чечевицы находится в Передней Азии, а не в Юго-Западной Азии. Е. И. Барулина в своей монографии (1930) все же делает вывод о том, что наибольшим эндемизмом отличается Юго-Западная Азия. Общее количество мелкосемянных разновидностей наиболь- шее и почти одинаковое — в Передней Азии и в Средиземно- морье, но эндемичных — больше в Передней Азии. Крупносемян- ных разновидностей больше в Средиземноморье, чем в Перед- ней Азии, а в Юго-Западной Азии их совсем нет (табл. 2). Эндемичных мелко семянных разновидностей больше всего в Юго-Западной Азии. Но с другой стороны в Юго-Западной Азии (см. выше) наблюдается большее экологическое разнооб- разие (Средняя Азия), и встречаются очень редкие эндемич- ные культурные формы с признаками дикорастущих форм (юго- восточный Афганистан, Индия), приближающиеся к абиссин- ским формам. Последнее можно объяснить предположением, что
Таблица 2 Распределение разновидностей культурной чечевицы по географическим областям Число мелко- семянных разновид- ностей Число крупно- семяннык разновид- ностей В том числеэндемич-нык В том числеэндемич-ных Область Передняя Азия (Закавказье, Малая Азия, Сирия, Палестина) . Собственно Средиземноморье, За- падная Европа, Эфиопия Юго-Западная Азия, Индия, 1(ашмир Эфиопия, Эритрея 46 44172 105 в Юго-Западной Азии (Афганистан, Индия) находится также первичный очаг развития вида Lens esculenta и вхождения его в культуру. Чтобы объяснить возможность существования этого второго первичного очага, надо допустить, что в прошлом ареал дикорастущего родоначальника (или родоначальников) L. escu- lenta был продвинут далее на восток, чем это имеет место в на- стоящее время. Возможно и другое предположение: вследствие примитивности земледелия в странах Юго-Западной Азии со- хранились примитивные культурные формы чечевицы, а раз- общенность здесь мелких земледельческих очагов привела к возникновению эндемичных примитивных форм с небольшим ареалом. В Эфиопию чечевица, вероятно, занесена пз Ин- ДИИ. UeHTp развития крупносемянных форм находится в Среди- земноморье п, главным образом, западном (Пиренейский полу- остров), но вся средиземноморская и европейская чечевица вышла из Передней Азии, а не из Юго-Западной Азии. В Юго- Западной Азии сохранились редкие, обособленные в пределах узких ареалов формы, которые не получили далекого распро- странения и не повлияли на состав европейско-средиземномор- ской чечевицы. Как видно из археологических данных, в глубо- кой древности всюду была распространена только мелкосемян- ная группа, а позднее в благоприятных условиях агросреды из нее вырабатывались среднесемянные и крупносемянные формы. Чина. Lathyrus sativus L. представляет собой вид с древне- средиземноморским культурным ареалом — от южной Европы до Индии. Археологические находки показывают, что в древ- ности (неолит) культивировалась только мелкосемянная чина. По данным Ф. Л. Залкинд, в настоящее время морфологическое и экологическое разнообразие мелкосемянных форм сосредото- чено в странах Юго-Западной Азии (Памир, Афганистан, северо- западная Индия, Иран). В этих странах совсем не имеется бело- цветковых и крупносемянных форм.
В Передней Азии сосредоточены, главным образом, промежу- точные по величине семян формы, но по окраске цветка и се- мени близкие к формам Юго-Западной Азии. Формы, похожие на переднеазиатские, встре1аются и в удаленных от Передней Азии странах (Испания, Северная Африка). Страны Западного Средиземноморья являюгся средоточием светлосемянных и круп- носемянных форм. Впечатление получается такое, что переднеазиатские и юго- западноазиатские чины — это как бы единый, более древний массив, чем чины Западного Средиземноморья. Но средне-юго- западноазиатские чины остановились в своем развитии — чело- гек над ними слабо работал, и они широко не распространялись, поэтому на востоке ареала образовался ряд обособленных очень своеобразных эндемичных форм. Из стран же Передней Азии чина распространилась далеко на запад. Издревле везде была (и на западе) только мелкосемянная и темноокрашенная чина, а позднее начали из них вырабатываться среднесемянные и крупносемянные формы — более молодые. Этот процесс далее всего пошел на западе культурного ареала, где формированию крупносемянных и светлоцветковых форм содействовали и климат, и более высокая культура земледелия. Расселение чины по Передней Азии на запад началось чрез- вычайно давно. Мелкосемянная и темноокрашенная чина из- вестна из слоев каменного века Венгрии (Lengyel и Aggtelelc). Ясно, что эта древняя чина не была заимствована из Среди- земноморья, а пришла в древности из стран Передней Азии через Балканы. История эта очень похожа на историю чечевицы и конских бобов, отчасти нута, вики и гороха. Все эти культуры распро- странились на север (на Русскую равнину) и на запад в Среди- земноморье и в страны Средней Европы в виде мелкосемянных и темноокрашенных форм, а позднее из них на западе вырабо- тались крупносемянные светлоцветковые формы. Это сходство в истории бобовых культур заставляет думать, что все они в глубокой древности распространились из Передней Азии на запад, и их распространение было, вероятно, более или менее синхронным. Однако между отдельными бобовыми культурами суще- ствуют и различия. Вика посевная не заходит так далеко на восток в страны Юго-Западной Азии, как остальные культуры. Яля конских бобов не имеется вполне достоверного дикора- стущего родоначальника. Наиболее близкая дикорастущая форма Faba pliniana находится на западе ареала, а на восточ- ном конце его имеются формы, близкие по конституции к дико- растущим (ssp. paucijuga и др.). Возможно, что вхождение в культуру конских бобов было политопным, в нескольких ме- стах древнесредиземноморского ареала, в том числе, вероятно, и в Передней Азии. В сорном состоянии бобы пока неизвестны. 107
У гороха, чечевицы, конских бобов и чины существование на восточном конце ареала в Юго-Западной Азии примитивных, близких к дикарям форм можно также объяснить и более широ- ким в древности, чем в настоящее время, распространением на восток их диких родоначальников (особенно для чечевицы, у которой дикий родоначальник Lens orientalis и теперь распро- странен в Средней Азии). Дикий родоначальник чины неизве- стен — он вымер. И совершенно неизвестно, имел ли он передне- азиатский ареал или распространился и далее на восток, а мо- жет быть, достиг восточного конца ареала не в дикорастущем, а в сорном состоянии. Чина как сорное растение широко распространена в Афга- нистане. Другой вид чины из той же секции чина нутовая — Lathyrus- cicera L. — имеется в культуре в более или менее значительных размерах только в Сибири, где употребляется на корм скоту п в пищу как зерновое растение. Этот вид чины дикой встречается на Кавказе, в (рыму и в Средней Азии. Вероятно, этот вид чины вошел в культуру в Передней Азии (Сирии?). Из кормовых видов чины заслуживает внимания в целях вве- дения в культуру эндемичная для 1(авказа чина киноваревая— L. miniatus М. В. Люцерна. Род Medicago L., возможно, и в целом имеет перед- неазиатское происхождение, что совершенно не вызывает сомнения в отношении подрода Falcago Grossh. — многолетних люцерн, который следовало бы, по существу, считать самостоя- тельным родом. Переднеазиатское происхождение имеют следующие виды.- 1) люцерна кавказская — М. praesativa Sinsk. — Кавказ, Малая Азия; 2) люцерна скальная — М. rupestris М. В. — Крым, Кав- каз; 3) люцерна клейкая — М. glutinosa sensu lato с включе- н«ем М. vires~cens Grossh., М. polychroa Grossh. и М. vardanis Vass. — область Главного I(авказского хребта, Дагестан; 4) лю- церна полуциклическая — М. hemicycla Grossh., полиморфный вид с несколькими подвидами — горные плато Армении, Грузии и Дагестана с достаточным увлажнением; 5) люцерна голубая— М. соеги1еа 1.ess. с несколькими подвидами — прикаспийские районы Дагестана и Азербайджана, Восточная Грузия; 6) лю- церна джавахетская — М. dzhawakhetica Bordz. — Закавказье Ахалкалакский район; 7) люцерна реснитчатая — М. ipapillosa Bioiss. — Малая Азия; 81 люцерна щебнистая — М. saxatilis M. В. — Каурым; 9) люцерна решетчатая — М. cancellata М. В.— Северный 1(авказ и юг европейской части СССР; 10) люцерна серпообразная — М. quasifal~cata Sinsk. — северо-западный 1(ав- каз; 11) люцерна диплоидная — М. difalcata Sinsk. ssp. daghe- stani~ca Sinsk.; по новейшим исследованиям эта форма оказа- лась самостоятельным видом, из которого следует исключить ssp. kazaichstani~ca Sinsk.; Дагестан; 12) люцерна железистая— 108
И. glandulosa David. — Причерноморье, окрестности Новорос- сийска и Геленджика, 1(рым. Характерно, что именно в странах Передней Азии (главным образом 1(авказ) распространены более древние диплоидные виды люцерны, судя по нашим наблюдениям последних лет, они еще и до сих пор не все выявлены. Значение Передней Азии, как важнейшей области видообра- зования люцерны, подчеркивает существование в ней древнего мезофильного интегрального вида — М. hemicycla, который об- наруживает переходные ряды ко многим другим родственным видам, объединяя в себе некоторые их черты. Из культурных видов люцерны для Передней Азии энде- мичны следующие: 1) эндемичная армянская люцерна — М. prae- sativa ss~p. armena Sinsl<. — Древняя Армения; 2) люцерна во- сточная — М. orientalis Vass. — Малая Азия; 3) люцерна сирий- ско-палестинская — -М. syria|co-palestinica Sinsl<. — Сирия и Па- лестина 4) люцерна йеменская — М. jemenensis Sinsk.— Иемен. Пажитник. Для северо-западного 1(авказа и Крыма эндемпчен пажитник меловой — Trigonella ~cretacea Gr~ossh., реликтовый вид, представляющий связующее звено между радами Tr! go- пеllа и Medicago. В Передней Азии имеется два эндемичных вида культурного пажитника: 1) пажитник йеменский — Т. jemenensis Sinsl<.— Иемен, 2) пажитник арабский — Т. агаЫса РеНе (возделы- вается кое-где в Палестине) Обыкновенный сенной культурный пажитник — Т. foenum- graecum L. — в Передней Азии представлен рядом передне- азиатских пажитников — series anatolicae Sinsk. В этот ряд вхо- дят два экотипа: 1) экотип Армении и Азербайджана, 2) экотип Малой Азии. Предполагаемый предок сенного пажитника — па- житник мечевидный — T. С1а<11а1а Stev. растет дико в Передней Азии и Средиземноморье. Пажитник гслубой — Т. соеги1еа (Desr.) Ser. растет в диком состоянии в Закавказье, в 1(рыму и на Северном 1(авказе. На Кавказе он поднимается в горы до 2000 м. Оттуда он занесен в центрально-черноземную полосу европейской части РСФСР II в Поволжье. Пажитник голубой считается большинством авто- ров культурным или одичалым растением, а за исходную дико- растущую форму последнего принимается распростертый пажит- ник — Т. pro~cumbens (Bess.) Rchb. Последний распространен E Передней Азии, на Балканах, в Восточной Сибири, Средней Европе и Средиземноморье. Пажитник голубой — древнее куль- турное растение (его прибавляют для запаха в хлеб и сыр). Вопрос о родине пажитника голубого еще недостаточно изучен. Донник. Донник желтый — Melilotus off icinalis Desr., по В. В. Суворову, ведет происхождение с 1(авказа и оттуда рас- пространился на север и на запад. На 1(авказе распространены два подвида М. officinalis: горный — ssp. montanus Suv. со 109
скальным, пастбищегорным и предгорным экотипом и 2) восточ- назакавказский — ssp. transcaucasicus Suv. (в Куринской низмен- п~>сти) . Лонник индийский — М. indicus Аll. — переднеазиатского происхождения. Встречается в Крыму, Туркмении и изредка на Северном Кавказе. Эспарцет. Весь род Onobrychis Adans. имеет переднеазиатское происхождение. Это же, в частности, относится и к подсекции Vulgatae Hand.-Макк., секции Eubrychis DC, к которой отно- сцгся большинство ценных в кормовом отношении видов. Передняя Азия является также и наиболее древним очагом ьхождения эспарцета в культуру. Переднеазиатскпе виды многолетнего эспарцета: . эспарцет высокий — О. altissima Grossh. — и эспарцет закавказский — О. transcaucasica Grossh.— стали разводиться на корм населением Закавказья и Ирана го- раздо раньше, чем начали сеять эспарцет в Западной Европе. В настоящее время в культуре еще встречаются оба эти вида в почти неизмененном состоянии, но в то же время, по мнению А. Г Хпнчук, формируется новый впд О. antasiatica Chintschul'. Этот молодой, создающийся уже всецело в условиях культуры вид конвергентного происхождения не вполне установился, хотя уже и имеет ядро популяций определенного состава. В. И. Андрукович не без основания считает, что О. antasia- tica — вид искусственный, и в Закавказье существует один круп- ный и очень полиморфный вид О. transcaucasica Grossh., а дико- растущий О. altissima Grossh. и культурный О. antasiati~ca Chintschuk входят в состав этого крупного вида. Такой взгляд подтверждает нашу мысль о значении крупных интегральных видов в первичных очагах происхождения куль- турных растений. Все кавказские ботаники согласны с тем, что в Закавказье находится вполне самостоятельный очаг вхождения в культуру эспарцета и этот очаг — более древний, чем западноевропейский. В подтверждение этого положения приводят многочисленные местные названия эспарцета: корнган, коринган, корингя, кулин- гар (Армения); куринга, хаша (Азербайджан); эсперес (Иран). Несомненно, что и многие другие закавказские виды эспар- цета ценны в кормовом отношении и заслуживают испытания в культуре и использования в селекционной работе. Это отно- сится прежде всего и эспарцетам секции Eubrychis, но не исклю- чена возможность, что и среди представителей других секций будут выявлены ценные виды. Степаняном произведены удачные опыты по введению в куль- туру эндемичного для Грузии засухоустойчивого эспарцета мес- хетского — О. meschetica Grossh. Клевер. Передняя Азия и раньше (А. Эйг) признавалась роди- ной клевера александрийского (берсима) — Trifolium alexandri- пит L. — и конкретно происхождение этого культурного вида 310
клевера велось из Палестины. Открытие Е. Г Бобровым более близкого, чем палестинские дикорастущие виды, родича клевера александрийского — Т. apertum Bobr. заставляет изменить преж- нюю точку зрения. Т. apertum — мезофит колхидской флоры. вероятно, является предком всей группы более ксерофильных клеверов, родственных берсиму. Клевер красный — Т. pratense L. sensu lato — образует боль- шие массивы в разных районах Кавказа. Существует ряд эко- типов дикорастущего клевера красного. Из них интересен эко- тип влажных нагорий Армении (Лорийское плато), отличаю- щийся очень высокой фертильностью. В пределах северо-запад- ного Кавказа и районов Кабарды с достаточным увлажнением сформировался особый местный сорт культурного двуукосного клевера (шунтукский красный). Представляют интерес дико- растущие клевера восточных районов Краснодарского края. Коровий горох. Коровий горох (или лобия китайская)— Vigna sinensis Endl. в странах Передней Азии, по А. М. Павло- вой, представлен следующими эндемичными агроэкологическпми группами: 1) азербайджанская — proles azerbaijanica А. Pavl.— Азербайджан, Армения, Средняя Азия; 2) закавказская — pro- les transcaucasica А. Pavl.— Абхазия, Азербайджан, Средняя Азия. В Закавказье распространены некоторые формы, которые Павловой отнесены к азиатской группе — proles asiatica А. Pavl. — Закавказье, Средняя Азия, Китай. Возможно, что коровий горох попал в Переднюю Азию из Китая (или Индии) через Среднюю Азию. Лен. В Передней Азии расположен один из трех мировых первичных очагов культурного льна, где имело место независи- мое вхождение в культуру (первый очаг — Индия, второй- Юго-Западная Азия, Афганистан и прилегающая к ним горная Индия, третий — Передняя Азия — Колхида). Наиболее древней реликтовой формой переднеазиатских льнов является озимью, склонный к простратности лен, который раньше имел широ- кое распространение, а теперь сохранился в культуре в Малой Азии и кое-где в Закавказье. Схема эволюции переднеазиатскпх льнов дана нами выше. Генетические исследования А. Q. Горгидзе (1955), проведен- ные в Институте ботаники АН Грузинской ССР под руковод- ством В. Л. Менабде, подтверждают нашу точку зрения. Колхидский реликтовый культурный лен оказался генети- чески очень близким к местным формам дикорастущего льна— L. angust~ifolium — и очень легко скрещивается с последним. Среди культурных форм льна по наиболее легкой скрещп- ваемости (90 — 100'/о удачных скрещиваний и высокая всхожесть гибридных семян) выделились именно переднеазиатские формы: сорта льна из Малой Азии и особенно горные грузинские куд- ряши. 111
Эти данные с убедительностью говорят о том, что колхид- ский стелющийся лен является родоначальным типом для всех других форм культурного льна, распространенных в Закавказье и вообще в Передней Азии (разумеется, мы говорим об энде- мичных местных, а не о завезенных сортах льна). Конопля. Местная культурная конопля — Cannabis sativa L.— представлена в Передней Азии тремя эндемичными типами (Т. Я. Серебрякова), не уступающими по ценности иностранной высокорослой южной конопле: 1) кавказским (Северный 1(ав- каз, Грузия, Армения); 2) кубинским (предгорные районы Азер- байджана); 3) сирийским (Сирия, Палестина, Малая Азия). Во многих районах Передней Азии распространена также и дикорастущая конопля. Культура масличных растений в Закавказье. теперь утратила былое значение и сильно сократилась в размерах. Многие мас- личные растения переходят в разряд реликтовых (масличный лен, ляллеманция, сурепица, рыжик). Кунжут. Древней масличной культурой является кунжут- '1езатит indicum L., который также постепенно теряет свое значение в Закавказье. Для Передней Азии В. М. Гильтебрандтом установлены сле- дующие хорошо различающиеся между собой региональные эко- типы: 1) закавказский (главным образом 1(уринская низмен- ность); 2) малоазиатский — из Баликессира; 3) малоазиат- ский — из Айдина; 4) малоазиатский — из Мерсины; 5) мало- азиатский — из Адалии и Денизли. Ляллеманция. Ляллеманция — Lallemantia iberica Fisch. et Меу. — в прежние времена возделывалась в Советской Армении на масло, которое шло в пищу и на освещение. В наше время культура ляллеманцин «ожила» и это растение опять «вошло в культуру» в Краснодарском крае ради очень ценного техни- ',åñê0ã0 масла с высоким иодным числом. В дикорастущем со- стоянии L. iberica нами нигде не обнаружена. Вероятно, этот вид возник в условиях культуры в Армении из какого-то дико- растущего родственного вида, а из какого именно — пока еще трудно определить. Таких близких видов имеется три: 1) лялле- манция седоватая — 1.. canescens Fisch. et Меу. — многолетний вид, редко встречающийся в горах Армении и Аджарии; 2) вновь открытый озимый скальный вид ляллеманция скзльная— L. rupestris Sinsk. et Voskr. — Армения, Азербайджан; 3) лялле- манция щитковидная — L. ~peltata Fisch. et Меу. — скальный и сорный вид, также озимый, но с меньшей степенью озимости, чем L. ruipestris — Армения, Азербайджан, Грузия. Ляллеманция скальная — L. rupestris — до известной сте- пени соединяет в себе черты L. ре11а1а (озимость, признаки се- мян) и L. iberica (ближе по форме прицветника). Этот вид и L. canescens являются наиболее древними видами и, возможно, родоначальными для двух остальных. 112
Для культуры представляет интерес не только L. iberica, но и 1 ..peltata и L. ru~pestris. Весь род Lallemantia Fisch. et Меу. переднеазиатского про- исхождения, как и культурный вид 1. iberica, и здесь же нахо- дится первичный очаг вхождения ляллеманции в культуру. Рыжик. Род Camelina Crantz также переднеазиатский. Куль- турный рыжик в Закавказье представляет собой эндемичный вид С. caucasica Sinsk., выделившийся из сорняков местного льна, этот рыжик стал уже культурным реликтом. Сурепица. Сурепица (Brassica campestris L.) — исчезающая культура, еще недавно ее сеяли в Ахалкалакском районе Грузии п кое-где в Армении. Как сорняк сурепица и теперь еще широко распространена. Для Передней Азии эндемичны следующие подвиды (ранее описанные нами как разновидности) яровой су- репицы: 1) В. camipestris ssp. ~саисаз1са Sinsk. — Грузия, Арме- ния, Малая Азия — сорняк, в культуре раньше возделывалась и в Закавказье; 2) В. campestris ssp. aserbaidjanica Sinsk.— сор- няк, Азербайджан. Горчица. Горчица черная — Brassica nigra Косй — представ- лена в странах Передней Азии и более примитивными формами (с мелкими, легко осыпающимися семенами, позднеспелая, с выраженной розеткой прикорневых листьев) и более окульту- ренными (скороспелыми, крупносемянными, с невыраженной при- корневой розеткой листьев) . Сюда относятся следующие под- виды (раньше описанные как разновидности): 1) В. nigra ssp. orientalis Sinsk. — культурная форма (Малая Азия); 2) В. nigra ssp. rigida Sinsk. — культурная форма (Сирия и Палестина); 3) В. nigra ssp. ipseudocampestris Sinsk. — сорная форма в по- севах сурепицы и имитирующая последнюю по ряду признаков (Малая Азия). Возможно, что Передняя Азия является первичным очагом культурной горчицы черной и родиной вида В. nigra. Однако решение этого вопроса затрудняется тем, что дикорастущий родоначальник горчицы черной пока не известен. Горчица белая — Sinaipis alba L. — представлена в Передней Азии особым экотипом — var. orientalis Sinsk. (Палестина). Индау. Индау — Егиса sativa Lam. — не так давно в неболь- ших размерах культивировался в Армении. В 1925 — 1926 гг. Е. А. Столетова встречала чистые посевы индау в Ленинакан- ском районе. Теперь это растение встречается изредка и только и сорном состоянии. Здесь, как и в Средиземноморье, эта куль- тура пользовалась в древности почетом, а теперь исчезла. Для Палестины описана эндемичная разновидность (скорее подвид) индау — Е. sativa var. orientalis Sinsk. Подсолнечник. Вид Helianthus annuus L. представлен в Арме- нии очень крупносемянными эндемичными грызовыми сортами, которые в настоящее время встречаются редко. Эти формы, возможно, являются самостоятельным видом.
Белямир. Вид Cephalaria syriaca Schrad. является эндемичной культурой Передней Азии (Малая Азия и Закавказье) . Упо- требляется как приправа к тесту и как масличное в Малой Азии. Как сорняк заходит в Среднеазиатские республики, Афга- нистан и Средиземноморские страны. В странах Передней Азии. имеются эндемичные формы этого вида. Здесь, по нашему мне- нию, имело место вхождение в культуру этого вида и здесь же- его родина. Мак. Вид Papaver somniferum L. представлен в Малой Азии эндемичными высокомо])фийными сортами, которые составляют подвид ssp. anatolicum М. Vesselov., заходящий в Палестину, Закавказье и на Балканы. Сафлор. Передняя Азия (Сирия, Армения) является областью развития рода сафлора — Carthamus L., так как здесь сосредо- точено наибольшее видовое разнообразие. Центр происхождения рода, однако, может быть приурочен к западной части ареала (Испания, Алжир и сопредельные страны) — здесь распространены многолетние и, вероятно, более примитивные виды С. arb~orescens L. и С. helenioides Desf. Диким родоначальником культурного сафлора считаются виды С. охуасапйа М.В., связанный переходами с культурным видом и, может быть, отчасти С. lanatus L. Ареал С. lanatus совпадает с ареалом рода — по всей Древнесредиземноморской области до Индии на востоке. С. ~oxya<cantha занимает восточную часть ареала рода. Культурный сафлор — С. tinctorius L.— и дикорастущий вид— С. охуа~сапйа — имеют одинаковое число хромосом (2п = 24) и хорошо скрещиваются. С. lanatus имеет число хромосом— 2n = — 64 и не скрещивается с культурным сафлором. Существо- вание переходных форм говорит о родстве первых двух видов. Поэтому, вероятно, С. охуасапйа и является родоначальником культурного сафлора или, во всяком случае, его ближайшим родичем. Обычно родиной сафлора считается Индия, но исследова- ниями А. И. 1(упцова выявлено, что разнообразие форм куль- турного сафлора в Передней Азии, Египте и Судане не меньше, чем в Индии. Перечисленные страны представляют собой также области древней культуры сафлора. В области Передней Азии А. И. 1(упцовым выявлены следующие экотипы: 1) экотпп За- кавказья (преобладающий); 2) экотип Армении; 3) экогпп Ви- финии (Малая Азия); 4) экотип окрестностей Наблуса; 5) эко- тпп районов Дамаска и Тель-Керама; 6) экотип окрестностей Назарета. В Закавказье культура сафлора известна издревле и здесь выработался наиболее культурный тип сафлора без колючек и с богатыми картамином цветками. Согласно исследованиям А. И. 1(упцова, в популяциях куль- турного сафлора из Закавказья, Ирана, Афганистана, Узбеки-
стана и Индии встречаются формы примитивного типа: очень позднеспелые с прикорневой розеткой и перисторассеченными нпжнимп листьями. Таких форм в коллекционных образцах пз Египта и Эфиопии не было обнаружено. Некоторые исследователи думают, что эти формы гибридного происхождения (с видом С. oxyacantha), но доказательств гиб- ридности мы не видим и считаем эти формы за примитивный тип С. tin~ctorius. Против происхождения С. tinctorius из Индии говорит то, что род Carthamus в целом древнесредиземноморскнй. А. Л. Гросс- гейм считает, что родина культурного сафлора — Восточное Средиземноморье. Египет почти не имеет самостоятельной, аборигенной куль- турной флоры. Таким образом, Передняя Азия — один из основных древних, первичных очагов культуры сафлора или, вероятно, основной очаг и родина этого вида. Клещевина. Вид Picinus microcarpus G. Pop. представлен в Азербайджане и Армении эндемичными формами (без воско- вого налета), относящимися к iproles persicus G. Рор. (область развития proles ipersicus — Иран и Афганистан). Существует мнение, что культура клещевины имеет единый центр происхождения — Африку. Но ряд фактов заставляет усомниться в этом. Мелкоплодные и при этом длиннодневные или нейтральные по длине дня формы сосредоточены в странах Передней Азии и по берегам Восточного Средиземноморья. Здесь же встречаются менее рослые, почти травянистые сорные формы мелкоплодной клещевины. Вероятно, имеются два очага вхождения клещевины в культуру: 1) Африка для крупноплод- ных, короткодневных и более многолетних форм; 2) Восточное Средиземноморье с Передней Азией — для мелкоплодных и длиннодневных форм. Маслина. K Передней Азии приурочивается и происхождение важнейшего масличного древесного растения — маслины. Чи- ферри (Ciferri, 1958) развивает гипотезу о происхождении куль- турной маслины — Olea europaea L. — от распространенной в Передней Азии дикорастущей маслины — О еа africana Mill. Он предполагает, что маслина была занесена в Европу через Египет и Малую Азию. Он же обращает внимание селекционе- ров по маслине на необходимость использования в качестве ис- ходного материала для селекции многообразия форм дикой переднеазиатской маслины. Кориандр. Вид Coriandrum sativum L. известен в странах Пе- редней Азии как весьма древняя культура, но используемая не как эфирномасличное, а как пряное и овощное растение. Ко- риандр обнаружен в диком состоянии в степи и полупустыне по реке Араксу. Вероятно, он вошел в культуру именно в Закав- казье и соседних странах Передней Азии. 115
По Е. А. Столетовой, для Передней Азии эндемичны следу.|о- щие типы кориандра: закавказский (Иран и Закавказье); гру- зинский, абхазский, малоазиатский. Фенхель. Вид Foenicuium vulgare Mill., по А. К.,Макашвили, обильно произрастает в диком виде на равнинах и в предгорьях Азербайджана. Вероятно, первичный очаг культуры этого расте- ния находится в Передней Азии, а оттуда оно распространилось на запад и на восток (древнесредиземноморский ареал}. Чабер. Вид Satureja spicigera (С. Косй) Boiss. эндемичен для Грузии, по А. К. Макашвили, произрастает в Грузии в диком и в культурном состоянии. Употребляется как приправа и сыру. Заменяет садовый чабер S. hortensis L. Шафран. Род Crocus L. — средиземноморский, а место древ- нейшей культуры одного из видов шафрана — С. sativus L.— Сирия и Палестина, где он возделывается со времен царя Соло- мона. В Закавказье шафран также древняя культура. Шафран возделывается в Иране, Малой Азии, Индии и в Среди- земноморье. Родиной считается большинством авторов Перед- няя Азия (Сирия и Иран, Малая Азия), но указывается также на древность культуры шафрана в Италии и Греции. Этот во- прос окончательно не выяснен. Тархун (эстрагон). Вид Artemisia dracunculus L. был очень давно известен в культуре в Сирии. Название Tarlchoum упо- требляется во многих восточных странах. В Европе его начали культивировать в средние века. Можно предполагать, что пер- вичной областью его культуры была Передняя Азия, но точно это не доказано. Анис. Вид Pimpinella anisum L. имеет средиземноморский ареал — от Канарских островов до Малой Азии включительно. Однако многие авторы считают, что анис здесь распространен не в диком; а в одичалом состоянии и настоящим диким родо- начальником аниса, по мнению многих, является Р. ~cretica Poir., распространенный в пределах того же ареала и очень близкий к обыкновенному анису. Л. В. Лузина считает родиной аниса Малую Азию и Египет. Однако культурная флора Египта в боль- шей части не аборигенна, и мы склонны считать основным оча- гом распространения аниса Малую Азию. Там сосредоточены описанные Л. В. Лузиной позднеспелые, мощно развитые, хо- рошо облиственные формы аниса, которые Лузина выделяет в proles turicicum. Подобные же позднеспелые и хорошо облист- венные формы описаны Лузиной для 1(ашгара, но по морфоло- гическим признакам кашгарские и малоазиатские мезофильные формы неодинаковы. Вероятно, предок современного аниса был мезофитом. Эволюция шла в сторону ксероморфизма и ранне- спелости (сокращение вегетационного периода). 1(ашгарские и малоазиатские позднеспелые формы не связаны непосредствен- ным родством, а представляют собой островки более древних по типу, некогда широко распространенных мезофпльных форм. 116
Интересно, что подобный экологический параллелизм между формами Кашгара и Малой Азии встречается и у других расте- ний. Например, по многим признакам сходны кашгарские и ма- лоазиатские мезофильные и высокорослые формы синей куль- турной люцерны, где Бардаков (1934) неверно предполагал су- ществование прямого родства. Мы же относим эти формы даже к различным видам (Mediicago orientalis Vass. — Малая Азия,. М. asiatioa ssp. sinensis Sinsk. — Кашгар). Известное сходство имеется между мезофильными формами кашгарского и мало- азиатского пажитника — Trigonella foenum-graecum L. Анизет. Родина и область первичного вхождения в культуру вида Pim~pinella anisetum Boiss. — Малая Азия (Каппадокия и Сивас). Разводился также в Древней Армении. 1(отовик (Nepeta L.). В Передней Азии находится центр рас- пространения цикла Mussinianae P~ojark. Эта группа эфирномас- личных растений представлена рядом слабо обособленных «ви- дов», связанных между собой непрерывными рядами переходных. форм, составляющих единый географический массив. Из них имеют значение как эфироносы: котовик Муссини — Nepeta mus- sinii Sipreng. Восточное Закавказье, Армения, 1(урдистан, ко- товик Рейхенбаха — N. reichenbaic~hiana Fisch. et Меу. — Южное Закавказье, Армения, Курдистан, котовик армянский — N. haja- stana Grossh. Южное Закавказье. 1(ритический вид — котовик закавказский — N. transcau~casiica Grossh.— Восточное и Южное Закавказье, Западный Иран — особенно ценен как эфиронос,. вводится в культуру, очень полиморфен, систематика его не уточнена. Вероятно, имеют значение для отбора и гибридизации и от- меченные выше родственные «виды», а также эндемичная, мало изученная форма котовика норашенского — N. noras~cheniica Grossh. (Южное Закавказье). дыня. По К. И. Пангало, для Передней Азии эндемичны сле- дующие виды дынь — Мело Adans. (некоторые из этих видов следует считать подвидами или региональными экотипами): 1) аджур — М. adzhur Pang. Малая Азия; 2) кассаба — М. icas- saba Pang. Западная Малая Азия; 3) М. adana Pang.— Малая Азия — 1(иликия; 4) канталупа — М. сап1а1ира Pang.— Армения; 5) М. figari Pang. сорные растения Сирии, Палестины и Восточной Африки. А. И. Филов (1960) дает более разработанную классифика- цию дынь, не выделяя их в отдельный род. Передняя Азия яв- ляется, по его данным, очагом развития дынь кассаб — ssp. iori- entalis Sageret, среди которых имеются летние сорта — var. zhu- kovskii (Pang.) Fil., осенне-зимние-- var hassanbey (Pang.) Fil. и гурбеки — var. gurbel< Fil. По П. М. 5Куковскому, в Армении существуют местные кан- талупы, но А. И. Филов считает основным очагом развития кан- талуп Индию. 117
Из стран Передней Азии дыни распространились в Европу. Огурец. По А. И. Филову (1960), основной очаг развития Cucumis salivous L. находится в странах Передней Азии (Малая Азия, Иран), а также Азербайджане, дагестане, Крыму, Ниж- нем Поволжье, где сформировался западноазиатский подвид огурца. Этот подвид заходит и в Средне-IOro-Западную Азшо (Афганистан, Средняя Азия) . К этому подвиду А. И. Филов относит шесть, по его мнению, хорошо выраженных разновидностей: 1) иранскую — var. irano- turanicus (Gab.) Fil., 2) анатолийскую — var. anatoliicus Ciab., 3) балканскую — чаг. bal~canicus Fil., 4) киликийскую — var. cili- cicus Gab., 5) астраханскую — var. astrachanicum Fil., 6) средне- азиатскую — var. turlcestanicus Fil. Последняя разновидность по ареалу должна быть отнесена к средне-юго-западной подобласти культурной флоры. Тыква. Род Cucurbita L. по сравнению с дынями мало распро- странен в странах Передней Азии. Тыква огромная — С. maxima Duch.— в Малой Азии пред- ставлена разновидностью var. ovalis Roem. Этот преимущест- венно кормовой сорт, вероятно, занесен и в нашу страну из Малой Азии. Репа. Brassica гароеигореа ssp. antasiatica Sinslc. составляет особый подвид в Малой Азии. Малоазиатские репы отличаются грубостью и примитивностью по сравнению со сходными евро- пейскими сортами. По нашему мненшо, репа отсюда проникла на север в Карелию и южную Европу (Италия) и дала начало многим европейским сортам. В Карелию малоазиатская репа, возможно, попала не прямо, а через Алтай и Урал, так как и там встречаются родственные ей стародавние сорта. Морковь. Род Вaucus I.. в дикорастущих популяциях Север- ного Кавказа и особенно Закавказья представлен обоими дико- растущими подвидами D. carota sensu lato, установленными М. К. Рубашевской (1931): ssp. orientalis и ssp. occidentalis. Пер- вый подвид дал начало бескаротиновым сортам азиатской куль- турной моркови, а второй, западный по ареалу, подвид дал начало группе каротиновых сортов европейской моркови (все подвиды могли бы быть названы и видами). Также очень пестры и стародавние популяции местной куль- турной моркови в Закавказье (теперь уступающие место чистым завозным сортам). П. Ф. Загородских (1939) выделил в составе переднеазиат- ской культурной моркови эндемичные формы: 1) очепь своеобразный эндемичный подвид сирийский — ssp. syriacus Zagor. (Сирия, Палестина, Иран, часто встречается и в Закав- казье); 2) эндемичный подвид киликийский — ssp. icilicicus Za- gor. (Малая Азия). Есть предположение, что в происхождении киликийской крупноцветковой моркови играла роль гибридиза- ция с дикорастущим видом D. maximus Desf. 118
Судя по сходству местных дикорастущих и местных культур- ных форм, морковь имела независимое вхождение в культуру во многих местах и в странах Передней Азии. Передняя Азия, во всяком случае, представляет собой очень важный очаг формообразования дикорастущей и культурной моркови. Здесь находится стык восточного и западного дикора- стущих подвидов. Может быть, здесь и возникли оба эти под- вида от общего предка. Свекла. Передняя Азия считается В. П. Зосимовичем (1939) родиной рода свеклы — Beta 1 . и дикорастущего вида В. maritima L., который является непосредственным родоначальником куль- турной свеклы — В. vulgaris L. Виды В. таг1 ima и В. vilgaris составляют самую древнюю секцию Vulgares Transch., которая дала начало двум более молодым секциям: sect. Corollinae Transch., развившейся в Передней Азии и заходящей на Бал- каны, и секции Patellares Transch., образовавшейся на запад- ном конце ареала (1~анарские острова). В. maritima, по мнению В. П. Зосимовича, возникла в засо- ленных степях Передней Азии п при движении на восток до- стигла Индии, а двигаясь на запад п на юг, достигла Канар- ских островов и на севере распространилась до берегов Дании и Швеции. При таком огромном приморском ареале дикого родоначаль- ника, естественно, что свекла могла входить в культуру неза- висимо в разных частях этого ареала, что, вероятно, и имело место. В. maritima в пределах Передней Азии отличается зна- чительным полиморфизмом. В. П. Зосимович выделяет здесь следующие ее формы: 1) proles syrio-mesopotamica (Сирия, Месопотамия); 2) proles irano transcaucasica (Закавказье, Иран). В. T. 1(расочкин (1960) пришел к выводу, что лиственная свекла впервые вошла в культуру на островах Сицилия, 1~рит и 1(ипр. Соответственно адриатический экотип ближе других экотипов В. таг10111а стоит к листовой свекле. По В. Т. 1(расочкпну, в Переднюю Азию (в Сирию и Закав- казье) листовая свекла была завезена из средиземноморских крупных островов, а из Сирии проникла в страны древнего Дву- 'речья, в противоположность мненшо В. П. Зосимовича, который полагает, что именно в Двуречье листовая свекла вошла в куль- туру. Закавказский экотип дикорастущей В. тагШта (этот экотип назван В. П. Зосимовичем ирано-закавказским, но В. T. Ера- сочкин не нашел его в Иране), согласно исследованиям В. Т. Красочкина, включает карликовые формы и, кроме того, более мощные и несколько более длинностадийные формы. Послед- ние, по В. П. 1~расочкину, представляют собой одичавшие ли- стовые свеклы. Эти формы не являются настоящими прпмор-
скими дикарями, а растут вблизи населенных пунктов (по реке Куре у Тбилиси, по реке Араксу у Еревана) и в прикаспийских степях, как сорняк в огородах, садах и по краям хлопковых плантаций. Листья этой сорной свеклы используются местным населением как овощное растение. Местные названия этих сор- ных форм сходны с древним названием листовой свеклы- силк, что указывает на сицилийское ее происхождение. Армяне называют ее «зилк», а азербайджанцы — «зылк». Таким обра- зом, по-видимому, названия этих сорных форм свеклы указы- вают на то, что она выходец из культуры. Однако в Передней Азии, по В. Т. Красочкину, имеются эндемичные группы сортов листовой свеклы, которые не происходят от вышеупомянутых сорных форм. В. Т. Красочкин различает здесь две группы форм: 1) анато- лийско-закавказскую и 2) полукорнеплодную. При высеве в се- верных широтах анатолпйско-закавказская группа дает 50— 100 /~ цветухи, в средних — 10 — 50 /~, а на tore — О. Образцы c rop особенно устойчивы к цветухе. Полукорнеплодная группа распространена в высокогорных районах Древней Армении (Эрзерум) — это особые формы, более длинностадийные с се- ребристой окраской и очень длинными черешками. Таким образом, хотя Передняя Азия и не является родиной листовой свеклы и первичным очагом вхождения ее в культуру, но в ней имеется очаг эндемичных форм — вторичный очаг фор- мообразования. В. Т. Красочкин считает Переднюю Азию первичным очагом развития корнеплодных форм культурной свеклы, но последние произошли не от приморских дикарей (В. maritima), а от форм листовой свеклы с мелкими зачаточными корнеплодами. Фор- мирование примитивных (первичных) корнеплодных форм из листовых происходило, по Красочкину, в условиях долинного жаркого климата (при орошении) и достаточно длинного веге- тационного периода. Такая первичная область создания корне- плодов приурочена и долинам Малой Азии и Закавказья, воз- можно — Ирана и Афганистана; короткая стадия яровизации примитивных форм и высокая их жароустойчивость говорят о южном их происхождении. Примитивные полукорнеплодные формы свеклы сохранились в западной Анатолии и Афганистане. В западной Анатолии имеется эндемичный сортотип Балыкесирская свекла и отчасти Алашехирская, в Афганистане — Бадахшанская; эти переходные к современным корнеплодам формы не могли образоваться в го- pax — там формируются более длинностадийные экотипы. У этих форм имеется склонность к однолетности, но высокая темпера- тура предохраняет их or цветушности. Ялинностадийные формы корнеплодов выработались, по В. Т. Красочкину, в прилегаю- щих к орошаемым долинам горах. Этот процесс имел место в высокогорных районах Малой Азии (Адана, Афьонкарахи- 120
сар) II также в Закавказье — позднее произошло расселение этих форм в более северные области. Балыкесирская полукорнеплодная свекла имеет ряд морфо- логических признаков, общих с Афьонкарахисарской свеклой, происходящей из расположенных более высоко над уровнем моря районов — это уже почти современная корнеплодная форма, сравнительно устойчивая к цветухе. Особняком стоит эндемичный сортотип длинночерешковой свеклы, распространенной в Закавказье, Иране, Афганистане и Средней Азии. По сообщению В. Т. Красочкина, в этих странах свеклу подсевали к хлопчатнику и люцерне и в борьбе с зате- няющими сопутствующими растениями выработалксь особые длинночерешковые формы, которые выбивались из нижнего яруса травостоя. В Малой Азии и Закавказье сконцентрировано и наибольшее разнообразие настоящих корнеплодных форм свеклы, среди ко- торых много эндемичных сортотипов. Например, в Иране выде- лены В. Т. Красочкиным эндемичные сортотипы красно-розовой п ярко-красной свеклы и сортотип свеклы с белым корнем; среди растений этого сортотипа встречаются оригинальные формы с плоским белым корнеплодом. К Передней Азии приурочено развитие секции дикорастущей горной свеклы Corollinae, ареал которой — Малая Азия, Закав- казье, северная Сирия, Крым, Балканы. Эволюция этои секции представляется нам, на основании данных В. Т. Красочкина, в следующем виде. Beta tntermedia (промежуточная) Ве ta lo m а to gona— Свекла каем чатоплодная Beta trigyna— Свекла трехстолбиковая Свекла промежуточная имеет общие признаки двух производ- ных от нее видов. Закавказская В. trigyna Wald. et Kit. имеет число хромосом 2n =54, а В. П. Зосимовичем выделены тетраплоидные (2n= =36) формы этого вида, которые им рассматриваются как са- мостоятельный вид — В. corolli lora Zoss. Это сорняк. Виды данной секции представляют интерес как исходный ма- териал для селекции. Таким образом, в Передней Азии сосредоточены: 1) очаг ви- дообразования и формообразования дикорастущей свеклы, в особенности секции Сого111пае; 2) вторичный очаг формообра- 121
зования листовой свеклы; 3) первичный основной очаг формооб- разования и расселения корнеплодной свеклы с переходными полукорнеплодными формами. Кервель. Вид Anthriscus cerefolium (L.) Hoffm., по А. К. Ма- кашвили, дико не встречается, а произошел от близкого дико- растущего вида А. trichosperma Sipreng., широко распростра- ненного на Кавказе. Капуста. Капуста кочанная — Brassica ica~pitata 1.1кр.— пред- ставлена в странах Передней Азии восточным подвидом — ssp. orientalis Lizg. Согласно новейшим исследованиям Т. В. Лизгуновой, сорто- типы кочанной капусты Малой Азии, Закавказья, Северного Кавказа имеют много общего с сортами Средней Азии и бал- канских стран. У многих видов растений, первичный очаг вхождения в куль- туру которых сосредоточен в Передней Азии, имеются вместе с тем формы первичного типа, заходящие в Среднюю Азию и на Балканы. Поэтому возможно, что именно Передняя Азия яв- .ляется первичным очагом формообразования и расселения ка- пусты кочанной. С этой точки зрения очень важно указание Т. В. Лизгуновой, что в Древней Армении и Абхазии имеются в культуре древние по типу формы листовой капусты. Такие формы есть также и на Пиренейском полуострове, но они удалены от основного ареала капусты кочанной, находящегося в Восточной Европе и Восточ- ном Средиземноморье. Среди кольраби (В. саи1ога~ра Pasq.) Т. В. Лизгунова (1959) выделяет азиатский подвид (зэр. asiatica Lisg.), приуроченный тлавным образом к Передней и Средней Азии. Этот подвид срав- нительно малокультурный, что указывает на относительную древность его возделывания в этих странах. Кресс-салат. Кресс-салат — Lepidium sativum L. — является в странах Передней Азии древней п распространенной культу- рой. Вопреки указаниям, имеющимся в литературе, нет осно- ваний считать его родиной Эфиопию. Там велико разнообразие форм этого растения по величине плодов и по окраске семян, а в странах Передней Азии открыто наибольшее разнообразие по форме листьев, их рассеченности, форме куста, по степени ме- зофильности, длине вегетационного периода. В Сирии, Пале- стине и реже в Иране описаны позднеспелые формы с ясно вы- раженной прижатой розеткой — var. tardif lorum Stchenli. Эти формы близки биологически к дикарю, имеют примитивную конституцию; подобных форм в Эфиопии нет. Кроме того, для Передней Азии характерны мезофильные формы с цельными нижними листьями — var. latifolium DC. Разновидностей кресс-салата, эндемичных для Передней Азии и имеющих там основное распространение, но заходящих в Афганистан и в Западную Европу, шесть: ~ аг. tardif lorum, за- 122
ходит в Афганистан; var. adpressum, эндемична для Передней Азии; var. intermedium, заходит в Афганистан; var. subpinnatum, заходит в Афганистан; var. ~pinnatum, заходит в Западную Ев- ропу; var. 1аНо1ш1т1, заходит в Западную Европу. Все эти раз- новидности распространены в культуре. Разновидностей, эндемичных для Эфиопии и Эритреи, три: var. nanum, сорная; var. 11пгсо1а, сорная; var. glau~cum, сорная и культурная. Разновидностей космополитических, распространенных по всему ареалу, всего одна — var. vulgare. Разновидностей, рас- пространенных на западном конце ареала, одна — var. сг1эрит (Тунис, Франция). Наиболее распространенная разновидность var. vulgare встречается по всему ареалу, она имеется также в странах Пе- редней Азии и вкраплена там почти во все популяции с другими разновидностями (кроме var. tardif lorum), характерна для Сре- диземноморья и Западной Европы, имеется в Эфиопии, Судане и Египте. В Эфиопии эта разновидность представлена двумя эндемич- ными формами: subvar. abyssinicum, отличающаяся мелкими признаками — окраской пыльников и окраской семян, ~i var. tri- valve с трехстворчатыми стручками (это уродливость). Следовательно, существует определенный общий костяк по- пуляций кресс-салата, связывающий все популяции всех стран. Разновидность var. vulgare проникла в Эфиопию, там несколько изменилась и дала эндемичные формы. В Эфиопию проникли из Передней Азии и разновидности с сильно рассеченными листь- ями, но в качестве сорняков, и из ННх иыработалпсь своеобраз- ные эфиопские сорные формы var. папит и var. linieola. Возможно, что кресс-салат был занесен в Эфиопию из Индии, где изредка разводится. В Индшо он занесен из Афганистана или Ирана. Таким образом, Эфиопия не только не может считаться пер- вичным очагом культуры кресс-салата, но и не является ни оча- гом наибольшего числа эндемичных форм, ни очагом раз- нообразия культурных форм. Состав ее форм бедный (если не принимать во внимание мелких признаков: окр".ñêè семян, окраски пыльников, окраски листьев), и она представляет собой небольшой вторичный очаг эндемичных сорных форм этого вида. Эти эндемичные формы строго замкнуты в своем очаге и не оказали никакого влияния на формы других стран. В Египет и Судан попала опять-таки космополитическая var. vulgare, пред- ставленная там крупноплодной формой subvar. тасгосагрит; но эта крупноплодная форма вовсе не оказалась эндемичной для собственно Средиземноморья, а распространена и в странах Передней Азии: Азербайджане, Армении и Иране. Наиболее примитивные формы или не выходят за пределы
Передней Азии (var. tardif lorum), или заходят только в Афгани- стан (var. айргеззит) и др. Две разновидности — var. pinnatum и var. latifolium — рас- пространились из стран Передней Азии на запад — в Западную .Европу, но в Передней Азии они гораздо сильнее варьируют и дают переходы к примитивныгл формам. Нежные салатные формы с неперистыми листьями (var. Iа- tifolium) возникли уже в Передней Азии и дают здесь все :переходы к перистым и к примитивным поздним разновидно- стям. Салатные «цельнолистные» формы распространились да- леко на запад, и здесь из них получились формы с курчавыми .листьями (var. сг1зрит — это та же var. latifolium, но по краю .сильно надрезанная, с курчаво загнутыми дольками). Наибольшее разнообразие примитивных и культурного типа форм кресс-салата сосредоточено, таким образом, в странах Пе- редней Азии. Оттуда пошло расселение на восток в Афгани- стан и Индию, где также сохранились примитивные формы (Аф- ганистан), но разнообразие форм меньше, чем в странах Перед- ней Азии. Разнообразие форм кресс-салата постепенно убывает к за- паду Европы, где выработалась наиболее культурная курчавая форма. В Эфиопии возник небольшой очаг сорных эндемич- ных форм. Такова картина истории и расселения форм кресс-салата, ко- торая может служить хорошим примером приложения ботанико- географического метода к вопросам происхождения культурных растений. Возможность нарисовать эту картину дает хорошо изученный и точно описанный М. С. IgeHKQBOH (1932) материал в монографии о кресс-салате. Лук. Некоторые авторы считают Переднюю Азию очагом про- исхождения лука-порея — А11шт iporrum L., так как здесь имеются близкие ему дикие виды (А. ampeloprasum L. и др.). Лук огородный, репчатый — А11шт сера L. представлен на 1~авказе и в Анатолии особой группой сортов — proles caucasi- cum Kuzn. Чеснок,— А11шт за1л ит 1 . — представлен в Передней Азии особой восточносредиземноморской (южноприморской) группой сортов (Черноморское побережье, I<psiM, Закавказье, Дагестан). Шпинат. Родиной шпината — Spinacia оlегасеа L. — возможно. является Передняя Азия. В Закавказье дико растет его ближай- ший родич — S. tetrandra Stev. Ревень. Ревень смородинный — Rheum ribes L. — распростра- нен дико в Малой Азии, Аравии, Закавказье, Сирии, Пале- стине, а также в Иране, Афганистане и в Белуджистане. В Си- рии — древняя культура. Салат-латук. Вид Lactuca sativa L. встречается в странах Пе- редней Азии дико и в культуре. Возможно, что здесь один из очагов вхождения в культуру этого растения, 124
Баклажан. Вид Solanum те1опдепа L. представлен, по А. И. Филову, следующими агроэкологическими группами или .экотипами: западноанатолийским, центральноанатолийским, смирнским, палестинским, аравийским, азербайджанским, гор- .ным. Горный экотип распространен в Малой Азии (Анатолии) и в Закавказье (Абхазия), а также в Индии. Филов (1958) счи- тает, что горный тип баклажана занесен из Передней Азии в Индию, а не наоборот. Палестинский тип поздний, но густо опушенный (признак, характерный для палестинских форм различных видов и родов), распространен широко — от Палестины до Южного 1~итая, но, судя по названию, А. И. Филов принимает за очаг его распро- странения Переднюю Азию (Палестину). Очаги первичной куль- .туры, по А. И. Филову, находятся в Индии и Передней Азии (Аравия). Дикие мелкоплодные родичи сохранились только .в Индии, а полукультурные формы имеются в Аравии. Из Ин- дии и Аравии одна ветвь развития пошла в Восточную Азию, .а другая возникла при распространении баклажана на запад. Предполагаемая схема эволюции и расселения баклажан: Индия Аравия Индокитай другие страны Передней Азии Япония — эЕвропа~ )25 Дикорастущие родственные культурные формы распростра- :нены в Индии, полукультурные — в Аравии. В Японии возделы- ваются зимой и часто не первой культурой, поэтому там вырабо- тались скороспелые формы. Портулак. Portulaca оlегаоеа L. чрезвычайно распространен в странах Передней Азии в диком виде. Вероятно, здесь имеет место и вхождение в культуру, как в других странах, где имеется его дикий родич. Груша. Передняя Азия сыграла особенно большую роль в раз- витии рода Pirus L., который, имея корни в тургайской флоре, возник в Восточной Азии, а во флоре Передней Азии является мезотермическим реликтом с продолжающимся видообразова- нием. Приводим список видов груши, распространенных в Закав- казье (по А. А. Федорову, 1951): груша иволистная — Р salici- folia Pall. (Дагестан и Предкавказье, Грузия, Армения; дико); груша Медведева — Р. medvedevii Rubtz. (Армения — Зангезур, Нахичеванская АССР; дико); груша Тахтаджяна — Р. takh-
tadzhianii An. Fed. (Южная Армения, Центральная Грузия:. дико, вероятно, прежде была в культуре); груша грузинская— Р georgica Kuth. (Центральная Грузия, Южная Армения; дико); груша сахокия — Р. sachokiana Kuth. (Центральная Гру- зия; дико); груша таохская — Р. taochia Woron. (Аджария, Ар- винский район Турции; дико); груша смешанная — Р сот~р1еха Rubtz. (Южная Армения — Зангезур; дико); груша остропильча- тая — Р o~xyprion %агап. (Южная Армения — Яаралагез, Ка- гызманский район Турции; дико); груша Федорова — Р. fedo- rovii Кий. (Центральная Грузия; дико); груша Сосновского- Р sosnovskyi An. Fed. (Южная Армения; дико); груша Дмит- рия — Р demetrii Kuth. (Центральная Грузия; дико); груша Всеволода — Р. vsevolodi Heidemann (Азербайджан: дико); груша Тамамшян — Р. tamamschianae An. Fed. (Южная Арме- ния; дико); груша Кецховели — Р ketzkhovelii Kuth. (Централь- ная Грузия; дико); груша эльдарская — Р. eldari~ca Cirossh. (Центральное Закавказье; дико); груша сирийская — Р syriaca Boiss. (Сирия, Южная Армения, Восточная Анатолия, Кипр; дико); груша зангезурская — Р. zangezura Maleev (Южная Ар- мения; дико); груша поникшая — Р. nutans Rubtz. (10жная Ар- мения, Зангезур; дико); груша Воронова — Р. voronovii Rubtz. (Южная Армения; дико); груша Радде — Р. raddeana Woron. (Южная Армения, Зангезур;; груша кавказская — Р. cau- ~casica An. Fed. (леса всего Кавказа). Кавказские популяции груши обыкновенной — Р communis выделены в самостоятельный вид. Возможно, что правильнее считать кавказские груши этого типа только подвидом илп региональным экотипом полиморфного вида Р. communis. Но для наших целей этот вопрос не имеет особого значения. Энде- мизм здесь во всяком случае имеется, хотя бы и по отношению к форме меньшего, чем вид, ранга. Груша эта представлена на Кавказе многими дикорастущими формами, формами переход- ными от диких к культурным и особыми местными стародавними культурными сортами. Груша Баланзы — Р. balansae Decne. (Лазистан; дико); груша Гроссгейма — Р. grossheimii An. Fed. (Талыш и, вероятно, Северный Иран; дико); есть предположе- ние, что эта груша вышла из культуры и теперь распространена в одичалом состоянии; груша гирканская — Р. hyucana An. Fed. (Талыш; дико); груша высокая — Р. elata Rubtz. (Южная Ар- мения; близка к китайским грушам; дико, или, может быть, одичавшая); груша Буассье — Р boissieriana Buhse (Северный Иран, Азербайджан, Туркмения — Копет-71аг; близка к гима-. лайско-китайской группе видов). Ан. А. Федоров (1951) указы- вает, что только для одного Кавказа известно 2б видов груш. Груши Ирана и Малой Азии изучены еще сравнительно слабо: груша туркменская — Р turcomani~ca Maleev (Туркме- ния — Копет-Даг; помещена в список, так как Туркмения включается нами в границы Передней Азии); груша лохоли- 126
стпая — P. elaeagrifolia Pall. (распространена в Крыму, Малой Азии и Турецкой Армении — это также вид переднеазиат- скпй). Прп стремлении многих систематиков к чрезмерному дробле- нию вида возможно, что некоторые из перечисленных видов на самом деле региональные экотипы. Тем не менее этот список дает представление об интенсивности видообразовательного и формообразовательного процессов в роде Pirus в Передней Азии, где имеется также много особых культурных местных сортов груши и можно встретить формы, переходные от культурных к диким — примитивным формам. Аборигенный для Северного Кавказа сорт груши Бергамот черкесский отличается очень хо- рошим вкусом и дает .лучшие дички для подвоя. Мы присоединяемся к мнению П. М. Жуковского, что не Сре- диземноморье, а именно Кавказ является ареной эволюции груши как дикой, так и культурной, основной ареной, ибо спо- радическое введение в культуру дикой груши и возникновение аборигенных местных сортов, вероятно, имело место и в других странах Средиземноморья и Западной Европы. Яблоня. Яблоня дикорастущая, в основном, представлена в Передней Азии полиморфным и еще недостаточно изученным видом Malus orientalis Uglitzk. (Кавказ, Малая Азия, Иран). Этот впд имеет переходные формы к другим видам и принимал участие в формировании эндемичных культурных сортов Перед- ней Азии. Культурная яблоня на Кавказе, видимо, часто дичает. Нередко смешиваются одичалые формы с дикорастущими. Культура яблони, очевидно, является более древней на Кав- казе, где имеется много форм, переходных от культурных и дикорастущим. Не легко бывает прп этом одичавшие формы отличить от действительно диких. Недавно среди куль- турных сортов яблони на Кавказе обнаружены формы, способ- ные к порослевому возобновлению. Эти формы будут иметь зна- чение в лесопосадке. Влияние кавказской восточной яблони сказывается на неко- торых средиземноморских культурных сортах (Ф. Q. Лихонос, 1954) . Заросли дикорастущей яблони и переходные к культурным формам имеются и в Малой Азии. По Д. Г. Редеру (1950), «...видно из хеттского кодекса законов и еще более раннего до- кумента — Тель-эль-Амарнского текста, повествующего о по- ходе Саргона Аккадского в Каппадокию и, наконец, из целого ряда упоминаний классических авторов, что яблоки были рас- пространенным плодом у хеттов». Г. Бушан считает, что впервые культурная яблоня появилась в Малой Азии (район Трапе- зунда). В Передней Азии имеется род Eriolobus DC. — род очень близкий к Malus. Сюда относится вид Е. trilobatus М. Poem, распространенный в Малой Азии и на Крите. 127
Айва. Айва (Cydonia М111.) — монотипный род. Вид С. oblonga Mill. растет в диком состоянии в Дагестане и Азербайджане, Иране и Туркмении (здесь возможны одичавшие растения). По Декандолю, родина айвы — север Ирана, берега Каспия п Анатолия. Отсюда айва в глубокой древности была занесена в северную Грецию и на остров Кипр. Иное название айвы- сидония — произошло от кипрского города Сидон, который сла- вился хорошими сортами айвы. Р. Я. Кордон (1953) выделяет три главных района распро- странения дикорастущей айвы в СССР: Талыш, Западный Ко- пет-Даг и 10жный Дагестан с прилегающими к нему районами Азербайджана. Наиболее древним районом распространения айвы Р Я. Кор- дон считает Талыш. За пределами СССР дикая айва распростра- нена по всему южному побережью Каспия — в Гиляне, Мазанде- ране и Астрабаде. Наибольшее разнообразие форм дикой айвы обнаружено в районе Каякента (Дагестан). Наряду с наиболее широко рас- пространенными разновидностями — грушевидной, яблоковид- ной и португальской, там встречаются особые формы дикой айвы: с обратнояйцевидными листьями — f. obovata, с округ- лыми листьями — f. rotundifolia. Р. Я. Кордон указывает на существование трех экотипов ди- кой айвы: 1) гигрофильный (в Талыше по зарастающим водое- мам); 2) ксерофильный (на сухих песках Апшерона); 3) соле- выносливый (на засоленных почвах). На Кавказе айва существовала еще в доисторическое время. У всех народов Кавказа имеются местные названия айвы: комши терокали (грузины); серкевели (армяне); абея (аб- хазцы); биа (имеретины, гурийцы, мингрелы); би (лезгины, ка- закутухи); хайва (кабардинцы). Введение в культуру айвы, а, может быть, и возникновение вида С. oblonga, произошло на Кавказе, где существуют хоро- шие местные сорта культурной айвы. Рябина. Род Sorbus L. имеет один из очагов видо- и формо- образования в странах Передней Азии, где растут следующие виды: рябина садовая — S. domestiica L. В диком состоянии рас- пространена в горных районах Малой Азии, Крыма и, может быть, северо-западного Кавказа (близ Новороссийска), а также на Балканах. Культивируется в садах Средней Европы, где иногда дичает. Очаг распространения — Передняя Азия и Бал- каны. Рябина обыкновенная — S. аисшр~~аг1а L. — бореальный евр- азиатский вид (по всей Европе, в Крыму и на Кавказе) . На Кав- казе вид S. аисираг1а представляет собой особую регио- нальную разновидность; рябина Буассье — S. boissieri Sneid. (Кавказ, Малая Азия); рябина буроватая — S. subfusca Boiss. (эндемична для Кавказа); рябина Альбова — S. albovll Zinserl. 128
(Кавказ); рябина полувойлочная — S. subtomentosa Zinserl. (эндемична для Кавказа); рябина колхидская — S. Icolchi~ca Zinserl. (эндемична для Кавказа); рябина бархатистая — S. че- lutina (Albov) Schneicl. (эндемична для Кавказа); р1бина Буша — S. buschiana Zinserl. (эндемична для Кавказа); рябина шемахинская — S. schema~chensis Zinserl. (эндемична для Кав- каза); рябина кавказская — S. caucasiica Zinserl. (эндемична для Кавказа); рябина Воронова — S. woronovii Zinserl. (эндемична для Кавказа); рябина армянская — S. armeniaca Hedl. (Закав- казье, Турецкая Армения); рябина Кузнецова — $. kusnetzovii Zinserl. (эндемична для Кавказа); рябина греческая — S. graeca Hedl. (Крым, Кавказ, Малая Азия, Балканы); рябина мигар- ская — S. migarica Zinserl. (эндемична для Кавказа); рябина Балдаччи — S. baldaecii Degen et Fritsch (Кавказ, Турецкая Ар- мения, Балканы, Азия); рябина крымская — S. taurica Zinserl. (Крым, KBBI<as); рябина тупозубчатая — S. obtusidentata Zin- serl. (эндемична для Кавказа); рябина двойная — S. dualis Zinserl. (Закавказье). Список дан главным образом для Кавказа. Рябины Малой Азии и Ирана изучены слабо. Некоторые из перечисленных ви- дов соответствуют скорее подвиду или региональному экотипу. Однако список дает представление о богатстве Передней Азии эндемичными формами этого рода. Боярышник. Род Crataegus L. представлен в Переднеи Азии большим количеством дикорастущих видов, из которых многие очень полиморфны. Плоды из дикорастущих зарослей иногда используются. Для плодоводства имеет значение боярышник понтийский— С. ipontica С. I<och (Иран, Турецкая Армения — по р. Чороху, Туркмения; заходит в Памиро-Алай). Родина — Передняя Азия. Дико и в культуре. Европейский культурный вид С. azarolus L. близок к С. pontica и переднеазиатскому виду С. aronia Возс и, вероятно, распространился в Европу из переднеазиатского очага. Боярыш- ник темно-красный — С. atrosanguinea Poj arly. (Армения, Иран, Турецкий Курдистан; дико и в культуре); боярышник Шовица- С. szowitsii Pojarlc. имеет съедобные плоды (Восточное Закав- казье). Мушмула. Род Mes~pilus L. монотипный, Кавказ является местом происхождения рода. Вид М. germanica L. растет дико на Кавказе, в Малой Азии, Крыму и в Северном Иране. Наибольшее разнообразие диких форм и культурных сортов в Азербайджане. Несомненно вошел в культуру на Кавказе. Малина и ежевика. Малина и ежевика (виды gubus L.) пред- ставлены на Кавказе большим количеством эндемичных дико- растущих видов и форм, которые являются интересным матери- алом для селекции. Синская Е. Н.
Слива. Род Prunus L. — мезотермический реликт с очагом со- временного видообразования в Передней Азии; этот очаг через Туркмению и Северный Иран сообщается со среднеазиатским очагом рода, имеющего свои первоначальные истоки в тургай- ской флоре Восточной Азии. В Передней Азии, в основном на Кавказе, находится основ- ной очаг интенсивного формообразования и, вероятно, родина алычи — Р. divaricata Ledeb. (разделена теперь на несколько ви- дов) Значительное разнообразие алычи имеется также в Сред- ней Азии. Н. В. Ковалев установил на Кавказе для алычи обыкновен- ной Р. divaricata следующие подвиды: 1) nairica I<ov. (Арме- .ния); 2) оо1сЫса Kov. (Колхида); 3) boreale-саисаз1са Kov. (Се- верный Кавказ); 4) naichitschevanica Коч. (Армения, район р. Аракса). Кроме того, Н. В. Ковалев различает следующие близкие к Р. divaricata (может быть тоже подвиды?) виды:1) Р. caspica Коч. (Каспийское побережье); 2) Р. ursina Kotschy (Ливан); 3) Р. spinosa L. (распространен по всему Средиземноморью и Европе, но нигде так не полиморфен, как на Кавказе; возможно, что именно здесь находится его родина, основной очаг его рас- пространения на запад и на север). Домашняя слива — Р. domestica L. — по теории, находящей много сторонников, произошла путем гибридизации терна и алычи. Родиной сливы, по этой теории, является Кавказ, где имеются целые рощи естественных гибридов между терном и алычей. Возможно, отдельные формы сливы произошли непо- средственно от алычи. Вишня, черешня. Из видов рода Cerasus Juss. многие ботаники приписывают переднеазиатское происхождение черешне — С. avium Moench, которая здесь отличается большим разнообра- зием дикорастущих форм и эндемичных местных культурных сортов. Существует предположение, что Передняя Азия — родина домашней вишни — С. vulgaris Mill, но этот вопрос неясен. Во всяком случае этот вид нередко встречается в одичалом со- стоянии на Кавказе и в Крыму. Происхождение домашней вишни — С. vulgaris связывают с двумя видами — С. avium и С. fruticosa Mill., которая распространена от Байкала на востоке до Пиренеев — на западе. С. fruticosa Mill. произошла от дикорастущей и широко рас- пространенной в СССР С. fruticosa (Pall.) Woron. (С. chamae- cerasus Loisel.) . В диком виде в Передней Азии встречаются антипка — С. mahaleb М111. (Закавказье, Крым); С. microcanpa (Boiss.) Mill. (Закавказье, Копет-Даг); С. araxina Piojark. (Армения); С. collina Lej. et Gourt. — имеется в одичалом состоянии в Ма- лой Азии. 130
Существует предположение, что дикорастущий впд С. со1Ипа Lej. et Court. дал начало культурной Владимирской вишне. Миндаль. Миндаль (Amygdalus L.) имеет главный очаг ви- дообразования в Передней Азии и представлен здесь несколь- кими видами — А. ioommunis L. — миндаль обыкновенный— в одичалом состоянии (некоторые думают, что в диком) распро- странен в Южной Армении, иногда в Азербайджане, Малой Азии, Аравии, Иране и Средней Азии. Имеются основания пред- полагать, что миндаль введен в культуру шумерами. В Средней Азии, может быть, вошел в культуру самостоятельно. Из дико- растущих видов ближе всех к культурному миндалю стоит А. fenzliana (Fritsch) Lipsky; представлен в Армении эндемич- ным подвидом (Зангезур), в целом этот вид переднеазиатский. Эндемичные для Передней Азии А. urartu Tamamsch. и А. nairica Fed. et Takht., по Ан. А. Федорову (1942), представляют собой «молодые прогрессивные виды»; миндаль Фенцля и мин- даль Урарту относятся к той же секции Euamygdalus Spach, что и культурный миндаль А. соттип1з. Эти виды очень поли- морфны. По мнению Ан. А. Федорова, миндаль Фенцля не яв- ляется непосредственным родоначальником современных армян- ских культурных миндалей (из которых некоторые занесены из Ирана и даже из Западной Европы), но принимал участие в про- исхождении культурного миндаля, хотя бы путем гибридизации с другими видами в условиях культуры. Вид А. oommunis, вероятно, и сформировался именно в усло- виях культуры в странах Передней Азии (включая Иран и Турк- мению), где сосредоточено наибольшее разнообразие видов этого рода (хотя истоки рода надо искать в Восточной Азии). Ан. А. Федоров, выявляя в роде Amygdalus различные ста- дии ксерофилизации, пришел к правильным, на наш взгляд, вы- водам, что «миндали развились не от каких-то дрсвних Pruno- ideae Передней Азии, а скорее от мигрантов третичных мезофиль- ных Prunoideae, проникших в Иран в незапамятное время и там ксерофилизировавшихся». Первоисточником распространения культурного миндаля на запад была Передняя Азия, отсюда он попал в Грецию. По сви- детельствам древних авторов (Атеней и др.), миндаль достав- лялся в Грецию жителями Малой Азии. Из Греции он проник в Рим и на Пиренейский полуостров. Лавровишня. Лавровишня — Laurocerasus officinalis Roem. в диком виде распространена на Кавказе, в Малой Азии, Иране и заходит на Балканы. Вид реликтовый, но и сейчас отличаю- щийся в Передней Азии значительным разнообразием. Абрикос. Род Armeniaca Mill. представлен в Передней Азии особой группой сортов — ирано-кавказской, относящейся к виду Armeniaca vulgaris L. Ирано-кавказская группа, составляющая вторичный очаг (культура абрикоса пришла сюда из Средней Азии) сортов аб- 131
р«Коса обыкновенного, по богатству форм уступает среднеазиат- ской, но превосходит европейскую. Особенно ценны беломясые формы с высокими вкусовыми качествами. Из Армении, где он культивировался уже в неолите, абрикос распространился в Грецию, затем в Италшо, а потом культура его в Европе про- двинулась и севернее. Поэтому долго в Европе родиной абрикоса считали Армению. Инжир. Ficus L. принадлежит к древнейшей ветви расти- тельного мира, вышедшей еще из мелового периода. В«д F. carica L. дико произрастает в Закавказье, Малой Азии и в Крыму. В Передней Азии имеются еще следующие дикора- стущие виды инжира: F. colchica Grossh. — эндемичный для Колхиды; F. hyrcana Grossh. — эндемичный для Талыша и F. serrata Forsk. — эндемичный для Аравии. Первые два вида, воз- можно, более правильно рассматривать как региональные эко- типы или подвиды F. carica, но тем не менее их открытие гово- рит о том, что Кавказ — один из очагов формообразования ин- жираа. Возможно, что в культуру инжир вошел независимо и в Пе- редней и в Средней Азии. По мнению А. А. Гроссгейма, дикорастущие кавказские виды F. colchiica и F. hyrcana отнюдь «не деградаты». Видообразова- ние в роде Ficus, которое стимулировалось в ксеротермические (третичный и послетретичный) периоды, продолжается и в ны- нешнюю эпоху. Типичный F. icarica, по А. А. Гроссгейму, встре- чается в культурном и одичалом, но не в подлинно дикорасту- щем состоянии. Гранат. Гранат (Ришса L.), как и инжир, является представи- телем древнейшего типа флоры, которая существовала раньше тургайской. Весь род представляет собой реликт, ибо известно всего 2 вида граната — один в Передней и Средней Азии, а дру- гой на острове Сокотра в Индийском океане у южных берегов Аравии. Вид Punica granaturn L., как и Р1сиз carica, а также род- ственные им виды, отнюдь не является деградирующим релик- том, но, напротив, отличается большим полиморфизмом. P. granatum произрастает дико в Закавказье, дагестане, Ма- лой Азии и горной Туркмении (Копет-Яаг). В Закавказье и Азербайджане имеется ряд местных сортов культурного граната; несомненно, что Передняя Азия является первичным очагом вхождения граната в культуру и родиной этого вида. В Азербайджане разнообразие сортов граната очень велико и наблюдаются все переходы от культурных форм к дикорасту- щим. Греки получили гранат из Малой Азии. Из Передней Азии гранат был в глубокой древности занесен в Индию (имеются санскритские названия его) . Гранат был известен и древним египтянам, но корни его на- званий не египетские, они сходны с арабскими, еврейскими, 132
коптскими, берберийскими. Гранат был занесен в Египет, воз- можно, пз Ассирии. Лорэ (V. Loret, 1892) и другие указывают, что древнеегипет- ские гранаты были мельче и проще по структуре, чем современ- ные: онп имели 4 — -6 отделений (гнезд, или камер), а совре- менные формы имеют 6 — 8 отделений. Кроме Р. granatum, в роде есть еще только один вид Р pro- tnipunica Balf. с удаленным островным ареалом (остров Сокетра в Индийском океане); этот вид не имеет отношения к происхож- дению современных культурных сортов. У Р. protopunica плоды также проще, чем у современного Р granatum, и гнезда у первого расположены в один ярус. Со- временные дикорастущие формы Передней Азии имеют такое же сложное строение плодов, как современные культурные сорта. Нам представляется правильным заключение А. Д.. Стребко- вой, что гранат с просто устроенными плодами представляет со- бои очень древний реликт, а в Передней Азии такие формы вы- мерли. А. g. Стребкова среди дикорастущих форм граната в Закав- казье находила кросс-стерильные с друп;ми близкими формами. Из этого, а также из свойственных здесь гранату чрезвычайного полиморфизма и пластичности А. Д. Стребкова заключает, что и в настоящее время в странах Передней Азии идут интенсив- ные процессы обособления форм и видообразования. Виноград. На Кавказе виноград произрастает дико; ближай- шии родич культурного винограда Vitis silvestris Gmel. обра- зует здесь два региональных экотипа. В Малой Азии и Закав- казье имеется много аборигенных сортов винограда. А. М. Негруль среди культурного винограда — V. vinifera зэр. sativa DC. — выделяет для Передней Азии следующие группы сортов: 1) proles georgica Negr. — сорта грузинского про- исхождения; 2) subproles сазрка Negr. — крупноплодные сорта Армении, Азербайджана, Дагестана, Средней Азии и Афгани- стана; 3) subiproles antasiatica Negr.— плотноягодные мясистые сорта Азербайджана, Дагестана и Туркмении. А. М. Негруль и многие другие исследователи производят культурный виноград V vinifera от дикорастущего очень поли- морфного вида V silvestris. Другие придерживаются мнения, что культурный виноград произошел не от одного, а от несколь- ких видов и что V vinifera и V. silvestris произошли от общего предка. Последнее нам представляется весьма вероятным. Хурма. Хурма кавказская — Diospyros lotus L. — произрастает в диком состоянии, иногда разводигся в Западной Грузии, в Талыше и в северном Азербайджане (Закаталы). Происходит пз 1~птая. Кизил. Comus mas L. — вид переднеазиатского происхожде- ния. В диком состоянии наиболее распространен на Кавказе 133
(Азербайджан), где имеется наибольшее разнообразие дикора- стущих форм и культурных сортов. Шелковица. Области древней культуры шелковицы — Китай, Япония, Западная Азия. Однако в Азербайджане и в Армении имеется большое разнообразие плодовых сортов Morus alba 1. и меньше — М. nigra L. Формы с удлиненной пластинкой листа сосредоточены в Южной Туркмении и имеют много общего с формами Закавказья, Украины и Балкан. Расселение на запад и север, вероятно, шло из Ирана. Границы между дикими и одичалыми формами в Передней Азии в некоторых случаях про- вести трудно. Фисташка. Фисташка (Pistacia L.) имеет истоки в древней (тургайской) флоре Восточной Азии, где обнаружен вид, очень близкий к широко распространенной в Средиземноморских странах Р. Ientis~cus 1. По Р. Я. Кордону, очаг возникновения новых видов, разновидностей и форм находится в центре ареала — в Малой Азии и Иране. Там как раз сталкиваются сре- диземноморские виды P. lentiscus, Р. terebinthus ~ ., Р. mutica Fisich. et Меу. со среднеазиатскими видами Р vera, Р integer- rima Stew. Ключ к познанию рода Pistacia во всем его видовом многообразии надо искать в Иране и Малой Азии. Фисташка настоящая — Р. vera L. — в диком состоянии рас- пространена в Малой Азии, Иране, Средней Азии и Афганистане, т. е. ее ареал распространяется и на Переднюю Азию, особенно, если включить в состав последней и Туркмению, где имеются значительные дикорастущие ее заросли. В Закавказье, Малой Азии и Туркмении имеются эндемичные сорта культурной фисташки. Передняя Азия во всяком случае входит в область первоначального распространения культурной фисташки, а может быть является и главным первичным очагом ее зарождения. Кевовое дерево — Р. mutica Fisch. et Меу. (из плодов добы- вают масло; используют и древесину) распространено в Азер- байджане, Иране, Анатолии, Аравии и, хотя на запад доходит до острова Родоса, а на востоке до Афганистана, в основном, в настоящее время имеет переднеазиатский ареал. Орешник. Орешник (Согу1из L.) мезотерм~ический реликт тур- гайской флоры, имеет вторичный центр развития в Передней Азии (Кавказ). Для Малой Азии эндемичен С. iponti~ca С. I(oich. Для Кавказа эндемпчны виды (возможно, что это не виды, а только региональные экотипы) С. oolchica Albov (Западная Грузия и Абхазия), древний вид С. iberica Wittm. et Kemular., С. cervorum Petr. (Талыш), С. imereti~ca Кети1аг-Nat. (Закав- казье). Кавказ и 1!!алая Азия составляют часть ареалов широко распространенных видов (в диком виде) С. со1пгпа L. и С. avel- ana L. На Кавказе имеется ряд нигде более не встречающихся куль- турных сортов орешника — С. avellana. Некоторые культурные 134
сорта орешника ведут евое происхождение от переднеазиатских ьидов С. maxima Mill. и С. pontica С. Koch. Каштан. Каштан (Castanea Mill.) имеет корни в древней флоре Восточной Азии. В Западном Закавказье и в причерноморской Малой Азии только один дикорастущий вид каштана — С. sativa Mill. (С. vesca Gaertn.), но на Кавказе имеется значительное разнообразие эндемпчных дикорастущих форм и свои культур- ные сорта каштана. Лекарственные растения. Передняя Азия изобилует видами и формами лекарственных растений. Из них упомянем следующие: белладонна кавказская — Atropa cau~casica Kreyer (Абхазия, Грузия, дагестан, Талыш; вводится в культуру); ландыш кав- казский — Convallaria transcau~casica Utkin (Закавказье; растет в диких зарослях); ромашка кавказская розовая представлена видамп: Pyrethrum roseurn M. В. (Кавказ, Малая Азия, Иран; дико и вводится в культуру); Р. сагпеит М. В. (Кавказ, Се- верный Иран, дико и вводится в культуру). Передняя Азия изобилует также дикорастущими видами ва- лерианы. Таким образом, пз всего изложенного выше следует, что Пе- редняя Азия (в широком понимании) является родиной или ос- новной областью развития следующих родов, к которым отно- сятся важные виды культурных растений. Из злаковых: ТгЖсит — пшеница; Secale — рожь; Aegilops, который не вошел в культуру, но имеет большие перспективы для использования в селекции. Возможно, здесь же находится первичная область развития рода Agrapyron. Таким образом, Передняя Азия — это важнейшая область развития всей трибы Hordeae сем. злаковых. Из бобовых: Cicer — нут; Lens — чечевица; Pisum — горох; Onobrychis — эспарцет; Medicago — люцерна (во всяком случае, самый древний и самый экономически ценный подрод Ра1саро); из губоцветных La 1lemantia — ляллеманция; из крестоцветных Camelina — рыжик; из маревых Beta — свекла (здесь важней- ший очаг вхождения в культуру корнеплодных форм, один из очагов видообразования, а может быть, и главный очаг). Из других родов следует отметить Mespilus — мушмула; Cydonia— айва; Carthamus — сафлор (область наибольшего развития рода, но, вероятно, не происхождения); Рипiica — гранат (очаг совре- менного развития и происхождения). Особенно велика роль Пе- редней Азии в происхождении и распространении культурных бобовых растений — она является родиной пяти важнейших ро- дов: нута, чечевицы, гороха, люцерны и эспарцета. К родам, содержащим в своем составе культурные расте- ния, имеющим в странах Передней Азии один из первичных очагов, либо более или менее крупный вторичный очаг своего
развития (видообразования), относятся: 1) овес — Анрепа (сор- ные; один из первичных очагов); 2) житняк и пырей — Agropy- ron (главным образом дикорастущие; один из первичных оча- гов); 3) вика — Vicia (очень крупный вторичный очаг или, мо- жет быть, один из первичных очагов; истоки рода, вероятно, в Восточной Азии); 4) клевер — Trifolium [один из вторичных очагов развития рода (дикорастущих видов), очаг развития груп- пы видов, близких к клеверу александрийскому]; 5) груша— Pirus (области интенсивного видообразования и формообразо- вания; вторичный крупный очаг развития рода; истоки рода — в Восточной Азии, как и Prunus); 6) слива — Prunus (имеет в Передней Азии вторичный очаг видообразования, но менее интенсивный, чем следующий род); 7) BpiIIIHH и черешня— Cerasus (вторичный очаг видо- и формообразования); 8) мин- даль — Amygdalus (один из первичных очагов видо- и формо- образования; истоки рода в Восточной Азии); 9) пнжир— Filcus (один из первичных очагов видо- и формообразования); 10) боярышник — Cra'aegus (один из вторичных очагов видо- и формообразования); 11) орешник — Согуlпз (вторичный очаг ви- дообразования); 12) рябина — $огЬиз (один из вторичных оча- гов видо- и формообразования); 13) фисташка — Pistacia (вто- ричный очаг видо- и формообразования). Следующие виды культурных растений строго эндемичны для Передней Азии, не выходят из ее пределов даже в куль- туре: 1) пшеница маха — Triticum macha (эндем для Грузии; в культуре); 2) колхидская полба — Т. paleooolchi~cum (эндем для Грузии; в культуре); 3) полба Тимофеева — Т. timepheevi (эндем для Грузии; в культуре); 4) полба Жуковского — Т. zhu- kovskyi (лучше рассматривать как подвид предыдущего аида); 5) пшеница дика — Т. persicum (эндем для Кавказа; в куль- туре); 6) пшеница ванская — Т. vavilovii (эндем для Армении; в культуре); 7) пшеница туранская — Т. turanicum (Месопота- мия, Иран, заходит в Туранскую низменность; в культуре); 8) полба-спельта — Т. spelta (вторичный очаг в Средней Европе); 9) пшеница исфаганская — Т. ispahanicum (Иран); 10) рыжик кавказский — Camelina icaucasica (сорное и в культуре, Закав- казье); 11) ляллеманция культурная — Lallemantia iberica (Древняя Армения; в культуре); 12) ляллеманция щитовид- ная — L. peltata (вводится в культуру); 13) ляллеманция скаль- ная — L. rupestris (перспективна для селекции и, может быть, для введения в культуру); 14) люцерна армянская — Medicago praesativa (дико и в культуре; Древняя Армения); 15) люцерна малоазиатская — М. orientalis (Малая Азия; культурная); 16) люцерна сирийско-палестинская — М. syriaco-palcstiniña (Си- рия и Палестина; культурная); 17) люцерна йеменская — М. jeme- nensis (Иемен, культурная); 18) люцерна месопотамская — М. mesopotamica (Месопотамия; культурная); 19) люцерна желтая майкопская предгорная — М. quasifalcata (вводится в куль- 136
туру); 20) люцерна железистая — М. glutinosa (перспективна для селекции, особенно подвид ssp. praefalcata — М. vardanis); 21) клевер открытый — ТгНо1шт apertum (вводится в куль- туру); 22) тригонелла йеменская — Trigonella jemenensis (Йемен; культурная); 23) эспарцет закавказский — Опobrychis transcaucasi~ca (Закавказье; дико и в культуре); 24) эспарцет высочайший — О. altissima (Закавказье; дико и в культуре; по мнению В. И. Андруковича, этот вид следует присоединить к предыдущему); 25) эспарцет куринский — О. cyri (интересен для селекции, местами вошел в популяцию О. antasiatica); 26) эспарцет Биберштейна — О. biebersteinii (чрезвычайно по- лиморфный вид, разбитый теперь на ряд мелких видов; перспек- тивен для селекции; местами принял участие в формировании популяций культурного эспарцета); 27) эспарцет переднеазиат- ский — О. antasiatica (Закавказье; формирующийся культур- ный вид, получающий теперь широкое распространение; пра- вильнее включить его в О. tranecau~casica); 28) эспарцет мсхет- ский — O. meschetica (ставятся опыты с введением в культуру); 29) вика изменчивая — Vicia variabilis (Кавказ, Малая Азия; вводится в культуру); 30) козлятник восточный — Galega orien- talis; 31) ромашка розовая — Pyrethrum roseum (Кавказ, Се- верный Иран; дико и вводится в культуру); 32) ромашка крас- ная — Р carneum (Кавказ, Северный Иран; дико и вводится культуру); 33) белладонна кавказская — Atropa caucasica (леса Кавказа; дико и вводится в культуру, может быть, это не самостоятельный вид, а подвид или региональный экотип); 34) Scopolia icaucasica Kolesn. (лекарственное; колхидские леса, возможно, что кавказская форма представляет собой только подвид S. caipnioliica Jacq.); 35) чабер колосоносный — Satureja s~pi~cigera (Грузия; дико и в культуре); 36) анизет — Р1т~р1пе11а anisetum (дико в горных районах Малой Азии и там же в куль- туре); 37) Pimpine la aromatica М. В. (Закавказье, дагестан; дико «38) роза дамасская — Rosa da- mas~ccna Mill. (Сирия; в культуре); 39) груша кавказская— Pirus >cau~casica (очень полиморфна; дико и дала начало куль- турным сортам; по нашему мнению, является подвидом Р. com- munis); 40) груша сирийская — Р. syria~ca (Сирия; дико, изредка в культуре); 41) груша Тахтаджана — Р. takhtadzhianii (Юж- ная Армения, UeHTpaJIbHBH Грузия; изредка в культуре); 42) яб- лоня восточная — Malus orientalis (Кавказ, Иран; дико и, ве- роятно, участвовала в происхождении некоторых культурных сортов); 43) Nolinia litoralis (Кавказ — Колхида; раньше куль- тивировалась как кровельный материал); 44) сростноплодник— Symiphyoloma graveolens С. А. Меу. (эндем Главного Кавказ- ского хребта). Виды, имеющие в странах Передней Азии один из первичных 'или интенсивный вторичный очаг своего развития, либо пред- ставленные эндемичными формами и сортами: 1) пшеница мяг-
кая — Triticum aestivum (главный первичный очаг); 2) пшеница твердая — Т. durum (один из первичных очагов); 3) полба-эм- мер — Т. Йсоасоп (главный первичный очаг); 4) ячмень — Hor- deum euroasiatiicum (крупный вторичный, а может быть, и вто- рой после восточноазиатского первичный очаг); 4) рожь куль- турная и сорная — $еса1е cereale (основной первичный очаг); 6) просо — Panicum miliaiceum (небольшой вторичный очаг), 7) чумиза — Setaria Иа11са (эндемичная сорная форма, которую В. Л. Менабде и А. А. Ерицян выделяют в вид S. ketzkhove ii); 8) ежа — Dactylis glomerata (один из очагов формирования и: развития дикорастущего вида); 9) костер прямой — Bromus ri- iparius (один из очагов развития дикорастущего вида); 10) рай- грас английский — 1 о1шт perenne (один из очагов распросгране- ния дикорастущего вида, эндемичные экотппы); 11) райграс вы- сокий — Arrhenatherum elatius (эндемичные дикие и культурные экотипы); 12) тимофеевка — Phleum pratense (эндемичные ди- кие и культурные экотипы); 13) житняк ширококолосый — Agro- pyron cristatum (один из очагов первичного развития, может быть, основной); 14) овсяница луговая — Festuca pratensis (один пз первичных очагов развития дикорастущего вида; эн- демичные экотипы); 15) овсяница красная — Festuca гиЬга (один из очагов развития дикорастущего вида; эндемичные эко- типы); 16) горох — Pisum sativum (главный первичный очаг}; 17) чечевица — Lens esculenta (главный первичный очаг); 18) нут — Ci~cer arietinum (главный первичный очаг); 19) кон- ские бобы — Faba vulgaris (один из первичных? очагов); 20} вика посевная — Vicia sativa (один из первичных очагов, вероятно, основной); 21) вика паннонская — Vilcia раппопьса (один из первичных очагов, вероятно, основной); 22) вика изменчивая— Vicia variabilis (основной первичный очаг); 23) чина посевная— Lath~yrus sativus (один из очагов первичного развития вместе с Юго-Западной Азией); 24) чина нутовая — Lathyrus екега (основной первичный очаг культуры — Сирия); 25) люцерна по- севная — Medicago sativa sensu lato (первичный очаг возникно- вения культуры и развития группы видов); 26) пажитник посев- ной — Trigonella foenum-graacum (эндемичные экотипы); 27) па- житник голубой — Trigonel la icoerulea (эндемичные формы) „ 28) донник желтый — Melilotus officinalis (главный первичный очаг развития вида); 29) клевер красный — Trifolium ðãàtåïse sensu lato (эндемичные дикорастущие и культурные экотипы); 30) клевер александрийский — Trifolium alezandrinum (важней- ший первичный очаг развития вида); 31) лен — Linum usitatis- simum sensu lato (один из трех первичных очагов культуры и формообразования льна; родина озимого льна и производных от него форм); 32) конопля — Cannabis satix а (эндемичные сорта и культурные экотипы); 33) кунжут — Sesamum indicum (ряд эндемичных экотипов); 34) ляллеманция обыкновенная— Lallemantia iberica (родина вида и первичный очаг введения 138
в культуру; только недавно начала распространяться за пре- делы Передней Азии и поэтому помещена в список эндемичных культурных растений); 35) сурепица — Brassica icampestris (~ряд эндемичных экотипов); 36) горчица черная — Brassica nigra (ряд эндемичных экотипов; возможно, что здесь первичный, мо- жет быть, и основной очаг развития вида); 37) горчица белая— Sinapis alba (эндемичный экотип; Палестина); 38) индау- Егиса satIva (эндемичный подвид — экотип; Малая Азия); 39) подсолнечник — ilelianthus annuus (эндемичные сорта круп- носемянного грызового подсолнечника); 40) белямир — Cephala- ria зуг1а~са (один из первичных очагов, скорее всего, основной; вторичный — в Средней Азии); 41) мак — Papaver somniferum (эндемичный подвид опийного мака — Малая Азия); 42) са- флор — Carthamus tinctorius (область современного развития, а может быть и родина рода, важнейший очаг развития вида, ряд эндемичных экотипов); 43) клещевина мелкоплодная— Ri~cinus т1сгосаприз, iproles persieus G. Рср. (эндемичные формы в Армении и Азербайджане); 44) маслина — 01~ca euroipaea (пер- вичный очаг); 45) кориандр — Coriandrum sativum (один из оча- гов развития вида; эндемичные экотипы; некоторые авторы счи- тают Переднюю Азию первичным очагом культуры); 46) фен- хель — Foeniculum vulgare (один из первичных очагов, может быть, основной); 47) огурец — Cucumis sativus (эндемичные под- виды и экотипы); 48) дыня — Melo (один из трех первичных оча- гов видообразования и развития рода); 49) репа — Brassi~ca ra- роеигореа (один из первичных очагов видообразования и введе- ния в культуру); 50) морковь — Daucus carota (один из первич- ных очагов введения в культуру и формообразования; имеет два эндемичных подвида); 51) свекла — Beta vulgaris (основной первичный очаг введения в культуру и формообразования; ряд примитивных эндемичных форм); 52) кервель — Anthriscus ce- refolium (один из основных или основной очаг вхождения в куль- туру); 53) капуста кочанная — Вгазз1са caipitata (эндемичный подвид); 54) кресс-салат — Lepidium sativum (очаг развития вида, вероятно, основной); 55) шпинат — Spinacea oleracea (ве- роятно, область происхождения или один из очагов вхождения в культуру); 56) ревень смородинный — Rheum ribes (древний очаг культуры и, может быть, основной); 58) салат-латук — Lac- tBca sativa (эндемичные формы, возможно, один из очагов вхож- дения в культуру); 58) баклажан — $о1апит melongena (один из двух первичных очагов, второй — в Индии; ряд эндемичных экотипов); 59) портулак — Portulaca оlегасеа (эндемичные формы); 60) алыча — Prunus divaricata (основной очаг вхожде- ния в культуру, происхождения и формообразования); 61) куль- турная вишня — Cerasus vulgaris (очаг формообразования, воз- можно, происхождения); 62) черешня — Cerasus avium (очаг формообразования и, вероятно, происхождения); 63) миндаль— Amygdalus ~communis (эндемичные формы; первичный очаг
вхождения в культуру); 64) абрикос — Armeniaca vulgaris (эп- демичная группа сортов); 65) инжир — Е1сиз саг1са (возможно, один из основных очагов первичной культуры, форма- и вндооо- разования); 66) гранат — Рипica granatum (первичный очаг культуры и формообразования; область развития рода); 67) ви- ноград — Vitis viniiera (эндемичные группы сортов); 68) муш- мула — Mes~pilus дегтапйса (первичный очаг вхождения в куль- туру и формообразования; область развития монотипного рода); 69) кизил — Comus mas (первичный очаг формообразования и вхождения в культуру, вероятно, родина вида); 70) шелковица белая — Могиз alba (эндемичные плодовые сорта); 71) шелко- вица черная — М. nigra (эндемичные плодовые сорта); 72) фи- сташка — Pistacia vera (один из первичных очагов культуры и родина эндемичных сортов); 73) лещина — Corylus ave lапа (эндемичные дикорастущие формы и культурные сорта); 74) каштан — Cas anea sativa (эндемичные дикорастущие формы и культурные сорта) . Богатство дикорастущей флоры стран Передней Азии выра- жается наиболее ярко прежде всего в том, что на этой террито- рии возникли или достигли интенсивного развития многие роды, из которых выделились виды культурных растений. Кроме того, страны Передней Азии имеют ряд эндемичных культурных видов (распространенных и в диком, и в культурном состоянии только здесь), которые в других областях не возделы- ваются. K таким культурным эндемикам относятся или релик- товые (исчезающие, местные и раньше не имевшие широкого распространения) культуры (Мо inia litoralis) или новые куль- туры (кавказская ромашка, кавказская белладонна). Многие культурные виды и даже целые роды (Triticum, Secale и др.) возникли здесь, а затем широко распространились в другие страны. Пришельцы из других стран в Передней Азии образовали эндемичные виды, подвиды, экотипы, формы и сорта. Степень вторичного видо- или формообразования зависит от длитель- ности и интенсивности воздействия новой агросреды на занесен- ные растительные формы. Высокая древность и высокий уровень культуры древних оби- тателей стран Передней Азии, историческая преемственность этой культуры были важнейшими факторами формирования бо- гатой культурной переднеазиатской флоры, ставшей крупней- шим очагом распространения культурных растений на запад и на север в Европу, а на восток — в Среднюю и Юго-Западную Азию.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕ РАСТЕНИЯ СОВЕТСКОЙ СРЕДНЕЙ АЗИИ И СОПРЕДЕЛЬНЫХ С НЕЮ СТРАН СРЕДНЕ-ЮГО-ЗАПАДНОЙ АЗИИ Средне-Юго-Западная Азия в нашем понимании включает Среднеазиатские советские республики, из которых в отноше- нии аборигенной культурной флоры особенно интересны Таджи- кистан и южные части Узбекистана и Туркмении. Сюда же от- носится северо-западная Индия и соседняя часть северо-запад- ного 1~итая. Аборигенные культурные растения Сибири рассмотрены ниже; сделано это для того, чтобы не отделять истории древ- него земледелия Сибири от истории земледелия Русской рав- нины. В то же время в Восточной Сибири на культурной и ди- корастущей растительности обнаруживается сильное влияние стран Восточной Азии. ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ СОВЕТСКОЙ СРЕДНЕЙ АЗИИ Следы древнейшего человека (мустьерский период) обна- ружены в 1938 г. в отрогах Гиссарского хребта на высоте 1500м над ур. м. в пещере Тешик-Таш близ городка Байсун (недалеко от Душанбе). Череп и кости скелета принадлежат ребенку б— 7 лет и отнесены к неандертальскому типу. Пещера Тешик-Таш, видимо, служила долговременной стоянкой первобытного чело- века — здесь найдены следы очагов и каменные орудия (скребла и клинки). Хотя палеолит Средней Азии изучен еще слабо, но доказано, что человек жил здесь в глубокой древности. В 1945 — 1950 гг. произведены были раскопки в городище На- мазга-Тепе (Б. А. Литвинский, 1952) эпохи энеолита. Это горо- дище находится в пределах Копет-Яага на реке Лоинсу в 7 км от железнодорожной станции Каахка в Туркменской ССР Тол- щина культурного слоя достигает здесь 30 — 40 м. Нижние остатки зданий относятся к III тысячелетию до н. э. Установ- лено, что основным занятием жителей было земледелие. Най- дены каменные зернотерки и зерна пшеницы, ячменя, нута и винограда. Исследователи выделяют в Намазга-Тепе 6 слоев. В пятом слое поселение достигает наибольшего размера. Зерно в это
время хранилось в специальных больших сосудах. Наступает бронзовый BEK. В это время, кроме пшеницы и ячменя, появ- ляется нут. В дальнейшем нападения врагов приводят поселение к упадку, и площадь его сокращается, но жизнь продолжалась на других соседних «холмах» (Н. Я. Мерперт и Д. Б. Шелов, 1961) . История поселения Намазга-Тепе особенно интересна тем, что здесь можно проследить постепенный переход от первобыт- ного лиманного земледелия к пашенному. Особенно много дали для изучения древнейшей человеческой культуры раскопки эпохи энеолита и позднейших эпох в пред- горьях Копет-Дага у станции Анау около Ашхабада, в 2 км к западу от развалин древнего города Анау, начатые еще B 1904 г. и продолжающиеся в настоящее время. Раскопаны два холма. Обнаружено несколько культурных слоев. А. А. Марущенко (1956) дает следующую периодизацию для Анау. Анау 1 и начало Анау II: мотыжное земледелие, ско- товодство; Анау II и начало Анау III: плужное земледелие, вы- сокоразвитое скотоводство, выделение ремесел; Анау III: высо- коразвитое земледелие и скотоводство; классовое рабовладель- ческое общество. Из орудий откопаны пластины, шаровидные каменные бу- лавы, а также каменные зернотерки и, что самое важное, най- дены отпечатки соломы и остатки зерен мягкой пшеницы и двурядного ячменя. Обнаружены также и кости диких живот- ных. Этот наиболее древний культурный слой относится к эпохе энеолита и датируется III тысячелетием до н. э. Поселение Анау представляет собой памятник древнейшей земледельческой культуры в Средней Азии. Однако эта куль- тура уже не относится к самым примитивным, на что указывает характер жилищ. Несомненно, что самое возникновение здесь земледелия имело место значительно раньше, когда — точно не- известно. В 30 км от Ашхабада в песчаной Каракумской пустыне ис- следовано древнейшее поселение Джейтун (Н. Я. Мерперт и Д. Б. Шелов, 1961). В обмазке полов джейтунских домов были обнаружены от- печатки зерен ячменя и пшеницы. По мнению археологов, жизнь в этом поселении прекратилась в V тысячелетии до н. э., воз- можно, вследствие нашествия неолитических племен, еще не знавших земледелия. Но в некоторых других древних туркменских поселениях жизнь длилась по нескольку тысячелетий и культурные слои здесь образовали холмы до 30 — 40 м высотой. Самое местоположение древнейших земледельческих посе- лений на tore Туркмении свидетельствует о том, что ранние земледельческие оазисы появились и здесь, в предгорьях, так как именно в таких местах возможен наиболее легкий и прими- 142
тивный способ орошения, так называемый болотный или лиман- ный. Этот способ орошения существовал в Средней Азии до са- мого недавнего времени, а кое-где его можно видеть и сейчас. Как и в Двуречье, здесь человек лишь в более поздние пе- риоды и постепенно переходил к земледелию вблизи больших рек, где требовались сложные ирригационные сооружения. Од- новременно шло и освоение человеком безводных равнин, но иным путем — при помощи системы колодцев. В Средней Азии, еще в глубокой древности, население выра- ботало надежные способы устройства колодцев для орошения и питья из доступного материала. Эти способы описаны В. Н. Ку- ниным (1950) . Археологические исследования в Анау выявили существова- ние связей энеолитической культуры Средней Азии с современ- ными им культурами Приуралья, Восточного Туркестана, Ирана и побережий Индийского океана. Более поздние слои — Анау II — относятся к энеолиту. Сле- дующие слои — Анау III — датируются концом III тысячелетия и II тысячелетием до н. э. и относятся к бронзовому веку. Среди бронзовых изделий здесь встречаются также медные и камен- ные. Анау III это значительно более высокая стадия развития. Керамика здесь уже тонкостенная, сделанная с применением гончарного круга. Имеются металлические серпы. Земледелие в этот период перемещается в долины рек и, по-видимому, на- ряду с богарным появляется также и поливное земледелие. Для среднеазиатских поселений бронзового века характерны связи их с аналогичными культурами Западной Сибири, По- волжья и Ирана. Что касается этнического состава населения в это время, то предполагается, что это были племена, говорив- шие на языках восточнояфетической ветви и на скифских язы- ках. В еще более поздних слоях — в Анау IV и также в городище Тали-Барзу, недалеко от Самарканда и в Гяур-Калы, в Старом Мерве, найдены памятники, очень сходные со скифскими памят- никами Восточной Европы. На рубеже II и 1 тысячелетий до н. э. в Средней Азии разви- ваются два типа хозяйств: в полупустынях — кочевое скотовод- ство, а в оазисах и в речных долинах — оседлое земледелие. В первой половине 1 тысячелетия до н. э. в Средней Азии уже существовали большие оросительные сооружения, что под- тверждает данные античных писателей о наличии в этот период здесь государства. Древнейшими из ранних государственных образований яв- ляются Бактрия (Пригиндукушье в Афганистане; центральный, южный и восточный Таджикистан и долина реки Сурхан-Дарьи в Узбекистане), Хорезм (часть северного Узбекистана и юго- западного Казахстана, по нижнему течению Аму-Дарьи) и Согд, или Сугда (долина Зеравшана и Кашка-Дарьи).
Современные таджики являются потомками земледельческих народов древности: согдийцев, бактрийцев, тохаров и др. Тад- жики сложились как народность 1000 с лишним лет тому назад. Имеются указания на связь бактрийской и других двух древ- них культур нашей Средней Азии — Хорезма и Согда с Месо- потамией (Ассирией) и Египтом, несомненны также Нх связи с Индией. Античный автор Квинт Курций пишет, что в Бактрии (перед походом в Азию Александра Македонского, 329 г. до н. э.) много винограда, плодородные почвы заняты посевами хлебных зла- ков, а другие — пастбищами, но много и бесплодных сухих про- странств. В период существования греко-бактрийского царства торго- вые связи Бактрии с различными странами усилились — через эту страну проходили дороги на юг и восток в Индию, Кашгар и на запад — через Парфию и по Аму-дарье. В последние столетия 1 тысячелетия до н. э. создалось пар- фянское царство на территории современной Туркмении и сосед- них южных областей, впоследствии сильно расширившее свои владения, включив в нпх Иран, Месопотамию и Армению, что п свою очередь способствовало расширению связей этих стран и взаимной ассимиляции их культур. Во II в. до н. э. была установлена связь Парфии и Китая. В китайских источниках сообщается о развитии земледелия Парфии, о возделывании там риса, пшеницы и винограда. По китайским источникам, переведенным Н. Я. Бичуриным (1950), в Аньси (Бактрия) вели оседлую жизнь и занимались земледелием, сеяли рис и пшеницу, делали вино из винограда. Китайские источники говорят об изобилии дикого ячменя р Западной Бактрии. Предполагается, что хлопчатник также был занесен в Западный Китай из Средней Азии. Большое влияние на соседние страны оказала и культура Хорезма. Обширнейшие работы по изучению древней хорезмской культуры, ведущиеся С. П. Толстовым с сотрудниками, воссоз- дали историю этого очага среднеазиатской культуры. Самые древние слои относятся здесь к кельтеминарской культуре и имеют общие черты с древними слоями Анау. Ме- стность, где на рубеже IV и III тысячелетий до н. э. развилась кельтеминарская культура, была болотистой, с озерами и напо- минала теперешнюю Барабинскую степь. Орудия изготовлялись из камня и кости. Уровень культуры был сравнительно высокий, так как обитатели жили в огромных родовых помещениях, по- строенных из камыша и тростника, рассчитанных человек на сто. Основное занятие кельтеминарцев — рыболовство и охота. Указаний на существование земледелия нет. Ирригационных сооружений не было. В поздних стадиях кельтеминарской куль- туры начинается скотоводство (2-я половина III тысячелетия до н. э.) . 144
Помимо связей с культурой позднейшего Анау, прослежи- ваются связи кельтеминарской культуры с древней дравидий- ской Индией, с неолитом Синьцзяна (Синьцзян-Уйгурский авто- номный район), с неолитической культурой «ямочка-гребенчатой керамики» северо-восточной Европы и афанасьевской культурой Сибири. Следующий период — тазабагъябская культура датируется серединой II тысячелетия до н. э. и относится уже и энеолиту. Медные изделия этих слоев напоминают андроновскую культуру Сибири. 5Кители тазабагъябского времени занимались скотовод- ством, так как кости домашних животных обнаружены там в большом количестве. Следов земледелия в этот период обнару- жено не было. Ирригационные сооружения в эту эпоху отсутст- вовали. По мнению С. П. Толстова (1948), там, где на речны; поймах высокий уровень почвенных вод обеспечивает достаточ- ную влажность для произрастания культурных растений, могло быть каирное земледелие. Следующая ступень развития культуры названа С. П. 1'ол- стовым суярганской. В суярганских стоянках, датируемых сере- диной II тысячелетия до н. э., уже не было микролитов, а мате- риалы напоминают средние слои Анау. К началу 1 тысячелетия до н. э. суярганская культура сме- няется амирабадской. Ирригация в это время еще не получила заметного развития. Амирабадские стоянки расположены в сто- роне от древних каналов и, видимо, базируются еще на каирах. В этом обществе земледелие еще, по-видимому, не вышло из за- чаточного состояния. И не удивительно. Выше уже отмечалось, что в Средней Азии земледелие раньше началось и развивалось более быстрыми темпами в предгорьях, чем на равнинах и доли- нах больших рек. Собственно хорезмская культура развивается с VIII — VII вв. до н. э. Первый ее период продолжается до IV в. до н. э. Глав- ной отраслью хозяйства является скотоводство (рогатый скот, лошади, верблюды). Земледелие имеет второстепенное значение, оно, в основном, уже поливное. В V в. до н. э. не только существовала грандиозная ороси- тельная система, но о ее возникновении уже складывались ле- генды. С. П. Толстов относит время ее сооружения к VIII— VI вв. до н. э., когда в Хорезме уже возникает сильное рабо- владельческое государство с пережитками матриархальной о6- щины и родового 6~|та. В IV — III вв. до н. э. население от ско- товодства постепенно переходит к земледелию, что связано с еще более высоким развитием ирригации. Ассортимент растений, возделывавшпхся в Хорезме, ве- роятно, в этот период был уже большим и близким по составу к культурам более позднего времени. В комнатах в откопанном замке Топрак-I(aza найдены се- мена пшеницы, ячменя, проса, дыни, косточки урюка, персиков, 145
винограда. Здесь же были обнаружены монеты III в. н. э., по ним можно датировать и найденные растения (хлопок и, веро- ятно, лен также были известны, найдены бумажные ткани). Со- вокупность всех данных по Средней Азии и прилегающим к ней странам говорит за то, что хлопчатник возделывался здесь в еще более ранние эпохи. Найденный бюст женщины с плодом гра- ната в руках свидетельствует о том, что уже тогда культивиро- вался и гранат. Изображения белых и красных лилий наводят на мысль о культуре орнаментальных растений в садах Хо- резма III в. н. э. В Х в. н. э., по персидским источникам, в перечислении пред- метов вывоза из Хорезма названы изюм, виноград, орехи (лес- ные) и кунжут. Несомненно, что начало культуры кунжута от- носится к гораздо более древнему периоду, ибо это растение было известно еще древним земледельцам Двуречья и позд- нее — Ур арту. В образцах А. М. Беленицкого, начальника отряда Таджик- ской археологической экспедиции АН СССР, группой сотрудни- ков ВИР были найдены просо, пшеница, ячмень, бобовые (среди них нут). Ячмень преобладал голозерный. Этот материал был датирован VI — VII вв. н. э. и найден в раскопках древнего го- родища Пянджикента в Таджикистане. Культура согдийцев и отчасти хорезмийцев была связана с культурой Киргизии. По всей северной Киргизии открыты следы скотоводческой киргизской культуры П тысячелетия до н. э. (бронзовый век). В южной Киргизии в этот период уже занимались земледелием: выращивали ячмень, пшеницу, урюк. В VII в. до н. э. Киргизия была заселена воинственным ко- чевым племенем саков (персидское название скифов), которые славились своими конями. Связи Киргизии с западом и восто- ком развились в период от III в. до н. э. до I в. н. э. Одно время 1~~~ргизия находилась под властью усуней. Усуни были скотоводами, но на зимовках занимались не- много и земледелием — сеяли рис и просо. Отсюда, с гор Тянь- Шаня, распространилось любимое среднеазиатское блюдо— плов. К югу от Ферганского хребта возвышается Памир — «крыша мира». Древнейшие из открытых на Памире памятников при- надлежат тем ранним кочевникам, которых по китайским источ- никам называли «Сэ» (V — III в. до н. э.) и «Усуни» (II в. до н. э. — 1 в. н. э.) .. Западный Памир имеет более благоприятный климат, там давно жили земледельцы. Восточный Памир с его очень суро- вым климатом был царством кочевников. На Западном Памире стояли грозные крепости против кочевников Восточного Па- мира. В это время в южной Киргизии и Фергане (в «царстве Да- вань») существовало интенсивное земледелие с разнообразным
.ассортиментом культур. Здесь было развито и орошение. Для орошения первоначально использовали небольшие горные речки и ручьи — освоить такие большие реки, как Кара-Дарья или Нарын, было не под силу земледельцам того времени. В Фергане Bo II тысячелетии до н. э. существовала своеоб- разная культура эпохи бронзы. Стоянка эпохи бронзы открыта в северной Фергане в Андижанской области в районе поселка Чуст. В горах Ферганского хребта имеется много наскальных изображений диких и домашних животных. Эти наскальные ри- сунки относятся к разным периодам; на скале имеются изобра- жения яка. По А. Н. Бернштаму (1942), китайцы научились возделывать виноград и люцерну именно у даваньцев. Усуньская кочевая культура столкнулась с более высокой культурой согдийцев около V в. н. э. В Чуйской долине тогда была широко развита ирригация на основе использования горных речек. Там разводили рис, вино- град, урюк, яблони, туту, ячмень, просо. Кочевники под напором согдийцев уходили в глубь гор. В XI в. в Чуйской долине было много городов с водопрово- дами. Почва обрабатывалась сохой и кетменем. Нашествие монголов в середине XIII в. прервало развитие киргизской культуры, как и хорезмской. Поселения были раз- граблены и сожжены, ирригационные сооружения разрушены пли сами собой пришли в упадок, так как оставшиеся в живых жители ушли. На месте пышной хорезмской культуры водвори- лась пустыня; о былой культуре и интенсивном земледелии в Чуйской долине можно судить лишь по данным археологиче- ских раскопок. Современные народы Средней Азии — казахи, киргизы, уз- беки, туркмены, кара-колпаки — почти все тюркоязычны. Тад- жики — ир аноязычны. На рубеже нашей эры европейский тип на территории Сред- ней Азии стал смешиваться с монголоидным. В это время из центральной Азии через горы Северной Монголии, Алтая и Тянь-Шаня двигались орды северогунн- ского союза. Народы Средней и Центральной Азии пришли в тесное взаимодействие. Победители усвоили культуру побеж- денных, а побежденные заимствовали у победителей язык. Для Семиречья проводником земледельческой культуры была согдийская колонизация. Согдийцы переселялись из до- лины Зеравшана в Семиречье после двух арабских нашествий: в V в. и в VI — VIII вв. н. э. Согдийцы стали сеять в Семиречье пшеницу, просо, ячмень, разводить абрикосы и виноград. К югу от Каратау между предгорьями и берегами Сыр-Дарьи жили пастухи — скотоводы одного из э владений государства Кангюй (Хорезм). Часть жителей осела здесь в I тысячелетии н. э, Первые посевы появились в болотистых местностях, при- 147
втыкающих к Сыр-дарье, и в каирах. Таким образом, здесь зем- леделие развилось раньше, чем в Тянь-Шане. После Закавказья наиболее богатым составом культурных растений отличаются в нашей стране Среднеазиатские респуб- лики (3-е место занимает Дальний Восток). Главное богатство видовых и сортовых ресурсов культурных растений сосредоточено здесь в горных районах Копет-para, Тянь-Шанд и Памиро-Алая. Земледелие в низменных районах поливное, приуроченное к оазисам. Казахстан и северная Киргизия по составу дикорастущей и культурной растительности стоят ближе к Сибири, чем к типич- ным для Средней Азии горным районам. Область наибольшего разнообразия и интенсивного формооб- разования культурных растений не совпадает с государствен- ными границами. Среднеазиатский советский очаг культурных растений основной территорией имеет горный Таджикистан. К нему примыкают Афганистан и горная северо-западная Ин- дия. Трудами советских исследователей доказано, что в отно- шении культурной флоры — это единая область. Обыкновенна сюда присоединяют и Пенджаб, отделяя последний от собст- венно индийской области, которая в основном имеет другие, ха- рактерные именно для нее, культурные растения. Пенджаб представляет собой переходную область от перед- неазиатской области распространения культурных растений к южноазиатской, охватывающей в широком понимании Индию, Индокитай и Индонезию с Филиппинами. Ассортимент культурных растений Ирана имеет смешанный характер. В северо-западной части он носит черты культурной флоры Передней Азии. Северная и восточная части имеют не- мало общих черт с Афганистаном и Туркменистаном. Туркмения — также переходная область от Передней Азии к Средне-Юго-Западной. Западный Китай, в котором лучше всего исследованы окрестности Кашгара, по составу культурной флоры также примыкает к рассматриваемой территории, хотя и имеет некоторые специфические черты. Пестрый состав популяций местных сортов культурных ра- стений и наличие полуокультуренных форм (морковь, репа, лю- церна и др.) особенно характерны для Афганистана. АБОРИГЕННЫЕ КУЛЪТУРНЫЕ РАСТЕНИЯСРЕДНЕ-ЮГО-ЗАПАДНОЙ АЗИИ В ЦЕЛОМ Ниже помещен краткий обзор аборигенных культурных ра- стений Средне-Юго-Западной Азии, куда включены горный Тад- жикистан, прилегающие районы других Среднеазиатских респуб- лик, Афганистан, северо-западная Индия и Западный Китай. П ш е н и ц а м я г к а я (Т. aestivum 1 .) представлена в Сред- не-Юго-Западной Азии необычайным разнообразием форм. Осо- 148
бенной пестротой отличаются афганские местные популяции. Од- нако, несмотря на широчайший диапазон изменчивости и иа большое число варьирующих признаков, это признаки мелких систематических единиц, отнюдь не видового ранга. Нет здесь и дикорастущих форм рода Triticum. Здесь перед нами явно вторичный очаг интенсивного формообразования и только одной ветви пшениц — мягкой пшеницы в широком объеме этого поня- тия. Очевидно, что культура пшеницы еще в глубочайшей древ- ности была принесена сюда из соседних стран Передней Азии. В Афганистане сохранились очаги древних форм мягкой пше- ницы — местных компактных карликовых пшениц Т. compactum Host, которые мн включаем в вид мягкой пшеницы Т. aestivum. Эти формы компактной пшеницы — промежуточный этап от древнейших пшениц Т. antiquorum и Т. protomacha к современ- ным мягким пшеницам. Инфлятные формы пшеницы, образую- щие очаги в горах Рошана, Шугнана и Афганистана, по нашему мнению, имеют конституцию древнего типа, обладают свойст- вом легко терять через ступень инфлятности ости (пшеницы древнего типа были короткоостистыми или безостыми). Некоторые древние черты (круглозерность, короткоости- стость или инфлятность) имеет и индийский вид — п ш е н и ц а кругл озер на я (Т. sphaerococcum Peric.). Эта форма обосо- билась в Пенджабе и с большим правом, чем другие (принимае- мые С. Н. Кудряшевым за «виды» инфлятные и компактные формы Афганистана и Памира), может считаться самостоятель- hbIM видом, хотя теснейшее родство ее с мягкой пшеницей не- сомненно. Вероятно, древнейшие примитивные пшеницы почти одновременно проникли и в Средне-Юго-Западную Азию и в Пенджаб, а отсюда в Китай и Монголию, а затем в Сибирь. Р аж ь (Sacale сегеа1е L.) в чистом посеве в Средне-Юго-За- падной Азии не культивируется, но имеется огромное разнооб- разие сорных форм ржи в посевах пшеницы. Однако это разно- образие не больше даже и по числу признаков, чем в Передней Азии, а эндемичных, дикорастущих или культурных видов ржи здесь совсем нет. Рожь явно пришла сюда из Передней Азии в качестве сорняка, и здесь находится вторичный центр поли- морфизма сорной ржи. Афганская сорная сильно ломкая рожь [S. afghanicum (Vav.) Poshev.], по нашим исследованиям, встре- чается и в Туркмении. По нашему мнению, эта рожь не яв- ляется самостоятельным видом, а только подвидом полиморф- ного вида S. cereale, включавшего и сорную рожь. Ячмень (Hordeum L.) хотя и является здесь, по-видимому, древней культурой, но у него обнаружено только интенсивное формообразование, видообразования нет. Однако в Средне-Юго- Западной Азии, особенно в Иране и Афганистане, имеются свои местные сорта и эндемичные ботанические формы. Ф. Х. Бахтеев (1948) выделяет иранскую (proles iranicum Vav. et ВасИ.) и ирано-туркестанскую (proles irano-turkestani-
цш Vav. et Bacht.) агроэкологические группы. Они по своему характеру не являются переднеазиатскими, хотя распростра- нены и в Иране. Все же, по сравнению с Передней Азией, разно- образие ячменей на рассматриваемой территории значительно меньше. Может быть, ячмени Юго-Западной Азии и не были привезены из стран Передней Азии, а представляют собой або- ригенные формы. Ближайший дикорастущий родич культурного ячменя, хотя и не прямой его родоначальник Н. s~pontaneum С. Koch, распространен в пределах Средне-Юго-Западной Азии — в Туркмении, Иране и Афганистане. Вопрос нуждается в дал ьнейшем детальном исследов ании. Ф. Х. Бахтеев (1959), изучающий дикие ячмени, нашел в долине Кельте-Чинар, в 2 — 3 км южнее поселка Калининского, колосья шестирядного ячменя, у которого боковые колоски были снабжены ножками, подобно ископаемому ячменю Н. lagunculi- forme ВасЫ. Р и с (Oryza sativa L.) только относительно древняя культура в Средней Азии. Во время походов Александра Македонского рис уже разводился в Бактрии. Здесь успели выработаться ме- стные сорта риса, ценные для селекции. По мнению Г Г Гу- щина (1938), рис занесен сюда из Китая, некоторые другие ав- торы предполагают, что из Индии. древнейший очаг культуры риса в СССР — Средняя Азия. Просо (Panicum milia~ceum L.) очень древняя культура в Средне-Юго-Западной Азии, особенно в советской Средней Азии. Отличается здесь довольно большим экологическим и мор- фологическим разнообразием. В. Н. Лысов (1949) установил су- ществование здесь двух экотипов ' развесистого проса (средне- азиатский горный и притяньшаньский) и экотип сжатого проса (среднеазиатский низинный). Последний, вероятно, отсюда рас- пространился и в Закавказье. Культура проса пришла сюда, ве- роятно, из Китая. С о р г о (Sorghum Moen~ch) также древнее культурное расте- ние Средне-Юго-Западной Азии, но распространилось сюда из Африки. Е. С. Якушевский (1949) относит сорго рассматриваемой территории к африкана-азиатской группе и в этой группе выде- ляет подвид азиатского сорго — Sorghum durra ssp. asiaticum Jakushev. Здесь встречается также и другой подвид — S. durra ssp. abyssinicum Jakushev. Эти подвиды представлены особыми ме- стными сортами. Флора Среднеазиатских республик и соседних стран богата видами и формами к о р м о в ы х з л а к о в. Большой интерес представляют виды Agro~pyron, Roegneria, Agrostis, Bromus ~ В книге «Просо», изданной в 1968 г., В. Н. Лысов называет их эколо- го-географическими группами; в дальнейшеч изложении агро-ботаническая классификация приводится по последнему изда|шю. 150
и др. Кормовые злаки этой географической области очень слабо изучены в культуре. Из к о р м о в ы х культурных б о б о в ы х растений наиболее древней является люцерна — Mediicago asiatica Ь1пзЕ. Весь род Medi~cago переднеазиатского происхождения, но возделываемая люцерна Средне-Юго-Западной Азии — культура автохтонная, первичная, ибо культурные сорта азиатской люцерны явно про- изошли от местных эндемичных дикорастущих видов. Последних имеется несколько на территории Среднеазиатских республик и Афганистана. Они далеко не так разнообразны, как кавказские люцерны того же подрода Falcago Grossh., границы между ними гораздо более сглаженные, и, несомненно, число основных азиат- ских видов должно быть значительно уменьшено против того, что имеется в монографии И. Т. Васильченко (1950). Тем не менее дикорастущие люцерны Средне-Юго-Западной Азии очень разнообразны и заслуживают основательного изучения в усло- виях культуры. Рассматриваемая географическая область яв- ляется вторым первичным очагом вхождения люцерны в куль- туру и вторым по значению очагом развития подрода многолет- них люцерн — Falcago. Первый очаг (более древний) развития подрода 1.'а1садо на- ходится в странах Передней Азии, это подтверждается тем, что: 1) даже близкие (нередко скрещивающиеся между собой) энде- мичные виды морфологически гораздо более обособлены и в смене популяций не связаны переходными формами, как в Средней Азии (например, такие виды, как М. g utinosa M. В. sensu laho, М. hemicycla Grossh., М. dzhawakhetiica Bordz. и др.); 2) в Передней Азии распространены интегральные, со- единяющие в организационном типе или в комплексе популяций черты других близких видов (М. hemicycla, М. glutinosa sensu lato); 3) Передняя Азия является главным очагом распростра- нения и формообразования более древних диплоидных видов синей и желтой люцерны. Диплоидные виды в Средне-Юго-За- падной Азии распространены только в прикаспийских районах и связаны переходными зонами популяций с близкими кавказ- скими формами, т. е. все они имеют переднеазиатский (большей частью закавказский) корень. Л ю ц е р н а а ф г а н с к а я культурная (Medicago asiatica Sinsk.) представлена в Средне-Юго-Западной Азии следующими эндемичными подвидами: 1) туркестанским — зэр. turkestanica Sinsk. (в этот подвид входят равнинно-туркестанский и семире- ченский экотипы); 2) туркменским — ssp. turkomaniica Sinsk.; 3) хивинским — ssp. khivinica Sinsk. 4) кашгарским — ssp. si- nensis Sinsk.; 5) кандагара-кабульским — ssp. kandagaro-kabu- lica ЯпзЕ; 6) гератско-хоросанским — ssp. geratica Sinsk. Тибетский гибридный экотип (ладак) имеет сложное гибрид- ное происхождение, где основой послужила, вероятно, люцерна афганская (кандагара-кабульского подвида). Эта совокупность
гибридных популяций, однако, весьма своеобразна, представляет собой характерный местный сорт, в настоящее время райониро- ванный в некоторых районах СССР Донни к бел ы й (Melilotus albus Desr.) представлен здесь предгорноазиатским экотипом (западный Тянь-Шань) и восточ- ностепным экотипом (главным образом южный Казахсган). П а ж и т н и к (Trigonella foenum-graecum 1 .) имеет в Сред- не-Юго-Западной Азии эндемичные экотипы: 1) западнокитай- ский и 2) иранский. Последний обнаруживает некоторые общие черты с западноевропейским экотипом и потому не связан с пе- реднеазиатским очагом пажитника. В Средне-Юго-Западной Азии идет интенсивное формообра- зование (не видообразование) зерновых бобовых культур. Полуди- кая чечевица (с растрескивающимися, очень мелкими бобами)— рго1ез subspontaneae Ваги1. вида Lens еэси1еп1а распространена в прииндийском районе Афганистана. Другие — мелкосемян- ные и близкие к дикорастущим формы iproles pilosae Ваги1. эндемичны для северо-западной Индии, входящей, по нашему мнению, в область Средне-Юго-Западной Азии, и встречаются также в Пенджабе, т. е. примыкают к окраинам и Переднеазиат- ской подобласти (эти пролесы должны быть выделены в подвиды, если не в виды) . Более близкие к культурным формам, но мелкосемянные и часто темносемянные, составляют proles asiati- cae Ваги1., распространенные в Передней и Юго-Западной Азии. В Передней Азии сосредоточены и промежуточные (iproles inter- mediae Ваги1.) формы, переходные к пролесам еигореае (Сред- няя Европа) и к средиземноморским, наиболее культурным, крупносемянным формам, которые начинают встречаться в Пе- редней Азии, восточнее их нет. Таким образом, разнообразие всех групп (пролесов) сосредоточено в Передней Азии. Из этого древнейшего первичного очага распространение чечевицы шло на восток и на запад. На востоке (Афганистан и прилегающий район северо-западной Индии) сохранились наиболее примитив- ные формы, а на западе интенсивно происходил процесс окуль- туривания. Подобная схема приложима и к куту, и к гороху, и отчасти к конским бобам. Средне-Юго-Западная Азия, хотя и не является первичным очагом нута, но имеет эндемичные группы: iproles turkestani- сит G. Рор., iproles kashgaricum G. Pop. и proles afghanicum G. Рор. Эти три пролеса отнесены к азиатскому подвиду ssp. asiati~cum G. Рор. Среди второго подвида ssp. orientale G. Pop. для данной геообласти эндемичны proles pamiricum G. Рор. и pro- les montanum G. Рор. Отсюда нут распространился в Индию и образовал proles indicum G. Рор., а из Индии — в Эфиопию. где образовался proles abyssinicum G. Рар. Маш золотистый [Phaseolus aureus (Roxb.) Pip.] расте- ние, несомненно, азиатского происхождения, но первичный очаг культуры его точно определить трудно. Наиболее близкий дико-
растущий вид — Ph. sublobatus Poxb. распространен в Индии и Индонезии. Если этот вид и не является непосредственным прародителем маша, то указывает на ареал их вымершего об- щего прародителя. Скорее всего первоначальный ареал этих родичей был приморский — по азиатским южным берегам Те- тиса вплоть до Японии. Соответственно растянут и ареал куль- турного маша — от советской Средней Азии до Японии. В Юго- Западной Азии имеется свой подвид sstp. iranicus G. Pop. Средне-Юго-Западную Азию можно рассматривать как вторич- ный очаг формообразования маша золотистого. Горох (Pisum sativum L.) представлен в Средне-Юго-За- падной Азии азиатским подвидом — ssp. asiaticum Gov., для которого особенно характерны агроэкологические группы форм: proles afghanicum Gov. ц proles persicum Gov. (Афганистан и Иранский Сейстан). В горных районах Средне-Юго-Западной Азии сосредоточены мелко- и темносемянные формы гороха, как и других бобовых. Конские бо бы (Viicia faba L. цли Faba vulgaris Moench) средиземноморского происхождения. В Юго-Западной Азии и отчасти в Индии сосредоточено разнообразие примитивных мелкосемянных форм этого вида, которые в древности были рас- пространены и в Европе. По В. С. Муратовой (1937), для рас- сматриваемой террцторцц характерна среднеазиатская горная агроэкологическая группа местных сортов — мелко- и темно- семянных. Ч и н а (Lathyrus sativus L.), по Ф. Л. Залкинд (1937), в Средне-Юго-Западной Азии представлена эндемичными груп- г.ами: 1) ~proles iranicus Zalk. (Афганистан, Таджикистан, Иран); ) некоторые эндемичные формы, входящие в состав proles indiicum Zalk. Классификация чины составлена очень за- путанно и разобраться в географии отдельных форм нелегко. go ХХ в. древняя для Юго-Западной Азии культура хл о п- ч а тн ц к а базировалась на местном виде с местным названием гуза (Gossypium herbaceum 1..). Этот вид дико растет в северо- западной Индии и в Восточной Африке по берегам Индийского океана, откуда распространился в страны Средне-Юго-Запад- ной Азии и отчасти в Переднюю Азию (зэр. euherbaiceum Nauer). Теперь здесь этот вид стал реликтовым. К о н о п л я растет дико в Азии к северу от Гималаев и Гин- дукуша, и первичных очагов вхождения ее в культуру, вероятно, несколько. Обыкновенная прядильная конопля Cannabis sativa L. пред- ставлена на рассматриваемой территории среднеазиатской груп- пой (Т. Я. Серебрякова, И. А. Сизов, 1940), похожей на средне- русский тип. На юге распространен эндемичный вид гашцшной конопли С. indiica (Lam.) Serebr. Культура индийской конопли занесена и в Малую Азию, Сирию, Палестину и Северную Аф- рику, но в диком виде распрос1ранена только в северо-западной 153
11ндии Восточном Афганистане и прилегающих к нему районах Иранд. Таким о~разом, здесь находится первичный очаг проис- хождения этого вида и вхождения его в культуру. Кенды рь представлен имеющими хозяйственное значение видами Apocynum scabrum Russ. (от Усть-Урта до Афганистана и Ирана) и А. lancifolium Russ. (бассейны Сыр-дарьи, Чу, Иртыша), Западный Китай (Кашгар). Л е н (Linum usitatissimum L.) в Средне-Юго-Западной Азии очень древняя культура. Он представлен масличными фор- мами. Здесь, по нашему мнению, находится первичный очаг (один из трех) — первоисточник вымерших форм дикорастущего льна L. angustifolium, некогда, вероятно, распространенного и в Азии по берегам исчезнувших морей. Наиболее древними из культурных форм являются поздние, медленно растущие яровые кудряши, в первый период вегетации имеющие простратный га- битус, слабее выраженный, чем у переднеазиатских озимых льнов. В условиях горной богары эти формы дали начало ран- ним кудряшам, а при продвижении в долины с умеренным увлажнением и на север — межеумкам, а затем — при дальней- шем движении на север через Урал — и долгунцам. Особое раз- нообразие льнов имеется в Таджикистане, а также и в Афгани- стане, где находится стык собственно индийских ранних крупно- коробочных льнов и аборигенных позднеспелых льнов Афгани- стана, К,ашмира и Таджикистана. Льны Афганистана, Таджикистана и Узбекистана дают в каждой стране сходные, но вместе с тем и несколько разли- чающиеся ряды форм от позднеспелых, склонных в начале жизни к простратному росту, через быстро растущие кудряши к еще быстрее растущим межеумкам. В горах выделяются также низкорослые кудряши, быстро растущие, но скоро прекращаю- щие рост. Схема филогенетического развития льна из этой обла- сти исторического развития культурной флоры приведена ниже. Лен в Хивинском оазисе представлен особенно поздними и медленно растущими формами, часто белоцветковыми. Лен в Кашгаре дает особую разновидность того же корня — от позд- него кудряша до позднего межеумка — часто белоцветковую и с особой звездчатой формой цветка. Среди этих льнов имеются формы с неплохим волокном. Кунжут (Sesamum indicum L., масличное растение) имеет в Средне-Юго-Западной Азии, по В. М. Гильтебрандту (1932), следующие экотипы: 1) хивинский (эндемичный для Хи- винского оазиса); 2) бухарский; 3) экотип Таджикистана, нося- щий черты сходства с бухарским и афганским; 4) афганский, заходящий и в Иран; 5) эндемичный экотип Ирана (район ва- рахша); 6) экотип Исфагана, обнаруживающий черты сходства с японским кунжутом. С а флор (Carthamus tinictorius L.) давно известен как кра- сильное растение и сравнительно недавно стал возделываться 154
Схема происхождения и распространения льна из индо-афганского очага Дикорастущий лен Linum angustifolium(вымершая форма, некогда существовавшаяпо берегам исчезнувших морей Средней Азии) Исходная культурная форма (медленно растущийкудряш со склонностью к простратностив первый период развития) Кашгарскиемедленно рас-тущие кудряши Хивинские мед- ленно растущие кудряши орные карликовыекудряши Кашгарскиесреднерослыекудряши Хивинскиесреднерослыекудряши Среднерослыекудряши Кашгарскиеполумежеумкии межеумки Казахстанские (и поволжские) лесостепные межеумки Северные русскиедол гунцы также на масло. В Средне-Юго-Западную Азию он давно при- шел из Индии или из Передней Азии; в Афганистане, Таджики- стане и соседних странах выделены А. И. Купцовым следующие экотипы: 1) экотип Кашмира; 2) экотип южного Афганистана; 3) экотипы Кабульской и Гератской провинций; 4) эндемичный экотип Герата; 5) экотип Памира; 6) экотип Туркестана; 7) эко- тип Чимкента; 8) экотип Семиречья, Бухары и Туркмении; 9) экотип северо-восточного Ирана. В направлении с юга на север экотипы становятся менее мощными. Захождения ареалов показывают, что экотипическая классификация не вполне доработана, однако обращает на себя внимание большой полиморфизм культурного сафлора в данной области исторического развития культурной флоры. Здесь на- ходится очаг интенсивного формообразования сафлора, сливаю- щийся с индийским и переднеазиатским очагами. В Афганистане встречаются примитивные формы культурного сафлора, имею- щие некоторые признаки дикорастущих родичей. 155
Кл ещеви н а (gicinus mi~crecarpus G. Pop.). В Азии, по Г М. Поповой, находится вторичный очаг распространения и формообразования мелкоплодной клещевины. В Афганистане встречаются дикие формы, но они распространены и в Малой Азии. Юго-Западной и Малой Азии свойственна общая группа сортов — proles persicum G. Pop. М а к (Papaver somniferum L.) представлен на рассматри- ваемой территории эндемичным подвидом, в состав которого вошли формы опийного назначения ssp. tianshanicum М. Vesse- lov. и ssp. centro-asiaticum М. Vesselov. (Киргизия, Узбекистан, Ь;ашгар). И н д а у (Eru~ca sativa Lam.) происходит из Северо-Запад- ной Африки. В Юго-Западной Азии нами открыто огромное раз- нообразие форм, засоряющих лен и другие культуры. Еще не- давно в горных районах возделывалась смесь индау и маслич- ного льна. К р е с с - с а л а т (Lepidium sativum L.) древняя культура в этой области. Но здесь распространены в основном такие же формы, как и в странах Передней Азии, где имеет место основ- ной очаг формообразования и расселения. Гop ч и ц а с а р еп тс к а я [Brassilca juncea (L.) Czern.] в культуре в Средне-Юго-Западной Азии, за исключением За- падного Китая (Кашгара), почти неизвестна, но имеется зна- чительное разнообразие сорных форм, относящихся к var. subsa- reptana Sinsk. (скорее — подвид). Район распространения дико- растущих форм — Джунгария, Монголия. Популяции H. juncea из Афганистана довольно полиморфны и недостаточно изу- чены. Кашгарские формы составляют особый кашгарский экотип, интересный для селекции. В Кашгаре и сопредельных районах находится основной очаг распространения желтосемянной са- рептской горчицы. С у р е п и ц а (Brassica campestris 1 .) имеет, по нашему мне- нию, первичный очаг формообразования в Юго-Западной Азии (Афганистан и Индия). На Памире и в Афганистане распро- странены эндемичные разновидности (скорее подвиды) маслич- ной сурепицы. Еще большее, чем в Афганистане и Таджикистане, разнообразие форм сурепицы свойственно Индии, где сформи- говались и близкие эндемичные виды. Нами описаны следую- щие разновидности (вернее, подвиды), соответствующие регио- нальным экотипам: 1) памирский — var. рат1гка Sinsl'. (высоко- масличный, близок к сурепзще Киргизии); 2) кабульский — var. 1<abuli~ca Sinsk.; 3) кандагара-кабульский — var. afghanica Sinslc. Ажгон (Carum ajowan Benth. et. Ноо!'.) довольно широко культивируется в Афганистане как эфирномасличная и лекар- ственная культура. Здесь он отличается большим полиморфиз- мом. Можно предполагать, что в Афганистане — первичный очаг культуры этого растения. Из него добывают тимол. 156
К о р и а н д р (Coriandrum sativum L.) распространен в Средне-Юго-Западной Азии как овощное и лекарственное ра- стение. Е. Н. Столетовой описан туркестанский «тип» этого растения, распространенный в Среднеазиатских республиках и в Западном Китае. А н и с (Pimipinella anisum L.) представлен здесь особой западнокитайской группой — iproles sinense Lusina. Ш ал фей му с к атн ый (Salvia эсlагеа L.) широко рас- пространен в горном и предгорном поясах Копет-Дага, Памиро- Алая, Тянь-Шаня. С. Н. Кудряшев считает Среднюю Азию одним из центров происхождения и формообразования мускат- ного шалфея, что не является вполне доказанным. Мускатный шалфей имеет огромный ареал — от западных до восточных гра- ниц Древнесредиземноморской области развития культурной флоры, и эколого-географическое разнообразие этого растения еще очень слабо изучено. Таран (Polygonum coriarium Grig.) ценное дубильное ра- стение. Введено в культуру в странах советской Средней Азии. Азиатские формы не вполне сходны с европейскими. Борщевик Л ем анн а (Нега~с1еит lehmannianum Bge.) эндемичен для Средней Азии (Памиро-Алай). Может быть использован для получения анетола. Т а у — с а г ы з (S~corzonera tau-saghyz Liipsch. et Bosse.) от- крыт в Каратау, а затем найден и в Таджикистане (Гиссарский хребет). Описан ряд видов, которые скорее являются формами или экотипами. К о к - с а г ы з (Taraxaicum kok-saghyz @odin) распростра- нен в отрогах Тянь-Е!аня в Казахстане. Возможно, имеется в Западном Китае. В Средне-Юго-Западной Азии находится первичная область развития эндемичных форм р е п ы, объединяемых в агроэко- логическую группу, которая может быть выделена в самостоя- тельный ботанический вид Brassica rapoasiatica Sinsk. Особенно велики разнообразие форм репы и пестрота популяций в Афга- нистане, где широко распространены примитивные формы реп с грубым, тонким, целиком погруженным в почву, бороздчатым корнеплодом. В советской Средней Азии и в Индии абориген- ные формы репы того же вида более окультурены и менее разно- образны. Дикорастущих родичей ни в Средне-Юго-Западной Азии, ни в собственно Индии не найдено. В этой области имеется третин первичный очаг видообразования репы и независимого вхождения ее в культуру. (первый очаг — в Средней и северной Европе, второй — в Малой Азии). Из Средне-Юго-Западной Азии репа попала в Японию, где дала начало вторичному очагу формообразования. Дикие родоначальники репы, вероятно, имели некогда широкий приморский ареал по берегам Тетиса, как и дикие родичи культурных редек и редисов. Родоначаль- ники европейских эндемичных форм репы, может быть, жили 157
где-нибудь по берегам Атлантического океана и Северного моря В целом, по нашему мнению, ареал исчезнувших диких родо- начальников репы сходен с известным нам огромным ареалом дикорастущей свеклы — от Индии до берегов Южной Швеции. P е д и с (Paphanus sativus 1 .) в Афганистане представлен особой группой местных сортов, близких к европейскому редису, но по признакам плодов, несколько приближающихся к азиат- ским (монгольско-китайским) формам. Средне-Юго-Западная Азия является, несомненно, первич- ным центром развития некаротиновой азиатской м о р ко в и (Dau~cus carota L.), которая заслуживает, по нашему мнению,, выделения в самостоятельный вид. П. Ф. Загородских (1939) выделяет эту полиморфную группу местных сортов моркови в подвид ssp. afghanicus Zagor. с двумя группами в его со- ставе: 1) флавоновой — желтой и 2) антоциановой — фиолетовой (последняя больше распространена в Афганистане и в Индии) Как и у репы, наиболее примитивные формы, приближающиеся к диким, сосредоточены в Афганистане. В советской Средней Азии распространены более окультуренные аборигенные формы желтой и белой моркови, а в Индии — антоциановой: фиолетовой и розовой. Дикорастущие формы моркови этого вида распростра- нены во всей области. По генеративным признакам они отли- чаются от европейской дикой и культурной моркови и сходны с азиатскими культурными формами некаротиновой моркови М. K. Рубашевская (1931) установила особый подвид дикора- стущей азиатской моркови — ss~p. orientalis Pubaschev., который. скорее следует считать видом. Основной очаг происхождения корнеплодной с в е к л ы (Beta vulgaris), по В. Т. Красочкину (1960), — Передняя Азия. Однако этот очаг частично как бы продолжается в Юго-Запад- ную Азию. В Афганистане обнаружен полукорнеплодный прими- тивный (переходный к типичным современным корнеплодным сортам столовой свеклы) сортотип — Бадахшанская. Д ы н я (Melo Adans.) имеет, по К. И. Пангало (1955), в Юго-Западной Азии следующие эндемичные виды (или под- виды?), распространенные в Среднеазиатских республиках, Аф- ганистане и Иране: 1) М. chandalalc Pang. 2) M. ameri Pang.; 3) М. zard Pang. 4) М. flexuosus Pang. По классификации среднеазиатских дынь существуют зна- чительные разногласия, однако, несомненно, что Средняя Азия— один из очагов формообразования сахаристых культурных дынь, относящихся, по А. И. Филову (1959, 1960), который не выде- ляет дыни в род Melo, к Cucumis те1о ssp. rigida (Pang.) Fil. Эти дыни представлены разновидностями: 1) хандалак — var. chandalal< (Pang.) Greb., 2) летние мягкие — var. bucharica (Pang.) IFil., 3) летние хрустящие — var. autumnalis Fil., 4) зим- ние — var. hibernus Fil. Дикий родич культурной дыни — М. ag- restis (Naud.) Pang. — засоряет посевы в советской Средней 158
Азии, в Афганистане и Иране. Юго-Западную Азию можно при- знать одним из первичных или, во всяком случае, очень древним центром культуры дыни (главный центр — Передняя Азия). Средне-Юго-Западная Азия, по А. И. Филову (1960), яв- ляется основной территорией интенсивного формообразования столового сахаристого а р б у з а (Citrullus Sichrad.) .. По А. И. Филову (1960), родоначальником столового арбуза является особая форма С. colocynthis L., широко распростра- ненная в Египте, по северным окраинам Сахары и в Алжире, т. е. по Средиземноморской Африке. Из африканской части Средиземноморья арбуз распространился в Средне-Юго-Запад- ную Азию, там происходило образование высококультурных сор- тов. По этой версии столовый арбуз перешел из Северной Аф- рики в Азию в виде дикаря или сорняка; в Азии он стал куль- .тивироваться; во всяком случае, крупнейший очаг формообра- зования арбуза находится в Средней Азии. Декандоль (1885) считает возможным, что первые шаги в окультуривании арбуза были сделаны в Индии — во влажных предгорьях Гималаев. Однако, по А. И. Филову (1960), окуль- туривание арбуза достигло высшей ступени не во влажном климате, а в сухом и жарком. В Средней Азии арбуз был издревле популярным растением. В местностях поливного земле- делия конкурентом его была и остается дыня. Наиболее древ- ние арабские авторы не упоминают о столовом арбузе, зато сведения о нем имеются у Авиценны (Ибн Сина), которыи жил в Х — XI вв. н. э. Ибн Байтар в XIII в. тоже пишет о столовом арбузе. Азиатское происхождение арбуза подтверждается существо- ванием многих местных названий его на таджикском, монголь- ском, татарском и других азиатских языках (по А. И. Филову, 1960): тарбиз — по-таджикски, хорпудж — по-бенгальски, тар- бах — по-монгольски, карбуз — по-татарски, тарбуз — по-уз- бекски, гармыз — по-азербайджански. О г у р е ц (Cuicumis sativus 1 .) в Западном Китае (Кашгар) представлен особой кашгарской группой рго1ез kasgharicus Fil., которая очень отличается от южно- и восточнокитайских сортов. В Средне-Юго-Западной Азии и в Передней Азии (Закав- казье, Иран) распространены тыквы вида Cucurbita moschata Duch., которые А. И. Филов (1960) включает в подвид turke- stani~ca Fil. последний, однако, не является эндемичным для Средней Азии. Сюда относится сорт Дербентская перехватка (разновидность пальцевпдная — longiicanpa Fil. и другая разно- видность перехватки — var. berberiica Naud.) . KpoMe Средней Азии и Закавказья, они встречаются в Иране, Западном Китае и Севернои Африке. Сюда относятся сорта Плов-Кеды, Ленин- абадская, Кашгарская, Бирючекутская 628, Перехватка 69 и др. Каким путем распространялись эти тыквы в древности? Наиболее вероятно, что путь их был следующим: из Северной 159
Африки в Переднюю Азию, затем в Среднюю Азию (через Иран) и из Средней Азии в Западный Китай (Кашгар). Лук р епч аты й (Allium сера L.) представлен в Средне- Юго-Западной Азии большим количеством эндемичных сортов. Здесь произрастают и его близкие дикорастущие родичи: А. pskemense В. Fedtsch. (Алатау), А. vavilovii М. Рор. et. Vved. и др. В этой области находится первичный очаг развития куль- туры репчатого лука. Для Среднеазиатских республик Н. Х. Тро- фимец (1939) выделяет среднеазиатскую группу местных сор- тов — proles turkestanicum Troph. По всей данной области в це- лом луки еще недостаточно изучены. Чеснок (А11шт sativum L.), по А. В. Кузнецову (1948), представлен главным образом стрелкующимися формами — ssp. sagittatum Кикп., в состав которых входит среднеазиатская группа местных сортов (Узбекистан, Таджикистан, Казахстан). Имеется также среднеазиатская группа местных сортов нестрел- кующегося подвида — ssp. vulgare Kuzn. По А. В. Кузнецову (1948), наибольшее разнообразие форм чеснока имеет место в северо-западной Индии, Афганистане, Таджикистане, Узбекистане, в западном Тянь-Шане. Здесь рас- пространен и дикий чеснок, следовательно, находится первич- ный очаг развития вида и вхождения его в культуру. В Юго-Западной Азии находится один из вторичных очагов видообразования и один из первичных очагов вхождения в куль- туру (очаг видообразования вторичный по отношению к Восточ- ной Азии) груши (Pirus L.), хотя далеко уступающий по раз- нообразию и значению переднеазиатскому очагу. Здесь распро- странены следующие дикорастущие виды, может быть принн- мавшие участие в эволюции культурного ассортимента и употребляющиеся как подвой: 1) Р. asiae-mediae М. Рор. (Тянь- Шань); 2) Р. liorshinskyi Litv. (Таджикистан); ) Р. regelii Rehd. (западный Памиро-Алай и Тянь-Шапь, Туркмения). Юго-Западная Азия имеет и свои виды дикорастущей я 6 л о н и, которые приняли участие в формообразовании куль- турных сортов и имеют значение, как исходный материал для селекции: 1) яблоня Сиверса — М. sieversii (Ledeb.) Roen>. (Памиро-Алай, Тянь-Шань; имеются формы, переходные к культурным; представляет большой хозяйственный интерес); 2) яблоня туркменская — М. turkmenorum Juz. et М. Рор. (Ко- пет-Даг); 3) яблоня Недзведцкого — М. niedzwetzl~yana Dieclc. (Тянь-Шань, Кашгар; большое разнообразие форм этого вида имеется и в Азербайджане). По Ф. Д. Лихоносу (1954), сорта, близкородственные яблоне Недзведцкого (с красными цветками и красной мякотью пло- дов), распространены в Казахской ССР под названием куль- джинок. По К. Ф. Костиной (1935), миндаль кул ьтур н ый — Ату- gdalus icommunis L. — распространен дико в горах Тянь-Шаня 160
и Копет-,Яага. Там же растет и близкий ему вид А. bucl>arica Korsh. На Кавказе произрастает другой близкий к культурному миндалю вид А. fenzliana Lipsky. Все остальные виды минда- лей Передней Азии, Афганистана, Ирана, все миндали Китая, а также миндали Северной Америки (Калифорния, Техас, Ка- нада) никакого участия в происхождении культурного ассорти- мента миндаля не принимали. Е1ентр видового разнообразия, по K. Ф. Костиной, находится в Иране, т. е. примыкает к Передней Азии и, следовательно, областью происхождения культурного миндаля приходится признать Переднюю Азию (куда включается Копет-,1~аг). Америка и Восточная Азия имеют по нескольку эндемичных видов Amygdalus. В западном Тянь-Шане и западном Памиро- Ллае произрастает А. ulmifolia M. Рор. Он интересен тем, что путем естесгвенной гибридизации его с алычой возник, по мне- нию ряда исследователей, новый вид сливы Prunus 1егganica Lincz. Алыча (Prunus divaricata Ledeb.) распространена в Сред- неазиатских республиках в дикорастущем и культурном состоя- нии, но не отличается таким полиморфизмом, как в Передней Азии, и особенно в Закавказье. Однако алыча очень разно- образна в южной Киргизии. М. Г Попов (1929) установил две разновидности среднеазиатская алычи: 1) var. orientalis М. Рор. (Копет-par); 2) var. turcomanica М. Рар. (Тянь-Шань, Па- миро-Алай) . С л и в а ф е р г а н с к а я (Prunus ferganica Lincz.) энде- мична для Тянь-Шаня и Памиро-Алая. Абрикос обыкновенный (Armeniaca vulgaris Lam) растет дико в Китае и в Средней Азии. Весь род происходит из Китая. K. Ф. Костина предполагает, что культура абрикоса в Средней Азии могла быть не только занесена, но и могла воз- никнуть самостоятельно, в результате окультуривания местных дикорастущих абрикосов Тянь-Шаня. Таким образом, в Средне- Юго-Западной Азии находится один из первичных очагов вхож- дения в культуру абрикоса обыкновенного. В среднеазиатской группе местных сортов К. Ф. Костина (1947) отмечает наиболь- шее разнообразие морфобиологических свойств абрикоса и вы- сокие качества сухих фруктов. В Средней Азии (и может быть отчасти в Передней Азии) создались эндемичные культурные виды абрикоса: 1) А. dasy- carpa Pers. возник путем гибридизации алычи и абрикоса обык- новенного; 2) А. leiocarpa Kost. включает формы, происшедшие от скрещивания алычи с голоплодными формами абрикоса обык- новенного. Пер си к обыкновенный (Persica vulgaris Mill.) проис- ходит из Китая. В Средней Азии имеется группа местных сор- тов. Однако Средне-Юго-Западная Азия имеет свой эндемичный вид культурного персика — п е р с и к ф е р г а н с к и й (Р. fer- Синская Е. Н. 163
~anensis Коч. et Kostina) . Имеются местные сорта персика в Средней Азии и Западном Китае. В и ног р ад (Vitis vinifera L.) представлен в странах Сред- не-Юго-Западной Азии и, особенно, в Среднеазиатских респуб- ликах огромным разнообразием местных винных и столовых сортов. В литературе имеется много указаний на произрастание здесь диких форм винограда, но П. А. Баранов и А. М. Негруль (по нашему мнению, правильно) считают эти заросли винограда не настоящими «дикими», а одичалыми (вопреки мнению Г П. Сумневича и И. Т. Васильченко, которые выделили даже ряд «видов» дикого винограда в районах западного Тянь-Шаня и Гиссарского хребта) . И. Андрашовский выделяет культурный виноград Западной Азии в особые виды: V. byzantina Andras., V. deliciosa Andras. и V. antiquorum Andras. А. М. Негруль (1946) относит среднеази- .атские и некоторые переднеазиатские сорта в одну rpynny— proles orientalis крупного вида — V. vinifera. Фисташка (Pistacia vera L.) растет дико в предгорьях Памиро-Алая, Тянь-Шаня, Копет-,дага и Бадхыза. Местом вхож- дения фисташки в культуру считают Переднюю Азию (Ассирия, Сирия, Малая Азия), но возможно, что она частично входила в культуру и в Средней Азии. В Средней и Малой Азии распро- странена одна и та же группа сортов — иранская (по P. Я. Кор- дону). Культура фисташки в разных странах, вероятно, распро- странялась и из Передней, и из Средней Азии. На это указы- вает и общий корень в названии фисташки в различных странах (Иран — |piste, pista; Туркмения — pista, psta, ipistac; арабское— fus ag; Грузия и Армения — pustugi; татарское в Крыму — fis- сЫа). В Западной Азии растет кинджак — Pistacia khinjuk Stocks, цз орехов которого добывают масло, а листья используют для дубления. Кевовое дерево (Pistacia mutica Fisch. et Меу.), веро- ятно, происходит из Передней Азии, но встречается и в Иране и Афганистане. В Афганистане растет кабульская фисташка — P. ca- bu 1 i ca Stocks. Гр ецкий орех (Juglans regia L.) распространен дико в советской Средней Азии, Афганистане и Западном Китае. Здесь находится один из первичных очагов вхождения в культуру этого дерева. Происходит из Восточной Азии. И н ж и р (Ficus carica L.) дико произрастает в Копет-Даге, западном Памиро-Алае, в Иране, северо-западной Индии и Бе- луджистане. Важнейшим первичным очагом более древнего вхождения в культуру и видообразования считается Передняя Азия (там, по-видимому, есть более древние по организации виды), но и вне ее имеются и другие виды инжира: в южном Иране — инжир персидский (F. persica Boiss.), геранелистный 362
(F. geraniifolia Miq.). На северных склонах Гималаев распрост- ранен F. roxburghii Wall. Несомненно, что и в Средне-Юго-За- падной Азии расположен один из первичных очагов вхождения инжира в культуру, его видо- и формообразования. Гранат (Punica granatum L.) имеет основной первичный очаг древней культуры и формообразования в странах Передней Азии, но, вероятно, входил в культуру и в ряде местностей Средне-Юго-Западной Азии, где он произрастает дико. Вместе с тем многие считают, что в диком виде он встречается, несом- ненно, только в Закавказье и Konez-Даге (а последний вклю- чается в Переднюю Азию), но такие утверждения еще недоста- точно проверены. Во всяком случае в Афганистане, Иране и других странах Средне-Юго-Западной Азии имеются оригиналь- ные местные сорта граната. Л ох (~Elaeagnus angustifolia L.) произрастает дико и в Пе- редней Азии и в Юго-Западной Азии, а также к северу от ос- новного ареала. культурный вид — Е. orientalis L. — имеет мест- ные сорта и в Закавказье, и в советской Средней Азии. Устано- вить место, где он впервые вошел в культуру, трудно. У н а б и (Zizyphus sativa Gaertn.) произрастает в Юго-За- падной Азии дико и в культуре. Вероятно, здесь находится один из очагов введения этого растения в культуру. Т у т а к р а с н а я (Morus гиЬга L.) представлена в Средне- Юго-Западной Азии многочисленными сортами, и здесь, веро- ятно, находится главный очаг развития и вхождения в культуру этого вида. Тута бел а я (Morus ~а1Ьа L.) и черная (М. nigra L.) не растут дико в Средне-Юго-Западной Азии, но везде разводятся, имеется много хороших местных сортов. Из Средне-Юго-Западной Азии вышли, по Н. А. Базилев- ской (1960), такие декоративные растения, как тюльпаны, ако- нит, мак восточный и др. Из всего сказанного выше следует, что Средне-Юго-Западная Азия не является областью возникновения или наиболее интен- сивного развития тех родов, к которым относятся важнейшие культурные растения. В Средне-Юго-Западной Азии, кроме двух каучуконосов, нет (или почти нет, — мы могли пропустить какие-либо малоизвест- ные культуры) эндемичных видов культурных растений, ко- торые здесь же впервые возникли, или эндемичных растений, которые здесь впервые вошли в культуру и далее не распрост- ранялисьь. В Средне-Юго-Западной Азии обнаружены первичные очаги вхождения в культуру следующих растений. 1. Эндемичные дикорастущие виды, культура которых рас- пространилась за пределы данной области исторического разви-
гия культурной флоры: кок-сагыз (Taraxacum koli-saghyz); тау- сагыз (Scorzonera tau-saghyz). II. Основной первичный очаг находится в Средне-Юго-За- падной Азии, а в Передней Азии только небольшие вторичные формообразовательные очаги: лук репчатый (Аllшт сера); чес- нок (А11шт sativum); индийская конопля (Cannabis indica); 'среднеазиатские дикорастущие виды кендыря. III. Крупные первичные очаги находятся и в Средне-Юго- Западной Азии, и в Передней Азии: люцерна азиатская (Medi- садо asiatica); лен (Linum usitatissimum sensu lato); репа (Bras- в1са гара sensu lato); дыня (Melo); некаротиновая морковь .(Daucus carota); сафлор (Carthamus tinctorius). Для лука репчатого и чеснока Средне-Юго-Западная Азия является единственной областью, где находится первичный очаг развития этих видов и вхождения их в культуру. В Передней Азии имеются лишь сравнительно слабые вторичные формо- образовательные очаги этих культур. У ряда культур — люцерны, льна, репы, дыни, сафлора имеются параллельные (викарные) крупные первичные очаги в 'Средне-Юго-Западной Азии и в Передней Азии. В Средне-Юго- Западной Азии находится и первичный очаг видообразования .дикорастущих люцерн, но более молодой и менее яркий, ем а Передней Азии. Культурная люцерна Средне-Юго-Западной Азии отнесена нами к одному виду со многими подвидами, связанными между собой переходными рядами промежуточных форм. Культурные люцерны Передней Азии и вышедшие оттуда иррадиации ха- рактеризуются большей обособленностью видов. Лен Средне-Юго-Западной Азии по существу (если прини- ъ~ать независимое происхождение и ряд особых свойств, прису- щих и исходным формам и иррадиациям) может считаться са- мостоятельной филогенетической ветвью. Судя по огромному полиморфизму и более далекому распространению в виде ко- ротких и длинных иррадиаций, первичный очаг льна в Средне- 20го-Западной Азии не менее важен, чем переднеазиатский, а по значению даже превосходит, ибо отсюда, вероятно, ведут происхождение северо-западные русские долгунцы. Дыня имеет первичные очаги форма- или видообразования (по К. И. Пангало, 1955) в Средне-Юго-Западной и в Передней Азии. Если считать, что все дыни составляют род Melo, то можно сказать, что обе подобласти являются очагами развития этого рода. Репа афгано-индийская, по нашему мнению, отпосптся к са- мостоятельному виду — Brassica rapoasiatica Sinsk., так как эта древнейшая форма характеризуется особым комплексом мор- фобиологических признаков и свойств. Развитие малоазиатских реп, составляющих подвид европей- ской репы — В. ràðoeuroðeà ssp. antasiatica Sinsk. пошло на 154
север и на запад — от них возникла карельская группа форм и некоторые сорта репы Южной и отчасти Средней Европы. Афганские наиболее примитивные формы азиатской репы дали начало более окультуренным формам Индии и Средне- азиатских республик (теперь там почти исчезнувшим) и высо- коокультуренной группе реп Японии. Примитивные формы репы Передней и Средне-Юго-Западной Азии дали далекие иррадиации, но в противоположную сторону, что подчеркивает, может быть, видовую самостоятельность этих реп. Первичный очаг вхождения в культуру имеет в Средне-Юго- Западной Азии некаротиновая морковь. Здесь произрастает и исходный дикорастущий вид азиатской моркови. Из Средне- Юго-Западной Азии морковь, относящаяся к этому виду, рас- пространилась на юг в Индию и на восток — в Японию. Первичный очаг некаротиновой моркови имеется и в Перед- ней Азии (Закавказье и Сирия). Имеется некоторое основание считать и Переднюю Азию первичным очагом образования всей моркови и очагом дивергенции, из которого вышла и азиатская некаротиновая морковь, а при расселении на запад формиро- вался полиморфный вид европейской (средиземноморской) ка- ротиновой моркови. Но эта теория пока не нашла полного под- тверждения, так как в Передней Азии не было найдено интег- ральных форм с признаками и той и другой моркови. Мы склонны считать Средне-Юго-Западную Азию основным пер- вичным очагом происхождения и расселения некаротиновой мор- кови. Первичный очаг формообразования сафлора простирается из Передней Азии в рассматриваемую область (Афганистан и се- веро-западная Индия). Область первичной культуры алычи, сливы и др., в основном приуроченная к Передней Азии, захо- дит и в Средне-Юго-Западную Азию, что видно из наших об- зоров. Мы нашли в странах Передней Азии 39 эндемичных куль- турных видов. Эта цифра в высшей степени ярко характеризует Переднюю Азию, как наиболее оригинальную и богатую об- ласть развития культурной флоры, по сравнению с Средне-Юго- Западной Азией. В странах Передней Азии гораздо больше обнаружено и та- ких видов, среди которых имеет место вторичное формообразо- вание, в разной степени интенсивное. Раньше Средне-Юго-Западная Азия переоценпвалась, как первичная область формирования культурной флоры. Эта пере- оценка имела место потому, что слишком большое значение приписывали количеству признаков каждого вида, хотя бы и эпдемичных, отмеченных в данной стране. Важны не отдельные признаки, а организм в целом — эндемичные формы: виды, эко- тнпы, сорта. Важна также не только эндемичность признаков 165
и форм — бывает и новый и древний эндемизм, а примитивность форм и признаков по отношению к дикорастущим родоначаль- никам. Роль последних в определении очага происхождения и распространения культурного вида явно недооценивалась. Роль скачкообразных изменений в процессе видообразования также недооценивалась или совершенно отрицалась, поэтому всякий, даже незначительный гиатус уже часто мешал призна- нию значения многих дикорастущих видов, как родоначальни- ков близких культурных видов и форм. Не обращалось также достаточного внимания на наличие древних интегральных ви- дов, как бы совмещающих до известной степени признаки дру- гих, более молодых родственных видов. По этим причинам Средне-Юго-Западная Азия считалась, например, первичным очагом видообразования пшениц, зерновых бобовых. С точки зрения принятых нами установок Средне-Юго-За- падная Азия — это совсем не область видообразования пшениц, а только громадный очаг вторичного формообразования (прав- да, очень интенсивного) мягкой пшеницы — Triticum aestivum. В такой же степени Средне-Юго-Западная Азия является вторичной областью формообразования сорной ржи, включаемой нами в один вид с культурной рожью — Secale сегеа1е. Что касается чечевицы, нута и конских бобов, то Средне- Юго-Западную Азию мы не считаем областью первичного видо- образования и первичного вхождения их в культуру; это скорее вторичный очаг, где в связи с отсталостью земледелия сохрани- лись более примитивные формы. Основным первичным очагом вхождения в культуру и рас- пространения на восток и на запад мы склонны считать страны Передней Азии. Здесь разобраны только наиболее характерные культурные растения. Провести сравнение в деталях состава культурных флор различных областей исторического развития культурной флоры легко по составленным нами обзорам. Небольшой объем обзора для Средне-Юго-Западной Азии не позволяет дать спи- сок всех культур, как это было сделано нами для Передней Азии. Несмотря на большое богатство культурной флоры Перед- ней Азии по сравнению с флорой Средне-Юго-Западной Азии и интенсивность видообразовательных процессов у многих родов и видов в Передней Азии, между обеими областями имеется поразительное сходство. Это сходство выражается в общности основного видового ассортимента, в викарности первичных оча- гов видообразования и формообразования у некоторых харак- терных для обеих областей культур. Kaz видно из обзоров по обеим областям, основным очагом расселения для многих культур была Передняя Азия, откуда распространение их шло на восток в Средне-Юго-Западную Азию, и на запад — в страны Европы и в Средиземноморье. !66
Наименьшее количество культур распространилось в Средне- Юго-Западную Азию с востока из Kmaa (просо, абрикос) или из Индии (кунжут). Передняя Азия и Средне-Юго-Западная Азия, в том объеме этих областей, как он нами принят, яв- .ляются достаточно монолитными территориями, имеющими свою собственную ботаника-географическую характеристику; в то же время они тесно связаны исторически и по существу являются частями единой более крупной географической области. Эта более крупная геофлористическая область и есть область древнего Средиземноморья, протянувшаяся от 1~анарских ост- ровов до Северных Гималаев. История земледелия третьей подобласти,1~ревнего Средизем- номорья (собственно Средиземноморья) изложена в разделе «Аборигенные культурные растения Средиземноморья».
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И ДРЕВНИЕ АССОРТИМЕНТЫ культурной Флоры русской рАвнины и сиБири Развитию славянской культуры вообще и земледельческой в частности предшествовали на юге Русской равнины триполь- ская и скифская культуры, между которыми существовала не- посредственная преемственность, и поэтому прежде всего кратко остановимся на более древних культурах Триполья и скифов. ТРИПОЛЬСКАЯ КУЛЬТУРА Выше неоднократно говорилось о том, что в Передней Азии и других горных странах земледелие начиналось в предгорьях— вблизи небольших горных речек и ручьев и уже в более позд- ний период своего развития оно спускалось на равнины, на поймы больших рек. Вопрос о том, как развивалось земледелие в равнинных уда- ленных от горных хребтов областях, разбирается Т. С. Пассек (1949) в большой работе по периодизации трипольских поселе- ний, в которых она (Пассек) в этом вопросе придерживается точки зрения Е. Ю. кричевского (1940). Они намечают два ос- новных типа перехода от раннего неолита к поздней его ста- дии; это связано с освоением различных частей ландшафта. Некоторые неолитические племена поселяются на нижних берего- вых террасах, на дюнах и даже на пойменных террасах. Основ- ным их промыслом очень долго продолжает оставаться рыбо- ловство, к земледелию они переходят позднее, и очень долго оно носит подсобный характер. Другие же неолитические пле- мена оставляют прежние места обитания вблизи рек, подни- маются на плато и выбирают места с подходящими для земле- делия почвами. Эти племена раньше переходят к земледелию. При дальнейшем развитии, еще в эпоху неолита, различия ме- жду этими двумч типами племен усугубляются. Охотничьи и ры- боловческие племена расселяются преимущественно в лесной и степной зонах, а земледельческие — в южной и западной лесо- степи Европы. По-другому шло развитие племен на свободных от леса лёс- сово-черноземных равнинах раннеземледельческой культуры, по- лучившей название дунайской, или культуры ленточной кера- мики. Эта культура распространилась на обширной территории от Дуная до северной Франции. Дальнейшее развитие ее про- слеживается в Дунайском бассейне, в Поднепровье и Подне- 158
стровье, по Пруту, частично в Побужье, на Балканах и даже в Македонии и Фессалии. Здесь, в восточных районах распро- странения культуры ленточной керамики, развилась культура крашеной керамики. По месту особенно продуктивных раскопок она получила название культуры Триполья и датируется III — II тысячелетиями до н. э. В Триполье прослежены слои от каменного до медного веков. Новейшими исследованиями трипольской культуры вскрыто, что ее связь с Малой Азией, греческими островами и Балканами прослеживается с III тысячелетия до н. э. и усиливается во I I тысячелетии. 1~ак пишет Т. С. Пассек, на основе «скрещивания» местных «дотрипольских» неолитических культур и интенсивных куль- турных взаимоотношений с Передней Азией и восточным Сре- диземноморьем (Египет) шло развитие земледельческих племен на территориях Поднепровья, Поднестровья, по Дунаю и на Бал- канах. В более поздней работе о раннеземледельческих племенах Поднестровья Т. С. Пассек (1961) указывает на вероятную связь трипольской культуры с домикенскими культурами. Она указывает на фракийский город как на носителя той культуры, которая главным образом лежит в основе развития ранне- греческой общины (на фракийскую культуру имеются многие указания у Гомера). Т. С. Пассек, вслед за Э. Р. Штерном, счи- тает балканские, дунайские и трипольские племена предками обширной группы фракийских и дако-фракийских народов, позд- нее поглощенных славянами. Установлены связи трипольской культуры с древневосточной культурой стран Двуречья (украшения, подобные трипольским и датируемые 2800 г. до н. э., обнаружены в Уре и в Шумере). Таким образом, культурные растения могли распространяться в область развития трипольской культуры и из восточносреди- земноморского очага древнего земледелия (включающего кри- та-микенскую культуру), а также из стран Древнего,Д вуречья, Малой Азии и Закавказья. У археологов и историков о происхождении трипольской культуры и ее жителей в основном существуют две гипотезы: 1) трипольская культура появилась на правобережной Украине в результате перемещения сюда племен из древневосточных зем- ледельческих цивилизаций (сторонники этой гипотезы пользуют- ся методом сопоставления керамики); 2) трипольская культура возникла в результате местного автохтонного развития дотри- польских племен. Сторонники второй гипотезы, однако, не отрицают очень сильного влияния востока на развитие трипольской культуры и в частности трипольского земледелия. Детально изучена трипольская культура С. Н. Бибиковым (1953) по материалам раскопок раннетрипольского поселения
Лука Врублевецкая. С. Н. Бибиков считает, что всякие попытка установить прямую связь между мезолитическими или неолити- ческими и трипольскими культурами пока не дают результатов и не подкреплены фактическим материалом. Археологические. материалы поселения Лука Врублевецкая привели С. Н. Биби- кова к следующим представлениям о возникновении и развитии культуры Триполья. В конце IV или на рубеже III тысячелетия до н. э. произошло перемещение племен, двигавшихся через Малую Азию. Одни племена шли в направлении к Средней Европе, возможно, по Дунайскому коридору, другие — на юго-восток Европы и заняли территорию междуречья Дуная и Днепра. Пришлые племена принесли с собой свою культуру, но сме- шались с местными охотничьими племенами и составили вместе- с ними обновленное, как бы аборигенное население. Охота стала постепенно терять свое значение. Собирательство сохранилось у трипольцев. В большом коли- честве использовались в пищу моллюски Unio, в раскопках об- наружены большие кучи раковин (и их обломков) этих моллюс- ков (Т. С. Пассек, 1961). Уже в раннем Триполье был вполне одомашнен рогатый скот. В раскопках обнаружено много костей быков, коз, овец, свиней. Возможно, и лошадь была в Триполье домашним животным. В. И. Громова установила, что в трипольское время в одо- машненном состоянии были две породы быка: одна порода мел- кая, похожая на торфяникового быка, а другая — крупная, по- хожая на черкасский скот. Таким образом, трипольская культура — результат смешения, или, точнее, слияния, двух куль- тур — местной охотничьей и пришлой — земледельческо-ското- водческой. С. Н. Бибиков полагает, что это слияние двух куль- тур явилось результатом постоянной инфильтрации, а не едино- временного нашествия. Трипольские поселения размещались главным образом на возвышенных участках, преимущественно на плато возвышен- ностей, огражденных оврагами и балками (такой рельеф харак- терен для лесостепи). Наиболее ранние трипольские поселения часто занимали низко расположенные приречные террасы, укры- тые возвышенностями и скалистыми обрывами. По С. Н. Бибикову (1953), в Триполье занимались не огород- ничеством, а полеводством. Под зерновыми хлебами, очевидно, находились значительные площади — это видно из того, что в поселениях были обнаружены большие запасы зерна. Значи- тельные площади посевов требовали тягловой силы. Вероятно, обработка почвы производилась при помощи быка, который тя- нул примитивную соху без лемеха. О возможности использова- ния быков как тягловой силы говорят археологические данные: были найдены глиняные модели саней. Очень часты в раскопках Триполья находки зернотерок, сосудов для хранения зерна. 170
Остатками земледельческого культа, вероятно, являются ста- туэтки женщин, вылепленные из глины. С. Н. Бибиков рассматривает трипольское земледелие как полевое мотыжное, возможно, что уже применялись соха и во- локуша. 1~ультурные растения были принесены сюда пришед- шими с востока племенами, ибо в этих местах не произрастает диких родичей культурных растений. О составе возделываемых растений в Триполье можно судить по данным раскопок и определениям ряда авторов, наиболее полно обобщенным С. Н. Бибиковым. Согласно описанию В. А. Петрова (1947), найден колосок полбы с равновеликими чешуями. Зерна этой полбы имели раз- меры 7,0;(4,3 и 6,2~~,'4,2 мм; они овальной формы, с округлен- ными окончаниями, глубокой и узкой бороздкой на брюшной пло- ской поверхности. Отсутствие на отпечатках следов толстых ки- левых жилок на острокилеватых колосковых чешуях, по мнению В. А. Петрова, позволяет предполагать, что боковой зубец был развит слабо или отсутствовал. Эти черты сближают рассмат- риваемый образец полбы с образцами полбы из неолига За- падной Европы. Обнаружены были также отпечатки зерен полбы на женских статуэтках — эти зерна имеют размеры более крупные, чем обугленные зерна, найденные вне статуэток (9,0;( 4,5 и 7,0;( 4,3 мм) . Интересно, что реликтовая культура полбы и теперь еще кое- где бытует в этих местах; Н. И. Вавилов впервые обнаружил в 1940 г. посевы полбы в Львовской и Черновицкой областях УССР. Уже на ранних этапах трипольской культуры сеяли пшеницу. Есть основания полагать, что разводили два вида пшеницы: твердую и мягкую. В раннетрипольском поселении Лука Вруб- левецкая отпечатки зерен пшеницы обнаружены на ряде глиня- ных статуэток — снаружи и внутри их. Отпечатки зерен пшеницы из Луки Врублевецкой были осмотрены М. М. Якубцинером. Он установил наличие мягкой и твердой пшеницы. Статуэтки также были обследованы В. Е. Писаревым — по его определению зерна относились к твердой пшенице. Хлебные зерна были найдены и в других поселениях Три- полья. С растительными остатками, собранными в Триполье, Веремье, Стайках и 1цербаковке, ознакомился Р. М. Окснер и определил среди них мягкую пшеницу, равно как и К. А. Флякс- бергер (1934). В. А. Петров (1945) изучал растительные остатки из глиняных обмазок, собранных Т. С. Пассек в 1~оломийщине и Владимировке, и отметил там присутствие чешуй твердой пше- ницы. Д. И. Персидский и О. Л. Липа (1935) изучили 40 образ- цов обмазки с отпечатками зерен из тех же стоянок в Веремье, Колодистом и Сушновке и установили там присутствие пшеницы. В Яербаковке и Триполье, по данным В. В. Хвойко (1907, 1913), были найдены поджаренные зерна и зерна пшеницы, ко- 171
торые находились в сосудах. С. С. Гальченко сообщаег о на- кодке зерен пшеницы в подольско-трипольских местонахожде- ниях — в Криничках и Корытном. Ячмень был обнаружен Ф. Х. Бахтеевым (1956) при осмотре. глиняных статуэток из Луки Врублевецкой. Ф. Х. Бахтеев счи- тает этот ячмень голозерным. Ячмень также был найден С. С. Гальченко в Криничках и Корытном; Д. И. Персидским и О. Липа (1935) — в Веремье, Колодистом и Сушновке; В. А. Петровым (1947) в глиняных обмазках в Коломийщине и у Владимировки (ячмень был определен В. А. Петровым как шестирядный) и др. Находки проса (Panicum miliaceum) были также довольно многочисленны в поселениях трипольской культуры. Остатки проса отмечал А. А. Спицин близ села Колодистое; С. С. Галь- ченко обнаружил просо в Криничках и Корытном; Д. И. Пер- сидский и О. Липа — в Веремье, Колодистом и Сушновке; В. А. Петров — в Коломийщине, Владимировке и в ряде других обследованных археологических пунктов. Д. Персидский и О. Липа (1935), которые изучали материалы из Веремья, Колодистого и Сушновки, изредка обнаруживали присутствие ржи. Очевидно, это была сорная рожь, а культуры ржи не было. Овса также не обнаружено, этой зерновой куль- туры в Триполье не было. При анализе отпечатков тканей на донышках сосудов из рас- копок в с. Стина и в других местах (М. М. Новицкая, 1948) установлено наличие двух видов ткани — из растительных воло- кон (лен или конопля) и пз шерсти. Некоторые авторы высказывают предположение, что в Три- полье разводили овощные растения. Древняя культура, получившая название трипольской, раз- вивалась и за пределами СССР в сопредельных странах. Указа- ния на эти местонахождения собраны С. Н. Бибиковым (1953), и здесь вкратце сообщается об этих данных. В Румынии, во Фрумушице, найдены обугленные зерна пше- ницы, которая определена как Triti~cum compactum. Мы предпо- лагаем, что это была не современная Т. compactum, а ископае- мый европейский, близкий к мягкой и карликовой пшеницам вид Т. antiquorum, который находят при раскопках, относящихся и к более ранней (соответствующей Триполью) эпохе, — в свайных постройках Швейцарии, Венгрии, Австрии, Дании и других ме- стностях. В Гумельнице, в Румынии, были также обнаружены зерна злаковых культур. На территории Болгарии в поселениях Юнадита, Свети-Кириллово, Коджа-Д,ермель и Банята находили зерна или оттиски пшеницы. В Банята, по определениям болгар- ских исследователей, были найдены полба (Т. с11сэасоп), тот же вид, что и в Триполье, и однозернянка Т. топососсит. Одно- зернянка пока не была обнаружена в раскопках поселений три- польской культуры на территории СССР. В Банята были най- 172
дены остатки ячменя, чечевицы EI вики. Бобовых в трипольских поселениях, расположенных на советской территории, пока не обнаружено, хотя возможно, что и эти культуры там имелись, но пока еще не удалось их найти. Во всяком случае, широкой куль- туры бобовых растений там, по-видимому, не было. На северо-востоке Балкан в Бутмире обнаружены обуглен- ные зерна двух видов пшеницы: однозернянки (Т. monococcum) и карликовой пшеницы (Т. eompactum), опять возникает во- прос — не является ли эта пшеница Т. antiquorum? 1(роме того„ в Бутмире найден ячмень. В Винче (Балканы) были обнару- жены зерна пшеницы. Таким образом, и в зарубежном Триполье представлен тот же в основном набор зерновых культур, а именно: пшеница, ячмень, просо с добавлением однозернянки и бобовых — вики и чечевицы. Трипольские земледельческие племена в свое время были представителями наиболее высокой культуры в Европе. В конце эпохи Триполья намечается переход к предскифским этническим образованиям. КУЛЬТУРА СКИФОВ Под общим именем скифов объединяют не один народ или племя. Скифы — это сложное этническое образование, объеди- няемое общностью языка и культуры. Племена скифской группы занимали обширные территории. Так, Б. Б. Пиотровский (1949) пишет, что в VII — VI вв. до и. э. в Закавказье три элемента сла- гали культуру: местный закавказский, древневосточный и скиф- ский. Скифы населяли в 1 тысячелетии до н. э. Среднюю Азию и являлись в числе других народностей древними предками таджиков (В. Г. Гафуров, 1949). В слоях Анау IV и городища Тали-Барзу близ Самарканда обнаружены изделия, очень похо- жие на скифские памятники Восточной Европы. Скифы насе- ляли Предкавказье, южные степи европейской части СССР по Волге, Дону и Днепру, а также Kpb>M. Материалов о древнем земледелии у скифов имеется мало. Геродот в Восточной Европе различает скифов-всадников (т. е. кочевников) и скифов-пахарей. Последние занимались мо- тыжным и плужным земледелием. Согласно новейшим изыска- ниям П. Д. Либерова (1951) основная территория, заки.;.аемая скифами-пахарями, находилась западнее Днепра и по течению Южного Буга. Скифы культивировали те же растения, которые, по-видимому, и до них сеяли жившие на этой же терр1пории земледельцы: пшеницу (и, вероятно, ячмень), просо, чечевицу, лук, чеснок, возможно, лен, коноплю (В. В. Мавродин, 1948). Геродот писал, что в Скифии конопля имеется и в дикорасту- щем, и в культурном состоянии. Описывая обряд погребения у скифов, он рассказывает следующее. При погребении скифских царей им разрезали живот, вычищали его и наполняли ладаном, 173
купырем, сельдереем, анисом, а затем снова зашивали. На ос- новании этого можно предположить, что скифы культивировали анис и сельдерей. Между прочим, сельдерей был связан с по- гребальными обрядами у древних египтян. В общем, ассортимент культур скифского земледелия был довольно богатым. Земледелие, хотя в основном имело подсоб- ный характер, было плужным. Во II в. до н. э. скифы были покорены савроматами, позднее сарматами и другими племенами. Наиболее долго существо- вали скифы в Крыму, а затем исчезли, растворившись в сме- нивших их этнических образованиях. На Северном Кавказе во II в. н. э. приобретает значение одно из сарматских племен— аланы, и постепенно это название вытесняет название сарматов. Существование земледелия у скифов доказывает преемствен- ность земледельческой культуры у народов, заселявших кикные степи нашей страны. ЗЕМЛЕДЕЛИЕ И КУЛЬТУРНЫЕ РАСТЕНИЯ У ДРЕВНИХПРОТОСЛАВЯН И СЛАВЯН В эпоху максимального (днепровского) оледенения обитатели Русской равнины не только умели пользоваться огнем, но уже жили в первобытных жилищах, охотились на крупных живот- ных. Они оставили следы своего изобразительного искусства (обычно рисунки зверей). В последующие эпохи также имели место надвигания льда; флора и фауна еще подвергались миграциям с юга на север и обратно, пока природа Русской равнины приняла характер, близкий к современному. Эта территория, по-видимому, в тече- ние всего четвертичного периода была ареной деятельности пер- вобытного человека. На пространствах в многочисленных раскопках на террито- рии нашей страны вскрыты следы пребывания древнего чело- века. Палеолитические находки мамонта обнаружены, например, на Украине по рекам Удаю, Орнеиде и Суле, в Гонцовской стоянке, где вскрыты несомненные следы деятельности и палео- литического человека. В Карачаровской стоянке по реке Оке, недалеко от г. Мурома, вместе с каменными орудиями найдень; кости мамонта (Elephas primigenius), носорога (rhinoceros ty- chorhinus) и первобытного быка (Bos primigenius). Мамонт был объектом массовой охоты. Кости мамонта широко использовали для изготовления раз- личных предметов, а также для постройки жилищ в качестве ос- новного строительного материала. Эти своеобразные «дома» имели вид шатров, в основании которых закладывались самые крупные кости, а поддерживающим каркасом служили жерди, Наверху шатер обычно украшался рогами северного оленя (Н. Я. Мерперт и g. В. Шелов, 1961). 174
В Костенковской стоянке, относимой к эпохе солютре (с. Кос- тенки на правом берегу реки Дон), было найдено много крем- невых орудий (целая их мастерская) и костей мамонта. Сход- ные палеолитические находки вскрыты и в Ковальской балке близ Кривого Рога. По берегам рек Десны,,Дона, Днестра, Днепра, Оки, Жиздры, на Южном Урале, в Крыму, на Северном Кавказе — везде жил палеолитический человек, и число извест- ных его стоянок растет в процессе развития археологической науки. Характерная черта южных стоянок — богатство остатками костей крупных диких животных: мамонта, носорога, первобыт- ного быка, северного оленя (Cervus tarandus), мускусного овце- быка (Ovibos moschatus), зубра (Bos priscus), ископаемого ро- дича лошади (Equus caballus fossilis), медведя, волка и др. Еще более широко распространены на необъятных просто- рах нашей родины известные в настоящее время памятники позднейших стадий каменного века — эпохи неолита. В Крымских горах с остатками скелета неолитического че- ловека найдены кости существующих и теперь ~кивотных: до- машней собаки, кабана и буйвола. Во многих неолитических стоянках найдены кости рогатого скота, овцы и др. В период нео- лита развиваются и скотоводство, и первобытное земледелие. Развитие последнего шло неравномерно в различных частях нашей страны. В киевских неолитических землянках были найдены зерно- вые терки, зерна проса или сорго (точно определить невоз- можно), а также зерна пшеницы или ячменя (распознать их трудно). Сходные находки были и в Петренах в Бессарабии (Ю. В. Готье, 1925) . Во многих приблизительно одновременных неолптических стоянках севера — Ладожской, Окской, Донецкой, Фатьяновской (в пределах нынешних областей: Московской, Ярославской, Ко- стромской, Владимирской, Калининской и соседних) и других следов земледелия или не обнаружено, или они слабые и часто не вполне достоверные, или их распространение носит островной характер. Однако новейшие археологические исследования вскрыли и здесь много неожиданного. Земледелие, вероятно, ос- тровное, уже во II тысячелетии до н. э. было местами сравни- тельно высоко развито и в нечерноземной зоне. Племенами лесных зон Восточной Европы (охотниками и рыболовами), жившими в землянках (зимой) и в шалашах (ле- том), создана культура ямочно-гребенчатой керамики (3000— 1000 лет до н. э.). Затем возникли острова с более высокой куль- турой — с текстильной керамикой и земледелием. В эпоху верхнего неолита и ранней бронзы у ряда племен зарождается земледелие, на юге — раньше, на севере — боль- шей частью позднее. На основе общей земледельческой куль- туры, а следовательно, близкого бытового уклада, этнической
близости складывалось постепенно древнее славянство, куда уходят корнями многие древние земледельческие племена. В лесостепи развилась земледельческая культура Триполья. Отсюда же распространилась на север и восток и земледельче- ская культура древнего славянства. Поэтому характер лесостеп- ного ландшафта и его история представляют большой интерес. Согласно современным взглядам большинства палеонтологов и ландшафтоведов (Ф. Н. Мильков, 1950) лесостепь рас- сматривается как древний зональный ландшафт, сформировав- шийся еще в доледниковое время, раньше степной зоны Русской равнины. Соответственно и земледелие сначала было более раз- вито в лесостепи, а позднее стало распространяться в степях. Для лесостепи характерен расчлененный рельеф — хорошо выраженные долины рек, много оврагов и балок. Такой рельеф был удобен для стоянок первобытного человека — он здесь легко мог найти защиту от ветров и нападения врагов. Лес доставлял топливо, материал для построек и поделок и пищу (плоды, ягоды и грибы). Лес служил убежищем при на- падении врагов, был обиталищем лесных птиц и зверей, кото- рые являлись объектами охоты. В то же время наличие в лесо- <'тепной зоне открытых, без древесной растительности, терри. 'IopHH давало широкие возможности для земледелия. Палеолитические стоянки в лесостепи располагались большей частью по крутым и расчлененным правобережьям рек. Здесь имелись всегда выходы коренных пород, которые доставляли материал для изготовления каменных изделий. Основным за- нятием древнейших обитателей лесостепи была охота. 1~ости ма- монта, носорога, северного оленя, песца, байбака, тушканчика в большом количестве встречаются во всех стоянках палеолита. Территория лесостепи стала и основной ареной развития сла- вянской земледельческой культуры. П. Н. Третьяков (1953) подразделяет древнее славянство на трп основные группы, которые в 1 тысячелетии до н. э. уже являлись славянами: 1) Лужицкую, 2) Янестровско-Средне- днепровскую (связанную со скифским миром) * и 3) Верхне- днепровскую. Образование этих трех славянских групп было не начальным, а заключительным этапом формирования восточ- ного славянства. Самое же зарождение этих групп относится ко П и III тысячелетиям до н. э. В последний, предславянский, период племена Средней и Восточной Европы составляли четыре локальные группы: 1) группа охотничье-рыболовческих племен лесной полосы Вос- точной Европы; 2) группа охотничье-рыболовческих и позднее скотоводческих племен степной полосы Восточной Европы; 3) группа дунайских и днепро-днестровских (трипольских) зем- ледельческих племен; 4) группа среднеевропейских охотничье- 176 ' Не непосредственно.
рыболовческих и примитивно земледельческих племен, занимав- ших бассейны рек Вислы, Одры, частично — Эльбы. Из четвертой группы выделились пастушеские племена, рас- пространившиеся на север в Ютландию, южную Скандинавию, юго-восточную Прибалтику (культура одиночных могил) и в Верхнее Поволжье (фатьяновская культура), где (в Прибалтике и Поволжье) они смешивались с местным населением. Именно на основе племен четвертой группы во II тысячелетии до н. э. сложились уже группы, представляющие собой непосредствен- ных предков указанных выше трех ранних славянских групп. Внутри четвертой группы отделились, вероятно, и литовские пле- мена, что было связано с нарастанием диалектных различий. Трипольцы в основном не были предками славян — здест~ имела место только преемственность культур и ассимиляция. Археологические исследования последних лет подтвердили наличие связей земледельческого населения скифского времени с предшествующими ему местными земледельческими племенами Среднего Поднепровья, Побужья и Поднестровья. В качестве непосредственных предков земледельческих племен западной Скифии (скифы-пахари) называют следующие три группы: 1) степные племена срубной культуры — это главные, основные предки собственно скифов; 2) племена, сохранившие в своей культуре трппольские черты; 3) некоторые племена, близкие к фракийской группе. Славяне, видимо, были преемниками земледельческой куль- туры Триполья, а в некоторой степени и скифской. Каково же было состояние земледелия у предков славян? Скотоводческие племена, заселявшие степные территории, про- исходили от более древних степных же племен и исторически были связаны со степными племенами Передней Азии и вообще Восточного Средиземноморья. С другой стороны, эти степные племена были связаны с земледельческо-скотоводческими пле- менами Восточной и Средней Европы. Все эти племена, по пред- положению ряда археологов, во II тысячелетии до н. э. были но- сителями индоевропейских языков. Среди этих племен возникли и предки славян. В исторических и археологических работах советского пе- риода стала выясняться история той группы пастушеских пле- мен, которая с конца III тысячелетия до н. э. обитала в бас- сейне реки Вислы, на Волыни и по Среднему Днепру. Племена этой группы в продолжение II тысячелетия до н. э. расселились по Верхнему Поднепровью и далее — по Десне и в Верхнем Поволжье. Эти племена и считаются протославянской группой, которая получила название балтославянской. В бронзовом веке в протославянской, или балтославянской, группе углубились различия между отдельными племенами. В середине II тысячелетия до н. э. существовала большая :группа тшенецких славян, обитавших в бассейнах Средней и ~77
Верхней Вислы; поселения этой группы доходили до верховьев- Вислы. По мнению ряда археологов, тшенецкие племена были основными или одними из основных предков протославянских лужицких племен. Последние возникли на рубеже II и ! тыся- челетий до н. э. в бассейнах рек Вислы и Одры. Лужицкие племена захватили также верховья левых при- токов Дуная — Моравы, Вага, Грана, может быть, доходили и до Дуная. Эти племена распространились и до верховьев При- пяти и Днепра, где вступили в соприкосновение с протославя- нами Днестра и Среднего Днепра. Имеются указания, что предками лужицких протославянских племен были также уне- тицкие и предлужицкие племена, обитавшие в начале и середине. II тысячелетия до н. э. по Верхней Эльбе, Одре и в междуречье Одры и Вислы. Советские ученые доказали, что лужицкие пле- мена были славянами, а не германцами, как утверждали неко- торые историки. Лужицкие племена имели развитое земледель- ческо-скотоводческое хозяйство. Особенно хорошо исследовано поселение на Бискупинском озере в Польше, которое датируется VI — V вв. до н. э. и даже ранее. Здесь были найдены роговые мотыги, бронзовые и железные серпы, а также пахотное орудие типа рала, что указывает на примитивное пашенное земледелие Были найдены колесницы, что свидетельствует о существэвании упряжного скота, который мог использоваться и для пахоты. В этом поселении найдены многочисленные остатки культурных растений: пшеницы, ячменя, проса, чечевицы, бобов, гороха,. мака. В середине ! тысячелетия до н. э. как большая политическая сила выдвинулись на арену истории акты и склавины, которые были, несомненно, настоящими славянами. Антские и склавин- ские племена рассматриваются историками как прямые наслед- ники предыдущей, более чем тысячелетней земледельческой культуры Среднего Поднепровья и Прикарпатья. Культура «полей погребальных урн», развивавшаяся ант- скими племенами в ! — IV вв. и. э., но восходящая к позднему неолиту, также послужила важным историческим звеном в про- цессе развития более поздней славянской земледельческой куль- туры. В Среднем Приднепровье культура «полей погребальных урн» исчезает к VI — VII вв. н. э., так как вследствие господства в ранний период подсечной системы примитивного землсделия акты постепенно передвигались, перенося заимки на новые ме- ста по мере истощения почвы. Однако от III до IX вв. н. э. по- степенно растет и техника земледелия. Соха «суковатка», ха- рактерная для эпохи подсечной системы земледелия, постепенно на юге сменяется плугом или ралом, а на севере — настоящей сохой. Настоящий серп с большим изгибом сменяет древний примитивный серп, похожий на кривой нож. «У них большое ко- личество разнообразного скота и плодов земных, особенно проса 178
и пшеницы»,— писал об актах Маврикий Стратег (VI — VII ».). Земледелие у актов было пашенное — в их поселениях находили железные лемехи, серпы, а также жернова. У актов, по свиде- тельству Маврикия, было очень много проса и пшеницы. В. В. Хвойко на Пастерском городище в бассейне реки Тясьми- ны (правый берег Среднего днепра) в жилище антского пле- мени нашел обугленные зерна проса и ячменя. У антов были коровы, лошади, мелкий рогатый скот, домашняя птица; об этом свидетельствуют большие скопления костей вокруг жилищ. За- нимались анты и рыболовством, но в основном это был земле- дельческий народ. К. А. Фляксбергер изучал семена культурных растений из раскопок 1928 г. под Минском (Банцеровское городище, дати- руемое IV — VIII вв.). Он нашел здесь много гороха, мелкосе- мянных конских бобов и проса. В горшках обнаружены семена вики посевной и зерна пшеницы мягкой. Здесь же .были най- .дены железные серпы. V,. А. Фляксоергер не обнаружил здесь ржи. Через два — два с половиной столетия после исчезновения ан- тов, возможно, и раньше, стало все шире распространяться в исторических памятниках другое название восточнославян- ских племен, которые фигурируют уже под именем росов (рос, росы, русь). Одновременно с распространением названия «рос» или «русь» оставалось и наименование «славяне», которое стало применяться уже в общем значении, а не только в применении к одним западным славянам. Несмотря на схематическое изложение, из сказанного выше видно, откуда пошла славянская земледельческая культура; как протославянами и затем славянами ассимилировались земле- дельческие навыки трипольцев, скифов и других народностей. Видно также, на какой значительной территории развивалась древняя славянская и ранее протославянская культура: от Бал- кан до Одры, Эльбы, Балтийского моря, Скандинавии и мест обитания угро-финских племен — на северо-западе. В ходе консолидации славянства в него включались и пле- мена с иными языками: даки, фракийцы, в какой-то мере кельты, собственно скифские (ираноязычные) племена, сарматские, ли- товские, угро-финские и др. Часть славянских племен в эту эпоху подверглась местной ассимиляции со стороны кельтов, скифов и других, позднее целиком ассимилированных другими народами. Эти исторические процессы протекали в конце 1 ты- сячелетия до н. э. и в 1 тысячелетии н. э., в эпоху ломки пер- вобытно-общинного строя, образования классового общества и государственности. В процессе консолидации славянства складывались славян- ские язык и культура. Общность славянства особенно ярко от- разилась именно То, что зем- леделие в экономике древних славян играло основную роль, до-
казывает существование множества общих для всех славян тер-- минов, относящихся к сельскому хозяйству. Особенно инте- ресны общеславянские названия культурных растений, указы- вающие на древнеславянский набор культур: пьшеница, льшено, овос. Общеславянскими наименованиями считаются также: лен, горох, сочевица (чечевица), лук, чеснок, репа, хмель, мак. Слово «жито» обозначает основную, главную культуру района, и так называли (и называют) и рожь, и пшеницу, и ячмень, а у ре- зьян — и кукурузу. Древнейшее славянское земледелие было в большей части подсечным. В период аятского земледелия у славян в основном опреде- лился состав культурных растений. В раскопках послеантского времени обнаружены те же основные зерновые культуры, что и у антов: пшеница, ячмень, просо, rooox. В 1~овшаровском городище Смоленской области (IX-- XIII BB.) К. А. Фляксбергер (1926) нашел больше всего яч- меня. А. Н. Лявданский нашел в этом городище две формы яч- меня (шестирядный и двурядный), овес, пшеницу и отпечатки льняной ткани. В Борщевском и 1~овшаровском городищах в верховьях Днепра, в остатках, датируемых IX — X вв., найдены зерна проса, ячменя, пшеницы, овса, гороха. Были известны тогда также и лен, конопля, рожь, репа, лук, чеснок. В киевских землянкаx XI в. в районе, где жила городская беднота, найдены зерна ржи, ячменя, проса, пшеницы. В. 1~. Гончаров (1950) в культурных слоях Райковецкого городища Житомирской области, относящихся к XI — XII и пер- вой половине XIII вв., нашел обугленные зерна пшеницы, яч- меня, ржи и овса. П. В. Дундулене (1950) сообщает, что, по данным раскопок в 5Кмуди (Литва), в слоях Х — XII вв. был найден овес. Данные о составе возделываемых культур имеются и из бо- лее западных районов. В раскопках на горе Бекеше (Вильнюс) в слоях XI, XII и XIV вв. в землянке была обнаружена смесь обуглившихся зерен ржи, пшеницы с ячменем и овсом. Там даже был найден хлеб, выпеченный из смеси зерен. В эту эпоху в Литве практиковались смешанные посевы (В. Голубович и Е. Голубович, 1945). Там в городищах Габриелшики и Вегоны в слоях XI, XII и XIII вв. найдены рожь, ячмень, просо, полба, овес и одно зерно гречихи. Интересны данные материалов Волго-Донской экспедиции (1958), относящихся к хазарскому и несколько более позднему времени. Население хазарской крепости Саркел, как и в соседних по- селениях салтовской культуры, обрабатывало землю тяжелым плугом с большими симметричными лемехами и массивным чреслам из железа и, по-видимому, с деревянным стволом. В плуг 180
запрягали несколько пар быков. Зерно хранилось в больших ямах. В этих ямах в культурном слое хазарского периода зерна были крупные и незасоренные. Здесь в то время сеяли просо, коноплю, двурядный ячмень, карликовую пшеницу. ~Кали же- лезными серпами. В каждом доме были легкие каменные жер- нова; в раскопках найдены косы и серпы, в одной из мусорных ям — остатки бахчевых и огородных культур, которые редко хо- рошо сохраняются и редко обнару>киваются в раскопках. Здесь найдены семена дынь и огурцов, косточки дикорастущих плодо- вых растений — садоводства тогда еще не было. Встречаемые изредка косточки абрикоса свидетельствуют о завозе в эти места фруктов с юга. В культурном слое городища встречалось боль- шое количество костей бобра, который теперь здесь не живет. Мясо бобров употребляли в пищу. В одном месте нашли даже кусок лепешки из муки грубого помола. Население города Саркела было стародавнее, местное, а позднее — в городе Бе- лой Веже (на месте Саркела) русское, пришлое. 1~ории и традиции русского земледелия в Подонье были иные, чем в хазарское время. Русское население принесло сюда с со- бой мягкую пшеницу, шестирядный ячмень, рожь, лен и маслич- ное растение — кользу (сурепицу). Общими с предыдущим пе- риодом культурами остались просо и конопля. В заполненном русском колодце в Белой Веже сохранилось деревянное трепало для обработки волокна льна и конопли. В Екимауцком городище (Г. Б. Федоров, 1953), существовав- шем на территории славян Поднестровья в XI в. и основанном славянским племенем — тиверцами, наряду с жерновами, сер- пами, лемехами и косами-горбушами было обнаружено при раскопке много зерна. В большинстве стоянок обнару>кены мяг- кая пшеница, рожь, просо, горох, овес; также найдены лен ц КОНОПЛЯ. В Яонецком городище, существовавшем до XII в. (около Харькова), найдены зерна проса, ржи, ячменя, гречихи, двух видов пшеницы (твердой и мягкой), мака, овса, нескольких форм льна. Рожь, возможно, была сорной. В районе Цимлянского водохранилища, по данным М. И. Ар- тамонова, были найдены зерна карликовой пшеницы и двуряд- ного ячменя, а позднее уже не карликовой, а мягкой пшеницы, ржи, льна, рыжика. Рыжик, вероятно, был сорный (во льне). Характерно, что здесь, как и в Западной Европе, карликовая пшеница была распространена раньше мягкой. Возможно, это была переходная форма от древнейшей ископаемой п;неницы типа Т. antiquorum и Т. protomacha. При анализе растительных остатков М. М. Якубцинер (1956) нашел здесь также зерна мягкой пшеницы и четырехрядного яч- меня. Набор культур был, таким образом, довольно большой. Зерна твердой пшеницы были более округлые, чем у совре- менных сортов твердой пшеницы. По сообщению Н. Н. Воро- l8l
нина, зерна, обнаруженные в раскопках городища в Гродненской области, существовавшего в Х — XII вв., определены А. С. Крото- вым как гречиха. Однако культура гречихи, по-видимому, древ- ней славянской эпохи и существовала еще у протославян и ски- фов. В Эрмитаже (Ленинград) в отделе «Древняя Русь» имеются плоды гречихи из Немировского городища. Они были найдены при раскопках скифского жилища, существовавшего в VII— VI вв. до н. 3. на территории Винницкой области. Семена этой ископаемой гречихи нескол ько мел ьче о бычных. Гречиха сравнительно рано вошла в культуру и на дону у сарматов, от которых могла перейти к соседним славянам. Миллер (1940) нашел плоды гречихи в раскопках сарматского погребенья, относящегося к 1 — III вв. н. э. (в районе Ростова- на-Яану). 1(ультура гречихи, вероятно, распространялась по Русской равнине с востока, но не татарами, как думает Е. А. Столетова (1940, 1952), а гораздо раньше. При археологических работах в районе 1(уйбышевской ГЭС (А. П. Смирнов и Н. Я. Мерперт, 1951) вскрыты слои от конца II тысячелетия до н. э. до золотоордынского времени. В двух более древних стоянках на границе Ульяновской области и Та- тарской АССР найдены каменные зернотерки и наконечники стрел, что указывает на существование в ту эпоху охоты и зем- ,леделия. Среди памятников бронзового века найдены серпы (1200 — 700 лет до н. х). Археологические изыскания и в этих местах говорят о непрерывной преемственности земледельческой культуры. В слоях золотоордынского периода в зерновых ямах най- дены запасы зерна с примесью семян сорных растении: гре- чишки, вьюнка полевого, мари белой, подмаренника и других сорняков, типичных для старопахотных почв. Археологи пред- полагают, что булгары здесь создали высокую земледельческую культуру, которая по технике и набору культур соответствовала таковым средневековой Европы. Земледелие булгарами было, вероятно, заимствовано от славян; материалы эти указывают также и на высокий уровень славянской земледельческой куль- туры. В раскопках в районе 1(уйбышевской ГЭС в запасах зерна первое место занимает пшеница, затем рожь, ячмень, овес, ro- рох, чечевица и просо. Интересные данные о булгарах и росах сообщает Ибн Фад- лан, посланник арабского халифа по торговым делам (Х в.). Из повествования Ибн Фадлана видно, что у русов имели большое значение пшеница, ячмень, просо, лук. Из дикорасту- щих зарослей в ту эпоху использовались орехи, яблоки (судя по описанию — сильнокислый вкус — это были именно дикие яблоки). Параллельно с эволюцией земледелия изменялся характер использования сельскохозяйственных животных. Еще в VIII и 182
IX вв. пользование рабочим скотом имело ограниченный ха- рактер по сравнению с последующими IX — XI вв. Когда утвер- дилось пашенное земледелие, вол и лошадь приобрели важное- значение, как тягловый скот. Однако на юге работали чаще на волах, а лошадь использовалась для верховой езды конными дружинниками. По мере роста поголовья лошадей увеличива- лось значение овса и ячменя. Кормовых трав в те времена не сеяли. Арабский автор Ибн Русте (начало Х в.) сообщал, что сла- вяне наиболее широко возделывают в качестве хлебного расте- ния просо. На основании многих археологических изысканий и письмен- ных памятников можно утверждать, что просо имело особое зна- чение у актов и настоящих славян и являлось здесь одной из- самых древних зерновых культур (если не самой древней). Про- со высевали и жители Триполья. На распространение проса могли оказать влияние и скифы. Как растение неприхотливое и скороспелое, просо является из- любленной культурой всех кочевых и полукочевых племен, где земледелие носит примитивный и подсобный к скотоводству ха- рактер. Просо было и в древние времена, и в еще сравнительно недавние, любимой культурой кочевников и полукочевников— скотоводов азиатских равнин, древних монголов. Культура проса к славянам пришла от трипольцев и ски- фов. Кроме того, просо здесь сеяли еще в неолите. Древность появления проса отмечена и в Сибири, и в Закавказье, и в Ме- сопотамии, и в странах Западной Европы, и в Индии и, осо- бенно, в Китае. Возможно, что эта культура предшествовала возделыванию голозерной пшеницы и ячменя. Кашное потребление древнее хлебного, и древние пшеницы — полба и однозернянки — тоже были кашными культурами. Но просо легче перерабатывалось на крупу, из него получали более разнообразные продукты (вспомним, что из проса в древности изготовляли пиво и коче- вые народы квасили его с молоком). Таким образом, к древ- нейшим пищевым культурам относятся просо, пшеница и ячмень. Овес и ро>кь появились позднее. Киевская Русь не является первым государством славян. Несомненно, до нее существовало государство росов на Днепре. Культура Киевской Руси непосредственно связана и с антской. Состав культурных растений КНеВсКОН Руси сложился в основ- ном раньше и отличался разнообразием. В Киевской Руси возделывались следующие полевые куль- туры: пшеница (мягкая и твердая), ячмень (шестирядный, дву-. рядный и голозерный), просо, овес, рожь, гречиха, лен, конопля, мак (для приготовления маковников), чечевица, горох, конские бобы, вика яровая (как пищевая зерновая культура). Из ого- родных растений разводились: репа, капуста, лук, чеснок, воз- 183
можно, морковь и сельдерей (он был у скифов) . Из пряных воз- делывались: горчица (?) и, может быть, сурепица, укроп, мята, анис, перец. Из плодовых деревьев были в культуре яблони, вишни и сливы. По исследованиям Н. Н. Воронина (1953), в Княжеском Де- тище в Гродно были найдены остатки сливы, вишни, а гакже большие запасы зерна пшеницы, ячменя, ржи, проса, овса, гре- чихи, гороха, мелких конских бобов. Яблоки были издревле из- вестны и в более северных областях. В первой Новгородской летописи говорится о необыкновен- ном граде: «зело страшно бысть, гром же и мълния, град же яко ябльков боле». Здесь сравнение с яблоками гGBopHT о том, что последние были обычным фруктом. В древнем Новгороде при раскопках Ярославова Дворища были найдены ягоды смо- родины. На юге в Тмутаракани разводился виноград. В станице Тамань экспедиция АН СССР добыла интересные сведения о столице древнерусского Тмутараканского княжества. Княжеские знаки Рюриковича здесь обнаружены на осколках корчаг, откопанных на месте, где, возможно, располагались кня- жеские кладовые. Ьыли найдены остатки огромных сосудов для хранения зерна и амфор — корчаг для виноградного вина. Воз- делывался также хмель. Славянский лен в большом количестве вывозился в Среднюю Азию через Дербент. О возможности культуры горчицы в древней Руси интерес- нейший документ приводится М. Тихомировым (1950). Послед- ний расшифровал древнюю надпись на разбитом глиняном со- суде, найденном археологом Авдусиным в кургане под Смолен- ском. Этот сосуд датирован Х в. Надпись содержала одно слово «горухща». Это слово (иначе «горуха» или «горушица») в древ- неславянских языках обозначало горчицу или горчичное семя. М. Тихомиров, впрочем, считает возможным, что под словом «горухща» следует понимать собственно не горчичное семя, а перец — одну из самых дорогих пряностей средневековья. Гор- чица, конечно, под Смоленском не сеялась, как и перец. «Нет ничего удивительного, — пишет М. Тихомиров, — что пряности в числе других товаров попали на север, в Гнездовский могиль- ник, и были погребены с покойником по старому языческому обычаю отправлять умерших в загробный мир с их любимыми и наиболее дорогими предметами». На юге горчица в это время, вероятно, была уже в культуре. Наиболее широко распространенное в наше время огородное растение — капуста кочанная — широко возделывалось в Киев- ской Руси. Упоминания о капусте имеются во многих древних рус- ских памятниках. Старославянское название капусты — «зеле»— сохранилось и теперь на Балканах. Документальные дан- ные о культуре капусты на Руси относятся к XI — XII вв. О ка- пусте упоминается в «Изборнике Святослава» (1073 г.), в «Ус- тавной грамоте Смоленского князя Ростислава Мстиславского». 484
В этом документе имеется такая фраза: «И даю на посвет свя- той богородицы из двора своего... на горе огород с капустни- ком». Очевидно, для культуры капусты отводились специальные места — «капустники». В сочинениях Эль Омара («Пути взоров по государствам разным») говорится о культивировании капусты в Золотой Орде, а также в городах черкесских, русских и ясских (см. сб.: «Ис- тория Татарии», 1937). О капусте имеются сведения и в поздней- ших древних памятниках: в сборнике «Мерило праведное~ (1350), в «Домострое» — в последнем говорится о необходимо- сти для тех, у кого нет поместья, пашни, сел и вотчины, де- лать путем закупок годовые запасы хлеба и «всякого жита на возах» и капусты. 1(апуста на Руси была продуктом широкого потребления. Щи и сейчас еще остаются национальным русским блюдом. Едва ли не самым древним огородным растением является репа. Это — типичное растение подсечкой культуры севера. Еще совсем недавно (в 1(арелии) репу сеяли только на гари, бросая семена в горячую золу. Мелкий пахотный слой способствовал формированию местных стародавних сортов репы с плоской формой корнеплода (Петровская, местные карельские сорта и др.). Подсечная система на юге Русской равнины раньше усту- пила место пашенной, чем на севере, где она сохранялась ме- стами до ХХ в. Поэтому на юге репа не имела такого большого хозяйственного значения, как на севере, а теперь совсем почти исчезла. На юге репа сохраняется в культуре только в некото- рых горных районах Закавказья, но эти репы имеют прямые бо- танически близкие связи не с северными репами, а с репами Малой Азии. В Новгородской Руси (Н. Я. Аристов, 1866) репу разводили на огородах и на полях. Поля, где сеяли репу, назы- вались «репищами» и постоянно упоминались в купчих XI в.. Несмотря на то, что примитивные формы земледельческой культуры на севере сохранились местами почти до наших дней, там существовали и более крупные очаги, где земледельческая культура достигала довольно высокого уровня еще в VII— XIII вв. и ранее. Приильменье было одним из крупных культурных центров Руси, о чем говорят раскопки В. И. Равдоникаса (1950) в Старой Ладоге, А. В. Арциховского (1949) в Новгороде. В этих районах пашенное земледелие существовало еще в те времена, когда на большей части территории севера господст- вовала подсечная система. Итоги археологических исследований Н. И. Репникова (1910 — 1913), В. И. Равдоникаса (1933, 1940 и др.), а затем и Г. П. Гроздилова (1938 — 1947) славянского поселения на Ла- доге в современном Волховском районе Ленинградской области вскрыли довольно высокий уровень сельского хозяйства в VIII— 185
1Х вв. на севере в лесной области. Важным является нахож- дение здесь сошников от многозубовой сохи, что указывает на высококультурное для того времени хлебопашество. В Старой Ладоге были найдены лесные орехи и зерна манника. Суще- ствование здесь культуры льна доказано остатками льняных тканей и семян. Были обнаружены также мелкие семена конопли, возможно, дикорастущей. В нижнем культурном слое VII в. об- наружены зерна ячменя. В довольно хорошем состоянии сохра- нились остатки проса обыкновенного — найдены зерна, чешуи и остатки стеблей. Найден также горох. Пшеница вначале не ,была обнаружена. Однако позднее, в 1953 г., М. М. Якубцинер (1955), исследуя один из образцов обугленных колосков пз по- горевшего амбара VII в., нашел хорошо сохранившиеся ко- .лоски полбы, которые, по его определению, напоминали по- волжскую полбу. Ладожская полба имела более мелкие ко- лоски, чем современная волжская (у ладожской длина чешуи 7 — 8 им, а у волжской — 10 — 12 мм~. В этом же образце .М. М. Якубцинер нашел обугленные зерна мягкой пшеницы. Здесь же Ф. Х. Бахтеев обнаружил мелкие зерна ячменя. В том же образце А. И. Мордвинкина нашла 3 зерна овса, а В. Ф. Ан- тропова — 2 зерна ржи. Таким образом, исследования говорят о значительном разно- образии культур на северо-западе Русской равнины в VIII в. Основными культурами, видимо, были полба, мягкая пшеница, просо; уже возделывались овес, рожь, лен, может быть, конопля. В это время граница культуры пшеницы и проса и граница па- шенного земледелия простиралась минимум до 60' с. ш. А. В. Арциховский (1951) открыл на территории древнего Новгорода мощный культурный слой. В Новгороде во время последних раскопок было обнаружено много амбаров, напол- ненных зерном: просом, мягкой пшеницей, ячменем и рожью раз- ных эпох (Х — XIV в.). Состав ископаемых растительных остатков в Новгороде ис- следовал также В. Л. Янин (1953). На Неревском конце были найдены зерна пшеницы, ячменя, ржи, бобовых. Есть данные о том, что в это время в Новгороде были развиты огородниче- ство и садоводство. В Новгороде найдены кости крупного рогатого скота, ".виньи, овцы, козы, лошади, собаки, кошки, а также кости диких жи- вотных, на которых охотились жители — кости лося, кабана, бобра, зайца, медведя, косули, барсука и тура (В. И. Цал- кин, 1952) . Не только в Старой Ладоге и на территории древнего Нов- города, но и в других местностях северной окраины расселения восточного славянства имеются следы очень древнего пашен- ного земледелия. С. А. Тараканова (1953), изучавшая раскопки на Псковской земле, производившиеся в 1948 — 1951 гг., обнару- жила следы пашенного земледелия даже на рубеже V — -VI вв. 186
В окрестностях Пскова возделывались в то время мягкая пше- ница, двурядный ячмень и горох. древнее земледелие в Новгороде основательно изучено А. В. Кирьяновым (1952), особенно подробно в его работе по истории земледелия новгородской земли Х — XV вв. (1959). В древнем Новгороде практиковалась очистка зерна. Осо- бенно большие скопления семян сорных растений были обна- ружены в Новгороде в слоях XI — XV вв. — это были отходы по- сле очистки зерна. Состав сорняков был разнообразен. Одна из находок включала следующие виды сорных растений: марь бе- лую, пикульник, гречишку птичью, бородавник, репу полевую, ярутку полевую, щавель кислый, щетинник сизый, бодяк по- левой, ромашку, куколь, костер ржаной и др. А. В. Кирьяков нашел (1952), что во ржи росли главным образом озимые сор- няки (бородавник, костер ржаной и др.), а мягкую пшеницу засоряли больше яровые сорные растения (гречишка развеси- стая, репа полевая, горчица полевая). Состав сорняков говорит о том, что они произрастали на старопахотных почвах. Есть предположение, что уже в XI в. в Новгороде существовала па- ровая система земледелия. Особую ценность для истории Новгородской земли в целом представляют материалы, найденные при раскопках Старой Ладоги. В основном ладожане возделывали полбу, в меньшей мере — мягкую пшеницу и овес. Вероятно, в это время возделы- валось и просо, хотя оно было найдено в более верхних слоях. Период полбяной культуры длился здесь сравнительно недолго. Как показали археологические материалы Старой Ладоги, в более поздних слоях никаких следов полбы не обнаружено. И в других районах Новгородской земли археологические ма- териалы не содержали остатков полбы. При раскопках городища Камна обнаружены были только зерна ячменя, пшеницы и го- роха. Зерна староладожского проса мелкие, продолговатой формы, с несколько заостренными концами, преобладает фиоле- товая окраска пленок, но встречаются также желтая и светло- серая. Сохранились и остатки просяного растения, отнесенного к просу метельчатому (Рап1сит miliaceum ssp. effusum). Старо- ладожское просо относится к северному экотипу, характерному для северного края ареала этой культуры, и уже этот факт до- казывает, что данная находка отнюдь не была случайной. А. В. Кирьяков считает, что мелкозерное и, вероятно, скороспе- лое староладожское просо было на севере вполне устойчивой культурой. Археологические материалы из раскопок Старой Ла- доги и Новгорода говорят о том, что просо возделывалось здесь в течение ряда столетий и занимало значительное место в зем- леделии новгородских славян вплоть до XII в. Овес в ранних раскопках Старой Ладоги был представлен только единичными зернами. Более поздние материалы из рас- копок в Старой Ладоге и Новгороде свидетельствуют о том, 187
что овес не был широко распространен в Приладожье и в бас- сейне Ильменя вплоть до XIII в. А. В. Кирьяков считает, что те немногие зерна овса, кото- рые были обнаружены в ранних слоях Старой Ладоги, относи- лись к сорному овсу, засорявшему полбу. Находка единичных зерен ржи в Старой Ладоге не свиде- тельствует о широком ее распространении здесь до XIII в. Ар- хеологическими изысканиями в смежных территориях не обна- ружено следов культуры ржи. Не найдена рожь и в ранних слоях городища Камна, по времени совпадающих с ранними слоями Старой Ладоги. Не обнаружена рожь и в городище Подгай бывшей Великолукской области, датируемом V — VII вв. (Я. В. Станкевич, 1952); не нашли также ржи при исследовании Банцеровского городища (около Минска), датируемого VI— VIII вв., и в Ковшаровском городище под Смоленском (K. А. Фляксбергер, 1926, 1934). А. В. Кирьяков считает, что единичная находка р?ки в ран- них слоях Старой Ладоги указывает на культуру яровой пжи, не получившей еще тогда широкого распространения. В зна~~~~тель- ных количествах зерна ржи были обнаружены в Старой Ладоге только к XIII — Х1Ч вв. K этому времени озимая рожь заняла ведущее положение в нечерноземной полосе Русской равнины. В ранний период земледелия набор культур в Старой Ла- доге состоял из зернобобовых и прядильных растений (лен и конопля). Железные наконечники сохи в Старой Ладоге по своей форме и размерам очень похожи на наральники первой поло- вины 1 тысячелетия н. э. из Среднего Поднепровья. Собран богатый археологический материал о пригородном земледелии древнего Новгорода. Некоторые участки, особенно с овощными культурами, находились в пределах городских уса- деб. Особенно часто встречалось там просо (но мелкие партии семян), в Новгороде возделывалось такое же метельчатое просо, как и в Старой Ладоге. В Новгороде было найдено мало ячменя и пока не встречено следов культуры пшеницы; А. В. Кирьяков объясняет это случайными обстоятельствами. Из зернобобовых культур здесь возделывался горох; найдено несколько зерен мелкосемянной чечевицы, по размерам и форме сходной с бул- гарской чечевицей в слоях XII в. Не было найдено в Новгороде ни ржи, ни овса. Рожь, возможно, и сеяли, но мало. В слоях XI — XII вв. уже были находки ржп, отсутствовавшей в более ранних слоях. В слоях XII в. рожь уже часто встреча- лась в раскопках Новгорода. Быстрое распространение ржи в Новгородской земле А. В. Кирьянов связывает с распростране- нием паровой системы земледелия. Уменьшение количества на- ходок проса по сравнению с X в., по А. В. Кирьянову, связано с распространением более устойчивой культуры ржи. В слое XI в. была только одна находка ячменя, а в слоях XII в. — четыре. Малый удельный вес ячменя в археологических 188
материалах по Новгороду не говорит о малом значении этой культуры вообще на севере. В XII в. несколько увеличилось значение пшеницы, о чем сви- детельствуют многочисленные находки ее в слое этого века. В слоях XI и XII вв. обнаружены семена конопли и небольшое количество стеблей льна. Рожь, широко распространенная в XI, XII и последующих веках (хотя в самом Новгороде она появилась позднее и ее сеяли не так много, как в соседних районах лесных областей), была озимой, о чем свидетельствует состав сорняков. А. В. Кирьяков считает, что рожь проникла в северо-запад- .ную часть лесной зоны из лесостепи. Свое мнение он подтвер- ждает следующими археологическими фактами: 1) при раскопке городища Екимауцы найдено много ржи в слоях IX — первой половины Х вв.; 2) при раскопках Новотроицкого городища .в Лебединском районе Сумской области найдено значительное количество ржи в наземном помещении в слоях VIII — Х вв. В другом жилище — в полуземлянке — найдена смесь пшеницы и ржи, последней в смеси было около 18о/о. Среди этой ржи най- ,дено 5 зерен костра ржаного. Находка костра ржаного говорит о том, что здесь сеяли рожь озимую. Таким образом, можно сделать вывод, что озимая рожь рас- пространилась на север из лесостепной полосы на рубеже X и XI вв. В самом Новгороде в слое X в. ржи не найдено, но в слоях XI — XII вв. она уже занимает главное место среди нахо- док зерна. Небольшой материал был собран Новгородским музеем в 1951 г. на городище Княжова Гора в 8 км от Яемянска. Горо- .дище это находится на территории бывшей Деревской пятины на р. Явонь, которая была одним из звеньев древнего селигер- ского пути, соединявшего Новгород с «низовскими землями». Здесь были зерна ржц, пшеницы, ячменя, гороха и единичные зерна овса. Находка датируется XII — XIII вв. Пшеничные зерна с городища Княжова Гора, по свидетель- ству А. В. Кирьянова, имеют поразительное сходство с зернами пшеницы, обнаруженными в Новгороде в слоях XI — XII вв. Со- став сорняков с Княжовой Горы был в общем такой же, как и в Новгороде, следовательно, и здесь имелись в XII в. старопа- хотные земли. Набор полевых растений на Новгородской земле XI II— XIV вв. в основном был тот же, что и в более раннее время. К этому времени увеличились площади посевов ржи, уменьши- лись посевы проса. Среди яровых зерновых культур занимал, вероятно, первое место ячмень. Продолжали сеять горох. В XV в. первое место принадлежало ржи и ячменю, пшеницы, видимо, было значительно меньше, чем ячменя. Просо к этому времени совершенно перестали возделывать. Овес в XV в. еще не приобрел большого значения как фуражная культура. В лесо-
степной полосе, пишет А. В. 1~ирьянов, овес интенсивно культи- вировался в IX — Х вв., что подтверждается данными раскопок в Екимауцах (Молдавия), где найдено много овса. В слое XV в. впервые были обнаружены гречишные пленки. На юге гречиха вошла в культуру раньше (раскопки Донецкого городища XIII — XIV вв., в Гродно в слоях XII — XIII вв. и др.). Вероятно, гречиха продвинулась на Новгородские земли с юга в XV» гречка часто упоми- наются в новгородских писцовых книгах. Археологические материалы проливают свет и на историю овощеводства на Новгородской земле. В летописях имелось только упоминание о репе. В слое Х в. были найдены 3 огуреч- ных семечка. Огурцы проникли сюда, вероятно, из Византии. Найдено было также семечко тыквы (Cucurbita реро). Широко возделывалась на Новгородской земле капуста. В небольшом количестве обнаружены семена огородных бобов в слое XIV в. В слоях XI — XII вв. найдены семена огородного укропа, может быть, он употреблялся при засолке огурцов. В Новгороде в слое Х в. были обнаружены в большом количестве косточки вишни, семена яблок и даже целое яблоко. «По-видимому, вишня куль- тивировалась новгородцами в очень отдаленное время», — пишет А. В. Кирьяков. Яблоко, найденное в слое Х в., и другое— в слое XIII в., относятся к виду Malus silvestris Mill. Ареал этого вида не доходит до Новгорода. Яблоня разводилась в са- дах Новгорода. К XV в. плодоводства в Новгороде достигло зна- чительных размеров, что можно проследить по писцовым книгам. В слое Х в. была найдена половинка яблока, определенного как сибирская яблоня Malus bactcata. Вероятно, в Новгороде был~г и свои культурные сорта яблони. В слое XI в. найдены две косточки сливы. Привозили ли их в Новгород в виде сухих фрук- тов или культивировали там — неизвестно. Из ягодных кустар- ников в Новгороде разводили черную смородину и малину. Все археологические материалы говорят о высоком уровне и самобытности Новгородской земледельческой культуры. Имеются археологические данные о древнем составе возде- лывавшихся культур в приокских районах, а также в Среднем Поволжье и в Прикамье. В старой Рязани (А. Л. Монгайт, 1955), в районах среднего течения Оки, куда к тому времени все- лились вятичи (XIV — XV вв.), были найдены рожь, пшеница, просо, меньшее значение имели ячмень, горох и овес. В раскопках Донды-Карского городища на р. Вычегде в Удмуртской АССР в слое Х в. были найдены овес, просо, яч- мень и пшеница (А. П. Смирнов, 1952). Данные археологии не только дают много сведений по со- ставу древних зерновых культур, но и проливают свет на исто- рию в нашей стране прядильных культур (лен и конопля), ко- торые с глубокой древности известны на севере Русской рав- нины. 190
На территории Финляндии и всей европейской таежной зоны .севера СССР в III — II тысячелетии до н. э. существовала куль- тура ямочка-гребенчатой керамики. Население в основном зани- малось охотой и рыболовством, явных указаний на занятие земледелием нет. Текстильная керамика там не была найдена. Ямочка-гребенчатый неолит сменяется новой культурой городищ, где камень остается основным материалом, но появляется и же- лезо, местами как бы минуя бронзовый век, хотя иногда памят- ники бронзового века и обнаруживают. При этом ямочка-гребен- чатая керамика сменяется текстильной. Именно к этой древней эпохе в центральной части Русской равнины и севернее расположенных местностях относятся за- чатки земледелия, о чем говорят также широко распространен- ные остатки текстильной, или сетчатой, керамики. Интересные материалы имеются в работе А. Я. Брюсова (1950). По собран- ным им данным, во многих местах СССР обнаружены черепки с отпечатками тканей, нитей, веревок, сетей, применявшихся при лепке или сушке глиняных сосудов. По А. Я. Брюсову, «...Самое -обилие подобных черепков и широкое распространение такой керамики свидетельствует о том, что отпечатавшаяся на них ткань была уже заурядным предметом домашнего обихода и использовалась нередко, может быть, в виде тряпок, на побоч- ные дом ашние нужды». Текстильные черепки найдены также близ г. Мурома, в стоянке у деревни Панфилово и отнесены к середине II тыся- челетия до н. э. На ряде черепков младшего Волосовского мо- гильника, датируемого 1300 — 1200 гг. до н. 3.. видно, что ткань изготовлялась посредством плетения и похожа на ткани из швейцарских свайных построек каменного века. В том же районе похожие ткани были найдены в стоянке на реке Ве- летьме и в стоянке Легалиноборской, а также у стыка волосов- екой культуры с рязанской культурой на стоянке 1~оренец, син- хронной с Волосовской, и на Владыченской стоянке, которая по находке в погребении бронзовой манжеты моравского типа (унетицкой культуры) датирована началом II тысячелетия до н. э. В пределах Рязанской культуры найдены также ткани на Борковской стоянке под г. Рязанью. Значительно севернее, на Языковской стоянке близ г. Ка- шина (первая половина II тысячелетия до н. э.), на Уницкой стоянке под Ростовом в Ярославской области, еще далее к се- веро-востоку — под Чухломой (Федоровская стоянка) и на стоянке по реке Юг (относится к федоровской культуре) най- дена сетчатая керамика. Таким образом, и на Верхней Волге ткани в раскопках появляются уже в начале II тысячелетия до н. э. Ткачество в ту эпоху было распространено и в более север- ных областях — на территории современных Вологодской, Архан- гельской областей и карельской АССР. На реке Шексне известны две стоянки с сетчатой керамикой: из деревень Ягорба и Бара-
чек. В последней найден отпечаток плотной ткани с четким диагональным переплетением. Эти остатки датируются второй половиной П тысячелетия до н. э. Сетчатая керамика найдена также значительно северпес- на правом берегу реки Модлоны, около впадения в нее реки Перечной. Здесь в верхнем культурном слое обнаружены отпс- чатки тканей из довольно толстых крученых нитей с переплете- ниями диагонального и полотняного типов. Эти находки дати- руются около 1600 — 1300 гг. до н. э. Сравнительно высокая ступень развития ткани на керамике этого верхнего слоя сви- детельствует о том, что искусство ткачества начало здесь раз- виваться задолго до этой даты. В нижнем культурном слое свайной постройки, относящейся к началу II тысячелетия до н. э., не было найдено сетчатой керамики. А. Я. Брюсов объяс- няет это тем, что в данную эпоху не употреблялся такой способ изготовления глиняной посуды, при котором на ней могла б» отпечатываться ткань.,Яоказательством правильности его сужде- ний служит факт нахождения здесь, в нижнем культурном слое свайного поселения, не только семян культурного льна, но и обломка деревянной прялки. Еще севернее, в окрестностях г. Петрозаводска, была отко- пана сетчатая керамика на месте древней бронзолитейной ма- стерской у устья реки Томицы. Эта мастерская датируется кон- цом I тысячелетия до н. э. Во всех перечисленных стоянках отпечатки показывают, что ткань была грубая с толстыми уточ- ными нитями и с небрежным, хотя и разнообразным переплете- нием. Эти ткани очень примитивны по сравнению с теми, отпе- чатки которых найдены в стоянках I тысячелетия до н. э. в более южных областях (например, на черепке из скифского поселения близ г. Никополя видна полотняная ткань, несомненно, выде- ланная на ткацком станке) . Однако в младшем Волосовском мо- гильнике оттиски на керамике соответствуют грубой ткани, а в погребении найден был кусочек полотна, что явно говорит о су- ществовании и здесь в это время ткацкого станка. «Следова- тельно, — заключает Брюсов, — для выделки посуды применя- лась грубая ткань, а для одежды — лучшая по качеству». Находка в свайном поселении на реке Модлоне семян куль- турного льна и находка в более позднем Волосовском погребе- нии полотняной ткани доказывают существование в это время земледелия на севере Русской равнины. Несомненно, кроме льна, сеяли и другие растения. Интересно, что культура пря- дильного льна была довольно широко распространена в это время в средней и северной Европе — от Швейцарии до Урала. Если в швейцарских свайных постройках (Робенгаузен) древ- нейшие находки льна и тканей датируются более ранним време- нем, то, по существу, неизвестно, как рано началась культура льна на нашем севере — может быть и раньше II тысячелетия до н. э. 192
Данные о наличии земледелия уже во II тысячелетии до н. э. у некоторых северных первобытных племен лесной зоны на се- вере Русской равнины идут вразрез с существовавшими раньше представлениями о чрезвычайно низком уровне культуры в этих местах во II — I тысячелетиях до н. э. Скорее всего, что земледе- лие в это время было островное и подсобное. Система земледе- лия, конечно, была подсечная. На современной территории СССР во II и I тысячелетиях до н. э. были два очага льняной культуры прядильного направ- ления. Южный центр находился в 1(олхиде, основывался на ози- мом льне местного происхождения и служил не только целям местного потребления, но давал лен для экспорта на запад. Такой же озимый лен, привнесенный, вероятно, из Колхиды, воз- делывался в период свайных построек в Швейцарии. Другой центр располагался в нечерноземной полосе Русской равнины и, вероятно, был издревле основан на культуре льна-долгунца. Этот лен не мог быть здесь в полном смысле первичной культу- рой, так как в этом районе никогда не было дикорастущих льнов. Русские северо-западные долгунцы представляли собой совер- шенно оригинальный морфобиологический тип, не похожий на долгунцы и межеумки других стран. Здесь был создан знамени- тый русский лен-долгунец, который формировался в течение тысячелетий из материала, занесенного, как можно судить на основании ботанических данных, из Юго-Западной Азии (гор- ные районы современного Таджикистана). Приведенные данные свидетельствуют о том, что на широ- ком пространстве нечерноземной полосы Русской равнины острова земледелия появились очень рано, в конце каменного века. Позднее древнейшее земледелие вступило в соприкосно- вение со славянским земледелием, стоящим на более высоком культурном уровне. Славянское влияние распространялось с юго-запада, и аван- посты его уже в V — VII вв. заходили далеко на север (Псков, Старая Ладога, Новгород). Культура древнейших обитателей севера была охотничье- рыболовческой, но и острова земледелия здесь имелись задолго до славянской эпохи. 1(ультура древних северных племен была связана с Уралом и Сибирью, о чем речь будет ниже. В соответствии с установленной теперь новейшими истори- ческими и археологическими изысканиями древностью земле- дельческой культуры на севере Русской равнины, здесь и ныне встречаются очень древние («стародавние») формы и сорта некоторых культурных растений. Таким «пережитком глубокой старины» являются, например, северные эндемичные сорта репы, низкорослая северная конопля — живые ископаемые севера. Сннская Е. Н. 193
КУЛЬТУРНЫЕ РАСТЕНИЯ, ВОЗНИКШИЕНА РУССКОЙ РАВНИНЕ Русская равнина была с древнейших времен ареной земле- дельческой культуры. В ледниковые эпохи Русская равнина была выпахана ледни- ком, многие дикорастущпе виды или вымерли, или отступили к tory. В настоящее время она по сравнению, например, с Кав- казом очень бедна видами высших растений. Растительные ре- сурсы Русской равнины относительно бедны. Однако помещен- ный ниже обзор показывает, что и здесь не так уже мало дико- растущих видов было взято в культуру, как этого можно было ожидать, и что ресурсы еще не исчерпаны. Северная т и м о ф е е в к а (РЫеит .pratense L.), несомненно, является аборигенной формой, так как культурный экотип во- логодской тимофеевки очень близок и биологически, и морфоло- гическц к местному дикорастущему экотипу. Русский север- зто область первичного введения в культуру тимофеевки. В Аме- рику она занесена была отсюда позднее. Эндемичные северные формы (экотипы) описаны также для других дикорастущих зла- ков: костра безостого, овсяницы луговой, овсяницы красной, лисохвоста, полевицы, ежи сборной, мятлика лугового и болот- ного, бекмании. Эти северные формы луговых злаков резко отличаются морфобиологическими и хозяйственными призна- ками. Богата формами (экотипами) луговых злаков центральная лесная зона и степные районы (главным образом поймы, балки и западины). дикорастущие экотипы луговых злаков уже по- служили исходным материалом для многих селекционных сор- тов, хотя использование этих ресурсов далеко еще не закон- чено. То же можно сказать и о дикорастущих бобовых расте- ниях. Местные дикорастущие формы к л е в е р о в (Trifolium pra- tense sensu lato, T. repens, Т. hybridum) красного, белого и гиб- ридного интересны для селекции, хотя еще далеко не полностью изучены. Об этом говорят, например, исследования М. С. пен- ковой в KOMH АССР, где местный дикорастущий печорский кле- вер оказался при сравнительном испытании перспективнее, чем сорта культурного клевера красного. Аборигенная л ю ц е р н а ж е л т а я (Medicago falcata sensu lato) послужила компонентом в создании ценных гибридных сортов посевной люцерны. Такими же компонентами в каждом районе были свои местные дикорастущие экотипы люцерны желтой: на севере — северный луговой экотип (марусинские сорта), на юге — степные экотипы (украинские, северокавказ- ские п другие местные и селекционные гибридные сорта). Эти сорта имеют большую хозяйственную ценность, многие из них распространены широко за пределамц первичного очага их 194
культуры и районированы даже в азиатской части СССР Се- верная дикорастущая люцерна желтая настолько своеобразна, что некоторыми ботаниками обоснованно считается самостоя- тельным видом — Medicago borealis Grossh. Сходные формы имеются в лесостепи Западной Сибири и в горной Киргизии. Ряд северных форм люцерны желтой (Дединовская, по рекс Мсте и др.) были изучены цитологически (В. А. Борковская, 1952; Е. Н. Синская и 3. П. Малеева, 1959). Они оказались ди- плоидами: имели число хромосом 2n = 16, как и древнейшие формы дикорастущей люцерны на Кавказе. Сходные по габптусу сибирские и киргизские формы еще цитологически не изучены. Если и они окажутся диплоидами, то можно предположить существование круговой миграции этих форм с Кавказа через Сибирь на север Русской равнины. Донник б ел ы й (Melilotus albus Desr.) представлен в не- черноземной полосе многими эндемичными экотипами северного подвида, а в степной зоне — эндемичными экотипами южного подвида. Ди ко р а стущи е э сп а р цеты [Onobrychis tanaitica S~preng., О. arenaria (КН.) Ser. и др.] имеют ряд экотипов и представляют интерес для использования в селекции. Эспарцет песчаный — О. arenaria (в широком понимании объема этого вида) начал входить в культуру в нашей стране сравнительно недавно. Первые опыты по введению этого растения в культуру, по-видимому, сделаны В. H. Черняевым. Интересно, что многие северные экотипы европейской части СССР не только люцерны желтой и донника, но и ряда других луговых злаковых и бобовых дикорастущих трав обнаруживают большее родство с западносибирскими и алтайскими экотипами тех же видов, чем с южными экотипами из пограничной полосы распространения (донник белый, люцерна желтая, костер без- остый). Такие факты заставляют предполагать, что в периоды отступления ледников миграция в нечерноземную полосу евро- пейской части СССР шла не только из южных убежищ, но и из Сибири, а в некоторых случаях с Алтая через Урал. Таким путем, по-видимому, могло идти расселение и растений, и человека. Помимо кормовых растений из дикарей, по-видимому, кое- где вошли в культуру местные дикорастущие формы тмина, хмеля, хрена. Хрен (Cochlearia armoracea L.) в дикорастущем состоянии распространен в Финляндии и по всей Русской равнине до Астра- хани. В Западной Европе — это одичалое, но не дикое растение. Родина его — Восточная Европа. Русское слово хрен — ichren имеет общеславянский корень. От этого корня произошли и не- которые немецкие и французские названия — kreen, cran, cran- son. Таким образом, хрен представляет собой аборигенное для Восточной Европы культурное растение.
К е н д ы р ь сибирский — Аросупит sibiricum (Pall.) Russ. (основной очаг — волжско-уральский) распространен на 1(авказе и в 1(рыму. Некоторые дикорастущие плодовые и ягод н ые р а сте- н и я имеют значение в селекции. Из них назовем виды дикора- стущей яблони (Malus silvestris Mill., М. ~praecox Вог1й.), дикую грушу (Pirus соттип1з L.), черную смородину (Ribes nig- rum L.), дикую вишню (Prunus fruticosa Pall.) и другие плодо- вые и ягодные деревья и кустарники. А. Д. Даниловым (1947) недавно были найдены в лесах Воронежской области не описанные раньше разновидности груши обыкновеннои: темно-бурая, светлокорая, темнолистная и светлолистная. По мнению А. Д. Данилова, эти разновидности представляют интерес для лесоводства и плодоводства. Состав популяций дикорастущих плодовых Русской равнины изучен еще недостаточно. Групповое разнообразие диких груш и яблонь следует детально изучить и в других районах европейской части СССР. По А. Н. Ипатьеву (1960), существуют в настоящее время следующие группы сортов культурных яблонь: 1) средне- или северорусская группа (Боровинка, Грушовка московская, Белый налив, Антоновка, Титовка, Скрижапель, Бабушкино и др.); '2) западнорусская группа (Суйслеппер, Штрейфлинг, Борсдорф- ское и др.); 3) волжская группа (Малы, Астраханское, Хоро- шавка и др.); 4) уральская группа близка к сибирским сортам. Бе р ескл ет 6 о род а вч а ты й (Euonymus verrucosa Scop.) имеет значение как гуттаперченос. Во флоре Русской равнины имеются также л е к а р ств е н- ные, ду бильные, медоносные и другие полезные расте- ния. Еще далеко не все из них достаточно исследованы. На се- веро-западе Русской равнины растет таран дубильный (Polygo- mum сог1аг1цт Grig.). Сделаны опыты по его выращиванию на Карельском перешейке (П. А. Якимов, Г В. Куршакова, А. Г Власова, 1954). Основной очаг происхождения точно не установлен. Азиатские формы отличаются от европейских. Для тех культурных растений, для которых в составе або- ригенной дикорастущей флоры не имеется диких родичей, источ- ник происхождения может быть двояким: 1) из местных (или завозных) сорняков; 2) из завозных семян культурных расте- ний. Кратко рассмотрим состав культурных растений европейской части СССР Из плодовых отметим холодостойкие абрикосы (Armeniaca vulgaris Lam.) — «жердели», распространенные в центральной и южной частях европейской части Союза. Здесь же и в сосед- них областях имеется много стародавних сортов яблони, груши, вишни, которые использованы широко в селекционной работе. Русские местные сорта яблони отличаются холодостойкостью. 195
Группа волжских сортов характеризуется засухоустойчивостью и ранним созреванием. Северорусские и украинские сорта груши характеризуются рядом полезных качеств: холодостой- костью, нетребовательностью к почве. Весь этот ассортимент местных сортов используется в селекции. Рассмотрим детальнее ассортимент полевых и огородных культур. Р ажь (Secale cereale 1.) появилась во всех районах ее куль- туры позднее ячменя и пшеницы. По археологическим данным, на территории современных Венгрии, Польши и других погранич- ных с СССР государств, а также ГДР и ФРГ рожь появилась не раньше железного века и в большинстве случаев обнаружена именно в раскопках, относящихся к славянской эпохе. На терри- тории СССР, по археологическим данным, рожь отмечена в Старой Ладоге в Ъ'11 — IX вв, в Новгороде — XI — XII вв., Лат- .вии — V, Х — XIII вв., Эстонии — XIII вв., Литве — XI — XII вв. Раньше XI B. она, однако, встречалась редко и в очень неболь- шом количестве. Как было сказано выше, в те же века появляется рожь и на юге славянской территории, в более ранних находках можно подозревать наличие сорной, а не культурной ржи. В древней- ших слоях она встречается только с пшеницей, а позднее и от- дельно. Смешанная культура ржи и пшеницы — суржа, еще недавно бытовала в южных районах, особенно в предкавказских степях. При продвижении на север условия становились все более благоприятными для ржи, и возникла уже чистая ее куль- тура, которая распространилась в лесной зоне СССР Рожь н основном была освоена нашими предками — протославянами и славянами. И в настоящее время основное мировое богатство сортов высококультурной ржи сосредоточено в нашей стране и, главным образом, в лесной зоне Русской равнины. Установлены следующие экологические группы сортов ржи (В. Ф. Антропова): 1) северорусская; 2) западноевропейская— также развилась, в основном, на базе древнеславянской куль- гуры; 3) степная, заходящая и в азиатские районы СССР. У типичной северной ржи больше общего с переднеазиат- ской сорной рожью, чем с сорной рожью Среднеазиатских рес- публик и Лфганистана, которая чаще имеет ригидное, более грубое строение колоса. О в е с (Avena sativa L.) также и по своим древнейшим оча- гам зарождения и по современному разнообразию сортов— культура славянская, отнюдь не западноевропейская. Сомнение вызывает только богатство и своеобразие китайских овсов. Там, может быть, имеет место самостоятельный очаг зарождения или, во всяком случае, развития этой культуры. Западная внеславянская Европа дала мировому земледелию совсем другой вид овса — Avena strigosa Schreb., который теперь не имеет широкого распространения и становится реликтовой
культурой. В местностях, примыкающих к границам СССР, в раскопках бронзового периода найдены только мелкозерный овес, по мнению многих авторов, не типичный для А. sativa L., относящийся к виду — А. strigosa. Позднее в тех же приблизи- тельно районах появляется овес посевной (и одновременно овсюг А. fatua L.), который получает широкое распространение в славянскую эпоху, примерно в тот же период, что и рожь, в VII — XIII; однако внедрение его шло медленнее. Обыкновенный овес — это культура географически восточно- европейско-азиатская, а исторически — славянско-китайская. Самые древние формы А. sativa, очевидно, полбяные сорные овсы этого вида. Очаги культуры полбы находились в Передней Азии (Закавказье), 1(амско-Волжских районах и на Балканах, т. е. окружали древнеславянские территории. Ячмень (Hordeum L.) представлен в европейской равнин- ной части Советского Союза северной группой — (proles boreale Vav. et ВасЫ., заходящей и в Скандинавию, и так называемой западноевропейской — proles aocidentali-еигореит Vav. et. ВасЫ., распространенной преимущественно в лесной и лесостеп- ной зонах европейской части СССР, а также в Чехословакии, ГДР, ФРГ и далее на запад до Англии и Франции включительно (Ф. X. Бахтеев, 1948) . В эти группы вошли и селекционные сорта, но в них не выделены систематиками ячменей наиболее древние корни. Тем не менее в обеих группах имеются, несом- ненно, стародавние местные сорта ячменя. Я хмени на террито- рии Русской равнины являются более древней культурой, чем рожь и овес, и на севере — более древней, чем пшеница. Древ- нейшим очагом, из которого распространился сюда ячмень, яв- ляется, вероятно, Передняя Азия. П ш е н и ц а могла попасть на Русскую равнину из Передней Азин (в основном) и из Юго-Западной Азии, в меньшей сте- пени — с Запада. В европейской части СССР мягкая пшеница (Triti~cum aestivum) представлена следующими агроэкологиче- скимц группами: северной лесной, лесостепной, степной волж- ской, степной. В эти группы входят и местные стародавние сорта, и селекционные, во многих случаях выведенные из местных. Твердая пшеница — Т. durum Desf. — представлена особой степной группой. К. А. Фляксбергер (1935) различал здесь две подгруппы: рыхлоколосую (кубанки) — proles laxiusculum и плотноколосую (белотурки) — proles densiusculum. Аборигенные твердые русские пшеницы пользуются мировой известностью благодаря высокому качеству зерна (высокое содержание белка) и широко используются в мировой селекции, особенно г, США и 1(анаде. Эта группа сортов формировалась в течение достаточно длительного времени, так как культура твердой пше- ницы на юге России известна со времен Триполья. Полба — Т. dicoccon Schrank — имела большой реликтовый очаг в Камско-Волжских районах (Татарская АССР, Чуваш-
ская АССР) и также в Югославии и Болгарии. Эта полба вы- делена К.А. Фляксбергером в proles tartaro-balcanicum. Из По- волжья полба распространилась далеко на север; ископаемая полба из Старой Ладоги похожа на волжскую. I(aK и в странах северо-западной Европы полба распространилась с юга на се- гер, а затем снова отступила к югу, где лишь кое-где до настоя- щего времени остались островки этой культуры (ныне уже почти исчезнувшие). Пришла полба на Волгу, вероятно, из Ма- лой Азии, через Балканы. Н. И. Вавилов предполагал, что в древности распростра- нение пшеницы шло в Европу через Переднюю Азию (1(авказ) и Балканы. Поэтому он искал в 1(арпатах следы реликтовых культур и действительно нашел там в культуре полбу-эммер в горном районе Путиля, в северо-восточной части Карпат (Ф. Х. Бахтеев, 1960). П р о с о — Panicum miliaceum L. — древнейшая культура Русской равнины. Просо было известно в Триполье и служило главным пищевым растением скифам. Нетребовательность куль- туры проса, его скороспелость и легкость изготовления из него пищи (каша, заквашенное зерно) сделали просо любимым ра- стением азиатско-европейских кочевников. Нуждаясь в корме для лошадей, скифы могли содействовать распространению овса, культивируя его вначале, вероятно, в смеси с полбой. Овес, по-видимому, вошел впервые в Европе в культуру именно на протославянской и скифской территориях, а просо скорее всего перешло к протославянам и скифам из Три- полья. В. Н. Лысовым установлены следующие региональные эколого-географические группы проса в пределах подвида effusum: северная и лесостепная; в пределах подвида contra- ctum: лесная, лесостепная (переходная к степкой), степная по- волжская и степная украинская. Г р е ч и х а ((Fago~pyrum sagittatum) в европейской части СССР представлена четырьмя агроэкологическими типами, от- носящимися к подвиду ssp. vulgare Яо1е1. В европейской части СССР распространены следующие группы (пролесы) и сорто- типы гречихи: 1) скороспелая (сортотипы Казанский, Терехов- ский); 2) среднеспелая — сортотипы Богатырь и сортотип Се- ребристый (А. С. Кротов, 1963). Гречиха пришла в Европу из Азии. Путь расселения ее по Русской равнине мог быть двояким: с востока (гречиха рано по- явилась у сарматов) или из Передней Азии. Местные сорта г о р о х а — Pisurn sativum L. — вытеснены почти всюду селекционными, но раньше они имели большое зна- чение, и некоторые из них использованы в селекции. Дольше сохранились местные сорта на севере (Приуралье и Коми АССР). В Поволжье встречается особая форма гороха, засо- ряющая вику — proles wolgense Gov., относящаяся к закавказ- скому подвиду — ззр. transcau~cas;cлп Соч.
На Русской равнине имеются стародавние сорта ч е ч е- в и ц ы — Lens esculenta Moench — и конских бобов — Faba vu1garis Моепсй. Последние большей частью относятся к бо- реальной агроэкологической группе. Можно отметить следующие эндемичные формы конских бобов: 1) f. minutissima Murat., год- ную для возделывания на корм (Ленинградская область); 2) f. mi~crosperma Murat. (Кировская область); 3) f. pumila Murat. (лесная зона СССР); 4) f. borealis Murat. (Предуралье, центральные области, заходит в Сибирь); 5) f. opaca Murat. (Предуралье); 6) f. ianthina Murat. (северные и центральные районы); 7) f. nigro-violacea Murat. (Тамбовская область). Эти ~рормы темносемянные и мелко- или среднесемянные (за исклю- чением последней). В и к а п о с е в н а я — Vicia sativa I . — представлена на тер- ритории европейской части Союза еще большим экологическим разнообразием, чем рассмотренные выше бобовые культуры. По А. Ю. Тупиковой, здесь имеются следующие агроэкологиче- ские группы: 1) юго-западная (Правобережная Украина, БССР); 2) восточная (Куйбышевская область, Мордовская АССР и Татарская АССР); 3) северо-восточная (Кировская и Свердловская области); 4) северная (северные области, север БССР). Бобовые культурные растения распространились на европей- скую часть СССР в основном из Передней Азии (древнейший очаг) и частично из Западной Европы (вторичный источник переднеазиатского же корня). На Русской равнине везде господствуют местные гибридные сорта люцерны или выведенные из них селекционные. Культурный русский к л е в е р (Trifolium:pratense I .) очень разнообразен и делится в основном на группы: 1) многолетний одноукосный (север); 2) малолетний двуукосный (южные степ- ные районы); 3) промежуточный — северный двуукосный (севе- ро-запад). П. Н. Лисицын считал его занесенным пз Среди- земноморья или южной Европы и в какой-то мере подвер- гавшимся влиянию дикорастущих местных клеверов. По Н. Г. Хорошайлову (1952), процесс формирования сортов куль- турного клевера протекал в различных районах по-разному: без участия завезенных западноевропейских раннеспелых сортов клевера, а также при слабом и значительном их участии. Во всяком случае, русские клевера настолько переработались в новой агроэкологической среде, что стали эндемичными фор- мами, сохраняя большой полиморфизм популяций. Русские кле- вера известны во всех странах умеренных зон земного шара. Переходя к техническим растениям, прежде всего следует отметить л е н — Linum usitatissimum L. Северо-Западная Русь дала лучшие в мире стародавние сорта прядильного льна, не- превзойденные русские долгунцы, которые во всех странах по- служили основой для выведения сортов прядильного льна. Куль- 200
тура льна в нечерноземной Руси существовала еще во II тыся- челетии до н. э. и даже, вероятно, раньше. К о н о п л я — Cannabis ваШа L. — также является древней культурой на Руси. В сорном и рудеральном состоянии (около жилищ и на «базах» — стойбищах скота) конопля широко рас- пространена в Поволжье, Южном Приуралье, Крыму, Предкав- казье, на Украине. Источником происхождения культурных форм могли быть местные сорные формы. На территории евро- пейской части СССР распространены две агроэкологические группы: северная и среднерусская. Особенно оригинальна край- няя северная разновидность (var. praecox Serebr.), низкорослая и раннеспелая, годная только для возделывания на масло. Из сравнительно недавно завезенных культур следует отме- тить русский масличный п о д с о л н е ч н и к — Helianthus annuus L. Он резко отличается от существующих дикорастущих, сорных и декоративных форм подсолнечника. Попал он в Сред- нюю Россию скорее всего из Армении лет 200 — 300 назад; за это время в условиях новой агросреды возникли его новые бота- нические формы. Использовать подсолнечник как масличную культуру начали в России. Эта важнейшая масличная культура аборигенна для степных и лесостепных районов Русской рав- нины. От каких именно видов и форм произошли сорта русского масличного подсолнечника в настоящее время решить трудно. Вероятно, в формировании его принимали участие Н. lenticula- ris Dougl. п грызовой подсолнечник из Армении. Армянские формы подсолнечника, эндемичные для Армении, своеобразны, имеют черты дикорастущих видов. В настоящее время выведен на основе местных сортов ряд высокомасличных селекционных сортов. Здесь мы видим при- мер сравнительно быстрого возникновения большой группы эндемичных сортов в результате внедрения нового способа ис- пользования растения. М а к — Paipaver somniferum L. — и, по-видимому, г о р- ч и ц а — Brassica jun~cea (L.) Czern. — были известны еще в древнеславянские времена, но не как масличные (мак — для пряников, а горчица — как пряное). Теперь сарептская горчица (В. juncea) имеет здесь один из самых больших очагов куль- туры этого вида на масло (второй очаг — Индия) и представ- лена здесь эндемичной разновидностью (вернее — подвидом) var. sareptana Sinsk. Русская поволжская горчица характери- зуется высокой урожайностью и масличностью и отличается большим полиморфизмом популяций. В культуру вошла из сор- няков льна, проса и зерновых культур и распространилась из Азии (Монголия, Казахстан, Средняя Азия). В Закавказье нет ни дикорастущей, ни сорной горчицы этого вида. На Русской равнине, особенно на Украине, распространены стародавние сорта (огородные) мака, относящиеся к евроазиат- скому подвиду. 201
К о р и а н д р — Coriandrum sativum L., не является древней культурой для Русской равнины. Воронежская область — ста- рейший очаг культуры этого растения. Завезен кориандр сюда в первой половине XIX в. Однако за это время успел вырабо- таться особый местный сорт воронежского кориандра. Р ы ж и к я р о в о й — Camelina sativa (L.) Crantz — также сравнительно новая культура. Богатство форм сорного рыжика еще недавно было огромным. Сорные и культурные яровые рыжики на Русской равнине делятся на 4 подвида: 1) рыжик льняной — зэр. linicola (Zing.) Sinsk., засоряет лен-долгунец, имеет высокое иодное число; 2) рыжик промежуточный — зэр. зиЬ11пйсо1а (Zing.) Sinsk., засоряет льны-межеумки; 3) ssp. glabrata (Zing.) Sinsk. засоряет изредка лен-кудряш, а также зерновые яровые культуры (ячмень); 4) ssp. subglabrata Sinsl'. засоряет лен в центральных районах и отчасти в Приуралье, имеет некоторое сходство с сибирским экотипом, вероятно, при- шел из Сибири (к нему относятся высокомасличный сорт ВНИИМК 17). Все эти эндемичные подвиды представляют ин- терес для селекции. Р ы ж и к аз им ы й — С. silvestris звр. pilosa Zing. — кое-где возделывался, но теперь почти не встречается в культуре. Воз- можно, что эта культура имеет ценность, и поэтому интересны как исходный материал те сорные формы озимого рыжика, ко- торые местами сохранились в посевах озимых хлебов. С у р е п и ц а я р о в а я — Brassica campestris 1 . — также- исчезла из культуры, хотя никогда не была широко распростра- нена. Представляет интерес как медонос и, может быть, как масличное. На территории европейской части СССР имеется эндемичная сорная форма — var. prostrata Sinsk., возможно, и другие. В ботанической литературе часто имеются указания на местонахождение одичалой черной горчицы — Brassica nigra Koch — в средней полосе по поймам рек. Мы лично видели целые заросли черной горчицы в районе заповедника Галичья Гора Липецкой области, где она растет в прирусловой песчаной по- лосе Дона вместе с ивняком, достигает 2 м высоты, обильно цветет и плодоносит. По общему габитусу эта горчица несколько напоминает горчицу из Малой Азии, возделываемую в чистых посевах, отличаясь от последней почти голыми нижними листьями. С. В. Голицын и другие воронежские ботаники сооб- щили, что эта форма черной горчицы широко распространена по Дону в Воронежской и Липецкой областях и неизменно при- урочена к одному типу местообитания — прирусловым пескам. С. В. Голицын определенно утверждает, что здесь черная гор- чица растет дико и что никогда никто ее здесь не сеял. Интересно, что такая же форма черной горчицы встре- чается значительно севернее — в Окском притеррасном заповед-- нике Московской области. Здесь черная горчица представлена. 202
той же формой, что и по верхнему Дону, и растет на местооби- тании этого же типа — на прирусловых песках в пойме Оки. В культуре в Московском отделении ВИР эта форма горчицы сохранила свою высокорослость. Обширные заросли ее по пой- мам Оки, Дона представляют очень интересный факт с точки зрения географического распространения растений. Если даже считать, что здесь Brassica nigra не является искони дико- растущим видом, во всяком случае следует признать, что это растение натурализовалось, стало как бы членом местной дикой .флоры с константным типом местообитания. Родина этого древ- него вида горчицы точно неизвестна. Условия той области, где она смогла натурализоваться, указывают на то, что по проис- хождению этот вид не обязательно должен быть южным. Из огородных растений едва ли не самым древним на Рус- ской равнине является р е п а — Brassi~ca rapaeuropaea Sinsk. Выявлено существование здесь двух эндемичных групп репы, очень различных по морфологии и по очагам происхождения. Первая — это петровская репа и примыкающие к ней сорта (ss~p. rossica Sinsl~.). Она имеет родство с примитивной группой тельтовских реп, сохранившихся кое-где в ГДР, ФРГ и Фран- пии. Эти две группы — дерпваты древнейшей европейской репы, и мы считаем, что в странах северного, более или менее уме- ренного климата Европы, находится один из первичных очагов происхождения репы. Группа реп петровского типа сложилась у финских и у славянских племен. Вторая группа эндемичных русских реп — это карельские репы, довольно разнообразные по внешним признакам. Но родство здесь совсем другое. Ряд об- щих признаков и свойств говорит о том, что репы эти были за- .несены из Передней Азии. В настоящее время очаг примитивных грубых форм этой линии родства сохранился в культуре в Ма- лой Азии. Б р ю к в а — В. napobrassi~ca Mill. — также имеет эндемич- ную группу желтомясых брюкв типа красносельской брюквы. ~Келтомясые брюквы, подобно желтомясым репам, возникли на севере Европы ц основной очаг их происхождения — это северо-восток Европы — Финляндия и север европейской части СССР Красносельская брюква использовалась в России и в Скандинавии для создания селекционных сортов. В европейской части СССР имеются стародавние сорта редьки. Редька пользовалась большой популярностью в Древ- ,ней Руси и в более поздние эпохи. Об этом говорит следующая поговорка, указывающая на разнообразие способов потребле- ния редьки: «редька — триха, редька — ломтиха, редька- с квасом, редька — с маслом, редька — так». Коч а н н а я капу ста — Brassica capitata (L.) Lizg.— .имеет эндемичные группы местных сортов. По Т. В. Лизгуновой (1959), в европейской части СССР имеются следующие геогра- :фические группы кочанной капусты: 1) сорта южной части
Украинской ССР, имеющие много общего с сортами Северного Кавказа, Закавказья и Средней Азии; возможно, что эти сорта распространились на север с Балкан и из стран Передней Азии;. 2) сорта центральной зоны, среди них есть эндемичные; 3) сорта юго-востока — Поволжье; 4) северные сорта. ,Лук — А11шт сера L. — на Русской равнине представлен сле- дующими подвидами, из которых каждый объединяет несколько групп местных сортотипов: 1) среднерусский — зэр. medio-rossi- cum Troph. 2) южный — ssp. australe Тгор1т. В северо-западных районах имеются своеобразные местные, вегетативно размножаемые сорта лука, биологически напоми- нающие шалот — A. ascalonicum L., но которые нельзя отожде- ствлять с последним. Лук дудчатый, или батун — А. fistulosum L. — представлен на Русской равнине особым подвидом — ssp. ruthenicum Troph Шнитт-лук — А. schoenoprasum L., — в европейской части СССР представлен особой среднерусской группой местных сор- тов (Н. Х. Трофимец). О г у р ц ы — Cucumis saiivus L. — представлены, по А. И. Фи- лову (1948, 1959), на европейской части СССР следующими эндемичными группами сортов: 1) клинской; 2) астраханской; 3) северной; 4) восточноевропейской (главным образом Украина). Первоначально огурцы были получены славянами из Визан- тии, а потом, возможно, они попали из стран Передней Азии. Родство с Передней Азией и Балканами обнаруживают разно- видности, выделенные A. И. Филовым — анатолийская, килий- ская, балканская н астраханская. Позднее на сортовом разно- сбразии сказалось влияние стран Западной Европы — Англии, Дании, Германии. Китайские огурцы входили в культуру в на- шей стране не непосредственно, а через страны Западной Ев- ропы. В северной европейской части СССР распространены мест- ные сорта огурцов, которые А. И. Филов (1960) относит к север- ной разновидности — var. borealis Fil. — европейско-американ- ского подвида. Сюда относятся сорта огурцов Муромский, Вяз- никовский, Павловский, Боровский, Кооператор, Неросимый,. Фоминский парниковый и др. К тому же подвиду относятся местные сорта разновидности var. eurapaeus Gab. — Степан Ра- зин, Хорошаевский, Берлизовский и другие, распространенные в центральной части черноземной полосы и южнее. Нежинские огурцы — var. nezhinensis Fil.— распространены на Украине и прилегающих с севера к ней районах. Имеются в СССР и сорта китайского подвида. Они занесены из Китая в Англию, Данию, ГДР и ФРГ, а оттуда — в нашу страну. Из этого завозного ма- териала у нас сформировались свои сорта: Телеграф, Сенсация и др. Из китайского подвида вышел клинский огурец — var. kli- nensis Ritov, разводится в теплицах. 204
Т ы к в а — Cucurbita maxima Duch. — в европейской части СССР распространена в Поволжье — тип Волжской серой тыквы или разновидности серых тыкв — var. viridis Ser. Этот тип тыквы культивируется также на Украине, в средней части Ка- захстана, в Закавказье, меньше — в Сибири. Разновидность бе- лых тыкв менее распространена (сюда относятся, например, Белая медовая, Безенчукская белая и др.). К разновидности мамонтовая — var. grossa броет.— относится сорт Шестипудо- вая. Тыква зимняя — var. hiberna Fil. — не так дано распро- странилась на Кавказе, занесена она из Испании. Относящиеся к другим разновидностям этого вида сорта встречаются редко. Из форм тыквы С. реро L. в нашей стране разводится раз- новидность цитрульная — var. Ыгош11а Naud., относящаяся к подвиду плетистому. Сюда относится сорт Мозолеевская. Рас- пространена эта разновидность на Украине и в Балканских странах, откуда, вероятно, занесена на территорию СССР. Зим- ние сахаристые тыквы занесены из Испании в предгорные и отчасти степные районы Северного Кавказа. Сюда относятся сорта вида — Cucurbita maxima — Испанская и Черкасская. Это — сравнительно новые здесь сорта. Ар 6 уз — Citrullus Schrad. — во второй половине I тысяче- летия н. э., по А. И. Филову (1960), стал распространяться на запад тюркско-монгольскими племенами и отчуда попал на Русь, например, в Подонье. В настоящее время в европейской части Союза имеется много превосходных местных и селекционных сортов арбуза. g ы н я — Cu~cumis melo L. — распространена в европейской части СССР сравнительно мало, но все-таки на Северном Кав- казе и еще кое-где на юге имеются и местные сорта дыни. Рус- ские дыни отнесены А. И. Филовым (1960) к европейскому под- виду — ssp. europaeus Fil., среди них имеются ранние разновид- ности — var. praecox Fil. и зимовки — var. hiemalis Fil. Интересные сведения о культуре дынь в XVII в. приводит Адам Олеарий в своей книге: «Описание путешествия в Моско- вию и через Московию в Персию и обратно» (данные взяты из заметки А. Васильева, 1960). Олеарий пишет, что дыня возде- лывается на Руси и там выработаны особые приемы культуры этого растения. Семя размягчается в парном молоке или в от- стоявшейся дождевой воде, в которую добавлено старого овечьего помета. Из соломы и навоза устраиваются грядки, по- крытые сверху хорошей почвой, в которой делают неглубокие ямы. Семена помещаются в середину ямы, чтобы они прогрева- лись снизу и сверху. Ночью и в холодные дни грядки прикры- ваются слюдяными крышками. Как мера ухода, применяется обрезка ветвей, а иногда и кончиков побегов. Свекл а (Beta vulgaris L.) на Руси стала довольно широко распространяться в XI — XIV вв. О ней говорится в «Домострое» (XVI в.), как об обычном огородном растении. Существует много 205
славянских названий для свеклы: русские — ботва, ботвинья; украинские — ботва, ботвина; польское — ботвина; сербское— битва, блитвина и др. Лингвисты ведут происхожение этих на- званий от лебеды — блитон. Другой ряд названий: церковносла- вянское — свекла; русское — свекла; древнеславянское — сеукл; польское — свикла; чешское — цвикла; сербское — цвекла; бол- гарское — цвекло и цекло; литовское — свиклое; албанское- сефкле, сфекле, сфеклия; венгерское — цекла — ведет начало от греческих и византийских названий свеклы (свекла, сеукала, сеутлои, сеуклон). За исключением немногих видов кориандра, подсолнечника, кормовых трав и некоторых других, вьппе были рассмотрены стародавние культуры, которые развились на территории европейской части СССР частично еще до славян и составляли костяк древнего славянского растениеводства. Лишь немногие пз перечисленных культур ведут происхождение от местных дикорастущих форм (хрен, ряд кормовых трав, бересклет). Еще недостаточно изучены дикорастущие растения Крайнего Севера — тундровой флоры и также пойм северных рек. Сравни- тельно большое внимание уделено дикорастущим северным ра- стениям, пригодным на корм оленям (В. Д. Александрова, 1940 и др.). Пищевые растения Севера изучены слабо (Н. А. Аврорин, 1933). Судя по работам американских исследо- вателей Арктики, в северной флоре имеются богатые ресурсы витаминоносных растений (Porsild А. Е., 1937; godahek, 1944, 1945, 1949, 1950) . В нашей Арктике эти природные богатства изучены еще недостаточно. Важнейшие русские культурные растения уходят своими кор- нямп в Переднюю Азию и распространились в основном оттуда (пшеница, рожь, зерновые бобовые, вероятно, ячмени, карель- ская репа, огурцы). Из Средне-Юго-Западной Азии пришел лен, сарептская горчица, вероятно, рыжик (в качестве сорняка), из Азии пришло просо. Америка внесла свой вклад значительно позднее — карто- фель, табак, томаты, перец, баклажаны, кукуруза, подсолнечник. Однако и по этим культурам сформировались местные сорта и выведены селекционные. Кукуруза относительно давно попала на славянскую терри- торию. В Молдавии и на западе Украины сформировались осо- бые эндемичные местные сорта кукурузы. Эти сорта отличаются мелкозерностью и плотностью расположения зерен в початке. Подобные сорта имеются и на Балканах. В еще более позднюю, современную эпоху в Молдавию и сопредельные районы попала соя и здесь отработались особые местные сорта, составившие западный подвид — ssp. slavonica Коч. et Pinz. Творческая переработка иноземного материала продол- жается. На наших глазах создаются новые, не существовавшие 206
прежде, ранние сорта хлопчатника, сои, риса и др. Страна ста- новится творческой лабораторией, где методами селекции соз- даются новые сорта и невиданные ботанические формы. ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ СИБИРИ Сибирь из-за своего сурового климата имеет относительно ограниченный набор культурных растений, почти равный набору культур Русской равнины. Так как географически Западно- Сибирская низменность является непосредственным продолже- нием Русской равнины, то ее земледельческая культура оказала несомненное влияние на историю земледелия и культурную флору Сибири. Климат Сибири не всегда был сходен с современным. В раз- ных местах Западной Сибири найдены остатки теплолюбивых форм нижнечетвертичной фауны (Elephas antiquus, Rhinoceros mercki и др.). С похолоданием, в связи с надвигающимся льдом, эта фауна исчезает и появляются животные холодного климата (мамонт — Elephas trogontherii, Bison priscus и др.). Ледники спускались на западе почти до 60' с. ш., и ими был прегражден сток Оби и Енисея. Теперешняя Западно-Сибирская низменность покрылась тогда водой, а арктическая фауна отступила к югу. Следов человека шелльской и ашельской эпох в Сибири пока не обнаружено. Остатки мустьерской культуры также недосто- верны. Наиболее древние из известных стоянокотносятся к концу ледникового периода — к верхнему палеолиту. Верхнепалеолити- ческих стоянок обнаружено много и они доходят до 61' с. ш. (А. П. Окладников, 1954). Эти стоянки сопоставляются со стоян- ками солютрейской или раннемадленской стадии Европы, хотя и отличаются от последних рядом особенностей. Существенной смены населения от палеолита к неолиту не замечено. Особенно это ясно для Прибайкалья, где видна пол- ная преемственность палеолитических и неолитических культур. Стоянки второй стадии верхнего палеолита на севере доходят до Томска, Красноярска и Киренска. Во второй период верхнего палеолита увеличились пространства, занятые степями. Стоянки третьей стадии верхнего палеолита, особенно многочисленны в районе Байкала и на Енисее. На селенгинских стоянках (при- близительно того же возраста) вместе с каменными орудиями нередки находки скорлупы страусовых яиц, часто просверлен- ных. Среди остатков фауны здесь же найдены и кости винторо- гой антилопы. Такие находки говорят о том, что климат здесь был тогда теплее, чем теперь. Эти данные очень интересны с точки зрения обширности ареала страуса в эту и в позднейшие эпохи. И. Бичурин (1884) на основании старинных китайских источников установил, что страус питался диким ячменем. Можно предположить, что и 207
ареал дикорастущего ячменя простирался в это время далеко вглубь Азии — вплоть до Южной Сибири. Эти факты по мере их накопления могут пролить свет на вопросы происхождения азиатских культурных ячменей. Селенгинские стоянки обнаруживают сходство со стоянками Северного Китая. По М. Г. Левину (1951), связи населения Во- сточной Сибири с Центральной и Восточной Азией отмечаются на всех стадиях верхнего палеолита. Археологические данные по неолиту и бронзовому веку Прибайкалья говорят о непрерывном развитии населения Прибайкалья в течение ряда тысячелетий (М. Г. Левин, 1951). К неолиту Прибайкалья примыкает и якут- ский неолит по Лене. Ниже Киренска неолит носит несколько иной характер. Неолит Енисея изучен слабо. Археологические исследования А. П. Окладникова (1951) в Бурятской АССР показывают, что древнейшая культура юж- ного Забайкалья обнаруживает общность с культурами лесного Забайкалья. Связи с Китаем установлены и здесь. В культурном слое Березовской стоянки, помимо обломков сосудов местного неолитического типа, оказались фрагменты глиняного сосуда- «трипода», повторявшего по форме и отделке поверхности древ- некитайские сосуды — «триподы ли». Отсюда следует, что ранне- скотоводческие племена Селенгинского края в конце II и в на- чале 1 тысячелетия до н. э. имели культурные связи с далеким юго-востоком, где издревле обитали древние земледельцы— предки современных китайцев. По А. П. Окладникову (1950), скотоводческие племена Забайкалья бронзового века имели связи также с западом (Средняя Азия) и с восточными ски- фами. Следующий этап в истории Забайкалья отражен в памятни- ках гуннского периода. В 1949 г. были проведены раскопки гуннского городища близ устья реки Нижней Иволги (гуннский этап — время, когда в степях Центральной Азии возникает клас- совое общество и первое государственное образование степных кочевников). Кочевники гунны, кроме мяса, молока и сыра, потребляли и земледельческие продукты. На Иволгинском горо- дище найдены зерна проса и наконечник мотыги. Этим подтвер- ждались сведения китайских источников о том, что гунны сеяли просо. Из изложенного выше становится ясным, что связи Сибири с Юго-Востоком Азии, с предками китайцев и затем с самими китайцами начались в доисторическую эпоху и продолжались вплоть до более поздних времен. В древние эпохи земледелие на обширной территории Сибири существовало только островами, да и там имело подсобный ха- рактер. Основным занятием жителей была охота и рыболовство. Собирательству также принадлежала видная роль. Основными объектами собирательства были характерные для местной флоры растения. Большое значение имели кедровые орехи, луковицы лилии, так называемой сараны (Lilium martagon L.), ягоды 208
(брусника, черемуха, смородина, облепиха и др.), грибы (осо- бенно грузди) и т. п. Что касается сараны, то в древности она имела довольно большое значение в питании населения. Об этом имеются ука- зания в китайских источниках, относящихся к эпохе от II в. до н. э. и вплоть до середины IX в. н. э. По этим источникам, в Прибайкалье было очень много сараны, жители «собирали ее коренья и приготовляли из них кашу». Н. Я. Бичурин добав- ляет при этом «и по горам от вершин Енисея на восток до Чикоя растет много саранки, иначе пунцовой сараны, корень которой и ныне буряты осенью собирают и запасают на зиму в большом количестве». Прядение и ткачество появились в Сибири раньше земледе- лия. Жители рыболовецких районов еще в очень древние вре- мена постигли искусство изготовления веревочных сетей. По А. П. Окладникову (1950), существование сетей вскрыто еще в неолите Прибайкалья. Там было найдено много каменных гру- зил. Сети делались из волокон дикой крапивы, заросли которой обильны в Сибири, и, возможно, из диких льна и конопли. Qn- кого льна (видов, далеких от культурного льна) много на Алтае, в степных и лесостепных районах Сибири и даже в Якутии, а конопли — на Алтае. Еще путешественники XVIII в. обратили внимание на пора- зившие их рыболовные сети и вообще текстильные изделия, распространенные у аборигенов Сибири и Приуралья. По сло- вам П. С. Палласа, башкиры, например, не зная еще земледелия, тем не менее умели изготовлять волокно для пряжи из дикора- стущей крапивы. История первых металлических культур южной Сибири про- работана С. В. Киселевым (1949). Первая металлическая куль- тура южной Сибири — это афанасьевская (названия культуры даются по ближайшему селу, где произведены первые раскопки) . Со времени палеолита таежная зона Сибири на север от ли- нии Сибирской железнодорожной магистрали и на восток от реки Енисей была заселена племенами монгольского типа, кото- рые были близки к тунгусам. Степная же зона Сибири (Мину- синская котловина, пространства Западной Сибири и Казах- стана) были с древности населены племенами, относящимися к палеоевропейскому типу, которые имели черты кроманьонцев палеолитической Европы. Среди таежных племен были и охотничьи, и рыболовческие. 5Кители степной полосы — афанасьевцы, вероятно, были самыми древними скотоводами в Сибири. Происхождение афанасьевцев не вполне установлено. Наиболее вероятно, что они произошли in situ от неолитических европеоидных предков. Связи афа- насьевцев были направлены на запад и на юго-запад (есть общ- ность со среднеазиатскими кельтеминарской и анауской куль- турами, а также с ямнокатакомбовыми слоями Триполья). Име- 209
лись у афанасьевцев коровы, овцы и лошади, но в небольшом количестве. Роль скотоводства была менее значительна, чем охоты, рыболовства и собирательства. Мотыг не найдено. Но может быть примитивное земледелие все-таки и существовало,. особенно на плодородных глинистых почвах по Енисею. В области распространения афанасьевской культуры позднее развилась андроновская культура. С антропологической точки зрения нет прямой преемственности между афанасьевской и андроновской культурами. Здесь предполагают «скрещение» с какими-то новыми этническими группами. Возможно, что из Средней Азии была перенесена в Сибирь культура пшеницы. В Хорезме с андроновской сопоставляется тазабагъябская куль- тура, и более ранняя кельтеминарская культура тех мест имеет общие черты с афанасьевской. Андроновцы знали бронзу и зо- лото, афанасьевцы — только медь и камень. В андроновских культурных слоях найдено много костей до- машних животных — скотоводство получило большее развитие, чем у афанасьевцев. Имелись кожаная обувь и шерстяные ткани. Андроновцы занимались и земледелием: в раскопках найдены каменные мотыги и зернотерки. Что сеяли — неизвестно. Земле- делие было распространено, по-видимому, неравномерно. В при- алтайских андроновских памятниках прямых указаний на земле- делие не обнаружено. В Андроновской стоянке по Тоболу (около села Алексеев- ского Кустанайского района) найдены две каменные мотыги. Форма их приближалась к треугольной с выемками по бокам. В выемках помещался ремень, которым мотыга привязывалась к глаголевидной рукоятке. Здесь же обнаружен серповидный секач, сделанный из бронзы, но, по мнению С. В. Киселева, та- кие орудия, не будучи серпами, служили орудиями для расчистки под посевы припойменных участков. Раньше, чем степь, должнь| были быть использованы для земледелия плодородные, доста-- точно влажные, илистые почвы по верхнему Тоболу. Найден был также серп в женском погребении № 3 андроновского могиль- ника, раскопанного Б. Н. Граковым у аула Тулайкина в 1935 г.. (С. В. Киселев, 1948). На жертвенном холме Алексеевского поселка были обнару- жены в специальных ямках остатки жертвоприношений. Ня дне ямок найдены пережженные зерна и стебли пшеницы. Во всех алексеевских землянках обнаружены каменные зерно- терки. В Приуралье открыты андроновские могильники, и здесь, как. и в Сибири, имеются признаки связи с западноевропейской культурой. В Нижнем Поволжье отмечена также преемствен- ность между древнеямной культурой, родственной афанасьев- ской, и позднейшей срубной культурой, которая родственна андроновской. Недалеко от Оренбурга была найдена каменная литейная форма, приспособленная для отлития разом трех сер- 210
пов, это говорит о большой потребности местных андроновцев в подобных орудиях. Андроновский быт в VIII в. до н. э. с приходом племен из Северной Монголии изменялся и заменялся карасукской его формой. На западе Сибири андроновская культура просущество- вала до окончания здесь бронзового века. Медные вещи, най- денные в Алексеевском поселке, относятся к концу бронзового века. На востоке этим памятникам современны карасукские, а на западе, в Нижнем Поволжье — позднесрубные (хвалынские). Яля карасукской эпохи земледелие было менее характерно. Выше указывалось на существование связей афанасьевской и андроновской культур с Уралом и Поволжьем. Остановимся подробнее на связях древнейших сибирских племен с западом. древние охотничье-рыболовческие племена северной и цент- ральной Европы были расселены широкой полосой на восток— в бассейн Верхней Волги и Оки и далее на Каму и Печору, доходя на западе до нижней Одры. Культура этих племен сло- жилась в III тысячелетии до н. э. и может быть еще раньше. Характерные черты их быта: хижины-землянки с коническими крышами; лодки однодеревки и сети; способ довольно искусной обработки костяных и каменных орудий; культ медведя, лося и водяной птицы. Эти особые черты культуры и быта сближали лесные племена Восточной Европы с древним населением При- уралья и таежного населения Сибири. В бассейне Камы (А. В. Збруева, 1954) остатки ананьинской культуры скифского времени на реке Чусовой дают возможность заключить, что основой этой культуры слу~кили земледелие и скотоводство. Найдены были каменные зернотерки, бронзовые серпы и мотыги. В одной из раскопанных землянок Луговской стоянки недалеко от ананьинского могильника обнаружены обугленные зерна проса. К этому времени охота и рыболовство постепенно утратили свою роль основных отраслей хозяйства, зато возросло значение земледелия и скотоводства. В ананьин- ской культуре земледелие и скотоводство выдвигаются на первое место. В связи с такими переменами в начале 1 тысячелетия до н. э. поселки из поймы переносились на возвышенные берега ближе к засеянным участкам. Земледелие у ананьинцев было подсечным и яровым. Выжигались небольшие участки леса, при- чем самые крупные деревья не вырубались. Почва разравнива- лась вручную деревянными мотыгами с костяными или бронзо- выми наконечниками. Посев производился прямо в золу. Затем участок бороновался. Ананьинцы держали лошадей, коров, сви- ней, овец, коз, собак. Лошадь была не только мясным животным, но употреблялась и для езды — в раскопках найдены удила. Ананьинская культура имела связи с севером и востоком— (Зауральем). Г. Ф. гребец (1954) утверждает, что срубная культура За- волжья была связана с Зауральем и с ананьинской культурой.
Об этом же говорят и исследования A. Я. Брюсова (!954) о предках народов Зауралья. Он пишет, что с озер Урала нача- лось, по-видимому, расселение древних племен по заволжскому северу вплоть до Балтики. В середине II тысячелетия до н. э. на основе уральских месторождений меди возник один из крупней- ших очагов древней металлургии, снабжавший металлом много- численные восточноевропейские племена бронзового века. Таким образом, в Приуралье и в Зауралье приходили в широкое сопри- косновение народы востока и запада. Близ нынешнего Тагила раскопан Горбуновский торфяник, представляющий собой осу- шенное озеро. Недалеко от Горбуновского торфяника находится Шигирский торфяник, который также раньше был обширным озером. Древнейшие вещи из Горбуновского торфяника отнесены к II I тысячелетию до н. э., а из Шигирского — к V и IV тысяче- летиям до н. э. Таким образом, человек заселял Зауралье еще в палеолите. На Урале раскопаны стоянки не менее древние, чем в Дании и Прибалтике. Север европейской части СССР, по за- ключению А. Я. Брюсова, мог заселяться не только с запада, но и с востока; земледельческие навыки и культурные растения также могли внедряться на севере Русской равнины с двух сто- рон — с запада и с востока. В древнейшие времена население в Прибалтике и Зауралье занималось только звероловством и рыболовством. Однако, по мнению А. Я. Брюсова, и земледелие здесь восходит к глубокой древности. Крайнее облесение местности не противоречит древ- нему развитию здесь мотыжного земледелия. Он считал, что нельзя исходить из современных представлений о земледелии. В древние времена земледелие было почти невозможно «на твердых берегах» даже в степных областях. Обрабатываться могли берега озер и поймы рек, освоение которых представляется делом более легким. На одной из стоянок Горбуновского торфяника, по сообще- нию А. Я. Брюсова, на пространстве в несколько квадратных метров было найдено 25 роговых мотыг. Все эти мотыги носят следы долговременного употребления. Такие же мотыги отко- паны и на Шигирском торфянике. Д. Н. Эдинг нашел здесь большой деревянный жом для выжимки растительного масла. Можно предположить, что этим жомом пользовались для вы- жимки масла из кедровых орехов. Но так как на реке Модлоне были найдены семена льна и части деревянной прялки, то воз- можно жом служил и для изготовления льняного масла. По раскопкам А. В. Арциховского в более поздние эпохи такими же жомами пользовались для отжимки масла и жители Великого Новгорода. Вероятно, конструкция этого жома была заимство- вана с востока. Для сообщения в эпоху свайных построек на описанных тор- фяниках пользовались лодками, лыжами, санями. 212
По А. Я. Брюсову, в последней четверти II тысячелетия до н. э. первое место в хозяйстве занимали охота и рыболовство. Роль земледелия возрастала постепенно. По В. Н. Чернецову (1953), на протяжении неолита и бронзо- вого века лесные районы Зауралья и Приобья имели культурные связи не только с западными лесными территориями Восточной Европы, но и с более южными лесостепными и степными обла- стями. Археологические изыскания последних лет уточнили об- ласть распространения кельтеминарской культуры и обнаружили даже на севере наличие близких ей аналогов. Аналогичные культурные связи, вероятно, распространялись и на более позднюю так называемую потчевашскую культуру, городища и курганы которой изучены В. И. Мошинской (1953). Наиболее изучено городище близ г. Тобольска. Эта часть берега Иртыша местным населением называется Потчеваш (возможно, это означает «Олений город»). Хозяйство этого городища было комплексным. Большое значение имело рыболовство, меньшее— охота и мотыжное земледелие. Мотыги были массивные, из оленьего рога. Здесь было довольно развито коневодства. Оби- татели Потчевашского поселения знали прядение ниток из ко- нопли, которая растет здесь в диком виде. Верхний предел суще- ствования Потчевашского городища относится В. И. Мошинской не позднее чем к 1 в. н. э. Потчевашская культура, согласно исследованиям В. H. Чер- нецова (1953), явилась исходной при формировании усть-полуй- ской культуры Приобья. Материалы последней датируются 1 в. до н. э. — II в. н. э. Культуры древнего Приобья были связаны с культурами южной Сибири и Китая. В. Н. Чернецов в той же работе пишет о связях древних культур Зауралья с Поволжьем. Медные изделия распространились в Поволжье из Приуралья и Зауралья. Первые иглы и шила из меди лишь к середине II ты- сячелетия до н. э. стали заменяться бронзовыми. Южные племена срубной и андроновской культур были, по В. Н. Чернецову, основными в формировании сарматов, а основ- ной территорией сарматских племен были современные Саратов- ская и Волгоградская области. Таким образом, можно просле- дить культурные и этнические связи древнего Зауралья и При- уралья с южным Поволжьем. В южной Сибири прослеживаются исторические связи с более западными и северными культурами и с земледельческим Китаем. Л. А. Евтюховой (1954) изучены в южной Сибири древности таштыкской эпохи. Таштыкское время относится к III в до н. э. — IV — V в. н. э. В Минусинской котловине в таштыкскую эпоху жили местные племена, одни из которых были кочевниками-скотоводами, другие же из этих пле- мен жили оседло и занимались земледелием. Культура ташты- ков была тесно связана с Китаем. В V в. н. э. в сельском хозяйстве приенисейских племен на- ступил перелом — мотыжное земледелие стало сменяться плуж- 213
ным. Древнехакасские мелиораторы с большим искусством строили оросительные сооружения, а древнехакасские мастера делали серпы. На смену примитивным зернотеркам пришли пар- ные вращающиеся жернова. Хакассы в то время (по китайским .источникам) сеяли просо, пленчатый ячмень, пшеницу и голый гималайский ячмень. У них были верблюды, коровы и овцы. На Алтае (С. В. 1(иселев, 1948) на территории раскопанных Куюмских курганов верхний слой носит определенные черты афанасьевской культуры, а нижние слои содержат более древ- ние остатки. 1(ерамика из этих нижних слоев похожа на иско- паемую посуду кельтеминарской культуры дометаллического Хорезма. Таким образом, на Алтае доафанасьевский неолитиче- ский период представляется весьма сложным. С одной стороны, его культура обладает чертами сибирского неолита (Чудацкая гора), а с другой — обнаруживаются связи со Средней Азией. Культурные растения могли распространяться на север и на запад через Алтай или из Средней Азии, или из 1(итая, или по обоим этим направлениям. Весьма вероятно и то, и другое, и третье, но детально выяснить эти вопросы пока еще трудно. На Алтае, а также в Прибайкалье по Енисею земледелие развивалось, видимо, неравномерно. Курган, раскопанный С. И. Руденко в Восточном Алтае на высоте 1600 м над уровнем моря, относится к середине I тысяче- летия до н. э. — к скифским временам. Вследствие вечной мерзлоты многое здесь хорошо сохранилось — утварь, ткани и т. д. В одном из курганов найдены семена конопли, что неуди- вительно, так как это растение на Алтае встречается как сорное, рудеральное и изредка даже как дикорастущее (Е. Н. Синская, 1935). Нахождение семян конопли отнюдь не указывает на нали- чие земледельческой культуры — население могло собрать ко- ноплю около своих жилищ, на местах становищ скота (на «6а- зах»), где конопля образует чистые заросли самосевом. В этом кургане найдены были также семена проса и кориандра, а последний, вероятно, был завезен сюда откуда-то с юга, воз- можно, из Средней Азии. Разнообразие найденной в этом кур- гане утвари говорит о больших торговых связях жителей с юго- западом. Кориандр не мог вызревать на семена в таком суро- вом климате. Вероятно, вследствие суровости климата, здесь в горах земледелия не было совсем. Саяно-алтайская киргизская культура VII — VIII вв. имела развитое земледелие и оказала влияние на сибирскую культуру. Советская археология много сделала и для выяснения исто- рии культуры в Восточной Сибири. А. П. Окладников (1949) в работе о прошлом Якутии до присоединения ее к Русскому государству успешно связывает историю палеолитических племен бассейна Лены с историей дру- гих областей — вплоть до бассейнов Волги и днепра. По его словам: «...древнейшие племена, проникшие к востоку от Урала 214
в конце ледникового периода сначала жили одинаковой жизнью с племенами Запада, не утрачивая связи с ними, и обладали об- щим в основе культурным достоянием». Лишь через много тыся- челетий «немноголюдные, широко рассеянные на колоссальных пространствах Сибири... племена эти утратили непосредственную связь с населением западных стран...» А. П. Окладников посвятил целую главу связи неолитических культур Якутии с западом и востоком, это же сделал он и для эпохи бронзы. Таким образом, Сибирь издревле не была изоли- рована от Зауралья. На это указывают и ботанические данные о распространении форм некоторых культурных растений. Среди сорных овсов Алтая имеются формы, сходные с ураль- скими и северорусскими овсами. У некоторых форм прядильного льна Тувинской АССР обнаружено сходство со стародавними уральскими популяциями льна и северными долгунцами (или полудолгунцами) Вологодской и Архангельской областей. По сообщению А. И. Мордвинкиной, привезенные нами с Южного Алтая сорные овсы сходны с некоторыми уральскими и север- ными овсами. Фиолетовые репы встречаются на Алтае, на Урале и распро- странены в Карелии. Эти репы явно происходят из Малой Азии и двигались на север Русской равнины, вероятно, кружным пу- тем через Алтай и Урал. По-видимому, теми же путями еще в неолите проникло из Китая на север современной русской территории и просо. По- следнее могло распространиться и посредством связей металли- ческих культур южной Сибири с Трипольем, а еще позднее— через сарматов и скифов. Многое сделано советскими археологами, особенно А. П. Ок- ладниковым, для изучения истории культуры народов Дальнего Востока. Окладников (1953) занимался памятниками древних государств дальнего Востока. Это были Бохайское царство и Чжурчжэньская золотая империя, существовавшие в VI I I— XII вв. н. э. Однако культуре этих государств предшествовали более древние примитивные культуры. Грубые рубящие камен- ные орудия были найдены у деревни Осиновка недалеко от г. Уссурийска. древняя стоянка открыта также близ станции Надеждинская около Владивостока. Там найдены орудия типа резца из обсидиана. Такие орудия употреблялись для обработки дерева, кости и рога. Этот резец отличался от тех, которые были найдены в раскопках Западной Сибири. Резцы, похожие на дальневосточные, были обнаружены в древних поселениях Мон- голии и пустыни Гоби. Еще более интересным является то, что сходные резцы открыты на Аляске — в одном из древнейших поселений человека на американском континенте близ г. Фер- бенкса. Большинство исследователей полагает, что человек про- ник в Америку из Азии около 10000 лет назад или еще раньше. Предполагается, что он перешел из Азии в Америку через терри- 215
торию, занятую теперь Беринговым проливом, возможно, в тс времена не существовавшим "' На Яальнем Востоке таким образом в отдаленнейшее время жил человек; тогда же создавалась особая культура первобыт- ных охотничьих племен, которая однако носит черты связей с культурами других географических областей и в то же время имеет свои особенности. Обследования поселений в районе Хабаровска показали, что древние обитатели этих мест не умели еще делать глиняной по' суды и шлифовать камни. Копья с каменными наконечниками, топоры и лодки были примитивны. Однако наличие лодок гово- рит о том, что еще в каменном веке возможны были морские путешествия и возможно, что человек переехал в Америку из Азии на лодке. В позднейших поселениях племен неолита уже находили керамику, хорошо шлифованные каменные орудия, луки и стрелы. Все эти предметы характерны именно для данной культуры и отличались от каменных изделий неолитических племен Западной Сибири и лесной области европейской части СССР. Особо важное значение в жизни таежных племен Восточной Сибири имела охота на лося, изюбря, косулю и других лесных зверей, а на ~альнем Востоке главным промыслом было рыбо- ловство. Одежды изготовлялись из рыбьих шкур и назывались «рыбьекожими». Черты рыболовческого жизненного уклада об- наружены на Амуре и в Приморье уже с III и II тысячелетий до н. э. Стоянки палеолита здесь сравнительно редки, а неолити- ческих стоянок очень много. В 1930 г. только на одном из япон- ских островов было насчитано около 11 тыс. стоянок и 617 рако- винных куч. Такое же обилие неолитических памятников открыто на Амуре, на Камчатке, на Сахалине, в Южной Маньчжурии и Корее. Собирательство морских моллюсков имело очень большое значение, что и доказывается большим количеством раковинных куч (в современной Японии моллюски и раковины, собираемые после отлива, являются важным подспорьем в питании населе- ния побережий). В более поздние времена возникли деревни свайных домиков и целые мастерские каменных орудий. Древний Тетюхинский поселок у реки Монастырки занимал площадь в 3 — 4 тыс. кв. м. Рыболовы переходили к оседлости. В глубокой древности они пользовались растительным волокном — найдены в раскопках ткацкие грузила и маховички для веретен в виде глиняных кружков. Появились большие сосуды — чаны. Для ры- боловческой культуры характерно обилие женских статуэток и криволинейно-ленточный стиль красивых спиральных узоров на керамике. дальневосточная культура рыболовов была выше син- хронной ей культуры лесных племен таежной Сибири. 216 '" Вследствие более низкого уровня воды в мировом океане в плейстоцене.
В легендах дальневосточных племен сохранились воспоми- нания о более жарких южных странах, о тех древних временах, когда «земля кипела» и шаману потребовалось «убить два солнца», чтобы стало прохладнее. Предки дальневосточных рыболовов может быть пришли с юга, а климат на Дальнем Востоке был более жарким (вторая половина лета и теперь там жаркая). Очевидна связь дальневосточной рыболовческой куль- туры с китайской культурой Яншао на реке Хуанхэ в III — II ты- сячелетиях до н. э. Промежуточные памятники известны из Маньчжурии. Таким образом, земледельцы неолитического Kz- тая были тесно связаны с рыболовами Приморья. Древние рыбо- ловческие племена Дальнего Востока земледелием не занима- лись, но продукты земледелия могли поступать к ним из 1~итая. Культурные и этнические связи древних племен Дальнего Во- стока (предков современных нивхов, гиляков, отчасти гольдов, нанайцев и др.) простирались и в другие страны. Наличие спи- рального и криволинейного орнамента говорит о связях древнего Дальнего Востока с Океанией, Индонезией и Японскими остро- вами в древней области расселения айнских племен. Последние, согласно исследованиям русского этнографа Е. М. Штернберга, имеют родственные связи с австралоидным населением По долине реки Амур, где был теплый и мягкий климат, жили южные неолитические племена, которые входили в сопри- косновение с охотничьими таежными племенами. Несмотря на более высокую культуру южных племен по сравнению с таеж- ными, у них не было земледелия. Единственным домашним жи- вотным была собака. Здесь также вскрыто множество раковин- ных куч, содержание которых постепенно менялось. В более древних слоях обнаружены грубые каменные орудия, которые постепенно совершенствовались. Если при раскопках в более древних кучах были обнаружены только мелководные раковины, то позднее в кучах стали появ- ляться и раковины с больших глубин, а в более поздних слоях найдены каменные зернотерки. Вместе с обломками каменных зернотерок в раковинных кучах имеются и каменные мотыги, фрагменты шиферных ножей особого типа. Эти ножи имели вид небольших пластинок с выпуклым односторонним лезвием и обычно с двумя отверстиями, просверленными в верхней части пластинок. Совершенно такие же шиферные ножи и особые мо- тыги с плечиками употребляли неолитические земледельцы древ- него 1~итая на реке Хуанхэ. Южные племена долины Амура были первым земледельческим населением Дальнего Востока, от них земледелие могло распространиться на соседние территории. В более поздние времена шиферные ножи и кинжалы стали из- готовляться и местными жителями Приморья по металлическим образцам других стран. Подобные кинжалы употреблялись пле- менами бронзового века, заселявшими степные пространства к востоку от Байкала — Забайкалье (верховья Амура) — вплоть 217
до Гоби и Ордоса на юге. Эти восточные племена бронзового века были соседями жителей Дальнего Востока. На их террито- рии в горах Восточного Забайкалья (по Онону и Аргуни) нахо- дились богатые залежи цветных металлов, в том числе и оловян- ного камня. Здесь издревле производилась обработка руд и выплавка металлов. Именно рудные богатства Восточного За- байкалья были той базой, на которой развивалась с половины II тысячелетия до н. э. бронзовая культура степных восточно- азиатских племен. Отсюда и попадали к берегам Амура и Тихого океана металлические изделия. В VIII в. н. э. на территории Приморья возникло Бохайское царство, которое было уже государством с довольно высокой общей и земледельческой культурой. На развитие бохайской культуры сильно влияла соседняя китайская земледельческая культура, здесь уже начали складываться классовые отношения. Приморье в эпоху Бохайского царства было густо заселено. Культура Бохайского царства была разрушена монголами. Страна стала пустынной и дикой. Новейшая история сибирского земледелия связана с раз- витием там русского земледелия в XVII в. (В. Н. Шунков, 1950). В связи с этим отметим следующее: 1) ни по территории, ни по преемственности трудовых навыков нельзя установить непосред- .ственной связи русского земледелия XVII в. с очагами древне- сибирского земледелия; 2) при установлении границ сибирского земледелия в XVII в. нельзя говорить о сплошном, хотя бы и редко заселенном земледельческом массиве; можно лишь наме- тить четыре русских земледельческих очага в Сибири XVII в.: Тобольский, Томский, Енисейска-Красноярский и Ангара-Лен- ский. Хотя развившееся в XVII в. русское земледелие и не имело «непосредственной» связи с древними аборигенными культурами Сибири, но стародавние сорта культурных растений, конечно, были переняты. Об этом говорит существование таких своеобраз- ных форм, которые несут на себе отпечаток многовекового при- способления к местным условиям, как монголо-бурятское просо, алтайская яровая рожь, стародавние сорта пшеницы и др. Культурная флора Сибири не богата по видовому составу, однако она имеет свои оригинальные черты и исторически сло- жилась из разных источников. Местная дикорастущая флора дала немного выходцев в культуру, хотя еще далеко не исчер- пана в этом отношении. Наиболее перспективна для дальнейшего освоения растительных ресурсов Восточная Сибирь, особенно Дальний Восток. Многие культурные растения пришли в Сибирь с юга, юго- востока и юго-запада. На составе культурных растений Восточ- ной Сибири особенно заметно сказывается влияние соседнего Китая и других сопредельных стран. Оттуда пришли просо, рис, позднее соя, перилла, некоторые сорта овощей (редька лаба, са- 218
латные горчицы и др.) . .В. Е. Писарев (1948) сближает эндемич- ные сибирские сорта пшеницы с китайскими. Возможно, что путь распространения пшеницы последовательно был таким: Перед- няя Азия (первичный очаг) — Средне-1Ого-Западная Азия— Китай и Монголия — Сибирь. Связи древних сибирских культур со Средней и Передней Азией несомненны. Кроме пшеницы, от- туда мог проникнуть кориандр. Не так давно на Алтае и на Урале существовали местные сорта репы, напоминающие мало- азиатские. Древняя культура Восточной Сибири несколько обособлена и имеет более тесную связь с Китаем, чем культура Западной Сибири. Последняя тесно связана с культурой Предуралья, За- уралья и далее на запад, на юге — с Поволжьем, а на севере- с культурой северных племен Восточной Европы до Прибалтики и ~анни. Из Сибири были возможны два пути распространения культурных растений: южный — по Поволжью (через сарматов и скифов) и северный через Урал — далеко на северо-запад. В новейший период истории сорта культурных растений пере- носились в Сибирь переселенцами из различных районов Рус- ской равнины. дикорастущая флора Сибири в целом значительно богаче, чем на Русской равнине, но в культуру вошло еще мало местных сибирских видов и форм. Особенно богата и интересна для освоения флора Алтая, но в древности на Алтае земледелие су- ществовало только островками. Лишь недавно начали осваи- ваться растительные ресурсы Алтая (Е. Н. Синская, 1925). Ниже дается краткий обзор видовых и сортовых раститель- ных ресурсов Сибири. На Алтае растет б ад а н (Bergenia сгазз11о11а Fritsch) пре- красное дубильное растение. Сибирь богата местными лекарственными растениями. UeHT- ром распространения (или одним из центров — так как китайско- тибетские Adonis L. слабо изучены) многолетних г о р и ц в е- т о в (Adonis L.) является Алтай. Однако общий ареал много- летних адонисов простирается от Пиренейского полуострова до Тихого океана. Это род — древнесредиземноморский. Б о~р щ е в и к с л а д к и й (Heracleum dulce Fish.) — едят стебли и черешки листьев (содрав кожицу). Прежде добывали сахар и гнали водку. Камчадальское название — учкуй или пнч-коу. Эндемичен для Камчатки. У борщевика р ассеченнолистн ого (Н. dissectum Ledeb.) молодые побеги и листья идут в пищу в маринован- ном виде. Хорошо поедается овцами. Б о р щ е в и к с и б и р с к и й (Н. sibiricum 1.) распростра- нен также в Северной и Средней Европе и на Кавказе. Хорошее кормовое растение, листья содержат до 19,5 л~г'/о каротина. Алтай богат формами д о н н и к о в (Melilotus 1 .) . В. В. Су- воровым (1950) описаны для Сибири следующие экотппы белого
донника: 1) сибирский равнинный (Западная Сибирь и Северный Алтай); 2) экотип песчаных почв (север Хакасской автономной области); 3) экотип засоленных почв (Хакасская и Горно-Алтай- ская автономные области); 4) экотип пониженных местообитаний (Западная Сибирь); 5) многостебельный (Восточный Алтай и север Западной Сибири); 6) пастбищный (Горно-Алтайская ав- тономная область). Североевропейский экотип очень близок к сибирскому равнинному экотипу. Многостебельный экотип дал начало сортам люцерновидного донника. Лесостепь Сибири и Алтай богаты формами л ю ц е р н ы желтой (Medicago falcata), которые послужили основной для вы- ведения холодостойких сортов люцерны. Сибирская люцерна желтая еще далеко не полностью изучена. Экотипы севера Рус- ской равнины, объединяемые в вид М. borealis, заходят и в Си- бирь. Некоторые из экотипов к о с т ра б е з о с т о г о (Bromus iner- mis Leyss.) Приуралья и севера Русской равнины близки к ал- тайским пойменным кострам, произрастающим по Иртышу. Возможно, что путь расселения многих луговых злаковых и бобовых трав проходил с Алтая через Западную Сибирь и Урал на север европейской части СССР. В Сибири имеются интересные для использования виды дико- растущих клеверов и местные формы культурного клевер а к р а с н о г o (Trifolium pratense L.). Н. Г Хорошайлов описал подвид дальневосточного культурного клевера — ssp. orientale СЬог., предком которого является местный дикорастущий клевер (возможно, что это одичавший клевер). Богаты формами дико- растущего клевера красного предгорные районы северо-запад- ного Алтая. Практический интерес представляют дикорастущий л юпинов идный кл евер (Т. lupinaster L.) и другие виды. Этот клевер еще не изучен достаточно как кормовое расте- ние. Заслуживают большого внимания и вводятся в культуру си- бирские э с п а р ц е т ы, типа песчаного эспарцета. Еще слабо ботанически и практически изучены заходящий далеко на во- сток Onobrychis яйг1са Turcz. и в западных районах О. tanai- tica Sprengel. Эти эспарцеты осваиваются в культуре как ис- ходный материал для селекции. В Сибири сосредоточено большое разнообразие форм и ви- дов вики. Из них Vicia amoena Fisch. вводится в культуру. Представляют интерес как кормовые л е с п е д е з а (Les~pedeza bicolor Turcz.) и ку.м мер о в и я (Kummerovia striata Schindl.). Заслуживают большого внимания и сибирские кормовые злаки. Введен в культуру в о л о с н е ц с и б и р с к и й (Cline- lymus sibiri~cus ценз!«); уже имеются селекционные сорта этого растения. Эндемичные формы костра безостого — Bromus iner- mis Leyss. (алтайский экотип) встречены нами в предгорьях Алтая. Заслуживают испытания в культуре и другие виды си- 220
сибирских костров, особенно В. sibiricus Drob. и В. richardsenii Link. Алтайская группа форм т и м о ф е е в к и (РЫеит pratense L.) состоит из нескольких экотипов. На Дальнем Востоке СССР в специфических условиях произрастания, по М. А. Шебалиной, сформировалась особая биологическая группа сортов. Л и с о х в о с т л у г о в о й (Alopacurus,pratensis L.) пред- ставлен алтайским предгорным и луговым западносибирским экотипами. Алтай, Саяны и другие районы Сибири изобилуют разно- образными формами ширококолосых житняков (Адго- ругоп cristatum sensu lato), а также житняков пустынного— А. desertorum Schult. и сибирского — А. sibiri~cum Beauv. Пырей бескорневищный — Poegneria tra~chycaulon Nevski — занесенный из Америки, нашел в Сибири вторую ро- .дину и послужил материалом для создания селекционных сор- тов. Дико в Сибири распространен близкий к нему вид регне- рия волокнистая — Я. fibrosa Nevsl<i. В Сибири имеется ряд дикорастущих экотипов еж и — Dacty- lis glomerata L. Алтайская ежа ближе к северным экотипам, распространенным на севере европейской части СССР и на За- падно-Сибирской низменности, чем к южнорусским и кавказ- -ским. Вероятно, расселение дикорастущей ежи шло на европей- ский север двумя путями: 1) с Кавказа на Украину и в Крым; 2) с Алтая на равнины Западной Сибири и оттуда на Русскую равнину. Представляют интерес алтайский экотип дикорастущей о в с я н и ц ы л у r о в о й (Festuca pratensis Huds.), дикорасту- шие формы мятликов, полевицы, бекмании и др. Что касается зерновых злаков, то на них несомненно лежит отпечаток древности сибирского земледелия. Ф. Х. Бахтеев выделяет две группы местных сортов я ч м е н я: 1) восточносибирскую — proles orientali-з1Ь|г1сит Vav. et ВасМ.; 2) дальневосточную — proles extremo-orientale Vav. et Bacht., заходящую и в Северо-Восточный Китай. Овес (Avena sativa), по А. И. Мордвинкиной (1936), пред- ставлен в Сибири 2 типами: 1) северный, распространен в гор- ном Алтае, Западной Сибири и на севере европейской части СССР; по нашему мнению, он распространился на северо-запад с Алтая; 2) восточный — в Восточной Сибири и на Дальнем Во- стоке; корни последней группы сортов надо искать в Монголии. Р о ж ь (Secale сегеаlе L.) представлена в Сибири, по В. Ф. Антроповой, особой восточносибирской группой местных сортов, озимых, но более короткостадийных, чем европейские северные сорта. Алтайский экотип яровой ржи, к сожалению, теперь почти исчезнувший, отличается быстрым ростом с весны и поэтому представляет большой интерес для весенней под- кормки скота.
В. Е. Писарев (1960) внутри п ш е н и ц сибирского типа об- наружил большое ботаническое разнообразие на юге Монголии и в Китае. Он считает, что группа восточносибирских пшениц является крайним северным отпрыском древней, некогда круп-- ной группы пшениц Китая, теперь сохранившейся в виде при- меси к современным более продуктивным сортам. Особенно ценны скороспелые местные холодостойкпе сорта пшеницы Triticum aestivum L. П р о со (Panicum miliaiceum L.) разнообразно в Сибири. В. Н. Лысов различает две эколого-географические группы: мон- голо-бурятскую и восточноазиатскую. На Дальнем Востоке СССР имеются местные сорта п а й з ы— Eehinochloa frumentacea (goxb.) Link, происходящей из Севе- ро-Восточного Китая. Гр еч и ха (Fagop~yrum sagittatum Gilib.), по А. С. Кротову (1955), представлена в Сибири двумя сортотипами: 1) сортотип тарский, распространенный в Сибири и на Урале, относится к северному скороспелому пролесу, который встречается и к западу от Урала и включает в себя сортотип,<азанскпй; распространение этого пролеса (вернее подвида) еще раз ука- зывает на связь земледельческой культуры Предуралья и За- уралья; 2) сортотип горношорский — распространен в лесостеп- HbIx районах Западной Сибири. Этот сортотип относится к южному среднеспелому пролесу. другой сортотип того же пролеса распространен в Башкирской АССР и Удмуртской АССР, т. е. в западном Предуралье. Распространение сортов, относящихся к этому пролесу, указывает также на связь с Си- бирью. Вероятно, сорта этого пролеса двигались (исторически) из Сибири через Урал в Предуралье и Поволжье, а уже отсюда на Украину и в Белоруссию. Е. А. Столетова (1940) выделила местные сорта гречихи Приморья в особый подвид — ssp. multifolium Stolet. представ- ленный, по A. С. Кротову, одним сортотипом — позднеспелым приморским. Гречиха татарская (кырлык) засоряет посевы пшеницы и гречихи обыкновенной в Сибири, особенно в районах, примы- кающих к Монголии. Раньше местные жители собирали кырлык ц употребляли его в пищу. Р ыжи к я р о во й (Camelina sativa ssp. glabrata) культи- вируется в Сибири. Сибирский рыжик представляет собой энде- мичный ценный в хозяйственном отношении экотип. Произошел он от местного сорного рыжика, распространенного там в посе- вах хлебов, а последний, вероятно, произошел от местного дико- растущего рыжика. На Алтае нами описана эндемичная крупно- плодная форма дикорастущего рыжика — С. micracanpa var. major Sinsk. На Алтае же среди рыжика ярового нами выде- лена ветвистая форма — С. sativa ssp. subliniIcola var. ramosa Sin sk. 222
Кое-где разводится г о р ч и ц а с а р е п т с к а я [Brassica 1ипсеа (L.) Czern.]. Особая форма с очень тонкими стручками— var. mongolica Sinsk. — занесена, вероятно, из Монголии. Кроме того, нами описан скороспелый алтайский экотип этого вида. На Алтае распространен эндемичный сорный экотип суре- н и ц ы я р о в о й — Brassica campestris var. altaica Sinsk. Формы этого экотипа обнаруживают родство с северными сор- ными формами нечерноземной полосы Русской равнины. На Яальнем Востоке СССР распространены две формы ш а н- др ы — Elsholtzia cristata Willd. и Е. serotina Кот. В семенах .этого растения содержится ценное техническое масло с высоким иодным числом. Удачные опыты поставлены с культурой шандры. Некоторые овощные растения также представлены в Сибири местными сортами. Сибирские и уральские сорта кочанной к а п у с т ы Т. В. Лизгунова объединяет в особую группу — си- -бирскую. Особые местные сорта р е д ь к и лаба занесены из Китая, как и встречающиеся на Дальнем Востоке СССР курчаволистные салатные формы сарептской горчицы. Ш н и т т - л у к — А111пт schoenoprasum L. — представлен и Сибири особой группой местных сортов — сибирской (Н. Х. Трофимец). Яикор астущие луки Сахалина пред- ставляют несомненный интерес для селекции репчатого лука и сортов лука, выращпваемых «на перо» (используемых в виде .листьев). Наибольшим богатством дикорастущих кустарников и де- ревьев отличается Дальний Восток. Род актинидия (Actini- dia Lindl.) происходит из Восточного Китая и Японии, но и у нас на Дальнем Востоке насчитывается 5 дикорастущих ви- .дов, из которых 2 введены теперь в культуру, а остальные пред- ставляют интерес для селекции. К первым двум видам отно- сятся: 1) А. ko omicta Maxim. — «изюм»; имеются хорошие сорта этого вида, выведенные И. В. Мичуриным; 2) А. arguta (Sieb. et Zu~cc.) Planch. И. В. Мичурин также вывел сорта, относя- щиеся и к этому виду актинидии. Следующие три вида с очень вкусными плодами еще слабо использованы в селекции: А. д raldii Diels, А.;polygama (Sieb. et Zucc.) Maxim. со съедобными ягодами; А. chinensis Plan~ch.— редкий у нас вид, распространенный больше в горах Китая. Восточноазиатское происхождение имеет также ценное ле- карственное и плодовое растение л и м о н н и к (Schizandra chinensis Ва111.), который растет дико на г(альнем Востоке. В и н о г р ад представлен во флоре дальнего Востока двумя видами: Vitis thunbergii $1еЬ. et Zuoc. и Ъ amurensis gupr. Последний вид введен в культуру. М а н ь ч ж у р с к и й о р е х (Juglans mar>shurica Maxim.) обитает в диком состоянии в СССР — в Приморском крае, но 223
основное разнообразие его форм приурочено к Северо-Восточ- ному 1~итаю. Разводится только как декоративное парковое дерево. Г р у ш а у с с у р и й с к а я (Pirus ussuriensis Maxim.) растет дико на Дальнем Востоке СССР в Северо-Восточном 1(итае п Корее. Введена в культуру и используется в селекции путем скрещивания с другими видами. Этот вид произошел, по-види- мому, из районов, расположенных южнее Дальнего Востока, у нас же имеются самые холодостойкие формы. Я б л о н я с и б и р с к а я (Malus pal asiana Jus.) распростра- нена от нашего Дальнего Востока по всей Восточной Сибири, включая Предбайкалье. Вид, возможно, аборигенного происхож- дения. И. В. Мичуриным эта яблоня широко использовалась в се- лекции и работа в этом направлении продолжается. IjpoMe того, на Дальнем Востоке встречаются дико следующие виды яблони: 1) М. mans!zuri~ca (Maxim.) Kom. — вид родственный предыду- щему, может быть подвид или экотип его; 2) М. sachalinensis (Kom.) Juz. близок к предыдущему виду (Сахалин). На Дальнем Востоке находится северная граница ареала холодостойкого сибирского а б р и к о с а IArmenia~ca sibirica (L.) Lam.]. Холодостойкие культурные сорта абрикоса являются здесь в основном гибридами абрикоса обыкновенного (А. vul- garis Lam.) с сибирским абрикосом и с завозимыми сюда маньч- журским [А. manshurica (Koehne) Skvortz.] и китайским [А. ansu (Kom.) I(~ost. абрикосами. В Сибири имеются богатые ресурсы видов и форм сморо- дины и кр ыжовни ка. Один из видов смородины красной (Ribes гиЬгит L.) введен в культуру как таковой, кроме того, используется для гибридизации. Смородина щетинистая (Ribes hispidulum Pojark.) тоже имеет значение в селекции. Группа ви- дов, близких к R. petraeum Wulf., также заслуживает внимания. А. Н. Пояркова выделила в этой группе 5 азиатских таежных видов, которые еще подобно своим близким европейским роди- чам почти не использованы в селекции, как и далекий от этой группы вид — R. paucif lorum Turcz., обитающий дико в Сибири. Смородина черная (Igibes nigrum L.) тоже распространена в Сибири. Таким образом, род Ribes L., имеющий широкое евроазиат- ское распространение, в Сибири, главным образом, в Восточной, представлен и формами широко распространенных видов и теми видами, которые свойственны Восточной Азии за пределами на- шей родины, но имеются также и эндемичные виды (во всяком случае, подвиды и экотипы) для Восточной Сибири и Дальнего Востока. Разнообразие крыжовника в этих областях сравнительно невелико и здесь нет эндемичных видов. Крыжовник буреин- ский — Grossularia burejensis Berger, растущий дико в бассейне рек Зеи и Буреи, а также в Приморском крае, и крыжовник 224
иглистый — G. aci~cularis Spach, растущий на Алтае и в Саянах, представляют некоторый интерес для селекции. Распространена в Сибири о б л е п и х а с и б и р с к а я (Hip- pqphae rhamnioides L. sap. еи-rhamnoides Servet.). Вводится в культуру как витаминоносное растение (Гатин, 1955). В лесах Дальнего Востока растут б е р е с к л е т — Euonymus maaickii Ruhr. — хороший каучуконос, и жимолость — Loniicera edulis Типск., плоды которой употребляются в пищу. Экономическое значение имеют в Сибири к е д р ы. Широко распространены кедр сибирский — Pinus sibiriea (Rupr.) Mayr и кедр-стланец — Р. ~pumila (Pall.) Rgl., которые дают большие сборы «орехов», используемых как лакомство и для изготовления масла. Кедр корейский — Р. koraiensis Sieb. et Zucc. растет по берегам Амура и уссури. Используется, как и предыдущие виды. Растения арктической части Сибири изучены слабо, хотя сибирская Арктика богаче ими, чем Арктика европейская. Еще А. Ф. Миддендорф (1867) довольно обстоятельно писал о «съедобных растениях глубоко северной флоры». Он отмечал хороший вкус некоторых северных растений Сибири и считал возможным окультуривание их для использования в качестве весенних и осенних овощей. Миддендорф заметил, что некото- рые растения на севере теряют свои ядовитые свойства. Напри- мер, «туруханский хрен» — корень чемерицы — Veratrum lobelia- пит Bernh. — употребляется как приправа к пище в Турухан- ком крае и там совсем не считается ядовитым. Б. А. Тихомиров (1958) также отмечает, что чемерица, растущая на севере Си- бири, не ядовита для лошадей. Эскимосы и чукчи используют в пищу и для других целей значительное количество растений из местной флоры. Б. А. Ти- хомиров (1958) составил список полезных растений, листья которых используются эскимосами Чукотки: А111пт sibiri- cum L.— лук сибирский (листья употребляются в сыром виде и хранятся в мешках из шкур нерпы); Salix speci~osa Hook.— ива (листья употребляются вместе с желудком оленя); свежие листья других видов ивы идут в пищу — вместе с жиром, кровью и мясом нерпы; травы с кислым вкусом — Охупа digyna (L.) Hill., Rumex graminifolius Lamb. — сибирский щавель (листья и корни употребляются в сыром виде). Листья арктического ща- веля — Rumex arctiicus L.— едят с топленым жиром и сахаром, с жиром и мясом употреблялась также надземная и подземная часть Polygonum elliipticum Willd. ex S)preng. В качестве при- правы к мясу используют свежие листья камнеломок — S Bxl- fraga punctata L. и S. nelsoniana D. Don. Идут в пищу листья Dnyas ~punictata Juz. и других видов этого рода. Листья кипрея — Chamaenerium angustif~olium (L.) Scop.— отваривают, отжимают и хранят в замороженном виде в мешках из шкур нерпы. Синская Е. Н. 225
Используются в пищу следующие растения: анемон сибир- ский — Anemone sibirica L. (корневища этого растения едят в сыром и вареном виде); Sedum а1паригригеит Hort., Асотайу- lis rossii Greene (корневища едят в сыром виде с жиром и мя- сом); Hedysarum obscurum L. и Oxytropis mertensiana Turcz. (корни едят в свежем виде); Nardosmia frigida (L.) Hook. (кор- невища едят в жареном виде), морошка — gubus ~chamaemo- rus L., брусника — Vaccinium vitis-idaea L., голубика — V. uligi- ловит L., водяники — Fm~petrum nigrum L., Е. hermaphroditum (Lange) Hagerup, Arictous alpina (L.) Nied. Ряд растений имеют лекарственное значение, например ко- рень Сое1ор1еигит Ledeb. Листья сабельника завариваются как чай. Многие растения используются на топливо. По данным Пор- сильда (Piorcild, 1950), эскимосы Гренландии также широко ис- .пользуют местную флору и очень многие дикорастущие расте- ния употребляют в пищу. Материалы о питании народов Арктики ясно свидетельствуют о том, что даже в странах со сравнительно бедными раститель- ными ресурсами местной флоры, далеко не исчерпаны все воз- можности введения в культуру новых пищевых, лекарственных si других растений.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ КИТАЯ, ИНДИИ И СОСЕДНИХ С НИМИ СТРАН дикорастущая флора Центральной, Юго-Восточной и Юж- ной Азии чрезвычайно богата. Китай и Индия вместе с Индокитаем являются родиной очень большого числа культурных растений. Из них многие имеют огромное хозяйственное значение и в нашей стране: рис, хлопчатник, чай, просо, гаолян, пайза и др. Начало земледель- ческой культуры в этих странах относится к глубочайшей древ- ности. С тех пор человек здесь ввел в культуру многие растения, которые затем распространились в соседние страны и за пре- делы Азии. Растительные ресурсы Китая и Индии еще далеко не исчер- паны. jIa и богатство культурных видов и сортов весьма слабо изучено; эти страны, в особенности Китай, представляютогром- ный интерес для интродукции новых культурных растений в СССР. ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ КИТАЯ И СОСЕДНИХ СТРАН ВОСТОЧНОЙ АЗИИ На территории Китая человек появился еще в конце третич- ного периода. Останки этого предка человека (синантроп) най- дены близ Пекина. Раскопками открыты стоянки первобытного человека, относящиеся к палеолиту, неолиту и энеолиту. Там же, где найдены останки синантропа, обнаружены п кости человека верхнего палеолита (пещера Чжоукоудянь неда- леко от Пекина) . Очень интересны верхнепалеолитические раскопки в районе Ордоса на севере Ганьсу и Шэньси. Находимые здесь предметы позволяют сблизить этот период с ориньяком, а позднее- с азильским периодом в Европе. Степные культуры бассейна Хуанхэ сходны с культурами верхнепалеолитических стоянок Селенги: и там, и тут были обнаружены остатки теплолюбивой фауны, в том числе и скорлупа страусовых яиц. Культура бас- сейна Хуанхэ и на Селенге синхронны с раскопками в Северо- Восточном Китае в районах Харбина и Чжалайнора. Их пред- положительно относят к плювиальному периоду В эту эпоху имело огромное значение собирательство, на что указывает большое количество в стоянках раковин улиток и скорлупы страусовых яиц. 227
В древнекитайских литературных памятниках и в легендах имеется много фантастических рассказов о далеком прошлом- о жизни первобытных людей на территории Китая. В труде «Хань Фэй-цзы» говорится, что в период Яо (1 ты- сячелетие до н. э.) люди жили в хижинах, крытых камышом и тростником, употребляли в пищу грубые сорта риса, ели и пили из глиняных сосудов, летом слегка прикрывали тело гру- 4ыми полотнами, а зимой одевались в оленьи шкуры. Из неолитических культур Китая лучше исследована куль- тура яншао, охватывающая северо-западные провинции: Ганьсу, Шэньси, Шаньси и западную — Хэнань. Жители раннего неолита Северного Китая занимались мотыжным земледелием, возде- лывали несколько сортов проса и гаоляна. Из домашних живот- ных были собаки и свиньи. В Южном Китае неолит изучен слабее. Южные неолитиче- скпе племена бассейна реки Янцзы занимались рыболовством и примитивным земледелием. Они возделывали корнеплоды, глав- ным образом, ямс. Обитатели этих стоянок жили не в землян- ках, как на севере, а, по-видимому, в свайных постройках. Юж- нокитайский неолит связан с Юго-Восточной Азией, на что ука- зывает и пестрота ее современного населения. Яанные по палеолиту в Японии мало достоверны. Археоло- гические сведения о древней Японии обобщены М. В. Воро- бьевым (1958). Им дается краткое изложение этого материала. 'Описания памятников каменного века появляются в самых древ- них письменных памятниках Японии. В «Описании земли Хи- тати» говорится: «в глубокой древности жили люди росту очень высокого, жили они на холмах. Собирали моллюсков и ели их, собирали ракушки и ели их. Складывали их и получались хо.1мы» (М. В. Воробьев, 1958). В раковинных кучах, относящихся к эпохе собирательства ракушек, находили также зернотерки. В древнейшие времена собиратели раковинных куч жили оседло в небольших землян- ках. Стоянки были редкие и небольшие — с 1 — 2 землянками. Раковинные кучи были небольших размеров. Позднее каменный инвентарь, находимый в раковинных кучах, становится разнообразнее и совершеннее. Близ городка Кикуна в раковинной куче был найден большой рыболовный крючок, сделан- ный из рога оленя, кости крупных глубоководных рыб и животных (тунец, акула), клык дикого кабана и кости некрупной домаш- ней собаки. В это время на керамике появляются веревочные оттиски. На более древних сосудах оттиски получались от свя- зывания сырого сосуда травяной веревкой. Позднее веревоч- ный узор покрывает всю поверхность сосуда и становится орна- ментом. Большую роль играло собирательство кореньев, кото- рые растирались в каменных ступах. «Расцветом раковинных куч» характеризовались неолит и энеолит; в меньшем количестве 228
их находят и в раннем железном веке. В позднем неолите встре- чаются целые слои каштановых и ореховых скорлупок. Еще позднее появляются кости домашней лошади и коровы. Землянки сменялись наземными жилищами. В эпоху позд- него неолита и энеолита, судя по археологическим данным, население было знакомо с разведением культурных растений: гречихи, бобов, конопли, позднее — риса. В конце неолита на Японию двинулись волны переселенцев. Аборигены в начале этого объединения двух культур еще только начинали знакомиться с металлом, делали первые шаги в возделывании культурных растений, они еще не знали ника- ких домашних животных, кроме собаки. Пришельцы изготов- ляли изделия из бронзы, возможно и из железа, возделывали рис. Население стало жить в больших поселках. По морским берегам сохранились раковинные кучи. Земледелие распростра- нялось на равнинах и на легко затопляемых заболоченных уча- стках, пригодных для рисосеяния. Население земледельческих деревень обитало в наземных жилищах, имевших небольшую подземную часть. Эти жилища делались из жердей, тростника и ветвей. В энеолите и бронзовом веке земледелие становится одной из отраслей хозяйства. На основании японских мифов, историчес- ких хроник и археологических памятников можно заключить, что в это время возделывали рис, ячмень, просо, бобовые и коноплю. Сначала земледелие было богарным, позднее развилось и по- ливное. Вследствие недостатка площадей, удобных для земледе- лия, оно долго не играло ведущей роли в хозяйстве японцев. Вопросы происхождения современных японцев и родства их с древнейшим населением архипелага решаются учеными раз- лично (М. В. Воробьев, 1958, и др.). Несмотря на различные волны этнических миграций форми- рование японцев было постепенным процессом. Интересна, по- видимому, весьма правдоподобная связь древнейшего населения Японии с сибирскими древними культурами, что нами было от- мечено выше в разделе об истории земледелия Сибири. Несмотря на то, что массовое переселение из Кореи и Китая имело место в сравнительно позднюю эпоху (в начале железного века), культурное влияние именно этих переселенцев было огромно. На это указывает состав культурной флоры Японии, где, безусловно, преобладают корейско-китайские элементы. Исторический период китайской истории начинается только ° с эпохи Инь. Иньская культура развилась во II тысячелетии до н. э. в долине по среднему течению реки Хуанхэ (на северо- востоке современной провинции Хэнань, в области, прилегающей к хребту Тайханшань). Основными занятиями иньцев были рыболовство и охота. Наряду с этими промыслами имели также значение скотоводство и земледелие. Скотоводство, видимо, иг- рало большую роль (скот приносился в жертву богам).
Земледелие было примитивного мотыжного типа, что, впро- чем, в Китае и Японии частично сохранилось и до самого послед- него времени. Земледельческие орудия изготовлялись из камня и дерева. Орошения не было, удобрения не применялись. Когда участок под пашней переставал родить, он забрасывался, и земледелец устраивал поле в другом месте (И. Клинген, 1898). В эпоху Инь возделывались: ячмень, пшеница, просо, рис, ту- товое дерево (Степугина, 1950). Родина риса — Южная Азия (Южный Китай, Индия, Бир- ма). Когда именно рис распространился в Китае, точно неиз- вестно. За 3000 лет до н. э. культура риса там уже была. Ко- личество местных сортов риса в Китае чрезвычайно велико. Экспедиция института археологии Академии наук Китая, про- изводившая (с 1955 г.) раскопки в долине реки Янцзы в местечке 1Яюйцзялинь уезда Цзиньшань, в 150 км западнее Учана, обна- ружила в красной обожженной глине много пленок от зерен риса. Эти находки отнесены к эпохе неолита. Затем такие же пленки были обнаружены и в местах стоянок неолита в Шиц- зяхэ (110 км западнее Учана), а также в Фаньинтае (Учан) . Там же было извлечено много каменных ножей, топоров, гончарных изделий и т. п. Все три места расположены в долине реки Ян- цзы на высотах 50 — 100 м над уровнем моря и пригодны для земледелия. Сравнивая найденные предметы, участники экспе- диции пришли к заключению, что находки относятся к периоду первобытнообщинного патриархального общества конца нео- лита — более позднему, чем памятники культуры яншго и куль- туры луншань. Новые находки насчитывают не более 4000 лет (Дин Ин, 1960) . В конце неолита красная глина служила основным строительным материалом, куда добавляли солому и мякину риса. Некоторая часть пленок сравнительно хорошо сохрани- лась. Кроме пленок, попадались обломки остей и измель- ченная солома. Ископаемые зерна риса имеют бледно-желтук~ или пепельно-желтую окраску, сходную с окраской зерен тех сортов, которые возделываются в настоящее время. На пленках обнаружено опушение. Судя по обломкам остей, они были очень толстые и длинные. От колосковой чешуи сохранился лишь от- печаток. Наибольшая длина зерна составляла 7,5 мм, наимень- шая — 6,8 мм, наибольшая ширина — 3,8 мм, наименьшая— 3,0 мл~; наибольшее отношение длины к ширине — 2,34, наимень- шее — 1,8. Иньская культура в основном относится к бронзовому веку„ но с пережитками неолита, особенно это касается земледельче- ских орудий. Древнее племя чжоу первоначально обитало вдоль реки Вэй в центре теперешней провинции Шэньси. Чжоу и иньцьг находились в сношениях друг с другом, но культура чжоу была, по-видимому, выше, Чжоу занимались скотоводством (играла 230
значительную роль лошадь) и земледелием, еще до своего пере- селения в долину реки Вэй, когда они вели еще полуоседлый образ жизни. Сеяли они пшеницу, ячмень и просо. Последнее имело особенно большое значение. Начало племени чжоу, со- гласно преданиям, идет от легендарного Хоу-1Язи, что означает «1(нязь — просо». П р о со — одно из древнейших культурных растений Китая. И в древности, и в настоящее время оно играло и играет огром- ную роль в питании китайцев (Ермашов, 1949). Ботанические данные указывают на большое разнообразие форм проса в Ки- тае, на эндемичность большинства из них и приводят к заклю- чению, что родина проса — это Китай. К древнейшим растениям Китая относятся также рис и соя, которые культивировались здесь еще за 2700 лет до н. э. древняя история Южного Китая изучена еще слабее. Во II и 1 тысячелетиях до н. э. на территории Чжэцзян, Фуцзянь и Гуандун обитали племена юэ, хозяйство и культура которых сильно отличались от быта племен Северного Китая и обнару- живали сходство с Индонезией и Океанией. Юэ занимались ры- боловством и земледелием. Из домашних животных имелась свинья. ~Кили юэ в свайных постройках, одежду частично приго- .товляли из луба. К западу от юэ в середине 1 тысячелетия до н. э. обитали племена, образовавшие княжества Чу и У. Культура их была близка к культуре юэ. Эти племена считаются предками таи. Существует предание, что плуг был изобретен около 2700 лет до н. э., но на самом деле — может быть и раньше. В это же время был составлен первый китайский травник, в котором го- ворилось о многих видах китайских аборигенных лекарственных растений. В Китае с древнейших времен культивируются апельсины. Почти для каждого вида и сорта этих плодов существует особое название на китайском языке, и о них упоминается во мно- гих древних памятниках. О мандаринах и помпельмусах упоминается в перечне пред- метов дани в 2300 г. до и. э. Лимон, вероятно, был завезен позднее, так как в древних документах он не упоминается. Многие исследователи считают, что чай начали культивиро- вать в Китае после VI в. н. э., но лингвистические данные ука- зывают, что он употреблялся, как напиток, а также и в меди- цине, гораздо раньше. В Китае чай растет и дико. По японским преданиям, чай происходит из Индии, но, возможно, что куль- тура его возникла йезависимо и в Южном Китае. Состав культур в Китае еще в древние времена был разно- образен даже на севере. По «повествованиям о восточных ино- земцах» (Н. Я. Бичурин, 1950), в начале нашей эры и позднее на территории современного Северо-Восточного Китая и южнее,
«родятся крупные каштаны», имеются «виды разных хлебов, пло- дов, огородного овоща и зелени, также сорта вин»... ~Государ- ственные подати вносят... шелком, пенькой и рисом»... «Хлеб вносят смотря по урожаю. Сеют просо и пшеницу, вино делают из риса». Значение проса очень велико, оно очень часто упоми- нается, подать собирается в некоторых местах, главным обра- зом, просом. Власти также иногда награждали подданных про- сом. 06 истории земледелия в Японии известно еще меньше, чем о собственно Китае, так как вообще история японцев слабо р азработана. Древняя керамика представлена в Японии высокими плоско- донными сосудами с «веревочным орнаментом». Эти сосуды по форме и размерам напоминают современные плетеные корзины Японии и Малайи. Широко было распространено плетение ци- новок и изготовление грубых тканей из крапивы. Интересно отметить и здесь значение крапивы, как древнейшего прядиль- ного растения, о чем упоминалось выше в разделе о Сибири. Неолитическая культура этого характера была распространенз и на юге Сахалина и на Курильских островах. К концу I тысячелетия до н. э. относится резкое изгленение хозяйственной жизни обитателей Японии. Новая культура яёй связана с началом земледелия. Обнаружены во многих местах каменные мотыги, зерна риса, кости лошади. Современные айны до самого последнего времени сохраняли многие черты жизни неолитического человека: занимались рыболовством и охотой,. жили в шалашах, пользовались каменными орудиями, приготов- ляли ткани из крапивы и т. п. Японская культурная флора имеет корни на материке Вос- точной Азии (в основном) и в Южной Азии (бамбуки, бенин- каза, таро и др.). Большинство культурных растений Японии заимствовано из Китая. Культурная флора этой страны в об- щем сходна с флорой Восточного Китая, хотя на Японских ост- ровах имеются и свои аборигенные сорта и даже виды. Многие древние классические источники, относящиеся к на-- чалу нашей эры, переведены Н. Я. Бичуриным. О Японии в них говорится, что там «земля произращает просо, рис, коиоплю.. Умеют ткать шелковые ткани и холсты... Климат теплый. Зимой и летом растет всякий овощ. Нет коров, лошадей, тигров, лео- пардов, овец и сорок» (Н. Я. Бичурин, 1950). Здесь говорится о зимнем и летнем земледелии, что способ- ствует разнообразию культур. Кроме упомянутых выше, есть имбирь, корица, померанцы, перец, мелисса. Земледелие в Япо- нии развивалось отдельно от скотоводства, которое начало раз- виваться лишь в последнее время. Исконные домашние живот- ные в Японии — куры и собаки. Черты примитивного земледельческого уклада сохранялись в Японии вплоть до нашего времени. До сих пор Япония — это
страна не плужного, а мотыжного земледелия,— поля большей частью малого размера и обрабатываются вручную; бытуют примитивные способы уборки и молотьбы. До войны японцы почти не употребляли пшеничного хлеба. Сейчас изделия из пшеничной муки все шире входят в быт населения Японии. Все эти особенности характерны и для Kmaa, особенно Вос- точного. В Западном Китае, по данным, относящимся к началу нашей эры и позднее (по переводам Н. Я. Бичурина, 1950), зем- леделие было интенсивно развито и набор культур был большой: рис, просо, кунжут, пшеница и другие колосовые хлеба, овощные культуры, виноград, плодовые, конопля, шелковичное дерево, гречиха и др. Луковые горы (хребет UyHJIHH) получили свое название от того, что на их вершинах в изобилии рос лук. Вследствие большой древности земледелия в Восточчом и Центральном китае там сложились системы вековых навыков и приемов обработки почвы и ухода за растениями, несмотря на примитивный инвентарь. Земледелие, благодаря высокой плотности населения и складывавшейся в течение тысячелетий системы земледелия имело интенсивный характер еще со времен глубочайшей древ- ности. Благодаря разнообразию условий бытовали различные системы земледелия. Н. Я. Бичурин (1844) перечисляет сле- дующие типы пахотных угодий: 1) гнездовые пашни; 2) плото- вые пашни — на плоты около берега рек и озер насыпают слой почвы и производят посев; 3) плотинные пашни — участки, обне- сенные земляными валами; 4) ящичные пашни — обнесенные крепкими насыпями; 5) лестничные пашни — расположенные уступами по склонам гор; 6) грязные пашни. «Такой способ по- севов производится вообще в местах, изобилующих водою и в привзморьях, где находятся земляные наносы, образуемые морскими приливами, в виде отмелей и островов. На послед- них местах вначале растут одни кустарники и солонцеватые травы, а затем засевают сии места диким просом, а когда солонцеватость совершенно испарится, то начинают сеять раз- ный хлеб». Здесь очень интересно сообщение о практикующихся посевах «дикого проса», — какой именно злак здесь имеется в виду, уста- новить пока трудно. Очень оригинальна культура на гнездовых пашнях. Гнездовой посев начал применяться за 1790 лет до н. э. и ныне употребляется преимущественно в гористых местностях, мало удобных для обыкновенного хлебопашества. Интересны смешанные посевы пшеницы и проса черезбороздно, а также проса с черными бобами (Н. Я. Бичурин, 1950). Некоторые исследователи думают, что территория между ре- ками Хуанхэ и Янцзы была родиной культурного риса, местом вхождения его в культру. Но в последнее время такое мнение
мало кем поддерживается. jIHH Ин (1960) считает родинок риса — Южный Китай. Многие районы Южного Китая, по нашему мнению, входяг в область развития культурной флоры Южной Азии, а не Вос- точной Азии, и с этой точки зрения взгляды тех исследовагелей, которые считают родиной риса Южной Китай, не противоречат нашей концепции о рисе, как аборигене Южной Азии. Однако более вероятно происхождение риса из южной Индии. Происходившая веками и тысячелетиями смена поколений культурных растений в условиях интенсивного возделывания,. естественного и искусственного отбора создавала высококуль- турные местные сорта из аборигенного и из завозного (из дру- гих стран) исходного материала. Поэтому-то многие культур- ные растения так оторвались от своих диких родоначальников, что их трудно опознать, или последние успели исчезнуть (просо и др.). Культуры, занесенные в Китай из других первичных оча- гов, здесь успели за огромный промежуток времени, в течение которого подвергались интенсивному воздействию человека, сильно измениться, приобрели ботаническую обособленность и стали казаться местными, аборигенными растениями (сорго и др.) .. Природа Китая богата и разнообразна. Северный Китай представляет собой естественное продолжение степей Забай- калья и Северной Монголии. Хребет Большой Хинган покрыт тайгой, и по нему распространились на юг многие представи- тели восточносибирской флоры, вплоть до окраины Ордоса. Здесь характерны леса из осины, березы, дуба и др. Южнее про- стираются степи и солонцеватые земли. На юг и юго-запад от Харбина расположены саванны. По мнению П. А. Баранова (1960), саванны в Китае носят первичный характер (например, по Красной реке) в проти- воположность саваннам Африки, где они почти везде вторич- ные. По словам К. К. Флерова (1959), в Ляодунской низменности из стеблей тростника и кукурузы делают плоты. Вода перека- чивается из каналов на поля силами мулов и ослов. Лесная растительность истреблялась в Китае в течение тыся- челетий. Леса сохранились главным образом в горных мест- ностях, заповедниках; однако состав древесной флоры чрезвы- чайно богат и разнообразен. По словам K. K. Флерова, ему приходилось насчитывать в лесу на площади величиной в четверть гектара до 69 древес- ных пород и до 30 видов лиан. По его данным, в Китае имеется 100 видов клена, 130 видов дуба. В подлеске имеют большое зна-- чение в лесах южных провинций рододендроны и бамбуки; и те, и другие представлены в Китае в количестве около 300 видов каждый. В лесах Южного Китая встречаются 50 видов фикусов, много видов пальм, магнолий, непентесов, камелий, некоторые из 234
этих видов при достаточном количестве влаги заходят довольно далеко на север. На стволах деревьев в изобилии растут эпифиты. Среди них имеется очень много красивых видов орхидей, из которых пока еще не так много используется в декоративных целях. В тропи- ческих лесах Южного Китая встречаются хлебное дерево, дре- вовидные молочаи и др. На плоскогорьях и горах Гуандуна и Гуанси растительность носит более ксероморфный характер. В лесах в изобилии растут разные хвойные: криптомерии, подокарпусы и др., реже встре- чаются саговники. Широко распространены саванны. В саван- нах, по данным П. А. Баранова (1960), в частности в окрест- ностях Юаньцзяна, встречаются деревья китайской фисташки— ближайшего родича средиземноморской фисташки. Здесь же П. А. Баранов встречал дерево из семейства молочайных из рода филлантус. Если плоды этого дерева взять в рот, они вначале не вкусны, а затем становятся сладкими. Китайцы называют этот филлантус «юканцзе», что значит «остается сладкий вкус». Из его плодов местное население приготовляет мармелад. В окрестностях биологической станции Юньнаньского универ- ситета П. А. Баранов встретил экземпляры гарцинии (Garcinia 1 .) — дикорастущего родственника мангустана. В Сишуанбанна П. А. Баранов посетил тропический лес. Одна из наиболее крупных древесных пород этого леса анчар (Antiaris toxicaria Lesch. из семейства тутовых). Млечный сок анчара содержит глюкозид антиарин, которым некогда смазы- вали наконечники стрел. Здесь также растет масса всевозмож- ных лиан и эпифитов, особенно замечательны из них так назы- ваемые фикусы-душители, плотно обвивающие древесные стволы. Хорошо запомнились П. А. Баранову леса на горе Нань- ношань, где живет древний народ айни, выращивающий очень ценный ассамский чай — Thea assamica Mast.— под пологом тро- пического леса. В одном из ущелий горы Наньношань Баранов видел деревья этого чая, достигавшие 6 м и имевшие, по сведе- ниям местных ботаников, возраст 500 — 600 лет. В горах илесах Китая можно встретитьтакиевиды растений, которые в литературе часто указываются, как вымершие. Так, по данным А. В. Гурского (1960), в лесном заповеднике на горе Темуса в 90 км от города Ханчжоу китайские лесоводы еще в 20-х годах обнаружили в дикорастущем виде реликтовую дре- весную породу гинкго — Ginkgo biloba L. Сведения о том, что это интересное дерево сохранилось только в культуре, прочно во- шли в научную литературу. На южных склонах горы Темуса по руслам временных потоков и на крутых обнажениях скал гинкго растет группами. Это светолюбивое дерево не может расти в сомкнутом лесу. Старые деревья гинкго все увешаны мхами и лишайниками. Они достигают высоты 25 — 30 м и тол- щины 60 — 80 см. Разновозрастность этих деревьев говорит 235
о том, что естественное семенное возобновление гинкго идет. успешно. В некоторых старых лесах недавно открыты заросли древ-- него представителя лесной флоры — метасеквойи. Чем южнее, тем богаче лесная флора и тем чаще встре- чаются редкие и мало известные растения. Замкнутое, не имеющее стока плоскогорье Центральной. Азии, входящее в состав Восточноазиатской флористической области (Е. В. Вульф, 1944), бедно растительностью и дало очень мало аборигенных культурных растений, поэтому нашим обзором оно, по-существу, не затрагивается. Смена дикорасту- щей флоры и состава культурных растений идет постепенно с востока на запад и здесь нельзя провести резкой границы. Од- нако самая западная часть 1~итая (Кашгар) имеет много общих. черт со Средне-Юго-Западной Азией и нами отнесена к послед- ней. Самая южная тропическая часть Китая относится к флори- стической области Южной Азии, куда входят Индия, Цейлон, Индокитай. Северную границу южноазиатской области также трудно наметить с полной определенностью. Восточноазиатская область развития культурной флоры за- нимает громадную территорию и подразделяется на ряд под- областей и провинций. Однако состав культурной флоры стран Восточной Азии еще недостаточно изучен, особенно очаги пер- вичного вхождения в культуру и географического распростра- нения культурных видов и форм и их родичей. Поэтому ниже: дается краткий обзор важнейших культурных растений всей Вос- точной Азии. ОБЗОР АБОРИГЕННЫХ РАСТЕНИЙ СТРАН восточной Азии Ячмень (Hordeum L.) в целом в Восточной Азии имеет эндемичные формы, выделенные Ф. Х. Бахтеевым в самостоя- тельный вид — Н. humile Vav. et Bacht. По внешнему облику эти: ячмени напоминают ископаемый ячмень свайных построек Швей- царии. )lo последнего времени из дикорастущих видов только один — Н. spontaneum С. Koch — включался в одну секцию с культурным ячменем — секцию Crithe Doell. Однако — это вид двурядный, т. е. с редуцированным колосом. Такое строение ко- лоса Н. spontaneum со всей определенностью говорит о том, что раньше существовал близкородственный дикорастущий вид ячменя с нередуцированным колосом, т. е шестирядный, кото- рый и был родоначальником как Н. Spiontaneum, так и совре- менных культурных ячменей. В подтверждение этих догадок были найдены на юго-вос- токе Тибетского нагорья, на границе с Южным Kmaeri, своеоб- разные дикорастущие шестирядные формы, близкие к культур- ному ячменю и получившие название Н. agriccrithon Aoberg. 236
Э. Оберг (Aberg, 1948) сообщает о находке культурного яч- меня, очень похожего на дикорастущий Н. agriocrithon»s Восгоч- ного Тибета. Это сообщение подкрепляет предположение о том, что Н. agriocrithon близок к родоначальнику культурного ячменя и что группа близких к нему форм имела раньше широкий ареал (да и в настоящее время этот ареал может оказаться не столь редуцированным, как это предс~ авляется теперь, вследствие пло- хой обследованности Тибета и прилегающих к нему территорий собственно Китая). В последнее время систематику Н. spontaneum изуча.1 Ф. Х. Бахтеев (1962). Он открыл в Туркмении особую разно- видность этого вида — чаг. lagunculiforme ВасЫ. Ячмени этой разновидности найдены и в ископаемом виде на Северном кав- казе. В состав этой разновидности входят двурядные и шести- рядные формы. По нашему мнению, подобная шестирядная форма, но только более мезоморфная, раньше имела более широкий ареал, прости- равшийся из Передней Азии далеко на восток, вплоть до Восточ- ной Азии, возможно, по берегам исчезнувшего Тетиса. Что касается Н. agriocrithon, то Ф. Х. Бахтеев приписывает ему гибридйое происхождение. Однако эти формы похожи на гибриды именно с какими-то дикарями. Вероятно, близкие ди- корастущие формы растут или росли вблизи местонахождения ячменя, описанного Э. Обергом. Возможно существование Н. ag- riocrithon указывает на прежнее простирание Н. spontaneum на восток. Если наши соображения имеют достаточно оснований, то ареал вымершего дикорастущего шестирядного ячменя с мезо- фильными чертами строения простирался от восточных окраин Тибета до Передней Азии (ареал Н. spontaneum — Иран, Афга- нистан, Туркменистан, юго-восточное Закавказье, Сирия, Па- лестина и Киренаика). Если это было так, то ячмень мог войти в культуру независимо в разных областях Восточной и UeH тральной Азии и в Передней Азии от одного полиморфного, ме- зофильного, шестирядного дикорастущего родоначальника. Условия влажного муссонного климата Восточной Азии осо- бенно благоприятны для сохранения древнего мезофильного шестирядного типа в культуре. Здесь меньше, чем в других ме- стах, возможно стимулирование редукционных процессов, веду- щих к образованию двурядных форм. Последних и нет среди эндемичных восточноазиатских ячменей Китая и Японии. По древним китайским источникам (переведенным И. Бичу- риным в 1851 г. и переизданным в 1950 г.), в Аньси (так назы- валась Бактрия) и в соседней стране Тяочжи, расположенной западнее, в большом количестве водился страус, который (по сведениям из старинной географии Гуан-чжи) «питается ячме- нем». С этими данными интересно сопоставить недавние на- ходки страусовых яиц, относящиеся к каменному веку, на севере Ганьсу и Шэньси в Китае, во Внутренней Монголии (в Мон- 237
гальской Народной Республике) и в СССР (в Сибири) в верх- непалеолитических стоянках по реке Селенге. Следовательно, ареал страуса, питавшегося в основном диким ячменем, был не- когда огромным и простирался от Аму-Дарьи до Селенги. Не указывают ли эти данные косвенно и на то, что ареал дикора- стущего ячменя (родоначальника мезофильной шестирядной формы, теперь исчезнувшей) простирался далеко на восток и север Азии? Ксерофилизированным дериватом этой вымершей ме- зофильной формы и является, по нашему мнению, Н. agriocri- thon, сохранившийся кое-где в Тибете. Овес (Avena L.) имеет самостоятельные очаги кульгуры г, Китае, главным образом Восточном, где находится область раз- вития крупносемянного голозерного овса, который некоторыми ботаниками считается самостоятельным видом — А. nuda L. Ди- ких родичей поблизости нет. Вероятно, А. nuda образовался в культурных условиях в области мусонного климата из каких-то сорных форм. Род пшеницы (Triticum L.) в Китае представлен немно- гими видами, но здесь много аборигенных сортов мягкой пше- ницы (Т. aestivum L.). Существует мнение, что пшеница сюда пришла из Индии: имеется ряд форм, сближающих китайские пшеницы с индийским видом — Т. sphaerocoocum Pere. Более ве- роятно, что пшеницы распространились и в Китай, и в Индию пз Средне-Западной Азии, а туда — из Передней Азии. В Китае наибольшее разнообразие сортов мягкой пшеницы встречается в провинции Юньнань. Из юньнаньских пшениц Цзин Шань-бао (1958) выделил новый тип пшеницы Т. yunnanense King. Скорее это не вид, а только подвид или сортотип мягкой пшеницы. Японские пшеницы (proles japonicum Flaksb.) как будто на первый взгляд отличаются некоторыми примитивными чертами (карликовость, плотноколосость), но они все же очень однооб- разны и представляют собой явную продукцию местной агро- среды: осенне-зимней культуры в условиях муссонного климата. Пшеницы Южного Китая и Тибета еще очень мало исследо- ваны. Существует точка зрения, что культура пшеницы в Китае восходит к столь глубокой древности, что не может не быть або- ригенной. Однако в пользу последней точки зрения нельзя при- вести никаких фактических доказательств: диких родичей по- близости нет; не обнаружено и таких примитивных интегральных типов, как в Закавказье. И. Бичурин (1844) сообщает следую- щие любопытные данные о китайской пшенице: «Северная пше- ница цветет днем; имеет шелуху тонкую, мучнистого вещества много и в пище здорова. Южная пшеница цвететночью и трудно варится в желудке, это происходит от почвы». Рожь (Secale L.) возделывается в горных местностях Цен- трального Китая, но слабо там изучена. Когда она распростра- нилась сюда из Передней Азии, неизвестно. 238
В Восточной Азии из занесенной (вероятно, очень давно, может быть, еще до колумба) из Америки куку р уз ы (Zea L.) развилась особая рецессивная эндемичная группа сортов вос- ковидной кукурузы — Zea мауз ssp. ceratina Kulesch. Эта свое- образная форма кукурузы, которую некоторые исследователи даже склонны считать самостоятельным видом, отличается, по- мимо воскообразного эндосперма, еще отсутствием антоциана и пониклой метелкой (похожа несколько на сорго). Интересно, что в приморской Восточной Азии найдены также восковидные формы проса и риса. Однако нельзя восточноазиатскую воско- видную кукурузу считать строгим эндемом Восточной Азии. Сходные формы открыты и в Южной Америке. П р о с о (Panicum miliaceum L.) по исключительной древ- ности культуры в Китае и большому количеству эндемичных форм, пестроте популяций, по мнению многих исследователей, имеет первичный очаг своего происхождения в Китае. Здесь же распространены сорные родичи — P. spontaneum Lyssov и P. mi- liare Lam. (в Юго-Восточном китае). В. Н. Лысов устанавливает следующие эколого-географиче- ские группы: у подвида развесистого проса — дальневосточную и восточноазиатскую с более широким ареалом, а у подвида сжатого проса — восточноазиатскую (маньчжурскую) . Многооб- разие китайского проса далеко еще не изучено. III,етини стае просо (Setaria italica P.В.) образует под- виды, связанные переходными формами, но часто считающиеся самостоятельными видами: 1) чумиза, или гоми, — Setaria italica ssp. maxima Alef. — имеет первичный очаг развития в Китае; 2) могар — S. italica ssp. moharium Alef., вероятно, происходит также из Китая. П а й з а (Echinochloa frumentacea Link), по мнению многих ботаников, происходит из Восточного 1~итая, где существуют пе- реходные формы к сорному куриному просу — Е. crus-galli (L.) Roem. et Schult. Возможно, что пайза распространилась из Юж- ной Азии. Гаолян [Sorhgum ~chinense Jakuschev.=S. ja~ponicum (Hack) Roshev.] представляет собой обособленную от других географических групп или видов культурного сорго группу энде- мичных сортов. диких родичей в Азии не найдено. В Китае, ве- роятно, имеет место вторичный очаг происхождения сорго. Е. С. Якушевский считает, что все сорго занесено в Азию из Африки. Перечисленные просовидные злаки имеют и кормовое зна- чение. Специально возделывающихся на корм животным злаков в Kurae до последнего времени не было, но в дикорастущей флоре в этом отношении перспективны многие виды. Из них упо- мянем о еж е (Dactylis L.). В лесных местностях Китая распро- странен особый эндемичный подвид древнего реликтового вида 239
~). aschersoniana зэр. hymalaensis Domin (Е. Н. Сннская, 1964) . Р и с (Oryza sativa L.) является очень древней культурой в Китае, но не здесь, а южнее — в Индии и прилегающих стра- нах Южной Азии — находится первичный очаг его происхожде- ния и вхождения в культуру. «Естественноисторические условия Китая лишены основных черт, которыми должен характеризо- ваться район первичного зарождения культуры риса. Мощные консерваторы влаги, подобные Гималаям, здесь отсутствуют, нет следов примитивной культуры риса, а самое главное, нет пред- ставителей дикорастущих видов, которые могли бы быть некогда вовлечены в культуру и показывали бы постепенный переход от диких форм к культурным сортам» (Г Г. Гущин, 1938). Воз- можно, что рис имеет гибридное и полифилитическое проис- хождение. Китайский, а также японский рис, по Г Г Гущину, в основ- ном представлен его обыкновенным подвидом — ssp. communis Gust. Этот подвид делится на две группы сортов: 1) proles indica Gust. 2) proles japonica Gust. Наибольшее разнообразие оба пролеса имеют на юге (Бенгалия, Ассам, Бирма), а по мере движения на восток и северо-восток все более доминируют формы, относящиеся к proles japonica; в Северном Китае, Япо- нии и Корее они доминируют окончательно. Несмотря на вторичное развитие, рис в Северном Китае и Японии представлен очень большим количеством местных сор- тов. В Японии имеются очень скороспелые формы. Большинство авторов считает родиной г р е ч и х и (Fa gopyrum sagittatum Gilib.) Индию (Гималаи), но и на север от Индии встречаются эндемичные местные сорта. В Северо-Восточном Китае и Восточной Сибири гречиха встречается только в одича- лом состоянии. Е. А. Столетова (1940) выделяет азиатскую куль- турную гречиху в особый подвид — ssp. ти1Яо1шт Stolet. Ареал этого подвида охватывает Индию, Китай, Японию, Корею, Даль- ний Восток СССР. Е. А. Столетова делит этот подвид на ряд типов: 1) индийский (Индия, занесен в Японию); 2) японский (Япония, Дальний Восгок СССР; 3) корейский (Корея); 4) ки- тайский (Китай). Соя (Glycine L.) — тропический род. Большинство видов сои обитает в тропической Африке, но эти виды не имеют отношения к происхождению культурной сои. Происхождение вида G. hispida Maxim., безусловно, приурочено к Юго-Восточной Азии. Здесь сосредоточено огромное разнообразие аборигенных сортов, имеются примитивные, с признаками дикарей, вьющиеся культурные формы, здесь же распространен родственный сорный вид — G. graci is Skworz.— и дикорастущий родич — G. ussu- riensis Rgl. et Maack. Некоторые авторы склоняются принять за родину культурной сои Индию, но, по многим соображениям, родиной этого расте- 240
ния является Китай. дикорастущий близкий вид — G. ussurien- sis — не заходит за Гималаи. Хотя вьющаяся индийская культур- ная соя и ближе к дикорастущей сое, чем вьющаяся китайская, но в Китае можно проследить все переходы от вьющейся до невьющейся культурной сои. В Северо-Восточном, Северном и Центральном Китае распространена сорная соя — G. gracilis, которая, вероятно, имеет вторичное происхождение от культур- ной сои — G. hispida — через приспособление к существованию в посевах мелкосемянных форм фасоли. В Восточноазиатской области сосредоточено основное богат- ство сортов и экотипов сои, объединенных В. Б. Енкеным (1959) в подвиды: 1) маньчжурский — ssp. manshurica Enken; 2) корей- ский — ssp. corajensis Enken; 3) китайский — ssp. chinensis Enken. В осточно аз иа тс ка я ф а соль адзуки [Phaseolus an- gularis (Willd.) Wight] ведет происхождение из Японии и Восточ- ного Китая, где имеется большое количество аборигенных форм и сортов. По данным Н. Р. Иванова (1960), восточноазиатские виды фасоли вошли в культуру на несколько тысячелетий раньше, чем в Америке. Фасоль адзуки не имеет диких форм. Автор думает, что этот вид вошел в Китае в культуру одновре- менно с соей, т. е. около 5000 лет до н. э. В Китае часто встре- чаются смешанные посевы сои и адзуки. Фасоль обыкновенная [Ph. vulgaris (L) Savi], веро- ятно, давно занесена сюда из Америки и образовала здесь группу эндемичных сортов, в особенности спаржевой фасоли. М а ш [Ph. aureus (Roxb.) Pip.] ведет происхождение из Ин- дии, но представлен в Восточной Азии особым подвидом — ssp. chinensis G. Рор., в котором Г. М. Попова (1937) различает сле- дующие агроэкологические группы: 1) маньчжурская — iproles mandchuricus G. Рор. (Дальний Восток СССР, Корея, Северо- Восточный Китай, Япония); 2) корейская~рго1ез korei~cus G. Pop. (в основном Корея); 3) японская — proles japonicus G. Pop. (Япония, Корея, Северо-Восточный Китай); 4) китайская— proles aksuensis G. Рор. (Китай, Корея); 5) западнокитайская— proles kashgaricus G. Рор. (Западный Китай, Корея). Н. Р Иванов считает, что маш происходит из Индии, но он стал широко возделываться в Китае в Х в. во времена династии Тан. Б а р х а т н ы е б о б ы [Stizolobium utile (Wall.) Ditm.], по мнению ряда авторов, имеют родоначальником дикорастущую лиану — S. иИе, а последняя произошла от тропической лианы Старого и Нового Света —,S. pruriens (L.) Medic. Вхождение дикорастущих форм в культуру могло произойти и независимо в разных странах, но первичным центром формообразования, вероятно, следует считать Южную Азию, где распространены два других культурных вида: S. hirsutum guntze и S. pruritum Kuntze. Вторичной областью развития считается Япония, где
имеется группа оригинальных местных сортов, объединенных ц подвид ssp. hassjoo (Piper et Tracy) Ditm. Кар о в ий гор ох (Vigna sinensis Endl.) имеет вторичный центр развития в Восточной Азии (Япония, Дальний Восток СССР, Корея, Китай), где образовалась группа аборигенных сор- тов из ~proles japonica А. Pavl. Формы коровьего гороха из Фи- липпин и Южного Китая выделяют в особый подвид — зэр. sesquipedalis Рйрег. (Китай, Япония, Филиппины, Индия). Вто- рой вторичный очаг культурного коровьего гороха находится в Индии, а первичный, вероятно, в Африке. Японская ло 6ия (Dolichos hassjoo Siebold) кое-где воз- делывается в Японии. Мало изученный вид. К о н с к и е б о б ы (Faba vulgaris Moench) представлены в Восточной Азии особой японо-китайской группой местных сортов. Японские местные сорта гороха (Pisum sati- vum L.) объединяются в особую японскую группу — proles japo nicum Gov. На севере области имеется монгольская группа мест- ных сортов — proles mongolicum iGov. Горох, наверно, давно рас- пространился на восток Азии и мало подвергался там селекции,. ибо восточноазиатские формы не лишены примитивности. Не- давно Оберг (Aberg, 1948) описал новую форму гороха из Во- сточного Тибета — Р. sativum tibetanum. Из бобовых сидератов интересен эндемичный для Японии и Восточного Китая культурный а с т р а г а л — генге (Astragalus sinicus L.) . .Этот астрагал представлен в Китае несколькими экотипами. Люцерн а синяя культу рн ая распространена в посе- вах в Китае. О кашгарском подвиде вида Medicago asiatica Sinsk. и о гибридной тибетской люцерне «ладак» говорилось выше в обзоре культурной флоры Средне-Юго-Западной Азии. Есть указания о посевах гибридной люцерны в окрестностях Пекина, где она находится в одичалом состоянии. Сорта лю- церны в Китае не изучены. И. Т. Васильченко (1949) без доста- точных на то оснований (мало материала и он экспериментально не изучен) выделяет дикорастущую люцерну из окрестностей Пе- кина в самостоятельный вид — М. beipinensis Vass. (по-види- мому, эта люцерна близка к М. соегиlеа Less.). Л е с п е д е з а (Lespedeza М1сЬх.) ведет свое происхождение из Восточной Азии. Следующие виды имеют экономическое зна- чение: 1) леспедеза двухцветная — L. bicolor Turcz. (Дальний Восток СССР, Северо-Восточный Китай, Корея, Восточная Мон- голия; заслуживает внимания как кормовое и декоративное ра- стение, кое-где культивируется); 2) леспедеза плотная — L. cyr- tobotrya Miq. (Япония, Корея, Северо-Восточный и Восточный Китай; культивируется в Западной Европе как декоративное); 3) L. hedysaroides (Pall.) Kitag. (Дальний Восток СССР, Япо- ния, Корея, Северный и Северо-Восточный Китай; имеет кормовое 242
значение на естественных оленьих пастбищах); 4) L. junicea Pers. (Корея, Япония, 1(итай, Индокитай, Малайский архипелаг, Ин- дия, Австралия; имеются опыты по возделыванию). К у м м е р о в и я (Kurnmerovia Schindl.) также восточно- .азиатского происхождения. В культуре известны виды под назва- нием корейского и японского клевера. 1(орейский клевер— 1(. stipulacea (Maxim.) Ма11по (Япония, 1(орея, Северо-Восточ- ный 1(итай) — возделывается в Южном Китае, Северной Аме- рике и Австралии в качестве сидерата и кормового растения; японский клевер — 1(. striata S~chindl. (Дальний Восток СССР, Япония, 1(орея, 1(итай) — разводится в качестве сидерата и кор- мового растения в Австралии и Северной Америке. Как кормовое растение в Китае разводится также с в и н о й с а л а т (Pistia stratiotes L.), который растет в прудах и неболь- ших водоемах. Из масличных, прядильных и технических растений иного назначения, кроме сои (последняя на родине своей является не масличным, а пищевым растением), для Восточной Азии харак- терны следующие. Судз а (Репllа ocymoides L.) доставляет ценное масло для лакокрасочной промышленности. Родиной всего рода является Восточная Азия. Следовательно, здесь находится первичный очаг вхождения периллы в культуру. Р. ocymoides образует две агроэкологические группы. 1) корейско-китайскую — proles ko- rejo-sinensis Bab., 2) более позднюю японскую — iproles japonica Bab. В Китайском ботаническом атласе имеются указания на культуры ый в ид п ер ил л ы [Perilla frutescens (L.) ВгЖ] и разновидность ее — var. crista Decne. Возможно, что эта пе- рилла представляет собой одну из форм вида Р. ocymoides L. По личному сообщению Н. Н. Глущенко (участницы экспедиции в 1(итай в 1956 г.), Р. frutescens употребляется для душистых курений в храмах, и, надо полагать, что культура этой периллы восходит к глубокой древности. Виды ш а н д р ы (Elsholtzia cristata Willd. и близкий ему вид Е. serotina Kom.) распространены в СССР на Дальнем Востоке и перспективны для возделывания ради технического масла с высоким иодным числом. Эти виды распространены также и в Северо-Восточном и Восточном 1(итае, как и декоративные виды этого рода. Необходима поисковая работа среди других близких родов растений семейства губоцветных для отыскания растений, даю- щих техническое масло (в Восточной Азии следует также про- смотреть с этой точки зрения виды змееголовника — Draicooapha- lum L.) . 1(унжут (Sesamum indicum L.) имеет вторичные очаги эн- демичных форм, очень ценных для селекции в Западном 1(итае и на востоке (Япония, 1(орея, Северо-Восточный и приморский 243
Китай). Кунжут появился в Китае в 1 тысячелетии н. э. (Haudri- court et Hedin, 1953). В. М. Гильтебрант (1932) различает маньчжурский и дальне- восточный экотипы. На этом скороспелом кунжуте строится вся селекция его в европейской части Советского Союза. Японский кунжут отличается очень большим полиморфизмом и еще слабо: изучен. ТаМ распространен и четырехплодолистиковый кунжут, неправильно выделенный в подвид; по остальным признакам ок не отличается от местного обычного кунжута с двумя плодоли- стиками и растет в одной популяции с последним. В Восточной Азии находится, вероятно, первичный очаг про- исхождения г о р ч и ц ы с а р е п т с к о й [Brassica juncea (L.) Czern.], наиболее экономически ценного вида горчицы. Разно- образие форм этой горчицы в Японии и Китае поразительно Здесь имеются ряд овощных (салатных) разновидностей и мас- личные сорта. Нами установлены следующие разновидности 1) var. crispifolia Sinsk. — курчаволистная салатная горчица; 2) var. subcrispifolia Sinsk. — слабокурчаволистная салатная горчица (в Японии листья этих двух разновидностей служат главным образом для украшения блюд); 3) var. integrifolia Sinsk. — цельнолистная салатная горчица; 4) var. subintegrifolia Sinsk. — полуцельнолистная салатная горчица; 5) var. subsarep- tana Sinsk. — масличные местные сорта с более или менее кисте- видным соцветием и явлениями «циклической стерильности»,. распространены в полевой культуре, в Кашгаре — поздние желто- семянные формы этой разновидности; 6) var. mongolica Sinsk.— c тонкими стручками (Монголия, Северный Китай); 7) var. sinensis Sinsk. — ранние масличные китайские формы, напоми- нающие индийскую раннюю сарептскую горчицу. В Китае встречается большое разнообразие раннеспелых и желтосемянных масличных форм сарептской горчицы и наиболь- шее разнообразие салатных ее сортов. Здесь же, на востоке Азии, произрастает и корнеплодная форма — В. naipiformis Bailey. Помимо способности образовы- вать съедобный корень (несколько напоминающий репу), эта овощная форма настолько сходна с обыкновенной горчицей— В. jun~cea, что могла бы быть включена с нею в один вид. В литературе имеются указания, что В. juncea встречается в дикорастущем состоянии кое-где в степях Монголии и Джун- гарии. Но эти указания сомнительны. Вероятно, были найдены только одичалые или сорные, но не настоящие дикорастущие растения. Экспериментальные генетические и цитологические исследо- вания японских и индийских ботаников доказали, что В. juncea— амфидиплоид, возникший путем скрещивания сурепицы— В. campestris L. — с черной горчицей — В. nigra (L.) Koch. Суре- пица (культурная и сорная) отличается в Восточной и Южной Азии огромным разнообразием форм, но в диком состоянии 244
неизвестна. Черная горчица менее распространена в культуре в азиатских странах, но нами была обнаружена в Индии энде- мичная форма — В. nigra — с дважды перисторассеченными листьями. Наличие эндемичной формы говорит о древности культуры черной горчицы в Индии, а, может быть, и в соседнем Южном Китае. Вероятно, где-то в этих широтах произошли те спонтанные скрещивания В. campestris с В. nigra, которые по- служили источником возникновения нового вида — В. juncea, получившего впоследствии очень большой ареал в культуре (но никогда не бывшего «диким», как и его прародительские виды). Может быть, происхождение В. juncea было политопным, и многообразие его форм в значительной степени объясняется огромным полиморфизмом сурепицы и близких к ней 20-хромо- сомных видов в Южной и Восточной Азии. М а с л и ч и а я с у р е п и ц а jBrassica campestris L. (abura-na)] отличается в Японии огромным полиморфизмом. Имеются пере- ходные формы сурепицы и к местной репе, и к китайским капу- стам — В. pekinensis Rupr. и В. chinensis L. и даже к В. nippo- sinica Bailey. Это разнообразие форм еще мало изучено бота- никами в Японии и особенно в Китае. В Китае местные сорта В. campestris используются двояко. При посеве весной их культивируют на масло, так как они дают обильный урожай семян. При посеве под осень они образуют корнеплоды и возделываются уже как овощное растение. Интересно, что именно в Восточной Азии распространены и многочисленные салатные сорта сурепицы и близких к ним видов китайской капусты и салатные горчицы, иногда несколько имитирующие по общему габитусу китайские капусты. М а с л и ч н а я к и т а й с к а я к а п у с т а (В. chinoleifera Ъ lehoever) очень близка к овощной китайской капусте(Тай-на)— В. с111пепз1з L. Р е д ь к а м а с л и ч н а я (gaphanus sativus var. oleiferus Metzg.) в настоящее время культивируется только в Китае. Воз- можно, занесена из,древнего Египта. Изменчивость ее не изучена. Некоторые формы редьки масличной имеют строение плодов, как у китайских редисов. Возможно, что редька масличная — это тонкокорневая форма редиса, в Китае — китайского, а в Европе — европейского про- исхождения. UeHTp развития рода тун г (Aleurites Forst.) в Юго-Восточ- ной Азии (Индокитай, Индонезия), но основные культурные виды приурочены к Китаю — А. fordii Hemsl. — и к Японии— А. cordata R. Br. дикорастущие формы распространены там же, где и культурные; следовательно, происхождение культурных форм аборигенное. Оба вида тунга отличаются большим поли- морфизмом. Виды к а м е л и и (Camellia sasanqua Thunb. и С. jaiponica L.) возделываются для получения парфюмерного масла. 245
Ма к (Papaver somniferum L.) хотя широко распространился в Китае только в историческое время, все же представлен там особой группой местных сортов опийного назначения, которая выделена в ssp. mongolicum М. Vesselov. и ssp. centro-asiaticum М. Vesselov. Кл е щ ев и н а к р у п н о п л о д н а я (gicinus macrocarpus G. Рор.) проникла из Африки, сначала распространилась в Индии, а оттуда в Китае и Японии. В Восточной Азии сформировался очаг скороспелых форм этого вида. Г. М. Попова выделяет здесь две агроэкологические группы форм: 1) японскую — proles japo- nicum (Япония); 2) китайскую — proles chinensis G. Pop. (Восточный, Северо-Восточный и Западный Китай, Дальний Восток СССР) . М о л о ч а й м а с л и ч н ы й (Euphorbia lathyris L.) встре- чается дико в горах на западе Китая. Растение длинного дня, вероятно, средиземноморского корня. Разводилось издревле в Японии и Китае; здесь, вероятно, первичный очаг его вхожде- ния в культуру. В Японии эта культура, по-видимому, исчезла. Необходима поисковая работа для отыскания новых растений с технически ценным маслом среди растений семейства молочай- ных в Восточной Азии. Р а м и fBoehmeria nivea (L.) Gaud.] растет дико в Японии и Восточном Китае и здесь же вошла в культуру. К а н а т н и к (Abutilon theophrastii Medic.) растетдико в СССР на Дальнем Востоке, в Японии, Северо-Восточном Китае, при- морской части Китая и I(opec. Здесь введен впервые в культуру. Т е м ед а (Themeda triandra var. japonica Makino) — злак, дает волокно. В Японии имеется особая разновидность, но во- обще вид более южного происхождения. Х л о п ч а т н и к (Gossypium L.) представлен в Восточной Азии эндемичными сортами, относящимися к виду G. агЬо- геит L. ss~p. nanling Mauer — Китай (Центральный, Северный и Северо-Восточный, Тайвань), юг Японии и Кореи. Родина этого вида — Южноазиатская область. Раньше в Китае, особенно в западной части, возделывался и G. herbaceum L. (гуза), занесенный из Средне-Юго-Западной Азии, но теперь эта культура становится реликтовой. По данным древних китайских источников, в Южном Китае на острове Хайнань хлопчатник возделывался до III — I в. до н. э., т. е. раньше завоевания этих областей китайцами. Широкое распространение культуры хлопчатника относится к Х — XIV вв. н. э., а на юге Японии — около VIII в. н. э. До развития хлопководства в Китае пользовались волокном конопли, рами и шелком. Ко н о п л я (C;annabis sativa L.) представлена в Японии (воз- можно, и в Китае) крупносемянными и высокорослыми сортами, выделенными в особые разновидности: var. japonica Serebr. и var. orientalis Serebr. 246
П р я д и л ь н а я п а л ь м а (Trachycarpus excelsus Makino), из волокна которой делаются веревки и канаты, имеет большое применение в Китае и на юге Японии. П у е р а р и я (Pueraria thunbergiana Benth.) растет дико в Японии и Восточном Китае, дает грубое волокно, из корней до- бывается крахмал. Р. edulis Thunb. употребляется в Китае в. пищу. С и т н и к п р я д и л ь н ы й (Juncus effusus чаг. decipiens ВисЬ.) называется «и» или «и-гуса», употребляется для изготов- ления циновок, имеются большие плантации в Японии. О с о к а п р я д и л ь н а я (Carex malae~censis Lam.) возделы- вается для той же цели на юге Японии (остров Кюсю). Б у м а ж н о е д е р е в о (Broussonetia papyrifera L'Her. и В. kasinoki Sieb.) растет дико и культивируется для изготовле- ния бумаги. K о р и а н д р (Coriandrum sativum L.) в Китае — не особенно старая культура, появившаяся в IV — Ч вв. н. э., но в своеобраз- ных условиях здесь создался особый экотип кориандра --мон- гольский (Е. А. Столетова, 1931). Анис 3 вез дч а тый (Illicium anisatum L., сем. Magnolia-- ceae) разводится в Китае и Японии как эфирномасличное. Г а р д е н и я (Gardenia florida L.) — эфирномасличное растение Китая; в Японии разводится также G. radicans Thunb. П р я ные р а стен ия (из рода Zanthoxylum L.) культиви- руются в Японии. Сюда относятся виды: Z. piperitum DC., Z. planispinum Sieb. et Zucc., Z. bungei Planish., Z. schinifolium Sieb. et Zucc. (Fagara schinifolia Engl.). Ш ел к о в и ц а (Morus L.) ведет происхождение из Восточ- ной Азии. Здесь в диком состоянии растет большинство ее видов, и здесь, вероятно, они впервые были введены в культуру. Пантропический род Morus L. имеет наиболее важный очаг культурных видов и форм в Восточноазиатской области. Основ- ные виды, давшие начало культурным сортам, — М. alba L., М. bombycis КоЫ7., М. multicaulis Perr., М. nigra L. Особо боль- шое значение имеют первые три. Наибольшее разнообразие сортов имеется в Китае и Японии. Отсюда культура шелковицы была перенесена в Средне-Юго-Западную Азию. Г у т т а п е р ч е в о е д е р е в о (Eucommia ulmoides Oliv.) про- исходит из Западного и Центрального Китая. Бе р ескл ет Ма а к а (Fuonymus maackii Ruipr.) — гуттапер- ченос, произрастающий на Яальнем Востоке СССР, в Японии и Корее. .Л а к о в о е д е р е в о (Rhus vernicifera DC. и другие виды,. в том числе R. succedanea L.) растет дико и культивируется в Японии и Восточном Китае. Восковое дерево, или з и м о ц в ет (Meratia praecox Rehd. et Wils.) — декоративное растение, родом из Восточной Азии, вероятно, из Японии. 247
Китайское сальное, или жирное, дерево (Sapium sebiferum (L.) goxb.] — в Китае и Индии дико и в культуре. Рас- пространено в Китае от острова Хайнань до Шэньси. Употреб- ляется для изготовления мыла и свечей. В Китае встречается около 10 видов растений этого рода, среди них есть эндемичные, но в целом вопросы происхождения видов и рода Sapium разра- ботаны еще недостаточно. М ы л ь н о е д е р е в о (Sapindus mukorossi Gaertn.) растет дико и в культуре. Восточная и Юго-Восточная Азия — родина многих к р а- сил ь ны х р а с те н и й: синюю краску дает Polygonum tincto- гшт Lour., красную — Strobilanthes flaccidifolius Nees, gubia cordifolia L., Lithosperrnum erythrorhizon Sieb. et Zuicc. Аконит (Aconitum wilsonii Stapf) разводится как лекарст- венное растение. Из лекарственных растений знаменит ж е н ь - ш е н ь (Рапах ginseng. С. А. Меу.), который растет дико в глухих девственных лесах на Дальнем Востоке в СССР и в Северо-Восточном Китае. Род Panax L., вероятно, происходит из Южной Азии. Ассортимент китайских и тибетских лекарственных растений чрезвычайно велик и нуждается в специальном изучении. Мята японская (Mentha сапайепз1з var. piperascens Briquet) — древняя культура. В Японии различают две разно- видности: Аомару и Акамару. Вероятно, те же разновидности культивируются и в Китае. К о п ы т е н ь в о с т о ч н ы й (Asarum sieboldii Miq.) — лекар- ственное растение. Называется диким жень-шенем, произрастает на Дальнем Востоке СССР и в Северо-Восточном Китае. Мел ия японская (Melia azedaraich L.) — лекарственное и декоративное (Южная Япония, Китай) растение. К а м ф а р н о е д е р е в о (Cinnamomum camphora Nees et ГЬегт.) растет дико и в культуре в Китае (Сычуань, Хубэй, Тайвань), Японии, Корее. К и т а й с к а я к о р и ц а [Cinnamomum icassia (L.) В1ите] растет дико и в культуре в Юго-Западном Китае. Ч а й (Thea sinensis L. sensu lato) — древнейшая культура 13 Китае. Вошел в культуру в Южном Китае и, возможно, неза- висимо от него в Индокитае. В горных районах Японии и Китая имеются наиболее холодостойкие формы. По В. И. Кречетовичу, в Восточной Азии произрастают сле- дующие культурные виды чая: 1) Thea ЬоЬеа L. (Япония, Ко- рея, смежное с ней побережье Китая); 2) Thea viridis L. (Юнь- нань, Юго-Восточный Китай); 3) Thea stricta Наупе (Средний Китай); 4) Thea cantoniensis Lour. (известняки Гуандуна). Рас- пространен последний вид также в другой геообласти — Южной Азии, где произрастает предполагаемый дикий прародитель пе- речисленных культурных видов — Thea mionticola V. Igrecz. (за- падный Юньнань и примыкающие районы Сычуани). 248
С а х а р н ы й т р о с т н и к (Saccharum sinense Roxb.) — энде- мичный для Юго-Западного Китая (Юньнань) вид сахарного тростника, растет дико и культивируется. Возможно, что он происходит из южноазиатской флористической области. Однако появление культурного тростника в Китае относится к 760— 750 гг. до н. э. В танскую эпоху (618 — 907 гг. н. э.) в Китае научились делать сахар. Своеобразную группу растений, растущих дико и культиви- руемых, составляют б а м б у к и (Phyllostachys Sieb. et Zuc~c., Arundinaria Michx., Bambusa Retz., Sasa Makino et Shibata и др.) . Употребляются они для изготовления мебели, циновок, разных поделок, водопроводных труб и т. д., а молодые ростки— в пищу. Распространены они вплоть до нашего Дальнего Во- стока и Сахалина, где сформировались северные травянистые виды, приспособленные к условиям сурового климата. Китай и Япония очень богаты овощными, вообще пищевыми растениями, среди которых имеются и эндемичные виды, под- виды, формы, сорта. Лук (А11шт L.) имеет в Китае и Японии первичный очаг развития следующих культурных видов: 1) лук китайский— А. ~chinense Don., цзюцао — особый вид многолетнего лука; 2) лук-татарка, или непахучий чеснок, — А. fistulosum L. (наи- более окультуренные сорта — в Японии); 3) лук крупнотычиноч- ный — А. maicrostemon I3ge., в Северном Китае; 4) лук пекин- ский — А. pekinense Prokh., вид, близкий к обыкновенному чес- ноку, возник в культуре (Корея и Япония) . Японская р едьк а, или да й кон [Raiphanus raphani- stroides (Makino) Sinsk.], представлена группой оригинальных аборигенных сортов — дайконов — в Японии, Корее и Восточ- ном Китае. Дайкопы составляют самостоятельный вид, энде- мичный для прибрежий Восточной Азии. Здесь же по берегам Тихого океана дико растет родоначальник культурного дай- кона. Таким образом, Восточная Азия — первичный очаг разви- тия редьки. Всего, включая и редис, четыре первичных очага: 1) Средиземноморьe и Европа; 2) Индия и Индонезия; 3) Япо- ния и Восточный Китай; 4) Северо-Восточный, Центральный 'и Западный Китай. Китайская группа р едисок и редек распростра- нена в Западном, Северном и Восточном Китае и в Монголии. Эту группу следует выделить в самостоятельный вид Raphanus sinensis Sinsk., ибо она имеет свой комплекс признаков, хотя по форме плодов носит промежуточные черты между европейским видом — Raphanus sativus — и японским — R. raphanistroides. Включает она три подгруппы: 1) монгольские, или восточно- китайские (лаба), белые и зеленоголовые редьки; 2) китайские редисы (Восточный и Центральный Китай; по вкусу менее острые, чем европейские); 3) розовые и красные китайские 249
.редьки и редисы (Западный Китай); к ним относится также дун- ганская редька. Восточная Азия является вторичным центром развития куль- турных салатных видов рода Brassi~ca L. диких родоначальни- ков здесь нет, произрастающие виды возникли в культуре от занесенных из Юго-Западной Азии культурных форм или сор- няков. Таким образом, это центр вторичный в том отношении, что родоначальниками не были аборигенные дикари, но виды возникли именно здесь, а не были занесены в виде какой-либо примитивной формы того же вида. Имеются в виду следующие виды: 1) В. pekinensis Rupr. 2) В. chinensis L.; 3) В. пйрроз1п1са Bailey; 4) В. dubiosa Bailey; 5) В. паг1поза Bailey. Эти виды связаны переходными формами между собой, с масличной капустой — В. tchinoleifera, сурепицей — В. сатре- stris (аЬигапа) и с японскими формами репы. Возможно, что исходными формами послужили репы, занесенные из Средне- Юго-Западной Азии. Имеются переходы от корнеплодных форм к тонкокорневым масличным и салатным видам Brassica. Листо- вая китайская однолетняя капуста — В. alboglabra Bailey— очень близка к В. о1егасеа. Происхождение не установлено. Встречается редко. P е п а (Brassica rapoasiatica за. jaipionica Sinsk.) пред- ставлена в Японии и в приморском Восточном Китае особой группой эндемичных сортов (особым подвидом), ведущей свое происхождение от реп Юго-Западной Азии (но по сравнению с исходными формами более мезофильных и нежных, окульту- ренных). Здесь имеется ряд переходных форм к масличной и салатной сурепице. M о р к о в ь (Daulcus sativus Zagor.) в Японии представлена двумя группами местных сортов: 1) кроваво-красной — ssp. japo- nica Zagor. 2) очень крупной (до 1 л~ длины) каротиновой морковью (остров Хоккайдо). Хрен я п о н с к и й (Wassabia japonica Matsum.) эндемичен для Японии. Ч истец к о р н е п л о д н ы й (Stachys sieboldii Miq.) энде- мичен для Японии (и, вероятно, приморского Китая) . Лопух к о р н е п л о д н ы й (La~ppa edulis Siebold ex Miq.) эндемичен для приморского очага Восточной Азии. Сп а р ж а к луб еньков а я (Asparagus 1исЫиз Lindl.) произрастает в Японии. Л и л и и со съедобными луковицами — Lilium tigrinur> Ker- Gawl., L. maximowiczii Rgl. — и другие виды. Из рода Ipomoea L. известны: Ipomoea aquatiica Forsl<. и 1. edu is Makino, а также батат. Б а т а т о б ы к н о в е н н ы й [Ipomoea batatas (L.) Poir.] широко возделывается в Китае, Японии и в других странах Восточной Азии. Имеется много эндемичных сортов, в том числе наиболее холодостойкие. Среди собственно японских сортов и 250
сортов с острова Тайвань имеются цветущие и дающие семена, что гораздо реже наблюдается у типичных американских сор- тов. Эти особенности заставляют некоторых исследователей культурной флоры подозревать политопное происхождение ба- тата: из Америки, Африки и Восточной Азии. Однако до сих пор все эти формы относят к одному виду, который должен иметь основным очагом происхождения одну какую-нибудь географи- ческую область, а таковой по всем данным является все же Америка. Для интродукции в наши субтропики особое значе- ние имеют восточноазиатские сорта батата. Из др угих овощных р а стен ий, произрастающих в Японии, назовем: Aralia cordata Thunb., Peucedanum jaiponi- сит Thunb., Actinostemma paniculatum Maxim. О г у р е ц (Cucumis sativus L.) представлен в Восточной Азии особыми группами эндемичных сортов: 1) японской— рг oles j aponi~cum 1 il. 2) южнокитайской — proles australi-ichi- nensis Fil.; 3) западнокитайской — iproles 1~азйдаг1сиз Fil.; 4) маньчжурской — ргоlез manshuricus Fil. Первая группа отнесена А. И. Филовым к индо-японскому подвиду — С. sativus ss~p. indo-japoni~cus Fil.; остальные три вхо- дят в подвид китайский — ssp. chinensis Fil. По данным А. И. Филова (1960), в Восточной Азии (Китае и Японии) имеется крупный очаг формообразования огурца. Здесь выработался особый подвид — ssp. chinensis Fil., который рас- пространился в страны Западной Европы, а затем и на террито- рию СССР. Д ы н я (Melo Adans.) представлена в Восточной Азии осо- быми группами сортов, которым К. И. Пангало (1955) дал сле- дующие видовые названия: 1) дыня китайская — М. chinensis Pang. (Китай, Япония); 2) дыня однополая — М. сопотоп Pang. (1(итай); 3) дыня гермафродитная — М. mono~clinus Pang. (Ки- тай, Монголия). Последние две группы сортов дыни раньше считались огур- цами. По-видимому, эти группы очень древние, заслуживающие детального изучения с точки зрения их филогении. А. И. Филов (1960) рассматривает историю и филогению бах- чевых с иной точки зрения. Он считает, что в Восточной Азии находится очаг формообразования китайских дынь, включаемых пм в подвид ssp. ciinensis Fil. Л ю ф ф а [Luffa cylindrica (L.) Poem.] используется как овощное растение и на мочалки. Основной областью распростра- нения считается Индия, но в Китае имеются, по-видимому, энде- мичные сорта. Т ы к в а (Cucurbita maxima Duch.), должно быть, очень давно попала в Китай, ибо там сформировалась оригинальная группа сортов с очень сплюснутой чалмовидной формой — ssp. turban (Spach) Fil., очаг этих эндемичных чалмовидных тыкв находится в Западном 1(итае. 251
8 1(итае чаше, по-видимому, распространена разновидность этого подвида с плодами средней величины — ча". feonstricta Alef., а также разновидность крупноплодная — var. тасго1иг- Lana Fil. Эти разновидности разводятся в Западном 1(итае и других местностях 1(итая, встречаются они и в Японии, где чаще культивируются мелкоплодные сорта. В Японии разводится также красная мелкоголовая — var. microcina Alef.— и зеленые мелкоголовые — var. appetit Naud.— декоративные тыквы, относящиеся к китайскому подвиду— С. maixima звр. turban (Spach) Fil. Тыквы этого подвида, ве- роятно, занесены в Японию из 1(итая. В равнинных районах 1(итая распространены эндемичные .сорта С. moschata Dutch. с очень тонкими высокосахаристыми плодами удлиненной формы, малосемянные, высококультурного типа. В Японии также имеется группа эндемичных сортов му- с к а т н о й т ы к в ы — С. moschata. Эти сорта мелкоплодные с деревянистой корой и глубоко сегментированной поверхностью. 1(огда и каким путем попала тыква в Японию, неизвестно. Здесь она представлена эндемичными сортами. А. И. Филов объединяет их в особый японский подвид — ssp. japonica (Zhi- ten.) Fil., который в меньшей степени распространен в прибреж- ных тихоокеанских районах Китая. Этот подвид довольно поли- морфен и содержит несколько разновидностей: 1) японская моз- говая — var. abbreviata Alef. 2) катушечная мозговал — var. trochlearis Fil.; 3) канталупообразная — var. melanaeformis Ва)1еу; 4) карликовая — var. ipygmaea Fil. По мнению А. И. Филова (1960), стол о в ы й а р б уз попал в 1(итай не с юга, а вероятно, с северо-запада, сначала он рас- пространился в Синьцзян-Уйгурском автономном районе, кото- рый включается нами в Средне-Юго-Западную подобласть куль- турной флоры. Первое упоминание об арбузе относится к 947 г. н. э., когда китаец Ни Kiao путешествовал по стране I(itan. )Кители 1(итака заимствовали арбуз у уйгуров. Виды х р и з а н т е м (Chrysanthemum coronarium L., Ch. morifolium Ram. var. sinense Mal<ino) используются как пище- вые растения (для салатов) . Ш и с о (виды Perilla L., идущие как овощные — P ocy- moides L., и отчасти P nanl<inensis Lour.). P. ocym~oides в Япо- нии имеет форму с темно-красными листьями, которые прибав- .ляются в маринады для придания плодам и овощам красивой окраски. В Восточной Азии (Северо-Восточный 1(итай, Дальний Во- сток CCCP) распространен съедобный вид ф и 3 а л и с а (Physa- lis glabripes Pojark.) .. P е в е н ь д л а н е в и д н ы й (Rheum palmatum L.) — овощное и лекарственное растение (Япония, К»тай, Сибирь). П о д б е л я п о н с к и й (Petasites 1aponicus F S~chmidt)— овощное растение (Япония). 252
Бубенчик к л у 6 н е н о с н ы й (Adenophora latifolia Fisch., А. vertioillata Fisch.) — овощное растение (Япония) . Камыш клубненосный, или водяной каштан (Eleocharis tuberosa S~chult.), распространен в Японии и Китае. Стр ел о лист кл у 6нено сны й (Sagittaria sagittifolia var. sinensis Makino) распространен в Японии. Il è ц а н и я ш ир о ко листвени а я (Zizania latifolia Tu- rcz.) родственна американскому водяному рису. Произрастает в Японии, Северо-Восточном Китае, на Дальнем Востоке в СССР В пищу употребляются стебли и влагалища листьев, пораженные головней. В о д я н ы е о р е х и (Trapa bicornis L., Т. bigpinosa Roxb.) и другие в о д я н ы е р а с т е н и я (Elatostema шпЬе11а1ит Blume var. convolucratum Makino) — овощные растения Японии. Я м с ы к и т а й с к и е (Dioscorea batatas Decne, D. japonica Thunb.) — корнеклубнеплоды. Вторичной областью развития яв- ляется Восточная Азия (происхождение рода — африканское). В 1(итае, особенно в южных провинциях, распространено клубнеплодное растение т а р о [Colocasia antiquorum (L.) Schottj, ~родина которого Малезия, но в Китае создались энде- мичные сорта, большей частью принадлежащие, по-видимому, к эндемичной для Южного Китая разновидности — var. globu- lifera Engl. et Krause. 1(о н ь я к (Amorphophallus konjak С. I(och) — клубненосное растение в Японии, 1(итае, 1(opec. Б а к л а ж а н (Solanum melongena L.), по А. И. Филову (1958), представлен в Восточной Азии особыми агроэкологиче- скими группами местных сортов: 1) японской скороспелой, 2) се- верокитайской. На юге Японии мы встречали также чрезвычайно крупно- ллодные и поздние сорта баклажана, занесенные сюда из Юж- ной Азии. Восточноазиатская флористическая область является пер- -вичной областью развития очень многих древесных лиственных пород, которые затем расселялись на запад. В этой области на- ходятся первичные очаги образования родов многих плодовых .деревьев. Г р у ш а (Pirus L.) имеет первичный очаг происхождения и развития в Восточноазиатской области. Здесь сосредоточено большое богатство видов этого рода, еще слабо изученных. bo- таники насчитывают здесь 15 — 20 эндемичных видов: Pirus ussuriensis Maxim., Р. ovoidea Rehd., Р serotina Rehd., Р. pashia Buich.-Íàò., Р. betulaefolia Bge., Р. aromatica Kikuchi et Nakai и многие другие. Наибольшее практическое значение для нас имеет груша уссурийская — Р. ussuriensis Maxim., распространенная дико и в культуре в Японии, Корее, Китае и у нас на Дальнем Востоке. Этот вид груши родствен груше обыкновенной европейской— 253
Р communis L. При расселении на запад уссурийская груш"- (или ее ближайший предок) дала начало европейской и очень. близкой к последней — кавказской — Р. cau~casi~ca А. Fed.— груше. Переднеазиатский вид — Р. boissieriana Buhse — обнару- живает родство с гималайской грушей — Р. pashia. Близки к ки- тайским предкам и некоторые другие переднеазиатские и средне- азиатские виды. Я б л о н я (Malus Mill.) также представлена в Восточной Азии разнообразием эндемичных видов, имеющих интерес для селекции: 1) М. baccata (L.) Borkh., культурный вид (Япония,. Китай); 2) М. manshurica (Maxim.) Кот., возможно, подвид пре- дыдущего вида, дико в Приморском крае СССР и Северо-Восточ- ном Китае; 3) М. halliana Koehne, в культуре в Сычуани (Китай); 4) М. floribunda Sieb., в культуре (Япония); 5) М. ringo Sieb.„ в культуре и дико (Япония); 6) М. formosana Kawak. et Koidz., дико (на острове Тайвань); 7) М. hupehensis Pehder, дико в Ки- тае; 8) ряд других слабо изученных видов. Согласно иссле- дованиям Юй Дэ-цзюнь, Ян Чжень-лун (1955), в Китае насчитывается около 20 видов яблони. Наибольшее количество видов имеется в провинциях Шэньси и Сычуань. Malus и Pirus — роды близкородственные, имеют, вероятно, одного общего предка и оба распространились в Средне-Юго- Западную Азию и Европу из Восточноазиатской области. Б о я р ы ш н и к (Crataegus L.) представляет собой также эле- мент реликтовой восточноазиатской флоры, где имеется многа эндемичных видов. Некоторые из них представляют интерес в ка- честве подвоя и для селекции. Р я б и н а (Sorbus L.), подобно видам Pirus, Crataegus, Ma- lus и многим другим, на запад распространялась из лесов Во- сточной Азии, где имеется много дикорастущих видов со съедоб- ными плодами, которые представляют известный интерес для селекции, особенно северные виды: 1) рябина бузинолистная— S. sambucifiolia Poem. (Дальний Восток); 2) рябина сибир- ская — S. sibirica Hedl. (Сибирь, Монголия); 3) рябина амур- ская — S. amurensis Koehne (Дальний Восток СССР, Япония, Ко- рея, Северо-Восточный Китай); 4) рябина смешанная — S. сот- mixta Hedl. (Дальний Восток СССР, Япония, Китай); 5) рябина анадырская — S. anadyrensis Кот. (Анадырь); 6) рябина кам- чатская — S. kamtsichatcensis Кот. (Дальний Восток СССР). Мушмул а в о сто ч н а я (Eriobotvya Lindl.) имеет первич- ной областью происхождения рода Восточную Азию, где пред- ставлена 10 — 12 видами. Мушмул а японская — би в а (Е. japonica Lindl.) возде- лывается в Японии, Корее, Китае. Хеномелес (Chaenomeles Lindl.) имеет в целом восточ- ноазиатское происхождение. Наибольшее значение имеют. 1) айва японская — Ch. japonica Lindl., дико и в культуре (Япо-
ния, Корея, Восточный Китай); 2) айва китайская — Ch. sinen- sis l<oehne, дико, изредка разводится (Китай). Ун а б и (Zizyphus vulgaris Lam.) широко распространен и :за пределами Восточной Азии. Род Zizyphus Mill., по-видимому, Босточноази атского происхождения. Вопрос о происхождении унаби спорный. Одни ботаники- географы культурных растений придерживаются той точки зре- ния, что родина унаби — Восточная Азия (Китай и Индия), а .другие считают первичным очагом его распространения Среди- земноморье. Род Prunus L. (sensu lato) подвергся разукрупнению. Теперь большинство ботаников рассматривает как отдельные роды аб- рикосы — Аггпе111аса Mill., черешни и вишни — Cerasus Juss., персики — Persica Mill., миндали — Amygda1 us L. Корни всех этих родов надо искать в реликтовой флоре Восточноазиатской области. Здесь встречаются интегральные типы с признаками различных родов Агтеп1аса holosericea (Batal.) Kost. и др. Таким образом, эта группа близких родов, взятых в целом, имеет главным первичным очагом Восточную Азию, второй пер- вичный очаг находится в Америке (главным образом, сливы и сливовишни), третий — в Передней Ами, где меньше дикорас- тущих видов, но откуда произошли важнейшиекультурные виды, а именно алыча, слива домашняя, вишня обыкновенная, че- решня. Род Amygdalus имеет область наибольшего современного развития в Передней и Средне-1Ого-Западной Азии. Собственно сливы — Prunus Mill. — представлены в Восточ- ной Азии не очень большим количеством видов. Из них имеют экономическое значение следующие: 1) Р. ussuriensis Коч. et Post., в культуре в Восточной Сибири и Северо-Восточном Ки- тае; 2) Р. simonii Carr., культивируется в Японии и Китае; 3) Р. заИс1па Lindl., дико и в культуре в Северном Китае. В и ш н я и ч е р е ш н я (Cerasus Juss.) не только происходят из Восточной Азии, но и в наше время представлены здесь ог- ромным количеством видов, из которых очень много эндемич- ных. Только в двух китайских провинциях Хубэй и Сычуань бо- таники насчитывают около 50 эндемичных видов Cerasus. Япо- ния имеет около 15 эндемичных видов. Японские виды являются в большинстве своем декоративными (знаменитые японские вишни с махровыми цветками), а не плодовыми растениями. В Гималаях и Тибете меньше эндемичных видов, однако эти области недостаточно изучены. Из восточноазиатских видов Cerasus отметим следующие: 1) С. tomentosa Wall. — холодо- стойкий вид с огромным ареалом — от Тихого океана до гра- ницы со Средней Азией; 2) С. jaiponi~ca Lois. (Япония); 3) С. glandulosa (Thunb.) Lois. (Корея, Китай); 4) С. maximo- wiczii (Puipr.) Кот., дико, употребляется как подвой (Дальний Восток СССР, Япония, Китай); 5) С. sachalinensis Кo1n. et Alviss., 255
дико (Приморский край, о. Сахалин); 6) С. glandulifolia Kom., дико (Приморский край). Род а 6 р и к о с (Armeniaca Mill.) сформировался в Китае. Абрикос обыкновенный — А. vulgaris Lam. — произрастает дико от северо-восточных до западных границ Китая и отсюда распространился в Среднюю Азию и далее на запад. Культурный китайский абрикос — А. апзи (Кот.) Kost.— распространен в Северо-Восточном Китае, Корее и Японии. Ди- кий родоначальник этого вида неизвестен. Муме — А. тите Sieb. — культивируется в Японии и Китае (больше декоративное, чем плодовое). На Дальнем Востоке СССР и в Северо-Восточном Китае культивируются и растут дико наиболее холодостойкие виды аб- рикосов: абрикос сибирский — А. sibirioa (L.) Lam., абрикос маньчжурский — А. manshuriea (Koehne) Skvortz. и абрикос Да- видиана — А. davidiana Carr. Все эти виды в той или иной сте- пени принимали участие в формировании культурных сортов. Дикорастущий вид — А. апота1а K!ost. — распространен в провинции Гуандун и соседних районах. Наиболее древним видом по своей организации является растущий в Восточном Тибете А. holoseriicea (Batal.) Kost. Этот интереснейший вид по своим признакам связывает род Prunus (слива) с родом Агте- nia~ca. Древним реликтовым видом К. Ф. Костина (1947) считает и А. тите, «ареал которого находится в самой южной части об- щего ареала рода и ближе всего подходит к центральнокитай- скому ареалу, наиболее богатому разнообразием видов и форм других родов подсемейства Prunoideae (Cerasus, Persiica)». Персик об ыкновенный (Persica vulgaris МШ.)— имеются указания на существование в диком состоянии в Китае, но это еще не установлено с полной достоверностью. Большин- ство авторов считает, что персик обыкновенный происходит из Китая, как и следующие виды: Р. davidiana Carr., дико на Се- верном Кавказе; Р. kansuensis Коч. et Kost., дико в Китае; Р. mira (Koehne) К~оч. et Kost., Р. potanini (Batalin) Kost. М и н д а л ь (Amygdalus L.) представлен в Восточной Азии небольшим числом видов, хотя зарождение его и произошло, вероятно, здесь, ибо именно в Восточной Азии зародилось вообще Prun~oideae и здесь существуют переходные формы между отдельными их родами. Область современного развития Amygdalus — Передняя и Средне-Юго-Западная Азия. Восточно- азиатские виды не имеют отношения к происхождению культур- ного миндаля — А. communis L. Г о в е н и я (Hovenia dulcis Thunb.) — в пищу употребляют сладкие плодоножки (Китай, Япония). Я п о н с к а я х у р м а (Dios~pynos kaki L.) широко распро- странена в культуре в Японии, Корее и Восточном Китае, где представлена многими местными и селекционными сортами. Происхождение этого плодового растения из Восточной Азии 256
несомненно, хотя весь род Diosipyros L. имеет очень большое число видов в Южной Азии, а также в Африке и Австралии— в тропических районах. В и н о г р а д (Vitis L.) растет дико в песках Восточной Азии. Из его видов отметим: ) виноград амурский — V. arnuren- sis Карг., дико и разводится (Дальний Восток СССР, Северо- Восточный Китай); 2) В. Тунберга — V thunbergii Sieb. et Zu~cc., декоративное (Дальний Восток СССР, Япония, Китай). Лох (Elaeagnus L.) растет в Восточной Азии (Япония, Ки- тай) дико и культивируется. В Японии наиболее известны сле- дующие виды: Е. multiflora var. hortensis Мах., Е. umbellata Thunb., Е. pungens Thunb. Род Elaeagnus имеет восточноазиат- ское происхождение. О б л е п и х а (Н1ррорйаё L.) — восточноазиатский род. Недавно описана новая засухоустойчивая форма облепихи обык- новенной — Н. rhamnoides ss~p. thibetiana Serv. Растет в Тибете на высоте 3000 — 5000 м над ур. м. К а шта н (Castanea Mill.) — растут дико и культивируются следующие виды: 1) С. crenata Sieb. et Zucc. (Япония, Корея); 2) С. mollissima Вl. (Восточный и Южный Китай). Другие виды также используются в диком состоянии. Весь род Castanea ве- дет происхождение из Восточной Азии и слабо там изучен. О р е х г р е ц к и й (Juglans L.) также происходит из Восточ- ной Азии. Культивируются: 1) J. manshurica Maxim:. (Северо- Восточный Китай, Дальний Восток СССР); 2) J. sinensis Dode (Япония); 3) J. sieboldiana Maxim. (Япония); 4) J. cordiformis Maxim. (Япония). Л ещи н а (Corylus L.) также происходит из Восточной Азии. К азиатским видам относятся: 1) С. heterophylla Fischer (Даурия, Корея, Япония, Восточный, Северо-Восточный и Цен- тральный Китаи), этот вид очень близок к ореху обыкновенному С. аче11апа L. 2) С. manshurica Мал1т. (СССР — Амурская об- ласть, Приморский край, Япония, Корея, Китай); 3) С. siebol- diana В1ите (Япония); 4) С. ferax Wall. (Центральный и Юж- ный Китай); 5) С. brevituba Кот. (Дальний Восток СССР, ареал недостаточно выяснен). О р ех м едв еж и й (С. ~colurna L.) имеет огромный ареал- от Балкан до Южного Китая и, по мнению некоторых авторов, происходит из Китая, а позднее распространился далеко на запад. Истоки всего рода Corylus находятся в Восточной Азии. Ф иста ш к а (Pistacia L.) имеет истоки в реликтовой (тур- гайской) восточноазиатской флоре. Близкий вид к средиземно- морской Р. lentiscus L., распространен в Китае (Юньнань, Сы- чуань, имеются представители этого рода на Тайване — Р. for- mosana Matsum.) . Очаг вторичного видообразования — Иран и Малая Азия, откуда вышла и фисташка настоящая— Р. vera L. Синская Е. Н. 257
К а р и я (Сагуа Nutt.) происходит из Америки, но в Китае и прилегающих странах распространен китайский пекан — С. cat- hayensis Sarg. Т о р р е я (Тоггеуа grandis Fort.) — Восточная Азия. М а л и н а и е ж е в и к а (Rubus L.) имеют одну из первич- ных областей развития в Восточной и Юго-Восточной Азии (вто- рая область первичного значения в Америке) и имеют в данное время большое хозяйственное значение. Из хозяйственно ценных видов отметим следующие: 1) R. phoenicolasius Maxim.— япон- ская «винная ягода» (Япония и Западный Китай), в культуре в Америке и Англии; 2) R. illecebrosus Foclce — земляничная малина (Япония), имеет значение для селекции; 3) R. innomina- tus Moore (Центральный и Западный Китай), участвовала в происхождении некоторых культурных сортов; 4) R. kuntzea- nus Hemsley (Центральный и Западный Китай), дико и введена в культуру. С м ар один а (Ribes L.) и кр ыж о в ни к (Grossularia Mill.) ведут свое происхождение из Восточной Азии, представ- лены там большим количеством видов, которые изучены больше в пределах Советского Союза и в Северо-Восточном Китае. А к т и н и д и я (Actinidia Lindl.) распространена в странах Восточной Азии. Для нас наиболее важны: 1) А. 1~olomicta Ma- xim. — «изюм», дико в Северном Китае, Корее, Японии, в СССР в Приморском крае, на Сахалине, Курильских островах; 2) А. ar- guta (Sieb. et Zucc.) Planch., дико (Северный Китай, Корея, Япония, СССР — Приморский край, Сахалин, Курильские ост- рова); 3) А. giraldii Diels. дико (тот же ареал); 4) А. ~polygama (Sieb. et ZDac.) Maxim., дико (тот же ареал); 5) А. chinensis Planch., дико и в культуре (Западный и Центральный Китай). Л и м о н н и к (Schizandra ichinensis Ва111.) — плодовое и ле- карственное растение (Дальний Восток СССР, Япония, Северный Китай) . Весь род восточноазиатский. С м и л а к с (Smilax china L.) — ценное лекарственное расте- ние, родом из Восточной Азии, занесен в Южную Азию (Япония, Китай, Корея) . М и р и к а, или н а г и (Myrica гиЬга Sieb. et Zuoc.) — плодо- вое дерево, дико и разводится (Япония и Китай). Г и н к г о (Ginkgo biloba L.) — реликтовое, некогда очень широко распространенное растение. Беднейшее население Япо- нии и Китая употребляет в пищу «ягоды» (семена с покровом) гинкго. В настоящее время встречается большей частью в куль- туре (часто около старинных храмов в Японии), но в последнее время найдено и в диком состоянии. Некоторые виды цитрусовых и многие культурные сорта тех же видов, которые вышли из стран Южной Азии, сформирова- лись в Восточноазиатской области развития культурной флоры. А пел ь сии сл ад к и й [Citrus sinensis (L.) Osbeck] вопреки видовому названию, вероятно, происходит из Восточной Бирмы,
Вьетнама (Бакбо и Намбо), но он распространен и в Китае (Юньнань). В давние времена его занесли в Средиземноморье. Ма н да р ин [C. reticulata Blanko (С. n~obilis Andrew., С. de- lieiosa Ten.)], вероятно, произошел из Юго-Восточного Китая. Морозостойкие сорта сосредоточены в Японии и Восточном ICu- тае. П о м п е л ь м у с, или ш е дд о к [C. grandis Osbeck (С. ma- xima Merrill)], по предположеншо одних авторов, происходит из Юго-Восточного Китая, других, — из Индонезии и Малайзии. Больше данных имеется в пользу южноазиатского происхожде- ния. В Восточной Азии, во всяком случае, культивируются более хозяйственно ценные (холодостойкие) эндемичные сорта. Лимон кантонский, или китайский, белый и кр а сиый лимоны (С. limonelloides Нау.) — Южный Китай (Гуандун). П а п ед а «ичанг» (С, ichangensis Swingle) — Центральный и Западныи Китай. Родина — провинции Хубэй, Сычуань, Гуан- чжоу. Самый холодостойкий из видов Citrus; В Восточной Азии произрастают только два дикорастущих вида из рода Citrus: С. 1асЫЬапа Тап. (юг Японии, Китай— Тайвань) и С. depressa Hayata (там же). Оба вида употреб- ЛЯ EOTCH ДЛ Я ПОДВОЯ. Таким образом, мы считаем основной областью первичного видообразования цитрусовых Южную, а не Восточную Азшо. Однако в Восточной Азии, несомненно, создалось большое сор- товое многообразие вследствие давности возделывания и широ- кого распространения в культуре. Из других померанцевых Восточноазиатской области разви- тия культурной флоры свойственны следующие: п о н ц и р у c— Powirus trifoliata (L.) Paf. — употребляется для подвоя, родом из Центрального и Северо-Западного Китая, монотипный род. К инка н — Fortunella Swingle, весь род происходит из Во- сточной Азии; F. margarita (1.оиг.) Swingle, в культуре (Юго- Восточный Китай); F. japonica (Thunb.) Swingle, дико и в куль- туре (Юго-Восточный Китай); F. hindsii (Cham~p.) Swingle, дико (Юго-Восточный Китай) . 1~итрусовые и другие померанцевые Китая, особенно южных провинций, еще ждут своего детального обследования и изуче- ния в культуре. В а м п и к и т а й с к и й (Clausena lansium Skeels) происхо- дит из Южного Китая, где разводится ради плодов. Употреб- ляется как подвой для цитрусовых. Л и ч и (I itchi Ichinensis Sonn.) разводится из-за вкусных пло- дов в Южной Японии и Китае. Родиной считается Южный Китай. Занесен в Южную Азию и Америку. Китай — родина ряда декоративных растений. По Н. А. Ба- зилевской ( 960), Китай и Япония дали 261 вид де ко р а т и в- н ы х р а с т е н и й. В Китае находится родина: пеона — Paeonia 9+
chinensis Vilm., P. albiflora Pall., астры — Callistephus 'chinensis (L.) Nees, глицинии — Wistaria chinensis DC., спаржи — Аврага- gus verticillatus L., астильбы — Astilbe davidii Henry, вьюнка— Calystegia chinensis Bailey, лилии — Lilium regale Wilson, ал- теи — Althaea rosea (L.) Cav., Diclytra spectabilis DC., садовой астры — Callistpphus з1пепз~з Bergmans — введена в культуру в Китае. Особой популярностью в Японии пользуются хризантемы (Chrysanthemum L.). Масса любителей занимается там выведе- нием новых декоративных сортов этого растения, разнообразие которых огромно. Восточная Азия — родина декоративных хри- зантем и астр. Восточноазиатская флористическая область является вели- чайшей ареной видообразования и формообразования не только дикорастущих, но и очень многих культурных растений. Восточ- ная Азия является родиной большинства наших лиственных по- род, которые распространились в Европу и в Северную Америку. Следующие роды плодовых растений всецело сформирова- лцсь в Восточноазиатской области: мушмула азиатская — Eri- obotrya, айва азиатская — C}>aenomeles, абрикос — Armeniaca, персик — Persica, кинкан — Fortunella, понцирус — Poncirus, ак- тинидия — AIctinidia, лимонник — Slchizandra. Здесь находится колыбель развития многих родов, и отсюда они распространились и в Переднюю Азию и в другие области Евразии, где в ряде случаев получили импульсы ко вторичному расцвету видообразования и формообразования. Из таких родов отметим следующие плодовые и орехоплодные: слива — Prunus, вишня и черешня — Cerasus, миндаль — Amygdalus. Восточная Азия является, таким образом, областью проис- хождения всего подсемейства РгипоЫеае, а также и других ро- дов: яблоня — Malus, груша — Pirus, рябина — S~~orbus, боярыш- ник — Crataegus, смородина — Jibes, крыжовник — Grossularia, унаби — Zizyphus, грецкий орех — Juglans, лещина — Corylus, фисташка — Pistacia, каштан — Castanea. Восточная Азия является родиной многих родов, представи- тели которых используются в качестве технических, лекарствен- ных и декоративных растений и т. д., например: дуб — Quericus, бук — Fa gus, бересклет — Euonymus, камфорное дерево — Cin- патотит, криптомерия — Cryptomeria, адамово дерево — Раи1о- wnia, сирень — Syringa, сосна — Pinus. Из родов, включающих в свой состав масличные растения, Восточная Азия является родиной периллы — Perilla и, ве- роятно, шандры — Elshol tzi a. Некоторые роды кормовых растений имеют свои истоки в Восточной Азии. Особенно заслуживают изучения следующие: леспедеза — Les~pedeza, куммеровия — gummerovia, вика — Vi- cia, чина — Lathyrus, ежа — Dactylis (предполагают, что этот 260
монотипный род также имеет свои истоки в лесах Юго-Восточ- ной Азии), регнерия — Roegneria. Многие роды, содержащие травянистые лекарственные расте- ния, ведут происхождение из Восточной Азии: аконит — Aconi- tum, ландыш — Convallaria. Многие роды из Восточной Азии являются источниками деко- ративных растений (лианы и травы), например: гортензия— Hydrangea, хризантема — Chrysanthemum, глициния — Wista- г1а. Шелковица — Могиз, по всей вероятности, имеет восточно- азиатское происхождение, водяной орех — Trapa — также ведет происхождение из Восточной Азии. Перечень таких родов отнюдь не является исчерпывающим. Здесь мы не приводим списка культурных и используемых в дикорастущем состоянии видов из тех родов, родиной которых является Восточная Азия. Многие культурные виды, относящиеся к родам, ро- дина которых находится вне Восточной Азии или еще с точ- ностью не установлена, имеют основную область происхождения и развития в Восточной Азии. Из них укажем следующие: просо — Panicum miliaiceum, пайза — Echinioichloa 1гитеп1асеа и чу- мнза — Setaria Йа1йса (первичный очаг), гаолян — Sorghum chi- nense (почти эндем, вторичный очаг), соя — Glyicine hislpida и фасоль адзуки — Phaseolus angularis (первнчный очаг), ячмень азиатский — Hordeum humile (эндем, первичный очаг), редька японская — Raphanus raphanistroides, редька китайская и ре- днс — R. sinensis (эндем, первичный очаг), хурма китайская— Diospyros kaki (первичный очаг), жень-шень — Panax ginseng, (первичный очаг), коньяк — ' АтогрМрйа11из 1<onjak (эндем), Brassica alboglabra (эндем, возник в культуре), виды В. chinen- sis и В. pekinensis (почти эндемы, возникли в культуре), виды В. narinosa, В. dubiosa, В. nipposinica (эндемы, возникли в куль- туре), горчица корнеплодная — В. na~piformis (эндем), горчица сарептская — В. juncea (первичный очаг), астрагал китайский— Astragalus sinicus (эндем), звездчатый анис — Illicium anisatum (эндем), гуттаперчевое дерево — Еисотт1а ulmoides (первичный очаг), виды луков: китайский — Аllшт chinense, пекинский— A. pekinense, крупнотычинковый — А. macrostemon (эндемы) . Эндемичны следуюшие виды: хрен японский — Wassabia ja~po- nica, чистец корнеплодный — Sta~chys sieboldii, лопух корнеплод- ный — Laip~pa edulis, клубневая спаржа — Asparagus lucidus, удо — Aralia ~cordata, горичник — Peucedanum 1a~ponicum, Acti- nostemma panicula turn, Chrysanthemum ~согопаг1ит, Ch. morifo- lium, дыня китайская — Меlо chinensis, дыня однополая — М. со- потоп, дыня гермафродитная — М. monoclinus, подбел япон- ский — Petasites japonic~ls, бубенчик широколистный — Adeno- phora latifolia, камыш клубненосный — Eleocharis tuberosa, стрелолист клубненосный — Sagittaria sagittifolia var. sinensis, Ipomoea aquatica, Elatostema umbellatum; говения — Hovenia 261
dulcis и карня — Carya cathayensis (почти эндемы), торрея— Тоггеуа grandis (эндем?), ситник прядильный — Juncus effusus var. decipiens и осока прядильная — Carex malaecensis (эн- демы); молочай масличный — Euphorbia lathyris (почти эндем, первичный очаг), цицания широколистная — Zizania latifolia (эндем), пальма прядильная — Trachycarpus ехсе1зиз и рами— Boehmeria пиеа (первичные очаги); канатник — Abutilon theophra- sti (основной первичный очаг); китайский сахарный тростник— Saccharum sinense (эндем); литчи — Litchi сЫпепз1з (первич- ный очаг), китайский вампи — Clausena lansium (первичный очаг, почти эндем); лобия японская — Dolichos hassjoo и м»- рика — Myrica rubra (эндемы); смилакс — Smilax china, тунг китайский — Aleurites fordii, тунг японский — А. cordata, пе- рилла — Perilla ocymoides (первичные очаги), P. frutescens (эндем, первичный очаг), мелия японская — Melia azedarach (первичный очаг), виноград амурский — Vitis amurensis (эндем), шелковица — Магия alba (основной первичный очаг); свиной салат — Pistia stratiotes (эндем). Список этот может быть значительно расширен. Одних энде- мичных видов из рода Cerasus в Восточной Азии имеется около 80, и все они так или иначе могут использоваться, хотя введенных в культуру не так много, а возможности применения их в селекции совсем не изучены. 1~роме того, имеется много мелких эндемичных культур второстепенного значения, которые не учтены нами. Тем не менее, и этот перечень дает ясное пред- ставление о богатстве культурной флоры Восточной Азии, об очень большом количестве распространившихся оттуда и энде- мичных для Восточной Азии культурных растений. Многие ценные широко распространенные культуры издревле были занесены в Восточную Азию, и здесь, в новых для них своеобразных условиях муссонного климата, под многовековым воздействием местной своеобразной интенсивной агротехники выработались новые группы форм, новые местные сорта. Из та- ких культур в особенности следует отметить следующие: 1) го- лозерные овсы, которые некоторыми ботаниками выделены в са- мостоятельный вид — Avena nuda L. (основной очаг их не выяс- нен); 2) особая группа (подвид) восточноазиатских реп (из Средне-Юго-Западной Азии); 3) особый подвид восточноазиат- ской флавоновой моркови (из Средне-Юго-Западной Азии); 4) особая группа (подвид) восточноазиатского хлопчатника (пз Индии); 5) особые агроэкологические группы крупноплодной кле- щевины (из Африки, вероятно, проникнувшие через Индию); 6) группа эндемичных разновидностей масличной сурепицы- Вгазз1са campestris (из Средне-Юго-Западной Азии); 7) особые, очень ценные ранние экотипы кунжута (из Индии); 8) самые раннеспелые сорта риса; 9) большое количество ценных местных сортов пшеницы (из Передней Азии); 10) большое количество местных сортов цитрусовых (которые в основном вышли из 262
Южной Азии); 11) особые агроэкологические группы сортов ба- клажана (из Индии); 12) эндемичные сорта огурца (из Индии). 1~ультурная флора Восточноазиатской области имеет свой собственный, особый характер и мало общего с культурной фло- рой Средне-Юго-Западной и особенно Передней Азии. Здесь сов- сем другой состав эндемичных растений, их гораздо больше, и они разнообразнее; другой состав родов, которые дали культур- ные астения. ревних культур, «заходящих» из других областей, не так много, как в двух рассмотренных выше подобластях (Передней и Средне-Юго-Западной Азии) Древнесредиземноморской 06- ласти. Роль Передней Азии как источника занесенных растений здесь сравнительно невелика, если вспомнить о том огромном влиянии, какое оказала культурная флора Передней Азии на флору Средне-Юго-Западной Азии, и взаимном влиянии, взаим- ных связях этих двух флор. Из Передней Азии Восточная Азия получила только пшеницу, горох, но, вероятно, не прямо, а через посредство Средне-Юго- Западной Азии. Из Средне-Юго-Западной Азии получены лю- церна, виноград, хлопчатник (гуза), репа, морковь (флавоно- вая). Влияние восточноазиатской культурной флоры на состав культурной растительности Передней и Средне-Юго-Западной Азии (обратная связь) очень велико. Это влияние двоякого порядка. Очень велико влияние не прямое, а через дикорасту- щую флору. Современная флора Древнесредиземноморской флористиче- ской области в очень большой мере сложилась на базе распро- странившихся из Восточной Азии элементов древней флоры ши- роколиственных лесов умеренного климата. Там надо искать корни представителей культурной флоры, относящихся к родам Ма1из, Pirus, Prunus, Vicia и многих других. Однако и прямое влияние — через распространение навыков земледелия и первичного набора культур — очень большое и древнейшее. Вспомним просо, которое, несомненно, пришло из Азии еще в каменном веке. В более позднее время наша страна получила из Восточной Азии сорта риса, чая, многих цитрусо- вых, хурмы, японской мушмулы, сои, кунжута, периллы, очень много декоративных растений. Очень важны связи Восточноазиатской области с Южной Азией: Индией, Индокитаем, Малезией. Тропическая раститель- ность постепенно переходит здесь в субтропическую и раститель- ность умеренных широт, и часто бывает трудно определить здесь границы Восточной и Южной Азии по отношению к ареалам ро- дов п видов дикорастущих и культурных растений. 1з Южной Азии Восточная Азия получила рис, кунжут, хлопчатник, многие цитрусовые, а также чай, баклажаны, огурцы, бананы и другие растения. 263
Короче говоря, Восточная Азия — это огромная самостоятель- ная географическая область культурной флоры. Наоборот, IIe- редняя и Средне-Юго-Западная Азия — это крупные подобласти единой и своеобразной области создания культурной флоры- Древнесредиземноморской. ЗЕМЛЕДЕЛИЕ В ДРЕВНЕЙ ИНДИИ И АБОРИГЕННЫЕКУЛЬТУРНЫЕ РАСТЕНИЯ ЮЖНОЙ АЗИИ Археологические изыскания в Индии показывают, что эта страна имеет очень древнюю историю. В 5 км к северо-западу от Кветты в нижней части насыпного холма Купе-Гулл-Мухаммед, у южной оконечности Боланского прохода открыта древнейшая докерамическая земледельческая культура. М. Уилер (Wheeler, 1953) указывает, что здесь были найдены кости овец или коз, и сопоставляет земледельческую культуру этого холма с культурой Джармо (в Северном Дву- речье), которая датируется примерно около 5000 лет до н. э. В северном Белуджистане известны раскопки в Рана-Гундай, нижний разрез здесь относится к неолиту и соответствует до- вольно ранней стадии развития земледелия (аналог Хассуна в Месопотамии). Нагорные культуры Ирана и соседствующие с Двуречьем в Месопотамии предшествовали культуре Хараппы и долго существовали одновременно с ней. По M. Уилеру, культуры месопотамская в Двуречье и прото- индийская развивались независимо, но между ними существо- вали сношения. И в Передней Азии, и в Индии земледелие на- чалось прежде в нагорьях, а потом спустилось в долины боль- ших рек и на равнины. В Южной Индии (А. М, Осипов, 1948) были открыты целые мастерские неолитических орудий, где изготовлялись и мотыги, служившие для примитивной обработки почвы. В раскопках неолита уже находили обрывки хлопчатобумажных тканей. Это говорит о чрезвычайной древности здесь культуры хлопчат- ника. На территории Пенджаба и Хайдарабада между IV и II тысячелетием до н. э. существовали четыре цивилизации. Самая древняя из них — амрийская — не знала еще металла. В III тысячелетии до н. э. развивались культуры Мохенджо- Даро и Харрапы и за ними культура Джхукары. Хотя культуры эти и стояли на высоком уровне, не уступая современной им культуре Двуречья, но следов письменности не было обнару- жено, и судить о жизни в те периоды можно только по археоло- гическим находкам. Нижние слои Мохенджо-swapo примерно со- ответствовали древнему египетскому царству. Люди этих куль- тур были членами раннего классового общества, занимались земледелием, охотой, рыболовством и скотоводством. Имело. место приручение слонов.
В настоящее время в Мохенджо-Даро климат сухой, не- благоприятный для земледелия. В III тысячелетии до н. э., судя по фауне, климат был более влажный. Теперь некоторые суще- ствовавшие тогда реки исчезли. В Мохенджо-Даро издревле применяли орошение. В эпоху Мохенджо-Даро уже пользова- лись металлами (олово, медь, бронза, золото; железа еще не было). В некоторых кремневых орудиях Э. Маккей усматривает лемехи. Нет определенных данных для суждения, было ли земледелие Мохенджо-Даро мотыжным или плужным. Были обнаружены зернотерки, найдены светильники; очевидно, поль- зовались растительным маслом, но каким — неизвестно, может быть, хлопковым. Земледельцы Мохенджо-swapo разводили хло- пок, ячмень, пшеницу, дыни, финики, лотос (Б. Грозный, 1940; Э. Маккей, 1951). Производство хлопка в ту эпоху было ограничено Индией, лишь в более позднюю эпоху хлопок стали возделывать на за- паде от Индии (Д. Неру, 1955). По Г. Чайльду (1956), жители долины Инда сеяли карли- ковую пшеницу (Triticum compactum). Возможно, что эта пше- ница сходна с круглозерной ископаемой европейской пшеницей Т. antiquorum Heer, которая многими авторами смешивается с Т. compactum. Здесь разводили также горох и кунжуг. Судя по распространенной единице веса, выращивался также и рис, но прямых указаний на эту культуру не имеется. Э. Маккей указывает на существование непосредственной связи между Индией эпохи Мохенджо-Qapo и Средне-Юго-За- падной Азией. Кельтеминарцы и суярганцы, по предположению ряда исследователей, относились к той группе народов, к ко- торой принадлежат современные кавказские народы, т. е. к восточной ветви народов кавказской системы языков. В нее входили протохетты, шумеры и древние обитатели Сирии. Б. Грозный усматривает в надписях на печатях из Мохенджо- Даро следы кавказского языка и думает, что носители культуры Харрапы относились к той же группе народов. Для всех этих земледельческих народов была характерной культура крашеной керамики. А. К. Струве надписи на печатях из Мохенджо-Даро признал хурритскими. Хурриты, или субарейцы, или иначе митанпийцы, образовали во I тысячелетии до н. э. большое государство, центр которого был в северном Двуречье. Между скульптурой из Мохенджо-Даро и скульптурой шумеров замечается некото- рое сходство. Северное Двуречье имело тесные связи с племе- нами Закавказья. Язык Урарту имеет родство с языками хур- ритских и сирийских памятников. Сухопутный караванный путь шел через Белуджистан. Воз- можно существование и морского пути в Шумер. Эти изыскания вскрывают связанность древнейших культур Передней Азии и Индии, Индии и Средне-Юго-Западной Азии. Древний мир в 235
эпоху культуры Мохенджо-Даро и расцвета культур Двуречья отнюдь не был разобщенным, а последнее означает и обмен культурными растениями. Древнее население долины Инда имело постоянные связи с Передней Азией. Рыба регулярно привозилась с берегов Ара- вийского моря в Мохенджо-Даро, товары из городов Индии рас- пространялись по рекам Тигру и Евфрату (Неру, 1955). Из-за отсутствия письменных источников нельзя последовательно про- следить историю земледелия в Индии после эпохи Мохенджо- Даро. Древнейшие письменные документы, в которых можно найти указание на характер земледельческой культуры Индии, относятся к ее ведической истории, которая началась более чем за тысячу лет до н. э. с вторжением сюда с северо-запада ариев. Арии, которые постепенно в доведическую эпоху продвигались на юг, тесня аборигенное население древнейшей Индии, были земледельцами. Само слово «ария» происходит от корня, озна- чающего «пахать, пашня» (Д. Неру, 1955). Профессор Винтермид (по Д. Неру, 1955) полагает, что истоки ведической литературы относятся к 2000 или даже к 2500 г. до н. э. Это весьма близко подводит нас к эпохе Мохен- джо-Даро. Однако большинство современных ученых относит гимны «Ригведы» (наиболее древняя и наиболее интере=ная в историческом отношении часть литературных памятников «Вед») к 1500 г. до н. э., а самые поздние из них — к I тысячелетию ДО Н. Э. Гимны «Ригведы» рассказывают о племенах, которые зани- мались скотоводством, рыболовством и охотой: «...с помощью бога поля мы обрабатываем эту землю. Впрягайте плуги, натя- гивайте ярмо, бросайте семя сюда в подготовленное лоно...» и далее «...они старательно вспахали этот ячмень упряжкой из восьми, упряжкой из шести». Мы видим, что это было время плужного земледелия и что в плуг впрягались быки. Ячмень по этим и другим данным был, вероятно, важнейшей полевой культурой в Северной Индии. В «Ведах» есть указания и на культуры риса, проса, кунжута, сахарного тростника. Ведическая Индия была знакома с производством хлопчато- бумажных тканей. Несомненно, что культура хлопчатника в Индии восходит к временам глубочайшей древности. Первые письменные указания о прядении и хлопковой пряже имеются в религиозных гимнах «Ригведы», относящихся к 1500 — 800 гг. до н. э. О священной хлопковой нити браминов говорится и в законах Ману (800 лет до н. э.). Древние греческие писатели Геродот (V в. до н. э.) Феофраст (IV — III вв. до н. э.) и Неарх (IV в. до н. э.) писали, что в Индии имеются обширные хлопко- вые поля. В Индокитае хлопководство также было распростра- нено в глубокой древности. Со льном вопрос обстоит несколько сложнее. Ботанические данные приводят к заключению, что Индия — это древнейший 266
очаг культуры льна, откуда он широко распространился на за- пад и юго-запад. Вместе с тем, имея большое разнообразие эндемичных, совершенно своеобразных форм масличного льна, современная Индия не знает культуры льна на волокно. Так, видимо, было и в древности. Поэма Тулси Даса «Рамаяна», созданная на основе древне- индийского эпоса, по существу отражает строй мыслей древней Индии, ее верования и предания. В «Рамаяне» лен (San) яв- ляется символом зла: «...он отдает свое волокно для того, чтобы связать узников, хотя это несет гибель и ему самому». Здесь видно прямое указание, что лен древней Индии шел именно на веревки или, может быть, из него еще изготовляли самые гру- бые мешочные ткани, которые, вероятно, и находили в древних р а скопках. В «Артхашастре» описана прядильно-ткацкая мастерская (Г Ф. Ильин, 1951). Здесь говорится о прядении шерсти, хлопка, конопли и льна. Вероятно, в отношении последнего речь шла опять только о грубых тканях. Таким образом, упоминание о находках льняных тканей в неолитических стоян- ках не стоит в противоречии с тем доказанным положением, что лен в Индии в древности, как и теперь, в основном был маслич- ным растением. Как сообщает Р. Пишель (1911), льняные ткани высокого качества были, однако, в употреблении у высших классов обще- ства в эпоху Будды (VI — Vвв. .до н. .э..). .Древние тексты сооб- щают, что Будда (Сиддхартха) был очень нежным ребенком и носил одежды «из тончайшего белоснежного льна». Скорее всего речь шла о льняных тканях, привозившихся из Египта или из Колхиды, где с древнейших времен возделывали лен на пряжу и где достигали замечательного искусства в производстве льня- HbIx тканей. Индия издревле имела связи с Западом, а Будда был сыном царя, и высшие классы, конечно, имели возможность пользоваться дорогими импортными изделиями. Лен в Индии в настоящее время представлен огромным ко- личеством эндемичных форм, которые, однако, объединены общими признаками. Крупнокоробочность, анатомические при- знаки и характер листьев указывают на близкое родство индий- ского льна со средиземноморскими крупнокоробочными льнами. Мы считаем несомненным, что лен из Индии был перенесен в Египет и другие страны Средиземноморья не через Закавказье (там господствовал в это время совсем другой лен — колхид- ский), а через Сирию. Через Сирию же и соседние страны Ара- вийского полуострова лен из Индии попал в Эфиопию. Это было в очень древние времена, почему и успел выработаться из зане- сенного из Индии своеобразный эфиопский лен специального использования (пищевой) . Своеобразное устройство цветка и особая гамма окраски лепестков с несомненностью выдают близкое родство эфиоп- 267
ского и индийского льна. Образование форм, похожих на эфиоп- ские, имело место еще в Индии, ибо и там в сухих местностях возделываются карликовые льны эфиопского типа. На прилагаемой схеме (стр. 280) показаны распространение и филогенетическое развитие индийского льна. Мы полагаем, что наиболее древними являются культурные растения, часто упоминаемые в древнем эпосе. Например, еще в «Ведах» говорится, что при добывании огня путем трения кусочков дерева искры обычно падали на хлопок. Упоминания о лотосе появляются в более поздних литературных памятниках, чем «Ригведа». В «Рамаяне» (перевод А. П. Баранникова, 1948) среди по- этических образов, связанных с растениями, часто упоминается о лотосе, хлопке и рисе. Эти культуры, несомненно, являются древнейшими в Индии. Приводим наиболее часто встречаю- щиеся в «Рамаяне» растения (всего упомянуто 80 растений): лотос, хлопчатник, рис, лен, кунжут (сезам), ячмень, кориандр, редька (вероятно, редис), бамбук, индиго, бхаич (индийская конопля, из листьев которой приготовляется крепкий спиртной напиток), манго, чампак (дерево с желтыми благоухающими цветами), лилия, пальма, платан, дуб, береза, жасмин, мальва, ашока (Joannesia asol<a — священное и лекарственное дерево), бет (лиана — Calamus rotang L. — лекарственное, корни и моло- дые побеги употребляются в пищу). Земледелие в Индии, как и в других странах, зародилось в горных местностях. Использование растений здесь начиналось с собирательства, где большую роль играли семена дикорасту- щего риса — Oryza fatua Prain, дагуссы — Eleusine indica (L.) Gaertn. — и разнообразные плоды. Большую древность имеет в Индии и сопредельных странах Южной и Юго-Восточной Азии культура клубнеплодов: таро, ямса и др. Введению клубнеплодов в культуру предшествовало их собирательство. Некоторые исследователи думают, что культуре риса поло- жили начало сорные формы последнего, растущие на плантациях таро и других клубнеплодов. В глухих углах горной Индии, и особенно в Бирме, еще и в настоящее время сохранилось первобытное земледелие на площадках из-под леса (подсечная система). При этом посев производится просто в лунки, сделанные палкой. Сначала рис культивировали в Бирме без орошения, со временем перешли к терраской культуре риса на склонах. Еще позднее земледелие спустилось в долины больших рек, где культура риса стала вестись путем затопления. И теперь еще во многих местах вспашка здесь производится первобытным плугом, который тащат буйволы. культура горного риса и риса поливного существовала из- давна в Малезии независимо от Индии. По А. Ходрикурту и 268
Л. Хедину (А. G. Haudricourt et L. Hedin, 1953), многие расте- ния распространялись в Малезию и Полинезию не через Индо- китай, а из Китая путь их распространения пролегал между Тайванем, Хайнанем и Бакбо. Об этом говорят лингвистические данные, полученные при изучении малайских и полинезийских языков. Их словарь указывает, что эти народности в очень дав- нюю эпоху культивировали поливной рис, пользуясь горными речками. Здесь орошение было связано с терраской культурой; в Индии вода для орошения держится в больших резервуарах— хранилищах. Как и в других географических областях, горное земледелие в Юго-Восточной Азии является более древней и примитивной фазой земледельческой культуры, чем земледелие на низмен- ностях. И примитивная культура риса также приурочена к горам. Исторические вехи рисовой культуры были следующие: 1) неорошаемый рис во влажных горных районах; 2) орошае- мый рис в условиях террасной культуры в горах и предгорьях; 3) орошаемый рис на низменностях при запасах воды в боль- ших водохранилищах. В глубокой древности из Индии и Бирмы культура риса была перенесена в Китай, а оттуда в Малезию. В Индии из- вестно около 8000 сортов риса. Не только рис, но и многие дру- гие культурные растения еще в очень древние времена распро- странялись на север в Китай (рис, цитрусовые, чай, таро и др.) и в Африку (лен, чечевица и др.). Из Африки в свою очередь культуры распространялись в Индию (кунжут, сорго и др.). Данные исторической этнографии говорят, что этнические связи между этими областями существовали, вероятно, еще в доземледельческие эпохи. Народы экваториальной (негро- австралоидной) большой расы заселяли некогда Южную Азию и Африку. В Южной Азии была распространена самая древняя ветвь — негро-австралоидная. В Малезии к этой расе принадле- жали меланезийцы, на юге Индии — дравидийские племена. По- следние отнесены к веддоидной (от цейлонского племени ведды), или южноиндийской расе. В более поздние эпохи в Индию устремились потоки евро- пеоидов, а в Индокитай и Индонезию — монголоидов. Передви- жения народов происходили и после возникновения земледелия, что имело огромное значение в отношении распространения культурных растений в пределах Евразии. Вне Индии по странам Южной Азии археологических данных значительно меньше. Приводим данные С. А. Семенова (1950) по Вьетнаму. Древнейшие следы человеческой деятельности в эпохи ран- него неолита и мезолита обнаружены в пещерах Хоа-биня и в горном массиве Бак-сон. Эти пещеры были, по-видимому, засе- лены человеком после окончания плювиального климатического 269
периода в Юго-Восточной Азии, после перехода от плейстоцена к современному геологическому периоду. Большинство пещер расположено на высоких берегах рек или среди выступов скал, откуда можно было обозревать нижерасположенные долины. Сходные пещеры служили обиталищем древнего человека также во Вьетнаме (Чунгбо, Намбо), Таиланде и в других об- ластях Индокитая и Индонезии. Эпоха собирательского хозяйства отражена здесь в скопле- ниях многочисленных раковин. Одновременно обитатели пещер занимались охотой на обезьян, диких свиней, оленей, тибетских медведей и буйволов. Первым прирученным здесь животным была, вероятно, свинья. Собирательское хозяйство постепенно сменялось земледелием — в очень древних пещерах находили зернотерки с глиняными пестами. Переход от неолита к бронзовому веку обнаружен в раскоп- ках в селении Сомронг-сен в Камбодже. Поселение древних земледельцев было расположено здесь на аллювиальных отло- жениях в дельте Меконга. Предполагают, что в течение недолго- срочного половодья жители покидали селение и пережидали до спада большой воды на бли>кайшем небольшом горном массиве. 5Кители древнего Сомронг-сена занимались рыболовством и земледелием. Плавали на специально приспособленных лодках по порожистой и быстрой реке. Эти лодки были снабжены аут- ригером — легким, заостренным на конце бревном, которое при- вязывалось на некотором расстоянии от лодки к длинному шесту. Аутригер придавал лодке устойчивость при ударах волн. Изготовлялся он из бамбука или связанных пучков камыша. Остатков культурных растений в Сомронг-сене не было найдено, но, по С. А. Семенову, можно предполагать, что земледельцы Сомронг-сена знали культуру риса, кунжута, диоскореи. Судя по найденным остаткам, з хозяйстве имелись свиньи и быки. Сахни (по gao Тхе Туан, 1960) обнаружил в Восточном Пенджабе ясные следы зерен риса на керамике племен Север- ной Индии, живших в конце I тысячелетия до н. э. В древнем тамильском произведении «Пуранас» имеется описание различ- ных сортов риса, которые употреблялись в религиозных цере- мониях в Южной Индии. Qao Тхе Туан предполагает, что на Цейлоне до 543 г. до н. э. рис возделывался как суходольная культура. На Цейлон рис, вероятно, распространился из Индии. Первые сведения об орошении риса на Цейлоне датируются 420 г. до н. э. Qao Тхе Туан (1960) пишет, что по литературным данным намечаются три очага, откуда, возможно, произошел рис: долина Инда, Бенгалия и Южная Индия. Однако, по мне- нию того же автора, следует исключить долину Инда, так как, согласно историческим данным, рис появился в этих районах не раньше 1 тысячелетия до н. э. Кроме того, климатические усло- вия, особенно в Хайдарабаде, не позволяли возделывать рис без орошения. 270
По К. Рамьяху и Р. Гошу (К. Ramiah and R. Ghose, 1951), родина риса — Южная Индия. Рис введен в культуру дравидий- скими племенами в очень древние времена; наибольшее разно- образие форм риса сосредоточено в Джайпуре. Однако, как уже упоминалось выше, вопрос о самостоятельном вхождении риса во Вьетнаме, в других странах Индокитайского полуострова и в Малезии еще не решен окончательно. Qao Тхе Туан (1960) пишет, приводя данные Семенова и комментируя их, что, по археологическим данным, самая древняя культура во Вьет- наме — культура Хоа-бинь на западе от Ханоя — эпохи верх- него палеолита. Следующей по времени культурой считается культура эпохи раннего неолита (Бак-сон — на север от Ханоя). Имеется пред- положение, что эта культура принадлежит индонезийским пле- менам (австроазиатским), которые под давлением арийцев пере- селились из Индии на полуостров Индокитай, там смешались с автохтонным меланезийским племенем и вместе с последним положили начало коренному населению этого полуострова. При миграции индонезийские племена, может быть, и не занесли с собой риса. Может быть, в Индии в ту эпоху еще и не было риса. Неизвестно, сами ли эти племена создали культуру риса или получили ее от племени юе из Южного Китая. По сведениям, приводимым gao Тхе Туаном, в 1 тысячеле- тии до н. э. некоторые племена юе ежегодно совершали стран- ствия на юг, и при этом часть людей оседала в новых местах в дельте Красной реки во Вьетнаме, на Филиппинских и Малай- ских островах, и этим путем, несомненно, могло распростра- ниться рисоводство, ибо земледелие в Восточном Китае тогда уже находилось на высоком уровне. Когда государство юе было разрушено (333 г. до н. э.), население племени юе в массе миг- рировало на юг. Населявшее дельту Красной реки племя лак-вьет создало бронзовую культуру. В раскопках этой культуры в Северном Вьетнаме в Данг-соне (провинция Тхань-хоа) был найден рису- нок, где изображались мужчина и женщина, обрушивающие рис пестиком в ступе, как это делают некоторые горные племена Северного Вьетнама еще и в настоящее время. В китайской записи «Гуанчжоу» (по тому же автору) ука- зывается, что в это время в дельте Красной реки уже существо- вала культура риса при орошении водой прилива. По другим источникам, тибета-бирманцы и индонезийцы являются родственными народами и имеют общее происхожде- ние. Тибета-бирманцы пришли из Тибета на полуостров Индо- китай, вероятно, в I тысячелетии до н. э. Они добывали средства к существованию по-разному, например лолоцы были скотово- дами, а тайцы земледельцами. Неизвестно, возделывали ли рис народы Индокитая до их прихода. Однако в Сомронг-сене (до- лине реки Меконг) открыты памятники позднего неолита, где 271
был найден пестик для растирания зерен, может быть, риса (С. А. Семенов, 1950). Интересны лингвистические данные, помещенные в книге Дао Тхе Туана (1960). Сначала приводятся те названия у раз- личных народов, которые происходят от трех санскритских кор- ней. При сравнении этого списка названий с названиями риса у народов Индокитая бросается в глаза резкое несходство по- следних с теми названиями риса, которые считаются происхо- дящими от санскритских корней. Лингвистические данные дают основание предполагать, что культура риса возникла у народов Индокитая самостоятельно или занесена туда была не из Индии. дикорастущая флора Южной Азии чрезвычайно богата ро- дами и видами растений. В густонаселенной Индии аборигенная растительность сохранилась хуже. Первобытная природа в большей степени имеет девственный характер во Вьетнаме и соседних странах. Для ландшафтов этих стран с их девствен- ными лесами и джунглями характерны заросли многих полез- ных растений и родичи культурных видов. Особенно типичны для этих мест бамбуки, дикие бананы, всевозможные лианы ит. п. География культурных видов Южной Азии еще требует де- тального изучения. В настоящее время ареалы многих культур- ных растений не уточнены, эндемичные культуры далеко не все выявлены и многие из них имеют только сугубо местное, для не- большого района, значение. Остановимся главным образом на тех культурных растениях, которые получили более широкое распространение. Поэтому здесь не дается подразделения Южноазиатской области исторического развития кульгурной флоры на подобласти, а дается обзор або ригенных и с та р од а в них кул ьтур ных растений для Южной Азии в целом. Из хлебных растений можно считать происходящими из Южной Азии следующие. П ш е н и ц а представлена в Индии эндемичным видом— пшеницей круглозерной — Triticum sphaerococcum Pere., кото- рая распространена главным образом в Пенджабе, а последний включается многими исследователями в область Средне-Юго- Западной или даже Передней Азии. Я чм ен ь (Hordeum euroasiaticum Vav. et ВасЫ.) представ- лен особой агроэкологической группой — proles indicum ВасЫ., ареал которой охватывает Пенджаб, Кашмир, Хайдарабад и Хайрпур. Недостаточно изучен. Вопрос о происхождении по сев ного р и с а остается спорным. Некоторые исследователи близкими к Oryza ваШа L. видами считали О. fatua Prain (2п =24), О. minuta Presl. (2n= =48) и О. officinalis Wall. (2n =48). Последние два вида много- летние, дикорастущие; родина О. minuta — Филиппины, а О. offi- 272
cinalis — Индия и Индокитай. Оба вида далеки от О. sativa» не могут быть его непосредственными предками. Кроме того, oáa вида являются тетраплоидами. Остается О. fatua, который, как и О. sativa, представляет собой однолетнее растение и имеет одинаковое с О. sativa число хромосом (2n=24), оба вида образуют гомологические ряды варьирующих признаков. Однако есть возражения против теории происхождения по- севного риса от О. fatua. С. Сампат и Нарасинга Pao (S. Sampath and Narasinga Rao, 1951) считают О. fatua сборной группой. Они указываюг, что О. fatua из Индии и О. fatua из Южного Китая не скрещи- в а ются между со бо й. Нам представляются заслуживающими внимания взгляды тех исследователей, которые считают предком культурного риса многолетний корневищный и плавающий рис — О. perennis Noench. Предполагается, что он является общим предком для двух видов культурного риса: О. sativa и африканского О. gla- berrima Steud. Эти три вида, хотя и затрудненно, скрещиваются между собой, но О. sativa и О. glaberrima образуют хорошие го- мологические ряды варьирующих признаков. Ареал О. perennis пантропический. Этот вид представлен различными типами в Африке и Америке (возможно, что эти типы следует считать подвидами или в некоторых случаях просто региональными экотипами). О. perennis представлен и более древними плавающими экотипами, и переходными фор- мами, и неплавающими (к последним относится и африканский тип, которому дано видовое название О. barthii А. Cheval.) .. В Южной Азии имеются разнообразные экотипы или физиологи- ческие расы с разными требованиями к влаге. Эволюция в этом отношении шла в следующем порядке (см. схему на стр. 274). По Р. Портеру (взято из книги Дао Тхе Туана, 1960), пла- вающий рис (О. sativa f. aquatica) распространен в Индии (Ассам), Бирме, Камбодже и Южном Вьетнаме. По мнению Qao Тхе Туана, его можно рассматривать как гредковую форму О. sativa и О. fatua» можно назвать его О. prosativa. Популяции О. prosativa при дальнейшей дивергенции дали начало, с одной стороны, О. fatua, с другой,— типичному О. sa- tiva. Некоторые формы О. fatua могли произойти и от типичного О. sativa подобно фатуоидным овсам, возникающим из Avena ~qtjyg. Таким образом, новейшие данные свидетельствуют о моно- филетическом происхождении О. sativa от одного вида — О. pe- rennis. Вместе с тем происхождение О. sativa могло быть поли- топным, т. е. истоки популяций этого вида находились в раз- личных местностях Южной Азии — из различных популяций азиатского О. perennis. Суходольный рис является по этой схеме 273
Этапы экологической эвол~оции риса Плавающие, глубоководные, многолетние Плавающие, многолетние, менее глубоководные Неплавающие многолетние(типа Q. barthit) Однолетние плавающие(О. sativa f. aquatica) Однолетние неплавающие, требующие более длительного затопления(О. sativa, О. fatua) Однолетние О. sativa, требующие менее длительного затопления Однолетние О. sativa, имеющие гпгрофитную конституцшо, но сравнительно менее требовательные к условиям увлажнения, в некоторых влажных районах способные произрастать без орошении (суходольный рис) последним звеном эволюции О. sativa, что подтверждается и историческими данными. Такое мнение о происхождении и позднем возникновении суходольного риса отнюдь не противоречит концепции Н. И. Ва- вилова о зарождении первичного земледелия и, в частности, культуры риса в горных и предгорных влажных районах, где для орошения пользовались горными речками, ручьями и клю- чами при помощи мелких запруд и небольших водных резервуа- ров. Там высевался не настоящий суходольный рис, а экотипы, требующие не столь длительного и глубокого затопления, как формы риса, культивируемые на низменностях. Настоящий сухо- дольный рис, также влаголюбивый, но при прочих благоприят- ных условиях способный возделываться и без орошения, возник гораздо позднее. В Южной Азии распространены и другие виды дикорасту- щего риса из различных секций рода, но они не имеют прямого отношения к происхождению культурного посевного риса. Рис лекарственный (О. officirtalis Wall.) — многолет- ний дикорастущий рис (Индия, Цейлон, Бирма, Малакка, Индо- незия). Р и с м е л к и й (О. mirtuta Presl.) — также многолетний вид. Некоторые авторы считают (неправильно), что этот вид 274
принимал участие в происхождении культурного риса. Исполь- зуются дикие заросли. Рис суходольный зернисты й (О. granulata Nees)— используется зерно дикорастущих растений (Индия, Цейлон, Индокитай, Индонезия, Филиппины). Д а гусс а [Eleusine coracana Gaertn и Е. indica (L.) Gaertn.] в древности имела большое значение как зерновое растение, в диком виде и как сорняк. Дикорастущие заросли и теперь ис- пользуются в неурожайные годы. П а с п а л у м я м ч а т ы й (Paspalum scrobiculatum L.) растет дико и культивируется (Индия, 1~ейлон, Индокитай). Родина этого растения — Индия, а отсюда оно широко распространи- лось, стало космополитом тропиков. Корм для птиц. С о р г о х л е б н о е [Sorghum durra (Forsk.) Stapf] отли- чается в Индии большим разнообразием и представлено здесь рядом эндемичных сортов. Однако в Индии вторичный очаг сорго — родина его в Африке. Р о с и ч к а (Digitaria sanguinalis Scop.) — в Индии, особенно в неурожайные годы, используется в пищу зерно сорных и дико- растущих растений. Раньше культивпровалась в Европе, ве- роятно, была занесена из Индии. П а й з а [Echinochloa frumentacea (Roxb.) Link] считается многими растением восточноазиатского происхождения, но этот вопрос спорный. Э. Верт (Е. Werth, 1937) пишет, что этот вид культивируется издавна в Индии и считается там «хлебом бед- няка», как и близкий вид (или подвид) Е. соlопа (L.) Beauv., который занесен в качестве сорняка в Южную Европу вплоть до Испании. Оба вида вошли в культуру из сорняков риса. К у р и н о е п р о с о [Echinochloa crus-galli (L.) Roem. et Schult.] широко распространено в качестве сорняка по субтропи- ческим и тропическим странам обоих полушарий и в умеренных широтах Европы и Азии. Местами в Индии и Африке разводится как как кормовое. Э. Верт предполагает, что очагом происхождения всех трех видов Echinochloa является Индия. Здесь возник единый поли- морфный вид, который затем дифференцировался и распростра- нился в разные страны на север и запад. К о и к с (Goix laicrima-jobi L.) — зерновое растение (Малезия и Индия). Употребляется также для изготовления четок. Из Ин- дии занесен в Грузию, где употребляется для изготовления оже- pea><A. Из растений, занесенных давно в Южную Азию, укажем на кукурузу. Хейне-Гельдерн (R. Heine-Geldern, 1958) считает, что кукуруза в Азию была завезена до нашей эры Гипотеза завоза кукурузы португальцами из Бразилии в Гоа неправильна, так как, по Хейне-Гельдерну, уже в 1579 — 1585 гг. кукуруза возделы- валась в Китае, проникла туда через Тибет и долгое время назы- валась тибетской пшеницей. Мало вероятно, чтобы к этому вре- 275
меня кукуруза акклимачизировалась в горных районах, сильно изменилась и продвинулась из Индии (Гоа) через Тибет в Ки- тай. В обширных списках возделываемых растений в Индии (А. Fuzl в конце XVI в. и Виппапп в конце XVIII в.) кукуруза не упоминается. Только Роксбург в 1800 г. встречал кукурузу в садах как редкое растение. Аналогичное предположение о завозе в Китай португальцами двух видов а м а р а н т а также неправильно, так как амарант возделывался в Китае еще в Х в. и. э. По археологическим данным установлено, что связи Китая с Мексикой начались с VIII — VII вв. до н. э. Считается возмож- ным (Heine-Geldern, 1958) завоз в Китай кукурузы и амаранта в 1 тысячелетии до н. э. Следовательно, кукуруза попала в Ки- тай не из Индии, а наоборот, в Индию кукуруза была завезена из Китая, а какими путями, установить трудно. Я и р и ц а (Amaranthus L.) представлена в Индии следую- щими культурными видами: зерновыми — А. Irumentaceus Huch.— Нат., А. speciosus Sims; овощными — А. anardana Bu~ch.-Нат., А. blitum var. oleraceus Watt., А. gangeti~cus L., А. tricolor L. Индия считается родиной важной крупяной культуры — г р е- ч и х и (Fagopyrum за@11а1ит Gilib.). Культурная гречиха и весь род Fagvpyrum Gaertn. азиат- ского происхождения — из Юго-Восточной Азии. В Бирме и Кашмире возделывают настоящую гречиху, а в Тибете и кое-где в Гималаях употребляется в пищу и ближайший родич обыкно- венной культурной гречихи — гречиха татарская (кырлык)— F. tatariicum (L.) Gaertn.; эндемичная высокогорная форма та- тарской гречихи, употребляемая в пищу, получила название гималайской — F. tataricum var. himalaica. В посевах обыкновенной гречихи и пшеницы татарская гре- чиха распространилась из горных районов Гималаев и Тибета на север в Сибирь и на запад. Татарская гречиха — это специа- лизированный сорняк, скорее всего она не является прародите- лем вида культурной гречихи — F. sagittatum, а оба представ- ляют собой пару близкородственных видов, происшедших от общего родоначальника. В Индии сосредоточены также богатейшие сортовые ресурсы бобовых растений, в особенности фасоли. Ф а сол ь (Phaseolus L.) представлена в Индии группой так называемого маша, которая, вероятно, оттуда и ведет свое про- исхождение. В пользу этого предположения говорит существо- вание в Индии близкородственного многолетнего дикорастущего вида Ph. sublobatus ЯохЬ., который является лекарственным и зерновым растением и иногда разводится. От этого вида, воз- можно, произошли оба вида маша. Первый — маш обыкновен- ный — Ph. aureus (Poxb.) Pip., который в Индии представлен особым подвидом ssp. indicus G. Pop. Внутри индийского под- вида выделяются следующие агроэкологические группы мест- 276
ных сортов: 1) proles punicus G. Po~p. (Пуза, Бомбей); 2) proles bengalicus G. Рор. (Бенгалия); 3) proles pundj abicus G. Pop. (Бирма, Индия — Пенджаб); 4) iproles lahorus G. Рор. (Индия, заходит в Таджикистан — дарваз?); 5) iproles nagpuricus G. Polyp. (Бирма, Индия — Ассам, Пенджаб); 6) iproles kashmiri~cus G. Рор. (Кашмир, Ассам, Северная Индия). Второй вид — маш индийский, или урд, — Ph. mungo 1, представлен в Индии многими эндемичными формами, еще не- достаточно изученными. По Н. Р. Иванову (1960), маш происходит из Индии (юго- западные отроги Гималаев), откуда он распространился в Аф- ганистан, Иран и Северо-Восточную Африку (Эфиопия). Фасоль аконитовая — Ph. aconitifolius Jacq., растет в Индии и на Цейлоне дико и культивируется. Может служить зерновым и кормовым растением, корень употребляется в пищу как корне- плод и на муку. Лекарственное (в Китае). Фасоль трехлопастная — Ph. trilobus Ait., близкий к Ph. асо- пНИо1шз вид, вошел в культуру из сорняка, сопутствующего Ph. aiconitifolius, возделывается на зеленый корм. Фасоль рисовая — Ph. са1сага1из Roxb., возделывается в Индии, Индокитае и Китае как зерновое, кормовое и сидерат,. встречается в диком виде. Таким образом, в 10жной Азии расположен один из первич- ных очагов культурных видов фасоли рода Phaseolus L. (основной первичный очаг — в Америке). Л о б ия, BJIJI ги а цинто вы е бобы (Doli~chos L), имеет один из очагов развития в Индии, а весь род — африканского происхождения. Лобия двухцветковая — D. biflorus L. — широко распростра- нена в диком состоянии и в культуре, аборигенна в Индии„ Бирме и Индонезии. Лобия обыкновенная — D. lablab L. очень широко распро- странена и представлена эндемичными формами в Южной Азии, но происходит, вероятно, из Африки. D. ablab попала в Азию давно, и здесь возникли многие эндемичные формы. В Индо- китае имеются полувосковидные формы. По Райвалсу (Р. Rivals, 1953), в Индии распространены сле- дующие эндемичные для Южной Азии подвиды D. 1аЬ1аЬ: 1) зэр. bengalensis Rivals (Индия и соседние страны Юго-Во- сточной Азии, заходит в Юньнань, где есть одна разновидность этого подвида); 2) ssp. ensiformjs Rivals (тот же ареал). В и г н а (Vigna Savi) в целом африканского происхождения, в Индии широко распространена и представлена эндемичными сортами коровьего гороха — V. sinensis Endl. Очаг развития этой культуры в Индии, вероятно, вторичный, а первичный находится в Африке. Монотипный род Cajanus DC. представлен видом С. indicus Sipreng.— голубиный горох. 27Т
Голубиный горох имеет в Индии большое количество энде- мпчных форм, возделываемых за 3000 лет до н. э. и имеющих санскритское название «ачрика». В Индии распространены двух- летние и многолетние формы, карликовые и гигантские, высоко- культурные и примитивные (мелкосемянные и темносемянные). Существует мнение, что голубиный горох происходит из Африки, но имеется много оснований считать родиной этого растения Индию. Отсюда он еще в древности проник в Египет (и в Uea- тральную Африку?). Вопрос о происхождении рода каянуса недоработан. Следует выяснить отношение этого рода к другим близким родам. С о я (61у cine 1 .) представлена в Индии эндемичным под- видом — G. hispida ssp. indica Enliven, по конституции прибли- жающимся к дикорастущим вьющимся видам. Основной очаг развития культурной сои — Китай. Чечевица (Lens еэси1еп1а Moench) представлена в Индии особой эндемичной группой мелкосемянной чечевицы — дгех pilosae Bar. Основной район распространения — Кашмир и от- части Пенджаб, примыкает к Средне-Юго-Западной Азии, а по- этому нами включена в ч1:сло элементов культурной флоры последней. Однако в Индии культура чечевицы отличается древ- ностью. Существует несколько санскритских названий чечевицы: renulce, masura, mangalya, отсюда иранское название manga. Го р ох (Pisum sativum L.) представлен в Индии группой местных сортов, объединенных в, proles ~пйсит Gov. довольно примитивного типа. Известно санскритское название ropoxa— hurense. Чин а (Lathyrus sativus L.) представлена в Индии особой агроэкологической группой местных сортов — proles indicus Zalk., входящей в состав азиатского подвида — ssp. asiaticus Zalk. Индийские чины родственны переднеазиатским и, вероятно, пришли в Индию из Передней и Средне-Юго-Западной Азии, а из Индии проникли в Эфиопию. Нут (Cicer arietinum L.) в Индии представлен эндемпчными сортами, которые составляют индо-абиссинский подвид — ssp. orientale G. Pop. Индийские эндемичные формы выделены в proles indicus G. Pap. Индию можно считать вторичным очагом развития обык- новенного нута. К о н с к и е б о б ы (Faba vulgaris Moench) представлены в Индии особым примитивным подвидом — ssp. paucijuga Murat., распространенным на северо-западе на границе с Средне-Юго- Западной Азией. П а жити и к (Trigonella foenum-graecum L.) также имеет вторичный очаг развития в Индии и представлен эндемичными формами — ssp. indica Sinsk. В составе этого подвида имеются поздние и ранние экотипы. 278
Род к а н а в а л и я (Canavalia Adans.) максимальное ви- довое разнообразие имеет в Южной Америке, но следующие виды происходят из Южной Азии: 1) С.;рос1осагра Dunn.— многолетник, больше B диком состоянии, иногда разводится (Цейлон, Южная Индия); 2) С. lineata DC. — многолетник, ра- стет дико, плоды сьедобны (Индокитай, Малакка, Индия, по песчаным морским берегам — на побережьях Китая, на о. Тай- вань, в Японии, Полинезии и Австралии); 3) С. gladiata РС.— возделывается в Индии, Индокитае, Индонезии и на Филиппи- нах, культура ее позднее из Индии распространилась (как овощное и зерновое) в Китай, Японию, Африку и Америку. Диким родоначальником считается С. virosa; 4) С. virosa W. Arn. — растет дико и возделывается, употребляются в пищу незрелые плоды (Индия, Бирма, Таиланд, Юго-Западный Китай, а также на Аравийском полуострове и в Эфиопии); 5) С. ensi- formis DC. (Индия, Индокитай, Япония, Индонезия, Америка, Африка), распространилась из Америки. По-видимому, имеются азиатские местные сорта. Б а р х а т н ы е б о б ы (Stizolobium P. Вго~чпе) — лианы тро- пиков Старого и 1-lового Света. Прародитель культурного вида S. pruriens (L.) Medi~c., имеющий лекарственное значение, рас- пространен в тропических лесах и как сорняк в Старом и Новом Свете. Виды S. ipruritum Kuntze и S. hirsutum Kuntze имеют зна- чение в Индии как сидераты. Возможно, что в разных странах имело место независимое вхождение в культуру. Для Южной Азии эндемичны следующие подвиды бархатных бобов — S. utile (Wall.) Ditm. 1) ssp. capitatum (Roxb.) Ditm. (Индия, Ява); 2) ssp. pachylobum (Piper et Tracy) Ditm. (Индия). Из бобовых растений в культуре Индии известны следующие клубненосные растения: Pachyrhizus angulatus Rich., Psophocar- pus tetragonolobus DC., Cyamopsis psoralioides DC., Мисипа utilis Wall. et Wight (Зондские острова) и другие виды. Китайский ка ртоф ель (Paichyrhizus angulatus Rich.) внешне похож на фасоль. Вьющееся многолетнее растение, но используется как однолетнее для получения клубней. Эти клубни едят сырыми. Они содержат около 80'/о воды, крахмал и сахар. По вкусу похожи на брюкву и репу. В жаркое время ими уто- ляют жажду. К р о т а л я р и я (Crotalaria 1 .) африканского происхожде- ния. Некоторые виды кроталярии используются в Африке и Индии как сидераты и кормовые растения. Кроталярия прядильная — Crotalaria jun~cea L. — дико рас- пространена в Индии, на острове Цейлон, Малайском архипе- лаге и в Австралии. Культивируется главным образом в Индии и Индокитае. Видами индийского происхождения считаются сле- дующие: С. verrucosa L., наиболее типичные формы которой представлены в Индии, и С. incana L., произрастающая в Индии и тропической Африке. 279
С ес6 а ни я (Sesbania aiculeata Poiret и S. cannabina Poi- те() — прядильное (грубое волокно) и сидерат. В Индонезии культивируется д р е в о в и д н о е 6 о 6 о в о е (Cynometra eauliflora L.). Лен (Linum usitatissimurn L.) в Индии представлен боль- шим количеством эндемичных форм. В Северо-Западной Индии, которая относится к Средне-Юго-Западной Азии, распростра- нены иные, хотя также древние по типу своей организации, формы льна. Индия — один из трех первичных очагов распро- странения льна. Схема происхождения и распространения льнов из индийского очагаДикорастущий лен L. angustifolium (исчезнувшая форма на восточном пределедревнего ареала) Исходная, исчезнувшая культурная форма с чертами абиссинского, в начале жизни про- стратного льна и, вероятно, с чертами медленно растущей, склонной к простратности афгано- индийской формы 1Карликовые индий-ские крупнокоробоч-ные кудряши Абиссинские мелкосе- мянные среднерослые кудряши со склонностью к простратности в нача- ле развития („гигрофиль- ная группа') Более высокие индий-ские крупнокоробочныекудряши и ранниемежеумки Абиссинские мелко-семянные карлики (,ксе-рофильная группа') Средиземноморские крупнокоробочные кудряши Средиземноморские крупнокоробочные полумежеумки Хлопчатник (Gossypium L.) имеет первичный очаг куль- туры в Индии и прилегающих странах. Яля Индии абориген- ным является древовидный хлопчатник — G. arboreum L. Его подвид — ss~p. nanking (Meyen) Маиег — распространился от- сюда в Китай, Японию и Корею, где получили развитие состав- ляющие этот подвид сорта. 10жной Азии свойственны следующие подвиды, относящиеся к виду G. arboreum: 1) ssp. obtusifolium (goxb.) Маиег (Запад- ная Индия, Цейлон); 2) ssp. neglectum (Tod.) Маиег (Индия, :280
Бирма, Индонезия); 3) ssp. регеппе (В1апоо) Mauer (Индия, Индокитай, Индонезия) . Ф. М. Мауер (1950) считает, что хлопчатник входил в куль- туру независимо в Южной Азии, Юго-Восточной Африке, Аме- рике и, может быть, на Мадагаскаре. Наиболее древним оча- гом является Северо-Западная Индия. Здесь можно наблюдать все переходы от дикорастущих форм к культурным. По Ф. М. Мауеру (1954), скороспелые сорта хлопчатника G. arboreum существовали с глубокой древности, что позволило этой культуре распространиться в северных и горных районах, а затем в Китай. Особый вид хлопчатника — G. stocl<sii Mast. — распространен в Индии и Восточной Африке. Бом ба кс, или в атно е дерево (Bombax malabaricum DC.) распространен в дикорастущем состоянии и иногда разво- дится. Шелковистое волокно его находится внутри плодов. При- меняется для набивки мебели, а также для изготовления спа- сательных кругов, так как обладает большой плавучестью. Капо к (Eriodendron pentandrum Harz.) относится к сем. мальвовых. Волокно употребляется как примесь к хлопку. Хорошее волокно дает Calotropis gigantea (L.) R. Br. Во- локно употребляется для набивания подушек. Джут (Cot>chorus L.) наибольшее распространение в куль- Tóðå имеет в Индии и в прилегающих странах и отличается в этой области большим разнообразием форм, составляющих виды С. caipsularis L. — джут короткоплодный и С. olitorius L.— джут длинноплодный. Предполагается, что отсюда и ведет про- исхождение эта прядильная культура. В Индии имеется очень большое экологическое и морфологическое разнообразие форм вида С. olitorius. Отсюда он распространился в Африку, Аме- рику и Австралию, а также в другие области Азии. С. capsularis — очень распространенная культура в Индии. Некоторые авторы считают родиной этого вида не Индию, а Китай. Р а м и [Boehmeria nivea (L.) Gaud.] имеет более северный ареал и происходит скорее из Восточной Азии. В. tenacissima (Roxb.) Gaud. — более теплолюбивый вид [по мнению некото- рых авторов, например А. Шевалье (Chevalier, 1952), только разновидность В. nivea, а не самостоятельный вид]. Из сем. Urtiica~ceae в Индокитае, кроме рами, имеются сле- дующие прядильные виды: Maoutia ipuya Wedeb. (= B~oehmeria puya Hoolc.) — Индия и Индокитай; Conocephalus mollis Merrill (лиана) — в лесах Вьетнама (Чунгбо). С и д а (Sida rhombifolia) — прядильное растение. Распро- странена в Индии в дикорастущем состоянии и в культуре. Кен а ф прядильный (Hibiscus cannabinus 1.) — Во- сточная Индия. Довольно широко распространен в культуре дру- 28k
rnx стран Азии, Европы, Африки и Америки. Некоторые авторы считают, что этот вид африканского происхождения. Розан кита й ски й (Н. rosa-sinensis L.) иногда исполь. зуется как прядильное. Широко распространенное декоративное растение. Родина — Малайский архипелаг. Розелла, или щавель яванский (Н.sabdariffa L),— овощное (употребляются в пищу цветочные почки), прядиль- ное и декоративное (Индия, Цейлон). К о н о п л я (Cannabis sativa L.) — очень возможно, что арийцы еще до своего переселения в Индию знали об употреб- лении конопли на веревки. Конопляное волокно называлось bhanga. Возможно также, что арийцы получали масло из семян конопли раньше, чем они познакомились с кунжутом. Индийские формы прядильной конопли слабо изучены. Прядильная ко- нопля, вероятно, родом из Восточной Азии (Япония, Восточный Китай). И з и н д и й с к о й к о н о п л и — Cannabis indiica (Lam.) Serebr. — получают сильный одуряющий наркотик — гашиш, ко- торый употребляют для курения. Вероятно, родина этого расте- ния Северо-Западная Индия. Из других прядильных отметим: а б р о м у — Abroma augusta L., с а н с е в и е р и ю — Sansevieria zeylani~ca Willd. и банан текстильный — Nusa textilis Nee (Филиппины, Индокитай) . Для к л е щ е в и н ы существует ведическое название eraud— это слово санскритского корня и перешло в индоевропейские языки. Р. Китель (по А. G. Наийггсоиг1 et 1. Hedin, 1953) счи- тает, что это название дравидическое и что культура клещевины имела место в Индии еще до прихода туда арийцев. Из масличных древесных растений следует отметить южный тунг — Aleurites то1иссапа Willd. (Зондские острова). Из других масличных растений Индии интересны следующие ВИДЫ. К у н ж у т (Sesamum indicum L.) представлен в Индии огром- ным количеством экологических и морфологических форм. Ин- дия является, несомненно, центром наибольшего развития этого вида, хотя и не первичным (первичный центр развития рода Sesamum приурочен к Южной Африке). Ведическое название кунжута — tila. Его семена в древности употребляли в пищу в виде каши вместе с овощами или хлебом. С а ф л о р (Carthamus tinctorius 1 .) нигде не выявляет та- кого морфологического разнообразия форм, как в Индии. В этой области предполагается один из первичных очагов вхождения в культуру сафлора. Страны Передней Азии, по-видимому, отли- чаются большим экологическим многообразием. Сурепицы индийские (виды, близкие к Brassi~ca сат- pestris L.) — Индия может быть названа вторичным центром группы 20-хромосомных Brassica, примыкающих к В. сатре- 282
stris. Сюда относится тория (В. campestris с черными семе- нами). Зафар Алам (Zafar А1ат, 1945) классифицирует индийскую торию как В. campestris ssp. oampestris var. dichotoma Waff. Мы не принимаем его деления В. campestris на два подвида: campestris и na~pus, считая В. napus самостоятельным видом. В.,'oamp~estris в Пенджабе называется кали. Она напоминает формы сурепицы из Афганистана, Таджикистана и из Восточ- ного Китая. Вероятно, в Индию и Китай она расселилась в ка- честве сорного растения, а может быть перешла в Индию непо- средственно из Восточного Китая, может быть, в истории имели место и тот и другой пути расселения. Из группы 20-хромосомных Brassi~ca в Индии распространен еще один вид, по-видимому, эндемичный для Индии — с а р з о н, и иди й с к а я с у р е п и ц а (В. glauca Wittm.) — вид очень свое- образный и интересный. Имеет общие черты с В. campestris, с В. naipus и с В. jun~cea. Вместе с тем сарзон, несомненно, само- стоятельный ботанический впд. Сарзон дает пищевое масло в противоположность торин, которая дает в основном техниче- ские масла. Зафар Алам (1945) классифицирует сарзон (мест- ное пенджабское название pili), как В. trilocularis Hook. et Thompson. Неудобство этого названия в том, что имеются и двухствор- чатые, и трехстворчатые, и четырехстворчатые формы этого ра- стения, а из названия вытекает, что она всегда трехстворчатая. Мы, однако, не можем вступать здесь в таксономическую дис- куссию, акцентируем лишь на видовой самостоятельности "ар- зона, включая сюда и наиболее распространенные желтосемян- ные формы и более редкие буросемянные, которые, однако, не следует смешивать с торией. По Зафар Аламу, ч е р н а я г о р ч и ц а (Brassica nigra Косй) называется в Индии Poorbirai и интродуцирована в Пенд- жаб с востока. Зафар Алам нашел, что индийская форма В. nigra похожа на описанную нами (Е. Н. Синская, 1928) мало- азиатскую разновидность var. orientalis. Этот факт указывает на древние связи Индии со странами Передней Азии. В коллекции ВИР был один образец В. nigra из Индии с сильно рассеченными листьями, привезенный В. В. Маркови- чем. Он получил название В. nigra var. indica Sinsk. По-види- мому, индийская горчица, относящаяся к виду В. nigra, до- вольно разнообразна, но ее формы еще недостаточно изу- чены. С а р е п т с к а я г о р ч и ц а (Brassica juncea Czern.), по За- фар Аламу, широко культивируется в Индии под названием raya. Этот автор думает, что сарептская горчица была интроду- цирована из Китая сначала в Северо-Восточную Индшо, а за- тем из Индии была занесен» в Афганистан, вероятно, через Пенджаб. 283
яестные популяции В. juncea из Индии, по нашим наблюде- ниям, очень полиморфны, но разнообразие форм в Индии меньше, чем в Восточном Китае. Эта горчица возделывается также в Пакистане и Белуджистане. Популяции в последних двух странах также пестры по составу и похожи на индийские. Там в ее популяциях как примесь встречается близкий к В. jun~cea вид корнеплодной горчицы — В. napiformis Bailey. Последняя образует довольно крупные реповидные корнеплоды, очень разнообразные по окраске: от фиолетовых до белых. Состав европейских популяций В. juncea (Поволжье, Украина) гораздо менее пестрый, чем азиатских, и компоненты этих популяций большей частью другие. Ясно, что расселение этого вида горчицы шло из Азии с востока на запад. В. junicea имеет в Индии вторичный очаг формообразования, а первичный находится в Восточном Китае и Японии. Б е л а я г о р ч и ц а (Sin atpis alba 1 .), по-видимому, в Индии почти не возделывается, встречается в ботанических садах. Све- жие молодые листья этого растения употребляются для приго- товления острого соуса. Экспедициями ВИР доставлены из Индии и Пакистана образцы культурной горчицы — Brassica carinata А. Braun, абиссинской горчицы, или капусты. В нашем распоряжении нет пока сведений о культуре этого вида в Индии, но судя по об- разцам коллекции, это местные сорта. В. lear inata возникла син- тетическим путем — соединением геномов В. nigra (2n = 16) и капусты (2n = 18) . Капуста имеет средиземноморское проис- хождение, и, вероятно, В. carinata занесена в Индию из Эфио- пии, может быть, еше в древние времена. В. carinata в Индии довольно полиморфна. По нашим наблюдениям над коллекцией культурных кресто- цветных ВИР, в Индии распространена форма и н д а у (Eruca sativa var. orientalis Ь|пз1 .) с розеткой нижних реповидных листьев и палевыми, довольно узкими лепестками. Раньше мы встречали подобную форму (в Палестине) в образцах коллек- ции ВИР, собранных Н. И. Вавиловым. Нахождение этой формы в Индии указывает на древние связи ее со странами Передней Азии, расположенными на Аравийском полуострове. В Индии (в Пенджабе) по краям полей распространено ра- стение из сем. крестоцветных с местным названием rai Brassica tournefortii Gouan. Особенно часто оно встречается в централь- ных районах, которые Зафар Алам считал центрами древнего земледелия. Он предполагает, что указанное растение в древ- ности было в течение долгого времени в культуре, а в настоящее время сохранилось только в сорном состоянии. Он приводит све- дения, будто бы оно культивируется в Западном Тибете и в за- падном Средиземноморье (Испания и Италия) . Мы не встре- чали нигде в других источниках указаний о культуре В. tourne- fortii в Средиземноморье. 284
Вообще этот вид переднеазиатский, распространенный в Иране, в Закавказье (Талыш) и изредка в пустынях Средней Азии. Л и м о н н о е с о р г о jCymbopogon citratus DC., а также С. martini (Roxb.) Stapf, С. pardus (L.) Rendle] дает эфирное масло, получаемое из листьев, применяется в медицине и мыло- варении. Этот злак используется также при приготовлении острых соусов и для изготовления щеток. С этой же целью употребляются и некоторые злаки из рода Chrysop~ogon Trin. и Eragrostis Host (Индокитай, Южная Индия). В Малайзии и Индии разводятся также эфирномасличные ра- стения Vetiveria zizanioides Stapf и иланг-иланг jCananga odo- rata (Lam.) Hook. et Thorns.]. П а ч у л и jPogostemon ~patchouli Pell. (= Pogostemon heyneanus Benth.) ] имеют ареал: Индию, Малакку, Южный Китай, Индонезию, Филиппины, Сингапур, в Америке— Парагвай и Антильские острова. Родина — Бенгалия или Фи- липпины. Заменителями пачули служат следующие растения, также из семейства губоцветных: Urena lobata L. (Индонезия), Hyptis suaveolens Poit. (Индонезия), Microtaena robusta Hemsl. (Китай — Сычуань), Nicrotaena icymoca Prain = PleIctranthus patchouli С. В. Clarke (Бакбо, Гуандун, введена в культуру в Ассаме). Кар и а ндр (Coriandrurn sativum L.), по Е. А. Столетовой (1930), в Индии представлен особым индийским типом. Расте- ние эфирномасличное, лекарственное и овощное. В Индии возделываются также ж а с м и н (Jasminum grandi- Eolium L), сант а ло в ое дерево (Santalum alatum L.). Эти растения занесены в 1(итай, по-видимому, из Индии. Следующие растения из рода Атотит Roxb. доставляют ароматические семена: А. aromaticum ЯохЬ. (Бирма), А. subula- tum Roxb. (Бенгалия, Гималаи), А. krervanh Pierre (Камбоджа), А. echinosphera К. S~chumacher (Ява, 1(амбоджа) и др. Виды рода Cinnamomum L. происходят из Юго-Восточной Азии. Используются как ароматические и лекарственные расте- ния. С. cassia В1ите происходит из Индокитая, С. zeylanicum Nees — из Индии. Гв аз ди ч но е де р ев о jEugenia caryophyllata Thunb. (Caryophyllus aromatiicus L.) ] — Малезия. Т м и н и н д и й с к и й (Cuminum oyminum 1 .) растет дико и в культуре (Индия). Южная Азия богата пряными и возбуждающими расте- ниями. Из них отметим следующие виды и м 6 и р я: Zingiber officinale Rose., Z. mioga Rose., Z. zerumbet Rose. — пряное и лекарственное (Индия, Индонезия); 2. mloga встречается также в Японии. Перечислим виды растений, у которых используются ароматические корни: ш а ф р а н и н д и й с к и й — Cuncuma longa L. (Индия), С. aromatica Salisb. (Е~ейлон), С. ipierreana 285
Qagnep. (Индокитай), С. хапйоггЫуа Roxb. — лекарственное растение, как и ко стус.— Costus sipeciosus Smith. Вероятно„ первичный очаг развития рода Сигсита L. — Южная Азия. Виды галанги — Languas galanga Stuntz, Каетр1ег1а langa L.— и другие виды кемпферии считаются магическими. Из пряных и ароматических имеют большое значение следую- щие. Перец черны й (Рйрег nigrum L.) ~распространен в диком состоянии и культивируется в Индии и Индокитае, как и Р. betle L., Р. longum L. и другие виды. К а рд а м о н (Elettaria cardamomum Maton и Е. major Sm.) растет дико на Зондских островах и в Индокитае. О р е х м у с к а т н ы й (Myristica fragrans Houtt.) растет дико и культивируется в Индии, Бирме, Малезии и других стра- нах Южной Азии. Из растений, используемых для приготовления возбуждаю- щих напитков, остановимся на ч а е. Наиболее глубоко вопрос происхождения и первичного расселения чая освещен В. И. Кре- четовичем. К подроду Theamorpha V. Krecz. относится только один декоративный вид — Thea amplexicaulis Pit. (Бакбо). Под- род Euthea V. I(recz. делится В. И. Кречетовичем на 2 цикла: ) цикл Theoidea — не содержащие кофеина; к этому циклу от- носится только один вид — Thea taliensis W. W. Sm. (Юньнань); 2) цикл чайдающих — Theifera V Krecz. Последний цикл вклю- чает две группы (или ряда) — китайскую и индокитайскую. Первая группа китайская — ряд Amblyophyl lac V. I(recz.: 1) Th. bohea L. (Япония, Корея, смежные побережья Китая); 2) Th. viridis 1. (Юго-Восточный Китай — Юньнань); 3) Th. stricta Наупе (Средний Китай); 4) Th. сап1оп1епз1з Lour. (из- вестняки Гуандуна). Все это культурные виды. Возможным их диким прародителем В. И. 1(речетович считает Th. monticola V. I(recz., распространенный в горах Западной Юньнани и смеж- ной Сычуани. Это растение обильно произрастает в горных ле- сах и верховьях рек Салуэна и Меконга. Отсюда это растение, по мнению В. И. Кречетовича, распространилось речным путем к востоку и к югу (по долинам рек Меконга и Янцзы идут древ- ние торговые пути в Индию). Вторая группа видов чая индокитайская — ряд Urephyllae V. I(recz. содержит следующие виды: 1) Th. assamiica Mast. (Ас- сам, Восточная Бенгалия); 2) Th. sinensis indica Coh. St. (Манипур, Лушан); 3) неизученная группа шанских чаев. По предположению В. И. 1(речетовича, диким прародителем группы является Th. theaster V. Кгесу, растущий в 1(итае (Юнь- нань), Индокитае и соседних странах. Популяции дикого чая, по 1(речетовичу, довольно однородны. В Восточной Бенгалии центры древней культуры чая были поглощены джунглями, и там вид Th. assamica выработался из одичавших чаев. Настоя- щий дикий чай распространен только на Шанском плата 286
(Бирма) и в Тибетских Гималаях. Южный Китай и Восточная Индия имеют только сортовое разнообразие культурного чая. Таким образом, вопрос о происхождении чая и первичных очагов введения его в культуру сложен. По-видимому все-таки первичным очагом следует считать 10жную Азию, с включением в него и некоторой лесистой части Южного Китая. Род Thea L. представляет большой практический и теоретический интерес с точки зрения систематики и географии. Необходимы деталь- ные исследования этого вопроса. Из технических растений отметим следующие. Б а м 6 у к и — распространены дико и культивируются очень многие виды: Dendrocalamus asper Ba~cker, Gigantochloa apus Kurz, G. atter Кигу,, G. verticillata Мипго (Зондские острова) и др. К каучуконо сам и гутта п е р ч ен о сам относятся сле- дующие виды: 1) хевея бразильская — Hevea brasiliensis Muell., происходит из Южной Америки, но основная область ее куль- туры теперь находится на Цейлоне, полуострове Малакка и Индонезии; распространилась хевея из ботанического сада на Цейлоне; 2) Ficus elastica Roxb. — прежде имел значение в Индии как каучуконос; 3) гуттаперчевое дерево — Palaquium gutta Burclc. (Индонезия и Малайзия). Многие красильные растения разводятся в Индии: Indigofera tinctoria L. (может быть, основной очаг вхождения в культуру в Китае), Rubia tinctorum L. — марена, Lawsonia alba Lam.— хенна (культивировалась также в Древнем Египте, в Индии растет и дико), Curcuma longa L. и многие другие. Из лекарственных растений отметим следующие. Кассия (Cassia angustifolia Vahl) растет дико и возделы- вается. О~р ех стр ихни но вый (Stryichnos пих-vomica '.), кр о- т о н (Croton tiglium I..) — Индия, Малайзия. Ч а й п о ч е ч н ы й (Orthosiphon stamineus Benth.) — важное лекарственное растение, в настоящее время вводится в культуру в Сухуми. Родина его Индонезия, встречается в культуре и в Индии, как II Oronylum indicum (L.) Vent. Род р а у в о л ь ф и я (Rauwolfia L.) представлен многими видами в тропиках Азии, Африки и Америки. Раувольфия— R. serpentina Benth. — растение Юго-Восточной Азии, упоми- нается в письменных памятниках Индии за 1000 — 800 лет до н. э. Ценное средство при лечении гипертонии. Наибольшей мощности достигает в Гималаях. Растет в Пригималайских частях Пакистана и Индии, в Индонезии, Малайзии. Из овощных, корнеклубнеплодных растений связаны своим происхождением с Южной Азией или представлены там энде- мпчными формами и сортами следующие. Огурец (Cucumis sativus L.) представлен в Индии энде- мичными культурными сортами и формами, переходными к ди- 287
корастущим видам. Здесь же распространен предполагаемый ближайший дикорастущий родич — огурец Хартвига (С. hard- v ickii Royle), который С. Г Габаев считал подвидом (ss~p. agrestis Gab.) обыкновенного огурца. Многие авторы считают Индию родиной огурца. За родо- начальника при этом принимается С. hardwickii, описанный Рой- лем. А. И. Филов не считает это растение настоящим огурцом. Он обращает внимание на интересные примитивные формы гор- ных огурцов гималайского подвида — ssp. himalaicus Fil., кото- рые представляют собой экотип, приспособленный к условиям неравномерного увлажнения, с двумя разновидностями: ран- ней — индийская богарная (var. aephemeri~cus Fil.) и более позд- ней — гималайская пустотелая (var. squamosus Gab.). Воз- можно, что эти формы, распространяясь на восток и запад, играли известную роль в эволюции огурца. Новейшие исследования Э. Т. Мещерова и А. А. Залькална проливают свет на вопрос происхождения огурца. Для богарной разновидности гималайского подвида характерны не загниваю- щие семенники, как у культурного огурца, а его зрелые плоды— шарики — перекатываются по земле, как плоды колоцинта. Эти огурцы вполне съедобны и могли войти в культуру. Огурец Хартвига очень горек, и вхождение его в культуру трудно пред- ставить. Форма огурца, названная Э. Т. Мещеровым и А. А. Залькал- ном индийская мелкоплодная, близка к богарной разновидности, но еще ближе последней к культурным формам — она вполне съедобна, вкус хороший. Эта форма образует хорошие плоды гри гибридизации с культурными огурцами. Авторы думают, что такие гибриды могли появиться в Индии издревле. А. А. Залькалн считает, что описанные примитивные формы ги- малайского огурца — сорнополевые (размножаются они самосе- вом на полях и огородах), а огурец Хартвига — это дикарь и не является предком, а только родичем культурного огурца. Интересны соображения Залькална о наиболее древней форме огурца. По его мнению, такой была гермафродитная форма. Сейчас гермафродитные сорта огурца (например, Ле- ман) известны только в США. Иногда такие формы возникают как мутанты в гибридных популяциях — это реверсивные мута- ции («выщепление» древних форм) . Очень важны наблюдения многих авторов, что первые цветки у различных сортов огурца часто гермафродитные. Оказалось, таким образом, что у всех типов огурца первоначальная фаза (этап) морфогенеза цветка гермафродитная. Этот древний при- знак теперь перешел в рецессив. «Выщепление» гермафродит- ных форм чаще происходит у образцов огурца из Индии и Кита я. Если древняя форма огурца представляла собой лиану, то это была не лесная лиана, приспособленная к лазаншо по де- 288
ревьям, а лиана, ползущая по земле. Усы у огурца слабые- они не могут поднять его на дерево. Огурец, по мнению Залькална, произошел в сравнительно сухих районах с муссонным климатом — в центральных и запад- ных районах Индии и на юго-западе 1(итая. Первичный очаг вхождения огурца в культуру находится в Индии. В японском подвиде — ssp. indo-japonicus Fil. — А. И. Филов выделяет группу индийскую — ~proles indicus Fil.— и считает, что из Индии огурцы распространились на восток и запад. В Индии, кроме того, по А. И. Филову, имеются растения из сем. Cucurbita~ceae, плоды которых в некоторой степени за- меняют огурец: Trichosanthes anquina L.. Т. dioica Poxb., Т. cu- cumeriana L., Т. ipalmata Poxb. Плоды первых двух видов едят свежими, а двух остальных — вареными. К. И. Пангало (1956) родиной всех а р б уз о в (Citrullus Schrad.) считает Индию, Иран и Малую Азию. Однако эта тео- рия пока никем не разделяется. Несомненно, более обоснована позиция сторонников африканского происхождения арбуза. К. И. Пангало описал в Индии новый род Praecitrullus Pang. (Citrullus fistulosus Stol<es) . Мо м о р ди к а (Мотогйса charantia L.), по-видимому, впервые вошла в культуру в Индии и оттуда распространилась в Китай и Японию. Дико и в культуре очень распространена. Дыня (Cucumis melo L.) в Индии культивируется с древ- нейших времен. Там имеются и дикорастущие формы, которые употребляются в пищу. И вместе с тем в Индии встречаются всевозможные переходные формы от совершенно диких, прими- тивных до высококультурных, к каковым относятся канталупы. По А. И. Филову (1959, 1960), индийские дикорастущие дыни— Cucumis melo ssp. agrestis (Naud.) Fil.— поздние. Затем в Ин- дии возникла скороспелая дыня — var. dudaim Naud. Последняя разновидность отнесена А. И. Филовым (1960) к подвиду полу- культурных дынь — ssp. subspontaneum Fil. Следующим шагом было укрупнение плода и возникновение разновидности var. chito Morren. Следующей ступенью на пути к получению крупной дыни было появление индийской разно- видности — var. indicus Fil. Близки к упомянутым трем формам и некоторые из индийских канталуп — это уже высококультур- ные и сахаристые сорта. В Индии имеются своеобразные местные сорта тыкв вида Cucurbita moschata Duch., отличающиеся округлой формой, мягкокоростью и позднеспелостью. А. И. Филов (1960) объеди- няет эндемичные сорта мускатной тыквы в подвид ssp. indica Гil., распространенный только в Индии. Г ар лянка, или тыква бутылочная (Lagenaria vul- garis Ser.), представлена в Южной Азии многими сортами. Культура горлянки в Америке имела место до прихода туда европейцев. Не совсем ясно, где находится первичный очаг куль- Синская Е. Н. 289
туры, но большинство ботаников считает родиной этого растения Юго-Восточную Азию. Л ю ф ф а [Luffa acutangula (1.) Roxb.] — родиной считается Индия. Б а х ч е в о е р а с т е н и е (Benincasa his~pida Cogn.) напоми- нает дыню HJIH тыкву, культивируется по всей Южной Азии и заходит на север вплоть до Японии. Большого значения не имеет. Р е п а довольно часто встречается в культуре в Индии и Пакистане. Относится к виду Brassi~ca ra~poasiati~ca Sinsk. М о р к о в ь (Daucus carota 1 .) занесена из Средне-Юго- Западной Азии, как и бескаротиновые формы моркови. Часто встречается фиолетовая морковь, похожая на таковую в Афга- нистане. Виды китайских к апуст (Brassica chinensis L. и В..pe- kinensis Ruhr.) занесены из 1(итая. Виды Raphanus, напротив, аборигенны. Сюда относятся: редис индийский — R. indicus (L.) Sinsk., эндемичный ин- дийский вид (Индия); р е д и с з м е е в и д н ы й — R. caudatus L., вид, родственный R. indicus; в пищу употребляются крупные не- зрелые стручки (Зондские острова, 1Яейлон). Т а р о IColocasia antiquorum (L.) S~chott] — древнейшая клубнеплодная культура Индии. Там и в сопредельных странах имеется много местных сортов. Отсюда таро распространилась на север в Китай и Японию. Некоторые авторы полагают, что культура таро в Индии и Индокитае древнее риса. Имеются указания, что в Индии таро встречается в диком состоянии. Существует теория, что рис вошел в культуру из сорняков таро. Дикое таро распространено на пониженных местах и часто встречается вдоль дорог. 1(лубни питательны, содержат много крахмала. Молодые листья и черешки также исполь- зуются в пищу (заменяют шпинат, обычно их варят с содой, чтобы понизить кислотность) . Род к о л е у с (Coleus Lour.) в целом африканского проис- хождения, но в Южной Азии имеются эндемичные клубненосные виды: С. barbatus Benth. (Индия), С. tuberosus Benth. (Южная Индия, Цейлон, Ява, Малайский архипелаг). Следующие декоративные виды Goleus с окрашенной листвой имеют южноазиатское происхождение: С. aromaticus Benth., С. btumei Benth., С. ratundifolius А. Chev. et Perrot. Последний происходит из Африки (Судан), но на острове Ява возделы- ваются эндемичные формы этого вида. М и г о (Plectranthus tuberosus Blume) — древнейшее куль- турное растение Индии из сем. губоцветных. Используются клубни и листья (ароматическое и инсектицидное). Я м сы (Dioscorea L.) при возделывании на почвах с глубо- ким пахотным слоем образуют очень длинные корнеклубне- ПЛОДЪ|. 290
Я м с п о с е в н о й (Dioscorea sativa Ь.) культивируется в Индии, Японии, I<opec и Западной Африке. В Индии имеются эндемичные сорта. Я м с к р ы л а т ы й (D. alata 1.) произрастает дико и в куль- туре (Индия, Малайзия). Возможно происходит из Малайзии. D. aculeata L. в тех же районах, как и D. alata; D. ЫэрЫа Roth. — на Зондских островах; D. ~pentaphylla L. — дико и в куль- туре на острове Ява; D. bulbifera L. — Малайзия. Ямсы устойчивы к засухе. Они имеют довольно большое значение как клубнекорневые пищевые растения. Корпе- клубнеплоды их, содержащие 14 — 21'/о крахмала, едят варе- НЫМИ. Сур ан, индийский канья к [Amonphophallus сатрапи- latus (Roxb.) Blumej культивируется и встречается в диком со- стоянии в Индии. Суран распространен в тропической Азии и Африке. В Индии больше всего известен в штате Бомбей. клубни весят до 10 кг. Возделывается под покровом люцерны, арахиса, редиса и других растений. Вопрос о происхождении сурана и первичном очаге его возникновения не ясен. Т а к к а (Тасса Ipinnatifida Forst) — клубненосное растение (Индия, Индокитай, острова Тихого океана) . Возможно, это растение аборигенно и в Африке. Ал о к а з и я [Alocasia maicrorrhiza (L.) Schott. (Arum тас- rarrhizum L.)j растет дико и культивируется (Индия, Цейлон, Малезия) . Свекл а индийская салатная (Beta orientalis Roth) эндемична для Индии и прилегающих стран. Имеет тонкий корень. Но совокупности признаков близка к дикорастущему р астению. Шпинат м ала 6 арский (Basella rubra L ) — Индия, Цейлон. У к р о п и н д и й с к и й [Anethum sowa Roxb. (Peucedanum graveolens Wats.) ] — Индия. Салат индийский (Lactuca indiica L.) — Индия. Т м и н и н д и й с к и й (Carum roxburghianum Benth. et EI ook.) — Индия. П л у х е я [Pluchea indica (L.) Less.] — овощное растение (Индия) . Б а к л а ж а н ы (Solanum melongena L.) отличаются в Индии большим экологическим и морфологическим разнообразием. Здесь распространены мелкоплодные дикорастущие (возможно, сорные формы, но по конституции близкие к дикорастущим) и переходные к культурным формам. В Индокитае встречаются очень поздние, многолетние сорта. Отсюда баклажаны распро- странились на Аравийский полуостров и страны Восточной Азии Из Индии баклажаны, вероятно, в очень древние времена по- пали в Переднюю Азию, судя по нахождению там полукультур- ных форм (А. И. Филов, 1958). Исторический путь распростране- 291
ния баклажан, по А. И. Филову, можно представить следующей схемой. Схема распространения баклажанИндия Аравийский полуостров Индокитай Япония Другие соседние страныПередней Азии Северные районы куль-туры этого растенияв Европе lЕвропа 292 Некоторые исследователи принимают два первичных центра вхождения баклажана в культуру: Индию и Аравийский полу- остров. Из плодовых культур аборигенными видами и сортами в Ин- дии являются следующие. U, и т р у с о в ы е в Индии и прилегающих странах отличаются огромным разнообразием видов и сортов. Некоторые виды и сорта аборигенны для Индонезии и Малезии. Вероятно, ареал видо- и формообразования цитрусовых очень широкий и, по- мимо первичных центров, имеется не один вторичный. Горные районы Индии, Вьетнама и Юго-Западного Китая изучены еще весьма недостаточно. Согласно многим авторам, из Южной Азии ведут свое про- исхождение следующие цитрусовые. U,и трон, или л а йм (Citrus птейса L.) распространился в Средиземноморье в древние времена (Непал, Ассам, Бихар, Верхняя Бирма). О нем упоминает Феофраст. Очень популярен л а й м к и с л ы й (С. тейса var. acida), заменяет лимон, упо- требляется как приправа и салат и для изготовления различных напитков. Сладкий лайм имеет кисло-сладкие и сладкие плоды. Л и м он [C. limon (L.) Вигт.], вероятно, произошел в усло- виях культуры от индийской разновидности цитрона. Б ига рдия, помер а нец (С. aurantium L ) происходит из Восточных Гималаев. А пел ьсин сл а дкий [C. sinensis (L.) Osbeck] предпола- гается, что происходит из Восточной Бирмы, Вьетнама, но рас- пространен до Китая (Юньнань). М а н д а р и н ди к и й (С. indica Tanaca) — Индия. Из подрода Papeda в Южной Азии растут: С. latipes (Hook. et Thorn.) Тап. — дико в Северо-Восточной Азии (Хази) и
Бирме; С. micrantha West. — дико, в Филиппинах; С. се1еЫса 1(oord.— дико, на острове Сулавеси (Целебес); С. ma~croptera Montr. — дико в Малезии; С. hyxtrix DC. — в Малезии. В Южной Азии произрастают дико различные виды цитру- совых, а в Восточной Азии — только два: С. ta~chibana Tan. и С. depressa Hayata. Южная Азия, несомненно,— основная пер- вичная область видообразования цитрусовых, хотя сортовое разнообразие очень велико и в Восточной Азии, но по направ- лению к 1(итаю число видов уменьшается. Настоящие цитрусовые появились в ту эпоху, когда Австра- лия отделилась от континента, в верхнемеловой период. По мнению Танака и других исследователей, эволюция совре- менных цитрусовых шла от группы Papeda (Hassk.) Tan. к типу Aurantium (Tourn.) Тап. через Limonellus (Rumph.) Tan. (лайм) и Cephalocitrus (Tan.) Luss (шеддок). Эволюция этих ветвей проходила на склонах Восточных Гималаев и в приле- гающих местностях. Тип Citrophorum (Reeker) Tan. постепенно формировался на западных склонах Гималаев, возможно, через 1 1топе1 1из. Род Fortunella Swingle был создан позднее в индомалай- ском и китайском очагах. Наиболее интенсивное развитие важ- нейших групп имеет место на индийских склонах, а не на китай- ской стороне. КНТВНсКНН очаг цитрусовых вторичный. Эгле м ар мел адны й (Aegle marmelos Gorr.) отличается сладкой и ароматичной мякотью, напоминающей тыкву. Дает смолу и краску. Лекарственное. Произрастает дико в лесах Юж- ной Азии. Родиной считается Индия, но распространен и в Индо- китае. Слоновое дерево (Feronia elaphantum Corr.) употреб- ляется как овощное растение (незрелые плоды). Зрелые плоды употребляются с сахаром. 1(ультивируется и растет дико в Ин- дии и Бирме. Манго, мангустан и дуриан дают наиболее вкусные в Южной Азии плоды. Манго (Mangifera L.) — одна из древнейших и популяр- нейших плодовых культу.р Южной Азии. Упоминания о ней встречаются в древнейших мифологических и религиозных па- мятниках. Вошло в культуру 4000 — 5000 лет назад. Род Mangifera (сем. Ana~cardiaceae) содержит более 40 ви- дов. Ареал рода — Индокитай и Малайский архипелаг. Наиболь- шая концентрация видов — на Малаккском полуострове. Наибо- лее примитивные виды — М. duperreana Pierre и М. сосЫпсЫ- nensis Engl. — распространены в Индокитае. Основной культурный вид — М. indica L. дикие формы этого вида растут в Бирме и Северо-Восточной Индии. Очень высо- кие деревья дикого манго обычны в лесах Ассама. На этом же ареале встречаются родственные виды — М. silvatica Roxb. и М. саlопеига Kurz. В Бирме и соседних странах встречаются
также виды М. foetida Lour., М. caesia Jaclc. Количество куль- турных и диких разновидностей манго в Индии очень велико. Дикорастущие виды дают плоды худшего качества, но ши- роКо используются местным населением, особенно в неурожай- ные годы. В пищу идут как незрелые, так и зрелые плоды манго. Незрелые используются для приготовления острого соуса, маринада и освежающих напитков. Зрелые едят сырыми или приготовляют из них пастилу. Род м а н г у с т а н (Garcinia L.) тропический, происходит из Южной Азии. Культурные виды — мангустан горький— G. mangostana L. и мангустан сладкий — G. dulcis (Roxb.) 1(urz — считаются вкуснейшими из всех фруктов. Мангустан ра- стет только в районах с влажным тропическим климатом, при- легающих к экватору. Родственные дикорастущие виды — G. cowa L. и G. xantho- chymus Anders.— дают съедобные плоды, но гораздо хуже по вкусу, используются чаще на джем. Основной очаг рода Garci- nia — Малайский архипелаг. Д у р и о (Durio rithinus DC.) возделывается в садах и ра- стет обильно в диком состоянии в лесах (Бирма, Индонезия). Большое дерево с вкусными плодами, но с очень неприятным запахом. Потребляется в большом количестве. Б а н а н (Musa L.) имеет огромное значение в тропических странах. В Южной Азии, по литературным данным, распростра- нены: М. balbisiana Colla и банан обыкновенный — М. sapien- tum L. дико и в культуре (Индия, 1Яейлон, Индокитай, Малезия); М. aicuminata Coll a — дико (Индокитай, Малайский архи- пелаг); М. paradisiaca L. — в культуре (Индия). Диким праро- дителем или близким родичем считается М. acurninata. М. ca- vendishii Lamb., вероятно, также ведет происхождение из Юж- ной Азии. Есть указание, что родина этого банана — Южный Китай. Основная область развития рода Musa и вхождения в культуру банана — Южная Азия. Отсюда банан распростра- нился в Восточную Азию. 1(итайский банан — М. cavendishii— своеобразен. Это карликовое растение с красными (позднее жел- теющими) плодами, менее вкусными, чем плоды лучших сортов банана обыкновенного. Наибольшее разнообразие сортов ба- нана, по-видимому, имеется в Индии. Все культурные сорта банана имеют бессемянные плоды. Напротив, дикорастущий банан — М. silvestris Colla — близок к М. acuminata, имеет мел- кие съедобные плоды с большим количеством семян, что пред- ставляет интерес для селекционеров. Для банана имеется два линнеевских эпитета: М. paradisiaca и М. sapientum. Многие авторы считают, что существует только один вид: N. sapientum ssp. ~paradisiaca Bal<er, или М. paradi- siaca зэр. sapientum Schman. Е. Чисман (Cheesman, 1948 — 1950) реконструировал род Musa, принимая во внимание число хромо- сом, и поделил его на 4 секции. Все съедобные культурные сорта 294
включены им в секцию Eumusa. Он изучил также разнообразие двух широко распространенных видов — М. acuminata и М. bal- bisiana. Все индо-малайские бананы он поделил на 3 группы. 1. Преобладают признаки М. acuminata — это и есть родо- начальное ядро всей группы. Этой группе он предложил дать имя М. paradisiaca. 2. Во второй группе преобладают признаки М. balbisiana— родоначальника группы, и эту группу он предложил называть М. balbisiana. 3. Яля форм этой группы характерна смесь признаков М. aicuminata и М. balbisiana. Группе в целом он дал назва- ние М. sapientum. В эту группу входит меньше сортов, чем в остальные две. Между тремя группами существует и частично описано боль- шое количество переходных форм. Банан, по этому автору, имеет основной очаг происхождения в Малайе, но встречается и на Индийском полуострове. М. balbisiana имеет ареал: LleA- лон, Индия, Бирма, Таиланд, Малайзия, Ява, Филиппины, Но- вая Гвинея. Литературные данные показывают, таким образом, огромное разнообразие форм у видов банана секции Eumusa и неотчет- ливость границ между видами, что и является характерным для первичных очагов развития культурной флоры. Род Artocarpus Forst. (сем. Moraceae) происходит из Индии, Малезии, Южного 1<итая и Океании, где имеется до 40 видов. Наибольшее значение имеет хлебное дерево джек — А. integri folia L. Распространено в культуре в Индии, Бирме и во влаж- ных районах сопредельных стран. 1(рупные плоды прикреплены непосредственно к центральному стволу дерева или к основным крупным ветвям. Имеются сорта со сладким и с кислым вку- сом. Мякоть спелых плодов (соплодий) едят сырой или варят, сушат. древесина идет на постройки и из нее приготовляют краску (одежда буддийских монахов обычно окрашивается этой желтой краской). Собственно хлебное дерево — А. incisa L. — имеет плоды на концах ветвей. Плоды крупные, до 30 — 35 см в диаметре (ве- личина с арбуз) . 1~енятся больше сорта с бессемянными пло- дами. Соплодие содержит внутри крахмалистую мякоть, в кото- рой располагаются семена. Эту мякоть нарезают ломтями, а затем варят в воде или поджаривают — вкус несколько напо- минает мягкий сладкий хлеб. Едят также с рисом, с соусами. Этот продукт очень сытный и питательный (с высоким содержа- нием крахмала и сахара). дикие виды хлебного дерева А. chaplasha goxb. и А. lak~oocha Roxb. растут в лесах и также имеют съедобные плоды. Кл у 6 н и ч н ое де р ев о ICornus capitata (Wall.) Nakai]— плодовое дерево [Индия, Непал, заходит в 1(итай (Юньнань}]. Весь род Comus L. восточноазиатский. 295
А в е р р о а ( Averrhoa сагатЬо1а L.) относится к семейству Geraniaceae. Невысокое дерево, со съедобными плодами, похо- жими на ребристый лимон. Имеются кислые и сладкие сорта Едят в свежем виде, идет для джемов и желе. Распространено в Индокитае и на Молуккских островах. Другой вид— А. bilimbi L. — имеет плоды худшего качества. Аверроа редко возделывается — обычно используется из дикорастущих или одичавших зарослей. М и р и а л (Bouea burmani~ca Griff.) относится к сем. Ana- cardiaceae. Входит в состав джунглей и не вышел в культуру (Бирма, Индия). Кислые плоды используют для приготовления маринада, джема, компота, иногда используются как овощи и т. д. Считается укрепляющим организм продуктом. Хур м а и иди й с к а я (Diospyros discolor Willd.) имеет съедобные плоды, но не вошла в широкое употребление. Рас- пространена в Индокитае (Бирма). Этот вид близок к китайской хурме D. l~aki L. Последняя происходит из Восточной Азии и в странах Южной Азии встречается редко. Дикорастущий родич D. lotus 1. растет дико и в Южной Азии употребляется как ПОДВОЙ. 1(а ш та н индийский I'Castanopsis indica DC. (Castanea 1пйса ЯохЬ.)] относится и сем. Fagaceae. Растет дико в лесах. В Передней Азии и Средиземноморье плоды жарят, как каш- таны. И н д и й с к и й м и н д а л ь (Terminalia catapipa L.) отно- сится к сем. Combretaceae. Распространено в Индокитае и Ин- дии. Сладкие плоды заменяют миндаль в кондитерских изде- лиях. декоративное. Родственный вид Т. chebula Ketz. Незрелые плоды потребляют как овощи. Индия считается родиной сахарного тростника (Sac- charum officinarum L.). Последний в диком состоянии не найден. Родственные дикорастущие виды того же рода почти все сосре- доточены в Индии: S. barberi Jesw. (Пенджаб), S. spontaneum L. (Бенгалия, Пенджаб, Средняя Азия), S. robustum L. (Новая Гвинея) и др. В Бирме и других странах Южной Азии дикорастущие виды сахарного тростника очень распространены по берегам рек, где эти берега не затапливаются. Сахарный тростник весьма харак- терен для ландшафта и быта Южной Азии. Из его сока получают особый сахар, который называется «джаггри». Способ приго- товления — самый примитивный — увариванием в котлах. Плитки этого сахара похожи на наш «постный сахар», кроме сахарозы, содержит глюкозу, белок и другие примеси. Очень питателен, но не может долго храниться. Из него приготовляется очень популярный напиток, который продается на улицах и ба- зарах во многих городах. Многообразное употребление сахар- ного тростника в странах Южной Азии говорит о большой древ- ности здесь его культуры.
Сравнительно небольшое значение имеет другое сахаронос- ное растение — п а л ь м а с а х а р н а я (Arenga saccharifera 1.abil ). Родина его, вероятно, Южная Азия (Индокитай, воз- можно, Индонезия). Пальмы Corypha utan Lam., С. umbraculifera L., С. laevis А. Cheval., происходящие из Индии и Индокитая, дают сладкий сок, из которого получают сахар и вино; листья раньше слу- жили вместо бумаги для писания. Пальма Ni~pa fru icans Thunb. имеет большое значение в Индокитае, редко возделывается, больше используется из естественных зарослей. Сладкий сок этой пальмы употребляется для изготовления особого напитка «tari». Из листьев делают маты, циновки и другие мелкие до- машние изделия. Из твердой части плодов делают пуговицы. Пальм а п а л ь м и р с к а я (Borassus flabellifer 1..) дает пальмовый сахар, который употребляется как пищевой продукт, а также в медицине. Плоды в большом количестве продаются на базарах. Молодые проростки употребляют в пищу. Листья используют как кровельный материал и на различные мелкие изделия (шляпы, веера, зонтики и т. д.). Из черешков делают мебель. Использование этой пальмы ведет начало из глубокой древности. Вообще разнообразные пальмы имели большое значение в истории культуры аборигенов стран Южной Азии. По Б. Рюри- кову (1963), в древние времена в Индонезии высушенные паль- мовые листья (Рюриков называет «лонтаровую пальму») слу- жили вместо бумаги, на тонких листьях острым металлическим стержнем выцарапывали буквы, а затем втирали сажу. Полу- чалось нечто вроде татуировки; такое письмо сложно и требует большого труда — зато и живут эти рукописи сотни лет. Рюри- ков лично знакомился с подобными рукописями в музеях Индо- незии. П альмы с а го вы е (Metroxylon sagu Rottb. и М. rumphii Mart.) доставляют саго, заменяющее рис. Растут на Молуккских островах, в Новой Гвинее, Индонезии и на Филиппинах. П а л ь м а а р е к а (Areca catechu L.) — дико и в культуре (Малакка, Малезия, Бирма и реже в Индии; в Индии насчиты- вают 14 видов этого рода). Имеются санскритские названия. У населения стран Южной Азии распространено жевание кусоч- ков плода пальмы ape«a — бетельного ореха, который служит наркотическим и стимулирующим средствм. Обычай жевать бе- тель очень древний. Бетелем всегда угощают гостей. Для жева- ния берется лист перца Piper betle L., в него завертывают очень немного (мазок) гашеной извести и кусочек бетельного ореха. П а л ь м а с а л а к (Zalacca edulis Blume) дает съедобные плоды (Индонезия) . Род Phoenix L. распространен в тропиках и субтропиках Аф- рики и Передней Индии.
П а л ь м а ф и н п к о в а я (Phoenix dactylifera L.) имеет Индии близкого дикорастущего родича Ph. silvestris Rovh., который распространен до Северной Африки включительно. Близок к нему вид Ph. reclinata Jacq., обитающий в Северной Африке. Вероятно, все три вида, включая и Ph. daictylifera, про- изошли от одного предкового вида, обитавшего на широкой тер- ритории в Индии и в Северной Африке. Вхождение в культуру, вероятно, происходило независимо в разных пунктах этого боль- шого ареала. Вид Ph. dactylifera создался в культуре. Из женских соцветий п а л ь м ы в и н н о й (Caryota urens L.) получают сахаристый сок. Из сока приготовляют напиток. Сахар винной пальмы даже экспортируется из Бирмы. Это дерево куль- тивируется и растет дико в Индии, Бирме и на Малайском архи- пелаге. Пальма кокосовая (Cocos nucifera L.) имеет большое значение в Индии, Индокитае, Малезии. Масличное растение. Один из важнейших очагов распространения находится в Юж- ной Азии. Вопрос о происхождении кокосовой пальмы спорен. Некото- рые исследователи считают, что она занесена морскими тече- ниями из Южной Америки. В настоящее время кокосовая пальма представляет собой весьма характерное растение для ландшафтов достаточно влажных стран Южной Азии. Кокосо- вое масло здесь часто употребляется в смеси с арахисовым. Большое распространение имеет потребление так называемого кокосового «молока». жмых употребляется на корм скоту. Волокно межплодника ореха идет на канаты, коврики и пр. Оно служит также для набивки мебели и матрацев. Из скорлупы выделываются разные мелкие предметы (бусы II т. п.). Древесина используется для строительства зданий и на столяр- ные поделки. Корень имеет лекарственное значение. Листьями кроют крыши. Из них же делают плащи, шляпы, из черешков- маты. Кокосовые орехи употребляются при всевозможных рели- гиозных церемониях. Последнее показывает на болыпую древ- ность культуры кокосовой пальмы в Индии и сопредельных странах. Та м ар и нд (Тamarindus indica L.) — родиной этого расте- ния некоторые считают Южную Азию, а другие думают, что оно было сюда занесено из Африки. Очень распространенное дерево в Индии, Бирме и соседних странах. Бобы кислого вкуса. Незре- лые плоды употребляются как овощи, листья идут в суп и на салат. Сухие растертые семена прибавляют в табак, при изготов- лении местных сигар. К распространенным в странах Южной Азии плодовым отно- сятся виды рода Eugenia L.: Е. aquea Burm., Е. jambos L.— дают вкусные плоды, которые едят сырыми и в консервированном виде, декоративные растения (Бирма, Ява и другие острова, Ма- лакка); Е. javanica Lam. — дает менее вкусные плоды (Малай- 298
ский архипелаг); Е. malaocensis L. — дает очень вкусные плоды (Малакка, Малайский архипелаг, встречается в Бирме). На юге 10жноазиатской области развития культурной флоры встречаются и другие плодовые деревья из этого древнего рода. Л и т ч и к и т а й с к и й (Litchi chinensis Sonn.-Nep~pelium litchi Camb.) культивируется в странах Южной Азии, но занесен сюда из Китая. Его дикорастущий родич L. longanum Camb. встречается B лесах, используется как подвой. В литературе встречаются упоминания и о других видах того же рода. Ю ю б а (Zizyphus jujuba Lam.) сем. Rhamnaceae растет в изобилии дико в Индокитае, хотя многие авторы считают ро- диной ее Китай. Из плодов приготовляют пастилу, напитки, упо- требляют их сушеными. В лесах Бирмы и соседних стран растет ююба лекарствен- ная — Z. rugosa Lam., которая также дает съедобные плоды. К Зондским островам приурочен один из первичных центров развития рода Rubus L., в особенности подродов Malachobatus Focke и Chamaebatus Focke. Из культурных видов отметим Rubus niveus Thunb. (Индия, Гималаи до Сиккима, Цейлон, Южный Китай, имеется в культуре в Америке) . Малина я ванская (R. rosaefolius Smith) возделывается на Яве. Янко на Зондских островах, Филиппинах, доходит до юга Японии. R. ellipticus Smith, R. lasiocarpus Smith, R. mallu- censis 1. и другие родственные, мало изученные виды растут дико, чаще в горных районах. Встречаются в Бирме и соседних странах. Плоды используются местным населением. Богатство Южной Азии дикорастущими и культурными пло- довыми очень велико и далеко не исчерпывается видами, упомя- нутыми в нашем обзоре, который все же дает достаточное пред- ставление о богатстве и своеобразии культурной флоры Южной Азии. Из тропиков Восточной Азии получены очень многие декора- тивные растения: пальмы — Areca ~catechu, Arenga saccharifera, Phoenix silvestris, Corypha umbraculifera и другие; виды орхи- деи, Begonia rex Putz. и другие и однолетники — Celosia cris- tata L., Imipatiens balsamina L., виды Coleus и др. Южная Азия дала мировому земледелию такие важные куль- туры, как рис, чай, цитрусовые, банан, хлопчатник (G. arbo- геит), а также особый масличный лен, распространившийся в Средиземноморье и Эфиопию, виды фасоли, большое разнооб- разие форм лобии — Doli~~chos — и коровьего гороха — Vigna. Здесь предполагается родина голубиного гороха — Cajanus, ви- дов Canavalia и некоторых других бобовых. В Южной Азии со- средоточено огромное разнообразие сортов кунжута, сафлора, масличных крестоцветных. Здесь родина пачули и многих дру- гих ароматических растений, родина сахарного тростника и са- харной пальмы, кардамона, мускатного ореха, черного перца, гвоздичного дерева и многих других пряных растений. Здесь 299
также, по-видимому, находится родина огурца и многих энде- мичных клубнекорнеплодов. В Южной Азии культивируются и произрастают в диком со- стоянии виды плодовых: цитрусовые, манго, мангустан, дуриан, хлебное дерево, финиковая пальма, кокосовая пальма, саговая пальма, пальма арека и многие другие. Культурная флора Южной Азии богата эндемичными родами и видами культурных растений. Тропическая флора в Азии непосредственно переходит по Южному Китаю в субтропическую и умеренную. Многие куль- турные растения, зародившиеся в Южной Азии, в тропиках, при расселении на север B Китае дали там формовое и сортовое разнообразие (чай, цитрусовые, банан и др.). Поэтому для на- шей страны гораздо важнее изучение культурной флоры Во- сточноазиатской флорпстической области, чем Южной Азии. Наиболее резко выявляется связь южноазиатской культурной флоры с восточноазиатской. На обширной территории Юго-Восточной Азии еще в доисто- рическое время шел обмен культурными растениями. Из Индии в Индонезию перешли улучшенные формы цитрусовых, Eugenia jambolana, Zizyphus jujuba, Michelia champaca, Mimusops elengi 1., вероятно, Cassia fistula 1, Elaeocarpus ganitrus Roxb., Mesua ferrea L. и др. Из Индии сюда же распространи- лись сафлор и hibiscus abelmoschus L., жасмины; манговое де- рево также перешло в Малезию из Индии. Из Малезип в Индию распространились многие растения, например пачули — Pogostemon heyneanus Henth. С другой сто- роны, интересно отметить связь Южноазиатской области с Аф- рикой. Из Африки 10жная Азия получила кунжут, нуг (гвизо- тию), сорго; до сих пор не выяснено, в Индии или в Африке родина лобии, коровьего гороха, арбуза и др. Что касается льна, то, по нашему мнению, здесь вопрос ясен: лен распространился в Африку (Эфиопию) из Азии, точнее из Индии. Еще точно не установлено, является ли уроженцем Ин- дии или Африки вид Tamarindus indica. То же можно сказать о6 Eleusine coracana, Gossypium herbaceum, Musa iparadisiaca. Эти растения переходили с одного континента на другой. Eleusine скорей всего распространился из Индии в Африку, а G. herba- ceum и Musa, вероятно, наоборот,— из Африки. Сахарная канна (Canna L.) и таро (Colvcasia Schott) имели в истории обратный путь распространения: из Индии в Африку. Яиоскорид указывает, что в счастливой Ара- вии (Йемен) умели изготовлять сахар в I в. н. э. Той же доро- гой через Аравийский полуостров пришли в Индию Coleus ro- tundifolius и Pennisetum ty~phoideum (Виг~ . f.) 1. С. Rich. Вне зависимости от того, где был первичный очаг расселения и как шло распространение того или иного культурного расте- ния (из Азии в Африку или обратно), видно, что обмен ассорти- 300
ментом культур имел место в глубочайшей древности, ибо общие роды и виды культурных растений представлены в Азии и Аф- рике различными формами или видами, а это видо- и формо- образование требует большого времени. В Индии, несомненно, имеется вторичный очаг формообразо- вания кунжута, но кунжут в Индии очень не похож на африкан- ский. Здесь развилось огромное количество эндемичных форм и сортов этого растения. Из занесенного из Эфиопии нуга в Индии сформировался совсем особый раннеспелый экотип и т. п. Эти факты следует учесть археологам и антропологам, кото- рые в дальнейших своих исследованиях могут помочь ботаникам решить вопрос о времени начала обмена культурными расте- ниями между Африкой и Азией (Индией) . Южной Азии и, в частности, Индии свойственны виды корот- кого дня. В горах формируются расы (экотипы), которые П. Райвалс (Р. Rivals, 1953) называет короткодневными, но наи- более благоприятной для этих рас длиной дня является 15— 16-часовой день. Такие расы Райвалс обнаружил у следующих короткодневных видов: Momordica charantia, Cyclan ther a pe- da а (L.) Schrad., Luffa cylindrica, Phaseolus mungo, Vigna catjang, (Burm.) Wal~p., Crotalaria juncea и др.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ АФРИКИ Прежде чем приступить к изложению специального раздела, посвященного Средиземноморью, MbI предпосылаем краткие об доры культурной флоры древнего Египта и Эфиопии, значение которой во многих работах сильно преувеличивается. Мы же рас- сматриваем Египет как часть Средиземноморья, хотя и характе- ризующуюся несколько специфическими условиями развития. Эфиопия — это стык влияния двух флор — средиземноморской и африканской, а не самостоятельная крупная область развития культурной флоры. Египет и Эфиопия рассматриваются нами раньше, чем Средиземноморье, чтобы оттенить значение и осо- бенности истории последнего. Культурная флора Африки изучена недостаточно и содержит в себе очень много элементов чисто местного значения. Поэтому аборигенные африканские возделываемые растения рассматри- ваются нами сравнительно кратко (основное внимание уделено культурам, получившим более широкое распространение и лучше изученным) . ЗЕМЛЕДЕЛИЕ И КУЛЬТУРНАЯ ФЛОРАДРЕВНЕГО ЕГИПТА Самая древняя доисторическая египетская цивилизация- это тазианская (по местечку Der Tasa). Здесь были найдены кремневые орудия, керамика, в том числе полированная крас- ная посуда. Тазианцы погребали своих умерших в земле. Эти погребения и были исследованы. Не имеется раскопок жилищ, что затрудняет изучение тазианской культуры. По Баумгертель (Е. Baumgartel, 1947), весь нубийский культурный комплекс— способ погребения, кремневые изделия, посуда — все это похоже на доисторическую культуру Египта. Это одно из оснований, которое заставляет некоторых современных ученых предпола- гать, что раннее население Египта пришло с юга. Тазианская культура развивалась в Верхнем Египте, а не в долине Нила. Нижний Египет не производил в это время такой посуды: север Египта отстал в культурном развитии от юга. По Чайльду (G. Childe, 1952), главными занятиями тазиан- цев, или тасийцев, были охота и рыболовство. Однако у них существовали скотоводство и земледелие, носившие подсобный характер. При раскопках тасийских поселений (Г Чайльд, 1956) 302
были найдены зерна пшеницы — двузернянки (полбы) и ячменя. Тасийцы уже пользовались для перемалывания зерен в муку ручными зернотерками, которые имели вид плоских камней, до- стигавших 50 сл~ в длину и 37 см в ширину. На них зерно расти- рали с помощью двигавшегося взад и вперед камня-куранта раз- мером до 25 )( 11,3 )(7,5 см. Самостоятельными культурными очагами были: район Ме- римде Бени-Салам на западной окраине Дельты, Файюм — не- сколько южнее и Эль-Омари — на юг от Каира. Все эти очаги заглохли в конце неолита. Файюмские поселения были расположены по краям обшир- ного озера. Жители этих поселений сеяли пшеницу двузернянку (полбу), ячмень и лен. Хлеба жали серпами из зазубренных крем- невых пластин, вставленных в деревянные рукоятки. Зерно хра- нили в специальных ямах, устланных соломенными циновками, и мололи на ручных зернотерках. Имеются остатки плетеной корзины и редкотканного полотна (Чайльд, 1956). Здесь особенно интересны указания на культуру льна, кото- рая (если эти указания подтвердятся) была известна в Египте в глубочайшей древности. Жители Меримде, по Чайльду (1956), сеяли пшеницу дву- зернянку (полбу) и ячмень и пользовались такими же серпами и зернохранилищами, как в Файюме. Поселение Эль-Омари было исследовано в 1942 г. Оно рас- положено близ Гелуана, восточнее Нила. По Чайльду (1956), жители этого поселения сеяли пшеницу однозернянку, ячмень и вику, употребляли в пищу финики. По сообщению Тэкгольм (V. Tackholm) на Международном конгрессе по генетике в 1948 г., в Эль-Омари была найдена однозернянка, но последующими исследованиями (Е. Schiemann, 1954) это сообщение было опровергнуто. Были исследованы колоски и зерновки эпох III, VI, XI, XII династий. Анатомиче- ские и морфологические данные показали, что однозернянки в Древнем Египте не было. Следующая стадия египетской культуры — это бадарийская (от местечка Badari). Бадарийская культура является непо- средственным продолжением тазианской. Бадарийцы жили также в Верхнем Египте. У них была в употреблении медь. Их кремневые орудия были совершеннее, чем у тазианцев, посуда их была великолепна. Очень хороши и изделия из кости. Их скульптура — женские статуэтки — указывает на связь с восточ- ным Средиземноморьем. Чайльд (1956) указывает, что бадарийцы одевались в шкуры животных, но, кроме этого, они умели изготовлять довольно грубую ткань из волокна, похожего на льняное. В погребении находили животных: овцу, козу, собаку или шакала, нечто вроде быка. Подобные же погребения животных были обнаружены и в нубийских ископаемых кладбищах, что
также указывает на глубокие связи египетской культуры с бо- лее южными странами Африки. У бадарийцев, несомненно, было земледелие, вероятно, оно было и у тазианцев. Бадарийцы возде- лывали полбу и ячмень (G. Вгип1оп, 1928). Остается неизвест- ным, какие другие зерновые и волокнистые растения они возделывали. Остатки были слишком разложившиеся. По-види- мому, волокнистые растения — лен. Употреблялось касторовое масло, что не удивительно. Его могли добывать с дикорастущих растений клещевины. Неизвестно, были ли бадарийцы вполне оседлыми или полу- номадами. Раса коз и овец не была определена. Однако ни коза, ни овца не являются аборигенными для Северной Аф- рики. Согласно некоторым авторам, древнейшая раса овцы в Северной Африке была короткохвостая — ее родина в Иране. Другая раса появилась в Северной Африке позднее — это была длинноногая овца — Ovis longipes, распространившаяся сюда из Западной Азии, может быть, и из Передней Азии. Последняя раса была обнаружена на посудном орнаменте в эпоху На- када П, а раньше следов ее существования не было найдено. Одно достоверно: ни одна из этих рас не могла распространиться из дельты Нила — эти животные очень чувствительны к сырости и не могут существовать в болотной местности. Значит, они распространились в дельту из Верхнего Египта, а не на- оборот. Бадарийские костяные изделия были сделаны из слоновой кости, которая могла быть местной или была занесена с юга. У бадарийцев была найдена бирюза, есть предположение, что она сюда занесена из Синайского полуострова. Раковины, слу- жившие для украшения, имели происхождение из Нила, Крас- ного моря, но не из Средиземного моря. Чайльд (1952) считает бадарийскую культуру непосредственным продолжением тази- анской. Многие исследователи считают, что родиной древних египет- ских земледельцев была Сахара, но это не доказано. Чайльд отмечает наличие сходных черт в неолитических культурах Судана, Сахары и Египта и придерживается мнения, что египетский неолит — африканского происхождения, хотя нельзя совсем отрицать наличия в египетском неолите и азиат- ских элементов. «Во всяком случае, — пишет Чайльд, — мнение о мировом приоритете Египта в области сельского хозяйства не оправды- вается историческими фактами». Чайльд считает, что месопо- тамская и египетская цивилизации были самостоятельными, но, несомненно, также и наличие взаимодействия между ними. За бадарийской последовала цивилизация Накада 1. Раз- вилась ли эта цивилизация из бадарийской или ее основатели пришли с юга, неизвестно. Идентифицировать египетские и ну- бийские доисторические памятники очень трудно, так как многие 304
их особенности, присущие только раннему Егигту, бытовали в Нубии очень долго. Баумгертель замечает, что приходизшие с юга народы не всегда были выходцами именно из Нубии и не обязательно направлялись именно к дельте Нила, они направля- лись также в различные оазисы Ливийской пустыни, где в не- скольких местах обнаружены остатки культуры, сходной с древ- ней египетской из Файюма. Сама нильская долина, вероятно, не была обитаема до времени Накада П. Здесь были открыты целые мастерские кремневых орудий. Уровень культуры на- много выше, чем у бадарийцев, но орошения еще не было— климат пустыни, окружающей теперь Нил, был в те времена более влажным и позволял выращивать растения без орошения. Охота, судя по орнаментам на посуде, была главным занятием. Имеется картина, относящаяся к протодинастическим временам, с изображением природы Ливии — деревьев и стад домашних животных. Наиболее часто изображались гиппопотамы и рога- тые олени (может быть, это были антилопы) . Найдены были ри- сунки, на которых представлена охота на оленя с собаками. По- рода собак была определена — она и ныне существует у хамити- ческих племен Центральной Африки (это опять указывает на связи с югом) . Следующая за цивилизацией Накада 1 была культура На- када II, которая настолько отличается от Накада I, что, по Баумгертель, не могла развиться непосредственно из Накада I. Разрисованная посуда и кремневые лезвия указывают на тес- нейшую связь с Азией. Предполагается, что первоначальная область обитания накадийцев II была недалеко от страны шу- меров. Накадийцы II ввели в Египте письменность и были осно- вателями царства в Нижнем Египте. Хотя культуры Накада I и Накада II были созданы разными народностями, но между ними была преемственность. Таким образом, древнеегипетская цивилизация имела двоя- кие истоки: 1) юг Африки — Нубия и соседние страны на юге и юго-востоке; 2) Переднюю Азию — древнюю культуру Дву- речья. Влияние собственно Средиземноморья возникло позд- нее. Развитие египетской культуры в дальнейшем протекало на небольшой территории — в условиях плодородной долины Ниж- него Нила и его дельты. Плодородная почва, периодические раз- ливы Нила, ежегодно удобряющие илом долину, теплый ровный климат — все содействовало здесь раннему развитию орошения и еще в глубокой древности высокому развитию ирригационных сооружений и оросительной техники. Уже в IV тысячелетии до н. э. в Египте существовало для того времени высокоразвитое земледельческое хозяйство, основанное на орошении. Сравни- тельно высокий уровень земледелия уже в эту отдаленную эпоху говорит о том, что начало земледелия здесь относится к еще более древним временам. 305
Имеются основания считать, что древние египтяне возделы- вали пшеницу еще до появления у них календаря, т. е. раньше IV тысячелетия до н. э. Один тот факт, что зерна пшеницы были в Египте обнару- жены под слоем торфа, говорит о том, что первые шаги земле- делия в долине Нила восходят к глубочайшей древности, может быть, с тех пор прошел десяток тысячелетий. Однако на самок заре земледелия первобытному человеку было трудно справ- ляться с организацией хотя бы и самого примитивного ороше- ния. Ему, казалось бы, было не под силу регулировать мощныс разливы могучей реки. Поэтому существует предположение, что египтяне пришли в древние времена откуда-то из горной мест- ности или из предгорий. Воздвижение величественных пира- мид — гигантских священных гробниц египетских царей (фарао- нов) — объясняется, как «далекое воспоминание о древнем культе гор и скал» в доисторическую эпоху (Авдиев, 1948). Не- которые историки предполагают, что египтяне пришли в долину Нила с востока (через Синайский полуостров). С наступлением пустыни и оскудением растительности древ- нее население Северной Африки переходило в оазисы и речные ДОЛИНЫ. ~очевые охотничьи племена, уже приобщавшиеся к зем- леделию, основались в долине Нила и здесь нашли благо- приятные условия для перехода от охотничьего уклада к вполне оседлому образу жизни и развитому земледелию. Новейшие исследования, основанные на документальных данных археологической науки, как изложено было в начале этого раздела, рисуют ход событий несколько иначе. По-види- мому, древние предшественники египтян и нередко соседних племен Северной Африки (Ливии) пришли с юга, из Нубии и более удаленных, расположенных на юге и на юго-западе мест- ностей Ливии, а жители позднейшей из доисторических цивили- заций Накада П пришли с востока, из Азии. Накада П и предшествующие цивилизации были земледель- ческими: общий уклад быта, религиозные верования складыва- лись постепенно еще у доисторических цивилизаций Верхнего Египта и перешли к позднейшим обитателям дельты Нила— настоящим древним египтянам. Самые древние исторические данные характеризуют египтян как народ земледельческий, и это отразилось на всем строе мыс- лей древних обитателей долины Нила, в содержании его куль- тов, верований и обычаев. ~ульт растений и животных имел место у египтян в глубокой древности. K очень давним временам восходит и почитание Ози- риса — бога воды и растительности. Впоследствии Озирис стал богом умирающей и воскресающей природы — эти верования нашли отражение в погребальном культе, где значительную роль играли растения. 306
В эпоху древнейшего классового общества египтяне жили .небольшими общинами. В связи с ростом численности населения и поднятия общего уровня культуры оросительная сеть посте- пенно усовершенствовалась. Оросительная система имела перво- степенное значение в жизни египетского общества, создалась необходимость централизованного управления ирригационной сетью, функции которого исполняли сначала общины, а потом взяла на себя государственная власть. Сохранилось изображе- ние царя архаической эпохи — покровителя земледелия, который держит в руках мотыгу. Несмотря на такое раннее развитие ирригации, земледельче- ская техника была примитивной. Мотыга бытовала в Египте с доисторических времен вплоть до позднейших исторических времен Нового царства. Примитивный плуг также известен в Египте еще в архаическую эпоху и был распространен наряду С МОТЫГОЙ. Имеются указания, что плуг и лошадь были заимствованы египтянами у шумеров. Посев производился на слегка разрых- ленной земле. Семена затем втаптывались в землю копытами животных. Молотили также при помощи скота. Аборигенные растения Египта относятся к береговой расти- тельности дельты и долины Нила. Эта растительность некогда, когда климат Египта был более влажным, чем теперь, отлича- лась большим разнообразием и более пышным развитием, чем в настоящее время. В гробу мумии (V Laurent-Ta~ckholm, 1940) были найдены полевица белая — Agrostis stolonifera L., совсем не встречающаяся теперь в Египте, и многолетний райграс— Lolium ~регеппе L., который теперь там встречается очень редко; найден был также ячмень мышиный — Hcrdeum mari- пит Huds. Имели ли эти злаки ритуальное значение, в точности неизвестно. другой дикорастущий злак — Eragrostis pilosa Beauv., похо- жий На мятлик, встречается в очень многих раскопках разных эпох. Семена его были найдены в древней пирамиде Дахшур (Dahschur, 3359 лет до н. э.). Некоторые исследователи думали, что эти находки можно определить, как тефф — Eragrostis abys- sinica Link, который является важным хлебным растением Эфиопии. Ф. 1~ёрнике, а за ним и другие высказали сомнение в этом определении и пришли к заключению, что в Египте ис- пользовался в пищу дикорастущий родоначальник теффа— Е. pilosa. Последний является обитателем легких песчаных почв и находит себе много подходящих местообитаний на песчаных наносах в дельте и долине Нила. По Г Бушану (Ci. Buschan, 1895), в P,ðåâíåì Египте находил применение и другой злак — Eragrostis icynosuroides Roem. et Sichult. (Leptochloa bipinnata Retz.), в настоящее время извест- ный под названием альфа. Это растение у древних египтян при- менялось для изготовления корзин и сит для веялок. 307
И> прибрежной флоры в Древнем Египте имели большое зна- чение чуфа и папирус. Чуф а (Cyperus esculentus L.) является аборигенным расте- нием долины и дельты Нила. Сначала ее собирали ради богатых маслом и сахаром клубеньков в дикорастущих зарослях, а затем возникла и культура чуфы, которая и сейчас имеет в Египте не- которое значение. Округло-овальные клубеньки чуфы были обна- ружены в могилах Джебелин (Gebelin, 2400 — 1000 лет до н. э, от XI до XXI династии), Дра-Абу-Негга (Dra-Abu-Negga, вблизи Фив, XII династия), Дейр-эль-Бахри (Deir-el-Bahri, также не- далеко от Фив, 1650 — 1100 лет до н. э.). Клубеньки чуфы из древних могил были более округлые и мелкие, чем у современной египетской чуфы. На употребле- ние этого растения в Египте указывает Геродот. Последний сооб- щает, что чуфа росла в Египте близ реки на песчаных отложе- ниях. Клубеньки употреблялись вареными в ячменном пиве как лакомство. П а п и р у с (Cyperus papirus 1.) также в изобилии рос по бе- регам Нила. Это высокое тростниковое растение было харак- терно для прибрежного египетского пейзажа. Найдено много древних изображений папируса. Дерева в Египте было мало, и поэтому лодки часто делались из связок папируса, скрепленных особой склеивающей замазкой. Стебли папируса давали также уголь. Крахмалистые корневища папируса употреблялись в пищу в вареном виде. Но самое важное применение папируса, имевшее большое значение для египетской культуры, — это изго- товление бумаги (пергамента) из его луба. Письмо на папирусе было средством общения для людей в течение нескольких тысяч лет. Из растений того же рода в пищу еще использовалось С. auricomus Sieber (по Феофрасту, «sari»). Из водных растений в Древнем Египте был широко распро- странен лотос (Nyrnphaea otus I.). Об этой крупной красивой кувшинке Геродот писал, что она растет в реке и напоминает розу, а плод ее похож на гнездо ос. Семена лотоса, похожие на оливковую косточку, египтяне употребляли в пищу. Красивое растение Nelumbium nuciferum Gaertn. теперь не встречается в Египте, но в древности было распространено в прибрежных водах Нила и употреблялось в пищу. Феофраст писал о нильских растениях следующее: «речных растений в Египте вообще так много, что перечислить их по от- дельности каждое невозможно: вообще говоря, они все съедобны и сладкого вкуса». Вышеперечисленные растения были аборигенами Египта и их использование (а также, вероятно, и других дикорастущих растений — обитателей прибрежных песков, болот и мелких вод, о которых не сохранилось следов в археологии и истории) имело особенно большое значение в самую раннюю эпоху египетской культуры, когда еще собирательство главенствовало над земле- 308
делием. С течением времени усилилась связь Египта с сосед- ними и отдаленными странами, и в основном земледелие Египта развилось не на аборигенных культурных растениях. Доставлявшие важнейший продукт питания хлебные зерно- вые растения пришли в Египет из других стран. Сведения, сообщаемые В. И. Авдиевым в «Истории Древнего Востока» (1948) о том, что в соседних с Египтом странах, напри- мер в Эфиопии, и предположительно даже в самой дельте, были распространены дикорастущие виды зерновых — «предшествен- ники ячменя и пшеницы», — неверны. Культурные растения были занесены в Египет из Азии, Сре- диземноморья и Северо-Восточной Африки. Из зерновых злаков в культуре Древнего Египта, по-види- мому, одним из первых хлебных растений появилась полба (Triti~cum diicoocon). Это растение имело в Древнем Египте боль- шое хозяйственное значение (Schulz, Buschan и др.). В настоя- щее время полба в Египте не возделывается. В Средиземномор- ские страны, в том числе и в Египте, а также в Центральную и Северную Европу полба проникла в глубочайшей древности из стран Передней Азпп, включая Двуречье. Полба была найдена археологами в доисторических слоях Египта во многих местах: Бадари (Badari), Мончагадда (Nontagadda), в пустыне Файюм (Fayoum) и других пунктах, где велись раскопки. Э. Верт (Е. Werth, 1939) сообщает о находках зерен полбы в доисторических раскопках, между Каиром и Александрией, относящихся приблизительно к V тысячелетию до н. э. Среди этого материала встречаются и нетипичные зерна, напоминаю- щие и Т. turgidum, и Т. aest!vum, но сохранившиесп чешуи по- казывают на характер сочленения, как у Т. йсосооп. В этих же раскопках встречался и шестирядный ячмень. По Г Швейн- фурту, иероглифическое название ЬоЫ означает, вероятно, полбу- эм мер. Но сообщению Д. Персиваля (J. Percival, 1936), Г Брантон (G. Brunton) нашел кусочки полбы и ячменя в Бадари. Эти ос- татки относятся к додинастическому периоду — 5000 — 3500 лет до н. э., возможно, что эти зерновки были еще более раннего происхождения. Такой же примерно древности и те образцы полбы, которые найдены Г. Кэтон-Томпсон (Caton-Thom~pson) в пустыне Файюм недалеко от озера Биркет-Карун (Birket- Qurun). Согласно исследованию Персиваля, зерновки эммера из ba- дари, найденные Г Брантоном, довольно разнообразны. На од- ном из рисунков в статье Персиваля (193б) изображено одно округлое зерно, а другие — более или менее удлиненные (до сильно удлиненных); по-видимому, они несколько деформиро- ваны. Зерна эммера из Файюма также додинастического пе- риода — очень короткие, округлые. В Египте, судя по этим дан- ным, была остистая полба.
По мнению 1(. А. Фляксбергера, египетская ископаемая полба близка к современной эфиопской полбе. В Египте в глубокой древности также существовала голо- зерная пшеница. Бушан указывает для неолита Египта плотно- колосую круглозерную пшеницу — Т. аотрас1ит var. globitorme Buachan. По словам этого же автора, эта древняя голозерная египетская пшеница очень похожа на неолитическую пшеницу из свайных построек Швейцарии — Т. antiquorum Heer. В. Л. Менабде (1948) высказывает предположение о широком распространении древней примитивной пшеницы типа колхид- ской — Т. prot~omaicha — и альпийской — Т. antiquorum; какая-то из них была завезена и в Египет. Таким образом, ряд авторов указывает, что в gpeBHeM Египте, кроме полбы-эммера, была еще какая-то пшеница, на- поминающая мягкую. Ф. Грозный (F. Hrozny, 1914) относит к этой голозерной пшенице древнеегипетское название «swot». Унгер (F. Unger, 1857 — 1859, взято из Н. S. Reed, 1942) нашел не- большое количество голозерной пшеницы в кирпичах из раско- пок, датируемых около 3357 г. до н. э., которую он определил, как Т. vulgare antiquorum, т. е. счел ее за сходную с ископаемой пшеницей из неолита Швейцарии. Может быть, это была форма, близкая к современной абиссинской пшенице — Т. aethiopicum Jakubz., вернее, к ее предковой форме — Т. protoaethiopicum, близкой к Т. antiquorum. Сначала полба, а затем и древнейшие голозерные пшеницы, вероятно, пришли в Египет не с севера, а с востока — через Си- рию — Синайский полуостров. Современные твердые пшеницы Египта — proles aegyptiacum Vav. — скороспелые, низкорослые. Много колосьев пирамидаль- ного строения. Эти пшеницы, по Н. И. Вавилову, являются свя- зующим звеном между Т. durum ssp. abyssinicum, т. е. между формами Т. aethiopicum, похожими на твердые пшеницы, и со- BpeMeHHbIMH типичными твердыми пшеницами — Т. durum ssp. expansum Vav. Мы полагаем, что первые зерновые культуры — полба, яч- мень, Т. protoaethiopicum и другие — древнейшее население Египта (тазианцы или др.) имело еще в то время, когда обитало на юге от Верхнего Египта, получив их через Сирию (и, может быть, ту часть Аравии, где теперь Иемен). Переселившись в Верхний Египет, эти пришедшие с юга племена принесли с собой и культуру древнейших зерновых — полбы, ячменя, при- митивной голозерной пшеницы. Таким образом, мы считаем, что из голозерных пшениц в Египте в древности была распространена Т. pnotoaethioyi- cum — прародитель современной абиссинской пшеницы — Т. ae- thiapicum, которая в Египте со временем исчезла, а сохранилась в Эфиопии вследствие низкого уровня ее земледельческой куль- туры. 310
Т. protoaethiopicum, по нашему мнению, близкородственна колхидскому интегральному виду Т. protomacha (включая и 28-хромосомную Т. yrotopaleocolchicum). Таким образом, древ- ние африканские и в том числе древние египетские пшеницы не аборигенны, а имеют свои истоки в древних земледельческих культурах востока. Смит и Нетолицкий (по Peake, 1927), Лаурент-Тэкгольм (Laurent-Tackholm, 1940) исследовали содержание несколь- ких мумий додинастического периода (времена неолита) и установили, что пищу древних египтян составляли ячмень, про- совидное растение — Echinochloa соlопа Beauv. и клубни чуфы. Ячмень находили в доисторических слоях погребений в ba- дари и Монтагадда, в пустыне Файюм (в Эль-Каб); в устье Нила, в слоях каменного века, были найдены зерна ячменя, пшеницы и проса (последнее точно не было определено). Персиваль (193б) считает, что ячмень возделывался в Египте' за 10000, даже, может быть, за 15000 лет до н. э. В статье Персиваля имеется фотография ископаемых зерен ячменя ид пустыни Файюм, они мало похожи на ископаемый ячмень свайных построек Швейцарии, описанный Геером. В работе Е. Шиманн (S~hiemann, 1954) ячмени из Эль-Омари рассмотрены более подробно. Здесь были обнаружены четырех- рядные и шестирядные пленчатые ячмени. Последние очень по- хожи на ископаемый ячмень из свайных построек Швейцарии— Hordeum hexastichum sanctum Heer. В небольшом количестве найдены симметричные зерна, что указывает на возможность существования здесь и двурядных ячменей. Кроме того, здесь же обнаружены очень мелкие ячмен- ные зерна. Неизвестно, были ли это зерна сорного ячменя или мелкие зерна вышеупомянутых культурных ячменей. Подобные мелкие зерновки ячменя обнаружены в раскопках в трех геогра- фических пунктах: Трое, Эль-Омари и Ленгиеле(Schiemann, 1954). Интересно обнаружение в египетском неолите ископаемого ячменя, похожего на ископаемый швейцарский ячмень Геера. Этот факт говорит о том, что ископаемый круглозерный яч- мень — Н. hexastichum sanctum, как и ископаемая круглозерная пшеница — Т. antiquorum Heer, был широко распространен в эту эпоху. Ячмень и пшеница были найдены в пирамиде Дахшур (V династия, т. е. 4000 — 3000 лет до н. э.), у древнего города Eileythia (также эпохи Ъ династии); ячмень был найден в мо- гиле XVIII династии (lб00 лет до н. э.) в Тель-эль-Амарна (Tell-el-Àòàãnà) и в других местах. Во времена Геродота из ячменя в Египте приготовляли пиво (booza). Ячмень был пищей низших классов. Установлено, что в древнем Египте тогда возделывался шестирядный ячмень. Бушан определил древнеегипетский ячмень, как Hordeum tetrastichum var. coerules~cens и Н. hexasti~chum var. pyramidale.
Не имеется никаких доводов ботанического характера в пользу аборигенного происхождения африканских и, в частности, еги- петских ячменей. Из просовых растений уже упоминали об Echinochloa colo- na Beauv., зерна которой были обнаружены в кишках мумии доисторического периода (неолита). В настоящее время этотвид встречается в Египте только как сорняк. Он распространен также в Южной Европе до Испании и иногда культивируется в Индии. Это растение является ближайшим родственником ку- риного проса — Echinochloa icrus-galli. Последнее растение имеет, по мнению большинства ботаников, азиатское происхож- дение, что, вероятно, относится и к Е~сЫпосЫоа соlопа, кото- рая, возможно, занесена была в Египет из Индии. Неизвестно, возделывалось ли это растение в Древнем Египте или же ис- пользовались заросли сорняка. Просо (Paniicum miliaceum) и м о г а р, или 6 o p (Setaria italica) — также растения азиатского происхождения, в Древнем Египте они не возделывались. Имеются указания на возделыва- ние в Древнем Египте негритянского проса (Pennisetum spicatum), но полной уверенности в этом нет. Относительно культуры в Египте с о р г о о б ы к н о в е н- и о г о [Sorghum vulgare Pers. (S. durra) ] существуют разногла- сия. Согласно Бушану, остатки листьев и стеблей сорго нахо- дили в древних погребениях. Имеются также изображения частей растения сорго, указывающие на то, что части этого рас- тения входили в состав жертвенных даров. На одном древнем рисунке из гробницы Бени-Хассан (Beni-Hassan) изображена сцена жатвы сорго — так истолковывал этот рисунок Ф. Вениг (Wonig, 1886). Многие считали, что сорго возделывалось в древнеегипет- ский период с середины IV тысячелетия до н. э., и указывали на изображение жатвы сорго в гробнице Атепепйа. Беккер-Дил- линген (J. Beclcer-Dillingen, 1929) и некоторые другие авторы утверждают, что указание Вёнига неправильно и изображения на стенах древнеегипетских построек относятся не к сорго, а к другим растениям. Имеется даже мнение, что это лен. Послед- нее предположение делается на том основании, что, как видно из древнего рисунка, растения при уборке выдергивались с кор- нем. Однако на рисунке ясно видно, что изображенные там со- цветия на лен совсем не похожи, а гораздо более похожи на сорго. Вёниг указывает, что на стенном изображении растения, принимаемые им за сорго, были высотой с человеческий рост, стебли были светло-зеленые или желтые, а соцветия красные. По Линденбейну (W. Lindenbein, 1941), сорго в Древнем Египте при уборке могли вырывать с корнем, если хотели использовать целые стебли — такой способ уборки сорго и теперь практикуется в Эфиопии. Однако Линденбейн указывает, что очень древних названий для сорго в Египте нет. 312
По всей вероятности, в Древнем Египте сорго возделывалось, но не являлось там растением аборигенным, а пришло туда или из Азии, или из Юго-Восточной Африки (последнее предположе- ние более вероятно) . В гробу египетской мумии, относящемся к доисторической эпохе (Laurent-Tackholm, 1940), найдены остатки клевера. Среди них был Trifolium medium, который теперь в Египте не встре- чается (или встречается чрезвычайно редко, и мы сомневаемся в точности определения этого вида по ископаемым остаткам). В этом же гробу был найден другой вид клевера — шабдар— Т. resupinatum L., который и теперь распространен в Египте в диком или, может быть, в одичалом состоянии. Бушан указы- вает на существование в Древнем Египте и клевера алексан- дрийского — Т. alexandrinum L. Это растение дико не растет в Египте, родом оно из Передней Азии и проникло в Египет от- туда. Какое было значение этих клеверов в Древнем Египте, неизвестно. Из пищевых бобовых растений как, возможно, аборигенную культуру отметим л ю п и н — Lupinus termis Forsk., который произрастает дико в Египте (Boissier, 1867), а также на остро- вах Сицилии, Сардинии и Корфу. Боб этого растения был най- ден в раскопках, относящихся к эпохе ХХ — XXVI династии, но определение нельзя считать вполне достоверным. На древне- еврейском языке нет названия для люпина. Отсюда Декандоль делает заключение, что это растение вошло в культуру в Египте после изгнания евреев. Швейнфурт (G. S~chweinfurth, 1884) ду- мает, что 1 . termis появился в Египте в греческую эпоху. По Бушану, в настоящее время семена этого вида люпина мочат в соленой воде и затем варят. Есть предположение, что он входит в число продуктов, из которых в Египте приготовлялось пиво. Швейнфурт нашел в гробнице XII династии семена люпина белого — L. albus L., который, возможно, также употреблялся в пищу древними египтянами. Ч е ч е в и ц а (Lens esculenta Moench), по свидетельству Ге- родота, Феофраста и других, широко употреблялась в Древнем Египте. Ее нашли в гробнице Среднего царства Дра-Абу-Негга (XII династия) . В памятниках Древнего царства зерновых бобовых не было найдено. Чечевицей питались строители пирамид — вообще это была пища низших классов общества. По Д. Г. Редеру (1950), чечевица в эпоху Нового царства упо- минается только три раза: в папирусе Анастаси IV (о запасах чечевицы) и два раза в отчете Унуамона о путешествии в Библ. Во времена Птоломея чечевица имела, несомненно, большое значение в Египте. В городе Арсиное (Файюмский оазис) была улица, носившая название «продавцов чечевицы», в дельте был город, носящий название Facusa (греческий корень — facus— чечевица). 313
В римскую эпоху чечевица экспортировалась в Рим из Египта; Плиний говорил о чечевице, как о египетском продукте, он писал о двух египетских сортах. Имеются древние изображе- ния корзин с чечевицей (Бушан). Существовал обычай варить чечевицу с маслом и чесноком. По Жибо (Gibault, 1912), древняя египетская чечевица не отличается от современной. В Лувре, по его данным, имеются три хорошо сохранившихся семени чечевицы из египетских гроб- ниц. Они «вполне аналогичны мелкосемянной разновидности, ко- торая и теперь сеется на севере и на востоке Франции под на- званием чечевицы красной (Lentille rouge)». Чечевица пришла в Египет с востока, из Передней Азии. 06 этом говорят все ботанические данные. Швейнфурт думает, что чечевица про- никла в Египет из стран, расположенных по Евфрату, хотя ассиро-вавилонское название ее пока неизвестно. Хотя чечевица и была распространенным и любимым пище- вым растением в Палестине, но это еще, конечно, не означает, что чечевица занесена в Египет евреями. Лоре (Loret, 1892) писал, что еврейское название чечевицы adasim не египетского корня. Чечевица пришла в Египет, если и из Палестины, то до заселения ее семитами, а может быть, она была занесена из дру- гой переднеазиатской страны, возможно, из Малой Азии. Имеются указания (G. Buachan, 1895) на возделывание в Древнем Египте ч и н ы (Lathyrus sativus L.) . .Швейнфурт опре- делил находку в древнем погребении эпохи ХХ династии как чину, Но Бушан считает сомнительным такое древнее существо- вание чины в Египте и указывает, как на более достоверные, на более поздние находки в @pa-Абу-Негга и Джебелин. Родина чины — Передняя Азия (области каспия, Армения, Причерно- морье). Отсюда и пришла чина в Египет. Э. Верт (Werth, 1939) указывает, что в раскопках между 1~аиром и Александрией, относящихся к V тысячелетию до н. э., найдены зерна го р ох а, который им определен как Pisum sa- tivum var. elatius Beck. Размеры этих зерен колебались между .следующими пределами: 3.)(3'/4, 3'/5 )(3'/з мм. Верт считал, что это была культурная (но приближающаяся к дикорастущей) форма гороха. У древних авторов указаний на нахождение го- роха в Древнем Египте нет. Это говорит о том, что горох там был мало распространен. В тех же раскопках были найдены семена вики (Vicia зр., но не V. sativa) и семена, похожие на мелкосемянную форму к о н с к и х 6 о 6 о в, в точности определения которой автор не уверен. Современные египетские (из Верхнего Египта) формы гороха отнесены Л. И. Говоровым к другому подвиду — ssp. asiaticum Gov., чем абиссинские и средиземноморские, oáe последние группы сортов отнесены к подвиду «обыкновенного» ropoxa— ssp. commune Gov. В составе азиатского подвида Л. И. Говоров 314
выделил египетский горох в особую агроэкологическую rpynny— proles aegyptiacum Gov., однако не обладающую особо резкими отл ичи ям и. К о н с к и е 6 о 6 ы (Faba vulgaris Moench) в настоящее- время в ОАР и Судане возделываются и широко употребляются местным населением в пищу. В древнеегипетских погребениях эпохи XII династии были найдены 2 семени конского боба без всякого сомнения в их оп- ределении. По Геродоту, египтяне питали отвращение к конским бобам. жрецы не позволяли себе даже смотреть на них. Редер (1950) подвергает сомнению вывод о повсеместном запрещении бобов в Египте, он полагает, что «табу, наложенное на бобы, ограничивалось определенной местностью и определенным куль- том», а Геродот частный случай распространил на всю страну. Конские бобы распространены по всему Средиземноморью, но вряд ли они в Египте являются первичной культурой, скорее культура бобов пришла сюда из стран Передней Азии. Голубиный горох (Cajanus indicus Spreng.) был в упо- треблении у древних египтян. Семена этого растения были най- дены в раскопках погребения XII династии в Дра-А6у-Негга. Родина этого растения, по одним авторам, Восточная, может быть, и Южная Африка, а по другим — Индия (последнее более вероятно) . К о р о в и й г о р о х (Vigna sinensis Endl ) был введен в культуру в Египте после эпохи фараонов. Таким образом, зерновые бобовые растения не являются або- ригенами Египта, как и зерновые злаки, и появились там значи- тельно позднее последних. Из технических растений древнего Египта, кроме папируса, о котором уже было сказано, заслуживает особого внимания лен. По сообщению Леггета (W. F. Legget, 1945), в египетской мифологии лен был первой вещью, сотворенной для себя богами перед их появлением на земле. Полотно было найдено в могиле Менеса — это погребение относится к 1 или ко II династии, т. е. к очень древней эпохе. Унгер (F. Unger, 1866) нашел семена и луб льна в пирамиде Яахшур (V династия) . По Леггету, раскопки древних погребений выявили хорошо развитую тех- нику изготовления ниток и тканей из льна в неолити- ческую эпоху. В египетских могилах, датируемых около 3400 лет до н. э., были найдены не только остатки льняной ткани высокого качества, но и каменный барельеф, изображающий примитивный горизонтальный ткацкий станок без челнока. Около 4000 лет до н. э. в Египте уже изготовляли прекрасное льняное полотно, похожее на тончайшую кисею. Его носили вы- сокопоставленные египтяне на торжественных церемониях. В очень древних могилах также эпохи 1 и II династий, в А6у- сир-эль-Мелек (Abusir-el-Melek) Меллер нашел коробочки льна 315
вместе C семенами. Такие же находки имели место и в раскоп- ках, отнесенных к позднейшим периодам. Например, Швейнфурт нашел коробочки с семенами и стебли льна в могиле XII дина- стии. Однако в древнейший период льняные ткани еще не были так широко распространены, как в последующие. В пирамиде Менкаура (IV династия) мумия еще одета в шерстяное покры- вало, а в более поздние времена мумии всегда завертывались в льняные ткани. Приблизительно через тысячу лет лен приоб- рел очень большое значение, о чем говорят многочисленные изо- бражения и упоминания о нем в надписях. В гробнице Сциум- неса (в Chum-el-Achmar) изображена уборка льна. В погребении Бени-Хассан показаны процессы обработки льна и изготовления тканей: мочка, сушка, трепание, прядение, тканье. Египтяне торговали льном с Индией, Персией, позднее с Гре- цией и Римом. Впоследствии фараоны организовали большие царские фабрики в Фивах, Мемфисе и в других городах, где при- менялся рабский труд. Для окрашивания льняной ткани упо- треблялись растительные краски, которые египтяне умели изго- товлять еще за 2000 лет до н. э. И вместе с тем, справедливо замечает Леггет: .<Египет нельзя назвать родиной льна, но можно назвать его колыбелью». По мнению некоторых историков, искусство изготовления льняных тканей пришло в Египет из стран Двуречья (Вавило- ния). По указаниям Бушана, в Древнем Египте возделывались две формы льна: 1) Linum usitatissimum vulgare Schiibl. et Маг1.; 2) L. humile Mill. = L. tcrepitans Вёеп. Лен-кудряш — L. humile — вовсе не соответствует льну-пры- гунцу — L. crepitans Dum. Сомнительно, чтобы лен-прыгунец действительно возделывался в Древнем Египте, этому нет ни- каких доказательств. Это растение приурочено более к западной части Средиземноморья. Вероятно, как и в настоящее время, в Древнем Египте возделывались льны низкорослые и ветвистые, или кудряши, а также формы более высокорослые и более при- годные для получения волокна — межеумки или полумежеумки (высокорослые и сравнительно слабо ветвящиеся кудряи1и). Ис- копаемые египетские льны из погребений Дра-Абу-Негга и Абд-эль-Курна (Abd el-Qurna) имели высоту коробочки 8,8 мм, а длину семени 5 мм. Следовательно, в древности в Египте воз- делывали, как и теперь, крупнокоробочные льны. Египетские современные льны не отличаются существенно от крупнокоро- бочных и крупносемянных средиземноморских льнов, распро- страненных не только в Египте, но и в Алжире, Марокко и дру- гих местностях Средиземноморья. Этот лен обнаруживает бли- жайшее родство со льном индийским, которому присуще боль- шое разнообразие и который, на основании многих соображений, является более древним. Несомненно, что в Египет и другие 316
страны Средиземноморья льны занесены из Индии. Однако в Индии лен возделывался как в древности, так и теперь только на масло, а льняные ткани привозились в Индию и Китай из Египта. Имеются указания, что и хлопчатник был известен в Древ- нем Египте. Семена его были найдены в одном сосуде из Фив и были определены как Gossypium religiosum L. Хлопчатник был, вероятно, мало употребителен в Древнем Египте. Геродот и Феофраст не упоминают о египетском хлопке. О дороговизне хлопка в Египте во время XIX династии (1400 летдон. э.) го- ворит следующий библейский эпизод. При возвышении Иосифа Прекрасного фараон пожаловал ему, как первому сановнику государства, хлопчатобумажное платье, а не льняное или шер- стяное. О растениях, используемых в Древнем Египте на масло, из- вестно следующее. К л е щ е в и н а (Ricinus miicrocarpus G. Pap.) возделывалась египтянами, о чем говорят неоднократные находки археологов и свидетельства древних авторов (Геродота, Феофраста и Диоско- рида). Масло употреблялось для освещения и, может быть, для медицинских целей. Мелкоплодная клещевина дико растет в Северной Африке и здесь входила в культуру, вероятно, также и в Египте, но все же это средиземноморское растение, а не специально египет- ское. Kу н ж у т (Sesamum 1пйсит L.), по предположению Бу- шана, попал в Египет в предгреческое время, а по Декандолю,— в греческую эпоху. Нет явных доказательств возделывания его во времена фараонов. Феофраст, Диоскорид и Плиний говорят, что кунжут возделывался в Египте, при этом Плиний указывает на Индию как на родину этого растения. По свидетельству Феофраста и Страбона, в Египте употреб- .лялось оливковое масло. Время начала культуры о л и в ы- Оlеа eurapaea L. — в Египте с точностью не установлено. Масперо (по Бушану) находил в текстах еще VIII династии слово tat и думает, что это было обозначение маслины и олив- кового масла. Швеинфурт считал, что олива была известна только, начиная с XI династии (3000 лет до н. э.). Лоре отодви- гает эту дату до XV111 династии (1500 г. до н. э.). От времен позднейших династий осталось много изображений ветвей и пло- дов оливы, но они не во всех случаях позволяют с достаточной "точностью определить в них изображение именно оливы. Археологические остатки более надежны. Косточки оливы найдены в одном из саркофагов Дра-Aбy-Негга (XII династия). В гробчице Тутанхамона были открыты гирлянды с вплетенными в них листьями оливы. Гирлянды из оливы были найдены в по- гребениях ХХ — XXVI династии. Много остатков открыто в слоях греко-римской эпохи.
Таким образом, можно считать установленным, что культура оливы существовала в Египте примерно за 1000 лет до н. э., может быть, и несколько раньше, но в Древнем царстве ее не было, и она, конечно, отнюдь не является в Египте аборигенным растением. Никаких доказательств последнего нельзя привести. Для решения вопроса о более или менее широком распро- странении оливы в Египте трудно пользоваться лингвистиче- скими указаниями. Слово tat или dt (соответствует коптскому djoit, еврейскому zait или zeit, арабскому zeitum) бесспорно является названием оливы и говорит о том, что олива пришла р Египет с востока — из Аравии, возможно, из Палестины (Ре- дер, 1950). Однако имеется еще одно и более древнее назва- ние — bak. Bak — было названием какого-То масличного расте- ния, которое имело большое экономическое значение и давало много масла. Но античные авторы указывали, что в Египте про- изводство оливкового масла невелико (Страбон, Плиний) . Согласно предположению Лоре (Loret, 1892), название bak относится к другому дереву — Moringa aptera Gaertn., которое росло в восточной части Египта и в Аравии (по Феофрасту и Диоскориду, — Balanos, по Плинию, — МугоЬа1апит) . Лоре указывает на факт нахождения одного зернышка из плода этого дерева в гробнице Дра-Абу-Негга. Миробалановое масло употреблялось для умащения и для лекарственных целей.. Д. Г. Редер считает, что «гипотезу Лоре нельзя игнорировать». Была ли или нет культура Moringa aptera в Египте, во вся- ком случае она не была там аборигенным растением, а была за- несена из Аравии (Иемен) или из Эфиопии. Для умащения в Египте употреблялась смола мирра. Мирру получали из Balsamea myrrha Baill. и других аравийских и африканских деревьев, относящихся к сем. Burseraiceae. Muppa доставлялась в Египет из южных областей, по Д. Г. Редеру,— главным образом из Пунта (нынешняя Эфиопия и Сомали) не- посредственно, а по Д. Г Брестеду, — через промежуточные племена вниз по Голубому Нилу, Атбаре и Белому Нилу. Реде- ром (1950) собраны сведения о попытках перевозки небольших мирровых деревьев из Пунта в Египет в кадках с землей. Однако и по Редеру остается неизвестным, насколько опыты с культурой мирры увенчались успехом. Тем не менее, эти сведения очень интересны, так как показывают любовь египтян к занятию ин- тродукцией чужеземных растений. Археологические данные определенно говорят о культуре в Древнем Египте с а ф л о р а (Carthamus tinctorius L.) . .В иеро- глифических текстах сафлор называется nas, nasi или nasti. Слово nas было встречено в надписи на пирамиде фараона Тета VI династии. В Древнем Египте знали сафлор больше как красильное, чем масличное растение. Из сафлора получали черно-коричневую краску. Мумия фараона Аменхотепа (XVIII династия около 1600 лет до н. э.) была украшена гир- 318
.ляндами из цветков сафлора, подобное же украшение было .найдено на другой могиле в Дра-Абу-Негга. Плиний упоминает и о сафлоровом масле, называя его «египетским». По исследованиям А. И. Купцова (1932), египетский сафлор и сафлор Судана очень разнообразны и по богатству форм не уступают сафлору Индии. А. И. 1(упцовым выявлены следующие экотипы сафлора: 1) экотип Верхнего Египта; 2) экотип Ниж- него Египта; 3) экотип Верхнего Египта и Судана. Таким образом, ботанические данные показывают, что в Древнем Египте сафлор мог быть и первичной культурой, так как там имеются и ero ближайшие дикорастущие родичи. По .Декандолю, родина сафлора находится на Аравийском полуост- рове между Индией и Египтом. Современная область развития рода Carthamus L. — Передняя Азия, а область наибольшего экологического разнообразия — Передняя Азия, Египет, Судан и, по-видимому, на втором месте — Индия, где больше выражено морфологическое разнообразие, чем экологическое. I(ар и андр (Coriandrum sativum L.) употреблялся в древ- неегипетской кулинарии, а также в медицине. Находимые архео- .логами изображения, начиная с V династии, показывают, чтоэто растение имело большое ритуальное значение и фигурировало в жертвенных приношениях и в погребальных обрядах. 0 куль- туре кориандра говорят и позднейшие находки в Дейр-эль-Бахри (XXVII династия) и в раскопках греко-римской эпохи. Сущест- вовали и иероглифические названия этого растения ounshavu или wenshivu и для семян — ounshi. 1(ориандр также широко употреблялся в египетской медицине при болезнях печени, ки- шок и др. Культура кориандра была в древности широко распростра- нена и не имеется оснований, чтобы считать ее первичной в Египте, как и а н и с а — Pimpinella anisum L. Дико ни анис, ни кориандр в Египте не растут. Лоре допускает, что анис появился в Египте ко времени XVIII династии. Существует египетское название этого растения — maquae или та1<и. 0 добывании эфирного масла из этих двух растений в Египте данных не имеется. Е. А. Столетова выделяет североафриканский экотип ко- риандра, который, впрочем, распространен не только в Египте, но и в Эфиопии (наряду с собственным, более позднеспелым экотипом Эфиопии и Эритреи) В папирусе Эберса упоминается также и ф е их ел ь (Foeni- си1ит vulgare Mill.) . Это растение средиземноморского или, может быть, переднеазиатского происхо>кдения. Семена г о р ч и ц ы п о л е в о й (Sin ap's arvens's L.) были най- дены в погребении Дра-Абy-Негга. Этот вид горчицы и в настоя- щее время широко распространен в OAP как сорное растение. Есть указание, что в Древнем Египте возделывался ш а ф- р а н (Crocus sativus L.).
В ранний период развития культуры Египет, по-видимому, не знал м а к а (Pa~paver somniferum L.) . .Во времена Плиния мак был у египтян лекарственным растением. Вероятно, он был занесен в Египет, как и в Грецию, из Малой Азии. А ни с также упоминается в папирусе Эберса как желудоч- ное средство. В египетской медицине употребляли многие из тех растений„ которые имели и другое назначение. В папирусе Эберса из Фив и в более древнем папирусе — травнике эпохи от XII до XVIII династии — упоминались как лекарственные ячмень, пше- ница, бобы, финики, смоковница, кедр, тамарикс и др. Использование многих растений имеет свою историю. Неко- торые овощные растения вначале потреблялись для других це- лей: как лекарственные, ритуальные, масличные и т. д. Редис (Ra~phanus sativus L.) использовался в стране фа- раонов и как масличное, и как овощное. Поэтому мы считаем, что в Египте возделывался именно редис (однолетняя форма), а не редька. Последняя дальше стоит от масличной редьки и вообще является овощем умеренного и холодного климата (японская редька не является исключением, ибо она возделы- вается зимой) . Нет оснований приписывать первичность куль- туры редиса именно Египту. Дикие родичи распространены по всем берегам Средиземного моря. Геродот сообщает, что строи- телей пирамиды Хеопса IV династии (около 2900 лет до н. э.) кормили редисом, луком и чесноком. Эти овощи были пищей низших классов. Изображения корней редиса имеются в рисун- ках храма в Карнаке (XII династия). В литературе имеются указания на возделывание в Древнем Египте с в е кл ы. При этом указывают на изображение какого-то растения в храме Бени-Хассан (XII династия, 2000 — 1788 лет до н. э.). Это изображение дано в работе Вёнига (Wonig, 1936); оно очень схематично и похоже как на корнеплод, так и на луко- вицу. Лоре, описывая культурную флору эпохи фараонов, неупо- минает о культуре корнеплодной свеклы, но у него имеется на- звание «гтит», что, может быть, соответствует арабскому назва- нию листовой свеклы — silk. Если свекла встречалась в Древнем Египте, то это была не корнеплодная, а листовая свекла, однако и существование последней только предположительно. О свекле упоминалось в папирусе III в., где рекомендовался свекольный сок для чистки золотых и серебряных вещей и для фальсификации красных растительных красок. Таким образом, тогда здесь была известна красная свекла, но это была не на- стоящая корнеплодная свекла в нашем понимании, а листовая свекла, вероятно, переходного типа с утолщенным грубым кор- нем. По В. Т. Красочкину (1955), культура корнеплодной свеклы в Египте относится к более поздней эпохе. Во время покорения Египта Александром Македонским свекла фигурировала 320
в списке доставлявшихся ему продуктов. В. Т. Красочкин отнюдь не считает свеклу первичной культурой в Древнем Египте. В коллекции Всесоюзного института растениеводства об- разцы свеклы из Египта относятся к тонкокорневой салатной свекле — к восточному ее виду — Beta orientalis Roth. Этот вид далек от культурной корнеплодной свеклы и не мог служить для создания корнеплодных форм. Вообще жаркий климат Египта неблагоприятен для корнеплодов, и они там быстро вырож- даются. Чеснок (А11шт sativum L.) и лук (А11шт ~сера L.) были распространенными овощными растениями в Древнем Египте, но не аборигенными. Древнеегипетское название лука — badjar, от этого корня выводятся и древнееврейские названия. Сохрани- лось много древних изображений лука, по Вёнигу, также чеснока и шалота. Такое изображение обнаружено в погребении IV ди- настии, а также в пирамиде V династии в Саккаре. Пучки лука находили в саркофагах вместе с мумиями. Чеснок 6ыл найден в могиле около Фив у Assasif и в Дра-А6у-Негга. Унгер считает, что изображение шалота (А11шт astcalonieurn L.) имеется на египетском памятнике Серабит-эль-Хадим на Си- найском полуострове. Вёниг также различал шалот на египет- ских рисунках. Пор ей (А11шт ~роггит L.) 6ыл в употреблении в Древнем Египте. Швейнфурт (1884) приводит как доказательство на- ходку в раскопках. Согласно Плинию, порей был известен в Египте. По Фолькенсу (из Бушана), этот порей из древних египетских погребений не совсем похож на какой-либо современ- ный вид лука, но представляет собой сочетание признаков нескольких различных видов. Такого мнения придерживался и Швейнфурт, считая, что в Древнем Египте была промежуточная форма между А. роггит L. и А. ampeloprasum L. Все эти расте- ния из рода Allium, кроме, может быть, порея, не являются або- ригенными для Египта. П о р тул а к (Piortulaca olera~cea L.) возделывался в Древнем Египте, но Лоре считает, что он использовался не как овощное, а как красильное растение. Г о р л я н к а (Lagenaria vulgaris Ser.) узнается в изображе- ниях, обнаруженных в некоторых древних египетских погребе- ниях, особенно в Дра-Абу-Негга. Находили и семена бутылочной тыквы, которая, вероятно, имеет основной родиной Америку. На древнеегипетских рисунках встречаются дыня и огу- р е ц. Не всегда легко решить, какое из этих двух растений нарисовано. По А. И. Филову (1959), древние египтяне не знали настоя- щего огурца. В Древнем Египте 6ыл распространен примитив- ный вид дыни — Cucumis chate (Forsk.) Hasselqui.', Встре- чается в долине Нила и дикорастущий вид дыни — С. agrestis. А р 6 у з (Citrullus vulgaris Schrad. var. coloeyntboides Синская Е. Н. 321
p~churf. = С. coloncyisthoides Pang.), согласно Бушану, часто встречался на египетских рисунках и употреблялся в «жертвен- ных приношениях и как освежающее средство». В саркофаге в Дейр-эль-Бахри между крышкой и мумией найдены листья арбуза. Семена были обнаружены и другом древнем погребении в Фивах. Швейнфурт определил это расте- ние как чаг. со!о"ynthoides (арбуз кормовой) и сообщает, что он в современном Египте возделывается как масличное. Следова- тельно, в Древнем Египте был известен кормовой арбуз— именно его остатки, а не столового арбуза археологи находили в гробницах. О распространении арбуза batti~ch в Египте писали Ф. Штульман (Ь[uhlmann, 1909) и др. Арбуз battich арабы от- личали от настоящего сахаристого столового арбуза. В. Браун (W. Brown, 1800) сообщал, что кормовой арбуз battich растет .на полях, т. е. как сорняк, и что этим арбузом кормят ослов, верблюдов и других домашних животных. Название кормового арбуза battiich сродно раннему копт- скому его названию betuke и относится к периоду, предшествую- щему завоеванию Египта арабами (VII в. до н. э.). Вышеприведенные названия имеют общий корень с древне- еврейским названием арбуза — «а66атихим». Последнее назва- ние упоминается в писании, где говорится о6 исходе евреев из Египта, из чего можно сделать вывод, что кормовой арбуз был в культуре раньше 1500 г. до н. э. По А. И. Филову (1960), собственно Средиземноморье является отчасти ареной формообразования сладкого арбуза— С. vulgaris, который в целом имеет африкана-азиатское проис- хождение. Одной из родоначальных форм сладкого арбуза, по А. И. Филову, была особая разновидность дикорастущего С. co- lacynthis. Ареал этой разновидности — Египет, северные окраины Са- хары, Алжир. Сельдерей (Аршт graveolens L.) — в погребении Шейк- Абд-эль-Курна (XXI династии), датируемом около 1200 г. до н. э., под изголовьем мумии лежал венок из цветков голубого лотоса и листьев сельдерея, которые были связаны нитками из папируса. Подобные гирлянды находились и в других погребе- ниях. Как овощное растение сельдерей в те времена не был известен, а употреблялся как ритуальное и лекарственное расте- ние. Швейнфурт пришел к заключению, что египтяне имели дело с дикорастуиц;м растением, которое в изобилии росло р устье Нила и в тени деревьев по садам и паркам, Таким обра- зом, возможно, что на сельдерей можно указать как на один из немногих примеров аборигенного египетского культурного рас- тения. Древние египтяне возделывали а р т и На фресках име- лись изображения корзин с артишоками в качестве жертвенных приношений. Унгер считал, что это был настоящий огородный 322
артишок — Супага scolymus L., а Вёниг думал, что у египтян был другой вид, близкий к С. сагйипсп1из L. Декандоль, Бушан и Лоре ставят под сомнение существование в Египте артишока, а на рисунках признают вместо Hero салат — Lactuca sativa L. Артишок и салат огородный — оба растения средиземноморского происхождения. Рид (Н. Reed, 1942) указывает, что в Древнем Египте разво- дилась и с п а р ж а, но другие исследователи сомневаются в точ- ности определения рисунков, где были изображены похожие на спаржу растения. В,древнем Египте, еще в ранние династические времена, наряду с зерновым хозяйством и огородничеством развивались садоводство и виноградарство. Во II тысячелетии до н. э. сель- ское хозяйство в крупных рабовладельческих поместьях дости- гало высокого развития. Инжир (Ficus сапса L.) — растение древнесредиземномор- ское с широким ареалом происхождения, занимало видное место среди плодовых деревьев )IpeBHero Египта. Изображение фиг- соплодий инжира — появляется со времени XII династии. В гроб- нице в Бени-Хассан найден рисунок, где изображен сбор инжира, причем ясность рисунка не внушает никаких сомнений. Хороший рисунок инжира найден также в гробнице в Дра-Абу-Herra. Существовало и иероглифическое изображение для инжира— tab. Поскольку среди памятников Древнего царства, следов ин- жира найдено пока не было, можно предполагать, что инжир был занесен в Египет из Аравии или из Двуречья. В Египте распространен был также си ком ар — Ficus syco- morus Roxb. — вид индийского происхождения. Гр анат (Punica granatum L.), родина которого находится в Передней Азии, изображен на рисунках во многих древних египетских храмах. Остатки цветков граната найдены в погребе- ниях ХХ — XXVI династий в Фивах, но, по Бушану, самое старое указание на гранат относится к XII династии. Существуют иероглифические названия — tet, teb, tep и arkmani. Судя по ископаемым остаткам, в )IpeBHeM Египте были примитивные формы граната, более мелкие и более простые по структуре, чем современные. Родиной граната является, вероятно, Передняя Азия (в широком объеме этого понятия). В Закавказье сосре- доточено большое разнообразие дикорастущих и культурных форм граната обыкновенного, а второй из двух видов рода рас- тет на острове Сокотра у южных берегов Аравии, т. е. его ареал примыкает к южным границам Переднеазиатской области. 1(этой области первичного развития рода Рипка примыкал, возможно, и Египет. Если это предположение верно, то культура граната в нем могла быть и аборигенной. За это говорит примитивность граната Древнего Египта. Однако более обоснованным является мнение, что культура граната перенесена в Египет с востока. Гранат не мог прийти в Египет с tora: его нет в Сомали, Эритрее
и Судане. Не найдено никаких следов граната в Древнем цар- стве, что говорит против аборигенности этого растения в Египте. 1~ эпохе Среднего царства (XII династия) относятся гранаты (примитивные формы), найденные в гробницах Яра-Абу-Негга (в районе Фпв). Гранаты упоминаются в папирусе Эберса времени XVIII ди- настии, но следует учитывать, конечно, что сообщаемые там све- дения относятся к более древним временам. От позднейших вре- мен сохранились гирлянды из цветов граната и изображения гранатового дерева на барельефах. jI,. Г. Редер (1950) указывает, что египетское название гра- ната ynrhm «безусловно чужеземного происхождения». Таким образом, совокупность ботанических и историко-ар- хеологических данных по гранату говорит за то, что: 1) это рас- тение не аборигенно в Египте; 2) гранат распространился в Египет из стран Передней Азии. Г Редер рисует следующий путь распространения гра- 'ната: «из южной Аравии в Иран, оттуда в двуречье, Палестину .и Египет». Против первичного вхождения граната в культуру а ИИрраанне е ггооввоорриит т ттоо, что там нет достоверных дикорастущих за- рослей граната, которые могли бы твердо считаться не одичав- .шими. В Египет гранат попал в глубокой древности, может быть, даже и раньше эпохи Среднего царства (хотя следы его найдены только в последнем). За это говорит примитивность структуры г,лодов ископаемого египетского граната. Вероятно, некогда везде, на тогдашнем ареале культуры, были распространены только примитивные формы граната, подобные ископаемым еги- петским и современным с острова Сокотра, а затем постепенно заменялись формами, по структуре плода близкими к современ- иым. Изображения, связанные с культурой в и н о г р ад а и виноде- лием, встречаются начиная с V династии. В старых погребениях находят ягоды, кисти и листья винограда. Египтяне разводили также ф и н и к о в у ю п а л ь м у. Финики и гранаты употреблялись и для медицинских целей. Персея (Mimusops schim~peri Hochst.), которую Страбон называл египетским миндалем, является плодовым деревом с крупными сладкими плодами; у египтян считалась священным деревом. Оно разводилось вблизи храмов и в садах знатных египтян. Многочисленные следы персеп найдены в памятниках Нового царства, но не в более древних. В Египет она была зане- сена в период Нового царства с юга, вероятно, из Эфиопии, но сейчас в ОАР не разводится — скорее всего, из-за сухости климата. В далекие, хотя и не в самые древние времена разводились в Египте и я о л о н и. Историю этого фруктового дерева под- робно рассматривает Д. Г Редер (1950). Упоминание о яблоках 324
имеется в памятниках XIX династии и в Большом папирусе Гар- рис (ХХ династия). В этих памятниках имеются указания, что яблоки заготовлялись и хранились в амбарах. В Большом папи- русе Гаррис сообщается, что Рамзес IV подарил на праздник в честь реки Нила храмам Гелиополя 848 мер яблок. Лоре также считает, что при XIX династии яблоня была обычным плодовым деревом в Египте и указывает, что арабское taffah, еврейское ta)ppuah и коптско-арабское djepeh означают яблоню. Согласно изысканиям Р,. Г. Редера (1950), яблоня культи- вировалась только в восточной части Яельты, откуда могла попадать и в Гелиополь, их нет в фиванском и мемфисском раз- делах папируса Гаррис. Феофраст писал, что в Египте яблоки плохого качества. Таким образом, яблони не были широко распространены в Египте в старину и теперь там также не возделы- ваются. В Египет яблони проникли из стран Передней Азии. Пра- вильны соображения Редера, что хотя «филологические данные указывают на Палестину», но «не здесь следует искать родину яблони». Правильно в общем то, что «именно хеттские садовники впервые в истории стали возделывать яблоню и они пере- несли ее в Палестину, а позднее в Европу». Трудно утверждать, что только хеттские садовники впервые ввели в культуру яб- лоню. Может быть, она одновременно входила в культуру у разных горных народов Передней Азии и, может быть, и еще раньше хеттов, но что яблоня из Малой Азии распространилась в Палестину, а оттуда и в Египет, и в Европу (что не исключает и аборигенности некоторых европейских сортов) — это, по всем соображениям, правильно. Среди историков, особенно зарубежных, существовало и от- части еще существует мнение, что Египет — это центр, колыбель развития культуры всего востока, в частности и земледельческой культуры. Вышеизложенный обзор составлен довольно подробно не только по той причине, что Египет долго был ареной интенсив- ных археологических исследований и по древним культурным растениям Египта имеется немало данных, но и для того, чтобы окончательно развенчать представление о Египте, как об области первичного происхождения многих культурных видов. Египет — это только часть огромной Средиземноморской области развития культурной флоры, и бесспорных аборигенныхкультур- ных растений Египет имеет очень немного. Они были указаны выше. Подобно Египту, была несколько преувеличена и роль Эфиопии как очага развития культурной флоры. О значении Эфиопии и других африканских стран будет сказано в после- .дующем изложении.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕ РАСТЕНИЯЭФИОПИИ И BHECPEДИЗЕМНОМОРСКОЙ АФРИКИ Дикорастущая флора Африки очень разнообразна. Флора Северной Африки носит средиземноморский характер, и в отно- шении культурных растений мы включаем ее в отличие от более широкой Древнесредиземноморской области в собственно Среди- земноморскую подобласть, в которую из африканских стран входят Египет, Ливия, Марокко, Алжир, Тунис. На юг от собст- венно Средиземноморья тянутся обширные пустынные террито- рии — сюда относятся пустыни Сахара, Ливийская и Нубийская. В целом же область пустынь северной средиземноморской Аф- рики относится к Голарктической флористической области, как и вся Европа и внетропическая Азия. На юг от Сахары характер флоры резко меняется, и отсюда простирается уже Палеотропическая флористическая область африканского континента, а юг и toro-запад Африки составляют Капскую флористическую область. Последняя, хотя по сравне- нию с Палеотропической областью африканского континента невелика, но очень своеобразна и мало изучена. Палеотропическая область на африканском континенте зани- мает огромную территорию и поэтому не может быть однород- ной и делится на ряд провинций: сахельская, суданская, эфиоп- ско-сомалийская, гвинейская лесная (Гвинея и Конго), южно- африканская тропических саванн и саванновых лесов, тропиче- ская высокогорная. На крайнем юге еще выделяется ряд менее обширных провинций (Калахари, Драконовы горы, Дамара и др). Из этих ботаника-географических провинций для нас особыи интерес представляет Эфиопско-сомалийская, так как она лучше других изучена по составу культурной флоры. На просторах африканского континента первобытный чело- век обитал издревле. Питекантроп жил на огромнейшей террито- рии — от Пекина и Явы до Британских островов и до крайнего юга Африки. Южная раса неандертальцев была расселена от Явы до Южной Африки. Связи неандертальцев с древнейшими расами современного человека не выяснены. Африканский континент был издревле заселен племенами африканской ветви экваториальной (или негро-австралоидной) большой р асы. E( ее негр оидной ветви относятся р л сы.- негрская в начале позднего палеолита занимала почти всю Африку; в ту эпоху бушменская раса занимала крайний юго- запад Африки, а негрилльская располагалась в центре конти- нента (в бассейне реки 1(онго). Африканские негры, согласно воззрениям антропологов (Ми- клухо-Маклай и др.), не произошли непосредственно от мелане- зийцев, с которыми сходны по многим общим внешним призна- кам, а сближаются с австралоидами и веддоидами. Эти две по- 326
следние расы считаются самыми древними представителями экваториальной большой расы. Экваториальная раса в лице древнейших австралоидов занимала Индонезию (откуда рассе- .лилась в Австралию и Африку, кроме северной ее части). Древнейшие представители европеоидной расы — крамань- онцы — занимали Европу (кроме севера, вероятно, незаселен- ного) и Северную Африку. На Востоке Африки образовались .смешанные расы в результате контакта племен африканской гетви экваториальной расы с древними европеоидами. Такой смешанный характер имеет сомалийско-абиссинская, или эфиоп- ская, раса. Древнейшие ископаемые скелеты эфиопского типа относятся к мезолиту, по данным других авторов, — уже к нео- литу. Таким образом, этническая связь Африки и, в частности, во- сточной ее части с Южной Азией прослеживается с древнейших времен, заходящих за грань современного нижнего «порога исто- рии». Интересен факт древнейших исторических связей Южной Азии, в частности Индии, с Восточной Африкой, ибо культурная флора Эфиопии и сопредельных стран имеет общие элементы с культурной флорой Южной Азии, особенно Индостана. Вряд ли для распространения культурных растений имели значение только первоначальные древние этапы расселения по Африке экваториальной расы. Вероятно, перенос культурных растений из Южной Азии в Африку и обратно совершался в основном в более поздние времена. Известно, что уже в металлическую эпоху индонезийцы переселяются на Мадагаскар и смеши- ваются там с древним негроидным населением. Такие же пере- селенческие волны могли двигаться из Азии в Восточную Африку уже в земледельческую эпоху и играть роль перенос- чиков культурных растений. Экскурс в область древнейшей этнической истории Евразии оправдывает наше присоединение Северной Африки к собст- венно Средиземноморской области культурной флоры — это область древней европеоидной культуры. Законно также, с исторической точки зрения, выделение Эфиопии и сопредельных стран, в первую очередь Сомали, в сме- шанную область культурной флоры, на которой сказалось влия- ние и европеоидного Средиземноморья и африканской абориген- ной области развития древнего негроидного земледелия. Огромную, слабо изученную палеотропическую область Африки с древним земледелием негроидных племен еще нельзя расчленить с достаточной ясностью на подобласти культурной флоры. Поэтому остановимся на культурной растительности тропической и Южной Африки лишь в самых общих чертах. Несомненно, и средиземноморская Африка и палеотропическая Африка в истории их материальной и, в частности, земледельче- ской культуры обнаружили взаимовлияние, но конкретные исто- 327
рические взаимосвязи этих областей еще слабо вскрыты. Лучше изучены историческая география и предыстория человеческого общества Северной Африки. Согласно новейшим изысканиям, Сахара никогда не была дном моря, как это многими предполагалось раньше. Не всегда Сахара отличалась сухим климатом. Существовали два влаж- ных периода в Сахаре, отдаленных друг от друга многими тыся- челетиями (А. Лот, 1962 и др.) . .Очень несовершенные орудия первого периода отнесены к раннему палеолиту. Орудия второго влажного периода датированы как неолитические. Первый влаж- ный период соответствует шелльскому и ашельскому европей- ским периодам. После первого влажного периода наступил длительный за- сушливый период, и пустыня тогда доходила до самых границ тропического леса. Во второй влажный период Сахару населяли охотники и рыболовы. Орудия второго влажного периода гораздо разнооб- разнее и совершеннее, чем орудия первого периода. Они встре- чаются не повсеместно, а обычно по берегам рек и озер. Среди орудий эпохи неолита, относящихся ко второму влажному пе- риоду, кроме стрел, скребков, примитивных топоров, обнару- жены были зернотерки и серпы. Трудно сказать, когда началось возделывание растений. ~Кать и дробить на зернотерках могли и зерно, собранное в ди- ких зарослях. Раскопки А. Аркелла (А. J. Arkell, 1949, 1955) около Хартума показали, что неолитический человек собирал дикорас- тущие злаки и перемалывал в муку их семена. Собирали также плоды с диких деревьев. Изучение скелетов обитателей неолита этих мест (поселение датируется примерно 3900 г. до н. э.) пока- зало, что по антропологическому типу они относились к негроид- кой расе. В наше время многие специалисты считают, ч1о неолит Са- хары не моложе неолитических культур Египта. В археологиче- ской и исторической литературе имеются высказывания, что неолит Файюма в Египте развивался под влиянием неолита Тибести и Судана. Судя по характеру неолитических орудий„ имеются предположения, что район Тибести был центром, из ко- торого распространилось культурное влияние на Файюм, Хар- тум и окрестные местности (А. J. Аг1<е11, 1955). По мере высыхания Сахары население отходило к водным источникам. Этих огромных водных потоков было много, боль- шинство их впадало в огромное озеро Чад. Сохранившиеся русла этих потоков называются вади. По склонам вади в огром- ном количестве находят кучи кухонных остатков (кухонные или раковинные кучи), а в них множество костей животных. Открыты целые скопления наскальных изображений животных. Эти изо- бражения приурочены к следующим периодам. 328
1. Древнейший период — так называемая «эфиопская фауна». Преобладают изображения носорогов, гиппопотамов, слонов, жирафов. Эти изображения датируются 5000 — 2000 лет до н. э. 2. Скотоводческий период (2500 — 1000 лет до н. э.). Был рас- пространен культ быка, что роднит эту культуру с древнеегипет- ской. Найдено было и изображение нильской лодки. Обнару- жены также изображения козы, собаки, слона, муфлона, анти- лопы, редко жирафа, в более верхних слоях — лошади, а в ку- чах пепла — жернова, зернотерки, скорлупа страусовых яиц. Все это указывает на степной характер местности. животные влажного периода — гиппопотамы, носороги — исчезли, слонов становится все меньше. Для скотоводческого периода харак- терны разнообразные одежды — от длинных туник до коротких передников, которые сплетались из травы. 3. Период колесниц и лошадей (1100 лет до н. э. — начало нашей эры). В это время часты изображения лошадей, встре- чаются наскальные рисунки колесниц. 4. Период верблюда. Верблюд распространился около III в. н. э. К этому периоду относятся изображения ливийско-бербер- ского (200 г. до н. э. — 700 г. н. э.) и арабо-берберского времени (700 лет н. э. и позднее). Причины усыхания Сахары до сих пор не выяснены. Что ка- сается смены травянистой степи или саванны пустыней, то очень убедительно мнение многих исследователей, что в опусты- нивании Сахары большая роль принадлежит крупному ро- гатому скоту, который уничтожал травянистую растительность этих мест. Следы существования земледелия намечаются с сере- дины или с конца скотоводческого периода. Для негроидного искусства характерно изображение «Белой дамы», что по пред- положению означает богиню земледелия — вблизи фигуры бо- гини изображено хлебное поле. Предполагается, что эта фреска относится к эпохе, когда начало сказываться влияние египет- ской культуры. В Сефаре найдены фрески с изображением ра- ботающих на поле женщин. Доисторические художники умели изготовлять очень стойкие минеральные краски, к которым при- ,мешивали казеин и растительный клей из акации. Древние связи аборигенов Сахары имели место не только с народами Южной Африки и с Египтом, но и с более северным Средиземноморьем. Вначале изображения колесниц на афри- канских наскальных фресках приписывались гарамантам, насе- лявшим (по Геродоту) земли современной Ливии, а затем при- шли к заключению, что стиль изображений более сходен с ми- кенским искусством на острове Крит. Около 1200 лет до н. э. племена острова Крит, имевшие намерение начать войну с Егип- том, высадились в Киренаике и смешались с местным ливийским населением. Фрески с рисунками колесниц подтверждают этот факт вторжения народов Моря, согласно некоторым рукописям Египта. 329
B данном очерке основное внимание уделяется Восточной Африке. По описаниям А. Брэма (по Г.. М. Штейнбергу, 1960) столеч" чому назад в районе Хартума по берегам Голубого Нила и рек Бахр-эль-Джебель и Бахр-эль-Араб росли «xopoIIIQ развитые галерейные леса, состоявшие из различных лиственных пород, перевитых лианами и поросших эпифитами». Е. М. Штейнберг (1960) на основании личных впечатлений до некоторой степени помогает читателям составить представление о зональной смене современных ландшафтов Африки. Теперь от лесов, описывае- мых Брэмом, сохранились жалкие остатки. По берегам много папируса. Североафриканские пустыни сменяются засушливыми са- ваннами, которые в Африке большей частью имеют вторичный характер. Чем дальше на юг, тем климат саванны становится влажнее и тем больше в ней древесной растительности. Саванна переходит в лесосаванну, а на границе с Конго начинается зона влажного тропического леса с годовым количеством осадков до 1500 мм в год, с 2 — 3 сухими зимними месяцами. У Эль- Обейда (12' сев. шир.) годовое количество осадков около 340 мм,, сухой период длится 5 месяцев. В северных районах Судана уже сказывается влияние ветров из Сахары и осадков очень мало, иногда их совсем не выпадает. В районе Хартума количество осадков около 150 мм в год. Растительность здесь скудная. Ха- рактерны кусты Acacia tortilis Hayne и безлистные колючие кусты Caip~paris decidua Edgew. В окрестностях Эль-Обейда встречается кустарник Acacia senegal (L.) Willd. Это растение дает камедь — гуммиарабик, которая добывается путем над- реза стволов. Особый колорит ландшафту засушливых районов придает баобаб с чрезвычайно толстым стволом. В горной части Судана в саванне имеется злаковый травя- нистый покров — Setaria incrassata Нас1~е1 — с отдельными де- ревцами Acacia scyal Balanifer. В предгорьях растет резиновое дерево, которое выделяет каучук — Boswe lia papyrifera Hochst. Из пищевых растений в Судане имеет большое значение бамия. Эфиопия отличается от Судана более богатой и разнообраз- ной растительностью. Это горная страна. Абиссинское нагорье имеет средние высоты 1800 — 2000 м, отдельные вершины дости- гают 4620 л. Климат более умеренный, чем в Судане, и очень разнообразен. В Аддис-Абебе температура редко спускается до 0', на больших высотах зима холоднее. Количество осадков во влажных местностях очень велико (до 2000 мл~), но есть и сухие районы. В древние времена большая часть Эфиопии была покрыта лесом, а теперь он лишь кое-где сохранился в виде небольших массивов. На месте бывших лесов сформировалась вторичная саванна с акациями. 330
Е. M. Штейнберг описывает сохранившийся лес в государ- ственном заповеднике Мадагаши, недалеко от Аддис-Абебы. Верхний ярус образован исполинскими можжевельниками, для второго яруса характерны тропические хвойные деревья— подокарпусы — Рос1осагриз gra~cilior Pilger. На высоте 2500 м растет древовидный вереск — - Eri~ca arborea L., в нижнем ярусе— шиповник. Е. М. Штейнберг описывает лес в toro-западной Эфиопии недалеко от Яжиммы. В нижнем ярусе много высоких папоротников и драцен. Здесь был встречен дикий банан. На бо- лее разреженных участках леса нижний ярус состоит, главным образом, из дикого кофейного дерева — СоПеа ara- bica L. Эти деревья были усыпаны цветками и плодами различ- ной степени зрелости и достигали высоты не более 4 лх. Они доставляют кофе, который по качеству ничем не отличается от культурного кофе. Местное население эксплуатирует эти за- росли. Если применить некоторый уход, возможно получать с диких зарослей довольно большие урожаи кофе. Кофейное дерево считается географическим элементом Во- сточной Африки (Эфиопии, Эритреи), но распространено и в Иемене. С другой стороны, несомненны родственные связи с тропиче- скими странами Африки, расположенными южнее. Весь род Coffea еще недостаточно изучен — недавно установлен новый подрод — Africoffen Moens — в Конго (Р Моепз, 1962). Эфиопия — страна древнего земледелия. Население в основ- ном амхары, основное занятие которых и теперь земледелие. Рассматривая основные черты культурной флоры Египта, (;у- дана и Эфиопии, прежде всего следует отметить земледельче- скую отсталость древних Нубии и Эфиопии по сравнению с Египтом. На основе археологических исследований Б. Б. Пиотровский (1962) пишет, что в конце IV тысячелетия до н. э. в Северной Нубии складывалась культура, которая легла в основу куль- туры древнего Египта. Позднее, когда центр древнеегипетской культуры был пере- несен на север, Нубия некоторое время развивалась обособ- ленно и отстала от Египта, а затем эти две культуры вступили во взаимодействие. Территорию Нубии во I I тысячелетии до н. э. заселили негроидные племена. При раскопках в районесооруженияАсуан- ской плотины были найдены в слое, соответствующем эпохе древнего египетского поселения, зернотерки, зерна пшеницы, плоды пальмы дум, вкладыши для серпов, скорлупа страусо- вых яиц, обломок браслета из слоновой кости. Поселение со- стояло из легких тростниковых построек и хозяйственных ям, покрывавшихся плетеными щитами. Экспедиция встречала здесь многочисленные наскальные изображения страусов, жирафов и .других животных. Эти изображения покрыты слоем темной 331
патины, что говорит о6 их глубочайшей древности. Наскальные изображения встречаются и в других местностях Африки. В Египте для развития земледелия были особо благоприят- ные условия, а в Нубии простора для полей было меньше и условия для развития орошения явно неблагоприятны. Дожди были редки. В Нильской Эфиопии не строилось дамб и кана- лов, развитие классового общества и образование государств там сильно задержалось. В южной части Верхнего Египта и в Северной Нубии в IV тысячелетии до н. э. обитатели Нильской долины занимались рыболовством, охотой, скотоводством и немного земледелием (где позволяли условия). Однако и в эти отдаленные эпохи здесь сеяли ячмень, просо (не Panicum milia- ceum, а другие виды), существовали огородничество и виноделие. Такое же замедленное развитие общественных отношений государственности и земледелия имело место в странах, распо- ложенных далее на юго-восток. В Эфиопии и ныне сельское хо- зяйство продолжает оставаться на низком уровне. И те- перь еще здесь бытуют примитивная техника земледелия и гру- бые несовершенные орудия для обработки почвы, обмолота и перетирания зерен. jjo самого недавнего времени в Эфиопии сохранялись примитивные каменные зернотерки, мотыги и дон- горы — колья для обработки почвы с насаженным сверху для утяжеления каменным кольцом. Поля обрабатываются в Эфиопии руками или примитивной деревянной сохой, запряженнои зебу. Еще и теперь сохранились своеобразные старинные способы использования некоторых культур. Например, лен употребляют в пищу в виде поджарен- ных семян. Растение, похожее на горчицу и дающее масло высо- кого качества — Brassica carinata, там используется не как мас- личное, а как салатное; кресс-салат — Lepidium sativum — воз- делывается ради семян, а не для зелени и т. п. Сознательному отбору, селекции культурные растения Эфиопии, видимо, не под- вергались, чем в значительной степени объясняется пестрота популяций абиссинских местных сортов. Эфиопия занимает нагорье с довольно ровным, теплым, в верхних зонах — с уме- ренно теплым климатом. Земледелие служит основой экономики страны. С древних времен связи с Египтом были постоянные,. взаимовлияние обеспечивалось караванами и военными похо- дами. культурные растения Эфиопии хорошо изучены. Для по- знания культурных растений Эфиопии больше всего дают мате- риалы путешествия Н. И. Вавилова и изучение собранных им коллекций. В противоположность Египту по Эфиопии археологического растительного материала почти не имеется, и о путях разви- тия эфиопской культурной флоры можно делать заключения, главным образом, на основе сравнительного ботанического изу- чения современных культурных растений, что и должен показать, ниже помещаемый краткий обзор. 332
Дикорастущих пшениц и представителей родственных им родов трибы Hordeae в Эфиопии нет. Эфиопская п ш е н и ц а в основном представлена энде,"личным, полиморфным, интеграль- ным (т. е. сочетающим в себе в нерезко выраженном проявле- BIBB признаки нескольких родственных видов пшеницы) видом— Triticum aeihiopicum Jal~ubz. Этот вид, по нашему мнению, близкородствен колхидским древним пшеницам — Т. ~pr~otoma~cha и Т. protopaleocolchicum — и занесен в Эфиопию из Закавказья в чрезвычайно древние времена, вероятно, через Сирию. В на- стоящее время эфиопский полиморфный вид пшеницы содержит в своем составе и формы примитивного типа, и сильно подви- нутые по пути специализированного развития в разных районах своей второй родины. В этом видно доказательство давности существования в Эфиопии пшеницы этого корня. Полба-эммер — Triiicum йсоссоп Schrank — представлена в Эфиопии особым подвидом — ssp. abyssinicum (Stol.) Vav., который имеет сходство с индийскими формами полбы. В Эфио- пию полба пришла или из Индии, или, вернее, непосредственно из Передней Азии (Закавказья) и уже из Эфиопии — в Индию. Так называемая польская пшеница — Т. ~polonicum L,. — пред- ставлена в Эфиопии эндемичной формой sslp. abyssinicum Vav., но, по нашему мнению, самостоятельного вида под этим назва- нием не существует, а абиссинские формы, причисляемые к Т. poloniicum, просто представляют собой изменения Т. aethio- р1сит, встречающиеся и у других видов пшеницы в гналогпчной каждому из них форме. Формы мягкой пшеницы — Т. aestivum L.— явно занесены в Эфиопию из других стран с востока и, может быть, частично с севера. Ареал распространения дикорастущего яч м ен я, так же как диких пшениц, очень удален от Эфиопии. Однако ячмени Эфио- пии своеобразны, полиморфны и ныне выделены Ф. Х. Б-1хтее- вым в самостоятельныи вид — Hordeum aethiopicum Vav. et Bacht. В составе эфиопских ячменей имеются и шестирядные, слабо редуцированные формы и, наоборот, проявляющие максималь- ную среди всех культурных ячменей степень редукции (у таких форм у боковых колосков редуцпроианы не только завязи, но и тычинки и пыльники). Сильно редуцированные формы являются более молодыми по сравнению с теми нередуцированными, от ко- торых они непосредственно произошлц. Область первичного про- исхождения культурных ячменей многие авторы видят в Перед- ней Азии. Сильное развитие редукционного процесса в Эфиопии говорит о том, что он протекал здесь в течение очень долгого периода и что ячмени впервые попали из Азии в Эфиопию в чрезвычайно давние времена. В Эфиопии распространен особый о в е с (Ачепа vaviloviana Nordv.) — сорняк, от которого произошел и эндемичный куль-
турный вид а б и с с и н с к о г о о в с а (А. abyssinica Iochst.), последний распространен и в йемене. Таким образом, Эфио- пия — это область развития двух эндемичных видов овса, род- ственных другим 28-хромосомным средиземноморским овсам. В Африке распространен прародитель к у л ь т у р и о г о р и с а — Oryza perennis Noench. Он представлен здесь несколь- кими региональными экотипамп. К последним относят и О. bart- hii А. Cheval. рис Барта. Он, по-видимому, не является само- стоятельным видом, а представляет собой тип (или подвид) древнего вида О. perennis. Рис Барта — это однолетний не- плавающий вид, довольно далеко филогенетически продвинутый от исходного О. perennis, который в своем типичном выражении является многолетним и плавающим. Но рис Барта и культуру не вошел и растет в тропической Африке только дико. От плавающего риса — О. perennis — произошел культурный африканский рис — О. glaberrima Steud. — однолетний, непла- вающий, родственный виду азиатского происхождения — О. sa- tiva ? . — и образующий гомологические ряды варьирующих признаков с последним. Однако О. sativa и О. glaberrima— самостоятельные виды, хотя и произошли от общего родона- чальника, но в различных местах и от разных популяций О. ~perennis. Имеются в Африке и одичалые формы О. sativa, занесенные из Азии, п гибриды О. sativa с О. glaberrima. По- следний растет и дико. В Африке распространены в диком со- стоянии еще следующие виды: О. ~рипс1а1а Kotschy — однолет- ний, распространен в диких зарослях (Судан), иногда исполь- зуется местным населением как объект собирательства; О. per- rieri А. Camus и О. tisseranti А. Chev.— дикорастущие виды. Также далек от видов культурного риса и О. stapfii Roshev., хотя и является однолетним. Еше филогенетически дальше одно- летний дикорастущий рис — О. brachyanta А. Chev. et Poehrich. Возможно, что кое-где и эти виды бывали или и теперь являются объектом собирательства. Что касается развития рода Oryza, то, несомненно, Африка является вторым по значению центром его развития (первый— Южная Азия, а третий — Америка). Д а г у с с а (Eleusine coracana Gaertn.) представляет собой весьма распространенное хлебное культурное растение в Эфио- пии и употребляется там для изготовления пива. Однако боль- шинство авторов, в том числе и мы, считают родиной этого ра- стения не Африку, а Индию (Е. Werth, 1937). В Эфиопии произрастает вид п р о с а (Panicum callosum Hochst.), близкий к обыкновенному к у л ь т у р н о м у п р о с у (Р miliaceum L.). Однако, как было показано выше, вся сово- купность ботанических и исторических фактов говорит за то, что родиной проса является Восточная Азия (Китай). Негр итянс кое п р о co [Pennisetum spicatum (L.) Poem. et Schult. (Р. typhoideum erich. = Pennicilaria s~pi~cata Willd.)] 334
также служит для производства пива и других пищевых продук- тов. Растение аборигенное для Африки и, возможно, именна для Эфиопии и смежных стран Восточной Африки. Некоторые авторы считают прародителем этого растения дикорастущий просовидный злак — Pennisetum villosum Brown, а другие пред- полагают, что происхождение этого растения полифилетическое; в нем принимали участие многие просовидные дикорастущие злаки: Penniseium ~perrottetii K. Schum., Р. violaceum (Lam.) Rich., Р. versicolor Schrad. (все три — пз Сенегала, Гамбии и Во- сточной Африки), Р mollissirnum Hochst. (Судан) и др. Верт (1937) считает определенно, что родина негритянского проса- это обласгь тем Iblx хамитов Северо-Восточной Африки, т. е. в основном Эфиопия. С о р г о х л е б н о е ISorghum vulgare Pers. (= S. durra Jakuschev.)] очень распространено в Эфиопии и других странах Восточной Африки. По мнению ряда авторов, ведет свое проис- хождение из Азии, но правильнее его считать африканским ра- стением, ибо Африка — родина всего рода Sorghum. Е. С. Яку- шевский объединяет африкана-среднеазиатское (не восточно- азиатское) сорго в один вид Sorghum durra с большим разно- образием в Африке, где, по Е. С. Якушевскому, имеются два подвида этого сорго: эфиопский — ssp. aethiopicum Jakuschev., нубийский — ssp. пиЫсит Jakuschev. Таким образом, Эфиопия составляет часть первичной области культуры Sorghu» durra. Т е ф ф (Eragrostis abyssinica Link) — это настоящая древ- няя эфиопская культура. О большом значении теффа в Эфио- пии писал Н. И. Вавилов. Это значение сохранилось и до на- стоящего времени. E. М. Штейнберг (1960) пишет: «Излюблен- ная зерновая культура здесь тефф, дающий очень мелкие се- мена и очень низкий валовой урожай. Из семян приготовляют муку и пекут кислые лепешки. Амхары настолько привязаны к этому напиональному блюду, что с большим трудом отказы- ваются от малопродуктивного теффа, хотя в тех же районах созревают и дают более обильный урожай ячмень и пшеница». Из него приготовляют также пиво — buza. Одно пз его туземных названии напоминает нашу кашу, но есть ли тут какая-нибудь связь или это случайное совпадение, неизвестно. Горох [Pisum sativum L. (с включением Р. arvense L.)]— растение, происходящее из Передней Азии. В Эфиопии Л. И. Го- воров (1937) выделяет особую группу proles aethiopi~cum мест- ных сортов. В Эфиопии и Йемене также произрастает в куль- туре и в диком состоянии вид Р. abyssinicum А. Вгаип, видовая самостоятельность которого спорна и скорее это только под- вид — Р. sativum. Н у т (Cicer arietinum L.) — растение переднеазиатского про- исхождения, представлен в Эфиопии особой группой местных форм — ~proles abyssinicum G. Pop. 333
>z о н с к и е б о б ы (Faba vulgaris Moench) имеют средизем- номорское происхождение (в рамках древнего Средиземья), в Эфиопии они представлены особой экологической группой- аравийско-абиссинской (В. С. Муратова, 1937). Чечевица (Lens esculenta Moench) — растение передне- азиатского происхождения (или шире — восточной части gpea- него Средиземноморья). В Эфиопии образовала в культуре осо- бую группу местных сортов — prol=s aethiopicae (Е. И. Бару- лина, 1937), распространенную также и в Иемене. Ч и н а (Lathyrus sativus L.) — растение средиземноморского происхождения. В Эфиопии представлена особой группой мест- ных сортов, распространенной также и в Египте — proles abyssi- nicum Zalk. (Ф. Л. Залькинд, 1937). В Эфиопии возделываются эфиопские и эритрейские п а- .ж и т н и к и, относящиеся к средиземноморскому подвиду обык- новенного пажитника — Trigonella foenum-graecum L. ssp. medi- terranea subseries aethiopicae Sinsk. (= grex aethiopieae Ser- pwch~ova). Возделываемые пажитники очень богаты формами, особенно по окраске семян, но в целом имеют признаки среди- земноморского подвида. Эфиопия, таким образом, не может счи- таться первичной областью происхождения культурного пажит- ника: здесь только вторичный очаг формообразования. В Эфио- .пии распространен в качестве сорняка возделываемый в чистом посеве в Иемене вид Т. jemenensis Sinsk. В и г н а (Vigna sinensis Endl.) представлена разнообраз- ными местными сортами в Эфиопии, Восточной Африке и ОАР Некоторые авторы считают это растение вышедшим из Африки, .а другие — из Индии или Юго-Западной Азии (F. Stuhlmann, 1909), что мало обосновано. Л о б ия, или ги а ци н тон ы е бобы (Doli~chos lablab L.)— древняя культура Африки, распространенная в Эфиопии, ОАР, Судане, 1хении и Уганде. Н. И. Вавилов считает именно Эфио- пию первичной областью вхождения лобии в культуру, но это вопрос спорный. Многие авторы находят, что культура лобии прежде возникла в Индии и Индокитае. По Н. Р. Иванову (1937), родиной лобии является тропическая Африка (в том числе и Эфиопия), из ко- торой она проникла в отдаленные времена в Индию, позднее— в Египет. В Сомали имеется эндемичный вид л ю п и н а (Lu~pinus somaiiensis Baker.). Л е н (Linum usitatissimum) возделывается в Эфиопии как зерновая культура, в пищу употребляются его семена. Эфиоп- ский лен — группа особых форм, обнаруживающая тесное родство со льнами Индии, где, по нашему мнению и дру- гих исследователей, имело место первичное вхождение льна в культуру во времена глубокой древности и откуда лен, также в очень отдаленные времена, занесен в Эфиопию. Для Эфиопии
эндемична горная гигрофильная подразновидность льна — sub- var. subprostratum Sinsk. В предгорьях и равнинах Эфиопии распространены карли- ковые формы льна, похожие на индийские — subvar. nanoere- ctum Sinsk. f. aethiopicum. Из Индии прослеживается постепенно изменяющийся ряд от крупнокоробочных индийских карликовых льнов к мелко- коробочным индийским карликам и далее к собственно эфиоп- ским карликовым кудряшам. В Эфиопии нет дикорастущих форм льна, близких к культурному льну. Туда лен не мог быть зане- сен из Средиземноморья, ибо связь между средиземноморскими и эфиопскими формами льна сравнительно небольшая — пере- ходные формы встречаются не так часто и только на границе ареала средиземноморских льнов. Типичные средиземноморские льны — самые крупноплодные, крупносемянные и крупнолистные, по некоторым признакам прямо противоположны эфиопским формам — мелкоплодным, мелкосемянным и мелколистным. Сами средиземноморские льны находятся в очень близком родстве с индийскими крупноплод- ными формами льна. Индия принимается нами за очаг рассе- ления, откуда тянутся ряды переходных форм льна и в Среди- земноморье, и в Эфиопию. Следовательно, именно Индия яв- ляется первичным очагом происхождения и расселения льна на юго-запад — в Эфиопию и на северо-запад в Средиземноморье. 'Однако в Индии не сохранилась исходная для индо-эфиопской ветви культурного льна (series indo-aethiopica Sinsk.) древней- шая форма со склонностью к простратному росту — она распро- странена только в высокогорьях Эфиопии с относительно влаж- ным климатом. По нашему мнению, это объясняется тем, что лен был занесен из Индии в Эфиопию в глубочайшей древности. Вследствие примитивного уровня земледелия в Эфиопии древ- няя форма культурного льна сохранилась до наших дней, а в Индии она исчезла. Н у г (Guizotia abyssini~ca Cass.) — масличное растение, аборигенное для Африки, которая является родиной всего рода Guizotia Cass. В Эфиопии распространены два близкородствен- ных вида — G. schultzii и G. bidentoides. Эфиопские названия: nuhk, nepuk, пЪо1~. Аборигенная в Эфиопии культура. Из Эфио- пии нуг занесен в Индию. Кл еще в и н а (Piicinus L.) — маслс.чное растение, возможно, африканского происхождения, но имс:.' ..e вторичные очаги в Индии и Восточном Средиземноморье. Мгл.'оплодный вид кле- щевины gi~cinus ппсгосагриз G. Pop. распространен в диком и одичалом состоянии в Северной Африке по берегам Средизем- ного моря, а также по берегам Красного моря и в Эфиопии, Сен- наре и Кордофане. Некоторые авторы считают Эфиопию важ- нейшим первичным очагом этой культуры, но по этому вопросу .существуют различные гипотезы. Интересна точка зрения
ф. Штульмана (F. Stuhlmann, 1909), что клещевина имела очень. широкии первоначальный ареал — от Индии через Аравийский полуостров до тропической Африки. Благодаря быстрому росту и способности семян долго сохранять всхожесть клещевина имеет способность к широкому распространению. Ф. Штульман предполагает, что клещевина была в отдаленные времена окуль- турена в двуречье и Южной Аравии и оттуда в глубокой древ- ности была перенесена в Египет и Индшо. Во всяком случае и Эфиопию с соседними странами надо признать если не основ- HbIM, T0 важным очагом древней культуры клещевины. К р е с с - с а л а т (LBDldium sativum L.) в Эфиопии назы- вается фета. Кресс используется здесь не ради листьев, а ради семян, дающих масло и употребляющихся в пищу непосред- ственно. Он распространен здесь как сорняк льна и в чистых посевах. Несомненно, он вышел в культуру из сорного состоя- ния. Первичным очагом происхождения этого растения Эфиопию назвать нельзя — поздние формы с розеткой листьев у основа- ния растения имеют, по нашему мнению, более древнюю кон- ституцию и распространены в Передней Азии, Афганистане и Иране. другое крестоцветное растение — к а п у с т а а б и с с и н с к а я (Brassica icarinata А. Вгаип.), хотя и дает прекрасное масло, в Эфиопии используется как салатное. Эта культура эндемична для Эфиопии, и имеются все основания предполагать, что именно здесь она и вошла в культуру. Родственные связи этот вид имеет с видами из Средиземноморья рода Brassica L. В. cari- nata возникла как амфпдиплоид от скрещивания двух среди- земноморских видов: черной горчицы — В. nigra (L.) Yoch и ого- родной капусты — В. оlегасеа L. с последующим удвоением числа хромосом. Как и все виды, связанные происхождением со Средиземноморьем, В. carinata — растение длинного дня. I есомненно, из Средиземноморья (в широком понимании) пришла в Эфиопию и г о р ч и ц а ч е р н а я (Brassica nigra), которая, однако, представлена в Эфиопии особой формой, ра- стущей в смеси с В. carinata и имитирующей последнюю. К р а м 6 e (Crambe abyssinica Hochst.) растет дико в Эфио- пии. Сейчас она вводится в культуру как масличное в СССР. Род Crambe L. древнесредпземноморскпй. Это растение длин- ного дня. Б а м и я (Hibiscus esculentus L.) возделывается и распро- странена в диком состоянии в Эфиопии. В Эфиопии имеются и другие овощные культуры. Эта группа слабо изучена. Из овощных местных растений назовем щ а в ел ь абиссинский (Яитех abyssini~cus Jacq), паслен эфиоп- с к и й (Solanum aethiopicum Tom., S. quineense Lam.), возможно, занесен из Гвинеи. Эфиопия составляет часть первичной области вхождения в культуру х л о п ч а т н и к а — вида Gossypium herbaceum 1 338
В Эфиопии, по Ф. М. Мауеру, распространены два подвида этого вида: «долговечный» (многолетнпй) рудеральный — ssp. pseu- йоагЬогеит (G. arboreum Forsl<.) и менее «долговечный» — ssp. frutescens Маиег, представленный сортами однолетней и много- летней культуры. Формы первого подвида распространены, кроме Эфиопии, также в Судане, Сомали и в Западной Африке, а второго — в Судане, Нигерии и оазисах Сахары. Е,о р и андр (Coriandrum sativurn L.), ч е р ну ш к а (Ni- gella sativa L.) и а ж г о н (Carum copticum Benth. et Hook.) имеют в Эфиопии область вторичного развития и представлены там эндемичными местными формами. Эфиопия считается одним из очагов введения в культуру м а сл и н ы (Olea europaea ? .), там распространен в диком со- стоянии ближайший дикорастущий родич — О. 'Chrysophylla Lam. (который, однако, имеет широкий ареал и вне Эфиопии: от Гималаев до Южной Африки и островов Макаронезии). Чат (Catha edulis Forslc.) — аборигенное культурное расте- ние Эфиопии и Иемена, где растет и дико. В пищу идут моло- .дые ростки. Аборигенным растением для Эфиопии является и кофей- н о е д е р е в o (Coffea arabiica L.) . .Здесь оно растет дико и от- сюда распространилось в Йемен и затем в другие страны. Б а н а н к у л ь т у р н ы й в отдаленную эпоху занесен вместе с таро, дагуссой и другими растениями из Южной Азии. В Эфио- пии имеется дикорастущий банан — Musa ensete J. F. Gmel. У Musa ensete употребляются на плоды, а крахмалистая серд- цевина междоузлий. Следующие виды Coleus Lour.— клубненосные растения из семейства губоцветных распространены в культуре в Эфиопии: С. edulis Vatke, С. penzigii Schweinf., в Эритрее — С. ingiar Sew., в Эфиопии и Эритрее — С. lanuginosus Hochst. Эти виды, ве- роятно, восточноафриканского происхождения. С. barbatus Benth. и С. sipicatus Benth., хотя и возделываются в Эфиопии, но, вероятно, занесены туда из Индии. Из рода Coleus эндемичен для Эфиопии и Анголы — С. insolitus Pobyns et Lebrun (вид, родственный С. floribundus Pobyns et Lebrun). Я м с а 6 и с с и н с к и й (Diosoorea abyssinica Hochst.) растет дико и местами разводится в Эфиопии и соседних странах Во- сточной Африки. М и р р о в о е д е р е в о (Balsamea myrrha Baill.) является древней культурой Иемена и стран древнего Пунта (Сомали, Эритрея), играло важную роль в парфюмерии древних (умаще- ния тела и воскурения). Родина — Восточная Африка. П е р с е я (Mimusops schimperi Hochst.) дает плоды, ценив- шиеся в древности в Египте, куда она была занесена, вероятно, из Судана, где растет дико (как, по-видимому, и в Эфиопии). К о р д и я (Cordia abyssinica Я. Br. из сем. Borraginaiceae) пмеет съедобные плоды. Растет дико.
из дикорастущих деревьев, распространенных в Эфиопии и соседних странах Восточной Африки, отметим: к о с о ( Iagenia abyssini~ca Wi1ld.) — лекарственное дерево; с т р о и т е л ь н ы й м о л о ч а й — Euphorbia candelabrum Trem. (для изгородей); Commiphora abyssinica Engler — дает душистую смолу и др. (иногда также называется миррой). Состав эфиопских культурных растений не очень велик и количество аборигенных культур в нем cpaBHHTeJIbHQ незначи- тельно — это тефф, нуг, абиссинский овес, абиссинская капуста, кофейное дерево, чат, эфиопский банан, возможно, негритянское просо. Территории окружающих Эфиопию стран изучены слабее, чем собственно Эфиопия, и поэтому по ряду культур трудно установить, HacKQJIbKQ они заслуживают названия эфиопских. Для ряда культур, хотя и не аборигенных, а нашедших себе в Эфиопии вторую родину, отмечается развитие здесь в усло- виях культуры совершенно своеобразных эндемичных видов, подвидов, форм и экотипов. Особенно примечательно вторичное развитие эндемичных форм льна, вид эфиопских пшениц, яч- меня, подвида абиссинской полбы, сорных форм кресс-салата, форм пажитника, черной горчицы, чечевицы и других бобовых. Таким образом, Эфиопия хотя и представляет собой срав- нительно большой очаг развития мировой культурной флоры, но не может в этом отношении стать в сравнение с такими геогра- фическими областями и подобластями, как Средиземноморье, Передняя Азия, Южноазиатская и Восточноазиатская области, .Мексика с прилегающими странами или Южноамериканская подобласть аборигенных культур. Несомненно, из Индии при- шли в Эфиопию лен, дагусса; из Передней Азии — пшеница, полба; из Е~ентральной или из Передней Азии — ячмень; из Сре- диземноморья (в широком смысле этого слова) — горох (может быть, и непосредственно из Передней Азии), чина, конские бобы, черная горчица, возможно, клещевина. С Индией выявляется связь и обратного порядка. Из Эфио- пии в Индию перешли Guizotia, сорго (может быль, не непосред- ственно из Эфиопии, но из Африки). Из африканских стран Эфиопия имеет с Иеменом не только общие культурные растения, но и растения, представленные там очень сходными формами; таковы африканские культуры: кофей- ное дерево, чат. 1~онские бобы через Иемен распространились в Эфиопии — существует общая аравпйско-эфиопская группа сортов. Чечевица представлена одинаковыми формами в Г1емене и Эфиопии. Разумеется, имеются общие культуры и в ОАР и Су- дане и с внесредиземноморскими африканскими странами. Интересным с точки зрения истории древнего человечества является тот факт, что в Эфиопии некоторые культурные расте- ния, имеющие здесь только вторичное развитие, все же сохра- няют очень примитивные черты, роднящие их с примитивными видами отдельных стран, например эфиопская пшеница, абис- 340
с инская полба, с несомненностью свидетельствующие о древней- шей связи Эфиопии с Передней Азией — Закавказьем; эфиоп- ский лен, указывающий на древние связи Индии и Эфиопии. Ячмень в Эфиопии заведомо не аборигенен. Вместе с тем не- сомненен необычайно долгий период исторического развития яч- меня в этой стране, результатом чего является формирование своеобразного эфиопского вида ячменя. Это заставляет нас предполагать существование древнейших связей Африки непо- средственно с Передней и LIeFITpBJIbHQA Азией, Южным 1~итаем, с предполагаемыми первичными очагами культуры ячменя. Историки пишут, что в доисторическое время имели место переселения народов из Азии в Африку. Изучение культурных растений Эфиопии дает факты, которые подтверждают выводы историков — такими переселениями можно объяснить родство эфиопского льна с индийским, родство эфиопской пшеницы с пшеницами Передней Азии примитивного типа и т. д. Из Азии культурные растения проникали через Аравию в об- ласти современных Судана и Эфиопии. Из этих или соседних областей двигались на север древние народы Египта, принося с собой и набор культурных растений. В дальнейшем земледельческая культура Египта достигла высокого уровня, а в Судане и Эфиопии осталась на низком уровне. Поэтому и сохранились здесь древние примитивные фор- мы пшеницы, ячменя и других культур, хотя и в измененном виде. Среди культурных растений Эфиопии явно различаются два основных географических элемента: 1) аборигенный африкан- ский (в широком понимании, но с выключением средиземно- морской Африки); 2) Средиземноморский (и древнесредиземно- морский), Эти два элемента относятся к географическим 06- ластям исторического развития культурной флоры и вместе с тем они связаны с геофлористическими областями — т. е. с флорой вообще. Древнесредиземноморскпй и средиземноморский (в уз- ком понимании) элементы входят в Голарктическую флористи- ческую область, а африканский элемент — Палеотропическую область. При анализе состава культурной флоры Эфиопии Н. И. Вави- лов обратил особое внпмачие на средиземноморские элементы. Последнее понятно — к этой группе относятся растения, имею- щие наибольшее хозяйственное значение. Большой интерес именно к этой группе растешш, вероятно, привел Н. И. Вави- лова к переоценке их значения, как особенно характерных ра- стений для Эфиопии, и к преувеличенному представлению об общем эндемизме культурной флоры этой страны. Для большей ясности дадим перечень растений средиземноморского корня, расп остраненных в Эфиопии. шеи ица э идем ична я эфиоп с к а я (Triti~cum aethio- рысит) ведет происхождение из Передней Азии (Закавказья). Полба-эммер — Т. йоассоп, хотя и представлена в Эфиопии
эндемичным подвидом, пришла в Африку из Передней Азии, Т. poloni~cum — польская пшеница, вероятно, представляет собой измененные формы Т. aethiopiicum. Кул ь тур ный я ч мень Эфиопии, хотя и выделен Ф. Х. Бахтеевым в самостоятельный вид, происходит от передне- азиатских предков. Ачепа vaviloviana — эндемичный сорный и А. abyssini~ca — эндемичный культурный о в е с Эфиопии явно родственны другим 28-хромосомным средиземноморским овсам. Горох, нут, чечевица, чина — это весьма распростра- ненные во всех странах бобовые, согласно Н. И. Вавилову, имеют первичный центр развития родов и вхождения в культуру в Передней Азии. По нашему мнению, весьма вероятно, что эти виды или их предки имели древнесредиземноморские ареалы. Б о б ы ко н с к и е имеют, по нашему мнению, именно древне- средиземноморский ареал. Основной центр развития п а ж и т- ника сенного в Передней Азии. Л ю п и н — 1.upinus termis— относится к группе собственно средиземноморских видов. К о р и а н д р имеет основной центр развития в Передней Азии, кресс-салат — в Г!ередней или Средне-Юго-Западной Азии, во всяком случае, в области Древнего Средиземноморья. Абис- синская к а пуст а — вид, хотя эндемичный для Эфиопии, про- изошел от скрещивания двух средиземноморских видов. Л ен распространился в Эфиопию из Индии, а дикий родич всего культурного льна имел древнесредиземноморское проис- хождение. Род Сгапйе также средиземноморский. Хотя Эфиопия и считается одним из древних очагов вхождения в культуру м а сл ин ы, однако дикорастущая близкая к культурному виду Оlеа chrysophylla имеет, по-видимому, древнесредиземноморский ареал. В Средне-Юго-Западной Азии находится первичный очаг развития а ж г о н а. Из приведенного перечня видно, что эндемичные виды и формы многих эфиопских растений имеют корни в Цревнесреди- земноморской области развития культурной флоры и в ряде случаев вышли из определенных ее подобластей. Внесредиземноморские страны Африки пока еще очень слабо изучены в отношении географии и истории культурных растений. Из новейших раскопок интересны данные эпохи раннего нео- лита в районе озера Тумба (западная часть Конго), позволяю- щие сделать заключение, что древнее население этой части Африки уже тогда было знакомо с земледелием (А. И. Собченко, 1952). Подобные находки сделаны и в Мали. Таких археологических данных, которые могли бы дать более детальную ориентировку в этом вопросе, по Африке почти не имеется. Поэтому ниже рассмотрены только некоторые абори- генные для внесредиземноморской Африки культурные растения. К у н ж у т (Sesamum L.) африканского происхождения. Дикорастущие виды кунжута распространены в Восточной Аф- рике, Судане, Гвинее, бассейне реки Конго, в Южной Африке.
В Африке используются 3 вида: 1) S. radiatum Schum. et Thonn. — дико растет и местами возделывается в Занзибаре, Су- дане, Западной Африке, употребляется как масличное; 2) S. an- gustifo!ium Engl.— растет дико в Восточной Африке и Гвинее, употребляются в пищу семена и все растение; 3) S. indi~curn var. integerrimum Engl.— по указаниям ряда авторов, распростра- нен в диком состоянии в Галла (Эфиопия) и в области 1~онго.. Однако вопрос, является ли здесь S. indi~curn действительно ди- ким или одичавшим, еще нуждается в выяснении. Возделы- вается S. indicum в Африке почти повсеместно. Кунжут в Аф- рике идет не только на масло, но используется как кормовое и лекарственное растение. Поедается всеми видами скота. В Нигерии S. indicum сопровождается двумя мимикрирую- щими сорными растениями того же семейства: Sesamum radia- tum Schum. et Thonn. и Ceratotheca sesarnoides Endl. Последнее растение является лекарственным, листья его съедобны. Нали- чие специализированных сорняков говорит о древности культуры кунжута в Африке. Согласно предположению Штульмана (F. Stuhlmann, 1909), путь распространения культурного кунжута S. indicum или его ближайшего родича (что вернее) шел из Центральной Африки через Аравию и Месопотамию, Иран и Афганистан в Северо-3а- падную Индию. В Индии этот вид имеет вторичную область ин- тенсивного формообразования. Здесь же растет дико S. ðãostãà- tum Retz. К л е щ е в и н а почти по всей Африке сопровождает человека как рудеральное растение. Африканское происхождение имеют следующие короткодневные виды клещевины: занзибарская— Ricinus zanzibarinus G. Рор. и крупноплодная — R. macro~carpus G. Рар. = R. communis var. sanguineus Mull.-Arg. Мелкоплод- ная клещевина — R. ioommunis L. sensu stricto — является длиннодневным растением, что связано с ее древнесредиземно- морским происхождением (произрастает дико го морским бере- гам Аравии и Средиземного моря). Tetracanpidium conophorum (Mu11.-Arg.) Hutch. et Dalz.— лиана, дающая масличные семена, растет в кустарниковых за- рослях Нигерии, Дагомеи, l:,г.меруна. Ма сл ичный и сто д (Ро1уда1а butyracea Нас!'el) — мас- личное растение Западной Африки. Следующие растения дают масла: Loiphira alata Banks — плоды содержат масло, употреб- ляются на мыло (дико, Yaxtepyz и другие места); On~coba glauca Planch.— в семенах содержится до 30'/о масла, масло употреб- ляется при лечении проказы (дико, Камерун, Берег Слоновой Кости). М а с л я н о е д е р е в о (Butyrospermum ~parkii Kotechy)— семена содержат масло, которое имеет техническое значение. Этот вид имеет местное название Каг11е, является единствен- ным представителем сем. Sapotaceae в Западной Африке. Он ха-
рактерен и для саванн Судана. Плоды его переносятся летучими мышами на значительные расстояния (P Jaeger, 1962). К сем. Cucurbitaceae принадлежат два африканских маслич- ных растения: Telfairia occidcntalis Ноо1<. и Cucumeropsis man- nii Naudin. Род с о р г о (Sorghum Moench) имеет около 40 — 50 видов, в целом африканского происхождения, но далеко не всецело абиссинского и даже не восточноафриканского; эволюция его, по-видимому, протекала на широком пространстве древнего африканского континента. Не вызывает сомнения африканское происхождение сорго центральноафриканского, распространен- ного также и в Восточной Африке — S. bantuorum Jakuschev., сорго южноафриканского — S. caffrorum Jakuschev., а также copro западноафриканского — S. guineense Jakus~chev. Спорным является вопрос о происхождении азиатско-афри- канской группы сортов зернового сорго — Sorghum durra (S. vul- gare Pers. sensu з1г1с1о). Кроме Эфиопии (очаг распространения подвида S. durra ssp. aethiopi~cum Jakuschev.), сорта этого вида распространены в Судане, где представлены особым под- видом S. durra ssp. nubicum Jal<uschev., и в Африке отличаются не меньшим, а скорее большим разнообразием, чем в Средней и Юго-Западной Азии (где распространен подвид S. durra ssp. arabicum Jakuschev.). Скорее всего и этот вид обыкновенного культурного сорго имеет родину в Африке и отсюда распростра- нился в Азию. Веских доводов за происхождение S. durra из Ин- дии не имеется. В Индии и вообще в Азии не имеется родствен- ных диких или переходных к культурным форм. Некоторые сто- ронники азиатского происхождения сорго указывают на сорное сорго — гумай — S. halepense (L.) Pers. как на родоначальника культурного сорго — дурро. Действительно, S. halepense, или Andropogon halepense Brot., очень широко распространен, однако этот вид в Китае и Индии вовсе не чувствует себя так хорошо, как в Африке и Средиземноморье. Кроме того, этот впд октоплоидный (2n =40) и не может быть родоначальником для тетраплоидных видов, к которым относится азиатско-афри- канское сорго — S. durra (2п =20). Тетраплоидные виды, как и диплоидные (2п=10, например, S. versicolor Anderss.), сосре- доточены в Африке. Там же возделываются метельчатые виды сорго, переходные к дикорастущим [суданская трава — S. suda- nense (Piper) Sta~pf], и другие родственные дикорастущие виды, которые выявлены Е. С. Якушевским. Китайское сорго — гао- лян — S. chinense Jal(uschev., хотя п отличается большим свое- образием, однако проникло в Китай в не столь глубокой древ- ности. Таким образом, все факты говорят за африканское про- исхождение всех видов сорго. Щ ет и н н п к (Setaria Beauv.) в целом, по-видимому, афри- канского происхождения, но культурные формы чумизы, или гоми — S. italica ssp. maxima Alf. и могар — S. italica ssp. то- 344
charium A1f. происходят из Восточной Азии (Китай), а отсюда распространились через Монголию, Переднюю Азию в Среди- земноморье и Африку (Северную и Южную). Р и с (Oryza L.) имеет область свое~о развития в Южной Азии (Индия, Индокитай), но также и в Африке. Из африкан- ских видов в культуре имеется О. glaberrima Steud. Местным населением, кроме эфиопского риса — О. punctata Kotschy (о котором уже упоминалось в разделе об Эфиопии), исполь- зуются О. breviligulata А. Chev. et Poehr. (дико и иногда разво- дится в Судане и Центральной Африке), О. glaberrima Steud. (дико и иногда разводится в Западной Африке). О. barthii ис- пользуется на зерно и как фуражное растение в странах тропи- ческой Африки. Из хлебных растений используются местным населением сле- дующие: Paspalum exile lfippist (Западный Судан); ибури— Digitaria iburua $1apf (Нигерия). Eleusine согасапа Ciaertner, вероятно, занесена из Индии, но. некоторые авторы считают ее африканским растением. Я м с (Diioscorea L.) широко распространен в тропиках и субтропиках. К используемым видам, возможно, африканского происхождения, относится ямс абиссинский — D. abyssinica H~ochst., о котором упоминалось выше. D. latifolia Renth. рас- пространен в диком состоянии по всей тропической Африке. Ядовит, но поедается местным населением в неурожайные годы. Этот вид, хотя и имеет луковички, отличается от D. bulbifera L., который растет дико в Индокитае и Малезии. Одна форма D. latifolia кое-где возделывается. D.,cayenensis Lam. с колю- чими и неколючими клубнями распространен в тропической Африке в диком, одичалом состоянии и в культуре. D. dumeto- гит Paix растет дико в кустарниковой зоне и по опушкам тропи- 1еских лесов, иногда возделывается. К о л е у с (Coleus 1.оиг.) широко распространен по всей Африке. Вероятно, весь род африканского происхождения, не- смотря на существование южноазиатских (Индия и южные острова) дикорастущих и культурных видов. Кроме видов, ра- стущих в Эфиопии, особенно распространены С. rotundifolius А. Chev. et Perr. (Судан, Мадагаскар, Наталь, Конго и др.), представленный в Африке разнообразными формами; С. flori- bundus Pobyns et J. 1 еЬгип (Наталь, Ангола, Конго, в Восточ- ной озерной области) и многие другие. К виду С. rotundifolius близки С. brazzavillensis А. Chev. (Республика Конго), С. escu- lentus (N. E. Brown) Taylor (Наталь) п С. mirabilis Briquet. В настоящее время культура Coleus почти исчезла. Согласно исследованиям Шевалье (А. Chevalier, 1953), клубненосные виды Coleus — все африканского происхождения, но их культура распространена и в Азии (Индия, Цейлон, Ява). Л о б и я (Dolichos L.) распространена в тропическом поясе Старого Света (немногие виды в Америке). 345
Гиацинтовые бобы (D. lablab L.) широко возделы- ваются в Африке и в Южной Азии. Выявлено большое разно- образие форм гиацинтовых бобов в Восточной горной Африке— в Восточной озерной области, а также в области Килиманджаро и на Мадагаскаре. В Восточной Африке встречаются гигантские, почти древовидные лианы до 20 м длиной, и здесь же обитают низковьющиеся мелколистные формы (Н. Р. Иванов). Здесь, по Н. Р. Иванову, находится родина этого вида, а не в Южной Азии — там вторичное разнообразие форм. Отметим еще следую- щие африканские виды: D. gibbosus ТйипЬ. (Капская провин- ция); D. ipratensis Taub. (Южная Африка). В и г н а (Vigna sinensis Endl.) отличается огромным поли- морфизмом не только в Восточной Африке (см. выше — o6 Эфио- пии), но также и в южной части континента. Род в целом афри- канского происхождения (более 40 видов обитает в Африке). ОсНоВНоН областью происхождения вигны (коровьего гороха) является Центральная и Южная Африка. Отсюда он распростра- нился через Аравию в Индию и Китай, где находится вторичный очаг формирования этого вида. Второй вторичный очаг нахо- дится в Закавказье. Vigna icatjang (Burm.) %а1р. культивируется в Южной Аф- рике. Отсюда распространился в Индию, Египет, Судан. Старейшее пищевое бобовое аборигенное растение Африки- это Voandzeia subterranea Thou., отличающееся геокарпией, рас- пространено по всей тропической Африке. В Западнои Африке растет другое бобовое, также геокарпное — Kerstingiella geoca- rpa Harms, а в районе озера Чад и соседних с ним растет клуб- неносное бобовое растение — Sphenostylis stenvcarpa Idarms. Второй по значению очаг вхождения в культуру хлопчатника, по Ф. М. Мауеру (1950), сосредоточен в Юго-Восточной Африке, где был введен в культуру вид Gossypium herbaiceum L., отсюда занесенный в Азию. В Африке сосредоточено огромное разно- образие форм и экотипов этого вида: от наиболее древних долго- летних форм — зэр. pseudoarboreum Маиег — до промежуточных, менее долговечных сортов. В Африке распространены следующие эндемпчные подвиды G. herbalceum: 1) ssp. africanum (Watt) Mauer — дикий долго- летнии кустарник (Юго-Восточная, Южная и Юго-Западная Африка), в Южной Африке имеются примитивные культурные многолетние формы этого подвида и переходные к ним; 2) ssp. frutescens (Deli е) Nauer — предшественник скороспелых суб- тропических форм ssp. euherbaceum Mauer, этот подвид распро- странен не только в Африке, но и в Индии; для Африки энде- мпчны следующие эколого-географические группы этого под- вида: западноафриканская, суданская и восточноафриканская; 3) ssp. ipseudoarboreum Mauer — примитивные рудеральные и культурные формы, используемые, по Мауеру, в приусадебной культуре как лекарственное п культовое (троппческая Африка, 346
Сомали, заходит в 10жную Аравию, Иран, изредка в Южную Индию). Вторичный центр развития вида G. herbaceum в Юго- Западной, Средней и Малой Азии. Африке свойственны еще следующие виды хлопчатника: G. soudanense Watt — происходит из Мадагаскара и Юго-Во- сточной Африки; G. somalense J. В. Hutch. — из Сомали и Во- сточного Судана; G. апота1ит Wawra et Peyr. — Судан, Ангола. Растет в тропических полупустынях и саваннах. Ф. М. Мауер (1954) считает эндемичными для Африки еще следующие дикорастущие виды хлопчатника: G. capitis-viridis Маиег — острова Зеленого Мыса; G. triiphyllum Hochr.— Юго- Западная Африка; G. ellenbeckii Mauer — Сомали, Галла, Кения (северо-восточная). Текстильное растение Hibiscus aspera Hook., по-видимому,. эндемично для Африки. Имеют значение следующие прядильные тропические дико- растущие растения. Sansevieria senegambiica Baker (S. liberi~ca Ger. et Labr.) дикорастущее в Камеруне, листья дают грубое волокно, орна- ментальное растение. Adansonia digitata L. (Baobab) растет в Камеруне и в дру- гих местах. Из коры получают волокно (для веревок); плоды и листья съедобные; медонос. Urena hobata L. культивируется в Конго на волокно. Заме- няет джут. Vigna unguiculata var. textilis Wal~p. — плодоножки очень длинные и дают белое прочное волокно. Растет дико в Каме- руне. Кон о пл я гви а иска я (hibiscus cannabinus L.) дает во- локно. Листья сьедобные. Растет дико в Камеруне и других местах. Б а м и я (Hibiscus es~culentus L.) имеет несколько разновид- ностей. Возделывается ради съедобных плодов. Едят также листья. Зола стеблей заменяет соль. Имеются и другие виды Hibiscus со съедобными плодами и листьями. В Судане и Сенегале возделывается текстильное растение Ceiba pentandra Gaertn. В опылении принимают участие мест- ные летучие мыши. Кол а (Cola Schott et Endl.) происходит из Западной эква- ториальной Африки, где имеется около 50 видов. Cola — лекар- ственные и возбуждающие растения. Из них особое значение имеют С. nitida Chev. (Нигерия, Гвинея, Камерун и соседние страны) и С. acuminata Schott et Endl. (Берег Слоновой Кости — дико, возделывается во многих странах экваториальной Африки) . П а л ь м а м а с л и ч н а я (Flaeis guineensis Jacq.) дает цен- ный растительный жир, происходит из лесов нижнего Конго. Отсюда в XIX в. занесена в Индонезию. 347
Род Cucumis L. богато представлен в Африке, и здесь его родина. Однако африканские виды не являются родоначальни- ками дыни и огурца. Африка богата «огуречноподобными» растениями из сем. тыквенных. К таковым А. И. Филов (1959 и 1960) относит, на- пример, следующие виды: Acanthosicyos horrida Welw. — растет дико в Юго-Западной Африке, важный продукт питания готтен- тотов и бушменов. Плоды едят свежими или вареными; Telfai- ria yedata Hook. — дает съедобные плоды, растет на Мадагас- каре, изредка возделывается Т. oocidentalis Hook. культиви- руется ради плодов (Западная Африка). Род Cucumis отличается в Африке большим разнообразием, многие виды имеют съедобные плоды, например: С. callosus Cogn., С. Iongipes Hook. (Ангола), С. hirsutus Sond., С. metuli- ferus Е. Меу. с кисловатыми плодами (почти по всей Африке). Некоторые виды африканских Cu~cumis заходят и в Азию: например, С. prophetarum L. с плодами без горечи растет в Се- верной Африке, Аравии, Афганистане и Передке"; С. fici- .fo ius А. erich. имеет съедобные плоды, распространен дико в Северной Африке, но заходит и в Аравию. Африканские представители Cucumis еще очень слабо изучены. Совсем не изучены виды этого рода из относительно увлажненных местообитаний, где, может быть, впоследствии найдутся непосредственные родоначальники настоящего огурца. В настоящее время схему происхождения и древнего рассе- ления огурца можно представлять себе следующим образом. 1~акие-то формы Cucumis, вероятно, из сравнительно болеевлаж- ных горных районов, распространились из Африки в Азию: с одной стороны, в Переднюю Азию (Аравию, Иран, Малую Азию) и оттуда гораздо позднее в Среднюю Азию и Европу; с другой — на восток в Индшо. В Индии сперва образовались ги- малайские сорта огурца, которые в древности были занесены в Восточную Азию (I(maA и Японию), где сформировалась осо- бая группа восточноазиатских сортов. Позднее эти сорта были занесены в Европу. По этой схеме очаг происхождения огурца— Северная Африка, а два центра интенсивного формообразова- ния — Передняя и Восточная Азия. Африка — очаг происхождения д ы и и. Дикие формы Cucu- mis, близкие к дыне (С. melo L.), распространены почти по всей Африке. Они имеются и в Южной Азии, на островах Тихого океана и в восточной части Америки, но они, по-видимому, рас- пространились на другие материки из Африки. Здесь можно предполагать некоторый параллелизм развития двух ветвей рода Cucumis — огуречной и дынной. По достаточно обоснованной теории А. И, Филова, основное разнообразие культурных форм того пли иного вида обыкновенно создается за пределами ареала распространения дикорастущих предков или на краях этого ареала (не в центре его). 348
Дикие формы родоначального вида — С. agrestis (Naud.) Рапо'. — в Северной Африке доходят до долины Нила включи- тельно, они заходят в Переднюю Азию (Аравию) и в Среднюю и Южную Азию — здесь-то и находятся области интенсивного формирования культурных сортов дыни. По А. И. Филову (1959, 960), культурные сахаристые дыни были созданы в Азии, их в Африке нет. Дыни (и арбузы) дали высококультурные формы только в Азии на северной границе распространения дикорасту-. щих форм. Здесь играет роль не только древность земледелия и многовековая работа человека, но и климат. Основными оча- гами эволюции дыни в культуре являются Индия (канталупы) и Передняя Азия, в особенности Западная Анатолия (кассабы). Африка является, несомненно, родиной рода Citrullus Schrad. Вопрос же о происхождении сладкого столового арбуза— С. vulgaris Schrad. — более сложен. Ко р м о в о й а р б уз (С. colocynthoides Pang.), -несомненно, имеет африканское происхождение. Ближайший дикий родствен- ник С. icolooynthis L., по А. И. Филову (1962), включает три группы форм. Группа форм с широкодольчатыми листьями рас- пространена только в Центральной и Южной Африке — это и есть, по А. И. Филову, родоначальник кормового арбуза. Кормовые арбузы имеются в Африке в культуре. Окультури- .вание связано с потерей горечи и увеличением размеров плода. Этот арбуз был знаком многим путешественникам по Африке: Брауну (W. Brown, 1800), Шрадеру (Н. А. Schrader, 1836, 1838) и другим под названием каффрского — С. caffer Sichrad. Ноден (Naudin, 1855) сообщал о каффрских арбузах, что имеется много их форм, нуждающихся в изучении. Встречались арбузы с раз- личной окраской семян и все имели желтую безвкусную мякоть. Ливингстон (D. Livingston, 1857) пишет, что арбузы С. caffer растут в диком состоянии в пустыне Калахари в большом коли- честве в сравнительно влажные годы, что бывает один раз в 10 или 11 лет. В эти благоприятные годы дикие арбузы утоляют жажду и голод людей и животных. Оказывается, однако, что не все ид плодов бывают съедобны, т. е. достаточно мягкие и соч- ные, без горечи, а сахаристых среди них совсем не бывает. В пустыне Калахари имеются маленькие оазисы, где растет трава и даже кустариики и деревья. Растительность также имеется по руслам древних рек. Эти русла заполняются водой в дождливое время. Здесь на рыхлой песчаной почве условия благоприятны для роста арбузов. Арбуз распространяется при посредстве людей и животных. Сок диких арбузов сохраняется в плодах в течение длительного времени, и этим пользуются путешественники по пустыне, запасая арбузы как источник необ- ходимой для жизни влаги. Семена из поедаемых арбузов, попа- дая на благоприятную почву, прорастают. Дальнейшее распро- странение арбузов происходит в качестве дикаря или сорного растения. Таково начало окультуривания кормового арбуза: он 349
постепенно изменяется вследствие приспособления к новым условиям и улучшается в результате искусственного отбора. Столовый арбуз, по А. И. Филову, имеет азиатско-африкан- ское происхождение. Исходной разновидностью Филов считает особую форму С. colocynthis с желто-серым венчиком и вырез- ными листьями, как у его азиатских форм, но в отличие от ази- атских С. соlоcynthis имеет не гладкие вздутые семена, а гло- ские с боковыми рубчиками, как у столового арбуза. Ареал этой разновидности: Египет, северные окраины Сахары, Алжир. Тыкв а в Африке является очень древней культурой. Воз- можно, что тыква была завезена в Африку португальцами (F. Stuhlmann, 1909). Тыква считается как бы туземным расте- нием по берегам Нигера в Гвинее, в Анголе и в других местах. В коллекции ВИР имеются тыквы, привезенные Н. И. Вавило- вым, они представляют собой своеобразные экотипы (А. И. Фи- лов, 1960). Н. И. Вавиловым была привезена с окраин Сахары также дикая мелкоплодная тыква, похожая на арбуз. По пред- положению А. И. Филова, плоды дикой тыквы из Америки пере- плывали через океан и таким образом достигали Африки. Тыквы в Эфиопии, по исследованиям А. И. Филова, весьма своеобразны, имеют сильно вытянутую форму, напоминают североамериканский сорт банана. Судя по материалу, приве- зенному экспедицией Ф. Ф. Сидорова и Т. Я. Зарубайло, эфиоп- ские тыквы относятся к виду С. maxima Duch. Каким путем и когда они сюда попали из Америки, неизвестно. За основную область первичной культуры б а н а н а (Musa L.) большинством принимается Азия (Индия и Индо- китай с островами, Китай, Япония), имеется второй очаг разви- тия в Африке. Кроме эфиопского банана — М. ensete, имеет зна- чение африканский дикорастущий текстильный банан — М. holstii K. Schum.— и родственные виды (тропическая Африка). Ьукудшт owariense Benth.— плодовое дерево родом из тро- п ичес к ой Аф рики. Anona L. — в основном американский род, но имеются и 2 — 3 африканских вида со съедобными плодами, из них наибо- лее распространен по всей тропической Африке А. senegaleiisis Pers. с ароматными плодами (дико и в культуре). Плодовых деревьев в Африке много. В парковых лесах Гам- бии и Казаманса (Сенегал) растет Lephira alata Banks со съедобными плодами. Важную роль в питании негров играют следующие виды: Маттеа africana G. Don., Sarcocepha us esculentus Afzelius (съедобные плоды с запахом персика), Treculia africana Decne. (крупные плоды содержат съедобные зерна), Antrocaryon micra- ster А. Chev. et А. Guill., Муг1апйиз arboreus Веаиъ. и др. Следующие растения содержат в семенах масло: Blighia sapida Koenig, Staudtia gabonensis Warburg, Pentadesma buty- racea Sabine, Раийа oleosa Pierre, Poga ioleosa Pierre и др. 350
Ficus asperifolia Miq. растет дико в Камеруне и других ме- стах, плоды и листья съедобны; F. capensis Thunb. растет дико в Камеруне и других местах, плоды съедобны. Следующие виды Zizyphus М111. распространены в Африке дико: Z. jujuba Lam. — с малосъедобными плодами Z. spina- christi AVilld. — плоды хорошего качества. Находят применение некоторые виды Euphorbia L. (местное название — Burowal тапка), например кактусовидный молочай. Цветущая верхушка этого растения, печеная вместе с головой змеи и плодами строфантуса, давала яд для стрел. В Африке имеется еще много древесных и травянистых або- ригенных растений, которые используются из диких зарослей или культивируются местным населением, но они не вошли широко в культуру и их филогения и география еще слабо изучены. де- коративными растениями особенно богата Капская провинция. Стол етн и к (Aloe L.) в целом африканского происхожде- ния. Из A. perfoliata Meyen добывается грубое текстильное во- .локно, идущее на маты, веревки, канаты; из сока А. afri~cana Mill. и А. ferox Mill.— лекарственное вещество сабир. Имеют .лекарственное значение и другие виды столетника, например, А. arborescens Mill. (чаще всего разводится в горшечной культуре) . Большинство видов алоэ распространено в Капской провин- иии (всего около 250 видов). Там же находится основной очаг развития рода Pelargoniunz L'Her. из сем. Geraniaceae. Многие виды имеют декоративное значение и разводятся под названием герани. Р. roseum Ш11д. дает эфирное масло (возделывается в Закавказье) . Африка дала много комнатных растений: виды Coleus Lour., спаржи, кливии, крестовника (Senecio L.), гемантуса, веничника и др. С Мадагаскара происходят декоративные виды цитруса, фикуса, каланхое и др. (М. Toivonen, 1958). Из лекарственных растений упомянем о р а у в о л ь ф и и р в о т- ной (Rauwolfia vomitoria Afzel.). Имеет следы серпентина, но есть другие полезные алкалоиды (резерпина вдвое больше, чем у R. senpentina Benth. ех Iyurz). Распространена в тропической Африке: в Западной — от Сенегала до Амбоина; в Восточной- до озер Виктория и Ньяса. Корни пользуются огромным спро- сом, главным образом в Африке, как лечебное средство. Африканская область культурной флоры, включая Восточную Африку без Северной — Средиземноморской Африки, дала не так мало мировому растениеводству. Можно отметить целые роды, включающие в свой состав культурные растения африканского происхождения: Guisotia — нуг, Sesamum — кунжут, Coleus— клубненосные растения, Pennisetum — негритянское просо, Sor- ghum — сорго, Vigna — коровий горох, Citrullus — арбуз, Cucn- mis (но виды не родственные огурцу и дыне), Cola — кола, Seta- ria (род, но не основные культурные виды), Indigofera — индиго- фера, Aloe — алоэ, Pelargonium — пеларгониум. 351
Часть родов имеет здесь одну из областей первичного разви- тия рода — хлопчатник, рис, банан. Многие растения имеют в Африке область вторичного разви- тия в культуре (не имеют здесь диких родичей, но представлены эндемичными видами или формами). Таким вторичным очагом видо- и формообразования культурных растений являются прежде всего Эфиопия с Эритреей (пшеница, ячмень, лен, кресс- салат, черная горчица, горох, нут, чина, конские бобы, триго- нелла, другие бобовые, дагусса, кориандр). Некоторые культурные растения представлены одиночными эндемичными видами, хотя род в целом не африканского проис- хождения, например абиссинская капуста — Brassica carinata. Некоторые культуры так давно занесены не только в Эфио- пию, но и в другие страны Африки, что производят впечатление аборигенных и представлены многими эндемичными формами, к каковым относятся: рис посевной — Oryza sativa L., таро — Colo- casia antiquorum Schott, неэндемичные виды банана Musa L., еще в глубокой древности проникшие в Африку из Азии (Индии и соседних стран) и др. Особенности состава африканских культурных растений вы- ражаются именно в том, что, помимо чисто аборигенных куль- тур, в Африке нашли вторую родину многие растения, очень давно пришедшие сюда из Азии (главным образом Индии, Ин- донезии), Передней Азии и Средиземноморья (в широком значе- нии этого понятия). Эфиопия и Эритрея не резко отграничены uG составу культурной флоры от африканских стран, располо- женных к югу и к западу. С другой стороны, Эфиопия не резко отграничена и от Средиземноморья, хотя средиземноморские растения (в широком объеме этого понятия) проникали в Эфио- пию чаще не с севера, а через Аравию. Таким образом, Эфиопию и Эритрею нельзя рассматривать как самостоятельную область культурной флоры, а можно считать ее областью влияния афри- канской и древнесредиземноморских культурных флор. Возможно, что эту часть Африки лучше выделить как Эфи- опско-Сомалийскую подобласть большой Африканской тропиче- ской области исторического развития культурной флоры, что согласуется и с представлениями о флористических подобластях. Чтобы окончательно решить этот вопрос, нужно дальнейшее на- копление материала. Нам представляется, что первым важным шагом к упорядо- чению вопроса об областях и подобластях культурной флоры на африканском континенте является отделение средиземноморской Африки от палеотропической, что мы и пытались сделать в дан- ном очерке.
АБОРИ ГЕННЫЕ КУЛЬТУРНЫЕ РАСТЕНИЯСРЕДИЗЕМНОМОРЬЯ В данном разделе под Средиземноморьем разумеются только страны, расположенные кругом современного Средиземного моря в отличие от установленной М. Г. Поповым Древнесредиземно- морской области, куда входят и Передняя и Средне-Юго-Запад- ная Азия с Афганистаном. Вопрос о значении Древнего Среди- земноморья в географии культурной флоры освещена нами выше. Под средиземноморскими странами подразумеваются Греция, Югославия, Италия, Южная Франция, Испания, Португалия и Северная Африка до южной границы Сахары. Сирия и Пале- стина и отчасти Иемен составляют область, переходную между Передней Азией и собственно Средиземноморьем (сведения о них имеются в разделе о Передней Азии). климат Средиземноморья характеризуется сухим и жарким летом и умеренной, обыкновенно дождливой зимой. Различают в Средиземноморье следующие климатические зоны: ) сухая — пустынная (пустыни Северной Африки); 2) полусухая (сухие степи) — часть Прованса во Франции, степные местности Греции и Италии, Испания, 1~рит, Сицилия, острова Эгейского моря; 3) умеренная — средний пояс южных склонов Альп; 4) влажная — слабо выражена (некоторые местности в Се- веннах, остров 1~ос); 5) высокогорная — верхние пояса Альп; 6) зона апельсина — приморская полоса (Ривьера) западной Италии и Южной Франции; некоторые местности в Испа- НИИ. Растительность (дикорастущая) в Средиземноморье в на- стоящее время преобладает ксерофильная, лесов очень мало, главным представителем лесной растительности является среди- земноморский дуб — Quercus ilex L. Прежде лесов было гораздо больше — они истреблены человеком. Средиземноморье издавна заселено европеоидами, древней- шими представителями которых являются кроманьонцы. В доисторическое время климат Средиземноморья был влаж- ным. В плювиальный период пустыни Африки изобиловали пресной водой и были в ббльшей своей части покрыты пышной 353 Синская Е. Н.
др~ддд растительностью. Обитатели этих мест были носителями единой палеолитической культуры Северной Африки и европей- ских средиземноморских стран, они занимались собиратель- ством, охотой и рыболовством; может быть, были и зачатки зем- леделия. Ледники отступили, климат становится суше и теплее. Люди спустились в долины рек и долины предгорий. В дельте и долине Нила осели предки египтян (египетская земледельческая куль- тур а является частью средиземноморской) . Неолитические стоянки в большом количестве открыты всюду в средиземно- морских странах, особенно в восточной части Средиземно- морья. Около первобьпных жилищ (пещер) появились растения, вы- росшие на почве, удобренной пищевыми отбросами. Затем воз- никли и опыты сознательной культуры. Возможно, что и в Средиземноморье, как и в других геообластях, нексторые пер- вые возделываемые растения имели не пищевое, а лекарственное п р иту альп ое зн ач ение. Археологические изыскания в средиземноморских и приле- гающих к ним странах очень многочисленны. Именно здесь впер- вые была разработана в основных чертах общая археологиче- ская периодизация, основы которой изложены выше. В дополне- ние приведем схему периодизация европейского земледелия по Р. Фюрону из его «Учебника общей предыстории» (Furon, 1932) . 1. Фландрский период начинается с максимума последнего большого оледенения (вюрмского), который соответствует эпохе влажного климата, пышной растительности и богатого населения животными и людьми той территории, где теперь находится огромная пустыня — Сахара. Эта эпоха заканчивается новым мустьерским периодом — 50000 лет до н. э. 2. Яалее последовала эпоха Солютре, когда уже существовал человек — кроманьонец. Эта =-noxa сменяется мадленской (верх- ний палеолит) — 15000 лет до н. э. В это время было положено начало приручению животных и, вероятно, еще не было куль- турных растений. 3. В эпохи мезолита (азильская, капсийская культуры) про- изошло погружение суши на месте пролива Па-де-1~але. 4. В начале неолита оседающее население уже возделывает хлеба в Передней Азии и занимается разведением домашнего скота — 8000 лет до н. э. 5. Расцвет первых больших азиатских цивилизаций и озер- ная свайная культура в Швейцарии — уже развитое земледелие v. Центральной Европе — 7000 — 3500 лет до н. э. 6. Окончательное высыхание Сахары. 1~ультура пшеницы и ячменя в неолитическом поселении в Файюмском оазисе в Егип- те, где зерно хранилось в ямах. Начало египетской цивилизации 5000 — 4000 лет до н. э. 354
7. Открытие меди и бронзы — в III тысячелетии до н. э. Древ- няя культура Крита 3800 — 2800 лет до н. э. Расцвет ее — 1700— 1500 лет до н. э. 8. Эпоха меди в Европе 2500 — 1900 лет до н. э. 9. Начало века железа в Передней Азии (Сирия) — 1300 лет до н. э. Первые железные плуги. 10. Расцвет карфагенской культуры — 1100 — 146 лет до н. э. 11. Расцвет Греции — 800 — 500 лет до н. э. (век Перикла). 12. Римская цивилизация 266 лет до н. э. — 350 лет н. э. Начиная с этой эпохи, земледелие в Галлии интенсивно раз- вивается, но впоследствии вторжение варваров вызвало продол- ж ител ьный р егр ес с. Разумеется, исчисления времени отдельных периодов здесь гипотетичны, и схема служит только общей придержкой. Важно, однако, то, что земледелие Средиземноморья и Центральной Европы за 10 000 — 8000 лет до н. э. уже было достаточно развито. До последнего времени историки были склонны преувеличи- вать значение переселений. Так, по Фюрону (R. Ригоп, 1932), и Европу земледелие было «занесено» из 1Ого-Западной Азии. Археологические материалы свидетельствуют о том, что земле- делие зарождалось спонтанно, независимо, во многих областях Земли. другое дело — культурные растения. Они вводились в куль- туру и из местных флор, а также (еще в чрезвычайно отдален- ные доисторические эпохи) переносились из одного о~ага древ- него земледелия в другой. Древнейшая цивилизация Восточного Средиземноморья, до- стигшая уже высокого уровня, — это крита-микенская, развитие которой изучено, начиная с неолита. Последний продолжался на Крите от начала VI до III тысячелетия до н. э. Земледелием на Крите занимались еще в эпоху неолита. В эпоху расцвета этой древней цивилизации, развившейся на базе рабовладельческого строя, скотоводство и земледелие до- стигли высокого уровня. Здесь возделывали пшениц~, ячмень, бобы, горох, чечевицу, лен, шафран, кунжут, мяту, виноград, маслину. Имелись и декоративные растения: ирисы, лилии, тюль- паны, гиацинты и др. Город Микены находился в южной части Балканского полу- острова, и жители его считаются греками — ахейцами, крестьян- ские же поселения — не греческие, существовали с XIV по XII век до н. э. Здесь было так же высоко развито земледелие, как и на Крите. Ахейцы, теснимые народами с севера, двигались на юг, вы- тесняя пеласгов и распространяя достижения своей культуры, в том числе и земледельческой. Ахейцы направлялись и в Севе- ро-Западную Малую Азию, стремясь утвердиться в Троаде и завоевать главный город — Трою. В Трое люди жили с отдален- нейших времен. 12"
При раскопках установлено 12 слоев поселений (деревень и городов), и гомеровская Троя является шестым городом, стояв- шим на холме Гиссарлык, считая снизу вверх. Раскопки в Трое показали, что здесь земледелие существовало задолго до по- хода ахейцев в XII в. до н. э., описанного Гомером в «Илиаде». Раскопками старой Трои (2000 лет до н. э.) установлено, что древние троянцы сеяли пшеницу (однозернянку), горох, чечевицу, бобы, разводили виноград. Таким образом, и на этом примере видно, что «переносилось» обыкновенно не зем- леделие, возникновение которого политопно (во многих ме- стах и часто одновременно), а навыки более совершенного зем- леделия и культурные растения. В результате военных походов и через караванные торговые сношения уже с древних времен Греция имела связи не только с Передней Азией, Египтом, но и с другими средиземноморскими и еще более отдаленными странами. Параллельно шел и обмен культурными растениями. В неолите Фессалии были найдены полба и пшеница, похо- жая на мягкую, ячмень, желуди — Quercus ~1ех, горький мин- даль, горох, конские бобы, смоква, груши (две формы), просо— PaniI.um miliaiceum. В «Илиаде» рисуется картина сельскохозяйственной жизни Греции в гомеровские времена. В поэме говорится о пашнях, засеваемых ячменем и пшеницей, о больших виноградниках, са- дах и огородах. Особенно большими пахотными угодьями владели гомеров- ские цари и другие представители военно-земледельческой:ари- стократии. В садах росли маслины, гранаты, смоковницы и другие пло- довые деревья. Техника земледелия в эту эпоху уже отошла от примитив- ного уровня. Для обработки почвы употреблялись плуг, кирка, колотушка и заступ. Нивы вспахивались три раза волами, за- пряженными в плуг. Жатва производилась серпами, за жницами шли вязаль|цики, которые связывали снопы, а за ними — дети, собиравшие ко- лосья. Здесь же присутствовал хозяин (басилей), ко1орый при- сматривал за работающими. В эпоху «великой» греческой цивилизации (VIII — VI в. до н. э.) в самой Греции уменьшилось возделывание хлебных зла- ков и получило дальнейшее развитие садоводство, виноградар- ство, огородничество и разведение маслин. Орошение, которое в примитивном виде существовало еще в гомеровскую эпоху, по- лучило дальнейшее развитие. Греческий поэт Гесиод (конца VIII в. до н. э.) говорит 06 ирригации и описывает некоторые сельскохозяйственные работы. Молотьба производилась волами на тщательно подготовленном току, имеются упоминания и о цепной молотьбе. Провеянное зерно собиралось в особые
сосуды и хранилось в специальных амбарах. Хранение зерна в специальных сосудах в эту эпоху практиковалось и в других странах (например, в Урарту). Гесиод составил сельскохозяй- ственный календарь. В VI — III вв. до н. э. развивается римская цивилизация, в об- ласти сельского хозяйства многое осваивается из стран, лежа- щих на восток, особенно из Греции. Эта эпоха для нас менее ин- тересна, так как видовой ассортимент средиземноморских расте- ний к этому времени уже в основном сложился. В Европе земледелие возникло очень рано не только в среди земноморских странах, но и в Центральной и даже в Северной Европе. Влияние Средиземноморья сказалось и на странах, рас- положенных намного севернее, и поэтому, чтобы впоследствии выделить костяк аборигенных или заносных, но очень древних культур в каждом крупном очаге древнего земледелия, удобнее дальше перейти к обзору по отдельным культурам. Род Triticum L. — п ш е н и ц а — довольно богато представлен и Средиземноморье. К реликтовым мало распространенным ви- дам относятся следующие. Д и к ар а стущ а я одноз ер н яика (Т. boeoticum Boiss ) распространена в Израиле, Сирии, Ливане и на Балканах, захо- дит на Карпаты, встречается в Испании, но, вероятно, зане- сена. Культурна я одноз ерн як к а (Т. топо~соссит L.) кое- где сохранилась в Испании, Южной Франции и в Марокко. gикор астуща я двузернянк а (Т. dicoccoides Коегп.) распространена в Сирии, Израиле, Ливане, т. е. в странах Пе- редней Азии, а не в собственно Средиземноморье. По л б а - э м м е р (Т. Йсоосоп Schrank) представлена в Сре- диземноморье особыми подвидами — ssp. maroocanum Flaksb. (Марокко). В Испанию (Наварра) заходит европейский подвид полбы — ssp. еигораеит Vav. Испанские формы выделены в осо- бую ветвь (proles) европейского подвида — iproles euscaldunense Flaksb. Т в е р д ы е п ш е н и ц ы (Т. durum Desf.) представлены в собственно Средиземноморье большой группой крупноколосых и крупнозерных сортов средней плотности (и рыхлых), большей частью позднеспелых. Эта группа распространена в Алжире, Тунисе, Марокко, Ливии (Триполитания), частично ОАР, в юж- ной Испании, южной Италии, на юге Балкан, в Сирии, Израиле. Твердые пшеницы с компактным колосом (группа duro<com- pactum F1aksb.) приурочены к Марокко, Алжиру, Кипру и встре- чаются в Египте, Сирии, Израиле. Пшеницы, относимые к Т. tur- gidum L., нами включены в вид Т. durum sensu lato, в Среди- земноморье они представлены высокорослыми и крупноколосымп формами — proles meridionale Flaksb. (особенно в Сицилии и Испании) . 357
В коллекции ВИР имеются ценные для селекции крупнозер- ные формы озимой м яг кой пшеницы — Т. aestivum L.— из приморской Югославии, выделенные М. М. Якубцинером (1956) в особый подвид звр. grandisemineum. М. М. Якубцинер считает возможным, что эти формы были использованы селек- ционерами Италии для выведения крупнозерных сортов. Европейская п о л б а - с п е л ь т а (Т. spelta 1 .) имеет вторич- ный очаг культуры в Южных Пиренеях. Я ч м е н ь (Hordeum sativum Jessen, точнее Н. eurnasiaticum Ча~. et ВасЫ.) представлен в Средиземноморье крупнозерным подвидом — ssp. mediterraneum ВасЫ., в составе которого для средиземноморских стран намечен ряд ветвей (пролесов): pro- les a~penninicum ВасЫ., proles aegyptiaicum Vav. et Bacht., proles boreali-africanum ВасЫ., ~proles ругепагсит ВасЫ., proles graeco- ma~oedoni~cum Vav. et ВасЫ., proles syraco-palestinicum Vav. et ВасЫ., proles jemeni~cum Vav. et ВасЫ. Последние два пролеса скорее относятся к Переднеазиатской подобласти. Овсы (Avena L.) в Западном Средиземноморье представ- лены большим разнообразием диких и сорных 14- и 28-хромо- сомных форм. Овес волосистый — А. hirtula Lag., распростра- ненный на Пиренейском полуострове, в Северо-Западной Аф- рике и Сицилии, является предком культурного песчаного овса— А. strigosa S~chreb. (2n = 14) . Песчаный овес вошел в куль- туру на Пиренейском полуострове, где имеется большое разнообразие сорных форм, и далее распространился на запад. Дикорастущие 28-хромосомные овсы — А. barbata Pott. и А. wiestii Steud.— имеют древнесредиземноморский ареал. Пер- BbIH доходит на востоке до Гималаев, а второй — до Передней Азии включительно. Византийский 42-хромосомный овес — А. byzantina С. Кос1т- возделывается по всей Средиземноморской области, здесь же распространен и считающийся его родоначальником А. sterilis L. sensu lato. Вид A. byzantina представлен в Средиземноморье (не считая Передней Азии) средиземноморским (ssp. eubyzan- tina Nordv.), марокканским (ssp. nodipubescens Nalz.) типами (А. И. Мордвинкина, 1960). Древнейшие народы и цивилизации не возделывали овса посевного — А. sativa L. Названия овса нет в санскритском языке и нет в более новых языках Индии. Об овсе не знали древние египтяне и евреи. О нем нет упоминаний в библии. Овес посевной не возделывался у древних греков и римлян. К наибо- лее ранним относятся сведения (Феофраст п др.) об овсе как сорняке. Плиний Старший (I в. н. э.) писал, что ячмень может перерождаться в овес, который сам становится культурным рас- тением, а германцы даже его сеют и употребляют в пищу в виде каши. Таким образом, Плиний прямо указывает, что овес вышел в культуру из сорного состояния вне пределов Греции и Рима, в более северных странах. Однако у Плиния имеются сведения 358
о культуре другого вида овса — «греческого» — он возделывался в смеси с бобовыми на зеленый корм. Гален сообщал о греческом овсе, что его сеют в Мизин на корм вьючным животным, но при нужде им питаются и люди. А. И. Мордвинкпна (1960) считает, что этот овес относился к виду А. byzantina С. Koch, который представлен особенно в большом разнообразии в Малой Азии и оттуда распростра- нился в Грецию. Этот вид овса древнее, чем А. sativa. В Западном Средиземноморье имеет первичную родину и а- н а р е е ч н и к (Phalaris сапаг1епз1з L.) .. Ал ь ф а (Stipa tenacissima L.) дает волокно, употребляемое главным образом на бумагу. Кормовое растение, ценное для сухих степей. Ареал — Западное Средиземноморье (юго-запад Испании, степи горных плато восточного Марокко, сухие степи Алжира и Ливии). С п а р т (Lygeum sparteum L.) — злак степных зон Запад- ного Средиземноморья. Имеет тот же ареал, что и альфа, но менее распространен. Употребление такое же, что и альфы. 3 о л о т о б о р о д н и к [Chrysoipogon gryllus (L.) Trin.]— средиземноморский впд. Возделывается в Северной Италии для приготовления щеток. Ampelodesma tenax Linl; — также средиземноморское расте- ние и идет на щетки (Северная Африка, Испания) . В Средиземноморье имеется большое разнообразие видов и сортов бобовых растений. Л ю п и н (Lupinus L.) главную область развития имеет в Америке, но в Средиземноморье находится второй очаг с не- большим числом видов, интересных в сельскохозяйственном от- ношении. В разделе, посвященном Египту, уже говорилось о древнем пищевом растении L. termis Forslc. Родиной этого вида счи- тается южноевропейское Средиземноморье (включая Балканы). Имеются указания на дикое произрастание его в ОАР, Сирии, Сицилии, Сардинии и Корфу. Употреблялся в пищу в Древней Греции (Феофраст) и в Риме (семена этого вида люпина были найдены при раскопках Помпеи). Средиземноморье является также родиной тех видов люпина, которые теперь имеют большое хозяйственное значение в нашей стране. Люпин синии (L. angustifolius L), люпин желтый (L. luteus L.), л ю пик в о лоси с ты й (L. pilosus L.) и б ел ы й (L. albus L.) распространены в Средиземноморье в диком со- стоянии от Греции до Италии и западнее. В Древней Греции и Риме использовали семена белого люпина на зеленое удобрение (Плиний), люпин в Греции имел также лекарственное зна- ЧЮНИЮ. Гор ох (Pisum заШшп L.), вероятно, ведет свое происхожде- ние пз Передней Азии, где и сейчас встречаются переходные 359
формы от дикорастущего родоначального вида Р elatius к куль- турному гороху. Но Р. elatius распространен по всему побе- режью Средиземноморья и на восток вплоть до Тибета, т. е. имеет древнесредиземноморский ареал. Поэтому не исключена возможность независимого вхождения в культуру гороха в раз- личных странах. Однако нужно принять во внимание и указа- ние Г Бушана (G. Buschan, 1895), что в собственно Средизем- номорье распространены не дикорастущие, а одичавшие формы. Если правильность этих указаний будет подтверждена, то область первичного вхождения гороха в культуру надо будет ограничить пределами Передней Азии и, может быть, несколько шире — далее на восток (тибетские дикорастущие формы недо- статочно изучены). достоверных указаний на культуру гороха в Египте нет. jIBa семени, найденные Унгером, были определены Швейнфуртом (G. Schweinfurth, 1884) как Vicia narbonensis; Яибо (G. Gi- bault, 1912) полагает, что зерна, найденные в погребечии XII династии, относятся к Р. elatius, который растет дико в Египте — они не походили на Р. sati> um; также нет ясных указаний и на культуру ropoxa у древних евреев. древние греки знали горох издревле. Римские агрономы 1холумелла и Палла- дий знали овощной горох, но не считали его ценным растением. У этих авторов чаще встречаются упоминания о нуте, люпине и чине. И в наше время овощной горох больше ценится на севере, чем на юге. Семена мелкосемянного гороха были найдены в нео- литическом слое при раскопках Трои II, то есть значительно раньше гомеровской эпохи. Горох обнаружен во многих свайных постройках Централь- ной и Южной Европы: в неолите — в Швейцарии, Западной Гер- мании (Вюртемберге), Верхней Австрии, Югославии (Хорва- тии); в слоях бронзового века — в Швейцарии, Савойе, Испании, Нижней Австрии; железного века — в Саксонии, Вестфалии и других местах Германии, Италии. Семена ископаемого гороха, найденные в неолите Швейцарии, имели от 3,5 до 4,4 мм в попе- речнике, а одно семя имело даже 5 л~л~. По Г Бушану (G. Bu- schan, 1895; с включением данных Геера), размеры ископаемых горохов из Центральной и Южной Европы колебались: в нео- лите — 4,0 — 4,4 мм в поперечнике, в бронзовом веке — 2,7— 3,6 мл~, в железном веке — 4,0 — 4,8 мл. Л. И. Говоров (1937) указывает, что размер семени совре- менного культурного гороха колеблется от 3,5 до 9,5 мл. Таким образом, ископаемые горохи Средней и Южной Европы и Малой Азии не отличаются по размерам от современных мелкосемян- ных культурных сортов. Бушан считает, что ископаемые евро- пейские горохи — все культурные, за исключением слишком мелкого гороха из Аргара (южная Испания). Мелкосемянные горохи распространены в настоящее время в горных районах Африки, Передней, Юго-Западной и Центральной Азии, т. е. 360
в странах с отсталой земледельческой культурой. По данным Бушана, нет существенных различий по размеру семян между горохами каменного, бронзового и железного веков, хотя в по- следнем встречаются семена несколько более крупные, чем в первые две эпохи. В настоящее время в Средиземноморье возделываются круп- носемянные формы гороха. Они, вероятно, выработались путем отбора из мелкосемянных форм, которые в древности были рас- пространены везде. Есть предположение, что в Среднюю Европу горох попал пз Южной России, а не с юга — из Средиземноморья. Горох был важным продуктом в средневековой Европе. Зеленый горошек вошел в употребление позднее — с XVIII в. Латинское название Pisum имеет родство с санскритским Pic, pis — быть разделен- ным, разложенным. Санскритское peci означает семена гороха, вынутые из боба. В и к а (Vicia L.) в Средиземноморье (в широком понима- нии) имеет область интенсивного развития, хотя истоки этого рода идут из Тургайской (арктотретичной) флоры. Подрод Euvicia Vis. средиземноморскии; к этому подроду от- носятся в и к а п о с е в н а я (V. sativa 1 .) и родственные ей виды. Посевная вика, вероятно, переднеазиатского происхожде- ния, но и в других странах собственно Средиземноморья имеются эндемичные формы этого вида. Балканские страны при- мыкают к первичному переднеазиатскому очагу формообразова- ния и, по-видимому, происхождения вики посевной. Оригинальные формы вики озимой обнаружены в Югосла- вии. Эти формы связаны некоторыми переходными чертами с видом V. angustifolia L., они интересны как исходный мате- риал для селекции озимых сортов вики посевной. V. ervilia (L.) XVilld. (известная под названием чечевица французская) — также вид, вероятно, переднеазиатского проис- хождения, но вхождение ее в культуру имело место и в Запад- ном Средиземноморье (Франция, Испания). V. ervilia была най- дена в неолитическом слое Трои II. Семена из Трои в попереч- нике 2,4 — 3,2 л~м, треугольно-округлые и имеют длинный узкий корешок, как это свойственно данному виду. В и к а одно ц в е т к о в а я (V. monanthos Desf.) разводится на Пиренейском полуострове как кормовое растение. Вид, по мнению ботаников, средиземноморского происхождения. Ч е ч е в и ц а (Lens Adans.) — также в широком смысле сре- диземноморский (вероятно, переднеазиатский) род. Ч е ч е в и ц а о б ы к н о в е н н а я культурная (1 . esculenta Moench) — вид переднеазиатского происхождения или из во- сточной части Древнего Средиземноморья. Для средиземномор- ских стран Западной Европы и Северной Африки в настоящее время характерны крупносемянные формы чечевицы — ssp. ma- crosperma Bar. 361
уже отмечалось, что чечевица возделывалась в Древнем Египте (см. выше). Популярность чечевицы у древних евреев иллюстрируется библейскими рассказамп. В библии чечевица называется adashum или adasim, отсюда арабское название adas. Это семитическое название перешло к берберам Северной Африки в измененном слове ades. У древних евреев полагалось есть чечевицу, когда начинался пост. В библии рассказывается, как Исав продал свое первородство Иакову за чечевичную похлебку. У восточных народов вареная чечевица с маслом и чесноком считается вкусным блюдом. Чечевица была найдена в старой Трое II и в неолите Греции. В Древней Греции она была широко распространена — это была пища народных масс. На Апеннинском полуострове чечевица была обнаружена в неолитическом поселении у Monte Lotfa (около Вероны). Катон дает указания о культуре чечевицы E употребляет названия Le]>tilus и Lens. В Центральной Европе- культура чечевицы является также очень древней — она найдена в неолите Швейцарии, Венгрии и Южной Германии; в бронзо- вом веке — во Франции, Швейцарии; в железном веке— в Швейцарии, Франции, Германии и Италии. Бушан (G. Busichan, 1895) приводит данные о размере семян ископаемой европейской чечевицы по десяти странам. Из данных Бушана можно заметить, что от неолита к железному веку раз- меры чечевицы постепенно увеличиваются. Очевидно, в древ- нейшие времена везде и на юге (Малая Азия, Италия) была распространена в основном мелкосемянная чечевица, хотя на севере (Германия) встречалась с более крупными семенами. От неолита к железному веку чечевица постепенно окультури- вается — семена становятся крупнее. В более новые времена, после железного века, в связи с повышенной интенсивностью роста земледельческой культуры крупность семян чечевицы сильно возросла — до 1 см в поперечнике. Крупносемянные формы в настоящее время встречаются и на севере, но особенно характерны для Средиземноморья, где отбор на крупность се- мян стимулировался экологическими условиями. В Средиземно- морье часто имеют место весенние засухи, поэтому растению выгодно иметь крупное семя (с большим запасом веществ), так как это обусловливает быстрый рост молодых растений. Конские бобы(РаЬа vulgaris Moen~ch=Viicia faba Ь)— средиземноморский род. Наличие особых мелкоссмянных форм, приближающихся к дикарям из Индии (ssp. ipaucijuga Murat.), говорит о том, что область развития рода большая, вмещается в границы Древнего Средиземноморья и даже, может быть, слегка выходит из них. Диким родоначальником конских бобов некоторыми ботани- ками считается Faba ~pliniana Trabut — мелкосемянный вид (мо- жет быть, только подвид), растущий дико в Алжире. Возможно, 362
ц полифилетическое вхождение конских бобов в культуру — не- которые дикие родоначальные формы могли с веками исчез- нуть. Как уже было отмечено, конские бобы были в употреблении в Древнем Египте. Большое количество конских бобов было обнаружено Г Шлиманом в раскопках старой Трои. Древние сказания говорят о культуре конских бобов в Древней Греции. В «Илиаде» упоминается о бобах с черной кожурой. Конские бобы были найдены и в раскопках на Крите. В Древней Греции бобами пользовались при подаче голосов. По данным Диоско- рида, бобы в Греции шли в пищу в виде этиолированных про- ростков, как спаржа. Некоторым богам приносили в жертву печенья из конских бобов. Во время празднеств, которые афи- няне справляли ежегодно в честь Аполлона, было в обычае есть конские бобы. Бобы обыкновенно употреблялись и на погребаль- ных угощениях. В Риме конские бобы находили широкое применение. Кан- дидаты на публичные должности раздавали бобы массам перед выборами. Один из древних родов Рима (gens) носил имя Fabia. У кельтов бобы были важным пищевым продуктом. Плиний писал, что кельтические племена по реке По широко употреб- ляли конские бобы, которые у них входили в состав каждого кушанья. Плиний сообщал также, что на некоторых островах Северного моря конские бобы растут дико, почему римские солдаты и назвали эти острова Fabariae — «бобовники». Вряд ли, впрочем, первоначальная область распространения дикорасту- щих конских бобов простиралась до Северного моря. В раскоп- ках около реки Эльбы, датируемых 100 г. до н. э., найдено много остатков конских бобов. Возможно, что в римское время бобы кое-где и дичали. В средние века в отношении конских бобов предрассудков не существовало. Бобы, чечевица и горох были в почете и слу- жили обычной пищей масс. В XIII в. конские бобы в виде пюре или в кожуре обильно продавались на улицах Парижа. Тогда же бобы были главной пищей крестьянства во Франции наряду с другими бобовыми, капустой, репой, луком и чесноком. Во всем Средиземноморье и Средней Европе бобы известны с глубокой древности — с нового каменного века. Бобы найдены в неолитических поселениях Египта, Малой Азии, Венгрии, Ита- лии и в Испании; в слоях бронзового века — во Франции, Греции, Италии, Швейцарии, Испании; в слоях железного века — в Гер- мании, Италии. Г. Бушан (G. Buschan, 1895) приводит данные по размерам семени конского боба из поселений эпохи неолита, бронзового и железного веков в разных странах.
В неолите бобы были и мелкие (El Garcel) и крупные (Cam- pos) в противоположность чечевице и гороху. Бушан объясняет это тем, что в доисторическое время уже существовали две (может быть, и больше) разновидности конских бобов. По его предположению, бобы из Малой Азии (Троя) и Греции (He- raclea) ~распространились затем в Венгрию (Lengyel и Aggte- lek — в Верхней Венгрии), Швейцарию (Petersinsel) и Италию (M~onte Gurazzo). Среди испанских находок эта первая (восточ- ного происхождения) — более мелкая и более округлая форма, по Бушану, неизвестна. Вторая разновидность имеет более ци- линдрическую форму, и только одна эта вторая разновидность встречается в Испании, т. е. на крайнем западе ареала. Вторая разновидность найдена также в раскопках во Фран- ции (Bourget), в Италии (Castione) и Австрии (Aquileja). Она обнаружена и севернее — в Саксонии (Koschiitz и $с1'.11еЬеп), в Западной Германии (Miischen), в Вестфалии (Klusenstein). Од- нако в Пруссии (Freiwalrle) в железном веке, т. е. сравнительно в позднюю эпоху, найдена очень мелкосемянная форма — мельче, чем даже из Малой Азии и 1(рита. Есть все основания думать, что в Среднюю и даже Северную Европу продвигалась куль- тура конских бобов и с востока (мелкие и более округлые), и с запада (крупные и более почковидно-цилиндрическпе). Но Декандолю и Бушану, продолговатая западная разно- видность происходит от нарбонской вики — Vi~cia narbonensis L. Швейнфурт (G. S~chweinfurt, 1884) поддерживает эту теорию. Он говорит, что в Эфиопии конские бобы называются Bacher (старосемитское — Bachar), а теперь такое название в Египте у феллахов относится к нарбонской вике. Этп авторы не прво- дят достаточно веских доводов за происхождение западной раз- новидности именно от V narbonensis, но вообще полифилетиче- ская точка зрения на происхождение конских бобов, вероятно, правильна. Однако родоначальником западной формы конских бобов скорее является алжирская Faba pliniana или, еще ве- роятнее, близкая к ней, теперь исчезнувшая форма, так как F. yliniana не очень похожа на западные формы конских бобов. Родоначальная исчезнувшая форма была, вероятно, предковой и для F. ~pliniana и для западных культурных бобов. Во всяком случае классификация конских бобов В. С. Муратовой носит формальный характер. Ее подвид Еи-faba слишком сборный. Внутри этого подвида следует выделять формы по совокупности признаков, в том числе и по форме и окраске бобов. Экологиче- ская классификация у В. С. Муратовой оторвана от общей си- стематики конских бобов. Интересна дагестанская эколого-географическая группа, описанная В. С. Муратовой, с мелкими шарообразными черными семенами. Не близка ли эта форма к той, которая двинулась с востока на запад в Европу и дала начало 1-й разновидности Бушана? В настоящее время имеются формы бобов с семенами 364
гораздо более крупными, чем самые крупносемянные из раско- пок железного века. В эту эпоху максимальная величина семени конских бобов была около 1 слх, а самые крупные из современ- ных форм достигают 4 см. В Средиземноморье и теперь рас- пространены преимущественно крупносемянные формы. Чина (Lathyrus L.), подобно роду Vicia L., имеет первич- ную область развития в умеренных странах (истоки рода были в древней лесной северной флоре Евразии) и вместе с тем в области вторичного интенсивного видообразования в Древнем Средиземноморье. Развитие секции Ciicercula Gren. et Godr. приурочивается к Древнему Средиземноморью. I< этой секцин относится чина посевная — L. sativus L., формы с наиболее примитивными признаками сосредоточены в Восточном Средиземноморье- Юго-Западной и Передней Азии. По направлению к западу растения приобретают все более окультуренный облик — с разви- тием признаков вторичного характера. Алжир, Испания, Порту- галия, Южная Франция и Италия являются областямн распро- странения наиболее молодых крупносемянных и светлопигмен- тированных форм чины, образующих proles mediterraneus Zall<. На Kpme распространены особые мелкосемянные формы — ssp. сург1сиз Zall~. В Средней Европе распространены формы ssp. europaeus Zalk., более близкие к чинам Юго-Западной и Передней Азии, чем к средиземноморским. Чина была известна в Древнем Египте, неолитические на- ходки ее в большом количестве имеются в Венгрии (Lengyel и Aggteleli), что указывает на возделывание ее здесь, а не на ис- пользование диких растений. Эти семена очень мелкие (длина 4,2 мм, ширина 3,2 мл, толщина 4 мм) . Чина была найдена в неолите Трои (Малая Азия) и была известна грекам под на- званием Lathyrus. Судя по географическому распределению форм, чина распространилась в Среднюю Европу из Передней Азии, а в Египет и Эфиопию — из Индии. К той же секции Cicercula, что и L. sativus L., принадлежит и L. cicera L., который считается средиземноморским видом и иногда разводится. В средиземноморских странах возделывается иногда и L. ochrus DC., растущий дико в Италии и Испании, и L. gorgonii Pari. Последний вид разводится в Сирии в каче- стве кормового растения. Родина популярного декоративного растения — д у ш и с т о г о г о р о ш к а (L. odoratus L.) — также Средиземноморье. Этот вид чины растет дико в Южной Италии, Сицилии и на Канар- ских островах. Нут (Cicer L.) переднеазиатского происхождения. По-види- мому, и для культурного нута — С. arietinum L. — первичная родина вида — Восточное Средиземноморье (Передняя Азия) . В собственно Средиземноморье в настоящее время распростра- 365
Ю невы крупносемянные сорта нута — proles hispanicum G. Pap. (Испания, Италия, Марокко, Алжир, Тунис). В Сирии имеется вид, очень близкий к культурному нуту,— С. judaicum Boiss. Точных сведений о культуре нута в Египте и у древних евреев не имеется. По-видимому, нут возделывался у древних греков под названием Fribinthus и Krios (Декандоль) . Римляне называли его Cicer, что напоминает алоанское — l~ikera, это до- пускает раннее появление нута — еще во времена пеласгов. В свайных постройках Средней Европы нут не был найден. Сулла, однолетний копеечнцк (Hedysarum icorona- гшт L.) разводится как кормовое растение в Южной Италии, Сицилии и Испании. Л ю ц е р н а (Wiedicago L.) переднеазиатского происхожде- ния, но ареалом ее развития в природе и в культуре служит вся Древнесредиземноморская область. В округах, переходных от Передней Азии к собственно Средиземноморью, сформировались следующие виды синей культурной люцерны — М. syriaco- palestinica Sinsk. (Сирия и Израиль), М. jemenensis Sinsk. (Йемен) . В собственно Средиземноморье сформировались М. tripoli- taniioa Sinsk. (Ливия — Триполи) и М. eusativa Sinsk. (Италия, Испания, юг Франции). П а жити и к (Trigonella L.) и дон н и к (Melilotus Adans )— древнесредиземноморские роды. В собственно Средиземноморье создался подвид пажит- ника — Т. foenum-graecum ssp. mediterranea Sinsk., имеющий в своем составе экотипы Туниса, Марокко, Египта и Ливии (Триполи). В собственно Средиземноморье сформировался и подрод дон- ника — Nicromelilotus Suv. с видами М. indicus All., М. itali- cus (L.) Lam., М. neaipolitanus Ten. и др. Кроме того, здесь сфор- мировался и средиземноморский подвид донника белого— М. albus ssp. maritimus Suv. (приморские районы Португалии, Испании, Франции и Италии). К л е в е р а л е к с а н д р и й с к и й (Trifolium alexandrinum 1 .) и целая группа родственных видов (Т. berytheum Boiss., Т. vavi- lovi Eig и др.) распространены в Сирии и Палестине. Хотя кле- вер александрийский и возделывался в Древнем Египте, но он там не чувствует себя, как на родине, и, по словам Делиля (Delile, 1923), часто в Египте «вырождается» — египтяне пред- почитают получать семена этого растения из Сирии, а не про- изводить их у себя. Ближайший дикий родич клевера алексан- дрийского — Т. apertum Bobr. — распространен в Колхиде (и в области влияния колхидской флоры). Клевер кр а сиый (Т. pratense L.) образует в Средизем- номорье особые региональные экотппы — быстрорастущие, ско- роспелые и малолетние. Сюда относится клевер итальянский. 366
Клевер белый (Т. repens Ь.) представлен в Средиземно- морье особой крупной формой — var. giganteum Vav. (Ломбар- дия). Этот клевер называют также лодийским. 1~л ев ер ин к ар и а тн ы й (Т. incarnatum L.) культиви- руется и растет дико в западном Средиземноморье (южная Италия, Сардиния, Балеарские острова, Алжир). С е р а д е л л а п о с е в н а я (Ornithopus sativus Broth.) имеет родину в Западном Средиземноморье. Вошла в культуру в Португалии. Д р о к (Ulex europaeus L.) — кормовое бобовое растение влажного приатлантического климата, распространено от Пор- тугалии до Англии. Используется также и из дикорастущих за- рослей. Возделывается кое-где в Португалии. Весь род Шех ~ . происходит из приатлантической Европы. Из прядильных растений древнейшим в Средиземноморье яв- ляется л ен (Linum usitatissimum L. ssp. mediterraneum Vav.). Средиземноморские формы льна, распространенные в Египте, Алжире, Марокко, Тунисе, южной Италии, представляют собой крупноплодные и крупносемянные формы, родственные индий- ским льнам, и были занесены в Средиземноморье пз Индии, где имеется также крупносемянный лен с переходами к средиземно- морской крупносемянной и абиссинской мелкосемянной формам льна. Однако лен распространился из Индии в Среди- земноморье в очень древние времена, ибо в погребениях Древ- него Египта были найдены семена уже крупносемянного льна. И н д а у (Eruca sativa Lam.) происходит из западного Сре- диземноморья (Алжир, южная Испания), где распространены все виды рода Eruca Adans. Отсюда сорный вид индау — Е. sa- tiva — распространился по всей области Средиземноморья и вышел даже за пределы ее (Индия). В собственно Средиземно- морье индау является древней культурой, которая была из- вестна в Греции и, особенно, в Риме в качестве овощного (са- латного) растения; была известна также во Франции и в Ита- лии. Греческое название этого растения означает «растение, даю- щее хорошую подливку, хороший соус». Сейчас это растение как овощное нигде не употребляется, а в Азии (Индия) исполь- зуется как масличное. Eruca tpinnatilida (Desf.) Роте1. — масличное и кормовое растение Северо-Западной Африки (местное название — harra). Частично к масличным, а также и к пищевым растениям средиземноморского происхождения относится ч у ф а (Cyperus esculentus L.). Средиземноморье является родиной г о р ч и цы б ел о й (Sinapis alba L.), которая встречается там как сорняк и, по-ви- димому, дико, а также в культуре; здесь же сосредоточено мор- фологическое разнообразие этого вида. К средиземноморским странам в общем приурочены более раннеспелые формы белой горчицы. Плиний писал, что масло получается из семян дико- 367
растущей горчицы. Возможно, что он говорил о г о р ч и ц е пол евой (S. arvensis L.), а не об S. alba. Г о р ч и ц а ч е р н а я (Вгаsslca п~дга K!Qch) в собственно Средиземноморье представлена разновидностью, точнее подви- дом var. occidentalis Sinsk. Она же распространена и в Средней Европе. В Сирии и Палестине распространена особая разновид- ность черной горчицы — var. rigida Sinsl<. Р а п с (Brassica napus ssp. oleifera Netzger) многими счи- тался средиземноморским видом, происходящим от каких-то форм капусты. Некоторые авторы считают, что средиземномор- ское происхождение имеет и наиболее близкии к капусте лист- венный раис — чаг. ~pabularia DC., который затем распростра- нился отсюда в ГДР, ФРГ и Данию, но, по нашему мнению, родина рапса вообще — приатлантическое и северное побережье Европы. М а к (Papaver somniferum L.) отличается в Средиземно- морье значительным полиморфизмом. Принадлежит к широко распространенному подвиду ssp. euroasiati~cum N. Vesselov., к которому относится мак всей Европы и Сибири. Греки умели возделывать мак и имели представление о его наркотических свойствах. Данные о маке имеются у Гомера, Феофраста, Диоскорида, последний упоминает о разновидности с белыми семенами. Греческое название Nelcon похоже на мак славянских народов. Может быть, греки получили мак с севера, с берегов Черного моря. Там был город Синоп (Sinope), который, по Гесиоду, имел более древнее название Nekon — город мака. Римляне были знакомы с маком. Семена этого растения были найдены в неолитической свайной стоянке в Lagozza. К у н ж у т (Sesamum indicum L.) имеет ряд форм в соб- ственно Средиземноморье (не считая Передней Азии) . В. N. Гильтебрандтом описаны экотипы острова Родос, экотип Крита и оригинальные формы из Италии. К р а м б е и с п а н с к а я (СгатЬе hispanica L.) растет дико в Западном Средиземноморье (Испания) и на Балканах на сол- нечных, щебнистых местах. Близка к С. abyssinica, которая испытывалась в СССР в культуре как масличное растение. Особенно богата культурная флора Средиземноморья овощ- ными растениями. Капуста (Brassica о1егасеа L.) распространена в диком состоянии по берегам Франции. Кроме того, по побережьям Средиземного моря растут очень близкие виды: В. montana Peur- rett — берега Южной Франции и Италии; В. rupestris Raf.— берега Италии; В. insularis Noris — Корсика, Сардиния; В. atlantica О. Е. Schulz — Алжир; В. cretica Lam. — Греция, Италия; В. incana Tenore — берега Италии, Сицилии; В. villosa Biv.-Bern. — Италия, Сицилия. Большое количество родственных дикорастущих видов соот- ветствует видовому и сортовому разнообразию культурных ка- 368
пуст. Феофраст различал три формы капусты: курчавую, с глад- кими листьями и капусту дикую. Т. В. Лизгунова (1959) приходит к заключеншо, что нельзя с точностью быть уверенным в том, что в Греции возделыва- лась настоящая огородная капуста — Brassica оlегасеа sensu lato. На основании описания Феофраста она полагает, что там в древности разводился один из видов Crambe: в пищу исполь- зуются обламываемые побеги, разводится он черенками, заго- товляемыми с кусочками корня. Другой формой, упоминаемой Феофрастом, была, вероятно, листовая капуста, а третьей (по мнению Т. В. Лизгуновой) — В. cretica Lam. Т. В. Лизгунова предполагает, что римлянам были известны несколько форм листовой и кочанной капусты и листовые брокколи. Катон очень высоко ценил капусту; он писал, что потребление капусты под- держивает здоровье и исцеляет разные болезни, даже мелан- XOJIHIO. В Средиземноморье капуста кочанная представлена особой группой сортов — ssp. mediterranea Lizg. В средиземноморских странах распространены формы, переходные от листовой ка- пусты к цветной. По-видимому, цветная капуста была известна в Греции. По описанию Плиния, эта капуста была похожа на брокколи. Появление цветной капусты во Франции, по )Кибо (G. Gibault, 1912), относится к середине XVI в. В старых книгах говорится о получении семян капусты с Мальты, Крита и из Италии. Раньше цветную капусту называли во Франции кипр- ской капустой. Ка пуста цвети а я (В. cauliflora Lizg.) в Средиземно- морье больше представлена подвидом ssp. simplex Lizg. (Ита- лия, Испания, острова Средиземного моря). По Т. В. Лизгуновой, в настоящее время более древние (близкие к дикорастущим) формы листовой капусты встре- чаются в культуре в Западном Средиземноморье (Испания), а также и в Восточном Средиземноморье (в Передней Азии— Древняя Армения и Абхазия) . На Пиренейском полуострове в наибольшем разнообразии сохранились в культуре примитивные эндемичные формь1 кочан- ной капусты. Древнейшие формы цветной капусты (ветвистые брокколи) до сих пор довольно широко распространены в культуре в Ита- лии и на островах Средиземного моря. Реп а (В. гара L.) была хорошо известна древним грекам и римлянам. Феофраст упоминает о репе как об огородном расте- нии. Грекам было известно несколько сортов репы. Римский писатель Колумелла употребляет два названия: naipus и гара; корнеплод под названием гара имел более широкие корни. Сле- довательно, название napus не соответствует современной брюкве — В. napus ssp. rapifera Metz~. == В. na~pobrassica. Оба названия относятся к различным разновидностям репы. Колу- 369
мелла приводит еще третье название для репы — Л!ursian gon- gy1is, греки называли репу geggulos пли ~одди11з. Плиний (Plinii, 1829) говорит о многих сортах репы. Он употребляет два названия: napus и гара. Под первым он, по-ви- димому, понимает мелкие, длинные, твердые репы. Сорта реп во времена Плиния различались по величине, окраске и форме корнеплода: известны были длинные, округлые и плоские репы. Римляне разводили репу не только на огородах, но и на полях. Культура репы занимала тогда одно пз первых мест после зер- HoBbIx хлебов и винограда. Репа достигала очень больших раз- меров — до 16 кг. Специально кормовых сортов тогда не было„ но уже были формы с крупными корнеплодами, из которых впо- следствии выработались кормовые репы (турнепсы). В настоящее время культура репы в средиземноморских странах имеет небольшое значение; на европейском побережье Средиземного моря культивируются раннеспелые сорта репы типа миланской. Культура репы пришла в собственно Среди- земноморье из Малой Азии (где общий характер реп похож на европейскую репу, но только более груб, примитивен) или, может быть, частично и из Средней Европы. Средиземноморье вместе со Средней и Северной Европой является областью сортового разнообразия р е д и с а е в р о п е й- с к о г о (Ra~phanus sativus L.) . Ближайший его дикорастущий родич — R. maritimus Smith — распространен по берегам Чер- ного, Средиземного морей и Атлантического океана. Некоторые европейские сорта редьки и некоторые сорта редиса (с черными и желтыми корнеплодами) в Средиземноморье не встречаются, а свойственны только странам Средней и Северной Европы. Возможно, что они произошли от приатлантических дикорасту- щих форм. С в е к л а (Beta L.) — древнесредпземноморский род, захо- дящий в Индию. Филогенетпчески наиболее древней считается секция Vulgares Tran., которая дала начало в Передней Азии секции Corollinae Tran., а в Западном Средиземноморье — сек- ции Patellares Tran. В Средиземноморье, по В. Н. Зосимовичу (1939), дикорастущая В. vulgaris L. представлена экотипами: proles mediterraneum и proles italio-adriatiicum. По В. Т. Красочкину (1960), в древнюю (исходную для рода) секцию Vulgares входят виды: В. vulgaris L. — культурная кор- неплодная свекла, В. maritima L. — дикорастущий ближайший предок культурной свеклы, В. orientalis Rath. — листовая свекла Индии и Египта и В. patula (Soland) W. Ait. (последняя яв- ляется связующим звеном между секцией обыкновенной свеклы и секцией канарской свеклы). Среди В. maritima В. Т. Красоч- кин выделяет следующие подвиды: скандинавский — ssp. danica I<rass., средиземноморский — ssp. mediterraneum Krass. с че- тырьмя разновидностями: var. ~prostrata Krass., var. erecta Кrass., var. atriplicifolia (Roy) Krass. и var. macrocarpa (Guss.) Krass. 370
Крупноклубочковая сицилийская разновидность — var. macro- carpa — выделяется многими исследователями в самостоятель- ный вид. Она сходна с полукорнеплодными формами Передней Азии (Анатолия и Афганистан) . Наиболее утолщенные и сахаристые корнеплоды имеет var. erecta, распространенная главным образом по побережью Адриатики и в долине реки По. По толщине корнеплода ее пре- восходит только скандинавский подвид. По В. Т. Красочкину, адриатические дикорастущие формы могли дать начало культур- ной листовой свекле. В. Т. Красочкин (1960) полагает, что культура листовой свеклы возникла не позднее чем во II тысячелетии до н. э. Он считает первичным очагом культуры листовой свеклы среди- земноморские острова: Сицилию, Кипр, Крит. Латинское название листовой свеклы (Beta ~cicla) Sicula beta происходит от названия острова Сицилия. Названия у древ- них восточных народов для свеклы очень сходны. Сходство названий говорит о том, что листовая свекла распространилась пз одного первичного очага. В Средиземноморье имеется большое разнообразие свеклы листовой. Она была в употреблении у древних греков и римлян. Аристофан упоминает о ней в V в. до н. э. Аристотелю за 300 лет до н. э. была известна листовая свекла с красными черешками. Феофраст называет ее sicula (отсюда видовое назва- ние cicla). Феофраст, Колумелла, Плиний и Палладиус знали белую и черную листовую свеклу. Черной называлась, ве- роятно, форма с интенсивно окрашенными антоцианом череш- ками. По показаниям Ювенала, Плиния и других древних писа- телей, листовая свекла была пищей ремесленников и бедного люда. В трудах греческих писателей V — IV вв. до н. э. имеются сведения об использовании свеклы. Древнегреческий врач Гип- пократ упоминает о свекле как о лекарственном средстве и о диетическом ее значении для больных. По этим данным, использовались не только листья, но и корни свеклы. Феофраст описывал корень свеклы как долговечный, стержневой, развет- вленный, но вместе с тем толстый и мясистый, приятного вкуса. Такие корни свойственны дикорастущим п многим формам ли- стовой свеклы. Указания на «долговечность» (многолетность?), разветвленность говорят о том, что в те времена настоящей корнеплодной свеклы еще не было, а употребляли в пищу дикие и культурные растения листовой свеклы, но имелись по призна- кам корня и переходные формы — толстые и относительно мяси- стые, хотя и разветвленные. У греческого писателя 111 в. н. э. Афинея (по В. Т. Красоч- кину, 960) перечислены дикие растения, которые употреб- ляются в пищу в вареном виде, в этом перечне упоминается и о свекле наряду с лебедой, мальвой, щавелем.
Q. Липпманн (Lippmann, 1925) считает, что свекла около 1000 г. до н. а. была известна в Сицилии, около 425 г. — в Гре- ции. В III в. до н. э. свекла была, согласно этому автору, хорошо известна в Греции, а около 200 г. — в Италии. Липпманн счи- тает, что свекла, прежде чем распространиться в Италию и Грецию, была известна в Сицилии, а в Сицилию была занесена с Востока финикиянами. Э,а версия предполагает, что первич- ное введение культуры свеклы имело место на востоке Среди- земноморья, но окончательно это не установлено. Возможно и полифилетическое зарождение культуры свеклы независимо в различных местах, так как ареал дикой свеклы — В. таг1- tima L. — тянется по всему побережью Средиземного моря и даже по берегам Атлантического океана. Тем не менее, страны Передней Азии и собственно Среди- земноморья являются областью развития многих форм столовой корнеплодной свеклы — там и теперь встречаются такие прими- тивные формы со склонностью к цветухе, в которых можно узнать «прообразы» современных европейских сортов (В. '1'. I(ра- сочкин, 1931) . древнее Средиземноморье — область развития и всего рода Daucus L. и моркови о г о р о д н о й (Dau~cus sativus Zagor., П. Ф. Загородских, 1939). Собственно Средиземноморье — o6- ласть развития каротиновой моркови — ssp. med!terraneum Za- gor., т. е. того подвида, к которому относятся наиболее распро- страненные у нас столовые сорта. Овощное растение средиземноморского происхождения п ет- р уш к а [Petroselinum crispum (Mill.) Nym.] была в культуре у древних греков и римлян. Греки называли ее Selinon, а рим- ляне — Anetum и Petroselinon (отличная Anetum ап1агц.п и А. viride от сельдерея). Петрушка была в употреблении как растение пряное и лекарственное. Колумелла знал сорта пет- рушки с крупными и курчавыми листьями. С ель де р ей (Apium graveolens L.) распространен в Среди- земноморье в культуре и дико, это аборигенная культура соб- ственно Средиземноморья. Сельдерей упоминается в «Одиссее» и у Феофраста, но, возможно, что в этих источниках упомина- лось о диком растении. Яиоскорид и Плиний говорили опреде- ленно и о диком, и о культурном сельдерее. Мы уже писали о ритуальном значении сельдерея в Древнем Египте. Этот обы- чай перешел и к грекам. При погребальных церемониях сель- дерей клали на головы мертвецам и сажали на могилах. У древних было еще одно растение из сем. зонтпчных, кото- рое заменяло сельдерей, — с м п р н и у м (Smyrnium olusat- гшп L.). Смирниум использовался в дикорастущем состоянии и возделывался на огородах. Теперь это растение совершенно вышло из употребления. Из растений, широко известных в древности, а в настоящее время совсем или почти утративших свое значение, следует 372
упомянуть о м а н д р а г о р е. Из-за причудливой формы под- земных органов, иногда напоминающих маленьких человечков, мандрагора считалась магическим растением и применялась, кроме того, в медицине. Два основных вида мандрагоры растут в прибрежных местностях Средиземноморья: Mandragora offici- narum L. и М. autumnalis Bertol. Один вид известен в Испании (М. hispanica Vierh.) и один — в Греции (М. Ьаизз!~пссМ11 Heldr.). Кроме того, по одному виду известно в Туркмении, Ги- малаях и Тибете. Род Mandragora L. интересен, таким образом, своим типичным древнесредиземноморским ареалом. Пастернак (Pastinaca sativa L.), овсяный корень (Tragoporon porrifolium 1.) и с к о р ц о н е р и с п а н с к и й (Sicorzonera hispanica L.) — аборигенные корнеплодные расте- ния собственно Средиземноморья, где распространены в диком состоянии и в небольших размерах возделываются, как и к е р- в е л ь л и с т о в о й [Ап11чг1ьсцз icerefolium (L.) Hoffm.]. Кер- вель употреблялся греками как приправа. Колумелла называет его Chaerophyllum. Средиземноморье считаетс» родиной пастер- нака (хотя самые древние находки его имеют место в Швейца- рии). Пастернак найден в раскопках в окрестностях Пармы и Меркураго (на переходе от бронзового века к железному) и в Фонтанеллато (железный век) . Пастернак — Р. sativa — очень близок к дикорастущему виду Р. silvestris Mill., который и считается непосредственным пра ю- дителем культурного пастернака. Ареал Р. silvestïs — вся Ев- ропа, Средиземноморье, Кавказ, Малая Азия. Возможно, пастер- нак входил в культуру в нескольких странах независимо. С к о р ц о н е р (Scorzonera delic~~oisa Guss.) распространен в диком состоянии в Испании и Сицилии, используется местным населением как овощное растение. U,èкорий (Cichorium intybus L.) распространен в Среди- земноморье в диком, сорном и культурном состоянии. Возможно, что культура его аборигенна не только в Средиземноморье— ареал распространения дикого растения очень большой. Средиземноморское происхождение имеет с а л а т - э н д и в и й (Ciichorium endivia L.), ближайший дикорастущий родоначаль- ник которого С. pumilum Jacq. растет в этой области. С а л а т - л а ту к (Lactuca sativa L.), возможно, аборигенен в Средиземноморье, хотя ареал его простирается и в Переднюю Азию; то же можно сказать и о п о р т у л а к е (Portulaca olera- cea L.). С п а р ж а — также аборигенная средиземноморская культура. Мелколистная спаржа — Asparagus tenuifolius Lam. — растет в лесистых и гористых местностях Европы, спаржа остролистная распространена в Южной Европе и Северной Африке. Эти виды спаржи употребляют в пищу в Италии, Испании и Алжире. Ареал спаржи обыкновенной огородной — А. offi~cinalis 1 . —. заходит в Средиземноморье (Южная Франция), и, возможно, здесь она
впервые вошла в культуру. Древние греки собирали побеги ди- кои остролистной спаржи; предполагают, что им была знакома и обыкновенная спаржа, но никаких указаний на культуру этого растения в Греции не имеется. Средиземноморье — родина а р т и ш о к а. Предок настоя- щего артишока — Супага cardunculus L. — растет дико на юге Франции, Испании, Италии, Греции, в Северной Африке и на островах Средиземного моря. культурный артишок — С. ьсо1у- mus L.— при одичании делается похожим на дикорастущий вид. По-видимому, артишок вошел в культуру в Испании и Северной Африке, но это была форма, близкая к С. cardunculus, получив- шая название «испанский кардан». Такой же примитивный арти- шок, вероятно, возделывался и в Греции. Феофраст упоминает этот артишок под именем ca~ctos — колючее растение, которое, по его словам, произошло из Сицилии. По сведениям, приводимым А. И. Филовым (1960), судить по названиям о существовании ты к в ы у древних греков очень ~рудно. Греческие писатели упоминали о Реро, Cucurbita. Мно- гие авторы (Декандоль, 1885; В. Ген, 1872; Н. 3. Умиков, 1953 и др.) считали, что эти названия относились к тыкве. Слово рероп означает древний плод, Cucurbita (по этимологическому словарю Эрнаута и Л!ейдеса) — «банка», что указывает на сход- ство с лагенарией. В. Ген считает, что Cucurbita происходит от corbis — голова, но corbis означает и корзину. А. И. Филов пишет, что слово зщиа следует понимать как растение, исполь- зуемое наподобие огурца. Все греческие термины, приписываемые теми или другими авторами тыкве, могли относиться к лагенарии, к полукультур- ной змеевидной дыне, к слабо окультуренным формам арбуза. В раскопках Старого Света, относящихся к доколумбий- скому периоду, достоверных остатков тыквы нет. Где и когда впервые тыква появилась в Европе, точно неизвестно. Во всяком случае она сначала распространилась в Средиземноморье, а за- тем отсюда к северу. Ты кв а (С. maxima Duch.) в настоящее время распростра- нена на западе Европы — в Испании, Италии, Франции. Ве- роятно, отсюда она распространилась севернее (ГДР, ФРГ, Голландия). Зимние сорта С. maxima являются эндемичными для Испании (испанская тыква). В странах Западной Европы и США возделываются мелкоплодные сорта (С. maxima) — мар- роны (var. marron Naud.) . В Западном Средиземноморье возделывается С. moschata var. brachicina Аlе1. Эта разновидность включена А. И. Фило- вым в туркестанский подвид мускатной тыквы — ssp. turkesta- nica Fil. 1~ак отмечает А. И. Филов, это самая крупноплодная разновидность данного подвида, что еще раз подчеркивает пред- ставление о Средиземноморье как об очаге крупноплодных и крупносемянных форм. 374
К р е с с - с а л а т (Lepidium sativum L.), вероятно, происхо- дит из Передней и Юго-Западной Азии, но и в Средиземноморье считается древней культурой. Предполагается, что Kardamon Феофраста — это кресс-салат. Римские авторы называют его Nasturtium. Детально вопрос о его происхождении и расселении рассмотрен в разделе о Передней Азии. Щ а вель (Rumex acetosa L. и другие виды) дико растет по всей Европе. Древнее население, по-видимому, культивировало его для кулинарного употребления, хотя он никогда не имел большого значения. Ч а бе р (Satureja hortensis L.), садов а я р у т а (Ruta graveolens L.) и блптум (виды Blitum rubrum С. А. Меу. В. virgatum L., В. ca~pitatum L.) считаются аборигенными куль- турами средиземноморских стран. Л у к -п о р е й (AI ium porrum L.) растение передне".çèàò- ского происхождения. Было известно в Древнем Египте и Гре- ции. О нем упоминает Гомер как о суповом овощном растешш. Ч е с н о к (Allium sativum L.) происходит из Средне-Юго- Западной Азии, имел большое значение в Древнем Египте. Рим- ляне употребляли его главным образом как лекарственное ра- стение. В Средиземноморье имеются особенно крупные формы чеснока, как и реп ч а того л ук а (Allium сера L.). Послед- ний представлен испанской группой сортов (с крупной лукови- цей) — ssp. australe Iproles hispanium Trophyme z. Многие эфирномаслпчные и пряные растения имеют среди- земноморское или древнесредиземноморское происхождение, но с образованием эндемичных форм в собственно Средиземно- морье, что еще недостаточно для ряда видов уточнено. А н и с (Pim~pinella anisum L.) в средиземноморских странах распространен дико и в культуре. Возможно, что это растение переднеазиатского происхождения. Анис был известен в Древ- нем Египте. Диоскорид и Плиний писали, что лучший анис про- исходит из Крита. Pim~pinella peregrina L.— эфирномасличное растение. Растет дико по всему Средиземноморью — от Испании до западного Тянь-Шаня. Тм ин (Сагит сагч1 L.) дико распространен в средиземно- морских странах (и в Средней Европе). К о р и а н д р (Coriandrum sativum L.) имел большое значе- ние в Древнем Египте. Он, вероятно, переднеазиатского проис- хождения. Издревле был известен в Сирии. Ф е н х е л ь (Foeniculum vulgare Mill.) считается средизем- номорским растением и дико растет в европейских и северо- африканских средиземноморских странах. Он также широко рас- пространен B полупустынных группировках Закавказья (Рагп- мов, 1950) и в других странах Передней и Юго-Западной Азии. Это растение имеет древнесредиземноморский ареал, и, воз- можно, основная область развития его — Передняя Азия. Вхо-
дить же в культуру он мог и независимо в разных частях ареала. Л а в а нд а (Lavandula L.) ведет происхождение из Запад- ного Средиземноморья. В культуре известны два вида: L. vera DC. (= L. officinalis Chaix.), ареал дикого растения — Южная Франция, Португалия, Корсика, Сардиния, Сицилия и L. spica L. (=L. latifolia Vill.), растет дико на Балеарских островах, в Испании, Тунисе, Алжире, Южной Франции, Италии, Далма- цпи. Ряд других видов лаванды, не вошедших в культуру, растет в Западном Средиземноморье. M е л и с с а (Melissa offiicinalis L.) — лекарственное, медонос- ное и эфироносное растение Средиземноморья (весь род среди- земноморский) . Ч абер (Satureja hortensis L.) введен в культуру, вероятно, в Южной Европе, род средиземноморский. И с с о п л е к а р с т в е н н ы й (Hyssopus officinalis L.) есте- ственно произрастает в горах Южной Европы. Известен в Древ- нем Египте. Род Нуззориз L. древнесредиземноморский. Род Najorana Moench — из Восточного Средиземноморья. М а й о р а н (M. hortensis Moench) возделывается в Египте, Гре- ции, Риме. Тимьян об ыкновенный (Thymus vulgaris L.) растет дико в средиземноморских странах от Португалии до Греции. Эфирномасличное растение. Раньше добывали из него тимол, который получают теперь из ажгона. У кр о п (Anethum graveolens L.) был известен древним гре- кам и евреям. Овощное и эфирномасличное (карвон). Средиземноморское происхождение приписывается следую- щим эфироносным и пряным растениям, из которых некоторые имеют скорее древнесредиземноморский ареал, и не для всех пока область первичного развития вполне ясно определена в пределах Древнего Средиземноморья: чернушка (tigella sativa L.), ~р о з м а р и н (Rosmarinus offiicinalis L.), и р и с (Iris pallida Lam.). Д а м а с с к а я р о з а (Rosa damascena Mill.) произошла из Сирии. Из красильных дико и в культуре распространена м а р ен а (Rubia tinctorum 1.). Многие лекарственные растения имеют средиземноморское происхождение; например, подрод однолетних г о р и ц в е т о в (Adonis L.) имеет центром распространения Древнее Средизем- номорье, включая Иран, Афганистан, Крым, Кавказ. Из древесных растений средиземноморского происхождения прежде всего следует отметить л а вр бл а го родный (Lau- rus nobilis L.), из дубителей — с у м а х (Rhus сопапа 1-.) (Испания и Италия) и из плодовых — о л и в у (Olea еиго- раеа L.), рожка вое дерево (Ceratonia siliqua L.). Оливковое дерево было издревле известно в Сирии и Пале- стине. В «Ветхом завете» QHQ носило название salt или zelt. Это
похоже на иранское seitum, арабское setum или zeitum, тюрк- ское и татарское seitum. Олива, как видно, распространилась из Сирии, откуда на запад она стала распространяться лишь в 1 тысячелетии до н. э. Древние греки были распространителями оливы в Италии. Остатки оливы найдены были в террамарах римского периода (железный Be«). На Пиренейском полу- острове, по мнению Бушана, олива имеет более древнее распро- странение. В собственно Средиземноморье сосредоточено сортовое и формовое разнообразие плодовых растений, но эта область не является ареной интенсивного видообразования. В и ног р ад (Vitis 1п11ега L.) — одно из древнейших куль- турных растений Европы. Семена найдены в свайных построй- ках Швейцарии. В произведениях классических авторов (1(а- така, Варрона, 1(олумеллы, Плиния и ряда других) сообщается много сведений о культуре винограда. Средиземноморье обла- дает богатым фондом форм и сортов культурного винограда. Сорта Балканского полуострова объединены А. М. Негрулем в proles balcanica Negr., а к proles oocidentalis Negr. относятся сорта Испании, Португалии, 1Лталпи, Франции, ГДР и ФРГ П е р с и к (Persica vulgaris М111.) был занесен в Европу из Ирана, на что указывают Феофраст и Колумелла. Греки и римляне называли его Persica. В Средиземноморье и Южной Европе создались особые сорта персика. Намечается следующий путь расселения персика из Азии в Европу: 1~итай (родина), отсюда в начале нашей эры — в Парфию, затем в область 1~ас- пия, в Переднюю Азию, из Ирана в Сирию и оттуда на запад, в Грецшо и Рим. А б р и к о с (Armenia~ca vulgaris Lam.) был занесен из Арме- нии в Грецию, а затем и в Рим. Поэтому абрикос назывался в течение долгого Бремени армянским яблоком и родиной его считали Армению. Тем не менее, европейская группа сортов абрикоса в настоящее время очень богата и содержит хорошие, крупноплодные и ароматные столовые сорта. Путь распростра- нения абрикоса был подобен тому, по которому продвигался на запад персик, а именно: впитай (родина) — древние страны Средней Азии — Передняя Азия (Иран и Закавказье) — из Ар- мении в Грецию и Рим. Родина о р е х а г р е ц к о г о (Juglans regia L.) — Передняя Азця и, может быть, Средняя Азия. В Европу пришел, вероятно, из Малой Азии, сначала в Грецию и Италию, а затем и в дру- гие страны. Это растение было популярно у древних греков и римлян. Упоминания о нем встречаются у Диоскорида, Фео- фраста, 11ицерона, Плиния, Овидия, Вергилия и Палладия. Римляне называли грецкий орех Jovis Glans — желудь или орех Юпитера, отсюда название рода; видовое название regia также римское. В Средиземноморье грецкий орех дико не ра- стет, но культурных сортов особенно много в Италии и 1Лспании. 377
О р ешни к ло м б а рдски й (Gorylus maxima Mill.) родом из восточной части,1~ревнесредиземноморской области, вероятно, пз Малой Азии. Растет дико в Балканских странах. В Италии имеются хорошие местные культурные сорта этого ореха; встре- чается одичавший. Доказано, что к а ш т а н н а с т о я щ и й (Castanea sativa L.) аборигенен в Малой Азии и на Кавказе. Из Малой Азии он уже в историческое время был занесен в Грецию, затем в Италию, а оттуда в Испанию. Образует леса, похожие на дикие заросли, во всех средиземноморских странах. Здесь трудно провести гра- нпцу между областями распространения культурного и дико- растущего каштана. В Средиземноморье имеется много местных сортов каштана. Уже Плиншо было известно 8 сортов. Ф иста ш к а культур н ая (Pistacia vera L.) в Средизем- номорье представлена эндемичными группами сортов: тунисской и сицилийсЕОЙ. Ун а б и (Zizyiphus vulgaris L.) был известен древним грекам и упоминается у Гомера. Родиной считается северная Африка Il средиземноморские острова. В Италию был ввезен с востока и начале нашей эры и очень быстро распространился по всему Средиземноморью. Унаби распространен также в Азии — Индии, Китае, Корее, Японии. Существует версия о происхождении этого растения из Восточной Азии. Многие чужеземные растения проникли в Средиземноморье в давние времена и нашли там как бы вторую родину. К тако- вым, теперь типичным для приморского средиземноморского ландшафта, южным растениям относятся некоторые виды цитру- совых. В странах собственно Средиземноморья имеется большое разнообразие форм и сортов цитрона, лимона, поме- р а н ц, а и а п е л ь с и н а. Цитрон известен в Греции со времен походов Александра Македонского, померанец появился в конце 1 тысячелетия до н. э., лимон — с XII — XIII, а апельсин — с XV— XVI вв. По Н. А. Базплевской, из Средиземноморья вышло 765 ви- дов декоративных растений, среди них н а р ц и с с (Narcissus L.), г и а цинт (Hya~cinthus L.), цикл ам ен (Cyclamen L.), л ев- и о й (Matthiola R. Br.), л а к ф и о л ь (Cheiranthus L.), ре- з еда (Reseda L.), ирис (Iris L.), анемо н (Anemone 1..), львиный з е в (Antirrhinum L.), гвоздика (Dianthus L.), ноготок (Calendula L.), кохия (Kochia Roth), душистый г о р о ш е к (Lathyrus odoratus Ь.), некоторые виды л и- лий идр. В Средиземноморье (в узком объеме этого понятия) сосре- доточено большое разнообразие форм и сортов культурных ра- стений. Но это не арена интенсивного видо- и родообразования культурных растений, а только подобласть основной области Лревнего Средиземноморья. 37S
В средиземноморских странах сосредоточены высококуль- турные, крупноплодные и крупносемянные сорта многих расте- ний, имеющих первичный очаг вхождения в культуру и распро- странения в Передней Азии и в меньшей степени в Средне-Юго- Западной Азии. Эндемичных культур в собственно Средиземноморье не так много, к ним относятся следующие: люлин термис — Lupinus termis Forsk., хотя он возделывается и в Сирии, примыкающей не к собственно Средиземноморью, а к Передней Азии; сулла— Hedysarum согопаг ит L. (Испания, Португалия, Италия, Си- цилия); дрок — Ulex еигоpaeus L. (Португалия, приатлантиче- ские районы Европы с влажным климатом); дрок прядильный— S~partium junceum L. (Португалия, Испания, Северная Африка); чина охряная — Lathyrus ochrus DC. (Италия, Испания); чина— Lathyrus gorgoni Pari., клевер инкарнатный — Trifolium incar- natum L. (Западное Средиземноморье, почти эндем; культура его вне Западного Средиземноморья незначительна); люцерна триполитанская — Medicago tripo itanica Sinsk., эрука пери- стая — Егиса pinnatifida P~omel, альфа — Sti~pa tenacissima L., AmIpelodesma tenax Link. Перечисленные 11 культур до сих пор имеют эндемичные ареалы — не возделываются в странах, расположенных вне соб- ственно Средиземноморья. Из родов, в составе которых имеются возделываемые виды, средиземноморское (в узком значении) происхождение имеют: Егиса Adans.— Западное Средиземноморье, Lavandula L. — За- падное Средиземноморье, возможно, Carthamus — Западное Сре- диземноморье, Sinapis L. — Средиземноморье, Cynara L.— Западное Средиземноморье, а также подрод донника — Micro- melilotus S uv. — Средиземноморье. Если же взять Яревнесредиземноморскую область в полном сбъеме, вместе с Передней и Средне-Юго-Западной Азией, то окажется, что отсюда происходит огромное число важнейших культурных растений. Собственно Средиземноморье является родиной капусты, европейского редиса, важнейших в сельскохозяйственном отно- шении видов люпина, 14- и 28-хромосомных овсов, лаванды, ар- тишока, средиземноморского подвида льна, видов синей лю- церны — М. tripoli tanica Sinsk., М. eusativa Sinsk., рожкового дерева и очень многих сортов зерновых, овощных и плодовых растений. Таким образом, растения, вышедшие из Древнесредиземно- морской области в полном ее объеме, могут иметь первичные очаги и в собственно Средиземноморье, и в Передней, и в Сред- не-Юго-Западной Азии. Некоторые виды распространены теперь в культуре во всех трех подобластях Древнесредиземноморской области или в двух из них. Часто очень трудно определить, где находится первичный очаг вхождения в культуру. Легче это 379
сделать в тех случаях, когда дикорастущие прародители сосре- доточены в определенном месте (например, дикие артишоки явно приурочены к западной части собственно Средиземноморья, весь род Eruca также имеет явно западное происхождение— Алжир, Испания). Труднее определить очаги вхождения в куль- туру отдельных родов, для которых можно предположить древ- несредиземноморское происхождение потому, что некогда такие роды были распространены по берегам исчезнувшего древнего средиземного моря метиса и далеко заходили на восток в Азию. Затем на большей части этого огромного прибрежного древнего ареала дикие родоначальники вымирали и сохранились только кое-где. 1~ таким родам можно (с различной степенью уверен- HocTII) отнести следующие. 1. Ra~phanus L. — редька и редис. Родоначальные дикорасту- щие виды сохранились на крайних западном и восточном кон- цах Евразии, а на крайнем юге приморского ареала древнейшие следы исчезнувших родоначальников обозначены теперь энде- иичнымп культурными видами (Е. Н. Синская, 1931). 2. Mandragora 1 . — местонахождение современных видов в разорванном ареале рода: Испания, Греция, Туркмения, Ти- бет, Гималаи. Этот ареал далеко простирается с запада на во- сток. 3. Daucus L. — морковь. Ареал рода прибрежный, проходит с крайнего западного до крайнего восточного конца Евразии, а эндемичные культурные виды [или подвиды (?) — это уже дело больше систематиков или географов] имеют такой же тип рас- пространения (подвиды западноевропейский, переднеазиатский, афгано-индийский) . 4. Beta L.— свекла. Ареал рода прибрежный, средиземно- морский и атлантический, но в Индии имеется тонкокорневой вид, близкий к дикорастущим свеклам. 5. Brassica L. — род в целом, возможно, также имел первич- ный ареал древнесредиземноморский. В частности, репа пред- ставлена двумя большими видами: европейским и азиатским с японским подвидом. Впрочем, восточноазиатский состав еще довольно слабо обособленных видов Brassica скорее возник в Восточной Азии в культуре из элементов, принесенных туда человеком из Средне-Юго-Западной Азии. Ареал дикорастущих древних, родоначальных представителей Brassiica мог не дохо- дить до Дальнего Востока Азии. Дикие исчезнувшие родоначаль- ные виды могли и не распространяться дальше определенной границы, например на широте Зондских островов и т. п. 6. Высказанные выше соображения относятся и к таким ро- дам из сем. бобовых, как чечевица, конские бобы, горох. При- митивные формы чечевицы обнаружены в Передней Азии и Ин- дии, а ранние (мелкосемянная чечевица) были широко распро- странены и в собственно Средиземноморье. Примитивные формы бобов нашлись в Алжире и Индии. Дикий родоначальник гороха 3SO
Pisum elatius распространен от Средиземноморья (в основ- ном в Передней Азии) до Тибета, а примитивные культурные срормы представлены и в Японии. 7. Подобные соображения относятся к культурному льну. Дикий родоначальник сосредоточен в собственно Средиземно- морье и Передней Азии, но эндемичные формы и очаг древней- шей культуры (Е. Н. Синская, 1954) открыты в Индии. 8. Предположение о древнесредиземноморском ареале диких родоначальников относится и к ячменю. Дикорастущие ро- дичи — Hordeum spontaneum — вместе с недавно открытыми Ф. Х. Бахтеевым новыми близкими к нему формами приурочены к подобластям Передней и, может быть, Средне-Юго-Западной Азии, но в Восточной Азии сохранились эндемичные формы очень древнего типа — Hordeum humile Vav. et Ba~cht. Роды и виды, имевшие первоначально древнесредиземномор- скпе ареалы, более древние, чем те, у которых исходные формы приурочены к отдельным подобластям Древнесредиземномор- ской области исторического развития культурной флоры. Дальнейшее накопление фактов должно многое выяснить и помочь в расшифровке уже имеющихся данных. Восстановление древнейших путей расселения исходных форм позволит уточнить историю путей распространения важнейших культурных расте- ний — в результате будет создана историческая ареалология культурной флоры.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕННЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ СРЕДНЕЙ И СЕВЕРНОЙ ЕВРОПЫ Во время возникновения и развития первых государств Qpejj- него Востока Европа представляла собой отсталую окраину. В эпоху неолита собственно Средиземноморье было ближай- шей периферией gpeBHejo Востока. Западная, Средняя и Юго- Восточная Европа была как бы периферией Средиземноморья ji далее — на север и восток — крайней зоной цивилизации. Однако и в этой зоне расселения рыболовческих и охотничьих племен земледелие возникло в более древние времена, чем ду- мали раньше. Ассортимент культурных растений в Средней и Северной Европе в основном складывался исторически под большим влия- нием средиземноморских стран в более узком значении этого понятия, а если взять Средиземноморье в широком аспекте N. Г Попова — древнего Средиземноморья, то можно сказать, что последнее было основным очагом, откуда древние обита- тели Средней и Северной Европы заимствовали культурные ра- стения. Однако история природы и народов носит черты свое- образия и разнообразия в разных частях Европы, а главное, земледелие здесь во многих местах оказалось очень древним Поэтому и принесенные сюда извне элементы культурной флоры имели здесь большие возможности для своего дальнейшего раз- «ития и преобразования в течение чрезвычайно длительных пе- риодов. КроМе того, аборигенная дикорастущая флора, хотя сильно обедненная во время ледникового периода, все же могла дать кое-что для введения в культуру. Богатый и хорошо изученный археологический материал вскрывает древний ассортимент культурных растений, начиная с каменного века. В южной Германии (Федерзее) останки палеолитическога человека были найдены в слоях, относящихся к кустарниковой тундре, когда еще не было леса. В тундре были распространены олени, и человек охотился на них. Здесь в нижних слоях была обнаружена пыльца полярных ив, карликовой березы, горной сосны; их последовательно сменяли береза, сосна обыкновен- ная; затем следует период дуба с берестом, липой и орешником в подлеске. Стоянки человека среднего палеолита, расположен- ные кругом озера Федер, относятся к периоду отступления сосны, времени наибольшего распространения орешника и к началу 382
° ° ° 4 ° ° С ° О ° е а ° ) а ° Э ° ° ° ° а Э ° ° sâ ! ° ° ° ° а ° ° 1 ° ! ° ° ° ° вв а а ° а ° ° а ° С r й ° ° а ° ° ° ° ° ° ° ° в ° ° ° ° ~ в е в ° 9 ° ° ] ф а ° а I 111 ° ° r ° ° ° а а ° ° в ° ° ° Э С ° С ° l ° С ° ° l Э С ° ° С ° ° ° ° а ° ° в f ° ° e ° 3 ° а ° а ° ° ° l 1i ° ° ° ° Э а ° ° ее r ° ° ° ° ° а С r ° ° С ° ° ° а ° ° ° й а ° С ° ° ° ° ° ° а J ° С ° ° ° ° ° ° ° ° а а ° ° а ° ° в 3 ° е ю ° а ° а ° ° ° в ° ° е а ° Э ! Э е 9 I в В ° ° и с ° Э ° ° ° ° r ° r а ° ° ° ° а ° ° 4 ° ° ° ° 3 ° ° 3 ° 9 ° ° е s ° 3 ° Э ° ° а a ° ° а ° ° ° а ° ° ° ° ° ° ° а а ° а ° ° с ° ° 3 ° ° ° а а ° ° а С Л ° в °а ° ° б ° ° б ° ° ° С ° ф ° ° ° Э а а
]94p) их было 5 (Skorhaek, Lindslcov, Brabrand, Signalbakl'еп, Bundse). Здесь хорошо выявляется последовательность появления хлебных злаков. Однозернянка культивировалась только в ка- менном веке — позднее она в раскопках уже не встречалась. Каменный век был эпохой наибольшего 'значения полбы, в брон- зовом веке полбы уже мало, а позднее она исчезает совершенно. Вероятно, однозернянка пришла вместе с полбой как засоритель последней. Век наибольшего распространения полбы был в то же время и веком культуры однозернянки. Когда полба на- чинает исчезать (бронзовый век), однозернянка исчезает совер- шенно. Неолит является эпохой и наиболыиего распространения голозерной пшеницы. Эта неолитическая пшеница не могла быть ничем иным, как той же самой пшеницей, которая характерна для свайных построек Швейцарии и названа была Геером (1865) T. vulgare antiquorum Heer. Эта пшеница в Дании встречалась и позднее, вплоть до римского периода, но уже в гораздо мень- шем количестве. В противополсжность трем неолитическим куль- турам Скандинавии — — однозернянке, полбе и Т. antiquorum— ячмень был там распространен, начиная с неолита, во все эпохи. Существование голозерного ячменя кажется нам сомнительным, вероятно, пленки у ископаемых ячменных зерновок просто истлели. Просо появляется здесь только в бронзовом веке и затем опять исчезает. Овес начинает появляться в бронзовом веке, но распростра- нение его значительно усиливается в ~келезном веке. В бронзо- вом веке был, вероятно, не обыкновенный овес — А. sativa, а песчаный — Л. strigosa. Эпохи появления и наибольшего рас- пространения овса и полбы в Данин не синхронны. Отсюда можно сделать заключение, что в Скандинавию овес попал не вместе с полбой как ее сорняк, а отдельно. Вероятно, полба и овес пришли в Скандинавию разными пу- тями: полба — из Передней Азии, точнее из Древней Колхиды, в виде древней формы, близкой к современной Т. ра1еосо1сЫсит, и овес — А. sativa — много позднее из Скифии и протославян- ской территории. Рожь появилась в Дании только в железном веке. Однозернянки из неолита Дании мелкозерные, мельче совре- менных дикорастущих. Размеры зерновок ископаемых датских однозернянок из стоянки Bundse, по даннь м К. Иессена (Е. Schiemann, 1940), колеблются: длина — 3,6 — 5,3 мм, ши- рина — 1,1 — 2,0, высота — 1,6 — 1,9 мм. K. А. Фляксбергер дает для современных однозернянок сле- дующие амплитуды размеров зерновок (в лм): дикорасту- щ и е о д н о з е р н я н к и — длина 6,0 — 7,5, ширина 1,0 — 1,5, вы- 384
сота — 20 — 26; культурные однозернянки — длина 7,0 — 8,5, ширина 1,8 — 3,0. QJIHHa зерен полбы — Т. dicoccon, по данным 1(. Иессена, из- меняется от 4,1 до 7,7 мм. Ископаемая датская полба мелкая, но максимальные размеры ее зерен крупнее минимальных у со- временных полб. Г о 1(. А. Фляксбергеру, длина зерновок совре- менной полбы колеблется от 6,5 до 10 мм. Тот же основной состав культур известен и из неолита Гер- мании. Неолитические слои из Trebus (Бранденбург), исследо- ванные Вертом (L. Werth, 1939), содержали полбу, шестиряд- ный ячмень и пшеницу, определенную как Т. antiquorum Heer; кроме того, была найдена пшеница, похожая на Т. ~compactum, но отличавшаяся от современной. Это была без сомнения также Т. antiquorum, которая характеризовалась значительным поли- морфизмом. Состав культур из слоев бронзового века обычно разнообразнее. Так, при раскопках в Dobenecl<, по Верту, были найдены: полб а — 'I'. Йсоссцт, пшеница, определенная как Т. com~paictum (но, скорее всего, это была еще Т. antiquorum), овес (одна зерновка, более короткая и узкая, чем у современ- ного овса, но не похожая на зерновку овсюга — А. fatua), шести- рядный ячмень, бобы, горох, чечевица. Зерновки ячменя имели длину 5,8 — 6,4 мл1, ширину 2,8 — 3,2 мм. По несимметричности зерен было определено, что это шестирядный ячмень. Чечевица была мелкосемянная со слегка треугольно-закруг- ленными зернами размером 3,0 )(2,8 ~ 1,8 л~м. Бобы также были мелкосемянные и определены Вергом, как Vi~cia faba var. icel- tica папа Heer, т. е. здесь найдена такая же форма, как и из свайных построек озерной Швейцарии. Были найдены также остатки плодов и семян дуба черешча- того, лещины и терна. Эти дикорастущие растения играли, не- сомненно, значительную роль в питании обитателей той эпохи и были обьектами собирательства. В других неолитических стоянках Германии находили также остатки древесины и семян вяза, липы и бука вместе с дубом. Эти стоянки относились к эпохе господства смешанного лиственного леса. Часто здесь же находили и семена дикого винограда — Vitis silvestris. Так, на- пример, в Хейльбронне на Неккаре, в слоях раннего каменного века, были найдены семена и кисти дикого винограда вместе с зернами полбы, однозернянки и шестирядного ячменя, семе- нами бузины и некоторых сорных трав: Chenopodium, Atriplex, Polygonum, Lyco~pus, Galium и Ча1ег1апе11а. Ископаемые остатки дикого винограда обнаружены и в дру- гих местах Германии, а также в Тироле, не говоря уже о более южных странах (Италия и др.). По-видимому, дикий виноград был представлен разными расами и широко распространен в древней Европе — вплоть до южной Швеции (Г'амс). Остатки его относятся главным обра- зом к постгляциальному теплому периоду. Синская Е. Н. 385
В другом пункте раскопки раннебронзового века около де- ревни в Lenzersilge были найдены остатки следующих расте- ний: ячмень шестирядный, полба-эммер, пшеница — Т. antiquo- rum, однозернянка, просо, конские бобы мелкосемянные, чече- вица мелкосемянная (около 3 мм в поперечнике), дуб, орешник. Ячмень состоял из смеси крупных и мелких зерен: максималь- ные размеры — 7,5~3,8 мм, минимальные 5)(2,7 мм. В этом месте однозернянка задержалась до бронзового века. Ясна преемственность культур в Германии от бронзового и железному веку. Например, в пробе из Plauen (ранний желез- ный век) Верт определил следующие растения: ячмень шести- рядный (размеры 6,)(4 и 3)(2мм), полба (5,5)(4,7 и 2,7;( Q 2,3 мл~), конские бобы (7,5)(6Р~ 6 и 6)(5,3)(5 мм). В железном веке для Германии, по-видимому, все еще харак- терны мелкосемянные формы зерновых и бобовых. Так, макси- мальные размеры зерен составляли (в мм): для ячменя шести- рядного — 6 )( 2,7, полбы-эммера — 5,5 )( 2,7, конских бобов— 7,5;(6,)(6; минимальные — соответственно 4 +2; 4,7;~;2,5; 6 ~; Q 5,2 )( 5. Согласно Кларку (Clark, 1952), человек постепенно изменял характер территории и умножал количество пригодных для земледелия угодий. Сначала имело место только мелкоостровное и придомное земледелие и скотоводство. Подсечная система спо- собствовала сокращению площадей под лесом, и все большая часть территории приобретала степной или лугостепной харак- тер. Кларк подкрепляет эти выводы пыльцевым анализом грун- тов в центральной Ютландии. Во время распространения там племен «боевых топоров» образовались участки степи — резуль- тат вырубки леса. На пыльцевой диаграмме видно, как восходя- щая линия древесных растений обрывается, а линия травянистых растений возрастает. В бронзовом веке здесь устанавливается вместо переложной системы земледелие на постоянных полях. Так, человек на всем пространстве, где он появлялся, изме- нял природную среду, и этот процесс имел место не только в Се- герной и Северо-Западной Европе, но и в центральной ее части. Особенно много для познания древнего ассортимента куль- турных растений Средней Европы дали археологические иссле- дования свайных построек Швейцарии, Франции, Венгрии и тер- рамар Италии. Особенно следует отметить заслуги исследова- теля свайных построек О. Геера, труд которого отличается тща- тельностью описания и снабжен хорошими рисунками. Е~енные сведения дало изучение материалов по ископаемой пшенице, приближающейся по типу к мягкой и компактной, но вместе с тем отличающейся от всех ныне существующих видов пшеницы и названной О. Геером Т. antiquorum Heer. Эта пше- ница представляет собой интегральный вид, т. е. как бы совме- щает в себе признаки нескольких современных видов. Раз- ными исследователями (Геер, Бушан, Верт, вейнингер, Вёниг,
Унгер и др.) описывались различные формы: близкие и кар- ликовой пшенице — Т. compactum, более близкие к современной мягкой — Т. aestivum, близкие к английской, или египетской, пшенице — Т. turgidum, и особые формы с мелкими грушевид- ными зерновками. Однако при тщательном сравнении описаний становится ясно, что все эти формы относились к одному поли- морфному виду — Т. antiquorum, а из современных пшениц имели общие черты только с реликтовыми видами Колхиды- Т. macha и Т. paleocolchicum — и с абиссинской эндемичной пше- ницей — Т. aethiopicum. Как отмечалось выше, в слоях, относя- щихся к неолиту и бронзовому веку Закавказья, были найдены аналогичные Т. antiquorum круглозерные и мелкозерные пше- ницы, дериваты которых сохранились в Грузии, а в Армении исчезли. Имеются веские основания предполагать, что древний прото- тип этих интегральных видов проник в Среднюю и даже Север- ную Европу (Дания) и дал там начало гееровской пшенице— Т. antiquorum, а также в Египет и Эфиопию. В Эфиопии суще- ствует дериват первоначального древнего прототипа, представ- ляющий собой своеобразный и полиморфный вид — Т. aethiopi- сит, который отличается от остальных современных ви- дов пшеницы и имеет общие черты только с колхидскими релик- товыми видами и исчезнувшей среднеевропейской Т. antiquorum. Последний вид пшеницы был широко распространен. Он от- крыт в неолите Швейцарии (gobenhausen, Wangen, Luscherz, Steckborn и др.), Германии (Federsee, Andreasthor, Metendorf, Ettersberg, Trebus и др.), Дании (Alsen и др.), Венгрии (Len- gyel, Felso-Dobsza, Aggtelel< и др.) .. По данным, собранным Бушаном, такая же круглозерная и мелкозерная пшеница — var. globiforme Buschan — обнаружена в слоях, относящихся к неолиту в Италии, Египте и к бронзо- гому веку в Австрии, Италии и Швейцарии. Одн озер н яика (Т. monoooccum), согласно сводке Е. Ши- манн (Schiemann, 1940), найдена в неолите Дании, южной Шве- ции, Боснии, Венгрии, Германии (Тюрингия, Саксония и др.), Швейцарии, французской Юре и Бельгии. В общем ее древний ареал находился в восточной части Западной Европы (без Рос- сии) . Пришла однозернянка в Среднюю и Северную Европу, несо- мненно, с востока. Она не имеет непосредственной родственной связи с дикорастущими однозернянками Балканского полуост- рова, ибо климат Северной Европы, даже и в более теплый ин- тергляциальный период, вряд ли был благоприятен для дикой однозернянки и, что особенно важно, сначала тундровые, а затем лесные фитоценозы Северной и Средней Европы вряд ли могли пропустить так далеко на север это горно-степное растение. Однозернянка появилась в Средней и Северной Европе, несо- мненно, уже в виде окультуренного растения. Сначала она рас- 387
прострянилась, вероятно, вместе с полбой-эм мерам — Т. dicoc- con — в качестве сорняка последней, а затем стала выходить и р чистые посевы на каменистых и более бедных почвах, где лучше мирилась с неблагоприятными условиями, чем полба. На связь однозернянки с полбой указывают многочисленные случаи r<x совместного нахождения в одной пробе при археологических раскопках. Описанные Геером полба-эммер и однозернянка (из одной и той же неолитической стоянки Wangen) не были похожи на современную полбу и однозернянку; они отличались более густым расположением колосков, полным отсутствием остей и мелкозерностью. Интересно, что и встречающаяся часто вместе с полбой и однозернянкой ископаемая пшеница типа мягкой— Т. antiquorum ~ai~a~~TepnaoBa~acb reMH»<e признаками. В эпоху неолита существовал, следовательно, общий морфологи- ческий тип, характе,')ный для всех культурных видов пшеницы того времени, ибо и неолитическая пшеница Закавказья отлича- лась плотным колосом, мелкозерностью. Закавказские, близкие к Т. antiquorum неолитические пшеницы (по крайней мере те, которые пока были найдены в раскопках) были остистые, но ости (орган испарения), вероятно, оказались ненужными в до- статочно влажном и прохладном климате приальпийской об- ласти и других местностей Средней и Северной Европы и были утрачены, или же (что вернее) в раскопках в Закавказье были найдены все же не самые древние формы, которые были без- остыми или короткоостистыми, как и предки пшеницы — пыреи из подрода El~ytrigia. Во всяком случае родство Т. antiquorum и древней закавказской пшеницы — Т. protomacha — не вызывает сомнений. Ngi полагаем поэтому, что все пшеницы — Т. anti- quorum, полба (в виде формы, близкой к Т. ipaleocolchiicum) и однозернянка — пришли в Среднюю Европу пз Передней Азии, или через Балканы, или через южную Россию, или двумя этими путями. Однозернянка была издревле в культуре в Малой Азии, что доказано находкой ее в старой Трое II. Т. antiquorum, как таковая, существовала еще в бронзовом веке, вероятно, и далее, постепенно переходя в формы, более близкие к современной карликовой пшенице — Т. compactum. Однозернянка до сих пор еще сохранилась в культуре в виде небольших островков на Балканах, Карпатах, в северных Аль- пах и в их северных отрогах. Культурные однозернянки из удаленных пунктов своего разорванного ареала имеют определенные различия, но эти раз- личия гораздо меньше, чем между географическими подвидами пол бы. Важно отметить, что основная разновидность однозернянки с красными чешуями и красными зернами — var. hornemanni С1ет. — распространена не только на Кавказе, но и в Крыму, на Балканах и в странах Средней Европы. Этот общий «костяк» 388
в популяциях однозернянки еще раз говорит о происхождении однозернянок из переднеазиатского очага (куда включается и Закавказье). Здесь она растет дико целыми массивами, имеются переходные формы от дикорастущих к культурным; здесь (За- кавказье) однозернянка недавно широко возделывалась, еще сохранились смешанные сообщества ее с более древними куль- турами — полбой-эммером и голбой Тимофеева, в лоне которых однозернянка прошла историческую стадию сорняка. Даром переднеазиатских древних народов является и полба Т. йсоссоп, которая, вероятно, была занесена в Европу в виде Т. protopaleocolchicum. Ботаническое изучение подвидов полбы-эммера позволяет наметить следующие пути ее распространения: 1) из Передней Азии по Балканскому полуострову, по Дунаю в Среднюю Европу и оттуда через Францию в Испанию; 2) с Балканского полуост- рова в область Волги и Камы; 3) из Передней Азии в Иран, Еги- пет и в Эфиопию, из нее в Индию (может быть, и-обратно — из Индии в Эфиопию). Подвид полбы Средней Европы является по своей организа- ции, по мезофильным свойствам наиболее древним, приближаю- щимся к исходной переднеазиатской форме типом. Европейская п ол б а - сп ел ь та (Т. spelta) представляет собой сравнительно молодой вид, возникший, по нашему предпо- ложению (Е. Н. Синская, 1955), из прототипа современной мяг- кой пшеницы — Т. antiquorum — в приальпийской области Сред- ней Европы. За такое ее происхождение говорят часто наблюдаю- щиеся у современной мягкой пшеницы уклонения в сторону признаков спельты, в резком своем выражении получившие название спельтоидов. Спельтоиды образуются большей частью под влиянием неблагоприятных для мягкой пшеницы условий: сухости почвы и воздуха, пониженных температур, поражения вредителями и болезнями и т. д. Подобным же образом могла возникнуть и спельта, не из мягкой пшеницы, а еще из более древнего типа — Т. antiquorum (или из ближайшей к последней производной формы), поэтому спельта имеет более древние черты (например, ломкость стержня), чем современные спель- тоиды. Ареал спельты в Европе в настоящее время ограниченный и таковым был и прежде: в археологических раскопках она най- дена только в слоях бронзового века в Швейцарии. К. А. Флякс- бергер предполагает, что римляне заимствовали это растение от швабов и алеманнов и занесли эту культуру в Астурию (Испа- ния}. Согласно такой точке зрения, спельта — это не столько реликт, а скорее эндемик приальпийской области, заселенный раньше древнегерманскими племенами. Теория европейского происхождения спельты изложена де- тально и, казалось бы, очень обоснованно К. А. Фляксбергером, который утверждал, что спельта в странах Передней и Средне-
~О„З~п~дно~~ Лзии не возделывается. Оказалось, что спельта имеет там определенный ареал. Куккук (Н. Kuckucl< und E. Schiemann, 1957) обнаружил, что спельта распространена в Западном Иране и представлена там большим разнообразием форм в посевах племени бахтиар. Мак-Фадден и Сирс (Е. S. МсРагЫеп and Е. R. Sears, 1946) в результате скрещивания Т. di~coocoides с Aegilops squarrosa получили форму, чрезвычайно сходную с Т. spelta. На основании этих фактов многие исследователи пшениц склонны считать первичным очагом происхождения Т. spelta не Европу, а Переднюю Азию. Куккук и Шиманн (1957), а также М. М. Якубцинер (1956) указывают на ряд древних местона- хождений спельты в странах Передней Азии и вне Ирана. Вероятным является первичное появление спельты в Закав- казье и прилегающих странах, где имеются и двузернянки, и виды Aegilops; позднейшее занесение этого вида в приальпий- ские районы Европы, возможно, через Балканы. История спельты в Европе, изложенная К. А. Фляксбергером, интересна тем, что подтверждает существование древнейших сношений переднеазиатских и европейских народов. Принятие переднеазиатского происхождения спельты лишний раз говорит о том, что Европа является лишь очагом формообразования у видов пшеницы, но не областью видообразования. Яч м ень, как и пшеница, является древнейшей культурой Европы. Он обнаружен в неолите Швейцарии (Stoore, Gortail- lod, Lusicherz, Blei~che-Arbon, Robenhausen, Wangen, Steel<born и др.), Венгрии (Aggtelek, Lengyel, Felso-Dobsza и др.), Герма- нии (Ettersberg, Andreasthor — Саксония и др.), Дании, южной Швеции, Франции, (Martres-de Чеуге, Bourget — Савойя), Верх- ней Италии, а также в слоях, относящихся к бронзовому веку Швейцарии, Германии, Дании, южной Швеции, Франции, Ита- лии, Испании, Греции. Местонахождения ячменя из неолита приурочены большей частью к горным местностям. I eepoM описан ячмень из неолита свайных построек Швей- царии (Robenhausen). Это был шестирядный ячмень с мелкими зернами, с мелкими и очень плотными безостыми колосьями (4,4 см длины) — 1lordeum hexastichum sanctum Heer. В мень- шем количестве была обнаружена более крупноколосая и круп- нозерная форма, но также с очень плотными колосьями — Н. he- xastilchum densum Heer. Длина зерен варьировала от 5 до 7 мм, а толщина — от 3 до 4 мм. Был найден один двурядный колос. Эта находка показывает, что в ту отдаленную эпоху имело место образование двурядных форм культурного ячменя, образующе- гося из шестирядного в результате редукции боковых колосков. Мелкозерный и круглозерный ячмень был также найден в Древнем Египте и в слоях, относящихся к бронзовому веку Закавказья (Шенгавит). Вероятно, ячмень еще в глубочайшей древности был занесен в Европу из Передней Азии.
Р о ж ь (Setcale сегеаlе L.) нигде в свайных постройках Швей- царии найдена не была. Геер нашел ее в слоях, относящихся к бронзовому веку в Австрии (Olmiitz), вместе с пшеницей, ячменем, просом и лещиной. Пшеница была мелкозерная, ве- роятно, Т. antiquorum. Геер думает, что эта рожь — наиболее древняя из находимых в раскопках. Скорее всего, это была не самостоятельная культура ржи, а сорная рожь из посевов пшеницы и ячменя, распространявшаяся вместе с ними. В Швей- царии рожь, по Гееру, возделывалась только в римское время, на что указывают находки ее в Buchs вместе с пшеницей, овсом и могаром. Геер указывает на находку ржи в руинах римской эпохи в Венгрии вместе с мелкосемянными конскими бобами. По словам Галена, Briza, возделывавшаяся во Фракии и Македо- нии и дававшая плохой черный хлеб, была, вероятно, рожью. Плиний называл ее Secale и считал культурой кельтов. В рас- копках Помпеи (70 лет до н. э.) были найдены рожь и овес. В Верхней Италии (Bor) в свайных постройках железного века рожь была найдена вместе с чечевицей, каштаном, остатками плодов оливы, винограда, черешни и видов Prunus. В раскопках близ г. Гута (Чехословакия) в слое, отнесенном ко времени римского владычества (около 400 г. н. э.), найдены семена пшеницы, проса, вики и других трав, а также семена ломкой ржи. Аналогичные семена были найдены в Восточных Альпах и Богемском лесу. Этн находки датируются 200 — 400 гг. н. э. Со- вершенно невероятно мнение о том, что некогда в этих местно- стях возделывались формы ломкой ржи. Несомненно, здесь была сорная рожь, а отнюдь не культурная. Распространение ломких форм ржи в Европе и, в частности, в Польше можно объяснить торговыми связями римлян с переднеазиатскими народами. В других странах рожь найдена в ряде раскопок железного века. В стоянке Niemitszsch (Бранденбург) нижние слои досла- вянские, а верхние — славянские. Рожь была обнаружена только в верхних славянских слоях, а в нижних найдены ячмень, просо, чечевица. Рожь вместе с овсом была найдена в Шлезвиг-Голь- штейне (Ahrensburg и др.), именно в славянских слоях, а также в славянских слоях около Вроцлава (с пшеницей и просом), в славянских слоях Пруссии (Striesow, Тогпо). Таким образом, культура ржи получила развитие только в железном веке и главным образом на востоке Европы в сла- вянский период. До этого времени рожь возделывалась мало, случайно, там, где условия были наиболее благоприятны для сорной ржи, она вытесняла культуру «хозяина» (пшеницу и ячмень) и появлялась в чистом посеве. Рожь сначала расселя- лась из Передней Азии (где возник самый род Secale и вид S. оегеаlе, составляющий один вид с сорной рожью) вместе с пше- ницей и ячменем, и в самостоятельную культуру вошла именно в земледелии славян. Нет никаких доказательств, что культура 391
р~и существовала в бронзовом веке. Рожь была только сорня- ком. В римское время рожь, по-видимому, была в культуре у кельтов, но не в большом масштабе, несравнимом с размерами ее культуры у славян. К кельтам (и на Балканы) она была также занесена вместе с пшеницей и ячменем из Передней Азии или, может быть, позднее из Скифии. Овес также нигде не обнаружен в раскопках раньше бронзо- вого века. Геер находил овес в свайных постройках Швейцарии, в бронзовых слоях из Petersinsel (400 лет до н. э.) и Montelier и из римских руин (Buchs). Зерна этого овса длиной 6,2 мл~ и шириной 2 мм имели на брюшной стороне только одну слабую бороздку, со спинной стороны — слабо выпуклые; у зародыша— заостренные, а на другом конце — закругленные. Зерна овса из слоев, относящихся к бронзовому веку Швейцарии, мельче, чем зерна всех современных сортов овса (А. ваШа), и сходны снай- денными в 1!!екленбурге. А. fatua был найден также в слоях, относящихся к бронзовому веку Австрии (Heidengebirge), Гер- мании (Р1аиеп), в Савойе (Bourget), Дании, в раскопках же- лезного века в Пруссии (Wittenberge), в римскую эпоху в Пом- пее, в гельвето-римское время в Швейцарии (Buchs) и на во- стоке во многих местах в славянское время (Ahrensburg— Шлезвиг-Гольштейн, Висмар и др.). Мы считаем, что мелкозерный овес из слоев бронзового века Швейцарии, описанный Геером, относился не к А. sativa, а кдру- гому виду — А. strigosa. Вероятно, и в других раскопках брон- зового века (в Мекленбурге, по Гееру, овес из бронзового века также мелкозерный) был именно А. strigosa, а не А. sativa. На- пример, Вертом (Werth, 1939) в раскопках в Dobeneck, относя- щихся к 1200 г. до н. э. (бронзовый век), найдено одно зерно овса, которое было короче и уже, чем у современного овса, и не походило, по словам Верта, на зерно овсюга— А. fatua. Верт исследовал также зерна овса из раннебронзовых слоев (900 — 700 гг. до н. э.) в раскопках у деревни Lenzersilge, кото- рые показались ему не совсем типичными для культурного овса Вероятно, в обоих случаях зерна относились к виду А. strigosa— песчаному овсу. А. strigosa древнее обыкновенного овса — А. sa- tiva — и в настоящее время кое-где сохранился в культуре на Пиренейском полуострове, в Англии (Уэльс и Шотландия), в западной и средней Франции (Овернь), Бельгии, в северной Германии (Мекленбург), а как сорняк — по всей Средней и Се- верной Европе (вплоть до западных областей CCCP). Песчаный овес сохраняется в культуре главным образом в гористых мест- ностях, где обыкновенный овес не дает устойчивых урожаев. Песчаный овес представляет собой реликтовую культуру, або- ригенную для Юго-Западной Европы (Испания, Португалия, от- куда был занесен во Францию и Англию). Близкий дикорасту- щий родич А. hirtula распространен в Испании, Португалии, 392
Северной Африке (Марокко, Алжире, Тунисе), Сицилии, Из- раилее. П р о с о (Panicum milia~ceum L.) почти не сеется в Швейца- рии, оно, по словам Геера, вытеснено теперь картофелем и зер- новыми яровыми хлебами, а в прежние времена оно было здесь широко распространено. Просо было найдено в свайных по- стройках Швейцарии (Pobenhausen) и в слоях бронзового века (Montelier) . В неолите оно обнаружено в немногих местах: в Ломбардии (Varese-See), в Венгрии (Lengyel) и Румынии. В слоях бронзового века просо найдено было, кроме Швейцарии, во Франции, в Италии, Австрии, Яании и Германии. Его нахо- дили в слоях бронзового века (алания), но гораздо чаще в ран- ний железный век и славянский период (Niemitszsch, I<onigs- walde) в Германии и др. Просо не было с достоверностью обнаружено в gpeBHeM Египте и вообще не является растением средиземноморского происхождения. Оно пришло в Европу из Азии, вероятно, по бе- регам Черного моря, а может быть, также и с севера. Кукуруза отнюдь не является аборигенной для Европы, а за- несена из Америки. Однако местные условия вызвали формооб- разование у этого вида. На Балканах имеются эндемичные мест- ные сорта мелкозерной и скороспелой кукурузы. Подобные формы обнаружены также в Молдавии и на западе Украины. Конские бобы в неолите найдены были в Венгрии (Lengyel и Aggtelek), но основное распространение их в каменном веке было более южное — Италия, Испания, Египет, Малая Азия. В слоях, относящихся к бронзовому веку Швейцарии (Pe- tersinsel и Moosseedorf), Геер нашел очень мелкие конские бобы, определенные им как Vicia faba:celtica nana Heer. Наиболее мелкосемянные бобы найдены в раскопках Малой Азии (неолит), Крита (бронзовый век), Верхней Венгрии (нео- лит), Германии (Dobeneci< — бронзовый век, Lenzersilge — ран- ний бронзовый век, Freiwalde — железный век и др.) . Эти во- сточные ископаемые формы отличались большей округлостью семян. Бобы крупные, более длинные, узкие и почковидные, свой- ственны западным раскопкам (Испания, Италия, Франция). Однако и в неолитических слоях на Западе (например, Испа- ния — EI Garcel) известны и очень мелкосемянные формы. Ве- роятно, сначала всюду были распространены только мелкосе- мянные формы. Из этих более примитивных форм на юге выра- ботались крупносемянные формы, которые позднее стали рас- пространяться на север. Может быть, под влиянием культуры этот процесс постепенного укрупнения шел и на севере, но го- раздо медленнее, чем на юге. Г о р о х (Pisum sativum) и ч е ч е в и ц а (Lens esculenta) из слоев бронзового века Швейцарии описаны Геером как очень мелкосемянные формы (Petersinsel, Moosseedorf). Семена гороха 393
имели в поперечнике 4,4 мм, а самые мелкие — 3,5 мм. В Pe- tersinsel найдены семена мелкосемянной чечевицы, которая Геером была названа Ervum Lens miicrospermum. Семена с обеих сторон плоско-выпуклые (в общем, п,поские) . Характеристика ископаемой европейской чечевицы по размерам дана выше. 1~ак конские бобы и горох, древняя чечевица была мелко- семянная; затем в условиях культуры началось постепенное укрупнение; быстрее этот процесс шел на юге, медленнее — на севере и в горах, где и теперь еще сохранились мелкосемянные формы. В Среднюю и Северную Европу чечевица распространилась из Передней Азии, вероятно, через Балканы, а может быть, от- части и по южной России. Из Передней Азии пришла в Европу и чина. В неолите Швей- царии и Италии чина найдена не была. В неолите Венгрии она обнаружена в большом количестве в Lengyel и Aggtelel<, что указывает на ее возделывание здесь. Семена неолитической венгерской чины мелкие, в 2 раза мельче современных: длина— 4,2 мм, ширина — 3,2 мм, толщина — 4 мм. Н у т (Cicer arietinum) не встречен в свайных постройках Швейцарии и Австрии. Он проник (из Передней Азии) в Сред- нюю Европу в более поздние времена, но все-таки здесь выра- боталась особая штамбовая форма — proles bohemi~cum G. Pop. Из кормовых бобовых прежде всего следует отметить эс- п ар цет в и к ол исти ы й (On~obryichis viciaefolia Scop.), кото- рый дико растет в Прибалтийских странах, Силезии, Моравии, Богемии, южной Австрии, Тироле. В ХЧ в. введен в культуру в Южной Франции и теперь широко распространен. Возделы- вается и в СССР, но в настоящее время уступает место абори- генным видам. У F. Medicus имеются сведения, что эспарцет возделывался в провинции Яофинэ ранее 1507 г. Из Франции культура эспарцета распространилась в начале XVII в. в Англию, а в XVIII в.— в Германию и Италию. С е р а д е л л а м елка я (Ornithopus ~perpusillus L.) рас.- пространена дико по всей Средней Европе и в юго-западных районах СССР. В Западной Европе возделывается местами в смеси с сераделлой посевной — О. sativus Broth. По всей Европе распространены сменяющие друг друга в по- рядке широтно-меридианальной и вертикальной зональнести «расы» (региональные экотипы) к л е в е р а к р а с н о г o— Trifolium pratense L. sensu lato. В составе европейского Т. pratense E. Г Бобров (1947) раз- личает следующие региональные разности: к л е в е р а т л а н- тический — proles atlanticum, клевер к онти не нта л ь- н ы й — proles continentale и клевер субальпийский — proles subal~pinum. В действительности подобных разностей (экотипов) имеется, конечно, больше. 394
Клевер венгер ский (Т. expansum Жа1йй. et КЯ.) воз- водится в ранг вида. Этот клевер растет дико в южной и юго- западной Венгрии. Согласно распространенному мнению, он оттуда завезен в США, где широко возделывается. По исследо- ваниям Н. Г Хорошайлова, американский опушенный клевер происходит не от венгерского Т. exlpansum, а от английского дикорастущего клевера, известного под названием «коровья трава» (Cow Grass). Среди культурных европейских клеверов Е. Г Бобровым различаются, кроме венгерского Т. expansum, следующие раз- ности Т. sativum: ssp. praecox Bobr., ssp. serotinum Bobr., ssp. mattenklee (горный). Шведский кл евер (Т. hybridum L.) имеет в основном западноевропейский ареал и представлен в различных странах особыми экологическими разностями. Во Франции, в Германии, Венгрии, Австрии, Скандинавских и соседних странах выработались многочисленные сорта гибрид- ной л ю ц е р н ы, полученные на основе естественных гибридов между дикорастущими местными формами желтой люцерны— Medicago falcata L.— и культурной европейской синей люцер- ной — М. eusativa Sinsk., с их последующим улучшением в усло- виях культуры. Из Германии эти гибридные люцерны были зане- сены в Северную Америку, где образовались под влиянием новых условий особые американские сорта. Некоторые гибрид- ные сорта (Венгерская и др.) были занесены в СССР и здесь подверглись переработке и образовали новые русские сорта. Так называемая французская люцерна, занесенная в СССР, путем гибридизации с местными желтыми люцернами и перера- ботки в новой агросреде дала ценные сорта — Украинская, Се- верокавказская (Славянская) и др. Подрод в и к и (Euvi~cia) считается древнесредиземномор- ским, а наиболее распространенный культурный вид — вика яро- вая (Vi~cia sativa L.), вероятно, переднеазиатского происхожде- ния, но получивший развитие в Средиземноморье, Средней и Се- верной Европе; об этом говорит наличие многих своеобразных местных европейских сортов. Вика была сначала пищевым рас- тением и известна в Европе с неолита, а затем стала фуражной и сидеральной культурой. В средние века вика была главным кормовым растением в Средней Европе (клевер, люцерна и эспарцет распространились позднее). Из кормовых злаков страны Средней и Северной Европы имеют многие экологические разности е ж и, к о с т р а п р я- м о г о, т и м о ф е е в к и и других злаков. Древний вид Dactylis aschersoniana Graebner введен в культуру из дикорастущих и лесных форм в Германии, Югославии и Чехословакии. На альпийских лугах Средней Европы распространены осо- бые формы костра прямого — Bromus erectus Huds., отличаю- щиеся от наших и, может быть, составляющие особый вид. 395
Долины южных Альп, вероятно, являются первичным очагом и родиной р а й г р а с а многолетнего — Lolium ~perenne. Этот вид близкородственен малолетнему виду — райграсу многоукосному, или итальянскому — L. multif lorum Lam., который вошел в куль- туру как кормовое растение в Ломбардии. Однолетняя .е разновидность итальянского райграса — райграс однолетний— var. westerwoldicum, зародилась во влажных странах Северной Европы (Голландии) и оттуда распространилась в Среднюю и Восточную Европу. Из технических растений следует остановиться на льне. Культура л ь н а в Швейцарии возникла в глубочайшей древ- ности. Геер находил в неолите свайных построек (Wangen, Robenhausen) разнообразные изделия из льна, что указывает на сравнительно высокий уровень культуры, несмотря на то, что эти остатки датируются 4000 — 3000 лет до н. э., а начало куль- туры льна здесь относится, несомненно, к еще более раннему периоду. В Robenhausen нашли смесь из семян и коробочек- это было, по-видимому, блюдо из льна. В Эфиопии и теперь едят жареные семена льна, а, по сообщению Плиния, лен употреб- лялся в пищу в Верхней Италии. Вероятно, и в неолитических свайных поселениях Швейцарии лен был не только прядильным, но и пищевым растением. Кроме Швейцарии (Robenhausen, Wangen, Moosseedorf, Steel<born), Австрии и Италии (Lagozza), лен был обнаружен в Испании — в слоях, относящихся к брон- зовому веку (Al gal ), в Германии — в слоях, относящихся к же- лезному веку и к славянскому времени. Какой же лен возделывали земледельцы каменного века? Ответ на этот вопрос дают исследования Геера в свайных по- стройках Швейцарии. Геер определил сначала этот лен как дикорастущий — Linum angustifolium, но позже сам отказался от такого определения и пришел к заключению, что в неолите Швейцарии возделывался особый озимый лен — L. hyemale. Этот озимый лен со склонностью к простратному росту и теперь Воа- делывается во влажных приморских районах Малой Азии и кое- где в Закавказье (Мегрелия, Ленкорань, Абхазия). Изредка он встречается в Баварии. Лен Геера имел такие же мелкие коро- бочки (даже еще мельче) и мелкие семена, как и современные формы колхидского озимого льна. Коробочки ископаемого швей- царского льна не раскрывались при созревании, что характерно именно для озимого льна, а не для L. angustifolium, у которого коробочки при созревании раскрываются. Реликтовый колхид- ский лен, несомненно, был занесен в приальпийскую область Средней Европы из Передней Азии (1~олхиды) в глубочайшей древности, вероятно, через Балканы. Среди образцов коллекции льна Всесоюзного института рас- тениеводства в образцах из Франции нами были обнаружены переходные формы от L. angustifolium к озимому многостебель- ному льну. Этот факт показывает, что и теперь еще в странах
Европы можно встретить интересные формы, могущие пролить свет на историю льняной культуры. Ма к (Pa~paver somniferum 1.) был известен неолитическим обитателям свайных построек Швейцарии (Robenhausen), Италии (Lagozza) и бронзового века Франции (Bourget). Не вполне достоверно, что это был культурный мак. Может быть, использовалось сорное или рудеральное растение. Геер писал, что семена мака, найденные в Robenhausen, имели сред- нюю величину между сорными и культурными маками. У куль- турного мака, по Гееру, на поверхности семян имеются 5— 6-угольные петли, которые не расположены определенными рядами. У Р. rhoeas L. эти петли мельче, 4 — 5-угольные и рас- полагаются более или менее рядами, особенно на спинке семени. По характеру петель семена швейцарского свайного мака по- хожи на культурный мак, так как петли у него крупнее, чем у Р. rhoeas, 5 — б-угольные, образуют хорошо выраженные поля — площадки. Площадки такой же величины, как у P. som- niferum, но семена мельче и сеть имеет меньше площадок, и это, вместе с мелкостью семян и 8-лучевым рыльцем, показывает, что ископаемый швейцарский мак не вполне сходен с современным культурным маком. Ископаемый мак по величине коробочек (длина — 12 мм, поперечник — 10 мм) сходен с Р. rhoeas, по форме коробочек с культурным — Р somniferum, по 8-лучевому рыльцу — с Р. setigerum DC. Таким образом, это был особенный мак, соединявшии признаки нескольких видов — интегральный вид. Этот-то теперь исчезнувший вид, очевидно, и был родона- чальником Р. somniferum и Р. setigerum. Р. setigerum многими считается родоначальником культур- ного мака — Р. somniferum (Р. setigerum растет дико в Испании, Алжире, Корсике, Сицилии, Греции и на острове Кипр). Однако Р. setigerum не может считаться предком Р somniferum, ибо Р. setigerum имеет число хромосом 2п =44, а Р somniferum- 2n = 22. Невероятно происхождение вида с меньшим числом хромосом от вида с числом хромосом вдвое большим. Вероятно, интегральный предок обоих видов сформировался южнее Швейцарии, где был край его ареала. Для разрешения вопроса нужно детальное исследование дикорастущих и сорных маков Средиземноморья. Г о р ч и ц а ч е р н а я (Brassica nigra Косй) в диком состоя- нии неизвестна, а в культуре имеет древнесредиземноморский ареал от Испании до Индии с захождением в Эфиопию. Исполь- зовалось в древности в медицине и как приправа. В средние века употреблялись в пищу и листья (в вареном виде). И н д а у (Егиса sativa) была распространена не только в Средиземноморье, но распространилась и в Среднюю Европу. Употреблялась как салат и в медицине. О з им а я с ур е пи ца (В. icampestris var. autumnalis DC.) культивируется только в ГДР и ФРГ и оттуда занесена 397
Б Польшу и Украину, где встречается в смеси с озимым рапсом и неправильно называется «мелкосемянным» рапсом. Озимая су- репица — теперь растение аборигенное для упомянутых стран; вероятно, первичная область ее происхождения приурочена и приморским областям (приатлантические и берега Северного моря) . Р еп а (Brassica гара 1.) имеет в Европе (ГДР, ФРГ, Фран- ция) особые эндемичные формы (тельтовская репа) примитив- ного характера с признаками дикорастущей конституции и с не- которыми чертами сходства с азиатскими примитивными репами (Афганистан). Нигде, кроме стран Средней Европы, не встре- чаются черные и серые репы, а также репы желтомясые. Группа желтомясых реп имеет родину на севере Русской рав- нины и в Финляндии. Разнообразие сортов репы в Западной, Средней и Северной Европе очень велико, и имеются все основа- ния предполагать, что здесь находится один из первичных оча- гов возникновения культуры репы (см. схему), которая, до вве- дения картофеля, имела очень большое значение в питании на- родных масс. Мы выделяем европейскую репу в особый вид — В. га- роеигареа Sinsk. Схема филогении культурной репы Д,икорастущие приморские азиатские тонко- корневые формы, близкие к В. гаро- 1 asiatica Репы индо-афган- ского подвида— ssp. indoafghanica дикорастущие малоазиатские при- морские тонкокор- невые формы дикорастущие европейскиесеверные тонкокорпевые1 формыРепы тельтовского типа—В. rapoeuropea ssp. teltovien-sis 1Репы малоазиатскогоподвида европейскоговида — В. rapoeuropeassp. antasiatica Репы петров- ского типа— ssp. rossica Сортотип чер- ных и серых реп — conc u1- ta t r is tie o1or ata астоящие европеиские репы — ssp. europea Репы японскогоподвида — ssp. japonica Схел~а филогении сурепииы и репы дикорастущие родоначальные формы, близкие к культурной озимой сурепице 1Культурная и сорнаяяровая сурепица 1Репа европейская 1Культурная озимаясурепица 398 Озимая сурепица и европейская репа связаны тесным род- ством (см. схему) .
диким предком и озимой сурепииы и европейских реп был, вероятно, общий, теперь исчезнувший предок, похожий на ози- мую сурепицу, но со склонностью к образованию зачаточного корнеплода. Предок озимой сурепицы и северных европейских реп был, вероятно, жителем прибрежий и островов Атлантиче- ского океана и Северного моря. Наши обычные корнеплодные растения — свекла, редис и отчасти морковь — имеют примор- ское происхождение. Б р ю к в а (Brassica napobrassica Mill.) не культивируется. на юге Европы, в Азии и Африке она неизвестна. Ьрюква- растение Средней и Северной Европы и из этих стран ведет свое происхождение. Вероятно, из Средней Европы происходят беломясые брюквы, а желтомясые — из Северной, точнее из Се- веро-Восточной Европы (Финляндия, Северо-Запад СССР). Вид возник, вероятно, как амфидиплоид в результате скрещивания озимой сурепицы — переходной формы к репе или древней формы репы с одной из форм капусты. Возможно, что это был дикорастущий вид теперь вымершей приатлантической капустьг из видов, близких к В. о1егасеа. Возможно, что переходным звеном к настоящей корнеплод- ной брюкве был лиственный рапс из подвида pabularia DC., среди популяций которого встречаются биотипы с утолщенным корнем. Зимостойкость лиственного рапса говорит о его северном, по нашему мнению, приатлантическом происхождении. По нашей теории, брюква и послужившие для ее образования исходные формы аборигенны для Северной Европы. Европейские страны — область развития сортового ассорти- мента европейского р е д и с а — Raphanus sativus, который ведет происхождение от средиземноморских прибрежных дикорасту- щих видов, но в Европе представлен многими эндемичными сор- тами. Только в Европе, например, встречаются редисы с жел- тыми и черными корнеплодами (в собственно Средиземноморье они не распространены). Европейские виды редьки и редиса приспособлены к усло- виям северного длинного дня, тогда как азиатские родственные виды в этих условиях стрелкуются, не дают хорошо развитых корнеплодов или развивают совсем тонкие корни. дикие их ро- доначальники распространены по морским побережьям, в том числе по берегам Атлантического океана и на севере по берегам Британских островов и Северного моря. Р ед ь к а (Raphanus sativus ssp. hybernus Alef.) — двулетняя форма европейского типа, которая должна быть выделена в са- мостоятельный вид; нигде, кроме Европы (включая и СССР), не встречается, отсюда и ведет свое происхождение, возможно, также от исчезнувших диких прибрежных форм северных морей. То же можно сказать и о сортах сахарной, столовой и кор- мовой с в е к л ы — Beta vulgaris. Родина ее и первичная область вхождения в культуру собственно Средиземноморье, но область 399
развития большого сортового ассортимента (столовая и кормо- вая свекла) — это Средняя и Северная Европа. Дикорастущий предок культурной свеклы — В. maritima, рас- пространенный по берегам Атлантического океана и Северного моря вплоть до берегов Дании и южной Швеции, образует осо- бый северный экотип. Возможно, что и эти северные дикорасту- щие формы послужили источником происхождения некоторых европейских культурных сортов. Листовая свекла в странах Передней Азии (Сирия, Пале- стина и др.), а также в Египте, Греции и Риме была в начале нашей эры широко распространена, но настоящая корнеплод- ная свекла, у которой мясистый корнеплод имеет правильную форму (и именно корнеплод, а не листья, используется в пищу), распространилась позднее. В средневековой Европе листовая свекла была еще широко распространена и как лечебное и как пищевое растение. Настоя- щие корнеплодные сорта усовершенствовались и постепенно вытесняли листовую свеклу, которая сделалась довольно редкой, почти любительской культурой. Европейская каротиновая м о р к о в ь (Daucus ~carota L. sensu lato) в основном средиземноморского происхождения, но ассор- тимент столовых и кормовых сортов получил особое развитие и в Средней Европе, где имеется много местных и селекционных сортов и где распространены дикорастущие формы. Остатки моркови найдены в неолите свайных построек Швейцарии (Ro- benhausen). Возможно, сначала использовали в пищу дико- растущую морковь, которая распространена в Европе повсе- местно. П а с т е р н а к (Pastinaca sativa L.) был известен жителям бронзового века Швейцарии, на что указывает находка этого растения в Maosseedorf, возможно, в этот период еще не было культурного пастернака, а использовалось дикорастущее расте- ние. Это растение входило в культуру не только в Средиземно- морье, но и севернее — в Средней Европе, где встречается и те- перь в диком состоянии. Еще в средние века пастернак пользо- взлся большой популярностью. Р а п с (Вгазйса naipus L.) — весьма полиморфное растение. Дикий предок неизвестен. Подобно брюкве, родственному корне- плодному виду, рапс — европейского происхождения. Возможно, что он возник в приатлантических странах из каких-то форм дикой капусты (2n = 18) путем удвоения хромосом или как амфндиплоид от скрещивания неизвестной формы капусты (2и = 18) с сурепицей (2n = 20) . Последний путь происхожде- ния вероятен для тех форм рапса, у которых число хромосом 2п = 38 (см. схему) . Наиболее древними формами, по нашему мнению, являются лиственные рапсы подвида раЬи1аг1а DC. Они очень холодо- стойки, возделываются преимущественно в Англии.
Схел~а происхождения рапса и брюквы приморской дикой капусты древнейшие приатлантические формы(2n = 18)тЛиственный раис подвида раЬи!аг!а (кори! 2n = 38)тРапс озимый с тонким корнем(2гг = 38 или 36)1Раис яровой (2n = 38 или 35) ! и часто со слабым утолщением,!Брюква беломясая (2n = 38)~Брюква желтомясая (2n = 38) Велая брюква часто имеет уродливые и твердые корнеплоды, 11з нее иногда выщепляются растения с тонким корнем, она бо- лее примитивна, чем брюква желтая. Ка пуста (Brassica о1егасеа sensu lato) как группа близ- ких видов происходит из Средиземноморья, но важная область развития культурного ассортимента кочанной, савойской и брюссельской капусты простирается на Среднюю и на Северную Европу. У кочанной капусты установлен особый подвид— ssp. еигараеа Lizg., включающий две группы сортов: западно- европейскую (Англия, Франция) и центральноевропейскую. Нам представляется, что брюссельская капуста и кольраби также произошли не в собственно Средиземноморье, а в Сред- ней и Северной Европе (Франция, Англия). Кочанная и савойская капуста, может быть, впервые появи- лась и в Средиземноморье, но это были примитивные рыхлоко- чанные формы. Крупно- и плотнокочанные формы выработались севернее — в Средней и Северной Европе. Основная область культуры белокочанной капусты — страны приатлантической Европы (Дания, Голландия, ГДР, ФРГ) и не- черноземная полоса СССР. Не исключено предположение, что эта группа сортов произошла от приатлантических, теперь вы- мерших форм дикорастущей капусты. Этот вид холодостоек и на юге не образует хороших плотных кочанов. 1~ветная капуста, безусловно, южного происхождения, более древними были формы подвида simplex Lizg. Позднее вырабо- тались более совершенные с точки зрения овощеводства формы цветной капусты подвида abortiva Lizg., и они сформировались севернее, а не в прибрежных странах Средиземноморья. К а п у с т а м о р с к а я (Crambe mar itima 1 .) дико растет по всем морским побережьям Западной Европы. Впервые начали ее культивировать в Англии. К а т р а н (Crambe tatarica Jacq.) в диком состоянии рас- пространен в Венгрии, Моравии, Валахии, в лесостепной и степ- ной полосе Русской равнины. Кювье и другие считают, что кат- ран был известен древним грекам и римлянам под названием chara, однако это требует уточнения. Во Франции это растение начали культивировать не так давно.
В неолите Швейцарии найдены семена т м и н а [Сагит carvi L. (gobenhausen)]. В дикорастущем состоянии тмин рас- пространен в странах Средней Европы. В средние века из тмина приготовляли дрожжи. О р и г а н (Origanum vulgare L.) — медонос, эфиронос, ле- карственное растение. В культуру вошел в Южной и Средней Европе, где имеется в дикорастущем состоянии. Возможно, что он самостоятельно вошел в культуру и в Передней Азии, и в соб- ственно Средиземноморье. Ареал рода Origanum L. средиземно- морский. Родиной м я т ы п е р е ч и о й (Mentha pi~perita L.) считается южная Англия, где известна с XVII столетия; была введена в культуру под названием белой и черной английской мяты. Брике считает, что перечная мята возникла в результате скре- щивания двух видов: М. spiicata 1 . р,' М. aqua tica I . Шургоф (Р. Н. Schurhoff) признает тригибридное происхожде- ние перечной мяты. М. silvestris (с 9 хромосомами), скрещенная с М. rotundifolia Huds. (с 27 хромосомами) дала н"~.àëo М. viri- dis (ic 18 хромосомами). Гибрид М. viridis р,' М. aquatica дал но- вый культурный вид — М. piperita (с18хромосомами). Боль|пин- ство форм перечной мяты стерильно, размножается черенкова- нием — это указывает на происхождение ее в условиях кульгуры. Мята курчавая — М. сг1эра L., вероятно, введена в культуру в Западной Европе. Возможно, гибридного происхождения. Мята длиннолистная — М. longifolia (L.) Huds. — пищевая приправа (для зеленого сыра). В Западной и Средней Европе имеется до 37 разновидностей. Там же, вероятно, и введена в культуру. Эфиронос. Мята колосовидная, или зеленая — М. s~picata L. (М. viri- dis L.) — эфиронос, введена в культуру в Европе. Мята круглолистная — М. rotundifolia (L.) Huds. — введена в культуру в Европе (Англия и Франция). Мята курчавовидная — М. icrispata Schrad. — введена в куль- туру в Западной Европе. Массовые находки семян м а р и б е л о й (Chenopodium al- Ьит 1 .) в свайных постройках Швейцарии (Pobenhausen) сви- детельствуют о том, что это растение не только собиралось из естественных зарослей, но и разводилось. Зимний салат «Витлуф», или салатный цико- рий (Cichorium inthybus L.) — в культуре имеются сорта этого вида, способные формировать в темноте крупные кочаны. Этц сорта выведены в Бельгии брюссельскими огородниками около 1876 г. из местных дикорастущих форм. Название «Витлуф» по- фламандски означает белый лист. При культуре в теплых поме- щениях (температура от 8 до 18') не требует света и дает про- дукт с октября по май. Содержит пиулин (рекомендуется диабе- тикам), глюкозу, полезные азотнокислые, сернокислые, соляно- кислые соли калия, интибин, оказывающие благоприятное 402
действие на кровеносную систему. Выращивается в некоторых хозяйствах под Москвой (Н. Г Василенко, 19бО). Медвежий лук, или черемш а (Allium ursinum L.)— западноевропейский вид, распространен по тенистым влажным лиственным лесам. Содержит витамины, служит противоцингот- ным средством. Молодые растения едят сырыми, жарят с яйцом, сушат на зиму (Г Л. Ремезова, 19бО). I(л у б н е н о с н ы е колокольчики, р а п у н ц е л ь (Сат- рапи1а raipunculoides 1. и некоторые другие виды) имеют сьедоб- ные корнеклубнеплоды, которые используются местным населе- нием. Имеются опыты введения в культуру. Виды щ а в е л я (Rumex acetosa L., R. montanus Desf. и scutatus L.) распространены по лугам Франции и других стран Средней Европы, особенно в горных местностях. Эти виды вводились и в культуру (первое упоминание относится к XII в.). По А. И. Филову (1960), о г у р е ц п о с е в н о й появился в Ев- ропе значительно позднее, чем считают многие авторы. Послед- ние часто смешивают древний огурец с «огуречноподобными» (по их использованию) растениями. А. И. Филов сомневается в существовании настоящего огурца в Древней Греции. Назва- ние «аджур» многие авторы принимают за название огурца, а по А. И. Филову, это название относится к дыне — Cucumis chate, которая была распространена и в Древнем Египте, где, по Филову, не знали настоящего огурца. Этот вид дыни и теперь культивируется в Малой Азии под названием аджур. Византийское название огурца Agguria или Aggurion, по А. И. Филову, не произошло от санскритского корня, как по- лагают некоторые исследователи. Огурец, возможно, был изве- стен в Византии в IV — V вв, н. э. Вполне же достоверные сведе- ния об огурце в Европе, по Филову, имеются только в VIII в. н. э. В это время огурец распространяется по разным странам Европы. 1( славянам он попал из Византии. А. И. Филов утверждает, что в свайных постройках Средней Европы не могло быть огурца, и относящиеся к этой эпохе све- дения об огурце ошибочны. В сравнительно позднее время в Европе выработались свои местные и селекционные сорта огурца. А. И. Филов (19бО) боль- шинство Нх относит к следующим разновидностям европейско- амерпканского подвида: 1) var. danicus Fil. (Дания, Франция, ГДР, ФРГ); 2) var. piculis Fil. (Западная Европа, Украина и Северный 1(авказ) . Из 1(итая занесены в Европу следующие разновидности, от- носящиеся к китайскому подвиду ssp. chinensis Fil. Из южной части 1(итая и Индии — var. anglicus Bailey, к ко- торой относятся тепличные сорта Чудо Прескотта, Ювель, Коп- ппц, Лондонский длинный зеленый и др.; в Германии выработа- лись сорта этого же подвида, относящиеся к var. germanicus Fil., — Немецкий длинный, Эрфуртский длинный и др. 403
~толовый а р б у з (Citrullus vulgaris), по А. И. Филову (1962), в страны Западной Европы был занесен в средние века из Византии — его первым названием было византийское слово ангурия. В европейскую часть Восточного Средиземноморья, на Балканы, арбуз пришел из Средней Азии. В европейских странах Восточного Средиземноморья, в Дрег- ней Греции были распространены змеевидные д ы н и, относя- щиеся к подвиду ssp. flexuo~us (L.) Greben. — тарра (var. tarra) и аджур — var. chate (Forsk.) Fil. Эги примитивные сорта яв- ляются предками настоящих сахаристых дынь и были еще в древности занесены в Восточное Средиземноморье из стран Передней Азии. Сорта настоящей культурной дыни были занесены в 'Европу много позднее, вероятно, также из Передней Азии (Анатолия). В Европе выработался свой подвид культурных дынь — ssp. euro- paeus Fil., куда относятся европейские летние дыни, европейские канталупы, роккифорды. Многие известные л е к а р с т в е н н ы е р а с т е н и я имеют европейское происхождение. К ним относятся: наперстянка крас- ная — Digitalis ригригеа L. (растет дико в Средней и приатлан- тической Европе, вошла в культуру во Франции), Sco~polia саг- пю1йса Ja~cq. (растет дико в Средней и Южной Еврспе, заходит в Предкавказье и европейские степи СССР) и др. Европейское происхождение имеет известное декоративное растение — л ь в и н ы й з е в (Antirrhinum majus L.). О в о щ н ы е р а с т е и и я, занесенные даже в сравнительно позднее время (томаты, перцы и др.), в настоящее время имеют европейские сорта, сильно отличающиеся от исходного мате- риала. В стоянках каменного и бронзового веков находили много косточек и семян дикорастущих древесных и кустарниковых по- род (лещины, дуба, яблони, груши, черемухи, бузины, различ- ных видов Ргипиз, винограда, малины и др.), некоторые из HiIx затем участвовали в создании европейских культурных сортов [яблоня — Malus з11vestris Mill., груша — Pirus communis 1 ., вишня — Cerasus fruticosa (Pall.) %огоп. С. prostrata Ser. и др.]. 1~ультурная яблоня — М. domesti~ca Borkh., по мнению ряда авторов, была занесена в Европу из Малой Азии, но, может быть, некоторые сорта, связанные происхождением с местными дикорастущими формами, и аборигенны. А. Н. Ипатьев (19бО) выделяет особую южноевропейскую группу сортов яблони (Ренеты, Кальвили, Бельфлеры и др.). Яблоне Сиверса — М. sieversii Roem. — родственны сорта Анис, Боровинка, Апорт, Малы, Орлеанский ренет и др.; яблоне ранней — М. ~praecox Borkh. — Белый налив, Грушовка московская, Белое астраханское; яблоне восточной — М. orien- talis Uglitzk. (кавказский вид) — Синап, Пепин, 1~альвиль, Роз- марин; это европейские сорта. 404
По Шевалье, в лесах Франции распространена дикая яб- лоня — Malus silvestris Mill., но сидровые сорта происходят не от нее, а от М. dasyphylia Borkh. Этот вид, по Шевалье, имеет «зиатское происхождение. По Ф. Д. Лихоносу, яблоня кавказская — М. orientalis— дала начало европейским сортам — Синапу, Пепину, 1(альвилю, Розмарину. Предки европейских культурных груш и яблонь распростра- нились из Восточной Азии по пути в Гималаи, Иран, Кавказ, Малую Азию и Кашмир, т. е. прежде в Восточное Средиземно- морье, везде уступая место порождаемым ими под действием иных условий новым видам. Культура яблони, груши и их сорта переходили из восточных стран в Европу. Но в Европе из при- шедших сюда с востока элементов сформировались свои дико- растущие виды, которые также приняли участие в формирова- нии культурного ассортимента. Эти дикие формы яблони и груши росли в лесах и маки Средней Европы, начиная от четвертичного периода. Во всяком случае они существовали в неолите, и плоды их служили пред- метом собирательства, о чем свидетельствуют многочисленные находки семечек в древних культурных слоях. По А. Шевалье (Chevalier, 1953), во Франции распростра- нены многочисленные формы Pirus icommunis L. Многие сорта груши в Бретани и Нормандии происходят от Р. cordata Desv. (очень близки к Р. communis). Некоторые французские сорта груши близки к Р. nivalis Jaicq. Один из подвидов этого вида — Р. salicifolia Pall. — рас- пространен во Франции, и к нему относятся некоторые высоко- культурные сорта. Дико во Франции растет Sorbus aria Crantz, которая рас- гространилась сюда из Азии через Восточную Европу. Из Во- сточной же Европы были занесены во Францию боярышники. Плоды боярышников — Crataegus oxya~cantha L. и С. monogyna Jaeq.— в довольно большом количестве находили местами при. раскопках неолитических стоянок. М у ш м у л а г е р м а н с к а я (Mespilus germanica 1 .)— переднеазиатский вид, аборигенный на Кавказе, в Иране и Гре- ции, был занесен во Францию, где в настоящее время имеются свои сорта этого плодового растения: крупноплодные и без ко- лючек. 3 е м л я н и к а в ы с о к а я (Fragariia elatior Ehrh.) распро- странена в диком виде почти по всей Европе, дала начало неко- торым культурным сортам (Миланская и Шпанка), которые имеются в СССР, встречаются в Италии, Франции и Англии. Европа — один из втори .ных центров развития рода Rubus, там имеется в диком состоянии большое разнообразие видов подрода Eubatus (ежевика). В культуру вышелевропейскийпод- вид м а л ин ы о бы кн о вен ной [R. idaeus звр. vulgatus Arrhen 405
(пз подрода Idaeobatus)], который является родичем многих культурных сортов. От европейского вича к р ы ж о в н и к а [Grossulria reclinata (L.) Mill.] п~роизошло очень много культурных сортов. Сборные виды с м ар один ы к р а си о й (Jibes rubrum) и ч ер н ой (Я. nigrurn) дико распространены почти по всей Европе, дали начало некоторым культурным сортам. В культурной флоре Средней и Северной Европы выявлены следующие географические элементы. 1. Переднеазиатский — из стран Передней Азии вышли пше- ница мягкая, ячмень, полба-эммер, полба-спельта, однозернянка, лен. Перечисленные выше растения (за исключением полбы- спельты) были занесены в Европу, вероятно, через Балканы, уже в неолите, и северная граница их культуры еще на заре земледе- лия доходила до Дании и южной Швеции включительно. Полба- спельта была занесена, вероятно, позднее. Еще недавно она счи- талась эндемом приальпийской области, но теперь ее древняя культура открыта в Иране, Азербайджане и Малой Азии, и стало ясно, что спельта — реликт, а не среднеевропейский эндем. 2. Восточноазиатский элемент сравнительно невелик. Сюда относится прежде всего просо, распространившееся в Европе в глубокой древности. Из 1(итая в Европу занесен, например, абрикос (через Армению). 3. Средиземноморский элемент включает растения, распро- странившиеся в Среднюю и Северную Европу из собственно Средиземноморья. Это артишок, лаванда и другие крупносемян- ные и крупноплодные сорта многих культурных растений. Наряду с растениями, занесенными в глубокой древности, несомненно, в культурной флоре Средней и Северной Европы аборигенный элемент представлен значительно большим коли- чеством видов и форм, чем ожидали сначала. Овощные растения занимают особенно заметное место среди культурных аборигенов Средней и Северной Европы. Из них отметим следующие: хрен — вошел в культуру в Северо-Восточ- ной Европе, озимая сурепица (почти реликт, мало распростра- нена), репа, европейские виды редьки и редиса, брюква, рапс, свекла (столовая и кормовая), пастернак, морковь. Таким образом, в Европе находится значительный очаг про- исхождения корнеплодов и родственных масличных растений (рапс, сурепица озимая). По нашему мнению, в Северной Европе произошли желтая брюква (а может быть, и вообще брюква), желтомясые русские репы и отсюда ведет начало (от тельтовской репы) почти вся европейская группа местных сортов беломясых реп. Здесь воз- никла и наиболее примитивная группа сортов рапса (листвен- ный рапс). Существует и северный приатлантический подвид дикорастущей свеклы, который, вероятно, принимал участие 1; формообразовании культурной европейской свеклы. Настоя- 406
щие двулетние формы редьки также по морфологическим при- знакам близки к северным экотипам других корнеплодных ра- стений и, по нашему мнению, произошли из стран Северной и Средней Европы с умеренным климатом. Сюда же относится хрен. Таким образом, Северо-Восточная и отчасти Средняя Ев- ропа (вместе с нечерноземной частью Русской равнины) яв- ляется одним из очагов первичного вхождения в культуру (во всяком случае, формообразования) многих овощных растений. Европейское происхождение имеют и следующие овощные растения: капуста кочанная, савойская, брюссельская, кольраби, катран морской, катран татарский, европейская черемша, цикор- ный салат «Витлуф» (распространен в Бельгии), некоторые виды клубненосных колокольчиков, виды щавеля. В итоге можно сде- лать вывод — в Средней и Северной Европе сформировался довольно большой ассортимент овощных растений. Из кормовых аборигенных европейских растений прежде ьсего следует назвать культурный эспарцет посевной, клевер розовый (шведский), клевер венгерский, тимофеевку луговую (вошла в культуру на территории Русской равнины), ежу Ашер- сона, райграс многолетний, райграс многоукосный и костер пря- мой. Из группы эфироносов и лекарственных растений в первую очередь следует отметить мяту, наперстянку лекарственную; из декоративных — львиный зев, нарцисс белый и др.; из дикора- стущих плодовых — яблони, груши, крыжовник, малину. Мы составили довольно подробный обзор культурной флоры Средней и Северной Европы, так как в нашей специальной ли- тературе до настоящего времени этого сделано не было. Оказа- лось, что Средняя и Северная Европа имела свои пути развития культурной флоры. Нельзя, конечно, отрицать, что Средиземно- морье оказало большое влияние на состав культурных растений всей Европы, но это влияние, с одной стороны, очень неравно- мерно сказалось на отдельных странах и было сильнее на за- паде (Франция, север Испании), чем в центре (Германия, Австрия, Венгрия, Швейцария) и на севере (Скандинавия). С другой стороны, культурный ассортимент Европы сложился под очень сильным влиянием земледельческой культуры Перед- ней Азии (связующей областью были Балканы). Средиземноморские (и позднее американские) овощные ра- стения в странах Средней и отчасти Северной Европы дали на- чало богатому ассортименту выработавшихся на месте европей- ских сортов. Занесенные элементы, особенно пришедшие с востока, до- стигли стран Средней и Северной Европы в глубочайшей древ- ности, поэтому они представлены в раскопках каменного и брон- зового веков примитивными формами и видами, которые некогда были широко распространены, а теперь в Европе исчезли. 407
Все привнесенные в Среднюю и Северную Европу элементы в новых условиях развили формообразование разного мас- штаба — от новых форм и сортов до вида. Таким образом, не- смотря на небольшой сравнительно со Средиземноморьем ви- довой состав, во флоре Средней Европы имеются реликты и сравнительно молодые эндемики. Некоторые культуры, хотя и не имеют в настоящее время в европейской флоре дикорастущих родичей, все же несомненно здесь вошли в культуру, т. е. для них Европа — первичный очаг. К таковым относятся брюква и европейская репа целиком как виды "". Характерны эндемичные репы типа тельтовской и пет- ровской, озимая сурепица, европейские редисы и редька, лист- венный и масличный рапс, может быть, и белокочанная капуста. Некоторые европейские культурные растения имеют в абори- генной дикой флоре своих непосредственных родоначальников. Из таковых наибольшее значение и распространение получил эспарцет виколистный — Onobrychis viciaefolia. История земледелия и культурных растений Средней и Се- верной Европы интересна тем, что в ней вскрываются, с одной стороны, древние связи с Закавказьем — Колхидой, а с дру- гой — более новые связи со славянским миром. Рожь и овес являются такими растениями, которые получили развитие и вошли в культуру у славянских племен, живших на территории СССР и на соседних территориях Западной Ев- ропы. На древней земледельческой культуре Европы следует оста- новить внимание еще и потому, что некоторые западноевропей- ские исследователи преувеличивают значение Западной Европы и, в частности, приальпийской области в развитии мирового земледелия и мирового ассортимента культурных растений (Bertsch, 1939 и др.). Спельта считается некоторыми зарубеж- ными авторами тем древним видом, от которого в приальпий- ской области произошли европейские мягкие пшеницы. Культур- ная однозернянка выводится из местных дикорастущих форм, которые будто бы раньше были более широко распространены к северу, чем в настоящее время. Берч (1939) говорит о самостоятельности и туземности древнегерманской земледельческой культуры в приальпийской области и о том, что больше всего реликтовых культурных ра- стений еще недавно можно было встретить в Швабии, Баварии, Вюртемберге и прилегающих районах. Земледелие там было, ве- роятно, аборигенным, т. е. с самого начала развивалось на базе культуры местных, аборигенных народностей, но все же оно с самого начала развивалось в основном на заносных (главным образом с востока) культурных растениях. ' По нашему мненшо, брюква составляет самостоятельный вид, который .не следует объед1~нять с рапсом.
ДРЕВНЕЕ ЗЕМЛЕДЕЛИЕ И АБОРИГЕН НЫЕ КУЛЬТУРНЫЕРАСТЕНИЯ НОВОГО СВЕТА Данные современной науки приводят к заключению, что че- ловек (протоамериканец) пришел в Америку из Азии северным путем, через Аляску, сначала в Северную Америку, а оттуда позднее — в Е~ентральную и Южную. Существуют еще три теории о появлении человека в Новом Свете. Одна из них предполагает существование исчезнувшего материка — Атландиты, откуда будто бы и явились первые оби- татели Америки. другая считает человека в Америке автохтонным. Третья предполагает расселение человека в Америку через Антарктиду. Вторая и третья теории в настоящее время почти не имеют сторонников. Не может иметь близкого отношения к данному вопросу тео- рия Вегенера о расхождении некогда соединенных материков, ибо это расхождение могло иметь место только очень давно, до появления на земле человека. Против этой теории теперь вы- двинуты веские возражения. Что касается существования Атлантиды, т. е. огромного острова, погрузившегося в Атлантический океан, то в последние годы эта теория получила некоторое обоснование. Обнаружено существование погрузившегося атлантического хребта. Когда образовался этот погрузившийся хребет, судить трудно, воз- можно, в палеозойскую эру, но переселение растений по атлан- тическому хребту было возможно и в четвертичный период (Л. С. Берг, 1949). Эти открытия могут многое разъяснить в географии дико- растущей флоры Нового Света, но в вопросе происхождения культурных растений Америки существование Атлантиды ничего не объясняет, так как древнее земледелие Америки основано почти исключительно на аборигенных растениях, взятых из местной растительности. Человек не перенес из Азии в Америку никаких культурных растений, а из домашних животных он захватил с собой только собаку. Переселение человека в Америку произошло в конце ледни- ковой эпохи, около 10 000 — 25 000 лет тому назад. 1(огда человек
переселился в Америку, он еще застал там ныне вымерших жи- вотных, в том числе мамонта. Древность человека в Америке не столь велика, как древность его в Старом Свете. Шелльская, ашельская п мустьерская культуры в Америке неизвестны. Древняя американская фолсомская культура напо- минает солютрейскую. Уисслер (Wissler, 1922) различает в Америке в период ее открытия следующие области. 1. Область Великих озер. Основные занятия жителей — охота на оленей и других животных, собирательство (ягоды, корни, плоды и т. п.). Здесь, к югу от озера Онтарио и реки Святого Лаврентия, ко времени прихода европейцев в дремучих лесах обитали полуоседлые племена, имеющие общее название ироке- зов. Домашних животных, кроме собаки, у ирокезов не было. Земледелие все же было им известно, но носило подсобный ха- рактер; занимались им женщины. Система земледелия была подсечная, вернее, система выжигания, ибо лес не срубали, а выжигали. Участки, удобренные золой сгоревшего леса, при- митивно обрабатывали палками и мотыгами, и здесь разводили кукурузу, фасоль, тыкву, подсолнечник, табак. Когда почва истощалась, участок забрасывали и выжигали в лесу другие площадки, куда переносили и свои легкие жилища, сделанные из древесной коры (Бломквист, 1951). Земледелию будто бы научил ирокезов их полулегендарный вождь Гайавата (живший около 1570 г. н. э.). 2. Область степей Северной Америки, где занимались также охотой и собирательством. 3. Северо-западная область к северу от Калифорнии. Основ- ное занятие рыболовство (сельди, лососи и др.), а также охота и собирательство (ягоды, грибы, орехи и т. п.). 4. Область к востоку от Миссисипи. Охота (олени, бизоны, индейки и др.) и собирательство, главным образом, использо- вание зарослей дикорастущего риса (Zizania aquatica L.), топи- намбура и подсолнечника были здесь основным источником су- ществования человека. 5. 1(алифорнийско-Техасская область. Источником питания являлся сбор дикорастущих плодов, корней и луковиц, отчасти рыболовство; использовались в пищу черепахи, змеи, насекомые, муравьи, термиты. б. Земледельческая субтропическая и тропическая область горной Мексики и Центральной Америки. Основной источник пи- тания — продукты земледелия, в основном кукуруза, а также батат, тыква и другие растения. 7. Вест-Индия. Основное занятие — охота и рыболовство, сбор моллюсков и только отчасти земледелие. 8. Тропическая Южная Америка бассейна реки Амазонки. Наряду с охотой и собирательством (дикпе плодовые и др.)
vìååò значение земледелие..Это была область возделывания ма- ниока — Manihot aiipi Ро111 и М. utilissima Pohl, земляного ореха — Arachis hyipogaea 1 ., канны — Canna edulis Ker-Gawl., ананаса и др. 9. Высокогорные районы Перу и Бразилии. Земледелие— основной источник пропитания. Область культуры картофеля и лебеды (киноа и каньява). 10. Остров Чилоэ и южные районы Чили (Араукания). Ос- новной источник пропитания — земледелие и собирательство. 1(ультура картофеля, вымершего теперь костра — Bromus mango Desv. и сбор семян араукарии и др. 11. Область Чака (Gran Chako) и прилегающие районы 10жной Америки к востоку от Аидов. Основное занятие — охота и собирательство (сбор диких плодов, ягод, орехов). Характе- рен напиток, приготовляемый из экстракта листьев дикого па- рагвайского чая (Ilex paraguariensis St. Hil.). Охота на гуанако. 12. Огненная Земля и примыкающие берега Тихого океана. Основные источники пропитания — рыболовство, сбор моллюс- ков, морских ежей и водорослей. С приходом европейцев аборигенные обитатели были разо- рены и большей частью вытеснены со своих исконных земель в районы малообитаемые и тяжелые для освоения. 1(ак пример, можно привести историю племен гуаяков (Н. Г Шпринцин, 1952), которые заселяли некогда значитель- ную часть лесистого плоскогорья восточного Парагвая в между- речье Параны и Мондая. Гуаяки были вытеснены из своей страны в так называемый Великий лес — тропические трудно- проходимые джунгли. В условиях постоянного преследования и систематического истребления их материальная и духовная куль- тура не могла подняться выше примитивного уровня. Они бро- дят с места на место в поисках пищи. Земледелия они не знают. Главное занятие — охота и собирательство. До сих пор бытуют каменные орудия: топоры, ножи, имеются луки и стрелы, для обтесывания употребляются раковины. Из верхушки ствола пал~мы пиндо они добывают сердцевину — «пальмовую ка- пусту», которую едят в сыром или жареном виде; из ствола этой пальмы приготовляется древесная мука. Из этой муки, замеши- вая ее на воде, изготовляют лепешки, которые едят сырыми или подсушивают их на костре. Из сердцевины пальмы пиндо также выжимают сладкий ароматный сок, служащий напитком. Собирательством занимаются главным образом женщины. С культурными растениями гуаяки познакомились поздно, и для них в языке гуаяков существуют только описательные назва- ния. Например, батат называется «корень, который кушается», маниок — «чуждая пища», «круглая пища», а бык получил длин- ное название: «тот, который имеет пару рогов и хорош для еды». Таким образом, из-за поистине варварского поведения «куль- турных» завоевателей Америки в ней еще сохранились отдален-
pb~e уголки, где можно наблюдать первобытный образ жизни- без земледелия — эпоху собирательства, через которую челове- чество в целом прошло в глубочайшей древности. 1(огда же европейцы появились в Америке, то они там вошли в соприкосновение не только с первобытными охотничьими и рыболовческими племенами, но и с племенами из крупных оча- гов древнего туземного земледелия, основанного на абориген- Hblx культурных растениях. После испанского завоевания большая часть литературных памятников индейской культуры была уничтожена. Из сохранив- шихся памятников замечательно произведение «Пополь Вух», содержащее предания и мифы, по которым можно судить о древ- ности земледелия у индейцев. 1(рупиейшим очагом древнего земледелия является Цен- тральная Америка (Гватемала, Гондурас, Сальвадор, Никара- гуа, Коста-Рика и Панама), включая Мексику. По Вавилову, большая часть Юкатана не входит в этот очаг эндемичной куль- турной флоры. Эти страны отличаются разнообразием экологических усло- вий: на уровне моря здесь расположены местности с влажным тропическим климатом, в горах климат суровый, и земледель- ческая культура поднимается здесь вверх до 3200 м над ур, м. Разные варианты сухого климата имеются в горных и равнинных полупустынях и пустынях. дикорастущая флора чрезвычайно богата, особенно в юж- ной Мексике, хотя еше сравнительно слабо изучена. Хемсли (W. Hemsley, 1879 — 1888) дает следующие сравни- тельные цифры видов цветковых растений: Северная Америка (Аляска, Канада, США) — 9403 вида; Центральная Америка и Мексика — 11 б2б видов (в том числе около 70~/о эндемичных). Земледелие стран Центральной Америки в основном неполив- ное, а в древности было исключительно богарным. Развитие земледелия и очаги возникновения и распростра- нения культурных растений в Америке, как и в Европе, при- урочены к субтропическим и тропическим горным зонам и сухим предгорьям и плато. Влажные лесные области тропиков мало- благоприятны для развития земледельческой культуры. Леса тропиков, хотя очень богаты видами, но это виды деревьев, важнейшие же культурные растения представляют собой травы. Богатство дикорастущей флоры такими видами, которые были пригодны для введения в культуру, способствовало раз- витию древних цивилизаций Центральной Америки и Мексики. Это была старотольтекская цивилизация северной Мексики, впо- следствии перешедшая к ацтекам, а в Центральной Америке- к народам майя. Эти цивилизации, несомненно, моложе древнейших цивилиза- ций Евразии. Хотя, по некоторым авторам, начало хронологии народов майя и тольтеков и относится к 111 — II тысячелетиям 412
до н. э., но главнейшие памятники этих культур датируются первыми веками новой эры, веками, которые были эпохой рас- цвета древних цивилизаций Центральной Америки. Знаменитые пирамиды вблизи Мехико были воздвигнуты около 300 г. н. э. Земледелие в этих странах, однако, возникло гораздо раньше, чем развились упомянутые цивилизации. Американские архео- логи указывают VI — V тысячелетия до н. э. как дату возникно- вения здесь земледелия, но, по ряду соображений, древность его может быть и значительно большая. Древние цивилизации Мексики и Центральной Америки до- стигли высокого уровня. В I в. до н. э. у майя была подсечная система земледелия. Основным орудием обработки почвы слу- жила заостренная деревянная палка. Тем не менее, благодаря олагоприягным климатическим и почвенным условиям получали высокие урожаи. Урожаи кукурузы снимали 4 раза в год. У народов майя существовала оригинальная письменность, наука. Имелись астрономические обсерватории. Искусство до- стигло высокой ступени развития. Народ этот был миролюби- вый, не знавший войн. Причины гибели культуры майя до сих пор недостаточно ясны. Одной из основных причин было ско- рее всего вторжение ицев-тольтеков с их высокоразвитой куль- турой. Тольтекская цивилизация в свою очередь перешла к ацте- кам. В настоящее время остатки племен майя сохранились в не- которых штатах Мексики (Юкатан, Чиапас и др.), в северной Гватемале и Британском Гондурасе. Огромные культурные достижения народов Мексики и U«- тральной Америки тем более удивительны, что они не знали железа и даже бронзы, хотя золото было в большом ходу. Не было у этих народов и домашних животных, кроме собаки, индейки и мускусной утки. Разводили съедобных муравьев. Из- вестны были пчелы и домашние осы. В Мексике разводилась также кошенильная тля для приготовления краски. Земледель- ческие орудия были примитивные. Землю обрабать.вали камен- ными мотыгами и своеобразными лопатами — медными на дере- вянной рукоятке и деревянными. Орошение начало применяться с древнего времени. Известны своеобразные «плавучие сады» — чинампа. Это были плоты из во- дяных растений, скрепленных древесными ветками. На эти пла- вучие островки насыпали почву и разводили кукурузу, фасоль и другие растения. Иногда здесь даже располагались хижины. Набор культур в Мексике и Центральной Америке был очень большой и своеобразный, Сведения, содержащиеся в «Пополь Вух», говорят о большом значении многих дикорастущих и воз- делываемых растений в обиходе древних жителей Центральной Америки. В «Пополь Вух» имеются упоминания о бобах, какао, тыкве (многократно), перце, табаке. Кукуруза являлась (и яв- ляется) важнейшей культурой этих стран. Она бь1ла основой жизни народа майя. В Мексике боги кукурузы (маиса) не менее
почитались, чем боги дождя или войны. Многие исследователи считают мексикано-центральноамериканский очаг родиной куку- рузы или трех ее групп: indurata Sturt., indentata Sturt. и everta S turt. Некоторые исследователи считают группу ату1асеа Sturt. более древней (Н. Н. 1(улешов, 1955), что далеко не доказано. Если это так, то тогда кукурузу можно выводить и из Южной Америки. Однако распространение ближайшего дикого родича кукурузы (не являющегося непосредственным ее родоначальни- ком) — теосинте — говорит о том, что кукуруза скорее происхо- дит из Мексики и Центральной Америки. Интересны археологические данные, касающиеся происхо- ждения кукурузы. Раскопками, проведенными в 1948 г. в Новой Мексике в покинутой пещере (Carton Gountry), были вскрыты слои отбросов в 6 футов в глубину. Эти слои датируются от 2500 г. до н. э. и до 500 — 1000 гг. н. э. Большинство образцов кукурузы хорошо сохранилось— уцелели даже нежные волоски на эпидермисе чешуй. Наблюда- лась эволюционная последовательность в характере початков из разных слоев. Початки из нижних древнейших слоев мелкие, гибкие, с вы- пуклыми пленками, зерна мелкие. От слоя к слою кверху по- чатки и зерна становились все крупнее, однако мелкие зерна встречались и в верхних слоях, но в меньшем количестве, и все же они оказались крупнее, чем в нижнем слое, где были только примитивные мелкозерные и мелкопочатковые формы. Форма зерен также изменяется, кверху они становятся шире. Таким образом, нижние слои были более однородными, а в верхних имело место значительное варьирование. Самое интересное, что в нижнем слое имелось до 20о/о настоящей пленчатой куку- рузы — Zea mays tuni~cata Sturt., в остальных слоях содержа- лись голозерные и со слабыми следами пленчатости формы. Эти данные рисуют процесс постепенного улучшения, окуль- туривания кукурузы, подтверждают ее аборигенное происхо- ждение, говорят против теории происхождения культурной ку- курузы от дикого родича — теосинте, так как растений при- знаками теосинте в нижних слоях здесь найдено не было. Согласно археологическим данным, древнейшая примитивная кукуруза была лопающейся (Z. т. everta), пленчатой и сравни- тельно с современной — мелкопочатковои '" ~ Недавно в районе реки Р»о-Гранде в Мексике археологи и биологи об- наружили в земной толще около 750 початков кукурузы, при этом установ- лено, что в различных геологических cJIQHx найдены разные сорта. В слоях, относящихся к глубокой древности, початки очень маленьких размеров; в слоях, относящихся к недавнему геологическому периоду, они значительно крупнее. С помощью радиоактивных изотопов установлено, что наиболее мо- лодые початки имеют возраст 1700 лет, наиболее древние — по крайней мере 3500 лет (см. «За рубежом», № 3, 1968). 414
Нужны разведки в еще более древних слоях, чтобы оконча- тельно выяснить вопрос. Более молодые слои ничего не дают. В раскопках, сделанных в Аризоне в каньоне Муэрто (F. Ander- son and F. Blanchard, 1942), найдена кукуруза, датируемая 500 — 700 гг. и. э. Эта ископаемая кукуруза очень похо>ка на ту, которую и ныне возделывает индейское племя папаго и род- ственные ему племена. По П. М. Жуковскому (1959), в самой столице Мексики об- наружена пыльца кукурузы, возраст которой исчисляется в 60000 лет. В это время земледелия в Мексике еще не было, II пыльца могла принадлежать только дикорастущей кукурузе. Наиболее древние формы кукурузы относятся к подвиду лопаю- щейся кукурузы — ssp. everta. При расселении на юг племен из центральной Америки вместе с ними в Мексику были занесены иноземные расы куку- pY3bI, и в дальнейшей истории имела место гибридизация с этими расами и с теосинте. Влияние теосинте- замечается только в самых верхних слоях. Эти данные приводятся довольно подробно как замечатель- ный пример, иллюстрирующий, какое значение может иметь археология в решении вопроса о происхождении культурных ра- стений. И. В. 1(ожухов установил следующие подвиды кукурузы: 1) ssp. montano-mexicana (Zea hirta Bonaf.) ет. Kozh.; 2) ssp. mexicana Kozh.; 3) ssp. peruviana (Wittm.) Kozh.; 4) ssp. arizo- nica Какй.; 5) ssp. everta (Sturt. repro parte) em. КокИ.; 6) ss~p. еигореа (Аl. pro parte) em. Kozh. 7) зэр. argentinica Kozh.; 8) ssip. indentata (Sturt.,pro ~parte) ет. Kozh.; 9) вар. asiatica I(ozh. Все подвиды И. В. 1(ожухова, за исключением европейского и азиатского, американского происхождения. Согласно исследованиям Уэлхаузена (Е. Ше11Ьаизеп, 1952) и Мангельсдорфа (P. Mangelsdorf, 1952), в Мексике сохрани- лись до настоящего времени 4 древнейшие расы лопающейся кукурузы, 4 доколумбийские экзотические расы, проникшие в Мексику с юга, и 13 доисторических метисных рас. Предполагается, что доколумбийские экзотические расы были привезены из Центральной или из Южной Америки — все они имеют своих представителей в Южной Америке. Согласно теории Мангельсдорфа, спонтанные скрещивания с теосинте — Eu~chlaena тех1сапа Schrad. — в той или иной сте- пени оказали влияние на все современные расы кукурузы, а сама теосинте возникла от скрещивания кукурузы с Tripsa- сит L. По новейшим литературным данным (Дао Тхе Туан, 1960; Я. Porteres, 1956 и др.), в Центральной Америке (1(уба) рас- пространен родоначальник культурных видов риса — Oryza pe- rennis Моепс1т. Этот вид представлен в Америке несколькими
экотипами. К нему присоединен в последнее время О. cubensis Ekman., распространенный на Кубе (2n =24). На зерно возделывается в Центральной Америке также хуаутли — Amaranthus pani~culatus L. var. leucocarpus Saif.— древнее эндемичное растение Мексиканского плоскогорья, ко- торое имело большое значение в культуре ацтеков как пищевое и ритуальное. Это растение, по мнению С. М. Букасова, древнее кукурузы. С. М. Букасов (1930) приводит мнение С. В. Юзепчука, что хуаутли характерно для Мексики как родины многих культур- ных растений. Из семян этого растения у ацтеков «...приготовля- лись изображения богов с глазами из фасоли и зубами из семян тыквы. Этим идолам приносились в жертву ладан, цветы, пульке и другие приношения. После церемоний пульке выпивали, съедобные жертвоприношения съедали. Поедали и самого идола, разламывая его на части. Больные хранили эти частицы для облегчения страданий». «Заслуживает внимания, — далее пишет С. М. Букасов, — что хуаутли сохранилось в северо-западной части Мексики (Сонора), откуда пришли и ацтеки. Самое сочетание в жертвенном идоле из хуаутли его с фасолью и тыквой наводит на мысль, что от- сутствие в этом сочетании кукурузы не случайно и в религиоз- ной церемонии сохранились наиболее древние растения Мекси- канского плоскогорья или иного исходного центра возникнове- ния ацтекской культуры». Эндемичными для этой области культурами являются также шпинатные и хлебные виды мари — Chenopodium nuttaliae Saff. и Ch. ambrosioides L. Эти растения также являются пережит- ками древнеацтекской культуры. Происхождение важнейших американских видов фасоли также связано с этим очагом. Имеются в виду фасоль много- иветковая — Phaseolus multiflorus Willd., фасоль лима — Ph. 1и- natus L., фасоль тепари — Ph. acutifolius А. Gray, Ph. retusus Ben th. Ph. multiflorus и Ph. luratus встречаются в диком состоянии. Существует, однако, указание, что фасоль тепари введена в культуру в Аризоне и Техасе. По П. М. Жуковскому (1959), лимская фасоль происходит из Перу. Что касается происхождения обыкновенной фасоли — Ph. vul- garis, то до сих пор считали ее родоначальником дикорастущий горный вид из Гватемалы — Ph. ma~crolepis Pip., однако и в Юж- ной Америке имеются дикорастущие виды, очень близкие и Ph. vulgaris. Н. P Иванов (1960) детально показал ход эволюции фасоли обыкновенной от многолетних лиан до однолетних кустовых форм. Совокупность фактов говорит за центральноамериканское происхождение фасоли обыкновенной. 416
Мексикано-центральноамериканский очаг считается и родиной среднеамериканского хлопчатника упланда — Gossyipium hirsu- tum L. История хлопчатника связана с цивилизацией майя. ~Лмеются здесь и дикие виды хлопчатника. По Ф. М. Мауеру, Мексике и Центральной Америке, а также Антильским островам и Колумбии, кроме G. hirsutum, свой- ственны следующие виды хлопчатника: 1) G. davidsonii 1(е11. (западная Мексика, нижняя Калифорния); 2) G. trilobuni Skov- sted (Мексика и Аризона); 3) G. aridum Skovsted (западная Мексика); 4) G. tricuspidatum Lam. (Центральная Америка, Вест-Индия, Бразилия). G. hirsutum представлен следующими подвидами: 1) дикие долговечные растения — ssp. maxiicanum Маиег (Центральная Америка); 2) формы многолетней культуры и рудеральные— ssp. punctatum Маиег (Центральная Америка, занесены в тро- пики Африки и Азии); 3) тропические формы однолетней и мало- летнеи культуры — звр. paniculatum Маиег (тропики Америки, Африки и Азии); 4) однолетние субтропические формы (упланд) — ssp. euhirsutum Маиег (субтропические и умеренные зоны земного шара) . G. tricusipidatum Lam. имеет следующие подвиды: 1) долго- вечные дикие формы — звр. rupestre Маиег (Колумбия, Антиль- ские острова); 2) формы многолетней культуры и рудераль- ные — ssp. purpurascens Маиег (Панама, Вест-Индия, 1(олум- бия); 3) менее долговечные тропические формы многолетней и малолетней культуры — sstp. glabrum Маиег (Вест-Индия, Ко- лумбия). На этих двух видах хорошо иллюстрируется вся эволюция хлопчатника: от многолетних — к малолетним и однолетним скороспелым формам. В настоящее время укрепилось мнение, что тетраплоидные виды хлопчатников Нового Света — G. hir- sutum, G. barbadense и G. tomentosum Nutt. ex Seem. — про- изошли в результате скрещивания старосветского диплоидного вида G. herbaceum c каким-то диплоидным американским хлоп- чатником. При этом в скрещивании участвовал не «зиатский, а африканский G. herbaiceum. Стеббинс (G. Stebbins, 1947) считает, что возникновение те- траплоидных хлопчатников в Северной Америке имело место в ранний третичный период — в это время популяции египетского хлопчатника входили в соприкосновение с американскими, что сделало возможным переопыление. Из эндемичных прядильных растений в Центральной Аме- рике культивируются агава-хенекен — Agave sisalana Perrine, агава икстли — Agave ixtli I(arw. — и лечигилья — Agave lechy- gilla Тогг. Из одного вида агавы — Agave atrovirens I(arw.— изготовляется любимый туземцами, известный с доисторических времен напиток пульке. Мексиканское нагорье считается цен- тром происхождения родэ Agave, а также рода юкка — Yuctca 1. 14 синская Е. н. 417
Yuoca baocata Torrey дает съедобные плоды. Юкка и другое ли- лейное растение — S amuel а Trel. — также используются как пря- дильные. Здесь же находится родина многих видов кактусов, некоторые (из рода Cereus Mill.) служат в качестве изгородей. 1(роме того, плоды кактусов из рода Opuntia Mill. под названием туна употребляются в пищу, разводигся также кошенильпый кактус — Nopalea coccineliiiera Salm-Рус1~. Особенно много кактусов в наиболее сухих районах тропиче- ской зоны. В Мексике насчитывается около 400 видов кактуса и около 150 видов агав. В Мексике растут злаки, идущие на щетки: Epicampes тасгоига Benth. и E. stricta Presl. (Мексика и Гватемала) . Таким же образом используется Agrostis tolu~censis Н. В. 1(. (Мексика) . Из Центральноамериканского очага, по Н. А. Базилевской (1960), вышло 547 видов декоративных растений. Здесь нахо- дится родина канн — Canna L., георгинов — Dahlia Cav., шал- фея — Salvia splendens Ker-Gawl., вьюнка — I~pomoea L., космеи— < osmea bipinnaia %111Й., бархатцев — Tagetes L., многих видов кактуса, орхидей из рода ОпсЫшт Sw., циннии — Zinnia L. 1( мексиканским эндемичным видам относятся: тубероза— Polyanthes tuberosa L., Dahlia variabilis Desf., Lobe ia fulgens Willd., Маигапйа barclaiana Lindl., Begonia gracilis Н. В. K., E~cheveria DC. и другие виды, устойчивые к низким температурам (Heuchera sanguinea Fngelm., Pentstemon campanulatus Willd., P. gentianoides Lindl. и др.). В Мексике растет дико пуансеттия — Euphorbia pulcherïmà (традиционный рождественский цветок в США). Из Гватемалы происходит известное декоративное растение Monstera deliciosa Liebm. (Philodendron ~pertusum) из сем. ароидных, которое на родине приносит сьедобные плоды с запа- хом ананаса. Цветы в изобилии разводили в Древней Мек- сике, HMH украшали храмы и жилища. Существовало древ- нее слово Xochitl, обозначавшее «цвет» (в декоративном значе- нии). Все виды тыквы американского происхождения. Cucurbita реро L. распространена от Канады до Гватемалы. В Южной Америке, по А. И. Филову, нет местных форм этого вида, но они имеются в Мексике. С. moschata Риса. — вид центральноамери- канского происхождения с многочисленными формами от Мек- сики до 1(олумбии и Венесуэлы. Горные ее формы — С. mixta Pang. — заходят в горы до 2600 м над ур. м. С. ficifolia Bouche— вид полудикий, короткого дня, по мнению большинства иссле- дователей, не является близким родственником культурных ви- дов настоящей тыквы. Этот вид распространен от Калифорнии до Аризоны, в 1(олумбии, Венесуэле, Эквадоре, Перу и Чили. Трудно сказать, где находится первичный очаг возникновения этого вида, возможно, что в Центральной Америке. 418
Археологические остатки тыквы были найдены в Гватемале в Петене (P. Vestal, 1938). Здесь были обнаружены плодоножки вида тыквы С. moschata. Остатки стеблей и семян видов С. реро и С. moschata, датируемые раньше чем 1000 лет до н. э., были обнаружены археологами в Пуэбло в штате 1(олорадо (А. Т. Er- win and Е. S. Haber, 1949; Т. Whitakeã and G. Carter, 1946). Семена С. moschata были найдены на юго-западе США в слоях, относящихся к 2000 — 1500 гг. до н. э. Стебли, кора и семена С. реро обнаружены в Мексике в Тамаулипас. Они датируются примерно 3000 г. до н. э. Эта одна из древнейших находок тыкв свидетельствует о большой древности культуры тыквы в Север- ной Америке. По данным Уайтекера и Бона (Т. Whitaker and G. Bohn, 1950), в начале второй половины ХХ в. археологи находили в Се- верной Америке тыкву: С. реро — 8 раз, С. mosichata — 4 раза. В 1Ожной Америке при раскопках тыква встречается реже, чем в Северной. Наличие индейских названий указывает также на древность культуры тыквы в Америке. Название тыквы — scvoch — анало- гично названию одного из племен; индейское название вида С. moschata — icushaw. Наличие диких родичей тыквы указывает также на ее аме- риканское происхождение. Диким родичем С. реро А. Грей счи- тал описанный им С. texana Gray, который сейчас большинство систематиков включает в вид С. реро. Как близкий родич С. maxima Duch. указывается С. andreana Naud., который также некоторыми систематиками включается в состав полиморфного вида С. maxima. Примитивной формой С. moschata А. И. Филов считает очень мелкую разновидность — var. seminoliensis Fil., которая разво- дится древнейшим непокоренным племенем индейцев — семино- лов, живущих в зарослях болотного кипариса во Флориде (А. Т. Erwin and E. Р. Lana, 1956). Древнейшим типом тыквы вообще А. И. Филов (1960) счи- тает тропический мезофит-лиану, распространенную в более влажной и теплой части Мексики и Флориды. Наиболее высокогорной формой дикорастущих тыкв, по А. И. Филову, является С. moorei Bailey, которая вьется по кустарникам и деревьям в Мексике на высотах 1750 — 2000 м над ур. м. Этот вид очень близок к С. ficifolia и является при- митивной формой последнего, вошедшего в культуру в высоко- горных районах. Уайтекер и Картер (Т. Whitaker and G. Carter, 1946) пишут, что тыква входила в культуру в Америке двумя путями. Плети- стые сорта, распространенные в полевой культуре, вышли из U,åíòðàëüíoé Америки (Мексики). Из восточной части Америки от дикорастущих форм типа texana произошли кустовые формы типа кабачков и патиссонов. 419
Эволюция форм вида тыквы С. moschata шла, по А. И. Фи- лову (1960), в нескольких направлениях. Одним из типов этого вида является тип сыра — с"еезе, с шаровидными некрупными плодами, напоминающими сыр. Н. Е. Житенева считает этот тип индейским, А. И. Филов также считает его местным и говорит, что такие тыквы индейцы разводили до открытия Америки 1~олумбом. Кроме того, имеются типы: 1~ушо — Cushaw, канадский, колумбийский и мексиканский. Эти данные говорят о пластичности вида С. то- schata, o чем свидетельствует его распространение от Канады до Мексики и южнее. Мексика с прилегающими странами считается родиной не- скольких видов тыквы. Существует мнение, что отсюда вышли культурные виды тыкв — С. реро 1., С. moschata Duch. и С. mixta Pang. Во всяком случае, эти виды тыкв и весь род Cucurbita 1. американского происхождения. Дикими родичами мускатной тыквы — С. mos~chata — яв- ляются С. gapa niata S. Wats., С. digitata А. Gray. Ч а й о т а (Sechium edule Sichwarz) — мексиканский огурец, заменитель нашего огурца. Плоды употребляются в пищу в све- жем, вареном и соленом виде. В Коста-Рике растет дико близкий к чайоте вид — такако— Polakowskia tacaco Pitt. Область первоначального распростране- ния чайоты — страны старотольтекской культуры и культуры майя. Аборигенное население издавна употребляло в пищу огур- цевидные плоды Gonolobus edulis Hemsl. (сем. Asclepiadaceae). Растением, по происхождению связанным с древней ацтекской культурой, считается и хикама — Pachyrhizus uberosus Spreng. Это оригинальный корнеплод из семейства бобовых. Б а т а т (Ipomoea batatas Poiret) в диком состоянии неиз- вестен. По А. Грею, предком батата является растущий дико на Антильских островах 1. fastigiata Sweet, клубни которого съедобны, как и у некоторых других видов I~pomoea. Батат был в культуре у майя и ацтеков. H. И. Вавилов считает его роди- ной Мексику и Центральную Америку, а С. М. Букасов полагает возможным возникновение его в Бразилии и Антильских остро- вах с последующим распространением в странах древних циви- .лизаций майя и ацтеков. Новейшие цитологические исследования проливают свет на происхождение культурного батата — 1. batatas. Следующие аме- риканские виды этого рода оказались диплоидными (2n= 15): 1. asarifolia (Desr.) Roem. et Sichult., 1. ~сагпеа Ja~cq., 1. arbo- rescens G. Don, 1. pendula Choisy и 1. quinquefolia L., а осталь- ные два из исследованных видов — тетраплоидами (2n = 30);. к этой последней группе относятся 1. gracilis R. Br. и 1. Ы1аcia (Wild.) Choisy. Авторы считают невозможным прямое проис- хождение гексаплоидного 1. batatas от тетраплоидного livia и предполагают, что культурный батат — 1. batatas— 420
образовался в результате скрещивания диплоидного и тетра- лоидного видов и представляет собой амфидиплоид. Последнее предположение делает понятным тот факт, что 1. batatas в ди- ком состоянии неизвестен. Родиной а р р о р у т а (Maranta агипйпасеа L.) Н. И. Ва- вилов считает Антильские острова, включая последние в терри- горию мексикано-центральноамериканского очага культурных растений. Но аррорут распространен в Южной Америке (Брази- лия, Венесуэла), поэтому вопрос о родине этого растения не решен. С. М. Букасов и С. В. Юзепчук обнаружили в Мексике и UeR- гральной Америке большое разнообразие дикорастущих видов к а рта ф ел я (Solanum L.), распределенных по следующим се- риям (С. М. Букасов, 1955). Серия горная (Demissa Buk.) включает виды: S. verrucosum S~chlechtd., S. demissum Lindl. и гибридогенные виды S. semide- missum Juz. н S. salan>anii Hawk. Вид S. demissum распространен дико — в тени пихтового леса по опушкам и полянам, нижняя граница его распространения 3000 м, засоряет поля картофеля. Серия Cardiophylla Buk. распространена на Мексиканском нагорье (на высоте от 1700 до 2400 м над ур. м.). Сюда отно- сятся виды: S. ~сагйорЬу11ит Lindl., S. соуосапит Buk., S. 1апс1- forme Rydb., S. ehrenbergii (Bitt.) Rydb. и др. Серия Bulbocastana Rydb. приурочена к южной Мексике и Гватемале (на высоте 1300 — 2500 м над ур. м.). К ней отно- сятся виды'. S. bulbocastanum Dun., S. longistylum Gorr., высоко- горный, S. с1агит Corr. (в Гватемале на высоте 3600 — 3800 м над ур. м.) и др. Серия Oxycarpa (Rydb.) Buk. включает виды: S. ox~zarpum Schiede (Мексика), S. longioonicum Bitt. (Коста-Рика), S. тапо- teranthum Bitt. (Панама), S. agrimonifolium Rydb. и др. Серия Polyadenia Buk. включает очень своеобразный вид S. polyadenium Greenm., обитающий в центральной Мексике на высоте 2100 — 2900 м над ур. м. Серия Pinnatisecta Rydb. распространена в горах (от 1500 до 2900 м над ур. м.): в Мексике, Никарагуа. К ней относятся: S. jamesii Torr., S. pinnatisectum Dun. и др. Серия Longi~pedicellata Buk. распространена в Мексике на высоте от 2200 до 3800 м над ур. м. К ней относятся виды: S. stoloniferum Schlec!ztr,, S. ajuscoense Bulc. S. vallis-mexici JUz. К серии Morelliformia Hawkes относится вид S. morelliforme Bitt. et Muench., эпифит, обитающий на деревьях и скалах во влажных лесах южной Мексики и Гватемалы (на высоте 2000— 2200 м над ур. м.) .. Некоторые зарубежные ботаники считают, что и обыкновен- ный картофель — S. tuberosum L. — происходит из Е~ентральной Америки, что опровергнуто советскими исследователями. 421
К эндемичным мексиканским растениям относится к а к о- M:ûòå — Tigridia pavonia Ker-Gawl. (сем. Iridaceae). Луковицы какомите (главным образом дикорастущих растений) местное население употребляет в пищу. Происходят из центральной Америки и отсюда распростра- нились в Бразилию виды американского таро (Taia или Yan- tia): Xanthosoma edule Schott, X. violaceum Schott, Х.brasiliense Engl., Х. Ьгlophyllum Кип111. К овощным эндемичным мексиканским растениям относится м е к с и к а н с к и й т о м а т, или мильтомат — Physalis aequata Jalcq. Имеются и другие культурные мексиканские виды этого рода — Ph. ixocarpa Brot., Ph. ~pubescens L. и др. В Мексике и прилегающих странах распространены ориги- нальные формы полудиких т о м а т о в. Вишневидный томат— Lyao~persi~con cerasiforme Dun. — является в Мексике широко распространенным рудеральным растением. Название томат происходит от индейского Tomatl (на наре- чии nahuatl) . Однако дикорастущие виды распространены в Перу, где они растут в низменных долинах и по берегам морей. По Д. Д. Брежневу, культура томата возникла в Южной Америке, а в Северной Америке распространилась значительна позднее. Мексика, по мнению многих исследователей, является также- родиной обыкновенного перца — CaIpsicum annuum 1 . (Са~рясит mexicanum Hasenbusch) — древнего мексиканского культурного растения. Здесь же распространен и дикорастущий многолетний перец — С. fruteacens L. По В. Л. Газенбушу, ко времени открытия Америки здесь уже существовали и крупноплодные культурные перцы, относя- щиеся к разновидности grossum Sendt. С. М. Букасовым были собраны в Мексике примитивные формы этой разновидности карликовые и сравнительно мелкоплодные, но по совокупности признаков типичные для современных сортов var. grossum. Однако существует мнение, что первичный очаг вхождения в культуру перцев находится в Южной Америке. Ч и а, м а с л и ч н ы й ш а л ф е й (Salvia chian 1 .), — одно пз интереснейших в историческом отношении растений Мексики. Здесь имеется много видов Salvia L., маслянистые семена неко- торых из них употребляются в пищу. «Судя по названию,— пи- шет С. М. Букасов, — и широкому употреблению чиа вплоть до Калифорнии, Аризоны и New Mexico, культура распространи- лась с очень давних времен, быть может, от самых древних оби- тателей Мексики — чиапанеков, предшествовавших толтекам» По сведениям, собранным С. М. Букасовым, различается форма чиа с крупными семенами, употребляющаяся для напит- ков (для приготовления напитков семена-орешки размешивают в воде, образуется слизистая жидкость), и форма с мелкими чер- ными семенами, которая идет и на масло. Масло чиа употребля- 422
лось в древней Мексике для живописи — краски долго сохра- няют свежесть; масло чиа по качеству выше льняного (иодное число 196,3). В СССР вызревает вблизи Баку. В Мексике имеется до 80 видов шалфея, многие из них со- держат в семенах ценное масло — S. tiliaefolia Vahl, S. lan- -ceolata Brouss. и др. Какао (Theobroma cacao L.) возделывается и встречается дико в Гватемале и Гондурасе. Само слово «какао» относится к наречию майя. В момент открытия Америки культура какао была известна от Панамы до Гватемалы. Хотя Декандоль придерживался мне- ния о южноамериканском происхождении какао, но многие авторы считают его родиной мексикано-центральноамериканский очаг. Th. ca~cao L. поделен теперь на более мелкие виды. В Цен- тральной Америке из них возделываются: Th. sativa Lignier е Le Веу, Th. leiooarpa Bernoulli; кроме того, в Мексике возде- .лывается особый вид Th. angustifolia Moc et Sesse, дающий продукт самого высокого качества, а в Коста-Рика — Th. simia- .гит Ропп. и др. По Шевалье (Chevalier, 1946), в Венесуэле распространены следующие виды какао: Th. leiocarpa, Th. cacao. Гибриды этих видов дали настоящее какао. Вероятно, центральноамериканское происхождение имеет и .древнее культурное растение, распространенное и в диком со- стоянии, ачиоте или биша — Bixa orel ana L. (сем. Bixaceae), красильное и плодовое. Из северной части пустынного Мексиканского нагорья про- исходит каучуконос — гваюла — Parthenium argentatum А. Gray. Она впервые вошла в культуру в Мексике и Калифорнии. Воз- делывалась в СССР (Азербайджан, Туркмения). Н. И. Вавилов считает, что махорка — Ni~cotiana rustiica L.— ведет свое происхождение из центральноамериканского очага древних земледельческих культур; можно предполагать более северное происхождение. Культура табака была широко распро- странена в доколумбовой Америке — от Канады до Патагонии. Табак был в обиходе у майя и ацтеков. У этих народов имеется много туземных названий для табака. Ацтеки культивировали в жаркой зоне N. tabacum, а на плоскогорье — N. rustica. По мнению Н. И. Вавилова, N. tabacum — вид южноамериканского происхождения. Другие авторы считают, что и махорка происхо- дит из Южной Америки. Р а увал ьфи я седоватая (Rauwolfia canescens L.) ис- г ользуется как средство против гипертонии. Распространена главным образом в Мексике, Гватемале, Никарагуа, Коста-Рика, Бразилии, на Антильских островах. Мексикано-центральноамериканскому очагу свойственны многие виды эндемпчных плодовых растений. Сюда относятся виды п а п а й и. В Мексике распространено главным образом 423
дынное дерево — Carica papaya L., используются в пищу и дру- гие виды папайи. Здесь же распространены техокоте — мексиканский боярыш- ник — Crataegus тех1сапа Моч. et Sesse, мексиканская слива— Stpondias mombin L. и S. ригригеа L., белая сапота — Casi- miroa edulis 1 а Llave, желтая сапата — Lucuma sali- cifolia Н. В. K., многие виды Anona L., восковое дерево — My- rica mexicana Willd. — и многие другие. Следует отметить, что этот же очаг дал и авокадо — Persea gratissima Gaertn., который начинает вводиться в культуру у нас на Черноморском побережье Кавказа. Авокадо ценился индей- цами как лекарственное растение. Ацтеки называли это растение Ahuacaquahuitl, испанцы — Ahuacate, а отсюда произошло назва- ние авокадо. В Мексике распространены следующие виды рода Carya. С. ovata var. mexicana Manning, С. palmeri Manning, С. ту- risticaeformis butt., С. illinoensis (Wang.) Кэсй. Но это все- не тропические виды. Тропические виды Carya имеются только в Азии. По Леруа (J. Leroi, 1953) большая часть видов Сагум распространена в восточной половине Северной Америки между 35 и 45' с. ш. На Антильских островах и в Гвиане имеется несколько видов в а н и л и (Vanilla Mill.), которые иногда культивируются: V guianensis Siplitg. (Гвиана), Ъ. ipoitaei Rei~chb. (Гаити), V lep- rieurii Porteres (Кайенна), V. phaeantha Rei~chb. (Куба). Характерная черта Центральной Америки и Мексики — бо- гатство дикими зарослями плодовых и других полезных расте- ний. При расчистке леса мексиканскую сливу, боярышник, гвайяву и другие оставляют, и они становятся растениями куль- турных угодий. Многие виды встречаются и в культурном, и и дикорастущем состоянии, причем дикорастущие формы свя- заны переходами с культурными (авокадо, сапата, гвайява и др.). Многие виды используются преимущественно в дико- растущем состоянии — лиана ваниль — Vanilla planifolia Andr.— u многие другие. Собирательство и теперь еще имеет очень большое экономи- ческое значение в Центральной Америке, тем более, велика была его роль в древности. Когда европейцы завоевали Мексику, они были худшими ботаниками по сравнению с туземцами. В Мек- сике был в это время устроен замечательный ботанический сад, подобного которому тогда не было еще в Европе (Н. S. Reed, 1942) . Вторым важнейшим очагом древней земледельческой куль- туры Нового Света является андийский горный очаг Южной Америки. Сюда относятся прежде всего горные районы Перу и Боливии. Растительность Анд вообще очень разнообразна. Анды яв- ляются не только дорогой миграций, но и мощным центром 424
видообразования. Насчитывается более 40 эндемичных аидий- ских родов. Возникли ли они после поднятия горной системы или произошли от форм, существовавших еще раньше, не вполне установлено. Однако монотипичность многих андийских энде- мичных родов говорит за их огромную древность. Наряду с этим Лиды продолжают оставаться и ареной интенсивного видообра- зования. Андийская Пуна Перу и Боливии — это высокогорная степь, для которой характерен 3 — 7-месячный засушливый период при 400 — 1000 мм годовых осадков. При уменьшении годовых осад- ков ниже 400 мл~ нормальная Пуна переходит в сухую Пуну, или толу, с растительностью полупустынного характера. Почвы Пуны плодородные, черноземовидные. Область Пуны с глубочай- шей древности была очагом земледельческой культуры, сначала доинкской, а затем инкской. Из домашних животных у аборигенов Пуны были приручены ~олько ламы и альпаки. В дикой флоре этой страны имеется много корнеплодов и клубнеплодов. Поэтому не удивительно, что к этой группе отно- сятся и очень многие культурные эндемы области. Андийским земледелием введено в культуру много видов к а р т о ф е л я. Советскими экспедициями (Н. В. Вавилов, С. М. Букасов, С. В. Юзепчук) в Южной Америке открыт целый мир видового и формового разнообразия картофеля, который раньше был почти неизвестен американцам. В настоящее время этот богатей- ший фонд широко используется в селекции. Классификация южноамериканского картофеля и география распространения его разработаны С. М. Букасовым. Серия Commersoniana Buk. распространена в Ла-Платской низменности. Сюда относятся So апит commersonii Dun., произ- растающий в Уругвае и прилегающих частях Аргентины и Бра- зилии, и близкие к нему виды. Серия Glabrescentia Buk. распространена в горах пампы, ок- ружающих Ла-Платскую низменность. К ней относятся S. gib- berulosum Juz. et Buk., S. cha~coense Bitt. и ряд других видов. Серия Cuneoalata Hawkes распространена на севере Арген- тины, Боливии и Чили. Сюда относятся S. xerophyllum Hawl'ås, S. unfundibuliforme Phil. и другие близкие виды. Серия Negistacroloba Card. et Hawkes произрастает в север- ных частях Аргентины и Боливии до 4300 м над ур. м. Сюда отно- сятся S. megistacrolobum Bitt. и другие клубненосные виды. Серия A~caulia Juz. — к видам этой серии, распространенным в Пуне северо-западной Аргентины, Боливии и Перу до высоты 4500 м над ур. м., относятся S. асаи1е Bitt., S. punae Juz. и дру- гие, а также культурные виды S. juzepczukii Buk., S. curti!obum Juz. et Buk. К серии Tuberosa (Rydb.) Buk. в узком смысле относятся виды, распространенные на тихоокеанском побережье Чили и 425
11еру: S. 1пЪегозип1 L., S. maglia Sichlechtd., S. molinae Juz.„. S. leptostigma Juz., и перуанские виды «ломы» — S. medians Bitt. и др. Серия Vaviloviana Ви1<. включает перуанские виды S. va- vilovii Juz. et Ви1~., S. wittma~ckii Bitt. Серия Etuherosa Juz. — виды этой серии не имеют с1олонов и клубней и распространены в Чили и на соседних островах. Сюда относятся S. etuberosum Lindl. и др. По С. М. Букасову, это далекое ответвление группы Tuberosa. Серия Transaequatorialia Buk. занимает обширный ареал (Перу, Боливия, Аргентина, Эквадор) с многообразием природ- ных условий. Высокогорный южный неморозостойкий сорный вид S. ага~сс-pampa Juz. произрастает на высоте до 3400 м над ур. м. Северный морозостойкий вид S. bukassovii Juz. доходит до 3800 м над ур. м. Самый южный вид этой серии S. macolae Buk. обитает в аргентинских Андах в окрестностях Мендоса. По количеству видов это самая многочисленная серия. Кроме упо- мянутых выше видов, в нее входят: S. berthaultii Haw<es, S. ca- nasense Hawkes, S. kurtzianum Bitt. et Wittm., S. microdontum Bitt., S. sparsipilum (Bitt.) Juz. et Buk., S. catarthrum Juz., S. vernei Bitt. et Wittm., S. sucrense Hawkes, S. brev>caule- Bitt., S. famatinae Bitt., S. multiinterruptum Bitt., S. virgul- torum (Bitt.) Card. et Hawkes и др. К серии Andigena Buk. относятся культурные виды карто- феля. В ее состав входят виды Боливии и Перу: S. stenotomum Juz. et Buk., S. ~phureja Juz. et Buk., S. goniocalyx Juz. et Buk., морозостойкий боливийский вид S.:ajanhuiri Juz. et Buk. и др. В Колумбии и Эквадоре распространены S. гуЪ1пй Juz. et Buk и другие виды. Наибольшим разнообразием форм, нередко очень урожайных, отличается S. andigenunl Juz. et Buk., широко культивируемый в Аргентине, Колумбии, Венесуэле, Гватемале, отчасти Мексике. дикорастущие эквадорские, колумбийские и венесуэльские виды Solanum далеки от культурных картофелей и включаются в большую сборную серию Coni~ciba~oca а Bitt. — S. santolallae Vargas, S. colombianum Dun. На север за экватор простирается также ареал серии Piurana Hawl<es — S. ipiurae Bitt. и др. К серии Circaeifolia Hawkes относятся виды S. capsicibalcca- tum Cardenas, S. cincaeifolium Bitt. (северная Боливия). Еще более далекие, большей частью не клубненосные виды серии Juglandifolia Rydb. представляют собой кустарниковые растения, распространенные в Колумбии и Эквадоре. Некоторые из этих видов имеют желтые цветки — S. Iyicepersicoides Dun. Эти виды интересны как прпмитивный тип Solanum. Таким образом, арена видообразования и формообразования клубненосных видов Solanum очень обширна и простирается 426
оТ южной Аргентины и Чили до южных штатов Северной Аме- рики. Наиболее же интересные дикорастущие и культурные виды сосредоточены в Перу, Боливии и Чили. Происхождение обыкновенного культурного картофеля— S. tuberosum — приурочено к тихоокеанскому побережью Чили. Дикие картофели тихоокеанского побережья Чили и Перу ботанически резко отличаются от видов других областей. Наи- более интересная местность здесь — это остров Чилоэ и Араука- ния — в дождливой части Чили с довольно прохладным клима- том. Здесь находится очаг гигрофильной чилийской группы роди- чей культурного картофеля и одновременно ареал аборигенной культуры S. tuberosum, давшего начало европейским сортам (С. М. Букасов и А. Я. Камераз, 1948). В прилегающей более сухой зоне теплого климата древней культуры картофеля не было. Здесь распространена ксерофиль- ная чилийская группа диких картофелей, представителем кото- рой является S. maglia. По данным, собранным I l. М. Жуковским (1959), обыкновен- ный картофель S. tuberosum — растение малых высот, длинного дня с диффузным освещением и пасмурного дождливого климата. П. М. Жуковский, побывав в Латинской Америке, убе- дился, что остров Чилоэ — родина тетраплоидного вида S. tube- гозит — имеет именно такой климат. П. М. ~Куковский пишет, что Чили до сих пор получает картофель с острова Чилоэ и из прилегающих к нему прибрежных районов материка, лежащих южнее г. Вальдивия, через порт Пуэрто-Монт. П. М. Жуковскому не удалось видеть S. tuberosum на о. Чи- лоэ в диком виде, и он считает, что вполне достоверных сведений о диком здесь произрастании картофеля не имеется; он полагает, что среди культурного картофеля на о. Чилоэ встречаются и диплоидные виды" Дикий вид S. maglia, по мнению П. М. Жуковского, не может быть прародителем S. tuberosum, ибо это вид триплоидный. Это почти псаммофит. Жуковский собирал его на крутых склонах побережья Тихого океана во влажных песчаных ложбинах. Второй по значению тетраплоидный культурный вид — S. an- digenum — короткодневный, высокогорный, светолюбивый. S. andigenum в больших количествах культивируется индейцами и креолами на северо-западе Аргентины в провинциях Жужуй, Сальта, Тукуман и Катамарка от 1500 м и выше. Характерное для него местообитание — оазис Тилькара, расположенный в глубокой долине реки Рио-Гранде. Индейцы селятся в боковых долинах по притокам Рио-Гранде и разводят картофель на не- "' В 1967 г. А. Г. Зыкиным в безлюдной лесистой ~асти острова Чилоэ собран, безусловно, дикий картофель, близкий к $. leptostigma Juz. 427
больших поливных участках. Здесь П. М. Жуковский наблюдал сложные популяции картофеля. Особые эфемерные виды картофеля распространены в Ломе — области низких холмов с малым количеством осадков. и увлажнением почвы посредством туманов. За Ламой тянется пустыня, изолирующая ареал чилийских тихоокеанских видов картофеля серии Tuberosa от Анд. Области происхождения S. andigenum и S. tuberosum, таким образом„ р азобщены. В Южной Америке, кроме видов картофеля, имеется ряд. клубнеплодов, которые за пределы Америки не распространи- ЛИСЬ. Очень своеобразны эндемичные клубнеплоды — ока — Oxa- lis tuberosa Molina и О. сгепа1а Jacq. Ока была известна в древ- ней культуре инков. Особенно большое распространение и разно- образие форм ока имеет в Перу, Боливии и Эквадоре. Значение ее в этих странах как продукта питания велико; отсюда она рас- пространилась в Колумбию и другие страны Америки. Большое пищевое значение имеет и имело у древних инков у л ь ю к о — Ullucus tuberosus Lozano, распространенный в Перу,. Боливии и отчасти в Колумбии. Америка дала Старому Свету широко распространенное цве- точное растение — н а сту р ци ю. К тому же роду Тгораео1ит L. относятся эндемичные виды со съедобными клубнеплодами. Са- мый известный из них а н ь ю — Тгораео1ит tuberosum Ruiz et Pavon, возделываемый подобно картофелю в Перу (откуда, по- видимому, происходит), а в Колумбии — Т. icubio, где, вероятно, был известен еще во времена древней колумбийской земледель- ческой культуры чибча. В Андах имеются и другие виды — Tro- paeolum со съедобными клубнями. Съедобные клубни дает также ачир а (Canna edulis Ker- Gawl.) . .Это древнее растение найдено в могильниках Перу. Мака (Lepidium meyenii Wal~p.) возделывается в Перу и Боливии„как своеобразная зерновая культура. Своеобразнейшими древними зерновыми культурами яв- ляются м а р ь кино а (Chenopodium quinoa Willd.) и к а н ь я у а (Ch. ра11Ы&саи1е Аеllеп) — старинные культуры Перу, Боливии, Чили и Колумбии. По П. М. 5Куковскому (1959), до настоящего времени еще сохранился в культуре в Перу памятник древнего растениевод- ства — марь киноа. Это растение возделывается индейцами в высокогорных ущельях с умеренным климатом в Пуне. Ин- дейцы нередко готовят пасту из смеси извести, киноа и листьев кокаиновых кустов (Erythroxylon соса и Е. raimondii), которые дико растут в провинции Куска и в долине Санта-Ана и воЬде- лываются там же и в других местах. Индейцы называют этот кустарник «кока-кока». Киноа служит заменителем кукурузы там, где последняя не растет. 428
В Перу также издревле культивируется амарант к и- ч у и ч а — Amarai>thus caudatus L. Л ю п и н и з м е н ч и в ы й (Lu~pinus mutabilis Sweet) дико растет в Колумбии и Перуанских Андах. Древнее инкское куль- турное растение (1 — 2-летнее), в пищу идут семена. Отличается высокой масличностью. По П. М. Жуковскому (1959), в Перу в провинции Амасонас, в Хунин и Пуно, в провинции Куска в долине Оропеса, Уасао на высотах 3200 м над ур. м. и выше и в настоящее время сохра- нилась культура крупносемянного люпина — Lupinus mutabilis. В национальном музее в г. Лиме имеется обелиск с отчетливым изображением растений и семян 1. mutabilis (изображенные се- мена более крупные, чем у современного L. mutabilis). Назы- вался этот вид люпина чочос, или таруй. Обелиск относится к древней доинкской культуре (VII — V вв. до н. э.). О быки ов ен н а я ф а сол ь (Phaseolus vulgaris L.), по Н. И. Вавилову, основную родину имеет в Центральной Америке; вторичный центр формообразования — в Южной Америке (Перу, Боливия) . Н. И. Вавилов считает, что первичный очаг развития вида л и м с к о й ф а с о л и (Ph. lunatus 1 .) — Центральная Америка, а в Южной Америке, в Перу находится только вторичный центр формообразования. Здесь возникли крупнозерные формы Ph. !u- natus gr. ma~crospermus. По П. М. Жуковскому (1959), лимская фасоль (у инков на- зывалась Pallar) происходит из долины реки Чиллон (Chillon) в Перу. Нижняя очень широкая часть этой долины находится недалеко от Лимы, а верхняя — в горах на высоте более4000м над ур. м., что соответствует большому разнообразию экологиче- ских условий в этой местности. Эта долина является родиной многих культурных растений, в том числе табака, видов тома- та — Lycopersicon peruvianum Mill. и L. hirsutum НитЬ. et Bonip. Земледельческая культура из высокогорных областей Перу и Боливии позднее распространилась в прилегающие прибрежные пустынные и полупустынные районы, где развилась и ирригация. Таким образом, и здесь, как и в Старом Свете, первичные очаги земледелия приурочены к горным местностям, а уже на следую- щем историческом этапе земледелие спустилось на равнины, где возникла ирригация. Здесь шло вторичное (но своеобразное) формообразование к у к у р у з ы, были созданы крупные формы кукурузы Куска. Перуанцы возделывали кукурузу на орошении, они удобряли поля гуано; на удобрение шла и рыба. Перуанцы не приготовляли муки из кукурузы, но употребляли ее целыми зернами, вареными или поджаренными. Поэтому здесь имел место отбор на крупность зерна. В нижней части этой долины выращивают крупнокоробчатый длинноволокнистый хлопчатник. П. М. Жуковский (1959) пишет, что в Перу эндемичные формы кукурузы с крупными крахмалистыми зернами, близкие 429
цо тцпу к древней крупнозерной кукурузе Куска, встречаются в культуре и в настоящее время. Перу — родина хлопчатника — Gossypium barbadense 1. ,дикорастущие, очень долговечные, многолетние формы G Ьаг- badense зэр. darwinii Mauer растут в Перу, Эквадоре, на Гала- пагосских островах. Многолетние культурные и рудеральные формы G. barbadense ssp. ruderale Mauer распространены и Южной Америке (заходят в Западную Африку). Менее долговечные тропические формы многолетней и одно- летней культуры хлопчатника — ssp. vitifolium (Lam.) Маиег- распространены в Перу, Бразилии, Колумбии, Эквадоре (также встречаются в Африке) . От этого подвида происходит подвид eubarbadense Mauer, включающий однолетние субтропические формы, распространенные в культуре вне Америки: в Египте, Алжире, южных районах СССР В Южной Америке имеется еще ряд дикорастущих видов хлопчатника: G. 1<lotzschianum Anderss. (Галапагосские ос- трова), С. raimondii Ulbr. (северное Перу). По П. М. 5Куковскому (1959), древнее название хлопчатника а Перу унтеку или кобо. Перуанский хлопчатник — G. ЬагЬа- с1епзе = G. peruvianum Cav. — и теперь встречается в Перу в ди- ком состоянии на бросовых землях. Особый подвид этого хлоп- чатника — ssp. darwinii — распространен в Перу и на Галапа- госских островах; коробочки и семена его обладают плавучестью и поэтому могли достигнуть архипелага. Культурные сорта пе- руанского хлопчатника относятся к ssp. vitifolium. В северном Перу в департаментах Кахамарка и Ла-Либертад встречается по побережьям 26-хромосомный дикий вид хлопчатника— G. га1топй1. Н. И. Вавилов считал, что некоторые виды растений вошли в культуру поливного земледелия из местной флоры достаточно увлажненных районов восточных Анд. К таковым он относил д ы н н у ю г р у ш у — Solanum muri~catum Ait. — и виды дикого и рудерального томата. Южная Америка является родиной подрода то м а т о в Erio- persiicon Ми11ег. Основные виды, составляющие этот подрод: Lycopersioon peruvianum (L.) Miller — распространенный от се- верного Перу до Чили и L. hirsutum НитЬ. et Вопр. — дико- растущее, большей частью многолетнее растение, распространен- ное в Перу и Эквадоре. Оба вида очень полиморфны; в их со- ставе выделяют теперь более мелкие виды. Из них L. glandulo- sum С. Н. Ми11ег, вероятно, является подвидом L. hirsutum, а недавно вновь открытый L. chilense Рип. примыкает к L. pe- ruvianum. Систематика этих мелких видов еще не уточнена. В Перу растет полудикая разновидность — L. езси1еп1ит ssp. subspontaneum Brezh. var. suocenturiatum (Pasq.) Brezh. По П. М. Жуковскому (1959), культурный томат L. езси1еп- 'turn происходит из Южной Америки. Имеются сведения, чтоэтот 430
вид, а также L. chilense растут дико на Галапагосских островах. П. М. 5Куковский сообщает, что первые крупноплодные сорта томата появились в Эквадоре. Там имели место процессы фас- циации и гибридизации L. esculentum и L. pimpinellifnlium Dun. Вид L. hirsutum довольно обычен в Перу в долине реки Чиллон. L peruvianum проник из Перу в северо-западную Аргентину. Вопрос, где вошел в культуру обыкновенный томат — 1. escu- 1еп(ит (здесь или в Центральной Америке), является спорным. Возможно и полифплетическое вхождение томата в культуру. По нашему мнению, более обоснована теория южноамерикан- ского происхождения культурного томата и всего рода. С восточных склонов Анд, может быть, вошла в культуру и т ы к в а — Cucurbita maxima Duch. Дикорастущий вид С. an- dreana Naudin распространен в Аргентине и Уругвае. Из мелко- плодных сортов этой тыквы приготовляют сосуды, из которых пьют парагвайский чай. Настоящая посудная тыква, или горл яика — Lage- nana vulgaris Ser. не всеми считается аборигенным для Южной Америки растением. Из предполагаемой своей родины — Юго- Восточной Азии — лагенария могла распространиться в Аф- рику, а затем проникнуть в Южную Америку. Позднее морскими течениями она могла быть перенесена в Бразилшо. Кемп (W. Camip, 1954) считает невозможным перенесение тыквы из Бразилии в Перу — преградой являются Анды. Он думает, что лагенария в Перу также перенесена 1ечениями. Уайтекер и Кар- тер (Т. Whital<er and (i. Carter, 1954) доказали возможность пре- бывания в морской воде этой тыквы в течение 224 дней без потери всхожести. Il. М. Жуковский (1959) предполагает, что м у с к а т н а я тыкв а — С. moschata Duch. — происходит из Южной, а не из Центральной Америки. Это древняя культура Колумбии и Перу. В раскопках Уала-Приета найдены остатки ее семян и чашели- стиков, давность которых определяется в 4000 лет. Из других видов тыквы С. ficifolia возделывается высоко в горах в Перу и Гондурасе. С. maxima Duch., по А. И. Филову (1960), имеет эндемичные формы в Перу, Боливии, Чили, воз- можно, в Бразилии и Аргентине. Все же большинство авторитет- ных исследователей считает, что этот вид тыквы (а также му- скатная тыква) вышел из Центральной Америки. По Смиту и Хейзеру (P. Smith, С. Heiser, 1957), существуют следующие виды п е р ц а: Capsiicum аппиит L., С. frutescens 1., С. pendulum Willd., С. pubescens Я. et Р. и С. sinense Jacq. (ero синонимы — С. luteum Lam. и С. umbili~catum Veil.). Ав- торы приводят ареалы этих видов в центральной и 10жной Аме- рике. Согласно классификации В. Л. Газенбуша (1958), С. umbi- li~catum Veil., а также С. odoriferum Veil. являются синонимами вида С. соп1сип1 Mayer — колумбийский перец. 431
J(poMe колумбийского перца, В. Л. Газенбуш включает в свою классификацию перуанскии перец — С. angulosum Mill. (Перу, Бразилия, Чили), С. pubescens (высокогорные районы Перу, Колумбии, Боливии и Гватемалы) и С. аппиит — мекси- канский перец (с включением в этот вид С. sinense и С. fru- tescens), сначала распространенный в Центральной Америке (главным образом Мексике) и Южной Америке, позднее очень широко распространившийся по всей Америке, Европе, в азиат- ских странах и в Африке. В Мексике, Гватемале и Кубе культи- вируются мелкоплодные примитивные формы С. аппиит, в стра- нах Центральной Америки встречаются и дикорастущие формы этого вида перца. Таким образом, вопрос о первичных очагах вхождения в культуру видов перца является спорным. Что касается наи- более распространенного вида — С. аппиит, то имеются сторон- ники той точки зрения, что родина этого вида Центральная Америка (Мексика), а в Южной Америке находится вторичный очаг его формообразования. Остальные виды перца, видимо, происходят из Южной Аме- рики. Вопрос может быть окончательно решен после более де- тального и всестороннего изучения дикорастущих перцев Америки. Н ар а их иль о (Solanum quitoense Lam.) считается плодо- вой культурой. На основании исследований Д. Костова и С. А. Эгиза предпо- лагается, что табак — Nicotiana tabacum L. возник путем скре- щивания андийских дикорастущих видов — N. silvestris Speg. et Comes и N. rusbyi Brit. В Перу и Боливии распространен вид табака N. paniculata L., в Чили — N. angustifo ia R. et Р., в Чили, Бразилии, Аргентине и Парагвае — N. langsdorfii %е1пт. и др. В Перу и Боливии обитают и виды, близкие к ма- хорке — N. rustica 1 . С восточных склонов Анд ведет, вероятно, свое происхожде- ние хинное дерево — Cinchona calisava Wedd., С. suocirubra Pav., используемое главным образом из дикорастущих зарослей, и кокаиновый куст — Егуйгоху1оп соса Lam. Небольшой очаг земледелия на Боготском плоскогорье AH- дийского хребта в 1(олумбии более или менее оторван от основ- ных очагов андийской земледельческой культуры в Перу, Боли- ВНН H Чили. Здесь, в области древней культуры чибча, распространены многие туземные виды картофеля — Solanum rybinii и др., а также арракача — Arracacia xanthorhiza Вапсг.— многолетнее корнеплодное растение из сем. зонтичных, распространенное в умеренно теплой и холодной зонах. Весь род Arracacia Bancr. происходит из северных Анд. Вероятно, в области древней куль- туры чибча был введен и колумбийский перец — Capsicum columbianum Hazenb. 432
Настоящая тропическая лесная флора Америки дала сравнительно мало культурных растений. Если принять во вни- мание богатство флоры Бразилии видами, то число культурных видов бразильского происхождения невелико сравнительно с су- хими и горными районами Лнд и Llen~paльной Америки. По Н. И. Вавилову, в Бразилии введено в культуру такое ценное растение, как арахис, или земляной орех — Arachis hy- родаеа L. Арахис получил широкое распространение на земном шаре, «озделывается и в нашей стране. По П. М. ~Куковскому (1959), аргентинскими ботаниками А. Краповицкас и В. Ригони в 1957 г. доказано, что культурный арахис — А. hy~pogaea — происходит не из Бразилии, как писал H. И. Вавилов (1935), а из северо-западной Аргентины и из южной Боливии. В провинциях 5Кужуй на высоте 1450 м, в ущелье Умауака на высоте от 1400 до 2800 м, в Сьерра де Сента на высоте 2300 м над ур. м. и в Кампо Дуран провинции Сальта распространен дикорастущий однолетний тетраплоидный вид — А. monticola b;raip. et Rig. Этот вид является, по-видимому, прародителем культурного А. hypogaea. Указанные два вида дают фертильные гибриды. Кроме этих видов, все остальные- диплоидные. Близкородственен А. hypogaea А. villosa Benth. Еще ближе стоит к культурному арахису другой дикорастущий вид — Л. зу1- westris А. Chev., с гладким односемянным бобом. В тропической Южной Америке имеется еще слабо изученный вид — А. rasteiro А. Chev. Это трехлетнее, хорошее кормовое растение. Все виды рода Arachis распространены в Южной Америке между 10 и 28' ю. ш. Отсюда вышли также маниак — Manihot utilissima Pohl и М. dulicis Рах, ананас — Ananas сотоза L., фейхоа — Feijoa selloviana Berg. и некоторые другие. Следует отметить, что, по Н. И. Вавилову, маниак, арахис, ананас и парагвайский чай приурочены к сравнительно сухим районам, а не к влажным тропическим лесам — гилеям. Андийским очагам Южной Америки свойственны многие виды Rubus L., из них отметим встречающиеся г, культуре: R. macro~carpus Benth. (Анды — Колумбия, Эквадор), R. glaucus Benth. (Эквадор, Перу, юг Мексики). В тропической части Южной Америки сосредоточено огром- ное разнообразие дикорастущих видов Rubus (один из первич- ных очагов видообразования). Своеобразныи вид грецкого ореха (Juglans neotroipica Diels) растет в Перу среди формации подокарпуса. В Лзии нет тропических видов из рода Juglans, а в Америке нет тропиче- ских видов орехов из рода Carya. Выше уже отмечалось, что наиболее древняя земледельче- ская культура была связана с андийскими районами, а позднее, но еще до Колумба, земледелие распространилось вниз до юж-
ного Чили. Здесь развилась небольшая, но оригинальная арау- канская культура. Климат южной части Чили и острова Чилоэ довольно влажный и умеренный. Здесь на 38 — 40' 10. ш. день достаточно длинный; здесь и возник наш обыкновенный карто- фель — Solanum tuberosum L. — растение длиннодневное, в про- тивоположность короткодневным выходцам из Перу и Боли- ВИИ. Из Араукании происходит также масличное растение м а д и я (Madia sativa Molina), которое в настоящее время мало распро- странено. В древности здесь имел большое значение как зерно- вая культура особый вид кастр а — Bromus mango Desv., ко- торый теперь вымер. Здесь в диком состоянии растет ч и л и й с к а я 3 е м л я н и к а (Fragaria chiloensis Duch.) .. Подспорьем в питании населения здесь с древних времен служат орехи хвойного дерева а р а у к а р и и — Агацсапа imbricata Pav., по имени которого получила наименование и вся культура. Это древнее дерево в диком состоянии имеет ра- зорванный ареал. Из тропической Америки происходит м ал а ига (Xantho- soma sagittifolium Sichott), которая найдена в перуанских мо- гильниках. В Бразилии в диком состоянии распространен а р р о р у т (Maranta агипйпасеа L.). Из Южной Америки это растение распространилось в Африку и Индию. Бразилия является родиной к а у ч у к о в о г о д е р е в а (Нечеа brasiliensis Muell. Arg.), распространенного по долине реки Амазонки. Несмотря на исключительное богатство амазонской флоры, она дала немного культурных представителей. Несколько видов Eugenia имеется в культуре в Бразилии и Парагвае. А м е р и к а н с к и й о р е х (Bertholletia excelsa НптЬ. et Bonpl.), крупное какао (Theobroma grandiflora К. Schum.) и другие виды какао используются в диком состоянии. Дико- растущие виды какао имеются также в Колумбии и тропических. районах Перу. Род Manill<ara Adans. из сем. Sapotaceae имеет пантропиче- ский ареал. Многие виды дают вкусные сладкие плоды, гутта- перчу, ценную древесину и т. д, Один из очагов распространения видов Manill'àãà — Бразилия (лесные области по Амазонке). Шевалье указывает на произрастание следующих видов: какао в Бразилии: Theobroma sipeiciosa Spreng. — дико и в культуре; Т. spruceanum Bern. — дико и в культуре, Т. miro- сапрпт Mart. — дико; T. grandiflora (Sipreng.) Ьс1шт. — дико и в культуре; Т. subincana Mart. — в культуре; Т. atrorubens. Hub. — дико; T. bicolor Н. В. К. — в культуре; Т. obovatum Bern. Из тропической Южной Америки происходит и ваниль— Vanilla aromatica Sw. (Ямайка, Колумбия). 434
В субтропических лесах Аргентины и Бразилии распростра- .нены в диком состоянии близкие родичи обыкновенной фасоли— Phaseolus vulgaris. По Буркарту (Burkart, 1953), родоначальником фасоли обык- новенной надо считать дикорастущую лиану — Ph. aborigineus Burk., которая распространена на северо-западе Аргентины. Обнаружение подобных видов не говорит с полной достовер- ностью о возможности происхождения обыкновенной фасоли из Южной Америки. Упомянутые виды недостаточно изучены. В Аргентине распространено бобовое дерево, являющееся пищевым растением,— Prosopis dulcis Кипй. По Н. А. Базплевской (1960), около 647 видов декоративных растении вывезено пз Южной Америки: Heliotropium peruvia- пит L. (Перу и Эквадор), Iresine lindenii Van Houtte (Эквадор), Alternanthera amoena Voss п А. versicolor Hort., Begonia sem- perflorens 11п1; (Бразилия). Из однолетников вывезены из Юж- ной Америки виды родов Тгорзео1ит, Petunia Juss., Verbena L., Portulaca L. Яля развития земледельческой культуры и создания фонда окультуренных растений мало одного богатства флоры. Для этого необходимо развитие человеческой культуры, а сырые тропические лесные дебри с их нездоровым климатом неблаго- приятны для жизни человека, поэтому здесь и не создалось крупных очагов земледелия. Нами охарактеризованы самые крупные очаги происхожде- ния культурных растений в Америке. Существование их, конечно, не исключает и случаев вхождения в культуру растений и на других территориях этого континента. Так, например, основная арена видо- и формообразования видов Solanum, близких карто- фелю, на севере заканчивается Мексикой. Но и севернее на юге США встречаются единичные виды се- рий Pinnatisecta (S. jamesii) и Longipedicellata (S. fendleri А. Gray и др.). Дикий п о д с о л н е ч н и к (Helianthus annuus L.) распро- странен в огромном количестве по краям полей, межам и пусты- рям в США и отчасти в южной Канаде. Н. И. Вавилов приурочивает введение в культуру подсолнеч- ника и родственного ему вида топинамбура, или земляной груши — Н. tuberosus L., к пограничной области СШЛ и 1ханады, т. е. вне двух основных очагов. Здесь же распро- странен близкий им родич культурного подсолнечника— Н. Ientiicularis Dougl. С. М. Букасов пишет, что ареал распространения сорныхпод- солнечников доходит до Мексики, где он заменяется внешне похожим на него сорняком кукурузы, но относящимся к другому роду — мексиканским подсолнечником — Tithonia tubaeformis &ass. 435
Среди культурных сортов подсолнечника — Н. annuus есть короткодневные и длиннодневные формы (Е. Н. Синская, 1955). Короткодневные, вероятно, возникли на юге ареала, а длинно- дневные выработались из них по мере распространения на север Растекающийся ареал имеет и т а б а к — виды Nicotiana. Кроме Южной Америки, распространены в Мексике N. герап4а Willd., в Центральнои и Северной Америке — N. torr yana: А. Nelson, только в Северной Америке — N. rnultivalvis Lindl, N. quadrivalvis Pursch и др. Все эти виды используются индей- цами для курения и жевания. В Северной Америке распространены дикорастущие виды кен дыр я, волокно которого использовалось индейцами в до- колумбовское время. Аросупит cannabinum L. распространен от Мексики до Канады, больше в центральных штатах; А. hyperi~ci- folium Ait. — в США и Канаде. Лекарственное растение Hydrastis сапа4епз1з L. распростра- нено в Канаде и северо-восточных штатах Северной Америки Александрийский лист, или сенна алексан- д р и й с к а я (Cassia aicutifolia Del.), распространен в восточной части Северной Америки (Мэриленд, Виргиния). Ценное лекарственное растение Podophyllum peltatum L (сем. барбарисовых), клубни которого содержат подофиллин, вывозится из Северной Америки и с побережья Мексиканского залива. Американский п о р т у л а к (Lewisia rediviva Pursch)— важное пищевое растение индейцев Северной Америки. В Северной Америке распространены аборигенные виды грецкого ореха (Juglans nigra L. и J. icinerea 1.).. Северная Америка является родиной американского каш- т а н а (Castanea dentata Borkh.) .. К л е й т о н и я (Claytonia perfoliata D~onn.) происходит из Се- верной Америки и разводится в Европе как салатное растение. Из Северной Америки происходят и некоторые виды п е- и а н а. Наибольший интерес из них представляет Carya oli- vaeformis Nutt., распространенный от Айовы до Техаса и дико произрастающий по берегам рек Миссисипи, Колорадо и их при- ТОКОВ. Североамериканские виды к р ы ж о в н и к а (Grossularia hirtella Spach., G. oxyanthoides Mill., G. icynosbati Mill., G. diva- ricata Соч. et Britt. G. nivea Spach и другие) участвовали в происхождении многих американских культурных сортов. Из североамериканских видов Rubus (малина и ежевика) назовем следующие: 1) R. idaeus зэр. strigosus Мс1тх.--дико от Ньюфаундленда до Орегона; 2) R. maicropetalus Dougl. от Бри- танской Колумбии (Канада) до Калифорнии; 3) R. loganobaocus Bailey — культурный вид тихоокеанской Северной Америки, про- исхождение не вполне выяснено; 4) R. titanus Bailey — культи- вируется в Калифорнии; 5) R. flagellaris Willd. — родоначальник 436
многих североамериканских культурных сортов; 6) R. velox Bailey — культурный вид Северной Америки; 7) R. trivialis. Michaux — дико и в культуре, от Виргинии до Флориды и Те- хаса; 8) R. pergatus Blanch. — дико и в культуре, Северная Аме- рика; 9) R. argutus Link — восточные штаты США; 10) R. ostryi- folius Rydb. — центральные штаты США. Имеются и другие виды этого рода. Из Северной Америки происходят следующие виды з ем л я- н и к и: 1) Fragaria platypetala Rydb. — вошла в культуру на Аляске; 2) F. virginiana Mill.— происходит из Северной Аме- рики, в прошлом культивировалась в Америке и Европе, в на- стоящее время не выращивается. К л ю к в а к р у п н о п л о д н а я [Oxycoocus п1асгосагриз (Ait.) P~ers.] — в диком состоянии в Северной Америке (Нью- фаундленд, Онтарио, Виргиния, Мичиган, Арканзас). Введена здесь в культуру. Имеются сорта. Новый Свет — родина многолетних люпинов. Интересен .ч ю- п ин м но го летний (Lupinus p~olyphyllus Lindl.), произрас- тающий дико по берегам рек в Северной Америке. Дикий «в о д я н о й р и с» (Zizania aquatiica 1 .) используется индейцами США и южной Канады. Виды Prunus распространены в основном в Северной Аме- рике и Канаде. Из них имеют селекционное значение: P. ame- ricana Marsch. (от Мексики до Канады), P. nigra Ait. (США и Канада), P. munsoniana Wight et Hedr. (южные штаты США), P. angustifolia Marsch. (южные штаты США). Эти виды слу- жили для выведения американских культурных сортов сливы. Северной Америке свойственны некоторые интересные для селекции дикорастущие холодостойкие сорта я б л о н и Malus rivularis Roem. из Аляски, а также группы эндемичных культур- ных сортов и их гибридных форм с дикорастущими формами. По А. Н. Ипатьеву (1960), существуют две группы американских сортов яблони: 1) американская — Уэлен, Джонатан, Мекин- тош, Бен Девис и др.; 2) американские крэбы — мелкоплодные сорта Северной Америки. Северные леса бедны фруктовыми деревьями, но и там встречаются Amelanchier alnifolia Nutt. Между Миссисипи и Атлантическим океаном в лесах растут Dios~pyros virginiana L., Asirnina triloba (L.) Dun. и некоторые орехоплодные. Многие дека р а ти в н ы е деревья происходят из Северной Америки. Сюда относятся виды магнолии — Magnolia grandi- Hora L., тюльпанное дерево — Liriodendron tulipifera L. Н. А. Базилевская (1960) пишет об особом североамерикан- ском центре происхождения декоративных растений. Она ука- зывает на происхождение отсюда следующих растений: Phlox L., Solidago L., Rudbecl<ia L., Coreopsis L., Не1епшт L., несколько видов Aquilegia L. и Lilium L, ,Оепойега biennis L., Ttadescantia ~.'irginiana L. Здесь находится родина однолетних декоративных 437
растений — Eschscholtzia californica Cham., С1аг11а Pursh, Go- detia Sipa~ch, Nemophilla Buccal. Сделанный нами обзор, несмотря на его сжатость, приводит к след ющим выводам. 1. ссортимент культурных растений Америки очень велик и своеобразен. Он базируется на аборпгенных видах и большей частью аборигенных родах. Лишь немногие роды имеют куль- турных представителей в Новом и Старом Свете; к ним отно- сятся следующие: Grossularia, Rubus, Fragaria, Gossypium, Pha- saolus, Solanum, Amaranthus, Chenapodium, Lupinus, Bromus, Prunus, Vitis (американские виды используются главным обра- зом в качестве подвоя), Malus, Crataegus, Diospyros, Eugenia, Juglans. Основные культурные растения Старого Света (пшеница, ячмень, овес, рожь, просо, горох, гречиха, рис, вика, чина, нут, бобы, лен, конопля, капуста, репа, брюква, люцерна, клевер, эспарцет и очень многие плодовые) появились в Америке лишь после прихода туда европейцев. По-видимому, только одно возделываемое растение проникло в Америку до Колумба — это горлянка — Lagenaria vulgaris, из плодов которой жители древних цивилизаций Центральной Аме- рики изготовляли посуду. Америка дала Старому Свету кукурузу, картофель, батат, табак, томат, дынную грушу, перец, какао, каучуковое дерево- гевею, гвайюлу, хинное дерево, новосветские виды фасоли и хлопчатника, некоторые сливы, многие декоративные и лекарст- венные, авокадо, фейхоа, ананас, подсолнечник, топинамбур, арахис, мадию, американские виды тыквы, американские виды кендыря и др. Таким образом, знакомство с фондом американских культур- ных растений отвергает теорию существования древнего исчез- нувшего материка Атлантиды, ибо через Атлантиду культурные растения Старого Света перебрались бы и в Новый Свет. Чело- век явился в Америку из Азии без культурных растений. 2. Происхождение огромного числа аборигенных культурных растений Америки в основном приурочено к двум подобластям: 1) Мексикано-Центральноамериканской и 2) Южноамерикан- ской, что не исключает случаев введения их в культуру и на ос- тальной территории нового континента. 3. Очаги первичной земледельческой культуры и происхож- дения большого количества культурных растений в Новом., как и в Старом Свете, приурочены в основном к горным районам и сухому климату. Поливное земледелие позднее развилось в при- легающих равнинах. Влажные лесные тропические области дали начало сравни- тельно немногим культурным растениям. 4. Своеобразие древнейших земледельческих цивилизаиий Америки состоит в том, что они почти не знали домашних живот- 438
ных, рабочего скота. Человек явился в Америку без домашних животных (за исключением собаки). 5. Древнейшие цивилизации Америки (принимая во внима- ние время их расцвета) моложе древних цивилизаций Старого Света, но земледелие все же в Америке является весьма древ- ним, что подтверждается археологическими и ботаническими данными. Д. Бранд (D. Brand, 1939) считает доказательством orpoM- ной древности аборигенного американского земледелия то, что многие культурные растения Нового Света в настоящее время не способны совсем или в слабой степени способны к размноже- нию семенами, а для утраты этой способности требуется очень много времени. К таковым относятся: маниак, картофель, батат„ аракача, ананас, ока, улыбка, анью, ксантозома — yahutia. Древ- ность этих культур доказана историческими, археологическими и лингвистическими данными.
ЗАКЛЮЧЕНИЕ Процессы форма- и видообразования идут на всей террито- рии земного шара, везде, где возможна жизнь растений. И в об- ластях тропических, и субтропических, и в областях с умеренным суровым климатом возникают новые виды или новые формы и экотипы, которые со временем могут дать начало видам. Однако видообразовательный процесс протекает не везде с одинаковой интенсивностью. Там, где имеется большое разно- образие экологических условий (как это обычно бывает в горных странах), видообразование носит особенно интенсивный харак- тер, а флора отличается наиболее богатым видовым составом и большим содержанием эндемичных, старых и молодых видов. В областях интенсивного видообразования человек находил (и находит) особенно много разнообразных видов и форм рас- тений, пригодных для использования и введения в культуру. Однако области массового окультуривания растений приурочены не ко всем областям интенсивного видообразования. Процесс эволюции культурных растений более сложен, чем у представителей дикорастущей флоры. Филогенетическая история культурного растения носит двой- ственный характер — оно представляет собой создание и при- роды и человека. В местностях, неблагоприятных для массового расселения первобытного человека, вошло в культуру сравнительно мало ви- дов растений (влажные тропики с их трудно освояемыми джунг- лями). Окультуривание растений происходило в большем масштабе .там, где развивались древние земледельческие цивилизации, где интенсивнее шел процесс развития общества, где раньше воз- никли государства. В нашей стране области древнейшей земледельческой куль- туры и раннего развития государственности приурочены к Закав- казью и предгорным областям Среднеазиатских республик. Именно для этих областей характерно особое богатство видами и формами дикорастущей флоры. Территории интенсивного окультуривания растений в нашей работе получили обобщенное название географических областей исторического развития культурной флоры. Они определены нами с учетом древности земледельческих цивилизаций, общ- -440
ности состава кульгурных и дикорастущих видов и роаов расте- ний, а также их ботанического разнообразия. Наибольшее внимание нами уделено странам, которые входят в огромную Древнесредиземноморскую область исторического развития и влияния культурной флоры. Именно по этой области имелось больше всего ботанического и археологического мате- оиала. Особое значение нами придавалось изучению Древнесреди- земноморской области, так как под ее непосредственным влия- нием происходило формирование культурной флоры на об- ширной территории Советского Союза. Влияние остальных областей на состав культурной флоры СССР сравнительно невелико. Влияние Восточноазиатской об- ласти незначительно, а влияние Южноазиатской области ничтожно. Влияние африканской культурной флоры не было непосредственным, а шло через Переднюю, Средне-Юго-Запад- ную и Восточную Азию. В Древнесредиземноморской области различаются три круп- ные подобласти: Передняя Азия, Средне-Юго-Западная Азия и собственно Средиземноморье. В состав первой входит Закав- казье, в состав второй — предгорные и горные районы Средне- азиатских республик. На примере этих двух подобластей можно ясно видеть большое различие между основными обла- стями и подобластями исчорического развития культурных флор. Если сравнить списки растений, эндемичных и имеющих пер- г.ичный очаг зарождения и вхождения в культуру в Древнесре- диземноморской и К)жноазиатской областях, то огромные раз- личия в отношении родов и видов сразу бросаются в глаза. На- против, подобласти Средне-Юго-Западная и Передняя Азия при их сравнении имеют общий костяк важных родов и параллель- ное развитие ряда культурных видов. Сравнение этих двух подобластей наглядно показывает вы-- сокую интенсивность видообразования в Передней Азии, и именно последняя является основным центром, откуда распро- страняется влияние на равнинные территории большей части нашей страны, а также Северной и Средней Европы. Примеры обзора таких территорий, которые по нашей терми- нологии обозначаются как области влияния, даны в разделах, посвященных Русской равнине и Сибири, Северной и Средней Европе, Эфиопии и Северной Америке. Для демонстрации того положения, что видообразователь- ный процесс у растений нигде не затухает и только различается по своей интенсивности, приведены краткие сведения о есте- ственных ресурсах дикорастущей флоры в основных областях и в областях влияния. Даже молодая (по геологическому счету времени) флора Русской равнины дала (и еще даст) растения для введения в культуру, перспективные для использования 441
в дальнейшем. То же можно сказать и о Сибири, Северной и Средней Европе. Особое внимание нами уделено Передней Азии и это сделано по двум причинам: 1) подобласть Передней Азии оказала влия- ние на культурную флору всей территории Советского Союза; 2) в нашем распоряжении по Передней Азии было больше всего материала. Пути распространения культурных растений СССР, Передне- азиатской подобласти и всей Древнесредиземноморскои области и связи их с другими крупными областями исторического раз- вития культурных флор земного шара в данной работе осве- щены, но не с той полнотой, которая является желательной. Мы надеемся, что эти вопросы найдут более полное освеще- ние в трудах других исследователей при дальнейшем накопле- нии фактов по исторической географии и филогенезу культур- ных растений.
ЛИТЕРАТУРА А б и 6 ул а ев О. А. Раскопки холма Кюль-Тапа. КСИИМК, вып. 51, 1953 А в д и е в В. И. История Древнего Востока. ОГИЗ, 1948. Агрикультура в памятниках Западного Средневековья. Переводы и ком- ментарии. Изд. АН СССР, 1936. А н ф и м о в Н. В. ~древние поселения Прикубанья. Краснодар, 1953. Ар а к ел я н Б. Н. Раскопки крепости Гарни. ВДИ, № 1, 1951. Ар и сто в Н. Я. Промышленность Древней Руси. Спб., 1866. А р и сто ф а н. Ахарияне. Спб., 1897. Ар т а м он о в М. И. Раскопки Саркела — Белой Веки в 1950г. «Вопросы.. истории», № 4, 1951. А р т а м о н о в М. И. Археологические исследования в зоне будущего Цимлянского водохранилища. Вести. ЛГУ, № 2, 1951. Ар т а мо н о в М. И. Белая Вежа. «Советская археология», т. 16,,1952.. Ар ц1и х о в ски й А. В. Введение в археологию. МГУ, 1940. Ар ци х о в ск,и й А. В. Раскопки восточной части дворища в Новгороде. Сб. «Материалы и исследования по археологии древнерусских городов>, т. 1,. 1949. Ар ци хо в ск,и й А. В. Раскопки на Славне в Новгороде. Сб. «Мате- риалы и ысследования по археологии древнерусских городов», т. 1, изд.. AH СССР, 1949. Ар ц их о в ск и й А. В. Археологические открытия в Новгороде. Вестн. АН СССР, Мо 12, 1951. Б а дер О. Н. Древнее городище на Верхней Волге. «Материалы и ис- следования по археологии СССР», № 13, 1950. Б а з ил е в ск а я Н. А. центры происхождения декоративных растений. В кн.." «Вопросы эволюции, биогеографии, генетики и селекции>, М. — Л, 1960. Б а й бур т я н Е. А. Культовый очаг из раскопок Шингавитского посс- ления в 1936 — 1937 гг. ВДИ, Ю~ 4, 1938. Б а р а н о в П. А. Под тропиками Рака. «Природа», № 7, 1960. Баранов,П. А., Негруль А. М. и Фролова К. И. Дикорасту- щий виногр ад Средней Азии. Сб. «Проблемы ботаники>, изд. АН СССР, 1955. Б а р ул и н а Е. И. Чечевица СССР и других стран. Тр. по прикладной ботанике, генетике, и селекции, приложение 40, 1930. Б а хт ее в Ф. Х. Эколого-географические основы филогении и селекции ячменей. Изд. АН СССР, 1948. Б а х т е е в Ф. Х. Ископаемая форма культурного ячменя Hordeum lagun- cu1iforme mihi. ДАН СССР, т. 110, М 1, 1956. Б а хт ее в Ф. Х. К истории культуры ячменя в СССР. Сб. «Материалы по истории земледелия СССР», Сб. II. М.— Л., 1956. Б а хт ее в Ф. Х. Открытие Hordeum lagunculiforme ВасЫ. на территории Туркменской ССР. ДАН СССР, т. 129, № 1, 1959. Б а х т е е в Ф. Х. Полба (Triticum dicoccum Schubl.), найденная Н. И. Ва- виловым в Карпатах. В кн.: «Вопросы эволюции, биогеографии, генетики и селекции», М.— Л., 1960. Б а х т ее в Ф. Х. Новое звено в дикорастущем виде ячменя. Ботаниче- ский журнал, Мо 6, 1962. 443
Б а хтеев Ф.Х. и Дар ев ск а я Е. М. Hordeum spontaneum С. Косй и Н. lagunculiforme ВасЫ. из Туркменской ССР. Ботанический журнал, № 2, 1962. Б е р г Л. С. Очерки по физической географии. Изд. АН СССР, 1949. Бердзенишвили Н., Джа ва хишвили И. и Джанаши я С. История Грузии. Т. 1, Тбилиси, 1946. Бе р н шта м А. Н. Культура древнего Киргизстана. Фрунзе, 1942. Б ер н ш та м А. Н. В, горах и долинах Памира и Тянь-Шаня. В кн.: <По следам древних культур», т. 3, М., 1954. Б и б,и к о в С. Н. Раннетрипольское поселение Лука Врублевецкая на .Днестре. «Материалы и исследования, по археологии СССР>, т. 38, 1953. Б и б и к о в С. Н. Пещерные палеол!итические местонахождения Южного Урала. «Советская археология», Мо 12, 1960. Б и ч у р,и н Иакинф. Земледелие в Китае. Спб., 1844. Б и ч ур.им Н. Я. (Иакинф). Собрание сведений о народах, обитающих в Средней Азии в древние времена. Т. 1 — 2, М.— Л., 1950. Б л ей х шт е й н ер Р. Субары древнего Востока в свете изучения яфе- тидов. «Язык и история», сб. 1, 1926. Бл о MKB и ст Е. Э. Союз ирокезов. «Вокруг света», № 4, 1951. Б о б р о:в Е. Г. Виды клеверов СССР. Тр. Ботанического ин-та :им. В. Л. Комарова, сер. 1, т. 6, 1947. Б о г а е в с к и й Б. Л. Очерк земледелия Афин. Пг., 1915. Б о г о л ю б с к и й С. Н. Происхождение и преобразование домашних .животных. М., 1959. Б о р д а ко в Л. П. Опыт систематического изучения вида Medicago sa- tix а L. Тр. по прикладнои ботанике, генетике и селекции, сер. 7, № 1, 1934. Б о р и с к о в с к и й П. И. Основные группы палеолитических местона- хождений. Среднего Поднестровья. КСИИМК, вып. 26, 1949. Бр ест ед Д. Г. История Египта с древнейших времен до персидского завоевания. Т. 1 — 2, М., 1915. Б р и в к а л и с Э. Древние поселения у городищ на территории Латвий- .ской ССР. М., 1951. Б р юсов А. Я. Сетчатая керамика. «Советская археология», т. 14, 1950. Б р ю с о в А. Я. Предки народов Зауралья. Сб. «По следам древних культур», т. 3, М., 1954. Бук асов С. М. Возделываемые растения Мексики, Гватемалы и Ко- лумбии. Тр. по прикладной ботанике, генетике и селекции, приложение 47, 1930. Б ук а со в С. М. Система видов картофеля. Сб. «Проблемы ботаники», М.— Л., 1955. Б ук а со в С. М. ~и К а м е р аз А. Я. Селекция картофеля. Сельхозгиз, 1948. Б у к,и н и ч Д. Д. История первобытного орошаемого зечледел!ия в Турк- .менской области в связи с вопросом о происхождении земледелия и ското. водства. «Хлопковое дело», № 3 — 4, 1924. В а в и л о в Н. И. О восточных центрах происхождения культурных ра- стеншь «Новый Восток>, 'K9 6, 1924. В а в ил ов Н. И. Центры происхождения культурных растений. Тр. по прикладной ботанике, генетике и селекции, т. 16, вып. 2, 1926. В а в и л о в Н. И. Ботанико-географичесиие основы селекции. В кн.: «Тео. ретические основы селекции растений», т. 1, М.— Л., 1935. В а в ил о в Н. И. Великие земледельческие культуры доколумбовой Амс. рики и их взаимоотношения. Известия Государственного географического об- щества, т. 71, № 10, 1939. В а в ил о в Н. И. Пять континентов. «География», М., 1962. В а силенка Н. Г. «Витлуф» — зимний салат. «Природа», № !О, 1960. В а сил ьев А. Дыня ча Руси в XVII веке. «Сельская жизнь», No 77 (8728), 1960. 444
В е село век а я М. А. Мак, его класоификация и значение как маслич- ной культуры. Тр. по прикладной ботанике, генетике и селекции, приложе- ние 56, 1933. В и н о г,р а д о в - Н и к,и т и н П. 3. Плодовые и пищевые деревья лесов .Закавказья. Тр. по прикладной ботанике, генетике и селекции, т. 22, вып. 3, 1929. В о е в од ски й М. В. Деснинская археолопическая экспедиция 1940 г. КСИИМК, вып. 13, 1946. В о р о н и н Н. Н. Пища и утварь. В кн.: «История культуры Древней Руси>, изд. АН СССР, 1951. В о,р о н и н Н. Н. На берегах Клязьмы и Немана. Сб. «По следам древ- них культур», т. 2, 1953. Вор о 6ьев М. В. Древняя Япония. М., 1958. В ул ь ф Е. В. Историческая география растений. Изд. АН СССР, 1944. Г а йду ке ви ч В. Ф. Некоторые итоги раскопок Тиритаки и Мирмекия. ВДИ, № 3, 1947. Г а йд у ке в и ч В. Ф. К истории древне;о земледелия в Средней Азии. ВДИ, № 3, 1948. ?' а т и н ~К. И. Биологические особенности облепихи и проблема введе- .ния ее в культуру для садоводства и лесных полос. Сб. «Проблемы бота- ники>, т. 2, М.— Л., 1955. Г а ф у р о в В. Г. История таджикского народа. М., 1949. Ге н. В. Культурные 1растения и домашние животные в их переходе из Азии в Грецию и в Италию, а также в остальную Европу; Спб., 1872. Г е н к о А. Н. О названиях плуга в северокавказских языках. ДАН СССР, .Ко 7, 1930. Ге ор ги е в ск и й Б. М. Южный Хорезм. Геологические и гидрологиче- ские исследован~ия. Ташкент, 1937. Гер одот. История в 9 книгах. Перевод Мищенко. М., 1888. Гибридная кукуруза. Сборник статей. Перевод М. Л. Бельговского и др. М., 1955. .Гильтебр а ндт В. М.,Кунжут. Тр. по прикладной ботанике, генетике и селекции, сер. IX, М 2, 1932. Г и и м е л ь Я. H. К проблеме археоботаники Закавказья. Сообщения Грузинского филиала АН СССР, 1, № 19, 1940. Г и м мел ь Я. И. Археологические очерки. Баку, 1940. Г и м м е л ь Я. И. К проблеме археоботаники Закавказья. Сообщения .АН Груз, инской 'ССР, т. 2, № 1 — 2, 1941. Г и н з б у р г В. В. Древние и современные антропологические типы Сред- ней Азии. Сб. «Происхождение человека и древнее расселение человечества», М., 1951. Г и п п ок р а т. Избранные книги. М., 1936. Г о л у б о в и ч В. и Г о л у б о в, и ч Е. Кривой город — Вильно. КСИИМК, вып. 11, 1945. Голуб о в ич В. и Голубов и ч Е. Хлебныс злаки из раскопок нз горе Бекеше. КСИИМК, вып. 11, 1945. Г о н ч а р о в В. К. Райковецкое городище. Киев, 1950. Г ар г и две А. g. Лен Грузии. Автореферат диссертации. Тбилиси, 1955. Г о т ь е Ю. В. Очерки по истории материальной культуры Восточной 'Европы. Л., 1925. Г р е к о в Б. Д. Образование Русского государства. В кн.: «220 лет АН СССР», т. 2, М.— Л., 1947. Греко в Б. Д. Киевская Русь. Учпедг,из, 1949. Г р ем я цки й М. А. и О кл а д н и к о в А. П. Тешик-Таш. МГУ, 1949. Г р и го р ье в М. Г Древняя Москва. В кн.: «По следам древних куль- тур», т. 2, Д,ревняя Русь, 1953. Г р и г о р ь е в Г. В. К вопросу о центрах происхождения культурных .растений. Известия Государственной Академии истории материальной куль- .туры, т. 13, вып. 9, 1932. 445
Гр игор ьев Г. В. Поселения древнего Согда. КСИИМК, вып. 6, 1940 Г р и чук В. П. Географические ландшафты Русской равнины леднико- вого периода. «Природа», № 2, 1952. Гр озд,ил о в Г. П. Раскопки в Старой Ладоге в 1948 г. «Советская археология», т. 14, 1950. Г р о з н ы й Б. Доисторические судьбы Передней Азии. ВДИ, т. 3 — 5,. 1940. Г р о з н ы й Б. Протоиндийские письмена и их расшифровка. ВДИ, № 2, 1940. Г р о м о в а В. И. Материалы к познанию фауны трипольской куль- туры. Ежегодник Зоологического музея, т. 28, вып. 1, 1927. Г р о ссгей м А. А. Реликты Восточного Закавказья. Баку, 1940. Г р ум м - Г р ж и м а йл о А. Г. В- поисках растительных ресурсов мира. Изд. АН СССР, 1962. Гур и н а Н. Н. Археологическое обследование реки Мсты. «Советская археология», т. 14, 1950. Г у ip с к и й А. В. В ботанических садах и заповедниках Китая. «При- рода», № 3, 1960. Г у р у А. Азия. Перевод с франц. К. М. Макаровой и А. В, Шмелева, М., 1956. Гущи н Г. Г. Р,ис. Сельхозгиз, 1938. Д а н и л о в А. Д. Разнообразие форм обыкновенной груши в Острогож- ских лесах Воронежской области. Зап. Воронежского с.-х. ин-та, т. 21, вып. 3, 1947. Д а о Тх е Т у а н. Происхождение, систематика и экология риса. Таш- кент, 1960. Д а Ip в и н Ч. Изменение животных и растений в домашнем состоянии. М., 1941. Де 6 ец Г. Ф. Палеоантропологические материалы из погребений срубнок культуры Среднего Заволжья. «Материалы и исследования по археологии СССР», № 42, 1954. Де 6ец Г. Ф. Происхождение коренного населения Америки. Сб. «Про- исхождение человека и древнее расселение человечества», М., 1951. Дебец Г Ф.,Трофимова Т. А. и Чебоксаров Н. И. Проблемы земледелия севера по антропологическим данным. Сб. «Происхождение че- ловека и древнее расселение человечества>, М., 1951. Д е б е ц Г. Ф. Заселение Южной и Передней Азии по данным антропо- логии. Сб. «Происхождение человека и древнее расселение человечества>, М., 1951. Де б е ц Г. Ф. Антропологические данные и заселение Африки. Сб. «Про- исхождвние человека и древнее расселение человечества», М., 1951. Д е к а н д о л ь А. Местопроисхождение возделываемых растений. Спб., 1885. Д е к а п р е л е в и ч Л. Л. Главнейшие полевые культуры эпохи Шота Руставели. Сб. «Материальные культуры эпохи Шота Руставели», Тбилиси, 1938. Де к а п р еле в ич Л. Л. Роль Грузии в происхождении пшениц. Сооб- щения АН ГрузССР, т. 1, № 10, 1940; т. 2, № 2, 1941; т. 3, № 4, 1942. Д е,к а п р е л е в и ч Л. Л. Грузинский очаг формообразования пшениц Сообщение АН ГрузССР, т. 3, № 7, 1942. Д е к а п р е л е в и ч Л. Л. О некоторых вопросах одомашнивания пше- ницы. В кн.: «Вопросы эволюции, биогеографии, генетики и селекции», М.— Л., 1960. Д е р ж а в и н Н. С. Происхождение русского народа: великорусского, украинского, белорусского. М., 1944. Д ж а в а х и ш в и л и И. Основные историко-этнологические проблемы истории Грузии, Кавказа и Ближнего Востока древнейшей эпохи. ВДИ, № 4, 1939. Д ж а н а ш в и л и М. Историческая справка по сельскому хозяйству на Кавказе. <Кавказское сельское хозяйство>, № 60, 1895.
Д и н И н. Исследование пленок риса, обнаруженных в долине р. Янцзы и красной обожженной глине, относящейся к эпохе неолита. «Агробиология», № 4, 1960. Дневник фиванского некрополя. ВДИ, № 4, 1950. Д о в т е и о к В. И. К вопросу о технике пашенного земледелия иа юге Древней Руси. Сб. «Археолог]я», т. 4, Киieв, 1950. Домострой по Конишенскому списку и подобным, под ред. А. Орлова, .кн. 1 — 2, М., 1908 — 1910. Древнейшая Индия в свете последних археологических раскопок. ВДИ, Мо 2, 1940. Дум а н Л. И. Очерк истории Китая. «Китай», сб. статей под ред. акад. В. М. Алексеева, Л., 1940. Д у н дуле н е П. В. Земледелие в дофеодальпой Литве. Краткие сооб- щения Института этнографии им. Н. Н. Миклухо-Маклая, т. 12, М., 1950. Дунин М. С. По Афганистану, Пакистану, Индии. Географгиз, М., 1952. Д ь я к о в М. М. Древняя Бактрия. Сб. «По следам древних культур», т. 3, М., 1954. Д ь я к о н о в И. М. Развитие земельных отношений в Ассирии. Л., 1949. Д ь я к о н о в И. М. Реформы Урукагины в Лагаше. ВДИ, № 1, 1951. Д ь я к о.н о в И. М. Последние годы Урартского государства. ВДИ, № 2, 1951. Е в т ю х о в а Л. А. Южная Сибирь в древности. Сб. «По следам древних культур», т. 3, М., 1954. Е н к е и В. Б. Соя. М., 1959. Е р м а ш е в И. И. Заря над Азией. <Октябрь», № 4, 1949. Е фа м е н к о П. П. Палеолитические стоянки Восточно-Европеи=кой рав- нины. В кн.: «Труды II Международной конференции Ассоциации по изуче- нию четвертичного периода Европы>, вып. 5, Москва — Ленинград, 1934. Е ф и м е н к о П. П. Первобытное общество. Киев, 1953. ;~К и т е н е в а Н. Е. Мировой сортимент культурных тыкв. Тр. по при- кладной ботанике, генетике и селекции, т. 23, вып. 3, 1930. ;~К ук о в ск и й П. М. Культурные растения и их сородичи. М., 1950. ~К у ко в скп~ й П. М. По центрам происхождения культурных растений Латинской Америки. Ботанический журнал, Мо 3, 1959. 3 а м я т н и н С. Н. О локальных различиях в культуре палеолитического периода. Сб. «Происхождение человека и древнее расселение человечества», М., 1951. 3 бр уе в а А. В. Население берегов Камы в далеком прошлом. Сб. <По следам древн1их культур», т. 3, М., 1954. 3 и м и н а Т. А. и К р юк о в а Т. Н. Дикорастущие луки Сахалина и перспективы введения их в культуру. Ботанический журнал, т. 47, № 6, 1960. 3 оз Н. Г. Резорпинсодержащие виды Pauvolfia L. Ботанический журнал, т. 45, Мо 8, 1960. 3 о л о т н и ц к и й. Наши садовые цветы, овощи и плоды. Их история, роль в жизни и вврованиях разных народов и родины. М., 1911. 3 о cfB м о в, и ч В. Н. Эколого-географическая характеристика диких видов свеклы (Beta L.). ДАН СССР, т. 24, № 1, 1939. И бы Си н а (Авиценна). Канон врачебной науки. Ташкент, 1956. И в а н о в Н. Р. Эволюция культурных видов рода Phaseolus L. В кн.: «Вопросы эволюции, биогеографии, генетики и селекции», М.— Л., 1960. И в а ш ен к о М. Материалы к 1изучению культуры колхов. Сб. «Мате- риалы по истории Г)рузии и Кавказа», вып. 11, изд. АН ГрузССР, 1941. И л b и н Г. Ф. Особенности рабства в Древней Индии. ВДИ, № 1, 1951. И п а ть е в А. Н. О дифференциальной географии культурных растении. В кн.: «Вопросы эволюции, биогеографии, генетики и селекции», М.— Л, 1960. История Татарии в материалах и документах. М., 1937. К а г а н к ат в а ц и Моисей. История агван. Перевод К. П. Патканов», Спб., 1861. 447
К аз а ко в а А. А. Местные луки северной зоны СССР Тр. по приклад- ной ботанике, генетике и селекции, т. 29, вып. 1, 1951. К а л а н д а д з е А. Н. Юго-Осетинская археологическая экспедиция. Научная сессия Института истории им. акад. И. А. Джавахишвили, посвя- щенная итогам полевых археологических исследований 1952 г. Тбилис~~, 1953. К а л л и сто в Д. П. Очерки по истории Северного Причерноморья антич- ной эпохи. ЛГУ, 1949. К а р м ен Р. Л. По странам трех континентов. «Молодая Гвардия», 1962. К а ц н е л ь с о н И. С. Проблемы исторического развития древнейшей Нубии. ВДИ, № 2, 1948. К и ip ь я н о в А. В. К вопросу о земледелии в Новгородской земле в XI — XII вв. КСИИМК, вып. 47, 1952. К(и р ь як о в А. В. История земледелия Новгородской земли Х — XV вв. «Материалы и исследования по археологии СССР>, № 65, 1959. К и с е л е в С. В. К вопросу о культуре древнейшего еэропеоидного насе- ления Сибири. ВДИ, № 1 (23), 1948. К и с е л е в С. В. Древняя история южной Сибири. «Материалы и иссле- дования по археологии СССР», № 9, 1949. К л и н г ен И. Среди патриархов земледелия народов Ближнего и Даль- него Востока (Египет, Индия, Цейлон, Китай IH Япония). Ч. 1, Спб., 1898. Ко в алев Н. В. Значение алычи в селекции косточковых плодовых по- род. В кн.: <Проблемы ботаники», М.— Л., 1955. К о в а л е в с к и й Г. В. История главнейших культурных растений. Гос- издат Украины, Одесса, 1926. ,К о в а л е в ски й Г. В. Человеческ)ие племена и растительные культуры. «Человек», № 2 — 4, 1928. Ко в алев ски й Г. В. Земледельческая Индия. Тр. по прикладной бо- танике, генетике и селекции, т. 21, вып. 5, 1929. К о в а л е вски й Г. В. К истории культуры арбуза и дыни в европей- ской части СССР. Тр. по пр(икладной ботанике, генетике и селекции, т. 23, вып. 3, 1929 — 1930. К о в а л е в с к и й Г. В. Культурно-.историческая и биологическая роль горных р а йо нов. «Пр ирода», № 2, 1931. К о в а р ск и й А. С. Пшеницы горной части Крыма. Тр. по прикладной ботанике, генетике и селекции, т. 22, вып. 2, 1929. Кол ч и н В. А. На раскопках Древнего Новгорода. «Наука и жизнь», № 4, 1952. К о м а р о в В. Л. Происхождение культурных растений. Сельхозгиз, 1931. К о н д р а т о в и ч К. Дикционер или Речениар по алфавиту российских слов о разных,произращениях, то-есть о древах, травах, цветах, семенах, ого- родных и полевых кореньях и о прочих былиях и минералах, собранный и сочиненный Имп. Академии наук Коллежским асессором К. Кондратовичем. Спб., 1780. К ар до н Р. Я. Айва СССР. Автореферат диссертации, Л., 1954. К о с т и н а К. Ф. Происхождение и эволюция культурного абрикоса. Тр. Никитского ботанического сада, т. 24, Мо 1, 1947. Кр а сочкин В. Т. Исходный ботанический материал свеклы и мор- кови и перспективы его использования в селекции. Сб. «Проблвмы ботаники», М.— Л., 1955. Кр а со чкин В. Т. Свекла. М.— Л.. 1960. Кр а сочкин В. Т. Из истории возделывавия свеклы. Материалы по истории сельского хозяйства и крестьянства СССР, сб. IV, 1960. Кр а со чк и н В. Т. и Узун о в В. Н. Свекла в странах ее древней культ ры. Тр. по прикладной ботанике, генетике и селекции, т. 26, 1931. р еч ето в ич В. И. Сем. Theaceae Mirb. и род Thea L. Культурная флора СССР. Рукопись. Кр и ч е в ски й Е. Ю. Мезолит и неолит Европы. КСИИМК, вып. 4, 1940. К р и ч е в с к и й Е. Ю. О роли межплеменных отношений в древнейшей истории. КСИИМК, вып. 13, 1946. К р и ш то ф он и ч А. Н. Палеоботаника. М.— Л., 1941. 448
Крото в А. С. Гречиха. М.— Л., 1963. К, р у гл о в А. П. Культовые места горного Дагестана. КСИИМ1~, вып. 12, 1946. Кр у п Ir о в Е. И. Краткий очерк археологии Кабардинской АССР На.~ь- чик, 1946. Кр у п н о в Е. И. Древняя история Северного Кавказа. М., 1940. К уд р я ш о в С. Н. Растительность Гузара. Ташкент, 1941. К у л е ш о в Н. Н. Географическое распределение сортового разнообразия кукурузы на земном шаре. Тр. по прикладной ботанике, генетике и селекции, т. 20, 1929. К у л е ш о в Н. Н. Ботаническое описание кукурузы (Zea may", L.). Записки Харьковского сельскохозяйственного института, т. 11 (48), 1955. Культурная флора СССР. Т. 1, 1935; т. 2, 1936; т. 4, 1937; т. 5, 1940; т. 6 и 7, 1941; т. 13, 1950; т. 16 и 17, 1936; т. 20, 1958; М. — Л. К у п ц о в А. И. Географическая изменчивость вида Carthamus tincto- rius L. Тр. по прикладной ботанике, генетике и селекции, сер. IX, № 1, 1932. К у п ц о в А. И. Географическое распределение культурной флоры в исто- рическом развитии. Известия Всесоюзного географического об-ва, т. 87, вып. 3, 1955. Ку ф т и н Б. А. Киргиз-казаки, культура и быт. М., 1926. К уф тик Б. А. К вопросу о древнейших корнях грузинской культуры на Кавказе по данным археологии. Вестн. государственного музея Грузии, т. 12-В, 1944. К у ф т и н Б. А. Три этапа истории культурного и этнического обособле- ния дофсодальной Абхазии. Тбилиси, 1949. К у ф т и н Б. А. Материалы к археологии Колхиды. Т. 1 — 2, Тбилиси, 1949 — 1950. Л а в р о в Л. И. Земледелие на северо-западном Кавказе. Сб. «Материалы по истории земледелия в СССР», М., 1952. Л а т ы н и н Б. Работы в районе проектируемой электростанции на р. На- рь1не в Фергане. В кн.: «Археологические работы Академии на новостройках в 1932 — 1933 гг.», М., 1935. Л е в а ше в а В. П. Из далекого прошлого южной части Красноярского края. Красноярск, 1939. Л е в и н М. Г. и Ч е б ок с а р о в Н. Н. Древнее расселение человече- ства в восточной и юго-восточной Азии. Сб. «Происхождение человека и древ- нее расселение человечества», М., 1951. Л е в и н М. Г. Древние переселения человека в северной Азии по данным антропологии. Сб. «Происхох дение человека и древнее расселение человече- ства», M., 1951 Л и в и н г с т о н Д. Путешествия и исследования в Южной Африке с 1840 по 1855 гг. Географгиз, 1955. Л е и у и а н Я. А. Дар Масиниссы (к вопросу о хлебной торговле Делоса в начале II в. до н. э.). ВДИ, г1 4, 1948. Л и б е р о в П. Д. К вопросу о скифах-пахарях. ВДИ, № 4, 195!. Л и з r у н о в а Т. В. История ботанического изучения огородной капусты. Тр. по прикладной ботанике, генетике и селекции, т. 32, вып. 3, 1959. Л и т в и н с к и й Б. А. Намазга-Тепе. По данным раскопок 1945 — 1950 гг. «Советская этнография», № 4, 1952. Л и х о н о с Ф. Д. Некоторые данные по систематике видов и культур- ных сортов яблони. Бюлл. Главного Ботанического сада, вып. 51, 1963. Л и Ill у - ч а п. Род Sapium в китайской флоре. ~1жиу фэнь-лэй сюэвао. Acta Phytotaxon, sinica, чо1. 5, № 2, 1956. Л он г ф ел л о Г У Песнь о Гайавате. Перевод Бунина. Пг., 1915. Л от А. В поисках фресок Тассили. Изд-во восточной литературы, М., 1962. Л уз и н а Л. В. Анис и анизет. Культурная флора СССР. Рукопись. Л ук и и A. Л. Нсолитическое селище близ Гудаут. «Советская археоло- гия», т. 12, 1950. Синская Е. Н. 449
Л ю б о м у д р о в Д. О культуре полбы в России до середины XVIII века. ~р. по прикладной ботанике, генетике и селекции, т. 18, вып. 1, 1928. Л я вда н ски й A. Н. Некоторые данные о городищах Смоленской гу- бернии. Научные известия Смоленского государственного ун-та, т. 3, вып. 3, 1926. Л ьв о в Н. А.,и Я к о вл е в а С. В. Изучение и селекция перечной мяты. Тр. по прикладной ботанике, генетике и селекции, т. 23, вып. 1, 1930. М а в р од и н В. В. Образование древнерусского государства. Л., 1945. М а к а ш в и л и А. К. Неизвестные культуры Колхиды. Автореферат диссертации, Тбилиси, 1951. .' 1 а к к е й Э. Древнейшая культура долины Инда. Перевод Граково- Свцр1'довой. М., 1951. М а к у шин а- Г о р о щек ко Е. Н. Новый вид пшеницы Triticum топ- tanum Makush. nom. поч. и его положение в системе рода Triticum. Ученые записки Ленинградского Государственного педагогического ин-та им. А. И. Гер- цена, 66, 1948. М ар ущен ко А. А. Анау. В кн.: «Архитектурные памятники Туркме- нии», вып. 1, Москва — Ашхабад, 1939. Марущенко А. А. Итоги полевых археологических работ 1953 г. Института истории, археологии и этнографии Академии наук Туркменской ССР. Тр. ИИАЭАН Туркм. ССР, т. 2, 1956. М ауер Ф. М. Происхождение культурных и дцких видов хлопчатника и основы их классификации. Тр. Среднеазиатского гос. ун-та, вып. 18, 1950. N а у е р Ф. М. Хлопчатник. Ташкент, 1954. Махабхарата Адипарва. Книга 1. Перевод Кальянова под ред. акад. А. П. Баранникова. М.— Л., 1950. М а ш к и н Н. А. Карфагенская держава до пунических войн. ВДИ, № 4, 1948. Ме н а 6 де В. Л. Ячмени Грузии. Тр. Тбилисского ботанического ин-та, т. 6, 1938. Ме н а бде В. Л. Ботанико-систематические данные о хлебных злаках древней Колхиды. Сообщения Грузинского филиала АН СССР, т. 1, № 9, 1940. Ме н а бд е В. Л. Пшеницы Грузии. Тбилиси, 1948. Мен а бде В. Л. и Ер иця н А. А. К изменив грузинской пшеницы зандури. Сообщения АН ГрузССР, т. 26, М 6, 1960. Мерило праведное. Сборник библиотеки Троице-Сергиевской лавры, около 1350. Мер п ер т Н. Я. ~и Ш ел о в Д. Б. Древности нашей земли. М., 1961. Мечников Л. И. Цивилизация и великие исторические реки. Киев— Харьков, 1899. М и з г и р е в а О. Ф. Мандрагора туркменская. Сб. «Проблемы бота- ники», М. — Л., 1955. М илл ер М. А. Сарматский сосуд с зерном из Нижне-Гниловского мо- гильника. ИРостОМК, вып. 2, 1940. Мил ько в Ф. Н. Лесостепь Русской равнины. М., 1950. Мо н г а йт А. Л. Старая Рязань. «Материалы и исследования по архео- логии СССР», Ю 49, 1955. Мо н га й т А. Л. Обзор полевых археологических, исследований ИИМК в 1947 г. ВДИ, Л% 2, 1948. Мордвин ки н а А. И. К истори1и культуры овса в СССР. «Материалы по истории сельского хозяйства и крестьянства СССР>, сб. IV, 1960. Мошинская В. И. и Смир н ов А. П. Археологические исследо- вания в РСФСР за 1947 г. КСИИМК, вып. 26, 1949. М о ш и н с к а я В. И. Городище и курганы Потчеиаш (К вопросу о пот- чевашской культуре), МИА, № 35, 1953. Не г р ул ь А. М. Происхождение культурного винограда и его классифи- кация, В кн.: «Ампелография СССР», т. 1, М., 1946. Н е р у Д. Открытие Индии. Изд-во иностранной литературы, 1955. Н и к о л ь ск а я 3. А. и Ш и л л и н г Е. М. Горное пахотное орудие тер- расовых полей Дагестана. «Советская этнография», № 4, 1952. 450
Н из а м и Г. Искандер-наме. Т. 1, перевод Е. Э. Бертельс, Баку, 1953 Н и к и шин И. И. Находка зерен в Керчи. КСИИМК, вып. 23, 1948 Н и кол ь ск и й В. К. Детство человечества. М., 1949. Н о ви цк а я М. К вопросу о текстиле Трипольской культуры. Сб. «Að хеология», изд. Ин-та археологии АН УССР, т. 2, Киев, 1948. О к л а д н,и к о в А. П. Прошлое Якутии до присоединсния к Русском~ государству. История Якутии, т. 1, Якутск, 1949. Окл ад нико в А. П. Неолит и бронзовый век Прибайкалья. Изд. АН СССР, 1950. О к л а д н и к о в А. П. Археологические исследования в Бурят-Монго- лии. Изв. АН СССР, сер. истории и философии, № 6, 1951. О кл ад недико в А. П. Реферат о работе Джона Марингера в разделе «Крлтика и библиография». ВДИ, № 5, 1951. О к л а д н и к о в А. П. У истоков культуры народов Дальнего Востока. Сб. «По следам древних культур от Волги до Тихого океана», М., 1954. О си п о в А. М. Краткий очерк истории Индии до Х века. М., 1948. Палеолит и неолит СССР. Под ред. Окладникова. «Материалы и иссле- дования по археологии СССР», М~ 39, 1953. П а н г а л о К. И. Дыни как самостоятельный род Melo. Ботанический журнал, т. 35, вып. 6, 1950. П а н г а л о К. И. Происхождение и эволюционный путь бахчевых куль- тур. В сб. «Проблемы ботаники>, М.— Л., 1955. П а с с е к Т. С. Периодизация трипольских поселений (111 — I I тысяче- летия до н. э.). «Материалы и исследования по археологии СССР», № 10, 1949. П а с с е к Т. С. Раннеземледельческие (трипольские) племена Поднест- ровья. Ы., 1961. П е в цо в М. В. Путешествие в Кашгарию и Кун-лунь. М., 1949. Пер с ид с ьк и й Д. и Л и п а О. Злаки з буд1вель трипольско1 куль- тури на Укра]н]. Науков] записки УИМК, кн. 5 — 6, Ки'1в, 1935. П етр о в В. А. Растительные остатки из культурного слоя Старой Ла- доги (IX — Х вв.). КСИИМК, вып. 11, 1945. П е т р о в В. А. Растительные остатки из глиняных обмазок раннетри- польского времени с Украины. Рефераты научно-исследовательских работ за 1945 г. (отд. истор. и илософ. АН СССР), М. — Л., 1947. H етр о в В. А. ищевые остатки из Пастырского городища. Сб. «Ар- хеолог1я», т. 2, Ки1в, 1948. Пио тр о в ск(и й Б. Б. Вишапы. Каменные статуи в горах Армении. Л., 1939. Пиотро в ски й Б. Б. История и культура Урарту. Ереван, 1944. П и отр о в ский Б. Б. Археологическое изучение древнего Закавказья. БДИ, Мо 3, 1947. П и о т р о в с к и й Б. Б. Город бога Тейшебы. ВДИ, Мо 4, 1948. П и о т р о в с к и й Б. Б. Археология Закавказья с древне~~ших времен до I тысячелетия до н. э. Л., 1949. П и о тр о в с к и й Б. Б. Кармир-Блур. Т. 1, Ереван, 1950. П и с а р е в В. Е. К вопросу о происхождении земледелия и полевых культур Восточной Сибири. Сб. «Материалы, по истории земледелия СССР», г. 2, М. — Л., 1956. П и с а р е в В. Е. Праисхождение яровых пшениц Восточной Сибири. В кнл «Вопросы эволюции, биогеографии, генетики и селекции», М.— Л., 1960. Пишель Р. Будда, его жизнь и учение. Перевод с немецкогоД. Н.Ану- чина. М., 1911. Повесть временных лет. Изд. АН СССР, 1950. Попов M. Г. Дикие плодовые Средней Азии. Тр. по прикладной бо- ганике, генетике и селекции, т. 22, вып. 3, 1929. П о п о в М. Г. Мировые фонды диких плодовых растений и их значение цля плодоводства. В кн.: «Достижения и перспективы в области прикладной ботаники, генетики и селекции>, Л., 1929.
П о п о в а Г. М. Очерк культурной растительности центрального Бухар- ского оазиса. Тр. по прикладной ботанике, генетике и селекции, т. 17, № 2, 1923. Пополь Вух. Родословная владык Тотоникапана. Перевод Кинжалова. Изд. АН СССР, 1959. П о я р ко в а А. И. Критический обзор дикорастущих видов смородины и крыжовника Советского Союза. Тр. Ботанического ин-та AH CCCP, сер. 1, вып. 2, М.— Л., 1936. Путешествие Ибн-Фадлана на Волгу. Под редакцией И. Ю. Крачков- ского. М. — Л., 1939. Р а в д о н и к а с В. И. Старая Ладога. КСИИМК, вып. 11, 1945. 1' а в до н и к а с В. И. Старая Ладога (Из итогов археологических иссле- дова:~ ш 1938 — 1947 гг.). Часть 11, «Советская археология», т, 12, 1950, Р а вд оп и к а с В. И. Древнейшая Ладога в свете археологических ис- следований 1938 — 1950 гг. КСИИМК, вып. 41, 1951. Р а г и м о в М. А. К биологии и экологии фенхеля. Тр. пн-та ботаники АН АЗССР, т. 15, 1950. Р аз ум о в В. И. Реакция на длину дня чумизы из различных провин- ций Китая. «Физиология растений>, т. 2, ¹ 7, 1955. Р а з у м о в с к и й Д. В. Обозрение растений, упоминаемых в священном писании. М., 1871. Р ед е р Д. Г. Из истории одного древнеегипетского города. ВДИ, № 2, 1948. Р ед е р Д. Г. Попытки акклиматизации чужеземных культурных расте- ний в Древнсм Египте. Учен. зап. Московского областного педагогического ин-та им. Н. КрупскоЙ, т. 14, вып. 1, 1950. Р е м е з о в а Г. Л. Медвежий лук. «Природа>, ¹ 8, 1960. Р о ж е в и ц Р. Ю. Злаки. М.— Л., 1937. Р уб а ш е в ск а я М. K. Дикая морковь в культуре и в природе. Тр. по прикладной ботанике, генетике и селекции, т. 2G, вып. 2, 1931. Р у б ц о в Н. И. Краткий обзор эндемов флоры Крыма. Тр. Государст- венного Никитского ботанического сада, № 29, 1959. Р у б ц о в Н. И. Понтида. «Природа», ¹ 8, 1960. Р уб ц о в Н. И. Опыт сопоставления флор горного Крыма п Западного Закавказья. Тр. Государственного Никитского ботанического сада, № 35, 1961. Р у"д е н к о С. И. К древней истории Горного Алтая (Горноалтайская экспедиция ИИМК АН СССР 1949 г.). ВИ, ¹ 2, 1950. Р у д е н к о С. И. Сокровища Пазырыкских курганов. Сб. «По следам древних культур», М., 1951. Р у д е н ко С. И. Горноалтайские находки и скифы. М.— Л., 1952. Руководство по апробации сельскохозяйственных культур: т. 1, 1947; т. 2, 1949; т. 3, 1949; т. 4, 1950; т. 5, 1948, М. — Л. Р ы б и н В. Гибриды терна и алычи и проблема происхождения куль- турной сливы. Тр. по прикладной ботанике, генетике и селекции, сер. 2, вып. 10, 1936. Р ю р и к о в Б. Встречи под созвездием Южного Креста. «Знамя», № 1, 1963. Р я б о в И. Н. и К. Ф. К о с т и н а Персики Ферганской долины. Тр. по прикладной ботанике, генетике и селекции, сер. 8, № 1, 1932. С е м е н о в С. А. Ранние археологические памятники Вьетнама. ВДИ, № 4, 1950. Сер г ее в В. С. История Древней Греции. ОГИЗ, 1948. С и н ск а я Е. Н. О полевых культурах Алтая. Тр. пл прикладной бота- нике, генетике и селекции, т. 14, вып. 5, 1925. С и н ск а я Е. Н. Масличные и корнеплоды сем. Cruciferae L. Тр. по прикладной ботанике, генетике и селекции, т. 19, 1928. C Ii H c к а я Е. Н. Краткий очерк сельскохозяйственного растениеводства в Японии. Тр. по прикладной ботанике, генетике и селекции, т. 22, вып. 5, 1930.
Си н ск а я Е. Н. К генезису культурных форм рода 1харйапиз. Тр. по прикладной ботанике, генетике и селекции, т. 26, вып. 2, 1931. С и н ск а я Е. Н. Динамика вида. М., 1948. Сии с к а я Е. Н. Классификация льна как исходного материала для се- лекции и его эволюция. Сб. работ по биологии развития и физпочогии льна, Сельхозвиз, 1954. С и н ск а я Е. Н. О составе сортовых популяций подсолнечнт~ка. Крат- кий отчет ВНИИМЭМК, Краснодар, 1955. С и н с к а я Е. Н. Происхождение пшеницы. Сб. «Проблемы ботаники», т. 2, М.— Л., 1955. С инск а я Е. Н. Видообразование и формообразование в роде Dacty- 1is. Ботанический журнал, т. 49, ЛЪ 2, 1964. С кр и п ч и н с к и й В. В. На новой родине. Ставрополь, 1954. С к р и п ч и п с к и й В. В. Драгоценные зерна. Ставрополь, 1954. С л о б од и и В. М. Развитие и смены систем земледелия до I тысячс- летия до н. э. «Материалы по истории земледелия СССР», М., 1952. С м и р п о в А. П. Очерки древней и средневековой истории пародов Среднего Поволжья и Прикамья. «Материалы и исследования по археоло- гии СССР», М~ 28, 1952. С м и р н о в А. П. и М е р п е р т Н. Я. Археологическис работы в pa i' оне строительства Куйбышевской ГЭС. Вестн. АН СССР, ~4 1, 1951. С м и р н о в А. П. и Ме р п е р т Н. Я. Из далекого прошлого народов Среднего Поволжья. Сб. «По следам древних культур от Волги до Тихого океана», М., 1954. С о б ч еи и о А. И. Литература по Западной Африке з1 !951 г. «Совет- ская этнография», № 4, 1952. С о к о л о в а И. I I. Коллекция риса в условиях дождевания. Тр. по при- кладной ботанике, генетике и селекции, т. 34, вып. 2, 1962. С ол о в ьев Л. Н. Селища с текстильной керамикой па побережье За- падной Грузии. «Советская археология», т. 14, 1950. С о л о в ь ев Л. Н. Энеолитическое селище у Очамчирского порта в Аб- хазии. Тр. Абхаз. НИИЯИ, вып. 15, 1939. С р е з и е в с к и й H. И. Материалы для словаря древнерусского языка по письменным памятникам. Т. 1, вып. 1 — 3, Спб., 1890 — 1S93. С т а н и к е в и ч Я. В. Итоги археологических работ в Великолукской об- ласти. КСИИМК, вып. 17, 1952. С т е п у г и н а Т. В. К вопросу о социально-экономических отношениях в Китае в XIV — XII вв. до н. э. БДИ, М 2, 1950. С т о л е т о в а Е. А. Полевые и огородные культуры Армении. Тр. по прикладной ботанике, генетике и селекции, т. 22, вып. 4, 1930. С т о л е т о в а Е. А. Кориа ндр. Л., 1931. С т о л е т о в а Е. А. Эколого-географическая классифпкация гречихи. до- клады ВАСХНИЛ, вып. 2, 1940. С то летов а Е. А. Гречиха. М.— Л., 1958. С т р е б к о в а А. Д. Гранат. Рукопись. С т р ж е л е ц к и й С. Ф. Отчет о раскопках раннетаврского поселения Уч-Баш, X — VIII вв. до н. э., в 1952 г., Архив ГХМ. Т а м а м ш я и С. Г. «Ископаемая» пшеница,из ССР Армении. Сб. «Рас- копки древнего Вагаршапата», Ереван, 1935. Т а н ф и л ь е в Г И. Очерк географии и истории главнейших культур- ных растений. Одесса, 1923. Т а р а к а н о в а С. А. Древности Псковской земли. Сб. «По слсдам древних культур», т. 2, М., 1953. Т и х о м и р о в N. Древнейшая русская надпись. «Литературная газета», Мо 41, 1950. Т их о м и р о в Б. А. Данные о полезных растениях эскимосов иго-восточ- ного побережья Чукотки. Ботанический журнал, т. 43, М 2, 1958. Тол сто в С. П. Древности Верхнсго Хорезма. ВДИ, М 1, 1941. Толстов С. П. Новые материалы по истории культуры древнего Хо- резма. ВДИ, М 1, 1946. 453
Т ол сто в С. П. По следам древнехорезмийской цивилизации. Изд. АН СССР, 1948. Т р е т ь я к о в П. И. Туруханский край, его пр; рода, житии. Спб 1871. Третьяков П. Н. Сельское хозяйство Ir промыыы. Сб. «История культуры Древней Руси>, М. — Л., 1948. Тр етьяко в П. Н. У истоков Древнеи Руси. Сб. «По следам древних культур», т. 2, Древняя Русь, М., 1953. Тр етья ко в П. Н. Восточно-славянские племена. М.— JI., 1953. Т р етья ко в П. Н. О происхождении славян. «Славяне», № 7, М., 1957. Т р у б н и к о в а Н. В. Племена Городецкой культуры. Тр. Государст- венного Исторического музея, вып. 22, М., 1953. Труды Волго-Донской археологической экспедиции. «Материалы и ис- следования по археологии СССР», № 62, 1958. Т у л с и Дас. Рамаяна. Перевод А. П. Баранникова. Изд. АН СССР, ? 948. Т у м а н я н М. Г. Культурные растения Урартского периода в Армян- ской ССР. Изв. АН АрмССР, № 1 — 2, 1944. Туп,и ко в а А. Ю. К изучению закавказского очага формирования по- севной вики (Vicia sativa L.). В кн.: <Вопросы эволюции, биогеографии, ге- нетики и селекции», М. — Л., 1960. Тур а е в Б. А. История Дрввнего Востока. Т. 1, Л., 1936. Т ю м е н е в А. И. Государственное хозяйство Древнего Шумера. Изд. АН СССР, 1956. Ум иков Н. 3. Плоды, ягоды, овощи, злаки и пряности. Из истории происхождения и врачебного применения их с древнейших времен. Тбилиси, 1953. Ф а н ь В э н ь-л а н ь. История Древнего Китая. Изд. АН СССР, 1958. Федор о в Ан. А. Дикие миндали Армении. Известия Армянского фи- лиала АН СССР, № 1 — 2, 1942. Ф едор о в Ан. А. Дикорастущие груши СССР и их значение для се- лекции )и гибридизации. В кн.: «Материалы Первого Всесоюзного совещания ботаников и селекционеров», вып. 1, М. — Л., 1951. Ф ед о о в Г. Б. Славяне Поднестровья. Сб. «По следам древних куль- тур», т. 2, ревняя Русь, М., 1953. Ф е д о р о в с ь к и й О. С. Археолог1чн1 розкопи в околицях Харкова. Хрон|ка археолог11 та мистецтва, ч. 1, 1930. Ф е о ф р а ст. Исследование о растениях. Перевод Сергеенко. Изд. АН СССР, 1951. Ф ил о в А. И. Огурцы мира. Сталинабад, 1948. Ф ил о в А. И. О происхождении огурца. Тр. по прикладной ботанике, генепике и селекции, т. 32, 1959. Ф и л о в А. И. Агроэкологическая изменчивость культурных тыквенных. Авто еферат диссертации, Л., 1960. и л о в А. И. К вопросу о систематике дыни. Вестн. с.-х. науки, № 1, 1960. Филов А. И. Баклажаны. <Культурная флора СССР», т. 20, )!!.— Л., 1958. Ф и л о Б А. И. Новые данные о происхождении и классификации ар- бузов (Citrullus Forsk.). Ботан~ический журнал, т. 47, № 7, 1962. Ф л ер о в К. К. По Китаю. «Природа», № 3, 1959. Ф лик с бергер К. А. Хлебные зерна из Ковшаровского городища Гри- невской волости Смоленского уезда. Научные известия Смоленского госу- дарственного ун-та, т. 3, вып. 3, 1926. Ф л я к сбе р гер К. А. Семена культурных растений VI — VIII века из- под Минска. «Природа», № 5, 1929. Ф л я к сб ер гер К. А. Древнеегипетская и современная полбы-эммеры. Тр. по прикладной ботанике, генетике и селекции, т. 19, вып. 1, 1929. Фл я кс бе р гер К. А. ТгШсит compactum antiquorum (Heer). Изве- стия Главного ботанического сада, т. 29, вып. 1 — 2, 1930.
Фл я к сбер гер К. А. Находки культурных растений доисторического периода. Тр. ин-та науки и техники, сер. 1, вып. 2, 1934. Ф л я к с б е р г е р К. А. Археологические находки хлебных растений в области, прилегающей к Черному морю. КСИИМК, вып. 8, 1940. Ф л я кс6 ер гер К. А. Зерновые хлеба в V X, ХШ вв. до н. э. на тер- ритории Латвийской ССР. Рига, 1941. Х в о й к о В. В. Начало земледелия и бронзовый век в Среднем По- днепровье. Тр. XIII археологического съезда, т. 1, 1907. Х в о й к о В. В. Древние обитатели Среднего Приднепровья и их куль- тура в доисторическое время. Киев, 1913. Хохлов С. С. Е~ентры происхождения или географическис фокусы ви- дообразования. Ботанический журнал, т. 26, № 1, 1947. Ц алкин В. И. Новая находка остатков первобытного быка в СССР. Бюлл. Московского об-ва испытателей природы, отделение биологии, т. 57, вып. 2, 1952. Lt 3 и н ь Ш а н ь - б а о. Виды китайской пшеницы и их распространение (па китайском языке с русским резюме). Сб. «Материалы юбилейной сессии ВАСХНИЛ», М., 1958. Ч а й л ьд Г. Древнейший Восток в свете новых раскопок. 5!., 1956. Чернецов В. Н., Мошинская В. И. и Талицкая И. А. Древ- няя история Нижнего Приобья. «Материалы и исследованил по археологии СССР», № 33, 1953. Ч и т а я Г. К вопросу о происхождении пахотных орудий. Сообщения ЛН ГрузССР, Мо 3 и 4, 1941. Ш а в р о в Н. Н. К познаншо огородничества и овощей в Туркестане и Закаспийской области. Спб., 1911. Ш а ф е р В. Основы общей географии растений. M., 1956. Ш ел о в Д. Б. Античный мир в Северном Причерноморье. М., 1956. Ш и ш к и н В. А. Археологические работы 1937 г. в западной части Бу- харского оазиса. Ташкент, 1940. Ш пр и п ц и н Н. Г. Индейцы гуаяки. «Советская этнография>, № 4, 1952. Ш р а де р О. Сравнительное языковедение и первобытная история. Линг- вистическо-исторические материалы для исследования индогерманской древ- ности. Спб., 1886. Шр адер О. Индоевропейцы. Перевод Ф. И. Павлова. Спб., 1913. Ш те й н бе р г Е. М. По Судану, и Эфиопии. «Природа», № 12, 1960. Ш у н к о в В. И. Географическое размещение сибирского земледелия в XVII веке. «Вопросы гвографии», сб. 20, М., 1950. Ig е н к о в а М. С. Географическая изменчивость и центры происхожде- ния кресс-салата. Тр. по прикладной ботанике, генетике и селекции, сер. IX, N~ 1, 1932. III, е н к о в а М. С. Дикорастущие многолетние кормовые травы Коми АССР в естественных условиях. М.— Л., 1961. Эди н г Д. Н. Береговая стоянка Горбуновского торфяника. Государст- венный историческии музей. Археологическая сессия Ученого совета 19— 22 марта 1941 г. Тезисы докладов. М., 1941. Э л л е н б е р г е р В. Трагический конец бушменов. Изд-во иностранной литературы, 1956. Ю дки н а О. Н. Вопросы древней истории на сессии АН СССР. ВДИ, № 2, 1948. Ю й Дэ-цз юн ь, Я нь Чже EI ь-лун. Изучение китайских видов рода Malus Mill. Чжиу феньлэй сюэвао. Acta Phytotaxon. Sinica, чо1. 5, М 2, 1956 (кит., рез. англ. Реферативный журнал, вып. Биология, Л' 40 — 537, 1957) . Я к о в л е в Н. Ф. Кавказская лингвистическая экспедиция. Вестн. АН СССР, № 11, 1949. Я к и м о в В. П. Ранние стадии антропогенеза. Сб. «Происхождение чсловека», изд. АН СССР, 1951. 455
Я к у бц и не р М. М. К познанию дикой пшеницы Закавказья. Тр. по при- клддиой ботанике, генетике и селекции, сер. 5, вып. 1, 1932. Я к у б ц и н е р М. М. О составе зерновых культур из Старой Ладоги. ЕСИИМ1~, вып. 57, 1955. Я к у б ц и н е р М. М. К истории культуры пшеницы в СССР. В кн.: «~Чатер«азы по истории земледелия СССР», т. 2, М. — Л., 1956. Я к у б ц и н е р М. М. Новые ботанические формы пшеницы, ценные для селекции. Бюлл. ВИР, № 2, 1956. Я ку бци нер М, М. Новые виды пшеницы. Вестн. с.-х. науки, № 12, 1959. Я EI и н В. Л. Великий Новгород. Сб. «По следам древних культур>, т. 2, Древняя Русь, М., 1253. А b e r g Е. Cereals and peas from Eastern Tibet and their importance for the knowledge of the origin of cultivated plants. Landbrukshogskolans аппа1ег (Annals of ihe Royal Agric. College of Sweden), чо1. 15, 1948. А n d e r s о п Е. and В l а п сh а r d F. D. Prehistoric maize from Canon del Muerto. Amer. Journ. Bot., vol. 29, No 1О, 1942. А п d er s s о п G. Die Geschichte der Vegetation Schwedens. Englers Bota- nische Jahrbucher, В. 22, 1897. Аn d e r s sо и G. und В i r ge r S. Die Geographische Verteilung und die Einwanderungsgeschichte der nordskandinavischen Flora. Englers Bot. Jahrb., Bd. 51, 1914. А r k e l l А. J. Early khartoum. Oxford, 1949. Ar k e l l А. J. А history of the Sudan from the earliest times to 1821. Lond on,1935. А i п а u d о w N. Pflanzenreste aus einer prahistorischen Siedlung in Sudbulgarien. Osterreichische Bot. Zeitschrift, Bd. 88, Н. 1, 1939. В а i l е у Ь. Н. Standart cyclopedia of horticulture. London, 1922. В а i l еу L. Н. The domesticated Cucurbitas. I. Gentes Herbarum, vol. 2, fasc. 2, Ithaca, 1929. В а i l е у L. Н. А medley of pumkins. Mem. hort. soc. of New Yorl<, 1920. В а k th е уе ч F. КЬ. А fossil form of cultivated barley Hordeum lagun- culiforme mihi. Annals of the royal agricultural college of Sweden, чо1. 2Э, 1957. В а и m g а r t e l Е. The cultures of prehistoric Egypt. London, 1947. В е с k e r - D i 11 i n g e n J. Handbuch des Getreidebaues. Berlin, 1929. В е r t s c h К. Die vorgeschichtlichen Wildreben-Funde Deutschlands. Berichte der Deutschen Botanischen Gesellschaft, Bd. 57, Heft 9, 1939. В е r t s c h F. Entwicklung und Herkunft unserer Getreide. Mannus, Bd. 31, 1939. В l а у d e s G. W. The romance of domesticateds plants. Annual Яер1. Board legends Smithsonian Instn. Washington, vol. 54, 1954. В о i s D. Les plantes alimentaires chez tous les peuples et а travers les ages. Paris, 1927. В о i s s i e r Е. Flora Orientalis. Basel, 1867 — 1884. В r а п d D. D. The origin and early distribution of New World cultivated plants. Agricultural history, April, 1939. В re t s п еi d er E. Botanicon sinicum. Part I — III, Londcn, 1882 — 1895. В r о w п Т. W. Cucurbitaceous fruits in Egypt. Journ. of Я. Horticultural Society, vol. 35, 1909 — 1910. В r о г п W. Peisen in Afrika, Egypten und Syrien. Weimar, 1800. В u r k а r t. Les Leguminosas Argentinas silvestris y cultivadas. Buenos- Ayres, 1953. В u s c h а n G. Vorgeschichtliche Botanik der Kultur- und Nutzpflanzen der alten Welt auf Grund prahistorischer Funde. Breslau, 1895. С а гп р W. Н. А possible source for American precolumbian gourds. Amer. Journ. Bot., vol. 41, No 9, 1954. С а r г i e r L. The beginnings of agriculture in America. London, 1923. С h а k r а ч а r t у Н. L. Studies on Indian Cucurbitaceae. Journ. of agri- cultural science, vol. 16, No 1, 1946. 456
Ch а ц gu1е В. А. and КЬ ot В. D. Four years with Suran. Indian Farm., vol. 7, No 9, 1957. С h е е s m а п Е. Е. Classification of йе bananas. Critical notes on species. Кебич Bulletin, N 1 — 3, 1948; N 1 — 4, 1949; N 1, 2, 195О, London. С h е ч а 1i e r А. Legumineuses fourageres cultivees dans ГInde anglaise. Revue intern. bot. appl., t. 4, 1924. Ch ev а l i er А. Les productions vegetables du Sachara et de ses con- fins N. et S. Passe, Present, Avenir. Revue bot. appl., 12 аппее, N 133 — 134, 1932. С h е ч а1i e r А. Les origines et Гечо1аЫоп de Гagriculture тей1егапее. Revue Bot. appl., 19е annee, N 217 — 218, 1939. С he vа1i e r А. Revision du genre Theobroma, сГapres ГНегЫег du Mu- seum d'Histoire naturelle de Paris. Revue intern. de bot. Appl. et d'agric. tropic., 26' аппее, N 285 — 286, 1946. С h еч а1i er А. D'îu viennent les Pommiers а bons fruits de nos jardin' et de nos vergers? Comptes rendus des seances de ГАсайеппе des Sciences, t. 234, N 22, Paris, 1952. С h е ч a l i e r А. L'origine. des poiriers et pommiers sauvages de nos forets et la part qu'ils ont prise dans la formation des varietes cultivees. Comptes rendus de ГАсаdemie des Sciences de Paris, 237, 1953. С h е ч а1 i e r А. Labiees du genre Coleus Loureiro cultivees dans les pays tropicaux de ГАпс1еп Monde. Revue intern. de bot. appl. et d'agric, tropic. Аппее 33п", N 369 — 370, 1953. С l а r k J. Prehistoric Europe. The economic basis. London, 1952. С о о k F. Peru as center of domestication. Journ. of Heredity, vol. 16, No 2, 1924. D ay G. М. The Indian as an ecological factor in the northeastern forest. «Ecology», vol. 34, No 2, 1953. D е 1 i 1 е А. Memoires botaniques extraits de la «Description de ГЕдур1е». П. Histoire des plantes cultivees en Egypte. Paris, 1813. Dictionnaire etymologique de la langue grecque. Heidelberg — Paris, 1916. D u c k е А. The genus Manilkara Adans. in Brazil. J. Linnean soc. Lon- don, vol. 55, No 362, 1957. D u ч е у r i e r Н. Les Touareg du Nord. Paris, 1864. Е i g А. Monographisch-Kritisch Ubersicht der Gattung Aegilops. Berlin, 1929. Е i g А. Ecological and phytogeographical observations on Palestine plants. Beihefte zum Botanischen Centralblatt. Bd. 50, Abt. 2, 1933. Е rm а n n А. Aegypten und aegyptisches Leben im Altertum. Tubingen, 1885. Е r w i n А. Т., L а п а Е. P. The seminola pumpkin. Economic Botany, vol. 10, No 1, 1956. F i е 1 d H. Field Museum-Oxford University Joint Expeditions to Pish from 1923 to 1929. Field Museum Anthropology Leaflet, N 28, Chicago, 1929. For sk а l P. Г1ога aegyptiaca-arabica sive descriptiones plantarum, quas per Aegyptum ш~епогет at Arabiam felicem detexit. Havniae, 1775. Г.ur о п R. Мanuel de prehistoire депега1. Paris, 1932. G а1е п о s. De alimentorum facultatibus liber secundus. Lipsiae et Bero- lini, 1923. G а 1i п а t %. С., М а n g e l s d о r f P. С., P i e r s о n L. Estimates of Teosinte introgression in archaeological maize. Bot. Museum Leaf 1., 17, No 4, 1955. G e а d w i п Н. S. Men out of Asia. New York, 1947. G i b а u l t G. L'histoire des legumes. Paris, 1912. G i 11 N. Т., P 1>. D. The origin of our garden crops, «The Gardeners' chronicle», vol. 4, third series, London, 1939. G i l m о r e M. К. Uses of plants of Indians. Ann. Report. Bur. of Amer. Ethnology, 1911 — 1912. G r а h 1е А. Fremdlandische Pflanzen im Haushalt. 6. Igaffe. I(osmos, Bd. 51, N б, Stuttgart, 1955. 457
Н а rl an I. Crops, weed and evolution. Sci. Monthly, No 5, 1955. H at t G. Landbrug i Danmarzs Oldtid. КвЬпйачп, 1937. Н au dricou r t А. G. et Н ed in L. Ь'Ьоп1п1е et les plantes cultivees. 7-me edition, Paris, 1943. Н au dr icou r t А. G., H ed in L. Pecherches recentes sur Г111я1о1ге des plantes cultivees. Яеч. internat. bot. appl. et agric. trop., аппее 33, N 373 — 37-, 1953. Н а wk e s I. G. Potato collecting expeditions in Mexico and South Аше- rica, Cambridge, 1944. К е d r i ck U. P. Sturtevant's notes on edible plants. Л1Ьапу, 1919. Н е е r О. Die Pflanzen der Pfahlbauten. ZQrich, 1865. Н ее r О. Uber den Flachs und Flachskultur im Altertum. ZQrich, 187 . Н е g i G. Illustrierte Flora von Mittel-Europa. MQnchen, 192'1' — 1923. Heh n V. Qulturpflanzen und Haustiere in ihrem Uebergang aus Asien nach Griechenland und Italien sowie das Qbrige Europa. 8. Auf1., Berlin, 1911. H e i п е - G e l d e r n Я. Qulturpflanzengeographie und das Problem vor- kolumbischer Kulturbeziehungen zwischen Alter und Neuer Же11. Anthropos, ~ »1. 53, fasc. 3 — 4, 1958. Н е l Ь а e k Н. Spelt (Triticum spelta L.) in Bronze age Denmark. Acta archaeologica. Bd. 23, 11952. H e m s 1 е у W. В. Biologia Centralia-America of contributions to Йе knowledge of the fauna and flora Mexico and Central America. Botany, vol. 1 — 4, London, 1879 — 1888. Hens1ow G. Origin and history of our garden vegetables and their dietetic values. Journ. Я. Hort. Soc., чоl. 36, 1911. Н r о z n y Fr. Sur quelques rapports entre Summer-Akkad et 1'Едур1е а IV millenaire avant. J. С. Arch. Orient., vol. 8, 1938. Н ro zny Fr. Das Getreide in alten Babylonien. Wien, 1914. Hutchinson J. В., Silow Я. А., Stephens S. G. The evolution of Gossypium. London, 1947. J а е g e r P. Note provisoire sur le role des Chauves-Souris dans la dissemination des fruits du КагИе (Butyrospermum parkii Ко1.), Ви11. Inst. 1гапс. Afrique noire. $ег1е А, t. 24, N 1, 1962. Ке] mer L. Die Gartenpflanzen im alten Agypten. Bd. 1, Berlin, 1924. pih а r а Н. Considerations on the evolution and distribution of Aegilops based on the analyser-method. Cytologia, vol. 19, 1954. К ih а r а Н. Japanese Expedition to the Hindukush (The native place of бх-ivheat). First International ~vheat Genetics Symposium. Canada, 1958. К i d d e r А. V., G u er n se у $. I. Archeological explorations in North- Eastern Arizona. Smithsonian Institute on Bureau of Am. Ethnology, Bull. 65, 11949. gris hnamurthi S., S esh а dr i Ч. S. Origin and evolution of cultivateds bananas. Proceed. of Internat. Symp. on Origin, Cytogenetics and Breeding of tropical fruits. Indian J. of Horticulture, vol. 15, No 3 — 4, 1958. К u ck u ck Н., S ch i em апп Е. Uber das Vorkommen von Spelz und Еттег (Triticum spelta Ь. und Тг. dicoccum Schiibl.) im Iran. Zeitschr. PflanzenzQchtung, Bd. 38, Н. 4, 1957. L а m о nt Р., В er bi g ie r А. Ь'alfa et 1'experimentation alfatiere en Algerie. gevue intern. bot. appl. et сГagric. trop., N 365 — 366, 1963. L а u re n t- T ac kh ol m V. А mummy coffin in the Egyptian museum, Stockholm, and its plant remains. Svensk Botanisk Tidskrift, Bd. 34, Н. 1940. Ь- eg ge J. The Chinese classics, vol. 4, Book XV. Second edition, Lon- don, 1871'. Le g get W. F. The story of linen. New York, 1945. L е г oi J. F. Notes geographiques виг les поуегв tropicaux (Juglans et Сагуа). Revue intern. bot. appl. et d'agric. trop., N 367 — 368, 1953. Ь i n d e n b e i n Ю. Ursprungszentrum, Igulturstromverbreitung und Ein- zelwanderung bei kultiyierten Andropogon — Arten. Bot. Jahrb., Bd. 71, Stutt- gart, 1941 °
L i р р т а п п Е. О. Geschichte der Rube (Beta) als gulturpflanze. Ber- 1in, 1925. L iч i n g st о п е D. Missionary travels and researches in South Africa, London, 1857. Lore t V. La Ноге pharaonique d'apres les documents hieroglyphiques et les specimens decouverts dans les tombes. Paris, 1892. L о w I. В. Arameischen Pflanzennamen. Leipzig, 1881. L un d m an В. The Problem of ancient oriental shipping in the North See. INES, No 2, 1957. Ма l zy P. Quelques plantes du nord Сатегип et leur utilizations. Journ. сГagric. appl. et de bot. арр1., N 5 — 6, 1954. М а п g е1s d о r f P. С,, S m i t h С. J. А discovery of remains of primi- tive maize in New Mexico. Journ. of Heredity, vol. 40, No 2, 1949. М а n g el s d о гf P. С., S m i t h С. J. New archaeological evidence on evolution in maize. Bot. Mus. Leaf 1. Harvard. Univ. 13, 1949. М а п g e l s d о г f P. С. Hybridization in the evolution of maize. «Hete- rosis», ed. J. Gowen, 1952. M а п g e l s d о r f P. С., Ьi ster Я. H. Archaeological evidence on the evolution of maize in Northwestern Mexico. Bot. Museum Leaf 1. 17, No 6, 1956. Mar quet -Кг au s e J. 1 а deuxieme campagne de fouilles а'Ау. В. Sy- ria, t. 16, Paris, 1935. М а s р е r о G. Histoire ancienne des peuples de 1'Orient. Paris, 1893. М а 1 1 а k о ч п а M. Weitere Untersuchungen uber mittelalterisches Get- reide aus Littauen. Acta Soc. Bot. Polon, vol. 6, N 4, 1929. N c F а d d е п Е. S., S еа rs E. К. The origin of Triticum spelta and its free-threshing hexaploid relatives. The Journ. of Heredity, чо1. 37, No Э, 4, 1946. М eissne r В. Babylonien und Assyrien. 1. Heidelberg, 1920. N o e n s P. La place systematique du Coffea Lebruniana Germain et Кеь- lег. Bull. Jardin bot. etat, vol. 32, N 1, 1962. М о l d е п h а w e r К. Nowa forma zyta kopalnego (Secale сегеа1е 1. ssp. ancestrale Zhuk. var. afghanicum Vav.) w wykopalisku na ziemiach Polski. Acta agrobot., N 7, 1958. М о r g а п J. Les premieres civilisations. Paris, 1909. Nu k h e r j е е S. К. The origin of Mango. Proceed. of Internat. Symp. ог Origin, Cytogenetics and Breeding of tropical fruits. Indian J. of Horticul- ture, чо1. 15, No 3~4, 1958'. Ми sс hlе r P. А manuel flora of Egypt. Vol. 1 — 2, Berlin, 1912. N au d i n Ch. Origin supposee des gourges et des potirons. gevue hor- icoIe, serie IV, t. 4, 1855. Р а r k e r А. С. Iroquois uses of maize and other food plants. St. Mus. Ви11., vol. 144, No 9, 191О. P а r o d i L. Pelaciones de la agricultura prehispanica con la agriculture Argentina actual. Annalas de la Academic national de agr., t. 1, Buenos-Aires, 1935. P е а ke Н. J. The beginnings and the early spread of agriculture. «Na- ture>, чо1. 1'19, No ЗО07, 1927. P е rci ч а l J. Cereals of Ancient Egypt and Mesopotamia. «Nature>, vol. 138, No 3485, 1936. P f e i f e r I. Die Werkzeuge des Steinzeit Menschen. Jena, 1920. P i р е r С. V. Studies in American Phaseolinae. Contr. U. S. Nat. Herb., чо1. 22, 1926. Р l i n i i С. Secundi. Historiae Naturalis. Libri XXXVII, чо1шпеп sextum, Parisii, 1829. Рorsi l d А. Е. Plant life in the Arctic. How plants survive in the Ear North. Plants and Gardens, Baltimore. Md., vol. 7, No 4, 1951. P о r s i l d А. E. I.~seful plants in the Canadian Arctic Archipelago. Eco- nomic Botany, vol. 13, No 1, New York, 1959. 459
P о r t er e s P. С. Taxonomic, agrobotanique des riz cultives О. sativa L. et О. glaberrima Steud. J. Agric. trop. bot. арр1., vol. 3, N 7 — 12, 1956. P и m р е11у R. Prehistoric civilization of Апац I, Washington, 1908. Я а m i а h К., Gh os е R. L. Origin and distribution of cultivated plants of South Asia. Rice. Indian J. Gen. and plant breeding, vol. 11, No 1, 1951'. Р е е d Н. S, А short history of the plant sciences. California, 1942. R i ч а l s P. Le dolique d'Egypte or Lablab (Dolichos lablab L,). Revue intern. bot. appl. et d'agric. trop., N 373 — 374, 1953. S а m р а t h S., N а r а s i n g а R. N. В. Interrelation between species in the genus Oryza. Indian J. genetica and plant-breeding, vol. 11, No 1, 1951. S а f f о r d W, Е. Our heritage from the American Indians. Ann. Smith- sonian Institution, 192б. S c h i e m а п и Е. Neue Probleme der Gerstenphylogenie. Der Zuchter, Bd. 11, Н. 6, 1939. S c h i e m а п п Е. Einkorn in Neolithicum Danemarks. Вег. Deutsch. Bot. Gesell., Bd. 57, Н. 10, 1940. S ch ie m а n n Е. Weizen, Roggen, Gerste. Jena, 1948. S сli i e in а пп Е. New results on the history of cultivated cereals. He- redity, vol, 5, 1951. S c h i e in а n n Е. Einkorn im alten Aegypten. Der Ziichter, Bd. 24, N 4 — 5, 1954. S c h r а d e r H. А. Enumeratio plantarum Africae australis extratropicae. Paris, 1836. S c h u1 z А. Der Emmer des alten «Aegyptens». Ber. Deutscl>. Bot. Ge- sell., Bd. 36, 1916. S c h w e i n f и r t h G. Vber Pflanzenreste aus altaegyptischen Grabern. Berichte deutsch. bot. Gesell., Berlin, 1884. S m i t h P. G., Н e i s e r Ch. В. Jr. Taxonomy of Capsicum sinense Jacq. and the geographic distribution of the cultivated Capsicum species. Bull. Torrey Bot. Club, vol. 84, No б, 1957. S о1ms - L а ub а с h Н. Weizen und Tulpe und deren Geschichte. Leip- zig, 1899. S t eb b i и s G. Types of polyploids: their classification and significance. Advances in Genetics, vol. 1, 1947. S t e h l е Н. Etude botanique et agronomique des Leguniineuses autochto- nes et exotiques des genres Сапача11а, СИопа et Cro.olaria aux Antilles Fran- caises. Revue intern. bot. appl. et d'argric. 39 annee, N 373 — 374, 1953. S t e h 1 е Н. Especes rares on spectaculaires de la Flora des Antilles frant„aises шепасеея de disparition et rnesures а envisager en vue de leur pro- tection. Bull. du Museum (Paris), 2' ser,, t. 28, N 1, 1956. S t u h l m а п n F. Beitrage zur I<ulturgeschichte чоп Ostafrika. Berlin, 1909. T а п а k а Т. The origin and dispersal of citrus fruits Iiaving their centre of origin in India. Proceed. of the Intern. Symp. on Origin, Cytogenetics and Breeding of tropicai fruits. Indian J. of Horticulture, vol. 15, No 3, 4, 1958. Т ch а1е п ko G. Villages antiques de la Syrie du Nord. Le massif de Belus а Гepoque romaine (lnstitut franglais d'archeologie de Beyrouth, Bib- liotheque archeologique et historique). Т. 1 —, 1953; t. 3, 1958, Paris. Т i n g I. С., К eh r А. Е., Mi l l е г I. C. А cytological study of Йе sweet potato plant lpomoea batatas (L.). Lam. and its rel~ated species. Amer. Naturalist, vol. 91, No 858, 1'957. T о i ч о п е п М. Samoilemme huonekasvien syntymaseutuja. Mista huo- nekswimme ovat Rotoisin. Puntarha, N 1, 1958. U n g e r F. Botanische Streif7uge аш сЬ1п Gebiete сапог g~ilturgeschichte. Sitz. Ber. Akad. Wien, 1866. V ac e J., Th о m p s о п. Prehistoric Thessaly, Cambridge, 1919. V e s t а1 P. А. Cucurbita moschata found ш pre-Columbian mounds i» Guatemala. Bot. Mus, Leaf 1. Harvard Univer., vol. 6, No 4, 1938. W e l l h а u s е п Е. Heterosis in а new popula'ions. <Hetervsis ed. J Gowen. Ames. Jowa, 1952.
Wer th Е. Emmer und Gerste aus dern 5 Jahrtausend ч. С. und andere vorgeschichtliche Kulturpflanzenfunde. Berichte Deutsch. bot. Gesell., Bd. 67, Н. 8, 1939. We r t h Е. Zur Geographic und Geschichte der Hirsen. Angewa»dte Bot., Bd. 19, Н. 1, 1937. W e r t h Е. Abstammung und Heimat des getiich. Angewandte Bot., Hd. 19, 1937. Wh е е1е r,М'. The Indus civilization. Cambridge, 1953. W h i t а 1' е r Th. Origin of cultivated Cucurbita. Amer, Na.uralist, vol. 90, No 852, 1956. Wh i t а k er Th., С а r t er G. Н. Critical notes of the origin and do- mestication of the cultivated species of Cucurbita. Amer. Journ. Botany, vol. ЗЗ, No 1', 1946. W И t а ke r Th. and В о h n G. The Taxonomy Genetics, Production and Uses of cultivated species of Cucurbita. Econ. Bot., vol. 4, No 1, 1950. Whi t а ker Т., С а r t er G. Oceanic drift of gourds experimental ob- servation. Amer. Journ. Botany, vol. 41, No 9, 1954. W i s s l e r С. The american Indians. 2nd edition. New York, 1922. Wi t t m а с k Ь. Our present knowledge of ancient plants. Trans. Akad. Sci. of St. Louis, vol. 15, No 1, 1905. Wo n i g Fr. Die Pflanzen im alten Aegypten. Leipzig, 1886. Z а f а г А. Nomenclature of o}eiferous Brassicas cultivated in the Punjab Indian Journal of agricultural Science, part 4, vol. 15, 1945.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ Abroma augusta Ь. 282 Abutilon theophrasti Medic. 246, 262 Acacia scyal Balanifer 330 — senegal (Ь.) Willd. 330 — tortilis Hayne 330 Acanthosicyos horrid a Welw. 348 Acomastylis rossii Greene 226 Aconitum L. 25, 261 — wilsonii Stapf 248 Actinidia Lindl. 223, 258, 260 — arguta (Sieb. et Zucc.) Planch. 223, 258 — chinensis Planch. 223, 258 — giraldii Diels 223, 258 — kolomicta Maxim. 223, 258 — polygama (Sieb. et Zucc.) Maxim. 223, 258 Actinostemma paniculatum Maxim. 250, 261 Adansonia digitata Ь. 347 Adenophora latifolia Fisch. 253, 261 — verticillata Fisch. 253 Adonis Ь. 219, 376 Aegilops Ь. 20, 21, 48, 79, 95, 135, 390 — biuncialis Vis. 96 — caudata Ь. 96 — columnaris Zhuk. 96 — comosa Sibtli. et Sm. 96 — cylindrica Host 79, 96 — mutica Boiss. 96 — ovata Ь. 96 — speltoides Tausch 96 — squarrosa Ь. 79, 96, 390 — triuncialis Ь. 79, 96 — uniaristata Vis. 96 — variabilis Eig 96 — ventricosa Tausch 96 Aegle marmelos Correa 293 Agave Ь. 417 — atrovirens Karw. 417 — ixtli garw. 417 — lechygilla Torr. 417 — sisalana Perrine 417 Agropyron Gaetn. 101, 135, 136, 150 — cristatum (L.) Gaertn. 101, 138,221 — dagnae Grossh. 101 — desertorum (Fisch.) Schult. 221 — pinifolium Nevski 72, 101 462 Agropyron ponticum Nevski 101 — риЬеги!ит (Boiss.) Prokud. 101 — sibiricum (Willd.) Beauv. 22! Agrostis Ь. 150 — stolonifera Ь. 307 — tolucensis Н. В. К. 418 Aleurites Forst. 245 — cordata R. Br. 245, 262 — fordii Hemsl. 245, 262 — moluccana Willd. 282 Allium L. 249, 321 — ampeloprasum Ь. 124, 321 — ascalonicum Ь. 204, 321 — сера L. 124, 160, 164, 204, 321, 375 — chinense Don. 249, 261 — fistulosum Ь. 204, 249 — macrostemon Bge. 249, 261 — pekinense Prokh. 249, 261 — porrum Ь. 124, 321, 375 — pskemense В. Fedtsch. 160 — sativum Ь. 124, 160, 164, 321, 375 — schoenoprasum Ь. 204, 223 — sibiricum Ь. 225 — ursinum L. 403 — vavilovii М. Pop. et Vved. 160 Alocasia macrorrhiza (Ь.) Schott 291 Aloe L. 351 — africana Mill. 351 — arborescens Mill. 351 — perfoliata Neyen 351 — ferox Mill. 351 Alopecurus pratensis L. 22I Althaea rosea (Ь.) Сач. 260 Alternanthera amoena Voss 435 — versicolor Hort. 435 Amaranthus Ь. 276, 438 — anardana ВысЬ.-Нат. 276 — ЬИшп Ь. 276 — caudatus Ь. 429 — frumentaceus Buch.-Ham. 276 — gangeticus Ь. 276 — paniculatus Ь. 416 — speciosus Sims 276 — tricolor Ь. 276 Amelanchier alnifolia Nutt. 437 Amomum Roxb. 285 — aromaticum Roxb. 285 — echinosphera К. S chum acher 285
Amomum krervanh Pierre 285 — sub ulàtum Roxb. 285 Amorphophallus konjak С. Koch 253, 261 — campanulatus (Roxb.) В!ите 291 Ampelodesma tenax Link 359, 379 Amygdalus Ь. 131, 136, 161, 255, 256, 260 — bucharica Korsh. 161 — communis Ь. 131, 139, 160, 256 — fenzliana (Fritsch) Lipsky 131, 161 — nairica Fed. et Takht. 131 — ulmifolia М. Рор. 161 — urartu Tamamsch. 131 Ananas comosa Ь. 433 Androsace L. 24 Anemone Ь. 378 — яЬЫса Ь. 226 Апейшп graveolens L. 376 — sowa Roxb. 291 Апопа Ь. 350, 424 — senegalensis Pers. 350 Anthriscus cerefolium (L.) Hoffim. 122, 139, 373 — trichosperma Spreng. 122 Antiaris toxicaria Lesch. 235 Antirrhinum Ь. 378 — majus L. 404 Ап1госагуоп micra ster А. Chev. et Guill. 350 Apium graveolens Ь. 322, 372 Аросупшп саппаЬ1пшп L. 436 — hypericifolium Ait. 436 — lancifolium Russan. 154 — scabrum Russan. 154 — sibiricum (Pall.) Russan. 195 Aquilegia Ь. 25, 437 Arachis Ь. 433 — hypogaea Ь. 411, 433 — monticola Кгар. et Rig. 433 — rasteiro А. Chev. 433 — sylvestris А. Chev. 433 — villosa Benth. 433 Aralia cordata Thunb. 251, 261 Araucaria imbricata Pav. 434 Arctous alpina (Ь.) Nied. 226 Areca catechu Ь. 297, 299. Arenga saccharifera Labill. 297, 299 Armeniaca Mill. 131, 255, 256, 260 — anomala Kostina 256 — ansu (Кот.) Kostina 224, 256 — dasycarpa (Ehrh.) Pers. 161 — davidiana Carr. 256 — hnlosericea (Batal.) Kostina 24, 255 256 — leiocarpa Kostina 161 — manshurica (Koehne) Skvortz. 224, 256 — mume Sieb. 24, 256 — sibirica (Ь.) Lam. 224, 256. Armeniaca vulgaris Lam. 131, 140, 161, 196, 224, 256, 377 Arracacia Bancroft 432 — esculenta DC. 46 — xanthorhiza Bancroft 432 Arrhenatherum elatius (L.) Nert. et Koch 100, 138 Artemisia dracunculus Ь. 116 Artocarpus Forst. 295 — chaplasha Roxb. 295 — incisa Ь. 295 — integrifolia Ь. 295 — lakoocha Roxb. 295 Arundinaria Michx. 249 Asarum sieb old ii Miq. 248 Asimina triloba (Ь.) Dun. 437 Asparagus lucidus Lindl. 250, 261 — officinalis L. 373 — tenuifolius Lam. 373 — verticillatus Ь. 260 Astilbe davidii Henry 260 Astragalus sinicus Ь. 242, 261 Atriplex Ь. 385 Atropa caucasica Кгеуег 135, 137 Avena Ь. 98, 136, 238, 358 — abyssinica Hochst. 43, 44, 334, 342 — barbata Pott. 98, 358 — byzantina С. КосЬ 76, 98, 99, 358, 359 — fatua Ь. 98, 198, 385, 392 — hirtulа Lag. 358, 392 — ludoviciana Dur. 98, 99 — nuda Ь. 238, 262 — sativa Ь. 39, 98, 99, 197, 198, 221, 273, 358, 359, 384, 392 — sterilis L. 76, 98, 358 — strigosa Schreb. 40, 197, 198, 358, 384, 392 — vaviloviana Mordv. 333, 342 — wiestii Steud. 358 Averrhoa carambola Ь. 296 — bilimbi Ь. 296 Balsamea myrrha Baill. 318, 339 Bambusa Retz. 249 Basella rubra Ь. 291 Begonia gracilis Н. В. К. 418 — semperflorens Link 435 — rex Putz. 299 Benincasa hispida Со п. 290 Bergenia crassifolia Ь.) Fritsch 219 Bertholletia Humb. et Bonpl. 46 — excelsa Humb. et Bonpl. 106, 434 Beta Ь. 36, 119, 135, 370, 380 — cicla Ь. 371 — corolliflora Zoss. 121 — intermedia Bunge 121 463
Beta lomatogona Fisch. et Mey. 121 — maritima L. 119, 120, 370, 372, 400 — orientalis goth 291, 321, 370 — patula (Soland) Ait. 370 — trigyna %. et К. 121 — vulgaris L. 77, 119, 139, 158, 205, 370, 399 Bixa orellana? . 423 Blighia sapida l<oenig 350 Blitum capitatum L. 375 — rubrum С. А. Меу. 375 — virgatum L. 375 Boehmeria nivea Gaud. 246, 262, 281 — tenacissima (Roxb.) Gaud. 281 Bombax malabaricum DC. 281 Borassus flabellifer Ь. 297 Boswellia papyrifera Hochst. 330 Bouea burmanica Griff. 296 Brassica L. 28, 41, 42, 250, 282, 338, 380 — alboglabra Bailey 250, 261 — atlantica О. Е. Schulz 368 — campestris L. 19, 78, 113, 139, 156, 202, 223, 244, 245, 250, 262, 282, 283, 398 — capitata (Ь.) Lizg. 122, 139, 203 — carinata А. Braun 43, 44, 284, 332, 338, 352 -- cauliflora Lizg. 369 — caulorapa Pasq. 122 — chinensis L. 245, 250, 261, 290 — сЫпо1еИега Viehoever 245, 250 — cretica Lam. 368, 369 — dubiosa Bailey 250, 261 — glauca Wittm. 283 — incana Tenore 368 — insularis Noris 368 — juncea (Ь.) Czern. 19, 22, 78, 156, 201, 223, 244, 245, 261, 283, 284 — montana Pourr. 368 — napiformis Bailey 244, 261, 284 — napobrassica Mill. 19, 203, 369, 399 — napus L. 19, 283, 368, 369, 400 — narinosa Bailey 250, 261 — nigra Косй 19, 76, 113, 139, 20?, 203, 244, 245, 283, 284, 338, 368, 397 — nipposinica Bailey 245, 250, 261 — oleracea Ь. 19, 250, 338, 368, 369, 399, 401 — pekinensis gupr. 245, 250, 261, 290 — гара Ь. 164, 369, 398 — rapoasiatica Sinsk. 157, 164, 250, 290, 398 — rapoeuropea Sinsk. 118, 139, 164, 203, 398 — rupestris Pafin. 368 — tournefortii Gouan 284 — trilocularis Hook. et Thompson 283 464 Brassica villosa Biv.-Bern. 368 Bromus L. 100, 150, 438 — erectus Huds. 395 — inermis 1.abyss. 220 — mango Desv. 411, 434 — richardяопИ Link 221 — riparius Pehm. 100, 138 — sibiricus Drob. 221 — variegatus М. В. 100 Broussonetia papyrifera Ь'Нег. 247 — kazinoki Sieb. 247 Вutyrosf ermum parkii potschy 343 Cajanus DC. 277, 299 — ш й cus S pren g. 277, 315 Calamus rotang L. 268 Calendula 1. 378 Call istephus chinensis (Ь.) Nees 260 — sinensis Bergmans 260 Calotropis gigantes (L.) R. Br. 281 Calystegia chinensis Bailey 260 Camelina Crantz 113, 135 — caucasica (Sinsk.) Vass. 64, 113, 136 — crepitans Sinsk. 22, 25 — linicola Schimp. et Sperm. 27 — microcarpa Andrz. 64, 222 — sativa (L.) Crantz 202, 222 — silvestris Wallr. 72, 202 Camellia j aponica L. 245 — sasanqua Thunb. 245 Campanula rapunculoides L. 403 Cananga odorata (Lam.) Hook. et Thorns. 285 Canavalia Adans. 279, 299 — ensiformis DC. 279 — gladiata DC. 279 — lineata DC. 279 — podocarpa Dunn. 279 — virosa W. Arn. 279 Canna L. 300, 418 — edulis Кег-Gawl. 411, 428 Cannabis indica (Ьат.) Serebr. 163, 164, 282 — sativa Ь. 112, 138, 153, 201, 246, 282 Capparis decidua Edgew. 330 Capsicum angulosum МИ1. 432 — annuum L. 422, 431, 432 — сошсшп Neyer 431 — co 1 umbi anu>n H a zenb. 432 — frutescens L. 422, 431, 432 — mexicanum Hazenb. 422 — pendulum Willd 431 — pubescens Quiz et Pav. 431, 432. — sinence Jacq. 431, 432 Carex malaecensis Lam. 247, 262 Carica papaya Ь. 424
Carthamus L. 114, 115, 135, 319, 379 — arborescens Ь. 114 — helenioides Desf. 114 — lanatus L. 114 — oxyacantha М. В. 114, 115 — tinctorius L. 114, 115, 139, 154, 164, 282, 318 Carum ajowan Вепй. et Hook. 156 — carvi L. 375, 402 — copticum Benth. et Hook. 339 — roxburghianum Benth. et Hook. 291 Сагуа Nutt. 257, 424, 433 — cathayensis Sarg. 258, 262 — illinoensis (Wang.) Koch 424 — myristicaeformis Nutt. 424 — olivaeformis Nutt. 436 — ovata (Mill.) Koch 424 — palmeri Manning 424 Caryota urens L. 298 Casimiroa edulis Ьа Llave 424 Cassia acutifolia ВеИе 436 — angustifolia Vahl 287 — fistula L. 300 Castanea МИ1. 135, 257, 260 — crenata Sieb. et Zucc. 257 — dentata Borkh. 436 — moll issimà В1. 257 — sativa Mill. 135, 140, 378 Castanopsis indica DC. 296 Catha edulis Forsk. 44, 339 Ceiba pentandra Gaertn. 347 Celosia cristata Ь. 299 Cepha1ana syriaca (L.) Schrad. 114, 139 Cerasus Juss. 130, 136, 255, 256, 260, 262 — araxina Pojark. 130 — avium (L.) Moench 130, 139 — chamaecerasus Lois. 139 — collina Lej. et Court. 130, 131 — fruticosa (Pall.) Woronow 130, 404 — grandulifolia Кот. 256 — glandulosa (Thunb.) Lois. 255 — japonica Lois. 255 — mahaleb (Ь.) Mill. 130 — maximowiczii (Яирг.) Кот. 255 — microcarpa (Boiss.) Mill. 130 — prostrata Ser. 404 — sachalinensis (Рг. Schmidt) Kom. et Aliss. 255 — tomentosa Wall. 255 — vulgaris Mill. 130, 139 Ceratonia siliqua Ь. 376 Ceratotheca sesamoides Endl. 343 Cereus Mill. 418 Chaenomeles Lindl. 40, 254, 260 — japonica Lindl. 254 — sinensis Кое1ше 255 Chamaenerium angustifolium (Ь.) $сор. 225 Cheiranthus Ь. 378 Cheno podium Ь. 385, 438 — album L. 402 — ambrosioides Ь. 416 — nuttalliae Saff. 416 — pallidicaule Aellen 428 — quinoa Willd. 428 Chrysanthemum Ь. 260, 261 — согопагшт L. 252, 261 — morifolium /am. 252, 261 Chrysopogop Trin. 285 — gryllus (L.) Тг!п. 359 Chrysosplenium Ь. 25 Cicer Ь. 102, 135, 365 — arietinum Ь. 75, 102, 138, 278, 335 365, 394 — cuneatum Hochst. 102 — judaicum Boiss. 75, 102, 366 — pinnatifidum Jaub. et Sp. 75, 102 Cichorium endivia Ь. 373 — intybus L, 373, 402 — pumilum Jacq. 373 Cinchona calisaya Wedd. 432 — succirubra Рач. 432 Cinnamomum L. 260, 285 — camphora Nees et Eberm. 248 — cassia (Ь.) В!ите 248, 285 — zeylanicum Nees 285 Citrullus Schrad. 159, 205, 289, 349, 351 — caffer Schrad. 349 — col ocynthis Ь. 159, 322, 349, 350 — colocynthoides Pang. 322, 349 — fistulosus Stockes 289 — vulgaris Schrad. 321, 322, 349, 404 Citrus L. 259 — aurantium Ь. 292 — celebica goord. 293 — depressa Hayata 259, 293 — hyxtrix DC. 293 — grandis Osbeck 259 — ichangensis Swingle 259 — indica Tanaca 292 — latipes Tanaca 292 — limonelloides Нау. 259 — limon (L.) Вигт. 292 — macroptera Nontr. 293 — medica L. 292 — micran(ha Wester. 293 — nobilis Andrews 259 — reticulata Blanco 259 — sinensis (Ь.) Osbeck 258, 292 — tachibana Tanaca 259, 293 C l arki a P ur sh 438 Clausena lansium Skeels 259, 262 Claytonia perfoliata Dorm 436 465
Clinelymus sibiricus (Ь.) Nevski 220 Cochlearia armoracea Ь. 195 Cocos nucifera Ь. 298 Coelopleurum Ledeb. 226 Coffea L. 331 — агаЫса L. 331, 339 Coix lacryma-jobi L. 275 Cola Schott et Endl. 348, 351 — acuminata Schott et Endl. 347 — nitida Chev. 347 Coleus Lour. 44, 290, 299, 339, 345, 351 — aromaticus Вепй. 290 — barbatus Вепй. 290, 339 — blumei Вепй. 290 — br azzavi1 1 ensis А. Chev. 345 — edulis Vatke 339 — esculentus (N. E. Brown) Taylor 345 — floribundus Pobyns et L~brun 339, 345 — ingiar Sew. 339 — insolitus Pobyns et Lebrun 339 — 1anuginosus Hochst. 339 — mirabilis Briquet 345 — penzigii Schweinf. 339 — rotundifolius А. Chev. et Perrot 290, 300, 345 — spicatus Benth. 339 — tuberosus Вепй. 290 Colocasia Schott 300 — antiquorum (L.) Schott 253, 290, 352 Commiphora abyssinica Engler 340 Conocephalus mollis Merrill 281 Convallaria L. 261 — transcaucasica Utkin 135 Corchorus L. 281 — capsularis Ь. 281 — о itorius L. 281 Cordia abyssinica Я. Br. 339 Coreopsis L. 437 Coriandrum sativum L. 115, 139, 157, 202, 247, 285, 319, 339, 375 Comus L. 295 — mas L. 133, 140, 295 — capitata (Wall.) Nakai 295 Corylus Ь. 134, 136, 257, 260 — avellana Ь. 134, 140, 257 — brevituba Кот. 257 — cervorum Petr. 134 — colchica Albov 134 — colurna L. 134, 257 — ferax Wall. 257 — heterophylla Fischer 257 — iberica Wittm. et Кепш1аг. 134 — imeretica Кепш1аг.-Nat. 134 — manshurica Maxim. 257 — maxima Mill. 135, 378 — pontica С. Koch 134, 135 466 Corylus sieboldiana В1шпе 257 Corypha laevis А. Chev. 297 — umbraculifera Ь. 297, 299 — utan Lam. 297 Cosmea bipinnata Willd. 418 Costus speciosus Smith 286 Crambe Ь. 338, 342, 369 — abyssinica Hochst. 43, 338, 368 — hispanica Ь. 368 — maritima L. 401 — tatarica Jacq. 401 Cratae gus L. 129, 136, 254, 260, 438 — aronia Bcsc 129 — atrosanguinea Poj ark. 129 — azarolus L. 129 — mexicana Moc. et Sesse 424 — monogyna Jacq. 405 — охуасапйа Ь. 405 — pontica С. Коей 129 — szowitsii Poj аг1'. 129 Crocus L. 116 — sativus L. 116, 319 Crotalaria L. 279 — incana L. 279 — juncea Ь. 279, 301 — verrucosa Ь. 279 Croton tiglium L. 287 Cryptomeria D. Don. 260 Cucumeropsis mannii Naudin 344 Cucumis L. 348, 349, 352 — agrestis (Naud.) Pang. 321, 349 — callosus Cogn. 348 — chate (Forsk.) Idasselquist 321, 403 — ficifolius А. erich. 348 — hardwickii Яоу1е 288 — hirsutus Sond. 348 — longipes Hook. 348 — melo L. 158, 205, 289, 348 — metuliferus E. Меу. 348 — prophetarum Ь. 348 — sativus L. 88, 118, 139, 159, 204, 251, 287 C ucurbi ta L. 118, 420 — andreana Naud. 419, 431 — digitata А. Gray 420 — ficifolia Bouche 418, 419, 431 — maxima Duch. 46, 118, 205, 251, 350, 374, 419, 431 — mixta Pang. 418, 420 — moorei Bailey 419 — moschata Duch. 159, 252, 289, 374, 418, 419, 420, 431 — palmata S. Wats. 420 — реро Ь. 190, 205, 418, 419, 420 — texana А. Gray 419 Cuminum супипшп Ь. 285 Curcuma L. 286 — aromatica Salisb. 285
Curcuma 1onga Ь. 285, 287 — pierreana Gagnep. 285 — zanthorrhiza goxb. 286 Cyamopsis psoralioides DC. 279 Cyclamen Ь. 378 Cyc1anthera pedata (Ь.) Schrad. 301 Cydonia Mill. 128, 135 — oblonga Mill. 128 Cymbopogon citratus (DC.} Stapf 285 — martini (Roxb.} Stapf 285 — nardus (L.} Rendle 285 Cynara L. 379 — cardunculus Ь. 323, 374 — scolymus Ь. 323, 374 Cynometra саиИ1ога Ь. 280 Cyperus auricomus Sieber 308 — esculentus Ь. 308, 367 — papyrus Ь. 308 Dactylis Ь. 100, 239, 260 — aschersoniana Graebn. 100, 240, 395 — glomerata Ь. 100, 138, 221 Dahlia Сач. 418 — variabilis Desf. 418 Daucus Ь. 118, 372, 380 — carota Ь. 118, 139, 158, 164, 290, 400 — m a xi mus Des f. 118 — sativus Zagor. 77, 250, 372 Delphinium Ь. 25 Dendrocal amus asper B acher 287 Dianthus Ь. 378 Diclytra spectabilis DC. 260 Digitalis purpurea Ь. 404 13igitaria Heist. 8 — iburua Stapf 345 — sanguinalis Scop. 275 Dioscorea Ь. 290, 345 — abyssinica Hochst. 339, 345 — aculeata Ь. 291 — alata Ь. 291 — batatas Decne. 253 — bulbifera Ь. 291, 345 — cayenensis Lam. 345 — dumetorum (Kunth.) Рах 345 — ЫярЫа Яой 291 — japonica Thunb. 253 — latifolia Benth. 345 — pentaphylla Ь. 291 — sativa Ь. 291 Diospyros L. 257, 438 — discolor Willd. 296 — kaki L. 256, 261, 296 — lotus Ь. 133, 296 — virginiana L. 437 Dolichos 1.. 277, 299, 345 — biflorus L. 277 Dolichos gibbosus Thunb. 346 — hassjoo Siebold 242, 262 — lablab Ь. 277, 336, 346 — pratensis Taub. 346 Dracocephalum L. 243 Dryas punctata Juz. 225 Durio rithinus DC. 294 Echeveria DC. 418 Echinochloa L. 8, 275 — сгия-galli (L.) Roem. et Schult. 239, 275, 312 — со1опа (L.) Веаич. 275, 311, 312 — frumentaceae (Roxb.) Link 222, 239, 261, 275, 285 Elaeagnus L. 257 — angustifolia Ь. 163 — пш1И1ога Thunb. 257 — orientalis Ь. 163 — pungens Thunb. 257 — umbellata Thunb. 257 Elaeis guineensis Jacq. 347 Elaeocarpus ganitrus goxb. 300 Elatostema umbella turn Blunie 253, 261 Eleocharis tuberosa (Roxb.) Schult. 253, 261 Elettaria cardamomum Naton 286 — п~а~ог Smith 286 Eleusine coracana Gaertn. 275, 300, 334, 345 — indica (Ь.) Gacrtn. 268, 275 Elsholtzia Willd. 260 — cristata Willd. 223, 243 — serotina Кот. 223, 243 Empetrum hermaphroditum (L ange) Hagerup 226 — nigrum L. 226 Epicampes macroura Benth. 418 — stricta Presl. 418 Eragrostis Host 285 — abyssinica Link 44, 307, 335 — cynosuroi des Poem. et Schult. 307 — pilosa (Ь.) Beauv. 307 Erica arborea L. 331 Eriobotrya Lindl. 40, 254, 260 — japonica Lindl. 254 Eriodendron pentandrum Harz. 281 Eriolobus DC. 127 — trilobatus М. poem. 127 Eruca Adans. 367, 379, 380 — pinnatifida (Desf.) Роте1. 367, 379 — sativa Lam. 113, 139, 156. 284, 367, 397 Ervum lens Ь. 394 Erythroxylon coca Lam. 428, 432 467
Erythrozylon raimondii О. E. Schulz 428 Eschscho1tzia califnrnica Cliani. 438 Euchlaena mexicana Schrad. 415 Eucommia ulmoides Oliv. 247, 261 Eu g equi a L. 298, 434, 438 — aquea Burm. 298 — ca ryo ply yl1~ t a Thu nh. 285 — j ambo1anа Lam. 300 — j ambos L. 298 — jачап1са I am. 298 — m ala ccens i s 1 . 299 Euonymus L. 260 — maackii Rupr. 225, 247 — verrucosa Scop. 196 Euphorbia L. 351 — candelabrum Trem. 340 — lathyris L. 246, 262 — pulcherrima Willd. 418 Faba pliniana Trab. 105, 107, 363 — vulgaris Moench (= Vicia faba L.) 75, 105, 138, 153, 200, 242, 278, 315, 336, 363, 364 Fagara schinifolia (Sieb. et Zucc.) Engl. 247 Fagopyrum Gaertn. 276 — sagittatum Gilib. 199, 222, 240, 276 — tataricum (Ь.) Gaertn. 276 Радия L. 260 Feijoa selloviana Berg. 433 Feronia eleph antum Corr. 293 Festuca pratensis L. 100, 138, 221 — rubra L. 138 Ficus Ь. 132, ] 36 — asperifolia Niq. 351 — capensis Thunb. 351 — carica L. 132, 140, 162, 323 — colchica Grossh. 132 — elastica Rozb. 287 — geraniifolia Niq. 163 — hyrcana Grossh. 132 — persica Boiss. 162 — roxburgl~ii Юа11. 163 — serrata Forsk. 132 — sycomorus Rozb. 323 Foeniculum vul gare Mill. 116, 139, 319, 375 Fortunella Swingle 259, 260, 293 — 1aponica (Thunb.) Swingle 259 — hindsii (Champ.) Swingle 259 — margarita (1.оиг.) Swingle 259 Fragaria L. 438 — chiloensis (L.) Duch. 434 — clatior Ehrh. 405 — platypetal а Rydb. 437 — virginiana Duch. 437 Fritillaria L. 24 468 Galena orient alis Lam. 137 Ля1шш 1.. 38~ Garci»ia I . 235, 294 — cowa L. 294 — du leis (Poxh.) Kurz 294 — mangostana 1 . 294 — ~anthochymus Hook. 294 Gardenia florida 1 . 247 — radicans Thunb. 247 Gigantoc1iloa apus (Schult.} Киги 287 — atter Кцгг 287 — verticillata (Willd.) Munro 287 Ginkgo ИоЬа L. 235, 258 Glycine L. 240, 278 — gracilis Skworz. 240, 241 — hispida (Moench) Maxim. 240, 241, 261, 278 — ussuriensis Rg1. et Maack 240, 24? Godetia Spach 438 Gonolobus edu1is Hemsl. 420 Gossypium Ь. 246, 280, 438 — апоп1а1шп Wavrra et Peyr. 347 — агЬогешп L. 246, 280, 281, 299 — aridum Skovsted 417 — barbadense L. 45, 417, 430 — capitis-viridis Mauer 347 — davidsonii Кеll. 417 — ellenbeckii (Gurke) Маиег 347 — herbaceum L. 42, 153, 246, 300, 338, 346, 347, 417 — hirsutum L. 45, 417 — klotzschianum Anderss. 430 — гаппопй1 Ulbr. 430 — religiosum L. 317 — вота!епяе (GQrke} J. В. Hutch. 347 — яошlanense Watt 347 — st ocksi i N a st. 281 — tomentosum Nutt. 417 — tricuspidatum Lam. 45, 417 — irilobum (Мое. et Sesse) Skovsted 4'1 7 — triphyllum (Harv.} Hochr. 347 Grossularia Mill. 258, 260, 438 — acicularis (Smith) Spach 225 — burejensis (Fr. Schmidt) Berger 2 4 — cynosbati i М111. 436 — divaricaia (Dougl.) Cov. et Britt. 436 — hirtella Spach 436 — nivea (Lindl.) Spach 436 — oxyanthoides (L.) Mill. 436 — reclinata (L.) Mill. 406 Guizotia Cass. 337, 340, 351 — abyssinica (L.) Cass. 44, 337 — bidentoides Oliver et Hiern. 337 — schultzii Hochst. 337 Hagenia abyssinica Willd. 340 Haynaldia Schur 25 Hedysarum coronarium Ь. 366, 379
Hedysarum obscurum Ь. 226 Helenium Ь. 437 Helianthus annuus L. 113, 139, 201, 435, 436 — lenticularis Dougl. 201, 435 — tuberosus Ь. 435 Heliotropium peruvianum Ь. 435 Негас1еит dissectum Ledeb. 219 — dulce Fisch 219 — lehmannianum Bge. 157 — ыЬЫсит L. 219 Heteranthelium Hochst. 25 Heuehera sanguinea Engelm. 418 Нечеа brasiliensis Muell. Arg. 287, 434 Hibiscus abelmoschus L. 300 — aspera Hook. 347 — cannabinus Ь. 281, 347 — esculentus 1.. 338, 347 — rosa-sinensis Ь. 282 — sabdariffa Ь. 282 Hippophae Ь. 257 — rhamnoides Ь. 225, 257 Hor deum Ь. 149, 198, 236 — aethiopicum Vav. et ВасЫ. 333 — agriocrithon Aoberg 236, 237, 238 — dIstichum Ь. 84 — euroasiaticum Vav. et ВасЫ. 96, 138, 272, 358 — hexastichum sanctum Heer 51, 311, 390 — hexastichum densum Heer 51, 390 — humile Vav. et ВасЫ. 41, 236, 261, 381 — lagunculiforme ВасИ. 150 — marinum Huds. 307 — заШит Jessen 358 — spontaneum (С. Косй) ВасЫ. 84, 150, 236, 237, 381 — tetrastichum Buschan 311 Hovenia dulcis Thunb. 256, 261 Hyacinthus? . 378 Hydrangea Ь. 261 Hydrastis canadensis L. 436 Hyptis suaveolens Poit. 285 Hyssopus L. 376 — officinalis Ь. 376 Ilex paraguariensis St. Hil. 411 Illicium anisatum Ь. 247, 261 Impatiens balsamina L. 299 Indigofera Ь. 351 — tinctoria Ь. 287 I pomoea L. 250, 418, 420 — aquatica Forsk. 250, 261 — arborescens Don. 420 — asarifolia poem. et Schult. 420 — batatas Poir. 20, 250, 261, 420, 421 — carnea Jacq. 420 — edUlis Makino 250 I pomoea f asti giata Street 420 — gracilis P. Br. 420 — pendula Choisy 420 — quinq<>efolia 1. 420 — iiliacia (%11!6.) Choisy 420 Iris L. 378 — р a 1 lid а Lam. 376 Iresine lindenii Чап Houtte 435 Jasminium gra»difolium L. 285 Joannesia asoka 268 Juglans L. 257, 260, 433, 438 — cinerea Ь. 436 — cordiformis Maxim. 257 — manshurica Мах~ш. 223, 257 — neotropica Diels 433 — nigra L. 436 — regia L. 162, 377 — sieboldiana Maxim. 257 — sinensis Dod e 257-Juncus e j f usus L. 247, 262 Kaempferia galanga L. 286 КегЫ~пд~е11а geokarpa Наппя 346 Kochia Яой 378 Kummeroivia Schindl. 243, 260 — stipul aces (Maxim.) МаЫпо 243 — striata (Thunh.) Schind l. 220, 243 Lactuca indica L. 291 — sativa L. 124, 139, 323, 373 Lagenaria vulgaris Ser. 289, 321, 431, 438 Languas galanga (L.) Stuntz 286 Lallemantia Fisch. et Меу. 113, 135 — canescens (L.) Fisch. et Mey. 112 — iberica Fisch. et Иеу. 112, 113, 136, 138 — peltata (L.) Fisch. ei Mey. 112, 113, 136 — rupestris Sinsk. et Voskr. 112, 113, 136 Ьарра edulis Siebold (=Arctium lappa L.) 250, 261 Lathyrus L. 260, 365 — cicera L. 75, 108, 138, 365 — gorgoni Pari. 75, 365, 379 — miniatus М. В. 108 — ochrus (L.) DC. 365, 379 — оdoratu L. 365, 378 — sativus L. 106, 138, 153, 278, 314, 336, 365 Laurocerasus officinalis Poem. 131 Laurus nobilis . 376 Lavandula L. 376, 379 — spica Ь. 376 — vera DC. 376 Lawsonia alba Lam. 287
Lens Adans. 105, 135, 361 — esculenta Noench 106, 138, 152, 200, 278, 313, 336, 361, 393 — kotschyana (Boiss.) Alef. 105 — lenticula (Schreb.) Alef. 105 — nigricans (М. В.) Godr. 105 — orientalis (Boiss.) Hand.-Макк. 105, 108 Lepidium meyenii Walp. 428 — sativum Ь. 76, 122, 139, 156, 332, 338, 375 Leptochloa bipinnata Retz. 307 Lespedeza Nichx. 242, 260 — bicolor Turcz. 220, 242 — cyrtobotrya Miq. 242 — hedysaroides (Pall.) Kitag. 242 — juncea Pers. 243 Lewisia rediviva Pursch 436 Lilium Ь. 24, 437 — martagon Ь. 208 — maximowiczii Regel 250 — regale Wilson 260 — tigrinum Кег-Gawl. 250 Linum angustifolium Hudsp. 36, 54, 56, 111, 154, 155, 280, 396, 397 — crepitans Dum. 22, 25, 316 — humile Mill. 316 — usitatissimum L. 18, 138, 154, 164, 200, 280, 316, 336, 367 Liriodendron tulipifera Ь. 437 Litchi chinensis Sonn. 259, 262, 299 — 1опдапшп Camb. 299 Lithospermum erythrorhizon Sieb. et Zucc. 248 Lobelia fulgens Willd. 418 Lolium multif lorum Lam. 396 — perenne L. 101, 138, 307, 396 Lonicera edulis Turcz. 225 Lophira alata Banks 344, 350 Lucuma salicifolia Н. В. К. 424 Luffa acutangula {1..) Roxb. 290 — cylindrica (L.) Roem. 251, 301 Lupinus Ь. 359, 438 — albus Ь. 313, 359 — angustifolius Ь. 359 — luteus Ь. 359 — mutabilis Sweet 429 — pilosus Ь. 359 — polyphyllus Lindl. 437 — somaliensis Baker 336 — termis Forsk. 75, 77, 313, 342, 359, 379 1 ycopersicon cerasiforme Dunal 422 — chilense Dunal 430, 431 — esculentum Mill. 430, 431 — glandulosum С. Н. Muller 430 — hirsutum Humb. et Вопр. 429, 430, 431 — peruvianum МИ1. 429, 430, 431 — pimpinellifolium Dunal 431 470 Lycopus Ь. 385 Lygeum sparteum Ь. 359 Майа sativa Mol. 434 Magnolia grandiflora Ь. 437 Najorana Moench 376 — hortensis Noench 376 Malus Mill. 127, 254, 260, 263, 438 — baccata (L.) Borkh. 190, 254 — dasyphylla Borkh. 404 — domestica Borkh. 405 — floribunda Sieb. 254 — formosana Kawak. et Koidz. 254 — halliana Коейпе 254 — hupehensis Rehder 254 — manshurica (Maxim.) Кот. 224, 254 — niedzwetzkyana Dieck 160 — orientalis Uglitzk. 127, 137, 404, 405 — pallasiana Juz. 224 — praecox (Pall.} Borkh. 196, 404 — ringo Sieb. 254 — rivularis Roem. 437 — sachalinensis (Кот.) Juz. 224 — sieversii (Ledeb.) Roem. 160, 404 — silvestris Mill. 190, 196, 404, 405 — turkmenorum Juz. et М. Рор. 160 Mammea africana G. Don. 350 Mandragora Ь. 373, 380 — autumnalis Bertol. 373 — haussknechtii Heldr. 373 — hispanica Vierh. 373 — officinarum Ь. 373 Nangifera Ь. 293 — caesia Jack 294 — caloneura Kurz 293 — cochinchinensis Engl. 293 — duperreana Pierre. 293 — foetida Lour. 294 — indica Ь. 293 — silvatica Roxb. 293 Nanihot aipi Pohl 411 — dulcis (J. F. Gmel.) Pax 433 — utilissima Pohl 411, 433 Nanilkara Adans. 434 Naranta arundinacea L. 421, 434 Maoutia риуа Wedeb. (= Boehmeria риуа Hook.} 281 Naranta arundinacea L. 421, 434 Natthiola R. Br. 378 Naurandia barclaiana Lindl. 418 Medicago Ь. 108, 109, 135, 151, 366 — asiatica Sinsk. 117, 151, 164, 242 — beipinensis Vass. 242 — borealis Grossh. 195, 220 — cancellata М. В. 108 — coerulea Less. 18, 108, 242 — difalcata Sinsk. 108 — dzhawakhetica Bordz. 108, 151
Nedicag0 eusativa Sinsk. 75, 366, 379, 395 — falcata Ь. 194, 220, 395 — glandulosa David. 72, 109 — glutinosa М. В. 18, 108, 137, 151 — hemicyc1a Grossh. 18, 108, 109, 151 — jemenensis Sinsk. 75, 78, 109, 136, 366 — mesopotamica Vass. 136 — orientalis Vass. 109, 117, 136 — papillosa Boiss. 108 — polia Vass. 78 — polychroa Grossh. 108 — praesativa Sinsk. 18, 78, 108, 109, 136 — quasifalcata Sinsk. 108, 136 — rupestris М. В. 108 — sativa L. 19, 138 — saxatilis М. В. 108 — syriaco-palestinica Sinsl'. 75, 109, 136, 366 — tripolitanica Sinsk. 366, 379 — vardanis Vass. 108, 136 — virescens Grossh. 108 Melia azedarach Ь. 248, 262 Nelilotus Adans. 219, 366 — albus Desr. 152, 195, 219, 366 — indicus All. 110, 366 — italicus Lam. 366 — neapolitanus Ten. 366 — off(etna(is (Ь.) Desr. 109, 138 Nelissa officinalis Ь. 376 Melo Adans. 117, 139, 158, 164, 251 — adana Pang. 117 — adzhur Pang. 117 — agrestis (Naud.) Pang. 158 — ameri Pang. 158 — cantalupa Pang. 117 — cassaba Pang. 117 — ehandalal»~ Pang. 158 — chinensis Pang. 251, 261 — conomon Pang. 251, 261 — figari Pang. 117 — flexuosus Pang. 158 — monoclinus Pang. 251, 261 — zard Pang. 158 Mentha aquatica Ь. 402 — canadensis Ь. 248 — crispa L. 402 — crispata Schrad. 402 — longifolia Huds. 402 — piperita Ь. 402 — rotundifolia (Ь.) Huds. 402 — silvestris Ь. 402 — spicata Ь. 402 — viridis Ь. 402 Meratia praecox Rehd. et Wils. 247 Mesgilus L 37, 129, 135 — germanica L. 129, 140, 405 Mesua ferrea Ь. 300 Metroxy1on rumphii (Will d.) Маг(. 297 — sagu Rottb. 297 Nichelia champaka L. 300 Nicrotaena cymosa Prain 285 — robusta Hemsl. 285 Nimusops elengi Ь. 300 — schimperi Hochst. 324, 339 Nolinia litoralis Host 101, 137, 140 Momordica charantia Ь. 289, 301 Nonstera deliciosa Liebm. 418 Noringa aptera Gaertn. 318 Norus Ь. 247, 261 — alba Ь. 134, 140, 163, 247, 26" — bombycis Koidz. 247 — multicaulis Perr. 247 — nigra L. 134, 140, 163, 247 — rubra Ь. 163 Nucuna utilis Ша11. et Wight 279 Nusa Ь. 294, 300, 350, 352 — acuminata Colla 294, 295 — balbisiana Colla 294, 295 — cavendishii Lamb. 294 — ensete J. F. Gmel. 339, 350 — holstii К. Schum. 350 — paradisiaca Ь. 294, 295, 300 — sapientum L. 294, 295 — silvestris СоИа 294 — textilis Nee 282 Myri anthu s arboreu s В еа пч. 350 Myrica mexicana AVilld. 424 — rubra Sieb. et Zucc. 258, 262 Myristica fragrans Houtt. 286 Narcissus Ь. 378 Nardosmia frigida (Ь.) Ноо(с. 228 Nelumbium nuciferum Gaertn. 308 Nemophilla Buckl. 438 Nepeta L. 117 — hajastana Grossh. 117 — mussinii Spreng. 117 — noraschenica Grossh. 117 — reichenbachiana Fisch. et Mey. 11Т — transcaucasica Grossh. 117 Nicotiana Ь. 436 — angustifolia R. et P. 432 — langsdorfii Weinm. 432 — multivalvis Lindl. 436 — paniculata Ь. 432 — quadrivalvis Pursh 436 — repanda Willd. 436 — rusbyi Brit. 432 — rustica Ь. 423, 432 — silvestris Speg. et Comes 43" — tabacum Ь. 423, 432 — torreyana А. Nelson 436 Nigella sativa Ь. 339, 376 Nipa fruticans Thunb. 297 Nomocharis Franch. 24 47I
Nopalea coccinellifera (L.) Salm- Dyck 418 Nymphaea lotus Ь. 308 Оепойега biennis Ь. 437 Olea africana Mill. 115 — chrysophylla Lam. 339, 342 — europaea L. 115, 139, 317, 339, 376 Oncidium Sw. 418 Опсооа glauca Planch. 343 Onobrychis Adans. 110, 135 — altissima Grossh. 110, 137 — antasiatica Chinschuk 110, )137 — arenaria (К11.) Ser. 195 — biebersteinii Sirj. 137 — cyri Grossh. 137 — jailae N. Tschern. 72 — meschetica Grossh. 110, 137 — miniata Stev. 72 — sibirica Turcz. 220 — tanaitica Spreng. 195, 220 — transcaucasi ca Grossh. 110, 137, 151 — viciaefolia Scop. 394, 408 Opuntia Mill. 418 Origanum L. 402 — vulgare L. 402 Ornithopus perpusillus Ь. 394 — sativus Broth. 367, 394 Огоху1ит indicum (L.) Vent. 287 Orthosiphon stamineus Benth. 287 Oryza Ь. 334, 345 — barthii А. Chev. 273, 274, 334, 345 — brachyanta А. Chev. et Poehr. 334 — breviligulata А. Chev. et goehr. 145 — cubensis Е1лпап. 416 — fatua Prain 268, 272, 273, 274 — glaberrima Steud. 273, 334, 345 — granulata Nees 275 — minu(a Presl. 272, 274 — officinalis Wall, 272, 274 — perennis Moench 273, 274, 334, 415 — perrieri А. Camus 334 — punctata Kotschy 334, 345 — sativa Ь. 150, 240, 272, 273, 274, 334, 352 — МарИ goshev. 334 — tisseranti А. Chev. 334 Oxalis crenata Jacq. 428 — tuberosa Nolina 428 Oxycoccus macrocarpus Pers. 437 Oxyria digyna (L.) Hill. 225 Oxytropis mertensiana Turcz. 226 Pachyrhizus a»gulatus erich. 279 — tuberosus (Lam.) Spreng. 420 472 Paeonia albiflora Pall. 260 — chinensis Vilm. 259 Palaquium дц11а Burck. 287 Panax L. 248 — ginseng С. А. Меу. 248, 261 Panda oleosa Pierre 350 Panicum Ь. 8, 99 — callosum Hochst. 334 — miliaceum Ь. 99, 138, 150, 172, 187, 199, 222, 239, 261, 312, 332, 334, 356, 393 — miliare Lam. 239 — spontaneum Lyssov 239 Papaver rhoeas L. 397 — setigerum DC. 24, 397 — somniferum Ь. 24, 114, 139, 156, 201, 246, 320, 368, 397 Parthenium argentatum А. Gray 423 Paspalum exile К~ррЫ 345 — scrobiculatum L. 275 Pastinaca sativa Ь. 373, 400 — silvestris Mill. 373 Paulownia Sieb. et Zucc. 260 Pelar gonium L'Herit. 351 — roseum Willd. 351 Pennisetum erich. 351 — molissimum Hochst. 335 — perrottetii (Klotzsch.) К. Schum. 335 — spicatum (L.) Roem. et Schult. 312, 334 — typhoideum (Вигт. f.) Ь. С. Rich. 300, 335 — versicolor Schrad. 335 — villosum Я. Br. 335 — violaceum (Lam.) Rich. 335 Pentadesma butyracea Sabine 350 Pentastemon campanulatus Willd. 418 — gentianoides Lindl. 418 Pepo Adans. 374 Perilla L. 252, 260 — frutescens (L.) Britt. 243, 262 — nankinensis (1.оиг.) Decne 252 — ocymoides L. 248, 252, 262 Persea gratissima Gaertn. 424 Persica Mill. 255, 256, 260, 377 — davidiana Carr. 256 — ferganensis Коч. et Kostina 161 — kansuensis (Rehd.) Kov. et Ков- tina 256 — mira (Igoehne) Коч. et Kostina 256 — potanini (Batalin) Kostina 256 — vulgaris МИ1. 161, 256, 377 Petasites japonicus F. Schmidt 252, 261 Petroselinum crispum (Mill.) Nym. 372 Petunia Juss. 435 Peucedanum graveolens S. Wats. 291
254, Peucedanum iaponicum Thunb. 251, 261 Phalaris canariensis L. 359 Phaseolus L. 276, 277, 438 — aborigineus Hurkart 435 — aconitifolius Jacq. 277 — acutifolius А. Gray 277, 416 — angutaris (Willd.) Wight 241, 261 — aureus (Roxb.) Piper 152, 241, 276 — calcara tu s Roxb. 277 — lunatus L. 416, 429 -- macrolepis Piper 416 — ты1И lorus Willd. 416 — mungo (L.) Piper 277, 301 — retusus Benth. 416 — sublobatus Roxb. 153, 276 — trilobus Ait. 277 — vulgaris (L.) Savi 241, 416, 429, 435 Philadelphus coronarius Ь. 80 Phleum pratense L. 100, 138, 194, 221 Phlox L. 437 Р11оепЫ . 297 — dactylifera L. 298 — reclinata Jacq. 298 — silvestris Roxb. 298, 299 Phyllostachys Sieb. et Zucc. 249 Physalis L. 422 — aequata Jacq. 45, 422 — glabripes Pojark. 252 — ixocarpa Brot. 422 — pubescens Ь. 422 Pimpinella anisetum Boiss. 117, 137 — anisum L. 116, 157, 319, 375 — aromatica М. В. 137 — eretica Poir. 116 — peregrina L. 375 Pinus L. 260 — koraiensis Sieb. et Zucc. 225 — pumila (Pall.) Rgl. 225 — sibirica (Rupr.) Mayr 225 Piper betle L. 286, 297 — longum L. 286 — nigrum L 286 Pirus Ь. 125, 127, 136, 160, 253, 254, 260, 263 — aromatica Е;йпсЫ et Nakai 253 — asiae-mediae М. Рор. 160 — balance e D ecne 126 — betulaefolia Bge. 253 — boissieriana Buhse 126, 254 — caucasica Fed. 126, 137, 254 — communis L. 126, 137, 196, 404, 405 — сот р1ека Rubtz. 126 — eor d a t a Des v. 405 — demetrii Kuthath. 126 — elaeagrifolia Pall. 127 — elata Rubtz. 126 — eldarica Grossh. 126 Pirus fedorovii Kuthatli. 126 — georgica Еыйай. 126 — grossheimii Гed. 126 — hyrcana Fed. 126 — ketzkhovelii Е,пйай. 126 — koishinskyi Litv. 160 — medvedevii Rubtz. 125 — nivalis Jacq. 405 — nutans Rubtz. 126 — ovoidea Rehd. 253 — oxyprion Woron. 126 — pashia Buch.-Ham. 253, 254 — raddeana Юогоп. 126 — regelii Relid. 160 — яасйоЫапа I<uthath. 126 — salicifolia Pall. 125, 405 — serotina Rehd. 253 — sosnovskii Fed. 126 — syriaca Boiss. 126, 137 — takhtadzhianii Fed. 125, 137 — t am amschi an ae Ге(1. 126 — taochia Woron. 126 — turcomanica Maleev 126 — ussuriensis Maxim. 224, 253 — voronovii Rubtz. 126 — vsevolodi Heidem. 126 — zangesura Ма1ееч 126 Pistacia Ь. 134, 136, 257, 260 — cabulica Stockes 162 — formosana Matsum. 257 — integerrima Stew. 134 — khinj uk $1ос1'es 162 — lentiscus L. 134, 257 — mutica Fisch. et Меу. 134, 162 — terebinthus L. 134 — vera Ь. 77, 134,140, 162, 257, 378 Pistia stratiotes L. 243, 262 Pisum Ь. 101, 102, 103, 135 — abyssinicum А. Braun 78, 103, 335 — arvense L. 102, 103, 335 — aucheri Jaub. et Spach 101 — elatius (Ы. В.) Stev. 101, 103, 360, 381 — iormosum (Stev.) Boiss. 101 — fulvum Sibth. et Sm. 101 — humile Boiss. et Noe 101 — sativum Ь. 75, 102, 103, 138, 153. 199, 242, 278, 314, 335, 359, 360, 393 Plectranthus tuberosus Blume 290 — patchouli С. В. Clarice 285 Pluchea indica Less. 291 Podocarpus gracilior Pilger 331 Podophyllum peltatum L. 436 Рода oleosa Pierre 351 Pogostemon patchouli Pell. 285 — heyneanus Benth. 285, 300 Polakowskia tacaco Pittier 420 Polianthes tuberosa Ь. 418 Polygala butyracea Heckel 343 473.
Polygonum Ь. 385 — coriarium Grig. 157, 196 — ellipticum Willd. ex Spreng. 225 — tinctorium Lour. 248 Po»cirus Rafin. 260 — trifo1iata (Ь.) Raf. 269, 260 Portulaca ? . 435 — oleracea Ь. 125, 139, 321, 373 Praecitrullus Pang. 289 Primula Ь. 24 Prosopis dulcis I<unth 435 Prunus Ь. 130, 136, 255, 256, 260, 263, 391, 404, 437, 438 — americana Narsch. 437 — angustifolia Narsch. 437 — caspica Коча1. et Ekim. 130 — divaricata Ledeb. 130, 139, 161 — domestica L. 19, 130 — ferganica Lincz. 161 — fruticosa Pall. 196 — munsoniana Wight et Hedr. 437 — nigra Ait. 437 — salicina Lindl. 255 — simonii Carr. 255 — spinosa L. 130 — ursina Ко1яс1~у 130 — ussuriensis Коч. et КояЫпа 255 Psophocarpus tetragonolobus DC. 279 Pueraria thunbergiana Benth. 247 — edulis Thunb. 247 Punica Ь. 132, 135, 323 — granatum L. 132, 133, 140, 163, 323 — protopunica Balf. 133 Pyrethrum carneum М. В. 135, 137 — roseum М. В. 135, 137 0uercus Ь. 260 — Иех Ь. 354, 356 Raphanus Ь. 26, 36, 380 — caudatus Ь. 26, 290 — indicns (L.) Sinsk. 26, 290 — maritimus Smith 370 — raphanistroides (МаЫпо) Sinsk. 18, 249, 261 — sativus Ь. 18, 158, 245, 249, 320, 370, 399, 400 — sinensis Sinsk. 249, 261 Rauwolfia Ь. 287 — canescens Ь. 423 — serpentina Benth. ex gurz 287, 351 — vomitoria Afzel. 351 Reseda L. 378 Rheum palmatum Ь. 252 — ribes Ь. 77, 124, 139 Rhus coriaria L. 376 — succedanea Ь. 247 — vernicifera DC. 247 Ribes L. 224, 258, 260 474 Ribes L. hispidulum Pojark. 224 — nigrum Ь. 196, 224, 406 — paucif lorum Turcz. 224 — petraeum Wulf. 224 — rubrum L. 224, 406 Ricinus communis Ь. 337, 343 — macrocarpus G. Pop. 246, 343 — microcarpus G. Pop. 115, 139, 156, 317, 337 — zanzibarinus G. Pop. 343 Roegneria С. Кос1~ 150, 261 — ftbrosa (Schrenk.) Nevski 221 — trachycaulon Nevski 221 Rosa damascena Mill. 137, 376 Rosmarinus officinalis Ь. 376 Rubia cordifolia Ь. 248 — tinctorum Ь. 287, 376 Rubus Ь. 129, 258, 299, 405, 433, 436 — argutus Link 437 — chamaemorus L. 226 — ellipticus Smith 299 — flagellaris Willd. 436 — glaucus Вепй. 433 — idaeus L. 405, 436 — illecebrosus Focke 258 — innominatus Moore 258 — kuntzeanus Непм1еу 258 — lasiocarpus Smith 299 — loganobaccus Bailey 436 — macrocarpus Benth. 433 — macropetalus Dougl. 436 — mallucensis Ь. 299 — niveus Thunb. 299 — ostryifolius Rydb. 437 — pergatus Blanch. 437 — phoenicolasius Maxim. 258 — rosaefolius Sm. 299 — trivialis Мк1ж. 437 — titanus Bailey 436 — velox Bailey 437 Rudbeckia Ь. 437 Rumex abyssinicus Jacq. 338 — acetosa Ь. 375, 403 — arcticus Ь. 225 — graminifolius Lamb. 225 — montanus Desf. 403 — scutatus L. 403 Ruta graveolens Ь. 375 Saccharum barberi Jesw. 296 — officinarum L. 296 — robustrum Ь. 296 — sinense Roxb. 249, 262 — spontaneum Ь. 296 Sagittaria sagittifolia Ь. 253, 261 Salix speciosa Hook. 225 Salvia Ь. 422 — chian Ь. 422 — lanceolata Brouss. 423
Salvia sclarea L. 157 — splendens Кег-Gawl. 418 — tiliaefolia Vahl 423 Samuela Trel. 418 Sansevieria liberica Ger. et Labr. 347 — senegambica Baker. 347 — zeylanica Willd. 282 Santalum alatum L. 285 Sapindus mukorossi Gaertn. 248 Sapium P. Br. 248 — яеЬИегит (Ь.) Roxb. 248 Sarcocephalus esculentus Afzelius 350 Sasa Nakino et Shibata 249 Satureja hortensis Ь. 116, 375, 376 — spicigera (С. Koch) Boiss. 116, 137 S axifra ga L. 25 — nelsoniana D. Don. 225 — punctata Ь. 225 Schizandra Nichx. 260 — chinensis Baill. 223, 258, 260 Scopolia carniolica Jacq. 137, 404 — caucasica Kolesn. 137 Scorzonera deliciosa Guss. 373 — hrspanica Ь. 373 — tau-saghyz Lipsch. et Bosse. 157, 164 Secale Ь. 96, 135, 140, 238, 391 — afghanicum (Vav.} Roshev. 97, 149 — africanum Stapf 98 — anatolicum Boiss. 80, 97, 98 — ancestrale Zhuk. 97 — cereale Ь. 97, 138, 149, 166, 197, 221, 391 — ciliatoglume (Boiss.) Grossh. 97 — dalmaticum Vis. 98 — daralagesi Thum. 97 — dighoricum (Vav.} Roshev. 97 — kuprijanovii Grossh. 97, 98 — montanum Guss. 97, 98 — segetale (Zhuk.} Roshev. 97, 98 — silvestre Host 98 — vavilovii Grossh. 80, 97 Sechium edule (Jacq.) Schwarz 420 Sedum atropurpureum Hort. 226 Senecio Ь. 351 Sesamum L. 282, 342, 351 — angustifolium Engl. 343 — indicum Ь. 77, 112, 138, 154, 243, 282, 317, 343, 368 — prostratum Petz. 343 — radiatum Schum. et Thonn. 343 Sesbania aculeata Poiret 280 — cannabina Poiret 280 Setaria Beauv. 8, 18, 344, 351 — incrassata Hackel 330 — italica (L.) Beauv. 62, 99, 138, 239, 261, 312, 344 — ketzkhovelii Men. et Eritz. 99, 138 Sida rhombifolia Ь. 281 Sinapis Ь. 379 — alba Ь. 76, 113, 139, 284, 367, 368 Sin Sm Sm Sol 426 253, 421 475 apis arvensis Ь. 319, 368 ilax china Ь. 258, 262 yrnium olusatrum Ь. 372 anum Ь. 46, 421, 426, 435, 438 асаы1е ВЖ 425 aethiopicum Tom. 338 agrimonifolium Pydb. 421 ajanhuiri Juz. et Buk. 426 ajuscoense Buk. 421 andigenum Juz. et Buk. 45, 427, 428 aracc-pal. a Juz. 426 berthaultii Hawzes 426 brevicaule Bitt. 426 bukassovii Juz. 426 bulbocastanum Dun. 421 canasense Hawkes 426 capsicibaccatum Cardenas 426 cardiophyllum Lindl. 421 catarthrum Juz. 426 chacoense Bitt. 425 circaeifolium Bitt. 426 clarum Corr. 421 colombia num Dun. 426 commersonii Dun. 425 соуосапшп Buk. 421 curtilobum Juz. et Buk. 425 demissum Ь1пй1. 421 ehrenbergii (Bitt.} Rydb. 421 etuberosum Lindl. 426 famаtinae Bit 426 fendleri А. Gray 435 gibberulosum Juz. et Buk. 425 goniocalix Juz. et Buk. 426 jamesii Torr. 421, 435 juzepczukii Buk. 425 kurtzianum Bitt. et Wittm. 426 lanciforme Pydb. 421 leptostigma Juz. 426, 427 longiconicum Bitt. 421 longistylum Corr. 421 lycopersicoides Dun. 426 macolae Buk. 426 maglia Schlechtd. 426, 427 manoteranthum Bitt. 421 medians Bitt. 426 megistacrolobum Bitt. 425 melongena Ь. 77, 78, 125, 139, 291 microdontum Bitt. 426 molinae Juz. 426 morelliforme Bitt. et Nuench. multiinterruptum Bitt. 426 muricatum Ait. 430 oxycarpum Schiede 421 phureja Juz. et Buk. 426 pinnatisectum Dun. 421 piurae Bitt. 426 polyadenium Greenm. 421 рыпае Juz. 425
Sol Sol Sor So 476 апшп quineense La:ï. 338 quitoense Lam. 432 rybinii Juz. et Buk. 426, 432 salamanii Hawkes 421 santolallae Vargas 426 semidemissum Juz. 421 sparsipilum (Bitt. ) Juz. et Buk. 426 stenotomum Juz. et Buk. 426 stoloniferum Schlechtd. 421 sucrense Hawkes 426 tuberosum L. 46, 421, 426, 427, 428, 434 unfundibuliforme Phil. 425 vallis-mexici Jus. 421 vavilovii Juz. et Huk. 426 vernei Bitt. et Wittm. 426 verrucosum Schlechtd. 421 virgultorum (Bitt.) Card. et На~ч- kes 426 wittmacl~ii Bitt.. 426 xerophyl1цгп Н awkes 425 idago L. 437 bus Ь. 128, 136, 254, 260 albovii Zinserl. 128 amurensis Кое1~пе 254 anadyrensis Кот. 254 aria Crantz 405 armeniaca Hedl. 129 aucuparia L. 128 baldaccii Degen et Fritsch 129 boissieri Schiieid. 128 buschiana Zinserl. 129 caucasica Zinserl. 129 colchica Zinserl. 129 commixta Hedl. 254 domestica L. 128 dualis Zinserl. 129 graeca (Spach) Hedl. 129 kamtschatce»sis Кот. 254 kusnetzovii Zinserl. 129 migarica Zinserl ° 129 obtusidentata Zinserl. 129 sambucifolia poem. 254 schemachensis Zinserl. 129 sibirica Hedl. 254 яиЫияса (Ledeb.} Boiss. 128 subtomentosa (А!Ьоч) Zinserl. 129 taurica Zinserl. 129 velutina (А1Ьоч) $сппеЫ. 129 worono~vii Zinserl. 129 rghum 5! oench 150, 335, 344, 351 b antuor um J akusch ev. 344 caffrorum (Beauv.} Jakuschev. 344 chinense Jakuschev. 239, 261, 344 durra (Forst<.) Stapf 150, 275, 335, 344 halepense (Ь.) Pers. 344 guineense (Stapf) Jakuschev. 344 sudanense (Piper) Stapf 344 Sorghum versicolor Anderss. 344 — vulgare Pers. 312, 335, 344 Spartium junceum L. 379 Sphenostylis stenocarpa Harms 346 Spinacia oleracea Ь. 124, 139 — tetrandra Stev. 124 Spondias mombin L. 424 — purpurea L. 424 Stachys sieboldii hliq. 250, 261 S(audtia gabonensis Warburg 350 Stipa tenacissimа Ь. 359, 379 Stizolobium P. Browne 279 — hirsutum (W.-А.) Kuntze 241, 279 — ргиг!епя (L.) Medic. 241, 279 — pruritum (Ноо1с.) Kuntze 241, 279 — utile (Wall.) Ditm. 241, 279 Strobilanthes flaccidifolius Nees 248 Strychnos nux-vomica Ь. 287 Symphyoloma graveolens С. А. Меу. 137 Syringa L. 260 — vulgaris L. 80 Syzygium owaricnse Henth. 350 Тасса pinnatifida Forst. 291 Tagetes L. 418 Tamarindus indica L. 298, 300 Taraxacum ко~-sa ghyz /odin 157, 164 Tetracarpidium conophorum Hutch. et Dalz. 343 Telfairia occidentalis Hook. ~4 1, 348 — pedata (Smith) Hook. 348 Terminalia catappa Ь. 296 — chebula I<etz. 296 Thea 1 . 287 — amplexicaulis Pit 286 — assamica Mast. 235, 286 — bohea L. 248, 286 — cantoniensis Lour. 248, 286 — monticola V. Кгеск. 248, 286 — morpha V. Krecz. 286 — sinensis L. 248 — sinensis indica Coh. St. 286 — stricta Hayne 248, 286 — taliensis W. W. Smith 286 — theaster V. Кгеск. 286 — viridis Ь. 248, 286 Themeda triandra Forsk. 246 Theobroma angustifolia Sesse et Nok. 423 — atrorubens Hub. 434 — bicolor Humb. et Bonpl. 434 — cacao Ь. 423 — grandiflora (Spr.) К. Schum. 434 — microcard um Mart. 434 — obovata Klotzsch. ex Bern. 434 — sativa (Ьат.) Lignier et Ье Веу 423
Theobroma simiarurn Bonn. 423 — speciosa S preng. 434 — spruceanum Bern. 434 — subincan a Mart. 434 Thymus vulgaris L. 376 Tigridia pavonia Кег-Gawl. 422 Tithonia tubaeformis Gass. 435 Torreya grandis Fort. 258, 262 Trachycarpus excelsus Nakino 247, 262 Tradescantia virginiana L. 437 Tragopogon porrifolium Ь. 373 Trapa L. 261 — bicornis L. 253 — bispinosa goxb. 253 Тгеси11а africana Decaisne 351 Trichosanthes anquina L. 289 — cucumeriana L. 289 — dioica Poxb. 289 — palmata goxb. 289 Trifolium L. 136 — alexandrinum L. 75, 110, 138, 313, 366 — apertum Bobr. 75, 111, 137, 366 ° — berytheum Boiss. 75, 366 — expansum Waldst. et Кй. 395 — hybridum L. 194, 395 — incarnatum L. 367, 379 — lupinaster Ь. 220 — medium Ь. 313 — pratense Ь. 111, 138, 194,200, 220, 336, 394 — repens L. 194, 367 — resupinatum L. 313 — sativum Crome 395 — vavilovii Eig 75, 366 Trigonella L. 109, 366 — arabica Delile 75, 109 — соеги1еа (Desr.) Ser. 109, 138 — cretacea (М. В.) Grossh. 72, 109 — foenum-graecum Ь. 109, 117, 138, 152, 278, 336, 366 — gladiata Stev. 109 — jemenensis (негр.) Sinsl~. 78, 109, 137, 336 — ргоситЬепв (Bess.) Rcht. 109 Tripsacum L. 415 Triticum L. 50, 93, 96, 135, 140, 149, 238, 357 — aestivum L. (Tr. vu1gare Host) 60, 94, 95, 138, 148, 149, 166, 198, 222, 238, 309, 333, 358, 387 — aethiop!curn Jakubz. 24, 51, 94, 95, 310, 333, 341, 342, 387 — antiquorum Heer (vu1gare anti- quorum Heer) 24, 39, 50, 61, 61, 85, 86, 94, 149, 172, 173, 181, 265, 310, 311, 383, 384, 385, 386, 387, 388, 389, 391 — araraticum Jakubz. 49, 76, 93 ticuin boeoticum Boiss. 49, 84, 94, 357 compaetum Host 85, 86, 94, 95, 149, 172, 173, 265, 310, 385, 387, 388 dicoccoides Коегп. 75, 79, 84, 93, 357, 390 dicoccon Schrank 78, 79, 81, 84, 85, 94, 138, 172, 198, 309, 333, 341, 357, 383, 385, 388, 389 durum Desf. 94, 95, 138, 198, 310, 357 ispahanicum Hes1ot 80, 94, 136 macha Dek. et Men. 24, 48, 51, 89, 86, 94, 136, 387 monocnccum L. 84, 94, 173, 357, 383, 387 ра1еосо1с111сшп Меп. 24, 48, 51, 86, 94, 136, 384, 387, 388 регясш.п Vav. 94, 136 polonicum L. 80, 94, 333, 342 рго(оасИпо1~1сшп 51, 94, 310, 311 protomacha 24, 48, 50, 51, 61, 86, 149, 181, 310, 311, 333, 388 protopa 1 eocolchicum 311, 333, 389 sphaerococcum Pere. 95, 149, 238, 272 spelta L. 25, 51, 79, 94, 96, 136, 358, 389, 390 timopheevi Zhuk. 49, 75, 94, 136 turanicum Jakubz. 94, 136 turgi~1!im Ь. 85, 86, 94, 95, 309, 357, 387 urart<> Thum. 93, 98 vavi1nvii ЛакцЬк. 94, 136 yunnanense IPing 238 zhukovskyi Men. et Eritz. 49, 94, 136 opaeolum L. 428, 435 tuberosum Quiz et Pavon 428 Ulex L. 367 — епгорасця ?.. 367, 379 Ullucus tuberosus Lozano 428 Urena lobata Ь. 285, 347 Vaccinium vitis-Маеа Ь. 226 — uli g i nnsum L. 226 VaIeriane11a М111. 385 Vanil1a N!11. 424 — aromatica Sw. 434 — guianensis $1~11~. 424 — leprieurii Рогteres 424 — phaeantha Peichb. 424 — planifo1ia Лш1г. 424 — poi(aei Яе1с1Й. 424 Veratrum lobelianum Bernh. 225 Verbena L. 435 Vetiveria zizanioides Stapf 285 477
Vicia Ь. 103, 136, 260, 263, 314, 361, 36з — amoena Fisch. 220 — angustifolia Ь. 104, 361 — erч111а (L.) Wi11d. 103, 104, 361 — faba Ь. 153, 362, 385, 393 — monanthos Desf. 361 — narbonensis Ь. 360, 364 — pannonica Crantz 104, 138 — sativa Ь. 101, 104, 138, 200, 314, 361, 395 — variabilis Freyn et Sint. 104, 137, 138 — villosa goth 104 Vigna Savi 87, 277, 299, 351 — catjang (Вигт.) Walp. 301, 346 — sinensis Endl. 111, 242, 277, 315, 336, 346 — unguiculata auct. 347 Vitis L. 257, 438 — amurensis gupr. 223, 257, 262 — antiquorum Andras. 162 — byzantina Andras. 162 — deliciosa Andras. 162 — silvestris Gmel. 133, 385 — thunbergii Sieb. et Zucc. 223, 257 — vinifera Ь. 133, 140, 162, 377 Voandzeia subterranea Thou. 346 ШаязаЫа japonica Matsum. 250, 261 Wistaria Nutt..261 — chinensis DC. 260 Yucca L. 417 — baccata Torrey 418 Zalacca edulis В1шпе 297 Zanthozylum L. 247 — hungei Planch. 247 — piperitum DC, 247 — planispinum Sieb. et 247 — schinifolium Sieb. et Zucc. 247 Zea L. 239 — mays L. 99, 239, 414, 429 Zingiber officinale pose. 285 — mioga (ТЬипЬ.} Rose. 285 — zerumbet (Ь.) Rose. 286 Zinnia Ь. 418 Zizania aquatica L. 8, 410, 437 — latifolia Turcz. 253, 262 Zizyphus Mill. 250, 260, 351 — jujuba Lam. 299, 300, 351 — rugosa Lam. 299 — sativa Gaertn. 163 — spina-christi Willd. 351 — vulgaris Lam. 255, 378 Zucc. Xanthosoma belophyllum Kunth 422 — brasiliense Engl. 422 — edule Schott 422 — sagittifolium (L.} Schott 434 — violaceum Schott 422
С ОДЕРЖАН ЙЕ Предисловие Принципы и методы установления историко-географических областей происхождения культурной флоры Археологические сведения в связи с историей древнего земледелия Факторы, имеющие решающее значение в процессе изменения при- роды растений при введении их в культуру Методы, применяемые при изучении происхождения культурных растений и путей их первичного распространения 11сторпко-географические области происхождения аборигенных культурных флор земного шара 17 29 Древнее земледелие и аборигенные культурные растения Передней 47 Азии ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 47 Земледелие и культурные растения в древнем Закавказье Развитие земледелия в ближайших областях влияния переднеазиат- ской культурной флоры в СССР 1~ультурная флора некоторых зарубежных стран, относящихся к Передней Азии Земледельческие культуры Месопотамии (древнего Двуречья) Обзор видов и форм аборигеппых культурных растений Передней Азии в целом 73 80 Древнее земледелие и аборигенные культурные растения советской Средней Азии и сопредельных с нею стран Средне-!Ого-Западной А зии 141 ,древнее земледелие и аборигенные культурные растения советской Средней Азии......................... 141 Лборигенные культурные растения Средне-Юго-Западной Азии в целом ........................... 148 древнее земледелие и древние ассортименты культурной флоры Рус- ской равнины и Сибири.................... 168 Трипольская культура...................... 168 Культура скифов ........................ 173 Земледелие и культурные растения у древних протославян и славян 174 Культурные растения, возникшие на Русской равнине...... 194 ,Л,ревнее земледелие и аборигенные культурные растения Сибири . 207 479 ,древнее земледелие и аборигенные культурные растения Китая, Ин- дии и соседних с ними стран.................. 227 ,древнее земледелие и аборигенные культурные растения Китая и соседних стран Восточной Азии................ 227 Обзор аборигенных растений стран Восточной Азии ..... 236 Земледелие в,древней Индии и аборигенные культурные растения южной Азии ......................... 264
Аборигенные культурные растения Средиземноморья Древнее земледелие и аборигенные культурные растения Северной Европы Средней и 382 Древнее земледелие и аборигенные С вета Нового 409 культурные растения 449 443 Заключение Литература Указатель латинских названий растений 46" Издано Л. О. издательства «Колос» по заказу Всесоюзного орденаЛенина научно-исследовательского института растениеводстваим. Н. И. Вавилова Синская Евгения Николаевна ИСТОРИЧЕСКАЯ ГЕОГРАФИЯКУЛЬТУРНОЙ ФЛОРЫ(НА ЗАРЕ ЗЕМЛЕДЕЛИЯ) НАУЧНЫЕ ТРУДЫПод ред. акад. ВАСХНИЛ Д. Д. Б р е ж н е в а Л., отделение издательства «Колос», 1969.480 стр. с илл. УДК 631.5/.9: 581.9.003 Редактор Г. А. П е и ь к о в аХудожественный редактор О. П. А н д р е е вТехнический редактор Л. Г. 5 а р а н о в аКорректоры Е. М. Носкова и Н. Г. Медви иска я Сдано в набор 21/Xt 1968 г. Подписано к печати 21/II 1969 г. М-20119. Формат 60Х90",1~. Печ. л. 30. Уч.-изд. л. 34,10. Byмага тип. № 3. Тираж 3000 экз. Цена 2 р. 76 к. Заказ № 1053. Отделение издательства «Колос», Ленинград, Д-186, Невский пр., 28. Типография им. Котлякова издательства «Финансы» Комитетэпо печати при Совете Министров СССР. Ленинград, Садовая, 21. Древнее земледелие и аборигенные культурные растения Африки .. 302 Земледелие и культурная флора Древнего Египта........ 302 древнее земледелие и аооригенные культурные растения Эфиопии и внесредиземноморской Африки ............... 326