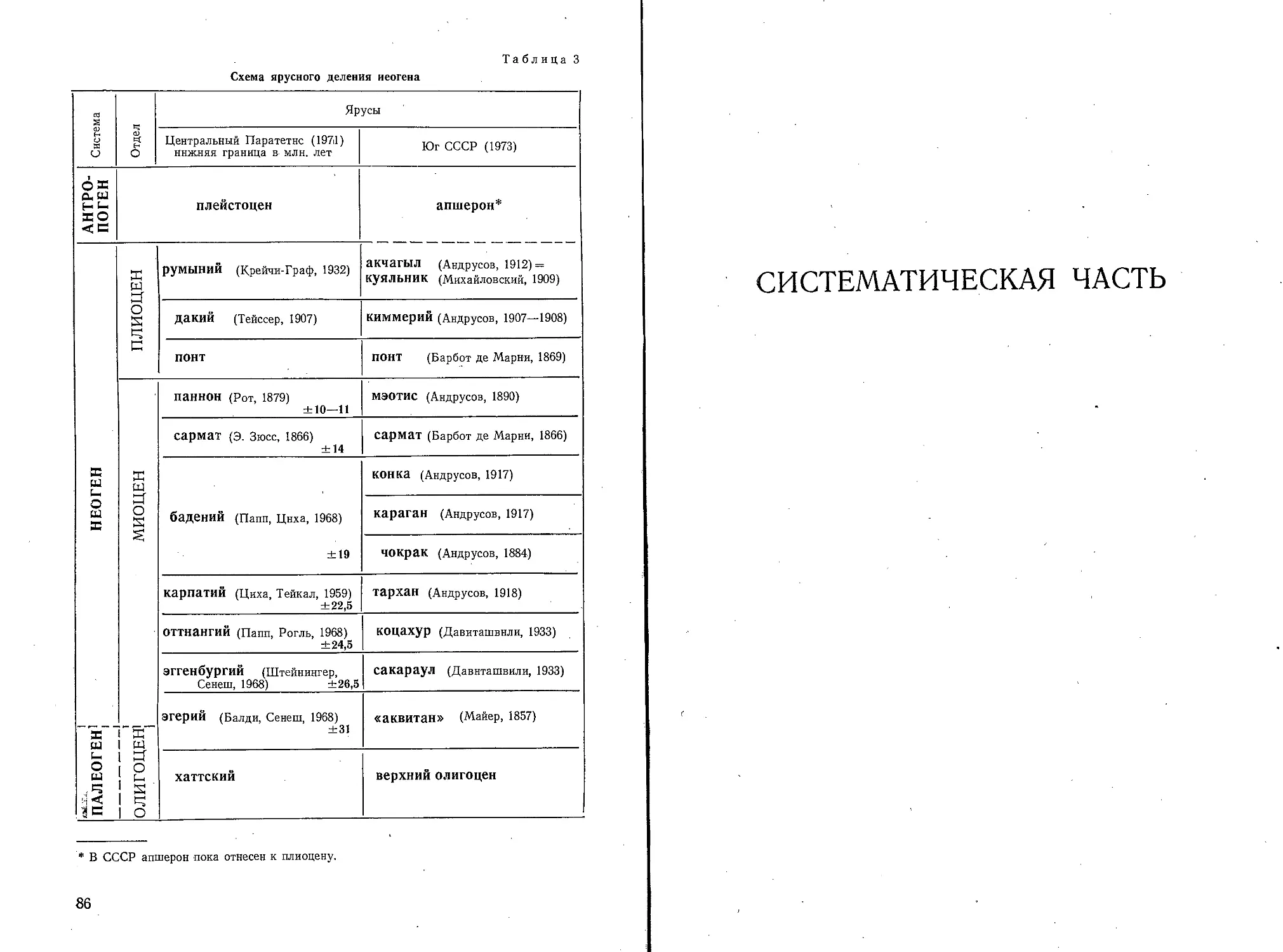
/





































































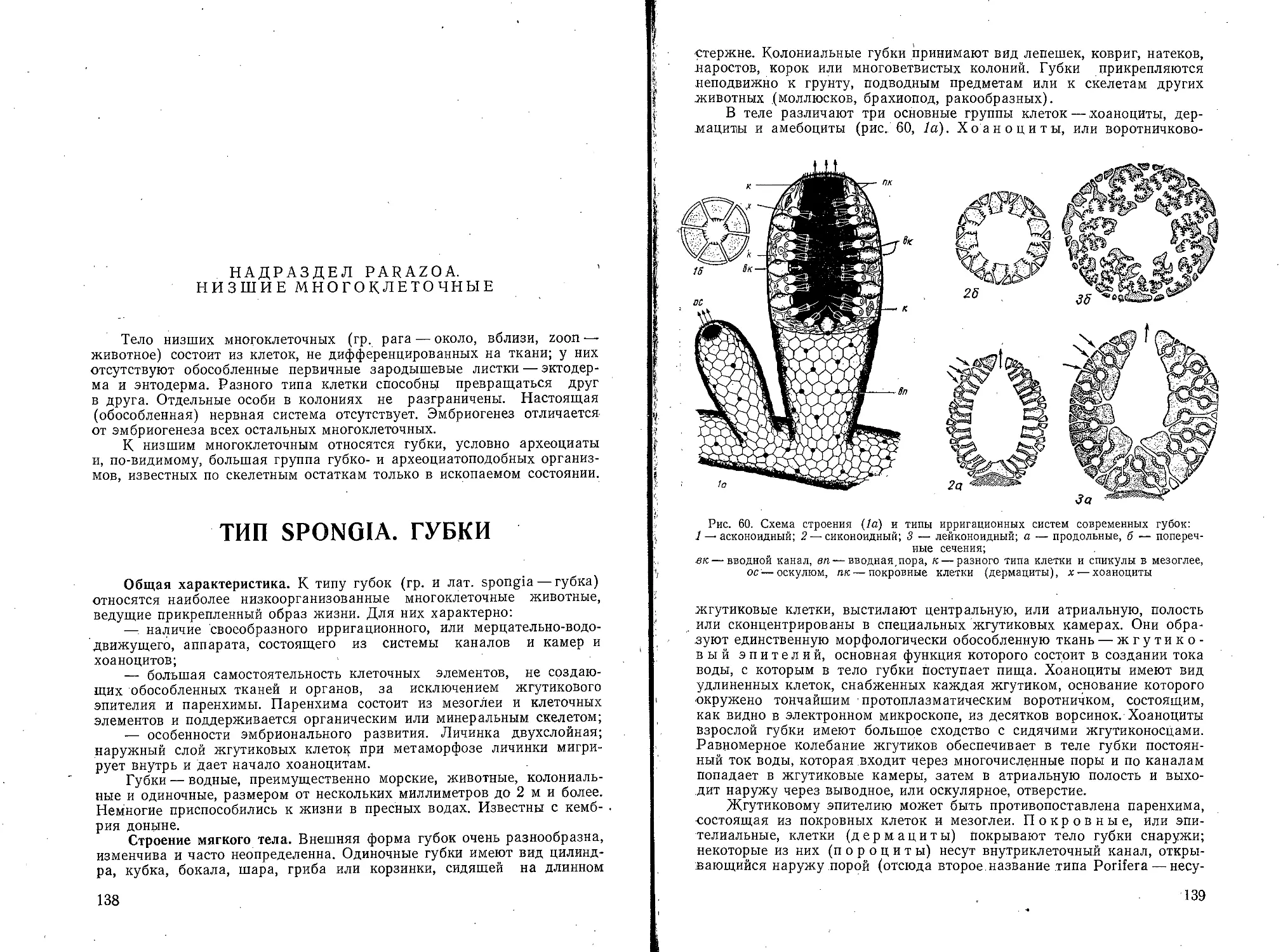



































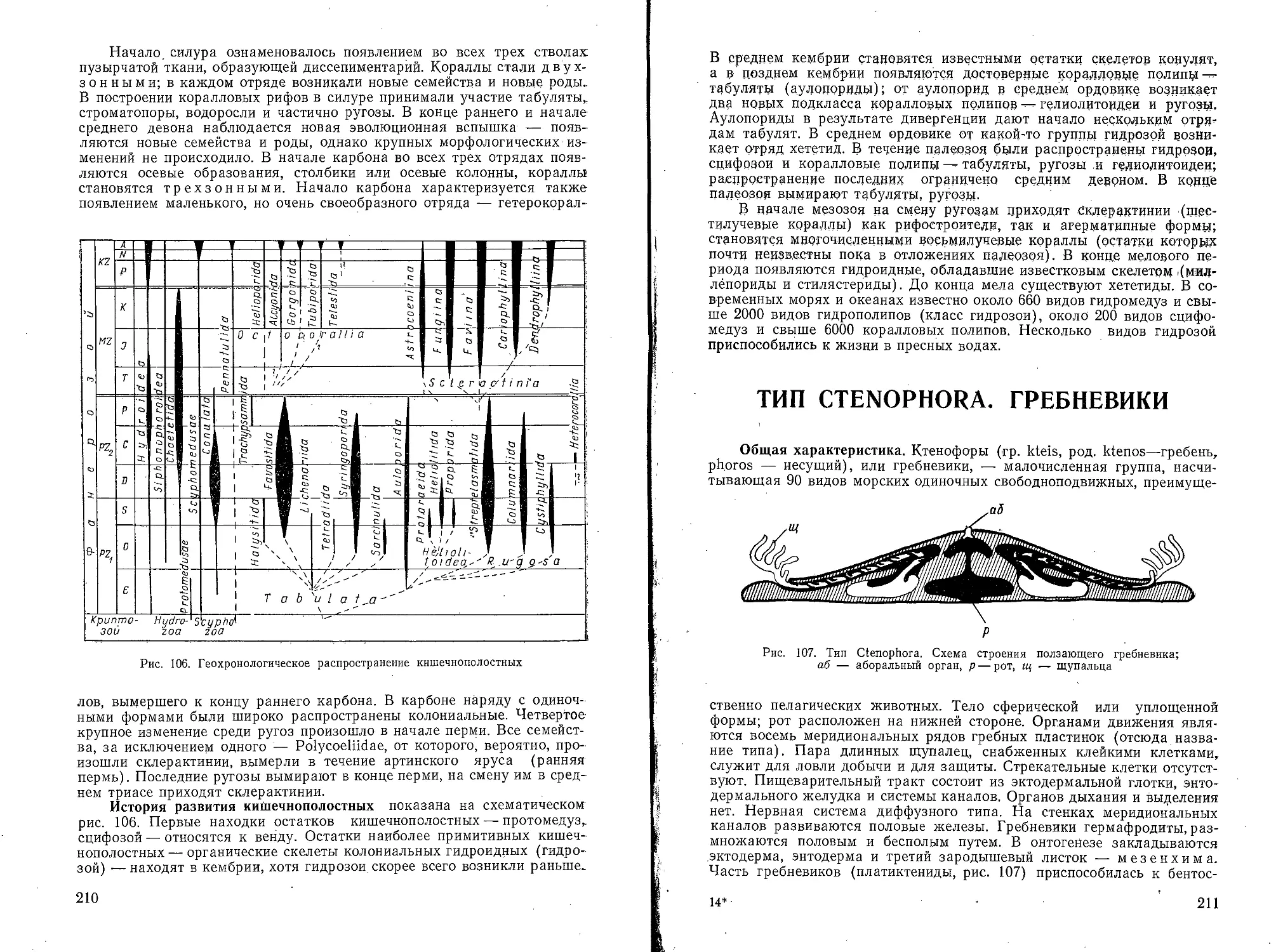



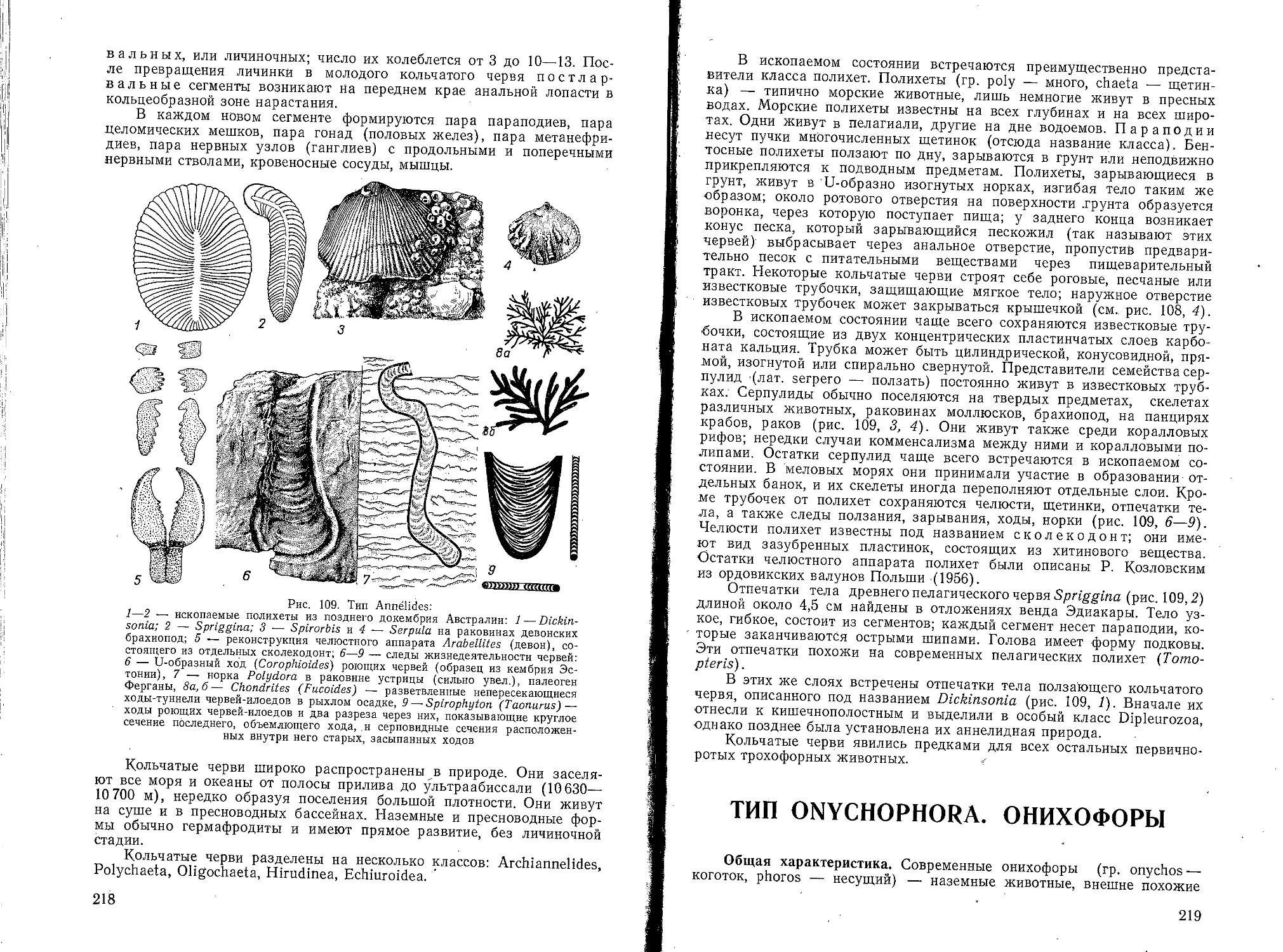





























































































































































Текст
ПАЛЕОНТОЛОГИЯ
БЕСПС8ЮНСЧНЫХ
В. В. ДРУЩИЦ
В. В. ДРУЩИЦ
ПАЛЕОНТОЛОГ
БЕСПОЗВОНОЧНЫХ
ДОПУЩЕНО МИНИСТЕРСТВОМ ВЫСШЕГО И СРЕДНЕГО
СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР
В КАЧЕСТВЕ УЧЕБНИКА ДЛЯ СТУДЕНТОВ
ГЕОЛОГИЧЕСКИХ СПЕЦИАЛЬНОСТЕЙ УНИВЕРСИТЕТОВ
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА
19 7 4
УДК 562
Учебник состоит из двух частей. В первой части рассматриваются предмет и зада-
чи палеонтологии, ее место среди естественных наук, ее подразделения, сохранность
беспозвоночных в ископаемом состоянии, история палеонтологии, основные принципы
систематики, прав,ила зоологической номенклатуры, значение палеонтологии для геоло-
гических наук. В систематической, основной, части описаны все таксоны — до отрядов,
включительно, с указанием характерных родов и иллюстрацией наиболее важных пред-
ставителей. Рассмотрен также ряд групп неясного таксономического значения: крибри-
циаты, соаниты, сфинктозои, ксенокоихии и др.
Рецензенты:
кафедра палеонтологии геологического факультета Ленинградского государственного
университета (зав. кафедрой профессор Д. Л. СТЕПАНОВ); доктор геолого-минерало-
гических наук, профессор Н. С. МОРОЗОВ
О Издательство Московского университета, 1974 г.
20801— 133
077(02)—74 175—74
Д
ПРЕДИСЛОВИЕ
Палеонтология, изучающая органический мир прошлых геологи-
ческих эпох, относится к циклу биологических наук, но всегда была тес-
но связана с геологическими науками — стратиграфией и исторической
геологией, литологией и палеогеографией. Успехи палеонтологии стиму-
лировали разработку детальной стратиграфии, изучение геологического
строения новых территорий доставляло палеонтологам новые материа-
лы о прошлом органического мира. Особенно много нового было сделано
в последние годы: были открыты остатки древнейших следов жизни
(2,7—3,0 млрд, лет), обнаружены молекулярные ископаемые в отло-
жениях протерозоя, остатки бесскелетных многоклеточных в венде, от-
крыты новые группы современных и вымерших организмов (погонофо-
ры, моноплакофоры, склероспонгии, петалонамы и многие другие).
Изучение процессов биоминерализации, разработка новой методики хи-
мического препарирования, исследование микро- и ультраструктур ске-
летов с помощью электронного микроскопа (трансмиссионного и ска-
нирующего) открыли новую страницу в истории палеонтологии. Все
это привело к пересмотру наших представлений о систематике многих
групп животного мира и разработке новых классификаций.
Тщательное изучение путей развития древних организмов, их сис-
тематики и филогении способствует восстановлению историй древних
бассейнов и разработке детальной стратиграфии. От уровня наших
палеонтологических знаний зависит детальность стратиграфических схем,
возможность региональных и межрегиональных корреляций. Палеонтоло-
гия относится к числу фундаментальных наук, без знания которой не
могут успешно развиваться ни биологические, ни геологические науки.
Учебник «Палеонтология беспозвоночных» служит источником для
получения первых, самых общих представлений о палеонтологии, ее
предмете и задачах, путях развития, особенностях сохранения живот-
ных в ископаемом состоянии, о взаимоотношениях животных между со-
бой и абиотическими факторами среды, о некоторых проблемах эволю-
ции, основах систематики и правилах зоологической номенклатуры, ис-
пользовании палеонтологических данных в различных разделах геоло-
гических наук. Учебник знакомит читателя со всеми типами и классами
животного мира, известными в наше время, и в ряде случаев с под-
классами и отрядами и сопровождается рисунками, раскрывающими осо-
бенности строения мягкого тела, различных элементов скелета и пока-
зывающими наиболее характерных представителей. Для многих групп
3
приведены таблицы геологического распространения отрядов и подклас-
сов, а для граптолитов указано распространение некоторых родов и
семейств.
Учебник рассчитан на студентов геологических и биологических фа-
культетов университетов, геологических факультетов геологоразведоч-
ных, горных и нефтяных институтов и составлен в соответствии с прог-
раммой по палеонтологии, утвержденной Министерством высшего и
среднего специального образования СССР.
Автор благодарен за особое внимание и доброжелательное отно-
шение зав. кафедрой палеонтологии академику В. В. Меннеру и своим
коллегам по кафедре: И. С. Барскову, О. Б. Бондаренко, Т. Н. Горба-
чик, Л. А. Догужаевой, Е. И. Кузьмичевой, Н. И. Маслаковой, И. А. Ми-
хайловой, О. П. Обручевой, Т. Н. Смирновой, М. Н. Шелеховой,
Б. Т. Янину, просмотревшим всю рукопись или ее отдельные главы.
Автор выражает большую благодарность своим официальным ре-
цензентам профессору Н. С. Морозову и доценту Г. Г. Пославской
(Саратовский университет), кафедре палеонтологии Ленинградского уни-
верситета, ее заведующему профессору Д. Л. Степанову и сотрудникам
кафедры 3. Г. Балашову, Е. А. Балашовой, В. И. Волгину, Л. В. Куш-
нарь, Р. С. Елтышевой, В. А. Сытовой, сделавшим полезные замечания
ко всей рукописи или ее отдельным главам.
Автор очень признателен докторам наук М. С. Зиновьеву, К. Б. Кор-
дэ, Г. П. Ляшенко, И. П. Морозовой, Л. А. Невесской, Я. И. Старобога-
тову, М. А. Семихатову, В. Н. Шиманскому, научным сотрудникам
О. В. Амитрову, Ю. Н. Арендту, А. В. Богоявленской, И. А. Гончаро-
вой,. А. Н. Иванову, Л. Б. Ильиной, Н. А. Заренкову, С. В. Попону,
А. Н. Соловьеву, В. А. Сысоеву, сделавшим ряд замечаний к отдельным
главам или передавшим оригинальные рисунки.
Большую помощь в подготовке рукописи к печати оказала
М. В. Кнорина, которая монтировала рисунки, В. Г. Куренкова, подго-
товившая ряд рисунков, и Т. В. Черноглазкина, корректировавшая
текст.
Автор будет весьма признателен всем, кто после знакомства с кни-
гой пришлет свои замечания и пожелания по адресу: 117234, Москва,
В-234, геологический факультет Московского университета.
ОСНОВНЫЕ ПОЛОЖЕНИЯ
И МЕТОДЫ
ПАЛЕОНТОЛОГИИ
ПРЕДМЕТ И ЗАДАЧИ ПАЛЕОНТОЛОГИИ
Палеонтология — наука биологического цикла, изучающая
органический мир прошлых геологических эпох и закономерности его
исторического развития. Материалом для палеонтологии служат лю-
бые сохранившиеся в слоях Земли остатки организмов или следы их
жизнедеятельности, называемые окаменелостями, или' фосси-
ли я м и (от лат.1 fossa — яма, ед. ч. — fossile — ископаемое, мн. ч.—
fossilia — ископаемые). Слово «палеонтология» в переводе с гречес-
кого языка означает учение, или науку, о древних организмах (гр. ра-
laios — древний, ontos — существо, logos — учение).
Остатки организмов сохраняются в осадочных породах только при
благоприятных условиях захоронения, при наличии прочного минераль-
ного или органического скелета. Очень редко сохраняются мягкие тка-
ни и органы: или при быстрой их минерализации сразу же после ги-
бели организма, или в форме отпечатков. В настоящее время на Зем-
ле существует свыше 1 млн. видов животных и растений, часто пред-
ставленных огромным ЧИСЛОМ индивидуумов, НО ОНИ обычно после ГИ"
бели исчезают совершенно бесследно, если не попадут в благоприятные
условия для их сохранения. Поэтому в ископаемом состоянии сохраняет-
ся лишь незначительная часть от брльшого числа ранее существовав-
ших организмов (рис. 1); это особенно наглядно видно на примере чле-
нистоногих, число видов которых в настоящее время приближается к
миллиону, причем одних жесткокрылых (жуков) в три раза больше, чем
всех моллюсков вместе взятых, а в ископаемом состоянии от большого
класса насекомых, вероятно, сохранилось не более 1%. И хотя палеон-
тологическая летопись неполна и несовершенна, она тем не менее слу-
жит единственным источником наших знаний о событиях, происходив-
ших на Земле в течение многих миллионов лет, а ископаемые остатки
являются своеобразным архивом, в котором хранятся молчаливые сви-
детели прошлой жизни.
Палеонтология связана с зоологией и ботаникой, изучающими мор-
фологию, анатомию современных организмов, их родственные связи и
взаимоотношение между собой и с окружающей средой, а также проб-
лемы формо- и видообразования, эволюционного развития. В то же время
1 В дальнейшем при переводе латинские слова будут сокращенно указываться
«лат.», а греческие — «гр.», если будет приводиться слово в родительном падеже —
«род.», в единственном числе — «ед. ч.», во множественном числе — «мн. ч.» '
7
палеонтология связана с геологическими науками, и в первую очередь
с биостратиграфией, исторической геологией, литологией, поскольку
изучение органических остатков, заключенных в осадочных по-
родах, с одной стороны, позволяет выяснять их происхождение (напри-
мер, морские или континентальные осадки), восстанавливать биономи-
ческие зоны моря, намечать очертания древних материков и океанов,
Простейшие
Рис. 1, Относительное количество известных современных и вымерших видов
животных.
Г равен 3500 современных и вымерших видов. Площадь сектора, покрытая точ-
ками, пропорциональна относительному количеству видов, известных в ископае-
мом состоянии
изучать геологическую историю данного региона, восстанавливать кар-
тины жизни прошлых геологических периодов, а с другой стороны,
позволяет использовать ископаемые остатки для выяснения относи-
тельного геологического возраста изучаемых слоев, развитых в регио-
нах, часто удаленных друг от друга на большие расстояния. Эволю-
ция органического мира, ее темпы, особенности и направление всегда
находились в тесной связи с особенностями развития Земли, измене-
ниями климатических условий, характером тектонических движений.
8
На основании изучения эволюции организмов, прошлого выделены ос-
новные этапы развития Земли и построены геохронологическая и стра-
тиграфическая шкалы, которые более подробно рассмотрены ниже.
Палеонтология состоит из двух крупных разделов: палеозооло-
гии и палеоботаники, которые по мере развития науки все, бо-
лее отдаляются друг от друга и в настоящее время представляют две
самостоятельные науки: первая из них связана с зоологией, вторая —
с ботаникой.
Палеозоология, изучающая, животный мир' геологического
Прошлого, разделена на палеозоологию беспозвоночных и палеозооло-
гию позвоночных. Палеозоология беспозвоночных, рассматриваемая в
этой книге, часто называется палеонтологией беспозвоночных; эти два
названия — синонимы. Провести границу между палеозоологией и
зоологией обычно легко, так как объектами изучения палеонтологов
являются вымершие организмы, существовавшие в геологическом прош-
лом, а зоологов — современные или недавно вымершие животные. Од-
нако в ряде случаев Многие ныне существующие организмы имеют
довольно длительную геологическую историю (например, некото-
рые моллюски, простейшие) и в этих случаях грань между зоологией
И палеонтологией становится условной; следует рекомендовать подобные
группы изучать совместными усилиями.
Перед палеонтологией стоят задачи, к числу которых относятся:
— изучение видового состава организмов по их ископаемым остат-
кам;
— изучение их морфологии, особенностей строения, изменчивости;
— изучение изменений отдельных признаков в онтогенезе;
— определение времени существования и ареала каждого изучае-
мого вида;
— на основании морф.офункционального анализа выяснение образа
жизни изучаемого вида и особенностей его взаимоотношения со средой;
— изучение конкретных филогенезов для видов, родов и более вы-
соких таксонов и разработка их систематики;
— восстановление основных путей развития изучаемой группы;
— выявление значения изученных видов для биостратиграфии, вос-
становления физико-географических, палеоклиматических условий, па-
леобиогеографии, абиотических и биотических особенностей геологичес-
кого прошлого.
Накопленный за последние годы большой палеонтологический ма-
териал привел к обособлению в палеонтологии специальных разделов.
Выделены: микропалеонтология, тафономия, палеоэкология, экогения,
палеобиогеография, палеоанатомия, палеофизиология, палеоэмбриоло-
гия, палеопатология, палеоихнология, палеотаксиология и ряд других
направлений.
Микропалеонтология. Объектом изучения служат остатки
мелких и микроскопических организмов, в первую очередь фораминифер,
радиолярий, остракод, тинтиннид. Особенно много специалистов работа-
ют в настоящее время над изучением фораминифер.
Тафономия представляет собой особый раздел палеонтологии,
выделенный И. А. Ефремовым (1940, 1950), занимающий пограничную
область между биологией и геологией. Тафономия (гр. taphos — захо-
ронить, nomos — закон) изучает закономерности захоронения и обра-
зования местонахождений ископаемых животных и растений. Образо-
вание местонахождений, связанное с переходом органических остатков
из биосферы в литосферу, осуществляется процессами, происходящи-
ми, с одной стороны, в биосфере, с другой — в литосфере. Каждое мес-
9
тонахождение при существующей неполноте геологической летописи,
впервые специально рассмотренной Ч. Дарвином, является важнейшим
документом палеонтологии; оно позволяет после детального исследова-
ния заглянуть в прошлое, восстановить хоть частично жизнь минувших
геологических эпох. В раздел тафономии можно включить а к ту о-
палеонтологию — особое направление, выделенное Р. Рихтером
(1928), содержанием которой является познание особенностей гибели
современных организмов и их постмортальных изменений до захороне-
ния в осадке (перенос после гибели, разрушение, рассеивание или, на-
оборот, концентрация в определенных местах). С актуопалеонтологией
тесно связано еще одно направление в тафономии — биостратоно-
мия (гр. bios -- жизнь, лат. stratum — слой), введенное в литературу
И. Вейгельтом (1919), задачей которой является выяснение особеннос-
тей пространственного расположения остатков организмов в слое и по
отношению друг к другу. Это направление тесно связано с литологией
и учением о фациях.
Палеоэкология, по определению Р. Ф. Геккера (1957), зани-
мается изучением взаимоотношений между миром организмов геоло-
гического прошлого и средой их обитания, т. е. решает те же задачи, ко-
торые ставит экология по отношению к ныне живущим организмам.
Палеоэкология выясняет образ жизни вымерших организмов —
способы питания, передвижения, нападения, защиты, особенности раз-
множения и жизненные циклы и на основании этих данных восстанав-
ливает условия жизни тех или иных форм или целых сообществ, свя-
занных с абиотической средой и биотическими взаимоотношениями.
К абиотическим факторам относятся: характер дна (для донных орга-
низмов), соленость воды, температура, газовый режим, степень подвиж;
ности воды. К биотическим факторам относятся те сложные взаимоот-
ношения, в которые вступают и вступали организмы между собой (ха-
рактер пищи, конкуренты, враги, мирные сожители).
Для успешного решения поставленных вопросов палеоэкологические
исследования должны быть тесно связаны с литологическими исследова-
ниями и экологией современных животных.
Изучение эволюции организмов невозможно без исследования адап-
таций и изменения их во времени и пространстве. Л. Ш. Давиташвили
предложил назвать экогенией учение о экогенезе, или историческом про-
цессе развития экологических отношений между организмами, с одной
стороны, и средой — с другой. Вопросы, которые рассматривает экоге-
ния, связаны с изучением процессов распространения и переселения жи-
вотных из одной жизненной области (океан, пресные воды, суша) в дру-
гую или в пределах одной области, обусловленные адаптивными измене-
ниями й строении, физиологии и поведении животных. Данные экогении
помимо теоретического имеют большое практическое значение, так как
помогают выяснять происхождение некоторых полезных ископаемых, в
первую очередь каустобиолитов.
Палеозоогеография составляет часть .палеогеографии, изуча-
ет закономерности распространения организмов геологического прошло-
го, изменение их ареалов и выясняет причины этих изменений. Палео-
биогеография тесно связана с экогенией, но отличается от послед-
ней по содержанию, задачам и методам исследований. Палеогеография
воссоздает физико-географические, биогеографические, климатические и
другие условия минувших эпох. •
Палео ан атомические исследования наиболее важны для
палеозоолога, но так как остатки мягких частей беспозвоночных сохра-
няются очень редко, то для их восстановления приходится прибегать к
10
сравнению с современными формами или использовать те уникальные
находки, которые иногда попадают в руки коллекционеров. К числу
таких уникальных находок относится сохранившийся пищеварительный
тракт трилобитов, двустворок, гастропод. В ряде случаев удается вос-
становить мускульную систему по мускульным отпечаткам, сохранив-
шимся на раковинах двустворчатых и головоногих моллюсков, брахио-
под, остракод. О форме мягкого тела можно судить, например, по строе-
нию жилой камеры головоногих моллюсков, по форме чашечки корал-
ловых полипов, по строению теки граптолитов и т., д.
Перед палеофизиологией стоят очень сложные задачи —
изучение у древних организмов жизненных отправлений, т. е. тех. про-
цессов, которые совершались в организме в течение всей его жизне-
деятельности — пищеварения, дыхания, выделения, кровообращения,
движения, образования скелета.
Изучение микро- и ультраструктур и химического состава скелетов
позволяет судить о палеофизиологических процессах, приводивших к
созданию различных типов скелета. Так, кремневые скелеты строили ра-
диолярии и низшие многоклеточные — губки; органические скелеты
известны у всех животных, но состав аминокислот, входивших в состав
белка этих скелетов, изучен еще недостаточно.
Палеоэмбриология изучает особенности эмбрионального
развития организмов прошлого. Развитие палеофизиологических и палео-
эмбриологических исследований стало возможным с внедрением в прак-
тику работ палеонтолога электронного сканирующего микроскопа.
Палеопатология изучает различные прижизненные поврежде-
ния и болезненные явления, приводящие к нарушению строения скелета.
Изучение повреждений, полученных животными и залеченных при жизни,
позволяет путем их анализа выяснить их природу. Так, сохранившиеся
на раковине одного из аммонитов следы укуса позволили высказать
предположение, что укус был сделан мозозавром, остатки скелета кото-
рого не были обнаружены в осадке вместе с раковинами аммонитов.
Следы повреждений на раковинах палеозойских брахиопрд вызваны уку-
сами головоногих моллюсков. Изучение прижизненных повреждений на
раковинах беспозвоночных интересно тем, что в некоторых случаях при
залечивании раковины возникает скульптура, характерная для предко-
вого вида, что позволяет намечать родственные связи. Паразитирующие
на теле и скелете хозяина организмы вызывают образование различной
формы опухолей, приводящих к патологическому изменению скелета, эти
изменения иногда принимались за систематические особенности и при-
водили к недоразумениям.
Палеоихнология представляет новый раздел палеонтологии,
посвященный следам жизнедеятельности организмов.
Палеотаксиология занимается изучением особенностей пове-
дения животных в зависимости от воздействия на них различных факто-
ров внешней среды: гравитации, фототаксиса, хемотаксиса, реотаксиса
(ориентировка животного против течения, с которым поступает пища);
однако восстановление этих особенностей у вымерших животных пред-
ставляет большие трудности.
СОХРАНЕНИЕ ОСТАТКОВ ЖИВОТНЫХ В ИСКОПАЕМОМ
СОСТОЯНИИ И МЕТОДЫ ИХ ОБРАБОТКИ
Любьщ остатки организмов или следы их жизнедеятельности назы-
ваются окаменелостями, или фосси лиями. Фоссилии встре-
чаются преимущественно в осадочных породах водного происхождения—
морских, лагунных, пресноводных — озерных или речных. Реже они мо-
гут быть встречены в метаморфических породах осадочного происхожде-
ния и в различных осадочно-вулканогенных породах (вулканических ту-
фах). Фоссилии распределены в осадочных породах крайне неравномер-
но: местами они образуют большие скопления, но часто на значитель-
ных участках отсутствуют. Очень немногие организмы сохраняются на
тех местах, где они обитали. К ним относятся преимущественно бентос-
ные организмы, прикрепленные, зарывающиеся, в том числе гидроид-
ные и коралловые полипы, прирастающие двустворки (рудисты, устри-
цы), брахиоподы (тецидные, рихтгофении), многие иглокожие, хотя
скелеты этих групп после их гибели могут отрываться от субстрата и
переноситься часто на значительные расстояния. Например, части скеле-
тов колониальных кораллов, особенно кустистых, могут волнами при-
боя отламываться и переноситься на несколько километров от места их
обитания.
ОБРАЗОВАНИЕ ОРИКТОЦЕНОЗОВ
Характер перехода остатков животных из биосферы в литосферу
и сохранение их в осадочных породах представляет србой очень слож-
ный и недостаточно изученный процесс. Выделяются три основных эта-
па перехода: 1) накопление органических остатков, 2) захоронение,
3) фоссилизация.
Накопление органических остатков. Образование каждого местона-
хождения начинается с гибели животных. Одни организмы погибают
естественной смертью, другие от различных неблагоприятных причин —
резкого изменения температуры или солености, внезапных заморозков
или заморов, связанных с недостатком кислорода, различных заболева-
ний, наводнений или засух, извержений вулканов и массового отравле-
ния. Значительную помощь в анализе захоронения могут дать наблю-
дения над накоплением остатков современных животных. При гибели
трупы животных либо остаются на месте, либо переносятся на различ-
ные расстояния и в каких-то местах накапливаются. При участии бакте-
рий и различных факторов среды происходит разложение мягких тка-
ней и органов и разрушение скелета. Органические остатки рассеива-
ются, переносятся в море течениями или волнами, на суше — реками и
различными временными потоками, ветром. Скелеты и остатки многих
прикрепленных организмов могут оставаться на месте обитания в при-
жизненном положении. Скопления остатков погибших организмов об-
разуют танатоценозы — сообщества смерти (гр. thanatos —
смерть). В танатоценоз попадают животные из различных биоценозов.
К скоплениям остатков бентосных морских организмов могут добав-
ляться погибшие представители планктона, нектона, псевдопланктона,
а также вынесенные реками остатки наземных организмов и погибшие в
воде различные насекомые. Количество сохранившихся в ископаемом
состоянии организмов зависит от очень многих причин, но в первую
очередь от наличия или отсутствия скелета. Бесскелетные животные
(медузы, осьминоги) имеют минимальные шансы на сохранение и, на-
12
оборот, формы с массивным скелетом (колониальные гидроидные, ко-
раллы, многие устрицы) сохраняются значительно чаще. Для построй-
ки скелета животные используют органические соединения — тектин,
спонгин, хитин и неорганические — водный кремнезем (опал), кальцит,
арагонит, фосфат кальция. Тектин известен у саркодовых, спонгин у
губок, хитин преимущественно у артропод, частично у моллюсков. Крем-
невый скелет строят радиолярии, солнечники, губки; кальцитовый —
фораминиферы, археоциаты,' губки, октокораллы, мшанки, брахиоподы,
полихеты, некоторые моллюски, ракообразные, иглокожие. Арагони-
товый скелет известен у гидроидных, склерактиний, большинства мол-
люсков; фосфат кальция используют брахиоподы, некоторые артропо-
ды и все позвоночные. Существенное значение для сохранения в
ископаемом состоянии имеет численность индивидов, плотность их посе-
ления, массовая гибель их на местах обитания йли расстояния, на
которые переносятся их остатки, а также посмертное накопление в опреде-
ленных местах в результате постоянного сноса в одном направлении.
При переносе формы с тонким и нежным скелетом обычно легко уничто-
жаются, а скелеты, перекатываемые волнами, подвергаются разруше-
нию, окатыванию, сглаживанию острых углов, обламыванию тонких кра-
ев. Разрозненность и редкость органических остатков, продолжительное
пребывание их в условиях свободного доступа кислорода, сильное меха-
ническое( разрушение, наличие трупоедов часто приводит к полному
уничтожению всех остатков. (
Захоронение. Второй стадией образования местонахождения являет-
ся захоронение, при котором остатки погибших организмов покрывают-
ся осадком, ограничивающим доступ кислорода и предохраняющим их от
механического повреждения. Однако процессы разрушения и разло-
жения продолжаются в рыхлом осадке, насыщенном водой. Быстрое за-
хоронение и прекращение доступа кислорода, захоронение в асфальте,
смоле, в водах, насыщенных растворами солей, благоприятствуют сохра-
нению скелетных остатков.
Захоронение органических остатков связано с процессами осадко-
накопления, уплотнения осадка, изменением его физико-химических
свойств. При быстром пропитывании мягких тканей минеральными
растворами возможны случаи захоронения мягких тканей и органов. На-
пример, и'з меловых отложений Англии известны остатки пищеваритель-
ного тракта гастропод, а из юрских отложений той же страны — такой
же тракт двустворок.
Скопления органических остатков, погребенные в жидком осадке и
еще мало измененные (нефоосилизированные), образуют тафоценоз,
или сообщество погребения (гр. taphos — могила). Тафоценозы при
этом становятся членами минеральных ассоциаций осадочных пород лито-
сферы и в качестве таковых подчиняются всем закономерностям процес-
сов, происходящих в литосфере. Тафоценоз никогда не соответствует
палеобиогеоценозу, а включает в себя представителей разных ценозов.
Только в очень редких случаях такое соответствие может наблюдаться
(например, при массовой гибели кораллового биогеоценоза в результате
резкого изменения солености воды). В процессе захоронения почти
полностью уничтожаются личинки, молодь, виды с органическим и внут-
ренним скелетом, хотя известен ряд случаев сохранения и тех, и других.
Одним из ярких примеров сохранения организмов в морском бассейне
служат окаменелости из позднеюрского бассейна Золенгофена (ФРГ),
в котором захоронены аммониты с устьем, закрытым аптихами, тонкие
скелеты десятиногих раков, бесстебельчатых морских лилий, гладиусы
кальмаров с сохранившимся чернильным мешком и многие другие
13
остатки уникальных организмов, в том числе и первой птицы — архео-
птерикса. Другим примером может служить верхнеюрское пресное озеро
в Каратау (Южный Казахстан) с прекрасно сохранившимися остатка-
ми насекомых, рыб, летающих ящеров, подробно описанное Р. Ф. Гек-
кером (1.948).
Фоссилизация. Третьим, завершающим этапом образования .место-
нахождения является превращение рыхлых осадков в горные породы
(литификация), сопровождающееся превращением органических остат-
ков в окаменелости (фоссилизация). Во время фоссилизации происходят
изменения органических остатков, связанные преимущественно с
различными химическими факторами, температурой, давлением. Фосси-
лизация протекает параллельно процессу литификации и подчиняется
сходным закономерностям. При фоссилизации может происходить даль-
нейшее уничтожение органических скелетов, а также частичное или
полное — минеральных. С другой стороны, наоборот, во время литифи-
кации могут идти процессы, способствующие Захоронению органичес-
ких остатков:
окаменение — заполнение минеральным веществом пор и
пустот, имеющихся в различных скелетах;
перекристаллизация — преобразование первичной структу-
ры скелета без изменения его химического состава. Например, менее
стойкий арагонит замещается более стойким кальцитом, опаловый ске-
лет радиолярий, губок может замещаться тонкокристаллическим квар-
цем; мелкие кристаллы кальцита скелетов многих. организмов заме-
щаются более грубыми кристаллами того же кальцита;
метасоматоз — замещение первичного вещества скелета хи-
мическим веществом иного состава. Наиболее широко распространены
процессы окремнения, карбонатизации, ожелезнения, доломитизации.
При окремнении первичное скелетное вещество замещается раз-
личными модификациями кремнезема; при карбонатизации кремневые
или фосфатные скелеты замещаются карбонатом кальция. В условиях
восстановительной среды происходит пиритизация — первичный ске-
лет замещается пиритом, марказитом; в условиях окислительной сре-
ды происходит замещение различными окислами железа. При доломи-
тизации первичное вещество замещается двойной солью карбоната
кальция и магния.
Все органические остатки, встреченные в ископаемом состоянии
в одном месте, в одном слое, образуют ориктоценоз (гр. oryk-
tos — ископаемое), или ископаемый комплекс остатков растений и
животных. Изучение ориктоценозов позволяет восстанавливать палео-
биогеоценозы прошлого. Задача эта крайне трудная и сложная. При
анализе состава морских ориктоценозов выделяются организмы бенто-
са, планктона и нектона; среди первых обособляются различные типы
бентоса: подвижный, прикрепленный или зарывающийся; выясняется
характер расположения органических остатков в породе (биостратоно-
мический анализ); выясняются тафономические особенности: дальность
переноса, степень разрушения, прижизненное положение, а также вос-
станавливается образ жизни и условия существования захороненных
организмов (палеоэкологический анализ). В заключение восстанавли-
ваются палеобиогеоценозы отдельных частей бассейна и, если возмож-
но, фауна всего бассейна. Фауна — это исторически сложившийся
комплекс животных, объединенных общностью области распростране-
ния, иначе говоря, комплекс животных, характерный для изучаемого от-
резка времени, для региона, биогеографической провинции.
14
ФОРМЫ СОХРАННОСТИ
Полностью организмы в ископаемом состоянии сохраняются крайне
редко. К идеальным формам сохранности можно отнести консервацию
насекомых, хелицеровых и других членистоногих в янтаре мелового и
палеогенового возрастов.
Скелеты или их части. Чаще всего в ориктоценозах сохраняются
минеральные скелеты — известковые, кремневые, реже органи-
Рис. 2. Формы сохранности:
1а, б — раковина; 2 — внутреннее ядро; 3 — сохранившийся пищеварительный
тракт мелового брюхоногого моллюска (Кейси, 1960); 4 — отпечаток мягкого те-
ла осьминога из верхнемеловых отложений Сирии; 5 — отпечатки мускулов на
ядре жилой камеры у Doleroceras (Cephalopoda, Oncoceratida); 6—9 — разроз-
ненные органические остатки: 6 — аптихи; 7 — ринхолит; 8 — членики стебля
морской лилии; 9 — иглы морских ежей
ческие — хитиновые, хитино-фосфатные, тектиновые. Скелеты сохра-
няются целиком или частично; к целым скелетам можно отнести рако-
вины фораминифер, моллюсков, брахиопод, скелеты кораллов, мшанок,
иглокожих и др. В виде разрозненных частей скелета- встречаются спи-
кулы губок, челюсти кольчатых червей и головоногих моллюсков, кры-
шечки, закрывающие устье раковины гастропод и аммонитов, членики
стеблей морских лилий, иглы морских ежей и морских звезд (рис. 2).
Внутренние и внешние ядра (рис. 3). Внутреннее ядро
представляет собой слепок внутренней полости раковины. Ядро обра-
зуется при заполнении полости раковины, освобожденной после разруше-
ния мягких частей тела, окружающим ее осадком; если же полость ра-
ковины изолирована от внешней среды, то возможно проникновение
водных растворов в ее полость через любые поры, отверстия с дальней-
шим выпадением из раствора кристаллов, образующих друзы или вы-
полняющих всю полость раковины. Внутреннее ядро отражает особен-
ности внутреннего строения раковины; у тонкостенных раковин, у ко-
торых внешняя скульптура отражается и на внутренней стороне (у мно-
15
гих двустворок), внутреннее ядро отражает особенности внешней
скульптуры. У головоногих моллюсков обычно полость жилой камеры
заполняется осадком, вместе с которым внутрь попадают мелкие рако-
винки других организмов, а полости гидростатических камер остаются
либо пустыми, либо инкрустируются кристаллами кальцита, кварца, пи-
рита, целестина, реже заполняются осадком, проникающим через сифо-
нальное отверстие в перегородках.
Рис. 3. Возможные формы сохранности на примере раковины двустворчатого
моллюска (Тениус, 1963):
1 — захоронение; 2 — отпечаток и псевдоморфоза; 3 — внутреннее ядро; 4 —
внешнее ядро; 5 — деформация; 6 — настоящая окаменелость; 7 — захоронение
и окаменение; 8 — химико-биологическое разрушение; 9 — механическое раз-
рушение; 10 — раскрытые створки; 11 — изоляция; 12 — перенос; 13 — вторич-
ное захоронение; 14 — неполная сохранность; 15 — вымывание из осадка; 16 —
перезахоронение
Внешнее ядро возникает после разрушения раковины, обра-
зования отпечатка (подобно форме, изготовляемой для гипсовых отли-
вок) и заполнения возникшей полости осадком, принесенным цирку-
лирующими подземными водами, или кристаллами, выпавшими из раст-
вора. Внешние ядра передают особенности внешней формы раковины;
они бывают трудно отличимы от внутренних ядер, возникших у тонко-
стенных раковин. Внешние ядра чаще всего бывают песчаными, из-
вестковыми, доломитовыми, ожелезненными, фосфатными.
Отпечатки. Кроме ядер в осадочных породах встречаются отпечатки
мягкого тела или скелетов. Отпечатки мягкого тела обычно редки, но
имеют большую ценность. Отпечатки скелетов или частей скелетов встре-
чаются значительно чаще. Они возникают в результате полного уничто-
жения скелетов. Обычно по отпечатку делается слепок из латекса, плас-
16
тилина, воска, гипса или какого-нибудь другого материала. Слепок в
таком случае представляет собой искусственное наружное ядро.
Наиболее древние отпечатки мягкого тела бесскелетных организ-
мов — медуз, восьмилучевых кораллов, кольчатых червей, членистоно-
гих и групп неясного систематического состава были обнаружены впер-
вые в Южной Австралии, в Эдиакаре, в отложениях венда, датируемых
от 0,6 до 1 млрд. лет. Из более молодых отложений известны отпечат-
Рис. 4. Типы следов жизни (Зейлахер, из Циглер, 1972)
ки мягкого тела многих групп животных — кольчатых червей, голово-
ногих (кальмаров, осьминогов) и многих других.
Следы жизни. К следам жизни, или биоглифам (гр. bios— жизнь,
glyphos — резать), относятся все проявления жизнедеятельности орга-
низмов (рис. 4); различают экзоглифы — следы, оставленные на
поверхности осадка, и эндоглифы — следы жизнедеятельности
внутри осадка. Форма следов жизни зависит от образа жизни и поведе-
ния животных. Одни и те же следы могут быть оставлены разными ор-
ганизмами и, наоборот, разные следы оставляются одним и тем же
животным. К экзоглифам относятся следы ползания при поисках пищи,
следы лежания, следы погружения в осадок в случае опасности; к эндо-
глифам относятся жилые постройки разных зарывающихся организ-
мов — норки сверлильщиков твердых известковых пород: сверлящих гу-
бок, полихет, мшанок, усоногих раков, двустворок; постройки обитателей
илистых и песчаных грунтов. Среди последних различают простые норки
(например, у лингул, двустворок), небольшие выемки в осадке (остав-
ленные раками, крабами, трилобитами) или U-рбразные трубки, в кото-
2
В. В. Друщиц
17
рых постоянно живут их обитатели; через один конец трубки они полу-
чают пищу и воду, через другой — удаляют отбросы (многие полихе-
ты); в достаточно пористом осадке выводная часть трубки может отсут-
ствовать.
Многие обитатели, живущие в осадке, относятся к детритофа-
г а м; они извлекают питательные вещества из осадка, пропуская его-
через свой пищеварительный тракт. Особенно много питательных ве-
ществ в поверхностной пленке, покрывающей осадок, здесь кроме орга-
нических остатков растительного и животного происхождения имеется
богатая микро- и макрофлора. Этой пленкой питаются как зарывающие-
ся организмы, так и передвигающиеся внутри осадка или ползающие по
поверхности, оставляя следы, известные под названием следов кормле-
ния; они отличаются от следов ползания обычно маленькой площадью и
меандрической формой. Такие следы оставляют гастроподы, многие чер-
ви, разнообразные членистоногие. Следы ползания обычно отличаются от
следов кормления прямолинейностью и зависят от способа передвиже-
ния: способ передвижения червей отличается от способа передвижения
гастропод, двустворок, членистоногих. Для каждой группы организ-
мов характерны свои следы ползания, следы кормления, свои норки.
К следам жизнедеятельности относятся также копролиты —
ископаемые экскременты животных. Копролиты отличаются большим
разнообразием форм и размеров. Изучение копролитов позволяет су-
дить о пище, которой питались организмы. Так, по копролитам ихтио-
завров было установлено, что их пищей служили белемниты.
Следы жизнедеятельности организмов позволяют восстанавливать
условия жизни и осадконакопления, жизнь тех групп, которые не сохра-
няются в ископаемом состоянии (например, черви), особенности биото-
пов, а также залегание слоев (нормальное или перевернутое), имеющее
значение для выяснения истинной последовательности слоев.
Хемофоссилии. В последние годы были обнаружены молекулярные
ископаемые, или хемофоссилии; они представляют собой наиболее устой-
чивые фрагменты крупных органических молекул (предельные углеводо-
роды — алканы, аминокислоты и жирные кислоты), ранее входившие в
состав организмов, а затем распавшиеся в процессе фоссилизации. По
ним нельзя определить, к каким организмам они принадлежали, но они
являются свидетелями ранней жизни; первые хемофоссилии обнаруже-
ны в породах, возраст которых определен около 2,7 млрд. лет. Иссле-
дования хемофоссилий только начаты.
СБОР И МЕТОДЫ ОБРАБОТКИ
ИСКОПАЕМЫХ ОСТАТКОВ
Предпосылкой для успешной научной обработки палеонтологичес-
кого материала и всестороннего использования его для целей био-
стратиграфии, палеогеографии и восстановления истории бассейна, а
также решения вопросов филогении и систематики служат послойные
сборы фауны из изучаемых разрезов, тщательные, по возможности мас-
совые, сборы ископаемых остатков, точная их привязка к слоям, изу-
чение пространственного изменения состава фауны по отдельным го-
ризонтам и подробное описание всех особенностей найденных органи-
ческих остатков.
Отбор образцов для изучения фораминифер, радиолярий, остракод.
производится через определенные интервалы, преимущественно из рых-
лых разностей пород. При исследовании разрезов, сложенных известня-
ками, тщательно осматриваются все слои и отбираются образцы для из-
18
готовления шлифов. Поиски ископаемых обычно начинаются с осмот-
ра осыпей, а затем исследуются обнажения и выбираются места, удоб-
ные для разработок. Особенно тщательно осматриваются ' выветрелые
поверхности, на которых можно иногда обнаружить окаменелости прек-
расной сохранности, отпрепарированные процессами выветривания (на-
пример, скелеты археоциат, фузулин). Основная масса ископаемых из-
влекается с помощью геологического молотка и зубила.
Мелкие формы удается отбирать из рыхлых отложений при просеи-
вании. Собранные фоссилии подвергаются в лаборатории предваритель-
ной технической обработке для очистки их от вмещающей породы.
Методика препарирования зависит от размеров, формы, степени сохран-
ности экземпляра и характера окружающей его породы. Обычно пре-
парирование требует осторожности, терпения и занимает много вре-
мени. Прежде чем приступить к освобождению образца из породы, сле-
дует получить представление об особенностях его строения.
Механическое препарирование. От глины и песка окаменелости очи-
щаются щетками и водой; лишние куски плотных пород удаляются от-
калыванием с помощью зубил, долот, игл и других инструментов. Обра-
зец при препарировании кладется на мешок с песком («подушку»).
Случайно отбитые кусочки подклеиваются клеем, специально приготов-
ленным для этих целей.
За последние годы находят применение виброинструменты, зубо-
врачебные боры и другие инструменты. При препарировании также при-
меняется ультразвук, позволяющий не только удалять «пустую» породу,
но и отрезать лишние куски.
Шлифы и пришлифовки. Скелеты всех вымерших животных изу-
чаются в шлифах и пришлифовках. Для изучения возрастных измене-
ний и деталей строения изготавливается серия последовательных шли-
фов и пришлифовок.
Метод реплик. С пришлифовки всегда можно изготовить ацетат-
ную реплику, на которой обычно хорошо бывают видны многие детали
изучаемой структуры. Для этой цели пришлифованная поверхность об-
разца полируется, подвергается травлению кислотой, промывается и су-
шится; затем на смоченную ацетоном поверхность накладывается от-
мытая от эмульсии фотопленка.
Методы обработки микрофоссилий. Для изучения микрофоссилий
(фораминифер, радиолярий, остракод) в каждом отдельном случае
требуются отдельные методы. Для извлечения микрофоссилий породу
предварительно дробят, измельчают, размачивают в воде, кипятят, ис-
пользуя при этом некоторые щелочи или кислоты. После многократной
промывки в осадке остаются микрофоссилии, которые после просушива-
ния осадка отбираются под бинокулярным микроскопом или лупой и
переносятся в специальные камеры (камеры Франке) для последую-
щего изучения.
Электронная микроскопия. В последние годы достигнуты большие
успехи в изучении микро- и ультраструктур скелетов коралловых по-
липов, мшанок, граптолитов, раковин фораминифер, моллюсков, брахио-
под с помощью электронного, трансмиссионного и сканирующего, микро-
скопов.
Химическое препарирование основано на применении различных
растворителей — щелочей и кислот, с помощью которых' растворяется
порода, окружающая окаменелость. Для освобождения из карбонатных
пород скелетов археоциат, кораллов, брахиопод, Моллюсков применяет-
ся уксусная, муравьиная или слабая соляная кислоты; для освобожде-
2:
19
ния скелетов из кремнистых пород основным реагентом является плави-
ковая кислота.
) Рентгеноскопия. Применение рентгеновских снимков палеонтологи-
ческих объектов, в том числе крупных плит, позволяет обнаружить
часто .скрытые в породе органические остатки со всеми деталями их
строения (например, остатки морских лилий, офиур, отпечатки щупалец
аммонитов) и наметить направление препарирования. В последние годы
с помощью рентгеноскопии были обнаружены следы окраски на рако-
винах головоногих и брюхоногих моллюсков.
Определение ископаемых. Обработанные и подготовленные к изуче-
нию ископаемые только тогда приобретают значение и ценность, когда
они будут определены, т. е. когда будет установлено их родовое и ви-
довое название, геологический возраст и географическое распростране-
ние.
При определении пользуются специальной литературой: атласами,
монографиями, справочниками, а также монографическими коллекция-
ми ранее описанных видов.
ОСНОВНЫЕ ЭТАПЫ РАЗВИТИЯ ПАЛЕОНТОЛОГИИ
Становлению палеонтологии как науки предшествовал длительный
период накопления фактического материала как биологического, так и
геологического и попыток создания естественнонаучных теорий. В разви-
тии представлений о природе, в том числе и древних организмах, насе-
лявших нашу планету, можно наметить следующие периоды: 1 — от
глубокой древности до начала XIX в. — период накопления биологи-
ческих и геологических фактов, 2 — возникновение палеонтологии и
ее додарвиновский период, 3 — период становления и развития эво-
люционной палеонтологии от Дарвина до 40-х годов нашего столе-
тия, 4 — развитие современной палеонтологии. Развитие палеонтоло-
гии шло параллельно с развитием всех естественных наук.
Первый период. В глубокой древности естествоиспытатели Индии,
Китая, Египта высказывали отдельные идеи о естественном происхож-
дении живых существ, о материальности мира, о естественном харак-
тере законов природы. Для познания природы и развития материалисти-
ческой философии много было сделано в античной Греции. Некоторые
греческие ученые и философы правильно понимали природу окамене-
лостей, рассматривая их как остатки организмов, некогда живших в
морских бассейнах. В древнегреческих мифах очень часто фантастика
переплеталась с правильными ' наблюдениями над жизнью природы.
Успехи естествознания древних греков были обобщены в трудах
Аристотеля (384—322 гг. до н. э.) —великого мыслителя своего време-
ни, который создал основы классификации животных, зачатки срав-
нительной анатомии и эмбриологии, сделал в естествознании ряд обоб-
щений: о постепенном переходе от более простых преобразований к
более сложным, о градациях в природе, высказал предположение о
корреляциях частей тела, ввел родовые и видовые понятия, охарактери-
зовал животных по строению их частей тела, образу жизни, поведе-
нию и привычкам. Аристотель различал животных с кровью (позво-
ночных) и животных без крови (беспозвоночных в современном пони-
мании). Последних он разделял на четыре «больших рода» — мягкоте-
лых (головоногие), мягкоскорлупых (высшие ракообразные), черепо-
кожих (моллюски, без головоногих) и насекомых. К насекомым Арис-
тотель относил многоножек, пауков, клещей, некоторых червей; медузы,
20
актинии, морские звезды и губки отождествлялись им с растениями и.
относились к зоофитам. Многие идеи Аристотеля — естественное возник-
новение живых существ путем самозарождения, представление о гра-
дации, т. е. о «иерархии» существ, учение о единстве и целостности
строения организмов, принцип коррелятивной связи органов —
сравнительно-анатомический метод, и представление о формообразова-
нии как о последовательном возникновении качественно нового в про-
цессе индивидуального развития — были использованы спустя только
несколько столетий при разработке теории развития органического ми-
ра. По Аристотелю, биологические процессы носили целесообразный ха-
рактер. Однако позднее представление Аристотеля о целесообразности
в природе было искажено в духе креационизма — источником целесо-
образности в природе считался бог — создатель мира. В таком ви-
де телеология Аристотеля (от гр. telos — цель) в течение более чем
двух тысяч лет служила опорой религиозно-метафизическому мировоз-
зрению и была сокрушена только в середине XIX в. Ч. Дарвином.
За периодом расцвета науки в Греции наступил тысячелетний период
средневековья — период феодально-церковного владычества, период
очень замедленного развития, а иногда и полного застоя естествозна-
ния. Только в эпоху Возрождения (со второй половины XV 'в.) на-
чалось новое развитие естественных наук: эта эпоха характеризова-
лась установлением метафизического взгляда' на природу, в соответст-
вии с которым отрицалось развитие, признавалась неизменность при-
роды и присущая ей изначальная целесообразность. Шло накопление и
описание фактического материала в зоологии и ботанике и делались
первые попытки классификации. 4
Леонардо да Винчи (1452—1519) — один из наиболее выдающих-
ся представителей естествознания итальянского Возрождения рассмат-
ривал окаменелости как остатки морских животных, которые сохрани-
лись в осадочных породах; развивал идеи единства природы, причин-
ности и закономерности жизненных процессов, необходимости изучать
организмы в динамике и развитии, от моментов зачатия до зрелого воз-
раста.
Те же взгляды на природу окаменелостей высказывали итальян-
ский профессор философии Фракасторо (1483—1553) и французский
ученый Б. Палисси (1510—1590).
Однако в течение XVII и первой половины XVIII в. многие уче-
ные продолжают считать окаменелости «игрой природы», рассматривая
их как «фигурные камни», только внешне похожие на животных, или
как результаты деятельности «пластической силы». В начале XVIII в.
распространяется представление, что ископаемые являются остатками
организмов, погибших во время потопа, на основании находок семян и
плодов устанавливалось даже время года, когда произошел «потоп».
Считалось, что Земля и ее обитатели существовали около 6000 лет.
Следуя учению о потопе, швейцарский ученый И. Шейхцер (1672—•
1733) описал ископаемые остатки крупной саламандры как человека, по-
гибшего во время потопа.
Среди представителей передовой науки XVIII в. одно из важных
мест принадлежит М. В. Ломоносову (1711—1765), который заслужен-
но считается основоположником естественноисторического материализма
в России. Ломоносов отвергал представление о «всемирном потопе» и
объяснял происхождение осадочных горных пород образованием их в
морских бассейнах. Ископаемые моллюски («черепокожие», по Аристо-
телю), встреченные в этих породах, обязаны своим происхождением
морям, существовавшим здесь в прошедшие геологические эпохи. Ло-
21
Моносов представлял себе смену различных периодов жизни на Земле
как последовательное чередование наступания и отступания морей,
объясняя эти явления медленными вековыми колебаниями суши. Ломо-
носов впервые высказал правильное представление о янтаре как
ископаемой смоле хвойных, образно описав в своей книге «О слоях зем-
ных» (1763), как попали и захоронились в янтаре разнообразные «чер-
Рис. 5. Михаил Васильевич Ломоносов
(1711—1765)
вяки и другие гадины». В отличие от Бюффона, он считал, что история
Земли насчитывает около 400 000 лет и, несмотря на преследование
церкви, не отказался от своих идей, как это сделал Бюффон.
В 1755 г. по инициативе Ломоносова был создан Московский уни-
верситет — один из первых научных центров подготовки русских уче-
ных. Первые палеонтологические сборы в России начались после ука-
зов Петра I о создании Кунсткамеры (1718) и открытия в Петербурге
Академии наук (8/II 1724). Из Кунсткамеры в дальнейшем развились
все естественноисторические музеи Академии наук, в том числе и па-
леонтологический.
В течение XVII и особенно XVIII в. был накоплен большой факти-
ческий материал по всем разделам естествознания. Первые классифи-
кации носили произвольный характер, это были алфавитные каталоги
либо классификации, основанные по произвольно избранным признакам.
Д. Рэй (1628—1705) в своей сводке «История растений» (1686—1703)
ввел понятие «вид», «род» и бинарную номенклатуру, однако получив-
шие признание только после работ К. Линнея. Применение первого
микроскопа (А. Ван Левенгук, 1632—1723) создавало новые возмож-
ности для изучения индивидуального развития.
22
В естествознании возникли два течения — преформизм и эпигенез.
В соответствии с идеями преформизма индивидуальное развитие
рассматривалось как простой рост, развертывание образованных ранее
зачатков — как их простое количественное увеличение. Один из пред-
ставителей преформизма Ш. Бонне (1720—1793) создал «лестницу су-
ществ», соединив известные в то время минералы и организмы в од-
ну общую цепь от низших к высшим. Главным в учении преформистов
были их представления о неизменности природы, о постоянстве видов.
В соответствии с идеями эпигенеза (У. Гарвей, 1578—1657)
развитие рассматривалось как возникновение нового, как последова-
тельное образование частей тела и органов в результате формообра-
зования; причем считалось, что разнородное развивалось из однород-
ного. Борьба эпигенетиков против преформистов создавала предпо-
сылки для формирования идей об изменяемости видов. Эпигенез в
XVIII в. стал передовым течением в естествознании, поскольку сто-
ронники эпигенеза боролись против догмы, против неизменяемости при-
роды, против ограниченности преформизма.
Большое значение в развитии науки имели труды шведского уче-
ного Карла Линнея (1707—1778), который построил систему всех из-
вестных в его время организмов, завоевавшую всеобщее признание.
Линней разделил всю природу на три царства: минералов, растений и
животных. Царство животных делилось на 6 классов: млекопитающих,
птиц, земноводных, рыб, насекомых и червей. Классы были разделены
на роды, роды на виды, а последние на разновидности. Каждому виду
присваивалось двойное латинское название (см. ниже) по правилам
бинарной номенклатуры. Линней описал свыше 4200 видов животных, в
том числе и многих вымерших, и около 10 000 видов растений. В конце
своей жизни Линней допускал возможность возникновения новых ви-
дов растений под влиянием внешней среды или в результате скрещива-
ния. Классические исследования Линнея в области систематики пред-
ставляют собой крупную веху в развитии естествознания, всей биологии
и палеонтологии, в частности.
Наряду с представлениями о постоянстве и неизменяемости видов
все чаще высказывались мысли об их изменяемости, о возможности пре-
вращения одних видов в другие. К числу сторонников идей трансформиз-
ма относятся член Петербургской академии наук К. Вольф (1733—1794),
предшественник Ламарка — французский ботаник А. Жюсье (1748—
1836), французский естествоиспытатель Ж. Бюффон.
Ж. Бюффон (1707—1788) —один из наиболее видных представителей
биологии эпохи Просвещения, автор 36-томной «Естественной истории»;
он рассматривал наиболее актуальные и трудные вопросы естествозна-
ния: историю Вселенной и Земли, происхождение и развитие жизни, ис-
торию животного и растительного мира и другие вопросы. Бюффон под-
черкивал отсутствие резкой границы между животным и раститель-
ным миром, говорил о едином плане строения животных, о наличии
промежуточных форм между разными группами животных, о длитель-
ности истории Земли (до 75 000 лет), защищал идеи эпигенеза.
Сторонником изменяемости видов в ранний период своей научной
деятельности был русский академик П. С. Паллас (1741—1811), пред-
лагавший изображать связь между видами в форме древовидных раз-
ветвлений, подчеркивая генетическую связь между вымершими и ныне
живущими организмами. Паллас был одним из зачинателей биогеогра-
фии как самостоятельной науки. Среди сторонников идей трансфор-
мизма можно отметить А. Н. Радищева (1749—1808), Э. Дарвина
23
(1721—1802), деда Ч. Дарвина, высказывавших идеи об историческом
развитии органических форм.
В геологии в XVIII в. шла борьба между плутонистами и нептунис-
тами. Одним из ярких выразителей идеи плутон истов (Плутон —
бог подземного царства у древних греков) был шотландский натуралист
Дж. Геттон (1726—1797), придававший большое значение внутреннему
теплу и давлению в образовании кристаллических пород, но он допус-
кал, что в образовании пород и формировании рельефа важную роль
играет вода. Геттон в общей форме сформулировал идеи унифор -
м и з м а — научное течение, основанное на идеях о непрерывности
изменений, происходивших на Земле в прошлом и происходящих в на-
стоящее время, о длительности происходящих событий и суммировании
незначительных отклонений. Геттон придавал первостепенное значе-
ние актуалистическому методу в геологических исследованиях; «настоя-
щее есть ключ к познанию прошлого» — это крылатое выражение
принадлежит Геттону.
Идейным вдохновителем нептунистов (Нептун — бог морей
у римлян) был А. Вернер (1750—1817), профессор Фрейбергской гор-
ной академии. Вернер придавал воде основное значение в образовании
горных пород. «Весь земной шар можно считать осадком из воды», —
писал он (из И. И. Шафрановского, «А. Г. Вернер — знаменитый ми-
нералог и геолог». М., «Наука», 1968, стр. 134). Вернер стремился к
созданию классификации горных пород, минералов, подчеркивал необ-
ходимость тщательных полевых наблюдений, стремился подойти к пост-
роению первой относительной геохронологии; он разделил горы по слага-
ющим их горным породам, «образовавшимся мокрым путем», на четы-
ре периода, соответствующих четырем последовательным стадиям об-
разования геологических формаций: первозданные горы («кристалличес-
кие осадки»); переходные горы с находками первых окаменелостей ор-
ганического происхождения; «флецовые», или пластовые, горы с боль-
шим количеством окаменелостей, образованные песчаниками, известня-
ками и базальтами, и «намывные горы», состоящие из песков, глин и
других типичных осадочных пород. В учении Вернера имелись элемен-
ты катастрофизма: признание необычайных сил, действовавших в прош-
лом, наличие резких границ между формациями и ограниченное время
существования Земли. Особенно резко критиковали взгляды Вернера
его талантливые ученики А. Бух (1774—1853) и А. Гумбольт (1769—
1859).
Второй период. К началу XIX в. накопился большой фактический
материал как в области биологии, так и в области геологии. Палеон-
тология как научная дисциплина выделилась из цикла естественных
наук. У ее истоков стояли английский геолог У. Смит и французские-
биологи Ж. Ламарк и Ж. Кювье.
Уильям Смит (1769—1839) — выдающийся английский ученый,
основатель геологии Англии, занимаясь геологическими и топографи-
ческими исследованиями, пришел к выводу о возможности использо-
вания органических остатков для определения относительного возраста
слоев, в которых они встречены. Смит полагал, что слои, развитые
в разных районах Англии и содержащие одинаковые органические ос-
татки, должны быть одновозрастными, а содержащие различные ис-
копаемые — разновозрастными. Смит составил для Англии первый
сводный стратиграфический разрез, в котором выделил 23 слоя. Каждо-
му слою он дал свое название, связанное либо с местом распространения
этого слоя (например, «Фуллерова земля», «лондонская глина») или
с какими-то внешними особенностями («Лесной мрамор», «Зеленый пес-
24
чаник», «Мел»). Выходы коренных пород на дневную поверхность Смит-
наносил на топографическую карту и для наглядности раскрашивал раз-
ными цветами. Так родилась не только первая геологическая карта
Англии и Шотландии, но и первая геологическая карта в мире.
200 экземпляров такой карты, раскрашенной вручную, сопровождаемой
объяснительной запиской к ней, были опубликованы в ' Англии
в 1815—1816 гг.
Рис. 6. Уильям Смит (1769—1839)
Идея составления геологической карты, стратиграфической колон-
ки и объяснительной записки к ней оказалась очень плодотворной. Гео-
логическая карта давала представление о последовательности слоев,
слагающих изучаемую территорию, о ее тектонике, геологической исто-
рии; она позволяла прогнозировать направление поисковых и геолого-
разведочных работ с целью выявления полезных ископаемых, пресных и
минеральных вод.
Вскоре геологические карты стали создаваться в других странах, в
первую очередь во Франции, Германии, России. Идеи Смита о выделе-
нии отдельных слоев, о названии этих слоев по месту их распростране-
ния и использование органических остатков для определения относи-
тельного возраста послужили основой для создания биостратиграфии,,
исторической геологии и палеонтологии как самостоятельных наук.
С первых своих шагов палеонтология развивалась параллельно с
развитием всех естественных наук и в первую очередь биологических и
геологических, оставаясь всегда биологической наукой.
25-
У. Бакленд (1784—1856), первый профессор геологии Оксфордского
-университета в Англии, предложил в 1818 г. классификацию стратигра-
фических подразделений в виде иерархической лестницы, подобно иерар-
хии биологических таксонов К- Линнея. Бакленд рекомендовал выделять
следующие стратиграфические подразделения: классы, порядки,
формации и слои/ Несколько позднее, в 1820 г., в Англии было
предложено формации объединить в системы. Палеонтологический
метод Смита и принципы классификации слоев Бакленда стимулирова-
ли геологов к выделению формаций и систем. Менее чем за двадцать
.лет (1818—1841) были выделены почти все системы, которые в дальней-
шем легли в основу существующей сейчас геохронологической шкалы
(об этом подробнее см. ниже).
Жан Батист Ламарк (1744—Г829), современник Смита, французский
натуралист, ботаник и зоолог, работал вместе с Сент-Илером и Кювье
в Национальном музее естественной истории в Париже. Он впервые раз-
работал последовательное, хотя и не лишенное недостатков учение о
развитии органического мира, которое изложил в своем труде «Филосо-
фия зоологии», опубликованном в год рождения Ч. Дарвина (1809).
Ламарк, находясь под влиянием идей Бюффона, отрицал метафизичес-
кие идеи о постоянстве и неизменяемости видов и утверждал, что виды
изменяются крайне медленно и незаметно, что между ними нет границ
и что они связаны между собой постепенными переходами. Живая при-
рода представлялась Ламарку в виде цепи непрерывно изменяющихся
-особей, которые объединяются в виды, существующие только в течение
•определенных отрезков времени.
Органический мир развивался, по Ламарку, от простейших форм к
более сложным. Животный мир позвоночных и беспозвоночных (послед-
нее название было предложено Ламарком) Ламарк разделил на
14 классов вместо 6 линнеевских, разместив их на шести ступенях: на
низшей — инфузории и полипы, на высшей — птицы и млекопитающие.
Постепенное усложнение организации живых существ в процессе эво-
люции Ламарк назвал градацией, считая при этом, что движущей
•силой развития является врожденное внутреннее стремление животных
к совершенствованию своей организации, к прогрессу как высшей цели,
изначально заложенной в них. Правильная градация, по Ламарку, нару-
шалась влиянием внешней среды, которая воздействовала либо прямо
(растения и низшие животные), либо косвенно через нервную систему
(высшие животные). Изменчивость связана с развитием и деятельностью
органов. Упражнение какого-либо органа развивает его, увеличивает и
придает ему силу, наоборот, неупражнение приводит к его ослаблению
и редукции (1-й закон Ламарка). Изменение внешних условий вызывает
изменение потребностей, а это влечет за собой изменение органов; все,
что было приобретено или изменилось в течение индивидуальной жиз-
ни, сохраняется благодаря наследственности и передается потомству
(2-й закон Ламарка).
Ламарк, создав теорию наследования приобретенных признаков и
изменчивости, соответствующей потребностям внешней среды, поставил
перед естествоиспытателями очень сложную проблему о взаимосвязи
изменчивости и наследственности. Он не смог ответить на вопрос,
как изменчивость становится достоянием последующих поколений.
Гениальная эволюционная идея основывалась на признании поступа-
тельного развития органического мира, однако отсутствие достаточного
материала и неразработанность многих проблем биологии не позволили
Ламарку правильно ответить на вопрос о движущих силах эволюции.
Признание в качестве движущей силы эволюции внутреннего стремления
.26
к прогрессу, якобы заложенного у организмов самой природой, приво-
дило к признанию каких-то сверхъестественных сил, а утверждение о
врожденной способности организмов изменяться адекватно изменениям
-среды позволяло признавать изначальную целесообразность, т. е.
признавать соответствие организмов той цели, которая якобы была с
самого начала поставлена для них «творцом». Ламарк не смог отве-
Рис. 7. Жан Батист Ламарк (1744—1829)
тить на вопросы, как можно объяснить все многообразие органического
мира и удивительные адаптации организмов к условиям окружающей
их среды и почему в процессе эволюции происходит повышение органи-
зации. Ламарк подвергал беспощадной критике катастрофизм и проти-
вопоставлял катастрофизму идеи униформизма, считая, что геологи-
ческие силы в истории Земли действовали ровно и постоянно в течение
длительного времени, а не 6000 лет, как утверждали церковники.
Ламарк опубликовал 32 статьи по ископаемым третичным моллюс-
кам Парижского бассейна, впервые стал применять термин «ископае-
мые» только к органическим остаткам (до Ламарка этим термином
обозначали также.и минералы и горные породы). Ламарк, используя
метод актуализма, допускал общность происхождения третичных мол-
люсков и современных, что шло в разрез с общепринятым мнением; на
основании анализа третичных моллюсков высказывал предположение
о глубине бассейна, в котором они существовали, и изменении климатов
прошлого.
27
Жорж Кювье'(1769—1832), ровесник Смита и современник Ламар-
ка и Сент-Илера, известен как выдающийся выразитель идеи неизменяе-
мости органического мира и одновременно как яростный противник
своих выдающихся современников — сторонников трансформизма — Ла-
марка, Сент-Илера. Свою теорию постоянства и неизменяемости видов
Кювье основывал на двух принципах: принципе корреляции и принци-
пе условий существования. В соответствии с первым принципом, или
Рис. 8. Жорж Кювье (1769—1832)
принципом соподчинения органов, всякое организованное
существо образует единую замкнутую систему, части которой соответ-
ствуют друг другу и содействуют путем взаимного влияния одной конеч-
ной цели. В соответствии со вторым принципом различные части тела со-
единены друг с другом таким образом, чтобы животное могло существо-
вать в его взаимоотношениях с окружающими существами. Оба принци-
па Кювье были теологичны, поскольку предполагалось, что все соз-
дано для определенной цели. Они, однако, послужили основой для созда-
ния целостного учения в области сравнительной анатомии.
Кювье обосновал четыре плана строения, в соответствии с
которыми все животные по строению нервной системы были разделе-
ны на четыре типа: позвоночные с нервной системой, состоящей
из головного и спинного мозга; моллюски, имеющие нервные узлы
и отходящие от них нервы; членистые с нервной системой в виде
двойной нервной цепочки; лучистые с радиальным расположением
нервных стволов. Принципы корреляции и условий существования были
28
установлены Кювье при сравнительно-анатомических исследованиях
нервной системы, кровообращения, дыхания и органов воспроизведе-
ния у различных классов моллюсков — брюхоногих, головоногих и. без-
головых (так Кювье называл двустворчатых), червей и насекомых. Эти-
ми работами Кювье заложил прочный фундамент сравнительной ана-
томии.
Изучая остатки третичных животных в Парижском бассейне, и в
первую очередь позвоночных, он установил, что существовала смена
органических форм во времени, что при переходе от древних геологи-
ческих отложений к более молодым возрастает сходство вымерших жи-
вотных с современными. Кювье во Франции одновременно со Смитом в
Англии, изучая биологические особенности развития организмов во вре-
мени и проверяя их в сочетании с геологическими, заложил основы па-
леонтологического метода в стратиграфии. Кювье пытался научно обос-
новать, что жизнь на Земле появилась на определенном этапе ее разви-
тия,. древние организмы принимали участие в осадконакоплении и од-
ни виды вымирали, другие появлялись; состав видов изменялся вместе
со слоями. Кювье наметил четыре фауны позвоночных во времени:
1) пресмыкающихся, 2) палеотериев, анаплотериев, 3) млекопитаю-
щих, ныне вымерших (мастодонты, мамонты, мегатерии), 4) совре-
менных видов и человека.
Кювье явился ярким выразителем идей катастрофизма, в соответст-
вии с которыми: 1) на Земле в органическом и неорганическом мире
происходили внезапные, скачкообразные изменения, 2) в истории Зем-
ли было три переворота, причем последний связывался с библейским по-
топом, 3) перевороты захватывали обширные участки, но не были, как
•сейчас выражаются, глобальными.
Заслуги Кювье, несмотря на допущенные им теоретические ошибки
(защита идеи постоянства видов, разработка теории катастроф), безус-
.ловно велики; его исследования по палеонтологии, сравнительной анато-
мии создали косвенные предпосылки для появления и развития дарви-
низма.
Жоффруа Сент-Илер (1772—1844), современник и противник Кювье,
резко выступал против теоретических построений Кювье, против идеи о
постоянстве видов и ратовал за создание философии зоологии. Сент-
Илер развивал идеи единства животного мира, изменяемости его форм
под влиянием'воздействия условий внешней-среды. Он сформулировал
два принципа: принцип гомологий и принцип равновесия, или компен-
сации. В соответствии с принципом гомологий, на основании
сравнительно-анатомических и эмбриологических исследований могут
быть установлены гомологичные органы, имеющие общее происхожде-
ние и сходное строение, но выполняющие разные функции. Этот принцип
гомологий позволил установить общие принципы морфологии; он имел
важное значение для решения вопросов систематики и происхождения
различных групп. Второй принцип — принцип компенсации
говорит о том, что сильное развитие одних органов совершается за счет
недоразвития других; отсутствие каких-то органов, например зубов у
китов на взрослой стадии, может быть результатом их недоразвития
или редукции на эмбриональной стадии, что заставляет с вниманием
относиться к изучению рудиментарных органов. На основании этих
двух принципов Сент-Илер устанавливал общность в строении всех клас-
сов позвоночных.
В нашумевшем научном споре с Кювье о планах строения, в кото-
ром столкнулись две противоположные точки зрения — идеалистичес-
кая и материалистическая, Сент-Илер потерпел поражение, потому что
29
вступил на путь сравнения не гомологичных, а аналогичных органов,,
выполняющих одинаковые функции, но различных по происхождению;
он пытался доказать единство всего животного мира, сравнивая строение
позвоночных и членистоногих или позвоночных и головоногих моллюс-
ков; в этом была его научная ошибка.
Первая половина XIX в. характеризуется большими успехами в раз-
витии геологии и биологии. Большую роль в развитии этих наук сыгра-
ли труды английского ученого Чарлза Лайеля (1797—1875), современни-
Рис. 9. Чарлз Лайель (1797—1875)
ка Ч. Дарвина. В своем капитальном труде «Основы геологии»
(1830—1833) Лайель на основании анализа большого фактического ма-
териала показал, что преобразование Земли в геологическом прошлом:
происходило не путем катастроф или внезапных революций, а при по-
степенном воздействии разнообразных процессов как внешних (экзо-
генных), так и внутренних (эндогенных). В соответствии с длительными
изменениями окружающей неорганической среды медленно и постепенно-
изменялся органический мир. Лайель в своих рассуждениях широко ис-
пользовал метод актуализма, т. е. метод сравнения процессов, протекав-
ших в прошлые геологические эпохи, с современными процессами.
Актуалистический метод Лайеля сыграл выдающуюся роль в формиро-
вании мировоззрения Дарвина.
Лайель явился главным выразителем униформизма — уче-
ния, основанного на трех принципах: а) силы, преобразующие земной:
30
шар, единообразны (принцип однообразия), б) эти силы дейст-
вуют непрерывно И без катастроф (принцип непрерывности),
в) малозаметные изменения суммируются и приводят к крупным изме-
нениям (принцип суммирования мелких отклонений). Униформис-
ты принимали, что геологические силы прошлого по их роду, энергии и
скорости тождественны с современными. В этом была их ошибка, но они.
наносили удар катастрофистам, идеи которых господствовали в геоло-
гии в первой четверти XIX в. Ф. Энгельс, критикуя воззрения Кювье,
высоко оценил заслуги Лайеля. Он писал: «Лишь Лайель внес здравый
смысл в геологию, заменив внезапные, вызванные капризом творца,,
революции постепенным действием медленного преобразования Земли»
(К. Маркс, Ф. Энгельс. Соч., т. 20, стр. 352).
К числу крупных открытий первой половины XIX в., оказавших,
влияние на развитие естественных наук, относятся создание клеточной
теории (Шлейден и Шванн, 1838—1839) и эмбриологические исследова-
ния X. Пандера и К. Бэра.
X. Пандер (1794—1865) был одним из основоположников палеонто-
логии в России, одним из ранних сторонников эволюционной идеи и
учения об изменчивости видов. Он разработал учение о зародышевых
листах, основанное на изучении эмбриогенеза цыпленка. Пандер издал
геологическое описание окрестностей Петербурга, снабдив его много-
численными рисунками ископаемых, обработал большие коллекции си-
лурийских и девонских рыб, впервые выделил и описал конодонты, —
зубы до сих пор не известных каких-то морских животных.
К числу крупных ученых, подготовивших почву для дарвинизма,
следует отнести русского академика К. Бэра (1792—1876), научные-
исследования которого проходили по трем направлениям — анатомии и
эмбриологии животных, физической географии и теоретическому ес-
тествознанию. Бэр заложил основы сравнительной эмбриологии. Изучая
зародыши позвоночных, он вывел закон, в соответствии с которым за-
родыши высших животных напоминают зародыши низших, а не их
взрослые формы, как предполагали ранее: у эмбрионов признаки круп-
ных таксонов закладываются раньше, чем специальные. Это открытие
имело важное значение для эволюционной теории; эмбриология совмест-
но с палеонтологией и сравнительной анатомией стала основным источ-
ником доказательства эволюции органических форм.
Современником Пандера и Бэра был профессор Московского универ-
ситет а К. Ф. Рулье (1814—1858), читавший первый курс палеонтологии
и развивавший лучшие традиции, заложенные Ломоносовым. Рулье при-
надлежит ряд интересных работ об экологическом направлении зооло-
гии, по стратиграфии юрских отложений Подмосковья, по палеоклима-
тологии и палеоэкологии. При решении геологических вопросов Рулье
пользовался сравнительно-историческим методом; он считал, что органи-
ческий мир развивался параллельно с неорганическим. Однако в учении
Рулье, так же как у Ламарка, оставались нерешенными две проблемы:
проблема возникновения видов и проблема целесообразности организ-
мов. Рулье подготовил своих учеников к восприятию дарвинизма.
В течение первой половины XIX в. были созданы все предпосылки
для появления эволюционной теории,Дарвина. В разных странах были
подготовлены и опубликованы крупные исследования по палеонтологии:
во Франции А. д’Орбиньи опубликовал «Палеонтологию Франции»
(1841—1850), в Англии отец и сыновья Соверби издали «Минеральную
конхиологию Великобритании» (1812—1846), в России Э. И. Эйхвальд
выпустил «Палеонтологию России» (1860—1868), в Чехословакии И. Бар-
ранд опубликовал свой труд в 24-х томах — «Силурийская система
31
Центральной Богемии» (1852—1888), швейцарский ученый Ф. Пикте на-
писал первый учебник по палеонтологии (1844—1846).
Третий период развития палеонтологии охватывает отрезок време-
ни от опубликования Дарвином книги «Происхождение видов» в 1859 г.
до 40-х годов нашего столетия. Появление книги Дарвина оказало ог-
ромное влияние на развитие всех биологических и геологических наук, в
том числе и палеонтологии; своим трудом Дарвин закрепил позиции
Рис. 10. Чарлз Дарвин (1809—1882) в год
опубликования книги «Происхождение видов»
(1859)
материалистического мировоззрения в естествознании, элементы которо-
го имелись в .трудах многих естествоиспытателей прошлого. Труд Дар-
вина настолько велик, затрагивает столько проблем, что изложить его
в кратком виде не представляется возможным. Базируясь на большом
числе фактов из практики выведения новых пород в животноводстве и
новых сортов в растениеводстве и данных живой природы, Дарвин при-
ходит к выводу, что размножение особей вида происходит в геометричес-
кой прогрессии, но до взрослого состояния доживает лишь небольшая
часть особей; большинство погибает в «борьбе за существование». Вы-
ражение «борьба за существование» Дарвин принимал в широком и
метафорическом смысле, включая сюда зависимость одного существа от
другого, успех одной особи в обеспечении себя потомством, во взаимоот-
ношении организмов с абиотической средой. Основываясь на опыте се-
32
лекционеров, Дарвин показал, что для всех Живых организмов характер-
на всеобщая изменчивость признаков и свойств; он выделил три типа
изменчивости: определенную, неопределенную и коррелятивную.
Под определенной, или групповой, изменчивостью
понималась массовая изменчивость, когда все виды или особи данного
вида или породы под влиянием определенной причины изменялись оди-
наковым образом и в одном направлении (например, рост организма
зависит от количества пищи). Определенная изменчивость не передается
по наследству. Неопределенная, или индивидуальная, измен-
чивость проявляется более часто в виде бесконечно разнообразных
слабых различий у отдельных особей вида или породы и не унаследо-
вана от своих предков. Неопределенная изменчивость, однажды появив-
шись, сохраняется в последующих поколениях, т. е. является наслед-
ственной. Коррелятивная, или соотносительная, изменчивость,
вытекает из принципа Кювье о корреляции частей тела или органов; в
соответствии с ней изменение одной части организма обычно обусловли-
вает изменение других его частей (например, белые кошки с голубыми
глазами обыкновенно глухи). Используя неопределенную и коррелятив-'
ную изменчивость, животноводы и растениеводы с помощью искусствен-
ного отбора выводят разнообразные породы домашних животных и
различные сорта культурных растений.
Дарвин различал две формы искусственного отбора: бессознатель-
ный и методический. В первом случае человек отбирал стихийно лучших
особей, не заботясь о конечных результатах отбора, во втором — отбор
проводился планомерно, методически, в заранее намеченном направле-
нии. Отбор всегда сопровождался выбраковкой (уничтожением) непод-
ходящих особей, скрещиванием форм, обладающих необходимыми приз-
наками для их последующего усиления от поколения к поколению.
Искусственный отбор являлся главным фактором в образо-
вании новых пород и сортов, которые выводились человеком от неболь-
шого числа диких предков. Эта мысль резко отличалась от господ-
ствовавшей в то время точки зрения, в соответствии с которой каждая
порода домашних животных имела своего дикого предка. Дарвин до-
казал, что все разнообразные породы голубей выведены из одного ви-
да — дикого сизого, или скалистого, голубя. Процесс, связанный с по-
лучением двух или нескольких новых пород путем усиления едва замет-
ных различий у потомков, Дарвин назвал дивергенцией, или рас-
хождением признаков (рис. 11).
В природе постоянно идет борьба за существование, которая всег-
да происходит при активной или пассивной конкуренции и сопровож-
дается избирательным уничтожением особей (элиминацией). Дарвин
различал три формы борьбы за существование: конституционную, меж-
видовую и внутривидовую.
Конституционная борьба, или борьба с факторами неорга-
нической природы, приводит к уничтожению особей, наименее приспо-
собленных к данным условиям среды. Межвидовая борьба, или
борьба с особями других видов, приводит к гибели от хищников и па-
разитов, при состязании за одни и те же пищевые ресурсы. Внутри-
видовая борьба, или конкуренция с особями одного и того же
вида, наиболее остра и является важнейшим фактором эволюции. Со-
поставляя два сделанных им вывода — о всеобщей изменчивости ц о
перепроизводстве потомства, Дарвин показал, что в процессе борь-
бы за существование особи уничтожаются избирательно; их выжива-
ние статистически в среднем зависит от тех незначительных изменений,
которыми любая особь отличается от множества других. Полезные из-
3
В. В. Друщиц
33
менения дают особям минимальное преимущество в .выживании по срав-
нению с теми особями, которые ими не обладают; вредные изменения
снижают плодовитость особей, уменьшают их выживаемость; безраз-
личные изменения не оказывают влияния на судьбу вида в целом.
Естественным отбором, или выживанием наиболее приспо-
собленных, Дарвин назвал «сохранение полезных индивидуальных раз-
o'4 о'4 р'4 У4 У4 о’4 е’4 mV'4 л’4 г'4 у’4 у’4 z'4
Д--Г---J--------1----7-----1--т--7--1-----------<--1--,---t-----г
—\ 1—1 1 / \ |_Д— XI/ \ / \ 1 / Н-/ \ 1 / \ 1 / vt,
\ 1 / VT7 \ 1 / ’ / \ ! / аю f’° т'° Е’° \ 1 / \ 1 / X'l Lit Щ—// F’° iv'0 z'°
IV’xp ' U'z’ /z
atty ty'fs ks Is tyrriS u! , w'tty -tyzs
а1 ц- Ч/77 к7tytn7 : . 1 Z vlii u7\i wV tyz7
'a6'# ''U* \/m6 \ " rll U6*i W 1/7
d5 kfy tym5 j tSty tyz5 ,/
‘ &d4 i4 "'tym4 i "'tyz4 /z
: 7-J • '
aty. s2'tym2 i /V if
(фф ДИ7' : ; Zz’ • 7
i 5 1 —
/ ---to bo /V Г С H I /< L II 1 ! II । /
Рис. 11. Схема дивергенции (Дарвин, 1859, с добавлением)
личий или изменение и уничтожение вредных». Изменения неполезные
и невредные не будут подчиняться действию естественного отбора. Ес-
тественный отбор, по Дарвину, «ежедневно, ежечасно расследует по
всему свету мельчайшие изменения, отбрасывая дурные, сохраняя и
слагая хорошие, работая неслышно, невидимо, где бы и когда бы толь-
ко ни представился к тому случай, над усовершенствованием каждого,
органического существа по отношению к условиям его жизни, органи-
ческим и неорганическим» (Ч. Дарвин. «Происхождение видов». М..,
1952. стр. 141).
Дарвин выделил естественный отбор в качестве главного механиз-
ма эволюции^ но в теории Дарвина существовало слабое место — неяс-
ное представление о. природе наследственности. Если взять за основу
это представление, то получится, что каждое возникшее наследствен-
ное изменение при новом скрещивании будет не накапливаться, а раз-
бавляться, поскольку при каждом скрещивании изменение, возникшее у
одного из родителей, будет разбавляться неизмененной наследствен-
ностью другого; в соответствии с арифметическими подсчетами вновь,
возникшее изменение будет уменьшаться вдвое (1/2, 1/4, 1/8 и т. д.).
В действительности эти изменения не уменьшаются, а сохраняются и
даже усиливаются.
34
Несмотря на нападки, которым подверглась теория происхожде-
ния видов, основанная на представлении об изменчивости, борьбе за
существование, наследственности и естественном отборе, эта теория ока-
залась единственно верной материалистической теорией, в соответствии
с которой органический мир развивался по естественным законам, не
нуждаясь в посторонней помощи, в помощи творца. В отличие от своих
предшественников Дарвин оценивал длительность жизни на Земле во
много миллионов лет, но подчеркивал, что неполнота геологической ле-
тописи и бедность наших палеонтологических коллекций являются при-
чиной отсутствия сведений о переходных промежуточных формах, свя-
зывающих одну группу с другой, и причиной кажущегося внезапным
появления многих групп. Резкие границы между формациями, отсутст-
вие переходных форм были источником представления о неизменяемости
видов и о наличии в истории Земли катастроф.
Рассматривая геологическую последовательность организмов, Дар-
вин пришел к очень важному для биостратиграфии закону о необра-
тимости эволюции. «Если вид однажды исчез с лица Земли, мы
не имеем основания думать, что та же форма когда-нибудь появится
вновь» (Ч. Дарвин, 1952, стр. 335). Этот закон вытекает из теории ес-
тественного отбора, причем вымирание древних форм является почти не-
избежным следствием возникновения новых; если какая-нибудь группа
совершенно исчезла, она не появится вновь, потому что цепь поколе-
ний прервана.
Господствующие формы, распространенные широко и дающие наи-
большее число разновидностей, стремятся населить мир близкими к
ним, но измененными потомками, и эти последние обыкновенно с ус-
пехом вытесняют группы, уступающие им в борьбе за существование.
Обитатели мира каждый последовательный период его истории побежда-
ли своих предшественников, — писал Дарвин (там же, стр. 355); в
этом смысле они выше своих предшественников и являются обыкновен-
но более специализированными в своей организации, в этом заключает-
ся объяснение того мнения, что организация в целом развивалась
прогрессивно. С позиций естественного отбора Дарвин объяснял причи-
ны возникновения у животных и растений целесообразных приспособ-
лений, которые до него Кювье рассматривал с креационистских позиций
«принципа условий существования».
Эволюционный процесс всегда носит адаптивный, или приспо-
собительный, характер; органическая целесообразность — это приспо-
собленность организма к существованию в определенных условиях сре-
ды; она всегда имеет относительный характер. Виды животных и рас-
тений приспособлены к разнообразным условиям среды и часто друг'к
другу. К числу приспособлений относятся различной формы защитные
окраски: покровительственная, предостерегающая, подражательная (ми:
микрия) и маскировочная; различные приспособления к перекрестному
опылению и многие другие.
Основываясь на дивергентном характере эволюционного процесса,
Дарвин развивал монофилетическую теорию происхождения
видов, доказывая, что каждая крупная группа животных или растений
происходит от общих предков и из одного центра. Теория Дарвина име-
ла огромное значение для дальнейшего развития биологии, для борьбы
с метафизикой и телеологией; она решала две крупные проблемы —-
проблему превращения одной органической формы в другую и проблему
целесообразности органических форм. Она опиралась на огромное ко-
личество фактов, взятых из различных биологических и геологических
дисциплин, и ставила перед ними новые задачи. Палеонтология из науки
3;
' 35
статической — собирать, описывать и классифицировать — постепен-
но стала превращаться в науку историческую — науку, изучающую кон-
кретные пути эволюционного процесса.
Как отмечалось выше, Дарвин не смог решить проблему наслед-
ственности. Эта задача была решена чешским ученым Грегором Мен-
делем (1822—1884) в 1865 г., но оставалась мало известной и была
переоткрыта в 1900 г. одновременно тремя исследователями: де Фризом
Рис. 12. Закономерности наследования признаков у Сераеа bortensis в Северном
море в соответствии с законами Менделя:
1 — правило доминирования, II — правило расщепления; А — доминантный ал-
лель, а — рецессивный аллель (Циглер, 1972, с добавлениями)
в Голландии, Корренсом в Германии и Чермаком в Австрии. 1900-й год
стал годом рождения новой науки — генетики. На основании опы-
тов по моногибридному скрещиванию сортов гороха, у которых роди-
тельские особи отличаются по одной паре противоположных признаков,
Мендель установил две закономерности. Первая закономерность за-
ключается в том, что у всех гибридов первого поколения проявляется
только один из парных признаков — правило доминирования.
Этот признак назван господствующим, или доминантным, а второй
скрытым, или рецессивным. Вторая закономерность относилась к ко-
личеству доминантных и рецессивных признаков во втором поколении,
соотношение которых равнялось 3: 1. Она получила название закона
расщепления (рис. 12). Дальнейшее изучение результатов различ-
ных типов скрещивания привело к открытию третьей закономерности, по-
лучившей название правила независимого распреде-
ления наследственных факторов. В соответствии с
этим правилом расщепление по каждой паре признаков идет незави-
симо от других пар признаков. Анализируя выведенные закономерности,
Мендель предложил гипотезу «чистоты гамет», согласно которой в по-
ловых клетках имеются какие-то наследственные факторы, или задатки,
которые определяют развитие отдельных признаков. Эти факторы, позд-
нее названные генами, всегда парные, один из них — доминант-
ный, другой — рецессивный; в дальнейшем они стали называться а л-
леломорфными, или аллельными, а отдельный ген — а л -
36
лелем. Доминантный аллель условно обозначают большой буквой (А),
а рецессивный — малой (а). Например, гаметы гороха желтого сорта
несут доминантный аллель (А), а зеленого — рецессивный (а). При
скрещивании одинаковых сортов в зиготах будут присутствовать одина-
ковые аллели (А + А=АА, а+а = аа); особи, развившиеся из подобных
зигот, названы гомозиготными. Если скрещивать два разных
сорта, то в зиготах объединятся доминантный и рецессивный аллели,
что даст комбинацию Аа; у особей, развивающихся из подобных зигот,
внешне будет выражен только доминантный признак А (правило доми-
нирования), являясь по своей наследственной основе гетерозиготным.
При скрещивании гетерозиготных особей, у которых одни половые клет-
ки содержат аллель (А), другие (а), возможны четыре разные комбина-
ции: АА, Аа, Аа, аа. Соотношение доминантных и рецессивных форм
в этом случае будет равно 3: 1 (правило расщепления); из них комби-
нации АА и аа дадут гомозиготных особей, а комбинации Аа, Аа — ге-
терозиготных. При дальнейшем скрещивании гетерозиготные особи
вновь дадут доминантные и рецессивные формы в соотношении 3:1.
Выведенные Менделем, таким, образом, три закономерности: 1 —
единообразие гибридов первого поколения, 2 — расщепление в про-
порции 3: 1, 3 — независимое расщепление наследственных факторов —
давали возможность представить механизм наследственности и опреде-
ляли значение полового процесса как важного фактора эволюции; они
связывали эволюцию и генетику между собой и отвечали на вопрос, на
который Дарвин так и не смог ответить. Наследственные признаки не
разбавляются от поколения к поколению, а являются дискретными
и сохраняются в новых поколениях. Дальнейшие работы в области ге-
нетики подтвердили выводы Менделя.
Идеи Дарвина вызвали бурный рост естествознания, способство-
вали ускорению прогресса науки, явились поворотным пунктом в раз-
витии всех естественных наук. Ч. Дарвин, писал Ленин, «...положил ко-
нец воззрению на виды животных и растений, как на ничем не связан-
ные, случайные, «богом созданные» и неизменяемые, и впервые поста-
вил биологию на вполне научную почву, установив изменяемость видов
и преемственность между ними...» (В. И. Ленин. Поли. собр. соч., т. 1,
стр. 139).
Большую роль в распространении дарвинизма сыграли Т. Г. Гекс-
ли, М. Неймайр, Э. Геккель, Л. Долло, братья В. О. и А. О. Ковалев-
ские, А. П. Карпинский, А. П. Павлов и М. В. Павлова, С. Н. Ники-
тин, Н. И. Андрусов, К- А. Тимирязев, А. А. Борисяк, А. Н. Северцов
и многие другие.
В кратком обзоре трудно рассмотреть роль многих ученых в раз-
витии эволюционной палеонтологии. Остановимся лишь на работах не-
которых отечественных и зарубежных исследователей. Идеи историчес-
кого развития органического мира лежат в основе почти всех работ это-
го периода.
Крупнейшим пропагандистом дарвинизма был английский натура-
лист Т. Г. Гексли (1825—1895), написавший ряд работ по белемнитам,
ракообразным, различным группам позвоночных. Гексли рассматри-
вал вопрос о геологической одновременности или одновозрастности, счи-
тая невозможным провести общую для всего земного шара периоди-
зацию геологической истории. Он предлагал заменить слово «геологи-
ческая одновременность» как неопределенное и двусмысленное на сло-
во «гомотаксис», или сходство порядка, которое определяло бы по-
рядок слоев в разрезах, удаленных друг от друга, так как точную синх-
37
ронизацию слоев установить трудно. Эти идеи подвергались критике
со стороны Ч. Дарвина, но они продолжают обсуждаться до сих пор.
В. О. Ковалевский (1842—1883)—один из талантливых пропаган-
дистов дарвинизма, основоположник биологического направления в па-
леонтологии. При изучении позвоночных (в первую очередь копытных)
В. О. Ковалевский прослеживал эволюционное изменение отдельных эле-
Рис. 13. Владимир Онуфриевич Ко-
валевский (1842—1883}
ментов скелета во времени (конечностей, черепа, зубов) в связи с из-
менением их функции, физиологии питания, обусловленной изменени-
ем условий существования. Важным достижением Ковалевского было
установление принципа адаптивной радиации (термин, предло-
женный позднее Осборном), уже намеченного Дарвином, который под-
черкивал широкую приспособительную дивергенцию в разнообразных
направлениях представителей господствующей группы. В качестве при-
мера Ковалевский приводил иррадиацию копытных. Эволюция лоша-
дей, прослеженная Ковалевским от палеогена до наших дней, вошла во
все учебники и учебные пособия по дарвинизму и биологии. На при-
мере парнопалых копытных Ковалевский, наметил два пути перестрой-
ки коррелятивных взаимоотношений конечностей скелета: первый,
названный им а д ап ти вн ы м, происходил при коренной перестройке
этих взаимоотношений, и второй — инадаптивный, не был связан с рез-
кой перестройкой. Эта закономерность позднее была распространена
на другие группы животных. Современники Ковалевского высоко оцени-
вали его палеонтологические работы.
А. О. Ковалевский (1840—1901) брат палеонтолога В. О. Ковалев-
ского, проделал большую работу по изучению эмбрионального развития
38
у многих современных животных: кишечнополостных, кольчатых чер-
вей, членистоногих, моллюсков, брахиопод, иглокожих и хордовых, уде-
ляя особое внимание онтогенезу у групп неясного систематического по-
ложения (ланцетнику, асцидиям, баляноглоссусу, брахиоподам).
Большой вклад в развитие дарвинизма внес И. И. Мечников (1845—
1916), разработавший теорию происхождения многоклеточных.
Исследования в области эмбриологии позволили Ф. Мюллеру и
Э. Геккелю сформулировать биогенетический закон: «Онтогенез является
коротким и быстрым повторением филогенеза, обусловленным физио-
логическими функциями наследственности (воспроизведения) и приспо-
собляемости (питания)». Биогенетический закон стал’широко использо-
ваться палеонтологами в установлении филогенетических связей. Гек-
кель, кроме того, создал свою теорию гастреи при рассмотрении про-
исхождения многоклеточных.
В Западной Европе идеи Дарвина в палеонтологии развивали
В. Вааген, М. Неймайр и многие другие.
В. Вааген (1841—1900), изучавший юрских аммонитов, установил
первый родословный ряд форм среди морских ископаемых животных
(1869) и ввел понятие «мутация», рассматривая ее в качестве разновид-
ности во времени. Он считал, что мутации, сменяющие друг друга во
времени, выражают развитие в определенном направлении, и время их
•существования отвечает зоне — единице геологического времени, уста-
новленной Оппелем (1865). Представление о мутациях подчеркивало
изменчивость форм во времени и служило для обозначения этапов раз-
вития разных групп. Впервые в систематику был введен генетический
принцип, сыгравший большую роль в развитии эволюционной палеонто-
-ЛОГИИ.
М. Неймайр (1845—1890)—одна из наиболее ярких фигур последар-
виновского периода; он считал дарвинизм основой науки о жизни и пред-
лолагал, что потребуется еще много времени для решения многих во-
просов, стоящих перед палеонтологией. Неймайр рассматривал естест-
венный отбор как главный фактор эволюции, но при этом значительную
роль отводил непосредственному воздействию среды. Он построил непре-
рывный филогенетический ряд неогеновых вивипарусов (гастропод) и
показал постепенное изменение этих моллюсков в зависимости от изме-
нений внешней среды в замкнутом внутреннем бассейне; для форм, жив-
ших в открытом море, можно построить только прерывистые ряды. Сос-
тавление подобных рядов, по мнению Неймайра, имеет большое значе-
ние для дальнейшей разработки эволюционного учения. В первой части
своей замечательной книги «Стволы царства животных» (1889) Неймайр
подробно рассматривает основные вопросы эволюционной палеонтоло-
гии и дает описание некоторых групп беспозвоночных: кишечнополост-
ных, иглокожих, червей, мшанок и брахиопод. Смерть помешала Ней-
майру закончить свою книгу.
Нельзя также не отметить большое значение первой сводки всего
палеонтологического материала, составленной немецким палеонтологом
К. Циттелем, — «Основы палеонтологии» (1876—1893), выпущенной в
нескольких томах. В дальнейшем это издание многократно перерабаты-
валось и издавалось в разных странах. В Советском Союзе оно было
переработано советскими палеонтологами, издано в 1934 г. и много лет
служило основным справочником для советских геологов и палеонто-
логов.
Американский ученый А. Хайэтт (1838—1902) на примере головоно-
гих моллюсков пытался выяснить закономерности их филогенетического
развития; он ввел в литературу большое число разнообразных терминов,
39
из которых отметим лишь некоторые: биопластология, изу-
чающая соотношение между онтогенетическим и филогенетическим
циклами; акселерация, или ускорение развития, — более раннее
воспроизведение разнообразных наследственных признаков у потомков;
ретардация — задержка в воспроизведении ряда наследственных
признаков у потомков на тех же стадиях, что у предков. Хайэтт создал
в Северной Америке школу палеонтологов, изучавших филогенез на ос-
новании онтогенетического закона. Подробные сведения о роли Хайэтта
Рис. 14. Александр Петрович Кар-
пинский (1847—1936)
и других зарубежных палеонтологов в развитии палеонтологии можно
найти в работе Л. Ш. Давиташвили «История эволюционной палеонто-
логии от Дарвина до наших дней» (1948).
Большую роль в популяризации идей Дарвина в России сыграли
лекции проф. С. С. Куторги в Петербургском университете, статья
Д. И. Писарева «Прогресс в мире животных» (1864).
Для развития эволюционной палеонтологии много сделали русские-
ученые А. П. Карпинский, С. Н. Никитин, А. П. Павлов.
А. П. Карпинский (1847—1936) вошел в историю отечественной нау-
ки как крупнейший геолог, палеонтолог и организатор науки (он был
первым выборным президентом Академии наук СССР в 1916—1936 гг.).
Карпинский внес ценный вклад в эволюционную палеонтологию, проведя
изучение онтогенетического развития ряда пермских (артинских) аммо-
ноидей и наметив их филогенетические связи с более древними, камен-
ноугольными аммоноидеями. Карпинский использовал онтогенетический
метод как палеонтолог-морфолог и как биостратиграф, изучая историю»
артинской фауны и закономерности ее распределения во времени и
40
пространстве. Он показал, какую важную роль играет филогения для
решения стратиграфических вопросов, вопросов изучения фаун, их про-
исхождения, миграции, изменений во времени.
А. П. Карпинский совместно с Ф. Н. Чернышевым и Г. П. Гельмер-
сеном был инициатором создания Геологического комитета (Геолком,.
1882), который руководил геологическими и палеонтологическими иссле-
дованиями в России. После Октябрьской революции до 1930 г. Геолком
возглавлял всю геологическую службу страны, позднее переданную Ми-
нистерству геологии.
С. Н. Никитин (1850—1909) подробно рассматривал проблему вида
в палеонтологии на основе дарвинской теории естественного отбора; он
считал, что границы между стратиграфическими подразделениями сле-
дует проводить не по исчезновению форм, господствующих в более древ-
них слоях, а по появлению новых. Он оставил в наследство хорошие
описания юрских отложений и аммонитов России.
Важную роль в развитии палеонтологии и геологии и в воспитания
молодых специалистов сыграл А. П. Павлов (1855—1929), воспитанник
Московского университета, начавший работу в университете еще тогда,,
когда лекции по геологии и палеонтологии в 1881—1883 гг. читал
В. О. Ковалевский; после его трагической смерти в 1883 г. А. П. Павлов,
занял кафедру геологии и возглавлял ее до последних дней своей жиз-
ни. Павлов высоко ценил труды Дарвина и Ковалевского; в статье о
геологической истории птиц (1884) он впервые применил идею Ковалев-
ского об адаптивных и неадаптивных изменениях к древним птицам,
подчеркнув, что эта идея выражает закономерность общего значения.
Большое влияние на работы Павлова оказал палеонтолог-дарвинист
М. Неймайр.
Большинство работ Павлова представляет сочетание палеонтологи-
ческих и стратиграфических исследований, посвященных изучению пре-
имущественно юрских и меловых отложений бореальной области. В сво-
их работах Павлов стремился наметить родственные связи между
различными родами аммонитов, белемнитов, двустворок и создать ес-
тественную классификацию, выявить палеогеографическое значение-
изученных групп. Он всегда подчеркивал, что особенно после Дарвина
палеонтология стала наукой биологической, задача которой состоит в-
доказательстве эволюции и выявлении основных факторов эволюции.
При изучении юрских аммонитов Павлов отметил интересное явле-
ние, названное им не совсем удачно, профетической, или пророческой,
фазой; на молодых оборотах у аммонитов появляются признаки, кото-
рые не рекапитулируют черты предков, а как бы предсказывают приз-
наки потомков. Впоследствии немецкий ученый Шиндевольф назвал это-
явление протерогенезом; правильное объяснение этому явлению-
дал советский ученый А. Н. Иванов, считавший, что описанное явление-
связано с задержкой, или ретардацией, развития. Это объяснение ли-
шало профетическую фазу того таинственного пророческого значения,
которое ему приписывалось некоторыми исследователями.
Интересны палеонтологические и стратиграфические работы, выпол-
ненные Н. И. Андрусовым (1861—1924), изучавшим неогеновые и чет-
вертичные бассейны Понто-Каспийской области; он был, бесспорно,
крупнейшим знатоком истории органического мира этой большой обла-
сти в неогеновое и раннечетвертичное время, впервые в истории науки
доказавшим зависимость эволюционного развития организмов от опре-
деленных изменений среды обитания. Андрусов по-дарвиновски объяс-
нял вымирание различных групп неогеновых моллюсков, и в первую»
очередь двустворчатых, выделяя среди них стойкие типы, отличающиеся
41.
продолжительным существованием, и типы, достигающие пышного рас-
цвета и быстро вымирающие; он установил, какие условия благоприят-
ны для адаптивной радиации моллюсков: соответствующая степень
солености, обширность бассейна, разнообразие в ней различных биото-
пов, длительность существования бассейна.
Особенно широкое развитие получили палеонтологические исследо-
вания в Советском Союзе после Великой Октябрьской революции. Боль-
Рис. 15. Алексей Алексеевич Борисяк
(1872—1944)
шой вклад в развитие отечественной палеонтологии внес академик
А. А. Борисяк (1872—1944), возглавлявший до 1930 г. палеонтологиче-
скую секцию в Геологическом комитете. В 1930 г. в Академии наук по
его инициативе был создан палеонтологический институт, который он
возглавлял до 1944 г. Борисяк много сделал для развития палеонтоло-
гии как биологической дисциплины. Он выделял в .ней три основные
проблемы: проблему формы и ее изменений (морфологию), проблему
вида (систематику) и проблему взаимоотношения организма и среды
(палеобиологию).
В 1919 г. в Московском и Ленинградском университетах были соз-
даны кафедры палеонтологии. Позднее такие кафедры были организо-
ваны в некоторых институтах и ряде университетов. В настоящее вре-
мя в качестве самостоятельных единиц кафедры палеонтологии сохрани-
лись в Ленинградском, Московском и Томском университетах.
Четвертый период в развитии палеонтологии начался после Великой
Отечественной войны (1941—1945 гг.). В палеонтологии обособилась
42
микропалеонтология, объектом изучения которой стали фораминиферы,
радиолярии и остракоды, палеоэкология, тафономия, палеофизиология,
палеоанатомия, палеопатология, палеоихнология и многие другие. Па-
леонтологические исследования развернулись на всей огромной терри-
тории Советского Союза; были открыты новые, ранее неизвестные груп-
пы древних организмов (археоциаты, табуляты, тентакулиты, хиолиты
и многие другие). В последние годы были сделаны фундаментальные
открытия в области генетики. Применение электронного микроскопа
при изучении клетки позволило установить тончайшие детали строения
цитоплазмы и ядра (некоторые детали строения клетки будут рассмот-
рены при описании простейших). Успехи в развитии экспериментальной
и теоретической генетики привели к установлению точных математико-
статистических законов наследования признаков, общей дискретной при-
роды наследственных факторов, относительно высокой их стабильности,
открытию структуры дезоксирибонуклеиновой (ДНК) и нуклеиновых
кислот, к открытию роли ДНК в синтезе новых молекул ДНК и в пе-
редаче наследственных свойств от материнской клетки к дочерним. Было
установлено, что ген представляет собой участок двойной спирали ДНК,
содержащий информацию о структуре белка, который будет создан.
Все это позволило по-новому оценить эволюционный процесс и опреде-
лить исходный материал, лежащий в его основе.
Современная наука ставит перед палеонтологами новые и крайне
важные задачи, к числу которых относится разработка совместно с гео-
логами проблемы среды и жизни минувших геологических эпох, особен-
но докембрия. Не менее важной для палеонтологии является проблема
факторов и-темпов эволюции, проблема этапности в органической эво-
люции и использование ее для целей биостратиграфии.
Новые современные методики исследований, применение электрон-
ного микроскопа как трансмиссионного, так и сканирующего, изучение
геологического строения дна морей и океанов и новых территорий суши
-открывают перед палеонтологией новые перспективы развития.
В целях объединения научных исследований палеонтологов всего
мира был создан Международный палеонтологический союз, позднее,
в 1969 г., переименованный в Международную ассоциацию палеонтоло-
гов, включенную в Международный союз теологических наук.
Для координации палеонтологических исследований в СССР при
отделении общей биологии АН СССР в 1964 г. был организован Научно-
консультативный 'совет по проблеме «Пути и закономерности историче-
ского развития животных и растительных организмов», при котором
были образованы постоянные комиссии по общим проблемам палеонто-
логии и по разным группам фауны и флоры. Большую роль для разви-
тия . палеонтологических исследований играет Всесоюзное палеонтоло-
гическое общество (ВПО) при АН СССР, созданное в 1916 г., которое
начиная с 1955 г. проводит в Ленинграде (во ВСЕГЕИ) свои ежегодные
•тематические палеонтологические сессии.
Накопление огромного палеонтологического материала привело к
необходимости его обобщения. В связи с этим были подготовлены и
опубликованы общие сводки по палеонтологии: во . Франции — «Traite
de Paleontologie» под редакцией проф. Пивето (1952—1961), в СССР —
«Основы палеонтологии» в 15 томах под редакцией акад. Ю. А. Орлова
(1958—1964), в США—«Treatise on Invertebrate Paleontology» под ре-
дакцией проф. Р. Мура и проф. К- Тейхерта (1953—1974, издание еще
незаконченное). Если в 80-х годах прошлого столетия с подобной рабо-
той справился один Циттель, то в составлении указанных выше сводок
принимали участие сотни палеонтологов.
43
ОРГАНИЗМ И СРЕДА
Область распространения живых существ на Земле образует осо-
бую оболочку, которую Ж.-Б. Ламарк назвал биосферой, но учение
о биосфере и ее роли в процессах, протекающих на поверхности Земли,,
было создано академиком В. И. Вернадским (1863—1945). Биосфера
возникла с появлением на Земле живых существ; она занимает всю по-
верхность суши от уровня моря до горных вершин; живые организмы
заселяют также все водоемы Земли — океаны, моря, озера, реки, общая
площадь которых составляет 361 млн. км2. Общая поверхность Земли:
равна 510 млн. км2; на поверхность суши приходится 149 млн. км2. Часть
организмов, населяющих сушу, проникает в атмосферу; большинство
поднимается в воздух более чем на 50—70 м; правда, многие птицы
взлетают значительно выше (например, альпийская галка была обнару-
жена на горе Эверест на высоте свыше 8 км). Споры бактерий и грибов^
заносятся на высоту до 22 км, не исключена возможность, что некото-
рые микроорганизмы достигают более высоких слоев стратосферы.
Жизнь проникает и в литосферу, где она концентрируется главным об-
разом в поверхностном слое на глубине до 6—8 м, но некоторые бакте-
рии найдены в слоях, залегающих на глубинах до 2—3 км.
Все живые существа, населяющие Землю, по наличию клеточной
организации делятся на две группы (иногда называемые империями):
доклеточных — вирусы и фаги и клеточных — все остальные ор-
ганизмы. Вирусы состоят из одной нуклеиновой кислоты, одетой бел-
ковой оболочкой; они могут вести только паразитический образ жизни..
Клеточные организмы разделены на две подимперии: прокариот,
или предъядерных, и эукариот, или ядерных форм. У прокари-
от (сине-зеленые водоросли и бактерии) цитоплазма не отделена от
ядра и ДНК распределена диффузно; по способу питания они относятся
преимущественно к автотрофам; они синтезируют сложные органические
молекулы — белки, углеводы и жиры — из простых неорганических со-
единений, используя солнечную энергию либо энергию, получаемую за
счет окислительных процессов. Эукариоты, или ядерные формы, име-
ют типичное клеточное строение с морфологическим и функциональным
расчленением клетки на ядро и цитоплазму. К эукариотам относятся
животные и растения; все животные — гетеротрофные'организмы; они
не способны синтезировать органические питательные вещества из не-
органических и питаются либо растительной (растительноядные), либо
животной пищей (плотоядные, или хищные животные).
Условия существования животных на Земле очень разнообразны и
определяются факторами как неорганического, так и органического по-
рядка. К неорганическим факторам относятся температура, влажность,,
соленость воды, глубина бассейна, давление; к органическим — много-
численные и разносторонние взаимоотношения, в которые организмы
вступают между собой. Эти взаимоотношения в первую очередь выра-
жаются пищевыми связями.
Всеобщей и характерной чертой жизни на Земле является дискрет-
ность живых форм и идентичное воспроизведение себе подобных форм1
по матричному принципу с определенным кодом, хранящимся в эле-
ментарных наследственных структурах — генах.-Жизнь всюду присутст-
вует в биосфере, что объясняется постоянно существующим «давлением
жизни», как писал Вернадский, вызываемым размножением всех орга-
низмов в геометрической прогрессии; если бы не было никаких препят-
ствий, то каждый вид произвел бы огромное потомство, заполнив всю
Землю.
44
Каждый вид обладает своим ареалом, занимая пространственно .
различные части земной поверхности. Изучением пространственного
распределения организмов на Земле занимается хорология. Хоро-
логическое изучение может вестись по двум направлениям: рассматри-
вать для каждого таксона его особенности ареала (географическая био-
логия и ареалогия) или каждый регион характеризовать населяющими
его организмами (биогеография).
Все организмы на Земле живут сообществами, называемыми био-
ценозами. Биоценоз представляет собой комплекс, или сообщество,
организмов, живущих совместно при определенном сочетании разнооб-
разных факторов среды. Биоценоз обычно получает название по наибо-
лее характерному для него количественно преобладающему организму.
Например, в биоценозе устричника преобладают устрицы, раковины ко-
торых прирастают ко дну или друг к другу. В нем живут многие орга-
низмы, которые используют раковины устриц в качестве субстрата для
поселения; к ним относятся губки, гидроидные полипы, мшанки, кольча-
тые черви — полихеты. Здесь же живут брюхоногие моллюски-хищники
и падалеяды, некоторые морские звезды. Другим примером может слу-
\жить биоценоз водоросли ламинарии, к которому приурочены преиму-
щественно растительноядные организмы, некоторые рыбы, брюхоногие
моллюски, фораминиферы, черви; кроме того, многие формы использу-
ют заросли ламинарий в качестве естественного укрытия. Каждый био-
ценоз занимает определенную территорию, называемую биотопом.
В результате сложных взаимоотношений между организмами и сре-
дой формируется определенная саморегулирующаяся подвижная эколо-
гическая система, которую В. Н. Сукачев назвал биогеоценозом.
Биогеоценоз является элементарной биохорологической единицей или
элементарным подразделением биосферы Земли. В нем протекает опре-
деленный тип биологического круговорота, который характеризует оп-
ределенную единицу биохимической работы, производимой живыми
организмами. В любом биогеоценозе каждый организм, входящий в его
-состав, занимает определенную экологическую нишу, под которой
понимается место, занимаемое определенным видом в биотической и
абиотической среде; с эволюцией вида может изменяться его экологи-
ческая ниша. Наука, изучающая образ жизни отдельных видов, назы-
вается этологией. Биогеоценозы имеют обычно прерывистое распро-
странение и распределяются по соответствующим биотопам.
Организмы, входящие в состав биогеоценоза, обычно по-разному ре-
агируют на колебания того или иного фактора среды — солености, тем-
пературы, давления, изменений климатических условий; одни могут су-
ществовать при широких колебаниях одного из факторов среды, другие,
наоборот, не переносят даже незначительных изменений этого фактора.
В первом случае прибавляется приставка э в р и- (гр. eurys—широкий),
во втором — ст ено- (гр. stenos — узкий).
Соответственно применяется следующая терминология:
фактор организм
биотоп (место жительства) эвритопный, стенотопный
глубина эврибатный, стенобатный
соленость эвригалинный, стеногалинный
температура эвритермный, стенотермный
Если желают выразить «любовь» или «боязнь» организма по от-
ношению к определенному фактору, то добавляют соответствующие суф-
фиксы, взятые из греческого языка-ф и л ь н ы й (fileo—люблю), -фоб-
45
н'ый (fobos — страх). Например, по отношению к солености различают
галофильные и галофобные организмы.
Способность организмов переносить малое, среднее или большое-
воздействие какого-либо фактора среды обозначается терминами с при-
ставками соответственно олиго-, мезо- и поли- (например, олиго-, мезо-
и полигалинные животные — животные, живущие при малой, средней и
большой солености).
Все виды в пределах своего ареала обычно распределены неравно-
мерно, и в каждом биотопе образуют более или менее обособленные
поселения — популяции. Каждая популяция реально существует во-
времени и пространстве как определенное единство, состоящее из сово-
купности особей, в течение длительного времени населяющих определен-
ную территорию, не ограниченных заметными изоляционными барьера-
ми и способных к воспроизведению потомства себе подобного. Соревно-
вание особей и естественный отбор протекают прежде всего внутри по-
пуляции. Популяция характеризуется различной плотностью — числом
особей на единицу площади, рождаемостью, смертностью, возрастным
составом, скоростью расселения, биотическим потенциалом — способ-
ностью к увеличению численности при стабильном возрастном составе-
к оптимальных условиях среды.
С генетической точки зрения популяция, даже если она фенотипи-
чески относительно однородна, представляет весьма гетерогенную смесь-
различных генотипов и отдельных редких мутаций, особенно рецессив-
ных, содержащихся в данной популяции в гетерозиготном состоянии в
различных количествах. При относительном постоянстве различных
факторов генотипический состав популяции может оставаться в течение-
более или менее длительного времени довольно стабильным; при про-
должительном одностороннем давлении одного из факторов генотипиче-
ский состав популяции изменяется, и это может привести к разделению-
популяции на две или несколько, между которыми могут возникнуть
изоляционные барьеры.
Условия обитания животных на суше. На суше, занимающей */з всей
поверхности земли, условия жизни крайне разнообразны. Распределе-
ние животных на суше в первую очередь зависит от климатических ус-
ловий. Каждый климатический пояс характеризуется своими темпера-
турными условиями, особенностями времен года, дневными и ночными-
колебаниями температуры, влажности, наличием или отсутствием рас-
тительного покрова, характером рельефа, высотой гор, рассеченностью
склонов, глубиной долин и многими другими чертами. На суше обита-
ют позвоночные и многие группы беспозвоночных. К последним относят-
ся черви, живущие в почве или ведущие паразитический образ жизни,
членистоногие, среди которых первое место занимают насекомые, осво-
ившие всевозможные ниши жизни. Не менее широко распространены
представители хелицеровых — паукообразные, скорпионы, фаланги и
многие другие; обитателями суши являются легочные брюхоногие мол-
люски, завоевавшие самые разнообразные биотопы, вплоть до высоко-
горных районов. К жизни в пресноводных водоемах суши приспособи-
лись жгутиковые, корненожки, солнечники, ресничные, губки, гидроид-
ные, низшие и высшие черви, двустворчатые и брюхоногие моллюски,
членистоногие (насекомые и ракообразные), мшанки.
Температурный режим определяет быстроту онтогенеза, продол-
жительность жизни. Температурные условия на суше очень измен-
чивы, но у животных в процессе исторического развития выработалисы
специальные приспособления, помогающие избегать вредного влияния
температур. Большинство животных обладает непостоянной температу-
46
рой тела, изменяющейся в определенных пределах в соответствии с из-
менением температуры среды.Только теплокровные животные (птицы и
млекопитающие) имеют механизмы терморегуляции, поддерживающие-
постоянную температуру тела. .Для сохранения воды в теле животных
имеются различные приспособления, предохраняющие их от избыточного-
испарения.
В соответствии с распространением различных групп животных
(главным образом млекопитающих) были установлены шесть главных,
биогеографических областей: палеарктическая (Европа, северная
часть Африки, Азия к северу от Гималаев), неарктическая (Грен-
ландия и Северная Америка на юг до северного плато Мексики), неот-
ропическая (Южная и Центральная Америка и южная часть Мекси-
ки), эфиопская (Африка без ее северной части, Мадагаскар), вос-
точная (Индия, Шри Ланка, Индокитай, Малайсций полуостров),,
австралийская (Австралия, Новая Зеландия, Новая Гвинея).
Условия обитания животных в океанах и морях. Водная среда
наиболее благоприятна для жизни. Вода имеет высокий показатель,
теплоемкости, является хорошим растворителем и легко проницаемой
средой для активно передвигающихся животных. Она допускает суще-
ствование во взвешенном состоянии многих организмов. Солнечный свет,
проникающий в толщу воды, обеспечивает существование разнообраз-
ных водорослей, для которых характерен быстрый рост, получение пи-
тательных минеральных веществ непосредственно из окружающей их.
воды. Водоросли считаются главным трансформатором световой энер-
гии, превращая ее в химическую; на океан приходится высокий процент
фотосинтеза, происходящего на всей планете. Кроме того, существова-
ние в воде водорослей и бактерий обеспечивает жизнь очень многих
животных.
Океан как среда обитания разделяется на две основные части: вод-
ную массу, или пелагиаль (гр. pelagos — открытое море), и дно океа-
на, или бенталь (гр. benthos — глубокий). Соответственно организмы,
населяющие толщу воды, называются пелагическими, а живущие
на дне — бентосными.
Распределение организмов в водной среде зависит в первую оче-
редь от солености, т. е. количества солей, растворенных в одном лит-
ре воды, которое измеряется единицами, называемыми промилле
(°/оо). Океаны и открытые моря, соленость которых составляет 33—
35%о, называются нормальносолеными. Моря, имеющие затруд-
ненную связь с океаном и большой приток пресных вод, имеют соленость
воды обычно ниже нормальной (Азовское море —11—12%0, Черное мо-
ре—16—22%о). Бассейны с соленостью воды от 24,5 до 5%о называют
солоноватоводными, а с соленостью меньше 5%о — пресно-
водными. Бассейны с повышенной соленостью, или ультра-
талинные, содержат свыше 35%о солей.
Другие факторы среды — температура, подвижность воды, количе-
ство растворенного в воде кислорода, характер грунта — также играют
важную роль в распределении водных организмов. Знание всех этих
факторов позволяет полнее изучать организмы прошлых геологических
эпох и восстанавливать физико-химические условия бассейнов прош-
лого.
В каждом морском бассейне выделяют три области бентали: нери-
товую, батиальную и абиссальную (рис. 16).
Неритовая область—это область подводной окраины материков,
или шельфа, представляющая часть суши, покрытую морем. Ширина
ее различна и может достигать нескольких сотен километров; на шель-
47
Рис. 16. Схематический профиль морского дна и биономические зоны моря
фё иногда располагаются огромные эпиконтинентальные бассейны (Ба-
ренцево море) со средней глубиной 200—500 м. Эта область разде-
ляется на зоны литорали и. сублиторали.
Литораль — узкая полоса вдоль берега, которая обнажается
при отливе и покрывается водой во время прилива. Организмы, живу-
щие в литорали, приспосабливаются к амфибиальному существованию.
Часть времени они проводят под водой, а другую — в условиях воздуш-
ной среды. Жизнь литорали обильна и разнообразна., Скопление орга-
нического детрита, поступающего с суши и намываемого морем, обеспе-
чивает обильное развитие водорослей (бурых, зеленых, багряных), ко-
торые в свою очередь создают благоприятные условия для жизни мно-
гих животных: фораминифер, червей, моллюсков, иглокожих, ракообраз-
ных. Особенно обильна жизнь в мангровых зарослях-—прибрежных
заболоченных тропических лесах. -Литораль являлась, по-видимому, для
многих организмов переходной ступенью от жизни в море к жизни на
суше.
.Сублитораль (до 200—500 м) —это та часть морского дна не-
ритовой области, которая постоянно покрыта водой. Сублитораль харак-
теризуется также обилием света (особенно в своих верхних частях),
изменчивой температурой, движением воды. В ней растут бурые, зеле-
ные, багряные водоросли и морская трава — небольшая группа цветко-
вых растений, среди которых преобладает зостера. Морские растения об-
разуют настоящие «подводные луга и леса», в которых живет множество
животных: моллюсков, червей, мшанок, гидроидных, ракообразных.
Глубже всех в сублиторали опускаются багряные водоросли (до 100—-
120 м).
Часть неритовой области, расположенную от 200 до 500 м, иногда
выделяют под названием п сев до а б и с с а л ь; в ней отсутствуют рас-
тения и животные имеют глубоководный облик,
Батиальная область (батиаль) — это область материкового скло-
на— от края шел'ьфа до ложа Мирового океана. Она отличается обыч-
но наиболее сложным рельефом и несколькими типами строения, от уз-
кого материкового склона до широкой зоны, состоящей из котловин и
порогов. В этой области температура воды постоянная, свет проникает
только в самые верхние части; жизнь довольно однообразна; преобла-
дают хищные и илоядные животные; растения отсутствуют.
Абиссальная ^область (абиссаль)—ложе Мирового океана, в кото-
ром выделяют .крупные океанические котловины, подводные хребты, воз-
вышенности, плато. Глубоководные впадины названы ультраабис-
салью. Для абиссальной зоны характерны полное отсутствие света,
постоянные температура воды (от —1,87 до 2°) и соленость, громадное
давление и повышенное количество углекислоты; известковые скелеты
развиты слабо — у моллюсков и брахиопод известковая раковина тон-
кая, морские ежи без твердого известкового панциря, у костистых рыб
понижено окостенение скелета; наблюдается замещение известкового
скелета кожистым, хитиновым или кремневым; очень много сидячих ор-
ганизмов с длинными стеблями (губки, восьмйлучевые кораллы).
В абиссали развиты осадки биогенного происхождения: птероподо-
вые, диатомовые, радиоляриевые илы и красная глубоководная глина.
По данным Л. А. Зенкевича, ложе современного океана, или абис-
саль, составляет 77,1%, материковый склон, или батиаль, —15,3%, а
материковая отмель, или неритовая область, — 7,6% всей поверхности
дна океанов.
Большая группа водных организмов ведет донный, или бентос-
ный, образ жизни от литорали до глубоководных впадин. Донная жизнь
4 в. В. Друщиц . • 49
более разнообразна, чем пелагическая, и, возможно, что первые в гео-
логической истории организмы развивались на плотном субстрате в мо-
ре; к пелагическому существованию они перешли позднее.
Донные организмы делятся на несколько биологических групп: сво-
бодно передвигающиеся по дну бассейна (многие простейшие, моллюс-
ки, иглокожие); свободно лежащие на дне (многие моллюски, некото-
рые древние брахиоподы); закапывающиеся в грунт временно; живу-
щие в иле или песке постоянно и способные передвигаться в самом
грунте (многие морские ежи, брюхоногие и двустворчатые, моллюски,,
кольчатые черви); живущие в грунте неподвижно и строящие себе
постоянное жилище; сверлящие известковые породы механически или
химически, выделяя своеобразный секрет (губки, двустворчатые мол-
люски) .
Многие донные организмы ведут прикрепленный образ жизни. При-
крепленные к субстрату животные имеются среди почти всех типов
животного мира. Они произошли, по-видимому, от подвижных форм;,
вся организация прикрепленных форм по сравнению с подвижными не-
сет' на себе черты упрощения. Они прикрепляются к различным под-
водным предметам или скелетам донных организмов, или к плавающим
предметам и животным. Прикрепленные животные часто образуют ко-
лонии (губки, кишечнополостные, мшанки), возникающие в результате-
бесполого размножения — почкования или деления. Способы прикреп-
ления крайне разнообразны: цементация к субстрату (губки, коралло-
вые полипы, двустворки — рудисты, устрицы), образование стебля (раз-
ные группы иглокожих — морские лилии, цистоидеи, бластоидеи) или
специальной ножки (брахиоподы); прикрепление при помощи биссус-
ных нитей (двустворки); образование особых корневидных выростов,,
или каблучка прирастания (губки, археоциаты, коралловые полипы,,
мшанки).
К обитателям пелагиали относятся планктонные и нектонные орга-
низмы. К планктонным принадлежат парящие или пассивно пере-
носимые вместе с водой организмы, не имеющие совсем или имеющие
очень слабые органы движения, организмы, не способные противостоять,
волнам и течениям. Различают зоопланктон и фитопланктон. К зоо-
планктону относятся радиолярии, некоторые фораминиферы, крыло-
ногие (класс брюхоногих), отдельные группы ракообразные, личинки
большинства водных беспозвоночных. Пищей для мнбгих планктонных:
животных служит фитопланктон, представленный различными
водорослями.
По величине тела среди планктона выделяют несколько размерных
категорий: ультр ацл ан кт он — менее 5 мк (главным образом бак-
терии); наннопланктон — от 5 до 50 мк (жгутиковые, кокколито-
фориды, диатомеи); микропланктон—от 50 мк до -1 мм (диато-
меи,-веслоногие рачки); мезопланктон — от1 до 5 мм; макро-
планктон— более 5 мм. Иногда выделяют мегапланктон с:
размерами тела более 5 см (сифонофоры, медузы, некоторые пелагиче-
ские моллюски). Однако в большинстве своем планктонные организмы,
характеризуются малыми размерами, прозрачным телом, обычно ради-
альной симметрией, большой поверхностью тела по сравнению с его
весом, особыми гидростатическими органами, наличием в цитоплазме-
включений, уменьшающих удельный вес тела.
Другую группу пелагических животных образуют активно плаваю-
щие организмы, обладающие различными органами движения, позво-
ляющими противостоять движению волн и течений. Они относятся к
нектонным животным (многие современные, головоногие, рыбы.,.
50
водные пресмыкающиеся и млекопитающие). Форма тела у них обтекае-
мая, двусторонне-симметричная; хорошо развита мускулатура, скелет и
органы чувств, позволяющие им ориентироваться в пространстве и пре-
следовать добычу. Среди них различают хватальщиков и фильтраторов.
Хв а та льщики преследуют добычу; обычно они хорошие пловцы, име-
ющие специальные приспособления для захвата добычи. Фильтрато-
ры (некоторые рыбы, беззубые киты) имеют специальный аппарат для
отцеживания воды, с помощью которого они собирают, пищу, состоящую
главным образом из планктона.
В последние годы установлено, что пданктонные организмы, так
же как и нектонные, встречаются во всех зонах пелагиали. В эпипе-
лагической зоне (до 50 м) процветают фитопланктон и очень раз-
нообразный зоопланктон. В мезопелагической зоне (50—200 м)
встречаются многие планктонные животные и хлорофиллоносные расте-
ния, которые сохраняют свою жизнеспособность, если периодически с
восходящими токами воды возвращаются в верхнюю зону. В инфра-
пелагической зоне (200—600 м) много разнообразных животных;
ниже, в батипелагической, или сумеречной, зоне (до
2000—2500 м) обитают настоящие глубоководные формы (рыбы,
медузы). В абиссопелагической зоне (до 6000—6500 м),
или зоне, полной тьмы, основу биомассы составляют мизиды,
десятиногие раки и щетинкочелюстные. В глубоководных
впадинах, наконец, наблюдается резкое обеднение состава фауны, в
которой преобладают ракообразные (бокоплавы, остракоды, веслоно-
гие рачки), изредка встречаются офиуры, брюхоногие моллюски, веду-
щие бентосный образ жизни. Установлено, что многие виды пелагиали
проводят всю свою жизнь в вертикальных миграциях, опускаясь и под-
нимаясь иногда на многие сотни метров. Предполагается, что причиной
миграции являются поиски пищи, бегство от преследователей и иные
причины.
К некропланктону (гр. necros — мертвый, plankton — блуж-
дающий) относятся раковины и, реже, тела мертвых организмов, пас-
сивно переносимые течениями, волнами (например, пустые раковины
головоногих моллюсков, фораминифер).
Псевдопланктон, или эпипланктон, образуют организмы,
прикрепляющиеся к какому-либо пассивно плавающему предмету или
организму (например, усоногие раки прикрепляются к различным пла-
вающим предметам — кускам древесины, пемзы, пустым раковинам).
Эпинектон составляют организмы, прикрепляющиеся к активно пла-
вающим, нектонным животным, неспособные сами к активному плава-
нию (например, усоногие раки, живущие на теле китов).
К триптону относят весь органический и неорганический детрит,
встречаемый в толще воды. Особо выделяют нейстон, состоящий из
организмов, населяющих поверхностную пленку воды (некоторые жгу-
тиковые) .
В зависимости от температуры поверхностных слоев воды в. нери-
товой зоне Мирового океана выделяют пять основных биогеографиче-
ских областей: одна тропическая, две, прилегающие к ней с севера
и юга, бореальная и нотальная, и две прилегающие к полюсам —
арктическая-на севере и антарктическая на юге. Для каждой
области характерен свой состав животного населения, причем одни виды'
приурочены только к одной области, другие.— космополиты, котсУ
рые могут жить в разных областях, независимо от температурных усло-
вий; третьи, так называемые биполярные, живут в бореальной и по-
тальной областях по обе стороны от тропической области.
4* . 51;
Распределение фауны батиали и абиссали, как известно, не зави-
сит от температуры воды (она постоянна), а зависит от многих других
факторов. Эта фауна формировалась в первую очередь за счет аркти-
ческой, а также за счет переселения из сублиторали в более глубокие
зоны моря.
Знание биогеографии прошлых геологических эпох имеет немало-
важное значение для биостратиграфии, палеогеографии и понимания
путей эволюции отдельных групп.
ПАЛЕОНТОЛОГИЯ И НЕКОТОРЫЕ
ПРОБЛЕМЫ ЭВОЛЮЦИИ
Основная задача, которая стоит перед палеонтологией, заключается
в изучении органического мира геологического прошлого и закономер-
ностей его исторического развития. Решение этой большой и сложной
задачи может быть осуществлено только путем изучения элементарных
факторов эволюции, ее движущих сил, форм и темпов эволюции, а так-
же разработки различных аспектов морфологических проблем эволю-
ции— корреляции в развитии организмов, соотношений онтогенеза и
филогенеза, направления эволюционного процесса, взаимоотношения
организма и среды, прогресса и вымирания.
В основе эволюционной теории Дарвина лежал синтез всех дости-
жений биологических и геологических наук того времени. Открытие дви-
жущей роли естественного отбора в эволюции на основе изменчивости,
наследственности и борьбы за существование явилось крупнейшим до-
стижением теории Дарвина. Вместе с тем такие понятия, как изменчи-
вость, наследственность, вид во времени, мутация, изоляция, популяция,
в те времена представлялись достаточно туманными. Достижения сов-
ременной. биологии, и в первую очередь генетики, переход от типологи-
ческого организмоцентрического мышления к популяционноцентричес-
кому, признание популяции, а не отдельных организмов элементарной
единицей эволюции подняло дарвиновскую теорию на новый уровень,
уровень диалектико-материалистической методологии. Современные до-
стижения биологических наук позволили выяснить природу многих ра-
нее неизвестных явлений..
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ
О НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
Одно из. возражений против теории Дарвина заключалось в-том,
что приобретённые свойства должны утрачиваться в .результате «раз-
бавления» при', скрещивании с особью, не имеющей этих изменений.
Это возражение было снято работами Менделя, доказавшего корпус-
кульный .характер .наследственности.
. Второе возражение состояло в том, что теория естественного отбо-
ра не может объяснить появление в организме структур, кажущихся
бесполезными. Сейчас установлено, что их появление связано с побоч-
ным проявлением эффекта генов.
1ч ’ Открытие законов Менделя и современные достижения генетики
позволили по-новому взглянуть на природу наследственности. В каж-
дой’ клетке. любого организма имее.тся определенное число хромосом,
которое у разных организмов колеблется от 2 до 1600 (у одного вида
круглого червя.их-2, у некоторых ракообразных до 200, у одной морской
32
радиолярии их около 1600). Хромосомы всегда парные, в каждой клет-
ке имеется по две хромосомы • каждого сорта; у человека 24 пары хро-
мосом, они отличаются друг от друга по форме, длине, очертанию. При
делении клетки (митозе) число хромосом удваивается, и в процессе миг
готического деления хромосомы равномерно расходятся в дочерние клет^
ки таким образом, что в каждой новой клетке сохраняется постоянное
диплоидное число хромосом. При образовании половых клеток, или га-
мет, происходит два клеточных деления (мейоз), в результате которых
Рис. 17. Типы деления ядра:
I — простое деление — митоз (А—Е — последовательные стадии); II — редук-
ционное дёление — мейоз при образовании половых клеток с гаплоидным набо-
ром хромосом (А—Д, Д')-,
1 — центросома, 2 — ядро, 3 — хромосома, 4 — центросома, 5 — веретено
число хромосом сокращается вдвое и каждая гамета содержит только
один набор хромосом (гаплоидный набор). При оплодотворении проис-
ходит соединение двух гамет, в результате чего оплодотворенное яйцо,
или зигота, и все клетки тела, развивающиеся из зиготы, содержат дип-
лоидное число хромосом (рис. 17).
В каждой парной хромосоме гены расположены в определенных
местах — локусах. Поэтому, когда в мейозе хромосомы расходятся, а
при оплодотворении перекомбинируются, то такое же явление происхо-
дит и с генами. Хромосомы состоят из белков и дезоксирибонуклеиновой
кислоты (сокращенно ДНК). ДНК ответственна за передачу генетиче;
ской информации от одного поколения к другому. Модель ДНК, соз-
данная в 1953 г. Дж. Уотсоном и Ф. Криком (см. Дж. Уотсон «Двойная
спираль». М., «Мир», 1969), состоит из двух парных цепей, скрученных
в спираль, похожую на винтовую лестницу, ступеньки которой образуют
53
соединенные между собой основания, или нуклеотиды (рис. 18, Л). Они
образуются соединением двух типов нуклеотидов: пуриновых (аде-
нин и гуанин) и пиримидиновых (тимин и цитозин); попарное сое-
динение носит закономерный характер и называется комплементар-
ным: пуриновое основание — аденин (А) всегда соединяется с пири-
мидиновым основанием — тимином (Т), а пуриновое основание —
гуанин (Г) — только с пиримидиновым — цитозином (Ц), т. е. по
форме А — Т, Т — А или Г — Ц, Ц — Г.
Рис. 18. Модель двойной спирали ДНК:
А — пары аденин (А)—тимин (Т) и гуанин (Г) — цитозин (Ц), входящие в
состав модели ДНК (пунктиром показаны водородные связи); Б — возможный
механизм репликации ДНК (Уотсон, 1969)
Модель Уотсона и Крика, подтвержденная многими химическими
и физическими экспериментами, показывает, что ДНК состоит из неод-
нородных участков — генов, слагаемых из определенного числа пар ос-
нований, и генетическая информация заключена в линейной последова-
тельности четырех оснований по длине молекул. Передачу генетической
информации, закодированной в ДНК, можно представить следующим
образом: в процессе деления клетки делится ядро и двойная спираль
ДНК частично раскручивается, ветви спирали расходятся, водородные
связи нарушаются; комплементарные основания каждой ветви форми-
руют недостающую половину молекулы (рис. 18, Б). Код, заключенный
в материнской ДНК, переходит из родительской клетки в дочернюю.
54
Информация передается от ДНК к рибонуклеиновой кислоте (РНК),
которая переносит ее к рибосомам — месту синтеза белка. РНК-посред-
ник формируется в ядре и переносит от ДНК к рибосоме генетическую
информацию, определяющую последовательность расположения амино-
кислот в белке. Транспортная РНК с помощью специальных ферментов
связывает аминокислоты и переносит их к рибосомам; для каждой ами-
нокислоты (их всего 20) имеется особая РНК-переносчик и ее положе-
ние в цепи молекулы белка определяется кодирующей единицей. Она
состоит из трех оснований нуклеотидов, образуя триплет (например,
АТГ, ЦАГ). В пределах хромосомы каждый ген занимает определенное
место — локус. Аллельные гены занимают одинаковые локусы. Каж-
дый организм имеет определенный генотип. Под генотипом понима-
ют сумму всех материальных структур клетки, обладающей функцией
наследственности. Генотип складывается из совокупности всех генов,
локализованных в хромосомах — хромотипа и совокупности всех
внеядерных компонентов клетки, обладающих генетической функцией —
ллазмотипа.
Каждая особь на любой стадии развития имеет определенный об-
лик, или фенотип. Фенотипические свойства формируются'в процессе
онтогенеза, в результате взаимодействия генотипа и среды.
Наследственная изменчивость. Неистощимым источником эволюци-
онных изменений организмов служит наследственная изменчивость.
Она возникает в результате генных и хромосомных мутаций. Мутации —
наследственные изменения, которые фенотипически проявляются силь-
но, слабо или вовсе не проявляются. Генные мутации связаны с
изменениями структуры гена, в результате чего изменяется синтез бел-
ка, контролируемый данным геномом. Хромосомные мутации
связаны со структурными изменениями хромосом; они возникают в ре-
зультате утраты какого-либо участка хромосомы, удвоения отдельных
участков (дупликация), поворота участка хромосомы на 180° (инверсия),
обмена участками разных хромосом (транслокация) или перекрестка
двух гомологичных хромосом с последующей рекомбинацией. Любая
мутация ведет к нарушению определенного течения синтеза белка и вы-
ражается в изменении норм реагирования организма На внешние усло-
вия среды. Если эти нарушения крупные, то они могут приводить к сни-
жению плодовитости и жизнеспособности мутанта, а иногда и к его
гибели.
Новая форма в популяции может оказаться жизнеспособной при
скрещивании с другими особями, приводящем к комбинативным изме-
нениям хромосом. Каждая популяция обычно содержит большой и скры-
тый резерв мутаций, накопленных в условиях данной среды. При неко-
торых изменениях внешних условий скрытые мутации могут оказаться
более жизненными и привести к изменении! фенотипа, или расширению
ареала вида, или в пределах ареала лучше перенести неблагоприятные
условия. Установлено, что для популяции неблагоприятны как чрезмер-
ная наследственная изменчивость, так и чрезмерное генетическое одно-
образие.
Фенотипическое однообразие смежных популяций на обширных про-
странствах обусловлено тем, что отбор благоприятствует сохранению
стабилизирующих генов, противостоящих вредному воздействию чуже-
родных генов. В создании новых генотипов важную роль играет обмен
генами (генный поток) между соседними популяциями, представляющий
главный источник наследственной изменчивости. В изменении генетиче-
ского состава популяции из поколения в поколение основная роль при-
надлежит отбору.
55
Наследственная изменчивость, таким образом, связана с возникно*-
вением мутаций, генным потоком и рекомбинацией и служит исходным
материалом для эволюционного процесса. Фенотипы создаются при вза-
имодействии генотипов со средой. Наследственная изменчивость прояв-
ляется в полиморфизме, под которым понимается существование внутри
единой популяции резко отличных (прерывистых) наследственно обу-
словленных фенотипов. Полиморфизм охватывает любые фенотипи-
ческие признаки — морфологические, физиологические или реакции по-
ведения, если они находятся под генетическим контролем. В качестве
примера можно привести диморфные фенотипы — мужской и женский
или полиморфные особи среди брюхоногих моллюсков, которые отлича-
ются числом полос на раковине или у насекомых числом пятен на теле
(у божьих коровок). Для особей, генетически почти идентичных, но от-
личающихся фенотипически, Майр предлагает термин «полифенизм».
Полифенизм — ненаследственные изменения фенотипа; ' например,
различные касты у общественных насекомых, определенные стадии жиз-
ненного цикла (личинки и взрослые), сезонные формы (летние и зим-
ние) .
Ненаследственная изменчивость. Наряду с наследственной изменчи-
востью существует изменчивость ненаследственная, или модификаци-
онная. Модификации возникают у всех организмов, и в них выра-
жается наследственная способность организмов отвечать на изменение
факторов внешней среды изменениями своих морфологических и физио-
логических свойств. Эти изменения обратимы и исчезают, если переста-
ют действовать вызвавшие их условия; они носят массовый адаптивный
характер. Модификации возникают под влиянием разнообразных внеш-
них факторов: температуры, влажности, света и многих других. Спо-
собность к модификациям наследственно обусловлена и является опре-
деленным видовым свойством. Одни организмы имеют стабильный фе-
нотип, который сохраняется при разных условиях внешней среды; дру-
гие отличаются большой пластичностью, что приводит к формированию,
разных фенотипов при существовании их в разных условиях внешней
среды. Пределы модификационной изменчивости называют нормой
реакции; норма реакции наследственна.
Различают следующие типы ненаследственной изменчивости: 1) ин-
дивидуальная изменчивость во времени, 2) экологическая изменчивость,
3) патологическая изменчивость.
Важное значение для палеонтолога имеют возрастные изменения
и связанная с ними возрастная изменчивость. У одних живот-
ных новорожденный потомок очень похож на своих родителей, у других —
резко отличается от взрослых особей (например, гусеница отличается
от бабочки, планктонные личинки от своих бентосных родителей). Воз-
растная изменчивость возникла в результате взаимодействия между от-
бором и генотипом вида. Те морфологические изменения, которые
наблюдаются на протяжении жизненного цикла особи, являются резуль-
татом взаимодействия двух тенденций: тенденции к все большей диффе-
ренциации, что приводит к возникновению различий у взрослых особей
двух родственных видов, и тенденции к приспособлению к условиям оби-
тания на всех стадиях жизненного цикла. Это может привести к тому,,
что различия между родственными взрослыми формами будут меньше,
чем между личинками.
У некоторых животных наблюдается сезонная изменчивость, связан-
ная с изменением фенотипа в разные сезоны; она хорошо выражена у
птиц и млекопитающих. У беспозвоночных, в частности у насекомых,
живущих непродолжительное время, наблюдается сезонная изменчи-
56
вость генераций: потомство, появившееся весной, отличается рядом приз-
наков от потомства, родившегося летом.
Экологическая изменчивость зависит от многих факторов.
Особи одного и того же вида в популяциях, живущих на разных биото-
пах, часто бывают довольно различны; среди таких локальных вариан-
тов могут быть экологические расы, или ненаследственные эко фено-
типы. Такие биотопические типы особенно часто встречаются среди
бентосных форм. Так, если устрица живет в условиях сильного течения
или приливно-отливной зоны, то ее створки становятся узкими и высо-
кими; если устрица растет в чистой воде на камне, то нижняя створка
углубляется и часто становится ребристой. Биотопические модификации
также хорошо выражены у кораллов: у особей, живущих в’зоне прибоя,
колония принимает массивную форму, у особей, живущих в спокойных
водах, колония имеет ветвистый характер. Скученность особей в попу-
ляции иногда порождает морфологическую изменчивость (например, у
многих двустворчатых моллюсков и брахиопод изменяется форма рако-
вины).
Особым типом изменчивости следует считать аллометрическую
изменчивость, возникающую при аллометрическом росте, когда некото-
рые структуры тела резко и непропорционально увеличены по сравнению
с другими частями тела, например голова муравьев, рога и грудь неко-
торых жуков. У. общественных насекомых — пчел, ос, муравьев, терми-
тов— имеются особые касты: царицы, рабочие и Солдаты, резко отли-
чающиеся друг от друга морфологически, но относящиеся к одному виду.
Патологическая изменчивость возникает у разных групп
животных в виде различного рода вздутий, деформаций скелета или не-
которых изменений органов, вызываемых различными паразитами или
травматизмом.
БОРЬБА ЗА СУЩЕСТВОВАНИЕ
И ЕСТЕСТВЕННЫЙ ОТБОР
Мутации, как было отмечено выше, могут выражаться по-разному
и включаться в жизнь популяции через скрещивание и в результате-
борьбы за существование. Формы и последствия борьбы за существо-
вание крайне разнообразны. Они определяют пути дальнейшей эволю-
ции; создание новых форм идет всегда исключительно через соревнова-
ние особей внутри данной популяции. Борьба за существование
включает в себя две стороны: с одной стороны, различные формы эли-
минации, т. е. уничтожение особей, с другой — активную или пассив-
ную конкуренцию между особями одного вида.
Типы элиминации крайне разнообразны. Различают общую эли-
минацию, или случайную истребляемость, носящую постоянный или
периодический характер (период дождей, засухи, размножение хищни-
ка), не зависящую от индивидуальных свойств организма. Гибель зави-
сит от слепого случая, а не определяется индивидуальными свойствами..
В этом случае могут выжить более плодовитые пары —естественный от-
бор будет происходить на максимальную плодовитость. Если общая
элиминация имеет возрастной характер, то при высокой истребляемости
взрослых происходит удлинение личиночной стадии, наоборот, при высо-
кой истребляемости личинок удлиняется взрослая фаза.
Индивидуальная элиминация, или избирательное устране-
ние, зависит от индивидуальных отличий особей и может иметь постоян-
ный или периодический характер; элиминация бывает прямой или кос-
57
венной. Прямая элиминация связана с действием физических фак-
торов, с неблагоприятными метеорологическими условиями, воздействи-
ем крайних температур, ветра, потоков воды, колебанием концентраций
солей. В этом случае переживают наиболее стойкие, наиболее защищен-
ные особи.
Биологические факторы (нападение хищников, паразитов) также
приводят к прямой элиминации, к отбору более активных особей с бо-
лее совершенными органами чувств, инстинктами, вооруженных средст-
вами активной защиты. Эта форма элиминации имеет значение в мор-
фофизиологическом совершенствовании организации, в прогрессивной
эволюции.
Косвенная элиминация вызывается физиологическими фак-
торами, приводящими к ослаблению организма вследствие голодания,
к снижению его плодовитости, к устранению от размножения; все это
соответственно способствует истреблению другими факторами (физичес-
кими и биологическими). Косвенная элиминация приводит к отбору на-
иболее успешно добывающих пищу, наиболее продуктивно ее исполь-
зующих, к совершенствованию органов чувств, средств нападения, пи-
щевой специализации.
Семейная элиминация приводит к уничтожению целых се-
мейств и вместе с тем вызывает отбор семейств, обладающих наиболее
защищенными кладками, наиболее совершенными средствами защиты
икры, приводит к улучшению способов заботы о потомстве (строитель-
ство гнезд, охрана потомства).
Групповая элиминация, основанная на генетической близос-
ти между особями одной естественной группы (популяции, подвида),
приводит к истреблению целых популяций или вытеснению их другими
формами. Она вместе с индивидуальной элиминацией способствует рас-
пределению форм по различным биотопам, служит источником дивер-
генции и обеспечивает эволюцию каждой группы.
. Элиминация приобретает индивидуальный характер только через
соревнование генетически разнородных особей внутри одной популяции.
Различают индивидуальное активное и пассивное соревнование между
отдельными особями одной популяции. Активное соревнование в пер-
вую очередь выражается в конкуренции за жизненные средства — пищу,
свет, воду, солнце и т. д. Пассивное соревнование проявляется в
борьбе с физическими и биологическими факторами за сохранение жиз-
ни и оставление потомства.
Различают две основные формы естественного отбора: стабилизи-
рующую и движущую. Стабилизирующая форма естественного
отбора направлена на поддержание установившегося «нормального»
строения и жизненных функций организма, его приспособленности на
всех стадиях развития к данным условиям существования и обеспечива-
ет установление оптимальной жизнеспособности и плодовитости; она реа-
лизуется на основе селекционного преимущества представителей средней
нормы перед всеми отклонениями от этой нормы и означает известное
динамическое равновесие между популяцией и внешней средой. Эта
•форма естественного отбора ведет к сохранению фенотипического одно-
образия популяций как генетических систем, к развитию регулирующих
механизмов, поддерживающих устойчивость развивающейся особи, по-
пуляции и вида к элиминации всех особей, отклоняющихся от феноти-
па (рис. 19).
Движущая, или творческая, форма естественного отбора наи-
более важная в эволюционном процессе. Накопленная в популяции из-
менчивость находится под контролем цитофизиологических механизмов
58
и экологических факторов. Все природные популяции обладают значи-
тельной наследственной изменчивостью, которая служит источником
эволюционных изменений. При изменении формы элиминации изменяется
направление действия естественного отбора; естественный отбор создает
и перестраивает аппарат ин-
дивидуального развития, на-
правленного на приобретение
новой нормы реакции, соот-
ветствующей видоизмененным
условиям жизни. Из относи-
тельно однородной популяции
возникают разнородные груп-
пы, и эти различия с течением
времени углубляются, группы
внутри популяций рее более
обособляются, увеличивается
разрыв внутри непрерывной
цепи изменчивости, и многооб-
разие приобретает прерывис-
тый характер. Две основные
группы факторов внешней
среды оказывают влияние на
направление и величину дав-
ления отбора: а) климатиче-
ские факторы, б) биотопиче-
ские и биотические факторы.
Они влияют на географиче-
скую изменчивость, т„ е. на те
Рис. 19. Схема стабилизирующего отбора
(Шмальгаузен, 1969).
Уклонения в онтогенетическом развитии, ве-
дущие к образованию крайних вариантов,
элиминируются. Сохраняются и свободно
накапливаются лишь варианты, находя-
щиеся в пределах нормы
различия, которые наблюда-
ются у пространственно раз-
деленных популяций. Не менее
важна хронологическая изменчивость организмов во времени, связан
ная с действием тех же факторов.
ИЗМЕНЕНИЕ ОРГАНОВ В ФИЛОГЕНЕЗЕ
В процессе онтогенеза происходит закладка и формирование раз-
личных органов. На ранних стадиях филогенеза каждый орган обычно
мультифункционален; кроме главной функции он выполняет ряд вто-
ростепенных. При изменении условий существования или приспособле-
нии к новым нишам жизни происходит изменение или перестройка орга-
нов без нарушения основных признаков организации. Под влиянием ес-
тественного отбора различные органы могут изменяться в определенных
адаптивных направлениях, при этом главная функция может терять
свое значение, а какая-либо из второстепенных может стать главной.
Например, первоначальная функция конечностей речного рака была
плавательной, а второстепенные — ходильная, хватательная и иногда
дыхательная. В процессе эволюции произошла дифференциация и сме-
на функций, головные и передние грудные конечности стали хвататель-
ными, и жевательными, задние грудные — ходильными, брюшные — пла-
вательными.
При прогрессивном развитии часто происходит дифференциация ор-
гана или его усиление, что приводит к расширению или усилению функ-
ций, сопровождаемому приобретением некоторых новых функций или
усилением прежних; при специализации решающее значение приобрс-
59
тает главная функция, а второстепенные теряют свое значение, что при-
водит к ограничению дальнейших преобразований. Смена функций про-
исходит всегда одновременно со сменой формы; в' процессе развития
форма и функция неразрывно связаны друг с другом. При смене функ-
ций происходит перестройка отдельных органов, что может приводить
к изменению всех корреляционных связей и соответственно аппарата
индивидуального развития.
АДАПТИВНАЯ РАДИАЦИЯ, КОНВЕРГЕНЦИЯ
И ПАРАЛЛЕЛИЗМ
Дарвин считал, что развитие организмов идет дивергентно: появ-
ление различий в строении организмов приводит к приспособлению к
местным географическим и экологическим условиям, к возникновению
географических изолятов, местных географических или экологических
форм. Дивергенция форм лежит в основе всего эволюционного процес-
Рис. 20. Конвергентное сходство у прикрепленных животных:
1 — четырехлучевой коралл Omphyma- 2 — брахиопода Richthofenia; 3 — дву-
створчатый моллюск из отряда рудистов; 4 — усоногий рак Pyrgoma
са. Она обусловлена борьбой за существование, естественным отбором
и мутационным процессом. Примером дивергентного развития может
служить класс двустворчатых моллюсков, который распался на ряд отря-
дов, приспособленных к разным местам обитания в морских и пресных
водах. Дивергенция любой группы животных приводит к специализации
ее отдельных представителей. Сходство форм объясняется общностью'
происхождения, а различия — приспособлением к разным условиям оби-
тания. Позднее, в 1905 г., X. Осборн предложил термин «адаптивная
радиация», который образнее слова «дивергенция» и полнее отражает
пути развития любой группы. Сущность адаптивной радиации заклю-
чается в том, что наибольшая сумма жизни достигается при наиболь-
шем разнообразии и что при дивергенции снижается давление внутри-
видовых и межвидовых противоречий и повышается коэффициент вы-
живаемости.
Конвергенция означает приобретение сходных признаков нерод-
ственными организмами в результате приспособления к сходным услови-
ям существования. Сходство объясняется приспособлением к сходным
условиям, а различия — отсутствием близкого родства. Это сходство за-
60
трагивает только внешнюю форму тела или отдельных органов, но не
затрагивает основной организации. Например, конвергентные формы:
акула (рыба), ихтиозавр (рептилия), дельфин (млекопитающее),
коралл, рихтгофения (брахиопода), рудист (двустворчатый моллюск),
усоногий рачок (рис. 20).
П а р а л л ел из м— независимое приобретение родственными орга-
низмами сходных признаков; в этом случае сходство объясняется частью
Рис. 21. Гомеоморфия у двустворчатых моллюсков:
/ — Cardium fittonl; 2 — Caladacna steindachneri; 3 — Caladacna escheri
общим происхождением, частью приспособлением к сходным условиям
существования. У двоякодышащих рыб и амфибий параллельно развива-
лись сердце и кровеносная система. Параллельность в развитии объяс-
няется близким родством двух групп, обладающих сходной наследствен-
ной основой, сходными нормами реакций и сходными взаимоотноше-
ниями со средой. Внешнее сходство одной или нескольких форм, принад-
лежащих к одной и той же систематической группе, приводит к гомео-
морфии (рис. 21).
МОНОФИЛИЯ и ПОЛИФИЛИЯ
Дарвин убедительно доказал, что виды одного и того же рода, какие
бы удаленные участки земного шара они не занимали, должны были
произойти из одного места, поскольку они произошли от общего праро-
дителя. Установление центров происхождения тех или иных групп жи-
вотных и их предков, установление путей и способов расселения, а также
изменения ареалов во времени и в пространстве представляет одну из
важнейших задач палеонтологии. Однако некоторые исследователи ста-
раются доказать, что большинство групп животного мира имеет поли-
филетичное происхождение, принимая конвергенцию, а не дивергенцию
за основной процесс в. эволюции. Развитие наших знаний действительно
приводит к открытиям гетерогенности ряда групп, ранее считавшихся
едиными, но это свидетельствует не о полифилетическом происхожде-
нии, а о недостатках наших знаний. Происхождение от одного предка
нельзя понимать как происхождение от одной пары особей; как было
показано выше, элементарной эволюционной единицей является популя-
ция. В процессе эволюции идет преобразование и дифференциация це-
лых популяций. Поскольку каждая особь происходит от двух родителей,
четырех прародителей и т. д., то родство особей в пределах популяции
и целого вида имеет сетчатый характер; вне вида родословные линии
расходятся, приобретают независимое, иногда параллельное течение.
Различные органы развиваются обычно с разными скоростями; в процес-
61
се перехода в новую адаптивную зону одна структура или комплекс,
структур подвергается более сильному давлению отбора и развивается
сильнее, наоборот, другие структуры могут задерживаться в развитии;
эволюционирующий тип или недостающее звено может представлять
мозаику из примитивных и продвинутых в своем развитии признаков, из
специализированных и неспециализированных признаков.
Каждую крупную систематическую единицу (тип) можно предста-
вить в виде родословного древа, от ствола которого отходят крупные
ветви (классы), по мере своего развития отделяются ветви второго по-
рядка (отряды), на них вырастают ветви третьего порядка (семейства),
затем ветви четвертого порядка (роды), которые распадаются на мель-
чайшие веточки (виды) (см. рис. 11). Рассмотренное древо наглядно-
показывает прогрессивное углубление разрывов по мере перехода от од-
ного таксона к другому. Эти разрывы невелики между видами и доста-
точно глубоки между отрядами и классами. В результате адаптивной
радиации создается все разнообразие данной группы, ее разные уров-
ни организации, приспособления к разным условиям среды, что обеспе-
чивает ей возможности развития, расселения и занятия разных экологи-
ческих ниш.
СООТНОШЕНИЕ МЕЖДУ ОНТОГЕНЕЗОМ
И ФИЛОГЕНЕЗОМ
В процессе индивидуального развития, или онтогенеза, организм
выступает как единое целое. Онтогенез начинается после оплодотворе-
ния с дробления яйца, которое идет под контролем геномных кор-
реляций, обусловливающих последовательную дифференциацию час-
тей тела и установление между ними на каждой стадии развития новых
соотношений. Внутри развивающегося организма существует сложней-
шая система связей, объединяющая в одно целое элементы наследствен-
ной субстанции и внутриклеточные биохимические процессы.
За геномными корреляциями вступают в действие морфогене-
тические корреляции, которые определяют последовательность
развития отдельных структур тела. Например, губа бластопора у поз-
воночных индуцирует образование хорды и нервной трубки. Каждый
предшествующий орган обеспечивает развитие нового последующего.
Организм развивается как единая целостная интегрированная система.
Каждая часть отвечает за все целое, а целое отвечает за развитие каж-
дой части, причем каждая стадия развития зародыша адаптивна и на-
ходится под контролем естественного отбора. Все корреляции отлича-
ются большой прочностью и нужны крупные сдвиги, чтобы нарушить эти
корреляции. Морфогенетические корреляции наблюдаются на всех ста-
диях морфогенеза и. захватывают все органы и признаки организма.
После завершения морфогенеза вступают в действие эргонтиче-
ские, или рабочие, корреляции, которые обусловливают окончатель-
ное формирование органов и отдельных частей организма. Например,,
в результате эргонтических корреляций происходит формирование ске-
летных частей и связанных с ними мышц.
Геномные корреляции, таким образом, служат той основой, на ко-
торой развиваются морфогенетические корреляции; последние являют-
ся основными факторами эмбрионального формообразования, опреде-
ляющими развитие исторически сложившихся взаимоотношений между
органами, охраняемыми высокими защитными порогами реакций. Все
эти корреляции обусловливают формирование нового организма, его>
62
дифференциацию и интеграцию. Под интеграцией понимается про-
цесс установления взаимных корреляций между частями организма по
мере его дифференциации.
Первый этап развития, происходящий под защитой яйцевых оболо-
чек или в материнском организме, называется эмбриональным;
после выхода из яйца или рождения — постэмбриональным. Если
из яйцевых оболочек выходит личинка, то такое развитие называется
непрямым, или развитием с метаморфозом; если новорожденный по-
хож на родителей, то развитие прямое, без метаморфоза, без личиноч-
ных стадий.
Филогенез, или филогенетическое развитие, представляет собой
историческое развитие данного животного или данного вида, рода, се-
мейства, отряда и т. д. Наиболее полную и всестороннюю теорию о вза-
имосвязи между индивидуальным и историческим развитием разработал
Дарвин в последней главе своей книги «Происхождение видов» — «Вза-
имное сродство между организмами». Он установил, что изменения у
потомков возникают на тех же возрастных стадиях, на которых они
наблюдались у предков, что зародыши разных видов одного и того же
класса весьма сходны (закон Бэра), но, достигнув взрослого состояния,
резко отличаются; однако часто на ранних.стадиях развития личинки
или молодые особи под влиянием естественного отбора могут отличать-
ся от родителей до любого предела. И в этом случае ранние стадии не
могут дать представления о строении взрослых предков. Геккель, как
было отмечено ранее, установив биогенетический закон, согласно кото-
рому онтогения есть краткое повторение филогении, упростил взгляды
Дарвина на соотношение индивидуального и исторического развития.
В процессе развития любой группы происходит перестройка онто-
генезов, поэтому филогенез следует рассматривать как исторический
ряд известных онтогенезов. Эволюция онтогенезов составляется из раз-
ных процессов: приспособлений зародыша или личинки к окружающей
среде, перестройки самого онтогенеза, усовершенствования факторов
индивидуального развития и ряда филэмбриогенезов.
Под филэмбриогенезами А. Н. Северцов понимал такие перестройки
эмбриональных формообразовательных изменений, которые связаны с
эволюцией взрослого организма. Различают три типа филэмбриогенезов:
анаболию, девиацию и архаллаксисы.
Анаболия, или надставка, —• наиболее распространенный спо-
соб онтогенеза — заключается в изменений конечных стадий формообра-
зования; законченный процесс формообразования дополняется дальней-
шими дифференцировками; в процессе онтогенеза рекапитулируются
(повторяются) исторические этапы развития предков. Девиация есть
уклонение в ходе формообразования; в этом случае у потомков сохра-
няется сходство только ранних стадий зародышевого развития с их
предками; поздние стадии отличаются от таковых у предков. В случае
архаллаксиса происходит изменение самих зачатков, онтогенез из-
меняется е первых стадий и историческая последовательность изменений
не отражается в онтогенезе. Эмбрионы потомков отличаются от эмбрио-
нов предков и использовать архаллаксисы для восстановления филоге-
нии оказывается невозможным.
Изменения зачатков могут быть выражены в изменении времени за-
кладки органа (гетерохрония) или положения (гетеротопия).
Девиации и архаллаксисы, таким образом, оказываются мало при-
годными для восстановления филогении. Рекапитуляция предковых при-
знаков является результатом изменения конечных стадий формообразо-
вания, результатом, эволюции посредством надставок, или анаболии.
63
Целый ряд вымерших групп позволяет изучать их онтогенез и'на
этом основании решать филогенетические вопросы. К числу таких групп
относятся аммониты, археоциаты, коралловые полипы и некоторые дру-
гие группы. Значительные трудности возникают при изучении онтогене-
зов вымерших членистоногих.
ПУТИ И ЗАКОНОМЕРНОСТИ
ЭВОЛЮЦИОННОГО ПРОЦЕССА
Эволюционный процесс на всех стадиях развития жизни на. Земле
носил приспособительный, или адаптивный, характер, т. е. шёл под зна-
ком приспособления организмов к окружающим их условиям. Наряду
с общим повышением организации, связанным с выработкой приспособ-
лений общего характера (например, появление жабр как органов водного
дыхания, развитие легких как органов воздушного дыхания), развива-
Рис. 22. Схема эволюционного процесса (Северцов,11967):
восходящие ленты (ар) — ароморфозы; горизонтальные плоскости
(ид) — идиоадаптации; снижающиеся плоскости (сЭег) — дегене-
рация
лись частные и специальные приспособления. При прогрессивном раз-
витии одних групп в других может наблюдаться застой или даже рег-
рессивное развитие. Процесс повышения организации идет обычно не-
равномерно, периоды бурного подъема организации сменялись перио-
дами спокойного приспособления. Прогрессивное развитие новых форм
жизни всегда сопровождалось биологическим регрессом или вымирани-
ем менее совершенных форм.
. . По А. Н. Северцову, биологический прогресс сопровождал-
ся возрастанием приспособленности организма к окружающей его среде,
ведущим к увеличению его численности и более широкому распростра-
нению во времени и пространстве. Биологическая стабилизация
шла при сохранении численности на одном уровне, организмы изменя-
лись соответственно изменениям окружающей их среды. Биологиче-
ский регресс приводил к снижению приспособленности организма,
отставанию от темпов эволюции близких форм, снижению численности,
возрастанию элиминации, приближению к вымиранию. А. Н. Северцов
выработал общую концепцию главных направлений эволюционного про-
цесса (рис. 22). Он выделил этапы поднятия морфофизиологической
организации на более высокий уровень; этот процесс он предложил на-
зывать ароморфозом.
г- Под а р о м ор ф оз о м, или морфофизиологическим прогрессом, по-
нимается получение организмом преимуществ общего порядка, поднятие
64
организации на более высокий уровень, дающее организму возможность
расширить использование • внешней среды. Отбор идет по линии
повышения жизнеспособности в разнообразных условиях среды.
Ароморфоз характеризуется тремя главными элементами: повышением
организации, развитием приспособлений широкого значения, расшире-
нием среды обитания. Эти три момента можно проследить на примере
организации млекопитающих: подъем жизнедеятельности их был связан
с приобретением постоянной температуры тела, волосяного покрова,
прогрессивного развития легких и кровеносной системы; усовершенст-
вование органов чувств, нервной системы и головного мозга обеспечили
млекопитающим господство на суше и жизнь в водных бассейнах.
Идиоадаптация. После процесса ароморфоза наступает, по Се-
верцову, период идиоадаптации — приспособления организмов к разно-
образным условиям среды, которое не сопровождается усложнением ор-
ганизации. Эти приспособления выражаются в занятии различных эко-
логических ниш, развитии средств для добычи питания и средств актив-
ной и пассивной защиты. Они могут происходить на всех стадиях жизни
организма. Приспособительные изменения на личиночной стадии носят
название ценогенозов. И. И. Шмальгаузен выделил несколько ти-
пов идиоадаптаций.
Теломорфоз означает частное приспособление к специфическим
условиям среды, при котором происходит одностороннее развитие неко-
торых органов и редукция других; организация в целом остается на
прежнем уровне или испытывает некоторое упрощение. К специализи-
рованным формам относятся хамелеоны, дятлы, муравьеды, саблезубые
тигры, усоногие раки, двустворки-сверлильщики и др.
Другой формой идиоадаптации является гиперморфоз. Этот тип
эволюции характеризуется процессами резкого увеличения тела, сопро-
вождаемыми непропорциональным увеличением отдельных частей тела,
нарушением координаций. Например, крупные динозавры имели очень
маленькую голову и маленький головной мозг; гигантский четвертичный
олень имел гигантские рога; многие слоны имели огромные бивни. Часть
этих структур, возможно, могла возникнуть в результате полового
отбора.
Катаморфоз связан с недоразвитием, упрощением организации
(дегенерацией, по А. Н. Северцову) и переходом к более простым соот-
ношениям со средой; при этом организм получает известные пре-
имущества в борьбе за существование, часть органов становится излиш-
ними и редуцируется, изменяются некоторые корреляционные связи.
Например, переход от активной жизни к пассивной, от подвижного об-
раза жизни к сидячему или малоподвижному. Однако в новых условиях
эта группа продолжает успешно развиваться. Например, мшанки, усоно-
гие раки, птеробранхии, оболочники.
Гипоморфоз представляет частную форму катаморфоза, связан-
ную с недоразвитием организма вследствие сохранения тех отношений
со средой, которые были характерны для личиночной стадии. Гипомор-
фоз, или неотения, приводит к недоразвитию тех органов, которые
развиваются позже всего (специальных органов), например аксолотль,
не пройдя стадии метаморфоза, становится половозрелым и остается на
всю жизнь водным животным с хвостом, жабрами. Некоторые авторы
предполагают, что явление неотении, или недоразвития, создает благо-
приятную основу для прогрессивного развития. У гипоморфных живот-
ных выпадают поздние стадии развития и поэтому ни эмбриологические,
ни сравнительно-анатомические, ни палеонтологические данные не дают
5
В. В. Друщиц
65
указаний на их происхождение. Предполагают, что насекомые произош-
ли путем неотении от личинок многоножек.
Развитие органического мира, таким образом, в силу естественного
отбора наиболее приспособленных и способных оставлять потомство,,
идет по пути смены приспособлений к меняющимся условиям внешней,
среды. Одни виды развиваются по пути биологического прогресса, уве-
личивая свою численность, расширяя ареалы, за которым следует био-
логическая стабилизация, сохранение постоянной численности, более-
или менее постоянных ареалов, она сменяется биологическим регрессом,,
который сопровождается уменьшением численности, сокращением ареа-
лов, появлением разорванных ареалов, что в конце концов завершается"
вымиранием.
Прогрессивное развитие связано с приобретением адаптаций общего*
значения, приводящим к ароморфозам; расширение ареалов ведет к раз-
витию по пути идиоадаптаций, которым и завершается ароморфоз про-
цветающей. формы. Дальнейшее развитие, специализация и приспособ-
ление к определенным условиям существования приводят к теломорфо-
зу — специализации, связанной с утерей пластичности, и в некоторых;,
случаях к гиперморфозу — явлению переразвитая. Однако рассмотрен-
ная смена направлений эволюционного прогресса не является един-
ственным путем эволюции в целом. Развитие по пути идиоадаптаций
может идти неограниченно долго; получение нового приобретения,,
дающего организму какие-то преимущества, может привести к прогрес-
сивной эволюции по пути ароморфоза и т. д.
ВЫМИРАНИЕ ОРГАНИЗМОВ
В соответствии с теорией Дарвина, развитие одних форм всегда^
сопровождается вымиранием менее приспособленных и менее совер-
шенных форм. В течение фанерозоя вымерли многочисленные крупные-
, и мелкие группы: археоциаты, ругозы, табуляты, строматопораты,
трилобиты, аммониты, белемниты, бактриты, многие классы иглокожих,,
граптолиты, хиолиты, тентакулиты и многие другие. Ветви, быстро*
расцветающие и быстро , специализирующиеся, вымирают обычно-
быстрее ветвей менее специализированных.
Вымирание и естественный отбор, по Дарвину, идут рука об руку,,
но увеличение численности вида постоянно задерживается различными*
враждебными причинами на всех стадиях индивидуального развития,,
что в конце концов может привести вначале к редкости, а затем обус-
ловить вымирание вида. Если какой-либо вид захватывает место, ранее.-
занимавшееся видом другой группы, и от него разовьются новые фор-
мы, то эти новые формы могут вытеснить формы старого вида. Внедре-
ние новых форм на новую территорию, обладающих некоторыми:
преимуществами перед местными, приведет к вытеснению этих местных
форм, но благодаря каким-нибудь особенностям одна из местных форм;
может сохраниться и существовать еще длительное время. Окончатель-
ное вымирание обычно более медленный процесс, чем видообразование...
Известно много примеров, показывающих, что вид сначала вымирает
на части своего ареала, затем ареал становится все меньше и меньше-
и в конце концов вид может окончательно вымереть. В некоторых
случаях реликтовые формы продолжают существовать очень длительное-
время. К числу подобных реликтов можно отнести наутилуса и триго-
нию, лингулу и кранию — представителей широко распространенных
групп в мезозое и палеозое. В основе вымирания лежат причины,,
связанные с изменениями соотношений организма с внешней средой..
66
Важное значение имеют различные абиотические факторы и их измене-
ние. Организм гибнет в борьбе за существование. Но следует учиты-
вать также, что специализация организмов ограничивает их возмож-
ности приспосабливаться к изменяющимся условиям среды, и даже
кратковременное изменение той среды, к которой они приспособлены,
ставит организм перед угрозой вымирания. В процессе теломорфоза,
рассмотренного выше, под влиянием внешних и внутренних факторов
происходит чаще вымирание специализированных групп. Специализа-
ция создает предпосылки для вымирания. Вымирание местных групп
организмов может быть вызвано внезапным изменением климата,
резким похолоданием или сильной засухой,' внезапным изменением
химизма морской воды в результате ливневых дождей или прорыва
перешейка, извержением вулканов, изменением солнечной радиации
и многими другими причинами.
НЕОБРАТИМОСТЬ ЭВОЛЮЦИИ
Закон необратимости эволюции был сформулирован Дарвином, но
получил известность благодаря бельгийскому палеонтологу Л. Долло.
В соответствии с этим законом вид, однажды исчезнувший, вновь не
может появиться, так как порвалась цепь развития. Этот закон лежит
в основе выделения всех стратиграфических единиц от эр до зон вклю-
чительно. Для каждой стратиграфической единицы характерны свой
группы организмов, которые после вымирания вновь не могут возник-
нуть, даже если возникнут такие условия внешней среды, которые были
благоприятны для существования вымерших групп. Эволюционный
процесс необратим, возможно только возникновение конвергентно
сходных форм, отличающихся особенностями своего строения и орга-
низации.
ПРИНЦИПЫ СИСТЕМАТИКИ ЖИВОТНЫХ И ПРАВИЛА
ЗООЛОГИЧЕСКОЙ НОМЕНКЛАТУРЫ
Принципы систематики животных. Систематика, или таксо-
номия,— наука о классификации организмов. Термин «систематика»
происходит от латинского слова systema, а таксономия — от сочетания
греческих слов «таксис» — порядок и «номос» — закон. Зоологическая
классификация представляет собой распределение животных по сопод-
чиненным группам на основании изучения их сходства и различий и
выявления родственных отношений. Основная цель систематики состоит
в построении системы животных, которая давала бы максимальное
количество информации о любой группе животных и имела бы большую
прогностическую ценность.
В истории таксономии можно выделить три периода.
Первый период, долиннеевский, был связан с изучением' местных
фаун, их описанием и наименованием всех животных на том языке,
на котором их описывали, созданием классификаций по каким-либо
отдельным признакам. Первую классификацию животных, основанную
на степени их совершенства (градации), дал Аристотель (см. раздел
«Основные этапы развития палеонтологии»), его система просущество-
вала почти 2000 лет.
5*
67
Второй период связан с именем К- Линнея (1707—1778) —основа-
теля систематики как науки. В десятом издании книги «Система при-
роды» (1758) К. Линней предложил систематику, основанную на
юоподчиненности таксономических единиц, или таксонов: царство, класс,
отряд, род, вид и вариетет. Он распределил все известные к тому вре-
мени группы животных среди соподчиненных таксонов и дал четкие и
ясные диагнозы. В течение ста лет до выхода в свет работы Ч. Дарвина
«Происхождение видов» (1859) основное внимание уделялось четкому
ограничению видов, основанному на признании их постоянства. Разра-
батывалась типологическая концепция вида, в соответствии
с которой каждый изучаемый экземпляр сравнивался с типовым
экземпляром, и по степени-морфологического сходства или отличия
определялся статут вида. Шло быстрое увеличение числа описываемых
видов как современных, так и вымерших, разрабатывались основные
методы и принципы классификации. Предложенная иерархия таксоно-
мических единиц за срок более чем 200 лет не испытала сколько-нибудь
существенных изменений, если не считать добавленных промежуточных
таксонов, указанных ниже.
По предложению К. Линнея, каждый вид получил двойное латини-
зированное название — родовое и видовое. Например, Homo sapiens L.,
Canis familiaris Carlo, Pinus silvestris L. Первое слово относится к на-
званию рода, который обычно объединяет несколько видов и является
основным; второе — к названию вида, которое является, как прави-
ло, определяющим, подчеркивающим какие-либо особенности
организма. В данном случае: Ното — человек, sapiens — мудрый; Ca-
ms — собака, familiaris — домашняя; Pinus — сосна, silvestris — лес-
ная. За видовым' названием всегда следует фамилия автора (или авто-
ров), впервые описавшего данный вид (L. — принятое сокращение
фамилий К. Линнея). Двойное название каждого вида послужило
основой для создания бинарной номенклатуры, а 1 января 1758 г. было
принято за дату, с которой действует закон приоритета. В соот-
ветствии с этим, законом первое название, данное любому виду или
роду, не; подлежит изменению.
С Линнея начинается период быстрого увеличения числа описывае-
мых современных.и вымерших/видов, разработка основных принципов
и методов классификации, введение единых латинизированных назва-
ний для всех изучаемых животных и растений. Основное внимание было
направлено на создание так называемой «естественной» системы, под
которой понималось раскрытие «плана творения». Поскольку эволю-
ционисты в понятие «естественная система» вкладывают совершенно
иной смысл, лучше от этого термина отказаться.
Существенный вклад в систематику животных был внесен работами
Кювье и Ламарка, выделивших из беспозвоночных («бескровных»)
животных ряд новых классов (головоногих, брюхоногих). Это по-
служило толчком для расформирования неестественных группировок
и выделения новых. Кювье на основании сравнительно-анатомических
исследований выделил четыре независимые группы животных: хордо-
вые, моллюски, членистые и лучистые, разработал принцип соподчине-
ния основных и второстепенных признаков.
Третий период в развитии систематики начался с появления книги
Ч. Дарвина «Происхождение видов» (1859). Дарвин отверг креационизм
и обосновал принципы филогенетической систематики, основанной на
выявлении реального родства по происхождению. Сущность «естествен-
ной» системы была выяснена — естественные группы существуют пото-
му, что они происходят от общего предка. Ч. Дарвин создал теоретиче-
68
ские основы естественной (без кавычек), или филогенетической, сис-
темы. Он установил, что в процессе филогенеза осуществляются два
процесса: ветвление и последующая дивергенция, или расхождение, воз-
никших ветвей. Разделение таксонов должно основываться на установ-
лении их ветвления (что указывает на общность происхождения) и на
степени различия. Степень изменений, испытанных в ходе развития
различными группами, выражается в размещении их по таксонам раз-
ных рангов: классов, отрядов, семейств.
Дарвин говорил о необходимости создания филогенетической систе-
матики, основанной на выделении признаков трех категорий: а) при-
знаки, выявляющие действительное родство (гомогёничное сходство);
б) признаки зачаточных или атрофированных органов, не имеющих
значения в жизни животных, но важных для систематики; в) особен-
ности эмбрионального строения сравниваемых форм, имеющие важное
значение для систематики.
Последарвиновский период—период борьбы за признание эволю-
ции— характеризовался созданием классификаций различных групп
животных и растений. Концепция типа стала постепенно замещаться
концепцией популяций, согласно которой виды состоят из
изменчивых популяций, причем даже в пределах крупных таксонов
возможны отклонения от «типа», характерного для данной категории.
Признание изменчивости вида заставило обратить серьезное внимание
на типы изменчивости и на оценку изменчивости методами популяцион-
ного анализа и статистическими методами. Создание классификации
поставило перед исследователем ряд сложных вопросов, например:
представляют ли изученные две формы один или два вида, чем обус-
ловлено их сходство — филогенетическим родством или только внеш-
ним, конвергентным сходством. Все эти вопросы представляют собой
третью задачу систематики, которая была поставлена Дарвином —
изучение и анализ внутривидовой изменчивости и выявление факторов
эволюции; при решении этих вопросов систематика соприкасается с ге-
нетикой, биогеографией, экологией, сравнительной анатомией и данными
палеонтологии.
Изучение любой группы с позиции систематики проходит три ста-
дии, именуемые: альфа-, бета- и гамма-систематикой (Майр, Линсли,
Юзингер, 1946). Первая — альфа-систематика — это аналитическая
стадия, в течение которой на современном научном уровне изучается
каждая группа и даются названия; вторая стадия — бета-системати-
ка — это систематическая стадия, включающая объединение элементов
группы в систему таксонов, и третья стадия, или гамм а-систематика,
является заключительной, теоретически обобщающей все полученные
результаты. Современная систематика сочетает в себе методы анализа
и синтеза.
В настоящее время различают филогенетическую и искусственную
систематики. Ф и л о г е н е т ич е с к а я систематика строится на
основании выяснения генетических взаимоотношений родственных групп
во времени и пространстве. Всех животных можно расположить в
систематическую иерархию, состоящую из таксонов, ранг которых
постепенно повышается. Филогения и систематика рассматриваются в
неразрывной связи как две стороны единого процесса познания факти-
ческой истории органического мира; причем если филогения изучает
родственные связи и выясняет общность отдельных таксонов, то систе-
матика стремится разделить выделенные филогенетические ветви на
отдельные соподчиненные таксоны. Поэтому нельзя отождествлять
филогению и систематику.
69
Искусственные систематики отличаются от филогенети-
ческой тем, что организмы группируются по внешним сходным призна-
кам, причем различают несколько разновидностей искусственных систе-
матик. К искусственной систематике по необходимости прибегают
тогда, когда классифицируются отдельные части организмов, например
ринхолиты, аптихи, конодонты, скелетные элементы голотурий; они
получают видовые и родовые названия и даже иногда объединяются в
более высокие таксоны. Для некоторых из них, например конодонт, до
сих пор неизвестно, к какой группе организмов они принадлежали.
Для таких групп предлагается выделить паратаксоны — особые ка-
тегории, которые не подчиняются правилам зоологической номенкла-
туры.
Имеются искусственные классификации по принципиальным сооб-
ражениям, когда авторы считают, что в действительности филогенети-
ческие системы не существуют, а имеются особые «естественные», или
типологические, систематики, основанные на изучении ряда признаков,
позволяющие находить черты сходства и различий, опираясь на стати-
стические обобщения. Они важны для наведения порядка в разнообра-
зии организмов. Хотя эти систематики и претендуют на «естествен-
ность», но они далеки от филогенетической систематики и представ-
ляют, как правило, искусственную группировку организмов. Многие
неродственные группы организмов, как известно, при сходных усло-
виях жизни часто приобретают внешне сходные черты и при искусст-
венной систематике часто оказывались в одном таксоне.
В настоящее время принимается следующее соподчинение таксоно-
мических единиц:
Царство — Regnum Animalia
Тип — Phylum Mollusca
Подтип — Subphylum —
Надкласс — Superclassis —
Класс— Classis Cephalopoda
Подкласс — Subclassis Ammonoidea
Надотряд — Superordo —
Отряд — Ordo (-Ida) 1 Lytoceratida
Подотряд — Subordo (-ina) Lytoceratina
Надсемейство — Superfamilia Lytoceratoidea
(-oidea) 1
Семейство — Familia (-idae) Lytoceratidae
Подсемейство — Subfamilia Lytoceratinae
(-inae)
Триба — Tribus (-ini) 1 —
Род — Genus Lytoceras
Подрод— Subgenus Biasaloceras
Вид— Species Lytoceras fimbriatum J. Sow.
Подвид — Subspecies L. fimbriatum fimbriatum J. Sow.
Для каждой группы имеются свои характерные признаки. Таксоно-
мические единицы реально существуют и важно найти критерии для
их выделения. Наиболее общими, приложимыми для всех групп, могут
служить критерии, рекомендованные В. Е. Руженцевым в качестве
принципов систематики. К их числу относятся следующие критерии,
1 В скобках указано рекомендуемое окончание. Иногда между подклассом и над-
отрядом вводятся инфракласс и когорта. Окончания для надсемейства и трибы рас-
сматриваются как рекомендации, но не являются обязательными.
70
31ли принципы: хронологический, гомологий, онтогенетический, актуализ-
ма, основного звена, хорологический.
Хронологический принцип — при выделении любой так-
сономической группы и решении вопросов филогении важно иметь по
возможности точные хронологические данные о изучаемой группе,
положении ее представителей в естественных разрезах; незнание хро-
нологии может привести к ошибкам и неправильным выводам.
Принцип гомологий основан на изучении гомологичных и
аналогичных структур, причем если гомологичное сходство проистекает
от общности происхождения, то аналогичное—от общности адаптаций
к сходным условиям. Сравнение на основе аналогичных образований
также приводит к ошибкам, к созданию искусственной систематики;
сравнение, основанное на гомологичных образованиях, позволяет выяв-
лять действительные родственные связи.
Онтогенетический принцип дает возможность прослежи-
вать развитие отдельных структур в процессе индивидуального разви-
тия, выявлять их сходство и различия. Ранние стадии онтогенеза ука-
зывают на родство более крупных таксонов и могут служить основой
для их выделения; более поздние стадии указывают на родственные
•связи низших таксонов (роды, виды). Весь ход онтогенеза показывает
направление филогенетического развития всей группы. Для выяснения
жонкретного филогенеза какой-либо группы следует особенно внима-
тельно изучать поздние онтогенетические стадии с целью отыскания
черт, характерных для ближайших предков. Для некоторых групп, на-
пример для насекомых, такой метод совсем не применим.
Принцип актуализма заключается в сравнении вымерших
животных с современными: на этом основании делаются попытки вос-
становить строение и адаптации вымерших форм. Принцип актуализма
может применяться с определенными оговорками.
Принцип основного звена базируется на выявлении тех
^изменений, которые возникают в данной группе в процессе развития и
приводят в дальнейшем к возникновению и - становлению новой груп-
пы, т. е. к выявлению узла дивергенции, места расхождения призна-
ков. Вначале возникшие отличия бывают выражены очень слабо,
затем они усиливаются и становятся ведущими. Установление основ-
ного звена — одна из наиболее сложных проблем систематики.
Хорологический принцип состоит в выявлении распреде-
.ления организмов в пространстве и их экологических особенностей, гео-
графической изменчивости.
Используя эти принципы, можно разрабатывать систематику и
•филогению исследуемой группы.
Низшей таксономической единицей является вид. Проблема опре-
деления вида всегда представляла предмет дискуссий. До выхода в свет
работы Дарвина (1859) основное внимание уделялось постоянству видов
и их четкому разграничению. Виды определялись по степени их морфо-
логического сходства и различия; однако при этом часто исследователь
сталкивался с различными затруднениями, связанными с морфологиче-
скими различиями полов (половой диморфизм), возрастными отличия-
ми — личинки часто резко отличаются от взрослых. Особые сложности
при морфологических критериях возникли при установлении с и м п а т -
рических естественных популяций, т. е. таких популяций, которые
распространены в одной и той же области, по морфологическим при-
знакам почти не отличаются друг от друга, но не скрещиваются; их
считают самостоятельными видами и называют видами-двой-
ликами.
71
В настоящее время принимается, что каждый вид состоит из грущ
пы популяций, особи которых фактически или потенциально скрещи-
ваются друг с другом и репродуктивно изолированы от других видов.
Вид представляет собой репродуктивное сообщество, объединенное эко-
логическим единством; котя он состоит из отдельных особей, но взаимо-
действует с другими видами как единое целое, обладающее генетиче-
ским единством и единым генетическим фондом. К палеонтологическому
материалу, безусловно, эти критерии непригодны и поэтому при выде-
лении видов приходится учитывать их ареалы, качественную обособ-
ленность, все морфологические признаки, проводить их морфо-
функциональный анализ и изучать изменения признаков во времени.
При достаточно больших палеонтологических коллекциях виды можно-
изучать в пространстве и во времени. О сложностях, связанных с опре-
делением видов-двойников, уже говорилось выше.
В природе довольно широко распространены политипические
виды — такие виды, которые состоят из двух или более подвидов. Если
виды не распадаются на подвиды, то их называют монотипиче-
скими. Подвид—составная часть вида, он ийеет свой ареал, кото-
рый составляет часть ареала всего вида. Подвиды всегда являются
аллопатрическими, т. е. не встречаются вместе на одной терри-
тории.
Виды объединяются в роды, роды в семейства и т. д. Большинство
исследователей считают, что высшие таксоны объективно существуют
в природе и соответствуют группам, качественно отличным друг от
друга. Это хорошо показал Дарвин, рассматривая вопросы диверген-
ции видов и вопросы систематики. Однако существует и такое мнение,
что высшие таксоны субъективны и созданы только для удобства. Эта
субъективистская точка зрения в настоящее время не находит сторон-
ников.
Правила зоологической номенклатуры. Номенклатура (лат. no-
men-— имя, calare—-называть) представляет собой систему названий
для всех таксонов. Основная задача номенклатуры состоит в создании
универсальности, стабильности и правильного однозначного понимания
одних и тех же научных названий животных. Правила зоологической
номенклатуры утверждаются на Международном зоологическом кон-
грессе и являются обязательными для всех занимающихся системати-
кой. Последний Международный кодекс зоологической номенклатуры
был утвержден в 1964 г. на Международном зоологическом конгрессе
в Лондоне. Зоологический и ботанический кодексы независимы. В ка-
честве научных названий используются латинские, греческие или лати-
низированные слова любого языка, отражающие какие-либо особен-
ности группы (например, класс Bivalvia — двустворки), ее географиче-
ское (Timanites) или стратиграфическое (neocomiensis) положение,
либо название, посвященное какому-либо лицу, реальному или мифоло-
гическому (например, orlovi, в честь академика Ю. А. Орлова или.
Neptunoceras — рог Нептуна).
Все таксономические единицы разделены на пять групп: 1) видо-
вая (вид, подвид); 2) родовая (род, подрод); 3) семействен-
ная (триба, подсемейство, семейство, надсемейство); 4) отрядно-
классная (подотряд, отряд, надотряд, подкласс, класс, надкласс);.
5) типовая (подтип, тип, надтип); Каждый таксон имеет свой тищ
своеобразный «стандарт» — справочный эталон, с которым сравнивают-
ся изучаемые формьц для устранения всяких сомнений и правильности
определений; Т и п является ядром таксона и основой его названия, он
объективен и неизменяем, но его границы или объем (кроме голотипа)
72
подвержены изменениям. Тип любого таксона не подлежит замене, за:
редким исключением. Для вида или подвида таким будет типовой-
экземпляр вида, впервые описываемый и называемый голотипо м,.
все остальные экземпляры будут считаться паратипами. При пале-
онтологических исследованиях в качестве голотипа обычно избирается
один из наиболее полных и хорошо сохранившихся экземпляров. Голо-
тип указывается при описании нового вида и не подлежит замене.
Термины «вариетет» и «форма» не подчиняются правилам номенкла-
туры и рассматриваются как категории инфраподвидовые.
Если при установлении нового вида голотип не был выделен, то-
все экземпляры типовойчсерии представляют собой синтипы, равно-
ценные в номенклатуре. Из этой серии любой систематик может выде-
лить один из синтипов в качестве лектотипа. Если голотип, лекто-
тип или синтипы утеряны или уничтожены, то любой другой экземпляр
может быть выделен в качестве не.отипа с соблюдением всех необ-
ходимых правил.
Для рода выбирается номинальный вид, именуемый типовым
видом, для семейства-—тот номинальный род, на котором основано-
название семейства. Все члены семейственной группы, основанные на
одном типичном роде, — триба, подсемейство, семейство, надсемейст-
во — пишутся с фамилией автора, давшего впервые название одному из
перечисленных таксонов. Например, семейство Hoplitidae было выделе-
но в 1890 г. Дувийе (Н. Douville), а в 1952 г. Райт (С. Wright) раз-
делил его на три подсемейства: Cleoniceratinae Whitehouse, 1926,
Hoplitinae Douville, Gastroplitinae Wright, 1952; автором номинального-
подсемейства остался Н. Douville.
Названия отрядно-классной и типовой групп отличаются тем, что-
они не привязаны к определенному типу, представляют собой отдель-
ные слова латинского или греческого происхождения, всегда стоят во
множественном числе (например, Primates—-приматы). Для названий
от трибы до отряда предложено к названию рода добавлять соответ-
ствующие окончания, указанные в скобках при перечислении таксонов,
(см. стр. 70).
Закон приоритета предусматривает признание только того назва-
ния вида или рода, которое было предложено первым по времени и
опубликовано с соблюдением всех правил, предусмотренных кодексом.
Все последующие названия считаются синонимами первого и не
употребляются в качестве самостоятельных. Если одно и то же назва-
ние дано двум разным видам внутри одного -рода или разным родам,
оно считается гомонимом; более позднее тождественное название
является недействительным и должно быть отброшено. Например,
Noctua—насекомое и Noctua — птица, одно из названий должно быть
изменено.
Если название не соответствует правилам кодекса, то оно считает-
ся недействительным и не имеет номенклатурного статута, т. е. пред-
ставляет собой по номенклатурным правилам nomen nuda (или nomina
nuda во множественном числе).
Все названия таксонов выше вида состоят из одного слова, т. е.
являются униноминальными; все названия видов, как было-
показано выше, состоят из двух слов, т. е. биноминальны; все
названия подвидов состоят из трех слов и являются триноминаль-
ными, Названия видов и подвидов пишутся со строчной буквы, на-
звания всех, вышестоящих таксонов — с прописной. В политипическом
виде один из .подвидов является но минальным, т. е. носителем
названия. Например, если вид назван album, то один из подвидов, дол1
73
жен называться album album, а название второго подвида будет со-
стоять из названия вида album и какого-нибудь слова, подчеркиваю-
щего особенности выделенного подвида.
В соответствии с кодексом названия вида и подвида должны быть
грамматически согласованы с названием рода. Прилагательное albus,
означающее белый, сохраняет окончание -us, если оно относится к муж-
скому роду, изменяет окончание на -а, если род женский, и на -ит,
если род средний. Подбор видовых названий и согласование их с на-
званием рода вызывают часто много затруднений, связанных с незна-
нием латинского языка, незнанием рода (Мужского, женского или
среднего), к которому относится название.
Если в процессе изучения будет установлено, что вид должен быть
отнесен к другому роду, то в этом случае фамилия автора ставится в
скобки. Однако это рекомендуется делать только в специальных
палеонтологических работах. Если род разделяется на подроды, то
подрод, содержащий типовой вид, становится типовым, или номиналь-
ным, подродом и сохраняет название рода, а второй и следующие
получают новые названия, при этом название подрода ставится в Скоб-
ки после названия рода. Например, род Hoplites разделен на два
подрода — Hoplites {Hoplites), Hoplites {Isohoplites).
Открытая номенклатура применяется в том случае, если
сохранность материала плохая и не позволяет дать точное видовое
определение. Название «открытая», - или «свободная», номенклатура
связано с тем, что описанные формы не попадают под охрану закона
приоритета и их названия при последующих исследованиях могут
Уточняться или изменяться. Существует много различных обозначений,
применяемых при описании или определении материала плохой сохран-
ности. Вот некоторые примеры: если принадлежность к роду недосто-
верна, то после названия рода ставится знак вопроса; если сохран-
ность не позволяет достоверно определить вид, то ставится cf. (сокра-
щение от слова conformis — сходный); если описываемый вид имеет
достаточно хорошую сохранность, но отличается от близкого вида
какими-то признаками, не позволяющими с уверенностью относить
экземпляр к данному виду, то между названием рода и вида ставится
знак aff. (сокращение от слова affinis — родственный, близкий). Если
исследователь не может точно определить вид, то он указывает только
на принадлежность описываемой формы к группе известных видов и в
этом случае между названием рода и вида ставит знак ex gr., что озна-
чает ex grege — из группы (дословно из стада). Например, Nautilus
ex gr. pompilius L. Если не может быть установлена принадлежность
изучаемых таксонов к семейству, отряду, классу, типу, то пишут соот-
ветственно incertae familiae, incertae ordinis, incertae classis, incettae
phylum (incertae — неизвестный).
ПАЛЕОНТОЛОГИЧЕСКИЙ МЕТОД И ОСНОВЫ
СТРАТИГРАФИЧЕСКОЙ КЛАССИФИКАЦИИ
Основным методом определения относительного возраста пород
является палеонтологический метод. Его применение базируется на
дивергентном поступательном развитии органического мира и принци-
пе необратимости эволюции, установленных Дарвином (1859).
Стратиграфическое расчленение осадочных пород, развитых в дан-
ном регионе, начинается со всестороннего изучения естественных обна-
74
жений и кернов скважин, вскрывающих эти отложения в закрытых
районах. Изучается литологический состав отдельных слоев, их взаимо-
отношения друг с другом, причем принимается, что при ненарушенном
залегании подстилающий слой является более древним, а покрываю-
щий более молодым (принцип Стенона). Если между слоями нет
«следов перерыва, то их формирование шло последовательно и непрерыв-
но; если же между ними наблюдается так называемое стратиграфиче-
ское несогласие, то предполагается наличие перерыва в осадконакопле-
нии, а также возможность размыва нижележащих слоев. В таких
случаях важно установить размеры перерыва. О следах перерыва сви-
детельствует неровная, эродированная поверхность нижнего -слоя, на-
личие в подошве вышележащего слоя грубообломочного материала —
гравия, гальки, желваков фосфоритов, остатков переотложенных скеле-
тов различных организмов. В кровле нижнего слоя наблюдаются следы
-зарывающихся и сверлящих организмов: норы ракообразных, сверля-
щих и зарывающихся Двустворок, заполненные осадком вышележащего
слоя. Если между слоями наблюдается угловое несогласие, то это
объясняется тем, что подстилающие породы до образования вышележа-
щих подвергались воздействию тектонических движений, в результате
которых их первоначальное горизонтальное залегание было изменено.
Из каждого слоя или группы слоев изучается систематический
•состав ориктоценозов. Для этого извлекаются органические остатки —
фоссилии, которые после технической обработки определяются. При
извлечении из слоев ископаемых отмечаются особенности их захороне-
ния, распределение в слое, количественные взаимоотношения между
отдельными видами, проводятся палеоэкологические наблюдения,
выясняется их прижизненное положение, принадлежность к различным
экологическим группам. Для выяснения наличия спор, пыльцы и микро-
фоссилий отбираются образцы для дальнейшей технической обработки.
Предварительный анализ фоссилий в полевых условиях позволяет
«отличать отложения, образованные в морских или континентальных
условиях, в лагунах или пресноводных бассейнах. Однако для «немых»
толщ, не заключающих органических остатков, решить этот вопрос
иногда бывает очень трудно.
На основании изучения естественных обнажений и прослеживания
по простиранию слоев или пачек слоев составляется сводный страти-
графический разрез изучаемого региона. Для обоснования возраста
-слоев, выделенных в разрезе,- определяются органические остатки:
выясняются их систематическая принадлежность, их ареалы и время
существования. Если сохранность материала не позволяет определить
«органические остатки до вида, то тогда, в зависимости от сохранности
материала и опытности палеонтолога, определение дается до рода, до
семейства и т. д. (в открытой номенклатуре). В тех случаях, когда
встречены совершенно новые формы, они описываются как новые виды,
роды. Крайне важным для установления возраста пород, из которых
извлечены органические остатки, является их анализ с точки зрения
биостратиграфического значения каждой из изученных форм. Отдельно
анализируются остатки растений (отпечатки листьев, остатки плодов,
семян, споры и пыльца), позвоночных и беспозвоночных. Среди морских
форм выделяют организмы бентоса, нектона и планктона, выявляют
«формы, широко распространенные (космополиты), и формы, развитые
только в данном регионе (эндемичные); формы, жившие на месте
•находок (автохтонные — кораллы, губки и другие прикрепленные
«формы) дли попавшие в ориктоценоз из других биоценозов (аллох-
тонные).
75
На основании определения состава организмов, живших в изучае-
мый отрезок времени, и выяснения их стратиграфического значения;
выделяются отдельные стратиграфические единицы, или стратоны,
и обосновывается их возраст. Дробность стратиграфических подразде-
лений зависит от масштаба геологосъемочных работ и масштаба карт,,
которые необходимо составить, от задач, стоящих перед тематическими
исследованиями, а также от обилия и сохранности палеонтологического
материала.
Прежде чем остановиться на значении некоторых групп беспозво-
ночных для целей стратиграфии, следует напомнить, что палеонтологи-
ческий метод впервые применил У. Смит при составлении шкалы
стратиграфических образований Англии (1799) и при составлении им
первой геологической карты Англии, Уэльса и юга Шотландии (1815)..
Эту возможность использования ископаемых органических остатков-
для выделения отдельных формаций вслед за Смитом применили
Броньяр и Кювье при изучении разрезов Парижского бассейна.
Важное значение для разработки основ стратиграфической и гео-
хронологической шкал имело предложение первого профессора
Оксфордского университета Англии Бакленда (1818) о выделении:
соподчиненных стратиграфических единиц. Бакленд, подобно Линнею,,
разделившему весь животный и растительный мир на соподчиненные
таксоны, предложил следующую иерархию стратиграфических единиц:
класс, порядок, формация, слой. Класс рассматривался как высшая
стратиграфическая единица, слой — как низшая. В частности, три слоя:
серый мел, нижний мел и верхний мел объединялись в формацию «мел»,
которая относилась к порядку «горизонтальному», т. е. пород, залегаю-
щих горизонтально, и к классу «вторичных пород», содержащих орга-
нические остатки и фрагменты более древних пород. Для каждого слоя
кроме названия указывались литологический состав, мощность и наи-
более типичные' места распространения. В дальнейшем эти названия
стратиграфических единиц изменились, но принцип выделения остался.
В это же время (1820) в Англии было предложено объединять форма-
ции в системы. За сравнительно короткий срок (1818—1841 гг.) в За-
падной Европе были выделены все системы, принятые в настоящее-
время.
В 1841 г. Д. Филлипс предложил объединить системы в такие
единицы, как палеозой, мезозой и кайнозой. Возникла новая отрасль
геологии — стратиграфия, развитие которой сыграло важную
роль для становления палеонтологии. Перед стратиграфией были
поставлены две взаимно связанные задачи: 1) расчленение разреза и.
выделение соподчиненных стратиграфических единиц, 2) внутрирегио-
нальная и межрегиональная корреляция выделенных стратонов. В соот-
ветствии с господствующим в первой половине XIX в. мнением о по-
стоянстве и неизменяемости видов предполагалось, что для каждой
формации и системы характерны свои фауна и флора, которые в конце
каждого периода уничтожались, а в начале каждого последующего-
создавались заново.
Известный французский палеонтолог и стратиграф д’Орбиньи
(1802—1857) в 1842 г. в своей крупной работе «Палеонтология Фран-
ции» предложил разделять систему на ярусы — стратиграфические
подразделения, объединяющие группы слоев, часто разного литологи-
ческого состава, но заключающие характерную для каждого яруса
фауну. Каждый ярус д’Орбиньи назвал, в соответствии с идеей Смита,
по местонахождению типичного разреза: например, оксфордский ярус
юрской системы был назван по месту его развития вблизи г. Оксфорда,
76
Англия; барремский и аптский ярусы меловой системы названы по
местам их развития в одноименных деревнях во Франции — Баррем и
Апт. Вначале ярусное деление было предложено для меловой и юрской
систем. В дальнейшем ярусы были выделены во всех системах, однако
для некоторых систем до сих пор нет общепринятого ярусного деления
(например, для кембрия,' палеогена, неогена).
Значительные трудности возникают при применении схемы ярус-
ного деления, разработанной для морских отложений при изучении
континентальных пород, в которых встречены наземные организмы. Так,
для морских девонских отложений выделение ярусов основано на эта-
пах развития аммоноидей и брахиопод, а для водных-континентальных
бассейнов — на этапах развития бесчелюстных — остракодерм и пан-
цирных рыб (плакодерм). Ярусная схема для морских отложений резко
-отличается от ярусной схемы континентальных пород и их сопоставле-
ние представляет обычно большие трудности.
В 1856—1858 гг. немецкий палеонтолог и стратиграф Оппель
(1831—1865) предложил ярусы разделять на более мелкие стратигра-
фические единицы — зоны, отмечая, что каждая зона содержит
характерные виды, отличные от видов, встречаемых в выше- и ниже-
лежащих слоях. Оппель предложил первое зональное деление для
юрских отложений Западной Европы и рекомендовал каждую зону
называть по карактерному виду, или «виду-индексу».
Палеонтологический метод казался в то время простым и практи-
чески удобным, так как расчленение и увязка, разрезов проводилась на
сравнительно небольшой территории Западной Европы. Для геологов
первой половины XIX в. ископаемые организмы были только свидете-
лями относительного возраста определяемых слоев и еще не стали
звеньями непрерывной цепи жизни,, которыми их сделала эволюцион-
ная теория Дарвина. Благодарен усилиям Дарвина и его последователей
палеонтология превратилась в эволюционную науку; органические
остатки стали звеньями цепи жизни на Земле. Однако Дарвин отметил,
что геологическая летопись несовершенна и отличается неполнотой;
значительная часть памятников истории жизни на Земле была разру-
шена и исчезла бесследно. Тем не менее имеющиеся палеонтологиче-
ские документы представляют большую ценность для расчленения и
корреляции разрезов. Фауна из отложений любого стратона по уровню
организации всегда будет промежуточной между фаунами предшест-
вующего и последующего стратонов. Это достаточно четко выявляется
по различным группам беспозвоночных, но особенно выразительно при
сравнении фаун позвоночных. Особое внимание Дарвин обратил на
кажущееся внезапным появление в кембрии разнообразных предста-
вителей животного мира, объясняя это не внезапностью, а метаморфиз-
мом докембрийских пород либо недоступностью их для наблюдений.
Не менее важной проблемой стала после открытия и освоения новых
материков и земель проблема геологической синхронизации отложений,
развитых на удаленных территориях, но относимых к одним и. тем же
стратиграфическим подразделениям.
Во второй половине XIX в. был накоплен большой фактический
материал по стратиграфии и фауне, и на первой сессии Международ-
ного геологического конгресса (МГК), состоявшейся в Париже в 1878 г.,
была создана комиссия по унификации стратиграфической номенкла-
туры. Результаты работы комиссии были рассмотрены второй сессией
МГК в Болонье (1881) и была принята схема стратиграфических под-
разделений и их хронологических эквивалентов. На восьмой сессии
МГК в Париже в 1900 г. в основу стратиграфической классификации
7.7
были положены подразделения геологического времени, а подразделе-
ния осадочных отложений рассматривались как эквиваленты этих вре-
менных подразделений. Принятая сессией схема двойной классификации
получила в дальнейшем всеобщее признание:
Геохронологические Стратиграфические
подразделения подразделения
Эра —
Период , Система
Эпоха Серия (Отдел)
Век Ярус '
— Зона
В этой схеме не было эквивалента эре. В нашей стране им стала
группа, а для эпохи — отдел. Для зоны не было принято геохронологи-
ческого эквивалента, была рекомендация употреблять момент, фаза,
но эти термины не получили распространения. Из классификации вид-
но, что для указания времени существования той или иной группы
организмов должны быть использованы временные категории (напри-
мер, трилобиты появились в раннем кембрии и вымерли в середине
пермского периода), но при указании о находках в соответствующих
слоях используются стратиграфические единицы («остатки трилобитов
встречены в отложениях нижнего кембрия»). Таким образом, эти две
шкалы взаимно связаны: одна указывает на время, другая — на отло-
жения, накопившиеся за соответствующий отрезок времени.
В областях и районах, где не могут быть выделены единицы
международной стратиграфической шкалы, устанавливают местные
стратиграфические подразделения: Серию, свиту, пачку. Свита
объединяет комплекс отложений либо однородных в литологическом
отношении, либо представляющих более или менее однообразное чере-
дование определенных типов пород, накопившихся в течение определен-
ного этапа развития данного региона. При характеристике свиты на
первое место выдвигаются литологические признаки, а затем, если
имеются органические остатки, дается палеонтологическая характерис-
тика. Свиты объединяются в серии и в свою очередь разделяются на
пачки. Серия рассматривается как крупное литолого-стратиграфиче-
ское подразделение, объединяющее несколько свит. Все местные лито-
стратиграфические единицы выделяются как определенные комплексы
осадочных, магматических и метаморфических пород, ясно отграни-
ченные от смежных комплексов, литологически легко опознаваемые в
поле, имеющие достаточно широкое площадное распространение.
Осложнения, возникающие при применении названий вспомога-
тельных единиц, связаны с тем, что в странах, говорящих на англий-
ском языке, для литостратиграфических единиц приняты: группа, фор-
мация, член (member) и слой. Термин «серия» используется в качестве
эквивалента русского понятия «отдел», а «группа» в качестве эквива-
лента «серии».
В последние годы в связи с накоплением нового фактического ма-
териала вновь значительно повысился интерес к вопросам стратиграфи-
ческой классификации и номенклатуры. Американские стратиграфы
предложили выделять три основные категории стратиграфических под-
разделений: литостратиграфические, биостратиграфические и хроно-
стратиграфические.
Литостратиграфические единицы выделяются по литологическим
признакам и закономерностям накопления осадков.
78
К биостратиграфическим единицам в настоящее время относится
только зона, остальные подразделения рассматриваются как просто
стратиграфические. Фактически к биостратиграфическим единицам
должны быть отнесены все стратиграфические единицы.
К хроностратиграфическим подразделениям относятся^такие, кото-
рые объединяют отложения, образованные в течение определенного
отрезка времени, и границы между ними представляют изохронные
(одновозрастные) поверхности, не зависящие от литологического
состава отложений и их палеонтологической характеристики, т. е. пред-
ставляют лишь отвлеченное понятие. Поэтому выделение хронострати-
графических подразделений как таковых излишне. . •
В соответствии с рекомендациями геологического конгресса жела-
тельно выделять следующие подразделения:
Геохронологические
эон фанерозой
эра мезозойская
период меловой
эпоха ранняя
век берриас
время dalmasi
Стр атигр а фические
эонотема фанерозой
эратема мезозойская
система меловая
отдел нижний
ярус берриас
зона Dalmasiceras:
dalmasi
При написании названия яруса в разных странах приняты следую-
щие окончания:
Язык Окончание Пример
русский нет баррем, Оксфорд
английский — ian barremian, - oxfordian
французский — ien barremien, oxfordien
немецкий — ium barremium, oxfordium
испанский — iense barremiense, oxfordiense
итальянский — iano barremiano, oxfordiano
В настоящее время выделены пять эр: архейская, протерозойская,
палеозойская, мезозойская и кайнозойская. Первые две образуют эон
криптозой, последующие три — эон фанерозой.
Архей и протерозой выделяются по данным абсолютного возраста
и по крупным тектоно-магматическим циклам. Для расчленения про-
терозоя используются строматолиты и онколиты — известковые продук-
ты жизнедеятельности сине-зеленых водорослей. Продолжительность
криптозоя составляет около 4 млрд, лет, в то время как длительность
фанерозоя не превышает 570 млн. лет. Радиологические методы (калий-
аргоновый, рубидий-стронциевый, уран-торий-свинцовый) дают цифры
абсолютного возраста пород. Однако к этим цифрам следует под-
ходить с большой осторожностью, так как допускаемые ошибки бывают
большими (табл. 1).
Существенная перестройка структуры земной коры, которая
произошла на рубеже 2600+100 млн. лет тому назад, служит границей
между археем и протерозоем. Отложения с возрастом от 2600±100 до
1700 + 50 млн. лет отвечают второму крупному этапу развития земной
коры, который относится к раннему и среднему протерозою; наконец,
79
последний этап в докембрии выделяется как поздний протерозой, или
р и ф е й, от 1700±50 до 570—600 млн. лет. Терминальная часть рифея
известна под названием венда, в отложениях которого были обнару-
жены первые бесскелетные отпечатки многоклеточных организмов.
Наиболее богатым местонахождением вендского комплекса фауны яв-
ляется в настоящее время местонахождение Эдиакара в Южной Австра-
лии, открытое в 40-х годах нашего столетия. В Эдиакаре .обнаружены
остатки губок, кишечнополостных, червей, членистоногих, иглокожих.
Фанерозойский эон раз-.
делен на три эры: палеозой-
скую, мезозойскую и кайно-
зойскую (см. табл. 2). В на-
стоящее время предлагается
палеозойскую эру разделить
на две самостоятельные:
собственно палеозойскую и
метазойскую (Друщиц, Ши-
манский, 1962). Эры были
выделены на основании из-
менения таких крупных так-
сонов, как отряды, подклас-
сы и даже классы; перио-
ды — на основании измене-
ний состава менее крупных
таксонов — отрядов, над-
семейств. Для расчленения
отложений каждого периода
используются наиболее бы-
стро эволюционирующие
группы организмов. Для
разделения кембрия важ-
ную роль играют трилоби-
ты, для ордовика и силу-
ра,— граптолиты, для дево-
на .и для мезозоя — аммо-
ниты, для палеогена и неоге-
Рис. 23. Возможные варианты распростране-
ния видов (родов) в соответствующих зонах
(Жемчужников, 1934, с изменениями):
п — появление, р — расцвет, в — вымирание
(объяснение см. в тексте)
на — фораминиферы, двустворки и брюхоногие моллюски.
На основании развития этих же групп животных выделяются
зоны — наиболее дробные стратиграфические единицы. Объем и гра-
ницы зоны устанавливаются по распространению одного или несколь-
ких характерных видов, определяющих название зоны. В отложениях
любой зоны могут встречаться остатки разных категорий организмов
(рис. 23): 1) жившие только в течение хронозоны, 2) появившиеся в
предыдущей хронозоне и испытавшие расцвет в изучаемый отрезок вре-
мени, 3) появившиеся в данной хронозоне, достигшие расцвета, но про-
должающие свое существование и дальше, 4) появившиеся в данной
хронозоне, но испытавшие расцвет позднее, .5) появившиеся ранее, ха-
рактерные для данного времени, но вымирающие позднее, 6) появляю-
щиеся на рубеже двух зон, 7) доживающие свой век в данной хронозо-
не, 8) транзитные, существующие в течение относительно длительного
времени, 9) вымершие в предыдущее время, 10) появляющиеся в сле-
дующей зоне. Для выделения зон наиболее важное значение имеют виды,
относимые к 1—3-й, 5-й группам (виды и роды).
Но кроме короткого времени существования для широких меж-
региональных корреляций важно, чтобы эти организмы обладали
80
и крупными ареалами; при малых ареалах выделяются только местные
зоны, или л о н ы (т. е. локальные зоны, название, предложенное не-
давно Г. Я. Крымгольцем). К числу организмов, имеющих большие
ареалы, относятся планктонные и нектонные организмы (например,
аммониты, белемниты, граптолиты, планктонные фораминиферы).
Ареалы многих видов обычно ограничены палеобиогеографическими
областями или провинциями. Поэтому зоны, выделенные в одной
палеозоогеографической области, обычно не могут быть распростра-
нены на другие области, и при корреляции одновозрастных зон, выде-
ленных в разных биогеографических областях, возникают значительные
трудности. В некоторых случаях помогают виды-космополиты. Еще
большие трудности возникают при корреляции морских и континен-
тальных отложений. Здесь на помощь приходит изучение спор и
пыльцы и наземных растений, часто заносимых в водные бассейны, изу-
чение разрезов в промежуточных зонах, в которых наблюдается чере-
дование морских и континентальных отложений.
Для одного и того же региона часто предлагают зональное деление
по разным группам. Естественно, что зоны, выделенные по одним груп-
пам, не будут совпадать с зонами, выделенными по другим, поскольку
скорости эволюции и закономерности развития у разных групп различ-
ны и обычно не совпадают. Все это служит основанием для дискуссии
практически по любой границе между стратонами. Чтобы достичь
однозначного решения о границах, правильнее будет проводить зональ-
ное деление по одной ведущей группе и рассматривать это деление как
эталонное, а развитие других групп животных контролировать и срав-
нивать с этапами развития эталонной группы. Для каждой зоны,
яруса избирается стратотипический разрез, или стратотип, который
должен быть наиболее полным, без перерывов, с верхней и нижней
границами и, самое главное, с остатками ведущей группы животных.
Наряду с зонами как стратиграфическими подразделениями выде-
ляются так называемые биозоны. Биозоны не представляют собой
единиц ни стратиграфических, ни геохронологических подразделений.
Биозоной называются отложения, отвечающие полному распростра^
нению какой-либо одной определенной систематической группы орга-
низмов (вида, рода, семейства, класса и т. д.-) (Степанов, 1958). Ин-
тервал времени, соответствующий биозоне и отвечающий полному
распространению любой группы, называют биохроном. Время
максимального развития группы носит название гемера (гр. heme-
ros — день, время); отложения, накопленные в течение гемеры, — эпи-
боль. Например, биозона аммонитов охватывает девон — мел, а их
гемера относится к юре и мелу. Предложено также выделять т е й л ь -
хрону — время распространения вида в данном районе — и тейль-
зо ну — слои, накопленные за это же время. Тейльзона и тейльхрона
в разных районах, как правило, не совпадают, так как существование
каждого вида в каждом регионе обусловлено определенным комплексом
условий и их изменение может приводить либо к полному исчезновению
вида, либо к его миграции в соседние районы. Виды бентоса особенно
чувствительны к изменениям фаций, в то время как нектонные и планк-
тонные формы от них не зависят. Представление о биозоне любого
таксона складывается на основании анализа данных о тейльзонах, а
представления о биохроне — на основании местных тейльхрон. Для
палеонтолога крайне важно знать как биохрону, так и тейльхрону
каждого таксона на всем ареале и на отдельных его участках.
Наиболее короткий период — четвертичный; его длительность оцени-
вается по последним данным в 1,7 млн. лет, т. е. не превышает продол-
6
В. В. Друщиц
81
жительности одной зоны остальных периодов; поэтому он не разде-
ляется на отделы и ярусы.
Применение палеонтологического метода в стратиграфии часто
осложняется по различным причинам: переотложение органических
остатков, эндемизм фаун, реликтовые и реккурентные формы.
Переотложение органических остатков и нахождение их во
вторичном залегании очень широко распространены в природе. Как
правило, переотложение возникает в результате размыва подстилаю-
щих пород, и переотложенные органические остатки чаще всего при-
урочены к базальному горизонту. В нем может встречаться галька,,
обломки и целые глыбы палеонтологически охарактеризованных пород,
из более древних отложений. Так, в базальном горизонте альба (ниж-
ний мел) центральной части Крыма встречены глыбы известняков с
фауной баррема и отдельные переотложенные раковины аммонитов,
нижнего и верхнего апта. Таких примеров можно привести довольно-
много.
Эндемичные формы обладают ограниченными ареалами и
могут быть использованы только для ограниченных районов; к ним
также относятся только что установленные новые виды, известные-'
вначале локально. Примером эндемичных фаун служит фауна Байка-
ла, состоящая на 75% из эндемичных форм, фауна ряда островов (на-
пример, Галапагосских островов, блестяще исследованных Ч. Дарви-
ном), долин рек, горных областей. Эндемизм чаще всего связан с изо-
ляцией. Следует помнить, что эндемизм может быть обусловлен не-
только изоляцией и древностью форм, но и молодостью форм, только
что возникших недавно в пределах области и не успевших распростра-
ниться.
К реликтовым, или суперститовым, формам относятся
древние формы, живущие среди более молодых; например, современ-
ные: неопилина галатея — последний представитель класса моноплако-
фор, кистеперая рыба — латимерия, последний представитель мезо-
зойских целекантид. Находки подобных форм в ископаемом состоянии
могут привести к ошибкам при определении возраста.
К реккурентным формам относятся одни и те же формы,,
которые появляются в различных стратиграфических горизонтах в связи:
с повторением в разрезе благоприятных для них фаций. Реккуренция
связана с миграцией фауны при возникновении неблагоприятных усло-
вий для ее обитания и возвращением ее при восстановлении условий*
близких к прежним. Реккурентные фауны чаще всего наблюдаются в.
условиях ритмичного чередования слоев, например в Донбассе морские
фауны повторяются в прослоях известняков, чередующихся с континен-
тальными угленосными отложениями. Фаунистические комплексы дома-
никовой фации (битуминозные известняки) повторяются в живетском,.
франском и фаменском ярусах девона на востоке Русской платформы
и в Приуралье, что вначале затрудняло в каждом отдельном случае
определение их возраста.
Таблица 1
Деление докембрия в СССР
Эон Эра Подэра, период нижняя граница в млн. лет Развитие жизни
ФАНЕ- РОЗОЙ ПАЛЕО- ЗОЙ ранняя, кембрий 570 ±30 «Взрыв ископаемых» — появление орга- низмов, способных строить скелет; увели- чение в атмосфере содержания кислорода и уменьшение углекислоты
венд=терми- нальный рифей Находки беескелетных многоклеточных: кишечнополостных, кольчатых червей, чле- нистоногих, первых иглокожих (фауна Эди- акары), погонофор, петалонам (формация Нама)
0R0-I-20
S О 00 рифей* поздний 950-4-50 Увеличение содержания кислорода в ат- мосфере; дивергенция многоклеточных; мик- рофлора с клеточной структурой из форма- ции Биттерспринг (Австралия)
ТЕРО средний 1350-1-50 Предполагаемое появление многоклеточ- ных (1 млрд.)
S о 00 о ПРО ранний Возникновение одноклеточных; развитие строматолитов и микрофитолитов
с S а. средняя* 1ОЛЛ_1_1ЛП • Появление кислорода в атмосфере (1,8 млрд.). Развитие бактерий и водорос- лей
ранняя 97ЛЛ-1-1ЛЛ Бескислородная атмосфера; молекулярные ископаемые; первые ископаемые остатки из формаций Ганфлинт (2,0 млрд.) и железо- рудной Соуден (2,7 млрд.)
щ X п. поздняя ЯЗЛЛ 3500 Химическая эволюция; молекулярные ис- копаемые и биогенные отложения из сис- темы Свазилэнд (Южная Африка, 3— 3,2 млрд.); возникновение первых прокари- от
ранняя (катархей) 4500 Химическая эволюция; химическое разно- образие, морфологическая простота
* М. А. Семихатов (1972) предлагает рассматривать в качестве эр рифей и афебий,
последний объединяет ранний и средний протерозой, а протерозой поднять до ранга эона,
противопоставляя его фанерозою и архею.
6!
83
Таблица 2
Основные геохронологические и стратиграфические подразделения
Эон Эоно- тема Эра Эра- тема Период Система Длитель- ность, млн. лет Эпоха Отдел Век ярус
ФАНЕРОЗОЙСКИЙ КАЙНОЗОЙСКАЯ KZ 67—70 млн. лет четвертичный Q 1,7 общепринятых ярусов нет хаттский p3h рупельский РзГ
неогеновый N (25±1) палеогеновый Р ZR7 । QA 25 41 плиоцен Na
миоцен Nt олигоцен Рз
эоцен ₽2 приабонский лютетский ипрский РгР ?2lt ?2i
палеоцен ₽х танетский Pit МОНСКИЙ Р1ГП '
МЕЗОЗОЙСКАЯ MZ 165—170 млн. лет ^0*' ‘ меловой к /197 i 70 поддняя Кг верхний датский K2d маастрихтский К2Ш кампанский К2ср сантонский K2st коньякский К2сп туронский K2t сеноманский К2ст
ранняя Kt нижний альбский Kjal аптскнй Kiap барремский Kib готеривский Kih. . валанжинский KjV берриасский Kjbs
Д1О / —т~О ) юрский J 55—58 поздняя J3 верхний титонский J3t волжский J3v кимериджский J3km оксфордский J3ox келловейский J3kl
средняя J2 средний батский J2bt байосский J2bj ааленский J2al
ранняя Л. нижний тоарский Jit плинсбахский Jip синемюрский JiS геттангский Jih
1 »JO 1— о у триасовый Т (230±10) 40—45 поздняя Т3 верхний рэтский Т3г норийский Т3п карнийский Т3к
средняя т2 средний ладинский Т21 анизийский Т2а
ранняя Т1 нижний оленекский Тхо индский Txi
84
Продолжение табл. 2
Эон Эоно- тема Эра Эра- тема Период Система Длитель- ность, млн. лет Эпоха Отдел Век Ярус
ФАНЕРОЗОЙСКИЙ ПАЛЕОЗОЙСКАЯ PZ 310—385 млн. лет РАННЯЯ 1 ПОЗДНЯЯ пермский Р 12R4-4-101 45 ПОЗДНЯЯ р2 верхний татарский P2t казанский Р2к уфимский Р2п
ранняя Pi нижний кунгурский Pikg артинский Рха сакмарский Pxs ассельский Pjas
каменно- угольный С 13.40+101 65—70 поздняя С3 верхний гжельский C3g касимовский С3к
средняя с2 средний московский С2ш башкирский С2Ь
ранняя Ci нижний намюрский С]П визейский C]V турнейский Cjt
девонский D :—(405±10)— силурийский S 144 0-4-1.41 55—60 30—35 поздняя D3 верхний фаменский D3fm франский D3fr
средняя d2 средний живетский D2g эйфельский D2e
ранняя Di нижний поздняя Sa верхний эмский D]em ; зигенский DiS жединский Dxg пржидольский S2p (даунтбнский) лудловский S2ld
ранняя Si нижний венлокский SjW лландоверийский SjJn
ордовикский 0 1400-1-1.41 60—70 поздняя Оз верхний ашгильский О3а карадокский ' О2_3с лландейльский О21 лланвирнский О21п
средняя о2 средний
ранняя Ох нижний аренигский О2а тремадокский Ojt
кембрийский е (570+30) 70—80 поздняя €з верхний общепринятых ярусов нет
средняя €2 средний майский 62т амгинский 62а ’
ранняя €1 нижний ленский 6J алданский 6ха
85’
Таблица 3
Схема ярусного деления неогена
Система Отдел 1 Ярусы
Центральный Паратетнс (197il) нижняя граница в млн. лет Юг СССР (1973)
АНТРО- ПОГЕН плейстоцен апшерон*
ПАЛЕОГЕН] НЕОГЕН 1 ПЛИОЦЕН Румынии (Крейчи-Граф, 1932) акчагыл (Андрусов, 1912) — куяльник (Михайловский, 1909)
ДИКИЙ (Тейссер, 1907) киммерий (Андрусов, 1907—1908)
ПОНТ ПОНТ (Барбот де Марии, 1869)
ОЛИГОЦЕН] МИОЦЕН паннон (Рот, 1879) ±10—11 мэотис (Андрусов, 1890)
сармат (э. Зюсс, 1866) ±14 сармат (Барбот де Марии, 1866)
бадений (Папп, Цнха, 1968) ±19 КОНКИ (Андрусов, 1917)
караган (Андрусов, 1917)
чокрак (Андрусов, 1884)
карпатий (Циха, Тейкал, 1959) ±22,5 тархан (Андрусов, 1918)
оттнангий (Папп, Рогль, 1968) ±24,5 КОцахур (Давиташвили, 1933)
эггенбургий (Штейнингер, Сенеш, 1968) ±26,5 сакараул (Давиташвили, 1933)
Эгерий (Балди, Сенеш, 1968) ±31 «аквитан» (Майер, 1857)
хаттский верхний олигоцен
* В СССР апшерон пока отнесен к плиоцену.
86
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТИП PROTOZOA. ПРОСТЕЙШИЕ
Общая характеристика. К типу простейших (гр. protos — первый,
zoon — животное)" относятся мельчайшие животные (протозооны),. тело
которых состоит' из одной клетки, обладающей либо одним простым
(моноэнергидная организация), либо сложным (полиплоидным) ядром,
либо многими простыми ядрами (полиэнергидная организация) при
неразделенной протоплазме. Некоторые одноклеточные образуют
колонии, возникающие вследствие нерасхождения моноэнергидных,
дочерних, клеток при бесполом размножении. И, наконец, ряд простей-
ших достигает состояния, близкого к многоклеточности: их тело со-
стоит из соматических и половых клеток.
Простейшие в физиологическом отношении представляют целост-
ный организм на клеточном уровне организации, приспособленный к
самостоятельной жизни. Простейшие обладают гетеротрофным типом
питания, ведут преимущественно подвижный образ жизни и размно-
жаются половым и бесполым путем. В ископаемом состоянии известны
начиная с кембрия.
Строение тела. Протозоон является совершенно самостоя-
тельным организмом, выполняющим все жизненные функции. Его тело
состоит из цитоплазмы и ядра, представляющих собой единую и неде-
лимую структурную и функциональную систему, В цитоплазме
различают три типа структур: основу, органоиды и включения
(рис. 24).
Основу образует эндоплазматическая сеть, состоящая из слож-
ной системы мембран — тончайших канальцев, заключенных в бес-
структурную массу. На мембранах расположены многочисленные гра-
нулы, содержащие рибонуклеиновую кислоту (РНК). На эндоплазмати-
ческой сети осуществляется синтез белков; она связана с наружной
оболочкой ядра и наружной мембраной клетки.
Органоиды, или органеллы, представляют собой отдельные
участки клетки, приспособленные к выполнению различных функций,
связанных с обменом веществ, синтезом белка, движением, размно-
жением. К органоидам относятся: митохондрии, лизосомы, аппарат
Гольджи, клеточный центр.
Митохондрии (гр. mitos — нить, chondros — зерно) представ-
ляют собой палочковидные структуры размером от 0,5 до 7 мк. Основ-
ная их функция заключается в освобождении энергии, поступающей в
89
клетку с питательными веществами, и преобразование этой энергии в
формы, пригодные для процессов синтеза и осуществления работы
клетки: механической — сокращение и вытягивание псевдоподий, хими-
ческой — осуществление синтеза и многих обменных реакций, электро-
химической— передача различных раздражений, осмотической — всасы-
вание или выделение разных веществ.
Лизосомы — группа внутриклеточных гранул, функционирую-
щих подобно пищеварительной системе многоклеточных. Они перевари-
вают вещества, захваченные клеткой извне, а также выделяют фермен-
Рис. 24. Схема строения клетки многоклеточных по данным электронно-
микроскопического исследования:
аг — аппарат Гольджи, л — лизосомы, м— митохондрии, ц — цитоплазма,
цс — центросома, ас — эндоплазматическая сеть, я — ядро, яд — ядрышко
ты на поверхность клетки для усвоения пищи, находящейся во внешнем
слое цитоплазмы.
Аппарат Гольджи — комплекс, состоящий из мембран, гра-
нул и вакуолей, в функцию которого входит накопление веществ, под-
лежащих удалению: продуктов жизнедеятельности, воды или ядовитых
веществ, попавших в клетку извне. Клеточный центр расположен
обычно вблизи ядра и аппарата Гольджи и выполняет важную роль
при делении клеток. К включениям относятся непостоянно присут-
ствующие запасные вещества, капли жира, белковые гранулы, вакуоли.
Каждая клетка содержит одно или несколько ядер, ограничен-
ных оболочкой, состоящей из двух мембран, пронизанных порами; на-
ружная мембрана связана с эндоплазматической сетью. Основным
структурным элементом ядра является хроматин (хромосомы),
представленный нитями и тяжами, расположенными в жидком ядерном
соке—кариолимфе. Хроматин состоит из нуклеопротеидов (белков),
химически связанных с дезоксирибонуклеиновой кислотой (ДНК).
Кроме ДНК в ядре имеется информационная РНК, которая опреде-
90
ляет специфику синтезируемого белка, снабжает цитоплазму РНК, под-
держивает в цитоплазме на определенном уровне многие обменные и
синтетические процессы. Ядро играет важную роль также в процессе
размножения, обеспечивая передачу наследственных признаков по-
томству.
Цитоплазма простейших представляет собой сложную систему
гидрофильных коллоидов, структура и агрегатное состояние которых
зависит от окружающих условий среды. Она дифференцирована на два
слоя, отличающихся друг от друга по физическому строению и, воз-
Рис. 25. Схема строения некоторых простейших:
1—4 — класс Sarcodina: 1 — Amoeba (подкласс Rhizopoda), 2 — Difflu-
gia (подкласс Rhizopoda), 3 — Acantharia (подкласс Radiolaria), 4 — Allo-
gromia (подкласс Foraminifera); 5 — класс Ciliophora, Paramaecium
можно, химическому составу. Наружный слой, или эктоплазма,
более прозрачная, более однородная, более плотная; внутреннее содер-
жимое, или эндоплазма, более жидкая, обладает более сложным
строением.
Общий план строения и сложность организации у простейших
очень разнообразны. Часть простейших имеет неправильную и не-
постоянную форму тела (амебы); тело других (радиолярии, солнечни-
ки) построено по радиальному плану, с большим числом осей и пло-
скостей симметрии или с одной главной продольной осью; ряд простей-
ших (часть саркодовых и инфузорий) двустсфонне-симметричны
(рис. 25).
Органами движения простейших служат: а) псевдоподии —
временные выросты цитоплазмы; б) жгутики — отдельные тонкие
нитевидные выросты эктоплазмы, расположенные на переднем конце
тела (от 1 до 8) или собранные в пучки; в) реснички — короткие
волосовидные выросты эктоплазмы размером 10—15 мк, число которых
может достигать 10 000—14 000.
Строение скелета. У большинства простейших имеются скелетные
образования, выполняющие опорные и защитные функции. Скелетные
элементы вырабатываются за счет деятельности цитоплазмы; в некото-
рых случаях это просто уплотненная эктоплазма, но чаще всего это
продукты ее выделения. Основой большинства скелетных образований
91
служит органическое вещество — тектин, в состав которого входят про-
теин и углеводы. Скелет может быть наружным и внутренним, органи-
ческим и минеральным. Наружный скелет бывает в виде сплошного
покрова — кутикулярной оболочки, окружающей все тело в виде рако-
винки, снабженной отверстием, через которое протоплазма сообщается с
внешней средой. Форма раковинок чрезвычайно разнообразна. Внут-
ренний скелет может быть сплошным, в виде центральной капсулы
(радиолярии), или состоять из отдельных опорных образований органи-
ческого или минерального состава. Обычно органический скелет укреп-
ляется включением в него минеральных примесей либо посторонних
минеральных частиц, либо выделяемым цитоплазмой углекислым каль-
цием ,или кремнеземом. В ископаемом состоянии сохраняется обычна
минеральный скелет, очень редко органический.
При неблагоприятных условиях многие простейшие переходят в со-
стояние анабиоза, при котором жизненные процессы замедляются,
тело приобретает шаровидную форму, выделяя вокруг защитную обо-
лочку — цисту. В таком состоянии простейшие очень стойки и способ-
ны переносить длительные неблагоприятные условия.
Размножение и развитие. Простейшие размножаются бесполым и
половым путем. При бесполом размножении новые особи возникают
путем деления или почкования. В первом случае клетка делится на-
двое или множественно, при этом вначале многократно делится ядро
(организм становится временно многоядерным) и затем на это же число
частей распадается цитоплазма, окружающая каждое ядрышко. При
почковании от материнской особи отделяется одна или несколько более
мелких дочерних почек, при этом крупной реорганизации материнского
организма не происходит; если дочерние почки сохраняют связь с мате-
ринским организмом, то возникают колонии. Колонии имеются среди
жгутиконосцев, солнечников, радиолярий. Каждая колония состоит из
моноэнергидных особей, некоторые из них отличаются высокой целост-
ностью и интеграцией — у них имеются соматические и половые клетки,
развиты стеблевые отделы и при раздражении вся колония реагирует
как единая особь.
При половом размножении вначале возникают половые клетки,
или гаметы; после слияния мужской и женской гамет образуется
зигота, из которой развивается новый организм. У некоторых про-
стейших наблюдается сложный жизненный цикл, который заключается
в чередовании бесполого и полового поколений.
Экология и тафономия простейших. Современные представители
простейших очень широко распространены в природе: они обитают в
морях и океанах (60%), в солоноватых и пресных озерах, в реках,
болотах, лужах, во влажной почве, в подземных водах, т. е. практи-
чески всюду, где есть влага (18%); 22% простейших ведет паразити-
ческий образ жизни. Простейшие входят во все сложные цепи пита-
ния, создавая возможности для существования и размножения всех
многоклеточных; они участвуют также в круговороте органических
веществ в природе.
Простейшие питаются готовыми органическими веществами и яв-
ляются гетеротрофными организмами. Автотрофные простейшие, синте-
зирующие органические соединения из неорганических, отнесены к рас-
тительным организмам.
Размер тела простейших обычно колеблется от 0,1 до 1 мм; «гигант-
ские» формы достигают 80—100 мм, а наиболее мелкие 2—4 мк.
Скелеты многих простейших покрывают дно морей и океанов, прини-
мают участие в образовании и накоплении разнообразных илов
92
(радиоляриевый, глобигериновый), Из скелетов образованы большие
толщи разнообразных горных пород — известняков, трепелов, опок,
яшм, радиоляритов.
В настоящее время известно около 25—30 тыс. видов современных
простейших и примерно столько же обнаружено в ископаемом состоя-
нии. Наиболее древние остатки простейших (радиолярии и форамини-
феры) описаны из пород кембрийского возраста, хотя ни у кого не вы-
зывает сомнения в том, что простейшие появились на заре развития
жизни на Земле и были одними из первых организмов. Докембрийские
простейшие, по-видимому, не обладали еще скелетом и поэтому не
'Сохранились в ископаемом состоянии. Но до появления способности
строить скелет простейшие прошли длительный путь развития, который
привел вначале к обособлению ядра от цитоплазмы, затем к усложне-
нию и совершенствованию внутриклеточных цитоплазматических
•структур.
Основы систематики и классификация. На основании особенностей
•строения двигательного аппарата, ядра и цитоплазмы, а также особен-
ностей размножения простейшие большинством исследователей разде-
ляются на пять классов: Sarcodina, Cnidosporidia, Sporozoa, Zoomasti-
jgophora, Ciliophora. Некоторые исследователи рассматривают их в
качестве типов.
КЛАСС SARCODINA. САРКОДОВЫЕ
Общая характеристика. Класс саркодовых (гр. sarx, род. sarkos—•
:мясо, тело, sarcodes — вещественный) объединяет большое количество
разнообразных по форме и строению наиболее просто устроенных про-
-стейших. У одних цитоплазматическое тело голое, у других снабжено
кутикулой, у третьих заключено в раковину (агглютинированную или
секреционную), четвертые имеют внутренний органический либо мине-
ральный скелет, кремневый или целестиновый. Характерной особен-
ностью всех саркодовых является способность их цитоплазмы в различ-
ных местах тела образовывать временные выросты — ложноножки,
или псевдоподии, которые служат для передвижения, захвата
пищи, газообмена и выделения непереваренных остатков пищи. Сарко-
довые питаются бактериями, водорослями, личинками, рачками и раз-
ными микроскопическими организмами. В морях известны на всех глу-
бинах, от поверхностных вод до больших глубин (3000—5000 м). Среди
них имеются бентосные и планктонные формы. Небольшая часть оби-
тает в пресных водах. Размножаются бесполым и половым путем; у
многих наблюдается чередование поколений. Представители саркодо-
вых наиболее часто встречаются в ископаемом состоянии, два под-
класса —- фораминиферы и радиолярии — имеют особенно важное зна-
чение для целей биостратиграфии.
На основании строения псевдоподий, состава и строения скелета и
особенностей размножения в классе саркодовых выделено 4 подкласса:
Rhizopoda, Foraminifera, Heliozoa, Radiolaria.
ПОДКЛАСС RHIZOPODA. КОРНЕНОЖКИ
В состав подкласса ризопод (гр. rhizo — корень, pous, род. podos—
нога) входят два отряда: голые амебы (Amoebina) и раковинные амебы
(Testacea). Первые (рис. 25, 7) не имеют постоянной формы тела и
твердого скелета; ложноножки могут быть лопастной, пальцевидной
или стрельчатой формы; в ископаемом состоянии неизвестны. У рако-
93
винных амеб (рис. 25, 2) цитоплазма заключена в однокамерную рако-
вину, имеющую одно устье, через которое выходят псевдоподии в виде
тонких прямых нитей или реже тупых лопастей. Раковина разнообраз-
ной формы, состоит обычно из тектинового вещества. У большинства
поверхность органического скелета инкрустируется различными мине-
ральными частицами — песчинками кварца и других минералов, спику-
лами губок, панцирями диатомей. Раковинные амебы обычно вначале
втягивают внутрь эти частицы, а затем выталкивают их в эктоплазму
и цементируют органическим веществом. Такая раковина называется
песчаной, или агглютинированной. Корненожки размнот
жаются делением надвое, при этом половина цитоплазмы выступает
из устця и вокруг нее образуется новая раковинка; одновременно с
этим ядро делится на два и одно из них переходит в дочернюю особь,
Затем цитоплазма между дочерней и материнской особями перешнуро-
вывается и особи переходят к самостоятельному существованию. При
наступлении неблагоприятных условий многие из них образуют цисты.
Корненожки живут в пресных водах, болотах, во влажных местах.
Раковинные амебы встречаются в ископаемом состоянии очень редко;
они известны с палеогена.
ПОДКЛАСС FORAMINIFERA. ФОРАМИНИФЕРЫ
Общая характеристика. К подклассу фораминифер (лат. foramen,
род. foraminis — отверстие, дыра, fero — носить) относится большая
группа саркодовых, насчитывающих до 20 000 современных и ископае-
мых видов, цитоплазма которых заключена в органическую, агглютини-
рованную или известковую раковину. Псевдоподии фораминифер со-
стоят из тонких, разветвленных, корневидных, соединяющихся между
собой (анастомозирующих) нитей, выходящих из раковины либо только
через устье, либо через устье и каналы, пронизывающие стенку рако-
вины, Фораминиферы в большинстве своем морские бентосные или
планктонные, свободноживущие или прикрепленные формы. Небольшая
часть фораминифер приспособилась к жизни в солоноватоводных бас-
сейнах и лишь немногие известны в пресных водоемах. В ископаемом
состоянии известны начиная с кембрия.
Строение тела. Цитоплазма фораминифер обычно бесцветная, иног-
да окрашена в розовый, оранжевый или желтый цвета. Эктоплазма,
довольно однородная по структуре, осуществляет обмен веществ с
внешней средой и служит местом образования. псевдоподий. Под
электронным микроскопом псевдоподии представляют собой пучок
волоконец разного диаметра; каждое волоконце окружено оболочкой.
Способность псевдоподий вытягиваться и втягиваться основана на
свойстве цитоплазмы изменять свое агрегатное состояние, переходя из
жидкого состояния (золь) в вязкое (гель). Псевдоподии, не связанные
с субстратом, разветвляются, соединяются перемычками и образуют
своеобразную ловчую сеть, в которую попадают личинки, различные
микроорганизмы и органический детрит (рис. 26). Переваривание
пищи нередко происходит вне раковины.
Эндоплазма заполняет внутреннюю полость раковины и отли-
чается от эктоплазмы более темной окраской и менее однородной
структурой. Эндоплазма содержит ядро, многочисленные ультрамикро-
скопические структуры и различные включения, к числу которых
относятся поступившие извне: пища, минеральные частицы, паразити-
ческие микроорганизмы, а также продукты жизнедеятельности клетки.
В эндоплазме многих фораминифер в симбиозе с ними живут микро^
94
Рис. 26. Отряд Allogromiida. Схема
строения Allogromia ovoidea Rhumb,
(совр.):
п — псевдоподии, р — раковина, у —
устье, эк — эктоплазма, эн — эндо-
плазма
скопические водоросли — зооксантеллы, которые в процессе фото-
синтеза поглощают углекислый газ, вырабатываемый фораминиферами
при дыхании, и выделяют кислород, необходимый последним для газо-
обмена. Перед размножением эндоплазма претерпевает глубокие из-
менения, она очищается от посторонних частиц и просветляется.' При
этом у многих фораминифер образуются временные оболочки — цисты,
состоящие из тектиновой основы,
посторонних частиц и продуктов
выделения, эндоплазмы.
Строение раковины. Подав-
ляющее число фораминифер име-
ет раковину, и лишь у неболь-
шой части цитоплазма окруже-
на утолщенной эластичной орга-
нической оболочкой — мембра-
ной. Раковина может быть отно-
сительно простой или достигать
большой сложности (рис. 27). Ее
размеры колеблются от 0,02 до
110—-120 мм. Стенка раковины
может быть органической, агглю-
тинированной и известковой. На-
иболее низкоорганизованные фо-
раминиферы (аллогромииды)
имеют стенку, состоящую из тек-
тина, представляющего комбина-
цию протеинов и углеводов. У
многих фораминифер в тектино-
вую стенку включены посторон-
ние частицы разного минераль-
ного и химического состава: зер-
на кварца, различных тяжелых
минералов, карбонатов, пластин-
ки слюды, спикулы губок, орга-
нический детрит (обломки спи-
кул губок, раковинки иных фора-
минифер, скелеты радиолярий,
обломки раковин моллюсков) и
другой «строительный материал».
При этом фораминиферы обычно
так же, как и раковинные аме-
бы, «заглатывают» во внутрь этот
«строительный материал». Через
некоторое время протоплазма
разбухает и «строительный мате-
риал» выталкивается на поверх-
ность, где цементируется текти-
ном, карбонатом кальция, окис-
лами или карбонатом железа.
Таким образом, возникают агглютинированные раковины.
Раньше предполагалось, что в редких случаях цементом у некото-
рых фораминифер мог быть кремнезем. Однако у современных фора-
минифер наличие кремневого цемента пока не установлено. Многие
исследователи считают, что наблюдаемый у ряда ископаемых форами-
нифер кремневый скелет является вторичным и развился в процессе
95
фоссилизации по карбонату кальция. Неясным остается также вопрос,
откуда берется железистый цемент, обладают ли фораминиферы спо-
собностью выделять железо из цитоплазмы или оно привносится извне
в виде обломков железистых минералов. Цитоплазма некоторых
фораминифер обладает своеобразной избирательной способностью —
для постройки скелета она «выбирает» материал только определенного
размера, цвета и даже состава, например только зерна кварца или
Рис. 27. Типы строения раковин фораминифер и предполагаемые пути их изменения
(Покорный, 1958)
спикулы кремневых губок, или листочки слюды. Но чаще всего исполь-
зуется всякий подходящий обломочный материал, рассеянный на дне
водоема. Цемент и агглютинированные частицы входят в состав рако-
вины в разной пропорции: у одних форм частицы плотно прилегают
друг к другу, у других они разделены участками цемента, иногда
цемент полностью преобладает. Микроструктура стенки агглютинирую-
щих фораминифер изучена недостаточно. У многих имеется внутрен-
няя органическая выстилка.
Большинство фораминифер имеет секреционную известковую рако-
вину, стенка которой состоит из тектиновой основы, пропитанной мине-
ральными солями; важную роль здесь играет углекислый кальций
(кальцит или арагонит) с различным количеством примеси углекислого
магния (до 18%) и фосфата кальция и магния. Стенка известковых
раковин по своему строению довольно разнообразна. Существует три
основных типа микроструктур стенки: микрогранулярный, фарфоровид-
ный и гиалиновый (стекловидный). В последнее время выделен еще
криптокристаллический. Употребляемые названия «фарфоровидный»
и «стекловидный» не очень подходящие, так как они отражают не спе-
цифику самой микроструктуры, а общий облик стенки, но эти названия
общеприняты и пока существуют в литературе.
96
Микрогр а нул я р н ый тип стенки наблюдается у палеозойских
эндотирид, фузулинид и у некоторых мезокайнозойских отрядов; он
характеризуется наличием зерен микрозернистого кальцита размером
от 1 до 5 мк, отсутствием цемента и непостоянной примесью агглюти-
нированных частиц. Раковина с таким типом микроструктуры стенки
не имеет скульптуры, дополнительных скелетных образований; внутрен-
ний скелет представлен в виде выростов стенки. Поверхность раковины
тусклая, светлого или серовато-желтого цвета.
Фарфоровидный тип стенки отличается беспорядочным рас-
положением кристаллов и их кристаллографических осей; кристаллы
имеют различную форму, их размеры от 0,5 до 5 мк. В отраженном
свете стенка белая, фарфоровидная, иногда блестящая. Стенка рако-
вины содержит органическую основу. Этот тип стенки характерен для
отряда милиолид.
Стекловидный, или гиалиновый, тип разделяется на два
подтипа: стекловато-зернистый и стекловато-радиальный. У первого
подтипа кристаллы кальцита или арагонита однообразной округлой
или угловатой формы, плотно прилегают друг к другу; размеры крис-
таллов 0,5—10 мк; оптические оси ориентированы беспорядочно или с
преобладанием определенной ориентировки с осью С под углом к по-
верхности стенки. У стекловато-радиального подтипа кристаллы каль-
цита или арагонита сильно удлиненные, расположены в основном пер-
пендикулярно к поверхности стенки; так же расположена оптическая
ось С.
Криптокристаллический тип микроструктуры стенки ха-
рактерей для палеозойских фораминифер; стенка состоит из кристал-
лов кальцита с нечеткими границами.
Нередко в процессе фоссилизации секреционных известковых ра-
ковин возникают вторичные микроструктуры, связанные с процессами
перекристаллизации. В одних случаях происходит укрупнение крис-
таллов, в других — распадение удлиненных кристаллов на мелкие суб-
изометрические зерна.
Макроструктуру стенки раковины образуют морфологически
обособленные слои, внутрикамерные выстилки, вторичные слои на
внешней поверхности раковины и на поверхности септ. .
Первичная стенка раковины может быть однослойной либо состоя-
щей из двух или нескольких слоев. Первично-однослойные
Стенки развиты преимущественно у представителей с фарфоровидной
микроструктурой, а также у многих агглютинированных и- тектиновых
раковин. У фораминифер со стекловидной и с микрогранулярной струк-
турой развиты как однослойные, так и многослойные .стенки; у много-
слойной стенки отдельные слои разделены тонкими' прослоями органи-
ческого вещества; слои, слагающие стенки, обычно .отличаются друг от
друга особенностями строения. Для некоторых групп (фузулиниды) эти
слои имеют специальные названия: первичная стенка называется
протекой; она состоит из наружного тонкого слоя — тектума и
основного внутреннего слоя, носящего различные названия. У шва-
герин он имеет ячеистое строение и назван кериотекой (см.
рис. 39). У стекловатых многослойных раковин трехслойную первич-
ную стенку предложено называть биламеллярной, поскольку пер-
вично в ней различали внутренний и наружный (или основной) слои.
Стенка раковины изнутри бывает выстлана тонкой органической
цленкой. На внешней поверхности раковины и на внутренних оборотах
развиты вторичные слои стенок раковины; они образуются после фор-
мирования новой камеры в виде последующих наслоений на наружной
7 В. В. Друщиц
97
или внутренней стороне ранее образовавшейся стенки (их называют
иногда слоями нарастания, или утолщения, или вторично-многослой-
ными) .
§ простейшем случае при образовании новой камеры вся открытая
часть раковины перекрывается новым раковинным веществом и ее-
старая часть значительно утолщается (рис. 28), в то время как вновь.
Рис. 28. Типы строения стенки:
1 — моноламеллярный, стенка первично-однослойная;.-2 — биламеллярный, стенка
первично-двуслойндя; 3 — роталоидный, стенка первично-однослойная (зачернено,
внутрисептальное пространство); а — общий вид, б — три последние камеры
образованная септа и все предшествующие септы остаются однослой-
ными (рис. 28, J); такой тип строения наблюдается у нодозариид,
булиминид и простейших семейств роталиид. Во втором случае при
образовании новой камеры раковинное вещество перекрывает всю от-
крытую часть раковины и налегает на предшествующую септу таким
образом, что она становится двойной, а вновь образованная апертурная
септа остается однослойной (рис. 28, 3). У подобных двойных септ &
полостях, остающихся между двумя слоями, развивается система сеп-
тальных каналов. Такой тип двойных септ с системой внутрисепталь-
ных каналов характерен для отряда роталиид и получил название-
роталоидных септ. В третьем случае вновь образованная каме-
ра с конечной апертурной септой является первично-двойной и по
способу образования напоминает первый случай (рис. 28, 2). Подобные
двойные септы, также снабженные системой каналов, характерны для
раковин некоторых групп отрядов булиминид и нуммулитид (орби-
тоиды). •
Пористость стенки. У многих фораминифер наблюдается
пористая стенка. Поры могут быть простыми и сложными. Простые-
поры представлены цилиндрическими канальцами диаметром 0,2—
0,5 мк; сложные поры характеризуются объединением мелких поровых
канальцев в более крупные (кериотекальная пористость у фузулинид).
У некоторых мезокайнозойских фораминифер наблюдается аль-
веолярное строение стенки, образованной различными выростами, со-
ставляющими дополнительные внутрикамерные скелетные образования.
Все поровые каналы обычно покрыты органической выстилкой. Форма
И частота пор на раковине в последние годы интенсивно изучаются с
применением электронного сканирующего микроскопа.
98
Форма раковины. Раковина фораминифер может быть одно-, двух-
и .многокамерной (рис. 29). При непрерывном росте образуется рако-
вина, не разделенная на камеры; такая раковина называется одно,
камерной. В простейшем случае однокамерная раковина имеет фор-
му шара или колбы, с одним устьем (Saccamtnina, Lagend) или с не-
сколькими отверстиями (Astrorhiza). Она может быть агглютинирован-
Рис. 29. Схема строения раковин фораминифер:
1—'Однокамерная; 2 — двухкамерная; 3—5 — многокамерные: 3.— однорядная, 4-—
спирально-плоскостная: 4а — сбоку, 46 — со стороны устья, 5 — спирально-кониче-
ская: 5а —-с дорзальной стороны, 56 — со стороны устья, 5в — с вентральной стороны;
АА — ось навивания, Д1 — большой диаметр, Д? — малый диаметр, к — кнль, рр —
плоскость симметрии, с — септальные швы, сп. — септальная поверхность, сш — спи-
ральный шов, Т — толщина раковины, у — устье
ной или известковой. При усиленном нарастании вдоль устьевого края
возникает раковина в форме трубки, открытой с одной стороны или
с обеих.
Двухкамерные раковины состоят из шаровидной начальной
камеры и второй, длинной, неразделенной, трубчатой, отделенной от пер-
вой одной перегородкой. Вторая, камера может быть прямой или раз-
ветвленной, или завитой в неправильную клубковидную, плоскую или
коническую спираль.
Раковина, у которой внутренняя'полость разделена перегородками,
или септами, на камеры, называется многокамерной (рис. 29,
3—5). Возникновение многокамерности связано с изменением характера
роста цитоплазмы и раковины. Рост из постоянного становится перио-
дическим, причем периоды усиленного роста отделяются друг от друга
периодами покоя. Каждому периоду роста отвечает образование новой
камеры, которая, как правило, бывает больше предыдущей; форма и
расположение новой камеры и апертурной септы,, отделяющей вновь
образованную камеру от внешней среды, зависят от физико-химических
свойств цитоплазмы, от величины краевых углов, образуемых расходя-
щимися псевдоподиями со стенками предшествующей камеры, и от
характера поверхности последней. Возникновение периодичности роста
имело большое значение в развитии фораминифер, так как освобожда-
ло их от необходимости непрерывно строить раковину. Следы такой
Периодичности можно уже наблюдать на некоторых одно- и двухкамер-
ных трубчатых раковинах, несущих легкие пережимы.
Наиболее простой формой многокамерной раковины можно счи-
тать одноосную или однорядную, когда каждая последующая камера,
имея форму шара как наиболее выгодную, обладающую наибольшим
объемом при наименьшей поверхности, наращивается над предыдущей.
Но у таких однорядных форм довольно велика опасность излома, осо-
7‘
99
бенно в местах пережимов, поэтому совершенствование формы приво-
дит к тому, что новая камера охватывает своей основной частью часть
предыдущей камеры, как бы надвигаясь на нее.
Другим способом укрепления раковины служит ее закручивание
в спираль. Наиболее примитивным типом будет неправильно-клубко-
йидный, при котором обороты навиваются беспорядочно в нескольких
направлениях. При упорядочивании такого навивания возникают
плектогирные раковины или раковины милиолинового типа. В первом
случае ось навивания последующего оборота отклоняется, на некоторый
угол от положения оси предыдущего оборота. Во втором случае камеры
образуют спирально-свернутый клубок, располагаясь в нескольких
взаимно пересекающихся плоскостях. Это объясняется тем, что направ-
ление оси навивания изменяется с ростом раковины на определенный
угол. Длина каждой камеры составляет обычно половину оборота.
У одних форм камеры отстоят друг от друга на 144° и располагаются
в . пяти плоскостях (Quinqueloculina), пересекающихся под углом 72°
(см. рис. 42), у других камеры расположены в трех плоскостях (Trilocu-
Нпа), взаимно пересекающихся под углом 120°, и, наконец, у третьих
каждая камера расположена от предшествующей на 180° (Pyrgo, иди
Biloculina).
Спирально-плоскостной тип рассматривается как видоизмененный
одноосный, у которого главная ось спирально завивается в одной
плоскости. Линии соприкосновения смежных оборотов спирали ракови-
ны называются спиральными швами. Воображаемая прямая
линия, вокруг которой происходит навивание оборотов раковины,
носит название оси навивания. По оси навивания у спирально-
плоскостных измеряется толщина раковины. Перпендикулярно к 4эси
навивания через начальную камеру проводится диаметр' раковины.
Сечение раковины перпендикулярное диаметру является эквато-
риальным. С экваториальным сечением совпадает плоскость сим-
метрии. Форма спирально-плоскостных раковин разнодбразна и зависит
от диаметра и толщины (см. рис. 41, 3). При диаметре, значительно
превышающем толщину, раковина имеет дисковидную или чечевице-
образную форму. При диаметре, почти равном толщине, раковина
.приобретает шаровидную форму. При толщине, значительно превышаю-
щей диаметр, возникает веретеновидная форма. Если при рассмотрении
^спиральной раковины сбоку .видны все обороты, она называется э во-
лю тн ой (см. рис. 35, /). Если последний оборот охватывает все пре-
дыдущие обороты, то раковина называется инволютной (см.
рцс. 48, 5). Между этими двумя крайними типами строения имеется
большое число форм, занимающих промежуточное положение (полу-
эволютные и полуинволютные).
' Степень возрастания оборотов' бывает различна. У большинства
спирально-плоскостных раковин возрастание оборотов происходит по-
степенно, но у некоторых форм обороты возрастают очень быстро и
раковина приобретает вид «рога изобилия» или даже становится вееро-
образной. Иногда быстрое возрастание оборотов может приводить к
смыканию противоположных концов веера и к возникновению цикличе-
ского типа раковины. У циклических раковин камеры расположены по
концентрическим окружностям в одной плоскости (см. рис. 49).
При спирально-коническом типе (роталиевый) камеры распола-
гаются по у л и т к о в и д н о й, или тр о хои д н о й, спирали (рис. 29,5).
Сторону, отвечающую основанию конуса, где виден обычно только
последний оборот, принято называть вентральной, или брюшной.
Сторона, отвечающая вершине конуса, где видны все обороты, назы-
100
вается* д Op с а л ьн о й, или спинной. Спиральный шов отделяет
друг от друга спиральные обороты.
Спирально-винтовой тип раковин отличается тем, что вы-
сота нарастания камер происходит по высокой спирали, которая зна-
чительно превосходит диаметр основания (см. рис. 37). Обычно у таких
раковин спиральное расположение камер выглядит как двух-, трех- или
многорядное расположение камер и поэтому для них чаще употреб-
ляются названия двухрядные, трехрядные или многорядные раковины.
Рис. 30. Типы строения устья:
1 — простые; 2 — устья с зубом; 3 — удлиненное; 4 — лучистое, с устьевой ка-
мерной; 5 — дендровидное; 6 — добавочные устья; 7 — сложные
У прикрепленных фораминифер раковина приобретает древовидную или
неправильно разветвленную форму (см. рис. 34, 4).
Форма камер отличается большим разнообразием. Различают
камеры: шаровидные, овальные, трубчатые, циклические, радиально
удлиненные, угловатые (конические, ромбовидные, усеченно-кониче-
ские), валикообразные.
©днако рассмотренными выше основными типами строения рако-
вин не исчерпывается все многообразие их форм.
Гетероморфизм. Нередко в процессе индивидуального развития
(онтогенеза) происходит изменение типа строения раковины, что при-
водит ее к гетероморфному строению. Например, начальная раковина
может быть спирально-плоскостной, следующий отдел может состоять
из двух редко расположенных камер и конечный отдел может быть
однорядным. Такая раковина называется триморфной. Если рако-
вина сочетает всего два типа строения, то она биморфна (см.
рис. 37, 26, в), и, наконец, если она однотипна по своему строению, то
ее называют мономорфной. Наиболее резко гетероморфное строе-
ние раковины бывает выражено у микросферических особей (шизон-
тов).
Апертура, или устье. Отверстие, при помощи которого цитоплазма
сообщается с внешней средой, расположенное в конце однокамерной
или в последней септе многокамерной раковины,, носит название
устья, или апертуры. Последняя септа образует септальную, или
устьевую, поверхность. При образовании новой камеры устье предыду-
101
щей камеры становится отверстием, соединяющим соседние камеры.
Это отверстие называется фораменом (отверстие, дыра); отсюда
весь подкласс получил название фораминиферы. Устье (рис. 30)
расположено в центре, эксцентрично или в основании апертурной пере-
городки; оно может быть простым, т. е. состоять из одного отверстия
разнообразной формы: округлой, овальной, щелевидной, крестообраз-
ной, ветвистой, радиально-лучистой. Сложное устье состоит из не-
скольких отверстий. Наиболее распространенным типом сложного устья
Рис. 31. Система каналов у роталиид:
1а — вид с вентральной стороны; 16 — внутренний
слепок по продольному сечению; вк — внутрисеп-
тальный канал, к — камеры, ск — спиральный канал,
у — устье, у' — устье спирального канала
является ситовидное устье, состоящее из многочисленных мелких от-
верстий. У многих фораминифер строение устья осложняется дополни-
тельными образованиями, к числу которых относятся особые выросты,
называемые зубными пластинками, или зубами. Они имеют важное
таксономическое значение и служат, по-видимому, для укрепления края
раковины и прикрепления пучка выходящих псевдоподий.
Кроме основного устья для выхода эктоплазмы наружу служат
различные отверстия, имеющиеся в раковине. К их числу относятся тон-
кие каналы, пронизывающие стенку некоторых агглютинированных и
известковистых микрозернистых и радиально-лучистых раковин; допол-
нительные устья расположены в разных местах: вдоль периферического
края, вдоль шва и т. д.
Система каналов. У наиболее высоко организованных фораминифер
(роталииды, нуммулитиды) имеется система каналов внутри раковины
(рис; 31). Основными элементами этой системы являются спираль-
ный и межсептальный каналы. Спиральный канал связан' с
брюшной лопастью каждой из камер; от него отходят межсептальные
каналы, расположенные в полостях двойных септ и открывающиеся
тонкими порами в швах. У некоторых роталиид система каналов очень
сложна: наблюдается не один, а два спиральных канала, от которых
отходят пупочные и межсептальные каналы.
Дополнительный скелет. К дополнительному скелету относятся те
образования, которые усложняют строение раковины и септ. Они
могут быть внутренними и наружными. К внутренним образованиям
относятся известковые отложения, расположенные у эндотирид и фузу-
линид по краям экваториального устья (хоматы) или по бокам допол-
нительных устьев (парахоматы), или прерывисто только около септ
1.02
((псевдохоматы). К ним относятся также столбики конической формы
$1уммулитид, пронизывающие раковину. На поверхности оборотов они
имеют вид бугорков — гранул и служат для укрепления раковины.
К числу наружных дополнительных скелетных образований отно-
сятся различные скульптурные элементы в виде ребрышек, ячеек, килей,
бугорков, игл, шипов и различных выростов на раковине.
У некоторых фораминифер, имеющих спиральную раковину, пу-
почная область бывает закрыта своеобразной втулкой или диском,
'Р
Рис. 32. Схема чередования поколений у фораминифер:
<а — микросферическая форма (шизонт В) с дочерними «эмбрионами»; б, б' —
мегасферические формы (гамонты Аь А2); г — гамета с гаплоидным (п) набором
хромосом, з — зигота с диплоидным (2п) набором хромосом, рр — редукционное
деление, 'э — дочерние «эмбрионы»
состоящим из стекловатого кальцита; нередко этот диск бывает прони-
цав канальцами, связанными с внутренней системой каналов. У многих
раковин планктонных фораминифер имеются тонкие длинные иглы,
значительно увеличивающие их общую поверхность и облегчающие
парение в толще воды.
Размножение и развитие. У фораминифер наблюдается сложный
жизненный цикл развития (рис. 32), сопровождаемый чередованием
бесполого и полового.поколений. При половом размножении на каком-
то этапе развития у особи, достигшей взрослого состояния, ядро
.делится на огромное число (тысячи) частиц, вокруг которых обособ-
ляется небольшая частица цитоплазмы. Таким путем возникают одно-
ядерные клетки, снабженные двумя жгутами. Это половые клетки,
или гаметы. По своему строению они совершенно одинаковы и бла-
годаря жгутикам обладают подвижностью. После слияния двух гамет
(оплодотворение), обычно происходящих от разных особей, возникает
•оплодотворенная клетка — зигота, имеющая диплоидный набор
хромосом. Вокруг зиготы выделяется первая (эмбриональная) извест-
ковая камера.. От нее, у многокамерных фораминифер, образуется вто-
рая, третья и т. д. камеры. Зигота дает начало микросферическому
поколению, или шизонту. Шизонт (форма В) сравнительно долгое
I
103
время остается одноядерным, но с диплоидным набором хромосом.
Затем на каком-то этапе роста происходит редукционное деление й
ядро становится гаплоидным (с одинарным набором 'хромосом). При
достижении шизонтом взрослого состояния ядро делится последователь-
но несколько раз и шизонт временно становится многоядерным; обра-
зуются десятки, а иногда свыше сотни маленьких ядер, вокруг которых
обособляется цитоплазма. В этом случае возникают так называемые
«э м б р и о н ы», или амебовидные зародыши. Вокруг каждого «эмбрио-
на» образуется довольно крупная эмбриональная камера. «Эмбрионы».
покидают материнскую раковину и переходят к самостоятельному
существованию. Этот процесс представляет собой бесполое размноже-
ние. Возникшие особи постепенно растут, строят новые камеры и дают
макросферическое поколение, полу-
Рис. 33. Отряд Allogromiida:
1 — Chitinosaccus (совр.); 2 — Chiti-
nodendron (поздний кембрий — си-
лур); 3 — Chitinolagena (поздний ор-
довик)
чивщее название гамонтов
(форма А).
Изучение онтогенеза форами-
нифер показало, что обычно на-
блюдается закономерное чередова-
ние гамонтов и шизонтов. Но иног-
да это закономерное чередование
нарушается, за одним шизонтом
(форма В) следует два поколения
гамонтов (формы Аь А2). В одних
случаях гамонты почти не отличи-
мы или несколько отличаются по
размерам,- в других •— гамонты
крупнее шизонтов и обладают
большим числом камер, в треть-
их — гамонты и шизонты отлича-
ются по размерам начальных ка-
мер. У макросферических особей
начальная камера обычно крупных
размеров, раковина сравнительно
небольшая и число камер меньше,
чем у микросферических особей.
Последние отличаются маленькими
размерами начальных камер, срав-
нительно большой раковиной и об-
щим большим числом камер. Явле-
фораминифер двух типов строения
Изучение диморфизма
ние, связанное с образованием у
раковины, получило название диморфизма.
(или триморфизма) важно не только с точки зрения систематики, но и
для изучения происхождения и родственных связей между форамини-
ферами. При этом более важное значение имеют особи, возникшие в
результате полового процесса и более полно отражающие онтогене-
тическое развитие.
Основы систематики и классификации. Важное значение для систе-
матики фораминифер имеют строение и состав стенки раковины, строе-
ние цитоплазмы и ядра, особенности чередования поколений й другие
признаки. На этом основании Д. М. Рауз ер-Черноусов а и А. В. Фур-
сенко (1959) выделили 13 отрядов. Американские исследователи
А. Леблик и Е. Таппан (1964) предложили разделять отряд форами-
нифер на пять подотрядов. В соответствии с принятым в учебнике
рангом фораминифер в качестве подкласса эти подотряды подняты до
уровня надотрядов. Подкласс фораминифер на основании строения
104
стенки раковины разделен на пять надотрядов: Allogromioidea, Textu-
larioidea, Fusulinoidea, Miliolidoidea, Rotalioidea.
НАДОТРЯД ALLOGROMIOIDEA.
АЛЛОГРОМИОИДЕИ
Стенка в виде тонкой органической мембраны или тектиновая, с
небольшой примесью агглютинированных частиц. Поздний кембрий —
ныне. Включает один отряд Allogromiida.
Отряд Allogromiida. Аллогромииды (гр. alios — различный, иной;
лат. groma = gruma— палочка). Раковина неправильной, трубчатой или
шаровидной формы с одним или несколькими устьями. Стенка в виде
тонкой органической мембраны, тектиновая, реже инкрустированная
агглютинированными частицами. При размножении образуют жгути-
ковые или амебовидные гаметы. Живут в пресных и морских бассейнах;
среди них имеются формы, сверлящие раковины других фораминифер.
О путях развития этого отряда почти ничего неизвестно. Поздний
кембрий—ныне. Около 14 родов (рис. 26, 33).
НАДОТРЯД TEXTULARIOIDEA.
ТЕКСТУЛЯРИОИДЕИ
Стенка раковины построена из разнообразных агглютинированных
частиц, сцементированных органическим или минеральным цементом.
Кембрий^—ныне. Объединяет следующие отряды: Astrorhizida, Ammo-
discida, Ataxophragmiida, Textulariida.
Отряд Astrorhizida. Астроризиды (гр. aster — звезда, rhiza, или
rhizoma,—корень). Раковина свободная или прикрепленная, одно- или
двухкамерная, шаровидная, эллипсовидная или трубчатая, прямая или
разветвленная, иногда с обособленной вздутой центральной частью;
перегородки отсутствуют; стенка агглютинированная. Устье простое
или о'тсутствует, тогда его заменяют отверстия между агглютинирован-
ными частицами. Развитие шло от звездчатой неправильной формы
раковины к шаровидной, трубчатой, а также к обособлению устья —
специального отверстия для выхода псевдоподий. Современные астро-
ризиды живут преимущественно в холодных морях, ведут бентосный,
прикрепленный или подвижный образ жизни. Кембрий — ныне. До
75 родов (рис. 34).
Отряд Ammodiscida. Аммодисциды (гр. Ammon — египетское бо-
жество, изображаемое в виде барана; discos — диск). Раковина двух-
камерная, состоит из начальной шаровидной камеры и. второй трубча-
той, клубкообразной, неправильно завитой или свернутой в плоскую
спираль; на поздней стадии иногда однорядная. У некоторых форм
раковина многокамерная. Стенка однослойная, реже сложная, диффе-
ренцированная на два слоя, агглютинированная, с секреционно-извест-
ковым цементом. Устье простое, реже сложное, ситовидное. Развитие
аммо'дисцид шло от наиболее примитивных двухкамерных форм, обла-
дающих второй трубчатой камерой, свернутой вначале клубкообразно,
а затем в плоскую спираль, через виды с зачаточными септами к видам
многокамерным, с полным разделением на камеры и образованием раз-
личного типа раковин. Силур — ныне. 55 родов (рис. 35).
Отряд Ataxophragmiida. Атаксофрагмииды (гр. а — отрицание,
taxo — приводить в порядок, phragma — изгородь, огораживать). Рако-
вина многокамерная, спирально-винтовая, реже спирально-коническая,
105
может быть би- или триморфной; стенка одно- или двухслойная, агглю-
тинированная, с известковым цементом. Устье простое щелевидное или
«сложное. Изменение скелета шло по пути сокращения спирально-вин-
Рис. 34. Отряд Astrorhizida:
1 — Astrorhiza (ордовик — ныне); 2 — Saccatnmina (силур — ныне); 3 — Rhab-
dammina (ордовик — ныне); 4 — Dendrophrya (плейстоцен — ныне)
Рис. 35. Отряд Ammodiscida:
1 а, б — Ammodiscus (силур — ныне); 2 — Glomospira (силур — ныне); 3 — Reophax
(карбон — ныне); 4 — Haplophragtnoides (карбон — ныне)
товой стадии развития и появления неправильно-однорядного или
правильно-однорядного расположения камер с усложнением их внут-
реннего строения (образование столбиков, перегородок, а также услож-
нение устья, появление дополнительных устьев, шейки и устьевых
зубов),
К этому отряду отнесены Орбитолиниды (Orbitolinidae), распрост-
раненные преимущественно в мелу, имеющие крупную (до 30 мм) кони-
106
ческую или дисковидную раковину. Камеры на разных стадиях серпо-
видные, затем дисковидные или кольцевые с дополнительными горизонт
тальными и вертикальными перегородками, особенно сложными в
Рис. 36. Отряд Ataxophragmiida:
1 — Verneuilina (юра — ныне); 1а — вид сбоку, 16 — со стороны устья; 2 —
Ataxophragmium (поздний мел); 3 — Arenobulimina (мел — ранний, палеоцен):
За — вид сбоку, 36 — ср стороны устья; 4 — Orbitolina (мел): 4а — конический
тип с тангенциальным срезом, 45 — поперечное сечение вдоль оси х—х, 4в —
увеличенная часть 46, видны поры;
. к — камеры, км — камерки, с — септы
Рис. 37. Отряд Textulariida:
1 а, б — Textularia (карбон — ныне); 2 — Bigenerina (юра—ныне): 2а — продольный
разрез; стадии роста: I — первая, II — вторая, у — устье, 26 — мегасферическая особь,
2в — микросферическая особь
периферической части раковины; связь между камерами осуществлялась
септальными или апертурными порами. Устье сложное, ситовидное.
Стенка двухслойная. Карбон — ныне; расцвет в меловое время. До
ВО родов (рис. 36).
107
Отряд Textulariida. Текстулярииды (лат. textus, textiles — плете-
ный). Раковина многокамерная, обычно двухрядная, на ранней стадии
иногда спирально-плоскостная, на поздней — однорядная. Развитие
заключалось в усложнении устья и переходе от мономорфной к би- и
триморфной раковине: спирально-плоскостная — двухрядная — одно-
рядная. Стенка агглютинированная, с большим количеством известково-
го цемента. Устье простое или сложное, состоящее из нескольких от-
верстий. Девон (?), карбон — ныне. До 20 родов (рис. 37).
НАДОТРЯД FUSULINOIDEA. ФУЗУЛИНОИДЕИ
Стенка известковая, состоит из микрогранулярного или волокнисто-
го кальцита, одно-, двух- или многослойная. Ордовик — триас. Объеди-
няет два отряда: Endothyrida и Fusulinida.
Рис. 38. Отряд Endothyrida:
1 — Bradyina (карбон): la — вид сбоку, 16 — со стороны устья; 2 — Endothyra
(карбон — пермь): 2а — сбоку, 26 — со стороны устья, 2в — поперечное сечение;
3 — Nodosinella (поздний девон—пермь), раковина с двухслойной стенкой: За —
продольное сечение, 36, в — поперечные сечения: 36 — через камеру, Зв — через
септу
Отряд Endothyrida. Эндотириды (гр. endon — внутри, thyra—дверь).
Раковина многокамерная, спирально-плоскостная или плектогирная,
т. е. ось навивания последующего оборота отклоняется на некоторый
угол от положения оси предыдущего оборота. Конечный отдел иногда
развернут. Имеются однорядные, однокамерные и шаровидные формы.
Устье обычно щелевидное, реже ситовидное. Септы плоские, выпуклые,
наклонные. Стенка известковая, секреционная, волокнистая или зер-
нистая, обычно двухслойная, иногда с агглютинированными известко-
выми частицами и тонкопористая. Развитие шло от форм с однокамер-
ной шаровидной раковиной через многокамерные однорядные, прямые
или изогнутые к многокамерным, вначале клубкообразным, а затем
спирально-плоскостным раковинам, с развитием пористости и много-
слойности стенки. Эндотириды представляют, по-видимому, одну из.
108
ветвей фораминифер, обособившихся от аллогромиид в связи с возник-
шей способностью цитоплазмы выделять микрозернистый кальцит.
У более поздних форм в основании камер развиты дополнительные
скелетные образования: хоматы, валики и гребни.
Эндотириды появляются в ордовике, но достигают расцвета в конце
девона и в раннем карбоне, заселяя моря Европы, Азии и Америки.
Последние представители вымирают в мелу. До 25 родов (рис. 38).
Отряд Fusulinida. Фузулиниды (лат. fusus, умен, fusulus — вере-
тено). Раковина крупных размеров, спирально-плоскостная, инволют-
Рис. 39. Строение стенки раковины у представителей отряда Fusulinida:
1 — эоштафеллы и новеллы; 2 — шубертеллы; 3 — профузулинеллы; 4 —
швагерины;
ал — альвеолы, к — кериотека, нт — наружный текториум, пр — протека,
т — тектум
пая, от дискоидальной или чечевицеобразной до веретенообразной и
шаровидной. Стенка раковины известковая, микрозернистая, состоит
из одного или нескольких слоев. Выделяют несколько типов стенки
(рис. 39): 1) однослойная, тонкая, без видимых пор, состоящая
Рис. 40. Отряд Fusulinida:
1 — схема продольного сечения раковины Tritiates; 2 — схематическое изобра-
жение складчатых септ; -
' г — туннель, хм — хоматы
из одного однородного слоя — протеки; 2) двухслойная, над
протекой развивается тонкий слой — тектум; 3) т р е х с л о й н а я, об-
стоящая из протеки, тектума и нового слоя, покрывающего его, — на-
ружного текториум а. У более высоко организованных фузулинид
на месте протеки развивается более светлый и широкий слой — д и а -
ф анотека и подстилающий её тонкий слой — внутренний тек-
ториум. У швагеринид (рис. 39, 4} стенка толстая, двухслойная, под
тектумом расположен толстый слой с крупными, ясно видимыми поро-
выми каналами,„образующими альвеолы; этот слой получил название
кериотеки.
Раковина начинается шаровидной камерой, размером от нескольких
микронов до 1 мм, имеющей одно отверстие. Вокруг начальной камеры
по спирали вдоль оси навивания располагаются новые камеры и- в за-
109
видимости от ее длины раковина приобретает форму от дисковидной
или шарообразной до веретеновидной. Наибольшая ширина камер
наблюдается в плоскости симметрии и почти сводится к нулю у полюсов...
Последняя • перегородка, отделяющая полость раковины от внешней
среды, пронизана отверстиями, через которые наружу выходила цито-
т
с
са
За 36 38
Рис. 41: Отряд Fusulinida:
1 — схема строения раковины; 2 — Schwagerina (пермь): 2а — поперечный, 26—про-
дольный разрезы; 3—форма раковин: За —' дисковидная {Staffella), 36 — шаровидная
{Schwagerina), Зв — веретеновидная {Fusulinella)',
D — диаметр, L — длина раковины вдоль оси, км — камерки, нк — начальная камера,
с — септы, са—септальные арки, сб — септальные борозды, сп— септальные поры, т —
туннель, у — устье
плазма. Устье одно или несколько, расположены в основании септ.
Последовательные отверстия в септах создавали своеобразный тун-
нель, который ограничивался с двух сторон небольшими валиками —
хоматами; парахоматы ограничивали туннели, проходящие
параллельно экваториальному туннелю. :
НО
У большинства фузулинид перегородки в нижней части были
складчатые (рис. 40), причем иногда соседние складки срастались,
вместе, образуя вторичные камерки, которые сообщались между собой
через' особые отверстия, возникавшие в результате ресорбции соседних
стенок. В продольных шлифах в случае складчатых септ видны харак-
терные арочки различной формы и очертаний. На наружной поверх-
ности раковины септам соответствуют септальные швы, разделяющие
эту поверхность на различные дольки.
С>узулиниды возникли в раннем карбоне от эндотирид, быстро-
эволюционировали и вымерли в конце перми. Развитие шло по пути
увеличения раковины (от диаметра 0,4—0,9 мм у первых форм до 16 мм
у последних при длине до 60 мм), изменения формы от дискоидальной
или чечевицеобразной до сферической, веретеновидной или удлиненно-
цилиндрической, усложнения строения стенки от однослойной до сложно-
построенной, четырехслойной, изменения формы септ — от прямых до
складчатых у более поздних. Параллельно с этими изменениями проис-
ходили: редукция хомат и усиление парахомат, заполнение осевой
части дополнительными отложениями извести, образование септул —
дополнительных выростов стенки камер.
Фузулиниды заселяли открытые пространства прибрежных частей
морей и океанов. Они были распространены в обоих полушариях, вос-
точном и западном; их остатки обнаружены на всех континентах, кроме
Австралии и Антарктиды. Скелетами фузулинид сложены многометро-
вые толщи известняков карбона -и перми. Быстрая изменчивость во вре-
мени, широкое распространение, четкие видовые отличия, массовые
количества сделали фузулинид одной из важнейших групп для расчле-
нения и корреляции позднепалеозойских отложений. Карбон — пермь.
Около 70 родов (рис. 41).
НАДОТРЯД MILIOLIDOIDEA. М И Л И О Л И Д О ИД Е И
Стенка раковины известковая, фарфоровидная, непористая. Кар-
бон— ныне. Включает один отряд Miliolida.
180° -®------------®----------------©----------@-о
А
Рис. 42. Отряд Miliolida.
Схема расположения камер: А—Spirolocu.lin.cr, Б— Quinqueloculina',
В — Triloculina; римские цифры — номера последовательных камер
Отряд Miliolida. Милиолиды (лат. milium — просо)/Раковина спи-
рально-плоскостная или клубкообразная, свернутая по милиолиновому
типу. Стенка непористая, фарфоровидная, состоящая из скрыто-
кристаллического кальцита. Спирально-плоскостная раковина имеет
шаровидную начальную камеру (Cyclogyra) и следующую за ней
трубчатую, эволютную или инволютную. У большинства милиолид
раковина клубкообразная; состоит из начальной камеры, следующей за
Рис. 43. Схема строения раковины Fasciolites (альвеолиниды)
ней трубчатой, занимающей от половины до 2—3 оборотов, и далее
трубчатых камер длиной в полоборота,, расположенных либо в пяти
плоскостях (квинкелокулиновый тип), либо в трех (трилокулиновый
тип), либо в двух (рис. 42), но пересекающихся под углом менее 180°
Рис. 44. Отряд Miliolida:
1— Ophthalmidium (поздний триас — ныне),'поперечное сечение, 1—5 .— номера от
начальной камеры до 5-й; 2 — Pyrgo (юра — ныне): 2а— вид сбоку, 26—со сторо-
ны устья; 3 — Triloculina (юра — ныне): За — срединное поперечное сечение, 36—
вид сбоку, Зв — со стороны устья; 4 —'Quinqueloculina (юра — ныне): 4а, б — вид
сбоку, 4в — со стороны устья
(сигмойлиновый тип). Устье простое, иногда с дополнительным зубом,
реже сложное, ситовидное. Ведут бентосный, преимущественно подвиж-
ный образ жизни, реже прикрепленный; в последнем случае .раковина
приобретает неправильную форму.
Условно к этому отряду отнесены альвеолиниды (ранний мел —
ныне), объединяющие фораминифер с крупной раковиной, внешне
112
иногда похожей на раковину фузулинид. На ранних стадиях камеры
располагаются неправильно-клубкообразно или по типу милиолид, на
последующих оборотах образуют плоскую спираль (рис. 43). Каждая
камера разделяется продольными перегородками на камерки; последние
снабжены отдельными устьями.
Исходной формой милиолид, по-видимому, была двухкамерная
фарфоровидная раковина (Cyclogyra), от которой одно направление1
-привело к развитию клубкообразного навивания, вначале беспорядоч-
ного, позднее закономерного с образованием пятирядного, затем трех-
рядного и, наконец, двухрядного расположения камер. Второе направ-
ление было связано с совершенствованием спирально-плоскостной
раковины, которое привело к возникновению сложнопостроенной рако-
вины альвеолинид.
Милиолиды представляют одну из слепых филогенетических ветвей,
которая возникла от каких-то позднепалеозойских аммодисцид, имев-
ших секреционную известковую раковину с беспорядочным расположе-
нием кристаллов. Милиолиды появляются в раннем карбоне, однако в
палеозое встречаются редко, гораздо шире они распространены в юре
и мелу, но особенного развития они достигли в неогене. До 65 родов
(рис.-44). I
НАДОТРЯД ROTALIOIDEA. РОТАЛИОИДЕИ
Стенка раковины известковая, пористая, часто снабженная кана
лами. Силур — ныне. Объединяет следующие отряды: Nodosariida, Ro
taliida. Heterohelicida. Nummuli-
tida, Buliminida.
Отряд Nodosariida. Нодоза-
рииды (лат. nodosus — узлова-
тый). Раковина известковая, тон-
копористая; стенка радиально-
лучистая, состоит из радиально
расположенных кристаллов каль-
цита; устье лучистое в виде
округлого отверстия и радиально
расходящихся щелей. Раковина
обычно многокамерная, различ-
ной формы; однорядная, прямая
или дугообразно изогнутая, спи-
рально-плоскостная, спирально-
винтовая или неправильной фор-
мы у прикрепленных,
Происхождение и пути эво-
люционного развития недоста-
точно ясны. Возможно, нодозари-
иды возникли в раннем силуре от
аллогромиид, у которых цито-
плазма приобрела способность
секретировать 'углекислый каль-
ций. Вначале раковина была од-
нокамерной, затем возникла мно-
Рис. 45. Отряд Nodosariida:
1 — Nodosaria (пермь — ныне); 2 —
Frondicularta (пермь — ныне); 3 — La-
gena (юра — ныне); 4 — Lenticulina
(триас — ныне): 4а — вид сбоку, 46 —
со стороны устья; 5 — лучистое устье
гокамерная однорядная, из кото-
рой позднее образовались спирально-плоскостные формы. В процессе
развития появились спирально-винтовые двухрядные и, возможно,
вновь однокамерные шаровидные. Нодозарииды ведут бентосный образ
8 В. В. Друщиц
113
жизни;- большинство из них подвижные, небольшая часть приспособи-
лась к прикрепленному образу жизни.
Прежнее название «лагениды» было предложено по ошибке, допу-
щенной м-икропалеонтологами. В 1854 г. Шульце на основании
современного рода Lagynis (отряд аллогромииды) предложил семейст-
во Lagynidae, которое затем ошибочно было заменено на Lagenidae,,
выделенное значительно позднее (1880) на основании рода Lagena.
В семейство Lagenidae были включены такие роды, как Nodosarict,.
Dentalina, Frondicularia, которые еще в 1838 г. Эренбергом были,
объединены в семейство Nodosariidae. По закону приоритета, когда-,
была обнаружена ошибка, название лагениды, как предложенное
позднее (1880), было заменено названием Nodosariidae, установленным,
хронологически раньше (1838). В соответствии с этим было изменено-
название отряда.
Нодозарииды появились, по-видимому, в силуре и существуют до*
настоящего времени. До 65 родов (рис. 45).
Отряд Rotaliida. Роталииды (лат. rota, умен, rotula — колесо). Ра-
ковина обычно спирально-коническая (трохоидная); на ранних стадиях
Рис. 46. Отряд Rotaliida:
1—2 — Globigerina (палеоцен-—ныне): 1 — современная, 2— из палеоге-
на: 2а— вид с дорзальной стороны, 26— с вентральной, 2в — сбоку; 3 —
Rotalia (поздний мел — ныне): За — вид с дорзальной стороны, 36— с
вентральной, Зв — сбоку; 4 — Orbulina (неоген — ныне), наружный шар
разрушен
у всех, а на поздних — иногда иной формы: двух- или однорядная, спи-
рально-плоскостная, циклическая или с неправильным ростом. Камеры
многочисленные, с простыми или двойными септами, иногда разделен-
ные на вторичные камефки. Стенка секреционная, известковая, порис-
тая, одно- или многослойная. Устье от простого до сложного; у некото-
рых вместо устья развиваются крупные поры. У более развитых
роталиид имеется сложная система каналов, состоящая из спираль-
ного канала, от которого отходят внутрисептальные каналы, откры-
вающиеся в швах, и умбональные, выходящие в умбональном диске-
(см. рис. 31). Среди роталиид известны бентосные, подвижные ц
прикрепленные формы, и. планктонные; у последних камеры шарооб-
1.14 .;
разные, иногда последняя камера- охватывает все предыдущие (Orbu-
Ипа}.
Развитие роталиид шло по пути приспособления, с одной стороны,
к бентосному, с другой — к планктонному образу жизни; среди первых
только небольшая часть перешла к прикрепленному способу обитания,
основная же часть вела и ведет в настоящее время подвижный образ
жизни; приспособление к планктонному образу жизни началось в юре.
У первых роталиид раковины были мелкопористые, неразделенные до-
полнительными перегородками на вторичные камерки, с однослойными
септами, без системы каналов. У последующих- роталиид появились
дополнительные перегородки, возникли внутрикамерные. форамены и
появилась система каналов, стенка стала многослойной, септы двойные;
устье изменилось от простого щелевидного до сложного, состоящего из
нескольких отверстий, или простого, снабженного дополнительными
скелетными элементами (зубная пластинка).
'От роталиид, вероятно, произошли гетерогелициды, булимдниды
и нуммулитиды. Первые представители отряда появились в юре, воз-
можно, в триасе, но широкого распространения достигают в мелу и
особенно в кайнозое. До 140 родов (рис. 46).
Отряд Heterohelicida. Гетерогелициды (гр. heteros — разный, helis,
род. helikos — спираль). Раковина в типичном случае двухрядная; на-
чальная часть спирально-плоскостная; во взрослой стадии иногда
Рис. 47. Отряд Heterohelicida:
1—2 — Heterohelix (мел): la — продольный разрез, 16, 2а, 26 — общий, вид; 3 —
Bolivina (поздний мел — ныне)
однорядная, неправильно циклическая или с веерообразным располо-
жением камер; очень редко — трех- и четырехрядная. Стенка пористая,
снаружи гладкая или ребристая. Устье простое, краевое, у однорядных
форм — терминальное. Среди гетерогелицид, которые возникли в нача-
ле мелового периода от каких-то до сих пор неизвестных предков
(роталиид?) со спирально-плоскостной раковиной, различают две группы.
Развитие первой группы было обусловлено приспособлением к бентос-
ному образу жизни и характеризовалось редукцией спирально-плоскост-
ной части раковины и развитием двух- или однорядного расположения
камер, обычно снабженных скульптурой. Развитие второй группы опре-
делялось приспособлением к планктонному образу жизни и шло по
пути возникновения и формирования шаровидных камер, расположен-
ных двух- или четырехрядно. Гетерогелициды были особенно широко
распространены в позднем: мелу и раннем палеогене, бентосные формы
дожили до наших дней, а планктонные вымерли в начале палеогена.
До 20 родов (рис. 47). ' .. ж • ... :
8* . 115
Отряд Nummulitida. Нуммулитиды (лат. nummulus — монетка).
Раковина многокамерная, спирально-плоскостная или циклическая, от
дисковидной, сжатой по оси навивания, до шаровидной, часто дости-
гающая крупных размеров (диаметром до 100—160 мм), обороты от
медленно' до быстро возрастающих, от эволютных до инволютных. Ра-
Рис. 48. Отряд Nummulitida:
1—5 — Nummulites (палеоцен — ныне): 1 — схема строения раковины, 2 — про-
дольное сечение, За, б, 5 — внешний вид, 4 — экваториальное сечение; 6—Орег-
culina (поздний мел — ныне), схема строения раковины (Голев, 1964);
вк — внутрисептальные каналы, к — камеры, с — септы, св — спиральный валик,
сг—септальные гранулы, сл — септальная линия, ср — стенка раковины, ис —
центральный столбик
ковина имеет вторичные скелетные образования в виде столбиков и
спирального валика и сложную систему каналов; стенка пористая.
Микросферические особи обычно отличаются от мегасферических
своими размерами и строением начальных чабтей. Начальная часть
раковины спирально-трохоидная или спирально-плоскостная. Нуммули-
.116
тиды появились в позднем мелу, получили широкое развитие в эоцене и
последние представители дожили до наших дней.
Среди нуммулитид различают несколько крупных стволов: нум-
мулиты, орбитоиды, дискоциклины и лепидоциклины, происхождение и
развитие которых не всегда достаточно ясно.
Нуммулиты имеют дисковидную или чечевицеобразную, дву-
сторонне-симметричную раковину, состоящую из эволютных или инво-
лютных оборотов (рис. 48). Септы, спиральный валик и столбики
пронизаны системой мелких канальцев. Камеры простые или разделен-
ные на более мелкие вторичные. Поздний мел — ныне.
Рис. 49.-Отряд Nummulitida:
1—4 —- Discocyclina (палеоцен — эоцен): 1а, б— внешний вид, 2—3 — схема строения
раковины: 2 — экваториальное (э) и продольное (п) сечения, 3 — экваториальное се-
чение, 4 — схема строения экваториального сечения микросферической особи; стадия
спирального навивания (камеры не заштрихованы), стадия циклического навивания
(камеры с точками); 5 — Asterocyclina (эоцен), начальные стадии развития (обозна-
чения те же, что для 4);
бк. — мелкие боковые камеры, к—канальцы, пв — первая и вторая камеры, эк — круп-
ные экваториальные камеры
Орбитоиды с дисковидной или чечевицеобразной раковиной,
в которой различают двух- или многокамерный эмбрион (начальная
стадия развития) — кольцо относительно крупных периэмбриональных
камер и экваториальные камеры, расположенные кольцеобразно; иногда
экваториальные камеры с обеих сторон покрываются боковыми камер-
ками. Поздний мел-—палеоцен (?).
Дискоциклины с дисковидной или звездчатой раковиной, со-
стоящей из экваториального слоя с боковыми камерками с каждой
стороны (рис. 49). Двухкамерный эмбрион окружен двумя кольцами
периэмбриональных камерок. Экваториальные камеры расположены
кольцами и разделены на камерки;. камеры соединяются столонами.
Развиты интерсептальные и внутристенные каналы. Распространены
в палеоцене и эоцене.
Лепидоциклины имеют дисковидные сжатые раковины; эква-
ториальные камеры разной формы с пористыми стенками и столонами,
117
без системы каналов. С обеих сторон экватора расположены или боко-
вые камерки или пластинчатый слой с вакуолями. Эмбрион двухкамер-
ный, окруженный периэмбриональными камерами, расположенными по
спирали. Распространены от эоцена до миоцена.
Отряд Buliminida. Булиминиды (лат. bulla — пузырь). Раковина
многокамерная, спирально-винтовая, много-, трех- и двухрядная, реже
однорядная, в Начальной стадии роталоидная (трохоидная). Камеры
округлые, у специализированных форм — угловатые; стенка пористая,
состоит из радиально расположенных кристаллов кальцита. Поверх-
ность раковины скульптированная или гладкая; скульптура в видешипи-
Рис. 50. Отряд Buliminida:
1 — Uvigerina (палеоген — ныне); 2 — Bulimina (палеоген — ныне); 3—4 — схе-
. ма строения устьевой части: 3 — Buliminella, 4—' Globobulimina-,
з — устьевой зуб, п — устьевая желобообразная пластина, yi — устье, у% —
устье второе
ков, ребер или тонких трубок, выдающихся над поверхностью. Устье
петлевидное, аркообразное, иногда с одним или несколькими зубами;
устья последовательных камер связаны между собой внутренними
желобками — особыми внутренними. устьевыми образованиями. Разви-
тие шло в сторону удлинения спирали, изменения положения и харак-
тера устья, изменения внутренней системы устьевых желобков и раз-
вития скульптуры.
Булиминиды ответвились, по-видимому, от роталиид в юре и полу-
чили широкое развитие в меловое и палеогеновое время. Они играют
значительную роль в бентосе мелководных зон современных умеренных
и тропических морей. До 60 родов (рис. 50).
История развития фораминифер (рис. 51). В развитии фораминифер
можно выделить следующие этапы: 1. Возникновение и образование
органической тектиновой однокамерной раковины. 2. Образование
агглютинированной и секреционной однокамерных раковин. 3. Возник-
новение среди агглютинированных двухкамерных вначале трубчатых,
а позднее клубко- и спирально-свернутых раковин. 4. Возникновение
многокамерных раковин, по-видимому, одновременно в разных фило-
генетических ветвях. О первом этапе нам ничего неизвестно. Наиболее
древними и примитивными представителями подкласса фораминифер
следует, по-видимому, рассматривать аллогромиид, первые ископаемые
остатки которых найдены в кембрийских отложениях. Целый ряд при-
знаков, такие как строение тектиновой однокамерной раковины с прос-
тым устьем и иногда с агглютинированной стенкой, позволяет связывать
фораминифер с корненожками.
118
Следующим этапом в развитии фораминифер было образование
агглютинированной раковины. Это событие в их жизни произошло, по-
видимому, еще в протерозое, так как в начале кембрия уже известны
первые астроризиды (батисифон). Аллогромииды и астроризиды были,
очевидно, двумя основными группами фораминифер в кембрии и ордо-
вике. В ордовике среди астроризид появляются двухкамерные формы,
Рис. 51. Геохронологическое распространение фораминифер
имевшие форму звезды или трубки; с этого же времени становятся
известными первые фораминиферы с известковой секреционной рако-
виной, о систематическом положении которых нет единого мнения:
«одни исследователи относят их к эндотиридам (см. характеристику
отряда), другие рассматривают их среди нодозариид.'В силуре астрори-
зиды дали начало отряду аммодисцид, представители которого имели
агглютинированную двухкамерную раковину, вначале с клубкообраз-
ным, а позднее со спирально-плоскостным навиванием второй труб-
чатой камеры, еще не имевшей перегородок. В начале силура (если
относить Saccamminopsis к эндотиридам) появляются первые нодо-
.зарииды.
Девон характеризуется развитием астроризид, аммодисцид и в
конце периода эндотирид. Последний отряд представлен большим
числом семейств, раковины которых имеют разнообразную форму, но
все являются многокамерными; стенка у них состоит из зернистого или
волокнистого кальцита, обычно двухслойная, тонкопористая. Важным
этапом в развитии фораминифер был конец девона и начало карбона.
119
Этот этап характеризовался развитием многокамерных аммодисцид
(реофакс, гаплофрагмоидес), появлением двух новых отрядов с агглю-
тинированной многокамерной раковиной — атаксофрагмиид и тексту л я-
риид; возникновением милиолид, имевших сплошную, непрободенную,
фарфоровидную стенку, состоящую из скрытокристаллического каль-
цита. В конце раннего карбона от эндотирид возникли фузулиниды —
один из важнейших отрядов позднего палеозоя. Фузулиниды быстро
эволюционировали, широко распространились в морях карбона и ран-
ней перми и вымерли в конце палеозоя. Раковина их достигла высшей
степени усложнения и дифференциации среди всех известных палеозой-
ских фораминифер. Эндотириды и фузулиниды были распространены в.
карбоне и перми в таких количествах, что впервые в истории развития
фораминифер явились важными породообразователями. Во многих
странах мира, особенно в северном полушарии, известны толщи из-
вестняков, образованных раковинами эндотирид и фузулинид (эндоти-
ровые, фузулиновые и швагериновые известняки).
В перми появляются многие роды нодозариид, имевшие пористую-
раковину, стенка которой построена из радиально ориентированных
кристаллов кальцита. На смену фузулинидам и эндотиридам в начале-
мезозоя приходят нодозарииды, которые постепенно становятся господ-
ствующей группой и сохраняют такое положение до позднего мела. Они
вели, бентосный, преимущественно подвижный, реже прикрепленный
образ жизни и отличались большим разнообразием раковин. В начале-
юры появляются первые представители отряда роталиид, имевшие спи-
рально-коническую (трохоидную) пористую раковину. Развитие этой
группы шло в основном по двум направлениям •— приспособлению к
бентосному и планктонному (пелагическому) образу жизни. Этот
отряд дал планктонных представителей, которые достигли особенно-
большого разнообразия в позднем мелу и в кайнозое.
Меловой период наряду.с широким развитием всех отрядов, имев-
ших агглютинированную раковину (астроризиды, аммодисциды, тек-
стуляриды и атаксофрагмииды), характеризовался появлением трех
новых отрядов: гетерогелицид, булиминид и нуммулитид, среди кото-
рых наиболее важное значение для мелового периода имели первые-
два отряда. Среди гетерогелицид одна группа приспособилась к планк-
тонному образу жизни и была широко распространена в позднем мелу.
/Меловой период отличался также образованием различных известковых
пород, в накоплении которых принимали участие скелеты различных
фораминифер: орбитолин (условно отнесенных к атаксофрагмиидам),.
альвеолин (отряд милиолид), орбитоидов (отряд нуммулитид); огром-
ные толщи писчего мела (поздний мел) были образованы раковинами
глобигеринид и гетерогелицид, а также микроскопическими водоросля-
ми — кокколитофоридами.
Палеоген, особенно эоцен (средний палеоген), отличается разви-
тием многих групп фораминифер. Широкое развитие получают нумму-
литиды, обитавшие в средиземноморской области, в связи с чем палео-
геновую систему иногда называют «нуммулитовой». Нуммулитиды
принимали участие в накоплении больших толщ нуммулитовых извест-
няков. Наряду с нуммулитами не менее широко были распространены
и многие другие отряды фораминифер — атаксофрагмииды, булимини-
ды, милиолиды. Среди роталиид появляется много новых родов планк-
тонных фораминифер; у бентосных роталиид внутри раковины разви-
вается сложная система каналов.
Состав неогеновых и плейстоценовых фораминифер приближается
к современному, причем число многих групп фораминифер, широко»
120
распространенных в палеогене, сокращается; это относится в первую
очередь к нуммулитидам, альвеолинидам и атаксофрагмиидам. С другой
стороны, в неогене широкое развитие получают милиолиды, продолжают
развиваться роталииды.
Экология и тафономия фораминифер. Большая часть современных
фораминифер живет в нормальносоленых бассейнах и не переносит
колебаний солености (стеногалинные формы). Некоторые виды
приспособились к жизни в бассейнах с повышенным содержанием
солей (Красное море), а небольшое число видов — к пониженной соле-
ности (Черное и Каспийское моря). Немногие фораминиферы переносят
значительные колебания солености (эвригалинные формы), оби-
тая в прибрежных частях морей, их заливах, эстуариях рек; понижение
солености у таких форм приводит к уменьшению размеров, изменению
формы раковины, к замене известковой раковины органической; имеют-
ся указания на присутствие фораминифер в грунтовых водах некоторых
районов Средней Азии; здесь их считают реликтовыми формами акча-
гыльского бассейна, заходившего на территорию Каракумов.
Большинство современных фораминифер ведет бентосный образ
жизни, и лишь немногие приспособились к планктонному существова-
нию и обитают в пелагической зоне морей. Среди бентосных форм,
связанных с жизнью на дне, небольшая группа фораминифер неподвиж-
но прикрепляется к подводном- предметам. У таких форм (некоторые
представители аммодисцид, астроризид, текстулярид) раковина агглю-
тинированная, в виде простой или разветвленной трубки либо состоит
из ряда соединенных между собой трубок. Раковина может прикре-
питься целиком или только начальной камерой, и часть ее приподни-
мается над субстратом или стелется по нему. Среди аллогромиид из-
вестны формы, ведущие сверлящий образ жизни.
Большая часть фораминифер относится к подвижному бентосу,
обитая на илистом или песчаном дне, среди коралловых рифов, а также
на скалистом грунте. Они медленно передвигаются с помощью псевдо-
подий. Распространение ряда групп связано с областями развития
водорослей. В неритовой зоне современных морей обитают форамини-
феры, отличающиеся большим разнообразием. Мелководные формы
теплых морей имеют толстые, прочны,е известковые раковины, снаб-
женные богатой скульптурой. В некоторых местах количество этих
раковин настолько велико, что они принимают участие в образовании
прибрежных отложений. В северных морях распространены преимущест-
венно фораминиферы с агглютинированной раковиной, хотя существуют
и известковые.
В батиальных и абиссальных глубинах различных областей бентос-
ная фауна фораминифер довольно близка по родовому составу; в этих
областях, характеризуемых сравнительно низкой, более или менее
постоянной температурой, распространены формы с агглютинированной
и в меньшей степени с известковой раковиной.
Различные глубинные илы (известковый, голубой) изобилуют рако-
винами фораминифер. В северных морях известны: увигериновый ил,
заключающий многочисленных фораминифер, относимых к роду Uvi-
gerina, рабдамминовая глина с Rhabdamtnina и билокулиновая глина
с характерной формой Biloculina (теперь Pyrgo). Некоторые глубинные
илы содержат до 85% извести, возникшей за счет раковин бентосных
и планктонных фораминифер. Планктонные фораминиферы обитают
в верхних слоях пелагиали (от 0 до 100 м), но встречаются и глубже.
Падающие на дно раковины принимают участие в образовании танато-
ценозов глобигеринового, птероподового и других илов. Глобигериновый
121
ил широко распространен в тропическом й умеренном поясах до изоба-
ты в 4000 м.
У планктонных форм раковина шаровидная, тонкая, известковая,
с различными шипами и другими скульптурными образованиями, уве-
личивающими ее поверхность. Важнейший экологический фактор, обус-
ловливающий распространение фораминифер, особенно планктонных, —•
температура. Известны формы холодноводные, формы умеренного пояса
и тепловодные. Известковые скелеты фораминифер обычно не дости-
гают дна больших глубин, растворяясь на пути.
Изучение ископаемых фораминифер показывает, что в геологиче-
ском прошлом также были распространены две основные группы фора-
минифер: бентосные и планктонные. К первым относятся вымершие
эндотириды и гетерогелициды, фузулиниды, нуммулитиды и многие
другие; ко вторым — глобигерины. Бентосные фораминиферы—хоро-
шие показатели среды обитания, поэтому их изучение можно исполь-
зовать для палеогеографических построений и фациального анализа.
Планктонные формы, не связанные с дном, используются при страти-
графическом сопоставлении разнофациальных толщ, часто удаленных
друг от друга на большие расстояния.
Мелкие размеры, большое количество, широкое географическое
распространение делают фораминифер важными ископаемыми при
изучении стратиграфии, особенно в нефтеносных районах, где геоло-
гические исследования ведутся с применением буровых работ.
В грубообломочных породах фораминиферы обычно отсутствуют,
в алевритово-песчаных они встречаются сравнительно редко; наиболее
благоприятны для захоронения фораминифер глинистые и мергелисто-
известковые породы, в которых они встречаются иногда в массовом
количестве, в ряде случаев являясь породообразующими.
ПОДКЛАСС HELIOZOA. СОЛНЕЧНИКИ
К подклассу солнечников (гр. helios •— солнце) относятся саркодо-
вые (несколько десятков видов), обитающие в пресной и морской воде.
Большинство солнечников — свободно плавающие организмы, лишенные
скелета; только некоторые прикрепляются к. субстрату с помощью тон-
кого стебелька. Тело шаровидной формы с многочисленными тонкими
радиальными псевдоподиями,, имеющими внутри прочную эластичную
нить; такие псевдоподии называются аксоподиями. Центральная
капсула в отличие от радиолярий отсутствует. Цитоплазма распадается
на эктоплазму (!/6—’/ю всего диаметра) и эндоплазму. В эктоплазме
имеются крупные вакуоли; в эндоплазме расположен ядерный аппарат,
состоящий из одного или иногда из большого числа ядер (до 200, поли-
энергидная организация). В эндоплазме осуществляется пищеварение.
Часть солнечников имеет наружный минеральный кремневый скелет в
виде отдельных игл, пластинок или решетчатых шаров. Размножаются
бесполым путем, делением надвое или отделением почйк. При половом
процессе образуются разной формы гаметы. В ископаемом состоянии
солнечники неизвестны.
ПОДКЛАСС RADIOLARIA. РАДИОЛЯРИИ
Общая .характеристика. К подклассу радиолярий (лат. radiolus —
лучик) относятся морские саркодовые, ведущие планктонный образ
жизни. Они обладают центральной капсулой, состоящей из органиче-
ского вещества, и внутренним минеральным скелетом (кремневым или
122
целестиновым), разнообразной формы и строения. Современные радио-
лярии одиночные или очень редко колониальные животные, обычно
размером от 40—50 мк до 1 мм и более. Колонии достигают нескольких
-сантиметров.
Рис. 52. Подкласс Radiolaria:
1—2 — схема строения; 3 — Cistidium princeps, современный вид из Индийского
океана;
вп— внутрикапсулярная протоплазма, ж — капли жира, з — зооксантеллы, и —
Иглы, иск— наружный скелет, п — псевдоподии, сд — саркодиктиум, ск—сту-
денистая калимма, см — саркоматрикс, цк — центральная капсула, я—ядро
Строение тела. Внешняя форма тела и размеры варьируют в очень
широких пределах (рис. 52). Чаще всего тело имеет шарообразную
форму и расходящиеся по радиусам скелетные элементы и псевдо-
подии. Центральная капсула, представляющая собой тонкую органиче-
скую мембрану, разделяет цитоплазму на две части: внутрикапсуляр-
123
ную (интракапсулярную) и внекапсулярную (экстракапсулярную).
Внутрикапсулярная цитоплазма имеет довольно однообразное
строение и содержит кроме ядерного аппарата запасные питательные
вещества (жиры, белковые гранулы), иногда,зерна пигмента, окраши-
вающие тело в различные цвета. Радиолярии имеют одно полиплоидное
(полиэнергидное) ядро (все спумеллярии и часть насселлярий), возни-
кающее в результате повторного деления, без разделения его оболочки,,
или множество простых (моноэнергидных) ядер, развивающихся в-
процессе онтогенеза из одного первичного ядра. Внекапсулярная
цитоплазма, окружающая центральную капсулу, состоит из трех слоев:
а) слой зернистой цитоплазмы, прилегающей к центральной капсуле, —
саркоматрикс, от него отходят псевдоподии (слой зарождения
псевдоподий); б) средний слой студенистой цитоплазмы — калиМма,
однородного или грубовакуолистого строения; у мелководных радиоля-
рий обычно в нем живут водоросли-симбионты зоохлореллы и зооксан-
теллы; в) тонкий сетчатый слой — саркодиктион — покрывает ка-
лимму; от него отходят короткие и тонкие псевдоподии, разветвленные,
анастомозирующие. Псевдоподии улавливают различные планктонные
организмы, переваривание которых начинается вне тела, в псевдопо-
диях. Радиолярии — плотоядные организмы;
Строение скелета. Радиолярии обладают сложным изящно пост-
роенным скелетом, который располагается в экстракапсулярной части
протоплазмы и по своей конструкции повторяет форму центральной
капсулы. В простейшем случае (у акантарий) центральная кап-
сула представляет собой слой уплотненной цитоплазмы, разделяющей
интра- и экстракапсулярные части цитоплазмы, у других форм она
имеет форму шара, стенка которого построена из белкового вещества —
тектина; у спумеллярий форма центральной капсулы варьирует, она
обычно многоосная — шаровидная, дисковидная или овальная с одно-
слойной стенкой, пронизанной мелкими порами; у насселлярий она одно-
осная, шлемообразная, радиально-симметричная с плотной однослойной
стенкой, имеющей поровое поле, пронизанное многочисленными мелкими
отверстиями. У феодарий центральная капсула шаровидная, сплюснутая
у полюсов, двусторонне-симметричная с двойной стенкой; стенка прони-
зана одним или тремя отверстиями: главное отверстие лежит на плоской
стороне капсулы, прикрыто крышечкой и заканчивается трубчатым
хоботком; два других, дополнительных отверстия расположены на про-
тивоположном полюсе по бокам и также снабжены трубочками. Через
поры в капсуле осуществлялась связь между интракапсулярной и
экстракапсулярной цитоплазмой.
У большинства радиолярий скелет состоит из водного кремнезема
с большей или меньшей примесью органического вещества. У одной
из групп (акантарии) установлено присутствие целестина (сернокислого
стронция) и сложного алюмокальциевого силиката, содержащего крем-
незем. В ископаемом состоянии обычно скелеты акантарий не сохра-
няются; в донных осадках современных морей они также не обнару-
жены.
В редких случаях, у наиболее примитивных групп, скелет отсутствует
и поверхность тела радиолярий защищена посторонними частицами —
панцирями диатомей, скелетами мелких радиолярий или их обломками,
прикрепленными к поверхности внекапсулярной цитоплазмы (подобно
агглютинированным раковинам фораминифер).
Скелет закладывается в эктоплазме в виде отдельных минеральных
иголочек. Различают несколько типов скелетов. В простейшем случае
скелет состоит из отдельных простых или двойных игл, лежащих в ка-
124
лимдое (рис. 53); вторым типом можно считать астроидный скелет,
представленный 20 иглами, радиально расходящимися от капсулы, но
не соединенными в центре (см. рис. 55); разновидностью астроидного
Рис. 53. Отряд Spumellaria. Современные представители:
/ — Cromyatractus, стенки концентрических эллипсоидов вскрыты; 2 — Xiphostylus;
3 — Heliodiscus-. За — общий вид, 36 — сбору, Зв — в разрезе; 4— распределение ске-
летных игл в калимме у Lamproxanthium\
цк — центральная капсула, я — ядро
скелета служит скелет, у которого противоположные иглы соединяются
в центре тела и скелет оказывается состоящим из 10 игл. При соедине-
нии боковых отростков, отходящих от радиальных игл, формируется
125
более сложный шаровидный скелет, или сфероидный; при наличии
нескольких ярусов таких ответвлений возникает скелет, состоящий из-
нескольких шаров, вложенных один в другой (рис. 53, 1).
У насселлярий скелет имеет форму шлема, башенки, суженный на
одном конце и расширенный на другом; исходным типом такого скелета
является внутренний каркас (рис. 54, 1—4), имеющий вид балки, соеди-
няющей два центра схождения лучей; лучи^вторично ветвятся, и между
ними развиваются дуговидные соединения. Устьевой край скелета
может закрываться решетчатой диафрагмой. При увеличении размеров
в области устья возникают пережимы, приводящие к образованию
многокамерного скелета; вершина таких скелетов получила название
цефалума, или головного отдела, следующая за ним камера — '
торакса, или грудного отдела, третья камера названа абдоме-
ном; эти названия заимствованы из терминологии специалистов ш>
артроподам и представляются малоудачными.
Скелет феодарий состоит из полых трубчатых игл, лежащих ра-
диально и тангенциально; радиальные иглы лежат довольно беспоря-
дочно и никогда не проникают в центральную капсулу. В более слож-
ных случаях скелет шаровидный, но одноосный с большим отверстием
на одном конце. Любопытным типом скелета являются двустворчатые
скелеты, состоящие из двух пористых створок, соединенных с помощью
замка.
Скелет радиолярии выполняет роль опоры для цитоплазмы, способ-
ствует «парению» в толще воды; он обычно отличается сочетанием
удивительной легкости с большой прочностью. Различные выросты тела
значительно увеличивают его поверхность; тело радиолярии хорошо
приспособлено для планктонного образа жизни.
Размножение и развитие. Процесс размножения у радиолярий изу-
чен недостаточно. При половом процессе образуются жгутиконосные
одинаковые гаметы, после копуляции которых формируется зигота.
При бесполом размножении наблюдается деление надвое, при этом
недостающие половины в дальнейшем достраиваются. У видов, обла-
дающих сложным скелетом, такой способ размножения не возможен, щ
по-видимому, у таких радиолярий формируются «эмбрионы» — одно-
ядерные зародыши, подобные «эмбрионам» фораминифер. Если цито-
плазма при бесполом размножении не разделяется, то возникают коло-
нии, у которых в цитоплазме заключено много отдельных центральных
капсул (спумеллярии).
Основы систематики и классификация. На основании особенностей
строения стенки и формы центральной капсулы, - разнообразного
устройства и состава скелета среди радиолярий выделяются 4 отряда:
Spumellaria, Nassellaria, Acantharia, Phaeodaria. Выделенный недавно
новый отряд Stycholonchia содержит один род, в ископаемом состоянии
пока неизвестен.
Отряд Spumellaria. Спумеллярии (лат. spuma — пена, spumella —
пеночка). Скелет кремневый, разнообразной формы: шаровидный^
дисковидный, овальный, цилиндрический с одной или несколькими
скелетными сферами. Радиальные иглы у современных форм .не доходяг
до центра тела; у древних иглы либо соединялись в одной точке в-
центре сферы, либо отходили от короткой срединной балки. Централь-
ная капсула шаровидная, ее стенка однослойная, пронизанная много-
численными порами. Имеются одиночные и колониальные формы. Раз-
витие шло от бесскелетных форм к спумелляриям, имеющим отдельные-
иглы, рассеянные в калимме, затем к возникновению сфер (до 10) за
счет разрастания и соединения отростков соседних радиальных игл;
.126
Дальнейшее усложнение привело к образованию дисковидных много-
слойных скелетов, цилиндрических, расчлененных «метамерно» вдоль,
главной оси, и эллипсоидальных форм, имеющих три взаимно перпен-
дикулярные оси неравной длины. В составе отряда 5 подотрядов и
около 35 семейств (см. рис. 53). Кембрий — ныне.
Отряд Nassellaria. Насселлярии (лат. nassa—носик, рыльце). Ске-
лет кремневый, одноосный, в форме шлема, башенки, суженный на
одном конце и расширенный на другом, или кольчатый. В начальной
Рис. 54. Отряд Nassellaria. Современные и вымершие представители:
1—4 —схема строения скелета: 1 — Artostrobtum, 2 — Plagiocar ра, 3 — Cortiniscusr
4 — Acanthodesmia; 5 — Anthocyrtium (совр.); 6 — Eucyrtidium (палеоген); 7 —
Dictyomitra (поздний мел)
камере имеется внутренний каркас, за счет разрастания боковых от-
ростков которого образуются разнообразной формы скелеты. Цент-
ральная капсула одноосная со сплошной однослойной стенкой и порами,
сконцентрированными на базальном полюсе. Насселлярии возникли,
по-видимому, от спумеллярий в девоне. Их предки имели эллипсовид-
ный, несколько удлиненный скелет, внутри которого находился каркас,
состоявший из соединенных игл и дуг. Каркас подвешивался на иглах,
проходивших через стенки наружного скелета. Увеличение цитоплазмы
приводило к созданию многосегментного скелета. У части насселлярий
наблюдается редукция скелета и его роль принимает на себя стенка
центральной капсулы. В составе отряда 3 подотряда и 6 семейств
(рис. 54). Девон — ныне.
Отряд Acantharia. Акантарии (гр. acantha — колючка). Радиолярии
с 20 радиальными не сходящимися в центре скелетными иглами,
пронизывающими центральную капсулу, или с 10 сходящимися в
центре. Иглы образуют 5 поясов по 4 иглы в каждом. Они прикреп-
ляются к наружному слою цитоплазмы с помощью особых волоконец,
способных сокращаться и изменять общий объем цитоплазмы в зави-
симости от колебаний температуры и солености. Скелет состоит из
целестина и алюмокальциевого силиката. Примитивные акантарии не
имеют центральную капсулу, но разделение цитоплазмы на экто- и
эндоплазму сохраняется. В процессе эволюции, по-видимому, услож-
127
нялся скелет, появилась центральная капсула, представляющая собой
уплотненный слой цитоплазмы без пор; у игл развиваются отростки,
которые разрастаются, соединяются между собой и образуют пористые
шарообразные скелеты. В составе отряда 4 подотряда, 17 семейств
(риС. 55). В ископаемом состоянии известна одна проблематичная
находка из отложений верхней юры Предкавказья (Кисловодск).
Рис. 55. Отряд Acantharia. Скелетные иглы:
1 — Icosaspis; 2 — Zygacantha; 3 — Heliolithium
Отряд Phaeodaria. Феодарии (гр. phaios, phaeo — темный, темное
тело). Радиолярии с двухслойной центральной капсулой, имеющей
одно — три отверстия. Кремневый скелет представлен полыми иглами и
перекладинами, иногда образующими сплошной панцирь. До сих пор
не известно, в какой форме содержится кремнезем. В современных дон-
ных осадках известно только 11 видов. В ископаемом состоянии фео-
дарии только недавно, в 1964, 1965 гг., были установлены в миоцене
румынским микропалеонтологом Думитрикрм.
История развития радиолярий. Древнейшие остатки радиолярий
были описаны из докембрийских отложений Франции, но некоторые
исследователи склонны относить эти породы к кембрию. Достоверные
находки радиолярий известны из кембрия. Наиболее широким рас-
пространением и обилием форм пользуются радиолярии ордовика, си-
лура и девона. Они представлены обычно сферическими формами
(спумеллярии). В девоне появились перйые насселлярии. Для мезозоя,
особенно для юры и позднего мела, характерно наличие радиолярий с
эллипсоидными, плоскими, дисковидными, а также двух-, трех- и много-
камерными скелетами. В кайнозое, особенно в среднем палеогене (эоце-
не) и раннем неогене (миоцене), радиолярии многочисленны и
разнообразны, и нередко среди них преобладают формы со сфериче-
ским гладким, плоским дисковидным и круглым скелетом, эллипсои-'
дальные и колпачковидные. Радиолярии используются для целей био-
сдратиграфии в значительно меньшей степени, чем фораминиферы, что
объясняется их сравнительно небольшим количеством и сравнительно
слабой изученностью.
Экология и тафономия. Современные радиолярии — исключительно
морские пелагические животные, не переносящие колебаний солености
и обитающие на разных глубинах; их нет в морях с пониженной соле-
ностью. Основная масса радиолярий ведет пелагический планктонный
образ жизни и связана с теплыми бассейнами. Форма тела приспособ-
лена к парению в воде. Выделяют стенобатные формы, приурочен-
ные к строго определенным глубинам (0—50, 50—200, 200—1000, 1000—
2000, 4000—8000 м), и эврибатные, приспособленные к жизни на
разных глубинах. Наибольшее видовое разнообразие радиолярий уста-
128
новлено на глубинах от 0 до 400 м. Количество радиолярий резко сокра-
щается в умеренных и холодных водах; имеются виды, приуроченные
только к арктическим и антарктическим районам.
В теплых водах у радиолярий развиты приспособления, увеличи-
вающие их плавучесть; радиолярии имеют маленькие размеры и
тонкостенные скелеты, снабженные иглами; шипами и разветвленными
радиальными отростками. В холодных водах распространены формы
более крупные и толстостенные со слабым развитием различных от-
ростков. Глубоководным радиоляриям свойственны толстостенные
мелкопористые скелеты; у феодарий развивается сплошной панцирь.
В осадках небольших глубин скелеты радиолярий смешиваются с
раковинами фораминифер, но на глубинах свыше 4000 м остатки
радиолярий становятся преобладающими и глобигериновый ил сме-
няется радиоляриевым, занимающим значительные площади дна
современных океанов, в составе которого диатомеи, радиолярии, губки
составляют до 40%.
/ В прежние геологические эпохи планктонные радиолярии прини-
мали участие в накоплении таких осадочных горных пород, как радио-
ляриты, трепела, опоки, диатомиты, фосфориты. Они присутствуют в
спонголитах, вулканических туфах, кремнисто-известковистых глинах,
кремнистых известняках, мергелях, писчем мелу. Они хорошо сохра-
няются также в алевролитах. В процессе фоссилизации кремневый
скелет может замещаться халцедоном, кварцем, кальцитом, окислами
и .сульфидами железа. В исключительных случаях сохраняются остатки
центральной капсулы, органическое вещество которой превращается в
углистый пигмент. Остатки скелетов феодарий и акантарий, отличаю-
щиеся особым минеральным составом, не обнаружены, хотя А. Е. Ферс-
ман указывал на находки скелетов акантарий в позднеюрских породах
около Кисловодска.
Радиолярии из рыхлых пород извлекаются дезинтегрированием
породы с последующей отмывкой, отмучиванием, просушиванием осадка
и отбором скелетов. Для просветления скелетов используются различ-
ные жидкости. Изучение радиолярий, заключенных в твердые породы,
ведется в шлифах.
КЛАСС CNIDOSPORIDIA. КНИДОСПОРИДИИ
К классу книдоспоридий (гр. knide — жгутик, spora — зерно) отно-
сятся простейшие, ведущие паразитический образ жизни (паразиты
морских и пресных рыб, насекомых). Для них характерны одноядерный
амебоидный зародыш и особые споры, обладающие стрекательными
капсулами. Они происходят от саркодовых. В ископаемом состоянии
неизвестны.
КЛАСС SPOROZOA. СПОРОВИКИ
Споровики — простейшие, паразитирующие в различных тканях и
органах беспозвоночных, позвоночных и человека. Наличие в жизнен-
ном цикле у большинства споровиков жгутиковых стадий и особен-
ности полового процесса (копуляция) позволяют предполагать их про-
исхождение от жгутиконосцев. В ископаемом состоянии неизвестны.
КЛАСС ZOOMASTIGOPHORA. ЗООМАСТИГОФОРЫ
К зоомастигофорам, или жгутиконосцам (гр. zoon—жизнь, masti-
gos — бич, жгут, phoros — несущий) относятся виды, живущие в мор-
9 В. В. Друщиц
129
ской и пресной воде, питающиеся : либо сапрофитно (осмотический®
путем), либо гетеротрофно (за счет готовых органических соединений)-
Часть жгутиконосцев перешла к паразитическому образу жизни в раз-
личных животных (трихомонады, трипаносомы) и растительных (лепто-
монасы1) организмах.
Тело зоомастигофор окружено плотной эластичной оболочкой и
снабжено одним или несколькими жгутиками. Жгутики представляют-
собой волосовидные выросты цитоплазмы и выполняют функцию орга-
нелл движения. Одна группа жгутиковых (ризомастигины) в своем,
строении сочетает признаки двух классов — саркодовых и жгутиконос-
цев. На переднем конце у них имеется жгутик, а на поверхности тела,
хорошо развиты псевдоподии. Цитоплазма разделена на эктоплазму и
эндоплазму. Ризомастигины занимают промежуточное положение между
двумя классами и указывают на родственные связи жгутиковых и.
саркодовых. В ископаемом состоянии неизвестны. Класс зоомастигофор
выделен из большого класса жгутиконосцев, который объединял про-
стейших с автотрофным и гетеротрофным способами питания, а также
виды, сочетающие в себе черты обоих типов обмена. Автотрофных жгу-
тиконосцев, по-видимому, справедливо относить к растительным орга-
низмам (Phytomastigophora), а гетеротрофных — к животным (Zoomas-
tigophora).
f
КЛАСС CILIOPHORA. РЕСНИЧНЫЕ
Ресничные (лат. cilium, мн. ч. cilia — ресница, photos — несу-
щий) — наиболее сложно устроенные организмы среди всех простей-
ших; они представляют собой полиэнергидных простейших. Движение
в течение всей жизни или только на молодых стадиях осуществляется
с помощью ресничек — коротких (10—15 мк) волосяных выростов на-
ружного слоя протоплазмы. Число ресничек может достигать 10 000—
14 000; Каждая ресничка отвечает жгуту жгутиконосцев. Ядерный ап-
парат состоит из двух ядер: большого — обычно полиплоидного макро-
нуклеуса и маленького — микронуклеуса, различных по своему строе-
нию и функциям. Бесполое размножение состоит в поперечном делении;,
половой процесс протекает по типу конъюгации — временному соеди-
нению двух особей, в течение которого они обмениваются своими поло-
выми ядрами (микронуклеусами). В настоящее время известно около
6000 видов. К этому классу относятся два подкласса: ресничные инфу-
зории (Ciliata) и сосущие инфузории (Suctoria). В' ископаемом состоя-
нии встречаются представители первого подкласса.
ПОДКЛАСС CILIATA. РЕСНИЧНЫЕ ИНФУЗОРИИ
Тело разделено на эктоплазму, имеющую сложное строение и вы-
деляющую тонкую оболочку пелликулу, и эндоплазму, содержа-
щую микро- и макронуклеус. Имеется своеобразное ротовое отверстие,,
ведущее в глотку. Тело покрыто ресничками полностью или частично.
Ресничные инфузории живут в пресных водах и морях; часть держится
в планктоне, часть ведет бентосный образ жизни. Некоторые морские-
пелагические ресничные инфузории имеют псевдохитиновый скелет-
(ло-рику), имеющий форму цилиндра, конуса или полусферы, поверх-
ность которого укреплена агглютинированными минеральными части-
цами. Передний конец тела, спрятанный в лорику, снабжен венчиком
мембранелл — сросшихся ресничек, которые высовываются наружу ж
служат для передвижения животного. Лорика обнаруживает ряд при-
130
способлений к активному движению; она узкая, вытянута в длину, снаб-
жена спирально расположенными килями, имеет заостренный задний
край и отогнутый назад , пе-
редний край; для увеличения
прочности лорика снабжена
сотообразной скульптурой.
Скелеты этих инфузорий от-
носимые к группе тинтиннин
(Tintinnina), встречаются в
ископаемом состоянии. Совре-
менные тинтиннины составля-
ют значительную часть планк-
тона морей; они встречаются
также в солоноватоводных
бассейнах (Каспийское море)
и в пресных озерах (Байкал,
Балатон). Размеры скелета
варьируют от 50 до 1000 мк,
составляя в среднем от 50 до
200 мк. В ископаемом состоя-
нии остатки известковых ске-
летов тинтиннин известны пре-
имущественно в известковых
породах, начиная с поздней
юры (рис. 56). Стенка имеет
волокнистую структуру, в ко-
торой волоконца ориентирова-
ны перпендикулярно поверхно-
сти. Предполагается, что при
Рис. 56. Класс Ciliophora:
1—2 — схема строения современного пред-
ставителя рода Tintinnopsis; 3—4 — скелет
Tintinnopsella carpathica Murg. et Filip,
(поздняя юра — ранний мел);
вд — воротничок домика, р — реснички,
хп — хвостовой придаток
захоронении происходило
обызвествление органического скелета.
ФИЛОГЕНИЯ ПРОСТЕЙШИХ
Простейшие одноклеточные с обособленным ядром, напоминающие
современных амебообразных и моноэнергидных существ, произошли,
по-видимому, в протерозое, а может быть и раньше от каких-то пер-
вичногетеротрофных организмов, еще не имевших клеточной организа-
ции (прокариот). От них не сохранилось никаких следов. Они дали,
с одной стороны, саркодовых, с другой — примитивных жгутиконосцев’
с очень мобильной организацией, легко переходящих в амебоидное
состояние (рис. 57).
Среди саркодовых сразу же наметилось два пути адаптации — к
ползающему образу жизни (корненожки и фораминиферы) и к плаваю-
щему,. планктонному, образу жизни (радиолярии и позднее солнеч-
ники).
Корненожки без раковины или с песчаной раковиной в ископаемом
состоянии почти неизвестны и их эволюцию проследить нельзя. Фора-
миниферы прошли сложный путь развития от форм, имеющих только
органический скелет или агглютинирующих различные посторонние
частицы, до форм с минеральным известковым скелетом. Они известны
начиная с кембрия и их развитие было рассмотрено выше. Радиолярии
также прошли длинный путь развития от бесскелетных форм к формам,
имеющим на начальной стадии органический внутренний скелет в виде
центральной капсулы, а затем минеральный, преимущественно крем-
9
131
невый скелет, состоящий из рассеянных в цитоплазме игл, а позднее
образующих разнообразный, иногда очень сложный минеральный ске-
лет. Время возникновения солнечников и их эволюция остаются не-
ясными.
Часть амебообразных гетеротрофных одноядерных организмов
перешла к паразитическому образу жизни и дала новый класс книдо-
споридий.
Metazoa
Царство
растений
t о z о а
и п
Procaryota
Рис. 57. Филогения ’простейших (Иванов, 1968, с: изменениями)
кл Phyto-
mostigo -
Zoomastigophor'a
к/i Sporozoa
п/кл. Rhizopoda
п/кл Foraminifera
п/кл. Radiolaria
п/кл. Heliozoa
к л. Sa reading
Cnidospondia
От примитивных жгутиконосцев (ризомастигин) произошли формы,
способные к фотосинтезу, — новое направление в развитии органическо-
го мира, которое привело к возникновению и развитию растительного
царства. От тех же примитивных жгутиконосцев, вероятно, возникли и
развились настоящие животные жгутиконосцы. От последних произо-
шли ресничные, или инфузории, представляющие боковую слепую ветвь
Специализированных простейших, приспособленных к самым разнооб-
разным средам обитания. Развитие ресничного аппарата у инфузорий
й изменение ядерного аппарата привели к значительному повышению
жизнедеятельности и усложнению форм взаимосвязей с внешней
средой. Такого рода преобразования могут быть отнесены к ароморфо-
зам. От жгутиконосцев, вероятно, произошли споровики, ведущие пара-
зитический образ жизни. Наконец, от колониальных форм животных
жгутиконосцев;— зоомастигофор — возникли многоклеточные, история
развития которых будет рассмотрена ниже.
METAZOA. МНОГОКЛЕТОЧНЫЕ
Происхождение и развитие многоклеточных. Существует достаточ-
но много гипотез о происхождении многоклеточных, недавно подробно
и критически рассмотренных А. В. Ивановым в работе «Происхождение
многоклеточных организмов» (1968). Большинством исследователей
принимаются гипотезы происхождения многоклеточных от клеточных
предков, и лишь немногие придерживаются мнения о независимом
происхождении клеточных структур простейших и многоклеточных от
доклеточных предков. Первая группа гипотез исходит из предположе-
ния о происхождении метазой от многоядерных или колониальных
простейших (протозойные гипотезы). Наибольшее распространение
получили теория г а с т р е и Геккеля (1872) и теория ф а гоците л л ы
Мечникова (1877), предполагающие, что первичные многоклеточные были
одиночными свободноподвижными животными.
Бионт простейшего представляет собой одну клетку, или колонию
клеток, или полиэнергидный комплекс. Бионт многоклеточных в отли-
чие от простейших состоит из большого числа клеток, дифференциро-
ванных на соматические (тканевые) и половые; соматические клетки
образуют различного типа ткани и органы, которые специализируются
на выполнении какой-либо одной или нескольких функций и подчи-
няются организму как единому целому. В то же время, если бионт
простейшего состоит из колоний клеток, то составляющие его отдель-
ные клетки дифференцированы слабо. Если бионт простейшего пред-
ставлен одной клеткой, подобной тканевой клетке, то эта клетка в от-
личие от последней физиологически и экологически является само-
стоятельным организмом. Простейшие и многоклеточные отличаются
строением ядерного аппарата и характером жизненного цикла. Если
строение ядра и жизненные циклы простейших очень разнообразны, то
для многоклеточных характерен специализированный способ деления
ядра (типичный кариокинез, см. стр. 53) и определенный тип
жизненного цикла, который для всех многоклеточных — от губок до
человека, за небольшим исключением, сохраняется единым. Жизненный
цикл состоит в следующем: мужская и женская гаплоидные гаметы
сливаются вместе и образуют зиготу; диплоидная зигота начинает
дробиться, распадаясь на ряд клеток, получивших название бласто-
мер. В результате последовательного дробления размеры бластомеров
уменьшаются и возникает шаровидной формы зародыш — морула без
полости или бластула с первичной полостью внутри (рис. 58, 1, 2).
В процессе дальнейшего развития, обусловленного морфогенетиче-
скими корреляциями, образуется двухслойный зародыш, или гастру-
133
л а. Образование гаструлы может происходить тремя путями: имми-
грацией, инвагинацией и эпиболией (рис. 58, За—в).
При иммиграции часть наружных клеток бластулы, размно-
жаясь, внедряется внутрь зародыша, где образует плотную внутреннюю
клеточную массу, дающую у кишечнополостных начало энтодерме.
Рис. 58. Эмбриональное развитие многоклеточных:
1 — морула гидроидного полипа; 2 — бластула ланцетника; 3— различные
способы образования гаструлы: За — инвагинация, 36 — иммиграция, Зв —
эпиболия; 4 — различные типы развития мезодермы: 4а — телобластиче-
ский — у первичноротых, 46 — энтероцельный — у вторичноротых;
мк — мезодермальные карманы, мп — мезодермальные полоски, пп —
первичная полость, т — телобласт, эк — эктодерма, эн — энтодерма
Подобный тип зародыша Мечников (1877) назвал фагоцителлой; на-
ружный слой ее клеток.он назвал кинобластом, а внутреннюю
клеточную массу — ф а г о ц и то б л а ст о м.
При инвагинации, стенки бластулы на одном полюсе впячи-
ваются внутрь и образуют энтодерм у, выстилающую первичную
пищеварительную полость, которая открывается наружу первичным
ртом, или б л а с т о п о р о м- Инвагинация наблюдается у сцифоидных
134
медуз и актиний. Геккель (1872) назвал описанный гаструлоподобный
«организм гастреей и разработал теорию, в соответствии с которой
«от гастреи произошли все многоклеточные животные. Геккель, на осно-
вании работ Вольфа, Пандера, Бэра, А. О. Ковалевского (см. стр. 38),
•обосновал свой биогенетический закон и идею о гомологии эктодермы
и энтодермы у всех многоклеточных. Геккель, приписывал абсолютное
рекапитуляционное значение начальным стадиям онтогенеза, считая,
что каждой стадии соответствует определенный предок. Особенно важ-
ное значение он придавал гастрее; от нее все многоклеточные унаследо-
вали первичный кишечник и два первичных зародышевых листка, из
которых развились все остальные ткани. Мезодерма в процессе эволю-
ции возникла позднее.
При эпиболии мелкие клетки (микромеры) обрастают более
крупные клетки (макромеры). Этот способ образования зародышевых
.листков наблюдается у моллюсков.
Развитие мезодермы характерно для так называемых трехслойных
многоклеточных. Мезодерма образуется двумя способами: телобласти-
ческим и энтероцельным (рис. 58, 4a, б).
При телобластическом способе в области бластопора
гаструлы обособляются две крупные клетки — телобласты. Они
делятся и образуют симметрично расположенные полоски клеток, кото-
рые в дальнейшем заполняют всю первичную полость гаструлы. Бласто-
лор превращается в ротовое отверстие взрослого животного. Этот
способ образования мезодермы характерен для большой группы трех-
слойных животных, объединяемых в подраздел первичноротых
(Protostomia).
При энтер о цельном способе образование мезодермы проис-
ходит иначе: от первичного кишечника отшнуровываются три пары
мешковидных впячиваний, которые разрастаются и заполняют первич-
ную полость гаструлы (рис. 58, 46). Бластопор замыкается, рот воз-
никает в другой части зародыша, а на месте бластопора образуется
•анальное отверстие. Этот способ формирования мезодермы характерен
.для второй группы трехслойных животных, объединяемых в подраздел
вторичноротых (Deuterostomia). У первичноротых и вторично-
ротых внутри мезодермы в дальнейшем образуется вторичная
яолость тела, или целом, выстланная слоем, эпителиеподобных
клеток.
Тело многоклеточных, таким образом, представляет собой в разной
степени интегрированную и индивидуализированную систему, ограни-
чивающую самостоятельность клеток, входящих в состав тела и подчи-
няющую себе их жизнедеятельность. Одной из важнейших черт про-
грессивной эволюции многоклеточных является усиление интеграции их
•организма; чем выше степень его интеграции, тем выше выражена его
собственная индивидуальность, тем более она подчиняет себе отдельные
клетки, тем более стирается их индивидуальность. Каждый тип много-
клеточных обладает определенным планом строения, определенным
уровнем организации, свойственным только.этому типу, и соответствую-
щими ступенями симметрии.
В развитии многоклеточных, по мнению А. В. Иванова, можно вы-
делить следующие стадии: 1) свободноплавающая шаровидная колония
с периферическим расположением, жгутиконосцев; все индивиды коло-
нии однородны; размножение бесполое; 2) колония с клетками, диф-
ференцированными на соматические и половые, с радиальной симмет-
рией; появление полового процесса; 3) возникновение свободноплаваю-
щего организма типа мечниковской фагоцителлы и разделение клеток
135
на наружный кинобласт и внутренний фагоцитобласт; разделение всего
онтогенеза на эмбриональный период, когда развивалась
бластулообразная личинка, и постэмбриональный, в течение ко-
торого осуществлялся рост, сопровождаемый делением клеток.
Дальнейшее развитие предположительно шло следующим путем
(рис. 59). Губки отделились от основного ствола многоклеточных, когда
у фагоцителлы еще не было ни рта, ни кишечника. Кинобласт у губок
сменил локомоторную функцию на гидрокинетическую, погрузившись
внутрь фагоцитобласта. В то же время, возможно, отделились архео-
циаты, крибрициаты и ряд групп неясного систематического положения.
Позднее, когда у фагоцителлы возникло ротовое отверстие, от основного
ствола возникли две новые группы многоклеточных. Одна дала начало
гастрееобразным первичным кишечнополостным, вторая — бескишеч-
ным сколецидам, у которых еще не обособился кишечник и пищеваре-
ние происходило в фагоцитах — блуждающих клетках — или в полостях
паренхимы.
От первичных кишечнополостных возникли настоящие кишечно-
полостные, ведущие сидячий образ жизни (гидроидные и коралловые
полипы) или перешедшие к подвижному образу жизни (сцифоидные
медузы), и гребневики, приспособленные к жизни в пелагиали. 'Фаго-
цитобласт этих групп превратился в первичный пищеварительный ор-
ган— энтодерму, достигшую сложного строения. За счет кинобласта
развилась эпителиальная ткань, покрывающая наружную поверхность
тела. Кишечнополостные и гребневики, достигшие в своем развитии
только стадии гаструлы (двухслойного мешка), объединяются в под-
раздел двухслойных многоклеточных. Как отмечалось выше, у
всех остальных многоклеточных закладывается третий зародышевый
листок—мезодерма и они образуют группу трехслойных живот-
136
ных. Среди первичноротых выделяют бесполостных и первичнополост-
ных, к которым относятся низшие черви. У них появилась кровеносная
система, выполняющая функцию газообмена и переноса питательных
веществ, обособились мышечная и нервная ткани, у части низших чер-
вей возник сквозной пищеварительный тракт, появились органы выде-
ления. Вторая группа первичноротых, наиболее разнообразная и много-
численная, представлена вторичнополостными, или целомическими,
организмами. Это — кольчатые черви, онихофоры, членистоногие, мол-
люски, мшанки, форониды. Развитие каждой группы шло своим путем
и будет рассмотрено ниже.
Развитие вторичноротых, к которым относятся иглокожие, геми-
хордовые и хордовые, шло по пути повышения организации, совершен-
ствования всех систем тканей и органов и привело в конце концов к
становлению человека. Иглокожие в своем развитии уклонились от
основного ствола вторичноротых и в связи с прикрепленным или мало-
подвижным образом жизни приобрели радиальную симметрию тела и
особую амбулакральную систему. Промежуточное положение между
первичноротыми и вторичноротыми занимают брахиоподы. До сих пор
неясным остается систематическое положение погонофор и хетогнат.
НАДРАЗДЕЛ PARAZOA.
НИЗШИЕ МНОГОКЛЕТОЧНЫЕ
Тело низших многоклеточных (гр. para — около, вблизи, zoon —
животное) состоит из клеток, не дифференцированных на ткани; у них
отсутствуют обособленные первичные зародышевые листки — эктодер-
ма и энтодерма. Разного типа клетки способна превращаться друг
в друга. Отдельные особи в колониях не разграничены. Настоящая
(обособленная) нервная система отсутствует. Эмбриогенез отличается
от эмбриогенеза всех остальных многоклеточных.
К низшим многоклеточным относятся губки, условно археоциаты
и, по-видимому, большая группа губко- и археоциатоподобных организ-
мов, известных по скелетным остаткам только в ископаемом состоянии.
ТИП SPONGIA. ГУБКИ
Общая характеристика. К типу губок (гр. и лат. spongia — губка)
относятся наиболее низкоорганизованные многоклеточные животные,
ведущие прикрепленный образ жизни. Для них характерно:
— наличие своеобразного ирригационного, или мерцательно-водо-
движущего, аппарата, состоящего из системы каналов и камер и
хоаноцитов;
— большая самостоятельность клеточных элементов, не создаю-
щих обособленных тканей и органов, за исключением жгутикового
эпителия и паренхимы. Паренхима состоит из мезоглеи и клеточных
элементов и поддерживается органическим или минеральным скелетом;
— особенности эмбрионального развития. Личинка двухслойная;
наружный слой жгутиковых клеток при метаморфозе личинки мигри-
рует внутрь и дает начало хоаноцитам.
Губки — водные, преимущественно морские, животные, колониаль-
ные и одиночные, размером от нескольких миллиметров до 2 м и более.
Немногие приспособились к жизни в пресных водах. Известны с кемб-
рия доныне.
Строение мягкого тела. Внешняя форма губок очень разнообразна,
изменчива и часто неопределенна. Одиночные губки имеют вид цилинд-
ра, кубка, бокала, шара, гриба или корзинки, сидящей на длинном
138
•стержне. Колониальные губки принимают вид лепешек, ковриг, натеков,
наростов, корок или многоветвистых колоний. Губки прикрепляются
неподвижно к грунту, подводным предметам или к скелетам других
животных (моллюсков, брахиопод, ракообразных).
В теле различают три основные группы клеток — хоаноциты, дер-
-мациты и амебоциты (рис. 60, 1а). Хоаноциты, или воротничково-
Рис. 60. Схема строения (1а) и типы ирригационных систем современных губок:
1 — асконоидный; 2— сиконоидный; 3 — лейконоидный; а — продольные, б — попереч-
ные сечения;
вк— вводной канал, вп—вводнаядора, к — разного типа клетки и спикулы в мезоглее,
ос'—оскулюм, пк — покровные клетки (дермациты), х— хоаноциты
жгутиковые клетки, выстилают центральную, или атриальную, полость
или сконцентрированы в специальных жгутиковых камерах. Они обра-
зуют единственную морфологически обособленную ткань — жгутико-
вый эпителий, основная функция которого состоит в создании тока
воды, с которым в тело губки поступает пища. Хоаноциты имеют вид
удлиненных клеток, снабженных каждая жгутиком, основание которого
-окружено тончайшим • протоплазматическим воротничком, состоящим,
как видно в электронном микроскопе, из десятков ворсинок. Хоаноциты
взрослой губки имеют большое сходство с сидячими жгутиконосцами.
Равномерное колебание жгутиков обеспечивает в теле губки постоян-
ный ток воды, которая входит через многочисленные поры и по каналам
попадает в жгутиковые камеры, затем в атриальную полость и выхо-
дит наружу через выводное, или оскулярное, отверстие.
Жгутиковому эпителию может быть противопоставлена паренхима,
состоящая из покровных клеток и мезоглеи. Покровные, или эпи-
телиальные, клетки (дермациты) покрывают тело губки снаружи;
некоторые из них (пороциты) несут внутриклеточный канал, откры-
вающийся наружу порой (отсюда второе название типа Porifera — несу-
139
щие поры); по каналу внутрь тела поступает вода и частицы пищи,
которые, прежде чем попасть в канал, подвергаются своеобразному
отбору по размерам и качеству. Если вода приносит «неприятные»
вещества, то пороцит сокращается и закрывает отверстие. Основу
паренхимы составляет мезоглея — бесструктурное студенистое ве-
щество, выделяемое особыми клетками. В мезоглее расположены раз-
личные клетки: археоциты — недифференцированные клетки не-
правильной формы, выполняющие разные функции и способные
превращаться во все прочие клетки, в том числе и половые; амебоци-
ты— осуществляющие функции внутриклеточного пищеварения, пере-
носа пищевых частиц, выделения; клетки-скелетообразователи и звезд-
чатые клетки — соединительно-тканные опорные элементы.
Характерной особенностью всех губок является система каналов и
камер, образующих так называемый ирригационный, или мерца-
тельно-вододвижущий, аппарат, выполняющий функции доставки пищи,
газообмена и выделения. Особенности его строения и различие в соста-
ве и конструкции скелета создают все многообразие губок. У губок
различают три основных типа строения ирригационного аппарата:
асконоидный, сиконоидный и лейконоидный (рис. 60). У асконоид-
ного типа (аскон)—наиболее примитивного, свойственного первым
стадиям развития губок и некоторым взрослым известковым губкам,,
хоаноциты выстилают всю центральную полость (рис. 60, 1). В следую-
щей по сложности стадии — сиконоидной (сикон) — жгутиковый
Впителий втягивается внутрь паренхимы и образует особые камеры или
трубчатые полости, выстланные хоаноцитами (рис. 60, 2). Жгутиковые-
камеры связаны с наружной средой системой вводных каналов, а в
центральную полость открываются большим отверстием. В лейко-
ноидном типе (лейкон)—-наиболее сложном и характерном, за не-
большим исключением, для всех взрослых стадий губок, жгутиковые
камеры обособлены, расположены в толще тела и снабжены одним
отводным каналом и системой вводных каналов (рис. 60, 3). У коло-
ниальных губок ирригационный аппарат достигает особой сложности.
Строение скелета. Стенки тела губок, образованные паренхимой,
поддерживаются внутренним скелетом, расположенным в мезоглее.
По составу скелет может быть органическим или минеральным. Орга-
нический, или роговой, скелет состоит из спонгиновых волокон, у кото-
рых можно различать более рыхлую спонгиновую сердцевину, окру-
женную концентрическими слоями плотного спонгина. Спонгин
представляет собой органическое вещество, пр составу близкое к белку
шелка, который является наиболее простым волокнистым белком, по-
строенным из линейных полипептидных цепей весьма характерного
аминокислотного состава и высокого молекулярного веса. Спонгин вы-
деляется особыми клетками — с по н го б л а ст а м и, которые строят
вначале сердцевидную часть, а затем на ней отлагают концентрические
слои органического вещества.
Минеральный скелет состоит из спикул. Спикулы могут быть
кремневыми и известковыми. Кремневые спикулы состоят из водного
кремнезема и закладываются внутри особых клеток — склеробласт.
Вначале образуется осевая нить из органического вещества, которая
ватем окружается кремнеземом. После достижения размеров склеро-
бласта молодая спикула освобождается и ее дальнейшее образование
осуществляется другими клетками, выделяющими на ее поверхности
концентрические слои аморфного кремнезема.
Среди кремневых спикул различают крупные спикулы — макро-
склеры и мелкие спикулы — микросклеры. Макросклеры образуют
140
«основу скелета -и бывают: одно-, трех-, четырех- и многоосными
(рис. 61). О д н о о с н ы е спикулы имеют вид стержня либо с одинако-
во, заостренными или притупленными концами, либо с различно по-
строенными концами. Трехосные спикулы состоят из шести лучей,
расположенных в трех взаимно перпендикулярных плоскостях. Отдель-
ные лучи могут редуцироваться и даже исчезать. Лучи могут быть
Рис. 61. Спикулы губок:
7—2 — известковые спикулы: 7 — фаретронный скелет, 2—известковые трех-
лучевые спикулы; 3—15 — кремневые спикулы: 3—4 — спикулы кембрийской
кремневой губки, 5—11 — микросклеры — спикулы трехосных кремневых губок
(5 —сигма, 6 — хела), 12—13 — макросклеры — спикулы трехосных кремневых
губок: 12 — диктиональный скелет, 13 — различные виды трехосных спикул,
14—15 — десмы — спикулы четырехосных кремневых губок: 14 — литистидный
скелет, 15 — четырехлучевые десмы
прямыми, изогнутыми, разветвленными, нести шипы или вздутия.
У многих губок лучи трехосных спикул могут соединяться вместе, обра-
зуя своеобразную пространственную решетку с кубическими петлями,
так называемый д и к т и о н а л ь н ы й скелет (рис. 61, 12). Ч етыр ех-
осные спикулы состоят из четырех лучей, расходящихся под разными
углами; чаще всего один из лучей бывает длиннее трех остальных и его
называют рукояткой; остальные три луча образуют ее ветви; они
могут быть направлены вперед или назад от рукоятки, перпендикуляр-
но к ней; их окончания могут заостряться или, наоборот, расширяться.
Грубые крупные спикулы, покрытые дополнительными вторичными
отложениями кремнезема, образующего неровности и бугры, называют-
ся десмами. Десмы имеют разнообразную форму и, плотно соеди-
няясь друг с другом, образуют так называемый литистидный
скелет (рис. 61, 14). Известны также многолучевые спикулы,
имеющие вид звездчатых тел и состоящие из 8—10 и больше лучей.
Микросклеры рассеяны в мезоглее. Каждая микросклера
создается внутри одной клетки. Внешняя форма их очень разнообраз-
на (рис. 61, 5—11): они могут быть построены по трехосному типу,
141
иметь вид многолучевых звездообразных тел, изогнутого стерженька,
снабженного на концах крыловидными придатками (хелы), или
S-образно изогнутых спикул (сигмы).
Известковый скелет состоит из углекислой извести (кальцита).
Различают три типа известковых спикул: одноосные, имеющие вид.
игл, обычно заостренных на обоих концах, трехлучевые, состоящие-
из трех лучей, лежащих в разных плоскостях (т р и р а д и а т ы), и
четырехлучевые, сходные с трирадиатами, но имеющие четвертый
луч, отличный по форме и размерам. Каждый луч этих спикул строится
парой клеток; утолщение и рост в длину осуществляется новыми клет-
ками, возникшими в результате деления первой пары. Основной формой
известковых спикул являются трирадиаты. Обычно спикулы распо-
лагаются в мезоглее несвязно, реже они образуют связный, или
фаретронный, скелет (рис. 61, 1). В последние годы обнаружен
известковый скелет, построенный из кристаллов арагонита. В основной
известковой массе встречены единичные кремневые спикулы.
Размножение и развитие. Губки размножаются бесполым и поло-
вым путем. При бесполом размножении различают наружное почкова-
ние и образование геммул. При наружном почковании, свиде-
тельствующем о низком уровне интеграции особей, на материнском
организме из амебоцитов возникает небольшой бугорок — почка, у кото-
рой позднее появляются поры и возникает устье. Иногда вновь образо-
вавшаяся почка отделяется от материнского организма и начинает
вести самостоятельный образ жизни. Одиночная особь приобретает
радиальносимметричную форму тела с одним устьем и единой иррига-
ционной системой. Однако чаще новые почки не отделяются от мате-
ринского организма и создают колонии. У наиболее примитивных коло-
ний отдельные особи — зооиды — соединяются своими основаниями и:
мало отличаются от одиночных форм. Но у большинства губок зооиды
входят в состав колоний, теряют свою индивидуальность и колония:
растет как единое целое. Образование такого типа колоний связано»
с простотой строения губок и отсутствием централизации функций. От
каждой особи, вошедшей в состав колонии, сохраняется только устье и
прилегающая к нему часть отводящей системы. В некоторых случаях не
только особи, возникающие почкованием, остаются слитыми с коло-
нией, но и особи, живущие рядом, соприкасающиеся друг с другом,,
могут сливаться в единое целое.
Образование геммул наблюдается у пресноводных и некоторых
морских губок. Геммулы имеют шаровидную форму и состоят из скоп-
лений амебоцитов, богатых питательными веществами, окруженных хи-
тиноидной оболочкой. Геммулы не только служат для размножения и
расселения, но и представляют покоящуюся стадию, способную пере-
носить неблагоприятные условия (засуха, замораживание). При благо-
приятных условиях из геммулы образуется новая губка.
Специальный половой аппарат у губок отсутствует, но среди них
есть раздельнополые и гермафродитные формы. Половые клетки — га-
меты — возникают из археоцитов в мезоглее и обслуживаются специаль-
ными питательными клетками; женские гаметы — яйцеклетки —
имеют вид амебы и образуются вблизи жгутиковых камер; мужские
гаметы — спермин — возникают преимущественно в наружных слоях
паренхимы. Зрелые спермин выводятся наружу по каналам иррига-
ционной системы. Каждая губка, фильтруя воду, улавливает спермин
Других особей своего вида. Оплодотворение зрелых яиц и их дробление1
осуществляется внутри материнского организма. Возникшая личинка —
паренхимула — имеет размер до 1 мм и состоит из внутренней
142
массы рыхло расположенных крупных зернистых клеток, окруженных
мелкими жгутиковыми клетками (рис. 62, 3). Личинка, поплавав не-
которое время, опускается на дно и прикрепляется к субстрату. На-
ружные жгутиковые клетки мигрируют внутрь и превращаются в хоано-
циты. В свою очередь крупные клетки перемещаются наружу и дают
мезоглее.
начало покровным клеткам и
У части известковых
губок личинка имеет вйд
пузырька (целобласту-
л а, рис. 62, /), затем она
превращается в амфи-
бластулу (рис. 62, 2).
У одного ее полюса распо-
ложены мелкие, высокие и
узкие клетки, несущие жгу-
тики; на другом полюсе —
крупные, зернистые. В та-
ком виде личинка выходит
из тела губки и начинает
Рис. 62. Личинки губок:
1 — целобластула; 2 — амфибластула; 3—
паренхимула
плавать с помощью жгути-
ков. Затем она опускается на дно и прикрепляется к субстрату. Мел-
кие клетки мигрируют внутрь и дают начало хоаноцитам. Крупные
клетки, так же как у паренхимулы, оказываются снаружи, создают
покровный слой, мезоглею и все остальные виды клеток.
Таким образом, у обеих личинок первично-наружные клетки ухо-
дят внутрь, образуя там жгутиковый эпителий, единственную морфо-
логически обособленную ткань. С момента ее обособления остальная
паренхима становится второй тканью, но чрезвычайно пестрой по свое-
му составу. Губки как бы выворачиваются наизнанку с тем, чтобы
создать ирригационный, или мерцательно-вододвижущий, аппарат.
Подобное «извращение зародышевых пластов» не наблюдается у других
многоклеточных.
В результате личиночного метаморфоза возникает послеличиночная
стадия — олинтус или рагон. Олинтус — это . мешковидная
губка асконоидного типа, из которой по мере роста формируются из-
вестковые губки. Рагон имеет вид сплюснутой губки сиконойдного
строения с обширной атриальной полостью; через некоторое время ра-
гон преобразуется в молодую губку с лейконоидным строением (класс
обыкновенных губок).
Основы систематики и классификации. Единой общепринятой сис-
тематики губок нет. В зависимости от строения и химического состава
скелета, формы спикул и особенностей эмбрионального развития в типе
губок большинство исследователей выделяет три класса: Demospongfa,
Hexactinellida, Calcispongia. В последние годы (1970) выделен новый
класс Sclerospongia.
КЛАСС DEMOSPONGIA. ОБЫКНОВЕННЫЕ ГУБКИ
Общая характеристика. К классу обыкновенных губок (гр.
demos — народ, множество) относятся морские, реже пресноводные,
формы. Скелет спонгиновый или кремневый, у ряда форм смешанный,
состоящий из сети спонгиновых волокон, к которым приклеиваются
кремневые спикулы. Иногда спонгин обволакивает пучки спикул. У не-
которых форм скелет литистидный. Макросклеры одно- или четырехос-
ные. Микросклеры встречаются редко и представлены сигмами или
143
хелами (см. рис. 61). Ирригационная система у взрослых форм лейконо-
идного типа. В ископаемом состоянии известны начиная с протерозоя.
Выделяется несколько отрядов, но общепринятого деления нет.
Среди обыкновенных губок известны роговые, кремнероговые, одно-
осные и каменистые губки. У роговых губок спонгиновый скелет
Рис. 63. Класс Demospongia. Отряды Heteractinellida (/), Lithistida (2—4)
и Cornacuspongia (5):
1 — Chancelloria (ранний кембрий), реконструкция (Журавлева, Кордэ,
1955); 2 — Jerea (мел); 3 — Siphonia (поздний мел—палеоген): За — об-
щий вид, 36 — продольный разрез; 4 — Astylospongia (ордовик — силур);
5 — Pemmatites (карбон—пермь)
состоит из древовидно ветвящихся волокон, либо не образующих в теле
губки сети, либо образующих сложную сеть. Обитают на небольших
глубинах, преимущественно в субтропических и тропических морях (на-
пример, туалетная или греческая губка). В ископаемом состоянии встре-
чаются крайне редко (карбон — ныне), обычно в виде отпечатков или
слепков каналов ирригационной системы. Кремнероговые губки
морские и пресноводные имеют органический кремневый или смешан-
ный скелет. Известны с кембрия (рис. 63, 5).
У одноосных губок скелет состоит из одноосных макросклер
без спонгинового волокна и микросклер звездчатой формы. К этой
группе относятся так называемые сверлящие губки (Clione),
живущие в известковых породах, раковинах моллюсков, скелетах ко-
ралловых полипов. По-видимому, определенные клетки этих губок спо-
собны растворять известь и, таким образом, протачивать разветвлен-
144
ные ходы внутри раковины. Эти ходы используются губкой в качест-
ве местообитания. Следы деятельности сверлящих губок обнаружены
на известковых породах начиная с силура; они представлены в виде
круглых отверстий диаметром около 1 мм.
У каменистых губок (отряд Lithistida) скелет состоит из
четырехосных десм, образующих связный литистидный скелет. Между
ними расположены микросклеры двух видов — S-образные сигмы и
звездчатые астры. Внешняя форма литистид разнообразна — от не-
правильно шарообразной до блюдцеобразной, иногда с ножкой, на
которой держится грушевидное или шаровидное тело. Например,
«кубок Нептуна» достигает в высоту 1,5 м. Современные представители
каменистых губок живут во всех морях, но преимущественно в теплых,
на глубинах от зоны прибоя до 4000 м. Шире всего они распростране-
ны на глубинах от 150 до 300 м. В ископаемом состоянии они известны
начиная с кембрия, но особенно многочисленны в ордовике и мелу
(рис. 63, 2—4).
Особый отряд представляют гетер актинеллиды — вымершие
губки с кремневым скелетом, состоящим из не связанных друг с дру-
гом многолучевых звездчатых спикул (рис. 63, 1). Гетерактинеллиды
существовали от кембрия до карбона.
КЛАСС HEXACTINELLIDA. ШЕСТИЛУЧЕВЫЕ ГУБКИ
Общая характеристика. У шестилучевых, или стеклянных, губок
(гр. hex — шесть, гр. hyaleos = hyalinos •—стеклянный) скелет состоит из
Рис. 64. Класс Hexactinellida (Hyalospongia):
1 — Myrmecioptychium (поздний мел): la — вид сбоку, 16 — вид снизу; 2 — Cr.ati-
cularia (поздняя юра — неоген); 3 — Ventriculites (мел)
Ю в в. Друшяи
трехосных кремневых спикул, имеющих три взаимно перпендикулярные
оси. Макросклеры свободные или спаянные. В последнем случае они
образуют диктиональный скелет, иногда снабженный фонарными спику-
лами. Микросклеры бывают двух типов: одни построены по трехосному
типу, причем концы шестилучевых игл могут заканчиваться пучком
тонких зубцов, изогнутых, остроконечных или с расширенными конца-
ми; другие имеют вид палочки, к которой прикреплены звездочки (ам-
фидиски). Жгутиковые камеры крупные, трубчатой формы. Внешняя
форма губок очень разнообразна; чаще они имеют вид цилиндра, бока-
ла или гриба. Сверху бокалы прикрыты крупноячеистой крышкой,,
предохраняющей атриальную полость от попадания в нее крупных
мертвых организмов. Нередко в полости поселяются креветки и равно-
ногие раки. У некоторых губок округлое комковатое тело сидит на
своеобразном корневом пучке. Современные представители живут глав-
ным образом на больших глубинах и известны под названием стеклян-
ных губок. В ископаемом состоянии появляются в кембрии, однако-
есть указания на нахождение отдельных спикул в протерозое. Класс,
очень многочислен. Известно 16 семейств (рис. 64).
КЛАСС CALCISPONGIA. ИЗВЕСТКОВЫЕ ГУБКИ
(лат. calx,
Общая характеристика. У известковых губок, или кальциспонгий
calcis — мел, известь), скелет состоит из микроскопи-
чески малых известковых спикул: одно-,,
трех- и четырехосных, чаще всего свобод-
ных, не спаянных между собой. У неболь-
шой группы спикулы сливаются вместе,,
образуя связанный, или фаретронный, ске-
лет. Ирригационная система устроена раз-
лично, от простого до наиболее сложного*
типа. Современные виды живут в нерито-
вой области моря и преимущественно в
затемненных местах. В ископаемом состоя-
нии встречаются только губки с фаретрон-
ным скелетом. Первые их достоверные •
находки описаны из девонских отложений.
Особенно широко они были распростране-
ны в юрском и меловом периодах. Около
70 родов (рис. 65).
род.
Рис. 65. Класс Calcispongia:
1 — Eudea (триас — мел);
2 — Peronidella (девон —
мел)
КЛАСС SCLEROSPONGIA.
СКЛЕРОСПОНГИИ
Общая характеристика. Новый класс
губок Sclerospongia (гр. scler — твердый) г
который установили Гартман и Горо-
(Hartman, Gor'eau, 1970) для шести родов-
(Ceratoporella, Astrosclera, Merlia, Stroma-
tospongia, Goreauella, Hispidopetra), из которых 3 новых и 3 были
известны ранее, отличается от всех губок рядом особенностей (рис. 66).
Скелет имеет массивное арагонитовое основание, от которого подни-
маются выступы, образующие неровную поверхность, покрытую тонким
слоем мягкой ткани. Арагонит в виде волокнистых кристаллов, веерооб-
разно -расходящихся от центров кристаллизации, образует структуру,,
внешне напоминающую фонтан. У одного из родов (Merlia) имеются
146
трубчатые образования с одним перфорированным днищем. Все склеро-
спонгии выделяют трабекулярный или волокнистый арагонит; кристал-
лы растут вверх и в стороны от оси, пока не встретятся с пучком со-
седних кристаллов. Их микроструктура подобна таковой у склерактиний
(см. ниже) и строматопор. На поверхности скелета некоторых склеро-
спонгий имеются ветвистые углубления, которые являются отражением
Рис. 66. Класс Sclerospongia:
1 — Ceratoporella (совр.): la — общий вид, 16 — кремневые спикулы, 1в —
выводные каналы на внешней поверхности
звездообразной системы выводных каналов в мягкой ткани губок. Эти
образования напоминают астроризы строматопор. У ряда форм они
отсутствуют. По мере роста мягких тканей вверх борозды на поверх-
ности скелета превращаются в каналы внутри скелета. Все роды совре-
менных склероспонгий выделяют простые кремневые спикулы, хотя у
некоторых видов они отсутствуют. Наличие известкового скелета и
кремневых спикул представляет одну из любопытных особенностей
нового класса губок.
Все современные склероспонгии обитают в глубоких водах (глуб-
же 70 м), избегают света и не могут соперничать с кораллами-рифо-
строителями.
Авторы, выделившие этот класс, считают, что склероспонгии близ-
ки к строматопорам и могут рассматриваться как их современные
представители. Возражения против подобного объединения будет
удобнее рассмотреть при описании строматопор.
Геологическая история класса не известна, но, по'-видимому, екле-
роспонгии появились в начале палеозоя, и ряд родов, относимых пока
к другим группам, после пересмотра будет переведен в рассматривае-
мый класс. К их числу принадлежат Dendrostroma из ордовика,
Lophiostroma из силура, относимые в настоящее время к стромато-
порам.
История развития губок. Губки представляют боковую слепую
ветвь в царстве многоклеточных животных. Они возникли, по-видимо-
му, от свободноплавающей фагоцителлы, не имевшей еще ротового от-
верстия, и приспособились к неподвижному, сидячему образу жйзни.
Первые губки, имели, вероятно, только органический скелет, состоящий
из древовидно разветвленных спонгиновых ветвей; позднее ’ возник
сетчатый органический скелет. В дальнейшем для придания скелету
10* . 14'7
большей прочности вокруг спонгиновых волоконец губки начали отла-
гаты кремнезем. Возникли первые кремневые губки с одноосными
спикулами. От них, по-видимому, вследствие естественного отбора
развились, с одной стороны, губки с трехосными, а с другой — с
четырехосными спикулами. Вначале спикулы располагались в теле
несвязно, а затем для укрепления скелета соединялись в пространст-
венную решетку. Весь этот процесс развития происходил в докембрий-
ское время. Не ясно, когда возникли гетерактинеллиды, скелет которых
состоит из звездчатых спикул.
В кембрии известны остатки четырех групп: одноосных, звездчатых,
каменистых и шестилучевых. В ордовике и силуре широкое распрост-
ранение получают представители последних двух групп. В девоне, по-
видимому, от демоспонгий возникли первые известковые губки. Первые
достоверные находки роговых губок известны с карбона, хотя они,
вероятно, являются предками всех губок и появились еще в раннем до-
кембрии. Все палеозойские остатки губок приурочены к мелководным
отложениям. В мезозое значительное распространение получают каме-
нистые, шестилучевые и известковые губки. Скопления их скелетов
иногда настолько многочисленны, что возникают настоящие губковые
слои. Мезозойские губки обитали как в мелководных, так и в глубоко-
водных зонах моря. В юре некоторые демоспонгии начинают заселять
пресноводные бассейны, проникают в озера и реки. Губки палеогена и
неогена в большинстве своем близки по составу к современным и в
ископаемом состоянии встречаются в самых разнообразных отложе-
ниях. История развития склероспонгий остается пока неясной.
Экология и тафономия. Современные губки обитают в морских бас-
сейнах на разных глубинах, но преимущественно в мелководье, в холод-
ных и теплых морях, но наиболее многочисленны в тропических морях.
Почти исключительно к тропическим морям приурочены роговые губки.
Одни губки прирастают к подводным предметам, другие свободно
лежат на дне и, наконец, третьи укрепляются в рыхлом грунте корне-
выми пучками длинных спикул. Форма тела губок хорошо приспособле-
на к условиям обитания. В литоральной зоне живут губки или с проч-
ным скелетом (известковым, кремневым) иди, наоборот, с эластичным,
роговым, способным изгибаться под действием волн. Каменистые губки
достигают глубины 200—300 м. На больших глубинах (до 6000 м) рас-
пространены стеклянные губки с изящным кремневым скелетом. Из-
вестковые губки обитают преимущественно в зоне мелководья. Только
около 20 видов губок живут в пресных водах.
В ископаемом состоянии чаще всего сохраняются губки с фарет-
ронным, диктиональным или литистидным скелетом. Несвязный скелет
после смерти губки обычно разрушается и отдельные спикулы рассеи-
ваются по дну. В некоторых случаях массовая гибель губок приводит
к накоплению на дне слоя спикул — «стеклянного войлока», который в
процессе диагенеза и фоссилизации превращается в спонгилит — гор-
ную породу, состоящую из спикул губок. Губки встречаются в виде
остатков> скелетов или отпечатков. В процессе фоссилизации скелет
может испытать перекристаллизацию или замещение другими минераль-
ными веществами. Кремнезем может быть перекристаллизован или за-
мещен карбонатом или фосфатом кальция, глауконитом, соединениями
железа. Известковый скелет, наоборот, иногда замещается кремнеземом.
Чаще всего сохраняются макросклеры; микросклеры обычно разру-
шаются, но иногда их находят в окружающих породах. Для изучения
губок в первую очередь необходимо получить препараты изолирован-
ных спикул или сделать, пришлифовки и шлифы.
148 --
ТИП ARCHAEOCYATHI.
АРХЕОЦИАТЫ
Общая характеристика. Археоциаты (гр. archaios — древний, kyat-
hos — кубок) — вымершие одиночные или колониальные животные,
обладавшие пористым органогенно-карбонатным скелетом. Скелет,
имеющий форму кубка, состоял из одной или двух стенок и различных
скелетных образований: стерженьков, перегородок, днищ. Внутренняя
стенка у примитивных форм, вероятно, необызвествлялась и поэтому
О ди ночные
Рис. 67. Внешние формы кубков и колоний археоциат:
к—каблучок прирастания
они считаются одностенными. Археоциаты вели бентосный образ,
жизни и обитали в мелководных морских бассейнах раннего кембрия’-
и, возможно, начала среднего.
Строение скелета. О строении мягкого тела можно судить только
предположительно на основании косвенных данных,в первую очередь
на основании морфофункционального анализа скелета. Среди археоциат
/ были как одиночные, так и колониальные: организмы. Одиночные
археоциаты имели известковый скелет различной формы: цилиндриче-
ского или конического бокала, от узко- до ширококонического, иногда
дисковидного, значительно реже полушаровидной или грибовидной
(рис. 67). Поверхность кубка была гладкой или с дополнительными про-
дольными складками или поперечными пережимами. Колониальные
археоциаты имели массивную, цепочковидную, ветвистую или иную
форму скелета.
Диаметр кубка,колеблется в больших пределах — от 3.—5 до 300—
500 мм; однако у большинства он не превышает 10—25 мм; высота
кубка пропорциональна диаметру и изменяется от 6—10 до 80—150 мм.
149
Среди археоциат были прикрепленные и свободно лежащие на
грунте. Прикрепление осуществлялось уплощенным основанием или при
помощи так называемого каблучка прирастания, массивного,
трубчатого или пластинчатого (рис. 67). Первые два типа каблучка
прирастания развивались на 2—5 мм, третий тип охватывал кубок до
13—15 мм (т. е. !/з—’/г высоты кубка). На взрослой стадии многие
археоциаты, по-видимому, свободно лежали на грунте. Археоциаты с
полушаровидным или мешковидным облегченным скелетом без всяких;
следов прирастания, вероятно, были неприкрепленными.
Скелет археоциат — собственно кубок — органогенно-карбонат-
ный, состоит из двух стенок: наружной и внутренней. У наиболее низко-
организованных археоциат внутренняя стенка либо отсутствовала, либо
не обызвествлялась, и в ископаемом состоянии сохраняется только
одна наружная; таких археоциат называют одностенными.
У двустенных археоциат в пространстве между двумя стенками —
интерваллюме — расположены различные скелетные элементы:
стерженьки, перегородки, тении, днища, либо в различных сочетаниях
эти скелетные элементы. Первичный скелет у археоциат состоял, в
отличие от губок, не из спикул, а из мелких (0,01—0,02 мм) зерен
кальцита, пропитанных органическим веществом; он достаточно моно-
литен и однороден.
Наружная стенка является основным элементом скелета; она про-
низана системой каналов, которые открываются наружу порами; тол-
щина, стенки колеблется от сотых долей миллиметра до 1—3 мм. Раз-
личают несколько типов строения наружной стенки (рис. 68, 1—6):
а) с простой пористостью, б) с дополнительной тонкопористой оболоч-
кой, в) с козырьками, г) с простыми и бугорчатыми тумулами.
Наиболее широко распространена стенка с простыми округлыми,
овальными, прямоугольными или шестигранными порами; они распо-
ложены рядами — с одним, двумя, четырьмя, четырьмя — восьмью ря-
дами пор в интерсептуме (пространство между двумя перегородками).
Наружная'стенка бывает пронизана крупными и мелкими каналами,
иногда коленообразно изогнутыми или ветвящимися. Некоторые поры
снабжены шипиками длиной 0,2—0,3 мм, выступающими наружу.
Усложнение стенки достигается разными путями. Между порами в ряде
случаев расположены вертикальные узкие пластины — внутренние реб-
ра, отходящие от наружной стенки в интерваллюм. Стенка имеет у ряда
форм полые вздутия — т у м у л ы, пронизанные одной или несколькими
порами (иногда до 20—25); тумулы с многими порами называются
бугорчатыми (рис. 68, 5—6). Усложнение наружной стенки наблю-
дается как у одностенных, так и двустенных археоциат. Обычно в про-
цессе эволюции' наблюдается уменьшение числа пор на 1 мм длины
как в наружной стенке, так и в перегородках, образование дополнитель-
ных оболочек, наложенных на основной каркас скелета, и переход от
простых тумул к гроздевидным.
Внутренняя стенка по строению более разнообразна, чем наруж-
ная (рис. 68, 7—10). Если внутренняя стенка отсутствовала или не
обызвествлялась, то внутренняя полость пересекалась пленками пузыр-
чатой ткани или со стороны наружной стенки укреплялась стерженька-
ми. При наличии внутренней стенки различают два ее типа: 1) воз-
никшая за счет втягивания внутрь верхней защитной оболочки —
пельты, 2) .образованная во внутренней полости. В первом случае
она представляет собой мешковидное образование, пронизанное тонки-
ми порами; во втором — она повторяет форму наружной стенки и
достигает большого разнообразия. Различают стенку с простой
150
пористостью, с округлыми или прямоугольными порами; чешуйчатую
•стенку, или стенку, построенную по типу «жалюзи», — у нее круглые
лоры прикрыты со стороны внутренней полости чешуями или козырь-
ками; стенку с плоскими кольцевыми козырьками, которые в поперечном
сечении могут иметь различную форму, вследствие чего форма лоро-
Рис. 68. Строение наружной (/—6) и внутренней (7—10) стенок археоциат:
вс — внутренняя стенка, гк — горизонтальные кольцевые пластины, ко — ко-
зырьки, мв — микропористая внутренняя оболочка, нс — наружная стенка,
с — септа, ту — тумулы
вых каналов бывает различна (рис. 68, 7—10). Поры внутренней стенки
могут быть усложнены различными защитными устройствами: шипи-
ками, пластинчатыми козырьками или пористой оболочкой
(рис. 68, 10).
Интерваллюм— пространство, ограниченное наружной и внутрен-
ней стенками (рис. 69). Он имеет вид усеченного полого конуса. У од-
них интерваллюм лишен скелетных элементов (рис. 69, 1), у других
в нем расположены стерженьки, ориентированные в трех направлениях:
радиальном, тангенциальном и вертикальном (рис. 69, 5); у третьих
интерваллюм заполнен перегородками, представляющими собой
радиальные вертикальные пластины, соединяющие наружную и внут-
реннюю стенки (рис. 69, 2, 3); они разделяют интерваллюм на интер-
септальные камеры и пронизаны порами округлой.или овальной формы.
Иногда поры отсутствуют. Перегородки, заполняющие интерваллюм,
могут быть расположены часто или редко; они могут быть равномерно
тонкими или утолщенными по краям. Новые перегородки обычно возни-
кают со стороны наружной стенки. У многих археоциат перегородки
укреплялись синаптикулами и пузырчатой тканью. Синаптикулы
представляют собой горизонтальные стерженьки, соединяющие пере-
городки. Пузырчатая ткань состоит из тонких изогнутых не-
пористых выпуклых известковых пластинок. Они возникали позднее
перегородок и расположены в интерсептальных камерах.
У большой группы неправильных, или гениальных, архео-
циат интерваллюм заполнен искривленными, продольно ориентирован-
ными пористыми пластинками, получившими название тении (см,
рис. 74). Различают сильно искривленные тении, которые вблизи на-
ружной стенки дихотомически расщепляются, и плоские тении, внешне
151
ничем не отличимые от перегородок (только изучение ранних стадий
онтогенеза позволяет их различать). Поры в тениях обычно угловатые
и располагаются наклонными рядами. Иногда тении соединяются плен-
ками пузырчатой ткани.
В интерваллюме часто присутствуют днища — выпуклые, реже
горизонтальные пористые пластины (рис. 69, 5). Они связывают между
собой наружную и внутреннюю стенки и располагаются через различ-
Рис. 69. Строение интерваллюма:
1 — без скелетных элементов; 2 — с перегородками; 3 — днища и перегородки с
круглыми порами; 4а, б — с перегородками и гребенчатыми днищами; 5 — систе-
ма стерженьков (наружная стенка слева); 6 — с перегородками и синаптикула-
ми; 7 — с перегородками и пузырчатой тканью; 8а, б — со стерженьками, ориен-
тированными радиально;
вс—внутренняя стенка, гр-—гребенчатое днище, дн — днище, нс — наружная
стенка, рс — радиальные стерженьки, с — септа,. ст — стерженьки
ные промежутки. Одни днища представляют единое целое с наружной
стенкой и возникали, по-видимому, одновременно с ней. Другие появ-
лялись вскоре после образования внутренней стенки, септ или тений
и были связаны с наружной и внутренней стенками. Днища имели
круглые или щелевидные поры. Особым типом днищ являются гре-
бенчатые днища — самостоятельные образования в каждой интер-
септальной камере, расположенные очень неравномерно. Они состоят
из кольцевого валика и многочисленных тонких стерженьков, лежащих
в одной плоскости (рис. 69, 4).
Центральная полость, ограниченная внутренней стенкой, обычно-
свободна от скелетных элементов. При жизни археоциат она заполня-
лась мягкой тканью, обызвествленная структура которой иногда сохра-
няется в ископаемом состоянии. У одностенных и неправильных архео-
циат она может быть заполнена пузырчатой тканью, ламинами или
внутренними краями тений. Центральная полость, ограниченная мешко-
видной внутренней стенкой, не гомологична полости всех остальных
археоциат. Она никогда не пересекается пленками пузырчатой ткани.
Верхний край кубка известен у немногих форм. У одностен-
ных археоциат верхняя часть кубка закрыта покровной оболочкой —
152
п е л ь т о й. Она может иметь вид плоской тонкопористой пластинки
с крупным центральным отверстием в центре или без него, либо вогну-
той пластинки, пористой или сплошной, также с крупным отверстием
в центре (рис. 70). Пельта предохраняла внутреннюю полость кубка
от попадания в нее посторонних частиц и, возможно, служила для филь-
трования воды. У двустенных археоциат все формы, имевшие днища,
заканчивали свое развитие образованием верхних днищ, пористых или
гребенчатых. У некоторых неправильных археоциат верхний край за-
канчивался пленками пузырчатой ткани.
Рис. 70. Схема строения пельты и верхнего края кубков (Журавлева, 1960):
1 — типы пельт; 2—5 — типы верхнего края кубков, образованные: 2— выпуклым
пористым днищем, 3 — гребенчатым днищем, 4 — ворсинчатыми концами, 5 — плен-
ками пузырчатой ткани
У многих археоциат наблюдаются вторичные утолщения скелетных
элементов— наружной стенки, перегородок, внутренней стенки. Все
вторичные утолщения отличаются слоистостью и различной насыщен-
ностью этих слоев органикой. Многие археоциаты имеют различные
выросты или разрастания. Трубчатые выросты носят название тер-
сии.- Они представляют собой молодые почки, которые повторяют
структуры начальной стадии развития кубка. Кроме терсиевидных вы-
ростов у археоциат наблюдаются разрастания интерваллюма, при
котором возникают дополнительные образования, отражающие строе-
ние интерваллюма основного кубка. Известны также различные случаи
прижизненных повреждений скелетов, залеченных при жизни.
Размножение и развитие. О размножении археоциат известно очень
мало. Колониальные формы размножались почкованием; широкое рас-
пространение археоциат указывает на половое воспроизведение, в ре-
зультате которого возникала личинка. После непродолжительного пре-
бывания в толще воды личинка, вероятно, опускалась на дно и при-
креплялась к подводным предметам или к дну. Послеличиночное раз-
витие археоциаты начинают постройкой непористого известкового кубка
с каблучком прирастания (рис. 71)-. Высота такого кубка была 0,4—
0,5 мм при диаметре до 0,1—0,12 мм. За стадией непористого кубка
следует стадия обособления наружной стенки и возникновения пор;
вначале поры располагаются неупорядоченно, далее намечается более
Закономерное расположение пор при соответствующем увеличении их
диаметра. Дальнейшее развитие скелета идет у разных археоциат по-
разному, что дает основание разделять их на два самостоятельных
класса: правильных и неправильных археоциат. У одностенных правиль-
ных археоциат морфогенез заканчивается на разных уровнях: у одних
на стадии, когда диаметр кубка равен 0,2—0,3 мм; у форм, имеющих
тумулы, при диаметре 0,6—0,8 мм; у форм с очень сложной стенкой
(пористостью ветвистого типа) — при диаметре около 3,0 мм; внутрен-
няя полость у них была свободной на всем протяжении. У одностенных
153
Продольные сечения
Диаметр кубка кл. Jrregulares кл. Regular es
1,8 мп и Более । *. - :&~7' /^/ К""** : *2 ‘ Co /
1,1 -1,7 мм IS? Я? tS S7 Е2 £1
v 0,8-1,0мм °&\ I'd cS* к? у i Co л iMl- '^**5 A w
0,5-0,7 мм К/ Г 2 | - ч MH M W 'в Л \ Ц Vi й 5^ %-У
0,22-0,45мм !Й> s2 t
«Г
0,13-0,2 мм м 1 1/ / ь2 Vе b1 \J~a
0,05 -0,12мп А Тип Metacyathus \J Тип Aj ac/cyathus .
Рис. 71. Схема онтогенеза правильных и неправильных археоциат (Журавлева, 1960):
А — общая зародышевая стадия; Bi—Ж1 — стадии развития представителей Re-
1 gulares; В2—Ж2 — стадии развития представителей- Irregulares;
а — форма с гребенчатыми днищами, 0—форма без днищ, у — форма с пористыми
днищами, х — форма с тениями, у — форма с тениями и днищами, г — форма с ше-
стигранными радиальными трубками;
а — наружная стенка, б—внутренняя стенка, в — радиальные стерженьки в интервал-
люме, г — перегородки, д — гребенчатые днища, е — пористые днища, ж— поро-
вые каналы наружной стенки, з — бугорчатые тумулы наружной стенки, и —
зачатки поровых каналов внутренней стенки, к — поровые каналы внутренней стенки,
л — ворсинки на концах поровых каналов внутренней стенки, м — стерженьки во
внутренней полости, н — пузырчатая ткань, о — тении, п — гениальные шипики, р —
выпуклые днища, с — шестигранные локулы
неправильных археоциат она была заполнена различно ориентирован-
ными стерженьками или ламинами и пузырчатой тканью. От одно-
стенных археоциат возникли двустенные. Появление двух известковых
стенок — важный этап в развитии археоциат. Все двустенные архео-
циаты в онтогенезе вначале проходят одностенную стадию, затем появ-
ляется простая внутренняя стенка.
У правильных археоциат внутренняя стенка с мелкими простыми
порами и радиальными стерженьками появляется при диаметре кубка
0*2 мм; несколько позднее (при диаметре 0,22—0,45 мм) образуются
перегородки, вначале крупнопористые, позднее с более мелкими пора-
ми; на этой же стадии появляются и днища, гребенчатые и пористые;
при диаметре 0,8—1,0 мм усложняется внутренняя стенка, а затем
происходит усложнение остальных скелетных элементов.
Развитие скелета у неправильных археоциат протекает несколько
иначе, с некоторым «запаздыванием» появления скелетных элементов
по сравнению с правильными. На стадии 0,2 мм у них еще не обособле-
на центральная полость, она заполнена редкими пленками пузырчатой
^гкани и беспорядочно ориентированными стерженьками; внутренняя
стенка появляется только при диаметре 0,5-—0,7 мм; позднее (0,8—
1,0 мм) из стерженьков, расположенных в трех плоскостях, начинают
формироваться днища и тении; еще позднее происходит усложнение
всех скелетных элементов.
Основы систематики и классификация. На основании особенностей
строения скелета и онтогенетического развития выделяются две основ-
ные группы археоциат, которые рассматриваются в качестве классов:
Regulates и Irregulares.
КЛАСС REGULARES. ПРАВИЛЬНЫЕ АРХЕОЦИАТЫ
К классу правильных археоциат (лат. regularis — правильный)
относятся одиночные или колониальные археоциаты, скелет которых
состоит из одной или двух пористых стенок. Диаметр кубков колеблется
от нескольких миллиметров до 30—50 см, но у большинства он не пре-
вышает 20—25 мм; высота кубков, как правило, пропорциональна диа-
метру и колеблется от 6—10 до 250—300 мм, в среднем составляя
80—150 мм. Внешняя форма кубков варьирует от узкоконических или
цилиндрических до грибообразных, полушаровидных или блюдцеобраз-
ных (см. рис. 67). Внешняя стенка с простыми или сложными порами
или с системой поровых каналов, у некоторых имеются тумулы. Внут-
ренняя стенка, возникшая онтогенетически несколько позже наружной,
обычно толще наружной и более разнообразна по своему строению:
от тонкой стенки с простыми округлыми порами до сложной стенки,
образованной кольцами, пластинами в сочетании с различного рода
каналами. Центральная полость ограничена внутренней стенкой. Ин-
Дерваллюм снабжен разного типа скелетными элементами: стержень-
ками, простыми или гребенчатыми днищами, перегородками или сочета-
нием перегородок и днищ, но никогда не имел тений.
Класс правильных археоциат разделен на три отряда: Monocyathi-
da, Ajacicyathida, Putapacyathida.
Отряд Monocyathida. Моноциатиды (гр. monos — один), или одно-
стенные археоциаты, объединяют археоциат с одностенным кубком.
Исходной формой принимают род Archaeolynthus, имевший маленький
кубок с простой пористой стенкой и массивным каблучком прираста-
ния. От этого рода возникли, с одной стороны, сферические и полусфе-
рические археоциаты с наружной стенкой, пронизанной каналами двух
155
f8
Рис. 72. Класс Regulares (ранний кембрий). Отряд Monocyathida:
la—в — Archaeolynthus-, 2 — Rhabdocyathella-, 3 — Tumuliolynthus: За — продоль-
ное сечение, 36—реконструкция (Журавлева, 1963)
Рис. 73. Класс Regulares (ранний кембрий). Отряд Ajacicyathida:
1 — Dokidocyathus с радиальными стерженьками (р): 1а— поперечное, 16— про-
дольное сечения; 2 — Ajacicyathus, поперечное сечение; 3 — Ethmophyllum: За —
поперечное сечение, внутренняя стенка с поровыми каналами (к), 36 — продоль-
ное сечение вдоль септы (с); 4 — Orbicyathus, реконструкция кубка; 5 — Cos-
cinocyathus, косой разрез кубка; 6 — Nochoroicyathus, поперечное сечение;
гд — гребенчатые днища, дн — днища, с — септы
-типов — широких и узких, сообщающихся друг с другом, с другой — две
ветви: первая дала формы с тумуловыми порами (Тumuliolynthus) и
вторая—-формы с горизонтальными стерженьками (Rhabdolynthus)-,
•от первых в дальнейшем развились археоциаты с ветвистыми каналами
(Rhabdocyathella) (рис. 72). Они были распространены в раннем кемб-
рии (томмотский — начало ленского яруса).
Отряд Ajacicyathida. Аяцициатиды (Ajax — род. Ajacis — один из
вождей греков в Троянской войне, по имени которого назван рудник
в Австралии, где найдены эти археоциаты) — одиночные, реже коло-
ниальные, двустенные археоциаты. Кубки разнообразной формы. Интер-
валлюм полый или заполнен скелетными элементами. На этом основа-
нии отряд разделен на подотряды: Dokidocyathina — с редкими радиаль-
ными горизонтальными стерженьками, Ajacicyathina — с радиальными
перегородками, но без днищ, Coscinocyathina — с пористыми днищами
.и перегородками, Nochoroicyathina — с гребенчатыми днищами и пере-
городками (только одиночные).
Аяцициатиды произошли от одностенных археоциат в начале алдан-
ского века и вымерли к концу раннего кембрия. Реликтовые формы
обнаружены в начале среднего кембрия. Развитие аяцициатид шло по
нескольким путям. После развития внутренней стенки у одних в ин-
Дерваллюме появились радиальные стерженьки (докидоциатины), у дру-
гих (аяцициатины) — перегородки, у третьих (косциноциатины) —
.днища и перегородки, у четвертых (нохороициатины) развились пере-
городки и гребенчатые днища, причем днища появились позднее пере-
городок. Параллельно у этих групп шло усложнение скелетных элемен-
тов, в первую очередь наружной и внутренней стенок. Известно свыше
150 родов (рис. 73).
Отряд Putapacyathida. Путапациатиды — одиночные или колони-
альные археоциаты маленьких размеров; кубки узкоконические или
цилиндрические. Кубок построен из двух сложных стенок, соединенных
пористыми днищами. В интерваллюме, кроме того, имеются тонкие
вертикальные пластинки и пузырчатая ткань. Ранний кембрий. Извест-
но 7 родов.
КЛАСС IRREGULARES. НЕПРАВИЛЬНЫЕ АРХЕОЦИАТЫ
К классу, неправильных, или гениальных, археоциат относятся
«одиночные или колониальные формы, обладавшие, кубками различных
очертаний, часто неправильной формы. Наружная стенка обычно про-
низана порами, внутренняя более сложная, с простыми порами или
норовыми каналами. Интерваллюм значительно шире, чем у правильных
.археоциат; в нем могут присутствовать различно ориентированные
стерженьки, пузырчатая ткань, прерывистые искривленные или прямые
тении, радиальные пористые трубки, пористые днища. В отличие от
правильных археоциат внутренняя стенка в онтогенезе появляется
довольно поздно, после заполнения внутренней полости различно
ориентированными стерженьками, ламинами и пузырчатой тканью.
У одной группы центральная полость заполнена призматическими труб-
ками (рис. 74).
Неправильные археоциаты включают также формы с одностенным
кубком, внутренняя полость которых заполнена пузырчатой тканью,
стерженьками или ламинами.
Неправильные археоциаты были распространены в раннем кембрии;
•единичные формы дожили до начала среднего кембрия.
157
В составе класса выделены три отряда: Thalassocyathida, Archaeo-
cyathida, Syringocnemidida.
Отряд Thalassocyathida. Талассоциатиды. Одностенные археоциаты,,
у которых во внутренней полости присутствуют пузырчатая ткань или
стерженьки, ориентированные в разных направлениях. К отряду отне-
Рис. 74. Класс Irregulares (ранний кембрий). Отряды Thalassocyathida (/), Аг-
chaeocyathida (2—4) и Syringocnemidida (5):
Г— Bacatocyathus, продольное сечение; 2 — онтогенез Pristnocyathus praesignis Fo-
nin (оригинал В. Д. Фонина); поперечные сечения при диаметре кубка:, а — 0,2 мм,
б — 0,5 мм, в — 0,7 мм, г—1,3 мм, д — 1,5 мм,'е—1,7 мм, ж — 2,5 мм, з — 3,5 мм;
стадии: а—б — напоминает Bacatocyathus, в—д — напоминает Rhizacyathus, е —
напоминает Dictyocyathus, ж—з — несет признаки рода; 3 — Archaeocyathus, по-
перечное сечение; 4 — Verthocyathus, реконструкция (оригинал В. Д. Фонина);
' 5 — Syringocnema, поперечное сечение
сены два рода: Bacatocyathus — с простой пористой стенкой и поло-
стью, заполненной пузырчатой тканью (рис. 74, 1) и Thalassocyathusr
имеющий в полости стерженьки и пузырчатую ткань.
К этому отряду условно отнесены археоциаты одиночные или
колониальные, имеющие внутреннюю стенку, расположенную эксцент-
рично, образованную путем впячивания части наружной стенки во
внутреннюю полость. Интерваллюм свободен от скелетных элементов.
Их выделяют в. подотряд Capsulocyathina.
158
Отряд. Archaeocyathida. Археоциатиды. Одиночные и колониальные;
кубки цилиндрические, конические или дискоидальные, с поперечными
пережимами или выростами. Наружная стенка с простыми порами или
частично замещена пузырчатой тканью, внутренняя стенка с одним ря-
дом крупных простых пор, реже отсутствует или состоит из отдельных
колец. В, интерваллюме развиты стерженьки, тении, пузырчатая ткань,
нередко днища. В центральной полости могут быть призматические
трубки. Отряд разделен на два подотряда: Archaeocyathina с тениями,
без днищ и Archaeosyconiina — с днищами и тениями. Распространен
в .раннем кембрии и начале среднего. Около 50 родов (рис. 74, 2—4).
Отряд Syringocnemidida. Сирингокнемидиды (гр. syrinx — трубка,
knemis — колено, фрагмент). Одиночные археоциаты, кубки кониче-
ской формы, наружная стенка простая, пористая, внутренняя с просты-
ми порами; интерваллюм заполнен призматическими пористыми
трубками; нет ни септ, ни днищ. На ранней стадии онтогенеза —
одностенного кубка — имеются стерженьки и пузырчатая ткань, позд-
нее появляется внутренняя стенка и пористые трубки в интерваллюме.
Ранний кембрий. Известно 6 родов (рис. 74, 5).
К археоциатам относится отряд Kazakhstanicyathida, основанный на
одном роде из раннего кембрия. Кубок одностенный, внутренняя полость
с пористыми днищами, и стерженьками. Систематическое положение
отряда неясно.
История развития археоциат. Вся геологическая история археоциат
прошла в течение раннего кембрия, т. е. примерно в течение 30—
40 млн. лет. За это время археоциаты возникли, широко распространи-
лись по всему свету и почти все, за исключением немногих реликтовых
групп, вымерли в конце раннего кембрия. Предки археоциат неизвестны
и пока недостаточно ясно, какие группы могли быть исходными для
археоциат. Возможно, предков археоциат нужно искать непосредствен-
но среди одноклеточных, и в первую очередь среди саркодовых. Пред-
полагается, что археоциаты возникли еще в позднем протерозое (венде),
однако настоящими находками это до сих пор не подтверждено. Пер-
вые известные археоциаты, относимые к одностенным, были распрост-
ранены в томмотском и в первой половине алданского, века в морских
бассейнах Сибири, северной части Монголии и Австралии. К концу
века они исчезают и в начале ленского века встречаются спорадиче-
ски только в Восточном Саяне и Забайкалье.
В первой половине алданского века появляются двустенные пра-
вильные и неправильные археоциаты. Они возникают в пределах Си-
бирско-Австралийского бассейна и очень быстро распространяются за
его пределы, достигают территории современной северо-западной Аф-
рики. Расцвет правильных и неправильных археоциат приходится на
конец алданского й начало ленского века. Они становятся известными
из морских бассейнов всех материков земного шара. К концу раннего
кембрия основные группы правильных археоциат вымирают. Единич-
ные правильные и неправильные археоциаты доживают до начала сред-
него кембрия на Сибирской платформе и в Северной Америке.
Экология и тафономия. Все археоциаты относятся к бентосным ор-
ганизмам (рис. 75, А). Большинство из них вело неподвижный образ
жизни, прикрепляясь к подводным предметам или к дну основанием
кубка, его боковой стороной или специальным каблучком прирастания.
Часть археоциат, имевшая дисковидные или блюдцеобразные скелеты,
по-видимому, свободно лежала на дне и, наконец, небольшая группа
археоциат с полушаровидным и мешковидным скелетом, возможно, пас-
сивно перекатывалась по дну. Большинство археоциат жило на неболь-
159
ших глубинах (20—50 м) в теплых нормальносоленых морях с неболь-
шим содержанием окислов магния. На глубине 20—30 м археоциаты
совместно с сине-зелеными водорослями строили биостромы, биогермы.
Биостромы — пластовые органогенные постройки, которые в
момент их образования не намного возвышались над дном; в их созда-
нии принимали'участие сине-зеленые водоросли и в меньшей степени
Рис. 75. А — распределение археоциат в зависимости от глубины бассейна: а — редкие
до глубины 10—12 м; б — биогермы на глубине 20—30 м; в — редкие дисковидные и
конические на глубинах от 30 до 50 м; г — редкие на глубине 50—60 м; д — единич-
ные на глубине около 100 м (Журавлева, 1960); Б — археоциатовый биогерм (Журав-
лева, Зеленов, 1955)
археоциаты. Биогермы — органогенные постройки, созданные мас-
совыми поселениями археоциат при участии сине-зеленых водорослей.
Биогермы возвышались над дном на 1—2 м в высоту (рис. 75, Б).
В таких условиях развивались колониальные формы, отдельные кубки
примыкали плотно друг к другу. Водоросли занимали пространство
между кубками и обычно их обрастали. Археоциатовые биогермы рас-
полагались на дне моря на значительном расстоянии друг от друга
(20—40 м); при благоприятных условиях они широко разрастались,
почти сливаясь друг с другом, образуя биогермные зоны и рифы боль-
шой протяженности. По мере углубления бассейна количество архео-
циат сокращалось и на максимальных для них глубинах (50—100 м)
их кубки становились меньших размеров, тонкостенными, с узким ин-
терваллюмом, поровая система усложнялась. Хотя иногда развивались
довольно крупные и ажурные скелеты. Вместе с археоциатами на этих
глубинах жили красные водоросли.
160
; ГРУППЫ НЕЯСНОГО СИСТЕМАТИЧЕСКОГО
ПОЛОЖЕНИЯ
В разные отрезки времени в палеозое были распространены от-
дельные группы многоклеточных, систематическое положение и таксо-
номический ранг которых до сих пор остаются неясными. От. этих групп
в ископаемом состоянии сохранились известковые скелеты. Эти скелет-
ные остатки принадлежали каким-то губко- или археоциатоподобным
организмам и относятся, по-видимому, к низшим многоклеточным. Они
представляют собой, вероятно, слепые, часто очень короткие, ветви в
эволюции и становлении многоклеточности. В связи с тем что они не
укладываются пока ни в один из известных типов или класбов, они
описываются в этом разделе под разными таксономическими рангами
неясного систематического положения. Ниже будут рассмотрены сле-
дующие группы: класс (?) Cribricyatha, класс (?) Sphinctozoa, класс (?)
Aphrosalpingoida-, класс (?) Receptaculita, семейство (?) Soanitidae,
класс (?) Stromatoporata, класс (?) Hydroconozoa.
КЛАСС (?) CRIBRICYATHA. КРИБРИЦИАТЫ
К новому классу крибрициат (лат.cribrum—сито,решето),выделен-
ному А. Г. Вологдиным (1964, 1966), отнесены ранне- и среднекембрий-
ские археоциатоподобные мелкие организмы (рис. 76). Известковый
скелет крибрициат имеет форму кубка высотой от 1,5 до 8 мм, при диа-
Рис. 76. Класс (?) Cribricyatha (ранний—средний кембрий):
1 — типы периптерат; 2—6 — продольные сечения: 2 •— Peripteratocyathus, 3 —
Pyxidocyathus, 4 — Thecicyathus, 5 — Lomaticyathus, 6 — Pubericyathus
метре от 0,8 до 2,0 мм. Кубки одно- или двустенные, различной фор-
мы — цилиндрической, конической, округлой, овальной. У крибрициат
имеется внутренняя полость цилиндрической, бочковидной или воронко-
видной формы, которая в отличие от всех других групп располагается
по отношению к оси кубка асимметрично, косо, обычно начинаясь внизу
сбоку и заканчиваясь вверху кубка. Наружная стенка построена очень
11 В. В. Друщиц 161
своеобразно: она слагается из системы двух скелетных элементов — так
называемых периптерат и бакулей. Периптераты (лат. periptera-
tus — окруженный крылышком) представляют собой тонкие крыловид-
ные образования, опоясывающие снаружи по горизонтали весь кубок.
Они разной формы и толщины: плоские, равномерно утолщенные или:
с утолщенным внутренним краем, слабо или сильно изогнутые. Б а т
кули (лат. baculae — палочка)—палочковидные скелетные элементы,,
которые соединяли между собой внутренние края периптерат. Внутрен-
няя стенка тонкая, с мелкими круглыми порами или ленточная, состоя-
щая из тонких лентовидных образований (толщиной около 0,01 мм)г
снабженных щелевидными порами. Сверху кубки могут иметь перекры-
тия, образованные периптератами.
Описанные выше скелеты принадлежали к каким-то организмам,,
распространенным в раннем и в начале среднего кембрия. Формы с
коническим кубком, вероятно, вели прикрепленный образ жизни, хотя>
следов прирастания на кубке нет; формы с изометрическими и непра-
вильными кубками вели, по мнению А. Г. Вологдина, планктонный,
образ жизни.
Особенности строения наружной стенки, наличие периптерат,.
форма и положение внутренней полости отличают крибрициат от на-
стоящих археоциат и поэтому их временно следует рассматривать как
класс неясного систематического положения, а не относить к археоциа-
там, как предлагает А. Г. Вологдин.
КЛАСС (?) SPHINCTOZOA. СФИНКТОЗОИ
К классу сфинктозой (гр. sphinktos — тесно связанный, zoon —
животное) относятся вымершие одиночные или колониальные много-
Рис. 77. Схема строения представителен класса Sphinctozoa (1—2) и класса (?) Aphro-
salpingoida (3):
1 — Barrosia (мел): la — внешний вид колонии, 16 — продольное сечение; 2 — АтЫу-
slphonella (карбон — триас), схема строения скелета; 3 — Aphrosalpinx (поздний си-
лур), поперечное сечение
клеточные организмы (рис. 77, 1—2), существовавшие на протяжении:
карбона — мела. Известковый скелет имеет кубкообразную, цилиндри-
ческую, коническую или неправильную форму, разделенную на отдель-
ные камеры горизонтальными перегородками, или днищами. Через;
162
камеры у некоторых сфинктозой проходит пористая трубка, обособ-
ляющая центральную полость. В центральной полости может быть
развита пузырчатая ткань. Сообщение между камерами осуществляется
через систему каналов в днищах. Пространство между днищами может
быть заполнено разветвленными стержневидными скелетными элемен-
тами или пузырчатой тканью. Наружная стенка у некоторых форм
имеет крупные отверстия, расположенные на бугорках. Наружная и
внутренняя стенки и днища пронизаны порами. Достоверно сфинктозой
известны начиная с карбона; их расцвет относится к перми и триасу;
в юре и мелу их очень мало, а из палеогена они пока неизвестны.
Систематическое положение сфинктозой неясно; они напоминают как
губок, так и археоциат. Некоторые исследователи считают, что скелет
сфинктозой состоит из спаянных трехлучевых спикул и поэтому относят
их к губкам. А. Г. Вологдин (1956, 1957) рассматривает сфинктозой в
составе типа археоциат. И. Т. Журавлева (1956, 1960), не исключая
возможности конвергентного сходства с археоциатами, рассматривает
их в качестве класса среди губок. В составе класса один отряд и около
30 родов.
КЛАСС (?) APHROSALPINGOIDA.
АФРОСАЛЬПИНГОИДЫ
К афросальпингойдам (гр. aphros — пена, salpinx — трубка) отно-
сится небольшая группа морских организмов, скелетные остатки кото-
рых были обнаружены в позднесилурийских отложениях Урала и выде-
лены Е. И. Мягковой (1955) в новый класс, который первоначально был
отнесен к археоциатам. По внешнему виду афросальпингоиды напоми-
нают, с одной стороны, археоциат, с другой — водоросли. Скелет из-
вестковистый, имеет коническую, цилиндрическую или неправильную
форму и состоит из двух стенок, из которых наружная стенка не яв-
ляется самостоятельной. Межстенное пространство заполнено верти-
кальными трубками — л оку л я м И'— различных типов диаметром
0,35—0,8 мм, округлого или полигонального сечения (рис. 77, 3).
Афросальпингоиды отличаются от археоциат строением интервал-
люма, состоящего из трубчатых локулей, характером центральной полос-
ти, заполненной рыхлой известковистой тканью, отсутствием перегоро-
док и самостоятельной стенки. Недавно Циглер и Ритшель (1970)
высказали предположение, что афросальпингоиды должны быть отне-
сены к водорослям, а не к археоциатам, как предполагалось раньше.
КЛАСС (?) RECEPTACULITA. РЕЦЕПТАКУЛИТЫ
К рецептакулитам (лат. receptaculum — резервуар) относятся мор-
ские организмы с известковым скелетом. Скелет имеет грушевидную,
чашевидную или кубкообразную форму и состоит из наружной и внут-
ренней стенок. Размер скелета колеблется от' нескольких миллимет-
ров до 20 см. Снаружи скелет покрыт ромбическими или шестиуголь-
ными табличками, под которыми расположены радиальные цилиндри-
ческие столбики — радиали.
Рецептакулиты были обитателями мелководных зон моря от позд-
него кембрия до начала карбона, среди них были одиночные и коло-
ниальные формы. В ископаемом состоянии они встречаются вместе с
остатками водорослей и моллюсков, реже с остатками строматопорат.
Наиболее широко они были распространены в силуре и девоне.
1Г
163
Высказывается предположение, что рецептакулиты представляют собой
остатки водорослей.
; 'У. СЕМЕЙСТВО (?) SOANITIDAE. СОАНИТЫ
К соанитам (soan — от СО АН — Сибирское отделение Академии
наук) относятся организмы неизвестного происхождения и неясного.
систематического положения, известковые скелетные остатки которых
были встречены в отложениях нижнего ордовика Сибирской платформы.
Скелет имеет вид кубка конической, рогообразной или субцилиндриче-
ской формы, диаметром до 45 мм, высотой до 50 мм, иногда с несколько
обособленной ножкоподобной нижней частью.
Соаниты относятся к глубоководным организмам. От губок их от-
личает отсутствие спикул и наличие двух стенок; от археоциат — строе-
ние первичного скелета, отсутствие в интерваллюме перегородок, днищ
и пузырчатой ткани. Соаниты, установленные Е. И. Мягковой в 1965 г.,
известны пока только из раннего ордовика Сибири. Не исключена воз-
можность, что они также относятся к водорослям. Класс и отряд не-
известны.
КЛАСС (?) STROMATOPORATA. СТРОМАТОПОРАТЫ
Общая характеристика. Строматопораты (гр. stroma — слой, poros—
отверстие, пора) — вымершие колониальные морские организмы, имев-
Рис. 78. Класс Stromatoporata:
/—3 — внешний вид колоний; 4—5 — Siromatopora (ордовик — пермь): 4 — часть
колонии, 5 — поперечное сечение (видны астроризы);
м — мамелоны, лл — латиламины
шие известковый скелет — ценостеум, отличающийся большим
разнообразием как внешней формы, так и размеров. Ценостеум может
быть пластинчатым, массивным, желвакообразным, цилиндрическим,
ветвистым или инкрустирующим (рис. 78). Ценостеум строился цено-
сарком, выделявшим различные скелетные элементы: цисты, лами-
ны, столбики, ценостелы, ценостромы, инфлексионные ламины и ин-
164
флексоны. Характерны астроризы — горизонтальные радиальные
каналы, расположенные в межламинарном пространстве и соединенные
вертикальным каналом. В основании скелета расположена морщинис-
тая эпитека; цилиндро-конические колонии часто имеют ножку при-
растания. Ордовик — мел.
Строение скелета. Ц е н о с т е у м строматопорат состоит из гори-
зонтальных и вертикальных элементов и астрориз. К горизонтальным
элементам относятся цисты, ламины и диссепименты. Цисты пред-
ставляют собой выпуклые пластинки, образующие серию пузырьков
чечевицеобразной формы (рис. 79, 16). Ламины — горизонтальные
пластинки, чаще всего параллельные друг другу. Ламины возникают
Рис. 79. Класс Stromatoporata. Строение скелета.
Сечения: тангенциальное (1а, 2а, 26, За, 4а, 6, 7а), продольное (16,2в, 36,46,5,76)-, ре-
конструкция Стерна, 1966 (4в, S);
1 — Labe'chia (ордовик — карбон); 2 — Actinostroma (кембрий — ранний карбон);
3 — Stromatopora (ордовик — пермь); 4 — Clathrodictyon (кембрий — девон); 5
Simplexodictyon (силур); 6 — Gerronostroma (силур — девон); 7— Trupetostroma (де-
вон); 8 — Syringostroma (силур — девон);
бс—большие столбики, ил — инфлексионная ламина, ин — инфлексон, к — колликулы, »
л — ламина, мс — маленькие столбики, пс — пролонгированные столбики, пт — псев-
дозооидная трубка, с — столбик, сс — сепаратные столбики, т — ткань, заполняющая
промежутки между столбиками, ц — цисты, цн — ценостелы, цс — цено.стром, ят —
ячеистая ткань
двумя способами. В первом случае от вертикальных элементов перио-
дически возникают горизонтальные отростки (к о л л и к у л ы;
рис. 79, 26); соединяясь друг с другом, они создают своеобразную
сетку, петли которой в дальнейшем зарастают особой тканью и обра-
зуют так называемые колликулятные ламины. Во втором
случае вертикальные элементы периодически разрастаются в горизон-
тальном направлении и, соединяясь друг с другом, образуют конти-
н у з н ы е, или сплошные, ламины (рис. 79, 6). Рост колонии проис-
ходил периодически, о чем свидетельствует закономерное сгущение
горизонтальных элементов, образующих так называемые л а т и л а -
мины.
К вертикальным элементам относятся столбики и ценостелы.
Столбики имеют правильную округлую форму и четко обособлены
друг от друга; при образовании ламин рост столбиков приостанавли-
вается, а при возобновлении роста колонии столбики образуются либо
на том же месте и пересекают несколько межламинарных промежутков
(пролонгированные столбики; рис. 79, 2в), либо разви-
ваются изолированно в каждом межламинарном промежутке (сепа-
ратные столбики, рис. 79, 5). Ценостелы представляют собой
вертикальные элементы, образованные при слиянии столбиков друг с
другом и имеющие в поперечном сечении неправильную форму
(рис. 79, 7а, 6). Ценостелы, как и столбики, бывают сепаратными и
пролонгированными. Особое место занимают инфлексионные
элементы, у которых переход от горизонтального направления к вер-
тикальному не всегда ясен. Вертикальные инфлексионные элементы
имеют форму полого конуса, обращенного основанием вверх (рис. 79, 4),
основания, соединяясь вместе, образуют неправильную изогнутую
«горизонтальную» пластинку. У рода Stromatopora горизонтальные й
вертикальные элементы сильно утолщены; их предлагается называть
ц е н о с т р о м а м и (рис. 79, 3).
Астроризы представляют собой систему звездообразно ветвя-
щихся каналов, не имеющих самостоятельных стенок и расположен-
ных на поверхности горизонтальных элементов в межламинарных про-
странствах (рис. 78, 5). Они возникают на ранних стадиях развития
неностеума. Если они расположены одна над другой, то их центры сое-
диняются вертикальным каналом. Форма и размеры астрориз разнооб-
разны и зависят от характера ветвления горизонтальных каналов и
способа их сочленения с вертикальными каналамйГ в вертикальных
астроризальных каналах развиты тонкие горизонтальные или слегка
выпуклые днища или диссепименты. Верхняя поверхность
ценостеума нередко несет конические возвышения — мамелоны,
через которые проходят вертикальные каналы астрориз (рис. 78, 4).
Микроструктура скелета сохраняется крайне редко. Скелет
состоит из мельчайших частиц неправильной формы размером от 1
до 5 мк. Установлено несколько типов микроструктур: компактная, по-
перечно- или перисто-волокнистая, ячеистая, пятнистая,, тонкорешетча-
тая.
Систематическое положение строматопорат. Систематическое поло-
жение строматопорат до настоящего времени является предметом
Острой дискуссии. Существуют две основные точки зрения: одни иссле-
дователи относят строматопорат К целентератам, выделяя их в качестве
самостоятельного класса или в качестве подкласса гидроидных; другие
относили и относят сейчас к пориферам. Решение' вопроса о губковой
и целентератной природе строматопорат зависит от выяснения способа
166
литания; если строматопораты были подобно губкам фильтраторами,
то их нужно отнести к этому типу; если они были хищниками и имели
стрекательные клетки, то они принадлежат целентератам. Однако об
этом можно судить, только анализируя строение скелета.
С новым классом губок — склероспонгиями — их сближает сходство
общего облика и микроструктуры скелета, особенности строения астро-
риз; к числу различий между этими двумя классами следует отнести
наличие .плотного арагонитового скелета и кремневых спикул у склеро-
спонгий и отсутствие последних у строматопорат, наличие у стромато-
тторат и отсутствие у склероспонгии ламин и диссепиментов. У склеро-
спонгий в мягкой ткани имеются каналы, которые . отражаются на
поверхности скелета в виде борозд, напоминающих астроризы. По мере
роста мягких тканей борозды сверху перекрываются скелетной тканью
и превращаются в каналы, расположенные внутри скелета. Предпола-
гается, что астроризы строматопорат имели такое же назначение и
происхождение. Отсутствие астроризальных каналов у ряда форм
•сторонники этой гипотезы объясняют тем, что их положение в мягкой
ткани не отражалось на строении скелета. Сторонники целентератной
природы строматопорат считают, что в астроризальных вертикальных
канадах или помещалось тело полипа, или жили какие-то симбиотиче-
ские или сверлящие организмы (последняя идея пока имеет ограни-
ченное число сторонников).
В связи с тем что еще недостаточно данных, подтверждающих губ-
ковую природу строматопорат, они рассматриваются пока в качестве
•самостоятельного класса неясного систематического положения. Есте-
ственно, что общепринятой систематики строматопорат также пока нет.
В последние годы О. В. Богоявленская (1969) по типу строения астрориз,
вертикальных и горизонтальных элементов скелета разделила ордо-
викско-девонских строматопорат на пять отрядов: Labechiida, Clathro-
dictyonida, Actinostromatida, Gerronostromatida, Syringostromatida.
История развития строматопорат. Вопрос о происхождении стро-
матопорат окончательно не решен. Кембрийские представители изуче-
ны недостаточно и их систематическое положение не ясно. Первые бес-
спорные строматопораты известны из среднего ордовика. Это лабехииды,
щеностеум которых состоит из налегающих друг на друга цист толщиной
.до 45 мк, и строматопораты с ценостеумом, имеющим ячеистую и тон-
корешетчатую микроструктуру, но почти не различимые столбики и ла-
мины; в позднем ордовике появляются строматопораты, имеющие скелет,
•образованный инфлексионными элементами и внешне морщинистыми
или волнистыми ламинами и короткими столбиками (клатродиктиони-
,ды). В силуре разнообразие строматопорат увеличивается: появляются
формы, скелет которых состоит из колликулятных ламин и разной дли-
ны столбиков, с компактной микроструктурой (актиностроматиды).
В конце силура возникают строматопораты с ценостеумом, состоящим
из континузных пористых ламин и столбиков с компактной микрострук-
турой (герроностроматиды). В позднем силуре существенно обновляет-
ся состав уже известных • строматопорат и появляются формы, скелет
которых построен из ценостромов, имеющих ячеистую или микрорешет-
чатую микроструктуру (сирингостроматиды). Все названные группы,
по-видимому, продолжали свое существование в девоне. Они жили на
•сравнительно небольшой глубине, вблизи берега и в открытом море.
В силуре и девоне они принимали участие в образовании рифовых по-
строек, биостромов и биогерм. Совместно с ними встречаются ругозы и
табуляты, аннелиды и брахиоподы. Очень часто наблюдается сожи-
тельство строматопорат и сирингопорид.
167
Строматопораты, таким образом, были очень широко распростра-
нены в силуре — девоне, но в то же время почти неизвестны в карбо-
не— триасе и вновь известны в юре и мелу. В конце мела они, по-види-
мому, вымерли окончательно. Степень изученности мезозойских пред-
ставителей значительно меньше, чем палеозойских; их выделяют в
особый таксон Sphaeractinoidea. Всего известно около 70 родов
(рис. 79).
КЛАСС (?) HYDROCONOZOA. ГИДРОКОНОЗОИ
К классу гидроконозой (гр. hydra—- гидра, лат. conus — конус, гр.
zoon — животное) относятся мелкие кишечнополостные, встреченные в
отложениях нижнего и среднего кембрия азиатской части СССР; в
у
Рис. 80. Класс Hydroconozoa:
1 — Hydroconus, 2 — Jakovleviella — из раннего кембрия Тувы (оригинал и реконст-
рукция К. Б. Кордэ);
аб — аборальный канал, б — прикрепившийся молодой бионт, н — ножка, нч— началь-
ная стадия скелета, ок — осевой канал, п — поры, рк— радиальный канал, ч— чашеч-
ка, щ — щупальца
последнее время обнаружены в отложениях ордовика, карбона, перми,
юры и мела. Они имеют наружный, эктодермический скелет конической
формы высотой до 30 мм, иногда с обособленной ножкоподобной частыО
'(рис. 80). В верхней части кубка находится ширококоническая чашеч-
168
ка, с почти отвесными стенками,' в которой помещалось тело бионта.
Снаружи скелет снабжен продольными ребрами или покрыт табличками
ромбической формы. Внутренний слой стенки обычно продольно-волок-
нистый или бесструктурный. В нижней части скелет бывает заполнен
либо септоподобными образованиями, либо горизонтально расположен-
ными днищами. В пристенной части обнаружены различной формы
полости. Самый низ скелета имеет вид подошвы или ножки; в ней
обнаружен так называемый аборальный канал. Кроме аборального
канала в центральной части скелета имеются один или несколько осе-
вых каналов, от которых отходят радиальные каналы; последние у
представителей подкласса Porata открываются . наружу порами
(рис. 80, 2). Эти каналы сравниваются с каналами гастро-васкулярной
системы медуз. О развитии и размножении пока известно мало, но
можно предполагать, что гидроконозои размножались бесполым и поло-
вым путем. Встречены кубки совсем юных форм, еще без каких-либо
следов внутренних скелетных элементов, с каналом на аборальном
конце; такие кубки, вероятно, возникали вскоре после прикрепления
личинки к субстрату.
Особенности строения скелета, продольных и радиальных каналов,
наличие или отсутствие пор позволили К. Б. Кордэ выделить описан-
ные формы в особый класс гидроконозои в составе двух подклассов:
Porata и Aporata (рис. 80). Вопрос о систематическом положении
гидроконозой недостаточно ясен.
НАДРАЗДЕЛ EUMETAZOA.
ВЫСШИЕ МНОГОКЛЕТОЧНЫЕ
К высшим многоклеточным (гр. ей — приставка, обозначающая на-
стоящий; meta — продолжающий; zoon — животное) относятся все ниже
рассматриваемые типы животных, для которых характерны дифферен-
цированные органы и ткани, развивающиеся из двух или трех зароды-
шевых листков: эктодермы, энтодермы и мезодермы. Из эктодермы
развиваются наружные покровы животного с различными кожными
образованиями, наружный скелет, центральная и периферическая
нервная система, в том числе и органы чувств, мерцательно-движущие »
приспособления, органы дыхания. Из энтодермы развиваются орга-
ны пищеварения, печень и ее железы, в том числе и пищеварительные
железы. Из.мез одер мы помимо выстилки тела образуется большая
часть внутренних органов и тканей: поперечнополосатая мускулатура,
выделительные органы, половые клетки и соединительная ткань —
мезенхима, дающая начало всей группе опорно-трофических тканей
от крови и лимфы до хряща и кости включительно.
Эволюция настоящих многоклеточных шла различными путями от
наиболее низкоорганизованных двухслойных до таких высокоорганизо-
ванных трехслойных животных, какими являются головоногие, членисто-
ногие и хордовые (здесь не рассматриваемые). Все многоклеточные раз-
деляются на две неравные группы: двухслойных, или лучистых
(Radialia), и трехслойных, или двусторонне-симметричных (Bilateralia).
РАЗДЕЛ RADIALIA. РАДИАЛЬНЫЕ
К радиальным (лат. radius—луч) относятся наиболее низкоорга-
низованные многоклеточные, стоящие в основании всего ствола много-
клеточных, ткани которых возникают из двух зародышевых листков
(отсюда второе название—двухслойные). Настоящий третий слой —
мезодерма у них отсутствует; но в постэмбриональный период за счет
эктодермальных и эктодермальных клеток формируется мезоглея —
неклеточный слой, состоящий из протеинов; мезоглея обычно студе-
ниста, но может достигать значительной плотности и быть похожей на
гиалиновый хрящ; в ней присутствуют клеточные элементы, в том
числе и свободные амебоциты. Для всех двухслойных характерна ра-
диальная симметрия. Возникновение радиальной симметрии обуслов-
170
лено прикрепленным или свободноподвижным, парящим в воде образом
жизни.
Среди двухслойных выделяют два типа: кишечнополостные (Coelen-
terata) и гребневики (Ctenophora).
ТИП COELENTERATA.
КИШЕЧНОПОЛОСТНЫЕ
Общая характеристика. Кишечнополостные, или целентераты (гр.
koilos —• полый, enteron — кишка, внутренность), — многоклеточные,
двухслойные организмы, обитающие преимущественно в морях и океа-
нах; немногие приспособились к жизни в пресных водах (гидра и
Рис. 81. Схема строения кишечнополостных:
1 — гидроидных, 2 — сцифоидных, 3 — коралловых полипов; сечения: а — продоль-
ное, б — поперечное;
кк — кольцевой канал, мз — мезоглея, од — оральный диск, р — рот, т — тека,
щ — щупальца, эк — эктодерма, эн — энтерон — гастральная полость
гидромедуза). Радиально-симметричное тело имеет одну гастральную
полость, часто разделенную перегородками на отдельные камеры. Из-
вестны две жизненные формы: полипы, ведущие прикрепленный образ
жизни, и медузы — свободноподвижные. Вокруг единственного рта рас-
положены щупальца, снабженные стрекательными клетками. Кишечно-
полостные возникли, по-видимому, в протерозое, но достоверные от-
печатки тела известны из отложений верхнего протерозоя (венда);
скелетные остатки появляются впервые в породах кембрия.
Строение тела. Тело кишечнополостных имеет вид открытого на
одном конце двухслойного мешка с внутренней полостью, называемой
кишечной, или гастральной (рис. 81). Стенки тела состоят
из производных эктодермы и энтодермы. Через тело можно провести
171
одну продольную ось, вдоль которой проходит несколько плоскостей
симметрии. На одном конце тела помещается ротовое отверстие, веду-
щее в гастральную полость. Другой конец замкнутый. Иногда эта
полость — целентерон имеет боковые слепые впячивания, образующие
кишечно-сосудистую систему; она выполняет функцию пищеварения
и распределения питательных веществ. Единственное ротовое отверстие
выполняет функцию вводного и выводного. Рот окружен венчиком
щупалец, расположенных чаще всего несколькими кольцами. В качестве
органов нападения и защиты служат стрекательные клетки,
разбросанные по всему телу, но чаще всего сосредоточенные в экто-
дерме щупалец. Они разнообразны по строению и функциям; они раз-
виваются из промежуточных клеток и состоят из хитиноидной овальной
капсулы, заполненной жидкостью и несущей спирально свернутую
стрекательную нить; у одних кишечнополостных эти нити пробивают
покровы добычи, у других обвивают, у третьих прилипают к телу
добычи, выпуская при этом ядовитое токсическое вещество, парали-
зующее и убивающее жертву. Добыча, захваченная щупальцами, посту-
пает через ротовое отверстие в гастральную полость, где и происходит
ее внеклеточное, полостное переваривание.
Кишечнополостные — хищники; они питаются личинками и яйцами
различных животных, мальками рыб, мелкими ракообразными,
двустворчатыми и брюхоногими моллюсками., В связи с переходом к
питанию крупной добычей у кишечнополостных впервые появляется
нервный аппарат, основная функция которого заключается в согласо-
вании работы всех клеток, участвующих в ловле и проглатывании
добычи. Он состоит из отдельных нервных клеток, соединенных между
собой отростками, и имеет диффузный характер; небольшие сгущения
нервных клеток наблюдаются только вокруг рта и на подошве. На при-
митивность нервной системы указывает ее положение в эктодерме.
Кишечнополостные размножаются половым или бесполым путем.
Медузы:— обычно свободноплавающие одиночные формы — имеют
вид колокола или зонтика с щупальцами по краям, рот обращен вниз.
У медуз хорошо развита мезоглея; нервная система состоит из двух
кольцевидных нервных сплетений, соединенных радиальными тяжами;
вдоль периферического края тела медузы развиты органы равновесия
и чувств. Полипы — прикрепленные организмы, их организация
проще медуз. Размножение полипов происходит бесполым путем —
почкованием или делением; при этом образуются колонии, состоящие
либо из однородных индивидов, — мономорфные колонии,
либо из разнородных специализированных особей — полиморфные
колонии. В полиморфной колонии одни особи выполняют только
функцию пищеварения, вторые — размножения, третьи — защиты коло-
нии от внешних врагов. В колонии все особи находятся в органической
связи друг с другом, осуществляя обмен веществ, направленный на
поддержание жизнедеятельности колонии. Полипы и медузы могут
существовать либо как самостоятельные организмы, либо как стадии
жизненного цикла одного организма.
Половые клетки развиваются за счет эктодермических или энто-
дермических зачатков. После оплодотворения и дробления яйца, кото-
рое осуществляется у разных групп по-разному, образуется двухслой-
ная личинка — планула, покрытая ресничками. Она не способна ни
питаться, ни размножаться и является, скорее всего, только рассели-
тельной формой. После прикрепления личинки к субстрату происходит
перестройка ее организации и образуется полип, при этом эктодерма
может выделять скелет — органический или минеральный.
172
На основании особенностей внутреннего строения, характера раз-
множения среди кишечнополостных выделяют три класса: Hydrozoa,
Scyphozoa и Anthozoa.
КЛАСС HVDROZOA. ГИДРОИДНЫЕ
Общая характеристика. К классу гидроидных (гр. hydor — вода,
hydra — водяная змея, гидра, zoon — животное) относятся наиболее
примитивные, большей частью мелкие кишечнополостные, колониаль-
ные, реже одиночные, обладающие радиальной или четырехлучевой сим-
Рис. 82. Подкласс Hydroidea:
1 — общий вид колонии Obelia; 2 — схема чередования поколений гидромедуз
(м) и полипов (п);
г — гонангий — ячейки, в которых созревают молодые гидромедузы, л — личин-
ка, мп,— молодой полип, с—сперматозоид (мужская гамета), я — яйцеклетка
(женская гамета)
метрией. Щупальца, окружающие рот, собраны в 1—3 венчика. Рот
переходит в простую гастральную полость, не разделенную перегород-
ками. У колониальных форм гастральная полость продолжается в ствол
и его ветви. Некоторые формы имеют органический (хитиноидный) или
минеральный (известковый) скелет, выделяемый эктодермой. Половые
продукты образуются в эктодерме. Гидроидные размножаются половым
и бесполым путем; характерно чередование поколений, имеются две
жизненные формы — полипоидная и медузоидная. В отличие
от других классов эти жизненные формы носят названия гидропо-
173
липов и гидромедуз; гидрополипов часто называют гидран-
тами, или просто зооидами. В ископаемом состоянии гидроидные
известны с кембрия.
Строение мягкого тела. Гидрополип и гидромедуза построены по
одному плану (рис. 82). Тело' гидрополипа представляет собой
двухслойный мешок, открытый с одного конца; между эктодермой и
энтодермой развит тонкий слой мезоглеи. Тело имеет бокаловидную-
или трубчатую форму диаметром до 2—3 мм с ножкой на конце, при
помощи которой оно прикрепляется к подводным предметам (рис. 83, 2).
Рис. 83. Подкласс Hydroidea:
1 — Hydractinia, часть современной колонии при большом увеличении; 2 — гидра, про-
дольный разрез;
газ — гастрозооиды, гз — гонозооиды, д — дактилозооиды, р—ротовое отверстие,
х — базальная хитиновая пластинка, щ— щупальце, экт— эктодерма, энт — энтодер-
ма, юп — юная почка
Рот окружен щупальцами (обычно 10—30), расположенными правиль-
ными венчиками или без строгого порядка. Гигантский одиночный полип
(до 1 м в высоту) имеет до 380 щупалец, а миниатюрный полип (в не-
сколько миллиметров) только одно щупальце. Щупальца несут много-
численные стрекательные клетки и служат для ловли добычи и осяза-
ния. За редким исключением (пресноводная гидра) все гидроидные
полипы — колониальные животные; отдельные зооиды располагаются на
вертикально стоящем стволе (см. рис. 82, /) или поднимаются над
горизонтально стелющимся корневищем — гидроризой. Все зооиды
колонии соединены друг с другом каналами, которые служат для
связи между членами колонии. Обычно колонии полиморфны; они
состоят из особей, выполняющих различные функции: питающие зооиды
снабжены ртом и щупальцами, другие зооиды лишены рта и выполняют
174
защитную функцию, третьи, лишенные рта и щупалец, отпочковывают
медуз — новых воспроизводительных особей.
Гидромедузы имеют вид диска, зонтика или колокола диа-
метром 2—3 мм и очень редко до 50 мм; между экто- и энтодермой
хорошо развита мезоглея (см. рис. 82, 2 м). На конце хоботообразно-
го выроста расположен рот, ведущий во внутреннюю полость, от кото-
рой отходят четыре радиальных канала. Они соединяются с кольцевым
каналом, проходящим по краю зонтика, вдоль которого расположены
щупальца и органы чувств. От свободного края зонтика по направ-
лению к центру отходит кольцевая краевая перепонка, или парус.
Гидромедуза имеет нервную систему более сложную, Нем у гидрополи-
па, состоящую из двух нервных колец. Они иннервируют органы чувств,
мускулатуру, глазки и органы равновесия — с т а т о ц и с т ы. Стато-
цисты представляют собой видоизмененные щупальца, имеющие вид
пузырька, стенки которого образованы чувствительными клетками. На
них лежит известковый камушек-—статолит, выделенный клетками
эктодермы, который при изменении положения тела медузы переме-
щается и давцт на соответствующие чувствительные клетки.
Строение скелета. Эктодерма выделяет тонкую органическую обо-
лочку, выполняющую роль наружного скелета. Скелет служит опорой
и защитой. Он развивается только у полипов, гидромедузы лишены
скелета. Органический скелет образуется вокруг ствола, ветвей и от-
дельных зооидов; в ископаемом состоянии он известен из ордовика
Скандинавии и из ордовикских валунов Польши (по данным Р. Коз-
ловского). Небольшая часть современных и большая группа древних
гидроидных обладали способностью выделять известковый скелет; он
имеет разную форму и строение и будет рассмотрен в каждом отряде
отдельно.
Размножение и развитие. Гидроидные размножаются бесполым и
половым путем (см. рис. 82). В первом случае новые особи возникают
в результате почкования, что свидетельствует о невысоком уровне
интеграции особей. При почковании в колонии развиваются как зоои-
ды, так и гидромедузы; последние возникают из почек; у них появ-
ляются четыре радиально расположенных канала и ротовой хоботок;
они, отделяясь от колонии, переходят к самостоятельному существова-
нию в толще воды. Гидромедузы раздельнополы и в свою очередь
размножаются только половым путем. В эктодерме у них обра-
зуются половые клетки. Оплодотворение яиц и развитие происходит в
воде. Из яйца выходит двухслойная личинка — планула, покрытая
снаружи ресничками. Поплавав некоторое время, личинка прикреп-
ляется к какому-либо подводному предмету. Происходит перестройка
ее организации, из двух зародышевых листков развивается эктодерма
и энтодерма. Формируется радиально-симметричный полип. За счет
эктодермы вокруг полипа развивается т ек а, или перисарк. Полип,
достигнув взрослой стадии, начинает размножаться почкованием, про-
дольным или поперечным делением, в результате чего возникает коло-
ния; у нее развиваются зооиды и гидромедузы и процесс развития
повторяется вновь. У некоторых гидроидных медузы отрываются от
колонии после созревания половых продуктов, а иногда они перестают
отрываться и сохраняют постоянную связь с колонией. В процессе
дальнейшего упрощения вместо медуз образуется эктодермический
мешочек — с п о р о с а к, в котором развиваются половые клетки.
Основы систематики и классификация. На основании особенностей
строения полипоидной и медузоидной стадий, степени полиморфизма,
175
особенностей строения скелета класс гидроидных разделяется начдва
подкласса: Hydroidea, Siphonophoroidea.
ПОДКЛАСС HyDROIDEA. ГИДРООБРАЗНЫЕ
Гидрополипы и гидромедузы как морские, так и пресноводные,
одиночные и колониальные организмы (см. рис. 83). У большинства
наблюдается чередование полового и бесполого поколений, причем из-
вестны все формы редукции медузоидной стадии от медузы до
споросака; у небольшой группы пресноводных форм (гидры) развита
только полипоидная стадия (см. рис. 83, 2); у морских трахилид
основной жизненной формой является медуза (диаметр колокола до
10 см); полипов нет, из планулы возникают не полипы, а медузы.
У многих колониальных форм эктодерма гидроризы (сетчатое осно-
вание) ствола и ветвей выделяет органический скелет — перидерму,
которая образует вокруг полипа бокаловидную ячейку. У представите-
лей двух отрядов Milleporida и Stylasterida, которые будут рассмотрены
ниже, развивается известковый скелет. Фоссилизированные остатки
гидроидных очень незначительны и не отвечают тому разнообразию
жизненных форм, которые существуют сейчас и жили, по-видимому, в
прошлом. Подкласс насчитывает около 26 семейств, включающих со-
временных и вымерших представителей. Очень редко встречаются отпе-
чатки гидромедуз, причем их палеозойские остатки определяются не
всегда надежно, но юрские и более молодые находки кажутся досто-
верными. Известковые скелеты колониальных гидроидных полипов
известны начиная с позднего мела, органические скелеты — с ордовика.
Отряд Milleporida. Миллепориды (лат. mille — тысяча, гр. porus —
пора, канал). Колониальные полипы с известковым скелетом разнооб-
разной формы; гидрополипы живут в известковых трубочках — ячейках,
выделяемых промежуточной тканью всей ' колонии — ценосарком.
Среди полипов различают гастрозооиды — питающие особи с
четырьмя очень короткими щупальцами и развитой гастральной
полостью и дактилозооиды — полипы-воины с короткими щупаль-
цами (5—7), без гастральной полости, добывающие пищу, и, наконец,
более крупные ячейки—ампулы, в которых развиваются гидро-
медузы. Колония обладает неограниченным ростом, и гидрополипы
периодически отделяются от своих более глубоких частей горизон-
тальными днищами, надстраивая вверх свой скелет. Гидрополипы зани-
мают только верхнюю часть скелета. На поверхности скелета после
гибели полипов видны круглые или полигональные отверстия —
гастропоры, принадлежащие гастрозооидам, и окружающие
более мелкие отверстия — дактилопоры, в которых жили дактияо-
зооиды; в промежутках между ячейками развит пористый известковый
скелет. Ампулы обычно имеют вид более крупных ячеек, прикрытых
пористым скелетом. Современные миллепориды живут среди коралло-
вых рифов, на глубине не более 30 м, принимают участие в постройке
рифов. В симбиозе с ними живут водоросли. В состав отряда входит
5 родов (рис. 84, 1). Поздний мел — ныне.
Отряд Stylasterida. Стилястериды (гр. stylos — столб, колонна,
aster — звезда). Внешняя форма колоний разнообразна: от инкрусти-
рующей до древовидной; промежуточная ткань — ценосарк — имеет
многочисленные каналы, которые соединяют между собой гастрозоои-
дов, дактилозооидов и ампулы; первые имеют цилиндрическую форму
с ротовым отверстием, расположенным на сводообразном возвыше-
нии— гипостоме; рот окружают тонкие нитевидные щупальца;
176
дактилозооиды имеют вид простых щупалец. У стилястерид отсутствует
стадия свободноплавающей медузы и половые клетки развиваются в
особых ячейках-—гонофорах, расположенных в ампулах. Скелет
стилястерид более плотный, с более мелкими порами, чем у милле-
порид. Гастрозооиды помещаются в трубчатых гастропорах, часто
Рис. 84. Отряды Milleporida (/) и Stylasterida (2—3):
- 1 — Millepora (поздний мел — ныне): 1а — общий вид современной колонии,
16 — продольное и 1в — тангенциальное сечения; 2 — Stylaster (палеоген —
ныне), деталь колонии (Х9); 3 — Stenohelia (совр.);
ам -т- ампула, гп — гастропора, гс — гастростиль, дп — дактилопора
снабженных по периферии небольшими известковыми перегородками и
осевой структурой (гастростиль), напоминающей столбик у коралло-
вых полипов; дактилозооиды (от 6 до 18) помещаются в более мелких
дактилопорах, окружающих гастрозооиды. Форма ампул и их располо-
жение различны. Современные стилястериды живут преимущественно
в тропических морях в зоне мелководья. Но известны представители
также на довольно больших глубинах. В состав отряда входит свыше
15 родов (рис. 84, 2, 3). Поздний мел — ныне.
ПОДКЛАСС SIPHONOPHOROIDEA. СИФОНОФОРЫ
К подклассу сифонофор (гр. siphon — сифон, phoros — несущий)
относятся свободноплавающие, пелагические, морские высокоспециали-
12 в. В. Друщиц
177
зированные гидроидные, образующие сложные колонии с полиморф-
ными особями, среди которых имеются как полипоидные, так и
медузоидные особи. Основу колонии составляет ствол, стенки кото-
рого состоят из эктодермы и энтодермы и бесструктурной пластинки,
(мезоглеи) между ними. Энтодерма выстилает гастральную полость,,
которая связывает все особи в единое целое. Медузоидные особи видо-
изменены. Одни превращены в плавательный пузырь (пневма-
Рис. 85. Подкласс Siphonophoroidea:
1—схема строения сифонофоры; 2 — Plectodiscus
(поздний девон), реконструкция (Кастер, 1942);
ар —. арканчики с дактйлозооидами (дз), газ —
гастрозооиды, гз — гонозооиды — половые особи,
к — крышечки, пк — плавательные колокола, пн —
пневматофор, с — ствол колонии
тофор), заполненный газом, другие — в плавательные колокола,
(не кт о форы), которые, последовательно сокращаясь и расширяясь,
приводят в движение всю колонию, третьи — в гонофоры, продуци-
рующие половые клетки, и т. д. Полипоидные особи выполняют функ-
цию питания (гастрозооиды), функцию защиты (дактилозооиды).
Сифонофоры размножаются бесполым и половым путем. Из оплодо-
творенного яйца развивается личинка; из последней путем почкования
возникает новая колония. Отдельные особи объединяются в особые-
группы — кор мидии. Почкование начинается от ствола. Стадия
прикрепленного полипа отсутствует. В ископаемом состоянии сифоно-
форы встречаются исключительно редко. Известны отпечатки 5 родов,
пневматофор из отложений ордовика, силура и девона Европы, Север-
ной Америки, Австралии (рис. 85).
178
Приложение к классу гидроидных.
ХЕТЕТИДЫ
Отряд (?) Chaetetida. Хететиды (гр. chaeta — щетинка). Полип-
няки сферические или пластинчатые; ячейки многоугольные, неправиль-
ной формы или меандрические размером от 0,15 до 1,2 мм. Отсутствуют
какие-либо септальные образования; развиты многочисленные днища.
10
Рис. 86. Группа Chaetetida:
1—3 — типы колоний; 4—6 — типы размножения хететид: 4а, б — размножение
делением (Chaetefes), 5а, б — межстенное почкование (Bauneia), 6 — базальное
почкование (Chaetetiporella, X 15); 7—9 — микроструктура: 7 — гранулезная, 8 —
ламеллярная, 9 — фиброрадиальная; 10—11 — меандрические типы колоний, кон-
вергентное сходство между родами Chaetetipora (девон — карбон) и Blasiochae-
tetes (средняя юра — мел), (Фишер, 1970);
дн — днища, по — поры, яч — ячейки
Микроструктура стенки фиброрадиальная, гранулярная или ламелляр-
ная. Связь между полипами, вероятно, отсутствовала. Размножались
продольным делением и базальным почкованием. Средний ордовик —
эоцен.
Строение скелета и мягкого тела. Хететиды — колониальные ор-
ганизмы, вели прикрепленный образ жизни. Колонии имеют различную
форму: шаровидную, желвакообразную, полусферическую, пластинча-
тую или коркообразную, размером от нескольких сантиметров до мно-
гих дециметров; колонии состоят из плотно прилегающих друг к другу
трубок призматической формы с полигональным сечением, расположен-
ных радиально-лучисто (рис. 86). Кроме полигональных трубок у
хететид встречаются меандрические формы, возникшие в результате
неполного деления (рис. 86, 10, И). Хететиды размножались, по-види-
мому, как половым, так и вегетативным путем. Последнее осуществля-
лось путем продольного деления и базального почкования. При деле-
12*
179
нии материнский полип расчленялся на две или несколько частей, а
вслед за этим происходило соответствующее деление скелета
(рис. 86, 4); если деление не доходило до конца, то возникала меандри-
ческая колония (рис. 86, 11). Кроме того, новые зооиды могли возни-
кать между старыми, в результате так называемого межстенного
почкования (рис. 86, 5). При базальном почковании,
которое наблюдается преимущественно у пластинчатых колоний, новые
полипы возникали в основании колонии (рис. 86, 6).
По мере роста колонии отдельные полипы отделялись от нижней
нежилой части горизонтальными днищами (рис. 86, 9), полными или не-'
полными; последние могли переходить в пузырчатую ткань. Сообщения
между кораллитами обычно отсутствуют, стенки сплошные, однако
иногда наблюдаются соединительные поры. Различают три основных
типа микроструктуры стенки: трабекулярную, гранулярную и пластин-
чатую, или ламеллярную (рис. 86, 7—9); трабекулярная состоит из от-
дельных склеродермитов, образующих структуру, напоминающую фон-
тан воды (рис. 86, 96); гранулярная — представлена мелкими кристал-
лами арагонита или кальцита; пластинчатая — состоит из тонких слабо
изогнутых пластин микроскопических размеров.
Хететиды появляются в среднем ордовике; от среднего девона до
конца перми известно свыше 10 родов, от триаса до эоцена—11. Наи-
более широко хететиды были распространены в карбоне. Они были
обитателями мелководных зон морей.
Систематическое положение хететид до сих пор вызывает споры.
Б. С. Соколов (1962), монографически изучавший палеозойских хететид,
и Ж. Фишер (1970) —мезозойских, сравнивали их с мшанками (трепо-
стоматами),.кораллами (табулятами) и гидроидными и пришли к вы-
воду, что хететиды ближе всего стоят к гидроидным.
КЛАСС SCyPHOZOA. СЦИФОЗОИ
\
К классу сцифозой (гр. skyphos — чаша) относятся морские, пре-
имущественно свободноплавающие, реже прикрепленные, кишечно-
полостные, обладающие, как правило, четырехлучевой симметрией.
Гастральная полость, выстланная • энтодермой, разделена четырьмя
перегородками на четыре камеры. Половые продукты закладываются
в энтодерме. Развитие сопровождается чередованием поколений, с пре-
обладанием медузоидной стадии; полипоидная стадия представлена
маленьким полипом — сцифостомой, дающим бесполым размно-
жением начало медузам. Скелет у большинства отсутствует или пред-
ставлен хитиново-фосфатной перидермой (у вымершей группы кону-
лят). Сцифозои в ископаемом состоянии встречаются очень редко начи-
ная с венда. Разделяются на три подкласса: Scyphomedusae, Conulata,
Protomedusae.
ПОДКЛАСС SCYPHOMEDUSAE. СЦИФОМЕДУЗЫ
Морские, преимущественно крупные медузы (гр. skyphos — чаша,
medusa — медуза), свободноплавающие или прикрепленные, обладаю-
щие хорошо развитыми органами чувств и кратковременной стадией
полипа — сцифостомы (рис. 87).
Медузы имеют зонтикообразную, колоколовидную или округло-ку-
бическую форму. Вдоль края зонтика расположены щупальца. В центре
нижней поверхности зонтика находится • четырехугольный рот; углы
рта вытянуты в четыре желобовидные лопасти, которые служат для
180
захвата пищи. Рот ведет в короткую глотку, выстланную изнутри экто-
дермой; глотка — в Пищеварительную полость, снабженную четырьмя
радиальными выростами. От пищеварительной полости отходят
4—16 радиальных каналов, впадающих в кольцевой канал, расположен-
ный у края зонтика медузы. По этим каналам, выстланным мерцатель-
ным эпителием, пища разносится по всему телу. Хорошо развита
мезоглея, содержащая свободные амебоидные клетки и выполняющая
Рис. 87. Подкласс Scyphomedusae. Жизненный цикл медузы Aurellia:
з — зигота, л — личинка, м— медуза, п — полип —nt — полипоидная стадия
с образованием эфир), с — сперматозоид, э — эфира, я — яйцеклетка
функцию скелета. Нервная система имеет вид кольца, лежащего по
краю зонтика; здесь же расположены органы равновесия и светочувст-
вительные глазки. Медузы, как правило, раздельнополы; четыре поло-
вые железы образуются в энтодерме желудка. Оплодотворение и разви-
тие яиц происходит в воде. Личинка, покрытая ресничками, после не-
продолжительного плавания оседает на дно и, прикрепляясь хитиноид-
ной ножкой, превращается в сцифостому, у которой в отличие от
гидроидных полипов образуются: эктодермическая глотка, последова-
тельно вокруг рта 4, 8, 16 щупалец и четыре перегородки в гастральной
полости. Далее сцифостома перешнуровывается поперечно на ряд
дисков, которые постепенно отрываются от материнского организма,
перевертываются и превращаются в маленьких медуз — эфир. Эфиры
звездообразной формы растут и превращаются в медуз. Таким образом,
181
у сцифомедуз существует чередование поколений, в котором основной
жизненной формой является медуза, а полип представляет только
короткую временную жизненную стадию (см. рис. 87).
Сцифоидные медузы отличаются от гидроидных медуз более слож-
но устроенной кишечно-сосудистой системой, большим развитием
нервной системы и органов чувств, расположением половых желез
в энтодерме, наличием эктодермической глотки, отсутствием паруса,
наличием эфир, а на взрослой стадии складчатым краем.
Современные медузы широко распространены в морях и океанах;
кроме свободноплавающих известны сидячие медузы, живущие на не-
больших глубинах, преимущественно в холодных морях. Известно около
70 родов. В ископаемом состоянии крайне редко встречаются в виде
отпечатков и слепков. Наиболее древние остатки медуз известны из
венда (местонахождение Эдиакара) и нижнего кембрия Австралии.
Несколько родов описано из верхнеюрских отложений Золенгофена
(ФРГ) и из более молодых отложений разных стран.
ПОДКЛАСС CONULATA. КОНУЛЯТЫ
Подкласс конулят (лат. conulus — маленький конус) состоит из
одного отряда конулярий. К конуляриям относятся вымершие морские
Рис. 88. Подкласс Conulata:
1 — скульптура наружной поверхности; 2—3 — форма скелетов конулярий; 4 —
юная особь прикрепленной конулярий Sphenothallus (средний ордовик); 5—6 — ре-
конструкции: 5 — Archaeoconularia (ордовик — силур), прикрепленная форма, 6 —
Exoconularla (ордовик), свободноплавающая форма; 7а, б — устье, закрытое четырь-
мя лопастями; 8—9 — поперечные сечения скелета
организмы, обладавшие тонким хитиноидным скелетом сигаровидной
или пирамидальной формы высотой до 10 см, как исключение до
30—40 см (рис. 88). Скелет представляет собой тонкую оболочку, или
182
.перидерму, эктодермического происхождения толщиной от 0,05 до
Ю,30 мм, реже 1 мм. Она состоит из тонких микроскопических слоев,
окрашенных в бурые цвета, расположенных параллельно поверхности;
Рис. 89. Строение мягкого тела и скелета Stephanoscyphus (Вернер, 1966):
1а — продольное сечение, 16, 1в— поперечные сечения; 2 — скелет (перидерма);
кк — кольцевой канал, п — перидерма, р — рот, рк — радиальный канал, щ — щу-
пальце, эс — эндодермальная септа
«слои слагаются хитином, хитиново-фосфатной основой с различным со-
держанием фосфора и фосфата кальция. Перидерма при жизни кону-
лярий была довольно эластична. На ранних стадиях развития конуля-
рии, возможно, прикреплялись к различным подводным предметам при
помощи хитиново-фосфатного диска; на его нижней поверхности видны
/тойкие радиальные струйки, на верхней — два кольца (рис. 88, 4).
У взрослых конулярий перидерма имела вид четырехгранной пира-
миды или коцуса с округлым, овальным, квадратным или ромбическим
сечением (рис. 88, 8, 9). По середине каждой стороны и по углам
перидермы, на наружной стороне, проходят более или менее отчетливо
выраженные бороздки, или пигментированные линии. На внутренней
•стороне им соответствуют внутренние гребни или внутренние септы с
раздвоенными краями. Снаружи перидерма украшена поперечной
•скульптурой — ребрышками или рядами бугорков; в промежутках
между ними расположены продольные гребешки, придающие скульпту-
ре сетчатый характер. Передний конец перидермы обычно прикрыт
Четырьмя треугольными или округленными лопастями и развитыми
между ними эластичными связками; лопасти служили для закрывания
устья, внутрь которого втягивались щупальца в случае опасности.
О наличии щупалец у конулярий свидетельствуют их отпечатки, обна-
руженные по периферическому краю перидермы, а также гребни или
183
перегородки на внутренней стороне, к которым, вероятно, прикрепля-
лась мускулатура щупалец. Задний конец закрыт поперечными пере-
городками, или диафрагмами (рис. 88, 7).
Систематическое положение ’конулярий до последнего времени
вызывало дискуссию. В течение долгого времени их относили к брюхо-
ногим моллюскам, но после обстоятельных исследований, проведенных
Кидерленом (Kiderlen, 1937), их стали относить к сцифоидным, рас-
сматривая их в качестве группы неясного таксономического ранга или
в качестве подкласса сцифоидных. Еще более подтвердилась их
медузоидная природа после исследований Вернера (Werner, 1966) со-
временного медузоидного полипа Stephanos су phus (рис. 89). Этот ма-
ленький полип живет в тонкой’ конической трубке размером 10—20,
редко до 50 мм. Рот окружен щупальцами (от 40 до 100),'которые втя-
гиваются в гастральную полость. Глотка отсутствует. Имеются 4 .экто-
дермальные септы, слегка вдающиеся в гастральную полость. В отли-
чие от всех полипов в головном отделе имеется кольцевой канал, от
которого отходят четыре радиальных канала. Эти каналы выполняют
функцию гидродинамической опорной системы головного отдела с его
кроной щупалец. Эта особенность строения, характерная для медуз,,
указывает на равноценность медуз и полипов, причем, по-видимому,,
медузы представляют собой полипов, приспособленных к пелагическому
образу жизни для расселения и размножения. В головном отделе
полипа расположены железы, выделяющие хитиновое вещество пери-
дермы. Этот признак связывает современных медуз с конулятами и под-
черкивает их медузоидную природу.
Конулярии размножались, по-видимому, бесполым и половым пу-
тем. Первые находки конулярий обнаружены в среднем кембрии; они
довольно разнообразны и многочисленны в силуре и девоне и редко-
встречаются в отложениях карбона—триаса. Описано 12 родов и
около 200 видов.
ПОДКЛАСС PROTOMEDUSAE. ПРОТОМЕДУЗЫ
К небольшому подклассу протомедуз (гр. protos — первый) отне-
сены отпечатки примитивных кишечнополостных, встреченные в поздне-
Рис. 90. Подкласс Protomedusae:
1—2 — Brooksella (докембрий — средний кембрий): 1 — реконструкция (Хар-
рингтон и Мур, 1956), 2а, б — слепок внутренней полости ископаемой медузы;
вп — вентральная полость, ж — желудок, ок — осевой канал, рк — ради-
альный . канал, щ — щупальце
протерозойских и кембрийских отложениях Северной Америки, в ран-
нем ордовике Швеции и Франции. Отпечатки имеют дискоидальнук?
184
или эллипсоидальную форму тела с четкими радиальными полостями,
широко выпуклыми на верхней стороне зонтика и узкими, превращен-
ными в валики, на нижней стороне; между главными лопастями
вклинены обычно дополнительные лопасти. От ротовой пластинки от-
ходят четыре — пять щупалец, внутри которых проходит осевой канал,
открывающийся наружу; центрального рта нет. Каналы рук впадают
в вентральные полости, расположенные над ротовой пластинкой; вент-
ральные полости связаны вертикальными осевыми каналами с диско-
образным желудком, расположенным в центре у верхней поверхности;
от него отходят радиальные каналы, по одному на каждую лопасть.
Отсутствие центрального рта, кольцевого канала, щупалец и органов
чувств позволяет рассматривать протомедуз в качестве наиболее при-
митивных медузоподобных кишечнополостных и выделять их в качестве
подкласса. Известно два рода. Докембрий — средний кембрий Северной
Америки (рис. 90).
КЛАСС ANTHOZOA. КОРАЛЛОВЫЕ ПОЛИПЫ
Общая характеристика. Коралловые полипы, или антозои (гр.
anthos — цветок) — исключительно морские, наиболее высоко органи-
зованные целентераты; одиночные или колониальные, имеющие только
полипоидную стадию. В отличие от гидроидных и сцифозой их гаст-
ральная 'полость разделена системой мягких мезентериальных пере-
городок на центральную пищеварительную полость и периферические
камеры. Глотка эктодермического происхождения, сплющена с боков и
несет один или два мерцательных желобка — сифоноглифа. На
ротовом диске расположены щупальца в количестве от 8 до нескольких
десятков. Радиальная симметрия нарушена щелевидной формой рта
и глотки, расположением мезентериев и мышечных утолщений на них.
Одни коралловые полипы лишены скелета, у других имеется наружный
известковый скелет, у третьих скелет расположен в мезоглее и состоит
из отдельных спикул, роговых или известковых; спикулы могут соеди-
няться вместе, образуя разных типов внутренний скелет. Обитатели
морей всех глубин от литорали до ультраабиссали. Первые коралловые
полипы известны из кембрия.
Строение мягкого тела. В отличие от гидроидных тело коралловых
полипов отличается большей величиной и более сложным внутренним
строением. У многих форм между отдельными полипами развита про-
межуточная ткань, или ценосарк, пронизанная каналами, соеди-
няющими отдельные особи. У современных представителей тело имеет
цилиндрическую или мешковидную форму; стенка тела состоит из
экто- и энтодермы и лежащей между ними мезоглеи, представляющей
собой мезенхиму. Ротовое отверстие щелевидной формы окружено
венчиком щупалец, полых или сплошных; щупальца расположены коль-
цами. Рот ведет в сжатую с боков глотку эктодермического происхож-
дения, вдоль узких краев которой развиты один или два желобка —
сифоноглифа, покрытых мерцательными ресничками или жгутиками.
Вдоль этих желобков вода поступает в гастральную полость, разде-
ленную на центральную и периферические части системой мезенте-
риальных перегородок. У восьмилучевых кораллов имеются восемь
мезентериев, несущих на свободных концах железистые утолщения —
гастральные нити, богатые пищеварительными и стрекательными
клетками; при пищеварении свободные края мезентериев сближаются
и отграничивают центральную полость кишечника от периферической,
таким образом временно возникает кишечник и полости вокруг него.
185
Пищеварение вначале полостное внеклеточное, а затем измельченные
частицы пищи захватываются эктодермальными клетками и перевари-
ваются внутриклеточно в пищеварительных вакуолях или в мезоглее.
Непереваренные остатки пищи и продукты обмена выбрасываются
наружу через рот.
В ротовом диске развита кольцевая мускулатура, образующая
своеобразный сфинктор, закрывающий устье.’ Вдоль мезентериальных
перегородок с одной стороны в определенном порядке, характерном
для каждой группы антозой, расположены продольные мускулы, играю-
щие роль при втягивании и выпячивании щупалец. В основании тела
полипов развита мускулатура, с помощью которой бесскелетные формы
(актинии) могут присасываться к субстрату или передвигаться.
Антозой являются хищниками, они питаются крупной добычей,
захватывая ее при помощи щупалец, снабженных стрекательными
клетками; добыча затем передается в рот; вокруг рта наблюдается
концентрация нервного сплетения.
Строение скелета. Среди коралловых полипов различают две
группы: у первой, сравнительно небольшой, отсутствует скелет (акти-
нии), у второй, преобладающей группы, имеется опорный минеральный
скелет двух типов: наружный или внутренний.
Наружный скелет образуется за счет клеток эпидермы, фор-
мирующих кальцитовые или арагонитовые фибры размером 0,01—
0,02 мм; из фибр развиваются два типа структур: пластинчатая, или
ламеллярная, и фиброзная. В ламеллярных структурах фибры
располагаются параллельно, плотно прилегая друг к другу и образуя
тонкие пластинки; в фиброзной — фибры образуют пучки с единым
центром, или скл ер о дер м иты. Склеродермиты и ламеллярные
структуры образуют так называемые трабекулы.
Внутренний скелет, в отличие от наружного, состоит из
отдельных спикул, роговых или известковых, образованных в мезо-
глее особыми клетками эктодермического происхождения — склеро-
бластами. Размеры спикул варьируют от десятков микрон до не-
скольких миллиметров. Спикулы обладают способностью сливаться
вместе, образуя разного рода внутренний скелет. У одних вокруг каж-
дого полипа образуется плотная трубочка, у других слияние спикул
приводит к образованию плотного осевого стержня, причем в его обра-
зовании принимают участие как известковые, так и роговые спикулы.
Внутренний скелет характерен для большинства восьмилучевых корал-
лов, наружный — для всех остальных современных и вымерших корал-
лов и двух отрядов восьмилучевых. Наружный скелет одного полипа
называется кораллитом, а колонии — по липняком. Детали
скелетов будут рассмотрены в каждом подклассе отдельно.
Размножение и развитие. Современные коралловые полипы раз-
множаются половым и бесполым путем. Половые клетки образуются
в энтодерме мезентериальных перегородок. Все развитие протекает вне
материнского организма или в гастральной полости матери. У боль-
шинства антозой вышедшая из яйцевых оболочек личинка после не-
продолжительного плавания прикрепляется ко дну и превращается в
полип. Одни полипы всю жизнь остаются одиночными, другие начи-
нают почковаться и . создают колонии. При почковании? новые особи
возникают либо в промежуточной ткани—ценосарке, лйбо путем про-
дольного деления тела полипа; в последнем случае при подобном деле-
нии образуются два новых нормальных полипа, либо, если деление
идет лишь частично и обособляется только ротовые отверстия и глот-
ки, возникают колонии с неразделенными полипами, с парными сим-
186
метричными рядами мезентериев, щупалец и известковых перегородок
(склеросепт) и многочисленными ртами, расположенными между
двумя рядами щупалец.
Основы систематики и классификация. Класс коралловых полипов
на основании количества и формы мезентериальных перегородок, на-
личия наружного или внутреннего скелета и особенностей его строения
разделен на пять подклассов: Octocorallia, Hexacorallia, Tabulata, Не-
liolitoidea, Tetracorallia (Rugosa). Первые два класса существуют в
настоящее время, остальные три известны только по скелетным остат-
кам преимущественно в палеозое; имеются указания на находки табу-
лят в более молодых отложениях.
ПОДКЛАСС OCTOCORALLIA. ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ
Общая характеристика. Октокораллы (гр. octo—восемь, corallion—
коралл), или альционарии (гр. .Alkyonion — имя собственное),—
колониальные прикрепленные кораллы; отдельные полипы не соприка-
Рис. 91. Подкласс Octocorallia. Схема строения колоний и отдельных спикул:
1 — Corallium rubrum — современный красный коралл, мягкое тело частично срезано;
2 —• Tubipora (неоген — ныне), общий вид современной колонии; 3 — часть осевого
скелета Isis (мел — ныне); 4—8 — спикулы некоторых современных восьмилучевых
кораллов;
о — ось скелета, ц — ценецхима, или внеячеистый скелет, щ — щупальца
саются друг с другом и соединены промежуточной тканью — цено-
сарком; щелевидный рот с одним сифоноглифом окружен венчиком
из восьми полых перистых щупалец. Пищеварительная полость
разделена восемью мягкими мезентериальными перегородками на
восемь камер. Скелет внутренний, формируется в мезоглее особыми
клетками— склеробластами и состоит из отдельных спикул,
известковых или роговых; спикулы могут сливаться вместе, образуя
различного типа внутренний скелет: рт осевых структур до массивного
фиброзно-волокнистого. У двух отрядов имеется наружный массивный
187
фиброзный скелет. Октокораллы широко распространены в современ-
ных морях; известностью пользуются кораллы: красный, голубой
(солнечник), черный, роговой и коралл-органчик. В ископаемом со-
стоянии встречаются очень редко; известны находки из отложений
перми и более частые начиная с юры.
Строение мягкого тела. Колония состоит из мягкой промежуточной
ткани — ценосарка и расположенных в ней маленьких полипов, размеры
тела которых обычно составляют несколько миллиметров, изредка под-
нимаясь до 5—10 см (рис. 91, 1, 2). Колония имеет разнообразный
облик — ветвистый, веерообразный, трубчатый, массивный, ее объем и
форма очень изменчивы; размеры отдельных колоний достигают
2—3 м; в некоторых колониях насчитывают до 35 000 особей.
В колонии различают обычно два типа полипов •— автозооиды и
сифонозооиды. Автозооиды имеют мешковидное тело, несущее на
одном конце щелевидной формы ротовое отверстие с одним сифоно-
глифом. Сифоноглиф покрыт ресничками, вызывающими ток воды в
гастральную полость. Ротовое отверстие окружено венчиком из восьми
полых перистых щупалец. Пищеварительная полость разделена 8 мяг-
кими перегородками на камеры, которые продолжаются в щупальца.
Перегородки прикреплены к глотке и свободно свешиваются вниз в
пищеварительную полость. Две перегородки, расположенные против
сифоноглифа, значительно длиннее остальных шести; они покрыты
ресничками, выводящими воду из тела полипа. К ротовому диску
прикреплен мускул-ретрактор, втягивающий щупальца внутрь полипа.
Стенка полипа состоит из экто- и энтодермы и развитой между ними
мезоглеи. Мезоглея наиболее полно развита в ценосарке; в ней имеются
амебоидные клетки, образующие стрекательные клетки и склеробласты.
Сифонозооиды представляют собой видоизмененные полипы,
у которых сохранились только сифоноглифы; они снабжают колонию
свежей водой и иногда продуцируют половые клетки.
Строение скелета. Склеробласты строят скелет для полипов и для
ценосарка. Скелет образуется из белкового рогового вещества —
горгонина (протеин с примесью серы, йода, брома) или из
известковых спикул, которые могут состоять из углекислого или
фосфорнокислого кальция (рис. 91, 4—8). Спикулы имеют разнообраз-
ную форму и расположены в колонии либо изолированно, либо взаимно
связаны известковым или роговым цементом. У одних кораллов (туби-
пориды) спикулы соединяются вместе, образуя вокруг полипа известко-
вую трубку; между полипами развиты своеобразные соединительные
платформы; у других — внутри ценосарка в результате объединения
спикул роговым или известковым цементом образуется плотная осевая
структура (горгониды, или роговые кораллы; пеннатулиды, или мор-
ские перья); у третьей группы — гелиопор ид — имеется массивный из-
вестковый скелет, в отдельных трубках которого сидят полипы, ценосарк
образует трубчатый'промежуточный скелет — цененхиму. Главные
Трубки — кораллиты — и трубочки цененхимы снабжены многочислен-
ными горизонтальными днищами.
Размножение и развитие. Восьмилучевые кораллы размножаются
половым и бесполым путем. Половые клетки образуются в мезенте-
риях; оплодотворение яйца и развитие обычно происходит внутри мате-
ринского организма. Из яйца выходит личинка — планула, которая
выносится в воду через ротовое отверстие; личинка, поплавав некоторое
время, прикрепляется к подводным предметам и превращается в поли-
па. От основания полипа в стороны начинают ответвляться полые вы-
росты— столоны, которые распространяются по поверхности субстра-
188
та, соединяясь друг с другом, формируя своеобразное сетчатое основа-
ние колонии. Новые полипы образуются брсполым путем от базальных
столонов или горизонтальных платформ из стенки старого полипа или
из мягкой промежуточной ткани ценосарка.
Основы систематики и классификация. Систематика октокораллов
•основана на строении мягкого тела и скелета. В зависимости от поло-
жения скелета подкласс разделен на два надотряда: Endosclerata
с внутренним положением скелета и Exosclerata с внешним скелетом.
В составе Endosclerata выделяются пять отрядов: Alcyonida (юра —
ныне), Gorgonida (мел — ныне), Pennatulida (триас — ныне), Telestida
(в ископаемом состоянии неизвестны), Tubiporida (мел — ныне). К над-
отряду Exosclerata отнесены два отряда: Trachypsammida (пермь) и
Helioporida (мел — ныне).
История развития и экология октокораллов. Геологическая история
и филогения восьмилучевых кораллов крайне скудны. В ископаемом со-
стоянии они встречаются очень редко, что обусловлено особенностями
строения внутреннего скелета. Сравнительно недавно из отложений
венда (Эдиакара, Южная Австралия) были описаны остатки, условно
отнесенные к восьмилучевым кораллам; отпечатки похожи на колонии
пеннатулид, однако полной уверенности в принадлежности их к корал-
лам нет. В палеозое остатки октокораллов практически отсутствуют.
В пермских отложениях о. Тимора были найдены остатки массивных
скелетов Trachypsammia, вначале отнесенного к табулятам, а позднее
выделенного в отдельный отряд октокораллов.
Начиная с триаса становятся достоверно известными пеннатулиды,
или «морские перья», скелет которых представлен роговой или извест-
ковой осью. В юре к ним присоединяются альциониды, известные по
остаткам спикул. В меловой период появляются горгониды, или рого-
вые кораллы, колонии которых поддерживаются цельной или членистой
известковой осью (Isis) и гелиопориды, имеющие наружный известко-
вый скелет и принимающие участие в образовании биогермов. В палео-
гене и неогене продолжается развитие всех известных в настоящее
время отрядов.
Современные восьмилучевые кораллы распространены преимущест-
венно в тропических морях и океанах, они встречаются на всех глуби-
нах от зоны сублиторали до абиссали; многие восьмилучевые кораллы
образуют большие поселения в области шельфа и на материковом
склоне. Большинство из них прочно прикрепляется к подводным пред-
метам с помощью корнеобразных выростов или столонов; небольшая
часть живет на мягком грунте, на илах или песке, закрепляясь с по-
мощью длинного стебля. Часть кораллов живет среди рифов, их
известковые спикулы после гибели входят в состав осадков, возникаю-
щих вокруг них. В симбиозе с кораллами живут самые разнообразные
организмы, к числу которых в первую очередь относятся водоросли
(зооксантеллы), разнообразные членистоногие (в частности, копеподы)
и аннелиды.
ПОДКЛАСС HEXACORALLIA. ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ
Общая характеристика. Шестилучевые коралловые полипы (гр.
hex— шесть, corallion — коралл)—современные и вымершие, коло-
ниальные и одиночные морские организмы, многие из которых имеют
хорошо развитый наружный известковый скелет.
Цилиндрическое мешковидное тело полипа имеет плоскую подошву
и на дистальном конце околоротовой диск, несущий полые щупальца.
189
Щелевидный рот у актиний с двумя сифоноглифами, из которых один
служит для ввода воды, второй для вывода; у склер актиний сифоно-
глифы отсутствуют. К глотке прикреплены шесть пар протомезен-
териев, ограничивающих центральную полость — физиологически:
обособленный кишечник; периферические камеры радиальные и меж-
Рис. 92. Подкласс Hexacbrallia. Схемы строения коралловых полипов, скелета и от-
дельных его деталей:
1 — взаимоотношение тела полипа со скелетом у склерактиний; 2 — общая схема
строения колонии; 3 — схема расположения мезентерий; 4 — пластинчатая септа, с
простыми трабекулами; 5 — сложная трабекула; 6 — взаимоотношение полипа и ко-
раллита; 7—10 — стенка: 7 — эпитекальная, 8 — септотекальная, 9 — паратекальная,.
10 — синаптикулотекальная;
бп — базальная пластинка, вк — верхний край септы, вщ — полип с втянутыми щу-
пальцами, гл — глотка, м — мезентерии (зачерненные — направляющие мезентерии),
мн — мезентериальные нити,,оя — основание полипа, пр — продольный разрез полипа,
пт — простая трабекула, р—рот, рщ — полипы с расправленными щупальцами, с —
септа, ск — стенка кораллита, ст—столбик, т — тека, тп—тело полипа, цн — ценен-
хима, щ — щупальце, эп — эпитека
радиальные. В процессе роста новые пары мезентериальных перегоро-
док — метамезентерии — возникают в межрадиальных камерах.
На основании особенностей строения мезентериев, наличия или
отсутствия скелета шестилучевые коралловые полипы разделяются на
несколько отрядов, среди которых наиболее важное значение имеет
190
отряд Scleractinia, насчитывающий до 450 родов; актинии, не имеющие
скелета (200 родов, 700 видов), в ископаемом состоянии не известны.
Отряд Scleractinia. Склерактинии (гр. skleros — твердый, жесткий,
actis, род. actinos — луч). Наиболее обширный отряд среди шестилуче-
вых коралловых полипов, обладающих наружным известковым скеле-
том. К этому отряду относится большинство мезокайнозойских корал-
лов, возникших в начале среднего триаса.
Строение мягкого тела. Тело полипа обычно размером от 1 мм до-
3 см, реже до 25 см, цилиндрическое, несет на одном конце оральный
диск с щелевидным ртом посередине; рот окружен одним или несколь-
кими венчиками щупалец. Глотка не имеет сифоноглифов. Гастральная
полость разделена шестью парами (или кратными шести) мезентериев,
на отдельные камеры (рис. 92, 1—3). Внутри пары мезентериев распо-
ложена известковая перегородка, или склеросепта; число склеро-
септ в полости равно или кратно шести. Полипы склерактиний раз-
дельнополы, реже гермафродиты или стерильны. Полипы помещаются
в верхней углубленной части кораллита, называемой чашечкой.
Строение скелета. Колонии склерактиний имеют различную форму
(массивные, кустистые, древовидные) и размеры. Особенно крупных
размеров достигают рифостроящие кораллы, обладающие к тому же
довольно интенсивным ростом. Отдельные кораллиты в колонии соеди-
няются вместе известковой промежуточной тканью — склеренхи-
мой, выделяемой мягкой промежуточной тканью — ценосарком.
Форма тела полипа и наружный скелет тесно связаны между собой,
и по форме скелета можно судить о форме тела полипа.
Чтобы рассмотреть особенности строения скелета, необходимо про-
следить онтогенез современного коралла. Незадолго до прикрепления
к субстрату у планулы закладывается вначале первая пара направ-
ляющих мезентериев, затем ротовая щель, оральный диск, глотка,
щупальца; позднее одна за другой возникают остальные пары мезен-
териев. Планула прикрепляется к субстрату и образует базальную
пластинку, которая в дальнейшем увеличивается в размерах и утол-
щается. Края пластинки растут вверх, и образуется своеобразная ча-
шечка, или прототека, внутри которой помещается тело полипа.
Затем подошва тела полипа сморщивается и образует серию радиаль-
ных складок (6), внутри которых возникают тонкие арагонитовые
пластинки :— склеросепты.
Строение склеросепт. Склеросепты строятся из склеродермитов
диаметром до 50—100 мк и ламеллярной склеренхимы. Склеродермиты
располагаются вертикальными рядами, образуя своеобразные ба доч-
ки, или трабекулы (см. рис. 92, 4, 5). Если трабекулы плотно-
прилегают одна к другой, то возникает пластинчатая сплошная септа;
если они располагаются на некотором расстоянии, то образуется по-
ристая, решетчатая септа. Различают три основных типа трабекул
(см. рис. 100): голаканты, монаканты и рабдаканты. Голаканты
представляют собой тонкую трабекулу бесструктурного облика, по-
груженную в ламеллярную склеренхиму; м о н а к а н ты, или простые,
трабекулы, построены из серии простых склеродермитов; рабдакан-
т ы, или сложные трабекулы, состоят из нескольких пучков склеро-
дермитов. Трабекулы редко расположены вертикально; обычно они
отклоняются от вертикали или расходятся веерообразно.
Вначале закладываются шесть склеросепт первого порядка, или
протосепт; затем образуются шесть склеросепт второго порядка,
12 склеросепт третьего, 24 склеросепты четвертого и т. д. Все склеро-
септы, кроме протосепт, называются метасептами. Вместе с ростом
191
полипа увеличиваются размеры скелета чашечки, стенки и возникают
новые скелетные элементы.
К производным септ относятся: столбик (колумелла), свайки
(пали) и балочки (синаптикулы)..
Столбик представляет собой структуру, развивающуюся в осе-
вой плоскости коралла вследствие схождения здесь септ. Если септы
не доходят до центра, то осевая область остается свободной и у корал-
литов столбик отсутствует. Столбик возникает как часть одной, двух
или нескольких септ, достигнувших центра кораллита. По форме раз-
личают: пластинчатый, грифелевидный, губчатый, пучковидный стол-
бики. Пластинчатый и грифелевидный столбики образуются за счет
удлинения протосепты, губчатый и пучковидный — за счет внутренних
концов трабекул нескольких протосепт или метасепт второго порядка;
если трабекулы септ переплетаются друг с другом, то образуется
компактный или губчатый столбик; если трабекулы располагаются
вертикально, то развивается пучковидный столбик.
Свайки, или пали, представляют собой свободные вертикаль-
ные пластинки, возникшие из трабекулярных элементов, отщепившихся
от внутреннего края септ и расположенных по периферии осевой
полости. Иногда свайки располагаются в несколько колец, соответст-
вующих разным порядкам септ.
Синаптикулы, или балочки, — межсептальные образова-
ния, напоминающие балочки и возникающие за счет соединения двух
шиповидных или зерновидных выступов на боковых поверхностях двух
смежных септ.
Полип занимает только верхнюю часть кораллита—чашечку, и
в процессе роста объем его тела часто увеличивается не пропорцио-
нально увеличению размеров скелета. Для устранения этого несоответ-
ствия образуются горизонтальные скелетные элементы: днища, или
табулы, диссепименты, или траверзы. Днища представляют собой
тонкие плоские или изогнутые известковые пластинки, пересекающие
центральную полость, отделяющие верхнюю, жилую часть кораллита,
занятую полипом, от нижней, нежилой. Диссепименты имеют
вид выпуклых известковых пластинок, развитых в межсептальном про-
странстве по периферии кораллита. Они располагаются обычно под
углом к вертикальной оси. Образование днищ и диссепиментов проис-
ходит следующим образом: по мере надстройки верхнего конца ко-
раллита полип перемещается вверх и от нижней части отделяется дни-
щем. По мнению некоторых специалистов, так как его перемещению
мешает жесткое прикрепление мезентериев к скелету, то, чтобы освобо-
дить полип от скелета, в нижней части полипа стенка тела впячивается
вовнутрь, выделяет днище и, таким образом, отсекает нижнюю часть
полипа, которая в дальнейшем дегенерирует. Рост- кораллита и полипа
идет периодически, и полип всегда занимает верхнюю часть коралли-
та. У склерактиний могут быть развиты только днища, либо только
диссепименты, либо те и другие, а также синаптикулы.
Строение стенки. По способу образования стенки различают: эпи-
текальную стенку, септотеку, паратеку и синаптикулотеку (см. рис. 92,7).
Эпитекальная стенка возникает за счет расширения перифери-
ческого края септы. Септотека создается утолщением перифериче-
ского края септ; паратека — за счет развития и утолщения по пери-
ферии диссепиментов (пузырчатой ткани). Синаптикулотека
формируется за счет развития по периферии септ синаптикул. У мно-
гих склерактиний стенка кораллита снаружи, от основания до чашечки,
покрывается тонким известковым чехлом, получившим название э п и -
192
'теки. Эпитека, окружающая не один кораллит, а целую колонию,
называется г о л о т е к о й.
Скелет склерактиний, таким образом, состоит из теки, эпитеки,
склеросепт, расположенных радиально, число которых чаще кратно
шести, хотя имеются различные отклонения, основной пластинки, стол-
бика, синаптикул, днищ, диссепиментов. Эти скелетные элементы при-
сутствуют в разном сочетании в кораллитах.
Рис. 93. Отряд Scleractinia.
Внешняя форма скелетов одиночных (А) и колониальных (5) кораллов
Форма кораллитов и колоний. Кораллиты одиночных представи-
телей имеют различную форму, которая зависит от формы тела полипа,
условий обитания, особенностей индивидуального развития. Наиболее
обычной является коническая форма кораллита с углом от 60 до 80°;
при угле 40° кораллит приобретает нормально коническую форму, при
угле 20° — узкоконическую. Встречаются и иные формы: дисковидные,
чашеобразные, цилиндрические, червеобразные (рис. 93, Л).
Форма колоний зависит от особенностей бесполого размножения и
условий обитания (рис. 93, Б). Бесполое размножение может осуществ-
ляться внутрищупальцевым или внещупальцевым почкованием и попе-
речным делением. При внутрищупальцевом почковании внутри
кольца щупалец материнского полипа возникают две, три или более
новых почек. В этих случаях могут возникать колонии: тамнасте-
роидного типа — кораллиты без ясно различимых границ и со сли-
вающимися септами в виде непрерывных пластинок, меандроид-
ного типа с чашечками, расположенными линейными, более или
менее изогнутыми рядами, при этом рты полипов сливаются в одну
общую щель либо располагаются посредине между щупальцами, ко-
раллиты утрачивают свою индивидуальность. Колония приобретает
вид мозга, отсюда один из кораллов называется коралл-моз-
г о в и к.
При внещупальцевом почковании новые почки возникают в
краевой зоне вне кольца щупалец материнской особи или в ценосарке.
В тех случаях, когда краевая зона имеет форму стелющегося побега
или нитей, то возникает стелющаяся колония. При обоих случаях
13 В. В. Друщиц
193
почкования могут возникать сотовидные, ветвистые или сноповидные
колонии.
Некоторые склерактинии размножаются делением; при этом
вначале ротовая щель, а затем ротовой диск делятся на две части;,
позднее материнский полип образует две новые дочерние особи, каж-
дая из которых строит свой скелет.
На основании микроскопического строения септ, типов строения-
стенки, синаптикул и строения мягкого тела полипов среди склеракти-
ний выделяется пять подотрядов (рис. 94).
Рис. 94. Отряд Scleractinia. Подотряды Astrocoeniina (1—3), Fungiina (4), Faviinai
(5—6), Caryophylliina (7), Dendrophylliina (8—9):
1 — Acropora (эоцен — ныне); 2 — Stylocoenia (эоцен — миоцен); 3 — Stylina (позд-
ний триас—ранний мел): За — поперечное сечение, 36 — продольное; 4 — Cyclolites:
(мел — эоцен); 5 — Leptoria (поздний мел — ныне); 56, 6 — Montlivaltia (средний три-
ас — мел); 7а,б,— Parasmilia (ранний мел — ныне); 8а, б — Thecopsammia (совр.);.
9 — Dendrophyllia (эоцен — ныне)
Подотряд Astrocoeniina. Астрокениины (гр. aster — звезда, koinos—
общий). Септы построены из относительно малого числа простых
трабекул (до 8); преимущественно колониальные с маленькими поли-
пами (1^-3 мм); полипы несут обычно 2 венчика щупалец. Средний
триас — ныне. Насчитывает до 50 родов (рис. 94, 1—3).
Подотряд Fungiina. Фунгиины (лат. fungus — гриб). Скелет порис-
тый; септы построены из многочисленных трабекул; одиночные или
колониальные; размеры полипов обычно более 2 мм, полипы несут
более 2-х венчиков щупалец, имеются синаптикулы. Средний триас —
ныне. Около ПО родов (рис. 94, 4).
Подотряд Faviina. Фавиины (лат. favus — пчелиные соты). Септы:
построены из одной или нескольких систем веерообразно расположен-
ных простых или сложных трабекул; одиночные или колониальные;,
стенка кораллитов эпитекальная, септотекальная или паратекальная..
Хорошо развиты диссепименты. Около 150 родов. Средний триас —
ныне (рис. 94, 5, 6).
Подотряд Caryophylliina. Кариофиллины (лат. carios—разрушение^
phyllum — лист). Скелеты преимущественно одиночные, реже коло-
194
ниальные; септы пластинчатые, с гладким краем, построены из простых
трабекул; стенка септотекальная или эпитекальная. Около 100 родов.
Юра — ныне (рис. 94, 7).
Подотряд Dendrophylliina. Дендрофиллиины (гр. dendron—дерево).
Одиночные или колониальные; септы первично-пластинчатые, но затем
вторично утолщены, более или менее пористые; стенка синаптикуло-
текальная. Поздний мел — ныне. Около 20 родов (рис. 94, 8, 9).
История развития и экология склерактиний. Первые склерактинии
появились в начале среднего триаса. Они произошли, по мнению одних
исследователей, от четырехлучевых кораллов, по мнению других —от
каких-то бесскелетных палеозойских коралловых полипов. В начале
среднего триаса они представлены двумя подотрядами (Astrocoeniina,
Fungiina), позднее к ним добавился третий—• Faviina. Первые
склерактинии строили небольшие биогермы; в конце триаса-число родов
значительно увеличилось; они распространились по всей средиземно-
морской области от Южной Европы до Малайзии и в пределах этой об-
ласти, распространявшейся от 60° с. ш. до 10° ю. ш;, в зоне мелководья
строили рифы, биогермы и рифовые банки. Кораллы-рифостроители
получили название герматипных кораллов. В юрский период
склерактинии начали проникать в более глубокие зоны моря, адаптиро-.
ваться к жизни при более низких температурах. В эти зоны моря
проникали преимущественно одиночные кораллы, не строящие рифов,—
агерматипные кораллы. Они известны преимущественно в под-
отряде Caryophylliina. В юрском периоде во всей средиземноморской
области вновь создаются благоприятные условия для образования
рифов; северная граница рифового пояса сместилась до 40° к югу и
проходила на 20° севернее линии распространения современных герма-
типных кораллов. Рифогенные постройки известны в Крыму, вдоль
южного склона Кавказа, в Грузии, Азербайджане. В середине раннего
мела (баррем — апт, ургонские фации) рифовые постройки известны на
Карпатах, Малом Кавказе, Малом Балхане, из многих стран Южной
Европы, Северной Африки, Японии, Мексики. Становятся также извест-
ны новые группы агерматипных кораллов из области развития нерито-
вых и батиальных отложений. В конце мела появляются новые группы
склерактиний (подотряд Dendrophylliina).
В начале эоцена возникает много новых родов склерактиний, до-
живающих до наших дней. В конце палеогена рифовые постройки
известны из Мексики, северной части Южной Америки, Западной
Индии, некоторых стран Южной Европы. Начиная с неогена обособ-
ляются две области развития герматипных кораллов: карибская и
индо-тихоокеанская. В настоящее время в Карибском море
обитает всего 36 видов, принадлежащих к 20 родам, в. то время как из
второй области известно до 500 видов. Выделяются две экологические
группировки: а) рифостроители, ограниченные глубинами 15—20 м, жи-
вущие при температуре воды от 25 до 29°, но переносящие колебания
температур от 15 до 36°; им необходимо достаточное количество света,
кислорода и нормальная соленость воды, однако они могут переносить
колебания солености в пределах от 27 до 4О°/оо; б) рифостроители спо-
койных лагун, живущие на глубинах до 90 м.
В симбиозе с герматипнымц кораллами живут зооксантеллы, кото-
рые используют продукты обмена, выделяемые кораллами. Вместе со
склерактиниями в постройке рифов принимают участие восьмилучевые
кораллы (альционарии), гидроидные (миллепорины и стилястерины),
мшанки, усоногие раки и обладающие известковым скелетом полихеты.
13:
195
Среди многообразия рифогенных построек различают: береговые
рифы, барьерные рифы, атоллы, коралловые банки и коралловые посе-
ления в лагунах и бухтах. Береговые рифы расположены вдоль
берегов, островов или материков; барьерные рифы отстоят от бе-
рега на некотором расстоянии, к ним относится Большой Барьерный
риф, который тянется вдоль восточного берега Австралии более чем на
1000 км, барьерный риф вдоль восточного берега Африки. Атоллы
Рис. 95. Образование атолла (3) из берегового рифа (Дарвин, 1842, из книги «Жизнь
животных», 1968, т. 1)
представляют собой кольцеобразные коралловые острова, способ обра-
зования которых был впервые изучен Ч. Дарвином (1842), подробно
описавшим различные типы коралловых построек. Дарвин проследил,
как один тип коралловых поселений — береговой риф, окаймляющий
остров, в условиях опускания морского дна переходит вначале в барьер-
ный риф и далее в атолл—кольцевой коралловый остров, заключаю-
щий внутри лагуну (рис. 95).
Современные агерматипные склер актинии живут на глубинах от 175
до 800 м, некоторые опускаются до глубин 6000 м и живут в полном
мраке, в этом случае без симбионтов. Они обитают при температуре от
4,5 до 10°С, перенося температуру около —1,1°С.
Находки рифостроителей в отложениях прошлых геологических
эпох указывают на условия их обитания, по-видимому, близкие к со-
временным. Это подтверждается также находками вместе с ними дру-
гих рифолюбов. В настоящее время вме'сте с кораллами живут разнооб-
разные фораминиферы, губки, кольчатые черви, прикрепленные и свер-
лящие двустворчатые моллюски (литофаги и фоласы), и прикреплен-
ные брюхоногие моллюски (верметус) или активно ползающие (трохус,
*гурбо), морские, ежи, морские лилии, брахиоподы, преимущественно
прирастающие, И разнообразные ракообразные. Если организмы, живу-
щие среди коралловых рифов, имеют прочные массивные скелеты, то
у обитателей спокойных вод — лагун скелеты тонкие и хрупкие; к ним
относятся морские звезды, голотурии, ядовитые морские ежи — диаде-
мы, снабженные полуметровыми тонкими иглами, и многие другие.
ПОДКЛАСС TABULATA. ТАБУЛЯТЫ
; ' Общая характеристика. Табуляты (лат. tabula —доска, пол) — вы-
'мершие колониальные кораллы; полипняк состоит из трубчатых корал-
литов разнообразной формы и небольших размеров (0,5—4 мм). Корал-
литы имеют стенку, снаружи покрытую эпитекой, Септальный аппарат
относительно простой, он представлен простыми сейтами, септальными
Щипиками и ребр'амй.' Очень хорошо развиты днища горизонтальные,
наклонные, воронковидные, полные или неполные, иногда сопровож-
дающиеся пузырчатой тканью. Скелет по происхождению наружный
•эктодермический, построен из фибр, образующих ламеллярйую склерен-
химу. Табуляты размножались: половым и вегетативным путем; в по-
196
следнем случае главным образом путем межстенного или промежуточ-
ного почкования. Различают два типа скелета: у одной группы — ком-
муникатной — различные кораллиты сообщаются друг с другом
соединительными образованиями: порами, трубочками или пластина-
ми; у другой —инкоммуникатной — сообщения между коралли-
тами отсутствовали. Табуляты появляются в позднем кембрии и
существуют до конца перми.
Строение мягкого тела и скелета. О строении полипов можно су-
дить только по форме кораллита, в котором помещалось тело. Внешняя
форма кораллитов крайне разнообразна и зависит от типа полипняка.
Различают три основных типа полипняков: массивный, кустистый и
стелющийся. В массивных полипняках кораллиты плотно прилегают
друг к другу и имеют призматическую форму с полигональным попе-
речным сечением. Внешняя форма полипняка может быть дисковидной,
массивно-ветвистой. В кустистых полипняках кораллиты всегда в
той или иной мере отделены друг от друга свободным пространством;
они могут состоять из параллельно расположенных кораллитов, связан-
ных соединительными трубками, или свободно расходиться одни от
других, или состоять из кораллитов, сросшихся друг с другом краями
(цепочечные кораллы). Кораллиты обычно круглого или овального
сечения, реже полигонального. У стелющихся полипняков корал-
литы образуют или стелющуюся цепочку, или, соединяясь друг с дру-
гом, сетку различной формы. Кораллиты обычно имеют рожкообраз-
ную форму с круглым поперечным сечением. Чашечка представляет
собой углубление в верхней части кораллита; ее форма отражает
форму основания полипа. Чашечки могут иметь призмо-коническую,
цилиндрическую или воронковидную форму.
Внутри кораллита имеются горизонтальные и вертикальные скелет-
ные элементы. К горизонтальным относятся днища и диссепименты.
Днища бывают полные и неполные, горизонтальные и наклонные,
воронкообразные. Некоторые формы совершенно лишены днищ. Непол-
ные днища обычно принимают характер пузырчатой ткани, которая
состоит из отдельных известковых пластинок — диссепиментов,
обычно развитых слабо. К вертикальным скелетным образованиям отно-
сятся септы, септальные шипики, возникавшие в радиальных склад-
ках тела полипа. Сплошные септы развиты сравнительно редко: их
число может быть равно 4, 6, 8; определенная закономерность в порядке
заложения септ для табулят не установлена,- Чаще всего септальный
аппарат состоит из ряда шипов, ребер, небольших выступов и чешуй.
Осевая часть кораллитов обычно лишена каких-либо скелетных эле-
ментов.
Стенки кораллитов образованы простой фибральной склеренхимой,
и трабекулярная структура наблюдается очень редко; в септах фибры
направлены перпендикулярно к плоскости септы, а в шипах—перпен-
дикулярно к оси шипа.
Отдельные кораллиты соединяются друг с другом при помощи
соединительных пор — отверстий, пронизывающих стенку и эпитеку,
служащих для связи между полипами; иногда они связаны при помощи
соединительных трубочек, представляющих собой радиальные выросты
стенок кораллитов. Более редко связь осуществлялась соединитель-
ными выростами стенки кораллитов, сопровождающимися развитием
стенных пор. У многих табулят связь между полипами отсутствовала.
Наличие или отсутствие связи между полипами послужило Б. С. Со-
колову основой для разделения табулят на две группы: и н к о м му-
ки кат и коммуникат.
197
Размножение и развитие. Табуляты размножались половым и бес-
полым путем. Данные о личиночной стадии и половом развитии табу-
лят отсутствуют. Постройка колонии начиналась с того момента, как
личинка садилась на дно, превращалась в полипа, а последний создавал
первую скелетную ячейку — п ротокораллит, имеющий кониче-
скую форму и по форме напоминающий кораллиты аулопор. Ауло-
пороидная стадия оказывается общей для всех табулят. Образова-
ние .колоний происходило в результате почкования; наиболее
широко было распространено межстенное или промежуточное
почкование, в результате которого новые кораллиты возникали в углах
между зрелыми кораллитами или между их стенками. Кустистые фор-
мы полипняков образовывались в результате бокового почкования
или из соединительных отростков и пластин (столонное почкование).
Своеобразно почкование у тетрадиид: оно выражается в том, что осе-
вые концы четырех септ сливаются друг с другом и материнский орга-
низм оказывается разделенным на четыре новые дочерние почки. Одна-
ко не всегда развивались все четыре почки.
Основы систематики и классификация. Важнейшими признаками
для систематики табулят являются: типы вегетативного размножения,
определявшие основное разнообразие колоний, строение ' септального
аппарата, структура периферической стереозоны и типы соединитель-
ных образований, связывавших гастральные полости зооидов. На осно-
вании этих признаков выделяются семь отрядов: Auloporida, Lichena-
riida, Tetradiida, Halysitida, Favositida, Syringoporida, Sarcinulida.
Каждый отряд от всех остальных четко отличается особенностями
морфогенеза скелетных образований, иными особенностями конструк-
ции скелета и принадлежностью к самостоятельной филогенетической
ветви..
Отряд Auloporida. Аулопориды (гр. aulos — трубка, porus — пора).
Полипняки небольших размеров, стелющиеся, пучковидные, сетчатые;
кораллиты маленькие, конической, рожкообразной или цилиндрической
форм; соединены друг с другом в местах' почкования. Стенки корал-
литов толстые, с пластинчатой микроструктурой, снаружи покрыты
морщинистой эпитекой; чашечки конические, воронковидные или цилин-
дрические; внутренняя полость с косыми или воронковидными днищами,
иногда отсутствующими; септальные образования в виде мелких шипи-
ков, бугорков, септальных бороздок; часто отсутствуют вообще. Раз-
множались боковым или базальным почкованием. Развитие шло по
линии разнообразия форм почкования, форм колониальных построек,
изменений в строении днищ и структуры стенок. Отличаются большой
устойчивостью признаков. Поздний кембрий — поздняя пермь. 28 родов
(рис. 96,/).
Отряд Lichenariida. Лихенарииды (гр. leichen—лишайник). Полип-
няки массивные, полусферические; кораллиты призматические, плотно
прилегающие друг к другу; сечение полигональное; чашечки глубокие;
днища чаще всего горизонтальные. Стенки кораллитов от тонких до
довольно толстых, трабекулярного строения. Септальный аппарат в
форме шипиков или сплошных септальных ребер. Размножение проме-
жуточным почкованием. Основное развитие связано с усложнением
структуры периферической стереозоны, созданием септального аппарата
в форме шипиков и ребер. Средний кембрий (?), ранний ордовик —ран-
ний силур. 12 родов (рис. 96, 2).
Отряд Tetradiida. Тетрадииды (гр. tetra—четыре). Колонии разно-
образной формы: массивные, цепочечные, кустистые или в виде одиноч-
ных трубок. Кораллиты призматические или округло-призматические,
198
длинные и узкие; стенки тонкие и плотные. Септальный аппарат состоит
из четырех септ первого порядка, расположенных крестообразно; кроме
них бывают септы второго и третьего порядков. При почковании концы
•септ соединяются в центре и делят материнский организм на четыре
части; делению септ предшествовало деление тела полипа; в соответст-
вии с подобным почкованием из одного полипа возникало четыре дочер-
Рис. 96. Отряды Auloporida (7), Lichenariida (2), Halysitida (3—4) и Tetra-
diida (5):
.1 — Aulopora (ордовик — пермь): la — общий вид колонии, 16— кораллит; 2 —
Lichenaria (ранний — средний ордовик); 2а, 26 — поперечные, 2в — продольное
•сечения; 3 — Tollina (ордовик), поперечное сечение; 4 — Halysites (силур); 4а —
поперечное и 46 — продольное сечения; 5—Tetradium (ордовик), поперечное
сечение;
дн — днища, с — шиповидные септы
них; встречается также боковое чашечное и обычное промежуточное
почкование. Днища тонкие и горизонтальные. Средний ордовик —
ранний силур. 5 родов (рис. 96, 5).
Отряд Halysitida. Хализитиды (гр. halysis—цепь). Полипняк свое-
образной кустистой и цепочечной формы; кораллиты длинные, эллипти-
ческого, круглого или полигонального сечения, срастаются друг с дру-
гом краями, образуя длинные цепочки, между которыми бывают пус-
тые пространства. Чашечки неглубокие; стенки толстые; септальный
аппарат в виде шипиков, иногда длинных, доходящих до центра корал-
лита и образующих здесь своеобразный столбик. Днища горизонталь-
ные; иногда развита пузырчатая ткань. Размножались боковым и
промежуточным почкованием. Средний ордовик — поздний силур. 6 ро-
дов (рис. 96, 3, 4").
Отряд Favositida. Фавозитиды (лат. favus — пчелиные соты). По-
лйпняки массивные, сферические или ветвистые; кораллиты полигональ-
199
ного сечения; сообщались соединительными порами, расположенными
по углам или по стенкам в один или несколько рядов. Стенка от мас-
сивной до тонкой с отчетливым швом; септальный аппарат в виде
сплошных или шиповатых ребер, шипиков. Днища горизонтальные,
воронкообразные, иногда неполные. Размножались промежуточным
почкованием. Поздний ордовик — поздняя пермь. 85 родов (рис. 97, 7).
Рис. 97. Отряды Favositida (1), Syringoporida (2) и Sarcinulida (5):
/ — Favosites (силур — пермь): la — схема строения полипняка, 16—поперечное
и 1в—'продольное сечения; 2 — Syringopora (ордовик — пермь): 2а — полипняк,
26 — поперечное и 2в — продольное сечения; 3 — Sarcinula (поздний ордовик —
ранний силур): За — поперечное, 36— продольное сечения;
дн — днища, п — поры, с — шиповидные септы, ст — соединительные трубки
Отряд Syringoporida. Сирингопориды (гр. syrinx, род. syringos—
Друбка, poros — отверстие, канал). Полипняки кустистые, образованы
несоприкасающимися кораллитами круглого сечения, соединенными тон-
кими трубками, ориентированными или расположенными беспорядочно..
Толщина стенок изменчива; структура волокнисто-пластинчатая; сеп-
тальный аппарат в виде шипиков; днища воронковидные и горизонталь-
ные, иногда переходящие в пузырчатую ткань. Размножение промежу-
точным и столонным почкованием. Развитие шло по пути отклонения
соединительных трубок от правильной ориентировки, последовательного
усложнения днищ, у одной ветви — утолщения стенки. Средний ордо-
вик— ранняя пермь. 18 родов (рис. 97, 2).
Отряд Sarcinulida. Сарцинулиды (лат. sarcina—узел). Полипняки.
Кустистые или массивно-кустистые, состоящие из кораллитов цилиндри-
ческой формы. Стенки пронизаны венчиками пор (до 20—24 пор в
каждом венчике). Кораллиты соединены своеобразными пластинками,
образованными сросшимися радиальными каналами, идущими от вен-,
200
чиков пор. Септальный аппарат в виде ребер или шипиков. Днища
горизонтальные, воронкообразные. Почкование из образований проме-
жуточного скелета. Средний ордовик—ранний силур. 3 рода (рис. 97,5).
История развития табулят. Первые табуляты становятся известными
в конце кембрия, но достаточно достоверные находки аулопорид и
лихенариид известны с начала ордовика. В среднем ордовике резко
увеличивается разнообразие табулят: появляются тетрадииды, хализи-
тиды, сирингопориды и фавозитиды. На границе ордовика — силура
заканчивается первый этап развития табулят, связанный с вымиранием
в начале силура лихенариид, тетрадиид и сарцинулид. В силуре и де-
воне наблюдается второй максимум развития табулят, широкое рас-
пространение хализитид, фавозитид и сирингопорид. Но первые выми-
рают в конце силура, а в конце девона исчезают многие группы фавози-
тид и сирингопорид. В девоне заканчивается второй этап развития
табулят. Третий этап начинается в карбоне обновлением состава фаво-
зитид, сирингопорид и аулопорид; в перми геологическая история табу-
лят заканчивается полным исчезновением всех отрядов.
ПОДКЛАСС HELIOLITOIDEA. ГЕЛИОЛИТОИДЕИ
Общая характеристика. Гелиолитоидеи (гр. helios — солнце,
lithos —камень) —палеозойские колониальные полипы, скелет которых
формировался полипами и ценосарком. Кораллиты круглого сечения
с хорошо развитыми днищами и постоянным числом септ; между ними
развит промежуточный скелет — цененхима, представленная бакулями,
пузырчатой тканью или мелкими трубочками. Появляются на границе
среднего и позднего ордовика и вымирают в середине девона.
Строение скелета и мягкого тела. Полипняки массивные, полусфе-
рические, лепешковидные, реже ветвистые. Скелет состоит из двух
частей: основной, созданной полипами, и промежуточной, созданной
ценосарком. Промежуточный скелет может быть представлен трубоч-
ками, или сифонопорами, полигонального сечения, разделенными
многочисленными тонкими днищами, или пузырчатой тканью,
состоящей из маленьких выпуклых известковых пластинок, несущих
иногда короткие шипики, или тесно сближенными бакулями. Чашечка
имеет звездчатое очертание, связанное с наличием 12 или кратных
12 септальных шипиков или септальных пластинок, и только у одного
рода их больше. В осевой зоне чашечки иногда наблюдаются сложные
столбики, возникающие за счет закручивания осевых концов септ.
Днища обычно горизонтальные и лишь иногда изогнутые. Микрострук-
тура стенки и других скелетных элементов трабекулярная.
Особенности размножения. Гелиолитоидеи размножались своеоб-.
разным ценосарковым почкованием, при котором новые полипы возни-
кали непосредственно из цецосарка и скелет строился за счет ценен-
химы. Реже новые почки возникали из материнского организма в его
чашечке.
Основы систематики и классификации. Гелиолитоидеи на основании
строения промежуточной цененхимы — бакулярной, трубчатой или
пузырчатой — разделены на три отряда: Protaraeida, Heliolitida, Рго-
porida.
Отряд Protaraeida. Протареиды (гр. protos—первый, araios —уз-
кий). Полипняки желвакообразные, инкрустирующие; цененхима губ-
чатая или пористая, построена из бакуль; последние оконтуривают;
чашечки кораллитов, слагают септальные и осевые образования. На
поверхности полипняка бакули выступают в виде мелких бугорков.
201
Днища развиты очень редко. Конец среднего ордовика—ранний венлок
(силур). 10 родов (рис. 98, 1).
Отряд Heliolitida. Гелиолитиды. Полипняки преимущественно жел-
вакообразной формы; промежуточный скелет трубчатый, каждая тру-
бочка с хорошо развитыми днищами. Кораллиты округлые или звездча-
тые, несут обычно 12 рядов септальных шипиков или пластинок; иногда
их насчитывается 36—48; днища горизонтальные; иногда имеется лож-
Рис. 98. Отряды Protaraeida (/), Heliolitida (2) и Proporida (3):
1 — Protaraea {средний—поздний ордовик); 2 — Heliolites (поздний ордо-
вик— средний девон); 3 — Ргорога (поздний ордовик — силур);
сечения: а — поперечное, б — продольное; бакули: ос — осевые, сеп — сеп-
тальные, дн — днища, к — кораллиты, пт — промежуточная ткань, цен — це-
ненхима
ный столбик. Гелиолитиды образуют основной отряд подкласса и
объединяют 10 родов (рис. 98, 2). Поздний ордовик — средний девон.
Отряд Proporida. Пропориды (гр. pros — передний, poros — канал).
Полипняки желвакообразные или корковидные. Кораллиты резко обо-
соблены и часто сближены; имеют 12 септальных выступов; днища
горизонтальные или пузырчатые. Цененхима состоит из пузырчатой
ткани, в которой могут быть заключены -трабекулы. Поздний ордо-
вик — поздний силур. 6 родов (рис. 98, 3).
История развития гелиолитоидей. Первые гелиолитоидеи появились
в конце среднего ордовика и, вероятно, произошли от аулопороидных
табулят, поскольку в основании у некоторых колоний удалось.обнару-
жить протокораллит, внешне похожий на аулопор. Развитие гелиолито-
идей шло по пути возникновения промежуточной ткани — ценосарка,
вначале выделявшей бакулярную цененхиму (отряд протареиды).
В позднем ордовике от протареид возникли гелиолитиды, цененхима
которых строилась из трубочек. Они просуществовали до среднего де-
вона.. Менее ясно происхождение пропорид, появившихся в середине
позднего ордовика и просуществовавших до конца силура. Все гелиоли-
202
тоидеи вели прикрепленный образ жизни и были широко распростра-
нены в зоне мелководья морей ордовика — девона. Они обитали обыч-
но на твердом субстрате совместно с табулятами, ругозами, стромато-
порами, брахиоподами и другими организмами.
ПОДКЛАСС TETRACORALLIA (RUGOSA).
ТЕТРАКОРАЛЛЫ (РУГОЗЫ)
Общая характеристика. Тетракораллы (гр. tetra — четыре)—оди-
ночные или колониальные палеозойские коралловые полипы, билате-
рально-симметричные, обладавшие эктодермальным скелетом, с хорошо
выраженными большими и малыми септами. У одиночных вначале
Закладывается шесть протосепт, новые метасепты развиваются только
в четырех секторах. Скелет ламеллярный, трабекулярно-ламеллярный
или трабекулярный. Краевая зона, или маргинарий, состоит из диссепи-
ментария, или пёриферической стереозоны, образованной утолщениями
септ, которые развиваются в зоне малых септ вокруг табулярия. Табу-
лярий построен из горизонтальных днищ или диссепиментов. В осевой
части могут быть развиты столбик или осевая колонна. Известны также
крышечные кораллы. Тетракораллы возникли от табулят в среднем
ордовике, испытали четыре крупных эволюционных изменения в силуре,
на рубеже раннего и среднего девона, в начале карбона и начале перми.
Последние представители вымерли в начале триаса.
Строение скелета и мягкого тела. О строении мягкого тела четырех-
лучевых кораллов можно судить на основании анализа скелетных
остатков. Внешняя форма одиночных кораллитов крайне разнообразна.
Чаще они конические или цилиндрические, прямые или изогнутые,
грибовидные или дисковидные; реже они имеют пирамидальную или
туфелькообразную форму и тогда снабжены одной или четырьмя кры-
шечками, которые закрывали устье кораллита, когда тело втягивалось
внутрь чашечки. Одиночные кораллиты прикреплялись к субстрату
каблучком прирастания или корнеобразными выростами. Колониальные
ругозы имеют две основные формы: кустистую и Пассивную. В первом
случае кораллиты неплотно прилегают друг к другу и имеют круглое
сечение; у вторых кораллиты прилегают тесно один к другому, имеют
полигональное сечение, общую стенку, охватывающую все кораллиты.
Колонии были полушаровидные, караваеобразные, пластинчатые.
Полип помещался в чашечке — углублении верхнего конца корал-
лита; форма и поверхность чашечки отражают детали строения ниж-
ней части полипа. Его верхняя часть, по-видимому, имела довольно
простое строение в виде кожистого мешка с одним ротовым отвер-
стием, окруженным кольцом. щупалец. Форма чашечки варьирует от
мелкой блюдцеобразной до глубокой бокаловидной. Эктодерма нижней
Части полипа формировала основные скелетные элементы как верти-
кальные, так и горизонтальные.
К вертикальным элементам скелета относятся септы. Они обра-
зуются на самых ранних этапах развития в складках базальной эпите-
ки (рис. 99, 1); вначале в плоскости симметрии закладываются глав-
ная и противоположная септы, затем появляются две боковые, обычно
под некоторым углом к главной, и две противоположно-боковые. Эти
6 первичных септ называются протосептами; они разделяют
полость кораллита на 6 секторов. Все последующие септы, именуемые
метасептами, закладываются в четырех секторах. Протосепты и
метасепты образуют основные, или большие, септы; появление большой
203
септы сопровождалось возникновением рядом с ней малой септы. Нали-
чие больших и малых септ— характерная черта ругоз.
Септы ругоз преимущественно пластинчатые, построенные из тесна
прилегающих трабекул, но чаще из трабекул и ламеллярной или
фиброзной склеренхимы. Пластинчатые септы построены из монакан-
тинных, рабдакантинных, голакантинных трабекул или сочетания раб-
дакантинных и голакантинных (рис. 100). Внешний край септы бывает
Рис. 99. Подкласс Tetracorallia:
1 — схема заложения и порядок септ; просепты: главная (гл), боковые (бс),
противоположная (п), метасепты (ж); 2—3 — типы чашечек; 4—5 — схема
строения кораллита;
вс — внутренняя стенка, ок — осевая колонна, пб — продольные борозды, пл —
поперечные линии нарастания, пт — пузырчатая ткань, ф — фоссула, ч— чашечка.
Рис. 100. Микроструктура септ.
Трабекулы: 1а—в — монакантинные, 2а—в — голакантинные, За, б — рабда-
кантинные, 4 — диморфакантинные;
гк — голокант, рк — рабдокант, тр — трабекулы
204
зубчатым, неровным. Септы бывают длинными и короткими, тонкими
и толстыми, клиновидными. Иногда вместо пластинчатых септ разви-.
Ваются только шиповидные или пластинч'ато-шиповидные. Главная и
противоположная септы могут быть расположены в особом углублении,
получившем название фо с су л ы. Фоссула бывает узкая или широ-
кая, мелкая или глубокая. Стенка, или т е к а, у ругоз имеет различ-
ное происхождение и строение. Она строилась из склеренхимы или
Рис. Г01. Типы днищ и осевых структур:
1а, б — осевая колонна; 2а, б — простой столбик; 3 — днища, 4—6 — диссепи-
ментарий (ди);
пт — пузырчатая ткань, тб — табулярий
периферическими концами септ, соединенными склеренхимой; у других
♦гека построена только за счет расширения периферических краев септ,
образуя так называемую септотеку, или за счет периферических
краев септ и диссепиментов, образуя паратёку. Снаружи тека по-
крыта эпитекой — известковой оболочкой, чаще всего морщинистой
(лат. rugosus— морщинистый, отсюда название кораллов—ругозы).
Эпитека, покрывающая снаружи колонию, называется . г о л о т е к о й.
Часто на границе табулярия — осевой части кораллита, занятой днища^-
ми —и диссепиментария развивается внутренняя стенка. Она может
возникнуть за счет расширения септальных пластин (ст е р е о т е к а),
при уплотнении смежных рядов диссепиментов (склеротека), либо
за счет загнутых окончаний септ, либо круто опущенными участками
днищ.
К горизонтальным скелетным элементам относятся днища
и диссепименты. Днища служили опорой полипу и отделяли чашечку,
в которой помещалось тело полипа, от нежилой части кораллита. Раз-
личают полные и неполные днища; последние не достигают стенки ко-
раллита и по периферии переходят в изогнутые мелкие пластинки,
образующие диссепименты. Днища и диссепименты построены
фибрами; ориентированными перпендикулярно отлагавшей их поверх-
ности полипа. Область кораллита, выполненная Диссепиментамщ назы-
вается диссепиментарием. Обычно диссепименты ’отлагались
после заложения ёепт. У лонсдалеоидных кораллов диссепименты рас-
положены по периферии; они отлагались раньше отходящих от них
гептальных пластин.
205
К осевым структурам относятся столбик и осевые колонны.
Столбик, или колумелла (рис. 101, 2), может быть истинным или
ложным. Истинный сто'лбик образуется за счет внутренних кон-
цов септ, в первую очередь за счет концов главной или противополож-
ной септ. Ложный столбик возникает за счет закрученных осевых
концов септ. Осевая колонна, или сложный столбик (рис. 101, 1),
построена из осевых концов протосепт и метасепт и тангенциальных
структур, горизонтальных и вертикальных. Осевые структуры у корал-
ловых полипов появляются начиная с карбона.
Рис. 102. Схема строения скелетов однозонных (/), двухзонных (2) и трех-
зонных (3) кораллов;
дн — днища, п—пузырчатая ткань, с — стенка, сп — септы, ст — столбик
По степени сложности внутренней полости ругозы подразделяются
на однозонные, имеющие только септы и днища, двухзонные —
имеющие еще пузырчатую ткань, и трех зонные—-имеющие кроме
перечисленных структур столбик или осевую колонну. Степень слож-
ности скелета отражает основное эволюционное направление ругоз, но
эти усложнения возникали параллельно в трех основных филогенетиче-
ских ветвях, которые намечаются среди ругоз (рис. 102).
Размножение и развитие. Как и все антозой, ругозы размножались
Половым и вегетативным путем. Половые клетки, вероятно, как у
склерактиний, формировались в энтодерме йезентериев, и эмбриональ-
ное развитие протекало в теле матери. Личинки являлись расселитель-
ной стадией. После оседания личинки на дно, на твердый субстрат, воз-*
никал полип; эктодерма полипа выделяла базальную пластинку и стен-
ку, окружавшую тело. В полость полипа закладывались парные
мезентерии. Базальная часть полипа сминалась в складки, внутри кото-
рых образовывались первые шесть септ, или протосепт; как отмечалось
выше, сначала формировалась главная и противоположная септы„
затем— боковые. Метасепты развивались только в четырех секторах:
в двух, прилегающих к главной септе, и в двух, заключенных между
боковыми септами. Дальнейшее развитие септ проходило по-разному:
у одних появление каждой большой септы сопровождалось появлением
соседней с ней малой, у других—-малые септы появляются сразу во-
всех промежутках между септами первого порядка и позднее, так же,
как метасепты, появляются только в четырех секторах. По мере роста
полипа возникают днища и пузырчатая ткань. Полип всегда занимал
только верхнюю часть кораллита.
Колонии возникают в результате вегетативного размножения —
почкования или деления. У ругоз известно боковое и межстенное прчко-
вание. Боковое почкование связано с возникновением новых
206
почек сбоку полипа и приводит к образованию ветвистых колоний.
Массивные колонии возникают при межстенном почковании,
при котором дочерние полипы образуются между соседними и при
своем появлении раздвигают стенки соседних кораллитов (рис. 103).
При осевом почковании, или делении, материнский организм
распадался на несколько дочерних и погибал либо новые почки раз-
вивались, не вызывая гибели материнского организма, а только откло-
няя его в сторону. При этом возникали ветвистые или букетовидные
Рис. 103. Типы почкования:
1 — «омоложение», 2а, б — осевое, За, б — периферическое, 4 — боковые, 5—6 —
межстенное
колонии. В некоторых случаях у одиночных кораллов наблюдаются
периодические резкие уменьшения и увеличения диаметра чашечки. Это
явление получило название омоложения (рис. 103, /).
Основы систематики и классификация. Систематика ругоз еще не-
достаточно разработана, однако на основании строения маргинария и
табулярия выделяются три отряда: Streptelasmatida, Columnariida, Су-
stiphyllida. Особое место среди ругоз занимает небольшой четвертый
отряд — Heterocorallida.
Отряд Streptelasmatida. Стрептелазматиды (гр. streptos — изогну-
тый, elasma — тонкая пластинка). Одиночные, реже колониальные,
слабо ветвящиеся кораллы; септы вначале толстые пластинчатые, пе-
ристо расположенные, позднее тонкие; имеется фоссула; маргинарий из
септальной стереозоны, или диссепиментар и.я, построенного из
маленьких интерсептальных диссепиментов; днища выпуклые. Стрепте-
лазматиды появились в середине ордовика, вначале имели только
днища и септы, а начиная с силура к скелетным элементам добав-
ляется пузырчатая ткань, образующая диссепиментарий. В карбоне
возникают осевые структуры. Средний ордовик — пермь. Около 220 ро-
дов (рис. 104, 1—4).
Отряд Columnariida. Колумнарииды (гр. columna — столбик). Пре-
имущественно колониальные, редко одиночные; у ранних форм марги-
нарий отсутствует, но позднее развивается в виде септальной стерео-
зоны; может замещаться лонсдалеоидным диссепиментарием. Септы
клиновидные и пластинчатые, перисто расположенные. Пузырчатая
ткань появляется у силурийских форм, а осевые структуры у каменно-
207
угольных. Днища полные и неполные. Колумнарииды появляются в
среднем ордовике и вымирают в ранней перми. Около 60 родов
(рис. 104, 5, 6).
Отряд Cystiphyllida. Цистифиллиды (гр. kystis — пузырь, phyl-
lum — лист). Одиночные, реже колониальные; немногие обладали од-
ной или четырьмя крышечками (так называемые крышечные корал-
6а
Рис. 104. Отряды Streptelasmatida (1—4) и Columnariida (5—6):
1 — Streptelasma (средний ордовик — средний силур); 2 — Lambeophyllwn (сред-
ний ордовик); 3 — Kodonophyllum (средний — поздний силур); 4 — Lithostrotionella
(карбон — ранняя пермь); 5 — Columnaria (средний девон); 6 — Lonsdaleia (ран-
ний—средний карбон); сечения: а — продольное, б — поперечное, в — общий вид;
дн — днища, п — пузырчатая ткань, сп — септы, ст — столбик
лы). Септальные трабекулы крупные; у первых представителей септаль-
ный аппарат в виде шипиков, днища отсутствовали, позднее (в конце
ордовика) появляются пластинчато-шиповидные септы, а в силуре осо-
бое развитие получает пузырчатая ткань, которая иногда заполняет всю
полость кораллита .(кроме чашечки), развиваются особые стереоплаз-
матические корки (ранее известные как септальные конусы), которые
откладывались периодически всей базальной поверхностью эктодермы
полипа (рис. 105, /). Предполагается, что образование подобных сте-
реоплазматических корок было связано с- сезонными изменениями или
изменением физиологических процессов, протекавших в полипах (на-
пример, связанных с размножением). Маргинарий в виде стереозоны,
в которой оси трабекул погружены в пластинчатую склеренхиму; диссе-
208
пйментарий состоит из шаровидных диссепиментов. Средний ордовик —
девон. Около 30 родов (рис. 105, 1—4).
Отряд Heterocorallida. Гетерокораллы. Кораллы одиночные, малень-
ких размеров, круглого или многоугольного сечения, диаметром 6—
16 мм; развиты днища и очень своеобразные септы. Вначале заклады-
ваются четыре протосепты, соединяющиеся в центре. Затем две из них
Рис. 105. Отряды Gystiphyllida (1—4) и Heterocorallida (5—6):
1 — Nardophyllum (ранний — средний девон): 1а — продольное, 16 — попереч-
ное сечения; 2 — Tryplasma (силур — ранний девон): 2а — продольное, 26 —
поперечное сечения; 3 — Goniophyllum (ранний — средний силур); 4 — Calceola
(ранний — средний девон): 4а.— продольное сечение, 46 — общий вид; 5 — схема
расположения септ у Heterocorallida; 6 — Heterophyllia (ранний карбон), раз-
витие септ;
гл — главная септа, кр — крышка, п — противоположная септа, пр — просепты,
ск — септальные конусы, I—V — метасепты в порядке появления
или все четыре у периферии дихотомически ветвятся, образуя четыре
квадранта между ветвями и четыре полости между каждым ветвлением
(«фоссулы»). Новые септы развиваются только в четырех квадрантах
и никогда не появляются в фоссулах (рис. 105, 5). Днища состоят из
отдельных сегментов, развитых в каждой полости между септами. На
периферии днища круто изгибаются и примыкают к толстой стенке.
Периферические концы септ утолщены и образуют стереозону (назван-
ную Шиндевольфом гетеротекой). Все скелетные элементы построе-
ны из фиброзной склеренхимы. К гетерокораллам относятся только два
рода, распространенные в раннем карбоне Европы и Азии, — Hetero-
phyllia, Hexophyllia.
История развития ругоз. Ругозы, вероятно, возникли от аулопоро-
идных табулят в начале среднего ордовика; они были маленьких раз-
меров, рогообразной формы, без днищ, с шиповидными септами. Сразу
же возникло три основных ствола ругоз: стрептелазматиды, колумна-
рииды и цистифиллиды, но в ордовике они были немногочисленны и не
играли существенной роли. Все они отличались отсутствием пузырчатой
ткани и получили название однозонных.
14 В. В. Друщиц . 209
Начало, силура ознаменовалось появлением во всех трех стволах
пузырчатой ткани, образующей диссепиментарий- Кораллы стали двух-
зонными; в каждом отряде возникали новые семейства и новые роды.
В построении коралловых рифов в силуре принимали участие табуляты,,
строматопоры, водоросли и частично ругозы. В конце раннего и начале
среднего девона наблюдается новая эволюционная вспышка — появ-
ляются новые семейства и роды, однако крупных морфологических из-
менений не происходило. В начале карбона во всех трех отрядах появ-
ляются осевые образования, столбики или осевые колонны, кораллы
становятся трехзонными. Начало карбона характеризуется также
появлением маленького, но очень своеобразного отряда — гетерокорал-
Рис. 106. Геохронологическое распространение кишечнополостных
лов, вымершего к концу раннего карбона. В карбоне наряду с одиноч-
ными формами были широко распространены колониальные. Четвертое-
крупное изменение среди ругоз произошло в начале перми. Все семейст-
ва, за исключением одного — Polycoeliidae, от которого, вероятно, про-
изошли склерактинии, вымерли в течение артинского яруса (ранняя
пермь). Последние ругозы вымирают в конце перми, на смену им в сред-
нем триасе приходят склерактинии.
История развития кишечнополостных показана на схематическом
рис. 106. Первые находки остатков кишечнополостных — протомедуз,,
сцифозой — относятся к венду. Остатки наиболее примитивных кишеч-
нополостных — органические скелеты колониальных гидроидных (гидро-
зой) — находят в кембрии, хотя гидрозон скорее всего возникли раньше.
210
В среднем кембрии становятся известными остатки скелетов конулят,
а в позднем кембрии появляются достоверные коралловые полипы —
табуляты (аулопориды); ОТ аулопорид в среднем ордовике возникает
два новых подкласса коралловых полипов — гелиолитоидеи и ругозы.
Аулопориды в результате дивергенции дают начало нескольким ртряг
дам табулят. В среднем ордовике от какой-то группы гидрозой возни-
кает отряд хететид. В течение палеозоя были распространены гидрозон,
сцифозои и коралловые полипы — табуляты, ругозы и гелиолитоидеи;
распространение последних ограничено средним девоном. В конце
палеозоя вымирают табуляты:, ругозы.
В начале мезозоя на смену ругозам приходят Склерактинии (щес-
тилучевые кораллы) как рифостроители, так и агерматипные формы;
становятся многочисленными восьмилучевые кораллы (остатки которых
почти неизвестны пока в отложениях палеозоя). В конце мелового пе-
риода появляются гидроидные, обладавшие известковым скелетом ((мил-
лёпориды и стилястериды). До конца мела существуют хететиды. В со-
временных морях и океанах известно около 660 видов гидромедуз и свы-
ше 2000 видов гидрополипов (класс гидрозон), около 200 видов сцифо-
медуз и свыше 6000 коралловых полипов. Несколько видов гидрозой
приспособились к жизни в пресных водах.
ТИП CTENOPHORA. ГРЕБНЕВИКИ
Общая характеристика. Ктенофоры (гр. kteis, род. ktenos—гребень,
ph.oros — несущий), или гребневики, — малочисленная группа, насчи-
тывающая 90 видов морских одиночных свободноподвижных, преимуще-
Рис. 107. Тип Ctenophora. Схема строения ползающего гребневика;
аб — аборальный орган, р — рот, щ — щупальца
ственно пелагических животных. Тело сферической или уплощенной
формы; рот расположен на нижней стороне. Органами движения явля-
ются восемь меридиональных рядов гребных пластинок (отсюда назва-
ние типа). Пара длинных щупалец, снабженных клейкими клетками,
служит для ловли добычи и для защиты. Стрекательные клетки отсутст-
вуют. Пищеварительный тракт состоит из эктодермальной глотки, экто-
дермального желудка и системы каналов. Органов дыхания и выделения
нет. Нервная система диффузного типа. На стенках меридиональных
каналов развиваются половые железы. Гребневики гермафродиты, раз-
множаются половым и бесполым путем. В онтогенезе закладываются
.эктодерма, энтодерма и третий зародышевый листок — мезенхима.
Часть гребневиков (платиктениды, рис. 107) приспособилась к бентос-
14*
211
ному образу жизни, в связи с чем их тело удлинилось в полости щупа-
лец; при ползании одно щупальце направлено вперед, другое назад;
гребные пластинки видоизменились, в области аборального органа (ор-
гана равновесия) наблюдается концентрация нервных узлов (ганглиоз-
ных скоплений).
Место гребневиков в системе животного мира остается недостаточ-
но ясным. Несомненно, что их эволюционный уровень выше уровня ки-
шечнополостных и они возникли позднее последних. А. О. Ковалев-
ский (1880)'считал платиктенид предками плоских червей (турбелля-
рий). Некоторые авторы рассматривали гребневиков как неотенические
формы, возникшие от личинок коралловых полипов, однако организа-
ция и тип онтогенеза гребневиков существенно отличаются от таковых
кишечнополостных. Гребневики образуют самостоятельный тип, стоя-
щий вблизи типа кишечнополостных. В ископаемом состоянии они не-
известны.
РАЗДЕЛ BILATERALIA.
ДВУСТОРОННЕ-СИММЕТРИЧНЫЕ
Все многоклеточные животные, стоящие выше кишечнополостных
и гребневиков, объединяются в раздел двусторонне-симметричных жи-
вотных (Bilateralia). Двусторонняя симметрия тела возникла в связи
с приспособлением к активному передвижению. Все ткани и органы те-
ла развиваются из трех зародышевых листков: эктодермы, эндодермы и
мезодермы, и поэтому'их называют также трехслойными (Triplas-
tica).
В процессе эволюции трехслойных выработались две основные вет-
ви — первичноротые и вторичноротые.
ПОДРАЗДЕЛ PROTOSTOMIA.
ПЕРВИЧНОРОТЫЕ
Первичноротые (гр. protos — первый, stoma — рот) характе-
ризуются следующими чертами: на месте личиночного рта, или бласто-
пора, развивается рот взрослого животного; анальное отверстие, если
оно есть, возникает на противоположном полюсе зародыша. Мезодерма
закладывается телобластическим путем: в области бластопора
между эктодермой и эндодермой обособляются клетки (телобласты),
которые после размножения заполняют всю первичную полость заро-
дыша. Скелет наружный, эктодермический. Нервная система состоит из
ганглиев, соединенных нервными стволами. К первичноротым относятся
низшие и кольчатые черви, членистоногие, моллюски, мшанки и неко-
торые другие группы.
Среди них различают бесполостных и первичнополостных (отсутст-
вует или имеется полость тела — низшие черви), объединяемых в про-
трохофорных животных (Protrochozoa), и вторичнополостных (разви-
та вторичная полость тела — высшие черви, членистоногие, моллюски,
мшанки), объединяемых в надтип трохофорных животных (от названия
личинки трохофоры).
ТИП SCOLECIDA. НИЗШИЕ ЧЕРВИ
К низшим червям относится большая группа протрохофорных жи-
вотных, объединяющая наиболее низкоорганизованных трехслойных мно-
213
гоклеточных животных. Они характеризуются малыми размерами, дву-
сторонне-симметричным строением тела, развитием кожно-мускульного
мешка, облекающего тело, отсутствием настоящей метамерии и плотных
покровов, защищающих тело. Низшие черви не способны к быстрому
движению. У них развивается сквозной пищеварительный тракт (круг-
лые черви, немертины), появляется обособленная кровеносная система
(немертины), примитивные органы выделения (протонефридии). Вопрос
о происхождении низших червей окончательно не решен. По мнению
А. В. Иванова (1968), они возникли от фагоцителлообразных предков,
обладавших ротовым отверстием; основное значение в выработке плана
строения плоских червей имел переход от плавающего к ползающему
образу жизни. В процессе развития у них выработалась двусторонняя
симметрия тела, обособился передний отдел, ротовое отверстие посте-
пенно переместилось вдоль брюшной стороны вперед.
Низшие черви населяют моря, пресные воды, почву. Многие из
них — паразиты животных, растений, человека. В ископаемом состоя-
нии известны с карбона как паразиты насекомых.
Среди сколецид выделяют 3 подтипа: Plathelminthes, Nemathelmin-
thes, Nemertini.
ПОДТИП PLATHELMINTHES. ПЛОСКИЕ ЧЕРВИ
Общая характеристика. Плоские черви, или платгельминты (гр.
platos — плоский, helmins, род. helminthos — червь), представляют наи-
более низкоорганизованный подтип среди червей. К ним относятся тур-
беллярии, ресничные и ленточные черви. Они имеют двусторонне-сим-
метричное, сплющенное в спинно-брюшном направлении, обычно удли-
ненное тело. Снаружи оно покрыто мерцательным эпителием, под
которым расположено несколько слоев гладких мышечных волокон,
создающих совместно с эпителием кожно-мускульный мешок, который
обеспечивает сохранение и изменение формы тела, его червеобразное
изгибание и прямолинейное плавание. Гладкие мышцы образуют прими-
тивную мышечную систему, чем они отличаются от гребневиков и
кйшечнополостных.
Пространство между покровами и кишкой заполнено соединитель-
ной тканью — паренхимой, вследствие чего плоских червей назы-
вают бесполостиыми, или паренхиматозными. Пищевари-
тельный канал состоит из передней эктодермической кишки, или глот-
ки, и эктодермической средней кишки, заканчивающейся слепо. Рот,
расположенный или на переднем конце тела или на брюшной стороне,
служит для приема пищи и удаления непереваренных остатков. Двой-
ная функция рта и отсутствие задней кишки с анальным отверстием
указывают на признаки низкой организации плоских червей. Нервную
систему составляют парный мозговой ганглий и идущие назад нервные
стволы. Кровеносная и дыхательная системы отсутствуют. Впервые по-
являются органы выделения, состоящие из двух или нескольких кана-
лов (протонефридий, гр. protos — первый, nephros — почка), откры-
вающихся наружу мелкими отверстиями.
Половая система гермафродитная, очень сложная. Развитие про-
текает с метаморфозом — из яйца выходит личинка, которая после
ряда превращений дает половозрелых особей. •
Плоские черви живут в морях, пресных водах и на суше. Многие
ведут паразитический образ жизни. В ископаемом состоянии известны
как паразиты некоторых насекомых карбона, перми и палеогена. Плос-
кие черви наиболее примитивные среди двусторонне-симметричных жи-
214
потных. На это указывают: ресничный покров, паренхиматозность, от-
сутствие полости тела, слабое развитие нервной системы, отсутствие
органов дыхания и кровообращения, сквозного кишечника, примитив-
ность органов выделения. В состав подтипа включают следующие клас-
сы: Turbellaria, Cestodes, Mesozoa, Trematodes и др.
ПОДТИП NEMATHELMINTHES. КРУГЛЫЕ ЧЕРВИ
Общая характеристика. Круглые черви, или нематгельминты
'(гр. пета — нить), имеют цилиндрическое, удлиненное, несегментиро-
ванное тело, состоящее из кутикулы, подстилающего ее эпителия и
мышц. Тело разделено на головной, туловищный и хвостовой отделы.
Круглые черви имеют полость тела — схизоцель, заполненную по-
лостной жидкостью, соответствующую полости гаструлы, поэтому эта
полость называется первичной, а сами черви первичнополостными жи-
вотными. Кожно-мускульный мешок имеется у нематод. Кровеносная и
дыхательная системы отсутствуют. Нервная система развита слабо.
-Органы выделения — протонефридии. Половая система сравнительно
простая; в отличие от плоских червей круглые черви раздельнополы.
Прямой трубкообразный кишечник разделяется на 3 отдела: передний,
средний и задний с анальным отверстием. Появление сквозного пище-
варительного тракта приводит к разделению функций: рот служит
только для приема пищи, а задняя кишка с анальным отверстием —
для удаления переваренных остатков. В пищеварительном тракте воз-
никает направленный ток пищевых частиц.
Круглые черви живут в морях и пресных водах. Часть из них ве-
дет паразитический образ жизни (аскариды, острицы, власоглавы).
В ископаемом состоянии известны как паразиты насекомых из карбона,
мела, палеогена. Они были также найдены в замороженных трупах жи-
вотных в четвертичных отложениях Сибири.
Круглые черви произошли от плоских червей, с которыми их свя-
зывает ряд общих признаков организации. В ходе эволюции у них раз-
вился сквозной пищеварительный тракт и прочный кутикулярный по-
кров тела, изменился способ передвижения: вместо передвижения с
помощью ресничек они стали двигаться, змеевидно изгибая тело. В со-
став подтипа включают следующие классы: Gastrotricha, Rotatoria,
Nematodes, Nematomorpha и др.
ПОДТИП NEMERTINI. НЕМЕРТИНЫ
Общая характеристика. К немертинам (гр. Nemertes—имя од-
ной из дочерей морского царя Нерея) относятся морские черви, имею-
щие удлиненное несегментированное тело, покрытое мерцательным эпи-
телием. Полость тела заполнена паренхимой, в которой лежат внутрен-
ние органы. Стенка тела с хорошо развитой мускулатурой. Впереди в
виде продольной щели расположен рот. Имеется выворачивающийся
•наружу хоботок. Нервная система в виде головного узла и двух про-
дольных стволов. В отличие от плоских червей у немертин имеется зад-
няя кишка с анальным отверстием и система кровеносных сосудов —
•один спинной и два боковых. Выделительная система устроена по типу
протонефридиев. В нее поступают накопляющиеся в крови конечные
продукты обмена веществ. Немертины раздельнополы; половая систе-
ма устроена просто. Они произошли от плоских червей и занимают про-
215
межуточное положение между ними и кольчатыми червями. В иско-
паемом состоянии неизвестны.
ТИП ANNELIDES. КОЛЬЧАТЫЕ
ЧЕРВИ
Общая характеристика. Кольчатые черви (гр. annulus — кольцо) —
наиболее высокоорганизованная группа среди огромного мира червей
(рис. 108). Они образуют основное ядро всех трохофорных животных.
, Рис. 108. Тип Annelides:
1 — личинка полихет — трохофора; 2 — отпечаток кольчатого червя из литографских
сланцев Золенгофена (поздняя юра); 3 — общий вид колонии современного многоще-
тинкового червя Serpula vermicularis L.; 4 — крышечка эоценовых серпулид;
м — личиночные мышцы, мз — мезодермальный зачаток, мп — мерцательный пояс,
пт — пищеварительный тракт, р — рот, тп. — теменная пластинка
К ним принадлежат первичные кольчецы, много- и малощетинковые
черви, пиявки и эхиуриды.
Тело аннелид разделено на голову, туловище и задний отдел. Го-
лова является производной верхнего. полушария трохофорной личинки;
216
она несет скопление нервных узлов («головной мозг») и рецепторы ося-
зания, зрения, обоняния и вкуса. Задний отдел тела, или пигидий, не-
членистый. Туловище состоит из большого числа (до 800) сегментов.
В каждом сегменте повторяются узлы нервной системы, органы вы-
деления, половые железы. Такое многократное повторение сегментов
тела и всех внутренних органов названо метамерией (гр. предлог
meta — вместе, с, meros — часть). Каждый сегмент несет боковые при-
датки, или параподии (в переводе означает «похожие на ноги»),
снабженные щетинками; параподии служат органами движения; они
действуют по принципу простого рычага. Метамерия возникла у коль-
чатых червей в связи с удлинением тела в процессе эволюции. Раз-
личают гомономную, или однородную, и гетерономную, или
неоднородную, метамерию.
- У кольчатых червей появляется вторичная полость тела, или це-
лом, возникающая в мезодерме. Она находится между кишечником и
стенкой тела и выстлана сплошным слоем эпителиальных клеток. По-
лость тела заполнена жидкостью, по составу близкой к морской воде.
Жидкость находится все время в движении; она выполняет функцию
своеобразного «гидравлического скелета» (подобно камере в баллоне
автомобильного колеса) и служит для переноса внутри тела питатель-
ных продуктов, кислорода, углекислого газа и продуктов выделения
желез внутренней секреции. Внутренняя полость тела разделена пере-
городками, отделяющими отдельные сегменты друг от друга; они защи-
щают тело при ранениях и позволяют изолировать раненый участок
от внешней среды.
Нервный аппарат слагается из «мозга» — совокупности нервных
узлов верхнего полушария трохофоры, пары брюшных стволов, соеди-
ненных с мозгом с помощью окологлоточных коннективов. У примитив-
ных аннелид брюшные стволы широко расставлены и соединены попе-
речными комиссурами, образуя «лестничную нервную систему». У боль-
шинства аннелид происходит сближение стволов и из «лестницы» об-
разуется брюшная цепочка.
Аннелиды имеют хорошо развитую кровеносную систему; кровь
движется по системе сосудов, стенки которых ритмично сокращаются.
Сердце отсутствует. Органами дыхания служат жабры, расположен-
ные на параподиях, голове или хвостовом отделе. Пищеварительный
тракт состоит из переднего отдела, средней и задней кишки. Рот, рас-
положенный на переднем конце, ведет в глотку; в ней расположены ро-
говые'челюсти и зубы, известные в ископаемом состоянии под назва-
нием сколекодонт. Глотка переходит в пищевод с мышечной стен-
кой; мышцы перистальтическими движениями проталкивают пищу в
среднюю кишку, выполняющую функцию желудка. Задняя кишка за-
канчивается анальным отверстием. Органы выделения —• метанефри-
дии — служат для выведения продуктов обмена и половых клеток; ме-
танефридии имеют форму воронки, снабженной длинным каналом; ка-
нал открывается в соседнем сегменте.
Кольчатые черви размножаются бесполым и половым путем. При
бесполом размножении длинное тело распадается на несколько час-
тей. При половом размножении из оплодотворенной яйцеклетки разви-
вается личинка — т р о х о ф о р а (рис. 108, /). Мезодерма разви-
вается телобластическим путем; за счет первичной полости развивают-
ся спинной и брюшной кровеносные сосуды. В мезодерме возникает
вторичная полость тела — целом. Из верхнего полушария личинки
развивается головной отдел, несущий нервные ганглии и органы чувств.
Сегменты тела, возникшие на стадии личинки, получили название л а р-
. 217
вальных, или личиночных; число их колеблется от 3 до 10—13. Пос-
ле превращения личинки в молодого кольчатого червя постлар-
вальные сегменты возникают на переднем крае анальной лопасти в
кольцеобразной зоне нарастания.
В каждом новом сегменте формируются пара параподиев, пара
целомических мешков, пара гонад (половых желез), пара метанефри-
диев, пара нервных узлов (ганглиев) с продольными и поперечными
нервными стволами, кровеносные сосуды, мышцы.
Рис. 109. Тип Annelides:
1—2 — ископаемые полихеты из позднего докембрия Австралии: 1 — Dickin-
sonia; 2 — Spriggina; 3 — Spirorbis и 4 — Serpula на раковинах девонских
брахиопод; 5 — реконструкция челюстного аппарата Arabellites (девон), со-
стоящего из отдельных сколекодонт; 6—9 — следы жизнедеятельности червей:
6 — U-образный ход (Corophioides) роющих червей (образец из кембрия Эс-
тонии), 7 — норка Polydora в раковине устрицы (сильно увел.), палеоген
Ферганы, 8а, б— Chondrites (Fucoides) — разветвленные непересекающиеся
ходы-туннели червей-илоедов в рыхлом осадке, 9 — Spirophyton. (Taonurus) —
ходы роющих червей-илоедов и два разреза через них, показывающие круглое
сечение последнего, объемлющего хода, .и серповидные сечения расположен-
ных внутри него старых, засыпанных ходов
Кольчатые черви широко распространены в природе. Они заселя-
ют все моря и океаны от полосы прилива до ультраабиссали (10 630—
10 700 м), нередко образуя поселения большой плотности. Они живут
на суше и в пресноводных бассейнах. Наземные и пресноводные фор-
мы обычно гермафродиты и имеют прямое развитие, без личиночной
стадии.
Кольчатые черви разделены на несколько классов: Archiannelides,
Polychaeta, Oligochaeta, Hirudinea, Echiuroidea. '
218
В ископаемом состоянии встречаются преимущественно предста-
вители класса полихет. Полихеты (гр. poly — много, chaeta — щетин-
ка) — типично морские животные, лишь немногие живут в пресных
водах. Морские полихеты известны на всех глубинах и на всех широ-
тах. Одни живут в пелагиали, другие на дне водоемов. Параподии
несут пучки многочисленных щетинок (отсюда название класса). Бен-
тосные полихеты ползают по дну, зарываются в грунт или неподвижно
прикрепляются к подводным предметам. Полихеты, зарывающиеся в
грунт, живут в U-образно изогнутых норках, изгибая тело таким же
образом; около ротового отверстия на поверхности .грунта образуется
воронка, через которую поступает пища; у заднего конца возникает
конус песка, который зарывающийся пескожил (так называют этих
червей) выбрасывает через анальное отверстие, пропустив предвари-
тельно песок с питательными веществами через пищеварительный
тракт. Некоторые кольчатые черви строят себе роговые, песчаные или
известковые трубочки, защищающие мягкое тело; наружное отверстие
известковых трубочек может закрываться крышечкой (см., рис. 108, 4).
В ископаемом состоянии чаще всего сохраняются известковые тру-
бочки, состоящие из двух концентрических пластинчатых слоев карбо-
ната кальция. Трубка может быть цилиндрической, конусовидной, пря-
мой, изогнутой или спирально свернутой. Представители семейства сер-
пулид (лат. serpero — ползать) постоянно живут в известковых труб-
ках. Серпулиды обычно поселяются на твердых предметах, скелетах
различных животных, раковинах моллюсков, брахиопод, на панцирях
крабов, раков (рис. 109, 3, 4). Они живут также среди коралловых
рифов; нередки случаи комменсализма между ними и коралловыми по-
липами. Остатки серпулид чаще всего встречаются в ископаемом со-
стоянии. В меловых морях они принимали участие в образовании от-
дельных банок, и их скелеты иногда переполняют отдельные слои. Кро-
ме трубочек от полихет сохраняются челюсти, щетинки, отпечатки те-
ла, а также следы ползания, зарывания, ходы, норки (рис. 109, 6—9).
Челюсти полихет известны под названием сколекодонт; они име-
ют вид зазубренных пластинок, состоящих из хитинового вещества.
Остатки челюстного аппарата полихет были описаны Р. Козловским
из ордовикских валунов Польши (1956).
Отпечатки тела древнего пелагического червя Spriggina (рис. 109,2)
длиной около 4,5 см найдены в отложениях венда Эдиакары. Тело уз-
кое, гибкое, состоит из сегментов; каждый сегмент несет параподии, ко-
торые заканчиваются острыми шипами. Голова имеет форму подковы.
Эти отпечатки похожи на современных пелагических полихет (Тото-
pteris).
В этих же слоях встречены отпечатки тела ползающего кольчатого
червя, описанного под названием Dickinsonia (рис. 109, 1). Вначале их
отнесли к кишечнополостным и выделили в особый класс Dipleurozoa,
однако позднее была установлена их аннелидная природа.
Кольчатые черви явились предками для всех остальных первично-
ротых трохофорных животных. <
ТИП ONYCHOPHORA. ОНИХОФОРЫ
Общая характеристика. Современные онихофоры (гр. onychos —
коготок, phoros — несущий) — наземные животные, внешне похожие
219
на кольчатых червей. Тело длинное, червеобразное, членистое, покры-
то мягкой кутикулой. Голова неясно обособлена; она несет три пары
придатков: длинные усики и две пары придатков около рта — и пару
бокаловидных глаз. Рот ведет в глотку; пища из глотки попадает в
короткий пищевод, длинную среднюю кишку; задняя кишка короткая,
заканчивается на заднем конце тела анальным отверстием. Кровенос-
ная система состоит из спинного трубчатого сосуда. Нервная система
Рис. НО. Класс Protracheata.
Реконструкция Aysheaia pendu.ncu.lata Walcott . (кембрий); голова и передняя
часть туловища увеличены (Шаров, 1966)
построена так же, как у аннелид, от надглоточных узлов тянутся вдоль
тела два нервных ствола. Онихофоры раздельнополы; многие способ-
ны к живорождению.
Тип онихофор представлен одним классом Protracheata с одним
отрядом того же названия. Современные протрахеаты живут в тропи-
ческих областях, во влажных затененных местах. Они медленно пере-
двигаются с помощью многочисленных ног; каждая нога представляет
вырост тела, заканчивающийся двумя коготками (отсюда название ти-
па). По наличию гомономной метамерии тела, кожно-мускульного меш-
ка, лестничному типу нервной системы, строению конечностей и отсут-
ствию наружного скелета онихофоры сходны с кольчатыми червями.
Они, вероятно, произошли от общих предков с ними, но приспособились
к жизни на суше. Онихофоры образуют самостоятельную филогенети-
ческую ветвь, занимающую промежуточное положение между аннели-
дами и членистоногими. В ископаемом состоянии известен один пред-
ставитель этого типа — Aysheaia, описанный Уолкоттом в 1911 г. из
морских отложений среднего кембрия Северной Америки (рис. ПО).
ТИП ARTHROPODA.
ЧЛЕНИСТОНОГИЕ
Общая характеристика. Членистоногие (гр. arthro—сочленять,
pous, род. podos — нога) образуют наиболее многочисленный тип
животного царства, насчитывающий около 1 млн. видов (по некоторым
данным, свыше 2 млн. видов), из которых на долю насекомых прихо-
дится 9/10 общего числа.
Членистоногие приспособились к обитанию в самых разнообразных
условиях. Они известны в морях и пресных водах, в почве, в пещерах,
в самых разнообразных местах на поверхности суши; большинство на-
земных форм приспособилось к полету, часть членистоногих — парази-
ты. Членистоногих можно встретить в любой части Земли — от тропи-
ков до северных широт, в морских и континентальных бассейнах —
от горячих источников до холодных вод.
К типу членистоногих относятся трилобиты и мечехвосты — оби-
татели морей, многочисленные ракообразные (раки и крабы, щитни и
дафнии, остракоды и усоногие раки) обитатели морских и пресных вод,
стрекозы и поденки, жуки и бабочки, термиты и муравьи, пауки и кле-
щи — обитатели суши и наземных водоемов (рис. 111).
Палеонтологические находки, хотя и известны начиная с конца про-
терозоя (венд Австралии), не дают отчетливых данных о происхожде-
нии членистоногих. Изучение эмбрионального развития различных ар-
тропод и сравнительно-анатомические исследования позволяют выска-
зать предположение, что этот древний тип возник от примитивных ан-
нелид, с которыми их сближают следующие черты: сегментированность
(метамерность) тела, строение кутикулы, покрывающей тело, располо-
жение внутренних органов по единому плану.
Для всех членистоногих характерно двусторонне-симметричное ге-
терономно сегментированное тело. Число сегментов различно и колеб-
лется от 8 до 180. В процессе эволюции наблюдается сокращение чис-
ла сегментов, объединение их в отдельные тагмы, или отделы, те-
ла. Обычно обособляется голова и туловище; в туловище обособляет-
ся грудь и брюшко; иногда голова срастается с грудью, образуя голо-
вогрудь; у клещей границы между отделами тела вообще утрачива-
ются.
Тело членистоногих покрыто плотной и прочной кутикулой. Ку-
тикула выделяется эпителием и состоит из хитина — сложного поли-
221
сахарида, содержащего азот и различные белковые вещества; у многих
членистоногих кутикула пропитывается углекислым кальцием или фос-
фатом кальция и достигает иногда большой прочности. В этом случае
она состоит из твердых участков — тергитов и склеритов и перепон-
чатых участков с очень тонкой кутикулой; такое строение обеспечива-
Рис. 111. Представители четырех основных подтипов членистоногих:
1—2 — подтип Trilobitomorpha, трилобиты; 3^-5 — подтип Cheljcerata: 5 — эв-
риптериды, 4 — арахниды, 5 — лимулиды; 6—9 — подтип Branchiate: 6 — выс-
шие раки, 7 — циррипедии, 8 — остракоды, 9 — конхостраки; 10—12 — подтип
Tracheata, насекомые
ет шарнирное сочленение, фиксацию суставов и подвижность отдель-
ных сегментов тела и конечностей относительно друг друга. Плотность
и прочность кутикулы ограничивает рост членистоногих, поэтому рост
тела у всех членистоногих сопровождается линькой. Когда старый
панцирь становится тесным, он лопается и сбрасывается; в очень ко-
роткий период, пока не образуется и не затвердеет новый панцирь,
происходит интенсивное увеличение размеров тела. Хитиновые покро-
вы сбрасываются не только с наружной поверхности тела, но и со всех
конечностей, передней и задней кишки. Таким образом, рост членисто-
ногих происходит прерывисто, ступенчато, размеры тела увеличиваются
скачкообразно после каждой линьки (речной рак в первый год своей
жизни линяет 6—10 раз).
В типичном случае каждый сегмент несет пару двуветвистых ко-
нечностей, которые подвижно сочленяются с сегментом тела и состоят
222
из отдельных члеников (отсюда название «артроподы», т. е. членисто-
ногие). Схема строения различных типов конечностей изображена на
рис. 112.
Рис. 112. Схема эволюции конечностей членистоногих
(Шаров, 1966):
А — примитивная лобоподная конечность; Б—Ж —
нога представителей: Б — Pycnogonid.es, В — Tetra-
cephalosomita, Г — Trilobita, Д — Crustacea, Е — Che-
licerata, Ж — Tracheata;
цифры соответствуют гомологичным частям; 1 — пре-
кокса, 2 — кокса; бд бедро, в —- вертлуг, г — го-
лень, л — лапка, пэ преэпиподит, т — тазик, тел —
телоподит, ж — экзоподит, эн — эндоподит
Голова состоит из нескольких слившихся сегментов (рис. ИЗ,
А—Г), которые несут основные органы чувств' — глаза, антенны, слу-
жащие для ориентировки в пространстве, и ротовые органы — челюстц,
предназначенные для захвата и размельчения пищи. У древних членис-
тоногих первично конечности выполняли функцию передвижения.
С дифференциацией тела предротовые конечности становятся носителя-
223
ми органов чувств (антенны), околоротовые превращаются в челюсти
(рис. 113, Д). При'срастании головы и груди передние ножки груди
изменяются в ногочелюсти и принимают участие в добыче пищи. Чис-
ло пар ног, которые служат для передвижения, у разных групп членис-
тоногих неодинаково. У насекомых их три пары, у паукообразных —
Рис. 113. Взаимное положение сегментов головы у Dicephalosomita (A), Tetracephaloso-
mita (Б), Chelicerata (В), Tracheata (В) и схема эволюции мандибул (Д);
римскими цифрами обозначены сегменты головы, арабскими—туловищные сегменты
(у Д членики телоподита);
пэ — преэпиподит, тел — телоподит
четыре, у некоторых многоножек — свыше сотни. Обычно брюшко ли-
шено конечностей либо они видоизменены в жабры или половые ор-
ганы.
Панцирь членистоногих является наружным скелетом и служит
для защиты внутренних органов от повреждения, а также местом при-
224
крепления мускулатуры и внутренних органов. Двигательная мускула-
тура обычно состоит из поперечнополосатых мышц; движение обеспе-
чивается сокращением мышц, прикрепленных к конечностям и к вы-
ростам панциря, направленным внутрь.
У всех крупных членистоногих развиваются специальные органы
дыхания, у водных — жабры — тонкие листовидные выросты брюш-
ных конечностей; у наземных — либо «легки е» — глубокие мешко-
видные впячивания, либо трахеи — тонкие простые или ветвящиеся
трубочки, которые открываются наружу небольшим отверстием. Мелкие
водные членистоногие дышат всей поверхностью тела. Строение крове-
носной системы зависит от степени развития органов дыхания; она не-
замкнутая; кровь, называемая у членистоногих гемолимфой, дви-
жется по системе полостей, или лакун, и приводится в движение пуль-
сирующим продолговатым сосудом — сердцем, расположенным в
спинной части тела. Перенос кислорода от жабр к внутренним органам
помимо кровеносной системы осуществляет полостная жидкость. Гемо-
лимфа при наличии железа в гемоглобине окрашена в красный цвет,
при наличии меди (гемоцианин) — в синеватый. Вторичная полость
тела, или целом, сливается с первичной и, образуя смешанную по-
лость тела, или миксоцель, перестаёт существовать как самостоя-
тельный аппарат. За счет мезодермы образуется цененхима, которая в
дальнейшем дает начало мускулатуре, жировому телу, кровяным клет-
кам, а также отдельным частям полового, выделительного и кровенос-
ного аппаратов.
Нервная система состоит из головного ганглия (мозга), около-
глоточных коллектив и брюшных стволов, соединенных комиссу-
рами. В каждом сегменте брюшной ствол утолщен (ганглионизиро-
ван); у многих членистоногих происходит слияние двух стволов в не-
парную цепочку, а также слияние последовательных метамерных ганг-
лиев. Характер и степень концентрации брюшной цепочки связаны с
характером и степенью гетерономности их метамерии и степенью ин-
теграции отдельных тагм и всего тела. Хорошо развиты органы чувств:
вкуса, обоняния, осязания, а также органы слуха и зрения; различают
простые и сложные глаза. Переход к активно-подвижному образу жиз-
ни привел к централизации нервной системы, развитию и усложнению
органов чувств. Совершенствование нервной системы привело к улуч-
шению средств защиты от врагов, развитию разнообразных способов
расселения, поисков и привлечения полового партнера, а также к
развитию способов заботы о потомстве, к выработке сложных форм
инстинктивного поведения, обмена информацией, строительства жилищ
(муравьи, термиты).
Пищеварительная система устроена разнообразно — это находит-
ся в прямой связи со способом питания. Она состоит из трех отделов:
эктодермической передней, энтодермической средней и эктодермиче-
ской задней кишки. Обычно передняя и задняя кишки выстланы хити-
новой кутикулой, которая меняется при линьке. Особенности строения
пищеварительной системы будут рассмотрены при характеристике под-
типов и классов.
Все членистоногие размножаются половым путем, реже партено-
генетически; большинство из них раздельнополые с ясно выраженным
половым диморфизмом. Типы оплодотворения разнообразны: наруж-
ное, наружно-внутреннее и внутреннее. Большинство членистоноги^ от-
кладывают яйца; изредка наблюдается живорождение. У многих раз-
витие проходит со сложным превращением. Те особи, которые выхо-
дят из яйца и резко, отличаются по строению от взрослых, носят на-
15 В. В. Друщиц
225
звание личинок. Очень часто личинки ведут иной, чем взрослые, об-
раз жизни и обладают рядом специальных личиночных органов. Пре-
вращение личинки во взрослый организм, наблюдаемый у первично-
водных членистоногих, сопровождается метаморфозом, во время кото-
рого личиночные органы распадаются и вместо них возникают и раз-
виваются органы взрослого.
Членистоногие — одна из древнейших групп; первые остатки из-
вестны из отложений позднего протерозоя — венда Австралии (место-
нахождение Эдиакара).
- Тип членистоногих разделен на пять подтипов: Proboscifera, Tri-
lobitomorpha, Chelicerata, Branchiata, Tracheata.
ПОДТИП PROBOSCIFERA. ПРОБОСЦИФЕРЫ
Подтип пробосцифер (лат. proboscis — хобот, fero — носить)
выделенный А. Г. Шаровым (1965), — наиболее примитивные членис-
тоногие, объединяющие два класса: Dicephalosomita и Pycnogonides.
Они характеризуются наличием хобота, унаследованного и гомологич-
ного хоботу некоторых полихет, многочленистыми конечностями, со-
стоящими из 11—13 члеников и отсутствием эндитов на базальной ча-
сти протоподита. Кембрий — ныне.
КЛАСС DICEPHALOSOMITA. ДИЦЕФАЛОСОМИТЫ
Общая характеристика. К дицефалосомитам (гр. di — два„
cephalon — голова, soma — тело, туловище) относится небольшая груп-
па членистоногих из среднего кембрия Канады, у которых голова со-
стоит из двух сегментов: верхнегубного, или лабрального, и глазного,,
или окулярного (рис. 114, 1). Тело состоит из головы и туловища; го-
лова была подвижной; первый сегмент представлен двухлопастной,
верхней губой; второй сегмент несет пару стебельчатых фасеточных
глаз; от второго сегмента отходит '«фронтальный отросток» (длиной
20—24 мм), представляющий выпячивающийся хоботок, гомологичный
подобному органу полихет; конец хоботка вооружен зубцами. Тулови-
ще длиной до 80 мм состоит из 16 гомономных сегментов и заканчи-
вается тельсоном. Каждый сегмент несет паратергальные (боковые)
выросты, под которыми расположены многочлениковые ножки с жа-
берными нитями. Известен только один род Opabinia (средний кемб-
рий).
КЛАСС PYCNOGONIDES. МНОГОКОЛЕНЧАТЫЕ
Общая характеристика. К классу многоколенчатых, или пикногонид.
(гр. pyknos — компактный, gon — колено, узел), относится один
отряд Pantopoda (гр. pan — целый), или морских пауков, объединяю-
щий около 500 видов очень своеобразных длинноногих артропод, оби-
тающих в морях на разных глубинах (до 7000 м). Туловище от 0,8 до>
250 мм, цилиндрическое или в виде бляшки, разделено на голово-
грудь, состоящую из 7—9 сегментов, и рудиментарное брюшко. Пер-
вые 4 сегмента, срастаясь, образуют голову, чаще называемую глазным
сегментом; остальные сегменты срастаются между собой и головой или.
остаются свободными. Спереди от глазного сегмента отходит цилинд-
226
Рис. 114. Классы Dicephalosomita (?) и Pycnogonides (2), реконструкции:
1 — Opabinia из среднего кембрия Канады: 1а — вид сбоку, 16— вид сверху;
2 — Palaeoisopus из девона Зап. Европы
Рис. 115. Класс Tetracephalosomita (средний кембрий Канады):
1 — Sidneyia inexpectans (реконструкция): la — общий,вид, 16 — ходильная ко-
нечность; 2 — Leanchoilia superlata, вид сбоку (реконструкция)
15*
рический хоботок, напоминающий таковой дицефалосомит и аннелид.
По бокам хоботка имеются три пары коротких конечностей: хе лиф о-
р ы с клешнями, многочлениковые пальпы и яйценосные ножки. За
ними расположены крупные ходильные ноги, обычно 4 пары, иног-
да; 5 и даже 6. Ноги настолько крупны, что кажется, будто бы тело
состоит только из одних конечностей (рис. 114, 2). На глазном сег-
менте на особом бугорке расположены две пары простых глаз. В ис-
копаемом состоянии известно два рода Palaeolsopus, Palaeopantopus
из нижнедевонских отложений Западной Европы. Наличие хоботка,
длинных боковых выростов кишечника, множественных парных гоно-
фор, расположенных в основании ходильных ног, указывает на родст-
во пикногонид с аннелидами, с одной стороны, и дицефалосомитами —
с другой, и позволяет объединять их с последними в один подтип.
ПОДТИП TRILOBITOMORPHA.
ТРИЛОБИТООБРАЗНЫЕ
Подтип трилобитообразных включает три неравных по объему
класса: очень'крупный, и разнообразный класс Trilobita и два неболь-
ших класса Tetracephalosomita и Trilobitida, объединяющих морских
среднекембрийских членистоногих, найденных Уоллкотом в штате Бри-
танская Колумбия (Канада). Трилобитоморфы возникли от хоботонос -
цев. Тело их покрыто спинным трехраздельным хитиновым панцирем.
Голова несет одну, пару усиков и 4 пары двуветвистых конечностей.
Первые трилобитоморфы известны из отложений венда Эдиакары
(Южная Австралия) .
КЛАСС TETRACEPHALOSOMITA. ТЕТРАЦЕФАЛОСОМИТЫ
Общая характеристика. Класс тетрацефалосомит (гр. tetra — четы-
ре), выделенный А. Г. Шаровым (1965), объединяет раннепалеозой-
ских трилобитоморф, которые произошли от дицефалосомит в резуль-
тате редукции хобота, перемещения верхней губы на нижнюю сторону,
добавления двух туловищных сегментов в головную тагму, развития
тактильных, органов — антенн, появления на предбазальных члениках
ног внутренних выростов. Все эти преобразования были связаны, по-
видимому, с переходом от активного хищного образа жизни к жизни
на илистом дне или в илу и соответственно с переходом к питанию
илистой органикой или мелкими организмами — обитателями ила
(рис. 115).
В состав класса включен отряд Limulava (средний кембрий) и
предположительно отряды Cheloniellonida (ранний девон) и 'Leancho-
iliida (средний кембрий).
КЛАСС TRILOBITIDA. ТРИЛОБИТИДЫ
К этому классу, неудачно названному трилобитидами, относятся
трилобитообразные членистоногие в составе 6 отрядов: Nectaspida,
Burgessida, Marrellida, Waptiida, Emeraldellida, Mimetasterida, состоя-
щие каждый из одного рода и одного вида и найденные все, за исклю-
чением последнего отряда, в отложениях среднего кембрия Канады.
Головной отдел состоит из семи сегментов, конечности устроены сходно
228
с конечностями трилобитов, но у многих видоизменены в связи с при-
способлением к различному образу жизни: активно-бегающему, планк-
тонному И Т. Д. '
Рис. 116. Класс Trilobitida. Реконструкция представителей отрядов:
1 — Nectaspida; 2 — Burgessida; 3 — Marrellida: За — вид сверху, 36 — первый
послеродовой придаток; 4 — Waptiida; 5 — Emeraldellida; все из среднего кем-
брия Канады; 6 — Mimetasterida, ранний девон;
ан — антенна, лаб — лабрум, пэп — преэпиподит, тп — телоподит
У Nectaspida (рис. 116, 1) тело состоит из головы и пигидия; у
Burgessida (рис. 116, 2) головной щит разросся и закрыл туловищные
сегменты, тельсон стал членистым; у Marrellida (рис. 116, 3) головной
щит снабжен четырьмя длинными отростками, направленными назад,
имеется вторая пара антенн; у Waptiida (рис. 116, 4) панцирь узкий,
сердцевидный, тельсон с двумя лопастями, глаза . стебельчатые; у
Emeraldellida (рис. 116, 5) тело удлиненное, напоминающее хелицеро-
вых, .вторая пара постантеннальных конечностей превратилась в орга-
ны захвата добычи. Представители отряда Mimetasterida (рис. 116, б)
229
из раннего девона вели, по-видимому, планктонный образ жизни, в
связи с чем размеры тела уменьшились, а некоторые, конечности пре-
вратились в длинные придатки, покрытые щетинками по бокам.
КЛАСС TRILOBITA. ТРИЛОБИТЫ
Общая характеристика. Трилобиты — морские палеозойские членис-
тоногие с удлиненным сегментированным телом, состоящим из голов-
ного щита, туловища и хвостового щита (рис. 117). Двумя продоль-
ными линиями тело разделено на три части: осевую и две боковых
(отсюда название класса, лат. tri — три, lobos — лопасть). Тело три-
лобитов покрыто со спинной стороны тонким (до 1 мм) хитиновым
обызвествленным панцирем. Голова несет одну пару антенн, сложные
глаза. Вентральная сторона закрыта тонкой мембраной, обычно не со-
храняющейся в ископаемом состоянии. Конечности неспециализирован-
ные, двухветвистые. Дыхание осуществлялось жабрами. Онтогенетиче-
ское развитие было связано с рядом личиночных стадий. Трилобиты
появились в раннем кембрии, достигли максимума в конце кембрия
и ордовике, к силуру число их существенно уменьшилось и продолжа-
ло уменьшаться вплоть до окончательного исчезновения в начале пер-
ми. Известно около 1500 родов и свыше 10 000 видов.
Строение скелета. Трилобиты имели эллиптическое тело длиной
3—10- см, однако у некоторых оно было не больше 5 мм и, наоборот,
у самых крупных представителей достигало 45 см (Paradoxides из
среднего кембрия) и даже'70 см (Uralichas из ордовика Португалии).
Тело четко разделено на три отдела, или тагмы: головной щит, или це-
фалон, туловище, или торакс, и хвостовой щит, или пигидий.
Наружный скелет состоит из сильно минерализованного покрова, или
интегумента, подобного хитиновой кутикуле современных членис-
тоногих; на спинной стороне он пропитан карбонатом или фосфатом
кальция и образует прочные покровы — тер гиты; хитиновый скелет
конечностей был меньше пропитан минеральными солями и поэтому
последние в ископаемом состоянии встречаются очень редко. Панцирь
состоит из трех слоев: тонкого пигментированного наружного слоя,
основного слоя, составляющего 0,7—0,8 всей толщины интегумента
(0,2—0,5 мм), и тонкого непигментированного внутреннего слоя. По-
видимому, эти три слоя соответствуют эндокутикуле современных ар-
тропод. Возможно, у трилобитов существовал и аналог эпикутикулы,
следы которой обнаружены у одного вида. Скелет трилобитов был
окрашен (следы окраски были обнаружены у нескольких экземпля-
ров). Наружный скелет по краям тела загибается на вентральную сто-
рону, образуя вдоль нее узкую кайму — дублюру, или заворот.
К дублюре прикрепляется мембрана — тонкий хитиновый необыз-
вествленный покров нижней брюшной части; она обычно не сохра-
няется в ископаемом состоянии.
Головной отдел, или цефалон. Покровы цефалона состоят из слив-
шихся тергитов, закрывающих акрон, преантеннальный, антенналь-
ный и четыре послёротовых сегмента, и по форме напоминают щит
(рис. 118, 1). Прямым задним краем он соединяется с туловищем. Осе-
вая, наиболее выпуклая часть головного >щита —- глабелла (лат.
glabellus — гладкий) — имеет различные размеры и форму. Она мо-
жет почти целиком составлять головной щит или быть очень узкой; мо-
жет доходить до переднего края или только до половины головного щи-
та; может быть гладкой или разделенной бороздами (1—4) на отдель-
230
иые лопасти. Задним затылочным кольцом глабелла отделяется от осе-
вой части туловища. Боковые части головного щита—щеки—лежат
Рис. 117. Класс Trilobita. Реконструкция трилобита Ceraurus из сред-
него ордовика:
1а — вид с брюшной стороны, 16 — вид со спинной стороны; 2 — про-
дольный разрез через тело трилобита;
ан — анальное отверстие, ант — антенна, вм — вентральная мембрана,
г — гипостома, гл — глаз, д — дублюра, ж — желудок, к — конеч-
ности, м—метастома, нц— нервная цепочка, п — пищеварительный-
тракт, р — рот, сп — спинной панцирь
по обе стороны глабеллы. На щеках расположены глаза. Глаза у три-
лобитов очень разнообразны по форме, размерам и строению (рис. 118,
231
2—4). Различают два основных типа глаз: голохроические, или
фасеточные, и шизохроические, или агрегатные. Первые со-
стоят из плотно прижатых многоугольных линз, число которых, как у
насекомых, может достигать 15000; вторые — из крупных двояковы-
пуклых линз, отделенных друг от друга толстой склеротической стен-
кой; число линз от 2 до 300—400. Особенно хорошо глаза сохраняются
Рис. 118, Класс Trilobita:
/ — схема строения головного щита; 2—3 —• голохроические глаза (продольное сече-
ние): 2— у Asaphus, 3 — у Dysplanu.s\ 4 — шизохроические глаза у Phacops: 4а —
продольное сечение, 46 — вид сверху; 5 — схема строения конечности; 6 — строение
головного щита (Шаров, 1966); 7 — схема строения панциря трилобита;
ан — антенны и места прикрепления их мускулов изнутри, гб — борозды на глабелле,
гк — глазная крышка, гл — глабелла, гщ — головной щит (цефалон), зп. — зритель-
ная поверхность глаза, зш — затылочный шип, к — кокса, л — лунки, или ямки, —
места прикрепления изнутри мускулов головных конечностей, лш — лицевой шов, нщ —
неподвижная щека, пк — прекокса, па — преэпиподит, сщ — свободная (подвижная)
щека, тел — телоподит, тул — туловищный отдел (торакс), хщ — хвостовой щит (пи-
гидий), щш — щечный шип
у трилобитов, скелеты которых подверглись окремнению. Среди трило-
битов встречаются первично-слепые формы и утратившие зрение вто-
рично. У трилобитов, имевших глаза, щеки разделялись лицевым швом
на две части: часть щеки, прилегающая к глабелле, получила назва-
ние неподвижной; часть щеки, лежащая кнаружи от лицевого шва,
называется свободной. Последняя соединяется с головным щитом
менее прочно, чем неподвижная щека, и после гибели животного легко
отделяется от головного щита и часто в ископаемом состоянии не со-
храняется. Глабелла вместе с неподвижными щеками образует единое
неразделимое целое — кранидий (от гр. kranion — череп).
232
Различают несколько типов лицевых швов (рис. 119):
о-п и сто п а р и е в ы й, или заднещечный, — задние ветви ли-,
цевых швов начинаются от заднего края головного щита, тянутся к
глазам, проходят мимо глаз с внутренней стороны и идут к переднему
краю щита, иногда соединяясь впереди глабеллы или у переднего края
головного щита либо переходя вниз на дублюру панциря;
Рис. 119. Типы лицевых швов:
м — метапариевый, пр — пропариевый (переднещечный), гон — гонатопариевый, on —
опистопариевый (заднещечный); А — вид головного щита снизу, Б — ъъщ сверху,
В — вид сбоку;
г — гипостома, р — ростр
гонатопариевый — задние ветви лицевых швов начинаются в
заднем углу головного щита;
пропариевый, или переднещечный, — задние ветви лице-
вых швов начинаются от боковых краев головного щита;
метапариевый — задние ветви, как у опистопариевых, а пе-
редние изгибаются назад и подходят к углу заднего края головного
щита. Эти лицевые швы, по-видимому, не функционировали. Они ха-
рактерны для наиболее древнего отряда трилобитов — оленеллид.
Лицевые швы, проходящие через глаза, облегчали сбрасывание
панциря при линьке и способствовали в первую очередь освобождению
глаз. При потере глаз лицевые швы на дорзальной стороне головного
щита исчезают. С боков, на неподвижных щеках, помещаются глаз-
ные крышки, к которым примыкает зрительная поверхность глаза. Они
представляют собой округленные или полулунные выступающие части
щек. От передних концов глазных крышек иногда вперед протягивают-
ся узкие глазные валики.
На брюшной стороне головного щита расположены три таблички;
ростр, или эпистома, гипостома и метастома (см. рис. 117). Ротовое
отверстие ограничено сзади небольшой выпуклой табличкой — мета-
стомой и спереди непарной табличкой — гипостомой, имеющей
разную форму и размеры. У гипостомы различают выпуклую, среднюю
часть и выпуклую или вогнутую кайму. Впереди к гипостоме примыка-
ет ростр — узкая непарная табличка, отделенная от всех сторон шва-
ми: от дублюры, от метастомы и с боков; у некоторых форм ростр от-
сутствует.
Туловище, или торакс, трилобитов состоит из коротких подвижно
сочлененных гомономных сегментов, покрытых с дорзальной стороны
тергитом. В каждом сегменте выделяется средняя часть, или осевое,
кольцо — рахис, и две боковые части, или плевры. Соседние сег-
233
менты либо просто перекрывают друг друга, либо сочленяются особым
замком; при жизни все сегменты туловища были подвижно соединены
друг с другом, что было важно при передвижении и особенно при свер-
тывании трилобитов. Количество туловищных сегментов варьировало у
трилобитов от 2 (у агносцитид) до 44 (у оленеллид).
Хвостовой отдел, или пигидий, за исключением оленеллид, имею-
щих один хвостовой сегмент, состоит из нескольких сегментов, число
которых может превышать 30 (тринуклеины). Часто тергиты хвостово-
го отдела сливаются вместе, образуя хвостовой щит. Осевая его
часть — рахис — обычно заостряется к заднему краю. Наружный край
хвостового щита может быть ровным или снабжен многочисленными
шипами, из которых бывает развит или один непарный, или одна пара
передних. Хвостовой отдел может быть больше или меньше головного
отдела либо быть равным ему. В соответствии с этим выделяют три-
лобитов крупно-, равно- и малохвостых (макропиги, изопиги и мик-
ропиги).
Конечности. Конечности у трилобитов, как отмечалось выше, были
одеты в хитиновый чехол, слабо пропитанный карбонатом, и поэтому в
ископаемом состоянии трилобиты с конёчностями встречаются сравни-
тельно редко. Тем не менее к настоящему времени известны конечности
у 19 видов (из 10000 известных!). На нижней стороне головного щита
около гипостомы к вентральной мембране прикрепляется пара антенн;
антенны многочлениковые (иногда до 40—50 члеников), одноветвис-
тые, разной длины, обычно направлены вперед. Все остальные конеч-
ности двухветвистые (рис. 118, 5). Каждая конечность состоит из ос-
нования (протоподита) и отходящих от него двух ветвей: наруж-
ной (преэпиподит) многочленистой оси, несущей жаберные листоч-
ки, и внутренней (телоподит), или ходильной ноги, состоящей из
восьми члеников. Последний членик имеет вид крючкообразного шипи-
ка; протоподит обычно состоит из двух члеников: прекоксы и коксы
(лат. соха — бедро). Преэпиподит прикреплялся к прекоксе и у живого
трилобита занимал дорзальное положение, был снабжен жаберными
листочками и выполнял функцию газообмена и, возможно, служил для
плавания. Жаберные листочки у разных видов были неодинаковой дли-
ны, но сходные по строению. Некоторые считают, что преэпиподит не
ветвь конечности, а жабра. Телоподит прикреплялся к коксе, был всег-
да длиннее преэпиподита и служил для ползания по дну моря. Следы
ползания трилобитов иногда сохраняются на тонких глинистых осадках.
Конечности головного щита служили для захвата пищи и ее из-
мельчения. Они приводились в движение системой мускулов, которые,
однако, в ископаемом состоянии не сохранились; но на глабелле и
около нее внутрь панциря вдаются отростки, к которым, по-видимому,
прикреплялись мышцы парных конечностей и антенн. Подобные от-
ростки, расположенные на осевой части, имеются на тергитах тулови-
ща и хвостового щита. У одного вида (Olenoides serratus) из среднего
кембрия Канады обнаружена одна пара неветвистых конечностей, при-
крепленных к последнему хвостовому сегменту (они называются
церки).
Строение мягкого тела. О строении мягкого тела и расположении
внутренних органов у трилобитов имеются весьма скудные сведения
(см. рис. 117). Выше уже говорилось о наличии глаз и системы муску-
лов, приводивших в движение конечности. Некоторые данные имеются
о пищеварительной системе. Рот располагался на вентральной сторо-
не головного щита и был ограничен спереди и сзади двумя табличка-
ми •— гипостомой и метастомой. Пища доставлялась с помощью двух-
234
ветвистых малоизмененных конечностей. Пища поступала через корот-
кий пищевод в желудок овальной формы, расположенный под глабел-
лой. Пиритизированные слепки полости желудка сохранились у двух
экземпляров трилобитов из ордовика Богемии и были впервые описаны
Баррандом (1852). От желудка в сторону щек отходили так называе-
мые «печеночные отростки», оставившие отпечатки в виде разветвлен-
ных тонких морщин. Желудок переходил в длинную кишку, которая
тянулась под рахисом туловища и хвостового щита. Описываемый в
литературе многочленистый орган, лежащий над пищеварительным
трактом и долгое время принимавшийся за многочленистое сердце, при
дополнительной проверке оказался частью членистой руки криноидеи,
каким-то образом попавшей под скелет трилобита. К сожалению, ни-
чего не известно ни о нервной, ни о половой, ни о выделительной си-
стемах трилобитов.
Размножение и развитие. Все трилобиты раздельнополые; в настоя-
щее время-'имеются данные о половом диморфизме; описаны пары «уз-
Рис. 120. Стадии онтогенетического развития трилобитов:
1 — Sao hirsuta: la, б — протаспис, 1в, г — мераспис; 2 — Paradoxides pinus:
2а — протаспис, 26, в — мераспис; 3 — Olenus, стадии протаспис
ких» и «широких» скелетов, которые предположительно принадлежали
соответственно самцам и самкам и отличались также некоторыми
особенностями скульптуры. Развитие трилобита начиналось в яйцевых
оболочках, и об этом эмбриональном развитии ничего не известно.
Правда, в слоях, заключающих скелеты трилобитов, иногда встречают-
ся известковые тельца размером 0,6—4 мм, которые, возможно, пред-
ставляют собой остатки яиц трилобитов.
. В постэмбриональном развитии различают три стадии (рис. 120):
1) протаспис, 2) мераспис и 3) голаспис. Первая личиночная стадия —
протаспис (гр. protos — первый, aspis — щит) охватывает отрезок
развития от выхода из яйца до обособления головного отдела от хвос-
тового. В головном отделе видны 5 сегментов. На переднем краю с
каждой новой линькой начинают обособляться глаза, появляются лице-
вые швы, щечные шипы; размер протасписа от 0,25 до 1 мм. В течение
следующей личиночной стадии — мераспис (гр. meros — часть) —
между головным и хвостовым отделами после каждой линьки возникал
235
новый туловищный сегмент (новая часть тела) и тело увеличивалось в
размерах в 6—12 раз, увеличивались размеры и очертания глабеллыг
изменялось положение глаз (от вентрального на дорзальное), глазных
валиков, лицевых швов, изменялись размеры шипов: одни увеличива-
лись, другие, наоборот, уменьшались. Стадия голаспис («полный
щит») начиналась после формирования последнего туловищного сег-
мента и продолжалась до конца жизни индивида, сопровождаясь линь-
кой и увеличением размеров тела. Число линек установить очень труд-
но; по данным некоторых исследователей, оно могло достигать 30.
Сходство личинки современных мечехвостов с трилобитами указывает
на возможность возникновения хелицеровых от трилобитоморф. Об
этом свидетельствуют также находки в отложениях венда Эдиакары ос-,
татков членистоногих, несущих черты сходства с трилобитами и хелице-
ровыми.
Основы систематики и классификация. На основании количества
туловищных сегментов трилобиты разделены на два подкласса: Miome-
га (2—3 туловищных сегмента) и Polymera (число туловищных сегмен-
тов от 5 и выше).
ПОДКЛАСС MIOMERA. МИОМЕРЫ
Подкласс миомер, или малочленистых (гр. melon -- меньше, мень-
ший), объединяет мелких трилобитов, у которых головной и хвостовой
Рис. 121. Отряды Eodiscida (У) и Agno-
stida (2):
1 — Serrodiscus (ранний кембрий); 2 —
Agnostus (поздний кембрий)
большое число родов (13) ранне- и
битов с 2—3 туловищными сегментами,
щиты почти равных размеров;
туловище состоит из 2 или 3 сег-
ментов (рис. 121). Глаза обыч-
но отсутствуют, однако у некото-
рых имеются глаза и лицевые
швы. Кембрий — ордовик. 2 от-
ряда: Agnostida и Eodiscida.
Отряд Agnostida. Агностиды
(гр. agndstos — непознаваемый)
объединяет мелких трилобитов,
у которых 2 туловищных сегмен-
та; головной и хвостовой щиты
почти равных размеров (изопи-
ги); глаза и лицевые швы от-
сутствуют. Рахис хвостового щи-
та с 3 или меньшим числом ко-
лец. Ранний кембрий — поздний
ордовик. Около 70 родов
(рис. 121, 2).
Отряд Eodiscida. Эодисциды
(rp..eos — восток, заря, утро,
discos — диск) объединяют не-
среднекембрийских мелких трило-
с головным щитом, равным
хвостовому (изопиги), со следами сегментации. Иногда имеются глаза:
с пропариевым лицевым швом (рис. 121, 1).
ПОДКЛАСС POLYMERA. ПОЛИМЕРЫ
К подклассу полимер, или многочленистых (гр. poly — много), от-
носятся все остальные трилобиты с числом туловищных сегментов от
5 и выше (трилобиты с 4 туловищными сегментами неизвестны). Под-
236
класс объединяет свыше 1400 родов. На основании особенностей строе-
ния головного и хвостового отделов, характера лицевых швов подкласс
разделен на 7 отрядов: Olenellida, Redlichiida, Corynexochida, Ptycho-
pariida, Phacopida, Lichida, Odontopleurida.
Отряд Olenellida. Оленеллиды (Olenus—один из героев греческой
мифологии, превращенный в камень) — наиболее примитивные много-
членистые трилобиты. Тело длинное, уплощенное; головной щит отно-
сительно крупный; щеки снабжены щечными шипами; глабелла с ясно
выраженными бороздами. Глаза, как правило, крупные, с глазными
Рис. 122. Отряды Olenellida (/), Redlichiida (2) и Corynexochida (3):
/ — Olenellus (ранний кембрий); 2 — Paradoxid.es (средний кембрий); 3 — Edel-
steinaspis (ранний кембрий)
крышками; лицевые швы метапариевого типа; передняя ветвь направ-
лена к щечному углу. Туловище состоит из непостоянного, но обычно
большого числа (12—44) сегментов, несущих сильные борозды и шипы,
а третий сегмент часто увеличен и несет особенно крупный шип. Хвос-
товой отдел очень маленький, нерасчлененный, почти целиком занят
осевой частью или шиповидным тельсоном. Ранний кембрий. Около
20 родов (рис. 122, 1}.
Отряд Redlichiida. Редлихииды (названы в честь К. Редлиха, опи-
савшего фауну кембрийских трилобитов Индии в 1901 г.) близки к
предыдущему отряду, отличаются крупными глазами с заднещечными
лицевыми швами. Глабелла четко отделена от щек; подвижные щеки
с длинными щечными шипами. Туловище состоит из многочисленных
сегментов (15—25); хвостовой отдел маленький. Ранний и средний
кембрий. Около 90 родов (рис. 122, 2).
Отряд Corynexochida. Коринексохиды (гр. koryne — булава, ехо-
chos — высокий, выдающийся). Тело удлиненное, овальное, головной
щит округленный, с хорошо развитыми щечными шипами; глабелла
длинная, с четкими бороздами, расширяется кпереди. Глаза меньших
размеров, чем у предыдущих отрядов, узкие, с глазными валиками.
Лицевые швы заднещечные. Ростральная пластинка соединена с гипо-
стомой или рудиментарна. Туловище состоит из небольшого числа
(5—12) сегментов. Хвостовой щит крупных (макропиги) или средних
размеров с краевыми шипами. Ранний — поздний кембрий. Около 75
родов (рис. 122, 3).
237
Отряд Ptychopariida. Птихопарииды (гр. ptyche — складка, pareia—
щека). Глабелла разнообразной формы; глаза от средних до малень-
ких, иногда отсутствуют; лицевые швы преимущественно опистопарие-
вые, реже пропариевые; у некоторых занимают краевое положение. Ги-
постома отделена от цефалона гипостомальным швом. Туловище состо-
ит из 6—25 сегментов, у многих групп их число постоянно. Хвостовой
щит хорошо обособлен и состоит из небольшого числа сегментов. По-
явились в раннем кембрии от общих предков с редлихиидами, достигли,
расцвета в позднем кембрии и ордовике и вымерли в перми. Отряд
объединяет до 800 родов, разделенных на пять подотрядов: Ptychopa-
riina, Asaphina, Illaenina, Harpina, Trinucleina.
У подотряда Ptychopariina простая, суживающаяся вперед глабел-
ла, разделена простыми параллельными бороздами (3—4); глаза сред-
ние или маленькие, лицевые швы заднещечные; относительно большое
туловище и маленький хвостовой отдел. Имеется фронтальный лимб.
Кембрий — ордовик. Около 500 родов (рис. 123, 1,2).
У подотряда Asaphina (гр. asaphus—неясный) головной и хвостовой
отделы почти одинаковых размеров (изопиги), туловище состоит из
6—9 сегментов. Глабелла слабо очерченная, почти без борозд; глаза на
стебельках или полулунные, иногда очень крупные; лицевые швы зад-
нещечные. Обладали способностью сворачиваться. Конец среднего кем-
брия — поздний ордовик, расцвет в раннем и среднем ордовике. Около,
110 родов (рис. 123, 3).
У подотряда Illaenina (лат. illaino — косоглазый) головной и хвос-
товой щиты почти равных размеров (изопиги); глаза полулунной фор-
мы; лицевые швы заднещечные; глабелла выражена слабо; имеется
крупная ростральная табличка. Туловище из 6—10 сегментов. Возник-
нув в ордовике, этот подотряд пережил всех своих конкурентов и вы-
мер в перми. Около 150 родов (рис. 123, 4, 5).
У подотряда Harpina (гр. harpe — серп) головной щит округленный,,
с большими удлинениями щек (щечными рогами); глабелла суживает-
ся вперед, с 1—3 парами борозд; глаза развиты слабо или отсутствуют;
лицевые швы краевые, реже заднещечные; туловище с 12—29 сегмен-
тами; хвостовой щит короткий. Поздний кембрий — поздний девон.
Около 20 родов (рис. 123, 7).
У подотряда Trinucleina (гр. tri—три, nucleus — ядро) головной
щит от округлой до треугольной формы, в несколько раз больше хвос-
тового, окружен широким лимбом или краевой каймой. Глабелла и ще-
ки выпуклые, с длинными щечными углами. Глаза имеются или отсут-
ствуют; лицевые швы заднещечные или краевые. Туловище из 5—7 сег-
ментов, редко до 30. Рахис узкий, плевры широкие. Хвостовой щит
треугольный, узкий, рахис достигает конца щита; иногда сохраняются
следы мест прикрепления мускулов. Ранний ордовик — поздний силур.
Около 50 родов (рис. 123, 6).
Отряд Phacopida. Факопиды (гр. phakos—чечевица, ops, род. opos—
глаз) — послекембрийские трилобиты мелких и средних размеров. Гог
ловной щит с крупной расширяющейся кпереди глабеллой; глаза ши-
зохроические. Лицевые швы пропариевые или гонатопариевые, реже
опистопариевые; туловище состоит из 8—19 сегментов. Хвостовой щит
различных очертаний, чаще средних размеров, несет срединные или
краевые шипы. Ордовик — девон. Около 180 родов (рис. 124).
Отряд Lichida. Лихиды (гр. Lichas — имя слуги Геракла, доставив,-
шего ему отравленный плащ) — заднещечные послекембрийские три-
лобиты от средних до очень крупных размеров. Головной и хвостовой
238
Рис. 123. Отряд Ptychopariida:
1—2 — подотряд Ptychopariina: 1 — Ptychoparia (средний кембрий), 2 — Olenus
(поздний кембрий); 3 — подотряд Asaphina, Asaphus (ранний — средний ордо-
вик); 4—5 — подотряд Illaenina: 4 — Illaenus (ордовик), 5 — Phillipsia (кар-
бон); 6 — подотряд Trinucleina, Опта (ордовик); 7 — подотряд Harpina, Har-
pes (средний девон)
Рис. 124. Отряд Phacopida: •
1 — Phacops (силур — девон); 2 — Cheirurus (поздний ордовик — средний девон);
3 — Deiphon (силур); 4 — Dalmanites (силур — ранний девон)
щиты крупные и очень своеобразные. Глабелла широкая; лицевые швы
опистопариевые. Хвостовой щит крупный, его плевральные части лис-
товидной формы или шиповидные. Дорзальная поверхность бугорчатая.
Ранний ордовик — поздний силур. 25 родов, (рис. 125, 1, 2).
Рис. 125. Отряды Lichida (/—2) и Odontopleurida (3):
1 — Dicranopeltis (поздний ордовик — силур); 2 — Terataspis (девон); 3 — Odonto-
pleura (средний силур)
Отряд Odontopleurida. Одонтоплевриды (гр- odous, род odontos —
зуб, pleura — бок). Мелкие трилобиты с выпуклым головным щитом,
глабелла конической формы, с расширенной частью у затылочного
кольца и часто с 2—3 парами боковых борозд. Глаза расположены по-
средине щек; лицевые швы опистопариевые. Туловище из 8—10 сег-
ментов, причем осевая часть вздутая, выпуклая; плевральные части
уплощенные. Хвостовой щит короткий, его рахис с 2—3 бороздами.
Панцирь этих трилобитов имеет большое количество шипов на окон-
чаниях почти всех сегментов (отсюда название отряда). Конец сред-
него кембрия — поздний девон, расцвет в конце силура — начале де-
вона. Около 25 родов (рис. 125, 3).
Образ жизни и условия захоронения. Все трилобиты были морски-
ми организмами, так как их остатки встречаются только в отложени-
ях морского происхождения совместно с кораллами, брахиоподами,
моллюсками, мшанками, граптолитами. Личинки трилобитов протас-
пис вели, скорее всего, планктонный образ жизни, что обеспечивало им
широкое географическое расселение и адаптацию к разным экологиче-
ским нишам. После превращения в личинку типа мераспис трилобиты,
по-видимому, переходили к бентосному образу жизни (рис. 126). На-
ходки трилобитов в осадках, характерных для мелководных участков
моря, — в тонких песках, глинах, глинистых и известковистых илах —
позволяют предполагать, что трилобиты ползали по дну с помощью
своих телоподитов — внутренних ветвей конечностей — и, возможно,
плавали у дна, используя наружные ветви конечностей — преэпиподи-
ты. Часть трилобитов зарывалась в ил, выставляя наружу глаза на
стебельках, а формы, вторично утратившие зрение, всю свою жизнь
проводили в илу, питаясь органическим детритом и мелкими организ-
мами, населявшими верхние части илистого осадка.
240
О приспособлений к жизни на дне свидетельствует уплощенное,
широкое тело, глаза на верхней стороне головного отдела, положение
рта на нижней стороне, тонкая мембрана, прикрывавшая брюшную
сторону, отсутствие специальных органов управления, возникающих у
Рис. 126. Предполагаемый образ жизни Трилобитов (1—5 — пелагический, 6—7 — бен-
тосный, 8—10 — типы свертывания):
1 — личинка Acantholoma-, 2 — Lonchodomas-, 3 — Symphysops; 4 — Staurocephalus;
5 — Ceratarges; 6 — Teratorhynchus; 7 — Paraharpes.
Типы свертывания: 8 — дискоидальный, 9 — двойной, 10 — сфероидальный; головной
Щит обозначен редкими точками, туловище — частыми, хвостовой щит — зачернен
всех быстро плавающих форм. Бентосный образ жизни подтверждает-
ся также приуроченностью трилобитов только к определенным фациям.
Дополнительными приспособлениями для жизни на илистом дне слу-
жили уплощения края головного щита — лимб и длинные шиповидные
отростки щек, а также различные шиповидные отростки различных
сегментов. Наличие шиповидного тельсона, подобного таковому мече-
хвостов, помогало этим трилобитам зарываться в осадок в поисках пи-
щи. Мелкие трилобиты с большим количеством шипов и различных-
выростов, возможно, могли вести пелагический образ жизни. Об этом
свидетельствуют также гипертрофированные глаза, иногда целиком за-
нимающие поверхность щек (рис. 126, 3).
Для защиты вентральной стороны у многих трилобитов выработа-
лась способность к свертыванию тела подобно хитонам или наземным
ежам. Различают три основных способа свертывания (рис. 126,8—10):
1) при сфероидальном свертывании, наиболее распрост-
раненном у трилобитов, происходило смыкание, хвостового и головного
отделов, причем плевры туловища накладывались друг на друга и за-
крывали тело с боков, защищая конечности. Приспособлением для
плотного смыкания служили так называемые («пандеровы органы». Для
прочного смыкания на дублюре переднего края головного щита разви-
валась либо борозда (факопиды), либо система зубчиков. В борозду
16 в. В. Друщиц
241
при свертывании упирался задний конец пигидия. «Пандеровы органы»
в действительности представляют структуры наружного скелета: бу-
горки или валики, расположенные на дублюре щечных углов и на дуб-
люре каждой плевры туловища. Они служили упорами для плевр при.
свертывании тела трилобита. Отверстия, расположенные за бугорками,,
относимые также к категории «пандеровых органов», служили, по-ви-
димому, для просачивания воды к жаберным ножкам, лежащим внутри,
свернутого панциря;
2) при двойном свертывании хвостовой отдел и несколько’
туловищных сегментов подгибались и затем происходило свертывание,,
при котором передний край головы соединялся с одним из тех туловищ-
ных сегментов, где происходил изгиб; пигидий и часть туловищных сег-
ментов оказывались.внутри свернувшегося трилобита;
3) при дискоидальном свертывании передний край голо-
вы соприкасался с задним краем хвостового отдела и трилобит оказы-
вался согнутым, как книга.
Способность к свертыванию возникала у разных групп трилобитов,
независимо, как защитная реакция от врагов, которых в морях палео-
зоя у трилобитов было более чем достаточно. К числу врагов могли от-
носиться остракодермы и рыбы, другие группы членистоногих (некото-
рые водные хелицеровые), головоногие моллюски: наутилоидеи, актино-
цератиты, эндоцератиты. О нападении врагов свидетельствуют различ-
ные виды травм на наружном скелете, залеченные при жизни.
В ископаемом состоянии сохраняются как скелеты погибших от раз-
ных причин трилобитов, так и скелеты, сброшенные при линьке. Боль-
шинство остатков трилобитов, найденных в ископаемом состоянии, при-
надлежит скелетам, сброшенным при линьке. Различные швы, в первую»
очередь лицевые швы, затем швы между головой и туловищем, облег-
чали трилобитам линьку. У трилобитов без лицевых швов панцирь раз-
рывался между головой й туловищем и в ископаемом состоянии йногда>
встречаются туловище с хвостом и лежащий рядом головной щит, часто»
перевернутый. При наличии лицевых швов чаще всего сохраняется кра-
нидий без подвижных щек. В бургасских сланцах среднего кембриж
Британской Колумбии (Канада) найдены остатки разнообразных чле-
нистоногих, в том числе трилобитид и трилобитов с сохранившимися Ос-
татками конечностей. Хорошо сохранились трилобиты в ордовикских из-
вестняках окрестностей Нью-Йорка и восточной части Канады.
История развития трилобитов (рис. 127). Первые трилобиты из-
вестны из алданского яруса нижнего кембрия. В отложениях самого»
нижнего яруса кембрия — томмотского — остатки трилобитов отсутству-
ют. Для расчленения кембрийских отложений используются почти ис-
ключительно трилобиты, достигавшие в кембрии большого разнообразия
и изобилия. Трилобиты используются для выделения биогеографических
областей и позволяют среди отложений кембрийской системы выделить-
отделы, ярусы и даже зоны. В раннем кембрии известны представители,
обоих подклассов: миомеры и полимеры. Среди первых главную роль,
играли Eodiscida, среди вторых Olenellida, Redlichiida, Corynexochida и
первые Ptychopariida. На рубеже раннего и среднего кембрия-полностью-
вымирают Olenellida и появляются Odontopleurida и в конце среднего»
кембрия первые Asaphina. В среднем кембрии становятся многочислен-
ными Agnostida, Corynexochida, Paradoxidacea из Redlichiida, ряд се-
мейств-из подотряда Ptychopariina.
На рубеже среднего и позднего кембрия вымирают Eodiscida, Red-
lichiida, появляются Harpina, в позднем кембрии становятся разнооб-
разными Agnostida, Ptychopariina.
242
На рубеже кембрия и ордовика весьма существенно изменился си-
стематический состав трилобитов, их значение для стратиграфии не-
сколько уменьшилось и ведущую роль стали играть граптолиты. Выми-
рают Corynexochida. От птихопариид возникают два новых специализи-
рованных отряда: Phacopida и Lichida. В пределах отряда птихопариид
обособляются два новых подотряда: Illaenina и Trinucleina, испытывают
расцвет Asaphina. В конце ордовика вымирают Agnostida и подотряды
Asaphina и Ptychopariina и резко сокращается общее число родов всех
трилобитов.
В силуре значение трилобитов для стратиграфии еще уменьшилось;
тем не менее среди трилобитов были распространены представители
Phacopida, Harpina, Odontopleurida. В конце силура вымирают Trinuc-
leina.
В девоне продолжали свое существование Harpina, Phacopida, Li-
chida и Odontopleurida, вымирающие в конце периода. Только последние
представители подотряда Illaenina продолжали существовать в морях
карбона и перми, причем до перми дожили только два семейства этого
подотряда. В середине пермского периода, насколько можно судить по
находкам, исчезают последние представители трилобитов.
В кембрийское время были обособлены три зоогеографические об-
ласти: Атлантическая (Прибалтика, Скандинавия, Англия, Марокко,
восточное побережье Канады), Тихоокеанская (вдоль западного побе-
режья Северной Америки от Аляски до Мексики), Восточно-Азиатская,
16;
243
или краевая Тихоокеанская (от Кореи до Австралии, захватывая Ин-
дию). В . каждой из областей были распространены свои характерные
трилобиты. .
ПОДТИП CHELICERATA. ХЕЛИЦЕРОВЫЕ
Общая характеристика. Хелицеровые, или клешнеусовые, — преиму-
щественно наземные членистоногие (свыше 35000 видов), населяют са-
мые разнообразные биотопы суши и дышат с помощью легких или тра-
хей. Они произошли от трилобитов и в раннем палеозое населяли моря,,
дышали жабрами; в девоне перешли к жизни на суше. Их немногочис-
ленные водные потомки (5 видов) — мечехвосты, настоящие «живые
ископаемые», сохранились в разорванных ареалах современных тропи-
ческих морей. К хелицеровым относятся кроме мечехвостов скорпионы,
фаланги, пауки, клещи (рис. 128). Тело хелицеровых отчетливо разделе-'
но на два отдела — просому и опистосому. Просома состоит из акрона
и шести сегментов, несущих 6 пар конечностей; антенны отсутствуют,
первая пара, конечностей — хелицеры, вторая — педипальпы, осталь-
ные 4 пары — ходильные ноги. Опистосома разделена на две части: ши-
рокую мезосому и узкую метасому; мезосома несет жаберные ножки,
преобразованные у наземных форм в органы воздушного дыхания, ме-
тасома лишена конечностей. В ископаемом состоянии морские хелице-
ровые известны с кембрия, наземные с девона.
Строение тела. Просома состоит из акрона и шести сегментов,
из. которых четыре отвечают четырем ларвальным сегментам личинки
(протаспис) трилобитов. Просома покрыта сверху дорзальным щитом,
а снизу — стернальным. Дорзальный щит состоит из слившихся тергитов
передних сегментов; он может быть разделен на две части — переднюю
и заднюю; у одних хелицеровых передняя часть гомологична головному
щиту трилобитов, у других (клещей) она состоит из двух сегментов и
негомологична голове. Задняя часть дорзального щита построена из раз-
личного числа сегментов. Опистосома у скорпионов и эвриптерид
образована 12 сегментами и анальной лопастью, или тельсоном; она
разделена на широкую мезосому, состоящую из 6 сегментов, и узкую
метасому, также включающую 6 сегментов; у многих представителей
метасома в той или иной степени редуцируется и может полностью от-
сутствовать (пауки, клещи), иногда она частично сливается с тельсоном
в длинную хвостовую иглу. Просома и опистосома соединяются с по-
мощью предполовогО .(седьмого) сегмента, лишенного конечностей и
обычно недоразвитого (рис. 128); соединение может происходить по всей
ширине сегментов или посредством узкого стебелька. У многих пауков и
клещей сегментация брюшка исчезает в результате слияния сегментов.
Акрон, или головная лопасть, гомологичен головной лопасти аннелид;
он несет глаза и подротовой вырост — эпистом с верхней губой. Ан-
тенны отсутствуют, отсюда второе название — безусиковые чле-
нистоногие. Первый сегмент просомы несет хелицеры, или щу-
пальцежвалы, снабженные клешнями или крючками, они служат для
захвата пищи; вторая пара конечностей'—педипальпы, или ного-
щупальца; каждое ногощупальце состоит из основного членика — кок-
сы, снабженной коксэндитом — жевательным отростком, и членис-
того щупальца, которое иногда также заканчивается клешней. Педи-
пальпы служат для захвата добычи, осязания или передвижения; их
•основные членики с эндитами способствуют приему пищи. Педипальпы—
гомологи мандибул ракообразных и насекомых.
244
Остальные четыре пары конечностей просомы — ходильные ноги;
они служат для передвижения, хотя иногда первая пара отличается от
остальных и выполняет функцию осязания. У клещей одна или две пары
задних ходильных'ног могут быть редуцированы. Обычно ходильные но-
ги состоят из тазика, или коксы, слитого с покровом тела, и свободной
конечности — эндоподита, который расчленяется на вертлуг, бедро, ко-
лено, голень и лапку; лапка заканчивается коготком (см. рис. 112). Чле-
ники ног часто несут органы чувств. Первый сегмент мезосомы (вось-
245
мой от головы) — половой; в нем расположено половое отверстие, а ко-
нечности преобразованы в половые крышечки. Конечности следующих
пяти сегментов мезосомы у первичноводных хелицеровых (мечехвосты,
эвриптериды) представлены жаберными ножками; у наземных — пар-
ными легочными мешками от четырех пар (скорпионы) до одной пары
(пауки), причем у пауков задние пары конечностей мезосомы преобра-
зованы в Две пары паутинных бородавок. Мезосома с брюшной сторо-
ны у наземных форм также имеет стернальный скелет, образованный за
счет слияния коксальных элементов конечностей.
Нервная система построена по общему типу брюшной цепочки,
характерной для членистоногих, но в процессе эволюции наблюдается
постепенное слияние ганглиев: у современных мечехвостов ганглии про-
сомы слиты в одну нервную массу; у скорпионов ганглии просомы сли-
ты с 4 ганглиями мезосомы; у остальных хелицеровых все ганглии сли-
ваются в единый «мозг».
Пищеварительная система сохраняет общий тип кифечника членис-
тоногих и имеет печень, в которой происходит внутриклеточное перева-
ривание пищи. • Имеются примитивные органы выделения, напоминаю-
щие целомодукты аннелид. ; :
Все хелицеровые раздельнополые, но половой диморфизм прояв-
ляется нерезко, в основном в размерах тела и строении отдельных орга-
нов. Среди хелицеровых есть живородящие, но, как правило, почти все
самки хелицеровых откладывают яйца, из которых появляются особи,
по своей организации сходные со взрослыми, так^как послезародыше-
вое развитие протекает без метаморфоза (за исключением клещей), со-
провождаясь ростом и линькой. !
Основы систематики й классификация. Хелицеровые до последнего
времени разделялись на два или три класса; обычно выделялись два
класса: водные хелицеровые — меростомовые и наземные хелицеровые—
паукообразные. Некоторые исследователи разделяют меростомат на два
самостоятельных класса: мечехвостовых и эвриптеридовых. В настоя-
щей работе принимается классификация хедицеровых, предложенная
В. Б. Дубининым (1959), основанная на эмбриологических и сравнитель-
но-анатомических данных, характере слияния сегментов тела и обособ-
лении соответствующих его отделов, особенностях строения Конечностей.
Хелицеровые разделены на пять классов: Merostomata, Scorpioiiomorpha,
Arachnida, Solifugomorpha, Acaromorpha.
КЛАСС MEROSTOMATA. МЕРОСТОМОВЫЕ
Общая характеристика. Меростомовые — древнейшие водные хели-
церовые, дышащие жабрами. Они объединяют вымерших палеозойских
эвриптерид и мечехвостов, известных с силура до наших дней. Тело от
1—3 до 10—20 см, иногда до 100—180 см. Просома округлого или шле-
мообразного очертания; на ней расположены простые и сложные глаза
и шесть пар конечностей: клешневидные хелицеры разной формы и
строения, педипальпы и четыре пары ходильных ног. Опистосома обыч-
но отделена от просомы сегментом, в той или иной степени редуциро-
ванным; опистосома состоит из разного числа сегментов. Тельсон имеет
вид шипа или плавника. В онтогенезе современные мечехвосты прохо-
дят стадию «трилобитовой» личинки, что указывает на их общность про-
исхождения. Обитатели морских, солоноватоводных и преснбвддных бас-
сейнов. В ископаемом состоянии известны Начиная с кембрия.
Класс мсростомовых разделяется на два подкласса: Xiphosura и
Euryplcridoidca. " .А ’ : ' .: 1
246
ПОДКЛАСС XIPHOSURA. МЕЧЕХВОСТОВЫЕ
К подклассу мечехвостовых (гр. xiphos — меч, ига — хвост) отно-
сятся водные хелицеровые с хорошо обособленными просомой и описто-
сомой, неодинакового строения; опистосома у древних представителей
могла состоять из отдельных сегментов, у современных лимулид обра-
зует сплошной щит. На основании особенностей строения тела, ко'личе-
Рис. 129. Подкласс Xiphosura:
1 — отряд Aglaspidida, Aglaspis из позднего кембрия Сев. Америки; 2 — отряд
Chasrriataspidida, Chasmataspis из ордовика Канады; 3 —- отряд Synziphosura,
Weltibergina из раннего девона Зап. Европы (реконструкция); 4 — отряд Li-
mulida: 4а, б — схема строения современного мечехвоста Limulus, 4в — при-
жизненное положение, вид сбоку;
окк — жабры и жаберные крышки, опис — опистосома, пргл простой, глаз,
прос — просома, pi — рот, слгл — сложный глаз, т тельсон, х — ^хелицеры,
хн —ходильные ноги
ства сегментов и степени их слияния друг с другом мечехвостовые раз-
делены на четыре отряда (рис. 129); Aglaspidida (кембрий), Chastna-
taspidida (ордовик), Synziphosura (поздний силур — ранний девон),
Limulida (девон — ныне).
Современные мечехвостовые живут в сублитораЛи тропических и
•субтропических морей, образуя разорванный ареалы; они известны у
берегов Юго-Восточной Азии и прилегающих к ней островов, у атланти-
ческих берегов Северной Америки.:
247
ПОДКЛАСС EURyPTERIDOIDEA. ЭвРИПТЕРИДЫ
Эвриптериды (гр. euris — широкий, pteron — крыло), или гиганто-
страки, объединяют большую группу водных хелицеровых, широко рас-
пространенных от ордовика до перми (рис. 130). Они имели удлиненное
Рис. 130. Отряд Eurypterida:
1 — Eurypterus из среднего силура Эстонии: 1а — вид с брюшной стороны, 16 —
вид со спинной стороны; 2 — Pterygotus из силура Сев. Америки (длина око-
ло 2 м);
м — метасома, нб — нога для балансирования, нп — нога для плавания, опис —
опистосома, пргл — простой глаз,- прос— просома, слгл — сложный глаз, т —
тельсон, х — хелицеры, хн — ходильные ноги, 1—12 — сегменты опистосомы
тело сравнительно небольших размеров (10—20 см); лишь отдельные
их представители достигали 100—180 см. Просома несет простые глаз-
ки, расположенные почти посередине на небольшой пластинке, и боль-
шие сложные боковые глаза. Снизу к просоме прикреплялись клешне-
видные хелицеры, выполнявшие функцию хватающих органов; педиг
пальпы и следующие 4 пары ног служили для передвижения. Ходильные
248
ноги состояли из цилиндрических члеников и обычно заканчивались ши-
пиками, шестая пара иногда видоизменялась в плавательные гребные
ноги (Eurypterus). Опистосома образована 12 подвижными сегментами и
разделена на мезосому и метасому; к последней присоединяется тельсон,
имеющий вид шипа или двух лопастей. Второй сегмент опистосомы нес
генитальные придатки.
Скелетные остатки эвриптерид встречены в континентальных отло-
жениях; возможно, они были обитателями пресных или солоноватовод-
ных бассейнов, преимущественно донными, сравнительно малоподвиж-
ными. Однако некоторые из них были хорошими пловцами. Эвриптери-
ды, по-видимому, произошли от агласпидовых; в свою очередь в силу-
ре от них возникли вначале водные, а позднее наземные скорпионы.
КЛАСС SCORPIONOMORPHA. СКОРПИОНООБРАЗНЫЕ
Общая характеристика. Скорпионообразные (гр. Scorpios — скор-
пион) •— древнейшие наземные хелицеровые, возникшие в начале силу-
ра, от общих предков с эвриптеридами. К ним относятся современные
Рис. 131. Отряды Scorpionida (У) и Pedipalpida (2—7):
1 — Palaeophonus из силура Зап. Европы; 2 — подотряд Amblypygi, Thelyphry-
nus из карбона Сев. Америки, вид с брюшной стороны; 3 — подотряд Pseudoscor-
pionidea, Pseudogarypus из палеогена Европы (балтийский янтарь); 4 — подотряд
Ricinulei, Ricinoides, современный, вид с брюшной стороны; 5 — подотряд Uropygi,
Prdthelgophonus из карбона Сев. Америки, вид с брюшной стороны; 6 — подотряд
P'alpigradi, Sternarthron из юры Зап. Европы; 7 — подотряд Kustarachnida, Kusta-
rachne из карбона Сев. Америки, вид с брюшной стороны
скорпионы и жгутоногие. Тело вытянутое или короткоовальное, разделе-
но на просому, мезосому и метасому; последняя состоит из 5 или 3 сег-
ментов и имеет иногда вид нити. Хелицеры клешневидные, педипальпы
крупные, клешневидные, хватательные или ноговидные-.- Ходильные ноги
тонкие, короткие или длинные. На 8-м сегменте расположено половое
отверстие с двумя генитальными крышками; 4 (скорпионы) или 2 (жгу-
249
тоногие) сегмента мезосомы несут по паре легочных мешков, гомологич-
ных жаберным ножкам эвриптерид. У первых силурийских скорпионов
наличие легких не установлено. Конечности на метасоме отсутствуют.
Брюшная поверхность тела покрыта своеобразными щитками — стер-
нит а м и, . которые, иногда сливаясь, образуют настоящий грудной
щит — стер.нум. Скорпионообразные в настоящее время распростра-
нены во всех частях света. Их первые представители появились в силуре.
Есть предположение, что они были уже обитателями суши, хотя досто-
верные наземные представители известны только с карбона. Класс раз-
делен на два отряда (рис. 131): Scorpionida (силур — ныне) и Pedi-
palpida (карбон •— ныне).
КЛАСС ARACHNIDA. ПАУКООБРАЗНЫЕ
Наземные хелицеровые, насчитывающие свыше 21 000 видов, имею-
щие тело размером от 0,8 до 10 см, реже до 20 см, расчлененное на
два отдела: просому и опистосому. Тело состоит из .16—18 сегментов,
причем в состав просомы входят акрон и 6 сегментов. Просома сверху
покрыта головогрудным, или просомным, а снизу стернальным щитом.
Глаза, если имеются, расположены на верхнем щите. Хелицеры клешне-
видные или крючковидные, часто с протоками ядовитых желез;- педи-
пальпы простые, похожи на ходильные ноги. Пауки имеют 4 пары хо-
дильных ног; ноги длинные или короткие, состоят из 6 основных члени-
ков (тазик, вертлуг, бедро, чашечка, голень и лапка) и заканчиваются
у современных двумя коготками. Опистосома, или брюшко, соединяет-
ся с просомой или по всей ширине сегмента (7-го), или узким стебель-
ком — видоизмененным предполовым сегментом. Опистосома состоит
из 9—12 сегментов, в разной степени слившихся друг с другом. На ней
расположены половые отверстия и половые крышки (сохраняются не
всегда), отверстия легочных мешков и паутинные бородавки.
Легочные мешки представляют собой преобразованные жабер-
ные ножки 8-го и 9-го сегментов водных форм. Примитивные пауки име-
ют две пары легочных мешков (четырехлегочные пауки); у боль-
шинства пауков вместо второй пары легочных мешков развиваются тра-
хеи (двулегочные пауки); у небольшой части пауков обе пары ле-
гочных мешков замещаются трахеями (безлегочные пауки). Трахеи —
новообразования у наземных форм — имеют вид простых или сложно
разветвленных трубочек, по которым воздух доставляется непосредст-
венно к органам или тканям. Трахеи дополняют или полностью заме-
щают легочные мешки; они открываются отверстиями — стигмами,
или дыхальцами, — на нижней поверхности брюшка. Стенки трахей об-
разованы продолжением наружного покрова и выстланы кутикулой. Как
известно, строение дыхательной системы связано с кровеносной; у форм
с легочным дыханием кровеносная система развита хорошо, у форм с
трахейным дыханием она менее развита.
Одной из характерных особенностей паукообразных является спо-
собность пауков вырабатывать паутину. Их брюшко превращено в свое-
образную фабрику паутины, вырабатываемой особыми паутинными же-
лезами. Паутина по химическому составу близка к шелку шелкопрядов;
ее основу составляет протеид — фиброин. Паутина выводится наружу по
тонким хитиновым трубочкам, расположенным внутри паутинных боро-
давок. Различают наружные паутинные бородавки, возникшие за счет
видоизменения брюшных ножек 10-го и 11-го сегментов, и внутренние,
образующиеся как выросты внутренней стенки тела. У современных па-
уков число паутинных бородавок варьирует от четырех пар до одной.
250
Выработка; паутины, сыграла важную роль в эволюции паукообразных,
поскольку современные паукообразные во всех своих жизненных отправ-
лениях — добывании пищи, размножении, расселении, переживании не-
благоприятных условий — пользуются паутиной.
Паукообразные раздельнополые, с хорошо выраженным половым
диморфизмом, внутренним оплодотворением, прямым развитием. Яйца
откладываются в особый кокон, сплетенный из паутины.
Основы систематики и классификация. По мнению большинства ис-
следователей, паукообразные произошли от общих предков со скорпио-
Рис. 132. Классы Arachnida (1—4), Solifugomorpha (5) „и Acaromorpha (t>):
/—2 — подкласс Araneae: 1 — Misumena из палеогена Европы (балтийский ян-
тарь), общий вид самки, 2 —. Eolathus из палеогена Европы (балтийский ян-
тарь), общий вид самки сбоку; 3 — подкласс Soluta, Cryptomartus из карбона
Зап. Европы; 4 — подкласс Opiliomorphae, Goniotarbus из карбона Зап. Европы;
5 — Protosolpuga из карбона Сев. Америки; 6 — Plategeocranus из палеогена
Европы (балтийский янтарь)
номорфами в девоне и получили широкое развитие в карбоне. По-види-
мому, сразу же возникли три/ветви, которые в настоящее время рас-
сматриваются в качестве отдельных подклассов (рис. 132): Opiliomor-
phae (карбон — ныне), Soluta (девон — пермь) и Araneae (карбон —
ныне).
251
КЛАСС SOLIFUGOMORPHA. СОЛЬПУГОПОДОБНЫЕ
К сольпугоподобным относится один отряд Solifugida (лат. sol —
солнце), объединяющий около 600 видов, современных и ископаемых^,
обитателей степных и пустынных областей тропического, субтропическо-
го и отчасти умеренного поясов. Сольпуги, или фаланги, — довольно
крупные хелицеровые (длина тела от 10 до 70 мм). Передний отдел про-
сомы, покрытый крупным щитом, несет клешневидные крупные хелице-
ры, педипальпы, приспособленные к захвату добычи, и одну пару хо-
дильных ног. Задний отдел просомы состоит из 3 сегментов, несущих
3 пары ходильных ног. Ноги снабжены коготками. Опистосома длинная,
лишенная конечностей, состоит из десяти сегментов с тергитом и стер-
нитом на каждом. Сольпуги дышат трахеями, пронизывающими все те-
ло. Основные трахейные стволы открываются стигмами на 9-м и 11-м
сегментах брюшка.
Класс сольпугоподобных возник от общих предков с педипальпида-
ми (жгутоногими) и образовал обособленную группу хелицеровых, имею-
щую некоторые черты сходства с акароморфами. В ископаемом состоя-
нии известны начиная с карбона (рис. 132, 5).
КЛАСС ACAROMORPHA. АКАРОМОРФЫ, ИЛИ КЛЕЩИ
К классу акароморф, или клещей (гр. акап=лат. acarus — клещ),
относится свыше 10000 видов мелких хелицеровых с размером тела ме-
нее 1 мм, живущих преимущественно в почве, на растениях или парази-
тирующих на животных; небольшая часть приспособилась к жизни в
пресных и морских водах. Сегментация тела выражена, как правило,
слабо. В отличие от всех хелицеровых передние три сегмента обособле-
ны в так называемую ложную головку (гнатосому), состоящую из
акрона и двух сегментов, несущих хелицеры и педипальпы. За ложной
головкой следуют два сегмента, несущие передние две пары ног; очень
часто эти сегменты сливаются с ложной головкой, образуя передний от-
дел тела (протеросому); за ней следуют два сегмента, несущих по-
следние две пары ног, и опистосома, или брюшко, состоящее из 6 слив-
шихся сегментов, реже 7—9 или 10—11. Два сегмента просомы и опис-
тосомы, сливаясь, образуют задний отдел тела (гистеросому). На
голове расположены пара боковых и один срединный глаз.
Опистосома несет особо устроенный половой аппарат, образованный
за счет срастания ножек 8—10-го сегментов. Клещи дышат трахеями
или всей поверхностью тела.. Хитиновые покровы развиты в разной сте-
пени; у одних они кожистые, у других образуют защитные щиты. По-
верхность тела покрыта щетинками, выполняющими функцию осязания,,
механической защиты, хеморецепторов и др.
Клещи раздельнополые, с внутренним оплодотворением. Дробление
яйца напоминает дробление яйца кольчатых червей; зародыш проходит
стадию, сходную с личинкой трилобитов (протаспис); выходящая из яй-
цевых оболочек личинка шестиногая, без трех последних сегментов, пос-
ле линьки приобретает четвертую пару ног и превращается в нимфу;
нимфа после трех линек (иногда одной — Двух), приобретая с каждой
линькой недостающие сегменты брюшка, становится половозрелым-
клещом.
В настоящее время класс акароморф разделен на три отряда; Аса-
riformes (девон — ныне), Parasitiformes (палеоген-—ныне), Opilioacari-
па (в ископаемом состоянии не известны). Первые акароморфы извест-
ны из девонских красных песчаников Шотландии; большое количество-
252
клещей известно из, олигоцена (балтийский янтарь); остатки клещей ча-
сто встречаются в бурых углях. Вероятно, акароморфы были очень ши-
роко распространены в геологическом прошлом, однако в ископаемом
состоянии сохраняются, как и все остальные хелицеровые, сравнительно
редко (рис. 132, 6).
Экология и история развития хелицеровых. Хелицеровые впервые
появляются в геологической истории в начале палеозоя. Они произошли
в раннем кембрии от примитивных трилобитов, на что указывают: го-
мология частей двухветвистых конечностей и жаберных ножек водных
хелицеровых; расположение рта в виде продольной щели между кок-
сальными эндитами ног; соответствие четырех ларвальных сегментов
просомы хелицеровых таковым трилобитов (см. рис. 128). В эмбриоге-
незе мечехвостов наблюдается трилобитоподобный зародыш. В процессе
эволюции у хелицеровых атрофировались органы чувств: исчезли пер-
вая и вторая пары антенн, сложные глаза, произошло объединение сег-
ментов тела в два крупных отдела — просому и опистосому. Однород-
ные конечности предков (трилобитов) изменились: первая пара конеч-
ностей превратилась в хватательные хелицеры, остальные 5 пар — в
двигательно-жевательные конечности. Антенны редуцировались. Конеч-
ности мезосомы сохранили дыхательную функцию, а конечности мета-
сомы атрофировались.
Слияние головы с грудью и образование просомы, по-видимому, бы-
ло связано с приспособлением к роющему образу жизни на мелковод-
ных участках моря и на дне пресноводных бассейнов. Это были крупные
хелицеровые с щитовидной головогрудью, покрытой тяжелым панцирем
(подобно современным мечехвостам), которые отыскивали добычу «на
ощупь» в осадке на дне, захватывали ее хелицерами и измельчали же-
вательными отростками ног. При таком образе жизни усики и глаза
были почти не нужны и становились даже помехой.
От водных меростомат возникли эвриптериды, от которых в начале
девона произошли их наземные родственники, объединяемые в класс
Scorpionomorpha — скорпионообразных, наиболее примитивных водных
и наземных хелицеровых; от скорпиономорф, по-видимому, возникли кле-
щи, пауки и сольпугоморфы. В конце силура и в начале девона клещи
обособились от скорпиономорф. У них первые два сегмента формируют
своеобразный отдел тела, получивший.название гнатосомы, обособ-
ленный от остального тела, которое либо бывает разделено на два или
три отдела, либо состоит из слитых воедино сегментов. Клещи приспо-
собились к самым разнообразным местообитаниям, к жизни в почве, на
разных растениях, вторично перешли к жизни в воде, часть паразити-
рует на животных.
Немного позднее, в середине или конце девона, возник еще один
класс наземных хелицеровых — паукообразные, перешедшие к хищному
способу питания. У паукообразных хорошо обособлена просома, жабер-
ное дыхание сменилось легочным, возникли паутинные бородавки, кото-
рые продуцируют паутину; наличие паутины сыграло важную роль в
эволюции этого класса; паутина служит для устройства коконов и вы-
стилания убежищ, для создания ловчей сети, для переноса ветром мо-
лодых паучков.
В карбоне, по-видимому, от тех же скорпиономорф отделились соль-
пугоморфы — небольшой класс хелицеровых с хорошо развитой трахей-
ной системой, у которых первые три сегмента просомы сливаются вме-
сте, оставляя остальные сегменты тела свободными; педипальпы похо-
жи на ходильные ноги. Сольпугоморфы приспособились к жизни в тро-
пических областях и ведут скрытый образ жизни.
253
ПОДТИП BRANCHIATA. ЖАБРОДЫШАЩИЕ
Подтип Branchiata (гр. branchia — жабры) состоит из одного клас-
са Crustacea (лат. crusta — скорлупа, корка), объединяющего преиму-^
щественно водных, морских и пресноводных членистоногих, которые ды-
шат жабрами, имеют- две пары антенн, личиночную стадию науплиус;
только небольшая часть ракообразных приспособилась к жизни на суше
и имеет органы воздушного дыхания. В ископаемом состоянии извест-
ны начиная с кембрия.
КЛАСС CRUSTACEA. РАКООБРАЗНЫЕ
Общая характеристика. Ракообразные — морские или пресновод-
ные, реже наземные членистоногие (раки, крабы, остракоды, усоногие
Рис. 133. Класс Crustacea:
1 — строение речного рака; 2 — схема строения конечности ракообразных;
3—4 — личинки: 3— науплиус, 4 — зоеа;
ан — анус, пл — плеоподы (конечности брюшка), пр — преэпиподит, про — про-
топодит, р — рот, т — тельсон, ур — уроподы (последние видоизмененные
брюшные конечности), экз — экзоподит, энд — эндоподит, эп —' эпиподит; рим-
ские цифры — номера видимых на рисунке конечностей
раки); дышат жабрами. Тело разделено на голову, грудь и брюшко
(рис. 133). Голова несет две пары усиков (у трилобитов — одну пару!)
и три пары челюстей; на голове помещаются пара сложных глаз и один
простой.
Задний сегмент головы образует складку тела — карапакс, кото-
рый охватывает сверху, и с боков головной и грудной отделы тела. Ка-
254
рапакс имеет форму полуцилиндра, щита или двустворчатой раковины;
у ряда представителей он отсутствует. Грудь и брюшко состоят из раз-
личного числа сегментов; иногда обособлена головогрудь. Брюшко за-
канчивается тельсоном. Грудные ножки выполняют различные функции:
передвижения, дыхания (несут жабры) или захвата пищи. Ракообраз-
ные раздельнополые и гермафродиты. Из яйца развивается личинка —-
науплиус. Кембрий — ныне.
Строение тела. Тело состоит из отдельных сегментов, частично срас-
тающихся между собой (рис. 133, 1). Каждый сегмент несет пару чле-
нистых конечностей. Сегменты тела образуют три отдела, или тагмы: го-
лову (цефалон), грудь (торакс) и брюшко (абдомен). Голова образо-
вана слиянием головной лопасти и пяти следующих за ней сегментов,,
несущих антеннулы и антенны, жвалы, или мандибулы, и две пары че-
люстей, или максилл. Антеннулы и антенны (иногда их называют пе-
редними и задними антеннами) выполняют функции обоняния, осяза-
ния и двигательную. Мандибулы размельчают пищу, максиллы подают
ее ко рту и фильтруют. Непосредственно на голове или на специальных
стебельках расположена пара сложных фасеточных глаз. У личинки —1
науплиуса — имеется один простой глаз, называемый науплиаль-
н ы м, у многих он сохраняется и на взрослой стадии. Каждая конеч-
ность, за исключением антенн, состоит из общего основания — прото-
подита и двух отходящих от него ветвей: наружной — экз оп о ди т а
и внутренней — эндоподита (рис. 133, 2).
Голова и часть грудного отдела у многих ракообразных покрыта
складкой, которая охватывает сверху и с боков головной и весь грудной,
отдел или только его часть. Эта складка пропитана хитином и образует
опорный и защитный скелет — карапакс. Грудной и брюшной отдельГ
состоят из разного числа сегментов; нередко один или несколько груд-
ных сегментов срастаются с головой и образуют головогрудь. Ко-
нечности этих сросшихся грудных сегментов видоизменяются тогда в но-
гочелюсти и принимают участие в захвате пищи. Остальные грудные
ножки выполняют двигательную и дыхательную функции. Каждая нож-
ка снабжена лопастями; некоторые наружные лопасти превращены в
жабры, другие создают ток воды ко рту, третьи служат веслами; у выс-
ших ракообразных эти функции выполняются различными конечностя-
ми. Брюшко состоит из 6—7 сегментов, из которых- обычно первые 5
несут плавательные ножки. Брюшко заканчивается тельсоном, гомоло-
гом анальной лопасти многощетинковых червей.
Нервная система построена по типу брюшной нервной цепочки. Пи-
щеварительная система состоит из кишечника, в котором различают пе-
реднюю, среднюю и заднюю кишки. Передняя состоит из короткого пи-
щевода, который переходит в желудок; у высших ракообразных он раз-
делен на два отдела: первый, снабженный хитиновыми зубчиками, в ко-
тором происходит перетирание пищи, и второй, где отцеживаются пище--
вые частицы и пища прессуется. У большинства ракообразных функции
желудка выполняет короткая средняя кишка, где пища переваривается
и всасывается; с нею связаны протоки печени. Задняя кишка, так же,
как и передняя, эктодермического происхождения, изнутри выстлана хи-
тиновой кутикулой.
Кровеносная система состоит из сердца и системы сосудов, обычно
она незамкнута, ее степень развития зависит от степени развития орга-
нов дыхания. У низших ракообразных кровеносная система отсутствует
или представлена спинным сосудом — сердцем, от которого посегментно1
отходят парные артерии. Кровь содержит либо гемоглобин, либо гемо-
цианин, в котором железо заменено медью (голубая кровь); она течет
255
по лакунам — особым полостям между внутренними органами. Органа-
ми дыхания служат у большинства жабры, представляющие собой вы-
росты грудных ног либо, возможно, видоизмененные брюшные конеч-
ности; у многих ракообразных жабр нет и дыхание осуществляется всей
поверхностью тела.
Наружный скелет ракообразных отличается большим разнообрази-
ем; обычно он хитиновый, но чаще всего пропитан карбонатом или фос-
фатом кальция. Скелет может состоять из двух боковых створок, соеди-
ненных на спинной стороне (остракоды, конхостраки, филлокариды),или
иметь вид щита или полуцилиндра, охватывающего голову и часть гру-
ди. У усоногих раков скелет состоит из нескольких известковых табли-
чек, образующих своеобразный домик. Скелет служит защитой для мяг-
кого тела, местом прикрепления различных мускулов и внутренних ор-
ганов. При линьке старый скелет сбрасывается, а затем образуется
вновь.
Размножение и развитие. Большинство ракообразных раздельно-
полые; самцы и самки диморфны; они различаются размерами тела и
строением конечностей; некоторые могут размножаться партеногенети-
чески, т. е. без оплодотворения яйца. Обычно самка вынашивает опло-
дотврренные яйца в специальной выводковой камере. Из яйца выходит
личинка науплиус с тремя парами конечностей: антеннулами, антенна-
ми и мандибулами; передняя часть тела науплиуса не сегментирована
(рис. 133, 3). После ряда линек, иногда сопровождаемых более или
менее сложнйм превращением, развивается молодой рачок. У ракооб-
разных с прямым развитием все личиночные стадии проходят в яйце.
Основы систематики и классификация. Систематика ракообразных
основана на характере сегментации тела, строении конечностей, их спе-
циализации, количестве на грудном и брюшном отделах, строении и раз-
витии органов дыхания, выделения, кровеносной системы, особенностях
постэмбрионального развития.
До последнего времени было принято деление ракообразных на два
подкласса: низших (Entomostraca) и высших (Malacostraca), осно-
ванное на непостоянном числе сегментов грудного и брюшного отделов
у первых и постоянном (8—7) — у вторых; но, как показали новые ис-
следования, если высшие ракообразные представляют собой вполне ес-
тественную группу, то в подкласс низших раков объединялись разнород-
ные группы. Поэтому в настоящее время предложено всех ракообраз-
ных разделять на 6 подклассов: Branchiopoda, Ostracoda, Cirripedia, Со-
pepodoidea, Cephalocarida, Malacostraca. .
ПОДКЛАСС BRANCHIOPODA. ЖАБРОНОГИЕ
Общая характеристика. Бранхиоподы, или жаброногие, отличают-
ся рядом примитивных признаков, к числу которых относятся: наличие
кожно-мускульного мешка, неустановившееся и значительное количе-
ство гомономных сегментов тела, следы сегментации головы, ясное раз-
деление тела на грудной и брюшной отделы, однообразно построенные
листовидные конечности, сравнительно простое строение нервной, пище-
варительной и половой систем, наличие хвостовой вилкй, или фурки;
развитие обычно с метаморфозом.
Жаброногие—обитатели морей и пресных вод; в ископаемом состоя-
нии встречаются начиная с кембрия. Подкласс жаброногих разделен на
пять отрядов: Lipostraca, Anostraca, Conchostraca, Cladocera, Notostraca.
Отряд Lipostraca. Липостраки (гр. leipo — отсутствие, ostrak —
панцирь, раковина). Отряд включает одно семейство и один род Lepido-
256
caris из среднего девона Шотландии. Тело длиной 3 мм, панцирь отсут-
ствует, туловище состоит из 23 сегментов и заканчивается раздвоенным
тельсоном; первый и последний сегменты туловища с рудиментарными
конечностями; 11 пар плавательных конечностей, из них 3 передние
сильно изменены, остальные — двухветвистые, напоминающие плава-
тельные ножки копепод. Голова несет две пары антенн, из которых вто-
рая пара — двухветвистая — служит приспособлением для плавания
(рис. 134, 1).
Рис. 134. Отряды Lipostraca (У), Notostraca (2) и Conchostraca (3—6):
1 — Lepidocaris из среднего девона Шотландии, вид сбоку; 2 — Triops (совр.);
3— Isaura (совр.), левая створка удалена; 4 — Pseudestheria из среднего дево-
на Тувы; 5 — Hemicycloleaia из позднего карбона Сев. Америки; 6 — скульпту-
ра раковины древних Limnadiidae
Отряд Anostraca. Жаброноги, или бесщитковые листоногие (ап —
отрицание), объединяют около 180 современных и несколько вымерших
видов ракообразных, обитателей пресных водоемов {Branchipus') и со-
леных материковых озер полупустынь и степей; некоторые живут при со-
лености от 40 до 23O°/oo (Artemia). Голова состоит из двух частей — пе-
редней с двумя парами антенн и глазами и задней, несущей мандибулы
и максиллы (челюсти). Обычно грудь состоит из 11 сегментов, у неко-
торых из 17—19; каждый сегмент несет пару листовидных ножек, вы-
полняющих функции дыхания, передвижения и создания тока воды с
пищевыми частицами ко рту. Длинное цилиндрическое брюшко состоит
из 8 сегментов, не имеющих ног; первые два брюшных Сегмента обыч-
но сливаются в один; с ним связаны органы размножения. Тело закан-
чивается двухветвистой фур кой. Из яйца выходит науплиус, который
после нескольких линек превращается в молодого рачка. Эоцен (?) —
ныне.
Отряд Conchostraca. Конхостраки, или филлоподы (гр. konche —
раковина, phyllon — лист), объединяют рачков, обитающих в пресно-
водных и солоноватоводных бассейнах. Тело заключено в двустворчатую
раковину, напоминающую раковину двустворок; обе створки соединены
17
Б. В. Друщиц
257
поперечным мускулом-замыкателем. Голова несет пару фасеточных глаз
и один науплиальный глаз; антеннулы служат для плавания; антенны
двухветвистые, обычно длиннее антеннул; туловище состоит из 10—32
сегментов, несущих соответствующее число пар листовидных ножек, вы-
полняющих функцию газообмена и подачи пищи ко рту; тело заканчи?
вается короткой фуркой. Створки охватывают тело полностью, но при;
линьке не сбрасываются, а увеличиваются в размерах за счет нараста-
ния новых полос роста, напоминающих кольца нарастания на ракови-
не двустворчатых моллюсков. Обычно поверхность полос роста несет'
разнообразную скульптуру: ячеистую, ребристую. Размножение половое
с оплодотворением или партеногенетическое. Яйца вынашиваются меж-
ду спинной стороной тела и раковиной. Они окружены плотной оболоч-
кой и имеют приспособления для переноса ветром, различными живот-
ными. Из яйца выходит науплиус, или метанауплиус; на стадии ме-
танауплиуса появляется раковина в виде плоского щита, который на
более поздних стадиях перегибается пополам и образует две створки.
По мере роста и линек происходит увеличение тела и раковины. Первые
конхостраки известны из девона; они были, по-видимому, обитателями
мелководных зон моря; из каменноугольных, юрских и меловых отложе-
ний известны остатки створок конхострак из пресноводных континен-
тальных бассейнов. Современные представители обитают в пресных и со-
лоноватых водах озер, прудов и луж. Девон — ныне. Около 70 родов.
Отряд Cladocera. Кладоцеры, или ветвистоусые (гр. klados — ветвь,,
ceras — рог), — обитатели пресных и солоноватоводных бассейнов, не-
сколько родов живут в морях. Тело заключено в двустворчатую ракови-
ну. Голова не покрыта раковиной; она несет пару сложных и пару про-
стых глаз и четыре пары конечностей: антеннулы, антенны, мандибулы
и максиллы. Антенны предназначены для плавания. Туловище имеет-
пять пар листовидных конечностей, служащих для газообмена. Размно-
жаются половым и партеногенетическим путем. Для кладоцер характе-
рен цикломорфоз — резкое изменение формы тела в зависимости от тем-
пературы воды — летние формы резко отличаются от зимних. Олиго-
цен — ныне. Около 20 родов.
Отряд Notostraca. Нотостраки (гр. notos — спина). Тело удлинен-
ное, покрыто со спинной стороны головогрудным щитом (поэто-
му нотострак называют щ и т н я м и). Голова несет пару сложных фасе-
точных глаз и один маленький науплиальный глаз. Антеннулы и антен-
ны развиты слабо; туловище состоит из 35—71 сегмента, из которых.
29—52 — постгенитальные; первая пара — очень длинная. Фурка мно-
гочленистая, длинная. Поздний карбон — ныне (рис. 134, 2).
ПОДКЛАСС OSTRACODO1DEA. ОСТРАКОДЫ,
ИЛИ РАКУШКОВЫЕ РАЧКИ
Общая характеристика. Остракоды (гр. ostracon — раковина, чере-
пок) — мелкие ракообразные, живущие в морских, пресноводных и со-
лоноватоводных бассейнах. Маленькое тело заключено в карапакс,
состоящий из двух створок. Большинство видов имеют микроскопические
размеры (от 0,4 до 1,5 мм), некоторые пресноводные формы бывают до
5 мм, но среди свободноплавающих морских форм встречаются особи,,
достигающие 30 мм. У большинства остракод хитиновый карапакс про-
питан солями кальция, гладкий или скульптированный, без следов на-
растания. Во время линьки (8—9 личиночных стадий) происходит рост-
и изменение тела и скелета. Тело без следов сегментации. Голова несет-
простой глазок и пару сложных фасеточных глаз. Туловище снабжено
258 '
от одной до трех пар ходильных ног; тело заканчивается обычно фуркой.
Кровеносная система и органы дыхания отсутствуют. Нервная система
состоит из двух окологлоточных ганглиев и брюшной цепочки. Раздель-
нополые; размножаются половым путем. В ископаемом состоянии из-
вестны с кембрия. Встречаются в морских, а начиная с карбона и в
пресноводных отложениях.
Строение мягкого тела. Тело короткое, сжатое с боков, без следов
сегментации, заключено в двустворчатую раковину, представляющую
обызвествленный карапакс (рис. 135). Голова несет четыре пары видо-
Рис.. 135. Подкласс Ostracoda:
1 — схема строения современного представителя; 2 — схема строения края раковины;
3 — схема сочленения створок (зачернены зубы)'; 4 — замок у Loxoconcha (мел —
ныне); 5 — внутреннее строение створки Cythereis (мел — ныне); 6— то же Proto-
cythere (юра — мел); 7 — форма отпечатков мускулов;
ан — антенна, ант — антеннула, вл — внутренний листок, гл — глаз фасеточный, глп —
глаз простой, зс — зона сращения наружного и внутреннего листков, азе — обызвест-
вленный слой, мз — мускул-замыкатель, мк — максилла, мн — мандибула, н — нож-
ки, нк — наружный край раковины,' нл — наружный листок, пк — поровые каналы,
пщ — пищеварительный тракт, ф — фурка, х — хитиновый слой, Щ — щетинки
измененных конечностей — антеннулы, антенны, мандибулы и максил-
лы. Антеннулы, или первые антенны, одноветвистые, в типичном случае
состоят из 8 члеников; выполняют локомоторную функцию (служат для
плавания, ползания или зарывания) или функцию органов чувств, либо
служат в качестве органов равновесия. Антенны также выполняют
функцию локомоторных органов и служат для плавания, хождения или
ползания. В отличие от антеннул они двухветвистые; прикрепляются по
бокам головы около верхней губы. Мандибулы расположены по обе сто-
роны’рта; они состоят из протоподита, экзоподита и эндоподита. Про-
17;
259
то по дит снабжен зубом, предназначенным для жевания, эндоподит
несет щетинки, которые служат для разрезания пищи и ее удержания.
Остракоды используют мандибулы при зарывании и ползании. Мансиллы
развиты в разных группах по-разному и взаимосвязаны с мандибулами.
Они служат для доставки пищи в рот, удаления несъедобных частиц и
создания тока, воды для газообмена около головного отдела.
Туловище несет от одной до трех пар конечностей. Первая пара ту-
ловищных ног прикреплена к телу около места соединения головы и ту-
ловища. Первая пара ходильных ног у одних остракод связана с челюст-
ным аппаратом и рассматривается в качестве вторых максилл, у других
выполняет функцию ходильных ног, причем часто ноги самцов отличают-
ся от ног самок и наблюдается асимметрия — правая нога отличается
от левой. Вторая пара туловищных ног, если присутствует, напоминает
первую пару; она выполняет, правда не всегда, локомоторную функцию.
Третья пара,,развитая у части остракод, служит для очистки внутренней
полости раковины или выполняет локомоторную функцию. Тело закан-
чивается фуркой разнообразной формы и строения, напоминающей по
своему строению конечности.
Пищеварительный тракт состоит из рта, пищевода, желудка с пище-
варительной железой и задней кишки, которая заканчивается анальным
отверстием. Рот образован шлемовидной верхней губой и примыкающей
к ней нижней, по бокам которой расположены мандибулы. Имеются
слюнные железы. В желудок вдается хитиновый аппарат, фильтрующий
пищу. Органы выделения представлены тремя парами выделительных
желез. Органы кровообращения отсутствуют; только у морских миодо-
копид имеется пульсирующий пузырек — сердце. Газообмен осущест-
вляется через покровы тела и внутренние листки створок, снабженных
жаберными придатками. Нервная система состоит из надглоточного и
подглоточного узлов (ганглиев) и брюшной нервной цепочки. Глаза
представлены одним простым и 1—2 личиночными глазками или двумя
фасеточными, расположенными по бокам головы, иногда отсутствуют.
Створки скреплены вдоль спинного края лигаментом и соединены мус-
кулом-замыкателем, который прикрепляется к внутренней поверхности
створок, оставляя след прикрепления.
Все конечности снабжены поперечнополосатыми мышцами, приводя-
щими их в движение. Следы прикрепления мышц к карапаксу также
иногда сохраняются на внутренней стороне. Тело снабжено многочис-
ленными щетинками, проходящими через поровую зону раковины. Кро-
ме того, многие конечности несут щетинки, выполняющие функцию ося-
зания.
Размножение и развитие. Остракоды раздельнополы, снабжены хо-
рошо развитыми половыми органами, размножаются часто партеногене-
тйчески, при этом развитие идет вне тела матери или в особой выводковой
камере. Из яйцевых оболочек выходит личинка науплиус (у морских
форм), снабженная двустворчатой раковиной, одним глазком и тремя
парами конечностей: антеннуламй, антеннами и мандибулами. В про-
цессе линьки сбрасывается не только карапакс, но и хитиновые покро-
вы ряда частей тела, в -том числе задней кишки; во время линек проис-
ходит последовательное развитие всех частей и органов тела. Размеры
карапакса и его пропорции изменяются аллометрически. На второй ста-
дии появляются максиллы, фурка и остальные конечности; на третьей —
порово-канальная зона с узким пояском поровых каналов; на пятой ста-
дии обычно появляется скульптура и начинает формироваться замок.
После седьмой линьки появляются признаки полового диморфизма.
260
Пресноводные остракоды достигают половозрелости обычно раньше
морских. У остракод это время колеблется от 30 дней до трех лет.
Строение скелета. Карапакс остракод состоит из двух створок раз-
нообразной формы и размеров (см. рис. 135, 2—6). Створки образованы
двойной складкой кожи, которая прикрепляется к телу на границе го-
ловы и туловища. Она состоит из двух листков — наружного и внутрен-
него. . Н а ру ж н ы й листок образован двумя хитиновыми и находя-
щимся между ними известковым слоями. Иногда хитиновые слои также
обызвествляются. Внутренний листок представляет собой тонкую
кутикулярную пластинцу, переходящую непосредственно в хитиновый по-
кров животного. Вдоль внешнего края пластинка обызвествляется, обра-
зуя краевую пластинку. Внутренняя и наружная пластинки срастаются
вдоль брюшного края, образуя так называемую зону сращения.
В наружной пластинке и в зоне сращения имеются поровыеканалы
(так называемая порово-канальная зона), через которые выходят щетин-
ки, выполняющие функции органов чувств.
В карапаксе различают дорзальную и вентральную части, передний
и задний концы. Вдоль спинной части расположен замок, вдоль брюш-
ного, переднего и заднего краев створки соприкасаются без сочленения.
Створки могут быть равными и неравными, симметричными и асиммет-
ричными; у неравностворчатых раковин одна створка охватывает дру-
гую. Створки могут быть гладкими или скульптированными. К скульпту-
ре относятся лопасти, бугры, ребра, шипы, борозды, ямки, ячейки. Ло-
пасть представляет собой выпуклую часть створки, ограниченную бо-
роздами; створка может иметь от двух до четырех лопастей, развитых
преимущественно у палеозойских остракод (особенно бейрихиид). Бу-
гор — высокий конический выступ на внешней части створки, располо-
женный чаще всего посредине створки. У многих форм выделяется так-
же глазной бугорок, расположенный в переднеспинной части створ-
ки против глазка остракод. Ребра представляют собой выступы створок,
расположенные в любой их части. Они могут быть продольными, попе-
речными и косыми. Иногда вдоль свободного края створки развивается
краевое ребро. Шипы представляют собой скульптурные образо-
вания; они могут быть сплошными, полыми, крупными, мелкими. К я м-
к а м относятся небольшие углубления на поверхности створок; к ячей-
кам — небольшие углубленные участки створок, окаймленные гранями.
Сочетание ямок, ячеек, ребер, бугорков и шипов создает различные типы
скульптуры, характерные для различных групп остракод.
На внутренней поверхности створок сохраняются (не всегда) следы
прикрепления различных мускулов: замыкающего, мандибулярных, ан-
теннальных и спинных, управляющих как конечностями, так и органами
задней части туловища. Чаще всего сохраняются следы прикрепления
мускула-замыкателя, расположенного ближе к переднему краю створок,
примерно посередине. Эти следы имеют форму, круглого пятна, состоя-
щего из отдельных бугорков (иногда до 200) микроскопических разме-
ров (см. рис. 135, 5—7). На внешней поверхности бугоркам обычно со-
ответствуют ямки- или бороздки. У мезокайнозойских остракод сохраня-
ются следы прикрепления мандибулярных мышц в виде двух отдельных
бугорков; в спинной части створок над мускулом-замыкателем наблю-
даются иногда следы прикрепления мускулов спинной группы, а впере-
ди них следы прикрепления антеннальных мускулов.
Вдоль спинной стороны створки соединяются эластичной связкой —
л и га ментом и замком. Замок состоит из выступов — валиков или
зубов и углублений — желобков, или ямок (см. рис. 135, 3—4). Строе-
ние замочного края различно. Он может иметь: а) гладкую замочную
261
площадку, б) прямой или изогнутый, гладкий или зазубренный валик
в одной створке и бороздку или желобок в другой створке, в) валик
и бороздку, сопровождаемые кардинальным зубом и соответствующей
ему ямкой, д) замок, состоящий из зубов в одной створке и ямок в дру-
Рис. 136. Подкласс Ostracoda:
1 — отряд Archaeocopida, Bradoria (ранний — средний кембрий); 2 — отряд Le-
perditicopida, Leperditia (силур — девон): 2а — левая створка, 26 — правая; 3—
4 — отряд Palaeocopida: 3 — Quadrilobella (ранний ордовик), 4 — Beyrichia
(силур — средний девон): 4а — женская особь с выводковой камерой (вк),
46 — мужская особь; 5—7 — отряд Podocopida: 5 — Cythere- (совр.), 6 —^Ваг-
dia (ордовик — ныне), 7 — Cypris (? юра, плейстоцен — ныне); 8—9 — отряд
Platycopida: 8 — Kloedenella (силур — карбон), поперечный разрез через перед-
нюю (а) и заднюю (б) части (левая створка зачернена), 9 — Cytherella (юра —
ныне); 10—12 — отряд Myodocopida: 10 — Cypridina (совр.), 11 — Cypridella
(карбон), 12 — Cyprella (карбон), вид спереди;
ин — инцизура; направление стрелки указывает передний край
гой либо чередования зубов и ямок в каждой из створок. Зубы могут
быть гладкими или мелко зазубренными — кренулированными. Строе-
ние замочного края имеет важное значение ддя диагностики остракод.
В последние годы выделено большое число типов замков,. получивших
соответствующие названия.;, ?.. , < о,
262
Половой диморфизм. Самцы обычно отличаются от самок не толь-
ко по строению половой системы, но и по строению раковин, хотя поло-
вые отличия на раковинах не всегда выражены достаточно четко. Обыч-
но раковины самцов более длинные и низкие, несут несколько отличную
•скульптуру. У древних остракод раковины самок выделяются по наличию
выпуклостей у заднего конца створок, наличием выводковых камер
(рис. 136, 4а). Интересно, что половые различия более четко выражены
на раковинах палеозойских форм, чем на скелетах современных.
Основы систематики и классификация. В основу выделения отрядов
среди вымерших остракод приняты особенности строения карапакса,
•форма и строение отпечатков мускула-замыкателя, строение замка. При
выделении отрядов среди ныне живущих представителей использованы
особенности строения мягкого тела. В настоящее время остракоды раз-
делены на 6 отрядов: Archaeocopida, Leperditicopida, Palaeocopida, Ро-
docopida, Platycopida, Myodocopida.
Отряд Archaeocopida. Археокопиды (гр. archaios — древний, коре —
весло). Карапакс обызвествлен, содержит большое количество хитина,
делающего его эластичным и гибким. Замочный край прямой и длинный.
Глазной бугорок обычно выступает. Рядом с глазным бугорком (немного
позади и ниже) расположен отпечаток мускула-замыкателя. Отряд
включает 12 родов (рис. 136, 1), распространенных в раннем и среднем
кембрии. Предположительно к этому отряду отнесен род Eremos из тре-
мадока Швеции.
Отряд Leperditicopida. Лепердитикопиды. Карапакс известковый,
толстый, крупных размеров; замочный край прямой. Отцечаток мускула-
замыкателя крупный, составляет до 30%' высоты створки и состоит из
многочисленных бугорков (до 200). Поверхность карапакса гладкая или
слабо скульптированная. Появились в конце кембрия, были широко рас-
пространены в ордовике и силуре и дожили до конца девона. Известно
15 родов (рис. 136, 2).
Отряд Palaeocopida. Палеокопиды (гр. palaios — древний, коре —
весло). Карапакс с прямым смычным краем, крупных размеров. Поверх-
ность гладкая, но чаще скульптированная; имеются лопасти, бугры, реб-
ра. Обызвествленная внутренняя пластинка отсутствует. Часто наблю-
дается половой диморфизм. Наиболее распространенный отряд палео-
зойских остракод, насчитывающий до 260 родов (рис. 136, 3—4). Пер-
вые представители появились в ордовике, максимального развития до-
стигли в силуре—девоне, но несколько родов существовало в карбоне—
перми; одно семейство существует в современных морях.
Отряд Podocopida. Подокопиды. Раковина от овальной до закруг-
ленно-прямоугольной, неравностворчатая, иногда с оттянутым задним
концом; на конце могут быть шипы; поверхность раковины гладкая или
скульптированная. У современных представителей антеннулы несут ког-
тевидные щетинки, мандибулы и максиллы имеют жаберные придатки.
Отпечаток мускула-замыкателя округленный, состоящий из многочис-
ленных бугорков у ранних представителей и небольшого числа у более
поздних. Широко распространены во всем мире и встречаются как в
морских, так и в пресноводных водоемах. Насчитывается около 450 ро-
дов (рис. 136,5—7).
Отряд Platycopida. Платикопиды (гр. platys = platos — плоский).
Отряд объединяет остракод с овальной неравностворчатой раковиной, с
необособленной порово-канальной зоной; у самок задняя часть раковины
вздутая; туловищный отдел несет две пары конечностей; отпечаток мус-
кула-замыкателя состоит из двух рядов бугорков, расположенных в пе-
263
редней трети раковины. Каждый ряд состоит из 5—9 бугорков. Спинной
край округленный. Ордовик—-ныне. Несколько родов (рис. 136, 8—9).
Отряд Myodocopida. Миодокопиды (гр. myodos — мускул, коре—-
весло). Карапакс состоит из двух сильно выпуклых створок, несущих на
переднем крае выступ — ростр и инцизуру — отверстие для выхода ан-
тенн; поверхность створок раковин гладкая, редко ребристая; у палео-
зойских .форм со срединной бороздой; раковина обычно слабо обызвест-
влена. Размеры колеблются от 20 до 30 мм. На последней стадии жиз-
ненного цикла ведут планктонный образ жизни. Первые представители
появились в ордовике. В палеозое существовало 30 родов; из мезозоя
и кайнозоя известно только 2 рода, в современных морях насчитывает-
ся около 25 родов (рис. 136, 10—12). ;
История развития и экология остракод. К наиболее древним остра-
кодам относятся представители отряда археокопид, появившиеся в на-
чале кембрия и имевшие слабо обызвествлённый карапакс с прямым
смычным краем, крупным отпечатком мускула-замыкателя, расположен-
ным непосредственно под глазным бугорком. Они имеют некоторое сход-
ство с конхостраками, к которым были вначале отнесены. Слабое обыз-
вествление карапакса отличает их от остальных остракод. По-видимому,
предки остракод, возможно существовавшие уже в конце протерозоя,
имели хитиновый покров. От археокопид в конце кембрия появились ле-
пердитикопиды. Они обладали крупным карапаксом, сильно обызвест-
вленным, сложным мускулом-замыкателем, имеющим форму шеврона.
Лепердитикопиды были широко распространены в ордовикских и силу-
рийских морях Европы, Азии и Северной Америки.
В начале ордовика возникают палеокопиды — широко распрост-
раненные в палеозое остракоды, среди которых наибольшее значение
имеют бейрихииДы, образующйе самостоятельный подотряд среди па-
леокопид. Бейрихииды отличались крупной равностворчатой ракови-
ной, обычно сильно расчлененной, снабженной шаровидными образова-
ниями; они были наиболее широко распространены в силуре и девоне.
Основная группа палеокопид вымирает в середине перми; в современ-
ных морях обнаружен один род.
В начале ордовика ответвилась новая ветвь остракод, выделенная
в отряд подокопид, разделившаяся с самого начала на две ветви —• соб-.
ственно подокопин и метакопин. Собственно подокопины представлены
большой группой бердиид (Bairdiacea), получивших наибольшее разви-
тие в карбоне (но доживших до наших дней), цитераций (Cytheracea),
развитых в мелу и кайнозое. Все они несут 7 пар конечностей, простые
глазки (иногда глаза отсутствуют). Тело заключено в округленную,
овальную раковину с хорошо дифференцированным замком и развитой
порово-канальной зоной. Цитерации кроме морских заселили также
пресноводные бассейны. Их представители известны из континентальных
отложений верхней перми Кузбасса и Русской платформы и из солоно-
ватоводных бассейнов раннего триаса Поволжья и Эмбы. Вторая ветвь,
подокопид — метакопины — отличается бобовидной или яйцевидной ра-
ковиной с крупным отпечатком мускула-замыкателя, состоящего из не-
многочисленных относительно круглых бугорков. Их надсемейство ци-
придации — обитатели пресноводных бассейнов начиная с триаса. В ор-
довике возникли платикопиды, у которых бугорки для мускула-замыка-
теля (13—14) расположены двурядно в передней трети раковины. Створ-
ки обычно неравные, с изогнутым замочным краем.
Особое место среди остракод занимают представители отряда мио-
докопид, появившиеся в начале ордовика. Они имели крупную ракови-
ну до 30 мм, снабжённую на переднем конце ростральным выступом а
264
особым отверстием -— инцизурой, служащей для постоянного выхода
антенн. Современные миодокопиды ведут планктонный образ жизни.
Раковина часто слабо обызвествлена.
До настоящего времени, таким образом, доживают представители
четырех отрядов: подокопид, платикопид, миодокопид и палеокопид.
Современные остракоды обитают в самых разнообразных бассейнах —
от морских до пресноводных. В морях планктонный образ жизни ведут
миодокопиды. Подркопиды встречены в морских и континентальных бас-
сейнах. Морские подокопиды ведут бентосный образ жизни, обитая
обычно на глубинах до 200 м, но чаще в более мелководных зонах и
преимущественно на водорослях. Многие остракоды' живут в илистых
осадках, ползая или зарываясь. Подокопиды континентальных водоемов
приспособились к жизни в разных условиях. Они Живут в пресных и
соленых озерах, реках, источниках, прудах, в торфяниках и даже в
подземных водах. Впервые подокопиды приспособились к жизни в кон-
тинентальных бассейнах в конце карбона (дарвинулиды). Они встрече-
ны вместе с остатками пресноводных моллюсков, червей и рыб.
ПОДКЛАСС CIRRIPEDIA. УСОНОГИЕ РАКИ
Циррипедии (лат. cirrus — усики, завивка), или усоногие раки, ве-
дут прикрепленный или паразитический образ жизни, что привело к
регрессивному изменению многих органов; тело и конечности заключены
в наружный кожный покров —• «мантию», покрытую снаружи или из-
вестковыми табличками или хитинизированным слоем (у паразитирую-
щих). Головной отдел редуцирован, ротовые конечности недоразвиты.
Туловище состоит из 6 сегментов, несущих пару двухветвистых конеч-
ностей, служащих для создания тока воды, с которым поступает пища
в мантийную полость. Развитие идет с метаморфозом; науплиус после
ряда линек превращается в циприсовидную личинку или «куколку», об-
ладающую двустворчатой хитиновой раковиной; после новых линек ли-
чинка прикрепляется головным отделом к субстрату, окружается «ман-
тией» и превращается в молодого рачка. Передний отдел тела превра-
щается в орган прикрепления. Шесть грудных ножек видоизменяются в
длинные усики, покрытые щетинками. «Мантия» охватывает все тело
рачка и на наружной поверхности образует вначале хитиновые, а затем
известковые таблички. Из них возникает прочный скелет, состоящий из
нескольких известковых табличек, среди которых выделяются: пар-
ные — скуты, терги, латералии и непарные — карина, ростр.
Систематическое положение усоногих раков решается неоднознач-
но. Одни исследователи объединяют их с копеподбидеями, другие рас-
сматривают в качестве самостоятельного подкласса, состоящего из од-
ного отряда и трех подотрядов. Подотряды выделяются по строению ске-
лета и образу жизни.
К подотряду торациковых (Thoracica) относятся морские желуди
(Balanomorpha), прикрепляющиеся к субстрату широкой подошвой, мор-
ские уточки (Lepadomorpha), сидящие на длинном мясистом стебельке,
или ножке, часто покрытом известковыми табличками (рис. 137). Авто-
ром описан случай гибели морских уточек, поселившихся на внешней
стороне раковины раннемелового аммонита и зажатых между двумя
оборотами спирали при последующем построении раковины. У одного
из погибших экземпляров морских уточек сохранились ножки с остат-
ками щетинок (рис. 137, <?). В отличие от большинства ракообразных
265
усоногие — гермафродиты с развитыми женскими и мужскими половы-
ми железами.
Второй подотряд—акроторациковых усоногих (Acrothoracica)
объединяет несколько родов, ведущих сверлящий образ жизни. Из яйца
выходит науплиус, который затем превращается в циприсовидную ли-
Рис. 137. Подкласс Cirripedia:
1 — схема строения скелета современного Balanus: la — общий вид, 16 — по- .
перечный разрез, 1в — вид скелета в плане; 2 — общий вид современного Lepas;
3 — Blastolepas из раннего мела Сев. Кавказа;
к — карина, кл — каринолатералия, л — латералия, нщ —.ножки с остатками
щетинок, рл — ростролатералия, ск — скут, ст — стебелек, т — терг
чинку; последняя садится на известковые раковины брюхоногих, пан-
цирных моллюсков, стволы кораллов, сбрасывает свою двустворчатую
раковинку и начинает сверлить, выделяя вещество, растворяющее из-
весть. Размеры тела самок не превышают 2 см, самцы обычно значи-
тельно меньше (0,4—2 мм). В ископаемом состоянии сохраняются следы
сверления на рострах белемнитов и на скелетах других организмов.
Третий подотряд усоногих (Rhizocephala) ведет паразитический об-
раз жизни, паразитируя в теле крабов, креветок, раков-отшельников.
В связи с паразитизмом тело превращено в тонкие ветвящиеся трубоч-
ки, которые оплетают кишечник, нервные стволы, половые железы,
внедряются в мускулатуру хозяина. Только наличие в постэмбриональ-
ной стадии науплиуса и циприсовидной личинки указывает на проис-
хождение этих паразитирующих рачков.
;.. В ископаемом состоянии, усоногие встречаются . начиная с силура.
266
ПОДКЛАСС COPEPODOIDEA. КОПЕПОДОВЫЕ
Копеподовые (гр. коре — весло), или веслоногие раки, имеют слит-
ную голову, срастающуюся с передним грудным сегментом, постоянное
количество^ грудных сегментов (6), не-
сущих двухветвистые конечности, лишен-
ные жаберных и жевательных придат-
ков; брюшные конечности отсутствуют,
органы дыхания редуцированы, дыхание
осуществляется всей поверхностью тела,
кровеносная система развита слабо;
часть копеподовых ведет паразитический
образ жизни. Развитие протекает с пре-
вращением, из яйца выходит личинка
науплиус. Девон — ныне. Копеподоидеи
включают 4 отряда: Copepoda, Branchiu-
ra, Mystacocorida, Ascothoracida. В ис-
копаемом состоянии известны предста-
вители только первого отряда. Известен
один род из триасовых отложений За-
падной Европы. Головогрудь прямоуголь-
ная, голова несет пару сложных глаз и
одну пару простых антенн; туловище
состоит из 6 сегментов, несущих 6 пар
конечностей (рис. 138, /). Тельсон с
хвостовой вилкой.
ПОДКЛАСС CEPHALOCARIDA.
ЦЕФАЛОКАРИДЫ
К подклассу цефалокарид (гр. cep-
halon — голова, лат. caris — креветка),
установленному несколько лет тому на-
зад, отнесены два рода, обитающие в
Тихом и Атлантическом океанах, и один
род, известный из карбона Америки.
В современных, морях цефалокариды ве-
дут донный образ жизни. Голова велика
и состоит из слившихся сегментов,
брюшные сегменты (более 7) лишены
конечностей. По строению двухветвистых
конечностей они напоминают листоногих,
Рис. 138. Подклассы Сореро-
doidea (/) и Cephalocarida
(2—3):
1 — Euthycarcinus из раннего
триаса Зап. Европы; 2 — Tes-
nusocaris из карбона Сев. Аме-
рики; 3 — Hutchinsoniella со-
временный вид из Атлантиче-
ского океана
по строению головы и заднего
отдела тела — копепод, по строению грудных конечностей и антенн
сходны с некоторыми высшими раками. Цефалокариды, таким образом,
несут ряд признаков, близких к предковым формам жаброногих, копе-
под и высших раков. Карбон — ныне. 3 рода (рис. 138, 2, 3).
ПОДКЛАСС MALACOSTRACA. МАЛАКОСТРАКИ
Малакостраки (гр. malacos — мягкий, ostracon — черепок, рако-
вина), или высшие раки, в большинстве своем водные обитатели, преи-
мущественно морские, бентосные и пелагические, населяющие все зоны
моря, , от литорали до абиссали. Небольшая часть живет в пресных во-
дах; немногие приспособились к жизни на суше и часть ведет парази-
тический образ жизни. К высшим ракам относятся широко известные
267
крабы, креветки, речные раки (см. рис. 133, 1), раки-отшельники, ома-
ры, лангусты, а также раки-богомолы, мизиды, кумовые раки, изоподы,
бокоплавы.
У всех высших раков тело состоит из постоянного числа сегментов:
голова — из пяти, грудь — из восьми, брюшко — из шести или семи.
Тело заканчивается анальной лопастью, или тельсоном, лишенной ко-
нечностей. Все остальные сегменты несут пару одно- или двухветвистых
Рис. 139. Надотряд Phyllocarida:
1 — отряд Leptostraca, схема строения современного представителя; 2—3 — от-
ряд Archaeostraca (реконструкции): 2 — Nahecaris из раннего девона Зап. Евро-
пы, 3 — Mesothyra из девона Сев. Америки
конечностей. Головные и часть грудных сегментов часто сливаются
вместе, образуя головогрудь. У многих малакострак головной и груд-
ной отделы покрыты хитиновым панцирем — карапаксом, имеющим вид
цилиндра, двустворчатой раковины; иногда карапакс отсутствует. На
голове расположены одна пара сложных фасеточных глаз , и пять пар-
конечностей: две пары усиков и три пары челюстей. Часть грудных но-
жек видоизменена в ногочелюсти, которые принимают участие в пе-
редаче пищи ко рту. Брюшные ножки служат для плавания, выполняют-
функции жабр и половых органов. Высшие раки достигают иногда зна-
чительных размеров: карапакс камчатского краба до 50 см в длину..
Кровеносная и дыхательная системы хорошо развиты. Желудок раз-
делен на жевательную и фильтрующую камеры; имеется хорошо раз-
витая пищеварительная железа. Развитие прямое или с метаморфозом..
Кембрий — ныне.
Систематика подкласса отличается большой сложностью. На ос-
новании строения придатков головы, степени слияния грудных сегмен-
тов с головными и способа размножения высшие раки разделены на
пять надотрядов: Phyllocarida, Syncarida, Peracarida, Eucarida, Hoplo-
carida.
Надотряд Phyllocarida. Филлокариды (гр. phyllon — лист, лат. caris,
род. caridis— креветка). К филлокаридам отнесены два отряда: Leptos-
traca и Archaeostraca (рис. 139).
268.
Отряд Leptostraca. Тонкопанцирные раки (гр. leptos— тонкий) оби-
тают в морских бассейнах, преимущественно в прибрежной зоне
(рис. 139, 1), только один вид ведет планктонный глубоководный образ
жизни. Карапакс имеет вид тонкой двустворчатой раковины, прикры-
вающей голову, весь грудной и часть брюшного отделов; спереди к ра-
ковине причленяется овальный пластинчатый ростр, прикрывающий
сверху стебельчатые глаза и основание антенн; антенны довольно длин-
Рис. 140. Надотряд Syncarida, отряд Anaspidacea:
1 — схема строения современного представителя; 2 — Uronectes из ранней перми
Зап. Европы
ные. Каждый из 8 сегментов грудного отдела несет пару листовидных
цожек; они служат для передвижения, дыхания, для передачи пищи
ко рту. 4 пары брюшных ножек снабжены щетинками и служат для
плавания;' следующие две пары ножек — короткие, редуцированные;
последний брюшной сегмент лишен конечностей; удлиненный тельсон
несет две длинные ветви — фурки; при зарывании тело может так из-
гибаться, что тельсон касается конца ростра. Развитие прямое, без ме-
таморфоза. В ископаемом состоянии известна одна находка из перм-
ских отложений Ирландии. Характерный род Nebalia (пермь — ныне).
Отряд Archaeostraca. Древнепанцирные объединяют вымерших
филлокарид (кембрий — триас), от которых сохраняются в ископаемом
состоянии только редкие скелеты. Скелет в виде двустворчатого щита
или единого целого щита, ростр свободный, расположен впереди щита
(рис. 139, 2—3). Тельсон несет одну—три пары придатков (фурка).
Характерные роды: Hymenocaris (кембрий — ордовик), Ceratiocaris
(поздний кембрий — пермь), Nahecaris (девон).
Надотряд Syncarida. Синкариды (гр. syn — вместе) полностью ли-
шены карапакса; грудные и брюшные сегменты имеют сходное строе-
ние, почти одинаковые размеры и не срастаются, только первый грудной
сегмент срастается с головой; грудные ножки двухветвистые; конечно-
сти последнего брюшка сегмента преобразованы в рулевые ноги-уропо-
ды. Развитие прямое, у взрослых сохраняется науплиальный глаз.
К синкаридам относится небольшое число видов (около 75) мелких и
микроскопических высших ракообразных, обитающих в пресных водах,
озерах, ручьях, подземных водах, пещерных водоемах, колодцах. Со-
временные синкариды представляют собой остатки древней, некогда
процветавшей группы. Синкариды девона и карбона жили в морях; в
дальнейшем они, по-видимому, перешли к жизни в пресных водах и
расселились по всем континентам. В состав надотряда входит три от-
ряда (рис. 140): Anaspidacea (девон — ныне), Bathynellacea (совр.),
Stygocaridacea (совр.).
269
Надотряд Peracarida. Перакариды (гр. рега — сумка). Один из об-
ширных надотрядов, включающий ракообразных, различных по внеш-
нему облику и образу жизни (рис. 141). Голова срастается с одним или
двумя передними сегментами груди; конечности груди превращаются в
Рис. 141. Надотряд Peracarida:
1 — отряд Mysidacea, Grangopsis из карбона Шотландии (реконструкция); 2 —
отряд Amphipoda, Gammarus из неогена Кавказа; 3 — отряд Tanaidacea, схема
строения современного представителя; 4—5 — отряд Isopoda: 4 — Cyclosphaero-
та из поздней юры, 5 — Cymodoce из неогена Керченского полуострова, вид сбоку-
ногочелюсти. Глаза.стебельчатые или сидячие; правая и левая мандибу-
лы асимметричные. Карапакс у ряда перакарид отсутствует. Самка под
грудью имеет выводковую сумку, образованную пластинчатыми вы-
ростами грудных ног; в сумке развиваются оплодотворенные яйца. На-
стоящих личинок нет, развитие прямое, без превращений. Перакариды
живут в морях, где они ведут бентосный и пелагический образ жизни,
в пресных водах и на суше; часть из них паразитирует на других жи-
вотных. В надотряд включены 7 отрядо^,: Mysidacea, Cumacea, Tanaida-
cea, Spelaeogryphacea, Thermosbaenacea, Isopoda, Amphipoda. В иско-
паемом состоянии известны представители 5 отрядов: Mysidacea (кар-
бон— ныне), Cumacea (юра — ныне), Tanaidacea (юра — ныне), Isopoda
(триас — ныне), Amphipoda (палеоген — ныне).
Надотряд Eucarida. Эукариды (гр. ей — приставка, означающая
настоящий, хороший). К эукаридам относятся морские и пресноводные
ракообразные, объединяющие свыше 9000 видов современных и иско-
паемых представителей (рис. 142). Карапакс срастается с головными
и грудными сегментами. Глаза стебельчатые; мандибулы симметрич-
ные. Выводковой камеры нет, самка носит яйца на брюшных ножках
или выметывает их в воду. Развитие обычно сопровождается сложным
метаморфозом, реже прямое. В ископаемом состоянии встречаются
сравнительно часто начиная с перми. Эукариды разделены на два не-
равных отряда: Euphausiacea (совр.) и Decapoda (пермь — ныне).
Надотряд Hoplocarida. Гоплокариды (гр. hopion —оружие) вклю-
чает только один отряд Stomatopoda (гр. stoma — устье, рот, pous, род.
podos — нога)—ротоногих ракообразных, или раков-богомолов
(рис. 143). Раки-богомолы —• хищные морские ракообразные, имеющие
удлиненное (до 35 см) тело. С головой срастаются пять грудных сег-
270
ментов, их конечности преобразованы в одноветвистые ногочелюсти (от-
сюда название отряда «ротоногие»), из которых особенно‘сильно раз-
вита вторая пара, напоминающая хищническую ногу богомола (отсюда
второе название «раки-богомолы»). Сегменты, несущие фасеточные гла-
Рис. 142: Надотряд Eucarida, отряд Decapoda:
1 — Pempix из триаса Зап. Европы; 2 — Егуоп из поздней юры Зап. Европы;
3 — Cancer из раннего олигоцена Мангышлака
Рис. 143. Надотряд Hoplocarida, отряд Stomatopoda:
1 — схема строения современного представителя; 2 — Sculda из поздней юры Зап.
Европы
271
за и первую пару антенн, свободные. Карапакс короткий, имеет вид
щита, покрывающего голову и пять грудных сегментов, впереди распо-
ложен ростр. Брюшко длинное, все его шесть сегментов хорошо разви-
ты; пять пар конечностей листовидной фбрмы, служат для- плавания и
газообмена. Самки вынашивают яйца под грудью. У самцов внутренние
ветви первой и второй пар брюшных ножек преобразованы в копулятив-
ные органы. Уроподы плоские, направлены назад; они расположены по
бокам лопатообразного тельсона.
Ротоногие раки живут в норах, в щелях, между ветвями кораллов.
Они роют норы с,помощью ростра и ногочелюстей, выбрасывая осадок
из норы тельсоном и уропода-
Рис. 144. Класс Myriopoda, подкласс Diplo-
poda:
1 — Acantherpestes из перми Чехословакии;
2 — Pleurojulus из перми Чехословакии;
ми. Норы могут достигать
глубины 1 м и обычно имеют
два выхода.
Из яйцевых оболочек вы-
ходят личинки (псевдозоэа,
синзоэа, антизоэа), ведущие
в основном планктонный об-
раз жизни; после ряда линек,
продолжающихся иногда 2—
3 месяца, раки-богомолы пе-
реходят к бентосному образу
жизни. Современные виды
(известно около 170) обитают
преимущественно в тропиче-
ских и субтропических морях
на небольших глубинах. В ис-
копаемом состоянии известны
начиная с карбона.
Филогенетические взаимо-
3 — Lithobius из олигоцена Европы (бал- отношения между надотряда-
тийский янтарь) ми высших ракообразных
(рис. 150). Восстановление
филогенетических взаимоотношений между надотрядами, входящими в
состав подкласса, возможно главным образом на основании сравни-
тельно-морфологических, а не палеонтологических данных. Последние
крайне скудны и не дают указаний ни о происхождении, ни о род-
ственных связях. Малакостраки образуют единый, подкласс, состоящий
из пяти надотрядов. По данным Я- А. Бирштейна (1960), предки выс-
ших ракообразных имели тело, состоящее из 20 сегментов (из них 5
головных, 8 грудных и 7 брюшных) и тельсона. Голова и грудь были
покрыты карапаксом, вытянутым спереди в ростр. Голова несла пару
фасеточных глаз и непарный науплиальный глаз, тельсон заканчивал-
ся фуркой. Сердце длинное, тянулось через все тело и имело боковые
сосуды. Гонады располагались по бокам сердца и открывались у са-
мок на 6-м, у самцов на 8-м грудном сегментах; конечности были двух-
вётвистыми.
Наиболее древними, но близкими к исходной предковой группе
рассматриваются филлокариды и гоплокариды, рано обособившиеся от
основного ствола; все они имеют трубкообразное сердце, длинные го-
нады, свободные грудные сегменты, но каждый из надотрядов несет чер-
ты специализации: у филлокарид возник двустворчатый карапакс, у гоп-
локарид редуцировался первый брюшной сегмент и передние 5 пар груд-
ных конечностей были преобразованы в ногочелюсти, а задние три па-
ры упростились, стали тонкими. Позднее от общего ствола ответвился
272
надотряд синкарид, сохранивший ряд примитивных черт организа-
ции — свободные, несросшиеся грудные сегменты, семь брюшных сег-
ментов (у некоторых), науплиальный глаз у взрослых, длинное сердце
и гонады; вместе с тем от других групп они отличаются рядом особен-
ностей, к числу которых относятся: редукция карапакса, прямое раз-
витие, тенденция к слиянию переднего грудного сегмента с головой и
другие особенности.
В карбоне и перми возникли два близкородственных надотряда —
перакариды и эукариды, имеющие сходное строение многих
внутренних органов, но резко отличающиеся в способах размножения.
У перакарид за счет пластинчатых отростков грудных конечностей об-
разовалась своеобразная выводковая сумка, в которой происходит пря-
мое развитие молоди; подобное приспособление в зачатке имеется у
филлокарид. У эукарид яйца откладываются в воду или прикрепляются
к брюшным ножкам самки и развитие происходит с метаморфозом;
наблюдается срастание карапакса с грудными сегментами.
ПОДТИП TRACHEATA. ТРАХЕЙНО ДЫШАЩИЕ
К подтипу трахейнодышащих (лат. trachia — дыхательное горло),
или неполноусых (Atelocerata, гр. ateleios — неполный, ceras — рог,
ус), относится самый крупный ствол членистоногих, ведущих назем-
ный образ жизни или вторично перешедших к жизни в водной среде.
К этому типу относятся многоножки и насекомые. У многоножек тело
состоит из головы и туловища; голова несет одну пару усиков и три
пары челюстей; у насекомых туловище разделено на грудь и брюшко.
Низшие представители, обитающие в почве, дышат всей поверхностью
тела, все наземные формы — с помощью трахей — системы разветвлен-
ных трубочек, оплетающих все внутренние органы. Трахеи открываются
стигмами в каждом сегменте туловища. Тело покрыто кутикулой, со-
стоящей из эндо- и экзокутикулы. Направление эволюции насекомых
связано с выработкой приспособлений для жизни на суше: развитием
водонепроницаемой эпйкутикулы, покрывающей тончайшим слоем кути-
кулу; совершенствованием трахейной системы; приспособлением выде-
лительной системы к минимальному выводу воды. Оплодотворение на-
ружно-внутреннее и внутреннее. Трахейнодышащие произошли, по-ви-
димому, от далёких предков, близких к кольчатым червям, перешедших
к жизни в почве, от которых возникли первые многоножки, а затем —
насекомые; последние отличаются сокращением числа туловищных сег-
ментов, обособлением груди и брюшка, возникновением крыльев. Под-
тип Tracheata разделен на 2 класса — Myriopoda и Insecta.
КЛАСС MYRIOPODA. МНОГОНОЖКИ
К классу многоножек (гр. myrios — бесчисленный) относится свы-
ше 10 000 современных и вымерших исключительно сухопутных видов
(рис. 144); среди них пользуются широкой известностью так называе-
мые тысяченожки (гломерисы, жервезии, кивсяки) и сколопендры. Те-
ло разделено на ясно обособленную голову и более или менее вытяну-
тое сегментированное туловище, почти все гомономные сегменты кото-
рого несут одну или даже две пары конечностей. Туловище не разде-
лено на грудной и брюшной отделы; каждый сегмент состоит из спин-
ной пластинки — т е р г и т а, брюшной — стер нита и двух боковых,
18 В. В. Друщиц
273
их соединяющих. Число сегментов туловища от 11—13 до 170. Разме-
ры тела варьируют от 0,3 мм до 30 см. На голове расположена одна
пара антенн и простые, реже сложные глаза. Во. рту различают верх-
нюю губу, пару мандибул и одну или две пары максилл (челюстей).
Нервная система в виде лестницеобразной брюшной цепочки; трахей-
Рис. 145. Класс Insecta. Схема внешнего строения
крылатого насекомого (/) и строение крыла (2):
ан —антенна,' бд — бедро, бр—брюшко, г — го-
лень, гол — голова, зг — заднегрудь, зк — заднее
крыло, л — лапка, пг — переднегрудь, пгл — про-
стые глазки, пк — переднее крыло, сг — средне- i
грудь, фгл — фасеточный глаз, ц — церка; жил-
ки крыла: С — костальная, SC — субкостальная, ।
R — радиальная, PS — радиальный сектор,
М — медиальная, Си — кубитальные, А — аналь-
ная
ная система в виде парных пучков в каждом сегменте туловища. Мно-
гоножки раздельнополые; из яйцевых оболочек выходят личинки, кото- ;
рые после каждой линьки приобретают новые сегменты (развитие с ана-
морфозом). С наступлением половозрелости анаморфоз прекращается.
По форме сегментов, строению конечностей, половых органов и ха-
рактеру питания многоножки разделены на шесть подклассов: Protochi-
lopoda (поздний силур — ранний девон), Chilopoda (карбон — ныне), "
Pauropoda (палеоген — ныне), Diplopoda (поздний карбон — ныне), j
Protosymphyla (поздний кембрий), Symphyla (палеоген — ныне). Мно- 1
гие исследователи рассматривают эти подклассы в качестве самостоя- ।
тельных классов. Но число различий между этими подклассами не пре-
вышает число сходных признаков, поэтому для изучающих палеонто-
логию сохраняется единый класс многоножек. Первые многоножки из-
274
вестны из позднего кембрия (Protosymphyla), из отложений верхнего
силура —нижнего девона (Protochilopoda); от последних, по мнению
1 А. Г. Шарова, возникли Chilopoda, Diplopoda, Pauropoda, хотя в иско-
паемом состоянии первые две группы известны только с середины
карбона, а третья группа появилась в конце палеогена. Пятый подкласс
Symphyla, представляющий, по-видимому, слепую ветвь, возник в кон-
це кембрия от Protosymphyla, однако в ископаемом состоянии до сих
пор известен только из палеогена. В протосимфилах А. Г. Шаров ви-
дит предков большого класса насекомых.
КЛАСС INSECTA. НАСЕКОМЫЕ
Насекомые (лат. insecare — надрезать)
— самый многочисленный класс членис-
млн. видов. Они живут в самых раз-
Общая характеристика,
или шестиножки (Hexapodа)
тоногих, насчитывающий свыше
нообразных условиях на су-
ше, завоевали воздух, прис-
пособились к жизни в прес-
ных водоемах и даже в мо-
ре. Тело разделено на три
отдела, или тагмы: голову,
грудь и брюшко (рис. 145),.
Голова несет глаза, одну
пару антенн и три пары че-
люстей. Грудь состоит из
трех сегментов и выполняет
локомоторную. функцию;:
каждый сегмент снабжен
парой ног (отсюда, название
«шестиножки»); у крылатых
к средне- и заднегрудным
сегментам . прикреплено по
паре крыльев. Брюшко со-
тоит из 11 сегментов, оно
лишено конечностей; в
брюшке сосредоточена ос-
1
Рис. 146. ’ Класс Insecta. Схема внутреннего
строения крылатого насекомого:
бц — брюшная цепочка нервной систему, ж —
желудок, зки — задняя кишка, м—мозг,
мо — мальпигиевы органы, пгг — подглоточ-
ный ганглий, по — половые-органы, р — рот,
с — сердце, сж — слюнные железы .
новная масса внутренних органов, включая желудок и заднюю кишку,
половые и выделительные органы. Дыхание с помощью трахей. Насе-
комые раздельнополые; постэмбриональное развитие часто сопровож-
дается метаморфозом. Девон — ныне.
Строение тела. Наружный покров тела насекомого состоит из хи-
тиновой оболочки, в которой различают твердые участки, или скле-
риты, и более тонкие, их соединяющие, — перепонки. Склериты
на спинной поверхности носят название тергитов, на брюшной —
стернитов, на боках — плевритов. Голова образована за счет
шести слившихся эмбриональных сегментов, всегда сильно склероти-
зирована, так как служит местом прикрепления сильной челюстной мус-
кулатуры. Голова несет членистые антенны, или усики, три пары челю-
стей разной формы и размеров; антенны гомологичны первой паре уси-
ков ракообразных. Верхняя челюсть, или мандибула, состоит из одного
массивного склерита; нижние челюсти, или первые максиллы, как пра-
вило, членистые, двухветвистые, с особым придатком — челюстным щу-
пиком; вторые максиллы срастаются вместе и образуют нижнюю губу
(лабрум), расположенную под ротовым отверстием. У многих насеко-
мых в состав ротового аппарата входит верхняя губа —: непарная склад-
18*
275
ка покровов, подвижно сочлененная с головной капсулой. Ротовые ор-
ганы насекомых в зависимости от способа добывания пищи могут раз-
лично видоизменяться; выделяют несколько типов ротового аппарата:
грызущий, лижущий, сосущий, колющий, колюще-сосущий и переходные
между ними. На голове расположены глаза простые и сложные, или фа-
сеточные. Фасеточные глаза всегда располагаются по бокам голо-
вы и могут достигать большой сложности (например, у стрекоз бывает
до 28 000 фасеток, у комнатной мухи — до 4000). У личинок на месте
сложных глаз вначале развиваются простые глазки (каждый простой
глазок имеет свою кутикулярную роговицу); у хорошо летающих на-
секомых простые глазки (2—3) расположены на лбу, между сложны-
ми глазами.
За головой следует грудь, состоящая из трех сегментов: передне-,
средне- и заднегрудь. У взрослых насекомых каждый сегмент несет па-
ру конечностей; исторически грудь взяла на себя функцию передви-
жения; в связи с этим усилилась мускулатура грудных сегментов,- их
наружный скелет усложнился. Нога насекомого состоит из пяти члени-
ков: тазик, вертлуг, бедро, голень и лапка. Три пары ног обычно сход-
ны между собой и служат для хождения или бегания; очень часто от-
дельные пары ног специализируются для выполнения особых функций
и в связи с этим видоизменяются в прыгательные (кузнечики), рою-
щие (медведки), хватательные (богомолы)., плавательные (жуки-пла-
вуны), При переходе к неподвижному образу жизни наблюдается ча-
стичная или полная редукция ног..
У летающих насекомых роль груди как локомоторного органа су.-,
щественно усиливается развитием двух пар крыльев, расположенных на
2-м и 3-м сегментах груди. Крылья развиваются при переходе насе-
комого во взрослое состояние; они закладываются, как плоские листо-
видные. выпячивания покровов, средне- и заднегруди.. По мере разрас-
тания этих выпячиваний нижняя и верхняя стороны каждого выпячи-
вания сближаются и образуют эластичную пластинку крыла, подвижно
соединенную с тергитами груди. У палеозойских насекомых передне-
грудь также несла крыловидные придатки,- однако они были соедине-
ны с нею неподвижно. Образования, подобные крыльям, в виде неболь-
ших складок боковых покровов сегментов брюшка (паранотальные вы-
ступы) наблюдаются у некоторых насекомых '(тараканов, личинок жу-
ков). Эти паранотальные выступьг брюшных сегментов рассматри- ;
вают в. качестве сериальных гомологов крыльев. Форма крыльев очень ।
разнообразна. У одних насекомых обе пары крыльев развиты одинако-
во (стрекозы), у других — передние крылья развиты сильнее (пчелы,
бабочки, цикады), что влечет за собой усиление среднегруди. Разви-
тие задних крыльев (многие прямокрылые,, жуки) усиливает задне-
грудь; иногда передняя пара крыльев превращена в твердые надкры-
лья, или элитры (жуки). Следует отметить, что одной из тенденций в
эволюции насекомых в связи с наличием ног и развитием крыльев было
создание монолитной груди.
Крыло укрепляется многочисленными хитиновыми трубочками —
жилками. После формирования крыла жилки становятся его механи-
ческой опорой; по ним проходит гемолимфа. Продольные жилки име-
ют важное значение для классификации насекомых (см. рис. 145, 2).
Передний край крыла укреплен костальной жилкой (С); за ней
следует субкостальная жилка (SC); сзади субкостальной жилки
находятся радиальные, состоящие из собственно радиальной жилки
(R) и так называемого радиального- сектора (RS). Посередине крыла
лежит--система-медиальных жилок (М), затем располагаются ку-
276
i
.3
битальные (Си) и анальные (А). У большинства насекомых про-
дольные жилки соединяются поперечными и крыло приобретает слож-
ную сетчатую форму. Вместе с тем в ряде групп сохраняется подвиж-
ность переднегруди, что обычно связано с особым развитием первой
пары ног (богомолы, медведки). Наряду с усилением крыльев у мно*
гих насекомых наблюдается противоположная тенденция — потеря спо-
собности к полету и связанная с этим редукция крыльев, вплоть до пол-
ного их исчезновения (вши, блохи и др.). Это в свою очередь приводит
к вторичному упрощению груди.
Брюшко у насекомых состоит максимально из 11 почти гомономных
сегментов, однако в процессе эволюции у многих групп наблюдается
укорочение брюшка за счет либо редукции сегментов, либо резкого
уменьшения диаметра сегментов и втягивания их внутрь брюшка. На
конце брюшка у примитивных форм сохраняются придатки 11-го сег-
мента — так называемые церки, представляющие собой видоизменен-
ные конечности. Возможно, что половые придатки, развитые на брюш-
ке ,(8—9-й сегменты), представляют собой также видоизмененные ко-
нечности.
У насекомых хорошо развит внутренний скелет, образованный за
счет складок хитинизированной кутикулы, вдающихся внутрь тела.
Различные выступы и выросты служат местом прикрепления мышц ко-
нечностей, опорой для узлов нервной системы и передней части кишеч-
ника. Наружные покровы насекомых также несут различные выросты и
придатки: бугорки, зубчики, щетинки, чешуи, волоски. Наружный по-
кров очень богат железами, расположенными во всех частях тела. Же-
лезы вырабатывают разнообразные по химическому составу и биоло-
гическому назначению вещества (например, шелк, воск, различные
яды).
Органы дыхания представлены трахейной системой, состоящей из
ряда трубочек — трахей, которые открываются по бокам тела от-
верстиями •— дыхальцами, или стигмами. Трахеи многократно
ветвятся и заканчиваются тончайшими трубочками — трахеолами.
У личинок ручейников, веснянок и поденок, живущих в воде, стигмы
закрыты и воздух поступает через тонкие покровы, образующие трахей-
ные жабры. Водные насекомые (водные жуки, клопы, многие личинки
комаров), несмотря на жизнь в воде, дышат свободным воздухом.
Кровеносная система незамкнутая; сердце — трубчатый спинной
сосуд — своим сокращением прогоняет по телу кровяную жидкость —
гемолимфу, которая разносит по телу питательные вещества, гормоны
и уносит продукты обмена, но не разносит кислород. Нервная система
построена по типу нервной системы низших членистоногих и напоми-
нает, таковую кольчатых червей: она состоит из надглоточного ганглия
и брюшной нервной цепочки, в последней различают подглоточный ган-
глий и 10 грудных и брюшных ганглиев. Органы чувств, или рецепторы,
разнообразны и часто имеют очень сложное строение. Покровы тела
несут множество разных рецепторов, выполняющих различные функ-
ции: вкуса, слуха, осязания, обоняния, химического чувства, улавли-
вания колебаний температуры, давления, скорости и направления по-
лета и т. д.
Пищеварительная система состоит из передней, средней и задней
кишки; ее строение зависит от способа питания (рис. 146). Передняя
кишка начинается ротовой полостью, в которую впадают слюнные же-
лезы, вырабатывающие в зависимости от характера пищи различные
ферменты. В средней кишке идет переваривание пищи; в слепых вы-
ростах кишки симбиотические организмы (бактерии, многожгутико-т
277
вне) способствуют разложению трудноразрушимых соединений (на-
пример, клетчатки). Переваренная пища и вода всасываются задней
кишкой. Органы выделения представлены тонкими трубчатыми вырос-
тами (от 2 до 150) кишечника, расположенными на границе средней и
задней кишки; они получили название мальпигиевых сосудов.
Выделение продуктов белкового обмена происходит в заднем отделе
Эпиморфоз
Г оло метаморфоз
к у колка
Рис. 147. Схема развития насекомых с неполным (эпиморфоз) и полным (голо-
метаморфоз) превращением
кишечника. Во внутренней полости расположено жировое тело, кото-
рое служит источником получения воды при неблагоприятных услови-
ях. Это достигается процессом окисления жира, при котором выделяет-
ся вода.
Размножение и развитие. Почти все насекомые раздельнополы,
только немногие гермафродиты; у многих хорошо выражен половой
диморфизм; у общественных насекомых нередко наблюдается резко
выраженный полиморфизм особей (пчелы, муравьи, термиты); в этом
случае самки, способные к размножению, сильно отличаются от других
особей — рабочих, солдат и т. д. Насекомые, как правило, размножают-
ся половым путем, иногда партеногенетически; оплодотворение наруж-
но-внутреннее у низших, живущих во влажной среде, в почве, и внут-
реннее — у более высокоорганизованных насекомых. В первом случае
самец откладывает сперматофор на землю, а самка подбирает его сво-
им половым отверстием. Эмбриональное развитие происходит, в яйце;
из яйцевых оболочек выходит личинка.
Различают три основных типа постэмбрионального развития
(рис. 147). У первичнобескрылых развитие идет без метаморфоза, ли-
чинка похожа на взрослых, но отличается меньшими размерами, не-
полным числом конечностей, недоразвитыми половыми органами. С каж-
дой линькой увеличиваются размеры тела, появляются новые пары ко-
нечностей (такой тип развития называется анаморфозом). У всех
крылатых развитие идет либо с неполным (эпиморфоз), либо с пол-
ным : метаморфозом-(г о л о м е т а м о р ф о з). При неполном метамор-
фозе личинка внешне похожа на взрослых (имаго), имеет фасеточные
278
глаза, три пары ног, но отличается отсутствием крыльев и обычно при-
способлена к иному образу жизни, чем имаго. У стрекоз, веснянок и
поденок личинки ведут водный образ жизни и отличаются наличием, ря-
да специальных личиночных органов (имеют трахейные жабры для ды-
хания в воде, иначе устроенные ротовые органы). С каждой линькой
размеры крыльев увеличиваются.
При полном метаморфозе кроме
личинки имеется стадия куколки.
Личинки резко отличаются от има^
го внешним видом, количеством ко-
нечностей и отсутствием фасеточ-
ных глаз. Личиночные формы край-
не разнообразны (рис. 147), их
строение и внешний вид зависят от
образа жизни и способа питания:
имеются личинки только с ротовы-
ми конечностями и зачатками груд-
ных, личинки с брюшными конечно-
стями и, наконец, личинки, лишен-
ные конечностей. За личиночной
стадией следует стадия куколки;
во время этой стадии происходит
разрушение специфических личи-
ночных органов и из особых клеток
возникают новые органы взросло-
го организма. Полное превращение
свойственно широко известным
жесткокрылым, двукрылым, пере-
пончатокрылым, сетчатокрылым
насекомым.
Основы систематики и класси-
фикация. Основой для классифика-
ции насекомых служат степень рас-
членения тела на три главных его
•отдела, наличие или отсутствие
крыльев, а при наличии крыльев
характер их жилкования, строение
и положение ротовых органов,
особенности эмбрионального и
постэмбрионального развития. По
наличию или отсутствию крыльев
насекомые разделяются на
Рис. 148. Подкласс Apterygota:
1 — отряд Monura, Dasyleptus из
ранней перми Кузнецкого бассейна
(реконструкция); 2 — отряд Thysa-
nura, Triassomachilis из триаса При-
уралья, вид с брюшной стороны
Apterygota и Pterygota
два подкласса:
ПОДКЛАСС APTERYGOTA. ПЕРВИЧНОБЕСКРЫЛЫЕ
К подклассу первичнобескрылых (гр. а — отрицание, pteron — кры-
ло) относится несколько тысяч видов, всегда бескрылых, очень мелких
насекомых, живущих в почве или затененных местах. Глаза простые,
очень редко сложные, ротовые органы грызущего типа, втянуты почти
у всех в ротовую капсулу; брюшко со следами конечностей, развитых
в разной степени, с двумя церками; развитие простое, без метаморфоза,
чйсло линек неопределенное. В состав подкласса входят пять отря-
дов: Collembola (девон — ныне), Monura (поздний карбон — ранняя
пермь), Protura (только совр.), Dlplura (палеоген — ныне), Thysanura
279
(триас —ныне). В ископаемом состоянии первичнобескрылые встре-
чаются крайне редко (рис. 148); первые находки известны из отложе-
ний верхнего девона (отряд Collembola). Наиболее многочисленны на-
Рис. 149. Подкласс Pterygota: инфраклассы Hemimetabola (1—4) и Holometa-
bola (5—S):
1 — отряд Odonata, Permepallage (крыло) из поздней перми Архангельской обл.;
2 — отряд Ephemeroptera, Mesoneta из ранней юры Приангарья- 3 — отряд Ortho-
ptera, Tholmanvissia из ранней перми Приуралья (реконструкция); 4 — отряд
Homoptera, Permocicada из поздней перми Архангельской обл. (реконструкция);
5—6 — отряд Coleoptera: 5 — Lithostoma из поздней юры Казахстана, 6 — Ne-
cromera из поздней юры Казахстана; 7 — отряд Mecoptera, Agetochorista из ран-
ней перми Приуралья (реконструкция); 8 — отряд Diptera, Mesoplecia из позд-
ней юры Казахстана
ходки первичнобескрылых в балтийском янтаре. Первичнобескрылые,
как предполагают, произошли от древних многоножек (Protosymphyla).
Ряд исследователей считают, что перечисленные выше отряды следует
280
Л//Л. TRACHEAE A
КЛ. MYRIOPODA
п/кл. Раис о ро da
(К-ныне)
п/кл. Diptopoda
(Су-ныне)
п/кл. Chilopoda
(С-ныне)
п/кл. ProtochHopoda
п /кл. Symphytta —-
s_______(Р- ныне)
кл. INSECTA
п/кл. Pteruqota
' (С - ныне)
„инфркл. Hofomefabo/a
(С/- ныне)
инфркл. Hemimetabola
(С-ныне)
уп/кл. Apt erуgota
n/m. BRANCHI ATA
КЛ. CRUSTACEA
n/m. C HE L I C ER AT A
CAROMORPHA
f п/кл. Malacostraca
' (С-ныне)
н/отр. Eucarida
' \(P-ныне)
н/отр. Syncarida
(с - ныне)
н/отр. Hoptocarida
_______(С - ныне)
''п/кл.Ostracodoidea
.________- ныне)
г п/кл.Copepodoidea
._______(Р-ныне)
п/кл. Cirripedia
_______($2-ныне)
п/кл. Branchiopoda
<______ (Д- ныне)
'п/кл. Cephalocarida
(С - ныне)
н lamp. Percarida
(С- ныне)
н/отр Phyllocarida
кл. ARACHNID А
п/кл. Dpiiiomorpha
(С-ныне)
п/кл. Sotuta
(D-P)
п/кл. Агапеае
(С- ныне)
КЛ. SOLIFUGOMORPHA
(С-ныне)
кл. SCORPIONOMORPHA
отр. Pedipatpida
(С- ныне)
отр. Scorpionida
(S- ныне)
n/m. PROBOSCIFE
КЛ. ME RO STOMATA
п/кл. Xiphosura
(Cj - ныне)
п/кл. Eury^teroidea
КЛ. PYCN060NIDES
(D, - ныне)
КЛ. DICEPRM.OSOMITA
(^1)
. TRI LOBITOMORPHA
кл. TRILOBITA
(^i~Pt)
кл. TRILOBITOIDEA
(Gt'О,)
Тип ANNELIDES
кл. TETRACEPHALOSOMITA
(GfDi)
Рис. 150. Предполагаемые взаимоотношения между типами и классами члени-
стоногих
рассматривать в качестве самостоятельных классов, которые, возникнув
от древних многоножек, развивались параллельно и независимо друг
от друга.
ПОДКЛАСС PTERYGOTA. КРЫЛАТЫЕ НАСЕКОМЫЕ
К подклассу крылатых насекомых относится основная масса пред-
ставителей класса, насчитывающая около 1 млн. видов, которые обла-
дают одной или двумя парами крыльев или утратили их вторично. Во
взрослом окрыленном состоянии они, за исключением поденок, не ли-
няют. Ротовые органы не втянуты в ротовую капсулу, поэтому иногда
их называют открыточелюстными, в отличие от скрыточелюстных (бес-
крылые без отряда Thysanura), у которых ротовые части втянуты в го-
ловную капсулу. Развитие происходит с неполным или полным мета-
морфозом и в соответствии с этим они разделены на два инфракласса:
Hemimetabola и Holometabola (рис. 149, 150),
ИНФРАКЛАСС HEMIMETABOLA.
НАСЕКОМЫЕ С НЕПОЛНЫМ ПРЕВРАЩЕНИЕМ
К этому инфраклассу отнесены 29 отрядов, из которых 15 известны
только в ископаемом состоянии (рис. 149, 1—4). В постэмбриональном
развитии они проходят стадию личинки, напоминающую имаго, но без
крыльев. Личинки некоторых отрядов живут в воде и имеют специаль-
ные трахейные жабры. Наиболее характерные отряды: Palaeodictyop-
,tera (карбон — пермь), Ephemeroptera — поденки (поздний карбон —
ныне), Odonata — стрекозы (карбон — ныне), Blattodea — таракановые
(карбон — ныне), Isoptera — термиты (палеоген — ныне), Manteptera—>
богомолы (палеоген — ныне), Plecoptera — веснянки (пермь — ныне),
Orthoptera — прямокрылые (карбон — ныне), Phasmatodea — палочники
(триас — ныне), Homopteraравнокрылые (ранний карбон — ныне),
Heteroptera — разнокрылые, или клопы (пермь — ныне).
И'НФРАКЛАСС HOLOMETABOLA.
НАСЕКОМЫЕ С ПОЛНЫМ ПРЕВРАЩЕНИЕМ
К насекомым с полным превращением относятся крылатые и вто-
рично бескрылые насекомые, постэмбриональное развитие которых про-
исходит через стадию личинки и куколки. К этому инфраклассу отнесе-
но 10 отрядов, доживших до наших дней. Первый представитель известен
только по одному отпечатку крыла из отложений нижнего карбона Се-
верной Америки. В отложениях нижней перми впервые встречаются до-
стоверные представители разных отрядов с полным превращением
(скорпионницы, жесткокрылые, сетчатокрылые). С триаса появляются
двукрылые, и с этого времени эта группа насекомых занимает главен-
ствующее место во всех палеоэнтомологических комплексах
(рис. 149, 5—8). Характерные отряды: Coleoptera — жесткокрылые, или
жуки (пермь — ныне), Neuroptera — сетчатокрылые (пермь — ныне),
Lepidoptera — бабочки, или чЬшуекрЫлые (палеоген — ныне), Diptera —
двукрылые, или мухи и комары (триас — ныне), Hymenoptera — пере-
пончатокрылые (триас — ныне).
282
ТИП MOLLUSCA. МОЛЛЮСКИ
Общая характеристика. Моллюски (лат. mollis, molluscus — мяг-
кий) образуют второй по величине после членистоногих тип беспозво-
ночных животных, насчитывающий около 115000 современных видов и
около 45 000 вымерших. К моллюскам, или мягкотелым, относятся мор-
ские, пресноводные и наземные организмы, достигающие большого раз-
нообразия в строении и размерах тела (рис. 151). Тело моллюсков не-
сегментированное, как правило двусторонне-симметричное (асимметрич-
ное только у брюхоногих моллюсков), разделяется на более или менее
обособленную голову (не обособлена у двустворок), туловище, за-
ключающее большинство внутренних органов, и ногу — мускулистый
:вырост брюшной части туловища. Промежутки между внутренними ор-
ганами заполнены паренхимой •— рыхлой соединительной тканью.
'Тело сверху и с боков одето мантией — особой складкой кожи, эпи-
телий которой выделяет известковую раковину. Между мантией и си-
стемой голова — нога обособлена мантийная полость с органами дыха-
ния, выводными каналами пищеварительного тракта, половых желез и
органов выделения. Пищеварительная система развита хорошо. В ро-
товой полости имеется радула, состоящая из органических (кутику-
лярных) зубчиков и предназначенная для перетирания пищи. Желудок
с парной печенью. Центральная нервная система у низших моллюсков
состоит из нервных стволов, у высших — из нескольких пар ганглиев,
соединенных друг с другом комиссурами и коннективами. Дыхание осу-
ществляется жабрами, легкими или поверхностью тела. Кровеносная
система незамкнутая. Кровь с дыхательными пигментами (гемоглоби-
ном или гемоцианином). Сердце находится в целомическом перикардии,
юно состоит из желудочка и предсердий, число которых различно у. раз-
ных моллюсков. Гонады расположены в задней части тела и заключены
в полость целомического происхождения, с ними обычно связаны орга-
ны выделения (почки). Полости тела как таковой нет. Раковина обыч-
но состоит из трех слоев: одного наружного органического и двух из-
вестковых. Первые два: наружный органический, или периострак, и на-
ружный известковый — выделяются краем мантии, внутренний извест-
ковый формируется передней частью мантии. Строение и форма рако-
вины очень разнообразны: раковины могут состоять как из двух створок,
охватывающих тело с боков, так и из нескольких пластин, прикры-
вающих тело сверху, могут быть в форме колпачка, также прикрывая
283
тело сверху, или в виде длинной конической трубки, открытой с двух
концов или только с одного, в которую заключено тело моллюсков;,
трубка может быть свернута в плоскую или коническую спираль или
иметь иную форму. Наконец, у ряда моллюсков раковина может быть-
внутренней либо может полностью отсутствовать.
Рис. 151. Схема строения основных классов моллюсков:
1 — Monoplacophora; 2 — Polyplacophora; 3 — Bivalvia; 4—5 — Gastro-
poda; 6 — Xenoconchia; 7 — Scaphopoda; 8—9 — Cephalopoda: 8 — Cole-
oidea, 9 —• Nautiloidea;
гол — голова, жб — жабры, н — нога, ом — отпечатки мускулов, пт —
пищеварительный тракт, ра — раковина, щ — щупальца
Моллюски раздельнополы или гермафродиты; развитие прямое или
с метаморфозом, с личинками — трохофорой и велигером.
Особенности раннего онтогенеза (трохофорная личинка) указывают на:
общность происхождения моллюсков и кольчатых червей от одного пред-
ка. Моллюски известны с начала кембрия. По особенностям строения
мягкого тела и раковины и особенностям онтогенеза моллюски разде-
лены на 10 классов: Aplacophora, Polyplacophora, Monoplacophora^
Gastropoda, Scaphopoda, Xenoconchia, Bivalvia, Cephalopoda, Tentaeuli-
ta, Hyolitha, из которых два последних известны только в ископаемом:
состоянии, а первый живет только в современным морях и в геологи-
ческом прошлом неизвестен.
284
КЛАСС APLACOPHORA. АПЛАКОФОРЫ
Аплакофоры (гр. а — отрицание, plax, род. placos — пластинка,
phoros — несущий), или соленогастры (гр. solen — борозда, gaster —
желудок), имеют цилиндрическое, вытянутое в длину тело, внешне на-
поминающее тело кольчатых червей (рис. 152, 1). На брюшной стороне
проходит узкая бороздка; в ней находится кожная складка, представ-
ляющая видоизмененную ногу моллюска (иногда и борозда и нога
•отсутствуют). Рот с малодифференцированной радулой, сидящей непо-
средственно на кутикуле глотки; желудок со слабодифференцированной
печенью или без нее.
Нервная система состоит из двух пар нервных стволов. Кровенос-
ная система несовершенная; кровь движется по лакунам; сердце —•
часть спинного кровеносного синуса; предсердия отсутствуют или имеет-
ся одна пара их. Жабры отсутствуют и газообмен происходит через
стенки тела или имеется одна пара. Целом разделен на перикардиаль-
ную и половую части. Аплакофоры раздельнополы и гермафродиты. Ли-
чинка напоминает таковую протобранхиальных бивальвйй. У некоторых
В ид изнутри
За
Рис. 152. Классы Aplacophora (1) и Polyplacophora (2—3):
1 — Neomenia (совр.); 2 — подкласс Paleoloricata, Septemchiton (ордовик),
реконструкция и отдельные створки; 3 —: подкласс Neoloricata, схема строе-
ния: За — вид сверху, 36 — вид сбоку;
ан — анус, бб — брюшная бороздка, ис — известковые спикулы, к — клоака,
н — нога, пн :— перинот с известковыми чешуями и гранулами, пт — пи-
щеварительный тракт, р — рот; створки: гс — головная, сс — срединные,
хс — хвостовая
форм закладываются 7 известковых щитков, налегающих друг на друга
черепицеобразно. В дальнейшем щитки атрофируются и в теле разви-
ваются известковые шипики в виде заостренных игл или чешуек.
Аплакофоры — обитатели морей, они встречаются почти во всех
морях земного шара на разных глубинах, до 3—4 тыс. м. Одни живут
на колониях кораллов, другие закапываются в илистый осадок. Апла-
кофоры не встречаются в ископаемом состоянии, хотя, наверное, суще-
ствовали в геологическом прошлом. Известковые шипики, следы полза-
ния при. известной целенаправленности поисков могут быть найдены.
Много споров вызывает систематическое положение и природа со-
леногастров. Одни считают их наиболее примитивными по строению, и
285
развитию моллюсками, другие, рассматривают их в качестве регрессив-
ной группы. Развитие моллюска из трохофорного типа личинки, нали-
чие в покровах известковых пластинок указывают на примитивность их
организации и на близость их к лорикатам, или хитонам. Из-за отсут-
ствия скелета их называют аплакофорами.
КЛАСС POLYPLACOPHORA. ПОЛИПЛАКОФОРЫ
Общая характеристика. Класс полиплакофор (подчеркивается боль-
шое число табличек, слагающих раковину, гр. poly — много, plax —
пластинка, phoros — несущий), или лорикат (лат. loricatus — одетый
в панцирь, lorica — кольчуга), объединяет около 1000 видов современ-
ных и ископаемых моллюсков, имеющих овально-эллиптическое двусто-'
ронне-симметричное тело, прикрытое сверху 7—8 известковыми щитка-
ми, налегающими друг на друга черепицеобразно; мантия образует по-
ясок (перинот) вокруг всего тела (см. рис. 152). На нижней стороне
обособлена нога с широкой подошвой. Между ногой и мантией распо-
ложены многочисленные жабры. Рот снабжен радулой; пищеваритель-
ный тракт прямой, с анальным отверстием на заднем конце тела. Серд-
це состоит из желудочка и двух предсердий. Нервная система — око-
логлоточное кольцо и четыре продольных нервных ствола. Единичные
находки скелетов известны начиная с позднего кембрия.
Строение мягкого тела. Тело полиплакофор, или чаще называемых
хитонами, удлиненно-эллиптическое, со спинной стороны прикрыто ман-
тией; утолщенный край мантии, пронизанный, мышцами и нависающий
крышеобразно над ногой, образует так называемый перинот (гр. pe-
ri — вокруг, notos — спина). Между перинотом и ногой остается узкое
пространство — м а нтийн а я б ор оз д а.
Голова в виде полукруга обособлена от ноги узкой бороздкой; на
ее нижней стороне расположен рот. Рот ведет в глотку, в которой рас-
положена радула, состоящая из нескольких хитиновых зубчиков (по 17
в каждом ряду). В желудок впадают протоки двух пищеварительных,
желез. Анальное отверстие расположено на заднем конце тела. Сердце
в перикардии (околосердечной сумке) лежит под последней пластинкой
скелета и имеет один желудочек и два предсердия. Половая железа
также расположена на спинной стороне; она отделена от перикардия
и почек; от нее отходят два канала, через которые наружу выводятся
половые продукты. Нервное кольцо окружает глотку, от него отходят
четыре нервных ствола, соединенных поперечными перемычками. Нога
имеет вид широкой подошвы, двумя продольными бороздами она отде-
ляется от боковых частей тела. Нога служит органом передвижения и
плотного прикрепления к различным подводным предметам. В продоль-
ных боковых бороздах с каждой стороны лежит ряд жабр, из которых,
только одна пара гомологична паре жабр других моллюсков, остальные
,(от 6 до 88 пар) возникли в результате полимеризации. В мантийную
полость впадают протоки половых и пары выделительных органов. Пан-
цирные моллюски — раздельнополые животные с наружным оплодотво-
рением, кладки яиц похожи на кладки гастропод. Из оплодотворенного
яйца развивается личинка — трохофора, с зачатками ноги и рако-
винных пластинок. После непродолжительного плавания личинка опус-
кается на дно и превращается в молодого хитона.
Строение раковины. В отличие от всех остальных моллюсков ске-
лет хитонов состоит из 7—8 пластинок, или створок, которые заклады-
ваются еще в эмбриональной стадии; они имеют вид щитков, черепи-
цеобразно налегающих пруг на друга. Подвижное сочленение щитков
286
позволяет хитонам в случае опасности свертываться. Передний и зад-
ний щитки отличаются своей формой от срединных (рис. 152). Каждый
щиток состоит из четырех слоев: 1) наружный — периострак, тон-
кий, органический, 2) тегмент (tegmen — крыша) — известковый
слой, пронизанный сетью каналов и лежащий непосредственно под пе-
риостраком, 3) артикуламент — известковый слой, образующий
основу каждого, щитка и выросты апофизы для соединения соседних
щитков, 4) внутренний известковый слой, или гипостра к. Иногда
между тегментом и артикуламентом обособляется еще один известко-
вый слой — мезоострак. Артикуламент вдоль края каждой створки
образует узкий выступ, погруженный в поясок мантйи, и два боковых
пластинчатых выступа (апофизы), направленных вперед под лежа-
щую впереди створку. Поверхность пластинок гладкая или скульпти-
рованная ребрами, гранулами. Иногда пластинки почти полностью за-
крыты пояском мантии. К краям створок прикрепляются мышцы. Ман-
тия на верхней стороне пояска выделяет цилиндрические известковые
шипики, или органические щетинки, а на нижней стороне пояска раз-
виваются плоские ромбовидные пластинки, которые служат для более
прочного прикрепления моллюска к подводным камням.
Классификация полиплакофор, или лорикат, основана на строении
скелета и перинота. В настоящее время выделяют два подкласса: Ра-
leoloricata (поздний кембрий — поздний мел); известно 7 родов
(рис. 152, 2) и Neoloricata (карбон — ныне); известно около 70 родов
(рис. 152, 3).
История развития. Хитоны встречаются в ископаемом состоянии
крайне редко. Это связано с тем, что после гибели моллюска его рако-
вина, состоящая из 7 или 8 щитков, распадается и плохо сохраняется.
Находки отдельных щитков известны в отложениях всех возрастов на-
чиная с позднего кембрия, но проследить подробно историю этого клас-
са до сих пор пока не удалось. Первые хитоны имели раковину, состоя-
щую( из 7 створок, без артикуламента, а поэтому створки не имели
апофизов. Один род (Septemchiton) из ордовика имел удлиненное чер-
веобразное тело, длина которого в несколько раз превышала его ши-
рину (рис. 152). В карбоне появляются хитоны подкласса Neoloricata,
у которых створки построены из нескольких слоев, в том числе из
артикуламента, дающего апофизы, направленные вперед, и боковую
кайму, входящую в перинот мантии. Большинство родов хитонов, жи-
вущих в палеозое, вымирает в конце палеозоя. В мезозое широкое рас-
пространение получают неолорикаты. В настоящее время известно око-
ло 360 ископаемых видов (80 из палеозоя, 30 из мезозоя, 250 из кай-
нозоя). Были найдены два отпечатка полных скелетов хитонов, один из
ордовика, второй из карбона; у последнего сохранилась радула с отпе-
чатком 8 рядов зубчиков.
Все современные хитоны — обитатели морей, населяющие главным
образом приливно-отливную зону. Они хорошо приспособлены к жизни
в условиях литорали и верхней сублиторали. Обтекаемая форма тела,
сильная нога, которой они присасываются к подводным предметам, по-
зволяют им противостоять ударам волн и сильным течениям. Длина
тела некоторых хитонов достигает 20—30 см. На раковине часто посе-
ляются усоногие раки, мшанки, кольчатые черви и другие прикреплен-
ные бентосные организмы.
КЛАСС MONOPLACOPHORA. МОНОПЛАКОФОРЫ
Общая характеристика. Моноплакофоры (гр. monos — один) имеют
двусторонне-симметричное тело и слабо обособленную голову; рот окру-
287
жен щупальцевыми придатками и снабжен радулой. Основные системы
внутренних органов расположены метамерно. Имеется пять пар жабр,
шесть пар целомодуктов; нервная система состоит из двух пар нервных
стволов. Нога округленная. Тело покрыто сверху колпачковидной рако-
виной; вершина сдвинута вперед; изнутри с несколькими парами от-
Рис. 153. Класс Monoplacophora:
1 — Neopilina (совр.): la — вид снизу, 16 — вид сверху; 2 — Trybliduim (си-
лур): 2а — вид снизу, 26 — вид сверху; 3 — Cyrtonella (силур — девон): За —
внутреннее ядро, 36 — вид сбоку;
ан — анус, жб — жабры, мб — мантийная бороздка, н — нога, ом — отпечатки
мускулов, пп. — пальцевидные придатки, р — рот, рщ — ротовые щупальца
печатков мускулов. Известно несколько десятков видов от раннего
кембрия до позднего девона и один современный род Neopilina с двумя
подродами.
Строение мягкого тела. Тело состоит из трех отделов: слабо обособ-
ленной головы, внутренностного мешка и ноги (рис. 153). Рот располо-
жен впереди, окружен щупальцевидными придатками, напоминающими
пальпы протобранхий (двустворки). Глотка снабжена радулой; в желу-
док впадают протоки двух симметричных долей печени; длинная свер-
288
нутая кишка открывается анальным отверстием на заднем конце тела.
Органы выделения представлены шестью парами каналов — цел о мо-
ду кто в, из которых две пары связаны с гонодуктами — каналами
половых желез. Имеются два отдельных сердца с двумя предсердиями,
лежащих в двух перикардиях в.задней части тела, по обе стороны от
пищеварительного тракта. Нервная система состоит из окологлоточного
кольца и двух пар нервных стволов, соединенных 10 парами коннекти-
вов. Имеется общая целомическая полость и под ней обособлены две
пары гонад. Все внутренние органы покрыты сверху мантией. Широкая
мантийная борозда окружает со всех сторон ногу, в ней по обе сторо-
ны тела помещаются 5 пар гребневидных жабр, или ктенидиев, и
открываются протоки 6 пар целомодуктов. Нога широкая, плоская, при-
крепляется несколькими парами мышц изнутри к раковине. С помощью
ноги моноплакофоры медленно ползают по субстрату и, вероятно, спо-
собны прочно присасываться подобно хитонам. Оплодотворение наруж-
ное. Эмбриональная раковина плоскоспиральная, экзогастрическая.
Строение раковины. Современные и древние моноплакофоры имеют
эмбриональную плоскоспиральную раковинку и коническую, колпачко-
видную — у взрослых. Вершина раковины приближена к переднему
краю и часто загнута вперед. Раковина гладкая или с сетчатой скульп-
турой, возникшей от пересечения тонких радиальных струек или реб-
рышек и концентрических линий нарастания.
История развития и систематическое положение моноплакофор.
Первые моноплакофоры известны начиная с кембрия. Они сравнитель-
но широко были распространены в ордовике — девоне. На внутренней
поверхности раковины видны мускульные отпечатки, обычно парные,
число которых у современных форм равно восьми парам, по числу мус-
кулов-ретракторов ноги; у раннепалеозойских форм это число колеба-
лось от 5 до 8 пар. Ископаемые остатки более молодых моноплакофор
до сих пор неизвестны. Раннепалеозойские моноплакофоры по форме
раковины и характеру мускульных отпечатков относились к брюхоногим
моллюскам и выделялись в отдельное семейство, затем в отдельный
подкласс (Венц, 1938; Найт, 1952).
После находок в Тихом океане Датской глубоководной экспедицией
на судне «Галатея» вблизи берегов Мексики на глубине около 3,5 км
современных реликтов этой группы и изучения анатомии мягкого тела
новой формы Neopilina galatheae было предложено объединить палео-
зойское надсемейство Triblidiacea и современных представителей в са-
мостоятельный класс Monoplacophora. Однако единого мнения о-таксо-
номическом ранге этой группы среди малакологов (специалистов, изу-
чающих моллюсков) до сих пор нет. Одни исследователи видят в мета-
мерии современной неопилины примитивные черты, указывающие
на родство с кольчатыми червями, другие считают эту метамерию вто-
ричной. Однако скорее всего вторая точка зрения ближ:е к истине.
У моллюска, имеющего одну колпачковидную раковину, парные внут-
ренние органы не образуют единых метамеров (5 пар жабр, 6 пар це-
ломодуктов, 8 пар мускулов-ретракторов, отсутствие парных метамер-
ных ганглиев нервной системы).
Вряд ли так же достаточно обосновано предложение рассматри-
вать моноплакофор в качестве подкласса гастропод — Orthoneura,
объединяющего низших гастропод, еще не испытавших торсионного
процесса. Моноплакофоры принадлежат к наиболее примитивным мол-
люскам, имеющим двусторонне-симметричное тело с симметричным
строением нервной системы и вторичным метамерным расположением
всех внутренних органов.
19 в. В. Друшии
289
В настоящее время предложено класс моноплакофор разделять
на два отряда: Tryblidiida (ранний кембрий — ныне) и Cyrtonellida
(ранний ордовик — девон). Всего известно около 25 родов (рис. 153)»
КЛАСС GASTROPODA. БРЮХОНОГИЕ
Общая характеристика. Брюхоногие, или гастроподы (гр. gaster —
желудок, pous, род. podos — нога) — группа моллюсков, впервые вы-
деленная Кювье, образует наиболее многочисленный (до 85000 совре-
с
/
Рис. 154. Схема строения современных представителей класса Gastropoda:
А—Б — подкласс Prosobranchia: А — Archaeogastropoda, Б — Neogastropoda;
В — подкласс Opisthobranchia; Г — подкласс Pulmonata;
ан — анальное отверстие, гл — глотка и пищевод, жб — жабры, ,ж — желудок,
кр — крышечка, л — легкие, н нервные узлы, р — рот, с — сердце, сф — сифон
менных и 15000 ископаемых видов) среди всех моллюсков и единствен-
ный класс, имеющий своих представителей как в водной среде, так и на
суше. У гастропод хорошо обособлена голова, туловище и нога, при-
способленная для передвижения (рис. 154). В отличие от всех осталь-
ных моллюсков, тело брюхоногих асимметричное, все внутренние ограны
расположены асимметрично; как правило, тело заключено в из-;
вестковую спирально-коническую раковину, к которой оно прикрепляет?
ся двумя или одним мускулом-ретрактором. Тело втягивается полно-
стью в полость раковины и закрывается подошвой ноги или крышечкой,
расположенной на ноге; однако у ряда форм раковина отсутствует.
Голова несет обычно 1—2 пары щупалец и парные глаза, лежа-
щие у основания или на вершине второй пары щупалец. Рот ведет ц
ротовую полость с парной или непарной челюстью и радулой. В рото?
вую полость впадают протоки слюнных желез. Ротовая полость пере-
290
ходит в тонкий пищевод и далее в желудок, в который открываются
протоки асимметричной печени или пищеварительной железы. От же-
лудка отходит различной длины кишка, которая открывается заднепро-
ходным отверстием, лежащим в мантийной полости над головой или
сбоку.
В передней части тела обособлена мантийная полость, в которой у
водных форм расположены одна или две неравные жабры, у назем-
ных — легкое. В мантийную полость кроме заднепроходного отверстия
впадают протоки выделительной и половой систем. Кровеносная систе-
ма незамкнутая, в соответствии с числом жабр у сердца имеются два
или одно (левое) предсердие. От желудочка отходит аорта, которая
разделяется на головной и внутренностный стволы, переходящие в ар-
терии и систему лакун; из них кровь попадает в синусы, из последних
венозная кровь собирается и возвращается к органам дыхания.
Выделяют три основных типа центральной нервной системы:
1) только нервные стволы, без обособленных ганглиев, но с утолщен-
ными участками, 2) пять пар ганглиев, соединенных коннективами, ли-
шенными нервных клеток, но иногда с сохранившимися педальными
нервными стволами, 3) семь пар ганглиев без нервных стволов, обычно
сливающихся вместе. У всех переднежаберных, низших заднежаберных
и легочных нервные тяжи образуют характерный для брюхоногих
нервный перекрест. Среди брюхоногих моллюсков имеются как
раздельнополые, так и гермафродитные формы. Оплодотворение внут-
реннее. У водных форм из яйца выходит личинка трохофорного типа —
велигер или эхиноспира; велигер имеет туловище, ногу и голов-
ной диск с ресничками. В личиночной стадии у многих гастропод про-
исходит так называемый торсионный процесс, в результате
которого внутренностный мешок и колпачковидная раковина поворачи-
ваются на 180°; нервные стволы, идущие от головы в туловище, ока-
зываются перекрещенными, а мантийная полость с жабрами и аналь-
ным отверстием, расположенная у личинки сзади, перемещается вперед
и занимает положение над головой (рис. 154). У наземных форм
и многих морских развитие проходит без личиночных стадий. На' осно-
вании строения органов дыхания, радулы, нервной системы, сердца, но-
ги класс брюхоногих моллюсков разделен на три подкласса: Prosobran-
chia, Opisthobranchia, Pulmonata.
ПОДКЛАСС PROSOBRANCHIA. ПЕРЕДНЕЖАБЕРНЫЕ
Общая характеристика. К прозобранхиям (гр. pros — передний,
branchia — жабры) относится преобладающее число брюхоногих с
асимметричным расположением внутренних органов; мантийная полость
находится впереди на спинной стороне моллюска. У примитивных форм
развиты две жабры, два осфрадия, два предсердия, две почки, две го-
нады. У вышестоящих наблюдается редукция органов, лежащих на
правой стороне, сохранение левого мантийного комплекса: левой жабры
и осфрадия, почки, предсердия, гонады, левой лопасти печени. Хорошо
развита нога, приспособленная к ползанию, закапыванию, прикрепле-
нию и даже к плаванию. Нога часто несет роговую или известковую
крышечку. У примитивных нервная система состоит из перекрещенных
стволов, у более развитых — из пяти пар ганглиев, соединённых кон-
нективами. Раковина спирально-плоскостная, спирально-коническая,
колпачковидная или червеобразная; у немногих форм отсутствует.
Переднежаберные — обитатели преимущественно морей, но часть
живет в пресных водах или приспособилась к жизни на суше. Расти-
19* ' 291
тельноядные и хищники. Раздельнополые, реже гермафродиты. Личин-
ка трохофора и велигер. Ранний кембрий — ныне.
Строение мягкого тела. Мягкое тело переднежаберных разделено
на голову, ногу и туловище. Голова несет основные органы чувств!7
пару щупалец, глаза; впереди на брюшной стороне расположен рот; у
некоторых хищных гастропод рот помещен на конце длинного хоботка,
способного выбрасываться наружу и затем втягиваться внутрь. Нога
тесно связана с головой, образуя с ней единую систему голова — но-
та, которая втягивается целиком в раковину сокращением мускула-
ретрактора, прикрепленного одним концом к ноге, другим к рако-
вине изнутри; у некоторых форм имеется два мускула-ретрактора.
У большинства прозобранхий нога имеет плоскую подошву и служит
для ползания по субстрату, иногда она четко разделяется на переднюю
и заднюю части. Передвижение происходит волнообразным сокраще-
нием продольных мускулов ноги, у пелагических прозобранхий нога име-
ет форму киля. На заднем конце ноги на дорзальной стороне имеется
железа, выделяющая роговую или известковую крышечку.
Все внутренние органы помещены внутри висцерального, или
внутренностного, мешка, занимающего всю полость раковины. Висце-
ральный мешок окружен мантией, которая прикрепляется вдоль устья
к краю раковины. Эпителиальные клетки мантии выделяют раковинное
вещество. У многих прозобранхий слева от головы край мантии вытяги-
вается, образуя вводный сифон, через который в мантийную по-
лость, обособленную в передней части тела, поступает свежая вода с
кислородом. Сифон наиболее хорошо развит у хищных гастропод,
имеющих хобот; выводной, обычно короткий сифон расположен в зад-
ней части тела (в верхнем крае устья) или в особой мантийной щели.
В мантийной полости расположены: жабры, осфрадии, гипобранхиаль-
ная железа; в мантийную полость открываются анальное отверстие и
выводные протоки почек, половых желез. У наиболее примитивных форм
имеются две жабры, или ктенидии, и сопровождающие их осфрадии,
лежащие у основания жабр и служащие для опробования воды (органы
химического чувства). У более совершенных форм сохраняются только
одна левая жабра й один оафрадий. Гипобранхиальная железа выде-
ляет секрет, который склеивает мелкие частицы, попадающие с током
воды в мантийную полость, в более крупные комки. Все эти частицы и
комки, а также продукты обмена, выходящие из анального отверстия,
выносятся наружу через особый сифон. У немногих прозобранхий
жабры редуцированы полностью и газообмен происходит при помощи
эпителия или вторичных жабр, возникших по периферии мантии (сем.
Patellidae с конической раковиной). Позади жабр лежит сердце, со-
стоящее из желудочка и одного или двух предсердий (число их нахо-
дится в коррелятивной связи с числом жабр). От желудочка отходит
главная аорта, которая затем разделяется на артерии, снабжающие
кровью через систему лакун мантию, ногу и все внутренние органы.
Отсюда кровь собирается в венозные синусы и через вены поступает в
почки, жабры и затем в предсердия. Кровь бесцветная или голубая от
наличия гемоцианина.
Пищеварительная система начинается ртом, ведущим в ротовую
полость и далее в глотку. Известна только одна находка раковины из
нижнего мела Англии, внутри которой сохранился обызвествленный пи-
щеварительный тракт (см. рис. 2, 3), описанный Кейси. В ротовой по-
лости лежат две челюсти, а в ее задней части язык — мускулистый
вырост, несущий терку, или радулу, с рядами мелких поперечных зуб-
чиков (рис. 155). Зубчики состоят из протеина с примесью железа, фос-
292
фора и других элементов. Число и форма зубчиков очень варьируют.
Общее их число колеблется от 1 до 750 000; число рядов зубчиков из-
меняется от одного до сотен. В зависимости от характера расположения
и формы зубчиков выделяют несколько типов радулы: рипидоглоссныщ
тэниоглоссный, стеноглоссный и т. д. Новые зубчики радулы возника-
ют в особом радулярном кармане. Вблизи языка открываются протоки
Рис. 155. Строение радулы у переднежаберных:
1 — схематический разрез через ротовую полость Charonia; 2 — вид на радулу
внутри глотки Natica; 3—6 — радулы: 3 — Gibbula, 4 — Buccinum, 5 — Scala,
6 — Patella; 7 — полый ядовитый зуб Conus, сильно увел.;
гл — глотка, р — рот, рад — радула, хр — хрящ, ч — челюсть
слюнных желез, способствующие перевариванию пищи; у некоторых
железы выделяют серную кислоту. За глоткой следует пищевод, пере-
ходящий в мешковидный желудок; с ним связана пищеварительная
железа, или печень, состоящая из двух неравных лопастей и занимаю-
щая обычно всю спиральную часть раковины. Печень у примитивных
форм двухлопастная и служит для переваривания пищевых частиц; у
вышестоящих она асимметрична, левая лопасть развита сильнее и яв-
ляется пищеварительной железой и местом абсорбции пищи.
Желудок имеет довольно сложное строение. В нем выделяются три
функционально различные области: сортировочное поле, состоя-
щее из системы складок и борозд, где происходит сортировка и распре-
293
деление пищевых частиц, а также их переваривание, кутикулярный
желудочный щит и мешок протостиля, отделенный от кишки про-
дольными складками. Протостиль — палочка, образованная из уп-
лотненной слизи; реснички мешка вращают ее и вытягивают из пище-
вода в желудок слизистый шнур с пищей, с которым он соединен. У ря-
да форм развит кристаллический стебелек, состоящий из бел-
ков и ферментов. Конец стебелька вдается в желудок, где его часть
растворяется и освобождает ферменты, способствующие перевариванию
углеводов. Желудок переходит в хорошо развитую кишку, которая слу-
жит для формирования фекалий и обволакивания их плотной слизью.
Это имеет приспособительное значение, так как анальное отверстие
открывается в мантийную полость. Пара почек или одна левая служат
органами выделения; они сообщаются с перикардием и мантийной по-
лостью.
Нервная система состоит у одних из нервных стволов, у других из
пяти пар нервных узлов, или ганглиев, соединенных коннективами, ли-
шенными нервных клеток. В голове расположены парные церебральные,
или мозговые, узлы, иннервирующие щупальца, глаза. Боковые ганглии
иннервируют стенки тела, сифон; ножные, или педальные, ганглии —
ногу. Две остальные пары расположены в висцеральном мешке и ин-
нервируют внутренние органы. Перекрест двух нервных стволов, идущих
от плеврального ганглия к висцеральному, связан с торсионным процес-
сом, который будет рассмотрен ниже. К органам равновесия относятся
статоцисты, представленные парой пузырьков, заполненных жидко-
стью, в которых лежат микроскопические известковые конкреции. Раз-
витие нервной системы у прозобранхий шло по пути усиления и уве-
личения ганглиев (ганглионизация).
Переднежаберные моллюски низшие — гермафродиты, высшие —
раздельнополые. У низших форм половые продукты выводятся в воду
и там оплодотворяются; у многих оплодотворение внутреннее, у них по-
ловые органы устроены довольно сложно. Оплодотворенные яйца окру-
жаются студнеобразными коконами; несколько коконов образуют раз-
личной формы кладки, характерные для каждой группы моллюсков.
У низших прозобранхий из яиц выходит личинка — трохофора,
у более развитых — в е л и г ер, или парусник, ведущий пелагический
образ жизни. На ранних стадиях развития велигер двусторонне-симмет-
ричен, имеет спиральную раковинку, ротовое отверстие впереди, аналь-
ное сзади, на более поздних стадиях происходит торсионный по-
ворот: висцеральный мешок и раковина поворачиваются на 180° про-
тив часовой стрелки. Стебелек, соединяющий туловище с ногой и
головой, при этом винтообразно закручивается. Мантийная полость
оказывается спереди, над головой моллюска. Вместе с ней перемещают-
ся задняя кишка с анальным отверстием, жабры, осфрадии, гипо-
брахиальные железы, отверстия почек, половые отверстия, сердце, при
этом все органы, лежащие до закручивания на правой стороне, после
закручивания оказываются на левой, и наоборот. Перекрещиваются вос-
ходящие и нисходящая части петли кишечника; нервные стволы, соеди-
няющие плевральные и париетальные ганглии, закручиваются в форме
восьмерки. Этот перекрест нервных стволов получил название х и а с то-
не в р и и.
Строение раковины. Первые обороты раковины образуются у личин-
ки; эта часть раковины носит название протоконха (рис. 156); про-
токонх обычно гладкий или покрыт тонкими струйками. Первый оборот
имеет вид округленного пузырька, переходящего в обороты геликоидной
спирали. Размеры протоконха и число оборотов сильно варьируют у
294
разных прозобранхий; переход в раковину, образованную после оседа-
ния личинки, бывает выражен довольно отчетливо типом скульптуры,
иногда изменением положения оси; если положение оси вращения спи-
рали протоконха отличается от положения оси вращения спирали по-
следующих оборотов раковины, или телеоконха, то такой протоконх
Рис. 156. Различные типы протоконхов:
1—5 — гомеострофный, 6 — гетерострофный
называется гетер остро |фным (рис. 156, 6), если эти оси совпа-
дают— гомеострофный (рис. 156, 1—5).
Стенка раковины построена из органического и известкового
вещества. Наружный слой, или периострак, строится, как у всех
моллюсков, передним краем мантии в особой мантийной складке и яв-
ляется органическим, предохраняющим известковый слой раковины от
растворяющего действия воды; в ископаемом состоянии этот слой со-
храняется редко; обычно он окрашен пигментом в разнообразные цве-
та, придающие раковине красоту и защитную окраску, позволяющую
моллюскам сливаться с окружающей средой. Следы окраски встречают-
ся у прозобранхий начиная с ордовика. Окраска лучше всего сохра-
няется у окремнелых раковин’. У всех прозобранхий наиболее полно
развит средний перекрещенно-пластинчатый слой, состоящий из па-
раллельных пластин первого порядка толщиной 0,02—0,04 мм, ориенти-
рованных перпендикулярно поверхности раковины. Каждая пластинка
построена из тончайших (до 0,001 мм) пластинок второго порядка, на-
клоненных под углом 41° к длинной оси пластинки первого порядка.
Внутренний слой перламутровый, образуется тончайшими пластинка-
ми (менее 0,001 мм) арагонита, расположенными параллельно поверх-
ности раковины; он развит преимущественно у археогастропод.
Форма раковины. Раковина несет защитную функцию и слу-
жит местом прикрепления мускулов. Различают три основных типа ра-
ковины: колпачковидную, планоспиральную, турбоспиральную (кони-
спиральную.). Колпачковидная раковина (рис. 157) имеет форму
конического двусторонне-симметричного колпачка, напоминающего ра-
ковину моноплакофор; открытое основание раковины называется усть-
ем, или апертурой, а начальная точка роста — вершиной, или
апексом. На внутренней поверхности раковины иногда сохраняется
подковообразный след прикрепления мускула-ретрактора, прерванный
впереди. Колпачковидная раковина, с одной стороны, возникла впервые
у самых примитивных прозобранхий, а с другой --- могла возникать
вторично в связи с приспособлением к малоподвижному образу жизни
в других группах; последнее подтверждается возникновением в онтоге-
незе спирально-конического протоконха.
Вторым и наиболее характерным типом раковины является спи-
ральная раковина, представляющая конической формы трубку, свер-
нутую в спираль, закрытую на одном, апикальном, конце и открытую
295
на другом. Тело моллюска занимает всю полость раковины, прикреп-
ляясь к ней одним или двумя мускулами, которые служат для втяги-
вания головы и ноги моллюска внутрь раковины. Различают два типа
спирали: планоспиральный и турбоспиральный. Планоспиральная
раковина (см. рис. 158, 1) имеет вид свернутой в одной плоскости труб-
ки и напоминает раковины многих головоногих моллюсков, но в отли-
Рис. 157. Схема строения и форма раковины и крышечки:
1 — спирально-коническая; 2 — удлиненно-яйцевидная (Oliva)-, 3 — коническая
(Conus); 4 — колпачковидная (Patella); 5 — незакономерно. свернутая (Verme-
tus); 6 — турителлоидная, с развернутыми последними оборотами (Nerinella);
7 — ушкообразная (Haliotis); 8 — спирально-конические (продольное сечение):
8а — с ложным пупком (лп) и 86 — с настоящим, или истинным, пупком (нп); 9—
10 — крышечки: 9 — Cirsotrema, 10 — Natica;
an —апертура, вг — внутренняя губа, вш — вершина, зв — завиток, кл — кал-
лус, нг — наружная губа, о — оборот, ос — отверстия заросшего синуса, осп —
основание последнего оборота, по — последний оборот, ст — столбик, ш — спи-
ральный шов
чие от последних ее внутренняя полость не разделена перегородками
на отдельные камеры и занята целиком висцеральным мешком моллюс-
ка. Этот тип раковины характерен для наиболее примитивных прозо-
бранхий. Раковина состоит из отдельных оборотов, разделенных швом;
обороты соприкасаются друг с другом, образуя так называемую эво-
лю тную раковину — раковину с необъемлющими оборотами. Два уг-
лубления, возникающие с боков в связи с увеличением оборотов, назьь
ваются пупком, или умбиликом. Если каждый последующий обо-
рот охватывает целиком или частично предыдущий оборот, то такая
раковина называется инволютной; у нее пупок либо отсутствует,ли-
бо представлен небольшим углублением (см. рис. 158,1,2). У прозобран-
хий, обладавших такого типа раковиной, все внутренние органы были
парными: две жабры, два осфрадия, одна пара целомодуктов, два пред-
сердия, две половые железы и т. д. В полости симметрии у внешнего
края устья проходит анальная выемка, через которую выводилась из.
мантийной полости вода с продуктами обмена и посторонними части-
296
Рис. 158. Отряд Archaeogastropoda:
1 — Bellerophon (силур — триас): la — со стороны устья, 16 — снаружи; 2 — Еиот-
phalus (сил^р — ранняя пермь): 2а — вид снизу, 26 — со стороны устья; 3 — Murchi-
sonia (ордовик — триас); 4 — Helcionella (кембрий); 5 — Ecculiomphalus (ордовик —
силур); 6 — Diodora (поздний мел — ныне); 7 — Pleurotomaria (юра — ранний мел);
8 — Theodoxus (олигоцен— ныне); 9—: Gibbula (поздний мел — ныне);
ащ — анальная щель, с — синус, мп •— мантийная полоска
цами. По мере роста анальная выемка частично зарастала, оставляя
на раковине мантийную полоску (рис. 158, J).
Турбоспиральная, или улитковидная (кониспиральная), ра-
ковина наиболее типична для прозобранхий (см. рис. 157, /). Она пред-
ставляет собой известковую трубку (геликон), свернутую в кониче-
скую спираль. Наблюдаются все переходы от планоспиральной к тур-
боспиральной раковине; вначале первая становилась несколько асим-
метричной и отличалась размерами пупков, асимметричным положе-
нием оборотов. Затем один из пупков увеличивался в размерах, а вто-
рой уменьшался, пока не исчезал совсем. Начальные обороты в этом
случае начинали выступать наружу. Раковина приобретала коническую
форму (рис. 158, 1, 2, 7—9).
Раковину ориентируют таким образом, чтобы ее замкнутый конец,
отвечающий начальной стадии роста, был обращен вверх, а устье на-
ходилось внизу и было обращено к наблюдателю. При подобной ори-
ентировке, если устье расположено справа, раковины называют право-
завитыми (их большинство), если устье расположено слева, —
левозавитыми. Правозавитая спираль закручена по ходу часовой
стрелки, если смотреть на раковину сверху, и против часовой стрелки—
у левозавитых. Один полный виток спирали образует оборот; самый
крупный оборот, несущий устье, называется последним; остальные
обороты, расположенные над ним, образуют завиток, или просто спи-
раль. У турбоспиральных раковин может быть только один умбилик.
Если он прослеживается до вершины спирали, то называется истин-
ным; если ограничен только последним оборотом — ложным. Если
обороты тесно соприкасаются друг с другом, то вместо умбилика воз-
никает известковая ось — с т о л б и к, или колумелла. В ряде слу-
чаев обороты спирали могут совсем не соприкасаться и раковина стано-
вится развернутой, или скаляридной. Подобные раковины встре-
чаются у форм, ведущих преимущественно прикрепленный образ жизни;
они похожи на известковые трубки серпул. Внешний облик турбоспи-
ральной раковины крайне разнообразен. Известны раковины шаровид-
ные, ушкообразные, конические, цилиндрические, башенкообразные,
винтообразные, веретенообразные. Часто при характеристике формы ра-
ковины пользуются определениями, производными от названий родов
(например, натикоформная, букциноформная, пателлиформная рако-
вина).
В зависимости от числа оборотов, слагающих спираль, различают
раковины малооборотные (от 2 до 4) и многооборотные
(от 5 и выше). Высота спирали зависит от количества оборотов, скорос-
ти их возрастания и взаимного их соотношения. Если последний оборот
охватывает все предшествующие обороты полностью, то такая раковина
называется конволютной, если не полностью с образованием апи-
кального пупка, то инволютной (см. рис. 157, 2, 3).
Устье, через которое моллюск высовывает наружу голову и ногу,
имеет разнообразную форму и размеры. В нем различают: наружный
край, или наружную губу, и внутренний край, или внутреннюю губу.
Устье может быть голостомным, или цельнокрайним, без си-
фональной вырезки или канала; оно характеризует более древних про-
зобранхий; и сифоностомным, снабженным в нижней части выем-
кой или более или менее вытянутой трубкой — сифонал ьным ка-
налом, в котором помещается вводной сифон. У археогастропод с тур-
боспиральной раковиной, так же как у форм с планоспиральной рако-
виной, наружный край устья снабжен узкой анальной выемкой, которая
по мере роста постепенно зарастает, оставляя анальную полоску (см.
298
рис. 158, 1, 7). По ней можно судить о положении выводного канала.
При прерывистом зарастании от полоски остается ряд щелей (род На-
liotis, рис. 157, 7). Анальная выемка так же, как и щели, указывает на
сохранение парных органов, при некоторой редукции правого комплекса
у форм, имевших турбоспиральную раковину. У прозобранхий с сифо-
.ностомным устьем сохраняется только левый мантийный комплекс. На-
ружная губа отличается большим разнообразием: она может быть тон-
кой или утолщенной, отвернутой и закругленной, с выемкой или без;
нее; иногда она приобретает форму крыла или пальцеобразных высту-
пов. Внутренняя губа также устроена по-разному, она может быть тон-
кой или толстой, узкой или широкой, только прилегать к столбику или
покрывать столбик тонким или, наоборот, толстым слоем, Образуя мас-
сивное мозолевидное утолщение, или каллус. Различают умбиликаль-
ный каллус, закрывающий умбилик, и париетальный столбик, располо-
женный в верхней части устья.
Скульптура. Раковины прозобранхий редко бывают гладкими.
Обычно они украшены различной скульптурой, в основном состоящей
из спиральных и аксиальных, или осевых, элементов. Спиральные
элементы расположены параллельно шву. Аксиальные элементы
ориентированы параллельно оси навивания оборотов. У колпачковидных
раковин различают концентрическую и радиальную скульптуру. Эле-
менты первой расположены параллельно контуру основания раковины,
а элементы второй лучеобразно расходятся от макушки. При сочета-
нии спиральных и аксиальных элементов возникает сложная скульптура.
Различают: сетчатую (ретикулятную) — спиральные и аксиальные
элементы резко развиты, декуссатную — преобладают элементы
спиральной скульптуры, канцеллятную — наоборот, преобладают
аксиальные элементы. К основным элементам скульптуры относятся
ребра, кили, бороздки, шипы, бугорки, ямочки. Ребра могут быть спи-
ральными и аксиальными, они различаются по величине и по форме.
Межреберные пространства также отличаются по форме и величине.
Часто скульптура последнего оборота отличается от таковой спирали.
Остальные элементы скульптуры также крайне разнообразны и измен-
чивы. Следы утолщенной наружной губы на предшествующих оборотах
образуют поперечные валики, или варикозные утолщения. Они отра-
жают периоды замедления дальнейшего роста раковины и связаны с
периодами размножения или изменения сезонных условий.
У большинства прозобранхий на задней части ноги имеется рого-
вая или известковая крышечка, служащая для закрывания устья
при втягивании тела внутрь раковины. Крышечка образуется за счет
особой железы, расположенной в ноге. Иногда она используется для
закапывания и как средство защиты или нападения; в этом случае у
нее имеется острый край. В зависимости от характера нарастания раз-
личают крышечки: спиральные, концентрические и сферические. Боль-
шинство крышечек плоские, но известны и конические. В ископаемом
состоянии встречаются только известковые крышечки. Отдельные на-
ходки были описаны из отложений начиная с силура.
Большинство прозобранхий раздельнополы, и половой диморфизм
проявляется в размерах и пропорциях раковины. У женских особей ра-
ковина обычно шире и крупнее, чем у мужских. У современных форм
(род Alcithoe) известны яйца с известковой скорлупой, но в ископаемом
состоянии они еще не найдены.
Внутреннее строение раковины. К сожалению, раковина прозобран-
хий не сохраняет никаких особенных внутренних структур. Строение
внутренних органов не находит отражения на раковине. У колпачковид-
299
ных форм видны отпечатки одного или двух мускулов в форме подковы;
у планоспиральных раковин иногда сохраняются отпечатки двух муску-
лов-ретракторов позади устья на стенке последнего оборота. Отпечатки
мускула-ретрактора у турбоспиральных раковин не наблюдаются. У ря-
да прозобранхий на наружной и внутренней губах развиты складки —
особые валики, уходящие внутрь устья. Остатки подобных складок со-
храняются на внутренних оборотах; они видны на пришлифовках, сде-
ланных, вдоль вертикальной оси раковины. Рассматривая оборот за
оборотом, можно проследить характер изменения и поведения складок
в онтогенезе (см. рис. 159, 4).
Основы систематики и классификация. Как было отмечено, ракови-
на несет очень мало информации о строении мягкого тела и внутрен-
них органов, поэтому классификация прозобранхий основана на особен-
ностях строения радулы, органов дыхания, нервной системы, орга-
нов размножения. Единой принятой системы прозобранхий до сих пор'
нет. В палеонтологической литературе принято разделять прозобранхий
на три отряда: Archaeogastropoda, Mesogastropoda и Neogastropoda.
Отряд Archaeogastropoda. Археогастроподы. Наиболее примитив-
ные прозобранхий с мантийной полостью впереди, перекрестом нерв-
ных тяжей, слабой ганглионизацией; у низших форм две перистые жаб-
ры, два осфрадия, два предсердия, у более развитых — редуцированы
правая жабра и осфрадий, но сохраняются два предсердия (одно руди-
ментарное). Большинство ведут малоподвижный образ жизни, питают-
ся растительной пищей; хоботок и сифон отсутствуют; пищеварение
внутриклеточное, желудок разделен на отделы, имеется хрустальный
столбик. У самых древних археогастропод раковина колпачковидная,,
затем спирально-плоскостная, но у большинства турбоспиральная, или
вторично-колпачковидная. Устье голостомное, часто с анальной вырез-
кой для выхода анального отверстия. Крышечка известковая, роговая
или отсутствует.
Большинство археогастропод—обитатели прибрежной полосы моря,
некоторые живут в зоне сублиторали и даже опускаются на большие
глубины (Pleurotomaria). Небольшая группа археогастропод приспо-
собилась к жизни в пресных водах (Neritidae) и даже на суше (Heli-
cinidae). В связи с этим все развитие у них проходит внутри яйцевых
капсул.
Отряд Mesogastropoda. Мезогастроподы. Мантийный комплекс
представлен одной жаброй, одним предсердием. Низшие мезогастропо-
ды питаются растительной пищей, в связи с чем желудок имеет сор-
тировочный отдел, хрустальный столбик; пищеварение внутриклеточ-
ное; радула тэниоглоссная, в каждом ряду 7 зубчиков. У вышестоящих
форм, перешедших к плотоядному питанию, эволюция нервной системы
привела к увеличению ганглиев, увеличению размеров левой жабры и
осфрадия, совершенствованию органов чувств, особенно глаз, и засе-
лению новых биотопов. Одни мезогастроподы сохранили ползающий об-
раз жизни и турбоспиральную раковину, другие перешли к неподвиж-
ному, прикрепленному образу жизни и приобрели развернутую черве-
образную раковину, третьи приспособились к зарыванию в осадок, чет-
вертые — к сверлению раковин двустворок, пятые перешли к пелагиче-
скому образу жизни и очень сильно изменились (отряд Heteropoda).
Тело у последних стекловидно-прозрачное, раковина спирально-плос-
костная, спирально-коническая, колпачковидная или отсутствует. Не-
большая группа паразитирует на иглокожих, голотуриях, при этом у
них редуцировалась раковина и ряд внутренних органов и резко изме-
нился общий облик тела. Небольшая часть мезогастропод приспособи-
300
и Neogastropoda (8—//):
Рис 159 Отояды Mesogastropoda (/—7) и Neogastropoda (8 11). ....
1 Vivinarus (юоа-ныне)- 2 - Cerithium (поздний мел-ныне); 3 - Tumtela
1‘Тне) &еХа Тюра-мел); 5 - Aporrhais Vermetus
(S-ныне) '7 -
»"ыГн.СнТ«(П"н.л“ыТн.нал, и, - « - уееь. „
складками
лась к жизни в пресных водах, заселив пресные водоемы почти всего
земного шара (Viviparidae, Melanidae), немногие перешли к жизни на
суше (роды Pterocyclus, Cyclostoma). Объединяет свыше 700 родов,
(см. рис. 159, 1—7).
Отряд Neogastropoda. Неогастроподы. Высшие прозобранхий, пре-
имущественно хищники, с сильно концентрированной нервной системой,
левым мантийным комплексом органов, с узкой радулой стеноглоссно-
го типа, с 1—3 зубчиками в каждом ряду. Радула приспособлена к пло-
тоядному способу питания. Хорошо развит сифон; рот расположен на
конце хоботка. Раковина турбоспиральная с хорошо развитым сифоно-
стомным устьем. Раздельнополы. Проходят стадию личинки велигера.
Некоторые сверлят раковины двустворок, другие давят их как грецкие
орехи, зажимая краем крышечки и специальным зубовидным отростком,
развитым в наружной губе. Неогастроподы появляются в мелу и су-
ществуют до настоящего времени. Объединяют около 400 родов (см.
рис. 159, 8—11). <
ПОДКЛАСС OPISTHOBRANCHIA. ЗАДНЕЖАБЕРНЫЕ
Общая характеристика. У опистобранхий (гр. opisthen — задний,
branchia — жабры) раковина от спирально-конической у примитивных до^
уплощенной с сокращением числа оборотов, расширением устья, до ох-
вата ее мантией, превращением ее во внутреннюю и редукцией до прос-
той'пластинки и, наконец, полным исчезновением. Мантийная полость
на правом боку тела, жабры концом направлены назад, хиастонев-
р и я известна у наиболее древних форм, обычно отсутствует, пищевари-
тельный тракт выпрямлен; у одной группы (голожаберные) развиты
вторичные жабры; предсердие позади желудочка; нервные узлы кон-
центрированы вокруг глотки; гермафродиты. Занимают разнообраз-
ные биотопы морей и суши; питаются растительной пищей или хищники,
иногда паразитируют. Известны начиная с раннего карбона.
Строение мягкого тела. Опистобранхии произошли от прозобран-
хий, поэтому более примитивные их представители несут ряд черт,
близких к последним. Мантийная полость смещена назад, сохраняются
одна жабра и одно предсердие и хиастоневрия. У вышестоящих форм
происходит раскручивание (деторсионный процесс): нервные тя-
жи становятся параллельными, ганглии концентрируются вокруг около-
глоточного кольца и у голожаберных все ганглии сливаются в одну об-
щую массу, тем самым пищеварительный тракт выпрямляется, тело
вытягивается и приобретает вторично-двустороннюю симметрию (см.
рис. 154, В). Жабра редуцируется и вместо нее на заднем конце тела
около анального отверстия возникают вторичные жабры. За счет пище-
вода развивается вторичный желудок — так называемый мышечный
зоб, вооруженный известковыми пластинками (зубами), предназначен-
ными для перетирания пищи или разламывания раковин двустворчатых
моллюсков. Половая система устроена сложно; все опистобранхии гер-
мафродиты; морские формы проходят личиночную стадию.
Строение раковины. У наиболее примитивных, стоящих близко к
прозобранхиям, раковина спирально-коническая с крышечкой. В связи
с освоением новых биотопов на разных глубинах и приспособлением к
пелагическому образу жизни раковина облегчается, теряет свою функ-
цию как защитное образование, обрастает мантией и постепенно реду-
цируется. Вначале последний оборот становится широким, раковина
приобретает форму колпачка, затем изогнутой пластинки, почти целиком
закрытой мантией. У одной группы опистобранхий (Berthelinia) разви-
302
валась двустворчатая раковина, конвергентно сходная с раковиной дву-
створок. У нее протоконх спирально-конический, а телеокойх состоит из
двух створок (рис. 160, 3), соединенных связкой, или лигаментом, вдоль
внешнего дорзального края, имеется один мускул-замыкатель. У мно-
гих заднежаберных раковина отсутствует полностью и на дорзальной
стороне развиты разнообразные выросты мантии, обычно окрашенные
в различные цвета.
Рис. 160. Подкласс Opisthobranchia:
1 — Spiratella (неоген — ныне, увел.); 2 — Acteocina (юра — ныне, увел.); 3 —
Berthelinia (эоцен — ныне, длина створок до 7 мм)
Основы систематики. Опистобранхии на основании строения внут-
ренних органов, наличия или отсутствия раковины разделены на не-
сколько отрядов, из которых будет рассмотрен один отряд Pteropoda.
Отряд Pteropoda. Птероподы (гр. pteron—крыло, pous, род. podos—
нога), или крылоногие, объединяют опистобранхий, перешедших к пе-
лагическому образу жизни; нога редуцирована; параподий превращен
в двухлопастной плавник; хорошо развиты органы равновесия — ста-
тоцисты; одни питаются микропланктоном, имеют раковинку малень-
ких размеров, спирально-коническую, лево- или правозавернутую, сла-
боразвитую радулу, известковые пластинки в желудке; другие —
хищники, утратили раковину, известковые пластинки в желудке, име-
ют хорошо развитые челюсти, радулу; у личинок возникает протоконх,
который потом сбрасывается. Крылоногие живут большими скопления-
ми, служат пищей для многих животных (китов); их скелеты прини-
мают участие в накоплении птероподовых илов, распространенных на
глубинах от 1000 до 2700 м. В глинах миоцена Керченского полуостро-
ва Крыма наблюдается массовое захоронение раковинок спириалисов
(отсюда название спириалисовые глины), накопленных на дне бассей-
на, в котором существовали неблагоприятные условия для жизни дон-
ных организмов.
Первые достоверные находки известны начиная с эоцена. Недосто-
верные находки указываются из позднего мела (рис. 160, 1).
ПОДКЛАСС PULMONATA. ЛЕГОЧНИКИ
Общая характеристика. Подкласс пульмонат (лат. pulmo, род.
pulmonis — легкое) объединяет наземных и пресноводных гастропод, у
которых возник орган воздушного дыхания — легкое, й небольшое чис-
ло вторично-морских. Голова несет одну или две пары щупалец, хоро-
шо развиты глаза, радула снабжена длинными заостренными зубчика-
303
ми, имеется непарная подковообразная челюсть, сердце состоит из од-
ного желудочка и одного предсердия, нервные ганглии сконцентриро-
ваны в окологлоточном кольце. Раковина турбоспиральная, планоспи-
ральная, колпачковидная или отсутствует (слизни). Растительноядные,
всеядные и хищники. Гермафродиты; развитие без стадии личинки (жи-
ворождение). Первые пульмонаты известны с карбона; более разнооб-
разны начиная с позднего мела.
Строение мягкого тела. Хорошо обособлена голова, несущая одну
или две пары щупалец; глаза в основании щупалец или на их концах;
хорошо развита радула, снабженная острыми удлиненными зубчиками.
В полости тела развивается легкое с большой сетью кровеносных со-
судов (см. рис. 154, Г); легкое открывается наружу особым отверсти-
ем — пневмостомом, сокращение которого обеспечивает газообмен
или наполнение легкого водой. Сердце лежит позади легкого, причем
предсердие впереди, желудочек сзади. Рядом расположена правая поч-
ка, которая одним отверстием открывается в перикардий, другим в ман-
тийную полость. Пищеварительный тракт приспособлен в основном к
растительной пище; отсутствуют сортировочный отдел и протостиль.
Печень выполняет функцию выделения пищеварительного сока и пере-
вариванйя. Нервные тяжи укорочены и раскручены (как правило, от-
сутствует перекрест), ганглии концентрируются вокруг пищевода. Все
пульмонаты гермафродиты, половой аппарат сложный. Продуцируют
немного яиц, чаще всего откладывая их подобно черепахам в землю
(наземные) и на твердый субстрат (водные). Яйца у некоторых назем-
ных форм достигают диаметра 45 мм и имеют известковую скорлупу.
В ископаемом состоянии в отложениях олигоцена Англии и миоцена
Уганды найдена скорлупа яйца диаметром до 30 мм. Индивидуальное
развитие у большинства без личиночной стадии, у некоторых водных
с личинкой велигер.
Строение раковины. Большинство пульмонат имеют известковую
раковину разнообразной формы: турбоспиральную, планоспиральную,
колпачковидную (у обитателей быстрых рек). Раковина выполняет
функции защиты от врагов, испарения, механических повреждений мяг-
кого тела. У форм, живущих в условиях сухого климата, раковина с
толстыми стенками, у обитателей влажного Климата, пресных вод —
тонкостенная. Раковина обычно гладкая, у форм, живущих на скалах,
часто ребристая; ребра придают раковине прочность, и при строитель-
стве раковины достигается экономия в материале (рис. 161). Обычно
при наступлении неблагоприятных условий устье закрывается эпи-
фрагмой — особой пленкой слизи, содержащей известь, затвердева-
ющей после ее выделения. У одной группы имеется крышечка, не го-
мологичная крышечке прозобранхий, представляющая видоизмененный
край наружной губы, подвижно сочленяющаяся с краем раковины и
плотно закрывающая устье. Подобная крышечка получила специаль-
ное название — клаузилий. Одна группа пульмонат полностью ли-
шена раковины (слизни).'
Основы систематики. На основании строения внутренних органов,
наличия одной или двух пар щупалец пульмонаты разделены на два
отряда: Basommatophora и Stylommatophora.
Отряд Basommatophora. Базомматофоры (гр. basis — основание,
дно, omma — глаз, phoros — несущий), или сидячеглазые, объединяют
пульмонат, ведущих преимущественно амфибиотический образ жизни
в опресненных участках моря, в пресных водах рек, озер, прудов, бо-
лот, карстовых пещер и лишь редко живущих на суше. Голова несет
одну пару щупалец; глаза расположены у их основания. У обитателей
304
моря сохраняется одна жабра или жабры отсутствуют, вся мантийная
полость наполняется водой, из которой извлекается кислород. Все ос-
тальные дышат легкими. Раковина имеет форму колпачка, конической
спирали (семейство прудовиков Lymnaeidae) или симметричной спира-
ли (семейство катушек Planorbidae). Устье голостомное или несет на
Рис. 161. Подкласс Pulmonata:
1 — Helix, (поздний мел — ныне): 1а,— положение раковины при движении мол-
люска, 16 — вид со стороны устья; ,2 — Lymnaea (поздняя юра — ныне); 3 —
Planorbis (поздняя юра — ныне): За — вид со стороны устья, 36 — вид со сто-
роны пупка
внутренней губе утолщенную складку. Сидячеглазые пульмонаты
появляются в поздней юре и существуют доныне, встречаясь в ископае-
мом состоянии преимущественно в континентальных пресноводных от-
ложениях (рис. 161, 2, 3).
Отряд Stylommatophora. Стиломматофоры (гр. stylos — палочка),
или стебельчатоглазые, объединяют наиболее высокоорганизованных
пульмонат, ведущих преимущественно наземный образ жизни, частично
земноводный, населяя зону прибоя, частично полуземноводный, оби-
тая вблизи пресноводных водоемов. Голова нерет две пары щупалец —
одну пару втягивающихся щупалец, на концах которых расположены
глаза, и одну пару более коротких тактильных щупалец. У большинст-
ва имеется хорошо развитая раковина, защищающая тело от избыточ-
ной потери влаги и от' многочисленных врагов, в первую очередь от на-
секомых. Раковина обычно массивная, иногда достигающая длины 14 см.
Устье часто снабжено с внутренней стороны различными зубцами или
особой крышечкой — замочком (клаузилий). Многие стиломматофоры
закрывают устье специально выделяемой эпифрагмой. У большой груп-
пы (слизни) раковина полностью отсутствует или имеет вид пластинки
на спине, прикрытой мантией. В ископаемом состоянии стиломматофоры
встречаются начиная с карбона, однако они наиболее разнообразны и
представительны в кайнозое, особенно в континентальных отложениях
неогена и антропогена. Современные представители отряда заселили все
участки суши от Арктики до тропиков, от равнин до горных цепей, оби-
тая на всех высотах, вплоть до 5000 м; они приспособились к перене-
сению жаркого, засушливого и холодного сезонов и других неблаго-
приятных условий, переходя в спячку иногда на несколько месяцев
(рис. 161, 1).
20 в. В. Друщии
305
История развития гастропод и экология. Наиболее древние брюхо-
ногие, относимые к отряду археогастропод, известны из кембрия..
Вероятно, они произошли от общих предков с моноплакофорами. Раз-
витие шло по пути превращения низкоконической раковины в высоко-
коническую, а затем в спирально-плоскостную (беллерофонтины). У по-
следних мантийный комплекс органов находился впереди над головой;,
они имели парные жабры, два предсердия и, наверное, обладали плос-
кой ногой, приспособленной для ползания. В позднем кембрии появи-
лись представители плеуротомариин, обладающие асимметричной,
турбоспиральной раковиной. Наличие у них мантийного синуса указы-
вает на наличие парных жабр, переднее положение мантийного комп-
лекса; нервная система обладала хиастоневрией, т. е. нервные стволы;
были перекрещены. От плеуротомариин возникают другие группы гаст-
ропод, сохраняющие две жабры, два предсердия и две почки; они ве-
дут малоподвижный образ жизни (пател л ины), питаются раститель-
ной пищей, обладают примитивным строением желудка — имеют сор-
тирующий отдел, хрустальный столбик, отличаются внутриклеточным
пищеварением. Часть археогастропод приспосабливается к жизни в-
приливно-отливной зоне моря, в солоноватоводных и пресных бас-
сейнах.
В карбоне от археогастропод возникают мезогастроподы, до-
стигающие расцвета в настоящее время. Они обладают одной жаброй,,
одним предсердием, одной почкой и характерной лентовидной радулой,
с 7 зубами в каждом ряду. Низшие мезогастроподы ведут также мало-
подвижный образ жизни, а некоторые превращаются в неподвижных,,
прикрепляющихся к подводным предметам (верметиды); они являются,
растительноядными микрофагами, или планктоноядными; они сохра-
няют сортировочный отдел желудка, внутриклеточное пищеварение, хру-
стальный столбик. Часть мезогастропод переходит к жизни в пелагиалш
(киленогие), другие, наоборот, — к зарыванию в осадок.
От мезогастропод в меловой период возникает новый отряд нео-
гастропод, представители которого приспосабливаются к плотояд-
ному питанию, в связи с чем увеличивается их подвижность, активность;
переход к активному хищничеству приводит к совершенствованию пище-
варительного тракта, увеличению размера жабры, развитию ганглиев,,
совершенствованию органов чувств, особенно глаз; развивается узкая,
радула с 1—3 зубами в каждом ряду.
В карбоне от археогастропод возникли два новых подкласса гаст-
ропод: заднежаберные и легочники. Они претерпели раскру-
чивание нервных стволов, утратили хиастоневрию; это вызвало у зад-
нежаберных смещение направо и назад мантийной полости и ктенидия,,
лежащего позади сердца, раскручивание петли кишечника. Организация
заднежаберных моллюсков оказалась очень пластичной. Заднежаберные-
моллюски освоили разные биотопы моря, в том числе и пелагиаль, и
эволюционировали по пути редукции раковины. Раковина вначале умень-
шается, становится легче, постепенно обрастает мантией, становится
внутренней и затем полностью исчезает. В связи с подобными измене-
ниями тело становится двусторонне-симметричным, червеобразным,,
уменьшается мантийная полость и жабра, развивается кожно-мускуль-
ный мешок, на заднем конце тела появляются вторичные жабры; у не-
большой группы развивается двустворчатая раковина. Хотя заднежа-
берные являются типичными морскими обитателями, часть из них при-
спосабливается к жизни в пресных водах.
Легочные моллюски на ранних этапах эволюции жили в при-
брежной зоне моря, обладали перекрещенной нервной системой, нодлж
306
Natica, 14 — Aporrhais, 15 — Nassarius на трупе рыбы, 16 — Buccinum,
17 — глубоководный зарывающийся моллюск; 8 — подкласс Opisthobran-
chia, отряд Pteropoda, Spiratella
20*
дыхания использовали атмосферный воздух. Из прибрежной зоны мо-
рей легочные моллюски проникли в пресноводные бассейны, а затем
распространились по всей суше. Они остаются в основном раститель-
ноядными организмами, но часть из них становится хищниками, пое-
дающими червей и разных наземных животных. Большинство назем-
ных моллюсков сохраняет раковину.
Палеонтологическая летопись заднежаберных и легочных моллюс-
ков очень бедна и пока еще не может быть прочитана.
Современные брюхоногие моллюски распространены очень широко
(рис. 162). Они встречаются на всех глубинах морей — от зоны при-
боя до абиссальных глубин. Они заселили все материковые воды. Их
можно встретить в соленых, солоноватых и пресных водоемах, в реках,
болотах и источниках. Все участки суши также заселены брюхоноги-
ми — от тропиков до полярных стран.
Морские гастроподы ведут преимущественно бентосный образ жиз-
ни, лишь немногие приспособились к жизни в пелагиали (киленогие,
крылоногие, голые заднежаберники). Бентосные гастроподы ползают
по поверхности грунта, по водорослям, питаясь растительной пищей;
в зоне литорали живут малоподвижные морские блюдечки (Patella),
обладающие колпачковидной раковиной. Некоторые в поисках пищи за-
рываются в илистый осадок (Aporrhais); немногие прикрепляются к
камням или водорослям при помощи ноги или цементирующего ве-
щества.
Хищные гастроподы нападают на малоподвижных двустворчатых
моллюсков, на кораллы; они -либо раскрывают раковину двустворок с
помощью ноги (Rapana), либо выделяют свободную кислоту, которая
размягчает раковину, а затем радулой высверливают круглое отверстие,
сквозь которое просовывают хоботок внутрь раковины (Naticd). Следы
сверления натик часто встречаются на раковинах вымерших двуство-
рок. Гастроподы, ведущие прикрепленный образ жизни,-являются филь-
траторами. Некоторые фильтраторы живут в симбиозе с морскими ли-
лиями (Platyceras). Тафономический анализ захоронений раковин гаст-
ропод позволяет восстанавливать условия обитания гастропод, условия
осадконакопления, степень подвижности воды и другие особенности
древних бассейнов.
КЛАСС SCAPHOPODA. ЛОПАТОНОГИЕ
Общая характеристика; Скафоподы (гр. scaphis — лопата, pous, род.
podos — нога), или лопатоногие, — небольшой класс морских моллюс-
ков, ведущих зарывающийся образ жизни. Удлиненное двусторонне-сим-
метричное тело заключено в известковую раковину, имеющую вид слег-
ка изогнутой конической трубки, открытой с обеих сторон. Голова не-
сет длинные щупальца — каптакулы. Нога приспособлена для зары-
вания. Раковина гладкая или ребристая. Ордовик — ныне.
Строение мягкого тела. Мягкое тело удлиненной формы окружено
мантией, сросшейся в сплошную трубку с двумя отверстиями на кон-
цах: через переднее отверстие высовывается и втягивается голова и но-
га, через заднее — в тело вводится свежая вода для газообмена и вы-
водятся продукты обмена и половые продукты. Голова слабо обособле-
на. Ротовое отверстие ведет в глотку, в которой имеется подковообраз-
ной формы челюсть и язык, снабженный радулой. Рот у одних окружен
венчиком листовидных придатков, у других они отсутствуют. Над голо-
вой расположены две кожные складки, от края которых отходят много-
308
численные длинные щупальца — каптакулы, вздутые на конце. Кап-
такулы служат для сбора пищи и осязания.
Пищеварительный тракт коленообразно изогнут, состоит из пище-
вода, желудка и задней кишки, которая на брюшной стороне откры-
вается анусом. Вогнутая сторона тела и раковины рассматривается в
качестве спинной, а выпуклая — в качестве брюшной. Ниже головы
расположена нога. У современных форм она имеет форму тупого кону-
Рис. 163. Класс Scaphopoda:
.1 — Denialium (триас — ныне): 1а — из неогена Италии, 16.— схема располо-
жения внутренних органов, 1в — передний отдел тела, увел., 1г — прижизненное
положение; 2 — Cadulus (мел — ныне): 2а — из эоцена Сев. Кавказа, 26 —
современный; 3 — Laevidentalium (триас — ныне);
»ан— анальное отверстие, м — мантия, мп — мантийная полость, н— нога,
нс — нервная система, п—печень, р—рот, ра — раковина, с — сердце,
щ — щупальца
са или червеобразная, с круглым диском на конце. С помощью ноги
моллюск зарывается в песчанистый или илистый грунт в наклонном
положении, оставляя на поверхности грунта небольшую часть заднего
конца раковины (рис. 163). Втягивание ноги внутрь раковины проис-
ходит с помощью пары мускулов-ретракторов. Нервная система состо-
ит из 6 пар ганглиев, соединенных между собой нервными стволами.
Сердце рудиментарное, с одним желудочком, без предсердия; кровенос-
ная система незамкнутая, состоит из системы лакун. Циркуляция кро-
ви осуществляется благодаря ритмичному сокращению мышц тела и
сердца. Жабры отсутствуют. Дыхание осуществляется с помощью тон-
ких складок мантии в стенке мантийной полости. Почки и печень пар-
ные. Скафоподы — раздельнополые животные; оплодотворение проис-
ходит в воде. Из оплодотворенного яйца развивается личинка трохо-
форного типа; у нее на спинной стороне вначале возникают зачатки
309
мантии, выделяющие тонкую двустворчатую раковину, позднее мантия
срастается в трубку и выделяет трубчатую раковину. Личинка перехо-
дит к бентосному образу жизни.
Строение раковины. Раковина известковая, обычно напоминающая
по форме бивень слона, открытая с обоих концов, небольших размеров
(до 20—50 мм, реже до 20—30 см). Раковина может быть почти пра-
вильно цилиндрической, длинной или короткой, веретеновидной со взду-
тием в средней или передней части. Переднее отверстие, через которое
высовывается нога и голова с щупальцами, простое и шире заднего.
Заднее отверстие в раковине либо простое, либо с выемкой или щелью;
щель может быть расположена или на брюшной стороне, или на спин-
ной; иногда щель частично зарастает и тогда образуется ряд отверстий,
похожих на отверстия в раковинах некоторых брюхоногих ^(например, у
Haliotis). Раковина бывает гладкой, со струйками нарастания, или реб-
ристой, с продольными ребрами. Она образуется за счет деятельности
мантии и состоит из четырех слоев: одного органического и трех из-
вестковых: призматического, перекрещенно-пластинчатого и перекре-
щенно-призматического.
Систематика и филогения. На основании особенностей эмбриональ-
ного развития, наличия двусторонней симметрии, строения мантии, но-
ги и ряда внутренних органов (нервной системы, почек, половых желез)
скафоподы занимают промежуточное положение между брюхоногими
и двустворчатыми моллюсками. Они обособлены в отдельный класс,
разделенный на семейства: Dentaliidae и Siphonodentaliidae. Эволюция
и филогения представителей этого класса и их геологическая летопись
известны недостаточно.
История развития и экология скафопод. Один род найден в ордо-
вике, данных о силурийских формах нет; из девона известно три рода,
принадлежащие к сем. Dentaliidae, их более многочисленные предста-
вители известны из меловых отложений; в мелу появляются представи-
тели и второго семейства. Современные скафоподы обитают во всех
морях обоих полушарий, но более разнообразны в тропических и суб-
тропических морях. Скафоподы — фильтраторы, питаются детритом и
мелкими фораминиферами, личинками моллюсков, а сами являются
объектами нападения хищных брюхоногих. В ископаемом состоянии
они обычно встречаются в виде отдельных разрозненных раковин. Из-
вестно около 350 современных видов, в ископаемом состоянии встрече-
но около 300 видов (рис. 163).
КЛАСС (?) XENOCONCHIA. КСЕНОКОНХИЙ
Класс ксеноконхий (гр. xenos — чужеродный, concha — ракови-
на), выделенный В. Н. Шиманским (1963), объединяет морских моллюс-
ков с мягким телом, заключенным в высококоническую раковину, за-
крытую на узком конце. Раковина прямая, с круглым или овальным
поперечным сечением длиной до 100 мм. На вершине раковины иногда
сохраняется своеобразный к<рубчик», возникший, вероятно, в результате
прикрепления мягкого тела к внутренней поверхности раковины. Стенка
раковины состоит, как показали результаты исследований в электрон-
ном сканирующем микроскопе, из трех слоев: наружного и внутренне-
го призматического и лежащего между ними перламутрового. Стенка
раковины ксеноконхий подобна стенке аммоноидей. Полость раковины
единая, без каких-либо продольных или поперечных перегородок. На
границе основной и апикальной частей наблюдается пережим, иногда
ограниченный небольшим валиком, удаленный от вершины на вентраль-
310
пой стороне, приближенный на дорзальной и косо направленный на
«боках. У одного из экземпляров обнаружен второй пережим, располо-
женный несколько дальше первого. Вершина раковины у одних закан-
чивается небольшим шипиком, у других равномерно закруглена.
О строении мягких частей тела можно только предполагать. По-
жидимому, висцеральный мешок, судя по форме раковины, был доста-
точно высокий и объемистый, пищевари-
тельный тракт был петлеобразно -изогнут,
мантийная полость с жабрами помещалась
в передней части раковины; тело, вероятно,
втягивалось внутрь, хотя отпечатки муску-
лов-ретракторов не обнаружены. Выделен-
ный новый класс отличается от остальных
классов моллюсков рядом существенных
признаков и, по-видимому, представляет
боковую ветвь, возникшую от общего с
гастроподами или хиолитами предка. В со-
став класса В. Н. Шиманский включил
один отряд Toxeumophorida( гр. toxeuma —
-стрела, phoros .— несущий) известный из
карбона и перми (рис. 151, 6; 164), с двумя
родами Toxeumophora, Pseudotoxeuma.
КЛАСС BIVALVIA. БИВАЛЬВИИ, ИЛИ
ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ
Общая характеристика. Бивальвии (гр.
bi — два, valva — створка) — морские,
Рис. 164. Класс (?) Хепосоп-
chia (по В. Н. Шиманскому):
1 — Pseudotoxeuma (ранняя
пермь), внешний вид ядра с
частично сохранившимся внут-
ренним (в«) и внешним (вш)
слоями раковины; в примаку-
шечной части пережим (п);
2 — Toxeumophora (карбон —
ранняя пермь), внешний вид
•солоноватоводные или пресноводные, мало-
подвижные или неподвижные моллюски
бентоса; тело заключено в двустворчатую
.известковую раковину, состоящую из двух
равных или неравных створок, соединен-
ных на спинной стороне лигаментом, слу-
жащим для их открывания, и одним или
двумя мускулами-аддукторами — закрыва-
ния створок (рис. 165). Для прочного сое-
динения двух створок вдоль смычного
края может быть развит замок. Лопасти мантии, сросшейся на спинной
стороне, соединяются с краем раковины на внутренней стороне пал-
лиальными мускулами, оставляющими след на нем в виде мантийной,
или паллиальной, линии. Нога служит для ползания, зарывания и
прикрепления, имеет биссусную железу (у Autobranchia и Septibran-
chia). В мантийной полости, сзади, расположены парные жабры, вы-
полняющие функцию газообмена.
Голова не обособлена от туловища; отсутствуют челюсти, радула,
-слюнные железы. Пищеварительный тракт состоит из короткого пище-
вода, объемистого желудка, печени из трех долей, открывающихся в
желудок с вентральной стороны, и длинной задней кишки. Кровеносная
система незамкнутая; сердце состоит из желудочка и двух предсердий.
У протобранхий имеются нервные стволы, у остальных ганглии распо-
ложены ниже рта, в ноге и позади заднего мускула. Имеется одна пара
щеломодуктов (почки связаны с гонодуктами). Двустворчатые моллюс-
ки гермафродиты и раздельнополые. В верхней части ноги расположе-
ны парные гонады. Половой диморфизм выражен слабо. Оплодотворе-
ние внешнее, в воде. Индивидуальное развитие сопровождается личи-
ночными стадиями — трохофорой и велигером; у пресноводных наблю-
дается живорождение или развитие особого типа личинок — глохидий,
лазидий или гаусториальных личинок, временно паразитирующих на
Рис. 165. Класс Bivalvia:
А — продольный разрез через тело пресноводной Anodonta; Б — схематический попе-
речный разрез; В — схема взаимодействия мускула-замыкателя (лз) и связки (лига-
мента); Г — строение раковины Venus;
ан — анальное отверстие, вес — вводной сифон, во — выводной сифон, вс — волокни-
стая связка, гн — гонады, ж — желудок, жб — жабры, же — желудочек сердца, зб —
зубы, зм — задний мускул, зя— зубная ямка, лн — линии нарастания, м — мантия,
ма — макушка, мз — мускул-замыкатель, мл — мантийная линия, мп — мантийная по-
лость, н — нога, ну — нервные узлы, п — почка, пер —периострак, пж — пищевари-
тельная железа, пк — перикардий, плс — перламутровый слой, пм — передний мускул,
пре — призматический слой, пс — пластинчатая связка, пер — предсердие, р — рот,
ра — раковина, рт — ретрактор ноги, с — связка, смл — синус мантийной линии
рыбах. В ископаемом состоянии известны со среднего кембрия, но су-
щественную роль в фауне бентоса начинают играть с конца триаса,
занимая разнообразные экологические ниши, занятые в палеозое бра-
хиоподами.
312
Рис. 166. Класс Bivalvia:
1—5 — схема строения мантии и сифона (см.
текст); 6 — рот и губы Mitilus edulis, вид спе-
реди; 7 — схема строения нервной системы;
ганглии: вг — висцеральный, гг — головной,
нг — ножной; р '— рот; стрелки указывают
ввод и вывод воды
Строение мягкого тела. Тело двусторонне-симметричное, несегмен-
тированное, покрыто с боков двумя лопастями мантии, сросшейся на
спинной стороне. Голова необособлена от туловища, сжатого с боков,
поэтому бивальвий иногда называют Acephala — безголовые (а — от-
рицание, cephalon — голова); на брюшной стороне тела развита муску-
листая нога, имеющая удлиненную, червеобразную или клиновидную
форму (отсюда еще одно название Pelecypoda — секироногие, от гр.
peles — секира), у примитивных представителей с уплощенной подош-
вой. В ноге находится бис-
сусная железа, выделяю-
щая клейкие биссусные ни-
ти, служащие для прикреп-
ления моллюска к подвод-
ным предметам. Нога выхо-
дит между створок и слу-
, жит для ползания, зарыва-
ния в рьихлый осадок, помо-
гает при сверлении твердых
известковых пород; у при-
крепленных форм бывает
недоразвита или редуциро-
вана полностью.
Все внутренние органы
заключены в раковину, вы-
деляемую мантией. Мантия
двумя лопастями охватыва-
ет тело с боков. В задней
части обособлена мантий-
ная полость. Лопасти ман-
тии соединены вдоль спин-
ного края и у одних оста-
ются с брюшной стороны
свободными (рис. 166); у других срастаются сзади, оставляя отверстие
для ввода и вывода воды и продуктов обмена из мантийной полости;
у третьих срастаются спереди и снизу, оставляя отверстие для ноги.
У многих форм задние части мантии вытягиваются в трубки разной
длины, образуя вводной и выводной сифоны. Сифоны могут быть раз-
дельными либо срастаются вместе по всей длине или только частично.
У одних двустворок при закрывании раковины сифоны с помощью мус-
кулов втягиваются внутрь раковины, у других остаются в течение всей
жизни выставленными наружу. Сифоны развиты у двустворок, ведущих
зарывающийся образ жизни.
Край мантии имеет три складки, выполняющие различные функции
(рис. 167). Наружная складка выделяет периострак и внешний извест-
ковый слой; средняя складка несет небольшие щупальца и другие ор-
ганы чувств; внутренняя складка снабжена многочисленными мелкими
мускулами (паллиальными), при помощи которых мантия прикрепляет-
ся изнутри к раковине, образуя мантийную, или паллиальную,
линию, проходящую почти параллельно внешнему краю раковины.
У двустворок, имеющих сифоны, в задней части тела обособлено место
для втягиваемых внутрь раковины сифонов, ограниченное мантийным,
или п а_л л и альным, синусом. По глубине мантийного синуса су-
дят о размерах сифонов. У форм, не втягивающих сифоны, раковина ос-
тается сзади зияющей. Двустворки, живущие на поверхности дна, си-
фонов не имеют.
313.
Раковина состоит из двух створок, соединенных связкой, или
лигаментом, и замыкающими мускулами, или аддукторами.
Мускулы, один или два, являются производными мантийной мускула-
туры. Они служат для замыкания створок, в то время как связка — для
открывания. Парные мускулы обычно расположены ближе к спинной
стороне; непарные — ближе к центру. Можно проследить все стадии ре-
дукции переднего мускула и усиление заднего. Поэтому среди двуствор-
Рис. 167. Схема строения переднего края мантии Anadonta cygnea (Тейлор, Кен-
неди, Холл, 1969):
бп — борозда периострака, вс — внутренняя складка, всл — внутренний слой,
кэе — клетки эпителия внешней складки мантии, ли — линии нарастания, мм —
мантийный мускул, п — периострак, пм — палиальный миострак, пел — призма-
тический слой, ск — слизистые клетки, сс — срединная складка, сел — средний
слой, эвп — эпителий внешней поверхности мантии
чатых различают: равномускульных, неравномускульных и одномус-
кульных.
Основными органами дыхания служат жабры, расположенные сим-
метрично с боков в мантийной полости в задней части тела (рис. 168).
У примитивных двустворок жабры имеют вид лепестков или пластинок,
расположенных в два ряда с каждой стороны под углом друг к другу
(гребенчатые жабры). Пластинки соединяются между собой со-
единительной мембраной, вдоль которой в нижней и верхней частях
проходят кровеносные сосуды. Нижняя и боковые поверхности пласти-
нок покрыты мерцательным эпителием, создающим ток воды к рото-
вому отверстию. У большинства двустворок имеются нитчатые жаб-
р ы, состоящие из коленообразно изогнутых нитей, свисающих в ман-
тийную полость. Нити наружного ряда изгибаются наружу, внутренне-
го — внутрь. Дальнейшее усложнение жабр связано с образованием
перемычек между нитями и превращением их в пластинчатые ре-
шетки (отсюда название класса Lamellibranchiata, или пластинчато-
жаберные). Оба типа жабр служат не только для газообмена, но и для
сортировки и транспортировки питательных частиц к ротовому отвер-
стию, являющемуся своеобразным фильтром-ситом.
Особым типом строения жабр отличается небольшая группа преи-
мущественно глубоководных двустворок, у которых мантийная полость
314
разделена горизонтальной или изогнутой перегородкой, или септой, на
верхний и нижний отделы (септальные жабры); в септе для про-
хождения воды имеются ситовидные пластинки с мелкими отверстиями
или несколько более крупных отверстий (рис. 168, 4). С помощью мышц
септа приводится в движение, образуя своеобразный мембранный насос
для создания тока воды. У этих бивальвий жабры служат только для
добывания пищи.
Рис. 168. Схема строения жабр:
1 — гребенчатые, 2 — нитевидные, 3 — пластинчатые, 4 — септальные;
вп — висцеральная полость, жб — жабры, м. — мантия, « — нога
Ротовое отверстие имеет форму щели. Вокруг рта расположены две
пары ротовых лопастей, покрытых, мерцательным эпителием. У нуку-
лид ротовые лопасти имеют длинные хоботовидные придатки с желоб-
ком посредине; они проходят в отверстие между створками позади но-
ги и служат для сбора пищи (см. рис. 175). По желобкам лопастей пи-
ща поступает в рот.
Пищеварительная система состоит из короткого пищевода, удлиг
ненного желудка, средней и задней кишки, печени и кристаллического
стебелька. Челюсти, радула и слюнные железы отсутствуют. Пища пе-
редвигается мерцательным ресничным эпителием. Кристалличес-
кий стебелек, как и у гастропод, перемешивает пищу, растворяясь,
выделяет ферменты, способствующие внеклеточному перевариванию.
Стенка желудка напротив стебелька превращена в своеобразный желу-
дочный щит. Мелкие пищевые частицы поступают в печень, состоящую
из слепых выростов — дивертикул, в которых происходит внутри-
клеточное переваривание и всасывание; в этом процессе важную роль
играют блуждающие клетки — амебоциты. Основной пищей двустворок
служат фитопланктон, зоопланктон, бактерии и органический детрит,
исключение составляют цептибранхии — хищники, и тередины, пита-
ющиеся древесиной.
По особенностям внутреннего строения различают несколько типов
желудков — митилоидный, пектиноидный, астартоидный и др. Из же-
лудка остатки пищи попадают в среднюю кишку, проходящую через
стенку перикардия, желудочек сердца (создается впечатление протыка-
ния сердца кишечником), далее в заднюю кишку, и фекалии выводятся
через анальное отверстие наружу. Фекалии, продукты обмена и непере-
варенные частицы облекаются слизью и превращаются в плотные мас-
сы шаровидной, овальной или лентовидной формы, часто снабженные
бороздами, отражающими соответствующие выступы тифлозоля. Из-
вестен один случай находки в ископаемом состоянии остатков пищева-
315
рительного тракта, сохранившегося в раковинах у Nuculana, из ранней
юры Англии вместе с ожелезненными остатками фекалий. По-видимому,
захоронение и ожелезнение произошли очень быстро.
Кровеносная система незамкнута; сердце состоит из желудочка и.
двух предсердий, кровь сокращением сердца гонится к органам тела
по артериям и венам, по системе лакун и синусов, расположенных в со-
единительной ткани. У низших двустворок имеются два желудочка, ле-
жащие по бокам кишки, или непарный желудочек, лежащий над или
под кишкой. У более развитых двустворок два желудочка срослись в
один и кишка оказалась внутри сердца. Кровь обеспечивает внутренние
ткани и органы кислородом и питательными веществами, уносит про-
дукты обмена, поддерживает постоянство внутренней среды и способст-
вует созданию в ноге гидравлического давления, необходимого при пол-
зании и особенно при закапывании и регулируемого особым клапаном
(кеберов орган). У септибранхий внутренняя поверхность мантии
принимает на себя дыхательные функции, обогащая кровь кислородом.
Лакуны и синусы представляют собой первичную полость тела, они не
имеют собственных стенок.
Вторичная полость тела редуцирована; она представлена двумя не-
большими полостями: перикардием, или сердечной сумкой, и полостью
гонад. С перикардием связаны перикардиальные железы, выполняющие
наряду с почками выделительную функцию. Почки имеют вид двух тру-
бочек или мешков, которые сообщаются с одной стороны с перикарди-
ем, а с другой — через почечные отверстия с мантийной полостью..
У низших двустворок почки имеют вид простых трубоцек, покрытых вы-
делительным эпителием; они связаны также с половыми желеаами..
У большинства, почки отделены от половых протоков и устроены слож-
нее.
Нервная система развита слабо, что обусловлено малоподвижным
образом жизни (см. рис. 166, 7); она симметрична и состоит из трех
пар ганглиев, соединенных длинными тяжами (коннективами); одна па-
ра лежит над ртом и иннервирует ротовые лопасти, передний мускул-
замыкатель и 'переднюю часть мантии, вторая пара расположена глубо-
ко в ноге, третья — позади заднего мускула-замыкателя, иннервируя:
все внутренние органы, задний мускул и заднюю часть мантии; каждая,
пара ганглиев соединена поперечными перемычками (комиссурами)..
Органы чувств развиты слабо; к ним относятся: органы кожного чув-
ства '— чувствительные клетки, выполняющие осязательные функции и
расположенные на щупальцах, по краю мантии и сифонов, по краю но-
ги, на бугорках около анального отверстия; осфрадии — жаберные
органы чувств, сходно устроенные с таковыми примитивных гастропод;
статоцисты — органы равновесия, устроенные также сходно с подоб-
ными органами других моллюсков, они расположены около педального'
ганглия; глаза, или фоторецепторы, расположенные по краю мантии,
на первых жаберных нитях, вокруг отверстий сифонов; они представ-
ляют собой не мозговые глаза в обычном смысле, а новообразования
от простых эпителиальных до довольно сложных глаз с хрусталиком и
сетчаткой.
Двустворчатые моллюски раздельнополы, очень редко гермафро-
диты; имеют парные гонады, расположенные внутри целома; протоки
гонад связаны с протоками почек или чаще имеют самостоятельные
отверстия. Оплодотворение происходит в воде. Развитие морских и
пресноводных форм происходит разными путями. У морских форм раз-
вивается личинка трохофора, похожая на трохофору полихет. Ра-
ковина, выделяемая раковинной железой, лежит на спинной стороне
316
а(О.Н,мм) 6 /0,30 мм) в (0.1,0 мм)
г ( 0,Ь8 мм) 0//00 мм)
Рис. 169. Развитие Cardium edule L.:
а —- личинка-велигер, б—д — послели-
чиночные стадии
личинки. Вначале это непарная кутикулярная пластинка, которая, по-
степенно разрастаясь, складывается вдоль, образуя две створки. Затем
вся верхняя часть ноги личинки превращается в парус, или в е л у м,—
диск, покрытый ресничками; трохофора превращается в типичную для
бивальвий личинку— велигер (рис. 469). Первичная раковинка ве-
лигера, или продиссоконх (гр. pro — префикс, означающий вна-
чале, впереди, dissos — парный, konche — раковина), состоит из двух
створок, соединенных вдоль спин-
ного края тонкой конхиолиновой
пленкой (лигамент еще не раз-
вит), личиночными зубами и од-
ним, а позднее двумя мускулами-
замыкателями. В течение планк-
тонной стадии идет процесс фор-
мирования многих органов (орга-
ногенез), при этом часть личи-
ночных органов исчезает, напри-
мер велум. Затем личинка
опускается на дно и превращает-
ся в молодого моллюска. Начи-
нает интенсивно расти нога, слу-
жащая для передвижения или
закапывания, начинает действо-
вать биссусная железа, выделяю-
щая клейкие нити.. У прирастаю-
щих форм (устриц) биссусная железа выделяет вещество, прикреп-
ляющее створку к подводным предметам. Постларвальная раковина
(диссоконх) быстро приобретает облик взрослых форм; у нее начи-
нают формироваться зубной аппарат и связка.
Эмбриональное развитие у пресноводных форм протекает иначе.
У всех, за исключением дрейссенид, имеющих личиночные стадии, и
унионид, эмбрионы развиваются в мантийной полости материнского
организма и оттуда выходит вполне сформировавшийся моллюск (жи-
ворождение). У унионид в мантийной полости развивается личинка —
глохидия, тело которой заключено в треугольную двустворчатую
раковину, имеющую один мускул-замыкатель. Через выводной сифон
глохидии выбрасываются в воду и прикрепляются особым зазубренным
шипом раковины и биссусными нитями к телу рыб, превращаясь посте-
пенно в эндопаразитов, которые питаются осмотическим путем с по-
мощью личиночной мантии. Через два-три месяца глохидии, претерпев
соответствующий метаморфоз и органогенез, покидают тело хозяина и
молодые моллюски переходят к самостоятельному образу жизни. Остает-
ся неясным, когда у унионид впервые развилось подобное приспособле-
ние к паразитическому существованию, играющему важную роль в рас-
селении пресноводных моллюсков. У южноамериканских пресноводных
форм развивается личинка-л а зи д ий, у африканских — гаусториаль-
ная личинка.
Строение раковины. Раковина бивальвий в типичном случае состоит
из двух равных створок, охватывающих тело с правой и левой
сторон, в соответствии с чем различают правую и левую створки. Обе
створки соединяются вдоль смычного края лигаментом и замком. За-
мок расположен на спинной стороне и вместе с выступающей макуш-
кой позволяет определить верхний (замочный), или спинной, край ство-
рок; противоположный край называется брюшным, или нижним; поло-
жение рта определяет передний край, а положение анального отверстия
317
и сифонов — задний. Если макушка занимает центральное положение,,
то такие створки называются, равносторонними; если макушка
сдвинута вперед или назад — неравносторонними. Раковина
с загнутой вперед макушкой называется прозогирной (гр. gyros —
округленный), с загнутой назад — опистогирной иортогирной,
если макушки двух створок направлены друг к другу. У прирастающих
форм чаще всего раковина асимметричная, неравностворчатая: одна
створка может иметь форму глубокой тарелки или конуса, а вторая —
плоской крышки. Конусовидные раковины конвергентно сходны с рихт-
гофениями (из брахиопод) или скелетами одиночных кораллов; при-,
растающая створка отличается от свободной своей толщиной и массив-
ностью.
В замкнутом состоянии створки обычно плотно прилегают друг к
Другу; У зарывающихся и сверлящих форм на переднем крае бывают
развиты зияния для выхода ноги, а на заднем — для выхода сифонов.
Такие раковины называют зияющими; у некоторых сверлильщиков
(тередо) для защиты длинных сифонов в процессе эволюции выработа-
лась ложная раковина в виде длинной трубки. Редукция раковин
наблюдается у тех бивальвий, у которых створки обрастают мантией;
раковина в этом случае становится тонкостенной, зияющей, с редуциро-
ванным замком и мускулами-замыкателями.
Внешняя форма раковины бивальвии достигает большого разнооб-
разия и отражает черты приспособления этих моллюсков к различным
биотопам и условиям обитания (см. рис. 188).
В последние годы в связи с применением электронного микроскопа
получены новые данные о структуре стенки раковины у бивальвий.
Стенка, как у всех моллюсков, состоит из органического и известково-
го слоев. Наружный органический слой, или периострак, образуется
клетками эпителия внешней складки мантии; он постепенно утолщается
•от макушки к брюшному краю. Известковая стенка раковины обычно
состоит из трех слоев — наружного, среднего и внутреннего; они обра-
зуются разными участками эпителия переднего края мантии (см.
рис. 167).
Различают следующие микроструктуры: простая призматическая,
сложная призматическая, перламутровая, перекрещенно-пластинчатая,
сложная перекрещенно-пластинчатая, листоватая, гомогенная, грануляр-
ная (рис. 170). Призматическая микроструктура построена из
призм арагонита или кальцита с кристаллической осью С, расположен-
ной вдоль длинной оси призмы; у некоторых наблюдается сложная
призматическая микроструктура (нукулиды, венериды) — приз-
мы первого порядка построены из призм второго порядка. Перламут-
ровая микроструктура состоит из многочисленных многоугольных лис-
точков или пластин арагонита, всегда параллельных внутренней поверх-
ности раковины, налегающих друг на друга. Ось С всегда перпендику-
лярна плоскости таблички; ось В ориентирована в направлении роста.
Наиболее распространенным типом микроструктуры является арагони-
товая перекрещенно-пластинчатая, у которой отдельные пла-
стины построены иглообразными удлиненными кристаллами, причем в
соседних двух пластинах первого порядка пластины второго порядка
имеют противоположное направление (рис. 170, 5). Сложная пе-
рекрещенно-пластинчатая микроструктура возникла из про-
стой и отличается от нее более разнообразной и неправильной ориенти-
ровкой блоков таких пластин (рис. 170, 6). Листоватая микрострук-
тура состоит из более или менее правильно расположенных листочков
кальцита, лежащих рядом или черепицеобразно налегающих друг на
318
друга. К гомогенной микроструктуре относится такая тонкокрис-
таллическая структура, в которой отдельные кристаллы неразличимы;
Рис. 170. Основные типы микроструктур стенки раковины:
1 — простая призматическая; 2 — сложная призматическая (стрелка — направ-
ление к макушке); 3 — листоватая; 4 — перламутровая; 5—6 — простая (5) и
сложная (б) перекрещенно-пластинчатая (55 — пластина 2-го порядка, 5в — плас-
тина 3-го порядка) (Тейлор, Кеннеди, Холл, 1969; 5 — оригинал Попова, 1974);
вм — внутрикристаллическая органическая матрица, лн — линии нарастания,
мкс — межпризматические конхиолиновые стенки, мм — межкристаллическая ор-
ганическая матрица, мпм — межпластинчатая органическая матрица, пвп —
призмы второго порядка
ее происхождение до сих пор не выяснено. Г ранулярная микро-
структура всегда арагонитовая, состоит из мелких карбонатных гра-
нул, обычно с одинаковой кристаллографической ориентировкой.
6 7 8 9 Ю
Рис. 171. Микроструктура стенки раковины некоторых групп бивальвий:
1 — Pteriacea, Pinnacea; 2 — Nuculacea; 3 — Lucinacea, Tellinacea, Veneracea;
4 — большинство гетеродонт; 5 — Zirfaea, Myidae; 6 — Solemya; 7 — Pectina-
cea, Anomiacea, Limacea; 8 — Ostreacea; 9 — Cuspidariidae, Thraciidae; 10 —
Mytilacea;
микроструктуры: гом — гомогенная, л — листоватая, м — миострак, пер — пер-
ламутровая, пп — простая призматическая, ппл —• перекрещенно-пластинчатая,
сп — сложнопризматическая, ст — сложно перекрещенно-пластинчатая
Особую структуру (миострак) образуют слои в местах прикреп-
ления мускулов-замыкателей, мускулов ножных и паллиальных (рис.
319
171). Она построена из видоизмененных арагонитовых кристаллов, ко-
торые приобретают форму призм разных размеров, ориентированных
нормально к поверхности слоя. Все кристаллы арагонита- и кальцита,
принимающие участие в образовании стенки раковины, окружены ор-
ганической оболочкой, толщина которой очень варьирует. Известковая
стенка раковины, как было отмечено выше, состоит обычно из трех
слоев: наружного, среднего и внутреннего, но у многих можно различить
только два слоя. По мнению Тэйлора (1973), исходным типом строения
стенки раковины (рис. 171) у всех бивальвий была трехслойная стенка,
состоящая из простого призматического слоя (наружного) и двух пер-
ламутровых (среднего и внутреннего). Она установлена у моноплако-
фор и у некоторых групп бивальвий (фоладомиид, унионин и тригониин).
Пока еще не установлено таксономическое значение отдельных ти-
пов микроструктур, которые наблюдаются в разных группах бивальвий,
однако первые наметки уже можно сделать. Призматическая и перла-
мутровая микроструктуры обнаружены у протобранхий, митилид; приз-
матическая — у иноцерамид, гиппуритид; листоватая кальцитовая —
у остреин (устриц), некоторых пектенид; гомогенная — у нукулан (про-
тобранхий), хиателл; гранулярная и перекрещенно-пластинчатая — у
миид (рис. 171); перекрещенно-пластинчатая простая и сложная —
у большинства бивальвий. с гетеродонтным замком. В строении рако-
вины в малых количествах участвуют и такие элементы, как стронций,
барий, магний. Концентрация элементов зависит от многих факторов и
в первую очередь от химического состава, солености и температуры
воды.
Скульптура. Поверхность раковины может быть гладкой, с линиями
нарастания или скульптированной. Различают два типа скульптуры:
радиальную и концентрическую. Радиальная представлена ребрами,
расходящимися от макушки к брюшному краю; по размерам различа-
ют ребра первого, второго порядков, с широкими или узкими межре-
берными пространствами. Кроме ребер бывают развиты крупные склад-
ки (Lopha, см. рис. 185, 4), отражающиеся на строении всей раковины,
а также сильные перегибы, или кили. Обычно имеется один хорошо вы-
раженный киль, отделяющий переднее большое поле от малого зад-
него (тригонии). Концентрическая скульптура выражена концентриче-
скими ребрами, идущими параллельно линиям нарастания; она отра-
жает ритмическое изменение количества минерального вещества, отла-
гаемого краем мантии. При пересечении радиальных и концентричес-
ких ребер возникает канцеллятная скульптура. Часто на пересечении
ребер развиваются бугорки, шипы и различной формы выросты, играю-
щие защитную роль. У некоторых бивальвий сзади и спереди вдоль
спинного края обособляются две площадки: передняя — луночка,
задняя — щиток, отделенные от остальной поверхности раковины реб-
рышком или валиком. На щитке обычно расположен лигамент (см.
рис. 180, 3).
Наружный слой, или периострак, часто окрашен в различные цве-
та, причем наиболее яркой и сочной окраской отличаются тропические
бивальвии. Окраска иногда пропитывает и внешний известковый слой.
У ископаемых бивальвий окраска сохраняется реже, чем у гастропод,
тем не менее следы окраски обнаружены в ультрафиолетовых лучах
при просмотре раковин девонских, каменноугольных и более молодых
моллюсков. При этом также были видны паллиальная линия и отпечат-
ки мускулов.
Внутреннее строение. На внутренней поверхности створок различа-
ют структуры, частично отражающие строение тела и расположение
320
некоторых органов. Вдоль утолщенного спинного края протягивается
замочная площадка, на которой расположены более или менее разви-
тые зубы, разделенные зубными ямками, в которые входят зубы проти-
воположной створки. Зубы и зубные ямки образуют замок, или з а-
мочный аппарат (рис. 172). Замок служит для прочного соедине-
ния створок; он корректирует угол раскрывания створок, препятствует
Рис. 172. Некоторые типы прегетеродонтных (Л 2, 4), гетеродонтных (5—7)
и ктенодонтного (3) замков:
1 — актинодонтный, 2 — лиродесмоидный, 3 — ктенодонтный, 4 — схизо-
донтный, 5 — циреноидный, 6 — люциноидный, 7 — астартоидный;
створки: л — левая, п — правая
их боковому сдвигу при закрытой раковине. Первичный замок, или п р о-
В инку л, закладывается на стадии продиссоконха; он состоит из ряда
небольших первичных зубчиков, которые образуются из радиальных ре-
бер створок; иногда эта стадия первичных зубчиков выпадает из онто-
генеза. В дальнейшем, при переходе к стадии диссоконха, на замочном
крае образуется утолщенная замочная площадка, на которой заклады-
ваются по четыре первичных зубных пластинки; из них затем разви-
ваются различные типы замков, среди которых можно выделить три
основных типа: ктенодонтный, прегетеродонтный и гетеродонтный.
Ктенодонтный (гр. ctenion — маленький гребень) замок рас-
сматривается как наиболее примитивный тип замка (характерен для
Nuculacea); он состоит из ряда зубов, расположенных по обе стороны
от макушки й один прямой или изогнутый ряд. Для замка характерно
увеличение числа радиусов по мере роста раковины. Он был ранее из-
вестен под названием т аж с о д о н т н о г о, или рядозубого (гр. ta-
xis — ряд). Однако подобный тип замка позднее независимо развился
у другой группы двустворок (Arcina). Поэтому рекомендовано первич-
ный таксодонтный замок называть ктенодонтный, а вторично воз-
никший— псевдоктенодонтным; развившийся независимо рядо-
21 В. В. Друщиц • S21
зубый замок у некоторых пресноводных бивальвий предлагается назы-
вать псевдотаксодбитным (пермский Palaeomutela и современ-
ный Pleiodon).
К прегетеродонтному замку ,(рис. 172, 1, 2, 4) отнесены та-
кие, у которых зубы представляют собой неизмененные или слабо изме-
ненные по форме и положению первичные зубные пластинки, радиально
расходящиеся от макушки. Различают замки: лиродесмоидный, актино-
донтный, схизодонтный, изодонтный, птеринеоидный, циртодонтный, ди-
зодонтный. Лиродесмоидный замок состоит из равных или почти
равных зубов, веерообразно расходящихся от макушки; этот тип зам-
ка характерен для ранних бивальвий (Lyrodesma, ордовик). А к тин о-
донтный замок отличается от лиродесмоидного укороченными зуба-
ми под макушкой. Схизодонтный замок (гр. schisma — расщеп-
лять) характеризуется развитием зубов, идущих параллельно спинно-
му краю, в левой створке имеет срединный крупный раздвоенный зуб,,
входящий в крупную зубную ямку правой створки, ограниченную дву-
мя зубами; каждый зуб с боков снабжен насечками. По-видимому, опи-
санный тип -замка развился из актинодонтного и независимо возник у
морских (тригониины) и пресноводных (унионины) бивальвий. И з о-
донтный замок (гр. isos — равный); в наиболее развитом виде со-
стоит из двух равных зубов, симметрично расположенных по обе сто-
роны от лигаментной ямки (Spondylus)-, у пектинид — узкие пластин-
чатые зубы, от одного до трех, расходящиеся радиально от макушки
по обе стороны от лигаментной ямки; эти зубы назван^ кардиналь-
ными, или замочными, крурами (лат. crus, мн. ч. crura — нога).
У птеринеоидного замка имеются лишь задние краевые зубы не-
равной длины. Циртодонтный замок развился из схизодонтного
путем редукции примакушечных участков зубов; он характеризуется на-
личием только краевых зубов, идущих параллельно краю. Дизодонт-
ный замок (гр. префикс dys — означает плохой, неэффективный) со-
стоит из слабо или совсем неразвитых зубов. ' . .
Гетеродонтные, или разнозубые, замки (гр. heteros —
разный) характеризуются дифференциацией зубов на кардиальные,
расположенные под макушкой, и латеральные, лежащие парал-
лельно краям (рис. 172, 5—7); различают передние и задние латераль-
ные; их предлагают обозначать римскими цифрами, а кардинальные
зубы — арабскими: четными для левой, нечетными для правой створок.
Обычно бывает развито не более трех кардинальных и не более двух
латеральных передних и задних зубов. Различают следующие типы зам-
ков: астартоидный, циреноидный, люциноиДный, кардитоидный, пахио-
донтный, десмодонтный, возникших, вероятно, от актинодонтного замка;
Астартоидный замок (характерен для рода Astarte) состоит из хо-
рошо развитых кардинальных зубов и переднего бокового; циреноид-
ный, или корбикулоидный, замок отличается присутствием в
каждой створке трех кардинальных зубов и парных передних и задних
латеральных, причем на левой створке нижний задний латеральный зуб
Ьбычно не развит; люциноидный отличается наличием двух карди-
нальных и таких же, как-в корбикулоидном, латеральных; кардито-
ид:ныи отличается удлиненными косо направленными кардинальными
зубами, отсутствием латеральных. П а х и о д о нт н ы й, или толстой
зу б ы й,'закок (гр: pachys—г толстый) состоит из двух-трех толстых,
неправильной формы зубов, возникших из кардинальных или, возможно,
латеральных. Десмодон'тнЫй - Замок отличается редукцией или. пол-
ным исчезновением зубов у форм, ведущих зарывающийся образ жизни;
и•'зн'ачйтельн’ым развитием-внутреннёй’связки,-расположенной иногда на
322
особом выступе—хондрофоре. Подобно право- и левозавернутым
раковинам гастропод у бивальвий иногда наблюдается так называемая
инверсия замка, при которой зубы, свойственные левой створке, раз-
виваются на правой, и наоборот. Это явление наблюдается у некоторых
морских семейств (Astartidae, Crassatellidae) и некоторых пресноводных
(Unionidae) и приводит к осложнениям при цифровых обозначениях
зубов.
Рис. 173. Типы связок:
1—2 — амфидетная; 3—4 — сложная связка: 3 — у Isognomon, 4а, б — у Area;
5—6 — опистодетная: 5а, б — у Cardium, 6 — у Mytilus;
зачернен пластинчатый слой связки, штриховка — волокнистый слой, точки —
раковинный слой; ш — шевроны
Лигамент, иди связка, расположенный вдоль спинного края,
служит для соединения створок и для их открывания, когда ослабевают
мускулы-замыкатели (рис. 173). Лигамент представляет собой упругий
эластичный тяж, состоящий из периострака, наружного пластинчатого
и внутреннего волокнистого слоев. Пластинчатый слой слагается
из параллельно расположенных пластинок протеина, а волокнистый
имеет волокнистую структуру, с фибрами, ориентированными длинной
осью к поверхности роста в местах их образования. Конхиолин, из ко-
торого строится лигамент, состоит в основном из протеинов; волокнис-
тый слой может частично обызвествляться с образованием плотной оси
(литодесмы). К трем описанным слоям иногда еще добавляется
четвертый — соединительный слой, развитый между периостраком .и
пластинчатым слоем. Связка, расположенная по обе стороны от макуш-
ки вдоль замочного края, называется, а м ф.идетно й .(гр. amphi —
двойной, с обеих сторон; detic — Связанный); лежащая позади макуш-
21* • .323
ки, иногда на особой подпорке (нимфе) — опистодегной (гр. opis-
thefi •— задний). Отношение толщины лигамента к его длине может со-
ставлять 1 : 10. У большинства прикрепленных цементацией бивальвий
(устриц) связка расположена между створками в особых ямках на свя-
зочной площадке, и здесь отношение длины лигамента в передне-зад-
нем направлении к толщине в дор.зо-вентральном направлении состав'-
ляет 3 : 1 или 4 : 1. У форм, имеющих ктенодонтный или псевдоктено-
донтный замок, обычно под макушкой развита площадка, или а рея, на
Рис. 174. Схема расположения мускулов-замыкателей (мз), ноги и педальных
мускулов (пдм):
1 — Yoldia, 2 — Area, 3 — Cardium, 4 — Тellina, 5 — Zirfaea, 6 — Chlamys,
7 — Isagnomon.-,
an — апофиз, 6 — биссус, зб — зубы, мс — мантийный синус, н — нога, пм —
передний мускул, с — связка
которой к бороздкам, сходящимся под углом к макушке (шевронам),
прикрепляется амфидетная связка. У пектинид лигамент разделен на
две части — пластинчатый слой расположен вдоль ушек, а волокнис-
тый — в особой ямке под макушкой. В сложной, или мультивинку-
лярной, связке (лат. multus •— много, vinculum — тяж), развитой у
иноцерамид и бакевеллид, волокнистый слой распадается на ряд час-
тей — резилий, каждая из которых лежит в отдельной ямке на свя-
зочной площадке; резилии соединяются между собой пластинчатым
слоем (рис. 173). У ряда бивальвий, ведущих сверлящий образ жизни,
связка редуцируется или полностью исчезает; ее функцию принимает
на себя передний мускул (см. рис. 181, 26).
На внутренней поверхности створок сохраняются места прикреп-
ления разнообразных мускулов, оставляющие на раковине отпечатки
(рис. 174). К ним относятся мускулы-замыкатели (аддукторы), мантий-
ные, или паллиальные, мускулы, мускулы ноги и некоторые другие.
Мускулы-замыкатели, два или один, служат для быстрого за-
324
крывания створок и действуют как антагонисты лигамента, стремящего-
ся в силу своей эластичности открыть створки. В этом антагонизме за-,
ключено большое приспособительное значение подобного взаимодей-
ствия. В случае опасности Моллюску необходимо быстро захлопнуть
створки; после того как опасность прошла, можно понемногу ослабить-,
напряжение мускулов и приоткрыть створки. После гибели моллюска и
разрушения мягкого тела створки под действием лигамента приоткры-
ваются. В зависимости от числа и размеров мускулов различают дву-
створок с двумя равными или неравными мускулами или с одним круп-
ным задним мускулом. Бивальвии с почти равными мускулами назы-
ваются изомиарными, гомо м парным и, или равномускуль-
ными. У рудистов мускулы прикрепляются к особым подставкам,
называемым миофорами. Обычно оба аддуктора действуют
одновременно, но у сверлящих моллюсков (фоладины) передний
вынесен на отворот переднего края створки и его действия противо-
положны действию заднего мускула, лежащего внутри створок; при
сжимании переднего мускула расширяется задний край (у фоладин
раковина зияющая); при сокращении заднего мускула расширяется
переднее отверстие.
У большой группы бивальвий — мономиарий, или одномускуль-
ных, имеется один крупный задний мускул, расположенный почти в
центре створки и обычно оставляющий большой отпечаток. Между изо-
миариями и мономиариями существует ряд двустворок, обладающих
двумя мускулами разных размеров — гетеромиарии, и между ними
можно найти все переходы; при этом наблюдается постепенное умень-
шение переднего мускула и усиление и смещение к центру заднего.
Каждый мускул состоит из двух типов пучков волокон; один пучок
«стекловидный» (быстрый) служит для быстрого закрывания створок,
а другой — «перламутровый», длительно удерживает створки в сомкну-
том состоянии.
Мантия прикрепляется на некотором расстоянии от края к внут-
ренней поверхности раковины с помощью мышечных волокон, назначе-
ние которых состоит в быстром втягивании края мантии внутрь ракови-
ны перед ее закрытием. Внутри раковины эти мускулы образуют ман-
тийную, или паллиальную, линию: у изомиарий она начинается
у переднего мускула и идет почти параллельно краю до заднего аддук-
тора; у мономиарий мантийная линия следует параллельно краю рако-
вины. У форм, обладающих сифонами, последние полностью или час-
тично втягиваются внутрь раковины при помощи особых сифональных
мускулов-ретракторов, оставляющих след прикрепления в задней части'
раковины в виде выемки •— мантийного, или паллиального, синуса. Би-
вальвий с мантийным синусом называют синупаллиатными, без
него — интегрипаллиат н ы м и.
Как было сказано выше, нога выполняет разнообразные функции
и у большинства бивальвий довольно подвижна. Она приводится в;
движение с 'помощью системы мышц, среди которых различают парные-
передние и задние ретракторы — мускулы, втягивающие ногу в
раковину, м у с к у л ы - п р о т р а к т о р ы—способствующие выдвиже-
нию ноги из раковины. У форм, обладающих биссусом, задний ретрак-
тор превращается в сильно развитую мышцу, которая выдвигается
вперед и иногда распадается на несколько пучков. Все мышцы одним
концом прикрепляются к раковине, другим—входят в основание ноги.
Отпечатки этих мускулов Можно видеть вдоль спинного края и около
мускулов-ретракторов (рис. 174). У форм мало или совсем неподвиж-
ных мускулатура ноги сильно упрощена или редуцирована. Очейь'
325:
редко сохраняются мускулы, прикрепляющие висцеральный мешок к
раковине (у некоторых тригоний, унионин), и жаберные мышцы-ретрак-
торы (у некоторых устриц и пектинид).
Основы систематики и классификация. До последних лет единой
общепринятой систематики двустворчатых моллюсков не было. Палеон-
тологи использовали для выделения отрядов особенности строения
замка, лигамента и аддукторов, но, как оказалось, близкие типы
замков могут конвергентно возникать в разных эволюционных стволах,
часто далеко отстоящих друг от друга. Зоологи для выделения отрядов
использовали строение внутренних органов и в первую очередь жабр.
В последние годы были предприняты попытки выделить более высокие
категории, такие, как подклассы или надотряды. В американских «Осно-
вах палеонтологии» (1969) класс бивальвий разделен на 6 подклассов:
Palaeotaxodonta, Cryptodonta, Pteriomorpha, Palaeoheterodonta, Hetero-
donta, Anomalodesmata. Советские малакологи (Невесская, Старобога-
тов, Скарлато и Эберзин, 1971) предложили для выделения надотрядов
и отрядов использовать анатомические и конхиологические данные с
учетом экологической дифференциации. Они предложили разделять
всех бивальвий на три надотряда, в пределах которых строение жабер-
ного аппарата определяет возможности адаптивной специализации:
Protobranchia, Autobranchia и Septibranchia.
НАДОТРЯД PROTOBRANCHIA. ПРОТОБРАНХИИ
К протобранхиям (гр. protos — первый,, branchia — жабры) отно-
сятся наиболее примитивные бивальвии, обладающие двоякоперистыми
гребенчатыми жабрами, выполняющими только функцию газообмена;
жабры не используются для отфильтровывания пищевых частиц, как
это происходит у аутобранхий. Околоротовые лопасти несут придатки,.
Которые служат для сбора пищи; желудок довольно примитивный, с
1—3 протоками печени, состоящей из сильно разветвленных дивер-
тикул —слепых отростков. Малый и большой тифлозоли в желу-
док не вдаются; сердце лежит над кишечником; нога с плоской подош-
вой, без биссуса.
Раковина равностворчатая, внутренний слой перламутровый, пере-
крещенно-пластинчатый или гомогенный. Два почти равных мускула-
замыкателя или неравных, из которых передний мускул крупнее зад-
него; замок ктенодонтный или криптодонтный; мантийная линия без
синуса или с синусом. Все протобранхии — обитатели моря, ведут
подвижный или малоподвижный образ жизни, ползают на широкой'
подошве ноги, частично или полностью- погружаясь в осадок, и соби-
рают питательный детрит с помощью придатков околоротовых лопас-
тей; часть ведет зарывающийся малоподвижный образ жизни, причем
у одних (Nuculana) задний конец раковины оттянут, у других (Sole-
mya), наоборот, передний конец удлинен, а задний укорочен.
Надотряд состоит из трех отрядов и насчитывает около 100 родов
(рис. 175). Отряд Ctenodontida (кембрий?, ордовик—ныне) с харак-
терными родами: Ctenodonta (средний ордовик), Nucula (мел-—ныне),
Malletia (мезозой — ныне), Nuculana (триас — ныне). Отряд Solemyida
(девон — ныне) с характерными родами: Solemya (девон—ныне),
Nucinella (юра — ныне), Manzanella (пермь), Huxleya (совр.). Отряд
Praecardiida (поздний кембрий?, ордовик— ранний карбон) с харак-
терными родами: Praecardium (силур — девон), Cardiola (поздний си-
лур— девон), Antipleura (силур). Первые представители надотряда
326
протобранхии становятся известными в конце кембрия — начале ордо-
вика, они относятся, к отрядам Ctenodontida и Praecardiida; в девоне
возникает отряд Solemyida. До настоящего времени доживают предста?
•ви.телй .двух отрядов; отряд Praecardiida вымирает в начале карбона.
Рис. 175. Надотряд Protobranchia; отряды Ctenodontida (/—2), Praecardiida
(3), Solemyida (4):
1 — Nucula (мел — ныне), схема расположения внутренних Органов и, внеш-
ний вид раковины; 2 — Leda (силур — ныне); 3 — Praecardium (силур —
девон).; 4 — Solemya (девон — ныне);
.жб — жабры, зб — зубы, зм — задний мускул,1 л — лунка, ма — макушка,
мл — мантийная линия, н — нога, пдм—...педальный мускул, пм — передний
мускул, прл — придаток ротовой лопасти, рл — ротовая лопасть, смл —; си-
. нус мантийной линии, щ — щиток
НАДОТРЯД AUTOBRANCHIA. АУТОБРАНХИЙ
К надотряду аутобранхий (гр. autos—самостоятельный) отнесена
основная масса бивальвий, у которых жаберный аппарат преобразован
в фильтр-сито, т. е. выполняет не только функцию газообмена, но и
отбор мелких пищевых частиц и доставку их к ротовому отверстию.
В желудке развиты сортирующие аппараты, в него глубоко вдается
тифлозоль с сопровождающей его кишечной бороздой;’ развит кристал-
лический стебелек; печень открывается большим числом самостоятель-
ных отверстий и имеет несколько дивертикул; выделяются различные
типы желудков, среди которых наиболее примитивным считается
униоидный, имеющий большой тифлозоль и общие протоки печени;
такой тип желудка, вероятно, был характерен для всех актинодонтид.
Митилоидный тип желудка имеет сложный тифлозоль; он характе-
рен для устриц, митилин, арцин, птериин и позволяет объединять их
в один отряд циртодонтид. Для' остальных групп аутобранхий характе-
рен свой тип желудка, что позволяет выделять несколько естественных
групп, которым придается ранг отрядов. В пределах всего надотряда
встречаются все типы раковин, замков и лигаментов, рассмотренные
в общей части. По строению замка, желудка и- жабр надотряд Auto-
branchia, распадается на восемь отрядов: Actinodontida, Pectinida,.
327
Astartida, Carditida, Venerida, Hippiiritida, Cyrtodontida, Pholadomytda.
Отряд Actihodontida. Актинодонтиды (гр. actis, род. actinos—луч,
радиус) объединяют как морских, так и пресноводных бивальвий,
имеющих равностворчатую раковину с перламутровым или перекре-
Рис. 176. Отряд Actinodontida; подотряды Trigoniina (/—3), Unioniina (4—5)
и Actinodontina (6—8):
1 — Neotrigoriia (совр.), схема строения; 2 — Trigonia. (средний триас —
мел); 3 — Lyrodesma (ордовик); 4 — Unio (триас — ныне); 5 — Anodonta
(поздний мел — ныне); 6 — Actinodonta (ордовик); 7а, б — Cycloconcha (ор-
, довик); 8а, б — Modiomorpha (силур—пермь);
зб— зубы, зм— задний мускул, зп—‘заднее поле, к — киль, н — нога,
пдм — педальный мускул, пм — передний мускул, пп — переднее поле, мл —
мантийная линия
щенно-пластинчатым слоем, лиродесмоидный, актинодонтный, схизо-
донтный или дизодонтный замки. Жабры образованы свободными или
сросшимися нитями — филаментами. Желудок униоидного типа.
Большой тифлозоль имеет вид дуги, проходящей через весь желудок.
Представители отряда известны е ордовика и существуют до наших
дней, причем если морские формы в настоящее время немногочислен-
ны, то пресноводные представляют процветающую группу. Отряд
328
объединяет три подотряда: Trigoniina, Unioniina, Actinodontina
(рис. 176).
Подотряд Trigoniina. Тригониины (гр. trias—три, gonia— угол)
объединяют большое семейство тригонийд, распространенное широко в
мезозое и представленное сейчас реликтовым родом Neotrigonia, оби-
тающим у берегов Австралии, и два палеозойских семейства. Раковина
имеет характерный схизодонтный замок, хорошо развитую скульптуру,
состоящую из радиальных ребер или бугорков и киля, идущего от
макушки к заднему краю и отделяющего большое переднее поле от
небольшого заднего. К подотряду отнесены также семейство лиродесма-
тид (1 род), развитое в среднем и позднем ордовике, имевшее раковину
с лиродесмоидным замком, состоящим из радиально расходящихся от
макушки нескольких зубов, и семейство миофорид (13 родов), развитое
от девона до конца триаса (рис. 176, 1—3). Подотряд существует с
ордовика.
Подотряд Unioniina. Униониины (лат. unio — объединение, а также
одна жемчужина) объединяют пресноводных бивальвий, имеющих
равностворчатую раковину до 30 см длиной, с хорошо развитым
периостраком, опистодетным лигаментом, двумя псевдокардинальными
и одним боковым зубами в левой створке (иногда полностью редуциро-
ванными). У некоторых представителей имеется псевдотакеодонтный
замок. Не исключена возможность, что в подотряд объединены нерод-
ственные группы, возникшие от разных предков и в разное время при-
способившиеся к жизни в пресных водах. У современных унионин в
онтогенезе имеются личинки — глохидий, лазидий или гаусториальная.
Подотряд появился в конце девона, дал довольно много представителей
в карбоне и перми (надсемейство Anthracosiacea, около 20 родов) и
получил широкое распространение начиная с триаса (надсемейство
Unioniacea свыше 150 родов) (рис. 176, 4—5).
Подотряд Actinodontina. Актинодонтины объединяют морских, пре-
имущественно палеозойских, бивальвий, раковина которых имеет
актинодонтный замок, несколько напоминающий лиродесмоидный, но
отличающийся от него развитием зубов, идущих параллельно спинному
краю раковины и укороченных под макушкой. Актинодонтины появ-
ляются в среднем кембрии и представляют собой наиболее древних
бивальвий, от которых возникли все остальные группы аутобранхий.
Они объединяют около 40 родов; основные представители вымирают к
концу перми, а их потомки, если включить в этот подотряд современ-
ный род Gastrochaena, существуют до наших дней (рис. 176, б—8).
Отряд Pectinida. Пектиниды (лат. pecten — гребешок). Раковина
округленная или вытянутоовальная, иногда неправильной формы,,
равно- и неравностворчатая, вдоль спинного края развиты ушки тре-
угольной формы; преобладает'радиальная скульптура, ребра от слабых
до очень сильных; концентрическая скульптура иногда сопровождается
выростами, шипами;- структура внутреннего слоя перекрещенно-
пластинчатая или листоватая. Замок изодонтный или с кардинальными
крурами; иногда отсутствует. Лигамент наружный и; внутренний. Ман-
тия: с несросшимися утолщенными краями, которые загнуты внутрь,,
образуя своеобразный «парус». Вдоль края мантии развиты фоторецеп-
торы-глазки (иногда до 100), короткие щупальца и другие органы
чувств. Имеется один большой мускул, занимающий почти центральное
положение, разделенный на переднюю часть, состоящую из поперечных
мышц, способную к быстрым сокращениям, и заднюю с гладкими
мышцами, приспособленную для удержания створок в закрытом со-
стоянии. Жабры состоят из свободных филаментов (нитей), образую-
329
щих две ветви: .нисходящую и ’после коденообразного изгиба — восхо-
дящую. Желудок пектиноидного типа, большой тифлозоль сравнительно
короткий. Отверстия дивертикул многочисленные. Пектиниды — филь-
траторы, обитают во всех морях и океанах, на самых разнообразных
глубинах, от мелководья до ультра абиссали (до 8000 м). На ранних
стадиях они прикрепляются с помощью биссуса; одни всю жизнь
остаются прикрепленными, к субстрату при помощи биссусного
пучка, превращенного в особый известковый стебелек, выходящий через
,ся
Рис. 177. Отряд Pectinida:
J — Chlamys (триас — ныне), правая створка снаружи и изнутри; 2 — Spondylus-
(юра — ныне), схема строения замка правой (вверху) и левой (внизу) створок; 3 —’
Anotnia (мел—ныне);
•бв — биссусный вырез, зб — зубы, зм — задний мускул, зя — зубные ямки, мл • —
мантийная линия, ся — ямка для внутренней связки, уш — ушки, щнс — щель для
наружной связки
отверстие в нижней створке (Anomia), или прочно цементируются
нижней створкой к подводным предметам (Spondylus); другие из бис-
сусных нитей строят себе гнездо, скрепляя биссусом мелкие камешки,
обломки раковин и другой строительный материал, выстилая «гнездо»
мягкими биссусными нитями (Lima). Преобладающее же число свобод-
но лежит на дне и способно в случае опасности передвигаться скач-
ками, хлопая створками (Pecten, Lima)-, в этом случае при захлопы-
вании створок края «паруса» плотно смыкаются и вода с силой вытал-
кивается через отверстия в основании ушек. К лучшим «пловцам» от-
носятся формы с гладкой раковиной.
Пектиниды, объединяющие свыше 100 родов, появились в ордо-
вике, но широкое развитие получили начиная с мезозоя (рис. 177).
Отряд Astartida. Астартиды (гр. Astarte — финикийская богиня
любви и плодородия). Раковина равностворчатая, иногда зияющая или
33Q
окруженная со всех сторон мантией (становится внутренней с редук-
цией аддукторов); преобладает концентрическая скульптура, имеются
.луночка и щиток. Замок астартоидный и люциноидный, иногда пол-
ностью редуцированный; два аддуктора,лигамент преимущественно
юпистодетный. Структура внутреннего слоя перекрещенно-пластинчатая,
реже гомогенная или призматическая. Жабры пластинчатые, состоят из
филаментов, соединенных перемычками; мантия с несросшимися края-
ми или срастается, оставляя отверстия для ноги и сзади для сифонов.
Рис. 178. Отряд Astartida:
1 — Lucina (поздний мел — ныне); 2 — Donax (эоцен — ныне); 3 — Hiatella
(поздняя юра — ныне); 4 — Crassatella (поздний мел — неоген); 5 — Astarte
(юра — ныне);
ли — мускул-замыкатель, мл — мантийная линия, мс — мантийный синус
Астартиды — обитатели морей, преимущественно умеренных и при-
полярных; живут в зоне мелководья, обычно на поверхности грунта;
немногие зарывающиеся обладают вытянутыми сифонами и соответст-
венно имеют мантийный синус. Первые астартиды появляются в ордо-
вике, но многочисленными и разнообразными становятся начиная с
мела. Они объединяют свыше 270 родов (рис. 178).
Отряд Carditida. Кардитиды (гр. cardia — сердце). Раковина равно-
створчатая, сердцевидная, тригональная, трапециевидная; замок карди-
тоидный, сходный с астартоидным по числу кардинальных зубов (2—3),
:но отличающийся от него сильной скошенностью зубов; имеется по
одному латеральному зубу с каждой стороны. Преобладает радиальная
скульптура; макушка прозогирная. Лигамент наружный, поддержи-
вается нимфами, иногда внутренний; имеются два мускула-замыкателя;
мантийная линия интегрипаллиатная. Жабры пластинчатые. Желудок
кардитоидного типа, большой тифлозоль очень длинный, закрученный
на конце в многорборотную спираль; дивертикулы печени открываются
тремя — четырьмя группами отверстий. По типу питания — инфаунные
фильтраторы. Первые кардитиды появляются в девоне, становятся
331
более разнообразными начиная с мела; современные кардитиды живут
преимущественно в тропических и субтропических морях, немногие
встречены в мелководье умеренных морей. Известно около 80 родов..
Отряд Venerida. Венериды (лат. Venus — богиня любви). Один из
наиболее крупных отрядов аутобранхий. Объединяет свыше 400 родов,
из них большинство ранее относимых к отряду гетеродонт (без астар-
Рис. 179. Отряд Carditida:
1 — Myoconcha (юра — мел); 2 — Venericardia (палеоцен — ныне); 3 —
Cardita (палеоцен — ныне);
мз — мускул-замыкатель, мл — мантийная линия
тид и кардитид) и почти все роды, относимые к отряду десмодонт. Жаб-
ры пластинчатые, желудок венероидного типа, с одним или двумя сле-
пыми впячиваниями; большой тифлозоль напоминает цифру 7. Рако-
вина равностворчатая у большинства; у прирастающих (подотряд ха-
мина) резко неравностворчатая; замок циреноидный, люциноидный или
десмодонтный. Обитатели морей, солоноватоводных и пресноводных бас-
сейнов, по типу питания—инфаунные фильтраторы или собиратели-грун-
тоеды; среди них: живущие на поверхности осадка, частично и пол-
ностью зарывающиеся, сверлильщики известковых пород (фоладины) и
древесины (тередины), прикрепленные биссусом. Венериды, вероятно,
возникли в позднем триасе от астартид. Основной ствол, выделенный в
подотряд венерина, — обитатели дна моря, ползающие, частично погру-
женные в осадок или зарывающиеся, с хорошо развитыми сифонами;
в юре от венерин обособилась группа сверлильщиков известковых пород,
плотных илов и глин, а начиная с палеоцена или конца мела — группа
сверлильщиков древесины, выделенная в подотряд фоладина. В палео-
цене часть венерид перешла к прикрепленному образу жизни, цементи-
руясь одной из створок, и в связи с этим претерпела резкие изменения
внешней формы раковины (подотряд хамина) .
Подотряд Venerina. Венерины объединяют равностворчатых би-
вальвий с люциноидным и циреноидным замками или без зубов, с Кон-
центрической и радиальной скульптурой, от слабо выраженной до
очень сильной; имеют раковины от маленьких до гигантских размеров,
достигающих длины 1,4 м и веса до 200 кг (Tridacna). Структура внут-
реннего слоя перекрещенно-пластинчатая или гомогенная. Имеют два
мускула-аддуктора, паллиальную линию с синусом и без синуса; среди
332
зарывающихся форм обособились две группы: с длинной, почти ци-
линдрической раковиной и короткими сифонами (сем. Solemdae) и с
овальной, зияющей на конце раковиной и очень длинными сифонами
(сем. Myidae). Обитатели морей, солоноватоводных и пресноводных
бассейнов. К жизни в пресных водах перешли дрейссенсиды, прикреп-
ляющиеся биссусом, с двумя неравными мускулами, внешне напоми-
Рис. 180. Отряд Venerida; подотряд Venerina:
1 — Муа (олигоцен—ныне); 2 — Mactra (эоцен — ныне); 3 — Venus (олиго-
цен — ныне); 4 — Cardium (неоген — ныне); 5 — Solen (миоцен — ныне);
бз — боковые зубы, зм — задний мускул, кз — кардинальный зуб, л — ложечка,
лг/ — лунка, ма — макушка, мл — мантийная линия, нф — нимфа, пм — перед-
ний мускул, смл — синус мантийной линии, ся — ямка для внутренней связки,
щ — щиток
нающие митилид из отряда циртодонтид. Венерины известны с конца
триаса и существуют до наших дней. Объединяют около 300 родов
(рис. 180).
Подотряд Chamina. Хамины (гр. chamai — живущие на грунте)
объединяют три рода, представители которых прикрепляются к под-
водным камням путем цементации; створки неравные, неправильных
•очертаний, с прозогирной макушкой, концентрической или радиальной
333
скульптурой с шипами; замок редуцирован; сохранилось по одному
кардинальному зубу в каждой створке и у некоторых по два карди-
нальных и одному латеральному; по внешней форме зубы несколько'
напоминают зубы гиппуритид. Хамины появились в конце мела (дат-
ском ярусе) и в настоящее время являются обитателями теплых морей
(рис. 181, 2).
Рис.-,' 181. Отряд Venerida; подотряды Chamina (/) и Pholadina (2—5):
la, б — Chama (палеоцен — ныне); 2— схема строения раковины сем. Pholadi- .
dae: 2а — вид снаружи, 26 — изнутри; 3—4 — сем. Teredinidae, Bankia-. 3 —
общий вид, 4а, б — раковина, увел.; 5 — схема взаимоотношения сифона и па-
летки: 5а — сифон выставлен, 56 — сифон втянут;
ап — апофиз, дп — дополнительные пластинки, зм — задний мускул, к — кал-
лум, мп — мускулы палеток, мс — мантийный синус, пал — палетки, пк — пе-
редний край раковины, пм — передний мускул, гас — пластинки, покрывающие
сифон, см — сифональный мускул, сф — сифон
Подотряд Pholadina. Фоладины (рр. pholas — моллюски, сверля-
щие породы) —труппа, приспособившаяся к сверлению известковых
. пород или древесины. В соответствии с подобной адаптацией \ выде-
ляют морских сверлильщиков — фо ладид и древоточцев — тер еди-
ни д; первые объединяют 28 родов, вторые — 21. Сверлящий образ
жизни привел к существенным изменениям раковины "и отдельных ее
структур.
Раковина у фоладид равностворчатая, без лигамента и замка;,
передний край образует макушечную складку — отворот, к которому
прикрепляется передний аддуктор; задний аддуктор крупный; сущест-
вует еще один дополнительный аддуктор на брюшной стороне; мускулы
передний и задний действуют поочередно. Передняя часть раковины
-вооружена резкими шиповатыми ребрами, играющими основную роль-
при сверлении. Каждая створка имеет внутри раковины ножевидный
апофиз, служащий для прикрепления мускулов ноги. Сифоны сильные,.
334
сросшиеся, покрытые хитинизйрованным. периостраком; нога с подош-
вой служит для прикрепления к стенкам норки. Фоладиды сверлят
известковые породы, сжимая поочередно передний и задний аддукторы
и частично поворачивая раковину, проделывают ходы в плотном иле,
глине; один род (Xylophaga), сверлящий древесину, недавно был
встречен на глубине около 7 тыс. м. Раковины фоладид снабжены до-
полнительными пластинками, покрывающими раковину на спинной
стороне спереди и сзади и с брюшной стороны. У некоторых форм
(мартезий) передний зияющий край бывает закрыт полностью или
частично особой известковой оболочкой—каллумом (рис. 181, 26).
Терединиды, приспособленные к сверлению древесины, имеют
очень маленькую раковину, зияющую спереди и сзади, с зазубренной
передней частью; тело длинное, червеобразное, с двумя сифонами на
заднем конце, расположено вне раковины; у основания сифонов имеется
пара известковых пластинок — палеток, которые при втягивании
сифонов внутрь норки закрывают входное отверстие; у одних палетки
простые, ложкообразные, у других напоминают колосок, состоят из
мелких, входящих друг в друга воронковидных зазубренных конусов
(рис. 181, 5). Сверление древесины, как и у фоладид, происходит с по-
мощью переднего зазубренного края раковины при поочередном сокра-
щении то переднего, то заднего мускулов и легкого поворачивания тела
при помощи ноги, играющей роль присоски. Ходы древоточцев изнутри
выстланы тонким слоем известковых отложений. Тередины питаются
мелкими планктонными организмами и дополнительно древесными опил-
ками, которые перевариваются внутриклеточно в дивертикулах печени.
В отличие от фоладид они гермафродиты.
В ископаемом состоянии представители подотряда встречаются
начиная с палеоцена. От фоладин кроме раковин сохраняются просвер-
ленные ходы, дополнительные пластинки, палетки..
Отряд Hippuritida. Гиппуритиды (названы по роду Hippurites, име-
ющему сходство с рогом) объединяют свыше 130 родов крупных
двустворчатых моллюсков, впервые появившихся в позднем силуре
(надсем. Megalodontacea). Палеозойские гиппуритиды имели равно-
створчатую гладкую раковину, с прозогирной макушкой, толстыми
кардинальными зубами (от одного до нескольких), с наружным, описто-
детным лигаментом и с глубоко сидящим передним аддуктором, рас-
положенным в передней части дорзального края. В начале поздней
юры (Оксфорде) от мегалодонтаций возникли рудисты.
У рудистов две створки неравные; различают три основных
варианта: 1) прикрепленная створка — спирально закрученная, свобод-
ная — в форме крышечки; 2) прикрепленная створка длинная, боль-
шая, коническая, свободная — плоская, коническая или закрученная;
3) прикрепленная — маленькая, коническая, свободная — значительно
крупнее первой, закрученная. Прикрепление осуществлялось обычно
левой или инверсно-правой створкой. У позднеюрских и раннемеловых
форм створки отличались закрученностью (Diceras), у позднемеловых
преобладали формы с конической раковиной, внешне напоминающей
скелеты одиночных кораллов (Radiolites, Hippurites). Конические рако-
вины некоторых рудистов достигали 2 м в высоту и до 0,6 м в диаметре.
Обычно имелись два крупных зуба в крышке и один в прикрепленной
створке, или наоборот. Лигамент наружный у ранних форм, внутрен-
ний— у поздних и, возможно, у некоторых отсутствовал.
У рудистов стенка обычно толстая, .до 10 см толщиной, ячеисто-
призматическая, . с толстым призматическим наружным слоем, состоя-
щим из полых призм; у ранних рудистов стенка ячеистая. В стенке на-
335
блюдаются различные каналы, вертикальные и радиальные, простые и
разветвленные. У ряда форм отмечаются поперечные днища, отделяв-
шие тело от нижней нежилой части раковины. Кроме основной внутри
раковины ' имелись еще дополнительные полости. Створки соединялись
двумя мускулами-аддукторами, которые прикреплялись к особым вы-
ступам— мио форам. Наличие мускулов и лигамента позволяв?
предполагать, что створки у основных групп рудистов открывались и
Рис. 182. Отряд Hippuritida:
1 — Hippurites (поздний мел): 1а — внешний вид, 16 — продольный разрез,
1в — поперечный разрез; 2 — Radiolites (поздний мел), поперечный разрез; 3 —
Requienia (мел); 4 — Diceras (поздняя юра);
вс — верхняя створка, д — днища, жб — жабры, зб — зубы, зм — задний мус-
кул, зя — зубные ямки, нс — нижняя створка, пб — продольные борозды, пм —
передний мускул, св — связка; стрелки указывают направление тока воды
закрывались, так же как у всех ординарных бивальвий. Однако у
радиолитов и гиппуритов с плоской крышкой и вертикально стоящими
зубами и соответствующими зубными ямками верхняя свободная створ-
ка могла, скорее всего, двигаться только вертикально. Возможно, дви-
жение осуществлялось вследствие изменения объема ноги и тела. Для
обеспечения питанием и газообмена существовало два сифона, структу-
ры которых выражены углублениями, складками в наружной стенке,
особыми столбиками; один сифон был вводным, другой — выводным
(рис. 182). Первые рудисты, как уже было отмечено, появились в начале
Поздней юры, ответвившись от протодицераса (надсем. Megalodonta-
сеа). Рудисты были обитателями тропических морей, главным образом
коралловых рифов, жившими большими поселениями; в раннемеловое
336
.время они были сравнительно немногочисленны (около 25 родов), но
.стали очень разнообразными и многочисленными в позднемеловое
время (до 80 родов) (рис. 182). Однако в конце Маастрихта все рудис-
ты вымерли. Невесская и др. (1971) присоединяет к гиппуритидам одно
современное семейство Arcinellidae, которое большинство малакологов
относит к хаминам.
Рис. 183. Отряд Cyrtodontida; подотряды Arcina (/—2) и Pteriina (3—5)'.
1 — Area (средняя юра — ныне); 2 — Cucullaea (юра — ныне); 3 — Inoceratnus
(юра — мел); 4 — Isognomon, (трас — ныне); 5 — Pteria (триас — ныне);
зб — зубы, вм — задний мускул, ма — макушка, мл — мантийная линия, пм —
передний мускул, сс — сложная связка, ш — шевроны
Отряд Cyrtodontida. Циртодонтиды (гр. kyrtos—изогнутый) имеют
нитчатые жабры со свободными филаментами, желудок митилоидного
типа; большой тифлозоль S-образно изогнут, дивертикулы печени от-
крываются рядом отверстий. Отряд возник от актинодонтид в начале
ордовика и сразу же дал ветви, выделенные в подотряды Arcina и
Pteriina, отличающиеся строением замка и раковины. В девоне возник
подотряд устриц, а позднее, в триасе, ответвился подотряд митилин.
22 В. В. Друщиц
337
Подотряд Arcina. Арцины (лат. area — коробка). Раковины оваль-
ные, трапецеидальные; замок циртодонтный и псевдоктенодонтный, со-
стоящий из ряда зубов, расположенных в один ряд вдоль замочного
края. Под макушкой развита арея, на которой расположена сложная
связка в шевронах. Нога развита слабо; имеются биссусная железа, два
Мускула-аддуктора; появляются в ордовике и в настоящее время живут
преимущественно в тропических морях. Объединяют до 80 родов
(рис. 183, 1, 2).
Подотряд Pteriina. Птериины (гр. pteron — крыло). Раковины рав-
но- и неравностворчатые, с крыловидными ушками, тригональные,.
Рис. 184. Отряд Cyrtodontida; подотряд Mytilina:
1 — Mytilus (поздняя юра — ныне): 1а — левая створка изнутри, 16 — при-
жизненное положение; 2 — Modiolus (девон — ныне); 3 — Pinna (ранний кар-
бон — ныне);
б — биссус, за — зубовидные выступы, зм — задний мускул, ма — макушка,
мл — мантийная линия, нф — нимфа, пм — передний мускул ,
квадратные или трапецеидальные; нога с биссусом; спинной край
прямой и несет многочисленные ямки для сложной связки. Скульптура
концентрическая и радиальная. Замок птериноидный, с задними крае-
выми зубами неравной длины или лишенный зубов. Птериины появ-
ляются в ордовике, в карбоне приспосабливаются к жизни в солоновато-
водных бассейнах. В юрское и меловое время получает широкое рас-
пространение семейство иноцерамид, имевших разнообразной формы
раковину, сложную связку и хорошо развитый призматический слой.
Современные птериины — обитатели мелководья, преимущественно теп-
лых морей. Объединяют до 150 родов (рис. 183, 3—5).
Подотряд Mytilina. Митилины (гр. mitos — нить). Раковины тре-
угольной формы, суженные спереди и расширенные сзади, образуют
значительные поселения в прибрежной части моря; мускулы неравные:
передний — маленький, задний — большой; нога пальцевидная, у моло-
дых служит для переползания, у взрослых выделяет биссусные нити:
для прочного прикрепления к подводным скалам или к песчинкам
при частичном погружении в осадок; связка опистодетная; хороша
развиты сифоны; замок без зубов или со слабо развитыми зубчиками.
338
в примакушечной части. Первые митилины появляются в девоне, в кон-
це перми часть из них приспосабливается к сверлению известковых
пород, однако сверление в отличие от фоладид осуществляется кислотой,,
выделяемой особой железой и растворяющей известковое вещество (ли-
'гофаги, или морские финики); поэтому раковина у них тонкая, более
или менее удлиненная, почти цилиндрическая. Подотряд объединяет
около 60 родов (рис. 184).
Подотряд Ostreina. Остреины, или устрицы (лат. ostrea, гр.
ostreon —устрица); ведут прирастающий образ жизни, имеют неравно-
створчатую раковину: левая створка блюдцевидная, правая — плоская,
Рис. 185. Отряд Cyrtodontida; подотряд Ostreina:
1 — Gryphaea (юра); 2 — Ostrea (мел —ныне): 2а — левая створка изнутри,
26 — схема внутреннего строения; 3 — Exogyra (мел); 4 — Lopha (триас —
ныне);
жб — жабры, зм — задний мускул, ся — ямка для связки
в форме крышечки. Макушка прирастающей створки может быть
клювовидной, нависающей (род Gryphaea) или завернута в форме
плоской спирали (Exogyra). После цементации нога и биссус атрофи-
руются, структура внутреннего слоя листоватая. Имеют открытую
мантию и свободные нитчатые жабры. Замок без зубов или с рудимен-
тами таксодонтных зубчиков. Лигамент внутри ямки на примакушечной
арее. Обитатели мелководья, живут на плотных грунтах и скалах на
глубине до 50—70 м, образуя устричные банки. Могут выносить значи-
тельное опреснение. Появляются в триасе и существуют до наших дней
преимущественно в теплых морях. Объединяют свыше 50 родов
(рис. 185).
Отряд Pholadomyida. Фоладомииды. Раковины равностворчатые,
с двумя мускулами, опистодетным лигаментом, у некоторых (Pandora)
лигамент с литодесмой. Скульптура концентрическая и радиальная;
22*
339
замок десмодрндный, внутренний слой перламутровый; жабры пластин-
чатые, желудок пандороидного типа— большой тифлозоль сравнитель-
но короткий; ведут зарывающийся образ жизни; хорошо развиты сифо-
ны. Фоладомииды возникли в девоне, по-видимому, от отряда актино-
донтид. В связи с зарывающимся образом жизни у одной группы
фоладомиид (Clavagelld) раковины полностью или частично редуциро-
Рис. 186. Отряд Pholadomyida:
1 — Clavagella (поздний мел — ныне); 2 — Pholadomya (поздний триас —
ныне); 3 — Allorisma (девон — пермь): За — вид сбоку, 36 — вид сверху;
ит — известковая трубка, ра — раковина
вались и тело заключено в известковую трубку (обособлены в подотряд
клавагеллин). Объединяет около 90 родов (рис. 186).
НАДОТРЯД SEPTIBRANCHIA. СЕПТИБРАНХИИ
К надотряду Septibranchia (лат. septum, мн. ч. septa •— перегород-
ка), или перегородчатожаберным, относится небольшая группа би-
Вальвий, преимущественно глубоководных, у которых жабры преобразо-
ваны в мускулистую поперечную перегородку, разделяющую мантий-
ную полость на две части — нижнюю и верхнюю,, отсюда название
надотряда; в перегородке, или септе, имеются отверстия; за счет
мышечных сокращений септа приводится в колебательное движение и
играет роль своеобразного насоса, перекачивающего воду; поэтому
жабры не используются, подобно аутобранхиям, в качестве фильтра для
улавливания пищевых частиц. Септибранхии по способу питания отно-
сятся к хищникам, питаясь зоопланктоном, в частности копеподами,
остракодами и личинками разных групп. В связи с этим хитиноидная
выстилка желудка, способствующая измельчению пищи, сильно увели-
чена при общем примитивном строении желудка. Раковина равноствор-
чатая с задним оттянутым в виде ростра концом для сифонов.; хорошо
развит перламутровый слой; замок рудиментарен, неясного происхож-
дения, со слабо развитыми кардинальными и латеральными зубами,
иногда отсутствующими. Связка опистодетная; волокнистый слой с из-
вестковой сердцевиной (литодесмой). Скульптура в виде радиальных
ребер, иногда кренуллированных. Надотряд разделен на три отряда
(рис. 187): отряд Verticordiida (палеоген — ныне) с родами Verticordia
(палеоцен — ныне), Euciroa и Lyonsiella (совр.); отряд Poromyida
(мел — ныне) с родами Poromya (мел — ныне), Cetoconcha (совр.);
отряд Cuspidariida (ордовик — пермь, мел — ныне) с родами Cuspida-
340
ria (мел — ныне) и Conocardium (?) (средний ордовик—поздняя пермь,
возможно триас) . Последний род в американских «Основах палеонтоло-
гии» возведен в ранг самостоятельного отряда Conocardioida, однако
без указания, к какому подклассу его следует относить.
,История развития надотряда недостаточно ясна; но, по-видимому,
его представители ответвились от основного ствола аутобранХий до-
Рис. 187. Надотряд Septibranchia; отряды Cuspidariida (/—2), Verticordiida (3)
и Pbromyida (4):
la, б — Conocardium (средний ордовик — пермь?, триас); 2а, б — Cuspidaria
(мел — ныне); 3 — Verticordia (палеоцен — ныне),; 4 — Роготуа- (мел — ныне)
вольно рано, возможно в ордовике,, и позднее дали три основных отря-
да, существующих до настоящего времени в виде небольшой специали-
зированной группы глубоководных бивальвий.
История развития бивальвий. Филогенетические взаимоотношения
между рассмотренными выше тремя над отрядами недостаточно ясны.
По-видимому, предками для всех бивальвий послужили какие-то при-
митивные протобранхии, от которых возникли аутобранхии с
жабрами, преобразованными в фильтр-сито, а позднее от них вследствие
преобразования пластинчатых йсабр возникли септибранхии — не-
большой специализированный надотряд глубоководных бивальвий.
Среди протобранхий как наиболее древние рассматриваются ктено-
донтиды, появившиеся в самом конце кембрия или в начале ордовика,
от них возникли прекардииды, просуществовавшие до начала карбона.
По-видимому, представители обоих отрядов входили в состав инфауны
и питались детритом, который они собирали с помощью приротовых
придатков. В девоне от прекардиид возникли солемииды — протобран-
хии, перешедшие к зарывающемуся образу жизни, но, как и все прото-
бранхии, питающиеся детритом. В среднем кембрии возникли первые
аутобранхии, представленные отрядом актинодонтид. В начале
ордовика они разделились на два подотряда: актинодонтин и триго-
ниин; первый из них был развит преимущественно в палеозое, а второй
получил особенно широкое распространение в мезозое. В среднем дево-
341
не стали известны первые униониды, возникшие от каких-то актино-
донтид и перешедшие к жизни в пресных водах, а начиная с триаса
широко расселившиеся практически по всему свету. От актинодонтид,
по-видимому, в ордовике возникли первые пектиниды, составляющие
существенную часть свободноживущей эпифауны; пектиниды частично
приспособились к плаванию на небольшие расстояния, прикреплению с
помощью биссуса к подводным предметам, а также прочному соедине-
нию с субстратом цементацией одной из створок.
От актинодонтид в ордовике, вероятно, возник отряд астартид,
входящих в состав свободноживущей эпифауны мелководья, некоторые
представители отряда приспособились к зарыванию.. Астартиды яви-
лись одним из важных отрядов аутобранхий; от них в позднем силуре
Ьозник отряд гиппуритид, первые представители которых, по-видимому,
прикреплялись к субстрату с помощью биссуса, а начиная с поздней
юры перешли к прикрепленному образу жизни, цементируясь одной из
створок; обитая среди коралловых рифов, они очень быстро достигли
расцвета, но и столь же быстро угасли в конце мела. В раннем девоне
от астартид произошли кардитиды, по типу питания относящиеся к ин-
фаунным фильтраторам. В позднем триасе от астартид возник очень
большой отряд аутобранхий — отряд венерид. обитателей дна, свободно-
живущих или зарывающихся, частично приспособившихся к жизни
в солоноватоводных бассейнах и даже пресных водах. В юре от основ-
ного ствола венерид ответвился подотряд фоладин, представители ко-
торого приспособились к сверлению известковых пород, а позднее, в
конце мела, к сверлению древесины. В конце мела часть венерид, обо-
собленная в подотряд хамин, перешла к прочному прикреплению путем
цементации к подводным предметам, подобно рудистам.
В девоне от актинодонтид возник отряд фоладомиид, приспособив-
шийся к зарыванию внутрь осадка, возможно возникший от актинодон-
тид; подотряд клавагеллин, появившийся в конце мела, характеризо-
вался частичной редукцией раковины и возникновением вместо нее на-
ружной известковой трубки (см. рис. 186, У). В ордовике от актино-
донтид возник отряд циртодонтид, две ветви которого — арцины и пте-
риины — существуют до наших дней и обитают в зоне мелководья,
прикрепляясь биссусом к различным понижениям дна, к веткам корал-
лов, к морским водорослям. В девоне от арцин ответвился подотряд ми-
тилин, перешедший к прикреплению биссусом к подводным скалам, об-
разуя массивные поселения, или к частичному погружению в осадок
и прикреплению с помощью биссуса, как якорем, к частицам осадка
(Pinna, некоторые митилиды). В триасе от птериин возник подотряд
устриц, цементирующихся одной из створок, чаще левой,' к твердым
подводным предметам; устрицы — обитатели мелководья, где они часто
образуют банки (рис. 188).
КЛАСС CEPHALOPODA. ГОЛОВОНОГИЕ МОЛЛЮСКИ
Головоногие моллюски, или цефалоподы (гр. kephale — голова,
pous, род. podos — нога), — наиболее высокоорганизованные билате-
рально-симметричные моллюски, своего рода «приматы» моря среди
всех беспозвоночных. К ним относятся современные кальмары, кара-
катицы, осьминоги, аргонавты, «жемчужный кораблик» — наутилус и
большое число вымерших форм, среди которых наибольшей извест-
ностью пользуются аммониты, белемниты. Число современных видов
составляет около 650 видов, число вымерших превышает 10 000.
343
Рис. 189. Геохронологическое распространение двустворчатых моллюсков
Голова, как правило, хорошо обособлена от туловища; вокруг рта
развиты или короткие, но многочисленные щупальца (до 90), способ-
ные втягиваться во влагалище (у наутилуса), или длинные, но немно-
гочисленные ногощупальца, или руки (8 или 10); последние всегда
снабжены присосками или крючочками; руки используются для удер-
жания добычи или для хождения по дну (некоторые осьминоги). Щу-
пальца в эмбриргенезе развиваются из того же зачатка, из которого
у всех остальных моллюсков развивается нога, и иннервируются пе-
дальным гднглием; из этого же зачатка формируется воронка —
очень z характерный орган всех современных головоногих, служащий
дйя реактивного движения: вода через широкую щель всасывается в;
мантийную полость и оттуда, после того как щель замыкается, с силой
выталкивается через воронку; реактивная сила струи воды толкает
головоногого моллюска задним концом тела вперед.
Удлиненное тело головоногих заключено в мантию, прирастающую'
на спинной стороне к висцеральному мешку. Снизу, с брюшной сторо-
ны, Мантия прикрывает глубокую мантийную полость; в ней лежат 2‘
или 4 симметрично расположенные жабры, анальное отверстие, распо-
ложенное медиально, отверстия почек, половых органов и нидаменталь-
ная железа. Нервная система и органы чувств, особенно глаза, хорошо-
развиты; нервные узлы, или ганглии, обычно сконцентрированы в сплош-
ную нервную массу, называемую иногда «головным мозгом», за-
ключенную в хрящевую капсулу.
344
Рис. 190. Подкласс Nautiloidea. Схема строения современного наутилуса:
1а — строение раковины и мягкого тела, 16 — начало спирали, 1в, г — располо-
жение щупалец у самки (7s) и самца (Уг), вид спереди;
вен — вентральная сторона, впс — внутренний призматический слой, вр — во-
ронка, вч — верхняя челюсть, га — ганглии, гск — гидростатическая камера,
дор — дорзальная сторона, жб — жабры, кап — капюшон, кон — конхиолино-
вый слой, нпс — наружный призматический слой, нч — нижняя челюсть, 1 об,
2 об — первый, второй обороты, п — почка, пер — перламутровый слой, пж —
половая железа, пс — призматический слой, пт — пищеварительный тракт, ра —
радула, с — сердце, 1с — первая септа, сп — спадикс, сф — сифон, уо — умби-
ликальное отверстие, ц — целом, ч — челюсть, щ — щупальца (сечения — ок-
руглой или треугольной формы)
Пищеварительная система развита достаточно сложно. Рот, окру-
женный щупальцами, снабжен сильными роговыми или обызвествлен-
ными челюстями; в глотке на поверхности мускулистого языка лежит
радула, несущая 9 или 13 рядов хитиновых зубчиков; за глоткой сле-
дует пищевод, имеющий особое выпячивание — зоб; за ним лежит
мускулистый желудок, выстланный кутикулой; в нем происходит пере-
тирание пищи; иногда обособлен еще железистый желудок; с задней
частью желудка соединен слепой вырост, служащий для всасывания
пищи. Протоки печени впадают в слепой вырост; печень служит не
только для переваривания пищи, но и участвует в процессах резорби-
рования пищи и накопления питательных веществ. Желудок переходит
в тонкую кишку, в которой пища окончательно переваривается и вса-
сывается; далее следует толстая, или прямая, кишка, в которой форми-
руются экскременты. У большинства современных и ряда вымерших
головоногих имеется очень характерный для этого класса чернильный
мешок, чернильная железа которого вырабатывает особую жидкость,
выбрасываемую через воронку (отсюда название современных голо-
воногих Tintenfische —• «чернильные рыбы»); жидкость образует свое-
образную «дымовую завесу» или парализует обонятельные нервы на-
падающих рыб, или образует в воде темную каплю, некоторое время
плавающую в толще воды и по форме напоминающую тело выбросив-
шего ее моллюска.
Кровеносная система высокоразвита, она почти замкнута, лишь
в некоторых местах сохраняются лакуны; сердце состоит из желудочка
и 2 или 4 предсердий (в соответствии с числом жабр); кроме основного
сердца имеются добавочные пульсирующие органы — венозные, или
жаберные, сердца, проталкивающие кровь через жабры. Кровь голу-
бого цвета, так как вместо гемоглобина содержит гемоцианин, в состав
которого входит медь. От вторичной полости тела сохранился перикар-
дий, заключающий сердце и полость, в которой лежит желудок, часть
кишечника и половая железа; у осьминогов вторичная полость реду-
цирована до полости, в которой лежит только половая железа; даже
сердце находится вне целома, что является исключением среди всех
моллюсков. Органами выделения служат почки, построенные по типу,
характерному для всех моллюсков; у наутилуса их четыре, у остальных
современных двужаберных — две.
Все современные головоногие раздельнополы. Непарная половая
железа занимает самый задний конец тела и открывается протоком в
мантийную полость. Сперматофоры переносятся в мантийную полость
самки при помощи одной из видоизмененных рук. У некоторых голово-
ногих эта рука резко отличается от всех остальных рук своей удлинен-
ной червеобразной формой (гектокотиль); гектокотиль может отры-
ваться от тела и самостоятельно плавать. У наутилуса видоизмененные
щупальца (с п а д и к с) выполняют половую функцию. У всех современ-
ных форм яйца крупные, богаты желтком, развитие прямое, т. е. це-
ликом проходит внутри яйцевых оболочек. Только что вылупившаяся
молодь отличается от взрослых величиной и формой тела, недоразви-
тием рук и некоторых органов .(половых). Иногда наблюдается живо-
рождение.
Мягкое тело современного наутилуса и большинства вымерших го-
ловоногих заключено в известковую раковину разнообразной формы,
представляющую собой трубку, замкнутую, подобно раковинам гастро-
под, на одном конце и открытую на другом. Тело моллюска помещает-
ся в жилой камере, занимающей переднюю часть раковины, и прикреп-
ляется изнутри при помощи мускулов-ретракторов. Задняя часть ра-
346
ковины, в отличие от таковой большинства брюхоногих, разделена пе-
регородками, или септами, на отдельные гидростатические камеры, за-
полненные газом и жидкостью. Через все камеры, через особые отвер-
стия в септах проходит сифон, представляющий собой вырост задней
части тела и состоящий из кутикулярной оболочки и соединительной тка-
ни, внутри которой проходят кровеносные сосуды — артериальный и
венозный; сифон служит для регулирования содержания жидкости и
газа в камерах. Сифон может быть узким или широким. Камерная
часть раковины называется фрагмоконом. Форма раковины отли-
чается большим разнообразием, но превалируют два основных типа:
прямая и спирально-плоскостная. У современного наутилуса раковина
второго типа;-у вымерших разнообразие форм раковины будет рассмот-
рено при описании соответствующих подклассов. У двужаберных голо-
воногих эволюция шла вначале по пути превращения раковины во
внутреннюю (белемниты), а затем по пути редукции ее вплоть до пол-
ного исчезновения; у каракатицы она превратилась в толстую удли-
ненно-овальную пластинку,, прикрывающую под мантией спинную часть
тела; у кальмаров — в тонкую роговую нластинку, напоминающую рим-
ский меч-гладйус; у осьминогов •— редуцировалась до двух маленьких
конхиолиновых палочек.
Головоногие моллюски появились в кембрии, прошли сложный путь
развития, дали много разнообразных и широко распространенных групп,
очень важных для биостратиграфических целей. Современные голово-
ногие распространены преимущественно в теплых морях, составляя су-
щественную часть нектона и нектобентоса. На основании особенностей
строения раковины и внутренних органов все головоногие разделены
на семь подклассов: Nautiloidea, Endocertoidea, Actinoceratoidea, Ortho-
ceratoidea, Bactritoidea, Ammonoidea, Coleoidea. Первые шесть подклас-
сов объединяли ранее в группу наружнораковинных, или Ectocochlia, и
противопоставляли внутреннераковинным, или Endocochlia, к которым
относили последний подкласс. В связи с тем что современный наутилус
с наружной раковиной имеет четыре жабры, а все остальные совре-
менные головоногие с внутренней раковиной — только две жабры, го-
ловоногих разделяли на два подкласса: четырехжаберных, или Tetra-
branchiata, и двужаберных, или Dibranchiata. Полученная новая ин-
формация о строении внутренних органов у аммонитов, близких к тако-
вым современных дибранхиат, заставляет отказаться от подобного де-
ления.
ПОДКЛАСС NAUTILOIDEA. НАУТИЛОИДЕИ
Общая характеристика. У наутилоидей (гр. nautes — поэтически,
nautilos — моряк) мягкое тело заключено в известковую раковину:
прямую, изогнутую, спирально-плоскостную или реже спирально-кони-
ческую; жилая камера занимает у спирально свернутых раковин около
7з оборота; септы фрагмокона прикрепляются изнутри к раковине по
почти прямой, слабо волнистой и редко очень извилистой перегородоч-
ной линии. Сифон проходит через отверстия в септе и охватывается
септальными трубками и соединительными кольцами. Строение сифо-
на и его положение очень разнообразны. Тело современного наутилуса
короткое, мешковидное; рот окружен многочисленными тонкими щупаль-
цами, которые втягиваются внутрь влагалища; имеются роговые челю-
сти с обызвествленным клювом и радула. Воронка состоит из двух не-
сросшихся лопастей; в мантийной полости расположены две пары жабр,
347
около них 4 выводных отверстия почек, два половых отверстия и два*
отверстия нидаментальной железы. Чернильная железа отсутствует;
имеются осфрадии и статоцисты. Центральная нервная система пред-
ставлена тремя дугообразными нервными тяжами, расположенными во-
круг пищевода. Раздельнополы; эмбриональное развитие изучено недо-
статочно. Распространены от кембрия доныне. Известно около 6501
родов.
Строение мягкого тела. О строении мягкого тела наутилоидей мож-
но судить на основании рассмотрения анатомии современного наутилу-
са (рис. 190); о его вымерших родственниках можно высказывать лишь
отдельные предположения. Тело наутилоидей окружено мантией, склад-
ки которой на вентральной стороне ограничивают большую мантийную
полость. Голова слабо обособлена от туловища, она несет глаза, капю-
шон, щупальца и воронку. В жилой камере расположен висцеральный:
мешок, заключающий все внутренние органы. От задней части тела,
отходит сифон, проходящий через отверстия в септах до начальной ка-
меры. В голове расположен Н-образный хрящ, к которому прикреп-
ляются два ствола мускулов-ретракторов, втягивающих голову с щу-
пальцами внутрь раковины. Около глаз расположёно по паре щупалец,,
возможно гомологичных головным щупальцам гастропод.
Остальные образуют два кольца щупалец — внешнее и внутреннее
(рис. 190, 1в, 1г); внешнее несет 38 щупалец, одинаковых у самцов иг
самок, из которых два дорзальных срастаются своими влагалищами и
образуют капюшон, закрывающий устье при втягивании головы
внутрь раковины; влагалища двух соседних с ними щупалец также*
участвуют в замыкании устья. Влагалища остальных срастаются вместе-
и образуют внешнее кольцо. Внутреннее кольцо состоит из трех групп
щупалец, различных у самцов и самок. У первых часть щупалец видо-
изменена в особый копулятивный орган — спадикс, у вторых кольцо»
распадается на три лопасти: нижнюю, или вентральную, несущую
28 щупалец, и две верхние — правую и Левую по 12 щупалец на каж-
дой. Внешнее кольцо гомологично кольцу рук современных колеоидей, а?
внутреннее — буккальной мембране современных десятиногих колео-
идей.
На вентральной стороне лежит воронка, состоящая из двух ло-
пастей. Вода засасывается через щель между складками мантии в ман-
тийную полость, а затем выбрасывается с силой через воронку, при
этом щель плотно закрывается. Мантийный комплекс составляют две
пары жабр, расположенные по обе стороны от анального отверстия,,
четыре отверстия почек, лежащие у основания каждой жабры, два по-
ловых отверстия, из которых функционируют только правые, два осфра-
дия и в самом конце мантийной полости лежат протоки нидаменталь-
ной железы, секрет которой образует внешние яйцевые оболочки.
Центральная нервная система имеет вид трех дугообразных нерв-
ных тяжей, образованных несколькими ганглиями и огибающих пище-
вод; от них отходят нервные стволы, иннервирующие все внутренние-
органы. Педальные ганглии дают начало нервам воронки и всех щупа-
лец. Н-образный хрящ поддерживает снизу центральную часть нервной
системы; к нему прикрепляется- ряд мускулов, в том числе мускулы-
ретракторы. Органами обоняния служат два глазных щупальца — ри-
нофоры, органами химического чувства — осфрадии, органами
равновесия и слуха — статоцисты, органами зрения —• глаза, по-
строенные по тому же типу, что и глаза у гастропод; глаз устроен до-
вольно примитивно — нет ни роговицы, ни хрусталика, ни стекловид-
ного тела.
348
• Рот расположен в центре кольца щупалец;.в глотке имеются радула
ж челюсти. Радула несет по 13 рядов мелких роговых зубчиков, пред-
назначенных для перетирания пищи и превращения ее в кашицу. Че-
люсти обызвествлены на к’онцах, предназначены для прокусывания до-
бычи. У наутилуса имеются пищевод, зоб и мускулистый желудок,
выстланные кутикулой, железистый желудок, участвующий в перевари-
вании пищи, слепой вырост для ее всасывания; в него впадают прото-
ки объемистой печени. От желудка отходит тонкая кишка, переходящая
в толстую, которая заканчивается в мантийной полости анальным от-
верстием. Пищеварительный тракт у наутилуса U-образно изогнут. Серд-
це состоит из желудочка и четырех предсердий. Целом разделен на
.два отдела, в первом лежит сердце, во втором —’желудок, часть ки-
шечника и половая железа. Перикардий двумя отверстиями сообщается
<с мантийной полостью. НауТилоидеи раздельнополы; самка откладыва-
ет крупные яйца до 45 мм в диаметре. К сожалению, до сих пор эмбрио-
жальное развитие наутилуса неизвестно.
Строение раковины. Раковина современного наутилуса спирально-
плоскостная, состоит из 2,5—3 оборотов. Она начинается эмбриональ-
ной раковинкой, построенной из одного оборота, диаметром 25—27 мм,
.покрытой сетчатой, так называемой юношеской, скульптурой. Эмб-
риональная раковинка состоит из жилой камеры и нескольких перего-
родок (к сожалению, точное число их не установлено). В первом обо-
роте спираль свернута не плотно, имеется пупковое, или умбили-
к а л ь н о е, сквозное отверстие (рис. 190, 16). В первой гидростатиче-
ской камере, имеющей форму полумесяца, начинается сифон, который из-
нутри прикрепляется к раковине. Во второй перегородке возникает от-
верстие для прохождения сифона. Сифон представляет собой тонкий
тяж, отходящий от задней части тела, окруженный конхиолиновой и
‘Спикульной оболочкой; внутри сифона в соединительной ткани вдоль
вентральной стороны проходит артериальный сосуд, а венозная кровь
собирается в многочисленных лакунах и поступает в крупную вену,
проходящую в центре сифона. Оболочка образуется за счет эпителия,
окружающего сифон. Край перегородки, через которую проходит сифон,
отогнут назад й образует так называемую септальную трубку,
или некк.
Раковина современного наутилуса почти инволютная, умбилик от
широкого до очень узкого; у одного из современных видов умбилик
.закрыт умбиликальной пробкой.
Наружная выпуклая сторона условно названа вентральной;
вогнутая внутренняя — дорзальной; а соединяющие их две стен-
ки — латеральными. Стенка раковины построена из органиче-
ского конхиолинового слоя и сложно построенного известкового слоя,
-состоящего из двух призматических и одного пластинчатого между
ними. У ископаемых наутилоидей различают два основных типа ра-
ковин: прямую, или ортоконовую, и планоспиральную, или нау-
тилоконовую. Между ними можно найти все переходы. Разли-
чают: о р т о к о н ы, или прямые раковины, тонкие и длинные или
•толстые и короткие; циртоконы— раковины, частично согнутые;
потериоконы — согнутые в начальной части, а затем боченкооб-
разно вздутые; гироконы — свернутые в свободную спираль, со-
стоящую из несоприкасающихся оборотов; серпентиконы —
состоящие из едва соприкасающихся оборотов; тарфиконы —
с соприкасающимися оборотами; наутилоконы — обороты, в той
или иной степени охватывающие друг друга; среди последних разли-
чают эволютные, с малообъемлющими оборотами и широким
349
умбиликом, и инволют ны е, с объемлющими оборотами, с уз-
ким или замкнутым умбиликом.' Поперечное сечение ортоконов и
циртоконов обычно овальное или круглое; у спирально свернутых —
вентральные и латеральные стороны образуют эволютную часть обо-
рота, а дорзальная, закрытая последующим оборотом — инволютную
(рис. 191, 8—10).
Форма поперечного сечения спирально свернутых раковин крайне
разнообразна. Редко встречаются трохоконы — спирально-кониче-
ские раковины. Различают также э кз о г а с тр и ч ес к и е раковины,
у которых наружная выпуклая сторона является вентральной, и эн-
до гастрические, если таковой является внутренняя, вогнутая
сторона. Положение вентральной стороны определяется наличием
синуса для воронки. Поверхность раковины бывает гладкой, только,
с линиями нарастания, или скульптировэнной; различают скульптуру
продольную и поперечную. Некоторые формы были снабжены бугор-
ками, шипами, иглами. Известно около 40 экземпляров с сохранив-
шимися следами окраски в виде поперечных, волнистых, зигзагооб-
разных, узких полосок, обнаруженных преимущественно у ортоконо-
вых раковин.
Жилая камера занимает место от устья до последней пере-
городки. Устьевой край отличается большим разнообразием — от ши-
рокого открытого до щелевидного многодольчатого. Обычно за
устьевым краем наблюдается утолщение раковинного слоя, а с вен-
тральной стороны выемка для воронки. Мягкое тело соединяется с
раковиной двумя сильными мускулами, передние концы которых
прикрепляются к Н-образному хрящу, а задние — к стенке раковины
изнутри; с дорзальной и вентральной стороны они соединяются
полосой фибр, образующих так называемый аннулюс. Мускулы-
ретракторы втягивали тело наутилоидей внутрь раковины. У древних
наутилоидей мускулы-ретракторы прикреплялись на вентральной или
дорзальной сторонах раковины и также соединялись аннулярным
кольцом. Отпечатки мускулов и аннулюса получили название пери-
фра к т а (гр. peri — вокруг, phractos — заключенный).
Фрагмокон — задняя часть раковины, разделенная септами на
отдельные гидростатические камеры. Септы, периодически образуемые
по мере роста моллюска задней частью тела, состоят из свободной и
пристенной, или муральной, части; свободная часть имеет фор-
му блюдца, прогнутого в сторону начальной части раковины; в ее
центре эксцентрично или у края расположено отверстие для сифона
круглой или овальной формы.
Муральной частью перегородка прикрепляется к стенке ракови-
ны по прямой линии либо, если ее внешний край изогнут, по вол-
нистой; изгибы, направленные вперед, получили название седел,
а направленные назад — лопастей. После удаления раковинного
слоя на внутренних ядрах видна перегородочная линия —
линия прикрепления перегородки к раковине. Обычно ее изображают
в развернутом виде в одной плоскости. Иногда она приобретает до-
вольно сложные изгибы, однако не достигающие той сложности,
которая характерна для подобных линий аммонитов. Лопасти пере-
городочной линии получают названия в зависимости от их положе-
ния; выделяют вентральную и дорзальную лопасти, лежа-
щие в плоскости симметрии на соответствующих сторонах раковины—
вентральной и дорзальной; латеральную—лежащую сбоку,
умбиликальную — лежащую на умбиликальной стенке.
350
Различают несколько типов септальных трубок (рис. 191, 11—14):
1) ахоанитовый — имеется только отверстие со слабо округлен-
ными краями; 2) ор тохоа нитовы й — септальная трубка корот-
кая цилиндрическая; 3) ц и р т о х о а н и то в ы й — септальная трубка
отгибается внутрь камер; 4) г о л о х о а н и то в ы й — септальная
трубка длинная, достигает предшествующую септу; 5) макрохоа-
нитовый — длина септальной трубки превышает длину одной ка-
меры. Между септами сифон окружается соединительными кольцами,
длина и строение которых бывают различными.
Камерные отложения. У многих наутилоидей внутри гидростати-
ческих камер обнаружены так называемые внутрикамерные отложе-
ния, возникшие при жизни животного; они отличаются от вторичных
отложений, образовавшихся при фоссилизации. У современного нау-
тилуса в переднем углу гидростатических камер камерные отложения
представлены конхиолином и мелкими кристаллами арагонита.
Внутрисифонные, или эндосифонные, отложения обнаруже-
ны у наутилоидей с широким сифоном. Они представлены: попереч-
ными диафрагмами, которые изредка встречаются у эллесмероцера-
тид; продольными пластинами (напоминающими в поперечном сече-
нии септы кораллов) у онкоцератид; воронкообразными структурами
(эндоконами), вставленными одна в другую, с эндосифональным
каналом в центре (наиболее развитыми у эндоцератоидей); аннуло-
сифонатными отложениями, развитыми около септальных трубок.
История развития. Наутилоидеи появляются в раннем кембрии,
достигают расцвета в ордовике и силуре, несколько сокращаются в
девоне, еще больше в конце палеозоя, в мезозое продолжает сущест-
вовать только один отряд наутилид, обладающий спирально-плоско-
стной раковиной, последний представитель которого в составе одного
рода и пяти видов доживает до наших дней. Современный наутилус
живет в тропических морях в юго-западной части Тихого океана, у
берегов Австралии и Юго-Восточной Азии. Пустые раковины заносятся
к берегам Японии, Новой Зеландии, Мадагаскара. Наутилусы по спо-
собу питания являются хищниками, они питаются рыбой, крабами,
раками. Гидростатические камеры заполнены газом (азот, аргон) и
жидкостью; в последних камерах жидкости больше, чем в первых.
Газовое давление в камерах и количество жидкости регулируются
при помощи кровеносных сосудов сифона.
Основы систематики и классификация. Подкласс наутилоидей на
основании строения раковины и сифона разделен на следующие отря-
ды: Volborthellida, Ellesmeroceratida, Discosorida, Ascoceratida,
Tarphyceratida, Oncoceratida, Nautilida.
Отряд Volborthellida. Фольбортеллиды (по имени А. Ф. Фоль-
борта). Раковина маленькая, ортоконовая, округлая или сжатая
дорзовентрально, сифон центральный, узкий, без септальных трубок
(ахоанитовый). Систематическое положение отряда и принадлеж-
ность его к наутилоидеям рядом исследователей оспаривается; неиз-
вестно строение стенки, сифона (рис. 192, 1).
Отряд Ellesmeroceratida. Эллесмероцератиды. Раковины ортоко-
новые, маленьких и средних размеров, но более часты циртоконовые,
экзо- и эндогастрические, многие короткие, ширококонические, глад-
кие или кольчатые. Септы сближены; сифон сравнительно широкий,
занимает краевое или почти краевое положение, внутри сифона ино-
гда наблюдаются поперечные диафрагмы; септальные трубки обычно
короткие, но иногда голохоанитовые, соединительные кольца толстые.
Первые эллесмероцератиды известны из позднего кембрия, наиболее
351
.широко распространены в ордовике, но к концу ордовика вымирают.
Эллесмероцератиды рассматриваются в качестве основного ствола
наутилоидей, от которого произошли актиноцератиты, . эндоцератиты
и многие отряды наутилоидей. Поздний кембрий— ордовик. Около
80 родов (рис. 192, 2, 3).
нмш
Рис. 191. Подкласс Nautiloidea. Форма и строение раковины (1—10) и типы
септальных трубок (И—14):
1 — ортоцераконовая; 2 — аскоцераконовая; 3 — циртоцераконовая; 4 — ли-
туитиконовая; 5 — ширококоническая с Т-образным устьем: 5а — продольное
сечение, 56 — вид с вентральной стороны, 5в — вид со стороны устья; 6 — тро-
хоцераконовая; 7—10 — планоспиральные: 7 — гироцераконовая, 8 — эволют-
ная, 9 — полуинволютная, 10 — инволютная; септальные трубки: И — ахоани-
товые, 12 — ортохоанитовые, 13 — циртохоанитовые, 14 — голохоанитовые;
вен — вентральная сторона, дор — дорзальная сторона, жк — жилая камера,
сф — сифон, фк — фрагмокон
Отряд Discosorida. Дискосориды (гр. diskos — диск, soros —
куча). Раковины ширококонические, циртоконовые, реже ортоконовые,
с открытым или сжатым устьем, обычно эндогастрические; сифон
довольно широкий от краевого до центрального, септальные трубки
циртохоанитовые. Соединительные кольца обычно выпуклые, толстые
и сложные; у некоторых форм имеются внутрисифонные отложения,
вначале в форме небольших колец на внутренней поверхности сеп-
тальной трубки, а затем в форме своеобразных эндоконов; наблю-
даются небольшие внутрикамерные отложения. Начальная камера
чашевидная, цекум — шаровидный. Дискосориды возникли от эллесме-
роцератид, от которых они отличаются более сложно дифференциро-
ванными соединительными ' кольцами. Развитие дискосорид шло по
352
пути увеличения раковины, усложнения структуры соединительных
колец, смещения сифона к краю, усложнения устьевого края. Диско-
сориды появились в среднем ордовике и достигали расцвета два раза,
в ордовике и в середине силура. Средний ордовик — средний девон.
Около 45 родов (рис. 192, 4—7).
Рис. 192. Отряды Volborthellida (/), Ellesmeroceratida (2—3) и Discosorida (4—7):
1 — схема строения раковины Volborthella (ранний кембрий); 2 — реконструк-
ция вентральной половины сифона с внутрисифонными образованиями у Bathmo-
ceras (ордовик); 3 — Ellesmeroceras, схема (ранний ордовик.); 4 — полиптихо-
коны; 5 —• схема строения соединительного кольца; 6 — Phragmoceras (средний
силур): 6а — вид со стороны устья, 66 — сбоку, 6в — схема продольного се-
чения; 7 — Discosorus (средний силур), ядро жилой камеры (жк) и сифона;
по — полиптихоконы, ст — септальные трубки, ск — соединительные кольца,
сф — сифон
Отряд Ascoceratida. Аскоцератиды (гр. ascos — мешок). Ракови-
на состоит из длинноконической (до 20 см) циртоконовой начальной
части и сильно вздутой бревиконовой, или аскоцероидной, взрослой
части (до 15 см), очень редко находимых вместе. Септы у начальной
части с простой перегородочной линией; первая камера в два раза
длинней второй; сифон узкий, в начальной камере лежит на вент:
ральной стенке, затем между осью раковины и вентральной стенкой;
септальные трубки короткие, ортохоанитовые. Вздутая взрослая часть
имеет довольно широкий четковидный сифон и отделена от начальной
части очень толстой септой, толщина которой равна толщине стенки
взрослой части. В широкой части обнаружено от 1 до 12 септ, чаще
всего 5; септы обычно параллельны друг другу; но у многих они
образуют на дорзальной стороне очень высокое седло в виде сигмо-
идального изгиба (рис. 193, /). Не исключена возможность, что
23 в. В. Друщиц
353
взрослые формы при жизни отбрасывали свою начальную часть рако-
вины. Асконератиды возникли в среднем ордовике от ортоцератид,
были распространены в ордовике и силуре Европы и Северной Аме-
рики и вымерли в конпе силура. Наличие гидростатических камер на
дорзальной стороне жилой камеры позволяет предполагать, что аско-
цератиды были хорошими пловцами. Ордовик — силур. Известно око-
ло 15 родов (рис. 193, /).
Отряд Oncoceratida. Онкоцератиды (гр. onkos — вздутый). Рако-
вины ортоконовые, циртоконовые, наутилоконовые и трохоидные; но-
преобладают короткоконические, с утолщенной жилой камерой и су-
женной или сложно разветвленной апертурой. Перегородочная линия
простая или слегка волнистая; сифон обычно краевой, узкий или
более широкий с продольными радиальными пластинами, которые
образуются за счет утолщений соединительных колец, отходящих
радиально к центру сифона; септальные трубки циртохоанитовые
короткие, реже длинные; соединительные кольца от выпуклых до во-
гнутых. Сифон начинается в первой камере шаровидным цекумом и
расположен вблизи вентральной стенки. Эмбриональная раковина
состояла из жилой камеры и одной или двух гидростатических камер.
Онкоцератиды возникли (от эллесмероцератид) в среднем ордовике,
достигли расцвета в середине силура (известно свыше 40 родов) и в.
середине девона (около 40 родов), после чего их число з-аметно сокра-
тилось и последние представители вымерли в начале среднего кар-
бона. Средний ордовик — начало среднего карбона. Общее числом
родов около 150 (рис. 193, 2—6).
Отряд Tarphyceratida. Тарфицератиды (гр. tarphis — тесный, тол-
стый) объединяют наиболее древних наутилоидей, обладавших ма-
ленькой планоспиральной раковиной, состоящей из 3—6 оборотов с
умбиликальным отверстием. Часто планоспиральная раковина пере-
ходит в прямой ствол до 30 см длиной; обороты обычно эволютные,
едва соприкасающиеся; длина жилой камеры составляет около-
’/г оборота или соответствующей части ствола; встречаются трохоид-
ные раковины. Положение сифона изредка изменяется от вентраль-
ного до дорзального; сифон начинается удлиненным цекумом, лежа-
щим на вентральной стенке первой камеры; септальные трубки орто-
хоанитовые; соединительные кольца от толстых у ранних форм до
тонких у более поздних. На внутренних ядрах обнаружены отпечатки
мускулов-ретракторов на вентральной стороне у спирально сверну-
тых и на дорзальной — у прямых. Имеются внутрикамерные и сифон-
ные известковые отложения. Первые тарфицератиды появляются в
начале ордовика, по-видимому они происходят от эллесмероцератид
и эволюционируют по пути приобретения спирально-плоскостной ра-
ковины с простым устьем и синусом для воронки (название рода и
отряда связано с наличием спирально свернутой' раковины), а позд-
нее с суженным устьем; в силуре и девоне появляются тарфицера-
тиды с трохоидной раковиной; последние представители отряда вы-
мерли в начале позднего девона. Около 90 родов (рис. 194, 1—4).
Отряд Nautilida. Наутилиды (гр. nautes — поэтически, nautilos —
моряк). Раковины преимущественно наутилоконовые, у палеозойских
представителей могут быть прямые, циртоконовые и гироконовые-
(с несоприкасающимися оборотами); поперечное сечение оборотов-
округленное, овальное, четырехугольное, многоугольное, сжатое с
боков или, наоборот, широкое. Поверхность раковины гладкая или
скульптированная. Перегородочная линия от простой до довольно-
сложной с отчетливыми лопастями и седлами (рис. 195); положение.-
354
сифона изменчиво —от почти центрального до краевого; сифон начи-
нается в первой камере неясно выраженным цекумом, прикреплен-
Рис. 193. Отряды Ascoceratida (У) и Oncoceratida (2—6):
1 — Ascoceras (силур): la — продольный разрез, 16 — жилая камера; 2 —
начальная часть раковины; 3 — Poterioceras (ранний карбон); 4 — Octame-
rella (средний силур): 4а — вид сбоку, 46 — вид со стороны устья; 5 — Суг-
thoceratites (средний девон); 6 — актиносифонатная структура сифона (Х4);
жк — жилая камера, сф — сифон, фк — фрагмокон
Рис. 194. Отряд Tarphyceratida:
1 — начальная часть раковины Curtoceras; 2 — Ophioceras (поздний силур);
3 — Estonioceras (ранний ордовик); 4 — Lituites (средний ордовик);
сф — сифон, уо — умбиликальное отверстие
ным к стенке раковины, иногда сохраняется рубец прикрепления.
Септальные трубки ретрохоанитовые, короткие; соединительные коль-
23*
355
ным отверстием, наутилиды возникли в
Рис. 195. Отряд Nautilida. Типы перегоро-
дочных линий:
1 — Nautilus (совр.); 2 — Siberionautilus;
3 — Aturia; 4 — Т ainoceras-, 5 — Cimomia-,
:. 6 — Neothrincoceras;
ао — аннулярный отросток, вс — вентраль-
ное седло; лопасти: вл — вентральная,
дл — дорзальная, лл — латеральная, ул —
умбиликальная; уш — умбиликальный шов
вины гладкая, у ряда форм развита
ца узкие. Внутрисифонные и камерные отложения не обнаружены.
Эмбриональная раковина довольно крупная с характерной «юноше-
ской» скульптурой, представленной радиальными и продольными
ребрышками; с узкой чашевидной начальной камерой и умбиликаль-
начале девона, вероятно от
онкоцератид. Наиболее раз-
нообразны и многочисленны
палеозойские и триасовые
представители, обладавшие
разного типа раковинами;
начиная с конца триаса со-
храняются формы с плано-
спиральной раковиной. От-
ряд включает свыше горо-
дов (рис. 196).
ПОДКЛАСС ENDOCERATOIDEA.
ЭНДОДЕРАТИТЫ
Эндоцератиты (гр. еп-
don — внутри, keras — рог)
имели прямые длинно- и
короткоконические ракови-
ны, реже эндогастрически
согнутые; в среднем ордови-
ке известны раковины до
9 м длиной, принадлежав-
шие наиболее крупным из
всех известных раковинных
беспозвоночных. Попереч-
ное сечение круглое, широ-
коовальное или эллиптиче-
ское; устьевой край ровный,
иногда с неглубоким вен-
тральным синусом. У боль-
шинства поверхность рако-
[еречная кольчатость, тонкая
штриховка. Отличительной чертой эндоцератит является широкий си-
фон, диаметр которого иногда достигает 2/з диаметра раковины, хотя
у ранних представителей он был довольно узким. Сифон, по-видимому,
заключал значительную часть внутренних органов мягкого тела мол-
люска. Перегородки вогнутые; септальные трубки от очень коротких,
ахоанитовых, до очень длинных, макрохоанитовых; соединительные
кольца от простых до довольно сложных.
Внутрисифонные отложения в типичном • виде представлены эндо-
конами и радиальными пластинками. Эндоконы — известковые
конические воронки, вставленные одна в другую. В вершине каждого
эндокона имеется отверстие, через которое проходит тонкая эндоси-
фонная трубка круглого или овального сечения. Форма и размеры
эндоконов изменяются в онтогенезе и у разных представителей раз-
личны. Недавно было обнаружено в одном сифоне два эндосифона,
разделенных перегородкой. У небольшой группы эндоцератит уста-
новлены внутри сифона своеобразные структуры, представленные
продольно-радиальными пластинами (структуры, описанные 3. Г. Ба-
лашовым); в поперечном разрезе они напоминают строение одиноч-
356
ных кораллов. Радиальные пластины, подобно септам кораллов, мог-
ли образоваться за счет складок оболочки сифона.
Установлено два типа апикального конца раковины. Первый
имеет сравнительно тонкий сифон, окруженный с самого начала гид-
ростатическими камерами (тип Proterocameroceras, рис. 197, 5);
у второго типа сифон занимает почти всю полость начальной камеры,
длина которой от вершины до первой перегородки равна- 38—50 мм,
Рис. 196. Отряд Nautilida:
1 — Rutoceras (средний девон); 2 — Ephippioceras (карбон — ранняя пермь); За, б —
Aturia (палеоцен — миоцен); 4а, б — Metacoceras (поздний карбон — пермь); 5 —
ринхолит;
к — капюшон, рук — рукоятка, сф — сифон
а диаметр ее переднего конца изменяется от 16 до 20 мм (Endoceras);
сифон расширен и несимметричен, гидростатические камеры распо-
ложены в полости сифона на значительном расстоянии от вершины
раковины (рис. 197, 2).
Эндоцератиты возникли от эллесмероцератид в раннем ордовике.
По строению сифона эндоцератиты резко отличаются от всех осталь-
ных подклассов головоногих; по-видимому, основная функция сифо-
на, который, может быть, стоит называть иначе, сводилась к образо-
ванию эндоконов или радиальных пластин, служивших для стабили-
зации раковины и придания ей горизонтального положения, что позво-
ляло этим моллюскам не только ползать, но и плавать, используя
для этих целей воронку. Однако они были, скорее всего, придонными
нектобентосными животными, подобно современным каракатицам.
О горизонтальном положении раковины свидетельствуют также следы
357
окраски дорзальной стороны и следы волочения на вентральной.
Небольшая группа эндоцератит с эндогастрической раковиной вела,
скорее всего, нектонный образ жизни, у нее были небольшие рако-
вины, относительно узкий сифон и слабо развитые эндоконы.
На основании строения сифона выделены два отряда: Endocera-
tida, Intejoceratida.
Отряд Endoceratida. Эндоцератиды. Внутри сифона имеются эн-
доконы. Развитие отряда началось в раннем ордовике (тремадоке)
и закончилось в позднем ордовике (ашгиле). Первые представители,
выделенные в подотряд Proterocameroceratina, имели ряд черт, сбли-
жающих их с эллесмероцератидами: маленькую раковину, широкий
сифон, но снабженный уже эндоконами простого строения, с апикаль-
ным конусом первого типа. Большинство родов этого подотряда
существовало в раннем ордовике (25 и 337); второй подотряд Endo-
ceratina объединяет более специализированных эндоцератит, которые
имеют второй тип строения апикального конца и различные типы
эндоконов, от простых до сложных; большинство родов этого под-
Рис. 197. Отряды Endoceratida [1—4) и Intejoceratida (5):
1 — Endoceras (ордовик): la — общий вид, 16 — основные структурные элементы,
1в — взаимоотношение мягкого тела с раковиной; 2—3 — типы апикального конца
раковины: 2 — у Endoceras, 3 — у Proierocameroceras, 4 — Cyrtendoceras (ордовик):
4а — продольное, 46 — поперечное сечения; 5 — Padunoceras (средний ордовик), ос-
новные структурные элементы раковины и сифона;
гск — гидростатическая камера, пт — перегородка с септальными трубками, сф — сифон
отряда существовало в среднем и позднем ордовике. Не исключена
возможность, что последние представители подкласса дожили до
середины силура, но они до сих пор известны слабо (род Нитеосе-
ras). Всего отряд объединяет около 70 родов (рис. 197, 1—4).
358
Отряд Intejoceratida. Интейоцератиды. Внутри сифона имеются
радиальные пластины, конвергентно сходные с подобными структу-
рами отряда онкоцератид; диаметр сифона равен */г диаметра рако-
вины. Интейоцератиды появились во второй половине раннего ордо-
вика (арениге) и вымерли в. начале среднего ордовика (лланвирне).
Они были распространены на территории Сибирской платформы.
Пока известно 3 рода (рис. 197, 5).
ПОДКЛАСС ACTINOCERATOIDEA. АКТИНОДЕРАТИТЫ
Подкласс актиноцератит (гр. aktis, род. aktinos—луч, ось) объ-
единяет палеозойских цефалопод, которые отличаются прямой или
•слабо изогнутой (циртоконовой) раковиной, средних и больших раз-
меров (до 6 м) и наличием Широкого сифона, заключающего слож-
ную сифонно-сосудистую систему. Подобно эндоцератитам, актино-
цератиты' были наиболее крупными цефалоподами палеозоя. Попе-
речное сечение круглое или овальное; поверхность раковины гладкая
или слабо скульптированная. Жилая камера несколько вздутая,
устье простое. Перегородки обычно вогнутые; перегородочная линия
простая и только у одной группы (Ellinoceras) состоит из правильно
чередующихся 14 лопастей и седел.
Рис. 198. Подкласс Actinoceratoidea. Схема строения раковины и сифона:
У — основные структурные элементы раковины и сифона: la — общий вид фраг-
мокона, 16 — строение одной гидростатической камеры, 1в — реконструкция си-
фонной системы; 2 — Armenoceras (ордовик — силур): 2a — продольное и 26 —
поперечное сечения сифона;
всо — внутрисифонные отложения, ко — камерные отложения, пк — продоль-
ный канал, пс — периспат, ст — септальные трубки, рк — радиальный канал
Сифон занимает субцентральное или краевое положение и обыч-
но имеет четковидную форму, очень расширяясь в камерах. Сеп-
тальные трубки циртохоанитовые; соединительные кольца имеют
веретеновидную, почти сферическую или дисковидную форму, причем
359
диаметр соединительных колец всегда превосходит диаметр септаль-
ной трубки. Ширина сифона у ранних форм больше его высоты (рас-
стояния между септами) в 5—6 раз, у более поздних — не более чем
в 2 раза; с возрастом, в онтогенезе, также наблюдается уменьшение
относительной ширины сифона. Внутри сифона расположена сложно>
устроенная сифонно-сосудистая система, состоящая из продольного
канала, от которого напротив каждой камеры отходят радиальные
трубки, доходящие до периспата — полости, возникшей между
соединительным кольцом и кольцом внутрисифонных отложений.
Радиальные каналы редкие или частые, простые или сложные; про-
стые отходят от продольного канала прямо или изгибаясь серпооб-
разно или аркообразно; сложные каналы перед стенкой периспата
многократно ветвятся или образуют сложно разветвленную сетчатую
систему. Строение сифонно-сосудистой системы отражает определен-
ные черты строения мягкого тела актиноцератит. Остается неясным,,
каким образом регулировалось содержание жидкости и газа в гидро-
статических камерах, так как радиальные каналы заканчивались в.
периспате и в соединительных кольцах нет никаких отверстий. Воз-
можно, соединительные кольца были тонкопористыми или губчаты-
ми. Внутри сифона и внутри камер наблюдаются внутрикамерные и
внутрисифонные отложения.
О строении мягкого тела можно судить по форме жилой камеры,
строению сифонно-сосудистой системы и отпечаткам щупалец, остав-
ленным на поверхности дна при ползании. По этим отпечаткам мож-
но предположить, что число щупалец, которыми обладали их вла-
дельцы, было небольшими: (до 10). Данные об онтогенезе довольно'
скудные. Первая камера 'сохранилась у нескольких форм, она широ-
коконическая или блюдцеобразная, значительно короче всех после-
дующих камер, у ранних форм сифон относительно большой, зани-
мает почти всю полость первой камеры, у поздних относительные
размеры сифона меньше. Данные о мускулах отсутствуют.
Актиноцератиты, по-видимому, так же как эндоцератиты, возник-
ли от эллесмероцератид в начале ордовика, но их развитие пошло по,
пути создания иного, чем у эндоцератит, типа сифонно-сосудистой
системы; они испытали расцвет в позднем ордовике и раннем силуре;
к началу девона число родов значительно сократилось; последние
представители вымерли в среднем карбоне. Они были распространены
в морях Сибири, Урала, Прибалтики, Западной Европы, Северной
Америки и Австралии.
Развитие актиноцератит шло по пути постепенного увеличения
размеров раковины, упрощения сифонно-сосудистой системы, от дре-
вовидно разветвленных радиальных каналов у ранних форм до про-
стых прямых у поздних и относительного уменьшения ширины сег-
ментов сифона — изменения соотношения ширины и длины сифона
от 6 : 1 до 2 : 1. Известно около 50 родов (рис. 198).
ПОДКЛАСС ORTHOCERATOIDEA. ОРТОЦЕРАТИТЫ
К подклассу ортоцератит (гр. orthos — прямой, ceras — рог) от-
носятся головоногие с ортоцераконовой, реже слабо циртоцераконовой
раковиной, иногда достигающей длины 1,5 м. Жилая камера относи-
360
тельно короткая, фрагмокон длинный; сечение округлое, овальное.
Сифон занимает положение от центрального до краевого. Он начи-
нается цекумом, который многочисленными тяжами прикрепляет-
ся к стенке раковины (рис. 199, 9). Септальные трубки короткие,
прямые, реже с отогнутым краем (субциртохоанитовые); соедини-
тельные кольца тонкие, сегменты сифона обычно цилиндрические.
Септы вогнутые, блюдцеобразные; перегородочная линия прямая,
наклонная или слабо изогнутая. Внутри сифона наблюдаются аннуло-
сифонатные отложения различного строения. Среди камерных отло-
жений различают: эписептальные на вогнутой, или передней, стороне
септы, гипосептальные на выпуклой, или задней, стороне септы и му-
ральные на стенке раковины. Происхождение камерных отложений
до сих пор остается недостаточно ясным; предполагают, что они от-
кладывались особой камерной мантией, которая оставалась в камере
после того как моллюск Подтягивался и начинал строить новую
септу, или же их происхождение связывают с присутствием камерной
жидкости.
Поверхность раковины гладкая, с хорошо выраженными линиями
нарастания или скульптированная. На дорзальной стороне иногда
удается обнаружить отпечатки мускулов-ретракторов в виде двух
более или менее различимых углублений. Начальная камера колпач-
ковидная, короткая или субсферическая, отделенная от фрагмокона
пережимом.
Ортоцератиты были широко распространены от раннего ордовика
до карбона; постдевонские ортоцератиты немногочисленны и изучены
недостаточно. Подкласс ортоцератит на основании особенностей
строения сифона разделен на два отряда: Orthoceratida, Pseudortho-
ceratida.
Отряд Orthoceratida. Ортоцератиды. Раковина ортоконовая, реже
слабо циртоконовая, с узким ортохоанитовым сифоном, с тонкими
соединительными кольцами, иногда с аннулосифонатными отложе-
ниями. Хорошо развиты камерные отложения. Начальная камера суб-
сферическая, отделенная пережимом от фрагмокона, реже длинная
колпачковидная. Ордовик — поздний триас. Около 70 родов
(рис. 199, 8—9).
Отряд Pseudorthoceratida. Псевдортоцератиды (гр. pseudos —
ложный). Раковина ортоконовая или циртоконовая; сифон от суб-
центрального до краевого, относительно широкий, субциртохоанито-
вый, с внутрисифонными и камерными отложениями. Начальная ка-
мера короткая колпачковидная, последующие камеры постепенно
увеличиваются в длину до третьей — шестой, четвертая — седьмая
короче предыдущих, затем длина камер снова увеличивается. По мне-
нию ряда исследователей (И. С. Барсков), отряд ближе стоит к акта-
ноцератитам, чем к орторератитам. Средний ордовик — пермь
(рис. 199, 7).
ПОДКЛАСС BACTRITOIDEA. БАКТРИТЫ
Подкласс бактритов (гр. bactron — палка) объединяет палеозой-
ских цефалопод, обладающих небольшой прямой, ортоконовой или
слабо согнутой раковиной, с краевым положением узкого сифона
(рис. 199, 1—6). Первая камера, или протоконх, полусферическая, яйце-
361
видная или колпачковидная. Эмбриональная раковина прямая, отде-
лена от последующих пережимом и первичным валиком; она, воз-
можно, состояла из жилой и одной-двух гидростатических камер.
Жилая камера заключала мягкое тело, которое, по-видимому, у не-
которых довольно далеко выступало из нее; о наличии воронки сви-
детельствует вентральный синус в приустьевом крае у некоторых
форм; внутри раковины имеются отпечатки мускулов-ретракторов
подковообразной формы. Фрагмокон состоял из гидростатических ка-
мер, разделенных вогнутыми перегородками; у узкоконических рако-
вин длина камер составляла до х/г диаметра, у ширококонических
до '/io диаметра. Прижизненные камерные отложения не обнаружены.
Перегородочная линия в большинстве случаев прямая или состоит из
небольшого числа лопастей и седел; различают следующие лопасти:
вентральную, о м н и л а т е р а л ьн у ю и дорзальную. Ом-
нилатеральная лопасть широкая и расположена на боковой стороне.
Если сифон прилегает тесно к стенке раковины, то в вентральной
лопасти образуются прямые стенки с незамкнутым основанием.
Такая лопасть называется вентральной тубулярной.
Септальные, трубки короткие, более или менее цилиндрические,
иногда бокаловидные или циртохоанитовые, ретрохоанитовые. Сифон
узкий, всегда расположен у вентральной стенки; соединительные
кольца имели, по-видимому, только органическую оболочку и сохра-
няются очень редко. Стенка раковины многослойная, состоит из не-
скольких слоев, возможно образованных снаружи за счет отворотов
мантии, которые закрывали приустьевую часть раковины.
Систематическое положение бактритов до последнего времени
вызывает дискуссию. Одни исследователи объединяют их с наутило-
идеями, другие— по строению и положению узкого сифона относят их
к аммонитам; В. Н. Шиманский (1951) на основании всестороннего
изучения раковины пришел к выводу о самостоятельности изучаемой
группы головоногих, равноценной подклассу наутилоидей и аммоно-
идей. В последние годы высказывается предположение о близости
бактритов и ортоцератит. Не исключена возможность, что они
представляют собой две родственные ветви головоногих моллюсков,
различающиеся только положением сифона и некоторыми другими
особенностями и их следует объединять в один таксон.
Бактриты, вероятно, возникли в ордовике от ортоцератид с шаро-
видным протоконхом и эксцентричным положением сифона. Древней-
ший представитель из среднего ордовика (лландейло) отнесен к роду
Eobactrites. Но только в силуре становится известным Bactrites —
основной род подкласса. Широкое развитие бактритов относится к
девону, карбону и ранней перми. В конце перми или триасе выми-
рают последние представители подкласса. Намечаются две основные
ветви: собственно бактриты, с расцветом в среднем и позднем девоне,
и парабактриты, с расцветом в ранней перми. Первые имели узкоко-
ническую раковину с почти прямой перегородочной линией. Они, по
мнению Эрбена, вероятно, были предками подкласса аммоноидей.
У некоторых форм имеется устьевой край, снабженный большими
синусами: вентральным, латеральным и дорзальным, указывающими
на значительную дифференциацию головного отдела. Вторая груп-
па— парабактриты — обладали ширококонической раковиной с бока-
ловидными септальными трубками, с многослойной стенкой; они, ве-
роятно, явились предками подкласса колеоидей и его самого древ-
него отряда — белемнитов. Известно 16 родов (рис. 199, 1—6).
362
ПОДКЛАСС AMMONOIDEA. АММОНИТЫ
Общая характеристика. Раковина преимущественно планоспи-
ральная, но может быть иного типа: прямой, согнутой, крючкообраз-
ной, спирально-конической. Жилая камера’ от 7г до 1,5—2 оборотов;
протоконх шаровидной или валиковидной формы, диаметром' от 0,3
до 0,8 мм. Сифон начинается шаровидным вздутием — цекумом
и прикрепляется изнутри к протоконху просифоном; на первых
оборотах положение сифона может варьировать от центрального до
краевого; начиная со второго — третьего оборота занимает вентраль-
нокраевое положение; исключение составляет позднедевонский отряд
климений с дорзальным положением сифона. Сифон’тонкий; септаль-
ные трубки от коротких до длинных, у палеозойских форм направле-
ны назад (как у наутилоидей) —ретрохоанитовые. у мезозойских
направлены вперед — прохоанитовые.
Периферические края септ изогнуты, волнисты и соответственно
перегородочная, или лопастная, линия состоит из значительного
числа лопастей и седел; по особенностям развития различают четыре
основных типа лопастных линий: агониатитовый, гониатитовый, цера-
титовый и аммонитовый. Устье закрывалось известковыми крышеч-
ками— аптихами. Челюсти и-радула по своему типу строения
напоминают таковые колеоидей; имелся чернильный мешок. Раковина
гладкая или скульптированная, до 2 м в диаметре.
Развитие, по-видимому, прямое, без личинок. Из яйца выходила
аммонителла, имевшая протоконх и первый оборот, состоявший
из жилой и одной или двух гидростатических камер. Яйца были малень-
ких размеров и довольно многочисленные. Раковина аммонителлы и
первые две септы построены из призматического слоя; все последую-
щие обороты состоят из трех слоев: двух призматических и лежащего
между -ними пластинчатого; все септы, кроме двух первых, построе-
ны только из пластинчатого слоя. Аммониты произошли от бактри-
тоидей в начале девона и вымерли в конце мелового периода.
. Строение мягкого тела. О строении мягкого тела аммонитов за
последние годы получены новые интересные данные. О форме и раз-
мерах мягкого тела можно судить по форме и размерам жилой ка-
меры, а также по изменению формы поперечного сечения в онто-
генезе. На ранних стадиях тело было сжато в дорзовентральном
направлении, затем после третьего — четвертого оборота принимало
округленную форму и далее в зависимости от типа раковины — сжа-
тое с боков, узкое и высокое или низкое и широкое, с малообъемлю-
щими краями или, наоборот, с сильнообъемлющими. В зависимости
от длины жилой камеры тело было толстым и коротким или, наобо-
рот, длинным и червеобразным и занимало до 1,5—2 оборотов.
Тело было окружено мантией, которая прикреплялась к послед-
ней перегородке и к устьевому краю; по-видимому, двумя мускулами-
ретракторами оно прикреплялось к бокам раковины; их отпечатки
обнаружены в последнее время на ядрах. По неясным отпечаткам,
обнаруженным с помощью рентгеновских лучей, установлены пирити-
зирбванные остатки мягкого тела — головной комплекс с. небольшим
числом (8—10) коротких рук, воронка и остатки пищеварительного
тракта. Сравнительно недавно обнаружены остатки радулы,
челюстного аппарата и чернильного мешка. Радула состоит и,з
7 рядов хитиновых зубчиков, как у современных колеоидей. Чер-
нильный мешок, обнаруженный у юрских форм, в задней части жилой
камеры имеет вид мешка грушевидной формы, обращенный сужен-
363
ной частью к устью. Его размеры составляют ’/4 длины жилой каме-
ры, что соответствует относительным размерам чернильного мешка
современных кальмаров. По-видимому, у аммонитов имелся капю-
шон, выполнявший ту же функцию, которую он выполняет у наути-
Рис. 199. Подклассы Bactritoidea (1—6) и Orthoceratoidea. (7—10):
1 — перегородочная линия: 1а — Bactrites, 16 — Lobobactrites; 2 — Bactrites (силур—>-
пермь): 2а — фрагмокон, 26 — начальная часть раковины (увел.); 3 — Ctenobactrites
(ранняя пермь), начальная часть раковины; 4 — Parabact'rites (ранняя пермь); 5 —
Lobobactrites (девон): 5а — вид с вентральной стороны, 56 — вид сбоку; 6 — Cteno-
bactrltes (ранняя пермь); 7 — Pseudocyrtoceras (карбон); 8 — Mictielinoceras (ордо-
вик — триас); 9 — начальная часть Orthoceras (Бёмерс, 1936); 10 — реконструкция
представителя ортоцератид (10а) и отпечатки щупалец (106) (Флауер, 1955);
вл — вентральная лопасть, всо — внутрисифонные отложения, ко — камерные отло-
жения, ом — омнилатеральная лопасть, пр — протоконх, пс — просифон, сф — сифон
луса, т. е. закрывал устье при втягивании тела внутрь раковины.
У многих аммонитов капюшон выделял известковую крышку, состоя-
щую из двух пластинок — а п т и х о в. Аптихи кальцитовые, плотно
закрывали устье раковины. В ископаемом состоянии они встречаются
обычно отдельно от раковины, так как после гибели тело аммонита
вываливалось из жилой камеры и пустые раковины, вероятно, разно-
сились на большие расстояния. Аммониты подобно всем головоногим
364
были раздельнополы. Удается установить по раковинам половой
диморфизм — раковины самок отличаются от раковин самцов формой
и размерами, а также числом оборотов. В жилой камере самки были
•обнаружены оболочки яиц. В нижнемеловых отложениях (апт) около
Ульяновска была найдена кладка яиц с эмбрионами аммонитов, у ко-
торых была раковина в 3/4 или полный оборот, имевшая 1—2 перего-
родки либо без перегородок.
Строение раковины. Форма раковины крайне разнообразна
{рис. 200). Раковина состоит из протоконха, фрагмокона и жилой
Рис. 200. Подкласс Ammonoidea. Форма раковин:
У—4 — планоспиральные, 5—15 — гетероморфные: 1 — кадикон (Cadoceras}-, 2 —
•сферокон (Phyllopachyceras)-, 3 — оксикон (Timanitesy, 4 — платикон (Haploce-
ras)\ 5 — криокон (Crioceras); 6—7 —турикон (6 — Hyphantoceras, 7 — Ostlin-
goceras); 8,14 — гетероконы (8 — Anaclinoceras, 14 — Heteroceras); 9—10 —
анцилокон (9 — Ancyloceras, 10 — Worthoceras)-, 11 — бакуликон (Baculitesy,
12, 13— птихоцераконы (12 Ptychoceras, 13 — Hamalina)-, 15 — скафитикон
(Scaphites)
камеры. Любая раковина аммоноидей начинается с протоконха, или
начальной камеры, шаровидной или бочонковидной формы диаметром
от 0,3 до 0,8 мм. Вокруг протоконха происходит навивание всех
остальных оборотов у планоспиральных раковин. Форма фрагмокона
определяет основные черты строения раковины. Среди планоспираль-
ных различают эволютные и инволютны'е формы, У первых
умбилик широкий, у вторых узкий.
В зависимости от формы поперечного сечения эти раковины мо-
гут иметь различное очертание: кадикон ы — эволютные,
сжатые в дорзовентральном направлении, с ф ер окон ы — инволют-
ные, шаровидные, оксикон ы — инволютные, с заостренным вент-
ральным краем, платиконы — с прямоугольным поперечным сече-
нием и плоскими боками, серпентиконы — эволютные, с медленно
возрастающими оборотами. Все иные типы раковин выделяются в
группу гетероморф: криоконы — планоспиральные раковины, со-
стоящие из несоприкасающихся оборотов, анцилокон ьр— обла-
дающие спирально-плоскостной частью и длинной выпрямленной, за-
365
канчивающейся крючкообразным изгибом, гетер о коны — имеющие
спирально-коническую и длинную выпрямленную часть, как у анцило-
цератид, турриконы — спирально-конические, иногда с разобщен-
ным последним оборотом, б а ку л и кон ы — напоминают литуити-
коны, состоящие из одного—двух спирально свернутых оборотов и
прямой или почти прямой трубки, тортиконы — раковины, свер-
нутые в запутанный клубок.
Жилая камера отличается от фрагмокона отсутствием пе-
регородок; у большинства ее длина составляет от ’/г до одного обо-
рота; однако у эволютных многооборотных раковин встречаются
жилые камеры, достигающие 1,5—2 оборотов; короткие жилые каме-
ры наблюдаются у форм с инволютной .раковиной. Устье может быть
с простым и сложным краем. Во втором случае оно может иметь
боковые ушки различной формы и длины или вентральный
выступ, широкий или узкий или, наоборот, может быть развит
гипономический синус. Наличие последнего Позволяет предполагать
присутствие у данного аммонита воронки; наличие выступа ставит
под сомнение присутствие у этих аммонитов воронки. У некоторых
форм наблюдается сильно суженное устье.
Раковина может быть гладкой или скульптированной. На глад-
ких раковинах наблюдаются струйки роста, отражающие форму
устьевого края. Они состоят из выступов и синусов. Как у всех мол-
люсков, скульптура бывает радиальной и продольной или спиральной.
Радиальная скульптура представлена ребрами различных
размеров и формы, главных и промежуточных, несущих бугорки и без
них; продольная — выражена преимущественно продольными
ребрами. Сетчатая скульптура возникает при сочетании двух
первых типов. Вдоль вентральной стороны бывают развиты бороз-
ды и кили (1—3). На раковинах часто различают пережимы;
они связаны с утолщением раковинного слоя и чаще выражены на
внутренних ядрах. Значительный интерес представляют воротни-
ки— своеобразные приустьевые отвороты, возникавшие периодически
и наблюдаемые только у мезозойских литоцератид. Параболические
линии и бугорки — остатки промежуточных устьев, имевших фигурное
очертание, в процессе роста заполнялись новым раковинным веще-
ством.
Окраска раковины наблюдается очень редко и описана у немно-
гих экземпляров; она представлена темными или светлыми полосами,
параллельными устьевому краю.
Септы и лопастные линии. Протоконх отделяется от фрагмокона-
первой септой, или просептой. Как и все последующие септы, про-
септа прикреплялась к раковине изнутри; линия прикрепления ее
образует первую лопастную линию, или просутуру. У первых ран-
недевонских аммонитов просутура была простой линией, подобной
таковой наутилоидей. В. Бранко (1879—1880) назвал ее а с е л л а т-
й о й, т. е. бесседельной, в ней выделяется маленькая умбиликальная
лопасть, расположенная на шве (рис. 201). Из аселлатной линии у
всех палеозойских и части триасовых аммонитов развивается лати-
селлатная, или широкоседельная, просутура, состоящая из ши-
рокого седла, развитого на вентральной стороне, и соответствующего
ему дорзального седла; между двумя седлами с каждой стороны рас-
положены одна или две лопасти: одна, лежащая на шве, получила
название умбиликальной, а вторая, лежащая рядом с вентраль-
ным седлом, — латеральной. Из латиселлатной просутуры в-
триасе возникла ангустиселлатная, или узкоседельная, с уз-
366
ким вентральным седлом, дорзальным седлом и двумя или тремя
лопастями между ними; третья лопасть получила название в нут-
Рис. 201. Строение лопастной линии:
J — филогения просутуры: а — аселлатная (ранний девон), б — латиселлатная
(девон — триас), в, в' — ангустиселлатная (триас — мел); 2 — возникновение
адвентивной лопасти (Л) и 3 — умбиликальной (С/); 4 — типы лопастных ли-
ний: а — агониатитовая, б — гониатитовая, в — цератитовая, г — аммонитовая;
лопасти: А — адвентивная,'О — дорзальная, / — внутренняя, L — латеральная,
U — умбиликальиая, V — вентральная
Рис. 202. Основные типы протоконха:
1 — аселлатный, 2 — латиселлатный, 3— ангустиселлатный; а — вид со стороны
первой перегородки, б — вид сбоку, в — вид с вентральной стороны
ренней боковой. В процессе филогенеза аммоноидей наблюдает-
ся развитие просутуры от аселлатной к ангустиселлатной. В соответ-
367
ствии с формой просутуры различают три типа протоконхов: аселлат-
ный, латиселлатный и ангустиселлатный (рис. 202). Вторая перего-
родка отличается от просепты возникновением на месте вентрального
и дорзального седел двух новых лопастей — вентральной (V) и дор-
зальной (D); соответственно вторая лопастная линия, или прима-
с у тур а, имеет вентральную и дорзальную лопасти и расположен-
ные между ними одну — три лопасти; число этих лопастей зависит от
Рис. 203. Внутреннее строение филлоцератид:
1—2 — Phyllopachyceras-. la — протоконх и начало первого оборота, 16 — уве-
личенная часть 1а; 2 — остатки сифона в жилой камере; 3—4 — Euphylloceras:
3 — переход от ретрахоанитовой (14) к прохоанитовой (18) септальной трубке,
4 — взаимоотношение органической оболочки сифона, манжеты и септальной
трубки: 4а — стенка сифона, 46 — сифональная трубка;
ан — аннулярные образования, гк — гидростатическая камера, Dit Dt — боль-
шой и малый диаметры, жщ — жилая камера, ио — известковая оболочка це-
кума, кр — кромка, м — манжета, оо — органическая оболочка цекума, осс —
органический сегмент сифона, пр — просифон, прот — протоконх, Ci, Cz...Cn —
первая, вторая и последующие септы, сиф — сифон, ст — септальная трубка,
стр — стенка раковины, ц — цекум
их числа в просутуре. Дальнейшее развитие перегородок и лопаст-
ных линий у разных аммонитов протекало по-разному. Различают
три основных типа лопастных линий (рис. 201, 4): гониатитовый, це-
ратитовый и аммонитовый.
Г ониатитовый тип — лопастная линия состоит из простых
нерасчлененных лопастей и седел, причем, как правило, лопасти за-
остренные, а седла округленные.
Цератитовый тип — лопастная линия состоит из округлен-
ных седел и мелкозазубренных лопастей.
Аммонитовый тип — лопастная линия состоит из расчле-
ненных лопастей и седел.
Иногда у девонских аммонитов выделяют агониа титов ый
тип — лопастная линия отличается от гониатитовой наличием очень
368
Широкой боковой (омнилатеральной) лопасти и небольшим числом
элементов.
Каждая лопасть получает буквенное обозначение и индекс; с по-
мощью обозначений и индексов можно составить формулу лопастной
линии каждого аммонита. У мезозойских литоцератид наблюдается
особый тип строения дорзальной лопасти; от нее отходили назад две
ветви задней части мантии, которые прикреплялись к предыдущей
перегородке, оставляя на ее дорзальной части отпечатки, названные
септальными крыльями (см. рис. 209). Изучение развития
лопастной линии в онтогенезе позволяет выявлять родственные связи
между аммонитами.
Сифон начинается в протоконхе небольшим шаровидным взду-
тием, названным цекумом (рис. 203). После того как тело аммо-
нита подтянулось и освободило полость протоконха, задняя часть
тела аммонита оказалась прикрепленной к стенке протоконха с по-
мощью связки, образованной за счет эпителия мантии. Эта связка
состоит из бокала, охватывающего цекум, и длинного или корот-
кого тяжа, или ленты, или нескольких тяжей. Эта связка полу-
чила название просифона, так как исследователь, предложивший
этот термин, предполагал, что просифон выполнял в протоконхе 'функ-
цию сифона; в действительности он служил только для прикрепления
задней части тела. После того как полость протоконха была осво-
бождена, задняя часть мантии строила просепту, которая прикрепля-
лась с дорзальной стороны к дистальному концу протоконха, остав-
ляя небольшой его конец, названный кромкой; вокруг цекума
выделялась тонкая известковая оболочка. Наличие в просепте двух
седел — вентрального и дорзального — обусловлено существованием
цекума, который служил препятствием для изгиба септы назад и об-
разования лопасти. Но когда тело подтянулось дальше и мантия
начала строить вторую перегородку, или примасепту, то между
просептой и примасептой образовался тяж сифона. В примасепте
для сифона возникло отверстие, что, по-видимому, обусловило появ-
ление на внутренней и наружной сторонах септы соответствующих
изгибов — дорзальной и вентральной лопастей. Они сохраняются у
всех последующих септ. В септах, вокруг отверстия сифона, образо-
вались септальные трубки; у палеозойских аммонитов они направле-
ны назад и называются ретрохоанитовыми, а у мезозойских
они на взрослых оборотах направлены вперед. Септальные трубки,
направленные вперед, называют прохоанитовыми. Сифон
обычно снабжен соединительными кольцами, которые, по-видимому,
выделялись его эпителием и служили для прочного соединения сифо-
на с септами. Соединение было необходимо для создания герметич-
ности камер. Сифон у аммонитов на первых оборотах может занимать
любое положение — от центрального до краевого, но на всех взрос-
лых оборотах — вентральнокраевое и только у одного отряда —
позднедевонских климений — он смещен к дорзальной стенке. Отно-
сительные размеры сифона уменьшаются с возрастом: если на пер-
вых оборотах его диаметр составляет 7з—’/4 высоты оборота, то на
последних может составлять только '/16-
Стенка раковины. Стенки протоконха и первого оборота, а также
двух первых септ построены из призматического слоя. Приблизитель-
но в конце первого оборота у всех аммонитов наблюдается резкое
увеличение раковинного слоя, получившего название первичного
валика; он расположен от устья протоконха на расстоянии от 270
до 360° и даже 380° (т. е. больше одного оборота). Валик обычно
24 в. в. Друщиц
369
сопровождается первичным пережимом. Его возникновение связано
с появлением перламутрового слоя. По-видимому, после того как
молодой аммонит — аммонителла — вышел из яйцевых оболочек и:
попал в новую для него среду, ему потребовалось некоторое время
для адаптации к новым условиям. Раковина аммонителлы состояла
из протоконха и первого оборота. Аммонителла имела одну-две-
гидростатических камеры, цекум с просифоном и первый сегмент
сифона. За время адаптации мантия аммонителлы приобретала спо-
собность выделять перламутровый слой, за счет которого строился
первичный валик. Затем наступал новый этап в жизни аммонита, он.
продолжал расти, строить новые части раковины, новые перегородки..
Все септы, начиная с третьей, строились за счет перламутрового-
слоя. После первичного валика стенка раковины всех последующих
оборотов построена из трех слоев — двух призматических и заклю-
ченного между ними перламутрового. Пережимы, или валики, кото-
рые наблюдаются на раковинах, всегда образуются за счет утолщения!
перламутрового слоя. Дорзальная стенка строится обычно очень ^эко-
номно: только за счет внутреннего призматического слоя, остальные
слои выклиниваются у боковой стенки предшествующего оборота.
Устье раковины аммонитов после втягивания тела внутрь рако-
вины закрывалось известковой или роговой крышкой, гомологичной:
таковой гастропод. Известковые крышечки, известные под названием:
ап тих о в, образовывались эпителием капюшона, гомолога подоб-
ного образования наутилуса. Если раковина строилась из арагонита,,
то аптихи всегда из кальцита. Обычно аптихи встречаются отдельно»
от раковины и довольно часто бывает трудно установить, каким
группам аммонитов они принадлежат. Поэтому их систематика, так.
же как систематика ринхолитов, является искусственной. Аптихи со-
стоят из двух створок; крышечка, состоящая из одной створки, назы-
вается анаптихом. Анаптихи встречаются в ископаемом состоянии
в течение всего времени существования аммонитов, т. е. с девонского»
по меловой периоды, в то время как аптихи известны преимущественно»
из юрских и меловых отложений.
Основы систематики и классификация. На основании изучения
изменений в онтогенезе лопастной линии, формы поперечного сечения
оборотов, сифона и септальных трубок подкласс аммоноидей разде-
лен на семь отрядов: Agoniatitida, Goniatitida, Clymeniida, Ceratitida,.
Phylloceratida, Lytocerati.da, Ammonitida.
Отряд Agoniatitida. Агониатиты (гр. jgonia — угол, a — отрица-
ние). Раковина преимущественно спирально-плоскостная, дисковидная,
или линзовидная, у ранних форм криоконная с несоприкасающимися:
оборотами, позднее с соприкасающимися, но с умбиликальным отвер-
стием и, наконец, у большинства свернутая в плотную спираль.
Сифон на всех стадиях вентральнокраевой, протоконх с аселлатной и
латиселлатной просутурой. Лопастная линия агониатитовая, гониати-
товая и у поздних форм — цератитовая. Примасутура у ранних форм.:
с широкой омнилатеральной лопастью, позднее замещающейся лате-
ральной; новые лопасти — умбиликальная и внутренняя — возникали
за счет умбиликального седла. Омнилатеральная лопасть была уна-
следована от бактритов, от которых возникли агониатиты в середине-
раннего девона. Развитие отряда шло по пути увеличения размеров-
раковины, возникновения разнообразных морфологических типов от
полностью инволютных до эволютных, усложнения лопастной линии и:
возникновения новых лопастей и седел, число которых у некоторых
форм достигало 50, превращения простой вентральной лопасти в трех-
370
раздельную, простой дорзальной — в двузубчатую, сложного расчле-
нения наружного седла и лопастей у пермских и триасовых форм.
Девон—триас. Около 100 родов (рис. 204).
Рис. 204. Отряд Agoniatitida:
1 — Agoniatites (средний девон); 2 — Manticoceras (поздний девон); 3 — Timanites
(поздний девон); 4 — Medlicottia (пермь)
Отряд Goniatitida. Гониатиты (гр. gonia — угол). Раковина раз-
нообразной формы от эволютной до инволютной, от субсферической
до дисковидной, гладкая или скульптированная, с продольной попе-
речной и сетчатой скульптурой. Протоконх с латиселлатной просуту-
рой. Лопастная линия преимущественно гониатитовая, у поздних
форм — цератитовая. Примасутура у ранних форм состояла из вент-
24;
371
ральной, латеральной и дорзальной лопастей. Сифон на ранних ста-
диях от центрального до дорзального, на всех взрослых — вентраль-
нокраевой. Септальные трубки ретрохоанитовые, от коротких до длин-
ных. Развитие шло по пути превращения простой вентральной лопа-
сти в двураздельную, возникновения новой адвентивной лопасти, рас-
положенной рядом с вентральной и внутренней боковой. Дорзальная
лопасть оставалась простой или становилась двух-, реже трехзубча-
Рис. 205. Отряд Goniatitida:
1 — Tomoceras (девон); 2 — Imitoceras (поздний девон — пермь); 3 — Paragas-
trioceras (пермь); 4 — Goniatites (ранний карбон)
той. Новые лопасти и седла возникали в результате многократного
деления адвентивной, умбиликальной и внутренней боковой лопастей.
Гониатиты возникли от агониатитов в среднем девоне, получили ши-
рокое развитие в карбоне и перми и в конце перми вымерли, не
оставив потомков. Около 200 родов (рис. 205, 211, 4).
Отряд Clymeniida. Климении (гр. klimax — лестница, подчеркну-
то ступенчатое строение пупка) отличаются от всех аммоноидей дор-
зальнокраевым положением сифона. Протоконх латиселлатный; .сифон
у ранних форм на первом обороте занимал вентральнокраевое поло-
372
жение, а затем смещался к дорзальной стороне. Сифональные трубки
ретрохоанитовые, длинные. Раковины преимущественно эволютные
с широким умбиликом, в одном случае с треугольным навиванием;
имеются полностью инволютные, шаровидные раковины. Просутура
латиселлатная; лопастная линия гониатитового типа; в отличие от
всех аммоноидей у надсемейства Clymeniacea в филогенезе вначале
вентральная лопасть заменяется вентральным седлом, а позднее то
Рис. 206. Отряд Clymeniida:
1 — Gonioclymenia (поздний девон); 2 — Clymenia (поздний девон)
же происходит с дорзальной лопастью. Климении возникли в начале
позднего девона в зоне manticoceras от агониатитов (подотряд
Anarcestina), стали особенно разнообразными во второй половине
позднего девона (фаменском ярусе) и вымерли на границе с карбо-
ном. Известно около 30 родов (рис. 206).
Отряд Ceratitida. Цератиты (гр. keros — рог) объединяют около
400 родов, главное распространение которых относится к триасу.
Первые цератиты появились в конце ранней перми, в артинском веке,
отделившись от агониатитов. Протоконх с латиселлатной просутурой,
имеет одну или две лопасти; на взрослых стадиях лопастная линия
цератитовая или аммонитовая. Сифон в начале первого оборота за-
нимает любое положение — от центрального до дорзально- или вент-
ральнокраевого; на последующих оборотах смещается к вентральной
стенке; септальные трубки вначале ретрохоанитовые, на поздних
оборотах прохоанитовые. Раковины разнообразной формы — от эво-
лютных до инволютных, гладкие или скульптированные, с ребрами,
бугорками, шипами, часто на вентральной стороне наблюдаются
кили; в конце триаса появляются немногочисленные цератиты с гете-
роморфной раковиной — турриконовой, криоконовой или бакулико-
новой. Ранняя пермь — триас (рис. 207).
Отряд Phylloceratida. Филлоцератиды (гр. phyllon — лист) воз-
никли в конце перми или начале триаса от цератитов и представляют
373
Рис. 207. Отряд Ceratitida:
1 _ I „ - \ис- м'- итряд Ceratitida:
Ceratites (средний триас); 2 — Pinacoceras (поздний
триас): 2а — общий вид,
•26 — лопастная линия
— Phutlncprni i ^ИС' ,208. Отряд Phylloceratida:
Phylloceras (юра-мел); 2 - Sowerbyceras (поздняя юра)- 3 - лопает
ная линия Salfeldiella (мел) ? лопает
собой небольшой отряд (около 30 родов), существовавший без зна-
чительных изменений до конца мела. Протоконх шаровидный или
.валикообразный с ангустиселлатной просутурой. Лопастная линия
взрослых форм аммонитовая, с большим количеством лопастей и
-седел; седла заканчиваются округленными листочками и никогда
•не бывают двураздельными; вентральная лопасть разделена средин-
ным бутылеобразным седлом на две ветви, боковая — трехраздель-
ная, дорзальная — с прямыми нерассеченными стенками. Сифон на-
чинается шаровидным цекумом, который прикрепляется к стенке про-
токонха многочисленными, но короткими тяжами (сложным проси-
фоном); на первых двух оборотах сифон занимает почти центральное
положение, а затем постепенно смещается к вентральной стенке;
’септальные трубки на первых оборотах ретрохоанитовые, а затем
прохоанитовые, короткие, сопровождаются манжетами — особыми
•соединительными кольцами, направленными назад. Раковины преиму-
щественно инволютные, тонкоребристые, редко груборебристые.
.Жилая камера занимает от */г до 3Д оборота. Пермь (?), триас—мел
(рис. 203, 208).
Отряд Lytoceratida. Литоцератйды (гр- lytos — неплотный, keras —
рог) возникли в начале юры от филлоцератид и дали большое коли-
Рис. 209. Отряд Lytoceratida:
1 — Lytoceras. (юра): la — раковина с воротниками, 16 — лопастная линия,
1в — септальные крылья; 2 — Pictetia (ранний мел); 3 — Hamulina (ранний
мел); 4 — Turrilites (мел); 5 — Anlsoceras (мел); 6 — Nipponites (поздний мел)
-чество родов и видов в юрском и меловом периодах. Протоконх вали-
жовидный с ангустиселлатной лопастной линией. Просутура с двумя
или тремя лопастями, расположенными между вентральным и дор-
зальным седлами. Лопастная линия взрослых форм с ограниченным
числом лопастей и седел. Вентральная и боковая лопасти, а также
•седла — двураздельные; дорзальная лопасть крестообразная, с сеп-
тальными крыльями; в вентральной лопасти вторичное седло с пря-
мыми стенками. Сифон начинается шаровидным цекумом, который
прикрепляется к протоконху, подобно таковому филлоцератид, много-
численными, но короткими тяжами и почти сразу, в отличие от фил-
лоцератид, занимает краевое положение. Септальные трубки вначале
375
ретрохоанитовые, а после второго оборота прохоанитовые; септаль-
ные трубки длинные, манжеты расположены внутри септальных! тру-
бок и лишь немного выступают назад. Раковины преимущественно
эволютные или полуэволютные и многие гетероморфные; среди по-
следних-—• криоконовые, турриконовые, крючкообразные. У всех гетет
роморф лопастная линия состоит только из четырех лопастей: вент-
ральной, дорзальной и двух, лежащих между ними; все лопасти и
седла двураздельные. Раковины гладкие или тонкоребристые, часто
с воротниками различной конструкции и размеров, от очень больших
до маленьких. Юра —мел. Около 100 родов (рис. 209, 211, 1).
Отряд Ammonitida. Аммонитиды (Ammon — египетский бог Солн-
ца) объединяют всех юрских и меловых аммоноидей, кроме филло-
цератид и литоцератид (свыше 800 родов), онтогенез которых изучен
во многих случаях еще недостаточно. Протоконх валикообразный с
ангустиселлатной просутурой, в последней две или три лопасти.
Лопастная линия взрослых форм отличается большим разнообра-
зием— от сложно расчлененной аммонитовой до упрощенной псевдо-
цератитовой у некоторых позднемеловых форм. Вентральная лопасть
двураздельная, латеральная трехраздельная, дорзальная сложно рас-
сеченная. Новые лопасти возникают в области умбилика за счет рас-
членения умбиликального седла или лопасти. Раковины преимущест-
венно спирально-плоскостные, от эволютных до полностью инволют-
ных, гладкие и ребристые, с килями и без них, с бугорками, шипами;
устье простое или сложное, с ушками или вентральным ростром.
В юре и особенно в мелу возникает ряд аммонитид с гетероморфной
раковиной — криоконовой, анцилоконовой, гетероконовой, скафиконовой
и многие другие формы. Аммонитиды возникли от литоцератид в
начале юры, достигли расцвета в юрском периоде (известно до 650 ро-
дов), были разнообразны и многочисленны в меловом периоде и
вымерли в конце мела (в Маастрихте). Этапы развития аммонитид
йспользуются для зонального деления ярусов юры и мела (рис. 210,
211,2, 5).
История развития аммоноидей. Аммоноидеи возникли в начале
раннего девона, по-видимому, от бактритов, имевших прямую или
слабосогнутую раковину. Наличие такой раковины чрезвычайно су-
жало эволюционные возможности этого подкласса. Возникновение
аммоноидей было связано с образованием вначале согнутой, а затем
криоконовой раковины, состоявшей из несоприкасающихся оборотов,
свернутых в одной плоскости. Появление планоспиральной раковины
имело большие эволюционные последствия; раковина стала более
компактной, удобной для маневрирования, тонкая апикальная часть
трубки оказалась защищенной последующими оборотами — все это-
значительно расширило эволюционные возможности аммоноидей и
привело их к адаптациям к самым разнообразным условиям сущест-
вования, подобно тому как возникновение планоспиральной раковины
у гастропод обеспечило. их эволюцию. Первые агониатиты имели ра-
ковину криоконового типа, с протоконхом аселлатного типа. Затем
возникли планоспиральные раковины вначале с умбиликальным
отверстием, потом без него, с протоконхом латиселлатного типа. По
особенностям развития лопастной линии наметились две параллельно
развивающиеся группы—агониатиты и гониатиты (рис. 217). Оснрв-
йое различие между ними сводится к развитию латеральной лопасти,
которая у агониатитов развивается за счет омнилатеральной, унасле-
дованной от бактритов и вначале сохраняющейся у первых агониати-
тов; новые лопасти развиваются в области умбилика за счет расчле-
376
1 — Stephanoceras (средняя юр ah / r P__ Cardloceras (поздняя юра); 6 —
ceras (поздняя юра); 4 - Sca^s ^ceras (поздний мел); 8 - Tissotia (позд-
Virgatttes (иозДняя^аЦ-А^» (ранни& мед)
яения лопасти или седла. У гониатитов, возникших от агониатитов в
среднем девоне, латеральная лопасть остается маленькой в области
умбилика, а новые лопасти, так называемые адвентивные, воз-
никают за счет седла, лежащего между латеральной и вентральной
лопастями, внешний вид которых на взрослой стадии не отличим от
настоящей латеральной. От агониатитов в начале позднего девона
Рис. 211. Внутреннее строение раковины литоцератид (7), аммонитид (2—3)
и гониатитов (4):
1 — Tetragonites: la — взаимоотношение органической оболочки сифона, ман-
жеты и септальной трубки, 76 — протоконх и начало первого оборота; 2 —
Sphaeroceras, протоконх и начало первого оборота; 3 — Parkinsonia, прото-
конх и первая септа; 4 — Agathiceras, часть продольного сечения;
в — валик, няС], впс2 — внутренний призматический слой одного оборота и
следующего, нпр — наружный призматический слой, пл — пластинчатый, или
перламутровый, слой. Остальные обозначения те же, что на рис. 203
возникает небольшой отряд климений, отличающийся дорзальным
положением сифона и развитием иногда вместо вентральной и дор-
зальной лопастей соответствующих седел. Климении вымирают на
границе девона и карбона.
В конце ранней перми от агониатитов развивается основной ствол
триасовых аммонитов — цератиты, имевшие протоконх с латиселлат-
ной, а в конце их существования — с ангустиселлатной просутурой.
• От них в конце перми или начале триаса возникают филлоцератиды —
378
консервативный ствол медленно эволюционирующих аммоноидей, но
чрезвычайно важный в филогении аммоноидей: от него в начале юры
возникли литоцератиды, а от последних — собственно аммониты
(отряд аммонитид). Вначале цератиты были представлены только
одним родом, в поздней перми значение этой группы постепенно воз-
растало и в конце перми она заняла ведущее положение, полностью
вытеснив гониатитов и частично агониатитов. В раннем триасе цера-
титы широко распространяются по всем морям земного шара, возни-
кают новые группы разного таксономического ранга. В конце триаса
произошло вымирание цератит; перед их массовым вымиранием
появились немногие цератиты с гетероморфной раковиной (криоко-
новой, турриконовой и бакуликоновой); часть цератйтов приспособи-
лась к новым условиям жизни. Общее число родов аммонитов, оби-
тавших в триасе, приближается к 400. Среди этой цифры несколько
родов филлоцератид. В конце триаса (рэтском ярусе) от большого
отряда цератит сохраняется немногим более 5 родов. Раннеюрский
этап характеризуется полным обновлением систематического состава;
от филлоцератид возникают литоцератиды и от последних аммони-
тиды. Если филлоцератиды и литоцератиды в юрское время были
распространены только в средиземноморской области, то аммонитиды
образуют характерные группировки для средиземноморского и бо-
реального поясов, часто по составу совершенно различные и неповто-
римые. Аммонитиды дают в юре небольшое число родов с гетеро-
морфной раковиной (надсемейство Spirocerataceae).
Многие аммонитиды в юрском и меловом периодах были очень
широко распространены, быстро эволюционировали и используются
для зонального деления юрских и меловых отложений. В конце юры
происходит почти полное обновление родового состава среди аммо-
нитид. Но филлоцератиды продолжают свое существование без осо-
бенно крупных изменений, в то же время наблюдается адаптивная
радиация среди литоцератид и аммонитид. Среди последних двух
отрядов в меловом периоде возникали многочисленные роды с гете-
роморфной раковиной, указывающие на приспособление к самым
разнообразным условиям жизни. Литоцератиды с турриконовой ра-
ковиной (туррилитиды), по-видимому, приспособились к бентосному
подвижному образу жизни, с клубкообразной раковиной (Nipponites)
были, вероятно, малоподвижны, с крючкообразной раковиной (Pty-
choceras, Hamites] вели нектонно-бентосный образ жизни. Кроме
перечисленных известны формы с криоконовой, анцилоконовой, ска-
фиконовой, бакуликоновой и другими типами раковин. Особенный
интерес представляют гетероцератиды, которые на ранней стадии
имели турриконовую раковину и, вероятно, ползали по дну, а затем
изменяли образ жизни и приобретали выпрямленную раковину с
крючком на конце. В конце мела, в Маастрихте, все три отряда аммо-
ноидей (филлоцератиды, литоцератиды и аммонитиды) полностью
вымирают.
ПОДКЛАСС COLEOIDEA. КОЛЕОИДЕИ
К подклассу колеоидей (гр. koleos •— футляр, ножны) относятся
все, за исключением наутилуса, современные головоногие (рис. 212),
насчитывающие до 650 видов и большое число вымерших родов и
видов; среди них можно назвать белемнитов, кальмаров, каракатиц,
осьминогов. Все они пелагические или придонные хищники, ведущие
нектонный, нектонно-бентосный и частично планктонный образ жиз-
379
ни. Тело мешковидное или веретеновидное, самых разнообразных раз-
меров— от нескольких сантиметров до 18 м, часто снабженное плавни-,
ками. В отличие от современного наутилуса колеоидеи имеют
только две жабры (отсюда название, двужаберные, или дибранхиа-
ты), две почки, главное сердце, состоящее из желудочка и двух пред-,
сердий, и два венозных, или жаберных, сердца, накачивающих кровь
Рис. 212. Подкласс Coleoidea. Схема строения современных Spirilla (/) и Se-
pia (2):
вр — воронка, гл — глаза, жб — жабры, лр — ловчие руки, р — руки, ра —
раковина, ро — ротовое отверстие,- се — сепион, ч — челюсти
в жаберные сосуды, два женских половых отверстия или одно муж-
ское, парные протоки нидаментальной железы. Голова несет восемь
или десять крупных щупалец, или рук, снабженных роговыми крю-
чочками или присосками; у десятиногих две руки длиннее остальных
восьми, они называются ловчими. Внутри кольца рук имеется,
буккальная мембрана; состоящая из 7-—10 своеобразных выростов.
Рот вооружен парой роговых челюстей и радулой, несущей 7—9 ря-
дов хитиновых зубчиков. В задней части тела у большинства имеется
чернильный мешок, сохраняющийся в ископаемом состоянии. Лопасти
воронки срослись в цельную трубку, и мантийная щель замыкается
особым замком, построенным по типу запонок. Нервные узлы скон-
центрированы в единую компактную массу, заключенную в хрящевую
капсулу, напоминающую хрящевой череп позвоночных. Высоко раз-
виты органы чувств, особенно глаза, максимальные размеры которых
достигают 30 см. Хрящевые образования имеются около глаз, у осно-
вания рук, около плавников и в других местах. Под кожей развиты
особые клетки — хроматофоры, содержащие различные пигменты,
окрашивающие кожу в защитные цвета.
Колеоидеи отличаются от всех вышерассмотренных подклассов
наличием внутренней раковины, с различной степенью редукции,
вплоть до полного ее отсутствия (отсюда еще одно название этого
подкласса — эндокохлии, или внутреннераковинные). Раковина может
состоять: а) из фрагмокона, проострака и ростра, б) из фрагмокона,
жилой камеры и телума, в) из видоизмененного проострака, или гла-
380
заключены в сравнительно
Рис. 213. Отряд Aulacoceratida. Схема строе-
ния скелета (Елецкий, 1966):
1 — Metabelemnites (поздний триас, Канада):
la — продольный разрез, 16 — строение сеп-
ты и стенки конотеки; 2 — Aulacoceras (позд-
няя пермь), общий вид;
жк — жилая камера, пр — протоконх, с —
септа, ск — стенка конотеки, сф — сифон,
тел — телум
диуса, г) из видоизмененного фрагмокона — сепиона, д) полностью
отсутствовать. Наконец, у одной группы колеоидеи (аргонавты) сам-
ки за счет эпителия рук образуют специальную раковину для вына-
шивания оплодотворенных яиц, очень напоминающую раковину аммо-
нитов, но без перегородок. Развитие всех современных колеоидеи
прямое, без личиночных ст
толстую капсулу и кладки
яиц обычно прикрепляются
самкой к различным под-
водным предметам.
Колеоидеи, как отмеча-
лось выше, возникли от
бактритоидей в раннем кар-
боне и вначале развивались
сравнительно медленно, но
после того как раковина
превратилась полностью во
внутренний скелет и тело
приобрело торпедовидное
очертание и плавники, ко-
леоидеи образовали одну
из наиболее распространен-
ных и высокоорганизован-
ных групп среди всех бес-
позвоночных.
На основании особен-
ностей строения скелета и
мягкого тела подкласс ко-
леоидей разделен на пять
-отрядов: Aulacoceratida,
Belemnitida, Teuthida, Sepi-
ida и Octopodida.
Отряд Aulacoceratida.
Аулакоцератиды (гр. aulax,
род. aulaxos — борозда).
Скелет состоит из фрагмо-
кона, жилой камеры и телу-
ма. Жилая камера длинная,
трубчатая, с перистомом,
имеющим на вентральной и
дорзальной сторонах по
языкообразному выступу, подобно таковому у одного рода бактритов
(Pseudobactrites). Жилая камера переходит в узкоконический фрагмо-
кон (угол от 5 до 12°, как исключение 15—20°), состоящий из много-
численных гидростатических камер, разделенных септами с прямой
перегородочной линией. Протоконх шаровидной формы, отделен от
фрагмокона тонкой мембраной, цекум й просйфон не установлены.
Сифон занимает краевое положение; септальные трубки прохоанито-
вые; соединительные кольца связаны с внутренней поверхностью сеп-
тальной трубки и направлены вперед и назад. Фрагмокон заключен в
телум, по форме напоминающий ростр белемнитов, но отличающийся от
последнего органическим составом; телум построен из слоев органи-
ческого вещества (конхиолина), чередующихся с прослоями карбона-
та кальция; у одной группы (хитинотеутид) телум целиком состоит из
конхиолина. Аулакоцератиды появились в раннем Карбоне' (возможно,
381
в позднем девоне) и вымерли в конце юры. Отряд включает около
10 родов (рис. 213).
Отряд Belemnitida. Белемниты (гр. belemnon — дротик, копье)
возникший в карбоне,.
образуют наиболее важный отряд колеоидей,
Рис. 214. Отряд Belemnitida:
1 — Belemnoteuthis syriaca из позднего мела
Сирии, реконструкция (Роже, 1944); 2 — про-
дольный разрез ростра; 3 — схема строения
скелета;
ал — альвеола, ач — альвеолярная часть ро-
стра, вр — воронка, гл — глаза, мп — мантий-
ная полость, ол — осевая линия ростра, печ—
печень, пж — половая железа, пл — плавник,
поч — постальвеолярная часть ростра, пр —
проострак, пт — пищеварительный тракт,
р — ростр, ру — руки с крючочками, сп —<
спайка, сф — сифон, фк — фрагмокон, ч —
челюсть, чм — чернильный мешок
давший большое число ро-
дов и видов в юре и мелуй
почти полностью вымерший
к концу мела, за исключе-
нием одной небольшой ре-
ликтовой группы, дожившей
до эоцена. В отличие от
аулакоцератид белемниты
имели скелет; состоявший
из фрагмокона, проострака
и известкового ростра, рас-
положенного внутри тела на
его дорзальной стороне
(рис. 214). Тело, судя по'
отпечаткам, имело удлинен-
ную цилиндрическую форму
и обладало, как минимум,
8, а скорее 10 руками, снаб-
женными двумя рядами ро-
говых крючков каждая; у
них не установлены ловчие
руки, известные у современ-
ных кальмаров; по-видимо-
му, они не обладали еще
подобными дифференциро-
ванными руками, фрагмо-
кон с апикальным углом от
12 до 32° по строению напо-
минает раковину бактритов.
и имеет коническую форму;
он разделен септами на от-
дельные гидростатические
камеры. Стенка фрагмокона
носит название конотеки.
Протоконх полусферической
формы, отделен резким пе-
режимом от остальных ка-
мер; устьевой край закрыт
сплошной вогнутой мембра-
ной, которая, по-видимому,
является просептой; стенка
протоконха состоит из двух
тонких призматических сло-
ев; снаружи протоконх ок-
ружен перламутровым сло-
ем, переходящим в наруж-
ный слой конотеки.
В отличие от аммоноидей, у которых после освобождения поло-
сти протоконха возникал просифон и соответственно цекум, у белем-
нитов тело прикреплялось вдоль замыкающей мембраны и поэтому
сифон начинается в первой камере грибообразным расширением,
382
которым он прикрепляется к замыкающей мембране; это расширение
сифона гомологично цекуму; вторая септа без септальных трубок,,
ахоанитовая, через ее отверстие проходит сифон, окруженный обо-
лочкой. В третьей и остальных септах септальные трубки ретрохо-
анитовые, короткие; септы трехслойные, состоят из перламутрового
слоя и двух органических. Вначале сифон расположен эксцентрично,
но уже между 4—6 камерами смещается к краю и между 25—30 при-
легает к вентральной стенке. Конотека, гомологичная стенке ракови-
ны остальных цефалопод, состоит из двух слоев, из которых наруж-
ный, по-видимому, перламутровый, содержит большое количество-
органики, а внутренний призматический. Поверхность конотеки глад-
кая или ребристая, иногда волнистая. Дорзальный край конотеки у
белемнитов образует лопатовидной формы выступ, или проострак,
который очень редко сохраняется в ископаемом состоянии. Проострак
гомологичен гладиусу современных кальмаров. Ряд исследователей
предполагают, что проострак представляет собой дорзальную стенку
жилой камеры, брюшная часть которой редуцировалась. У белемни-
тов она, вероятно, выполняла защитную функцию и служила местом
прикрепления ряда мускулов.
В гидростатических камерах, начиная с третьей и кончая 13—20,.
у преимущественно юрских белемнитов наблюдаются камерные отло-
жения, коррелятивно связанные с формой тонкого, узкоконического,,
относительно легкого ювенильного ростра. У геологически более мо-
лодых белемнитов имелся воронковидный более тяжелый эмбриональ-
ный ростр и камерные отложения отсутствовали.
Ростр — часть скелета белемнитов, гомологичная телуму аула-
коцератид, чаще всего сохраняется в ископаемом состоянии. Ростр
построен из концентрических слоев кальцита, лежащих среди органи-
ческой основы. Поэтому ростры обычно окрашены в коричневые цве-
та различных оттенков. В онтогенезе вначале возникает эмбриональ-
ный ростр, имеющий разную форму: тонкую, узкоконическую, стрело-
видную или клиновидную, воронковидную; не исключена возмож-
ность, что молодой белемнит, только что покинувший яйцевые обо-
лочки, имел протоконх и эмбриональный ростр; последний играл роль
своеобразного органа равновесия у активно плавающего молодого
белемнита. Ростр взрослого белемнита имеет очень разнообразную
форму и состоит из чередующихся призматических и перламутровых
слоев. В передней части ростра находится углубление — альвеола,,
в которой помещался фрагмокон; размеры и глубина альвеолы очень
различны и зависят от размеров фрагмокона. Часто края альвеолы
разрушаются и не отвечают первичному строению. От конца альвеолы
через весь ростр к его заднему концу проходит осевая линия. Попе-
речное сечение ростра также очень разнообразно: от круглого до-
угловатого, но преобладает круглое и овальное; на поперечных сре-
зах хорошо видны концентрические кольца, отражающие форму сече-
ния ростра на разных возрастных стадиях; изменение формы ростра,
с возрастом можно также проследить на продольных разрезах.
На наружной поверхности ростра имеются продольные борозды,
а также наблюдаются отпечатки кровеносных сосудов. Две борозды,
расположенные по бокам, соответствуют местам прикрепления боко-
вых плавников. Бороздки на конце ростра — дорзальная и вентраль-
ная— могут быть короткими и длинными. В приальвеолярной части
также бывают бороздки — вентральная или дорзальная. Чаще всего-
ростры раскалываются вдоль вентральной борозды; у некоторых
форм, между ней и альвеолой существует спайка — гладкая плоскость
383
вдоль линии раскола, напоминающая спайность кристаллов; у поздне-
меловых белемнителл на месте спайки развивается вентральная щель.
Спайка и щель отражают какие-то особенности строения мягкого
тела белемнитов.
Ростр выполнял, по-видимому, три функции: служил противо-
весом подъемной силе фрагмокона, защищал фрагмокон и служил
для усиления прочности тела.
Рис. 215. Отряд Belemnitida. Ростры (1—6) и онихиты (7):
1 — Paleoceras (ранний карбон); 2 — Cylindroteuthis (юра); 3 — Pachyteuthis (позд-
няя юра); 4а, б — Duvalia (поздняя юра — ранний мел); 5 — Neohibolites (мел); 6.—
Belemnitella (поздний мел); 7 — онихиты — конхиолиновые крючочки
В основу выделения подотрядов положены такие признаки, как
наличие или отсутствие продольного альвеолярного канала и связан-
ных с ним спайки или альвеолярной щели: у подотряда Belemnitina
они отсутствуют, у подотряда Belemnopseina имеются либо спайка,
либо альвеолярная щель; по особенностям строения ростра и проост-
рака выделен подотряд Diplobelina с необычно узким проостраком,
толстым, коротким ростром, без радиальной призматической струк-
туры, столь характерной для двух предыдущих подотрядов.
Белемниты появились в начале карбона, до сих пор неизвестны
в перми, редки в триасе, очень многочисленны и разнообразны в юре
и мелу; редкие находки известны из эоцена. Отряд объединяет около
2500 видов (рис. 215).
384
Отряд Teuthida. Теутиды, или кальмары. К теутидам (гр. teut-
his — кальмар) относятся современные кальмары, обладающие 10 ру-
ками, из которых одна пара ловчих, и небольшая группа глубоко-
водных вампироморф, ранее относившихся к октоподам. По мнению
Ю. А. Елецкого (1966), несмотря на восемь рук, эта группа обладает
многими признаками, сближающими их с теутидами. Руки несут
два—четыре продольных ряда присосок на стебельках, снабженных
опорными роговыми кольцами, которые превращены у многих в крю-
' Рис. 216. Отряды Sepiida (1—2), Teuthida (3—5) и Octopodida (6):
1 — Sepia, сепион (совр.); 2 — Spirula, раковина (совр.);. 3 — онихиты — крю-
чочки рук ископаемых кальмаров; 4 — Geopeltis, гладиус (юра); 5 — Parabelo-
peltis с сохранившимся чернильным мешком (юра Поволжья); 6 — Argonauta
argo L., самка (совр.);
бп — боковое поле, п — перегородки, пк — первая камера, рр — рудимент рост-
ра, сп — срецинное поле, сф — сифон, чм — чернильный мешок
чочки (онихиты); у вампироморф имеется один ряд присосок и спра-
ва и слева от них по чувствительному усику; щупальца у них соеди-
нены перепонкой. Тело кальмаров удлиненное, торпедовидное, с ром-
бовидными, весловидными плавниками по бокам. Раковина имеет
вид тонкой пластинки — г л а д.и уса, гомологичного проостраку
белемнитов (рис. 216,4—5). Гладиус — роговой или частично обыз-
вествленный, разделен на три части — осевую и две боковых. У пер-
вых представителей на заднем конце сохраняется короткий фрагмо-
кон с сифоном и маленьким ростром. У последующих фрагмокон и
ростр редуцируются и сохраняются лишь остатки конотеки в. виде:
25 В. в. Друщи!
385
конической или ложкообразной воронки, без перегородок. Осевая
часть гладиуса продольными линиями разделена на несколько полей;
латеральные части очень варьируют по размерам, форме и обычно-
отделяются от срединного поля (рис. 216, 4). У ископаемых форм ино-
гда сохраняется чернильный мешок, столь характерный для всех
современных кальмаров (рис. 216,5).
В отряд теутид включены фрагмотеутиды (рассматриваемые;
Елецким в качестве самостоятельного отряда), существовавшие от-
перми до триаса и обладавшие фрагмоконом и редуцированным рост-
ром. Они рассматриваются в качестве боковой ветви теутид. Настоя-
щие теутиды известны в ископаемом состоянии с поздней юры. Зна-
чительный интерес представляет один из современных кальмаров (род.
Moroteuthis), у которого имеется рудимент ростра, напоминающий
по форме ростр белемнитов, но построенный из эластичного хряще-
видного вещества, образующего концентрические слои; строение этого-
ростра напоминает велум аулакоцератид. Это послужило основанием
для предположения, высказанного Ю. К. Кабановым, о возможной
прижизненной эластичности ростра белемнитов; ростр, по мнению-
этого исследователя, обызвествлялся посмертно. Однако это предпо-
ложение. не является пока общепринятым. Известно свыше 20 родов-
(рис. 216,3—5).
Отряд Sepiida. К сепиидам (гр. и лат. Sepia — каракатица) отно-
сятся каракатицы, спирулы, аргонавты й ряд вымерших форм, извест-
ных в ископаемом состоянии. Тело современных сепиид сжато в дор-
зовентральном направлении, окаймлено плавниками; вокруг рта рас-
положено кольцо из 10 коротких щупалец-рук, из которых одна пара
ловчих, обычно втянутых около головы в особые карманы; присоски
на стебельках с роговыми кольцами, крючочков нет. Скелет современ-
ных сепиид трех типов. У одних (спирул) он представлен маленькой
спирально эндогастрически свернутой раковиной, расположенной
внутри тела на спинной стороне. Раковина имеет протоконх, просифон
и цекум; спиральная часть (2,5 оборота) простыми перегородками раз-
делена на 30—40 гидростатических камер; сифон расположен
вдоль внутренней стороны оборотов; ростр и проострак отсутствуют..
У других (собственно сепиид) раковина имеет вид широкой и доволь-
но толстой овальной, пористой пластинки, целиком скрытой под спин-
ной частью мантии, выполняющей защитную функцию. Эта пластин-
ка, или с е п и о н, является видоизмененным фрагмоконом предков;
сепиид. Третий тип скелета, установленный у позднемеловых сепиид.
(семейство Groenlandibelidae, выделенное Елецким), имел прямой
длинноконический фрагмокон, начинающийся протоконхом, и длин-
ный узкий проострак; сифон начинается цекумом, который прикреп-
лен к протоконху просифоном; сифон занимает вентральнокраевое-
положение; септальные трубки от ахоанитовых до коротких ортохоа-
нитовых. От подобного типа раковины можно проследить переходные-
формы к двум вышеописанным типам. В первом случае (спирулы)
наблюдается постепенное эндогастрическое закручивание фрагмокона
(рис. 216, 2), постепенное уменьшение ростра и редукция проострака,.
приводящее в конце концов к их полной редукции и развитию пла-
носпиральной раковины. Во втором случае, при образовании сепиона^.
редукции подвергались ростр и брюшная часть фрагмокона
(рис. 216, 1); септы на дорзальной стороне постепенно удлинялись и
приняли наклонное положение, фрагмокон был уплощен в дорзовент-
ральном направлении, а брюшная часть септ уменьшалась и, нако-
нец, полностью исчезла; возник сепион, внутренняя полость которого
386
разделена косонаправленными сближенными септами, представляю-
щими собой тонкие, слегка обызвествленные, конхиолиновые пла-
стинки, соединенные поперечными перекладинами; сзади в виде ма-
ленького шипика иногда сохраняется маленький ростр. Сепииды
встречаются в ископаемом состоянии редко, начиная с юры. Известно
свыше 20 родов (рис. 216, 7—2).
Отряд Octopodida. Октоподиды, или осьминоги (лат. octo —
восемь). К октоподидам относятся осьминоги, имеющие в отличие от
сепиид восемь щупалец. Тело мешковидное, короткое, щупальца с
присосками; раковина обычно отсутствует; у современного рода
Argonauta самка значительно крупнее самр,а, она несет тонкостенную
экзогастрически свернутую известковую раковину, лишенную камер,,
образованную особой железой, расположенной на дорзальной стороне
обеих щупалец. Раковина служит для вынашивания оплодотворен-
ных яиц и молоди, она не гомологична таковой остальных цефало-
под. Чернильный мешок у современных форм отсутствует, у поздне-
мелового осьминога (см. рис. 2, 4), отпечаток тела которого (длиной
4,5 см без щупалец) обнаружен в верхнемеловых отложениях Сирии,
сохранились остатки чернильного мешка, челюстей и рудиментарной
раковины. Мел — ныне. Около десяти родов (рйс. 216,6).
История развития колеоидей. Геологическая история 1 колеоидей
до последних лет представлялась в следующем виде: от бактритоидей
в конце девона или начале карбона возникли первые белемнитиды,
имевшие фрагмокон, проострак и ростр; от них в процессе эволюции,
которая шла по пути редукции раковины, возникли три дивергентные
группы: теутиды, сепииды и октоподиды; у первых сохранился проост-
рак и редуцировались фрагмокон и ростр, у вторых сохранился видо-
измененный фрагмокон и отсутствуют ростр и проострак и, наконец,
у третьих скелет редуцировался почти полностью.
В последние годы, благодаря исследованиям канадского палеон-
толога Ю. А. Елецкого (1966), в филогению и систематику колеоидей
внесены существенные изменения. По мнению этого исследователя:,
от бактритоидей возникли сразу две ветви: одна ветвь дала аулако-
цератид с велумом, фрагмоконом и жилой камерой, вторая — белем-
нитид- с известковым ростром, фрагмоконом и проостраком. От обще-
го ствола с белемнитидами возникли теутиды, сепииды и октоподиды
(рис. 217).
Сепииды и октоподиды, вероятно, ответвились от общего ствола
с теутидами в начале мезозоя. У теутид и сепиид имеется много
общих черт, говорящих о их общем- происхождении: наличие ловчих
щупалец и буккальной мембраны, присоски на стебельках с роговыми
кольцами, замок по типу хрящевых' кнопок для замыкания- мантий-
ной щели, нидаментальные железы, практически замкнутая кровенос-
ная система, без лакун, сходное строение жабр и сердца. Однако
сепииды отличаются от теутид по строению скелета. Особый интерес
представляет строение раковины современной Spirula и мелового рода
Groenlandibelus, имеющих протоконх с просифоном и цекумом, напо-
минающим такие же структуры у аммоноидей; как уже отмечалось,
подобные структуры неизвестны ни у аулакоцератид, ни у белемни-
тид, ни у теутид. Вероятно, это может служить основанием для пред-
положения о более раннем возникновении сепиид от общего предка—
бактритоидей. Последний род отнесен к отдельному семейству
Groenlandibelidae (поздний мел). Особенности строения ростра и про-
острака послужили вначале основанием к отнесению этих колеоидей
к отряду белемнитид; но по особенностям строения скелета их рас-
25* 387
сматривают в качестве боковой ветви сепиид с сохранившимися чер-
тами предков. Возможно, от общих предков с ними развились две
ветви сепиид, рассмотренные выше.
Октоподиды представляют особую ветвь колеоидей, которая воз-
никла от общего ствола с теутидами; их развитие пошло по пути
Рис. 217. Геохронологическое распространение головоногих
полной редукции раковины, с сохранением иногда двух хрящевых
палочек, лежащих под мантией, прогрессивного развития нервной
системы, концентрации ганглиев в компактную массу («мозг»), с одно-
временным наличием ряда примитивных черт, таких как веслообраз-
ные плавники, зоб, наличие присосок без стебельков, без роговых
колец и крючков, одно- и двухрядное расположение присосок. В со-
временной фауне колеоидей, таким образом, представлены тремя
отрядами —теутидами, сепиидами и октоподидами — обитателями
преимущественно теплых морей всех глубин, от зоны мелководья до
ультраабиссали, общее число видов которых составляет около 750.
РИНХОЛИТЫ
Ринхолиты (гр. rhynchos — клюв, lithos — камень) представляют
собой обызвествленные кончики челюстей вымерших головоногих, си-
стематическая принадлейсность которых обычно устанавливается с
большим трудом.’ Ринхолит (см. рис. 196, 5) состоит из передней ча-
сти.—капюшона • и задней —рукоятки. ' Капюшон — треу голь-
388
ный, ромбовидный или стреловидный; рукоятка—либо почти прямо-
угольная с выпуклой верхней поверхностью, либо треугольная, с бо-
роздкой посредине. Рукоятка, по-видимому, была скрыта в челюсти,
а капюшон выставлялся из роговой челюсти наружу и служил голо-
воногим моллюскам для прокусывания добычи, отрыва кусков или
раздавливания твердых скелетов.
В ископаемом состоянии ринхолиты известны в отложениях от
конца карбона до середины палеогена (эоцена). Особенно часто
остатки ринхолитов встречаются в отложениях среднего триаса, юры
и мела средиземноморской области. Сравнительно недавно ринхоли-
ты были впервые описаны из меловых и эоценовых отложений Север-
ной Америки. Из керна скважины, пробуренной на дне Атлантиче-
ского океана из нижнего мела с глубины 5323 м, были извлечены три
маленьких ринхолита длиной до 4 мм. И, наконец, из нерасчленен-
ных отложений карбона — перми Южной Америки был описан кон-
хиолиновый ринхолит, найденный в жилой камере аммонита Eoasi-
anites.
Ринхолиты, как правило, встречаются отдельно от раковины и
установить их принадлежность к соответствующей группе головоно-
гих моллюсков — аммоноидеям, наутилоидеям или колеоидеям- —
очень трудно. Различают две основные группы ринхолитов: Phyncho-
lithes (5 родов) и Rhynchoteuthis (7 родов). Представители первой
группы, вероятно, представляли челюсти наутилоидей. Челюсти со-
временного наутилуса принадлежат роду Rhyncolite. Ринхолиты груп-
пы Rhynchoteuthis принадлежали другим головоногим (возможно,,
аммоноидеям), они более многочисленны и разнообразны и могут
быть использованы для стратиграфических целей при изучении
юрских и меловых отложений средиземноморской области; за пре-
делами этой области они пока неизвестны.
При описании ринхолитов, так же как и других частей скелета
(например, аптихов), пользуются искусственной систематикой, выде-
ляя формальные роды и виды, основанные на особенностях строения
и формы капюшона и рукоятки, на их взаимоотношениях и- относи-
тельных размерах; известно около 15 родов.
КЛАСС TENTACULITA. ТЕНТАКУЛИТЫ
Общая характеристика. Тентакулиты (лат. tentaiculum — щупаль-
це) имеют коническую раковину с округленным сечением длиной от
долей миллиметра до 30—50, реже 70 мм, гладкую, с крупными пе-
режимами или покрытую скульптурой в виде колец разных разме-
ров. В раковине выделяются начальная камера, юношеская и взрос-
лая части; внутренняя полость свободна от перегородок или отделена
от начальной камеры одной перегородкой или разделена многими:
перегородками на отдельные камеры; в центре септ имеется узкое'
отверстие. Обитатели морей вели, по-видимому, бентосно-планктонный:
и планктонный образ жизни. Тентакулиты появились в силуре (вен-
локе), испытали расцвет в среднем девоне и вымерли в конце девона
(в начале фамена).
; Строение тела и раковины. О строении мягкого тела почти ничего
не известно. Если судить по форме раковины, то тело было удлинен-
ным, узкоконическим, ротовое отверстие, возможно, окружалось вен-
чиком щупалец; в случае опасности тело втягивалось внутрь ракови-
ны и устье закрывалось крышечкой, которая иногда сохраняется в
ископаемом состоянии.
389-
Раковина узкоконической формы, известковая, с апикальным
углом от 2 до . 10°, в очень редких случаях до 25° (рис. 218). Стенка
раковины состоит из трех слоев: наружного и внутреннего органиче-
ских и лежащего между ними известкового. Наружный органический
слой представляет собой тонкую мембрану; внутренний слой, более
Рис. 218. Класс Tentaculita. Схема строения раковины:
1—3 — отряд Tentaculitida: 1—2 — продольные разрезы, 3 — строение колец;
4 — отряд Homoctenida; 5 — отряд Nowakiida; 6— отряд Styliolinida; (1,3—6 —
оригиналы Г. П. Ляшенко, 2 Блинд, 1969);
жк жилая камера, я — начальная раковина, п — часть раковины с перего-
родками
толстый, состоит из нескольких лент, разделенных тонкими прослой-
ками кальцита; известковый слой построен из таблитчатых кристал-
лов кальцита, лежащих в стенке эмбриональной камеры и юношеской
части параллельно стенке, В переходной зоне от юношеской к взрос-
лой части в стенке раковины появляются так называемые псевдо-
п о р ы; при малых увеличениях они имеют вид темных линий и оши-
бочно принимались за поры; они достигают значительной длины в
крупных кольцах. Стенка при больших увеличениях, представляет
собой волнообразные структуры, у которых тесно лежащие друг к
зэо
другу кристаллы описывают дугу, направленную наружу. Псевдопоры
возникали в области устья и, вероятно, были связаны с прикрепле-
нием мускулатуры кольца щупалец, предположительно существовав-
ших на голове вокруг ротового отверстия.
В раковине выделяют три части: начальную камеру, юношескую
и взрослую. Начальная, или эмбриональная, камера имеет кони-
ческую, каплевидную или шаровидную форму, иногда с тонким ост-
рием на конце; ее поверхность гладкая или тонкоребристая, длина
составляет 0,2—0,4 мм, диаметр устья 0,05—0,1 мм, толщина стенки
в среднем 12 мк; от следующей, юношеской, части эмбриональ-
ная камера отделена пережимом или первым кольцом; у многих в
конце начальной камеры располагается первая и иногда единственная
септа. Юношеская, или задняя, часть раковины соответствует, по-
видимому, первому этапу постэмбрионального развития; ее длина
составляет в среднем ’/2—Уз общей длины раковины; на этой стадии
лоявляется скульптура, представленная либо одинаковыми кольцами,
либо изгибами раковины, напоминающими гофрировку. Кольца рас-
полагаются на одинаковом расстоянии друг от друга. В полости этой
части раковины расположены тонкие септы. Взрослая часть обыч-
но субцилиндрической формы; ее длина равна длине юношеской
части; скульптура этой части отличается большим разнообразием:
у кольцевых форм она представлена кольцами четырех порядков.
Взрослая заканчивается приустьевой частью; у толстостенных форм
она отличается утоньшением стенки раковины, у тонкостенных, наобо-
рот, утолщением, что, возможно, связано с наличием крышечки,
закрывавшей устье.
Внутренняя полость раковины разделена на две части: жилую
камеру и перегородочную. У основной группы тентакулитов (отряд
Tentaculitida) перегородки расположены в юношеской части рако-
вины и иногда частично занимают взрослую часть. Септа состоит из
свободной и муральной частей; свободная слегка прогнута в сторону
лротоконха; муральная может быть очень короткой или, наоборот,
очень длинной. В центре каждой перегородки имеется небольшое
углубление или сквозное отверстие. Через отверстия осуществлялась
связь между камерами; имеются также указания на остатки сохра-
нившегося сифона (Б линд, 1969).
Раковина большинства тентакулит с'кульптирована; различают
четыре основных типа скульптуры: поперечную, продольную, бугорча-
тую и ячеистую. Поперечная скульптура представлена
кольцами разных размеров; различают кольца первого — четвертого
порядков (рис. 218, 1—4). Кольца первого порядка наиболее крупные,
развиты у представителей только одного отряда Tentaculitida; они
высокие, симметричные или асимметричные, с округленной или заост-
ренной вершиной. Кольца второго порядка, или средние, меньших
размеров, расположены между крупными; кольца третьего порядка,
или мелкие, наблюдаются между кольцами второго порядка, и, на-
конец, кольца четвертого порядка, самые маленькие, покрывают
кольца первого порядка и промежутки между ними. Размеры крупных
колец, как правило, меньше, чем их промежутки. Кольца сла-
гаются и образуются за счет изгибов отдельных микрослоев, но, как
правило, эти изгибы не затрагивают всей стенки раковины. У одной
группы тентакулит (Nowakiida) изгибы охватывают все слои и стен-
ка приобретает своеобразную гофрировку, она как бы состоит из
пережимов и вздутий, размеры которых закономерно или незаконо-
мерно увеличиваются. Продольная скульптура представлена
391
продольными ребрами или бороздками, которые тянутся вдоль всей
или только части раковины. Бугорчатая скульптура выраже-
на мелкими бугорками, а ячеистая состоит из своеобразных
ячеек, наблюдаемых на поверхности некоторых раковин.
Основы систематики и классификация. Большинство исследова-
телей относят тентакулит к моллюскам. Г. П. Ляшенко (1955, 1958)
включает их в состав моллюсков под названием Coniconchia, Фишер
(1962)—под названием Cricoconarida, Боучек (1964)—под назва-
нием Tentaculita, Блинд (1969) рассматривал тентакулит в качестве
подкласса головоногих моллюсков. Многие исследователи сравнивали
тентакулит с птероподами, заднежаберными брюхоногими, иногда
объединяли их с другими группами, имевшими также коническую ра-
ковину (например, с хиолитами, конуляриями).
На основании строения и формы начальной камеры, структуры
стенки и характера скульптуры тентакулиты разделены на 4 отряда:
Tentaculitida, Homoctenida, Nowakiida, Styliolinida.
Отряд Tentaculitida. Тентакулитиды (лат. tentaculum — щупаль-
це) объединяют формы, имеющие раковину размером до 70 мм, с ко-
нической эмбриональной раковиной, псевдопористой структурой стен-
ки и скульптурой, представленной кольцами различных размеров,
между крупными кольцами первого порядка расположены кольца
второго — третьего порядка, кольца четвертого порядка покрывают и
кольца первого порядка и промежутки между ними. Силур (венлок) —
девон (рис. 218, 1—3).
Отряд Homoctenida. Гомоктениды (гр. homoios — равный, kteis,
род. ktenos — гребень). Раковина тонкоконическая длиной до 10 мм,
покрыта мелкими одинаковыми кольцами; внутренняя поверхность,
гладкая или кольчатая, стенка микрослоистая, псевдопоры отсутству-
ют, имеется одна септа. Распространены только в девоне (рис. 218, 4).
Отряд Nowakiida. Новакииды (отряд назван по роду Nowakia в.
честь чешского палеонтолога Новака). Раковина маленькая — от до-
лей миллиметра до 10 мм, конической формы, состоит из узких или
широких кольцеобразных пережимов и вздутий; начальная камера
каплевидная или шаровидная, иногда с острием на вершине, отделе-
на септой от остальной раковины. Внутренняя поверхность гладкая
или повторяющая внешнюю скульптуру. Девон (рис. 218, 5).
Отряд Styliolinida. Стилиолиниды (гр. stylos — палка). Раковина
маленькая — от долей миллиметра до нескольких миллиметров, тон-
костенная, гладкая или с тонкой продольной ребристостью. Эмбрио-
нальная камера неотчетливо каплевидная, притупленная, иногда с
коротким шипом на вершине. Септы отсутствуют. Девон (рис. 218, 6~)~
История развития и экология тентакулит. Первые тентакулиты,.
относимые к отряду Tentaculitida, известны из венлока (силура)
Западной Европы, В раннем девоне становятся известными предста-
вители трех других отрядов. О родственных связях между четырьмя
отрядами говорить трудно, хотя, скорее всего, отряд Homoctenida,
у которого раковина покрыта равномерными кольцами, произошел от
тентакулитид. Два других отряда — Nowakiida и Styliolinida, по-види-
мому, возникли от общих предков с тентакулитами, но сразу же обо-
собились в связи с приспособлением к планктонному образу жизни.
Их маленькие тонкостенные раковины, а также массовые скопления
в отдельных местонахождениях /свидетельствуют о их пелагическом
образе жизни. Тентакулитиды и гомоктениды, обладавшие довольно-
крупной и скульптированной раковинкой, вели, скорее всего, придон-
ный образ жизни. Камеры .в задней . части раковины, заполненные
392
газом, облегчали ее удельный вес. В ископаемом состоянии тентаку-
литы встречаются в разнообразных морских фациях совместно с
раковинами и скелетами фораминифер, бивальвий, гастропод, цефа-
лопод, остракод, трилобитов, криноидей, из которых иглокожие и це-
фалоподы указывают на нормальную соленость древних бассейнов.
Находки скелетов тентакулит в битуминозных породах доманика, в
которых не встречены бентосные организмы, подтверждают их пела-
гический образ жизни. Последние представители тентакулит известны
из фаменских отложений позднего девона европейской части СССР и
Средней Азии.
КЛАСС HYOLITHA. ХИОЛИТЫ
К классу хиолитов (гр. hyos — побег, lithos — камень) относятся
палеозойские моллюски, обладавшие небольшой конической или пира-
Рис. 219. Подклассы Orthothecimorpha (2—3) и Hyolithomorpha (/, 4—7):
1 — схема строения раковины; 2 — Spinulitheca (ранний кембрий);-3 — Tcharatheca
(ранний кембрий); 4а, б — Hyolithes (кембрий—пермь); 5 — Phragmotheca (силур);
6—7 — реконструкции {6 — ТЛжсаржевсшЪ, 1969; 7 — оригинал В. А. Сысоева);
ан — анальное отверстие, вг — вентральная губа, вк — воздушные камеры, вс — вент-
ральная сторона, дс — дорзальная сторона, жб — жабры, жк — жилая камера, кр —
крышечка, н — нога, перегородки, р — рот, ра — раковина, эм. — эмбриональная
часть
мидальной раковиной, прямой или изогнутой (рис. 219, У). Раковина
длиной от 1 до 150 .мм, в среднем 10—40 мм, как и у гастропод, была
замкнута с одного, апикального, конца и открыта с другого, дисталь-
ного, конца. По обе стороны от плоскости симметрии проходят обычно
хорошо различимые две продольные линии, отделяющие вентральную
393
сторону от дорзальной; у некоторых форм с круглым сечением эти сто-
роны почти не отличимы; однако у большинства дорзальная сторона
отличается от вентральной скульптурой и некоторыми особенностями
строения. На вентральной стороне у многих форм около устья имеется
вентральная губа. Поперечное сечение раковины круглое, оваль-
ное, треугольное, четырехугольное или округленно-пятиугольное.
В раковине различают три части: эмбриональную, полость воздуш-
ных камер и жилую камеру. Эмбриональная часть состоит из
эмбриональной камеры конической, цилиндрической или шаровидной
формы; п о л о сть воздушных камер состоит из отдельных камер,
отделенных друг от друга перегородками, плоскими или вогнутыми.
Перегородки сплошные, без каких-либо следов отверстий. У некоторых
хиолитов перегородки не установлены, хотя в макушечной части на-
блюдается утолщение, не перекрывающее всю внутреннюю полость ра-
ковины. Жилая камера, в которой при жизни помещалось тело
моллюска, занимает основную часть раковины; внутри раковины, в ее
средней части, с брюшной стороны или по бокам расположены мускуль-
ные отпечатки — следы прикрепления мускулов-ретракторов.
Устье закрывалось крышечкой, простой или состоявшей из двух
неравных половинок (лимбов), расположенных под углом друг к другу
(рис. 219, /). Простая крышечка имеет вид пластинки или низкого ко-
нуса. На внутренней стороне крышечки видны валики, число которых
(от 1 до 4, иногда больше) имеет таксономическое значение. Хиолиты
обладали парой тонких и длинных придатков, дугообразно изогнутых,
отходящих в стороны от устья, обычно 'Слабо обызвествленных. Роль
этих придатков трактуется по-разному: предполагается, что они вы-
полняли функцию поддержек мантии или играли роль своеобразных
стабилизаторов, или служили для поддержки рук, предназначенных
для сбора пищи. Стенка раковины кальцитовая, одно- или двухслойная,
призматическая; у одной группы дорзальная сторона состоит из двух
стенок, пространство между которыми разделено поперечными перего-
родками.
На основании строения раковины и крышечки В. А. Сысоев (1968)
разделил хиолитов на два подкласса: Orthothecimorpha и Hyolitho-
morpha.
Подкласс Orthothecimorpha. Ортотециморфы (гр. orthos — прямой,
thece — чашечка, morphe — форма, вид). Раковина коническая или пи-
рамидальная, с круглым, овальным, треугольным, четырехугольным
или многоугольным поперечным сечением; прямая или отчасти изогну-
тая (рис. 219, 2—3). Вентральная сторона обычно вогнутая, реже пло-
ская, без губы. Мускульные отпечатки отсутствуют. Крышечка плоская
или низкоконическая. Устьевые придатки не установлены. Внутренняя
полость с перегородками или без них. Кембрий—девон. В. А. Сысоев
включает три отряда: Circothecida (кембрий — ордовик) с круглым по-
перечным сечением, ровным устьевым краем или одним синусом;
Orthothecida (кембрий — девон) с дифференцированной вентральной и
дорзальной сторонами, многочисленными перегородками; Exilithecida
(ранний кембрий) без перегородок,, вентральная сторона отличается от
дорзальной.
Подкласс Hyolithomorpha. Хиолитоморфы (основная группа хиоли-
тов) имеют коническую или пирамидальную раковину; вентральная
сторона снабжена удлиненной губой и отличается по строению от дор-
зальной (рис. 219, 4—7). Крышечка разделена на две части (лимбы),
расположенные под углом друг к другу. На внутренней стороне несет
' один—четыре валика, их бывает и больше. Внутри жилой камеры
394
Рис. 220. Отряд Hyolithel-
mintida:
1 — Hyolithellus (кембрий),
реконструкция раковины в
прижизненном положении с
закрытой крышечкой (Фи-
шер, 1962); 2 — Torellella
(кембрий — ордовик)
наблюдаются мускульные отпечатки. У ряда форм обнаружены
устьевые придатки. Внутренняя полость разделена 1—-6 септами. У рода
Matthevia имеется одна септа, муральная часть которой простирается
от апикального конца почти на 2/3 длины всей раковины, удваивая тем
самым толщину стенки. Кембрий — карбон (пермь —триас ?). Подкласс,
объединяет отряды: Hyolithida (кембрий — карбон) с прямой рако-
виной, воздушными камерами, отпечатками мускулов по бокам рако-
вины; Diplothecida (кембрий) с прямой раковиной, воздушными каме-
рами и двойной дорзальной стенкой, полость которой разделена пере-
городками; Globorilida (кембрий) —раковина прямая, с шаровидной
эмбриональной камерой; Pterygothecida (девон — пермь?) — по бокам
и на дорзальной стороне раковины имеются крылообразные выросты;
Ceratothecida (силур — девон)—раковина широкая, изогнутая вбок, с
вырезанным спинным краем устья.
История развития хиолитов изучена недостаточно. Момент их по-
явления и первые этапы эволюции неизвестны. В самом начале кембрия
(томмотском ярусе) появляются представи-
тели всех трех отрядов ортотециморф, оби-
тавших в эпиконтинентальных бассейнах.
Несколько позднее появляются хиолито-
морфы. Они быстро распространились, ин-
тенсивно дивергировали; одни группы хио-
литов сменяли другие, что позволило
В. А. Сысоеву использовать их этапы раз-
вития для разделения нижнего кембрия на
зоны. Хиолиты были широко распростране-
ны в кембрии, ордовике и силуре, число
родов существенно сокращается в девоне.
В карбоне и перми известны единичные на-
ходки. Лучшие находки хиолитов с сохра-
нившимися боковыми придатками известны
из бургесских глин (средний и верхний
кембрий) Британской Колумбии (Канада),
где они найдены вместе с прекрасно сохра-
нившимися остатками трилобитов и других
членистоногих.
Систематическое положение и образ
жизни хиолитов пока неясны. , Хиолиты,
возможно, произошли от общего с гастро-
подами предка, но в отличие от последних
эволюционировали по пути образования
воздушных камер; они представляют собой,
вероятно, инадаптивную слепую ветвь. Подобно. всем моллюскам, их
мягкое тело было окружено мантией, пищеварительный тракт, как у
скафопод, был петлеобразно изогнут; рот и анальное отверстие были
сближены; тело с помощью мускулов втягивалось внутрь раковины, и
устье закрывалось крышечкой.
Класс неизвестен.
ХИ ОЛ ИТЕЛ ьми нтиды
Отряд Hyolithelmintida. Хиолительминтиды (hyolithes — производ-
ное от хиолитов, см. выше, helmins — червь). Раковины в форме узко-
конических трубочек размером от 5 до 15 мм, апикальный угол 1—4°;
начальная часть раковины слепка согнута; сечение овальное или круг-
395
лое. Устье закрывалось крышечкой круглого сечения, многослойной,-
на ее нижней поверхности сохраняются отпечатки лары мускулов. Стен-
ка раковины кальцитофосфатная, слоистая, обычно относительно тол-
стая, внутренняя поверхность гладкая; полость единая, без перегородок.
Представители отряда известны в ископаемом состоянии из отложений
верхнего кембрия — ордовика. 7 родов (рис. 220).
Отряд по строению и химическому составу раковины отличается
от всех моллюсков и, вероятно, ближе стоит к кольчатым червям или
форонидам. В. А. Сысоев (1958) отнес его к хиолитам, Д. В. Фишер
(1962) выделил в качестве отряда неясного систематического поло-
жения.
ТИП BRYOZOA. МШАНКИ
Общая характеристика. Мшанки, или бриозои (гр. Ьгуоп — мох,
zoon — животное), — водные, преимущественно морские, колониальные
вторичнополостные животные, ведущие прикрепленный образ жизни.
Колонии разнообразной формы и размеров состоят из мелких зооидов,
у пресноводных мшанок мономорфных, у морских — полиморфных; сре-
ди последних различают автозооиды и гетерозооиды. У автозооида зад-
няя часть тела превращена в цистид, защищенный толстой 'кутикулой,
а передняя — видоизменена в нежный лолипид, несущий венчик щупа-
лец, втягивающихся внутрь цистида при помощи мускула-ретрактора.
У многих форм кутикула обызвествлена и колония имеет прочный из-
вестковый скелет. В центре лофофора расположен рот. Пищеваритель-
ный тракт петлеобразно изогнут, анальное отверстие расположено на
спинной стороне, вне венчика щупалец. Полость тела разделена диа-
фрагмой на два отдела — туловищный и лофофоральный. Надглоточ-
ный ганглий иннервирует внутренние органы и щупальца лофофора.
Целомическая полость двух- или трехраздельн'ая. Органы кровообра-
щения, дыхания и часто выделения отсутствуют. Мшанки — гермафро-
диты; эмбриональное развитие происходит в овицеллах, или гонозоо-
идах. Свободноплавающая личинка после прикрепления претерпевает
некробиотический метаморфоз и дает начало первичному зооиду, кото-
рый почкованием образует зооиды новой колонии. Мшанки появились
в ордовике и существуют до настоящего времени.
Строение мягкого тела. Все мшанки — колониальные организмы.
Колонии бывают похожи на мох (отсюда название типа — моховидные
животные) или на водоросли; иногда имеют форму корок, обрастаю-
щих подводные камни, водоросли или раковины моллюсков, брахиопод,
скелеты членистоногих, кораллов; некоторые имеют вид сетки, состоя-
щей из прутьев и петель разных размеров и формы. Часто колонии
Приобретают вид. шаровидных или узловатых тел. Форма и размеры
колонии зависят от условий существования: солености и температуры
воды, характера субстрата, наличия течений или волнений. Один и тот
же вид может иметь разную форму колонии: при обитании в зоне при-
боя — массивную, в условиях спокойной воды — ветвистую,. кустистую.
Колония слагается из отдельных зооидов (рис. 221). микроскопических
размеров (обычно до 1 мм). У пресноводных мшанок, колония состоит
из мономорфных зооидов, у морских — из полиморфных особей. Среди
397
последних различают две группы особей: автозооиды, или питаю-
щие зооиды, и гетеро зооиды — видоизмененные особи, утратившие
функцию питания и полового размножения и выполняющие в колониях
функции защиты, прикрепления и т. д.
Автозооид состоит из цистида — задней части зооида, имеющего
прочную, часто обызвествленную, кутикулу, и по липида — перед-
Рис. 221. Схема строения автозооидов (/—2), ранние стадии почкования кругло-
ротых мшанок (3), строение гонозооида (4), лунария (5) и вибракулярия (3);
ав — авикулярий, ан — анальное отверстие, ац — анцеструля (первая ячейка),
гз — гонозооид, жг — жгут, и — известковый слой, к — кутикула, кр — крыш-
ка, м — мускулы-ретракторы, нг — нервный узел, ов — овицелла, п — прото-
эциум- (диск прикрепления), пк — пищеварительный канал, пол — полипид,
пп — первая почка, пт — полость тела, р — рот, см — семенник, ц —• цистид,
щ — щупальца, эк — эктодерма, яи — яичник
ней выпячивающейся части зооида. Рот находится на переднем Конце
полипида и окружен венчиком щупалец, расположенных на лофофоре
кольцеобразно или в форме подковы. Щупальца полые, покрыты рес-
ничками, создающими Вводной и выводной токи водьг^с первым посту-
пает пища, со вторым выводятся продукты обмена. Основной, пищей
мшанок служат различные мелкие организмы, водоросли, простейшие
и органический детрит. Рот ведет й петлеобразно изогнутый Пищевари-
тельный тракт, состоящий из глотки, пищевода, средней кишки, или же-
лудка, и задней кишки, открывающейся на спинной стороне за преде-
лами кольца щупалец анальным отверстием. Пищеварительный тракт
прикреплен к стенке тела при помощи мускула-ретрактора, предназна-
ченного для втягивания, полипида внутрь цистида в случае опасности.
Выпячивание тела и распрямление щупалец происходит гидростатиче-
ским путем у разных групп по-разному. Кроме того, средняя кишка
прикреплена к стенке циСтиДа особым тяжом — канатиком. У пресно-
водных мшанок над ртом расположен особый вырост —эпи сто м (от-
сюда название класса пресноводных мшанок — покрыторотые), отсут-
ствующий у морских представителей. Стенка тела зооида у первых со-
398
стоит из эпителия и слоя кольцевых и продольных мышц и целомиче-
ского эпителия; она похожа на стенку кожно-мускульного мешка чер-
вей. У морских форм она лишена мышц и состоит из эпителия и
кутикулы органического происхождения, очень часто пропитанной
карбонатом кальция, вследствие чего стенка цистида приобретает
прочный минеральный известковый скелет.
Нервная система состоит из надглоточного ганглия, окологлоточного
кольца и нервов — чувствительных и двигательных, идущих к щупаль-
цам, пищеварительному тракту и другим органам. Полость, лежащая
между стенкой зооида и пищеварительным трактом, заполнена цело-
мической жидкостью, выполняющей функцию кровеносной системы.
У пресноводных мшанок целом разделен на три части, у морских — на
две. Целомическая полость продолжается в эпистом, щупальца. У ряда
морских мшанок полости тела соседних зооидов сообщаются между
собой. Кровеносная система отсутствует; газообмен, движение полост-
ной жидкости, удаление продуктов обмена, по-видимому, происходят
в процессе втягивания и высовывания маленького тела полипида. Поло-
вая система состоит из семенников и яичников; первые расположены
в стенке переднего отдела, вторые возникают на канатике, и в стенке
заднего отдела тела.
К гетер о зооидам, предназначенным для выполнения специали-
зированных функций, относятся гонозооиды, авикулярии, вибракулярии,
кенозооиды и наннозооиды. Гонозооиды — своеобразные зооиды,
возникающие из автозооидов (отряд циклостомат), в которых разви-
ваются оплодотворенные яйцеклетки (рис. 221, 4). Гонозооид возникает
при дегенерации полипида и часто имеет кувшинообразную форму.
Внутри гонозооида идет развитие зародыша, дающего в свою очередь
начало многочисленным вторичным зародышам (полиэмбриония),
которые питаются' за счет расположенной здесь ткани.
Для защиты колонии от непрошенных поселенцев у многих Совре-
менных мшанок развиты авикулярии и вибракулярии. Пер-
вые напоминают птичью головку (лат. avis — птица), у них редуциро-
ван полипид и крышечка, прикрывающая устье цистида, превращена в
хватательный орган (рис. 221, 2). Вибракулярии несут длинный подвиж-
ный жгут, представляющий собой также видоизмененную крышечку
(рис. 221, 6). Кенозооиды образуют корневые нити и стелющиеся
столоны — выросты колонии, выполняющие прикрепительную и опор-
ную функции. Наннозооиды —- тоненькие трубочки (в три раза
тоньше цистидов автозооидов); сидящий в них полипид имеет рудимен-
тарный кишечник, одно щупальце, но нормально развитую мускулатуру;
роль их и функции недостаточно ясны. Особые структуры у автозооидов
хейлостомат представляют ооеции, или о виц ел л ы. Они располо-
жены над устьем каждого зооида. В них протекает эмбриогенез и раз-
вивается личинка; до сих пор неясно, представляет ли она упрощенную
особь или продукт дифференцировки цистида автозооида (см. рис. 229).
Строение скелета. По химическому составу колонии, или зоарии,
могут быть органические и известковые. У одних стенки зооидов, абсор-
бируя большое количество воды (до 95%), образуют защитную пере-
понку, не способную сохраняться в ископаемом состоянии; у других
кутикула цистида построена из белкового вещества, содержащего муко-
полисахарид и стереоизомер настоящего хитина (В-полимер, N-ацетил-
глюкозамина). Средний слой кутикулы имеет волокнистое строение и
представляет собой матрицу, на которой формируются кристаллы кар-
боната кальция в форме кальцита или арагонита, образующие мине-
ральный скелет. Некоторые мшанки в качестве примеси содержат
399
стронций (<0—6% SrCO3) или магний (>7—4% MgCO3), У хейло-
стомат скелет построен из кальцита.
Размеры колонии современных мшанок иногда достигают 1—2 м,
среди ископаемых известны до 60 см, но обычно же величина их незна-
чительна. Зооеции, или ячейки, составляющие колонию, имеют харак-
тер длинных цилиндрических трубок или призматических коротких
камер; иногда они приобретают вид груши с широким основанием и вы-
Рис. 222. Формы ячеек Cystoporata
(/), Cryptostomata (2) и Treposto-
mata (3):
/ т- трубчатая, 2 — грушевидная
(7—2 — продольные сечения); 3
тангенциальное сечение колонии;
ап — акантопоры, в — вестибюль,
вг — верхняя гемисепта, дф — диаф-
рагмы, м — мезопоры, нг — нижняя
гемисепта, оя — основание ячейки,
у — устье,, цф — цистифрагмы
тянутым трубчатым вестибюлем.
В сечении ячейки могут иметь
форму параллелипипеда с прямо-
угольным, овальным, многоуголь-
ным основанием. Каждая ячейка
открывается устьем, имеющим раз-
ное очертание: округлое, овальное,
звездчатое, многоугольное. Устье
расположено на вершине ячейки,
на переднем ее конце, или на верх-
ней части передней стенки. Вокруг
устья может быть развит пери-
стом (приподнятый валик) или
л у н а р и й (полулунный выступ
около устья). Устье бывает снаб-
жено крышечкой. У циклостомат
крышечка отсутствует и устье за-
крывается тонкой мембраной с тер-
минальной порой в центре.
У многих трубчатых ячеек
внутренняя полость разделена пере-
городками, или диафрагмами,
отражавшими определенные эта-
пы жизнедеятельности зооида
(рис. 222). Диафрагмы бывают
сплошные и с отверстием в центре.
Известны также особые полупере-
городки — гемисепты, отделяющие вестибюль от основания ячей-
ки (у криптостомат), и цистифрагмы — особые пузыреобразные
структуры, развитые в ячейках; их образование,, возможно, связано с
дегенерацией полипида. Стенка ячеек обычно пронизана порами, через
которые осуществляется связь между полипидами зоарии. У крипто-
стомат утолщенные части скелета были пронизаны капиллярными тру-
бочками, служившими для газообмена.
Кроме ячеек нормальных размеров у ископаемых мшанок имеются
ячейки меньшего диаметра, представляющие собой остатки скелета ге-
терозооидов; среди них выделяют мезопоры и акантопоры. Мезопоры
имеют круглое или угловатое сечение и снабжены диафрагмами., Акан-
топоры — трубчатые образования малого диаметра с узкой полостью
и толстыми стенками; над поверхностью колонии они поднимаются в
виде шипов. У ряда морских мшанок около устья и вдоль киля у сетча-
тых зоарий развиты шипы, которые у современных хейлостомат пред-
ставляют собой скелеты редуцированных зооидов. Различные известко-
вые опорные и механические элементы колоний (кенозооиды), располо-
женные на нижней стороне, также развиваются за счет видоизменения
зооидов.. Пластичность организации зооидов мшанок, таким образом,
очень велика.
Размножение и развитие. Почти все мшанки — гермафродиты; в
400
верхней части цистида расположены семенники, в нижней — яичники.
Яйца оплодотворяются мужскими половыми клетками других особей,
попадающими в полость тела с током поды. Первые стадии эмбрио-
нального развития проходят в гонозооидах, в овицеллах или в полостях
тела материнского организма. Г а ст рул а образуется впячиванием;
клетки мезодермы развиваются телобластическим путем после замы-
кания бластопора. Личинка свободноплавающая. Поплавав в толще во-
ды некоторое время, она опускается на дно и прикрепляется к твердым
предметам. Кишечник и другие личиночные органы распадаются, и
после некробиотического метаморфоза формируется первая ячейка —
анцеструля с уплощенной начальной частью —п р от о е ц и е м
(см. рис. 221, 3). Индивидуальное развитие у пресноводных и морских
мшанок протекает различно и будет рассмотрено при характеристике
классов.
Основы систематики и классификации. На основании особенностей
астогенеза (онтогенеза колонии) и строения колоний тип мшанок разде-
лен на три класса: Phylactolaemata, Stenolaemata и Gymnolaemata.
КЛАСС PHYLACTOLAEMATA. ФИЛАКТОЛ ЕМАТЫ
К классу филактолемат (гр. phylactos — защищенный, laimos —
глотка), или покрыторотых мшанок, относится небольшое число видов
(до 100) современных пресноводных мшанок, практически неизвестных
Рис. 223. Класс Phylactolaemata. Схема строения колонии (1—2) и статобласт
(3-5):
1 — поперечный разрез на уровне рта;- 2 — продольный разрез колонии Crista-
tella mucedo-, 3—5 — статобласты: 3 — спинобласты, 4 — флотобласты, 5 —
сессобласты;
ан — анальное отверстие, бт — бурые тела, втп — втянутый поли'пид, зп — зо-
на почкования, лф — лофофор, по — подошва, р — рот, сем—семенник, ст —
статобласты, эп — эпистом, яи — яичник
в ископаемом состоянии, за исключением одного рода из пресноводных
меловых отложений Богемии (Чехословакия). Колония состоит из моно-
морфных зооидов и отличается значительной степенью слияния цисти-
лов. Полипид имеет эпистом над ротовым отверстием; под эпителием
тела лежит слой кольцевых и продольных мышц, имеются мускулы-
ретракторы,. протракторы и мускулы, связанные с лофофором; лофофор
в. отличие от гимнолемат имеет подковообразную форму (рис. 223) и
26 в. В Друщиц 401
несет от 20, до, 100 щупалец. Надглоточный ганглий и нервные стволы
лофофора находятся в передней части полости целома. Пищеваритель-
ный тракт устроен проще, чем у голоротых мшанок. Пищеварение це-
ликом полостное. Целомическая полость тела разделена на три части..
Продукты обмена выводятся через два ресничных .канала.
Филактолематы, как и все .мшанки, гермафродиты; у них наблю-
дается живорождение — все развитие, вплоть до образования зародыша»,
протекает внутри .материнского организма. Из зародыша без стадии
личинки развивается дочерняя колония — цистид с двумя полипида-
ми, — которая после непродолжительной свободной жизни оседает на.
дно и дает начало колонии. При наступлении неблагоприятных условий:
(низкие или, наоборот, высокие, температуры, высыхание или вымерза-
ние водоемов), зооиды образуют статобласты, или внутренние поч-
ки (рис. 223, 3—5).-Они обладают плотной оболочкой и бывают при-
способлены к плаванию (фл ото б л асты), лежанию на дне (сессо-
бласты) или имеют крючочки для прикрепления к посторонним пред-
метам или животным (спи но б л асты). Известны случаи, когда птицы,
из Африки заносили в дельту Волги африканские .виды мшанок. В даль-
нейшем из статобластов весной развивается новая колония. У филакто-
лемат широко распространены дегенерация и регенерация. Колония
одного из представителей филактолемат (Cristatella) ведет подвижный
образ жизни; эта колония представляет компактное целое с двусторон-
ней симметрией, имеет единую мускулатуру с общеколониальным нерв-
ным сплетением, общий целом, который обеспечивает единство внутрен-
ней среды; цистиды колонии утратили свою индивидуальность, вошли;
в состав общеколониальной стенки тела.
Филактолематы существуют по всему земному шару в чистых про-
точных и спокойных водах, встречаются в озерах на глубине до 170 м
на разных широтах и на разной высоте над уровнем моря (до 4000 м)..
Предки филактолемат были, по-видимому, морские подвижные черве-
образные животные с трехчленистым целомом и кожно-мускульным
мешком. Однако нам неизвестна их история развития и происхож-
дение.
КЛАСС STENOLAEMATA. СТЕНОЛЕМАТЫ
Класс стенолемат, или узкоротых мшанок (гр. stenos — узкий, lai—
mos — глотка), объединяет один отряд современных и четыре отряда
вымерших морских мшанок. Колония состоит из автозооидов и гетеро-
зооидов. Среди последних различают гонозооиды, кенозооиды, нанно-
зооиды, акантопоры, мезопоры. Автозооиды — длинные трубчатые (от-
сюда название класса); стенка цистида обызвествлена. Передний конец,
тела у современных форм закрыт мембраной с терминальной порой
посредине; ниже ее расположены вестибюль и влагалище для втягивае-
мых щупалец. Полипиды упрощены, лишены эпистома, несут на конце-
кольцо щупалец. Развитие оплодотворенных яиц происходит в особых
гонозооидах. Личинка дает начало одному зооиду; от него почкованием
возникают два новых; последующие зооиды образуются в передней
общеколониальной зоне роста — области, не разделенной на цистиды.
Полости зооидов сообщаются только порами. Существуют с конца кем-
брия до наших дней.
Класс разделен на пять отрядов: Cyclostomata, Trepostomata».
Rhabdomesonata, Cystoporata, Cryptostomata.
Отряд Cyclostomata. Циклостоматы (гр. kyklbs — круг, stoma —
рот). Колонии кустистые, ветвистые, инкрустирующие, пластинчатые..
402
Зооиды, или ячейки, трубчатые цилиндрические или призматические,
устье терминальное, без крышечки. Наружные стенки пронизаны псев-
допорами для газообмена; внутренние стенки, разделяющие соседние
зооиды, соединены порами, которые служат для прохождения целоми-
ческой жидкости. Стенки полипида лишены мускулатуры; на переднем
конце имеется терминальная перепонка, или мембрана, с терминальной
порой в центре, ведущая в предверье, за которым следует щупальцевое
Рис. 224. Отряд Cyclostomata. Начальная стадия астогенеза (/) и форма ко-
лоний (2—5):
1 — астогенез Berenicea patina {1 — начальный диск, 2—4 — зооиды в по-
рядке возникновения, 5 — общеколониальная зона роста); 2 — Mitoclema
(ордовик — силур); 3 — Ногпега (эоцен — ныне), передняя сторона колонии;
4 — Terbellaria (юра); 5 — Stomatopora (юра — ныне); у — устье
влагалище. При расширении, поры и уменьшении полости зооида про-
исходит выпячивание щупалец; этому помогает перепончатый мешок,
заполненный целомической жидкостью. Кроме автозооидов в колонии
имеются гетерозооиды: гонозооиды и кенозооиды. В гонозооидах
по типу полиэмбрионии происходит развитие зародышей; из каж-
дого зародыша развивается свободноплавающая личинка. После при-
крепления образуются протоеций и анцеструля с полипидом
цилиндрической формы (см? рис. 221). В процессе дальнейшего почкова-
ния колония имеет общую зону роста; рост идет по краю терминальной
мембраны, под которой находится общий целом (рис. 224, 1}. Полости
зрелых зооидов отделены друг от друга известковыми стенками, но со-
общаются между собой порами. Промежутки между зооидами иногда
заполнены пузырчатой тканью или плотным известковым веществом.
Кенозооиды, развитые в основании колоний, выполняют опорную и
прикрепительную функции. Имеются также наннозооиды, функция кото-
рых неясна.
Циклостоматы появляются в начале ордовика, немногочисленны в
палеозое, но разнообразны в юрском и меловом периодах. В кайнозое
их число сокращается и в настоящее время они представлены неболь-
шим числом родов. Известно около 250 родов (рис. 224).
Отряд Trepostomata. Трепостоматы (гр. trepo — поворачивать). Ко-
лонии массивные, полусферические, пластинчатые, ветвистые. Различат
26*
403.
ют две зоны: незрелую — осевой участок колонии с тонкостенными
призматическими трубками ячеек — и зрелую — периферическую.
В зрелой зоне трубки отогнуты наружу, стенки утолщены, развиты бо-
лее многочисленные диафрагмы и межзоециальные гетерозооиды — ме-
зопоры и акантопоры; мезопоры •— трубчатые полости угловатого
сечения с многочисленными диафрагмами; акантопоры и микро-
Рис. 225. Отряд Trepostomata. Схема строения колоний (/—6) и реконструкция обра-
за жизни (7):
I —• продольное и поперечное сечение цилиндрической колонии; 2 — Diplotrypa (ордо-
вик), тангенциальное сечение; 3 — пятно (макула) в колонии, состоящее из скоплений
Мезопор и крупных устьев ячеек; 4 — дистальная часть акантопоры; 5 — Esthoniopora
(ордовик), массивная колония; 6 — Dittopora (ранний ордовик), ветвистая колония;
7 — реконструкция ордовикской мшанки Dianulites petropolitanus (Pander) на во-
доросли;
ап — акантопоры, ди — диафрагмы, зз — зрелая зона колонии, мп — мезопоры, нз —
незрелая Йона колонии, я — ячейки
акантопоры развиты только в зрелой зоне. Стенки ячеек пронизаны
системой капилляров диаметром от 5 до 20 мк. Протоеций большой, вы-
пуклый; от него развивался первичный зооид, дававший одну средин-
ную и две боковые почки; от последних возникали по две новые почки.,
Ордовик — триас. Отряд включает около 120 родов (рис. 225).
Отряд Rhabdomesonata. Рабдомезонаты (гр. rhabdos — палка, гпе-
sos — средний). Колонии ветвистые, иногда сочлененные у основания;
ячейки удлиненные, трубчатые, призматического сечения, с диафрагма-
ми и гемисептами. В колонии выделяется осевая зона с трубчатой по-
лостью внутри или с осевым пучком параллельных трубочек; стенки
ячеек в осевой зоне тонкие, в периферической — утолщены. Устья оваль-
ные, круглые или с выемкой. Среди гетерозооидов выделяют: мет а по-
ры— трубчатые образования угловатого сечения диаметром 0,02—
404
0,08 мм; акантопоры, имеющие узкий осевой канал, диаметром до
0,1 мм; бугорки — шиповидные выросты на поверхности зоарий.
Ордовик — пермь. Известно около 40 родов (рис. 226, 1—2).
Отряд Cystoporata. Цистопораты (гр. kystis — пузырь, poros — от-
верстие). Колонии обрастающие, массивные, одно- или многослойные,,
или двухслойные, двусторонне-симметричные. Ячейки цилиндрические,
Рис. 226. Отряды Rhabdomesonata (/—2) и Cystoporata (3):
1 — Rhabdomeson. (карбон — пермь); 2 — Streblascopora (пермь); 3 — Fistull-
рога (ордовик — пермь); сечения: а — тангенциальное, б — продольное, в — по-
перечное; г — общий вид колонии;
л — лунарий, я — ячейки
призматические или коленчатообразно изогнутые; стенки ячеек тонкие
или слабо и неравномерно утолщены, пористые. Устье округлой или
многоугольной формы с крышечками или с лунариями; в ячейках раз-
виты диафрагмы. Между стенками соседних ячеек развиты неправиль-
ной формы пузыревидные структуры — цис топоры (отсюда название
отряда). Имеются также минутопоры — иглоподобные образования
диаметром 0,01—0,02 мм и капилляры — тонкие волосовидные тру-
бочки диаметром менее 0,01 мм. На поверхности зоарии выделяются
бугорчатые возвышенные площадки — пятна, сложенные скоплением
цистопор, реже более крупных или более мелких ячеек; их функция и
происхождение неясны. Ордовик — пермь. Включает 60 родов
(рис. 226, 3).
Отряд Cryptostomata. Криптостоматы (гр. cryptos — скрытый). Ко-
лонии сетчатые, состоят из прямых прутьев, соединенных через правиль-
ные промежутки неячеистыми перекладинами. Ячейки расположены на
одной стороне прутьев в два или несколько рядов; сетки могут быть
веерообразными, воронковидными, лирообразными, реже ветвистыми и
•пластинчатыми. Ячейки грушевидной формы, состоят из расширенного
основания и трубчатого вестибюля (см. рис. 222).. Основание бывает в
сечении овальным, четырех-, пяти- или шестиугольным. Внутри вести-
бюля обычно^ обособлены две геми септы — верхняя и нижняя. Устья
ячеек круглые или овальные; у них иногда развиты лунарии и крышеч-
ки, устьевые бугорки, и шипы. В стенке ячеек различают три слоя: внут-
4(У5
ренний тонковолокнистый, средний, промежуточный, зернистый и на-
ружный толстый, пронизанный многочисленными 'капиллярами. Наруж-
ный слой одевает все элементы колонии, перекладины, соединительные
выросты. У прутьев некоторых форм развит киль, вдоль которого рас-
положены шипы и бугорки, разрастание бугорков у некоторых родов
приводило к образованию сетки, защищающей полипидов сверху.
Рис. 227. Отряд Cryptostomata. Схема строения колоний:
1 — поперечное сечение прута колонии; 2 — общий вид и различные сечения
сетчатой колонии; 3 — Phaenopora, (ордовик —, девон), продольное сечение; 4 —
Penestella (ордовик — триас), тангенциальное сечение; 5 — Polypora (ордовик
ранний триас), тангенциальное сечение; 6 — Archimedes (карбон — пермь), вин-
тообразная колония, общий вид;
вг — верхняя гемисепта, вс — внутренняя стенка, кб — килевые бугорки, кл —
киль, кп — капилляры, м — мезопора, нг — нижняя гемисепта, нс — наружная
Стенка, п —; перекладина, пр — прут, пря — продольное .сечение ячейки, сп —
срединная пластина, у — устье, я — ячейка
К числу скелетных остатков полиморфных особей относятся ови-
целлы, килевые бугорки и шипы, различные корневые выросты, выпол-
няющие прикрепительную функцию. При изучении астогении колонии
иногда удается наблюдать большой протоеций, отделенный от суб-
страта тонкой базальной пластинкой, увенчанной трубчатой анце-
струлей; от нее отходят почки первой, затем второй и т. д. генераций.
Характер последующего почкования определяет форму колонии и у
разных групп протекает своим путем. Ордовик — триас. В отряде насчи-
тывается около 150 родов (рис. 227).
КЛАСС GYMNOLAEMATA. ГИМНОЛЕМАТЫ
Класс гимнолемат (гр. gymnos— голый, laimos-^-глотка), или по-
лоротых,объединяет дв-а отряда морских мшанок,- Зооиды цилиндриче-
ские или коробчатые. Крлонця состоит Из полиморфных -особей^ авто-
.406
зооидов и гетерозооидов. Зооиды имеют лофофор, поддерживающий
венчик щупалец; эпистом отсутствует (отсюда название «голоротые»).
Скелет перепончатый или известковый. Развитие известкового скелета
и прикрепленный образ жизни привели к утрате в стенках тела зооидов
мускулатуры и к развитию специализированных .мышц, предназначен-
ных, в случае опасности, длн быстрого втягивания полипида внутрь
.цистида, для высовывания
лофофора с щупальцами,
предназначенными для сбо-
ра пищи; в последней' опе-
рации принимает участие
гидростатический аппарат.
-Устье цистида при втягива-
нии полипида внутрь у мно-
гих гимнолемат закрывает-
ся особыми приспособле-
ниями в виде крышечки или
воротничка с венчиком щу-
палец, приводимыми в дви-
жение с помощью соответ-
ствующих мышц. Пищева-
рительный тракт имеет пет-
леобразную форму; перева-
ривание белков и углеводов
является полостным, а жи-
ров — внутриклеточным.
Целомическая полость раз-
делена на два, а не на три
отдела. Целом лофофора
продолжается в щупальцьг
л
Рис. 228. Отряд Ctenostomata. Поперечный
разрез полипида (/) и форма колонии (2):
1 — поперечный разрез на уровне рта; 2 —
Vinella (ордовик — мел): 2а — колония внут-
ри брахиоподы, 26 — участок той же коло-
нии (Х14);
ан — анальное отверстие, л — лофофор, р —
’Среди гетерозооидов вы'де- рот
ляются авикулярии, вибра-
кулярии, кенозооиды. Оплодотворенные яйца развиваются в ови'Целлах.
Личинка дает начало одному зооиду. Колония развивается фронталь-
ным или боковым почкованием. Класс гимнолемат в настоящее время
разделен на два отряда; Ctenostomata и Cheilostomata.
Отряд Ctenostomata. Ктеностоматы (гр. kteis, роД. ktenos — гре-
бень). Колонии состоят из тонких трубчатых образований—с т о л о-
н о в; зооиды самостоятельны. Эти особенности указывают на прими-
тивность отряда. В отряде имеются формы, перешедшие к одиночному
•существованию (Monobryozoon, два вида). Полиморфизм развит сла-
бо— гетерозооиды представлены столонами и ризоидами; последние
•служат для прикрепления колонии к субстрату. Стенки ячеек перепон-
чатые, студнеобразные или состоят из органического вещества, поэтому
ктеностоматы в ископаемом состоянии встречаются крайне редко;
обычно сохраняются частично обызвествленные столоны или отпечатки
ячеек. Для закрывания устья зооида служит крышечка, напоминающая
гребешок, состоящий из щетинок. Колонии растут боковым почкованием
от стелющихся столонов, расходящихся радиально или беспорядочно.
Ордовик — ныне. Среди современных ктеностомат выделяют четыре
подотряда; в ископаемом состоянии встречаются представители под-
отряда Stolonifera (лат. stolon —побег, ветвь, fего —-нести). Около
40 родов (рис. 228).
Отряд Cheilostomata. Хейлостоматы (гр. cheilos — губа, край)—
наиболее широко распространенный и высокоорганизованный отряд
407
мшанок. Колонии разнообразной формы: обрастающие или прикреп-
ленные одним 'концом; в последнем случае ветвистые, сетчатые, члени-
стые, дисковидные. Зооеции в виде коротких камер различной формы
(рис. 229). Передняя стенка может быть обызвествлена полностью или
только по краям. Устье, расположено в верхней части передней стенки;
его форма круглая, полукруглая или в виде выемки, оно закрывается
крышечкой. Сообщение между зооидами осуществляется системой пор.
Стенки ячеек отличаются большой 'сложностью и обычно многослойные.
Рис. 229. Отряд Cheilostomata. Внешняя форма и схема строения зооидов:
1—2 — строение зооида у Anasca: 1 — зооид с расправленными щупальцами,,
общий вид (Рилэнд, 1970); 2 — схема взаимоотношения фронтальной мембраны
и. париетальных мускулов: 2а — продольное и 26 — поперечное сечения; 3 —
взаимоотношение компенсационного мешка и париетальных мускулов у Ascopho-
га (полипид не изображен); 4—7 — общий вид ячеек современных Anasca (4—5)
и Ascophora (6—7): 4 — Bugula, 5 — Cribrilina, 6 — Schizoporella, 7 — Hlppo-
thoa\
к — крышечку, км — компенсационный мешок, мк — мускул крышечки, мр —
мускул-ретрактор, о — овицелла, п — пора, пм — париетальные мышцы, с — се-
менники, фм — фронтальная мембрана, щ — щупальца, э — эмбрион, я —
яичники
Полипид втягивается в цистид с помощью мускула-ретрактора, а высо-
вывается с помощью особой гидростатической системы. У одной группы
в полости цистида имеется. особый компенсационный мешок,,
заполненный целомической жидкостью, при втянутом положении поли-
ли да его объем минимальный; при высовывании полипида объем.мешка
.увеличиваете^*.цКомпенсациояный мешок открывается наружу особой.
4Q8
порой или в устье цистида. У другой группы с помощью специальных
мускулов передняя стенка цистида прогибается в полость и полипид
высовывает наружу свои щупальца (рис. 221, Z).
У хейлостомат развиты разнообразные гетерозооиды. К их числу
относятся авикулярии и вибракулярии, выполняющие защитную функ-
цию (см. рис. 221, 2, 6), кенозооиды, служащие для прикрепления коло-
нии, шипы — для защиты. О вицелл ы представляют выросты стенки
автозооидов. В овицеллах проходит эмбриональное развитие оплодо-
Рис. 230. Геохронологическое распространение мшанок
творенных яиц. Свободноплавающая личинка — ци фон аута — несет
двустворчатую кутикулярную раковину, имеет теменную пластинку,
кольцо ресничек, на заднем конце присоску, петлеобразно изогнутый
пищеварительный тракт, развитую мускулатуру и нервную систему. По-
сле прикрепления, как и у остальных гимнолемат, происходит некро-
биотический метаморфоз и возникает первый цистид — анцеструля с
первичным полипидом, дающим путем почкования три новые почки;
второе поколение состоит уже из шести Цочек, третье — из одиннадца-
ти. Скорость роста новой колонии выше, чем у остальных отрядов,
Хейлостоматы появились в юре и существуют доныне. По наличию или
отсутствию компенсационного мешка отряд разделен на два подотряда:
Anasca (гр. ап — отрицание, askos'—мешок, сумка) и Ascophora
(гр. phoros — несущий). Отряд включает около 600 родов (рис. 229).
История развития и экология мшанок (рис. 230). Первые мшанки
достоверно известны с начала ордовика, хотя имеются указания на на-
ходки их в верхнекембрийских отложениях. В ордовике довольно много-
численны находки стенолемат, среди которых, установлены представи-
тели всех отрядов. Редко встречаются цтенострматы., Предками .м.щанок
.409
были морские червеобразные животные с трехчленистым целомом и
личинкой типа трохофоры. Мшанки возникли, по-видимому, в кембрии,
в отложениях которого можно ожидать встретить первых представите-
лей типа. Они перешли к прикрепленному образу жизни, созданию коло-
ний, что обусловило специфику организации всей группы, ее специали-
зацию и некоторое упрощение организации.
Мшанки дивергировали на две ветви: основателями первой ветви —
стенолемат — были циклостоматы, основателями второй ветви —
гимнолемат — ктеностоматы. Циклостоматы имеют простые цисти-
ды цилиндрической формы с пористыми стенками и округленным терми-
нальным устьем без крышечки. От них ответвился отряд цистопор ат,
обладающий цистопорами. В ордовике от циклостомат возник отряд
трепостомат, у представителей которого также имелось терминальное
устье, сходные с циклостоматами ранние стадии астогении колоний, но
с более сложной их последующей дифференциацией, появлением мезо-
пор и акантопор, более сложной структурой стенок. Трепостоматы были
широко распространены в ордовике и силуре, являясь в это время гос-
подствующей группой среди мшанок. От них, по-видимому, также еще
в кембрии возникли рабдомезонаты и криптостоматы, которые были
широко распространены уже в ордовике. В силуре их число возрастает.
Девон становится временем расцвета четырех отрядов: криптостомат,
рабдомезонат, цистопорат и .трепостомат. В карбоне господствовали
криптостоматы (подотряд фенестеллин) и рабдомезонаты. Остальные
отряды играли подчиненную роль. В пермский период происходит общее
уменьшение числа родов и видов среди всех отрядов; к концу перми
полностью вымирают цистопораты и рабдомезонаты, резко уменьшают-
ся криптостоматы и трепостоматы, последние представители которых
доживают до триаса. В мезозое существенно изменяется таксономиче-
ский состав мшанок (триасовые мшанки известны недостаточно). В юре
и мелу господство принадлежит циклостоматам.
Вторая филогенетическая ветвь — гимнолематы — состоит из
Двух отрядов: ктеностомат и хейлостомат. Ктеностоматы наиболее при-
митивные, без известкового скелета, их колония состоит из обособлен-
ных самостоятельных столонов и отходящих от них зооидов, сидящих
в ячейках с органическим скелетом. Редкие находки ктеностомат извест-
ны из ордовика Прибалтики. Хейлостоматы появляются в средней юре;
они берут начало, по-видимому, от 'ктеностомат. Их первые представи-
тели относятся к подотряду анаска с более простой внутренней органи-
зацией — без компенсационного мешка. Несколько позднее, в мелу, воз-
никают аскофоры с компенсационным мешком. В палеогене и неогене
хейлостоматы очень разнообразны и многочисленны, хотя с ними до-
вольно успешно конкурируют циклостоматы. К концу неогена число
родов циклостомат постепенно сокращается; в современных морях пре-
обладают хейлостоматы; вместе с ними живут циклостоматы и ктено-
стоматы. В пресноводных водоемах широко распространены филакто-
лематы.
Современные морские мшанки известны во всех морях мира, на
всех широтах. Они отличаются хорошей адаптацией к самым разнооб-
разным условиям существования. Мшанки живут в условиях сублито-
рали, некоторые опускаются на значительные глубины. Многие из них
являются эвритермными; некоторые относятся к стенотермным формам,
живущим только При небольших колебаниях температур (например,
среди коралловых рифов). Одни мшанки хорошо переносят колебания
солености (эвригалинные формы) и способны'жить даже в условиях
опреснения, вблизи устьев рек, хотя не переносят, наоборот, повышения
410
солености; другие являются стеногалинными. Мшанки принимают уча-
стие в образовании коралловых рифов. Позднепалеозойские рифы При-
уралья построены с участием крупных сетчатых колоний криптостомат;
неогеновые рифы Керченского и Таманского полуостровов построены из
скелетов хейлостомат. Многие, особенно палеозойские, мшанки играют
существенную роль при определении возраста пород.
ТИП PHORONO1DEA. ФОРОНИДЫ
К типу Phoronoidea относится небольшая реликтовая группа, вклю-
чающая два современных рода и несколько -видов. Форониды— мор-
ские бентосные небольшие (0,6—15 см) животные, обитающие в зоне
литорали и верхней сублиторали (до глубины 50 м) тропической и уме-
ренной зон всех океанов. Они живут в трубочках, построенных эпидер-
мисом из органического вещества, иногда агглютинированных, обычно
погруженных в осадок; некоторые форониды сверлят известковые по-
роды или пустые раковины моллюсков (двустворок и гастропод). Тело
форонид червеобразное, состоит из трех отделов: эпистома, лофофора
с щупальцами и туловища. Э п и с т о м прикрывает рот. Щупальцевый
аппарат состоит из подковообразного лофофора, несущего большое
число полых щупалец, свернутых спиралью и расположенных в два
ряда. Щупальца служат для сбора пищи (наннопланктона), -газообмена
и -создания выводного тока воды. Туловище представляет собой кож-
но-мускульный мешок, одетый тонкой -кутикулой, выделяемой эпидер-
мисом. Пищеварительный тракт петлеобразно изогнут; ротовое и аналь-
ное отверстия сближены, но последнее лежит вне лофофора. Пищева-
рение в полости желудка внеклеточное, в выпячиваниях желудка —
внутриклеточное. Целом трехраздельный; состоит из полости щупальце-
вого аппарата, полости эпистома и общей- полости туловища. Кровенос-
ная система почти замкнутая. Движение крови достигается сокращением
продольных сосудов: спинного, бокового и сосудов лофофора. Нервная
система мало дифференцирована, состоит из сплетения волокон и нерв-
ных клеток -с нервной регуляцией по типу диффузной нервной сети ки-
шечнополостных. Форониды в большинстве гермафродиты и лишь не-
многие раздельнополые. Размножение преимущественно половое. Ли-
чинка — актинотроха (1—5 мм в длину) ведет пелагический образ
жизни. При оседании на дно одни личинки прикрепляются к мягкому
осадку и делают в нем норки, другие садятся на твердый субстрат,
высверливая норки в известковых породах или раковинах моллюсков.
При превращении актинотрохи в молодого форониса не происходит зна-
чительных изменений во внутренней- организации, которое наблюдается
у мшанок. Норки, в которых живут форониды, обйчно прямые и рас-
положены на небольшом расстоянии друг от друга (от 1 см и более).
Трубки форонид достигают в длину от 1 до 15 см; их верхний конец
выступает над уровнем дна или находится на несколько миллиметров
ниже, образуя небольшое углубление.
В ископаемом состоянии достоверные остатки форонид неизвестны.
Возможно, вертикальные трубочки и ходы, описанные в древних песча-
никах под названием Scolithus, могут принадлежать форонидам. Форо-
ниды, по-видимому, являются древней группой. По строению личинок,
I ряду морфологических признаков они близки к мшанкам и объединяют-
ся вместе с ними в один надтип Actinotrochozoa. В организации взрос-
лых форм у форонид имеются как примитивные, так и вторичные черты,
К числу примитивных черт относятся: червеобразная фор-ма тела с тре-
411
мя отделами, .кожно-мускульный мешок, эпидермальное . положение?
нервной системы, внутриклеточное пищеварение, черты сходства актино-
трохи с трохофорой. К вторичным чертам можно отнести: сильное раз-
витие щупальцевого аппарата, почти замкнутый характер кровеносной
системы, сближенное положение ротового -и анального отверстий. По-
совокупности признаков внутренней организации форониды более при-
митивны, чем мшанки. Последние, перейдя к колониальному образу
жизни, утратили ряд черт, свойственных их червеобразным предкам, и.
приобрели некоторые преимущества — колониальный образ жизни, по-
лиморфизм зооидов, выполняющих различные функции, органический
или минеральный скелет колонии, что позволило им приспособиться,
к самым разнообразным условиям существования.
ТИП BRACHIOPODA. БРАХИОПОДЫ
к типу брахиопод (гр. brachis — плечо, pous, род. podos — нога)
относятся только морские одиночные бентосные животные, занимающие
промежуточное положение между первичноротыми и вторичноротыми.
Мягкое тело заключено в двустворчатую раковину, известковую
или хитиново-фосфатную, состоящую из двух створок: педальной и бра-
хиальной; при жизни тело ориентировано передним концом вверх
(рис. 231). Плоскость симметрии, в отличие от плоскости симметрии
двустворчатых моллюсков, проходит посередине створки и делит ее на
две половины. Створки соединяются замком, состоящим из двух зубов
в брюшной створке и двух зубных ’ямок в спинной (замковые брахио-
поды), или только системой мускулов (беззамковые брахиоподы). Тело
состоит из туловищного отдела, спирально свернутого лофофора и нож-
ки. Туловищный отдел занимает заднюю треть полости раковины и от-
делен от передней, или мантийной, полости мягкой перегородкой. Ман-
тийная полость выстлана двойной складкой тела, состоящей из двух
лопастей, называемых не очень удачно мантийными (мантия брахиопод
не гомологична мантии моллюсков). В перегородке расположено рото-
вое отверстие щелевидной формы. По бокам рта прикрепляются две
руки, или лофофоры, лежащие в обширной мантийной полости и. снаб-
женные щупальцами. У большинства брахиопод лофофор поддерживает-
ся известковым скелетом — брахиальным аппаратом, состоящим из
крючков, петли или конусов. Брахиоподы, подобно двустворчатым мол-
люскам, являются биофильтраторами; они питаются частицами органи-
ческого детрита и микроорганизмами. Рот ведет в пищеварительный
тракт, состоящий из короткой глотки, относительно крупного желудка
и кишечника. Последний у замковых брахиопод заканчивается слепо,
у беззамковых — он длинный с анальным отверстием на конце. В желу-
док впадают протоки парной пищеварительной железы. Выделительная
система представлена метанефридиями. Нервная система состоит из
одного или двух ганглиев, окологлоточного кольца и нервных стволов,
иннервирующих внутренние органы, лофофор, мускулы и ножку. Крове-
носная система незамкнутая, развита слабо; от сердца отходят крове-
носные сосуды, несущие бесцветную жидкость — кровь в мантию и во
все внутренние органы. Большинство брахиопод раздельнополы, но сре-
ди них имеются и гермафродиты. Половые железы расположены в кана-
лах мантии. Брахиоподы размножаются половым путем. Развитие пря-
413
мое или с личиночной стадией. Проплавав некоторое время, личинка
опускается на дно, испытывает метаморфоз и превращается в молодую
особь. Все брахиоподы как современные, так и вымершие — обитатели
морского дна. Они прикрепляются к субстрату при помощи ножки или
прирастанием одной из створок. Во взрослом состоянии некоторые бра-
хиоподы могут лежать свободно на дне, частично погружаясь в ил.
Немногие зарываются в рыхлый осадок при помощи длинной и подвиж-
Рис. 231. Схема строения раковины и мягкого тела брахиопод:
1 — беззамковых, 2—3 — замковых, 4 •— схема расположения мантийных кана-
лов у Terebrcdulina;
бс — брахиальная створка, гп — отпечатки генитальной полости, ж — желудок,.
л — лофофор, ма — мантия, мз — мускул-замыкатель, мо — мускул-отмыка-
тель, мп. — мантийная полость, н,—ножка, ос — отпечатки основных каналов,
печ — пе.чень, пж — половая железа, пл — поддержка лофофора, пс — педаль-
ная створка, пел. — поперечное сечение лофофора, р — рот, ск — слепая кишка,
цп — целомическая полость
ной ножки или закрепляются ею в иле как якорем. Некоторые формы
с возрастом- изменяют свой образ жизни, переходя от прикрепленного
к свободнолежащему.
В геологической истории брахиоподы появляются очень рано. Их
первые представители известны с начала кембрия. Расцвет брахиопод
относится к палеозою, в течение которого они занимали господствую-
щее положение среди бентосной фауны, но в конце палеозоя многие
группы брахиопод вымирают; в мезозое их роль в составе бентоса зна-
чительно сокращается, уступая место двустворчатым моллюскам. В со-
временных морях известно около 70 родов.
На основании особенностей эмбрионального развития, строения
внутренних органов, способа образования ножки, химического состава
414
раковины, характера сочленения створок тип брахиопод разделен на
два. класса: Inarticulata, Articulata.
КЛАСС INARTICULATA. ИНАРТИКУЛЯТЫ,
или БЕЗЗАМКОВЫЕ БРАХИОПОДЫ
Общая характеристика. К классу инартикулят (лат. in —отрица-
ние, articulatus — сочлененный), или Ecardines, относятся наиболее
примитивные брахиоподы (около 130 родов), имеющие хитиново-фос-
фатную, реже известковую, раковину, пористую или непористую. Соеди-
нение створок осуществляется сложной системой мускулов; замковое
сочленение отсутствует. Лофофор не имеет известковых поддержек.
Желудок переходит в длинную сквозную кишку, которая открывается
в мантийной полости анальным отверстием. Ножка образуется брюш-
ной лопастью мантии и проходит между створками по специальному
желобку в педальной створке или на взрослой стадии отсутствует. «Ли-
чинка» ведет планктонный образ жизни, обладая почти всеми чертами
взрослой формы; взрослые инартикуляты ведут бентосный прикреплен-
ный или зарывающийся образ жизни. Известны с кембрия доныне.
Строение мягкого тела. Мантия состоит из двух частей: вентраль-
ной и дорзальной, выстилающих полость раковины. Тонкая перегородка
отделяет большую мантийную полость от висцеральной; в первой рас-
положен лофофор, ко второй приурочены основные внутренние органы.
В перегородке имеется щелевидный рот, ведущий в короткий пищевод,
который переходит в желудок, состоящий из передней и задней частей.
Над ртом лежит особый вырост тела — эпистом. В желудок впадают
протоки пищеварительной железы (2—4). От желудка тянется тонкая
петлеобразно изогнутая кишка, которая открывается анальным отвер-
стием в мантийную полость. Полость тела пересекается большим числом
мезентериев, поддерживающих пищеварительный тракт. Для закрыва-
ния и открывания створок существует сложная система мускулов, среди
которых выделяются пара центральных и пара умбональных, или при-
макушечных, и четыре пары косо направленных мускулов (рис. 232).
В мантийной полости расположен лофофор, состоящий из двух ветвей;
лофофор имеет форму трубки, вдоль которой проходит пищевой жело-
бок, окаймленный парой валиков; на одном валике сидят в один или
два ряда щупальца; на другом, более узком, имеющем вид губы, рас-
положены реснички. Внутри лофофора находятся два неравных канала,
заполненных целомической жидкостью. Щупальца создают ток воды, с
которым приносятся вода и пищевые частицы. Движением ресничек
мантии создается выводной ток воды.
Нервная система состоит из- кольца вокруг пищевода с подглоточ-
ным ганглием, от которого отходят нервные стволы, идущие в мантию,
лофофоры, ножку и другие части тела. Кровеносная система развита
слабо, она не замкнутая, состоит из сердца, сосудов и лакун. От не-
большого пульсирующего пузырька — сердца отходит спинной канал,
идущий в лофофор, который, разветвляясь, образует сосуды в брюшной
и спинной складках мантии. Кровь бесцветная. Целом сохраняет свое
трехчленистое строение; целомической жидкостью заполнены эпистом
(первый отдел целома — процель), большой и .малый каналы лофо-
фора, полость тела вокруг пищевода (второй отдел целома — мезо-
цель), полость в ножке лопасти мантии (третий отдел — метацель),
образующие мантийные синусы, с которыми связаны половые железы.
Органы выделения в виде одной пары метанефридиев служат не только
для выделения, но и для выведения половых клеток. В целомической
415
Жидкости имеются различные клетки: амебоцисты, гемэритрин с крас-
ным дыхательным пигментом, близкий к гемоглобину позвоночных.
Брахиоподы раздельнополы. Половой диморфизм не выражен; оплодо-
творение происходит вне тела матери, в воде. Весь эмбриогенез (изучен
у современных лингул) протекает в яйцевых оболочках; здесь идет фор-
мирование мантии, ресничек вдоль ее края, лофофора с щупальцами,
раковинки, ножки, всех внутренних органов (рис. 232, 6). Ножка раз-
Рис. 232. Класс Inarticulata:
1—2 — система мускулов у Lingula-, 3—4 — система мантийных каналов у: 3 —
Lingula, 4 — Crania-, 5 — педальная створка Тrimeriella-, 6 — личинка Lingula,
педальная створка изнутри;
бс — брахиальная створка, ж — желудок, к — кишка, н — ножка, нф — нефри-
дии, пл — платформа, пс — педальная створка, см — система мускулов, ф —
филоменты, щ — щетинки, э — эпистом
вивается у заднего конца вентральной части мантии и вначале лежит
внутри двух тонких хитиновых створок. Щупальца лофофора вначале
расположены так же, как у личинки актинотрохи (мшанки), а затем
их положение изменяется. Из яйцевых оболочек выходит «личинка»,
внешне похожая на своих родителей (скорее ее можно сравнивать с
нимфой насекомых), и поэтому ее нельзя называть личинкой, скорее
можно говорить о прямом развитии. «Личинка» ведет вначале планктон-
ный образ жизни, плавая с помощью ресничек лофофора, которые перио-
дически высовываются наружу при открывании створок. После оседа-
ния на дно ножка молодой лингулы выходит между створками и выде-
ляет желатинообразное вещество, твердеющее в воде, прикрепляя моло-
дую особь к субстрату.
Строение раковины. Раковина органически-фосфатная или извест-
ковая, снаружи покрытая периостраком. Органически-фосфатная рако-
вина состоит преимущественно из фосфата кальция (у лингулид от 74
416
до 94%).; органическое вещество представлено хитином и протеином.
Органическое и неорганическое вещество образует тонкие слои
(рис. 233), лежащие параллельно поверхности раковины; они форми-
руются передним краем мантии. Периострак построен из чере-
дования прослоек хитина и- протеина и по своей структуре напоминает
кутикулу насекомых. Известковая раковина построена из карбоната
кальция (около 88%). У краниид кроме периострака, в состав которого
Рис. 233. Схема строения раковины инартикулят:
1 — Crania (совр.): 1а — продольный разрез, 16, в — строение брахиальной
створки, 1г — участок места прикрепления аддуктора, 1д — краевая часть бра-
хиальной створки, 1е — часть мантии и педальной створки; 2 — Lingula (совр.),
строение стенки раковины: 2а — поперечный и 26—продольный разрезы; 3—4 —
схема строения стенки раковины представителей Kutorginida (5) и Obolellida (4);
бс — брахиальная створка, вс — внутренний слой, вэ — внутренний эпителий,
к — каналы, мз — мускул-замыкатель, нс — наружный слой, нэ — наружный
эпителий, ос — органический слой, пер — периострак, пс — педальная створка,
сб — субстрат, ст — соединительная ткань, фс — фосфатный слой, хс — хити-
новый слой
входит преимущественно протеин, а не хитин, различают еще два слоя:
тонкий пористый наружный и внутренний пластинчатый, обычно доволь-
но толстый, построенный из известковых фибр и органической основы.
Оба слоя более хорошо выражены в брахиальной створке.
Первичная раковина, или п роте гул, возникшая в эмбриональной
стадии, часто сохраняется у ископаемых форм. Дальнейший рост рако-
вины может происходить по-разному; у одних новые части раковины
образуются вокруг протегула концентрическими полосами (голове-
риферический рост), в этом случае возникает створка конической
формы и протегул оказывается на ее вершине (у дисцинид); у других
раковинное вещество образуется по бокам и особенно интенсивно у пе-
реднего края (г ем и п ер и фер ический рост), при этом раковина
27 в. В. Друщиц
417
приобретает лопатовидную удлиненную форму (лингулиды, рис. 234, <?);
протегул располагается на заднем донце раковины; у третьих, облада-
ющих так называемым миксопериферическим ростом1, новое ра-
ковинное вещество отлагается так же, как у первых (т. е. голоперифе-
рически), но задний край наклонен вперед (рис. 234, 2), образуя по-
верхность, называемую палинтролом; раковина приобретает непра-
вильно-коническую форму. Форма раковин, таким образом, у беззамко-
Рис. 234. Типы роста раковин:
1а, б — голопериферический (Orbiculoidea), 2а, б — миксопериферический
(Apsotreta), За, б — гемипериферический (Lingula)
вых брахиопод зависит от способа роста; она может быть округленной^
удлиненно-овальной, лопатовидной, конической.
Внутреннее строение. Створки у беззамковых брахиопод сочленя-
ются при помощи системы' мускулов, число которых непостоянно у раз-
ных групп. В местах прикрепления мускулов образуются мускульные-
отпечатки; здесь эпителий мантии внутриклеточно выделяет раковинное-
вещество. На внутренней поверхности раковин бывают видны отпечатки:
двух пар аддукторов, расположенных симметрично относительно-
медиальной линии, в том числе одной пары около внутренней стенки,,
другой около макушки, и отпечатки косых мускулов, различных у раз-
ных групп: четырех пар у лингулид, трех у дисцинид, двух у кранйид.
(рис. 232, /). iK их функции относится помощь аддукторам при откры-
вании и закрывании створок и при относительном боковом или продоль-
ном их смещении. У одной группы вымерших инартикулят (тримерел-
лид) для прикрепления мускулов были развиты особые платформы,,
сплошные или с полостью внутри, образуемые за счет внешнего эпи-
телия. Замочное сочленение между створками отсутствует, поэтому
инартикулят еще называют беззамковыми (Ecardines, лат. е — означает-
отрицание, cardo —крюк двери).
В области, где расположена мантийная полость, часто видны отпе-
чатки мантийных каналов; на брюшной створке видна одна пара основ-
ных каналов, на спинной — две пары; от них обычно отходят боковые-
ветви (рис. 232, 3—4). У краниид с ними связаны гонады, у остальных,
беззамковых брахиопод эти каналы заполнены целомической жидкостью..
Основы систематики и классификация. На основании строения и-
формы раковины, характера ножки, класс беззамковых брахиопод раз-
делен на пять отрядов: Lingulida (кембрий — ныне), Acrotretida (кемб-
рий— ныне), Craniida (ордовик — ныне), Obolellida (ранний и средний:
418
кембрий), Paterinida (ранний кембрий — средний ордовик). В качестве
отряда неясного систематического положения выделяется Kutorginida
(кембрий — ранний ордовик).
Отряд Lingulida. Лингулиды (лат. lingula — язычок). Раковина
хитиново-фосфатная, округленная или удлиненно-овальная, равноствор-
чатая, слабо выпуклая. Поверхность гладкая, с концентрическими ли-
ниями нарастания (гемипериферический рост). Ножка проходит между
Рис. 235. Отряды Lingulida (/—2), Craniida (3) и Acrotretida (4—5):
Г— Lingula (силур — ныне) в прижизненном положении; 2 — Obolus (средний кем-
брий— ранний ордовик): 2а, б — педальная створка снаружи и изнутри, 2в — бра-
хиальная створка изнутри; 3— Crania (мел — ныне): За — брахиальная створка сна-
ружи, 36 — педальная створка изнутри, Зв — прижизненное положение; 4 — Acrotreta
(ордовик): 4а, б — педальная створка изнутри и сбоку, 4в — брахиальная створка;
5 — Discinisca (юра — ныне): 5a — вид сбоку, 56 — педальная створка снаружи;
жн — желобок для ножки, лб — лимб, ом — отпечатки мускулов, ос — отпечатки ос-
новных мантийных каналов
створками, часто по желобку в педальной створке; она в несколько раз
длиннее раковины; с помощью своей мускулистой ножки лингулиды
выкапывают в илистом осадке глубокие норки и сидят в них, выставив
наружу передний край раковины. В случае опасности лингулиды втяги-
вают раковину внутрь норки. Некоторые лингулиды прикрепляются в
мягком осадке своей ножкой как якорем, выделяя на ее дистальном
конце клейкое вещество. Современные лингулиды приспособились к
жизни в зоне литорали, в солоноватых и опресненных водах эстуариев.
Лингулиды появляются в раннем кембрии, достигают расцвета в ордо-
вике и доживают до наших дней (2 рода). В состав отряда включают
около 50 родов (рис. 235, 1—2). Надсемейство Trimerellacea, относимое
многими исследователями к этому отряду, перенесено в отряд обо-
леллид.
27;
419
Отряд Acrotretida. Акротретиды (гр. akron — вершина, tretos —
продырявленный). Раковина хитиново-фосфатная, округленная; одна из
створок коническая, рост створок голопериферичеокий или миксопери-
ферический, Отверстие для ножки (форимен) округлое или треугольное,
расположено позади макушки. Брахиальная створка имеет срединную
септу, два крупных мускульных отпечатка и два маленьких. Хорошо
выражены отпечатки двух .мантийных каналов на обеих створках. Акро-
третиды появляются в раннем кембрии, достигают расцвета в позднем
кембрии, ордовике и силуре, затем число родов резко сокращается; в
настоящее время известно 3—4 рода (рис. 235, 4—5).
Объем этого отряда еще недостаточно определен, так как степень
изученности его невелика. Одни исследователи (Rowell, 1965) рассмат-
ривают их в составе двух подотрядов: акротретидин и краниидин, дру-
гие (Горянский, 1960) разделяют их на три самостоятельных отряда:
акротретид, сифонотретид и краниид. В настоящем учебнике кранииды
рассматриваются в качестве самостоятельного отряда, а сифонотретиды
включены в отряд акротретид.
Отряд Craniida. Кранииды (гр. cranion — череп). Раковина извест-
ковая, неравностворчатая, пористая, трехслойная, брахиальная створка
обычно коническая, педальная субконическая или выпуклая, состоит из
органического периострака и одного известкового слоя. На внутренней
поверхности отчетливы отпечатки двух пар аддукторов, двух пар косых
мускулов и двух разветвленных мантийных каналов. Ножка отсутствует.
Кранииды ведут прикрепленный образ жизни, прирастая педальной
створкой. Для прочного смыкания створок вдоль внешнего края развита
широкая площадка — лимб. Кранииды появляются в раннем ордовике
и существуют до наших дней. В состав отряда включают 15 родов
(рис. 235, 3).
Отряд Obolellida. Оболеллиды (гр. obolos — маленькая монета).
Отряд объединяет небольшое количество раннепалеозойских беззамко-
вых брахиопод, имеющих известковую двояковыпуклую раковину, округ-
ленную или овальную; у одних ножка выходит между створками, у дру-
гих— через отверстие в педальной створке. На внешней поверхности
хорошо выражены линии нарастания и радиальные струйки. Внутри, у
заднего края брахиальной створки, развита платформа и две пары ман-
тийных каналов. Кембрий — силур. Около 10 родов (рис. 236, 1^—2).
Отряд Paterinida. Патериниды (лат. patera — блюдце) объединяют
беззамковых брахиопод, имеющих фосфатную раковину округленного
или эллиптического очертания; в педальной створке треугольное отвер-
стие (дельтирий) для выхода ножки, прикрытое одной пластинкой
(псевдодельтидием). В брахиальной створке имеется треугольное отвер-
стие (нототирий), частично прикрытое одной пластинкой (гомеохили-
дием). Кембрий —ордовик. Около 10 родов (рис. 236, 3).
Отряд Kutorginida. Куторгиниды (в честь С. С. Куторги, геолога
и палеонтолога, профессора Петербургского университета) объединяют
три раннекем|брийских рода, имеющих известковую раковину неболь-
ших размеров с дельтирием на педальной створке и нототирием на бра-
хиальной; оба отверстия прикрыты соответствующими пластинками
(рис. 236, 4). По два основных канала васкулярной системы в каждой
створке, похожих на каналы остальных беззамковых брахиопод (род
Rustella, как показали новые исследования, возможно, является сино-
нимом рода Kutorgina и не может служить основой для отряда рустел-
лида). Систематическое положение отряда остается недостаточно ясным.
Он занимает промежуточное положение между беззамковыми и замко-
выми брахиоподами, по строению раковины стоит ближе к последним.
420
История развития и экология беззамковых брахиопод. В раннем
кембрии уже известны представители четырех отрядов: Paterinida,
Obolellida, Acrotretida и Lingulida и проблематичного отряда Rutor-
ginida. Представители первого отряда, имеющие в отличие от трех
остальных отрядов известковую раковину, вымирают в среднем ордо-
вике. Второй отряд, хотя и имеет фосфатную раковину, но по типу
Рис. 236. Отряды Obolellida (У—2), Paterinida (3) и Kutorginida (4):
1 — Obolella (ранний кембрий): 1а, б — педальная створка, 1в — брахиальная
створка изнутри; 2 — Trimerella (силур), педальная створка изнутри; За—в —
Paterina (кембрий), За, б — общий вид, Зв — педальная створка; 4 — Kutorgina
(ранний кембрий): 4а — педальная створка снаружи, 46 — вид сбоку;
бс — брахиальная створка, пс — педальная створка
строения и отпечатков мускулов резко отличается от всех остальных
беззамковых брахиопод. Он вымирает в силуре. Представители двух
других отрядов имели ножку, выходившую через отверстие в педальной
створке, и вели прикрепленный образ жизни. Акротретиды представ-
лены в кембрии 5 семействами, из которых одно было развито в раннем
кембрии, два появились в позднем, а два существовали в течение всего
кембрия. Лингулиды были представлены в кембрии одним семейством
(Obolidae). Ордовик и силур можно рассматривать как период расцвета
лингулид и акротретид. В то же время в ордовике появляются первые
кранииды и тримереллиды (группа, условно отнесенная к оболеллидам).
Кранииды и тримереллиды вели прирастающий образ жизни, прикреп-
ляясь педальной створкой к подводным предметам. В силуре роль без-
замковых брахиопод в бентосе остается довольно существенной, но в
девоне она значительно уменьшается. В дальнейшем продолжают суще-
421
ствовать представители только трех отрядов: акротретид, краниид и лин-
гулид, которые доживают до наших дней.
КЛАСС ARTICULATA. АРТИКУЛЯТЫ,
или ЗАМКОВЫЕ БРАХИОПОДЫ
Общая характеристика. К классу артикулят, или замковых бра-
хиопод (Testicardines), относится около 1700 родов. Раковина извест-
ковая, створки соединяются системой 'мускулов: аддукторов и дидукто-
ров. Замок состоит из двух зубов, развитых в педальной створке, и двух
ямок в брахиальной створке. Зубы поддерживаются зубными пластин-
ками. Лофофор у части форм имеет известковые ручные поддержки в
виде крючков, петли или спирали. Пищеварительный тракт заканчи-
вается слепо. Личинка состоит из трех отделов. Развитие с метаморфо-
зом. Ножка развивается за счет хвостового отдела; лопасти мантии
формируют две известковые створки. Для выхода ножки имеется дель-
тирий или форамен. Взрослые артикуляты ведут бентосный образ жиз-
ни. Известны с кембрия доныне.
Строение мягкого тела. Внутренний эпителий мантии образует
срединную перегородку, отделяющую висцеральную полость от мантий-
ной. Узкий щелевидный рот ведет в относительно длинную глотку и
пищевод, переходящий в небольшой желудочек; от него отходит корот-
кий кишечник, с заостренным или вздутым концом, заканчивающимся
слепо. В желудок впадают обычно четыре пищеварительные железы, как
у лингулид. Органы выделения — метанефридии — имеют вид длинных
трубок конической формы, которые открываются в мантийную полость
и выводят в нее не только продукты обмена, но и половые клетки. По-
добно. инартикулятам целомическая жидкость у артикулят заполняет
каналы лофофора, мантийные синусы, висцеральную полость, но в от-
личие от первых не проникает в ножку. В каждой лопасти мантии
имеется одна или две пары основных мантийных каналов, от которых
в стороны отходят тонкие ветви; характер ветвления мантийных кана-
лов используется в целях систематики.
У артикулят имеется достоянное число, мускулов: одна пара а д -
дукторов, или мускулов-замыкателей, и одна пара ди д у кто р о в,
или мускулов-отмыкателей (рис. 237). Аддукторы прикрепляются около
середины педальной створки, отсюда они, раздваиваясь, идут к брахи-
альной створке, образуя на ней четыре отпечатка. Дидукторы прикреп-
ляются одним концом к брахиальной створке, к ее краю или замочному
отростку, а другим концом—к педальной створке по краям аддукторов.
Кроме аддукторов и дидукторов имеются ножные мускулы, которые
обеспечивают незначительное движение раковины — повороты ее во-
круг оси, наклон к субстрату, втягивание ножки внутрь раковины и ее
обратное выталкивание.
Лофофор расположен около рта, прикреплен к брахиальной ло-
пасти мантии и внутренней стенке. В онтогенезе лофофор проходит
несколько стадий: вначале он окружает рот и эпистом над ним (или
брахиальную губу) в виде венчика щупалец (трохолофусная стадия),
затем срединной септой он разделяется на две ветви (схизолофусная
стадия), или две руки; позднее у ринхонеллид развиваются две спирали
(спиролофусная стадия), а у терабратулид ветви вытягиваются в длин-
ные петли, которые закручиваются на концах в спирали, лежащие в
плоскости симметрии (зиголофусная стадия). Развитие завершается
плектолофуснбй стадией — образованием двух петель у теребратулид.
422
У многих артикулят лофофор поддерживается известковым ручным ап-
паратом, строение которого будет рассмотрено ниже.
Нервная система артикулят состоит из двух ганглиев: надглоточ-
ного и подглоточного, соединенных окологлоточным кольцом. Ганглии
иннервируют лофофор, лопасти мантии, мускулы, ножку и другие части
тела.
Рис. 237. Схема расположения и взаимодействия мускулов у современной зам-
ковой брахиоподы (Magellania):
la — общий вид, 16 — фрагмент брюшной створки (вид изнутри); 2a — закры-
тые створки, 26 — открытые створки;
•6с — брахиальная створка, зб — зубы, зо — замочный отросток, мз — мускул-
замыкатель и его отпечатки на раковине, мн — мускулы ножки, мо — мускул-от-
мыкатель и его отпечатки на раковине, « — ножка, пс — педальная створка,
pa —ручной аппарат в виде петли
Кровеносная система изучена слабо; предполагается, что она в от-
личие от инартикулят замкнутая. Трехчленный характер целома не
выражен. Целомической жидкостью заполнены полость висцерального
мешка, каналы лофофора, полости в брюшной и спинной лопастях
мантии.
Артикуляты размножаются половым путем. В процессе эмбриогене-
за целом возникает энтер сдельным путем, как у всех вторичноротых,
а бластопор зародыша переходит в ротовое отверстие, как у первично-
ротых. Из яйцевых оболочек выходит трехчленистая личинка с голов-
ным, туловищным и хвостовым отделами; головной отдел покрыт рес-
ничками, круговая складка туловищного отдела образует лопасти ман-
тии (рис. 238). У личинки нет ни щупалец, ни ротового отверстия. После
•оседания личинки на. дно и прикрепления, ее хвостовым сегментом к
подводным предметам она испытывает глубокий метаморфоз, при кото-
ром; лопасти мантии поворачиваются на 180°, охватывая головной и
туловищный отделы; за счет хвостового сегмента развивается ножка;
эпителий лопастей мантии выделяет известковую раковину, развивается
лофофор с' щупальцами, открывается ротовое отверстие, развиваются
все внутренние органы и .мускулы.
Строение раковины. Раковина состоит из двух неравных створок,
обычно двусторонне-симметричных, с плоскостью симметрии, проходящей
посредине (рис. 239). Более крупная створка с отверстием для выхода
ножкц называется педальной (гр. pous, род. podos — нога), или
вентральной, вторая створка меньших размеров, к которой изнутри
прикреплен лофофор, или ручной аппарат, называется брахиальной
423
(гр. brachion— плечо, рука), или дорзальной. В каждой створке
имеется макушка — наиболее выпуклая часть створки и в то же время
наиболее старая. Периферические части створок называют краями:
передним и боковыми; примакушечная часть является задней, или за-
мочной; вдоль заднего края в брюшной створке расположены два зуба,
входящие в зубные ямки спинной створки (отсюда второе название
класса — Testicardines, или замковые брахиоподы (лат. testa — рако-
вина, cardo — крюк двери). Зубы создают жесткое соединение для двух
створок. Передний, или лоб-
Рис. 238. Схема эмбрионального развития со-
временной замковой брахиоподы (а — общий
вид, б — в разрезе):
1 — свободноплавающая личинка; 2 — ста-
дия прикрепившейся личинки; 3 — молодая
брахиопода с первичной раковиной;
бл —; брюшная лопасть мантии, бс — брахи-
альная створка, гс — головной сегмент, зк —
замочный край раковины, мр — место заклад-
ки створок раковины, н — ножка, пс — пе-
дальная створка, сл — спинная лопасть ман-
тип, т — туловищный сегмент, х — хвостовой
сегмент
ный, край является проти-
воположной частью ракови-
ны, при жизни брахиоподы
он ориентирован вверх; при
раскрывании створок через
передний край поступает
пища в желобки лофофо-
ров. Вдоль переднего края
осуществляется основной
рост раковины. Передний и
задний края соединяются
боковыми краями.
Раковины всех артику-
лят известковые,' состоящие
из карбоната кальция (от
94,6 до 98,6%) с небольшой
примесью карбоната маг-
ния, окиси алюминия, орга-
нического вещества. По-
верхность раковин всех ар-
тикулят покрыта тонким
органическим слоем — пе-
риостраком, состоящим из
•протеина, без хитина; в со-
став протеина входят: ар-
гинин, глицин и другие
аминокислоты. Периострак
образуется в складке ман-
тии (рис. 240). Под органи-
ческим слоем выделяются два известковых слоя: наружный первичный
и внутренний вторичный. Наружный состоит из призматических кри-
сталлов кальцита, ориентированных перпендикулярно наружной по-
верхности. Непосредственно за клетками, строящими первичный слой,
расположены клетки, выделяющие вторичный слой, состоящий из тон-
ких фибр кальцита, ориентированных под небольшим углом, к кри-
сталлам первичного слоя. Каждый кристалл окружен тонкой органи-
ческой. оболочкой, поскольку его образование происходит внутрикле-
точно. Толщина вторичного слоя очень различна и достигает наиболь-
шей величины в задней части раковины.
Различают три типа строения раковины: сплошное, пористое и
ложнопористое. В первом случае оба известковых слоя сплошные, во
втором случае стенка раковины пронизана тонкими канальцами, имею-
щими конусовидную форму; они открываются наружу порами
(рис. 240, 1) и характерны для некоторых групп артикулят (в частнос-
ти, теребратулид). Возникновение канальцев связано с образованием
у переднего края мантии отростков эпителиального слоя. Ложнопорис-
424.
тые раковины содержат во вторичном слое особые стерженьки — т а -
л е о л ы, около которых происходит резкий изгиб известковых слойков;
на внутренней поверхности раковины талеолам соответствуют неболь-
шие бугорки или ямки, создающие ложное впечатление пористости
(рис. 240, 9).
Форма раковины. Наиболее распространенная форма раковины —
двояковыпуклая, причем- педальная створка всегда крупнее брахиаль-
ной и отличается более выпуклой макушкой (рис. 241). Различают так-
же; плоско-выпуклые, выпукло-плоские, вогнуто-выпуклые, выпукло-
Рис. 239. Внутреннее строение раковин:
1 — схема замкового сочленения створок; 2 — внутреннее строение педальной
створки Choristites-, 3 — то же Magellania;
ар — арея, бс — брахиальная створка, г — генитальные отпечатки, др — дель-
тирий, дп — дельтидиальные пластинки, зб — зубы, зп — зубные пластинки,
зч — зубчики, зя — зубные ямки, м — мускульное поле, мз — отпечатки муску-
лов-замыкателей, мн — то же мускулов ножки, мо — то же мускулов-отмы-
кателей, пс — педальная створка, ф — форамен, я — ямки для зубчиков
' вогнутые, при этом первое слово указывает на изгиб брахиальной
створки, а второе—на изгиб педальной. При выпуклой педальной
створке брахиальная может быть выпуклой, плоской или вогнутой и,
наоборот, при выпуклой брахиальной створке педальная может быть
выпуклой, плоской или вогнутой. Передний край створок может иметь
глубокие складки, идущие почти от макушки. Срединное продольное
углубление от макушки к переднему краю в педальной створке назы-
вается синусом, соответствующее ему срединное возвышение на бра-
хиальной створке носит название седла. Синус может быть осложнен
вторичными складками. У прирастающих артикулят педальная створка
может иметь форму бокала, напоминая кубок одиночного коралла, а
брахиальная — форму невысокой вогнутой или плоской крышки. У не-
которых палеозойских брахиопод передние части створок вытягивают-
ся, образуя так называемый шлейф.
Размеры раковин брахиопод очень различны: от 0,1 до 40 см в длину,
однако преобладают размеры 3—7 см.
У многих артикулят под макушкой развита площадка—арея; она
бывает как на педальной, так и на брахиальной створках, но чаще на
первой. На арее, под макушкой, имеется треугольное отверстие —
дельтирий (А — буква греческого алфавита в виде треугольника,
thyra — дверь), служащее для выхода ножки. Подобное отверстие на
спинной створке носит название нототирий (гр. notos — спина).
Дельтирий может прикрываться одной или двумя пластинками, умень-
шающими размеры отверстия. Две пластинки, прикрывающие дельти-
рий, образуют вокруг ножки отверстие круглой формы — форамен:
Нототирий ;прикрывается только одной пластинкой — хи л ид и ем
(рис. 241,0.
425
Скульптура (рис. 242). Поверхность раковины редко бывает глад-
кой; на ней наблюдаются линии нарастания и разнообразные элементы
Рис. 240. Схема строения раковин артикулят:
1—7 — отряд Terebratulida: la — передний край мантии й раковины, 16 —
строение верхней части канала, 1в — Podichnus — след прикрепления ножки бра-
хиоподы к ростру белемнита (Х25); 2—7 — типы ножек у современных: 2 —
Cryptopora, 3 — Gwynia, 4 — Clindonophora incerta, 5 — Macandrevia, 6 —Euca-
lathis, 7 — Clindonophora chuni; 8 — отряд Rhynchonellida: 8a — передний край
мантии и раковины, 86 — схема строения стенки; 9 — ложнопористая стенка;
10 — строение стенки (отр. Strophomenida);
гз — генеративная зона, лп — ложные поры (талеолы), мпс — мукополисахарид,
ом — органическая мембрана; остальные обозначения см. рис. 233
скульптуры: радиальные и концентрические. Различают макро- и микро-
скульптуру; последняя различима только при увеличениях. Радиальная
скульптура представлена ребрами, складками. Ребра образуются на пе-
реднем крае створок, благодаря мелкой складчатости лопастей мантии.
Ребра имеют различное очертание — округленное, треугольное; расстоя-
426
ния между ребрами и межреберными промежутками бывают различны-
ми. Возникновение ребер, возможно, связано с наличием щетинок вдоль
Рис. 241. Форма раковин (1—6, 10) и типы роста (7—9):
1 — двояковыпуклая, 2 — плоско-выпуклая, 3 — выпукло-плоская, 4 .— вогну-
то-выпуклая, 5 — обратно-изогнутая; 6 — схема замочного края с двумя арея-
ми на створках; 7а, б — голопериферический (Plesiomys)-, 8а, б — миксоперифе-
рический (Dinorthls); 9а, б — гемипериферический (Productus); 10 — бокаловид-
ная педальная створка Prorichthofenia;
ар — арея, бс — брахиальная створка, др — дельтирий, закрытый дельтидием,
ла — корневые выросты, к — нототирий, закрытый хилидием, пс — педальная
створка
Рис. 242. Типы скульптуры:
1 — Atrypa, справа раковина с сохранившимися концентрическими пластинами нара-
стания, слева — без них; 2 — Echinoconchus, педальная створка с концентрическими
полосами и многочисленными тонкими иглами; 3 — Leptaena, педальная створка с
морщинами на висцеральной части; 4 — схема расположения крупных игл на педаль-
ной створке (отр. Productida)
лобного края мантии. Кроме ребер бывают развиты крупные складки —
сравнительно широкие изгибы створок. Ребра значительно увеличивают
427
прочность раковины, не утяжеляя ее, препятствуют боковому смещению
створок. Концентрическая скульптура представлена линиями нараста-
ния и поперечными морщинами, возникающими вследствие прерыви-
стого и 'неравномерного роста раковины. Кроме этих видов скульптуры,
на раковинах артикулят можно встретить различные бугорки, иглы и
другие образования. Иглы — полые трубчатые выросты мантии, вокруг
которых отлагались концентрические слои кальцита. Иглы бывают ко-
роткие конусовидные или длинные цилиндрические и достигают не-
скольких сантиметров в длину. Иглы расположены по всей поверхности
раковины или приурочены только к определенным местам: вдоль карди-
нального края, в области ушек, только в брюшной створке
(рис. 242,2,-4).
Внутреннее строение. Сочленение двух створок у артикулят про-
исходит с помощью двух зубов в брюшной створке, расположенных по-
краям дельтирия. Зубы представляют собой булавовидные выступы,,
часто с мелкими зубчиками или мелкими насечками. Они образуются:
за счет деятельности внутреннего эпителия. Зубы входят в соответст-
вующие ямки спинной створки, расположенные по бокам, замочного-
отростка. Зубы обычно поддерживаются особыми зубными пластинками;
они тянутся вдоль боковых краев дельтирия и могут опираться на дно>
створки. Зубные пластинки могут срастаться имеете обычно на некото-
ром расстоянии от дна педальной створки и чаще всего поддержива-
ются срединной септой; иногда септа отсутствует и тогда зубные пла-
стинки находятся в висячем положении или опираются на дно створки..
Все эти структуры зубных пластинок называются спондилием.
Валики, ограничивающие зубные ямки спинной створки и сопро-
вождающие их структуры, образуют кардиналий, который отли-
чается большим разнообразием у разных родов. В центре кардиналия
расположен замочный отросток, который служит местом прикрепления
муокулов-отмыкателей (дидукторов). От замочного отростка дидукторы
протягиваются к -брюшной створке, раздваиваются и прикрепляются по
краям мускулов-замыкателей. Отпечатки мускулов-замыкателей (аддук-
торов) расположены почти посередине каждой створки, образуя на дне
педальной створки два отпечатка, а на дне брахиальной — четыре
(см. рис. 237).
Важную роль для систематики артикулят имеет строение, структур,
связанных с лофофором. Различают (несколько типов структур. У одних
(продуктиды) лофофор не имел известкового скелета и прикреплялся ко-
Дну брахиальной створки, оставляя брахиальные отпечатки. Простей-
шими структурами являются брахиофоры — известковые короткие-
выступы, расположенные в области кардиналия на внутреннем крае зуб-
ных ямок; основание лофофора опиралось на брахиофоры. Следующей
стадией усложнения структур явилось развитие отростков брахиофор,
получивших название крур. У современных форм круры- соприкасают-
ся по обе стороны рта с лофофором.' Круры характерны для отряда
ринхонеллид. У теребратулид с кардиналием связана известковая пет-
ля различной формы и размеров, которая служит для поддержания
лофофора (рис. 243, 3, 4). Более сложной структурой является спи-
ральный ручной аппарат, развитый у спириферид (рис. 243, 5—6):
Он представляет собой ленты, свернутые в две спирали, вершины кото-
рых могут.-быть направлены в разные стороны. По мере роста раковины
петля и спирали резорбируются и выделяются вновь внутренним эпите-
лием, но уже более крупных размеров. На внутренней поверхности
обеих створок бывают видны отпечатки двух или двух, пар основных '
мантийных каналов и их ответвлений. У большой группы палеозойских
428
брахиопод—лродуктид— эта система 'каналов до сих пор неизвестна.
Около мускульного поля и каналов васкулярной системы наблюдаются
так называемые генитальные отпечатки, представляющие следы при-
крепления гонад (см. рис. 239, 2).
В спинной, реже в брюшной, створке имеется срединная септа, рас-
положенная в плоскости симметрии раковины.
Рис. 243. Схема строения ручного аппарата и следы прикрепления лофофора:
1 — следы прикрепления лофофора у отр. Productida; 2 — известковые поддерж-
ки лофофора (брахиофоры) у отр. Orthida; 3 — простая и 4 — сложная петля
у отр. Terebratulida; 5 — спиральный ручной аппарат у отр. Atrypida; 6 — то же
у отр. Spiriferida
ар — арея, бо — брахиальные отпечатки (следы прикрепления лофофора), бр —
брахиофоры, зо — замочный отросток, ая — зубные ямки, к — круры, ом — от-
печатки мускулов, п — петля, с — срединная септа, сп — спиральные конусы,
юо — югальный отросток
Основы систематики и классификация. На основании строения опор-
ных элементов лофофора, характера отверстия для ножки и приурочен-
ных к нему образований, особенностей структуры раковины и других
.признаков класс артикулят разделен на 8 отрядов: Orthida, Strophome-
nida, Productida, Pentamerida, Rhynchonellida, Spiriferida, Atrypida,
Terebratulida.
Отряд Orthida. Ортиды (гр. orthos — прямой). Наиболее древние
артикуляты, появившиеся в раннем кембрии. Раковины обычно двояко-
выпуклые радиально-ребристые, реже плоско- или вогнуто-выпуклые.
У первых представителей стенка сплошная, у ордовикских — пористая
(энтелетации) и ложнопористая, (гонамбонитации). Обе створки несут
. ареи с дельтирием и нототирием. Отверстия для ножки открытые или
429
прикрыты соответственно псевдодельтидием и хилидием. В вершине
дельтидиальной пластинки может быть развит форамен для выхода
маленькой ножки. В педальной створке два простых зуба, они поддер-
живаются зубными пластинками, которые иногда срастаются, образуя
ложкообразный спондилий. Замочный отросток брахиальной створки
простой или лопастной, -служил для прикрепления дидукторов. Зубные
Ямки ограничены валиками, от которых отходят брахиофоры, иногда
несущие отростки для поддержки лофофора; последний, по-видимому,
Рис. 244. Отряд Orthida:
1 — Orthis (ранний ордовик): 1а, 1в — педальная створка снаружи и изнут-
ри, 16 — вид раковины сбоку; 2 — Clitambonites (ордовик): 2а — педальная
и 26 — брахиальная створки изнутри, 2в, 2г — предполагаемое прижизненное
положение на дне на более ранней (2в) и более поздней (2г) стадиях роста;
ар — арея, др — дельтирий, дт — дельтидиальная пластинка, зб — зубы,
зо — замочный отросток, зя — зубные ямки, « — ножка, сп — спондилий,
ср — срединная септа
был схизолофусного и спиролофусного типов. Хорошо выражено мус-
кульное поле на вентральной створке, к которому прикреплялись аддук-
торы и дидукторы. На обеих створках имеются отпечатки мантийных
синусов и половых желез. Первые ортиды появляются в раннем кембрии
(биллингселлиды), испытывают расцвет в ордовике (известно до 125
родов); в силуре число родов сокращается почти в три раза (около 45),
до перми доживают представители только одного надсемейства (энтеле-
тации), вымирающего в начале триаса. Всего в отряде насчитывается
около 200 родов (рис. 244).
Отряд Strophomenida. Строфомениды (гр. strophus — согнутый,
тепе, род. menos — месяц). Раковины двояковыпуклые или вогнуто-
выпуклые с длинным замочным краем. Обе створки имеют хорошо раз-
витые, но низкие ареи. Дельтирий прикрыт псевдодельтидием, а ното-
тирий—хилидием; последний часто крупнее первого и, вероятно, при-
крывал место прикрепления дидукторов к замочному отростку. Зубы
простые, часто с дополнительными зубчиками; поддерживаются иногда
зубными пластинками. Крупные валики ограничивают зубные ямки;
замочный отросток двухлопастной. У многих форм имеется апикальный
430
форамен (в вершине псевдодельтидия). Ножка функционировала толь-
ко на ранних стадиях онтогенеза, с возрастом она редуцировалась,
отверстие зарастало известковым слоем, и взрослые строфомениды
лежали свободно в осадке или цементировались макушкой к субстрату.
Стенка раковины ложнопористая, реже сплошная. На брахиальной
створке иногда развиты платформы для опоры лофофора. Строфоме-
ниды появляются в ордовике, достигают расцвета в ордовике и силуре,
в девоне число родов сокращается и до триаса доживает только один
Рис. 245. Отряд Strophomenida:
1 — Strophomena (ордовик): la — педальная створка, 16 — вид со сто-
роны замочного края; 2 — Orthotetes (карбон), педальная створка;
ар — арея, бс — брахиальная створка, др — дельтирий, пс — педаль-
ная створка
род. Всего насчитывается около 180 родов (рис. 245). Включают два
подотряда Strophomenina (ордовик — пермь, 150 родов) и Chonetina
(ордовик —пермь, 30 родов).
Отряд Productida. Продуктиды (лат, productus— удлиненный).
Раковины вогнуто- или плоско-выпуклые, у прикрепленных — кониче-
ской формы, причем педальная створка в форме бокала, брахиальная —
в виде крышечки. У взрослых бывает развит шлейф — длинное про-
должение переднего края обеих створок. Смычный край наиболее ши-
рокий, развиты ушки. У самых примитивных представителей на каждой
створке были развиты ареи, дельтирий и нототирий, прикрытые псевдо-
дельтидием и хилидием, в брюшной створке зубы с зубными пластин-
ками. У большинства продуктид нет ни ареи, ни зубов; зубы замещают-
ся мелкими зубчиками, развитыми вдоль смычного края. Раковины обычно
ребристые и морщинистые, чаще всего покрыты иглами, среди которых
одни служили для прикрепления, другие — для сохранения равновесия
в осадке, третьи —для защиты от поселения личинок. Иглы обычно
полые и могут достигать большой длины (до 25 см). На внутренней
поверхности створок бывают также развиты мелкие иглы. На внутрен-
ней поверхности педальной створки почти всю среднюю часть раковины
занимают мускульные отпечатки; аддукторы образуют два или четыре
отпечатка, они расположены внутри или спереди от дидукторов; послед-
ние образуют два крупных отпечатка. На брахиальной створке хорошо'
выражен замочный, или кардинальный, отросток различных размеров
и формы; к нему прикреплялись мускулы-дидукторы. В средней части
проходит срединная септа, по обе стороны которой видны две пары
431
отпечатков мускулов-аддукторов. Вокруг них расположены два спираль-
ных изогнутых брахиальных валика, служивших для поддержки лофо-
фора. Мантийные каналы сохраняются плохо, по-видимому, потому что
на внутренней поверхности бывают развиты мелкие иглы. У прирастаю-
щих форм на дне конической' раковины развиты горизонтальные пла-
стинки— своеобразные днища, возникающие по мере роста раковины.
Рис. 246. Отряд Productida:
1 — Productus (карбон): la — педальная створка, 16 — схема соотношения
створок, 1в — брахиальная створка изнутри; 2 — Richthofenia (пермь), продоль-
ное сечение; За, б — Strophalosia (пермь); 4 — Oldhamina (пермь), брахиальная
створка снаружи; 5 — изменение образа жизни с возрастом: 5а — прикрепление
короткой ножкой, 56 — прирастание макушкой и опорными иглами, 5в — сво-
бодное лежание в осадке;
ар — арея, бр — брахиальные отпечатки, бс — брахиальная створка, др —
дельтирий, ао — замочный отросток, иг — иглы, ом — отпечатки мускулов, пс —
педальная створка, с — срединная септа, ш — шлейф
Продуктиды были очень широко распространены. Первые предста-
вители отряда известны из отложений раннего девона, они, вероятно,
произошли от хонетид; в позднем девоне и особенно в раннем карбоне
продуктиды становятся одной из самых распространенных групп арти-
кулят, населяя моря всех континентов, достигая наиболее крупных раз-
432
меров (Gigantoproductus до 35 см). В позднем карбоне число родов
уменьшается, но в перми вновь наблюдается широкое дивергентное
развитие продуктид, появляются рихтгофениды, ведущие прикрепленный
образ жизни. Большинство продуктид на взрослой стадии не имело
ножки и свободно лежало на дне, сохраняя равновесие при помощи игл;
одни прирастали макушечной частью брюшной створки, на которой со-
Рис. 247. Отряд Pentamerida: • ’
la, б — Pentamerus (силур); 2а, б — Conchidium (поздний ордовик — ран-
ний девон); За—г — схема развития спондилия и брахиофория;
бп — брахиальные пластинки,, бр .— брахиофорий, бс —' брахиальная
створка, др — дельтирий, зп — зубные пластинки, пс — педальная створ-
ка, сп — спондилий, ср — срединная брюшная септа
храняется след прикрепления. У лежащих форм развивался шлейф.
Последние продуктиды дожили до конца перми. В отряде известно сей-
час около 180 родов (рис. 246).
Особое место среди продуктид занимает подотряд Oldhaminidina
(поздний карбон — поздний триас), у представителей которого ракови-
на неправильной формы, состоящая из конической’ ложнопористой
брюшной створки и спинной створки, имеющей вид крышечки с паль-
цеобразными краями.
Отряд Pentamerida. Пентамериды (гр. penta — пять, meros —
часть). Раковины двояковыпуклые, до шаровидных, с хорошо развитыми
седлом и синусом, гладкие или ребристые. Дельтирий прикрыт дельти-
диальной пластинкой. Ареи развиты слабо. Зу^ы . поддерживаются
крупными зубными пластинками, которые, срастаясь, образуют ложко-'
образный спондилий, опирающийся на срединную септу и занимающий'
более половины длины раковины. Спондилий служил для прикрепления
мускулов. В спинной, или брахиальной, створке для поддержки лофо-
фора развит брахиальный аппарат, состоящий из брахиального отроет-:
28 в. В. Друщиц
433
ка и двух довольно длинных пластин, наружной и внутренней; наруж-
ные пластинки иногда, срастаясь, образуют круралий (рис. 247).
Пентамериды появляются в среднем кембрии, довольно разнообразны
в ордовике, наибольшего развития достигают в силуре и раннем девоне.
В начале позднего девона они вымирают.
Отряд Rhynchonellida. Ринхонеллиды (гр. rhynchos — клюв, rhyn-
chonella — клювик). Раковины двояковыпуклые, гладкие, но чаще реб-
ристые, с хорошо развитым синусом и седлам. Замочный край короткий,.
Рис. 248. Отряд Rhynchonellida:
la—в — Camarotoechia (средний девон); 2а—в — Hypothyridina (девон);
3—4 — Rhynchonella (поздняя юра — ранний мел): За—в — общий вид
раковины, 4 — изменение образа жизни с возрастом: 4а, 46 — прикрепле-
ние ножкой, 4в — прикрепление ножкой с опорой на края створок;
бс — брахиальная створка, пс — педальная створка, сд — серла, сн —
синус
арея узкая; дельтирий прикрыт двумя дельтидиальными пластинками,
ограничивающими небольшой форамен в центре; нототирий отсутствует.
Спондилий обычно отсутствует. Хорошо развиты зубы; у мезозойских
форм вдоль смычного края имеются мелкие зубчики. Зубные пластинки
развиты в разной степени. В спинной створке между зубными ямками
расположена замочная пластинка для прикрепления ножных мускулов.
От нее отходят парные круры; строению крур придается систематическое-
значение и среди мезокайнозойоких ринхонеллид выделяется их восемь-
различных типов, имеющих особые названия (радулиферовый, септи-
феровый и др.). В каждой лопасти мантии имеется по одному главному
мантийному каналу, от которого отходят многочисленные ответвления.
Срединная септа брахиальной створки поддерживает септалий, состоя-
щий из двух сросшихся пластинок. Стенка раковины сплошная, у не-
434
которых внутренний слой пористый. Для многих мезокайнозойских
ринхонеллид характерна своеобразная чешуйчатая структура раковины,
возникающая при соединении отдельных фибр кальцита в чешуйки.
У современных ринхонеллид имеются две пары метанефридиев, спиро-
лофусный лофофор, который поддерживается двумя крурами; вершины
конусов лофофора направлены дорзовентрально.
Ринхонеллиды появляются в среднем ордовике, довольно разнооб-
разны в позднем палеозое. Общее число ринхонеллид — 270 родов, в
Рис. 249. Отряды Spiriferida (1—4) и Atrypida (5):
la—г — Choristites (карбон — ранняя пермь), серия поперечных разрезов педаль-
ной створки; 2—3 — Cyrtospirifer (поздний девон — ранний карбон): 2а—в —
серия поперечных разрезов, 3 — общий вид; 4—Aihyris (девон — триас), об-
щий вид; 5 — Atrypa (силур — девон): 5а — вид с- переднего края, 56 — схема
строения ручного аппарата, 5в — схема расположения генитальных и мантийных
отпечатков на педальной створке;
ар — арея, бс — брахиальная створка, го — генитальные отпечатки, дп — дель-
тидиальная пластина, др — дельтирий, зб — зубы, зо — замочный отросток,
зп — зубные пластины, зя — зубные ямки, мо — мантийные отпечатки, мп —
мускульное поле, пс — педальная створка, у — ушки, ф — форамен,
палеозое известно около половины, в мезозое около 100 родов, а в кай-
нозое только 20 родов. В современных морях обитает немногим более
10 родов. Ринхонеллиды относятся к довольно консервативному стволу
артикулят и за время эволюции претерпели сравнительно небольшие
изменения. Современные ринхонеллиды распространены в Атлантиче-
ском и Тихом океанах, ведут прикрепленный образ жизни, прикрепляясь
с помощью ножки; на взрослой стадии дополнительно опираются на
края педальной створки (рис. 248).
Отряд Spiriferida. Спирифериды (лат. spira — спираль, fero — не-
сти). Раковины двуяковыпуклые, плоско-выпуклые, округленные или
удлиненные вдоль смычного края. Ареи обычно хорошо развиты, осо-
бенно в педальной створке. Дельтирий прикрыт двумя дельтидиаль-
ными пластинками, у атирид развит форамен. Зубы поддерживаются
28’
435
двумя раздельными или сросшимися пластинками. Замочный1 отросток
построен из тонких пластинок. Ручной аппарат состоит из двух извест-
ковых спиралей (отсюда название отряда) с вершинами, обращенными
в стороны. Обычно развиты синус и седло. Стенка раковины сплошная
или пористая. Отряд спириферид разделен на два подотряда: Spirife-
ridina и Athyrididina (рис. 249). Первый, наиболее многочисленный
подотряд Spiriferidina (средний ордовик — ранняя юра) объединяет
около 190 родов, второй — Athyrididina (поздний ордовик — юра)! объ-
единяет около 50 родов.
Рис. 250. Отряд Terebratulida:
1 •— Terebrdtula (неоген)': la — вид сбоку, 16 — со стороны брахиальной створ-
ки, 1в—: прижизненное- ’положение; 2 — Pygope (юра — ранний мел); 3 —
Stringocephalus (средний девон): За — продольный разрез, Зб — брахиальная
створка изнутри, Зв — вид сбоку;
бс — брахиальная створка, дт — дельтидиальные пластинки, зо — замочный от-
росток, к круры, м— мускульное поле, мк — мантийные каналы, п — петля,
. . пс — педальная, створка, с — срединная септа, ф — форамен
Отряд Atrypida. Атрипиды (гр. а — отрицание, trype— отверстие;
при установлении рода автор не заметил форамена и присвоил имя
Atrypa). Раковины округленные, двояковыпуклые; арея слабо выражена
или отсутствует; стенка сплошная. Скульптура выражена разного типа
ребрами. Форамен круглый, расположен под макушкой, ограничен дву-
мя дельтидиальными пластинками. Ручной аппарат состоит из двух
спиралей, вершины конусов которых направлены в сторону брахиальной
створки (см. рис. 243, 5), поэтому она чаще более выпуклая, чем пе-
436
Рис. 251. Подот-
ряд (?) Thecidei-
dina:
Thecidea (мел),
вид со стороны
брахиальной
створки
дальная; последняя с синусом. Средний ордовик — поздний девон. Около
60 родов, (рис. 249, 5).
Отряд Terebratulida. Теребратулиды (лат. terebratus — просвер-
ленный). Раковины двояковыпуклые с синусом и седлом, обычно, пори-
стые (отсюда название отряда). Замочный край короткий, изогнутый;
макушка брюшной створки нависает над спинной. Зубы хорошо развиты,
зубные пластины могут отсутствовать. Дельтирий прикрыт двумя дель-
тидиальными пластинками, ограничивающими снизу круглый форамен.
Ручной аппарат развит в виде петли, которая прикрепляется к прима-
кушечной части брахиальной створки и служит для поддержания лофо-
фора; петли могут быть простыми и сложными.
Теребратулиды прикрепляются в течение всей
своей жизни к субстрату с помощью ножки, кото-
рая может быть короткой или длинной, в послед-
нем случае она с корневидными отростками. Пер-
вые теребратулиды появляются в раннем девоне и
продолжают свое существование до наших дней,
достигая максимума развития в мезозое. Отряд
объединяет около 290 родов (рис. 250).
Отряд неизвестен (Ordinis incertae sedis).
Особое место среди артикулят занимает подотряд
Thecideidina — тецидные артикуляты, распростра-
ненные от триаса до наших дней, объединяющие
25 родов. Очертание раковины неправильное,
ножка отсутствует; створки соединяются хорошо
развитыми зубами, имеется бпондилий на.
срединной септе или прикрепленный ко дну створ-
ки. Стенка раковины ложнопористая Или по-
ристая, толстая. Педальная створка коническая, с ареей и псевдодель-
тидием. Брахиальная створда в форме крышечки, без ареи, с карди-
нальным отростком и развитыми брахиальными валиками. Лофофор
схизолофусный или птихолофусный. Тецидные артикуляты ведут, при-
крепленный образ жизни (рис. 251). Вопрос о систематическом поло-
жении тецидных артикулят остается не достаточно ясным. Возможно,
их следует объединять с отрядом строфоменид.
Экология артикулят. Личинки артикулят ведут подвижный планк-
тонный образ жизни. Морские течения могут разносить их на значи-
тельные пространства, что способствует их расселению. После непро-
должительного плавания личинки оседают на дно и прикрепляются к
субстрату ножкой или цементацией брюшной створки. Все артикуляты
являются обитателями морского дна и входят в состав эпйбентоса. Они
образуют иногда значительные поселения на разных участках дна. Све-
дения об экологии древних брахиопод довольно скудны; об образе
жизни многих родов сведения полностью отсутствуют. В настоящее
время, можно выделить три основных экологических типа:
1. Якорный. Прикрепление, к субстрату короткой или длинной
разветвленной ножкой; формы в течение жизни могут несколько менять
место своего обитания. Некоторые формы помимо ножки прикрепля-
лись еще с помощью игл, развитых вдоль замочного края. В течение
жизни некоторые формы меняли свой образ жизни. Большинство ринхо-
неллид, вероятно, лежало на' педальной створке, опираясь на ее края
и поднимая седло брахиальной створки над дном. Другие артикуляты
могли переходить к свободнолежащему образу жизни.
2. Свободнолежащ-ий. Брахиоподы свободно лежали на дне,
частично погружаясь в осадок. Ножка, у них редуцировалась за пена-
437
добностью (большинство продуктид). Развитые на педальной створке
многочисленные иглы (строфомениды, продуктиды) препятствовали
слишком глубокому погружению в илистый осадок. Для поднятия над
дном! лофофора и предохранения переднего края от загрязнения у неко-
торых продуктид развивался шлейф.
3. Прирастающий. Раковина прирастает педальной створкой к
твердым подводным предметам. Прирастание происходит путем цемен-
Рис. 252. Геохронологическое распространение брахиопод
тации; при этом форма педальной створки становится конусовидной
(рихтгофении) или приобретает неправильные очертания (ольдгамини-
тины и тецидные артикуляты); брахиальная створка превращается в
крышечку, часто с лопастными краями. Для поддержания конической
раковины в вертикальном положении у рихтгофений развивались допол-
нительные баковые выросты, у других артикулят — опорные иглы.
История развития артикулят (рис. 252). К наиболее древним арти-
кулятам относятся ортиды, первые представители которых известны в
раннем кембрии. В конце среднего кембрия появляются пентамериды.
Ордовик характеризуется расцветом ортид, появлением в начале перио-
да строфоменид, возникновением в среднем ордовике трех новых отря-
дов: спириферид, атрипид и ринхонеллид. Строфомениды были наиболее
широко распространены в ордовике и силуре. В девоне господство
принадлежит спириферидам и атрипидам; существенно уменьшается
число ортид, строфоменид, к концу периода вымирают атрипиды и пен-
тамериды,; но в начале девона появляются два новых отряда: продукти-
438
ды и теребратулиды; первый становится господствующим в карбоне и
частично перми. К концу палеозоя вымирают ортиды, строфомениды и
почти все продуктиды, резко сокращается число спириферид, теребра-
тулид и ринхонеллид. В перми особое развитие приобретают прирастаю-
щие продуктиды (надсемейство Richthofeniacea), имеющие кораллопо-
добную раковину. Не менее своеобразны представители подотряда
•Oldhaminidina, относимые к продуктидам, имевшие неправильной фор-
мы раковину — они доживают до конца триаса. Начиная с триаса гос-
подствуют представители двух отрядов: теребратулид и ринхонеллид.
В начале юрского периода исчезают последние спирифериды. В триасе
появляются так называемые тецидные брахиоподы, ведущие прирастаю-
щий образ жизни. До сих пор остается неясным, к какому отряду они
относятся. К началу кайнозоя значительно сокращается число родов
теребратулид и ринхонеллид, доживающих до наших дней. В современ-
ных 'морях известно небольшое число родов и видов этих артикулят и
два рода тецидных брахиопод.
ПОДРАЗДЕЛ DEUTEROSTOMIA.
ВТОРИЧНОРОТЫЕ
Вторичноротые (гр. deuteros — вторичный, stoma — рот) характе-
ризуются следующими чертами: первичный рот, или бластопор, зароды-
ша замыкается и рот возникает вторично в другой его части, а
в области бластопора образуется анальное отверстие. Мезодерма закла-
дывается энтероцельным путем, т. е. первичный кишечник образует
симметрично расположенные мешковидные выпячивания, которые от-
шнуровываются от кишечника, разрастаются и заполняют первичную
полость. Скелет, как правило, внутренний мезенхимного происхожде-
ния. Все вторичноротые имеют целом.
К вторичноротым относятся следующие типы: Echinodermata,
Hemichordata, Chaetognatha, Chordata и условно Pogonophora и Petalo-
narhae.
ТИП ECHINODERMATA.
ИГЛОКОЖИЕ
Общая характеристика. Иглокожие (гр. echinos — еж, derma, род.
dermatos — кожа) — очень своеобразный, хорошо обособленный тип
вторичноротых животных, для которых характерны: 1) радиальная, пя-
тилучевая симметрия; 2) внутренний скелет, состоящий из табличек
кальцита или отдельных разрозненных известковых телец; 3) амбулак-
ральная система целомического происхождения, состоящая из кольце-
вого и радиальных каналов и полостей; 4) многочисленные щупальца,
или ножки, отходящие от радиальных каналов. Современные иглоко-
жие: морские ежи, морские звезды, офиуры, голотурии и морские ли-
лии, объединяемые в пять классов, — морские, стеногалинные, одиноч-
ные организмы; живут в нормальносоленых морях на всех широтах и
на всех глубинах: от литорали до дна глубоководных впадин. Они ведут
прикрепленный или подвижный бентосный, реже пелагический или
псевдопланктонный образ жизни. Геологическая история иглокожих на-
чинается с венда, в палеозое известно 16 классов вымерших иглокожих.
Строение тела. Форма и размеры тела иглокожих очень разнооб-
разны, они могут иметь форму звезды, мешка, шара, бутона, чашечки
440
со стеблем и руками. Самые мелкие и самые крупные представители
наблюдаются среди морских лилий; длина достигает более 10 м; чашеч-
ка, вмещающая все внутренние органы, может не превышать 1—3 мм.
У многих форм, ведущих прикрепленный образ жизни, рот расположен
на верхней стороне, называемой оральной, противоположная ей сто-
Рис. 253. Схема строения тела иглокожих:
1 — морской звезды; 2 — морского ежа; 3 — кожи голотурии;
амн — амбулакральные ножки, ан — анальное отверстие, еэ — внутренний эпи-‘
телий, ж — желудок, состоящий из двух отделов, и — иглы, из — известковые
тельца, кк — каменистый канал, клк — кольцевой канал, км — кольцевые мыш-
цы, крс — кровеносная система, м — мадрепорит, нс — нервная система, нэ —
наружный эпителий, печ — печеночный придаток (слепой радиальный отросток
желудка), пж — половая железа, пт — пищеварительный тракт, р — рот, рк —
радиальный канал, с — соединительная ткань, ст —-стенка тела, цел — целом,
ча — челюстной аппарат («аристотелев фонарь»), ш — шипы
рона носит название а б о р а л ь н ой. У подвижных иглокожих (голоту-
рии) рот может быть расположен на переднем конце, а анус—на-заднем,
либо рот лежит на нижней стороне, обращенной к субстрату (морские
звезды, морские ежи), а анальное отверстие наверху, или сзади. В ти-
пичном случае тело иглокожих разделено на пять зон, или лучей. Вдоль
лучей проходят радиальные каналы, от которых по бокам ответвля-
ются щупальца, или ножки. Эти ножки располагаются вдоль радиаль-
ных зон, названных амбулакрами (лат. ambulacrum — аллея, огра-
ниченная рядами деревьев), или радиусами, в два ряда; промежу-
точные зоны соответственно названы интерамбулакрам и, или
интеррадиусами.
Для многих взрослых иглокожих характерна пятилучевая симмет-
рия, хотя личинки первично, и некоторые взрослые вторично имеют
441
двустороннюю симметрию. Пятилучевая симметрия часто нарушается
расположением отдельных органов или скелетных табличек (рис. 253).
Стенка тела состоит из трех слоев: наружного эпителия, или эпи-
дермиса, среднего слоя и внутреннего эпителия. Наружный эпителий
покрывает все тело, несет в отдельных местах реснички. Средний слой,
или собственно кожа, состоит из мышечной ткани, мезенхимы и клеток,
вырабатывающих известковый скелет. Внутренний эпителий выстилает
целомическую полость.
Нервная система радиально-симметричная; имеются три взаимосвя-
занные системы, расположенные на разных уровнях внутри тела: экто-
невральная, гипоневральная и энтоневральная. Каждая из них состоит
из околоротового кольца и почти радиальных нервных стволов, имею-
щих характер лент или тяжей. В эпидермисе расположена э кто нев-
ральная (оральная), или чувствительная, система эктодермического
происхождения. Она связана с чувствительными клетками, амбулак-
ральными ножками и иглами. У современных представителей от ротово-
го кольца отходят нервы к пищеварительной системе. Гипоневраль-
ная система лежит глубже эктоневральной и выполняет основную
моторную, или двигательную, функцию. Энтоневральная или
аборальная, система выполняет основную двигательную функцию, она
хорошо развита у морских лилий.
У иглокожих имеются разнообразные органы чувств: органы-рецеп-
торы, чувствительные к свету, запаху, качеству и составу воды, органы
ориентации.
П ищеварительная система. У большинства свободноживущих
иглокожих (морские звезды, офиуры, правильные морские ежи) рот
расположен на нижней стороне, анальное отверстие находится на верх-
ней стороне. У голотурий и некоторых неправильных морских ежей
(спатангоидов) рот смещен к переднему краю, а анальное отверстие —
к заднему. У прикрепленных форм рот и анальное отверстие располо-
жены на верхней стороне, причем рот в центре, а анальное отверстие
всегда в заднем интеррадиусе. Пищеварительный тракт, развивающий-
ся из энтодермы, у одних иглокожих (морские лилии, морские ежи)
имеет форму узкой трубки, свернутой петлеобразно по часовой стрелке,
у других (морские звезды, офиуры) является мешковидным. Пищева-
рительный тракт обычно разделяется на пищевод, желудок, кишечник
и заднюю кишку.
Амбулакральная, или водно-сосудистая, система представ-
ляет собой обособленную часть вторичной полости тела — целома и со-
держит жидкость, близкую по составу к морской воде. Наличие этой
системы, свойственной только иглокожим, представляет для этого типа
одну из наиболее важных и характерных черт.
Амбулакральная система состоит из следующих структур: 1) коль-
цевого, или орального, канала; 2) пяти радиальных каналов; 3) камени-
стого канала с гидропорой или мадрепоритом/ Кольцевой, или
оральный, канал окружает глотку; от него по радиусам отходят пять
радиальных каналов. Между ними расположены (у морских звезд)
пять полиевых пузырей, содержащих запас жидкости. У морских лилий
и морских ежей этих пузырей нет, а у голотурий их число может дости-
гать 50. От радиальных каналов отходят боковые ответвления с амбу-
лакральными ножками, которые у морских лилий образуют пучки,
состоящие из трех остроконечных ножек, или щупалец, у остальных
отходят по одной ножке, которые заканчиваются подошвой. Амбулак-
ральные ножки выполняют различные функции: газообмена, доставки
пищи, передвижения или органов чувств. У морских ежей, голотурий и
442
морских звезд каждая ножка снабжена ампулами. Каждый радиальный
канал заканчивается чувствительным к свету концевым щупальцем. От
кольцевого канала отходит так называемый каменистый канал, в
стенках которого имеются известковые спикулы. Канал открывается
наружу одним отверстием — г и др о п о р о й- или связан с перфориро-
ванной табличкой, называемой мадрепоровой, или мадрепо-
ритом. Кроме амбулакральной системы в теле иглокожих имеется
ряд полостей целомического происхождения, выстланных внутренним
эпителием.
С амбулакральной системой связана характерная для иглокожих
радиальная симметрия. Она обычно является пятилучевой, но нару-
шается присутствием одной плоскости симметрии, в которой расположен
осевой комплекс, установленный у современных морских звезд, офиур и
морских ежей. Осевой комплекс тесно морфологически и функционально
связан с кровеносной системой; он представляет собой обособленную
структуру, в состав которой входят осевой синус кровеносной системы,
половой тяж, каменистый канал и поровый канал с гидропорой или
мадрепоритом, окруженные общей оболочкор. Гомологичная структура
наблюдается у морских лилий.
Все современные иглокожие в личиночной стадии имеют двусторон-
нюю симметрию, что указывает на их происхождение от двусторонне-
симметричных животных. Пятилучевая симметрия взрослых форм рас-
пространяется почти на все системы: нервную, кровеносную, целомиче-
скую, амбулакральную,_ а также на расположение скелетных элементов.
Она возникла параллельно в разных группах иглокожих.
Кровеносная система состоит из системы лакун, связанных
сетью каналов и синусов. Выделяется околоротовое гемальное кольцо,
от которого отходят пять радиальных лакун; кроме того, имеется або-
ральное гемальное кольцо и связанные с ним пять генитальных гемаль-
ных лакун и гемальная лакуна, связанная с пищеварительным трактом.
Органы дыхания развиты слабо. Функцию газообмена выпол-
няет обычно амбулакральная система и покровный эпителий. У совре-
менных морских звезд и морских ежей существуют дополнительные
вторичные кожные жабры.
Строение скелета. Известковый скелет образуется в среднем слое,
или собственно коже. Он состоит из известковых табличек, или пласти-
нок, телец, спикул, микроскопических гранул и различного рода игл.
Скелет строится на органической основе (строме), которая иногда со-
ставляет до 60% всего объема; она обычно аморфная, реже волокнис-
тая. В строме возникают мелкие, едва заметные зернышки (гранулы)
кальцита, которые затем вытягиваются в трех направлениях, образуя
трехлучевую известковую спикулу. Каждый луч такой спикулы много-
кратно дихотомически ветвится под углом 120°; в дальнейшем концы
лучей сливаются и образуются решетчатые таблички с шестиугольными
ячейками (рис. 254). Подобным образом создается решетчатый скелет,
или стереома.
У голотурий весь скелет ограничивается разбросанными в коже
микроскопическими известковыми тельцами. У остальных иглокожих
эти тельца сливаются своими краями и образуют скелетные элементы.
Они имеют грубоячеистое строение; лишь в иглах морских ежей и мор-
ских звезд, в стеблях и руках морских лилий происходит вторичная
минерализация и губчатое строение может почти исчезать.
Образование игл происходит несколько иначе, чем табличек и спи-
кул. В типичном случае образование иглы морского ежа или морской
звезды начинается с возникновения известковой гранулы (рис. 254),
443
развивающейся в шестилучевую звездочку, лучи которой на концах
раздваиваются, разрастаются и, соединяясь между собой, образуют
кольцо — основу иглы. От центра звездочки вверх растет основной
отросток, который дает три ветви, разрастающиеся вверх и наружу и.
образующие на определенном уровне новое шестиугольное кольцо, па-
раллельное основному кольцу. От него вверх растут шесть отростков,,
которые, соединяясь между собой, образуют шестиугольный каркас
иглы, напоминающий ствол шахты. В дальнейшем этот ствол запол-
Рис. 254. Схема образования внутреннего скелета иглокожих:
1а—г — стадии развития пластинки молодой офиуры; 2а—в — последова-
тельные стадии развития иглы морского ежа; 3 — схема развития решетчатой
пластинки
няется кальцитом и игла может стать сплошной. Челюсти морских
ежей, в/ отличие от решетчатого скелета, характеризуются особыми
структурами — конус в конус (con-in-con),
По своим оптическим свойствам скелетные элементы иглокожих,
состоящие из кальцита, очень сходны между собой. К кальциту приме-
шивается в небольших количествах карбонат магния (от 3 до 15%),
количество которого возрастает с увеличением температуры воды. В ма-
лых количествах присутствуют фосфат и сульфат кальция; обнаружены
следы алюминия, железа, стронция, бария, меди, цинка, никеля и других
элементов. При фоссилизации органическое вещество замещается вто-
ричным кальцитом и скелетные элементы иглокожих легко распознают-
ся по. спайности, сохранению решетчатой структуры и первичного поло-
жения оптических осей кристаллов кальцита. В последние годы установ-
лено, что кристаллографическая ориентировка кристаллов у разных
групп иглокожих варьирует, но ориентировка оси С постоянна для раз-
ных таксонов. Иногда скелет иглокожих сравнивают с таковым позво-
ночных, хотя это сходство только поверхностное, у тех и других скелет
внутренний. В отличие от позвоночных скелет у иглокожих: 1) всегда
известковый й никогда не состоит из фосфата кальция; 2) спикулы об-
разуются внутри клеток; 3) отличается характерными кристаллографи-
ческими особенностями.
Размножение и развитие. Иглокожие размножаются половым и
очень редко бесполым путем; в последнем случае тело делится надвое
(некоторые морские звезды, офиуры). При половом размножении из
яйца ' развивается двусторонне-симметричная личинка — д и плевру-
444
ла, ведущая планктонный образ жизни (рис. 255). У некоторых игло-
кожих наблюдается прямое развитие, без личиночной стадии. Диплев-
рула имеет венчик ресничек вокруг рта, прямой пищеварительный тракт,
анальное отверстие, возникшее на месте бластопора (вторичноротость).
Дальнейшее превращение диплеврулы протекает в разных классах по-
разному: у морских ежей она превращается в личинку — ехиноплутеус,
у офиур — офиоплутеус, у морских звезд — бипиннарию, у голотурии—
аурикулярию, у'морских лилий — вителлярию. Личинки морских ежей и
Рис. 255. Основные типы личинок иглокожих (1—4) и личинка гемихордовых' (5):
1 — диплеврула, 2 — аурикулярия голотурии, 3 — бипиннария морских звезд,
4 — вителлярия морских лилий, 5 — торнария
офиур, с одной стороны, голотурий и морских звезд — с другой, имеют
много общих черт, что может быть обусловлено конвергенцией. Даль-
нейшее развитие иглокожих сопровождается сложным метаморфозом
личинок. Личинка иглокожих похожа на личинку гемихордовых. Это
сходство позволяет предполагать, что иглокожие, гемихордовые и хордо-
вые произошли от общего двусторонне-симметричного предка, имевшего
три пары целомических мешков, причем если иглокожие перешли к ма-
лоподвижному или прикрепленному образу жизни, то хордовые пере-
шли к активному движению и в связи с этим обе группы эволюциони-
ровали по-разному. У иглокожих в связи с малоподвижным образом
жизни возникла пятилучевая радиальная симметрия, а хордовые в связи
с активным образом жизни эволюционировали с развитием двусторонне-
симметричного тела.
Радиальная симметрия иглокожих нарушается расположением гид-
ропоры или мадрепорита и некоторых внутренних органов, в частности
каменистого канала. В процессе эволюции всего типа у некоторых групп
(голотурии, морские ежи) возникла вторично-двусторонняя симметрия.
Основы систематики и классификация. На основании особенностей
эмбрионального развития (у современных форм), типов роста тела,
характера и плана симметрии, особенности строения скелета иглокожие
разделены на четыре подтипа: Carpozoa, Crinozoa,. Asterozoa и Echino-
zoa, в отличие от прежнего деления на Pelmatozoa и Eleutherozoa, осно-
ванного на экологических особенностях типа. Иногда выделяют еще пя-
тый подтип Haplozoa, однако он многими исследователями отвергается:
ПОДТИП CARPOZOA. КАРПОЗОИ
К подтипу Carpozoa (гр. karpos — плод), или Homalozoa (гр. ho-
malos — плоский, zoon — животное), относятся раннепалеозойские
445
иглокожие, имеющие мешковидной или уплощенной формы асимметрич-
ное тело без каких-либо следов радиальной симметрии. Скелет состоит
из чашечки, брахиолы и у некоторых — из стебля.
В состав подтипа включены три класса: Carpoidea, Homostelea и
Homoiostelea.
КЛАСС CARPOIDEA. КАРПОИДЕИ
Общая характеристика. К классу карпоидей относятся карпозои с
плоским асимметричным телом — текой, построенной из многочислен-
ных разнородных табличек, и отходящего от нее длинного отростка —
аулакофора, или брахиолы. Рот расположен около аулакофора, анус —
на противоположном конце (рис. 256). Средний кембрий — средний
девон.
Рис. 256. Класс Carpoidea:
1 — Ceratocystis (средний кембрий): 1а — вид сверху, 16 — вид снизу, 1в —
увеличенная часть аулакофора у Cothurnocystis-, 2 — Mitrocystites (средний —
поздний ордовик): 2а — вид снизу, 26 — вид сверху, 2в — вид сбоку;
ау — .аулакофор, щп — щелевидные поры
Строение скелета. Скелет состоит из теки и единственного отростка*
отходящего от нее, — аулакофора. Тека уплощенная, овального
или округленно-прямоугольного очертания, обычно асимметричная. Она
построена только из крупных краевых табличек или из крупных
(10—12) и многочисленных мелких, образующих верхнюю и нижнюю
поверхности. Нижняя поверхность обычно плоская или вогнутая, верх-
няя — выпуклая.
446
От переднего конца теки отходит отросток — аулакофор (гр.
aulakos — борозда, phoros — несущий), или б р ахи о л а; различают
три части •— проксимальную, ближайшую к теке, состоящую из
кольцевых табличек, полых внутри, среднюю, построенную из кони-
ческой массивной известковой таблички — стилокона, и длинную
узкую дистальную, состоящую из отдельных табличек с желобком,,
прикрытых парой кроющих табличек (рис. 256, 1). По-видимому, аула-
кофор обладал подвижностью; внутри него находились нервный ствол,
гемальный синус и целомическая полость. Вдоль срединного желобка
проходил амбулакральный канал с щупальцами.
Аулакофор, таким образом, не выполнял функцию стебля, как пред-
полагалось ранее, а служил для сбора пищи. Рот, по-видимому, был
расположен внутри теки. Пища по желобку аулакофора поступала в.
ротовое отверстие, скрытое внутри теки, около проксимальной части
аулакофора. Анальное отверстие расположено на противоположном
конце от места прикрепления аулакофора и прикрыто набором табличек
(таблички перипрокта), окружающих его. На верхней поверхности око-
ло аулакофора, справа в теке, имеется несколько отверстий разного'
типа и строения, по-видимому, выполнявших разные функции. У некото-
рых родов на верхней поверхности в пластинках теки имеются тонкие
щелевидные поры, переходящие в полость теки.
Внутренняя полость теки срединной септой или своеобразным вали-
ком разделена на две части. Септа косо направлена от переднего края
к заднему. Во внутренней полости теки, по-видимому, помещался пище-
варительный тракт, который мезентериальной мембраной прикреплял-
ся к септе..
Экология и тафономия. Карпоидеи, по-видимому, лежали свободно
на дне моря; об этом свидетельствует отсутствие стебля или каких-либо
отростков, служивших для прикрепления тела к субстрату. Аулакофор
выполнял функцию сбора пищи, но также, возможно, ориентировки тела
в пространстве и некоторого передвижения. Передвижение, возможно,
осуществлялось также путем выбрасывания воды через анальное отвер-
стие, как это делают некоторые современные голотурии.
Скелетные остатки карпоидей встречаются преимущественно в гли-
нистых осадках, часто в конкрециях, реже в известковых осадках.
История развития карпоидей. Первые карпоидеи известны из сред-
него кембрия Богемии. Наиболее многочисленные находки сделаны из
отложений нижнего ордовика юга Франции и среднего ордовика Боге-
мии; последние находки обнаружены в среднем девоне Богемии.
Класс Carpoidea разделен на два отряда — Cornuta и Mitrata
(рис. 256, 1—2).
В последние годы было предложено выделить род Ceratocystis в
новый класс Calcichordata и рассматривать его в качестве промежуточ-
ного класса между типами иглокожих и хордовых. Для класса Carpo-
idea предложено также название Stylophora.
КЛАСС HOMOSTELEA. ГОМОСТЕЛЕИ
Класс гомостелей (гр. homos — равный, подобный, stele — стебель)
объединяет карпозой, у которых тека овальной и округлой формы, упло-
щенная, с вогнутой нижней и выпуклой верхней сторонами; она построе-
на из крупных краевых табличек и многочисленных мелких, покрываю-
щих верхнюю и нижнюю стороны (рис. 257). На переднем конце теки
расположены два отверстия: одно большое, другое малое. Малое отвер-
стие расположено между двумя краевыми табличками; от него направо
447
и налево отходят два желобка, покрытых мелкими табличками, у одних
коротких, у других длинных. Большое отверстие ведет в своеобразную,
конической формы камеру, называемую дистальной; она прикрыта
особой табличкой — крышечкой. Предполагается, что малое отверстие
Рис. 257. Класс Homostelea:
1 — Тrochocystites (средний кембрий): 1а — вид сверху, 16—
вид снизу, 1в — вид спереди;
дк — дистальная камера, ж — желобок, кр — крышечка, сп —
суборальная пластинка, ст — стебель
выполняло функцию рта, по желобкам доставлялась пища, а большое
отверстие служило клоакой, через которую выбрасывались не только
шлаки, но и половые продукты. Стебель, состоящий из двух рядов таб-
личек, имел небольшой осевой канал, который открывался в полость
теки. Он, по-видимому, не выполнял настоящую, функцию стебля, а
служил для более стабильного положения гомостелей на дне моря, бу-
дучи погруженным в осадок.
Происхождение гомостелей остается пока неясным.
В ископаемом состоянии известны 4 рода из среднего кембрия.
КЛАСС HOMOIOSTELEA. ГОМОЙОСТЕЛЕИ
Тело гомбйостелей (гр. homoios •— подобный) состоит из трех час-
тей: теки, двурядной руки и стебля. Тека неправильной асимметричной
формы, овальная, округленная, треугольная, сжатая, покрыта много-
численными табличками, тесно лежащими около друг друга, что указы-
вает на то, что при жизни скелет был прочным и жестким. От переднего
конца теки отходила одна двурядная рука, полость которой служила
продолжением полости теки. Срединный желобок руки был покрыт дву-
мя рядами брахиальных табличек. У основания руки должен был быть
448
расположен рот, однако его следов не сохранилось; возможно, как и
у представителей класса карпоидей, он был скрыт внутри теки; около
основания руки в теке имеются два отверстия, одно из которых выпол-
няло функции гидропоры, другое — гонопоры. Анальное отверстие, при-
крытое анальными таблич-
ками, расположено на зад-
нем конце теки на левой
стороне и обычно достигает
довольно крупных размеров.
От середины задней части
теки отходит стебель, со-
стоящий из трех частей. У
проксимальной части он по-
лый, построен из четырех
рядов табличек, образующих
круг, и по своему строению
подобен аулакофору карпо-
идей. Средняя часть пере-
ходная от тетрамерной к
двухрядной дистальной ча-
сти. Последняя часть двух-
рядная, длинная, напоми-
нает настоящий стебель.
В состав класса вклю-
чено 10 родов (рис. 258).
Первые гомойостелеи из-
вестны из отложений верх-
него кембрия; наиболее
многочисленные находки
сделаны в полимиктовых
обломочных осадках ордо-
вика Чехословакии, Австра-
лии. В силуре их остатки
пока не обнаружены. Не-
многочисленные находки
известны из нижнего девона
Австралии, ФРГ.
Рис. 258. Класс Homoiostelea:
1 — Dendrocystoid.es (поздний ордовик), ре-
конструкция (Батер, 1913); 2 — lowacystis
(поздний ордовик), реконструкция; 3 — Syrin-
gocrinus (средний ордовик), реконструкция;
ан — анальное отверстие, гоп — гонопора,
гп — гидропора, др — двурядная рука, ст —
стебель
ПОДТИП CRINOZOA. КРИНОЗОИ
К подтипу Crinozoa (гр. crinon — лилия) отнесены все иглокожие,
за исключением карпозой и эдриоастероидей, ранее объединявшихся в
подтип Pelmatozoa. Для всех классов, входящих в состав подтипа, ха-
рактерны следующие особенности: 1) все внутренние органы заключены
в известковый скелет — теку, или чашечку, имеющую преимущест-
венно пятилучевую симметрию, за исключением некоторых примитивных
представителей; 2) тека имеет чашевидную, грушевидную или шаровид-
ную форму; 3) все кринозои постоянно или временно (часть морских
лилий) прикрепляются к подводным предметам стеблем или основанием
чашечки; 4) рот расположен на верхней стороне; пищеварительный
тракт петлеобразно изогнут и заканчивается анальным отверстием,
расположенным в интеррадиусе; 5) для сбора пищи имеются особые
отростки — брахиолы или выросты тела — руки (морские лилии,
паракриноидеи), в последние продолжаются целом и ряд внутренних
29 в. В. Друщиц
449
органов. Посредине брахиол и рук проходят пищевые желобки, по кото-
рым пища доставляется ко рту. Ток воды создается колебанием ресни-
чек или щупалец — видоизмененных амбулакральных ножек. Рост теки,
происходит в меридиональном направлении, а рук или брахиол в ра-
диальном.
В состав подтипа включены 7 классов: Cystoidea, Blastoidea, Crino-
idea, Eocrinoidea, Paracrinoidea, Edrioblastoidea, Parablastoidear
существовавших в палеозое; только один класс Crinoidea дожил
до наших дней. Условно в подтип включен класс Lepidocystoidea.
КЛАСС CYSTOIDEA. ЦИСТОИДЕИ
Общая характеристика. К классу цистоидей (гр. kystis — пузырь^
относятся раннепалеозойские кринозои, прикрепленные или свободна
лежащие на дне, с шаровидной, удлиненно-овальной текой, образован--
Рис. 259. Класс Cystoidea:
7—2 — схема строения: 1 — внешний вид, 2 — продольный разрез; 3—5 —
отряд Diploporida: 3 — Glyptosphaerites (ордовик); 4 — строение парных
пор: 4а — в плане, 46 — в разрезе, 4в — расположение пор на табличке; 5 —
Holocystites (поздний ордовик — ранний силур);
аж — амбулакральные (пищевые) желобки, амс — амбулакральная система,
ан — анальное отверстие, бра — брахиолы, брп — места прикрепления бра-
хиол, гп. — гидропора, по — половое отверстие, р — рот
ной многоугольными, многочисленными или немногочисленными прочна
соединенными табличками. Таблички пронизаны двойными или ромбо-
выми порами. Рот расположен на верхней поверхности, от него (Отходят
пищевые желобки, по краям которых находятся двухрядные брахиолы.
Анальное отверстие прикрыто/анальными табличками; у некоторых име-
ются гидропора и гонопора. Одни имели стебель, другие прирастали к.
субстрату текой. Средний кембрий (?), ордовик — средний девон.
450
*
Строение скелета. Скелет состоит из теки, стебля (иногда отсутст-
вует) и брахиол (рис. 259). Тека имеет разнообразную форму, шаровид-
ную; бокаловидную, мешковидную или другую, построенную многоуголь-
ными табличками, в количестве от многих сотен до 13—15; таблички
часто расположены поясками. В последнем случае различают пояски:
базальных, латеральных (инфра- и медиолатеральных), радиальных и
дельтоидных, или оральных, табличек. Перистом расположен на верхней
оральной поверхности, в центре которой находится рот; перистом по-
крыт мелкими табличками. Форма рта разнообразна — щелевидная или
субквадратная. От рта отходит 2—5 простых или разветвленных пище-
вых желобков, также прикрытых парными табличками; эти желобки
иногда называют амбулакральными. Вдоль пищеварительных
желобков расположены брахиолы — длинные, неразветвленные вы-
росты теки, с продольным каналом, прикрытым двумя рядами табличек.
Брахиолы служили для сбора и доставки пищи к ротовому отверстию.
Число брахиол варьирует от 2 до 400. Перипрокт с анальным отверсти-
ем расположен между пищевыми желобками, на оральной, реже на
боковой, стороне. Анальное отверстие, как правило, прикрыто несколь-
кими табличками, образующими анальную пирамидку.
У многих цистоидей между перистомом и перипроктом расположены
два отверстия — гидропора, связанная с амбулакральной системой, и
гонопора — отверстие для вывода половых продуктов. Иногда оба от-
верстия сливаются в одно. Предполагается, что вокруг глотки распола-
гался кольцевой канал, от которого к гидропоре отходил каменистый
канал.
Таблички, слагающие теку, состоят из трех слоев: тонкого наруж-
ного — «эпидермиса», толстого среднего и тонкого внутреннего слоев.
Средний слой многих табличек бывает пронизан каналами, которые
заканчиваются порами под эпидермисом.
Различают два основных типа пор: парные и ромбовые. Дипло-
поры, или парные поры; представляют собой отверстия U-образного
канала, пронизывающего средний слой таблички. Обычно пара пор ле-
жит в овальном углублении (рис. 259, 4). Диплопоры расположены в
пределах табличек по-разному, с определенной зональностью. Непра-
вильной формы поры получили название га пл опор; они бывают
одинарными или незакономерно разветвленными (в этом случае их на-
зывают перипорами). Ромбовые поры представляют собой
каналы щелевидной формы, пронизывающие две соседние таблички
таким образом, что одна половина канала одной таблички служит про-
должением второй половины другой таблички. Наиболее длинные кана-
лы расположены посредине шва, соединяющего две таблички, а отсюда
в оба противоположных направления длина щелей постепенно убывает.
В плане площадь, занятая щелями, имеет форму ромба. Ромбовые поры,
таким образом, представляют собой щели глубиной в несколько милли-
метров. Чаще всего средняя часть щели закрыта известковой табличкой
и в этом случае по краям остается по одной краевой поре, ведущей в
полость щели; иногда эта табличка бывает пронизана несколькими по-
рами (рис. 260, 6в). У немногих цистоидей под эпитекой от одной поры
к другой проходят три канала (рис. 260, 6г).
Число и положение ромбовых пор у цистоидей очень варьирует:
у одних они имеются на каждой табличке, у других — только на боко-
вых табличках теки или на табличках около перистома. Парные и ром-
бовые поры служили скорее всего для газообмена.
Стебель. У одних цистоидей стебель отсутствовал и тека, по-ви-
димому, лежала на субстрате свободно; у других — маленький стебель
29
451
существовал только на ранних стадиях развития и отсутствовал у взрос-
лых, у третьих — развивался довольно длинный стебель, с большой
полостью у теки и узкой на дистальном конце. Возможно, эти цистоидеи
прикреплялись стеблем к субстрату, как якорем, или стебель мог обви-
ваться и закрепляться вокруг каких-либо других животных (мшанок,
кораллов).
Рис. 260. Класс Cystoidea, отряд Rhombifera:
1 — Echinoencrinites (ранний ордовик); 2 — Echinosphaerites (ордовик); 3 —
Pleurocystites (ордовик); 4 — Staurocystis (поздний силур); 5—6 — ромбовые'
поры: 5 — схема расположения, 6 — продольные разрезы; 6а — Leptocystis,
66 — Cariocrinites, 6в — Stichocystis, 6г — Echinosphaerites-,
ан — анальное отверстие, бра — брахиолы, вс — внутренняя складка, гп —
гидропора, п — поры, р — рот, рп — ромбовые поры, ст — стебель, ш — шов
Экология и тафономия. Цистоидеи свободно лежали на дне, либо
прирастали нижней частью теки, либо прикреплялись к субстрату при
помощи стебля. Цистоидеи собирали пищу с помощью брахиол. Некото-
рые цистоидеи заселяли значительные участки дна, образуя своеобраз-
ные подводные луга. Расселению цистоидей способствовали свободно-
плавающие личинки. Многие цистоидеи были широко распространены
в морях ордовика — среднего девона.
История развития цистоидей. Сомнительные остатки цистоидей с
парными порами указываются из среднего кембрия. Из раннего ордови-
ка известны многочисленные представители класса. Ордовик — время
расцвета цистоидей. Известно около 90 родов, в среднем ордовике
существовало 40, в среднем девоне только 2, а в позднем девоне —
1 род.
452
КЛАСС BLASTOIDEA. БЛАСТОИДЕИ
• Общая характеристика. К классу бластоидей (гр. blastos — бутон,
побег) относятся палеозойские кринозои, для которых характерны: бу-
тонообразная тека с хорошо развитой пятилучевой симметрией, постро-
енная из 18—21 крупных табличек, расположенных поясками; от рта,
лежащего в центре теки, отходят пять радиальных амбулакров, вдоль
которых расположены в два ряда многочисленные брахиолы; своеобраз-
ная система гидроспир, представляющих собой известковые складки
стенки теки, открывается порами вдоль амбулакров и отверстиями на
дельтоидных табличках. Бластоидеи распространены от силура до
перми.
Рис. 261. Класс Blastoidea, отряд Spiraculata:
1 — схема строения чашечки Pentremites (карбон): 1а— в плане, 16 — сверху,
1в — снизу, 1г — сбоку; 2 — реконструкция чашечки, продольный разрез; 3 —
разрез через амбулакральный желобок и брахиолу;
аж — амбулакральный желобок, ан — анальное отверстие, б аж — боковой ам-
булакральный желобок, бз — базальные таблички, бра — брахиолы, гс — гидро-
спира, дт — дельтоидные таблички, кп — краевая /пера, кт — кроющие таблич-
ки, р — рот, раж — радиальный амбулакральный желобок, рд — радиальные таб-
лички, с — спиракулы
Строение скелета. Тека, или чашечка, имеет бутоновидную, шаро-
видную, удлиненно-шаровидную или вазообразную форму (рис. 261).
Она образована у большинства из 18—21 крупных табличек, располо-
женных поясками. Три нижние таблички — базальные, или основ-
ные,— лежат непосредственно над стеблем; одна из них узкая, две —
вдвое шире, образованы за счет слияния двух маленьких..Выше следует
поясок из пяти радиальных табличек, чередующихся с базальными;
каждая радиальная табличка имеет глубокую выемку, получившую
название радиального синуса, в котором расположен пищевой
желобок. Размеры радиальных табличек и радиальных синусов различ-
ны; обычно их длина составляет от 1/2 до 2/3 высоты теки. Вдоль края
радиальных табличек расположены поры или щели, ведущие в гидро-
453
спиры. Третий поясок образован пятью дельтоидными табличками
треугольной или неправильно ромбической формы. У одного из отря-
дов бластоидей на каждой из четырех дельтоидных табличек было по
два отверстия — спиракул, а на пятой — три, третье отверстие —
анальное, поэтому и табличка называется анальной. В процессе
эволюции произошло слияние двух отверстий в одно, а на анальной
табличке — трех в одно. •
На верхней, оральной, стороне теки находится пятиугольный п е-
ристом, в центре которого помещается ротовое отверстие, покрытое
Рис. 262. Класс Blastoidea, отряд Fissiculata (1—7) и класс Parablastoidea (S):
1,5~6 — Orophocrinus (ранний карбон): 7 — реконструкция; 2 — Ceratoblastus
(пермь); За, б — Thaumatoblastus (пермь); 4 — Pleuroschtsma (девон); 7 :—Ро-
lydeltoldeus (силур); 8 — Blastoidocrinus (ордовик): 8а — вид снизу, 86 — вид
сверху;
амп — амбулакральное поле, ббр — бибрахиальные таблички, бра — брахиолы,
дт — дельтоидные таблички, ибр — интербрахиальные таблички, к — «корни»,
кр — крыловидные таблички, р — рот, рд — радиальные таблички, ст — стебель,
! сщ — спиракульные щели, 'ч — чашечка
всегда мелкими неподвижно сочлененными табличками. От рта отходят
пять радиальных амбулакральных полей, на которых расположены пи-
щевые желобки; средняя часть пищевого желобка занята узкой ланце-
товидной табличкой с осевым каналом посредине, от которого в обе
стороны отходят боковые желобки. Они переходят на небольшие бра-
хиолы, расположенные по краям амбулакрального поля. Боковые
части амбулакрального поля прикрыты так называемыми боковыми
(поровыми) и наружнобоковыми табличками. Все желобки сверху так-
же закрыты покровными табличками. В боковых (поровых) табличках
имеются краевые поры; они ведут в гидроспиры — особые складча-
тые каналы, расположенные по обе стороны под пищевым желобком
(см. рис. 261). Гидроспиры подвешены к стенкам полости теки и при-
крепляются к ланцетовидной табличке или отдельным гидроспирным
табличкам. По гидроспирам, вероятно, циркулировала вода для газооб-
мена, входившая через краевые, поры и выходившая через спир аку-
лы, расположенные на дельтоидных табличках, У многих блаетоидей
454
«спиракулы имеют вид щели; эти спиракульные щели расположены
параллельно амбулакральным полям; по краям последних вместо круг-
лых пор наблюдаются гидроспирные щели; с. каждой стороны амбулак-
ра насчитывается до 18 щелей (рис. 262).
Вокруг рта, под текой, у бластоидей обнаружен кольцевой канал,
•от которого отходят пять радиальных каналов; они расположены под
ланцетовидными табличками, поэтому пищевые желобки соответствуют
радиальным амбулакральным полям и называются амбулакрами. Вдоль
краев амбулакров расположены двухрядные брахиолы; они прикрепля-
ются к небольшим бугоркам. Каждая .брахиола состоит из члеников,
подвижно соединенных друг с другом. При помощи брахиол вода с пи-
щей попадала на боковые каналы, а затем по пяти срединным каналам
амбулакров доставлялась ко рту. Пищей, скорее всего, служили раз-
личные планктонные организмы. Длина брахиол обычно была в два—
три раза больше длины теки. Их число очень различно; у молодых
насчитывается до 200, у взрослых — до 500.
Стебель бластоидей состоит из коротких цилиндрических члени-
ков, пронизанных осевым каналом круглого сечения. Отдельные членики
неотличимы от члеников стебля криноидей и цистоидей. Длина стебля
не превышала 20 см; у некоторых бластоидей стебель, по-видимому,
•отсутствовал (во всяком случае на взрослой стадии).
Основы систематики и классификация. Класс бластоидей на осно-
вании строения гидроспир, формы спиракул разделен на два отряда
Fissiculata и Spiraculata.
Для отряда Fissiculata' (силур—пермь) характерно наличиегидро-
спирных щелей и спиракул в виде щелей. Известно около 35 родов
(рис. 262). Для отряда Spiraculata (силур — пермь) характерны
скрытые гидроспирные щели; около 45 родов, (см. рис. 261).
История развития бластоидей. Происхождение бластоидей неизвест-
но. Одни исследователи предполагают, что они возникли от цистоидей
•с ромбовыми порами, другие убедительно аргументируют происхожде-
ние бластоидей от небольшого класса эдриобластоидей. Возможно, что
вначале возникли фиссикуляты, а затем спиракуляты. Первые бласто-
идеи из силура, два рода принадлежат отряду фиссикулят и один —
•отряду спиракулят. В девоне бластоидеи становятся более многочислен-
ными (15 родов); в раннем карбоне они достигают расцвета (около
•30 родов, из них 20 — спиракулят). В перми сохраняются 3 рода спира-
кулят, но более разнообразными (16 родов) оказываются фиссикуляты.
В конце перми все бластоидеи вымирают.
КЛАСС CRINOIDEA. КРИНОИДЕИ,
или МОРСКИЕ ЛИЛИИ
Общая характеристика. Криноидеи (гр. krinon — лилия), или мор-
ские лилии, — наиболее крупный класс кринозой, тело которых состоит
из чашечки, заключающей внутренние органы, пяти обычно хорошо раз-
витых рук, служащих для сбора пищи, и стебля или системы усиков,
предназначенных для прикрепления к подводным предметам. Чашечка
радиально-симметричная, построена из пояска радиальных табличек и
•одного или двух поясков основных табличек. Чашечка сверху прикрыта
крышечкой, или тегменом, в которой находятся амбулакральные желоб-
ки, переходящие на руки и далее на пиннулы. Ордовик — ныне.
Строение тела. Внутренние органы морской лилии заключены в
чашечку, в центре которой на верхней стороне расположено ротовое от-
455
верстие (рис. 263). Рот ведет в пищеварительный тракт, делающий
один или несколько петлеобразных изгибов и открывающийся анальным
отверстием в заднем интеррадиусе. Пищеварительный тракт лежит во
вторичной полости тела и подвешен к стенкам тела при помощи мезен-
Рис. 263. Схема строения морской лилии:
1а, б — моноциклическая чашечка; 2а, б — дициклическая чашечка;
3 — схематический разрез через чашечку; 4 — общий вид прикреплен-
ной морской лилии;
амк — амбулакральный канал, ан — анальное отверстие, к — «корни»,
кр — крона, пи — пиннулы, р — рот, рук — руки, ст — стебель, ч —
чашечка, таблички: бз — базальные, бр — брахиальные, иб — инфра-
базальные, рд — радиальные
териальных перепонок. От чашечки отходят пять неразветвленных или
ветвящихся рук. Чашечка вместе с руками образует крону. Вокруг
пищеварительного тракта расположен кольцевой канал амбулакральной
системы; от него в руки отходят пять радиальных каналов, вдоль кото-
рых расположены амбулакральные ножки; у морских лилий они остро-
456
конечны, лишены ампул, присасывательных дисков и выполняют пище-
сборную, дыхательную и чувствительную функции. Пищей криноидеям
служат планктонные организмы и мелкие частицы детрита. Пища до-
ставляется ко рту по желобкам на руках с помощью амбулакральных
Ножек и ресничек покровного эпителия. Количество пищи, добываемой
морской лилией, зависит от степени разветвления рук и соответственно
длины бороздок, или желобков. У одной тропической лилии с 68 ветвя-
ми рук общая длина пищевых борозд достигает 100 м. Вокруг рта
расположено нервное кольцо, от которого по пяти радиусам в руки отхо-
дят нервные стволы, обеспечивающие их движение.
Скелет чашечки. Чашечка, или тека, разнообразной формы,
коническая, бокаловидная, дисковидная или шаровидная (рис. 263).
Часть чашечки ниже мест прикрепления рук называется спинной, или
дорзальнрй, а верхняя часть — крышечкой, или тегменом.
Дорзальная часть теки образована двумя илй тремя поясками табли-
чек. Различают пояски: радиальных (RR), базальных (ВВ) и инфра-
базальных (IB) табличек; каждый из поясков состоит из пяти табличек.
От основания чашечки отходит стебель или у бесстебельчатых
форм — усики, или цирри; к радиальным табличкам прикрепля-
ются руки. Чашечка, дорзальная часть которой кроме пояска радиаль-
ных табличек имеет поясок базальных, называется моноцикличе-
ской; если она имеет поясок базальных и инфрабазальных табличек —
дициклической. Дорзальная часть теки иногда бывает построена
исключительно из радиальных табличек, реже' только из базальных.
Часто в строении дорзальной части принимает участие ряд других таб-
личек, среди которых выделяются анальная- (одна или несколько), рас-
положенная в заднем интеррадиусе, радианальная и др. В процессе
эволюции у морских лилий наблюдается увеличение размеров чашечки.
Это увеличение происходит за счет включения в состав чашечки члени-
ков нижних частей рук и развития новых, так называемых интерради-
альных и интербрахиальных табличек (см. рис. 271,6—8).
Скелет руки. От радиальных табличек чашечки отходят руки.
Они редко остаются простыми, большей частью раздваиваются один
или несколько раз. Руки состоят из отдельных члеников, напоминающих
по форме позвонки, которые соединяются друг с другом с помощью
мышц или эластичной связки. Как правило, они снабжены короткими
членистыми придатками — пиннулами. Членики рук сочленяются
также при помощи особых площадок, часто имеющих один или два ва-
лика. Руки обладают гибкостью и большой подвижностью. При небла-
гоприятных условиях (высокой температуре, недостатке кислорода,
нападении врагов) морские лилии способны обламывать свои руки, при
этом утраченные части позднее восстанавливаются. Руки и пиннулы
снабжены довольно глубокими желобками, выстланными у современных
форм ресничным эпителием. Вдоль желобка проходит радиальный
амбулакральный канал, от которого отходят пучками (по 3) остроко-
нечные амбулакральные ножки без ампул; они выполняют функцию
осязания и дыхания. Боковые ветви радиальных каналов заходят также
в пиннулы.
Руки предназначены для сбора пищи. В руки продолжаются вто-
ричная полость тела, нервные стволы, сосуды кровеносной системы. По
пищевым желобкам-рук пища попадает в ротовое отверстие, располо-
женное в центре тегмена. В процессе эволюции увеличивается длина и
степень разветвленности рук. Однорядная рука у примитивных форм
заменяется двухрядной (рис. 264,2); двухрядная рука позволяет мор-
ской лилии собирать больше пищи. Увеличение длины рук происходит
457
при их дихотомическом ветвлении или при образовании перистой руки
(рис. 264,/). Однако в процессе эволюции возникали морские лилии,
у которых руки частично или полностью редуцировались. При редукции
Рис. 264. Схема строения рук:
1 — развитие перистой руки из дихотомически разветвленной; 2 — разви-
тие двухрядной руки из однорядной; 3—часть руки (4 членика) с амбула-.
кральным каналом и кроющими табличками; 4 — два членика руки Dia-
menocrinus (девон)
рук могли исчезать и, поддерживающие их
шечки.
радиальные таблички ча-
Рис. 265. Типы строения крышки (тегмена):
1 — развиты только оральные таблички; 2 — крышка с амбулакральными по-
лями; 3 — оральные таблички редуцированы (сохранились около рта); 4 — пи-
щевые желобки расположены под прочной крышкой;
ан — анальное отверстие, м — мадрепорит, таблички: ам — амбулакральные,
ат — анальная, иам — интерамбулакральные, ор — оральные
Т е г м е н у большинства современных форм почти совершенно ли-
шен крупных скелетных элементов. Он пронизан многочисленными пора-
ми, ведущими в полость тела; через , поры амбулакральная система
пополняется водой. Амбулакральные ножки, расположенные около рта,
458
бывают видоизменены в околоротовые щупальца. У древних морских
лилий тегмен был прикрыт пятью ор а л ьными, или ротовыми, таб-
личками, расположенными интеррадиально (рис. 265). Оральные таб-
лички развиты в разной степени: у одних форм они известны только в
личиночной стадии и отсутствуют у взрослых; у других — хорршо раз-
виты и плотно соединяются друг с другом; у третьих крышка состоит из
многочисленных мелких табличек, среди которых различают таблички,
Рис. 266. Типы стеблей морских лилий:
1 — стебель Eifelocrinus, прикрепленный к колонии мшанок (реконструкция); 2—
«якорь» у Ahcyrocrinus; 3 — двусторонне-симметричный стебель с усиками (ус)
у Myelodactylus, окружающий крону (кр); 4 — стебель, спирально свернутый
вокруг чашечки у Ammonicrinus
покрывающие пищевые желобки, и расположенные между ними интер-
амбулакральные таблички. Эти таблички, соединяясь друг с другом, об-
разуют своеобразный свод над чашечкой; под таким сводом расположен
рот, и пища поступает по пищевым желобкам, лежащим под крышкой.
Анальное отверстие размещается на верхней стороне орального
диска чашечки интеррадиально, ближе к ее краю. У морских лилий,
живших в спокойных, малоподвижных водах, возникала анальная труб-
ка, покрытая мелкими табличками. Подобная трубка позволяла живот-
ному удалять экскременты на значительном расстоянии от рта..
Стебель. К нижней стороне чашечки, к ее центральной табличке,
прикреплен гибкий стебель, состоящий из члеников различной формы;
круглой, эллиптической, четырехугольной, пятиугольной и очень редко
треугольной и шестиугольной (рис. 266). У одних родов стебель до-
стигает длины нескольких метров, у других остается коротким или со-
вершенно атрофируется. У некоторых форм чашечка прирастала своим
459
основанием. Через весь стебель проходит осевой канал, имеющий раз-
личное поперечное сечение. У древних морских лилий стебель состоял
из пяти рядов табличек, расположен-
ных в чередующемся порядке. В про-
цессе эволюции наблюдается переход
к циклическому расположению и к
Рис. 267. Эволюция стебля от пя-
тирядного к циклическому
слиянию при этом каждых пяти сосед-
них табличек в один членик стебля
(рис. 267). Нередко среди одинаковых
члеников встречаются более крупные
так называемые узловыечленики,.
несущие усики. Морские лилии при-
крепляются к субстрату разными пу-
тями: прирастанием стебля к каме-
нистому дну путем выделения вокруг
основных члеников значительного ко-
личества извести и образования при-
крепительного диска, развитием на
конце стебля разветвленных корневидных ветвей, наличием 'своеоб-
разного якоря, предназначенного
У некоторых морских лилий длин-
ный тонкий стебель обвивал водо-
росли или полипняки кораллов и
служил для временного прикреп-
ления, у других — закручивался
вокруг чашечки в плоскую спи-
раль и, возможно, служил для
медленного и недалекого пере-
движения по дну с помощью дву-
рядно расположенных усиков (см.
рис. 266, 3). Известно также раз-
витие на нижнем конце стебля
шаровидного вздутия, разделен-
ного перегородками на отдельные
камеры и служившего, по-видимо-
му, плавательным пузырем при
планктонном образе жизни. Нако-
нец, у ряда форм стебель отсут-
ствовал и отсутствует на взрос-
лой стадии у многих современных
криноидей. У таких бесстебель-
чатых морских лилий стебель су-
ществует только на первых ста-
диях развития в течение месяца—
полутора, после чего их чашечка
самопроизвольно отламывается
от стебля и молодая морская ли-
лия переходит к свободному об-
разу жизни. В основании чашеч-
ки развиваются усики, или
цирри. Движение таких лилий
закрепления в илистом грунте..
Рис. 268. Онтогенез Antedon bipida:
1—2 — свободноплавающие личинки (те-
менная пластинка обращена вниз); 3 —
прикрепленная стадия (напоминающая
цистоидей); 4 — пентакринусовая ста-
дия с обособленными руками;
пр — присоска, таблички: бз — базаль-
ные, ор — оральные, рд — радиальные
происходит при помощи рук, однако за один прием они проплывают не-
большое расстояние (до 3 м), совершая при этом до 100 ударов в ми-
нуту. Число, размеры, длина и внешний вид усиков зависят от условий
обитания: морские лилии, живущие на мягких илах, имеют тонкие,
460
длинные, почти прямые усики; лилии, обитающие на камнях, снабжены
короткими изогнутыми усиками.
Размножение и развитие. Наиболее подробно изучено размножение
и развитие современной бесстебельчатой морской лилии, принадлежа-
щей к роду Antedon (рис. 268). Морские лилии раздельнополые. Поло-
вые клетки созревают в пиннулах рук; выброс половых продуктов про-
исходит обычно одновременно, и оплодотворение яиц осуществляется в
воде. Оплодотворенные яйца заключены в оболочку, часто снабженную
различными шипами, иглами. В этих оболочках происходит развитие
яиц до стадии личинки. Вначале личинка лишена рта и питается только
желтком. На брюшной стороне у нее имеется прикрепительная присоска.
Поплавав некоторое время в воде, личинка опускается на дно, прикреп-
ляется к субстрату передней частью тела. Узкая передняя часть превра-
щается в стебель, а широкая задняя — в чашечку. Реснички, покры-
вающие тело личинки, исчезают, происходит вращение комплекса внут-
ренних органов на 90°. Возникают пять оральных табличек, образующих
на верхней стороне пирамиду, снизу развиваются пять базальных таб-
личек. Между ними и началом стебля появляются 3—5 инфрабазальных
табличек. В это время скелет молодой морской лилии несколько напо-
минает скелет’ некоторых палеозойских цистоидей. Вскоре между
Рис. 269. Подкласс Camerata. Схема строения (/—3 — в плане, 4 — сбоку):
1 — Cleiocrinus (средний ордовик); 2 — Glyptocrinus (поздний ордовик); 3 —
Platycrinites (девон — пермь); 4 — Acrocrinus (карбон)
базальными и оральными табличками развивается поясок, состоящий
из пяти радиальных табличек, и возникают руки. На границе между
чашечкой и стеблем формируются новые членики стебля. Через пять
недель после оседания личинки на стебельке раскачивается миниатюр-
ная морская лилия высотой около 4 мм. В дальнейшем руки постепенно
удлиняются, каждая рука делится на две ветви; вдоль руки возникают
пиннулы, которые чередуются друг с другом. На этой стадии морская
лилия похожа на представителей стебельчатой морской лилии рода
Pentacrinus. Спустя некоторое время оральные таблички редуцируются,
и на верхней стороне развивается кожный покров — тегмен. Базальные
461
таблички также редуцируются. Затем чашечка самопроизвольно отла-
мывается от стебля, и молодая лилия, превратившись в бесстебельча-
тую, начинает вести подвижный образ жизни, передвигаясь с помощью
рук. Для временного прикрепления в основании чашечки развиваются
цирри. Изучение онтогенеза современных морских лилий указывает на
возникновение бесстебельчатых представителей от прикрепленных.
Рис. 270. Подкласс Inadunata. Схема строения:
1 — эволюция чашечки у инадунатных криноидей; 2 — Cornucrinus (ордовик):
2а — вид сзади, 26 — вид сверху; 3 — locrinus (средний — поздний ордовик);
4 — Cupressocrinites (средний девон): 4а — чашечка с руками, 46 — вид чашеч-
ки сверху; 5 — Cromyocrinus (карбон);
дк — дорзальный канал, таблички: ан — анальная, ор— оральная (остальные
обозначения см. рис. 263)
Основы систематики и классификация. Систематика морских лилий
основывается на строении чашечки в целом, на строении ее дорзальной
части, крышечки (тегмена), рук и стебля, на количестве и характере
расположения анальных, интеррадиальных и интербрахиальных табли-
чек. В состав класса включены четыре подкласса: Camerata, Inadunata,.
Flexibilia, Articulata, из которых первые три существовали от ордовика
до перми, а представители четвертого, появившись в начале триаса, про-
должают 'свое существование в современных морях (рис. 269—272).
Экология и тафономия. Морские лилии в палеозое и мезозое были
обитателями преимущественно мелководных зон моря. Часть из них
жила под защитой коралловых рифов, часть опускалась на небольшие
глубины. Современные морские лилии живут на всех глубинах: от суб-
литорали до абиссали (до 9700 м), от тропиков до полярных широт.
Иногда они образуют большие поселения — «луга», состоящие из одно-
го вида. Вероятно, такие «луга» морских лилий существовали в прош-
лые геологические времена, так как известны слои криноидных извест-
няков, состоящих из члеников стеблей, реже рук й остатков чашечек..
Глубоководные морские лилии имеют длинные ветвистые руки, тонкие
стебли и небольших размеров чашечку; на конце стебля развиты раз-
462
личной длины корешки. У морских лилий, живущих в теплых водах,
чашечки более массивные, иногда снабженные шипами; стебель, как
правило, укороченный, в некоторых случаях полностью отсутствует, и
морская лилия прикрепляется к субстрату непосредственно чашечкой.
Большинство современных морских лилий ведет свободный образ жизни,
медленно плавая около дна при помощи рук. Подобные свободнопла-
Рис. 271. Подкласс Flexibilia:
1 —. чашечка Taxocrinus из девона с раковиной брюхоногого моллюска Platy-
ceras (Pl)', 2а, б — часть стебля из карбона, поврежденного паразитирующими
мизостомидами (Annelides)-, 3,8 — Sagenocrinites (силур), в состав чашечки
входят брахиальные (бр) и интербрахиальные (ибр) таблички; 4 — Protaxocri-
nus (средний ордовик — силур); 5—6 — увеличение размеров чашечки за счет
брахиальных табличек; 7 — Ichihyocrinus (силур — ранний девон), в состав ча-
шечки входят брахиальные таблички; (обозначения см. рис. 263)
вающие криноидеи встречались уже в палеозое (силуре, перми), не-
большая часть морских лилий, по-видимому, вела планктонный образ
жизни. К ним относится силурийская форма (Scyphocrinites), у которой
на конце стебля возник шаровидный плавательный аппарат (пневмато-
фор). Меловая бесстебельчатая морская лилия (Saccocoma) плавала
благодаря наличию маленькой чашечки и длинных рук. Малая подвиж-
ность морских лилий делает и делала их раньше доступными для посе-
ления на них других организмов, для различного рода сожительства,
вплоть до настоящего паразитизма. Черви семейства Myzostomidae
часто поселяются на морских лилиях, одни из них ползают по стеблю
и чашечке, другие образуют на пиннулах для своего жилья особые
вздутия, третьи паразитируют внутри тела. Некоторые брюхоногие мол-
463
люски, вероятно, были комменсалистами морских лилий. На брюшной
стороне чашечки некоторых палеозойских криноидей находят раковины
брюхоногих моллюсков (платиноцератид), расположенные таким обра-
зом, что устьевой край был приближен к анальному отверстию морской
лилии; не исключена возможность, что эти брюхоногие моллюски пита-
лись экскрементами лилии.
Рис. 272. Подкласс Articulata:
1 — Uintacrinus (поздний мел); 2а, б — Marsupites (поздний мел); 3 — Saccocoma
(поздняя юра — мел);
таблички: ибр — интербрахиальные, по — пятиугольная табличка основания (остальные
обозначения см. рис. 263)
История развития криноидей. Происхождение криноидей до сих пор
не выяснено. Предполагают, что они обособились в кембрии от общего
предка с цистоидеями и их развитие было связано с возникновением
радиальных выростов тела — рук, предназначенных для сбора пищи.
Руки не гомологичны брахиолам цистоидей и бластоидей. В раннем ор-
довике становятся известными представители двух подклассов: камерат
и инадунат, а начиная со среднего ордовика •— подкласс флексибилий.
Если первые два подкласса образуют дивергирующие группы, то под-
класс флексибилий остается на протяжении палеозоя небольшой груп-
пой, вымирающей в середине перми. Особенно многочисленны и разно-
образны были камераты и инадунаты в девоне и раннем карбоне. Число
камерат резко сокращается к концу карбона, и последние представители
этого подкласса вымирают в середине перми. Инадунаты, наоборот, в
перми дают новую вспышку и характеризуются довольно широким рас-
пространением. Один из подотрядов инадунат (энкринина) сохраняется
в триасовое время, но и он к концу триаса вымирает. Первые предста-
вители подкласса артикулят появляются в триасе; в юре и мелу они
становятся многочисленными; среди них наряду с прикрепленными сте-
бельчатыми формами появляются бесстебельчатые подвижные крино-
идеи. В современных морях артикуляты стебельчатые (75 видов) и бес-
стебельчатые (свыше 500 видов) являются единственными представи-
телями некогда обширного не только класса криноидей, но и всего
подтипа кринозой.
КЛАСС EOCRINOIDEA. ЭОКРИНОИДЕИ
К классу Eocrinoidea (гр. eos — восток, krinon — лилия) относится
небольшое число р.аннепалеозойских прикрепленных иглокожих, внешне
несколько напоминающих криноидей. Скелет состоит из теки, брахиол
464
из пяти табличек. На оральной стороне
Рис. 273. Классы Paracrinoidea (/) и Eocrinoi-
dea (2—4):
1 Comarocystites (средний ордовик); 2 —
Rhipidocystis (ранний — средний ордовик);
3 — Bockia (ранний — средний ордовик); 4 —
Rhopalocystis (ранний ордовик); (2—4 — ре-
конструкции) .
и стебля (последний иногда может отсутствовать). Тека имеет разно-
образную форму: неправильно-сферическую, овально-удлиненную, кони-
ческую, субцилиндрическую, уплощенную, обычно с пятилучевой симмет-
рией, правда не всегда четко выраженной (рис. 273,2—4). Тека пост-
роена из различного числа табличек (иногда до 500); у более прогрес-
сивных форм число табличек ограничено и, как правило, они располо-
жены поясками. Поясок сост
в центре перистома распо-
ложено ротовое отверстие.
Вокруг перистома лежат
пять оральных табличек и
многочисленные таблички
между ними, несущие бра-
хиолы. От ротового поля
отходят пять пищевых же-
лобков, простых или раз-
ветвленных, причем чаще
всего два желобка противо-
поставлены трем другим.
На концах желобков рас-
положены брахиолы. Число
брахиол варьировало от 8
до 44. Недалеко от пери-
стома, на оральной поверх-
ности, расположено аналь-
ное поле, или перипрокт,
с анальным отверстием,
прикрытым мелкими тре-
угольными табличками, об-
разующими анальную пи-
рамидку. У некоторых ро-
дов наблюдается гидро-
пора и, возможно, го но-
по р а — небольшое отвер-
стие, связанное с половыми
железами. У многих эокри-
ноидей между швами текальных табличек имеются небольшие (0,15—
0,5 мм) сутуральные поры эллипсовидной формы, иногда сложного
строения.
Стебель построен по-разному: у наиболее примитивных форм
он представляет собой удлиненный вырост аборальной части теки, по-
крытый многочисленными табличками; у более развитых стебель закан-
чивается вздутым диском, покрытым табличками, или состоит из от-
дельных члеников с большим каналом в центре; диаметр члеников по-
степенно уменьшается к дистальному концу.
Эокриноидеи по своему строению, положению рта и анального отвер-
стия на верхней поверхности теки, наличию брахиол и стебля ближе
всего стоят к цистоидеям; однако отсутствие текальных пор не позво-
ляет их объединять с последними. Сходство с криноидеями достаточно
поверхностное, и отличия заключаются прежде всего в отсутствии
у эокриноидей рук, представляющих у морских лилий продолжение
полости тела. Эокриноидеи, по-видимому, являются самостоятель-
ным классом, существовавшим от раннего кембрия до силура вклю-
чительно.
Класс объединяет около 30 родов (рис. 273,2—4).
30 В. В. Друщиц
465
КЛАСС PARACRINOIDEA. ПАРАКРИНОИДЕИ
К паракриноидеям (гр. para — около, вблизи, krinon — лилия>
относится небольшой класс кринозой, тека которых построена из раз-
личного числа табличек (от 30 до 175), расположенных без особой
закономерности; тека недифференцирована на дорзальную чашечку ш
тегмен; следы пятилучевой симметрии не обнаружены. Таблички глад-
кие или с ребристой скульптурой. Рот в виде щели, расположен на
верхней стороне, несколько сбоку, в центре перистома, прикрытого мно-
гочисленными мелкими табличками. От рта отходят четыре амбула-
кральных бороздки (или пищевых желобка). На конце бороздок у од-
ного отряда (Branchiata) расположены четыре так называемые сво-
бодные руки, соединенные с текой своими проксимальными концами.
У второго отряда (Varicata) часть проксимального конца рук прочно-
прикреплена к табличкам теки. У всех паракриноидей руки однорядные,
снабжены однородными отростками, или пиннулами. Руки простые или
ветвящиеся. Анальное отверстие с анальной пирамидкой расположено*
между пищевыми желобками почти в центре верхней стороны теки, про-
тив места прикрепления стебля.. Около рта имеется узкое, щелевидное-
отверстие гидропоры; наличие гонопоры не установлено. Таблички теки
двухслойные, состоят из эпитеки и внутреннего слоя — стереоте-
ки, обычно пронизанного порами. От аборальной стороны теки отходит-
стебель, состоящий из чдеников равного диаметра, с каналом в центре;,
стебель служил для неподвижного прикрепления к субстрату.
Паракриноидеи образуют небольшой класс, который отличается от
криноидей строением теки, отсутствием пятилучевой, симметрии, от
бластоидей — отсутствием гидроспир, от цистоидей — наличием одно-
рядных пиннулбносных рук. 8 родов из среднего ордовика (рис. 273).
КЛАСС EDRIOBLASTOIDEA. ЭДРИОБЛАСТОИДЕИ
К эдриобластойдеям (гр. hedra —- сидячий) -относится один род
Astrocystites из среднего ордовика Канады. Тека высотой 21 мм состоит
из 20 расположенных четырьмя поясками табличек; каждый поясок
состоит из пяти табличек. От рта, расположённого в центре верхней
поверхности, отходят пять пищевых желобков, прикрытых двумя рядами
табличек. На одной из дельтоидных табличек расположено анальное-
отверстие с анальной пирамидкой; около него находится гидропора.
Между швами табличек, образующих основание пищевых желобков;
имеются поры. В отличие от бластоидей и цистоидей у эдриобластоидей
отсутствуют брахиолы. От теки отходит стебель, состоящий из полиго-
нальных табличек. -
Вначале описанный род (рис. 274, Г) был отнесен к классу бласто-
идей; сходство с ними -оказалось чисто внешним; отсутствие брахиол^
спиракул не позволяет относить этот род к последним, и он послужил7
основой для нового класса — эдриобластоидей, названного так за внеш-
нее сходство с бластоидеями.
КЛАСС PARABLASTOIDEA. ПАРАБЛАСТОИДЕИ
К классу парабластоидей относятся стебельчатые кринозои, у ко-
торых тека прикреплялась к субстрату с помощью стебля. Тека состоит
из- многочисленных табличек; оральная и аборальная части довольно-
хорошо обособлены. Аборальная часть построена из нескольких поясков-
табличек. К пояску базальных прикреплен стебель, состоящий из от-
466 ; - . - ” 'г
дельных члеников круглого сечения. Оральная сторона построена из
пяти крупных дельтоидных табличек и пяти амбулакральных полей,,
состоящих из многочисленных табличек, посредине которых расположе-
но по три так называемых крыловидных таблички. Каждое поле имеет:
пищевой желобок, прикрытый рядами парных табличек. Вдоль пище-
вого желобка расположены адамбулакральные таблички, несущие па
/ длинной брахиоле. Ротовое отверстие
находится в центре; оно плотно при-
крыто пятилучевой табличкой и, по-
видимому, было связано с пятью пище-
выми желобками, идущими под по-
кровными табличками. Под каждой
Рис. 274. Класс Edrioblastoidea: Рис. 275. Класс Lepidocystoidea:
Astrocystites (средний ордовик)': Lepidocystis (ранний кембрий), реконструкция"*
1а — вид сбоку,/б— вид сверху; (Дарэм, -1967); '
амб — часть амбулакра без по- ан— анальное отверстие, др — двухрядная ру-щ
кровных табличек с краевыми. по- . .. ка, ор — основание руки, сп — сутуральные .
рами, ан — анальное отверстие, поры,' чт — чешуевидные таблички ’ ;
гп.— (?) гидропора, ст — стебель, ... Г/
таблички: бз — базальные, дт — . :
дельтоидные, 'ор— оральные, рд— : '
радиальные
дельтоидной табличкой лежат серии параллельных складок, называе-
мых к а т а сп и р а м и. Они связаны с порами, расположенными вДоль
края амбулакрального поля и с порами на адоральном крае каждой
дельтоидной таблички. По-видимому, катаспиры служили для газооб-
мена; вода поступала в них через одни поры и выходила через поры
дельтоидных табличек. Катаспиры сверху были прикрыты скелетной
тканью — стереомой.
Парабластоидеи отличаются от бластоидей большим числом таб-
личек теки, строением амбулакров. Они, вероятно, представляли собой'
слепую ветвь кринозой. Известно два рода — Blasiocystis и Blastoidoo
rinus. (рис,- 262, Sj; из ордовика СССР и Северной Америки.
30* 46Г
КЛАСС LEPIDOCySTOIDEA. ЛЕПИДОЦИСТОИДЕИ
К классу Lepidocystoidea (гр., lepis — чешуя, cystis — пузырь)
относится только один род Lepidocystis, остатки скелета которого
(рис. 275) были обнаружены в отложениях нижнего кембрия (зоне
Olenellus) в штате Пенсильвания (США), вместе с остатками медузо*
подобных камптостроматоидей.
Тело заключено в известковый скелет конусовидной формы, в кото-
ром различают оральную и аборальную части. Оральная поверхность
покрыта многочисленными табличками, несущими еутуральные поры.
Они, по-видимому, служили для газообмена. На оральной поверхности
циклично, по трем пояскам, расположены двухрядные руки, по пять
рук в каждом круге. В центре, под табличками, вероятно, располага-
лось ротовое отверстие, следы которого не сохранились. Пища по же-
лобкам рук поступала по специальным каналам, которые должны были
быть под оральной крышкой. Около периферического края оральной
поверхности, между основаниями двух рук, расположено анальное от-
верстие, прикрытое табличками. Аборальная часть имеет удлиненную
коническую форму; ее верхняя часть покрыта крупными табличками,
налегающими друг на друга чешуевидно, а нижняя — многочисленными
мелкими (рис. 275).
Лепидоцистоидеи образуют обособленный класс среди типа иглоко-
жих, отличающийся по своему строению от эокриноидей, эдриоастеро-
идей и цистоидей. Класс лепидоцистоидей условно отнесен к подтипу
Crinozoa.
ПОДТИП ASTEROZOA. АСТЕРОЗОИ
К астерозоям (гр. aster — звезда, zoon — животное) относятся
подвижные иглокожие с пятилучевой симметрией тела, у которых раз-
личают центральный диск и отходящие от него радиальные лучи, или
руки, причем у одних лучи 'обособлены нерезко, у других довольно рез-
fco, Рот расположен внизу; скелет рук защищает радиальные каналы
а'йбулакральной системы, вдоль которых в два ряда расположены амбу-
ййкральные ножки. В ископаемом состоянии астерозои известны начи-
ная с ордовика. По особенностям строения центрального диска и лучей
астерозои подразделятся на три класса: Somasteroidea, Asteroidea,
Ophiuroidea (рис. 276).
КЛАСС SOMASTEROIDEA. СОМАСТЕРОИДЕИ
К классу сомастероидей (гр. soma — тело, aster — звезда) отнесе-
нд-йеболыпая группа довольно примитивных астерозои. Тело пятиуголь-
ной формы, лучи почти не обособлены от центральной части
(рис. 276,7). Верхняя сторона покрыта сетью известковых трех- и че-
тырехлучевых спикул. На нижней стороне расположен крупный рот
пятиугольной формы, снабженный челюстями — видоизмененными пар-
ными амбулакральными табличками. От рта по радиусам отходят пять
амбулакров, состоящих из двух рядов полуцилиндрических амбула-
кральных табличек, расположенных в чередующемся порядке и охваты-
вающих полностью или частично амбулакральные каналы. Между
двумя табличками проходят поры для выхода амбулакральных ножек. ।
Пищевые желобки, идущие ко рту вдоль амбулакров, прикрыты таб-
личками или защищены шипами. Под углом от амбулакральных табли-
468
чек к периферии лучей диагональными рядами отходят виргалии—
палочковидные таблички.
Сомастероидеи появляются в раннем ордовике (тремадоке) и пред-
ставляют наиболее примитивных астерозои, существовавших до девона.
Высказывается предположение, что сомастероидеи явились предковой
Рис. 276. Классы Somasteroidea (1), Asteroidea (2—8) и Ophiuroidea (9—10):
1 — Villebrunaster (ранний ордовик); 2 — реконструкция морской звезды с больбо-
поритами (Елтышева, 1955); 3 — Bolboporites из ордовика: За — общий вид, 36 — в
разрезе; 4 — Protopalaeaster (ордовик); 5 — Stenaster (ордовик); 6—8 — скелетные
элементы морских звезд: 6 — из ордовика, 7 — из силура, 8 — из девона; 9 — Aploco-
та (ранняя юра); 10 — Stephanoura (поздний девон);
амп — амбулакральное поле, вир — виргалии, пер — перистом
группой для морских звезд и офиур. Класс объединяет 6 родов
(рис. 276, 7). В последнее время высказано, не лишенное основания;
предположение, что современный род Platasterias, возможно, может
быть отнесен к классу сомастероидей.
КЛАСС ASTEROIDEA. МОРСКИЕ ЗВЕЗДА
Общая характеристика. Тело уплощенное, звездообразное или пяти-
угольное, центральный диск нерезко обособлен от пяти лучей, внутрь
которых продолжаются полость тела, отростки желудка и ряд внутрен-
них органов..Амбулакральные ножки снабжены подошвой и ампулами.
Мадрепорит крупный, лежит на верхней стороне. На этой стороне име-
ются многочисленные шипы и педицеллярии (у более поздних предста-
вителей) . Ордовик — ныне.
Строение тела. Морские звезды „имеют уплощенное тело с цент-
ральным диском (рис. 276, 2—5), постепенно переходящим в ради-
469
альнб расходящиеся пять лучей, или рук; у морских звезд, обитающих
в арктических водах, число лучей иногда достигает 50. Для характерис-
тики формы тела морских звезд пользуются соотношением между рас-
стоянием от центра диска до конца луча и таковым от центра диска до
края диска в промежутке между лучами. У обитателей коралловых
рифов тело может иметь форму круглой подушки, или булки; недавно
описана морская звезда в форме шара диаметром около 1 см. Размеры
тела морских звезд варьируют в больших пределах от 1 до 80—90 см,
в среднем преобладает диаметр 10—20 см. В руки продолжаются по-
лость тела, половые железы, слепые-выросты кишечника, радиальные
каналы амбулакральной системы, стволы нервной системы. Рот распо-
ложен на нижней стороне, на верхней стороне лежат анальное отвер-
стие, мадрепорит и мелкие отверстия половых желез. От рта отходят
открытые амбулакральные желобки, образованные двумя рядами плос-
ких амбулакральных табличек, к которым с каждой стороны примыкает
по ряду адамбулакральных табличек; к последним присоединяются по
два ряда краевых, образующих боковую стенку луча. Амбулакральные
ножки выходят через специальные отверстия, расположенные между
амбулакральными табличками. Ножки имеют на своих концах присоски,
а у основания сократимые пузырьки — ампулы. Предполагается, что
амбулакральные ножки у палеозойских морских звезд Не имели ампул.
Вокруг рта имеется скелетное неподвижное кольцо, образованное
двумя парами амбулакральных и одной парой адамбулакральных таб-
личек. Рот ведет в большой мешковидный желудок; у морских звезд с
длинными и гибкими руками желудок может выворачиваться наружу и
обволакивать добычу, переваривая ее без заглатывания. ‘Звезды с ко-
роткими руками обычно заглатывают свою добычу и затем неперева-
ренные остатки выбрасываются через рот. Пищей морских звезд слу-
жат двустворчатые моллюски, брахиоподы, губки, коралловые полипы.
Нервная система морских звезд довольно примитивна. Чувствительнйе
нервные клетки имеются в присосках амбулакральных ножек, у основа-
ний игл и педицеллярий. Последняя непарная амбулакральная ножка
каждого луча несет глазок, состоящий из отдельных глазных бокальчи-
ков; глазок реагирует на свет.
Верхняя сторона у одних морских звезд покрыта правильной систе-
мой известковых табличек, у других — она более гибкая, имеет сетча-
тый скелет, состоящий из известковых балочек; у третьих известковый
скелет исчезает совсем и верхняя сторона покрыта только кожей. Очень
часто на верхней стороне развиваются защитные образования — извест-
ковые иглы, шипы или микроскопические «пинцеты» — педицеллярии.
В отложениях ордовика Прибалтики были встречены известковые шипы,
описанные под родовым названием Bolboporites, принадлежащие, по-
видимому, морским звездам.
Большинство морских звезд раздельнополы, но нередки случаи
гермафродитизма. По внешнему виду обычно мужские и женские особи
неразличимы. Эмбриональное развитие может протекать со стадией
свободноплавающей личинки или может быть прямым. Двусторонне-сим-
метричная личинка бипиннария и брахиолярия, после непродолжитель-
ной жизни в толще воды оседает на дно, иногда прикрепляется к суб-
страту и, претерпевая метаморфоз, превращается в молодую звездочку.
При прямом развитии из яйцевых оболочек выходят молодые звезды,
внешне напоминающие взрослых. В редких случаях морские звезды раз-
множаются бесполым путем; при этом диск делится на две части, таким
образом, что лучи остаются неповрежденными; обе части начинают
вести самостоятельный образ жизни, восстанавливая недостающие лучи
470
31 части диска. У морских звезд высоко развита способность к регене-
рации; они способны восстанавливать утраченные лучи или при опас-
ности самопроизвольно их отбрасывать.
Основы систематики и классификация. На основании особенностей
строения скелета, наличия или отсутствия и характера педицеллярий
класс морских звезд разделен на пять отрядов: Platyasterida (средний
ордовик — ныне), Paxillosida (ранний ордовик — ныне), Valvalitida
(ранний ордовик — ныне), Spinulosida (средний ордовик — ныне),
Forcipulatida (ранний ордовик — ныне);
История развития астероидей. Морские звезды, по-видимому, про-
изошли от сомастероидей еще в кембрии, хотя ископаемые остатки ни
тех, ни других в отложениях этого возраста до сих пор не известны.
В раннем ордовике появляются представители трех отрядов: Paxillosida,
Forcipulatida и Valvalitida. В среднем ордовике возникают отряды
Platyasterida и Spinulosida. Если представители Platyasterida несут
довольно много черт сходства с сомастероидеями, то первые три ранне-
ордовикских отряда резко отличаются друг от. друга и от сомастеро-
идей.
Развитие каждого отряда шло своим путем, однако сведения об
.этих путях развития довольно скудны, так как остатки морских звезд
в ископаемом состоянии встречаются редко. В раннеюрское время су-
щественно обновляется состав морских звезд; .появляются новые подот-
ряды, существующие до наших дней. Всего известно около 120 родов
морских звезд. В современных морях и океанах обитают до 1700 видов.
КЛАСС OPHIUROIDEA. ОФИУРЫ, или ЗМЕЕХВОСТКИ
Общая характеристика. У офйур (гр. ophis — змея, oura — хвост)
центральный диск резко обособлен-от тонких удлиненных лучей. Рот
и мадрепорит внизу; анальное отверстие отсутствует. Скелет рта под-
вижный. Желудок мешковидный, не выворачивается. Отростки желудка
в лучи не заходят. Скелет лучей состоит из цельных, подвижно соеди-
ненных табличек (позвонков). Движение осуществляется при помощи
лучей и частично амбулакральных ножек. Первые офиуры известны на-
чиная с ордовика (аренига).
Строение тела. Центральный диск округло-пятиугольного сечения,
резко обособлен от длинных очень подвижных членистых лучей
(см. рис. 276, 9—10), напоминающих хвосты змей (отсюда' Название
змеехвостки). От морских звезд офиуры отличаются отсутствием от-
крытых амбулакральных борозд; .отростки, желудка не заходят в лучи.
Диаметр диска 1—2 см, но иногда достигает 10 см; длина рук, число
которых обычно равно 5, реже 4, 6, 7, 8—9, в несколько раз превышает
диаметр диска. Внутри лучей развит сильный внутренний скелет, со-
стоящий из отдельных позвонков; позвонки соединены друг с другом
специальными мышцами и связками; у одних поверхность' сочленения
позвонков гладкая, у других она состоит из ямок и бугорков. Снаружи
позвонки покрыты четырьмя рядами тонких табличек (брюшных и
спинных) и двумя рядами боковых. Амбулакральные ножки выходят
через отверстия в брюшных табличках; они лишены присосок и ампул
и выполняют функцию газообмена или осязания.
Центральный диск сверху прикрыт мелкими табличками, имеющими
вид чешуек; у основания лучей выделяется по паре более крупных
радиальных табличек — радиальные щитки. Таблички диска иногда
'бывают снабжены иголочками, бугорками, гранулами. Рот звездооб-
разной формы помещается. в центре диска на нижней стороне; в него
471
вдаются пять челюстей треугольной формы; они служат для захвата и
удержания пищи. Рядом с челюстными табличками расположены адо-
ральные щитки, охватывающие с боков оральный щиток; один из
оральных щитков пронизан порами и выполняет функцию мадрепорита.
Желудок большой, мешковидный, занимает почти весь диск, анальное
отверстие отсутствует.
Большинство змеехвосток раздельнополы, однако известны случаи
гермафродитизма. Из оплодотворенных в воде яиц развивается личинка
(офиоплутеус), ведущая планктонный образ жизни. После сложных
изменений маленькая офиура опускается на дно и начинает вести бен-
тосный образ жизни. Офиуры растут несколько лет и достигают макси-
мальных размеров на 4-6-м году жизни. Среди офиур также известны
случаи бесполого размножения — делением поперечного диска. Чаще
всего делятся офиуры, диск которых имеет 6 лучей; недостающие части
диска и лучей затем восстанавливаются. У офиур широко развито
явление регенерации.
Офиуры передвигаются при помощи двух пар лучей, при этом они
ведут себя как двусторонне-симметричные животные, направляя пятый
луч вперед или назад. При передвижении, особенно при закапывании,
частично используются амбулакральные ножки. Они же принимают
участие при захвате пищи, которой служат различные мелкие живот-
ные, водоросли, донный детрит. Офиуры обитают на различных грун-
тах, на глубинах от прибрежной зоны до абиссали; многие мелководные
виды живут на губках, кораллах, среди водорослей, между иглами
морских ежей; более глубоководные формы часто зарываются в ил
или песок, выставляя на поверхность свои лучи.
Первые офиуры появились в раннем ордовике (отряд Stenurida),
несколько позднее, но также в раннем ордовике возник отряд Oegophiu-
rida, имевший уже характерную структуру лучей. Оба других отряда
появились в конце силура —начале девона (Phrynophiuridfa, Ophiurida);
у них хорошо развиты в лучах позвонки. В начале юрского периода в
составе последнего отряда появляются два новых подотряда. Класс
офиур насчитывает окодо 70 родов и около 200 видов.
ПОДТИП ECHINOZOA. ЭХИНОЗОИ
К подтипу Echinozoa (гр. echinos — еж, zoon — животное) отно-
сятся преимущественно подвижные иглокожие, обладающие шаровид-
ным или дисковидным телом, без рук, брахиол или иного вида выдаю-
щихся лучей, свойственных астерозоям и кринозоям. Тело, как правило,,
радиально-симметричное, с ростом в меридиональном направлении.
Амбулакральная, нервная система и органы размножения расположены
по первичной пятилучевой симметрии, на которую у некоторых групп
накладывается адаптивная двусторонняя симметрия. Эхинозои появля-
ются в ранном кембрии (зоне Olenellus) и существуют доныне. В со-
став подтипа включены следующие классы: Echinoidea, Edrioasteroidea,
Holothurioidea, Ophiocistioidea, Helicoplacoidea, Cyclocystoidea, Campto-
stromatoidea (рис. 277).
КЛАСС ECHINOIDEA. МОРСКИЕ ЕЖИ
Общая характеристика. Морские ежи — свободноподвижные игло-
кожие, имеющие субсф.ерической, конусовидной или сердцевидной
формы скелет, внутри которого заключены основные органы. Скелет
472
Рис. 277. Предполагаемые родственные связи между классами и отрядами подтипа
Echinozoa (Фелл и Мур, 1966)
состоит из известковых табличек, расположенных меридиональными ря-
дами по пяти амбулакральным (А) и пяти интерамбулакральным (IA)
полям. Под каждым амбулакральным полем проходит радиальный ка-
кал, от которого отходят .амбулакральные ножки; для каждой ножки
в амбулакральной пластинке имеется пара пор. Перистом с ротовым
отверстием обращен к субстрату; у многих рот снабжен челюстным
аппаратом; перипрокт с анальным полем лежит на верхней стороне
Рис. 278. Схема строения панциря «правильных» (Z) и «неправильных» (2)
морских ежей (а — вид сверху, б — вид снизу);
поля: А — амбулакральное, IA — интерамбулакральное; ан — анальное
отверстие, би — бугорки для игл; вщ — вершинный щиток, гл — глазные
таблички, м — мадрепорит, п — половые таблички, па — поры амбула-
кров, пл — пластрон, пп — перипрокт, пс — перистом, пт — петалоиды,
р — рот, ф — фасциола
внутри вершинного щитка или в заднем интеррадиусе вне его. Вершин-
ный щиток, расположенный на верхней (аборальной) стороне панциря,
построен из пяти окулярных и четырех-пяти генитальных табличек.
Ордовик — ныне.
Строение тела. Тело морского ежа имеет разнообразную форму
(рис. 278). По форме тела, расположению рта и анального отверстия
различают две группы морских ежей: «правильных» и «неправильных».
У «правильных» морских ежей рот расположен в центре нижней по-
верхности, а анальное отверстие — на верхней стороне в центре вер-
1 шинного щитка; у двусторонне-симметричных, или «неправильных»,
474
морских ежей рот расположен в центре или у переднего края, а аналь-
ное отверстие — в заднем интеррадиусе вверху, сзади или на нижней
стороне вне вершинного щитка. У «правильных» морских ежей панцирь
бывает шаровидным, сплющенным у полюсов, уплощенным, дисковидным.
У «неправильных» морских ежей форма панциря более разнообразна
и может быть овальной, конусовидной, сердцевидной. Наиболее широ-
кая часть скелета называется амбитусом.
Тело снаружи одето тонкой кожей. Таблички панциря, лежащие в
средней части кожи, несут подвижно соединенные иглы различных раз-
меров и формы, от крупных до очень мелких (миллиарных). На нижней
стороне тела расположен перистом, или ротовое поле, затянутое
кожистой перепонкой, в центре которой расположен рот (см. рис. 278,/) ,
у некоторых снабженный челюстным аппаратом. Рот ведет в длинный
пищеварительный канал, делающий внутри тела несколько петель и за-
канчивающийся анальным отверстием, которое находится внутри пери-
прокта, наверху, в области вершинного щитка или вне его, сзади, или
на нижней поверхности.
Амбулакральная система построена так же, как у морских звезд.
Вокруг рта1 над челюстным аппаратом расположен кольцевой канал, от
которого отходят пять радиальных каналов; они проходят под амбу-
лакральными полями скелета и заканчиваются в окулярных табличках
вершинного щитка светочувствительным щупальцем. От радиальных
каналов через каждую пару пор, расположенных на известковых амбу-
лакральных табличках, выходят амбулакральные ножки, снабженные
ампулами и присосками; Ножки на нижней, или оральной, стороне тела
предназначены для движения, на верхней стороне выполняют функции
газообмена И хеморецепторов. :.
От кольцевого Канала вверх отходит каменистый канал, входящий
в состав осевого комплекса; каменистый канал заканчивается много-
численными порами на апикальном поле, или вершинном щитке, на од-
ной из генитальных табличек. Эта табличка называется мадрепо-
ритом.
Параллельно с амбулакральной расположена кровеносная система.
Ее кольцевой канал лежит над аристотелевым фонарем; от кольцевого
канала отходят стволы кровеносной системы, идущие к ротовому от-
верстию, вдоль . каменистого • канала, к аборальному кольцу, половым
железам и другим внутренним органам. Осевой комплекс, в состав
которого входит каменистый канал, целомическая полость, кровеносный
сосуд и нервный ствол, окружен единой оболочкой; он представляет
собой сложный орган, иногда называемый сердцем. Нервная система
также построена по пятилучевой системе; она. состоит из кольца и
расходящихся по радиусам радиальных нервных стволов, иннервирую-
щих амбулакральные ножки, иглы панциря, половые железы, челюстной
аппарат и все остальные органы. Половые железы расположены интер-
радиально в адапикальных частях панциря (у аборального полюса) и
открываются наружу через особые отверстия в генитальных табличках,
входящих в состав вершинного щитка. У «правильных» морских ежей
число гонад равно пяти, у «неправильных» может быть сокращено до
четырех, трех и редко двух.
Строение панциря. Все внутренние органы морских ежей заключены
в известковый скелет, или панцирь, построенный из известковых таб-
личек. Каждая табличка представляет собой единый кристалл каль-
цита, ось С которого ориентирована или перпендикулярно поверхности
тела, или тангенциально. Панцирь состоит из четырех систем табли-
чек: корональной, вершинного щитка, перипрокта и перистома.
475
Корональная система табличек образует основу скелета
любого морского ежа. У современных морских ежей она состоит из
20 меридиональных рядов табличек, образующих пять амбулакраль-
ных и пять интерамбулакральных полей. Каждое поле состоит из двух
рядов табличек. Амбулакральное поле расположено над радиальным,
каналом амбулакральной системы. Для_выхода одной амбулакральной
ножки в каждой табличке имеются две сближенные ' поры
(см. рис. 283). Интерамбулакральные таблички не несут никаких пор, а
обычно снабжены крупными и мелкими бугорками, к которым при-
крепляются иглы; этими особенностями они легко отличаются от амбу-
лакральных табличек. У палеозойских морских ежей, в отличие от
мезокайнозойских, амбулакры состояли из 2—20 рядов, а интерамбу-
лакры из 1—14 рядов табличек.
Амбулакральные таблички могут быть простыми и сложными;
простая табличка имеет вытянутую пятиугольную форму и несет одну
пару пор; сложная табличка состоит из нескольких сросшихся простых
табличек и несет несколько пар пор. Часто швы между слившимися:
табличками незаметны и об их числе судят по числу пар отверстий..
Различают несколько типов сложных табличек, развитых у .«правиль-
ных» морских ежей начиная с мезозоя; у палеозойских форм известны
только простые амбулакральные таблички. В сложной табличке раз-
личают первичные таблички, если они протягиваются вдоль всей таб-
лички, и полутаблички, если они слагают только ее краевые части..
У «неправильных» морских ежей сложные амбулакральные таблички
известны у представителей отряда Holectypoida. У большинства других
представителей этой группы пластинки остаются простыми, но внеш-
няя форма пор может существенно изменяться; они могут иметь оваль-
ную или каплевидную форму; в каждой паре поры могут либо соеди-
няться узкой бороздкой, либо отделяться небольшим бугорком. У «не-
правильных» морских ежей с четким разделением панциря на верх-
нюю и нижнюю поверхности амбулакры верхней части часто расши-
ряются посредине и приобретают форму лепестков, или петалоидов.
В таком случае говорят о петалОидной части амбулакра. В петало-
идах таблички удлинены, поры имеют овальную или щелевидную-
форму. Петалоидные части амбулакров могут быть выпуклыми (отр.
Clypeasteroida) или углубленными (отр. Spatangoida). В приротовых
частях у некоторых представителей отряда Cassiduloida амбулакры
могут принимать листовидную форму, образуя так называемые ф и л-
лодии. Поры филлодий остаются круглыми, но каждая пара поме-
щается в особом углублении.
Интерамбулакры состоят из относительно крупных табли-
чек, несущих большие бугорки, или туберкулы. Новые интерамбу-
лакральные таблички, как и амбулакральные, возникают у края оку-
лярных, входящих в состав апикального поля. Как было отмечено
выше, у палеозойских морских ежей в состав интерамбулакров могло-
входить до 14 рядов табличек, у всех постпалеозойских — только два
(за исключением некоторых аберрантных форм). Интерамбулакры,
построены сравнительно просто и претерпевают существенный измене-
ния лишь у сердцевидных морских ежей на оральной стороне. У по-
следних интерамбулакр, примыкающий сзади к ротовому отверстию,,
расширен и имеет вид щита, или пластрона (рис. 279). Приротовая
табличка интерамбулакра, примыкающая к ротовому отверстию, вытя-
гивается и иногда образует губу, или л а б р у м. В зависимости от рас-
положения следующих за ней интерамбулакральных (стернальных и
амфистернальных) табличек различают четыре типа пластрона:
476
а) амфистернальный — за лабрадьной табличкой расположена
пара крупных стернальных табличек, протягивающихся почти до амби-
туса скелета и образующих стернум, или вентральный щит; б) мери-
.достернальный — за одной табличкой,, прилегающей к ротовой,
«следуют два ряда клиновидных табличек, связанных зигзагообразным
швом; в) ортостернальный — состоит из одной крупной стер-
Рис. 279. Типы пластронов (1—5) и фасциол (6):
1—2 — амфистернальный: 1 — без стернума, 2 — со стернумом; 3 — меридостерналь-
ный; 4 — ортостернальный; 5 — метастернальный; 6 — типы фасциол: а—анальная,
лт — латеральная, м — маргинальная, пп — перипетальная, са — субанальная
калькой таблички, примыкающей к лабруму, и двух следующих за ней
симметрично расположенных табличек;' г) метастернальный —
за лабрумом следуют три или четыре таблички, образующие один ряд.
Почти на всех табличках панциря имеются бугорки, или туберку-
лы, к которым прикрепляются иглы (рис. 280). Туберкулы имеют
вид усеченного конуса, который заканчивается круглой головкой, пред-
ставляющей поверхность для сочленения с иглой. Сочленение осу-
ществляется при помощи связки и мышц. Бугорки на днтерамбула-
жральных табличках (IA) достигают большой величины и расположе-
ны на округлой или овальной площадке — ареоле, или дворике.
.Ареола бывает окаймлена мелкими бугорками, или гранулами. Голов-
ка туберкула может иметь ямку для связки, прикрепляющей иглу;
такой туберкул называют перфорированным; если его основной
конус рассечен насечками, то его называют к рену л и р о в а иным.
У «неправильных» сердцевидных морских ежей на оральной стороне, в
.задней части панциря и некоторых других местах, обособляются участ-
ки с более крупными бугорками, что связано с прикреплением крупных
специализированных игл.
У многих' сердцевидных морских ежей (спатангоидов) имеются
•очень мелкие булавовидные иглы, расположенные на мельчайших бу-
горках, образующих узкие ленты, или фасциолы (лат. fasciola —
.лента). Различают несколько типов фасциол (рис. 279, 6):
а) субанальная — эллиптической формы, расположена ниже пери-
прокта (у рода Micraster), б) маргинальная — протягивается
вдоль амбитуса в основании верхней поверхности (род Cardiaster),
в) перипетальная — окружает все петалоиды (род Hemiaster),
г) внутренняя — окружает вершинный щиток, д) латеральная
(латероанальная) — отходит от перипетальной, е) анальная —
окружает перипрокт. Функциональное значение игл (клавул), рас-
477
положенных на фасциолах, связано с выделением слизи, скрепляющей
стенки норы у спатангоидов, созданием токов воды в определенном
направлении и санитарных условий в норке.
Иглы у морских ежей отличаются.большим разнообразием форм
и размеров, от микроскопических миллиарных до гигантских (до 30 см),
превышающих в пять раз диаметр панциря. Иглы бывают: цилиндри-
ческие, заостренные, булавовидные, лопатовидные (рис. 280).
В каждой игле различают: тело иглы, или стержень,
Рис. 280. Форма игл (1) и педицеллярий (2); 2а — змееголовая, 26 — шаронос-
ная, 2в, г — трезубая;
гл — головка, ст — стержень, ш — шейка
удлиненную шейку, с кольцевым валиком и головку. Головка
представляет собой утолщенный конец иглы с углублением, служащим
для сочленения с бугорком. Игла прикрепляется к бугорку с помощыо>
связки, расположенной в ее центре, и мускулов, идущих от кольцевого
валика шейки к ореоле. Сверху шейка и головка иглы покрыты кожей.
Иглы служат для разных целей: защиты, осязания, у части «правиль-
ных» морских ежей (отр. Cidaroida) для передвижения; у некоторых,
сердцевидных морских ежей для закапывания (в этом случае они
приобретают лопатовидную форму).
На теле морского ежа между крупными иглами имеются видоиз-
мененные иглы — педицеллярии (рис. 280, 2), состоящие из сте-
белька с известковой палочкой внутри и головки, построенной из трех,
известковых челюстеобразных створок; последние при помощи муску-
лов могут открываться и закрываться. Различают четыре основных
типа педицеллярий: а) шароносные — головка расширена и створ-
ки заканчиваются острым зубом; у некоторых морских ежей они несут
ядовитые железы; б) трезубые, обладающие тремя створками дли-
ной 2—3 мм; в) змееголовые — наиболее часто расположены в
приротовой области; концы их створок закруглены и напоминают
голову змеи; г) трилистные, имеющие створки в виде листочка с
тонкозазубренными краями. После гибели морскйх ежей педицелля-
рии легко отделяются от панциря и поэтому почти неизвестны в иско-
паемом состоянии. Педицеллярии служат для захвата добычи, защи-
ты от личинок, стремящихся поселиться на твердом панцире, очистки
панциря от инородных тел.
478
Вершинный щиток, или апикальное поле, представляет собой вто-
рую систему табличек панциря (рис. 281). Он расположен на его
верхней стороне и построен из окулярных и генитальных табличек..
У правильных морских ежей он состоит из пяти окулярных и пяти
генитальных табличек. Окулярные, или глазные, таблички несут
по одному отверстию, через которое выходит щупальце радиального
канала, заканчивающееся светочувствительным глазком; поэтому их
и называют окулярными. От них начинаются .амбулакральные поля*
Рис. 281. Типы вершинных щитков; •
1 — моноциклический (Melonechinus); 2—3 — дициклический (2 — Strongylocen- '
trotus, 3 — Pseudodtadema); 4^-5 — компактный (4 — Conulus, 5 — Holectypus); ;
6 — удлиненный (Echinocoris); 7 — монобазальный (Clypeaster)-, 8 — разор- -
ванный (Collyrites)',
поля: А — амбулакральные (/—V), IA — интерамбулакральные (1—5); пп —
перипрокт, таблички: гл — глазные, м — доадрепоровая. п — половая
и около них возникают новые таблички корональной системы как ам-
булакральные, так и интерамбулакральные. Генитальные, или
половые, таблички обычно крупнее окулярных и могут нести не только
одно отверстие, но иногда и несколько; через эти отверстия в воду
выводятся половые продукты. Одна из генитальных табличек пронизана
многочисленными гидропорами, которыми наружу открывается камени-
стый канал. Эта мадрепоровая табличка крупнее остальных. Оку-
лярные и генитальные таблички образуют вокруг перипрокта оДно
или два кольца. Если все окулярные и генитальные таблички сопри-
касаются с перипроктом, то такая апикальная система называется
однорядной, или моноциклической (рис. 281,/) если с пери-
проктом контактируют только генитальные таблички, образующие
одно кольцо, а окулярные лежат между ними и образуют второе
кольцо, то такая система называется двухрядной, или ди цикли-
ческой (рис. 281, 2). .
Однорядный тип вершинного щитка характерен для большинства
палеозойских морских ежей, а двухрядный — для мезокайнозойских.
В центре апикальной системы у правильных морских ежей расположен
перипрокт с анальным отверстием. Вершинный щиток с перипроктом в
479
центре называется эндоциклическим. Перипрокт затянут кожи-
стой пленкой и обычно покрыт табличками. Число и размеры этих таб-
личек у разных групп морских ежей довольно сильно варьируют.
В ископаемом состоянии они обычно не сохраняются.
У «неправильных» морских ежей, названных так в связи со сме-
щением перипрокта из области вершинного щитка по пятому интер-
амбулакру назад, изменяется строение вершинного щитка. Вершинный
щиток с перипроктом, лежащим вне апикального поля, называется
экзоциклически м. Впервые такое смещение наблюдается у ран-
неюрского рода Pygaster-, перипрокт еще примыкает к апикальному
полю, но при этом редуцируется одна гонада и соответственно одна
генитальная табличка. Оставшиеся четыре генитальные таблички зани-
мают место перипрокта, смыкаются вместе и образуют компактный
вершинный щиток (см. рис. 281, 4—5). При дальнейшем перемещении
перипрокта назад вдоль интерамбулакра вершинный щиток удлиняется,
передняя и задняя пары генитальных табличек оказываются разделен-
ными парой сближенных окулярных табличек; в этом случае на перед-
нем конце вершинного щитка расположена одна, а на заднем две оку-
лярные таблички; такой вершинный щиток получил название удли-
ненного, или интеркалярного (см. рис. 281, 6). Если скелет
морского ежа удлиняется, становится двусторонне-симметричным, то
задняя пара глазных табличек может отделиться от основной части
щитка и тогда образуется разорванный вершинный щиток (см.
рис. 281, 8). У некоторых морских ежей имеется так называемый
монобазальный щиток, когда все генитальные таблички сраста-
ются в один диск, по краям которого располагаются окулярные таб-
лички; мадрепорит лежит в центре, а генитальные поры по углам
диска (рис. 281, 7).
Перипрокт представляет собой отверстие в панцире для выхода
ануса. Обычно оно затянуто эластичной мембраной, через которую
проходит задний конец пищеварительного тракта. Как было отмечено
выше, у «правильных» морских ежей перипрокт расположен внутри
вершинного щитка, у «неправильных» — вне его, на заднем интерра-
диусе. Перипрокт у большинства морских ежей прикрыт различным
числом табличек (от 4 до нескольких десятков), чаще всего располо-
женных концентрически. В ископаемом состоянии таблички перипрокта
сохраняются редко.
Перистом — приротовое поле различной формы и величины, в
центре которого расположено ротовое отверстие; перистом затянут ко-
жистой мембраной, обычно покрытой известковыми табличками, ле-
жащими чешуеобразно друг на друге. У «правильных» морских ежей
вокруг рта расположены пять пар крупных букальных табличек, несу-
щих по одной паре пор для амбулакральной ножки. У современных
«неправильных» морских ежей перистом закрыт чешуйчатыми таблич-
ками без пор. У «правильных» морских ежей амбулакральные и интер-
амбулакральные таблички вокруг перистома несколько приподняты и
загнуты внутрь, образуя выросты; эти выросты на амбулакральных
табличках называются ушками, или аурикулами, а на интер-
амбулакральных — апофизами. Ушки могут быть незамкнутыми,
ёсли их края свободны, или замкнутыми, когда концы аурикул соеди-
няются вместе. Аурикулы и апофизы образуют околочелюстной,
или перигнатический, поясок (рис. 282, 1); он служит для при-
крепления мускулов аристотелева фонаря.
Аристотелев фонарь состоит из 40 известковых скелетных эле-
ментов: пяти зубов, пяти пар апофизов, пяти двойных пирамидок, пяти
480
дужек (состоящих из двух сросшихся половинок) и пяти скобок. Зубы
имеют форму зубила; у палеозойских форм и некоторых «правильных»
морских ежей они снабжены желобками, у остальных «правильных» и
челюстных «неправильных» морских ежей они имеют внутренний про-
дольный киль. Зубы заключены в пирамидки, составляющие главную
часть челюстного аппарата; сверху пирамидки горизонтально лежат
пять скобок и пять двойных дужек. Весь челюстной аппарат приводится
Рис. 282. Схема строения челюстного аппарата:
1 — реконструкция; 2—5 — типы .пирамид и сечения зубов: 2 — цидароидный,
3 — аулодонтный, 4 — стиродонтный, 5 — камародонтный;
поля: А — амбулакральное, IA — интерамбулакральное; ап — апофизы, ау —
аурикулы
в движение 60 мускулами; он расположен либо вертикально, либо го-
ризонтально (у форм с уплощенным панцирем). Предполагается, что
некоторые части челюстного аппарата являются измененными амбула-
: кральными и интерамбулакральными табличками. У части «неправиль-
ных» морских ежей челюстной аппарат редуцирован и следы его обна-
• ружены в онтогенезе некоторых представителей. Строение зубов и пи-
s'; рамидок имеет таксономическое значение. У цидароид зуб снабжен
желобком, отростки, или эпифизы, пирамидок маленькие; у аулодонт-
ных зубов имеется желобок, эпифизы крупные, но друг с другом не
• соединяются; у стиродонтных зубов тянется продольный киль, эпифизы
крупные; у камародонтных зубов развит киль, эпифизы соединены,
образуя дужку пирамидки (рис. 282).
У части «неправильных» морских ежей сохраняется центральное
/ положение перистома и челюстной аппарат, при этом иногда реду-
) цируются отдельные его элементы и изменяется их положение из вер-
у тикального на наклонное и даже почти горизонтальное. При переме-
•' щении вперед перистома у «неправильных» морских ежей челюстной
аппарат редуцируется и исчезает. Форма ротового отверстия изменяет-
ся и изменяется также ориентировка перистома по отношению к суб-
страту (см. рис. 290). Перед перистомом на переднем крае панциря
развивается борозда для амбулакра. Она может быть различной глу-
бины и длины.
Кроме рассмотренных выше частей скелета у морских ежей в мяг-
, ких тканях имеются отдельные скелетные элементы — спикулы и пла-
стинки. Они располагаются в стенках амбулакральных ножек, в пе-
ристомальной мембране, в стенке пищеварительного тракта.
31 В В. Друщии
481
Размножение и развитие. Морские ежи раздельнополы, однако-
половой диморфизм выражен редко. Оплодотворение, как й у морских
звезд, происходит в воде. Из оплодотворенного яйца развивается ли-
чинка — вначале диплеврула, затем эхиноплутеус; из левой части
личинки выпочковывается зародыш ежа, который на этой стадии (диа-^
метр тела около 1 мм) своей пятиугольной формой с равной оральной и
аборальной сторонами похож на молодую морскую звезду. Первона-
чально все скелетные элементы расположены в коже в виде отдельных,
табличек, затем они срастаются и образуют радиально-симметричный
панцирь. В дальнейшем новые таблички возникают под глазными таб-
личками вершинного щитка и постепенно выталкиваются наружу и
старые таблички постепенно отодвигаются книзу. Число рядов табли-
чек, слагающих амбулакральные и интерамбулакральные поля, остает-
ся постоянным, и изменяются только их размеры, толщина и форма..
На табличках сохраняются линии нарастания. В онтогенезе «непра-
вильный» морской еж проходит радиально-симметричную стадию, свой-
ственную «правильным» морским ежам. Перестройка панциря по дву-
сторонне-симметричному плану осуществляется за счет неравномерного
роста в различных зонах. Наличие в онтогенезе «неправильных» мор-
ских ежей радиально-симметричной стадии позволяет предполагать,
что они возникли от «правильных» морских ежей.
Основы систематики и классификация. На основании строения
панциря, вершинного щитка, околочелюстного пояса класс морских
ежей в настоящее время предложено разделять на два подкласса:
Perischoechinoidea и Euechinoidea.
ПОДКЛАСС PERISCHOECHINOIDEA. ПЕРИШЭХИНОИДЕИ
К подклассу перищэхиноидей относятся все палеозойские морские
ежи и один отряд Cidaroida, доживший до наших дней. Панцирь по-
строен из амбулакральных и интерамбулакральных полей. Амбула-
кральное поле состоит из 2—20 рядов табличек, интерамбулакральное
из 1—14 рядов. Наружные перистомальные жабры отсутствуют; пери-
гнатический, или околочелюстной, поясок отсутствует или представлен
только апофизами; имеется челюстной аппарат, амбулакральные таб-
лички простые, несут одну пару пор каждая. Вершинный щиток эндо-
циклическйй однорядный или двухрядный. Ордойик — ныне. В состав
подкласса входят четыре отряда: Bothriocidaroida, Palaechinoida,.
Echinocystitoida и Cidaroida.
Отряд Bothriocidaroida. Ботриоцидароиды (гр. bothrion — ма-
ленькая ямка, cidaris — диадема, тюрбан персидских царей) обра-
зуют небольшой отряд очень своеобразных «правильных» морских
ежей, отличающихся от всех остальных наличием пяти однорядных
полей, неперфорированных табличек и пяти двухрядных.полей, каждая
табличка которых несет пару пор и перфорированный бугорок на краю-
таблички (рис. 283). Радиальный канал проходит внутри неперфориро-
ванных табличек, от него под углом расходятся латеральные каналы,
соединяющиеся с амбулакральными ножками. Вершинный щиток со-
стоит из пяти, по-видимому генитальных, а не окулярных, как предпо-
лагалось раньше, табличек, из которых одна выполняла роль мадре-
порита. Перистом пятиугольной формы; челюстной аппарат несколько-
напоминает «аристотелев фонарь» более поздних морских ежей, снаб-
жен пятью зубами. Известно два рода из ордовика.
Отряд Palaechinoida. Палеэхиноиды (гр. palaios — древний) —
небольшой отряд палеозойских морских ежей, обладавших жес-тким
482
панцирем. Амбулакральные таблички налегают на лежащие рядом
интерамбулакральные. Околочелюстной поясок отсутствует, перистом
покрыт амбулакральными и интерамбулакральными табличками;
вершинный щиток эндоциклический. Амбулакры состоят из двух и
более рядов простых табличек, интерамбулакры — из одного и более
рядов. Силур — пермь. В состав отряда входит 10 родов (рис. 284, 3),
Рис. 283. Отряд Bothriocidaroida:
1 — Bothriocidaris (средний — поздний ордовик): 1а — общий вид сбоку,
16 — схема челюстного аппарата (Мяннилы, 1962); 2 — Neobothriocidaris (ор-
довик): 2а — схематический разрез через радиальный канал и реконструкция
амбулакральной ножки (Поль, 1967), 26 — деталь строения амбулакра и
интерамбулакра, 2в — реконструкция части амбулакральной системы;
ам — ампула, би —бугорок для иглы, бк — боковые каналы/н — амбула-
кральная ножка, рк — радиальный канал; таблички: нпт — неперфорирован-
ные (М), пт — перфорированные (Л); ча — челюстной аппарат
Отряд Echinocystitoida. Эхиноциститоиды (гр. kystes—пузырь).
Панцирь нежесткий, йзменчивой формы: шарообразный у ордовик-
ских, сплющенный у карбоновых и пермских; таблички черепицеоб-
разно налегают друг на друга. Каждое амбулакральное поле состоит
из 2—20 рядов простых табличек, несущих одну пару пор каждая.
Интер амбулакр альное поле состоит из 2—14 рядов табличек. Вершин-
ный щиток эндоциклический, однорядный. Рот в центре перистома, по-
крытого только амбулакральными табличками. Перигнатический поясок
отсутствует. Пирамидки и зубы челюстного аппарата расположены на-
клонно. Ордовик — пермь. Отряд включает около 25 родов (рис. 284,
1—2). Род Eothuria (ордовик) относился некоторыми исследователями
к голотуриям, однако строение амбулакральных и интерамбулакраль-
ных полей, наличие генитальных табличек указывают на его близость
к морским ежам, а не к голотуриям.
Отряд Cidaroida. Цидароиды (гр. cidaris — диадема, тюрбан пер-
сидских царей), или копьеносные ежи, имеют шаровидный панцирь,
уплощенный сверху и снизу. Амбулакральные поля узкие, лентовидные,
31
483
состоят из двух низких простых табличек с одной парой пор каждая.
Интерамбулакральные поля состоят из двух рядов (у палеозойских от
4 до 8) широких табличек. Вершинный щиток, как правило, двухряд-
ный, эндоциклический. Перистом сплошь покрыт табличками. Жабры
отсутствуют. Аристотелев фонарь прикрепляется к апофизам около-
челюстного пояска. Аурикул нет. Интерамбулакральные таблички не-
Рис. 284. Отряды Echinocystitoida (1—2), Palaechinoida (3) и Cidaroida
(4-5):
1.— Lepidesth.es (девон — карбон); 2 — Lepidocentrus (девон — ранний
карбон); 3 — Melonechinus (ранний карбон); 4 — Archaeocidaris (ран-
ний карбон), вид снизу; 5 — Cidaris (совр.), вид сверху;
поля: А — амбулакральное, IA — интерамбулакральное; ча — челюст-
ной аппарат
сут многочисленные гранулы и по одному крупному, часто кренулиро-
ванному, бугорку с ямкой в центре. Иглы массивные, длинные
(I —10 см), по 2—9 в каждом ряду; шароносные трезубые педицелля-
рии прикрепляются к мелким бугоркам. Они выполняют очистительную
и хватательную функции.
Первые находки, относимые к копьеносным морским ежам, извест-
ны из отложений силура и позднего девона; они представлены только
остатками игл, панцирь не обнаружен; достоверные находки панцирей
установлены начиная с карбона, причем. скелет состоял из налегающих
друг на друга табличек и поэтому, как правило, после гибели распа-
дался на части. Жесткий панцирь появляется у триасовых цидароид.
Отряд включает 5 семейств и 65 родов (рис. 284, 4—5). Современные
цидароиды объединяют около 30 родов, распространенных по всем
океанам й морям; они живут как в зоне литорали, так и на больших
глубинах (до 4000 м), но чаще всего на глубинах менее 500 м.
484
ПОДКЛАСС EUECHINOIDEA. ЭУЭХИНОИДЕИ,
или НАСТОЯЩИЕ ЕЖИ
Подкласс эуэхиноидей (гр. ей — настоящий) объединяет «правиль-
ных» и «неправильных» морских ежей, обладающих жестким панцирем,
построенным из пяти меридиональных амбулакральных и интерамбу-
лакральных полей, чередующихся между собой; каждое поле в отли-
чие от перишэхиноидей состоит из двух рядов табличек. Перипрокт
эндоциклический («правильные») и экзоциклический («неправильные»
морские ежи). Челюстной аппарат имеется у всех «правильных» и час-
ти «неправильных» ежей. Жабры и жаберные щели имеются у неко-
торых представителей. Педицеллярии всех типов. Подкласс разделен
на 3 надотряда: Diadematacea, Gnathostomata, Atelostomata.
НАДОТРЯД DIADEMATACEA.
ДИАДЕМОВЫЕ МОРСКИЕ ЕЖИ
Надотряд объединяет большое число «правильных» морских ежей,
напоминающих цидароид, имеющих жесткий полушаровидный панцирь.
Амбулакральные таблички обычно сложные, образованы из не-
скольких простых. Вершинный щиток моно- и дициклический; пери-
Рис. 285. Надотряд Diadematacea:
1 — схема строения амбулакральных (А) и интерамбулакральных (/А) табли-
чек; 2 — развитие сложных амбулакральных табличек; 3 — отряд Diadematoida,
Lissodiadema (совр.): За — вид сверху, Зб — вид снизу; 4 — отряд Echinothu-
riCida, Pelanechinus (поздняя' юра): 4а — вид снизу, 46 — амбулакральные
таблички
прокт всегда эндоциклический. Вокруг перистома развиты вторичные
жабры; челюстной аппарат с зубами аулодонтного, .стиродонтного и
камародонтного типа, прикрепляется к апофизам и аурикулам пери-
гнатического пояска. Иглы разнообразной формы и размеров, преиму-
щественно полые; у современных диадемовых ежей иглы могут дости-
гать длины 30 см; железистые клетки, расположенные в коже, окру-
485'
жающей иглы, способны выделять ядовитый секрет. Диадемовые ежи
передвигаются на амбулакральных ножках; большинство живет в глу-
боких водах, но имеются обитатели литорали и верхней сублиторали,
особенно в тропиках и субтропиках. Надотряд насчитывает свыше
250 родов (рис. 285, 286).
гл
Рис. 286. Надотряд Diadematacea:
1 — Salenia (мел — ныне): 1а — вид сверху, 16 — вид сбоку, 1в — вид сни-
зу; 2 — Hemicidaris (средняя юра — мел), вид сверху;
поля; А — амбулакральное, IA — интерамбулакральное; пп — перипрокт,
таблички: гл — глазные, п — половые
Описанный надотряд американские эхинологи разделяют на два
надотряда: Diadematacea и Echinacea, считая, что они представляют
две ветви одного общего ствола, берущего начало от цидароид. Однако
черт сходства между этими надотрядами значительно больше, чем
различий, и поэтому они объединены в один надотряд.
НАДОТРЯД GNATHOSTOMATA.
Г Н АТ О С Т О МАТ Ы, или Ч Е Л Ю С Т Н О Р О Т Ы Е
Челюстноротые морские ежи (гр. gnathos — челюсть, stoma —
рот) обладают жестким панцирем, челюстным аппаратом с килеватыми
зубами; перипрокт'экзоциклический, расположен вне апикального поля;
последнее состоит из 5 окулярных и 4—5 генитальных табличек. Амбу-
лакральные таблички пррстые; бугоркй для игл перфорированные и
кренулированные. В состав надотряда включено два отряда: Holecty-
poida и Clypeasteroida.
Отряд Holectypoida. Голектипоиды (гр. holos — целый, ectypos —
вырезанный). Панцирь с уплощенной или даже вогнутой нижней сто-
роной и полушаровидной или конусовидной верхней. В амбулакраль-
ных и интерамбулакральных полях — по два ряда табличек. Амбулак-
ры лентовидные или петалоидные; амбулакральные таблички простые,
соединены по три, без образования сложной таблички. Вершинный
щиток компактный, состоит из 5 окулярных и 4—5 генитальных таб-
личек. Перипрокт расположен вне апикального поля, в заднем интер-
радиусе на верхней или даже на нижней стороне скелета. Перистом
центральный, с выемкой для жабр. Околочелюстной пояс с аурикулами
и апофизами. Аристотелев фонарь у одних (юрских и меловых) сохра-
няется в течение всей жизни, у других (юрских — современных)
имеется только на ранних стадиях онтогенеза. Интерамбулакральные
таблички несут по одному бугорку, кренулированному или гладкому.
Бугорки расположены меридиональными и диагональными рядами.
Представители отряда были хорошо распространены в юрских и мело-
вых морях. В настоящее время известны только два рода: род Echino-
neus живет в тропических морях от зоны литорали до глубины 120 м,
486
Micropetalon известен около Гавайских островов и в Ю.жно-Китайском
море. Отряд насчитывает всего 30 родов (рис. 287, 1).
Дарэм и Мелвий (1957, 1962) выделили род Pyga&ter и еще два
.рода в отдельный отряд Pygasteroida (юра — мел) и отнесли его к
диадематоидным морским ежам.
Рис. 287. Надотряд Gnathostomata:
1 — отряд Holectypoida, Conulus (поздний мел): 1а — вид сзади, 16 —
вид снизу; 2 — отряд Clypeasteroida, Clypeaster (эоцен — ныне): 2а —
вид сверху, 26 — вид сбоку, 2в — вид снизу;
поля: А — амбулакральное, 1А — интерамбулакральное; ан — анальное
отверстие, р — рот
Отряд Clypeasteroida. Клипеастероиды (гр. clypeus — щит, aster —
звезда). Панцирь уплощенный, дисковидный, реже шлемообразный или
конический. Амбулакры на верхней стороне обычно петалоидные, раз-
личной формы и очертаний, амбулакральные таблички простые, в пе-
лалоидах — сложные; кроме обычных пор имеются мелкие вторичные
поры, расположенные на амбулакральных и интерамбулакральных
табличках. Вершинный щиток монобазальный, с центральной
табличкой, образованной за счет слияния генитальных. Перистом рас-
положен в центре, кожистый, без перистомальных жабр. На оральной
поверхности у многих представителей развиты пищевые желобки; че-
люстной аппарат прикрепляется к аурикулам, а у уплощенных форм
пирамидки занимают наклонное положение. Внутри панциря обычно
развиты дополнительные скелетные образования. Иглы полые, за-
остренные на концах или тупоголовые, небольших размеров, располо-
жены на поверхности панциря обычно очень густо. У современных
форм в тканях внутренних органов, в стенках амбулакральных ножек
имеются известковые спикулы, до сих пор не обнаруженные в ископае-
мом состоянии. Педицеллярии разных типов. Поздний мел — ныне.
'Отряд насчитывает до 75 родов (рис. 287, 2).
НАДОТРЯД ATELOSTOMATA. АТЕЛОСТОМАТЫ
К ателосто'матам (гр. atelos — незаконченный, stoma — рот) отно-
сятся морские ежи, обладающие жестким панцирем, построенным из
20 рядов табличек; перипрокт расположен вне вершинного щитка в
заднем интерамбулакре. Вершинный щиток экзоциклический. Перистом
в центре или смещен к переднему краю. Челюстной аппарат и жабер-
487
ные щели на взрослой стадии отсутствуют. Амбулакральные таблички
всегда простые, интерамбулакральные — обычно крупнее амбулакраль-
ных; первичные бугорки перфорированные и кренулированные; иглы
полые. Юра — ныне. В состав надотряда входят два отряда: Cassidu-
loida и Spatangoida.
Отряд Cassiduloida. Кассидулоиды (лат. cassis — шлем). Панцирь
от округленного у древних форм до удлиненного у послеюрских.
Амбулакры простые, лентовидные или петалоидные; у юрских форм
Рис. 288. Надотряд Atelostomata, отряд Cassiduloida:
1 — схема строения панциря: 1а — вид сверху, 16 — вид снизу; 2 — Cassi-
dulus (эоцен — ныне): 2 а — вид сверху, 26 — вид снизу;
, поля: А — амбулакральное, 1А — интерамбулакральное
петалоиды простираются до амбитуса, позднее их размеры уменьша-
ются и их расположение ограничивается только верхней частью скеле-
та, что, по-видимому, связано с полузарывающимся образом жизни,
при котором петалоиды всегда остаются над осадком. На нижней
оральной стороне амбулакры образуют листовидной формы расшире-
ния, называемые филлодиями; в процессе эволюции филлодии рас-
ширяются и число пар пор сокращается, у некоторых послесеноман-
ских кассидулоид иногда вне петалоидов развиваются одинарные по-
ры. Перипрокт экзоциклический, у одних он почти соприка-
сается с вершинным щитком, у других занимает краевое нижнее или
почти краевое положение. Вершинный щиток у юрских и раннемело-
вых форм тетрабазальный, начиная с позднего мела становится моно-
базальным. Перистом занимает почти центральное положение или
несколько смещен вперед. Челюстной аппарат и жабры на взрослой
стадии отсутствуют. Бугорки без ямок; иглы мелкие. Ранняя юра —
ныне. Отряд насчитывает до 60 родов; в современных морях живет
7 родов (рис. 288), ведущих полузарывающийся образ жизни. Род.
Neolampas (совр.) вместе с шестью кайнозойскими родами Дарэм и.
Вагнер (1966) выделяют в самостоятельный отряд Neolampoida; у неп>
не развиты петалоидные амбулакры. Отряд изучен недостаточно. Воз-
можно, он объединяет неотенические формы кассидулоид.
Отряд Spatangoida. Спатангоиды, или сердцевидные морские ежи
(гр. spatanges — один из видов морских ежей), в большинстве своем
ведут зарывающийся образ жизни. Панцирь удлиненно-овальный, на
чаще сердцевидный, с бороздкой впереди для амбулакрального поля.
488
Вершиный щиток состоит из четырех генитальных и пяти окулярных
табличек (рис. 289); у одних он удлиненный, у других — разорванный,
у третьих — компактный этмофрактный или этмолический; у э т м о-
фрактного четыре генитальные таблички соприкасаются между
собой, у этмолического — мадрепоровая табличка вытянутая и
разделяет задние окулярные таблички.
Перипрокт расположен на заднем конце панциря, на особой плсг-
щадке. Рот обычно смещен к переднему краю, иногда имеет полулун-
Рис. 289. Отряд Spatangoida:
Г — схема строения панциря Collyrites, вид сверху; 2 — Echinocorys (поздний:
мел): 2 а вид снизу, .26 — вид сбоку;
поля: А — амбулакральное, /А — интерамбулакральное; ан — анальное от-
’ верстие, р — рог
ную форму. Передний амбулакр расположен в углублении. За ртом ле-
жит приротовая табличка, или ла б рум, за которой находится
пластрон. На верхней части панциря амбулакры обычно располо-
жены в петалоидах. Хорошо развиты фасциолы. Иглы, покрывающие-
панцирь, отличаются разнообразием: на фасциолах расположены мел-
кие иголочки (клавули), снабженные ресничками; на пластроне —
крупные веслоподобные иглы, помогающие при передвижении; малень-
кие иголочки вокруг перистома и вдоль петалоидных амбулакров слу-
жат для защиты амбулакральных ножек, и, наконец, имеются длинные-
иглы, расположенные на верхней и задней поверхностях панциря, дей-
ствия которых связаны с респираторным и санитарным каналами.
Спатангоиды ведут зарывающийся образ жизни, передвигаются в-
осадке или неподвижно сидят в специально устроенных норках. Каж-
дая норка (рис. 290) имеет два канала — один, идущий вверх, —
дыхательный, или респираторный, другой — расположенный сзади,,
является санитарным. Ножки и иглы принимают участие в постройке-
каналов; ножки, расположенные вокруг перистома, помогают в сборе-
пищи.
В настоящее время спатангоиды встречаются в морях с нормаль-
ной соленостью на всех глубинах, от зоны литорали до больших глубин
(до 5000 м). Спатангоиды зарываются на разную глубину (от 5 до
20 см) в песчаный осадок с помощью боковых игл; норка и каналы —
респираторный и санитарный — укрепляются слизистыми выделения-
ми; через респираторный канал поступает свежая вода, через него
наружу могут высовываться сильно удлиненные амбулакральные нож-
ки. Некоторые спатангоиды медленно передвигаются внутри осадка,,
делая заново дыхательный канал.
489-
На основании особенности строения панциря, в первую очередь
пластрона, отряд разделен на два подотряда: Holasterina и Spatangina.
У первых пластрон меридостернальный, у вторых — амфистернальный
(американские эхинологи рассматривают их в качестве отрядов). >Спа-
тангоиды появляются в ранней юре, многочисленны и разнообразны в
Рис. 290. Отряд Spatangoida:
7 — Toxaster (мел): la — вид сверху, 16 — вид снизу; 2 — Micraster (поздний мел):
2а — вид снизу, 26 — вид сбоку; 3—4 — прижизненное положение Spatangus (3) и
Echinocardium (4);
поля: А — амбулакральное, IA — интерамбулакральное; амн — амбулакральная нож-
ка (видоизмененная), ан — анальное отверстие, р — рот, рк — респираторный канал,
ск — санитарный канал
торе и мелу, достигают расцвета в эоцене, после чего их число сокра-
щается. Всего известно около 250 родов.
КЛАСС EDRIOASTEROIDEA. ЭДРИОАСТЕРОИДЕИ
К классу Edrioasteroidea (гр. edraios — устойчивый, aster — звез-
да), или текоидей, отнесены прикрепленные или свободно лежащие
эхинозои, ранее рассматриваемые среди пельматозой, но по строению
скелета, амбулакров и отсутствию брахиол или рук ближе всего стоя-
щие к эхинозоям. Тека дисковидная, шаровидная или башенковидная
размером до 60 мм; рот, анальное отверстие и гидропора расположены
на верхней стороне. От перистома, прикрытого пластинками, отходят
5 амбулакральных полей, прямых или серповидно по часовой или про-
тив часовой стрелки изогнутых; вдоль них лежат пищевые желобки,
прикрытые покровными табличками; амбулакральные таблички распо-
ложены в один или два ряда, обычно пронизаны порами, через кото-
рые, вероятно, выходили амбулакральные ножки (рис. 291, 7);
последние имели ампулы и были, по-видимому, связаны с радиальными
каналами, проходившими под амбулакрами (правда, следы этих кана-
лов не сохранились); у однорядных амбулакров поры не обнаружены
за исключением одного рода (Anglidiscus), у двухрядных имеются по-
ры в амбулакральных табличках.
490
Рот находится в центре перистома, окруженного кольцом табличек,
к нему подходят пять пищевых желобков, идущих по амбулакральным
полям. На одной из интеррадиальных табличек около перистома рас-
положено небольшое отверстие, которое рассматривается в качестве
гидропоры или, возможно, гидрогонопоры; оно связывало каменис-
Рис. 291. Класс Edrioasteroidea:
1 —• схематический разрез: 1а — через теку, 16 — через амбулакральный
Желобок; 2 — Lepidodiscus (средний девон — ранний карбон), реконст-
рукция, вид сверху; 3 — Cyathocystis (средний — поздний ордовик): За —
вид сбоку, Зб —' вид сверху;
поля: А — амбулакральное, IA — интерамбулакральное; амс — амбула-
кральная система, ан — анальное отверстие (и пирамидка), м — мадре-
порит, п — поры для амбулакральных ножек, пт — покровные таблички,
р — рот
тый канал с внешней средой и, возможно, служило также отверстием
для вывода половых продуктов. В одном из интеррадиусов лежит пери-
прокт, прикрытый несколькими табличками (от 4 до 15), образующими
анальную пирамидку. Не исключена возможность, что анальное отвер-
стие у эдриоастероидей играло, так же как у голотурий, некоторую
роль при газообмене.
Первые эдриоастероидеи появились в раннем кембрии (известен
один род Stromatocystites), достигли расцвета в среднем ордовике
(известно, 14 родов) и вымерли в конце раннего карбона. До сих пор
не найдены эдриоастероидеи в позднем кембрии и неясна причина рез-
кого их сокращения (до 1 рода) в конце силура. Эдриоастероидеи вхо-
дили в состав эпифауны нормальносоленых морей, обитали преиму-
щественно в зоне мелководья; для прикрепления предпочитали плотный
491
субстрат, прирастая к раковинам брахиопод, бивальвий, скелетам губок,,
кораллов, мшанок, конулярий. Некоторые формы, по-видимому, могли
изменять свое положение или свободно лежали на дне. Класс насчи-
тывает 17 родов (см. рис. 291).
Систематическое положение эдриоастероидей остается недостаточ-
но ясным. С эхинозоями их сближает строение пор, пронизывающих
амбулакральные таблички, отсутствие стебля и каких-либо экзотекаль-
ных выростов, или рук, предназначенных у всех кринозой для сбора
пищи. От типичных эхинозои они отличаются неподвижным образом
жизни, положением на верхней стороне теки рта, ануса и гонопоры,.
способом доставки пищи по ресничным пищевым желобкам.
КЛАСС HOLOTHURIOIDEA. ГОЛОТУРИИ
Класс голотурий (гр. holothurion — морской огурец), морских огур-
цов, или кубышек, объединяет донных, подвижных, зарывающихся,,
прикрепленных, а также пелагических эхинозой, обитателей всех глу-
Рис. 292. Класс Holothurioidea:
1 — схема строения тела; 2 — общий вид Stichapus (совр.); 3 — склериты из
карбона (За), юры (36) и палеогена (Зе);
амн — амбулакральные ножки, ан — анальное отверстие, вл — водные легкие,
пт — пищеварительный тракт, р — рот
бин — от литорали до ультраабиссали (до 10 240 м). Тело самой раз-
нообразной формы, от удлиненной червеообразной до шаровидной, бу-
тылеобразной, . у пелагических — медузообразной, радиально-симмет-
ричной или билатеральной; рот, расположенный обычно на переднем;
конце, окружен щупальцами — видоизмененными амбулакральными
ножками; задняя кишка видоизменена в клоаку, в нее открываются два
протока водных легких, представляющих два сильно разветвленных
ствола, выполняющих функцию газообмена. У сидячих и зарывающих-
ся форм, рот и анус обычно расположены близко друг от друга на
верхней стороне. Ротовые щупальца в количестве от 10 до 30 бывают
простые, пальцевидные, перистые, щитовидные или древовидные
492
•(рис. 292); они служат для сбора пищи или помогают при зарывании,
частично выполняют функции осязания и газообмена. Вокруг глотки
расположен кольцевой канал амбулакральной системы; от него вдоль
спинной стороны проходят два. (бивиум), а вдоль брюшной — три
(тривиум) радиальных канала. Амбулакральные ножки бивиума
•служат для осязания и газообмена; ножки тривиума снабжены при-
сосками и служат для передвижения. У ряда форм (безногих голоту-
рий) ножки отсутствуют. Вокруг глотки у голотурий расположено из-
вестковое кольцо, состоящее из 5 радиальных и 5 (или более) интерра-
диальных табличек, от которых назад отходят известковые отростки.
Не исключена возможность, что известковое кольцо голотурий гомоло-
гично перистомальной системе табличек эдриоастероидей. К извест-
ковому кольцу прикрепляются продольные мышцы, ретракторы и про-
тракторы, служащие для втягивания и расправления передней части
тела. Имеется одна гонада и мадрепорит. Отличительной чертой боль-
шинства голотурий являются мягкие кожные покровы; считают, что у
наиболее примитивных, а возможно, и наболеее древних, имелся на-
ружный скелет, состоящий из табличек, чешуеобразно налегавших друг
на друга; панцирь покрывал все тело или только отдельные его участ-
ки — спинную сторону или область выводковых камер. У ротового и
анального отверстий в табличках имелись поры для прохождения
амбулакральных ножек. Однако у большинства скелет состоит из мик-
роскопических известковых телец — склеритов (рис. 292, <?),
рассеяных в соединительной ткани, число которых у одной современной
голотурии достигает 2 млн. штук. Скелет, как у всех иглокожих, раз-
вивается из гранул, которые превращаются вначале в миниатюрные
палочки, затем, их концы, раздваиваются под углом 120° и образуются
-склериты различной формы. Они располагаются в коже. После гибели
голотурий тело разрушается, склериты могут рассеиваться по дну или
накапливаться в отдельных местах. До сих пор известно только 3 на-
ходки трех видов целиком сохранившихся голотурий — один из ран-
него девона ФРГ (Palaeocucumaria) и два других из верхнеюрских
золенгофских сланцев ФРГ (Protholothuria, Pseudocaudina). Много-
численные другие находки, описанные ранее, ошибочно относились к
голотуриям (в частности, Eothuria}.
Голотурии, как правило, раздельнополы, лишь некоторые холодно-
водные формы—гермафродиты; оплодотворение внешнее; после дробле-
ния оплодотворенного яйца развивается личинка — аурикулярия;
у холодноводных наблюдается забота о потомстве и молодь вынаши-
вается в особых выводковых камерах в теле матери.
На основании строения внутренних органов класс голотурий раз-
делен на три подкласса: Dendrochirota, Aspidochirota и Apoda, пред-
ставителей которых в ископаемом состоянии по склеритам почти нель-
зя выделить.
История развития голотурий известна мало. Наиболее древние го-
лотурии, вероятно, имели сплошной известковый панцирь, подобно не-
которым современным дендрохиротам; они возникли, очевидно, от
общих предков с эдриоастероидеями и геликоплакоидеями; у послед-
них было веретенообразное тело, покрытое многочисленными таблич-
ками, рот спереди, анус сзади. Не исключена возможность, что первые
голотурии подобно древним морским ежам имели челюстной аппарат,
..за счет которого у голотурий развивалось вокруг глотки известковое
кольцо. Дендрохироты, ведущие прикрепленный образ жизни, внешне
очень напоминают эдриоастероидей; это может быть конвергентное
•сходство или сходство, связанное с близким родством. О последнем
493
говорит строение околоротового кольца эдриоастероидей и известкового
кольца дендрохирот.
От голотурий в основном сохраняются отдельные склериты. По
внешней форме склериты разбиты на отдельные семейства, роды и ви-
ды и описываются в соответствии с правилами биноминальной номен-
клатуры; такая систематика является, как и для других подобных
групп, искусственной, но она позволяет разобраться в различных типах
склеритов. Выделяют семейства, объединяющие склериты в форме па-
лочек, пластинок, колесиков, якорей и т. д.
КЛАСС OPHIOCISTIOIDEA. ОФИОЦИСТИИ
К классу офиоцистий (гр. ophis — змея, kiste — ящик, коробка)
относится небольшая группа (5 родов) подвижных вымерших иглоко-
Рис. 293. Классы Helicoplacoidea (1), Ophiocistioidea (2—3) и Cyclocystoidea
(4), реконструкции:
1 — Helicoplacus (ранний кембрий), Дарэм, Кастер, 1966; 2 — Volchovia (ор-
довик), Геккер, 1940; 3 — Sollasina (поздний силур), вид снизу; 4 — Cyclo-
cystoid.es (средний ордовик — средний девон): 4а — вид. снизу, 46 — то же,
амбулакральные таблички сняты и показаны амбулакральные борозды, 4в —
общий вид (Никольс, 1972);
поля: А — амбулакральное, IA — интерамбулакральное; амн — амбулакраль-
ные ножки, ан — анальное отверстие (и пирамидка), вк — внутреннее коль-
цо, нк — наружное кольцо, о — отверстие половое (?) или гидропора (?),
од — оральный диск, on — оральный полюс, пп — перипрокт, сам — скеле-
тизированные амбулакральные ножки
жих, распространенных от раннего ордовика до позднего силура (воз-
можно, среднего девона). Тело дисковидное, овальное, в форме колпа-
ка или шлема, с ясно выраженной нижней и верхней сторонами. Руки,.
494
или лучи, отсутствуют. Рот, расположенный на нижней поверхности,,
снабжен пятью челюстями, каждая из которых состоит из двух табли-
чек. От рта отходят пять полей радиальных и 5 полей интеррадиаль-
ных табличек. Каждое радиальное поле обычно широкое, состоит из
трех рядов: среднего ряда перрадиальных и двух боковых адрадиаль-
ных табличек; вдоль краев перрадиальных табличек расположены пар-
ные поры для выхода амбулакральных ножек. Интеррадиальное поле
узкое, состоит из одного ряда табличек. Верхняя поверхность скелета
покрыта крупными табличками у одного рода (Volchovia) (рис. 293,2)
и неправильно-полигональными у другого (Sollasina). На верхней
поверхности выделяют дорзоцентральную табличку и перипрокт, лежа-
щий около нее, прикрытый клапанной пирамидкой; -кольцо из пяти
радиальных табличек, лежащих вокруг него, и 5 полей радиальных и
интеррадиальных табличек, отличающихся размерами, формой и чис-
лом. Размеры тела офиоцистий достигали 90 мм. На нижней поверх-
ности сохранились остатки амбулакральных ножек, расположенных
вдоль радиальных полей. Они, в отличие от всех известных современ-
ных иглокожих, были покрыты мелкими чешуеобразными известковыми
табличками. Максимальная длина амбулакральной ножки достигала
40 мм при диаметре 6 мм. Самые маленькие ножки были расположены
около рта. Недалеко от края перистома обнаружена мадрепоровая таб-
личка диаметром до 4 мм (Eucladia). На верхней поверхности, вблизи
клапанной анальной пирамидки, у рода Volchovia, обнаружено отвер-
стие, рассматриваемое как гонопора или гидропора. Офиоцистии,
названные так за некоторое внешнее сходство с офиурами, с которыми
их вначале сблцжали, вели, по-видимому, подвижный образ жизни.
Одни из них (Volchovia) были малоподвижными, другие, вероятно’
передвигались довольно быстро с помощью скелетизированных амбула-
кральных ножек.
В ископаемом состоянии остатки офиоцистий встречаются крайне
редко; они известны из отложений нижнего ордовика — верхнего
силура. Остатки, описанные из среднего девона как скелетизированные
ножки, отнесены к роду Rhenosquama. Возможно, они принадлежат
также к классу офиоцистий, тогда его распространение будет расши-
рено до среднего девона.
КЛАСС HELICOPLACOIDEA. ГЕЛИКОПЛАКОИДЕИ
К классу гелико'плакоидей (гр. helix, род. helicos — спираль,
plax — пластинка), установленному в 1963 г. Дарэмом и Кастером,
относится один род Helicoplacus (см. рис. 293, /), скелетные остатки
которого были найдены в отложениях нижнего кембрия (зоны Olenellus)
США. Скелет размером до 3 см, веретеновидной формы, состоит из-
нескольких рядов известковых табличек, расположенных по спирали.
На одном из узких концов лежит ротовое отверстие, размеры которого,
по-видимому, не превышали 1 мм; следы анального и генитального
отверстий не обнаружены. Структура перистома также неизвестна. От
перистома по спирали расположены ряды амбулакральных и интер-
амбулакральных табличек, неплотно соприкасающихся друг с другом,
что приводило к распаду скелета после гибели на отдельные таблички.
Первый ряд амбулакральных табличек начинается от перистома и
образует два полных оборота спирали, но не достигает апикального
края; второй ряд начинается через полоборота и отделен от первого
двумя рядами интерамбулакральных табличек. Каждый ряд построен
из четырех и более мелких табличек, и, хотя следы пор для амбула-
495
кральных ножек не обнаружены, возможно, эти ножки выходили меж-
ду табличками. Интерамбулакральные поля состоят из трех рядов таб-
личек — из одного центрального и двух латеральных; на некотором
расстоянии от апикального края каждый ряд утраивается. Малые
размеры амбулакральных полей позволяют предполагать, что амбу-
лакральные ножки выполняли только функцию газообмена. Helicopla-
cus относится, вероятно, к детритоядам, однако образ жизни гелико-
ллакоидей не достаточно ясен. Возможно, они были частично погруже-
ны в мягкий осадок или свободно лежали на дне. В связи с наличием
гибкого скелета они, вероятно, почти в два раза могли изменять объем
тела. Скелетные остатку Нelicoplacus встречены вместе с остатками
трилобитов, эдриоастероидей, эокриноидей. Эти находки, по-видимому,
представляют наиболее ранние остатки иглокожих. Не исключена воз-
можность, что геликоплакоидеи были связаны родственными связями
с эдриоастероидеями и голотуриями и что иглокожие возникли еще в
докембрии; их предки, вероятно, вели подвижный образ жизни.
КЛАСС CyCLOCySTOIDEA. ЦИКЛОЦИСТОИДЕИ
К классу циклоцистоидей (гр. kykos — круг) отнесен один род
Cyclocystoides и несколько его видов, известных от среднего ордовика
до среднего девона Европы и Северной Америки, имеющих скелет в
форме маленького диска (см. рйс. 293, 4). В диске различают два коль-
ца: внешнее,- или маргинальное, состоящее из многочисленных
мелких чешуевидных табличек, частично погруженных в гибкий инте-
гумент, и внутреннее, или субмаргинальное, прилегающее к
внешнему, построенное из крупных табличек, вдоль которых проходит
кольцевой канал. К табличкам субмаргинального кольца с верхней
стороны прилегает оральный диск, а с нижней — аборальный. В центре
орального диска находится рот, от которого отходят пять амбулакраль-
ных желобков, раздваивающихся несколько раз по мере приближения
к субмаргинальному кольцу; они прикрыты кроющими табличками.
В каждой табличке субмаргинального кольца имеется по два отвер-
стия, через которые выходили амбулакральные ножки, вероятно видо-
измененные в щупальца, подобно таковым голотурий. Анальное отвер-
стие располагалось в одном из интеррадиусов, построенных из одного
ряда мелких табличек. К субмаргинальному кольцу присоединяется
также аборальный диск, покрытый многочисленными табличками; он
был, по-видимому, более прочным и жестким, чем более гибкий ораль-
ный диск. Предполагается, что циклоцистоидеи были ориентированы
оральной стороной вверх, а аборальной — вниз. Не исключено, что по-
добно современным голотуриям они плавали или вели бентосный образ
жизни, прикрепляясь или свободно располагаясь на субстрате. Цикло-
цистбидеи представляют еще довольно слабо изученную группу иглоко-
жих, условно относимую к подтипу эхинозой.
КЛАСС CAMPTOSTROMATOIDEA. КАМПТОСТРОМАТОИДЕИ
К классу Camptostromatoidea (гр. kampto — гибкий, stroma —
слой) относится только один род (рис. 294), известный из отложений
нижнего кембрия (зоны Olenellus) штата Пенсильвания (США). Тело
колоколообразной формы, радиально-симметричное, рот расположен на
нижней стороне, вдоль края «колокола» лежат подвижно сочлененные
руки (4—8), покрытые табличками. На оральной стороне вокруг круп-
ных табличек по периферии находятся мелкие таблички, несущие так
496
называемые сутуральные поры. Подобные поры имеются также по пе-
риферии верхней поверхности. В центре этой поверхности на централь-
ном диске расположено анальное отверстие, прикрытое анальной пира-
мидкой, состоящей из 7 табличек. За центральным диском следует
периферическая зона, покрытая ребрами, число которых превышает 40.
Скелетные таблички, вероятно, были погружены в толстую кожу и не
образовывали сплошного скелета; после смерти животного они рассеи-
Рис. 294. Класс Camptostromatoidea:
1 — Camptostroma (ранний кембрий),
реконструкция (Рюдеман, 1933): 1а—
вид сбоку, 16 — вид сверху
Рис. 295. Классы Cyamoidea (/) и
Cycloidea (2):
1 — Peridionites (средний кембрий):
1а — реконструкция, 16, в — части
скелета; 2 — Cymbionites (средний
кембрий): 2а — полный скелет, по-
строенный из 5 табличек, 26 — одна
из табличек
вались по дну и поэтому целиком в ископаемом состоянии камптостро-
матоидеи почти не сохраняются.
Образ жизни этих животных не очень ясен, хотя колоколообразная
форма тела позволяет предполагать, что они вели пелагический образ
жизни, подобно современной пелагической голотурии (Рelagothuria}.
Форма тела камптостромы вначале ввела исследователей в заблужде-
ние, и род Camptostroma, послуживший основой для целого класса, был
отнесен к типу кишечнополостных. Систематическое положение этого
класса в типе иглокожих недостаточно ясно; условно они отнесены к
подтипу Echinozoa.
ПОДТИП (?) HAPLOZOA. ГАПЛОЗОИ
К подтипу гаплозой отнесено два класса — Cycloidea и Cyamoidea,
основанные каждый только на одном роде, остатки которых были най-
дены в среднем кембрии Квинсленда (Австралия).
КЛАСС (?) CVCLOIDEA. ЦИКЛОИДЕИ
К классу циклоидей отнесен только один род Cymbionites. Его ске-
лет имеет полусферическую форму или форму, напоминающую напер-
сток, диаметром 12 мм (рис. 295, 2). В типичном случае скелет постро-
32 в, в. друщии
497
ен из пяти плотно соединенных табличек. Углубление в середине ске-
лета может быть разной величины. Периферический край чашечки’
фестончатый; чашечка несет многочисленные борозды.
КЛАСС (?) СУАМОЮЕА. ЦИАМОИДЕИ
К этому классу'также отнесен только один род Peridionites, ске-
лет которого напоминает одну половину двояковыпуклой линзы оваль-
ного очертания, диаметром, превышающим 10 мм (рис. 295, /). Скелет
Рис. 296.’ Геохронологическое распространение иглокожих
построен из пяти известковых табличек — одна срединная апикальная
соединяет две большие терминальные таблички и с противоположной:
стороны расположены две маленькие медиолатеральные. В середине-
скелёта имеется центральная полость конической формы.
Строение скелета обоих родов не вызывает сомнения о их принад-
лежности к иглокожим. Однако практически ничего не известно о-
строении мягких частей описанных животных. Остается также не яс-
ным, являются ли эти остатки целыми скелетами или только их частя-
ми. Все это' заставляет с осторожностью подходить к выделению этих
двух классов и подтипа гаплозой. Американские палеонтологи указан-
ные два рода относят к иглокожим неясного систематического поло-
жения.
Распространение основных групп иглокожих показано на рис. 296„
ТИП HEMICHORDATA.
ГЕМИХОРДОВЫЕ
Гемихордовые (гр. hemi — приставка, означающая половину,
chorde — струна, хорда) объединяют три класса: кишечнодышащих,
крыложаберных и граптолитов, которые характеризуются общностью
начальных стадий эмбрионального развития (два первых класса), на-
личием нотохорды — спинного выроста кишечника — и особенностями
строения вторичной полости тела.
КЛАСС ENTEROPNEUSTA. КИШЕЧНОДЫШАЩИЕ
Класс Enteropneusta (гр. enteron — кишка, рпео — дыхание) —
энтеропнейсты, или кишечнодышащие, объединяет небольшое число
(64 вида, 12 родов) малоподвижных животных, имеющих длинное чер-
веобразное тело. По форме тела и зарывающемуся образу жизни они
похожи на червей. Кишечнодышащие являются в основной своей массе
грунтоедами, живущими в прибрежной зоне моря, небольшая часть.
их — хищники — живет среди коралловых рифов и немногие, по-ви-
димому, питаются планктонными водорослями. В последнее время
энтеропнейсты были обнаружены в других зонах моря, вплоть до глу-
бин 4500 м. -
Червеобразное тело энтеропнейст имеет размеры в среднем от 10 до
15 см, но иногда достигает 2 и даже 2,5 м и четко разделено на три
отдела: хоботок, воротничок и туловище (рис. 297). Передний отдел
тела — хоботок — удлиненное мускулистое образование, служащее
для зарывания. Полость хоботка заполнена целомической жидкостью,
которая придает упругость хоботку и играет существенную роль при
зарывании животного. За хоботком расположен воротничок —
короткий отдел тела, имеющий полость, также заполненную целомиче-
ской жидкостью. За воротничком следует длинное червеобразное ту-
ловище, одетое однослойным эпителием и снабженное вторичной
полостью тела. Целом образуется энтероцельным путем.
В основании хоботка на брюшной стороне расположен рот, веду-
щий в короткую глотку. Спинная часть глотки образует вырост в по-
лость хоботка, получивший название «нотохорда» (или ст омо-
хор да). Нотохорда поддерживается хрящеобразной пластинкой и
считается гомологом хорды позвоночных. Глотка переходит в пищевод,
32'
499
боковые стенки которого пронизаны двумя рядами подковообразных
жаберных щелей. К жаберным щелям подходят кровеносные сосуды, и
кровь насыщается кислородом, приносимым водой. Такой тип жабер-
ных щелей известен только у представителей типа хордовых. За жа-
берным следует печеночный отдел кишечника, снабженный печеноч-
ными придатками, служащими для переваривания пищи. Этот отдел
переходит вначале в отдел средней кишки, а затем задней. Последняя
Рис. 297. Класс Enteropneusta:
1 — Balanoglossus (совр.): la — общий вид, 16 — схематический разрез через
головной отдел, 1в — в прижизненном положении;
в — воротник, жщ — жаберные щели, крс— кровеносные сосуды, нс — нервные
стволы, нх — нотохорда, р— рот, т— туловище, х— хоботок
открывается наружу анальным отверстием. Над нотохордой располо-
жено сердце — полый мускулистый мешок; сокращаясь, он протал-
кивает кровь из центральной кровеносной лакуны (полости) по двум
окологлоточным сосудам в брюшной сосуд, по которому кровь на-
правляется к заднему концу тела и через сеть лакун поступает в спин-
ной кровеносный сосуд, достигая центральной лакуны, расположенной
в области хоботка.
Нервная система развита хорошо. Она состоит из двух стволов —
брюшного и спинного, идущих параллельно кровеносным сосудам.
В области воротничка имеется нервное кольцо, которое соединяет оба
ствола; спинной ствол образует здесь цилиндрическое утолщение,
своеобразное подобие спинного мозга.
Энтеропнейсты — раздельнополые животные, размножаются по-
ловым путем и очень редко бесполым; половые железы в количестве
от 16 до 100 пар расположены по бокам кишечника. Половые продук-
ты выводятся в воду через короткие протоки, отходящие от каждой
гонады. Из оплодотворенных яиц у одних кишечнодышащих выходит
личинка — торн ария, названная так за свое постоянное вращение,
очень похожая на личинку морской звезды (см. рис. 255, 5). По мере
роста у личинки вначале закладываются жаберные щели (вначале
5—6), затем развивается хоботок, появляется воротничок; из нижней
части личинки формируется туловище. У других представителей личин-
ка, выходящая из яйца, состоит уже из трех отделов, соответствующих
хоботку, воротничку и туловищу, но имеет еще некоторые временные
личиночные органы. Наконец, среди энтеропнейст есть один вид живо-
500
родящий и один вид, способный поперечным делением тела давать
новые особи, ,т. е. размножаться бесполым путем; бесполые особи
внешне отличаются от полового поколения. Таким образом, среди вто-
ричноротых кишечнодышащих наблюдается чередование поколений —
бесполого и полового.
Энтеропнейсты строят в песчаном осадке U-ббразные норы, стен-
ки которых сцементированы слизью; при помощи хоботка животные
зарываются в грунт, заглатывая его при этом. Переваривание пита-
тельных веществ происходит в печеночном отделе кишечника. Фекалии
выбрасываются через анальное отверстие в виде спирально закручен-
ных экскрементов (рис. 297, 1в).
Для энтеропнейст характерна высокая способность к регенерации,
которая осуществляется за счет задней части тела.
В ископаемом состоянии кишечнодышащие не сохраняются. Не
исключена возможность, что могут быть встречены остатки U-образных
норок,
Кишечнодышащие занимают промежуточное положение между
двумя основными ветвями вторичноротых — типом иглокожих и типом
хордовых. Первые перешли к малоподвижному или сидячему образу
жизни; этот переход сопровождался кардинальной перестройкой всего
плана строения тела своих предков. Вторые перешли к активному пе-
редвижению, активному питанию, что привело к прогрессивному раз-
витию всех систем органов и в первую очередь нервной системы. О на-
личии общих предков с иглокожими свидетельствует сходство личинок
кишечнодышащих (торнария) и морских звезд (бипиннария). Строение
жаберного аппарата, нотохорды, нервной системы указывает на про-
исхождение кишечнодышащих, иглокожих и хордовых от одного обще-
го предка.
КЛАСС PTEROBRANCHIA. КРЫЛОЖАБЕРНЫЕ
Класс Pterobranchia (гр. pteron — крыло, branchia — жабры) объе-
диняет 3 рода современных и 4 рода вымерших колониальных или
псевдоколониальных морских животных, ведущих сидячий образ жизни.
Птеробранхии, в отличие от.энтеропнейст, живут на глубинах преиму-
щественно от 100 до 650 м и отличаются маленькими размерами тела
от 0,5 до 2—7 мм, реже до 14 мм. Колонии могут достигать 25 см в
поперечнике. Строение тела напоминает строение тела энтеропнейст;
оно состоит из трех отделов — хоботка, воротничка и туловища
(рис. 298, 299). Хоботок имеет форму голов н.о го диска, или щи-
та, содержащего полость целома. Воротничок несет 1—8 пар рук,
снабженных щупальцами. Такой воротничок у птеробранхий назван
лофофором. Туловище короткое, мешковидное, заканчивается
стебельком или ножкой. . На границе между головным щитом и лофо-
фором лежит рот, ведущий в широкую глотку; по бокам глотки распо-
ложены жаберные щели, рудиментарные у рабдоплеврид. Пищевод
переходит в желудок, от которого вверх поднимается кишечник, закан-
чивающийся анальной порой. Пищеварительный тракт, таким образом,
у крыложаберных петлеобразно изогнут, ротовое и анальное отверстия
сближены. Нотохорда — вырост дорзальной стенки кишечника — го-
мологична нотохорде энтеропнейст. Впереди нотохорды лежит сердце.
Кровеносная система вследствие маленьких размеров тела построена
сравнительно просто. Она состоит из системы лакун, по которым цир-
кулирует кровь; спинной, центральной, брюшной и пары околоротовых;
брюшная продолжается в стебелек. Нервная система состоит из двух
501
стволов — спинного и брюшного, соединенных нервным кольцом во-
ротничка. От него же отходят нервные тяжи в руки. Брюшной нерв-
ный тяж продолжается в стебелек. Органы чувств не развиты.
Рис. 298. Отряд Rhabdopleurida. Схема строения колонии Rhab do pleura:
а — автотеки с сократившимися зооидами, б —с выступающим зооидом, в —
боковая иг — концевая ветви колонии; 1 — замкнутый конец растущей ветви,
2. — концевая почка, 3 — юная почка, 4 — зимовочная почка, 5 — столотеки,
6 — перегородки между столотеками, 7 — столон, покрытый черной кутикулой,
8 — то же, молодая часть без кутикулы, 9 — стебелек, 10 — часть автотеки, сте-
лющаяся по субстрату, 11 — свободно поднимающаяся часть автотеки
Рис. 299. Отряды Rhabdopleurida (/—2) и Cephalodiscida (3—4):
1 — Rhabdopleura, схема строения зооида; 2а, б — три фрагмента Rhabdopleura
sp. из позднего мела; 3 — Cephalodiscus, схема строения зооида; 4 — Cephalo-
discus (Idiothecia), общий вид колонии (совр.);
ки — кишка, лф — лофофор, п— почки, под — приоральный диск, р — рот, ст —
столон, сте — стебелек, тз — трубка зооида, те — теки, ц — цисты
Отдельные особи птеробранхий живут в ячейках, которые они
строят при помощи выделяемого железами головного диска органиче-
ского белкового вещества. Строение этих ячеек отличается большим
разнообразием. Представители рода Atubaria, живущие только у по-
бережья Японии, ячеек не строят; они живут на колониях, гидроидных
502
31 ведут подвижный образ жизни, передвигаясь при помощи рук и сте-
белька. Виды рода Rhabdopleura образуют колонии; их отдельные зоо-
иды соединены между собой столонами. Виды третьего рода Cephalo-
discus* известные только в морях южного полушария, собираются
.вместе, образуя псевдоколонии; их отдельные особи независимы друг
от друга.
Птеробранхии раздельнополые, но половые различия могут быть
и не выражены; имеются гермафродитные особи. Гонады, одна или
пара, расположены в передней части целома туловища. Личинка очень
похожа на личинку кишечнодышащих; она несет пару зачатков рук.
Целом образуется энтероцельным путем. Рот расположен на переднем
конце, анальное отверстие — на заднем. После оседания на дно и
прикрепления строится органический скелет, дальнейшее развитие про-
исходит бесполым путем — почкованием. Новые почки возникают на
-свободных верхних концах столона.
На основании особенностей строения тела и формы скелета птеро-
бранхии разделены на два отряда: Rhabdopleurida, Cephalodiscida.
Отряд Rhabdopleurida. Рабдоплевриды (гр. rhabdos — прямой,
pleura — бок). К отряду относятся три рода: один современный
{Rhabdopleura') и два рода из ордовика (рис. 299, 1—2). Рабдоплев-
риды образуют маленькие разветвленные колонии, скелет которых,
или ценоций, состоит из стелющихся трубочек и поднимающихся от
них вертикальных трубочек, или тек; в т е к а х на стебельках сидят
..зооиды крошечных размеров (меньше 1 мм). При помощи стебельков
зооиды соединены со столоном, проходящим внутри основной горизон-
тальной трубки. Столон покрыт хитиноидной кутикулой черного цвета.
На концах столонов находятся незрелые концевые зооиды. Новые поч-
ки, из которых потом развиваются зооиды, возникают на столоне на
некотором расстоянии от концевых зооидов. Возникшие зооиды выде-
ляют поперечные перегородки и вся горизонтальная трубка оказывает-
ся разделенной на отдельные столотеки. Затем зооид резорбирует
(растворяет) стенку трубки (так называемое перфорирующее
почкование) и над этим отверстием строит вертикальную трубку
(теку). Трубка строится при помощи органического белкового вещест-
ва, выделяемого железами головного щита. Зооид, поворачиваясь в
трубке на полоборота то в одну, то в другую сторону, откладывает
полукольцо, или веретенце; полукольца соединяются друг с другом
зигзагообразным швом и образуют цилиндрическую трубку. Иногда
одна из новых почек превращается в концевой зооид и ее столон на-
чинает расти в длину; так возникает ветвление столона и соответствен-
но колонии. Среди зооидов имеются самцы, самки и бесполые особи,
но чаще самцы и самки строят отдельные колонии. Зооиды и их сте-
бельки снабжены системой мускулов, которые обеспечивают втягива-
ние зооида внутрь теки, втягивание рук лофофора, полувращательное
.движение зооида внутри теки. Выпячивание рук происходит за счет на-
гнетания в них целомической жидкости из области воротника; послед-
ний сообщается порами с внешней средой.
; Современные рабдоплевриды живут' в Атлантическом и Тихом
океанах и у берегов Антарктиды. Они прикрепляются к подводным
предметам. В ископаемом состоянии известны из ордовика (в ледни-
ковых валунах, найденных в Польше), позднего мела Польши и эоцена
Англии.
Отряд Cephalodiscida. Цефалодисциды (гр. kephale — голова,
disc — диск). Зооиды живут небольшими сообществами, но настоящих
колоний не образуют. В отличие от рабдоплеврид цефалодисциды
503
имеют несколько пар рук (до 8), парные гонады и пару жаберных пор.
Форма скелета отличается большим разнообразием. Ценоций может
состоять из толстых ветвей, в которых имеются специальные помеще-
ния для зооидов. Эти помещения изолированы или сообщаются друг
с другом каналами. В отдельных помещениях могут сидеть или отдель-
ные особи, или группы их. Зооиды присасываются и удерживаются
при помощи стебельков. Они способны вылезать из домика, передви-
гаться по поверхности скелета при помощи головного щита, выделяя
при этом дополнительный кортикальный слой, состоящий из белкового'
вещества. В составе сообщества живут самцы и самки. Размножение
происходит половым и бесполым путем. В первом случае из опло-
дотворенного яйца развивается червеобразная личинка, во втором —
новые почки возникают на конце стебелька зооидов. Представители
рода Atubaria скелета не имеют. Отряд объединяет 4 рода, из которых
два известны из ордовика и два (Atubaria и Cephalodiscus) обитают в.
современных морях (рис. 299, 5, 4).
КЛАСС GRAPTOLITHINA. ГРАПТОЛИТЫ
Граптолиты (гр. graptos — начертанный, lithos — камень) —-
палеозойские морские колониальные животные, прикрепленные, планк-
тонные или псевдопланктонные. Колония, или рабдосома, имела
разнообразную форму: древовидную, кустистую, сетевидную, инкру-
стирующую, решетчатую или состоящую из одной или нескольких вет-
вей. Колония начиналась сикулозооидом, строившим первичную
ячейку, или сикулу. Все остальные зооиды, моно- или полиморфные,
находились в ячейках, или теках. Зооиды были связаны друг с другом
столонами, которые не всегда сохранялись в ископаемом состоянии.
Скелет наружный, органический, склеропротеиновый, состоял из двух
слоев — фюзел л ярного , или веретенчатого, и кортикального,,
или пластинчатого. Прикрепленные и псевдопланктонные граптолиты
прикреплялись сикулой и: специальными выростами кортикального-
слоя, а планктонные имели пневматофор или поплавок. Грапто-
литы размножались половым и бесполым путем. В результате беспо-
лого размножения возникали колонии, иногда очень сложные. Класс
объединяет около 180 родов. Первые граптолиты известны из среднего
кембрия; расцвет относится к ордовику и силуру; последние грапто-
литы вымирают в конце раннего карбона.
Класс граптолитов на основании особенностей строения теки ко-
лоний, наличия столонов разделен на 8 отрядов, причем одни исследо-
ватели объединяют эти отряды в два подкласса (А. М. Обут): стерео-
столонат и собственно граптолитов, другие специалисты (Р. Козлов-
ский, О. Вульман) не объединяют отряды в более высокие таксоны..
В книге принимается первая точка зрения.
ПОДКЛАСС STEREOSTOLONATA. СТЕРЕОСТОЛОНАТЫ
Общая характеристика. Стереостолонаты (гр. stereos — твердый,,
солидный, лат. stolo, род. stolonis — побег, ветвь) — бентосные и псев-
допланктонные граптолиты, образующие кустистые, древовидные, сет-
чатые и инкрустирующие колонии. Каждая колония имела три типа
ячеек — автотеки, битеки и столотеки (рис. 300). Внутри них прохо-
дили столоны, заключенные в плотную органическую оболочку, обра-
зованную протеинами. Стереостолонаты (до 70 родов) появились в.
среднем кембрии и дожили до раннего карбона.
504
Строение колонии. Колония, или рабдосома, в типичном виде
кустистой, древовидной формы или похожа на сеть, состоящую из от-
дельных ветвей, соединенных перемычками. У инкрустирующих форм
рабдосома имеет форму пластины — т е к о р и з ы, над которой воз-
вышаются автотеки или битеки.
На основании химических анализов, проведенных в последние го-
ды, установлено, что скелет рабдосомы состоит из различных' склеро-
Рис. 300. Подкласс Stereostolonata. Детали строения колонии:
1 —• начальная часть колонии Dendrograptus; 2а, б — характер почкования
у Dendrida-, 3—4 — строение перидермы: 3 — поперечный разрез через
ветвь Koremagraptus, 4 — взаимоотношение кортикального (кс) и фюзел-
лярного (фс) слоев;’
а — автотека, б — битека, вс — внутренняя и нс — наружная части пер-
вичной столотеки, мп — место перфорации стенки просикулы первичным
столозооидом, мс — метасикула, оме — отверстие метасикулы, пд — при-
крепительный диск, пс — просикула, с — столотека, ст — столон,, у — устье
протеинов (серин, аланин, глицин и др.), а хитин, как предполагалось
раньше, полностью отсутствует у граптолитов. Стенка каждой теки
состоит из двух слоев: фюзеллярного и кортикального (рис. 300, 4}.
Фюзеллярный слой построен из отдельных узких полосок — вере-
тенец длиной в полоборота, соединенных в теке двумя зигзагообраз-
ными швами. По типу строения и химическому составу теки напоми-
нают строение трубок птеробранхий. Кортикальный, или плас-
тинчатый, слой покрывает снаружи теки и принимает участие в обра-
зовании соединительных перемычек.
Автотека — наиболее крупная зооидная трубка, обычно прямая,
слегка расширяющаяся к дистальному концу; ее устьевой край часто
снабжен парными отростками, или шипами. К автотеке подходит отно-
сительно длинный столон. У части стереостолонат автотеки принимают
участие в образовании текориз. В этом случае автотеки состоят из двух
частей: горизонтальной, входящей в состав текоризы, и вертикальной—
трубчатой, возвышающейся над текоризой.
Б и т е к а значительно меньше автотеки, имеет трубчатую форму
с округлым устьевым краем без шипов; столон короткий; битека всегда
находится рядом с автотекой.
505
Столотеки ячейки таких же размеров, как и битеки, распола-
гаются обычно вдоль одной из сторон ветви, и их устьевой край за-
канчивается у основания автотеки. Внутри каждой столотеки проходит
основной столон, от которого отходят три столона: два коротких, иду-
щих к битеке и новой столотеке, и один длинный, идущий к автотеке.
Каждая столотека представляет собой как бы основание автотеки.
В результате очередного почкования от столотеки развивались три но-
вых теки — автотека, битека и столотека (триадное почкование)
или две новых теки (ди ад ное почкование). По’мере'роста колонии
отдельные ветви могли раздваиваться. В этом случае в местах ветв-
ления возникали две новые автотеки, дававшие начало новым ветвям.
Для укрепления рабдосомы отдельные ветви могли соединяться особы-
ми полыми поперечными перемычками, образованными кортикальной
тканью. Иногда эти перемычки называют д и ссеп и м ент а ми.
Строение тела. О строении мягкого тела, расположения органов
у стереостолонат можно только предполагать, анализируя строение и
форму скелета. Зооиды помещались в теках и имели короткое мешко-
видное тело. Все особи были связаны столонами, подобно тому, как
•они связаны между собой у птеробранхий. В автотеках, наиболее круп-
ных ячейках, как предполагает Р. Козловский (1949), находились жен-
ские особи; по-видимому, они были снабжены хорошо развитым ло-
фофором; косвенным подтверждением этому могут служить различные
околоустьевые образования. В битеках сидели, скорее всего, мужские
особи, более мелкие, с менее развитым лофофором; не исключено, что
.лофофор у них вообще отсутствовал. Неясной остается роль столотек.
По мнению Обута, в них сидели почкующиеся особи, дававшие начало
новой триаде или диаде особей. Вульман, анализируя строение столо-
тек, предполагает, что поскольку непосредственным продолжением
столотеки служит автотека, то места для почкующихся зооидов в сто-
лотеке не было. Каждая столотека представляла собой раннюю стадию
развития автозооида.
Скелет рабдосомы строился так же, как у современных крыложа-
берных, за счет выделения склеропротеиновых веществ железами го-
ловного щита. Зооиды, по-видимому, строили веретейца фюзеллярного
слоя, поворачиваясь в теке на 180° и соединяя каждое • веретенце
друг с другом, зигзагообразным швом. Менее ясен способ образования
кортикального слоя, покрывающего снаружи теки. Этот слой, по-види-
мому, служил для укрепления скелета.
По плану строения зооиды граптолитов, вероятно, очень напоми-
нали зооидов птеробранхий, что позволило В. Н. Беклемишеву (1951)
объединить их в один класс.
Размножение и развитие. Астогенез колоний стереостолонат наи-
более подробно изучен Р. Козловским (1949) у представителей отрядов
.дендроидей и тубоидей. Личинка бентосных граптолитов, относимых
к отряду дендроидей, после плавания в толще воды опускалась на дно
и прикреплялась к подводным предметам при помощи прикрепитель-
ного диска. Возможно, она имела первичный скелет — просикулу,
представляющую собой тонкостенную трубку, усиленную спирально
расположенными нитями. Личинка превращалась в первичного зооида,
который начинал строить вторую часть своего скелета —- метасику-
Л у; метасикула состоит из отдельных полуколец, или веретенец, сое-
диненных на противоположных сторонах зигзагообразными швами, и
по своему строению резко отличается от просикулы, напоминая строе-
ние всех остальных тек. Таким образом, у первичного зооида скелет
состоял из просикулы, по-видимому, эмбрионального образования и
.506
метасикулы постэмбрионального скелета. Обе эти части образуют ске-
лет первого зооида колонии — сикулозооида, получившего назва-
ние сикулы (рис. 300, 1). Сикула стереостолонат всегда прикрепля-
лась к подводным предметам.
От сикулозооида в результате бесполого размножения возникал
столон, дававший первую почку. Эта почка, подобно почкам рабдоплев-
рид, растворяла часть скелетного вещества просикулы и через воз-
никшее отверстие давала начало первому зооиду. Этот зооид, по Вуль-
ману (1972), в дальнейшем превращался в автозооид и одновременно
давал начало двум другим: мужскому зооиду, строившему битеку, и
почке, строившей вначале столотеку, переходящую в дальнейшем в
новую автотеку. Таким образом, первый зооид давал начало первой
триаде новых особей. Такой тип почкования получил название триад-
ного почкования. Вслед за образованием первой триады зооидов
возникали одна за другой следующие 4—5 триад. После образования
четвертой или пятой триады возникали две новые автотеки, которые
приводили к образованию двух ветвей; для укрепления колонии ветви
соединялись перемычками. Разветвление могло происходить неодно-
кратно; в результате многократных ветвлений возникала сложная
огромная колония, в которой насчитывалось до 20 000—30 000 особей.
Астогенез представителей других отрядов, за исключением тубо-
идей, практически не изучен. Астогенез тубоидей отличался тем, что
первичный столон, достигнув стенки просикулы, делился на два столона
и в стенке для их прохождения возникало два отверстия. Первая воз-
никшая тека была битекой. В дальнейшем один из столонов вновь де-
лился дважды (диадное почкование), образуя новую теку и
новый столон. Новые теки располагались по спирали, право- или лево-
завитой (см, рис. 302, 2). Способ образования текоризы у них пока еще
не изучен.
Основы систематики и классификации. На основании особенно-
стей строения рабдосомы и тек стереостолонаты разделены на 7 отря-
дов: Dithecida, Dendrida, Inocaulida, Tubida, Camarida, Crustida,
Stolonida.
Отряд Dithecida. Дитециды (гр. di — два, theca — ячейка). К от-
ряду дитецид отнесены стереостолонаты, имевшие кустообразные или
древовидные рабдосомы, с незакономерным ветвлением. Колония со-
стоит из автотек и столотек, возникавших при очередном (диадном)
почковании; Наличие столонов у них еще не установлено. Обут предпо-
лагает, что дитециды образуют наиболее древний отряд граптолитов,
появившийся в конце среднего кембрия и просуществовавший до
начала девона. Около 10 родов (рис. 301, 1—4). Характерные роды:
Dithecodendrum (средний кембрий), Siberiograptus (поздний кембрий).
От дитецид в позднем кембрии произошел отряд дендрид.
Отряд Dendrida. Дендриды (гр. dendron — дерево, ствол) — наи-
более важный и широко распространенный отряд подкласса стереосто-
лонат. Колония древовидной формы прикреплялась к субстрату по-
средством прикрепительного диска или корневых выростов, образо-
ванных кортикальным слоем. На каждой ветви располагались три типа
ячеек: автотеки, битеки и столотеки. При очередном почковании воз-
никала триада новых тек. Первые дендриды имели кустообразные или
древовидные колонии, состоявшие из прямых или изогнутых ветвей,
вначале не соединенных перемычками. У последующих дендрид ветви
были соединены диссепиментами и колония приняла вид сети. В начале
ордовика среди дендрид возникли колонии (род Anisograptus), у ко-
торых ветви, несущие теки, были направлены вниз от сикулы, как у
507
типичных граптолоидей. Эти граптолиты занимали промежуточное поло-
жение между стереостолонатами и граптолоидеями и явились,,
по-видимому, исходной родоначальной группой для последних. В нача-
ле ордовика дендриды дали начало нескольким сравнительно коротким
ветвям стереостолонатных граптолитов — тубидам, камаридам и сто-
Рис. 301. Отряды Dithecida (1—4) и Dendrida (5):
1 — Siberiograptus (поздний кембрий); 2 — Dithecodendrum (сред-
ний кембрий); 3 — Siberiodendrum (средний — поздний кембрий);
4 — Aellograptus (поздний кембрий); 5 — Dictyonema (поздний
кембрий — карбон): 5а—г — различные стадии развития колонии,
5д — общий вид колонии, 5е — часть колонии в крупном плане;
а — автотека, б — битека, не — нема, пд — прикрепительный диск,
си — сикула
лонидам. Дендриды были широко распространены также в ордовике;
многие роды существовали длительное время. В конце намюра (ранний
карбон, по данным советских стратиграфов, или средний карбон, по
данным зарубежных специалистов) последние дендриды вымерли. Все
дендриды вели прикрепленный образ жизни, образуя в морях ордо-
вика подводные «луга», прикрепляясь к различным подводным предме-
там. Возможно, некоторые представители прикреплялись к талломам
плавающих водорослей и вели, таким образом, эпипланктонный образ
жизни. Поздний кембрий — ранний карбон. Около 20 родов (рис. 301,
5а—5е).
Отряд Inocaulida. Инокаулиды (гр. ino — волокно, лат. caulis —
ствол дерева). Рабдосома состоит из толстого ствола и отходящих от
него крупных ветвей, на которых расположены тонкие трубчатые ав-
тотеки, собранные на концах ветвей в пучки. Отряд был выделен Бу-
чеком (Boucek, 1957), но изучен еще недостаточно. Систематическое
положение многих родов остается неясным. Ордовик — силур. Около
10 родов (рис. 302, 1).
508
столотек в колонии имеются коноте-
3 тер
Рис. 302. Отряды Inocaulida (/) и Tubida
(2-3):
1 — Inocaulis (силур): la — общий вид
колонии, 16 — фрагмент (Х5); 2 — схема
астогении Dendrotubus (1—4 — последова-
тельные стадии образования тек); 3— Idio-
tubus (ранний ордовик), общий вид коло-
нии, реконструкция (Козловский, 1948);
а — автотека, б — битека, сс — столон си-
кулы, тер — текориза
Отряд Tubida. Тубиды (лат. tubus — трубка). Рабдосома древо-
видная, кустистая или сетчатая, иногда инкрустирующая; в последнем
случае состоит из текоризы, от которой поднимаются вверх трубчатые
автотеки и битеки. Имеется три типа тек с хорошо развитыми столо-
нами. Кроме автотек, битек и
к и — конической формы ячей-
ки с маленьким отверстием в
вершине конуса, функциональ-
ное. значение которых изучено
еще недостаточно. Обычно на-
блюдается диадное почкова-
ние. Автотеки длинные, труб-
чатые, вдоль устьевого края
снабжены отростками, или
шипами. У Tubidenrum авто-
теки свернуты в плотную спи-
раль. У колоний, имеющих те-
коризу, автотеки состоят из
горизонтальной части, входя-
щей в состав текоризы, и вер-
тикальной, возвышающейся
над ней. В древовидных коло-
ниях битеки встречаются в два
раза чаще, . чем автотеки, и
расположены довольно неза-
кономерно. Ордовик — силур.
Известно 12 родов (рис. 302,
. 2—3).
Отряд Camarida. Камари-
ды (гр. kamara — камера).
Колонии имеют текоризу, со-
стоящую из автотек и столо-
тек. Битеки установлены не у
всех представителей. Часть
автотеки, входящая в состав
текоризы, имеет форму взду-
той, удлиненной камеры, по-
груженной в своеобразную
межкамерную ткань. От каме-
ры вверх отходит трубчатая
часть, по-видимому соответ-
ствующая трубчатой части ав-
тотек предыдущего отряда.
Иногда трубчатая часть от-
сутствует и заменяется отвер;
стием в верхней части камеры.
Битеки имеют форму трубок и расположены незакономерно. Столотеки
представлены раздваивающимися трубками, внутри которых проходят
столоны, идущие к автотекам. Ранний ордовик. Пять родов (рис. 303).
Отряд Crustida. Крустиды (лат. crusta — раковина). Рабдосома
состояла из автотек, битек и столотек и обычно имела, насколько мож-
но судить по фрагментам, форму обрастающей корки, несколько напо-
минающей колонии камарид. Автотеки крупные, овальной формы,
состоят из горизонтально расположенной части, над которой возвы-
шается узкая, в виде горла, трубка (некк), переходящая в широкое
509
устье, снабженное двумя складками,
с устьями, расположенными рядом с
Битеки трубчатые, разной длины,,
устьем автотеки либо позади или
впереди него. Столотеки
Рис. 303. Отряд Camarida.
Общий вид колонии Bithecocamera (ранний ор-
довик), реконструкция (Козловский, 1948);
б — битека, к— камера, с — столотека, тр —
трубчатая часть камеры
также длинные, изогнутые,
на дистальном конце пере-
ходят в основание дочерней
автотеки. Столоны, дости-
гающие толщины 20—35 мк,
хорошо сохранились. Вну-
три автотек наблюдаются
особые вздутия, получившие
название цист. Природа
их не совсем ясна. Возмож-
но, цисты каким-то образом
были связаны со столонами
или представляли дегенери-
ровавших зооидов. Полости
автотек иногда заполнены
так называемым крипто-
Рис. 304. Отряды Crustida (7—2) и Stolonida (3):
1 — Bulmanicrusta -(ранний ордовик), реконструкция части колонии
(пунктиром показана внутренняя часть столотеки и битеки, справа —
поперечные сечения); 2а, б — Graptoblasius (ранний ордовик), ре-
конструкция; 3 — Stolonodendrum (ранний ордовик), фрагмент
колонии;
а —автотека, б — битека, зк — задняя камера, кп — криптопиле,
пк — передняя камера, с — столотека, ст — столон
бластом, который представляет собой овальной формы тело, несущее-
на одном конце маленькое отверстие — криптопиле. Внутренняя
полость криптобласта может быть разделена перегородкой на две ча-
сти; стенка построена из двух слоев; наружного — тонкого, бесструк-
510
турного и внутреннего — толстого, фюзеллярного. Криптобласты обна-
ружены также отдельно, вне автотек крустид в отложениях нижнего»
ордовика. Ордовик. Известно 7 родов (рис. 304, 1—2).
Отряд Stolonida. Столониды (лат. stolo, род. stolonis — побег,,
ветвь). Колония состоит из неправильно ветвящихся трубок, червеоб-
разной формы, диаметром от 50 до 350 мк. Стенки трубок толстые,
полости заполнены вторичным веществом. Теки однотипные, отходят
непосредственно от столотек, в которых имеются столоны. В этот отряд
Р. Козловский включил два рода (рис. 304, 3), представители которых:
были встречены в ордовике.
ПОДКЛАСС GRAPTOLOIDEA. ГРАПТОЛОИДЕИ
Общая характеристика. Планктонные и псевдопланктонные грап-
толоидеи, рабдосома которых состояла из пневматофора или поплавка
и отходящих от него одной, или нескольких ветвей. На ветвях распо-
лагались обычно в один — два ряда мономорфные теки. Устье тек
было направлено либо в одну сторону с устьем сикулы, либо в диамет-
рально противоположную. Наличие склеротизированных столонов не
установлено. Граптолоидеи произошли от дендридных стереостолонаг
в начале ордовика и просуществовали до начала девона, дав около»
ПО быстро эволюционировавших родов.
Рис. 305. Подкласс Graptoloidea. Детали строения колонии:
1а — сикула, 16 — просикула; 2 — сикула; 3 — сикула с тремя первыми
ячейками; 4 — схема строения теки; 5—6 — схема строения части колонии;
а — автотека, вг — виргелла, ей — виргула, кс — кортикальный слой,
мт — метатека, мс — метасикула, пре — просикула, пт — протека, си — си-
кула, ст — столон, фс — фюзеллярный слой
Строение колонии. Форма и особенности строения колонии завися-
ли от типа ее развития (астогенеза), порядка почкования и способа
образования новых тек. Рабдосома граптолоидей может рассмат-
риваться как упрощенная форма колонии дендрид, у которой сокра-
511
тилось число ветвей, исчезли перемычки между ними, рёдуцировались
битеки, сохранились одни автотеки, исчезли склеротизированные сто-
лоны. Рабдосома состоит из пневматофора или поплавка и ветвей, на
которых в один — два ряда располагались мономорфные, т. е.
одинаковой формы, теки. Сикула, так же как и у стереостолонат, со-
стоит из просикулы и метасикулы. В отличие от просикулы стереосто-
лонат она снабжена не только спиральными нитями, но и несколькими
продольными, 3 или 4 из которых доходят до начала немы (рис. 305).
Метасикула по строению’резко отличается от просикулы; она построе-
Рис. 306. Основные типы расположения ветвей:
1—3 — пендентное, 4 — горизонтальное, 5 — наклонное, 6—7 — скандентное
на так же, как и все остальные теки, но часто имеет ряд внутренних
кольцевидных утолщений стенки, причем первое утолщение наблю-
дается в конце просикулы. У некоторых форм этих колец нет. Возмож-
но, кольца служили для укрепления сикулы. Вдоль шва одной из сто-
рон сикулы проходит виргелла — утолщение стенки метасикулы;
вдоль устьевого края сикулы виргелла заканчивается ложкообразным
выступом или удлиненным шипом. У Linograptus виргелла заканчива-
лась зонтикообразной структурой — приспособлением для парения
(рис. 305, 2). Общие размеры сикулы варьируют в довольно больших
пределах; в среднем ее длина составляет 1,5—2,5 мм, иногда 5—6 мм
у некоторых родов (Cystograptus} может достигать 10 мм и более.
У отдельных форм (Retiolites) сикула иногда не сохраняется; возмож-
но, она была слабо склеротизирована. В просикуле у ранних предста-
вителей и в метасикуле у более поздних для первой почки или столона
путем перфорации возникало отверстие.
Нема служила для прикрепления рабдосомы к плавательному
пузырю. У двухрядных и однорядных граптолитов вместо немы разви-
валось опорное образование, получившее название виргула, в отли-
чие от виргеллы, развитой только у сикулы.
Теки. У граптолоидей, в отличие от стереостолонат, все теки оди-
наковы и состоят из двух частей: протеки, гомологичной столотеке
дендрид, и метатеки, отвечающей автотеке (рис. 305, 6). В простейшем
случае теки имеют форму цилиндрической трубки. Кроме трубчатых
встречаются сигмоидально изогнутые теки и так называемые лопаст-
ные теки. У последних метатека обычно изогнута крючкообразно. Теки
расположены вдоль ветвей рабдосомы в один, два, реже четыре ряда.
Различают четыре основных типа расположения ветвей по отно-
шению к Сикуле: пендентное, горизонтальное, наклонное и скандентное
(рис. 306). В первом случае ветви рабдосомы направлены вниз от
сикулы, во втором — в стороны; в третьем — косо вверх и в последнем
случае — вверх вдоль немы. Скандентный тип расположения ветвей,
512
по-видимому, создавал для зооидов, сидевших в теках, наиболее благо-
приятные условия для сбора пищи.
Многие граптолоидеи имели пневматофор, или плавательный
пузырь, к которому прикреплялась нема или виргула. Наличие пнев-
матофора указывает на планктонный образ жизни. Для этой же цели
служил поплавок — центральное двухслойное тело, растянутое
между начальными частями ветвей. По-видимому, часть граптолоидеи
вела псевдопланктонный образ жизни, прикрепляясь с помощью корот-
кой немы с диском на конце к плавающим подводным предметам.
Рис. 307. Типы развития колоний. Схема расположения ячеек у представителей
отрядов Axonolipa (1—2) и Axonophora (3—5):
1 — дихограптидный; 2 — лептограптидный; 3—4 — диплограптидный; 5 —
монограптидный;
Р, I2 и т. д. — порядок образования тек, сикула зачернена
Размножение и развитие. За последние годы астогенез грапто-
лоидей был изучен у многих представителей этого подкласса. Обычно
он начинался с формирования просикулы (длиной 400—500 мк), обра-
зования на ней спиральных, продольных нитей и немы (эмбриональная
стадия). Начало формирования метасикулы соответствует новому
этапу развития, возможно личиночному; метасикула строилась из по-
луколец (веретенец), соединенных на противоположных концах зигза-
гообразным швом. Вдоль вентральной стороны скелет утолщался~'и
строилась виргелла, конец ее вытягивался в длинный шип; на перио-
дические остановки роста метасикулы, возможно, указывают. кольца,
вдоль которых стенка метасикулы значительно утолщена. Однако у не-
которых ранних граптолоидеи (дихограптиды) виргелла отсутствова-
ла. После завершения образования сикулы в ней.при развитии первого
зооида возникала пора. У ранних граптолоидеи она образовывалась
в просикуле, у более молодых — в метасикуле. У них это отверстие
могло возникать вначале как глубокое углубление края метасикулы
(своеобразный синус), которое позднее ограничивалось снизу новыми
полукольцами нарастания. По-видимому, через это отверстие столон
давал начало первой почке, из которой развивался первый зооид.
33 в. В. Друщии
513
В зависимости от характера дальнейшего почкования выделяют
четыре основных типа развития и соответственно четыре типа коло-
ний: дихограптидный, лептограптидный, диплограптидный и монограп-
тидный (рис. 307). У дихогр аптидного типа, наблюдаемого у наи-
более ранних граптолоидей, первый зооид давал начало сразу двум
новым зооидам; от одного из них путем дальнейшего почкования раз-
вивалась ветвь, на которой в один ряд в теках располагались зооиды;
от второго, пересекающего сикулу, шло образование второй ветви, на-
правленной в противоположную сторону.. У лептограптидного
Рис. 308. Схема строения синрабдосомы Orthograptus
типа ветвление начиналось со второго зооида, причем ветви рабдосомы
располагались горизонтально или даже несколько изгибаясь вверх,
В диплограптидном типе вначале один за другим возникали три
новых зооида с устьями, направленными в сторону, противоположную
устью сикулы (скандентный тип), и только от третьего зооида возни-
кали четвертый, пятый и все дальнейшие. В монограптидном
типе устье всех тек направлено вверх и все зооиды сидели в один ряд.
Кроме этих основных четырех типов колоний имеется еще несколько-
производных.
Синрабдосома. Среди диплограптидных встречены сложно»
построенные колонии, состоящие из нескольких рабдосом и получившие
название синрабдосом (рис. 308). Они представляют скопление двух-
рядных ветвей, которые прикреплены к шаровидным телам, располо-
женным вокруг центральной капсулы. Существует два объяснения:
строения синрабдосом. Р. Рюдеман (Kuedemann, 1908) считал, что цен-
тральная капсула представляла собой пневматофор, а расположенные»
вокруг него Шаровидные тела — гонотеки, или гонангии, от которых
шло образование сикул.’ Каждая сикула давала двухрядную ветвь.
По мнению Р. Козловского, синрабдосома — сложная колония, состоя-’
щая из многих рабдосом диплограптидного типа, прикрепленных к
пневматофору.
514
В ©тличие от стереостолонатных граптолитов колонии собственно
граптолоидей насчитывали значительно меньше зооидов, у одних их
было от 2000 до 3000, у других не превышало 10—20.
Основы систематики и классификация. Систематика граптолоидей
основывается на наличии или отсутствии виргулы, типе аетогенеза ко-
лоний, особенностях строения тек и колоний. В настоящее время одни
исследователи выделяют два отряда — Axonolipa и Axonophora, дру-
гие обосновывают выделение только подотрядов, рассматривая грап-
толоидей в ранге отряда.
Рис. 309. Отряды Axonolipa (7—6) и Axonophora (7—12):
1. — Dichograptus (ранний ордовик).; 2 — Tetragraptus (ранний ордовик);
3 — Didymograptus (ранний — поздний ордовик); 4—5 — Dicellograptus
(поздний ордовик); 6 — Phyllograptus (ранний ордовик); 7 — Diplograp-
fus. (ордовик—ранний силур); 8 — Monograptus (силур — ранний девон);
9 — Spirograptus (силур); 10 — Rastrites (ранний силур); И — Cyrto-
graptus (средний силур); 12 — Retiolites (ранний силур);
а'в — антивиргула, ей— виргула
Отряд Axonolipa (гр. axon — ось, leipo — покидать, оставлять).
Аксонолипы, или безосные граптолоидей, объединяют два подотряда
Dichograptina (или Didymograptima) и Leptograptina. Они характери-
зуются наличием немы, отсутствием виргулы, дихограптидным или
лептограптидным способом образования рабдосомы. Наиболее древние
представители ответвились от дендрид в начале ордовика; у них отсут-
ствуют диссептименты, сократилось число ветвей и исчезли битеки..
Эти изменения были, по-видимому, связаны с переходом части грапто-
33:
515
ордовике от основного ствола
Рис. 310. Типы тек:
1 — Rastrites; 2 — Leptograptus; 3 —
Glyptograptus; 4 — Climacograptus;
5—6 — Monograptus
литов к пелагическому образу жизни. Колония состояла из многочис-
ленных ветвей, дихотомически ветвящихся (рис. 309, 1—6)-, поплавок
служил для поддержания колонии в толще воды. У первых представи-
телей число ветвей могло превышать 64 (Bryograptidae), затем в тече-
ние раннего ордовика произошло сокращение ветвей до 16—8 (Logano-
graptus), затем до 8 (Dichograptus) и 4 (Tetragraptus) , к началу сред-
него ордовика сократилось до одной ветви (Azygograptus). В раннем
лип ответвилась боковая ветвь
филлограптид (Phyllograptidae),
рабдосомы которых состоят из 4
или 2 сросшихся дорзальными
сторонами ветвей скандентного
типа. На смену дихограптинам в
среднем и позднем ордовике
пришли лептограптины, у кото-
рых рабдосома состояла из двух
ветвей, отходящих от второй или
третьей теки. Ветви несли сигмо-
идально изогнутые теки и были
направлены вверх; у некоторых
форм начальные части ветвей
срастались, образуя на некото-
ром расстоянии двухрядную раб-
досому (Dicranograptidae).
В конце ордовика все безосные
граптолиты вымерли. К отряду
относится свыше 50 родов
(рис. 309, 1—6).
Отряд Axonophora (гр.
axon — ось, phoros — несущий).
Аксонофоры, или осеносные
граптолиты, объединяют ордо-
викских и силурийских грапто-
лоидей, у которых рабсодома со-
стояла из одной или нескольких
прямых или в различной степени изогнутых ветвей, вначале с двумя
рядами, а позднее с одним рядом тек (рис. 309, 7—12). Устья тек и
устье сикулы ориентированы в диаметрально противоположных направ-
лениях. Вследствие этих особенностей астогенеза внутри рабдосомы
располагалась опорная ось — виргула. Они характеризовались дипло-
граптоидным и монограптоидным способом образования колоний. На
этом основании выделяют подотряды. , ,
Первые аксонофоры появились в середине ордовика (лланвирне)
и были представлены преимущественно двухрядными формами, причем
у одних аксонофор ветви срастались своими дорзальными частями
(подотряд Diplograptina), у других — боковыми (подотряд Glosso-
graptina). В последнем случае боковые стороны тек одной ветви срас-
тались с боковыми сторонами тек другой ветви. Теки имели различную
форму (рис. 310). В силуре от двухрядных граптолоидей возникли
формы, имевшие, сложнопостроенный скелет, состоящий из решетчатого
остова — кл атрия и покрывающей его сетки — ретикулюма
(сем. Retiolitidae). Обычно у этих форм сикула и теки были склероти.-
зированы только частично и почти не сохранились в ископаемом со-
стоянии. В скелете ретиолитид имеется" виргула и расположенная про-
тив нее опорная вторая ось, получившая название антивиргула.
516
sajiJispU'-A J
— sn^dojbosjdA iq
snidojdouow
snydpjfypH^
51700/173011074
sn^dojdoipiJd
sn 70^00 oqajo/ш iq
S32?7O?^'^
| | sn^dojooisoi
sn?dojfioi/:pjq J
snidDjdoidiq
sn? ^dOJ^oboUJl 70
t/7 \doj6oUDJ3'/q\^r
sn jdojfo 77ЭЗ 7q [^T^
tn/dDjfioyjfipifl .,
sn 7^0 J6077^(7
\sn2dDj6DJ73J
dDpndDjdoL/siq
________ZPPU'ddjflQfijQ
____\-\Xl- —
^piu0707S dujo
op77snjo~diub '
DpUDLUDJ dUJO
~op тдпj ~dwo
diuo\
Сэ
с:
Сз
§
сз
•=4
7Ъ>'
с:
I
§>
орттпозоис
~“ орторизд dwo
ортээцра Ошо
Рис. 311. Геохронологическое распространение граптолитов
5з 5: <ъ <\э евин унт он !длоб нлок Л" 'Ъ §
& «3 И чз 1
шгиль предок ландейло ланбирн рениг ремадок оздний
к
Пневматофор отсутствовал, и, возможно, в подобном сетчатом скелете
были какие-то приспособления для плавания в толще воды
(рис. 309, 12).
Если в начале силура еще встречаются двухрядные аксонофоры, то
во второй половине силура и начале девона в морях безраздельно гос-
подствовали монограптидные граптолоидеи (подотряд Monograptina)
с однорядными рабдосомами, причем наряду с прямыми ветвями име-
лись колонии, у которых ветви были свернуты в плоскую или гелико-
идальную спираль. В конце силура среди монограптид появились фор-
мы со сложными колониями, состоящими из главной ветви и боковых;,
боковые ветви возникали вследствие того, что один из зооидов давал
начало не одной, а двум новым почкам (и, соответственно, текам).
Одна из новых почек служила продолжением главной ветви, а вторая
давала начало новой, боковой ветви; последняя получила название
«кл адий» (гр. klados — побег), или псевдоветвь.
Аксонофоры произошли от аксонолип в середине ордовика, про-
шли быстрый путь эволюции от двухрядных до однорядных'рабдосом
и вымерли в начале девона. К аксонофорам относится около 60 родов.
Граптолиты имеют очень важное стратиграфическое значение
(рис. 311). На основании эволюции представителей подкласса грапто-
лоидей в ордовике и силуре выделено свыше 40 зон.
ТИП POGONOPHORA. ПОГОНОФОРЫ
К типу погонофор (гр. pogon — борода, phoros — несущий) отно-
сится около 100 видов современных морских, в большинстве своем глу-
боководных, животных и несколько видов, встреченных в ископаемом
состоянии. Тело современных погонофор состоит из трех отделов: двух
передних коротких и третьего необычайно длинного туловищного. Тело
их нитевидное, очень длинное (до нескольких десятков сантиметров
при толщине до 3 мм); помещается в хитиновых цилиндрических тру-
бочках диаметром 0,06—2,8 мм и длиной от нескольких сантиметров до
1,5 м. На переднем конце расположен сложный щупальцевый аппарат,
•состоящий от 4—6 до 250 щупалец. Щупальца выполняют функцию
дыхания, служат для сбора пищи, ее переваривания и всасывания про-
дуктов пищеварения. Ротовое отверстие и кишечник отсутствуют. Пого-
нофоры раздельнополы. Оплодотворенные яйца развиваются в перед-
ней части трубки самки. Дробление идет по радиальному типу с обра-
зованием мезодермы энтероцельным путем. Молодое животное, выйдя
из яйцевых оболочек, после некоторого периода плавания, оседает на
дно и переходит к бентосному образу жизни, выделяя свою собствен-
ную трубку.
Современные погонофоры — обитатели бентоса в морях и океанах
преимущественно абиссальной области, но некоторые виды живут на
глубинах от 90 до 200 м. На твердых трубках погонофор поселяются
фораминиферы, губки, гидроиды, мшанки и многие другие прикреплен-
ные организмы. До сих пор остается неясным принадлежность погоно-
фор к вторичноротым. Не исключена возможность, что они занимают
особое место в системе животного мира.
В отложениях венда и нижнего кембрия встречены тонкие эла-
стичные цилиндрические хитиновые трубочки длиной до 70—120 мм
при диаметре 0,5—3 мм. Они отнесены к нескольким родам (Sabel-
lidites, Saarina, Sokoloviina), объединенным Б. С. Соколовым в отряд
Sabelliditida. Предположительно к погонофорам отнесены некоторые
посткембрийские находки из валунов ордовикских пород на территории
Польши, отложений силура Таймыра, олигоцена Орегона (США),
конкреций раннего плиоцена Бельгии. Древние погонофоры, появив-
шись в конце протерозоя, были приурочены к мелководным зонам
шельфа; позднее были оттеснены в более глубоководные зоны моря
Мирового океана и сохранились в глубоководных океанических впади-
нах до настоящего времени.
519
ТИП CHAETOGNATHA. ЩЕТИНКОЧЕЛЮСТНЫЕ
Хетогнаты (гр. chaeta — щетинки, gnathos — челюсть) — неболь-
шая группа (до 40 видов) морских планктонных организмов, имеющих
удлиненное стреловидное тело (длиной 1—9 см), покрытое кутикулой.
По бокам расположены одна — две пары плавников, а на.заднем кон-
це — непарный хвостовой плавник. Голова вооружена сильным ловчим
аппаратом, состоящим из пучков сильных крючьев, приспособленных
для захвата добычи. Рот расположен на переднем конце брюшной сто-
роны. На голове имеются два глаза. Хорошо развита нервная система.
Органы дыхания, выделения и кровеносная система отсутствуют. Хето-
гнаты — гермафродиты. Развитие прямое, без личиночной стадии. Они
распространены во всех морях и океанах.
Хетогнаты занимают обособленное положение среди всех билате-
ралий. Они относятся к вторичноротым; гаструла образуется путем ин-
вагинации, целом — энтероцельным путем; он распадается на три
части — головной, туловищный и целом половых желез. В области
бластопора образуется задний конец тела, рот возникает на противо-
положном конце,
В ископаемом состоянии (средний кембрий Канады) известны отпе-
чатки, относимые к хетогнатам и описанные под названием Amiskwia.
Других находок до сих пор не обнаружено.
ТИП PETALONAMAE. ПЕТАЛОНАМЫ
За последние 7 лет из кварцитов Кибис формации. Нама, развитых
в Юго-Западной Африке, найдены скелетные остатки организмов, кото-
рые палеонтолог Пфлюг (Pflug, 1970, 1972) выделил в самостоятель-
ный тип Petalonamae (гр. petalon — лист, Nama — название форма-
ции). Тело петалонам чашевидной, валиковидной или веретеновидной
формы, состоит из нескольких равных листовидных (петалоидных)
частей (рис. 312). Каждый петалоид разделен на три части: ра-
дикс (корень), рамус (ветвь) и петалон (листообразная пластин-
ка). Переход от радикса к рамусу всегда характеризуется коленооб-
разным изгибом. Внутри петалоида проходит осевой канал, от которого
отходят в стороны каналы второго порядка. Каждый петалон имеет
вид веерообразной пластинки, состоящей из многочисленных тонких
трубочек, соединенных промежуточной тканью. Шесть петалоидов, сое-
диненных вместе, образуют единую разветвленную часть — ф л а б е л-
лий, от которого отходит стебель (трунк). Стебель служил для заяко-
ривания тела. Два флабеллия срастаются вместе, образуя тело пета-
лонам. Внутри тела обособлена одна или две полости: центрарий и
контрарий. В местах соединения двух флабеллий расположены два
желобка: верхний и нижний. На этих желобках обнаружены три отвер-
стия: посредине верхнего желобка (це нт р о сто м), на трункальном
конце (трункостом) и на терминальном конце нижнего желобка
(термистом). Тонкие поры на повёрхности верхнего желобка поз-
воляют предполагать наличие ресничек или усиков для сбора пищи
(вероятно, планктона). Пища собиралась вдоль желобка и через трун-
костом поступала в центрарий, где происходило ее переваривание. Не-
переваренные остатки выделялись через термистом.
На' основании особенностей строения скелета, формы и характера
разветвления петалоидов Пфлюг предложил тип петалонам разделить
на три класса: Rangeomorpha, Pteridinomorpha, Erniettomorpha.
520
Систематическое положение петалонам остается недостаточно яс-
ным. По мнению Пфлюга, остатки Pteridinium из Эдиакары, которые
были отнесены к восьмилучевым кораллам, принадлежат петалонамам.
Рис. 312. Тип Petalonamae. Реконструкция трех разных представителей (Пфлюг, 1972).
Пи, Пи' — интерпеталоидные сутуры, тр— трунк, Фц Фа — флабеллии; части скелета:
а, — трунк, б — мезосома, в — терминальный конец (т)
Они не представляют собой колонии, а являются остатком части ске-
лета одной особи. По особенностям строения скелета, петалоидам пета-
лонамы ближе всего стоят к иглокожим. Однако в строении петалонам
многое остается еще неясным и требует дополнительных исследований.
ЛИТЕРАТУРА‘
Общая
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных, т. 1,
Проморфология; т. 2, Органология. Изд. 3. М., «Наука», 1964.
Берман 3. И. и др. Современные проблемы эволюционной теории. Л., «Нау-
ка», 1967.
Бондаренко О. Б., Михайлова И. А. Краткий определитель ископаемых
беспозвоночных. М., «Недра», 1969.
Борисяк А. А. Избранные труды. К столетию со дня рождения. М., «Наука»,
1973.
Вялов О. С.‘ Следы жизнедеятельности организмов и их палеонтологическое зна-
чение. Киев, 1966.
Геккер Р. Ф. Ископаемое юрское озеро в хребте Каратау. «Тр. ПИН», 1948,
т. 15.
Геккер Р. Ф. Наставление для исследований по палеоэкологии. М., Изд-во
АН СССР, 1954.
Геккер Р. Ф. Введение в палеоэкологию. М., Госгеолтехиздат, 1957.
Геология в Московском университете за 50 лет Советской власти. Под ред.
Д. И. Гордеева. Изд-во МГУ, 1967.
Давиташвили Л. Ш. История эволюционной палеонтологии от Дарвина до
наших дней. М., Изд-во АН СССР, 1948.
Давиташвили Л. Ш. Теория полового отбора. М., Изд-во АН СССР, 1961.
Давиташвили Л. Ш. Современное состояние эволюционного учения на За-
паде. М., «Наука», 1966.
Давиташвили Л. Ш. Причины вымирания организмов. М., «Наука», 1969.
Давиташвили Л. Ш. Изменчивость организмов в геологическом прошлом.
Тбилиси, 1970.
Давиташвили Л. Ш. Учение об эволюционном прогрессе. Теория ароморфо-
за. Тбилиси, 1972.
Д а р в и н Ч. Происхождение видов. М., Сельхозгиз, 1952.
Друщиц В. В., Обручева О. П. Палеонтология, изд. 2. Изд-во МГУ, 1971.
Друщиц В. В., Шиманский В. Н. Об объеме палеозойской эры. ДАН
СССР, 1962, т. 144, № 5.
Ефремов И. А. Тафономия и геологическая летопись, кн. 1. «Тр. ПИН»,
1950, т. 24. .
Ж а м о й д а А. И., Ковалевский О. П., Моисеева А. И. Обзор зару-
бежных стратиграфических кодексов. М., «Наука», 1969.
Жемчужников Ю. А. Курс палеофаунистики. М., 1934.
Жизнь животных. Под ред. Л. А. Зенкевича. М„ «Просвещение», т. 1, 2, 1968;
т. 3, 1969.
Завадский К. М. Вид и видообразование. Л., «Наука», 1968.
Зенкевич Л. А. Биология морей СССР. М., Изд-во АН СССР; 1963.
Зернов С. А. Общая гидробиология. М.—Л., Изд-во АН СССР, 1949.
Зубкович М. Е. Методы палеонтолого-стратиграфических исследований. Осно-
вы биостратиграфии. М., «Высшая школа», 1968.
Иванов А. В. Происхождение многоклеточных животных. Л., «Наука», 1968.
Коробков И. А. Палеонтологические описания, изд. 2. Л., «Недра», 1971.
Крымгольц Г. Я- Методика сбора и обработки палеонтолого-стратиграфиче-
ского материала. Изд-во ЛГУ, 1954.
522
Леонов Г. П. Основы стратиграфии. Изд-во МГУ, т. 1,- 1973, т. 2, 1974.
Майр Э. Систематика и происхождение видов. М., ИЛ, 1947.
М а й р Э. Зоологический вид и эволюция. М., «Мир», 1968.
Майр Э. Принципы зоологической систематики. М., «Мир», 1971.
МайрЭ., Л и нс ли Э., ЮзингерР. Методы и принципы зоологической си-
стематики. М., ИЛ, 1956.
Международный кодекс зоологической номенклатуры, принятый XV Международ-
ным зоологическим конгрессом. М., «Наука», 1966.
Методика палеонтологических исследований. Под ред. Б. Каммела и Д. Раупа.
М., «Мир», 1973.
Найдин Д. П. Морфология и палеобиология верхнемеловых белемнитов. Изд-во
МГУ, 1969.
НевесскаяЛ. А. и др. Новые представления о системе двустворчатых мол-
люсков. «Палеонтол. ж.», 1971, № 2.
Основы палеонтологии. Под ред. Ю. А. Орлова. М., Изд-во АН СССР — Госгеол-
техиздат, 1958—1964;
Палеонтологический словарь. Под ред. Г. А. Безносовой, Ф. А. Журавлевой. М.,
«Наука», 1965.
Палеонтология беспозвоночных. Под ред. Ю. А. Орлова. Изд-во МГУ, 1962.
Равикович А. И. Развитие основных теоретических направлений в геологии
XIX века. «Тр. ГИН», вып. 189. М., «Наука», 1969.
Рауп Д., Стэнли С. Основы палеонтологии. М., «Мир», 1974.
Руженцев В. Е. Принципы систематики, система и филогения палеозойских
аммоноидей. «Тр. ПИН», т. 83. М., «Наука», 1960.
Р у т т е н М. Происхождение жизни (естественным путем). М., «Мир», 1973.
Северцов А. Н. Главные направления эволюционного процесса. Морфобиоло-
гическая теория эволюции, изд. 3. Изд-во МГУ, 1967.
Симпсон Д. Г. Темпы и формы эволюции. М., ИЛ, 1948.
Степанов Д. Л. Принципы и методы биостратиграфических исследований. «Тр.
ВНИГРИ», вып. 113. Л., Гостоптехиздат, 1958.
Стратиграфическая классификация, терминология и номенклатура. Под ред.
А. И. Жамойды. Л., «Недра», 1965.
Тимофеев - Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Крат-
кий очерк теории эволюции. М., «Наука», 1969. ’
У о т с о н Д. Д. Двойная спираль. М., «Мир», 1969.
Федотов Д. М. Эволюция и филогения беспозвоночных животных. М., «Нау-
ка», 1966.
Ходалевич А. Н., Торбакова А. Ф. Палеонтология. М., «Высшая шко-
ла», 1965.
Шмальгаузен И. И. Проблемы дарвинизма, изд. 2. Л., «Наука», 1969.
A g е г D. Principles of paleoecology. N.-Y., London, 1963.
Babin C. Elements de Paleontologie. Ser. «Geologie», Collection U. Paris, 1971.
Eckman S. Zoogeography of the Sea. London, 1967.
Geyer O. F. Grundziige der Stratigraphie und Fazieskunde. Bd. 1. Stuttgart, 1973.
E a s t о n W. H. Invertebrate Paleontology. N. Y„ 1960.
Jeletzky J. A. Comparative morphology, phylogeny and classification of fossil
Coleoidea. «Univ. Kans. Paleontol. Contribs», 1966, No. 7.
Land H. S. Treatise of marine Ecology and Paleoecology. N. Y., 1957.
Moore R. C., L a 1 i c k e r C. G., Fischer A. G. Invertebrate Fossils. N.Y.—
Toronto — London, 1952.
Muller A. H. Grundlagen der Biostratonomie. Abh. Deutsch.. Akad. Wiss.,. Nr. 3.
Berlin, 1950.
Muller A. H. Lehrbuch der Palaozoologie. Bd. 1—3. Jena, 1957—1966.
P f 1 u g H. D. Systematik der jung-prakambrischen Petalonamae Pflug 1970. «Pa-
laont. Z.», 1972, Bd. 46, Nr. 1/2.
Raup D. M., Stanley S. M. Principles of Paleontology. San Francisco,. 1971.
Ry 1 a n d J. S. Bryozoans. London, 1970.
ScharovA. G. Basic Arthropodan Stock with Special Reference to Insects. Per-
gamon Press, 1966, vol. 30.
Schaffer W. Actuo-Palaontologie nach Studien in der Nordsee. Frankfurt, 1962,
Schindewolf О. H. Grundfragen der Paiaontologie. Stuttgart, 1950.
Shrock R., Twenhof el W. Principles of Invertebrate Paleontology. N. Y. —
Toronto — London, 1953.
S p i n a r Z. Zaklady paleontologie bezobratlych. Praha, 1960.
Tasnadi-Kubacska A. Palaopathologie. Pathologie der Vorzeitlichen Tiere.
Jena, 1962.
Taylor J. D. The structural evolution of the Bivalve shell. «Palaeontology», 1973,
vol. 16, Pt. 3.
523
T h e n i u s E. Versteinerte Urkunden. Berlin, 1963.
T er mi er H. et G. Paleontologie Stratigraphique. Paris, 1960.
T e r m i e r H. et G. Biologie et ecologie des premiers fossiles. Paris, 1968.
Traite de Paleontologie (Publie sous la direction de J. Piveteau), T. 1—3. Paris,
1952—1953.
Treatise on Invertebrate Paleontology. (First Editions directed and edited
R. C. Moore, second — by C. Teichert). Geol. Soc. of America and University of Kansas
Press. 1953.
Ziegler B. Allgemeine Palaontologie Einfiihrung in die Palaobiologie, Teil 1.
Stuttgart, 1972.
Палеонтологические журналы
«Палеонтологический журнал». М., Изд-во АН СССР.
«Acta Paleontologia Polonica». Warszawa.
Lethaia. Universitetvorlaget. Oslo.
«Palaontologische Zeitschrift». Stuttgart.
«Palaeontology». London.
«Journal of Paleontology». USA.
Атласы, монографии и труды
Атлас руководящих форм ископаемых фаун СССР, тт. 1—13. М., Госгеолтехиз-
дат, 1939—1949.
Ежегодник Всесоюзного палеонтологического общества. М., Госгеолтехиздат.
Палеонтология СССР. Монографии. М., Изд-во АН СССР.
Труды Палеонтологического института. М., Изд-во АН СССР.
ОГЛАВЛЕНИЕ
Предисловие ................................................................ 3
ОСНОВНЫЕ ПОЛОЖЕНИЯ И МЕТОДЫ ПАЛЕОНТОЛОГИИ
Предметы и задачи палеонтологии ......................................... 7
Сохранение остатков животных в ископаемом состоянии и методы их обработки 12
Образование ориктоценозов . . .... 12
Формы сохранности....................................................... 15
Сбор и методы обработки ископаемых остатков............................ 18
Основные этапы развития палеонтологии . . .'.........................20
'Организм и среда...........................................................44
Палеонтология и некоторые проблемы эволюции.................................52
Современные представления о наследственности и изменчивости ..... 52
Борьба за существование и естественный отбор............................57
Изменение органов в филогенезе..........................................59
Адаптивная радиация, конвергенция и параллелизм.........................60
Монофилия и полифилия...................................................61
Соотношение между онтогенезом и филогенезом.............................62
Пути и закономерности эволюционного процесса............................64
Вымирание организмов.................................................. 66
Необратимость эволюции . . .................'.....................67
Принципы систематики животных и правила зоологической номенклатуры . . 67
Палеонтологический метод и основы стратиграфической классификации ... 74
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Тип Protozoa. Простейшие..............................................................89
Класс Sarcodina. Саркодовые..................................................93
Подкласс Rhizopoda. Корненожки.............................................93
Подкласс Foraminifera. Фораминиферы . . . ..... 94
Надотряд Allogromioidea. Аллогромиоидеи . . ....................105
Надотряд Textularioidea. Текстуляриоидеи . . ....................105
Надотряд Fusulinoidea. Фузулиноидеи ...................................108
Надотряд Miliolidoidea. Милиолидоидеи..................................111
Надотряд Rotalioidea. Роталиоидеи.......................... . . . 113
Подкласс Heliozoa. Солнечники.............................................122
Подкласс Radiolaria. Радиолярии....................................... 122
Класс Cnidosporidia. Книдоспоридии . . 129
Класс Sporozoa. Споровики.................................................129
Класс Zoomastigophora. Зоомастигофоры.....................................129
Класс Ciliophora. Ресничные...............................................130
Подкласс Ciliata. Ресничные инфузории ..................................130
Филогения простейших............................................................ 131
Metazoa. Многоклеточные.
Надраздел Parazoa. Низшие многоклеточные
Тип Spongia. Губки..........................................................138
Класс Demospongia. Обыкновенные губки . . 143
Класс Hexactinellida. Шестилучевые губки......................................145
Класс Calcispongia. Известковые губки.........................................146
Класс Sclerospongia. Склероспонгии............................................146
Тип Archaeocyathi. Археоциаты....................................................... 149
Класс Regulares. Правильные археоциаты........................................155
Класс Irregulares. Неправильные археоциаты....................................157
Группы неясного систематического положения.......................................... 161
Класс (?) Cribricyatha. Крибрициаты........................................161
Класс (?) Sphinctozoa. Сфинктозои ............................................162
Класс (?) Aphrosalpingoida. Афросальпингоиды..................................163
Класс (?) Receptaculita. Рецептакулиты........................................163
Семейство (?) Soanitidae. Соаниты...........................................164
Класс (?) Stromatoporata. Строматопораты......................................164
Класс (?) Hydroconozoa. Гидроконозои..........................................168
Надраздел Eumetazoa. Высшие многоклеточные
Раздел Radialia. Радиальные............................... . 170
Тип Coelenterata. Кишечнополостные............................................ 171
Класс Hydrozoa. Гидроидные .................................173
Подкласс Hydroidea. Гидрообразные.....................................176
Подкласс Siphonophoroidea. Сифонофоры ........ 177
Приложение к классу гидроидных. Хететиды..................... . . 179
Класс Scyphozoa. Сцифозои . . . . .........................180
Подкласс Scyphomedusae. Сцифомедузы .*..............................180
Подкласс Conulata. Конуляты.........................................182
Подкласс Protomedusae. Протомедузы..................................184
Класс Anthozoa. Коралловые полипы ......................................185
Подкласс Octocorallia. Восьмилучевые кораллы..........................187
Подкласс НехасогаШа. Шестилучевые кораллы..........................189
Подкласс Tabulate. Табуляты.........................................196
Подкласс Heliolitoidea. Гелиолитоидеи.................................201
Подкласс Tetracorallia (Rugosa). Тетракораллы (ругозы) .... 203
Тип Ctenophora. Гребневики................................................... 211
Раздел Bilateralia. Двусторонне-симметричные . . .. 213
Подраздел Protostomia. Первичноротые
Тип Scolecida. Низшие черви...................................................213
Подтип Plathelminthes. Плоские черви......................................214
Подтип Nemathelminthes. Круглые черви................................... 215
Подтип Nemertini. Немертины.............................................. 215
Тип Annelides. Кольчатые черви................................................216
Тип Onychophora. Онихофоры................................................... 219
Тип Arthropoda. Членистоногие.................................................221
Подтип Proboscifera. Пробосциферы.........................................226
Класс Dicephalosomita. Дицефалосомиты...................................226
Класс Pycnogonides. Многоколенчатые................................... 226
Подтип Trilobitomorpha. Трилобитообразные.................................228
. Класс Tetracephalosomita. Тетрацефалосомиты . 228
Класс Trilobitida. Трилобитиды..........................................228
Класс Trilobita. Трилобиты..............................................230
Подкласс Miomera. Миомеры.............................................236
Подкласс Polymera. Полимеры...........................................236
Подтип Chelicerata. Хелицеровые . 244
Класс Merostomata. Меростомовые........................................ 246
Подкласс Xiphosura. Мечехвостовые.....................................247
Подкласс Eurypteridoidea. Эвриптериды...................., . . . 248-
Класс Scorpionomorpha. Скорпионообразные................................249
Класс Arachnida. Паукообразные..........................................250
Класс Solifugomorpha. Сольпугоподобные..................................252
Класс Acaromorpha. Акароморфы, или клещи................................252
Подтип Branchiata. Жабродышащие...........................................254
Класс Crustacea. Ракообразные...........................................254
Подкласс Branchiopoda. Жаброногие . 256
Подкласс Ostracodoidea. Остракоды, или ракушковые рачки . . . 258
Подкласс Ci^ripedia. Усоногие раки.................................. 265
Подкласс Copepodoidea. Копеподовые....................................267
Подкласс Cephalocarida. Цефалокариды..................................267
526
Подкласс Malacostraca. Малакостраки................................
Подтип Tracheata. Трахейнодышащие . . . '. ..................
Класс Myriopoda. Многоножки............................................
Класс Insecta. Насекомые...............................................
Подкласс Apterygota. Первичнобескрылые...............................
Подкласс Pterygota. Крылатые насекомые...............................
Инфракласс Hemimetabola. Насекомые с неполным превращением
Инфракласс Homometabola. Насекомые с полным превращением
Тип Mollusca. Моллюски.........................................................
Класс Aplacophora. Аплакофоры..........................................
Класс Polyplacophora. Полиплакофоры....................................
Класс Monoplacophora. Моноплакофоры....................................
Класс Gastropoda. Брюхоногие...........................................
• Подкласс Prosobranchia. Переднежаберные..............................
Подкласс Opisthobranchia. Заднежаберные . ..................
Подкласс Pulmonata. Легочники........................................
Класс Scaphopoda. Лопатоногие..........................................
Класс (?) Xenoconchia. Ксеноконхий.....................................
Класс Bivalvia. Бивальвии, или двустворчатые моллюски..................
Надотряд Protobranchia. Протобранхии.................................
Надотряд Autobranchia. Аутобранхии ..................................
Надотряд Septibranchia. Септибранхии.................................
Класс Cephalopoda. Головоногие моллюски................................
Подкласс Nautiloidea. Наутилоидеи....................................
Подкласс Endoceratoidea. Эндоцератиты................................
Подкласс Actinoceratoidea. Актиноцератиты............................
Подкласс Orthoceratoidea. Ортоцератиты . . . .
Подкласс Bactritoidea. Бактриты......................................
Подкласс Ammonoidea. Аммониты........................................
Подкласс Coleoidea. Колеоидей . . ’..................................
Класс Tentaculita. Тентакулиты.........................................
Класс Hyolitha. Хиолиты................................................
Класс неизвестен. Хиолительминтиды.....................................
Тип Bryozoa. Мшанки.................................................
Класс Phylactolaemata. Филактолематы...................................
Класс Stenolaemata. Стенолематы........................................
Класс Gymnolaemata. Гимнолематы........................................
Тип Phoronoidea. Форониды . . . . ....................... . . .
Тип Brachiopoda. Брахиоподы.................................................'.
Класс Inarticulate. Инартикуляты, или беззамковые брахиоподы .
Класс Articulate. Артикуляты, или замковые брахиоподы . . . .
267
273
273-
275
279
282'
282
282
283
285
286'
287
290
291
302
303
308-
310
311
326
327
340
343
347
356
359
360
362
363
379
389
393
395
397
401
402
406.
411
413
415
422
Подраздел Deuterostomia. Вторичноротые
Тип Echinodermata. Иглокожие....................................................... 440'
Подтип Carpozoa. Карпозои.......................................................445
Класс Carpoidea. Карпоидеи....................................................446
Класс Homostelea. Гомостелей................................................. 447
Класс Homoiostelea. Гомойостелеи............................................ 448.
Подтип Crinozoa. Кринозои..................................................... 449
Класс Cystoidea. Цистоидеи . . . 450
Класс Blastoidea. Бластоидеи................................................ 453
Класс Crinoidea. Криноидеи, или морские лилии.................................455
Класс Eocrinoidea. Эокриноидеи................................................464
Класс Paracrinoidea. Паракриноидеи............................................466
Класс Edrioblastoidea. Эдриобластоидей........................................466
Класс Parablastoidea. Парабластоидеи..........................................466
Класс LepidocystOidea. Лепидоцистоидеи........................................468
Подтип Asterozoa. Астерозои.....................................................468
Класс Somasteroidea. Сомастероидеи............................................468
Класс Asteroidea. Морские звезды ............................................ 469
Класс Ophiuroidea. Офиуры, или змеехвостки . .'........................471
Подтип Echinozoa. Эхинозои .....................................................472
Класс. Echinoidea. Морские ежи................................................472
Подкласс Perischoecninoidea. Перишэхиноидеи.............................. 482’
Подкласс Euechinoidea. Эуэхиноидеи, или настоящие ежи .... 485
Надотряд Diadematacea. Диадемовые морские ежи............................485
Надотряд Gnathostomata. Гнатостоматы, или челюстноротые . . . 486
Надотряд Atelostomata. Ателостоматы . . ...... 487
527
Класс Edrioasteroidea. Эдриоастероидей................................
Класс Holothurioidea. Голотурии . .................................
Класс Ophiocistioidea. Офиоцистии.....................................
Класс Helicoplacoidea. Геликоплакоидеи.............................• .
Класс Cyclocystoidea. Циклоцистоидеи .................................
Класс Camptostromatoidea. Камптостроматоидеи. . . ' .
Подтип (?) Haplozoa. Гаплозои...........................................
Класс (?) Cycloidea. Циклоидеи........................................
Класс (?) Cyamoidea. Циамоидеи........................................
Тип Hemichordata. Гемихордовые................................................
Класс Enteropneusta. Кишечнодышащие...................................
Класс Pterobranchia. Крыложаберные
Класс Graptolithina. Граптолиты ......................................
Подкласс Stereostolonata. Стереостолонаты...........................
Подкласс Graptoloidea. Граптолоидеи ........
Тип Pogonophora. Погонофоры...................................................
Тип Chaetognatha. Щетинкочелюстные................................, . . . .
Тил Petalonamae. Петалонамы........................................ . . .
Литература....................................................................
490
492
494
495
496
496
497
497
498
499
499
501
504
'504
511
519
520
520
522
ВЛАДИМИР ВАСИЛЬЕВИЧ ДРУЩИЦ
ПАЛЕОНТОЛОГИЯ БЕСПОЗВОНОЧНЫХ
Тематический план 1974 г. № 175
Редактор Н. В. Баринова
Переплет художника
Е. А. Михельсона
Технический редактор .
В. И. Овчинникова
Корректоры М. И. Эльмус, С. Ф. Будаева, Н. И. Коновалова, И. С. Хлыстова,
Л. А. Айдарбекова
Сдано в набор 29/VI I 1974 г. Подписано к печати 20/XI 1974 г.
Л-50540 Формат 70X1081/i6 Бумага тип. № 2
Физ. печ. л. 33,0 Уел. печ. л. 46,2 Уч.-изд. л. 45,01
Изд. №’ 2226 Заказ 600 Тираж 3600 экз. Цена 1 р. 93 к.
Издательство
Московского университета.
Москва, К-9, ул. Герцена, 5/7.
Типография Изд-ва МГУ. Москва, Ленинские горы