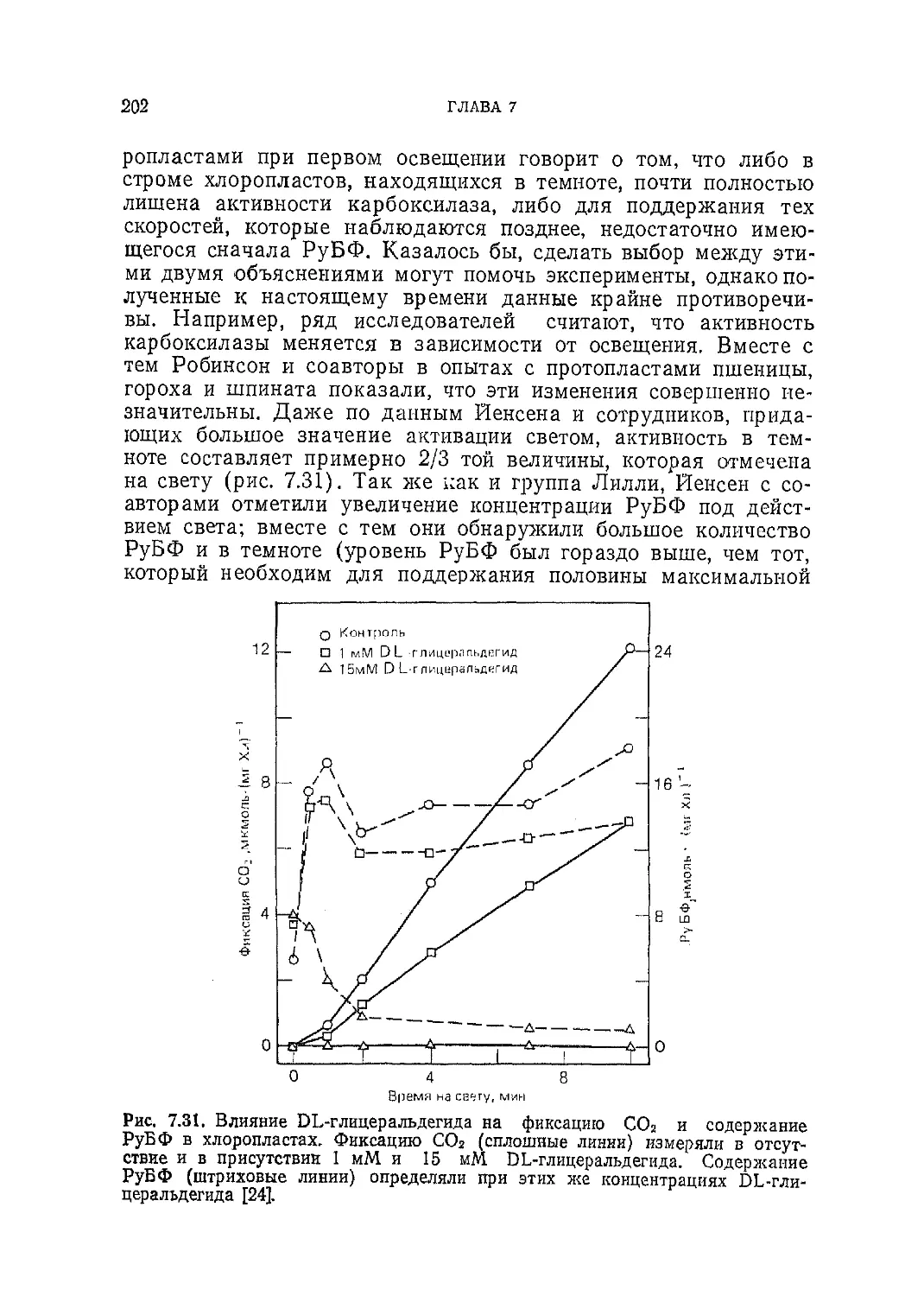
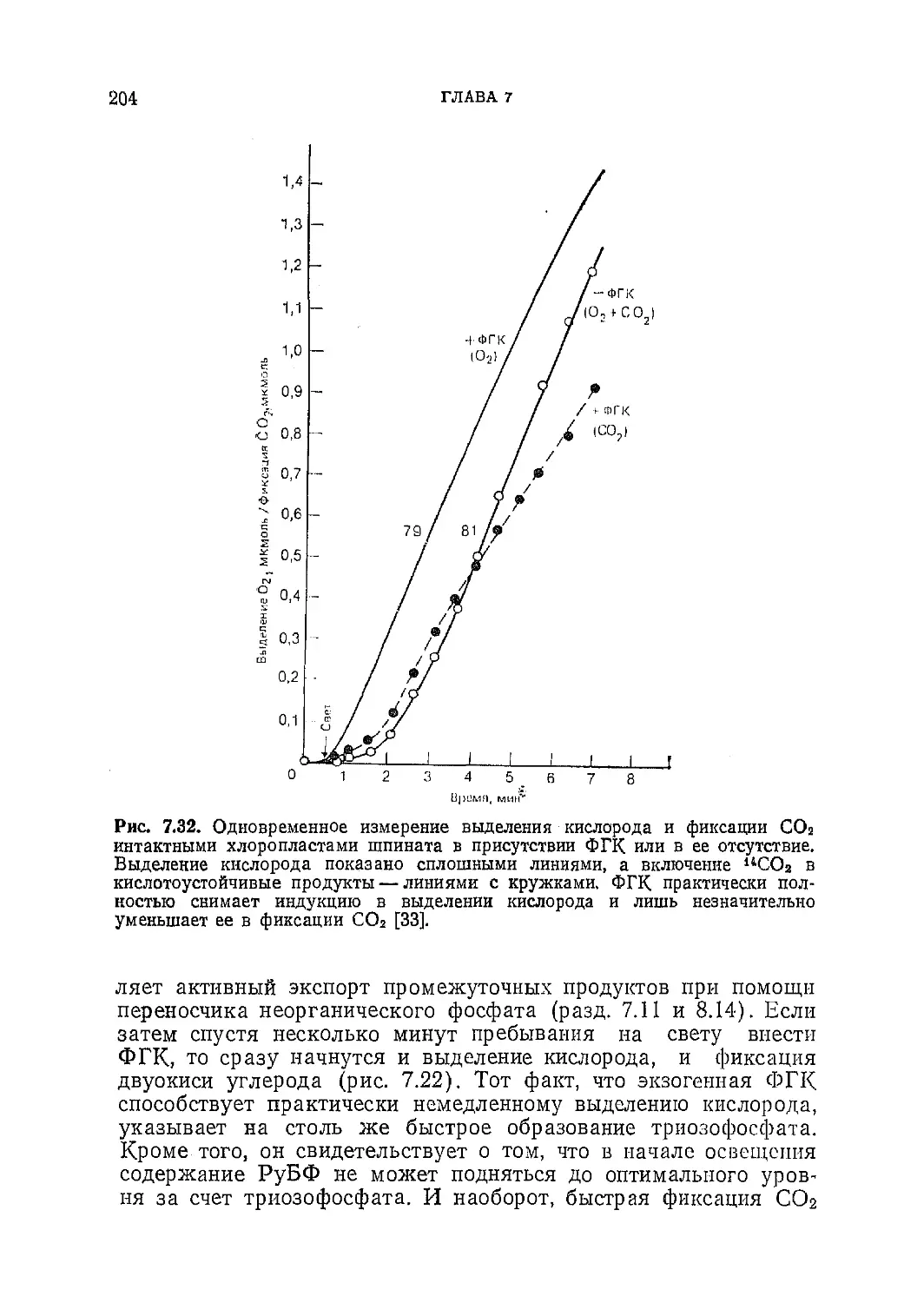
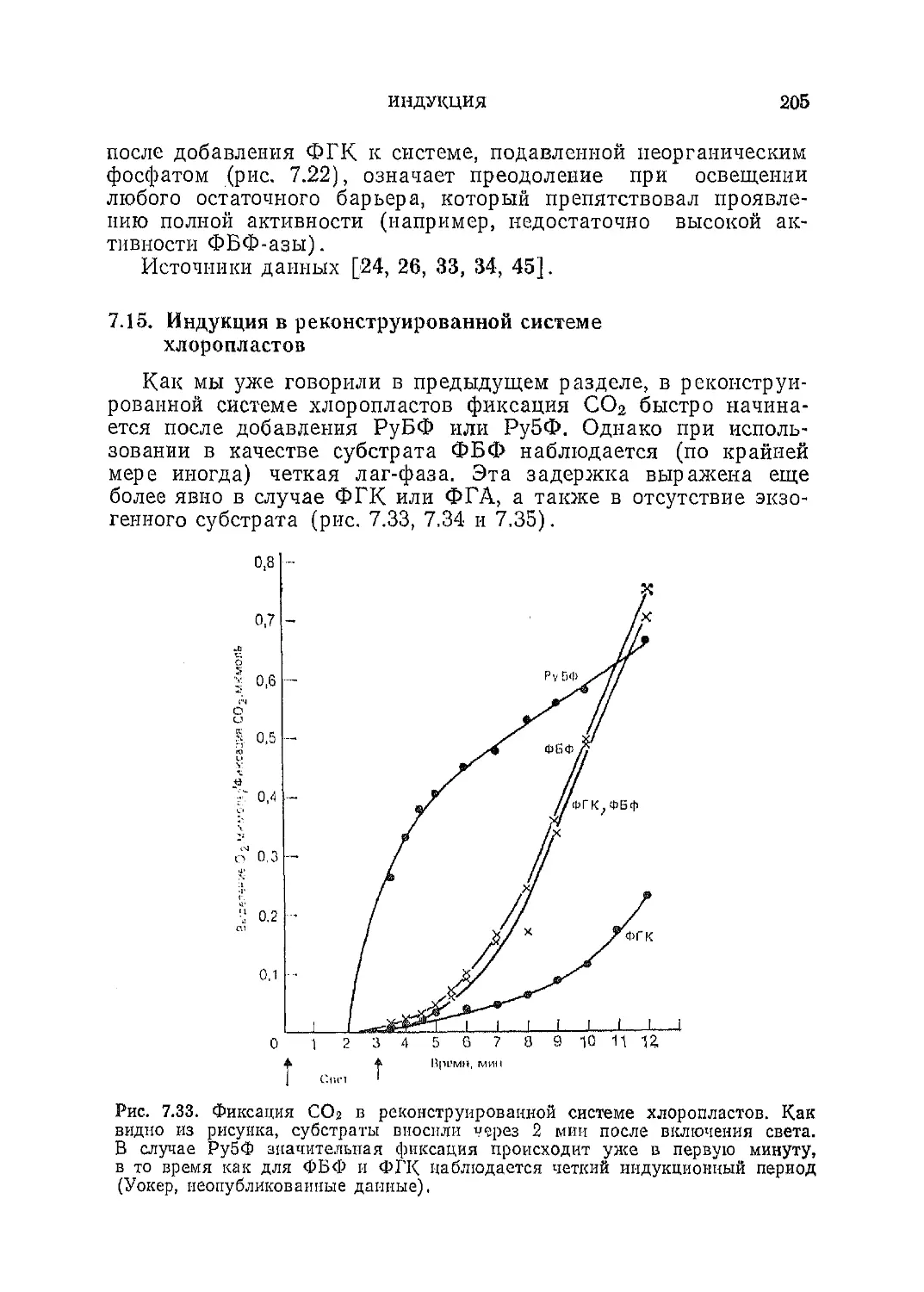
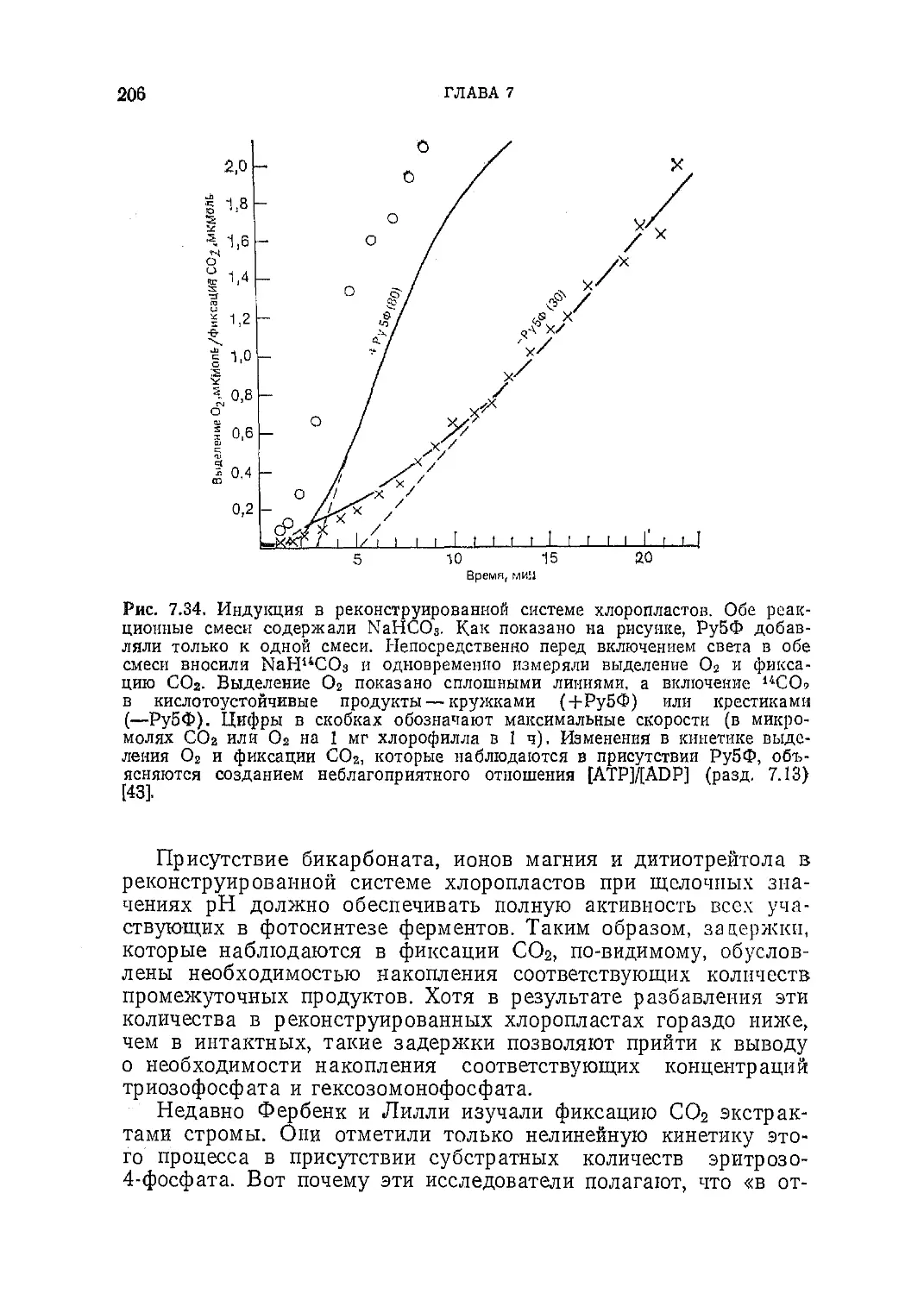
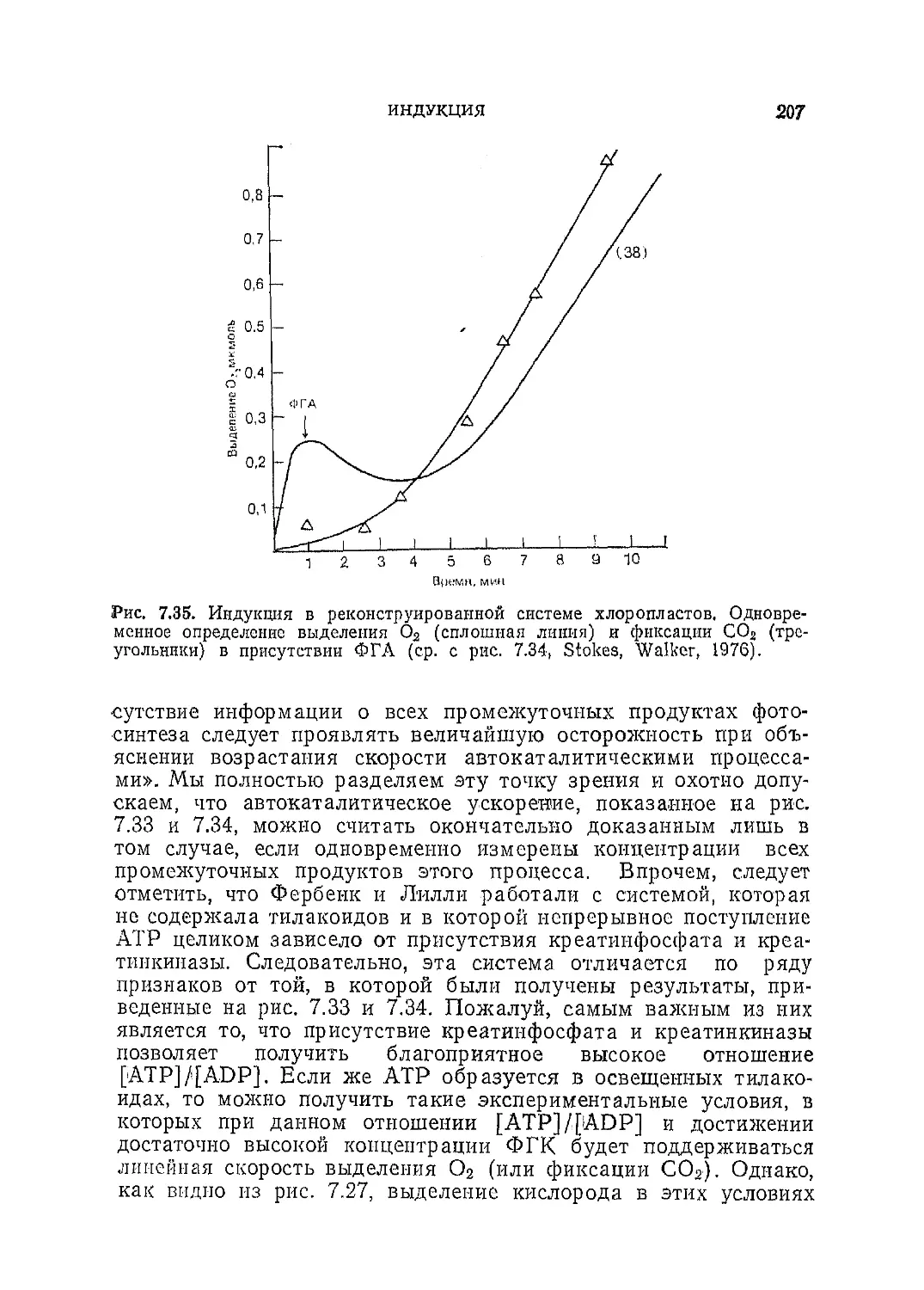
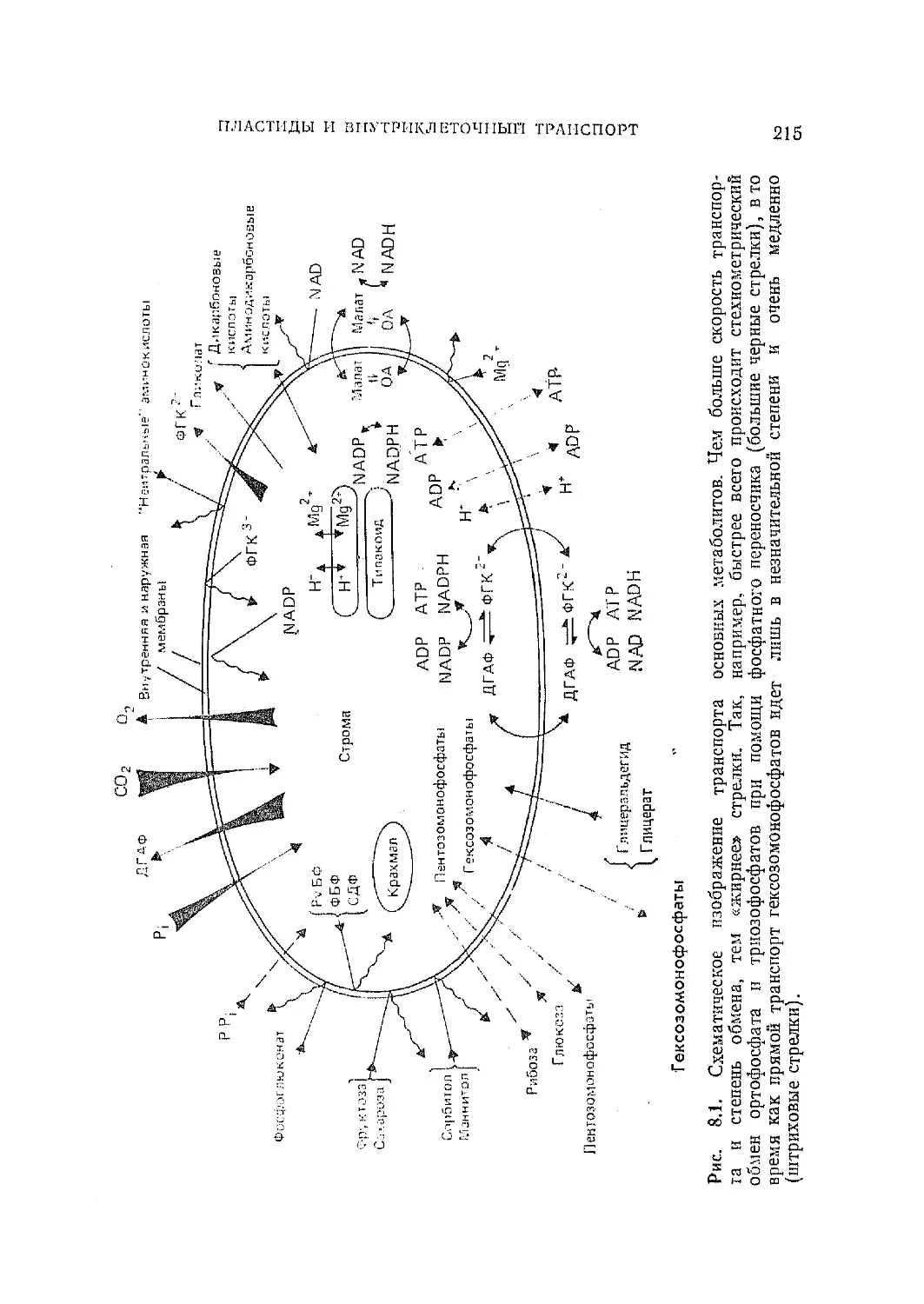
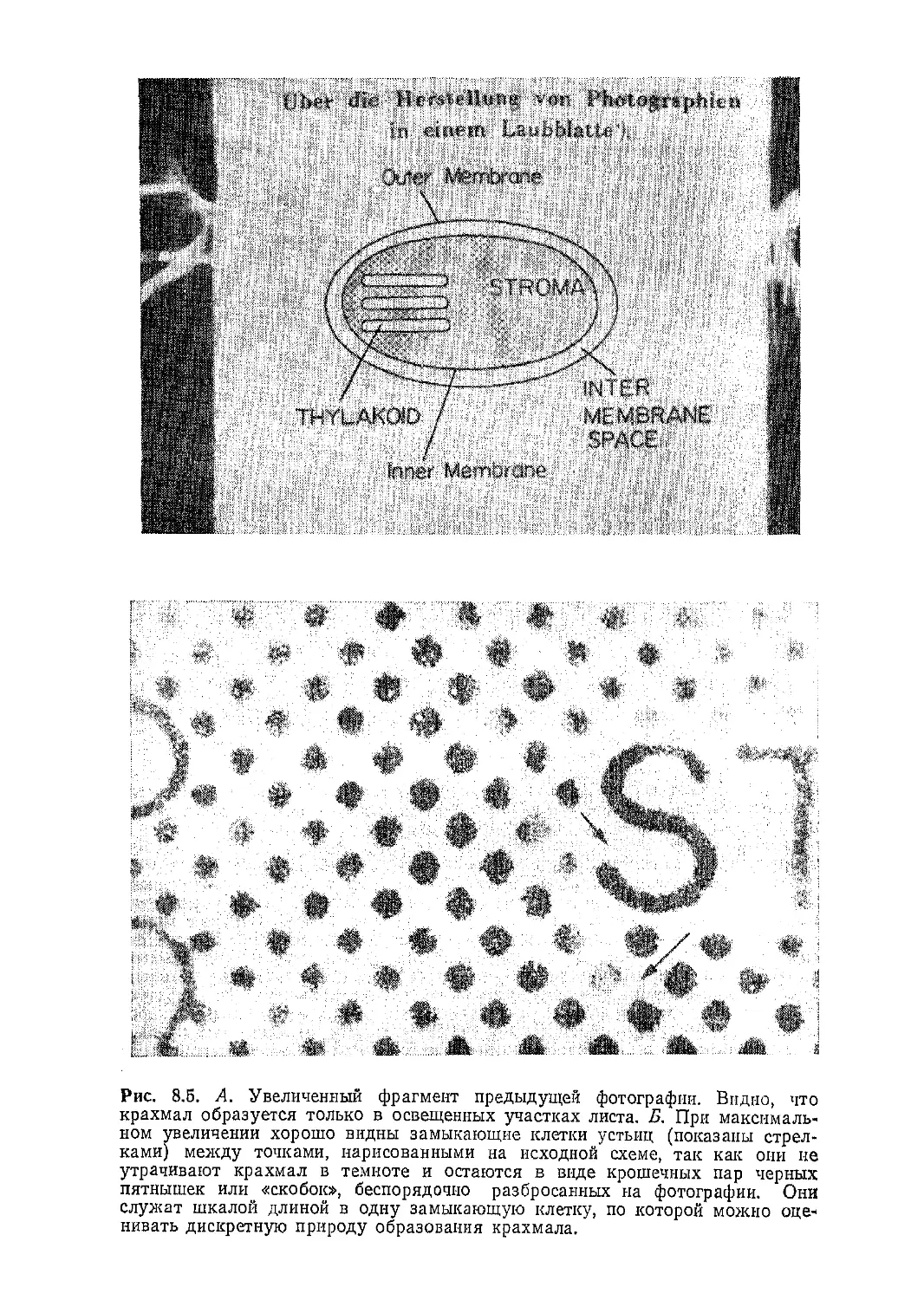
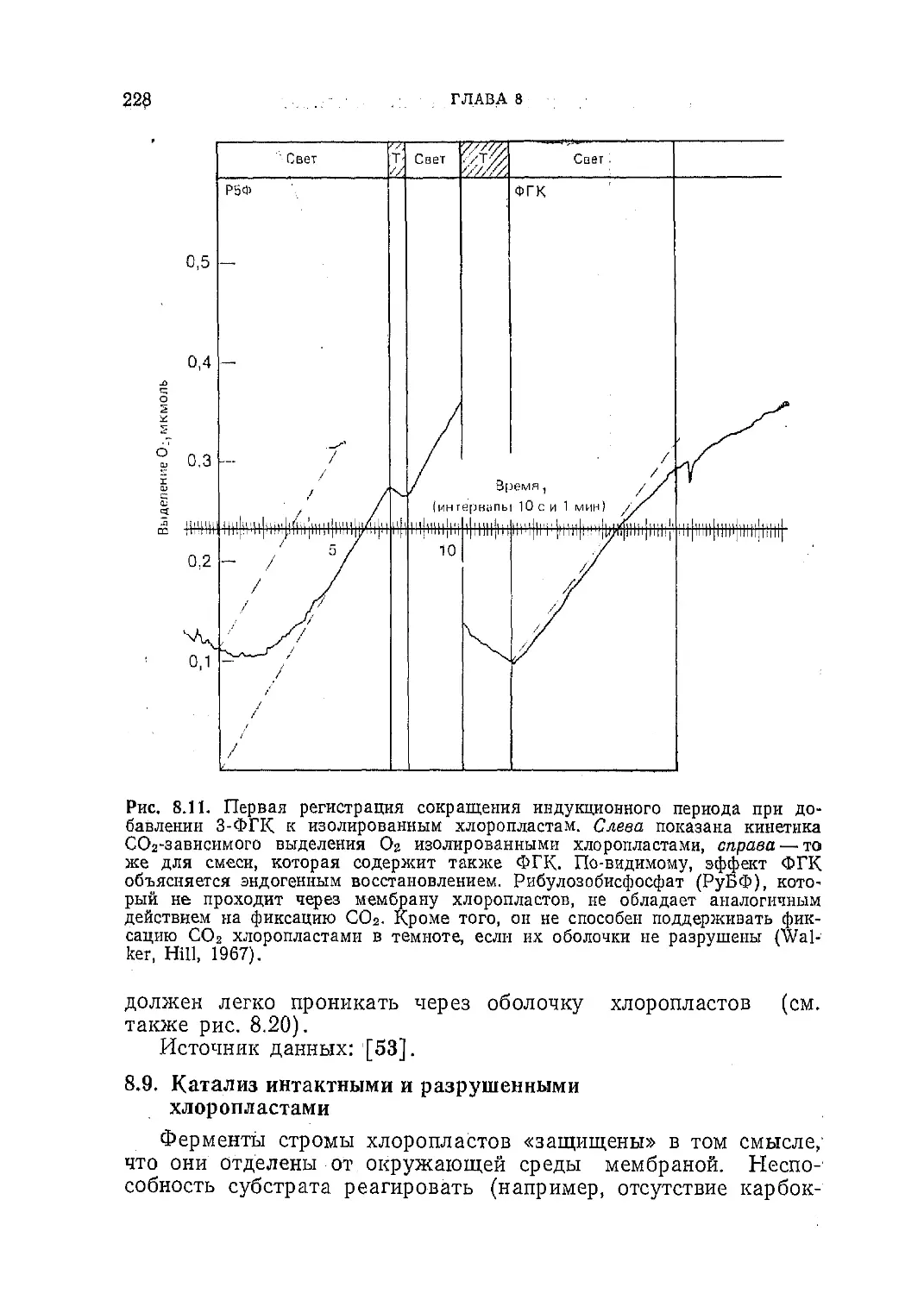
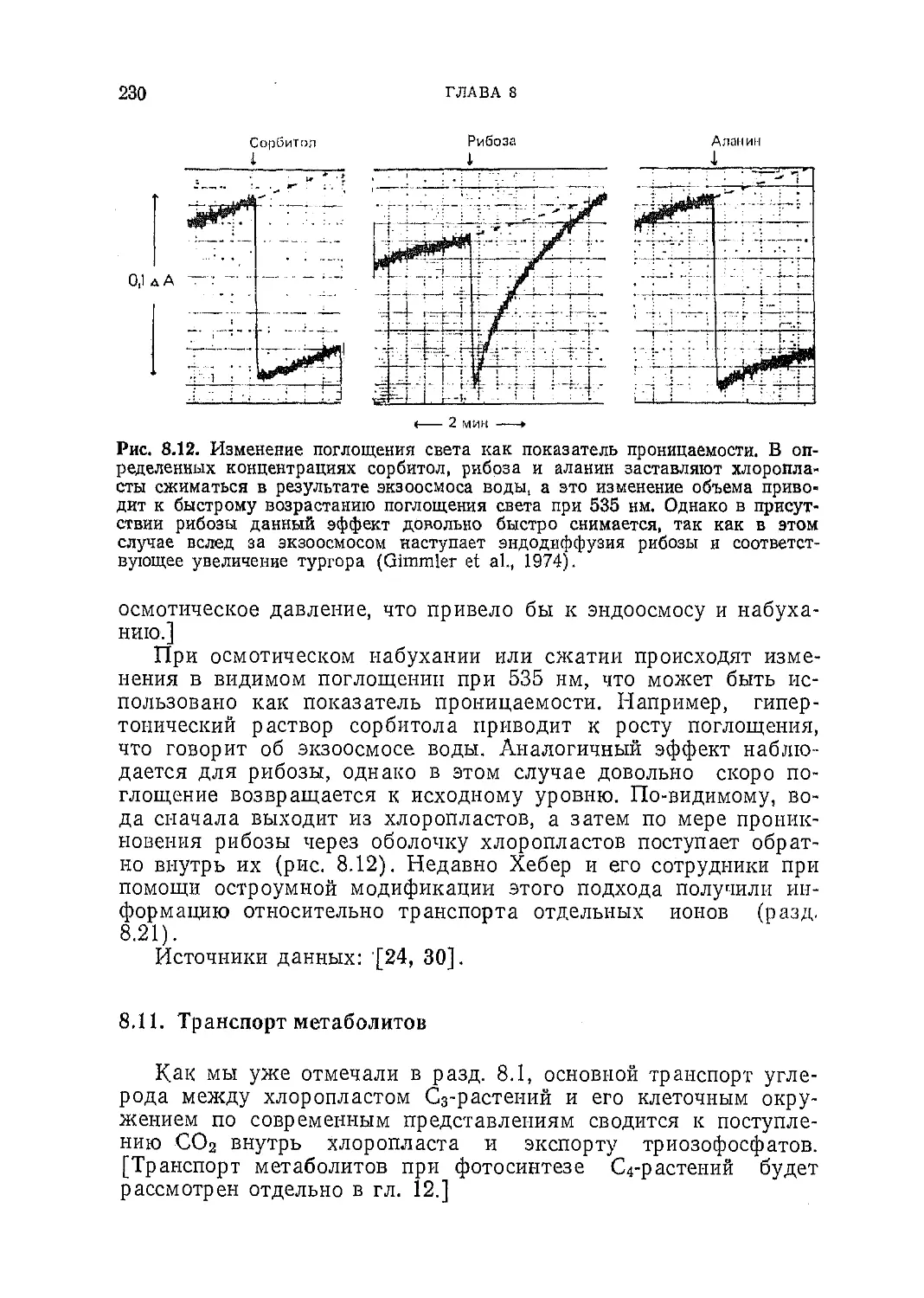
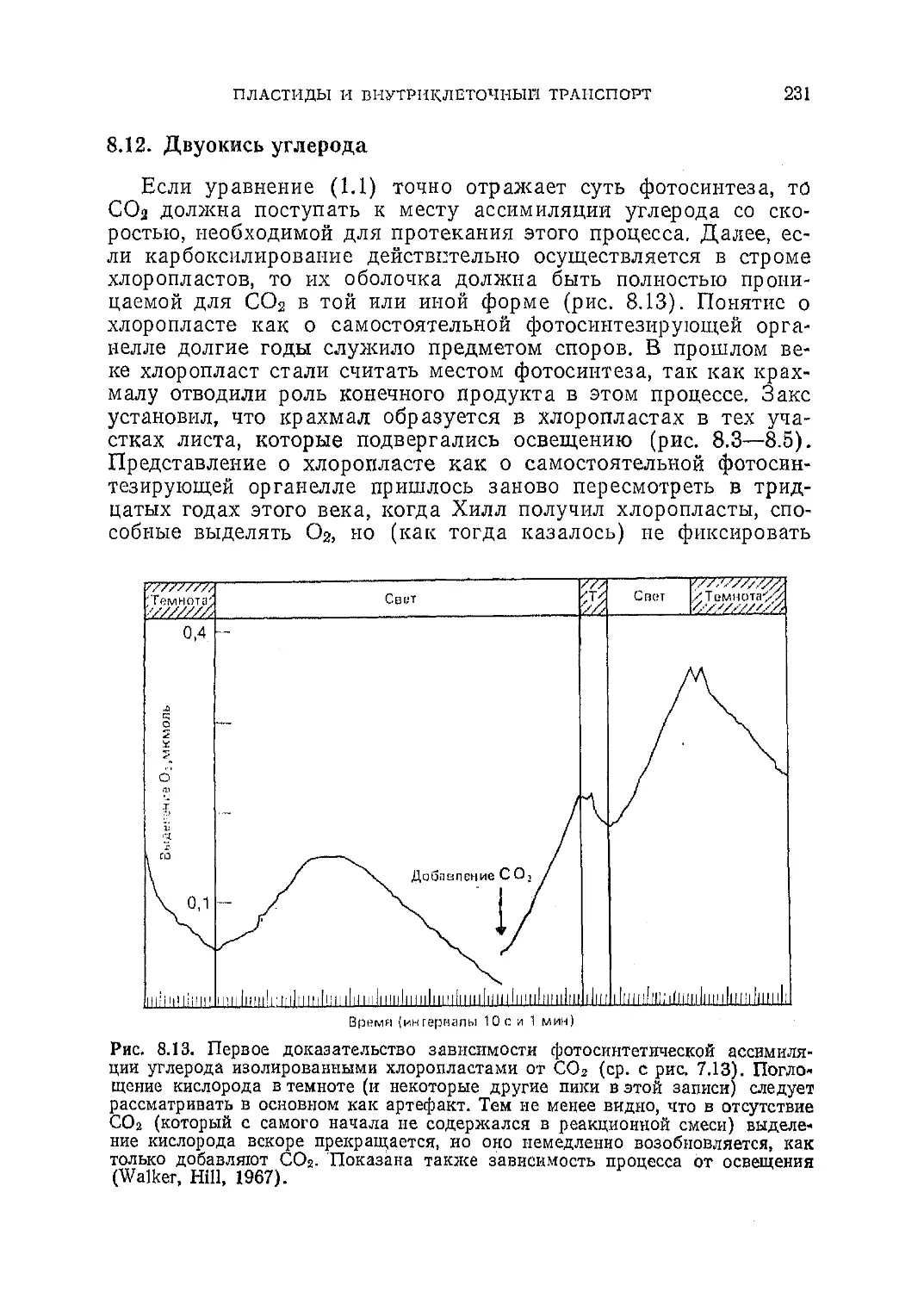
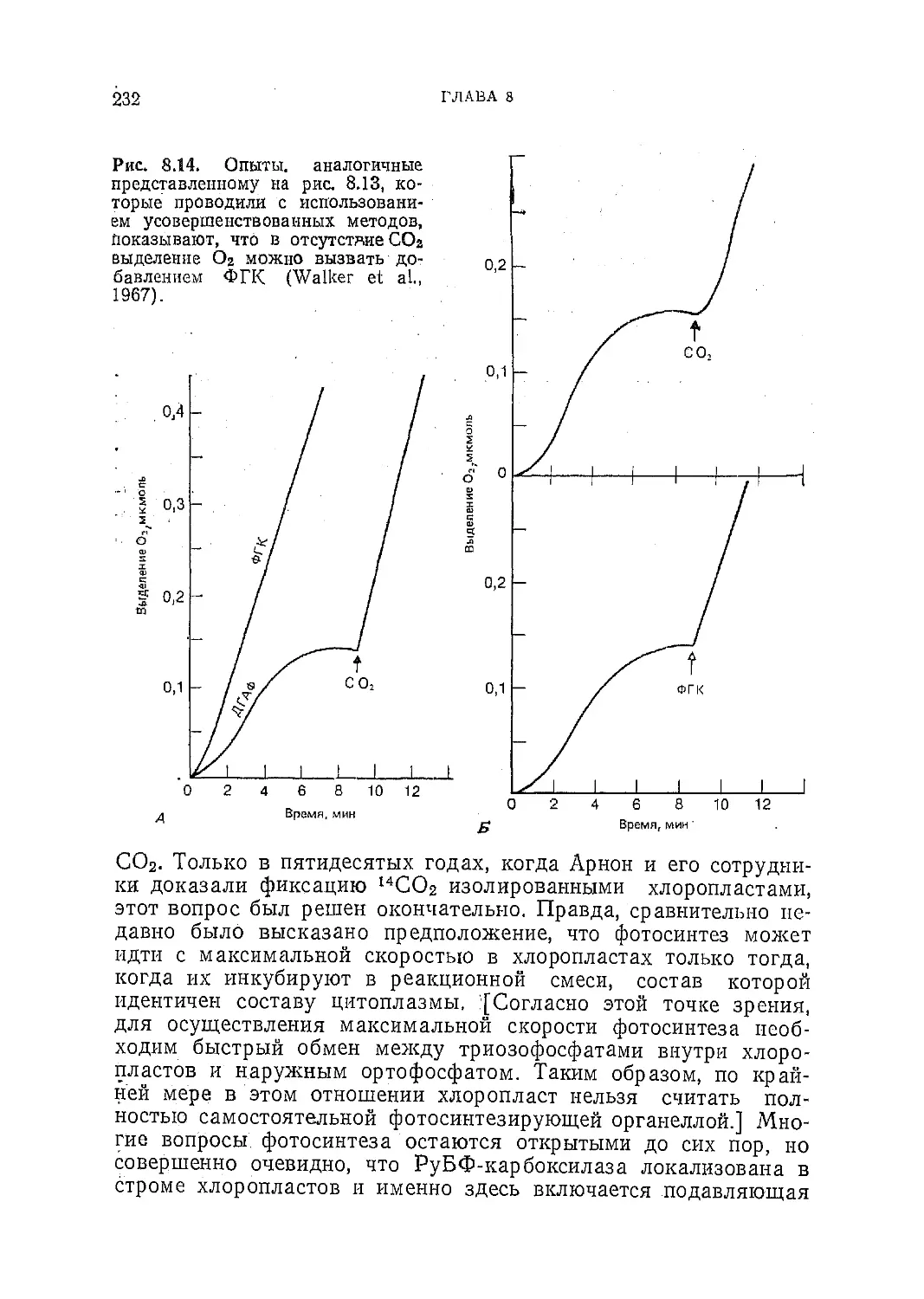
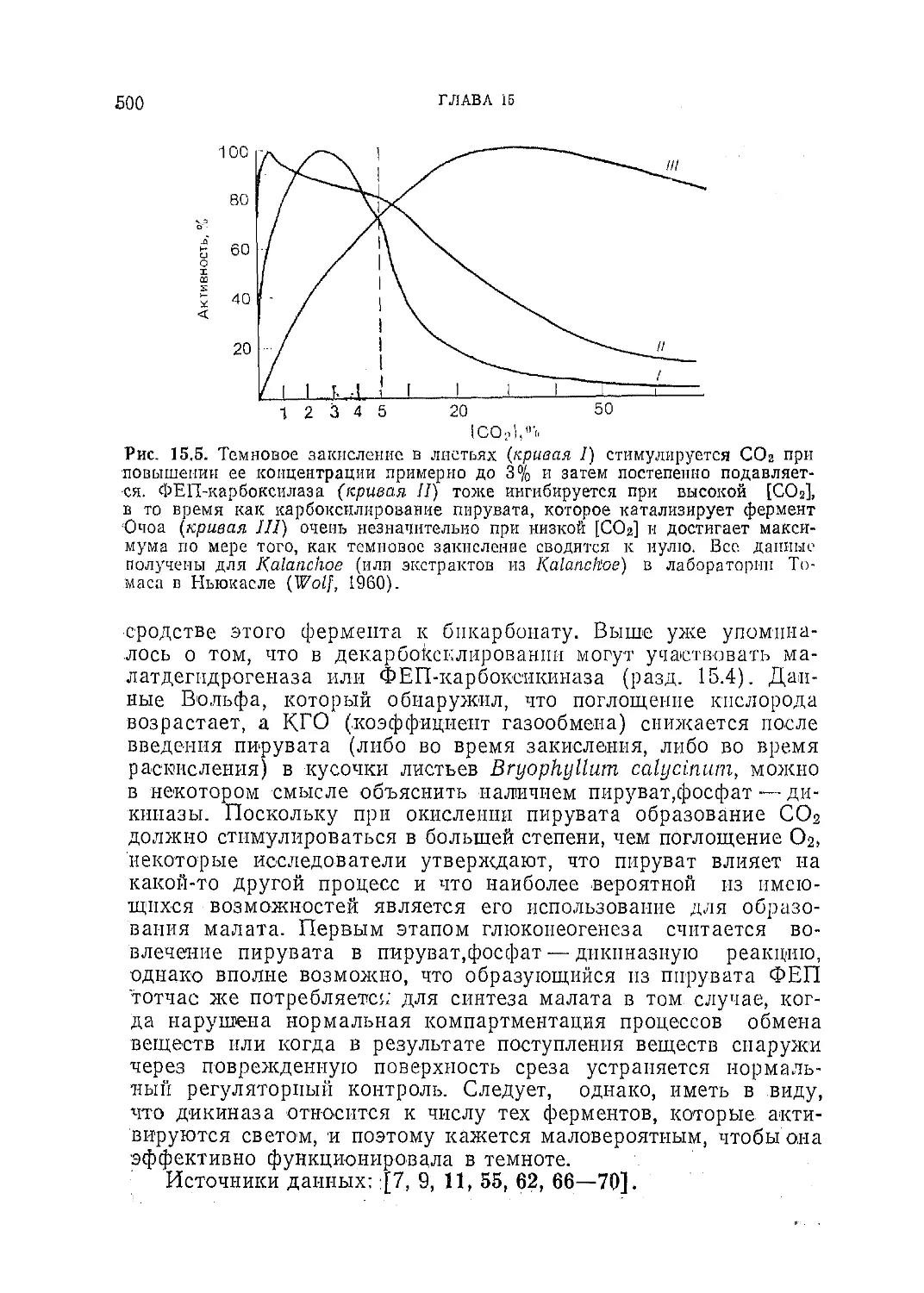
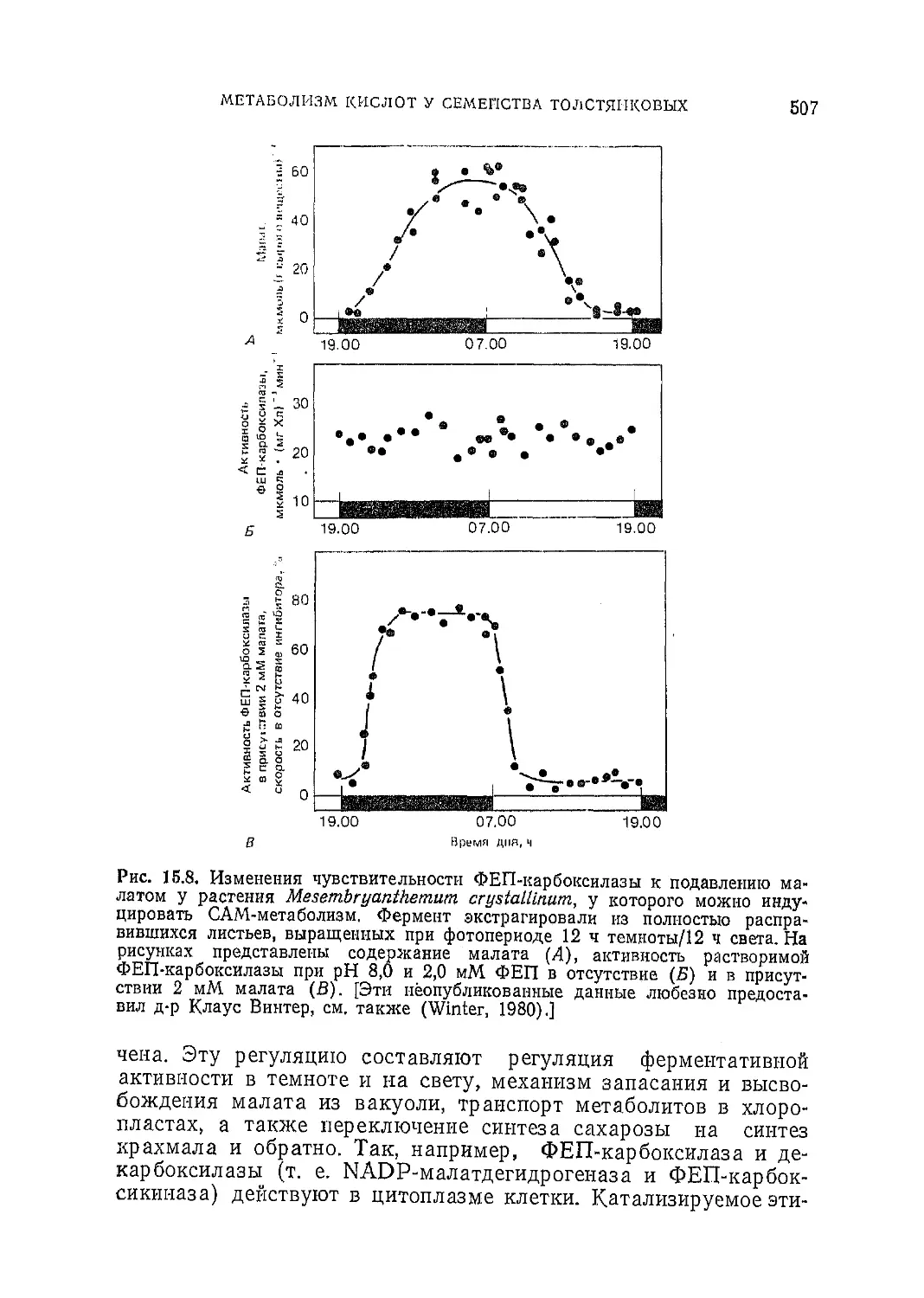
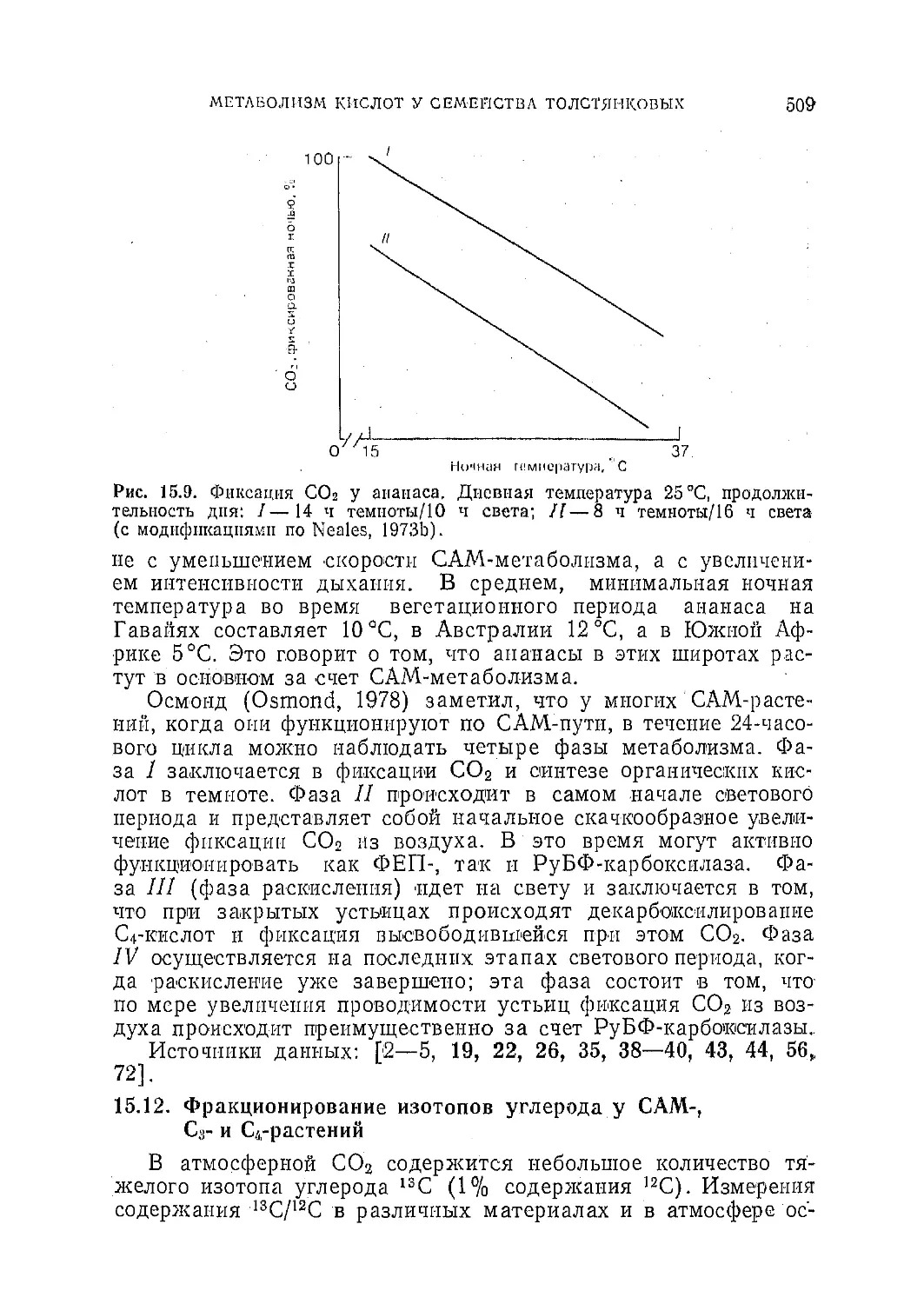
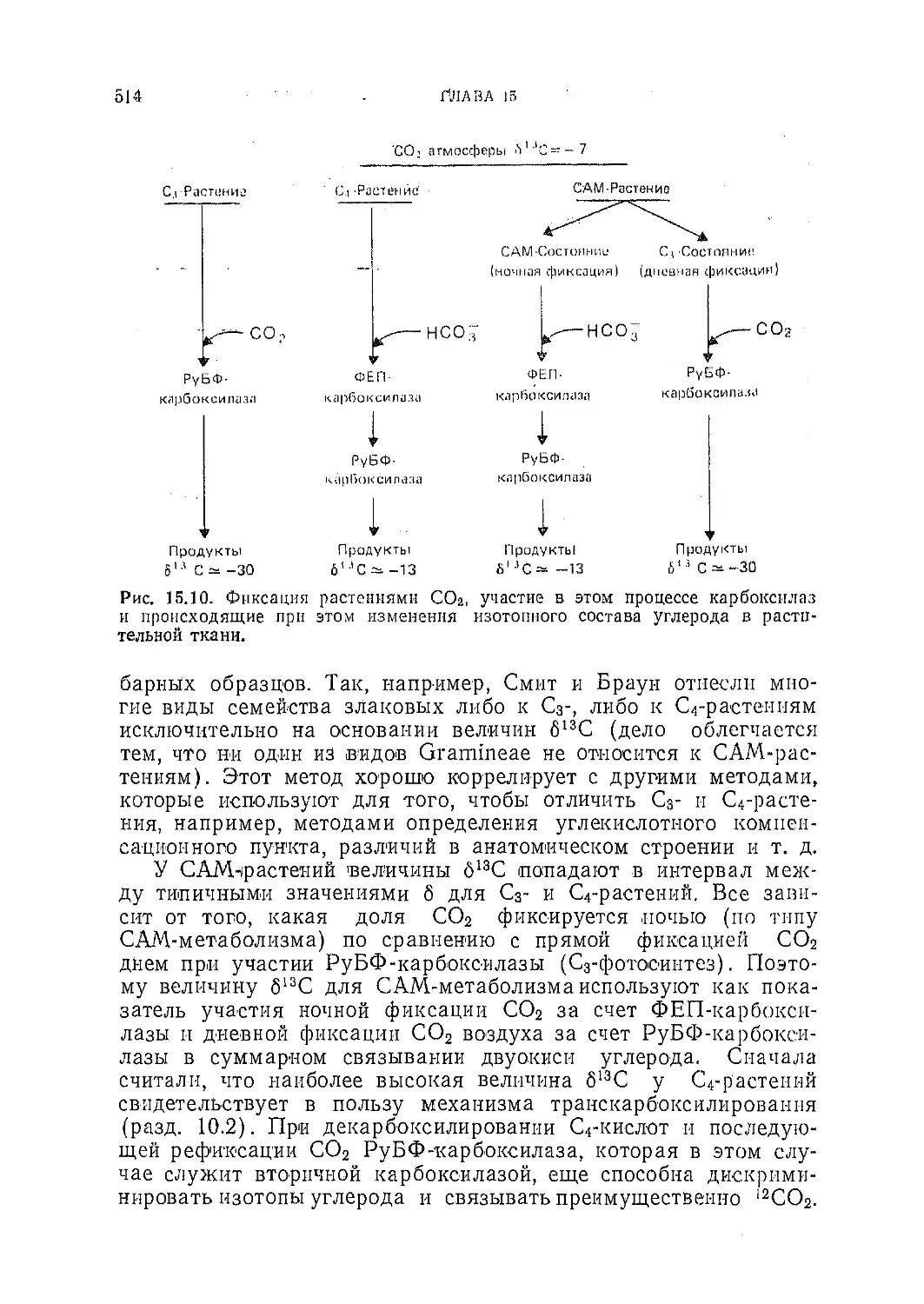
/















































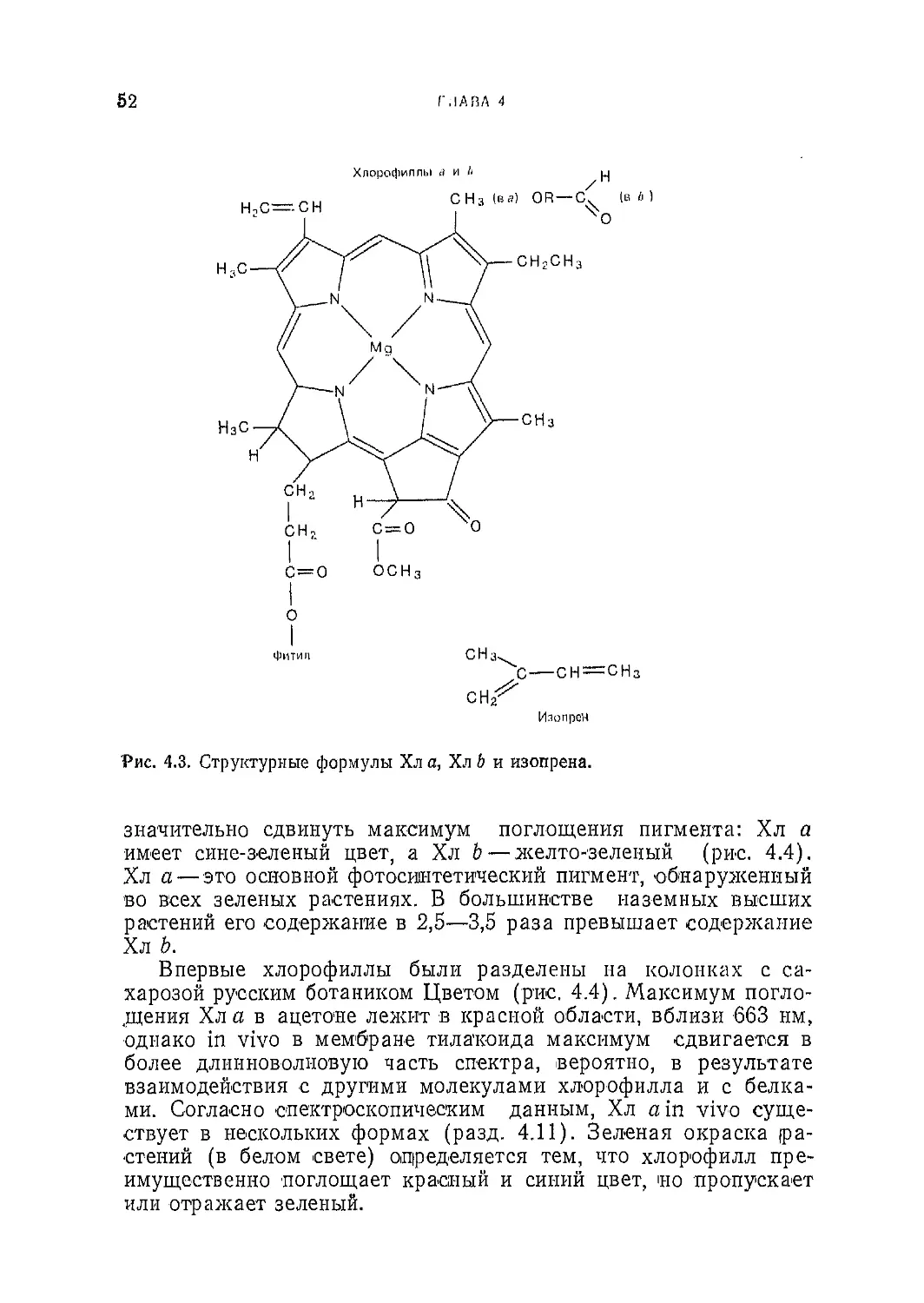






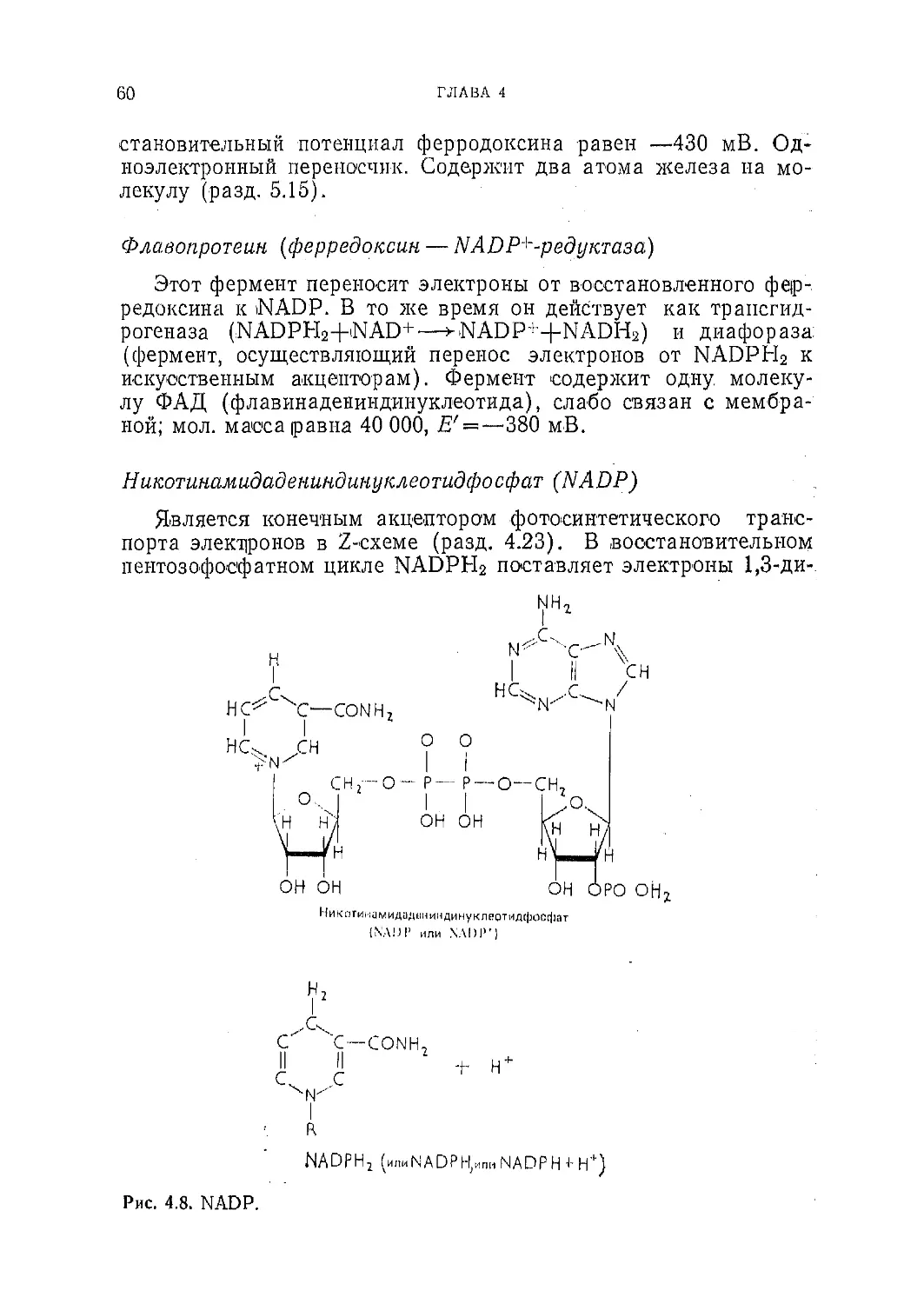
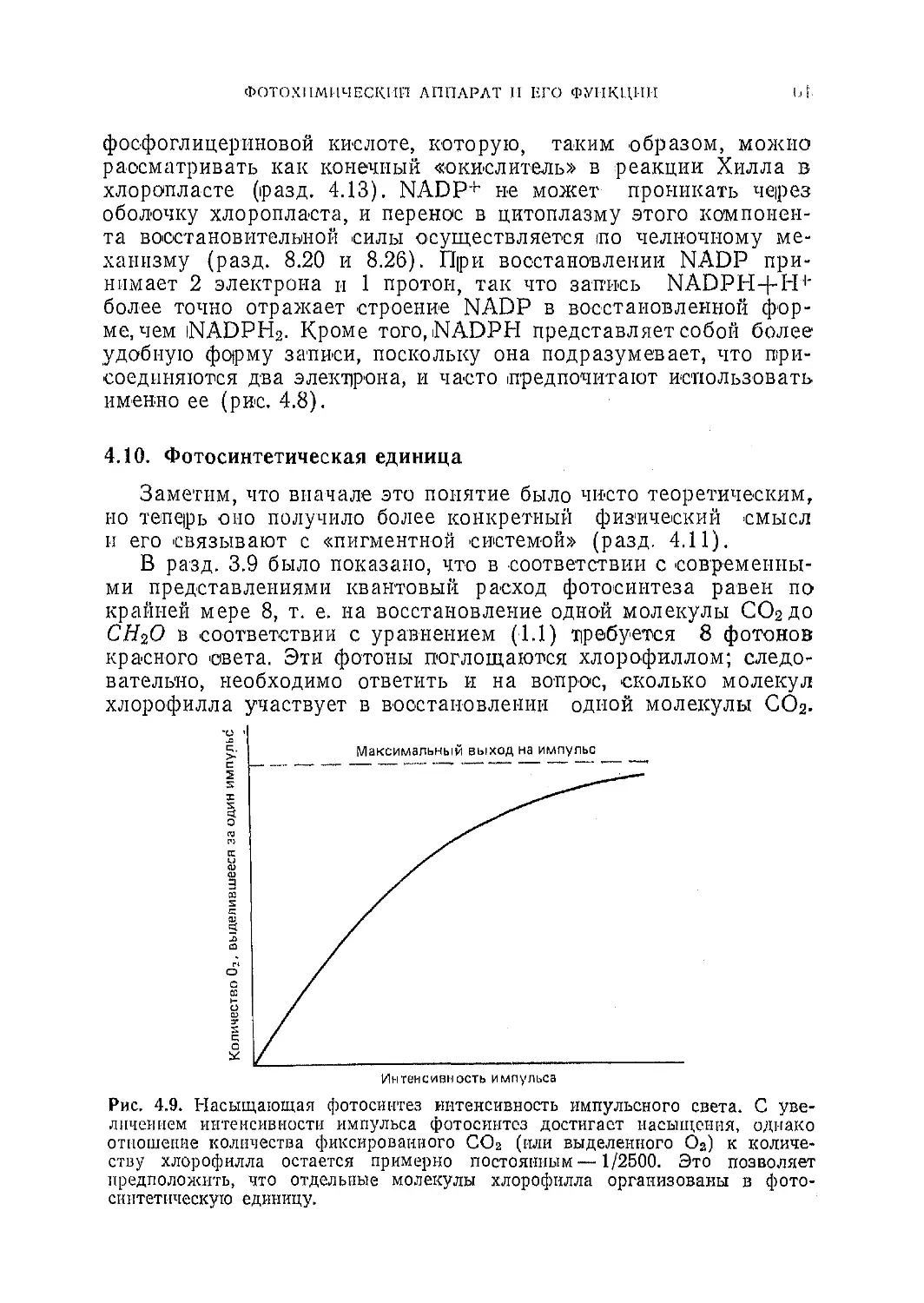













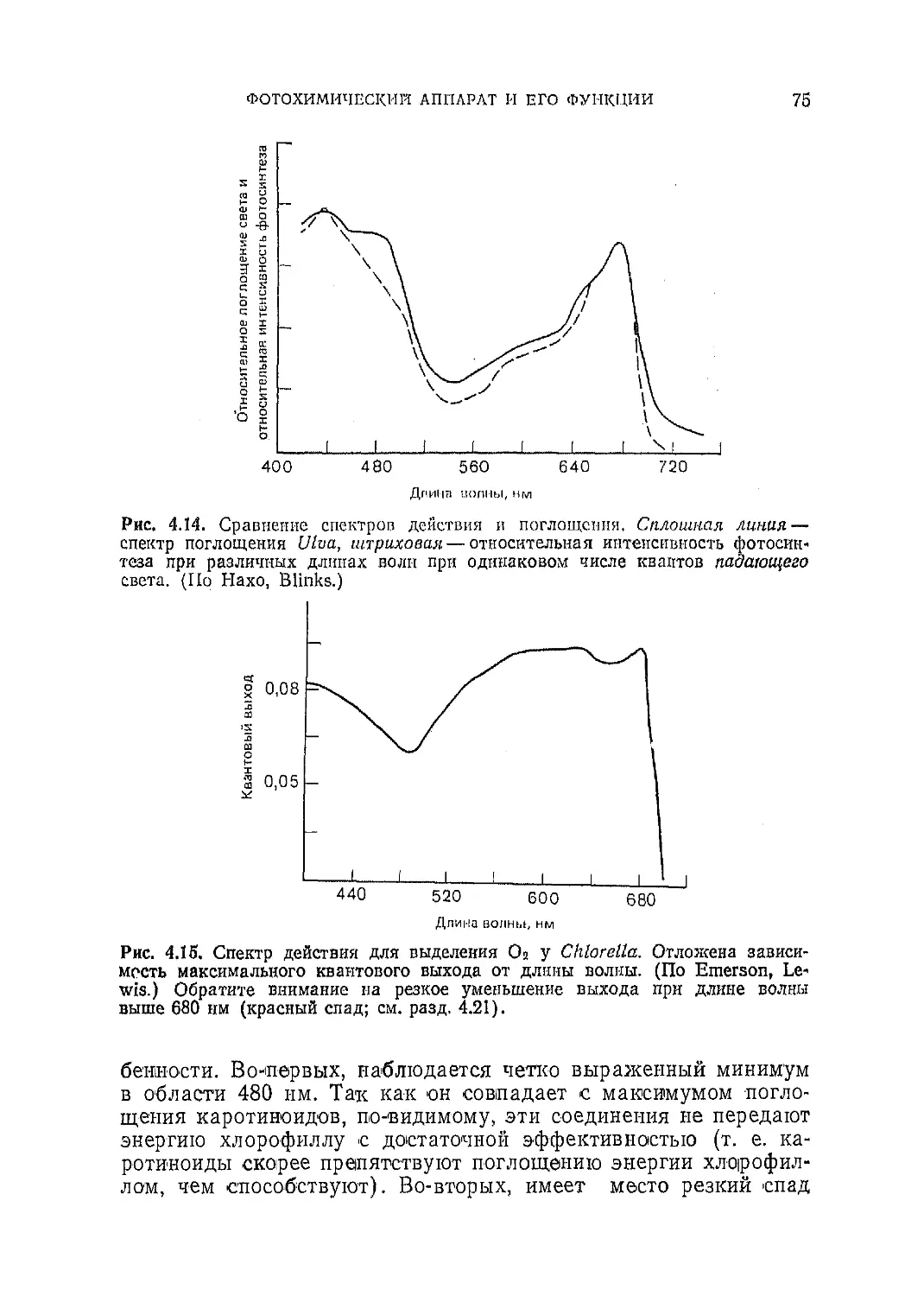
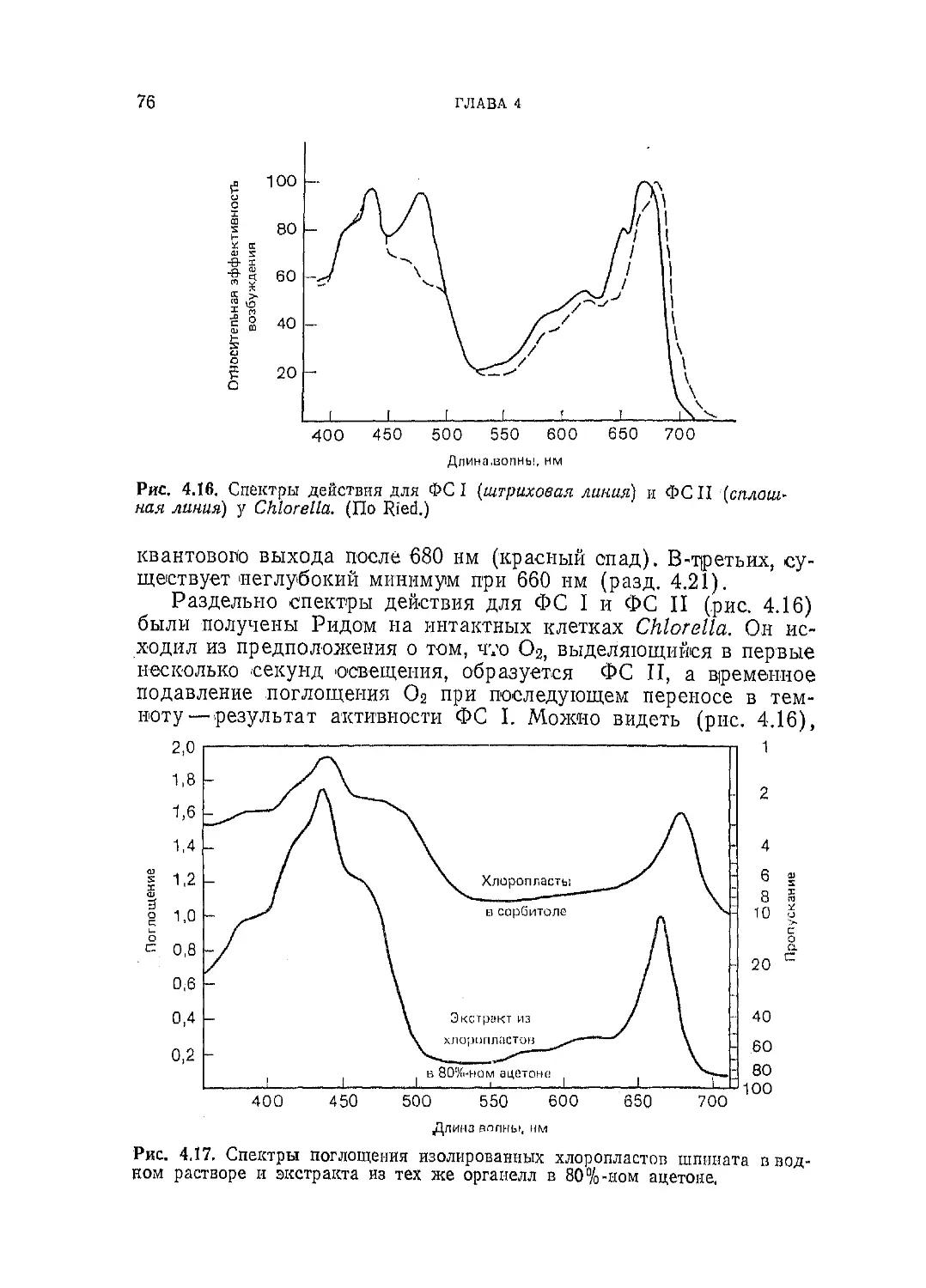


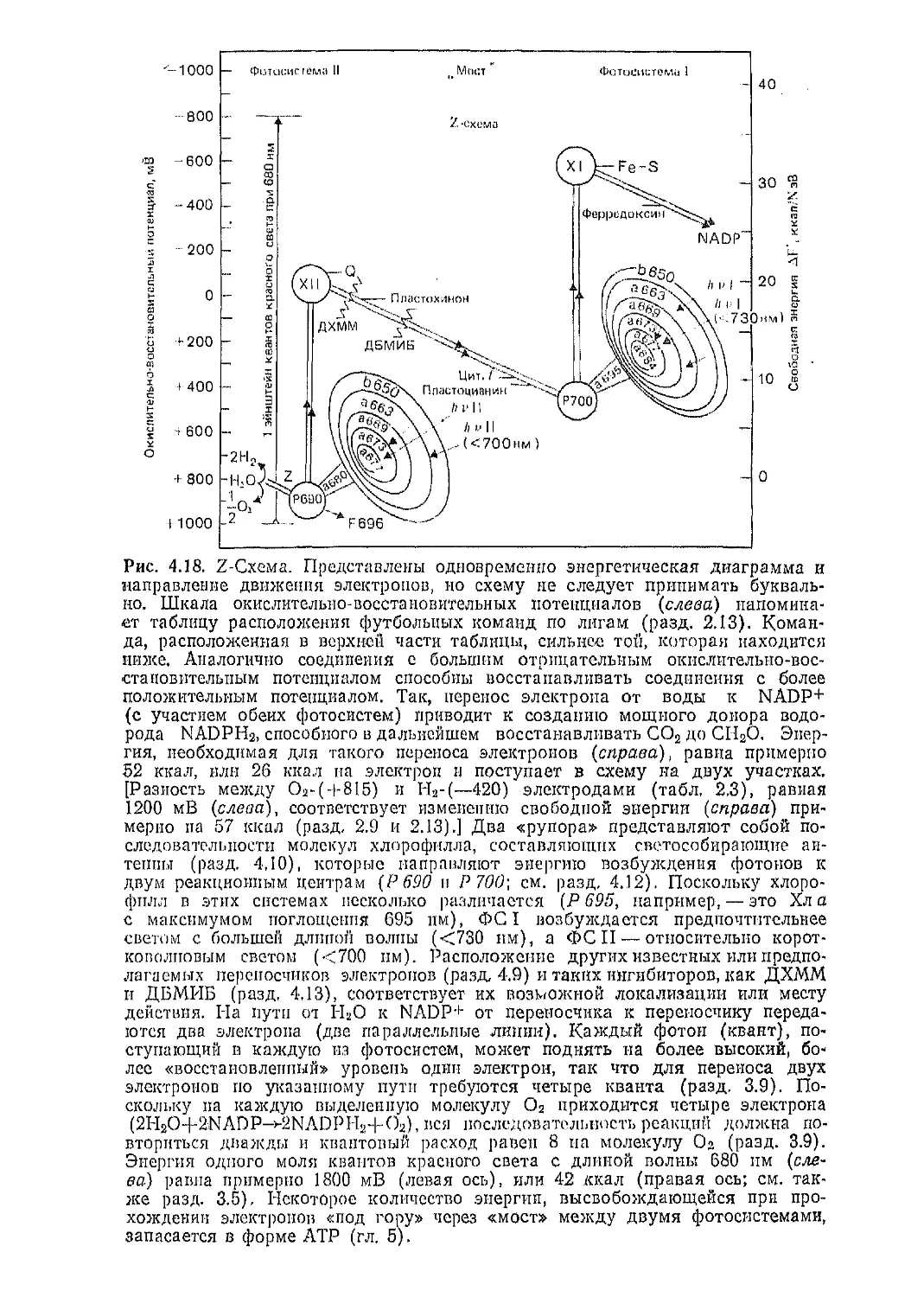










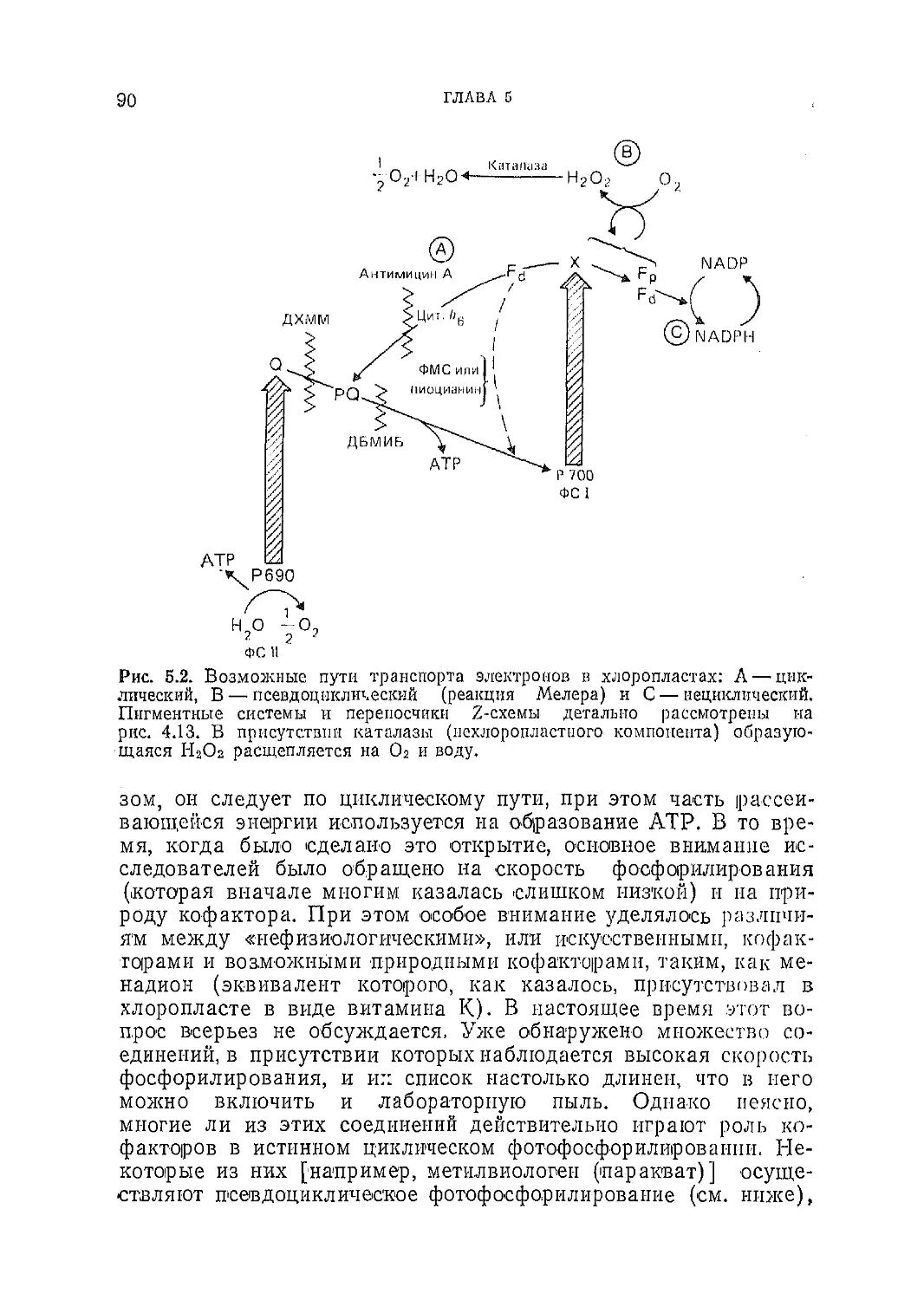




























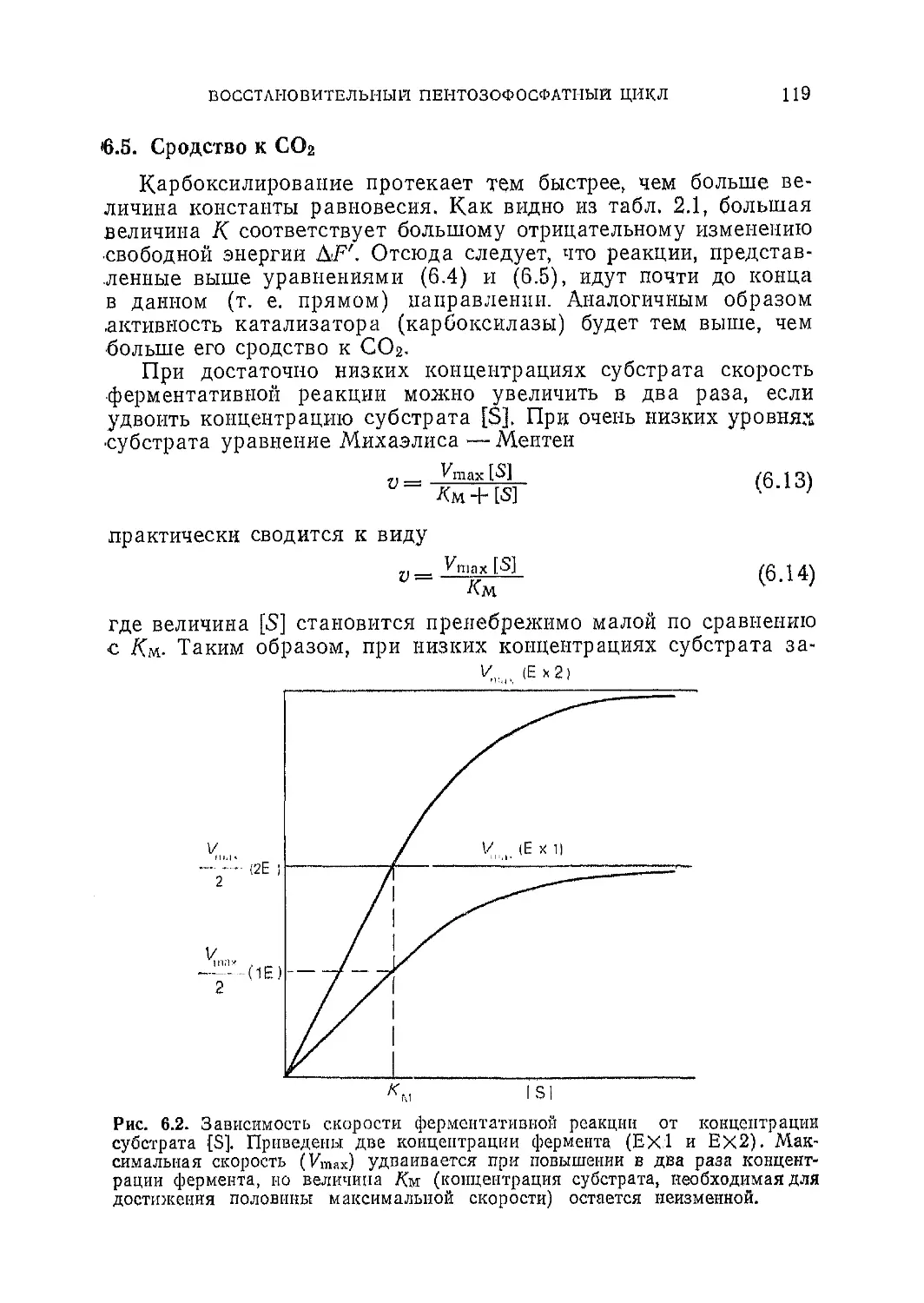




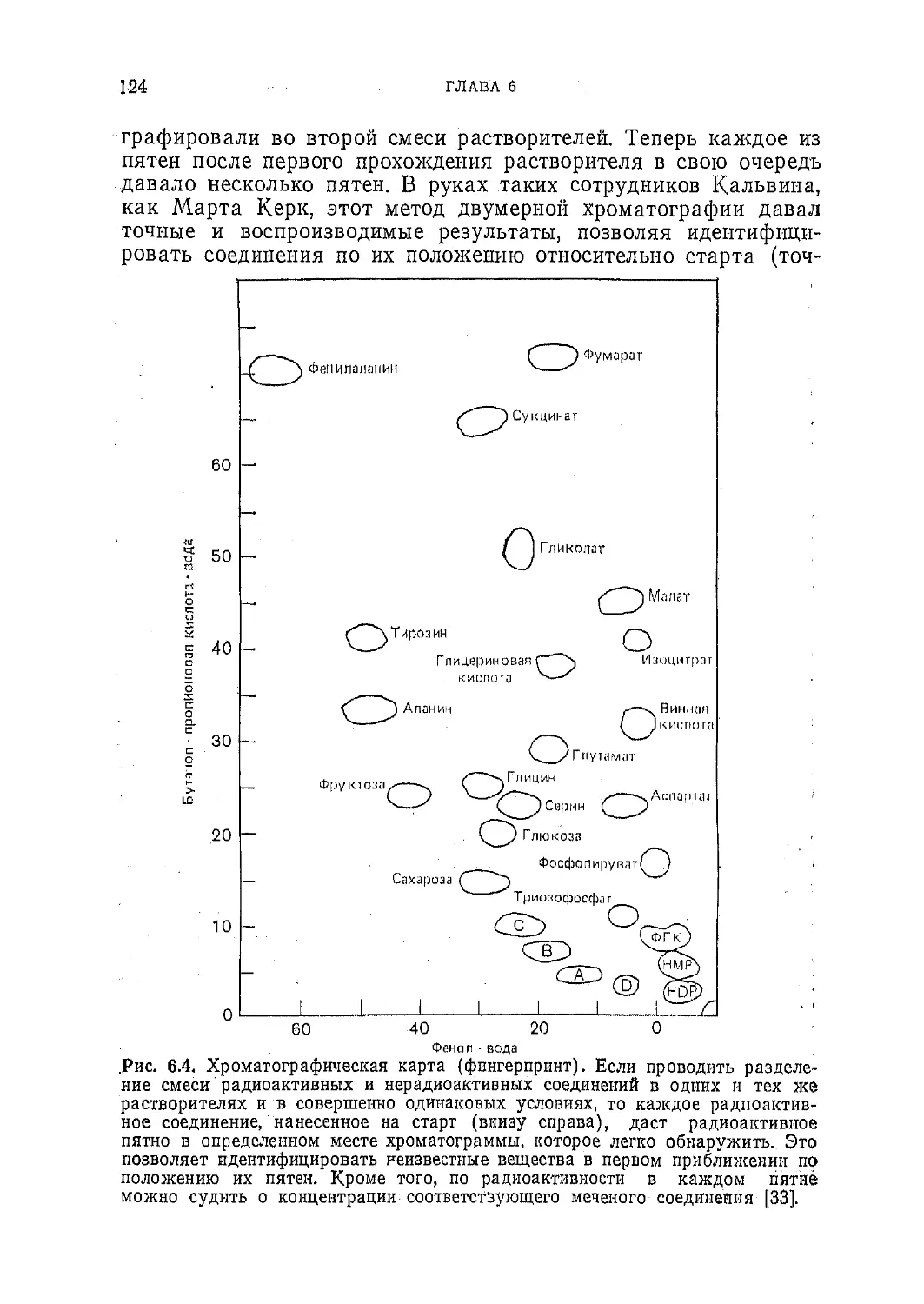
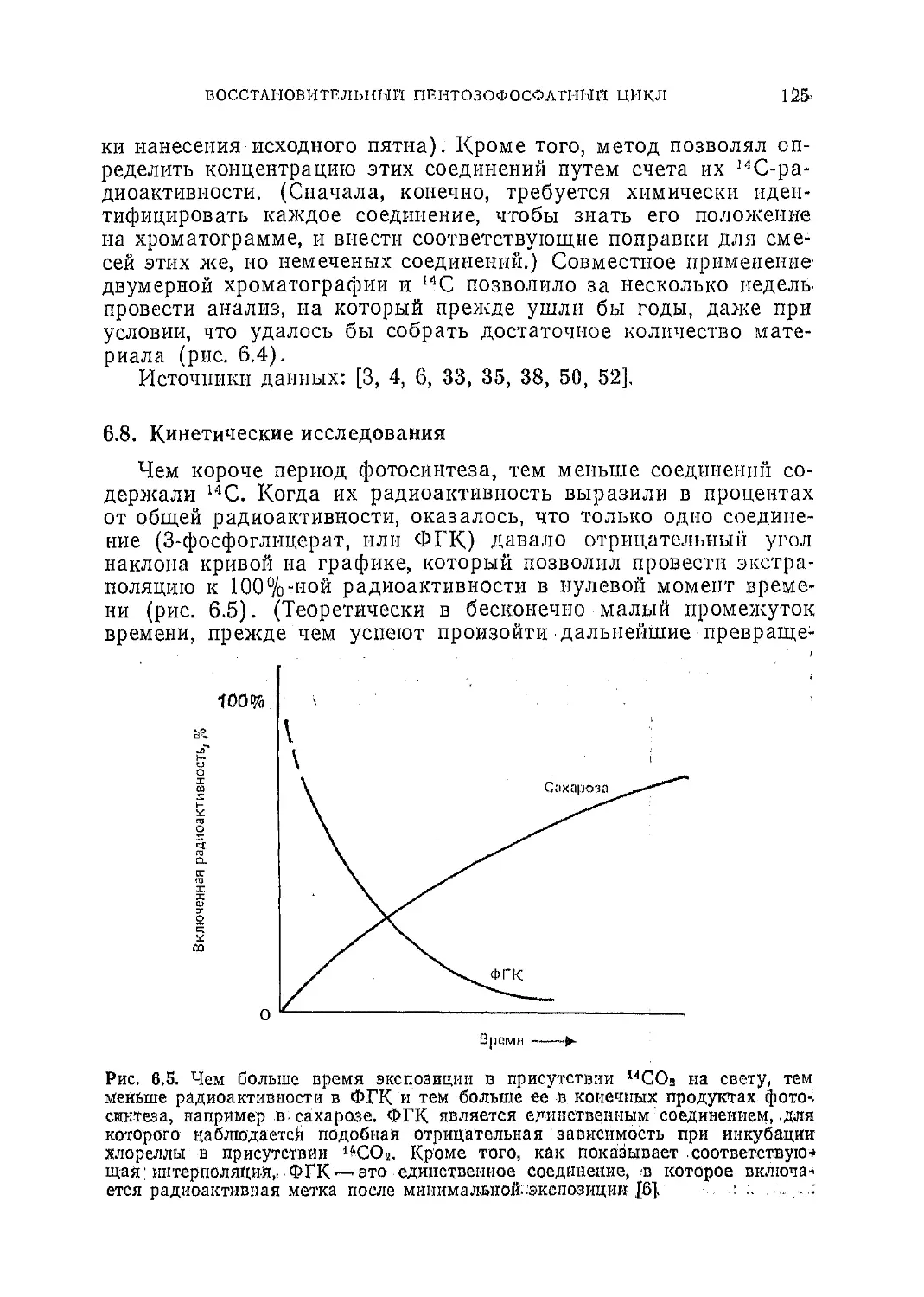
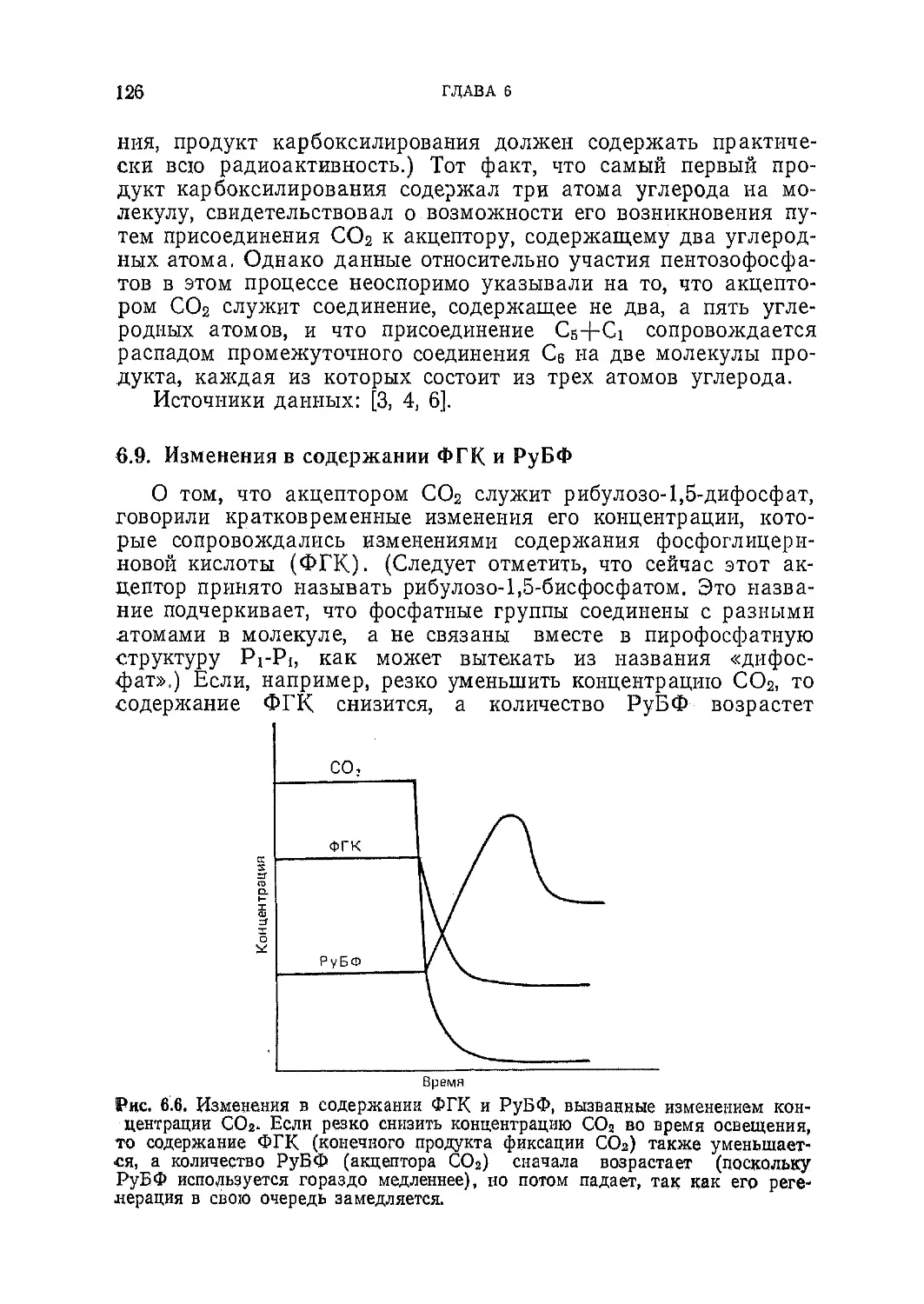
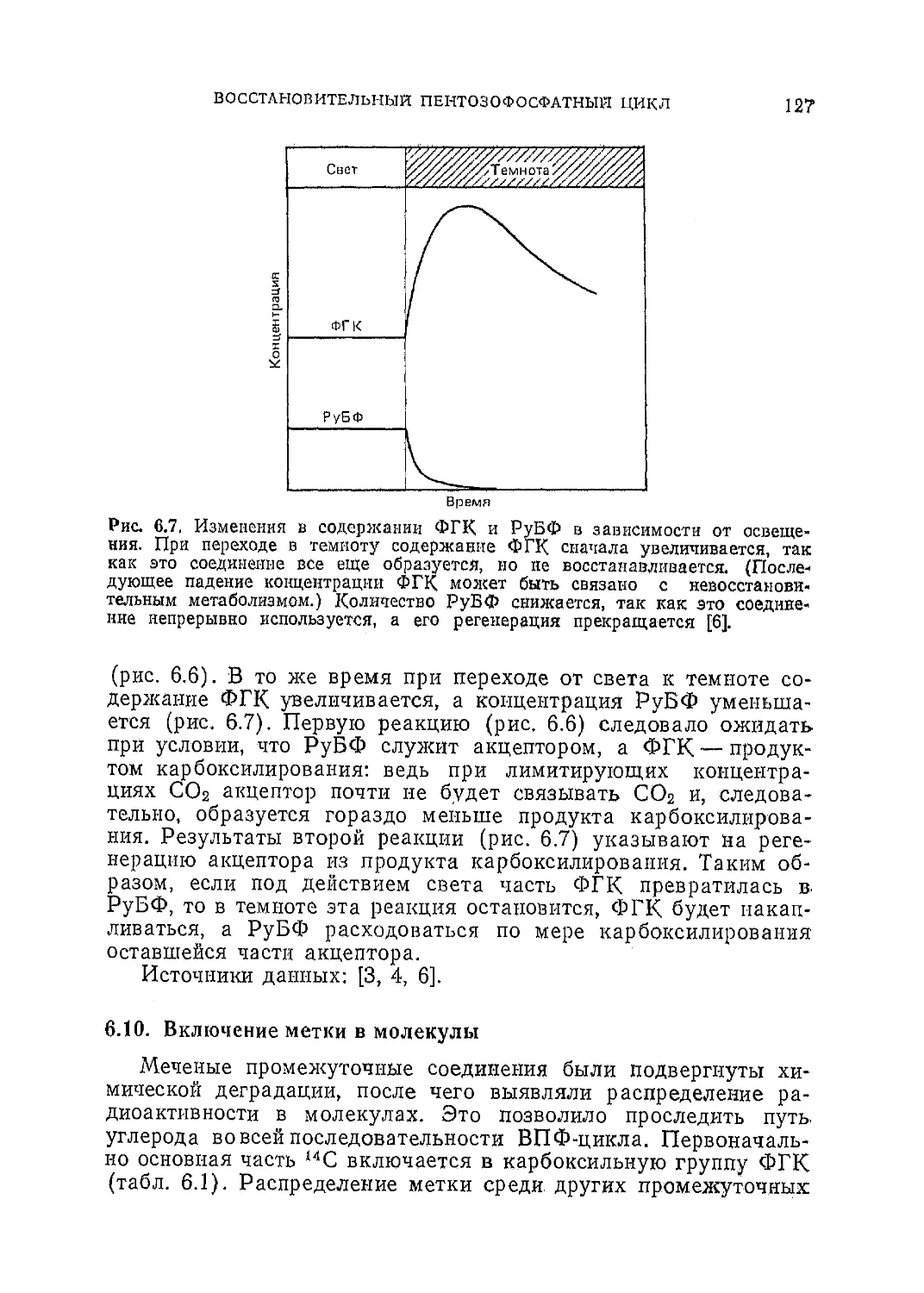

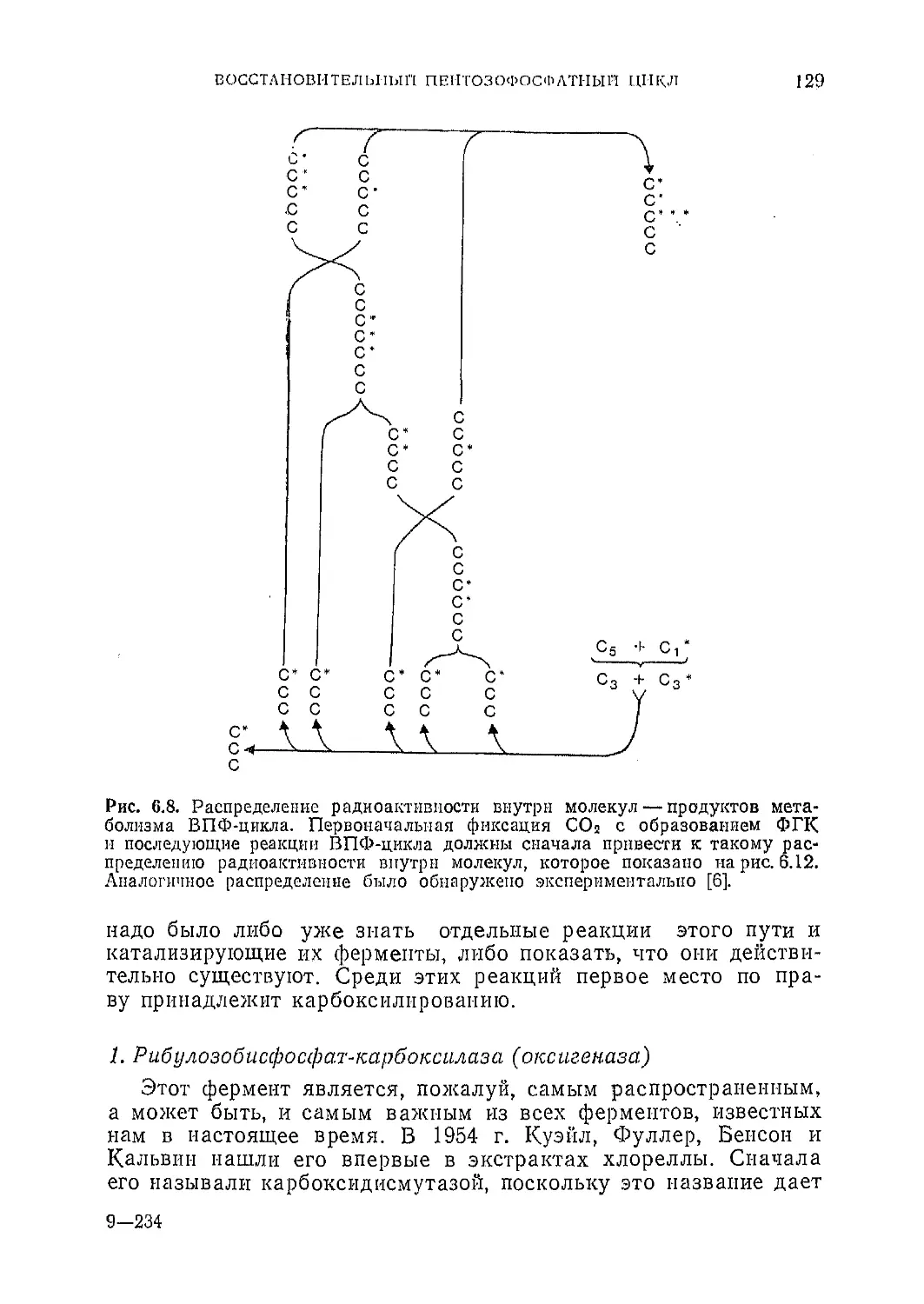










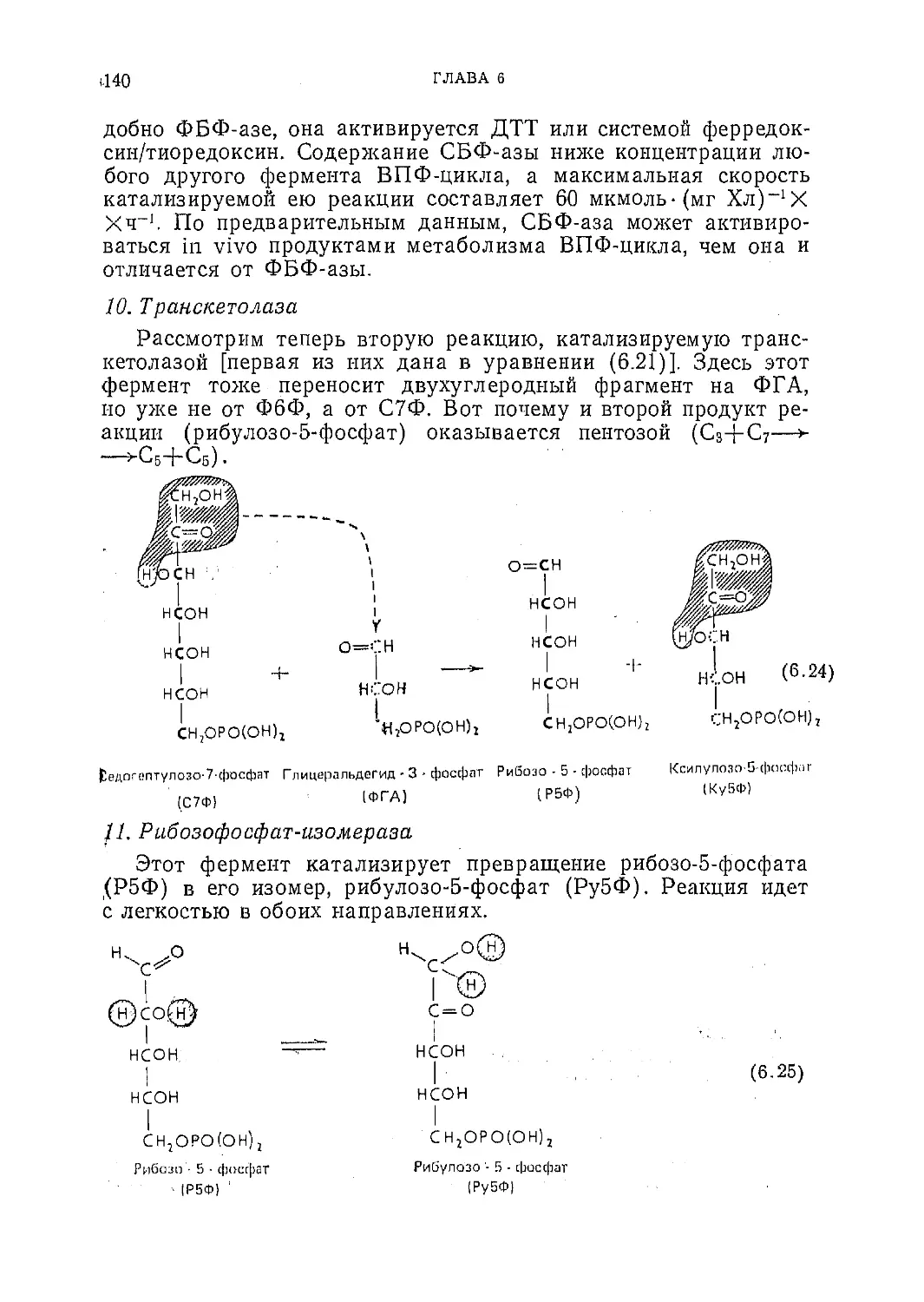









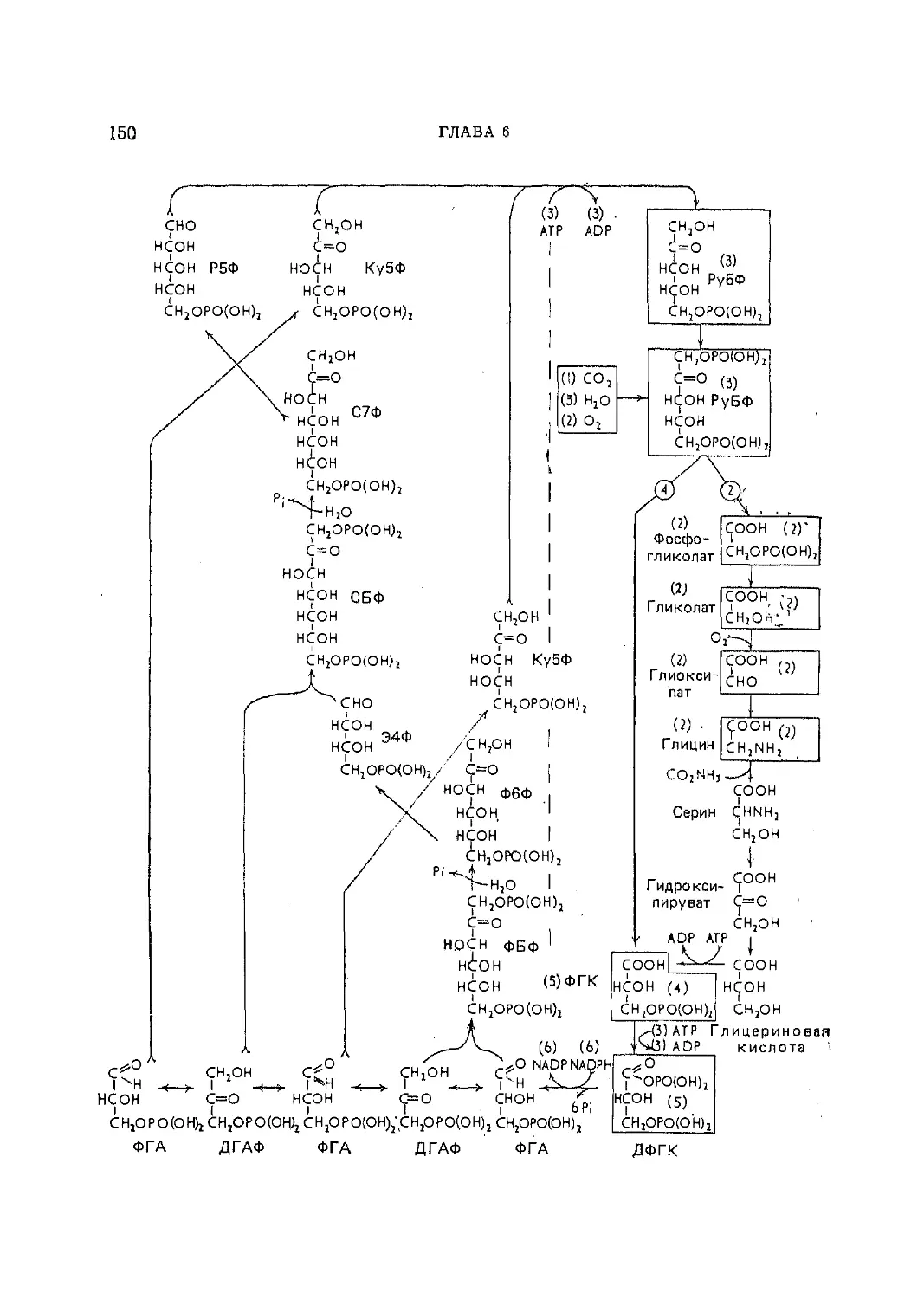




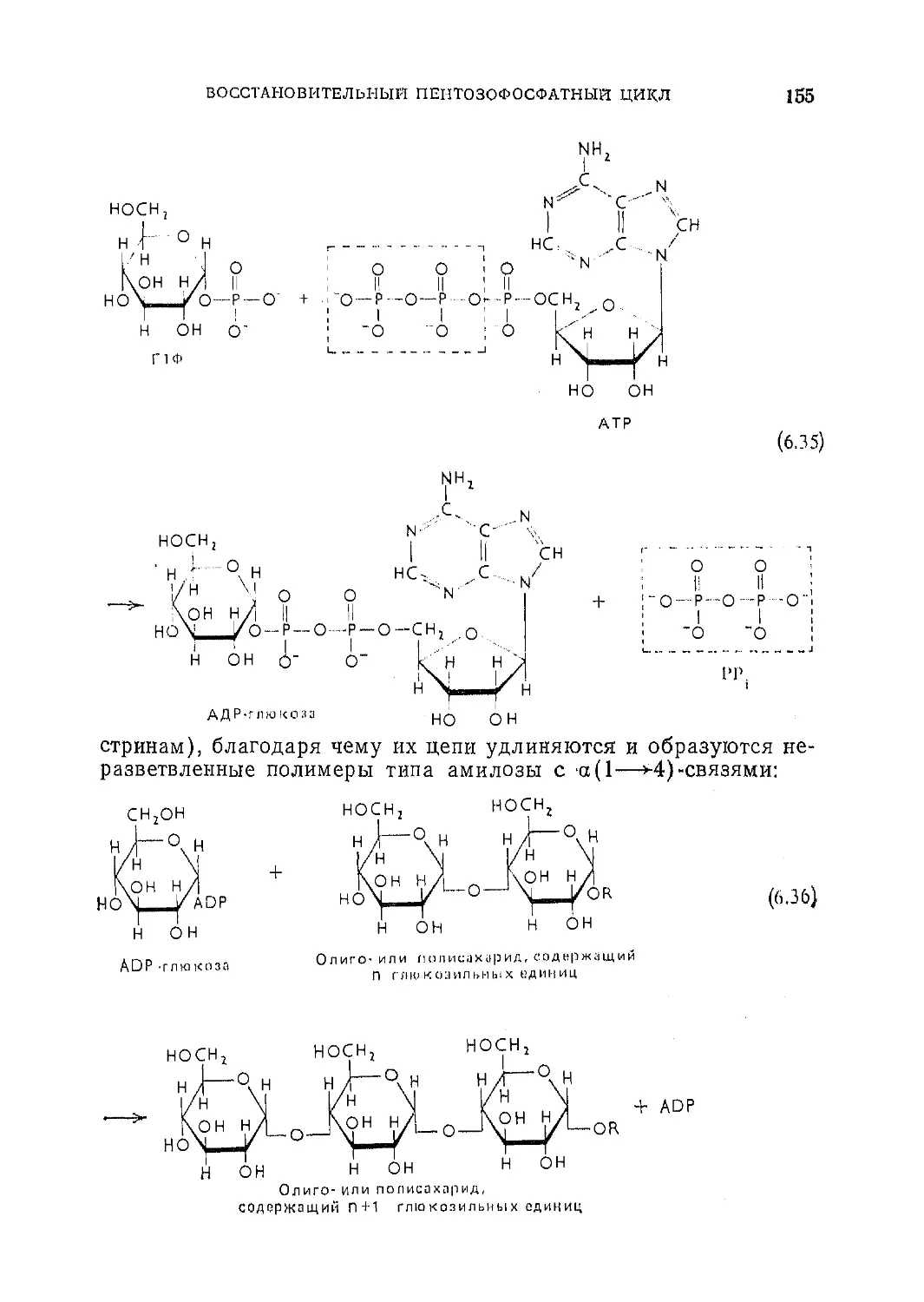

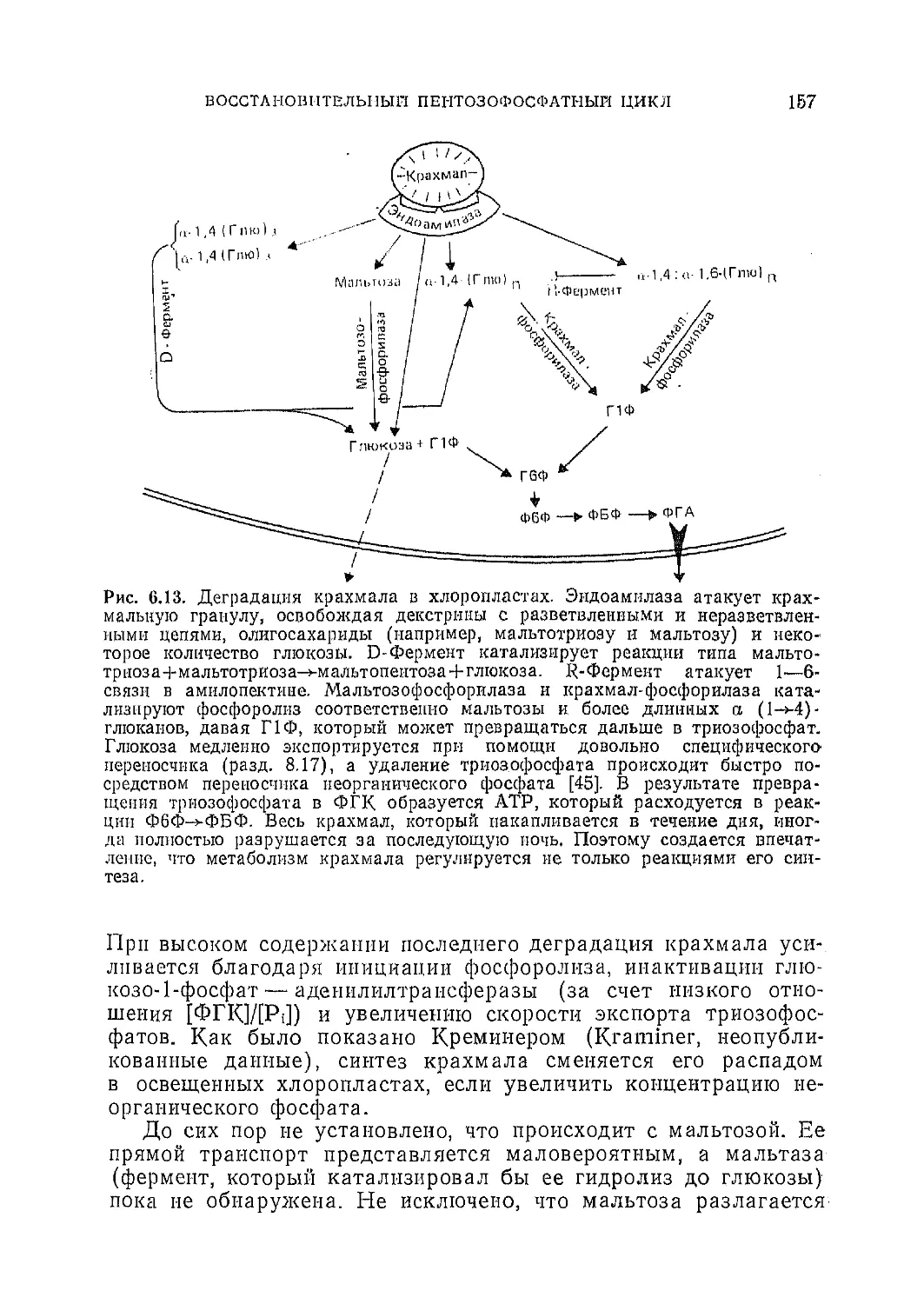

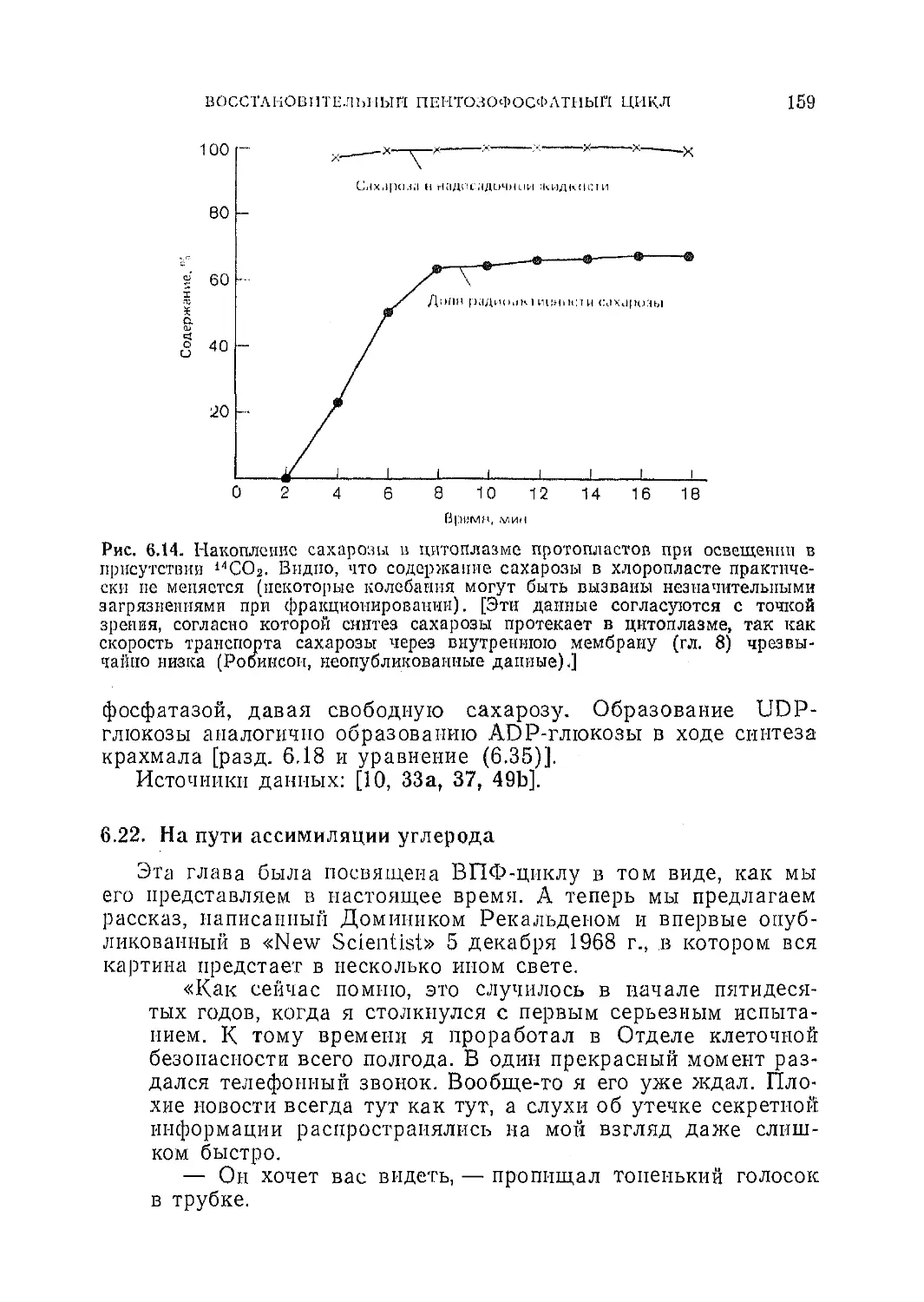








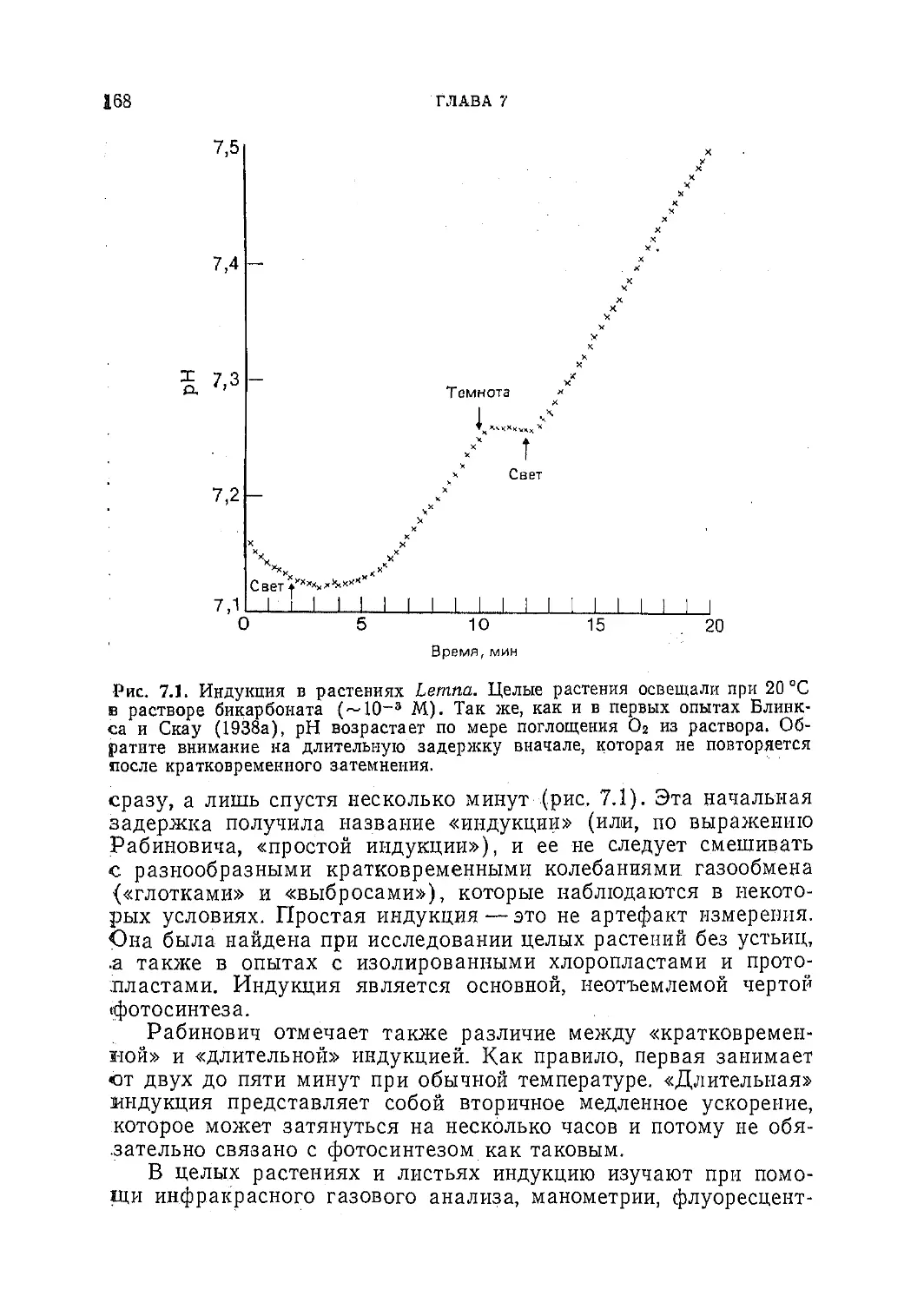
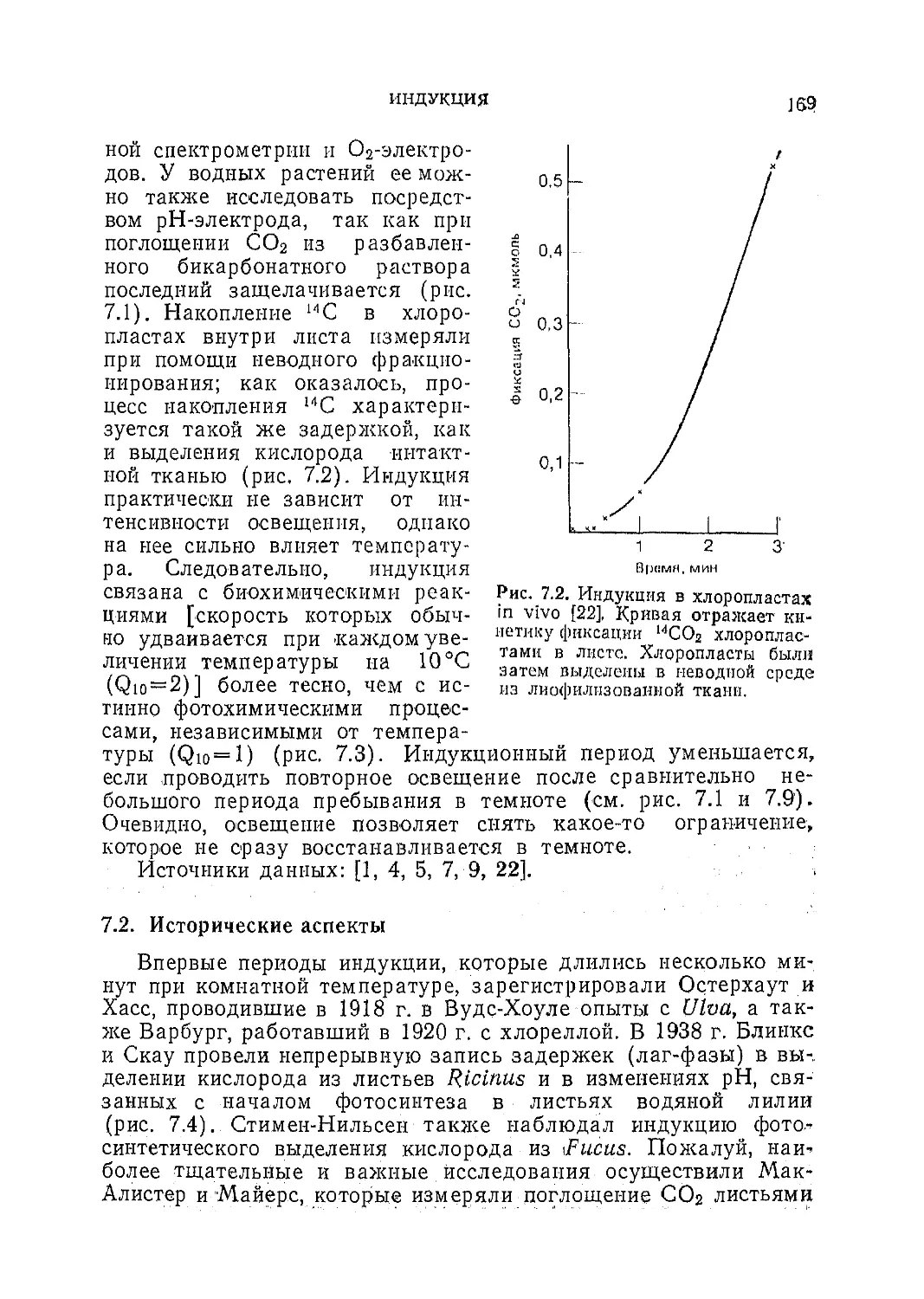
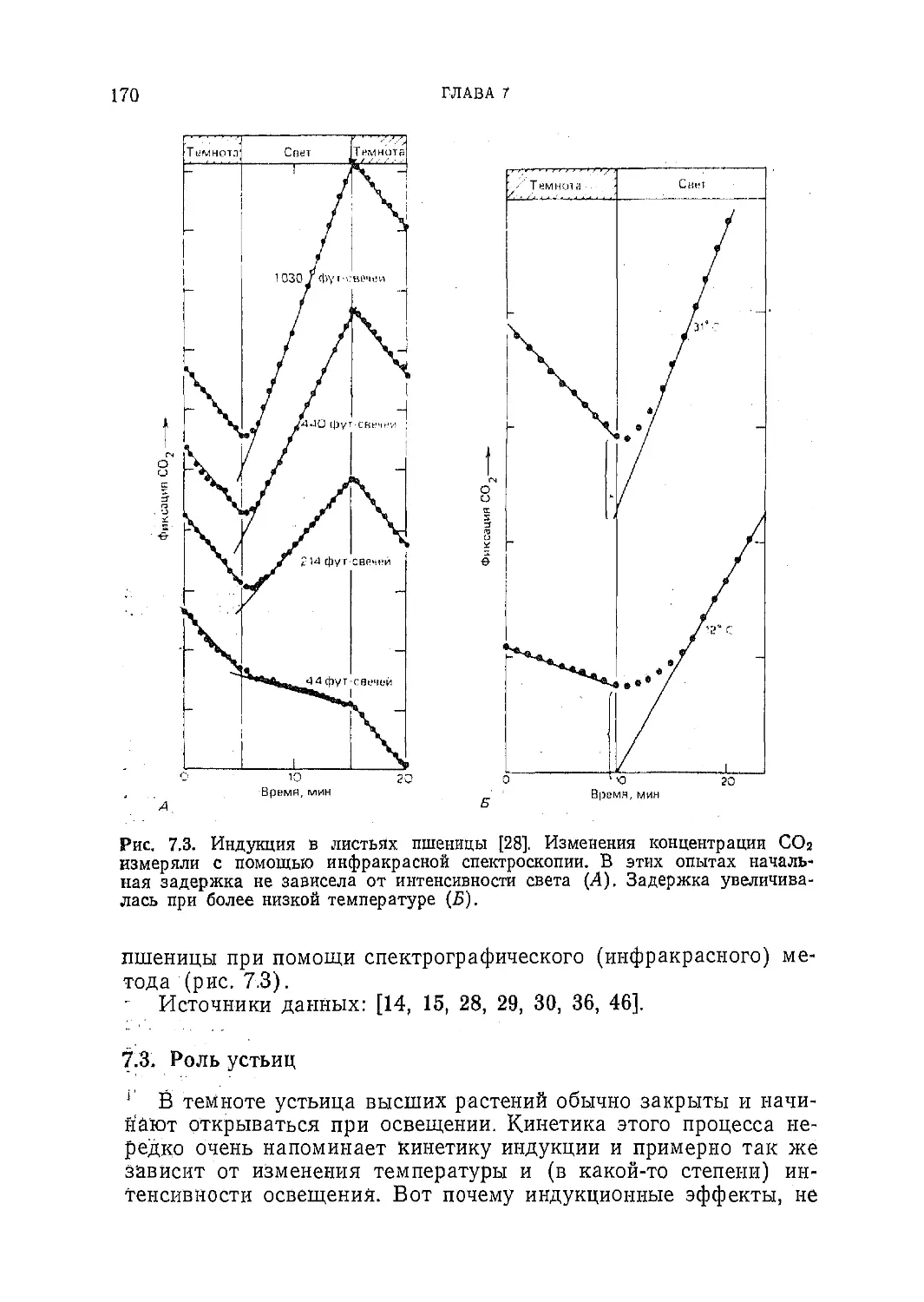
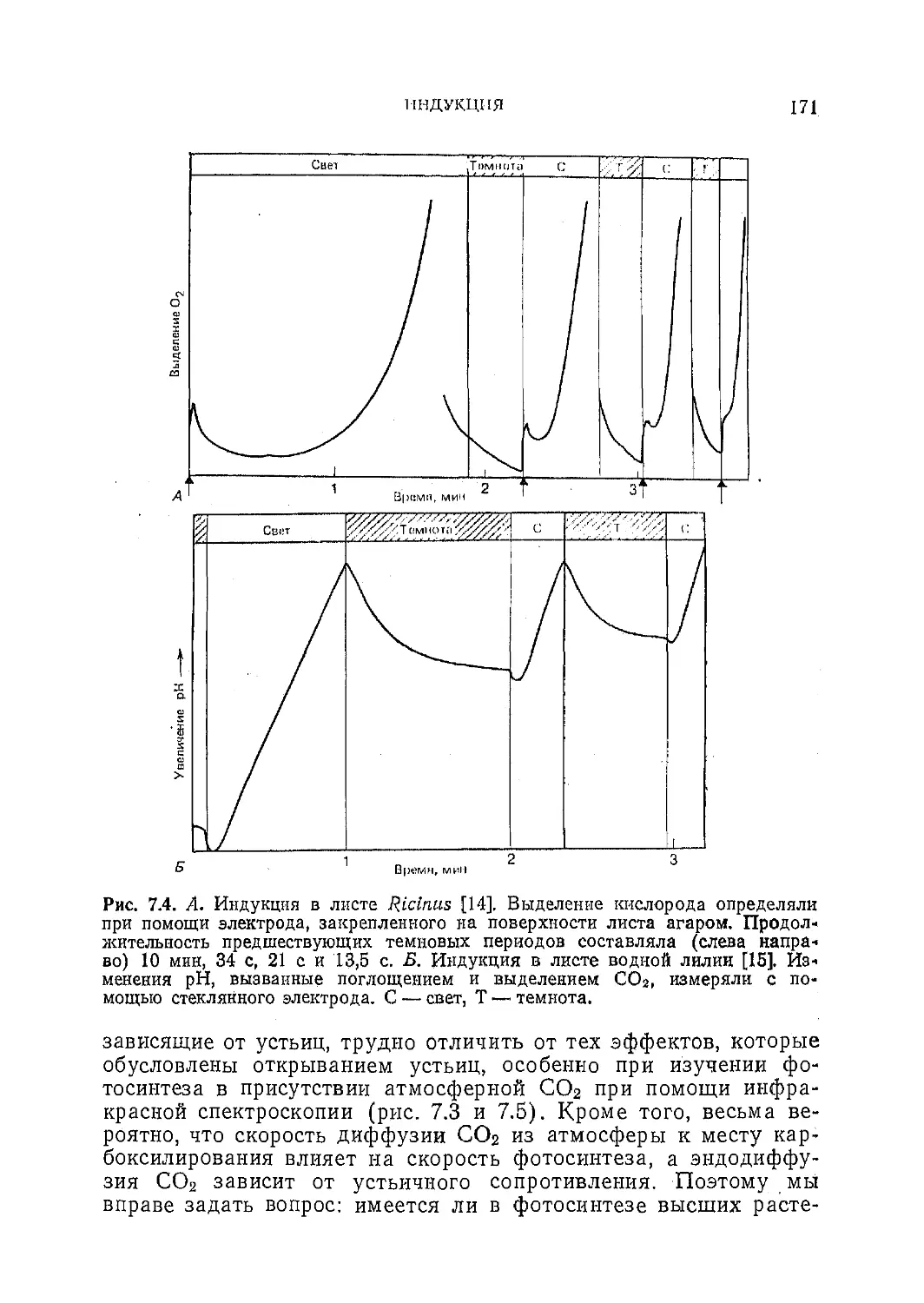

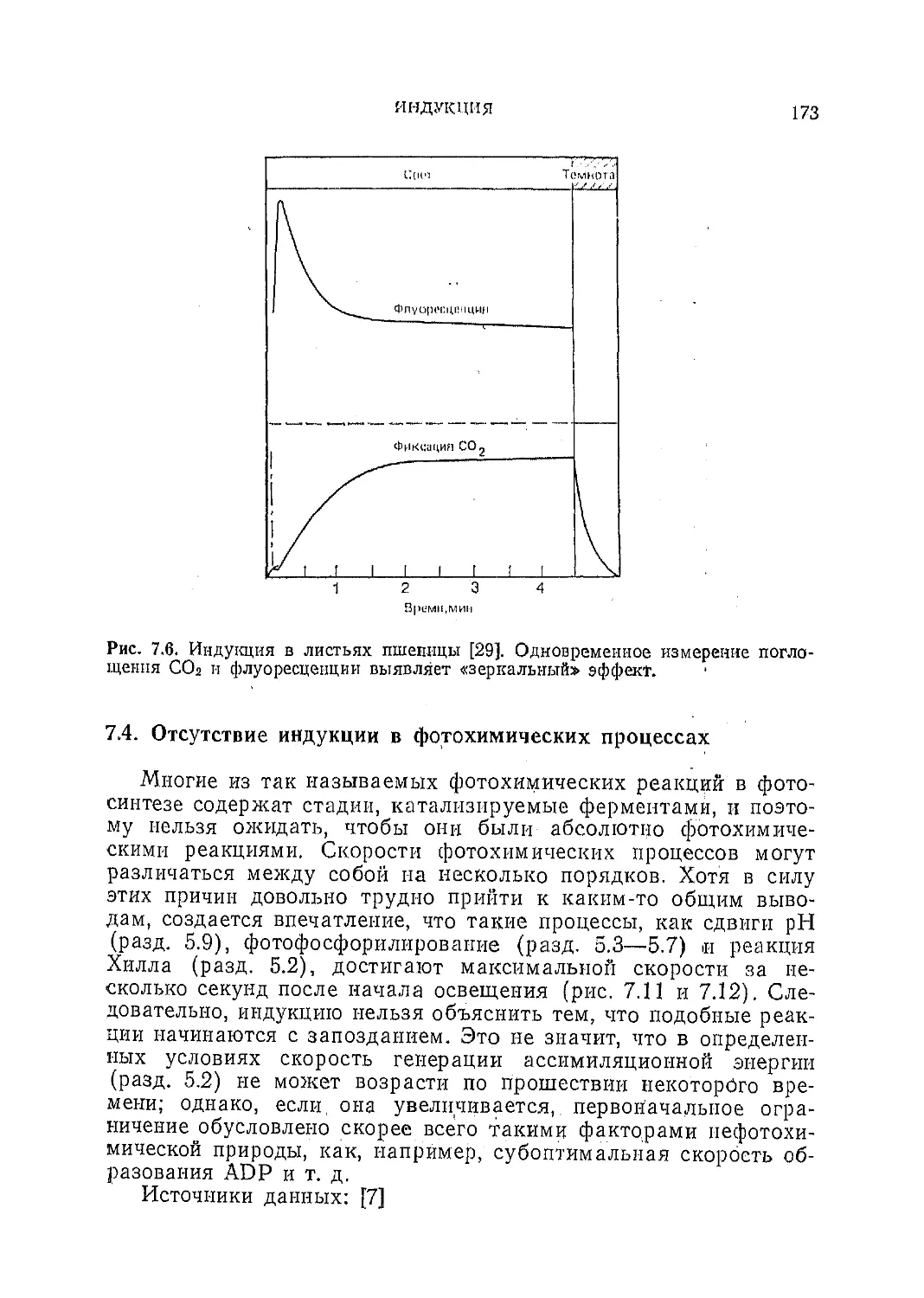
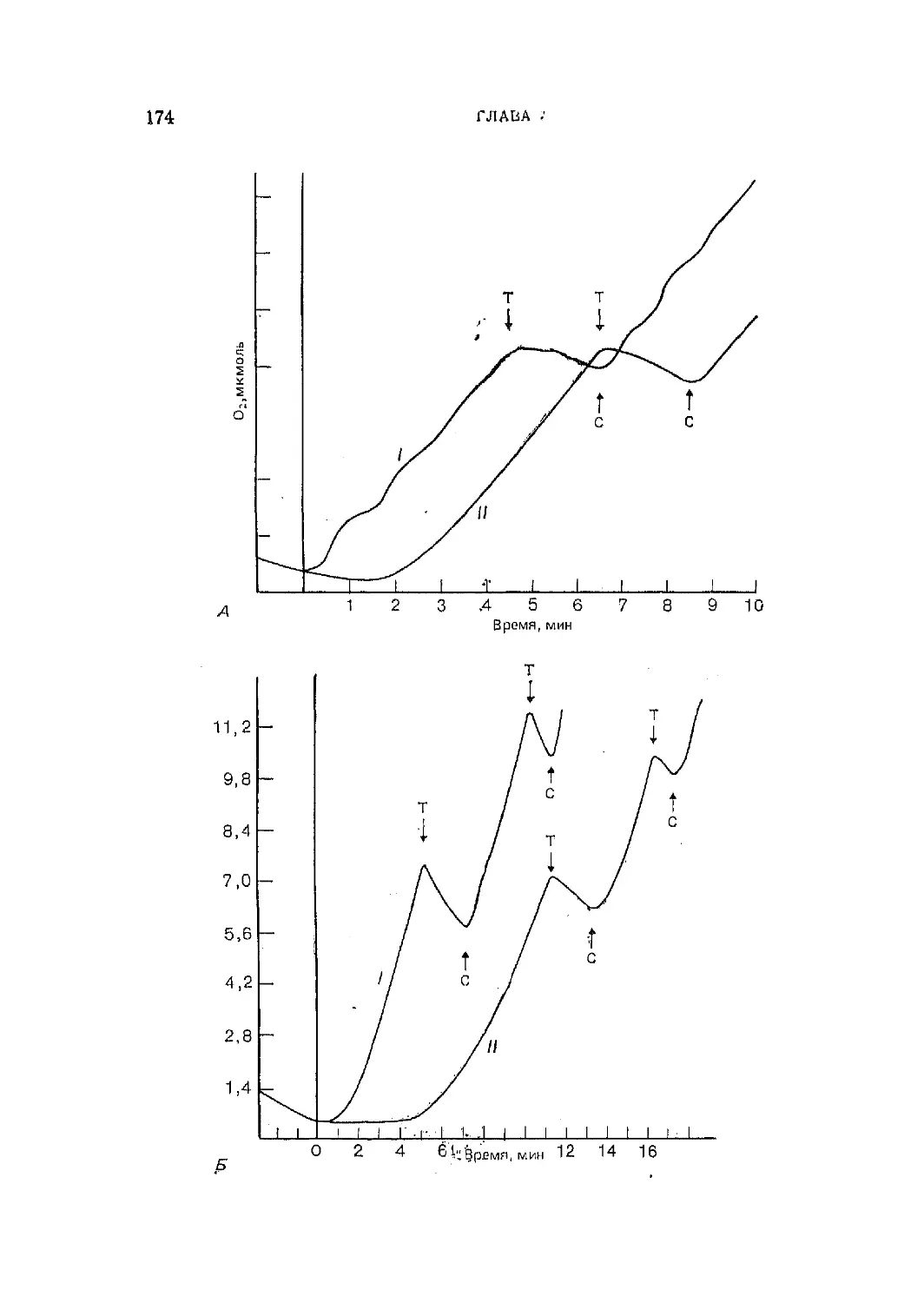
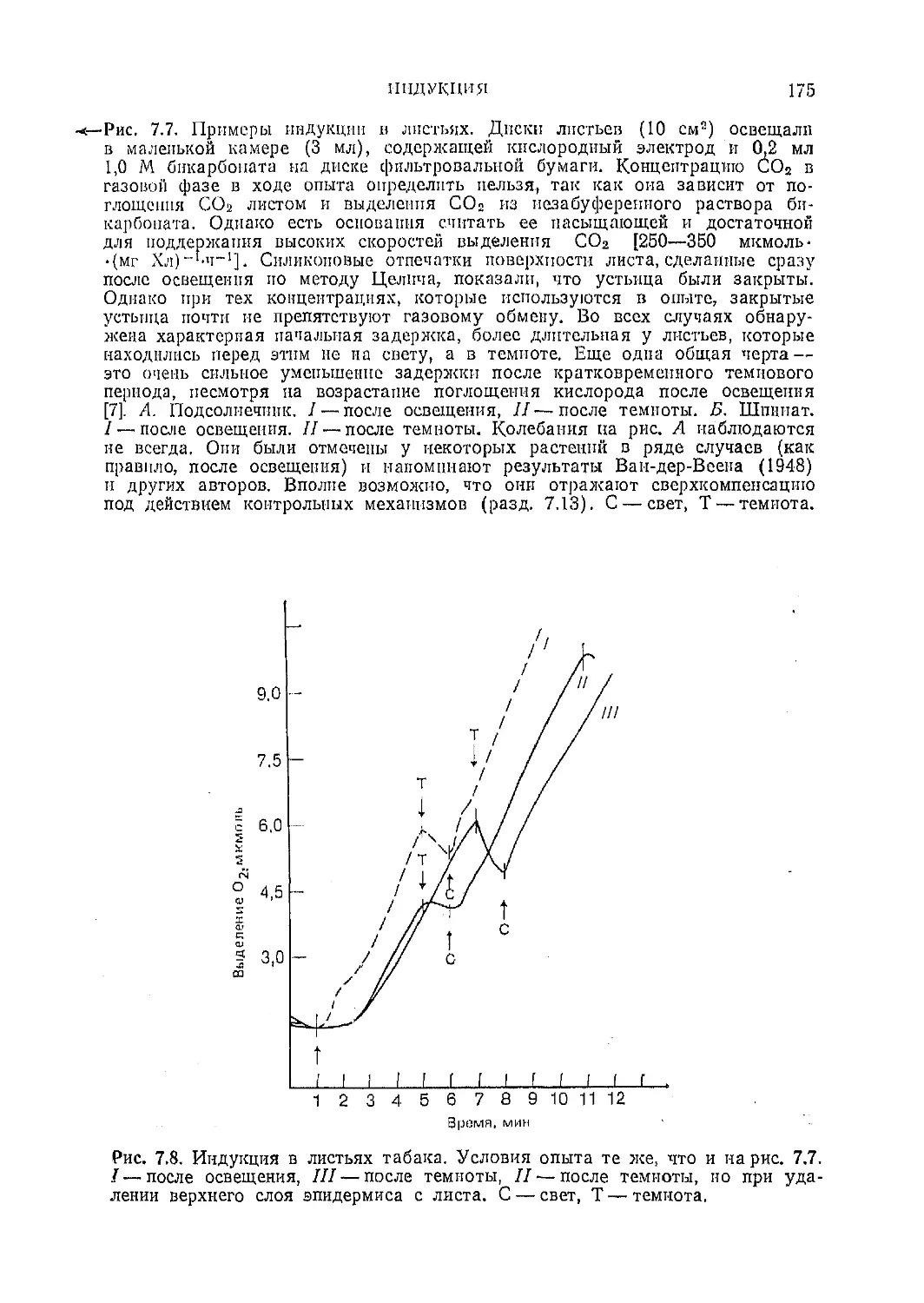
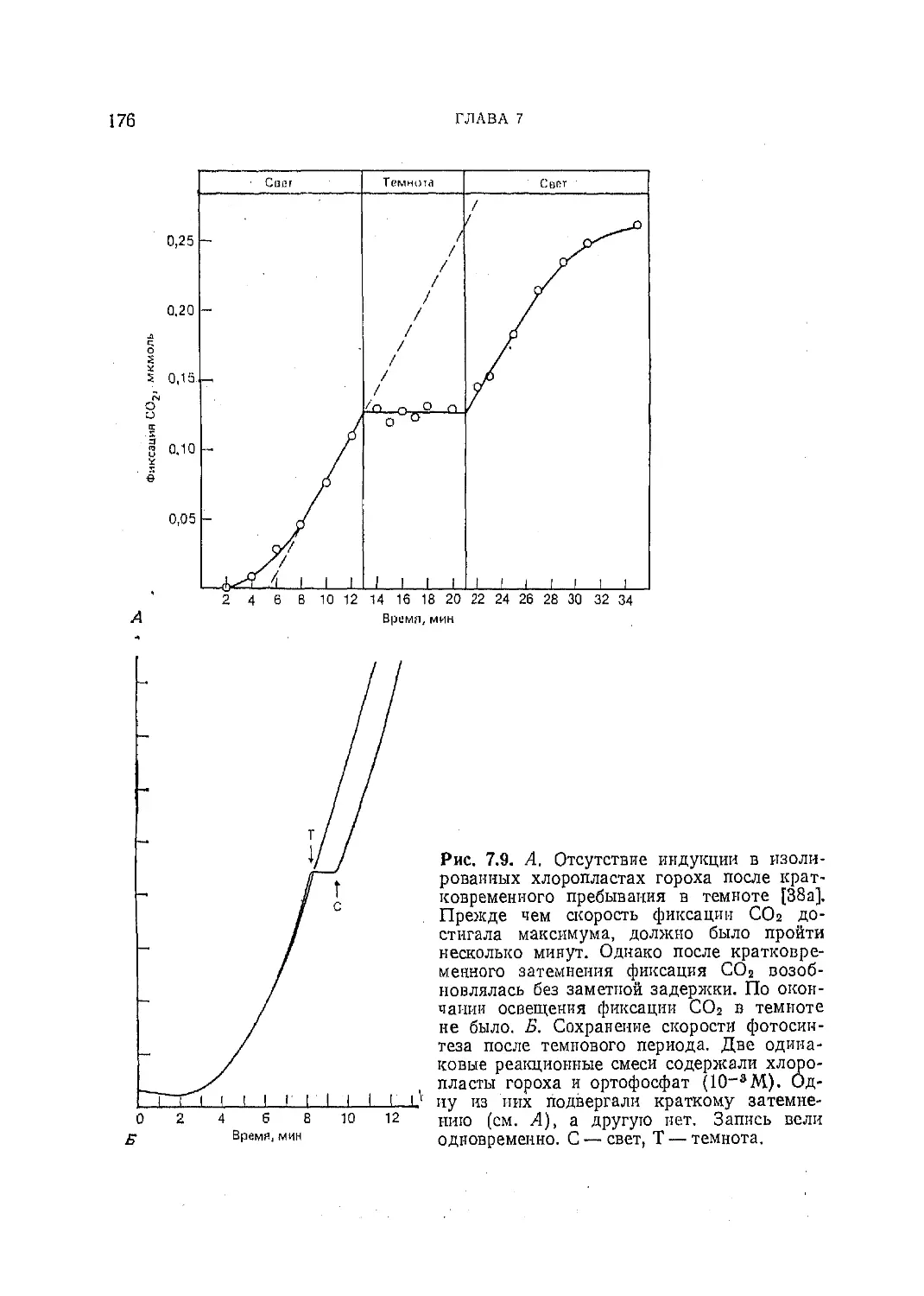
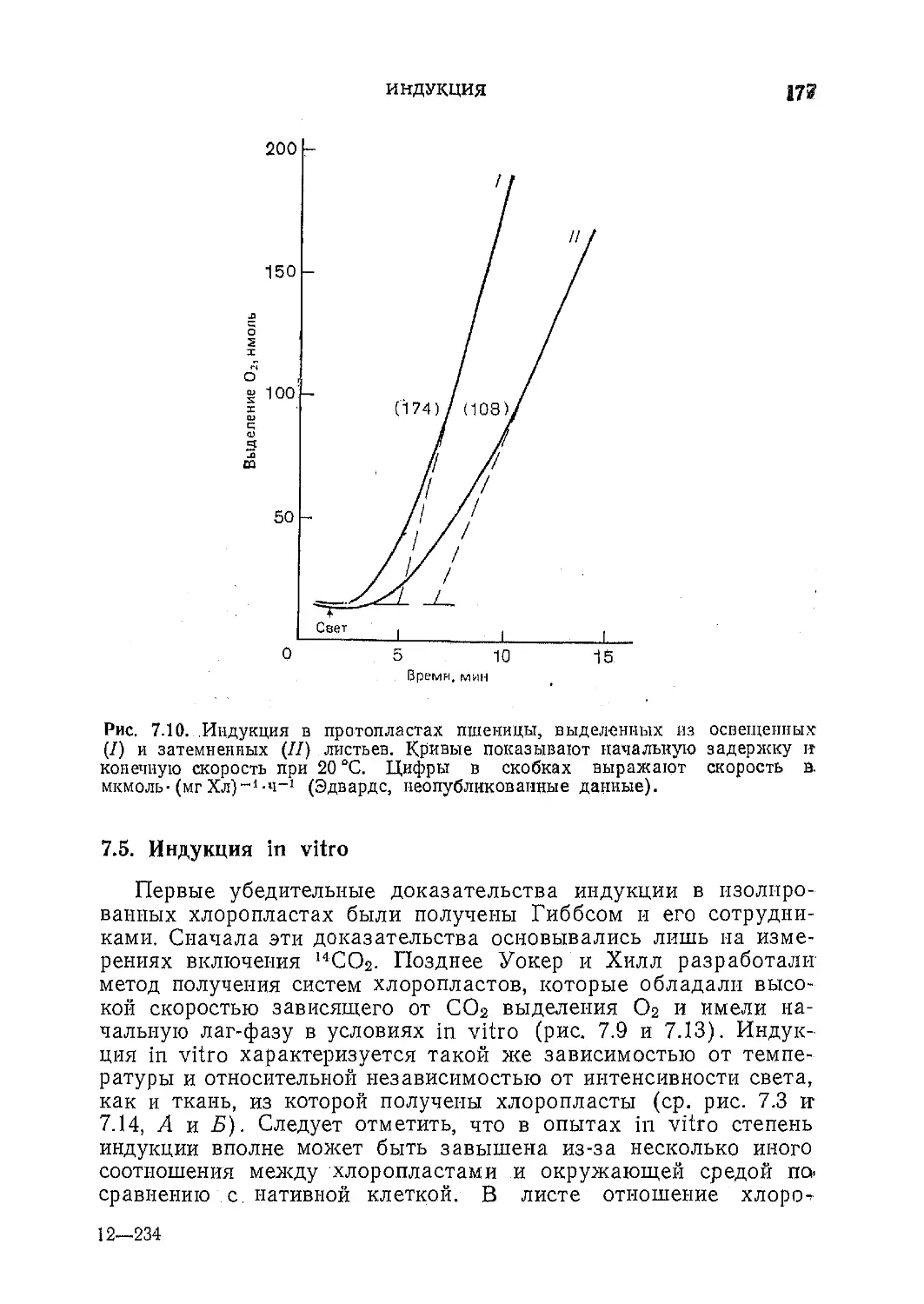
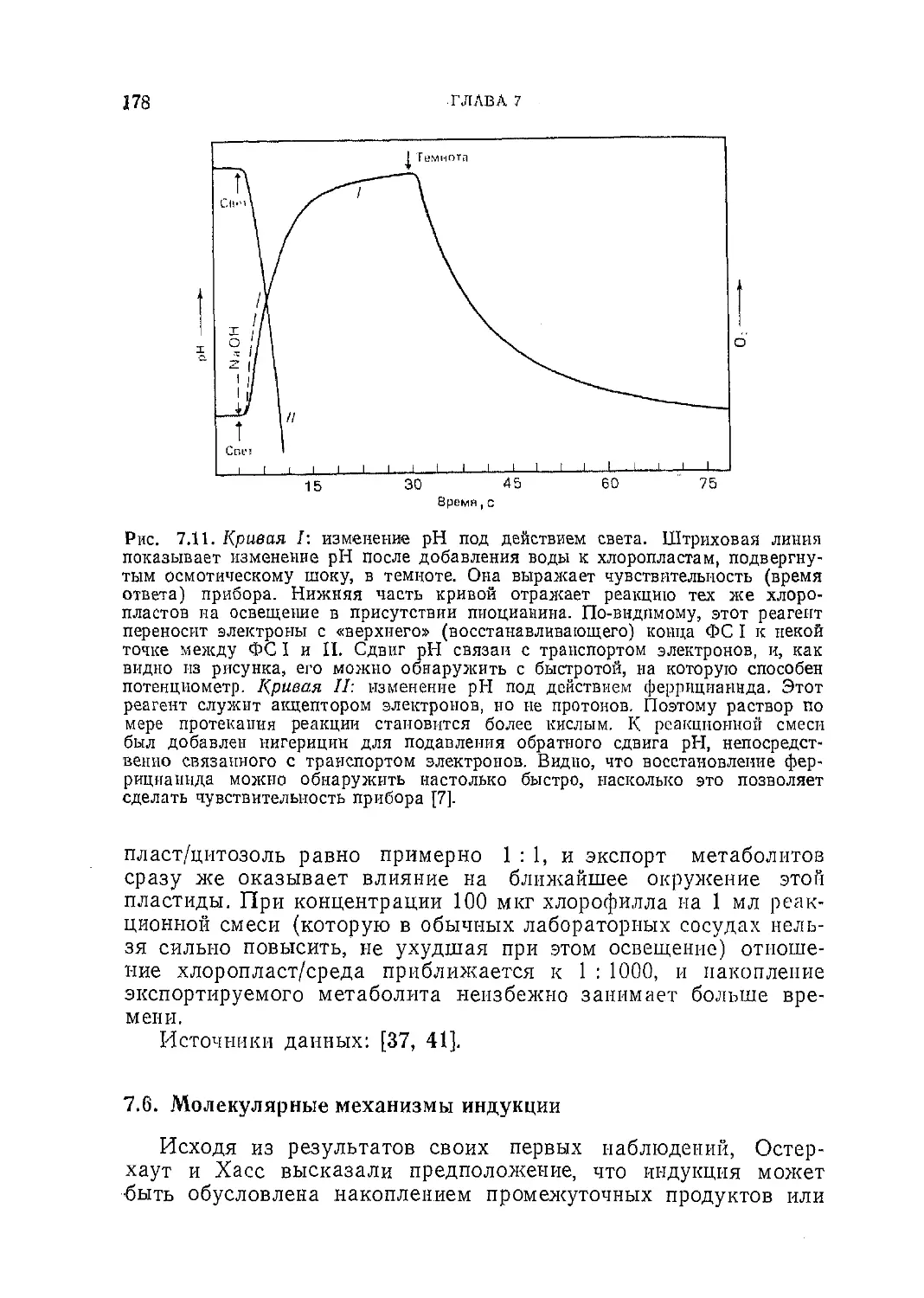
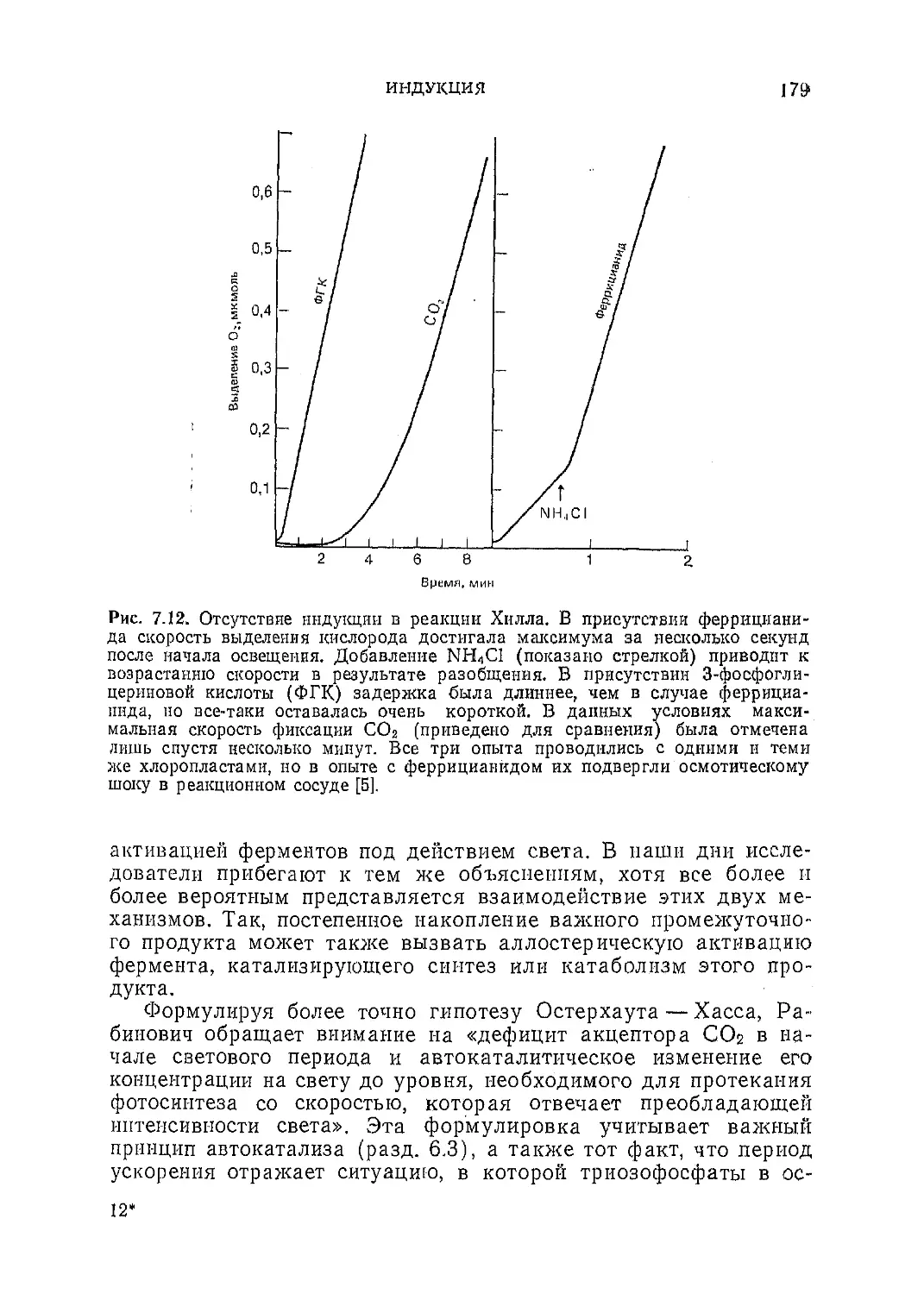
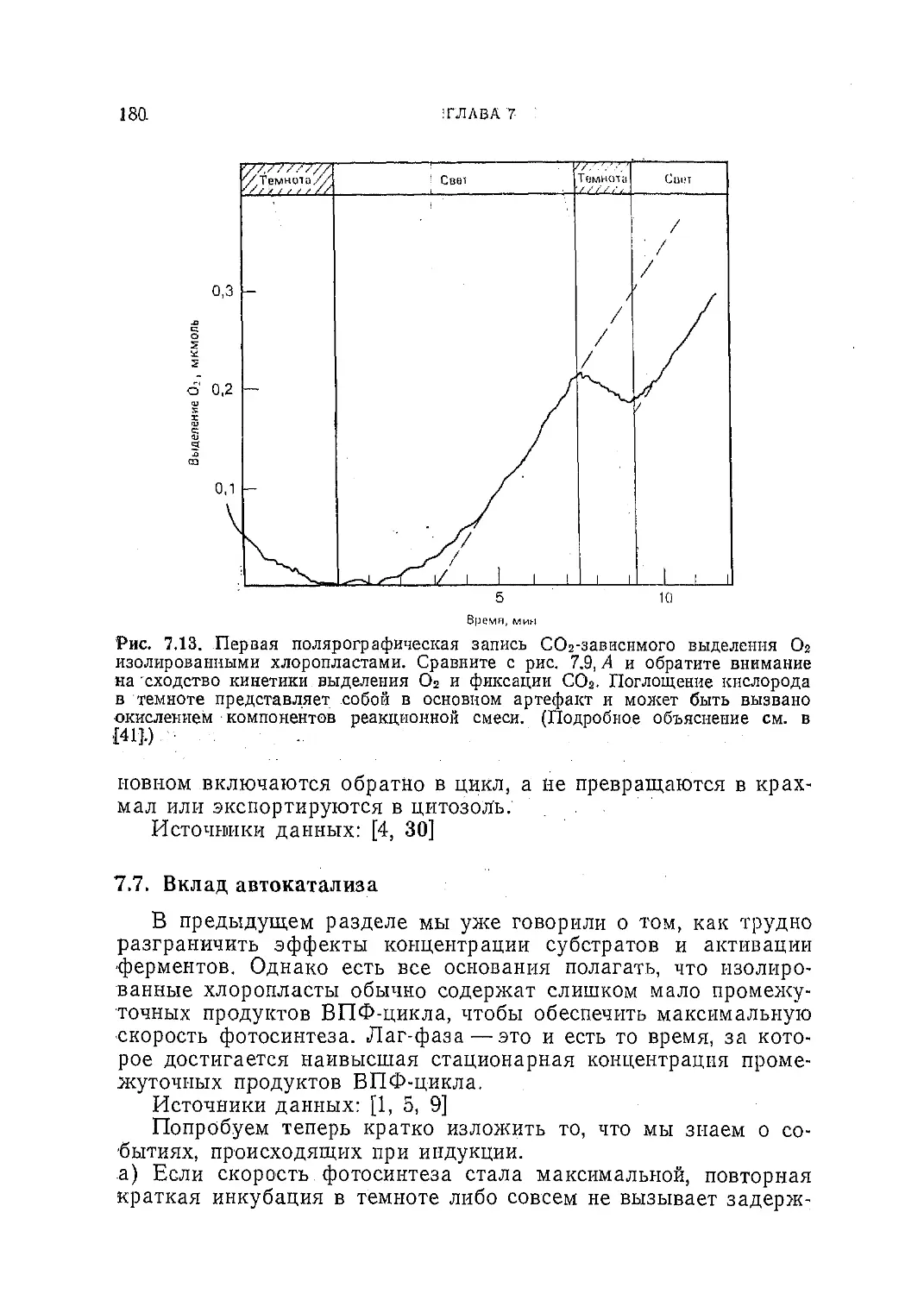
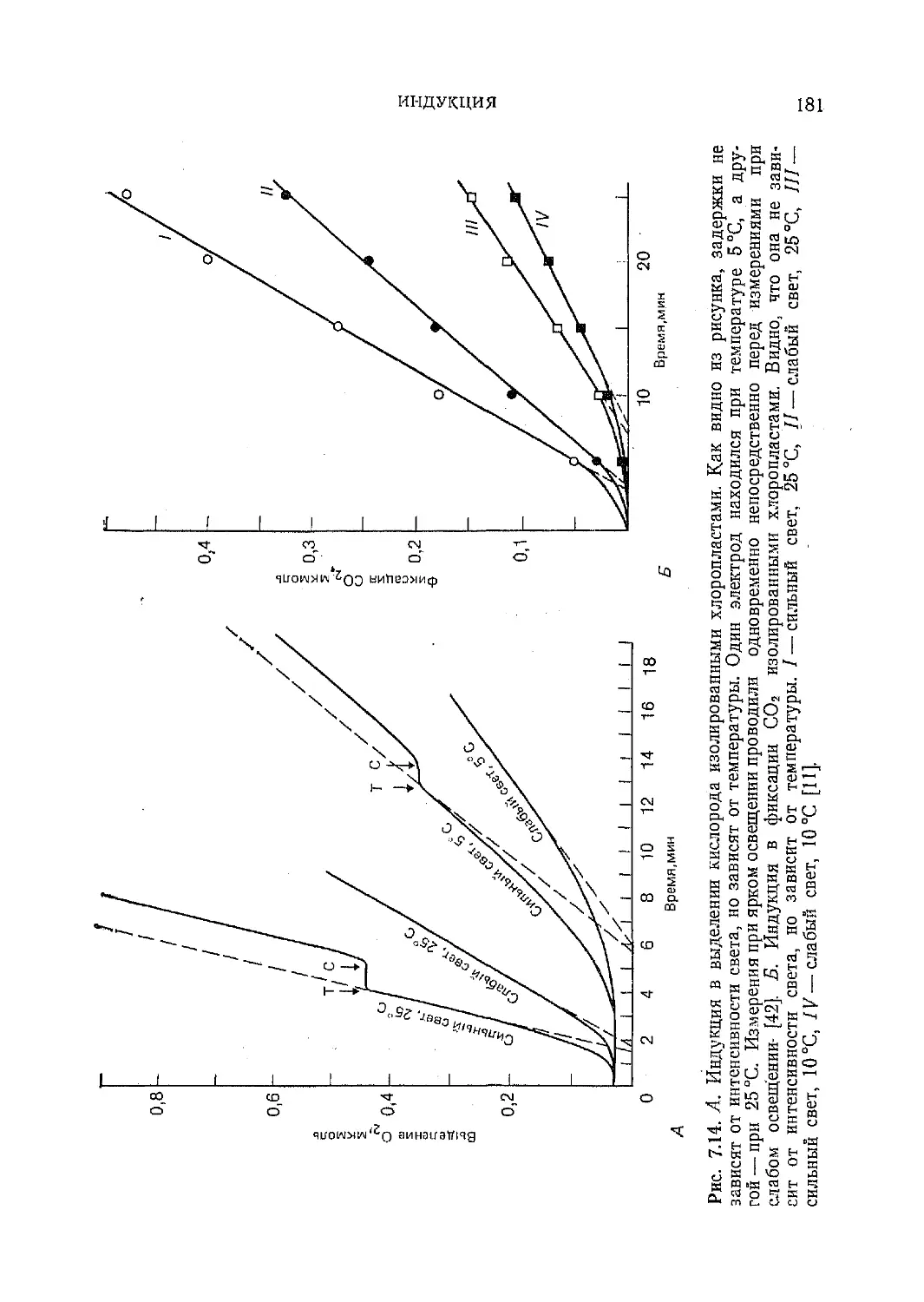



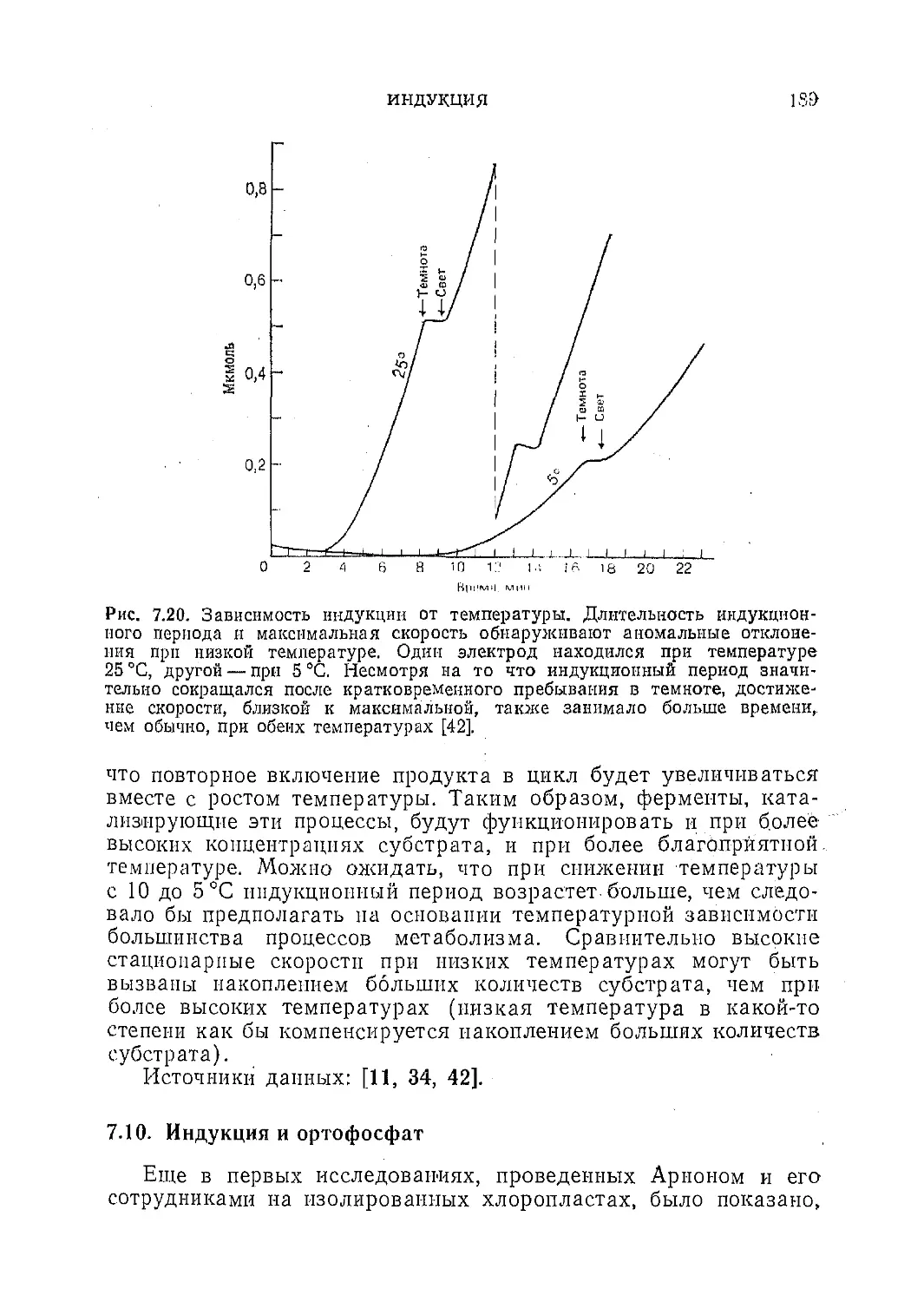
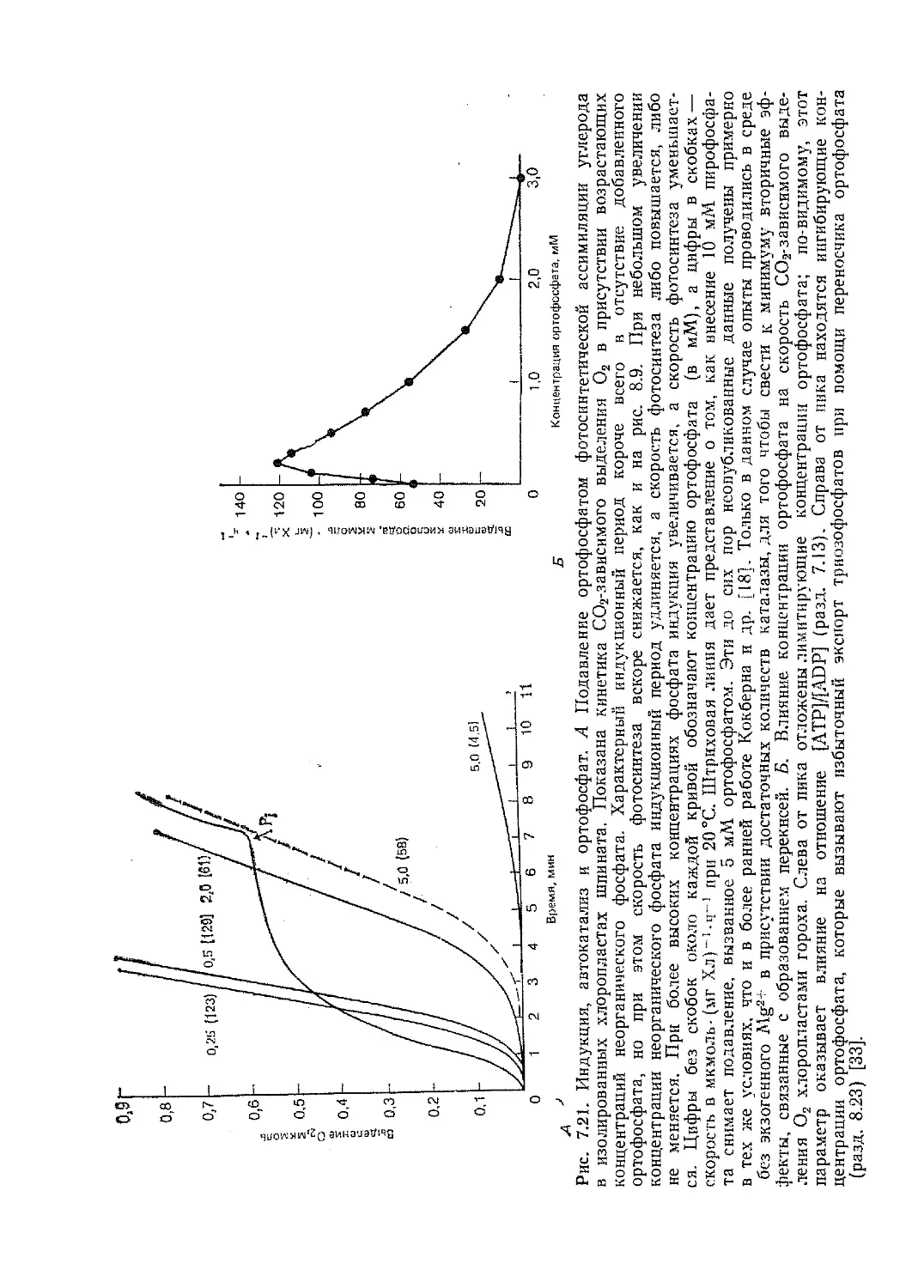

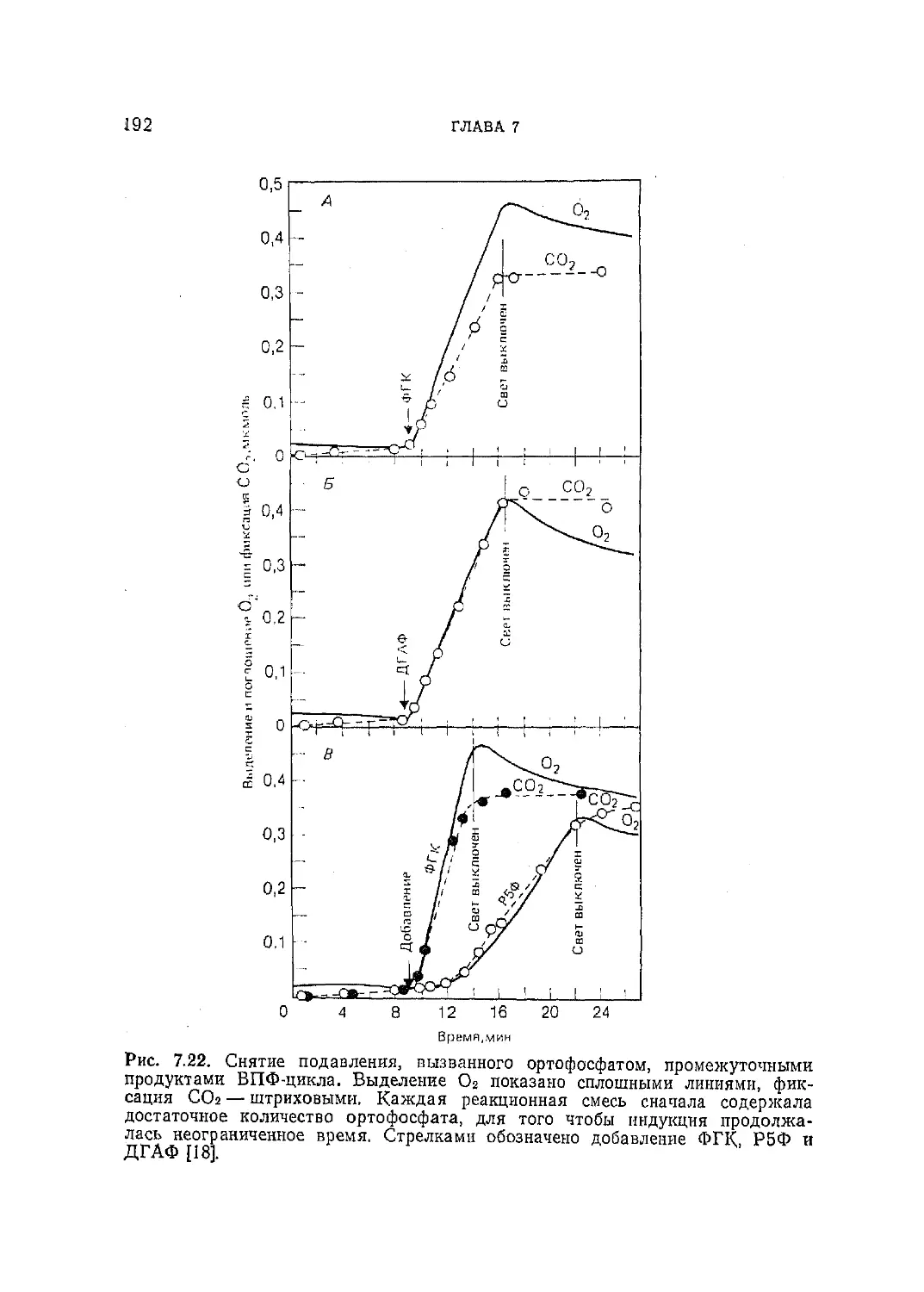




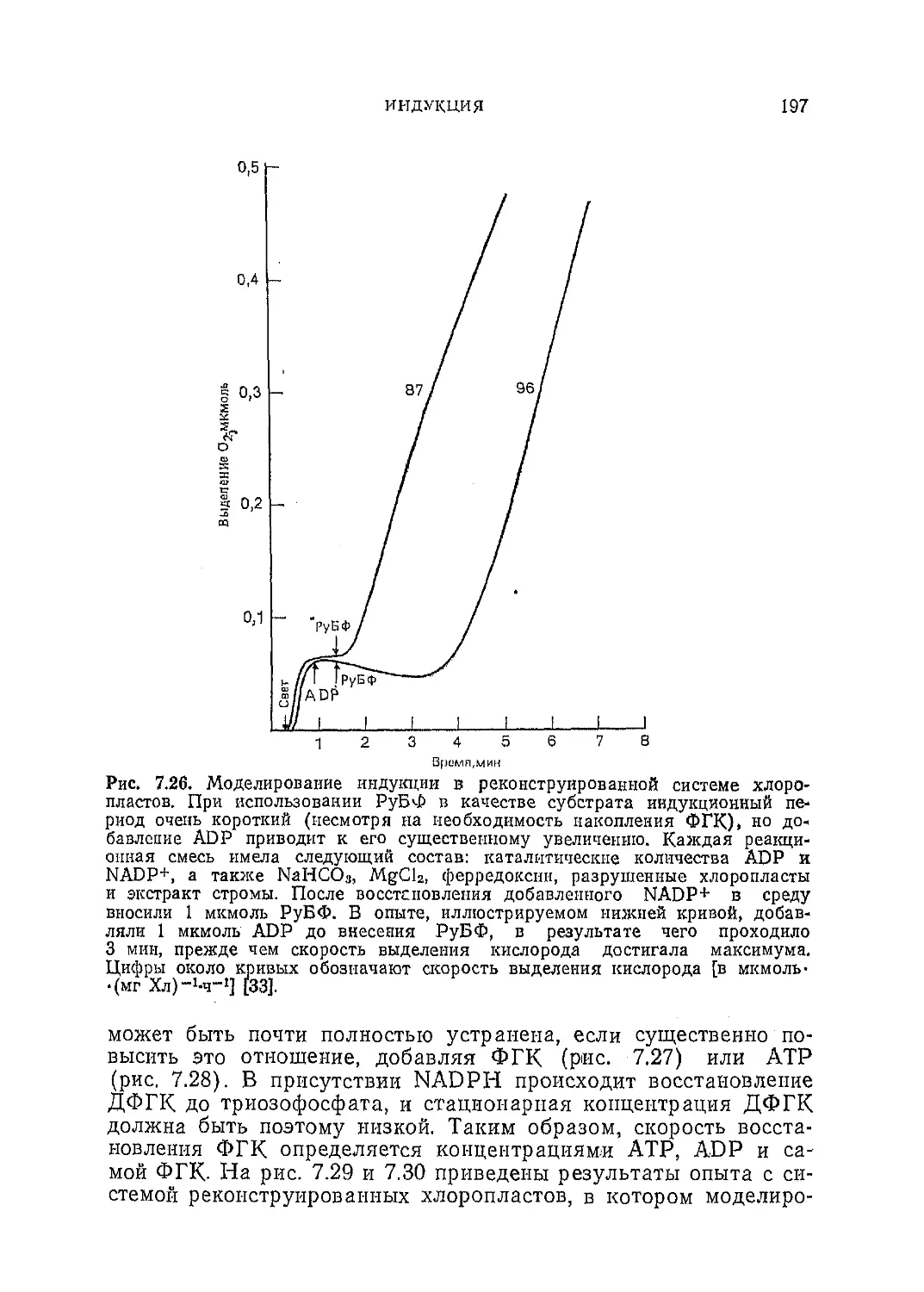
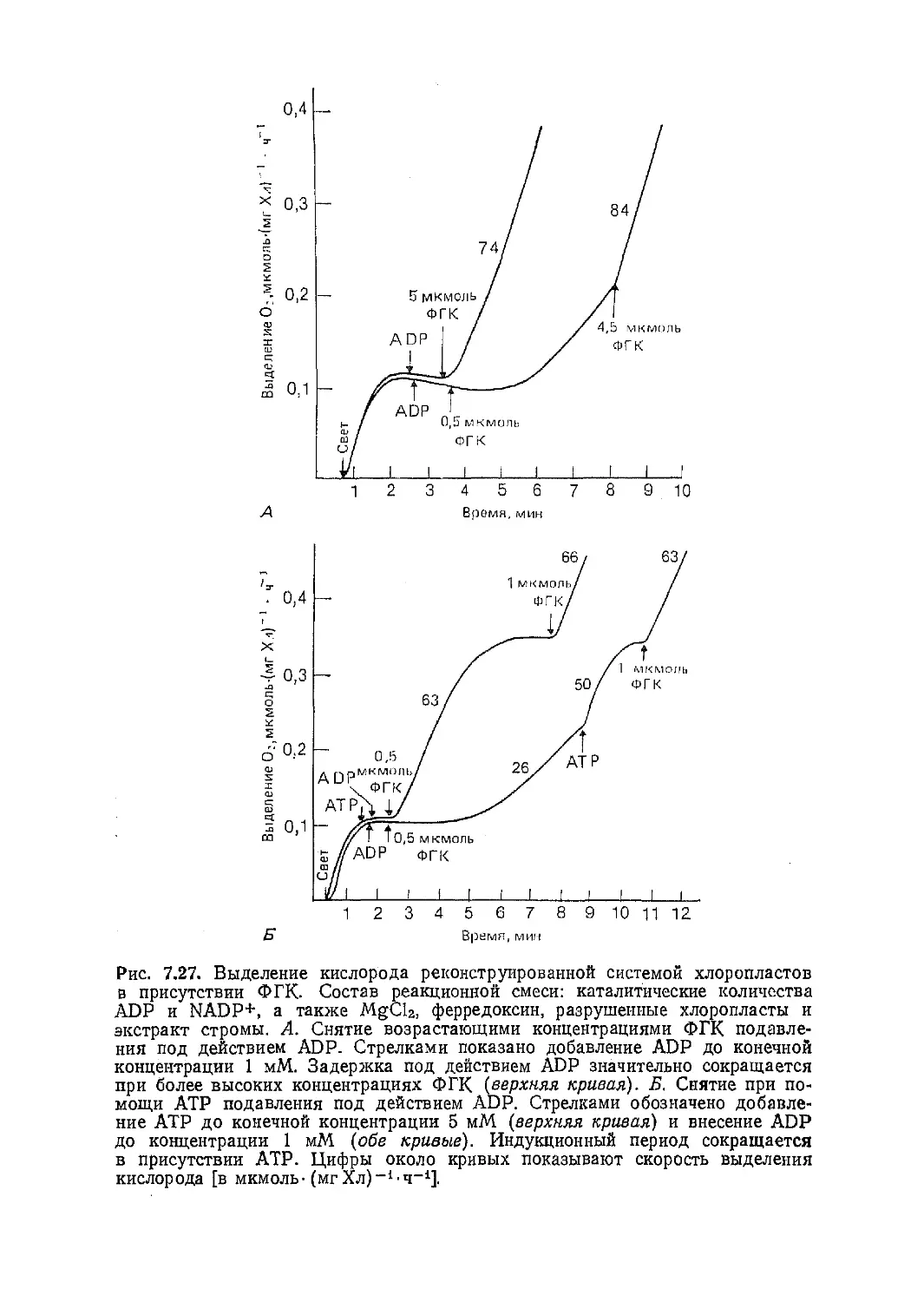
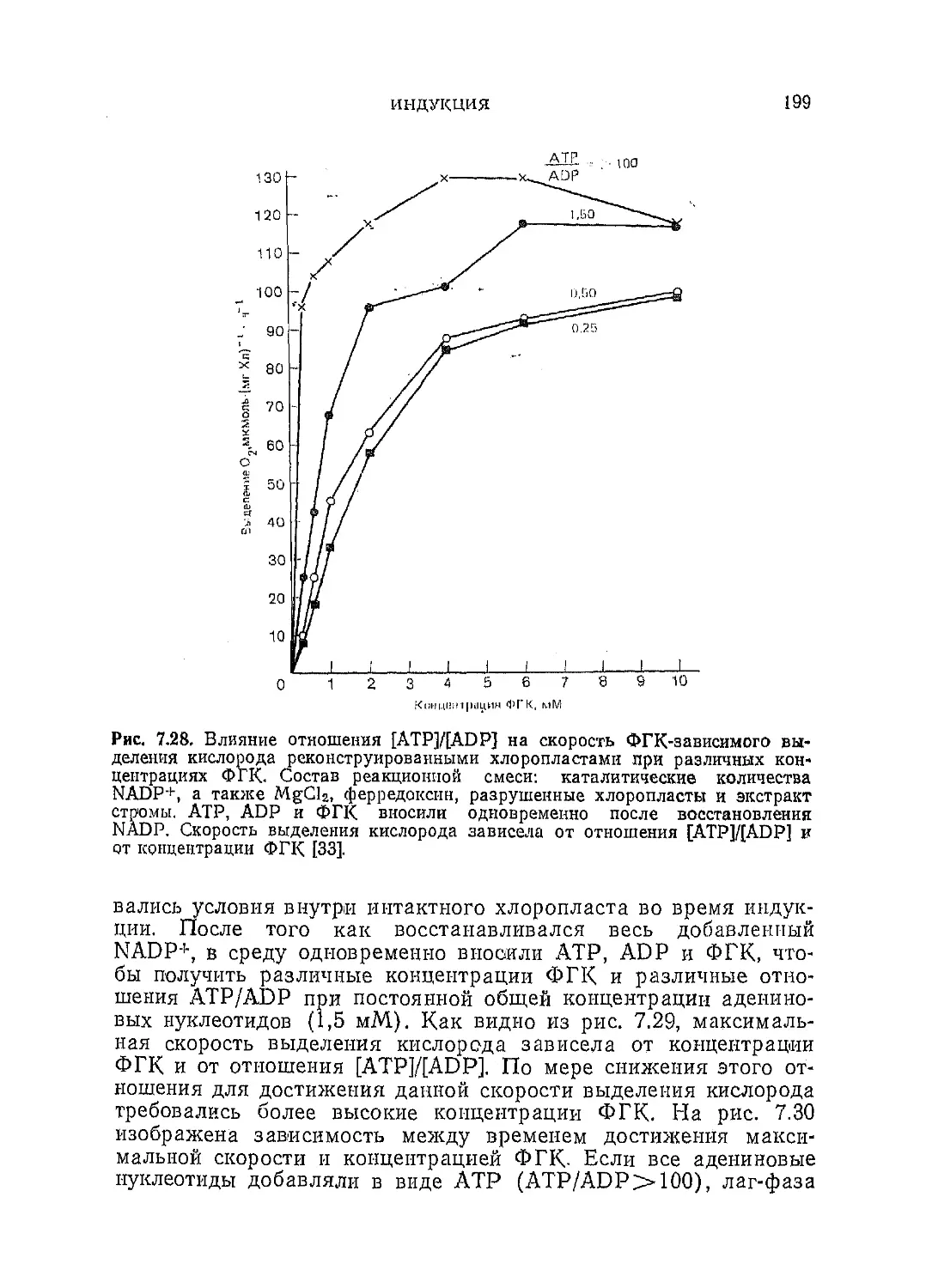
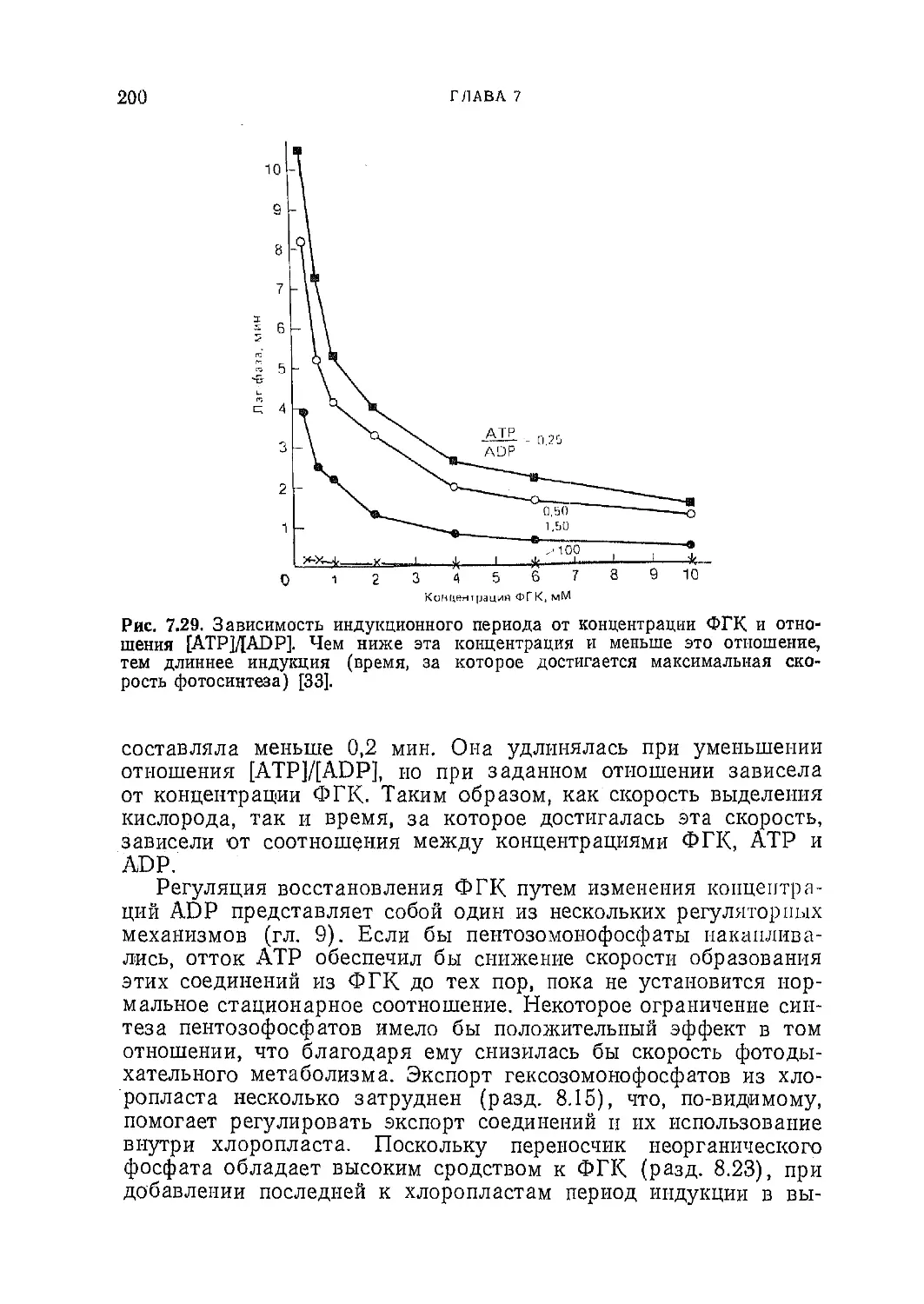
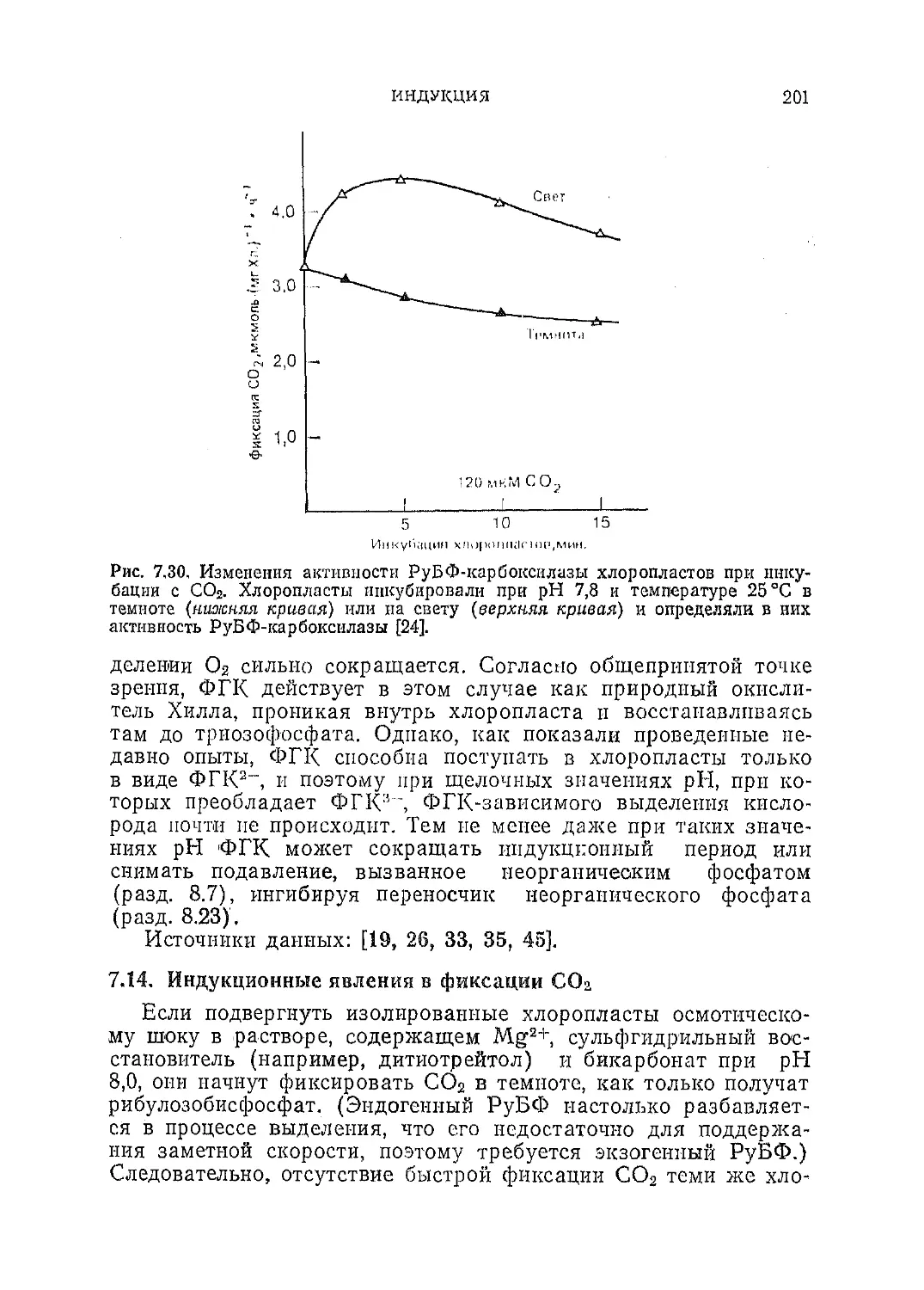













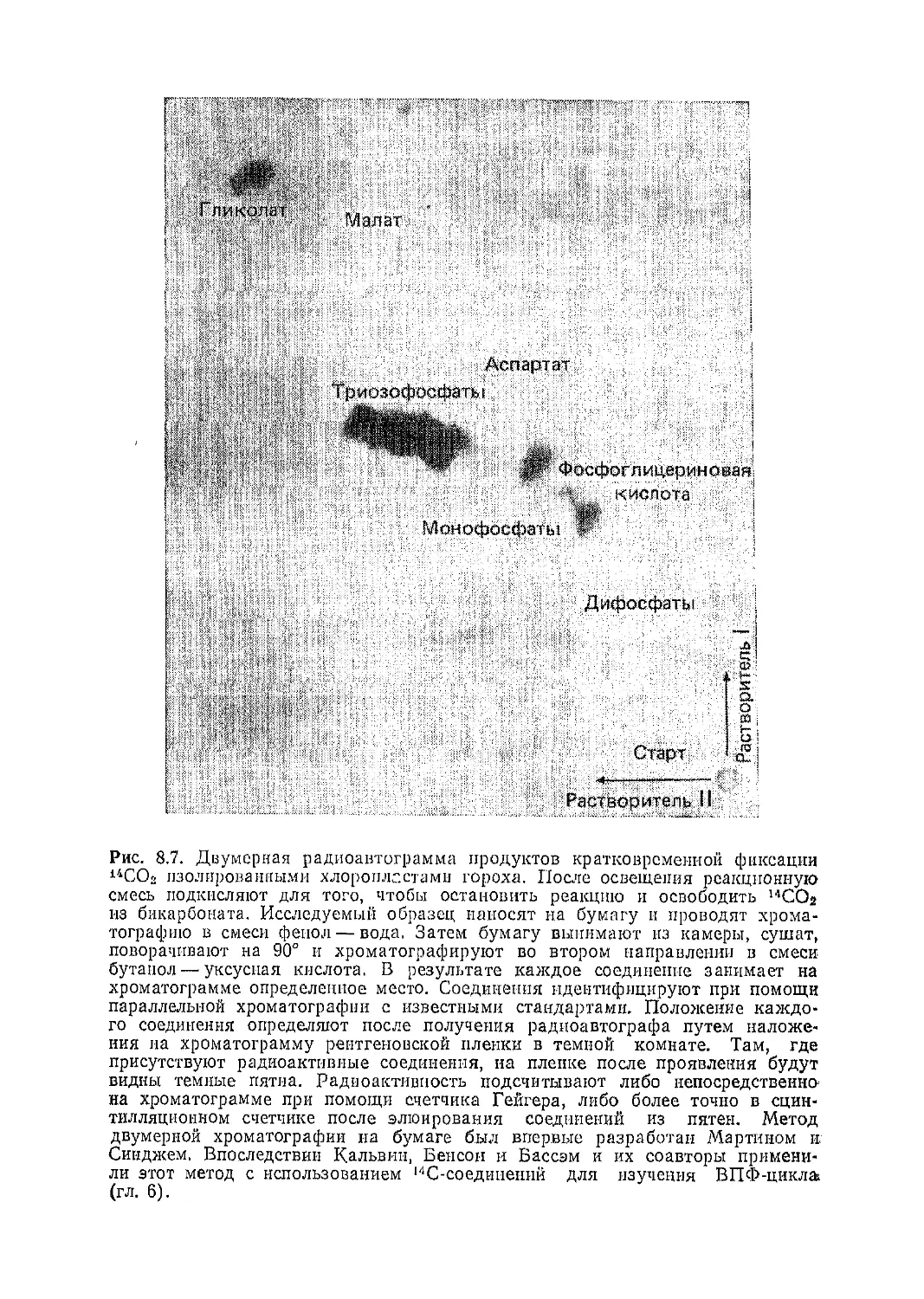

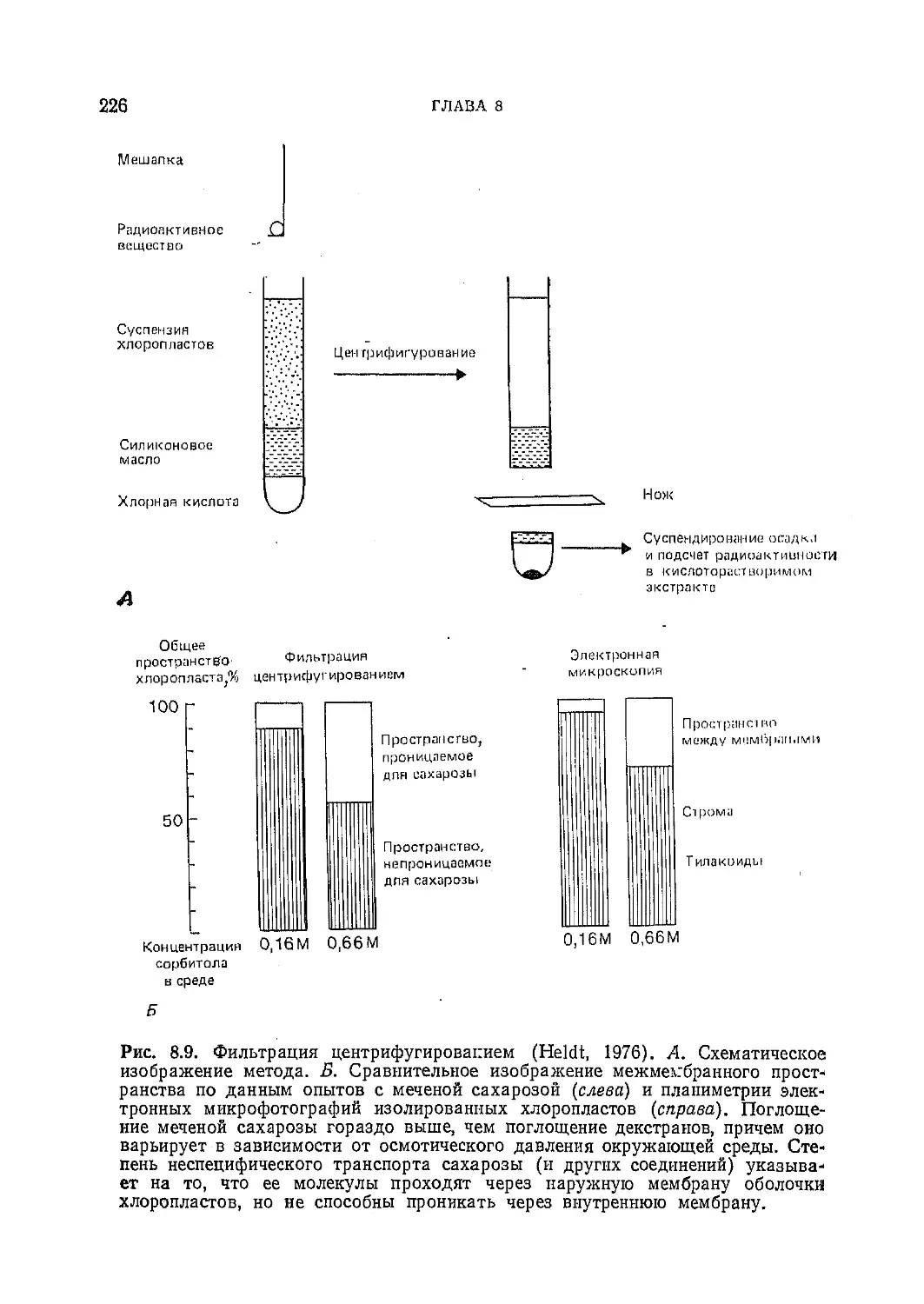
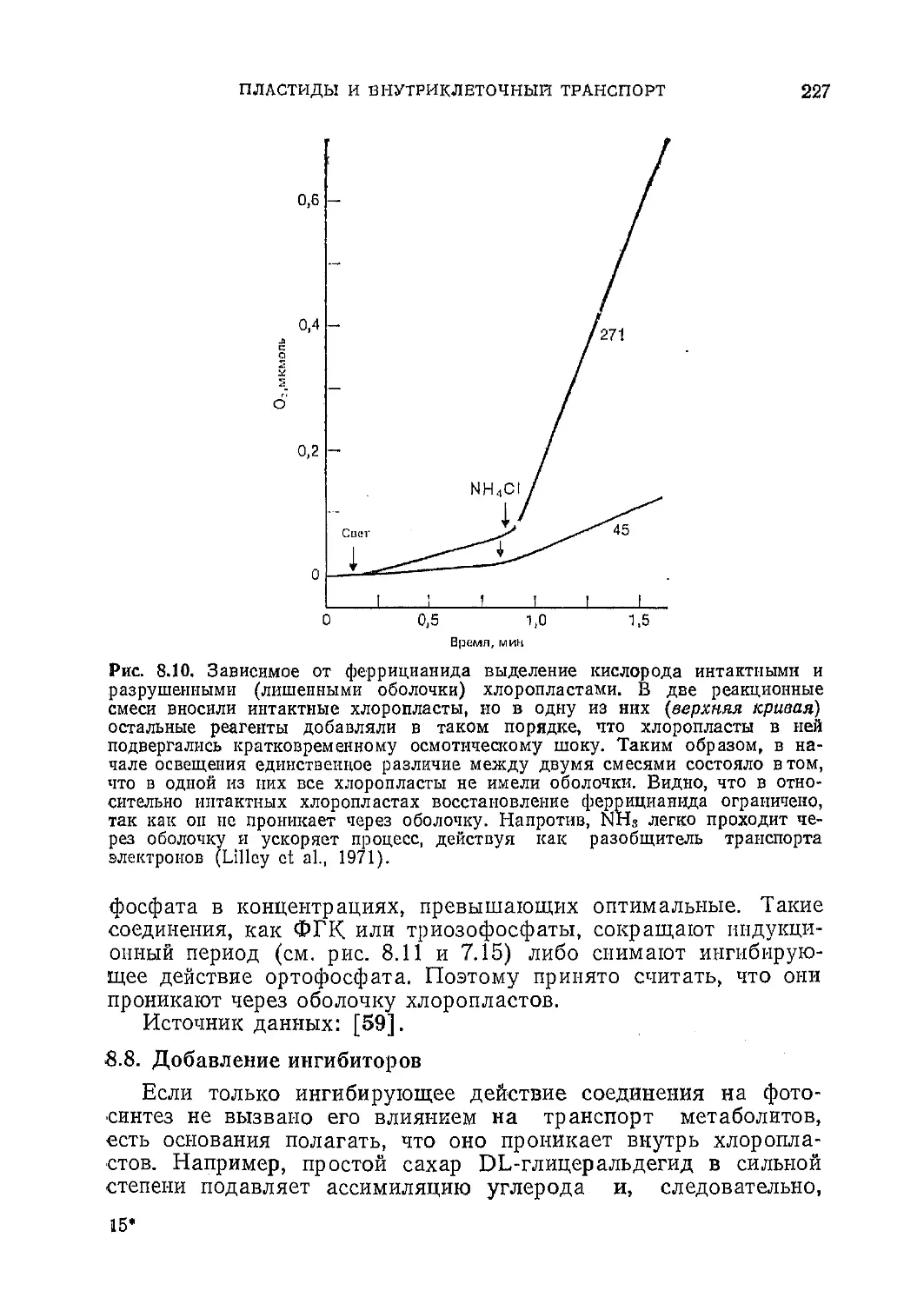



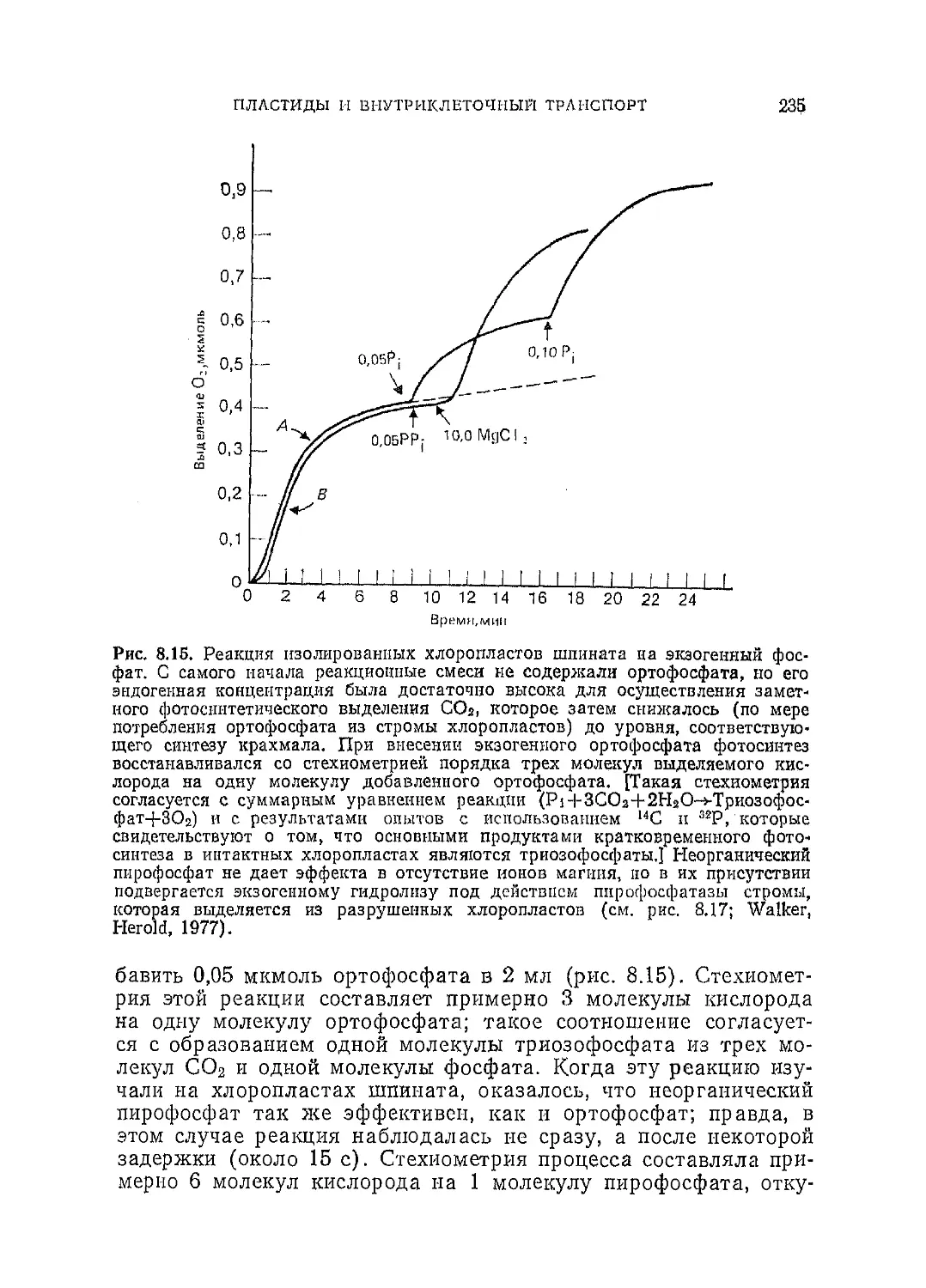
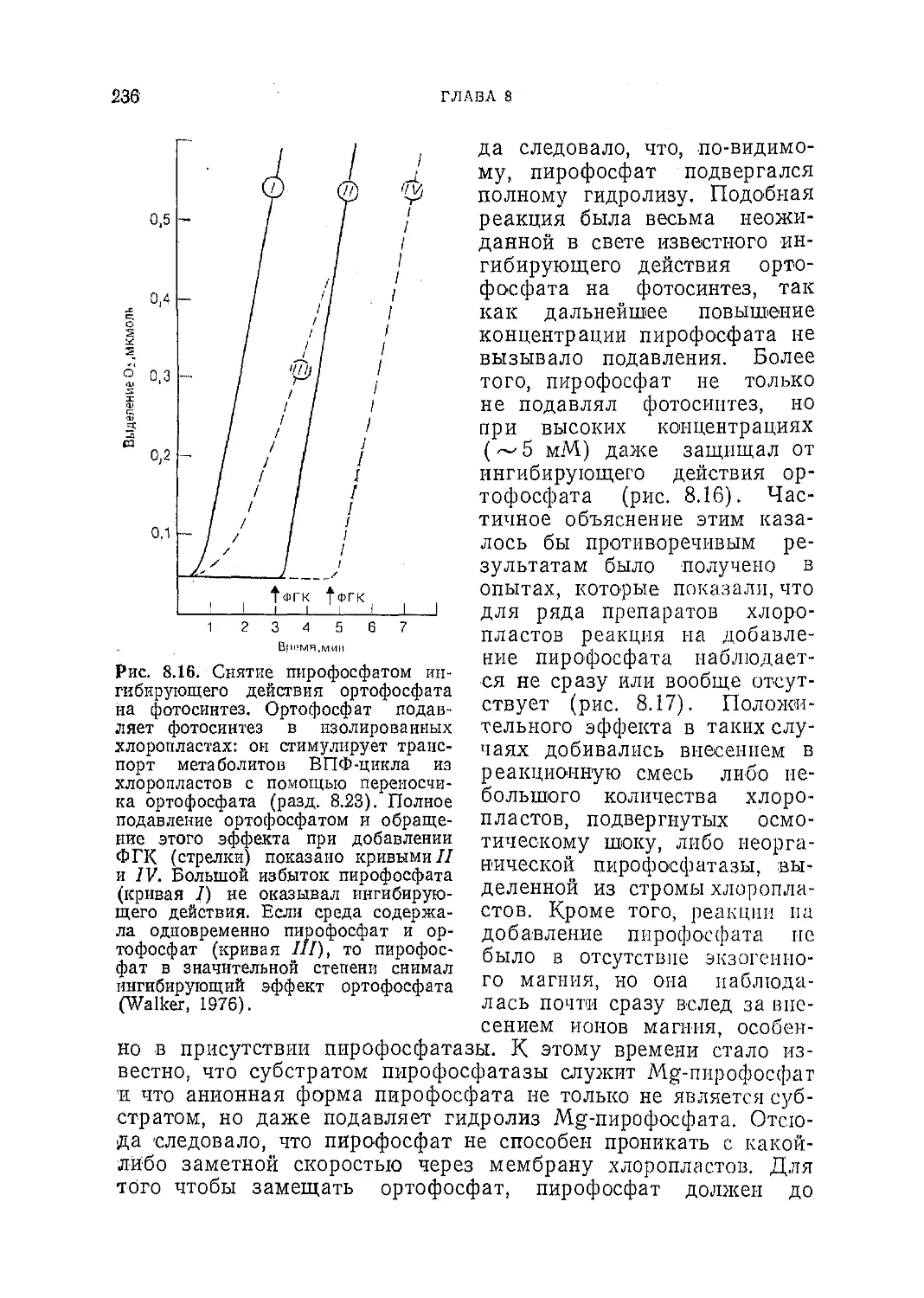
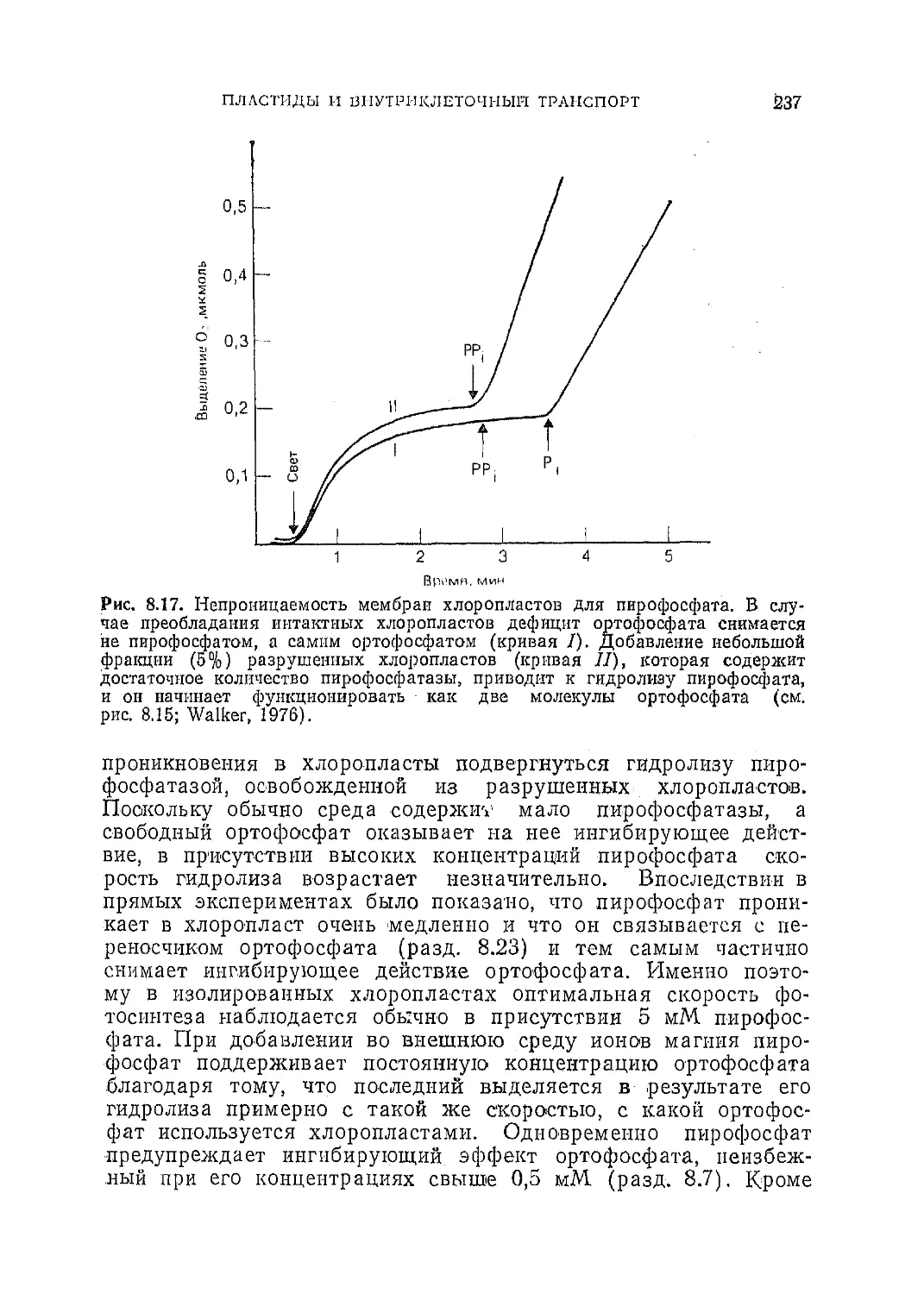
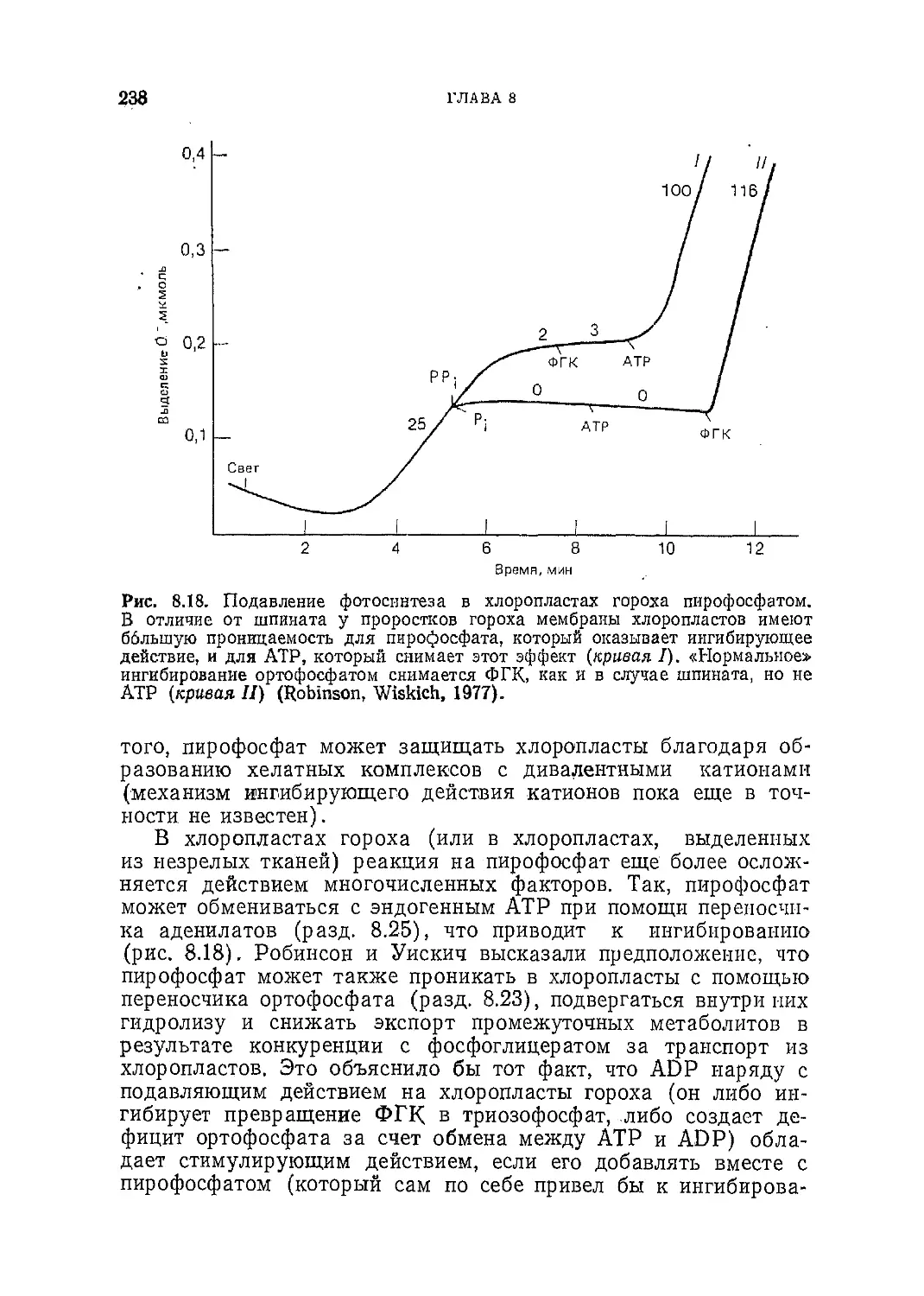


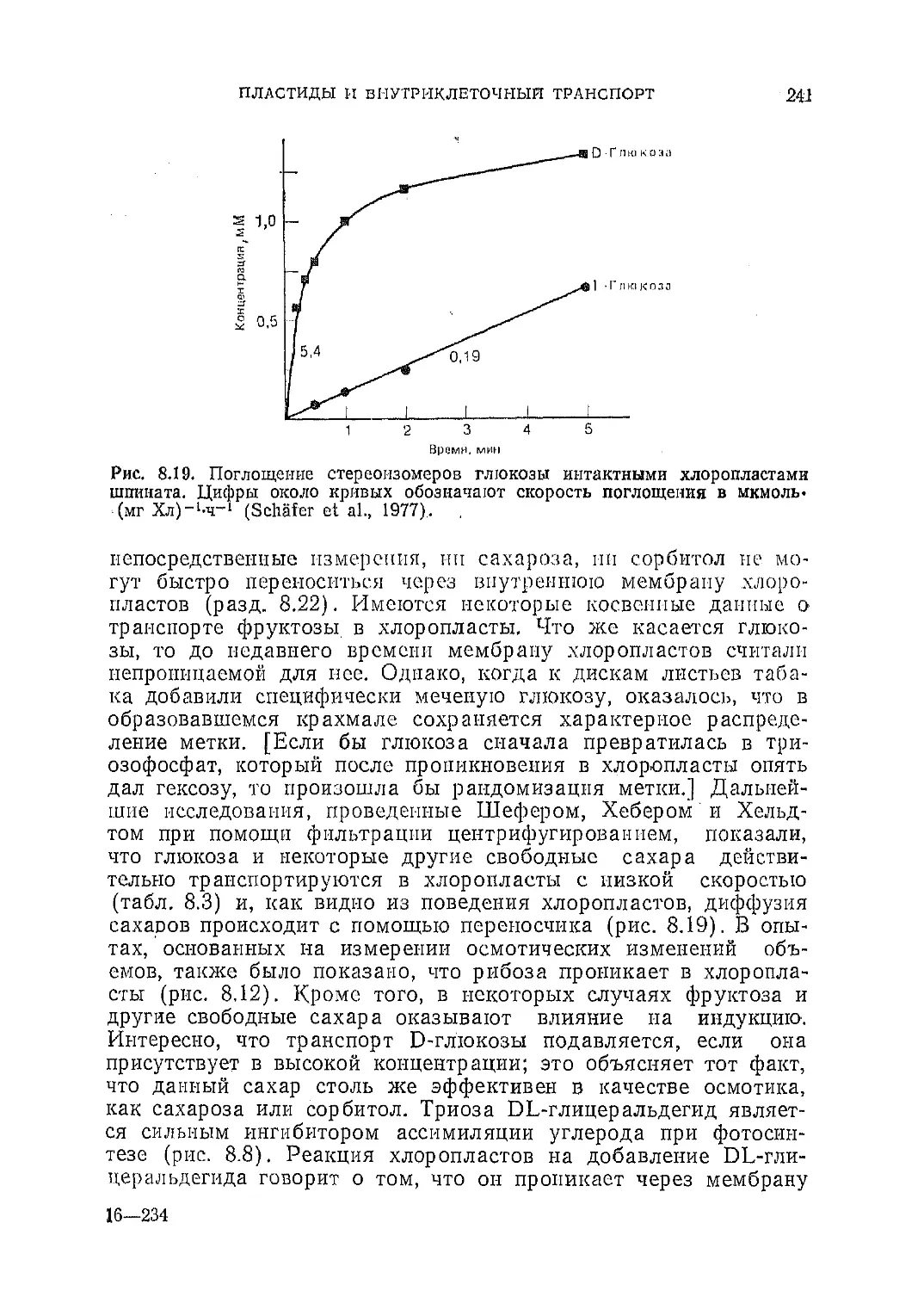
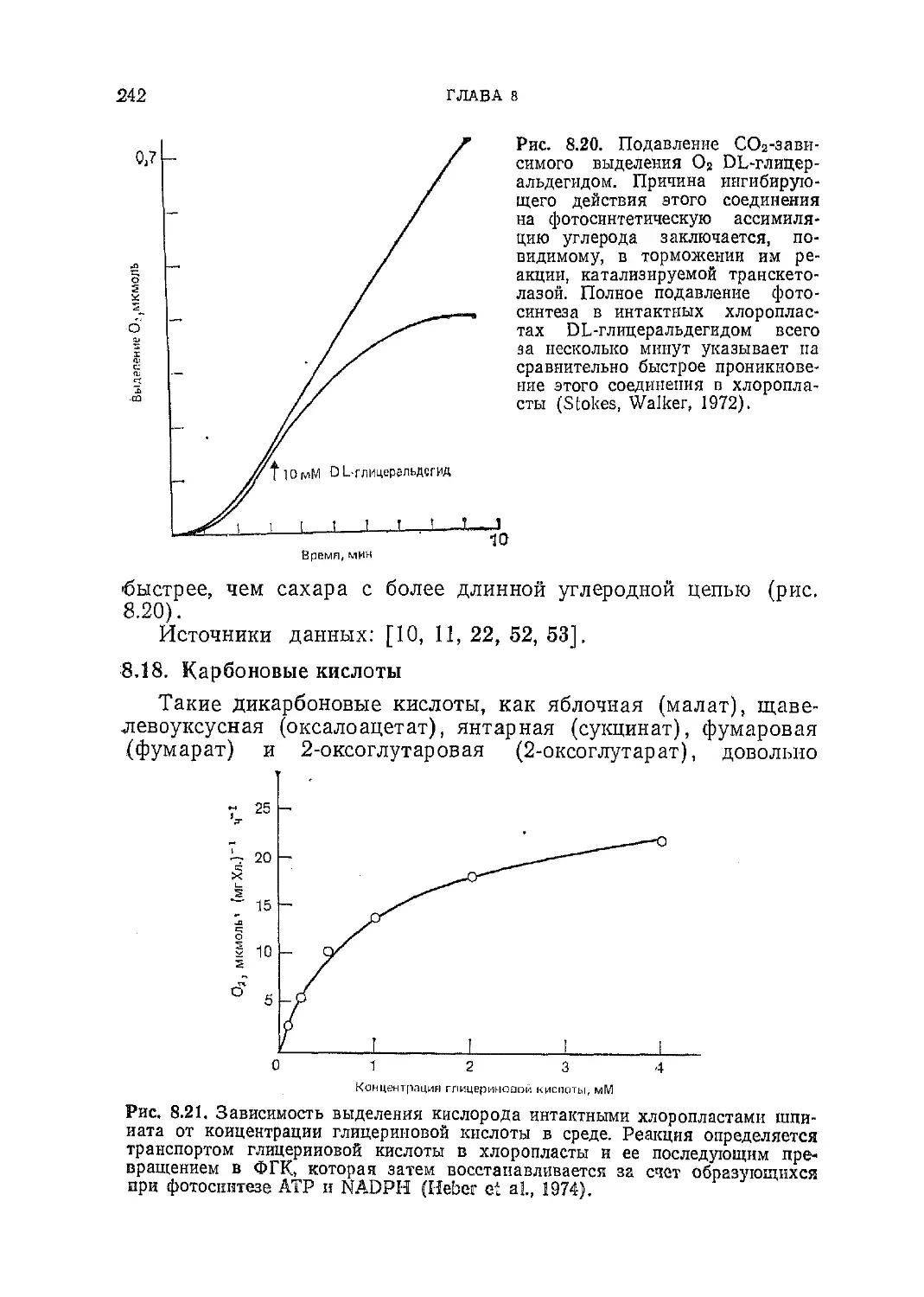

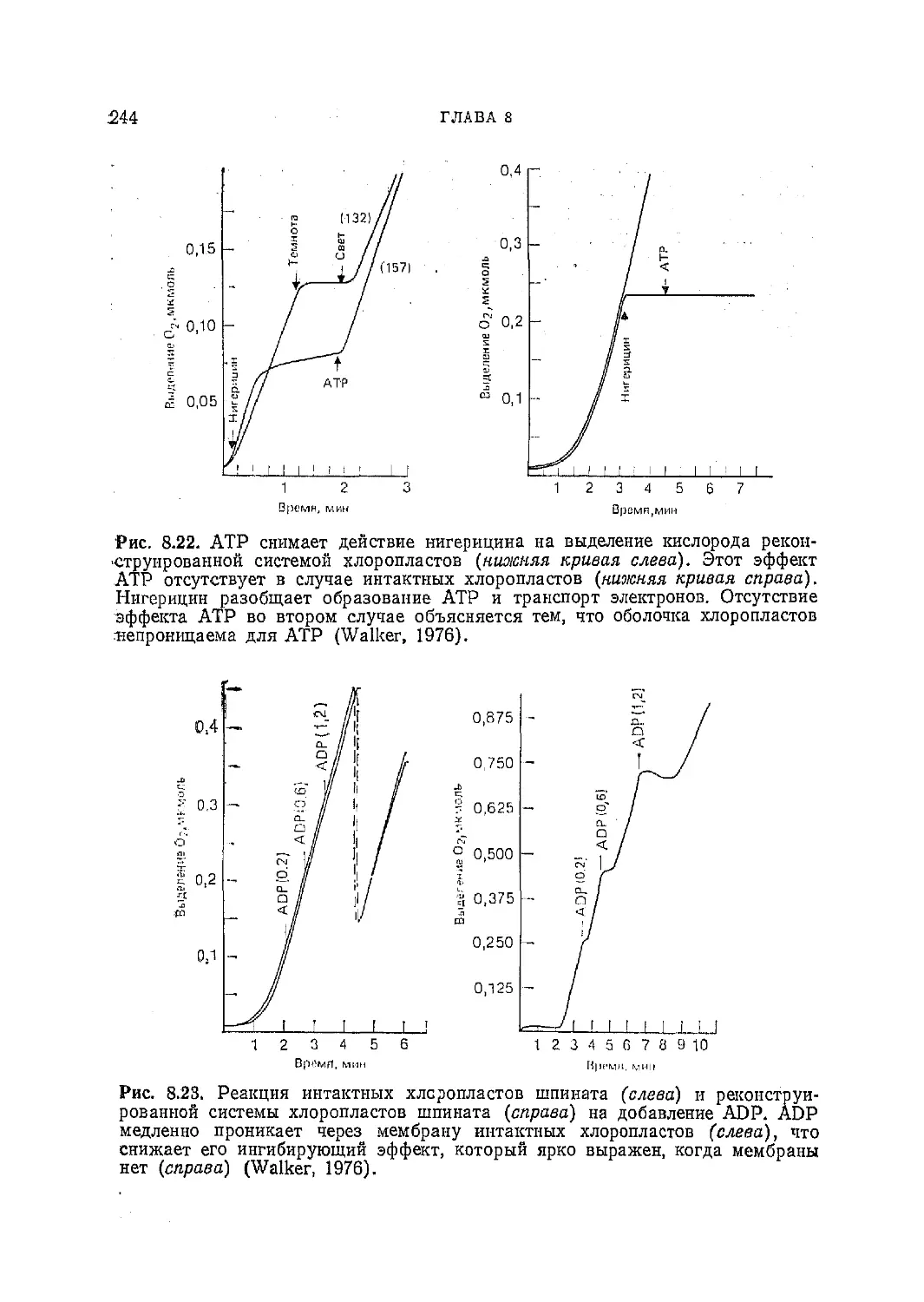




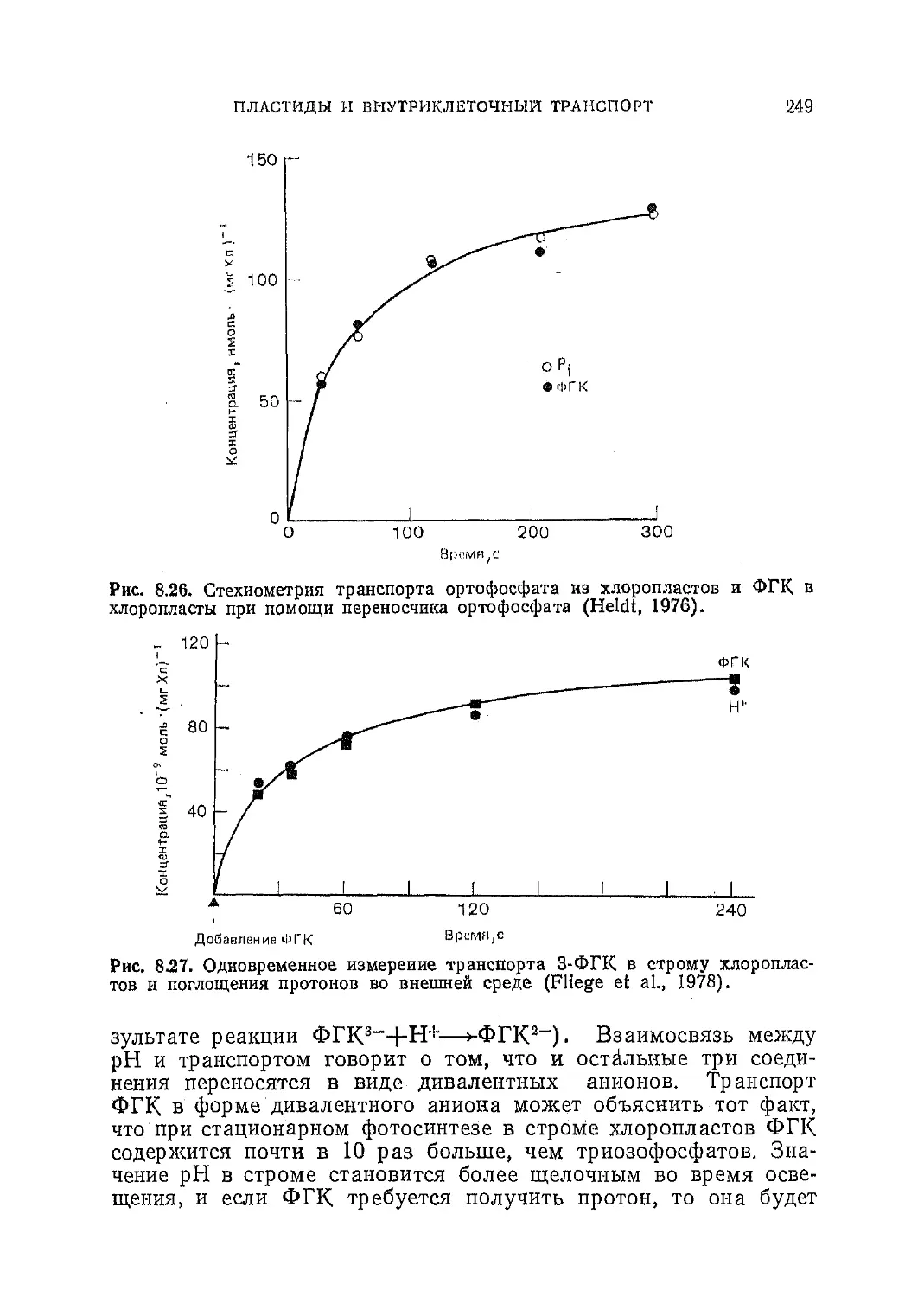

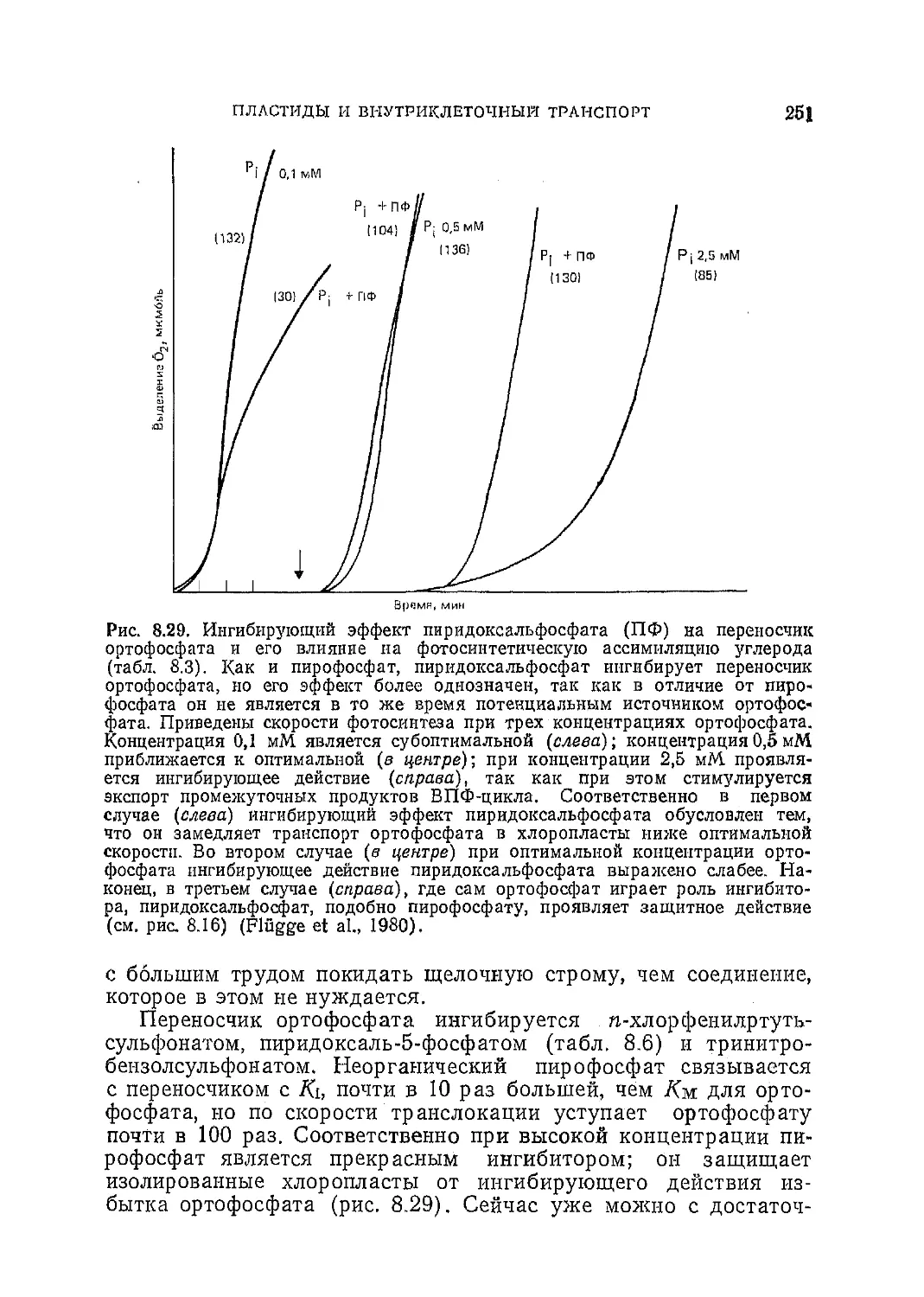
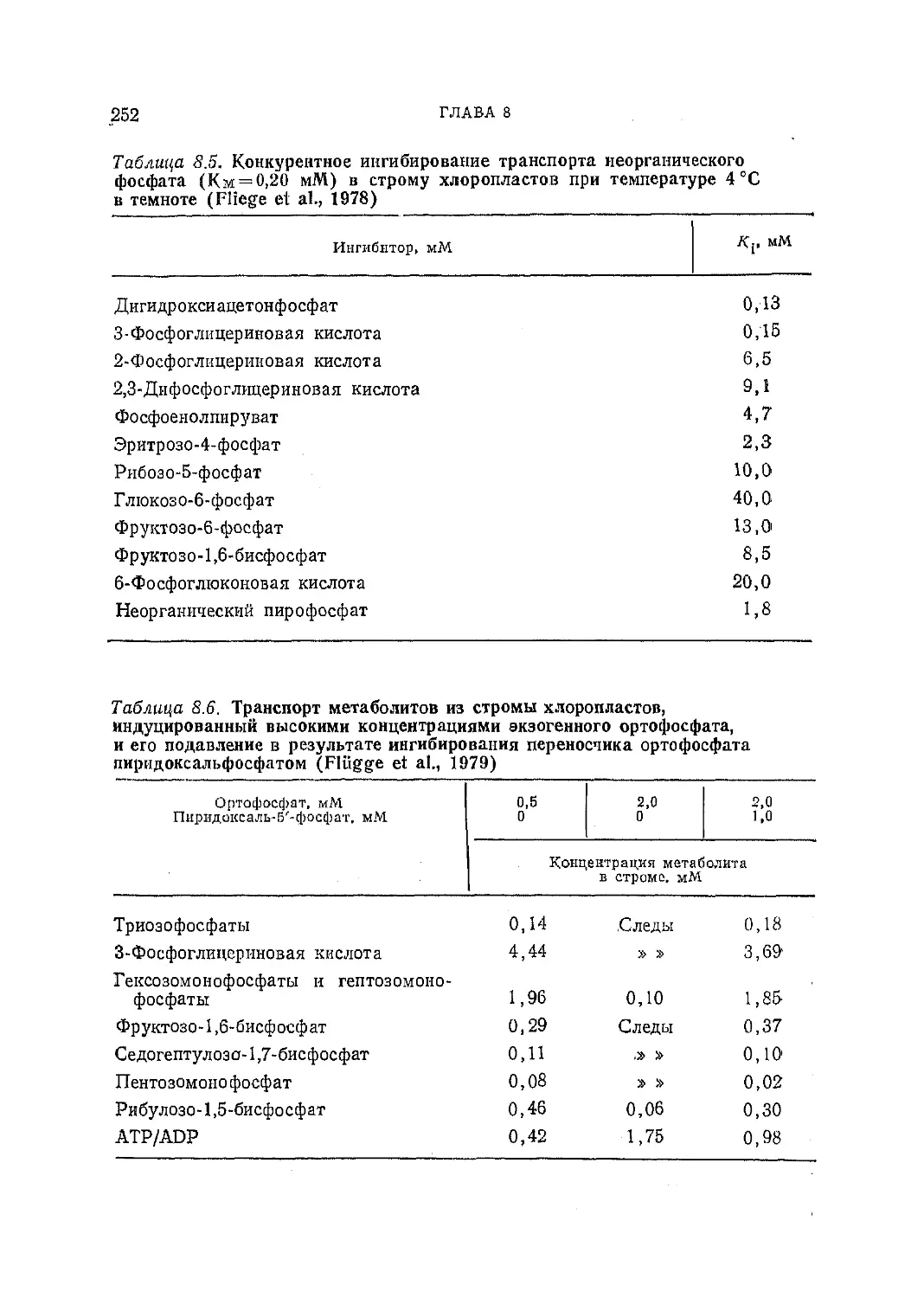
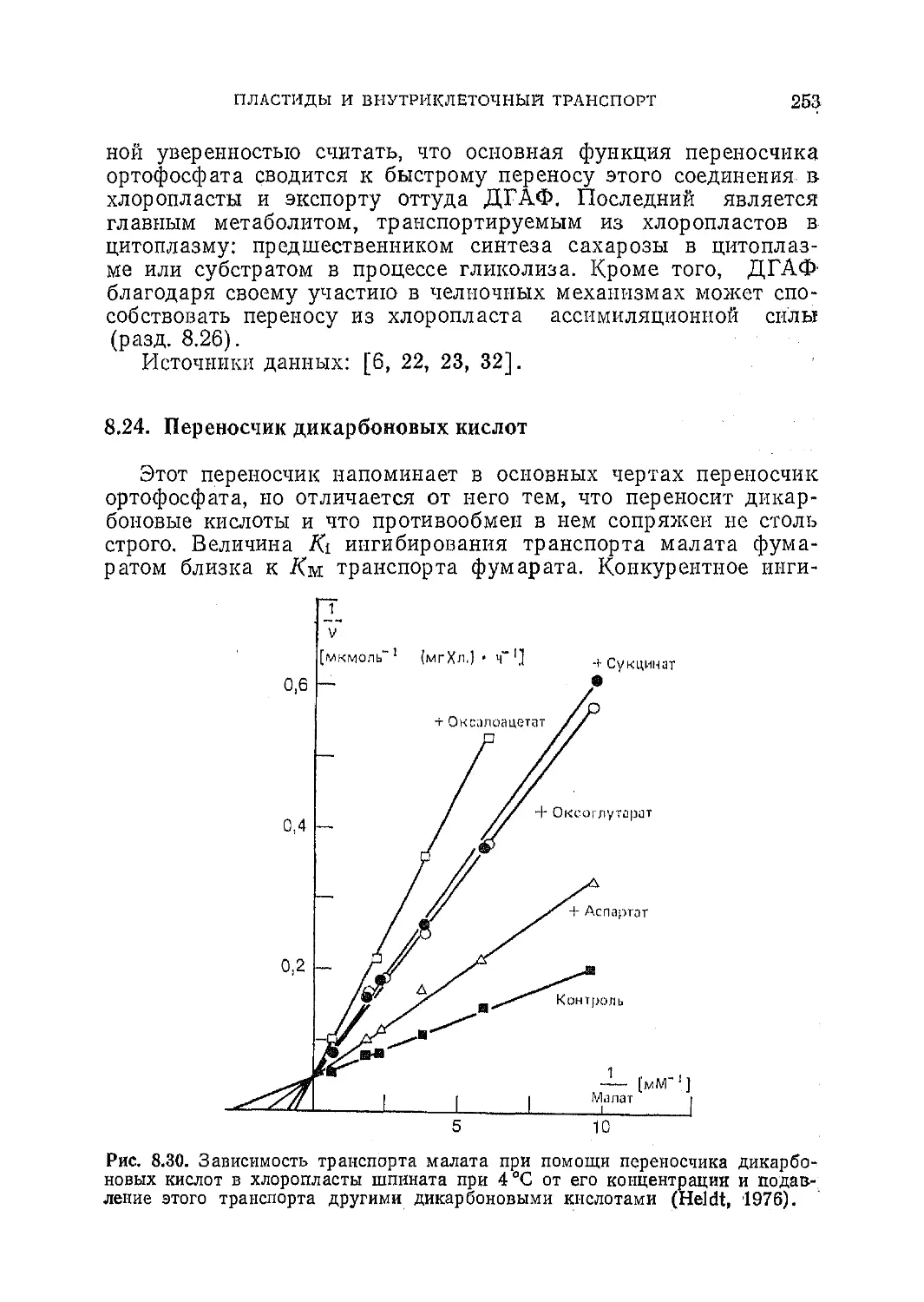









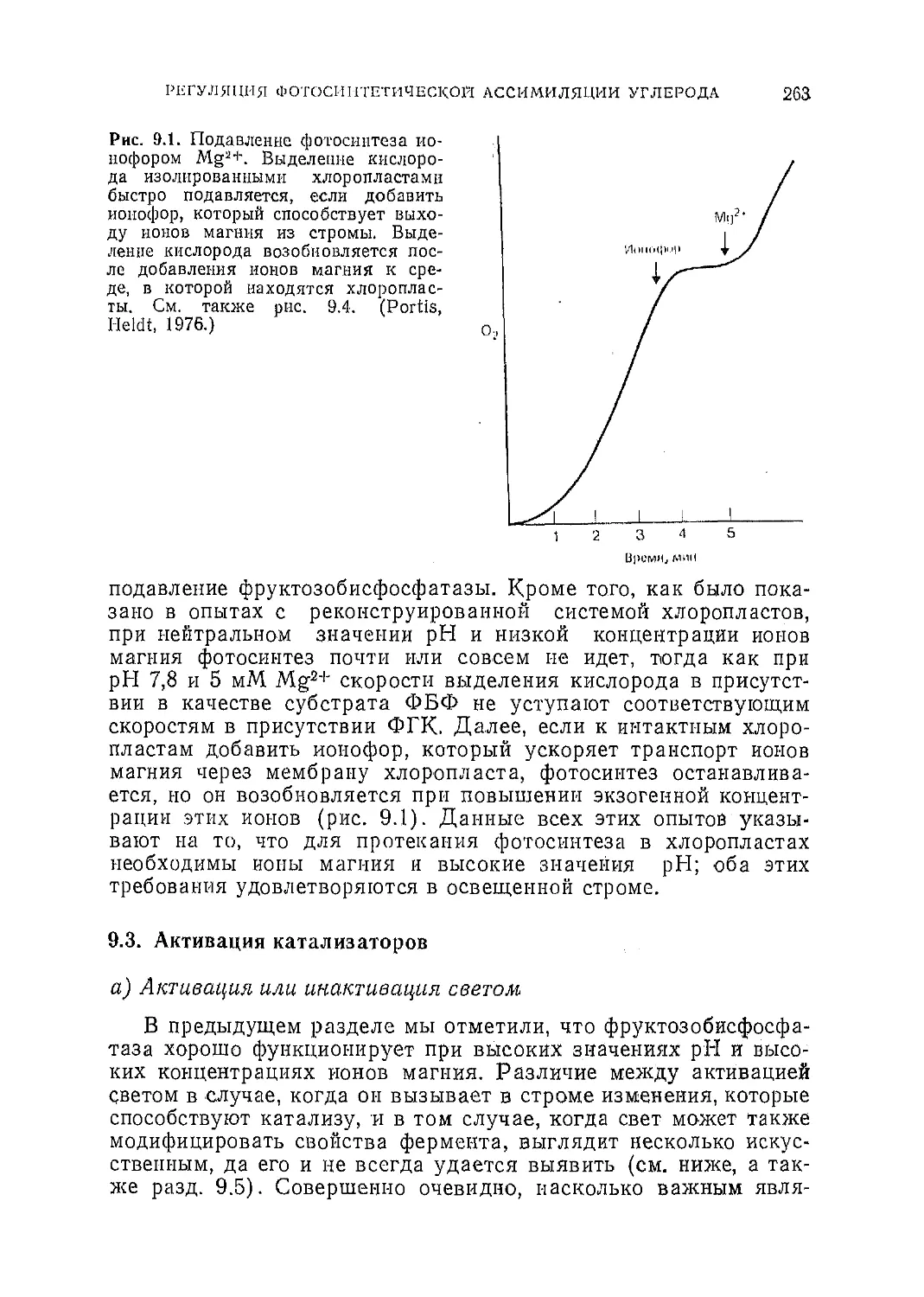
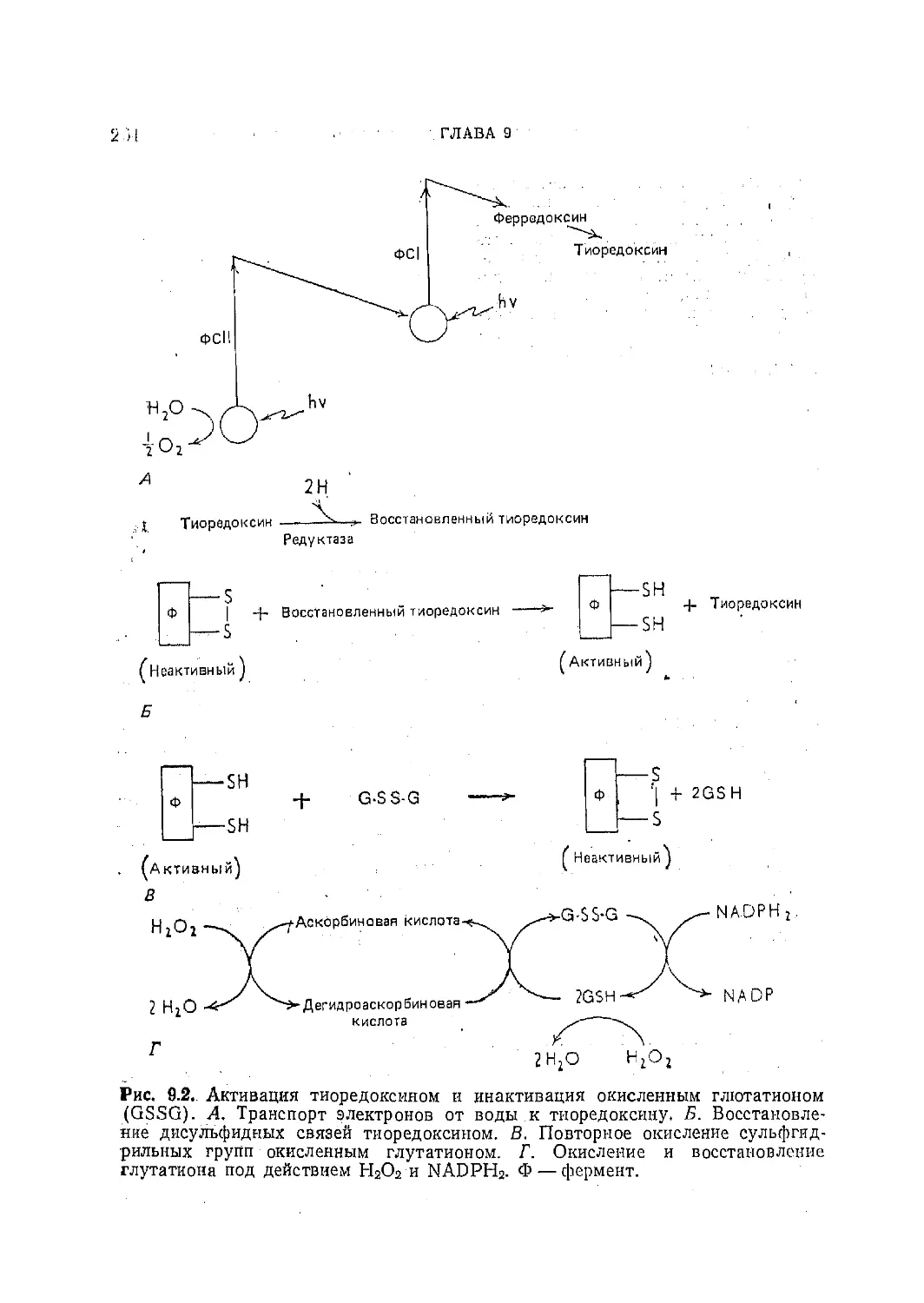
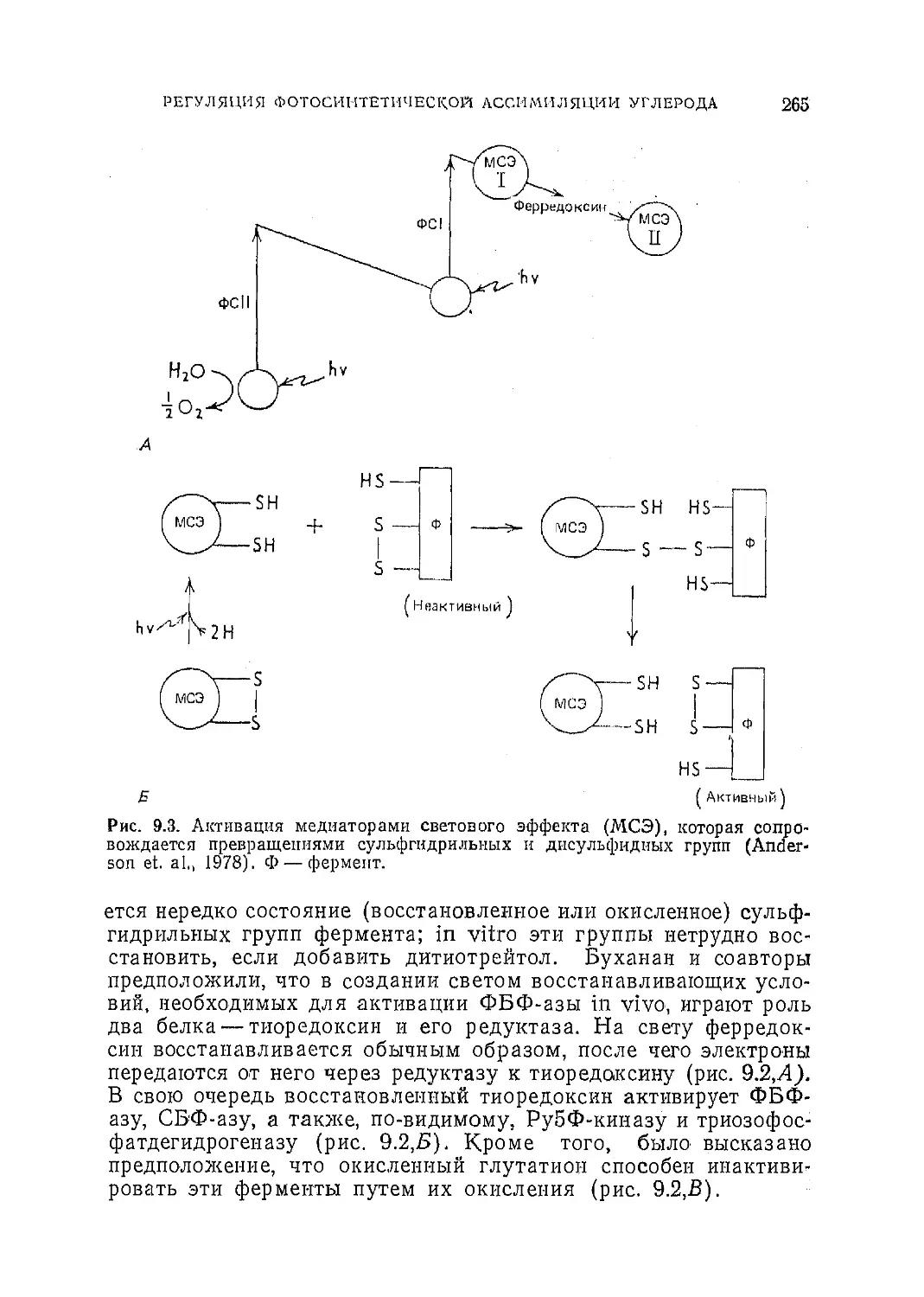











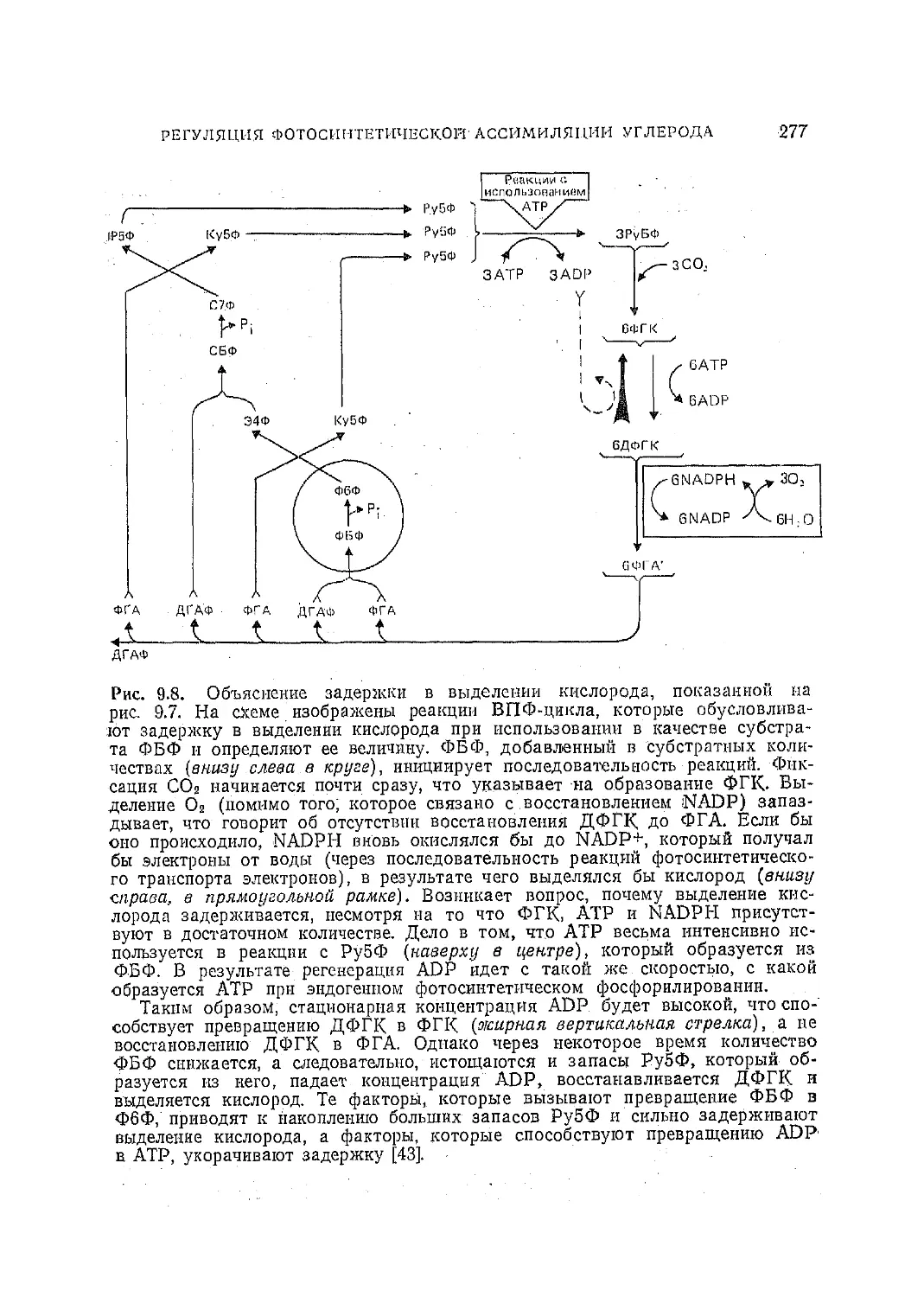

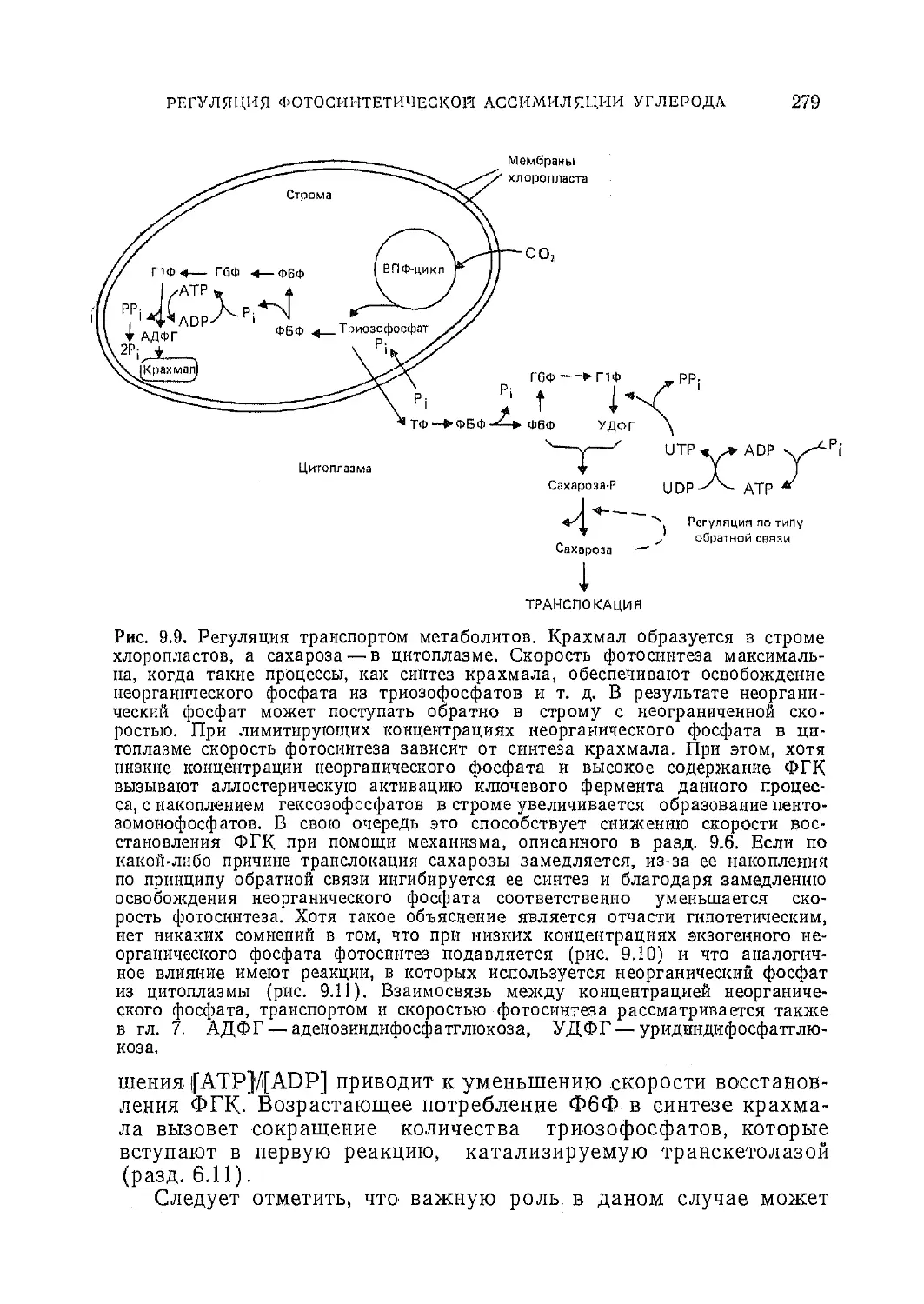
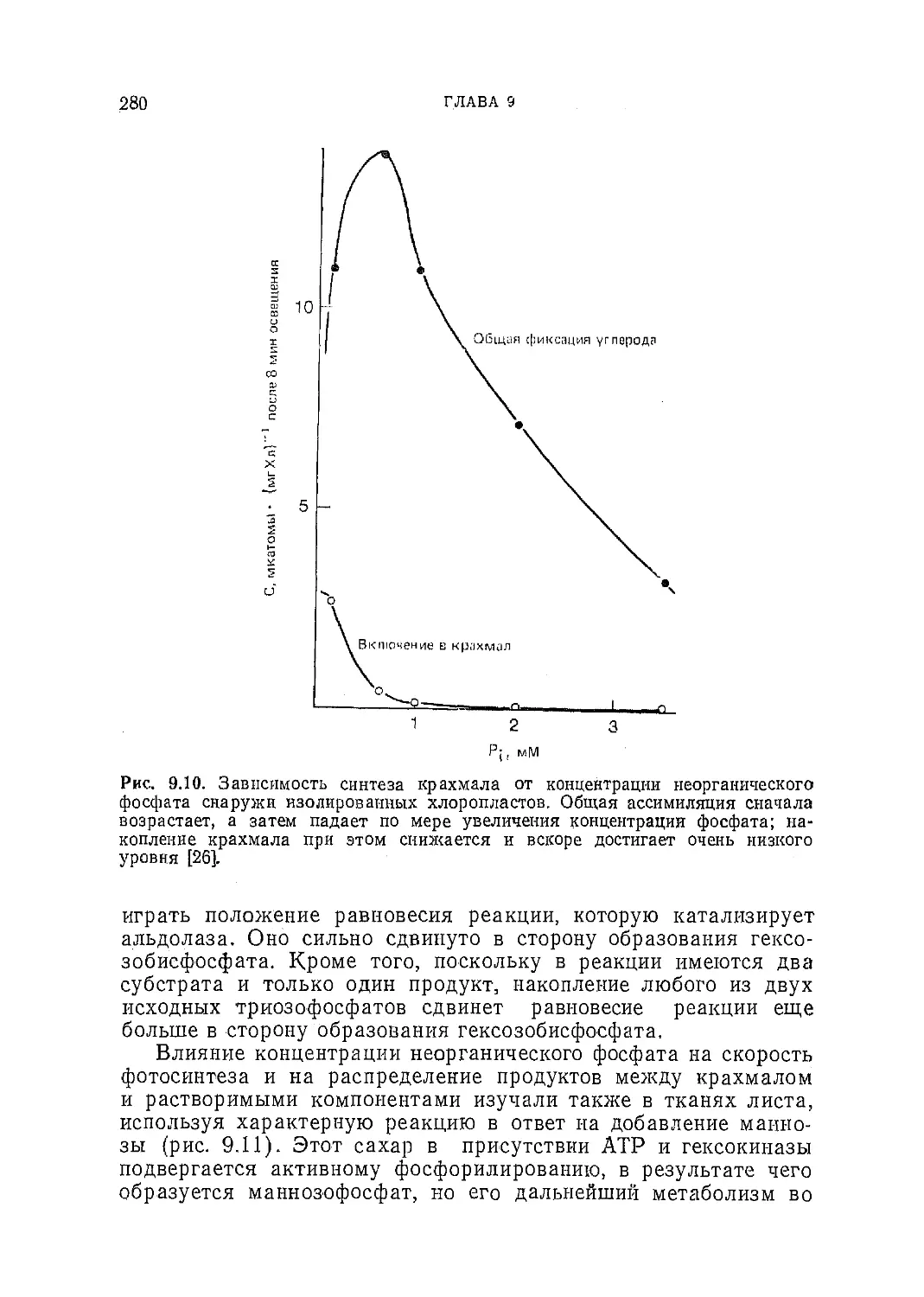
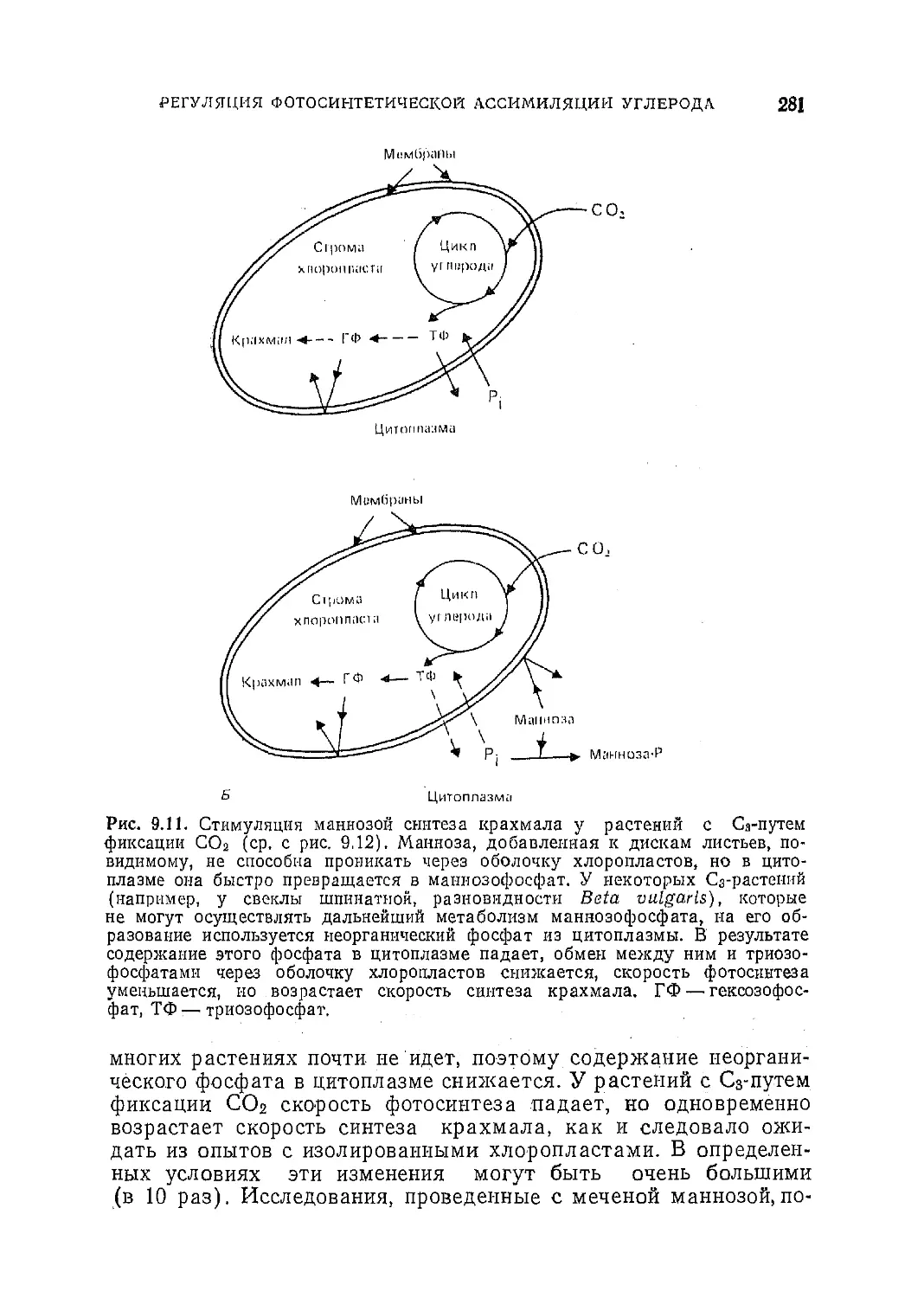
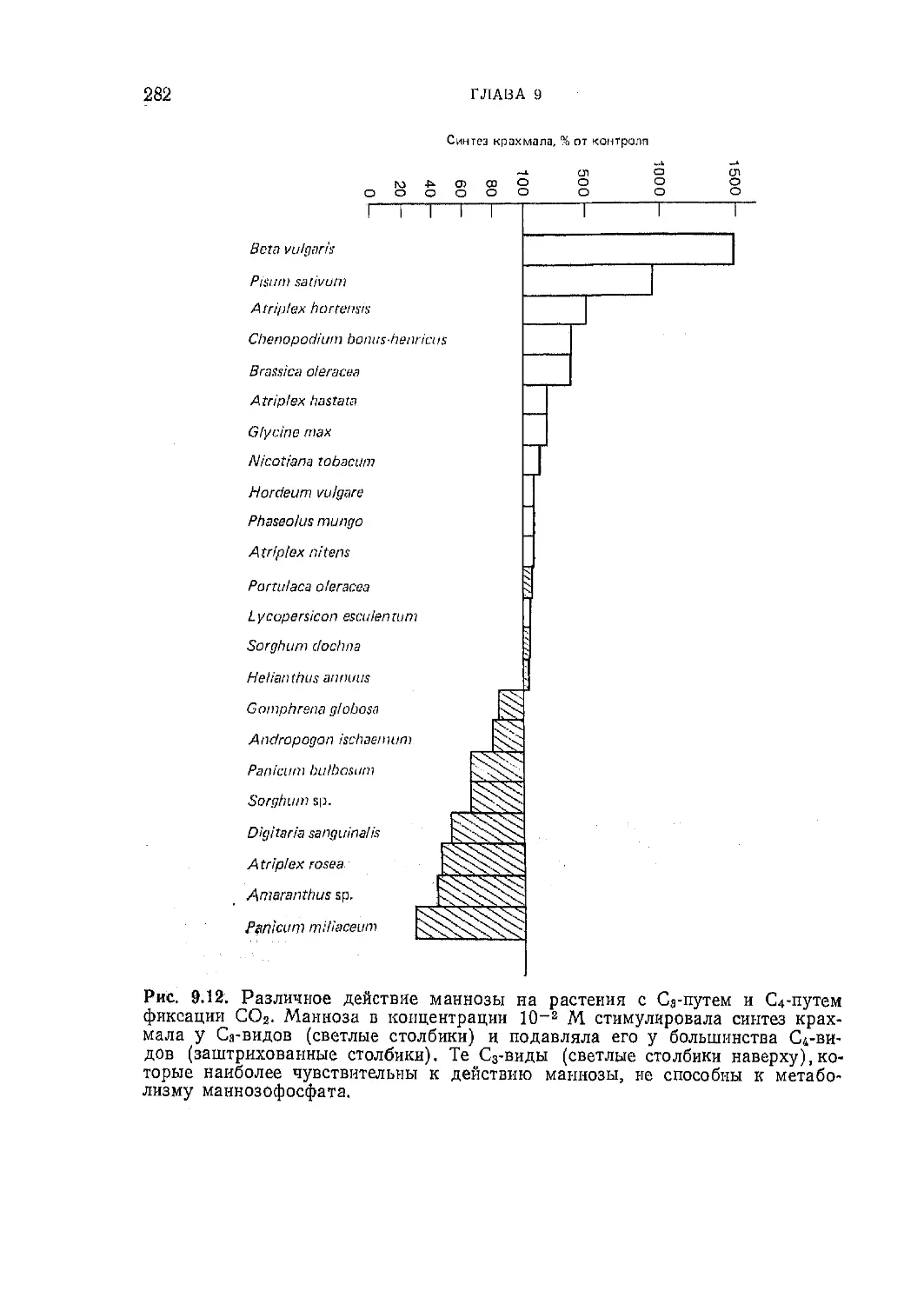




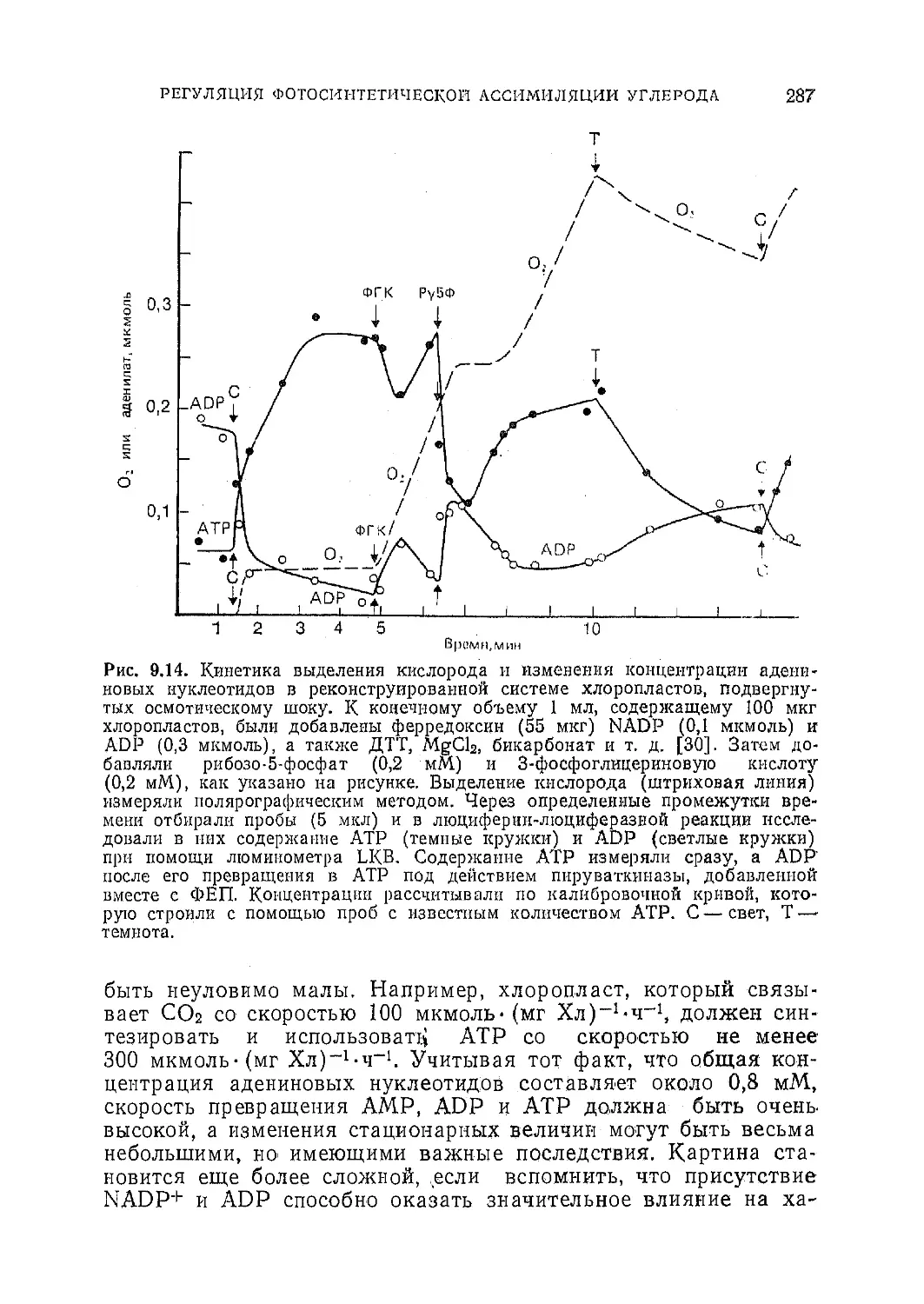









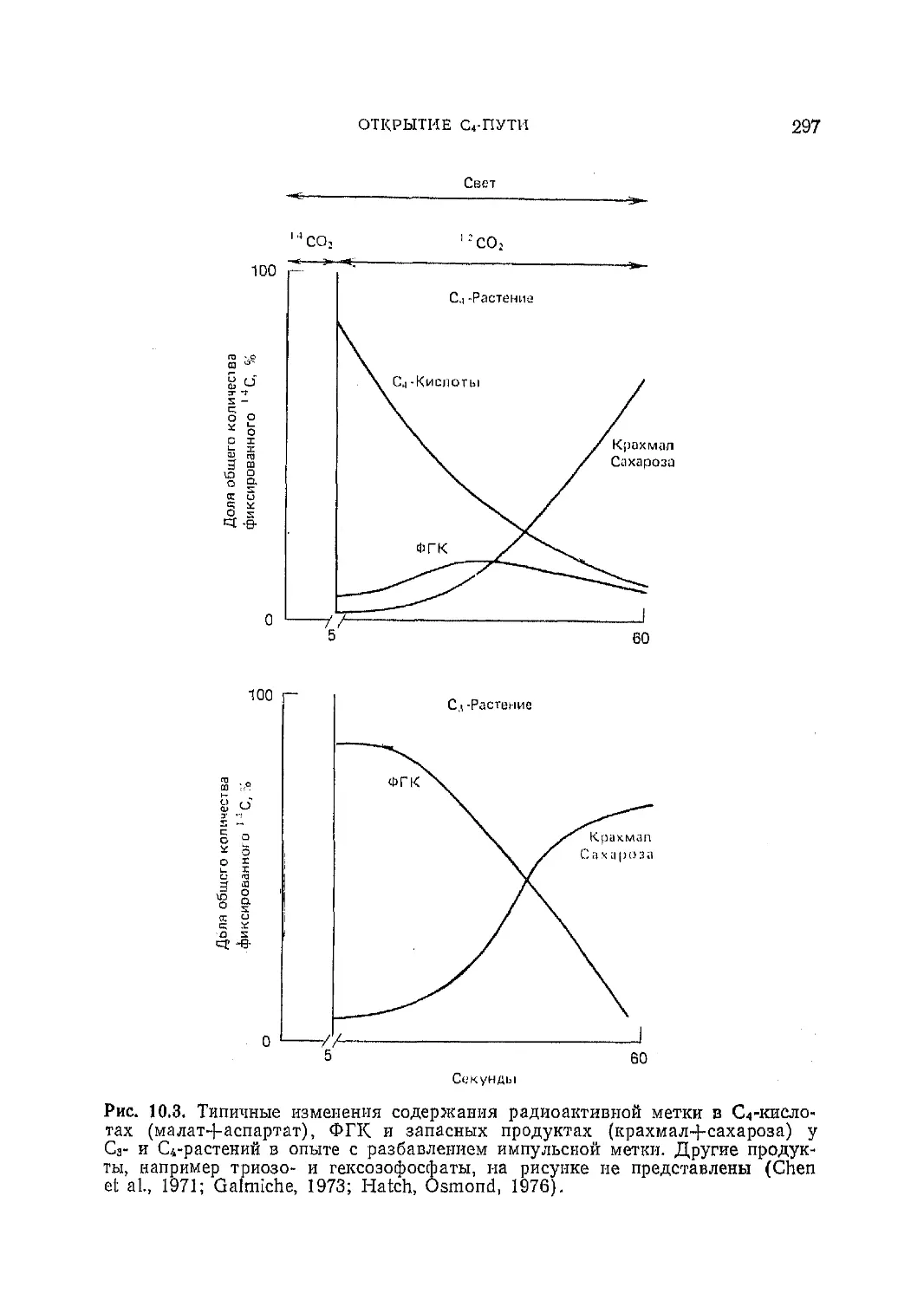








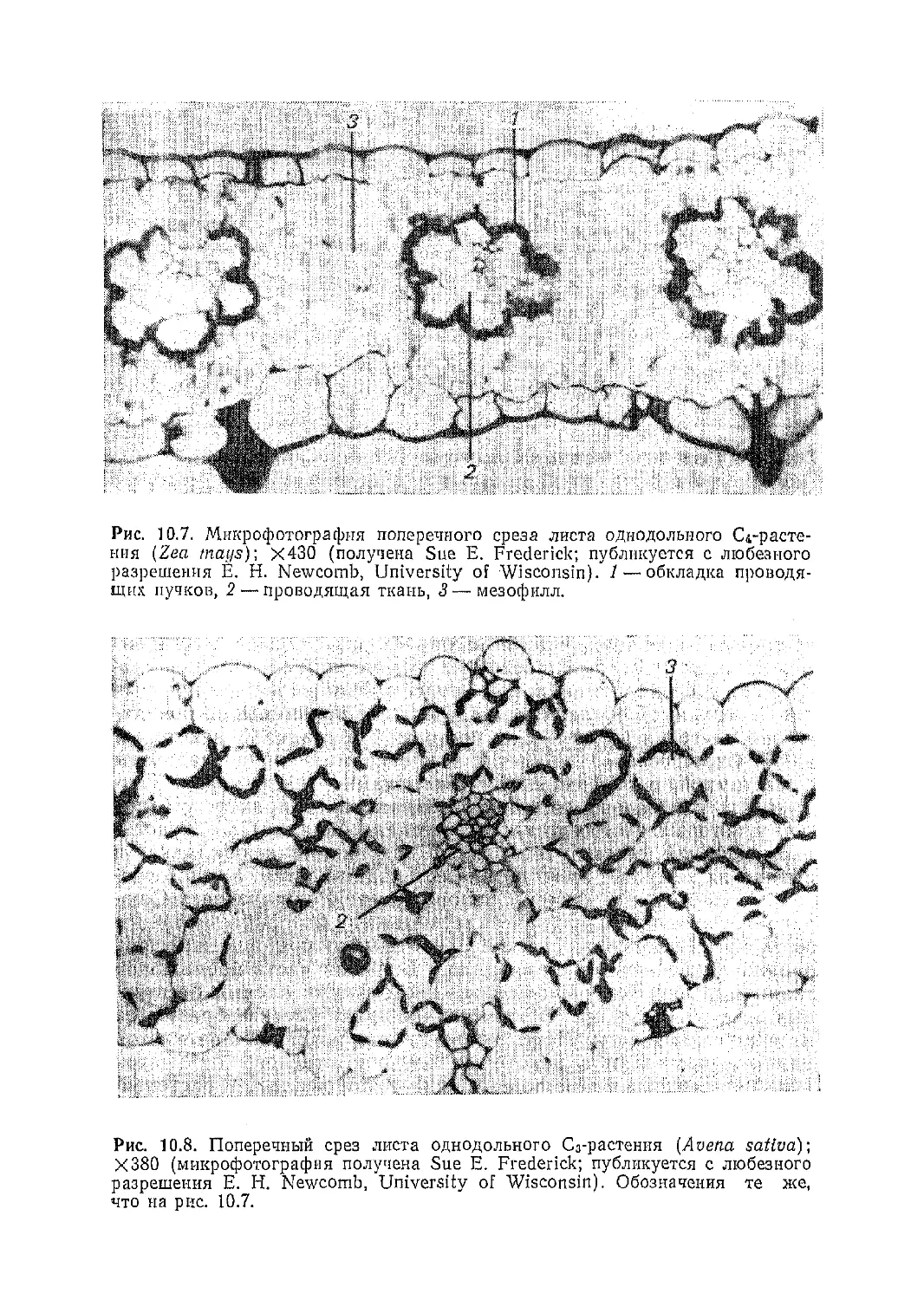
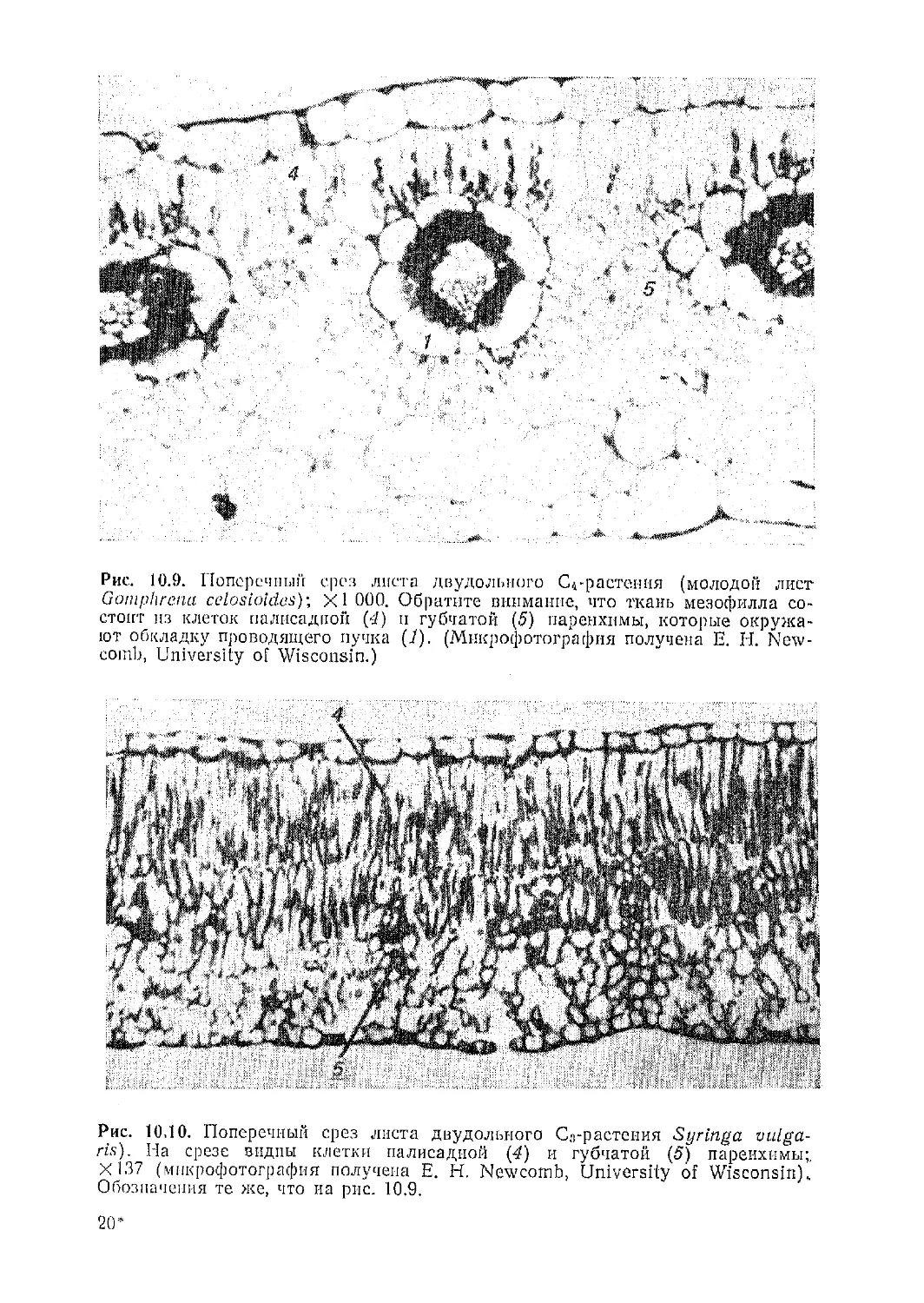
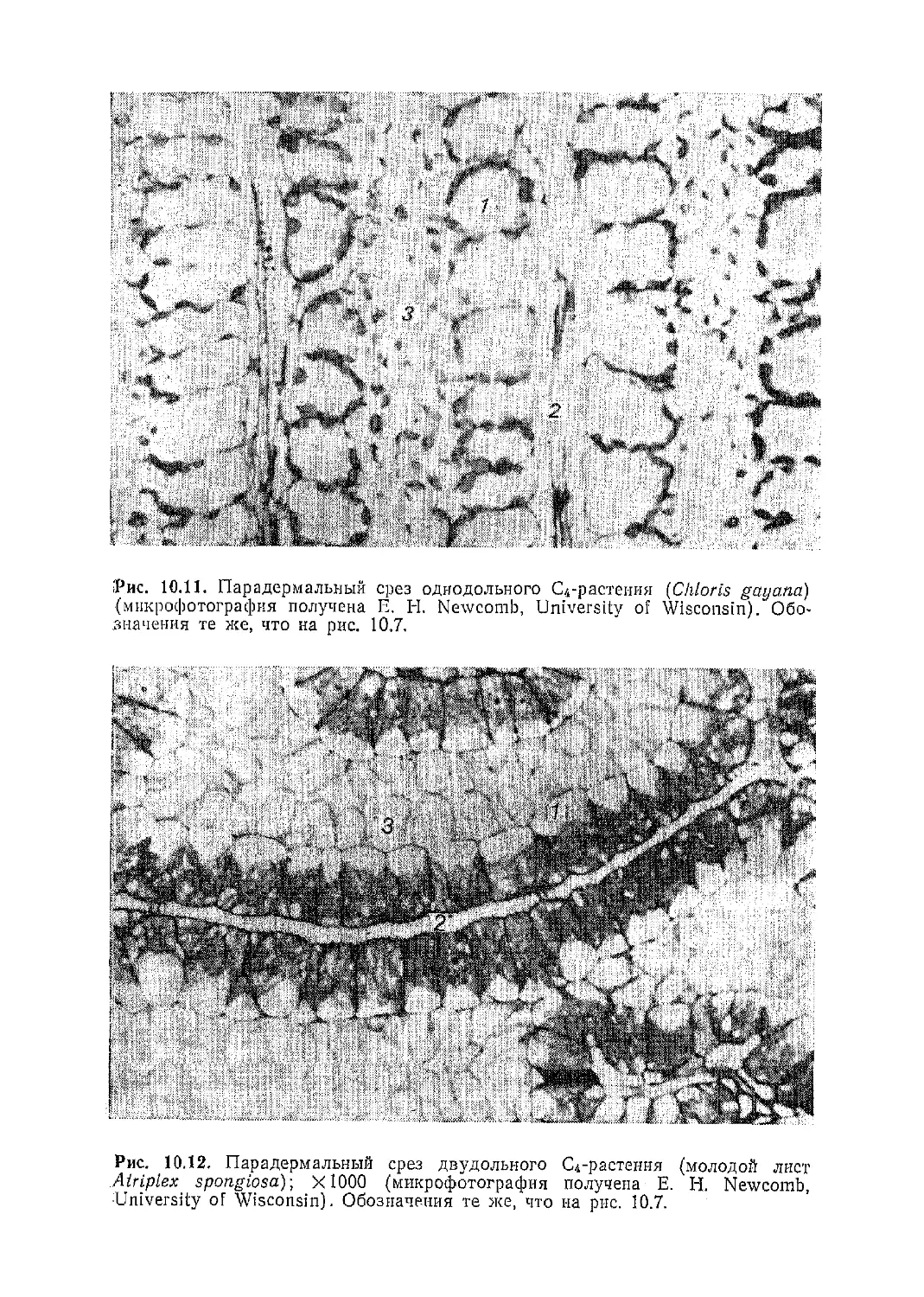


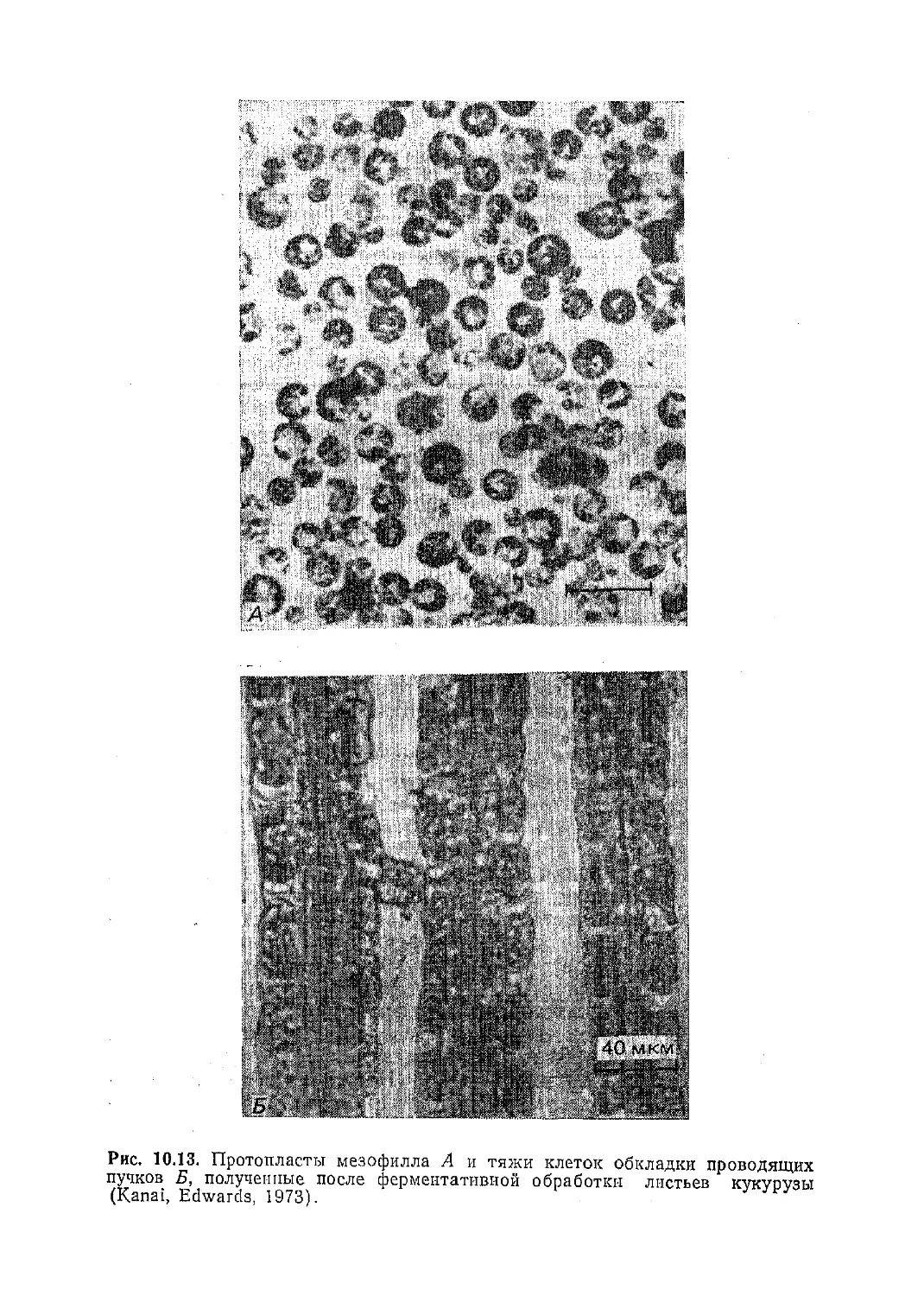















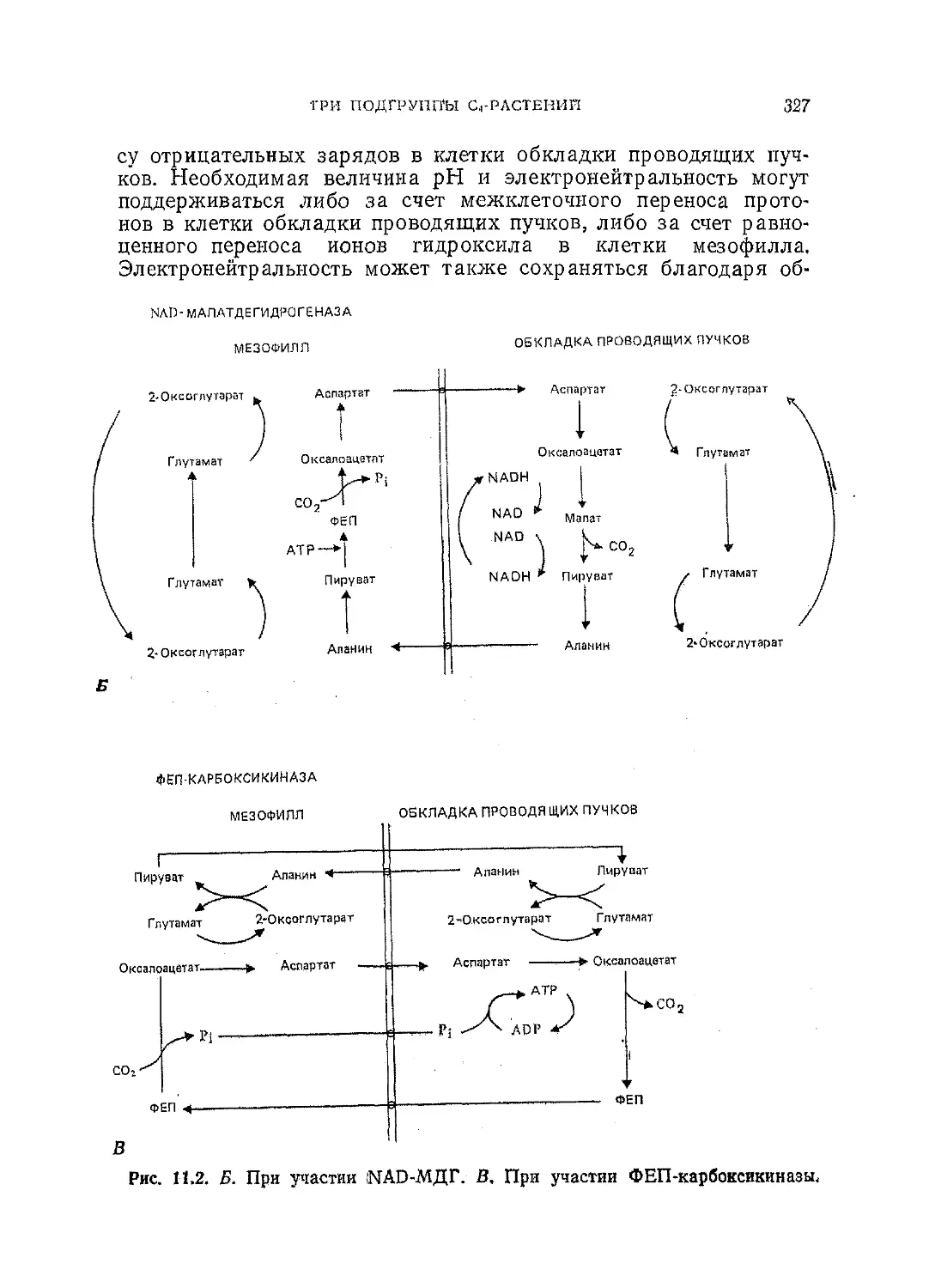








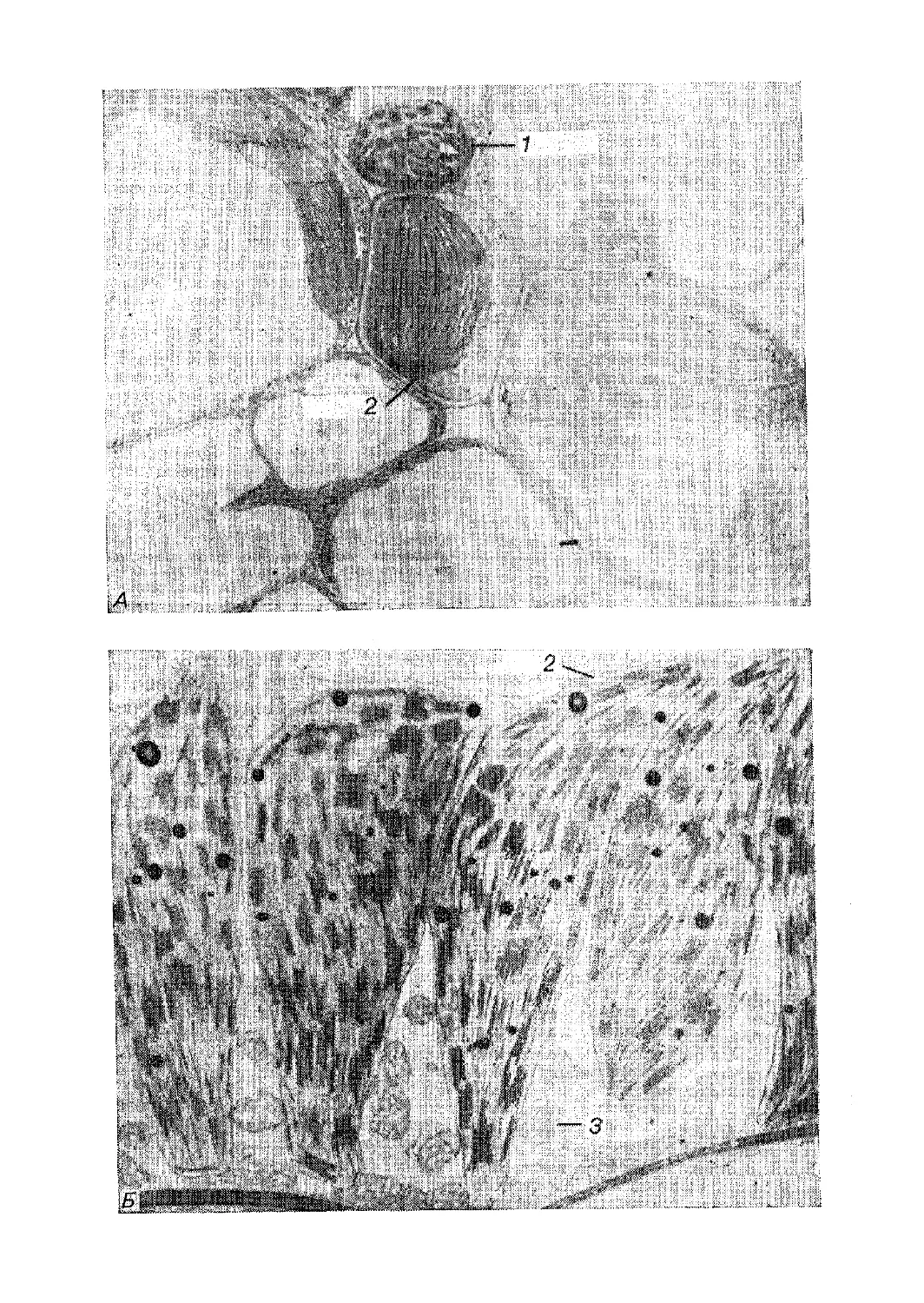

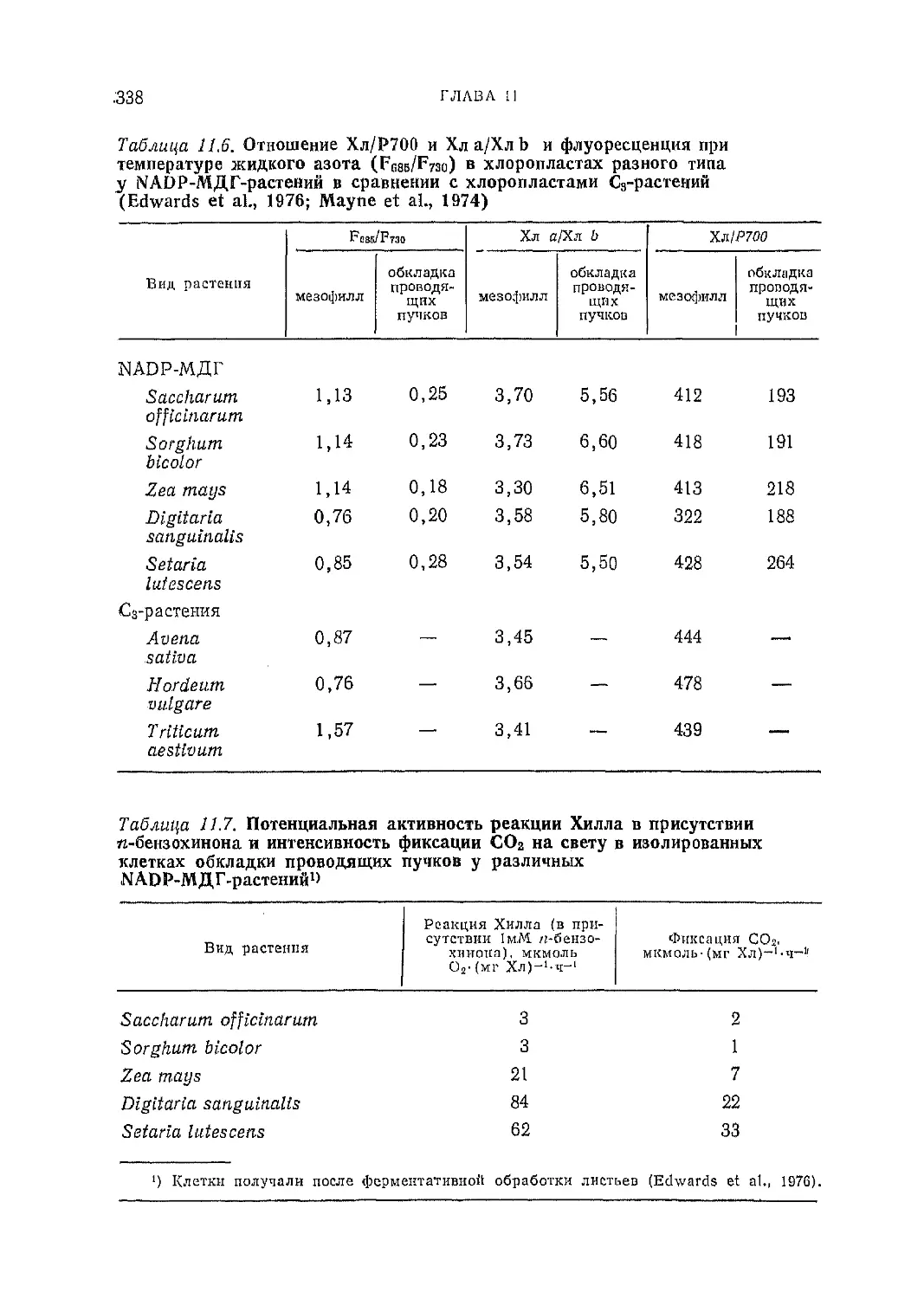


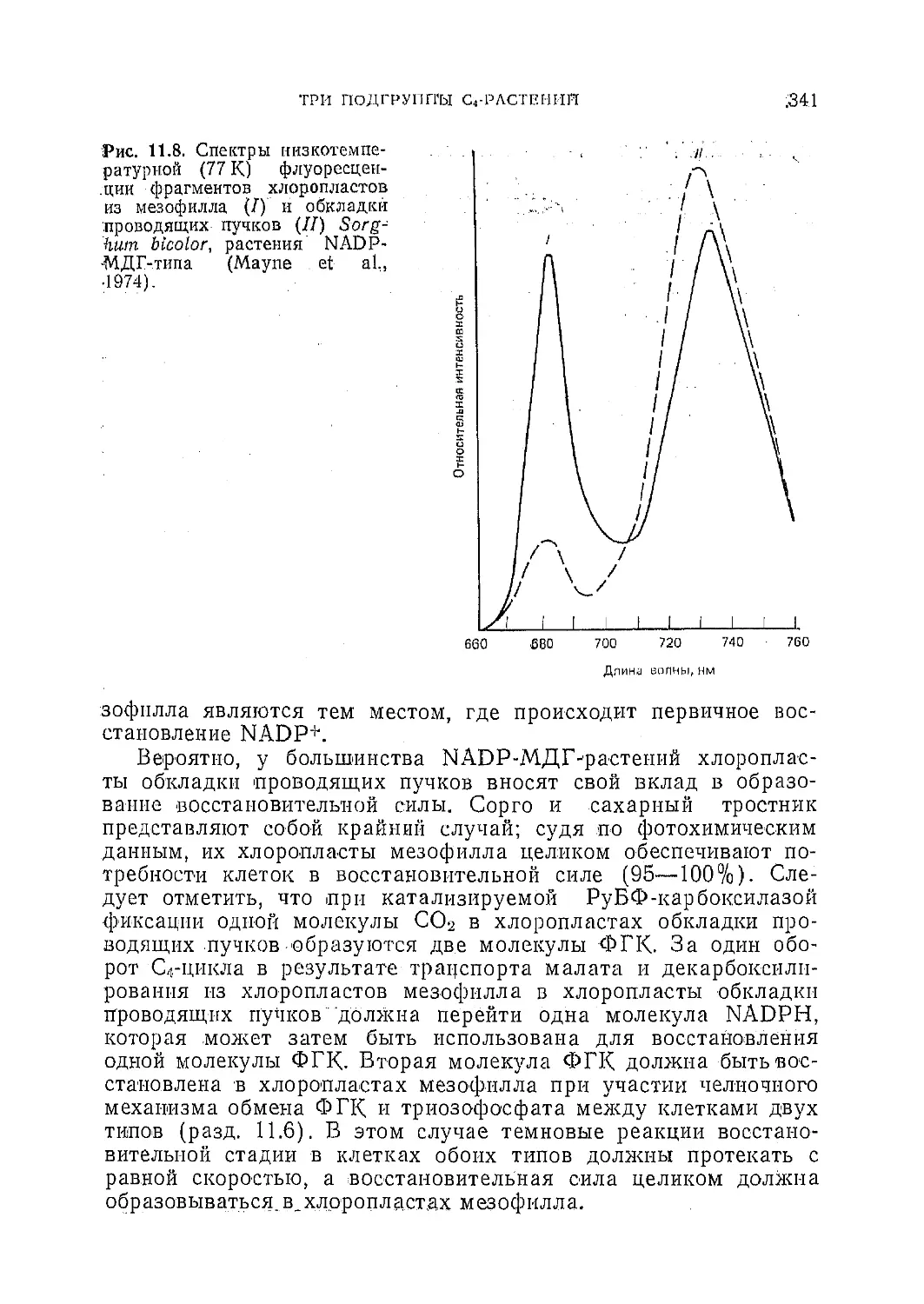














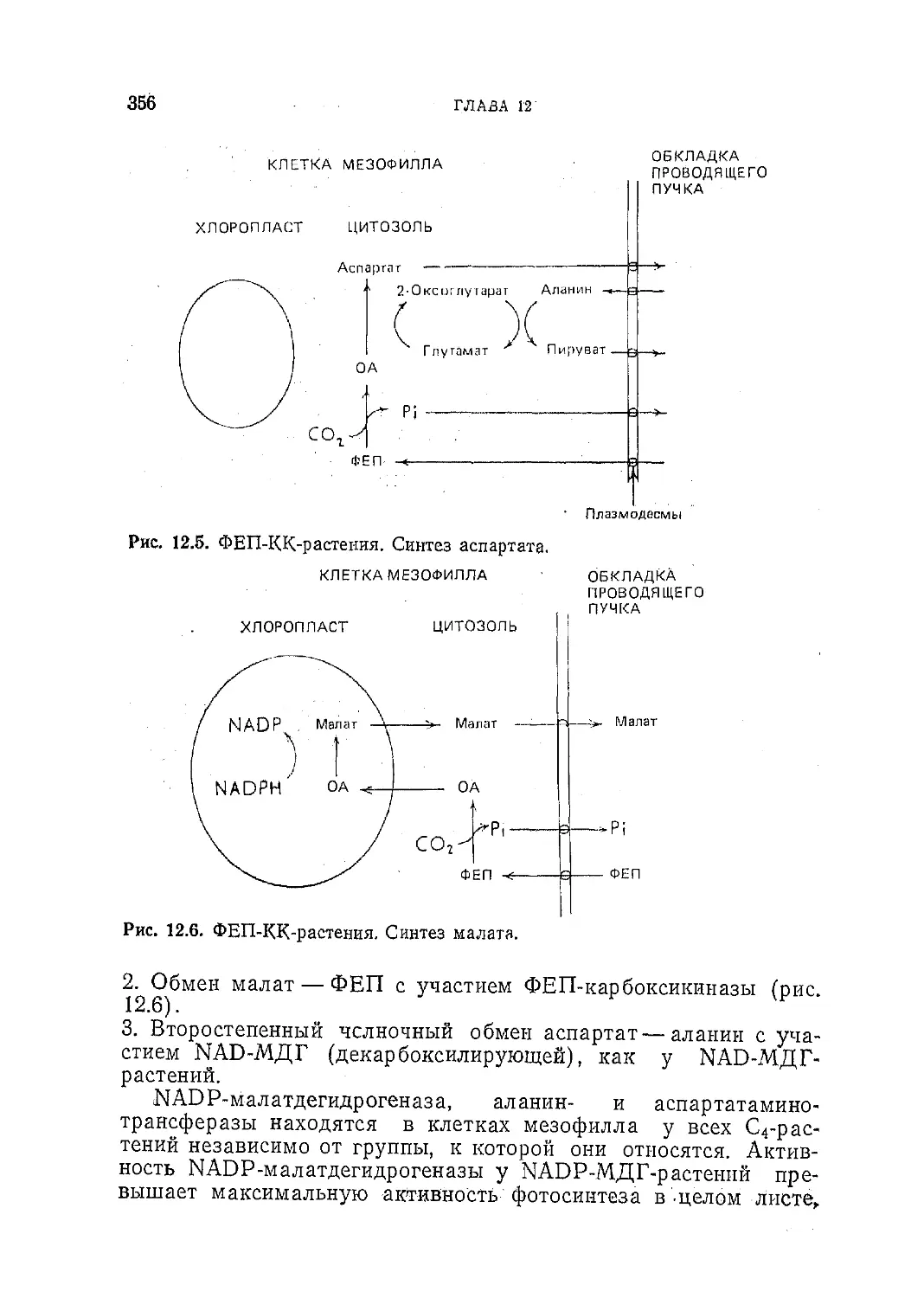

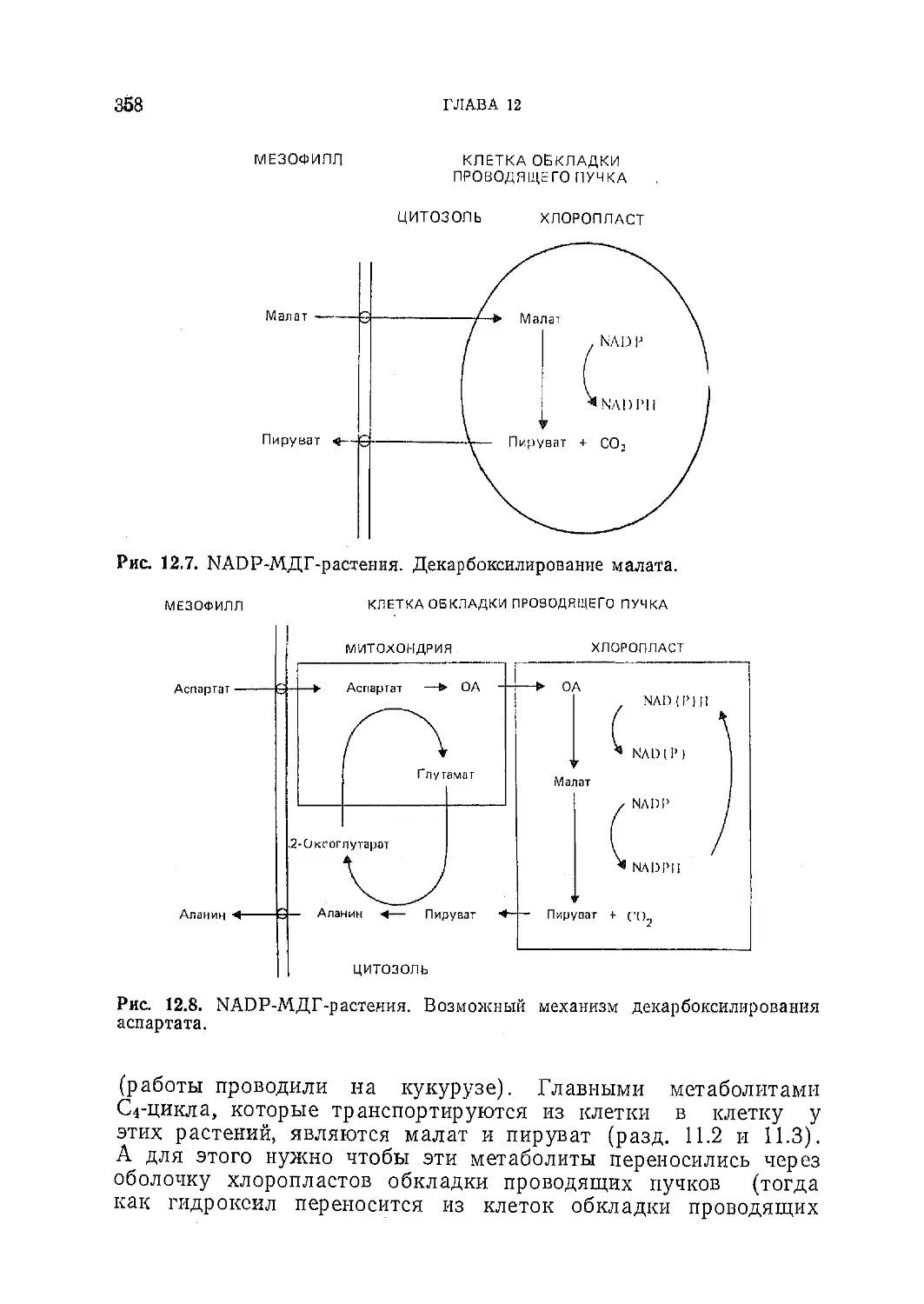

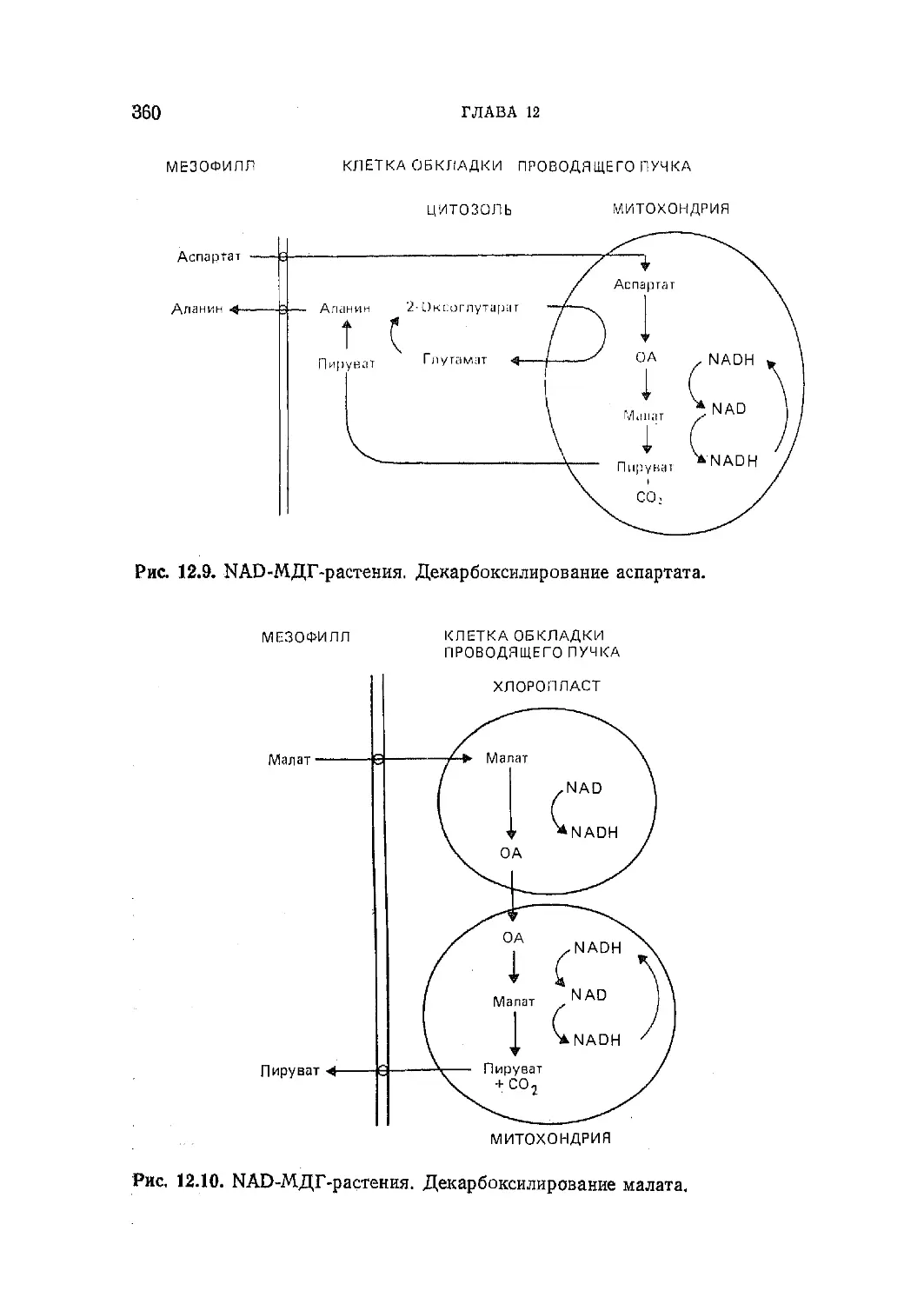

























































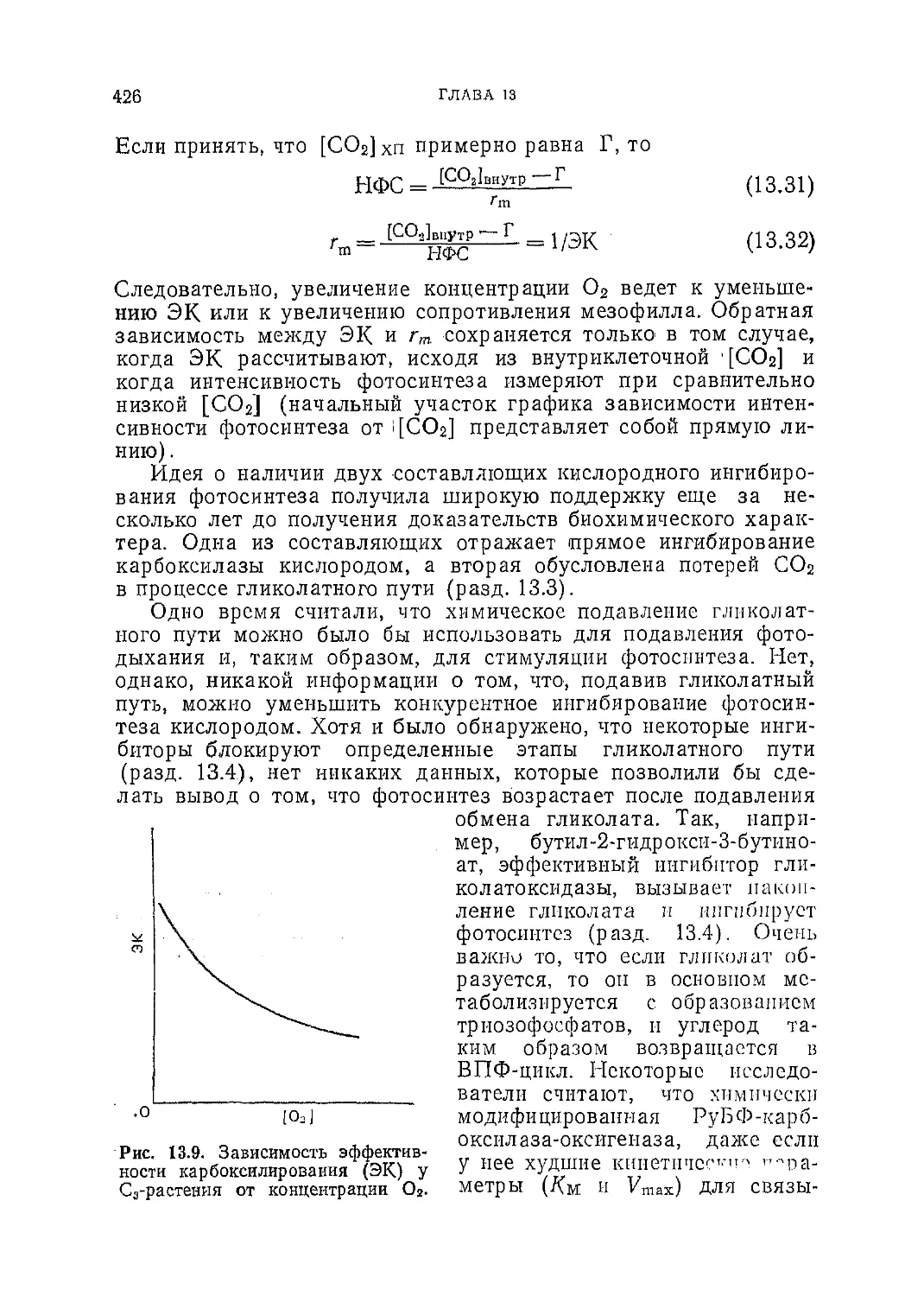





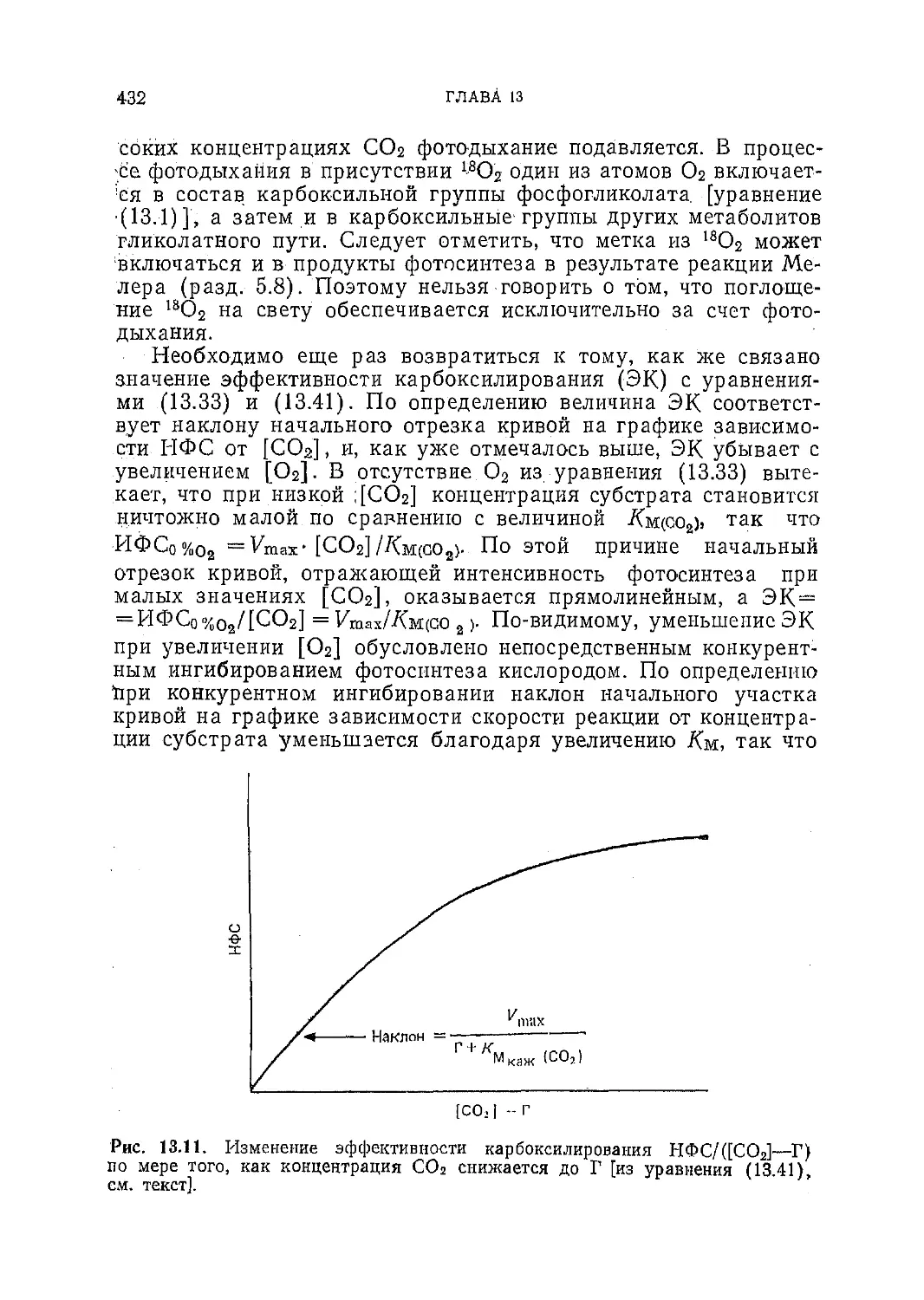
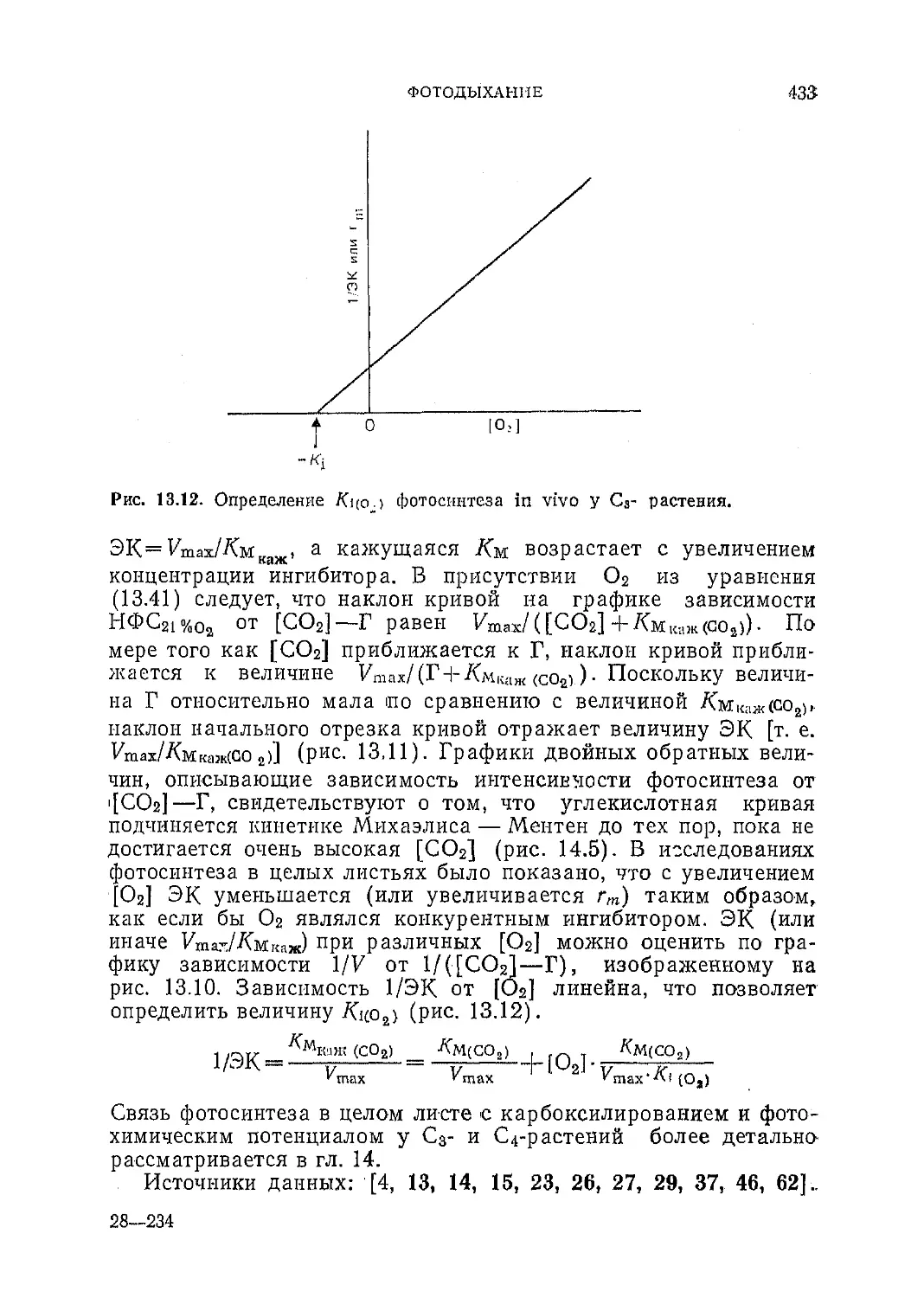

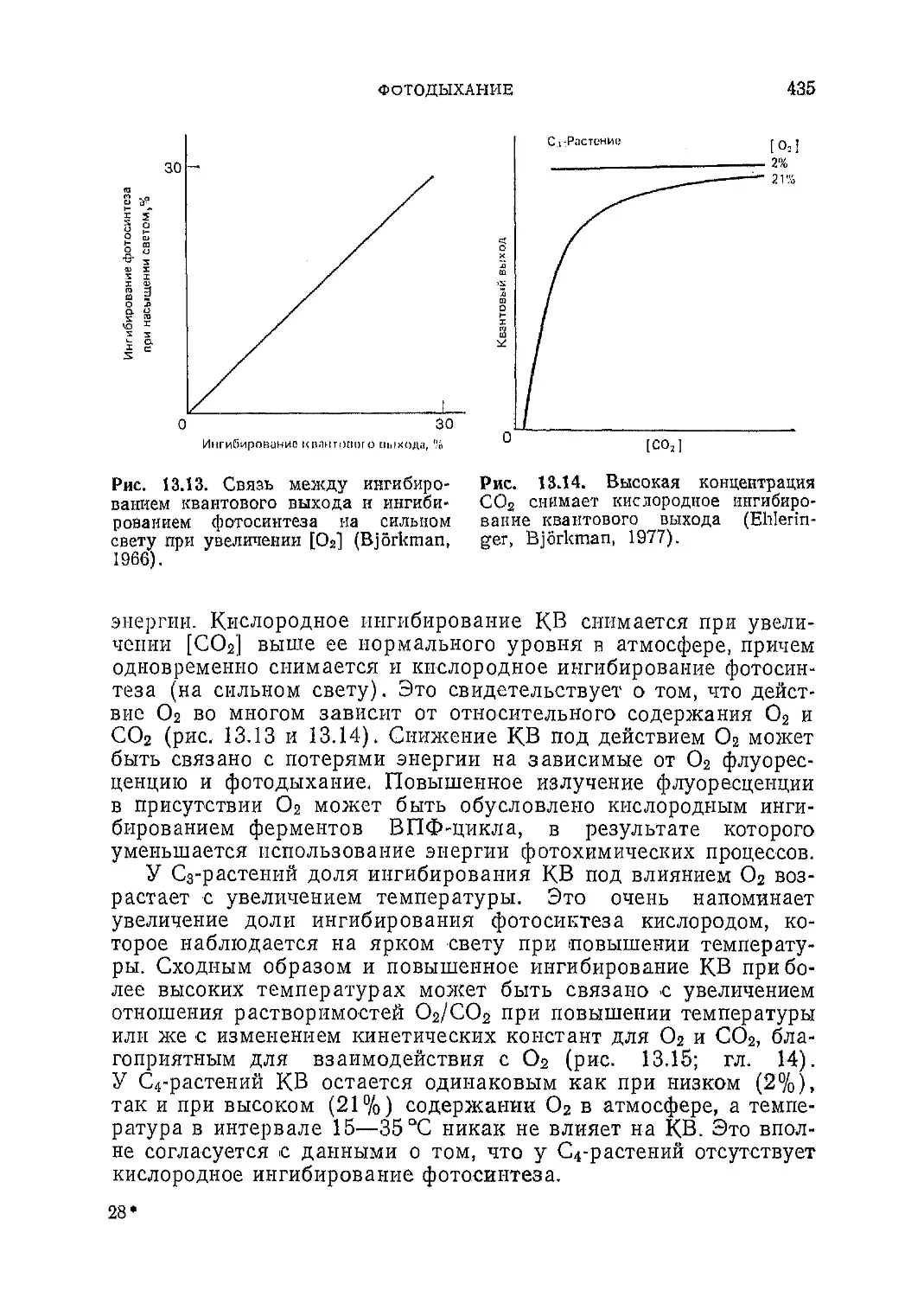






























































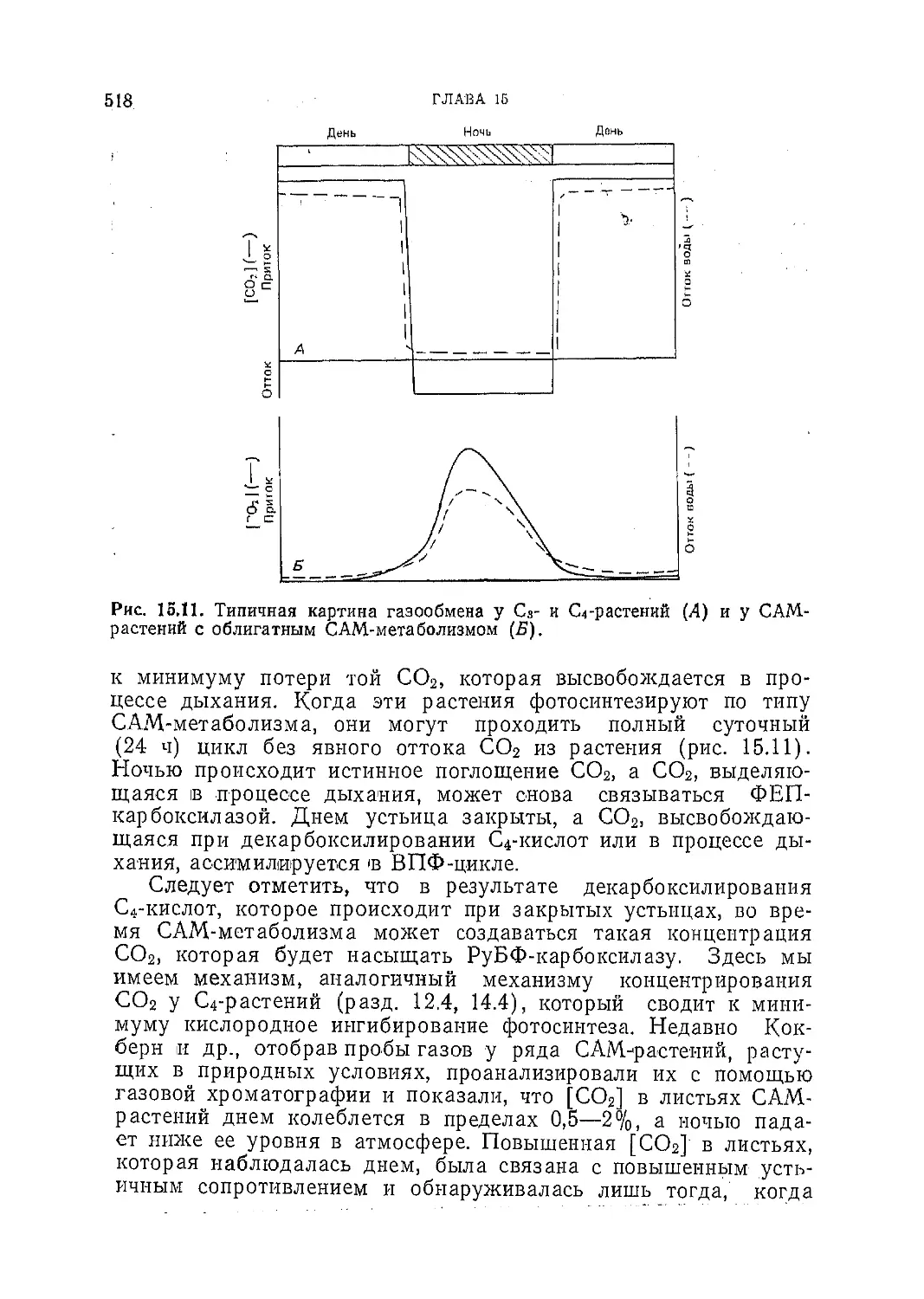














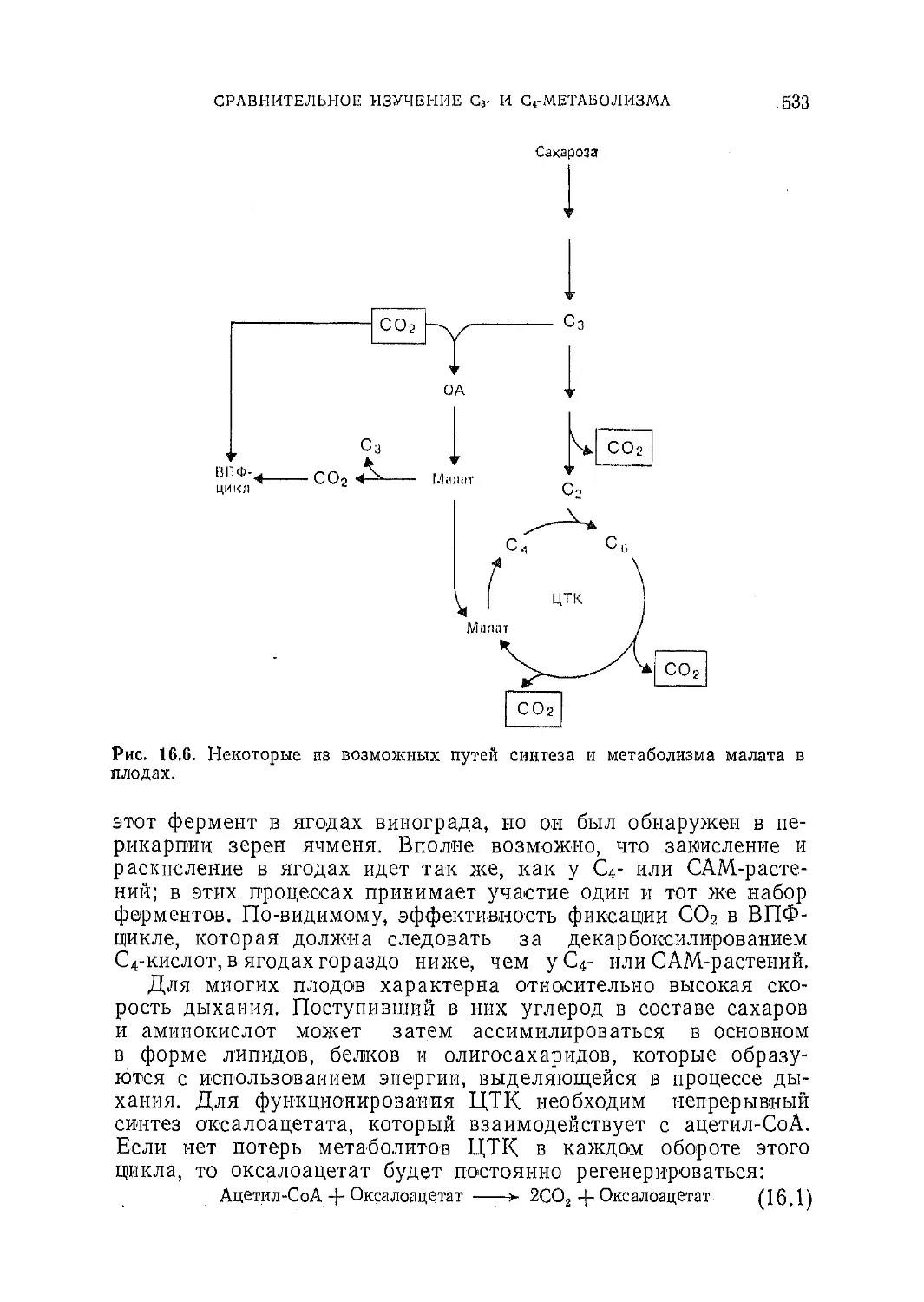

























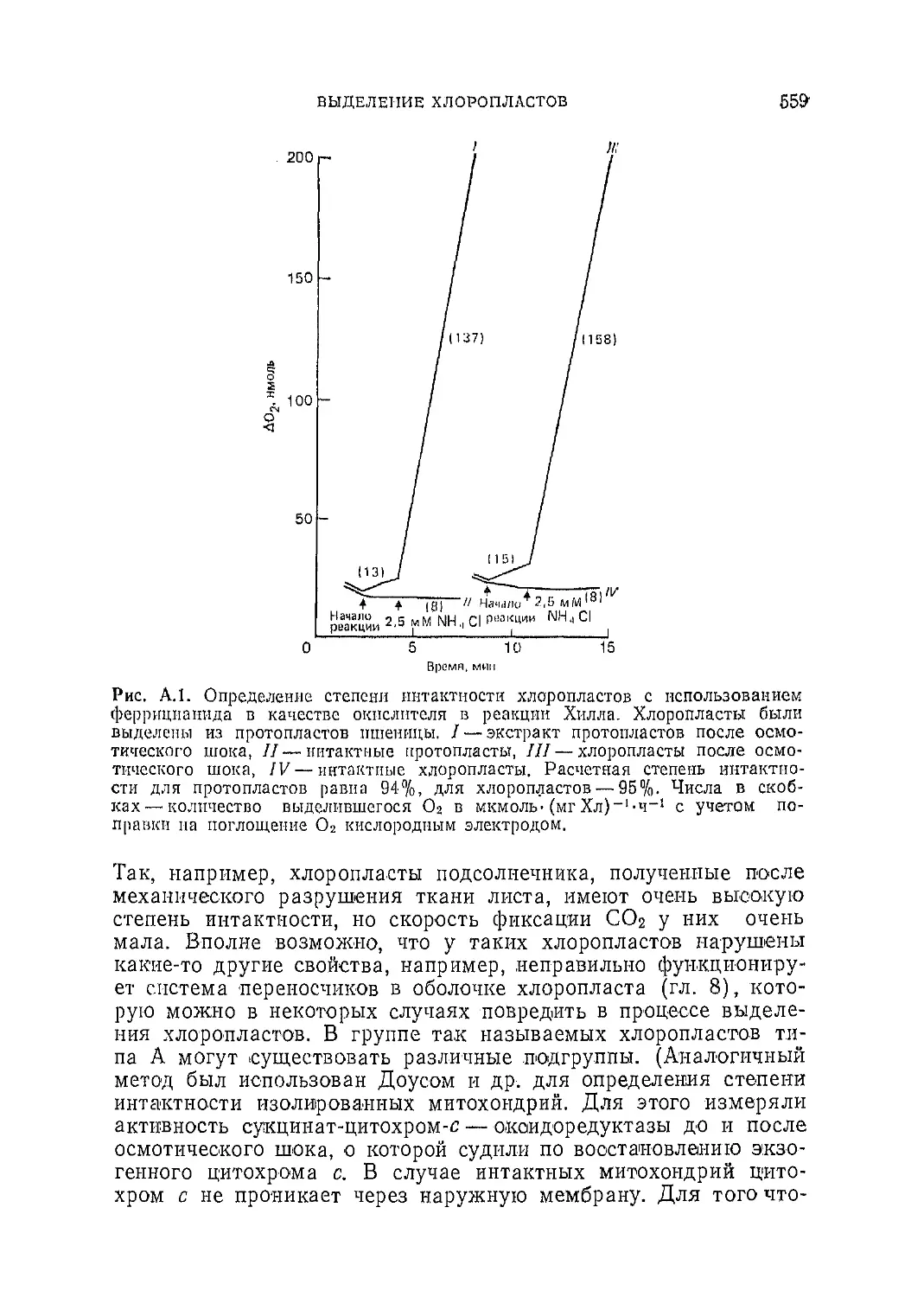













































Текст
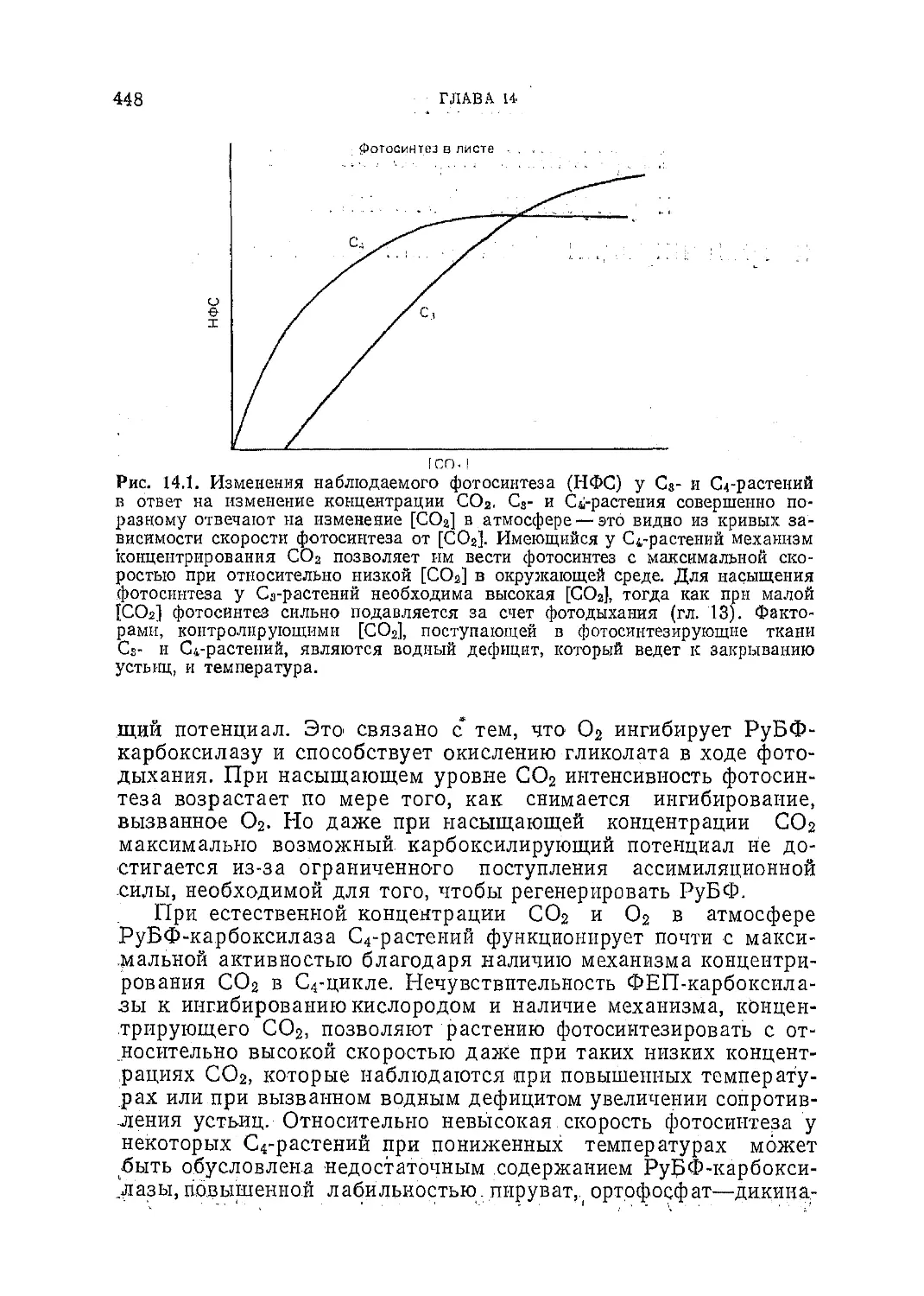
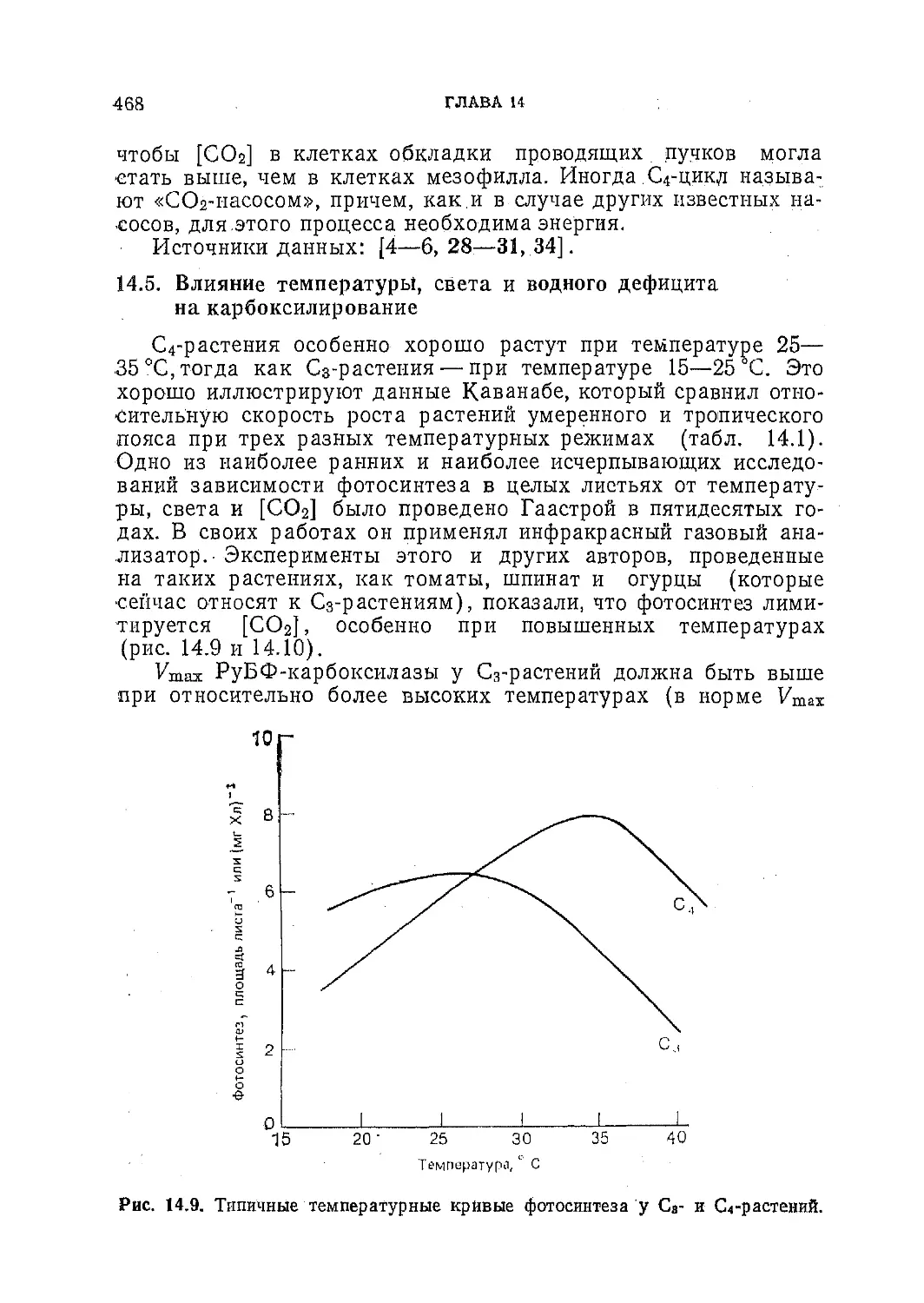
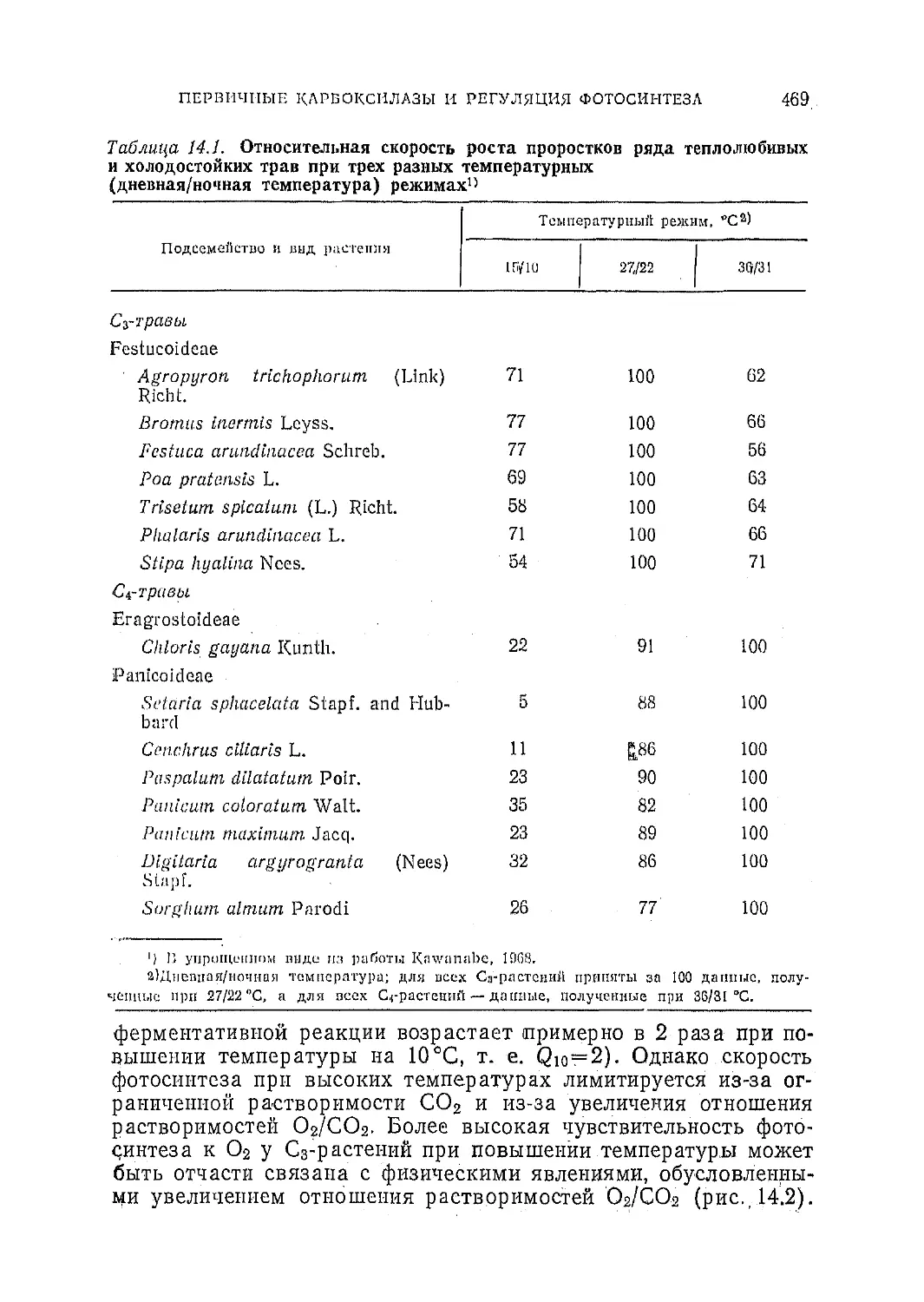
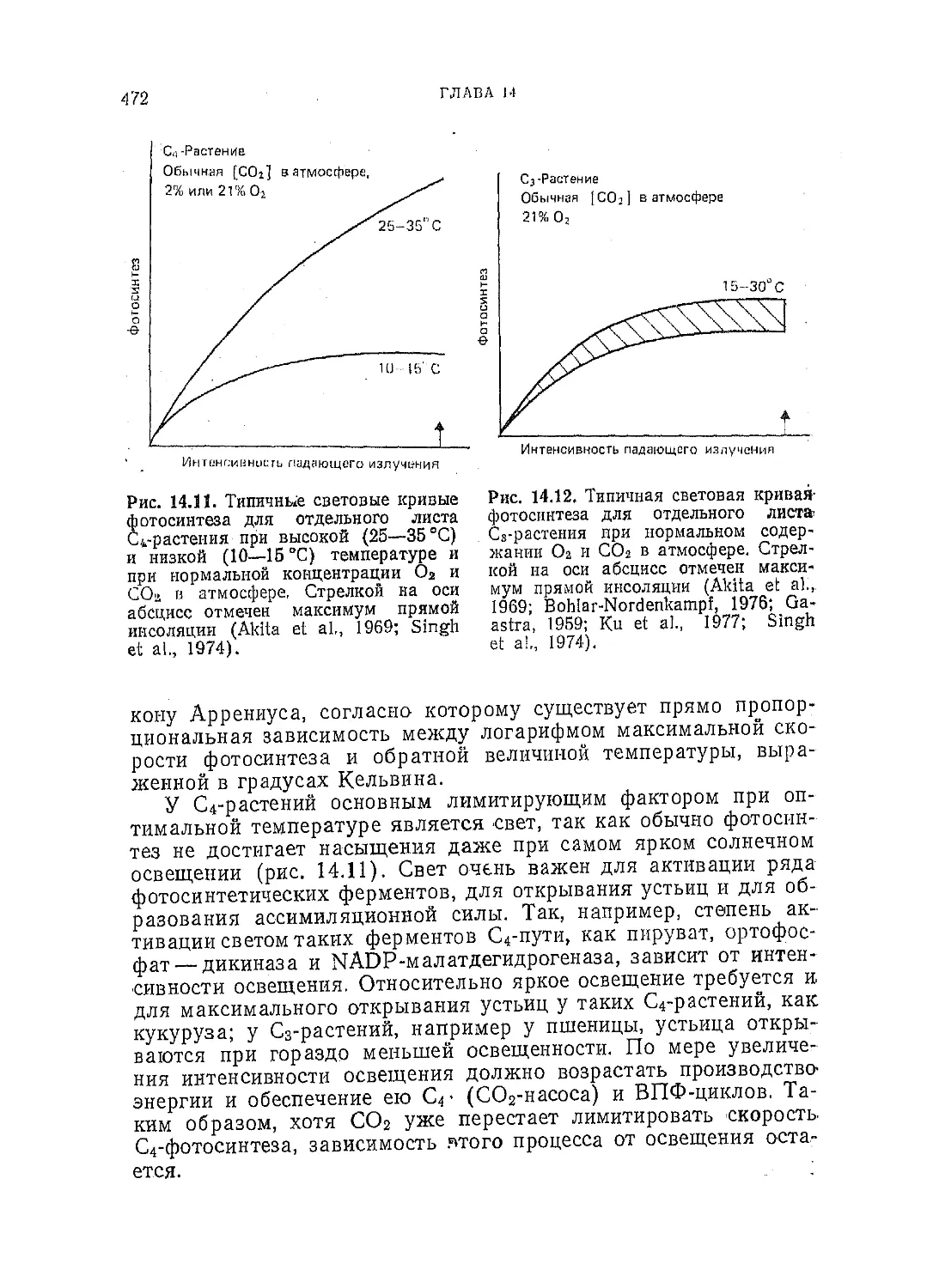
Фотосинтез С3- и С4-р астений:
механизмы и регуляция
С3, С< ; mechanisms,
and cellular and environmental regulation,
of photosynthesis
GERRY EDWARDS
Professor, Department of Botany,
Washington State University
DAVID WALKER
Professor of Biology,
Department of Botany University of Sheffield
BLACKWELL SCIENTIFIC PUBLICATIONS
OXFORD LONDON
EDINBURGH BOSTON MELBOURNE
Дж. Эдвардс
Д. Уокер
Фотосинтез
Сз"
растении:
механизмы
регуляция
Перевод с английского
канд. хим. наук М. И. Верховцевой,
канд. биол. наук Г. Л. Клячко-Гурвич,
канд. биол. наук. В. И. Мельгунова
под редакцией
чл.-корр. АН СССР А. Т. Мокропосова
Москва «Мир» 1986
lib К 28.07
318
УДК 581.132
Эдвардс Дж., Уокер Д.
318 Фотосинтез С»- н Сграетеиий: механизмы и регуляция:
Пер. с англ. — М..: Мир, 1986. — 590 с., пл.
11 книге американского и английского авторов...........извествых снсцналистоя в
оЛласти физиологии п биохимии растений — рассмотрены механизмы фотосинте-
тической ассимиляции углерода, организация фотохимического аттрита хлоро-
пласта. и также факторы, определяющие первичную продуктивность растения;
имеегея приложение с описанием применяемых при изучении i|n>ioeiiirre.in м,-
годон.
Для физиологов и биохимиков растений, биофизикой, пгроиомоа. дли студен-
тов, ueiiupiiiiTOB и преподавателей биологических вузов.
„ 2004000000—397
3 . — 137—86, ч. 1
ББК 28.07
Редакция литературы по биологии
(С) 1988 by Blackwell Scientific Publications
© перевод на русский язык, «Мир», 1986
1986
Предисловие редактора перевода
Издательство «Мир» предлагает вниманию читателей перевод книги из-
вестных исследователей фотосинтеза Дж. Эдвардса и Д. Уокера, посвящен-
ной физико-химической организации и регуляции фотосинтеза у растений с
Сд- и С^-типами метаболизма.
Как известно, уникальная биосферная роль фотосинтеза, определяющая
космическую роль зеленого растения и его вклад в создание жизненных ре-
сурсов человечества давно привлекает внимание исследователей. Начиная с
40-х годов работы в области физико-химических основ фотосинтеза привели
к столь значительным успехам, что они превзошли по значению все ранее
известное о природе этого замечательного явления.
Основные результаты послевоенных исследований фотосинтеза относят-;
ся к нескольким направлениям: 1. Исследована структура фотосинтетическо-
го аппарата иа субклеточном, мембранном и молекулярном уровнях. 2. Изу-
чены механизмы трансформации энергии света в хлоропластах, ведущие к
эиергизации мембран п накоплению ассимиляционной силы — АТР и
NADPHS. 3. Открыт фотосинтетический пеитозофосфатный цикл как основа
фиксации и восстановления СО;> при фотосинтезе. Обнаружены специфиче-
ские механизмы концентрирования и подачи СО2 в фотосинтетический цикл
у растений с С<- и САМ-тнпами метаболизма, , 4, Открыт геном хлороплас-
тов и исследовано его взаимодействие с геномом ядра, составляющее основу
генетической регуляции фотоавтотрофной эукариотической клетки.
Если к этому добавить еще работы в области физиологии и экологии
фотосинтеза, разработку теории фотосинтетической продуктивности фитоце-
нозов, создание высокопродуктивных биотехнологических систем с использо-
ванием фототрофных организмов, то станет очевидным, что теория фотосин-
теза сформировалась как одна из самых значительных проблем естествозна-
ния XX века.
Появление па русском языке книги Эдвардса и Уокера существенно до-
полнит отечественную литературу по фотосинтезу. Эта обширная сводка,
включившая в себя как хорошо обоснованные, ставшие хрестоматийными,
результаты исследований, так н новейшие факты и гипотезы, еще не про-
шедшие испытание временем. Основу книги составляют материалы о первич-
ных механизмах трансформации энергии в хлоропластах и о фотосинтети-
ческом метаболизме углерода. Вместе с тем, особенно в последних главах,
авторы рассматривают многие эколого-физиологические аспекты фотосинтеза,
Можно не сомневаться, что книга вызовет живой интерес у всех, кто
изучает фотосинтез. Исследователи этого процесса найдут в ней богатый
справочный материал, а для студентов и аспирантов монография послужит
первоклассным учебным пособием.
Представляя книгу читателю, отметим удачную ее композицию, которая
позволила авторам обобщить и аранжировать в логической последователь-
ности огромный экспериментальный и теоретический материал. Первая часть
книги (гл. J—5) посвящена энергетике фотосинтеза и включает общие све-
дения о законах термодинамики, о природе света, о первичных фотофизиче-
ских и фотохимических реакциях хлорофилла и т. п. В двух главах пред-
ставлены молекулярная организация фотосинтетического аппарата, функцио-
нирование двух фотосистем и процессы фотофосфорилирования (накопление
ассимиляционной силы) при циклическом и нециклическом транспорте элек-
тронов. Читателю, который пожелает более широко познакомиться с энер-
гетикой фотосинтеза, представленной в книге Эдвардса и Уокера довольно
6
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
лаконично, можно рекомендовать монографию Л. Н. Белла «Энергетика фо-
тосинтезирующей растительной клетки» (М., Наука, 1980).
Вторая, основная часть книги (гл. 6—16) представляет новейшую свод-
ку по организации и регуляции фотосинтетического метаболизма углерода.
Дается детальный анализ восстановительного пентозофосфатиого цикла
(ВПФ-цикла, или цикла Бенсона — Кальвина). Рассмотрены механизмы регу-
ляции ВПФ-цикла с помощью активации ферментов, изменения концентрации
или транспорта метаболитов в системе хлоропласт — цитоплазма. Представ-
лены данные о биохимии и энергетике фотодыхания. Пожалуй, впервые пос-
ле известной монографии Е. Рабиновича «Фотосинтез», Эдвардс и Уокер вве-
ли в свою книгу главу об индукционных явлениях фотосинтеза в переход-
ных условиях свет —темнота или темнота — свет. Значительную часть книги
составляет материал о фотосинтезе растений, имеющих С$- или САМ-типы
метаболизма. Хотя число видов с Ci-фотосинтезом, по-видимому, не превы-
шает 1—2% общего числа видов высших растений, интерес к ним все рас-
тет. Во-первых, Ci-тип метаболизма отражает одно из основных направле-
ний эволюции фотосинтеза в новейшее время, когда увеличивается арндпость
и ксерофильность климата и установилось новое соотношение СО2 и О2 в
атмосфере; во-вторых, основу фотосинтетического метаболизма у Ct- и
•CAM-растений, так же как и у С3-видов составляет фотосинтетический цикл
Бенсона — Кальвина. На новом этапе эволюции возникли лишь метаболитные
надстройки (ароморфозы, арохимозы), которые обеспечивают фиксацию и
концентрирование СО2 для подачи в цикл Бенсона — Кальвина в условиях вы-
сокой температуры, дефицита воды и низкой концентрации СО2.
Несомненным достоинством биохимических глав книги является стремле-
ние авторов во всех случаях, когда это возможно, показать стехиометрию ре-
акций и определить их энергетический баланс. Хотелось бы также обратить
внимание читателей на композиционную структуру глав; каждая глава на-
чинается с краткой аннотации представленных в ней материалов. Далее в
тексте почти не цитируются литературные источники, но в конце каждой гла-
вы дается перечень наиболее значительных источников. Введена четкая руб-
рикация глав. Такая композиция оптимальна для учебного пособия.
В книге есть ценное дополнение — приложение. В нем приведены мето-
дические указания для получения изолированных интактных хлоропластов с
высокой функциональной активностью. Во второй части приложения приве-
дена номенклатура и основные характеристики ферментов фотосинтетиче-
ского метаболизма.
К сожалению, нужно отметить, что в этой капитальной сводке не нашли
•отражения работы советских исследователей. Работы в области фотосинтеза,
начиная с исследований К. А. Тимирязева, являются традиционными для
отечественной науки, и с именами Т. Н. Годнева, А. А. Красновского, А. А. Ни-
чипоровича, О. В. Заленского, Н. П. Воскресенской и других связаны крупные
достижения в изучении фотосинтеза.
В заключение хочу с благодарностью отметить добросовестный и квали-
фицированный труд переводчиков. Справедливость требует упомянуть о том,
что многие разделы оригинала представляли серьзную трудность для пере-
вода иа русский язык. Гл. 1—5 переведены Г. Л. Клячко-Гурвич, гл. 6—9 —
М. И. Верховцевой, а гл. 10—16, приложение А и Б —В. И. Мельгуновым.
А. Т. Мокроносов
Предисловие
Эта книга посвящена рассмотрению основных аспектов фиксации угле-
рода в процессе фотосинтеза у Са- и С4-видов, а также у растений с мета-
болизмом кислот по типу толстянковых (САМ). Книга рассчитана на сту-
дентов университетов и на специалистов — физиологов и биохимиков расте-
ний. Она содержит подробное описание фотохимического аппарата и фото-
химических реакций, в результате которых генерируется ассимиляционная
сила (ATP+NADPH) [Часть А, гл. 1—5].
В части Б обсуждаются важнейшие проблемы фотосинтетической асси-
миляции углерода, в том числе ряд вопросов, которые были изучены за по-
следнее десятилетне и поэтому не получили достаточно полного освещения
во многих современных изданиях. Например, в гл. 6—9, в которых говорится
о Са-пути фотосинтеза, рассмотрены не только основные черты восстанови-
тельного пеитозофосфатиого цикла, ио и способы его регуляции, а также
взаимосвязь с транспортом метаболитов между хлоропластами и цитоплаз-
мой. В гл. 13 подробно изложены процессы фотодыхания, а в гл. 10—12 —
относительно новая проблема: фотосинтез у С4-растений, где читатель найдет
описание основных этапов функционирования С4-цикла, компартментации и
регуляции ферментов, транспорта метаболитов, фотохимических реакций и
их потребностей в АТР и NADPH, таксономических различий и физиологи-
ческих эффектов.
Обсуждение метаболизма кислот по типу толстянковых в гл. 15 сосре-
доточено па истории его открытия, а также па биохимических и экологиче-
ских аспектах САМ. Гл. 16 касается фотосинтеза в органах размножения и
различных путей метаболизма малата у растений. В приложении А описаны
методики выделения хлоропластов, а в приложении Б дана номенклатура
ферментов, участвующих в С3-, С4-метаболизме и CAM-путях ассимиляции
углерода при фотосинтезе.
Мы изложили пашу точку зрения на современное состояние проблемы
фотосинтетической ассимиляции углерода. Эту книгу ие следует рассматри-
вать как обзор, так как обилие литературы, имеющейся по этому вопросу,
делает подобную задачу вообще непосильной. Как правило, после каждого
раздела внутри главы приведены номера ссылок, в самом же тексте мы ста-
рались их нс давать. В конце каждой главы читатель найдет список реко-
мендуемой литературы, который позволит ему получить более подробную ин-
формацию по тому или иному вопросу. Номера ссылок на узкоспециальную'
литературу в конце разделов гл. 6—16 выделены жирным шрифтом.
Мы стремились излагать материал как можно проще и ориентировались
на читателя, располагающего знаниями в области биологии и физики в объ-
еме высшего учебного заведения. Хотя главной темой книги является асси-
миляция СО2 и мы уделили ей основное внимание, наряду с этим подробно
8 ПРЕДИСЛОВИЕ
’рассмотрены многие другие аспекты фотосинтеза и книга может быть ре-
комендована при изучении биохимии данного процесса в широком смысле
этого слова.
Неоценимую помощь при обсуждении и подготовке рукописи нам оказали
Alice Herold и Joyce Foster. Мы также благодарны всем, кто высказал свои
критические замечания и предложения по прочитанным главам: Randy Alberte,
Mordhay Avron, Jim Barber, Dong Doehlert, Christine Foyer, Garry Harris,
Ulrich Heber, Hans Heldt, Robert Hill, Peter Horton, Steve Huber, Maurice Ku,
Richard Lccgood, Joseph Neumann, Barry Osmond, Simon Robinson, Mark
Schmitt, Marly Spalding, Hideaki Usuda, Klaus Winter. Мы хотели бы выразить
признательность Mark Schmitt за подготовку некоторых микрофотографий для
^публикации и Kathryn Oldman, которая перепечатала большую часть рукописи
и вместе с Shirley Walker помогала нам сохранять присутствие духа. Мы бла-
годарны Richard Walker за его оригинальные иллюстрации к части А.
Дж. Эдвардс
Д. Уокер
Часть А
ГЛАВА 1
Введение
1.1. Что такое фотосинтез
Фотосинтез — это процесс, в ходе которого световая энергия
используется для превращения двуокиси углерода в сахара.
Этот процесс часто представляют с помощью уравнения (1.1)'
или сходных уравнений, отражающих те же основные особенно-
сти:
nitv
со2 + н2о--->CZ/20 + 02. (1.1)
В этом уравнении световая энергия обозначена буквами nhv',
смысл которых будет разъяснен ниже (разд. 3.5), а СН2О нэпе-,
чатаио курсивом, чтобы подчеркнуть, что это не какое-то реаль-
ное соединение, а нечто с такой же общей структурой, как уг-
леводы. Иногда все уравнение умножают на 6, в результате че-
го несуществующее СН2О превращается в C8Hi2OB. Привлекав
телыю, что СН2О становится при этом реальным сахаром: та-
кую эмпирическую формулу имеет, например, глюкоза. Однако
глюкозу также нельзя считать основным продуктом фотосинте-
за, и надо иметь в виду, что в этом смысле СаН^Ов — такой же
символ или удобное обозначение, как и СН2О, и что в ходе вто-
ричных реакций образуются другие органические соединения
(разд. 12.9). Существенно, что СО2 превращается в один из уг-
леводов в результате реакции восстановления (разд. 1.3), в ко-
торой донором водорода служит Н2О и в ходе которой выделя-
ется О2. Это в свою очередь неизбежно приводит нас к рассмот-
рению двух фундаментальных аспектов физической химии, t
1.2. Разрыв химических связей
Чтобы превратить СО2 в СН2О, согласно уравнению (1.1),
необходимо удалить Н2 из Н2О, а для этого нужно, разорвать
связь Н—О. Для разрыва химических связей всегда необходима
энергия, а при их образовании энергия всегда высвобождается^
В ходе фотосинтеза энергия света используется для осуществ-
ления фотолиза (расщепления под действием света) воды; вы-
10
ГЛАВА 1
!.н —о, —[н; + [о]
Под действием света разрываются связи Н — О.
свобождающийся водород присоединяется к СОг, восстанавли-
вая последний до уровня углеводов.
1.3. Окисление и восстановление
Окисление — процесс, противоположный восстановлению.
В быту под окислением понимают процесс соединения кислоро-
да с каким-либо веществом. Когда топливо — нефть, газ или
уголь—сгорает, оно окисляется. В большинстве случаев при-
родное топливо имеет сложный состав, но древесный уголь со-
стоит преимущественно из углерода, и его окисление можно
представить уравнением
С + О2 >СО2. (1.2)
Однако химики применяют термин «окисление» и к таким реак-
циям, как (1.3), в ходе которой соединение ХН2 окисляется пу-
тем передачи На акцептору (А), что сопровождается его вос-
становлением:
ХН2 + А ---------> АН, + X. (1.3)
(донор (акцептор (восстанов- (окисленный
водорода) водорода) ленный доньр)
акцептор)
Некоторые соединения, не содержащие водорода, отдают акцеп-
тору только электроны. Уравнение (1.4) описывает процесс окис-
ления иона железа, который приобретает при этом еще один
положительный заряд, отдавая электрон акцептору, приобрета-
ющему отрицательный заряд:
Fe2+ 4- А ---> Fe2+ 4- А". (1.4)
(акцептор (восстановлен-
электрона) ный акцептор)
Иногда передача электрона осуществляется после перехода мо-
лекулы в возбужденное состояние, например после возбужде-
ния хлорофилла под действием света:
1 IlV
Хл ---> Хл* -—>Хл+4-е-.
ВВЕДЕНИЕ
11
Процесс передачи можно изобразить таким образом, как будто
после удаления электрона остается положительно заряженная
дырка:
Ф -+ е
(1-5)
Все представленные выше, а также другие окислительно-восста-
новительные реакции обладают общими особенностями (табл.
1.1). Во всех таких случаях электрон (или электроны) переда-
ется от окисляемого соединения к восстанавливаемому. Атом
водорода представляет собой протон плюс электрон:
Н = Н* + е~
При окислении происходит перенос электрона.
Следовательно, перенос водорода всегда сопровождается пере-
носом электрона. Перенос электрона в реакции типа представ-
ленной уравнением (1.2) менее очевиден. Он обусловлен тем,
что кислород притягивает электроны сильнее, чем углерод, и в
СОа электроны немного смещаются в сторону атома кислорода,
Таблица 1.1, Окисление и восстановление.
Общие положения
Окисление Восстановление
Присоединение кислорода
Удаление водорода
Удаление электрона
Смещение электрона (от
окисляемой группировки)
Удаление кислорода
Присоединение водорода
Присоединение электрона
Смещение электрона
(в сторону восстанавливав'
мой группировки)
который при этом восстанавливается, и удаляются от атома
углерода, который окисляется:
х* * * ХС£
XX XX * \ *
X X X Г X X X Q X X р ххгхх
xUxxUx^xUxx U xxUx
XX XX
(1.7)
Можно видеть, что во всех рассмотренных случаях окисление
(потеря электрона) сопровождается восстановлением (присо-
единением электрона), даже если это происходит в пределах
одной молекулы.
13
ГЛАВА I
1.4. Фотосинтез
как окислительно-восстановительный процесс
Основное уравнение фотосинтеза [уравнение (1.1)] очень
полезно как простое обобщение, но в силу своей простоты оно
весьма неточно. Глядя на него, можно подумать, что часть вы-
деляемого С>2 происходит из СС>2- В действительности же весь
С>2 образуется из воды в ходе фотолиза [уравнение ('1.8)]. Про-
цесс включает перенос И на акцептор, который затем передает
его молекуле СО2 [уравнение (1.9)]:
I
2Н2О + 2А 2АН2 + О2, (1.8)
СО2 2АН2 -----> CUfi + НгО 2А. (1.9)
В сумме эти два уравнения дают уравнение
СО, + 2Н2О СН2О + О2 + Н2О, (1.10)
которое описывает процесс более точно, чем уравнение фотосин-
теза (1.1) но лишние молекулы воды в правой и левей частях
уравнения обычно сокращают, чтобы запись была как можно
короче. Согласно этим уравнениям, фотосинтез представляет
собой окислительно-восстановительную реакцию. Вода окисля-
ется в результате удаления водорода и выделения кислорода,
а двуокись углерода восстанавливается до уровня углевода.
1.5. Как заводится главная биологическая пружина
Чтобы определить фотосинтез полнее, чем это можно сделать
с помощью уравнения (1.1), следует рассмотреть концепцию
окисления — восстановления л тот факт, что для расщепления
химических связей необходима энергия. Это позволяет увидеть,
что процесс фотосинтеза не просто поставляет органический уг-
лерод (хотя, как будет показано дальше, это очень существен-
Свет «заводит» главную биологическую пружину.
ВВЕДЕНИЕ
13
пая его особенность): в ходе фотосинтеза световая энергия с
помощью реакции расщепления связей Н—О используется для
завода главной биологической пружины. Почти .все другие био-
логические процессы в конце концов ведут к восстановлению
этих связей. Фотосинтез связан с расщеплением связей Н—О,
а дыхание—-с их образованием (рис. 1.1). С этой точки зрения
человек занимает особую экологическую нишу, так как в отли-
чие от многих других видов он сжигает ископаемое топливо,
Фотосинтез
Дых»..
Рис. 1.1. При фотосинтезе И—0-связи разрываются, О2 выделяется и про-
исходит восстановление СО2 до СН10. При дыхании CHt,0 окисляется, СО2
выделяется и связи Н—О восстанавливаются.
чтобы удовлетворить свои энергетические потребности, что при-
водит к образованию связей Н—О, в свое время разорванных в
процессе фотосинтеза прошлых геологических эпох.
1.6. Фотосинтез
как источник органического углерода
Существует мнение, что мы неправильно поступаем, сжигая
нефть, так как это слишком ценный источник органического уг-
лерода, чтобы растрачивать его таким образом. Это замечание
вполне справедливо. Мы часто склонны забывать, что, в то вре-
мя как наши топливно-энергетические проблемы можно решить,
используя силу морского прилива или термоядерные реакции,
проблема обеспечения органическим углеродом остается нере-
шенной. Почти все, что мы видим вокруг себя, в той или иной
мере создано с участием фотосинтеза. Садовник обычно гово-
рит, что он «кормит» растение, применяя удобрения; бытует мне-
ние, что растение поглощает питательные вещества из почвы. Но
это не так. Растения получают из почвы минеральные соли, но
основное «питание» — из воздуха. То, что растения развиваются
не за счет почвы, показал еще в XVII в. Ван Гельмопт, который
•посадил черенок ивы в тщательно взвешенную почву и 'поливал
его дождевой водой (рис. 1,2). Через 5 лет растение весило
74,4 кг, а вес почвы уменьшился на 56,7 г. Будучи честнейшим
14
ГЛАВА 1
человеком, Ван Гельмонт приписал последнее эксперименталь-
ной ошибке и сделал вывод, что ива росла целиком за счет по-
требления воды. Теперь мы можем уточнить эту интерпретацию.
Большую часть сырой массы растения составляет вода, погло-
щенная из почвы. Его сухую массу составляют преимуществен-
но углерод и кислород, происходящие из атмосферного СОг
(табл. 1.2). Очень небольшая часть, обычно около 1%, прихо-
дится на долю минеральных солей. Это, несомненно, необходи-
Рис. 1.2, Ван Гельмонт за поливкой дерева.
мые элементы, но они дают лишь малый вклад в массу расте-
ния.
Все живые организмы — продукты фотосинтеза. Почти все,
что окружает человека, также или является продуктом фотосин-
теза, или создано с использованием ископаемого топлива. Это
относится к натуральным и искусственным волокнам, пласти-
кам, стали, керамике и т. д. В настоящее время считают, что
мировых запасов нефти хватит на 50 лет, а угля — на 200—
300 лет. Очевидно, за это время человечество неизбежно долж-
но превратиться в общество, которое использует углерод, со-
здаваемый в ходе современного, а не протекавшего в прошлом
Таблица 1.2. Химический состав листьев
(% от сухой массы)
С
О
н
N
Р, S, К, Na, Са, Mg
и микроэлементы
(вместе с N)
-45%
-45%
-5%
Часто 1—3%
Обычно 1—5%
ВВЕДЕНИЕ
15
фотосинтеза, оставившего нам углерод в виде запасов ископае-
мого топлива. Для достижения этой цели едва ли будет до-
статочно сельского хозяйства в его современной форме, по-
скольку оно тесно связано с использованием ископаемого топ-
лива п, как правило, расходует больше энергии для получения
урожая, чем то количество энергии, которое содержится в этом
урожае в виде химической энергии.
1.7. Некоторые аналогии и заключение
Основные особенности фотосинтеза можно проиллюстриро-
вать с помощью аналогий. Почти каждый, кто изучал элемен-
тарную физику, наблюдал опыт по электролизу, в ходе которо-
го вода расщепляется на водород и кислород. В основе этого
процесса лежит слабая диссоциация воды на ион водорода Н+
и гидроксильный ион ОН”. При диссоциации образуются и бо-
лее сложные ионы, но принципиальный ход реакции может быть
представлен уравнением (1.11). Реакция продолжается до тех
пор, пока нс образуется один ион Н+ и один ион ОН” на каж-
дые 554 млн. молекул Н2О. [На этом основано понятие pH как
меры кислотности среды. Знак «р» обозначает отрицательный
логарифм дайной величины, и поскольку на 1 л воды приходит-
ся 10~7 молей Н+, то отрицательный логарифм концентрации
ионов водорода равен 7. В кислом растворе находится больше
ионов водорода и pH меньше 7.]
Н2О ч=* Н+ + ОН-. (1.11)
Если продукты реакции удаляются, диссоциация воды в соот-
ветствии с уравнением (1.11) может продолжаться и дальше. На
Практике для этого в сосуд с водой помещают два платиновых
электрода (рис. 1.3), которые соединяют с помощью проводни-
ков с химическим элементом или батареей. Окислительно-вос-
становительная реакция приводит к переносу электронов в этой
ячейке. По проводникам электроны перемещаются от анода
(оставляя положительно заряженные дырки) к катоду, который
становится отрицательно заряженным. (Соответствующие изме-
нения происходят и в самой батарее.) Поскольку противопо-
ложные заряды притягиваются, водородный и гидроксильный
ионы перемещаются соответственно к катоду и аноду. На като-
де протоны Н+ присоединяют электроны и выделяется Н2:
2Н+-|-2е~---*- На. (1.12
На аноде гидроксильные ионы отдают электроны положитель-
но заряженным дыркам и образуется О2:
2О1-Г —> Н2О ф 4- О2 + 2е-. (1.13)
16
ГЛАВА I
Таким образом, за счет электрической энергии образуется
«сток» для Н+ и ОН-, и в результате неблагоприятное для раз-
рыва связей Н—О равновесие смещается в нужную сторону.
Рис. 1.3. Фотолиз воды в небиологической системе. При электролизе место
фотоэлектрического элемента занимает химический элемент или батарея.
Можно видеть, что фотоэлектрический элемент (солнечная батарея) играет
такую же роль, что и хлоропласт при фотосинтезе.
Если через смесь газов О2 и Н2 пропустить искру, энергия,
которая очень долго, в течение, быть может, нескольких часов,
накапливалась в системе, мгновенно в виде взрыва выделится в
ходе реакции образования связей между кислородом и водоро-
дом:
Н2 + -g- О2
----> Н2О Энергия.
(1.14)
Космические исследования, получившие широкое развитие в
последние годы, способствовали созданию солнечных (фото-
электрических) батарей, в которых электроны освобождаются
в достаточном количестве и с достаточно высоким потенциа-
лом, чтобы можно было осуществлять электролиз воды (рис.
1.3). Основной принцип работы остается тем же, что и раньше,.
ВВЕДЕНИЕ
17
но «пусковой» реакцией является возбуждение с помощью све-
та. Это заставляет электроны перетекать из ячейки солнечной
батареи к вакантным дыркам, образовавшимся при возбужде-
нии, Полная последовательность событий включает превраще-
Восстановление связи между водородом и кислородом.
ние энергии света в электрическую, а затем и в химическую
энергию. Как и при фотосинтезе, световая энергия инициирует
перенос электронов, что приводит к диссоциации связей Н—О.
Очевидно, что «фотоэлектролиз» (рис. 1.3), как можно было'
бы назвать этот процесс, во многих отношениях сходен с фото-
синтезом. В обоих случаях световое возбуждение инициирует
перенос электронов, что сопровождается диссоциацией связей
Н—О. В большинстве случаев фотосинтез не приводит к непо-
средственному выделению в свободном виде О2 и Н2, хотя в.
хлоропластах в присутствии соответствующих доноров или ак-
цепторов может выделяться О2 или Н2. Вместо этого «водород»
передается через ряд посредников к СО2, восстанавливая его
(разд. 5.9). Образующийся СН2О одновременно представляет
собой депо химической энергии и материал, из которого строят-
ся живые организмы. При дыхании из этого депо высвобожда-
ется энергия в результате воссоединения водорода с кислоро-
дом, ио в данном случае этот процесс протекает постепенно,,
а не взрывообразно, как при прямом их взаимодействии. В хо-
де этого процесса происходит запасание энергии в форме
АТР.
2—234
18
ГЛАВА I
В то же время при дыхании СО2 возвращается в атмосферу,
что создает возможность повторения цикла. Действительно, весь
углерод, содержащийся в тканях нашего тела, многократно про-
шел через фотосинтетическую ассимиляцию и со временем по-
вторит этот путь снова.
Вес обращается в прах.
ГЛАВА 2
Энергия и законы физики
и химии
2.1. Законы термодинамики
В гл. 1 мы говорили о том, что фотосинтез заводит главную
биологическую пружину, что энергия поступает от Солнца и что
благодаря фотосинтезу эта энергия становится доступной живым
организмам. Так как в процессе фотосинтеза идет превращение
энергии, с одной стороны, и превращение углерода—с другой,
следует рассмотреть и тот и другой процессы более детально.
Это требует некоторого знакомства с законами термодинамики,
а так как каждый образованный человек должен быть знаком
с этими законами, вероятно, следует остановиться на них более
подробно. К счастью, законы природы такого рода представля-
ют собой простое обобщение человеческого опыта, и поэтому
мы можем сформулировать их в более или менее произвольной
форме. Здесь мы представим их в следующем виде (более стро-
гие определения даны в схеме 2.1):
Первый закон: вы не можете выиграть.
Второй закон: вы не можете остаться «при своих».
Третий закон: вы не можете не участвовать в игре.
В такой формулировке эти законы приобретают философское
звучание и могут быть распространены не только на энергети-
ческие явления, но и иа человеческие отношения. В более при-
вычном выражении, применяющемся в физике, эти законы со-
ставляют основу для множества полезных расчетов и предсказа-
ний (схема 2.1). Первый закон утверждает, что энергия (как и
материя) не может ни создаваться, ни исчезать. Происходит
Схема 2.1
Первый закон Энергия не может ни создаваться, пи исчезать. Энергия Вселенной постоянна
Второй закон Энтропия Вселенной всегда возрастает. Сущест- вует общая тенденция к увеличению хаоса и беспорядка
Третий закон Энтропия равна нулю лишь для совершенного кристалла при температуре абсолютного пуля
лишь обмен энергией между системой и ее окружением, пере-
ход различных форм энергии (тепловой, электрической, меха-
2
20
ГЛАВА 2
нической и т. д.) одна в другую, но полная энергия остается
постоянной. При постоянных температуре,‘ давлении и объеме
второй закон может быть представлен в виде
A# = A£ = AF4-TAS, (2.1)
В этом уравнении знак А обозначает «изменение», Н — энталь-
пия, Е—полная энергия системы, F — свободная энергия, Т —
абсолютная температура (в градусах К, равных °С-|-273), S —
энтропия (разд. 2.3).
2.2. Что такое система
Система, о которой упоминалось выше (разд. 2.1), — это лю-
бой объект, выбранный исследователем (хлоропласт, клетка,
организм, сообщество) и отграниченный от окружающей его
среды, которая простирается «плоть до пределов известной нам
области Вселенной.
Строго говоря, принципы термодинамики применимы только к
замкнутым системам, которые не обмениваются материей с
окружающей средой. Совершенно очевидно, что хлоропласт,
клетка или интактное растение представляют собой открытые
системы; они могут находиться в стационарном состоянии, ког-
да синтез (анаболизм) полностью компенсирует распад (ката-
болизм), но никогда не достигают подлинного термодинамиче-
ского равновесия. Тем не менее при анализе биологических про-
цессов классическую равновесную термодинамику можно ис-
пользовать как упрощенную идеализированную основу, вполне
пригодную во многих случаях в отсутствие необратимой, или
неравновесной, термодинамики.
2.3. Энтропия (5)
Это весьма гибкое и несколько мистическое понятие. Энтро-
пия (обозначаемая буквой S) описывает состояние материи с
точки зрения возможности случайных движений атомов и моле-
кул, из которых она состоит. Например, при повышении темпе-
ратуры воды или при ее переходе из твердого состояния (лед)
в жидкое или из жидкого состояния в газообразное движение
молекул становится все более быстрым и все меиее упорядочен-
ным. Таким образом, энтропия — это мера беспорядка, и второй
закон полагает, что существует общая тенденция к увеличению
беспорядка системы. Это и есть концепция космической обре-
ченности, или энтропийной смерти. Только в кристаллах при
температуре, равной абсолютному нулю, царит высший поря-
док и отсутствует тепловое движение. Живые организмы спо-
собны на какое-то время обращать эту общую тенденцию к
ЭНЕРГИЯ И ЗАКОНЫ ФИЗИКИ II ХИМИИ
2 1
•установлению статистически равномерного состояния, увеличи-
вая степень упорядоченности за счет окружающей их материи.
Однако в конце концов всегда наступают смерть и распад, все
обращается в гграх п пыль.
В уравнении (2.1), предполагающем постоянство температу-
ры, объема и давления, изменение энтропии AS для любого ре-
ального процесса всегда положительно (разд. 2.4), и T&.S со-
ставляет ту часть общей энергии, которая не может быть ис-
пользована для совершения полезной химической работы.
2.4. Свободная энергия
Биологические процессы часто протекают при постоянных
температуре и давлении, и, следовательно, А/70 (изменение стан-
дартной свободной энергии), входящая в уравнение (2.1),очень
важна. В этом смысле F° (иногда обозначаемая так, как впер-
вые предложил Гиббс —буквой G) представляет ту долю об-
щей энергии Е, которая может быть использована для соверше-
ния полезной химической работы. Изменение стандартной сво-
бодной энергии (А/70 или ДО”) равно увеличению или уменьше-
нию свободной энергии при превращении 1 моля исходного ве-
щества в 1 моль продукта реакции, протекающей при 25 °C и
pH 7,0, Эту величину нельзя путать с реальным изменением АТ7,
или AG, которое зависит от условий реакции. Следует отметить,
что биологическая стандартная свободная энергия, обычно обо-
значаемая А/7' или Дбб, отличается от соответствующей физи-
ческой величины AF0, или AG0. Проводя расчеты, физики всег-
да, в том числе и в случае попов водорода,, используют моляр-
ные концентрации; биологи же в качестве точки отсчета предпо-
читают нейтральную величину pH, т. е. концентрацию И4’, рав-
ную 10“7 г-ион-л-1. Точно так же при выводе стандартных вели-
чии они часто основываются на атмосферной концентрации СО2
(0,03%), выражаемой не в молях, а в процентах. Необходимо
также подчеркнуть, что используемые величины приблизитель-
ны, основаны на многих предположениях и на данных, не всег-
да отличающихся большой точностью.
Член АТ7 в уравнении (2.1) представляет то изменение общей
энергии А£, с помощью которого может выполняться полезная
химическая работа. В отличие от отдельных эндергонических,
т. е. требующих энергии реакций, которые не могут протекать
изолированно, ио связаны с другими реакциями — экзергониче-
скими, т. е. поставляющими энергию, в реальных процессах
свободная энергия системы по мере приближения к равновесию
всегда уменьшается, а энтропия возрастает. Преимущество ис-
пользования AF по сравнению с AS при попытках предсказать
22
ГЛАВА 2
протекание той или иной химической реакции заключается в
том, что в первом случае достаточно знать изменение для си-
стемы, в то время как во втором — для системы и ее окружения.
2.5. Свободная энергия и равновесие
Величина и знак изменения свободной энергии позволяют
нам предсказать глубину протекания химических реакций и их
направление. Здесь вступает в силу закон действующих масс,,
который утверждает, что если в результате реакции между со-
единениями А и В образуются С и D, то скорость прямой ре-
акции в любых условиях максимальна в начале процесса и
связана с концентрацией соответствующих соединений, обозна-
чаемой [А], [В] и т. д., так, как это представлено на схеме 2.2.
Схема 2,2
At ,
Дана реакция А+В C-|-D
Скорость прямой реакции=У1=[А1[В1А1
Скорость обратной peaKUHH=lZ2=[C][D]fa
(где fa и fa —константы скорррти реакции).
Аналогичным образом, скорость обратной реакции в начальный момент
времени равна 0, но по мере образования С и © опа увеличивается, в то
время как скорость прямой реакции уменьшается, пока не будет достигнуто
равновесие, когда обе скорости становятся равными по величине и противо-
положными по знаку. Тогда
[AJ[B]fa=[C][D]fa,
откуда
„ [С1[Р]
fa [А] [В] ’
где А — это константа равновесия.
Из схемы 2.2 можно видеть, что если в результате прямой
реакции [А] и [В] сильно уменьшаются, то К. является очень
большой величиной. Если же в равновесии [А]-'[В] =1[С]
то а если равновесие сдвинуто влево, то К<1. Константа
равновесия К связана с рассмотренной выше свободной энер-
гией AF соотношением
АК' = —/?Т1пК, (2.2)
где R— газовая постоянная, Т — абсолютная температура (°С+
+273), In К —натуральный логарифм (т. е. логарифм по осно-
ванию е). Подставив в это уравнение значения 7? и Т и заме-
нив loge на Igio, получим (для температуры 25 °C)
AF = —1,361g К. (2.3)
Эта стандартная свободная энергия является мерой различий
между стандартными условиями и состоянием равновесия, ког-
да ДА=0. Обычно ее измеряют в килокалориях или джоулях
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ II ХИМИИ
23
Таблица 2.1. Связь между К и А/7' (из соотношения А/''= —RT In К)
ЛЕ', ккал к Характеристика реакции
-|-6,81 0,00001 Идет с большим трудом 1-5,45 0,0001 Идет с трудом, неблагоприятна -1-4,09 0,001 1 0 Qi J Может начаться довольно легко +1,36 0,1 । 0,00 1,0 } Легко обратима —1,36 10 J —2,73 100 1 _4 од 1 000 1 От УмеРенио благоприятной до очень ’ [ благоприятной —5,45 10 000 1 —6,81 100 000 Идет практически до конца Биологический реакции (не фотохимические), характеризующиеся величиной LF{>7— <8 ккал, по существу необратимы.
(разд. 2.6). Например, если &F'=—1,36 ккал, то 1g К= =
=4, и, поскольку антилогарифм 1 равен 10, К будет равно 10.
Аналогичным образом, если ДЕ'=—2,72, то 1g К=2 и /\ =
= 100. Из табл. 2.1 можно видеть, что большим отрицательным
значением ДР отвечают большие положительные значения К и
при этом равновесие реакции сильно сдвинуто вправо. Однако
сами эти величины ничего не говорят о вероятности протекания
реакции. Эта вероятность определяется другими факторами,
в частности наличием или отсутствием катализатора. Напри-
мер, энергия, высвобождающаяся при соединении водорода и
кислорода:
Н2 + -*-О2 —> Н2О, (2.4)
составляет около —56 ккал (—235 кДж|), но соответствующая
смесь газов может сохраняться сколь угодно долго без видимых
изменений. Однако, если пропустить через эту смесь искру, про-
изойдет мгновенная реакция.
Теоретически все реакции, даже такие, как соединение во-
дорода и кислорода, обратимы. Если бы это было не так, фото-
синтез, зависящий от фотолиза воды, был бы невозможен. По-
добным же образом неблагоприятная реакция может иногда
идти в прямом направлении, если имеется «сток», с помощью
которого удаляются один или несколько продуктов реакции
24
ГЛАВА 2
При наличии «стока» равновесие сдвигаемая вправо,
А + В ч С а
\
П1
, . Сток
(если идет удаление их в буквальном смысле или путем исполь-
зования в последующей реакции). Для осуществления многих
биохимических реакций, как правило, необходимо не более
8 ккал (33 кДж). Следует также отметить, что реакции с высо-
ким положительным значением ДЕ (эндергонические, пли по-
треблющие энергию, в противоположность экзергоническим
реакциям, в ходе которых энергия выделяется) практически со-
всем не идут, если они не сопряжены с реакциями, характеризу-
ющимися достаточно большой отрицательной величиной ДД так
что суммарное изменение свободной энергии становится отри-
цательным (разд. 2,4).
2.6. Единицы энергии
В последнее время для измерения энергии, света, расстояния
и т. д. все большеепримеиение находит система СИ (Междуна-
родная система единиц). Она привлекательна прежде всего тем,
что в пей используются величины, кратные 1000, с общими для
всех них приставками (кило='Ю3, милли=10~3, микро='10-®,
нано=10-9 и т. п.). Однако СИ не является единственной си-
стемой. Во-первых, во всех ранних работах используются ста-
рые единицы, и поэтому современный читатель должен быть
знаком и с ними. Во-вторых, в старых системах использовались
иногда весьма наглядные единицы. Может быть, старомодно из-
мерять расстояние в футах, но у нас есть две ноги (игра слов:
по-английски foot — это нога и одновременно мера длины), что
позволяет сразу представить себе эту единицу. Такой же аргу-
мент можно привести и в отношении единицы энергии. В си-
стеме СИ энергия измеряется в джоулях. В свою очередь эту
величину можно вывести из дин и эргов (дина — это сила, сооб-
щающая телу массой 1 г ускорение 1 см-с-2; эрг — это работа,
совершаемая силой в 1 дину на расстоянии 1 см. Десять мил-
лионов эрг равны 1 джоулю, т. е. 1 эрг= 10“7 Дж). Джоуль~
очень распространенная единица, из нее легко получить ватты
(1 Вт=1 Дж-с-1), ио все же в ряде случаев его пользуются не
так охотно, как калориями. Большая калория (когда диетологи
говорят о калориях, они имеют в виду «большие калории», или
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ И ХИМИИ
25
ккал) — все еще наиболее привычная единица во многих обла-
стях биоэнергетики. Одна калория, или грамм-калория, — это
количество тепла, необходимое для того, чтобы поднять темпе-
ратуру 1 г воды на 1 °C (от 14,5 до 15,5°). Калория соответ-
ствует примерно 4,2 Дж (4,1855 Дж).
2.7. Энергия образования углеводов
При сжигании в калориметре одного моля ('180 г) глюкозы
выделяется количество тепла ДЯ=673 ккал. По оценкам при
постоянной температуре изменение стандартной свободной энер-
гии ДД при окислении глюкозы составляет —686 ккал. В этих
условиях &Н=ЛЕ, и, следовательно, в соответствии с уравне-
нием (2.1) ГД5 = 4-13 ккал.
Для этой и других сходных реакций, протекающих при по-
стоянной температуре, величина TES— разность между точно
измеренной величиной Д/7 и вычисленной величиной ЕЕ'— рав-
на примерно 112 ккал (673-^6) при образовании гипотетическо-
го углевода СН2О из СО2 и Н2О в соответствии с уравнением
(1.1). Если, однако, учитывать изменение реальной, а не стан-
дартной свободной энергии, то эта величина возрастет до 114—
115 ккал (основано на 686-1-6), а в определенных условиях до
120 ккал. Тот факт, что предпочитают использовать все-таки бо-
лее низкое значение, определяется тем, что оно получено про-
стым калориметрическим измерением и является весьма точ-
ным, в то время как более высокое значение, основанное на
ряде вычислений и предположений, менее точно; в то же время
во многих случаях различие между тем и другим значениями
не очень существенно.
2.8. Энергия связи
Сходные оценки величин, упомянутые выше, можно получить
из эмпирических значений энергии связей. Огги основаны на
теплоте образования различных соединений. Однако здесь воз-
никает одно затруднение: энергия связи не абсолютная величи-
на, она зависит от того, в каком месте в молекуле эта связь на-
ходится, Например, для разрыва первой связи Н—О в молеку-
ле воды требуется меньше энергии, чем для разрыва второй.
Подобным же образом двойная связь С = О в какой-либо кисло-
те прочнее, чем две одинарные связи С—О, а С = О в СО2 проч-
нее, чем в альдегиде —-С (табл. 2.2). Поэтому значение
\О
энергии связи, полученное для одного какого-то соединения,
нельзя безоговорочно использовать для другого; однако из на-
бора известных значений можно выбрать то, которое наиболее
подходит к данному случаю. Таким образом, хотя выбор оцен-
26
ГЛАВА 2
Таблица 2.2.
Эмпирические значения анергии связей, ккал-моль-1 Образование углеводов
[уравнение
с-ы с—с С—0 с=о с=о Н-0 0 = 0 н—н 99 яч Разрыв связи Образование связи
(спирт) (кислота) (СО2) 77 180 190 НО 118 104 -1-190 4-190 + 110 +110 +600 Разность= + 113 ккал -83 -99 —77 -НО -118 —487
ки не •совсем объективен, зато мы приходим к одному и тому
же результату независимо от того, используем ли мы значения
энергии связи, полученные непосредственно из данных по окис-
лению глюкозы (см. выше) или из данных по совершенно дру-
гим реакциям. Так, взяв значения энергии связей, приведенные
в табл. 2.2. для гипотетической единицы СН^О, и учитывая чис-
ло таких единиц, соединенных связью С—С в глюкозе, сахаро-
зе или крахмале, мы вновь получим для энергии их образова-
ния величину, равную примерно 112 ккал (470 кДж).
190 190 НО НО
о == = С ===== О + Н -------- О —------ н
н
99
-—— с —---О •-----Н + о ===== о
S3 77 HO Н8
2.9. Расщепление воды
Расчеты подобного рода позволяют оценить полное измене-
ние энергии при образовании и диссоциации воды. Как мы уже
говорили, связь Н—О в молекуле воды разрывается при фото-
синтезе и вновь образуется при дыхании, так что можно оце-
нить полное изменение энергии, связанное с переносом водоро-
да (транспортом электронов) в этих процессах:
Н-Н + 4- (о^=о) —> н — о-111 н, (2.6)
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ и химии
27
Разрыв связей Образование связей
4" (118) — 59 2(110) = -220 Разность = —57 ккал
104 -1-163 (—240 кДж)
Для протекания реакции в обратном направлении, т. е. для рас-
щепления воды (в виде пара) до Н2 и Ог, требуется +57 ккал
(рис. 4.13). Во многих случаях, однако, процесс фотосинтеза не
приводит к образованию газообразных водорода и кислорода.
Вместо этого происходит перенос водорода от воды на кофер-
мент NADP+ — конечный окислитель в фотосинтетической си-
стеме транспорта электронов (разд. 4.9).
Сходным образом при дыхании водород передается от
NADPH (или от родственного кофермента NADH) к кислороду
с образованием воды. Изменение свободной энергии при восста-
новлении NADP (или при окислении NADPH) составляет около
52 ккал.
2.10. Резонанс
Как мы уже говорили, энергия одной и той же связи может
иметь разное значение в зависимости от условий; иначе гово-
ря, она изменяется в соответствии с положением связи в мо-
лекуле. Отчасти это связано с так называемым'резонансом. На-
пример, в уравнении (1.7) молекула СО2 представлена таким
образом, чтобы подчеркнуть способность атомов кислорода от-
тягивать на себя электроны. Такое изображение позволяет так-
же увидеть распределение электронов между С и О; в сфере
притяжения каждого ядра находится по 8 электронов:
хх
([Кислороду необходимы два электрона, чтобы довести их
число до восьми, и он является двухвалентным; углероду необ-
ходимы четыре электрона, и он четырехвалентен.] Подобное
распределение двух пар электронов между С и О порождает
двойные связи, и, если хотят указать структуру связи, СО2 обыч-
но представляют в виде О = С = О. Сходным образом бензольное
кольцо, содержащее шесть атомов углерода, можно представить
так, как это сделано на рис. 2.1. Видно, что в формах А и Б все
требования к структуре с точки зрения валентности (Н однова-
лентен, С четырехвалентен) выполнены и четко зафиксировано
28
ГЛАВА 2
положение одинарных и двойных связей в кольце. Однако и бо-
лее условные формы В и Г также отвечают этим требованиям.
В последнее время отдают предпочтение форме Д, в которой
положение двойных связей не фиксируется. Фактически В и Г'
представляют собой крайние формы, а Д — промежуточную. На
самом деле форма Д лучше отражает реальную ситуацию, чем
В или Г,поскольку измерение расстояния между атомами угле-
рода показывает, что в беизолы-юм кольце длина связей не со-
Рис. 2,1. Различные способы изображения бензольного кольца.
ответствует ни одинарным, ни двойным связям, а представляет
что-то среднее между ними. Другими словами, реальная струк-
тура, которую лучше всего отражает форма Д, лежит между
А и Б (или В и Г). [В квантовой механике атомные орбитали
обозначают буквами s, р и т. д., которым отвечают определен-
ные квантовые числа. Связи С—С и С—Н в молекуле бензола
образуются из гибридных «рЕ-орбиталей. При этом у каждого
атома углерода остается неспаренный электрон на З^-орбитали,
которая перекрывается с соответствующими орбиталями сосед-
них атомов углерода. Соответственно комбинация пар атомных
рх-орбиталей приводит не к образованию л-орбиталей, локали-
зованных между двумя атомами углерода, а к полной делока-
лизации л-О|рбиталей, охватывающих все шесть атомов углеро-
да.] Более того, по существу, благодаря резонансу бензольное
кольцо становится стабильной структурой. В данном случае
имеет место не чередование относительно прочных (двойных) и
относительно слабых (одинарных) связей, а одинаковое взаимо-
действие между всеми составляющими молекулу атомами. Та-
ким образом, СО2 можно изобразить в виде
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ II ХИМИИ
29
X X Г\ X X ГЛ XX
X ХГ X а также в виде х U «
XX XX
XX
или * Q £
XX
или
Благодаря резонансу расстояние между атомами углерода и
кислорода уменьшается, и СО2 становится особенно стабильной
п компактной молекулой.
[С точки зрения квантовой механики гибридные орбитали
spx и п-связи также могут участвовать в формировании СО2,
что и подразумевается в изображенных выше структурах, ио,
даже если это так, аксиальную симметрию бензольного кольца
и его высокую стабильность можно объяснить лишь с помощью
резонанса.].
2.11. Свободная энергия гидролиза
аденозинтрифосфата (АТР)
Перенос электронов в ходе фотосинтеза в норме приводит
к образованию восстановительной силы в форме АТР и NADPH2.
Об АТР обычно говорят как о соединении, богатом энергией,
и в этой •связи уместно разобраться — почему? Как мы уже
видели, для разрыва химических связей всегда необходима
энергия, а если в ходе какой-то реакции энергия выделяется, то
в некоторых случаях ее можно определить простым вычитанием
суммы энергий, затрачиваемых на разрыв связей, из общей
энергии, высвобождающейся при образовании связей. При при-
менении этого подхода к реакции гидролиза АТР (рис. 2.2)
можно ожидать, что полная энергия останется без изменений,
так как разрушается и образуется равное число связей соот-
ветственно О—Р и О—Н.
Однако измерения показывают, что изменение стандартной
свободной энергии составляет примерно 7 ккал (AF' = —7 ккал),
Это обусловлено тем (табл. 2.2), что энергия связи меняется в
зависимости от того, в каком окружении опа находится в моле-
куле. Как п СО5, молекула неорганического фосфата (ортофос-
фата, или Pi) очень стабильна благодаря резонансу. У АТР воз-
можность появления резонанса ограничена, поскольку электро-
ны одновременно и в равной степени отталкиваются в двух про-
тивоположных направлениях. Кроме того, в результате иониза-
ции и появления локальных положительных и отрицательных
зарядов в полифосфатной части молекулы возникает противо-
действие одноименных зарядов (рис. 2.2)."Здесь можно провес
30
ГЛАВА 2
О" О' О'
* f t
А—о— р*-о"*он + нго
О' О' о~
О' о~ О'
t I t
А—0 — он -+- ~о— р+— он + н +
О' О' О'
Рис. 2.2. Гидролиз АТР до ADP и Pi.
сти механистическую аналогию с системой из трех магнитных
брусков, связанных нитью друг с другом и с деревянной опорой
(рис. 2.3). Если они располагаются на плоскости таким обра-
зом, что их северные (и южные) полюсы находятся по одну
сторону, то они будут отталкиваться друг от друга, натягивая
нить. Если нить, соединяющую первый и второй брусок, пере-
резать, то первый брусок резко переместится вправо. То же са-
мое произойдет и со вторым бруском, если перерезать вторую
нить. Однако если перерезать третью нить, то магнит, связан-
ный с деревянной опорой, останется неподвижным. Природа
связи (нить) одинакова во всех случаях, однако их «магнитное»
окружение различно. АТР тоже можно рассматривать как со-
•Рис. 2.3. Гидролиз АТР. Аналогия.
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ II ХИМИИ
31
единение компонентов, постоянно находящихся в противодейст-
вии. Расщепление молекул (гидролиз) приводит к выделению
тепла (освобождению энергии) и образованию более устойчивых
соединений (у продуктов реакции резонанс более вероятен).
При гидролизе АТР до ADP А/7' равно примерно —7 ккал
(—29 кДж), и примерно такое же количество энергии освобож-
дается при гидролизе ADP до АМР. Однако при дальнейшем
гидролизе АМР выделяется значительно меньше энергии, по-
скольку при этом разрывается такая же связь, но ее окружение
в молекуле другое из-за предшествующего отщепления фосфат-
ных групп.
2.12. АТР как компонент ассимиляционной силы1
Фотохимические события, происходящие при фотосинтезе,
приводят к генерации ассимиляционной силы (ATP-j-NADPH),
которая, как показали Арион с сотрудниками, необходима для
Рис. 2,4. Экспериментальное разделение фотосинтеза на две фазы. Слева: на
свету в тилакоидах образуются АТР и NADPHj. Справа: в темноте фермен-
ты стромы хлоропласта используют АТР и NADPH2 для восстановления СО2
до СН2О.
превращения СО2 в СН2О (рис. 2.4). При расчетах можно при-
нять, что в реакциях типа (2.7) вклад энергии от АТР равен
количеству энергии, высвобождающейся при его гидролизе:
ФГК -|- АТР —> ДФГК + ADP. (2.7}
Но это не значит, что энергия существует как нечто независи-
мое и передается без участия реагирующих соединений (даль-
нейшие детали этой реакции см. в разд. 6.11 и рис. 9.6). В вос-
1 Ассимиляционная сила, или ассимиляционная энергия — понятие, вве-
денное Д. Арионом, обозначает совокупный продукт фотобиохимической фа-
зы фотосинтеза, используемый в последующих восстановительных фермента-
тивных реакциях. Он включает АТР, NADPH, трансмембранный электрохи-
мический потенциал Дцп+ — Прим. ред.
•32
ГЛАВА 2
•становитель'ном пентозофосфатном цикле (ВПФ-цикле), или в
•цикле Бенсона — Кальвина (гл. 6), АТР расходуется в двух
реакциях, одна из которых представлена уравнением (2.7); кро-
ме того, существует восстановительный этап, на котором
NADPH2 служит донором водорода, или восстановителем. Для
восстановления одной молекулы СО2 используются три молеку-
лы АТР и две молекулы NADPH2. Стандартная свободная энер-
гия гидролиза АТР равна примерно—7 ккал (реальное измене-
ние свободной энергии может достигать —12 ккал из-за присут-
ствия Mg, который связывает АТР и ADP в разной степени).
Поэтому вклад энергии от АТР, равный 3-7=21 ккал, в действи-
тельности значительно меньше, чем от NADPH2. Последнее со-
единение служит донором водорода, в то время как АТР — до-
нором фосфата (Р1). Изменение свободной энергии, связанное с
переносом водорода от NADPH2 к ДФГК [восстановительный
этап ВПФ-цикла; уравнение (6.17)], значительно меньше из-
менения, связанного с переносом водорода от NADPH2 к кис-
лороду (—52 ккал; разд. 2.9). Однако, если учесть, что процесс
переноса может продолжаться вплоть до соединения водорода
с О2 (при дыхании), то потенциальный вклад энергии составит
2-52 ккал, так как для восстановления СО2 используются две
молекулы NADPH2. Следовательно, 3ATP+2NADPH2 в сумме
«стоят» 214-104 = 125 ккал, и если учесть, что многие использо-
ванные здесь величины весьма приближенны, можно сделать вы-
вод, что реальный процесс ассимиляции углерода, требующий
112 ккал, протекает со значительно более высокой эффективно-
стью, чем образование ассимиляционной силы (гл. 5).
2.13. Окислительно-восстановительный потенциал
Как мы уже говорили, зная величину и знак изменения сво-
бодной энергии данной реакции, можно попытаться предсказать
глубину ее протекания и направление. Соединения, способные к
окислению/восстановлению, можно расположить в порядке из-
менения их потенциальной способности реагировать друг с дру-
гом. Некоторые примеры окислительно-восстановительных по-
тенциалов приведены в табл. 2.3. В таких таблицах в верхней
части помещают соединения с наиболее высоким отрицатель-
ным потенциалом. Эти соединения способны восстанавливать
соединения, лежащие ниже и характеризующиеся менее отрица-
тельным или положительным потенциалом. В некоторых случа-
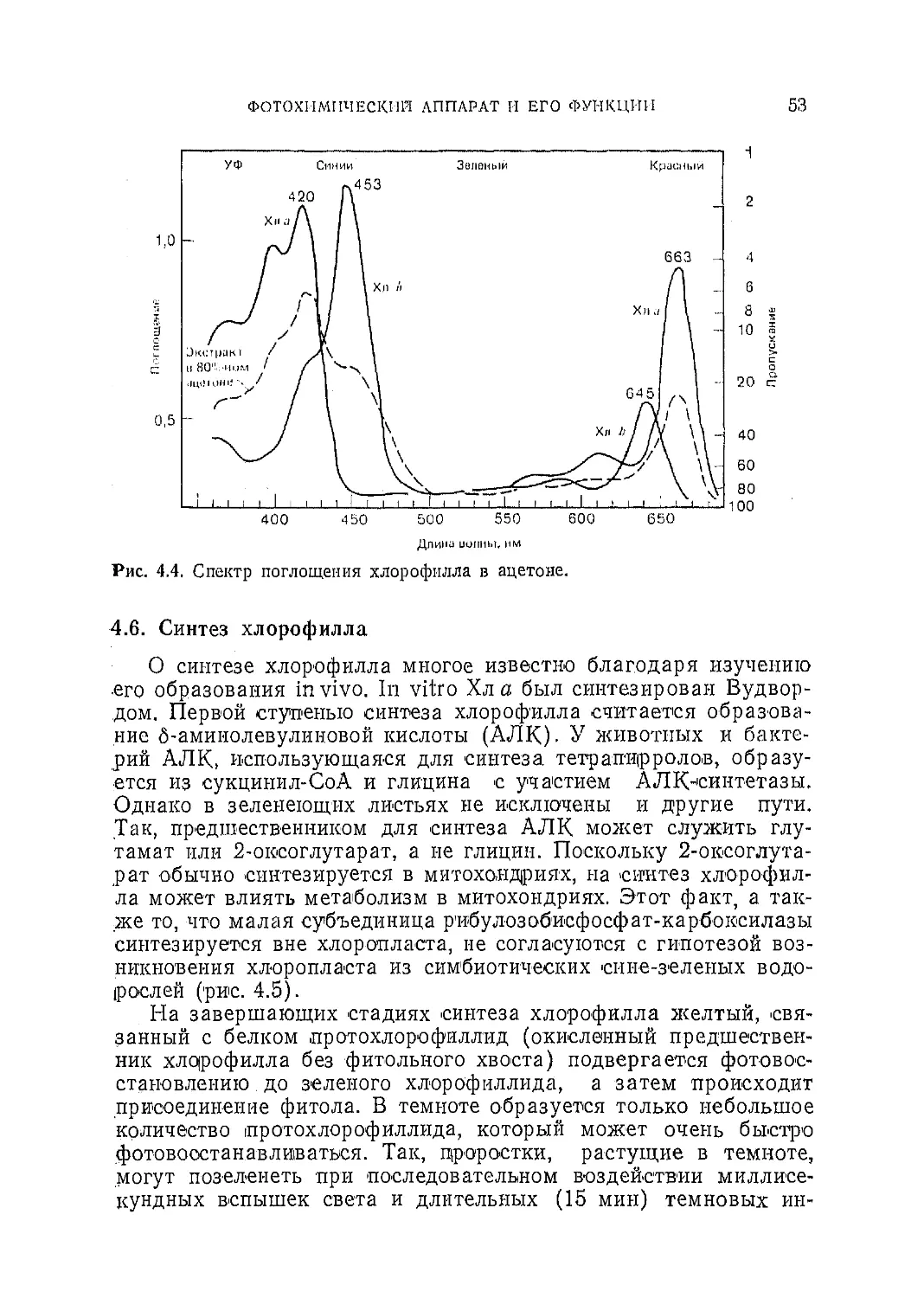
ях значение потенциала зависит от pH, и, хотя его всегда соот-
носят с соответствующей величиной для стандартного водород-
ного электрода, определяемой при рН = 0, он обычно связан с
pH следующей зависимостью:
Е' = Е°—0,06 pH, (2.8)
ЭНЕРГИЯ II ЗАКОНЫ ФИЗИКИ и химии
33
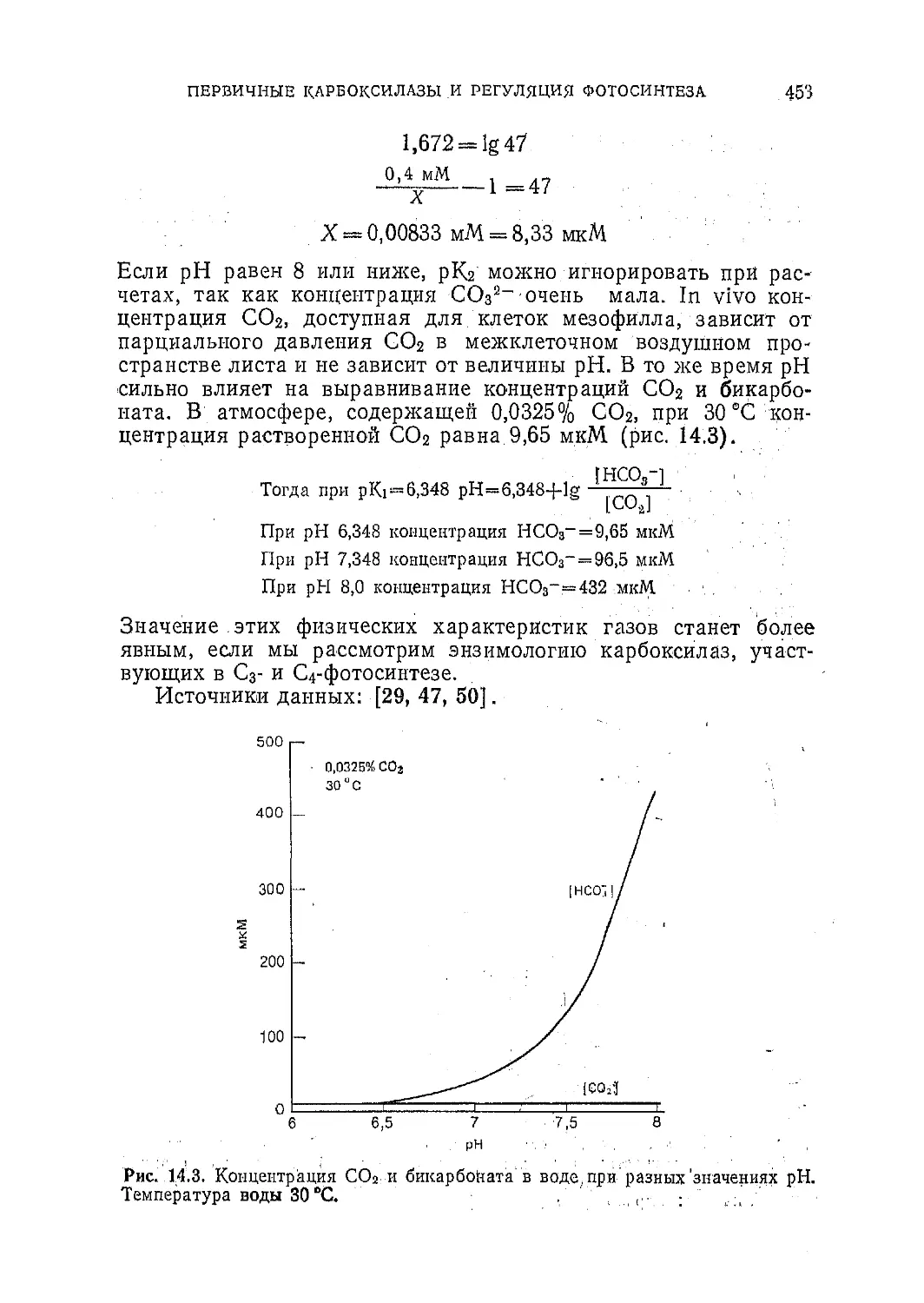
Таблица 2.3. Некоторые окислительно-восстановительные потенциалы1
Пара Окислительно-восстанови- тельный потенциал Е', мВ
1?е<1 Ох
Fe2+—S F3+—S —6004—700
Ферредоксин2* Ферредоксин-'1* —430
н2 2Н+ —420
NADPH+H+ NADP+ —320
Малат Оксалоацетат -165
Qred Qox —100
Цитохром /2+ Цитохром р+ +3004—1-370
Пластоциаинн* Пластоциаинн2* +390
Р700 Р700+ -I-430+-I 530
Н2О 1 2 °2 4-815
1 Болес детально некоторые из указанных здесь переносчиков электронов описаны в
гл. 4.
где Е°— значение потенциала при рН=0, а Е'-—его значение
•п|р'и данном pH. Так, при pH = 7,0 значение потенциала для
На/2Н+ становится равным —0,42 (7-0,06). При необходимости
изменения свободной энергии &F' можно получить из окисли-
тельно-восстановительного потенциала с помощью следующего
уравнения:
EF'~nfEE', (2.9)
где п-—число переносимых электронов, f—число Фарадея
( = 23). Например, из табл. 2.3 видно, что Н2 может восстанав-
ливать О2 (Нг+у Оз—>-Н2О); подставив в уравнение (2.9)
разность между соответствующими окислительно-восстанови-
тельными потенциалами (—0,420 и 4-0,815) и значения п и f,
мы получим
AF' = —2.23 ккал/В-[0,815В—(—0,420В)] = — 56,8 ккал (2.10)
Следует отметить, что энергия поступает в ВПФ-цикл в ходе
только трех реакций, в остальных десяти реакциях энергия рас-
ходуется. Этот цикл вращается подобно обручу, подгоняемому
палкой, Большинство «ударов» (все «удары» от NADPH2 и две
трети от АТР) обеспечивают восстановление ФГК до цриозо-
фосфата. 5/6 этого триозофосфата вновь окисляется до ФГК
(гл. 6), поддерживая тем самым «вращающий момент» цикла.
3—234
ГЛАВА 3
Энергия и свет
3.1. С чего все начинается
В конечном счете источником энергии при фотосинтезе слу-
жит Солнце, где благодаря чрезвычайно высокой темпера-
туре происходит термоядерный синтез водорода, сопровождаю-
щийся выделением энергии. Часть этой энергии достигающей
поверхности Земли, поступает с так называемым «видимым»
светом (длина волны 400—700 нм). Эти пределы (рис. 3.1) оп-
ределяются свойствами глаза человека, который лучше всего
видит в зеленой области спектра и не способен воспринимать
свет с длиной волны ниже 400 нм (верхняя граница ультрафио-
летовой части спектра) и выше 700 им (нижняя граница ближ-
него инфракрасного света). [1 нм = 1 нанометр = 1/1 000 000 000
метра = 10-9 метра. В системе СИ (разд. 2.6) обычно предпочи-
тают использовать именно эту единицу, а не миллимикроны
(ммк) или ангстремы (А), которые часто применялись ранее
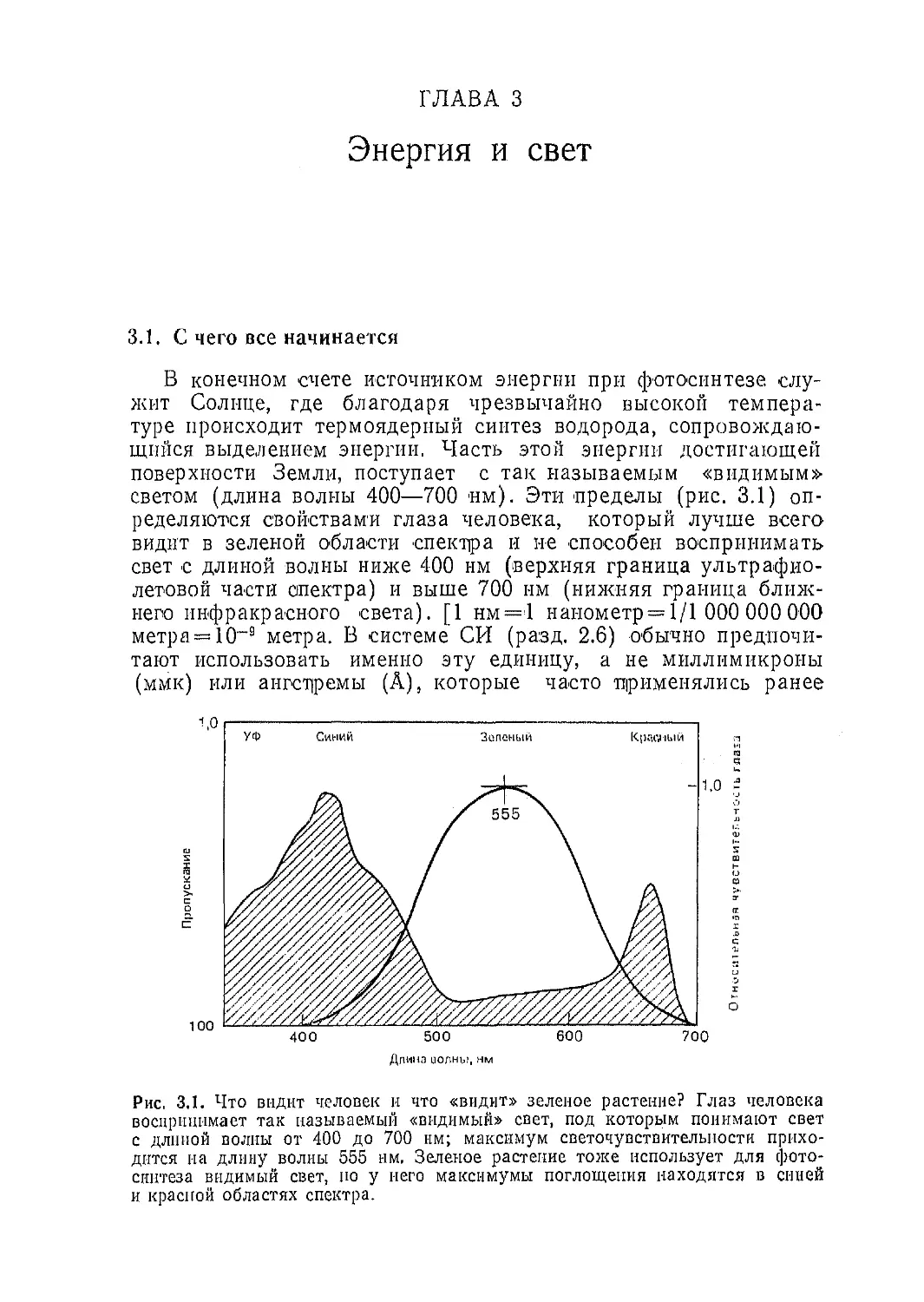
Рис. 3.1. Что видит человек и что «видит» зеленое растение? Глаз человека
воспринимает так называемый «видимый» свет, под которым понимают свет
с длиной волны от 400 до 700 нм; максимум светочувствительности прихо-
дится на длину волны 555 нм, Зеленое растение тоже использует для фото-
синтеза видимый свет, по у него максимумы поглощения находятся в синей
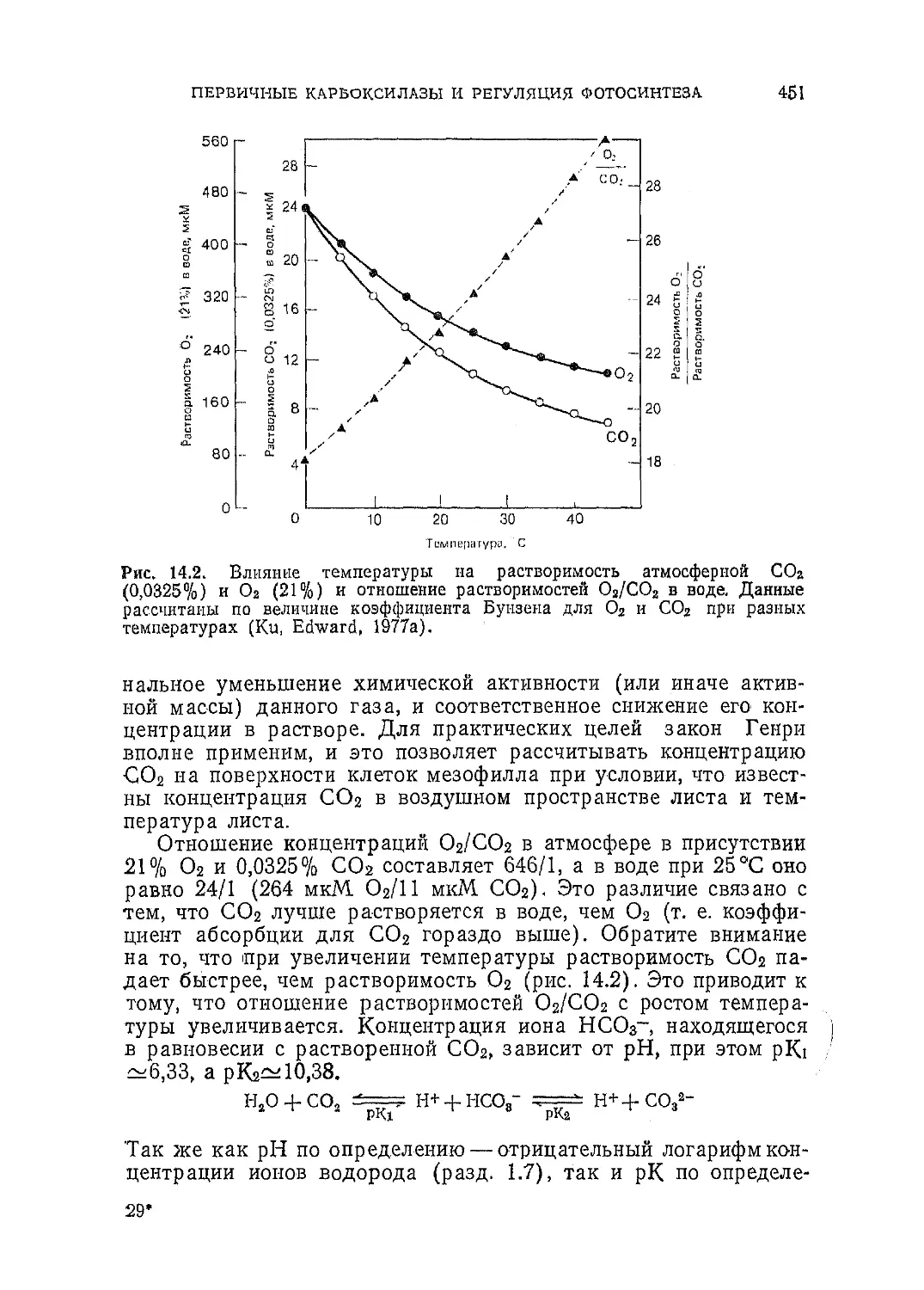
и красной областях спектра.
ЭНЕРГИЯ И СВЕТ
35
для обозначения такого порядка величин. 1 нм = 1 ммк=10 А.]
В этом отношении зеленые растения напоминают человека,
поскольку они также неспособны использовать световую энер-
гию вне указанного диапазона. Однако в отличие от глаза че-
ловека растения менее чувствительны к зеленому свету.
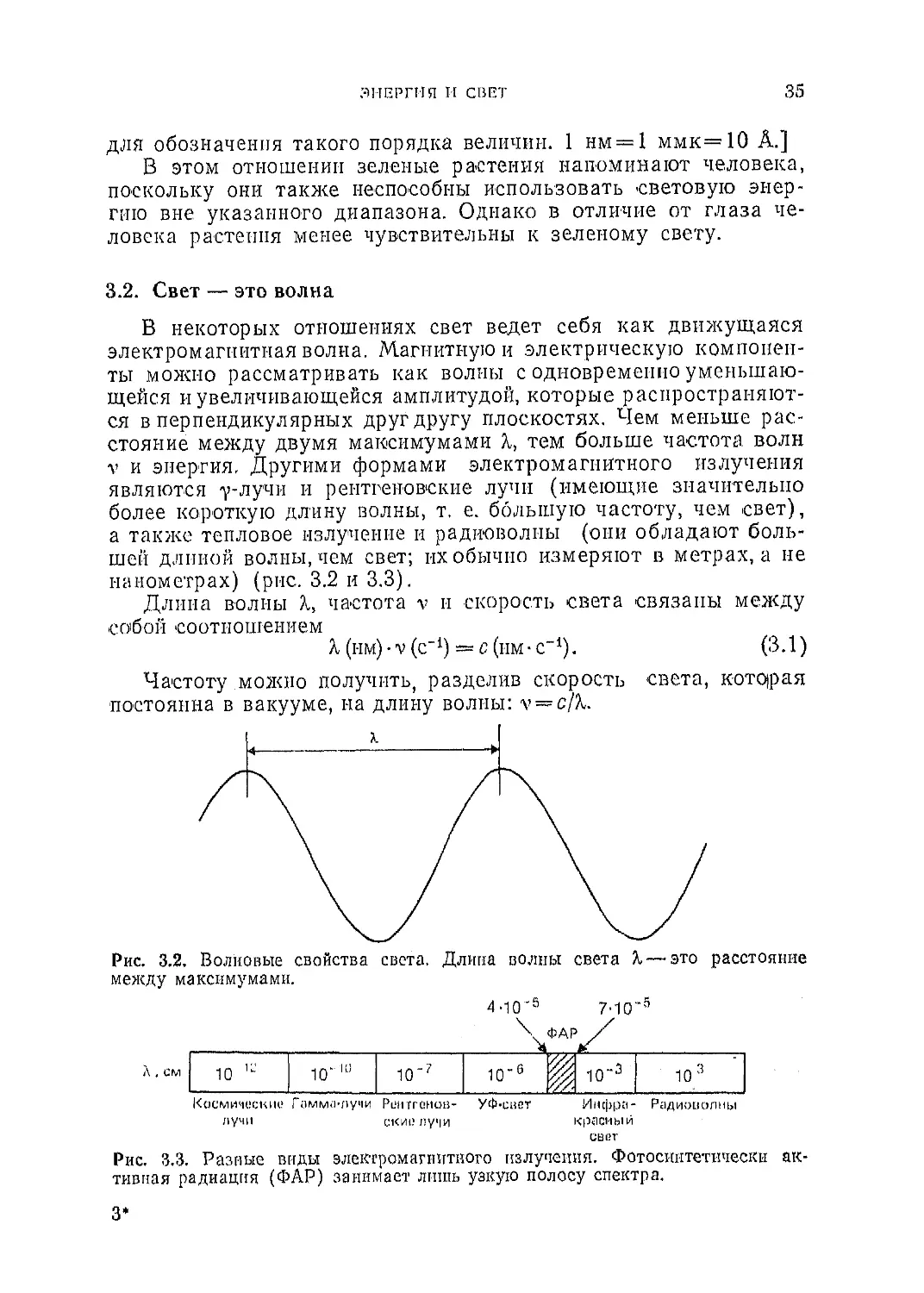
3.2. Свет — это волна
В некоторых отношениях свет ведет себя как движущаяся
электромагнитная волна. Магнитную и электрическую компонен-
ты можно рассматривать как волны с одновременно уменьшаю-
щейся и увеличивающейся амплитудой, которые распространяют-
ся в перпендикулярных друг другу плоскостях. Чем меньше рас-
стояние между двумя максимумами X, тем больше частота волн
v и энергия, Другими формами электромагнитного излучения
являются у-лучи и рентгеновские лучи (имеющие значительно
более короткую длину волны, т. е. большую частоту, чем свет),
а также тепловое излучение и радиоволны (они обладают боль-
шей длиной волны, чем свет; их обычно измеряют в метрах, а не
нанометрах) (рис. 3.2 и 3.3).
Длина волны X, частота v и скорость света связаны между
собой соотношением
X (нм) • v (с-1) = с (им- с-1).
(3-1)
Частоту можно получить, разделив скорость света, которая
постоянна в вакууме, на длину волны: v —с/Х.
Рис. 3.2. Волновые свойства света. Длина волны света X — это расстояние
между максимумами.
Л. см
Рис. 3.3. Разные виды электромагнитного излучения. Фотосинтетически ак-
тивная радиация (ФАР) занимает лишь узкую полосу спектра.
3*
36
ГЛАВА 3
3.3. Свет — это поток частиц
Вместе с тем свет ведет себя как поток частиц. Ньютон и
Эйнштейн придерживались именно такого представления, и мы
могли бы описать свет только с этих позиций, одиако, оставаясь
в рамках данной концепции, мы совсем ие отказываемся от рас-
смотренной выше. Когда достаточно надежные эксперименталь-
ные данные одновременно свидетельствуют в пользу двух про-
тивоположных концепций, часто оказывается, что обе они nipa-
впльиы. Поэтому, по крайней мере в настоящее время, мы мо-
жем считать, что ие до конца понимаем природу света, и ис-
пользовать в каждом случае наиболее приемлемую из этих двух
концепций.
3.4. Кванты
Частицы света называют квантами или фотонами. В соответ-
ствии с концепцией о волновой природе света их энергию свя-
зывают с длиной волны: чем ближе максимумы волн друг к
другу, тем больше энергия. Следовательно, синий свет с длиной
волны 450 им обладает большей энергией, чем красный свет с
длиной волны 650 нм.
3.5. Энергия света
Энергию одного фотона света можно определить, умножив
его частоту у на так называемую постоянную Планка;
E — hv, (3.2)
и именно поэтому энергию кванта часто представляют в виде
произведения hv, а энергию /г квантов — как nhy рем., например,
уравнение (1.1)]. Так как v равно скорости света с, деленной
на длину волны X, это уравнение можно записать также в виде
£ = р (3.3)
Хотя хлорофилл поглощает как «синие», так и «красные» фото-
ны, обычно для вычисления максимального количества энергии,
доступной для фотосинтеза, выбирают длину волны, близкую к
максимуму поглощения хлорофилла в красной области (напри-
мер, 680 им). Это связано с тем, что избыточная энергия фото-
на синего света ие может эффективно использоваться (разд.
4.16). Так как скорость света в вакууме равна З-Ю10 см-с~г
(или З-Ю17 нм-с-1), а постоянная Планка = 6,6-10~27 эрг-с, для
красного света с Х=680 нм имеем
Е = = 6,6-10~27 эрг-с- -3-' _ 2 9 • 1.0~1а эрг. (3.4)
А, ’ 1 с 680 нм ’
ЭНЕРГИЯ И СВЕТ
37
Как показано в разд. 2.6, 107 эрг=1 Дж, а 4,2 Дж = 1 кал.
Следовательно, Е, выраженная в калориях, равна
, Е ЭРГ i = 6 940-20 Кал. (3.5}
4,2-107 эрг-кал 1 \ /
Это очень малая величина. Масса одной молекулы кислорода,
выраженная в граммах, также очень мала и для удобства ее
можно умножить на число Авогадро (число ..атомов в грамм-мо-
лекуле). Масса одной грамм-молекулы, или одного моля, Оа
равна 32 г. Следовательно, энергия фотонов, число которых рав-
но числу .Авогадро (6-Ю23), для красного света составляет
6,9-10~В * * * * * * * * * * * 20 кал-6-1023 ~ 42 ккал. (3.6)
.{Число фотонов N, равное числу Авогадро (моль фотонов),
иногда обозначают как «эйнштейн», ио, так как последнюю
Таблица 3.1. Различные характеристики солнечного света
Освещенность Фотосинтетически активная радиация
па 1 № в 1 с
Люкс Фут-свечн Вт-м-2 Эрг Дж Кал Фотоны 1 мкмоль
квантов
100000 10 000 500 5-10» 500 120 1,3 10й1 2200
Фотометрические из- Радиометрические изме- Число квантов (400—
мереиия рения (400—700 нм) 700 нм)
В таблице сопоставляется полный солнечный свет (слева) и фотосинтетиче-
ски активная радиация при средней длине волны 575’.нм (справа). Внизу
указан способ измерения.
N. В. Все приведенные выше данные характеризуют плотность светового по-
тока. Поток — это скорость поступления вещества, выраженная в еди-
ницах количество/время. Плотность светового потока, или поток на
единицу поверхности, — это величина, выраженная в единицах коли-
чество-время-1-площадь-1. Мерой потока является ватт (1 Дж-с-1),
а мерой его плотности (облученности) Вт-м-2, или Дж-с-’-м-2. Та-
ким образом, фотосинтетически активная радиация (ФАР)—это про-
чего радиация в области 400—700 нм, а фотосинтетическая облучен-
ность (ФО) = ФАР-(единица времени)-1-(единица площади)-1. [Хотя
в опытах па растениях эти единицы применяются весьма широко, их
нельзя считать корректной характеристикой интенсивности света, пада-
ющего на единицу поверхности. Интенсивность — свойство источника
света. Например, стандартная свеча испускает световой поток силой
в 1 свечу, или 4л люмена (лм). Соответствующая мера плотности по-
тока света, падающего (разд. 3.6) на поверхность сферы радиусом
1 фут от стандартной свечи, находящейся в ее центре, составляет-
1 лм-фут-3, или 1 фут-свечу; рис. 3.4. 1 лк=.1 лм-м-2; 1 фут-свеч-а =
= 1 лм-фут-2; 1 фот=1 лм-см-2; следовательно, 1 лк=1-10-4, фот=
= 0,1 миллифот=0,0929 фут-свечи. 1 фут-свеча = 1 лм-фут-2 =
= 1,0764 миллифот= 10,764 лм/м2=10,764 лк.]
38
ГЛАВА 3
единицу нередко используют применительно к энергии моля фо-
тонов, мы будем говорить просто о числе фотонов (квантов) в
расчете на моль. Например, в табл. 3.1 использованы микромо-
ли квантов, а не микроэйнштейны, чтобы избежать путаницы.]
В некоторых случаях целесообразно перевести эрги в элект-
ронвольты, разделив первые на 1,6-10~12:
2,9-Ю-12 2,9 1 о о
-^=1,8 эВ.
Отсюда видно, что энергия фотона с длиной волны 680 нм соот-
ветствует энергии, которую приобретает электрон под действи-
ем разности потенциалов 1,8 В.
(3.7)
1,6-ю-12
3.6. Интенсивность света
Количество энергии, поглощаемой листом, зависит не толь-
ко от «качества» падающего света, но и от его интенсивности.
Если рассматривать свет как волну, то «качество» света будет
Рис, 3.4. Определение интенсивности света в свечах. Если стандартную свечу
поместить в центр сферы радиусом 1 фут, то плотность потока, падающего
на ее внутреннюю поверхность, составит 1 лм.фут-2, что равно одной фут-
свече. При радиусе в 1 м поток будет равен 1 лм>м~2= 1 м-свече=1 лк =
= I • 10-4 фот.
ЭНЕРГИЯ II СВЕТ
391
определяться расстоянием между двумя последовательными
максимумами X, а интенсивность — амплитудой волны. Если же
считать свет потоком частиц, то энергия падающего света будет
определяться энергией каждой частицы (фотона или кванта) и
количеством частиц, падающих на поверхность листа (разд.
3.2).
Человеческий глаз'—очень эффективный инструмент при
сравнении низких освещенностей, но он очень плохо определяет
абсолютную интенсивность света в отсутствие стандартных об-
разцов, которые он мог бы •использовать для сравнения. Это
связано с тем,что для защиты сетчатки от яркого света радуж-
ная оболочка, действуя как ирисовая диафрагма, суживает зра-
чок, и мозг очень приблизительно, основываясь на опыте, может
оценить, сколько света поступает в глаз в этих условиях. Веро-
ятно по этой причине мы бываем очень удивлены, впервые узнав,,
что в хорошо освещенной комнате интенсивность света может
быть в 50 раз ниже полной интенсивности солнечного света.
Вначале силу света определяли, сравнивая данный источник
с так называемой «стандартной свечой» (рис. 3.4). Такая свеча
испускает световой поток в 4л лм (4л — телесный угол). Если
такую свечу поместить в центре сферы радиусом 1 фут, то плот-
ность потока, падающего на ее внутреннюю поверхность, т. е.
освещенность, будет равна 1 люмену на 1 фут2 (1 лм-фут~2)
или 1 фут-свече. Если радиус сферы увеличить до I м, освещен-
ность уменьшится до 1 лм-м"2 [или 1 метр-свеча, имеющая так-
же специальное название «люкс» (сокращенно лк)]. Так как
поток уменьшается пропорционально квадрату расстояния от
источника света и 1 м = 3,2808 фут, то 1 фут-свеча = 3,28082 лк =
=40,8 лк.
3.7. Солнечный свет и свеча
Измерение интенсивности света в метр-овечах (или люксах)
полезно для повседневного пользования, но недостаточно точ-
но при определении потребляемой энергии, поскольку другие ис-
точники света (не свеча) испускают свет с другим спектраль-
ным составом. Учитывая все это, мы можем произвести при-
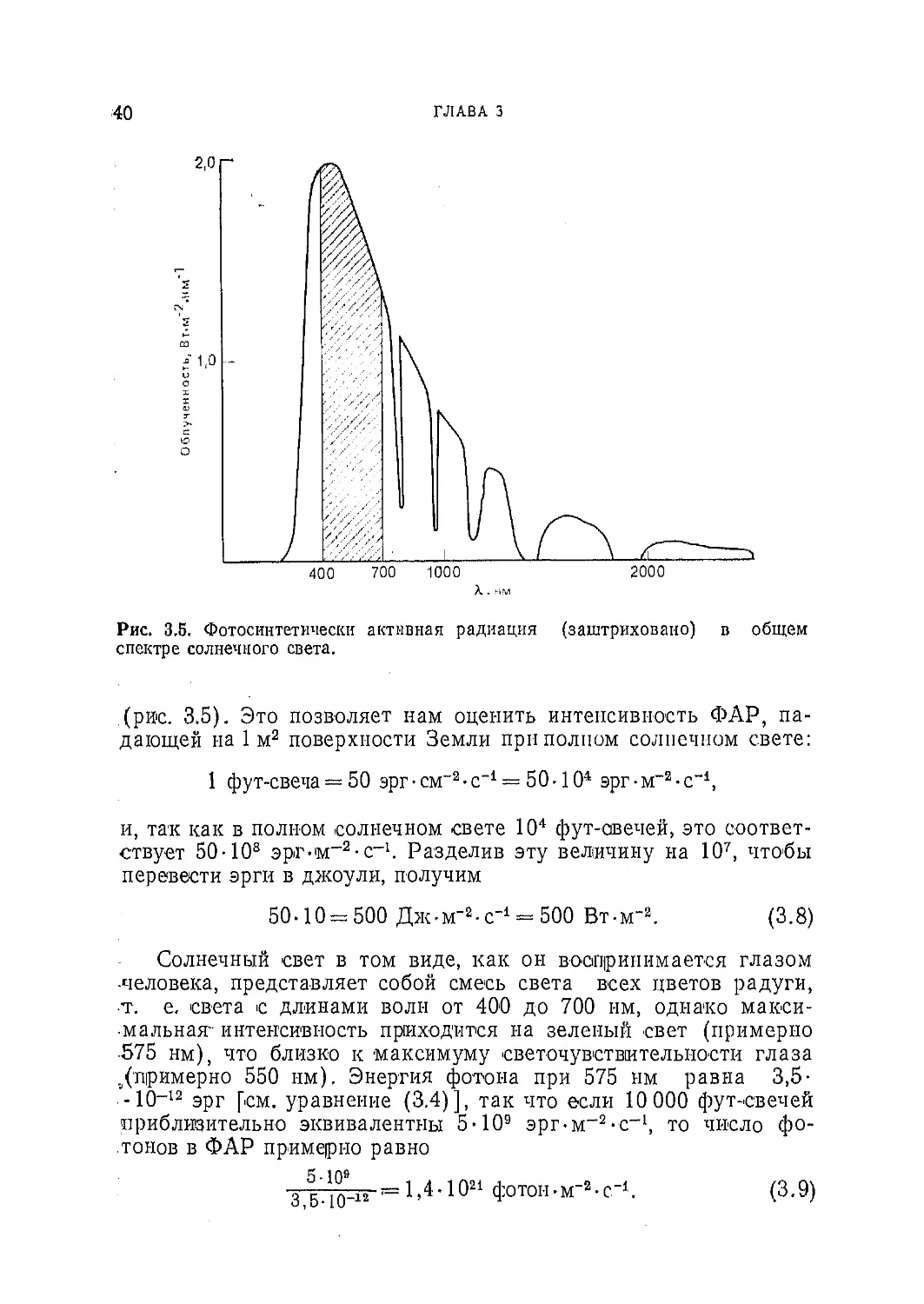
близительные расчеты. Плотность потока солнечного света у по-
верхности моря (прямой солнечный свет в полдень и рассеян-
ный синий свет неба) эквивалентна примерно 10 000 фут-свечам..
Для солнечного света (включая ультрафиолетовое и инфракрас-
ное излучение) 1 фут-свеча соответствует примерно 100 эрг-
•см-2-с-1. Так как па видимую часть спектра, т. е. на область
фотосинтетически активной радиации (ФАР; см. табл. 3.1),
приходится лишь 50% полной энергии солнечного излучения, то
для ФАР 1 фут-свеча соответствует примерно 50 эрг-см"2 -с"1
40
ГЛАВА 3
Рис. 3.5. Фотосинтетически активная радиация (заштриховано) в общем
спектре солнечного света.
(рис. 3.5). Это позволяет нам оценить интенсивность ФАР, па-
дающей на 1 м2 поверхности Земли при полном солнечном свете:
1 фут-свеча — 50 эрг • см~2 • с-1 — 50 -104 эрг • м“а • с-1,
и, так как в полном солнечном свете 104 фут-авечей, это соответ-
ствует 50-108 эрг-м-2-с_1. Разделив эту величину на 107, чтобы
перевести эрги в джоули, получим
50-10 = 500 Дж-м"2-с-1 = 500 Вт-мЛ (3.8)
Солнечный свет в том виде, как он воспринимается глазом
человека, представляет собой смесь света всех цветов радуги,
•т. е. света с длинами волн от 400 до 700 им, однако макси-
мальная- интенсивность приходится на зеленый свет (примерно
575 нм), что близко к максимуму светочувствительности глаза
./примерно 550 нм). Энергия фотона при 575 нм равна 3,5-
-10-12 эрг [см. уравнение (3.4)], так что если 10 000 фут-свечей
приблизительно эквивалентны 5 • 109 эрг-м-2-с-1, то число фо-
тонов в ФАР примерно равно
1,4 • 1021 ф0Т0н. м-а. с-1 (3.9)
0,0*10 л 4 z
ЭНЕРГИЯ II СВЕТ
41
Средняя годовая облученность для Англии составляет около 100 Вт-м-2.
Как мы видели, моль квантов часто оказывается более удоб-
ной единицей, чем одиночный квант, или фотон. Так,
1,4- 1021 фотонов соответствуют 3 моль квантов =
== 2,3-103 мкмоль квантов. (3.10)
Таким образом, интенсивность ФАР (400—700 нм) равна при-
мерно 2200 мкмоль квантов (или микроэйнштейнов) на 1 мг
в 1с. Характеристики различных параметров ФАР можно полу-
чить с помощью различных приборов.
а. Путем фотометрических измерений определяют световой
поток на единицу площади, или освещенность, в данном случае
составляющую 10 000 фут-свечей.
б. При радиометрических измерениях можно определить по-
ток излучения, или облученность, равную 500 Вт-м~2.
в. При измерении по числу квантов (например, с помощью
датчика, чувствительного в области 400—700 нм) можно опре-
делить поверхностную плотность потока квантов, в данном слу-
чае равную 2300 мкмоль-м~2-с"1.
Каждую из этих величии можно с каким-то приближением
nqpeBecTH в другую. Например, как показано выше, ФАР от
солнечного света (40 000 фут-свечей) ~500 Вт -M-2csi2300 мкмоль
квантов на 1 м2 в 1 с. Однако, проводя опыты на растениях, ис-
пользуют различные источники света, и без учета спектрального
состава напускаемого ими света подобный перевод делать нель-
зя. Так, если величины, указанные в какой-то работе, выраже-
ны в фут-свечах, их нельзя перевести в Вт-м-2 или (мкмоль
квантов) -м“2-с-1, не зная спектра излучения использованного
42
ГЛАЗА 3
источника света и нельзя количественно оценить ФАР различ-
ных источников в фут-свечах или Вт-м~2. Ясно, что если значи-
тельная часть энергии, испускаемой источником, лежит вне диа-
пазона 400—700 нм (лампа накаливания или ксеноновая лам-
па), то при использовании единицы Вт-м~2 мы получим завы-
шенную оценку ФАР. Аналогичным образом для ламп, испус-
кающих свет преимущественно в синей или красной области
спектра, мы получим заниженную оценку ФАР, если будем про-
водить измерения в фут-свечах. Правильнее сравнивать источ-
ники света по числу квантов или по облученности в области
400—700 нм, что дает гораздо более точные оценки ФАР. Но да-
же в этом случае существуют некоторые ограничения. Напри-
мер, натриевая лампа имеет интенсивную полосу испускания
при 560—620 нм; это дает высокую величину ФАР, однако ра-
стения предпочитают синий и красный свет.
3.8. «Зеленый человек»
Потребность в энергии у мужчины, ведущего малоподвиж-
ный образ жизни, составляет около 100 ккал-ч"1 (немногим
больше, чем электрическая энергия, потребляемая лампой
100 Вт). Средняя годичная облученность в разных участках
земной поверхности различается меньше, чем это можно пред-
положить. В районе Красного моря она составляет примерно
300 Вт-м-2, в Австралии — 200, в Соединенных Штатах—185,
в Англии — около 105 Вт-м-2. Таким образом, даже в Англии
человек мог бы получить всю необходимую ему энергию с 1 м2,
если бы умел со 100%-ной эффективностью превращать сол-
нечную энергию в энергию химических связей. Так как зеленые
растения используют энергию Солнца с эффективностью, не пре-
вышающей 5%, ясно, что «зеленому человеку», способному осу-
ществлять фотосинтез, все же пришлось бы «добывать» осталь-
ные 95% обычным путем, пока ему не удалось бы увеличить
поверхность своего тела в 10 раз. Площадь земной поверхно-
сти, необходимая для обеспечения годичных потребностей чело-
века сельскохозяйственными продуктами, может быть очень
разной в зависимости от внешних условий и характера питания,
однако она вряд ли будет меньше 400 м2.
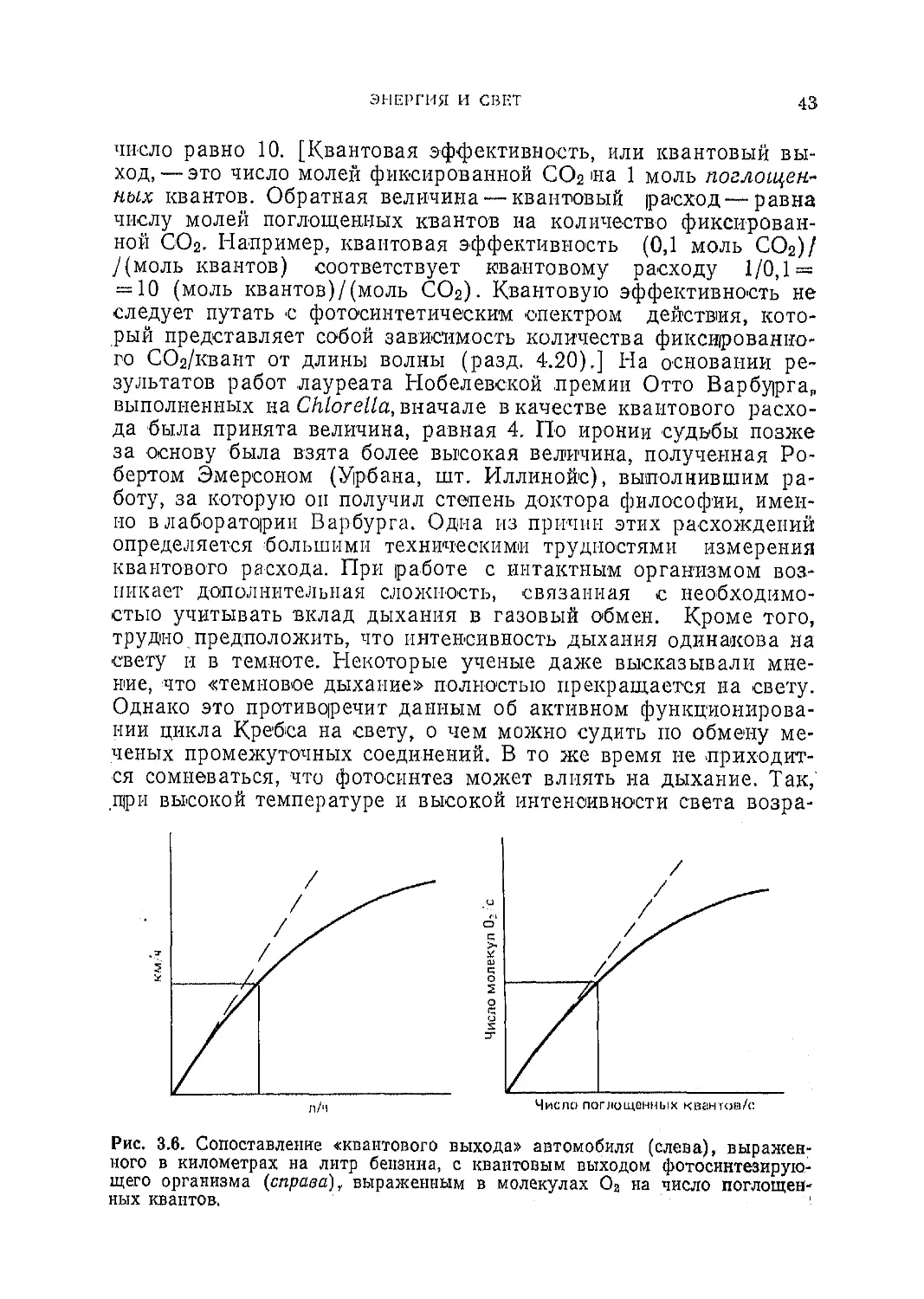
3.9. Квантовая эффективность
Так как энергия поступает от Солнца определенными пор-
циями (в виде фотонов) и довольно точно известно, какое ко-
личество энергии нужно для восстановления СО2 до СН2О,
можно, поставить вопрос, сколько фотонов необходимо для осу-
ществления этого процесса. В настоящее время принято счи-
тать, что требуется как минимум 8 фотонов, а скорее всего их
ЭНЕРГИЯ И СВЕТ
43
число равно 10. [Квантовая эффективность, или квантовый вы-
ход,— это число молей фиксированной СО2 на 1 моль поглощен-
ных квантов. Обратная величина — квантовый расход—равна
числу молей поглощенных квантов на количество фиксирован-
ной СО2. Например, квантовая эффективность (0,1 моль СО2)/
/(моль квантов) соответствует квантовому расходу 1/0,1 =
= 10 (моль квантов)/(моль СО2). Квантовую эффективность не
следует путать с фотосинтетическим спектром действия, кото-
рый представляет собой зависимость количества фиксированно-
го СОг/квант от длины волны (разд. 4.20).] На основании ре-
зультатов работ лауреата Нобелевской премия Отто Варбурга»
выполненных на Chlorella, вначале в качестве квантового расхо-
да была принята величина, равная 4. По иронии судьбы позже
за основу была взята более высокая величина, полученная Ро-
бертом Эмерсоном (Урбана, шт. Иллинойс), выполнившим ра-
боту, за которую он получил степень доктора философии, имен-
но в лаборатории Варбурга. Одна из причин этих расхождений
определяется большими техническими трудностями измерения
квантового расхода. При работе с интактным организмом воз-
никает дополнительная сложность, связанная с необходимо-
стью учитывать вклад дыхания в газовый обмен. Кроме того,
трудно предположить, что интенсивность дыхания одинакова на
свету и в темноте. Некоторые ученые даже высказывали мне-
ние, что «темновое дыхание» полностью прекращается на свету.
Однако это противчречит данным об активном функционирова-
нии цикла Кребса на свету, о чем можно судить по обмену ме-
ченых промежуточных соединений. В то же время не приходит-
ся сомневаться, что фотосинтез может влиять на дыхание. Так,'
,п|ри высокой температуре и высокой интенсивности света возра-
Рис. 3.6. Сопоставление «квантового выхода» автомобиля (слева), выражен-
ного в километрах на литр бензина, с квантовым выходом фотосинтезирую-
щего организма (трава), выраженным в молекулах Оа на число поглощен-
ных квантов.
Число поглощенных квантав/с
44
ГЛАВА 3
стает роль «фотодыхания» (гл. 13). При оценке влияния света
•на дыхание в ранних экспериментах с использованием мано-
метрической техники ситуация осложнялась различными «вы-
бросами» и «глотками» при переходе от света к темноте и на-
оборот.
Квантовый выход можно оценить из кривой зависимости ско-
рости фотосинтеза от интенсивности света (рис. 3.6). В усло-
виях, когда интенсивность света очень низкая и концентрация
СОг не лимитирует фотосинтез, эта зависимость является линей-
ной. Здесь уместна аналогия с автомобилем: можно определить,
.какое расстояние он проходит, используя 1 л бензина, и вычис-
лить таким образом эффективность использования бензина по-
добно тому, как оценивалась эффективность фотосинтеза. Для
машины она выражается в км-л"1, а для растения — в коли-
честве О2 на квант (рис. 3.6).
3.10. Эффективность фотосинтеза
на молекулярном уровне
Энергия восьми «молей квантов» фотонов красного света при
680 нм равна 8-42 = 336 ккал. При сжатии в калориметре одной
грамм-молекулы СН2О выделяется 672/6=112 ккал. Следова-
тельно, превращение энергии света в тепловую энергию осуще-
ствляется с эффективностью 112/336, или 33%. Это максималь-
но возможная эффективность фотосинтеза на молекулярном
уровне. Наибольшее количество энергии теряется при превраще-
нии световой энергии в ассимиляционную силу (ATP-j-NADPHs).
Можно считать, что при возбуждении хлорофилла энергия све-
та не теряется или почти не теряется (разд. 4.16), и, как было
показано выше (разд. 2.12), полагать, что на генерацию асси-
миляционной силы расходуется 125 ккал. Реальная эффектив-
ность фотосинтеза на молекулярном уровне значительно ниже,
чем приведенная максимальная величина. Квантовый расход
может быть равен восьми, однако скорее всего он равен все-та-
ки 10. Кроме того, квантовый расход примерно одинаков для
разных участков спектра, что связано с малым временем жизни
хлорофилла в возбужденном состоянии при освещении его синим
светом (разд. 4.16). Соответственно фотосинтез при синем све-
те с большой энергией квантов значительно менее эффективен.
Даже если принять среднее значение энергии фотона в видимой
области спектра равным 3,5-10~12 эрг-фотон-1 (==50 ккал) (ср.
.разд. 3.5) и квантовый расход равным 10, то эффективность
уменьшается от 33 до 22%. Если считать, что на долю ФАР при-
ходится 50% всего солнечного света (видимый свет-|-Уф-|-ИК
и т. д.), то эта величина составит лишь 11%.
ЭНЕРГИЯ И СВЕТ
45
3.11. Максимальная эффективность фотосинтеза
на уровне растения в целом
Для растения в целом эффективность использования энергии
света (т. е. его фотосинтетическая эффективность) существен-
но ниже, чем на молекулярном уровне. Все факторы, чье совме-
стное действие снижает эффективность фотосинтеза, можно объ-
единить в основном в две группы. Во-первых, это факторы, огра-
ничивающие эффективное поглощение света, во-вторых, фак-
торы, связанные с диссипацией энергии, которая могла бы быть
использована для фотосинтеза.
Растения имеют зеленую окраску, так как пропускают и рас-
сеивают зеленый свет (рис. 3.1). Потери энергии при этих и
других процессах оценить очень трудно. Однако, основываясь
на расчетах,, выполненных для процессов на молекулярном уров-
не с учетом потерь при облучении синим светом (разд. 4.16),
можно предположить, что эффективность уменьшится еще на
25%, т. е. от 11 до 8%. (При этом предполагается, что расте-
ния занимают всю освещаемую поверхность, хотя обычно это
далеко.не так, особенно в начале вегетационного периода).
Диюсипационные потерн обычно сводятся к потерям при ды-
хании и фотодыхании. В этом случае тоже можно дать лишь са-
мую приближенную оценку, согласно которой около 40% угле-
рода, ассимилированного при фотосинтезе, теряется в ходе ме-
таболизма, а не накапливается в виде запасных продуктов или
новых тканей. Итак, 'эффективность фотосинтеза составляет
около 5%, т. е. примерно 5% общей энергии падающего света
выделяется в виде тепла при сжигании в калориметре растения,
поглотившего свет. Это согласуется с большим числом экспери-
ментальных данных, указывающих на величину 4—6%. К сход-
ной величине можно прийти и другим путем. Например, для Реп-
nisetum typhoides, выращиваемого в Катарине (Восточная Авст-
ралия) и получающего в сутки 5100 ккал-м-2 энергии, приво-
дится величина 54 г-м~2-сут_1. Если принять среднюю калорий-
ность растительного материала равной 4,25 ккал-г-1 (что пре-
вышает калорийность углеводов 3,7 ккал-г-1), можно рассчи-
тать эффективность использования энергии:
Л^-100 = 4,5%.
3.12. Максимальная эффективность
Если у растений 5% энергии света превращаются в хими-
ческую энергию, можно оценить максимальную эффективность,
исходя из данных о средней облученности (т. е. энергии, усред-
46
ГЛАВА 3
ценной за 365 сут. при 24-часовом периоде освещения) и кало-
рийности 4,25 ккал на 1 г сухой массы.
Для Англии средняя годовая облученность составляет
105Вт-м2 (примерно 25 кал-м“2/с_‘, или 90 ккал•м'”2/ч1). 5%
от этой величины (4,5 ккал) могли бы быть эквивалентны при-
мерно 1 г-м~2*ч~1 (около 9 кг-м"2-год-1 — 90 т-га-1, год-1). Со-
ответствующая величина для Австралии и США была бы при-
мерно в два раза выше.
Однако в реальных полевых условиях такой урожай редко
удается получить. Так, в холодных климатических зонах боль-
шая часть света поступает при температурах, значительно более
низких, чем те, что требуются для фотосинтеза; листовая по-
верхность не покрывает всю площадь земли и т. д. Вместе с тем
для большинства С3-растений интенсивность света слишком ве-
лика, насыщение наступает при 1/5 полного солнечного света,
и на ярком свету при высокой температуре может быть избы-
точным фотодыхание. Тем не менее, как мы уже указывали,
Pennisetum typhoides в Австралии дает 54 г сухой массы на
1 м2/сут, что несколько выше величины, полученной из средней
годовой облученности. Даже в Англии урожай картофеля может
достигать 88 т сырой массы на 1 га, что составляет 30% или
более от теоретически возможного максимума, если учитывать
всю «биомассу» растения. Картофель дает особенно высокий
урожай в условиях умеренного климата, что отчасти определя-
ется его Длительным вегетационным периодом и способностью
расти при температуре ниже жизненного минимума для многих
видов Ci-растений, обладающих потенциально большими воз-
можностями.
ГЛАВА 4
Фотохимический аппарат
и его функции
Фотохимический аппарат
В классическом эксперименте Требса, Тсудзимото и Ариона
было установлено, что фотосинтез можно подразделить на две
стадии: световую и темновую (рис, 2.4). Они показали, что в
простейшем виде восстановление CO2 до СЯ2О (гл. 1) может
происходить в темноте при наличии восстановительной силы
(АТР и NADPH2), образованной независимо от этого процес-
са на свету. Темновая стадия катализируется растворимыми
белками стромы хлоропласта, а световая — осуществляется с
помощью фотохимического аппарата.
4.1. Структура хлоропласта
У человека главный красный пигмент локализован в эритро-
цитах. У растений преобладающий зеленый пигмент — хлоро-
филл—сконцентрирован в органеллах, называемых хлоропла-
стами (рис. 4.1), которые по размеру мало отличаются от эрит-
роцитов. Хлорофилл распределен по хлоропласту не равномер-
но, но сосредоточен в мембранах тилакоидов (разд. 4.2), со-
ставляющих фотосинтетический аппарат. Слово «тилакоиды»
греческого происхождения. Когда-то так называли мешковатые
шаровары, которые носили турки, а к мембранам этот термин
применили потому, что они также напоминают «мешки». Обыч-
но эти «мешки» уплощены, и в хлоропластах растений мембра-
ны представляются сдвоенными. Часто тилакоиды собраны в
стопки, которые в световом микроскопе выглядят как «граны»,
или зерна. Фотохимический аппарат погружен в строму — ге-
леобразную массу растворимых белков, в число которых входят
все ферменты, участвующие в ассимиляции углерода. Хлоро-
пласт окружен двумя мембранами; внутренняя мембрана играет
большую роль в переносе веществ между хлоропластом и окру-
жающим его цитозолем (гл. 8). [Хлоропласт, как и целая ра-
стительная клетка, реагирует на осмотическое давление. Если
полоску эпидермиса лука поместить в концентрированный рас-
твор сахарозы, будет наблюдаться плазмолиз: содержимое клет-
ки сжимается, отходит от клеточных стенок по мере перехода
воды в окружающий раствор. Это связано с выходом воды на-
48
ГЛАВА 4
Рис. 4.1. Строение хлоропласта: схематическое изображение основных компо-
нентов.
ружу через клеточную мембрану— плазмалемму, в результате
чего наружный раствор разбавляется (экзосмос) и концентра-
ция -раствора по обе стороны мембраны выравнивается. Подоб-
ным же образом вода переходит в мешочек, образованный по-
лупроницаемой мембраной и содержащий молярный раствор са-
харозы до тех пор, пока гидростатическое давление не достигнет
величины, эквивалентной 22,4 атм. Осмотическое давление рас-
твора определяется числом растворенных частиц независимо от
того, молекулы это, атомы или ионы. Так, в эквимолярном рас-
творе соли создается более высокое осмотическое давление,
чем в растворе сахара, из-за диссоциации: NaCl—»-Na++Cl-.]
Чтобы выделить из листа неповрежденный хлоропласт, необхо-
димо разрушить ткань в изотоническом растворе, т. е. в раство-
ре с таким же осмотическим давлением, как в органеллах, це-
лостность которых необходимо сохранить. Для этой цели часто
используют растворы сахаров, которые не могут проникать че-
рез внутреннюю мембрану (гл. 8), в результате чего поддержи-
вается осмотическое равновесие по обе ее стороны. Однако пос-
ле выделения хлоропласты легко разрушаются под действием
осмотического шока; если их поместить в дистиллированную
воду, они быстро разбухают в результате эндосмоса. Разрушен-
ные хлоропласты при этом теряют строму, ио нерастворимые
мембраны тилакоидов остаются относительно неповрежденными.
Тилакоиды в свою очередь разбухают, если их -оставить в воде,
но их наружная мембрана разрывается примерно через 30 с и,
если за это время осмотический потенциал наружного раствора
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
49
восстановить путем добавления сахара, то мембраны тилакоида
остаются целыми. Выделенный из хлоропласта фотосинтетиче-
ский аппарат или даже его фрагменты сохраняют способность
при освещении генерировать ассимиляционную силу (восстано-
вительную силу, ATP-j-NADPIT2). Такие препараты из изоли-
рованных мембран тилакоидов иногда называют разрушенны-
ми, фрагментированным1!-! хлоропластами или хлоропластами
без оболочек, иногда — свободными ламеллярными телами
(имея в виду, что ламелла— одиночный мембранный слой).
4.2. Тилакоидный компартмент
Как мы уже говорили, мембраны тилакоидов в хлоропласте
имеют вид уплощенных мешков или двойного мембранного
слоя. Представляется вероятным, что в хлоропласте все они
составляют единую систему (рис. 4.1) и что этот мембранный
слой разветвлен, уложен складками (иногда образует граны —
структуры, напоминающие стопки монет) или формирует поло-
сти (фреты). Совокупность этих мембран образует единый ком-
партмент — фотохимический аппарат.
4.3. Стромальный компартмент
Если все это верно, то отсюда следует, что единый непре-
рывный тилакоидный компартмент располагается внутри еди-
ного стромального компартмепта, который в свою очередь окру-
жен внутренней оболочкой хлоропласта (рис. 4.1). Строма —
это белковый тель, главный каталитический компонент которого
представлен рибулозобисфосфат-карбоксилазой (белок фрак-
ции I). Полагают, что в хлоропластах клеток мезофилла С4-рас-
тений эта карбоксилаза отсутствует (табл. 11.2). Если растение
синтезирует крахмал (такой способностью обладают не все рас-
тения: классический пример — подснежник Galanthus nivalis),
то он локализуется в строме в виде зерен, не связанных с какой-
либо специализированной мембранной системой. Кроме того, в.
строме находят другие включения, такие, как пластоглобулы
(липидные глобулы) и 705-рибосомы (частицы, участвующие в
синтезе белка). В большинстве случаев в строме содержатся
также ионы (например, Mg2+), коферменты, такие как АТР и
NADPy растворимые белки, участвующие в транспорте электро-
нов (например, ферредоксин) и ферменты, ответственные за ас-
симиляцию углерода.
4.4. Структура мембран
Тонкая внутренняя структура мембран тилакоида пока до>
конца не установлена. Известно, что они состоят из примерно'
одинакового количества липидов (жиров) и белков, и есть осно-
4- 4
50
ГЛАВА 4
Рис. 4.2. Схематическое изображение мембраны тилакоида; переносчики элек-
тронов расположены в соответствии с представлениями об их локализации
в цепи переноса электронов. Белки тилакоида и переносчики электронов по-
гружены в полужидкий матрикс. Световая энергия, улавливаемая пигмента-
ми светособирающего комплекса (Хл а/Ь) — белок, направляется к фотохими-
чески активным молекулам Хл а реакционного центра ФС II — Р680 (Р690).
При возбуждении Р680 теряет электрон, и в реакционном центре образуется
сильно окисленный Р680+. Это приводит к окислению первичного донора
электронов ФС II, Z, что сопровождается потерей электрона в системе рас-
щепления воды Y. Полное окисление воды достигается при удалении из Y
четырех электронов. Электроны от Р680 улавливаются первичным акцептором
электронов ФС II — пластохиноном, а затем передаются к вторичному ак-
цептору В и далее к пулу молекул пластохинона (PQ). Пул пластохинонов
участвует также в переносе протонов через мембрану. Железосерусодержа-
щий белок переносит электроны от PQ к цитохрому f (цит. f), а затем к
пластоцнанину (PC), связанному с хлорофиллом реакционного центра
ФС1 — Р700. При возбуждении светом Р700 теряет электрон, место которого
занимает электрон, получаемый от PC. Первичный акцептор электронов в
ФС I, принимающий электрон от Р700, возможно, представляет собой димер
хлорофилла, который передает электроны на цепь железосерусодержащих
центров, обозначенных как X, В и А. От последнего электрон поступает к
молекуле ферредоксина и далее к ферредокснн-NADP — редуктазе -- флаво-
протеину, способному использовать электрон, полученный от ферредоксина,
для восстановления NADP+ с образованием NADPH. Через цитохром
(цит. bs), переносящий электрон обратно к PQ, может осуществляться цик-
лический транспорт электрона. Перенос электронов через мембрану тилако-
ида создает протонный градиент, что способствует образованию АТР из ADP
с помощью сопрягающего фактора, который состоит из двух компонентов
CF, и CF(i. CF0 погружен в мембрану тилакоида и представляет собой про-
теолипидпый протонный канал, CF, проникает в строму и обладает АТРаз-
ной активностью.
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
51
вания полагать, что эти компоненты находятся в жидкокрис-
таллическом состоянии. Некоторые факты говорят в пользу то-
го, что эта структура представляет собой элементарную мембра-
ну, в которой липидный бислой (и 'связанные с ним пигменты)
находятся между двумя слоями белка. Молекулы липидов со-
стоят из полярной (гидрофильной) головки, обычно содержа-
щей одну или больше молекул шестиуглеродного углевода — га-
лактозы, и длинных неполярных углеводородных «хвостов».
Кроме того, в мембране присутствует также небольшое коли-
чество более полярных липидов, содержащих фосфат (фосфо-
липиды) .
Позже было показано, что белки не образуют сплошной слой,
а находятся в форме глобул или гранул или пронизывают ли-
пидный бислой в определенных местах (рис. 4.2). Во всяком
случае, образуется более или менее непрерывная гидрофобная
структура, благодаря которой мембрана не растворяется в воде
п включает пигменты, растворимые в липидах, и в то же время
ие препятствует взаимодействию мембранных белков (разд.
5.12) с водорастворимыми компонентами стромы. Наряду с хло-
рофиллом в тилакоидах находятся также компоненты электрои-
траиспортной системы (разд, 4.9).
Оболочка хлоропласта также состоит из смеси белков и ли-
пидов; из них на долю наиболее изученного в настоящее время
фосфолипида —- фосфатидилхолина приходится около 20%, Дус
и Бенсон показали, что мембраны лишены хлорофилла (они
окрашены в желтый цвет) и содержат уникальный набор каро-
тиноидов (разд. 4.7), которые, очевидно, могут выполнять роль
фильтра, поглощающего синий свет и защищающего хлорофилл
тилакоидов от разрушительного фотоокисления. Эти мембраны
существенно отличаются от мембран тилакоидов ji, судя по про-
ницаемости (гл. 8), состав и/илн строение наружной и внут-
ренней оболочек существенно различаются.
4.5. Хлорофиллы
Хлорофиллы (Хл) встречаются у высших растений в двух хи-
мически различающихся формах — а и b (рис. 4.3). Как и гем,
который ответствен за красный цвет крови, хлорофиллы пред-
ставляют собой порфирины, однако четыре пиррольных кольца
порфиринового ядра связаны не с Fe, как в геме, а с Mg. По-
мимо тетра'пирролы-юго макроцикла хлорофилл имеет длинный
фитольный «хвост», состоящий из четырех изопреновых единиц,
что придает молекуле гидрофобный характер, Хл а незначитель-
но отличается от Хл b (одна из его боковых цепей представля-
ет собой СН3-, а не СНО-группу), но этого достаточно, чтобы
4*
52
Г.1АВЛ 4
Рис. 4.3. Структурные формулы Хл а, Хл b и изопрена.
значительно сдвинуть максимум поглощения пигмента: Хл а
имеет сине-зеленый цвет, а Хл b — желто-зеленый (рис. 4.4).
Хл а — это основной фотосинтетический пигмент, обнаруженный
во всех зеленых растениях. В большинстве наземных высших
растений его содержание в 2,5—3,5 раза превышает содержание
Хл Ь.
Впервые хлорофиллы были разделены на колонках с са-
харозой русским ботаником Цветом (рис. 4.4). Максимум погло-
щения Хл а в ацетоне лежит в красной области, вблизи 663 нм,
однако in vivo в мембране тилакоида максимум сдвигается в
более длинноволновую часть спектра, вероятно, в результате
взаимодействия с другими молекулами хлорофилла и с белка-
ми. Согласно спектроскопическим данным, Хл a in vivo суще-
ствует в нескольких формах (разд. 4.11). Зеленая окраска ра-
стений (в белом свете) определяется тем, что хлорофилл пре-
имущественно поглощает красный и синий цвет, но пропускает
или отражает зеленый.
ФОТОХИМИЧЕСКИЙ АППАРАТ И ЕГО ФУНКЦИИ
53
Рис. 4.4. Спектр поглощения хлорофилла в ацетоне.
4.6. Синтез хлорофилла
О синтезе хлорофилла многое известно благодаря изучению
его образования in vivo. In vitro Хл а был синтезирован Вудвор-
дом. Первой ступенью синтеза хлорофилла считается образова-
ние 5-амииолевулиновой кислоты (АЛК). У животных и бакте-
рий АЛК, использующаяся для синтеза тетрапирролов, образу-
ется из сукцинил-СоА и глицина с участием АЛК-еинтетазы.
Однако в зеленеющих листьях не исключены и другие пути.
Так, предшественником для синтеза АЛК может служить глу-
тамат или 2-оксоглутарат, а не глицин. Поскольку 2-оксоглута-
рат обычно синтезируется в митохондриях, на синтез хлорофил-
ла может влиять метаболизм в митохондриях. Этот факт, а так-
же то, что малая субъединица р'ибулозобисфосфат-карбоксилазы
синтезируется вне хлоропласта, не согласуются с гипотезой воз-
никновения хлоропласта из симбиотических сине-зеленых водо-
рослей (рис. 4.5).
На завершающих стадиях синтеза хлорофилла желтый, свя-
занный с белком протохлорофиллид (окисленный предшествен-
ник хлорофилла без фитольного хвоста) подвергается фотовос-
становлению. до зеленого хлорофиллида, а затем происходит
присоединение фитола. В темноте образуется только небольшое
количество протохлорофиллида, который может очень быстро
фотовоостанавливаться. Так, проростки, растущие в темноте,
могут позеленеть при последовательном воздействии миллисе-
кундных вспышек света и длительных (15 мин) темновых ин-
54
ГЛАВА 4
соон соон
сн, ___
CH, + NH3-CH2 ch; + СО, + СоА
I 2 I I
C-S-CoA СООН С —CH,-—NH3
II
О
Сукцииил-CoA Глицин 6- Аминопевулиновап кислота
Рис. 4.5. Синтез хлорофилла: образование амниолевулииовой кислоты из сук-
цинил-СоА.
тервалов, в ходе которых снова образуется протохлорофпллид.
Хвойные, некоторые зеленые водоросли, а также многие папо-
ротники способны синтезировать хлорофилл в полной темноте.
Однако не исключено, что позеленение семян цитрусовых обус-
ловлено светом, проникающим через плод.
4.7. Дополнительные пигменты
Наряду с хлорофиллами а и b хлоропласты содержат каро-
тиноиды, главным образом p-каротин, лютеин, виолаксаптин и
неоксантин (табл. 4.1). Это изопреновые (полиеновые) высоко-
полимерные соединения с циклогексановыми кольцами на кон-
цах. Считают, что каротиноиды играют роль дополнительных
рецепторов света, увеличивающих тот спектральный диапазон,
который может использоваться для фотосинтеза, и, кроме того,
защищающих хлорофилл от окисления.
4.8. Курица или яйцо
При изучении свойств фотосинтетического аппарата мы стал-
киваемся с проблемой, заключающейся в том, что нельзя уста-
новить его тонкую структуру, не зная функциональной роли
отдельных элементов, а это в свою очередь нельзя до конца по-
нять, не зная структуры. Будем пока исходить из предпосылки,
что первична функция фотосинтетического аппарата, состоя-
щая в трансформации энергии, т. е. превращении энергии света
сначала в электрическую энергию, а затем в энергию химиче-
ских связей (ассимиляционную силу; разд. 5.2). По современ-
ным представлениям это достигается с помощью такой его
структурной и функциональной организации, которая позволяет
фотонам «стекать» в две фотосистемы (рис. 4.6). Эти системы
соединены друг с другом через ряд переносчиков электронов,
образующих своеобразный мост между ними. Совокупность двух
55
фотохимический аппарат и его функции
Рис. 4,6. Фотохимический аппарат: основные функции. Две фотосистемы со-
единены «мостом» из переносчиков электронов (разд. 4.9). Чтобы «поднять»
два электрона от молекулы воды через реакционный центр ФС II — Р690
(разд. 4.12) к первичному акцептору электронов, необходимо два фотона. От-
сюда электроны «сбегают» вниз по термодинамической «лестнице» «моста» и
заполняют дырки, образовавшиеся при возбуждении Р700 ФС I. Далее элек-
троны проходят через X к NADP+; количество фотонов, необходимое для
переноса 2е от воды к NADP+, равно четырем (см. рис. 4.2 и 4.18).
фотосистем (разд. 4.13) и соединяющего их «моста» составляет
Z-схему (разд. 4.23). Чтобы попасть от воды к NADP+ элект-
роны должны пройти весь этот путь. Ниже мы рассмотрим бо-
лее подробно структуру и механизм действия Z-схемы.
4.9. Компоненты электрон-транспортной цепи
Переносчики электронов, ассоциированные с двумя фотоси-
стемами, объединяют их в единую Z-схему (разд. 4.13, 4.23).
К числу переносчиков относятся все промежуточные соединения
на пути электронов от Н2О до NADP+, однако их локализация,
количество и природа установлены далеко не во всех случаях.
Некоторые из этих соединений, а также их окислительно-восста-
новительные потенциалы Ег (см. разд. 2.13) приведены ниже в
порядке их предполагаемого места в цепи переноса электронов.
Белок, содержащий марганец (Y)
Полагают, что в состав ФС II входит марганецсодержащий
белок У, который катализирует первые стадии выделения О2.
[При слабом нагревании и под действием гидроксиламина или
56
ГЛАВА 4
Таблица 4.1. Основные и дополнительные пигменты фотосинтетического
аппарата высших растений и их максимумы поглощения (приблизит.)1
в нанометрах
В органических растворителях In vivo
Хлорофилл а 420, 663 (в 80%-ном ацетоне) 435, 670-680
Хлорофилл b 453, 645 (в 80%-ном ацетоне) 480, 650
[1-Каротин 425, 450, 480 (в гексане) 445, 470, 500
Лютеин 425, 445, 475 (в этаноле) 445, 465, 495
Виол аксаи тин 425, 450, 475 (в этаноле) 445, 470, 595
1 Максимумы поглощения в органических растворителях определены точно. Значения,
полученные in vivo, менее точны, особенно для каротиноидов (0-каротии, лютеин, вио-
лаксатин), полосы поглощения которых перекрываются, а также для синей полосы хло-
рофиллов.
трис-буфера с высоким pH, когда хлоропласт теряет марганец,
выделение О2 подавляется, хотя цепь переноса электронов мо-
жет функционировать при наличии доноров электронов, отлич-
ных от Н2О. Водоросли, растущие на среде, дефицитной по Мп,
теряют способность выделять О2, ио могут осуществлять фото-
восстановление СО2 с помощью Н2. То, что Мп в хлоропластах
плохо поддается определению с помощью ЭПР, наводит на
мысль, что он связан с белками.] На 400 молекул хлорофилла
приходится около 5—8 атомов марганца. Очевидно, минималь-
ное количество Мп, необходимое для полного выделения О2, со-
ставляют 4 атома иа реакционный центр (Е' для Мп3+/Мп2'-
равно примерно +1100 мВ).
Ион хлора
Хлор необходим для осуществления окислительной функции
ФС II. При отмывке от С1~ хлоропласт теряет способность ис-
пользовать Ы2О в качестве донора электронов, хотя по-прежие-
му может использовать в качестве допоров аскорбат или дифе-
нилкарбазид.
Z
Это первичный донор электронов для ФС II. Обнаружен с
помощью ЭПР; ничего другого о нем неизвестно.
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
57
Цитохром 6559
Бендалл описал форму цит. Ьй с высоким потенциалом (Е' =
= 370 мВ) и с максимумом поглощения при 559 нм, которая,
возможно, ассоциирована с Y (см. выше). Об этом свидетель-
ствует то, что цит. &559 при низкой температуре окисляется с по-
мощью ФС II, но не ФС I и ие восстанавливается ФС II. Воз-
можное объяснение этого эффекта было предложено Батлером.
Р 690
Этот переносчик обозначают также через Р 680 или Р 683.
Он является реакционным центром (РЦП) фотосистемы II
(ФСП). Представляет собой молекулу (или димер) Хл а (разд.
4.12). В ФС II служит ловушкой фотонов, поглощаемых после-
довательным рядом молекул хлорофилла, составляющих антен-
ну (рис. 4.18), что приводит к изменению спектра прш 690 нм
(отсюда название пигмента). Может существовать в виде комп-
лекса Z-РЦ II-Q, где электрон поступает от Z и следует к Q.
Q (тушитель флуоресценции)
В окисленном состоянии этот компонент тушит флуоресцен-
цию молекул Хл а, передающих энергию на РЦ II (или Р 690),
и служит первичным акцептором электронов ФСП. С измене-
нием окислительно-восстановительного состояния Q связано из-
менение поглощения при 550 им некоего компонента С-550, но
нельзя сказать, что этот компонент и есть соединение Q. Флуо-
ресценция хлорофилла сильнее всего проявляется, когда блоки-
рован «нормальный» путь диссипации энергии. Тушение флуо-
ресценции может осуществляться многочисленными окислами,
которые восстанавливаются по мере перехода хлорофилла в
окисленное состояние. Таким образом, «нормальный» путь сня-
тия возбуждения, по-видимому, играет роль эффективного ту-
шителя флуоресценции. Содержание Q по отношению к хлоро-
филлу составляет 1/400, —35 мВ. Q представляет собой свя-
занный пластохинон, подвергающийся одноэлектронному восста-
новлению с образованием семихинона QH. (Согласно Краусу и
др., тушение флуоресценции может определяться кроме того,
высокоэнергизованным состоянием мембран тилакоида.)
Пластохинон
Впервые этот компонент охарактеризован Кофлером; он пред-
ставляет собой диметилбензохинон с боковой цепью из девяти
изопреновых единиц (и, следовательно, сходен с убихиноном
58
ГЛАВА 4
сн5
CHj
(CH2 -CH -C CHj), н
СНз-.П|
CH3-J
сн’
(СН2- сн с сн2)9 н
[Зпестаноцленнын ।ныстоХИНОН
Рис. 4.7. Пластохинон.
митохондрий, а по структуре «хвоста»—с каротиноидами и хло-
рофиллами). Существует ряд его аналогов и изомеров. Этот пе-
реносчик электронов содержится в хлоропласте в самом боль-
шом количестве (1 молекула на 10 молекул хлорофилла) и на-
ходится в виде мобилы-юго пула в мембране. Окислительно-вос-
становительный потенциал его равен нулю, и поэтому, а также
по ряду других причин пластохинон наиболее тесно ассоцииро-
ван с ФС II. Он может присоединить два атома водорода. Окис-
лительно-восстановительная реакция между Q- и пластохино-
ном требует участия связанного хинона В, восстанавливающе-
гося до В~ первым фотоном и до В2- — вторым, т. е. перенося-
щим оба эквивалента к пулу пластохинона.
Цитохром f
Это цитохром с-типа (из листьев L. frons), обнаруженный
Хиллом и Скарисбриком, с максимумом поглощения в интерва-
ле 550—555 нм (в зависимости от источника) и окислительно-
восстановительным потенциалом 300—370 мВ. На 400 молекул
хлорофилла приходится примерно 1 молекула этого соединения.
Ассоциирован с ФС I. Переносит один электрон.
Белок, содержащий железо и серу
Этот белок (£' = 290 мВ) расположен между пластохиноном
и цитохромом f.
ФОТОХИМИЧЕСКИЙ АППАРАТ И ЕГО ФУНКЦИИ
59
Пластоцианин
Медьсодержащий белок (два атома Си на молекулу белка),
впервые обнаруженный Като. Синий в окисленной форме, бес-
цветный—'в восстановленной; Z?/ = -|-370 мВ. На 400 молекул
хлорофилла приходится примерно 1 молекула этого соединения.
Ассоциирован с ФС I, а также выполняет функции связующего
звена между двумя фотосистемами (разд. 4.13). Переносит один
электрон.
Пигмент 700 (Р 700)
По данным Вессела Кока, Р 700 представляет собой реакци-
онный центр ФС I. Это одна из форм Хл а; существует в виде
димера с максимумом поглощения в красной области около
700 им. Обесцвечивается при окислении. На 400 молекул хло-
рофилла приходится примерно 1 молекула Р 700. Окислительно-
восстановительный потенциал £,/ = 430—530 мВ. Переносит один
электрон.
X (Р430)
Р 430 является первичным акцептором электронов в ФС I
(разд. 4.13). Это соединение неизвестной природы, с более от-
рицательным потенциалом, чем у ферредоксина, по-видимому,
равным 600—700 мВ (это значение получено на основании спо-
собности хлоропласта восстанавливать виологены с низким по-
тенциалом). Возможно, представляет собой комплекс железосе-
русодержащего белка (мембраносвязанный ферредоксин?) с ка-
ким-то другим соединением. Выявляется по изменению спектра
поглощения при 430 нм при освещении, а также методом ЭПР.
Возможно, существуют два (Fe—S)-компонента В и А, кото-
рые служат посредниками между X и ферредоксином.
Ферредоксин
Это соединение вместе с флавопротеином (см. ниже) впер-
вые упоминается в литературе Давенпортом, Хиллом и Уэтли
как фактор, восстанавливающий метгемоглобин, а затем Сан
Пиетро и Лангом как фотосинтетическая пиридиннуклеотидре-
дуктаза. Позже Мортенсоном из бактерий был выделен некий
железосерусодержащий белок. Сходное соединение из шпината
выделили Арнон и Тагава, которым удалось с его помощью в
присутствии флавопротеина провести каталитическое восстанов-
ление NADP+ в освещенных хлоропластах. Окислительно-вос-
60
ГЛАВА 4
становительный потенциал ферродоксина равен —430 мВ. Од-
ноэлектронный переносчик. Содержит два атома железа на мо-
лекулу (разд. 5.15).
Флавопротеин (ферредоксин — NADP^-редуктаза)
Этот фермент переносит электроны от восстановленного фер-
редоксина к 1NADP. В то же время он действует как трапсгид-
рогеназа (NADPH2-HNAD+—>-NADP++NADH2) и диафораза:
(фермент, осуществляющий перенос электронов от NADPH2 к
искусственным акцепторам). Фермент содержит одну молеку-
лу ФАД (флавинадениндинуклеотида), слабо связан с мембра-
ной; мол. масса равна 40 000, Е' =—380 мВ.
Никотинамидадениндинуклеотидфосфат (NADP)
Является конечным акцептором фотосинтетического транс-
порта электронов в Z-схеме (разд. 4.23). В восстановительном
пентозофосфатном цикле NADPH2 поставляет электроны 1,3-ди-
п 2 ,Сх Н N С" У 1 II , нсч НС- с—CO\’HZ N HCx. <:н ° ° | | | СН2--0 Р--Р -о сн, °М 1 1 /°х \Н Н/ ОН ОН Си , W нгН он он он с Никогинамидадининдинуклвотидфосфат ОЛ’ или Т2 .„.С.„ С 'С—CONH, II II 2 -н Н + С. С С|- 1 R. blADPHj (ипиМАОРНрти NADPH г н4 Л СН / "N )РО OHZ
Рис. 4.8. NADP.
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ и!
фосфоглицериновой кислоте, которую, таким образом, можно
рассматривать как конечный «окислитель» в реакции Хилла в
хлоропласте (разд. 4.13). NADP+ не может проникать через
оболочку хлоропласта, и перенос в цитоплазму этого компонен-
та восстановительной силы осуществляется то челночному ме-
ханизму (разд. 8.20 и 8.26). При восстановлении NADP при-
нимает 2 электрона и 1 протон, так что запись NADPH4-H+
более точно отражает строение NADP в восстановленной фор-
ме, чем iNADPH2. Кроме Toro.iNADPH представляет собой более
удобную форму записи, поскольку она подразумевает, что при-
соединяются два электрона, и часто предпочитают использовать
именно ее (рис. 4.8).
4.10. Фотосинтетическая единица
Заметим, что вначале это понятие было чисто теоретическим,
ио теперь оно получило более конкретный физический смысл
и его связывают с «пигментной системой» (разд. 4.11).
В разд. 3.9 было показано, что в соответствии с современны-
ми представлениями квантовый расход фотосинтеза равен по
крайней мере 8, т. е. на восстановление одной молекулы СО2до
СНцО в соответствии с уравнением (1.1) требуется 8 фотонов
красного света. Эти фотоны поглощаются хлорофиллом; следо-
вательно, необходимо ответить и на вопрос, сколько молекул
хлорофилла участвует в восстановлении одной молекулы СО2.
Рис. 4.9. Насыщающая фотосинтез интенсивность импульсного света. С уве-
личением интенсивности импульса фотосинтез достигает насыщения, однако
отношение количества фиксированного СОа (или выделенного О2) к количе-
ству хлорофилла остается примерно постоянным— 1/2500. Это позволяет
предположить, что отдельные молекулы хлорофилла организованы в фото-
синтетическую единицу.
62
ГЛАВА 4
Чтобы ответить на этот вопрос, следует убедиться, что все или
большинство молекул хлорофилла в данной системе действи-
тельно поглотили фотон в ходе опыта (иначе «несработавшие»
молекулы будут учитываться наряду с действующими и ответ
будет неправильным). Эта необходимость обеспечить фотонасы-
щение привела к использованию импульсного света (рис. 4.9).
При непрерывном освещении скорость фотосинтетической асси-
миляции СО2 увеличивается с возрастанием интенсивности све-
та, но у многих растений уже при 1/5 интенсивности солнечного
света достигается насыщение (хотя эффективность многих дру-
гих фотохимических процессов продолжает увеличиваться при
дальнейшем возрастании интенсивности света). Это означает,
что ассимиляцию углерода лимитирует не фотохимическая ре-
акция. В 1932 г. Эмерсон и Арнольд показали, что при импульс-
ном освещении Chlorella интервал между вспышками, за кото-
рый может завершиться темновая реакция, составляет 0,02 с
при 25 °C. Эмерсон и Арнольд провели опыты, в которых была
установлена интенсивность света, необходимая для фотонасы-
щения, при импульсах длительностью 10~в с с интервалами
около 0,09 или 0,05 с. Определяя «единицу» как ту часть фото-
синтетической системы, которая участвует в фотохимической
реакции при восстановлении одной молекулы СО2, они пришли
к заключению, что при фотонасыщении число фотосинтетических
единиц должно быть равно числу молекул СО2, восстановлен-
ных за один импульс. Так как время, необходимое для осуществ-
ления темновой реакции, равно 0,02 с, единица не может осу-
ществить работу дважды во время импульса длительностью
10~5 с. Опыты такого рода проводились при различной кон-
центрации хлорофилла, для чего или брали различное количе-
ство клеток Chlorella, или использовали водоросли, выращен-
ные в таких условиях, при которых они содержали разное ко-
личество хлорофилла. Как и предполагалось, выход на один им-
пульс увеличивался с возрастанием количества хлорофилла, но
отношение восстановленный СО2/Хл оставалось примерно оди-
наковым— 1/2480 (рис. 4.9). Эмерсон и Арнольд ие смогли дать
удовлетворительного объяснения, почему их единица содержит
так много хлорофилла. Позже, однако, Гаффрои и Воль вы-
двинули концепцию «фотосинтетической единицы», в которой
фотон может мигрировать через плотно упакованные молекулы
хлорофилла, пока не достигнет центра, способного осуществлять
фотохимическую реакцию, которая ведет к выделению О2. Если
2500 молекул хлорофилла кооперирование участвуют в выделе-
нии одной молекулы О2 и этот процесс протекает с квантовым
расходом равным 8, то, следовательно, фотосинтетическая еди-
ница может состоять из восьми субъединиц, каждая из которых
содержит 300 молекул хлорофилла и один реакционный центр,
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
63
или имеет такое 'строение, что 2500 молекул улавливающих свет
пигментов, направляют фотоны к 2—8 реакционным центрам.
Отличительной особенностью фотосинтетической единицы яв-
ляется то, что, хотя в 'возбужденное состояние переходят по су-
ществу все молекулы, энергия возбуждения большинства из них
не реализуется, так как занят реакционный центр, поскольку
единица не может использовать больше восьми фотонов за им-
пульс. Таким образом, максимальный выход можно определить
только в том случае, если мы уверены, что в возбужденное со-
стояние переходят вое молекулы хлорофилла. Напротив, мак-
симальный выход в расчете на квант (при этом подразумева-
ется, что энергия ие теряется) достигается только тогда, когда
в возбужденное состояние переходят 8 из 2500 молекул, хотя
это возбуждение может мигрировать через значительно большее
число молекул хлорофилла, возможно даже через все 2500, на
пути к реакционному центру.
4.11. Пигментные системы
В настоящее время ие делается почти никакого различия
между фотосинтетической единицей (разд. 4.10) и пигментной
системой. Пигментная система рассматривается как совокуп-
ность улавливающих свет молекул (антенна), которые поглоща-
ют фотоны и передают энергию возбуждения к реакционному
центру (разд. 4.12). Как будет показано на рис. 4.18 и в разд.
4.23, в настоящее время полагают, что хлорофилл включается
в общую последовательность фотосинтетических реакций в двух
участках и что в действительности существуют две фотосистемы
(обозначенные Дюйсенсом ФС I и ФС II; см. разд. 4.13), каж-
дая со своей собственной пигментной системой. О составе двух
пигментных систем судят прежде всего по спектрофотометриче-
ским измерениям, но не по данным .прямого анализа. Это скорее
умозрительное, чем функциональное и точное описание. Пола-
гают, что пигментная система II содержит основную массу до-
полнительных пигментов и большую часть, а возможно, и весь
Хл Ь. Кроме того, считают, что в ней содержатся некоторые
мембраносвязанные формы Хла с максимумами поглощения
650, 663, 673, 677 и 680 нм. Энергия возбуждения направляется
от этих светособирающих пигментов в «ловушку» реакционного
центра (Р 690).
Пигментная система I практически не содержит Хл Ь, но в
ней находятся четыре главные спектральные формы Хл а (а 663,
669, 677, 684) и «длинноволновая» форма хлорофилла (а 695),
полностью отсутствующая в пигментной системе II. Этот ряд
пигментов направляет энергию в ловушку, или реакционный
64
ГЛАВА 4
центр, обозначаемый как Р 700. В обеих системах энергия пе-
редается от «коротковолновых» пигментов к «длинноволновым».
Методом гель-электрофореза белковых комплексов, выделенных
с помощью додецилсульфата, были обнаружены четыре основ-
ных хлорофилл-белковых комплекса. Один из них, комплекс
Р 700—Хл а —белок, рассматривается как ядро фотосистемы I
и содержит до 15% суммарного количества хлорофилла. Осталь-
ные три комплекса содержат Хл а и Хл b и все вместе составля-
ют светособирающий (Хл а/Ь) — белковый комплекс. В Сз-хло-
ропластах в его состав входит до 50% хлорофилла. [Различия
между ’С8- и С4-хлоропластами и растениями, в которых они
встречаются, подробнее будут рассмотрены в части Б этой кни-
ги.] Остальной хлорофилл при экстракции с помощью доде-
цилсульфата оказывается в свободной форме. Считают, что этот
хлорофилл образуется в результате частичного разрушения хло-
рофилл-белковых комплексов антенны ФС I и реакционного
центра ФС II, неустойчивых к действию додецилсульфата. При-
нято, что энергия от светособирающего (Хл а/b) —белкового
комплекса поступает прежде всего в ФС II. (Существуют ука-
зания на то, что хлорофилл-белковый комплекс реакционного
центра ФС II, содержащий Р 690, имеет мол. массу около
32 000.)1 Однако этот комплекс расположен в хлоропласте та-
ким образом, что в некоторых случаях возможен перенос (спи-
ловер) энергии к ФС I, например когда пул акцепторов ФС II
сильно восстановлен или недостаточно велик. Таким образом,
поглощающие свет молекулы хлорофилла образуют единую по-
следовательность, направляющую энергию в обе фотосистемы,
а не распределяются строго между ФС I и ФС II. Представля-
ет ли вся пигментная система в целом (I-f-II) одну единицу с
2400 молекулами хлорофилла (см. выше о фотосинтетической
единице), состоит ли она из двух субъединиц по 1200 молекул
хлорофилла в каждой или же из четырех субъединиц по 600 мо-
лекул, пока не установлено. Джолио, измерявший выделение Оя
при коротких интенсивных импульсах света, установил, что при
первом импульсе О2 не выделяется, а затем выход его постепен-
но возрастает и достигает максимума при третьей, седьмой,
одиннадцатой и т. д. вспышках. Это согласуется с современны-
ми представлениями о том, что при выделении одной молекулы
О2 пигментная система II возбуждается четыре раза (разд.
4.18), т, е. для того чтобы накопить заряд и энергию для этого
акта, необходимы четыре связанных между собой реакционных
центра или четыре субъединицы.
1 Белок, содержащий Р690, имеет мол. массу 47 000—51 000. Наряду с
ним ФС II содержит белки с мол, массой 32 000. — Прим. ред.
фотохимический аппарат и его функции
б:'
4.12. Реакционные центры
Реакционный центр, или ловушка энергии, пигментной си-
стемы I (Р 700) изучен значительно лучше, чем реакционный
центр ФС II (Р 690). Полагают, что он представляет собой мо-
дифицированную форму Хл а (димер в специфическом окруже-
нии); ого обозначают как Р 700, поскольку его максимум погло-
щения лежит при длине волны 700 нм. Беосел Кок, открывший
Р 700, показал, что его окислительно-восстановительный потен-
циал (разд. 2.13) равен примерно -J-400 мВ. На долю этого
пигмента приходится около 0,3% от общего количества Хла.
Р 700 обесцвечивается при окислении на красном свету и вос-
станавливает свой цвет в темноте.
Реакционный центр II (Р 690) изучен в меньшей степени,
чем Р 700] хотя он также может быть одной из форм Хл а, от-
сюда еще не следует, что он во всем подобен Р 700. Возможно,
это более сложный комплекс, и, несомненно, что выделение Ог
в котором Р 690 играет ключевую роль, сильнее подавляется
механическими и тепловыми факторами, чем большинство реак-
ций, протекающих в ФС I. В то же время в других реакциях он
не менее стабилен, чем ФС I.
4.13. Фотосистемы (ФС I и ФС II)
Эти две системы составляют основу Z-схемы (разд. 4.23 и
гл. 5), с помощью которой осуществляется перенос электронов
от воды к NADP+ (рис. 4.18). Каждая фотосистема содержит
свою пигментную систему (разд. 4.11) и ассоциированные с ней
переносчики электронов. Природа, количество и локализация
этих компонентов электрон-траиспортной цепи точно не установ-
лены, но в разд. 4.9 подведены итоги некоторым современным
представлениям в этой области. Первоначально Дюйзенс и дру-
гие исследователи называли восстановление NADP+ «первой»
фотореакцией, а выделение О2— «второй», поскольку хемотроф-
иые фотосинтезирующие бактерии не обладали способностью
выделять О2 и в эволюционном плане восстановление NADP-1’
можно было рассматривать как «первую» реакцию. Такое на-
именование фотосистем сохранилось, хотя на пути от Н2О к
NADP+ электроны поступают сначала в ФС II, а затем в ФС I.
Некоторая путаница связана также с терминами «окислитель»
и «восстановитель». «Окислитель» — это акцептор электронов.
Множество соединений, например феррицианид, действуют как
«окислители в реакции Хилла», т. е. принимают электроны от
молекул воды в реакции, осуществляемой освещенными хлоро-
пластами.
5—234
66
ГЛАВА 4
Эти терминологические трудности связаны с тем, что пере-
носчики электронов составляют единую цепь. Так, соединение,
играющее роль окислителя (акцептора электронов) в одной ре-
акции функционирует как восстановитель (донор электронов)
в следующей по цепи переноса электронов реакции. Например,
кофермент NADP+ часто рассматривают как «природный» окис-
литель в реакции Хилла, связанный с фотохимическим аппара-
том, хотя это лишь один из многих компонентов, принимающих
электроны на разных этапах восстановления СОг. Точно так
же Р 700 является окислителем пластоцианина и восстановите-
лем NADP+, a NADPH осуществляет восстановление СО2.
Пигментные системы, а следовательно, и соответствующие
фотосистемы, можно частично разделить при механическом раз-
рушении хлоропласта (с помощью пресса Френча или под дей-
ствием ультразвука) или путем обработки детергентами (напри-
мер, дигитонином или тритоном Х-100). Частицы, обогащенные
ФС I, содержат главным образом Хл а и реакционный центр
Р 700, в то время как частицы, обогащенные ФС II, содержат
Хл а и Хл Ь. В хлоропластах Cg-растений соотношение между
общим количеством хлорофилла и количеством Р 700 составля-
ет 300/14-500/1. В частицах ФС I это соотношение равно 200/1.
При возбуждении коротковолновым светом (435 нм) хлоро-
пласта и частиц, обогащенных ФС II, в их спектре испускания
при комнатной температуре появляется полоса Хтах = 685 нм,
интенсивность которой возрастает при температуре жидкого
азота (77 К). Частицы ФС I при комнатной температуре почти
не флуоресцируют, но при температуре жидкого азота у них
появляется интенсивная полоса при 730 нм. Флуоресценция при
730 нм определяется Хл а антенны ФС I. Флуоресценция при
685 нм, очевидно, обусловлена светособирающим комплексом
(Хл а/Ь) — белок, связанным прежде всего с ФС II,
Кроме того, при комнатной температуре наблюдается замед-
ленная флуоресценция (испускание света за время от несколь-
ких секунд до нескольких минут после прекращения освещения),
специфически связанная с ФС II. Дихлорфепилдиметилмочеви-
на (ДХММ), блокируя поток электронов между Q и пластохи-
ноном, подавляет эту замедленную флуоресценцию, а дибромме-
тилизопропилбензохинон (ДБМИБ) —антагонист пластохино-
на— стимулирует ее. На замедленную флуоресценцию влияет
концентрация как восстановленных акцепторов, так и окислен-
ных доноров ФС II. Она связана также с диссипацией протои-
движущей силы, возможно возникающей при участии пласто-
хинона. Если ни функционирование реакционного центра, ни
размер пула связанных с ним акцепторов электронов не лими-
тируют процесс, замедленная флуоресценция восстанавливается.
Таким образом, ее можно рассматривать как показатель актив-
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
67
ности ФС II. У ФС I замедленной флуоресценции не проявляет-
ся, так как по неизвестным причинам квантовый выход флуо-
ресценции ФС I очень мал.
Функции
Как показано в разд. 4.8, строение фотохимического аппара-
та тесно связано с его функцией, и отделить одно от другого
невозможно. Тем ие менее в предыдущем разделе основное
внимание было уделено строению фотохимического аппарата,
а в данном разделе будут преимущественно рассмотрены спосо-
бы и основные принципы функционирования хлоропласта (разд.
4.14).
4.14. Поглощение света атомами
Простейший атом — атом водорода-—состоит из ядра (про-
тона Н+) и электрона е~, вращающегося по определенной орби-
те вокруг него, подобно спутнику, вращающемуся вокруг Зем-
ли. Чтобы вывести спутник на определенную орбиту, необходи-
мо затратить энергию, причем тем' большую, чем больше уда-
лена эта орбита от Земли. Электрой, вращающийся вокруг яд-
ра, занимает определенную орбиталь, и наиболее устойчивым
считается такое состояние атома, при котором электрон нахо-
дится на ближайшей к ядру орбитали (состояние с наименьшей
энергией, или основное состояние). В результате поглощения
кванта света с соответствующей энергией электрон может пе-
рейти из основного состояния в возбужденное, т. е. перейти на
более удаленную орбиталь (рис. 4.10). Поскольку разрешенны-
ми являются лишь строго определенные орбитали, такой пере-
ход возможен только при поглощении кванта со строго опре-
деленной энергией. Квант с более высокой или более низкой
энергией будет в этом смысле
неэффективен. Разность меж-
ду энергией основного и воз-
бужденного состояний весьма
велика, тем ие менее энергии
кванта ультрафиолетового или
видимого света достаточно
для осуществления такого пе-
рехода, и в спектре поглоще-
ния соответствующего атома
имеется черная линия, отве-
чающая длине волны погло-
щенного кванта. Положение
этой линии можно определить
5!
68
ГЛАВА 4
с помощью уравнения k=hc/AE, где А£ — энергия перехода из
основного состояния в возбужденное. Вероятность такого пере-
хода зависит от интенсивности падающего света, так как с уве-
личением интенсивности возрастает вероятность поглощения
кванта необходимой длины волны.
Возбуждение под действием света осуществляется практиче-
ски мгновенно (за время порядка 10~15 с) и сопровождается пе-
ремещением электрона с очень малой массой в атоме или мо-
лекуле, обладающих несравненно большей массой.
4.15. Поглощение света молекулами
Молекула содержит несколько ядер, которые участвуют в
колебательном и вращательном движении. Электроны распреде-
ляются между ядрами таким образом, что занимают самые низ-
кие молекулярные орбитали, т. е. обладают наименьшей энерги-
ей. При образовании из атомов молекулы выделяется энергия
(гл 2). В основном и возбужденном состояниях электрон в мо-
лекуле может находиться на нескольких разных колебательных
подуровнях. И в этом случае энергия кванта должна точно со-
ответствовать энергии, необходимой для перевода электрона с
одного из основных подуровней на один из возбужденных, но,
поскольку существует много различных подуровней, диапазон
квантов, способных осуществлять такой переход, расширяется.
В связи с этим вместо четких линий, характерных для спект-
ров поглощения атомов, в спектрах поглощения молекул из-
бирается набор линий, которые сливаются, образуя относитель-
но широкие полосы поглощения. Каждая индивидуальная линия
в полосе соответствует поглощению кванта с определенной дли-
ной волны, а вся совокупность линий — распределению энер-
гии вблизи максимума поглощения. (Эта картина еще больше
усложняется из-за того, что помимо электронных переходов
происходят колебательные движения в самих молекулах, и это
приводит к размыванию энергетических уровней. При очень низ-
ких температурах наблюдаются более узкие полосы поглоще-
ния, так как уменьшается число подуровней, на которых может
находиться электрон.)
4.16. Переход хлорофилла
в возбужденное состояние
Хлорофилл может находиться в двух возбужденных состоя-
ниях, соответствующих полосам поглощения в красной и синей
областях. Эти полосы частично перекрываются, что свидетель-
ствует о перекрывании колебательных подуровней. Возбужден-
ное состояние 1 является низкоэнергетическим, переход в него
фотохимический аппарат и его функции
69
Безызлучательный
переход
Возбужденное
состояние 2
Возбужденное
состояние 1
Химические реакции
„Синий”
'Ху'Х/-* Флуоресценции
Основное состяние
Рис, 4.11. Электронные переходы в молекуле хлорофилла. Параллельные ли-
нии отвечают разным энергетическим подуровням, или электронным орбита-
лям. Энергия, полученная системой при поглощении «синего» фотона (сле-
ва}, достаточна для перехода электрона в возбужденное состояние 2, из ко-
торого он быстро спускается путем безызлучательных переходов с одного
подуровня на другой до возбужденного состояния 1. Фотон красного света
(в центре) обладает энергией, достаточной только для перехода электрона
в возбужденное состояние 1, но это состояние достаточно стабильно для со-
вершения полезной химической работы и служит исходной точкой для всех
остальных событий фотосинтеза. В органических растворителях энергия
электрона, находящегося в состоянии 1, может быть израсходована па флуо-
ресценцию в дальней красной области.
осуществляется при поглощении «красного» фотона с энергией
около 40 ккал/эйнштейн (рис. 4.11). Возбужденное состояние 2
более высокоэпергетическое, переход в него связан с поглоще-
нием «синего» фотона с энергией около 50 ккал/эйнштейн. Так
как колебательные подуровни перекрываются, может происхо-
дить быстрый переход из состояния 2 в состояние 1 (за время
порядка 10~13 с), сопровождающийся тепловой диссипацией
энергии (безызлучательной потерей возбуждения); он состоит
из ряда последовательных переходов с одного колебательного
подуровня на другой. Этот процесс осуществляется так быст-
ро, что избыточная энергия состояния 2 ие успевает израсхо-
доваться на совершение полезной работы (фотохимической ре-
акции) и даже на испускание света (флуоресценцию), которое
происходит за время порядка 10-9 с. Таким образом, отправной
точкой для фотосинтетического процесса служит возбужденное
состояние 1; даже если хлорофилл переходит в него непрямым
путем, после поглощения «синего» фотона, избыточная энергия
теряется прежде, чем она успевает использоваться на соверше-
ние полезной работы.
70
ГЛАВА 4
4.17. Снятие возбуждения
Итак, энергия возбуждения может рассеиваться в виде теп-
ла, причем это рассеяние происходит очень быстро путем по-
следовательных переходов с одного колебательного уровня на
другой. Энергия может быть израсходована и <на испускание
кванта видимого света, хотя хлорофилл, как и многие другие
молекулы, флуоресцирует только при переходе в основное со-
стояние из более стабильного возбужденного состояния 1. Од-
нако, даже находясь в состоянии 1, молекула хлорофилла ча-
стично теряет энергию в результате тепловой диссипации, так
как флуоресценция происходит при переходе с самого низко-
энергетического подуровня, которому соответствует большая
(более «красная») длина волны (стоксов сдвиг). Экстрагиро-
ванный из листьев хлорофилл (в водном растворе ацетона) на
просвет выглядит зеленым, поскольку он поглощает «красные»
и «синие» фотоны. «Зеленые» фотоны проходят через раствор,
а кроме того, человеческий глаз к ним более чувствителен.
В органических растворителях снятие возбуждения хлорофилла
не может осуществляться с помощью тех же механизмов, что и
в хлоропласте. Возбужденное состояние 2 легко переходит в со-
стояние 1, и значительная часть энергии возбуждения высвечи-
вается в виде флуоресценции. Из-за стоксова сдвига этот свет
более длинноволновый, чем падающий. Если смотреть на осве-
щенный сосуд с раствором хлорофилла под прямым углом к
направлению падающего пучка света, он будет выглядеть кро-
ваво-красным.
Происходящая в этих условиях сильная флуоресценция —
недостаточно эффективный механизм снятия возбуждения и не
защищает молекулу хлорофилла от фотоокисления; раствор в
проходящем свете вскоре становится желтым в результате раз-
рушения хлорофилла.
4.18. Фотохимические реакции и выделение О2
Помимо тепловой диссипации и флуоресценции существует
еще один механизм снятия возбуждения— совершение полезной
работы (фотохимическая реакция). In vivo интенсивность флуо-
ресценции хлорофилла гораздо меньше, поскольку здесь воз-
можности для протекания «полезного» фотохимического процес-
са значительно выше, чем в освещенном растворе. Соответст-
венно в интактном листе хлорофилл гораздо меньше разруша-
ется при длительном освещении, хотя может происходить и
обесцвечивание, особенно если блокирован главный канал фо-
тохимического пути (т. е. если в окружающей атмосфере отсут-
ствует СО2).
фотохимический аппарат и его функции
71
В Итоге: 2Н2р + 4 hv + 4 Fe3*- 4Н1 I О.,+4 Fe2 *'
Рис. 4.12. Фотохимия реакции Хилла с феррицианидом в качестве акцептора.
В центре диаграммы изображена молекула хлорофилла ФС II (Р 690), кото-
рая при поглощении фотона переходит па достаточно высокий энергетиче-
ский уровень, чтобы восстановить акцептор Q до Q-. Этот процесс повторя-
ется четыре раза, и в Y (Mn-содержащем расщепляющем воду белке) накап-
ливаются четыре окислительных эквивалента. Эти эквиваленты используют-
ся одновременно при расщеплении двух молекул воды на О2 и 4Н+ н по-
ступлении в цепь четырех электронов. Акцептор электронов Q передает их
затем на другие переносчики Z-схсмы. In vivo эти электроны передаются на
NADP+ и в конечном счете на СО2. В реакции Хилла СО2 не восстанавлива-
ется, так как электрон передается искусственному акцептору, в данном слу-
чае феррицианиду. «Точка пересечения» определяется окислительно-восстано-
вительным потенциалом окислителя, и, как предполагает диаграмма, донором
служит не обязательно Q~. Хотя эта схема очень упрощена, опа показыва-
ет, как происходит накопление четырех окислительных эквивалентов в Y на
каждую молекулу выделяющегося О2 (.слева) и каким образом фотохими-
ческое возбуждение приводит к подъему электронов выше барьера энергии
активации (справа), препятствующего восстановлению молекулы искусствен-
ного или природного акцептора.
Чтобы между двумя стабильными молекулами произошла
реакция, необходимо преодолеть энергетический барьер. Ясно,
что для осуществления реакции молекулы должны столкнуть-
ся, и чем сильнее это столкновение, тем больше вероятность,
что барьер будет преодолен и за столкновением последует ре-
акция. Обычно химические реакции протекают быстрее при бо-
лее высокой температуре, так как увеличивается скорость дви-
жения молекул и соответственно чаще и с большей силой про-
исходят столкновения. Реакции протекают легче в присутствии
катализаторов благодаря снижению барьера (уменьшения
энергии активации) .
В некоторых фотохимических реакциях барьер не снижается,
а преодолевается. Именно так |Л|роисходит в случае реакций с
участием хлорофилла. Как мы уже говорили (разд. 4.16), об-
лучение хлорофилла как красным, так и синим светом приводит
72
ГЛАВА 4
к переходу его в возбужденное состояние 1, попав в которое,
электрон преодолевает тем самым энергетический барьер и мо-
жет перейти к акцептору электрона, или окислителю. При био-
логических температурах даже в присутствии соответствующего
катализатора энергия активации остается слишком большой,
чтобы электрон в иевозбуждснном состоянии мог ее преодо-
леть.
Одним из примеров такого процесса служит реакция Хилла
с феррицианидом в качестве окислителя (акцептора электро-
нов). Вначале в результате возбуждения под действием света
электроны в хлорофилле переходят из основного состояния в
возбужденное (рис. 4.12, в центре). Для выделения одной моле-
кулы кислорода необходимы две молекулы воды, выполняющие
роль доноров:
2Н2О --> Os-|-4H+ + 4e-. (4.1)
Четыре молекулы феррицианида (Fe3+) восстанавливаются до
ферроцианида (Fe2+), принимая четыре электрона, и четыре
иона водорода (Н1') выделяются в раствор. За реакцией Хил-
ла можно следить, регистрируя повышение кислотности раство-
ра, сопровождающее этот процесс.
Как видно из рис. 4.12, для каждого акта возбуждения хло-
рофилла требуется один фотон; электрон молекулы хлорофилла
передается затем тушителю флуоресценции Q, при этом об-
разуется одна молекула Хл+ (или Р 690+) и одна молекула Q-
(разд. 4.9). Для выделения одной молекулы О2 окисляющий во-
ду комплекс Y должен накопить четыре окислительных эквива-
лента. На рис. 4.12 последние представлены четырьмя ионами
окисленного марганца (Мп3+), образующимися при отдаче че-
тырех электронов четырьмя ионами Мп2+ при образовании че-
тырех положительных дырок у Р 690 под действием света. В ра-
боте Жолио и Кока было показано, что у адаптированных к
темноте водорослей или хлоропластов выделяется одна моле-
кула О2 на каждый четвертый импульс света при насыщающих
импульсах длительностью 1 мкс. [В действительности первая
молекула О2 выделяется после третьего импульса, последую-
щие— после 7-го, 11-го, 15-го и т. д. В рамках схемы, представ-
ленной на рис. 4.12, это означает, что в темноте уже имеется
один ион Мп3'1' и три иона Мп2+. Соответственно необходимы че-
тыре импульса для образования четырех ионов Мп3'1', которые
принимают 4е~ от двух молекул Н20 с выделением одной
молекулы 02.] Это также означает, что квантовый расход равен
4 (на 02). На практике феррицианид принимает электроны от
ФС I, если используются интактные тилакоиды (с квантовым
расходом 8), и от обеих фотосистем в случае поврежденных
тилакоидов.
фотохимический аппарат и его функции
73
4.19. Перенос энергии в пигментных системах
Прежде чем реакционный центр сможет передать электроны
переносчикам соответствующей фотосистемы, происходит погло-
щение фотонов светоообирающим комплексом и передача энер-
гии возбуждения через молекулы хлорофилла антенны к этому
реакционному центру.
Как уже отмечалось, снятие возбуждения некоего пигмента
А может происходить путем испускания кзанта света — флуо-
ресценции (разд. 4.17 и рис. 4.11). Если длина волны испускае-
мого света такова, что этот свет может быть поглощен вторым
пигментом В, то энергия будет передаваться от А к В путем
последовательных возбуждений, снятия возбуждения и повтор-
ного поглощения. В действительности эффективность переноса
энергии столь велика, что описанный выше механизм практи-
чески исключается, а энергия передается путем резонанса. При
этом энергия, которая могла бы высвободиться при переходе
электрона в основное состояние (флуоресценция), более или
менее непосредственно используется для возбуждения соседней
молекулы, находящейся в основном состоянии (рис. 4.13). Эф-
фективность этого процесса зависит от расстояния между мо-
лекулами. Чтобы передача энергии между двумя молекулами
хлорофилла была возможна, расстояние между ними должно
Хл а 667
Рис. 4.13. Перенос энергии от одной молекулы хлорофилла к другой путем
одновременно протекающих возбуждения и снятия возбуждения.
74
ГЛАВА 4
быть меньше 7 нм, а при расстоянии 2,5 нм эффективность
переноса возрастает до 100%. Доказательством реализации ре-
зонансного механизма в фотосинтезирующих организмах служит
наличие сенсибилизированной флуоресценции; например, Хла
может флуоресцировать при возбуждении Хл Ь. В случае оди-
наковых плотно упакованных молекул энергия с большой эф-
фективностью переносится при случайном блуждании экситона
(пары электрон — дырка). Этот процесс можно себе предста-
вить как обмен электронами между орбиталями с близкими
уровнями возбуждения, причем соответствующее движение
электрона между орбиталями основного состояния обеспечивает
согласованное движение электрона и «дырки».
Однако не исключено, что правильнее было бы считать хло-
рофилл, находящийся в агрегированном состоянии в пигментной
системе, сверхмолекулой, в которой возбуждение по существу
делокализовано. В процессе переноса энергии к реакционному
центру по всей последовательности молекул проходит волна
снятия возбуждения.
4.20. Спектр действия
Как было показано в разд. 4.16, хлорофилл поглощает свет
в синей и красной областях спектра, причем «красное» воз-
бужденное состояние хлорофилла является той отправной точ-
кой, с которой начинаются все остальные события фотосинтеза.
Учитывая это, можно ожидать, что при освещении хлоропластов
синим или красным светом будет наблюдаться интенсивный
фотосинтез, в то время как освещение зеленым светом даст ми-
нимальный результат. Этот вывод следует из формы спектра
действия (рис, 4.14): интенсивность света, вызывающего дан-
ный фотосинтетический ответ, обратно пропорциональна эффек-
тивности поглощения (т. е. для фиксации данного количества
СО2 требуется значительно меньше красного света, чем зеле-
ного). Спектр действия и спектр поглощения хлорофилла очень
близки по форме. Это позволяет считать хлорофилл главным
фотосинтетическим пигментом. Более полезную информацию
можно получить из распределения квантового выхода в види-
мой области ,(рис. 4.15). Из разд. 4.16 следует, что «синие» и
«красные» фотоны должны приводить к одинаковой ответной
реакции, так как избыточная энергия «синего» кванта не мо-
жет быть использована для совершения полезной работы. На
первый взгляд зеленый свет значительно менее эффективен,
однако, хотя при этом интенсивность фотосинтеза мала, мало и
количество поглощенных фотонов, так что квантовый выход
при освещении зеленым светом мало отличается от такового
для красного и синего света. Спектр действия имеет три осо-
фотохимический аппарат и его функции
75
Рис. 4.14. Сравнение спектров действия и поглощения. Сплошная линия —
спектр поглощения Ulva, штриховая — относительная интенсивность фотосин-
теза при различных длинах волн при одинаковом числе квантов падающего
света. (По Нахо, Blinks.)
Рис. 4.15. Спектр действия для выделения Оа у Chlorella. Отложена зависи-
мость максимального квантового выхода от длины волны. (По Emerson, Le-
wis.) Обратите внимание па резкое уменьшение выхода при длине волны
выше 680 нм (красный спад; см. разд. 4.21).
бенности. Во-первых, наблюдается четко выраженный минимум
в области 480 нм. Так как он совпадает с максимумом погло-
щения каротиноидов, по-видимому, эти соединения не передают
энергию хлорофиллу с достаточной эффективностью (т. е. ка-
ротиноиды скорее препятствуют поглощению энергии хлорофил-
лом, чем способствуют). Во-вторых, имеет место резкий спад
76
ГЛАВА 4
Рис. 4.16. Спектры действия для ФС I (штриховая линия) и ФСП (сплош-
ная линия) у Chlorella. (По Ried.)
квантового выхода после 680 нм (красный опад). В-третьих, су-
ществует неглубокий минимум при 660 им (разд. 4.21).
Раздельно спектры действия для ФС I и ФС II (рис. 4.16)
были получены Ридом на интактных клетках Chlorella. Он ис-
ходил из предположения о том, что О2, выделяющийся в первые
несколько секунд освещения, образуется ФС II, а временное
подавление поглощения О2 при последующем переносе в тем-
ноту— результат активности ФС I. Можно видеть (рис. 4.16),
Рис. 4.17. Спектры поглощения изолированных хлоропластов шпината в вод-
ном растворе и экстракта из тех же органелл в 80 %-ном ацетоне.
фотохимический аппарат и его ФУНКЦИИ
77
что в спектре действия ФС II проявляется дополнительный пик
в синей области, связанный с присутствием каротиноида и Хл Ь,
и что спектр поглощения Хл а ФС I сдвинут в красную область
примерно на 10 нм (см. разд. 4.11 и рис. 4.16).
4.21. Красный спад
In vivo хлорофилл продолжает поглощать свет и при дли-
нах волн, превышающих 680 нм (рис. 4.14, 4.17); в то же время
в спектре действия в этой области наблюдается резкий спад
(рис. 4.15). Впервые на то, что длинноволновый красный свет
не столь эффективен при фотосинтезе, как более коротковолно-
вый, обратил внимание Эмерсон (разд. 4.20). (Более коротко-
волновые фотоны обладают большей энергией, ио наблюдаемое
различие в эффективности только этим объяснить нельзя).
4.22. Эффект усиления
Обнаружение красного спада (разд. 4.21) привело Эмерсона
к мысли исследовать эффект примешивания к длинноволновому
красному свету относительно слабого света с более короткой
длиной волны. При таком освещении интенсивность фотосинте-
за оказывалась больше, чем сумма интенсивностей при раз-
дельном действии составляющих. Такое увеличение эффективно-
сти красного света с большой длиной волны (примерно 700—
730 нм) получило название «эффекта усиления». Это явление
объясняется в рамках Z-схемы, для обоснования которой оно в
свою очередь привлекалось; например, при 700—730 нм реак-
ционный центр ФС I (Р 700) может окисляться быстрее, чем
его ‘восстанавливают электроны из ФС II. Напротив, при более
коротковолновом облучении интермедиаты в ФС II (например,
восстановленный пластохинон) могут образоваться быстрее,
чем ФС I способна их снова окислять. Подобным рассогласова-
нием можно объяснить минимум в спектре действия при 660 нм
(непонятный «синий спад»),
(Как видно из рис. 4.16, свет с длиной волны 650—680 нм
активирует обе фотосистемы, хотя более эффективен он в от-
ношении ФС II. Напротив, свет с длиной волны больше 680 нм,
очень эффективный для ФС I, ие способен активировать ни один
из компонентов ФС II в их максимуме поглощения. Далее, в то
время как «коротковолновые» формы хлорофилла могут пере-
давать энергию «длинноволновым», обратный переход, «в гору»,
невозможен. Именно по этим причинам уменьшение эффектив-
ности с увеличением длины волны (красный спад) проявляется
гораздо сильнее, чем соответствующее уменьшение в синей об-
ласти спектра.)
78
ГЛАВА 4
Спектр действия для эффекта усиления можно получить, из-
меряя степень усиления в результате примешивания коротковол-
нового света с разной длиной волны к основному свету с фик-
сированной (большей) длиной волны. Наличие максимума при
670 нм в таком спектре подтверждает представление об отно-
сительном обогащении пигментной системы II Хла 670. Жолио
и др. в опытах с хлоропластами шпината получили раздельные
спектры действия для двух фотосистем. Для этого они измеря-
ли а) количество О2 при фиксированной длине волны 720 нм
с примесью света с переменной длиной волны и б) восстанов-
ление метилвиологена при фиксированной длине волны 650 нм
с примесью света с переменной длиной волны. Для изолирован-
ных хлоропластов были получены спектры действия (разд, 4 20)
для системы Н2О—»NADP+ и Н2О—’-дифлорфенолиндофенол,
а также (в присутствии ДХММ как ингибитора ФС II) для
транспорта электронов от системы аскорбат/дихлорфеиолиндо-
фенол, поставляющей электроны к ФС I. В соответствии с
Z-схемой для двух первых систем наблюдался красный спад,
а для последней, где функционировала только ФС I, при боль-
шей длине волны эффективность возрастала.
4.23. Z-Схема
Как говорилось в разд. 4.8 и будет обсуждаться в гл. 5,
Z-схема Хилла и Бендалла основана на предположении о на-
личии двух световых реакций при фотосинтезе. В результате их
совместного действия электроны «поднимаются» на двух участ-
ках пути от воды (исходный донор) до NADP+ (конечный ак-
цептор). Две рассматриваемые фотосистемы (разд. 4.13) связа-
ны друг с другом последовательностью переносчиков, образую-
щих «мост» между ними, идущий «под гору», так что электроны
проделывают Z-юбразный путь. На рис. 4.18 представлена схе-
ма, где указано направление движения электронов и промежу-
точные соединения, лежащие на пути электронов от Н2О до
NADP+ (разд. 4.4, 4.9). На этом рисунке приведена также энер-
гетическая диаграмма, где указаны разность окислительно-вос-
становительных потенциалов и количество высвободившейся
или затраченной энергии на отдельных участках пути. Кисло-
род выделяется в ходе реакций, протекающих в ФС II, которые
осуществляются при участии более коротковолнового света. На-
против, NADP'1' восстанавливается в ФС I с использованием
более длинноволнового света. Светособирающая антенна на-
правляет энергию возбуждения к реакционным центрам (разд.
4.12). Электроны, выбитые при возбуждении первичного донора
реакционного центра II, замещаются электронами молекул во-
ды, по-видимому, при участии фермента, содержащего марта-
Рис. 4.18. Z-Схема. Представлены одновременно энергетическая диаграмма и
направление движения электронов, но схему не следует принимать букваль-
но. Шкала окислительно-восстановительных потенциалов {слева) напомина-
ет таблицу расположения футбольных команд по лигам (разд. 2.13). Коман-
да, расположенная в верхней части таблицы, сильнее той, которая находится
ниже. Аналогично соединения е большим отрицательным окислительно-вос-
становительным потенциалом способны восстанавливать соединения с более
положительным потенциалом. Так, перенос электрона от воды к NADP+
(с участием обеих фотосистем) приводит к созданию мощного донора водо-
рода NADPH2, способного в дальнейшем восстанавливать СО2 до СН2О. Энер-
гия, необходимая для такого переноса электронов {справа), равна примерно
52 ккал, или 26 ккал па электрон и поступает в схему на двух участках.
[Разность между О2-(-|-815) и Н2-(—420) электродами (табл, 2,3), равная
1200 мВ {слева), соответствует изменению свободной энергии {справа) при-
мерно па 57 ккал (разд. 2.9 и 2.13).] Два «рупора» представляют собой по-
следовательности молекул хлорофилла, составляющих светособнраюпиге ан-
тенны (разд. 4,10), которые направляют энергию возбуждения фотонов к
двум реакционным центрам (Р 690 и Р700-, см. разд, 4.12). Поскольку хлоро-
филл п этих системах несколько различается {Р 695, например, — это Хл а
с максимумом поглощения 695 нм), ФС I возбуждается предпочтительнее
светом с большей длиной волны (<730 нм), а ФС II — относительно корот-
коволновым светом (<700 им). Расположение других известных или предпо-
лагаемых переносчиков электронов (разд, 4.9) и таких ингибиторов, как ДХММ
и ДВМИБ (разд, 4,13), соответствует их возможной локализации или месту
действия. На пути от Г12О к NADP+ от переносчика к переносчику переда-
ются два электрона (две параллельные линии). Каждый фотон (квант), по-
ступающий в каждую из фотосистем, может поднять на более высокий, бо-
лее «восстановленный» уровень один электрон, так что для переноса двух
электронов по указанному пути требуются четыре кванта (разд, 3.9). По-
скольку па каждую выделенную молекулу О2 приходится четыре электрона
(2H2O+2NADP->-2NADPH2-|-C2), вся последовательность реакций должна по-
вториться дважды и квантовый расход равен 8 па молекулу О2 (разд. 3.9).
Энергия одного моля квантов красного света с длиной волны 680 им {сле-
ва) равна примерно 1800 мВ (левая ось), или 42 ккал (правая ось; см. так-
же разд. 3.5). Некоторое количество энергии, высвобождающейся при про-
хождении электронов «под гору» через «мост» между двумя фотосистемами,
запасается в форме АТР (гл. 5).
80
ГЛАВА 4
иец (разд, 4,9). Они передаются через Q и пластохинон цито-
хрому f и пластоцианину, ассоциированным с ФС I, Здесь они
заполняют положительно заряженные дырки, образовавшиеся в
результате возбуждения (окисления) Р700. Электроны из реак-
ционного центра ФС I через X, ферредоксин и флавопротеин
(разд. 4.9) перемещаются к NADP+, Некоторое количество энер-
гии, выделяющейся при движении электронов «под гору», запа-
сается в форме химической энергии в АТР. АТР образуется в
двух центрах фосфорилирования, что подтверждают опыты с
ингибитярами, блокирующими окисление пластохинона, и с раз-
личными донорами и акцепторами, взаимодействующими с пе-
реносчиками, которые образуют «мост» между фотосистемами.
Один участок сопряжения переноса электронов с фотофосфо-
рилированием расположен у «моста», другой —между Г12О и
реакционным центром ФС II.
Используя слово «участок», мы подразумеваем, что энергия
запасается при прохождении электрона между вполне опреде-
ленными акцепторами. Если принять, что в образовании АТР
принимает участие протондвижущая сила (ПДС) (разд. 5.9,
5.10), то этот термин потеряет свою определенность: фосфори-
лирование осуществляется в некоем компартменте после обра-
зования ПДС. Локализован не сам процесс образования ПДС,
а лишь его механизм. Например, показано, что электроны мо-
гут возвращаться от вершины ФС I к Р 700 через пиоциапин,
минуя обычный путь через «мост» между двумя фотосистема-
ми. Этот искусственный кофактор образует свой собственный
участок в том смысле, что он переносит протон с наружной сто-
роны мембраны (после получения электрона от X в ФС I) во
внутренний, компартмент (после окисления под действием Р700).
Таким образом, при световом возбуждении ФС I протонный
градиент образуется даже в том случае, когда такие переносчи-
ки, как пластохинон, цитохром f и пластоцианин, не участвуют
в процессе.] По, современным представлениям расщепляющий
воду фермент (Мп-белок), а также Р690, Р 700, цитохром f и
пластоцианин расположены с внутренней стороны мембраны, и
там же происходит выделение О2. Напротив, Q, ферредоксин и
флавопротеин расположены с наружной стороны.
Концепция Z-схемы была предложена Хиллом и Беидаллом
в 1960 г. Она опиралась (по крайней мере отчасти) на опыты'
Эмерсона, проведенные в 1958 г. и показавшие, что, хотя даль-
ний красный свет сам по себе не индуцирует фотосинтез (крас-
ный спад, разд. 4.24), он увеличивает (разд. 4.22) его эффектив-
ность при освещении более коротковолновым светом. Эта кон-
цепция получила экспериментальное подтверждение в работах
Дюйзенса и его сотрудников в Лейдене, показавших, что даль-
ний красный свет возбуждает преимущественно ФС I (разд.
ФОТОХИМИЧЕСКИ» АППАРАТ II ЕГО ФУНКЦИИ
81
Рис. 4.19. Аналогия Z-схемы.
4.13) и при этом цитохром f 'Окисляется, в то время как при
возбуждении красным более коротковолновым светом преиму-
щественно функционирует фотосистема II и цитохром f восста-
навливается. Кроме того, в 1963 г. Хоху удалось эксперимен-
тально (но не физически) разделить две системы и показать,
что дальний красный свет индуцирует интенсивные реакции в
ФС I, но не в ФС И, в то время как красный свет оказывает
противоположное действие.
На рис. 4.19 'Представлена механическая аналогия Z-схемы.
Некий джентльмен слева «нагружает» фотоном коротковолно-
вого света ФС II. Частица, подскакивающая при этом вверх,—
это электрон, который направляется к первичному акцептору
ФС II. Частицы, скатывающиеся вниз, запускают генератор
АТР (в центре рисунка). Затем некий джентльмен справа «на-
гружает» фотоном ФС I, и на этот раз частица проделывает
путь через X (справа вверху) в корзину с NADP+.
4.24. Отношение Р/ег
Точное количество АТР, образующегося при прохождении
двух электронов по 2-схеме (отношение Р/е2), определить труд-
но. Это связано с наличием «основного потока» электронов без
6—234
82
ГЛАВА 4
фотофосфорилирования, который следует учитывать в расчетах.
К сожалению, сам этот «основной поток» ингибируется АТР,
поэтому неясно, каким образом его учитывать. «Основной по-
ток» может возникать из-за неполного сопряжения в результате
утечки протонов из компартмеита тилакоида (разд. 4.2), ско-
рость которой зависит в свою очередь от градиента pH через
мембрану тилакоида и, следовательно, от интенсивности света
(который повышает этот градиент) и синтеза АТР (который его
уменьшает).
In vivo или в интактных изолированных хлоропластах фак-
торы, уменьшающие потребление АТР, увеличивают утечку
протона и скорость «основного потока» электронов. Хебер рас-
сматривает это как некий механизм контроля, в результате
«подвижного сопряжения» изменяющий отношение Р/е2 от 1 до
2. Вообще говоря, эта величина, по-видимому, близка к единице,
хотя для того, чтобы имело место соответствие с ВПФ-циклом,
в ходе которого расходуются три молекулы АТР на каждые две
молекулы окисляющегося NADP, это отношение должно быть
равно 1,5. На величину отношения Р/е2 могут оказывать влия-
ние также циклическое фотофосфорилирование (разд. 5.4) и фо-
тофосфорилирование, связанное с восстановлением кислорода
(разд. 5.6).
4.25. Запасание энергии в Z-схеме
Для переноса четырех электронов от двух молекул воды с
выделением молекулярного О2 и восстановлением двух молекул
NADP+ в Z-схеме необходимо 8 квантов. Часть этой энергии
при сопряжении с транспортом электронов запасается в форме
АТР.
Как отмечалось выше (разд. 4.24), отношение Р/е2 нельзя
точно определить и in vivo его значение варьирует. Тем не менее
представляется маловероятным, чтобы при нормальных усло-
виях сопряжения в хлоропласте (разд. 5.11) это отношение бы-
ло меньше единицы или больше двух. Если принять, что сво-
бодная энергия окисления NADPH (AFZ) равна —52 ккал,
а свободная энергия гидролиза АТР (до ADP и Р,) —10,8 ккал,
то максимальное количество запасенной энергии для каждой
грамм-молекулы выделенного О2 составит около 148 ккал ((2-
•52)-(-(4-11)], а минимальное 126 [(2 52) + (2 11)]. При длине
волны 680 нм, близкой к максимуму поглощения хлорофилла в
красной области, на 8 эйнштейн при 42 ккал-эйнштейн-1 при-
ходится 336 ккал, т. е. запасается от 37,5% (126/336) до 44%
(148/336) энергии. Если в результате включения циклического
(разд. 5.5) и псевдоциклического (разд. 5.6) путей переноса
электронов квантовый расход повышается до 10, то эта вели-
ФОТОХИМИЧЕСКИЙ АППАРАТ II ЕГО ФУНКЦИИ
83
чина уменьшается до 35% (148/420). Следовательно, с энергети-
ческой точки зрения Z-схема представляется вполне правдопо-
добной, так как количество поступающей в нее энергии в три
раза больше количества высвобождающейся энергии.
Поскольку для обеспечения энергией пентозофосфатного цик-
ла требуется 2 NADPH24-3 АТР молекул (при расходе 112—
120 ккал на СН2О), AF7 цикла составляет 17—25 ккал, а «эф-
фективность» 87% (другие термодинамические аспекты будут
рассмотрены в разд. 5.14). Следует отметить, что реальную по-
требность в энергии на молекулярном уровне следует увеличить
на 50%, поскольку необходимо учесть подавление фотосинтеза
О2 (гл. 13).
4.26. Квантовый расход для Z-схемы
Как показано выше, в Z-схеме на каждый электрон, пере-
мещенный в каждой фотосистеме, приходится один фотон (один
квант). Результаты измерений хорошо согласуются с этими
оценками, т. е. для выделения одной молекулы О2 требуется
восемь фотонов. Квантовый расход, равный единице, зарегист-
рирован для транспорта электронов к Р 700, для фотоокисления
ферроцитохрома с препаратами ФС I при 710 нм, для перено-
са электрона от воды к дихлорфенолиндофенолу при 630—
660 нм или от воды к феррицианиду при 640 нм, для переноса
электрона от аскорбата к NADP'1' при 700 нм. Два кванта необ-
ходимы также для переноса двух электронов от Н2О к NADP4*
при 640—678 нм.
6*
ГЛАВА 5
Образование АТР;
генерация ассимиляционной силы
Краткое содержание
Фотохимическая стадия фотосинтеза завершается генераци-
ей ассимиляционной силы (АТР+восстановлениый NADP), не-
обходимой для осуществления -процесса ассимиляции углерода.
В освещенном хлоропласте электроны через ряд переносчиков
(гл. 4) движутся от Н2О к NADP+, восстанавливая его. Транс-
порт электронов приводит к образованию протонного градиента
и разделению электрических зарядов, что в свою очередь стиму-
лирует образование АТР. К процессам, с которыми сопряжено
фотофосфорилирование, относятся:
а. Нециклический транспорт электронов (от воды к NADP+
или — в нефизиологических системах — от различных доноров
к различным акцепторам). При возбуждении хлорофилла под
действием света электроны переходят на более высокие энерге-
тические уровни и затем передаются от одного переносчика к
другому вниз по термодинамической лестнице («под гору»),
б. Псевдоциклический транспорт электронов (электроны, доно-
ром которых служит вода, проходя через последовательность
соответствующих переносчиков, вновь соединяются с Os с обра-
зованием воды).
в. Циклический транспорт электронов (электроны фотосистемы I
вследствие возбуждения хлорофилла под действием света пе-
реходят на более высокий энергетический уровень и затем воз-
вращаются в этой же системе на более низкий энергетический
уровень).
Во всех этих случаях движение электронов «под гору» и со-
пряженное с ним фосфорилирование по существу аналогичны
фосфорилированию в дыхательной цепи. При фотофосфорилиро-
вании вследствие возбуждения хлорофилла электроны поднима-
ются «;в гору». При дыхании электроны движутся вниз по тер-
модинамическому градиенту (если при фотосинтезе электроны
отщепляются от воды, то при дыхании они возвращаются к Ог,
и в итоге образуется Н2О), но эти два события — первичные фо-
тохимические реакции и дыхание у животных — могут быть раз-
делены длинной пищевой цепью.
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
85
5.1. Окислительное фосфорилирование
При дыхании связь Н—О, разрываемая в процессе фотосин-
теза, образуется вновь и при этом высвобождается энергия. Как
и в случае фотосинтеза, важную роль здесь играют кофермен-
ты ряда никотинамида. При фотосинтезе электроны перемеща-
ются от воды к NADP+, и затем NADPH2 восстанавливает СО2
до СН2О. При дыхании электроны движутся от СН2О к NAD,
а затем NADH2 через цитохромы передает электроны к О2, в ре-
зультате чего вновь образуется вода. Несомненно, существует
несколько путей, ведущих в конечном счете к полному окисле-
нию углеводов до СО2 и Н2О, и при этом не во всех случаях
участвует NAD+. Тем не менее основной принцип остается тем
же самым, т. е. электроны, поступая к какому-то переносчику,
восстанавливают его, затем 1восстановленный переносчик вновь
окисляется, а конечным акцептором электронов является моле-
кулярный кислород.
На промежуточных этапах метаболизма восстановленные
формы коферментов поставляют водород для целого ряда вос-
становительных процессов, так что не весь кислород от СН2О
незамедлительно связывается с О2, часть его используется по-
степенно.
Таким образом, NADH2, NADPH2 и т. п. могут служить до-
норами электронов и энергии как при дыхании, так и при фото-
синтезе. Аналогичным образом АТР сначала образуется, а за-
тем используется в самых разнообразных реакциях обмена и
процессах жизнедеятельности. Образование АТР представляет
собой механизм запасания энергии: часть энергии, которая вы-
свобождается при переносе электронов от субстрата к кислоро-
ду, используется для синтеза АТР из ADP и Рь Установлено,
что процесс запасания энергии дыхания происходит в основном
в субклеточной органелле, называемой митохондрией, при пере-
носе водорода от NADII2 к О2. [И здесь в некоторых реакциях
участвуют иные, чем NADH2 восстановленные переносчики, од-
нако механизм переноса остается тем же.], Процесс окислитель-
ного фосфорилирования (или дыхательная цепь) можно пред-
ставить в общем виде следующим образом:
NADH2 4- Оа----->- NAD -|- НаО (5.1)
3ADP -|- ЗР( —-> ЗАТР -|- 3HSO (5.2)
Следовательно, если транспорт электронов полностью сопряжен
с образованием АТР, то при переносе двух электронов otNADH2
к О2 должны образоваться по меньшей маре три молекулы
АТР. Если принять, что ДА' для окисления NADH2 равна
86
ГЛАВА 5
3-7
—52 ккал, а для гидролиза АТР — 7 ккал, то мы получим •
•100, т. е. запасается примерно 40% энергии.
Когда в 1954 г. Арион, Аллеи, Уотли и независимо от них
Френкель открыли фотофосфорилирование, биохимики уже бы-
ли знакомы с изложенными выше концепциями. Однако, как
мы увидим ниже, этот факт лишь затруднил, а не облегчил по-
нимание механизма фотофосфорилирования. Полная ясность в
этом вопросе была достигнута лишь с построением Z-схемы.
5.2. Зачем нужна ассимиляционная сила
До 1930 г. существовало общее представление, что восста-
новление СО2 прямо связано с возбуждением хлорофилла. Впер-
вые это положение было подвергнуто сомнению в работах Ван-
Ниля на фотосинтезирующих бактериях. Им было показано, что
зеленые серобактерии, например, могут расти на смеси серово-
дорода (H2S) и СС2 и при этом способны к фотосинтезу. Этот
процесс можно описать с помощью следующего уравнения:
hv ,е о\
СО2 + 2HjS --> СНгО + Н2О + 2S (5.3)
При этом Ван-Ниль отметил поразительное сходство между
этим уравнением и уравнением, описывающим фотосинтез зеле-
ных растений:
, Г л ч
СО2 + 21-1,0 —СНг0 + Н2О + О2 (5.4)
Кроме того, известны также хемосинтезирующие бактерии,
которые обладают способностью восстанавливать в темноте СО2
и использовать при этом водород, выделяющийся при окислении
того или иного субстрата в таких условиях (как было показа-
но Лебедевым в 1921 г.), когда возбуждение хлорофилла ис-
ключено. Все это позволило Ван-Нилю постулировать обоб-
щенное для бактерий и высших растений уравнение фотосинтеза
AV /г-
СО2+2Н2А ----> СЯ2О + 2А +Н2о (5.5)
В таком обобщенном процессе возбуждение хлорофилла под
действием света приводит к генерации восстанавливающей еди-
ницы Н и окисляющей единицы ОН:
4Н2О 4/7 + 4ОЯ (5.6)
Затем как у растений, так и у бактерий Н восстанавливает СО2:
со± + 47/ --------------------> СИгО + Н2О (5.7)
ОБРАЗОВАНИЕ АП’; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
87
При этом у высших растений ОН используется для образования
кислорода и воды:
40Я —>Ог + 2Н3О (5.8)
а у бактерий ОН окисляет субстрат:
2H2S -|- 40Я -> 4Н2О -|- 2S (5.9)
Эта схема получила всеобщее признание, так как находилась в
полном соответствии с представлениями Варбурга о необходи-
мости четырех квантов для восстановления молекул СОг. Одна-
ко положение о том, что возбуждение хлорофилла больше свя-
зано с фотолизом воды, чем с восстановлением СОг, представ-
лялось не столь очевидным.
Примерно в это же время была опубликована первая работа
Роберта Хилла, которой суждено было окончательно изменить
представления о фотосинтезе. В ней сообщалось, что изолиро-
ванные хлоропласты могут на свету выделять кислород, вос-
станавливая соединения (искусственные окислители), иные,
чем СОа,. как, например,
2Fe»+ -|- НаО ->- 2Fea+ -|- 2Н+ + 4' (5.10)
Таким образом, гипотеза о фотолизе воды получила экспери-
ментальное подтверждение. Однако природа естественного окис-
лителя (акцептора водорода) оставалась неизвестной.
В 1943 г. Рубен обобщил некоторые более ранние представ-
ления Тимана и Липмана и высказал гипотезу, которая впослед-
ствии получила четкое подтверждение. В ней предполагалось,
что для фотосинтеза необходимы как АТР, так и восстанови-
тель. При этом считалось, что АТР может использоваться дву-
мя путями — поставлять энергию для карбоксилирования и уча-
ствовать в восстановлении карбоксильных групп с образованием
вначале более легко восстанавливаемого карбонилфосфата. Со-
гласно этой гипотезе, фотосинтетический восстановитель /Одол-
жен обладать таким же восстановительным потенциалом, какой
имеют коферменты ряда никотинамида, неспособные самостоя-
тельно осуществлять восстановление карбоксила до альдегида.
Макроэргичсские фосфатные эфиры (АТР) должны регенери-
роваться при прямой или непрямой рекомбинации Н и ОН. Эта
гипотеза, которую Рабинович, лишь однажды утратив свою
обычную проницательность, нашел неправдоподобной, обрела
вскоре твердую экспериментальную основу в работах по изуче-
нию цикла Кальвина — Бенсона (восстановительного пентозо-
фосфатного пути метаболизма), в которых доказывалось нали-
чие точно такого же пути использования NADPH2 и АТР, какой
предполагала гипотеза Рубена. Эти данные, а также выявлен-
88
ГЛАВА 5
ная Арионом и его коллегами способность хлоропластов восста-
навливать в темноте СОг до СН%0 с использованием при этом
ассимиляционной силы (ATP4-NADPH2) подтвердили гипоте-
зу Рубена,
Однако в одном отношении гипотеза Рубена не выдержива-
ла критики. Она, как и многие более поздние схемы, предпола-
гала, что образование АТР связано с реокислением не прямо,
а опосредованно, причем преимуществом такого пути может
быть использование метаболитического аппарата митохондрий
при поступлении в них Н в форме NADPH2, Однако, когда Ар-
ион, Аллен и Уотли в 1954 г. открыли циклическое фосфорили-
рование, они сделали это на препаратах хлоропластов, практи-
чески свободных от митохондрий. Более того, этот феномен на-
блюдался и в анаэробных условиях, не поддерживающих окис-
лительного фотофосфорилирования. Таким образом, можно бы-
ло предположить, что фотофосфорилирование связано с прямой
рекомбинацией Н и ОН; это положение получило подтвержде-
ние в рамках современной концепции циклического фотофосфо-
рилирования (см. ниже). Однако в августе 1957 г. Арион и его
сотрудники перевернули все представления о фотосинтезе, объ-
явив о существовании нециклического фотофосфорилирования:
NADP+ + ADP + Pj + Н2О NADPH, -|- АТР -|- Н2О + Ог (5.11 >
Совершенно очевидно, что ситуация, описываемая этим уравне-
нием, обратна окислительному фосфорилированию, при котором
АТР образуется по мере окисления NADH2, сопровождающегося
поглощением О2 и образованием Н2О. [Согласно уравнению
(5.11), вода (в левой части уравнения) поставляет электроны
NADP+, а вода, которая вновь образуется (в правой части урав-
нения), высвобождается при конденсации ADP и Pi.]
Если к безоговорочному принятию концепции циклического
фотофосфорилирования и восстановления NADP+ весь фотосин-
тетический мир оказался полностью подготовленным, то к идее
о том, что образование АТР связано с восстановлением NADPА
а не с его окислением, он готов не был. На первый взгляд эта
концепция была диаметрально противоположна фосфорилиро-
ванию в дыхательной цепи, и даже осознание того, что движу-
щей силой фотофосфорилирования является свет, а окислитель-
ного—субстрат, не прояснило этот вопрос. Труднее всего ока-
залось объяснить тот факт, что восстановление NADP'1’ происхо-
дит быстрее, когда оно сопряжено с образованием АТР, чем ког-
да ADP и Pi отсутствуют. Как указывали Хилл и Боннер, такое
сопряжение должно было бы, казалось, исключить возмож-
ность того, что образование АТР целиком зависит от рекомби-
нации Н и ОН; ведь если такая рекомбинация происходит не-
ОБРАЗОВАНИЕ ЛТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
89
зависимо от фосфорилирования, то трудно понять, почему при-
сутствие ADP и Pi должно было ускорять этот процесс. [Заме-
тим, что хемиосмотическая гипотеза еще не была сформулиро-
вана.] Если рекомбинация II и ОН зависит от присутствия ADP
и Р|, то скорость образования Н и ОН должна была бы умень-
шаться параллельно их воссоединению в процессе фосфорили-
рования.
Эта загадка была разрешена только после того, как Хилл
и Бенделл предложили свою Z-схему. Согласно этой схеме, фо-
тофосфорилирование не является «восстановительным» процес-
сом, каким оно представлялось вначале, а образование АТР
происходит, как и при окислительном фосфорилировании, при
движении электронов «под гору» (окислительным путем) через
ряд переносчиков, включая цитохромы.
5.3. Циклическое фотофосфорилирование
Процесс, названный вначале «фотосинтетическое фосфори-
лирование», позже получил название «циклическое фотофосфо-
рилирование», чтобы отличить его от открытого позже вариан-
та, в котором в качестве акцептора электронов выступают
NADP+ или феррицианид.
Циклическое фотофосфорилирование (рис. 5.1) ие связано с
поглощением или выделением О2 и может быть представлено
просто как
ADP р, АТР + Н,0 (5.12)
Термин «циклический», относящийся к потоку электронов, был
введен Арионом и его коллегами и основывался на идее Льюи-
са и Липкииа о том, что возбуждение хлорофилла ведет к по-
тере электрона. В своей схеме Арион и его коллеги считают фо-
тоокисление хлорофилла первичным событием. Затем электрон
передается на кофактор и — через ряд носителей — обратно,
чтобы заполнить дырку в окисленном хлорофилле. Таким обра-
Рис. 5.1. Схема циклического фотофосфорилирования (или транспорта элек-
тронов) в ее первоначальном виде (по Ариону и др.).
90
ГЛАВА 5
'К^РбЭО
Н,0 -о,
2 2 '
ФС II
Рис. 5.2. Возможные пути транспорта электронов в хлоропластах: А — цик-
лический, В — псевдоциклическнй (реакция Мелера) и С — нециклический.
Пигментные системы и переносчики Z-схемы детально рассмотрены на
рис. 4.13. В присутствии каталазы (нсхлоропластного компонента) образую-
щаяся Н2О2 расщепляется на О2 и воду.
зом, он следует по циклическому пути, при этом часть (рассеи-
вающейся энергии используется на образование АТР. В то вре-
мя, когда было сделано это открытие, основное внимание ис-
следователей было обращено на скорость фосфорилирования
(которая вначале многим казалась слишком низкой) и на при-
роду кофактора. При этом особое внимание уделялось различи-
ям между «нефизиологическими», или искусственными, кофак-
торами и возможными природными кофакторами, таким, как ме-
надион (эквивалент которого, как казалось, присутствовал в
хлоропласте в виде витамина К). В настоящее время этот во-
прос всерьез не обсуждается, Уже обнаружено множество со-
единений, в присутствии которых наблюдается высокая скорость
фосфорилирования, и их список настолько длинен, что в него
можно включить и лабораторную пыль. Однако неясно,
многие ли из этих соединений действительно играют роль ко-
факторов в истинном циклическом фотофосфорилировании. Не-
которые из них [например, метилвиологен (паракват)] осуще-
ствляют псевдоциклическое фотофосфорилирование (см. ниже),
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
91
роль других до конца не выяснена. Лишь о двух нефизиологиче-
ских кофакторах известно, что они не участвуют в поевдоцикли-
ческом фотофосфорилировании: это феназинметосульфат (ФМС)
и пиоцианин (один из продуктов его фотоокисления). [Однако
даже относительно этих соединений имеются некоторые сомне-
ния. Чистый пиоцианин катализирует фотофосфорилирование в
хлоропластах высших растений, но менее активен в хлоропла-
стах водорослей. ФМС широко используется в экспериментах
с хлоропластами высших растений, однако может терять ак-
тивность, если создать условия, предотвращающие его фотоокис-
ление в пиоцианин.] В соответствии с Z-схемой пиоцианин «за-
корачивает» поток электронов где-то вблизи места расположе-
ния связанного ферредоксина, возвращая их к участку около
Р700 (рис. 5.2). Таким образом, циклическое фотофосфорилиро-
вание прежде всего ассоциировано с ФС I, и катализируемая
пиоцпашшом реакция может идти в присутствии ДХММ (инги-
битора ФС II), если созданы условия для предотвращения чрез-
мерного восстановления. [В этом и подобных случаях очень ва-
жен механизм поддержания «равновесия» в окислительно-вос-
становительной системе, так как переносчики играют двойную
роль: они и акцепторы электронов, и их доноры. Если ФМС
становится слишком восстановленным, его способность прини-
мать электроны ослабевает —факт, с помощью которого можно
было бы дать альтернативное объяснение неспособности соеди-
нения «работать» при строго анаэробных условиях; см. выше.]
5.4. Циклическое фотофосфорилирование,
катализируемое ферредоксином
Ферредоксин способен катализировать циклическое фотофос-
форилирование и в аэробных, и в анаэробных условиях. Как и
в случае пиоцианина, при анаэробных условиях существует
проблема «равновесия». Ясно, однако, что ферредоксин спосо-
бен функционировать в цикле только три условии, что большая
часть его находится в восстановленной форме. Соответственно
в начале циклического фотофосфорилирования наблюдается
лаг-период, который уменьшается под действием факторов, уси-
ливающих начальное восстановление (например, в результате
поступления электронов от воды или от системы аскорбат/ди-
хлор фенол индофенол). Однако, как и в случае пиоцианина, су-
ществует опасность чрезмерного восстановления (электроны от
воды могут конкурировать с электронами циклического пути
при восстановлении пластохинона). По этой причине ХММ и
ДХММ (ингибиторы фотосистемы II) при низких концентраци-
ях стимулируют циклическое фотофосфорилирование, катализи-
руемое ферредоксином, а при высоких подавляют. (Если бы
92
ГЛАВА 5
проблемы «равновесия» не существовало, циклическое фотофос-
форилирование могло бы осуществляться при полном ингибиро-
вании фотосистемы II.
В случае использования ферредоксина в качестве катализа-
тора циклическое фотофосфорилирование стимулируется длин-
новолновым красным светом, оптимальным для фотосистемы I,
и избирательно подавляется антимицином А (антибиотиком, по-
давляющим перенос электронов между цитохромами b и с в
дыхательной цепи фосфорилирования). Последнее говорит о
том, что электроны от ферредоксина могут вновь переходить на
«мост» между двумя фотосистемами через звено (цитохром Ь
563), более чувствительное к антимицину А, чем тот путь, по
которому поступают электроны от воды. Тот факт, что ДБМ.ИБ
ингибирует как циклическое, так и нециклическое фотофосфо-
рилирование, говорит о том, что пластохинон участвует в функ-
ционировании обоих путей (рис. 5.2).
5.5. Циклическое фотофосфорилирование in vivo
Осуществляется ли in vivo при нормальном функционирова-
нии нециклического фотофосфорилирования циклическое фото-
фосфорилирование, пока неизвестно. Арнон высказал предполо-
жение, что NADPH2 может «уравновешивать» систему подобно
ДХММ. Так, если по каким-либо причинам уменьшается окис-
ление NADPHj под действием СО2, то накопление NADPH2 мо-
жет способствовать циклическому фосфорилированию. Однако
трудно предположить, что восстановленный ферредоксин может
быть вовлечен во взаимодействие с другими переносчиками в
цепи; более вероятным представляется прямое окисление его мо-
лекулярным кислородом (см. ниже). Такой процесс мог бы спо-
собствовать образованию АТР в большей степени, чем процесс,
связанный с восстановлением NADP+.
5.6. Псевдоциклическое фотофосфорилирование
Как отмечалось выше, термин «циклический» относится к
процессу, при котором электроны, следуя по циклическому пути
через фотосистему I, попадают обратно в реакционный центр,
из которого они начали свое движение. В этом случае цикличе-
ская система не потребляет и не выделяет кислорода, и транс-
порт электронов внутри ее не может быть выявлен поэтому кри-
терию. Методы измерения количества О2 не могут быть приме-
нены и в том случае, когда в системе одновременно поглощается
и выделяется одинаковое количество кислорода. В связи с этим
такой процесс был назван «псевдоциклическим», поскольку его
трудно отличить от «настоящего» цикла, в котором О2 не погло-
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
9®
Рис. 5.3. Псевдоцпклический транспорт электронов. Хотя в этом процессе
не происходит выделения О2, это не истинно циклический транспорт, подоб-
ный представленному на рис. 5.1. Выделение О2 замаскировано в данном
случае эквимолярным его поглощением; тем не менее электроны перемеща-
ются через обе фотосистемы, восстанавливая О2, а не NADP+.
вдается и не выделяется. В разд. 5.8 рассматривается случай
нециклического транспорта электронов, в котором молекуляр-
ный кислород является акцептором электронов (реакция Ме-
лера) (рис. 5.3).
5.7. Нециклическое фотофосфорилирование
Как видно уже из самого названия, эта форма фотофосфо-
рилирования связана с переносом электронов от донора (напри-
мер, НаО) к акцептору (такому, как NADP+ или феррицианид).
Если в роли донора и акцептора выступают Н2О и NADP+, то в-
фотофосфорилирование такого типа включаются обе фотосисте-
мы. (Более детально нециклический перенос электронов рас-
смотрен в разд. 4.13 и па рис. 5.2.) Однако в случае блокирова-
ния ФС II с помощью ДХМ.М. этот процесс может осуществлять-
ся только одной ФС I, если электроны поступают от восстанови-
теля с достаточно низким потенциалом непосредственно в Z-cxe-
му между ФС II и ФС1. Таким восстановителем является голу-
бой краситель 2,'6-дихлорфеиолиидофенол (ДХФИФ), который
часто используется для визуальной демонстрации реакции Хил-
ла (рис. 5.4):
Краситель (голубой, окисленный) -|- Н2О ->•
---> Краситель Н2 (бесцветный, восстановленный)-!--^- О2 (5.13)
94
ГЛАВА 5
Рис. 5.4. Нециклический транспорт электронов с участием одной фотосистемы.
Транспорт электронов из ФС II блокирован ДХММ; донором электронов
служит система аскорбат/дихлорфенолиидофеиол (ДХФИФ)
Если его добавить в каталитическом количестве вместе с фер-
рицианидом (другой окислитель, который сам по себе мо-
жет поддерживать нециклическое фотофосфорилирование), то
ДХФИФ будет захватывать электроны и передавать их ферри-
цианиду в обход некоторых переносчиков «моста» между двумя
фотосистемами, в связи с чем уменьшается количество образу-
ющегося АТР. Если, однако, использовать ДХФИФ вместе с ас-
корбатом (как донором электронов) и если при этом ФС II
блокировать с помощью ДХММ, то электроны перейдут от ас-
корбата через «мост» и ФС I к NADP+. Поэтому АТР будет об-
разовываться даже в том случае, когда ФС II не работает. [Этот
подход был впервые использован Лозадой, Уотли и Арноном
для демонстрации того факта, что нециклическое фотофосфори-
лирование можно разделить с помощью химических манипуля-
ций на два этапа: фотолиз воды и последующий транспорт элект-
ронов через АТР-генерирующую систему к NADP+. В настоящее
время полагают, что расщепление воды также сопряжено с фос-
форилированием (разд. 5.9). В случае системы аскорбат —
ДХФИФ поток электронов минует пластохиноновый участок и
электроны поступают в цепь иа уровне пластоцианина. KCN
(блокирует процесс у пластоцианина) ингибирует эту систему
доноров, а ДБМИБ (антагонист пластохинона) или ДХММ не
оказывают на нее никакого влияния. ДХФИФЩ служит, по-ви-
димому, искусственным переносчиком протонов через мембрану,
генерируя протондвижущую силу, необходимую для фосфорили-
рования (разд. 5.9). Эффективным искусственным донором для
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
95
ФС I является система аскорбат — ТМФД (н-тетраметил-п-фе-
нилендиамин), однако эта система не способна к фосфорилиро-
ванию, так как ТМФД только поставляет электроны, но не
переносит протоны через мембрану.
5.8. Реакция Мелера
В 1951 г. Мелер сообщил об обнаруженном им варианте ре-
акции Хилла, в которой молекулярный кислород является ко-
нечным окислителем. В ходе реакции Хилла на каждую молеку-
лу воды выделяется молекулы кислорода:
hv 1 1 лч
Н2О + А --> АН2 + у О2 (5.14)
в то время как в реакции Мелера происходит поглощение О2:
Н2О + О2 Ы2О2+4- О2 (5.15)
что эквивалентно
Н2О +~О2 -----> Н2О2 (5.16)
В присутствии избытка каталазы (фермента, который способст-
вует превращению перекиси водорода в воду и О2) не наблюда-
ется никакого обмена О2 (рис. 5.3):
Н,0 + О, ---> Н2О2 4- 4- О2
(5.17)
Н2О2----> Н2О + -у- 02
(5.18)
И действительно, если мы сложим уравнения (5.17) и (5.18),
то никакого результирующего изменения не обнаружим, хотя
электроны проходят через Z-схему.
Свойствами реагентов Мелера обладает большое число при-
родных и искусственных акцепторов электронов, например ме-
тилвиологен (паракват) и флавиновые коферменты, такие, как
иизкопотенциальные и легко самоокисляющиеся акцепторы в.
реакции Хилла:
Н2О А -----> АЫ2 -р "у О2
АН2+О2 ----> А + Н2О2
Сумма: Н2О + О2--> Н2О2 + -уг О2
(5.19)
(5.20)
(5.21)
96
ГЛАВА 5
Хотя эти уравнения отражают основные черты реакции Медера,
было бы неверно думать, что перенос На к О2 представляет со-
бой простой процесс, который протекает всегда одинаково не-
зависимо от природы окислителя. Так, например, приведенные
выше уравнения предполагают, что два электрона и два про-
тона (т. е. два атома водорода) переносятся к Oj с образова-
нием пероксида. Однако обычно одиночный электрон может
присоединяться к О2 с образованием супероксидного радикала
О~2:
Oa+e- ----у о3- (5.22)
В присутствии фермента супероксиддисмутазы О_2 превраща-
ется в пероксидный радикал О2~2 и перекись водорода:
20,-—>о?~ + о, (5.23)
0ЛЧ-2Н+------>Н2О2 (5.24)
но супероксидный радикал может окислять такие соединения,
как аскорбат:
АН, + 20,- -у А + 20?- + 2Н+ (5.25)
Таким образом, перенос двух электронов от воды может одно-
временно приводить к окислению аскорбата и поглощению 02
в количестве (2—~) = 1 j молекул:
Н20 + 20, у 2Н+ + 20,- + -у о2 (5.26)
АН, + 20," -у А + 2Н+ + 20?- (5.27)
20?- + 4I-I+-у 2Н,0, (5.28)
Сумма: И,О + АН, +1 02 у 2Н,0, А (5.29)
|[В присутствии избытка каталазы эта реакция могла бы
протекать с поглощением 4 02 на молекулу окисленного ас-
корбата.]
5.9. Хемоосмос и установление электрохимической
разности потенциалов, или протондвижущей силы
Под действием света хлорофилл переходит в более высоко-
энергетическое, возбужденное состояние, и при этом индуциру-
ется перенос электронов (разд. 4.16). От воды, первичного до-
нора электронов, электроны поступают через фотосистемы и
связанные с ними переносчики к NADP (разд. 4.9). Каждая
окислительно-восстановительная ступень этой последовательно-
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
$7
Рис. 5.5. Последовательность событий, происходящих в хлоропласте после
освещения. Пигменты и компоненты цепи транспорта электронов распола-
гаются в мембране тилакоида. Возбужденный хлорофилл в фотосистеме II
(ФС II) получает электроны. Для переноса электронов к ФС I необходимо,
чтобы протоны поглощались из тилакоидного компартмента и освобож-
дались в строму. pH стромы возрастает, и в нее поступает противо-
ион Mg2+. По мере прохождения Н+ через АТРазу в процессе фото-
фосфорилирования концентрация АТР увеличивается, a ’[ADP] и, [Pi]
уменьшается. При получении электронов из ФС I образуется восстанови-
тель, способный к прямому или непрямому восстановлению ферредоксина,
NADP и дисульфидных групп. Постепенно повышается концентрация мета-
болитов (ВПФ-цикл).
сти реакций обладает своими особенностями. Такие соединения,
как пластохинон, могут принимать и отдавать два протона,.
Другие, например цитохром и ферредоксин, могут акцепти-
ровать только один электрон, Конечный акцептор NADP
может принять два электрона и один протон. Таким обра-
зом, перенос электронов осуществляется с помощью после-,
довательиых реакций, в ходе которых протоны Либо выделяют-
ся в окружающую среду, либо поглощаются из нее, Как пока-
зали Хинд и Ягендорф, слабо забуференная среда, в которой
суспендированы лишенные оболочек хлоропласты, при освещен
иии становится более щелочной, а при затемнении подкисляется.
С помощью хемиосмотической гипотезы Митчелла в приложе:
7—234
98
ГЛАВА 5
Рис. 5.6. Другой аспект того механизма, который представлен на рис. 5.5.
Показано также распределение в мембране основных переносчиков и место
действия таких ингибиторов, как ДХММ.
нии к фотосинтезу сделана попытка сопоставить все эти факты
и отвести им центральную роль в синтезе АТР. Предполагается,
что протоны, поглощаемые в определенном участке электрон-
транспортной цепи, поступают только из наружного простран-
ства и высвобождаются только во внутритилакоидиое простран-
ство. В результате при транспорте электронов через серию пере-
носчиков на мембране устанавливается протонный градиент.
В этом смысле протоны не проходят прямо сквозь мембрану, а
лишь поступают на ее внутреннюю поверхность и забираются
с ее наружной поверхности, что является следствием транспорта
электронов. В результате внутритилакоидиое пространство под-
кисляется, а наружное становится более щелочным (рис. 5.5 и
5.6).
Предполагается, что при нециклическом транспорте электро-
нов существуют два участка, на которых создается протонный
градиент: при расщеплении воды внутри тилакоидного компарт-
мента и при окислении-восстановлении пластогидрохинона —
пластохинона. Например, опыты с гидрофильными и гидрофоб-
ными акцепторами и донорами электронов для ФС I и ФС II (а
также опыты с антителами против специфических белков-пере-
носчиков) указывают на то, что ФС II расположена около
внутренней поверхности мембраны тилакоида, а ФС I — около
наружной. Пластохинон, мобильный переносчик протонов через
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
99
мембрану, по-видимому, принимает по мере своего восстановле-
ния протоны из пространства стромы и отдает их внутрь тила-
коида по мере окисления пластогидрохинона (рис. 5.6). Фото-
фосфорилирование, связанное только с ФС II, можно обнару-
жить, блокируя поток электронов между ФС II и ФС I с по-
мощью KCN (на уровне пластоциаиина) или ДБМИБ (на уров-
не пластохинона) и добавляя бензохинон или феиилендиамин
в качестве акцепторов электронов. Можно думать, что цикли-
ческий поток электронов в отсутствие искусственных катализа-
торов также включает транспорт электронов через пластохинон
(рис. 5.2 и разд. 5.4).
При освещении ДрН между тилакоидным компартментом и
стромой составляет около 3—3,5 единиц и может быть измере-
но по поглощению слабого основания. Изменения pH в суспен-
зии тилакоидов при смене света и темноты настолько значитель-
ны, что их можно без труда зарегистрировать обычным стек-
лянным электродом. В слабозабуференной реакционной смеси,
содержащей 50 мкг Хл на 1 мл, при облучении светом, интен-
сивность которого близка к насыщающей, быстро обнаружива-
ется сдвиг pH на 0,2—0,3 единицы. При затемнении наблюдается
медленный возврат pH к величине, близкой к начальной, и эти
подъемы и падения pH в ответ на смену света и темноты могут
продолжаться сколь угодно долго. Величина сдвига pH является
функцией транспорта электронов. Ее можно увеличить, добавив
кофакторы, которые усиливают транспорт электронов без пря-
мого (вследствие восстановления) воздействия на pH. По мере
уменьшения освещенности pH среды становится более кислым
и возвращается к исходной величине при повторном включении
полного света. Обработка детергентами и другие воздействия, в
результате которых, как полагают, мембрана становится более
проницаемой для протонов (и поэтому, по-видимому, ускоряет-
ся обратный поток), в большинстве случаев уменьшают сдвиг
pH. Тот факт, что pH при освещении не возрастает до бесконеч-
ности, а быстро достигает насыщения, говорит об усилении об-
ратного потока протонов через мембрану по мере увеличения
их концентрации с внутренней стороны. Обратное давление,
обусловленное увеличением концентрации протонов с одной
стороны мембраны, будет подавлять транспорт электронов, т. е.
высвобождение протонов в среду с высокой их концентрацией
(внутри) или изъятие из среды с низкой концентрацией (снару-
жи) будет затруднено. Усиление потока электронов под дейст-
вием разобщителей является ответной реакцией па уменьшение
обратного давления при снижении протонного градиента.
Градиент, образующийся в результате переноса электронов,
имеет электрический и химический компоненты. Накопление по-
ложительно заряженных протонов приводит к неравенству элек-
7*
100
ГЛАВА! Б
трического заряда, которое может в той или иной мере компен-
сироваться перемещением внутрь или наружу соответственно
анионов или катионов. Протондвижущая сила (ПДС) представ-
ляет собой сумму электрического (Атр — величина положитель-
ного заряда внутри тилакоида) и химического (Z\pH) компо-
нентов, где Z—RT/f >=60 мВ:
ПДС = Дф-|-КАрН (5.30)
Один из вариантов этого аспекта гипотезы Митчелла был
предложен Витом и его коллегами, которые пришли к выводу,
что первичным событием в фотохимическом акте является раз-
деление зарядов. Так, при длительности импульсов 20 нс мемб-
раны тилакоидов становятся электрически заряженными. Таким
образом, при возбуждении хлорофилла создается некое силовое
поле, и отрицательные заряды (электроны) локализуются на
наружной поверхности мембраны, а положительные заряды
(дырки) — на внутренней, создавая на мембране разность по-
тенциалов Дф. Затем на наружной поверхности мембраны два
электрона от фотосистемы II восстанавливают пластохинон и
два — от фотосистемы I — восстанавливают NADP+ (рис. 5.5);
Каждый из этих процессов восстановления сопровождается по-
глощением протонов из наружного пространства. На внутренней
поверхности по мере окисления PQ выделяются протоны. Хотя
Внутри
Тилакоид
Си аружи
О
о
A_Q_p_0 —Р —(ОН
О'
А— О- - Р — О — Р~~ О—Р—О Н
О”
О"
0~
Рис. 5,7. Упрощенная схема синтеза АТР. В результате транспорта электро-
нов формируется протонный градиент (рис. 5.6), который выполняет функ-
цию стока для протонов и гидроксильных ионов, освобождающихся при
Этерификации, и таким образом смещает равновесие в сторону образования
АТР. .
>Н+->НгО
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
101
за это время исходное разделение зарядов исчезает, электриче-
ский потенциал, или неравенство зарядов, остается, .так как по-
ложительно заряженные протоны перемещаются с наружной
стороны мембраны (которая при этом становится отрицатель-
ной) к ее внутренней стороне (которая поэтому приобретает по-
ложительный заряд).
5.10. Механизм образования АТР
Работы Ягендорфа, Хинда, Неймана и Юрайба привели к
выводу, что роль индуцируемого светом высокоэнергетического
промежуточного продукта — предшественника АТР —может иг-
рать просто градиент pH. Ягеидорф и Юрайб в своем знамени-
том эксперименте показали,что искусственно созданный гради-
ент pH может приводить к синтезу АТР в темноте. Стимулом к
этой работе послужила хемиосмотическая гипотеза Митчелла,
которую она и подтвердила: согласно этой гипотезе, синтез АТР
ассоциирован со снятием протонного градиента через обрати-
мую АТРазу. В упрощенном виде имеет место следующая по-
следовательность событий:
а) в результате транспорта электрона через мембрану создает-
ся трансмембранный градиент протонов (разд. 5.9);
б) путем обращения гидролиза АТР с участием локализован-
ной в мембране обратимой и действующей направленно
ATP-азы происходит синтез АТР;
в) в процессе этерификации при синтезе АТР происходит на-
правленное расщепление молекул воды, так что с одной сторо-
ны мембраны концентрируются протоны, а с другой — гид-
роксильные ионы; это снимает протонный градиент, создаваемый
при переносе электронов (рис. 5.7). С этой точки .зрения про-
тонный градиент служит «стоком» для водородного и гидрок-
сильного ионов, сдвигая равновесие в сторону образования
АТР. Даже в таком упрощенном виде процесс , перемещения
ионов включает как электрический, так и химический компонен-
ты, и исчезновение химического компонента при рекомбинации
IT+ и ОН- с наружной стороны мембраны сопровождается со-
ответствующим изменением электрического дисбаланса. Со вре-
мени проведения этих первых экспериментов все большее рас-
пространение приобретало представление о том, что фосфори-
лирование осуществляется с помощью иротоидвижущей силы
(т, е. с помощью протонного градиента, разделения зарядов
или совместного действия обоих факторов). Так, известны слу-
чай, когда фосфорилирование ие было связано с градиентом'pH
или’ протекало, при "условиях, когда-неадекватный градиент pH
дополняется разделением, зарядов,, обусловленным, замещением
Н+ на К+/И наоборот, известны'случаи разобщения (разд. 541)
без снижения градиента pH. ’’: '
102
ГЛАВА 5
Хемиосмотическая гипотеза, особенно ее более поздние ва-
рианты, хорошо объясняет многие экспериментальные данные.
Так, например, не вызывает сомнения, что на свету в хлоро-
пластах быстро образуется градиент pH и что снижение этого
градиента связано с образованием АТР. Ускорение транспорта
электронов с помощью разобщителей или ADP-ф-Рь (фотосинте-
тический контроль) можно объяснить уменьшением обратного
давления, обусловленного градиентом и связанным с ним раз-
делением зарядов. Довольно давно Аврон выделил из хлоро-
пластов сопрягающий фактор (обозначаемый теперь как CFi;
разд. 5.12). Если этот фактор удалить путем промывания хло-
ропластов раствором ЭДТА, то произойдет разобщение транс-
порта электронов и фосфорилирования. При добавлении сопря-
гающего фактора фосфорилирование возобновляется, а также
восстанавливается способность хлоропласта изменять транспорт
электронов под действием ADP и Р(-. Рэкер показал, что CFi
обладает АТР-азной активностью. Сопрягающие факторы мито-
хондрий, бактерий и хлоропластов очень похожи. Сопрягающий
фактор состоит из двух компонентов—АТР-азного (обозначае-
мого Fi у бактерий и митохондрий и CFi у хлоропластов) и
Рис. 5.8. Накопление протонов во внутритилакондном пространстве. Транс-
мембранный протонный градиент может рассеиваться путем переноса прото-
нов с помощью сопрягающего фактора, что способствует синтезу АТР, или
под действием искусственных разобщителей. CFi — АТР-азный компонент
сопрягающего фактора, CFo — внутримембранный канал, через который про-
ходят протоны.
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
103
о
•I
О“—-Р—О~
I
он
о
I
О“—Р—он
I
он
+
н+
о о
I 4
Д—О —р—Q _. р — О-
I I
О' О“
н+
н+
Рис. 5.9. Возможная последовательность событий при синтезе АТР (сверху
вниз). Поступление Pi в компартмент фосфорилирования (в виде HiPO<2-)
облегчается в результате протонирования: Н1РО?- + Н+-*-Н2РО4-‘. Находя-
щийся снаружи ADPS- замещается внутренним АТР3-, диссоциация которо-
го снаружи (АТР3“-+-АТР'---|-Н+) в свою очередь облегчает этот обмен.
мембранного (обозначаемого CFo). В норме мембраны непро-
ницаемы для протонов. Компонент CFo создает канал (рис. 5.8),
по которому протоны проходят сквозь мембрану через АТРазу
в процессе фосфорилирования. Весь сопрягающий комплекс хло-
ропласта в целом обозначают CFt—CF0 (разд. 5.12 и рис. 5.8).
На самом деле механизм синтеза АТР может быть гораздо
более сложным, чем это показано на рис. 5.7. Так, например,
Диксон предположил, что Р/ поступает в компартмент, где про-
исходит фосфорилирование в виде ГДРСД2-. Протоны, концент-
рирующиеся в этом компартменте вследствие транспорта элек-
тронов, облегчают этот перенос внутрь компартмента как путем
создания положительного заряда, так и за счет «стока», при
котором Н1РО?- частично превращается в Н2РО4~ (рис. 5.9).
«Внешний» ADP3“ превращается в сопрягающем комплексе в
мембране тилакоида в АТР3~. Затем АТР3~ диссоциирует сна-
ружи тилакоидной мембраны на Н+ и АТР4-, что способствует
протеканию реакции. Таким образом, накопление протонов при-
водит к увеличению концентрации ADP и Р; в центре фосфори-
лирования. Более того, фосфат может перемещаться после ко-
валентного связывания с носителем, который в результате про-
'104
ГЛАВА 5'
тонирования становится более сильным фосфорилирующим аген-
том, чём Н1РО42-. [В любом случае между стромой и наружной
поверхностью тилакоида должен возникать-диффузионный гра-
диент АТР и ADP, чтобы мог осуществляться перенос АТР от
места синтеза к месту использования в реакции фотосинтетиче-
ской ассимиляции углерода.] ;
АТР — макроэргическое соединение, так как в его молекуле
имеются расположенные рядом фосфатные группы, между кото-
рыми существует электростатическое отталкивание. В результа-
те синтез АТР из ADP и Р, легче протекает в таких условиях,
когда йерекрывающиеся электростатические взаимодействия об-
легчают конденсацию реагирующих веществ. Бённан предполо-
жил, что образование электростатического комплекса с участи-
ем CFi (разд. 5.12) может способствовать накоплению энергии
(см. также гл. 2). •
5.11. Сопряжение, разобщение и фотосинтетический
контроль
In vivo транспорт электронов сопряжен с фосфорилирова-
нием, т. е. два этих процесса взаимосвязаны. При полном со-
пряжении в отсутствие образования АТР не должно было бы
происходить никакого переноса электронов. In vitro скорость
переноса электронов можно увеличить в 4—6 раз в присутствии
ADP и Р(-, но всегда существует основной, или не связанный
с фосфорилированием транспорт электронов, вероятно, в ре-
зультате утечки протонов (разд. 5.9—5.10). Известно большое
количество соединений, которые действуют как разобщители
фотофосфорилирования, уменьшая отношение АТР/2е и увели-
чивая скорость не связанного с фосфорилированием транспорта
электронов. С позиции теории Митчелла и ее модификаций «об-
ратное» действие протонного градиента подавляет транспорт
электронов, и любые факторы, устраняющие это подавление,
должны ускорить перенос электронов. Так, детергенты, способ-
ствующие образованию пор в мембране, снижают градиент, соз-
давая условия для быстрой обратной диффузии. Арсенат и тио-
фосфат в присутствии ADP действуют как разобщители (конку-
рируя с Рг); отсюда следует, что они образуют нестабильные
легко разрушающиеся комплексы с ADP. Такие соединения,
как NH3, вызывающие отток протонов (NH3+H-'-—s^NH^-), и
антибиотики — нигериции и грамицидин D, — которые, как
предполагают, облегчают замену Н+ на К+, очевидно, также
уменьшают химический компонент градиента Н+ и, таким обра-
зом, усиливают транспорт электронов и снижают фотофосфори-
лирование. Механизм действия этих соединений не всегда бы-
вает таким простым, как может показаться на первый взгляд,
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ |Q5
и во многом остается . непонятным.. Например,: NH3 создает
«сток» для протонов в соответствии ,с реакцией НН'з+Н+—>
—»NH,4+, однако, несмотря на снижение [Н |_], вклад заряда.в
ПДС не уменьшается. С другой, стороны, если" происходит диф-
фузия N.H3 в тилакоидный компартмент, a NH4+ — наружу,
уменьшаются как химический, так и электрический компоненты
градиента Н+. Некоторые разобщители действуют именно таким
образом. В нормальных условиях происходит также движение
ионов, компенсирующих заряды ’ (такие* ионы, как С1~, перемеща-
ются внутрь, а,такие катионы, как Mg21',наружу), посколь-
ку в отсутствие этого движения электрический компонент гра-
диента протонов .достиг бы слишком большой величины. Осто-
рожное нагревание хлоропластов ускоряет перенос протонов , и
внутрь, и наружу, так что градиент не изменяется или увели;
иивается, но фотофосфорилирование при этом подавляется. Эм-
мет и Уокер объясняют это явление увеличением проницаемо-
сти мембран под действием нагревания, что обеспечивает быст-
рый перенос ионов, компенсирующих заряд, так что уменьшает-
ся электрический компонент градиента Н1'.
В присутствии ADP и Р,- градиент Н+ тоже уменьшается в
результате функционирования обратимой АТРазы и транспорт
электронов усиливается. По мере расходования ADP поток
электронов замедляется. Это усиление и последующая задерж-
ка транспорта электронов представляют собой «фотоспитетиче;
ский контроль», подобный «дыхательному контролю» в мито-
хондриях.
Б.12. Сопрягающий фактор
Как уже отмечалось (разд. 5.10), Аврон в 1963 г. сделал
очень важное открытие, показав, что в лишенных оболочки
хлоропластах происходит разобщение транспорта электронов и
фосфорилирования и в то же время они теряют «сопрягающий
фактор» CFj. Если добавить сопрягающий фактор, то в при-
сутствии Mg2'1' сопряжение синтеза АТР н транспорта электро-
нов восстановится. Впоследствии сопрягающие факторы ак-
тивно изучались Авроном, Рэкером, Мак-Карти, Нелсоном и
другими. Непродолжительное нагревание, инкубация с трипси-
ном или восстановление в присутствии высокой концентрации
сульфгидрильных реагентов приводят к превращению свободно-
го CFi в высокоактивную Са2+-зависпмую АТРазу, указывая
на то, что CFi играет какую-то роль в фотофосфорилировании.
CFi встраивается в мембрану тилакоида в виде комплекса со
специфическим гидрофобным белком CF0, составляя сложный
АТР-азный комплекс (CF0 — CFi), который подвергается, кон-
формационным изменениям на свету, а также в условиях новы,-
106
ГЛАВА S
шения градиента pH. С позиций хемиосмотической гипотезы
Митчелла (разд. 5.11) CF0 можно рассматривать как канал,
направляющий протоны к CFi — катализатору синтеза АТР. Бу-
дучи встроенным в искусственные фосфолипидные липосомы,
комплекс CFo — CFt способен катализировать обмен АТР —Р,-
и синтез АТР при наличии трансмембранного градиента.
5.13. Термодинамика синтеза АТР
Стандартная свободная энергия гидролиза АТР до ADP и
Рг (разд. 2.4, 2.11) в какой-то степени зависит от присутствия
Mg2+, и обычно ее считают равной примерно —7 ккал. Это от-
носится к случаю, когда АТР, ADP и Р, находятся в эквимоляр-
ных концентрациях; на самом деле потенциал фосфорилирова-
ния (свободная энергия образования АТР) зависит от концент-
рации компонентов реакции следующим образом:
лг=ду+ет1п.1А[^Ц (6.31)
и имеет максимальное значение, когда большинство аденилатов
присутствует в форме АТР. Так как в лишенных оболочки хло-
ропластах почти весь добавленный ADP легко превращается в
АТР, потенциал фосфорилирования, полученный исходя из на-
блюдающегося соотношения между аденилатами в системе, со-
ответствует реальному изменению свободной энергии. Подобные
измерения, проведенные Крайепгофом, дали величину около
14 ккал. Соответствующий электрический потенциал можно
рассчитать (разд. 2.13) по формуле
АД'
AF = —nfE', откуда Е' =-
Для реакции, протекающей с участием двух электронов, и при
&F'=—14 ккал имеем
£'=-1£-~300 мВ (5.32)
что сравнимо по величине с уменьшением потенциала на уча-
стке между пластохиноном и цитохромом f (рис. 4.18). [Полное
уменьшение потенциала между двумя фотосистемами составля-
ет около 450 мВ для двух электронов.] Таким образом, синтез
АТР в этом центре фосфорилирования энергетически вполне
возможен. Соответствующие оценки для второго центра
(разд. 4.23) весьма приблизительны, поскольку окислительно-
восстановительные потенциалы для Y и Z (разд. 4.23) и реак-
ционного центра II (разд. 4.12) точно неизвестны. Однако если
принять, что только 70% энергии (1800 мВ/моль квантов) фото-
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ Ц)7
нов при 680 нм поступает в цепь транспорта электронов и раз-
ность потенциалов между водой и первичным акцептором элек-
тронов в фотосистеме II равна примерно 800 мВ, то первичное
падение потенциала в этом центре составит около 460 мВ. Та-
ким образом, и в данном случае этой величины более чем до-
статочно, чтобы покрыть те затраты (падение потенциала на
300 мВ), которые необходимы при потенциале фосфорилирова-
ния 14 ккал.
5.14. Связь между транспортом протонов,
транспортом электронов и энергией
Согласно хемиосмотической теории, на каждый электрон,
который проходит через электрон-транспортиую цепь, внутрь
входит один протон и на каждую синтезируемую молекулу
АТР выходят наружу два протона. Это должно было бы
соответствовать отношению Р/2е~, равному 1,0 для каждого из
участков. Измерения показали, что на одни электрон действи-
тельно приходится один протон, но отношение Н'7АТР лежит
в диапазоне 2—4. Предполагают, что в Z-схеме имеются два
центра фосфорилирования (разд. 5.9), откуда следует, что па
два электрона должно приходиться 4 протона (рис. 5.5). Для
получения реального отношения Н+/АТР выход молекул АТР
соответственно должен был бы составить 1, 1,33 или 2. Как
было показано в разд. 5.13, потенциал фосфорилирования, рав-
ный 14 ккал, эквивалентен 300 мВ для процесса, протекающе-
го с участием двух электронов, и, в свою очередь, соответствует
ДрН = 5,0 (если принять, что единица рНвябО мВ). Наблюдае-
мые на опыте величины ДрН лежат в интервале 2,5—3,5 и это
наводит на мысль о вкладе в этот потенциал как протонного
градиента, так и мембранного потенциала [т. е. ДрН=3,3
( = 200 мВ) и мембранного потенциала около 100 мВ]. Сущест-
венным моментом в подобных расчетах является то, что доступ-
ная энергия зависит не только от разности окислительно-восста-
новительных потенциалов, по и от числа участвующих в про-
цессе электронов. Так, три электрона, а следовательно и три
протона (если принять, что 1 е~ эквивалентен 1 Н+) дадут
в 1,5 раза больше энергии, чем два электрона при такой же
разности потенциалов. Если для синтеза одной молекулы АТР
необходимы три протона, то ДрН = 3,3 (=200 мВ) будет доста-
точно и без вклада мембранного потенциала. Однако при нали-
чии мембранного потенциала, равного 100 мВ, необходимая
разность pH составит лишь 1,7 единицы. [Хотя величина мемб-
ранного потенциала точно неизвестна, есть основания полагать,
что она может быть равна 100 мВ.] При транспорте в Z-схеме
двух электронов на четыре кванта образуются четыре Н+
j 08 . ГЛАВА' б
(рис. 5.6). Это может дать выход 2 АТР/4 кванта. Для транс-
порта электронов в Z-схеме требуется строго определенное ко-
личество квантов. Так, для каждого фотохимического акта
необходим один электрон и, следовательно, требуется один
квант. NADP принимает от воды электроны, прошедшие после-
довательно через всю цепь переносчиков; следовательно, для
восстановления NADP в этом процессе требуются четыре кван-
та, так как два электрона проходят через две фотосистемы.
Однако квантовый расход для синтеза АТР не установлен, так
как он зависит от того, сколько протонов поступает в тилакоид
при транспорте каждого электрона и как много молекул АТР
генерируется этим протонным градиентом. Если с внутренней
стороны мембраны тилакоида освобождается два протона при
поступлении в ФС II от воды двух электронов (и еще два про-
тона при поступлении двух электронов от пластохинона к ФС I),
то перенос в Z-схеме двух электронов приведет к освобождению
внутрь тилакоидного компартмеита четырех протонов. Соотно-
шение между процессом транспорта электронов и образовани-
ем АТР зависит, таким образом, от того, сколько протонов
должно пройти через «генератор» АТР (АТРаза Митчелла),
чтобы образовалась одна молекула АТР из ADP и Pi.
Для ассимиляции одной молекулы СОа в ВПФ-цикле необ-
ходимы три молекулы АТР. Как видно нз табл. 5.1, для син-
теза этих трех молекул АТР необходимо 6, 9 и 12 квантов при
соотношении между Н1' и АТР, равном соответственно 2, 3 и 4.
Но поскольку, кроме этого, требуются еще две молекулы
NADPHa, минимальный квантовый расход должен быть не
меньше восьми. Именно отсюда получены цифры 8, 9 и 12, при-
веденные в последней колонке табл. 5.1. При отношении
Н+/АТР, равном 2, на каждые две молекулы образующегося
NADPH должно образоваться на одну молекулу АТР больше
по сравнению с минимальной потребностью в ВПФ-цикле, При
Таблица 5.1. Минимальный квантовый расход (КР) при образовании
ассимиляционной силы и ассимиляции СО2 в ВПФ-цикле1)
Расход 11+ при синтезе АТР Н+ или кван- ты/3 АТР КРи Н+/2 NADPH Минимальный
2 6 8 8
3 9 8 9
4 12 8 12
’) Расчет сделан исходя из условия, что для переноса одного протона необходимо
поглощение одного кванта света. Отношение Н+/АТР изменяется от 2 до *1; для фикса-
ции 1 СОа требуется минимум 3 АТР и 2 NADPH.
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ
109
более высоком квантовом расходе не имеет значения, покрыва-
ется ли этот дефицит АТР за счет циклического или. нецикли-
ческого транспорта электронов. Например, при Н+/АТР, рав-
ном 4, возникает дефицит в одну молекулу АТР на каждые две
молекулы восстановленного NADP. Образование дополнитель-
ных молекул АТР потребовало бы четырех протонов и такой
градиент мог бы быть достигнут при прохождении двух электро-
нов через 2 центра фосфорилирования (при нециклическом
транспорте электронов с расходом 4 кванта) или четырех элек-
тронов через 1 центр (для циклического транспорта, также
требующего 4 кванта). Однако, если циклический транспорт
электронов вносит больший вклад в образование АТР, чем не-
циклический (т. е. если циклический транспорт электронов при-
водит к переносу четырех протонов на 2 электрона с использо-
ванием двух квантов), то общий квантовый расход на фикса-
цию СОг достигает 10.
5.15. Восстановление никотинамидадениндинуклеотид-
фосфата (NADP)
Ферредоксин
Еще в ранних работах Роберта Хилла было показано, что
при наличии искусственного окислителя или в присутствии
водного экстракта из листьев, предварительно обработанных
ацетоном, изолированные хлоропласты выделяют Ог. Это позво-
ляло предположить существование «растворимого акцептора
водорода для процесса фотосинтеза в самих листьях». Хилл и
Скарисбрик добавляли к системе хлоропластов метгемоглобин
й оксалат железа. Хлоропласт восстанавливал оксалат железа,
который в свою очередь восстанавливал метгемоглобин; обра-
зующийся при этом гемоглобин взаимодействовал с кислородом.
Появление окисленного гемоглобина позволяло контролировать
реакцию спектроскопически. Эти наблюдения привели Хилла и
его соавторов к постановке опытов, с помощью которых в экст-
ракте, полученном из ацетонового порошка листа, удалось об-
наружить белковую фракцию, способную восстанавливать мет-
гемоглобин (разд. 4.9).
Однако способность хлоропластов восстанавливать также
NAD+ и NADP+ при участии белкового фактора, была установ-
лена лишь десять лет спустя несколькими авторами, независи-
мо друг от друга. В 1957 г. Арион, Уотли и Аллен выделили
ПАОР+-восстанавливающий фактор, а в 1958 г. Саи Пиетро и
Ланг частично очистили «фотосинтетическую пиридиииуклео-
тидредуктазу». Немногим позже, в 60-е годы уже ие было
сомнений, что все эти экстракты содержат один и тот же белок.
по
ГЛАВА 5
Когда было показано, что он близок к бактериальному фер-
редоксину, Арнон счел возможным назвать хлоропластный бе-
лок «ферредоксином».
Ферредоксин — NADP-редуктаза
При дальнейшей очистке стало ясно, что сам по себе ферредок-
син не способен восстанавливать NADP+, скорее он представлял
собой просто носитель электронов с низким потенциалом. [Спо-
собность хлоропласта восстанавливать искусственные перено-
счики электронов с низким потенциалом, такие как метилвиоло-
ген (паракват), была установлена независимо друг от друга
Ягендорфом и Авроном, а также Хиллом и Уокером. Потенциал
метилвиологена ~ —0,450 В, потенциал ферредоксина
~—0,430 В, потенциал NADP+~—0,320 В.] Ферредоксин, обыч-
но связанный с тилакоидами, взаимодействует также с молеку-
лярным кислородом (поддерживая псевдоциклическое фото-
фосфорилирование) и с другими переносчиками электронов
Z-схемы, возможно, цитрохромом Ъ. При освещении смеси от-
мытых тилакоидов, ферредоксина и NADP интенсивно протека-
ет реакция Хилла.
Редуктаза представляет собой типичный флавопротеин с
максимумами абсорбции при 275, 385 и 456 им. Сродство редук-
тазы kNADP+ (/См=9,8- 10-еМ) настолько больше,чем kNAD+
(Лм=3,7-10~3 М), что ее можно рассматривать как NADP'h-cne-
цифический фермент в физиологических условиях. Однако фер-
мент может функционировать как трансгидрогеи аз a (NADPH-f-
NAD+4=*:NADH-|-NADP+) и диафораза (окисление NADPH2 ря-
дом акцепторов электронов, таких как феррицианид и хиноны).
Механизм восстановления NADP
Обычно электроны проходят от воды через переносчики Z-схе-
мы к ферредоксину. Последний поставляет электроны флавопро-
теиновой редуктазе, которая в свою очередь восстанавливает
NADP+ (NADP+ при восстановлении принимает 2 электрона и
1 протон). Чтобы перенести электроны от ферредоксина к
NADP+, требуется специфический флавопротеин — ферредок-
син— ЫАОР+-редуктаза. Этот фермент может участвовать так-
же в циклическом фотофосфорилировании. Физиологическая
роль редуктазы в псевдоциклическом транспорте электронов и
окислении NADPH2 пока не установлена, но, очевидно, в хлоро-
пласте имеется механизм диссипации энергии, который иногда
объясняют фотодыханием (гл. 13).
ОБРАЗОВАНИЕ АТР; ГЕНЕРАЦИЯ АССИМИЛЯЦИОННОЙ СИЛЫ Щ
ОБЩАЯ ЛИТЕРАТУРА
1. Arnon D. I. Role of ferredoxin in photosynthesis, Naturwissenschaften., 56,
295—305 (1969).
2. Avron M. Energy transduction in chloroplasts, Ann. Rev. Biochem., 46, 143—
155 (1977).
3. Beale S. 1. 6-Aminolevulinic acid in plants: Its biosynthesis, regulation, and
role in plastid development, Ann. Rev. Plant Physiol., 29, 95—120 (1978).
4. Bickford E. D. and Dunn S. Lighting for Plant Growth, The Kent State Uni-
versity Press, 221 pp., 1972.
5. Bishop N. I. Photosynthesis: the electron transport system of green plants,
Ann Rev. Biochem., 40, 197—226 (1971).
6. Black С. C. Photosynthetic carbon fixation in relation to net CO2 uptake,
Ann. Rev. Plant Physiol., 24, 253—286 (1973).
7. Blinks L. R. The photosynthetic function of pigments other than chlorophyll,
Ann. Rev. Plant Physiol., 5, 93—111 (1954).
8. Boardman N. K. Physical separation of the photosynthetic photochemical
systems, Ann. Rev. Plant Physiol., 21, 1 15—140 (1970).
9. Boardman N. K. Comparative photosynthesis of sun and shade plants, Ann.
Rev. Plant Physiol., 28, 355—377 (1977).
10. Boyer P. D. et al. Oxidative phosphorylation and photophosphorylation, Ann.
Rev. Biochem., 46, 955—1026 (1977).
11. Brown J. S. Forms of chlorophyll in vivo, Ann. Rev. Plant Physiol., 23,
73—86 (1972).
12. Butler IV. L. Energy distribution in the photochemical apparatus of photo-
synthesis, Ann. Rev. Plant Physiol., 29, 345—378 (1978).
13. Cheniae G. M. Photosystem II and O2 evolution, Ann. Rev. Plant Physiol.,
21, 467—498 (1970).
14. Cramer IF. A. and Whitmarsh J. Photosynthetic cytochromes, Ann. Rev. Plant
Physiol., 28, 133—172 (1977).
15. Goedheer J. C. Fluorescence in relation to photosynthesis, Ann. Rev. Plant
Physiol., 23, 87—112 (1972).
16. Hall D. O. The coupling of photophosphorylation to electron transport in
isolated chloroplasts. In: The Intact Chloroplast, ed. J. Barber, Elsevier/
North-Holland Biomedical Press, The Netherlands, pp. 135—170, 1976.
17. Hatch M. D. and Slack C. R. Photosynthetic CO2-fixation pathways, Ann.
Rev. Plant Physiol., 21, 141—162 (1970).
18. Heber U. Metabolite exchange between chloroplasts and cytoplasm, Ann.
Rev. Plant Physiol., 25, 393—421 (1974).
19. Hill R. The biochemists’ green mansions. The photosynthetic electron-trans-
port chain in plants, Essays Biochem., 1. 121 —151 (1965).
20. Hinkle P. C. and McCarty R. E. How cells make ATP, Sci. Am., 238, 104—
123 (1978).
21. Jackson W. A. and Volk R. J. Photorespiration, Ann. Rev. Plant Physiol., 21,
385—432 (1970).
22. Joliot P. Cinetiques des reactions liees a Remission d’oxygene photo-
synthetique, Biochim. Biophys. Acta, 102, 116-134- (1965).
23. Junge IF. Membrane potentials in photosynthesis, Ann. Rev. Plant Physiol.,
28, 503—536 (1977).
24. Kelly G. J. and Latzko E. Regulatory aspects of photosynthetic carbon meta-
bolism, Ann. Rev. Plant Physiol., 27, 181—205 (1976).
25. Kirk J. T. O. Chloroplast structure and biogenesis, Ann. Rev. Biochem., 40,
161—196 (1971).
26. Myers J. Enhancement studies in photosynthesis, Ann. Rev. Plant Physiol.,
22, 289—312 (1971).
27. Raschke K- Stomatai Action, Rev. Plant Physiol., 26, 309—340 (1975).
28. Rebeiz C. A. Protochlorophyll and chlorophyll biosynthesis in cell-free sys-
ГЛАВА 5.
и 2
terns from higher plants, Ann. Rev. Plant Physiol., 24, 129—172 (1973).
29. Ried A. Improved action spectra of light reaction .1 and II, Proc, of the
. 2nd International-Congress on PhofosyntheSis Research Stresa-1971, Vol. I.
Eds. G. Forti, M. Avron and A. Melandri, Dr. W. Junk, N. V, The Hague,
• PP..763—772, 1972. . i'
30. Schwartz M. The relation of ion transport to phosphorylation, Ann. Rev.
. - Plant Physiol., 22, 469—484 (1971).
31. Simonis Й7. and Urbach IF. Photophosphorylation in vivo, Ann. Rev. Plant
Physiol., 24, 89—114 (1973).
32. Thornber J. P. Chlorophyll-proteins: Light-harvesting and reaction center
, : cdmponents of plants, Ann. Rev. Plant Physiol., 26, 127—158 (1975).
33. Trebst A. Energy conservation in photosynthetic electron transport of chlo-
roplasts, Ann. Rev. Plant Physiol., 25, 423—458 (1974).
34. Walker D. A. and Crofts A. R. Photosynthesis, Ann. Rev. Biochem., 39, 389—
; 4?8 (1970.).
35. Zelltch I. Pathways of carbon fixation in green plants, Anu. Rev. Biochcin.,
; 44,123—145 (1975).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
36. Barber J. (ed.) .The Intact Chloroplast — Topics in photosynthesis, vol. 1,
' feisevier/North-I-iolland: -Biomedical Press, The Netherlands, 1976. '
37. Clayton R. /<. Light and Living Matter vol. 1, McGraw-Hill, New York,
1970.
38. Douce R. and Jot/ard I. Structure and function of the plastid envelope, Adv.
‘ Bot. Res., 7, 1—117 (1979).
39. Gibbs M. (ed.) Structure and Function of Chloroplasts, Springer-Verlag,
Berlin, Heidelberg, New York, 1971.
40. Giese A. C. Photophysiology, Academic Press, New York, 1964.
41. Govindjee (ed.) Bioenergetics of Photosynthesis, Academic Press, New York,
1975.
42. Hath M. D. arid Boardman N. R. (eds.) The Biochemistry of Plants. A Com-
prehensive Treatise, vol. 8, Academic Press, New York, 1981.
43. Hill R. and Bendall F. Function oi the two cytochrome components in chlo-
roplasts: a working hypothesis, Nature, London, 186, 136—137, 1960.
44. Lavorel J. and Enienne A. L. In vivo chlorophyll fluorescence, In: Primary
Processes of Photosynthesis (ed. J. Barber), pp. 203—268, Elsevier/North-
Holland Biomedical Press, The Netherlands, 1977.
45. 'McCarty R. E. Roles of a coupling factor for photophosphorylation in chlo-
roplasts, Ann. Rev. Plant Physiol., 30, 79—104 (1979).
46. McElroy IF. D. and Glass M. (eds.) Light and Life, The John Hopkins Press,
Baltimore, 1961.
47. Mitchell P. Chemiosmotic coupling in oxidative and photosynthetic phospho-
rylation, Biol. Rev. Cam. Philps. Soc., 41, 445—502 (1966).
48. Olson J. H. and Hind G. (eds.) Chlorophyll-proteins, reaction centres and
photosynthetic membranes, Brookhsaven Symposia in Biol. no. 28, Brookhaven
Nat. Lab. Assoc. Universities Inc. Upton, New York, 1976.
49. Rabinowitch E. I. Photosynthesis and Related Processes, vol. I, II, Wiley
(Interscience), New York, 1945, 1951, 1956.
50. San Pietro A. et al. (eds.) Haevestihg the Sun, Academic Press, New York,
1967.
51. Trebst A. and Avron M. (eds.) Photosynthesis I. Photosynthetic Electron
Transport and Photophosphorylation, Encyclopedia of Plant Physiology (New
Series), vol. 5, Springer-Verlag, Berlin, Heidelberg, New York, 1977.
52. Walker D. A. Energy, Plants and Man, Packard Publishing Ltd., Chichester,
1979.
53. Witt H. T. Coupling of quanta, electrons, fields, ions and phosphorylation in
the functional membrane of photosynthesis, Quarterly Rev. Biophys., 4, 365—
477 (1971).
Часть Б
ГЛАВА 6
Восстановительный пентозофосфатный цикл
и связанные с ним реакции
Краткое содержание
Живые организмы обладают только одним первичным меха-
низмом карбоксилирования. Первичным в том смысле, что он
порождает все остальное. Это путь биосинтеза всех углеродсо-
держащих органических соединений из двуокиси углерода, Он
известен под названиями цикла Кальвина, цикла Бенсона —
Кальвина, фотосинтетического углеродного (восстановительно-
го) цикла или, как мы и будем называть его в дальнейшем,
восстановительного пеитозофосфатного цикла (ВПФ-цикла,
рис. 6,1). Он характерен для всех растений: Ся-, С4- и тех, чей
кислотный метаболизм идет по типу толстянковых; впрочем,
двум последним присущи также дополнительные вспомогатель-
ные или предварительные механизмы фиксации.
Восстановительный пентозофосфатный цикл складывается из.
четырех основных этапов.
1. Карбоксилирование. На этом этапе СО2 соединяется с ак-
цептором рйбулозо-1,5-бисфосфатом (РуБФ), давая две моле-
кулы 3-фосфоглицернновой кислоты (ФГК). В этой реакции-
равновесие сильно сдвинуто в сторону ФГК, Реакцию катали-
зирует РуБФ-карбоксилаза, сложный фермент с высоким срод-
ством к СО2.
2. Восстановление. ФГК восстанавливается за счет ассими-
ляционной энергии до триозофосфата.
3. Регенерация. Пять молекул триозофосфата подвергаются
перегруппировке, в результате которой регенерируются три мо-
лекулы акцептора СО2 (5Сз—>ЗСд).
4. Автокатализ. На каждые три молекулы СО2, вовлеченные
в ВПФ-цикл, образуется одна молекула продукта—триозофос-
фата С3. Этот продукт способен превращаться в крахмал, саха-
розу и т. д., он может также повторно подвергаться регенера-
ции, приводя тем самым к автокаталитическому образованию
промежуточных соединений. Таким образом, цикл может функ-
ционировать в качестве «производительной» реакции, в которой
8—234
a i4
ГЛАВА В
г~г
Ру5Ф Ру5Ф
t t
Р5Ф КубФ
Продукт в цикл
АТР^-ЗН-О
ЗАОГЗР, >»С0;
ЗРубФ < ЗСО; 1-3 Н;О
ВФГК
ГВФ^ПФ (бАТР^бН.О
S' 6ДФГК6АОР 6Р,
Крахмал 6р. 6N ADPH +
6ФГА 6NADP
'6Н:О
ЗСО- 1- 2Н.0 + Pj - СН > ОН СО СН; ОРО(ОН)г + ЗОД
Рис. 6.1. Восстановительный пеитозофосфатный цикл (ВПФ-цикл). Справа
три молекулы РуБФ соединяются с тремя молекулами СОг и тремя молеку-
лами воды, давая шесть молекул ФГК. Эти молекулы фосфорилируются за
счет АТР с образованием ДФГК, которая восстанавливается NADPIT до
ФГА. Часть ФГА превращается в свой изомер ДГАФ. Альдольная конденса-
ция этих двух триозофосфатов дает молекулу ФБФ, которая подвергается
гидролизу до Ф6Ф. Этот гексозофосфат служит также предшественником
Г6Ф и ПФ; последний после дальнейших превращений дает начало крахма-
лу. Кроме того, Ф6Ф участвует в первой реакции, катализируемой транске-
толазой, давая двухуглеродный фрагмент ФГА, что приводит к образованию
ксилулозо-5-фосфата (КубФ) и эритрозо-4-фосфата (Э4Ф). Процесс конден-
сации, фосфорилирования и переноса двухуг лер одного фрагмента повторяет-
ся, в результате чего образуются соответственно СБФ, С7Ф и еще две пеи-
тозофосфатные молекулы. В конечном счете все три молекулы пентозомоно-
фосфата превращаются в РуБФ, который фосфорилируется до РуБФ, Весь
цикл можно подразделить на три фазы. Сначала происходит карбоксилиро-
вание (I фаза), а затем восстановление (II фаза) до триозофосфатов. Пять
Сз-молекул подвергаются перегруппировке (III фаза), в результате которой
регенерируются три Cs-молекулы акцептора СО2. Следует отметить, что на
образование одного триозофосфатного продукта (который может по-
вторно включаться в цикл, обеспечивая автокаталитическое ускорение) в
цикле используется 9 молекул АТР и 6 молекул NADPI-I. Суммарно в самом
цикле используются пять молекул воды, и три молекулы воды выделяются в
процессе генерации ассимиляционной энергии. Если бы образующийся три-
озофосфат подвергался гидролизу до свободной триозы в реакции с потреб-
лением одной молекулы воды, неорганический фосфат совсем бы не исполь-
зовался, и вся последовательность реакций свелась бы к классическому сум-
марному уравнению фотосинтеза [уравнение (1.1)].
восстановительный пентозофосфатный цикл
115
молекул СОг-акцептора образуется больше, чем потребляется.
Эта уникальная автокаталитическая способность лежит в осно-
ве роста растений.
Хотя сахарозу считают иногда конечным углеводным про-
дуктом фотосинтеза, на самом деле она служит основным
транспортируемым метаболитом во многих растениях и синте-
зируется в цитоплазме из поступающего туда триозофосфата.
В отличие от сахарозы крахмал образуется внутри хлоропласта
и встречается в большинстве листьев высших растений в каче-
стве временного запасного продукта. Его содержание наиболее
велико тогда, когда фотосинтез преобладает над потреблением
ассимилированных соединений в процессе метаболизма, т. е.
лист запасает крахмал днем и использует его ночью.
6.1. Фотосинтетическая фиксация углерода
Фотосинтез сводится к «фиксации», или включению, атмо-
сферной двуокиси углерода в органическую углеродную моле-
кулу в тканях растения. Все известные процессы фиксации при
фотосинтезе — это реакции карбоксилирования, т. е. СО2 соеди-
няется с уже имеющимся акцептором R таким образом, что об-
разуется новая карбоксильная группа R-COO':
R + СО2 —R-COO" (6.1)
6.2. Регенерация
Для того чтобы фотосинтетическая фиксация СО2 протекала
без остановки, акцептор R должен образовываться с той же
скоростью, с какой он потребляется. Таким образом, должен
существовать некий сопряженный механизм, обеспечивающий
регенерацию акцептора СО2 (разд. 6.13).
6.3. Автокатализ
Большинство процессов в высших растениях (и большинство
живых организмов) прямо или косвенно зависит от соединений,
возникающих в процессе фотосинтетической ассимиляции угле-
рода. Однако исходно должен существовать один основной са-
мостоятельный процесс, обеспечивающий образование и рост
содержания собственного субстрата. Реакция, которая дает
больше субстрата, чем его потребляет (производительная реак-
ция), является автокаталитической, поскольку в определенных
пределах ее скорость должна возрастать с увеличением кон-
центрации субстрата. Хотя необходимость в автокатализе явля-
ется самоочевидной, о ней нередко забывают, и поэтому нам
хотелось бы подчеркнуть ее важность. Если бы с помощью ме-
8*
J16
ГЛАВА 6
ханизма карбоксилирования (т. е. карбоксилирование плюс свя-
занные с ним реакции синтеза) регенерировалось лишь точно
такое же количество субстрата, которое потребляется, то рас-
тение не способно было бы расти и приспосабливать свою ско-
рость фотосинтеза к более благоприятным условиям (например,
к возрастанию интенсивности освещения и температур в течение
дня). Следовательно, зеленое растение должно обладать по
крайней мере одним механизмом, который обеспечивал бы пре-
вращение фотосинтетического продукта в дополнительный
субстрат. Единственным известным механизмом, который удов-
летворяет этому важному требованию, является цикл Бенсона —
Кальвина, или восстановительный пентозофосф атный цикл
(ВПФ-цикл) (разд. 6.13).
6.4. Энергетика
В реакции карбоксилирования затрачивается примерно
столько же энергии (-|-7 ккал), сколько при образовании АТР
из ADP и неорганического фосфата Рг-. Изменение энтальпии,
.рассчитанное из полученных эмпирическим путем значений энер-
гии связи и резонанса, составляет ----1-3 ккал/моль. При рас-
чете ДА' необходимо также учитывать изменение энтропии
(разд. 2.3), которое достаточно велико, если СО2 находится в
газообразном состоянии, но в любом случае точность рассчи-
танной величины лежит в пределах ±5 ккал. Молекула дву-
окиси углерода весьма стабильная, с большим числом возмож-
ных резонансных структур, и энергия, затрачиваемая па ее раз-
рушение, почти па 7 ккал превышает энергию, которая выделя-
ется при образовании новой карбоксильной группы. По этой
причине некоторые реакции карбоксилирования, например по-
казанное ниже образование оксалоацетата из пирувата и СО2:
СН3• СО. СООН + СО2 > НОО С СНа СО СООН (6.2)
настолько невыгодны с энергетической точки зрения, что их
можно не принимать во внимание в качестве возможных меха-
низмов биологического карбоксилирования. Другие реакции,
например восстановительное карбоксилирование пирувата до
малата, приведенное ниже:
СНз • СО СООН + NADPHj + СО2 ->
------------------> НООС СНОН СН2 СООН + NADP+ (6.3)
можно считать более выгодными энергетически, ио и тут ве-
роятность возникновения новых карбоксильных групп весьма
мала. Дело в том, что в атмосфере содержится очень мало СО2
(около 300 м. д. или 0,03%), и эта СО2 должна еще диффунди-
ровать в лист, где ее концентрация будет составлять, очевидно,
восстановительный пентозофосфатный цикл
117
7 мкмоль или даже меньше в отсутствие СО2-концентрирующих
механизмов (гл. 14). Основной механизм (или механизмы) фо-
тосинтетического карбоксилирования должен функционировать
в. рамках этих ограничений, и, конечно, ему будут благоприят-
ствовать такие константы равновесия, при которых накапли-
вающийся продукт не сразу замедляет скорость реакции. В на-
стоящее время четко установлены только две основные реакции
фотосинтетического карбоксилирования [уравнения (6.4) и (6.5)].
Обе они сопровождаются снижением свободной энергии пример-
но на 7—8 ккал.
ФЕП 4- СО„ -]-1IjjO ->- ОксалоацетатР1 [ДР ——7 ккал] (6.4)
РуБФ4-С02 4-НаО ---
--->- 23-Фосфоглицерииовая кислота [ДР =—8 ккал] (6.5)
Оба субстрата, фосфоеиолпируват (ФЕП) и рибулозо-1,5-бис-
фосфат (РуБФ), можно рассматривать как макроэргическиесо-
единения, которые обладают достаточным количеством энергии,
чтобы довести данную реакцию карбоксилирования практиче-
ски до конца.
Следует остановиться еще на двух аспектах энергетики кар-
боксилирования. Возникает вопрос, почему в ВПФ-цикле кар-
боксилируется РуБФ, а не Ру5Ф. Ведь обе эти молекулы бога-
ты энергией в том смысле, что они ие образуют пятичлепиые
кольца, которые сделали бы их более стабильными. (Кроме того,
карбоксилирование Ру5Ф привело бы к значительному умень-
шению свободной энергии, а образование глицериновой кисло-
ты в качестве одного из продуктов реакции могло бы быть ис-
правлено последующим фосфорилированием.) С точки зрения
энергетики ответ па этот вопрос заключается в том, что образо-
вание РуБФ способствует протеканию реакции регенерации в
нужную сторону. Реакция с участием Ру5Ф-кипазы также игра-
ет важную роль в регуляции (гл. 9).
ФЕП служит примером того, как одно и то же макроэрги-
ческое соединение может быть использовано в различных реак-
циях. Так, при прямом гидролизе ФЕП высвобождается
~14 ккал:
ФЕП + Н2О ------> Пируват-l-Pj [ДР =—14 ккал] (6.6)
При гликолизе около 7 ккал из этих 14 сохраняются в фор-
ме АТР [уравнение (6.7)]:
ФЕП -|- ADP -----> Пируват -|- АТР [ДF' = —7 ккал] (6.7)
Однако эта реакция все еще идет с изменением свободной энер-
гии ДЕ' приблизительно —7 ккал, что сильно затрудняет ее об-
ратимость.
118
ГЛАВА 6
Альтернативно около 7 ккал энергетического потенциал;®
ФЕП может быть использовано для «запуска» столь же энерге-
тически выгодной реакции карбоксилирования с AF' —7 ккал:
ФЕП 4- СО2 -|- Н2О -> Оксалоацетат 4-Pi [AF'=— 7 ккал] (6,8}
Если происходит и образование АТР, и карбоксилирование,
например в реакции, катализируемой ФЕП-карбоксикиназой
[уравнение (6.9)], то изменение свободной энергии приближает-
ся к нулю. Такая реакция увеличивает вероятность участия фос-
фоенолпирувата в процессе обратного гликолиза (глюконеоге-
неза):
Оксалоацетат 4-АТР .»’ ФЕП 4-СО3 4~ ADP [A/?'=0] (6.9)
При Ci-фотосинтезе регенерация ФЕП может (теоретически)
быть достигнута за счет обеих макроэргических групп АТР
Пируват 4- АТР -> ФЕП + AMP -|- Pi [АД' = 0] (6.10)
Благодаря этому было бы преодолено энергетически невы-
годное равновесие реакции, катализируемой пируваткиназой
[противоположной той, которая дана в уравнении (6.7)]. Дейст-
вительно, при изучении этой реакции (которая известна у ряда
бактерий) в результате целенаправленного поиска была обна-
ружена пируват,ортофосфат—дикиназа. В реакции используют-
ся обе макроэргические группы АТР:
Пируват 4-Р( 4-АТР ---> ФЕП4-АМР4-РР( [AE'=+7 ккал] (6.11)
Сама по себе эта реакция столь же энергетически невыгодна,
как и реакция, обратная той, которая дана в уравнении (6.7)
[AF'=-[-7 ккал в направлении синтеза ФЕП]. Однако она свя-
зана с гидролизом пирофосфата РР,-:
PPi 4- Н2О --> Р, 4- Pi [АТ' = -7 ккал] (6.12)
Таким образом, суммарное уравнение этих двух реакций
имеет ДЕ' около нуля, что обеспечивает полную обратимость
процесса (разд. 2.5). Для того чтобы процесс стал возможен
не только с энергетической, но и с биологической точки зрения,
необходимы два условия: достаточно высокое содержание пиро-
фосфатазы и очень низкая величина ее К-л (большое сродство
к РР,). В противном случае в первой реакции (уравнение
(6.11) не образовывалось бы быстро столько продукта, сколько,
его требуется для смещения ее энергетически невыгодного рав-
новесия за счет функционирования пирофосфатазы [уравнение
(6.12)].
Источники данных: [5, 7, 17, 18, 19, 20, 22, 28, 32, 53].
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ ЦИКЛ
119
6.5. Сродство к СО2
Карбоксилирование протекает тем быстрее, чем больше ве-
личина константы равновесия. Как видно из табл. 2.1, большая
величина К соответствует большому отрицательному изменению
свободной энергии AF'. Отсюда следует, что реакции, представ-
ленные выше уравнениями (6.4) и (6.5), идут почти до конца
в данном (т. е. прямом) направлении. Аналогичным образом
активность катализатора (карбоксилазы) будет тем выше, чем
больше его сродство к СО2-
При достаточно низких концентрациях субстрата скорость
ферментативной реакции можно увеличить в два раза, если
удвоить концентрацию субстрата [S]. При очень низких уровнях
субстрата уравнение Михаэлиса — Ментен
.. _ Утах [*SJ
Am + [S]
(6.13)
практически сводится к виду
v
(6.14)
Утах [5]
Км
где величина [S] становится пренебрежимо малой по сравнению
с Км- Таким образом, при низких концентрациях субстрата за-
Рис. 6.2. Зависимость скорости ферментативной реакции от концентрации
субстрата (S). Приведены две концентрации фермента (ЕХ.1 и ЕХ2). Мак-
симальная скорость (Vmax) удваивается при повышении в два раза концент-
рации фермента, ио величина Ам (концентрация субстрата, необходимая для
достижения половины максимальной скорости) остается неизменной.
120
ГЛАВА.6-
висимость между v и [S] приближается к линейной,, хотя никог-
да и не достигает ее. При увеличении концентрации субстрата,
в конце Концов все активные центры фермент-а оказываются
насыщенными. Если выразить графически зависимость между
скоростью реакции и концентрацией субстрата, то сначала прй
очень низких концентрациях она будет носить линейный харак-
тер. Однако'по мере приближения к насыщению зависимость,
начнет отклоняться от линейной (рис. 6.2).
Таким образом, эта кривая является частью прямоугольной
гиперболы, которая приближается к теоретическому максимуму
Vmax (максимальная скорость), но никогда не достигает его.
(или достигает при бесконечно высоких концентрациях субстра-
та) . Величина' Утах характерна для данного фермента, взятого
в определенной концентрации. Фермент характеризуется также
константой Михаэлиса (/См), равной концентрации субстрата,
при которой достигается .половина максимальной скорости.
Дополнительное преимущество константы Михаэлиса со-
стоит в том, что она не зависит -от концентрации фермента
;(рис. 6.2).
(На практике максимальную скорость не определяют путем
повышения концентрации субстрата до бесконечности, так как
это нецелесообразно. Вместо этого часто пользуются следующим
приемом: в качестве координат откладывают обратные величи-
ны скорости и концентрации субстрата 1/ц и 1/[S]; при этом
вместо гиперболы получают прямую линию. При бесконечно,
большой [S] ее обратная величина 1/[S] бесконечно мала, т. е.
равна нулю, и точка пересечения прямой с вертикальной осью
(рис. 6.3) соответствует l/Vmax- Аналогичным образом 1/Км па
этом графике соответствует отрицательному значению величи-
ны, при которой экстраполированная кривая пересекает гори-
зонтальную ось.)
Константа Михаэлиса обратно пропорциональна сродству
фермента к субстрату. В нескольких словах это можно сформу-
лировать так: если ферменту требуется много субстрата для
того, чтобы он мог работать с высокой скоростью (большая ве-
личина Км), следовательно, он обладает низким сродством к
субстрату. Очевидно, карбоксилаза активно функционирует в
присутствии низких концентраций СО2 благодаря высокому
сродству к этому субстрату (низкая величина Км). Ферменты,
катализирующие реакции, представленные уравнениями (6.4)
и (6.5), удовлетворяют этому требованию. Фосфоенолпируват-
карбоксилаза [катализатор реакции (6.4)] была впервые описа-
на в 1954 г., и уже тогда отмечалось ее высокое сродство к
СО2/бикарбонату. Открытая примерно в это же время РуБФ-
карбоксилаза породила значительные трудности, которые не
удавалось разрешить долгие годы. Дело в том, что для работы
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ цикл
П21
Рис. 6.3. Преобразование зависимости, приведенной на рис. 6.2, с использо-
ванием обратных величин скорости реакции и концентрации позволяет ги-
перболические кривые перевести в более удобную для расчетов форму,-—
в прямые. При [S]=oo 1/[S]=O и точки пересечения с вертикальной осью да-
дут величины l/Vmax для каждой концентрации фермента. (Теоретически мак-
симальная скорость достигается в присутствии бесконечно высоких концент-
раций субстрата.) Величину Лм можно получить так же, как и на рис. 6.2,
проведя вертикальную линию из точек, соответствующих обратным величи-
нам половины максимальных скоростей. Однако проще провести экстрапо-
ляцию к точке пересечения с отрицательной горизонтальной осью, что даст
ту же числовую величину 1//(м, но со знаком минус.
с — Vmax этому ферменту требовалось 6% со2, а это противоре-
чило представлению о его предполагаемой роли in vivo. Впо-
следствии было установлено, что опыты проводились с частично
инактивированным ферментом; при условии же соответствую-
щей активации РуБФ-карбоксилаза по сродству к СО2 не усту-
лала ФЕП-карбоксилазе.
(Теоретически величина Утах так же важна для фермента,
как и сродство к субстрату. Иными словами, все равно, сколько
субстрата требуется ферменту для функционирования с нужной
скоростью, лишь бы организм содержал достаточное количество
фермента. Однако на практике возможны пространственные
ограничения в отношении данного катализатора. Во всяком слу-
чае, РуБФ-карбоксилаза является основным белком стромы и
может составлять до половины всех белков в тканях листа.
Столь высокое содержание этого фермента отражает выработав-
шуюся в процессе эволюции способность сохранять фотосинте-
тическую активность вопреки падению концентрации СОо и воз'г
122
ГЛАВА 6
растанию концентрации О2, которые были вызваны этой же ак-
тивностью.)
Источники данных: [7, 17, 18, 19, 26, 34, 39, 42, 44, 53, 54].
6.6. Основные характеристики карбоксилирования
Главный механизм карбоксилирования при фотосинтезе обя-
зан отвечать следующим четырем требованиям:
1. Должен обеспечивать регенерацию акцептора СО2.
2. Должен функционировать автокаталитически в качестве
производительной реакции, в ходе которой субстрата образует-
ся больше, чем используется.
3. Равновесие реакции карбоксилирования должно быть
энергетически очень выгодным, чтобы компенсировать низкую
концентрацию СО2 в окружающей среде.
4. Катализирующая реакцию карбоксилаза должна обладать
высоким сродством к СО2, чтобы ее функционирование при низ-
кой концентрации СО2 было эффективным.
Всем этим четырем требованиям удовлетворяет только вос-
становительный пентозофосфатный цикл.
Источник данных: [55].
6.7. Изучение восстановительного пентозофосфатного
цикла
Восстановительный пентозофосфатный цикл (или путь) был
исследован в пятидесятых годах группой ученых во главе с
Кальвином и Бенсоном из Университета Беркли, штат Калифор-
ния. Важность этих исследований была отмечена присуждением
Кальвину Нобелевской премии. Полную последовательность
изученных реакций часто называют циклом Кальвина или цик-
лом Бенсона — Кальвина (чтобы отдать долг вкладу в работу
Бенсона). Сейчас, правда, это название употребляется редко,
да и другим путям метаболизма ие принято присваивать имена
их исследователей, список которых выглядел бы слишком длин-
ным. Более приемлемы такие нейтральные названия, как «фото-
синтетический углеродный цикл», но они страдают некоторой
неопределенностью, поскольку могут быть также отнесены ко
всему С4-пути (гл. 10). Вот почему мы остановились на назва-
нии «восстановительный пентозофосфатный цикл (ВПФ-цикл)»,
которое отличается точностью и отражает суть предмета.
Кальвин и его сотрудники воспользовались двумя новыми
для того времени методами — применением радиоактивных изо-
топов и хроматографией на бумаге. Долгоживущий радиоактив-
ный изотоп углерода ИС был обнаружен Сэмюэлем Рубеном и
Мартином Кейменом в 1940 г. и стал доступен в больших коли-
восстановительный пентозофосфатный цикл
123
чествах в 1945 г. в качестве побочного продукта ядерных реак-
торов. Мартин и Синдж разработали метод хроматографии, за
который в 1952 г. они были удостоены Нобелевской премии,
.а хроматография на бумаге стала применяться для разделения
аминокислот еще в 1944 г. (Первым хроматографию на колон-
ках с различными сорбентами применил русский ботаник Цвет
для разделения хлорофиллов и близких к ним пигментов из экс-
трактов листьев. Его имя на редкость подходит для первооткры-
вателя процесса разделения окрашенных соединений, названно-
го хроматографией1. Впрочем, сейчас термин «хроматография»
используется и при разделении неокрашенных соединений.)
Совместное применение 14С и бумажной хроматографии позво-
лило разработать новый успешный подход к решению весьма
сложной проблемы фотосинтетической ассимиляции углерода.
Кальвин и его сотрудники ввели радиоактивную метку 14С
в СО2 и затем с относительной легкостью проследили за появ-
лением радиоактивности во вновь синтезированных соединениях,
снабжая 14СО2 на свету зеленую водоросль хлореллу. Говорят,
что Кальвин выбрал хлореллу (ставшую теперь самым прослав-
ленным из фотосинтезирующих организмов, как видно, напри-
мер, из разд. 3.9 и 4.10) потому, что ему как химику она напо-
минала зеленый раствор и поэтому импонировала больше, чем,
скажем, капуста. Как бы то ни было, хлорелла оказалась иде-
альным объектом исследований, так как ее можно было под-
вергать равномерному и интенсивному освещению, а затем
быстро переносить в горячий этанол, чтобы сразу останавли-
вать ферментативные реакции. (Аналогичным подходом вос-
пользовались Гаффрон и его сотрудники из Чикаго, которые ра-
ботали с другой водорослью — Scenedesmus. Вообще, подобные
эксперименты лучше проводить сначала с очень крупными по-
пуляциями растущих в одинаковых условиях водорослей, а ие
с небольшими изменчивыми образцами листьев высших расте-
ний.)
После кратковременной экспозиции на свету в присутствии
14СО2 и последующей экстракции горячим 80%-ным этанолом
порции экстракта наносили в виде небольшого пятна в верхнем
углу большого листа фильтровальной бумаги. Когда пятно вы-
сыхало, верхний край бумаги опускали в хроматографическую
«лодочку», в которую заливали смесь растворителей в опреде-
ленном соотношении, и помещали всю систему в плотно закры-
тую термостатированную хроматографическую камеру. Раство-
ритель постепенно стекал по свешивающейся из лодочки бума-
ге, и исходное пятно разделялось при этом на несколько пятен.
Лист вынимали, высушивали, поворачивали на 90° и хромато-
1 От грёч. chroma, т. е. «цвет». — Прим, персе.
124
ГЛАВА 6
графировали во второй смеси растворителей. Теперь каждое из
пятен после первого прохождения растворителя в свою очередь
давало несколько пятен. В руках-таких сотрудников Кальвина,
как Марта Керк, этот метод двумерной хроматографии давал
точные и воспроизводимые результаты, позволяя идентифици-
ровать соединения по их положению относительно старта (тон-
Гис. 6.4. Хроматографическая карта (фингерпринт). Если проводить разделе-
ние смеси радиоактивных и нерадиоактивных соединений в одних и тех же
растворителях и в совершенно одинаковых условиях, то каждое радиоактив-
ное соединение, нанесенное на старт (внизу справа), даст радиоактивное
пятно в определенном месте хроматограммы, которое легко обнаружить. Это
позволяет идентифицировать неизвестные вещества в первом приближении по
положению их пятен. Кроме того, по радиоактивности в каждом пятне
можно судить о концентрации: соответствующего меченого соединения [33].
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ никл
125-
ки нанесения исходного пятна). Кроме того, метод позволял оп-
ределить концентрацию этих соединений путем счета их 14С-ра-
диоактивности. (Сначала, конечно, требуется химически иден-
тифицировать каждое соединение, чтобы знать его положение
на хроматограмме, и внести соответствующие поправки для сме-
сей этих же, но немеченых соединений.) Совместное применение’
двумерной хроматографии и ИС позволило за несколько недель-
провести анализ, на который прежде ушли бы годы, даже при
условии, что удалось бы собрать достаточное количество мате-
риала (рис. 6.4).
Источники данных: [3, 4, 6, 33, 35, 38, 50, 52],
6.8. Кинетические исследования
Чем короче период фотосинтеза, тем меньше соединений со-
держали 14С. Когда их радиоактивность выразили в процентах
от общей радиоактивности, оказалось, что только одно соедине-
ние (3-фосфоглицерат, или ФГК) давало отрицательный угол
наклона кривой на графике, который позволил провести экстра-
поляцию к 100%-ной радиоактивности в нулевой момент време-
ни (рис. 6.5). (Теоретически в бесконечно малый промежуток
времени, прежде чем успеют произойти дальнейшие превраще-
Рис. 6.5. Чем больше время экспозиции в присутствии 1‘|СО2 на свету, тем
меньше радиоактивности в ФГК и тем больше ее в конечных продуктах фото*
синтеза, например в-сахарозе. ФГК является единственным соединением, .для
которого наблюдается подобная отрицательная зависимость при инкубации
хлореллы в присутствии 14СО2. Кроме того, как показывает соответствую*
щая; интерполяция,- ФГК —это единственное соединение, -в которое включа-
ется радиоактивная метка после минимальной--экспозиции ,[6]. : ..
126
ГЛАВА 6
ния, продукт карбоксилирования должен содержать практиче-
ски всю радиоактивность.) Тот факт, что самый первый про-
дукт карбоксилирования содержал три атома углерода на мо-
лекулу, свидетельствовал о возможности его возникновения пу-
тем присоединения СО2 к акцептору, содержащему два углерод-
ных атома. Однако данные относительно участия пентозофосфа-
тов в этом процессе неоспоримо указывали на то, что акцепто-
ром СО2 служит соединение, содержащее не два, а пять угле-
родных атомов, и что присоединение С5+С1 сопровождается
распадом промежуточного соединения С6 на две молекулы про-
дукта, каждая из которых состоит из трех атомов углерода.
Источники данных: [3, 4, 6].
6.9. Изменения в содержании ФГК и РуБФ
О том, что акцептором СО2 служит рибулозо-1,5-дифосф ат,
говорили кратковременные изменения его концентрации, кото-
рые сопровождались изменениями содержания фосфоглицери-
новой кислоты (ФГК). (Следует отметить, что сейчас этот ак-
цептор принято называть рибулозо-1,5-бисфосфатом. Это назва-
ние подчеркивает, что фосфатные группы соединены с разными
атомами в молекуле, а не связаны вместе в пирофосфатную
структуру Pi-Pi, как может вытекать из названия «дифос-
фат».) Если, например, резко уменьшить концентрацию СО2, то
содержание ФГК снизится, а количество РуБФ возрастет
Рис. 6.6. Изменения в содержании ФГК и РуБФ, вызванные изменением кон-
центрации COj. Если резко снизить концентрацию СО2 во время освещения,
то содержание ФГК (конечного продукта фиксации СОа) также уменьшает-
ся, а количество РуБФ (акцептора СО2) сначала возрастает (поскольку
РуБФ используется гораздо медленнее), ио потом падает, так как его реге-
нерация в свою очередь замедляется.
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ
127
Рис. 6.7, Изменения в содержании ФГК и РуБФ в зависимости от освеще-
ния. При переходе в темноту содержание ФГК сначала увеличивается, так
как это соединение все еще образуется, ио не восстанавливается. (После-
дующее падение концентрации ФГК может быть связано с невосстанови-
тельным метаболизмом.) Количество РуБФ снижается, так как это соедине-
ние непрерывно используется, а его регенерация прекращается [6].
(рис. 6.6). В то же время при переходе от света к темноте со-
держание ФГК. увеличивается, а концентрация РуБФ уменьша-
ется (рис. 6.7). Первую реакцию (рис. 6.6) следовало ожидать
при условии, что РуБФ служит акцептором, а ФГК — продук-
том карбоксилирования: ведь при лимитирующих концентра-
циях СО2 акцептор почти ие будет связывать СО2 и, следова-
тельно, образуется гораздо меньше продукта карбоксилирова-
ния. Результаты второй реакции (рис. 6.7) указывают на реге-
нерацию акцептора из продукта карбоксилирования. Таким об-
разом, если под действием света часть ФГК превратилась в.
РуБФ, то в темноте эта реакция остановится, ФГК будет накап-
ливаться, а РуБФ расходоваться по мере карбоксилирования
оставшейся части акцептора.
Источники данных: [3, 4, 6].
6.10. Включение метки в молекулы
Меченые промежуточные соединения были Подвергнуты хи-
мической деградации, после чего выявляли распределение ра-
диоактивности в молекулах. Это позволило проследить путь
углерода во всей последовательности ВПФ-цикла. Первоначаль-
но основная часть ИС включается в карбоксильную группу ФГК
(табл. 6.1). Распределение метки среди других промежуточных
128 :: глава 6
Таблица 6.1. Распределение ИС среди промежуточных продуктов ВПФ-цикла
Углерод ФГК Гексоза Гептоза; 1 Пентоза
1 : 82* 3 ; 2 ; 11*
2 6 3 2 10*
3 ' 6 43* 28* 69* **
4 42* 24* 5
5 3 27* 3
6 з 2
7 2
соединений, перечисленных в табл. 6.1, проще всего объяснить
перегруппировкой пяти молекул, содержащих по три углерод-
ных атома, как это изображено на рис. 6.8. В последовательно-
сти, которая является частью ВПФ-цикла (разд. 6.11 и рис. 6,1),
две С3-молекулы (справа на рис. 6.8) соединяются особым об-
разом — «голова-к-голове», как это обусловлено их структу-
рой,— формируя соединение из. шести углеродных атомов. Это
соединение отдает С2-фрагмент С3-акцептору, что приводит
к возникновению молекулы из пяти атомов углерода. Оставшая-
ся часть Се-соединения содержит четыре углеродных атома и
соединяется с другой молекулой С3, образуя С7. Перенос С2-
фрагмента от С7-молекулы к С3-акцептору дает еще два проме-
жуточных соединения из пят*! углеродных атомов. Если теперь
суммировать радиоактивность этих соединений, окажется, что
средние углеродные атомы гексозы С0 и гептулозы С7 мечены
одинаково, в то время как центральный углеродный атом пенто-
зы С5 содержит гораздо большую метку. Напомним, что данная
последовательность входит в цикл, который функционировал
в этом опыте в течение короткого промежутка времени. В силу
циклической природы всего процесса метка затем перейдет от
пентозных производных к триозным. Полученные данные не сов-
сем совпадают с распределением радиоактивности, предсказан-
ным схемой на рис. 6.8. Тем не менее они достаточно хорошо
согласуются с ним, чтобы исключить любое другое объяснение
на основании известных реакций.
Источники данных: [3, 4, 6].
'6.11. Ферменты
Совместное, применение радиоактивного углерода и двумер-
ной хроматографии сыграло решающую роль в исследовании
пути углерода при фотосинтезе. Однако для полноты картины
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ цикл
129
Рис. 6.8. Распределение радиоактивности внутри молекул — продуктов мета-
болизма ВПФ-цикла. Первоначальная фиксация СОа с образованием ФГК
н последующие реакции ВПФ-цикла должны сначала привести к такому рас-
пределению радиоактивности внутри молекул, которое показано на рис. 6.12.
Аналогичное распределение было обнаружено экспериментально [6].
надо было либо уже знать отдельные реакции этого пути и
катализирующие их ферменты, либо показать, что они действи-
тельно существуют. Среди этих реакций первое место по пра-
ву принадлежит карбоксилированию.
1. Рибулозобисфосфат-карбоксилаза (оксигеназа)
Этот фермент является, пожалуй, самым распространенным,
а может быть, и самым важным из всех ферментов, известных
нам в настоящее время. В 1954 г. Куэйл, Фуллер, Бенсон и
Кальвин нашли его впервые в экстрактах хлореллы. Сначала
его называли карбоксидисмутазой, поскольку это название дает
9—234
130
ГЛАВА fi
представление о характере катализируемой реакции [уравнения
(6.15)].
CHjOPOIOHI; СН/ЭРО(ОН1? сн/жооно
’СО; 0 = 0 СК . 1 ЧС- СОН 1 И С ОН * 1 -
неон —>- НО' С —О ' Н2О ' h О
неон 1 Н СОИ х но’с--о 16.15а> ।
сн?оро(он).. СНЮРОЮиО г 1С ОН 1
Рибу 11030 1 ,5бисфосф.п 11 ромежУ ТОЧНЫЙ ироду 1' снл,ороюнО .1 2 Фосфит fiHiu'I’OHiiBdH
А (РуБФ) кисло 1 а ( ФГ К 1
о 4
CHjOPOtCHi,, ( <1 1 56 У
1 но’о’сон СН;ОРО1ОН|2 но’С =о 1 .
1 + Н,0‘ с=о >. A 1 О сон ь неон + н-'°
1 неон СНгОРО(ОНС,
CH,,OPOIOH)2
Промежуточный продукг 2 - Фосфси jinK.on.il 3 • Фисфог лицер.н
Б
К. Ф. 4.1.1.39
/<м (СО2) = 12 мкМ
Л’м (О2) = 250 мкМ.
Л'м (РуБФ) = 40 мкМ. Концентрация РуБФ в хлоропластах 0,2—-0,6 мМ.
ДБ' = —8,4 ккал (карбоксилазная реакция)
Мол. масса = 550 000; 8 больших субъединиц (мол. масса по 55 000) и
8 малых субъединиц (мол. масса по 15 000)1.
1 Размер белков обычно выражают в дальтонах. Один дальтон — это масса одного атома
водорода, а общая масса числа Авогадро водородных атомов почти точно равна
1 г. Одна грамм-молекула (моль)• РуБф-карбоксилазы весит 550 кг.
Реакция, катализируемая этим ферментом, сводится к одно-
временному присоединению СО2 и воды и внутримолекулярному
окислению — восстановлению (дисмутации). Таким образом,
присоединение одной Ci-молекулы (СО2) к С5-акцептору дает
два С3-продукта (3-ФГК), один из которых содержит ион водо-
рода, а второй — ион гидроксила из молекулы воды [уравнение
(6.15а)]. Раньше предполагали, что (по крайней мере in vivo)
одно из двух Сз-соединений может стать более восстановленным
в результате какого-либо прямого восстановительного процесса,
но это предположение ие подтвердилось. Сейчас уже точно уста-
новлено, что в реакции in vitro образуются только две молеку-
лы 3-фосфоглицериновой кислоты, одна из которых содержит
в своей карбоксильной группе только что связанный углерод-
ВОССТАНОВИТЕЛЬ! 1ЫП ПЕНТОЗОФОСФАТИЫЙ ЦИКЛ
131
ный атом из СО2 (углеродный атом номер один в отличие от
углеродного атома номер три, соединенного с фосфатом).
Впоследствии оказалось, что кислород способен конкуриро-
вать с СО2 и что карбоксилаза может также функционировать
в качестве оксигеназы (разд. 6.16). В этом случае одна моле-
кула 3-ФГК образуется так же, как и в предыдущем, а остав-
шийся С2-фрагмент становится 2-фосфогликолевой кислотой
[уравнение (6.156)]. Именно реакция, катализируемая РуБФ-
оксигеиазой, лежит в основе фотодыхателыгого цикла окисления
углерода. Ее связь с восстановительным пентозофосфатным
циклом показана на рис. 6.11 и 6.12, а подробно этот вопрос
обсуждается в гл. 13. Поскольку СО2 и О2 конкурируют за
один и тот же центр связывания на молекуле фермента, скоро-
сти этих двух реакций определяются концентрациями обоих
газов.
При высокой концентрации СО2 и низкой концентрации О2
будет преобладать карбоксилазная реакция. Истинные ско-
рости этих двух реакций зависят также от других факторов,
например от pH и температуры. Тем не менее в грубом прибли-
жении в условиях in vivo (т. е. при 250 мкм О2 и 10 мкм СО2)
карбоксилазная активность в 3—5 раз больше оксигеназной.
Как мы уже отмечали, карбоксилаза характеризуется очень
большой величиной Км по отношению к СО2 (разд. 6.5): для
достижения половины максимальной скорости реакции ей тре-
буется приблизительно 6% СО2 в газовой фазе. Это обстоятель-
ство довольно долго оставалось своего рода загадкой, так как
вполне естественно возникал вопрос: каким образом карбокси-
лаза со столь низким сродством к СО2 может эффективно функ-
ционировать при тех незначительных концентрациях СО2, кото-
рые должны присутствовать в месте карбоксилирования? А ведь
эти концентрации будут еще меньше, если СО2 поступает в ор-
ганизм из атмосферы (воздуха), содержащей всего 0,03%
(300 м. д.) этого газа, путем простой диффузии. Вот почему
некоторые авторы предположили существование механизмов,
концентрирующих СО2. Впоследствии оказалось, что величина
Км для СО2 резко снижается в результате активации, если
карбоксилазу преинкубируют с ионами магния и бикарбоната
при высоких значениях pH.
Процессы активации и карбоксилирования представлены
уравнениями (6.15в) и (6.15г). (Е— фермент, от англ, enzyme.)
АКТИВАЦИЯ:
Медленно Быстро ,
E-J-CO. ----> Е—СО,Mg2+ 3==t Е—СО2—Mg (6.15в)
(неакт.) (неакт.) (акт.)
9
132
ГЛАВА 6
КАРБОКСИЛИРОВАНИЕ:
Е—СО2—Mg + РуБФ + *СО2
,=fc Е—СО2—-Mg + ’ФГК + ФГК (6.15г)
Неактивная форма фермента сначала соединяется с моле-
кулой СО2 в ходе медленной обратимой реакции. Затем этот
фермент-субстратный комплекс реагирует с Mg2+, и на этой вто-
рой обратимой стадии быстро образуется комплекс фермент —
СО2— Mg, в котором фермент и проявляет полную активность
[уравнения (6.15в) и (6.15г)]. Mg2+ требуется для активации
фермента, а не для карбоксилазной реакции. Карбоксилирова-
ние РуБФ как таковое осуществляется тогда, когда с фермен-
том реагирует вторая молекула СО2 [уравнение (6.15г)]. Пер-
вая молекула СО2, которая связывается с ферментом па стадии
активации [уравнение (6.15в)], не участвует в карбоксилазной
реакции. Вполне вероятно, что эти две молекулы СО2 присоеди-
няются к разным центрам связывания на ферменте. Было уста-
новлено, что именно СО2, а не НСО3_ отвечает и за процесс
активации, и за саму реакцию карбоксилирования. Большая
часть углерода в щелочной среде стромы освещенного хлоро-
пласта существует в форме бикарбонатного иона. Равновесие
между СО2 и НСО3~ поддерживается ферментом карбопат-де-
гидратазой, который катализирует реакцию Н+-]-НСОз"ч=ьСО2-[-
+Н2О. Благодаря этому молекулы СО2, израсходованные
в процессе карбоксилирования, быстро пополняются за счет би-
карбоната.
Удаление СО2 и Mg2+ приводит к диссоциации активного
комплекса и появлению неактивной формы фермента. Таким
образом, концентрации СО2 и Mg2+ определяют активированное
состояние фермента. Процесс активации зависит от pH и имеет
щелочной оптимум, что говорит о возможном образовании при
активации в процессе формирования комплекса фермент — СО2
карбамат-иоиа. В результате активации увеличивается макси-
мальная скорость реакции и, как мы уже отмечали, повышается
сродство фермента к СО2. В итоге значительно возрастает ак-
тивность карбоксилазы при концентрации СО2, характерной для
атмосферы. По мере изменения активированного состояния
фермента активность оксигеназы увеличивается или уменьша-
ется так же, как и активность карбоксилазы. Было установле-
но, что высокие концентрации РуБФ приводят к субстратному
ингибированию фермента. Оказалось, однако, что это ингиби-
рование вызвано продуктами химического разложения РуБФ.
Ряд других продуктов метаболизма хлоропластов, особенно
NADPH, ферредоксин и 6-фосфоглюконат, увеличивают актив-
ность РуБФ-карбоксилазы или изменяют степень ее активации
(гл. 9). Роль этих метаболитов in vivo выяснить довольно труд-
восстановительный пентозофосфатный цикл
133
но, поскольку концентрация многих из них относительно коли-
чества фермента может быть гораздо ниже, чем в условиях
in vitro, где был отмечен этот эффект. Концентрация активных
центров связывания на РуБФ-карбоксилазе в хлоропластах мо-
жет достигать 3—4 мМ. Столь большая цифра обусловлена вы-
соким содержанием этого белка в хлоропластах и наличие^
восьми центров связывания на каждой его молекуле. Концент-
рация же многих метаболитов в хлоропластах гораздо ниже
этой цифры, и поэтому установить точно их действие на карбок-
силазу весьма трудно.
Источники данных: [34, 42, 47, 49].
2. Фосфоглицераткиназа
Этот фермент был уже хорошо известен по гликолизу, где
он катализирует реакцию [уравнение (6.16)], приводящую к об-
разованию ФГК. При фотосинтезе этот фермент катализирует
синтез ДФГК (1,3-дифосфоглицериновой кислоты, или глице-
рат-1,3-бисфосфата) из ФГК и АТР.
| "''он
НСОН + АТР
СН2ОРО(ОН)2
3 • Фосфоглицериновая
кислота (ФГК)
ро?о(он)2
НСОН
СН2ОРО(ОН)г
4- ADP
(6.16)
1,3 * Дифосфоглицериновая
кислота (ДФГК)
К. Ф. 2.7.2.3
Ли (ФГК)даО,5 мМ. Концентрация ФГК. в хлоропластах 3—5 мМ
К-щ (АТР) =0,1 мМ. Концентрация AMP+ADP+ATP в хлоропластах 1 —
3 мМ
ДГ/=+4,5 ккал
Мол. масса=47 000
Это единственная реакция в ВПФ-цикле с большой положи-
тельной величиной АГ', указывающей на то, что равновесие
реакции сдвинуто в сторону образования ФГК- Фосфоглицерат-
киназа не активируется под действием света в хлоропластах и,
по-видимому, не подвергается аллостерическим изменениям в
присутствии других метаболитов. Для того чтобы реакция про-
текала в направлении синтеза ДФГК, как это требуется для
фотосинтеза, необходимо высокое значение отношения [ФГК]
[АТР]/[ДФГК][АОР]. Такое отношение действительно наблюдает-
ся в хлоропластах, где содержание ДФГК очень мало, в основ-
ном из-за ее использования в реакции, катализируемой глице-
134
ГЛАВА G
ральдегидфосфатдегидрогеназой {уравнение (6.17)]. Напротив,
концентрация ФГК в хлоропластах довольно высокая, что дает
очень большую величину отношения [ФГК]/[ДФГК]. Кроме того,
как видно из приведенного выше соотношения между субстра-
тами и продуктами реакции, последняя чувствительна к кон-
центрациям АТР и ADP. Поэтому при любом уменьшении
[АТР] или увеличении [ADP] скорость образования ДФГК умень-
шается (гл. 9).
Для того чтобы фотосинтез все время шел с высокой ско-
ростью, содержание промежуточных продуктов в хлоропласте
в процессе функционирования ВПФ-цикла не должно падать
ниже необходимого уровня. В случае ФГК использование про-
межуточных продуктов постоянно компенсируется, так как этот
метаболит быстро поступает в хлоропласт при помощи фосфат-
ной транспортной системы и его концентрация довольно высо-
ка. Утечке ФГК в цитоплазму препятствует высокое значение
pH в строме хлоропласта, благодаря чему большая часть ФГК
присутствует в форме трехвалентного аниона ФГК3- (в цито-
плазму может переходить лишь двухвалентный анион ФГК2~)-
В результате концентрация ФГК внутри хлоропласта остается
довольно высокой, что способствует ее восстановлению, а в ци-
топлазму ФГК не поступает (разд. 8.23 и табл. 8.6).
3. Триозофосфатдегидрогеназа [AADP+]
При гликолизе NAD-специфический фермент катализирует
окисление глицеральдегид-3-фосфата до ДФГК. При фотосин-
тезе довольно близкая к этой реакция [уравнение (6.17)] ката-
лизируется NADP-зависимым ферментом, однако она идет с
восстановлением, а не с окислением, и неорганический фосфат
при этом высвобождается, а не включается.
..•°
Т'-ОРО(ОН)2
ИСОН +- NADP(hJ)
CHjOPOlOHh
1,3 ' Дифосфоглицериновая
кислота (ДФГК)
.О
I"®
НСОН + ®ОРО(ОН)!
I + NADP (6.17)
CHjOPOtOH);
Глицеральдегид • 3 - фосфат (ГЗФ,или ФГА)
КФ. 1.2.1.13
Км (ДФГК) = 1 мкМ. Концентрация ДФГК в хлоропластах очень низкая
(точные данные отсутствуют)
Км (NADPH) =4 мкМ. Концентрация NADPH+NADP в хлоропластах 0,1—
0,5 мМ
AF'==— ‘ 1,5 ккал
Мол. масса=600 000
восстановительный пентозофосфатный цикл
135.
Фермент активируется светом при помощи системы ферре-
доксин/тиоредоксин (гл. 9), а также in vitro дитиотрейтолом.
Благодаря присутствию NADPH происходит возрастание актив-
ности, которое носит сигмоидный характер; максимальные ско-
рости наблюдаются в интервале концентраций NADPH от 0,5
до 1,5 мМ, АТР оказывает аналогичное действие в диапазоне
от 1 до 6 мМ.
4. Триозофосфатизомераза
Это еще один фермент, известный по гликолизу, который
катализирует [уравнение (6.18)] превращение глицеральдегид-3-
фосфата (ГЗФ, или ФГА) в его изомер, дигидроксиацетонфос-
фат (ДГАФ).
ФГА и ДГАФ известны также под общим названием триозо-
фосфатов. В ВПФ-цикле две пятых триозофосфатов используют-
ся в форме ДГАФ. Равновесие реакций [уравнение (6.18)] сдви-
нуто вправо, и в состоянии равновесия 95,6% всех триозофосфа-
тов находятся в виде ДГАФ. Фермент ингибируется низкими
концентрациями фосфогликолата.
Нк ...о
С
(Н)со(н) —
СН7ОРО(ОН)7
Гпицв|1;>1>ьд|’г ид 3 - фодфаг
(ФГА'
(6.18)
С=О
CHjOPOlOHi,
Д иг идри к <: на цегон фосфат
(ДГАФ)
К. Ф. 5.3.1.1.
Дм (ДГАФ) = 1,1 мМ
Хм (ФГА) =0,3 мМ. Концентрация ДГАФ-|-ФГА в хлоропластах 0,3—0,4 мМ.
&F'=—1,8 ккал
Мол, масса=53 000
5. Альдолаза (ФБФ-альдолаза)
Этот фермент также известен по гликолизу. При фотосинте-
зе он катализирует реакцию [уравнение (6.19)] альдольной (от
слов «альдегид» и «алькоголь») конденсации, откуда и проис-
ходит его название, и обратную реакцию расщепления фрук-
тозо-1, 6-бисфосфата (ФБФ) на составляющие его триозофосфа-
ты (ФГА и ДГАФ).
136
ГЛАВА б
•ФГА
CHjOPOIOHJj
с=?о
н
СГ' _ :===
НО''' (Н)
нг.он
:'.Н2ОРО(ОН),
ДГАФ
CHjOPOtOHJi
с^-о
носн
J.o®
нгон
'.Н2ОРО(ОН)2
Фруктозе • 1,6 бисфосфат
(ФБФ1
(6.19)
К. Ф. 4.1.2.13
Хм (ФБФ) =20 мкМ
Хм (ФГА) = 0,3 мМ
Хм (ДГАФ) =0,4 мМ
ДГ'=—5,5 ккал
Мол. масса=150 000
Сильно отрицательная величина &.F' для образования ФБФ
из ДГАФ и ФГА свидетельствует о том, что равновесие этой
реакции сильно сдвинуто в сторону образования ФБФ. Однако,
поскольку в этой реакции имеются два субстрата и один про-
дукт, на положение равновесия значительно влияют их кон-
центрации. Например, если исходная суммарная концентрация
ФГА и ДГАФ составляет 1 мМ, то, прежде чем будет достигну-
то равновесие, в ФБФ превратится 76% этого количества. Если
же исходная концентрация этих соединений равна 0,1 мМ, то
в ФБФ превратится лишь 42% триозофосфатов. Это различие
продиктовано тем, что ФГА и ДГАФ присутствуют не в равных
количествах, так как равновесие триозофосфатизомеразной
реакции [уравнение (6.18)] сдвинуто в сторону ДГАФ при со-
отношении 22 : 1. Поэтому равновесие альдольной конденсации
-[уравнение (6.19)] сильно сдвигается в сторону ФГА и ДГАФ:
например, если исходная общая концентрация этих соединений
равна 0,1 мМ, то они превратятся в ФБФ только иа 33% при
условии, что ФГА и ДГАФ также находятся в равновесии, обус-
ловленном триозофосфатизомеразной реакцией.
б. Фруктозобисфосфатаза (ФБФ-аза)
Этот фермент катализирует гидролиз ФБФ, в результате ко-
торого образуются фруктозо-6-фосфат (Ф6Ф) и свободный не-
органический фосфат [уравнение (6.20)].
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ цикл
137
CH7OPO(OH)i
с=о
I
НОСН
I +
неон ~
I
неон
CH2OPO(OH)i
Фруктозо • 1,6 бисфосфат
(ФБФ)
СН2О®
с=о
I
НОСН
| ф fHOlPOlOH);
неон
I
неон
I
СН2ОРО(ОН)2
Фруктозо 6 • фосф<1Г (ФбФ)
(6.20)
К. Ф. 3.1.3.11
Км (Mg2+)=3 мМ
Км (ФБФ) =0,2 мМ. Концентрация ФБФ в хлоропластах 0,1—0,3 мМ
ДК'=—4,0 ккал
Мол. масса—160 000
Этот гидролиз является второй реакцией в ВПФ-цикле, про-
текающей с образованием свободного неорганического фосфата.
Реакцию катализирует специфическая ФБФ-аза хлоропластов,
которая отличается от соответствующего фермента из цитоплаз-
мы. Большая отрицательная величина &Р' указывает на то, что
равновесие реакции сдвинуто в сторону образования Ф6Ф. При
удалении СО2 как акцептора электронов активность фермента
увеличивается от 100 до 250 мкмоль-(мг Хл)“1-ч~1. Как пока-
зали опыты с изолированными хлоропластами, протопластами
и экстрактами целых листьев, активность ФБФ-азы хлороплас-
тов, измеренная при pH около 8,0, значительно повышается пос-
ле освещения. Можно также активировать очищенный фермент,
преинкубируя его с такими восстанавливающими агентами, как
дитиотрейтол (ДТТ) или восстановленный ферредоксин. В этих
условиях активация сопровождается увеличением числа доступ-
ных сульфгидрильных групп, т. е. белок при этом действительно
восстанавливается. Механизм активации под действием света
обсуждается в гл. 9. В результате активации понижается опти-
мальная величина pH и возрастает сродство к ФБФ и Mg2+.
Изменения степени активации фермента, а также изменения,
происходящие в строме хлоропластов, указывают на то, что
в темноте активность фермента должна быть очень низкой.
7. Транскетолаза
Этот фермент участвует также в дыхании (в окислительном
пентозофосфатном цикле). При- фотосинтезе ои катализирует
138
ГЛАВА 6
две реакции. В одной из них [уравнение (6.21)] транскетолаза
переносит «верхнюю» часть молекулы Ф6Ф из двух углеродных
атомов (комплекс транскетолазы и ФГА известен под назва-
нием дигидроксиэтилтиаминпирофосфат) на первый углеродный
атом ФГА. В результате реакции образуется ксилулозо-5-фос-
фат и эритрозо-4-фосфат (С6+С3—^С3+С4).
Фруктозе 6 - фосфат
(ФВФ)
о=сн —>-
неон
I
•ГН^ОРО^ОН),
о=сн
I
неон
неон
I
CHjOPOfOHJi
неон
I
CHjOPOfOH),
Глицерапьдвгид 3 фосфат Эритрозо 4 • фосфат Ксилулозо • 5 фосфат
(ФГА)
(Э4Ф) (КуВФ)
К. Ф. 2.2.1.1
Км (Ф6Ф) не известна. Концентрация Ф6Ф в хлоропластах 0,6—-1,5 мМ
Км (ФГА) не известна. Общая концентрация ДГАФ и ФГА в хлоропластах
0,3—0,4 мМ
ДР'=1,47 ккал
Мол. масса= 140 000
Одно время считали, что переносимый транскетолазой фраг-
мент из двух атомов углерода может служить важным источ-
ником гликолата при фотодыхании (гл. 13). Сейчас, впрочем,
эта точка зрения не пользуется популярностью, и образование
фосфогликолата из РуБФ более не подвергается сомнению.
8. Альдолаза
Вторая реакция альдольной конденсации [первая дана в
уравнении (6.19)] сводится к присоединению ДГАФ к эритро-
зо-4-фосфату, который образуется в результате первой из двух
реакций, катализируемых транскетолазой [уравнение (6.21)].
Продуктом этой альдольной конденсации является седогептуло-
зо-1,7-бисфосфат (С4-[-Сз—>-Су).
ВОССТАНОВИТЕЛЬНЫ!"! ПЕНТОЗОФОСФАТИЫЙ ЦИКЛ
139
СН2ОРО(ОН)2
НО
неон
'I
неон
I
енгоро(он)г
СН,ОРО(ОН)2
I
с--о
НОСН
нео®
н-еон
неон
I
СНгОРО(ОН)г
(6.22)
Эритроза • 4 - фосфат ДГАФ
(Э4Ф)
Седогепту лозо-1,7-бисфосфат
(СБФ)
9. Седогептулозобисфосфатаза (СБФ-аза)
Это вторая, катализируемая фосфатазой реакция гидролиза
[первая дана в уравнении (6.20)], в процессе которой фосфат от-
щепляется от седьмого углеродного атома седогептулозо-1,7-бис-
фосфата (СБФ) и образуется седогептулозо-7-фосфат (С7Ф).
СНгОРО(ОН)2
I
с^-о
НОСН
неон + (Н’2)
I
неон
I
неон
I
СН2ОРО(ОН)2
Седогептулозе-1,7-бисфосфат
(СБФ)
СН2О®
С^О
НОСН
неон + (но) ро:.он)2
неон
неон
<!:н2оро(он)2
Содогоптулозо-7-фосфат
(С7Ф)
(6.23)
К. Ф. 3.1.3.37
Хм (СБФ) =0,24 мМ. Концентрация СБФ в хлоропластах 0,2—1,0 мМ
Д1?/=—4,0 ккал [по аналогии с реакцией (6.20)]
Мол. масса~50 000
СБФ-аза, выделенная из листьев шпината, высоко специфич-
на по отношению к СБФ и для проявления своей активности
нуждается в двухвалентных катионах (Mg2+ или Мп2+). По-
1-140
ГЛАВА 6
добно ФБФ-азе, она активируется ДТТ или системой ферредок-
син/тиоредоксин. Содержание СБФ-азы ниже концентрации лю-
бого другого фермента ВПФ-цикла, а максимальная скорость
катализируемой ею реакции составляет 60 мкмоль-(мг Хл)-1Х
Хч-1. По предварительным данным, СБФ-аза может активиро-
ваться in vivo продуктами метаболизма ВПФ-цикла, чем она и
отличается от ФБФ-азы.
10. Транскетолаза
Рассмотрим теперь вторую реакцию, катализируемую транс-
кетолазой [первая из них дана в уравнении (6.21)]. Здесь этот
фермент тоже переносит двухуглеродный фрагмент на ФГА,
ио уже не от Ф6Ф, а от С7Ф. Вот почему и второй продукт ре-
акции (рибулозо-5-фосфат) оказывается пентозой (Сз+С?—>-
—^Сз+Сб) .
о=сн
неон
НСОН
I -1-
нсон
СН2ОРО(ОН)г
СН2ОРО(ОН)г
£ш.1пгептулозо-7-фосфат Глицеральдегид - 3 • фосфат РиВоао • 5 • фосфат Ксилулозо-В-фосф.и
(С7Ф) 1ФГА) (Р5Ф) ||<у5Ф)
11. Рибозофосфат-изомераза
Этот фермент катализирует превращение рибозо-5-фосфата
(Р5Ф) в его изомер, рибулозо-5-фосфат (Ру5Ф). Реакция идет
с легкостью в обоих направлениях.
н, .о
с^
неон
1
I
НСОН
/О®
СН,ОРО(ОН);
Рибоза • 5 • фосфат
' (Р5Ф) '
С = О
НСОН
I
НСОН
I
сн2оро(он)2
Рибуло'зо • 5 - фосфат
(Ру5Ф)
(6.25)
восстановительный пентозофосфатный цикл
141
К. Ф. 5.3.1.6
л'м (Р5Ф)=2,0 мМ
Afz = 0,64 ккал
Мол. масса = 54 000
12. Рибулозофосфат — 3-эпимераза
Данный фермент катализирует реакцию, которая, как и пре-
дыдущая [уравнение (6.25)], является изомеризацией, но ее суб-
страт и продукт представляют собой эпимеры, отличающиеся
друг от друга лишь ориентацией водородного атома и гидр-
оксильной группы при третьем углеродном атоме. Субстратом
служит ксилулозо-5-фосфат (Ку5Ф), который образуется в ре-
зультате двух реакций, катализируемых транскетолазой [урав-
нения (6.21) и (6.24)], а продуктом — тоже рибулозо-5-фосфат.
снгон
с=о
-
неон
СН2ОРО(ОН)2
Ксилулозо-5-фосфат
(Ку6Ф|
СН2ОН
с —о
. (6,26)
неон
CH2OPO(OHJ2
Рибулозо•5 • фосфат
(Ру5Ф)
к. ф. 5.1.3.1
/<м (Ку5Ф) =0,5 мМ
AF'=—0,13 ккал
Мол. масса=46 000
Реакция полностью обратима, о чем говорит низкая величи-
на &F', и в состоянии равновесия отношение Ку5Ф/Ру5Ф варьи-
рует от 1 до 3.
13. Фосфорибулокиназа
Этот фермент катализирует последнюю реакцию ВПФ-цик-
ла — превращение Ру5Ф, который образуется в реакциях
(6.25) и (6.26), в рибулозо-1,5-бисфосфат (РуБФ) за счет АТР.
142
ГЛАВА о
сн,о СН7ОРО(ОН)г
с —о с -о
1 НСОН + АТР -— 1 НСОН +ADP 1
1 НСОН 1 НСОН
сн,оро(0н)г СН,ОРО(ОН)2
Рибуло:ю • 5 - фосфат Рибулозо-1.5 бисфосфаТ
(РуБФ) (РуБФ)
(6.27)
К. Ф. 2.7.1.19
/<м (РуБФ) =0,2 мМ. Суммарная концентрация пентозомонофосфатов в хло-
ропластах 0,1 мМ
Ам (АТР) =0,1 мМ.. Концентрация AMP-j-ADP-f-ATP в хлоропластах 1—3 мМ
A.F'——5,2 ккал
Мол. масса=240 ООО
В этой реакции используется одна треть всего количества
АТР, которое требуется для фиксации СОг- Как показывает ве-
личина ДА7, равновесие реакции сильно сдвинуто в сторону об-
разования РуБФ. Получены убедительные доказательства в
пользу того, что in vivo фермент активируется под действием
света. Активация фермента дитиотрейтолом in vitro свидетель-
ствует о том, что возрастание активности фермента in vivo при
освещении, по-видимому, вызывается восстанавливающим аген-
том, который образуется в результате фотосинтеза. Фосфорибу-
локиназа регулируется также энергетическим зарядом аденилат-
ной системы, т. е. концентрациями АТР, ADP и АМР.
6.12. Свободные энергии
Обычные в биохимических реакциях изменения свободной
энергии ДА7 рассчитывают с учетом особенностей биологиче-
ских систем, и потому они отличаются от изменений свободной
энергии, которыми пользуются в физической химии (ДА0). На-
пример, ДА7 рассматривают для pH 7,0, а ДА0 — для pH 0.
Опубликованные в литературе значения нередко сильно разли-
чаются между собой. Эти различия вызваны трудностями, кото-
рые встречаются при измерениях, неравнозначными допущения-
ми, а подчас и ошибками в расчетах. Еще большие расхожде-
ния порождаются тем, как используются величины ДА7 и какие
выводы делаются при этом. Не следует забывать, что любое из-
менение свободной энергии (которое само по себе есть не что
иное, как слегка завуалированная константа равновесия) гово-
рит лишь о возможности протекания реакции, но не о ее скоро-
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТНЫП ЦИКЛ
143
сти. Как мы уже отмечали в разд. 2.9, окисление Н2 молекуляр-
ным О2 идет с уменьшением энергии примерно на 56 ккал. В оп-
ределенных условиях эти газы бурно реагируют друг с другом,
ио обычно их смесь может стоять бесконечно долго, и никакой
реакции при этом не происходит. Хотя сама по себе величина
ДР ничего не говорит о скорости реакции, тем не менее опа не-
сет определенную информацию, и в этом можно убедиться па
следующем примере. Малатдегидрогеназа является активным и
широко распространенным ферментом, ио ее ДД' имеет очень
большое положительное значение в направлении малат—>-окса-
лоацетат. Поэтому нельзя представить себе данную реакцию
протекающей в клетке именно в этом направлении, разве лишь
как звено в цепи реакций, например в цикле трикарбоновых кис-
лот, где это неблагоприятное равновесие сдвинуто за счет не-
прерывного синтеза субстратов и удаления конечных продук-
тов реакции. Вместе с тем, если можно так сместить столь не-
благоприятное равновесие, возникает вопрос: дают ли нам хоть
что-то значения ДД, а если дают, то что именно? Ответ па этот
вопрос заключается в том, что они позволяют сформулировать
проблемы, связанные с изучением метаболизма, даже если такой
подход будет несколько субъективным. Одно время предполага-
ли, что такие ферменты, как фермент Очоа, катализирующий
восстановительное карбоксилирование пирувата с образо-
ванием малата [уравнение (6.3)], могут участвовать в фотосин-
тетической фиксации углерода. Однако относительно неблаго-
приятные значения ДД реакций, катализируемых этими фермен-
тами, послужили отправной точкой в поисках иных механиз-
мов и помогли, например, установить роль фосфоенолпируват-
карбоксилазы [уравнение (6.8)] в метаболизме кислот по типу
растений из семейства толстянковых (гл. 15).
Бассэм уделял большое внимание величинам AF как показа-
телям «регуляторных ферментов». Однако при этом он подчер-
кивал, что следует соблюдать осторожность при интерпретации,
Действительно, реакция с большим значением ДД пойдет почти
до конца, если катализирующий ее фермент не подвержен топ
или иной регуляции. Это не означает, что реакция с относитель-
но низким значением ДД7 не существенна для путей использова-
ния углерода. В данном случае фермент, катализирующий реак-
цию, может и ие находиться под контролем регуляции, но зато
определенную роль будет играть само реакционное равновесие,
закон действия масс.
В табл. 6.2 приведены значения ДД7 по данным Бассэма и
Краузе. Необычное положение равновесия реакции, катализи-
руемых альдолазой, объясняется наличием двух субстратов
и одного продукта реакции. По мере увеличения концентрации
триозофосфатов равновесие постепенно сдвигается в сторону
144
ГЛАВА 6
Таблица 6.2. Изменения свободной энергии в реакциях ВПФ-цикла [32]
Уравнение AF'1, ккал
(6.15) РуБФ—Э-ФГК —8,4
(6.16)+ (6.17) ФГК—Э-ДФГК—>ФГА -г 4,3
(6.18) ФГА—> ДГАФ —1,8
(6.19) ФГА+ДГАФ—»-ФБФ -5,2
(6.20) ФБФ—>Ф6Ф -3,4
(6.21) Ф6Ф+ФГА—>Э4Ф+Ку5Ф +1,5
(6,22) Э4Ф+ФГА—>СБФ —5,6
(6.23) СБФ—>С7Ф —3,4
(6.24) С7Ф+ФГА—>Р5Ф+Ку5Ф +0,1
(6.25) Р5Ф—>Ру5Ф +0,5
(6.26) Ку5Ф—>Ру5Ф +0,2
(6.27) Ру5Ф—>РуБФ —5,2
1 А/7'— стандартное «биологическое» изменение свободной энергии (pH 7,0).
конденсации. Большая отрицательная величина &F' для РуБФ-
карбоксилазы (—8,4 ккал) согласуется с ее ролью в ассимиля-
ции углерода из атмосферы, где содержание СО2 составляет
всего ~0,03% (300 м. д.). Эта величина указывает также на
то, что карбоксилаза является регуляторным ферментом. Одна-
ко, хотя влияние многих метаболитов на ее активность можно
считать бесспорным, инактивация карбоксилазы в темноте не
столь очевидна, как это предполагали раньше (гл. 9). Бисфос-
фатазы также отличаются сравнительно большими значениями
&F' (от —3 до —4 ккал), и можно с уверенностью полагать, что
фруктозо-бисфосфатаза активируется светом (гл. 9). Значитель-
но более высокая активность этого фермента в щелочной среде
стромы под действием света говорит о том, что он может быть
совершенно неактивным в темноте (гл. 9).
Приведенная в табл. 6.2 величина А/7' для уравнений (6.16)
и (6.17) (ФГК—>ДФГК—>ФГА) равна +4,3 ккал; она гораз-
до менее выгодна по сравнению с довольно широко используе-
мой величиной —3,0 ккал (на рис. 9.6 AF' равно 1,2 ккал). Тем
не менее сейчас принято считать, что AF' для реакции (6.16)
ФГК—>ДФГК составляет около +4,5 ккал; в таком случае для
всей реакции восстановления ФГК до ФГА необходимо от 1,2
до 4,3 ккал. Отсюда вытекает, что реакция (6.16) является са-
мой термодинамически неблагоприятной во всем ВПФ-цикле и
поэтому больше других зависит от изменений в концентрациях
восстановительный пентозофосфатный цикл
145
ATP.ADP и ФГК (разд. 9.6). За исключением реакции Ру5Ф—>
—>-РуБФ, представленной уравнением (6.27), равновесие в ко-
торой сильно смещено в сторону образования РуБФ, все осталь-
ные реакции полностью обратимы. Даже относительно неболь-
шие величины ДА' могут быть достаточно велики,чтобы играть
существенную роль, а, например, смещение равновесия между
двумя триозофосфатами в сторону ДГАФ указывает, вероятно,
на преобладание этого соединения среди прочих удаляемых про-
дуктов метаболизма (гл. 8).
Источники данных: [7, 18, 19, 32].
6.13. Функционирование ВПФ-цикла
По характеру функционирования ВПФ-цикл можно подраз-
делить на несколько этапов, Сначала происходит карбоксили-
рование РуБФ. На рис. 6.1 в уравнении этой реакции простав-
лены коэффициенты «3» перед РуБФ, СО2 и Н2О для того, чтобы
установить правильное количественное соотношение между все-
ми продуктами, возникающими на следующих стадиях цикла.
Кроме того, это позволяет ясно увидеть, что на каждые три
молекулы СО2, вовлеченные в цикл, образуется одна молекула
продукта — триозофосфата (ЗХС1—>1ХС3). Затем происходит
восстановление. Это единственная реакция восстановления на
всем основном пути фотосинтетической ассимиляции углерода.
Поэтому в ней используется весь NADPH2, который потребляет-
ся в ВПФ-цикле. Теоретически восстановление ФГК до триозо-
фосфата может идти в одну стадию, ио в действительности об-
щий энергетический барьер снижается за счет использования
АТР в реакции, катализируемой фосфоглицераткиназой [урав-
нение (6.16); рис. 6.9].
После образования триозофосфатов в результате восстанов-
ления ФГК [уравнения (6.16) — (6.18)] все остальные десять,
реакций (так называемый углеводно-фосфатный шунт) сводятся
к регенерации РуБФ, акцептора СО2, причем сумма этих реак-
ций есть не что иное, как механизм, посредством которого пять
Л*
Фотохимии ЗРуБФ’ "'•ц
.-------- ---’ / V
ь н,0 -t I ?• .bNADPiaPj . ЬФГА 5Н3О + ЗСОг
t 9ADP V V
ЗО >/ ж ь NADPH, -'X 'ЬФГК I ФГА
1 т QATP \ '---------------------'
9 Н "Темновая"
биохимия
Рис. 0.9. Использование ассимиляционной энергии (NADPHj-l-ATP) и-
ВПФ-цикле.
10-234
146
ГЛАВА S
триозофосфатов подвергаются «перегруппировке» с образова-
нием трех пентозофосфатов (5хС3—>-ЗХСб).
Благодаря реакциям, представленным уравнениями (6.25) и
(6.26), все регенерированные пентозофосфаты оказываются в
форме рибулозо-5-фосфата. Наконец, фосфорибулокиназа [урав-
нение (6.27)] катализирует превращение РубФ в РуБФ при
участии АТР, и цикл завершается.
Источники данных: [3, 6, 55].
6.14. Использование ассимиляционной энергии
Как мы уже говорили в предыдущем разделе (см. также
рис. 6.1), ассимиляционная энергия используется в трех точках
ВПФ-цикла. Основная часть АТР (шесть молекул на каждые
три фиксированные молекулы СОг) идет на фосфорилирование
ФГК [уравнение (6.16)]. Эта энергия будет затем использована
при восстановлении ДФГК до триозофосфата [уравнение (6.17)],
на которое затрачивается шесть молекул NADPH2 (по одной
на каждый триозофосфат). Таким образом, основная часть об-
щей ассимиляционной энергии идет на восстановление ФГК, и
в принципе большая часть (5/6) возникающих триозофосфатов
затем вновь окисляется до ФГК (через цикл) в процессе кар-
боксилирования. Лишь сравнительно малая доля АТР (три мо-
лекулы или 5% ассимиляционной энергии) включается в цикл
на стадии, представленной уравнением (6.27). Благодаря этому
одновременно вводится энергия, необходимая для работы цикла,
и поддерживается соответствующая рабочая концентрация
РуБФ. [Свободная энергия образования РуБФ из РубФ ~
~—5 ккал (табл. 6.2), что указывает на сильное смещение
равновесия реакции (6.27) в сторону образования РуБФ
(разд. 2.5).] Иначе, пожалуй, трудно понять, для чего вторая
фосфатная группа вводится именно на этой стадии. [РуБФ — это
макроэргическое (богатое энергией) соединение в том смысле,
что фосфатная группа при пятом углеродном атоме препятст-
вует замыканию кольца, которое повысило бы резонансную
устойчивость. Именно этим объясняется тот факт, что карбокси-
лирование РуБФ, которое само по себе является процессом,
нуждающимся в энергии (разд. 6.4), приводит к уменьшению
свободной энергии (выделению энергии) примерно на 8 ккал.
Однако присоединение второго фосфата к первому углеродно-
му атому не дает такого эффекта. Создается впечатление, что
фосфорилирование, за которым следует карбоксилирование, по
сравнению с обратным порядком этих реакций (сначала кар-
боксилирование, а потом уже фосфорилирование) не имеет ка-
кого-либо преимущества в термодинамическом отношении по-
мимо того, о котором мы говорили выше (разд. 6.4).]
Источники данных: [5, 22].
восстановительный пентозофосфатный цикл
147
6.15. Повторное включение в цикл
Пять из шести молекул триозофосфата, образующихся в
ВПФ-цикле (рис. 6.1), идут на регенерацию акцептора СО2
(РуБФ), тогда как оставшаяся молекула представляет собой
продукт ассимиляции СО2. Ее дальнейшая судьба может быть
различной: например, она может транспортироваться в цито-
золь (гл. 8) или участвовать в синтезе крахмала внутри хлоро-
пласта. Однако в некоторых случаях эта молекула способна
опять включаться в основной ВПФ-цикл (разд. 6.3). Теоретиче-
ски это позволило бы повысить вдвое концентрацию РуБФ за
каждые пять оборотов цикла, если рассматривать цикл так, как
он изображен на рис. 6.10. [Из трех молекул РуБФ путем кар-
боксилирования образуются шесть молекул ФГК. В свою оче-
редь из этих шести молекул пять идут на регенерацию акцепто-
ра СС>2, а одна остается. После пяти оборотов цикла это даст
уже пять дополнительных молекул С3. Если эти пять молекул
не будут удалены в качестве продукта реакции, а вернутся об-
ратно в цикл, то появятся еще три молекулы РуБФ.]
Источник данных: [55].
6.16. Связь между ВПФ-циклом
и фотодыхательным циклом окисления углерода
Подробно фотодыхание будет рассматриваться в гл. 13. Сей-
час есть все основания полагать, что этот процесс представляет
собой циклическую последовательность реакций, которые состав-
со,
со,
со,
АТР ADP
6ADP ЬАТР
6NADP 6NADPH
Рис. 6.10. Восстановительный пентозофосфатный цикл. Более упрощенная мо-
дификация схемы, приведенной па рис. 6.1; показаны превращения углерода
в цикле.
10*
J 48
ГЛАВА 6
bADP ЬАТР
bNADP bNADPH
Рис. 6.11. Связь между ВПФ-циклом и фотодыхательным циклом в точке
компенсации СО2. Две из каждых трех молекул РуБФ окисляются, причем
каждая из них дает одну молекулу ФГК и одну молекулу фосфогликолата
(двухуглеродного соединения). На диаграмме отсутствует большая часть ре-
акций, в которых участвуют вышеназванные двухуглеродиые соединения.
Конденсация двух соединений, содержащих два атома углерода, дает новое
трехуглеродное соединение, которое опять вступает в цикл. Следует отме-
тить, что реально конечный продукт в этих условиях ие образуется. (Более
подробно эти реакции показаны на рис. 6.12).
ляют единое целое с ВПФ-циклом (рис. 6.11 и 6.12). Согласно
этой гипотезе, РуБФ-карбоксилаза функционирует также как
оксигеназа. Карбоксилазная активность этого фермента преоб-
ладает при высоких концентрациях СО2 или при низком содер-
жании О2. Однако в аэробных условиях часть РуБФ расщепля-
ется в реакции, идущей с потреблением Ог, давая одну молекулу
фосфоглицериновой кислоты и одну молекулу фосфогликола-
та НООС-СН2ОРО(ОН)2 вместо двух молекул фосфоглицери-
новой кислоты. Фосфогликолат подвергается гидролизу до гли-
колата, который переходит из хлоропласта в пероксисому, окис-
ляется там до глиоксилата СНО-СООН, а затем путем транса-
минирования превращается в глицин CH2NH2COOH. Глицин
поступает в митохондрии, и там из каждых двух его молекул
-образуется одна молекула серина CH2OH-CHNH2COOH:
2CH2NH2COOH + Н2О-->
Г лицин
---> СНгОН CHNHjCOOH + СО2 4- NH3 4- 2Н+ + 2е~ (6.28)
Серин
Эта реакция служит источником СО2 и NH3, которые выде-
ляются при фотодыхании, в то время как кислород использует-
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ
149
ся в исходной реакции, катализируемой оксигеназой, и при
последующем окислении гликолата. Серин переходит в перокси-
сомы, где из него возникает гидроксипируват СН2-СО-СООН,
который восстанавливается до глицериновой кислоты СН2ОН-
•СНОН-СООН. Последняя переносится в хлоропласт и фосфо-
рилируется до фосфоглицериновой кислоты, приходя в равнове-
сие с ФГК, которая образовалась раньше и находилась в стро-
ме (рис. 6.12).
Эта последовательность реакций хорошо согласуется с кар-
тиной включения метки 14СО2, указывающей на возникновение
гликолата из какого-то промежуточного соединения ВПФ-цикла
при помощи механизма, который приводит к включению одной
молекулы 18О2 в карбоксильную группу гликолата. Кроме того,
данная схема позволяет объяснить, почему фотодыхание подав-
ляется высокими концентрациями СО2 или низкими концентра-
циями О2 и каким образом 18О2, включившийся в гликолат, по-
является затем в глиоксилате, глицине, серине, гидроксипирува-
те н фосфоглицериновой кислоте. Наконец, схема согласуется
с тем фактом, что синтез гликолата из монофосфатов сахаров
ВПФ-цикла нуждается в АТР и что в присутствии 18О2 обога-
щение ФГК (в точке компенсации, или углекислотном компен-
сационном пункте) почти в 5 раз меньше обогащения гликола-
том (как видно из рис. 6.11 и 6.12, лишь одна из каждых пяти
молекул ФГК, образующихся в точке компенсации, возникает
: из гликолата и серина).
: Количество аммиака, который выделяется в этой цепи реак-
ций, может в несколько раз превосходить его количество, участ-
вующее в первичной ассимиляции. Поэтому растение должно
опять ассимилировать его, чтобы не лишиться всего своего азо-
та. Действительно, как показали исследования с применением
15N, растение вновь усваивает этот аммиак. Было высказано
предположение, что глутамин HOOC-CHNH2(CH2)-CONH2'об-
разуется в цитоплазме из глутаминовой кислоты и аммиака в
реакции, катализируемой глутаминсиитетазой:
CONH2 i
NH3 + АТР —(СНг)2 + ADP + Р; (б.29)
CHNHj
I
соон
соон
(сн2)2 +
chnh2
I
соон
Затем глутамин поступает в хлоропласт и в результате реак-
ции, представленной уравнением (6.30), образуются две новые
150
ГЛАВА 6
СНО
неон
неон Р5Ф
неон
СН2ОРО(ОН).
сн.он
с=о
НОСН Ку5Ф
неон
СН2ОРО(ОН>
CHjOH
г
носн
неон С7Ф
неон
н<!:он
СН2ОРО(ОН)2
Р'"Ч-Н1О
СН,ОРО(ОН),
I *
с=о
но<5:н
неон сбф
неон
неон
СН2ОРО(ОН)2
НОСН Ку5Ф
носн
СН2ОРО(ОН)
СНО
неон
I Э4Ф
неон
СН2ОРО(ОН)2 / С=О |
х / носн ф6ф
неон I
к НС^ОН I
CHjOPO(OH)
pi ^4 .
r-HjO I
'СН.ОН
СН3ОРО(ОН)2
с=о
НОСН ФБФ I
HcfzOH
НСОН (5)ФГК
СН2ОРО(ОН)2
соон
Серин CHNH;
CHjOH
Гидрокси-
пируват С=О
соон| —
неон (4)
СН2ОН
соон
Н(^ОН
ёндОРО(ОН)2 CHjOH
>3)АТР Глицериновая
уХЗ) АРР кислота '
. (6) (6)
^О NADPNAQPH
снон
С^° CHjOH С*°
I ХН I INH
НСОН 0=0 НСОН '.пип ,
I I I Т I 6Р.
С HjO Р О (О H)j CH jOP О (ОH)j С н jO Р О(ОН); С Н2О Р 0(0 Н) 2 СН2ОРО(ОН) 2
CHjOH
r°
се°
l'-OPO{OH)j
НСОН (5)
CH2OPO(OH)j
ФГА ДГАФ ФГА
ДГАФ ФГА
ДФГК
ВОССТАНОВИТЕ ЛЫ1ЫП ПЕНТОЗОФОСФАТИЫЙ ЦИКЛ
151
молекулы глутаминовой кислоты НООС-CHNIH^CHb^COOH:
СООН CONH2 1 СООН 1 СООН 1
'(СНг)г + (сн2)2 и- 2Н44- 2а-- -(сн2)г н- (СНг) г
1 со CHNH, CHNHj CHNHZ 1
1 СООН СООН СООН СООН
(6.30)
В качестве восстановителя в этой реакции участвует ферредок-
син. Образовавшаяся глутаминовая кислота способна опять
вступать в реакцию, катализируемую глутаминсиитетазой (см.
рис. 13.6), или давать 2-оксоглутаровую кислоту,
НООС-СО(CHshCOOH в одной из реакций переамииирования
[уравнение (6.31)], в ходе которой гликолат превращается
в глиции:
Глутаминовая кислота Глирксилат —
--->• Глиции + 2-Оксоглутаровая кислота (6.31)
Исходя из соображений стехиометрии, вторая молекула гли-
цина в этой последовательности реакций должна происходить
из серина в результате еще одного переамииирования:
Глиоксилат + Серии -->- Глиции-]-Гидроксипируват (6.32)
В этой схеме все сходится очень хорошо: из нее, например,
вытекает, что окисление глицина до серина сопряжено с появ-
лением трех молекул АТР. Этого количества АТР более чем
Рис. 6.12. Подробное изображение ВПФ-цикла и фотодыхательного цикла.
Реакции слева от штриховой линии — это те же реакции, которые схемати-
чески представлены на рис. 6.1 или 6.10 (фаза регенерации). Точно так же
карбоксилирование и восстановление ФГК до триозофосфатов не отличают-
ся от реакций, показанных на рис. 6.1 и 6.10 (т. е. от реакций собственно
ВПФ-цикла), ио здесь дополнительно показаны превращения углерода в
этих реакциях. Вместо трех молекул РуБФ карбоксилированию подвергает-
ся лишь одна, так что на этой стадии образуются только четыре молекулы
ФГК. Две молекулы фосфогликолата, которые появляются вместо этого в ре-
акции, катализируемой оксигеназой, претерпевают следующее превращение:
гликолат->-глиоксилат-->глици11-^серин->гидроксипируват-э-глицери11овая кисло-
та. Наконец, глицериновая кислота фосфорилируется, давая пятую молекулу
ФГК, так что общее количество молекул триозофосфата равно пяти. Это мо-
лекулы способны регенерировать акцептор СО2 (5CS—>ЗС5), но шестое трехуг-
леродное соединение (которое могло бы стать продуктом) теряется. Иными
словами, два из трех углеродных атомов, которые должны были бы вклю-
читься в продукт, вообще не фиксируются, так как вытесняются кислородом,
а третий освобождается в результате превращения глицина в серин
(2С2-»-С3 + СО2). Номерами в скобках обозначено число каждой из заклю-
ченных в рамки молекул, участвующих в цикле реакций, изображенных на
рис. 6.11.
152
глава е
достаточно для повторной ассимиляции аммиака с включением
его в глутамин, благодаря чему растение ие теряет свой азот.
[Другой путь использования восстановительной энергии от окис-
ления глицина заключается в том, что часть ее может быть пе-
редана пероксисоме и использована для восстановления гидр-
оксипирувата в глицериновую кислоту (см. рис. 13.6).]
Источники данных: [41, 43, 56].
6.17. Участие углерода в ВПФ-цикле
и в фотодыхательном цикле
Вопрос о том, сколько именно молекул РуБФ подвергается
окислению, остается спорным. На каждые три фиксированные
в ВПФ-цикле молекулы СОг образуется шесть молекул ФГК
(рис. 6.10). Пять из них в ходе реакций цикла превращаются
опять в РуБФ и только одна служит конечным продуктом в
форме триозофосфата. Если бы в результате окисления возни-
кали одна молекула фосфогликолата и пять молекул ФГК
(т. е. если бы общая фиксация сократилась иа одну треть), то
на каждые 15 атомов углерода основного цикла из фотоды-
хательного цикла поступало бы лишь полтора атома углерода
(иными словами, вклад фотодыхателыюго цикла составлял бы
10%)- При окислении двух молекул РуБФ на каждую карб-
оксилироваиную молекулу РуБФ (как это показано на
рис. 6.11) одна пятая общего количества синтезированной ФГК
(20% углеродных атомов в цикле) происходила бы из серина.
Это можно рассматривать как основную последовательность со-
бытий в точке компенсации СО2, когда конечный продукт сов-
сем ие образуется. Если бы иа долю серина приходилось более
20% общего количества углерода, который проходит через оба
цикла, то это можно было бы объяснить только полным исполь-
зованием уже синтезированных продуктов или метаболитов.
В этих условиях были бы исчерпаны все промежуточные про-
дукты ВПФ-цикла, и статус-кво мог бы сохраняться лишь бла-
годаря запасным веществам (например, крахмалу).
Источники данных: [43, 56].
6.18. Синтез крахмала
Крахмал нередко называют типичным конечным продуктом
фотосинтеза, и он действительно часто накапливается в хлоро-
пластах в течение дня и потребляется ночью (разд. 8.1 и 9.7).
Однако количество накапливающегося крахмала варьирует в
широких пределах. Некоторые виды растений, например под-
снежник (Qalanthus nivalis), совсем его ие образуют, а листья
ряда основных сельскохозяйственных культур, скажем пшени-
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТНЫЙ 11ИКЛ
153
цы, накапливают крахмал лишь в небольших количествах. При-
нято считать, что растения табака синтезируют много крахмала,
а между тем некоторые его сорта при выращивании их в опти-
мальных условиях накапливают очень мало этого полисахарида.
Чрезмерное накопление крахмала и снижение способности
разрушать весь крахмал, синтезированный в течение предыду-
щего дня, нередко служат симптомом неблагоприятных условий
выращивания или нарушения какой-либо функции, например
связанной с инициацией цветения. Многие растения совсем не
накапливают крахмал (за исключением отложений в замыкаю-
щих клетках), если их выращивать при слабом, пусть даже не-
прерывном освещении. По этим и подобным им причинам труд-
но делать какие-либо общие выводы, но одно кажется во вся-
ком случае бесспорным: если максимальный фотосинтез мета-
болитов идет быстрее, чем их использование, то избыток про-
дукта реакции может во многих случаях сохраняться в виде
временного запасного вещества — крахмала — в строме хлоро-
пластов. Следует отметить, что, как правило, крахмал образу-
ется быстрее и разрушается медленнее в изолированных листь-
ях, откуда растворимые продукты уже не могут быть перенесе-
ны в другие части растения. Роль неорганического фосфата в
регуляции синтеза крахмала рассматривается подробно в гл. 9.
Принято считать, что крахмал, находящийся в строме хлоро-
пластов, образуется исходно из триозофосфатов, которые слу-
жат конечным продуктом ВПФ-цикла. Большая часть этих
триозофосфатов возникает путем прямого восстановления ФГК,
но небольшое их количество образуется из углерода, который
возвращается из фотодыхательного цикла обратно в ВПФ-цикл,
пополняя уже имеющиеся там запасы ФГК. Мы еще раз напо-
минаем, что триозофосфаты, являющиеся конечным продуктом
ВПФ-цикла, могут либо экспортироваться из хлоропласта, либо
использоваться для регенерации акцептора СО2 (РуБФ) этого
цикла, либо, наконец, использоваться в синтезе крахмала. Если
триозофосфат остается внутри хлоропласта, он вступает в те
сн.он сно
I I
С=О НСОН
I I
носн носн
I I
НСОН НСОН
I | (6.33)
НСОН НСОН
I I
СН2ОРО(ОН)2 СН2ОРО(ОН)2
Фруктозо-6-фосфат Г люкозо-б-фосфат
(ФбФ) (Г6Ф)
154
ГЛАВА 6
же реакции, которые протекают в ходе синтеза сахарозы в ци-
топлазме. Иными словами, часть триозофосфатов подвергается,
альдольной конденсации, в результате которой образуется ФБФ.
В ходе гидролиза ФБФ возникает Ф6Ф, который затем превра-
щается в свой изомер Г6Ф в результате реакции, катализируе-
мой гексозофосфат-изомеразой.
В реакции, катализируемой фосфоглюкомутазой, Г6Ф пре-
вращается в ПФ.
Г6Ф
носн2
н он
Г1 Ф
(6.34 ).
В состоянии равновесия в присутствии вышеназванных фер-
ментов образуется смесь трех гексозофосфатов примерно в сле-
дующем соотношении:
Ф6Ф 4==*: Г6Ф ПФ
10 20 1
Хотя эти реакции полностью обратимы, общее равновесие
играет немаловажную роль в распределении атомов углерода
между. крахмалом и пентозомонофосфатами в освещенном хло-
ропласте. Как Ф6Ф, так и ТФ (триозофосфат) служат субстра-
тами первой транскетолазной реакции [уравнение (6.21)].
От этого в свою очередь зависит, сколько ТФ вступит во вто-
рую реакцию конденсации, катализируемую альдолазой [урав-
нение (6.22)], и во вторую транскетолазную реакцию [уравне-
ние (6.24)]. Таким образом, при активном удалении ПФ
углерод будет направляться на синтез крахмала. Весьма вероят-
но, что удаление ПФ обеспечивается глюкозо-1-фосфат — аде-
нилилтрансферазой, которая катализирует реакцию, представ-
ленную уравнением (6.35), и активируется при высоком соотно-
шении между концентрациями ФГК и Pi.
Это соотношение будет максимальным, когда при низкой
наружной концентрации неорганического фосфата замедляется
экспорт триозофосфатов и ФГК. Следовательно, максимальное
использование ПФ должно теоретически совпадать с макси-
мальным поступлением предшественника крахмала. Наконец,
под действием крахмал — синтазы глюкозильные остатки пере-
носятся от ADP-глюкозы к соответствующим затравкам (напри-
мер, к мальтозе, мальтотриозе и другим олигосахаридам и дек-
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ цикл
155
но он
АТР
стринам), благодаря чему их цепи удлиняются и образуются не-
разветвлеиные полимеры типа амилозы с а(1—>4)-связями:
сн2он
н он
ADP -глю
носн2 носнг
.О- О.
нон н о н
(6.36)
Олиго- или полисахарид, содержащий
П глю к оз ил ьн ы х единиц
НОСН2 НОСН; НОСН;
...........................
н он н он н он
Олиго- или полисахарид,
содержащий П+1 глюкозильных единиц
ГЛАВА 6
156:
Синтез амилозы может также катализироваться крахмал-
фосфорилазой [уравнение (6.37)]. Однако эта возможность яв-
ляется скорее теоретической, так как реакция полностью обрати-
ма, и более вероятно, что данный фермент катализирует реак-
цию деградации крахмала.
Источники данных: [21, 48].
6.19. Деградация крахмала
Крахмал, который находится в запасающих тканях, разру-
шается в результате совместного действия ряда ферментов.
Крахмал-фосфорилаза катализирует фосфоролиз а(1—>-4)-
глюкозильных цепей с образованием Г1Ф:
а-1,4 (Глюкозил)п+1Pj 4=^ а-1,4 (Глюкозил)п-|-ПФ (6.37)>
а-Амилаза атакует а(1—>-4)-связи амилозы (которая состо-
ит из неразветвленных цепей и дает характерную синюю окрас-
ку с иодом) или амилопектина [который имеет а(1—>-6)-связи
и окрашивается иодом в фиолетовый цвет]. В качестве конеч-
ных продуктов образуются короткие цепи декстринов, а также
мальтотриоза, мальтоза и некоторое количество глюкозы.
[3-Амилаза действует только на невосстанавливающие концы це-
пей, отщепляя пары глюкозильных остатков в виде мальтозы.
Так называемый Д-фермеит атакует 1—>6-связи, которые явля-
ются точками ветвления в молекуле амилопектина, делая этот
компонент крахмала доступным для расщепления |3-амилазой
или фосфорилазой. Наконец, D-фермент катализирует перенос
глюкозильных остатков между декстринами с короткими цепя-
ми, в результате чего образуется смесь глюкозы и декстринов
с цепями разной длины.
Весьма вероятно, что в листьях ф-амилаза содержится в ци-
топлазме и может разлагать крахмал только в старых хлоро-
пластах или в хлоропластах, которые приобрели свойства ами-
лопластов. Очевидно, деградация крахмала в хлоропластах на-'
чинается под действием эндоамилазы, которая содержится в
строме и несколько отличается от других а-амилаз растительно-
го происхождения (не нуждается в ионах Са2+ и термолабиль-
на) (рис. 6.13). Под действием этого, а также R- и D-фермеитов
образуются продукты, которые легко подвергаются фосфороли-
зу. Затем ПФ превращается в триозофосфаты через ряд проме-
жуточных соединений при участии фосфофруктокиназы;
*ФГА
ПФ ----> Г6Ф ---> Ф6Ф---> ФБФХ (6.38)
ДГАФ
Эти триозофосфаты могут экспортироваться из хлоропласта
при помощи переносчика неорганического фосфата (разд. 8.23).
восстановительный пентозофосфатный цикл
157
Рис. 6.13. Деградация крахмала в хлоропластах. Эядоамилаза атакует крах-
мальную гранулу, освобождая декстрины с разветвленными и иеразветвлен-
ными цепями, олигосахариды (например, мальтотриозу и мальтозу) и неко-
торое количество глюкозы. D-Фермент катализирует реакции типа мальто-
триоза+мальтотриоза->-мальтопентоза+глюкоза. R-Фермент атакует 1—6-
связи в амилопектине. Мальтозофосфорилаза и крахмал-фосфорилаза ката-
лизируют фосфоролиз соответственно мальтозы и более длинных а (1->-4)-
глюканов, давая Г1Ф, который может превращаться дальше в триозофосфат.
Глюкоза медленно экспортируется при помощи довольно специфического'
переносчика (разд. 8.17), а удаление триозофосфата происходит быстро по-
средством переносчика неорганического фосфата [45]. В результате превра-
щения триозофосфата в ФГК образуется АТР, который расходуется в реак-
ции Ф6Ф->ФБФ. Весь крахмал, который накапливается в течение дня, иног-
да полностью разрушается за последующую ночь. Поэтому создается впечат-
ление, что метаболизм крахмала регулируется ие только реакциями его син-
теза.
При высоком содержании последнего деградация крахмала уси-
ливается благодаря инициации фосфоролиза, инактивации глю-
козо-1-фосфат — аденилилтрансферазы (за счет низкого отно-
шения [ФГК]/[Р[]) и увеличению скорости экспорта триозофос-
фатов. Как было показано Кремииером (Kraminer, неопубли-
кованные данные), синтез крахмала сменяется его распадом
в освещенных хлоропластах, если увеличить концентрацию не-
органического фосфата.
До сих пор не установлено, что происходит с мальтозой. Ее
прямой транспорт представляется маловероятным, а мальтаза
(фермент, который катализировал бы ее гидролиз до глюкозы)
пока не обнаружена. Не исключено, что мальтоза разлагается
158
ГЛАВА 6
до глюкозы и ПФ под действием фермента мальтозофосфори-
лазы.
Источники данных: [36, 40, 45, 45а, 46, 48а, 511.
6.20. Регуляция синтеза и деградации крахмала
Подробно мы рассмотрим этот процесс в разд. 9.7, а пока
нам хотелось бы отметить, что многое в нем остается невыяснен-
ным. Мы не знаем, например, насколько крахмал разлагается
при освещении, в то время как совершенно очевидно, что какая-
то часть его должна разлагаться в этих условиях, если только
активность эндоамилазы не подавлена полностью. Де Фекит и
Выоег высказали предположение, что мальтоза ингибирует ак-
тивность амилазы. Кроме того, мальтоза является хорошей за-
травкой для крахмал — синтазы. Поэтому, по мнению этих
авторов, мальтоза в большой концентрации ие только непосред-
ственно стимулирует синтез крахмала, но и настолько подавля-
ют амилазную активность, что это приводит благодаря повышен-
ному содержанию субстратов крахмал — синтазы к увеличению
скорости его синтеза.
Фруктозобисфосфатаза в темноте инактивируется (разд. 9.2),
•благодаря чему предотвращается бесполезная работа цикла
[уравнение (6.39)], которая наблюдалась бы в присутствии ак-
тивной фосфофруктокиназы:
/,ФБФ\ (6.39)
Фосфофруктокиназа АТР Р: ФБФ-аза
Ф6Ф *
.Аналогичным образом в присутствии высокоактивной фосфо-
фруктокиназы ВПФ-цикл на свету не функционирует, и любая
•активность в освещенной строме привела бы к затратам АТР на
образование ненужного продукта.
Источники данных: [10, 36].
6.21. Синтез сахарозы
Хотя одно время хлоропласты считали местом синтеза саха-
розы, в продуктах фиксации 14СС>2 изолированных хлоропластов
юна обычно отсутствует (рис. 6.14). В проведенных в последнее
время исследованиях по распределению ферментов и образова-
нию продуктов было убедительно показано, что синтез сахаро-
зы сосредоточен в цитоплазме (рис. 9.9). Предпоследняя реак-
ция в последовательности, показанной на этом рисунке, катали-
зируется сахарозофосфат-синтазой, которая облегчает перенос
глюкозного остатка от UDP-глюкозы к Ф6Ф [уравнение (6.36)].
•Образовавшийся фосфат сахарозы гидролизуется специфической
восстановительный пентозофосфатный ЦИКЛ
159
Рис. 6.14. Накопление сахарозы в цитоплазме протопластов при освещении в
присутствии 14СО2. Видно, что содержание сахарозы в хлоропласте практиче-
ски ие меняется (некоторые колебания могут быть вызваны незначительными
загрязнениями при фракционировании). [Эти данные согласуются с точкой
зрения, согласно которой синтез сахарозы протекает в цитоплазме, так как
скорость транспорта сахарозы через внутреннюю мембрану (гл. 8) чрезвы-
чайно низка (Робинсон, неопубликованные данные).]
фосфатазой, давая свободную сахарозу. Образование UDP-
глюкозы аналогично образованию ADP-глюкозы в ходе синтеза
крахмала [разд. 6.18 и уравнение (6.35)].
Источники данных: [10, 33а, 37, 49Ъ].
6.22. На пути ассимиляции углерода
Эта глава была посвящена ВПФ-циклу в том виде, как мы
его представляем в настоящее время. А теперь мы предлагаем
рассказ, написанный Домиником Рекальденом и впервые опуб-
ликованный в «New Scientist» 5 декабря 1968 г., в котором вся
картина предстает в несколько ином свете.
«Как сейчас помню, это случилось в начале пятидеся-
тых годов, когда я столкнулся с первым серьезным испыта-
нием. К тому времени я проработал в Отделе клеточной
безопасности всего полгода. В один прекрасный момент раз-
дался телефонный звонок. Вообще-то я его уже ждал. Пло-
хие новости всегда тут как тут, а слухи об утечке секретной
информации распространялись на мой взгляд даже слиш-
ком быстро.
— Он хочет вас видеть, — пропищал тоненький голосок
в трубке.
160
ГЛАВА 6
— Я буду готов через пятнадцать минут, — ответил я.
Выскочив из комнаты, я ухватился за пустую молекулу,
плавающую в растворе. Мы проехали по ретикулуму до
ядерной мембраны. Я вынул свой пропуск и сунул его под
нос вахтеру, который читал комикс и с наслаждением ко-
вырял в зубах.
— Вас ждут, — буркнул он. Он даже не поднял глаз.
Мне стало неловко, что я посмел побеспокоить его.
Шеф уныло разглядывал открытую папку, когда я во-
шел к нему в кабинет. По-моему, она была пустая, а впро-
чем, я не уверен.
— Какие меры вы приняли в отношении утечки инфор-
мации?
— Четкий вопрос, — подумал я, — но...
— Боюсь, что пока не за что ухватиться. Может быть,
у вас есть какие-нибудь идеи?
— Клеточная безопасность — это ваша работа. И, судя
по всему, вы ее скоро потеряете. Да и мы все с вами за-
одно. Враг уже совсем подобрался к сравнительной биохи-
мии. Если он узнает наши секреты, он узнает все. И о сце-
недесмусе, и о хлорелле. Даже о самом главном, вроде
шпината. Все теперь под угрозой.
В мрачном настроении я покинул кабинет шефа и на-
правился на завод, чтобы еще раз бегло все осмотреть.
Я перебирал в уме то немногое, что нам стало известно.
Уже почти год мы страдали от утечки жизненно важной
информации о нашем фотосинтетическом механизме для
изготовления сахаров. Ясно, что дело тут было в простом
воровстве. Они, наверно, нашли способ выкрадывать проме-
жуточные продукты в разных местах конвейера, а потом
соединяли их по порядку. Но вскоре я отбросил эту мысль.
Ведь уже шесть недель я ие покидал завода, проверял по
списку весь инвентарь, подсчитывал все глюкозные молеку-
лы, которые мы изготавливали. Все было на месте.
Правда, время от времени Они устраивали истребление.
Нас, как скот, загоняли небольшими партиями в крошечный
контейнер под ослепительный свет, а потом бросали в
кипящий спирт — жуткая процедура, хотя это, пожалуй, еще
не самое страшное, на что Они способны. Ясно, Они нас
анализировали. Может быть, даже применяли двумерную
хроматографию — важнейший только что открытый в то
время прием. Но какая им от этого польза? Они ие знали,
в каком порядке расположить промежуточные продукты.
Нам же хорошо известно, что Они пытались вытянуть хоть
что-нибудь и из наших ферментов. А ведь всем известно, что
ферменты с детства приучены не выдавать ценной информа-
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТИЫЙ цикл
161
ции, если они окажутся вне клетки. «Имя, номер, звание» —
и больше фермент ничего не скажет. И вот, несмотря на
все это, Они получали от нас информацию, а каким обра-
зом — мы не могли понять.
Я заметил, что уже некоторое время стою с унылым
видом в упаковочном отделе и смотрю на старика
Джорджа.
— Привет, Джордж. Как дела?
Джордж поднял дюжину ящиков с крахмальными гра-
нулами и поставил их на подъемник.
— Старею, видать, — заметил он. — Бывало, по четыр-
надцать поднимал зараз. А сейчас только двенадцать.
— Может, они стали тяжелее, — предположил я.
— Как-то ие подумал об этом, —сказал он.
Я тоже об этом раньше не думал. А ведь это, пожалуй,
мысль.
— У нас еще осталась старая продукция, Джордж? От
прошлого года?
— Наверху.
— Дай-ка мне один из этих ящиков, — сказал я.
Я отнес его наверх и поставил на весы. Потом я взвесил
ящик с продукцией прошлого года. Ои был легче. Не намно-
го, но легче. На первый взгляд это могло показаться неваж-
ным, ио я чувствовал, что напал на след. Только я еще ие
мог понять, в чем тут дело. Как сейчас помню, я уже хотел
плюнуть на все это, как у меня в мозгу вроде что-то щелк-
нуло. Меня как громом поразило. Я был так взволнован, что
мне пришлось три раза набрать номер госпиталя, пока мои
дрожащие пальцы справились с добавочным номером.
— Госпиталь?
— Говорит Отдел безопасности. Откуда у вас больше
всего поступило больных в прошлом году?
— Наконец-то кто-то догадался спросить. Тут что-то
странное. Очень много больных из Отдела поглощения
двуокиси углерода. Никак ле пойму, в чем тут дело.
Я чуть сознание ие потерял.
— А они жалуются на какую-нибудь определенную бо-
лезнь? Есть какие-то четкие симптомы?
— Тут тоже странная вещь. Почти все они обнаружи-
вают симптомы — это звучит глупоно они все как будто
страдают от облучения. Это ерунда, я знаю, но симптомы...
Я бросил трубку и высоко подпрыгнул. Потом я сбежал
вниз, схватил Джорджа в объятия и закружился с ним
в вальсе, распевая «Веселую вдову». После тридцати двух
тактов я остановился перевести дух, а потом опять бегом
поднялся наверх и позвонил в Отдел контроля.
11—234
162
ГЛАВА 6
— Шеф? Говорит Отдел безопасности. Вы можете себе
представить... метка радиоактивного углерода!
— О чем вы говорите?
— Я понял, как Они все узнали. Они дают нам СОг
с меченым углеродом. Потом Они анализируют наши про-
дукты и следят за разбавлением радиоактивности. Дальше
Им остается только расположить промежуточные продукты
в порядке увеличения разбавления, и Они все узнают.
— Ничего не понимаю.
— Шеф? Могу я предложить кое-что в плане контрраз-
ведки?
— Подайте рапорт.
Moii рапорт был шедевром, хоть это я вам сам говорю.
И я, правда, горжусь тем, что мы потом сделали. Короче
говоря, мы выпустили целый набор бессмысленных фермен-
тов, которые катализировали совершенно нелепые превра-
щения молекул сахаров. У нас 3-углеродные сахара превра-
щались в 5-углеродные, 4-углеродиые — в 7-углеродные и
обратно в 5-углеродпые. Получился полный кавардак. Но>
самое главное в моем плане заключалось в том, что полная
схема была представлена необыкновенно сложным циклом
(Оии ведь обожают циклы!), в одной точке которого про-
исходил захват двуокиси углерода, а в другой получался
(иакоиец-то!) сахар. Все пошло как по маслу. Через полго-
да Они были совершенно убеждены, что выяснили весь путь
ассимиляции углерода. Они хлопали друг друга по плечу
и получали Нобелевские премии. После этого нас никто уже
не беспокоил. Опп думают, что раскрыли тайну клетки, и
поэтому счастливы. Мы же вернулись к нашему старому
простому способу фиксации углерода и тоже счастливы.
При таком обилии счастья в этом мире нам остается только’
улыбаться!»
Хотя этот рассказ представляет собой антропоморфическую
фантазию, в нем заложено предостережение. Когда был выяс-
нен ВПФ-цикл, казалось, что мы узнали о фотосинтетической
ассимиляции углерода почти все и ничего нового ожидать ие
приходится. Говорят, что один почтенный ученый заявил: «Ко-
нечно, в фотосинтезе еще остаются нерешенные вопросы, но они
не связаны с путем ассимиляции углерода». Однако такой опти-
мизм оказался преждевременным, о чем говорят исследования
фотодыхания и С4-фотосиитеза, а также установление экологи-
ческой роли метаболизма кислот по типу растений из семейства
толстянковых. Сейчас довольно трудно представить, что еще бу-
дут обнаружены целиком новые пути метаболизма. Однако со-
вершенно очевидно, что от полного понимания взаимосвязи
ВОССТАНОВИТЕ/!Ы1ЫП ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ
163
между хлоропластом и его клеточным окружением нас отделяют
еще долгие годы. Весьма удачным нам кажется утверждение,
что «ферменты с детства приучены не выдавать ценной инфор-
мации, если они окажутся вне клетки». Бесспорно, что целый
ряд исследований можно проводить только с высокоочищенны-
ми ферментами. Однако не следует забывать, что выделение
РуБФ-карбокснлазы в высокоочищенном виде не помогло
объяснить, почему ее активность столь низка in vitro, и только
выполненная раньше работа Пона подсказала, что этот фер-
мент нуждается в ионах Mg:;+. В настоящее время кажется не-
понятно низкой активность сахарозофосфат-синтазы. По-внди-
мому, решение этого вопроса потребует совместных усилий уче-
ных, работающих в области клеточной биологии и энзимологии.
Источники данных: [47, 54].
ОБЩАЯ ЛИТЕРАТУРА
1. Анит D. I. Cell-free plwlosynliesis and the energy conversion process. In:
Light and Life. (McElroy W. D., Glass B. (eds.), 489—569. The John Hopkins
Press, Baltimore, 1961.
2. Artion D. I. Photosynthetic activity of isolated chloroplasts. Physiol. Rev.,
47, 317—368 (1967).
3. Bassham J. A., Calvin M. The Path of Carbon in Photosynthesis. 1—104.
Englwood Cliffs, N. J., Prentice-Hall Inc. (1957).
4. Benson A. A., Calvin M. Carbon dioxide fixlion by green plants. Ann. Rev.
Plant Physiol., 1, 25—40 (1950).
5. Calvin M„ Pon N. G. Carboxylations and decarboxylations. J. Cell. Comp.
Physiol., 54, 51—74 (1959).
6. Calvin M., Bassham J. A. The Photosynthesis of Carbon Compouds, 1—127.
Benjamin, New York, 1962.
7. Conn E. E„ Stumpf P. K- Outlines of Biochemistry, 1—629. John Wiley, Sons,
Inc., New York, 1976.
8. Franck J., Loomis W. E. (eds.). Photosynthesis in Plants, 381—401. Iowa
State College Press, Ames, 1949.
9. Gibbs M. Photosynthesis. Ann, Rev. of Biochem., 36, Para II, 757—84- (1967).
10. Gibbs M. (ed.). Structure, and Function of Chloroplasts, L—286. Springer-
Verlag, Berlin, 1971.
11. Goodwin T. W. (ed.). Biochemistry of Chloroplasts. Vol. II. Proceedings
NATO Advanced Study Institute, Aberystwyth, 1965, 1—706. Academic Press,
London, 1967.
12. Hatch M. D„ Slack C. R. Photosynthetic COa-fixtion pathways. Ann. Rev.
Plant Physiol., 21, 141—62 (1970).
13. Hatch M. D„ Osmond С. B., Staiger R. O. (eds.). Photosynthesis and Photo-
respiration. Proceedings of the Conference at A.N.U., Canberra. Wiley Inter-
science, New York, 1970.
14. Jensen R. G., Bahr J. I. Ribulose 1,5-bisphosphate carboxylase/oxygenase,
Ann. Rev. Plant Physiol., 28, 379—400 (1977).
15. Kok B., Lagendorf A. T. (eds.). Photosynthetic Mechanisms in Green Plants.
Washington D. C.: Nat. Acad. Sci. 1145, 1963.
16. Kelly G. J., Latzko E. Regulatory aspects of photosynthetic carbon metabo-
lism. Gibbs M. (ed.). Ann. Rev. Plant Physiol., 27, 181—205 (1976).
17. Lehnlnger A. L. Biochemistry, 1—833. New York: Worth Pulishers, Inc.,
1970.
11
164
ГЛАВА 6
18. Lelininger A L. Bioenergetics, 1—245. Philippines: W. A. Benjamin, Inc.,
1971.
19. Nellands J. B., Stumpf P. K. Outlines of Enzyme Chemistry, 1—411. John
Wiley, Sons Inc., New York, 1958.
20. Pauling L. The Nature of the Chemical Bond, 1—644. Cornell University
Press, Ithaca, New York, 1960.
21. Pridhain J. B. (ed.). Phytochemical Society Symposium on Plant Carbohyd-
rate Biochemistry, Edinburgh, 1972. Academic Press, London, 1974.
22. Quayle J. R., Ferenci T. Evolutionary aspects of autotrophy, Microbiological
Reviews 251—73 (1978).
23. Rubinowitch E. !. Photosynthesis and Related Processes. Vol. 2, Part I,
537 et seq. Wiley Intcrscience, New York, 1956.
24. Robinson S. P., Walker D. A. Photosynthetic carbon reduction cycle (Calvin
cycle) (including fluxes and regulation). In The Biochemistry of Plants.
A comprehensive treatise, (eds. P., K. Stumpf and E. E. Conn.) Vol. 8
(Photosynthesis). Academic Press, London, 1981.
25. San Pietro A., Greer F. A., Army T. J. (eds.). Harvesting the Sun, 1—342.
Academic Press, London, 1967.
26. Siegelman H. IE., Hind G. (eds.). In: Photosynthetic Carbon Assimilation.
Proceedings of a Symposium held at Brookhaven National Labratory, 1—445.
Upton, New York, 1978. Plenum Press, New York, 1978.
27. Stiller M. The path of carbon in photosynthesis, Ann. Rev. Plant Physiol.,
13, 151—70 (1962).
28. Symposia of the Soc. for Exp. Biol. V. Carbon Dioxide Fixation and Photo-
syntesis (19'51).
29. Walker D. A., Crofts A. R. Photosynthesis, Ann. Rev. Biochem., 39, 389—
428 (1970).
30. Zelltch J. Photosynthesis, Photorespiration and Plant Productivity. 1—347.
Adademic Press, London, 1971.
31. Zelltch I. Pathways of carbon fixation in green plants, Ann. Rev. Biochem.,
44,123—45 (1975).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
32. Bassham J. A., Krause G. H. Free energy changes and metabolic regulation
in steady-stale photosynthetic carbon reduction, Biochim. Blophys. Acta, 189,
207—221 (1969).
33. Benson A. A., Bassham J. A., Calvin M., Goodale T. C„ Haas V. A., Stepka 117.
The path of carbon photosynthesis. V. Paper chromatography and radioauto-
graphy of the products, J. Amer. Chem. Soc., 72, 1710—18 (1950).
33a. Bird I. F., Cornelius M. J., Keys A. J., Whittingham С. P. Intracellular site
of sucrose synthesis in leaves, Phytochemistry, 13, 59—64 (1974).
34. Calvin M., Pon N. G. Carboxylations and decarboxylations, J. Cell. Comp.
Physiol., 54, 51—74 (1959).
35. Consden R., Gorden A. H., Martin A. J. P. Qualitative analysis of proteins:
A partition chromatography method using paper, Biochem. J., 38, 224—32
(1944).
36. De Fekete M. A. R., Vleweg G. H. Starch metabolism: Synthesis versus
degradation pathways, In Plant Carbohydrate Biochemistry, Proc. Phyto-
chem. Soc. Symp. (ed. J. B. Pridham), 127—44, Edinburgh, 1973, Academic
Press, London, 1974.
37. Everson R. G., Cockburn W., Gibbs M. Sucrose as a product of photosynthe-
sis in isolated spinach chloroplasts. Plant Physiol., 42, 840—4 (1967).
38. Gaffron H„ Fager E. IF., Rosenberg J. L. Intermediates in photosynthesis.
Formation and transformation of phosphoglyceric acid. In: Symposia of the
Soc. for Exp. Biol. V. Carbon Dioxide Fixation and Photosynthesis, 261—
283 (1951).
ВОССТАНОВИТЕЛЬНЫЙ ПЕНТОЗОФОСФАТНЫЙ цикл 165
39. Heath О. V. S, Assimilation by green leaves with stomatai control elimina-
ted, In: Symposia of the Soc, for Exp. Biol. 'V, Carbon Dioxide Fixation and
Photosynthesis, 94—115 (1951).
40, Heldt H, IF,, Chon C. J., Maronde E„ Herold A., Stankovic Z. S., Wal-
ker D. A., Kramlner A, Kirk M. A., Heber U, Role of orthophospsate and
other factors in the regulation of starch formation in leaves and isolated
chloroplast, Plant Physiol,, 59, 1146—1155 (1977),
41, Keys А Л, Bird 1. F„ Cornelius M. J., Lea P, Wallsgrove R. M„ Mlf-
lin B. J. Photorespiratory nitrogen cycle, Nature, 275, 741—3 (1978),
42. Lorimer G. H„ Badger M. R„ Andrews T. J. The activatlion of ribulose-1,5-
bisphosphate carboxylase by carbon dioxide and magnesium ions. Equilibria.,
kinetics, a suggested mechanism and physiological implications, Biochemistry,
15, 529—36 (1976),
43, Lorimer G. H,, Wo /(, C„ Berry J. A, Osmond С. B. The C2 photorespiratory
carbon oxidation cycle in leaves' of higher plants; Pathway and consequences.
In; Photosynthesis 77, Proceedings of the Fourth International Congress im
Photosynthesis, September 1977 (Hall D. O„ Coombs J,, Goodwin T. W„
eds,), 311—322, Biochemistry Society, London, 1978. -
44. Poincelot R, P. Uptake of bicarbonate ion in darkness by isolated chloro-
plast envelope membrane and intact chloroplast of spinach, Plant Physiol.,
54, 520—6 (1974),
45, Okita T. 1F„ Greenberg E,, Kuhn D, N„ Preiss J, Stibcellular localization of
the starch degradativc and biosynthetic enzymes of spinach leaves, Plant
Physiol,, 64, 187—92 (1979).
46, Peavey D. G., Steup M., Gibbs M. Characterization of starch breakdown in
the intact spinach chloroplast, Plant Physiol,, 60, 305—308 (1977).
47. Pon N. G. Studies , on the carboxydismutase system and related materials.
Ph, D, Thesis, University of California, Berkeley, 1959.
48. Preiss J., Levi C, Regulation of a-l,4-glucan metabolism in photosynthetic
systems. In; Photosynthesis 77. Proceedings of the Fourth International
Congress in Photosynthesis, September 1977, Hall D. O„ Coombs J., Good-
win T, W. (eds.) 457—68, Biochem, Soc., London, 1978.
49, Quayle I. R„ Fuller R, C., Benson A A, Calvin M, Enzymatic carboxylation
of ribulose diphosphate, J, Amer, Chem, Soc,, 76, 3610—1 (1954).
49a, Robinson S, P., McNeil P. H„ Walker D. A. Ribulose bisphosphate carbo-
xylase— lack of dark inactivation of the enzyme in experiments wits proto-
plasts, FEBS Letters, 97, 290—300 (1979),
49b, Robinson S. P„ Walker D, A, The site of sucrose synthesis in isolated leaf
protoplasts, FEBS Letters, 107, 295—299 (1979),
50, Ruben S„ Kamen M. D, Radioactive carbon of long half-life, Phys. Rev,, 57,
549 (1940).
5. 1. Thomas M,, Ranson S. L„ Richardson J. A. Plant Physiology, Longman, Lon-
don, 1973.
52. Tswett M„ 1906, См.; Rabinowitch E. I. Photosynthesis and Related Proces-
ses, Vol, 1, 402, Interscience, New York, 1945.
53, Walker D. A. Pyruvate carboxylation and plant metabolism, Biol, Rev., 37,
215—56 (1962).
54, Walker D, A. Photosynthetic induction phenomena and the light activation
of ribulose diphosphate carboxylase, New Phytol,, 72, 209—35 (1973),
55. Walker D, A. Some characteristics of a primary carboxylating mechanism.
Phytochemical Sociaty Symposium on Plant Carbohydrate Biochemistry,
Edinburgh, 1972 (ed. J. B. Pridham) 7—26. Academic Press, London, 1974.
56, Walker D. A Report of Symposium 7 (Regulation of Metabolism), In; Pho-
tosynthesis 1977, «Proceedings of the Fourth International Congress on
Photosynthesis», September 1977. D. O. Hall, J. Coombs, and.T. W, Good-
win (eds,), 489—493, Biochemistry Society, London, 1978,
ГЛАВА 7
Индукция
Краткое содержание
Индукция — это неотъемлемая особенность фотосинтеза.
Если перенести целые листья растений, протопласты или хлоро-
пласты из темноты на яркий свет, то фотосинтетическая ассими-
ляция углерода начинается не сразу, а лишь по истечении неко-
торого индукционного периода. Может пройти несколько минут,
прежде чем скорость станет максимальной. Напротив, большин-
ство «фотохимических» процессов, например транспорт электро-
нов, идет с более или менее максимальной скоростью с самого
начала. Следовательно, индукция связана непосредственно с
восстановительным пентозофосфатный циклом. Она часто встре-
чается в тех случаях, когда ие требуется участия устьиц.
Гипотеза Остерхаута — Хасса объясняет индукцию либо ак-
тивацией катализаторов под действием света, либо накоплением
субстратов. Первое объяснение означает, что некоторые фер-
менты, катализирующие ассимиляцию углерода, неактивны в
в темноте и «включаются» на свету. Второе объяснение связа-
но с автокатализом (разд. 6.3), т. е, способностью цикла обра-
зовывать больше промежуточных продуктов, чем он их ис-
пользует; в темноте количество одного или нескольких из этих
продуктов уменьшается и не сразу возрастает на свету. Фото-
синтетический транспорт электронов и фотофосфорилирование
прекращаются в темноте немедленно. Поэтому реакции, завися-
щие от ассимиляционной энергии, тоже останавливаются сразу
или почти сразу. В то же время другие реакции, например кар-
боксилирование, могут протекать еще некоторое время и в тем-
ноте, что приводит к истощению накопившихся промежуточных
продуктов.
Возможность аллостерических взаимодействий предполагает
существование сложных связей между активацией ферментов
и количеством различных промежуточных продуктов. В совре-
менных исследованиях изучаются оба аспекта гипотезы Остер-
хаута—-Хасса и считается, что такие взаимодействия вполне
возможны. Следует также отметить, что скорость катализа мо-
жет повыситься либо в результате активации ферментов (т. е.
перехода ферментов из неактивной формы, которая существует
в темноте, в активную форму на свету), либо благодаря более
ИНДУКЦИЯ
167
благоприятным условиям (фермент, уже обладающий полной
активностью, может функционировать еще лучше в относитель-
но щелочной среде освещенной стромы). Кроме того, оба эффек-
та способны дополнять друг друга.
Мы считаем, что индукция тесно связана с двумя особеннос-
тями ВПФ-цикла. Превращение ФГК в ДФГК представляет со-
бой полностью обратимую, но энергетически невыгодную реак-
цию, которая при фотосинтезе осуществляется в присутствии
высоких концентраций ФГК и весьма чувствительна к падению
отношения [ATP]/[ADP], Таким образом, лаг-фаза (индукцион-
ный период, задержка) в выделении кислорода вызвана преж-
де всего возрастанием концентрации ФГК, которое происходит
в течение первых минут освещения. (Выделение кислорода со-
пряжено с переносом электронов от Н2О к ЫАЭР+-акцептору,
который регенерируется путем восстановления ФГК через
ДФГК до триозофосфата.) Аналогичным образом задержка,
которая наблюдается в фиксации СО2, объясняется, ио-видимо-
му, в первую очередь увеличением концентрации гексозомоно-
фосфата. В фазе регенерации ВПФ-цикла триозофосфат исполь-
зуется одновременно в четырех реакциях (две из них катализи-
руются транскетолазой, а две — альдолазой). Кроме того, из
всех промежуточных продуктов цикла триозофосфат быстрее
всего экспортируется из хлоропласта. Вот почему превращение
пяти молекул триозофосфата в три молекулы пентозомонофос-
фата связано со значительными изменениями в количественном
содержании этих соединений под действием света, так как об-
разование триозофосфата из ФГК немедленно прекращается в
темноте и даже, по всей вероятности, этот процесс идет в обрат-
ном направлении. Необходимые концентрации этих соединений
могут обеспечиваться смещением равновесия катализируемой
альдолазой реакции, в которой продукт образуется в результа-
те конденсации двух реагирующих соединений, а также благо-
даря первоначально низкой активности бисфосфатаз.
Индукция представляет собой в основном искусственный
процесс в том отношении, что более или менее внезапный пере-
ход растений или листьев после довольно длительных периодов
темноты в условия яркого освещения в природе встречается
чрезвычайно редко. Этот процесс весьма интересен для исследо-
вателей, поскольку он позволяет гораздо лучше понять механиз-
мы регуляции и взаимосвязь между хлоропластом и клеткой,
в которой он находится.
7.1. Индукция в целых растениях и листьях
Если ярко осветить зеленые листья, которые до этого нахо-
дились в полной темноте, фотосинтетические процессы выделе-
ния О2 и фиксации СО2 достигают максимума, как правило, не
168
ГЛАВА 7
Рис. 7.1. Индукция в растениях Lemna. Целые растения освещали при 20 °C
в растворе бикарбоната ( — 10-» М). Так же, как и в первых опытах Блинк-
са и Скау (1938а), pH возрастает по мере поглощения О2 из раствора. Об-
ратите внимание на длительную задержку вначале, которая ие повторяется
после кратковременного затемнения.
сразу, а лишь спустя несколько минут (рис. 7.1). Эта начальная
задержка получила название «индукции» (или, по выражению
Рабиновича, «простой индукции»), и ее не следует смешивать
с разнообразными кратковременными колебаниями газообмена
(«глотками» и «выбросами»), которые наблюдаются в некото-
рых условиях. Простая индукция — это не артефакт измерения.
Она была найдена при исследовании целых растений без устьиц,
а также в опытах с изолированными хлоропластами и прото-
пластами. Индукция является основной, неотъемлемой чертой
<фотосинтеза.
Рабинович отмечает также различие между «кратковремен-
ной» и «длительной» индукцией. Как правило, первая занимает
ют двух до пяти минут при обычной температуре. «Длительная»
индукция представляет собой вторичное медленное ускорение,
которое может затянуться на несколько часов и потому не обя-
зательно связано с фотосинтезом как таковым.
В целых растениях и листьях индукцию изучают при помо-
щи инфракрасного газового анализа, манометрии, флуоресцент-
ИНДУКЦИЯ
159
ной спектрометрии и О2-электро-
дов. У водных растений ее мож-
но также исследовать посредст-
вом pH-электрода, так как при
поглощении СО2 из разбавлен-
ного бикарбонатного раствора
последний защелачивается (рис.
7.1). Накопление 14С в хлоро-
пластах внутри листа измеряли
при помощи неводного фракцио-
нирования; как оказалось, про-
цесс накопления 14С характери-
зуется такой же задержкой, как
и выделения кислорода интакт-
ной тканью (рис. 7.2). Индукция
практически не зависит от ин-
тенсивности освещения, однако
на нее сильно влияет температу-
ра. Следовательно, индукция
связана с биохимическими реак-
циями [скорость которых обыч-
но удваивается при каждом уве-
личении температуры иа 10 °C
(Qio=2)] более тесно, чем с ис-
тинно фотохимическими процес-
Время, мин
Рис. 7.2. Индукция в хлоропластах
in vivo [22]. Кривая отражает ки-
нетику фиксации 14СО2 хлороплас-
тами в листе. Хлоропласты были
затем выделены в неводной среде
из лиофилизованной ткани.
сами, независимыми от темпера-
туры (Qio = l) (рис. 7.3). Индукционный период уменьшается,
если проводить повторное освещение после сравнительно не-
большого периода пребывания в темноте (см. рис. 7.1 и 7.9).
Очевидно, освещение позволяет снять какое-то ограничение,
которое ие сразу восстанавливается в темноте.
Источники данных: [1, 4, 5, 7, 9, 22].
7.2. Исторические аспекты
Впервые периоды индукции, которые длились несколько ми-
нут при комнатной температуре, зарегистрировали Остерхаут и
Хасс, проводившие в 1918 г. в Вудс-Хоуле опыты с Ulva, а так-
же Варбург, работавший в 1920 г. с хлореллой. В 1938 г. Блинке
и Скау провели непрерывную запись задержек (лаг-фазы) в вы-,
делении кислорода из листьев Ricinus и в изменениях pH, свя-
занных с началом фотосинтеза в листьях водяной лилии
(рис. 7.4). Стимен-Нильсен также наблюдал индукцию фото-
синтетического выделения кислорода из \Fucus. Пожалуй, наи-
более тщательные и важные исследования осуществили Мак-
Алистер и Майерс, которые измеряли поглощение СО2 листьями
170
ГЛАВА 7
Рис. 7.3. Индукция в листьях пшеницы [28]. Изменения концентрации СО2
измеряли с помощью инфракрасной спектроскопии. В этих опытах началь-
ная задержка не зависела от интенсивности света (Д). Задержка увеличива-
лась при более низкой температуре (5).
пшеницы при помощи спектрографического (инфракрасного) ме-
тода (рис. 7.3).
Источники данных: [14, 15, 28, 29, 30, 36, 46].
7.3. Роль устьиц
В темноте устьица высших растений обычно закрыты и начи-
нают открываться при освещении. Кинетика этого процесса не-
редко очень напоминает кинетику индукции и примерно так же
зависит от изменения температуры и (в какой-то степени) ин-
тенсивности освещения. Вот почему индукционные эффекты, не
ИНДУКЦИЯ
171
Рис. 7.4. А. Индукция в листе Ricinus [14]. Выделение кислорода определяли
при помощи электрода, закрепленного на поверхности листа агаром. Продол-
жительность предшествующих темновых периодов составляла (слева напра-
во) 10 мин, 34 с, 21 с и 13,5 с. Б. Индукция в листе водной лилии [16]. Из-
менения pH, вызванные поглощением и выделением СОг, измеряли с по-
мощью стеклянного электрода. С — свет, Т — темнота.
зависящие от устьиц, трудно отличить от тех эффектов, которые
обусловлены открыванием устьиц, особенно при изучении фо-
тосинтеза в присутствии атмосферной СО2 при помощи инфра-
красной спектроскопии (рис. 7.3 и 7.5). Кроме того, весьма ве-
роятно, что скорость диффузии СО2 из атмосферы к месту кар-
боксилирования влияет на скорость фотосинтеза, а эндодиффу-
зия СО2 зависит от устьичного сопротивления. Поэтому мы
вправе задать вопрос: имеется ли в фотосинтезе высших расте-
1.72
ГЛАВА 7
Рис. 7.5. Поглощение ССЬ листьями шпината, измеренное при помощи инфра-
красной спектроскопии (Делени, неопубликованные данные). Диски листа
(10 см2) подвергали сильному освещению (~6000 фут-свечей) при 20 °C на
воздухе, содержащем 300 м. д. СО2 (скорость потока 1 л/мин). Кривая Д;
диск листа, предварительно подвергнутого сильному освещению, в течение
1 ч. Кривая Б: диск листа, находившегося при комнатном , освещении
(~150 фут-свечей). Так же, как на рис. 7.1—7.4, в данном случае проходит
несколько минут, прежде чем скорость становится максимальной. Кроме
того, видно, что 'скорость возрастает быстрее в диске листа, который подвер-
гался более, сильному освещению. Повторная задержка, которая следует
после кратковременного затемнения, гораздо короче. В данных эксперимен-
тальных условиях первоначальная задержка может быть отчасти вызвана
сравнительно медленным открыванием устыщ.
ний индукционная лаг-фаза помимо той, которая вызвана от-
крыванием уртьиц? На основании сравнительных данных мож-
но сделать вывод о том, что индукция — это неотъемлемое свой-
ство фотосинтеза. Это. положение подтверждается следующими
фактами. Много опытов по изучению индукции было проведено
на водорослях, которые вообще лишены устьиц. При умеренном
освещении устьица открыты довольно широко, и тем не менее
при..переходе от умеренного освещения к насыщающему свету
некоторый индукционный эффект все-таки наблюдается. Не ис-
ключено; что закрытые устьица могут помешать исследованиям
индукции при помощи измерения газового обмена, однако вряд
ли при этом задерживается начало фотосинтеза в листе. Вот по-
чему представляет интерес тот факт, что флуоресценция целых
листьев тоже имеет в самом начале подъем, который может
служить указанием на подавление фотосинтеза (рис. 7.6). На-
конец, индукция была обнаружена при насыщающих концент-
рациях СОг (рис. 7.7), а также в дисках листьев с удаленным
эпидермисом (рис. 7.8) и в опытах с изолированными хлоро-
пластами (рис. 7.9) и протопластами (рис. 7.10).
Источники данных: [10, 29, 36, 38а, 46, 47].
ИНДУКЦИЯ
173
Рис. 7.6. Индукция в листьях пшеницы [29]. Одновременное измерение погло-
щения СОа и флуоресценции выявляет «зеркальный» эффект.
7.4. Отсутствие индукции в фотохимических процессах
Многие из так называемых фотохимических реакций в фото-
синтезе содержат стадии, катализируемые ферментами, и поэто-
му нельзя ожидать, чтобы они были абсолютно фотохимиче-
скими реакциями. Скорости фотохимических процессов могут
различаться между собой на несколько порядков. Хотя в силу
этих причин довольно трудно прийти к каким-то общим выво-
дам, создается впечатление, что такие процессы, как сдвиги pH
(разд. 5.9), фотофосфорилирование (разд. 5.3—5.7) п реакция
Хилла (разд. 5.2), достигают максимальной скорости за не-
сколько секунд после начала освещения (рис. 7.11 и 7.12). Сле-
довательно, индукцию нельзя объяснить тем, что подобные реак-
ции начинаются с запозданием. Это не значит, что в определен-
ных условиях скорость генерации ассимиляционной энергии
(разд. 5.2) не может возрасти по прошествии некоторого вре-
мени; однако, если, она увеличивается, первоначальное огра-
ничение обусловлено скорее всего такими факторами нефотохи-
мической природы, как, например, субоптимальная скорбеть об-
разования ADP и т. д.
Источники данных: [7]
1, мин 12 14 16
0:,мкмоль
о
ГЛАВА
ИНДУКЦИЯ
175
Рис. 7.7. Примеры индукции в листьях. Диски листьев (10 см°) освещали
в маленькой камере (3 мл), содержащей кислородный электрод и 0,2 мл
1,0 М бикарбоната на диске фильтровальной бумаги. Концентрацию СО2 в
газовой фазе в ходе опыта определить нельзя, так как оиа зависит от по-
глощения СОа листом и выделения СО2 из иезабуферепного раствора би-
карбоната. Однако есть основания считать ее насыщающей и достаточной
для поддержания высоких скоростей выделения СО2 [250—350 мкмоль-
•(мг Хл)“*'Ч-1]. Силиконовые отпечатки поверхности листа, сделанные сразу
после освещения по методу Целина, показали, что устьица были закрыты.
Однако при тех концентрациях, которые используются в опыте, закрытые
устьица почти ие препятствуют газовому обмену. Во всех случаях обнару-
жена характерная начальная задержка, более длительная у листьев, которые
находились перед этим не на свету, а в темноте. Еще одна общая черта —
это очень сильное уменьшение задержки после кратковременного темнового
периода, несмотря па возрастание поглощения кислорода после освещения
[7]. /1. Подсолнечник. I— после освещения, II—после темноты. Б. Шпинат.
I — после освещения. II — после темноты. Колебания на рис. А наблюдаются
не всегда. Они были отмечены у некоторых растений в ряде случаев (как
правило, после освещения) и напоминают результаты Ван-дер-Веена (1948)
и других авторов. Вполне возможно, что они отражают сверхкомпенсацпю
под действием контрольных механизмов (разд. 7.13). С — свет, Т — темнота.
Время, мин
Рис. 7.8. Индукция в листьях табака. Условия опыта те же, что и на рис. 7.7.
I — после освещения, III — после темноты, 17 —после темноты, ио при уда-
лении верхнего слоя эпидермиса с листа. С — свет, Т — темнота.
176
ГЛАВА 7
Фиксация СО,, мкмоль
Рис. 7.9. А, Отсутствие индукции в изоли-
рованных хлоропластах гороха после крат-
ковременного пребывания в темноте [38а].
Прежде чем скорость фиксации СОа до-
стигала максимума, должно было пройти
несколько минут. Однако после кратковре-
менного затемнения фиксация СО2 возоб-
новлялась без заметной задержки. По окон-
чании освещения фиксации СО2 в темноте
не было. Б. Сохранение скорости фотосин-
теза после темнового периода. Две одина-
ковые реакционные смеси содержали хлоро-
пласты гороха и ортофосфат (10_®М). Од-
ну из них подвергали краткому затемне-
нию (см. Л), а другую нет. Запись вели
одновременно. С — свет, Т — темнота.
ИНДУКЦИЯ
17?
Рис. 7.10. Индукция в протопластах пшеницы, выделенных из освещенных
(/) и затемненных (II) листьев. Кривые показывают начальную задержку и
конечную скорость при 20 °C. Цифры в скобках выражают скорость в.
мкмоль-(мг Хл)-1-ч-1 (Эдвардс, неопубликованные данные).
7.5. Индукция in vitro
Первые убедительные доказательства индукции в изолиро-
ванных хлоропластах были получены Гиббсом и его сотрудни-
ками. Сначала эти доказательства основывались лишь иа изме-
рениях включения 14СО2. Позднее Уокер и Хилл разработали
метод получения систем хлоропластов, которые обладали высо-
кой скоростью зависящего от СО2 выделения О2 и имели на-
чальную лаг-фазу в условиях in vitro (рис. 7.9 и 7.13). Индук-
ция in vitro характеризуется такой же зависимостью от темпе-
ратуры и относительной независимостью от интенсивности света,
как и ткань, из которой получены хлоропласты (ср. рис. 7.3 и
7.14, А и Б). Следует отметить, что в опытах in vitro степень
индукции вполне может быть завышена из-за несколько иного
соотношения между хлоропластами и окружающей средой по.
сравнению с. нативной клеткой. В листе отношение хлоро-
12—234
178
ГЛАВА 7
Рис. 7.11. Кривая I: изменение pH под действием света. Штриховая линия
показывает изменение pH после добавления воды к хлоропластам, подвергну-
тым осмотическому шоку, в темноте. Она выражает чувствительность (время
ответа) прибора. Нижняя часть кривой отражает реакцию тех же хлоро-
пластов на освещение в присутствии пиоцианина. По-вндпмому, этот реагент
переносит электроны с «верхнего» (восстанавливающего) конца ФС I к некой
точке между ФС I и II. Сдвиг pH связан с транспортом электронов, и, как
видно из рисунка, его можно обнаружить с быстротой, на которую способен
потенциометр. Кривая II: изменение pH под действием феррицианида. Этот
реагент служит акцептором электронов, но не протонов. Поэтому раствор по
мере протекания реакции становится более кислым. К реакционной смеси
был добавлен нигерицин для подавления обратного сдвига pH, непосредст-
венно связанного с транспортом электронов. Видно, что восстановление фер-
рицианида можно обнаружить настолько быстро, насколько это позволяет
сделать чувствительность прибора [7].
пласт/цитозоль равно примерно 1 : 1, и экспорт метаболитов
сразу же оказывает влияние на ближайшее окружение этой
пластиды. При концентрации 100 мкг хлорофилла на 1 мл реак-
ционной смеси (которую в обычных лабораторных сосудах нель-
зя сильно повысить, не ухудшая при этом освещение) отноше-
ние хлоропласт/среда приближается к 1 : 1000, и накопление
экспортируемого метаболита неизбежно занимает больше вре-
мени.
Источники данных: [37, 41].
7.6. Молекулярные механизмы индукции
Исходя из результатов своих первых наблюдений, Остер-
хаут и Хасс высказали предположение, что индукция может
быть обусловлена накоплением промежуточных продуктов или
ИНДУКЦИЯ
170
Рис. 7.12. Отсутствие индукции и реакции Хилла. В присутствии феррициани-
да скорость выделения кислорода достигала максимума за несколько секунд
после начала освещения. Добавление NH<C1 (показано стрелкой) приводит к
возрастанию скорости в результате разобщения. В присутствии 3-фосфогли-
цериновой кислоты (ФГК) задержка была длиннее, чем в случае феррициа-
нида, по все-таки оставалась очень короткой. В данных условиях макси-
мальная скорость фиксации СО2 (приведено для сравнения) была отмечена
лишь спустя несколько минут. Все три опыта проводились с одними н теми
же хлоропластами, но в опыте с феррицианидом их подвергли осмотическому
шоку в реакционном сосуде [5].
активацией ферментов под действием света. В паши дни иссле-
дователи прибегают к тем же объяснениям, хотя все более и
более вероятным представляется взаимодействие этих двух ме-
ханизмов. Так, постепенное накопление важного промежуточно-
го продукта может также вызвать аллостерическую активацию
фермента, катализирующего синтез или катаболизм этого про-
дукта.
Формулируя более точно гипотезу Остерхаута — Хасса, Ра-
бинович обращает внимание иа «дефицит акцептора СОе в на-
чале светового периода и автокаталитическое изменение его
концентрации на свету до уровня, необходимого для протекания
фотосинтеза со скоростью, которая отвечает преобладающей
интенсивности света». Эта формулировка учитывает важный
принцип автокатализа (разд. 6.3), а также тот факт, что период
ускорения отражает ситуацию, в которой триозофосфаты в ос-
12*
180.
'ГЛАВА 7
Время, мин
Рис. 7.13. Первая полярографическая запись СО2-зависимого выделения О2
изолированными хлоропластами. Сравните с рис. 7.9, А и обратите внимание
на сходство кинетики выделения О2 и фиксации СО2. Поглощение кислорода
в темноте представляет собой в основном артефакт и может быть вызвано
окислением компонентов реакционной смеси. (Подробное объяснение см. в
14Ц.) ..
новном включаются обратно в цикл, а не превращаются в крах-
мал или экспортируются в цитозоль.
Источники данных: [4, 30]
7.7. Вклад автокатализа
В предыдущем разделе мы уже говорили о том, как трудно
разграничить эффекты концентрации субстратов и активации
ферментов. Однако есть все основания полагать, что изолиро-
ванные хлоропласты обычно содержат слишком мало промежу-
точных продуктов ВПФ-цикла, чтобы обеспечить максимальную
скорость фотосинтеза. Лаг-фаза — это и есть то время, за кото-
рое достигается наивысшая стационарная концентрация проме-
жуточных продуктов ВПФ-цикла.
Источники данных: [1, 5, 9]
Попробуем теперь кратко изложить то, что мы знаем о со-
бытиях, происходящих при индукции.
а) Если скорость , фотосинтеза стала максимальной, повторная
краткая инкубация в темноте либо совсем не вызывает задерж-
Рис. 7.14. Л. Индукция в выделении кислорода изолированными хлоропластами. Как видно из рисунка, задержки не
зависят от интенсивности света, но зависят от температуры. Один электрод находился при температуре 5 °C, а дру-
гой — при 25 °C. Измерения при ярком освещении проводили одновременно непосредственно перед измерениями при
слабом освещении- [42]. Б. Индукция в фиксации СО2 изолированными хлоропластами. Видно, что она не зави-
сит от интенсивности света, но зависит от температуры. I-—сильный свет, 25 °C, II-—слабый свет, 25 °C, III-—
сильный свет, 10 °C, IV-—слабый свет, 10 °C [11].
Время,мин
ИНДУКЦИЯ
00
o
ИНДУКЦИЯ
183
ки, либо приводит к гораздо меньшей лаг-фазе, чем в первый
раз (рис. 7.1, 7.4, 7.7, 7.8, 7.9) [37—39, 41, 42]. То же самое про-
изошло бы при активации катализаторов светом, если бы ие
следующее обстоятельство.
б) Если поместить хлоропласты в среду, в которой до этого
осуществляли фотосинтез другие хлоропласты (а потом были
удалены), лаг-фаза сильно сократится (рис. 7.15). Очевидно,
в среде накопилось какое-то вещество, способное проникать в
новые хлоропласты и сокращать их лаг-фазу. Конечно, не ис-
ключено, что оно воздействует на ферменты, связанные с фото-
синтезом, однако в данном случае подобный эффект должен
быть вторичным (т. е. активацию вызывает накапливаемый про-
дукт) [12].
в) Как показывает анализ, в среде, содержащей освещенные
хлоропласты, появляются различные промежуточные продукты
ВПФ-цикла (главным образом триозофосфаты). Если вносить
каждое из этих соединений в чистом виде в среду со свежими
хлоропластами, период задержки сократится (рис. 7.15). Эффек-
тивность этих соединений различна, по-видимому, из-за того,
что они с разной скоростью проникают внутрь хлоропласта
(гл. 8). Однако те соединения, которые проникают легче всего
(триозофосфаты и ФГК), наиболее эффективно уменьшают за-
держку. Эффективность таких медленно поступающих в хлоро-
пласт соединений, как пеитозомонофосфаты, можно повысить,
инкубируя их с хлоропластами перед освещением. Все соедине-
ния обладают каталитическим действием (рис. 7.15,5) [6, 12,
13, 16, 38, 40, 41]
Рис. 7.15. А. Сокращение индукционного периода под действием соединений,
выделяемых освещенными хлоропластами, а также при добавлении проме-
жуточных продуктов цикла. Кривая IV: СОа-зависимое выделение Сц хлоро-
пластами, полученными из листьев, которые перед этим находились 12 ч в
темноте. Кривая III: то же, что и IV, с одним отличием: хлоропласты нахо-
дились в среде, из которой до этого были удалены центрифугированием дру-
гие хлоропласты, инкубированные в ней 30 мин на свету. Кривая I: то же,
что и IV, но в присутствии РубФ. Кривая II: то же, что и IV, но при добав-
лении ФГК. Обратите внимание на то, что соединения, выделяемые освещен-
ными хлоропластами, сокращают индукционный период почти так же, как
и добавленная ФГК. Следовательно, этот продукт играет важную роль в
сокращении индукционного периода в хлоропластах. РубФ также уменьшает
индукцию, ио кинетика процесса в его присутствии больше напоминает ки-
нетику в случае хлоропластов, полученных из предварительно освещенных
листьев. Добавление ФГК в самом начале подавляет индукционные процес-
сы [7]. Б. Сокращение индукционного периода в присутствии каталитических
количеств ДГАФ. Горизонтальные черточки на кривых соответствуют кисло-
роду, который образовался бы в результате количественного превращения
ДГАФ в ФГК (в отдельных реакциях ВПФ-цикла) и последующего восста-
новления обратно до триозофосфата [25а].
ср
Рис. 7,16. Изменение содержания АТР и возрастание
концентрации РуБФ во время индукции [26].
ГЛАВА 7
Свет бремя, мин
Рис, 7,17. Возрастание концентрации ФГК и гексозо-
монофосфатов (ГМФ) в строме хлоропластов во время
индукции [26]. (В этом опыте и на рис. 7.16 длитель-
ность индукции составляла около 3 мин.) ПМФ — пен-
тозомонофосфаты.
ИНДУКЦИЯ
186
Рис. 7.18. Сокращение индукционного периода в хлоропластах в результате
предварительного освещения листьев, из которых они были получены (ср. с
рис. 7.8 и 7.10). Кривая I: СО2-зависимое выделение О2 хлоропластами, по-
лученными из листьев шпината, которые перед этим освещались 30 мин бе-
лым. светом (2000 фут-свечей). Кривая II: то же, что и I, но для хлоро-
пластов, которые были выделены одновременно из неосвещенных листьев.
Все листья собирали после пребывания в темноте в течение 12 ч и смеши-
вали перед экстракцией [7].
г) Оказалось, что некоторые соединения, прежде всего ФГК,
а также гексозофосфаты (рис. 7.17) и РуБФ (рис. 7.16), снача-
ла присутствуют в хлоропластах в небольших количествах. По
окончании индукции их содержание возрастает [26].
д) Ортофосфат, способствующий экспорту метаболитов, по-види-
мому, не вызывает существенного подавления активации фер-
ментов цикла под действием света (разд. 7.8) [2, 8, 17].
е) Так же как и в целых тканях, индукция удлиняется при низ-
кой температуре, ио почти не зависит от интенсивности света
[12, 42].
ж) Кроме того, индукция занимает гораздо больше времени в
случае хлоропластов, выделенных из ткани, которая находилась
в темноте (рис. 7.18). Этот факт можно объяснить как деграда-
цией важных промежуточных продуктов цикла, так и инактива-
цией ферментов в темноте [7, 17, 38, 41].
186
ГЛ AHA 7
7.8. Активация ферментов под действием света
Итак, мы убедились в том, что индукция связана с автока-
талитическим накоплением промежуточных продуктов ВПФ-
И'нкла (разд. 7.7). Это не означает, что активация ферментов,
участвующих в фотосинтезе, под действием света ие играет ни-
какой роли. Более подробно мы рассмотрим этот вопрос в гл. 9.
Хотелось бы обратить внимание на то, что не следует смеши-
вать активацию ферментов светом с появлением под действием
освещения условий, благоприятных для катализа. Например, на
свету pH стромы будет более щелочным, чем в темноте, что
способствует действию ферментов с более щелочными оптиму-
мами pH. Кроме того, при высоких значениях pH ферменты мо-
гут переходить из менее активной формы в более активную.
Хотя эти эффекты по своей природе совершенно различны, они
могут приводить к одному и тому же конечному результату.
Если же оба проявляются одновременно, их действие может
стать аддитивным. Однако на этом основании довольно трудно
объяснить практически полное отсутствие фотосинтетической
ассимиляции углерода сразу (в течение 1—2 мин) после начала
освещения. При прочих равных условиях нулевая скорость
должна говорить о том, что ферменты полностью лишены ак-
тивности. Тем не менее, хотя активность может сильно меняться
при освещении, некоторые ферменты (например, РуБФ-карб-
Таблица 7.1. Скорости СОг-зависимого выделения О2 и влияние освещения на
активность РуБФ-карбоксилазы из протопластов пшеницы1
Номер ‘опыта Выделение кислорода Активность РуБФ-карбо- ксилазы Отношение между актив- ностями РуБФ-карбокеи- лазы на свету и в темноте
в темноте на свету
1 139 182 223 1,25
2 152 161 186 1,16
3 167 174 206 1,18
4 136 150 207 1,38
5 117 165 166 1,01
6 115 144 141 0,98
1 Активность РуБФ-карбоксилазы определяли в протопластах поело 6-ммиутной ин-
кубации в темноте или на свету в тех же условиях, что и при измерении выделения
кислорода (2,5 мМ NaHCO3, pH 7,6). В опытах 5 и 6 растения пшеницы перед тем,
как из них выделяли протопласты, находились в темноте в течение 24 ч. Скорости вы-
ражены в мкмоль-(мг Хл)-1-*!— Заметной активации не было и в опыте, в котором кон-
центрация СО2 была такая же, как в воздухе. Начальные скорости выделения кислорода
протопластами пшеницы во время индукции чрезвычайно низкие. Скорости, которые на-
блюдаются в присутствии препарата РуБФ-карбоксилазы, соизмеримы с конечными ско-
ростями фотосинтеза. Однако они намного превышают скорости, которые потребовались
бы для карбоксилирования во время индукции [32].
индукция
187
140
100
во
60
40
20
140
7 120
X 100
3 80
;; 60
S 40
i 20
800
600
400
200
1200
1000
800
600
400
200
1200 < »7
St 2 з
юоо £ *_•
2400
140
120
Л 100
* 80
~ 60
§ 40
20
2000
- 400
6 o 1 2 3 4 5 6
Время, мин
1600
1200
800
a £
э 5
A *
Время, мик
РуБФ Kap69KSIVIa?S, '|мг Хи)”1'
s
Рис. 7.19. А— Д. Этот опыт показывает, что между проявлением полной ак-
тивности ферментов (в результате активации светом) и максимальной ско-
ростью фотосинтеза (вследствие автокатализа) существует вполне опреде-
ленный промежуток времени. Приведены зависимости скоростей фотосинтеза
в хлоропластах пшеницы и активации под действием света следующих фер-
ментов: NADP-зависимой глицеральдегидфосфатдегидрогеназы (А), фосфо-
рибулокиназы (5), фосфоглицераткиназы (В), РуБФ-карбоксилазы (Г) и
ФБФ-азы (Д). Обозначения: кружки — выделение О2, квадраты—активность
ферментов, светлые кружки и квадраты — в присутствии 10 мМ NaHCO3,
темные кружки и квадраты, а также штриховая линия (фиксация ИСО2 на
рис. Б и Д) —в присутствии 10 мМ ФГК и 10 мМ NaHCO3. При измерении
активации светом одновременно определяли активность в пробах в отсутст-
вие дитиотрейтола. Каждая кривая показывает, что скорости фиксации СО2
или выделения О2 достигают максимума в присутствии любого из ферментов
раньше, чем произойдет заметное выделение О2 в смесях, содержащих толь-
ко СО2 [25а].
оксилаза) проявляют гораздо более высокую активность в тем-
ноте, чем та, которая требуется для поддержания низкой скоро-
сти фотосинтеза во время индукции (табл. 7.1).
Совершенно ясно, что in vivo ряд других ферментов в темно-
те инактивируется. Поэтому следует ожидать, что их реактива-
ция на свету влияет иа индукцию. Остаточная индукция (30 с
или даже меньше), которая наблюдается в присутствии высоких
188
ГЛАВА 7
концентраций триозофосфата (рис. 7.15, Б), может быть вызва-
на именно этой причиной. И все-таки активация под действием
света, например ФБФ-азы, происходит слишком быстро'
(рис. 7.19), чтобы ею можно было объяснить более длительные-
задержки в начале фотосинтеза, которые наблюдаются во мно-
гих случаях. За 30 с освещения в подобного рода эксперимен-
тах активность ФБФ-азы и других ферментов ВПФ-цикла
достигает достаточно высокого уровня, чтобы обеспечивать мак-
симальный фотосинтез. Тем не менее этот потенциал не реали-
зуется еще 2—3 мни, если не добавить такие экзогенные мета-
болиты, как ФГК или ДГАФ.
Источники данных: [20, 21, 23, 24, 25а, Ь, с, 27, 32, 33, а, Ь].
7.9. Влияние интенсивности света и температуры
Раньше мы уже говорили о том (разд. 7.1 и 7.5), что индук-
ция почти не зависит от интенсивности света, ио начальная за-
держка удлиняется по мере понижения температуры (рис. 7.3 и
7.14). С чисто теоретической'точки зрения этот факт полностью
согласуется с важной ролью, которую играет концентрация про-
межуточных продуктов ВПФ-цикла. Так, например, если бы ско-
рость фотосинтеза падала при уменьшении интенсивности света,
это привело бы к снижению стационарной концентрации суб-
стратов, необходимой для поддержания новой, более низкой
скорости; соответственно сократилось бы и время, за которое
достигается эта новая скорость. Точно так же, если бы скорость
была прямо пропорциональна концентрации субстратов, то дли-
тельность лаг-фазы ие изменялась бы. И наоборот, при более
низкой температуре понадобилось бы больше субстрата для
поддержания данной скорости, и время, за которое достигался
бы этот более высокий уровень субстрата, увеличивалось бы.
Предположим, например, что в стационарном состоянии паде-
ние температуры на 10 °C снижает вдвое скорость функциони-
рования ВПФ-цикла. По-видимому, концентрация промежуточ-
ных продуктов, обеспечивающая эту более низкую скорость, ос-
танется неизменной, но время, необходимое для создания такой
концентрации в смеси, освещенной с самого начала при более
низкой температуре, возрастет. Действительно, удлинение лаг-
фазы при низких температурах порой бывает удивительно боль-
шим, а конечная скорость фотосинтеза очень высокой (рис. 7.20).
Первое явление можно отчасти объяснить высокими значения-
ми Qio, которые характерны для фотосинтеза при низких тем-
пературах. Так, для температур ниже 15 °C при повышении тем-
пературы на 10 °C скорость может возрасти гораздо больше, чем
вдвое (величина Q10=2 типична для многих процессов метабо-
лизма). Предполагают, что это явление обусловлено автоката-
литическим характером ВПФ-цикла (разд. 6.3), а также тем;
ИНДУКЦИЯ
Рис. 7.20. Зависимость индукции от температуры. Длительность индукцион-
ного периода и максимальная скорость обнаруживают аномальные отклоне-
ния при низкой температуре. Один электрод находился при температуре
25 °C, другой — при 5 °C. Несмотря на то что индукционный период значи-
тельно сокращался после кратковременного пребывания в темноте, достиже-
ние скорости, близкой к максимальной, также занимало больше времени,
чем обычно, при обеих температурах [42].
что повторное включение продукта в цикл будет увеличиваться
вместе с ростом температуры. Таким образом, ферменты, ката-
лизирующие эти процессы, будут функционировать и при б.олеё
высоких концентрациях субстрата, и при более благдпрйятной.
температуре. Можно ожидать, что при снижении температуры
с 10 до 5 °C индукционный период возрастет-больше, чем следо-
вало бы предполагать на основании температурной зависимости
большинства процессов метаболизма. Сравнительно высокие
стационарные скорости при низких температурах могут быть
вызваны накоплением больших количеств субстрата, чем при
более высоких температурах (низкая температура в какой-то
степени как бы компенсируется накоплением больших количеств
субстрата).
Источники данных: [11, 34, 42].
7.10. Индукция и ортофосфат
Еще в первых исследованиях, проведенных Арионом и его
сотрудниками на изолированных хлоропластах, было показано,
Рис. 7.21. Индукция, автокатализ и ортофосфат. А Подавление ортофосфатом фотосинтетической ассимиляции углерода
в изолированных хлоропластах шпината. Показана кинетика СОа-зависимого выделения О2 в присутствии возрастающих
концентраций неорганического фосфата. Характерный индукционный период короче всего в отсутствие добавленного
ортофосфата, но при этом скорость фотосинтеза вскоре снижается, как и на рис. 8.9. При небольшом увеличении
концентрации неорганического фосфата индукционный период удлиняется, а скорость фотосинтеза либо повышается, либо
не меняется. При более высоких концентрациях фосфата индукция увеличивается, а скорость фотосинтеза уменьшает-
ся. Цифры без скобок около каждой кривой обозначают концентрацию ортофосфата (в мМ), а цифры в скобках —
скорость в мкмоль-(мг Хл)-’-ч“’ при 20°С. Штриховая линия дает представление о том, как внесение 10 мМ пирофосфа-
та снимает подавление, вызванное 5 мМ ортофосфатом. Эти до сих пор неопубликованные данные получены примерно
в тех же условиях, что и в более ранней работе Кокберна и др. [181. Только в данном случае опыты проводились в среде
без экзогенного Alg2 f в присутствии достаточных количеств каталазы, для того чтобы свести к минимуму вторичные эф-
фекты, связанные с образованием перекисей. 5. Влияние концентрации ортофосфата на скорость СО2-зависимого выде-
ления О2 х.торопластами гороха. Слева от пика отложены лимитирующие концентрации ортофосфата; по-видимому, этот
параметр оказывает влияние на отношение [ATP]/[ADP] (разд. 7.13). Справа от пика находятся ингибирующие кон-
центрации ортофосфата, которые вызывают избыточный экспорт триозофосфатов при помощи переносчика ортофосфата
(разд. 8.23) [33].
ИНДУКЦИЯ
191
что фотосинтез in vitro подавляется высокими концентрациями
ортофосфата. Позднее стало ясно, что кинетика подавления не-
органическим фосфатом имеет сложный характер и по крайней
мере отчасти связана с удлинением индукционного периода
(разд. 8.14). Так, если повысить концентрацию неорганического
фосфата, начальная задержка тоже увеличится (рис. 7.21). При
более или менее низких концентрациях неорганического фосфа-
та удлинение индукционного периода не сопровождается сниже-
нием скорости фотосинтеза. Однако по мере дальнейшего повы-
шения концентрации Pi и возрастания задержки конечная ско-
рость фотосинтеза падает.
Источники данных: [7, 17].
7.11. Снятие подавления, вызванного ортофосфатом,
промежуточными продуктами ВПФ-цикла
При концентрациях неорганического фосфата приблизитель-
но 10 мМ удлинение индукционного периода и снижение скоро-
сти могут стать настолько большими, что фотосинтез почти пол-
ностью прекратится (разд. 8.14). Однако это подавление можно
затем в значительной степени спять, если добавить некоторые
промежуточные соединения ВПФ-цикла. Оба триозофосфата и
ФП< почти сразу снимают подавление, в то время как действие
Р5Ф проявляется лишь после довольно длинной лаг-фазы.
Фруктозобисфосфат также снимает подавление не сразу, по это
вызвано, вероятно, тем, что он должен сначала (еще до про-
никновения в хлоропласт) превратиться в триозофосфат. В при-
сутствии трпозофосфатов и Р5Ф кинетика снятия подавления
одна и та же как для фиксации СО2, так и для выделения О2
(рис. 7.22). Следовательно, в данном случае ВПФ-цикл функ-
ционирует полностью. Если с самого начала добавить одновре-
менно ФГК и неорганический фосфат в высокой концентрации,
то О2 выделяется немедленно, а фиксация СО2 увеличивается
постепенно. Это свидетельствует о том, что некоторое количе-
ство ФГК может восстанавливаться независимо от фиксации
СО2. Если же добавить ФГК не сразу, то скорость выделения
О2 после добавления ФГК увеличится и немедленно начнется
фиксация СО2 (рис. 7.22). Очевидно, высокая концентрация не-
органического фосфата каким-то образом нарушает нормальное
функционирование цикла, а ФГК снимает этот эффект
(разд. 7.14). Такие соединения, как РуБФ, неспособные прони-
кать через мембрану хлоропласта, не снимают подавления, выз-
ванного неорганическим фосфатом (гл. 8).
Источники данных: [18, 33].
192
ГЛАВА 7
Время,мин
Рис. 7.22. Снятие подавления, вызванного ортофосфатом, промежуточными
продуктами ВПФ-цикла. Выделение О2 показано сплошными линиями, фик-
сация СОз — штриховыми. Каждая реакционная смесь сначала содержала
достаточное количество ортофосфата, для того чтобы индукция продолжа-
лась неограниченное время. Стрелками обозначено добавление ФГК, Р5Ф и
ДГАФ [18].
ИНДУКЦИЯ
193
7.12. Природа подавления под действием ортофосфата
Ортофосфат является конкурентным ингибитором РуБФ-кар-
боксилазы, чем отчасти и может объясняться подавление им
фотосинтеза. Напомним, что индукция отражает время, за кото-
рое происходит автокаталитическое накопление промежуточных
продуктов, и что добавление промежуточных соединений, спо-
собных проникнуть внутрь хлоропласта, сокращает индукцион-
ный период (разд. 7.7). Если это так, то мы вправе предполо-
жить, что неорганический фосфат удлиняет индукцию, затруд-
няя автокатализ (гл. 6). На основании этого предположения
было сформулировано понятие о переносчике неорганического
фосфата и его свойствах (разд. 8.23). Согласно современным
представлениям, действие неорганического фосфата сводится к
тому, что ои обеспечивает активный экспорт триозофосфатов из
цикла. Взаимосвязь между концентрацией неорганического
фосфата, транспортом метаболитов и индукцией показана па
рис. 7.23. Так, во время индукции триозофосфаты почти не уда-
ляются из цикла и обеспечивают автокаталитическое ускорение.
В стационарном состоянии происходит экспорт триозофосфатов
в обмен иа неорганический фосфат снаружи хлоропласта. Если
повысить концентрацию неорганического фосфата, то благода-
ря активному транспорту индукционный период удлиняется, и
поэтому неорганический фосфат окажет гораздо более сильное
подавление, чем в том случае, когда его добавляют после до-
стижения стационарного состояния. [Следует отметить, что
предполагаемые взаимосвязи между автокатализом, транспор-
том л переносчиком неорганического фосфата многочисленны
и разнообразны. Изучение каждого из этих явлений способству-
ет выяснению природы двух других. Важную роль здесь играет
многолетнее сотрудничество между лабораториями Хебера
(Дюссельдорф), Хельдта (Мюнхен) и Уокера (Лондон и Шеф-
филд). Однако не следует считать, что представление о контро-
лируемой проницаемости основывалось па прямых измерениях.
В одном из обзоров, опубликованных в 1970 г., говорится о том,
что «непосредственный активный обмен между ортофосфатом
(снаружи хлоропласта) и фосфатом сахара (внутри хлоропла-
ста) мог бы объяснить подавление фотосинтеза ортофосфатом
и снятие этого подавления под действием фосфата сахара».]
Как мы увидим позднее, взаимосвязь между неорганическим
фосфатом, триозофосфатом и скоростью фотосинтеза может
быть гораздо сложнее, чем это дано в приведенной выше схеме.
Так, неорганический фосфат, находящийся снаружи хлоропла-
ста, способен не только ускорять экспорт триозофосфатов из
хлоропласта, ио и препятствовать их проникновению в эту орга-
неллу. Нередко между процессами внутреннего метаболизма и
13—234
194
ГЛАВА 7
МимбранЫ
Рис. 7.23. Схематическое изображение взаимосвязи между индукцией, кон-
центрацией ортофосфата и транспортом метаболитов. При нормальных кон-
центрациях экзогенного ортофосфата триозофосфаты, образующиеся в ВПФ-
цикле, могут: а) включаться обратно в цикл, б) превращаться далее в крах-
мал и другие соединения в строме, в) экспортироваться из хлоропласта. При
включении света наблюдающийся индукционный период будет отражать
время, за которое благодаря автокаталитическому включению концентрация
промежуточных продуктов достигнет стационарного уровня, определяемого
преобладающей интенсивностью света. В течение этого периода быстрое ис-
пользование промежуточных продуктов в цикле будет способствовать их
включению в цикл. По мере достижения стационарного уровня относитель-
ный избыток триозофосфатов приведет к тому, что они все больше и больше
будут экспортироваться из хлоропласта, а также запасаться внутри его в
виде крахмала. При низких концентрациях экзогенного ортофосфата
(в экспериментальных условиях этого можно добиться в некоторых тканях
листа, давая им маннозу [33]) экспорт трнозофосфатов снижается, а их из-
быток идет на синтез крахмала. При высоких концентрациях экзогенного
ортофосфата усиленный экспорт приведет к уменьшению количества триозо-
фосфатов, которые могут вновь включаться в цикл, и удлинению индукцион-
ного периода свыше обычных 1—3 мин (рис. 7.21). Существующие in vivo
менее резкие колебания концентраций ортофосфата регулируют равновесие
между накоплением продуктов внутри хлоропласта и их экспортом в цито-
плазму.
экспорта наблюдается конкуренция за триозофосфат. Сначала
количество триозофосфата, предназначенного для экспорта, об-
ратно пропорционально эффективности альдольной конденсации
(в которой триозофосфат превращается в фруктозобисфосфат,
о чем уже было сказано в разд. 6.11). Позднее, однако, триозо-
ИНДУКЦИЯ
195
фосфат начинает использоваться также в реакциях, катализи-
руемых транскетолазой (разд. 6.11). В некоторых случаях как
трнозофосфат, так и ФГК, находящиеся снаружи хлоропласта,
способны подавлять фотосинтез в изолированных хлоропластах.
Если концентрация триозофосфата в окружающей среде доста-
точно высока, то он будет подавлять выделение Ог, а не фикса-
цию СОг. По-видимому, подобное действие триозофосфата обус-
ловлено его обменом с ФГК, которая в противном случае вос-
станавливалась бы внутри хлоропласта. Как ФГК, так и трио-
зофосфат вызывают снижение концентрации неорганического
фосфата в хлоропласте.
Источники данных: [3, 9, 20, 25, 31].
7.13. Индукция в выделении кислорода
и роль ФГК
Одно время принято было считать, что в процессе индукции
накапливаются пентозомонофосфаты. Как было показано Лилли
и сотр. [26], во время лаг-фазы возрастает содержание рибуло-
зобисфосфата, но особенно сильно увеличивается количество
ФГК и гексозомонофосфатов (рис. 7.17). В среде триозофосфа-
та накапливается гораздо больше, чем ФГК, но его уровень в
строме остается низким. Как показали опыты с реконструиро-
ванными хлоропластами, ФГК совсем или почти совсем не вос-
Рис. 7.24. Добавление РубФ в реконструи-
рованную систему хлоропластов приво-
дит к выделению кислорода в результате
последовательности реакций, показанных на
рис. 7.25. Однако в этом процессе большое
количество АТР уходит на превращение
РубФ в РуБФ, а возникающий при этом
ADP блокирует превращение ФГК в ДФГК
(рис. 7.25). Данный опыт показывает, что
в присутствии добавленного источника
АТР (который способствует эндогенному
фотофосфорилированию) выделение кисло-
рода происходит, несмотря на присутствие
РубФ или добавление ADP. Напротив,
само по себе фотофосфорилирование не
может компенсировать неблагоприятное
(слишком низкое) отношение [ATP]/[ADP],
которое обусловлено сначала присутствием
РубФ, а затем добавлением ADP (Уокер,
неопубликованные данные).
13*
196
ГЛАВА 7
Ру ЗФ
—7^^'РуБТ ' СО;
АТР ADP i
тгк
©ФГК
ЯРГЛ
Рис. 7.25. Схема, отражающая подавление образования кислорода в результа-
те использования АТР в реакции превращения Ру5Ф->-РуБФ и ингибиро-
вания реакции ФГК->-ДФГК образующимся в первой реакции ADP.
станавливается до триозофосфата в присутствии пентозомопо-
фосфата или фруктозобисфосфата. Это вызвано тем, что при
превращении рибулозо-5-фосфата под действием своей киназы
в РуБФ [уравнение (6.27) в разд. 6.11] используется очень боль-
шое количество АТР (разд. 9.6). При этом в свою очередь соз-
дается неблагоприятное соотношение между концентрациями
АТР и ADP, за счет чего подавляется восстановление ФГК
(рис. 7.24) отчасти, вероятно, путем аллостерической регуля-
ции, а отчасти путем воздействия на равновесие ДФГК^ФГК.
Следовательно, восстановление ФГК и связанное с ним выделе-
ние кислорода будут подавлены до тех пор, пока содержание
ФГК остается низким, а концентрация РуБФ высокой (рис. 7.25).
Накоплению ФГК, которое требуется для преодоления неблаго-
приятного соотношения между АТР и ADP, должно способство-
вать следующее обстоятельство. При щелочных значениях pH,
которые характерны для освещенной стромы, ФГК песет на
один отрицательный заряд больше, чем триозофосфат, и по-
этому ее экспорт происходит с меньшей скоростью по сравне-
нию с триозофосфатом. Действительно, в строме во время ин-
дукции содержание триозофосфата почти не возрастает, хотя
он и накапливается в окружающей среде. Кроме того, концент-
рация ФГК внутри хлоропласта почти в 10 раз выше, чем сна-
ружи.
Скорость восстановления ФГК в экстрактах хлоропластов
зависит от отношения [ФГК][АТР]/[АЮР] при физиологических
значениях концентраций этих метаболитов. В процессе РуБФ-за-
висимого выделения кислорода реконструированными хлоропла-
стами задержка, которую вызывает добавление ADP (рис, 7.26),
ИНДУКЦИЯ
197
Рис. 7.26. Моделирование индукции в реконструированной системе хлоро-
пластов. При использовании РуБФ в качестве субстрата индукционный пе-
риод очень короткий (несмотря на необходимость накопления ФГК), но до-
бавление ADP приводит к его существенному увеличению. Каждая реакци-
онная смесь имела следующий состав: каталитические количества ADP и
NADP+, а также NaHCO3, MgCl2, ферредоксин, разрушенные хлоропласта
и экстракт стромы. После восстановления добавленного NADP+ в среду
вносили 1 мкмоль РуБФ. В опыте, иллюстрируемом нижней кривой, добав-
ляли 1 мкмоль ADP до внесения РуБФ, в результате чего проходило
3 мин, прежде чем скорость выделения кислорода достигала максимума.
Цифры около кривых обозначают скорость выделения кислорода [в мкмоль-
.(МГ Хл)-1-!-"] [33].
может быть почти полностью устранена, если существенно по-
высить это отношение, добавляя ФГК (рис. 7.27) или АТР
(рис. 7.28). В присутствии NADPH происходит восстановление
ДФГК до триозофосфата, и стационарная концентрация ДФГК
должна быть поэтому низкой. Таким образом, скорость восста-
новления ФГК определяется концентрациями АТР, ADP и са-
мой ФГК- На рис. 7.29 и 7.30 приведены результаты опыта с си-
стемой реконструированных хлоропластов, в котором моделиро-
Б
Время, мин
Рис. 7.27. Выделение кислорода реконструированной системой хлоропластов
в присутствии ФГК- Состав реакционной смеси: каталитические количества
ADP и NADP+, а также MgCl2, ферредоксин, разрушенные хлоропласта и
экстракт стромы. А. Снятие возрастающими концентрациями ФГК подавле-
ния под действием ADP. Стрелками показано добавление ADP до конечной
концентрации 1 мМ. Задержка под действием ADP значительно сокращается
при более высоких концентрациях ФГК (верхняя кривая). Б. Снятие при по-
мощи АТР подавления под действием ADP. Стрелками обозначено добавле-
ние АТР до конечной концентрации 5 мМ (верхняя кривая) и внесение ADP
до концентрации 1 мМ (обе кривые). Индукционный период сокращается
в присутствии АТР. Цифры около кривых показывают скорость выделения
кислорода [в мкмоль. (мгХл)-1'Ч-1].
ИНДУКЦИЯ
199
Рис. 7.28. Влияние отношения [ATP]/[ADP] на скорость ФГК-зависимого вы-
деления кислорода реконструированными хлоропластами при различных кон-
центрациях ФГК. Состав реакционной смеси: каталитические количества
NADP+, а также MgCl2, ферредоксин, разрушенные хлоропласты и экстракт
стромы. АТР, ADP и ФГК вносили одновременно после восстановления
NADP. Скорость выделения кислорода зависела от отношения [ATP]/[ADP] и
от концентрации ФГК [33].
вались условия внутри интактного хлоропласта во время индук-
ции. После того как восстанавливался весь добавленный
NADP+, в среду одновременно вносили АТР, ADP и ФГК, что-
бы получить различные концентрации ФГК. и различные отно-
шения ATP/ADP при постоянной общей концентрации аденино-
вых нуклеотидов (1,5 мМ). Как видно из рис. 7.29, максималь-
ная скорость выделения кислорода зависела от концентрации
ФГК и от отношения [ATP]/[ADP], По мере снижения этого от-
ношения для достижения данной скорости выделения кислорода
требовались более высокие концентрации ФГК. На рис. 7.30
изображена зависимость между временем достижения макси-
мальной скорости и концентрацией ФГК- Если все адениновые
нуклеотиды добавляли в виде АТР (ATP/ADP>100), лаг-фаза
200
ГЛАВА 7
Рис. 7.29. Зависимость индукционного периода от концентрации ФГК и отно-
шения [ATP]/[ADP], Чем ниже эта концентрация и меньше это отношение,
тем длиннее индукция (время, за которое достигается максимальная ско-
рость фотосинтеза) [33].
составляла меньше 0,2 мин. Она удлинялась при уменьшении
отношения [ATP]/[ADP], но при заданном отношении зависела
от концентрации ФГК. Таким образом, как скорость выделения
кислорода, так и время, за которое достигалась эта скорость,
зависели от соотношения между концентрациями ФГК, АТР и
ADP.
Регуляция восстановления ФГК путем изменения концентра-
ций ADP представляет собой один из нескольких регуляторных
механизмов (гл. 9). Если бы пентозомонофосфаты накаплива-
лись, отток АТР обеспечил бы снижение скорости образования
этих соединений из ФГК до тех пор, пока не установится нор-
мальное стационарное соотношение. Некоторое ограничение син-
теза пентозофосфатов имело бы положительный эффект в том
отношении, что благодаря ему снизилась бы скорость фотоды-
хательного метаболизма. Экспорт гексозомонофосфатов из хло-
ропласта несколько затруднен (разд. 8.15), что, по-видимому,
помогает регулировать экспорт соединений н их использование
внутри хлоропласта. Поскольку переносчик неорганического
фосфата обладает высоким сродством к ФГК (разд. 8.23), при
добавлении последней к хлоропластам период индукции в вы-
ИНДУКЦИЯ
201
Рис. 7.30. Изменения активности РуБФ-карбоксилазы хлоропластов при инку-
бации с СО2. Хлоропласты инкубировали при pH 7,8 и температуре 25 °C в
темноте (яияскяя кривая) или на свету (верхняя кривая) и определяли в них
активность РуБФ-карбоксилазы [24].
делении О2 сильно сокращается. Согласно общепринятой точке
зрения, ФГК действует в этом случае как природный окисли-
тель Хилла, проникая внутрь хлоропласта и восстанавливаясь
там до триозофосфата. Однако, как показали проведенные не-
давно опыты, ФГК способна поступать в хлоропласты только
в виде ФГК2“, и поэтому при щелочных значениях pH, при ко-
торых преобладает ФГК:,_, ФГК-зависимого выделения кисло-
рода почти ие происходит. Тем не менее даже при таких значе-
ниях pH ФГК может сокращать индукционный период или
снимать подавление, вызванное неорганическим фосфатом
(разд. 8.7), ингибируя переносчик неорганического фосфата
(разд. 8.23) .
Источники данных: [19, 26, 33, 35, 45].
7.14. Индукционные явления в фиксации СОа
Если подвергнуть изолированные хлоропласты осмотическо-
му шоку в растворе, содержащем Mg2+, сульфгидрильный вос-
становитель (например, дитиотрейтол) и бикарбонат при pH
8,0, они начнут фиксировать СО2 в темноте, как только получат
рибулозобисфосфат. (Эндогенный РуБФ настолько разбавляет-
ся в процессе выделения, что его недостаточно для поддержа-
ния заметной скорости, поэтому требуется экзогенный РуБФ.)
Следовательно, отсутствие быстрой фиксации СО2 теми же хло-
202
ГЛАВА 7
ропластами при первом освещении говорит о том, что либо в
строме хлоропластов, находящихся в темноте, почти полностью
лишена активности карбоксилаза, либо для поддержания тех
скоростей, которые наблюдаются позднее, недостаточно имею-
щегося сначала РуБФ. Казалось бы, сделать выбор между эти-
ми двумя объяснениями могут помочь эксперименты, однако по-
лученные к настоящему времени данные крайне противоречи-
вы. Например, ряд исследователей считают, что активность
карбоксилазы меняется в зависимости от освещения. Вместе с
тем Робинсон и соавторы в опытах с протопластами пшеницы,
гороха и шпината показали, что эти изменения совершенно не-
значительны. Даже по данным Йенсена и сотрудников, прида-
ющих большое значение активации светом, активность в тем-
ноте составляет примерно 2/3 той величины, которая отмечена
на свету (рис. 7.31). Так же как и группа Лилли, Йенсен с со-
авторами отметили увеличение концентрации РуБФ под дейст-
вием света; вместе с тем они обнаружили большое количество
РуБФ и в темноте (уровень РуБФ был гораздо выше, чем тот,
который необходим для поддержания половины максимальной
Рис. 7.31. Влияние DL-глицеральдегида на фиксацию СО2 и содержание
РуБФ в хлоропластах. Фиксацию СО2 (сплошные линии) измеряли в отсут-
ствие и в присутствии 1 мМ и 15 мМ. DL-глицеральдегида. Содержание
РуБФ (штриховые линии) определяли при этих же концентрациях DL-гли-
церальдегида [24].
ИНДУКЦИЯ
203
активности выделенной карбоксилазы). Теоретически одновре-
менное существование больших концентраций РуБФ и значи-
тельной активности карбоксилазы невозможно из-за большой
отрицательной величины ДР' карбоксилирования, которая ука-
зывает на то, что реакция должна идти до конца. Даже если
карбоксилаза обладает полной активностью (разд. 7.8), она
может не проявляться из-за неблагоприятных для нее условий
в строме хлоропластов, находящихся в темноте. Это нельзя
объяснить влиянием ионов магния, так как они действуют на
активацию, а ие на катализ. Как показали последние исследо-
вания Краузе и Беи-Хайима (разд. 9.3), фиксация СО2 не пре-
терпевает существенных изменений в изолированных хлоропла-
стах, в которых вызванное светом возрастание концентрации
ионов магния в строме снимается под действием аммиака. Ма-
ловероятно, наконец, чтобы в относительно кислой среде за-
темненной стромы активность карбоксилазы полностью подав-
лялась или чтобы реакция на щелочные значения pH при ос-
вещении была очень медленной и вносила существенный вклад
в индукционную задержку порядка нескольких минут.
Итак, напрашивается вывод, что если Йенсен и соавторы
правильно определили содержание РуБФ, то большая часть
его не сразу доступна карбоксилированию. [Величина 7<м
РуБФ равна 20—30 мкМ, что при объеме стромы в 25 мкл па
1 мг хлорофилла составляет 0,50—0,75 нмоль на 1 мг хлоро-
филла. На рис. 7.31, где, кстати, показано подавление фиксации
СО2 D.L-глицер альдегидом, первоначальная концентрация
РуБФ на порядок меньше исходной концентрации РуБФ.] Та-
ким образом, повышение содержания РуБФ, которое Йенсен и
Лилли с сотрудниками (1977) отметили при освещении, согла-
суется с тем, что задержка в фиксации СО2 отражает время,
за которое концентрация акцептора СО2 достигает своего ста-
ционарного рабочего уровня. Если активация ферментов играет
важную роль в накоплении РуБФ, то это относится скорее все-
го к таким ферментам, как бисфосфатазы и рнбулозо-фосфоки-
наза. Например, ФБФ-аза проявляет низкую активность в тем-
ноте, а кинетика ее активации при 20°C иногда напоминает ин-
дукцию в фиксации СО2. Разграничить выделение кислорода и
фиксацию СО2 в интактных хлоропластах не так-то просто.
Однако если добавить ФГК с самого начала, то выделение О2
наступает почти сразу, в то время как задержка в фиксации
СО2 при этом не сокращается (рис. 7.32).
Вместе с тем при увеличении лаг-фазы с помощью высоких
концентраций неорганического фосфата кинетика ФГК-зависи-
мого выделения кислорода н активация ФБФ-азы остаются в
целом неизменными (рис. 7.19). Это согласуется с современ-
ным представлением о том, что неорганический фосфат подав-
204
ГЛАВА 7
Рис. 7.32. Одновременное измерение выделения кислорода и фиксации СО2
интактными хлоропластами шпината в присутствии ФГК или в ее отсутствие.
Выделение кислорода показано сплошными линиями, а включение 14СО2 в
кислотоустойчивые продукты — линиями с кружками. ФГК практически пол-
ностью снимает индукцию в выделении кислорода и лишь незначительно
уменьшает ее в фиксации СО2 [33].
ляет активный экспорт промежуточных продуктов при помощи
переносчика неорганического фосфата (разд. 7.11 и 8.14). Если
затем спустя несколько минут пребывания на свету внести
ФГК, то сразу начнутся и выделение кислорода, и фиксация
двуокиси углерода (рис. 7.22). Тот факт, что экзогенная ФГК
способствует практически немедленному выделению кислорода,
указывает на столь же быстрое образование триозофосфата.
Кроме того, он свидетельствует о том, что в начале освещения
содержание РуБФ не может подняться до оптимального уров-
ня за счет триозофосфата. И наоборот, быстрая фиксация СОа
ИНДУКЦИЯ
205
после добавления ФГК к системе, подавленной неорганическим
фосфатом (рис. 7.22), означает преодоление при освещении
любого остаточного барьера, который препятствовал проявле-
нию полной активности (например, недостаточно высокой ак-
тивности ФБФ-азы).
Источники данных [24, 26, 33, 34, 45].
7.15. Индукция в реконструированной системе
хлоропластов
Как мы уже говорили в предыдущем разделе, в реконструи-
рованной системе хлоропластов фиксация СО2 быстро начина-
ется после добавления РуБФ или РубФ. Однако при исполь-
зовании в качестве субстрата ФБФ наблюдается (по крайней
мере иногда) четкая лаг-фаза. Эта задержка выражена еще
более явно в случае ФГК или ФГА, а также в отсутствие экзо-
генного субстрата (рис. 7.33, 7.34 и 7.35).
Рис. 7.33. Фиксация СОг в реконструированной системе хлоропластов. Как
видно из рисунка, субстраты вносили через 2 мин после включения света.
В случае РубФ значительная фиксация происходит уже в первую минуту,
в то время как для ФБФ и ФГК. наблюдается четкий индукционный период
(Уокер, неопубликованные данные).
206
ГЛАВА 7
Рис. 7.34, Индукция в реконструированной системе хлоропластов. Обе реак-
ционные смеси содержали NaHCO3. Как показано на рисунке, РубФ добав-
ляли только к одной смеси. Непосредственно перед включением света в обе
смеси вносили NaH14CO3 и одновременно измеряли выделение О2 и фикса-
цию СО2. Выделение О2 показано сплошными линиями, а включение “СО?
в кислотоустойчивые продукты — кружками (+Ру5Ф) или крестиками
(—РубФ). Цифры в скобках обозначают максимальные скорости (в микро-
молях СОа или Os на 1 мг хлорофилла в 1 ч). Изменения в кинетике выде-
ления О2 и фиксации COs, которые наблюдаются в присутствии РубФ, объ-
ясняются созданием неблагоприятного отношения [ATP]/[ADP] (разд. 7.13)
Присутствие бикарбоната, ионов магния и дитиотрейтола в
реконструированной системе хлоропластов при щелочных зна-
чениях pH должно обеспечивать полную активность всех уча-
ствующих в фотосинтезе ферментов. Таким образом, зацержки,
которые наблюдаются в фиксации СОз, по-видимому, обуслов-
лены необходимостью накопления соответствующих количеств
промежуточных продуктов. Хотя в результате разбавления эти
количества в реконструированных хлоропластах гораздо ниже,
чем в интактных, такие задержки позволяют прийти к выводу
о необходимости накопления соответствующих концентраций
триозофосфата и гексозомонофосфата.
Недавно Фербенк и Лилли изучали фиксацию СОз экстрак-
тами стромы. Они отметили только нелинейную кинетику это-
го процесса в присутствии субстратных количеств эритрозо-
4-фосфата. Вот почему эти исследователи полагают, что «в от-
индукция
207
Рис. 7.35. Индукция в реконструированной системе хлоропластов. Одновре-
менное определенно выделения О2 (сплошная линия) и фиксации СО2 (тре-
угольники) в присутствии ФГА (ср. с рис. 7.34, Stokes, Walker, 1976).
сутствие информации о всех промежуточных продуктах фото-
синтеза следует проявлять величайшую осторожность при объ-
яснении возрастания скорости автокаталитическими процесса-
ми». Мы полностью разделяем эту точку зрения и охотно допу-
скаем, что автокаталитическое ускорение, показанное на рис.
7.33 и 7.34, можно считать окончательно доказанным лишь в
том случае, если одновременно измерены концентрации всех
промежуточных продуктов этого процесса. Впрочем, следует
отметить, что Фербенк и Лилли работали с системой, которая
не содержала тилакоидов и в которой непрерывное поступление
АТР целиком зависело от присутствия креатинфосфата и креа-
тинкиназы. Следовательно, эта система отличается по ряду
признаков от той, в которой были получены результаты, при-
веденные на рис. 7.33 и 7.34. Пожалуй, самым важным из них
является то, что присутствие креатиифосфата и креатинкиназы
позволяет получить благоприятное высокое отношение
[ATP]/’[ADP], Если же АТР образуется в освещенных тилако-
идах, то можно получить такие экспериментальные условия, в
которых при данном отношении [ATP]/pADP] и достижении
достаточно высокой концентрации ФГК будет поддерживаться
линейная скорость выделения О2 (или фиксации СО2). Однако,
как видно из рис. 7.27, выделение кислорода в этих условиях
208
ГЛАВА 7
зависит как от концентраций АТР и ADP, так и от концентра-
ции ФГК. Короче говоря, накопление ФГК даст ощутимые ре-
зультаты даже тогда, когда отношение [АТР] /([ADP] не будет
сохраняться на высоком уровне. Исследования подобного рода
позволяют сделать вывод, что реакция, катализируемая ФГК-
киназой (разд. 6.11), играет ключевую роль как в индукции,
так и во внутренней регуляции ВПФ-цикла (гл. 9).
Источники данных: [20, 33, 43, 44].
7.16. Последовательность событий
во время индукции
а) Под действием света сразу начинается быстрый транспорт
электронов. В результате связанного с ним направленного дви-
жения протонов из стромы в тилакоиды строма становится бо-
лее щелочной (pH меняется с -~7 на ~8).
б) Чтобы восстановить равновесие зарядов, нарушенное в ре-
зультате оттока протонов, внутрь тилакоидов начинают посту-
пать такие анионы, как С1-, а из тилакоидов в строму — катио-
ны, например Mg2+. Одно время принято было считать, что в
активации ферментов важную роль играет транспорт попов
магния. Однако, по последним данным, концентрация попов
Mg2+ в строме всегда достаточно высока, чтобы обеспечивать
активацию ферментов и катализ.
в) В мембране хлоропласта находится протонный насос, кото-
рый облегчает обмен Н+ и К+. Насос помогает сохранить гра-
диент pH между стромой и цитозолем, который иначе, по мере
того как протоны медленно диффундировали в хлоропласт, ис-
чезал бы. На работу насоса влияет концентрация ионов маг-
ния снаружи хлоропласта. Кроме того, насос способен взаимо-
действовать с переносчиком неорганического фосфата.
г) Отношение [ATP]/[ADP] в строме начинает быстро расти,
так как фотофосфорилирование идет с максимальной ско-
ростью, а потребление АТР в первое время лимитируется недо-
статком субстратов для реакций, катализируемых киназой.
д) Отношение [NADPH]/[NADP+] увеличивается, так как сна-
чала восстановление NADP преобладает над его использовани-
ем. До того как будет достигнута полная активность реакции,
в которой используется NADPH (т. е. максимальная скорость
восстановления ФГК), электроны направляются на восстанов-
ление тиоредоксина и т. д. В результате вся среда стромы при-
обретает восстанавливающие свойства.
е) При щелочных значениях pH, а также благодаря действию
тиоредоксина и подобных ему соединений восстанавливаются
дисульфидные группы ферментов. Это приводит к активации
ИНДУКЦИЯ
209
таких ферментов, как Ру5Ф-киназа, триозофосфатдегидрогена-
за и бисфосфатазы. Пока еще неизвестно, какое именно влия-
ние оказывает такая активация на индукцию. Впрочем, ФБФ-
аза и СБФ-аза отличаются весьма низкой активностью в тем-
ноте; а если низкая активность обусловлена малой концентра-
цией субстратов, то активация этих ферментов может играть
важную роль на ранних стадиях индукции. Одно время
РуБФ-карбоксилазу также относили к ферментам, которые ак-
тивируются светом; кроме того, полагали, что она способна не-
посредственно активироваться под действием NADPH. Однако
эта точка зрения была опровергнута результатами опытов с
протопластами, в которых было показано, что в темноте
РуБФ-карбоксилаза почти столь же активна, как и на свету,
ж) Некоторые ферменты ВПФ-цикла (включая ФБФ-азу)
функционируют лучше при щелочных значениях pH освещенной
стромы. Это связано не с их активацией (см. предыдущий
пункт), а с оптимумом pH.
з) Концентрация РуБФ, ФГК и гексозомоиофосфатов возраста-
ет. Создается впечатление, что в некоторых случаях накопле-
ние гексозомоиофосфатов может быть обусловлено деградаци-
ей крахмала. По мере, увеличения содержания Ру5Ф АТР идет
на его превращение в РуБФ, и отношение [АТР]/\[ADP] сни-
жается, что затрудняет восстановление ФГК. Накапливаемая
ФГК удерживается в хлоропласте из-за щелочной среды стро-
мы. Как видно из рис, 7.26, даже если РуБФ-карбоксилаза об-
ладает полной активностью и не испытывает недостатка в со-
ответствующем субстрате, при неблагоприятном отношении
[ATP]/[,ADP] кислород все равно выделяется после некоторой
задержки. Такое подавление под действием ADP можно ком-
пенсировать высокой концентрацией ФГК (рис. 7.27). В ре-
конструированной системе хлоропластов при добавлении ФБФ
фиксация СО2 наступает очень быстро, тогда как выделение О2
может задерживаться па неограниченное время. В таких систе-
мах индукция в фиксации СО2 наблюдается лишь в отсутствие
добавленного субстрата или в присутствии промежуточных
продуктов — предшественников ФБФ. По-видимому, для них
время, за которое накапливаются необходимые количества раз-
личных промежуточных продуктов, значительно больше време-
ни, характерного для интактного хлоропласта, что объясняет-
ся разбавлением реакционной смеси (объем реконструирован-
ной системы почти в 1000 раз больше в пересчете на 1 мг хло-
рофилла, чем объем интактного хлоропласта). Тем не менее
ферменты, катализирующие эти процессы, полностью активиро-
ваны. Следовательно, по крайней мере в данных условиях, ин-
дукция вызвана главным образом необходимостью накопления
субстратов.
14—234
210
ГЛАВА 7
Как только ферменты цикла полностью активируются и на-
копится достаточное количество их субстратов, фотосинтез
вступает в стационарную фазу. В присутствии насыщающих
концентраций СОг и соответствующих количеств неорганичес-
кого фосфата скорость определяется интенсивностью света и
температурой.
Максимальный фотосинтез идет беспрепятственно лишь в
присутствии экзогенного неорганического фосфата. Если его
концентрация слишком снижается, то затрудняется экспорт
триозофосфатов. Большая величина отношения [ФГК]/[Pi]
способствует аллостерической активации глюкозо-1-фосфат-
аденилилтрансферазы (разд. 9.7). Если бы Ф6Ф мог превра-
щаться в крахмал со скоростью, достаточной для того, чтобы
воспрепятствовать дальнейшему падению концентрации неор-
ганического фосфата в строме, то теоретически было бы воз-
можно переключение с экспорта триозофосфатов на накопление
их внутри хлоропласта в виде крахмала (скорость фотосинте-
за при этом не меняется). Однако в действительности, по-види-
мому, содержание Ф6Ф возрастает и в присутствии достаточ-
ных количеств триозофосфатов приводит к ускоренному потреб-
лению углерода в первой реакции, катализируемой транскето-
лазой.
Таким образом, всякая тенденция к накоплению пентозо-
фосфатов неизбежно повлечет за собой дальнейшее уменьше-
ние отношения [АТР] /'[ADP], так как АТР расходуется в ре-
акции РубФ—>РуБФ. В свою очередь это вызовет подавление
восстановления ФГК, а регенерация РубФ снизится из-за его
превращения в РуБФ. Короче говоря, в фазе регенерации
ВПФ-цикла нет дополнительных возможностей для использо-
вания имеющегося АТР помимо тех, которыми он располагает.
Только при оптимальной концентрации неорганического фосфа-
та можно быть уверенным в том, что скорость регенерации до-
статочна и ассимиляционная энергия используется оптимально.
7.17. Восстановление лаг-фазы
Если на короткое время выключить свет, а затем опять
включить его, то лаг-фазы практически нет. Однако она опять
наблюдается после более длительного пребывания в темноте.
В связи с этим представляет интерес тот факт, что лаг-фаза
восстанавливается быстрее в целых листьях и протопластах,
чем в изолированных хлоропластах. Лигуд установил, что в
пшенице ФБФ-аза быстро инактивируется и реактивируется как
в хлоропластах, так и в протопластах. Отсюда следует, что раз-
личие в поведении протопластов и хлоропластов обусловлено,
ИНДУКЦИЯ
211
по-видимому, содержанием метаболитов, которое в первых сни-
жается быстрее, чем в последних.
Источник данных: .[25с, ЗЗЬ].
ОБЩАЯ ЛИТЕРАТУРА
1. Gibbs AL Carbohydrate metabolism by chloroplasts. In Structure and Func-
tion of Chloroplasts (Gibbs M., cd.), 169—214, Springer-Verlag, Berlin,
1971.
2. Heidi H. IF. Metabolite carriers of chloroplasts. In Encyclopedia of Plant
Physiology. Transport in Plant III (C. R. Stocking, U. Heber eds.), New
Series, Volume 3, 137—43. Springer-Verlag, Berlin, 1976.
3. Herold A., Walker D. A. Transport across chloroplast envelopes — the role
of phosphate, In: Membrane Transport in Biology (G, Giebisch, D. C. Tos-
teson, J. J. Ussing eds.), Vol. II, 412—439. Springer-Verlag, Heidelberg,
1979.
4. Rabinowitch E. I. Photosynthesis and Related Processes, Vol. П, Pt. 2,
537 et seq., Interscience, New York, 1956.
5. Walker D. A. Psotosynthetic induction phenomena and the light activation
of ribulose diphosphate carboxylase, New Phytol., 72, 209—35 (1973).
6. Walker D. A. Chloroplast and Cell-Concerning the movement of certain key
metabolites etc. across the chloroplast envelope, Med. Tech. Publ. Ini. Rev.
Sci. Biochem. Ser. 1, (D. H. Northcote, cd.) Vol. 11, 1—49, Butterworths,
London, 1974.
7. Walker D. A. COa-fixation by Intact Chloroplasts: Photosynthetic induction
and its relation to transport phenomena and control mechanisms, In: The
Intact Chloroplast. (J. Barber, ed.), Chapter 7, 235—78, Elsevier, Amster-
dam, 1976a.
8. Walker D. A. Plastids and Intracellular Transport. In: Encyclopedia of Plant
Physiology. Transport in Plants III, (C. R. Stocking, U. Heber, eds.), New
Series, Volume 3, 85—136. Springer-Verlag, Berlin, 1967b.
9. Walker D. A., Crofts A. R. Photosynthesis, Ann. Rev. Biochem., 39, 389—
428 (1970).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
10. Balasubratnanlam S., Willis A. J. Stomatai movements and rates of gaseous
exchange in excised leaves of Vicia faba, New Phytol., 68, 663—75 (1969).
11. Baldry С. IF., Bucke C., Walker D. A. Some effects of temperature on carbon
dioxide fixation by isolated chloroplasts, Biochim. Biophys. Acta, 126, 207—
13 (1966).
12. Baldry C. IF,, Walker D. A., Bucke C, Calvin'—cycle intermediates in rela-
tion to induction phenomenon in photosynthetic carbon dioxide fixation by
isolated chloroplasts, Biochem. J., 101, 641—6 (1966).
13. Bamberger E. S., Gibbs M. Studies on photosynthetic carbon dioxide fixation
by whole spinach chloroplasts, Plant Physiol., Suppl. 38, (1963).
14. Blinks L. R., Skew R. K. The time course of photosynthesis as shown by a
rapid electrode method for oxygen, Proc. Natn. Acad. Sci. U.S.A., 24, 420—7
(1938a).
15. Blinks L. R., Skow R. K. The time course of photosynthesis as shown by the
glass electrode, with anomalies in the acidity changes, Proc. Natn. Acad, Sci.
U.S.A., 24, 413—9 (1938b).
16. Bucke C., Walker D. A., Baldry C. IF. Some effects of sugars and sugar
phosphates on carbon dioxide fixation by isolated chloroplasts, Biochem. J.,
101, 636—41 (1966).
14*
212
ГЛАВА 7
17. Cockburn. W., Baldry C. W., Walker D. A. Some effects of inorganic phospha-
te on O2 evolution by isolated chloroplasts, Biochim. Biophys. Acta., 143,
614—24 (1967).
18. Cockburn W., Walker D. A., Baldry- C. IF. Photosynthesis by isolated chlo-
roplasts. Reversal of orthophosphate inhibition by Calvin cycle intermediates,
Biochem. J., 107, 891—95 (1968).
19. Fllege R., Flugge U.-L, Werdan K., Heldt H. W. Specific transport of inor-
ganic phosphate, 3-phosphoglycerate and triose phosphates across the inner
membrane of the envelope in spinach chloroplasts, Biochim. Biophys. Acta.,
502, 232—47 (1973).
20. Furbank R. T., Lilley R. M. C. Effects of inorganic phosphate on the photo-
synthetic carbon reduction cycle in extracts from the stroma of pea chloro-
plasts, Biochim. Biophys. Acta, 592, 65—75 (1980).
21. Heber U., Perceld P. Substrate and product fluxes across the chloroplast
envelope during bicarbonate and nitrite reduction, In: Proceedings of the
Fourth International Congress on Photosynthesis (D. 0. Hall, J. Coombs,
T. W. Goodwin, eds.), 107—18, Biochem. Soc., London, 1977.
22. Heber U., Willenbrink J. Site of synthesis and transport of photosynthetic
products within the leaf cell, Biochim. Biophys. Acta., 82, 313—24 (1964).
23. Jagendorf A. T., Hind G. Studies on the mechanism of photophosphorylation.
In: Photosynthetic Mechanism of Green Plants, 599—610, National Academy
of Sciences, National Research Council Publications, 1145 (1963).
24. Jensen R. G., Slcher R. C. Jr., Bahr J. T. Regulation of ribulose 1,5-bisphos-
phate carboxylase in the chloroplast, In: Photosynthetic Carbon Assimilation,
Basic Life Sciences Vol. Il, «Proceedings of a Symposium held at Brookha-
ven National Laboratory, New York, May 31 —June 2, 1978» (H. W. Siegel-
inan, G. Hind, eds.), 95—112. Plenum Press, New York, 1978.
25. Kaiser IF., Urbach W. Rates and properties of endogenous cyclic photophos-
phorylation of isolated intact chloroplasts measured by CO2 fixation in the
presence of dihydroxyaeetone phosphate, Biochim. Biophys. Acta, 423, 91—
102 (1976).
25a. Leegood R. C., Walker D. A. Autocatalysis and light activation of enzymes
in relation to photosynthetic induction in wheat chloroplasts, Arch. Biochem.
Biophys., 200, 575—582 (1980).
25b. Leegoad R. C., Walker D. Л. Modulation of fructose bisphosphatase activity
in intact chloroplasts, FEBS Letters, 116, 21—4 (1980).
25c. Leegood R. C., Walker D. A. Photosynthetic induction in wheat protoplasts
and chloroplasts. Autocatalysis and light activation of enzymes, Plant, Cell
and Environment, 4, 59—66 (1981).
26. Lilley R. McC., Chon C. J., Mosbach A., Heidi H. IF. The distribution of
metabolites between spinach chloroplasts and medium during psotosynthesis
in vitro, Biochim. Biophys. Acta, 460, 259—72 (1977).
27. Lorimer G. H., Badger M. R-, Heldt H. W. The activation of ribulose 1,5-bis-
phosphate carbxylase/oxygenase. In: Photosynthetic Carbon Assimilation, Ba-
sic Life Sciences Vol. II, Proceedings of a Symposium held at Brookhaven
National Laboratory, Upton, New York, May 31—June 2, 1978 (H. W. Sie-
gelman, G. Hind, eds.), 43—59. Plenum Press, New York, 1978.
28. McAlister E. D. Time course of photosynthesis for a higner plant, Smithson
Misc. Coll., 95, 24 (1937).
29. McAlister E. D., Myers J. The time course of photosynthesis and fluorescence,
observed simultaneously, Smithson, Misc. Coll., 99, 6 (1940),
30. Osterhout W. J. V., Hass A. R. C. On the dynamics of photosynthesis, J. Gen.
Physiol., 1, 1—16 (1919).
31. Peavey D. G„ Gibbs M. Photosynthetic enhancement studied in intact spinach
chloroplasts, Plant Physiol, 55, 799—802 (1975).
ИНДУКЦИЯ
213
32. Robinson S. P„ McNeil P. II., Walker D. A, Ribulose bisphosphate carboxyla-
se— lack of dark inactivation of the enzyme in experiments with protoplasts,
FEBS Letters, 97, 260—300 (1979).
33. Robinson S. P., Walker D. A. Regulation of photosynthetic carbon assimila-
tion. In: Photosynthetic Carbon Assimilation, Basic Life Sciences Vol. II,
Proceedings of a Symposium held at Brookhaven National Laboratory, Upton,
New York, May 31 —June 2, 1978 (I-I. W. Siegehnan, G. Hind, eds.), 43—59.
Plenum Press, New York, 1978.
33a. Robinson S. P„ Walker D. A. The distribution of metabolites between chlo-
roplast and cytoplasm during the induction phase of photosynthesis in leaf
protoplasts, Plant Physiol., 65, 902—90.5 (1980).
33b. Robinson S. P., Walker D. A. The significance of light activation of enzy-
mes during the induction phase of photosynthesis in isolated chloroplasts,
Arch, Biochem. Biophys., 202, 617—23 (1980).
34. Selwi/n M. J. Temperature and photosynthesis II. A mechanism for the effects
of temperature on carbon dioxide fixation, Biochim. Biophys. Acta, 126,
214—24 (1966).
35. Slabas A. R., Walker D. A. Transient inhibition by ribose-5-phosphale of
photosynthetic O2 evolution in a reconstituted chloroplast system, Biochim.
Biophys. Acta, 430, 154—64 (1976).
36. Steeman-Nielsen E. Der Mechanisinus der Photosynthesc, Dansk hot. Ark.,
11, 2 (1942).
37. Turner J. F., Black С. C., Gibbs M. The effect of light intensity on lhe tri-
phosphopyridine nucleotide reduction, adenosine triphosphate formation and
carbon dioxide assimilation in spinach chloroplasts, J. Biol. Chem., 237, 577—
9 (1962).
38. Walker D. A. Improved rates of carbon dioxide fixation by illuminated chlo-
roplasts, Biochem. J., 92, 22c—23c (1964).
38a. Walker D. A. Photosynthetic activity of isolated pea chloroplasts. In: Bio-
chemistry of the Chloroplast (1965) NATO Adv. Study Inst. Aberystwyth
(T. W. Goodwin ed.), Vol. 2, 53—69, Academic Press, New York, 1967.
39. Walker D. A., Baldry C. W., Cockburn IF. Photosynthesis by isolated chloro-
plasts, simultaneous measurement of carbon assimilation and oxygen evo-
lution, Plant Physiol., 43, 1419—22 (1968).
40. Walker D. A., Cockburn IF., Baldry C. W. Photosynthetic oxygen evolution
by isolated chloroplasts in the presence of carbon cycle intermediates, Natu-
re, 216, 597—9 (1967).
41. Walker D. A., Hill R. The relation of oxygen evolution to carbon assimila-
tion with isolated chloroplasts, Biochim. Biophys. Acta., 131, 330—8 (1967).
42. Walker D. A., Kosclukiewicz K., Case C. Photosynthesis by isolated chloro-
plasts: some factors affecting induction in CO2-dependent oxygen evolution,
New Phytol., 72, 237—47 (1973).
43. Walker D. A., Lilley R. McC. Autocatalysis in a reconstituted chloroplast
system, New Phytol.; 73, 657—662 (1974).
44. Walker D. A., Slabas A. R. Stepwise generation of the natural oxidant in a
reconstituted chloroplast system, Plant Physiol., 57, 203—6 (1976).
45. Walker D. A., Slabas A. R., Fitzgerald M. P. Photosynthesis in a reconstitu-
ted chloroplast system from spinach. Some factors affecting CO2-dependent
oxygen evolution with fructose-l,6-bisphosphate as substrate, Biochim. Bio-
phys. Acta,, 440, 147—62 (1976).
46. Warburg O. Uber die Gesclrwindigkeit der photochemischen Kohlensaitrezer-
setzmig in lebenden Zellen II, Biochem. Z., 103, 188—217 (1920).
47. Willis A. J„ Balasubramaniam S. Stomatai behaviour in relation to rates of
photosynthesis and transpiration in Pelargonium, New Phytol., 67, 265—85
(1968).
ГЛАВА 8
Пластиды
и внутриклеточный транспорт
Краткое содержание
В общем хлоропласты Сз-растений можно рассматривать
как органеллы, которые с высокой скоростью превращают по-
ступающие в них СО2 и ортофосфат в триозофосфат, а затем
экспортируют последний. Поэтому, несмотря на то что в хло-
ропластах протекает множество метаболических процессов, ос-
новные функции этих органелл в анаболизме и транспорте
можно представить следующим уравнением:
ЗСО2 -ф 2Н2О -|- Pj -s- Триозофосфат ЗО2
Прямой транспорт ассимиляционной силы (АТР и NADPH)
не может осуществляться с большой скоростью, так как внут-
ренняя мембрана (оболочка) хлоропласта относительно непро-
ницаема для аденилатов и совершенно непроницаема для пи-
котинамидиуклеотидов. Однако эти соединения способны об-
мениваться благодаря своему участию в челночных механиз-
мах (рис. 8.1).
8.1. Хлоропласт как транспортирующая органелла
Главная функция хлоропластов заключается в образовании
ассимиляционной силы и ее использовании на восстановление
СО2 до углеводов. Вместе с тем растение в целом выиграет от
этой функции лишь в том случае, если образующиеся продук-
ты будут экспортироваться из хлоропласта. А это в свою оче-
редь возможно только при условии, что в него будут посту-
пать субстраты для их синтеза. В конечном счете должно ус-
тановиться равновесие между поступлением одних соединений
в хлоропласт и экспортом других из него, и в каждый данный
момент транспорт ключевых метаболитов должен соответство-
вать скорости фотосинтеза. Вместе с тем необходимо также,
чтобы хлоропласт непрерывно регенерировал свой акцептор
СО2 и чтобы ВПФ-цикл мог функционировать как автокатали-
тическая последовательность, или «производительная» реак-
ция (гл. 6) . Эти требования в корне противоречат друг другу
и поэтому транспорт метаболитов, коферментов и ионов меж-
ду хлоропластом и его клеточным окружением должен
Г ексозомонофосфаты
Рис. 8.1. Схематическое изображение транспорта основных метаболитов. Чем больше скорость транспор-
та и степень обмена, тем «жирнее» стрелки. Так, например, быстрее всего происходит стехиометрический
обмен ортофосфата и триозофосфатов при помощи фосфатного переносчика (большие черные стрелки), в то
время как прямой транспорт гексозомонофосфатов идет лишь в незначительной степени и очень медленно ю
(штриховые стрелки). Si
ПЛАСТИДЫ и внутриклеточный транспорт
216
ГЛАВА 8
Рис. 8.2. Транспорт основных метаболитов через оболочку хлоропластов шпи-
ната. Двуокись углерода и ортофосфат Р( поступают в хлоропласт, а три-
озофосфаты (ТФ) и кислород выходят из него. Восстановление NADP+ и
синтеза АТР из ADP+ осуществляются в тилакоидах, в которых находятся
пигменты и другие компоненты фотохимического аппарата. ВПФ-цикл про-
текает в строме. В этом цикле образуются трнозофосфаты, которые могут
транспортироваться, опять подвергаться превращениям в ВПФ-цикле и через
гексозофосфаты (ГФ) участвовать в синтезе крахмала.
каким-то образом контролироваться. Данные, накопив-
шиеся за последнее десятилетие, свидетельствуют о том,
что основными соединениями, которые поступают внутрь хло-
ропласта, являются неорганический фосфат и СО2, а экспорти-
руются в основном триозофосфаты (рис. 8.2). В следующих
разделах мы рассмотрим, как изучали транспорт этих, а также
некоторых других соединений. (Методы, при помощи которых
были получены эти данные, приведены также в табл. 8.1.)
8.2. Экспериментальная основа
исследований транспорта — фракционирование
целых тканей
Для изучения путей метаболизма широко используется ра-
диоактивный углерод. Казалось бы, самый простой способ ис-
следовать транспорт — это вводить ИСО2 в интактный лист, за-
тем гомогенизировать его и определять, какой продукт фото-
синтеза выделяется из хлоропласта. Однако осуществить это
в эксперименте довольно трудно в силу нескольких причин. Во-
первых, транспортируемый метаболит должен в конечном сче-
те подвергаться дальнейшему метаболизму, а такие процессы,
как ВПФ-цикл и гликолиз, используют одни и те же промежу-
ПЛАСТИДЫ и внутриклеточный транспорт
217
Таблица 8.1. Методы изучения транспорта метаболитов в хлоропластах
А. Информация о распределении и транспорте метаболитов in vivo
I. Фракционирование клеток в неводной среде
а) Кинетические измерения концентрации и распределения субстратов
б) Кинетические измерения распределения метки
II. Фракционирование клеток в водной среде
Определение содержания метаболитов в интактных хлоропластах после
«быстрого» выделения
Б, Информация о распределении и транспорте метаболитов in vitro
I. Прямые методы изучения хлоропластов, выделенных в водной среде
а) Изучение распределения метаболитов или метки между хлороплас-
тами и средой
б) Определение концентрации субстратов после центрифугирования
хлоропластов через слой силиконового масла
II. Косвенные методы изучения хлоропластов, выделенных в водной среде
а) Изучение влияния добавленных метаболитов на метаболизм хлоро-
пластов
б) Определение активности «защищенных» ферментов
в) Изучение осмотической реакции хлоропластов на внесение метабо-
литов
В. Изучение распределения метаболитов или метки между хлоропластом и
клеточной средой с использованием изолированных протопластов
точные продукты. Поэтому, естественно, не так-то просто уста-
новить, переносится ли соединение X из хлоропласта в цито-
плазму в неизменном виде или — для облегчения транспорта —
оно сначала превращается в Y, а затем снова в X на первой
стадии последующего метаболизма. Во-вторых, хлоропласты и
другие субклеточные органеллы, как правило, очень малы и
весьма хрупки. Представьте себе, что вы пропустили через мя-
сорубку сумку с покупками; если вы не знали, какие именно
покупки были сделаны, и не можете прочитать, что написано
на пакетах, вряд ли вы сумеете точно установить содержимое
каждого пакета. Многие методы фракционирования клеток яв-
ляются столь же грубыми. Если же позволить листьям осу-
ществлять фотосинтез в присутствии 14СО2, затем быстро за-
морозить их в жидком азоте, а потом уже проводить фракцио-
нирование в неводных растворителях, тогда у нас будут основа-
ния считать, что соединения, находившиеся внутри хлороплас-
тов во время фотосинтеза, останутся там и при фракциониро-
вании. Правда, при использовании данного метода некоторое
количество крахмала обнаруживается не только во фракции
хлоропластов, т. е. этот метод тоже несовершенен. Тем не ме-
нее он позволяет получить ценную информацию о распределе-
нии метаболитов. ^Следует отметить, что крахмал относится к
тем немногим соединениям, локализацию которых можно лег-
218
ГЛАВА 3
Рис. 8.3. Крахмал, Зимой 1864 г. знаменитый немецкий физиолог растений
Закс подверг повторному освещению частично закрытый фольгой лист без
крахмала и показал при помощи реакции крахмала с иодом, что крахмал
вновь образовался только в освещенных хлоропластах. [В подобного рода
опытах крахмал из листьев удаляется после инкубации их в темноте в тече-
ние 24—28 ч. Затем их освещают и кипятят в 80 %-ном этаноле, где экстра-
гируется хлорофилл, а обесцвеченную ткань листа отмывают водой и окра-
шивают раствором иода в иодистом калии. Комплекс крахмала с подом на
самом деле имеет синий цвет (амилаза окрашивается в синий цвет, а амило-
пектин— в фиолетовый), но при большой концентрации выглядит черным.}
В двадцатых годах этого века Молпш повторил опыты Закса, но он освещал
лист через фотографический негатив и получил па нем позитивное изобра-
жение. Этим методом можно добиться очень красивых картинок из крахма-
ла. Здесь вы видите репродукцию картины «Невинность», которую Поль-
Пьер Прудон (1758—1823 гг.) в свое время нарисовал мелом. В данном
случае она выполнена крахмалом на листе Pelargonium и, пожалуй, нисколь-
ко не проиграла от этого.
ко установить невооруженным глазом (рис. 8.3—8.5). Если
экстрагировать хлорофилл из листа горячим этанолом и окра-
сить обесцвеченную ткань иодом, то крахмал благодаря реак-
ции амилозы с иодом четко выступит в виде сине-черных гра-
нул. В интактном листе эти гранулы связаны с пластидами.]
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
219
Рис. 8.4. Схематическое изображение хлоропласта по Хельдту и факсимиле
заголовка первой статьи Молиша о получении фотографий на листьях расте-
ний. Сейчас пас особенно интересует достигнутая здесь степень разрешения,
так как главным метаболитом, который выносится из хлоропластов, служит
триозофосфат, а основным метаболитом, поступающим в результате транс-
порта в проводящие ткани, является сахароза. Если инкубировать диски
листьев в растворе сахарозы в темноте, то крахмал образуется равномерно
по всей ткани, т. е. для синтеза крахмала спет нс требуется, он нужен только
для фотосинтеза. Возникает вопрос, почему образующаяся при фотосинтезе
сахароза, которая в опытах по получению изображений на крахмале долж-
на на своем пути к проводящим тканям проходить через непосредственно
прилегающие затемненные клетки, не размывает изображения. Сама сахаро-
за не может повторно проникать в хлоропласты, по если экзогенная сахаро-
за способна инициировать синтез крахмала в темноте, то туда должны про-
никать продукты ее деградации. Не исключено, что все это просто вопрос
концентрации. Возможно также, что сахароза, образующаяся при фотосин-
тезе, и экзогенная сахароза поступают в разные отделы клетки. В чем бы ни
заключалась причина этого явления, она должна быть связана с транспор-
том метаболитов. Очевидно, что транспорт метаболита будет наиболее эф-
фективным, если этот метаболит оказывается каким-то образом защищенным
от деградации на пути к проводящим тканям. Как именно достигается эта
защита в листьях, мы узнаем лишь тогда, когда поймем, почему изображе-
ния иа крахмале отличаются столь замечательной четкостью.
* » ф * ♦ е ‘>
*
а * Ф «
♦ • ♦ *
Рис. 8.5. А. Увеличенный фрагмент предыдущей фотографии. Видно, что
крахмал образуется только в освещенных участках листа. Б. При максималь-
ном увеличении хорошо видны замыкающие клетки устьиц (показаны стрел-
ками) между точками, нарисованными на исходной схеме, так как они не
утрачивают крахмал в темноте и остаются в виде крошечных пар черных
пятнышек или «скобок», беспорядочно разбросанных на фотографии. Они
служат шкалой длиной в одну замыкающую клетку, по которой можно оце-
нивать дискретную природу образования крахмала.
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ транспорт
221
Быстрое фракционирование в водных растворителях применя-
ется реже, но и оно проливает свет на прижизненную локали-
зацию тех метаболитов, которые не сразу выделяются из раз-
рушенных органелл.
Источники данных: [10, И, 42, 50, 51, 62, 64].
8.3. Экспериментальная основа исследований
транспорта in vitro
Мы рассмотрели сейчас фракционирование целых тканей,
ио возможен и другой подход к изучению транспорта метаболи-
тов. Он заключается в выделении функционально активных ор-
ганелл и выяснении того, что они образуют in vitro и что вы-
деляют в окружающую среду. Выделение функционально ак-
тивных хлоропластов — далеко не простая задача (приложе-
ние А). Прошло почти тридцать лет, прежде чем Роберту Хил-
лу удалось продемонстрировать СО2-зависимое выделение кис-
50 мкл 0,4 М сорбитола
100 мкл силиконового масла
•50 мкл 0,4 М сахарозы
Нейлоновая сетка _________
(диаметр отверстий 20 мкм)
___ 150 мкл _______
суспензии протопластов
Рис. 8.6. Новый метод, разработанный Робинсоном для быстрого отделения
цитоплазмы и хлоропластов от изолированных протопластов. Суспензию про-
топластов помещают в верхний контейнер, где их можно освещать или под-
вергать иной обработке. В нулевой момент времени их разрушают центрифу-
гированием через сетку с диаметром отверстий 20 мкм. Цитоплазма остается
над слоем силиконового масла, а хлоропласты проходят через него в денату-
рирующий раствор. Данный метод позволяет быстро определять распределе-
ние метаболитов между этими двумя фракциями при фотосинтезе или ды-
хании.
222
ГЛАВА 8
лорода изолированными хлоропластами, скорость которого бы-
ла сравнима с выделением О2 в присутствии искусственных ак-
цепторов водорода. Кроме того, хлоропласты — очень хрупкие
органеллы, и, поскольку вероятность присутствия хлоропластов
с разрушенными оболочками и дальнейшего их разрушения при
выделении достаточно велика, проведение эксперимента и ин-
терпретация его результатов неизбежно усложняются. Предло-
женный нами в последние годы способ получения изолирован-
ных протопластов (приложение А) и разработанный Робинсо-
ном метод быстрого отделения хлоропластов от остальных кле-
точных фракций значительно повысили ценность исследований
in vitro (рис. 8.6). В двух нижеследующих разделах мы рас-
смотрим прямые методы, применяемые в таких исследованиях.
Источники данных: ([34, 35, 45, 48, 49, 63].
8.4. Хроматографический анализ
Изучение ВПФ-цикла было основано на хроматографичес-
ком анализе водорослей, которые получали 14СО2 (разд. 6.7).
Бассэм и его сотрудники из Беркли в Калифорнии (прежде
всего Марта Керк, которой также принадлежат большие за-
слуги в более ранних исследованиях) применили этот подход
к изолированным хлоропластам (рис. 8.7), Как и в случае мно-
гих других методов, используемых для изучения изолированных
хлоропластов, главный недостаток этого подхода (помимо его
трудоемкости) состоит в том, что он не позволяет четко отли-
чать процессы внутри хлоропластов от тех явлений, которые
происходят во внешней среде при повреждении оболочки этих
пластид.
Источники данных: [16, 17].
8.5. Фильтрация центрифугированием
Этот метод был впервые предложен Веркхайзером и Барт-
ли, а затем Клингенберг и его сотрудники стали широко при-
менять его для изучения митохондрий. Хельдт весьма успешно
использовал этот метод в своих опытах с хлоропластами (разд.
8.23). Сущность метода сводится к следующему: хлоропласты,
содержащие радиоактивный метаболит, пропускают под дейст-
вием центрифужной силы через фильтрующий слой силиконо-
вого масла, под которым находится слой денатурирующего
агента, например хлорной кислоты. Затем отрезают дно цент-
рифужной пробирки с осадком хлоропластов и хлорной кисло-
той и определяют распределение метки между ними (рис. 8.9).
„Добавление различных немеченых метаболитов позволяет так-
же изучать степень обратного обмена (например, между мече-
пиколат
Малат
Аспартат
Гриозофосфаты
Фосфоглицериновая’
X кислота ;
Монофосфаты •
Дифосфаты
Старт
*..........
Растворитель II
□а I
е;5
а?
н
S
сх
о
£0 ,
га ’
О- 1
Рис. 8.7. Двумерная радиоавтограмма продуктов кратковременной фиксации
14СО2 изолированными хлороплс.стами гороха. После освещения реакционную
смесь подкисляют для того, чтобы остановить реакцию и освободить ’’СОг
из бикарбоната. Исследуемый образец наносят на бумагу и проводят хрома-
тографию в смеси фенол — вода. Затем бумагу вынимают из камеры, сушат,
поворачивают на 90° и хроматографируют во втором направлении в смеси
бутанол — уксусная кислота. В результате каждое соединение занимает на
хроматограмме определенное место. Соединения идентифицируют при помощи
параллельной хроматографии с известными стандартами. Положение каждо-
го соединения определяют после получения радиоавтографа путем наложе-
ния иа хроматограмму рентгеновской пленки в темной комнате. Там, где
присутствуют радиоактивные соединения, иа плепке после проявления будут
видны темные пятна. Радиоактивность подсчитывают либо непосредственно'
на хроматограмме при помощи счетчика Гейгера, либо более точно в сцин-
тилляционном счетчике после элюирования соединений из пятен. Метод
двумерной хроматографии иа бумаге был впервые разработай Мартином и
Синджем. Впоследствии Кальвин, Бенсон и Бассэм и их соавторы примени-
ли этот метод с использованием |4С-соедииений для изучения ВПФ-цикла
(гл. 6).
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
225.
ным неорганическим фосфатом в хлоропластах и триозофосфа-'
том на переносчике фосфатов). В тех случаях, когда меченое
соединение способно подвергаться метаболизму (например, при
добавлении к хлоропластам меченого неорганического фосфата
на свету при комнатной температуре), распределение продук-
тов можно изучать при щомощи колоночной хроматографии.
Поскольку небольшое количество среды остается связанным с
хлоропластами даже после фильтрации через силиконовое мас-
ло, следует ставить параллельные опыты с мечеными соедине-
ниями, например декстранами, которые не проходят через мем-
браны (разд. 8.22).
Источники данных: |[6, 32, 37, 66].
8.6. Косвенные методы изучения транспорта
метаболитов
Эти методы основаны на том, что можно воздействовать на
поведение или функциональную активность хлоропластов, до-
бавляя во внешнюю среду различные реагенты. Например, ес-
ли какое-либо соединение стимулирует или подавляет фотосин^
тез, это достаточно убедительно говорит о том, что оно способ-
но проникать через мембрану хлоропластов. Данное утвержде-
ние относится прежде всего к тем соединениям, которые могут
влиять на процессы, протекающие в строме или тилакоидах
хлоропластов (рис. 8.10). Недостатки подобного подхода про-
истекают из того, что полученные результаты не могут быть
однозначно истолкованы. Ведь некоторые из добавленных со-
единений способны воздействовать на фотосинтез и не проникая
внутрь хлоропластов, а другие могут сначала подвергаться из-
менениям во внешней среде и только после этого проникают че-
рез мембрану хлоропластов.
Источник данных: {41].
8.7. Сокращение индукционного периода
и снятие ингибирующего действия ортофосфата
Как мы уже отмечали в гл. 7, фотосинтез начинается не
сразу после освещения, а лишь спустя некоторое время (индук-
ционный период). Это время увеличивается в присутствии орто-.
Рис. 8.8. Хлоропласт транспортирует метаболиты в окружающую его клеточ-
ную среду. Эти электронные микрофотографии клеток мезофилла шпината
дают хорошее представление об относительных объемах хлоропласта, цитозо-
ля и большой вакуоли; Х6400. (Фотографии любезно предоставлены
А. Д. Гринвудом из Империал-колледжа в Лондоне.)
15—234
226
ГЛАВА 8
Мешалка
Радиоактивное С.
вещество
Суспензия
хлоропластов
Силиконовое
масло
Хлорная кислота
Л
Нож
Суспендирование осадки
и подсчет радиоактивности
в кислоторастворимом
экстракте
Общее
пространство- Фильтрация
хлоропласта % центрифугированием
100
50
Концентрация
сорбитола
в среде
Электронная
микроскопия
Строма
Т илакоиды
Простран ci во
между мембранами
Рис. 8.9. Фильтрация центрифугированием (Heldt, 1976). А. Схематическое
изображение метода. Б. Сравнительное изображение межмембранного прост-
ранства по данным опытов с меченой сахарозой (слева) и планиметрии элек-
тронных микрофотографий изолированных хлоропластов (справа). Поглоще-
ние меченой сахарозы гораздо выше, чем поглощение декстранов, причем оно
варьирует в зависимости от осмотического давления окружающей среды. Сте-
пень неспецифического транспорта сахарозы (и других соединений) указыва-
ет на то, что ее молекулы проходят через наружную мембрану оболочки
хлоропластов, но не способны проникать через внутреннюю мембрану.
Б
ПЛАСТИДЫ и внутриклеточный транспорт
227
Рис. 8.10. Зависимое от феррицианида выделение кислорода интактными и
разрушенными (лишенными оболочки) хлоропластами. В две реакционные
смеси вносили интактные хлоропласты, ио в одну из них (верхняя кривая)
остальные реагенты добавляли в таком порядке, что хлоропласта в ней
подвергались кратковременному осмотическому шоку. Таким образом, в на-
чале освещения единственное различие между двумя смесями состояло в том,
что в одной из них все хлоропласта не имели оболочки. Видно, что в отно-
сительно интактных хлоропластах восстановление феррицианида ограничено,
так как он не проникает через оболочку. Напротив, NH3 легко проходит че-
рез оболочку и ускоряет процесс, действуя как разобщитель транспорта
электронов (Lilley ct al., 1971).
фосфата в концентрациях, превышающих оптимальные. Такие
соединения, как ФГК или триозофосфаты, сокращают индукци-
онный период (см. рис. 8.11 и 7.15) либо снимают ингибирую-
щее действие ортофосфата. Поэтому принято считать, что они
проникают через оболочку хлоропластов.
Источник данных: [59].
8.8. Добавление ингибиторов
Если только ингибирующее действие соединения на фото-
синтез не вызвано его влиянием на транспорт метаболитов,
есть основания полагать, что оно проникает внутрь хлоропла-
стов. Например, простой сахар DL-глицеральдегид в сильной
степени подавляет ассимиляцию углерода и, следовательно,
15*
228
ГЛАВА 8
Рис. 8.11. Первая регистрация сокращения индукционного периода при до-
бавлении 3-ФГК к изолированным хлоропластам. Слева показана кинетика
СОа-зависимого выделения Ог изолированными хлоропластами, справа — то
же для смеси, которая содержит также ФГК. По-видимому, эффект ФГК
объясняется эндогенным восстановлением. Рибулозобисфосфат (РуБФ), кото-
рый не проходит через мембрану хлоропластов, не обладает аналогичным
действием на фиксацию С02. Кроме того, он не способен поддерживать фик-
сацию СО2 хлоропластами в темноте, если их оболочки не разрушены (Wal-
ker, Hill, 1967).
должен легко проникать через оболочку хлоропластов (см.
также рис. 8.20).
Источник данных: [53].
8.9. Катализ интактными и разрушенными
хлоропластами
Ферменты стромы хлоропластов «защищены» в том смысле,'
что они отделены от окружающей среды мембраной. Неспо-
собность субстрата реагировать (например, отсутствие карбок-
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ 229
силирования при добавлении рибулозобисфосфата) указывает
на то, что он не проникает в хлоропласты. Однако могут быть
и другие причины. Так, реконструированная система хлоропла-
стов (разрушенные хлоропласты с добавленным к ним ферредок-
сином) легко выделяет кислород в присутствии ФГК и 1 мМ
MgCl2. Следовательно, все необходимые фотохимические про-
цессы и ферменты, катализирующие восстановление ФГК до
триозофосфата, эффективно функционируют в этих условиях
(рис. 9.4). Если же субстратами служат РуБФ и СО2, кислород
почти совсем ие выделяется до тех пор, пока концентрацию
MgCl2 не повысят до 2 мМ. В этом случае разрушается обо-
лочка хлоропластов и РуБФ становится доступным для дейст-
вия карбоксилазы. Однако одновременно концентрация Mg2+ в
строме хлоропластов настолько уменьшается, что в данных, экс-
периментальных условиях РуБФ-карбоксилаза не активирует-
ся. [Фотосинтез в интактных хлоропластах подавляется также
ионофором, который делает оболочку проницаемой для Mg2+;
процесс восстанавливается при повышении концентрации
Mg2+ во внешнем растворе.]
Источники данных: [41а, 45, 58].
8.10. Осмотические изменения объема
В некоторых случаях хлоропласты ведут себя как почти
идеальные осмометры. Теоретически, если какое-то пространство
окружено истинной полупроницаемой мембраной (которая про-
пускает молекулы воды, но не растворенные в ней вещества),
оно будет сжиматься или расширяться в зависимости от отно-
сительной концентрации внутреннего и наружного растворов.
Так, если концентрация веществ выше внутри, чем снаружи, во-
да будет стремиться внутрь (эндоосмос), и более концентриро-
ванный внутренний раствор будет разбавляться. Этот приток
воды вызовет увеличение внутреннего пространства (набуха-
ние). Наоборот, если наружный раствор является более кон-
центрированным и поэтому отличается более высоким осмоти-
ческим давлением, то произойдет экзоосмос воды и внутреннее
пространство уменьшится (сжатие) (разд. 4.1). Известно, од-
нако, что хлоропласты можно успешно выделять в растворах,
содержащих в качестве осмотиков сахарозу или сахарный
спирт, например сорбитол. Следовательно, мембрана хлоро-
пластов почти непроницаема для этих соединений. [Осмотика-
ми называют относительно нереакциоиноспособные (предполо-
жительно) соединения, которые добавляют с целью предот-
вращения осмоса, Соединение, проникающее в хлоропласт под
действием своего градиента диффузии, повысило бы внутреннее
230
ГЛАВА 8
0,1 д А
Рис. 8.12. Изменение поглощения света как показатель проницаемости. В оп-
ределенных концентрациях сорбитол, рибоза и аланин заставляют хлоропла-
ста сжиматься в результате экзоосмоса воды, а это изменение объема приво-
дит к быстрому возрастанию поглощения света при 535 нм. Однако в присут-
ствии рибозы данный эффект довольно быстро снимается, так как в этом
случае вслед за акзоосмосом наступает эндодиффузия рибозы и соответст-
вующее увеличение тургора (Gimmler et al., 1974).
осмотическое давление, что привело бы к эндоосмосу и набуха-
нию.]
При осмотическом набухании или сжатии происходят изме-
нения в видимом поглощении при 535 нм, что может быть ис-
пользовано как показатель проницаемости. Например, гипер-
тонический раствор сорбитола приводит к росту поглощения,
что говорит об экзоосмосе воды. Аналогичный эффект наблю-
дается для рибозы, однако в этом случае довольно скоро по-
глощение возвращается к исходному уровню. По-видимому, во-
да сначала выходит из хлоропластов, а затем по мере проник-
новения рибозы через оболочку хлоропластов поступает обрат-
но внутрь их (рис. 8.12). Недавно Хебер и его сотрудники при
помощи остроумной модификации этого подхода получили ин-
формацию относительно транспорта отдельных ионов (разд.
8.21).
Источники данных: [24, 30].
8.11. Транспорт метаболитов
Как мы уже отмечали в разд. 8.1, основной транспорт угле-
рода между хлоропластом Cg-растений и его клеточным окру-
жением по современным представлениям сводится к поступле-
нию СО2 внутрь хлоропласта и экспорту триозофосфатов.
[Транспорт метаболитов при фотосинтезе Сграстений будет
рассмотрен отдельно в гл. 12.]
ПЛАСТИДЫ и внутриклеточный транспорт
231
8.12. Двуокись углерода
Если уравнение (1.1) точно отражает суть фотосинтеза, то
СОз Должна поступать к месту ассимиляции углерода со ско-
ростью, необходимой для протекания этого процесса, Далее, ес-
ли карбоксилирование действительно осуществляется в строме
хлоропластов, то их оболочка должна быть полностью прони-
цаемой для СО2 в той или иной форме (рис. 8.13). Понятие о
хлоропласте как о самостоятельной фотосинтезирующей орга-
нелле долгие годы служило предметом споров. В прошлом ве-
ке хлоропласт стали считать местом фотосинтеза, так как крах-
малу отводили роль конечного продукта в этом процессе. Закс
установил, что крахмал образуется в хлоропластах в тех уча-
стках листа, которые подвергались освещению (рис. 8.3—8.5).
Представление о хлоропласте как о самостоятельной фотосин-
тезирующей органелле пришлось заново пересмотреть в трид-
цатых годах этого века, когда Хилл получил хлоропласты, спо-
собные выделять О2, но (как тогда казалось) ие фиксировать
Рис. 8.13. Первое доказательство зависимости фотосинтетической ассимиля-
ции углерода изолированными хлоропластами от СО2 (ср. с рис. 7.13). Погло-
щение кислорода в темноте (и некоторые другие пики в этой записи) следует
рассматривать в основном как артефакт. Тем не менее видно, что в отсутствие
СО2 (который с самого начала не содержался в реакционной смеси) выделе-
ние кислорода вскоре прекращается, но оно немедленно возобновляется, как
только добавляют СОг. Показана также зависимость процесса от освещения
(Walker, Hill, 1967).
232
ГЛАВА 8
Рис. 8.14. Опыты, аналогичные
представленному на рис. 8.13, ко-
торые проводили с использовани-
ем усовершенствованных методов,
Показывают, что в отсутствие СОа
выделение Оз можно вызвать до-
бавлением ФГК (Walker et al.,
1967).
СО2. Только в пятидесятых годах, когда Арион и его сотрудни-
ки доказали фиксацию 14СО2 изолированными хлоропластами,
этот вопрос был решен окончательно. Правда, сравнительно не-
давно было высказано предположение, что фотосинтез может
идти с максимальной скоростью в хлоропластах только тогда,
когда их инкубируют в реакционной смеси, состав которой
идентичен составу цитоплазмы, '[Согласно этой точке зрения,
для осуществления максимальной скорости фотосинтеза необ-
ходим быстрый обмен между триозофосфатами внутри хлоро-
пластов и наружным ортофосфатом. Таким образом, по край-
ней мере в этом отношении хлоропласт нельзя считать пол-
ностью самостоятельной фотосинтезирующей органеллой.] Мно-
гие вопросы фотосинтеза остаются открытыми до сих пор, ио
совершенно очевидно, что РуБФ-карбоксилаза локализована в
строме хлоропластов и именно здесь включается подавляющая
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ 233
часть всей двуокиси углерода, фиксируемой Сд-р астениями на
свету.
В растворе СОг подвергается гидратации с образованием
угольной кислоты:
СО3 + НаО «=> HSCO3 I-P 4- НСО3- (8.1)
Эта реакция катализируется различными неорганическими ио-
нами, например ортофосфатом, и ферментом карбоангидразой
(карбоиат-дегидратазой). В какой именно молекулярной форме
используется СО2, определить не просто. Купер с. сотрудника-
ми проводили опыты с РуБФ-карбоксилазой, в которых они
сравнивали фиксацию СО2 в реакционных смесях, содержащих
либо НиСОз“+12СО2, либо Н12СО3“+14СО2. Они пришли к вы-
воду, что в этой реакции используется именно СОг, а не бикар-
бонат. [Напротив, в реакции, катализируемой ФЕП-карбоксила-
зой, используется бикарбонат, а не СО2 (разд. 14.4).] Пуансело
установил, что интактные хлоропласты используют НСО3~ зна-
чительно быстрее, чем СО2. Вместе с тем Вердан и Хельдт об-
наружили, что накопление бикарбоната внутри хлоропластов
согласуется с градиентом pH в их оболочке и с уравнением
Гендерсона — Хассельбаха, которое выражает зависимость рас-
пределения ионных форм СО2 от преобладающего значения pH.
Эти авторы пришли к заключению, что подобное поведение
обусловлено быстрым и свободным движением СО2 (тогда как
движение ионов бикарбоната ограничено), как видно из урав-
нения (8.2):
Оболочка
Среда Хлоропласт
НСОз +Н' ^Н2О +со2 —- СО2 + Н7.ОгЖСО;+Н* (8.2)
Далее, Хебер и Пурцзельд показали, что бикарбонат вызы-
вает необратимое осмотическое сжатие хлоропластов (а это го-
ворит об отсутствии свободного проникновения ионов). Кроме
того, им удалось подтвердить данные Вердена и его сотрудни-
ков относительно того, что карбоангидраза ведет себя как «за-
щищенный» фермент (интактные хлоропласты почти ие ускоря-
ли гидратацию СОз, тогда как ее скорость резко возрастала в
смесях, содержащих хлоропласты с разрушенными оболочка-
ми).
Роль, которую карбоангидраза играет в строме хлороплас-
тов, остается непонятной. Нс исключено, что этот фермент об-
легчает транспорт СОз внутрь самой стромы, благодаря чему
там накапливается большое количество бикарбоната [при pH
234
ГЛАВА 8
8,0, т. е. при pH стромы освещенных хлоропластов (разд. 5.9)
98% общего количества СО2 находится в форме бикарбоната]:
СО2(2%) XZ>CO2(2%)
(8.3)
нсо; (98%)' ХХт'* нс°з (98%)
СО2 и бикарбонат диффундируют со скоростью, пропорцио-
нальной их концентрации, и, хотя РуБФ-карбоксилаза исполь-
зует только СО2, высокое содержание бикарбоната в строме
ускоряет транспорт двуокиси углерода.
Источники данных: [13, 14, 21, 30, 34, 44, 51, 54, 61, 65].
8.13. Триозофосфат и 3-фосфоглицерат (ФГК)
Интактные хлоропласты начинают быстро выделять кисло-
род, если добавить ФГК в отсутствие СО2 (рис. 8.14). Следо-
вательно, ФГК может играть роль окислителя Хилла в суммар-
ной реакции:
Н,ОЧ .NADP
ФГАЗ-АОР + Р,
(8.4)
ФГК+АТР
Кроме того, она должна для этого поступать в хлоропласт со
скоростью, соизмеримой со скоростью выделения кислорода.
ФГК и оба триозофосфата [фосфоглицериновый альдегид (ФГА)
и дигидроксиацетонфосфат] сокращают индукционный период
и снимают ингибирующее действие ортофосфата (разд. 7.11 и
7.12). На основании этого наблюдения Хельдт и его сотрудни-
ки провели прямые измерения (разд. 8.5), которые позволили
сформулировать гипотезу о переносчике ортофосфата в ее со-
временном виде (разд. 7.12 и 8.23). Транспорт всех трех соеди-
нений осуществляется при помощи переносчика со скоростями,
не уступающими максимальной скорости фотосинтеза или даже
превосходящими ее.
Источники данных: [6, 32, 55, 56].
8.14. Ортофосфат и неорганический пирофосфат
Если скорость фотосинтеза в изолированных хлоропластах
падает в результате истощения запасов ортофосфата, то ее
можно быстро восстановить за счет экзогенного ортофосфата.
При этом для того, чтобы С02-зависимое выделение кислорода
вновь достигло относительно высокого уровня, достаточно до-
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ транспорт
235
Рис. 8.15. Реакция изолированных хлоропластов шпината на экзогенный фос-
фат. С самого начала реакционные смеси не содержали ортофосфата, но его
эндогенная концентрация была достаточно высока для осуществления замет-
ного фотосинтетического выделения СОз, которое затем снижалось (по мере
потребления ортофосфата из стромы хлоропластов) до уровня, соответствую-
щего синтезу крахмала. При внесении экзогенного ортофосфата фотосинтез
восстанавливался со стехиометрией порядка трех молекул выделяемого кис-
лорода на одну молекулу добавленного ортофосфата. [Такая стехиометрия
согласуется с суммарным уравнением реакции (Р1-|-ЗСО2-|-2Н2О-->Триозофос-
фат-(-ЗО2) и с результатами опытов с использованием 14С и 32Р, которые
свидетельствуют о том, что основными продуктами кратковременного фото-
синтеза в интактных хлоропластах являются триозофосфаты.] Неорганический
пирофосфат не дает эффекта в отсутствие ионов магния, но в их присутствии
подвергается экзогенному гидролизу под действием пирофосфатазы стромы,
которая выделяется из разрушенных хлоропластов (см. рис. 8.17; Walker,
Herold, 1977).
бавить 0,05 мкмоль ортофосфата в 2 мл (рис. 8.15). Стехиомет-
рия этой реакции составляет примерно 3 молекулы кислорода
на одну молекулу ортофосфата; такое соотношение согласует-
ся с образованием одной молекулы триозофосфата из трех мо-
лекул СО2 и одной молекулы фосфата. Когда эту реакцию изу-
чали на хлоропластах шпината, оказалось, что неорганический
пирофосфат так же эффективен, как и ортофосфат; правда, в
этом случае реакция наблюдалась не сразу, а после некоторой
задержки (около 15 с). Стехиометрия процесса составляла при-
мерно 6 молекул кислорода иа 1 молекулу пирофосфата, отку-
236
ГЛАВА 8
Рис. 8.16. Снятие пирофосфатом ин-
гибирующего действия ортофосфата
на фотосинтез. Ортофосфат подав-
ляет фотосинтез в изолированных
хлоропластах: он стимулирует транс-
порт метаболитов ВПФ-цикла из
хлоропластов с помощью переносчи-
ка ортофосфата (разд. 8.23). Полное
подавление ортофосфатом и обраще-
ние этого эффекта при добавлении
ФГК (стрелки) показано кривыми II
и IV. Большой избыток пирофосфата
(кривая I) не оказывал ингибирую-
щего действия. Если среда содержа-
ла одновременно пирофосфат и ор-
тофосфат (кривая III), то пирофос-
фат в значительной степени снимал
ингибирующий эффект ортофосфата
(Walker, 1976).
но в присутствии пирофосфатазы. К этому времени стало из-
вестно, что субстратом пирофосфатазы служит Mg-пирофосфат
и что анионная форма пирофосфата не только не является суб-
стратом, но даже подавляет гидролиз Mg-пирофосфата. Отсю-
да следовало, что пирофосфат не способен проникать с какой-
либо заметной скоростью через мембрану хлоропластов. Для
того чтобы замещать ортофосфат, пирофосфат должен до
да следовало, что, по-видимо-
му, пирофосфат подвергался
полному гидролизу. Подобная
реакция была весьма неожи-
данной в свете известного ин-
гибирующего действия орто-
фосфата на фотосинтез, так
как дальнейшее повышение
концентрации пирофосфата не
вызывало подавления. Более
того, пирофосфат не только
не подавлял фотосинтез, но
при высоких концентрациях
(—' 5 мМ) даже защищал от
ингибирующего действия ор-
тофосфата (рис. 8.16). Час-
тичное объяснение этим каза-
лось бы противоречивым ре-
зультатам было получено в
опытах, которые показали, что
для ряда препаратов хлоро-
пластов реакция на добавле-
ние пирофосфата наблюдает-
ся не сразу или вообще отсут-
ствует (рис. 8.17). Положи-
тельного эффекта в таких слу-
чаях добивались внесением в
реакционную смесь либо не-
большого количества хлоро-
пластов, подвергнутых осмо-
тическому шоку, либо неорга-
нической пирофосфатазы, вы-
деленной из стромы хлоропла-
стов. Кроме того, реакции па
добавление пирофосфата не
было в отсутствие экзогенно-
го магния, но она наблюда-
лась почта сразу вслед за вне-
сением ионов магния, особен-
ПЛАСТИДЫ и внутриклеточный транспорт
237
Время. мин
Рис, 8.17. Непроницаемость мембран хлоропластов для пирофосфата. В слу-
чае преобладания интактных хлоропластов дефицит ортофосфата снимается
не пирофосфатом, а самим ортофосфатом (кривая /). Добавление небольшой
фракции (5%) разрушенных хлоропластов (кривая II), которая содержит
достаточное количество пирофосфатазы, приводит к гидролизу пирофосфата,
и он начинает функционировать как две молекулы ортофосфата (см.
рис. 8.15; Walker, 1976).
проникновения в хлоропласты подвергнуться гидролизу пиро-
фосфатазой, освобожденной из разрушенных хлоропластов.
Поскольку обычно среда содержит' мало пирофосфатазы, а
свободный ортофосфат оказывает на нее ингибирующее дейст-
вие, в присутствии высоких концентраций пирофосфата ско-
рость гидролиза возрастает незначительно. Впоследствии в
прямых экспериментах было показано, что пирофосфат прони-
кает в хлоропласт очень медленно и что он связывается с пе-
реносчиком ортофосфата (разд. 8.23) и тем самым частично
снимает ингибирующее действие ортофосфата. Именно поэто-
му в изолированных хлоропластах оптимальная скорость фо-
тосинтеза наблюдается обычно в присутствии 5 мМ пирофос-
фата. При добавлении во внешнюю среду ионов магния пиро-
фосфат поддерживает постоянную концентрацию ортофосфата
благодаря тому, что последний выделяется в результате его
гидролиза примерно с такой же скоростью, с какой ортофос-
фат используется хлоропластами. Одновременно пирофосфат
предупреждает ингибирующий эффект ортофосфата, неизбеж-
ный при его концентрациях свыше 0,5 мМ. (разд. 8.7). Кроме
238 ГЛАВА 8
Рис. 8.18. Подавление фотосинтеза в хлоропластах гороха пирофосфатом.
В отличие от шпината у проростков гороха мембраны хлоропластов имеют
ббльшую проницаемость для пирофосфата, который оказывает ингибирующее
действие, и для АТР, который снимает этот эффект (кривая I). «Нормальное»
ингибирование ортофосфатом снимается ФГК, как и в случае шпината, но не
АТР (кривая II) (Robinson, Wiskich, 1977).
того, пирофосфат может защищать хлоропласты благодаря об-
разованию хелатных комплексов с дивалентными катионами
(механизм ингибирующего действия катионов пока еще в точ-
ности не известен).
В хлоропластах гороха (или в хлоропластах, выделенных
из незрелых тканей) реакция на пирофосфат еще более ослож-
няется действием многочисленных факторов. Так, пирофосфат
может обмениваться с эндогенным АТР при помощи переносчи-
ка аденилатов (разд. 8.25), что приводит к ингибированию
(рис. 8.18). Робинсон и Уискич высказали предположение, что
пирофосфат может также проникать в хлоропласты с помощью
переносчика ортофосфата (разд. 8.23), подвергаться внутри них
гидролизу и снижать экспорт промежуточных метаболитов в
результате конкуренции с фосфоглицератом за транспорт из
хлоропластов. Это объяснило бы тот факт, что ADP наряду с
подавляющим действием на хлоропласты гороха (он либо ин-
гибирует превращение ФГК в триозофосфат, либо создает де-
фицит ортофосфата за счет обмена между АТР и ADP) обла-
дает стимулирующим действием, если его добавлять вместе с
пирофосфатом (который сам по себе привел бы к ингибирова-
ПЛАСТИДЫ и внутриклеточный транспорт
239
нию). По-видимому, если ADP и пирофосфат присутствуют од-
новременно, они конкурируют друг с другом за транспорт
внутрь хлоропластов. В результате наблюдается стимулирую-
щий эффект под действием пирофосфата либо по вышеуказан-
ным причинам, либо потому, что пирофосфат приостанавлива-
ет транспорт метаболитов, вызванный ADP-индуцированным
экспортом АТР.
Источники данных; [18, 19, 20, 33, 40, 46, 47, 60, 61].
8.15. Пентозомонофосфаты и гексозомонофосфаты
Все три пентозомоиофосфата ВПФ-цикла снимают ингиби-
рующее действие ортофосфата, но не сразу, а после некоторой
задержки (см. рис. 7.22), что можно расценивать как доказа-
тельство их медленного проникновения в хлоропласты. Кроме
того, Хельдт и соавторы изучали транспорт, непосредственно
измеряя поглощение Р5Ф, подавляемое ортофосфатом, что
указывает на медленный перенос (около 1—2 мкмоль па 1 мг
хлорофилла в 1 ч) этого пентозомоиофосфата через мембрану
хлоропластов при помощи переносчика ортофосфата (табл.
8.2).
Таблица 8.2. Транспорт фосфорилированных метаболитов в строму
хлоропластов шпината при температуре 20 °C в темноте (Fliege et al., 1978)
Метаболит (2 мМ) Скорость транспорта мкмольЦмгХл)-1-1!-1
1-1- неоргани- ческий фос- фат (0,2 мМ) -1- D-глюкоза 1(20 мМ)
Неорганический пирофосфат 1,3 0,4 Не опреде- ляли
Рибозо-5-фосфат 1,7 1,1 1,9
Г люкозо-1 -фосфат 0,07 0,04 0,08
Глюкозо-6-фосфат 0,43 0,44 0,24
Фруктозо-6-фосфат 0,32 0,20 0,34
Фруктозо-1,6-бисфосфат 0,85 0,53 0,86
Глюкоиат-6-фосфат 0,23 0,22 0,28
Прямыми измерениями продемонстрирован также транспорт
Ф6Ф и Г6Ф в хлоропласты со скоростями, почти в четыре раза
меньшими, чем в случае Р5Ф. Поглощение Ф6Ф и Г6Ф подав-
ляется соответственно ортофосфатом и глюкозой, что говорит
об участии в транспорте Ф6Ф переносчика ортофосфата (разд.
8.23), а в транспорте Г6Ф — переносчика глюкозы (разд. 8.17).
Имеются данные, что Ф6Ф снимает ингибирующее действие ор-
240
ГЛАВА 8
тофосфата, но этот эффект не всегда удается воспроизвести.
Ф6Ф и Г6Ф довольно быстро появляются в цитоплазме листь-
ев, освещенных в присутствии 14СОг, но это может быть и ре-
зультатом их экзогенного образования из триозофосфатов.
Источники данных: ,[18, 22].
8.16. Бисфосфаты сахаров
Рибулозобисфосфат (РуБФ) не появляется в цитоплазме ос-
вещенных листьев или в среде, в которой хлоропласты инкуби-
руют на свету. Он не снимает ингибирующего действия орто-
фосфата и не сокращает индукционный период. Если добавить
РуБФ в темноте к разрушенным хлоропластам, он стимулирует
фиксацию МСО2 в гораздо большей степени, чем в случае пре-
обладания интактных хлоропластов. Короче говоря, все экспе-
риментальные данные указывают на то, что мембрана хлоро-
пластов в основном непроницаема для этого соединения. Пред-
полагают, что СБФ и ФБФ ведут себя аналогично РуБФ. Впро-
чем, следует остерегаться однозначного толкования результа-
тов, полученных для ФБФ, поскольку это соединение очень
легко образуется из триозофосфатов в присутствии альдолазы.
Источники данных: [3, 10, 11].
8.17. Свободные сахара
Хлоропласты можно выделять в растворах, содержащих в
качестве осмотиков сахарозу, фруктозу или глюкозу (а также
сорбитол или маннитол, которые чаще используются с этой
целью). Уже сам по себе этот факт говорит об их неспособно-
сти быстро проникать в хлоропласт (разд. 8.10). Как показали
Таблица 8.3. Скорость транспорта сахаров
в хлоропласты (время измерения 20 с;
Schafer et al., 1977)
Сахар (5 мМ) Скорость транспор- та, МКМОЛЬ'(МГ Хлу-'-ч-1
D-глюкоза 7,4
L-глюкоза 0,3
D-манноз а 8,4
D-фруктоз а 2,8
D-ксилоза 9,3
D-рибоза 6,0
L-арабиноза 7,7
D-арабиноза 1,6
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ транспорт
241
Время, мин
Рис. 8.19. Поглощение стереоизомеров глюкозы интактными хлоропластами
шпината. Цифры около кривых обозначают скорость поглощения в мкмоль*
(мг Хл)-1-ч~‘ (Schafer et al., 1977).
непосредственные измерения, ни сахароза, пи сорбитол не мо-
гут быстро переноситься через внутреннюю мембрану хлоро-
пластов (разд. 8.22). Имеются некоторые косвенные данные о
транспорте фруктозы в хлоропласты. Что же касается глюко-
зы, то до недавнего времени мембрану хлоропластов считали
непроницаемой для нее. Однако, когда к дискам листьев таба-
ка добавили специфически меченую глюкозу, оказалось, что в
образовавшемся крахмале сохраняется характерное распреде-
ление метки. [Если бы глюкоза сначала превратилась в три-
озофосфат, который после проникновения в хлоропласты опять
дал гексозу, то произошла бы рандомизация метки.] Дальней-
шие исследования, проведенные Шефером, Хебером и Хельд-
том при помощи фильтрации центрифугированием, показали,
что глюкоза и некоторые другие свободные сахара действи-
тельно транспортируются в хлоропласты с низкой скоростью
(табл. 8.3) и, как видно из поведения хлоропластов, диффузия
сахаров происходит с помощью переносчика (рис. 8.19). В опы-
тах, основанных на измерении осмотических изменений объ-
емов, также было показано, что рибоза проникает в хлоропла-
сты (рис. 8,12). Кроме того, в некоторых случаях фруктоза и
другие свободные сахара оказывают влияние на индукцию.
Интересно, что транспорт D-глюкозы подавляется, если она
присутствует в высокой концентрации; это объясняет тот факт,
что данный сахар столь же эффективен в качестве осмотика,
как сахароза или сорбитол. Триоза DL-глицеральдегид являет-
ся сильным ингибитором ассимиляции углерода при фотосин-
тезе (рис. 8.8). Реакция хлоропластов на добавление DL-гли-
церальдегида говорит о том, что он проникает через мембрану
16—234
242
ГЛАВА 8
Рис. 8.20. Подавление СО2-зави-
симого выделения О2 DL-глицер-
альдегидом. Причина ингибирую-
щего действия этого соединения
на фотосинтетическую ассимиля-
цию углерода заключается, по-
видимому, в торможении им ре-
акции, катализируемой транскето-
лазой. Полное подавление фото-
синтеза в интактных хлороплас-
тах DL-глицеральдегидом всего
за несколько минут указывает па
сравнительно быстрое проникнове-
ние этого соединения в хлоропла-
сты (Stokes, Walker, 1972).
быстрее, чем сахара с более длинной углеродной цепью (рис.
8.20).
Источники данных: [10, 11, 22, 52, 53].
8,18. Карбоновые кислоты
Такие дикарбоновые кислоты, как яблочная (малат), щаве-
левоуксусная (оксалоацетат), янтарная (сукцинат), фумаровая
(фумарат) и 2-оксоглутаровая (2-оксоглутарат), довольно
Концентрация глицериноаой кислоты, мМ
Рис. 8.21. Зависимость выделения кислорода интактными хлоропластами шпи-
ната от концентрации глицериновой кислоты в среде. Реакция определяется
транспортом глицериновой кислоты в хлоропласты и ее последующим пре-
вращением в ФГК, которая затем восстанавливается за счет образующихся
при фотосинтезе АТР и NADPH (Heber ei al., 1974).
ПЛАСТИДЫ й ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
243
быстро проникают в хлоропласты при помощи переносчика ди-
карбоновых кислот. Столь же быстро должна транспортиро-
ваться глицериновая кислота, судя по тому, что ее добавление
К интактным хлоропластам почти сразу приводит к выделению
кислорода (рис, 8,21). Данные сравнительной биохимии, а так-
же роль гликолата и фосфогликолата в фотодыхании (гл. 14)
позволяют предполагать относительно свободное передвижение
этих соединений (или хотя бы одного из них) через мембрану
хлоропластов, однако прямые доказательства их транспорта,
как это ни странно, до сих пор отсутствуют.
Источники данных: <['29, 39].
8.19. Аминокислоты
Аспартат и глутамат переносятся через мембрану хлоро-
пластов при помощи переносчика дикарбоновых кислот почти
с той же скоростью, что и малат, а аспарагин и глутамин не-
сколько медленнее. Однако, как следует из имеющихся экспе-
риментальных данных, транспорт нейтральных аминокислот ие
может происходить быстрее, чем это необходимо для поддержа-
ния белкового синтеза [~0,1 мкмоль-(мгХл)"1^"1], Резуль-
таты исследования транспорта глицина и серина указывают на
то, что эти соединения нс могут непосредственно переноситься
со скоростью, которая позволила бы им играть существенную
роль в транспорте углеродных соединений из хлоропластов.
Источники данных: [3, И, 24].
8.20. АТР и NADP
NADP действует как окислитель Хилла в присутствии фер-
редоксина и хлоропластов, подвергнутых осмотическому шоку,
но ие интактных хлоропластов. По-видимому, прямого транс-
порта пикотинамлдиуклеотидов почти не происходит. Вопрос о
прямом транспорте адениновых нуклеотидов (помимо медлен-
ного переноса при помощи переносчика аденилатов) остается
открытым, но нет сомнения, что хлоропласты, подвергнутые ос-
мотическому шоку, фосфорилируют экзогенный ADP гораздо
быстрее, чем интактные хлоропласты. Кроме того, АТР не сни-
мает подавление фотосинтеза, вызванное такими разобщающи-
ми агентами, как нигерицин или аммиак, либо флоридзином
(который, как известно, подавляет конечную стадию синтеза
АТР). Если бы все эти соединения лишь препятствовали обра-
зованию АТР и если бы АТР свободно проникал через мембра-
ну хлоропластов (рис. 8.22), то он снимал бы это подавление.
В зрелых растениях шпината, например, аденилаты не способ-
ны к свободному прохождению через мембрану (рис. 8.23). Од-
16*
.244
ГЛАВА 8
Рис. 8.22. АТР снимает действие нигерицина на выделение кислорода рекон-
струированной системой хлоропластов (ниокняя кривая слева). Этот эффект
АТР отсутствует в случае интактных хлоропластов (нижняя кривая справа).
Ннгерицнн разобщает образование АТР и транспорт электронов. Отсутствие
эффекта АТР во втором случае объясняется тем, что оболочка хлоропластов
непроницаема для АТР (Walker, 1976).
Время,мин
Рис. 8.23. Реакция интактных хлсропластов шпината (слева) и реконструи-
рованной системы хлоропластов шпината (справа) на добавление ADP. ADP
медленно проникает через мембрану интактных хлоропластов (слева), что
снижает его ингибирующий эффект, который ярко выражен, когда мембраны
нет (справа) (Walker, 1976).
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
245
нако, как показали последние исследования, в хлоропластах го-
роха переносчик аделанитов (разд. 8.25), гораздо активнее
(рис. 8.18). Эта разница в результатах, по-видимому, объясня-
ется не межвидовыми различиями, а возрастными, поскольку
в опытах с более старыми проростками гороха (по сравнению
с обычно используемыми 10—15-дневными) аденилаты не про-
никали через мембрану. Если проницаемость мембраны хло-
ропластов действительно меняется с возрастом, это в первую
очередь свидетельствует о том, что роль хлоропласта в общей
экономике метаболизма тоже при этом меняется.
Источники данных: [3, 4, 5, 11].
8.21. Транспорт ионов
Если осветить хлоропласты, подвергнутые осмотическому
шоку, они захватывают протоны из окружающей среды, и pH
тилакоидов снижается примерно на 3 единицы (разд. 5.14).
В интактных хлоропластах строма становится более щелочной
по сравнению с внешней средой. Сохранение относительно ще-
лочных значений pH в строме (гл. 5.9) свидетельствует о том,
что мембрана хлоропластов ие отличается большой проницае-
мостью для ионов ОН" или Н+.
Добавление KNO2, NaNO2 и NH4C1 к суспензиям интактных
хлоропластов приводит к осмотическому сжатию, которое сни-
мается внесением валиномицина в смеси, содержащие соли К+
и NH4+ (валиномицин делает мембрану проницаемой для ионов
калия и аммония). Создается впечатление, что мембрана про-
ницаема для NO2" и С1~, так как сам по себе облегченный
транспорт катионов не вызывает обращения осмотического эф-
фекта из-за ингибирования нарушенного равновесия зарядов.
NaNO3 также не снимает осмотический эффект, что подтверж-
дает приведенное выше объяснение и указывает па непроницае-
мость мембраны для Na+. Непроницаемость мембраны для
NH4+ следует из того, что при освещении интактных хлоропла-
стов в присутствии NH4C1 внешняя среда подкисляется. Это
объясняется диссоциацией:
NH4+---->NHS + H+ (8.5)
после которой NH3 поступает внутрь хлоропластов. Добавление
(NH4)2SO4 К суспензии хлоропластов в KNO2 приводит к быст-
рому осмотическому набуханию, которое объясняется взаимо-
действием ионов:
NJV + NO2- ---> NH3 + HNO3 (8.6)
и проникновением NH3 и HNO2 в хлоропласты. Валиномицин
почти ие влияет на осмотическую реакцию хлоропластов в
246
ГЛАВА 8
(NOJ +ЗНгО ► N Н 4 + 20Н~ + 1,5O2)
Рис. 8-24. Транспорт ионов через оболочку хлоропласта. Обычно оболочка
сравнительно непроницаема для таких дивалентных катионов, как Mg2+,
и для таких моновалентных катионов, как Na+ и Н+, хотя такие монова-
лентные анионы, как С1~ и КЮ2~, проходят через нее довольно легко. Неко-
торые ионофоры и другие соединения, показанные на рисунке, способны из-
менять проницаемость оболочки. Приведены также уравнения реакции вос-
становления NOa- (разд. 12.9).
K2SO4 или (NH4)2SO4, что указывает на низкую проницаемость
мембраны хлоропластов для сульфат-иона. Вместе с тем еще
с 1968 г. известно, что сульфат подавляет фотосинтез в интакт-
ных хлоропластах (возможно, замещая ортофосфат при фото-
фосфорилировании) и что это подавление легко снимается ор-
тофосфатом. Столь же хорошо известно, что в хлоропластах
происходит ассимиляционное восстановление сульфата. Недав-
но Мурью и Дус на основании своих исследований с помощью
фильтрации центрифугированием сформулировали понятие о
переносчике сульфата, который осуществляет обмен между ор-
тофосфатом и сульфатом со скоростью ~20 мкмоль-(мгХл)-1-
•ч-1. На рис. 8.24 представлены многие из найденных взаимо-
связей, в том числе восстановление нитрита в аммиак (разд.
12.9).
Валиномицин не снимает осмотического сжатия в присутст-
вии фосфата калия. Следовательно, поглощение фосфата мо-
жет происходить только благодаря противообмену триозофос-
фатов или ФГД при помощи переносчика ортофосфата.
Источники данных: [15, 21а, 25, 28, 30, 43].
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
247
8.22. Специфическая проницаемость
внутренней мембраны
Мембрана хлоропластов непроницаема для меченого декст-
рана, но, как было установлено, при помощи фильтрации цент-
рифугированием, через нее легко проходят все испытанные со-
единения сравнительно низкой молекулярной массы. Опыты с
использованием меченой сахарозы позволили обнаружить кор-
реляцию между степенью проницаемости для этих соединений
и объемом между двумя мембранами оболочки, который в ги-
пертонической среде увеличивается (разд. 8.10). Таким обра-
зом, это проницаемое для сахарозы пространство можно рас-
сматривать как межмембранное (рис. 8.9,Б), причем из двух
мембран, ограничивающих его, специфической проницаемостью
обладает только внутренняя мембрана. Селективность прони-
цаемости этой мембраны на редкость удивительна: так, напри-
мер, транспорт ортофосфата через мембрану хлоропластов
шпината осуществляется с очень большой скоростью, в то вре-
мя как пирофосфат почти не проходит через нее (рис. 8.15,
8.17 и 8.21), а D-глюкоза проникает в 25 раз быстрее, чем
L-глюкоза.
8.23. Переносчик ортофосфата
Как мы уже отмечали в разд. 7.12, косвенные данные позво-
ляют предполагать, что ортофосфат обменивается с фосфатами
сахаров. Хельдт и его соавторы обнаружили такой обмен меж-
ду ортофосфатом и триозофосфатами при помощи прямых из-
мерений (разд. 8.5), благодаря чему понятие о переносчике
ортофосфата обрело реальность (рис. 8.25). Количество орто-
фосфата, которое выделяется из хлороплг.стов, преинкубирован-
ных с 32Р-ортофосфатом, эквивалентно количеству использован-
ной 14С-ФГК, что говорит о строгом сопряжении экспорта и
импорта метаболитов (рис. 8.26, 8.27). Ортофосфат не способен
свободно диффундировать из хлоропластов; выходить оттуда он
может лишь в присутствии экзогенной ФГК или триозофосфа-
тов. Кроме того, переносчики транспортируют арсенат и отчас-
ти вещества типа 1-глицерофосфата. Транспорт рибозо-5-фосфа-
та идет медленно, а соединения, подобные фруктозо-6-фосфату,
практически совсем не переносятся. Транспорт ортофосфата,
ФГК и триозофосфатов осуществляется очень быстро-----200—
300 мкмоль- (мгХл)-1 • ч-1 при 20°C. Уже через 240 с концент-
рация ортофосфата или ФГК в строме становилась в 20 раз
больше, чем во внешней среде. С точки зрения кинетики пере-
носчик ведет себя как фермент, и зависимость 1/v от 1/[3] вы-
ражается прямой линией (рис. 8.28). Четыре основных соеди-
248
ГЛАВА 8
Рис. 8.25. Переносчик ортофосфата (ПОФ) находится во внутренней мембра-
не и способствует обмену между экзогенным ортофосфатом и эндогенным
ДГАФ в строго стехиометрическом соотношении. Другие соединения, напри-
мер ФГК, также обмениваются с ортофосфатом или ДГАФ. В зависимости
от условий транспорт может идти в обоих направлениях (Heldt, 1976; Wal-
ker, 1974; Walker, 1976).
нения, которые переносятся при помощи переносчика, конкури-
руют друг с другом, и величины их Ki (концентрация одного из
соединений, необходимая для того, чтобы снизить вдвое ско-
рость транспорта другого соединения) сходны с величинами их
Км (концентрация, требующаяся для достижения половины
максимальной скорости транспорта). Величины /(i, Км и Ктах
приведены в табл. 8.4 и 8.5.
При одновременном измерении концентрации протонов в
среде и их транспорта в хлоропласты было показано, что перенос
ФГК сопровождается транспортом протона (возможно, в ре-
Таблица 8.4. Кинетические константы транспорта фосфатов в строму
хлоропластов, полученные при изучении противообмена при 4 °C
в темноте (Fliege et al.. 1978)
Фосфат Дм, мМ V max’ мкмоль«(мг Хл)—1«ч—1
Неорганический фосфат 0,30 (0,15—0,50) 57 (40-91)
Дигидроксиацетонфосфат 0,13 (0,06-0,22) 51 (36-74)
D-Г лидер альдегидфосф ат 0,08 (0,03-0,17) 41 (37-44)
З-Фосфоглицериновая кислота 0,14 (0,09—0,19) 36 (29-45)
2-Фосфоглицериновая кислота 2,8 (1,8-3,7) 24 (20-26)
1-Фосфоглицерол 1,1 (0,7-1,4) 59 (38-71)
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
249
Рис. 8.26. Стехиометрия транспорта ортофосфата из хлоропластов и ФГК в
хлоропласты при помощи переносчика ортофосфата (Heldt, 1976).
Рис. 8.27. Одновременное измерение транспорта 3-ФГК в строму хлороплас-
тов и поглощения протонов во внешней среде (Fliege et al., 1978).
зультате реакции ФГК3“+Н+—>-ФГК2~)- Взаимосвязь между
pH и транспортом говорит о том, что и остальные три соеди-
нения переносятся в виде дивалентных анионов. Транспорт
ФГК в форме дивалентного аниона может объяснить тот факт,
что при стационарном фотосинтезе в строме хлоропластов ФГК
содержится почти в 10 раз больше, чем триозофосфатов. Зна-
чение pH в строме становится более щелочным во время осве-
щения, и если ФГК требуется получить протон, то она будет
250
ГЛАВА 8
Транспорт Р; Транспорт ФГК
Ингибирование ФГК Ингибипоеакий Pj
К-, (ФГК) ~ O.SOmM Xj (Pj ) = 1,23 мМ
Транспорт Pj Транспорт ФГК
ХМ|Р, | = 0,44 мМ
X; (ДГАФ) = 0,19 мМ
хм (ФГК) --- 0,33 мМ
X; (ДГАФ) =0,21 мМ
Рис. 8.28. Перенос ортофосфата и' ФГК с помощью переносчика ортофосфата.
Каждый переносимый метаболит способен конкурировать с другими транс-
портируемыми соединениями за этот переносчик (Werdan, Heidi, 1971).
ПЛАСТИДЫ и внутриклеточный транспорт
261
Рис. 8.29. Ингибирующий эффект пиридоксальфосфата (ПФ) на переносчик
ортофосфата и его влияние на фотосинтетическую ассимиляцию углерода
(табл. 8.3). Как и пирофосфат, пиридоксальфосфат ингибирует переносчик
ортофосфата, но его эффект более однозначен, так как в отличие от пиро-
фосфата он не является в то же время потенциальным источником ортофос-
фата. Приведены скорости фотосинтеза при трех концентрациях ортофосфата.
Концентрация 0,1 мМ является субоптимальной (слева); концентрация 0,5 мМ
приближается к оптимальной (в центре); при концентрации 2,5 мМ. проявля-
ется ингибирующее действие (справа), так как при этом стимулируется
экспорт промежуточных продуктов ВПФ-цикла. Соответственно в первом
случае (слева) ингибирующий эффект пиридоксальфосфата обусловлен тем,
что он замедляет транспорт ортофосфата в хлоропласты ниже оптимальной
скорости. Во втором случае (в центре) при оптимальной концентрации орто-
фосфата ингибирующее действие пиридоксальфосфата выражено слабее. На-
конец, в третьем случае (справа), где сам ортофосфат играет роль ингибито-
ра, пиридоксальфосфат, подобно пирофосфату, проявляет защитное действие
(см. рис. 8.16) (Flflgge et al., 1980).
с большим трудом покидать щелочную строму, чем соединение,
которое в этом не нуждается.
Переносчик ортофосфата ингибируется п-хлорфенилртуть-
сульфонатом, пиридоксаль-5-фосфатом (табл. 8.6) и тринитро-
бензолсульфонатом. Неорганический пирофосфат связывается
с переносчиком с Ль почти в 10 раз большей, чем Лм для орто-
фосфата, но по скорости транслокации уступает ортофосфату
почти в 100 раз. Соответственно при высокой концентрации пи-
рофосфат является прекрасным ингибитором; он защищает
изолированные хлоропласты от ингибирующего действия из-
бытка ортофосфата (рис. 8.29). Сейчас уже можно с достаточ-
252
ГЛАВА 8
Таблица 8.5. Конкурентное ингибирование транспорта неорганического
фосфата (Км = 0,20 мМ) в строму хлоропластов при температуре 4 °C
в темноте (Fliege et al., 1978)
Ингибитор, мМ мМ
Дигидрокси ацетонфосфат 0,13
З-Фосфоглицериновая кислота 0,15
2-Фосфоглицериновая кислота 6,5
2,3-Днфосфоглицериновая кислота 9,1
Фосфоенолпируват 4,7
Эритрозо-4-фосфат 2,3
Рибозо-5-фосфат 10,0
Глюкозо-6-фосфат 40,0
Фруктозо-6-фосфат 13,0'
Фруктозо-1,6-бисфосфат 8,5
6-Фосфоглюконовая кислота 20,0
Неорганический пирофосфат 1,8
Таблица 8.6. Транспорт метаболитов из стромы хлоропластов,
индуцированный высокими концентрациями экзогенного ортофосфата,
и его подавление в результате ингибирования переносчика ортофосфата
пиридоксальфосфатом (Fliigge et al., 1979)
Ортофосфат, мМ Пиридоксаль-5'-фосфат, мМ 0,5 0 2,0 0 2,0 1.0
Концентрация метаболита в строме, мМ
Триозофосфаты 0,14 .Следы 0,18
З-Фосфоглицериновая кислота 4,44 3,69
Гексозомоиофосфаты и гептозомоно- фосфаты 1,96 0,10 1,85
Фруктозо-1,6-бисфосфат 0,29 Следы 0,37
Седогептулозо-1,7-бисфосфат 0,11 » 0,10'
Пентозомоно фосфат 0,08 » » 0,02
Рибулозо-1,5-бисфосфат 0,46 0,06 0,30
ATP/ADP 0,42 1,75 0,98
ПЛАСТИДЫ и внутриклеточный транспорт
253
ной уверенностью считать, что основная функция переносчика
ортофосфата сводится к быстрому переносу этого соединения в
хлоропласты и экспорту оттуда ДГАФ. Последний является
главным метаболитом, транспортируемым из хлоропластов в
цитоплазму: предшественником синтеза сахарозы в цитоплаз-
ме или субстратом в процессе гликолиза. Кроме того, ДГАФ'
благодаря своему участию в челночных механизмах может спо-
собствовать переносу из хлоропласта ассимиляционной силы
(разд. 8.26).
Источники данных: [6, 22, 23, 32].
8.24. Переносчик дикарбоновых кислот
Этот переносчик напоминает в основных чертах переносчик
ортофосфата, но отличается от него тем, что переносит дикар-
боновые кислоты и что противообмеи в нем сопряжен не столь
строго. Величина Ki ингибирования транспорта малата фума-
ратом близка к Км транспорта фумарата. Конкурентное инги-
Рис. 8.30. Зависимость транспорта малата при помощи переносчика дикарбо-
новых кислот в хлоропласты шпината при 4 °C от его концентрации и подав-'
ление этого транспорта другими дикарбоновыми кислотами (Heldt, 1976).
254
ГЛАВА S
Таблица 8.7. Кинетические константы транспорта дикарбоновых кислот
в непроницаемое для сорбитола межмембранное пространство
хлоропластов шпината при температуре 4 °C (Lehner, Heldt, 1978)
Дикарбоновая кислота ^гпах ‘ мкмоль-(мг Хл)-|-ч-1 Км . мМ
L-Малат 18,6 0,39
Сукцинат 14,0 0,26
Фумарат 18,6 0,21
L-Аспартат 31,1 0,72
а-Кетоглутарат 26,4 0,19
L-Глутамат 7,9 1,17
Для хлоропластов водной культуры шпината были получены величины Vmax в Б раз превышающие значения, приведенные выше.
бирование транспорта малата было продемонстрировано и в
присутствии других дикарбоновых кислот (рис. 8.30). При 4 °C
величина Ктах равна 19 мкмоль-(мгХл)-1-ч-1 для малата и
31 мкмоль-(мгХл)-1-ч-1 для аспартата (табл. 8.7). [Эти вели-
чины составляют примерно от одной второй до одной третьей
значений Ктах при 4°C для переносчика ортофосфата.]
Переносчик дикарбоновых кислот играет, по-видимому, бо-
лее важную роль в функционировании челночных механизмов
(разд. 8.26), чем в транспорте углеродных соединений.
Источники данных: [6, 23, 32, 39].
8.25. Переносчик аденилатов
Переносчик аденилатов облегчает противообмен адениновых
нуклеотидов. В хлоропластах шпината самые высокие скоро-
сти [~5 мкмоль-(мгХл)-1-ч-1 при 20°С] наблюдаются в при-
сутствии экзогенного АТР. В хлоропластах молодых листьев
гороха скорость транспорта аденилатов еще выше (в 5 или 10
раз); пирофосфат, по-видимому, также переносится со значи-
тельной скоростью и вызывает ингибирование, которое снима-
ется ADP или АТР (разд. 8.14). Какова бы ни была роль пе-
реносчика аденилатов, низкая скорость его функционирования
не позволяет предполагать, что он в значительной мере способ-
ствует экспорту ассимиляционной силы из зрелых хлороплас-
тов. Правда, в молодых тканях он может играть более важную
роль.
Источники данных: [6, 31, 32, 39].
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ транспорт
255
8.26. Челночные механизмы
Как только стало ясно, что ассимиляционная сила не мо-
жет быстро переноситься из хлоропластов путем прямого транс-
порта, было высказано следующее чисто теоретическое сооб-
Рис. 8.31. Челночный механизм транспорта ФГК и ДГАФ. Внутри хлоропла-
ста ФГК восстанавливается до триозофосфата за счет NADPH и АТР. Во
внешней среде образуются iNADH и АТР из триозофосфата ДГАФ, который
экспортируется из хлоропласта. ФГК может опять поступать в хлоропласт,
что обеспечивает беспрерывное действие челночного механизма (Heber, 1974}
Walker, 1974).
Рис. 8.32. Челночный механизм транспорта малата и оксалоацетата. Внутри
хлоропласта оксалоацетат восстанавливается до малата за счет NADPH. В окру-
жающей цитоплазме малат опять окисляется под действием малатдегидроген-
азы. Таким образом при помощи этого челночного механизма через мембрану
хлоропласта переносятся восстановительные эквиваленты (Heber, 1974; Wal-
ker, 1974).
256
ГЛАВА 8
ражение. Не исключено, что триозофосфаты транспортируются
из хлоропластов в цитоплазму и уже там окисляются до ФГК.
При этом синтез АТР и NADPH также протекает в цитоплазме
(рис. 8.31), и если бы ФГК опять проникала в хлоропласты и
подвергалась в них восстановлению, то этот процесс мог про-
должаться до бесконечности. Впоследствии Стокинг и Ларсон
действительно зарегистрировали такой транспорт. Хебер и
Краузе наблюдали скорости до 50 мкмоль-(мг Хл)-1 -ч-1. Эк-
зогенное окисление триозофосфатов может также катализиро-
вать необратимая NADP-специфическая триозофосфатдегидро-
геназная система, описанная Келли и Гиббсом. Эта реакция не
сопряжена с синтезом АТР, и она обеспечивает экспорт только
NADPH. Восстанавливающие эквиваленты могут также перено-
ситься через мембрану хлоропластов при помощи челночного
механизма дикарбоновых кислот (рис. 8.32). Первоначально
такие челночные механизмы рассматривали как способ экс-
порта дикарбоновых кислот, вместе с тем они могли бы, на-
пример, участвовать в синтезе АТР внутри хлоропластов ночью.
[Так, с помощью челночного механизма триозофосфатов и
ФГК могли бы переноситься в хлоропласты АТР и восстанав-
ливающая сила, а с помощью челночного механизма дикарбо-
новых кислот могла бы транспортироваться восстанавливаю-
щая сила из хлоропластов, что приводило бы к синтезу АТР
внутри хлоропластов.]
Источники данных: '[126, 27, 36, 38, 57].
ОБЩАЯ ЛИТЕРАТУРА
1. Douce R., Joyard J. Structure and function of the plastid envelope. In: Adv.
Bot. Res., 7, 1—116 (1979').
2. Gibbs M. Carbohydrate metabolism by chloroplasts. In: Structure and Func-
tion of Chloroplasts, (Gibbs M., ed.), 169—214, Springer-Verlag, Berlin,
1971.
3. Heber U. Flow of metabolites and compartmentation phenomena in chloro-
plasts. In: Transport and Distribution of Matter in Cells of Higher Plants
(Mothes K., Muller E., Nelles A., Neumann D. eds.), 152—84. Akademie-Ver-
lag, Berlin, 1970.
4. Heber U. Metabolite exchange between chloroplasts and cytoplasm, Ann. Rev.
Plant Physiol., 25, 393—421 (19'74).
5. Heber U., Krause Q. H. Transfer of carbon, phosphate energy and reducing
equivalents across the chloroplast envelope. In: Photosynthesis and Photo-
respiration (Hatch M. D., Osmond С. B., Slatyer R. O. eds.), 218—25 (Proc.
Conf. Australian Nat. Univ. Canberra, 1970), Wiley Interscience, New York,
1971.
5a. Heber U., Heldt H. W. The chloroplast envelope: structure, function and
role in leaf metabolism, Ann. Rev. Plant Physiol., 32, 139—68 (1981).
6. Heber U., Walker D. A. The Chloroplast Envelope, Barrier or Bridge?,
T.I.B.S, 4, 11 (1979).
7. Heidi H. IT. Metabolite transport in intact spinach chloroplasts. In: Topics
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ ТРАНСПОРТ
257
in Photosynthesis, Vol. 1: The Intact Chloroplast (Barber J. ed.), 215—34.
Elsevier, Amsterdam, 1976.
8. Herold A., Walker D. A. Transport across chloroplast envelopes — the role
of phosphate. In: Membrane Transport in Biology, (Giebisch G., Toste-
son D.. C;, Ussing J. J. eds.), Vol. II, 412—39, Springer-Verlag, Heidelberg,
New York, 1979.
9. Kelly G. J., Latzko E., Gibbs M. Regulatory aspects of photosynthetic car-
bon metabolism, Ann. Rev. Plant Physiol., 27, 181—205 (1976).
10. Kelly G. Latzko E. The Cytosol. In: The Biochemistry of Plants: A Com-
prehensive Treatise. (Stumpf P. K., Conn E. E., eds.) Volume 1, The Plant
Cell (Tolbert N. E., ed.), Chapter 4, Academic Press, New York, 1979.
10a. Krause G. H„ Heber U. Energetics of intact chloroplasts. In: The Intact
Chloroplast. (Barber J., ed.), Chapter 5, 171—214. Elsevier, Amsterdam, 1976.
11. Walker D. A. Chloroplast and Cell. Concerning the movement of certain key
metabolites etc. across the chloroplast envelope. In: Med. Tech. Publ. Int.
Rev. Sci. Biochem., Ser. I (Northcote D. H. ed.), Vol. XI, 1—49, Butterworths,
London, 1974.
12. Walker D. A. Plastids and Intracellular Transport. In: Encyclopedia of
Plant Physiology. Transport in Plants III (Stocking C. R., Heber U., eds.)
New Series, Volume 3, 85—136. Springer-Verlag, Berlin, 1976.
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
13. Allen M. B., Arm 1. B., Capindale J. B., Whatley F. R., Durham L. J.
Photosynthesis by isolated chloroplasts. III. Evidence for complete photo-
synthesis, J. Am. Chem. Soc, 77, 4149—55 (1955).
14. Ar non D. I., Allen M. B., Whatley F. R. Photosynthesis by isolated chloro-
plasts, Nature, 176, 394—6 (1954).
15. Baldry C. W., Cockburn W., Walker D. A. Inhibition by sulphate of the oxy-
gen evolution associated with photosynthetic carbon assimilation, Biochim.
Biophys. Acta, 153, 476—83 (1968).
16. Bassham I. A., Calvin M. The Path of Carbon in Photosynthesis, 1—104,
Prentice-Hall Inc., Englewood Cliffs, N. J., 1957.
17. Bassham J. A., Jensen R. G. Photosynthesis of carbon compounds. In: Har-
vesting the Sun (San Pietro A., Greer F. A., Army T. J. eds.), 79—110,
Academic Press, New York, 1967.
18. Cockburn W., Baldry C. W., Walker D. A. Oxygen evolution by isolated chlo-
roplasts with carbon dioxide as the hydrogen acceptor. A requirement for
orthophosphate or pyrophosphate, Biochim. Biophys. Acta, 131, 594—6
(1967).
19. Cockburn IV., Baldry C. W., Walker D. A. Some effects of inorganic phos-
phate on O2 evolution by isolated chloroplasts, Biochim. Biophys. Acta, 143,
614—24 (1967).
20. Cockburn W., Walker D. A., Baldry C. W. Photosynthesis by isolated chlo-
roplasts. Reversal of orthophosphate inhibition by Calvin cycle intermediates,
Biochem. J., 107, 89—95 (1968).
21. Cooper T. G., Fllmer D., Wishnick M., Lane M. D. The active species of
. «СО2» utilized by ribulose diphosphate carboxylase, J. Biol. Chem., 244,
1081—3 (1969).
21a. Enser LL, Heber H. Metabolic regulation by pH gradients. Inhibition of
photosynthesis by indirect proton transport across the chloroplast envelope,
Biochim. Biophys. Acta, 592, 577—91 (1980).
22. Fliege R., FlUgge LL I., Werdan K-, Heldt H. W. Specific transport of inorga-
nic phosphate, 3-phosphoglycerate and triosephosphates across the inner mem-
brane of the envelope in spinach chloroplasts, Biochim. Biophys. Acta, 502,
232—47 (1978).
17—234
258
ГЛАВА 8
23. Flilgge и. Freisl М., Heldt Н. IV. Balance between metabolite accumula-
tion and transport in relation to photosynthesis by isolated spinach chloro-
plasts, Plant Physiol., 65, 574—7 (1980).
24. Ginunler H., Schafer G., Kraminer A., Heber U. Amino acid permeability of
the chloroplast envelope as measured by light scattering, volumetry and ami-
no acid uptake, Planta, 120, 47—61 (1974).
25. Hampp R., Ziegler I. Sulfate and sulfite translocation via the phosphate
translocator of the inner envelope membrane of chloroplasts, Planta, 137,
309—12 (1977).
26. Heber U., Santarlus K. A. Direct and indirect transport of ATP and ADP
across the chloroplast envelope, Z. Naturforsch., 25b, 718—78 (1970).
27. Heber U., Krause G. H. Transfer of carbon, phosphate energy and reducing
equivalents across the chloroplast envelope. In: Photosynthesis and Photo-
respiration (Hatch M. D., Osmond. С. B., Slatyer R. O., eds.), 218—25, Proc.
Conf. Australian Nat. Univ. Canberra, 1970, Wiley Interscience, New York,
1971.
28. Heber U., Krause G. H. Hydrogen and proton transfer across the chloroplast
envelope. In: Progress in Photosynthesis (Forti G., Avron M., Melandri A.,
eds.), Vol. II, 1023—1033. (Proc. 2nd Intern. Congr. Photosyn. Res. 1971),
N. V. Junk, The Hague 1972.
29. Heber U., Kirk M. R., Ginunler H., Schafer F. Uptake and reduction of gly-
cerate by isolated chloroplasts, Planta, 120, 31—46 (1974).
30. Heber U„ Purczeld P. Substrate and product fluxes across the chloroplast
envelope during bicarbonate and nitrate reduction, Proceedings of the Fourth
International Congress on Photosynthesis 1977 (Hall D. 0., Coombs J., Good-
win T. W., eds.), 107—18, The Biochemical Society, London, 1978.
31. Heidi H. IV. Adenine nucleotide translocation in spinach chloroplasts, FEBS
Letters, 5, 1 1—4 (1969).
32. Heldt H. IV. Metabolite carriers of chloroplasts, Encyclopedia of Plant Phy-
siology New Series, Vol. 3. Transport in Plants III. (Stocking C. R., He-
ber U., eds.), 137—43, Springer-Verlag, Berlin, Heidelberg, New York, 1976.
33. Herold A., IValker D. A. Transport across chloroplast envelopes — the role
of phosphate. In: Membrane Transport in Biology, Vol. II (Giebisch G., Tos-
teson D. C., J. J. Ussing eds.), Vol. II, 412—39. Springer-Verlag, Heidelberg,
New York, 1979.
34. Hill R. Oxygen evolved by isolated chloroplasts, Nature, 139, 881—2 (1937).
35. Hill R. The biochemists’ green mansions. The photosynthetic electron-trans-
port chain in plants, Essays Biochem., 1, 121—51 (1965).
36. Kelly G. J., Gibbs M. Nonreversible D-glyceraldehyde 3-phosphate dehydro-
genase in plant tissue, Plant Physiol., 52, 111—18 (1973).
37. KHngenberg M., Pfaff E. Means of terminating reactions, Methods Enzymol.,
10, 680—4 (1967). _
38. Krause G. H. Indirekter ATP-Transport zwischen Chloroplasten mid Zyto-
plasma wahrend der Photosynthcse, Z. Pflanzenphysiol. 65, 13—23 (1971).
39. Lehner K-, Heldt H. IV. Dicarboxylatc transport across the inner membrane
of the chloroplast envelope, Biochim. Biophys. Acta, 501, 531—44 (1978).
40. Lilley R. McC., Fitzgerald M. P., Rienits K- G., Walker D. A. Criteria of
intactness and the photosynthetic activity of spinach chloroplast preparations,
New Phytol., 75, 1—10 (1975).
41. Lilley R. McC., Schwenn I., Walker D. A. Inorganic pyrophosphatase and
photosynthesis by isolated chloroplasts. II. The controlling influence of ortho-
phosphate, Biochem. Biophys. Acta, 325, 596—604 (1973).
41а. Lilley R. McC., Walker D. A. Studies with the reconstituted chloroplast sys-
tem, Encyclopedia of Plant Physiology (New Series) — Photosynthesis,
Vol. II, 41—53. Regulation of photosynthetic carbon metabolism and rela-
ted processes, (Gibbs M., Latzko E., eds.), Springer-Verlag, 1979.
42. Molisch H. Populace Biologische-Vortrage, 1—306. G. Fischer, Jena, 1922.
ПЛАСТИДЫ И ВНУТРИКЛЕТОЧНЫЙ транспорт
25
43. Mourioux G., Douce R. Transport du sulfate a travers la double membrane
tumitanlc, ou enveloppe, des chloroplasts d’epinard, Biochimie, 61, 1283—92
(1979).
44. Poincelot R. P. Uptake of bicarbonate ion in darkness by isolated chloro-
plast envelope membrane and intact chloroplast of spinach, Plant Physiol.,
54, 520—6 (1974).
45. Portis A. R. Jr., Heldt H. IF. Light-dependent changes of the Mg2+ concen-
tration in the stroma in relation to the Mg24' dependency of CO2 fixation in
intact chloroplasts, Biochim. Biophys. Acta, 449, 434—46 (1976).
46. Robinson S. P., Wiskich J. T. Pyrophosphate inhibition of carbon dioxide fixa-
tion in isolated pea chloroplasts by uptake in exchange for endogenous ade-
nine nucleotides, Plant Physiol., 59, 422—7 (1977).
47. Robinson S. P., Wiskich J. T. Inhibition of CO2 fixation by adenosine 5'-di-
phosphate and the role of phosphate transport in isolated pea chloroplasts,
Archs. Biochem. Biophys., 184, 546—54 (1977).
48. Robinson S. P., Edwards G. E., Walker D. A. Established methods for the
isolation of intact chloroplasts, In: Methodological Surveys in Biochemistry,
Ch. 9, 13—24 (Reid E., ed.), Ellis Horwood, Chichester, 1979.
49. Robinson S. P., Walker D. A. Rapid separation of the chloroplast and cyto-
plasmic fractions from intact leaf protoplasts, Arch. Biochem., 196, 319—23
(1979).
50. Sachs J. Uber den Einflusz des Lichtes auf die Bilduug des Ainylums in den
Chlorophyllkornern, Bolan. Z., 20, 365—73 (1862).
51. Sachs J. Lectures on the Physiology of Plants. Translated by H. M. Ward,
309 seq. Clarendon Press, Oxford, 1887.
52. Schafer G-, Heber U., Heldt H. IF, Glucose transport into spinach chloro-
plasts, Plant Physiol., 60, 286—9 (1977).
53. Stokes D. M., Walker D. A. Photosynthesis by isolated chloroplasts. Inhibi-
tion by DL-glyceraldchydc of carbon dioxide assimilation, Biochem. J., 128,
1147—57 (1972).
54. Urnbreit TV7. W., Burris R. H., Stauffer J. F. Manometric Techniques and
Related Methods for the Study of Tissue Metabolism, 1—387, Burgess, Min-
neapolis, Minnessota, 1945.
55. Walker D. A., Cockburn IF., Baldry C. IF. Photosynthetic oxygen evolution
by isolated chloroplasts in the presence of carbon cycle intermediates, Natu-
re, 216, 597—9 (1967).
56. Walker D. A., Hill R. The relation of oxygen evolution to carbon assimilation
with isolated chloroplasts, Biochem. Biophys. Acta, 131, 330—8 (1967).
57. Walker D. A., Crofts A. R. Photosynthesis, Ann. Rev. Biochem., 39, 389—428
(1970).
58. Walker D, A. Photosynthetic induction phenomena and the light activation
of photosynthetic carbon assimilation, Vorgetragen auf der Botaniker-Tagung
59. Walker D. A. Photosynthetic induction and its relation to transport pheno-
mena and control mechanisms in chloroplasts, In: The Intact Chloroplast.
(Barber J., ed.), Ch. 7, 235—78, Elsevier, Amsterdam, 1976.
60. Walker D. A„ Slankovic Z. S. Photosynthesis by isolated chloroplasts. Some
effects of adenylates and inorganic pyrophosphate, Plant Physiol., 59, 428—-
61. Walker D. A., Herold A. Can the chloroplast support photosynthesis unaided?,
In: Photosynthetic Organelles: Structure and Function (Fujita Y., Katoh S.,
Shinata I<., Miyachi S., eds.), Special Issue of Plant and Cell Physiol., 295—-
340 (1977).
62. Walker D. A„ Robinson S. P. Chloroplast and Cell. A contemporary view
of photosynthetic carbon assimilation, Vorgetragen auf der Botaniker-Tagung
in Marburg am 13 September 1978, Ber. Deutsch. Bot. Ges. Bd., 91, 513—
26 (1978).
17*
260
ГЛАВА 8
63. Walker D. A. Preparation of higher plant chloroplasts. Photosynthesis and
nitrogen fixation, Methods in Enzymology, 94—104, Academic Press, New
York, 1979.
64. Walker D, A. Energy, Plants and Man. (A booklet for first year students in
Environmental Sciences etc.), Packard Publishing Ltd., Chichester, 1978.
65. IFerdun K., Heldt H. IV<, Geller G. Accumulation of bicarbonate in intact
chloroplasts following a pH gradient, Biochim. Biophys. Acta, 283, 430—41
(1972).
65a. Werdau K„ Heldt //. UY The phosphate translocator of spinach chloroplasts.
In: Progress in Photosynthesis (Forti G., Avron M., Melandri A., eds.),
Vol. II, 1137—44 (Proc. 2tid. Intern. Congr. Photosynthesis Res. 1971),
N. V. Junk, The Hague, 1971.
66. Werkhelser 117. C., Bartley IF. The study of steady-state concentrations of
internal solutes of mitochondria by rapid centrifugal transfer to a fixation
medium, Biochem. J., 66, 79—91 (1957).
ГЛАВА 9
Регуляция фотосинтетической
ассимиляции углерода
Краткое содержание
Так же как и другие процессы метаболизма, ассимиляция
углерода при фотосинтезе подвержена контролю пли регуля-
ции, хотя до сих пор мы ие имеем ясного представления ни о
детальных механизмах этой регуляции, ин о всех ее эффектах.
В основе регуляции лежат катализ (изменение ферментативной
активности), действие масс (влияние концентрации метаболи-
тов и т. п.) и транспорт (передвижение метаболитов между
хлоропластом и его клеточным окружением). Как правило, ре-
гулируемыми являются те ферменты, которые катализируют
«необратимые» реакции (т. е. реакции с большой величиной
отношения между скоростями прямой и обратной реакций, на-
пример Ру5Ф—нРуБФ'—>-ФГК и ФБФ—>-Ф6Ф). Последова-
тельность превращений ФГК—>ДФГК—КТриозофосфат в
большой степени зависит от действия масс, и на ее первую ста-
дию оказывают сильное влияние концентрации ADP, АТР и
ФГК. Транспорт через мембраны хлоропластов определяется
доступностью ортофосфата, от чего в первую очередь и зави-
сит, какая часть продукта фотосинтеза подвергнется дальней-
шему метаболизму внутри хлоропласта, а какая сохранится в
нем (в виде крахмала) или будет транспортирована из хлоро-
пласта в окружающую его клеточную среду.
9.1. Основные принципы
Если мы представим себе завод, производящий сложные ма-
шины, скажем самолеты, нам сразу бросится в глаза, как пре-
красно скоординирована там работа. Естественно, для беспере-
бойного функционирования завода необходимо, чтобы все мно-
гочисленные материалы и детали поступали на конвейер для
сборки своевременно и в нужном количестве. Избыток шасси
на заводе не восполнит недостаток оборудования для радара’.
Примерно с такими же проблемами сталкивается и живой ор-
ганизм, который должен чрезвычайно четко регулировать свой
метаболизм. Это относится прежде всего к высшим растениям,
которые в отличие от некоторых организмов ие способны сами
искать воду или избегать ее. Кроме того, высшие растения в це-
262
ГЛАВА S
лом отличаются весьма ограниченной способностью доставать
необходимые им соединения и избавляться от тех веществ, ко-
торые им не нужны. Как и животные, растения регулируют раз-
личные процессы своего роста и развития при помощи гормо-
нов. Помимо этого растения располагают и более прямыми спо-
собами контроля, которые в широком смысле можно подразде-
лить на три основные категории: регуляция катализа, регуляция
действием масс и регуляция транспортом.
9.2. Регуляция катализа
Степень протекания любой реакции зависит от ее константы
равновесия, изменения свободной энергии и, в ряде случаев,
от ее окислительно-восстановительного потенциала. Как мы уже
отмечали в разд. 2.5 и 2.13, все эти величины взаимосвязаны,
так что, например, большое отрицательное изменение свободной
энергии ДК' эквивалентно большому значению константы рав-
новесия К. В данном случае значения и ДК', и К говорят о том,
что реакция пойдет практически до конца, если прочие факторы
позволят ей начаться. Самым важным из этих факторов с точ-
ки зрения биологии является наличие фермента, катализирую-
щего данную реакцию. Следовательно, если реакция А—ЯБ ха-
рактеризуется большим отрицательным значением ДР и ката-
лизируется ферментом с высокой активностью, А будет превра-
щаться в Б с той же скоростью, с какой образуется А, при ус-
ловии, что этот каталитический процесс ие регулируется.
В ВПФ-цикле скорость реакции, катализируемой фруктозобис-
фосфатазой, увеличивается при высоких значениях pH и боль-
шой концентрации ионов магния, т. е. данная реакция является
ярким примером того, как изменения в строме способны моду-
лировать ферментативную активность. В темноте в более кислой
среде (в строме) катализ подавляется; по этой причине, а так-
же из-за того, что сам фермент находится в основном в неак-
тивном состоянии (разд. 9.3), в этих условиях фруктозобисфос-
фат почти не подвергается гидролизу. На свету протоны начи-
нают быстро переходить из стромы в тилакоиды, и ее pH увели-
чивается от 7,0 до почти 8,0. Выход протонов из стромы сопро-
вождается поступлением в нее ионов магния из тилакоидов, и,
как показали Байер и Лацко, подобные качественные и количе-
ственные изменения приводят к значительной активации бис-
фосфатазы. Как установил в 1979 г. Ульрик Эйсер в лаборатории
в Йорке, в присутствии высоких концентраций солей слабых
кислот (например, ацетата или пропионата) Д pH на мембране
хлоропластов меняется и строма становится по сравнению с ок-
ружающей средой более кислой. При этом накапливается ФБФ,
а содержание гексозомонофосфата падает, что указывает на
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ углерода
263
Рис. 9.1. Подавление фотосинтеза ио-
нофором Mg2+. Выделение кислоро-
да изолированными хлоропластами
быстро подавляется, если добавить
ионофор, который способствует выхо-
ду ионов магния из стромы. Выде-
ление кислорода возобновляется пос-
ле добавления ионов магния к сре-
де, в которой находятся хлороплас-
ты. См. также рис. 9.4. (Portis,
Heldt, 1976.)
подавление фруктозобисфосфатазы. Кроме того, как было пока-
зано в опытах с реконструированной системой хлоропластов,
при нейтральном значении pH и низкой концентрации ионов
магния фотосинтез почти или совсем не идет, тогда как при
pH 7,8 и 5 мМ Mg2+ скорости выделения кислорода в присутст-
вии в качестве субстрата ФБФ не уступают соответствующим
скоростям в присутствии ФГК. Далее, если к интактным хлоро-
пластам добавить ионофор, который ускоряет транспорт ионов
магния через мембрану хлоропласта, фотосинтез останавлива-
ется, но он возобновляется при повышении экзогенной концент-
рации этих ионов (рис. 9.1). Данные всех этих опытов указы-
вают на то, что для протекания фотосинтеза в хлоропластах
необходимы ионы магния и высокие значения pH; оба этих
требования удовлетворяются в освещенной строме.
9.3. Активация катализаторов
а) Активация или инактивация светом
В предыдущем разделе мы отметили, что фруктозобисфосфа-
таза хорошо функционирует при высоких значениях pH и высо-
ких концентрациях ионов магния. Различие между активацией
светом в случае, когда он вызывает в строме изменения, которые
способствуют катализу, и в том случае, когда свет может также
модифицировать свойства фермента, выглядит несколько искус-
ственным, да его и не всегда удается выявить (см. ниже, а так-
же разд. 9.5). Совершенно очевидно, насколько важным явля-
ГЛАВА 9
Редуктаза
(Неактивный )
Восстановленный тиоредоксин
ф
— SH
Тиоредоксин
— SH
(Активный)
Б
——SH
—-SH
(активный)
4- G-SS-G
Ф
— S
+ 2GSH
;--S
{ Неактивный)
В
Рис. 9.2,. Активация тиоредоксином и инактивация окисленным глютатионом
(GSSG). А. Транспорт электронов от воды к тиоредоксину. Б. Восстановле-
ние дисульфидных связей тиоредоксином. В. Повторное окисление сульфгид-
рильных групп окисленным глутатионом. Г. Окисление и восстановление
глутатиона под действием Н2О2 и NADPH2. Ф —фермент.
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ УГЛЕРОДА
265
HS —
(Неактивный
( Активный)
Рис. 9.3. Активация медиаторами светового эффекта (МСЭ), которая сопро-
вождается превращениями сульфгидрильных и дисульфидных групп (Ander-
son et. al., 1978). Ф — фермент.
ется нередко состояние (восстановленное или окисленное) сульф-
гидрильных групп фермента; in vitro эти группы нетрудно вос-
становить, если добавить дитиотрейтол. Буханан и соавторы
предположили, что в создании светом восстанавливающих усло-
вий, необходимых для активации ФБФ-азы in vivo, играют роль
два белка — тиоредоксин и его редуктаза. На свету ферредок-
син восстанавливается обычным образом, после чего электроны
передаются от него через редуктазу к тиоредоксину (рис. 9.2,А).
В свою очередь восстановленный тиоредоксин активирует ФБФ-
азу, СБФ-азу, а также, по-видимому, Ру5Ф-киназу и триозофос-
фатдегидрогеназу (рис. 9.2,В). Кроме того, было' высказано
предположение, что окисленный глутатион способен инактиви-
ровать эти ферменты путем их окисления (рис. 9.2,В).
266
ГЛАВА 9
Впоследствии Буханан и соавторы установили, что в актива-
ции участвуют два тиоредоксина, больший из которых активи-
рует фруктозобисфосфатазу, рибулозокиназу и триозофосфатде-
гидрогеназу, а меньший — только ЫАБР+-специфическую малат-
дегидрогеназу (декарбоксилирующую). Оба тиоредоксина вос-
станавливаются либо ферредоксин/ферредоксин-тиоредоксинре-
дуктазой, либо дитиотрейтолом.
Луиза Андерсон и сотр. из Чикаго обнаружили два медиато-
ра светового эффекта (МСЭ), прочно связанных с тилакоидны-
ми мембранами. Оба медиатора служат акцепторами электро-
нов на восстанавливающем участке ФС1 (рис. 9.3,А): один пе-
ред ферредоксином, а другой после него. По-видимому, как и
тиоредоксины, эти медиаторы активируют ферменты ВПФ-цикла
(или инактивируют глюкозофосфатдегидрогеназу) путем восста-
новления дисульфидных связей; тем: самым они модифицируют
потенциально, активные центры этих ферментов. Впрочем, дан-
ные Андерсон наводят на мысль о том, что механизм процесса
представляет собой не прямое восстановление, а тиодисульфид-
ный обмен (рис. 9.3,5). Хотя медиаторы светового эффекта не-
сколько напоминают тиоредоксиновую систему, они в то. же вре-
мя отличаются от нее тем, что связаны с тилакоидами, а не на-
ходятся в свободном виде в растворе. Не исключено, что между
тиоредоксиновой системой и М.СЭ существует некая взаимо-
связь, но в чем именно она заключается, мы пока еще не можем
сказать.
б) Активация ионами магния
Ионы магния активируют РуБФ-карбоксилазу (разд. 6.11).
В течение многих, лет принято было считать, что этот фермент
ие способен играть ту роль, которую сначала ему отводили in
vivo, так как его каталитическая активность представлялась
незначительной. Столь низкую активность объясняли в основ-
ном слишком малым сродством этого фермента кСО2. [Согласно
сделанным при расчетах допущениям, для достижения половины
максимальной скорости этому ферменту может понадобиться
8% СО2 в газовой фазе, в то время как концентрация СО2 в
строме хлоропластов С3-растений не превышает 0,01%.] Как
оказалось, большинство измерений проводили с ферментом, на-
ходившимся в более или менее инактивированном состоянии.
Лоример и соавторы установили, что РуБФ-карбоксилаза обра-
зует комплекс с СО2, который затем быстро активируется иона-
ми магния при щелочных значениях pH. В этом случае сродство
фермента к СО2 составляет '—10 мкМ СО2, что соответствует
(при pH 8,0 и температуре 20°C) примерно 0,03% СО2. Вполне
возможно, что при изменении концентрации ионов магния и
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ УГЛЕРОДА
267
Рис. 9.4. Активация РуБФ-зависим ого вы-
деления кислорода ионами магния в ре-
конструированной системе хлоропластов.
Нижняя кривая: концентрация ионов маг-
ния достаточна для того, чтобы обеспечить
транспорт электронов к NADP+ (первое
выделение кислорода — слева от плато) или
к триозофосфатам, которые нуждаются в
образовании АТР (второе выделение кис-
лорода— справа от плато, после добавле-
ния ФГК). Верхняя кривая: РуБФ присут-
ствует с самого начала, но выделение кис-
лорода замедляется после первоначального
восстановления NADP-1- в течение первой
минуты. Последующее добавление ионов
магния приводит к тому, что РуБФ пре-
вращается в ФГК и опять начинает быст-
ро выделяться кислород [29aJ.
Время, мин
протонов in vivo РуБФ-карбоксилаза активируется так же, как
и ФБФ-аза. Такая активация была воспроизведена в реконст-
руированной системе хлоропластов в присутствии 1 мМ MgCl2;
этой концентрации ионов магния было достаточно для обеспе-
чения 90% максимального транспорта [электронов от Н2О к
ДФГК (рис. 9.4). Однако фиксация СО2 оставалась на низком
уровне до тех пор, пока концентрацию ионов магния не увели-
чили почти до 2,5 мМ.. [Эти изменения концентрации ионов маг-
ния близки к тем, которые, как предполагают, происходят в
строме хлоропластов при освещении. На симпозиуме, состояв-
шемся в Йорке в апреле 1979 г., Краузе и Бен-Хайим доложили
о результатах своих исследований, проведенных с металлохром-
ным индикатором эриохромом синим (Biochim. Biophys. Acta
460, 500—510, 1977), в которых они обнаружили, что при низ-
ких концентрациях NH4O1 индуцированное светом увеличение
содержания Mg2+ в строме изолированных хлоропластов снима-
ется, НО' фиксация СО2 при этом почти не изменяется. Эти авто-
ры пришли к выводу, что концентрация ионов магния, которая
существует в темноте в строме хлоропластов, достаточна для
поддержания высокой активности «регуляторных ферментов».]
Бар и Йенсен определяли активность карбоксилазы в освещен-
ных хлоропластах и установили, что она составляет 40—60%'
полной активности этого фермента. Лоример и Хельдт отвели
некую роль в активации ортофосфату и обнаружили, что осве-
щение интактных хлоропластов приводит к полной активации
этого фермента. При низкой концентрации СО2 активация кар-
268
ГЛАВА 9
Таблица 9.1. Активация светом ферментов ВПФ-цикла (Leegood,
неопубликованные данные)
Фермент Кратность возраста- ния активности 'Конечная скорость, . мкмоль-(мг Хл)-“''Ч*1
ФБФ-аза 5,0 120
РуБФ-карбоксилаза 1J 200
NADP — ФГА-дегидрогепаза 1,4 1000
Ру5Ф-киназа 1,5 1100
Фосфоглицераткиназа 1,0 2200
боксилазы может также зависеть от присутствия таких метабо-
литов и коферментов, как 6-фосфоглюконовая кислота, фрукто-
зо-1, 6-бисфосфат, ФГК и NADPH. Как мы уже отмечали, РуБФ-
карбоксилаза может утратить свою активность, если она ока-
жется вне хлоропластов (особенно в процессе выделения). Ак-
тивность фермента восстанавливается после инкубации с иона-
ми магния и СОг при слабощелочных значениях pH. Впрочем,
В,.настоящее время появились серьезные сомнения в том, что в
темноте происходит полная или хотя бы почти полная инакти-
вация, Первоначальное представление об инактивации в темно-
те (возникло на основании следующих наблюдений: содержание
РуБФ, падает в темноте медленнее, чем этого следовало бы ожи-
дать, исходя из скорости фиксации СО2 на свету и известного
благоприятного положения равновесия реакции. Объяснение
этих, наблюдений осложняется трудностями, связанными с точ-
иым. измерением небольших количеств РуБФ. Отчасти эти ре-
зультаты могут быть обусловлены замедлением катализа и
присоединением РуБФ к центрам, где невозможно карбоксили-
рование. Большая работа в данной области была проведена в
Последнее время в нашей лаборатории в Шеффилде. Получен-
ии.^, результаты указывают иа очень высокую активность карбок-
силазы в хлоропластах и протопластах, которые были выделе-
нье'.ид. ткани,(.хранившейся в темноте. В том случае, когда эта
активность превышает скорость фотосинтеза, совершенно непо-
нятно, каким образом «темновая инактивация» может давать
какое-либо преимущество в метаболических процессах. По-види-
мому, инактивация является отчасти артефактом процесса вы-
д'елеция, и просто катализ идет быстрее в условиях освещенной
стромы (разд. 9.5). Действительно, как показали исследования,
проведенные в последнее время в Шеффилде, ферменты ВПФ-
цикла почти не подвергались инактивации в темноте, за исклю-
чением ФБФ-азы (табл. 9.1). Следовательно, медиаторы свето-
вого эффекта или ферредоксии-тиоредоксиновые системы
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ УГЛЕРОДА
269
(разд. 9.3) можно рассматривать скорее как. репарационные
системы, которые исправляют повреждения, нанесенные пере-
кисными радикалами и подобными им соединениями (разд. 5.8),
а не как регуляторы в строгом смысле этого слова.
(Следует отметить, что раньше определять активность РуБФ-
карбоксилазы было весьма трудно (разд. 6.11), так как, во-пер-
вых, этот фермент нуждается в определенных факторах актива-
ции, а во-вторых, его ингибируют, продукты деградации РуБФ
(данные Толберта и соавторов). Поэтому к опубликованным до
1975 г. результатам исследований максимальной скорости реак-
ций, катализируемых РуБФ-карбоксилазой, ее сродства к СОг
и т.. д. необходимо относиться с особой осторожностью.] В по-
следние годы широко дискутируется роль аскорбиновой кисло-
ты и прочих восстанавливающих агентов в хлоропластах. Содер-
жание аскорбиновой кислоты в растениях может быть очень
высоким, особенно летом, и ее часто используют в качестве за-
щитного агента или донора электронов при работе с изолиро-
ванными хлоропластами. Функция аскорбиновой кислоты сво-
дится, по-видимому, к тому, что она выступает в роли антиокис-
лителя. Холлиуэлл и Фойер уточнили эту роль, высказав пред-
положение, что аскорбиновая кислота может способствовать
диссипации Н2О2. Хлоропласты ие содержат каталазы, однако
они могут образовывать I-I2O2 за счет переноса электронов от
таких доноров, как ферредоксин, к кислороду. Восстановлен-
ный глутатион способен окисляться под действием Н2О2 либо
непосредственно при помощи глутатиоипероксидазы, либо кос-
венным путем, за счет гидро аскорбиновой кислоты. Последую-
щее восстановление окисленного . глутатиона посредством.
NADPH позволяет сохранять статус-кво и ферменты в окислен-
ном состоянии, которые в противном случае подвергались бы
инактивации под действием Н2О2 (рис. 9.2,Г).
Источники данных: [16—23, 25, 30—34, 39, 41—45]
9.4. Различие между регуляцией катализа
и активацией, катализаторов
В предыдущих разделах (9.2 и 9.3) мы попытались провести
различие между действием света на катализаторы и катализ.
Во многих отношениях подобное различие носит чисто акаде-
мический характер, так как оно сводится в разграничению меж-
ду эффектами на фермент и на реакцию, которую этот фермент
катализирует. Тем не менее не следует полностью пренебрегать
этим различием даже тогда, когда такие параметры, как pH,
оказывают одновременное воздействие и на катализатор, и на
катализ. Так, например, если вы обратитесь к учебнику по эн-
зимологии, написанному лет двадцать назад, вы вряд ли .встре-
титесь там с понятием «регуляция». Вместе с тем уже давно
70
ГЛАВА 9
известно, что большинство' ферментов проявляет активность в
узком диапазоне pH (будучи белками, ферменты представляют
собой многовалентные электролиты). Поэтому ионизационный
статус составляющих их аминокислот, а следовательно, и
каталитическая активность этих ферментов зависят от концент-
рации водородных ионов. Аналогичным образом карбокси-ионы
аминокислот будут взаимодействовать с такими катионами, как
Mg21’. Действие pH связано в основном с изменением величин
Vmax или Км либо со стабильностью самого фермента, хотя сле-
дует отметить, что в реакциях, в которых принимают непосред-
ственное участие протоны, важную роль может играть влияние
концентрации водородных ионов на саму реакцию. Например,
реакция, катализируемая малатдегидрогеназой (декарбоксили-
рующей), характеризуется величиной ДР, неблагоприятной
для протекания реакции в направлении окисления малата
(разд, 6.12). .'Вот почему реакция пойдет в этом направлении
только в том случае, если сдвинуть равновесие путем удаления
конечных продуктов реакции: цианид (связывающий оксалоаце-
тат) и высокие значения pH (связывание Н+) будут способство-
вать окислению малата. В течение многих лет принято было
определять активность ферментов при оптимальных значениях
pH и очень часто (когда это казалось необходимым) добавлять
цистеин или восстановленный глутатион для защиты сульфгид-
рильных групп. Сам по себе тот факт, что фермент «работает»
при pH 8,0 (как в освещенных хлоропластах), а ие при pH 7,0
(как в затемненных хлоропластах), еще не говорит о достаточ-
но глубокой физической модификации самого фермента, приво-
дящей к его «инактивации». Вместе с тем выделенный фермент,
например РуБФ-карбоксилаза, может легко инактивироваться
in vitro и нуждаться в щелочном значении pH для завершения
процесса реактивации. Аналогичным образом ионы магния мо-
гут требоваться для активации фермента (также, например, для
РБФ-карбоксилазы), но не для катализа (как в реакциях, ката-
лизируемых киназами). Далее, фермент типа ФБФ-азы, кото-
рый, по всей очевидности, находится в сильной зависимости от
своего восстановительного статуса и шочти не проявляет актив-
ности в темноте, будет тем не менее обнаруживать некоторую
активность, если ее определять в присутствии 10—20 мМ MgClz
и при pH 8,0. Эти данные позволяют заключить, что практиче-
ски полное прекращение реакции в темноте обусловлено соче-
танием окисления этого фермента и неблагоприятных условий в
затемненной строме.
9.5. Темновая инактивация
Как мы уже отмечали (разд. 9.3), системы световой актива-
ции способны функционировать в качестве репарационных си-
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ углерода
271
стем. Столь же вероятно, что они принимают участие в регу-
ляции, т. е. составляют часть системы, которая «включает» фер-
мент в освещенном хлоропласте и «выключает» его в темноте.
Различие между репарацией и регуляцией, если оно действи-
тельно существует, может быть вопросом степени. Оно прояв-
ляется не в способности фермента активироваться под дейст-
вием света, а в том, насколько световая активация необходима,
т. е. в степени темновой инактивации, без обращения которой
невозможно нормальное функционирование фермента. Например,
если при освещении хлоропластов в соответствующей среде ак-
тивность фермента возрастет в десять раз, то можно считать,
что этот фермент активируется светом. Если же фермент обна-
руживает почти полную активность сразу после выделения из
затемненного хлоропласта, то он, по-видимому, не нуждается
в световой активации. Напротив, если сразу после темноты фер-
мент проявляет очень низкую активность, это свидетельствует
о необходимости его «включения». В табл. 9.1 можно видеть
такие различия у ферментов. Как мы уже отмечали, вполне
возможно, что они отражают артефакты в экспериментальных
методиках. Тем не менее совершенно ясно, что только один фер-
мент (ФБФ-аза) действительно выключается в этих условиях.
Следует заметить, что даже в случае ФБФ-азы некоторая ос-
таточная активность была обнаружена и в темноте. Однако
нельзя забывать, что в подобного рода экспериментах актив-
ность ферментов определяют при оптимальных значениях pH и
других параметров (за исключением света). В темноте же эти
параметры стромы 'Перестают быть оптимальными, что приво-
дит к полной инактивации ферментов. Интересно также, что
активность ФБФ-азы была довольно низкой, когда ее сравнивали
с активностью Ру5Ф-киназы. Это согласуется с регуляторной
ролью фермента. Однако вместе с тем действительная скорость,
которая сохраняется in vivo, зависит от сродства фермента к
его субстрату или субстратам и от их стационарных концентра-
ций. В этом отношении концентрацию Ру5Ф можно с достаточ-
ной уверенностью считать очень низкой, и поэтому следует с
осторожностью относиться к результатам, полученным при из-
мерении скоростей, в более благоприятных условиях.]
Если принять, что различные ферменты подвергаются инак-
тивации в темноте в разной степени, возникает вопрос, как это
происходит. Вполне возможно, что для проявления своей актив-
ности фермент нуждается в постоянном восстановлении, и если
удалить восстановитель, то он переходит в неактивное окислен-
ное состояние. Не исключено, впрочем, что здесь действует со-
вершенно иной механизм. Например, было высказано предпо-
ложение, что' глутатион окисляется в темноте под действием
перекиси водорода, образовавшейся на свету, и глутатионперок-
272
ГЛАВА 3
сидазы, а затем окисляет сульфгидрильные группы чувствитель-
ных ферментов. Наблюдения Холлиуэлла и Фойера (которые
установили, что глутатион существует исходно в восстановлен-
ном состоянии как в темноте, так и на свету) не подтвердили это
предположение, хотя сами по себе их результаты не опроверга-
ют окончательно такой возможности (рис. 9.2 и разд. 9.3).
Данные Лигуда говорят в пользу первой возможности (посто-
янное восстановление) и указывают на потребность в молеку-
лярном кислороде или акцепторе электронов. Лигуд обнаружил,
что ;ФБФ-аза не только удивительно чувствительна к потоку
электронов, но и реагирует на присутствие (или отсутствие) це-
лого ряда акцепторов электронов. Например, СОг препятствует
полной активации, по-видимому действуя в качестве эффектив-
ного альтернативного акцептора электронов. Точно так же, если
исключить из реакционной смеси кислород, то световая актива-
ция не исчезает в темноте, но оксалоацетат (который способен
служить акцептором электронов подобно СОа или Оа) инакти-
вирует фермент в отсутствие кислорода и на свету, и в темно-
те. Антимицин А, ингибитор циклического- транспорта электро-
нов, почти не влияет на активность ФБФ-азы при высокой ско-
рости электронного потока. Однако этот антибиотик вызывает
заметную дополнительную активацию, когда транспорт элект-
ронов замедляют добавлением диурона (ДХММ, англ. DCMU)
(в концентрации, достаточной для подавления этого процесса
иа 50%) или уменьшением интенсивности света. Напротив, пио-
цианин, стимулятор циклического транспорта электронов, сни-
жает активность ФБФ-азы. Эти зависимости схематически пока-
заны иа рис. 9.5. Как видно из данной схемы, соединения, заби-
рающие электроны, которые должны идти на восстановление
ФБФ-азы, будут снижать активность этого фермента на свету,
и наоборот. В темноте транспорт части этих электронов будет
происходить в обратном направлении; окисление под действием
молекулярного кислорода, его производного или какого-либо
иного адекватного соединения приведет к окислению сульфгид-
рильных групп и, следовательно, к инактивации фермента. Во-
прос о том, принимает ли непосредственное участие в окисли-
тельной инактивации и восстановительной активации ферментов
тиоредоксин, остается открытым.
Источники данных: [28а, Ь].
Как (мы уже отмечали выше, сам по себе факт активации,
которой может подвергаться фермент под действием света, еще
не служит доказательством того, что регуляция этого процесса
осуществляется в природе. В настоящее время хорошо извест-
но, что РуБФ-карбоксилаза легко инактивируется во время экс-
тракции или быстро реактивируется в присутствии тех концент-
раций Н+ и Mg2+, которые характерны для освещенного хлоро-"
РЕГУЛЯЦИЯ фотосинтетической ассимиляции углерода
27Л
Рис. 9.5. Активация ФБФ-азы на свету и ее инактивация в темноте. Электро-
ны, которые передаются к ФБФ-азе через тиоредоксин, ферредоксин и фото-
системы, восстанавливают дитиоловые группы и повышают активность. Дру-
гие акцепторы электронов (СОа, О2, оксалоацетат, нитрит), забирающие их
от ФБФ-азы, и циклический транспорт электронов способствуют ее инакти-
вации в темноте и снижению активации на свету [28Ъ].
пласта. Как показано в проведенных недавно опытах, если
хлоропласт сразу после освещения подвергнуть лизису в реак-
ционной смеси, то в нем не проявляется полная потенциальная
активность карбоксилазы, которую следовало бы ожидать судя
по предварительной инкубации с высокими концентрациями
ионов магния при щелочных значениях pH. Тем не менее эта
активность будет несколько больше активности аналогичных
хлоропластов, которые перед этим находились в темноте. В дан-
ном случае наблюдается некоторое несоответствие между отно-
сительной величиной этих изменений и их значением. При
Vmax~1000 мкмоль-(мг Хл)_1-ч-1 возрастание активности фер-
мента в 1,5 раза под действием света может и не играть особой
роли, в то время как четырехкратная активация безусловно
приобретает огромное значение. Тем не менее, хотя четырех-
кратная активация действительно наблюдалась, не следует ис-
ключать возможность инактивации фермента в процессе экспе-
риментов и пренебрегать тем фактом, что некоторые исследо-
ватели обнаружили изменения активности в пределах 1 —
1,5 раза.
9.6. Действие масс
Если реакция характеризуется большой величиной Д77' (т. е.
большим значением /<), то она практически не испытывает
влияния со (Стороны своих конечных продуктов (разд. 2.5).
18--2.34
274
ГЛАВА 9
Рис. 9.6. Примерные соотношения величии свободной энергии при восстанов-
лении ФГК. Для прямого восстановления ФГК до триозофосфатов требуется
приблизительно 8 ккал энергии, но использование АТР снижает этот барьер
почти на 7 ккал. В результате суммарное превращение ФГК в ФГА пред-
ставляет собой легко обратимую реакцию с величиной AK'~ 1 ккал. Тем не
менее первая стадия этой последовательности реакций характеризуется
Д/?,~4,5 ккал. Это означает, что протекание реакции ФГК+АТР->ДФГК+
+ADP особенно чувствительно к снижению отношения [ATP]/[ADP], а следо-
вательно, к скорости фосфорилирования, с одной стороны, и к активности
реакций, в которых используется АТР, — с другой (объяснение см. в
тексте).
В данном случае регуляция будет осуществляться лишь за счет
модуляции фермента, катализирующего эту реакцию (разд. 9.2).
И наоборот, если ДГ' имеет положительное или малое значение
(величина К небольшая или близка к единице), то протекание
реакции будет в очень сильной степени зависеть от накопления
субстрата и удаления продукта.
Прекрасным примером вышесказанного может служить вос-
-становление ФГК до триозофосфата (разд. 6.11) —единственная
восстановительная стадия в цепи фотосинтетической ассимиля-
ции углерода. Как видно из рис. 9.6, суммарная реакция идет в
две стадии. Первая стадия невыгодна с точки зрения термоди-
намики и нуждается в АТР:
ФГК + АТР---->ДФГК-|~АЭР (9.1)
Вторая стадия является термодинамически выгодной, в ней
в качестве донора электронов используется NADPIT:
ДФГК + NADPH ----> ФГА + Р[ + NADP+ (9.2)
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ углерода
275.
Эти две стадии составляют вместе легко обратимую окисли-
тельно-восстановительную реакцию, которая отличается от ре-
акции гликолиза лишь тем, что коферментом в ней служит
NADP+, а не (NAD+ При гликолизе окисление представляет со-
бой сопряженное с субстратом фосфорилирование, на которое1
в целом затрачивается 8,2 ккал, причем 7 ккал из них сохраня-
ется в виде АТР. При фотосинтезе восстановление ФГК до трио-
зофосфата становится энергетически возможным благодаря ис-
пользованию АТР. Тем не менее термодинамически невыгодное-
положение равновесия первой стадии приводит к тому, что реак-
ция легко идет в обратную сторону в присутствии ADP. Как
показали опыты с реконструированной системой хлоропластов,
ФГК-зависимое выделение кислорода быстро подавляется в ре-
зультате добавления небольшого количества ADP (рис, 8.17),
Реконструированная система хлоропластов состоит из хло-
ропластов, подвергнутых осмотическому шоку, к которым после
этой обработки добавляют коферменты и белки стромы. Хлоро-
пласты в такой системе лишены интактных оболочек, и поэтому
ничто не препятствует свободному поступлению соединений
внутрь хлоропластов и выходу из них. Во многих отношениях
такую систему можно рассматривать как один большой хлоро-
пласт, ограниченный стенками реакционного сосуда. Как мы уже.
отмечали, реконструированная система с ФГК в качестве суб-
страта будет выделять кислород до- тех пор, пока к ней не до-
бавят ADP. Однако фотофосфорилирование (мы пока еще не.
знаем, является ли оно циклическим, псевдоциклическим или и
тем и другим) продолжается и после того, как прекращается,
выделение кислорода. По мере превращения ADP в АТР вре-
менное ингибирование снимается. Аналогичное подавление могут
вызывать Р5Ф и другие соединения* использующие энергию АТР:
АТР ADP
Р5ф----------->Ру5Ф----------з-РуЬФ (9.3)
Таким соединением является также глюкоза, добавляемая
вместе с ферментом гексокиназой:
АТР ADP
. (9.4)
Глюкоза—*-Глюкозо-6-фосфат
Во всех этих случаях подавление полностью снимается, если
одновременно добавить к системе соединения, которые с высо-
1S*
276
ГЛАВА !)
кой эффективностью превращают ADP в АТР, например креа-
тинфосфат и его киназу:
г Креатинфосфат.-|- ADP -> Креатин-у АТР (9-5)
Регуляторная природа подобного взаимодействия особенно
четко проявляется в том случае, когда субстратом служит ФБФ.
Ассимиляция углерода начинается почти сразу после добавления
этого метаболита, однако выделение кислорода наблюдается
лишь после некоторой лаг-фазы (рис. 9.7). Как мы уже говори-
ли об этом раньше (разд. 9.3), ФБФ-аза активируется в присут-
ствии ряда восстановительных агентов или ионов магния в
больших концентрациях, а также при высоких значениях pH. Эти
факторы удлиняют задержку в выделении кислорода, которую
можно снять добавлением креатинфосфата и его киназы. За-
держка увеличивается также при внесении дополнительного
количества ФБФ-азы либо при использовании части фруктозо-
фосфата и форме Ф6Ф, а не ФБФ (доля первой формы эквива-
лентна гидролизуемой части ФБФ до внесения). Таким образом,
здесь имеется кажущееся противоречие, которое заключается в
том, что любая из стадий, ускоряющих одну реакцию ВПФ-цик-
ла, будет автоматически' подавлять другую реакцию. Причина
во всех случаях одна и та же ;(рис. 9.8). Повышенный уровень
активности ФБФ-азы приводит (через ФБФ и т. д.) к большей
стационарной концентрации РубФ, который для своего превра-
щения в РуБФ использует энергию АТР. В результате снижается
отношение [ATP]/[ADP], благодаря чему подавляется восстанов-
ление ФГК и связанное с ним выделение кислорода. При ста-
ционарном фотосинтезе в строме сохраняется высокая концепт-
Bpi'Wi, мни
Рис. 9.7. Задержка в выделении кисло-
рода реконструированной системой хло-
ропластов при использовании в качест-
ве субстрата ФБФ [43]. См. объяснение
на рис. 9.8.
РЕГУЛЯЦИЯ фотосинтетической ассимиляции углерода
277
Рис. 9.8. Объяснение задержки в выделении кислорода, показанной на
рис. 9.7. На схеме изображены реакции ВПФ-Цикла, которые обусловлива-
ют задержку в выделении кислорода при использовании в качестве субстра-
та ФБФ и определяют ее величину. ФБФ, добавленный в Субстратных коли-
чествах (внизу слева в круге), инициирует последовательность реакций. Фик-
сация СО2 начинается почти сразу, что указывает на образование ФГК. Вы-
деление О2 (помимо того, которое связано с восстановлением NADP) запаз-
дывает, что говорит об отсутствии восстановления ДФГК до ФГА. Если бы
оно происходило, NADPH вновь окислялся бы до NADP+, который получал
бы электроны от воды (через последовательность реакций фотосинтетическо-
го транспорта электронов), в результате чего выделялся бы кислород (внизу
справа, в прямоугольной рамке). Возникает вопрос, почему выделение кис-
лорода задерживается, несмотря на то что ФГК, АТР и NADPH присутст-
вуют в достаточном количестве. Дело в том, что АТР весьма интенсивно ис-
пользуется в реакции с Ру5Ф (наверху в центре), который образуется из
ФБФ. В результате регенерация ADP идет с такой же скоростью, с какой
образуется АТР при эндогенном фотосинтетическом фосфорилировании.
Таким образом, стационарная концентрация ADP будет высокой, что спо-
собствует превращению ДФГК в ФГК (мирная вертикальная стрелка), а не
восстановлению ДФГК в ФГА. Однако через некоторое время количество
ФБФ снижается, а следовательно, истощаются и запасы Ру5Ф, который об-
разуется из него, падает концентрация ADP, восстанавливается ДФГК и
выделяется кислород. Те факторы, которые вызывают превращение ФБФ в
Ф6Ф, приводят к Накоплению больших запасов Ру5Ф и сильно задерживают
выделение кислорода, а факторы, которые способствуют превращению ADP
в АТР, укорачивают задержку [43].
278
ГЛАВА 9
рация ФГК. и низкое содержание пентозомонофосфатов. Теперь
нам ясно, почему это происходит: при любом увеличении уров-
ня Ру5Ф ингибируется восстановление ФГК и, следовательно,
образование Ру5Ф. Поэтому в стадии регенерации ВПФ-цикла
может участвовать лишь такое количество триозофосфатов, ко-
торое соответствует имеющейся скорости синтеза АТР. Чем сла-
бее освещение, тем меньше молекул триозофосфатов будет во-
влечено в регенерацию акцептора COs, и наоборот. Точно так
же любая тенденция к увеличению количества триозофосфатов,
которые подвергаются перестройке на стадии регенерации, в
стационарном состоянии повлечет за собой (соответствующее
снижение восстановления ФГК и, по-видимому, изменение соот-
ношении между ФГК и триозофосфатами, экспортируемыми из
хлоропластов в цитоплазму (гл. 7).
Источники данных: [29, 36, 37, 38, 40]
9.7. Регуляция транспортом
Бесперебойный экспорт метаболитов из хлоропласта (гл. 8)
нуждается в том, чтобы ортофосфат, который транспортируется
из стромы в виде органического фосфата, поступал туда обрат-
но в форме неорганического фосфата (рис. 9.9). Ограниченное
снабжение хлоропласта ортофосфатом приведет к уменьшению
отношения l[ATP]/[ADP], подавлению восстановления ФГК и
замедлению стадии регенерации ВПФ-цикла. Максимальный
фотосинтез немыслим без максимального экспорта, В этом от-
ношении следует отметить, что триозофосфаты используются в
двух реакциях: либо в реакции конденсации с образованием
гексозобисфосфата, либо в транскетолазной реакции. В свою
очередь участие триозофосфатов в транскетолазной реакции за-
висит от имеющегося количества Ф6Ф и С7Ф, которые служат
донорами двухуглеродного фрагмента [см. уравнения (6.21) и
(6.24)].
Прейс и соавторы обнаружили, что при низкой концентра-
ции ортофосфата и высокой концентрации ФГК в строме хло-
ропластов (когда скорость экспорта незначительна) активность
глюкозо-1-фосфат— аденилилтрансферазы сильно возрастает.
Этот фермент играет важную роль в синтезе крахмала, и его
аллостерическая активация при высоких значениях [ФГК]/[Р1]
должна способствовать тому, чтобы он имел самую высокую
активность в наиболее благоприятных для синтеза крахмала
условиях в строме. Действительно, как было показано рядом
исследователей, этот фермент имеет наибольшую активность в
том случае, когда изолированные хлоропласты не получают
достаточного количества ортофосфата (рис. 9.10). Как и следо-
вало ожидать, экспорт при этом замедляется, а снижение отно-
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ УГЛЕРОДА
279
Регуляция по типу
обратной связи
Сахароза
I
ТРАНСЛОКАЦИЯ
Рис. 9.9. Регуляция транспортом метаболитов. Крахмал образуется в строме
хлоропластов, а сахароза — в цитоплазме. Скорость фотосинтеза максималь-
на, когда такие процессы, как синтез крахмала, обеспечивают освобождение
неорганического фосфата из триозофосфатов и т. д. В результате неоргани-
ческий фосфат может поступать обратно в строму с неограниченной ско-
ростью. При лимитирующих концентрациях неорганического фосфата в ци-
топлазме скорость фотосинтеза зависит от синтеза крахмала. При этом, хотя
низкие концентрации неорганического фосфата и высокое содержание ФГК
вызывают аллостерическую активацию ключевого фермента данного процес-
са, с накоплением гексозофосфатов в строме увеличивается образование пенто-
зомонофосфатов. В свою очередь это способствует снижению скорости вос-
становления ФГК при помощи механизма, описанного в разд. 9.6. Если по
какой-либо причине транслокация сахарозы замедляется, из-за ее накопления
по принципу обратной связи ингибируется ее синтез и благодаря замедлению
освобождения неорганического фосфата соответственно уменьшается ско-
рость фотосинтеза. Хотя такое объяснение является отчасти гипотетическим,
нет никаких сомнений в том, что при низких концентрациях экзогенного не-
органического фосфата фотосинтез подавляется (рис. 9.10) и что аналогич-
ное влияние имеют реакции, в которых используется неорганический фосфат
из цитоплазмы (рис. 9.11). Взаимосвязь между концентрацией неорганиче-
ского фосфата, транспортом и скоростью фотосинтеза рассматривается также
в гл. 7. АДФГ — аденозиндифосфатглюкоза, УДФГ — уридиндифосфатглю-
коза.
шения ([ATP]/i[ADP] приводит к уменьшению скорости восстанов-
ления ФГК. Возрастающее потребление Ф6Ф в синтезе крахма-
ла вызовет сокращение количества триозофосфатов, которые
вступают в первую реакцию, катализируемую транскетолазой
(разд. 6.11).
Следует отметить, что важную роль в даном случае может
280
ГЛАВА 9
Рис. 9.10. Зависимость синтеза крахмала от концентрации неорганического
фосфата снаружи изолированных хлоропластов. Общая ассимиляция сначала
возрастает, а затем падает по мере увеличения концентрации фосфата; на-
копленне крахмала при этом снижается и вскоре достигает очень низкого
уровня [26].
играть положение равновесия реакции, которую катализирует
альдолаза. Оно сильно сдвинуто в сторону образования гексо-
зобисфосфата. Кроме того, поскольку в реакции имеются два
субстрата и только один продукт, накопление любого из двух
исходных триозофосфатов сдвинет равновесие реакции еще
больше в сторону образования гексозобисфосфата.
Влияние концентрации неорганического фосфата на скорость
фотосинтеза и на распределение продуктов между крахмалом
и растворимыми компонентами изучали также в тканях листа,
используя характерную реакцию в ответ на добавление манно-
зы (рис. 9.11). Этот сахар в присутствии АТР и гексокиназы
подвергается активному фосфорилированию, в результате чего
образуется маннозофосфат, но его дальнейший метаболизм во
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ углерода
281
Рис. 9.11. Стимуляция маннозой синтеза крахмала у растений с Сз-путем
фиксации СОа (ср. с рис. 9,12). Манноза, добавленная к дискам листьев, по-
видимому, не способна проникать через оболочку хлоропластов, но в цито-
плазме она быстро превращается в маннозофосфат. У некоторых Сз-растений
(например, у свеклы шпинатной, разновидности Beta vulgaris), которые
не могут осуществлять дальнейший метаболизм маннозофосфата, на его об-
разование используется неорганический фосфат из цитоплазмы. В результате
содержание этого фосфата в цитоплазме падает, обмен между ним и триозо-
фосфатамн через оболочку хлоропластов снижается, скорость фотосинтеза
уменьшается, но возрастает скорость синтеза крахмала. ГФ — гексозофос-
фат, ТФ—триозофосфат.
многих растениях почти не идет, поэтому содержание неоргани-
ческого фосфата в цитоплазме снижается. У растений с Сз-путем
фиксации СО2 скорость фотосинтеза падает, но одновременно
возрастает скорость синтеза крахмала, как и следовало ожи-
дать из опытов с изолированными хлоропластами. В определен-
ных условиях эти изменения могут быть очень большими
(в 10 раз). Исследования, проведенные с меченой маннозой,по-
282
ГЛАВА 9
Рис. 9.12. Различное действие маннозы на растения с Са-путем и Сл-путем
фиксации СО2. Манноза в концентрации 10~2 М стимулировала синтез крах-
мала у Са-видов (светлые столбики) и подавляла его у большинства Севи-
дов (заштрихованные столбики). Те С3-виды (светлые столбики наверху), ко-
торые наиболее чувствительны к действию маннозы, не способны к метабо-
лизму маннозофосфата.
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ ассимиляции углерода
2
казали, что ее углеродные атомы ие включаются прямо в моле-
кулы крахмала; обе радиоактивные метки (углерода и фосфора)
накапливаются во фракции, которая по своим хроматографиче-
ским характеристикам соответствует маннозофосфату. У расте-
ний с С4-путем фиксации СО2 манноза не стимулирует синтез
крахмала, а подавляет его (рис. 9.12). Это происходит, по-види-
мому, в результате снижения концентрации неорганического
фосфата в цитоплазме. Не исключено также, что манноза за-
трудняет регенерацию ФЕП в реакции, катализируемой пиру-
ват, фосфат — дикиназой; в результате скорость фиксации СО2
снижается настолько, что это перевешивает любую возможную
стимуляцию синтеза крахмала (разд. 12.10).
Мы хотели бы обратить особое внимание на потенциальную
важность регуляции транспортом метаболитов и концентрацией
неорганического фосфата, а также иа следующий факт. Если
хлоропласту приходится функционировать в клетке или в ис-
кусственной среде с низким содержанием неорганического фос-
фата, то суммарная скорость его- фотосинтеза снижается, в то
время как доля и общее количество углеродных атомов, кото-
рые включаются в молекулы крахмала, возрастают. В клетке
большая часть триозофосфатов, которые экспортируются из
хлоропласта, превращается в сахарозу, основной транспорти-
руемый метаболит. В литературе встречаются доволы-ю противо-
речивые данные относительно влияния иа фотосинтез различных
процессов, протекающих в растениях. Нилес и Ииколл вообще
считают, что подавление фотосинтеза его- продуктами, накапли-
ваемыми в клетке, до сих пор однозначно* ие доказано. Тем не
менее вряд ли можно сомневаться в том, что по крайней мере
в ряде случаев скорость ассимиляции углерода при фотосинтезе
меняется под действием отдаленных реакций, в которых исполь-
зуются те или иные соединения. Результаты, полученные в этих
случаях, можно объяснить регуляцией неорганическим фосфа-
том (рис. 9.9). Например, как показал Хокер, сахароза ингиби-
рует свою фосфатазу. Это одна из стадий, на которой неоргани-
ческий фосфат, экспортируемый из хлоропласто-в, поступает об-
ратно р эти пластиды благодаря процессам, идущим в цито-
плазме. Таким образом, вполне возможно, что пониженная
активность реакции, в которой используется сахароза, приведет
к снижению скорости ее транспорта, а это- в свою очередь за-
медлит освобождение неорганического фосфата в цитоплазме и,
следовательно, экспорт метаболитов из хлоропластов. Точно
так же любое/накопление триозофосфатов или ФГК будет
уменьшать скорость экспорта, и эффективная концентрация
этих соединений в цитоплазме также будет зависеть от кон-
центрации неорганического фосфата. Именно так можно объяс-
нить результаты, полученные в опытах с использованием манно-
284
ГЛАВА 9
зы. Растения, испытывавшие недостаток в неорганическом
фосфате, накапливали крахмал. Например, Каки установил, что-
содержание крахмала возрастает в растениях табака с низкой
концентрацией неорганического фосфата, хотя скорость фото-
синтеза у них была невысокой, <а Александер обнаружил на-
копление крахмала и пониженное содержание сахарозы в расте-
ниях сахарного тростника, испытывавших недостаток в неоргани-
ческом фосфате.
Источники данных: [13, 16а, 24, 27, 28, 35].
При стационарном фотосинтезе регуляция неорганическим
фосфатом неотделима от его роли в индукции (разд. (7.10—
7.12 и рис. 7.23). Вот почему мы хотели бы здесь еще раз оста-
новиться на основных общих чертах этих двух процессов. Преж-
де всего следует отметить, что часто на кривой, выражающей
зависимость скорости фотосинтеза в изолированных хлоропла-
стах от концентрации неорганического фосфата, наблюдается
очень четкий оптимум (рис. 7.21).
Попытаемся проанализировать, что происходит по обе сто-
роны этого оптимума. При относительно небольшом содержа-
нии неорганического фосфата низкое отношение [ATP]/[ADPJ
обусловит замедление восстановления ФГК (разд. 9.6 и рис. 9.8).
В то же время в отсутствие неорганического фосфата снаружи
хлоропластов уменьшится скорость экспорта из них триозофос-
фатов при помощи переносчика неорганического фосфата
(разд. 8.23). В результате большая часть триозофосфатов (со-
держание которых очень невелико) пойдет на синтез крахмала.
Этому способствует возросшая активность гЛюкозо-1-фосфат —
аденилилтрансферазы, так как этот фермент подвержен алло-
стерической регуляции, индуцируемой высоким отношением
[ФГК]ДР1]. При оптимальном содержании неорганического фос-
фата его количества будет достаточно для обеспечения настоль-
ко. большой скорости транспорта в хлоропласты этого- фосфата,
чтобы удовлетворялось максимальное требование—1 молеку-
ла Pi на 3 молекулы СО2 — в реакции 3CO2-|-2H2O-|-Pi—-Трио-
зофосфат-|-ЗО2. Это определяет, какие именно реакции пойдут
при. избыточном образовании продуктов фотосинтеза. Иными
словами, при оптимальных и субоптимальных концентрациях
неорганического фосфата скорость фиксации СО2 может превы-
шать скорость поступления неорганического фосфата в хлоро-
пласты лишь настолько, чтобы обеспечивался синтез крахмала
согласно- классическому суммарному уравнению фотосинтеза
[уравнение (1.1)], т. е. это процесс, в конечном счете не нужда-
ющийся в неорганическом фосфате. Соответственно если коли-
чество триозофосфатов, образующихся при фотосинтезе, превы-
шает непосредственную потребность в них метаболических про-
цессов в. цитоплазме, то избыточные продукты фиксации СО2
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ углерода
285
могут накапливаться в виде крахмала. Вместе с тем если уро-
вень неорганического фосфата в цитоплазме высок (например,
если метаболизм в цитоплазме и отдаленные реакции, в кото-
рых используются продукты, имеют такую большую активность,
что концентрация неорганического фосфата в цитозоле дости-
гает и даже превосходит оптимум, необходимый для фотосинте-
за), то те триозофосфаты, которые в противном случае могли
бы пойти на синтез крахмала, будут экспортироваться из хлоро-
пластов. В результате рециркуляция неорганического фосфата
будет все больше и больше обусловливаться процессами, проте-
кающими в цитоплазме. В этих условиях с понижением отноше-
ния [ФГК]</1[Р1] будет уменьшаться активность глюкозо-1-фос-
фат—аденилилтраисферазы и реакции, обеспечивающие по-
ступление субстратов для ВПФ-цикла, будут преобладать над
реакциями синтеза крахмала. Однако в конце концов экспорт
триозофосфатов станет конкурйровать с регенерацией РуБФ,
и скорость фотосинтеза упадет.
9.8. Значение регуляции
Как мы уже подчеркивали (разд. 9.1), регуляция необходима
для эффективной фотосинтетической ассимиляции углерода,
как, впрочем, и для осуществления любой другой важной реак-
ции метаболизма. В ряде случаев (например, при распределе-
нии продукта между его использованием внутри хлоропластов,
экспортом из них и запасанием в виде таких соединений, как
крахмал; разд. 9.7) картина более или менее ясна. Мы уже
имеем достаточно четкое представление о вероятной регуляции
метаболических процессов у растений и о тех преимуществах,
которые они получают от функционирования того или иного
контрольного механизма. Это утверждение справедливо и для
инактивации глюкозо-6-фосфатдегидрогеназы под действием
света. Бассэм и соавторы изучали механизмы активации и инак-
тивации этого фермента (отличающиеся от тех, которые были
рассмотрены в разд. 9.3) и нашли, что между ВПФ-циклом,
функционирующим на свету,, и окислительным пеитозофосфат-
ным циклом, действующим в темноте, возможны переключения.
Во многих листьях, например, весь или почти весь крахмал, ко-
торый накапливается в течение дня, используется ночью. Весьма
вероятно, что синтез крахмала и его распад не могут идти одно-
временно. [В этом отношении важную роль должна играть ре-
гуляция глюкозо-1-фосфат — аденилилтраисферазы (разд. 9.7) и
ФБФ-азы (разд. 9.3). Хотя здесь не все еще ясно, имеются дос-
таточно убедительные доказательства в пользу того, что триозо-
фосфат служит важным продуктом распада крахмала в хлоро-
пластах. Если процесс идет через ФБФ, то очевидно', например,
что при одинаковой активности фруктозо-6-фбсфаткиназы и
286
ГЛАВА 9
Рис. 9.13. Холостой цикл функционирования фруктозо-6-фосфаткииазы и
фруктозо-1,6-бисфосфатазы.
фруктозо-1,6-бисфосфатазы в одном и том же хлоропласте цикл
будет совершенно бесполезным (рис. 9.13).] В других случаях
смысл регуляции менее ясен. Телеология (т. е. тенденция иссле-
дователя во всем усматривать смысл) еще бытует в науке (см.
Krebs, 1954), ио вряд ли следует полагать, что каждая потенци-
альная регуляторная реакция предоставляет организму какое-то
преимущество. Не удивительно, что ферменты ВПФ-цикла про-
являют высокую активность при тех значениях pH, концентра-
циях ионов магния и уровнях окислителы-ю-восстаиовителыюго
потенциала, которые характерны для стромы освещенного хло-
ропласта. Не исключено, однако, что снижение каталитической
.активности этих ферментов в результате изменений, происходя-
щих в строме хлоропластов в темноте, дает какой-то выигрыш.
Вместе с тем отсюда [еще не следует, что некоторые из этих
ответных реакций являются неотъемлемыми компонентами фо-
тосинтеза.
Ряд дополнительных вопросов, связанных с регуляцией асси-
миляции углерода при фотосинтезе Ссрастений, мы рассмотрим
в гл. 12.
'9.9. Роль адениновых нуклеотидов в регуляции
В разд. 7.13 и 9.7 мы обращали внимание на то, каким об-
разом хлоропласты (.«понимают», что они находятся в среде с
низкой концентрацией неорганического фосфата. Было высказа-
но предположение, что один или несколько фотосинтетических
процессов регулируются адениновыми нуклеотидами, может
быть, при помощи энергетического заряда, действия масс или
отношения l[ATP]/[ADP]. Действительно, контроль подобного
рода вполне возможен, если в какой-то момент общее содержа-
ние свободного неорганического фосфата внутри хлоропласта
падает гораздо ниже того уровня, который необходим для пре-
вращения всех имеющихся молекул АМР и ADP в АТР. Для
того чтобы подтвердить это предположение, необходимо прове-
сти точные измерения адениновых нуклеотидов, что на первый
взгляд кажется вполне выполнимым благодаря современному
уровню исследований. Однако следует отдавать себе отчет в
том, что изменения в состоянии адениновых нуклеотидов могут
РЕГУЛЯЦИЯ фотосинтетической АССИМИЛЯЦИИ углерода
287
Рис. 9.14. Кинетика выделения кислорода и изменения концентрации адени-
новых нуклеотидов в реконструированной системе хлоропластов, подвергну-
тых осмотическому шоку. К конечному объему 1 мл, содержащему 100 мкг
хлоропластов, были добавлены ферредоксин (55 мкг) NADP (0,1 мкмоль) и
ADP (0,3 мкмоль), а также ДТТ, MgCl2, бикарбонат и т. д. [30]. Затем до-
бавляли рибозо-5-фосфат (0,2 мМ) и 3-фосфоглицериновую кислоту
(0,2 мМ), как указано на рисунке. Выделение кислорода (штриховая линия)
измеряли полярографическим методом. Через определенные промежутки вре-
мени отбирали пробы (5 мкл) и в люцифернн-люциферазной реакции иссле-
довали в них содержание АТР (темные кружки) и ADP (светлые кружки)
при помощи люмииометра LKB. Содержание АТР измеряли сразу, a ADP
после его превращения в АТР под действием пируваткиназы, добавленной
вместе с ФЕП. Концентрации рассчитывали по калибровочной кривой, кото-
рую строили с помощью проб с известным количеством АТР. С—свет, Т —•
темнота.
быть неуловимо малы. Например, хлоропласт, который связы-
вает СО2 со скоростью 100 мкмоль-(мг Хл)“1-ч~1, должен син-
тезировать и использован» АТР со скоростью не менее
300 мкмоль-(мг Хл)-1-ч-1. Учитывая тот факт, что общая кон-
центрация адениновых нуклеотидов составляет около 0,8 мМ,
скорость превращения AMP, ADP и АТР должна быть очень
высокой, а изменения стационарных величий могут быть весьма
небольшими, но, имеющими важные последствия. Картина ста-
новится еще более сложной, .если вспомнить, что присутствие
NADP+ и ADP способно оказать значительное влияние на ха-
•288
ГЛАВА 9
рактер и скорость, транспорта электронов. Тем не менее, как
показали опыты с реконструированной системой хлоропластов,
которые Хоуп, Карвер и Уокер провели недавно в Шеффилде,
изменения, вызываемые адениновыми нуклеотидами, согласуют-
ся с предположениями, высказанными в разд. 9.6. На рис. 9.14
видно, что сначала происходит быстрое выделение Ог, причем
стехиометрия процесса согласуется с восстановлением добавлен-
ного NADP+. После этого нециклический транспорт электронов
прекращается из-за недостатка окислителя до тех пор, пока до-
бавление ФГК не вызовет регенерацию NADP+ из NADPH. За-
тем восстановление ФГК связанное с ним выделение О2 оста-
навливают добавлением Ру5Ф, который взаимодействует с
АТР (разд. 9.6). Спустя короткий промежуток времени выделе-
ние О2 возобновляется, после чего оно .прекращается в темноте
(Т) и опять начинается в результате включения света (С). Как
мы сейчас увидим, соотношения адениновых нуклеотидов ;и их
изменения можно в основном предсказать. Некоторое количест-
во АТР и АМР (концентрацию АМР в данном опыте ,не опреде-
ляли) образуется в темноте, так как присутствующая аденилат-
киназа (миокиназа) катализирует превращение 2ADP—*-АТР-(-
4-АМР. Свет вызывает быстрый рост содержания АТР и паде-
ние концентрации ADP, причем эти процессы вскоре замедляют-
ся по мере того, как восстанавливается весь добавленный
NADP, а нециклический транспорт электронов заменяется цик-
лическим (или псевдоциклическим) транспортом. Как и следо-
вало' ожидать, добавление ФГК на короткое время опять меняет
ситуацию, поскольку при этом представляется дополнительное
количество ADP и NADP, но теперь на каждую молекулу выде-
ляющегося кислорода требуются две молекулы АТР:
2NADP+ + 2Н2О 4- 2ADP + 2Рг ->- 2NADPH2 + 2АТР + О2 (9.6)
2ФГК + 2АТР 4-2NADPHZ —> 2ФГА + 2Р; ф-2ADP + 2NADP+ (9.7)
• АТР, который необходим во второй реакции (9.7), быстро
компенсируется нециклическим фотофосфорилированием в пер-
вой реакции (9.6), и примерно через минуту концентрации АТР
и ADP возвращаются к исходным стационарным значениям
(рис. 9.14).
Сходное, но более сильное отклонение от стационарного
уровня адениновых нуклеотидов наблюдается при добавлении
Ру5Ф. Однако' в этом случае в результате первоначального па-
дения отношения [ATP]/[ADP] прекращается нециклический
транспорт электронов и связанное с ним выделение О2. Термо-
динамически выгодное отношение может быть вновь обретено
лишь посредством той или иной формы циклического фотофос-
форилирования. Интересно, что в данном опыте стационарное
отношение [ATP]/[ADP] после добавления ФГК составляло при-
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ ассимиляции углерода
289
мерно 5, а выделение О2 прекращалось, когда это отношение
достигало 1, возобновлялось при отношении, несколько меньшем
чем 3, и повое стационарное состояние (как для Оа, так и для
адениновых нуклеотидов) достигалось при отношении ~4. Это
новое значение ниже, по-видимому, чем первоначальное, так как
АТР непрерывно используется в реакции РубФ, чем может так-
же объясняться и зарегистрированная теперь меньшая ско-
рость выделения кислорода. Впрочем, не следует забывать, что
данная реакционная смесь будет иметь теперь очень сложный
состав, поскольку из РубФ все время возникают новые молеку-
лы ФГК, и наоборот.
При выключении света отношение l[ATP]/[ADP]| снижается,
а затем опять возрастает, если /снова включить свет. Первая
стадия этого последнего возрастания будет, очевидно', отражать
присутствие достаточных количеств NADP, а не немедленное
возобновление восстановления ФГК.
Результаты этого и аналогичных опытов подтверждают пред-
ставление о том, что в регуляции фотосинтеза важную роль мо-
гут играть соотношения между концентрациями АТР и ADP, а
также их изменения в зависимости от содержания неорганиче-
ского фосфата. Кроме того, они содержат в себе важное предо-
стережение. Дело в том, что большие изменения скорости по-
требления АТР могут происходить без изменений стационарной
концентрации АТР (эти изменения могут быть лишь временны-
ми), Следует также подчеркнуть, что значение имеет не столько
П1ТП1/ГЛПП1 [АТР]/[ФГК]
отношение [ATPj/[ADP], сколько отношение~[двр]ддфГд]~
Таким образом, выделение кислорода при фотосинтезе про-
текает в интактных хлоропластах, в которых отношение
[ATP]/[ADP] меньше единицы, но оно достигается в присутст-
вии относительно высоких концентраций ФГК-
Источники данных: [36, 37, 40],
ОБЩАЯ ЛИТЕРАТУРА
1. Ainsworth S, Steady-state Enzyme Kinetics, Macmillan, London, 1977.
2. Anderson L. E. Interaction between photochemistry and activity of enzymes,
Encyclopedia of Plant Physiology (New Series), Vol. VI, 271—278, Springer-
Verlag, Berlin, 1970.
3. The Intact Chloroplast, Topics Photosynthesis, Vol. I (Barber J., ed.), Else-
vier, Amsterdam, 1976.
4. Bassham J. A, The control of photosynthetic carbon metabolism, Science, 172,
526—534 (1974).
5. Gibbs M., Latzko E. Regulation of photosynthetic carbon metabolism and
related processes, Encyclopedia of Plant Physiology (New Series) Photo-
synthesis, Vol. II, 1—578. Springer-Verlag, Berlin, 1978.
6. Halliwell B. The chloroplast at work—a review of modern development on
our understanding of chloroplast metabolism, Prog. Biophys. Molec. Biol.,
33, 1—54 (1978).
19—234
290
ГЛАВА 9
7. Herold. A., Walker D. A. Transport across chloroplast envelopes — the role
ol phosphate, In: Handbook on Transport across Biological Membranes (Gie-
bisch G., Tostcson D. C., Ussing J. J., eds.), 412—439. Springer-Verlag, Hei-
delberg, 1978.
8. Jensen R. G., Bahr J. T. Ribulose 1,5-blsphosphate carboxylase oxygenase,
Ann. Rev. Plant Physiol., 28, 379—400 (1977).
9. Kelly Q. K., Latzko £., Gibbs M. Regulatory aspects of photosynthetic car-
bon metabolism, Ann. Rev. Plant Physiology, 27, 181—205 (1976).
10. Krebs H. A. Excursion into the borderland of biochemistry and philosophy,
Bull. John Hopkins Hosp., 95, 45—51 (1954).
11. Krebs H. A. The role of equilibria in the regulation of metabolism, Curr. To-
pics hi Cellular Reg., 1, 45—55 (1969).
12. Lehninger A. L. Biochemistry, Worth Pub, Inc., New York, 1970.
13. Preiss J., Kosnge T. Regulation of enzyme activity in photosynthesis systems,
Ann. Rev. Plant Physiol., 21, 433—466 (1970).
14. Walker D. A. Regulatory mechanisms in photosynthetic carbon metabolism,
In; Current Topics in Cellular Regulation (Horecker B. L., Stadtman E.,
eds.), 11, 203-24-1 (1976).
15. Walker D. A., Herold A. Can the chloroplast support photosynthesis unai-
ded? Im. Photosynthetic Organelles: Structure and Function (Miyachi S.,
Katoh S., Fujita Y., Shibata K., eds.), Special Issue of Plant and Cell Phy-
siol., 295—310 (1977).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
16. A/iazania T. Ribulose-l,5-bisphosphate carboxylase, New Encyc. Plant Phy-
siol., Vol. II, 208—25 (Gibbs M., Latzko E., eds.) Springer'-Verlag, Berlin,
1979.
16а. Alexander A. G. Sugar Cane Physiology, Elsevier, New York, 1977.
17. Anderson L. E., Advani V. R. Chloroplast and cytoplasmic enzymes: three
distinct enzymes associated with the reductive pentose phosphate cycle.,
Plant Physiol., 45, 583—585 (1970).
18. Anderson L. E., Nehrlich S. C., Champigny M.-L. Light modulation of enzyme
activity. Activation of the light effect mediators by reduction and modulation
of enzyme activity by thioldisulfide exchange? Plant Physiol,, 61, 601—
605 (1978).
19. Bahr J. T., Jensen R. G. Activation of ribulose bisphosphate carboxylase in
intact chloroplast by CO2 and light, Archs. Biochem. Biophys., 185, 39—48
(1978).
19a. Buchanan В. B. Role of light in the regulation of chloroplast enzymes, Ann.
Rev. Plant Physiol., 31, 341—374 (1980).
20. Baier D., Latzko E. Properties and regulation of C-l-fructose-l,6-diphosphate
from spinach chloroplasts, Biochim. Biophys. Acta, 396, 141—8 (1975).
21. Buchanan В. B., Kalberer P. P., Amon D. I. Ferredoxin-activated fructose
diphosphatase in isolated chloroplasts, Biochem. Biophys. Res. Comtnun,, 29,
74—79 (1967).
22. Buchanan В. B., Schurmann P., Kalberer P. P. Ferredoxinactivated fructose
diphosphaiase of spinach chloroplasts, J. Biol. Chem., 246, 5952—59 (1971).
23. Buchanan В. B., Wolosluk R. A. Photosynthetic regulatory protein found in
animal and bacterial cells, Nature, 264, 669—670 (1976).
24. Chen-she S. H., Lewis D. H., Walker D. A. Stimulation of photosynthetic
starch formation by sequestration of cytoplasmic orthophosphate, New Phy-
tol., 74, 383—392 (1975).
24a. Halliwell B., Foyer С. H. Properties and physiological function of a gluta-
thione reductase purified from spinach leaves by affinity chromatography,
Planta, 139, 9—17 (1978).
РЕГУЛЯЦИЯ ФОТОСИНТЕТИЧЕСКОЙ АССИМИЛЯЦИИ УГЛЕРОДА
291
25. Heldt Н. IF., Chon С. J., Lorimer G. Н. Phosphate requirement for the light
activation of ribulose-l,5-bisphosphate carboxylase in intact spinach chloro-
plasts, FEBS Letters, 92, 234—240 (1978).
26. Heldt H. HL, Chon C. J., Maronde D., Herold A., Stankovic Z. S„ Wal-
ker D. A., Kramlner A., Kirk M. A., Heber U. The role of orthophosphate and
other factors in the regulation of starh formation in leaves and isolated chlo-
roplasts, Plant Physioh, Б9, 1146—1155 (1977).
27. Herold A., Lewis D. H., Walker D. Л. Sequestration of cytoplasmic orthophos-
phate by mannose and ils differential effect on photosynthetic starch synthe-
sis in C3 and C4 species, New Phytol., 76, 397—407 (1976).
28. Kakle T. Effects of phosphorous deficiency on the photosynthetic carbon dio-
xide fixation products in tobacco plants, Soil Sci. Plant Nutr., 15, 245—251
(1969).
28a. Leegood R. C., Walker D. A. Modulation of fructose bisphosphatase activity
in intact chloroplasts, FEBS Letters, 116, 21—24 (1980).
28b. Leegood R. C.. Walker D. Л. Regulation of fructose 1,6-bisphosphatase acti-
vity in intact chloroplasts. Studies of the mechanism of inactivation, Biochim.
Biophys. Acta, 593, 362—370 (1980).
29. Lilley R. McC., Walker D. A. The reduction of 3-pbosphoglycerale by recon-
stituted chloroplasts and by chloroplast extracts, Biochim. Biophys. Acta,
368, 269—278 (1974).
29a. Lilley R. McC., Holboraw K-, Walker D. A. Magnesium activation of pho-
tosynthetic COs fixation in a reconstituted chloroplast system, New Phytol.,
73,'657—662 (1974).
30. Lilley R. McC., Walker D. A. Studies with the reconstituted chloroplast sys-
tem, Encyclopedia of Plant Physiology (New Series), Photosynthesis Vol. II.
Regulation of photosynthetic carbon metabolism and related processes
(Gibbs M., Latzko E., eds.), 41—52. Springer-Verlag, Berlin, 1979.
31. Lorimer G. H., Badger Ml. R., Andrews T. I. The activation of ribulose-1,5-
bisphosphate carboxylase by carbon dioxide and magnesium ions. Equilibria,
kinetics, a suggested mechanism and physiological implication, Biochemistry,
15,529—536 (1976).
32. Lorimer G. H„ Badger M. R„ Heldt H. IF. The activation of ribulose 1,5-bis-
phosphate earboxylase/oxygenase, In: Photosynthetic Carbon Assimilation.
Ribulose 1,5-bisphosphate Carboxylase/Oxygenase, Brookhaven Symposium
in Biology, Vol. 30 (Siegelman I-I. W., Hind G., eds.), 283—306. Brookhaven
Nat. Lab., Upton, New York, 1978.
33. Neales T, F., Incoll L. D. The control of leaf photosynthetic rate by the level
of assimilate concentration in the leaf. A review of the hypothesis, Bot. Rev.,
34, 107—125 (1968).
34. Portis A. R., Heldt H. IF. Light-dependent changes of the Mg2+ concentration
in lhe stroma in relation to the Mg?+ dependency of CO2 fixation in intact
chloroplasts, Biochim. Biophys. Acta, 449, 434—446 (-1976).
35. Priess I. The regulation of the biosynthesis of a-l,4-glucans in bacteria and
plants, Curr. Topics in Cell Reg., 1, 125—160 (1969).
36. Robinson S. P„ Walker D. A. The control of PGA reduction in isolated chlo-
roplasts by the concentrations of ATP, ADP and RGA, Biochim. Biophys.
Acta, 545, 528—536 (1979).
37. Walker D. A., Robinson-S. P. Regulation of photosynthetic carbon assimila-
tion, In: Photosynthetic Carbon Assimilation. Ribulose. 1,5-Bisphosphate Car-
boxylase/Oxygenase, Brookhaven Symposium in Biology, Vol. 30 (Siegel-
man H. W., Hind G., eds.), 43—49. Brookhaven Nat. Lab., Upton, New York,
1978.
38. Robinson S. P„ McNeil P. H., Walker D. A. Ribulose bisphosphate carboxy-
lase — lack of dark inactivation of the enzyme in experiments with proto-
plasts, FEBS Letters, 97, 296—300 (1979).
19*
292
ГЛАВА V
39. Scluirniann Р., Wolosiuk R. A., Breazeale V. D., Buchanan В. B. Two pro-
teins [unction in the regulation of photosynthetic CO2 assimilation in chloro-
plasts, Nature, 263, 257—258 (1976).
40. Slabas A. R., Walker D. A. Localization of inhibition by adenosine diphos-
phate of phosphoglycerate-depeiident oxygen evolution in a reconstituted chlo-
roplast system, Biochem. J., 154, 185—192 (1976).
41. Paech C., McCurry S. D., Pierce J., Tolbert N. E. Active site of ribulose
1,5-bisphosphate carboxylase/oxygcnase, In: Photosynthetic Carbon Assimi-
lation. Ribulose 1,5-Bisphosphale Carboxylase/oxygenase, Brookhaven Sympo-
sium in Biology, Vol. 30 (Siegelman H. W., Hind G., eds.) 227—43, Brookha-
ven Nat. Lab., Upton, New York, 1978.
42. Walker D. A., Lilley R. McC. Ribulose bisphosphate carboxylase — an enigma
resolved, Proceedings of the 50th Anniv. Meeting. SEB Cambridge (ed. Sun-
derland N.), 189—198, Pergamon Press, Oxford, 1974.
43. Walker D. A., Slabas A. R., Fitzgerald M. P. Photosynthesis in a reconsti-
tuted chloroplast system from spinach. Some factors affecting COa-dcpendent
oxygen evolution with fruclose-l,6-bisphosphate as substrate, Biochim. Bio-
phys. Acta, 440, 147—162 (1976).
44. Wolosiuk R., Buchanan В. B. Thioredoxin and glutathione regulate photo-
synthesis in chloroplasts, Nature, 266, 565—567 (1977).
45. Wolosiuk R., Buchanan В. B. Activation of chloroplast NADP-linked giyce-
raldehyde-3-phosphate dehydrogenase by the ferredoxin/thioredoxin system,
Plant Physiol., 61, 669—671 (1978).
ГЛАВА 10
Открытие С4 -пути
Краткое содержание
В этой главе мы бегло рассмотрим историю открытия С4-фото-
синтеза и обсудим некоторые особенности этого метаболическо-
го пути. Основу для биохимических исследований фотосинтеза
у Сграстений заложили в своих ранних экспериментах Хэтч и
Слэк, Затем потребовалось больше 10 лет научных поисков для
того, чтобы выяснить, что механизм фотосинтеза у подобного
рода растений связан с декарбоксилированием С4-кислот и по-
следующей передачей СО2 в ВПФ-цикл и что в этом механизме
ие участвует транскарбоксилаза, как предполагали вначале, За
этот же промежуток времени были достигнуты значительные ус-
пехи в разработке методов выделения клеток, протопластов и
хлоропластов из С4-растений, что способствовало прогрессу био-
химических исследований. Не известно ни одного исключения
из общего правила: Сгфотосинтез присущ только таким покры-
тосеменным растениям (это единственная группа, где встреча-
ются С^растения), у которых есть фотосинтезирующие клетки
двух типов. На свету С4-растеиия осуществляют- перенос атмо-
сферной СО2 в ВПФ-путь через цикл С4-кислот. Все эти факты
можно использовать в качестве критериев при оценке, насколько
обмен С4-кислот в той или иной фотосинтезирующей ткани (от
водорослей до покрытосеменных растений) близок к метаболиз-
му у С4-растеиий.
10.1. С4-дикарбоновые кислоты — ранние продукты
фотосинтеза
История открытия С4-пути фотосинтеза у некоторых высших
растений ведет свое начало от исследований Коршака и сотруд-
ников, которые в конце пятидесятых годов работали в Гонолулу
на Experimental Station of Hawaiian Sugar Planters' Association.
Они заметили, что листья сахарного тростника после кратковре-
менной (в течение нескольких секунд) инкубации в атмосфере
с 14СОг на свету включают значительную часть радиоактивной
метки (70—80%) в малат и аспартат. В темноте количество
фиксируемой СО2 не превышало' 1% количества СО2, фиксируе-
мой на свету. Малат и аспартат содержат по 4 атома углерода,
294
ГЛАВА 10
Рис. 10.1. COs-насос (С4-фотосиитез). Для того чтобы обеспечить поступле-
ние СОг в высокой концентрации, гарантирующей насыщение восстанови-
тельного пентозофосфатного пути субстратами, некоторые растения в ходе
эволюции приобрели своего рода метаболический С02-насос, Это и есть
С4-цикл, для работы которого нужна энергия (АТР). Функция этого цикла
заключается в улавливании и захвате атмосферной СО2 фотосинтезирующи-
ми клетками наружного слоя листа (мезофилл) и ее передаче за счет челноч-
ного механизма во внутренний слой листа (кранц-клеткн), где она и ассими-
лируется.
поэтому этот путь фиксации СОг стали называть С^-путем, чтобы
отличить его от Сз-пути (или, ВПФ-пути) (гл. 6), в котором
первоначальный продукт фотосинтеза содержит только 3 атома
углерода. Почти одновременно с исследованиями, проводивши-
мися иа Гавайях, сходные результаты были получены на куку-
рузе Ю. Карпиловым и сотрудниками .-в СССР. Но данные со-
ветских исследователей стали известны более широкому кругу
ученых только в середине шестидесятых годов после публика-
ции работы Коршака 4и др. (Kortschack et al., 1965). Таким об-
разом, открытие высокого (в процентном отношении) включения
метки в С4-кислоты позволило выявить глубокие различия в
составе начальных продуктов фотосинтеза у сахарного тростни-
ка и кукурузы, с одной стороны, и у ранее изученных водорос-
лей и высших растений — с другой.
В работах-Кальвина и Бенсона, выполненных в сороковые
годы, было показано, что основным начальным продуктом фото-
синтеза у Chlorella и Scenedesmus и ряда высших растений яв-
ляется ФГК (гл. 6). Еще в ранних исследованиях фотосинтеза
у водорослей было обнаружено, что малат тоже относится к
ранним продуктам фотосинтеза. Когда Chlorella и Scenedesmas
помещали на несколько секунд в атмосферу 14СО2, то основным
меченым продуктом была ФГК, но при этом небольшое количе-
ство метки регистрировали и в составе малата и гликолата. Эти
наблюдения позволили в 50-х годах Бенсону и Кальвину, а так-
же Гаффрону и Фагеру и Очоа предложить ряд схем, описываю-
ОТКРЫТИЕ СгПУТИ
295
Рис. 10.2. Гипотетическая схема двойного карбоксилирования, предлагавшая-
ся в пятидесятые годы ([3J; см. также Bassham et al., 1950).
щих циклы фиксации СО2, которые включали в себя С2- и С4-
промежуточные продукты и, как неизбежное следствие этого,
два отдельных акта карбоксилирования. В них рассматривалась
также возможность непосредственного участия в данных цик-
лах таких Сгкислот, как оксалоацетат, малат и «сукцинат
(рис. 10.2). Наиболее важной особенностью всех этих схем бы-
ли две реакции карбоксилирования в ходе цикла, каждая из ко-
торых вносила свою лепту в суммарную фиксацию СО2. Как
будет рассмотрено ниже, у С^-растений действительно имеются
два этапа карбоксилирования, но к суммарной, или реальной,
фиксации СО2 приводит только та реакция карбоксилирования,
которая идет через РуБФ. В ранних работах со Scenedesmus
Бассэм и др. (Bassham et al., 1950) установили, что отравление
растений малоиатом влечет за собой сильное ингибирование об-
разования малата, ио слабо влияет на включение 14СО2 в ФГК.
Именно поэтому малат и рассматривали не как промежуточный
продукт данного первичного пути ассимиляции углерода, а как
вторичный продукт фиксации СО2. Механизм действия малоната
был неизвестен, однако было показано, что малонат ингибирует
сукцинатдегидрогеназу, а еще позднее, что таким же образом
он действует на ФЕП-карбоксилазу. В отличие от этих данных,
полученных на водорослях, Коршак и др. обнаружили, что у
растений сахарного тростника малонат сильно ингибирует фото-
синтез, образование малата и синтез ФГК. Это заставляло пред-
полагать, что для синтеза ФГК необходимо предварительное об-
разование малата.
Кальвин и др., работая со Scenedesmus, тоже заметили, что
при очень низкой интенсивности света (45 фут-свечей) основным
первичным продуктом фиксации СО2 является малат, а основ-
ным вторичным продуктом— сахароза. К этому времени уже
было доказано, что малат — это первичный продукт темновой
фиксации СО2. Поэтому реальная фиксация СО2 у водорослей
при очень низкой интенсивности света могла быть результатом
296
ГЛАВА 10
наложения двух процессов: световой и темновой фиксации.
В это же время Фагер и Гаффрои отметили, что такой процесс
может быть вторичным и ие связанным с механизмом фотосин-
теза.
У Сз-растений первичное включение атмосферной СОг про-
исходит с участием РуБФ-карбоксилазы (гл. 6). В листьях Сз-
растений в С4-КИСЛОТЫ включается только небольшая доля (ие
более 5%) связанной СО2. Малат может образовываться в ре-
зультате вторичного карбоксилирования, следующего за реак-
цией, катализируемой РуБФ-карбоксилазой '[уравнения (10.1—
10.5)], или из ФЁП, появляющегося в ходе гликолиза.
РуБФ + СО2 -----> 2(3-ФГК) (РуБФ-карбоксилаза) (Ю.1)
3-ФГК ----> 2-ФГК (3-фосфоглицеромутаза) (10.2)
2-ФГК -----> ФЕП (енолаза) (10.3)
ФЕП НСО3_ --------> Оксалоацетат Р, (ФЕП-карбоксилааа) (10.4)
Оксалоацетат -|- NAD(P)H -р Н+--->
-----------> Малат -|- NAD(P) + [МАО(Р)Н-малатдегидрогеназа] (10.5)
Источники данных; [4, 12, 25, 36].
10.2. Вклад Хэтча и Слэка
в проблему С4-фотосинтеза
Необычайно высокое включение меченой 14СОг в ,малат и
аспартат в листьях сахарного тростника очень заинтриговало
Коршака, и он убедил австралийских ученых Хэтча и Слэка
заняться изучением биохимических аспектов этого явления. За
4 года работы Хэтч, Слэк и Джонсон внесли очень большой
вклад в исследуемую проблему. Они, во-первых, провели опыты
по разбавлению импульсной метки у сахарного тростника, во-
вторых, идентифицировали ряд ферментов С4-пути, в-третьих,
получили предварительные доказательства того, что у кукурузы
некоторые ферменты по-разному распределены между хлоро-
пластами /мезофилла и хлоропластами обкладки проводящих
пучков, и, в-четвертых, выявили несколько других видов расте-
ний, у которых характер включения метки был точно таким же,
как у сахарного тростника.
В опытах по разбавлению импульсной метки создавали такое
освещение, которое позволяло растениям достичь равновесного
состояния фотосинтеза в нормальных условиях (т, е. в присутст-
вии атмосферной 12СОг). Затем в течение определенного проме-
жутка времени (в данном случае в течение нескольких секунд)
проводили импульсное включение 14СО2, после чего растение
немедленно переносили в нормальные условия (атмосферная
12СО2). Через определенные интервалы после инкубации с 14СО2,
т. е. в процессе разбавления метки в атмосфере 12СО2, отбирали
ОТКРЫТИЕ Cj-ПУТИ
297
Свет
Рис. 10.3. Типичные изменения содержания радиоактивной метки в С4-кисло-
тах (малат+аспартат), ФГК и запасных продуктах (крахмал+сахароза) у
С3- и Сд-растений в опыте с разбавлением импульсной метки. Другие продук-
ты, например триозо- и гексозофосфаты, на рисунке не представлены (Chen
et al., 1971; Galmiche, 1973; Hatch, Osmond, 1976).
298
ГЛАВА 10
пробы листьев. Листья фиксировали (например, в кипящем эта-
ноле или в жидком азоте) и затем при помощи хроматографии
анализировали состав !4С-продуктов. Эксперименты подобного
рода, в которых общее количество включенной радиоактивности
во время разбавления метки не меняется, позволяют выявлять
самые ранние из первичных продуктов фотосинтеза и 'их даль-
нейшие превращения. Как видно из рис. 10.3, начальными про-
дуктами фотосинтеза у С4-растений оказались малат и аспартат.
После переноса в атмосферу 12СОг содержание радиоактивной
метки в аспартате и малате уменьшается, а количество метки,
включенной в ФГК, сахарозу и крахмал, возрастает. Большая
часть включенной радиоактивности обнаруживается в С-4-поло-
жении малата, а в ходе разбавления метки сначала больше
всего метится С-1-атом ФГК- Опыты такого рода позволили
Хэтчу и Слэку получить первые доказательства, что у С4-расте-
ний углерод С4-кислот быстро переходит в другие соединения и
что С4-кислоты ие являются конечными продуктами.
В отличие от С^растений, у которых первичные продукты
фотосинтеза представлены малатом и аспартатом, у Сз-растений
начальным продуктом фиксации СО2 является ФГК. При сход-
ной с представленной на рис. 10.3 постановке опыта по мере
разбавления импульсной метки радиоактивность ФГК убывает,
а радиоактивность сахарозы и крахмала возрастает. Такое по-
ведение метки наблюдалось и в более ранних исследованиях
фотосинтетического метаболизма углерода; в настоящее время
оно рассматривается как типичное для Сз-растений.
Чтобы быть уверенным в достоверности данных, отражаю-
щих метаболические превращения в процессе фотосинтеза in vi-
vo, исследователи должны правильно ставить эксперименты по-
добного рода. Здесь прежде всего следует упомянуть такие
факторы, как равноценное освещение, внесение 14СОг только в
условиях равновесного состояния фотосинтеза и поддержание
относительно низкой суммарной концентрации СОг во время им-
пульсного включения метки (т. е. ще более 0,04—0,05% уже
после добавления меченой СОг с высокой удельной активностью
к атмосферной СОг, концентрация которой в воздухе составляет
~0,03%). Отклонения от этой общей процедуры могут иметь
определенный интерес, однако полученные данные следует ин-
терпретировать только исходя из нормальных условий окру-
жающей среды.
. .Начальными продуктами С^фотосинтеза обычно принято
считать и малат, и аспартат. Оксалоацетат рассматривают как
самый ранний, но не самый главный продукт включения метки,
Даже при минимально возможном времени включения (~1 с).
Если в некоей последовательности реакций образуется продукт,
содержание которого относительно мало' по сравнению с количе-
ОТКРЫТИИ С.4-ПУТИ
299
ством других продуктов того же пути, то выявить его довольно
трудно. Именно это наблюдается, по-видимому, в случае окса-
лоацетата у Ст-растений.
Рассмотрим последовательность реакций А+СО2—>В—^С.
Если содержание вещества В очень мало по сравнению с содер-
жанием вещества С, то в состав В войдет относительно малая
доля включенных импульсов (общее число включенных импуль-
сов равно произведению удельной радиоактивности и общего со-
держания вещества). Однако при максимально коротком вре-
мени инкубации с 14СО2 удельная радиоактивность исходного
продукта (число ими./мин, приходящихся на 1 мкмоль радиоак-
тивного вещества) будет наивысшей. После кратковременной
(менее 1 с) инкубации листьев С<-растений в атмосфере 14СО2 и
мгновенной фиксации радиоактивность оксалоацетата крайне
мала, а радиоактивность аспартата и малата очень высока. Это
говорит о том, что содержание меченого оксалоацетата очень
мало, а скорость его круговорота очень велика.
Если во время равновесного стационарного фотосинтеза при
близком к атмосферному уровне СО2 (0,03%) в С4-Р астения
внезапно ввести импульсную метку (по концентрацию ,14СО?
сделать в 10 раз больше) и через несколько секунд определить
состав исходных продуктов, то 50—60% метки обнаруживается
в оксалоацетате (Huber, Edwards, 1975). Скорее всего, внезап-
ное увеличение уровня субстрата при равновесных условиях
приводит к временному увеличению метки в конечном продукте
даже в том случае, когда содержание последнего в нормальных
условиях невелико.
Ко всему прочему оксалоацетат неустойчив и декарбоксили-
руется за счет неферментативных реакций с образованием пиру-
вата и СО2, особенно в присутствии двухвалентных катионов и
при повышенной температуре. Этим можно объяснить относи-
тельно низкое содержание оксалоацетата в биологических систе-
мах. При фиксации растительного материала после включения
14СО2 необходимо создать такие условия, которые бы способст-
вовали сохранению оксалоацетата, например обработать его
динитрофеиилгидразином, чтобы получить стабильное производ-
ное фенил гидр азо и а. Иначе в результате неферментативного
декарбоксилирования оксалоацетата, особенно, в том случае,
когда метка находится в С-4-положении, 14С полностью исчезнет
из этого соединения.
С-1 СООН СООН
С-2 i=O + ldCO2
С-2 (kl2 * il-Lj
С-4 14СООЫ
300
ГЛАВА 10
Само по себе включение метки из ИСО2 в малат и аспартат у
С4-растений ничего не говорит об относительной скорости синте-
за этих дикарбоновых кислот. Так, например, можно очень бы-
стро пометить аспартат прямо в листе в результате перехода
метки из радиоактивного оксалоацетата (при этом переамини-
рование идет намного быстрее, чем обычная реакция). В то же
время содержание аспартата остается неизменным. Суммарный
эффект можно записать так:
14С-Оксалоацетат Аспартат ——> Оксалоацетат -|- 14С-Аспартат
Реакция идет с участием аспартатаминотрансферазы (рис. 10.4).
В процессе разбавления 12СО2 метка в аспартате должна
вновь уменьшиться вследствие изотопного обмена с немеченым
оксалоацетатом, но первичный поток углерода может проходить
и через малат. Хотя и малат, и аспартат могут непосредственно
передавать свой углерод другим соединениям, очевидно, относи-
тельная радиоактивность этих кислот после нескольких секунд
пребывания на свету не отражает степени их непосредственного
участия в Сгдикле.
С4-Кислоты не относятся к числу конечных продуктов фото-
синтеза у С4-растений. Ими являются гексозофосфаты, сахароза
и крахмал, для синтеза которых требуются определенные трех-
и шестиуглеродные предшественники. Поэтому было необходи-
мо рассмотреть механизм возможного переноса углерода С4-кис-
лот в трех- или шестиуглеродные соединения и способы непре-
рывной регенерации акцептора СО2, т. е. ФЕП, необходимого
для первоначальной фиксации СО2 (разд. 10.3). Как уже отме-
чалось выше, Хэтч и Слэк идентифицировали ряд ферментов
Сгпути, правда сначала только у сахарного тростника. У С4-рас-
тений такие ферменты, как пируват, ортофосфат — дикиназа,
ФЕП-карбоксилаза и NADP-малатдегидрогеназа, участвуют в
карбоксилирующей стадии С4-пути и локализованы в клетках
ФЕП - + z ,4СО3
14С*Оксалоацвтат ---------Л [Аспартат]
NADPH
NADP
14 ОМалат
Г лутамат
2-Оксоглутарат
Рис. 10.4. Включение 14С в аспартат в отсутствие суммарного (реального)
синтеза.
ОТКРЫТИЕ С„-ПУТИ
30!
мезофилла. На этом этапе пируват и СО2 превращаются в ма-
лат (подробно см. в разд. 11.1).
Пируват + Pf + АТР ->
---> ФЕП + АМР-|-РР| (пируват, ортофосфат—дикияаза) (10.6)
ФЕП + НСОц- -------------------------------------> ОксалоацетатPj (ФЕП-карбокснлаза) (Ю.4)
Оксалоацетат N ADPH -ф- Н+ ->
——> Малат -ф NADP+ (NADP-малатдегидрогеиаза) (10.7)
Обычно в листьях Сз-растений активность пируват,ортофосфат—
дикиназы вообще ие выявляется, в листьях С4-растепий актив-
ность ФЕП-карбоксилазы раз в 50 выше, чем в листьях С3-рас-
теиин, а уровень NADP-малатдегидрогеназы достаточно высок и
у тех и у других.
Применив неводные среды, состоящие из смеси гексана и
четыреххлористого углерода (в таком соотношении, чтобы плот-
ность среды составляла 1,3—1,4 г/мл), Слэк и др. (Slack et al.,
1969) получили первые доказательства, что некоторые фотосин-
тетические ферменты в листьях С4-растений по-разному распре-
делены между хлоропластами мезофилла и хлоропластами об-
кладки проводящих пучков. Основанием для разделения хлоро-
пластов (в этой работе, из кукурузы) на два типа послужил тот
факт, что хлоропласты мезофилла ие содержат крахмала и
имеют меньшую плотность, чем содержащие крахмал .хлоро-
пласты обкладки проводящих пучков. Пируват,ортофосфат —
дикиназа и NADP-малатдегидрогеиаза, участвующие в реакциях
С4-пути, локализованы в хлоропластах мезофилла. Точное на-
хождение ФЕП-карбоксилазы не было установлено, так как ре-
зультаты опытов, в которых применяли неводные среды, сильно
варьировали. Ограниченность используемого метода разделения
хлоропластов (низкий выход хлоропластов обкладки проводя-
щих пучков, недостаточная чистота получаемых фракций и воз-
можное загрязнение хлоропластов ферментами цитозоля) и от-
сутствие определенных сведений относительно энзимологии С4-
фотосиитеза не позволяли сделать вполне определенных выво-
дов. Точная локализация ферментов была установлена только
в последующих экспериментах, проведенных на изолированных
клетках и протопластах (гл. 11).
Источники данных: [8, 9, 28, 40].
10.3. Метаболизм Са-дикарбоновых кислот
Установлению механизма передачи углерода С4-кислот в
ФГК и другие продукты фотосинтеза у С4-растений во многом
способствовали исследования, проведенные в начале семидеся-
тых годов во многих лабораториях. Сначала Слэк и Хэтч (Slack,
302
ГЛАВА 10
Рис. 10.5. Гипотетические реакции транскарбоксилирования после фиксации
СО;! с участием ФЕГГ-карбоксилазы (Hatch, Slack, 1966; 1970).
Hatch, 1967) обнаружили, что активность РуБФ-карбоксилазы в
листьях С4-растеиип очень мала и составляет приблизительно
20—35 мкмоль-(мгХл)-1 ч-1, тогда как скорость фотосинтеза
у Сграстений близка к величине 200 мкмоль СО2- (мг Хл)-1 я-1.
Поэтому в то время С^путь рассматривали как путь, альтерна-
тивный ВПФ-пути. Чтобы логически объяснить наблюдения,
сделанные в опытах по разбавлению импульсной метки в целых
листьях, Хэтч и Слэк предположили наличие в клетках специ-
ального фермента транскарбоксилазы. Такая транскарбоксила-
за должна была катализировать прямой перенос карбоксильной
группы Сгкислоты либо иа С2-, либо на Cs-соединение, что
должно было затем приводить к образованию ФГК. Поскольку
для истинной фиксации СО2 (гл. 6) требуется автокатализ, в
дополнение к траискарбоксилазе в реакциях должны были уча-
ствовать ферменты, регенерирующие С2- или Cs-акцептор.
На роль возможного С2-акцептора был выдвинут гликоаль-
дегидфосфат, который в процессе транскарбоксилирования дол-
жен был превращаться в фосфогидроксипируват. Продуктом
восстановительного трапскарбоксилирования была бы при этом
ФГК. Если бы С2-акцептором служил гликолевый альдегид, то
продуктом транскарбоксилирования был бы гидроксипируват,
который далее мог бы превращаться в глицерат и затем в ФГК-
Предлагалась и такая схема транскарбоксилирования, когда
возможным акцептором был РуБФ. При транскарбоксилирова-
нии фермент переносил бы карбоксильную группу от С4-кислот
ОТКРЫТИЕ Cj-ПУТИ
303
прямо на пятиуглеродный акцептор, что устраняло бы потреб-
ность в РуБФ-карбоксилазе (рис. 10.5). Гипотеза, предполагав-
шая декарбоксилирование С^кислот и последующее рекарбок-
силирование с участием РуБФ-карбоксилазы, первоначально не
привлекала особого внимания. Тому было несколько причин.
1. Активность РуБФ-карбоксилазы и NADP-малатдегидрогеназы
(декарбоксилирующей; или малик-энзима, или яблочного фер-
мента) в полученных из листьев экстрактах была очень мала и
могла обеспечить не более 10% фотосинтеза в листьях. 2. В том
случае, когда листья метили, инкубируя с 14СО2, а затем пере-
носили их в атмосферу без СО2, ие регистрировали никакой
потери метки из листьев. В случае декарбоксилирования С4-кис-
лот такая потеря действительно наблюдалась бы. К тому же и
разбавление, метки в атмосфере с 5% 12СО2 не влияло иа пере-
нос 14С из С4-кислот в триозо- и гексозофосфаты, тогда как
следовало бы ожидать сильного (разбавления метки даже в том
случае, если бы в газовую фазу, содержащую 5% 12СО2, высво-
бождалось хотя бы небольшое количество 14СО2. Сейчас уже
известно, что карбоксилирование, ведущее к образованию С4-
кислот, и декарбоксилирование этих кислот происходят в разных
компартментах листа. Очевидно, декарбоксилирование С^кислот
в клетках обкладки проводящих пучков, каким-то образом
ограничивает потери СО2 и препятствует свободному уравнове-
шиванию высвобождаемой СО2 с СО2, присутствующей в газо-
вой фазе, окружающей растение. 3. Из-за недостатка знаний ре-
акция транскарбоксилирования казалась в то время самым
простейшим из возможных объяснений. Вероятно, эта гипотеза
возникла еще и потому, что Хэтч до этого изучал транскарбок-
силазу вместе со Штумпфом (Hatch, Stumpf, 1961).
Хэтч и Слэк были вынуждены отказаться от своей гипотезы
транскарбоксилирования лишь после того, как были получены
убедительные доказательства декарбоксилирования Сгкпслот и
фиксации высвобождающейся СО2 в реакции, катализируемой
РуБФ-карбоксилазой. В 1969 г. Бьёркман и Гауль сообщили о
значительной РуБФ-карбоксилазиой активности в экстрактах,
полученных из листьев нескольких С4-растеиий. То, что Хэтчу и
Слэку не удалось обнаружить столь же высокую активность
РуБФ-карбоксилазы, очевидно', связано с причинами техниче-
ского характера. РуБФ-карбоксилаза у С4-растеиий локализова-
на в клетках обкладки проводящих пучков, а эти клетки имеют
толстую клеточную стенку и очень устойчивы к разрушению.
По-видимому, в ранних исследованиях преимущественно разру-
шались клетки мезофилла, и это занижало уровень РуБФ-кар-
боксилазы, особенно при расчете на единицу Хл. К тому же и
определение активности данного фермента могли проводить в
условиях, далеких от оптимальных (разд. 6.11). Для оптималь-
304
ГЛАВА 10
Таблица 10.1. Средняя скорость фотосинтеза в листьях и активность
карбоксилаз в экстрактах, полученных из листьев С3- и С4-растений
Скорость фотосинтеза А кт и в и ост ь фермента
При нор- мальном со- держании СО2 в атмо- сфере В атмосфе- ре, насы- щенной CO*j ФЕП-карбо- ксилаза РуБФ-кар боксилаза
мкмоль* (мг Хл)—‘«ч-1
Сз-Р астения
С4-Растения
100—200 200—400 20—40 150—500
200-400 200—400 800-1200 100-200
кого функционирования РуБФ-карбоксилазы ее необходимо
предварительно активировать магнием и бикарбонатом. Даже
во многих современных работах уровень РуБФ-карбоксилазы,
выявляемый в экстрактах из различных Сграстений, не соответ-
ствует скорости фотосинтеза в целом листе. В табл. 10.1 приве-
дены типичные значения скорости фотосинтеза в целых листьях
и величины активности ФЕП-карбоксилазы и РуБФ-карбокси-
лазы в экстрактах, полученных из листьев С3- и С4-растений.
Механизм С4-цикла стал гораздо понятнее в семидесятые
годы, когда была обнаружена значительная активность различ-
ных декарбоксилаз С4-кислот.
Малат 4- NADP+ ----> Пируват 4- СО2 4- NADPH 4-
+ Н+ [NADP-малатдегидрогеиаза (декарбоксилирующая)]
Малат NAD+---------> Пируват 4- СО2 4- NADH 4-
4- Н+ [NAD-малатдегидрогеназа (декарбоксилирующая)]
Оксалоацетат 4- АТР ---------------->
---->• ФЕП 4- СО2 4- ADP (ФЕП-карбоксикиназа)
(10.8)
(10.9)
(10.10)
В настоящее время С4-растения подразделяют на 3 основные
подгруппы в соответствии с механизмом первичного декарбокси-
лирования (разд. 11.2, 11.8).
10.4. Анатомия фотосинтезирующей ткани
у Ci-растений
С4-растения характеризуются кранц-анатомией. В конце
прошлого столетия в листьях некоторых видов Сурегасеае Ха-
берландт (Haberlandt, 1884) обнаружил слой несущих хлоро-
филл клеток, которые явно отличались от других клеток, содер-
жащих хлоропласты. Он назвал эти клетки кранц-клетками (по-
немецки Kranz — венец, корона), так как они как бы венцом
ОТКРЫТИЕ СгПУТИ
305
Основная паренхима
Окаймляющая
паренхима
Местомная обкладка
Проводящая ткань
Рис. 10.6. Возможное происхождение кранц-клеток у С4-растений.
окружали проводящую ткань. К настоящему времени в кранц-
анатомии и эволюции кранц-клеток у разных растений выявлены
значительные различия. Так, у многих видов сем. Gramineae и
Сурегасеае (с кранц-анатомией и без нее) обнаружены две об-
кладки вокруг проводящего пучка, а именно местомная обклад-
ка и .окружающая ее паренхимная обкладка. У Gramineae-
кранц-клетки возникают либо из паренхимной обкладки (напри-
мер, у подсем. Eragrostoideae) или из местомной обладки (на-
пример, у большинства родов подсем. Panicoideae, у которых,
нет паренхимной обкладки). У Сурегасеае краиц-клетки появля-
ются с внутренней стороны от местомной обкладки. У некоторых
видов Сурегасеае кранц-клетки и клетки мезофилла разделены:
и местомной, и паренхимной обкладками (например, у Fim-
bristylis) или только местомной обкладкой (например, у Суре-
rus, у которых отсутствует паренхимная обкладка). У двудоль-
ных растений обычно есть только паренхимная обкладка, и
именно из нее чаще всего возникают кранц-клетки. У многочис-
ленных Ci-растений из сем. Chenopodiaceae, Polygonaceae и Com-
positae, имеющих цилиндрические листья или фотосинтезирующие
ветви, кранц-клетки образуют непрерывный цилиндрический слой,,
окруженный клетками мезофилла. Большая часть проводящей
ткани отделена от кранц-клеток несколькими слоями бесцвет-
ных клеток паренхимы. В этом случае кранц-клетки образуются:
из ткани паренхимы. Итак, кранц-клетки возникают из клеток
различного типа (и в некоторых случаях отделены от клеток
мезофилла клетками, не содержащими хлорофилла. У растений,,
применявшихся в ранних исследованиях С4-фотосинтеза (злаки
и некоторые двудольные растения), венец клеток образуется из
клеток обкладки проводящего пучка.
Термин «клетки обкладки проводящего пучка» вполне при-
меним ко многим Ci-растениям, но сейчас общепризнано, что-
20 - 234
3
Рис. 10.7. Микрофотография поперечного среза листа однодольного Ci-расте-
ния (Zea mays)-, Х430 (получена Sue Е. Frederick; публикуется с любезного
разрешения Ё. Н. Newcomb, University of Wisconsin), / — обкладка проводя-
щих пучков, 2 — проводящая ткань, 3 — мезофилл.
Рис. 10.8. Поперечный срез листа однодольного С3-растения (Avena sativa)-,
Х380 (микрофотография получена Sue Е. Frederick; публикуется с любезного
разрешения Е. Н. Newcomb, University of Wisconsin). Обозначения те же,
что на рис. 10.7.
Рис. 10.9. Поперечный срез листа двудольного С4-растения (молодой лист
Gotiiplirena celosioldes)-, XI 000. Обратите внимание, что ткань мезофилла со-
стоит из клеток палисадной (4) и губчатой (.5) паренхимы, которые окружа-
ют обкладку проводящего пучка (7). (Микрофотография получена Е. И. New-
comb, University of Wisconsin.)
Рис. 10.10. Поперечный срез листа двудольного Сз-растения Syrlnga vulga-
ris). Па срезе видны клетки палисадной (4) и губчатой (5) паренхимы;,
Х1.37 (микрофотография получена Е. Н. Newcomb, University of Wisconsin).
Обозначения те же, что иа рис. 10.9.
20
Рис. 10.11. Парадермальный срез однодольного С/,-растения (Chloris gayana)
(микрофотография получена Е. Н. Newcomb, University of Wisconsin). Обо-
значения те же, что на рис. 10,7.
Рис. 10.12. Парадермальный срез двудольного Сл-растения (молодой лист
Airiplex spongiosay, X1000 (микрофотография получена Е. Н. Newcomb,
University of Wisconsin). Обозначения те же, что иа рис. 10.7.
ОТКРЫТИЕ С,-ПУТИ
309
краиц-клетки не всегда (хотя и достаточно часто) возникают
из клеток обкладки проводящего пучка, примыкающих к прово-
дящей ткани. Когда речь идет о С^растениях, правильнее ис-
пользовать термин «краиц-клетки». У Сз-растений часто выявля-
ется слой клеток обкладки вокруг проводящей ткани, в которых
лишь время от времени встречаются немногочисленные разроз-
ненные хлоропласты. Эти клетки, однако, явно отличаются от
кранц-клеток, которые крупнее, имеют толстую клеточную стен-
ку и многочисленные хлоропласты (рис. 10.6).
На рис. 10.7—10.12 показаны основные различия в анатоми-
ческом строении листа у однодольных и двудольных С3- и С4-
растений. Обратите внимание на характерные клетки обкладки
проводящих пучков у однодольных (рис. 10.7) и двудольных
(рис. 10.9) Сграстений и на отсутствие этих клеток у однодоль-
ных (рис. 10.8) и двудольных (рис. 10.10) С3-растений. Ужуку-
рузы хлоропласты клеток обкладки проводящих пучков локали-
зованы в максимально удаленном от центра 'положении
(рис. 10.7), тогда как у G. celosioides они находятся ближе к
центру (анатомия представителей различных подгрупп С4-расте-
ний рассматривается в разд. 11.8). У некоторых двудольных
Сграстений клетки обкладки проводящих пучков окружены па-
лисадной и губчатой паренхимой (как на рис. 10.9), а у осталь-
ных видов имеются клетки мезофилла обычного типа, такие же,
как у однодольных С4-растеиий. Парадермальиые срезы хорошо
иллюстрируют взаимоотношения между мезофиллом и клетками
обкладки проводящих пучков у однодольного С4-растения с
параллельным жилкованием (рис. 10.11) и у двудольного С4-
растения с разветвленным расположением жилок (рис. 10.12).
У некоторых Сграстеиий между клетками мезофилла и клетка-
ми обкладки проводящих пучков, а также между присутствую-
щими в них хлоропластами наблюдаются поразительные раз-
личия по размеру.
В сороковые годы Роадес и Карвальо исследовали клетки
мезофилла и клетки обкладки проводящих пучков у кукурузы
с помощью микроскопа. Они заметили, что зеленые клетки об-
кладки проводящих пучков светлее, чем клетки мезофилла. Днем
в пластидах накапливался крахмал, который в темноте исчезал из
пластид клеток обкладки и оставался в пластидах клеток мезо-
филла. После 78 ч инкубации срезанных листьев в среде с са-
харозой в пластидах мезофилла накапливалось большое коли-
чество крахмала. Роадес и Карвальо заметили также, что в
пестрых листьях кукурузы (испещренных зелеными и 'белыми
полосами) явно нормальные содержащие хлорофилл клетки об-
кладки проводящих пучков не запасали крахмал в том случае,
когда по соседству с ними находились клетки мезофилла, лишен-
ные хлорофилла. Отсюда исследователи сделали вывод: «Нам,
310
ГЛАВА 10
к счастью, удалось однозначно показать, что крахмал, образую-
щийся в этих пластидах (пластидах клеток обкладки проводя-
щих пучков), возникает из растворимых углеводов, образую-
щихся в пластидах мезофилла и затем поступающих в клетки
паренхимной обкладки, где и происходит синтез крахмала»,
Прошли годы, и стало ясно, что для обеспечения высокой ско-
рости фотосинтеза у С4-растений необходим С4-Цикл, который
функционирует как механизм, концентрирующий СОа, и который
зависит от функционально активного состояния фотосинтезирую-
щих клеток мезофилла. Клетки мезофилла у С4-растений не
могут самостоятельно синтезировать сахара из СОа, однако упо-
мянутая выше концепция продержалась достаточно долго — до
того момента, когда были проведены основополагающие биохи-
мические исследования (синтез сахарозы и крахмала у С4-расте-
пий обсуждается в разд. 12.10).
В 1964 г. Джонсон при подготовке диссертации провел фун-
даментальное исследование ультраструктуры клеток обкладки
проводящих пучков и мезофилла у многих злаков, включая и те
виды, для которых характерна кранц-анатомия. Часть этой ра-
боты была опубликована в 1973 г. после того, как было устано-
влено биохимическое значение кранц-анатомии (Johnson, Brown,.
1973). В середине шестидесятых годов Лэтш начал электронно-
микроскопические исследования сахарного тростника, а затем и
других С4-Растений. К 1968 г. несколько исследователей совер-
шенно независимо друг от друга (Downes, Hesketh, Downton,
Tregunna, Johson, Hatch, Laetsch) пришли к выводу, что у мно-
гих растений существует некая связь между анатомическим
строением листа и рядом характеристик С4-фотосинтеза (Black,.
1973).
Источники данных: [5, 6, 10, 16, 18, 27, 33, 38, 43].
10.5. Выделение хлоропластов,
протопластов и клеток из С4-растений
В наиболее ранних работах по разделению хлоропластов ме-
зофилла и обкладки проводящих пучков у сахарного тростника
и некоторых других С4-растений были испытаны центрифугиро-
вание в градиенте плотности (как в водных, так и в неводных
растворах) и избирательное механическое разрушение ткани
(Black, 1973). Все методы избирательного разрушения основаны
на том, что клетки мезофилла разрушаются раньше, чем клетки
обкладки проводящих пучков, так как у последних клеточная
стенка гораздо^ толще. После механического разрушения впер-
вые были разделены клетки мезофилла и тяжи обкладки прово-
дящих пучков (проводящие пучки с прикрепленными к ним
клетками) из Digitaria sanguinalis (Edwards et al., 1970). Для
Рис. 10.13. Протопласты мезофилла А и тяжи клеток обкладки проводящих
пучков Б, полученные после ферментативной обработки листьев kvkvdvsw
(Kanai, Edwards, 1973). *
312
ГЛАВА 10
Таблица 10.2. Активность ФЕП-карбоксилазы и РуБФ-карбоксилазы
в экстрактах кукурузы (Kanai, Edwards, 1973)
Фермент Ф е р м е нт а ти ан ы it экстракт из протопластов мезофилла Ферментативный экстракт из тяжей обкладки прово- дящих пучков
РуБФ-карбоксилаза мкмоль - (мгХл)-1 -ч-1 0 389
ФЕП-карбоксилаза 864 10
выделения клеток мезофилла после механического разрушения
ткани могут быть использованы только1 С4-растения из рода
Digitaria. У других видов растений клетки мезофилла легко, раз-
рушаются после механического растирания ткани листа. Рюзи
Капай впервые применил для разделения клеток у С4-растений
(кукуруза, сахарный тростник, сорго) ферменты и выделил про-
топласты мезофилла и тяжи обкладки проводящих пучков
(рис. 10.13). Позже ферментативный метод применяли в самых
разных исследованиях С4-фотосинтеза у ряда С4-растений
(Edwards, Huber, 1978). Приемы выделения протопластов и
хлоропластов из протопластов описаны в приложении А. Для
выделения клеток из некоторых растений использовали также и
различные сочетания ферментативных и механических методов
(Che,n et al., 1973).
Успехи в выделении и разделении клеток мезофилла и об-
кладки проводящих пучков повлекли за собой очень важные
последствия. Так, например, на выявленных препаратах было
показано, что активность ФЕП-карбоксилазы связана только с
клетками мезофилла, а активность РуБФ-карбоксилазы — толь-
ко с клетками обкладки проводящих пучков. В протопластах
мезофилла С4-растений, так же как и в хлоропластах, получен-
ных из этих протопластов, очень мало или совсем нет РуБФ-
карбоксилазы «1% содержания фермента в клетках обклад-
ки проводящих пучков), нет в них и белка [фракции I или со-
ставляющих его компонентов. И наоборот, активность РуБФ-
карбоксилазы и присутствие белка фракции I характерны для
изолированных клеток обкладки проводящих пучков (Edwards,
Huber, 1978). Для выяснения локализации белка фракции I в
интактных тканях Хаттерсли и др. (Hattersley et al., 1977) ис-
пользовали очень остроумный метод. Применив иммунофлуорес-
центное мечение белка фракции I, они обнаружили на попереч-
ных срезах различных С4-растений, что этот белок локализован
в клетках обкладки проводящих пучков. В табл. 10.2 представ-
лена активность ФЕП-карбоксилазы и РуБФ-карбоксилазы в
ОТКРЫТИЕ СгПУТИ
313
изолированных препаратах мезофилла и обкладки проводящих
пучков кукурузы, полученных после ферментативной обработки.
Источники данных: [4, 20, 23, 30, 32, 40].
10.6. Современная упрощенная схема С^фотосинтеза
Результаты интенсивных исследований на уровне ферментов
фотосинтеза и фотосинтетического метаболизма в изолирован-
ных клетках мезофилла и обкладки проводящих пучков и в хло-
ропластах различных растений (гл. 11 и 12) можно представить
в виде упрощенной схемы фотосинтеза у С4-растений (рис. 10.14).
Отметим наиболее важные особенности данной схемы.
а) С помощью челночного механизма (Д-цикла СО2 переносится
из атмосферы в ВПФ-цикл, происходящий в клетках обкладки
проводящих пучков.
б) Полагают, что этот механизм создает относительно высокую
концентрацию СО2, обеспечивающую протекание ВПФ-цикла в
клетках обкладки проводящих пучков. Эта концентрация пре-
вышает нормальную концентрацию СОг, которая устанавлива-
ется в результате диффузии СО2 в лист из окружающей среды
с низким содержанием СО2 (0,03%). В результате СО2 стано-
вится нелимитирующим субстратом фотосинтеза, а это приводит
к подавлению фотодыхания, о значении которого пойдет речь в
дальнейшем (гл. 14).
в) Для челночного механизма необходим быстрый перенос
определенных метаболитов между клетками мезофилла и клет-
ками обкладки проводящих пучков.
г) В клетках мезофилла в ходе Сгцикла не происходит ре-
альной фиксации СО2.
Рис. 10.14. Ci-Фотосинтез.
314
ГЛАВА 10
10.7. Какие растения можно считать Ci-растениями
Термины «С3- и С4-растения» возникли, когда было выяснено»
различие между основными первичными продуктами фотосинте-
за у определенных высших растений. Эта терминология была
создана сразу же, как только были открыты С4-растения, еще
задолго до того, как у них был установлен механизм фотосин-
теза. У С4-растений углерод, исходно' связавшийся в составе
С4-кислот, передается в ВПФ-путь фотосинтеза. Если С4-расте-
ния выдержать несколько секунд с 14СО2, то 70—80% радиоак-
тивности обнаруживается в составе С4-кислот — малата и аспар-
тата. Лишь небольшая часть (1—5%) включенной метки выяв-
ляется в составе оксалоацетата (разд. 10.2). После установления
пути фотосинтеза у С4-растеинй этому термину можно дать бо-
лее строгое определение, учитывающее следующие свойства,
а) Главными первичными продуктами фотосинтеза являются
С4-дикарбоновые кислоты — оксалоацетат, малат и аспартат,
б) Углерод передается от С4-кислот в ВПФ-путь фотосинтеза
(С-4-атом С4-кислот в С-1-положение, ФГК).
в) Эти реакции происходят иа свету (в отличие от реакций у
САМ-растеиий, см. гл. 15).
г) Как правило, имеются два типа фотосинтезирующих 'кле-
ток—клетки мезофилла и краиц-клетки (клетки обкладки).
В ряде работ были получены данные, поставившие под сом-
нение необходимость кранц-аиатомии для С4-фотосинтеза. На
сегодняшний день у всех видов растений, относимых к С4-расте-
нням, обнаружена краиц-анатомия, т. е. показано наличие явно
отличающихся друг от друга фотосинтезирующих клеток двух
типов, которые располагаются концентрическими кругами
(разд. 10.4). И действительно, изучение анатомического строе-
ния любого из наземных растений является одним из наиболее
простых ,и быстрых методов для выяснения того, относится ли
данное растение к С4-типу. Главными продуктами фиксации
14СО2 в культуре фотосинтезирующей ткани таких С4-растений
как, Amaranthus retroflexus (Usuda et al., 1971), Froelichia gra-
cilis (Laetsch, Kortschack, 1972) и Portulaca oleracea (Kennedy,
1976), являются С4-КИСЛОТЫ, однако кранц-анатомия у этих рас-
тений не обнаружена. Тот факт, что в культуре клеток Portulaca
oleracea снижено фотодыхание, указывает иа протекание от-
дельных реакций С4-фотосинтеза в культуре ткани. Не известно,
однако, в какой степени поток углерода при фотосинтезе в та-
ких культурах отражает события, происходящие в целом рас-
тении. Так, например, источником ФЕП для фиксации СО2
культурами ткани, поддерживаемыми в среде с сахарозой, мо-
жет отчасти служить гликолиз, а синтез С^-кислот может быть
сопряжен со сравнительно высокой активностью ФЕП-карбокси-
ОТКРЫТИИ СгПУТИ
315
лазы, которая присуща этим клеткам. Таким образом, опыты с
культурой ткани не могут однозначно свидетельствовать о том,
что высокая скорость С4-фотосинтеза достигается и без кранц-
апатомии.
Время от времени появлялись собщения о наличии С4-фото-
синтеза у водных растений. Однако опубликованные данные
были недостаточно убедительными; в некоторых случаях позже
было доказано наличие у них С3-фотосинтеза. Так, сообщалось,
что морская трава Thalassia testudinum имеет все фотосинтети-
ческие характеристики С4-растений, хотя у нее и нет кранц-ана-
томии (Benedict, Scott, 1976). Однако при изучении ранних ра-
диоактивных продуктов и в опытах по разбавлению импульсной
метки, проведенных совсем недавно, было показано, что и у
этой и у других морских трав (Thalassia. spp. и Halophila spp.)
фиксация СО2 происходит в результате типичного С3-фотосинте-
за (Andrews, Abel, 1979; Benedict et al., 1980). В отличие от на-
земных Сз-растений водные С3-растеиия практически не разли-
чают 13СО2 и 12СО2. Очевидно, морские травы характеризуются
высоким диффузионным сопротивлением для поглощения СО2 в
водных средах. Это может приводить к связыванию всей погло-
щенной СО2, в результате чего изотопный состав углерода СО2
в морской воде и в растительных тканях практически одинаков
(о разделении изотопов углерода у растений см. разд. 15.12).
Водное цветковое растение Hydrilla verticillata при летних тем-
пературах и летнем фотопериоде проявляет ряд свойств, близ-
ких к свойствам С4-растеиия, и в то же время не имеет кранц-
анатомии (Holaday, Bowes, 1980). Углекислотный компенсаци-
онный пункт у него находится достаточно низко, основными на-
чальными продуктами являются аспартат и малат, а активность
ферментов С4-пути (пируват, ортофосфат—дикиназы и ФЕП-кар-
боксилазы) довольно высока. Скорость фиксации ИСО2 на свету
относительно невелика по сравнению со скоростью темнового
дыхания. Известно, что в темноте в различных растительных
тканях (даже в условиях дыхания) происходит включение мет-
ки в С4-кислоты. Это может в значительной степени влиять на
распределение метки между продуктами фиксации ИСО2 при
низкой скорости ассимилляции, что затрудняет выяснение пути
углерода при фотосинтезе. Помимо этого в отдельных частях
клеток могут существовать сравнительно высокие внутренние
пулы СО2, что в еще большей степени осложняет -проведение
опытов по разбавлению импульсной метки и затрудняет после-
дующую их интерпретацию. У другого водного макрофита, Ege-
rla densa, СО2 непосредственно фиксируется в ВПФ-цикле при
тех условиях, которые существуют в природной среде обитания,
в то время как сравнительно низкие концентрации СО2> приме-
нявшиеся в ряде исследований водных цветковых растений, мо-
316
ГЛАСА 10
гут способствовать фиксации СО2 в составе С4-кислот (Browse
et al., 1977). Вместе с тем показано, что для водного цветкового-
растения Potamogeion peclinatus характерен С3-фотосинтез; это
следует из состава исходных продуктов фиксации СО2 в интер-
вале концентраций СО2, создающихся при различных значениях
pH (Winter, 1978).
Ни одна из водорослей не отнесена к С4-р астениям, хотя в
ряде случаев показано, что значительная часть исходных мече-
ных продуктов представлена С4-кислотами. Предполагают, что к
С4-фотосинтезу способны некоторые сине-зеленые водоросли
(сравнительно высокая активность ФЕП-карбоксилазы, С4-кис-
лоты как первичные продукты фиксации 14СО2 и ингибирование
фотосинтеза малонатом; см. Colman, Coleman, 1978).
Без строгого научного исследования трудно провести четкую
границу между специфической функцией С4-фотосинтеза, как это
уже было определено, и образованием С4-кислот за счет других
механизмов (т. е. вторичного карбоксилирования). Для класси-
фикации растения как растения С4-типа недостаточно, чтобы
большая часть метки, включившейся в ранние продукты фото-
синтеза, находилась в составе С4-кислот (малате и аспартате).
Изучение разбавления импульсной метки при постановке опы-
тов, подобной постановке Хэтча и Слэка (опыты на сахарном
тростнике, разд. 10.2), может помочь выявить среди С4-кислот
доноры углерода для ВПФ-цикла (или для других путей, таких,
как обращенный цикл трикарбоновых кислот у некоторых бак-
терий). Не исключено, что метка, включающаяся в С4-кпслоты,
может переходить в сахарозу, минуя ВПФ-цикл (разд. 16.6).
Поэтому для полноты анализа следует продемонстрировать так-
же и перенос метки из С-4-положения С4-кислот в С-1-положе-
ние ФГК. Фотосинтезирующие ткани, в которых С4-кислоты
(аспартат и малат) являются продуктами фотосинтеза, нельзя
назвать, строго говоря, тканями С4-типа, если углерод С4-кислот
(независимо от той доли, которую они составляют) не перено-
сится в ВПФ-путь. В таких случаях может наблюдаться С4-фо-
тосинтез другого- рода, выполняющий сам по себе уникальную
функцию. Исключительное связывание атмосферной СО2 в со-
ставе С4-дикарбоновых кислот на свету и высокая скорость пе-
редачи углерода в ВПФ-путь были обнаружены только у таких
растений, у которых реакции С4-карбоксилирования и декарбок-
силирования пространственно разделены и происходят в клетках
двух разных типов. Несколько видов наземных растений, имею-
щих промежуточные характеристики (и С3-, и С4-растеиий), бу-
дут рассмотрены ниже (разд. 11.9). Метаболизм С4-кислот у
растений, не относящихся к САМ- или С4-растениям, тоже об-
суждается ниже (гл. 16).
Источники данных: [11, 14, 16, 21, 22, 26, 35, 37, 41, 42].
ОТКРЫТИЕ С4-Fli-'T И
317
ОБЩАЯ ЛИТЕРАТУРА
Симпозиумы
1. СО2 Metabolism and Productivity of Plants. (R. H. Burris and С. C. Black,
eds.) University Park Press, Maryland, U.S.A. (1976).
2. Photosynthesis and Photorespiration. (M. D. Hatch, С. B. Osmond and
R. 0. Slatyer eds.) Wiley-Intersciencc, New York (1971).
3. Symposium on Chemical Transformation of Carbon in Photosynthesis, Fed-
Proc., 9, 524—553 (1950).
Обзоры
4. Black С. C. Photosynthetic carbon fixation in relation to net CO2 uptake,.
Ann. Rev. Plant Physiol., 24, 253—286 (1973).
5. Brown IF, V. The Kranz, syndrome and its subtypes in grass systematics,.
Memoirs of Torrey Botanical Club, 13, 1—97 (1977).
6. Coombs J. Interaction between chloroplasts and cytoplasm in C4 plants, In:
The Intact Chloroplast (ed. J. Barber), pp. 274—313. The Netherlands, Else-
vier, 1976.
7. Edwards G. E., Huber S. C. The C4 pathway, In: The Biochemistry of Plants,
A Comprehensive Treatise, Vol. 8, Photosynthesis (eds. M. D. Hatch and
N. K. Boardman), pp. 237—281, New York, Academic Press, 1981.
8. Hatch M. D„ Osmond С. B. Coinparlincnlation and transport in C4 photo-
synthesis, In: Encyclopedia of Plant Physiology (New Scries), Vol. Ill,
Transport in plants, (eds. C. R. Stocking and U. Heber), pp. 144—184, New
York, Springer-Verlag, 1976.
9. Hatch M. D., Slack C. R. The C4-carboxylic acid pathway of photosynthesis,
In: Progress in Photochemistry, (eds. L. Reinhold and Y. Liwschitz), pp, 35—
106. London, Wiley-Interscience, 1970.
10. Laetsch IF. M The C4 syndrome: a structural analysis, Ann. Rev. Plant Phy-
siol, 25, 27—52 (1974).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
11. Andrews T. J., Abel К. M. Photosynthetic carbon metabolism in sea grasses.
“C-labelling evidence for the. C3 pathway, Plant Physiol, 63, 650—656
(1979).
12. Bassham J, A., Benson A. A„ Calvin AL The path of carbon in photosynthesis.
VIII. The role of malic acid, J. Biol. Chcm, 185, 781—787 (1950).
13. Benedict C. R., Scott J. R. Photosynthetic carbon metabolism of a marine-
grass, Plant Physiol, 57, 876—880 (1976).
14. Benedict C. R., Wong W. IF. L., Wong J. H. H. Fractionation of the stable-
isotopes of inorganic carbon by seagrasses. Plant Physiol, 65, 512—517
(1980).
15. Bjorkman O., Gauhl E. Carboxydismutase activity in plants with and without
P-carboxylation photosynthesis, Planta, 88, 197—203 (1969).
16. Brown W. V. Variations in anatomy, associations, and origins of Kranz tis-
sue, Amer. J. Bot, 62, 395—402 (1975).
17. Browse J. A., Dromgoole F. I., Brown J. AL A. Photosynthesis in the aquatic-
macrophyte Egeria densa. I. '"’СОг fixation at natural CO2 concentrations,
Aust. J. Plant Physiol, 4, 169—176 (1977).
18. Carotin R. C., Jacobs S. W. L., Vesk M. Leaf structure in Chenopodiaceac,
Bot. Jahrb. Syst, 95, 226—255 (1975).
19. Chen T. M., Brown R. H., Black С. C. Photosynthetic CO2 fixation products
and activities of enzymes related to photosynthesis in bermuda grass and1
other plants, Plant Physiol, 47, 199—203 (1971).
318
ГЛАВА 10
20. Chen Т. М., Campbell IS7. И., Dittrich Р., Black С. С. Distribution of carbo-
xylation and decarboxylation enzymes in isolated mesophyll cells and bundle
sheath strands of C4 plants, Biochem. Biophys. Res. Cominun., 51, 461—
4G7 (1973).
21. Colman B., Coleman J. R. Inhibition of photosynthetic CCL fixation in blue-
green algae by malonate, Plant Sci. Lett., 12, 101—105 (1978).
22. DeGroote D., Kennedy R. A. Photosynthesis in Elodea canadensis Michx,
Plant Physiol., 59, 1133—1135 (1977).
23. Edwards G. E., Huber S. C. Usefulness of isolated cells and protoplasts for
photosynthetic studies. In: Proceedings of the Fourth International Congress
on Photosynthesis, (eds. D. O. Hall, J. Cootnhs, and D. W. Goodwin), pp. 95—
106. London, The Biochemical Society, 1978.
24. Edwards G. E., Lee S. S„ Chen T. AL, Black С. C. Carboxylation reactions
and photosynthesis of carbon compounds in isolated bundle sheath cells of
Digitaria sanguinalis (L.). Scop, Biochem. Biophys. Res. Commun., 39, 389—
395 (1970).
25. Gajfron H., Pager E. W. The kinetics and chemistry of photosynthesis, Ann.
Rev. of Plant Physiol., 2, 87—114 (1951).
26. Gaitniche I. M. Sludies on lhe mechanism of glycerate 3-phosphate synthe-
sis in tomato and maize leaves, Plant Physiol., 51, 512—519 (1973).
27. Haberlandt G. Physiological Plant Anatomy (Transl. M. Drummond), Lon-
don, Macmillan, 1884.
28. Hatch AL D., Slack C. R. Photosynthesis by sugarcane leaves. A new carbo-
xylation reaction and the pathway of sugar formation, Biochem. J., 101,
103—111 (1966).
29. Hatch M. D., Stumpf P. K. Fat metabolism in higher plants. XVI. Acetyl
coenzyme A carboxylase and acyl coenzyme A-malonyl coenzyme A trans-
carboxylase from wheat germ., j. Biol. Chem., 236, 2879—2885 (1961).
30. Hullerslcy P. W., Watson L.. Osmond С. B. In situ innminofluorescent labe-
ling of ribulose 1,5-bisphosphate carboxylase in leaves of C3 and C4 plants,
Aust. J. Plant Physiol., 4, 523—539 (1977).
31, Holaday A, S., Bowes G. C4 acid metabolism and dark CO2 fixation in a
submersed aquatic macrophyte (Ht/drilla verticillate.). Plant Physiol., 65,
331—335 (1980).
32. Huber S. C., Edwards G. E. Regulation of oxaloacetate, aspartate and malate
formation in mesophyll protoplast extracts of several C4 plants, Plant Phy-
siol., 56, 324—331 (1975).
33. Johnson C., Brown W. V. Grass leaf ultrastructure variations. Amer. J. Bot.,
60, 727—735 (1973).
34. Kauai R., Edwards G. E. Separation of mesophyll protoplasts and bundle
sheath cells of maize leaves for photosynthetic studies, Plant Physiol., 51,
1133—1137 (1973).
35. Kennedy R. A. Photorespiration in C3 and C4 plant tissue cultures. Signifi-
cance of Kranz anatomy to low photorespiration in C( plants, Plant Physiol.,
58, 573—575 (1976).
36. Kortschack H. P., Hartt С. E.. Burr G. O. Carbon dioxide fixation in sugar-
cance leaves, Plant Physiol., 40. 209—213 (1965).
37. Laetsch W. M.. Kortschack H. P. Chloroplast structure and function in tissue
cane leaves, Plant Physiol., 40, 209—213 (1965).
.38 . Rhoades M. M., Carvalho A. The function and structure of the parenchyma
sheath plastids of the maize leaf, Bull. Torrey Bot. Club, 71, 335—346
(1944).
39. Slack C. R., Hatch M. D. Comparative studies on the activity of carboxylases
and other enzymes in relation to the new pathway of photosynthetic carbon
dioxide fixation in tropical grasses. Biochem. J., 103, 660—665 (1967).
40. Slack C. R., Hatch M. D„ Goodchild D. J. Distribution of enzymes in meso-
phyll and parenchyma sheath chloroplasts of maize in relation to the C4-di-
ОТКРЫТИЕ О ПУТИ
319
carboxylic acid pathway of photosynthesis, Biochem. J., 114, 489—498
(1969).
41. Usiidu 11., Kanai It, Takeuchi M. Comparison of carbon dioxide fixation and
the fine structure in various assimilation tissue of Amarauihus retroflexus L,
Plant Cell Physiol., 12, 917—930 (1971).
42. Winter K,. Short-term fixation of 14carbon by the submerged aquatic angio-
sperm Potaniogeton pcciitialus, J. Exp. Bot., 29, 1169—1172 (1978).
43. Winter K„ Troughlon J. II. Photosynthetic pathways in plants of coastal
and inland habitats of Israel and the Sinai, Flora, 16, 1—34 (1978).
ГЛАВА 11
Три подгруппы С4 -растений;
их биохимия, фотохимия и систематика
Краткое содержание
С4-цикл можно разделить на две стадии: стадию карбокси-
лирования, происходящую в клетках мезофилла, и стадию де-
карбоксилирования, идущую в клетках обкладки проводящих
пучков. Общим для всех С4-растений является то, что СО2 на
•стадии карбоксилирования фиксируется при участии ФЕП-
карбоксилазы. По способу' декарбоксилирования при участии
NADP- или NAD-малатдегидрогеназы (декарбоксилирующей)
(МДГ, называемой также малик-энзимом и яблочным фермен-
том) или ФЕП-карбоксикиназы у С4-растений можно выделить
три группы: NADP-МДГ-, NAD-МДГ- и ФЕП-КК-типа соответ-
ственно. У NADP-МДГ-растений главными метаболитами, во-
влеченными в обмен между клетками, являются малат и пиру-
ват, у NAD-МДГ-растений— аспартат и аланин и у ФЕП-КК-
растений — аспартат и ФЕП. Фотохимические особенности хло-
ропластов мезофилла и обкладки проводящих пучков у растений
этих трех подгрупп соответствуют выявляемым различиям в
энергетическом обеспечении их С4-цикла. У NADP-МДГ-расте-
ний транспорт малата обеспечивает передачу восстановитель-
ной силы из мезофилла в клетки обкладки проводящих пучков.
В клетках обкладки отсутствует фотосистема II, и они могут
за счет челночного механизма обмена передавать свыше поло-
вины всей ФГК (продукта РуБФ-карбоксилазной реакции в
клетках обкладки проводящих пучков) в клетки мезофилла,
где этот метаболит восстанавливается. Таким образом, клетки
мезофилла у растений этой подгруппы являются тем местом,
где первично образуется восстановительная сила (NADPH).
Для синтеза аспартата в клетках мезофилла у ФЕП-КК- и
NAD-МДГ-растений нужен только АТР. У них местом первич-
ного образования NADPH, используемого для восстановления
ФГК, являются хлоропласты обкладки проводящих пучков. По-
требность в источнике энергии для связывания СО2 у растений
всех трех подгрупп одинакова. Источником дополнительной
энергии сверх той, что требуется для ВПФ-цикла, служит
АТР, необходимый для энергизации С4-цикла.
С4-Растения обнаружены среди представителей 16 семейств.
К ним относятся ряд главных зерновых культур и многие сор-
ТРИ ПОДГРУППЫ СгРАСТЕНИИ
321
Рис. 11.1. Разнообразие декарбоксилирующих систем при С4-фотосинтезе.
У разных Сл-растеиий имеются «СО2-иасосы» различного типа. После фикса-
ции атмосферной СО- и виде С4-кпслот образовавшиеся в клетках мезофилла
малат и аспартат переносятся в клетки обкладки проводящих пучков, где
СО- слона высвобождается в результате декарбоксилирования. У одних рас-
теши'! декарбоксилирующим ферментом служит ФЕП-карбоксикиназа, у дру-
гих—NADP- пли NAD-МДГ (декарбоксилирующая). Это разнообразие
Сд-цикла обусловлено различиями в фотохимии процесса, анатомическом
строении листа и систематическом положении.
ные травы. У злаков сем. Gramineae • выявлена корреляция
между особенностями строения клеток обкладки проводящих
пучков (и их хлоропластов) и биохимическими различиями в
обмене веществ у представителей вышеуказанных подгрупп.
11.1. Стадия карбоксилирования в С4-пути
Стадия карбоксилирования у С4-растений протекаете клет-
ках мезофилла с. образованием из С3-предшественникрв и ат-
мосферной СОа Сгкислоты. Известны три способа карбоксили-
рования, которые присущи большинству изученных С4-расте-
ний. Следует, отметить, однако, что соотношение метаболиче-
ских путей, ведущих к образованию малата или . аспартата,
варьирует. .
а) Пируват-|-СО2 --->;Малат (11.1)
б) Аланин СО2 -—s- Аспартат .... (11.2)
в) ФГК-СО2 —Малат + Рг. (11.3)
В превращении пирувата и СО2 в малат [уравнение (11.1)] участ-
вуют пять ферментов: .................... ..........
Пируват 4- Р;.+ АТР ——>
---> ФЕП АМР -|- РР; (пируват, ортофосфат—дикиназа) (11.4)
21-234
322
ГЛАВА 11
Пирофосфат ---->- 2 Ортофосфат (пирофосфатаза) (Ц.5)
АМР + АТР ------> 2ADP (аденилаткнназа) (Ц.6)
ФЕП + СО2 -----»- Оксалоацетат + Рг (ФЕП-карбоксилаза) (11.7)
Оксалоацетат -|- NADPH -|- Н+ -->
---->- NADP+ + Малат (NADP-малатдегидрогеназа) (11.8)
В итоге: Пируват 4- СО2 + 2АТР + NADPH 4- Н+ ------->
-----э- 2ADP + 2Рг + NADP+ + Малат (11 -9)
Можно задать вопрос: какова же роль хлоропластов в клет-
ках мезофилла у С4-р астений? Как было наглядно показано в
приведенной выше последовательности реакций, для фиксации
1 молекулы СО2 в ходе С4-цикла требуются 2 молекулы АТР и
1 молекула NADPH. Следовательно, С4-путь зависит от актив-
ности хлоропластов мезофилла, обеспечивающих образование
восстановительной силы (гл. 5).
Для превращения аланина и СО2 в аспартат [уравнение
(11.2)] требуется шесть ферментов:
Аланин 2-Оксоглутарат ------>-
----> Пируват + Глутамат (аланинаминотрансфераза) (11.10)
Пируват ф- 2АТР --------> 2ADP + ФЕП + Рг (11.4—11.6)
ФЕП СО2 ------> Оксалоацетат Р; (ФЕП-карбоксилаза) (11 - 7)
Оксалоацетат-ф-Глутамат ---->-
---->- Аспартат 4-2-Оксоглутарат (аспартатаминотрансфераза) (11.11)
В итоге: Аланин 4- СО2 4- 2АТР ---->- Аспартат 4- 2ADP 2Р; (11.12)
Обратите внимание, на то, что в конечном итоге нет необ-
ходимости в непрерывном синтезе глутамата и 2-оксоглутарат а,
так как эти два соединения регенерируются в цикле с участи-
ем аланин- и аспартатаминотрансфераз. NADPH также не тре-
буется, но для превращения аланина и СО2 в аспартат необхо-
димо затратить две молекулы АТР,
Для превращения 3-ФГК и СО2 в малат и Р, [уравнение
(11.3) ] требуются четыре фермента:
3-ФГК ------>- 2-ФГК (фосфоглицеромутаза) (11.13)
2-ФГК ---->- ФЕП (енолаза) (11.14)
ФЕП4-СО2 ------> ОксалоацетатР; (ФЕП-карбоксилаза) (Ц .7)
Оксалоацетат 4- NADPH 4- Н+ ----->-
----> Малат 4- NADP 4- (NADP-малатдегидрогеназа) (11.8)
В итоге: 3-ФГК 4~ СО2 4- NADPH 4- Н+ —-*
----> Малат 4- NADP+ Рг (11.15)
ТРИ ПОДГРУППЫ СгРАСТЕНИИ
323
Это превращение не требует АТР, но для фиксации СО2 необ-
ходима одна молекула NADPH.
Активности этих ферментов [уравнения (11.4—11.14)], пе-
реходящих в экстракт из клеток мезофилла С4-растений, впол-
не достаточно для того, чтобы поддерживать приемлемую ско-
рость фиксации СО2 в СгЦикле. Сообщалось, одиако, что уро-
вень пируват, ортофосфат — дикиназы в клетках варьирует,
возможно, в связи с различной степенью активации этого фер-
мента. К тому же существуют доказательства, полученные в
опытах иа препаратах мезофилла (разд. 12.3), что все три ме-
таболита (пируват, аланин и ФГК) могут служить предшест-
венниками на стадии карбоксилирования в С4-пути.
Для каждого из возможных вариантов стадии карбоксили-
рования в С4-пути необходима ассимиляционная энергия в той
или иной форме. Общий баланс АТР и NADPH, требующихся
для фиксации СО2 в составе С4-кислот, определяется относи-
тельной скоростью фиксации СО2 в каждом из возможных трех
путей. Как будет рассмотрено ниже, вариации потока углерода
в ходе стадии С4-карбоксилироваиия влияют иа перераспреде-
ление энергии между клетками мезофилла и клетками обклад-
ки проводящих пучков, но ие иа общее количество энергии, не-
обходимой для фиксации СО2 С4-растениями (разд. 11.6).
11.2. Стадия декарбоксилирования в С4-пути
Эта стадия данного цикла протекает в клетках обкладки
проводящих пучков при участии одного из трех ферментов де-
карбоксилирования С4-КИСЛОТ. В соответствии с первичным ме-
ханизмом декарбоксилирования С4-кислот (табл. 11.1, разд.
11.8) все С4-растения подразделяются на 3 подгруппы:
Таблица 11.1. Примеры видов, представляющих каждую из трех подгрупп
'Ссрастений, выделенных на основании различий в механизмах
декарбоксилирования
Сгподгругша Вид растения
NADP-МДГ Кукуруза Сахарный тростник Сорго Росичка кровяная Zea myas Saccharum officinarum Sorghum bicolor Digitaria sanguinalis
NAD-МДГ Щирица колосистая Портулак огородный Просо южное Amaranthus retroflexus Portulaca oleracea Panicum miliaceum
•ФЕП-КК Просо крупное Хлорис Panicum maximum Chloris gayana
2 1*
324
ГЛАВА '!
а) NADP-МДГ (декарбоксилирующая)
NADP-МДГ необходима в том случае, когда малат транс-
портируется из мезофилла в клетки, обкладки проводящих пуч-
ков, а. пируват возвращается обратно в клетки мезофилла:
Малат +' NADP+ >- Пируват + СО2 + NADPH + Н+ (11.16)
Этот механизм связывает декарбоксилирование малата с ВПФ-
циклом. Для ВПФ-цикла декарбоксилирование малата необхо-
димо, чтобы обеспечить этот цикл СО2, а для декарбоксилиро-
вания малата нужно, чтобы образующийся NADPH вновь окис-
лялся бы в реакциях ВПФ-цикла. NADP-зависимая МДГ ло-
кализована в хлоропластах обкладки проводящих пучков. Как
правило, малат у NADP-МДГ-растений является самым ран-
ним из первичных продуктов фотосинтеза. Возможна, однако,
й такая последовательность ферментативных реакций, когда
функционирование NADP-МДГ сопряжено с превращением ас-
партата в СО2 и аланин (разд. 12.2). Но наиболее типичными
участниками обмена метаболитами между клетками мезофилла
н клетками обкладки проводящих пучков в процессе декарбок-
силирования являются малат и пируват.
б) NAD-МДГ (декарбоксилирующая)
Для декарбоксилирования с участием NAD-МДГ требуются
четыре фермента, поскольку в этом случае главными метаболи-
тами, мигрирующими между клетками мезофилла и клетками
обкладки проводящих пучков, являются аспартат и аланин.
NAD-МДГ локализована в митохондриях клеток обкладки про-
водящих пучков. Растения, использующие в качестве первичной
декарбоксилазы NAD-МДГ, характеризуются наличием: хорошо
заметных, многочисленных митохондрий в клетках обкладки
проводящих пучков.’• (см? рис.' Г1.7, Б). У,таких растении в клет-
ках обоих типов обычно повышена активность аспартат- и
аланинамииотр.ансфераз,; а -радиоактивность раньше всего
включается в аспартат, который представляет собой типичный
первичный продукт фотосинтеза.
Аспартат-]-2-Оксоглутарат ->• Оксалоацетат + Глутамат (П.Ц)
. Оксалоацетат — NADII-j-Н+ •—->
——> Малат NAD+ [NAD-МДГ (недекарбоксилирующая)] (11.17)
... ... MaJiaT4-NAD+------> Пируват-р-СО2 + NADH-J- :•
+ Н+[NAD-МДГ (декарбоксилирующая)] (11.18)
ПируватГлутамат ----->- Аланин-|-2-Оксоглутарат (11.10).
-В итоге: Аспартат --> АланииСО, (11.19)
ТРИ ПОДГРУППЫ СгРАСТЕНИИ
325.
Обратите внимание 'на то, что при. такой, последовательности
реакций восстан&вительная сила' в .конечном итогё как бы не
генерируется и не используется. В . клетках, где декарбоксили-
рование малата идет с участием-’NAD-МД Г, участниками обме-
на могут быть, хотя-И в меньшей степени, малат и пируват.
в) ФЕП-карбоксикиназа (ФЕП-КК)
У ФЕП-КК’Растеиий’ самым ранним из первичных продук-
тов фотосинтеза обычно является аспартат, который к тому же
служйт и главным метаболитом, транспортируемым из клеток
'мезофилла, в клеТйтг обкладки- проводящих пучков. Поэтому на
стадии декарбоксилирования здесь используются два фермента:
Асцартат.-ф 2-Оксоглутарат -> Оксалоацетат-|-Глутамат (11.11)
Оксалоацетат + АТР —ФЕП СО2 -|- ADP (ФЕП-КК) (11.20) * В
В итоге; Аспартат 2-Оксоглутарат АТР -->
--> ФЕП -р Глутамат СОа ADP (11.21)
Поскольку нет никаких доказательств высокой активности пи-
руваткиназы (необходимой для превращения ФЕП в пируват)
в клетках обкладки проводящих пучков у таких растений, по-
лагают, что ФЕП возвращается в клетки мезофилла без како-
го-то бы ни было изменения. Это должно приводить к более
сложному челночному передвижению метаболитов между клет-
ками двух типов, как это будет вкратце изложено ниже (разд.
11.3) при рассмотрении трех возможных путей обмена. ФЕП-
КК локализована в цитоплазме клеток обкладки проводящих
пучков (разд. 12.2). Некоторые различия в регуляторных свой-
ствах декарбокендаз и их потребности в кофакторах обсужда-
ются в разд. 12.8.
Источники данных: [14, 16, 17, 25].
11.3. Краткое изложение основных предполагаемых
путей переноса углерода с участием трех
декарбоксилирующих механизмов
В том случае, когда главным исходным продуктом С4-фото-
сиитеза является аспартат, для протекания С4-цикла необходи-
мо наличие фондов аспартата и аланина. Такие фонды могут
поддерживаться за счет восстановления азота (восстановление
нитрата до солей аммония и образование аминокислот). При
этом восстановление азота1 не связано непосредственно с син-
тезом аспартата в ходе фиксации СО2 в С4-цикле, так как ами-
ногруппа - аспартата, постоянно обменивается в процессе'реа-к1
326
ГЛАВА 11
ций переаминирования, катализируемых аспартат- и аланин-
аминотрансферазами. У ФЕП-КК-растений для поддержания
азотного обмена используется челночный обмен пирувата и
аланина или 2-оксоглутарата и глутамата, в результате кото-
рого аминный азот переносится из клеток обкладки проводя-
щих пучков в клетки мезофилла. У растений данного типа де-
карбоксилирование аспартата в изолированных клетках об-
кладки проводящих пучков зависит от количества внесенного в
среду пирувата или 2-оксоглутарата. Поскольку концентрация
2-оксоглутарата, необходимая для челночного обмена метабо-
литов, на порядок величины выше, чем концентрация пирува-
та, челночный обмен пирувата и аланина более предпочтите-
лен. Для работы цикла не требуется наличия пируват .ортофос-
фат— дикиназы. У ФЕП-КК-растений в декарбоксилировании
может участвовать также и NAD-МДГ, которая в этом случае
использует пируват,ортофосфат — дикиназу клеток мезофилла
(разд. 12.1, в).
11.4. Сбалансированность зарядов при межклеточном
транспорте метаболитов в ходе С-рЦикла
В результате карбоксилирования, происходящего в Срцик-
ле, содержимое клеток мезофилла должно закисляться, а со-
держимое клеток обкладки проводящих пучков вследствие де-
карбоксилирования Сркислот, наоборот, должно защелачи-
ваться. Предполагаемый обмен метаболитами между клетками
этих двух типов должен в конечном итоге приводить к перено-
NADR -МАЛАТДЕГИДРОГЕНАЗА
МЕЗОФИЛЛ
Малат
NADP
NADPH '
Оксалоацетат
СОг
ОБКЛАДКА
ПРОВОДЯЩИХ
ПУЧКОВ
Малат
NADP
NADPH
СО2
Пируват
ФЕП
2 АТР
Пируват
II
А
Рис. 11.2. Декарбоксилирующие механизмы. А. При участии NADP- МДГ.
ТРИ ПОДГРУППЫ Ci-РАСТЕИИН
327
су отрицательных зарядов в клетки обкладки проводящих пуч-
ков. Необходимая величина pH и электронейтральность могут
поддерживаться либо за счет межклеточного переноса прото-
нов в клетки обкладки проводящих пучков, либо за счет равно-
ценного переноса ионов гидроксила в клетки мезофилла.
Электронейтральность может также сохраняться благодаря об-
ОБКЛАДКА ПРОВОДЯЩИХ ПУЧКОВ
NAD-МАЛАТДЕГИДРОГЕНАЗА
МЕЗОФИЛЛ
2-Оксоглутарат
Глутамат
V
Глутамат
2* Оксоглутарат
ФЕП-КАРБОКСИКИНАЗА
Рис. 11.2. Б. При участии INAD-МДГ. В, При участии ФЕП-карбоксикиназы
328
ГЛАВА 11
МЕЗОФИЛЛ
ОБКЛАДКА ПРОВОДЯЩИХ ПУЧКОВ
Малат2' -----
Пируват <
—> Мапат2'
--- Пируват
+
со2
NADP
NADPH'
ДГАФ2’
▲
ОН' ч—
ФГК3' <
♦- ДГАФ2"
ФГК3'
н*
Рис. 11.3. Сбалансированность зарядов при межклеточном транспорте ве-
ществ.
мену ФГК — ДГАФ (рис. 11.3) (челночный перенос ФГК и
ДГАФ между клетками этих двух типов рассматривается в
разд. 11.6). Вопрос о том, существует ли градиент pH между
клетками во время стационарного состояния фотосинтеза и ес-
ли существует, то как он влияет на транспорт метаболитов, ос-
тается открытым.
11.5. Межклеточная локализация ВПФ-цикла
у С4-р астений
Как видно из табл. 11.2, стадии регенерации и карбоксили-
рования ВПФ-цикла протекают только в клетках обкладки про-
водящих пучков, тогда как восстановительная стадия осуществ-
ляется в клетках обоих типов. Что касается энергетики про-
цесса, то соответствующее распределение ферментов между
клетками предполагает, что АТР, используемый на стадии ре-
генерации, образуется в хлоропластах обкладки проводящих
пучков, а восстановительную стадию фотосинтеза обеспечива-
ют энергией хлоропласты и тех и других клеток.
11.6. Фотохимические затраты при С4-фотосинтезе
В табл. 11.3 суммированы фотохимические затраты в ходе
С4-цикла и ВПФ-цикла у растений с разными декарбоксилиру-"
ющими механизмами. При высвобождении .одной молекулы. СО2
ТРИ ПОДГРУППЫ Сч-РЛСТЕНИП
329
Таблица 11.2. Сводка данных по локализации стадий ВПФ-цикла
в клетках мезофилла и в клетках обкладки проводящих пучков
Стадия Выявляемая активность Локализация
мезофилл обкладка проводящих пучков
Регенерация РуБФ Фосфорибулокиназа — +
Карбоксилирование РуБФ-карбоксилаза — +
Восстановление ФГК Фосфоглпцераткпназа .+ +
до триозофосфатов NADP-триозофосфатде- гидрогеиаза + +
Таблица 11.3. Фотохимические затраты при фиксации одной
молекулы СО2 в С4-цикле
Группа Источник энергии Затраты в СгЦПКЛ с1) Дополни- тельные за- траты в ВПФ-цжлс Общие за- траты (в рас- чете на I молекулу СОД
NADP-МДГ АТР 2 3 5
NADPH 1 1А 2
NAD-МДГ АТР 2 3 5
NADPH 0 2 2
ФЕП-КК АТР 1 3 4
NADPH 0 2 2
') С учетом челночного обмена метаболитами (разд. 11.3). .
В результате декарГюксилироиаппя с участием ' NADP-МДГ в конечном итоге об-
разуется одна молекула NADPH.
вследствие декарбоксилирования малата образуется половина
того количества восстановительной силы, которое необходимо
для фиксации одной молекулы СО2 в ВПФ-цикле.
Долю участия клеток разного типа в образовании восстано-
вительной силы оценить гораздо труднее. В настоящее время
при оценке степени разделения «труда» между клетками де-
лают ряд допущений. Во-первых, предполагают, что в декар-
боксилировании С4-кислот у исследуемого растения участвует
только одни из декарбоксилирующих ферментов (что, по-види-
мому, соответствует действительности).
Во-вторых, считают, что метаболиты, переносимые из клет-
ки в клетку у NADP-МДГ-растеиий, представлены только ма-
латом и пируватом, у NAD-МДГ-растений — только аспарта-
том и аланином, а у ФЕП-КК-растений — только аспартатом и
330
ГЛАВА II
Таблица 11.4. Расчет фотохимических затрат в клетках мезофилла
и в клетках обкладки проводящих пучков у С4-растенийс
Группа растений Источник энергии СгЦикл Сэ-Цмкл О|бщие затраты
мезофилл обкладка проводящих пучков восстанови- тельная стадия стадия ре- генерации
ч § 'Н- о со <ъ Я обкладка проводящих пучков мезофилл обкладка проводящих пучков мезофилл обкладка проводящих пучков
NADP-МДГ АТР NADPH 2 1 0 2) 1 1 1 2) 0 0 1 0 3 2 2 0
NAD-МД Г АТР 2 0 1 1 0 1 3 2
NADPH 0 0 1 1 0 0 1 1
ФЕП-КК АТР 0 1 1 1 0 1 1 3
NADPH 0 0 1 1 0 0 1 1
) Обратите внимание на допущения. сделанные в тексте, и на те причины. по :
торым эти данные нельзя использовать для точной оценки распределения энергии между
теми и другими клетками,
2) См. сноску 2) к табл. 11.3.
ФЕП (разд. 11.3). Соответствующие этим представлениям
энергетические затраты приведены в табл. 11.4.
В-третьих, принимают, что степень восстановления ФГК в
ходе восстановительной стадии фотосинтеза одинакова и в тех
и в других клетках (если исходить из примерно равной актив-
ности ферментов восстановительной стадии в клетках обоих ти-
пов). Это обеспечивает распределение энергетических затрат
в ВПФ-цикле, приведенное в табл. 11.4.
Обратите вимание на то, что представленные в этой табли-
це примеры свидетельствуют о высокой степени потребления
NADPH в клетках мезофилла у NADP-МДГ-р астений, о чуть
меньшей потребности этих клеток в АТР у NAD-МДГ-растений
и о гораздо большем расходе АТР в клетках обкладки проводя-
щих пучков у ФЕП-КК-растений. Об этом же свидетельствуют
и фотохимические особенности хлоропластов в клетках мезо-
филла и в клетках обкладки проводящих пучков (разд. 11.7).
По ряду причин данные, приведенные в табл. 11.4, нельзя
рассматривать как строго установленные и абсолютные фото-
химические показатели энергетических затрат в клетках обо-
их типов. Любое Сграстеиие скорее всего, хотя и не всегда,
использует для первичного декарбоксилирования С4-кислот
только один из трех известных декарбоксилирующих механиз-
ТРИ ПОДГРУППЫ СгРАСТЕНИИ
331
ХЛОРОПЛАСТ
МЕЗОФИЛЛА
ХЛОРОПЛАСТ ОБКЛАДКИ
ПРОВОДЯЩИХ ПУЧКОВ
Рис. 11.4. Челночный механизм обмена ФГК — ДГАФ.
мов. Хотя первичный механизм декарбоксилирования у ФЕП-
КК-растений достаточно ясен и о нем говорит само название
этой группы растений, однако эти растения могут использовать
и NAD-МДГ. В клетках мезофилла всех С4-растений, к какой
бы группе они ии относились, обнаружена активность пиру-
ват,ортофосфат— дикиназы. Этот фермент у ФЕП-КК-растений
может функционировать в цикле, включающем NAD-МДГ.
У С4-р астений всех трех подгрупп радиоактивная метка
прежде всего включается в аспартат и малат. Вполне вероят-
но, что в обмене метаболитами между клетками мезофилла и
клетками обкладки проводящих пучков участвуют обе эти
С4-кислоты. Транспорт малата связан с фотохимическими затра-
тами NADPH, тогда как для транспорта аспартата не требует-
ся фотохимического образования NADPH в С4-цикле.
По-видимому, в наибольшей степени приспособлен к изме-
нению энергетических затрат в клетках разного типа тот уча-
сток, где восстанавливается ФГК, образованная в ходе РуБФ-
карбоксилазной реакции в хлоропластах клеток обкладки про-
водящих пучков. ФГК либо сразу восстанавливается в клетках
обкладки проводящих пучков, либо транспортируется для вос-
становления в хлоропласты мезофилла. В последнем случае
функционирует челночный механизм обмена ФГК — ДГАФ
между хлоропластами тех и других клеток (рис. 11.4).
В мезофилле шпината (С3-растение) ФГК может погло-
щаться хлоропластами за счет обмена с ДГАФ на переносчике
фосфата (разд. 8.23). У С4-растений переносчик фосфата в хло-
ропластах мезофилла и обкладки проводящих пучков также
способен катализировать подобного рода обмен ФГК — ДГАФ.
Другая возможность, хотя и менее выгодная с точки зрения
энергетических затрат, заключается в транспорте ФГК в клет-
ки мезофилла и ее последующем превращении в глицерат с
332
ГЛАВА 11
Рис. 11.5. Другой возможный механизм челночного обмена ФГК — ДГАФ
(в упрощенном виде по Randall, Tolbert, 1971).
участием ФГК-фосфатазы цитозоля. После этого глицерат и Pi
могут снова поглощаться хлоропластами и превращаться в
ФГК под действием глицераткиназы. Такой челночный меха-
низм был предложен в связи с тем, что, согласно некоторым
данным, экстракты, полученные из клеток мезофилла С4-рас-
теннй, содержат сравнительно много. ФГК-фосфатазы и глице-
раткиназы (Randall, Tolbert, 1971) (рис. 11.5).
Почти половина ФГК, образующейся в хлоропластах об-
кладки проводящих пучков, может переноситься в хлоропла-
сты мезофилла и там восстанавливаться. Для постоянной реге-
нерации РуБФ в ВПФ-цикле необходимо расходовать 5/6 ФГК,
образующейся в ходе РуБФ-карбоксилазной реакции (гл. 6).
Поэтому если половина ФГК восстанавливается в клетках ме-
зофилла, то по меньшей мере 2/3 этого количества следует воз-
вратиться обратно в хлоропласты обкладки. проводящих пуч-
ков, где идет ВПФ-цикл.
Вместе с тем ие исключено, что ДГАФ превращается в са-
харозу прямо в клетках мезофилла, и в этом случае перенос
триозофосфатов назад в клетки обкладки проводящих пучков
не требуется (рис. 11.6). Такие ферменты, как глюкозо-1-фос-
фат — уридилилтрансфераза и сахарозофосфат-синтетаза, кото-
рые участвуют в синтезе сахарозы, обнаружены и в тех и в
других клетках у очень немногих из исследованных С4-расте-
ний. Даунтон и Хокер предполагают, что у кукурузы первич-
ный синтез сахарозы происходит в клетках мезофилла (разд.
12.10).
Еще раньше был рассмотрен другой способ метаболизма
ФГК в клетках мезофилла, согласно которому ФГК in vitro
служит предшественником на стадии карбоксилирования в
С4-цикле и используется со значительной скоростью. Если про-
дукты декарбоксилирования в клетках обкладки проводящих
ТРИ ПОДГРУППЫ Cj-РАСТЕНИИ
333
ХЛОРОПЛАСТ
МЕЗОФИЛЛА
ХЛОРОПЛАСТ ОБКЛАДКИ
ПРОВОДЯЩИХ ПУЧКОВ'
Рис. 11.6. Челночный механизм обмена, приводящий к синтезу сахарозы в
клетках мезофилла.
пучков (такие, как пируват или ФЕП) отчасти используются в
других путях метаболизма, то ФГК в этом случае может слу-
жить альтернативным предшественником в С^цикле.
Источники данных: [10, 14, 16].
11.7. Фотохимические различия хлоропластов
разных типов
Предполагаемые различия в затратах энергии при ассими-
ляции углерода у представителей трех подгрупп С4-р астений
подтверждаются тем, что их хлоропласты явно различаются фо-
тохимически.
а) Количественная оценка
Для определения относительного фотохимического потенци-
ала хлоропластов применяют несколько методов. На величину
светособирающего потенциала хлоропластов влияет распреде-
ление хлорофилла (Хл) листа между клетками мезофилла и
клетками обкладки проводящих пучков. Представления об этом
распределении можно составить, если полностью разделить
клетки этих двух типов при помощи интенсивной обработки
ферментами или же если рассчитать эту величину по отноше-
нию Хла/ХлЬ в целом листе (ТУ), хлоропластах мезофилла
(Л4) и хлоропластах обкладки проводящих пучков (В).
В последнем случае содержание хлорофилла в клетках ме-
зофилла (в процентах) определяют по формуле
100 (2W+ 1) (Г— В)
(IF + I) (М — В)
334
ГЛАВА il
Этот метод приемлем только тогда, когда отношение Хла/ХлЬ
в клетках мезофилла и в клетках обкладки проводящих пуч-
ков 'Очень сильно различается, как, например, у NADP-МДГ-
растений.
Для нециклического переноса электронов необходима фото-
система II. Это можно показать, определив скорость реакции
Хилла в присутствии различных окислителей и измерив за-
медленную флуоресценцию (разд. 4.13).
Отношение активностей фотосистем I и II служит показа-
телем интенсивности процессов циклического и нециклическо-
го переноса электронов (высокое отношение ФС1/ФСП свиде-
тельствует о высокой потенциальной возможности для проте-
кания процессов циклического переноса электронов и о низкой
потенциальной возможности для нециклического переноса
электронов и для восстановления NADP). Взятые в совокуп-
ности, такие показатели, как отношение Хла/ХлЬ, низкотемпе-
ратурная флуоресценция в жидком азоте и содержание Р700,
позволяют оцепить относительную активность ФС1 и ФСП
(разд. 4.13).
Главным светособирающим пигментом ФС I в хлоропла-
стах Сз-растений служит Хл а, а в ФСП эту роль выполняют
Хла и ХлЬ. Отсюда следует, что чем выше отношение Хла/ХлЬ,
тем выше и потенциальная активность ФС I. Для фотосистем I
и II характерна интенсивная флуоресценция в областях 730 и
685 им соответственно. Это было установлено в опытах с хло-
ропластами Сз-растений, у которых ФС1 и ФСП можно отде-
лить друг от друга (разд. 4.13).
Поскольку Р700 представляет собой Хл реакционного цент-
ра ФС1 (разд. 4.12), отношение Хл/Р700 может меняться в за-
висимости от относительного содержания ФС I и ФС II. Так,
например, если отношение Хл/Р700 в хлоропласте составляет
400/1 и Хл при этом равномерно распределен между фотоси-
стемами, то отношение Хл/Р700 для частиц ФС I будет равно
200 (общее правило для хлоропластов Сз-растений; разд. 4.13).
Тогда это отношение для хлоропластов, утративших активность
ФСП, будет ниже, а именно будет равно 200.
Низкое отношение Хл/Р700 не всегда может служить пока-
зателем пониженной потенциальной активности ФСП. Так, на-
пример, с уменьшением содержания Хл в светособирающем
комплексе, не затрагивающем Хл реакционного центра, свето-
вые затраты, необходимые для насыщения ФС II, увеличивают-
ся, но максимально возможная потенциальная активность фо-
тосистемы при этом не меняется. Однако при недостаточной
освещенности хлоропластов in vivo ее активность заметно
уменьшается. Содержание Р700 обычно выражают через со-
ТРИ ПОДГРУППЫ С<-РАСТЕНИИ
335
Таблица 11.5. Нециклический перенос электронов в изолированных
хлоропластах NADP-МД Г-растений
Вид растения Обычнее название Процент общей активности нециклического переноса электронов Степень раз вития гран в хлоро- пластах
хлоропласты мезофилла хлороцласты обкладки проводящих пучков
Saccharum officitiarum Сахарный тростник 100-97 0-3 Гран нет
Sorghum bicolor Сорго 100-95 0—5 Гран пет
Zea mays Кукуруза £5—70 15-30 Рудимен- тарные
Digitaria sangulnalis Росичка кровяная 70 30 Рудимен- тарные
1 Для того чтобы измерить количество кислорода, выделяющегося и процессе не-
циклического переноса электронов, применяли различные окислители: метилвиологеп,
феррицианид, л-беизохипоп и NADP.
держание Хл в реакционном центре ФС1, тан как количествен-
но определить Хл в реакционном центре ФСП очень трудно.
Впервые количество Р700 у С^растений было определено в
хлоропластах суммарного экстракта листьев. Полученные ре-
зультаты свидетельствовали о том, что для хлоропластов
Сграстений характерно более высокое, чем у Сз-растений, от-
ношение Р700/Хл (Black, Mayne, 1970). Это вполне согласует-
ся с данными о том, что Сграстения расходуют в большей сте-
пени АТР, чем NADPH (5ATP/2NADPH по сравнению с
ЗАТР/2 NADPH у Сз-растений), поскольку повышенное содер-
жание Р700 может быть связано с относительно более высоким
уровнем циклического фотофосфорилирования.
б) NADP-МДГ-растения
У представителей NADP-МДГ-растений процесс нецикличе-
ского переноса электронов, по-видимому, на 70—100% проте-
кает в хлоропластах мезофилла. Структура хлоропла-стов в
клетках обкладки проводящих пучков постепенно меняется, и
если у росички кровяной и кукурузы в хлоропластах видны ру-
диментарные граны, то у сорго (рис. 11.7,А) и сахарного
тростника граны полностью отсутствуют. В таких хлоропластах
нет ФС II, что связано с утратой способности к образованию
гран (табл. 11.5). Хлоропласты обкладки проводящих пучков
у NADP-МДГ-растений—это тот единственный известный слу-
ТРИ ПОДГРУППЫ Щ-РЛСТЕНИИ
337
чай, когда можно видеть, что способность к формированию гран
в значительной степени редуцирована.
До сих пор не выяснены полные потенциальные 'возможно-
сти для протекания нециклического транспорта электронов in
vivo в хлоропластах различного типа. Данные, полученные в
опытах с изолированными хлоропластами, мало что говорят об-
относительной интенсивности нециклического переноса электро-
нов в клетках этих двух типов. Так, например, скорость нецик-
лического переноса электронов в изолированных . хлоропластах,
обкладки проводящих пучков кукурузы сплы-ю варьирует в за-
висимости от того, какие окислители использованы для реак-
ции Хилла. Из некоторых данных следует, что потенциальная
активность нециклического транспорта электронов в хлоропла-
стах мезофилла и обкладки проводящих пучков кукурузы оди-
накова (в присутствии ряда окислителей получены величины,
порядка 75—150 мкмоль О2-(мг Хл)_1-ч-1, В других работах
указывается, что у кукурузы хлоропласты мезофилла являют-
ся тем местом, где первично происходит нециклический пере-
нос электронов, скорость которого в присутствии п-бензохино-
на (одного из окислителей, применяемых в реакции Хилла) до-
ходит до 600 мкмоль О2-(мгХл)_1-ч-1, что вполне достаточно
для. достижения скорости фотосинтеза в целом листе. Некото-
рая противоречивость полученных данных может быть связана,
с различиями в возрасте ткани (в ходе онтогенеза хлоропла-
сты обкладки проводящих пучков теряют граны), разной осве-
щенностью при выращивании растений, разной степенью поте-
ри активности в процессе выделения хлоропластов различного-
типа, обоюдным загрязнением отдельных фракций при их вы-
делении или различными условиями определения активности.
Размеры фотосинтетической единицы в хлоропластах обкладки
проводящих пучков у NADP-МДГ-растений меньше (понижен-
ное содержание светособирающего Хл в реакционном центре),
чем в хлоропластах мезофилла (табл. 11.6). Поэтому in vivo-
процесс нециклического переноса электронов в хлоропластах
Рис. 11.7. А. Электронная микрофотография агранальных хлоропластов (1)
в клетках обкладки проводящих пучков и содержащих граны хлоропластов
(5) в клетках мезофилла Sorghum sudanen.se, растения NADP-МДГ-типа.
Х6000. (Микрофотография получена S. Е. Frederick; публикуется с любез-
ного разрешения Е. Н. Newcomb, University of Wisconsin.) Б. Электронная
микрофотография содержащих граны хлоропластов (2) в клетках обкладки
проводящих пучков молодого листа Portulaca oleracea, растения NAD-МДГ-
типа. X16 300. Обратите внимание на хорошо заметные митохондрии (3), ко-
торые у Ci-растений этой подгруппы являются местом декарбоксилирования
малата (разд. 11.2). (Микрофотография получена Mary Е. Doohan; публи-
куется с любезного-разрешения Е. Н. Newcomb, University of Wisconsin.)
22-234
.338
ГЛАВА И
Таблица 11,6. Отношение Хл/Р7О0 и Хл а/Хл Ъ и флуоресценция при
температуре жидкого азота (Ровб/Гтзо) в хлоропластах разного типа
у NADP-МДГ-растений в сравнении с хлоропластами С3-растений
(Edwards et al., 1976; Mayne et al., 1974)
Вид растения Вваб/Ртзо Хл с/Хл b ЫР700
мезофилл обкладка проводя- щих пучков мезофилл обкладка проводя- щих пучков мезофилл обкладка проводя- щих пучков
NADP-МДГ
Saccharum o/'/ici/iarum 1,13 0,25 3,70 5,56 412 193
Sorghum bicolor 1,14 0,23 3,73 6,60 418 191
Zea mays 1,14 0,18 3,30 6,51 413 218
Digitaria sanguinalis 0,76 0,20 3,58 5,80 322 188
Setaria lutescens 0,85 0,28 3,54 5,50 428 264
Сз-растения
Avena sativa 0,87 — 3,45 — 444 —•
Hordeum vulgare 0,76 — 3,66 — 478 —
Triticum aestivum 1,57 — 3,41 — 439 —
Таблица 11.7. Потенциальная активность реакции Хилла в присутствии
n-бензохинона и интенсивность фиксации СО2 на свету в изолированных
клетках обкладки проводящих пучков у различных
КАОР-МДГ-растений1»
Вид растения Реакция Хилла (в при- сутствии 1мМ /г-бензо- хинопа). мкмоль О2- (МГ Хл)-1-!!-1 Фиксация СОг. мкмоль-(мг ал)-1.,!-1'
Saccharum officinarum 3 2
Sorghum bicolor 3 1
Zea mays 21 7
Digitaria sanguinalis 84 22
Setaria lutescens 62 33
) Клетки получали после ферментативной обработки листьев (Edwards et at., 1976).
ТРИ ПОДГРУППЫ С.-РАСТЕПИИ
339
мезофилла может достигать насыщения при сравнительно низ-
кой интенсивности освещения, тогда как в хлоропластах об-
кладки проводящих пучков — при гораздо более высокой.
Если -не забывать о том, что разные авторы приводят раз-
ные цифры, то величины, представленные в табл. 11.5, могут
служить показателем распределения активности нециклическо-
го переноса электронов между клетками двух рассматриваемых
типов у некоторых NADP-МДГ-растений.
Существует и другая корреляция — между зависимым от-
/г-бензохинона выделением кислорода и способностью к фикса-
ции СС>2 в изолированных клетках обкладки проводящих пуч-
ков у ряда NADP-МДГ-растений (табл. 11.7). При этом вели-
чины, характеризующие скорость реакции Хилла и скорость
фиксации СО2 в изолированных клетках сахарного тростника и
сорго, ие превышают 1 % скорости фотосинтеза в целом листе.
С4-растений. Возможны два объяснения этих данных. Они свиде-
тельствуют либо о том, что в хлоропластах (аграиальных) об-
кладки проводящих пучков у сорго и сахарного тростника in
vivo ие функционирует нециклический перенос электронов, ли-
бо о том, что изолированные клетки обкладки проводящих пуч-
ков очень легко теряют фотосистему II, особенно те из иих, ко-
торые имеют аграиальиые хлоропласты. Для оценки активно-
сти нециклического переноса электронов Даунтон п др. (Down-
ton et al., 1970) применили метод определения in vivo. Вместо,
реактива Хилла они взяли тетразолиевый синий, который тем-
неет после восстановления. Эти авторы сделали вывод, что ак-
тивность нециклического переноса электронов полностью отсут-
ствует в хлоропластах обкладки проводящих пучков у сорго,
крайне мала в хлоропластах кукурузы и сравнительно велика
в хлоропластах Pani.cu.tn miliaceum (в которых есть граны),.
В последующих экспериментах, связанных с изучением нецик-
лического переноса электронов в изолированных хлоропластах
и клетках обкладки проводящих пучков, данные Даунтона и
соавторов были подтверждены.
Хотя и существует определенная корреляция между струк-
турой и функцией хлоропластов данного типа у Страстен ий,.
тенденцию к появлению в фотосинтезирующей ткани аграналь-
ных хлоропластов не всегда можно рассматривать как доказа-
тельство отсутствия ФС II (так, например, в хлоропластах
красных водорослей встречаются только единичные ламеллы).
В среднем у NADP-МДГ-растений примерно 90% замедлен-
ной флуоресценции приходится на долю хлоропластов мезофил-
ла и 10%— на долю хлоропластов обкладки проводящих пуч-
ков. Это опять убедительно свидетельствует об отсутствии
ФСП в хлоропластах обкладки проводящих пучков. Обычна
22*
340
ГЛАВА II
у NADP-МДГ-растений содержание Хл в клетках мезофилла
слегка выше, чем в клетках обкладки проводящих пучков (при-
мерно 60%), как и должно быть в центре первичного восста-
новления .NADP+ (разд. 4.13).
И отношение Хла/Хлй, и спектры флуоресценции в жидком
азоте, и содержание P70Q свидетельствуют о том, что отно-
шение активностей ФС1/ФСП в хлоропластах обкладки про-
водящих пучков у NADP-МДГ-растений достаточно велико.
Отношение Хла/Хлб в хлоропластах мезофилла у этих расте-
ний близко к соответствующему отношению у Сз-растений.
В хлоропластах обкладки проводящих пучков это отношение
гораздо больше, что предполагает и более высокую активность
ФС I (табл. 11.6). В хлоропластах Сз-растений светособираю-
щий Хл ФС I представлен Хл а, а светособирающий Хл ФСП
состоит из Хла и Хл Ь, содержание которых в этом случае оди-
наково (разд. 4.13-)-. Основная часть Хл листа приходится на
долю светособирающего Хл, а содержание Хл в реакционных
центрах намного ниже. Таким образом, по величине отношения
Хла/Хлб можно оценить распределение светособирающего Хл
между этими двумя фотосистемами. Так, например, в хлоро-
пластах с отношением Хл а/Хл Ь, равным 3, в ФС I присутству-
ют 2 молекулы Хла, а в ФСП'—по одной молекуле Хла и
ХлЬ. Это обеспечивает примерно равное распределение Хл
между разными фотосистемами. При отношении Хла/ХлЬ,
равном 6, как в хлоропластах обкладки проводящих пучков у
некоторых NADP-МДГ-растений, в ФС1 присутствуют 5 моле-
кул Хла, а в ФСП — по одной молекуле Хла и Хл b (или ина-
че, около 70% Хл в ФС I и 30% Хл в ФСП). Если число ак-
цепторов в ФСП ограничено, то комплекс Хла/Ь — белок так-
же может передавать энергию в ФС I (разд. 4.11). Это позво-
ляет Хл клеток обкладки проводящих пучков улавливать энер-
гию, необходимую в первую очередь для работы ФС I.
Хлоропласты обкладки проводящих пучков у NADP-МДГ-
растений характеризуются относительно низким отношением
Fees/Fyao (табл. 11.6, рис. 11.8), что свидетельствует о высоком
отношении ФС I/ФС II. Отношение Хл/Р700 в хлоропластах об-
кладки проводящих пучков у растений этого типа в 2 раза ни-
же, чем в хлоропластах мезофилла, и близко по величине к
аналогичному показателю для частиц ФС I из хлоропластов
Сз-растений. Отношение Хл/Р700 в Сд-хлоропластах мезофилла
NADP-МДГ-растений и в хлоропластах С3-растений практичес-
ки одинаково (табл. 11.6). Рассматриваемые в совокупности
данные фотохимических исследований свидетельствуют о том,
что в хлоропластах обкладки проводящих пучков очень высок
уровень активности ФС I, которая, как предполагают, связана
с циклическим фотофосфорилированием, и что хлоропласты ме-
ТРИ ПОДГРУППЫ СгРЛСТЕНИИ
.341
Рис. 11.8. Спектры низкотемпе-
ратурной (77 К) флуоресцен-
ции фрагментов хлоропластов
из мезофилла (Г) и обкладки
.проводящих пучков (77) Sorg-
hum bicolor, растения NADP-
МДГ-типа (Mayne et al.,
4974).
зофплла являются тем местом, где происходит первичное вос-
становление NADPK
Вероятно, у большинства NADP-МДГ-растений хлороплас-
ты обкладки проводящих пучков вносят свой вклад в образо-
вание восстановительной силы. Сорго и сахарный тростник
представляют собой крайний случай; судя по фотохимическим
данным, их хлоропласты мезофилла целиком обеспечивают по-
требности клеток в восстановительной силе (95—100%). Сле-
дует отметить, что при катализируемой РуБФ-карбоксилазой
фиксации одной молекулы СО2 в хлоропластах обкладки про-
водящих пучков образуются две молекулы ФГК. За один обо-
рот С4-цикла в результате транспорта малата и декарбоксили-
рования из хлоропластов мезофилла в хлоропласты обкладки
проводящих пучков 'должна перейти одна молекула NADPH,
которая может затем быть использована для восстановления
одной молекулы ФГК. Вторая молекула ФГК должна быть вос-
становлена в хлоропластах мезофилла при участии челночного
механизма обмена ФГК и триозофосфата между клетками двух
типов (разд. 11.6). В этом случае темновые реакции восстано-
вительной стадии в клетках обоих типов должны протекать с
равной скоростью, а восстановительная сила целиком должна
образовываться.в_хл.оропласт.ах мезофилла.
342
ГЛАВА 11
в) NAD-МДГ- и ФЕП-КК-растения
У NAD-МДГ- и ФЕП-КК-растений граны есть и в хлоро-
пластах мезофилла, и в хлоропластах обкладки проводящих
пучков. Содержащие граны хлоропласты обкладки проводящих
пучков одного 'Из растений NAD-МДГ-типа (Portu.la.ca olera-
сеа) изображены на рис. 11.7, Б. У представителей названных
выше двух групп С4-растеиий от 2/3 до 3/4 Хл листа находит-
ся в хлоропластах обкладки проводящих пучков. Это отличает
их от NADP-МДГ-растений, у которых большая доля Хл ли-
ста находится в клетках мезофилла. Изучая анатомию расте-
ний, Уолтер Браун заметил, что у представителей Eragrostoi-
deae, одного из подсемейств сем. Gramineae (относящихся, по
современным представлениям, к растениям NAD-МДГ- и ФЕП-
КК-типа), основная часть Хл листа находится в клетках об-
кладки проводящих пучков.
Судя по активности реакции Хилла и замедленной флуорес-
ценции, у NAD-МДГ- и ФЕП-КК-растений ФСII активна и в
хлоропластах мезофилла, и в хлоропластах обкладки проводя-
щих пучков. Однако более высокое содержание восстанови-
тельной силы в клетках обкладки проводящих пучков может
быть связано и с большим содержанием Хл листа в этих -клет-
ках. Поэтому ФГК У этих растений скорее всего восстанавли-
вается в клетках обоих типов не в равной степени, как это
следует из данных табл. 11.4, а преимущественно (более поло-
вины всего количества ФГК) в хлоропластах обкладки прово-
дящих пучков.
В рассмотренных выше расчетах энергетических затрат у
NAD-МДГ-растений (табл. 11.4) указывалось, что отношение
Таблица 11.8. Отношение Хл/Р700 и замедленная флуоресценция
в хлоропластах мезофилла и в хлоропластах обкладки проводящих пучков
у NAD-МДГ- и ФЕП-КК-растений (Edwards et al., 1976)
Группа XnlP700 Доля замедленной флуоресценции, %
мазофялл обкладка проводящих пучков мезофилл об кладка проводящих пучков
NAD-МДГ
Panicum capillars 271 376 13 87
Panlcum miliaceum 207 292 17 83
ФЕП-КК
Panicum maximum 403 242 42 58
Panicum texanum 428 342 35 65
ТРИ ПОДГРУППЫ СгРАСТЕНИИ
343
немного боль-
пучков (2/1).
мезофилла у
ATP/NADPH в хлоропластах мезофилла (3/1)
ше, чем в хлоропластах обкладки проводящих
К тому же отношение Хп!Р700 в хлоропластах
этих растений меньше, чем в хлоропластах обкладки проводя-
щих пучков. Вместе с тем хлоропласты мезофилла у ФЕП-КК-
пастений характеризуются относительно большим отношением
Хл/Р700 (табл. 11.8), что согласуется с низкой величиной отно-
шения ATP/NADPH в этих хлоропластах (табл. 11.4).
В настоящее время фотохимические характеристики хлоро-
пластов разного типа можно сравнивать только в самых об-
щих чертах, исходя из предполагаемого пути переноса углеро-
да при фотосинтезе у данной группы С4-растений. Степень ис-
пользования энергии в клетках разного типа меняется в зави-
симости от энергетических затрат в данном С4-цикле, соотно-
шения процессов транспорта аспартата и малата и степени
восстановления ФГК в хлоропластах мезофилла. Следователь-
но, фотохимическое обеспечение энергией происходит коорди-
нированно с реакциями ассимиляции углерода.
Источники данных: [5, 9, 10, 15, 17, 21, 22, 28].
1.8. Систематика
С4-растения идентифицированы только среди наземных по-
крытосеменных растений (разд. 10.7). Известно, что примерно
из 300 семейств цветковых растений только шестнадцать вклю-
чают в себя С4-растения. Сюда относятся Gramineae (злаки),
Сурегасеае (осоки) и 14 семейств двудольных растений:
Acanthaceae, Aizoaceae, Amaranthaceae, Asteraceae, Boragina-
ceae, Capparidaceae, Caryophyllaceae, Chenopodiaceae, Euphor-
biaceae, Nyctaginaceae, Polygonaceae, Portulacaceae, Scrophula-
riacea и Zygophyllaceae. По-видимому, С4-фотосинтез возникал
в ходе эволюции во многих отдельных случаях. Ни одно из се-
мейств не состоит целиком из С4-растений, в то же время от-
дельные роды, трибы или подсемейства в пределах одного се-
мейства могут включать в себя только С4-растения. Так назы-
ваемый С4-сиидром особенно часто встречается у представите-
лей семейств Gramineae, Сурегасеае и Chenopodiaceae. Брюс
Смит и Уолтер Браун, применив для идентификации ^-расте-
ний методику разделения изотопов углерода (разд. 15.12), ус-
тановили, что свыше половины из обследованных ими более
500 видов злаков относятся к С4-растениям.
Большинство самых злостных на Земле сорняков является
С4-растениями (табл. 11.9), а большинство сельскохозяйствен-
ных культур относится к Сз-растениям. Из наиболее важных с
хозяйственной точки зрения сельскохозяйственных культур к
числу С4-растений принадлежат сорго, кукуруза и сахарный
344
ГЛАВА 11
Таблица 11.9. Распространение С4-растений среди сорных трав
Влд растения Обычное название Классификация по последним данным
А. Десять самых злостных на Земле сорняков
(Holm, 1969; Black et al., 1 969)
Cynodon daciylon Свинорой С4 (NAD-МДГ)
Cyperus rotundas Сыть округлая СДИАПР-МДГ)
Echinochloa colonum Ежовник крестьянский с4
Echinochloa crusgalli Ежовник (просо кури- ное) С4(МАПР-МДГ)
Eleusine indica Елеузина С4 (NAD-МДГ)
Im per at a cylindrica Императа цилиндричес- кая С4
Panicutn maximum Просо крупное СДФЕП-КК)
Sorghum halepense Гумай (сорго алепское) C4(NADP-Mflr)
Eichhornia crasslpes Речной гиацинт Сз
Lantana camara Лантана Сз
Б. Десять самых злостных сорняков полевых культур в США (Doresch, 1970)
Amaranthus spp. Щирица С4 (NAD-МДГ)
Cyperus spp. Сыть С4(ЫАПР-МДГ)
Echinochloa crusgalli Ежовник (просо кури- ное) С4(ЫАПР-МДГ)
Panicutn spp. С4(первично NAD-МДГ)
Setaria spp. Щетинник С4(МАПР-МДГ)
Sorghum halepense Гумай (сорго алепское) С4(ЫАПР-МДГ)
Agropyron. repens Пырей ползучий Сз
Chenopodium album Марь белая Сз
Ipotnoea spp. Ипомея С3
Xaniiutn spp. Дурнишник Сз
тростник (входящие в NADP-МДГ-подгруппу), просяные куль-
туры (Pennisetum typhoides, Panicutn miliaceum, чумизы из рода
Setaria и Eleusine согасапа) и ряд пастбищных трав. Многие
травы прерий также являются Сграстениями (табл. 11.10).
Сем. Gramineae изучено, пожалуй, лучше других. Это ка-
сается и выявления отдельных Сз- или Севидов, и классифи-
кации обнаруженных С4-растений по разным подгруппам.-
У Gramineae можно выделить три главных подсемейства, к ко-
ТРИ ПОДГРУППЫ с.-растений
345
-Таблица 11.10. Важнейшие С4-травы прерий
Вид растения Обычное название Подгруппа
Andropogon scoparlus Бородач веничный (NADP-МДГ)
Andropogon geradii Бородач (NADP-МДГ)
Botileloua gracilis Грамова трава (бутелоуа изящная) (NAD-МДГ)
Bouteloua curtlpendula Бутелоуа (ФЕП-КК)
Buchloe dactyloides Бизонова трава (NAD-МДГ)
Panicum virgatum Просо лозное (NAD-МДГ)
Sorghasirum nutans Соргаструм поникающий (NADP-МДГ) ;
Таблица 11.11. Основные подсемейства
сем. Gramineae (Gould, 1968)
Подсемейство Доля общего числа видов, встречаю* щихся в США, %
Panicoideae 33
Eragrostoideae 28,5
Festucoideae 36
Итого: 97,5
торым относится большая часть американских злаков (табл.
И-11).
К С4-растениям принадлежит большинство видов . подсем.
Panicoideae и все виды подсем. Eragrostoideae, тогда как все
представители подсем. Festucoideae относятся к С3-растениям.
Растения подсем. Panicoideae и Eragrostoideae в общем лучше
всего приспособлены к теплому климату, а некоторые из видов
Eragrostoideae устойчивы к засухе.
На рис. 11.9 приведена классификация родов сем. Grami-
neae согласно результатам обследования механизма декарбок-
силирования у типичных представителей. Растения NADP-МДГ-
типа найдены только среди видов подсем.' Panicoideae (в том
числе сорго и сахарный тростник из трибы Andropogoneae, ку-
куруза из трибы Maydeae) и Aristoideae. Представители каж-
дой из подгрупп С^злаков отличаются характерными особен-
ностями анатомического строения (рис. 11.10). Так, у NADP-
МДГ-растений хлоропласты в клетках обкладки проводящих
пучков смещены в направлении от проводящего пучка ц в ос-
новном не имеют граи (см. также рис. 10.7 11.7, Д)_. Исклю,-
346
ГЛАВА 11
NADP-МДГ
NAD-МДГ
ФЕП-КК
"J
ANDROPOGONEAE
CHLORIDEAE
CHLORIDEAE
Andropogon
Saceharum
Sorghastrum
Sorghum
MAYDEAE
Euchlaena
Zea
PANICEAE
Cenchrus
Digitaria
Echinochioa
Panicum
Paspa/um
Pennisetum
Setaria
ARISTIDEAE
Aristida
Boutefaua
Bucioe
Ch far is
E/eusine
Leptochloa
Boute/oua
Chfaris
ERAGROSTEAE
Eragrostis
SPOROBOLEAE
Muhtenberyia
Sporobolus
ZOYSIEAE
Zoysia
PANICEAE
Panicum
SPOROBOLEAE
Sporuoc/us
PANICEAE
Brachiaria
Erfachfaa
Urochfaa
| ARISTIDOIDEAEJ | PANICOIDEAE | |erAGROSTOIDEAe|
Рис. 11.9. Схема филогенетического родства С4-подгрупп у сем. Gramineae,
основанная на биохимических и цитологических исследованиях растений под-
сей. Aristidoideae, Eragrostoideae и Panicoideae (Gutierrez et al., 1974, 1976).
чая несколько видов Panicum, NAD-МДГ-растения обнаруже-
ны только среди видов подсем. Eragrostoideae. На самом деле
к группе NAD-МДГ-растений могут быть отнесены только ви-
ды, действительно принадлежащие к роду Panicum, например
Panicum miliaceum. Все другие виды, которые относят к Pani-
cum, скорее всего принадлежат к другим родам. У NAD-МДГ-
растений хлоропласты обкладки проводящих пучков имеют
оформленные граны и смещены в направлении к проводящему
пучку.
ФЕП-КК-растения найдены только среди видов, относящих-
ся к ряду родов подсем. Eragrostoideae. Исключение составля-
ют несколько родов подсем. Panicoideae (Urochloa, Eriochloa
и Brachiaria). Несколько видов Panicum, такие, как Panicum
maximum, принадлежащие к ФЕП-КК-группе, сейчас правиль-
нее относить к роду Brachiaria. У ФЕП-КК-видов Gramineae
хлоропласты обкладки проводящих пучков имеют граны и
первично смещены в направлении от проводящего пучка.
Как уже отмечалось выше, агранальные хлоропласты
NADP-МДГ-растений не способны к нециклическому транспор-
ТРИ ПОДГРУППЫ СеРАСТЕНИИ
34Г
NADP-МДГ-тила
Рис. 11.10. Схема поперечного среза проводящего пучка, иллюстрирующая
расположение хлоропластов обкладки проводящих пучков у растений раз-
личных С4-подгрупп сем. Gramineae. Обратите внимание на то, что растения
NADP-МДГ- и ФЕП-КК-типа отличаются отсутствием или наличием гран в
хлоропластах.
ту электронов (разд. 11.7). Но никаких других функционально-
структурных связей, основанных на тех различиях С4-подгрупп
сем. Gramineae, которые представлены на рис. 11.10, устано-
вить не удалось. Более того, у двудольных С4-растений не об-
наружено никакой связи между механизмом декарбоксилиро-
вания и расположением хлоропластов в клетках обкладки про-
водящих пучков. По меньшей мере у нескольких видов двудоль-
ных Сграстений положение хлоропластов в клетках обкладки
проводящих пучков одинаково (они находятся у внутренней по-
верхности обкладки) независимо от того, принадлежат расте-
ния к NAD-МДГ- или к NADP-МДГ-подгруппе (рис. 10.9).
Кранц-анатомия у разных семейств сильно различается, и для
выявления возможной корреляции между биохимическими .и
анатомическими характеристиками необходимы дополнитель-
ные исследования.
Источники данных: [3, 6, 13, 14, 16, 27].
11.9. Растения с промежуточными (С3/С4) характеристиками
Есть несколько видов растений, которые по общепринятым
критериям нельзя отнести ни к Сз-, ни к С4-р астениям, потому
что они обладают рядом свойств как той, так и другой груп-
пы. Впервые это было обнаружено при изучении гибридов Сз-
348
ГЛАВА! Г
и Севидов Atriplex spp. Активность фЕП-карбоксилазы у рас-
тений Fi- и Р2-поколений варьирует в широких пределах, а
кранц-анатомия развита в разной степени. Углекислотный ком-
пенсационный пункт у всех потомков ближе к соответствующе-
му пункту Сз-растений. Интересно, что были получены гибри-
ды, которые по уровню карбоксилазной активности и по ана-
томическому строению почти не отличались от С4-растений, ио-
биохимически их нельзя было к ним отнести. Обычно скорость
фотосинтеза у таких: гибридов была ниже, чем у Сз- или
С4-растений, использованных для скрещивания. Исследования
подобного рода показали, что С4-фотосинтез функционирует
только тогда, когда налицо все параметры С4-синдрома и ког-
да эти параметры надлежащим- образом скоординированы.
В род Mollugo (Aizoaceae) входят как С3-, так и С4-расте-
ния, а кроме того, и виды с промежуточными характеристика-
ми, К числу последних, в частности, принадлежит Mollugo ver-
ticillata. У этого растения среди самых ранних первичных про-
дуктов фиксации СО2 обнаруживаются как С3-, так и С4-кисло-
ты, фотодыхание находится на промежуточном уровне, а в
клетках обкладки проводящих пучков есть хлоропласты. Ткань
мезофилла у него состоит из палисадной и губчатой паренхи-
мы. Опыты по разбавлению импульсной метки (12СО2 после
14СО2; разд. 10.2) позволили выяснить пути обмена С4-кпслот.
Экотипы этого вида обнаруживают различную степень проме-
жуточности между Сз- и С4-растениями. Некоторые из видов
Panicum (из секции Саха) также отнесены к промежуточному
типу С3/С4-растений.- Для таких растений, например для Pani-
cum mltioides (независимо обнаруженного в 1975 г. Брауном и
Брауном, а также Канаи и Касиваги), углекислотный компен-
сационный пункт имеет значение ~20—25 м.д. (сравните с
величиной 40—60 м.д. у С3-растений), а-уровень ингибирова-
ния фотосинтеза кислородом довольно низок. Им присуща ана-
томия кранц-типа (явно выраженный слой содержащих хлоро?
пласты клеток обкладки проводящих пучков, которые окружа-
ют проводящую ткань, см. разд. 10.4); правда, между проводя-
щими пучками находится гораздо- большее число клеток мезо-
филла, чем у С4-р астений. • Они также обладают существенно
более высокой активностью ФЕП-карбоксилазы, чем С3-рас-
тения, и есть данные о том, что у них, как у Mollugo verticil-
lata, 14СО2 связывается в составе С4-кислот типа малата и ас-
партата. Судя по углекислотному компенсационному пункту
и ингибированию фотосинтеза кислородом, Moricandia arvensis
также -является видом с промежуточными С3/С4-хар актер исти-
нами.
У- Mollugo -verticillata, Р. milioides и Moricandia arvensis
нет такой строго дифференцированной компартментации ФЕП-
ТРИ ПОДГРУППЫ С.1-РЛГ.ТЕ11ПП
349
карбоксилазы и. РуБФ-карбоксилазы. в. клетках разного типа,,
как у Сграстений. В листьях Р. mllioides присутствует пиру-
ват,ортофосфат — дикиназа, откуда следует, что ФЕ1Г может
частично образовываться не из 3-фосфоглицерата, а из пиру-
вата. Остается, однако, неясным, фиксируется ли СОг в соста-
ве С4-КИСЛ0Т в. результате вторичного карбоксилирования
(о субстрате ФЕП-карбоксилазы, возникающем из ФГК, кото-
рая образуется с участием РуБФ-карбоксилазы, см. разд. 10.1)
или же у этого растения происходит первичное карбоксилиро-
вание. Следует отметить, что у всех растений, у которых обна-
ружены промежуточные Сз/Сгхарактеристики, изотопный сос-
тав углерода (выраженного как б13С, см. разд. 15.12) близок к
изотопному составу углерода у Сз-растений. Это указывает на
то, что в нормальных атмосферных условиях прямая фиксация
СО2 из окружающей среды с участием ФЕП-карбоксилазы идет
гораздо слабее, чем фиксация с участием РуБФ-карбоксилазы.
Итак, идентифицировано всего несколько растений, имеющих
промежуточные Сз/Сгхарактеристпкп процесса фотосинтеза, и
получены немногочисленные данные о наличии у этих растений
крапц-подобпых клеток обкладки проводящих пучков. В то же'
время не обнаружено пи одного растения с С4-фотосинтезом, у
которого не было бы пространственного разделения процессов;
метаболизма (разд. 10.7).
Источники данных: [1, 2, 7, 11, 19, 20, 24, 26].
Общая литература приведена в гл. 10.
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
1. Apel Р., Ohle II. COj-Kompensationspunkt und Blattanatomie bei Arten der
Gattung Moricandia DC. (Crticiferae), Biochem. Physiol. Pflanzen, 174,.
68—75 (1979). . . •
2. Bjorkman 0., Nobs M. A., Pearcy R. 117. Hybrids'between Atriplex species,
with and without B-carboxylalion photosynthesis, Carnegie Inst. Wash: Year-
Book, 69, 624—648 (1971). •
3. Black С. С4 Chen T: M., Brown R. II. Biochemical basis for plant competition,
Weed Sci., 17, 33S—344 (1969). '
4. Black С. C„ Mayne В. С. P700 activity and chlorophyll content of plants,
with-different photosynthetic carbon dioxide'fixation cycles, Plant Physiol.,
45, 738—741 (1970).
5. Brown IV. V. Leaf anatomy in grass systematics, Bot.-Gaz, 11.9, 170-i-178'.
(1958). '
6, Brown IE. V. The Kranz syndrome and its subtypes in grass systematics,
Memoirs Torrey Bot. Club., 23, 1—97 (1977). .
7. Brown R. II,, Brown IV. y, Photosynthetic characteristics of Panictim milioi-
des, a -species with reduced "photorespiration, Ci‘op. .Sti., 15,' 681—685'
(1975).
8. Doreseh R. The 10 worst weeds of field crops, Crops and Soils,-' 1, 14
. (1970). ' ' '
9. Downtoil IV. I. S., Berry I. A., Tregupna E. B. Crphot'osynthesis: non’cyclic-
electron.flow and grana development in bundle sheath chloroplasts, Z. Pflan-
zenphysioh, 63, 194—199 (1970), . ' . ' • ' • "
•350
ГЛАВА 11
10. Edwards G. E. et al. Variation in photochemical activities in C4 plants in
relation to CO2 fixation. In: Metabolism and Productivity of Plants (eds.
R. H. Burris and С. C. Black), pp. 83—112, Baltimore, Maryland, University
Park Press, 1976.
11. Golstein, L. D. et al. Biochemical characterisation of Panicum species which
are intermediate between Сз and C4 photosynthesis plants, Plant Sci. Lett.,
6, 85—90 (1976).
12. Gould F. IV. Grass Systematics, p. 97. New York, McGraw-Hill, 1968.
13. Gutierrez M., Edwards G. E., Brown IV. V. PEP carboxykinase containing
species in the Brachiaria group of the subfamily Panicoideae, Biochem.
System. Ecol., 4, 47—49 (1976).
14. Gutierrez M., Gracen V. E„ Edwards G. E. Biochemical and cytological
relationships in C4 plants. Planta, 119, 279—300 (1974).
15. Hardt H., Kok B. Comparison of photosynthetic activities of spinach chloro-
plasts with those of corn mesophyll and corn bundle sheath tissue, Plant
Physiol., 62, 59—63 (1978).
16. Hatch M. D., Kagawa T., Craig S. Subdivision of C4-pathway species based
on differing C4 acid decarboxylating systems and ultrastructural features,
Aust. J. Plant Physiol., 2, 111—128 (1975).
17. Hatch M. D., Osmond С. B. Compartmentation and transport in C4 photo-
synthesis. In: Transport in plant. Encyclopedia of Plant Physiology, Vol. Ill
(eds. C. R. Stocking and U. Heber), pp. 144—184, Berlin, Springer-Verlag,
1976.
18. Holm L. Weed problems in developing countries, Weed Sci., 17, 113—118
(1969).
19. Kanai R., Kashiwagi M. Panicum milioides, a Gramineae plant having a
Kranz anatomy without C4 photosynthesis, Plant Physiol., 16, 669—679
(1975).
20. Ku S. B., Edwards G. E., Kanai R. Distribution of enzymes related to Сз and
C4 pathway of photosynthesis between mesophyll and bundle sheath cells of
Panicum Mans and Panicum milioides, Plant Cell Physiol., 17, 615—620
(1976).
21. Ku S. B., Gutierrez M., Kanai R., Edwards G. E. Photosynthesis in mesophyll
protoplasts and bundle sheath cells of C4 plants. II. Chlorophyll and Hill
reaction studies, Z. Pflanzenphysiol., 72, 320—337 (1974).
22. Mayne В. C., Dee A. M., Edwards G. E. Photosynthesis in mesophyll pro-
toplasts and bundle sheath cells of various types of C4 plants. Fluorescence
emission spectra, delayed light emission and P700 content, Z. Pflanzen-
physiol., 74, 275—291 (1974).
23. Randall D. D., Tolbert N. E. Two phosphatases associated with photosynthe-
sis and the glycolate pathway. In: Photosynthesis and photorespiration, (eds.
M. D. Hatch, С. B. Osmond and R. O. Slayter), pp. 259—266, New York,
Wiley Interscience, 1971.
24. Rathnam С. К- M„ Chollet R. CO2 donation by malate and aspartate reduces
photorespiration in Panicum milioides, а Сз/С4 intermediate species, Biochem.
Biophys. Res. Commun., 83, 801—808 (1978).
25. Rathnam С. К. M., Edwards G. E. C4 acid decarboxylation and CO2 dona-
tion to photosynthesis in bundle sheath strands and chloroplasts from species
representing three groups of C4 plants, Arch. Biochem. Biophys., 182, 1—13
(1977).
26. Sayre R. T„ Kennedy R. A. Ecotypic differences in the C3 and C4 photo-
synthetic activity in Mollugo verticillata, а Сз/С4 intermediate, Planta, 134,
257—262 (1977).
27. Smith B. N., Brown W. V. The Kranz syndrome in the Gramineae as indi-
cated by carbon isotopic ratios, Amer. j. Bot., 60, 505—513 (1973).
28. Walker G. H„ Izawa S. Photosynthetic electron transport in isolated maize
bundle sheath cells, Plant Physiol., 63, 133—138 (1979).
ГЛАВА 12
Интеграция функций
при С4-фотосинтезе
Краткое содержание
Биохимия Сгфотосинтеза и внутриклеточная локализация
ферментов этого процесса изучены достаточно хорошо. Это по-
зволяет в общих чертах охарактеризовать путь углерода во
время фиксации СО2 внутри листа у каждой из известных трех
подгрупп Сграстений. Результаты исследований, проведенных
иа изолированных препаратах мезофилла или обкладки прово-
дящих пучков in vitro, вполне определенно свидетельствуют о
разделении «труда» между фотосинтезирующими клетками
различного типа. Для протекания С4-фотосинтеза необходимы
четкая координация С4- и ВПФ-циклов и интенсивный транс-
порт метаболитов. Именно эти вопросы рассматриваются в дан-
ной главе. Кроме того, мы в общих чертах обсудим в ней яв-
ные пробелы в современном понимании процесса. Другие тесно
связанные с фотосинтезом события, а именно синтез крахмала
и сахарозы и метаболизм нитрата, освещены здесь с точки зре-
ния их компартментации у С4-растеиий. В первой части главы
будут рассмотрены факты, свидетельствующие о транспорте
метаболитов С4-цикла внутри (внутриклеточный транспорт)
клеток мезофилла и клеток обкладки проводящих пучков, и
С4-метаболизм in vivo (разд. 12.1—12.5). При изучении внут-
риклеточного транспорта использовали следующие подходы:
выяснение внутриклеточной локализации ферментов, исследо-
вание метаболизма субстратов в изолированных органеллах,
оценка изменений объема при сжатии — набухании и постоян-
ная регистрация поглощения метаболитов (на входе) или их
обмена (иа выходе) между органеллами, например с примене-
нием центрифугирования в силиконовом масле (разд. 8.5),
12.1. Внутриклеточная локализация ферментов
С4-Цикла в клетках мезофилла
Для определения внутриклеточной локализации ферментов
С4-пути использовали хлоропласты мезофилла, полученные ли-
бо после механического разрушения ткани, либо из приготов-
ленных заранее протопластов. В ранних работах на кукурузе
Слэк и Хэтч, применив методы, основанные на использовании
352
ГЛАВА 12
Рис. 12.1. В функционировании С4-цикла в листе участвуют фотосинтезирую-
щие клетки двух типов. Поэтому должны существовать механизмы, позво-
ляющие переносить метаболиты из клетки в клетку и через оболочку хлоро-
пласта (ХП). Роль хлоропластов в С4-фотосинтезе можно изучать, выделяя
эти органеллы из клетки. Координация обмена веществ и в тех и в других
клетках не ограничивается С4- и ВПФ-циклами; она включает также синтез
сахарозы, крахмала и обмен аминного азота.
неводных сред, пришли к выводу, что активность пируват,орто-
фосфат — дикиназы, пирофосфатазы, аденилаткиназы и NADP-
малатдегидрогеназы связана с хлоропластами мезофилла. Ло-
кализацию ФЕП-карбоксилазы определить не удалось, посколь-
ку она иногда была ассоциирована с хлоропластами, а иногда
'обнаруживалась в цитозоле. .[Цитозоль — это растворимая
фракция клетки, в которую погружены органеллы.]
Использование протопластов при исследовании локализа-
ции ферментов имеет два преимущества: 1) при этом получа-
ют чистые препараты- мезофилла-и 2) в большом количестве
выделяют интактные органеллы (80—99%). Можно отдельно
выделять хлоропласты и митохондрии вместе с микрбтельцами.
Фракции хлоропластов или митохондрий с микротельцами от-
деляют: от цитозоля путем дифференциального центрифугирова-
ния при 400 или 10 000 g соответственно. Можно также наслаи-
вать суммарный экстракт протопластов на градиент концентра-
ции сахарозы и разделять эти органеллы, применяя метод изо-
пикнического центрифугирования, при котором органеллы рас-
пределяются в соответствии с их плавучей плотностью (разд.
13.7).
Когда хлоропласты мезофилла выделяют с применением
водных растворов, ФЕП-карбоксилаза выявляется главным об-
разом во фракции цитозоля, хотя небольшая доля активности
часто обнаруживается и во фракции хлоропластов. Так, на-
пример, в случае хлорбпластов, выделенных из протопластов
.мезофилла кукурузы, NADP-триозофосфатдегидрогсназу опре-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
3.53
Таблица 12.1. Локализация ФЕП-карбоксилазы в цитозоле
протопластов мезофилла кукурузы1’
NADP-триозофосфатдегидрогеназа, %
ФЕП-карбокснлаза, %
осадок после цент-
рифугирования
при 400g
надосадочная
жидкость
осадок после цент-
рифугирования
при 400g
надосадочная
жидкость
82 18
3
97
) Общая активность NADP-трцозофосфатдегидрогоиазы и ФЕП-карбоксилазы со-
ставляла соответственно 705 и 864 мкмоль-(мг Хл)_1-ч-‘ (Gutierrez et al., 1975).
деляют во фракции хлоропластов, а 97% активности ФЕГЪ
карбоксилазы обнаруживают в цитозоле (табл. 12.1).
Если хлоропласты, полученные из протопластов мезофилла
С4-р астений, фракционируют далее в градиенте концентрации
сахарозы, то свыше 80% NADP-триозофосфатдегидрогеназы
выявляют в зоне интактных хлоропластов, а вся активность
ФЕП-карбоксилазы остается в верхней части градиента, во
фракции цитозоля. Следовательно, ФЕП-карбоксилаза локали-
зована только в цитозоле клетки. Не исключено, что in vivo
этот фермент непрочно связан с наружной частью оболочки
хлоропластов. Если к тому же учесть и транспорт метаболитов,
то напрашивается вывод о том, что этот фермент должен нахо-
диться снаружи от хлоропласта.
Локализация ферментов, участвующих в превращении пиру-
вата и СО2 в малат, по-видимому, одинакова у всех С4-расте-
пий. Все ферменты этой части С4-пути, :за исключением ФЕП-
КЛЕТКА МЕЗОФИЛЛА
ХЛОРОПЛАСТ
ЦИТОЗОЛЬ
Рис. 12'.2. Синтез малата у С4-растений.
23-234
354
ГЛАВА 12
карбоксилазы, находятся в хлоропластах мезофилла. Локали-
зация ФЕП-карбоксилазы имеет решающее значение, посколь-
ку она определяет транспорт метаболитов, необходимых для
хлоропластов. Присутствие этого фермента в цитозоле говорит
о том, что к числу тех метаболитов, которые должны транс-
портироваться через оболочку хлоропласта относятся ФЕП, Р[
и оксалоацетат, а также пируват и малат (рис. 12.2).
a) NADP-МДГ-растения
У NADP-МДГ-растений первичными метаболитами С4-цик-
ла, участвующими в транспорте между клетками мезофилла и
клетками обкладки проводящих пучков, являются малат и пи-
руват (разд. 11.2 и 11.3). Поэтому главная функция клеток ме-
зофилла в С4-фотосинтезе заключается в превращении (СО2+
-фпируват) в малат.
Вторичный обмен метаболитами может осуществляться за
счет челночных передвижений аспартата и аланина. В хлоро-
пластах мезофилла у NADP-МДГ-растений обнаружена высо-
кая активность аспартатаминотрансферазы. В хлоропластах и
цитозоле росички кровяной выявляется значительная актив-
ность алапинаминотрансферазы. Поэтому остается неясным,
какая же из аминокислот (вероятнее всего, глутамат или ала-
нин) поглощается хлоропластами и служит донором амино-
Ю1ЕТКА МЕЗОФИЛЛА
Рис. 12.3. NADP-МДГ-растения. Синтез аспартата.
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
355
группы в синтезе аспартата. Если хлоропласты поглощают ала-
нин, то транспортируемыми метаболитами будут аланин, ас-
партат, Pi, ФЕП и оксалоацетат (рис. 12.3).
б) NAD-МДГ-растения
У этих растений главными метаболитами, транспортируемы-
ми в процессе С^-фотосинтеза из клетки в клетку, являются ас-
партат и аланин (разд. 11.2 и 11.3). В клетках мезофилла у
растений этой группы выявляется высокая активность аспар-
тат- и аланинаминотрансфераз, которые локализованы в цито-
золе. Поэтому в ходе превращения аланина и СОз в аспартат
через оболочку хлоропласта должны транспортироваться толь-
ко ФЕП, Р; и пируват. Челночный обмен малат — пируват у
этих растений незначительный и рассматривается как вторич-
ный процесс (рис. 12.4).
в) ФЕП-К.К.-растения
У ФЕП-КК-растений высокая активность аланин- и аспар-
татаминотрансфераз обнаружена в цитозоле клеток мезофил-
ла. У растений этой группы могут функционировать три чел-
ночных механизма обмена, из которых основным является об-
мен аспартат — ФЕП.
1. Обмен аспартат — ФЕП с участием ФЕП-карбоксикиназы.
При работе этого цикла ни один из метаболитов не переносит-
ся через оболочку хлоропласта (рис. 12.5). Межклеточный
транспорт веществ через плазмодесмы рассматривается в разд.
12.6.
КЛЕТКА МЕЗОФИЛЛА
ХЛОРОПЛАСТ ЦИТОЗОЛЬ
Рис. 12.4. NAD-МДГ-растения. Синтез аспартата.
23*
356
ГЛАВА 12
Рис. 12.5. ФЕП-КК-растения. Синтез аспартата.
КЛЕТКА МЕЗОФИЛЛА ОБКЛАДКА
ПРОВОДЯЩЕГО
Рис. 12.6. ФЕП-КК-растения. Синтез малата.
2. Обмен малат — ФЕП с участием ФЕП-карбоксикиназы (рис.
3. Второстепенный челночный обмен аспартат—аланин с уча-
стием NAD-МДГ (декарбоксилирующей), как у NAD-МДГ-
растений.
NADP-малатдегидрогеназа, аланин- и аспартатамино-
трансферазы находятся в клетках мезофилла у всех 04-рас-
тений независимо от группы, к которой они относятся. Актив-
ность NADP-малатдегидрогеназы у NADP-МДГ-растений пре-
вышает максимальную активность фотосинтеза в-целом листе,
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ Сн-ФОТОСИНТЕЗЕ
357
тогда как у NAD-МДГ- и ФЕП-КК-растений активность этого
фермента гораздо ниже и не превышает по величине скорость фо-
тосинтеза в листе. Активность аспартат- и аланинамииотрапс-
фераз максимальна в ткани мезофилла у NAD-МДГ-растений
и ФЕП-КК-растений; оиа выше, чем активность фотосинтеза в
листе, и обе эти активности превышают активность данных
ферментов у С3-растений. Эти результаты хорошо согласуются с
выводом о том, что первичным продуктом и транспортируемым
метаболитом у NADP-МДГ-растеиий является малат, а у NAD-
МДГ- и ФЕП-КК-растений — аспартат.
12.2. Локализация ферментов и внутриклеточный
транспорт метаболитов Ci-цикла
в клетках обкладки проводящих пучков
И аспартат, и малат участвуют в межклеточном транспорте
метаболитов С4-цикла (разд. 11.2, 11.3 и 11.6). Принято счи-
тать, что у всех трех подгрупп (Д-растеинй малат транспорти-
руется в хлоропласты обкладки проводящих пучков, а аспар-
тат — в митохондрии этих же клеток. Основой для этой гипо-
тезы послужил тот факт, что у всех Сграстений аспартатами-
иотраисфераза локализована в митохондриях клеток обкладки
проводящих пучков и совсем не выявляется в хлоропластах тех
же клеток. В хлоропластах же обкладки проводящих пучков
присутствуют ферменты, позволяющие метаболизировать ма-
лат. У NADP-МДГ-растений это NADP-МДГ (декарбоксилиру-
ющая), а у других видов — различные малатдегидрогеназы.
Исходное усвоение малата в хлоропластах позволяет использо-
вать восстановительную силу непосредственно в ВПФ-цикле.
Как будет показано ниже, исходное превращение аспартата в
митохондриях или малата в хлоропластах не всегда означает,
что и декарбоксилирование происходит в той же самой орга-
нелле. В настоящее время все схемы, связывающие С4-цикл с
внутриклеточным транспортом в клетках обкладки проводящих
пучков, основаны на результатах немногочисленных исследова-
ваиий, проведенных па нескольких видах растений. И поныне
выделение для таких исследований интактных органелл из кле-
ток обкладки проводящих пучков сЗязано со значительными
трудностями.
а) NADP-МДГ-растения
Внутриклеточный транспорт метаболитов С4-цикла в клет-
ках обкладки проводящих пучков проще всего, по-видимому,
устроен у NADP-МДГ-растеиий. В хлоропластах обкладки про-
водящих пучков находится NADP-МДГ (декарбоксилирующая)
358
ГЛАВА 12
МЕЗОФИЛЛ КЛЕТКА ОБКЛАДКИ
ПРОВОДЯЩЕГО ПУЧКА
Рис. 12,7. NADP-МДГ-растения. Декарбоксилирование малата.
Рис. 12.8. NADP-МДГ-р астения. Возможный механизм декарбоксилирования
аспартата.
(работы проводили на кукурузе). Главными метаболитами
Сц-Цикла, которые транспортируются из клетки в клетку у
этих растений, являются малат и пируват (разд. 11.2 и 11.3).
А для этого нужно чтобы эти метаболиты переносились через
оболочку хлоропластов обкладки проводящих пучков (тогда
как гидроксил переносится из клеток обкладки проводящих
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
359
пучков в клетки мезофилла; разд. 11.4). Механизм такого
транспорта неизвестен (рис. 12.7).
Транспорт аспартата и аланина может осуществляться за
счет вспомогательного челночного механизма, Предполагают,
что у Gomphrena globosa в этом процессе участвует NADP-МДГ
(декарбоксилирующая) (рис. 12.8). Сначала считали, что у
NADP-МДГ-растений имеется достаточно высокая активность
NAD-МДГ (декарбоксилирующей). Но скорее всего эта актив-
ность обусловлена тем, что при pH, близком к 7,5, NADP-МДГ
может, хотя и в незначительной степени, использовать в каче-
стве кофактора NAD.
б) NAD-МДГ-растения
У этих растений внутриклеточный связанный с С<-циклом
транспорт метаболитов в клетках обкладки проводящих пучков
включает в себя метаболизм соединений углерода и в цитозоле,
и в митохондриях. Главным механизмом транспорта является
челночный обмен аспартат—аланин. В этом случае необходим
транспорт аспартата, пирувата, 2-оксоглутарата и глутамата
в митохондрии клеток обкладки проводящих пучков (рис. 12.9).
Число митохондрий в этих клетках очень велико (рис. 11.7, Б).
У NAD-МДГ-растений может функционировать и вспомога-
тельный цикл, в результате которого обмениваются малат и
пируват. Для этого цикла необходим транспорт малат — окса-
лоацетат в хлоропластах по схеме, представленной на рис.
в) ФЕП-КД-растения
Поскольку сообщалось, что у нескольких видов растений,
принадлежащих к этой подгруппе, ФЕП-карбоксикиназа нахо-
дится в цитоплазме клеток обкладки проводящих пучков, по-
видимому, декарбоксилирование у них ограничено цитоплазмой
клеток. Главным механизмом транспорта метаболитов Сд-цикла
является челночный обмен аспартат—ФЕП. Если аспартат пре-
вращается в оксалоацетат в митохондриях обкладки проводя-
щих пучков (как у растений NAD-МДГ-типа), то необходим
транспорт митохондриями аспартата, оксалоацетата, 2-оксоглу-
тарата, и глутамата (рис. 12.11).
В челночном обмене малат—ФЕП между клетками различ-
ного типа декарбоксилирование, катализируемое ФЕП-карбок-
сикииазой, может целиком происходить в цитоплазме (рис.
12.12), хотя начальная стадия этого процесса (превращение
малата в оксалоацетат) может протекать внутри хлоропласта.
360
ГЛАВА 12
МЕЗОФИЛЛ КЛЕТКА ОБКЛАДКИ ПРОВОДЯЩЕГО ПУЧКА
Рис. 12.9. NAD-МДГ-растения. Декарбоксилирование аспартата.
МЕЗОФИЛЛ КЛЕТКА ОБКЛАДКИ
ПРОВОДЯЩЕГО ПУЧКА
Рис. 12.10. NAD-МДГ-растения. Декарбоксилирование малата.
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С,-ФОТОСИНТЕЗЕ
361
МЕЗОФИЛЛ КЛЕТКА ОБКЛАДКИ
ПРОВОДЯЩЕГО ПУЧКА
Рис. 12.11. ФЕП-КК-растения. Декарбоксилирование аспартата.
МЕЗОФИЛЛ КЛЕТКА ОБКЛАДКИ
ПРОВОДЯЩЕГО ПУЧКА
^NAO
*NADH
ATP
^ADp
Рис. 12.12. ФЕП-КК-растения. Возможный механизм декарбоксилирования
малата.
362
ГЛАВА 12
Кроме того, у ФЕП-КК-растений может осуществляться, хо-
тя и в ограниченной степени, челночный обмен аспартат — ала-
нин при участии NAD-МДГ (декарбоксилирующей).
Источник данных: [38].
12.3. Доказательства фотосинтетической
деятельности клеток мезофилла
Для того чтобы понять механизм фотосинтеза у С4-расте-
ний, прежде всего следует изучить те реакции, которые проте-
кают в изолированных клетках и субклеточных фракциях. При
сравнении таких препаратов с исходной фотосинтезирующей
тканью не следует забывать, что скорость фотосинтеза у
С4-растений равна ~200 мкмоль СО2-(мгХл)~*-ч-1. Поскольку
реальная фиксация ССЕ в конечном итоге идет только за счет
ВПФ-цикла, при почти равном распределении хлорофилла ли-
ста между клетками разного типа скорость С4-цикла в клетках
мезофилла будет составлять, как и скорость ВПФ-цикла в клет-
ках обкладки проводящих пучков, 400 мкмоль-(мгХл)“,-ч-1.
В первоначальных исследованиях было показано, что изоли-
рованные клетки С4-мезофилла росички кровяной (Digilaria
sanguinalis, растение NADP-МДГ-типа) фиксируют СОг на све-
ту [~15 мкмоль СО2-(мгХл)1 ч '] в том случае, когда их
постоянно снабжают пируватом. Это указывает на то, что фик-
сация СО2 в клетках мезофилла зависит от поступления такого
предшественника в С4-цнкл. Высокая скорость фиксации СО2
была достигнута и при добавлении ФЕП, что хорошо согласу-
ется с данными о локализации ФЕП-карбоксилазы в клетках
этого типа. В принципе скорость индуцируемой ФЕП фиксации
СО2 выше, чем активность ФЕП-карбоксилазы, выявляемая в
этих клетках, так как СО2 фиксируется и на свету, и в темно-
те, а первичным продуктом является оксалоацетат. Внесение
ФЕП позволяет оценить карбоксилирующий потенциал С4-цик-
ла и сравнить его со связыванием СО2, которое зависит от пи-
рувата или аланина. В последнем случае для ' фиксации СО2
нужна энергия, возникающая за счет фотохимических процес-
сов, и полная функциональная активность стадии С4-карбоксп-
лирования в клетках мезофилла.
Изолированные хлоропласты мезофилла С4-растений прак-
тически -неспособны фиксировать СО2.на свету. В них нет
РуБФ-карбоксилазы и других ферментов ВПФ-цикла, а ФЕП-
карбоксилаза находится в цитозоле. Поскольку в таких хлоро-
пластах отсутствуют кар боксилирующие ферменты, не удиви-
тельно, что они не могут фиксировать СО2. В присутствии ФЕП
скорость стимулируемой светом фиксации СО2 в хлоропластах
мезофилла С4-растений очень невелика. Это связано, по-внди-
ИНТЕГРАЦИЯ ФУНКЦИЙ ПРИ (^-ФОТОСИНТЕЗЕ
363
Таблица 12.2. Зависимая от света фиксация СО2 в экстрактах
протопластов мезофилла из разных растений (Edwards et al., 1976 b)
Группа Спет Темнота +5 мМ пируват
Свет Темнота
мкмоль-(мг XjiJ-’-h-1
NADP-МДГ Digitaria, sanguinalis NAD-МДГ 4 I 267 1
Eleusine indica 6 1 146 1
ФЕП-КК Urochloa panicoides c3 4 1 210 1
Triticum aestivum 168 1 148 1
мому, с тем, что внутри хлоропластов остается лишь незначи-
тельная доля ФЕП-карбоксплазиой активности клетки (разд.
12.1). Фиксация СО2 в этом случае может стимулироваться све-
том за счет образуемой в этих условиях восстановительной си-
лы, которая способствует превращению оксалоацетата (силь-
ного ингибитора ФЕП-карбоксилазы) в малат.
Существует несколько условий, необходимых для того, что-
бы хлоропласты мезофилла Сграстений стали способны фик-
сировать СО2 на свету с высокой скоростью. Во-первых, следу-
ет обеспечить поступление ортофосфата и трехуглеродного
предшественника (пирувата или аланина), а во-вторых, нужно
объединить фракцию хлоропластов с фракцией цитозоля. По-
следнее легче всего осуществить, получив бесклеточные экст-
ракты из протопластов. При достаточно мягком разрушении
протопластов хлоропласты остаются интактными и для экспе-
римента можно брать суммарный экстракт. Внесение пирувата
и СО2 в экстракт, полученный из протопластов, приводит к син-
тезу оксалоацетата и малата. При добавлении аланина и 2-ок-
соглутарата в экстракты протопластов NAD-МДГ- пли ФЕП-
КК-растений иа свету синтезируются оксалоацетат и аспартат.
Скорость включения СО2 в С4-ки-слоты в таких препаратах при
40°С доходит до 250—350 мкмоль-(мг Хл)-1-ч-1. Для фикса-
ции СО2 нужно, чтобы хлоропласты превращали пируват и ор-
тофосфат в ФЕП, используя энергию фотохимических процес-
сов, и чтобы в цитозоле находилась ФЕП-карбоксилаза. Зави-
симость фиксации СО2 от пирувата продемонстрирована в опы-
тах с экстрактами, полученными из протопластов мезофилла
Сграстений, относящихся ко всем трем подгруппам. Как и еле-
364
ГЛАВА 12
довало ожидать, фотосинтез в хлоропластах пшеницы (С3-рас-
теыие) не зависит от пирувата (табл. 12.2).
Если хлоропласты мезофилла С4-растений отделить от рас-
творимой фракции экстракта протопластов, то скорость зави-
симой от пирувата фиксации СО2 заметно снизится. Считают,
что остаточная активность связана с незначительной актив-
ностью ФЕП-карбоксилазы, остающейся в хлоропластах после
дифференциального центрифугирования экстрактов протопла-
стов (см. табл. 12.1). Как и следовало ожидать, у ячменя
(Сз-растения) скорость фотосинтеза в хлоропластах (свобод-
ных от фракции цитозоля) и в экстрактах протопластов одина-
кова, так как карбоксилаза, участвующая в ВПФ-цикле, оста-
ется внутри хлоропластов (табл. 12.3).
Таблица 12.3. Зависимая от света фиксация СОг в экстрактах протопластов
и в хлоропластах (Edwards et al., 1976а)1’
Вид растений
Экстракт
протопластов
Хлоропласты
NADP-МДГ
Digllarla sanguinalis
С3
Hordeum vulgare
мкмоль СОа-(мг Хл)-1-!!-1
325 Ю
126 114
') Хлоропласты выделяли из экстракта протопластов мезофилла после 3 мин цент-
рифугирования при 600g и последующей однократной отмывки. Для индукции фиксации
СО2 У A sangulnatis использовали 5 мМ пируват и 0,5 мМ оксалоацетат,
Скорость зависимой от пирувата фиксации СО2 в экстрак-
тах протопластов мезофилла С4-растений возрастает в несколь-
ко раз после добавления оксалоацетата в небольшой (0,5 мМ)
концентрации. Внесение оксалоацетата стимулирует процесс не-
циклического переноса электронов, поскольку восстановитель-
ная сила используется для превращения оксалоацетата в ма-
лат. Это позволяет обеспечить превращение пирувата в ФЕП
энергией, образующейся в ходе нециклического фотофосфори-
лирования. Если не добавлять оксалоацетат, то связывание
СО2 с пируватом будет зависеть от циклического и псевдоцик-
лического фотофосфорилирования до тех пор, пока не синте-
зируется достаточное количество оксалоацетата.
Скорость фиксации СО2 в экстрактах протопластов мезо-
филла С4-растений стимулируется {>200мкмоль-(мгХл)‘--ч-1]
ФГК как на свету, так и в темноте. На свету, когда субстра-
том служит ФГК, продуктами фиксации СО2 являются оксало-
ацетат, малат и аспартат. Возможно, что ФГК выполняет роль
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ Сч-ФОТОСИНТЕЗЕ
365
3-ФГК (истинное поступление)
4
2-ФГК
Рис. 12.13. Суммарное поступление углерода в С4-цикл из ФГК.
предшественника при образовании ФЕП под действием фосфо-
глицеромутазы и енолазы. Насколько активным предшествен-
ником служит ФГК в этом пути in vivo, неизвестно. Для того
чтобы С4-цикл эффективно функционировал in vivo, существен-
но необходимо, чтобы содержание метаболитов этого цикла
поддерживалось на определенном уровне. Считают, что С4-цикл
обеспечивает стехиометрический перенос углерода из атмосфе-
ры в ВПФ-цикл. При этом общий пул углерода в С4-пути ие
должен меняться. Если бы в этом цикле не происходило ника-
ких реальных потерь углерода, то новое поступление углерода
в цикл было бы необходимо только для того, чтобы обеспечить
рост растения. Но некоторая потеря углерода при работе 04-
цикла in vivo (например, при транслокации аминокислот и их
использовании как предшественников для синтеза макромоле-
кул, при дыхании или при изменении размеров пула в ответ па
колебания условий окружающей среды) вполне естественна.
Поскольку в конечном итоге COs усваивается с участием РуБФ-
карбоксилазы, поступление углерода в С4-цикл должно обес-
печиваться за счет метаболического превращения ФГК в ФЕП.
Скорость поступления углерода ФГК в.С4-цикл может быть ни-
же, чем скорость потока углерода в этом цикле, ио в отсутст-
вие поступления углерода из ФГК С4-цикл постепенно лишит-
ся всех промежуточных продуктов и потеряет способность конт-
ролировать увеличение их содержания в ответ на изменяющие-
ся потребности метаболизма (рис. 12.13).
Хэтч и Слэк проследили временные изменения в распреде-
лении метки между малатом и ФГК (табл. 12.4) после вклю-
366
ГЛАВА 12
Таблица 12.4. Распределение метки в малате и ФГК после инкубации
листьев сахарного тростника с 14СО2 (Hatch, Slack, 1970)
Время после экспозиции с * 14СО,2, с Распределение метки. %
малат ФГК
С-1 С-2 1-С-З С-4 С-1 С-21-С-З
3 10 0 90 98 2
45 29 4 67 61 39
150 33 25 42 45 55
чения радиоактивной метки в целые листья сахарного трост-
ника.
Если бы Сгцикл был полностью замкнутой системой, то
следовало бы ожидать, что метка включится только в С-4-по-
ложение малата. Если же частично предшественником в.
С4-цикле служит и ФГК, то малат должен метиться не только
по С-4, но и по другим положениям. В этом случае следовало
бы ожидать, что метка сначала появится в С-1-положении ма-
лата, а затем уже и в (C-2-J-C-3)-положениях. По С-1-поло-
жению ФГК метится за счет исходного карбоксилирования
14СО2, катализируемого РуБФ-карбоксилазой, По мере того
как некоторая часть 14С-ФГК поступает в реакции ВПФ-цпкла,
начинает метиться и РуБФ. Это приведет к тому, что ФГК ста-
нет меченой и по С-2-, и по С-З-положению. Если ФГК частич-
но используется как предшественник С4-цикла, то метка из
С-1-положения ФГК должна перейти в С-1-положение малата,
а при включении метки в (С-2-(-С-3)-положения ФГК она по-
явится в соответствии с характером включения метки (табл.
12,4) в (С-2-ф-С-З)-положениях малата. Существует и другая
возможность появления метки в С-1-положении малата в ре-
зультате частичной рандомизации метки за счет уравновешива-
ния с фумаратом под действием фумаразы (разд. 15.8), Метка
в С-1- и (С-2+С-3)-положениях малата может появляться и
вследствие уравновешивания ФГК с ФЕП под действием фос-
фоглицеромутазы и енолазы, когда углерод из ФГК в конеч-
ном итоге не поступает (рис. 12.14).
Хотя в отсутствие цитозоля хлоропласты мезофилла С4-рас-
тений почти не фиксируют СО2, они еще способны к выделе-
нию на свету О2, но только после внесения в среду определен-
ных субстратов, таких, как ФГК или пируват+оксалоацетат.
Тот факт, что выделение О2 зависит от ФГК, хорошо согласует-
ся с данными о локализации реакций восстановительной фазы
ВПФ-цикла в хлоропластах мезофилла и, кроме того, указыва-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С<-ФОТОСИНТЕЗЕ
367
ет на то, что ФГК легко пог-
лощается хлоропластами. Как
и в хлоропластах Сз-растений,
для восстановления ФГК до
триозофосфатов под действи-
ем фосфоглпцератктшазы и
NADP - триозофосфатдегндро-
геназы необходимы и АТР, и
NADPH.
Скорость выделения 02 и
скорость нециклического пере-
носа электронов в хлороплас-
Пируват
3-ФГК ^=5= 2-ФГК =£== ФЕП
Гсо’
ОА
'г
Фумарат Малат
Рис. 12,14.
тах мезофилла росички кровя-
ной резко возрастают после добавления в среду пирувата и не-
большого количества оксалоацетата и практически не увеличи-
ваются, если в среду вносят один оксалоацетат. Это указывает
на то, что нециклический перенос электронов сопряжен с син-
тезом АТР и что для максимальной скорости необходимо ис-
пользовать как АТР, так и NADPH (использование АТР для
превращения пирувата в ФЕП описано в разд. 11,1). В отли-
чие от фиксации СОг хлоропластами С4-мезофилла для зави-
симого от пирувата-|-оксалоацетат выделения О2 хлоропласта-
ми не нужна фракция цитозоля, так как участия ФЕП-карбок-
силазы не требуется (рис. 12.15).
Источники данных: [2, 11 — 14, 19, 24, 28].
12.4. Доказательства фотосинтетической
деятельности клеток обкладки проводящих пучков
В ранних опытах клетки обкладка проводящих пучков ро-
сички кровяной (растения NADP-МДГ-типа) получали после
механического разрушения ткани. Такие клетки фиксировали
ФЕП Пируват Малат
Рис. 12.15. Использование ассимиляционной силы, образующейся в процессе
нециклического переноса электронов в хлоропластах мезофилла, для Ci-фото-
сиитеза.
368
ГЛАВА 12
СОа с довольно большой скоростью, если в состав среды вхо-
дил рибозо-5-фосфат. Не исключено, что значительная стиму-
ляция фиксации СО2 под влиянием рибозо-5-фосфата была
обусловлена повреждениями хлоропластов в процессе их выде-
ления, в результате чего рибозо-5-фосфат быстро входил в та-
кие клетки. Для восстановления пула метаболитов внутри хло-
ропластов вполне достаточно даже незначительного поступле-
ния рибозо-5-фосфата в клетки. При этом уменьшается инкуба-
ционный период и достигается максимальная скорость фикса-
ции (разд. 7.7). Сейчас уже вполне очевидно, что фиксация
СО2 в изолированных клетках обкладки проводящих пучков у
NADP-МДГ-растений лимитируется на свету из-за отсутствия
в них фотосистемы II (ФСП; разд. 11.6). В более поздних ис-
следованиях скорость фиксации СО2 в клетках обкладки про-
водящих пучков, полученных из NAD-МДГ- и ФЕП-КК-расте-
ний, превышала 100 мкмоль СО2-(мгХл)-1 -ч-1 и без добавле-
ния органических субстратов.
Дихлорфенилдиметилмочевина (ДХММ) препятствует вос-
становлению NADP+ за счет нециклического переноса электро-
нов и таким образом ингибирует фиксацию СО2 в изолирован-
ных клетках обкладки проводящих пучков. Малат может час-
тично снимать ингибирование, вызываемое ДХММ в изолиро-
ванных клетках, полученных из С4-р астений всех трех групп.
Это указывает на то, что малат может участвовать в передаче
восстановительной силы в клетки обкладки проводящих пуч-
ков. У NADP-МДГ-растений в этом процессе, по-видимому,
участвует NADP-МДГ (декарбоксилирующая), а у NAD-МДГ-
и ФЕП-КК-растений это может быть NAD-малатдегидрогеназа.
Фиксация СО2 в клетках обкладки проводящих пучков подав-
ляется ингибитором ВПФ-цикла (гл. 8) глицеральдегидом, тог-
да как на зависимую от пирувата фиксацию СО2 в препаратах
мезофилла он влияет гораздо слабее.
Для исследования процесса передачи углерода С4-кислот в
ВПФ-цикл в изолированных клетках обкладки проводящих
пучков или выделенных из них хлоропластов применяют два
следующих метода.
1) Регистрация зависимого от Сц-кислот выделения О2 ла све-
ту в присутствии малата, аспартата или оксалоацетата. Окса-
лоацетат очень лабилен и либо медленно превращается в пиру-
ват и СО2 в результате неферментативного декарбоксилирова-
ния, либо быстро декарбоксилируется в присутствии двухва-
лентных катионов и некоторых белков. Поэтому, используя
этот субстрат in vitro, следует отчасти учитывать и такое неес-
тественное поступление СО2 в ВПФ-цикл.
2) Хроматографический анализ продуктов зависимого от света
включения метки из 14 С-4-атома малата или аспартата в мета*
ИНТЕГРАЦИЯ ФУНКЦИЙ ПРИ Сч-ФОТОСИНТЕЗЕ
369
Аспартат -------► ОА
2-Оксоглутарат Глутамат
Аланин »-—:— Пируват
Рис. 12.16.
болиты ВПФ-цикла (т. е. в ФГК., триозо- или гексозофосфаты).
Это дает прямые сведения о том, что углерод переходит в ВПФ-
цикл. С практической точки зрения второй метод более трудо-
емок, чем первый. Применив эти методы, можно получить ско-
рость поступления углерода С4-кислот в ВПФ-цикл, равную
100—300 мкмоль СО2 или Os- (мгХл)-1-ч~1.
Максимальная скорость зависимого от аспартата выделения
О2 клетками обкладки проводящих пучков NAD-МДГ- или ФЕП-
КК-растений достигается после внесения в среду пирувата
(1 мМ) или 2-оксоглутарата (10 мМ). 2-Оксоглутарат нужен
для начальной реакции стадии декарбоксилирования у этих
двух подгрупп С4-растений. Пируват может стимулировать де-
карбоксилирование аспартата в результате сопряжения реак-
ций с участием аспартат- и аланинаминотрансфераз (рис.
12.16).
Для исследования декарбоксилирования С4-кислот в изолн*-
рованных клетках обкладки проводящих пучков был применен
еще один метод. В среду вносили 14С-4-дикарбоновые кислоты,
и по количеству 14СО2, высвобождающейся на свету в присут-
ствии D.L-глицеральдегида, судили об интенсивности декарб-
оксилирования. Глицеральдегид препятствует повторной фикса-
ции ИСО2 в ВПФ-цикле. В его присутствии ФГК резко усили-
вает декарбоксилирование малата в изолированных клетках
обкладки проводящих пучков у С4-фастений всех трех групп.
Это говорит о том, что окисление малата сопряжено с восста-
новительной стадией ВПФ-цикла. Декарбоксилирование малата
в клетках обкладки проводящих пучков у ФЕП-КК-растений
ингибируется 0,1 мМ 3-меркаптопиколиновой кислотой (инги-
битор ФЕП-КК). Этот ингибитор ие влияет иа декарбоксилиро-
вание малата у NAD-МДГ- и NADP-МДГ-растений. Щавелевая
кислота (0,1 мМ) специфически ингибирует декарбоксилирова-
ние малата в клетках обкладки проводящих пучков у NADP-
МДГ-растений (росичка кровяная). Это свидетельствует о том,
что в декарбоксилировании малата у разных представителей
С4-растений участвуют разные механизмы. 3-меркаптопиколи-
новая и щавелевая кислоты, взятые в той же концентрации, не
влияют на фиксацию 14СО2 в изолированных клетках обкладкй
24—324
.370
ГЛАВА 12
КЛЕТКА ОБКЛАДКИ ПРОВОДЯЩЕГО ПУЧКА
ЦИТОЗОЛЬ
МИТОХОНДРИЯ
Рис. 12.17. Предполагаемый транспорт метаболитов в митохондриях во время
декарбоксилирования аспартата при участии NAD-МДГ (декарбоксилирую-
щей).
проводящих пучков, из чего можно сделать вывод о том, что
оба этих вещества специфически ингибируют декарбоксилиро-
вание С4-кислот.
Для того чтобы декарбоксилирование 14С-4-аспартата в
клетках обкладки проводящих пучков в присутствии D.L-глп-
церальдегида шло с максимальной скоростью, необходимо до-
бавить 2-оксоглутарат или пируват. З-Меркаптоппколшювая
кислота сильно ингибирует декарбоксилирование аспартата у
ФЕП-КК-растений (но не действует на растения NAD-МДГ-
или NADP-МДГ-типа). Таким образом, главный путь декарб-
оксилирования аспартата у этих растений, по-видимому, вклю-
чает в себя ФЕП-карбоксикиназу. Свет стимулирует декарбок-
силирование 14С-4-аспартата в клетках обкладки проводящих
пучков у ФЕП-КК-растений. Причина такой стимуляции непо-
нятна; возможно, что в результате фотохимических реакций об-
разуется АТР, необходимый для декарбоксилирования, ката-
лизируемого ФЕП-карбоксикиназой. У NAD-МДГ-растений
скорости декарбоксирования аспартата на свету и в темноте
одинаковы. Это вполне естественно, поскольку для декарбокси-
ИНТЕГРАЦИЯ ФУНКЦИЙ ПРИ Щ-ФОТОСИНТЕЗЕ
371
лироваиия аспартата под действием NAD-МДГ энергии не
требуется.
Скорость декарбоксилирования аспартата в митохондриях
обкладки проводящих пучков NAD-МДГ-растений возрастает
после внесения в среду малата, Pi и 2-оксоглутарата. Для того
чтобы аспартат превратился в митохондриях в пируват и СОг,
необходимо поступление аспартата и 2-оксоглутарата внутрь
органеллы в обмен на глутамат и пируват. Малат и Pi служат
своего рода катализаторами метаболического превращения ас-
партата в пируват и СО2.
В митохондриях млекопитающих и в митохондриях из не-
фотосиптезирующих растительных тканей поглощение малата
может быть связано с переносчиками фосфата. Высокое отно-
шение NADPH/NAD+ в митохондриях обкладки проводящих
пучков должно способствовать превращению аспартата в ма-
лат, так как для этой реакции необходим NADH. Поглощение
малата, сопряженное с функционированием переносчиков фос-
фата п дикарбоновых кислот, может влиять на окислительно-
восстановительное состояние митохондрий н таким образом
способствовать достижению максимальной скорости превраще-
ния аспартата в пируват и СО2 (рис. 12.17).
В некоторых митохондриях предполагают существование
переносчика, обменивающего малат на 2-оксоглутарат. Поэто-
му малат и Pi тоже могут способствовать облегченному погло-
щению 2-оксоглутарата. Механизм транспорта веществ в мито-
хондриях клеток обкладки проводящих пучков еще предстоит
выяснить.
Источники данных: (2, 11, 14, 24, 45, 46].
12.5. Механизмы внутриклеточного транспорта
метаболитов
Для фотосинтеза у С4-растений необходимо, чтобы опреде-
ленные метаболиты С4-цикла быстро переносились через обо-
лочку хлоропластов н митохондрий. О механизме транспорта
метаболитов, особенно в хлоропластах обкладки проводящих
пучков и в митохондриях, мало что известно. Связь транспорта
метаболитов с С4-циклом была изучена в опытах с хлоропласта-
ми мезофилла С4-растепий. Для этого регистрировали на спект-
рофотометре при 535 им осмотические изменения объема хлоро-
пластов под влиянием субстрата и оценивали методом центри-
фугирования в силиконовом масле поглощение метаболитов хло-
ропластами или их отток в окружающую среду.
Осмотическое набухание применяли в исследованиях транс-
порта метаболитов в митохондриях млекопитающих, в расти-
тельных митохондриях из нефотосинтезирующих тканей и в.
24*
372
ГЛАВА 12
хлоропластах С3- (шпинат) и С4- (росичка кровяная)-растений.
Этот метод позволяет выявить суммарное поглощение веществ,
но не позволяет оценить и измерить обмен метаболитами (за
исключением обмена гидроксил — анион при поглощении солей
аммония) через мембраны (гл. 8 и рис. 12.19).
Метод центрифугирования в силиконовом масле можно ис-
пользовать для выяснения механизма обмена веществ в ходе
транспорта и для изучения кинетики этого процесса. Если ис-
пользовать физиологические концентрации метаболитов, то
можно даже различить поглощение за счет диффузии и погло-
щение с участием переносчика. В отличие от диффузии транс-
порт, опосредуемый переносчиком, характеризуется кинетикой
с насыщением при 'высоких концентрациях субстрата (обычно
в пределах 10 мМ). Ему свойственна высокая чувствительность
к температуре, а метаболиты, транспортируемые одним и тем
же переносчиком, обычно конкурируют друг с другом. Погло-
щение, опосредуемое переносчиком, подавляется некоторыми
сульфгидрильными реагентами. [Сульфгидрильные реагенты
взаимодействуют с сульфгидрильными группами белков и та-
ким образом мешают белку выполнять его каталитическую
функцию.] Ингибирование поглощения, например специфиче-
скими аналогами субстрата, также служит доказательством
участия в этом процессе переносчика.
Переносчик пирувата в хлоропластах мезофилла
Сграстений
В хлоропластах мезофилла С4-растений (росичка кровяная)
присутствует электрогенный переносчик пирувата (транспорт
метаболитов на переносчике не электронейтрален). Интактные
хлоропласты, сохраняемые в сорбитоле (0,25 М; взят в качест-
ве осмотика), сразу же сжимаются после добавления 0,1 М
пирувата калия. Исходный объем хлоропластов снова восста-
новится, если произойдет поглощение растворенного вещества
как единого целого (т. е. если через мембрану пройдут и ка-
тион, и анион). Добавление валиномицина, который облегчает
транспорт К+, ведет к быстрому набуханию хлоропластов. Это
говорит о том, что проникающим ионом является анион пиру-
вата. В то же время валпиомицин не вызывает набухания хло-
ропластов шпината в присутствии пирувата калия, что свиде-
тельствует о том, что анион пирувата не проникает в хлоро-
пласты С3-р астений. Некоторое набухание хлоропластов мезо-
филла Сграстений наблюдается в присутствии 0,1 М пирувата
аммония, однако этот процесс идет не столь быстро, как по-
глощение пирувата калия в присутствии валиномицина. Отсю-
да следует, что обмен пируват — гидроксил чем-то лимитиру-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С4 ФОТОСИНТЕЗЕ
373
А"-------> д-
(Например, пируват")
х+-------► х +
Рис. 12.19. Поглощение солей аммо-
(Например, Н*) ния с участием обмена гидроксил-
аниона (А-).
Рис. 12.18. Три примера электроген-
ного транспорта.
(Например, ДГАФ'"
ФГК-’")
ется. [В случае солей аммония поглощение аниона в обмен на
ионы гидроксила можно оценить по набуханию хлоропластов.
NH3, который легко проникает через мембраны, поглощается
хлоропластами и уравновешивается с МНЦ. При наличии со-
ответствующего переносчика ионы гидроксила, которые обра-
зуются внутри хлоропласта, обмениваются на анноны, присут-
ствующие во внешней среде, что и приводит к поглощению дан-
ной соли аммония.] Центрифугирование в -силиконовом масле
показало, что градиент pH на оболочке хлоропласта практиче-
ски не влияет на поглощение 14С-пирувата. Это тоже говорит
о том, что поглощение пирувата в обмен на гидроксил огра-
ничено (рис. 12.18 и 12.19).
Изменения скорости поглощения иС-пирувата, выявленные
этим способом, описываются кинетической кривой Михаэлиса —
Ментен, а кажущаяся Км для пирувата составляет 0,7—1 мМ.
Это тоже свидетельствует о том, что пируват скорее всего по-
глощается не за счет диффузии, а с участием переносчика.
Поглощение 14С-пирувата хлороплэстамп мезофилла С4-рас-
тений подавляется мерсалилом, сульфгидрильным реагентом.
Мерсалил ингибирует также набухание хлоропластов в присут-
ствии пирувата калия и валииомнцина. Такое ингибирование
свидетельствует о том, что транспорт пирувата опосредован пе-
реносчиком.
Производные а-коричных кислот являются мощными инги-
биторами поглощения пирувата в митохондриях животных,
374
ГЛАВА 12
идущего с участием переносчика. Поглощение пирувата хлоро-
пластами мезофилла С4-растений тоже специфически ингиби-
руется рядом аналогов пирувата и различными производными
tt-коричной кислоты.
Обмен ФЕП—Pi на переносчике фосфата
в хлоропластах мезофилла (^-растений
В хлоропластах мезофилла С4-растений (росичка кровяная}
есть переносчик фосфата (локализованный в мембране пере-
носчик, обменивающий Pi и/или анионы органических фосфатов
точно так же, как и переносчик в хлоропластах Сз-растений,
гл. 8), который катализирует обмен ФЕП и ортофосфата.
В изолированных хлоропластах содержится пул ФЕП, который
может экспортироваться из хлоропласта в обмен на Pi или
ФГК. ФЕП не является проникающим анионом, так как хлоро-
пласты в растворе фосфоенолпирувата калия не набухают и в
присутствии валиномицина, и без него.
По данным, полученным путем центрифугирования в сили-
коновом масле, ФЕП конкурентно ингибирует (70 = 0,45 мМ)
поглощение 32Pi (Км = 0,2 мМ) в хлоропластах мезофилла
С4-растеннй. Это свидетельствует о том, что ФЕП обладает
сравнительно высоким сродством к переносчику фосфата в та-
ких хлоропластах. В хлоропластах шпината Kt для вызываемо-
го ФЕП ингибирования поглощения 32Р примерно в десять
раз выше, чем в хлоропластах С4-растений, а это указывает на
то, что ФЕП плохо транспортируется в хлоропласты Сз-расте-
ний.
Синтез ФЕП в хлоропластах С4-растений, инкубируемых на
свету в среде с пируватом, но без Pi, идет медленно, однако
концентрация ФЕП внутри хлоропластов достигает очень вы-
сокого уровня (отношение концентрации ФЕП внутри хлоро-
пластов к концентрации снаружи равно 30). Внесение в среду
Pt в небольшой концентрации (0,1 мМ) приводит к быстрому
снижению уровня ФЕП в хлоропластах (отношение концентра-
ций внутри и снаружи хлоропластов падает до 2). Следова-
тельно, Pi необходим и для синтеза ФЕП, и для его выхода из
хлоропластов. Транспорт может быть электрогенным и иа пе-
реносчике пирувата, и на переносчике фосфата. А как следст-
вие этого транспорт всех трех метаболитов (ФЕП, Р| и пиру-
вата) должен быть электроиейтральным (рис. 12.20).
В хлоропластах мезофилла С4-растений (росичка кровяная)
присутствует также переносчик дикарбоновых кислот, который
имеет Км для оксалоацетата 0,13 мМ. Поглощение оксалоаце-
тата конкурентно ингибируется малатом (Ki = 0,26 мМ) и ас-
партатом (Kt = 0,2 мМ), что свидетельствует об участии в
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
375
Рис. 12.20. Транспорт пирувата, ФЕП и Pi в хлоропластах мезофилла Q-
.растений.
1/О15щая концентрация осмотически активных ионов и молекул, М’
Рис. 12.21. Влияние концентрации сорбитола на объем межмембранного про-
странства и стромы в хлоропластах мезофилла росички кровяной. Объем
стромы определяли по разности величин общего объема хлоропласта (3Н2О)
И межмембранного пространства (иС-сахароза). В реакционную смесь в
каждом случае вносили по 6 мкг Хл (Huber, Edwards, 1977b).
376
ГЛАВА 12
транспорте этих дикарбоновых кислот одного и того же пере-
носчика.
Считают, что такие переносчики локализованы во внутрен-
ней мембране хлоропластов. Как следует из данных по погло-
щению 3Н2О и 14С-сахарозы, сахароза проникает через наруж-
ную мембрану хлоропластов мезофилла С^растений, но не мо-
жет проходить через внутреннюю. При повышении осмотичес-
кой силы раствора за счет изменения концентрации сорбито-
ла общий объем хлоропласта остается постоянным (судя по
поглощению 3Н2О), а межмембранное пространство (измерен-
ное по поглощению 14С-сахарозы) увеличивается. Следователь-
но, с увеличением концентрации сорбитола уменьшается объем
стромы и увеличивается межмембранное пространство, а это
говорит о том, что внутренняя мембрана малопроницаема для
ряда веществ (рис. 12.21).
Итак, есть данные о том, что во внутренней мембране хло-
ропластов мезофилла С4-растений имеются переносчики, кото-
рые катализируют обмен метаболитов по типу обмена, пред-
ложенного в разд. 12.1. Механизмы транспорта метаболитов в
хлоропластах обкладки проводящих пучков до сих пор еще не
исследованы.
Источники данных: [34, 35].
12.6. Механизм межклеточного транспорта
метаболитов
Для фотосинтеза у С4-растений необходим быстрый перенос
метаболитов из клеток мезофилла в клетки обкладки проводя-
щих пучков и обратно. Обычно считают, что такой перенос осу-
ществляется за счет симпл астич еского транспорта через плаз-
модесмы. Симпластический транспорт между клетками проис-
ходит через поры (плазмодесмы), которые обеспечивают непре-
рывность цитоплазмы, несмотря на наличие у клеток барьера
в виде мембран или клеточных стенок. Апопластический транс-
порт происходит через клеточную стенку и заключается в пе-
реносе вещества через плазматическую мембрану.
Есть несколько сообщений о том, что клетки мезофилла и
клетки обкладки проводящих пучков соединяются плазмодес-
мами. Так, например, приводятся доводы в пользу того, что
плазмодесмы у кукурузы находятся в области первичных по-
ровых полей между этими клетками. Большое количество пер-
вичных поровых полей между клетками мезофилла и клетками
обкладки проводящих пучков видно на производящих впечат-1
ление микрофотографиях Крэйга и Гудчайлда, полученных с
помощью сканирующего электронного микроскопа (рис. 12.22).
Одна из простейших гипотез заключается в том, что метаболи-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ ^ФОТОСИНТЕЗЕ
377
Рис. 12.22. Микрофотография ткани листа Triodia irritans после ее обработки
проиазой. На фотографии, полученной с помощью сканирующего электронно-
го микроскопа, первичные поровые поля в клеточных стенках клеток обклад-
ки проводящих пучков видны со стороны проводящего пучка. X1500. Стрел-
ками показаны первичные поровые поля в клеточной стенке. Каждое поровое
поле состоит из многочисленных плазмодесм (Craig, Goodchild, 1977).
ты передвигаются из клетки в клетку по градиенту концент-
рации за счет диффузии через плазмодесмы.
Трудности в выявлении путей транспорта между клетками
отчасти связаны с недостатком знаний о структуре плазмодесм.
Не ясно, являются ли плазмодесмы открытыми каналами или
же передвижение ионов и молекул между клетками строго ог-
раничено и избирательно. Робардс считает, что плазмодесма
состоит из тонкого тяжа десмотрубочки (измененный эндо-
плазматический ретикулум), проходящего сквозь пору. Оценка
действительных размеров открытой площади плазмодесм и их
общего числа — задача не из легких (рис. 12.23).
При рассмотрении модели Робардса возникает вопрос, что
же участвует в транспорте: эндоплазматический ретикулум
(ЭР), аннулярный слой цитоплазмы или оба этих компонента?
ЭР представляет собой сложную сеть липопротеиновых мем-
бран, образующих различного рода трубочки и полости, кото-
рые простираются от ядра, а возможно, и от других органелл
по всей цитоплазме клетки. Эта структура может выполнять
378
ГЛАВА 12
Плазмодесма
Рис. 12.23. Схематическое изображение структуры плазмодесм (в упрощен-
ном виде по Robards, 1975).
множество различных функций, таких, например, как транс-
порт и синтез белков, а также выступать в качестве предшест-
венника при образовании других мембран клетки. Изучив
электронные микрофотографии плазмодесм, соединяющих
клетки мезофилла и клетки обкладки проводящих пучков
у кукурузы, Эверт и др. предположили, что перенос веществ
через аннулярный слои цитоплазмы ограничен и что ско-
рее всего транспорт метаболитов идет через десмотрубочки,,
а следовательно, через ЭР. Если допустить, что ЭР представля-
ет собой сеть из трубочек, то для межклеточного транспорта
необходимо, чтобы метаболиты и в тех и в других клетках про-
ходили через мембраны ЭР. Кажется маловероятным, чтобы
обмен метаболитов через плазмодесмы был свободным или не-
избирательным. Время от времени Эверт и др. наблюдали при-
крепление ЭР к наружной оболочке хлоропластов, поэтому они
предположили, что хлоропласты мезофилла н хлоропласты об-
кладки проводящих пучков соединены друг с другом через ЭР.
Во всяком случае, наружная мембрана хлоропластов мезофил-
ла Сграстений легко проницаема для таких веществ, как сор-
битол и сахароза, тогда как внутренняя мембрана ограничива-
ет транспорт метаболитов (разд. 12.5). Отсюда следует, что
ЭР, соединенный с наружной мембраной хлоропласта, непо-
средственно контактирует с веществами, растворенными в ци-
тозоле, но не с веществами, находящимися в хлоропласте.
В ходе Сгцикла межклеточный транспорт метаболитов дол-
жен быть направлен от места их синтеза в одной клетке к ме-
сту их использования в соседней клетке. Если транспорт С^-кис-
лот определяется только скоростью их диффузии, то можно
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
379
рассчитать градиент концентрации между клетками мезофилла
и клетками обкладки проводящих пучков, необходимый для
обеспечения потока вещества в соответствии с наблюдаемой
скоростью фотосинтеза. Для этого, во-первых, надо знать число
плазмодесм па единицу поверхности клетки и размер пор,
сквозь которые проходят плазмодесмы, и, во-вторых, при рас-
чете надо исходить из допущения о том, что через поры осу-
ществляется свободное передвижение метаболитов. Используя
эти расчеты, пришли к заключению, что градиент концентрации
Сд-аминокислот между клетками мезофилла и обкладки прово-
дящих пучков должен составлять ~ 10—30 мМ. Расчеты
активного пула Сд-кислот тоже дают цифры ~40—60 мМ при
условии, что та или иная Сд-кислота полностью находится в ци-
топлазме клеток мезофилла. Вместе с тем малат и аспартат в
концентрации 1 —10 мМ ингибируют ФЕП-карбоксилазу и за-
висимую от пирувата фиксацию СО2 в препаратах клеток мезо-
филла Сд-растений (разд. 12.8). Остается неясным, так же чув-
ствительна ФЕП-карбоксилаза in vivo или же расчеты дают
завышенную величину пула Сд-кислот в цитозоле мезофилла.
Все рассмотренные выше соображения лишний раз подчерки-
вают, что современные представления о межклеточном транс-
порте метаболитов Сд-цикла в основном исходят из умозритель-
ных предпосылок.
Источники данных: i[16, 24, 26, 43, 50].
12.7. Сд-метаболизм, сопряженный с циклическим,
псевдоциклическим и нециклическим
фотофосфорилированием
В хлоропластах мезофилла С4-р астений содержится пиру-
ват,ортофосфат— дикиназа, которая вместе с адеиилаткиназой
использует две молекулы АТР для превращения пирувата в
ФЕП (разд. 11.1). При скорости фотосинтеза в листе
200 мкмоль-(мгХл)-1-ч-1 скорость превращения пирувата в
ФЕП в расчете па единицу Хл в ткани мезофилла составляет
~400 мкмоль - (мгХл)~1-ч~1. Это соответствует скорости по-
требления АТР в клетках мезофилла ~800 мкмоль - (мгХл)-1-
-ч~!. Очевидно, метаболизм пирувата в хлоропластах мезо-
филла предъявляет высокие требования к системе образования
АТР при фотофосфорилировании. Это позволяет создать иде-
альную систему для оценки потенциальных возможностей цик-
лического, псевдоциклического и нециклического фотофосфори-
лирования в интактных органеллах, так как расход АТР очень
велик, а нециклический перенос электронов можно контроли-
ровать, добавляя оксалоацетат или ФГК.
380
ГЛАВА 12
В экстрактах, полученных из протопластов мезофилла
С4-растений, продуктивность фотофосфорилирования можно»
оценить косвенным путем, измерив зависимое от пирувата свя-
зывание 14СО2. При фиксации СО2, сопряженной с активностью
пируват,ортофосфат —дикиназы, пирофосфатазы, аденилатки-
назы и ФЕП-карбоксилазы, на связывание каждой молекулы
СО2 затрачиваются две молекулы АТР (разд. 11.1). Такую сте-
хиометрию проверили экспериментально, измерив скорость зави-
симой от пирувата фиксации СО2 в темноте в присутствии
АТР [она составляет 40—70 мкмоль-(мгХл)-1 -ч-1]. АТР по-
глощается хлоропластами с помощью переносчика адениловых
нуклеотидов, причем на связывание каждой молекулы СО2 тра-
тится две молекулы АТР.
Потенциальные возможности циклического фотофосфорили-
рования можно оценить, измерив зависимую от пирувата фик-
сацию СО2 на свету в экстрактах протопластов на фоне 2% О2
(чтобы подавить псевдоциклическое фотофосфорилирование за
счет реакции Мелера). Концентрация Хл в пробе при этом
должна быть достаточно низкой, чтобы свести к минимуму на-
копление оксалоацетата. В противном случае оксалоацетат бу-
дет взаимодействовать с NADPH через NADP-малатдегидроге-
назу и таким образом индуцировать нециклический перенос
электронов. В присутствии 2% О2 скорость циклического фото-
фосфорилирования составляет ~60—80 мкмоль-(мгХл)-1 -ч-1.
Такое фотофосфорилирование чувствительно к антимицпну А
(10 мкМ) и ДБМИБ (6 мкМ), но ие чувствительно к ДХММ
(предполагаемые участки ингибирования представлены па
рис. 5.2). На самом деле ДХММ в низкой концентрации
(0,2 мкМ) сильно стимулирует циклическое фотофосфорилиро-
вание, очевидно потому что этот реагент предотвращает чрез-
мерное восстановление (разд. 5.4).
Псевдоциклический перенос электронов служит главным ис-
точником АТР тогда, когда концентрация О2 в. газовой фазе
очень велика, циклический перенос электронов заблокирован
антимицииом А, а содержание Хл в пробе достаточно мало,
чтобы свести к минимуму накопление оксалоацетата. Скорость
лсевдоциклического фотофосфорилирования в атмосфере с 21%
О2 близка по величине к скорости циклического фотофосфори-
лирования [60—80 мкмоль АТР-(мгХл)-1-ч-1]. Полумакси-
мальная скорость лсевдоциклического фотофосфорилирования
достигается при [О2] ~30% (рис. 5.2 и 5.3).
Нециклическое фотофосфорилирование служит главным ис-
точником АТР для зависимой от пирувата фиксации СО2 в при-
сутствии 0,5 мМ оксалоацетата. Скорость этого процесса нд
фоне 2°/о О2 (который подавляет псевдоциклическое фотофос-
форилирование) и антимицина А (который блокирует цикличе-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
381
ское фотофосфорилирование) составляет 500—700 мкмоль
АТР- (мгХл)~[• я-1. Как псевдоциклическое, так и нецикличе-
ское фотофосфорилирование ингибируется ДХММ и ДБМИБ
(рис. 5.2 и 5.3).
Поскольку С4-растения расходуют иа фиксацию одной мо-
лекулы СО2 5 молекул АТР и 2 молекулы NADPH (разд. 11.6),
следует ожидать, что некоторая часть АТР поступает в резуль-
тате циклического и псевдоциклического фотофосфорилирова-
ния. При отношении Р/2е-=1—2 иа 1 молекулу NADPH при-
ходится 2—4 молекулы АТР (разд. 5.14). Зная фотохимиче-
ские характеристики хлоропластов мезофилла и обкладки про-
водящих пучков (разд. 1L7), можно предсказать потенциаль-
ные возможности циклического фотофосфорилирования у
Сграстеиий, относящихся к разным подгруппам, ио ничего
нельзя сказать об относительном вкладе циклического и псев-
доциклического фотофосфорилирования. Оба этих процесса
могут служить важным источником АТР в клетках мезофилла,
особенно тогда, когда главным продуктом С4-фото синтез а яв-
ляется аспартат, так как для реакции Алаиип+СО2—^Аспар-
тат требуется АТР (разд. 11.1).
Источники данных: [15, 30—33].
12.8. Регуляция ферментов С4-цикла
а) Освещение этиолированной ткани
В этиолированной ткани активность ферментов, которые
участвуют в фотосинтезе, зачастую очень невелика, но она воз-
растает во много раз после позеленения такой ткани. Это-
справедливо прежде всего для таких ферментов С4-цикла, как
пируват,ортофосфат — дикиназа, адеиилаткиназа, ФЕП-карбок-
силаза, аланин- и аспартатаминотрансферазы и NAD-МДГ (де-
карбоксилирующая). У кукурузы и сорго такое индуцируемое
светом увеличение активности ФЕП-карбоксилазы, пируват,ор-
тофосфат— дикиназы и аденилаткииазы ингибируется и хлор-
амфениколом, и циклогексимидом (двумя ингибиторами белко-
вого синтеза). Это говорит о том, что увеличение активности
ферментов в процессе позеленения связано скорее с синтезом
белка, чем с активацией светом пр ед существующих белков.
Активность изоцитратдегидрогеназы и глюкозо-6-фосфат — де-
гидрогеназы, не принадлежащих к числу ферментов фотосин-
теза, не возрастает после освещения этиолированной ткани,
т. е. в С4-цикле участвуют вполне определенные ферменты. Так,
например, активность ФЕП-карбоксилазы в этиолированной
ткани невелика, а в ткани зеленого листа появляется другая
изоформа этого фермента. По-видимому, уровень изофермен-
382
ГЛАВА 12
тов С4-цикла довольно низок и в тех листьях, функция кото-
рых не связана с С4-фотосинтезом.
Активность ферментов С4-цикла в фотосинтезирующих
листьях регулируется несколькими способами. К ним относятся:
световая модуляция ферментативной активности; регуляция ал-
лостерических ферментов их эффекторами (разд. 12.8, в); регу-
ляция уровнем содержания субстрата, величиной отношения
NADPH/NADP+, адениловыми нуклеотидами, pH и двухвалент-
ными катионами.
•б) Световая активация фотосинтетических ферментов
Два фермента С4-цикла, пируват, ортофосфат — дикииаза и
NADP-малатдегидрогеназа, активируются, когда листья нахо-
дятся на свету, и инактивируются, когда листья попадают в тем-
ноту.
Пируват --->- ФЕП -> Оксалоацетат -> Малат
2АТР СОа NADPH---------> NADP+
‘Степень активации ферментов зависит от интенсивности света;
для полной активации необходимо сильное освещение. Чтобы
максимально активировать пируват,ортофосфат — дикиназу пе-
ред ее экстракцией из клеток, Хэтч и Слэк помещали листья
С4-растений на солнце или в эквивалентные условия освещения.
Полагают, что световая активация NADP-малатдегидрогеиа-
зы обусловлена восстановлением дисульфидных групп этого бел-
ка. Точная природа данного восстановления не выяснена, но
можно рассмотреть простейшую из возможностей. Восстанови-
тельная сила для такого превращения, вероятно, поступает из
•системы нециклического переноса электронов. Восстановитель
(модулятор), который образуется в терминальной части цепи
нециклического переноса электронов, затем передает свои элект-
роны на фермент (рис. 12.24).
Сходным образом за счет восстановления дисульфидных
групп белка активируются на свету и NADP-малатдегидрогена-
за, и ферменты ВПФ-цикла (NADP-триозофосфатдегидрогеназа,
•фруктозо-1,6-бисфосфатаза, седогептулозо-1,7-бисфосфатаза и
фосфорибулокиназа) у С3-растений (гл. 9). У этих растений в
восстановлении дисульфидных групп, по-видимому, участвует
растворимый низкомолекулярный белок, называемый тиоредок-
сином, который в свою очередь восстанавливается ферредоксин-
тиоредоксин-редуктазой. In vitro активацию можно вызывать
неферментативно, восстанавливая тиоредоксин дитиотрейтолом.
Для активации пируват,ортофосфат — дикиназы необходимы
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ Щ-ФОТОСИНТЕЗЕ
383
н«Т|«ый Акги..
Ьмкж В,;„ок
Рис. 12.24. Восстановление дисульфидных групп белков.
Pi и высокомолекулярный термолабильный белок. Этот процесс
ингибируется АМР и ADP. Для инактивации фермента требует-
ся ADP. Активация in vitro может происходить и в отсутствие
восстанавливающих агентов. Это свидетельствует о том, что для
активации этого белка не требуется восстановления дисульфид-
ных групп (Т. Stigiyama, М. D. Hatch, личное сообщение). In
vivo активация может быть отчасти опосредована превраще-
нием АМР и ADP в АТР в хлоропластах иа свету.
в) Регуляция ФЕП-карбоксилазы
Для некоторых ферментов С4-цикла характерны другие ти-
пы регуляции. Хорошо изучены кинетические свойства и регуля-
ция ФЕП-карбоксилазы у С4-растеиий. Как и для бактериально-
го фермента, для ФЕП-карбоксилазы С4-растений характерна
S-образная (сигмоидная) кинетическая кривая зависимости ак-
тивности фермента от [ФЕП], т. е. она обладает аллостериче-
скими свойствами, но в отличие от бактерий у растений несколь-
ко другие эффекторы этого фермента. У аллостерических фер-
ментов метаболиты связываются ие в центре связывания суб-
страта, а с другими участками молекулы. Это приводит к кон-
формационным изменениям белка, которые либо усиливают в
случае активатора, либо ослабляют в случае ингибитора срод-
ство фермента к одному или нескольким из его возможных суб-
стратов. Так, например, малат и аспартат являются отрицатель-
ными эффекторами ФЕП-карбоксилазы у С4-растеиий, они вы-
зывают увеличение Км для ФЕП. Глюкозо-6-фосфат действует
как аллостерический активатор, повышая сродство фермента к
ФЕП (величина Км уменьшается). Эффекторы практически по-
влияют на Vmax (рис. 12.25). Сильным ингибитором ФЕП-кар-
боксилазы у С4-растений является оксалоацетат.
S-Образная /форма кинетических кривых для какого-либо-
фермента свидетельствует о том, что у него не один центр свя-
384
ГЛАВА 12
Рис. 12.25. Кинетика ФЕП-карбоксилазной реакции у С*-растеиий без эффек-
тора (контроль) и в присутствии положительного (2 мМ Г6Ф) или отрица-
тельного (1 мМ аспартат) эффектора. Внизу справа изображен график в
координатах Хилла. Обратите внимание на то, что активность фермента
сильнее всего контролируется эффекторами при концентрациях субстрата,
далеких от насыщающих (Huber, Edwards, 1975b).
зывания субстрата, а несколько. По мере увеличения концентра-
ции субстрата, когда фермент (Взаимодействует более чем с
«одной молекулой субстрата, взаимодействие фермента со сле-
дующей молекулой намного облегчается. Для определения сте-
пени кооперативности при связывании молекул субстратов ис-
пользуют графики Хилла (зависимость lg(t»/V—v) от IgS, где
v — скорость, V=Vmax, 5 — концентрация субстрата]. Наклон
•кривой на графике Хилла (п) соответствует минимальному чис-
лу участков связывания молекул субстрата на белке (т. е. при
п=2 число таких участков равно по меньшей мере двум). При
.обычной кинетике Михаэлиса — Ментен п = 1.
Аллостерические ферменты с кооперативной кинетикой могут
терять свои регуляторные свойства (например, в процессе очи-
стки, при изменении температуры, pH и т. д.), но сохранять при
этом свои каталитические свойства и давать типичную кинетику
Михаэлиса — Ментен. Отсутствие аллостерического контроля
или «S-образной кинетической кривой не может служить строгим
доказательством того, что фермент полностью утратил способ-
ность к такого рода регуляции.
Сочетание аллостерического контроля с немихаэлисовой ки-
нетикой обеспечивает максимальную степень регуляции эффек-
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СЦ-ФОТОСИНТЕЗЕ
385
торами при лимитирующей концентрации субстрата. Vmax для
ФЕП-карбоксилазы у С4-растений в несколько раз выше, чем
наблюдаемая скорость фотосинтеза в листе, In vivo' содержание
ФЕП, вероятно, лимитирует скорость реакции, а это создает все
условия для оптимальной регуляции аллостерическими эффек-
торами.
В случае ФЕП-карбоксилазы из С4-растений максимальная
регуляция фермента аспартатом, малатом и глюкозо-6-фосфа-
том наблюдается при сравнительно низких значениях pH и при
низкой концентрации Mg2+ (столь же низкой, как концентрация
ФЕП). In vivo активность ФЕП-карбоксилазы контролируется
величиной pH, уровнем Mg2+, ФЕП, С4-кислот и глюкозо-6-фос-
фата. Скорость использования С4-кислот для передачи углерода
в ВПФ-цикл в свою очередь может влиять на карбоксилирова-
ние в С4-цикле, вызывая ингибирование ФЕП-карбоксилазы по
типу обратной связи. Глюкозо-6-фосфат является ключевым ме-
таболитом глюконеогенеза, ведущего к синтезу сахарозы, крах-
мала и других макромолекул. В клетках мезофилла С4-растений
ФЕП может использоваться как для синтеза глюкозо-6-фосфата,
так и для образования С4-кислот. Активация ФЕП-карбоксила-
зы под действием глюкозо-6-фосфата может препятствовать
утечке углерода из С4-цикла, обусловленной превращением
ФЕП в глюкозо-6-фосфат,
ФЕП-карбоксилаза обычно встречается у всех растений, ио
активность этого фермента в листьях С4- и САМ-растеыий во
много раз выше, чем у Сз-растений. Кинетические константы
этого фермента для ФЕП и Mg2+ варьируют в зависимости от
его источника (Сз-растения, С4-растения, CAM-растения, нефо-
тосинтезирующие ткани).
При рассмотрении различных карбоксилаз важно, во-первых,
в какой форме фиксируется неорганический углерод (НСО3~
или свободная СО2) и, во-вторых, какова величина Км для это-
го субстрата. Это имеет особое значение для тех работ, в кото-
рых сравнивают С3- и С4-фотосинтез или рассматривают роль
ФЕП-карбоксилазы и РуБФ-карбоксилазы в первичной фикса-
ции атмосферной СО2. Все эти вопросы обсуждаются в гл, 14.
г) Регуляция декарбоксилирующих ферментов
Для активации всех трех ферментов, которые декарбоксили-
руют С4-кислоты, необходимы двухвалентные катионы: для
NADP-МДГ Mg2+ .или Мп2+, для NAD-МДГ Мп2+ и для ФЕП-
карбоксикиназы Мп2+, а еще лучше Mn2+-|-Mg2+. Декарбоксили-
рование, катализируемое NADP- или NAD-МДГ, подавляется
бикарбонатом (не исключено, что активной формой является
25-234
386
ГЛАВА 12
СОг). Это позволяет регулировать активность Ci-цикла in vivo.
Высокая [СО2] в Ингибирует де- Увеличивает кои- Ингибирует
клетках обкладки-> карбоксилирова- ->цептрацию С<-кпс- ФЕП-кар-
проводящих пуч- ние и образование лот боксплазу
ков Сз-предшествен-
иика
Такой цикл должен функционировать со скоростью, доста-
точной для поддержания повышенной [СО2] в клетках обкладки
проводящих пучков. Излишне высокая скорость этого процесса
может привести к значительной потере СО2 и к непроизводитель-
ному расходу энергии. Дополнительный контроль цикла возмо-
жен на уровне NADP-МДГ, поскольку этот фермент ингибиру-
ется продуктами декарбоксилирования (пируватом и NADPH).
Относительная концентрация NADP+ и NADPH тоже регули-
рует активность этого фермента. Декарбоксилирование с уча-
стием NADP-МДГ зависит ,от восстановительной фазы фотосин-
теза, которая обеспечивает регенерацию образующегося NADP+.
Малат + NADP+ ---> Пируват 4- NADPH 4- Н+ -ф- СО2 (12.1)
РуБФ-1-СОз----> 2ФГК (12.2)
2ФГК ----> 2(1,3-ДФГК) --> 2 Триозофосфат (12.3)
2АТР ---> 2ADP 2NADPH + 2Н+ ----------> 2NADP+
Для восстановления ФГК в ВПФ-цикле необходимо в 2 раза
больше NADPH, чем его образуется в результате декарбоксили-
рования малата. Это гарантирует постоянную доступность
NADP1" для декарбоксилирующего фермента. Хлоропласты об-
кладки проводящих пучков у NADP-МДГ-растеиий характери-
зуются низким уровнем нециклического переноса электронов
(разд. 11.7). Чрезмерно большой фотохимический потенциал об-
разования NADPH в таких клетках был бы невыгоден из-за
конкуренции за NADP+ с МДГ (декарбоксилирующей) и в ко-
нечном итоге ограничения декарбоксилирования.
NAD-МДГ (декарбоксилирующая) (из Atriplex sponglosa и
Amaranthus edulis) обладает аллостерическими свойствами и
проявляет кинетическую кооперативность, при этом положитель-
ными эффекторами фермента служат ацетил-СоА, Со А и фрук-
тозо-1, 6-бисфосфат, а отрицательным эффектором является
НСО3 . В обычных условиях пируват метаболизируется до аце-
тил-СоА-ф-СОг в ЦТК. После этого ацетил-СоА и оксалоацетат
соединяются вместе с образованием изоцитрата. Это может при-
вести к тому, что метаболиты С^цикла полностью окислятся в
ходе дыхания. Здесь существенно то, что NAD-МДГ (декарбок-
силирующая) активируется ацетил-СоА, а это способствует об-
ращению цикла вспять (Оксалоацетат—>-Малат) и, кроме того,
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С.,-ФОТОСИНТЕЗЕ
387
обеспечивает циклическое поступление пирувата (продукта де-
карбоксилирования Сд-кислот) обратно в клетки мезофилла.
Декарбоксилирование, идущее с участием ФЕП-карбоксики-
назы, характеризуется высокой специфичностью в отношении
АТР по сравнению с другими нуклеозидтрифосфатами. Фрукто-
зо-1,6-бисфосфат, ФГК и ДГАФ (1—5 мМ) частично ингибируют
этот фермент.
д) Выводы
Ряд ферментов С4-цикла ,инактивируется в темноте. Это
препятствует бесполезному челночному обращению СОз в систе-
ме за счет энергии дыхания. На свету ферменты С4-цикла акти-
вируются промежуточными продуктами других метаболических
путей, в которые «оттекают» метаболиты С4-цикла [т. е. ФЕП-
карбО'Ксикиназа активируется глюкозо-6-фосфатом, а NAD-МДГ
(декарбоксилирующая)—ацетил-Со А]. ФЕП и пируват могут
метаболизироваться и Другими путями, участвуя в реакциях
глюконеогенеза и дыхания.
Некоторые метаболиты ВПФ-цикла влияют на активность
С4-цикла, а связанные с переходом свет — темнота изменения
[Mg2+] и pH в определенных компартментах клетки влияют на
активность определенных ферментов. Наши представления о
регуляции ферментов в цикле, который охватывает две клетки
и их многочисленные компартменты, весьма и весьма спорны.
Прежде всего следует быть очень осторожными при рассмотре-
нии возможной физиологической роли любого из факторов, ко-
торые влияют на активность фермента in vitro. Вполне возмож-
но, что некоторые из таких факторов вообще не имеют никакого
физио л огического значени я.
Источники данных: [2, 5, 17, 18, 21—23, 29, 36, 48, 51, 52, 55,
56].
12.9. Ассимиляция азота:
сравнение С4- и С,грастений
Для восстановления и включения азота нитрата в аминокис-
лоты листьев растений необходима энергия, поставляемая хло-
ропластами. Сначала нитрат должен превратиться в аммиак
под действием иитратредуктазы и нитритредуктазы [уравнения
(12.4) и (12.5)]. В течение многих лет считали, что в превраще-
нии аммиака в аминный азот участвует глутаматдегидрогеназа
[уравнение (12.6)]. В митохондриях присутствует NAD-глута-
матдегидрогеназа, а в хлоропластах — NAD (Р)-глутаматдегид-
рогеназа. Сравнительно недавно был открыт другой возможный
25*
388
ГЛАВА 12
механизм, который включает в себя глутаминсинтетазу — глу-
таматсинтазу [уравнения (12.7) и (12.8)], и были получены убе-
дительные доказательства тому, что эти ферменты участвуют в
основном пути ассимиляции азота. Очень часто' главным пер-
вичным продуктом ассимиляции 15NO3 или 15МЩ в фотосинтези-
рующей ткани является глутамин, после чего метка появляется
в глутамате в полном соответствии с той последовательностью
включения аммиака, которая описывается уравнениями (12.7) и
(12.8). Метионинсульфоксимин (МСО) ингибирует глутаминсин-
тетазу, а азасерин подавляет реакции переноса амидной группы,,
в том числе п глутаматсинтазную реакцию. В ряде работ дей-
ствие этих ингибиторов на превращение азота (у сине-зеленых
водорослей), нитрита или аммиака в глутамат рассматривалось
как доказательство того, что ассимиляция азота идет по глут-
аминсинтетаза-глутаматсинтазному пути. Так, например, мета-
болическое превращение аммиака или нитрата+2-оксоглутарат
в глутамат в изолированных хлоропластах гороха (С3-растение)
ингибируется МСО и азасерином. В присутствии азасерина ам-
миак или нитрат включается в глутамин, а не в глутамат, что
хорошо согласуется с действием ингибитора на глутаматсиитазу.
Км для аммиака у глутаматдегидрогеназы из высших растений
примерно на 2 порядка больше, чем аналогичная величина у
глутаминсинтетазы. При сравнительно низкой концентрации
аммиака это делает глутаминсинтетазную реакцию более веро-
ятной.
NO3 + NADI-I + Н+--------->- NO2 + NAD+ (нитратредуктаза) (12.4)
NO2~ + 6Фд (восстановленный) ------->
——> NH4+-|-6Фд (окисленный) (нитритредуктаза) (12.5)
NH3 NAD(P)H -|- Н+ + 2-Оксоглутарат ----------->-
----»- NAD(P)+ + Глутамат Н2О (глутаматдегидрогеназа) (1 2,6)
NH3 -р АТР + Глутамат ---------------------->-
----Глутамин -|- ADP + Р; (глутаминсинтетаза) (12.7)
Глутамин 2-Океоглутарат + 2Фд (восстановленный) —*•
—► 2 Глутамат 2Фд (окисленный) (глутаматсиитазя) (12.8)
Неизвестно, существует ли прямая связь между ассимиля-
цией азота и Сгфотосинтезом. Следует отметить, что для превра-
щения нитрата в аминный азот необходима энергия, а локали-
зация этих процессов в листьях С4-растений интересна в связи
с существованием фотосинтезирующих клеток двух типов. При
изучении представителей всех трех подгрупп Сграстений пока-
зано, что в клетках мезофилла локализовано свыше 80% актив-
ности ферментов, восстанавливающих нитрат до аммиака (нит-
ратредуктаза, нитритредуктаза). По данным ряда работ, глут-
ИНТЕГРАЦИЯ ФУНКЦИЙ ПРИ С,-Ф0Т0СИНТЕЗЕ
389
аминсинтетаза и глутаматсинтаза находятся главным образом'
в клетках мезофилла, тогда как, согласно другим работам, ак-
тивность этих ферментов достаточно велика в клетках обоих
типов. Но, даже если правы авторы последних работ, клетки
мезофилла следует рассматривать как место первичной ассими-
ляции азота у Си-растений.
Путь превращения нитрата в аминный азот в клетках мезо-
филла; по-видимому, одинаков и у Сз-, и у С^растений. Нитрат-
редуктаза локализована в цитозоле или в наружной мембране
хлоропласта. И в том и в другом случае восстановительная сила
должна поступать из хлоропласта за счет челночного механиз-
ма. Возможно несколько вариантов такого механизма
(рис. 12.26).
Первый (Л) из них заключается в том, что восстановитель-
ная сила переносится в цитозоль при участии малатдегидроге-
назы; при этом переносчик дикарбоновых (кислот облегчает
транспорт С4-кислот через оболочку хлоропластов (разд. 8.24 и
12.5). Второй (£) и третий (В) варианты состоят в том, что экс-
порт ДГАФ из хлоропластов сопряжен с поглощением ФГК при
участии переносчика фосфата (разд. 8.23). И в том и в .другом
случае ДГАФ окисляется до ФГК в цитоплазме, в результате
чего возникает восстановительная сила. В случае необратимой
реакции, катализируемой NADP-триозофосфатдегидрогеназой
(вариант Б) образуется ФГК. и возникает восстановительная си-
ла [уравнение (12.9)]:
ФГА NADP+ —> 3-ФГК NADPH + Н+ (12.9)
При обратимой реакции, катализируемой NAD-триозофосфат-
дегидрогеназой, продуктом окисления ФГА является 1,3-ДФГК.
Следующее за этим образование ФГК под действием фосфогли-
цераткипазы завершается синтезом АТР, который может исполь-
зоваться в других процессах метаболизма, идущих в цитозоле,
например в синтезе сахарозы (гл. 9),
Другие ферменты обмена нитрита и глутамата локализованы
в хлоропластах (рис, 12.27). [Некоторые глутаминсиитетазы ло-
кализованы в цитозоле, а в митохондриях присутствует NAD-
глутаматдегидрогеиаза. Функция этих ферментов в метаболиз-
ме соединений азота пока неизвестна,] Для восстановления NC>2~
до ИНД (эквивалентно 3 NADPH) под действием иитритредук-
тазы необходимы ферредоксин и 6 е~. Для превращения одной
молекулы нитрата в аминный азот под действием глутаминсин-
тетазы —глутаматсинтазы требуется 1 АТР и 5 NADPH (в экви-
валентах восстановительной силы). А иа обмен нитрата, связан-
ного с его1 превращением в аминный азот под действием глута-
матдегидрогеназы, необходимо потратить эквивалентное количе-
ство восстановительной силы, но совершенно не требуется АТР.
390
ГЛАВА 12
Рис. 12.26. Способы челночного переноса восстановительной силы из хлоро-
пластов для восстановления нитрата в цитозоле. А. Челночный механизм
обмена оксалоацетат — малат через малатдегидрогеназу. Б. Челночный меха-
низм обмена, включающий необратимую реакцию, катализируемую NADP-
триозофосфатдегидрогеназой. В. Челночный механизм обмена, включающий
обратимую реакцию, катализируемую NAD-триозофосфатдегидрогеназой.
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ СгФОТОСИНТЕЗЕ
391
Рис. 12,27. Возможная схема метаболизма нитрита в хлоропластах мезофил-
ла С3- н С^-растений. 1 — нитратредуктаза; 2— нитритредуктаза; 3—глута-
мипсиптетаза, ингибируемая метионинсульфоксимином (МСО); 4— глутамат»
синтаза, ингибируемая азасерииом; 5 — трансаминаза; 6 — NADP-глутамат-
дегидрогеназа (штрихами показан возможный вспомогательный путь);
2-ОГ — 2-оксоглутарат (Rathnam, Edwards, 1976; Lea, Miflin, 1978; Moore,
Black, 1979).
Рассмотрим энергетические затраты на ассимиляцию азота у
Сграстений в связи с энергетикой (^-фотосинтеза (для сравне-
ния с Сз-растениями см. разд. 13.6). При ассимиляции СО2 у С4-
растений иа .связывание одной молекулы СО2 расходуется
5ATP/2NADPH (разд. 11,6). Содержание азота в листьях со-
ставляет примерно 6% содержания углерода (при расчете
атом: атом), так что можно, допустить, что и скорость ассими-
ляции азота составляет 6% скорости фотосинтеза. Тогда соот-
ветственно расход энергии при фиксации одной молекулы СО2 в
Сд-цикле и ВПФ-пути плюс расход энергии при превращении
нитрата в аминный азот в листьях будет равен
(5АТР+ 2NADPI-I/CO,) +0,06 (1АТР + 5NADPH) = 5,1АТР/2,3NADPH
Полученный результат свидетельствует о том, что на ассимиля-
цию азота в процессе фотосинтетической ассимиляции углерода
392
ГЛАВА 12
расходуется — 13% восстановительной силы. Синтез белков из
аминокислот и синтез таких углеводов, как сахароза и крахмал,
требуют дополнительных затрат энергии.
Ассимиляция нитрата может быть еще более расточительной
.для углеродного баланса растений. В процессе поглощения и
восстановления нитрата образуются ионы гидроксила, которые
могут быть нейтрализованы органическими кислотами (обычно
это малат). Для этого требуется потратить по меньшей мере
одни эквивалент карбоксила на каждую молекулу нитрата, вос-
становленного’ до аммиака. При стехиометрии реакции Яблоч-
ная кислота+2КЫО3'—нК2Малат-|-2 (—-NHs) на каждую молеку-
лу восстановленного нитрата необходимо перевести в органиче-
ские соединения 2 атома углерода (разд. 16.5). Большая часть
малата, запасенного в растительных вакуолях, может использо-
ваться для компенсации защелачивания, связанного с превра-
щением нитрата в азот аминокислот. Смысл этого состоит в сле-
дующем: чем. больше белка синтезирует растение, используя
нитрат в качестве источника азота, тем больше углерода оно
должно превратить в анионы органических кислот.
В ряде исследований показано, что ферменты ассимиляции
азота у С^растений первично локализованы в клетках мезофил-
ла. Поэтому восстановительная сила, возникающая в клетках
мезофилла, может быть использована для синтеза малата, вос-
становления ФГК, поступившей из клеток обкладки проводящих
пучков, восстановления нитрата и его превращения в азот ами-
нокислот. По этой же причине восстановительная сила, возни-
кающая в клетках обкладки проводящих пучков (в результате
декарбоксилирования малата или фотохимических реакций),
первично используется в ВПФ-цикле.
Обнаружено, что около 80% активности NAD-глутаматде-
гидрогеназы в листьях С4-растений находится в митохондриях
клеток обкладки проводящих пучков. В каких реакциях (амини-
рования или дезаминирования) участвует этот фермент, пока не
выяснено; отмечено лишь, что для глутаматдегидрогеназ харак-
терны очень высокие значения Км для аммиака. У С3-растений
декарбоксилирование глицина (2 Глицин—^Серин+СОг+ПНз)
в гликолатном пути происходит в митохондриях. Считают, что
по меньшей мере часть реакций гликолатного пути у С4-расте-
ний протекает в клетках обкладки проводящих пучков
(разд. 13.9). Аммиак, высвобождающийся в ходе декарбоксили-
рования глицина, может вновь ассимилироваться либо за счет
глутаматдегидрогеназы митохондрий, либо за счет глутаминсии-
тетазы хлоропластов обкладки проводящих пучков.
Источники данных: {1, 20, 37, 39—42, 44].
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ Сд-ФОТОСИПТЕЗЕ
393
12.10. Синтез крахмала и сахарозы у С4-растений
Запасать крахмал могут хлоропласты и мезофилла, и об-
кладки проводящих пучков, ио первично этот процесс происхо-
дит в хлоропластах обкладки проводящих пучков. Ранее пред-
полагали, что эти хлоропласты выполняют функцию амилоплас-
тов, ио сейчас стало очевидно, что они участвуют в очень важ-
ном карбоксилировании при фотосинтезе. У некоторых растений,
таких, как кукуруза, крахмал обнаруживается и в хлоропластах
мезофилла, но только тогда, когда растения находятся при по-
стоянном освещении. У кукурузы и ряда других исследованных
Сграстеиий ферменты метаболизма крахмала (глюкозо-1-фос-
фат — аденилилтрансфераза, АОРглюкоза-крахмалсинтетаза и
фосфорилаза) в основном находятся в клетках обкладки прово-
дящих пучков. Способность хлоропластов мезофилла синтезиро-
вать крахмал может сильно различаться у разных видов. В ка-
ком процессе участвует фосфорилаза: в синтезе крахмала или в
его распаде, ие известно.
Де Фекет и Впвег обнаружили, что изолированные клетки
обкладки проводящих пучков кукурузы синтезируют крахмал
па свету в том случае, когда в среде присутствует глюкозо-1-
фосфат (ПФ), а не СОа. На этом основании они пришли к вы-
воду, что1 для синтеза крахмала in vivo необходимы клетки ме-
зофилла, которые обеспечивают поступление ПФ. /Однако от-
сюда отнюдь ие следует, что СОг превращается в гексозофосфа-
ты в клетках мезофилла. Изолированные клетки обкладки про-
водящих жучков у кукурузы включают 14СОа прежде всего в
ФГК, хотя некоторая часть метки появляется и в крахмале.
Скорость фиксации СО2 при этом сравнительно невелика, так
как из-за отсутствия фотосистемы II ограничено поступление
восстановительной силы. In vivo восстановительная сила для
синтеза триозофосфатов в этих клетках поступает от малата под
действием NADP-МДГ (декарбоксилирующей). Поэтому вполне
логично', что изолированные клетки обкладки проводящих пуч-
ков способны синтезировать крахмал из ДГАФ или ПФ, а не
из ФГК или СОа (разд. 11.7).
Ферменты синтеза сахарозы, глюкозо-1-фосфат — уридилил-
трансфераза, сахарозосиитетаза и сахарозофосфатфосфатаза, у
нескольких видов С4-растеиий были обнаружены как в клетках
мезофилла, так и в клетках обкладки проводящих пучков. У не-
которых растений, таких, как кукуруза или росичка кровяная,
большая часть этих ферментов находится в клетках мезофилла.
Даунтон и Хоукер предположили, что клетки мезофилла у куку-
рузы являются основным местом синтеза сахарозы.
Уровень фруктозобисфосфат-альдолазы (гл. 6) в клетках
обкладки проводящих пучков у некоторых С4-растений пример-
394
ГЛАВА 12
но на порядок выше, чем в клетках мезофилла. Если уровень
альдолазы в клетках мезофилла лимитирует метаболическое
превращение триозофосфатов в сахарозу, то гексозофосфаты
могут поступать из клеток обкладки проводящих пучков. На
каждую молекулу гексозофосфата, синтезированного под дей-
ствием альдолазы и далее превращенного в сахарозу, приходит-
ся, однако, по меньшей мере пять молекул гексозофосфата, син-
тезированного' в ходе регенерирующей стадии ВПФ-цикла, ко-
торая протекает в клетках обкладки проводящих пучков
(разд. 11.5). Поэтому не удивительно, что альдолаза в основном
обнаруживается в клетках юбкладки проводящих пучков. Гексо-
зофосфат, синтезируемый в таких клетках при участии фрукто-
зобисфосфат-альдолазы, далее используется либо в реакциях
восстановительной стадии ВПФ-цикла, либо для синтеза крах-
мала и отчасти сахарозы. Гексозофосфат в клетках мезофилла
используется в основном для синтеза сахарозы. К тому же вос-
становленный углерод может транспортироваться из мезофилла
в клетки обкладки проводящих пучков в виде гексозо- или трио-
зофосфата.
У Сз-растений концентрация Pi в цитозоле, по-видимому,
играет регуляторную роль в процессе передачи углерода на
синтез сахарозы (гл. 9). У С^-растений такие ферменты, участ-
вующие в последних стадиях синтеза сахарозы, как глюкозо-1-
фосфат — уридилилтрансфераза и сахарозофосфатсинтетаза, ло-
кализованы в цитоплазме. Поэтому синтез сахарозы, вероятно,
как и у Сз-растений, идет д цитоплазме. Уровень Pi в цитозоле
клеток обкладки проводящих пучков может влиять на скорость
выхода ФГК или триозофосфата из хлоропластов. В ходе даль-
нейших превращений ФГК или триозофосфатов в сахарозу, ко-
торые осуществляются в цитозоле клеток обоих типов, Pi снова
высвобождается и возвращается обратно в хлоропласты кле-
ток обкладки проводящих пучков (разд. 11.6). В хлоропластах
мезофилла. С4-растений Рг поглощается в обмен на ФЕП
(.разд. 12.5). У Сз-растений в первичном обмене на переносчике
фосфата участвуют Pi и ДГАФ, а выход ФГК в процессе фото-
синтеза незначителен. Следовательно, и содержание ФГК в ци-
тозоле может быть тоже относительно невелико. Предполагают,
что у Сграстений ФГК выходит да клеток обкладки проводя-
щих пучков и затем восстанавливается в хлоропластах мезофил-
ла: Поэтому уровень ФГК в цитозоле в процессе фотосинтеза
может быть сравнительно высок. Содержание ФГК в цитозоле
клеток мезофилла может частично контролировать скорость
синтеза сахарозы за счет обмена ФГК—ДГАФ в хлоропластах.
В отличие от условий, существующих в клетках С3-растений,
для цитозоля клеток мезофилла С4-растений характерен высо-
кий уровень Pj, что способствует обмену ФЕП и ингибированию
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С,-ФОТОСИНТЕЗЕ
395
синтеза сахарозы за счет конкурентного подавления поглоще-
ния ФГК хлоропластами.
Считают, что синтез сахарозы у С3-растений частично кон-
тролируется экспортом триозофосфатов из хлоропластов в обмен
на Pt, связанный с переносчиком фосфата. Поэтому в условиях
ограниченного содержания Pi в цитозоле может понижаться вы-
ход углерода из хлоропластов, уменьшаться скорость синтеза
сахарозы и усиливаться синтез крахмала. В некоторых тканях
манноза удерживает Pi в виде маннозофосфата и оказывает
стимулирующее действие на фотосинтетическое образование
крахмала у некоторых С3-растеиий (разд. 9.7). В отличие от
этого введение маннозы в ткань листа С4-растений на свету ин-
гибирует синтез крахмала (см. рис. 9.12). Хлоропласты обкладки
проводящих пучков обычно являются главным местом синтеза
крахмала у Сграстений. Можно ожидать, что снижение уровня
Pi в цитозоле клеток обкладки проводящих пучков (например,
в результате взаимодействия с маннозой) приведет к уменьше-
нию поступления ,ФГК или ДГАФ из хлоропластов и таким об-
разом вызовет стимуляцию синтеза крахмала, как и у С3-расте-
ний. Однако фотосинтез у Сграстений связан с Сд-циклом, а для
этого цикла необходимо, чтобы Pi поглощался хлоропластами
мезофилла в обмен на ФЕП (разд. 12.5). Поэтому захват Р; ци-
топлазмы маннозой в ткани листа С4-растений приводит к замет-
ному подавлению и фотосинтеза, и синтеза крахмала.
Источники данных: [3, 4, 7—10, 27, 53, 54].
Общая литература приведена в гл. 10.
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
1. Beevers L., Hageman R. Н. The role of light in nitrate metabolism in Higher
plants. In: Photophysiology (ed. A. C. Giese) 7, 85—113, New York, Acade-
mic Press, 1972.
2. Black С. C. Photosynthetic carbon fixation in relation to net CO2 uptake,
Ann. Rev. Plant Physiol., 24, 253—286 (1973).
3. Bucke C., Oliver I. R. Location of enzymes metabolizing sucrose and starcli
in the grasses Pennisetum purpureum and Muhlenbergla moniana, Planta,
122,45—52 (1975).
4. Chen T. M., Dittrich P., Campbell W. H., Black С. C. Metabolism of epider-
mal tissues, mesophyll cells, and bundle sheath strands resolved from matu-
re nutsedge leaves, Arch. Biochem. Biophys., 163, 246—262 (1974).
5. Coombs J. Interactions between chloroplasts and cytoplasm in C4 plants,
In: The Intact Chloroplast (ed. J. Barber), pp. 279—313, The Netherlands,
Elsevier, 1976.
6. Craig S., Goodchild. D. J. Leaf ultrastructure of Triodia irritans: A C4 grass
possessing an unusual arrangement of photosynthetic tissue, Aust. J. Bot.,
25, 277—290 (1977).
7. Davies D. R. Some aspects of sucrose metabolism. In Plant Carbohydrate
Biochemistry, (ed. J. B. Pridham), pp. 61—81, New York, Academic Press,
1974.
3 96 ГЛАВА 12
8. De Fekete M. A. R., Vieweg G. H. Starch metabolism: Synthesis versus
degradation pathways, in: Plant Carbohydrate Biochemistry (ed. J. B. Pri-
dham), pp. 127—144, New York, Academic Press, 1974.
9. Downton IF. J. S., Hawker J. S. Enzymes of starch and sucrose metabolism
in Zea mays leaves, Phytochemistry, 12, 1551—1556 (1973).
10. Edwards G. E., Black С. C. Photosynthesis in mesophyll cells and bundle
sheath cells isolated from Digitaria sanguinalis leaves. In: Photosynthesis
and Photorespiration, (eds. M. D. Hatch, С. B. Osmond, and R. 0. Slatyer),
pp. 153—168, New York, Wiley-Interscience, 1971.
11. Edwards G. E., Lee S. S., Chen T. M., Black С. C. Carboxylation reactions
and photosynthesis of carbon compounds in isolated bundle sheath cells of
Digitaria sanguinalis (L.) Scop, Biochem. Biophys. Res. Commun., 39,
389—395 (1970).
12. Edwards G. E., Huber S. C., Gutierrez M. Photosynthetic properties of plant
protoplasts. In: Microbial and Plant Protoplasts, (eds. J. F. Peberdy,
A. H. Rose, H. J. Rogers and E. C. Cocking), pp. 299—322, New York,
Academic Press, 1976a.
13. Edwards G. E., Huber S. C., Ku S. B., Gutierrez Л4., Raihnam С. К. M.,
Mayne В. C. Variation in photochemical activities in C4 plants in relation to
CO2 fixation. In: CO2 Metabolism and Productivity of Plants, (eds. R.FI. Bur-
ris and С. C. Black), pp. 83—112, Baltimore Maryland, University Park
Press, 1976b.
14. Edwards G. £., Huber S. C. C4 metabolism in isolated cells and protoplasts.
In: Encyclopedia of Plant Physiology, New series, Vol. 6, Photosynthesis II,
Photosynthetic carbon metabolism and related processes, (eds. M. Gibbs and
E. Latzko), pp. '102—112, New York, Springer-Verlag, 1979.
15. Edwards G. E., Huber S. C. Usefulness of isolated cells and protoplasts for
photosynthetic studies. In: Fourth International Congress on Photosyn-
thesis, (eds. D. 0. Hall, J. Coombs and T. W. Goodwin), pp. 95—106, London,
The Biochemical Society, 1978.
16. Evert R. F., Eschrich IF., Heyser IF. Distribution and structure of the plas-
modesmata in mesophyll and bundle sheath cells of Zea mays L, Planta,
136, 77—89 (1977).
17. Goatley M. B., Smith H. Differential properties of phosphoenolpyruvate car-
boxylase from etiolated and green sugarcane, Planta, 117, 67—73 (1974).
18. Graham D„ Hatch M. D„ Slack C. R., Smillle R. M. Light-induced formation
of enzymes of the C4 dicarboxylic acid pathway of photosynthesis in deta-
ched leaves, Phytochemistry, 9, 521—532 (1970),
19. Gutierrez M., Huber S. C„ Ku S. B., Ranai R., Edwards G. E. Intracellular
carbon metabolism in mesophyll cells of C4 plants. In: Third International
Congress on Photosynthesis Research, (ed. M, Avron), pp. 1219—1230, The
Netherlands, Elsevier, 1975.
20. Harel E., Lea P. J., Miflin B. J. The localization of enzymes of nitrogen
assimilation in maize leaves and their activities during greening, Planta,
134, 195—200, 1977.
21. Hatch M. D. Regulation of enzymes in C4 photosynthesis. In: Current Topics
in Cellular Regulation, (eds. B. L. Horecker and E. K. Stadtman), Ch. 14,
pp. 1—28, New York, Academic Press, 1978.
22. Hatch M. D., Mau S. Properties of phosphoenolpyruvate carboxylase in C4
pathway photosynthesis, Aust. J. Plant Physiol., 4, 207—216 (1977).
23. Hatch M. D., Mau S., Kagawa T. Properties of leaf NAD malic enzyme from
plants with C4 pathway photosynthesis, Arch. Biochem. Biophys., 165, 188—
200 (1974).
24. Hatch M. D., Osmond С. B. Compartmentation and transport in C4 photo-
synthesis. In: Encyclopedia of Plant Physiology, New series, Vol. Ill, Trans-
port in plants, (eds. C. R. Stocking and U. Heber), pp. 144—184. New York,
Springer-Verlag, 1976.
ИНТЕГРАЦИЯ ФУНКЦИИ ПРИ С4ФОТОСИНТЕЗЕ
397
25. Hutch М. D., Slack С. R. The С4 carboxylic acid pathway of photosynthesis.
In: Progress in phytochemistry, (eds. L. Reinhold and Liwschitz), pp. 35—
106, Loudon, Wiley Interseience, 1970.
26. Haltersley P. IV., Watson I.., Osmond С. B. Metabolite transport in leaves
of C4 plants: specification and speculation. In: Transport and Transfer Pro-
cesses in Plants, (eds. 1, F. Wardlaw and J. B. Passioura), pp. 191—201,
New York, Academic Press, 1976.
27. Herold A., Lewis D. II., Walker I). A. Sequestration of cytoplasmic ortho-
phosphate by inaiuiose and its differential effect oil photosynthetic starch
synthesis in Ca and C4 species, New Phytol., 76, 397—407 (1976),
.28 . Hither S. C„ Edwards G. E. Regulation of oxaloacctate, aspartate and inalale
formation in mesophyll protoplast extracts o( several C4 plants, Plant Phy-
siol., 56, 324—331 (1975a).
29. Huber S. C., Edwards G. E. Inhibition of phosphoeiiolpyruvate carboxylase
from C4 plants’ by inalale and aspartate, Canad. J. I3ot., 53, 1925—1933,
1975b,
30. Huber S. C., Edwards G. E. Effect of DBM1B, DCMU and anthnycin A on
cyclic and uoncyclic electron flow in C4 mesophyll chloroplasts, FEBS Let-
ters, 58, 211—214 (1975c). .
31. Huber S. C., Edwards G, E. The effect of oxygen on CO2 fixation by meso-
phyll protoplast extracts of Сз and C4 plaids, Biochem. Biophys. Res. Com-
mon., 67, 28—34 (1975d).
32. Huber S. C., Edwards G. E, Studies on cyclic photophosphorylation in C4
mesophyll chlorophisls, Bioebini. Biopliys. Acta, 449, 420—433 (1976).
33. Huber S. C„ Edwards G, E. The effect of reducing conditions on inhibition
of cyclic photophosphorylation by DBM1B, EDAC and anlimycln A by C4
mesophyll chloroplasts, FEBS Letters, 77, 207—211 (1977a).
34, Huber S, C., Edwards G. E. Transport in C4 mesophyll chloroplasts. Charac-
terization of the pyi'uvate carrier, Biochim. Biophys, Acta, 462, 583—602
(1977b),
35, Huber S. C., Edwards G. E. Transport in C4 mesophyll chloroplasts. Evidence
for an exchange of inorganic phosphate and phosphoenolpyruvate, Biochim.
Biophys. Acta, 462, 603—612 (1977c).
36, Kagawa T., Hatch M. D. Regulation of C4 photosynthesis: characterization
of a protein factor mediating the activation and inactivation of NADP-malate
dehydrogenase, Arch. Biochem. Biophys., 184, 290—297 (1977).
37. Kelly G. J., Gibbs M. A mechanism for the indirect transfer of photosynthe-
tically reduced nicotinamide adenine dinucleotide. phosphate from chloro-
plasts to the cytoplasm, Plant Physiol., 52, 674—676 (1973).
38. Ku M. S. B., Spalding м. H., Edwards G. E, Intracellular localization of
phosphoeiiolpyruvate carhoxykinasc in leaves of C4 and CAM plants, Plant
Sci. Letters, 19, 1—8 (1980).
39. Lea P. Miflin B. J. Photosynthetic ammonia assimilation. In: Encyclope-
dia of Plant Physiology, New series, Vol. 6, Photosynthesis II, Photosynthetic
carbon metabolism and related processes, (eds. M. Gibbs and E. Latzko),
pp. 445—455, New York, Springer-Verlag, 1979.
40. Leech R. M., Murphi/ D, /. The cooperative function of chloroplasts in the
biosynthesis of small molecules. In: The Intact Chloroplasts, (ed. J. Barber),
pp. 365—401, Amsterdam, Elscvier/North-Holland Biomedical Press, 1976.
41. Miflin B. J., Lea P. J. Amino acid metabolism, Ann. Rev. Plant Physiol,, 28,
299—332 (1977).
42. Moore R., Black С. C. Nitrogen assimilation pathways in leaf mesophyll and
bundle sheath cells of C4 photosynthesis plants formulated from comparative
studies with Digitaria sangulnalis (L), Scop, Plant Physiol., 64, 309—313
(1979).
43. Osmond С. B., Smith. F. A. Symplaslic transport of metabolites during C4
photosynthesis. In: Intercellular Communication in Plants: Studies on Plas-
398
ГЛАВА 1Й
modesmata, (eds. В. Е. S. Gunning and A. W. Robards), pp. 229—240, New
York, Springer-Verlag, 1976.
44. Rathnam С. K. Al., Edwards G. E. Distribution of nitrate assimilating enzy-
mes between mesophyll protoplasts and bundle sheath cells in leaves of
three groups of C4 plants, Plant Physiol., 57, 730—733 (1976).
45. Rathnam. С. K, M„ Edwards G, E, C4 acid decarboxylation and CO2 dona-
tion to photosynthesis in bundle sheath strands and chloroplasts from spe-
cies representing three groups of C4 plants, Arch, Biochem, Biophys., 183,
1—13 (1977),
46, Rathnam С. K. Al., Edwards G. E, C4-dicarboxylic—acid metabolism in
bundle sheath chloroplast, mitochondria and strands of Eriochloa borumensis,
a PEP-carboxykinase type C4 species, Planta, 133, 135—144 (1977).
47. Ray T. B., Black С, C. Inhibition of oxaloaeetate decarboxylation during C4
photosynthesis by 3-mercaplopicolinic acid, J. Biol. Chem., 251, 5824—5826
(1976a).
48. Ray T. B„ Black С. C. Characterization of phosphoenolpyruvate carboxyki-
iiase from Punicuni maximum, Plant Physiol., 58, 603—607 (1976b).
49. Repo E., Hatch AL D. Photosynthesis in Gomphrena celosioides and its clas-
sification amongst C4-pathway plants, Aust. J. Plant Physiol., 3, 863—876
(1976).
50. Robards A, IV". Plasmodesmata, Ann. Rev. Plant Physiol., 26, 13—29
(1975),
51. Sugiyama T. Proteinaceous factor reactivating an inactive form of pyruvate,
Pi dikinase isolated from dark treated maize leaves, Plant Cell Physiol.,
15, 723—726 (1974),
52. Uedan K-, Sugiyama T. Purification and characterization of phosphoenolpyru-
vate carboxylase from maize leaves, Plant Physiol,, 57, 906—910 (1976),
53. Usuda H., Edwards G. E. Localization of glycerate Kinase and some enzy-
mes for sucrose synthesis in C3 and C4 plants, Plant Physiol,, 65, 1017—
1022 (1980).
54. Usuda H., Kanai R., Miyachi S. Carbon dioxide assimilation and photosys-
tem II deficiency in bundle sheath strands isolated from C4 plants, Plant
Cell Physiol., 16, 485—494 (1975),
55, Utter Al. E., Kolen-brander H, M, Formation of oxaloaeetate by CO2 fixation
on phosphoenolpyruvate. In: The Enzymes, Vol, VI (ed. P. D, Boyer),
pp, 117—168, New York, Academic Press, 1972.
56. Walker D. A. Physiological studies on acid metabolism 7, Malic enzyme from
Kalanchoe crenata; effects of carbon dioxide concentration, Biochem. J,, 74,
216—223 (1960).
ГЛАВА 13
Фотодыхание
Краткое содержание
Фотодыхание, как следует уже из самого названия, — это ды-
хание на свету. У Сз-растений его можно обнаружить самыми
различными способами, и все результаты, полученные с помощью
этих способов, свидетельствуют о том, что у С^растеиий фото-
дыхание скорее всего отсутствует. Точно измерить абсолютную
интенсивность фотодыхания очень трудно, так как в стационар-
ных условиях иа свету фотосинтез и фотодыхание идут одновре-
менно. Считают, что исходным субстратом для «светового дыха-
ния» служит гликолат. Место разветвления процессов фотоды-
хания и фотосинтеза находится па уровне РуБФ-карбоксилазы-
оксигепазы. Кислород, взаимодействуя с РуБФ, обеспечивает и
синтез гликолата, и фотодыхание, а СО2, вступая в реакцию с
РуБФ, обеспечивает фотосинтез. Если отношение О2/СО2 ста-
новится достаточно высоким, растения начинают на свету ды-
шать п фотосинтезировать с одинаковой скоростью (это проис-
ходит, например, в компенсационном пункте). И наоборот, ког-
да отношение О2/СО2 невелико, как, например, в клетках обклад-
ки проводящих пучков во время С4-фотосинтеза, активность фо-
тодыхания сравнительно мала. Кислород и СОг ведут себя как
субстраты, конкурирующие друг с другом за взаимодействие с
РуБФ-карбоксилазой-оксигспазой (т. е. О2 ингибирует карбок-
силазу, а СОг—оксигеназу). Поэтому ингибирование фотосин-
теза кислородом у Сз-растений обусловлено подавлением кар-
боксилирования под действием Оз и синтезом гликолата, что в
итоге ведет к стимуляции фотодыхания.
Количество энергии, расходуемой на фиксацию СО2 у Сз-рас-
тенпй, возрастает с увеличением концентрации О2 (т. с. О2 сни-
жает квантовый выход). Независимо от того, что (О2 или СО2)
служит субстратом для РуБФ-карбоксилазы-оксигеиазы, конеч-
ным продуктом является ФГК, которая восстанавливается и ис-
пользуется в ВПФ-цикле для регенерации РуБФ. Кислород
шунтирует этап карбоксилирования, и поэтому чем больше Ог
вступает в реакцию, тем больше энергии тратится иа фиксацию
СО2. Первопричиной снижения квантового выхода фикса-
ции СО2 в присутствии О2 является дополнительный приток уг-
лерода из ВПФ-цикла, а ие расход энергии в гликолатном пути.
400
ГЛАВА 13
Рис. 13,1. Фотодыхание и кислородное ингибирование фотосинтеза. О2 и С02
конкурируют друг с другом за РуБФ в ВПФ-цикле. Это создает условия для
метаболизма углерода по двум путям. Реакция РуБФ с СО2 ведет к ассими-
ляции углерода, а реакция РуБФ с О2 шунтирует реакцию карбоксилирова-
ния и приводит к потере СО2 на свету (фотодыхание). Интенсивность фото-
синтеза и доля кислородного ингибирования этого процесса зависят от отно-
шения СО2/О2 з листе и от относительного сродства указанных выше ре-
акций к этим двум газам. О2 ингибирует фотосинтез, конкурируя с СО2 за
РуБФ или же способствуя потере СО2 в результате фотодыхания. У Ct-pac-
тений увеличение концентрации СО2 в клетках обкладки проводящих пучков
приводит к стимуляции фотосинтеза и к подавлению фотодыхания.
Квантовый выход фиксации СО2 при нормальной концент-
рации О2 в атмосфере и 30 °C у С3- и С4-растений одинаков. Из-
за кислородного ингибирования С3-растения тратят больше
энергии на фиксацию СО2, а С4-растения потребляют больше
энергии в Сд-цикле.
Предполагают, что в ходе связанных с фотодыханием про-
цессов метаболизма осуществляется массовый обмен метаболи-
тами внутри клетки между хлоропластами, пероксисомами и
митохондриями. Механизм такого транспорта изучен длохо, и
роль фотодыхания пока непонятна. Ясно лишь, что С3-растения
явно благоденствуют тогда, когда в условиях атмосферы, обога-
щенной СО2, фотодыхание искусственно подавлено.
13.1. Введение
В темноте большинство зеленых растений поглощает О2 и
выделяет СО2. У CAM-растений такой суммарный обмен в неко-
торой степени маскируется из-за темновой фиксации СО2. По-
глощение О2 и выделение СО2 являются результатом потребле-
ФОТОДЫХАНИЕ
401:
ния метаболитов в ходе дыхания. Главным субстратом дыхания:
в типичном зеленом листе, вероятно, служит гексозофосфат, об-
разующийся из крахмала, однако в семенах и других органах,,
предназначенных для длительного хранения веществ, для дыха-
ния могут расходоваться липиды и белки. Фосфаты сахаров и-
подобные им соединения разрушаются в ходе гликолиза, окис-
лительного пентозофосфатного пути и цикла трикарбоновых,
кислот. Восстановленные коферменты вновь окисляются и, но-
мере того как электроны проходят к кислороду по цепи перено-
са, состоящей из ряда цитохромов и других переносчиков, энер-
гия окисления запасается в виде АТР и становится доступной:
для других процессов обмена веществ. Сходным образом отте-
кают и метаболиты других путей, чтобы затем принять участие'
в синтезе новых компонентов клетки. Несомненно, что некото-
рые из этих процессов продолжают идти и на свету, хотя ка-
жется маловероятным, чтобы они протекали в совершенно неиз-
менном виде и никак бы не зависели от фотосинтетических,
процессов, которые в это же самое время происходят в сосед-
них компартментах клетки. К. тому же выявлены и новые ре-
акции, в ходе которых поглощается О2 и выделяется СО2. Эти
реакции протекают только на свету и в совокупности их обычно-
называют «фотодыханием», чтобы отличить от реакций «темно-
вого дыхания». Термин «фотодыхание» иногда применяют и для
того, чтобы охватить все связанные с дыханием явления, кото-
рые происходят на свету. Точно так же, как газообмен, обуслов-
ленный темновым дыханием, может изменяться или маскиро-
ваться за счет других процессов метаболизма, так и выделение
Ог н реальная фиксация СО2 при фотосинтезе могут снижаться
под влиянием газообмена, связанного с фотодыханием. Если
газообмен оценивают без поправки на дыхание, то употребляют
термин «наблюдаемый фотосинтез». В первую очередь это отно-
сится к целым растениям, клеткам и протопластам. В том слу-
чае, когда учитывают поправку па дыхание, употребляют тер-
мин «истинный фотосинтез».
13.2. Распознавание фотодыхания
а) Скачкообразное увеличение концентрации СО2
Из-за маскирующего действия .-фотосинтеза фотодыхание
распознается гораздо труднее, чем «темповое дыхание». Проще
всего, прервать фотосинтез в точке максимума и посмотреть, как
возобновляется «темновое дыхание», ,т. е. остается ли оно неиз-
менным или же происходит перестройка этого процесса за счет
переходных эффектов. Когда это было проделано Декером, а
впоследствии и другими .исследователями, все они наблюдали
26-2.34
402
ГЛАВА 13
скачкообразное увеличение концентрации СОэ. Таким образом
если растение поместить в закрытый сосуд и позволить ему фо-
тосинтезировать до тех пор, пока концентрация СОа в сосуде не
достигнет стационарного уровня, а затем быстро выключить ос-
вещение, то в темноте СО2 сначала будет выделяться с высокой
скоростью, и лишь затем, по истечении определенного времени,
ее содержание снизится до уровня, характерного для нормаль-
ного «темнового» дыхания. Это ясно говорит о том, что на свету
происходит какой-то процесс, который не сразу останавливается
после выключения освещения. Правда, это явление одинаково
легко объяснить как влиянием света на «темновое» дыхание,
так и временным последействием «светового» процесса. Послед-
нее объяснение более убедительно, так как оно подтверждается
данными о том, что наблюдаемый эффект резко уменьшается
при понижении концентрации О2 в окружающей среде до вели-
чины ~2%. Темновое дыхание по существу нечувствительно к
такому снижению концентрации О2, так как цитохромоксидаза,
которая играет главную роль в «темновом» дыхании, обладает
очень высоким сродством к Ог. Следует, однако, отметить,' что в
опытах подобного рода часто используют условия (например,
низкую концентрацию СО2), благоприятствующие фотодыханию.
При высокой концентрации СОг, которая, как полагают, подав-
ляет фотодыхание (и уменьшает скачкообразное увеличение
концентрации СОг), все еще можно наблюдать неоднократное
кратковременное «заглатывание» О2 при переходе свет — темно-
та. Причина этого явления до сих (пор не установлена, но а
priori можно предположить, что оно отражает либо переходные
эффекты, связанные с регуляцией размеров пула метаболитов,
либо действие света на «темновое» дыхание, либо и то и другое
вместе. Этот пример еще раз подчеркивает, насколько осторож-
но надо употреблять термин «фотодыхание» для объяснения ка-
ких-либо переходных эффектов.
б) Выделение СО2
Если поместить растение в условия, способствующие фикса-
ции СО2 на свету, а затем заменить газовую фазу на воздух,
лишенный СО2, то в атмосферу выделится некоторое количест-
во СО2. Хотя такое выделение СО2 через несколько минут может
пойти на убыль, обычно оно- идет быстрее, чем в темноте. Такие
опыты можно провести и с б) * * * * * * * 14СОг. При этом удельная активность
выделяющейся ИСО2 будет выше, чем в темноте. Отсюда сле-
дует, что субстратом служит другое гораздо позже образующее-
ся соединение (если исходить из допущения о том, что метка
будет все больше и больше разбавляться по мере продвижения
из фотосинтетического цикла в существующие пулы запасных
ФОТОДЫХАНИЕ
403.
веществ). Этот метод, ,'как и большинство других, позволяет
только качественно охарактеризовать фотодыхание, так как пе-
ренос растений в среду с низким содержанием СО2 на свету бо-
лее благоприятствует фотодыханию, чем фотосинтезу.
в) Углекислотный компенсационный пункт (Г)
При освещении в небольшом замкнутом пространстве С3-рас-
тения могут снижать концентрацию СО2 до величины ~50 м.д.
(ср. с 326 м.д., содержащимися обычно в атмосфере). Принято
считать, что при такой концентрации СО2 в углекислотном ком-
пенсационном пункте поглощение СО2 в процессе фотосинтеза
строго сбалансировано или «скомпенсировано» с утратой СОг в
результате дыхания. При низкой концентрации О2 (2% и ме-
нее) содержание СОг снижается почти до дуля. Разность между
числовыми значениями’ компенсационного пункта в этих двух
разных условиях можно использовать для оценки фотодыхания.
При этом предполагают, что связанное с фотодыханием выделе-
ние СО2 при высокой .концентрации О2 в среде достаточно-
мощно, чтобы сместить компенсационный пункт в сторону более
высоких концентраций, в то время как «темповое» дыхание, ко-
торое менее чувствительно к Ог, может почти целиком маски-
роваться даже в том случае, когда фотосинтез полностью на-
правлен только на фиксацию той СОг, которая иначе выделялась
бы наружу. СгРастения, у которых, по-видимому, нет фотоды-
хания, имеют очень низкие числовые значения углекислотных
компенсационных пунктов (обычно около 0—5 м.д.), и эти вели-
чины по существу не зависят от концентрации Ог во внешней
среде.
г) Поглощение Ой
В ранних исследованиях поглощения О2 у водорослей и
высших растений часто использовали относительно высокие (на-
много превышающие атмосферное содержание) концентрации
СОг и низкие (ниже уровня О2 в атмосфере) концентрации О2.
Сейчас известно, что такие условия неблагоприятны для связан-
ного с фотодыханием обмена веществ. Не так давно для изуче-
ния механизма и путей фотодыхания был применен 18О2. Этот
изотоп кислорода включается в карбоксильную группу гликола-
та, после чего 18О можно обнаружить в составе карбоксильной
группы глицина и серина. Скорость поглощения 1аО2 на .свету
намного выше, чем в темноте. По мере увеличения ![СО2] коли-
чество 18О2, поглощаемого Сз-растеииями в процессе фотосинте-
за, уменьшается, что хорошо согласуется с предположением о
том, что СО2 подавляет фотодыхание, снижая скорость этого
26*
404
ГЛАВА 13
процесса (разд. 13.10). В поглощении 18О2 в ходе фотодыхания
участвуют РуБФ-оксигеиаза, гликолатоксидаза и глициноксида-
за; при этом реакция, катализируемая последним ферментом,
сопряжена с митохондриальным фосфорилированием. Значитель-
ное поглощение 1аО2 на свету наблюдается также и у таких
С4-растений, как кукуруза, хотя это, вероятно, и не связано с
обменом веществ в ходе фотодыхания. Радмер и Кок обнаружи-
ли, что во время индукционного периода фотосинтеза поглоще-
ние 18О2 и выделение 16О2 у водорослей идут линейно, с одина-
ковой скоростью (что отражается в отсутствии видимого обмена
О2) и без фиксации СО2. После того как .'фиксация СО2 достиг-
нет стационарной скорости, включение 18О2 подавляется. Эти
данные рассматривали как доказательство псевдоциклического
транспорта электронов, когда 'О2 и СО2 конкурируют за восста-
новительную силу. Трудно связать математически скорость выде-
ления СО2 за счет фотодыхания со скоростью поглощения О2 на
свету, так как поглощение 18О2 может быть обусловлено и реак-
циями транспорта электронов (реакция Мелера, см. разд. 5.8),
и дыханием, протекающим в митохондриях (и ие связанным с
гликолатным путем).
д) Наблюдаемый фотосинтез как функция [СО2]
При концентрации СО2 выше компенсационного пункта и при
других благоприятных условиях скорость фотосинтеза возрас-
тает с увеличением [СО2]. Сначала наблюдается почти линейная
зависимость между этими двумя величинами. Если прямую,
отражающую эту зависимость, экстраполировать к нулю, то она
пересечет ось ординат в области отрицательных значений. Это
и указывает на наличие фотодыхания. При концентрациях СО2,
превышающих ее концентрацию в воздухе, скорость фотосинте-
за у Сз-растений продолжает возрастать до тех пор, пока не
наступит насыщение СО2. Фотосинтез у С4-растений в гораздо
меньшей степени зависит от [СО2] даже при концентрации
>300 м.д.; в атмосфере азота различия между С3- п С4-ра-
стениями выражены гораздо слабее, чем в обычных усло-
виях (разд. 14.3 и 14.4). Что касается светового насыщения
фотосинтеза, то у многих Сз-растений оно наблюдается при ин-
тенсивности освещения, близкой к 30% прямой инсоляции, в то
время как скорость фотосинтеза у С4-растений продолжает бо-
лее или менее возрастать с увеличением освещенности. И здесь
наблюдается тенденция к стиранию различий между этими дву-
мя группами растений при их инкубации в атмосфере азота, и
опять-таки все эти эффекты можно истолковать как результат
.фотодыхания, свойственного Сз-растениям.
Таким образом, существует целый ряд приемов, которые,
ФОТОДЫХАНИЕ
405
«если ими воспользоваться в совокупности, позволяют выявить
фотодыхание. Количественная оценка этого процесса в условиях
стационарного фотосинтеза крайне затруднена (разд. 13,'Ю).
Переходные эффекты, связанные с обменом газов в нестацио-
нарных условиях или при нефизиологических концентрациях
СО2 и О2, могут завышать или, наоборот, занижать величину
фотодыхания, свойственного растениям в условиях нормального
фотосинтеза,
Источники данных: [1, 2, 4, 13, 14, 21, 23, 56]
13.3. Происхождение гликолата
То, что гликолат является одним из продуктов фотосинтеза,
было впервые установлено в пятидесятых годах Кальвином и
его сотрудниками. Это соединение синтезируется на свету и
представляет собой продукт ассимиляции углерода в ВПФ-цик-
ле. Выдвигались самые различные предположения о том, как
синтезируется и как затем используется гликолат в процессе
фотодыхания (см. рекомендуемую литературу в конце главы).
Многие из известных аспектов фотодыхания у высших растений
можно, однако, понять и объяснить, рассмотрев только один ме-
ханизм синтеза гликолата и всего один путь его дальнейшего
метаболизма (разд. 13.4). Так, Бовес, Огрен и Хагеман обнару-
жили, что РуБФ-карбоксилаза может служить также и оксиге-
назой, в результате чего наряду с фосфоглицератом образуется
и фосфогликолат. Затем фосфогликолат должен превратиться в
гликолат под действием присутствующего в хлоропластах фер-
мента фосфогликолатфосфатазы.
О2 РуБФ ----->- Фосфогликолат-|-ФГК (РуБФ-оксигеназа) (13.1)
Фосфогликолат —-> Pj Гликолат (фосфогликолатфосфатаза) (13.2)
СО2-|-Н2О-|-РуБФ --э- 2ФГК (РуБФ-карбоксилаза) (13.3)
Очень важно отметить, что реакции (13.1) и (13.3) катализиру-
ются одним и тем же ферментом. О2 является при этом и конку-
рентным ингибитором РуБФ-карбоксилазы, и субстратом этого
фермента, когда он проявляет свойства РуБФ-оксигеназы. Точно
так же и СО2, с одной стороны, конкурентно ингибирует оксиге-
назную реакцию, а с другой, служит субстратом для этого фер-
мента, когда он проявляет свойства РуБФ-карбоксилазы. Наи-
меньшие из известных в литературе величин Км для О2, когда
он служит субстратом для оксигеназы, и Ki, когда он ингибирует
карбоксилазу, находятся в пределах 200—400 мкМ О2. Анало-
гичные наименьшие величины Км (для карбоксилазы) и Ki (для
оксигеназы) для СО2 варьируют в пределах.6—15 мкМ. [Km-
это константа Михаэлиса. Она равна той концентрации субстра-
406
ГЛАВА 13
та, при которой скорость реакции составляет 1/2 Vmax. Ki —кон-
станта диссоциации фермент-ингибиторного комплекса EI.
El Е-\-1
К tgl W
~ [£/]
Таким образом, чем выше сродство ингибитора к ферменту, тем
ниже величина Ль] Концентрации СО2 и Ог, соответствующие
указанным выше величинам Км, находятся в пределах раство-
римости атмосферных СОг и Ог в воде (разд. 14,2). Относитель-
ная величина Ктах для оксигеназы, по-видимому, ие превышает
1/4 величины Ктах для карбоксилазы. Поэтому при концентра-
циях О2 и СОг, близких по величине к соответствующим Км,
скорость реакции карбоксилирования должна быть примерно в
4 раза выше, чем скорость оксигеназной реакции. Известно, что
высокая концентрация Ог и сравнительно низкая концентрация
СО2 способствуют синтезу гликолата. Эти же условия наиболее
благоприятны и для фотодыхания, и для вызываемого О2 инги-
бирования фотосинтеза.
Из-за того, что синтез гликолата связан с активностью
РуБФ-карбоксилазы-оксигеназы, он зависит от присутствия и
СО2 и О2. Фиксация определенного количества СО2 необходима
для того, чтобы регенерировать РуБФ, субстрат оксигеназной
реакции. Без такой фиксации СО2 сопряженное функционирова-
ние гликолатиого пути и ВПФ-цикла приводило бы только к
фотодыханию и к оттоку метаболитов из этих двух путей
(рис. 13.2). (Подробно гликолатиый путь рассматривается в
разд. 13.6).
В условиях обезвоженности, когда может снижаться внутри-
клеточная концентрация СО2, растения, чтобы избежать исто-
щения субстратов, могут метаболизировать в ВПФ-цикле и гли-
колатиом пути некоторую часть углерода, находящегося в соста-
ве запасных веществ. Это ведет к утрате углерода эндогенных
веществ растения, обусловленной их .деградацией в процессе
фотодыхания. Если в закрытую камеру одновременно поместить
С3- и С4-растения и затем держать их на свету, то Сз-растение
погибнет от потери углерода, обусловленной фотодыханием, а
С4-растение будет захватывать «отработанную» СО2 и останет-
ся живым и здоровым по меньшей мере в течение некоторого,
хотя и ограниченного, промежутка времени.
В случае хлоропластов, выделенных из Сз-растений, главным
продуктом фотосинтеза в присутствии 21% О2 при пониженной
концентрации СО2 (например, в присутствии 0,4 мМ НСО3_, что
при pH 8,0 соответствует примерно. 8 мкМ СО2) является глико-
лат. И РуБФ-оксигеназа, и фосфогликолатфосфатаза находятся
ргуЮДЫХАНИЕ
407
10
Фас фогли колат
5СО;
5 ФГК
10 РуБФ + 10 О.
10 ФГК -----к 15ФГК «-----
9 РуБФ
Рис. 13.2. Потеря углерода из ВПФ-цикла в ходе фотодыхания.
в хлоропластах. Другие ферменты, участвующие в дальнейших
превращениях гликолата, локализованы вне хлоропласта, в пер-
оксисомах и митохондриях (разд. 13.7).
Источники'данных: [1—8, 11, 12, 18—20, 22, 32, 40, 48].
13.4. Гликолатный путь у С3-р астений
Гликолатный называется метаболический путь, в ходе кото-
рого гликолат превращается в триозофосфат. Современные пред-
ставления об этом пути сформировались после того, как в ран-
них работах был применен такой прием, как инкубация водо-
рослей или ткани целого листа с мечеными субстратами. Резуль-
таты, которые легли в основу этих представлений, были получе-
ны в пятидесятых и шестидесятых годах Толбертом, Зелитчем,
Вейгудом, Уиттиигемом и их сотрудниками. В ткань листа вво-
дили меченный 14С гликолат, глицин или серин и затем анализи-
ровали состав полученных продуктов. Среди продуктов метабо-
лизма 14С-гликолата, .образующихся па свету, были выявлены
глицин, серии, глицерат и сахароза. Продуктами аналогичных
превращений 1‘1С-глицина были серии, глицерат и сахароза. Все
три изученных меченых метаболита быстро метаболизировались
с образованием сахарозы и крахмала. Эти данные привели ис-
следователей к заключению, что гликолатный путь имеет глюко-
неогенетическую функцию. |[Глюкоиеогенез — это синтез сахаров,
например глюкозы, в результате обращения реакций гликолиза.]
В других работах исследовали продукты фиксации 14СС>2, об-
разующиеся в присутствии возможных ингибиторов метаболиз-
ма гликолата и без них. Так, например, Зелитч обнаружил за-
висимое от света накопление гликолевой кислоты в тканях листа
табака, которое вызывалось добавлением а-гидроксисульфоиата,
ингибирующего гликолатоксидазу. Уиттингем и др. показали,
408
ГЛАВА 13
что включение 14СОг в гликолат и глицин у Chlorella возрастает
в восемь раз после добавления изоникотииилгидразина (ИНГ, ин-
гибитора трансаминаз), а скорость фиксации СОг при этом не
меняется. Сейчас известно, что ИНГ сильно ингибирует превра-
щение глицина в серин [уравнения (13.8) и (13.9)]. Аминоаце-
тонитрил конкурентно ингибирует окисление глицина в изолиро-
ванных митохондриях. При низких концентрациях Н14СО3_он по-
давляет фотосинтез в протопластах пшеницы и стимулирует вклю-
чение метки в глицин, а при высоких концентрациях НСО3’ этот
ингибитор не влияет на фотосинтез. Бутил-2-гидрокси-З-бутиноат
необратимо ингибирует гликолатоксидазу, что в присутствии
21% Ог приводит к накоплению гликолата и подавлению фото-
синтеза, тогда как в атмосфере азота он не влияет на эти про-
цессы. Иногда оказывается, что ингибитор, про который сначала
думали, что он является относительно, специфичным, на самом
деле обладает множественным действием на самые различные
участки, и очень часто приходится тратить много усилий на вы-
яснение побочных эффектов.
Более поздние исследования показали, что 18Ог включается
в карбоксильную группу гликолата, но не в ФГК и что в этой
реакции участвует РуБФ-оксигеназа. 18Ог, введенный на свету в.
целые листья, быстро включается в карбоксильные группы гли-
цина п серина. Эти данные хорошо согласуются с предложенным
механизмом синтеза и последующими метаболическими превра-
щениями гликолата (разд. 13.5 и 13.6).
Источники данных: [1—8, 45, 52, 58, 63, 66, 69].
13.5. Упрощенная схема пути углерода
в гликолатном цикле
Результаты исследований интактных организмов с помощью
меченых метаболитов и изучение ферментов гликолатного пути
позволяют схематически, в сжатом виде, представить путь угле-
рода в этом цикле у высших растений (рис. 13.3) (более под-
робно вопросы, связанные с ферментами цикла, рассматривают-
ся в разд. 13.6). Можно назвать следующие основные особенно-
сти этого, пути:
1. СО2 образуется во время превращения двух молекул глицина
в серин.
2. Кислород расходуется как при синтезе гликолата, так и в
процессе его дальнейшего метаболизма (т. е. с участием РуБФ-
оксигеназы и гликолатоксидазы). Кроме того, если окисляется и
NADH, образующийся при декарбоксилировании глицина в ми-
тохондриях, то потребление О2 возрастает.
3. Утрата СОг и потребление О2 на свету в процессе метаболиз-
ма гликолата и называется фотодыханием.
ФОТОДЫХАНИЕ
409
2 ФГК
+
2 Фосфогликолат
2 0.-^
2РуБФ
2Н2О + О2
t
2Р, 20. 2Н:О2
—► 2 Гликопат —-► 2Глиоксипат
<— 2 (-№;)
2 Г лицин
NAD |
NADU*7 „
Серин +• МЦ 4- С02
ФГК ------- Глицерат 4-- Гидроксипируват
ADP АТР NAD NADH
Рис. 13.3. Гликолатный путь.
4. В ходе этого метаболического цикла образуется свободный
аммиак.
5. Гликолатный путь является глюконеогенетическим. ФГК
представляет собой продукт, который может использоваться для
синтеза сахарозы и крахмала.
13.6. Реакции гликолатиого пути
и энергетические затраты
Гликолатный путь включает в себя следующие реакции, ко-
торые в конечном итоге ведут к превращению двух .молекул
гликолата в ФГК, предшественник синтеза сахарозы у высших
растений:
2 Гликолат 2О2 ------>• 2 Глиоксилат -ф- 2Н2О2 (глико чатоксидаза)
21-12О2 --> 2Г12О О2 (каталаза)
Г лиоксилат + Глутамат —►
--->- Глицин 2-оксоглутарат (глиципаминотрансфераза)
Глиоксилат Серии ....................>
---> Глицин + Гидроксипируват (серин-глиокснлат—аминотрансфераза)
Глицин -|- ТГФ1> NAD+ --------------------------->
---> NADH + I-I+ + Метилеи-ТГФ СО2 NH3 (глицинсинтаза) (13.8)
Метилен-ТГФ -|- Глицин Н2О -------------------->-
------------->- Серин 4-ТГФ (серин-гидроксиметилтрансфераза) (13.9)
1) тгф—тетрагидрофолат.
(13.4)
(13.5)
(13-6)
(13-7)
410
ГЛАВА 13
Гидроксипируват NADH Н+ ——>
----->- NAD+ -|- Глицерат (глиоксилатредуктаза)11 (13.10)'
Глицерат + АТР------> ФГК + ADP(глицераткппаза) (13.11)
В итоге: 2 Гликолат Ог + Глутамат -р АТР -----
--->- ФГК + ADP-|- С.О2 + NHS + 2-оксоглутарат + Н2О (13.12)
Лм(о2), гликолатоксидазы из Pisum sativum очень велика —
170 мкМ. РуБФ-оксигеназа также характеризуется высокой Км
для Ог (разд. 13.3). Поэтому для синтеза гликолата и его даль-
нейшего метаболизма требуется более высокая |[Ог], например
21% (см. разд. 14.2, где приводится растворимость Ог), чем для
темнового дыхания, которое достигает насыщения при низкой
концентрации Ог, т. е. при концентрации ~2%. На последую-
щих этапах гликолатного пути |NH4+ должен включиться в глу-
тамат. Это включение обеспечивается либо за счет глутаматде-
гидрогеназы, либо в результате сопряжения реакций глутамии-
синтетазы и глутаматсинтазы. И /в том и в другом случае тре-
буется один эквивалент NADPH. Поскольку глутаматдегидроге-
наза имеет очень высокую /<м для аммиака (разд. 12.9), более
вероятно участие в гликолатиом цикле глутаминсиитетазы —
глутаматсинтазы.
Глутамат -|- АТР NH3 ----->-
---->- Глутамин -р ADP + Р( (глутаминсинтетаза) (13.13)
Глутамин -р 2-Оксоглутарат + 2Фд (восстановленный) -->
-------> 2 Глутамат 2Фд (окисленный) (глутаматсинтаза) (13.14)
Восстанавливаясь, ферредоксин (Фд) захватывает один электрон.
Поэтому для превращения аммиака в азот аминной группы требуется 2 электро-
на или две молекулы восстановленного Фд.
В итоге: 2-Оксоглутарат + NH3 + 2Фд (восстановленный) + АТР --->
----э- Глутамат 2Фд (окисленный) + ADP + Р; (13.15)
Если рассматривать две молекулы Фд (восстановленного) как
энергетический эквивалент одной молекулы NADPH, то уравне-
ния (13.12) и (13.15) можно суммировать следующим образом:
2 Гликолат 4~ О2 -|- 2АТР + NADPH Н+ ------->
---->- ФГК + Н2О + 2ADP + Р, + СО2 + NADP+ (13.16)
Таким образом, для превращения двух молекул гликолата в
ФГК в результате указанных выше реакций требуется одна мо-
>) Этот фермент, зависящий от NAD, может катализировать восстанов-
ление либо глиоксилата, либо гидроксипирувата. В соответствии с номенкла-
турой ферментов он назван глиоксилатредуктазой.
ФОТОДЫХАНИЕ
411
лекула NADPH и две молекулы АТР. Однако при оценке дей-
ствительного расхода энергии в гликолатном цикле следует учи-
тывать его влияние па энергетику фиксации СО2 в ВПФ-цикле.
Рассмотрим следующий пример: при атмосферных условиях от-
ношение активностей карбоксилазы и оксигеназы равно 3,5,
т. е. с РуБФ взаимодействует 3,5 СО2/1 О2 или 7 СО2/2 О2. (Учти-
те, что в углекислотном компенсационном пункте это отношение
равно 1 СО2/2 О2; см. рис. 6.11.)
7СОа + 7РуБФ I- 7Н3О --> 14ФГК (13.3)
2О„ + 2РуБФ --->- 2ФГК 2 Фосфогликолат (13.1)
2 Фосфогликолат -|- 2Н2О ->- 2Pj 2 Гликолат (13.2)
2 Гликолат -|- О2 2АТР NADPH + Н+------>
---> ФГК + Н2О -I- 2ADP + + СО,2 + NADP+ (13.16)
В итоге: 6СО, + 9РуБФ 8НгО -j- 3OS -|- 2АТР -|- NADPH Н+ ->•
---> 17ФГК 4-2ADP+NADP+4-3Pi (13.17)
17ФГК+ 17АТР+ 17NADPH 17Н+ -------->
---> 17ТФ-1- 17ADP+ 17NADP+4- 17Р, (13.18)
15ТФ-|-9АТР4-бН<.0 ----> ЭРуБФ9ADP-|-6Pj (13.19)
Сумма уравнений (13.17—13.19) равна 6СО2 -|- ЗО2 +
-|- 28АТР + 18NADPH -|- 18Н+ 14Н2О ---->
---> 2ТФ 28ADP + 26Р, 18NADP+ (13.20)
18NADP++18Н2О -----> 18NADPH-j-18Н+SO2 (13.21)
28ADP-|-28Р|--->- 28АТР + 28НаО (13.22)
Сумма уравнений (13.20—13.22) равна 6"О2 + 2Pj 4- 4Н2О ->
---> 2ТФ + 6О2 (13.23)
На рис, 13.4 суммированы энергетические затраты для рассмот-
ренного примера, которые в этом случае составляют 28 АТР,
18NADPH на 6 СО2, или, иначе, 4,67 АТР, 3 NADPH на одну мо-
лекулу ассимилированной СО2. Интересно сравнить, как связа-
ны между собой скорости ассимиляции аммиака и углерода и
каковы энергетические затраты в том и другом случае. Если
вернуться к рассмотренному выше примеру, то при фиксации
каждых шести молекул СО2 -(реальная фиксация, или иетто-
фиксация) в результате фотодыхания должна высвобождаться
одна молекула аммиака. Для реассимиляции такого аммиака и
его превращения в аминную группу глутамата необходимо из-
расходовать энергию одной молекулы АТР и одного эквивалента
NADPH; при расчете на одну молекулу связанной СО2 [если в
реассимиляции аммиака участвует глутаминсинтетаза — глута-
412
ГЛАВА J3
2 АТР, 2 Фд (вс>сстании||1’||пый) 1\АГ)1’11
Рис. 13.4. Теоретически рассчитанные затраты энергии в процессе фотосинте-
за у Cs-растения (при условии, что стехиометрия реакции с РуБФ-карбокси-
лазой-оксигеиазой составляет 7СОз/2О2, а концентрации СО2 и О2 в атмо-
сфере не отличаются от нормальных). Подробности см. в тексте.
матсинтаза в соответствии с уравнениями (13.13) и (13.14)] это
составляет 0,17 АТР и 0,17 NADPH. Это достаточно малая ве-
личина, если сравнить ее с той энергией, которая необходима
для фиксации СОг (табл. 13.1). Поэтому главной причиной рез-
кого увеличения расхода энергии при Сз-фотосинтезе скорее все-
го является не затрата энергии на реассимиляцию аммиака, а
конкурентное ингибирование РуБФ-карбоскилазы кислородом и
потеря СОг в ходе фотодыхания.
Повторное циклическое использование аммиака при фотоды-
хании приводит к тому, что растение в конечном итоге не полу-
чает аминного азота извне. Растения обычно получают амин-
ный азот для синтеза белков в результате превращения нитра-
та, для которого необходима дополнительная энергия. Содер-
жание азота в органических соединениях листа составляет
примерно 6% суммарного, содержания в них углерода (при
расчете атом/атом). Для превращения нитрата в аминокислоты,
в котором участвуют нитратредуктаза, нитритредуктаза и глу-
таминоинтетаза—глутаматсинтаза, требуется израсходовать од-
ну молекулу АТР и 5 эквивалентов NADPH (разд. 12.9). При
отношении азот/углерод в листе, равном 0,06/1, энергетические
затраты на Са-фотосинтез и обмен нитрата составляют
(4,67АТР + 3NADPH/CO2) + 0,06(1 АТР + 5NADPH/NO3~) =
= 4,73ATP/3,3NADPH на одну молекулу связанной СО2. Таким
ФОТОДЫХАНИЕ
413
Таблица 13.1. Суммарная сводка теоретически возможных затрат
энергии в ВПФ-цикле и в ходе ассимиляции нитрата у Сз-растений1’
Метаболический путь Расход в расчете на одну молекулу связанной СО2
АТР NADPH (или его эквиналсит)
ВПФ-цпкл (в отсутствие О2) .3 2
ВПФ-цикл (в присутствии 21% О2) 4,67 3
ВПФ-цикл+превращсние нитрата в глутамат (в присутствии 21% О2) 4,73 3,3
ЧаЬть, обусловленная рсасспмпляци- сй ’ аммиака при фотодыхании (в присутствии 21% Оа) 0,17 (3,6%) 0,17 (3,2%)
Часть, обусловленная превращением нитрата в глутамат (в присутствии 21% О2) 0,06 (1,3%) 0,30 (9,1%)
') Цифры и скобках означают, какая доля суммарной энергия расходуется для ре-
асеимиляции аммиака и для превращений нитрата.
образом, расход энергии при превращении нитрата в азот ами-
нокислот сравнительно мал; о и гораздо меньше, чем затраты
энергии, связапиы-е с фиксацией СОг (табл. 13.1). Из-за цик-
лического повторного использования аммиака, высвобождающе-
гося в процессе фотодыхания, скорость превращения аммиака в
аминный азот в листьях Сз-растений может быть выше, чем в
листьях С4-растеиий. Для приведенного выше случая общая
скорость ассимиляции аммиака у Сз-растений должна состав-
лять ~23°/о скорости фиксации СОг (при условии, что количест-
во выделяющегося аммиака составляет 1/6 или, иначе, 17% ко-
личества связываемой СОг, а скорость восстановления нитрата
равна 6% скорости фиксации СОг). При скорости фотосинтеза-
в листьях, равной 100—200 мкмоль СО2-(мгХл)-1 -ч-1, скорость
ассимиляции аммиака у С3-растений соответственно равна 23—
46 мкмоль- (мгХл)“1-ч-1. При отношении активностей карбо-
ксилазы и оксигеназы, равном 7/2, в присутствии О2 фиксиру-
ется 6 молекул СО2 (рис. 13.4), а в отсутствие О2 примерно-
9 молекул СО2. Эта величина вполне приемлема для оценки-
увеличения карбоксилирования после удаления О2 из атмосфе-
ры. Число молекул СО2, которые дополнительно вступают в ре-
акцию в отсутствие О2, ие обязательно точно соответствует сни-
жению активности оксигеназы. Относительная скорость карбок-
силазной реакции в присутствии О2 и без него, как и скорость
оксигеназной реакции, определяется концентрацией этих двух
414
ГЛАВА 13
газов и кинетическими константами (Итак и Л’м) [разд. 13.10,
уравнения (13.33), (13,36) и (13.39)]. В разбираемом выше
примере доля вызываемого Ог ингибирования фотосинтеза со-
ставляет
±=1(100) = 33%
Эта величина очень близка к величине ингибирования фотосин-
теза в листе и к величине, характеризующей снижение кванто-
вого выхода фотосинтеза при переходе от 2 к 21% О2
(разд. 13.11 и 13.12). В этом случае в нормальных атмосферных
условиях иа долю фотодыхания приходится 1/3, а на долю кон-
курентного ингибирования — 2/3 суммарного ингибирования фо-
тосинтеза кислородом. Это вполне разумное приближение к ис-
тинным значениям этих двух составляющих, которые in vivo
оценить не так просто (разд. 13.10).
При поглощении листом энергии О2 снижает фиксацию СО2.
Это приводит к увеличению энергетических затрат иа фиксацию
СО2. Если для фиксации СО2 в ВПФ-цикле в присутствии 2% О2
требуется 3 молекулы АТР и 2 эквивалента NADPH, то сниже-
ние квантового выхода на 33% (или ингибирование фотосинте-
за кислородом, см. разд. 13.12) в атмосфере, содержащей
21% О2, приведет к тому, что энергетические затраты возрас-
тут до
3 ATP/2NADPH па 0,67 молекулы фиксированной СО2 или 4,48
ATP/3NADPH иа одну молекулу фиксированной СО2
В этом случае 1/3 энергии, вероятно, расходуется на обмен ве-
ществ, связанный с кислородным ингибированием фотосинтеза.
Итак, между величинами, которые получают при оценке
энергетических затрат двумя различными способами, наблюда-
ется полное соответствие, даже несмотря на то, что не совсем
понятно, как сбалансированы использование и образование
энергии в гликолатном пути (разд. 13.7). Ясно, что О2 увеличи-
вает расход энергии при фиксации СО2 у Сз-растений
(разд. 13.12), но остается непонятным, куда тратится эта энер-
гия. Убыль поглощенной энергии в присутствии О2 может быть
связана с увеличением флуоресценции (вероятно, из-за конку-
рентного ингибирования фотосинтеза кислородом) или с исполь-
зованием энергии фотохимических процессов для фотодыхания.
Оценка квантового выхода в целых листьях и измерение его
снижения под действием О2 не позволяют сделать определен-
ный выбор между этими двумя возможностями.
Источники данных: ([5, 26, 33, 40, 41]
ФОТОДЫХАНИЕ
415
13.7. Внутриклеточная локализация ферментов
гликолатного пути
Гликолатный путь у растений был обнаружен в начале шес-
тидесятых годов. В 1968 г. Толберт и его коллеги выделили из
листьев органеллы, которые они назвали пероксисомами. Было
установлено, что в пероксисомах находятся некоторые ключе-
вые ферменты гликолатного пути, а именно глпколатоксидаза,
каталаза и глиоксилатрсдуктаза (функция последнего в перок-
сисоме заключается в том, что он восстанавливает гндроксипи-
руват до глицерата, КФ 1.1.1.26). Эти органеллы обычно рас-
сматривают как своеобразные микротельца. Диаметр перокси-
сом варьирует от 0,5 до 1,5 мкм. В отличие от митохондрии и
хлоропластов эти органеллы окружены не двойной, а одинар-
ной мембраной (рис. 13.5). Для разделения митохондрий, хло-
ропластов н пероксисом обычно используют методы центрифу-
гирования.
Хлоропласты, которые отличаются более крупными размера-
ми, имеют большую скорость седиментации, чем митохондрии
Рис. 13.5. Электронная микрофотография, на которой видны пероксисомы
листа (7), митохондрия (2) и часть двух хлоропластов (5) в клетке мезо-
филла молодого листа табака (Nicotiana tabacum L.). Х48 000. Большой кри-
сталл внутри пероксисомы, по-видимому, каталаза. (Микрофотография полу-
чена Sue Е. Frederick, любезно предоставлена Е. Н. Newcomb, University of
Wisconsin.)
416
ГЛАВА 13
или пероксисомы. Поэтому при помощи дифференциального
центрифугирования (т. е. последовательного центрифугирования
экстракта, полученного из листьев или протопластов, при раз-
личных ускорениях) можно получить фракцию (500—1000 g в
течение нескольких минут), обогащенную хлоропластами. Ана-
логично, если экстракт, полученный из листьев или протоплас-
тов, наслоить на градиент концентрации сахарозы и отцентрифу-
гировать достаточно долго (например, 15 мин), то можно отде-
лить интактные хлоропласты от смешанной фракции митохонд-
рий и пероксисом. Из-за более высокой скорости седиментации
хлоропласты продвинутся по градиенту дальше, чем другие ор-
ганеллы (например, в таком градиенте, как у Лича илиМифли-
на и Биверса). Этот метод, однако, ие годится для отделения
пероксисом от митохондрий, так как скорость седиментации
этих органелл одинакова.
Еще один метод разделения органелл основан на равновес-
ном (изопикническом) центрифугировании в градиенте плотно-
сти. Полученный из листьев экстракт наслаивают иа градиент
концентрации сахарозы и затем центрифугируют до тех пор,
пока органеллы не достигнут равновесного положения в гради-
енте в соответствии с их плавучей плотностью. Разделение ор-
ганелл в данном случае основано ие на различии в скорости их
седиментации, а на различии в их плотности. Пероксисомы име-
ют более высокую плавучую плотность, чем интактные хлоро-
пласты, а интактные хлоропласты имеют более высокую плаву-
чую плотность, чем интактные митохондрии. Обычно полученные
из листьев экстракты наслаивают на градиент концентрации са-
харозы (примерно 20—60% по весу), который затем центрифу-
гируют в роторе с подвесными стаканами в течение 3—4 ч при
100 000 g. Равновесное положение пероксисом достигается при
шлотности — 1,25, интактных хлоропластов— при плотности
1,20—1,22, а митохондрий — при плотности 1,18—1,20. Бо-
лее высокая плавучая плотность пероксисом может быть
обусловлена тем, что они частично проницаемы для са-
харозы. После центрифугирования градиент делят иа фракции
(объемом около 1 мл; весь объем стакана обычно составляет
— 40 мл), в которых определяют активность исследуемых фер-
ментов. Для каждой органеллы характерны определенные мар-
керные ферменты; так, например, для митохондрий это может
•быть цитохром с — оксидаза, для пероксисом — гликолатоксида-
за, а для интактных хлоропластов — NADP-триозофосфатдегид-
.рогеназа. Равновесное центрифугирование в градиенте плотно-
сти использовали Брейденбах и Биверс, которым удалось с его
помощью открыть глиоксисомы (микротельца, которые находят-
ся в семенах, содержащих много жиров). Толберт и др. приме-
нили этот метод для выделения и очистки пероксисом. Цеитри-
ФОТОДЫХАНИЕ
417
фугирование в градиенте концентрации сахарозы очень удобно
для разделения органелл и исследования состава входящих в
них ферментов, однако число методов выделения и очистки
функционально активных (т. е. активно метаболизирующих) пе-
роксйсом, митохондрий и хлоропластов из ткани листа неве-
лико.
Митохондрии играют ключевую роль в метаболизме гликола-
та, так как в них находятся те ферменты, которые осуществля-
ют декарбоксилирование глицина, ведущее к образованию сери-
на и СО2. Поэтому выделение СОг, которое связано с фотоды-
ханием, происходит в этой органелле. Синтез гликолата и его
дальнейший метаболизм определяются отдельными реакциями
гликолатного пути, которые происходят в хлоропластах, перок-
сисомах и митохондриях, и, следовательно, для него необходим
интенсивный перенос метаболитов из органеллы в органеллу.
В пероксисомах содержатся те ферменты, которые нужны
для превращения гликолата в глицин [уравнения (13.4) —
(13.6)], а серина в гидроксипируват и глицерат [уравнения
(13.7)— (13.10)]. Глицераткиназа локализована в хлоропластах,
где, очевидно, и происходит превращение глицерата в ФГК-
В гликолатном пути гликолат синтезируется в хлоропластах,
переносится в пероксисомы и затем превращается в глицин. Пос-
ле этого глицин транспортируется в митохондрии и декарбокси-
лируется с образованием серина, СОг и NH3. В этом превраще-.
нии участвуют глицинсиитаза и серин-гидроксиметилтрансфера-'
за [уравнения (13.8) и (13.9)]. В дальнейшем серин метаболи-
зируется до глицерата в пероксисомах, а глицерат метаболизи-
руется до триозофосфата в хлоропластах. Предполагают, что в
пероксисомах происходят обмен и транспорт гликолата, глици-
на, серина и глицерата, а в митохондриях осуществляется
транспорт глицина, серина и СОг (рис. 13.6). До сих пор не
выяснено, как происходит транспорт метаболитов между перок-
сисомами и митохондриями, т. е. участвует ли в этом процессе
переносчик или же он идет за счет диффузии. Конечным резуль-
татом функционирования гликолатного пути является суммар-
ный перенос азота из пероксисом в митохондрии. Поэтому азот
может повторно использоваться в пероксисомах в виде амино-
кислоты. Для аминирования и превращения NH3 в глутамат
требуется один эквивалент NAD(P)H. Одна молекула NADH
образуется в митохондриях в результате декарбоксилирования
глицина, и одна молекула NADH используется в пероксисоме
в ходе глиоксилатредуктазной реакции. Для : метаболического
превращения'двух молекул гликолата в глицерат необходимо
реальное поступление восстановительной силы (один эквива,-
леит NADPH), которая, как предполагают, происходит;, из хло-
ропластов. Если бы дополнительная восстановительная силадля
27—234
418
ГЛАВА 13
Рис. 13.6. Возможный челночный перенос метаболитов между пероксисома-
ми, митохондриями и хлоропластами в ходе гликолатного цикла (полный
список ферментов, участвующих в этом процессе, см, в разд. 13.6).
реакций гликолатного пути возникала в ходе дыхания в мито-
хондриях, это приводило бы к дальнейшим потерям СО2. По-
ступление восстановительной силы из хлоропластов необходи-
мо либо для превращения гидроксипирувата в глицерат, либо
для превращения аммиака в азот аминокислот.
ФОТОДЫХАНИЕ
419
В митохондриях растений имеется NAD-глутаматдегидроге-
наза, которая может переводить аммиак обратно в состояние
.аминного азота. Тогда для реассимиляции аммиака требуется
восстановительная сила, возникающая в результате декарбок-
силирования глицина:
2 Глицин NAD+ Н2О ----->
----> NADH+H+ +Серин 4-СОг + МНз (13.8 и 13.9)
NADH Н+ 4- 2-Оксоглутарат NH3 —->•
---> NAD+ Глутамат -]-Н2О (13.24)
Более вероятно, что путь ассимиляции аммиака лежит через
глутамиисинтетазу— глутаматсинтазу (разд. 12.9), поскольку
/<м для аммиака у NAD-глутаматдегидрогеназы очень велика.
Известны данные о том, что глутаминсинтетаза локализована и
в хлоропластах, и в цитоплазме, а глутаматсинтаза находится
только в хлороиластах. Поэтому, как показано на рис. 13.6, ам-
миак может реассимилироваться в цитоплазме, а глутамат мо-
жет синтезироваться в хлоропластах. Перенос ассимилированно-
го азота может в этом случае происходить за счет челночного
механизма транспорта глутамата и 2-оксоглутарата между хло-
ропластами и пероксисомами. Восстановительная сила, которая
возникает в митохондриях в ходе декарбоксилирования глицина,
может переноситься в пероксисомы за счет челночного обмена
оксалоацетат — малат (рис. 13.6) и использоваться там для вос-
становления гидроксипирувата.
В опытах с препаратами митохондрий были получены дан-
ные о том, что образование восстановительной силы в ходе
декарбоксилирования глицина может быть связано с окислитель-
ным фосфорилированием. Остается неясным, используется ли
восстановительная сила, возникающая при декарбоксилирова-
нии глицина, непосредственно для энергизации других реакций
этого пути in vivo или же она служит источником АТР для аль-
тернативных метаболических процессов. В последнем случае
хлоропласты могут обеспечивать восстановительной силой и ас-
симиляцию аммиака, и превращение гидроксипирувата в гли-
церат.
Было выдвинуто предположение о том, что гликолатный
путь является составной частью главного пути синтеза сахаро-
зы. Считают, что синтез сахарозы у Сз-растений происходит в
цитозоле. Метаболическое превращение триозофосфатов в саха-
розу в цитозоле приводит к высвобождению Pi, который может
поглощаться хлоропластами в результате обмена на триозофос-
фат, происходящего на переносчике фосфата (разд. 8.23). При-
нято думать, что глицерат — продукт гликолатного пути — мета-
болизируется до триозофосфата в хлоропластах. Триозофосфат,
27*
420
ГЛАВА 13
синтезирующийся таким" способом, входит в: общий пул трио-
зофосфатов, синтезирующихся непосредственно в хлоропластах
при участии РуБФ-карбоксилазы. Следовательно, триозофосфат,
возникающий в ходе метаболизма гликолата,. будет время от
времени использоваться в реакциях восстановительной фазы
ВПФ-цикла для синтеза крахмала или сахарозы. По это вовсе
не означает, что существует прямой или альтернативный путь
синтеза сахарозы из ФГК, возникающей в ходе метаболических
превращений гликолата.
Источники данных: [4, 5, 28, 34, 41, 43, 47, 60, 61, 65, 68].
13.8. Метаболизм в изолированных органеллах
и его связь с гликолатным путем
Для того чтобы оценить роль хлоропластов, митохондрий и
пероксисом- в метаболизме гликолата, а также выяснить меха-:
низм обмена метаболитами и особенности регуляции отдельных
реакций этого пути, можно использовать изолированные орга-
неллы. В большинстве работ, в которых исследовали метабо-
лизм в изолированных органеллах, были, использованы лишь
частично очищенные препараты (технические трудности, связан-
ные с выделением органелл, описаны в разд. 13.7).
Хлоропласты
Хлоропласты синтезируют гликолат и метаболизируют гли-
церат до триозофосфата. При низких концентрациях НСОз-
(0,4—1,0 мМ) и высоких концентрациях О2 (21-—100%) глав-
ным продуктом фотосинтеза в хлоропластах является гликолат.
Нет никаких данных о том, что гликолат окисляется в резуль-
тате фотодыхания и далее метаболизируется именно в хлоро-
пластах. Если изолированные, хлоропласты инкубировать иа
свету в среде с 14С-гликолатом, то метка лишь иногда, и то в
следовых количествах,, появляется в составе глицина и серина.
Это может быть обусловлено загрязнением хлоропластов цито-
плазмой. Кроме того, О2 ингибирует фотосинтез в изолирован-
ных хлоропластах при низкой концентрации НСОз-, а высокая
концентрация НСОз~ препятствует ингибированию. Таким об-
разом, . кислородное ингибирование фотосинтеза проявляется . в
изолированных, хлоропластах так же,, как и в фотосинтезирую-
щих клетках или в ткани листа, несмотря, на то.что в хлоро-
цластах отсутствует фотодыхание (см. такрке,разд, 13.10).
- .. Изолированные хлоропласты шпината- (Сз-раетение) в.ка:
кой-то степени способны к превращению глицерата. На .свету
хл.оропласты способны к зависимому .от глицерата выделению
(Э2; 'это. видно, если все определения проводят в отсутствие
ФОТОДЫХАНИЕ 421
НСОз~, что устраняет выделение Ог, зависимое от СО2. ; Для
превращения глицерата в триозофосфат требуются две молеку-
лы АТР и одна молекула NADPH:
Глицерат-)-АТР-->ФГК + АОР (13.11)
ФГК + АТР ---> ДФГК + ADP (13.25)
ДФГК + NADPH-J-Н+ --->- NADP+-|-ТФ + Р; (13.26)
Глицерат 2АТР + NADPH + II+—> ТФ Р; 2ADP + NADP+ (13.27)
Нет никаких данных о том, что гликолат или глицерат транс-
портируются через оболочку хлоропласта путем, отличным от
простой диффузии в виде протонированной кислоты (гл. 8).
Пероксисомы.
Хрупкость изолированных пероксисом и использование для
их очистки изопикнического центрифугирования при высокой
концентрации осмотически активных молекул сахарозы затруд-
няет их выделение в интактном и пригодном для метаболиче-
ских исследований виде. Есть данные о том, что изолированные
пероксисомы способны метаболизировать 140-гликолат до глиок-
силата и глицина. Когда донорами аминогруппы служат глута-
мат, аланин или серин, главным продуктом метаболических
превращений гликолата становится глицин.
Митохондрии
Митохондрии, выделенные из листьев шпината,' декарбокси-
лируют глицин в соответствии со стехиометрией уравнений
(13.8) и (13.9), когда из двух молекул глицина образуется 1 мо-
лекула СО2+1 молекула серииа + 1 молекула NH3. Эта реакция
может быть связана с транспортом электронов [при этом сте-
хиометрическое соотношение СО2 (выделение) и О2 (поглоще-
ние) составляет 2/1] и с окислительным фосфорилированием
(в этом случае соотношение Р/2е“ приближается к 3, что ука-
зывает на связь декарбоксилирования глицина с восстановлени-
ем NAD+). Декарбоксилирование глицина в митохондриях сти-
мулируется, если добавить оксалоацетат; при этом реакция де-
карбоксилирования ие сопряжена с транспортом электронов.
Очевидно, эта активность связана с повторным ’ окислением
NADH при участии малатдегидрогеназы. In vivo восстанови-
тельная сила может переноситься из митохондрий в пероксисо-
мы за счет челночйого обмена дикарбоновыми кислотами
(рис. 13.6): . . ’
2 Глицин 4-NAD+ ->-
.----> NADH+Н++Серин+ CO2 + NW3- (13,8 и. 13.9)
Оксалоацетат-j-NADH-р Н,|_NAD+ .Малат (13.28)
422
ГЛАВА 13
В настоящее время нет никаких данных о том, что восстанови-
тельная сила, образующаяся в процессе дыхательного окисле-
ния глицина, может использоваться для реассимиляции аммиа-
ка в митохондриях с участием глутаматдегидрогеназы.
Источники данных: [4, 10, 24, 64, 68].
13.9. Гликолатный путь у С4-растений
Все методы, применяемые для обнаружения фотодыхания
(разд. 13.2) в целых листьях, говорят о том, что у С4-растений
фотодыхание или вообще отсутствует, или же его активность
очень невелика. Это, однако, не связано с отсутствием фермен-
тов гликолатного пути. Вероятнее всего, обмен веществ, связан-
ный с функционированием гликолатного пути, просто подавлен
и не функционирует. Предполагают, что в результате работы
С4-цикла в клетках обкладки проводящих пучков в процессе
фотосинтеза поддерживается относительно высокий уровень СО2
(разд. 12.4). В результате этого отношение СО2/О2 у них выше,
чем у Сз-растений, а это благоприятствует фотосинтезу и спо-
собствует подавлению синтеза гликолата. Однако не исключено,
что какая-то часть гликолата синтезируется за счет РуБФ-ок-
сигеназы, так как следует ожидать, что в обкладке проводящих
пучков должна поддерживаться определенная концентрация О2.
Это особенно справедливо для С4-растений, у которых очень вы-
сока активность фотосистемы II, а хлоропласты обкладки прово-
дящих пучков характеризуются повышенной способностью к вы-
делению О2 (разд. 11.7). Кислород ингибирует зависимую от
света фиксацию СО2 в изолированных клетках обкладки прово-
дящих пучков С4-растений. Поэтому, когда С4-цикл не функцио-
нирует, фотосинтез в хлоропластах обкладки проводящих пуч-
ков становится столь же чувствительным к О2, как и фотосин-
тез в хлоропластах С3-растений.
У самых разных исследованных С4-растений было показано,
что более 9О°/о активности гликолатоксидазы и глиоксилатре-
дуктазы локализовано в клетках обкладки проводящих пучков.
Активность фосфогликолатфосфатазы и каталазы также в ос-
новном присуща клеткам обкладки проводящих пучков. При
расчете на единицу хлорофилла активность этих ферментов у
Сз-растений в несколько раз выше, чем в клетках обкладки про-
водящих пучков у С4-растений. Вся СО2, которая высвобожда-
ется в гликолатном пути у С4-растений, обнаруживается глав-
ным образом в клетках обкладки проводящих пучков. Эта СО2
может вновь фиксироваться при участии РуБФ-карбоксилазы в
клетках обкладки проводящих пучков или за счет ФЕП-карбок-
силазной активности в клетках мезофилла. Глицераткиназа, од-
нако, локализована у С4-растений в клетках мезофилла. По-ви-
ФОТОДЫХАНИЕ
423
димому, весь глицерат, который синтезируется за счет гликолат-
ного пути в клетках обкладки проводящих пучков, должен
транспортироваться в клетки мезофилла, чтобы там превра-
титься в триозофосфаты. У Сч-растений нет такой непосредствен-
ной связи между ВПФ-циклом и гликолатным путем, как у
Сз-растений.
Экстракты, полученные из листьев Сз-растений, характери-
зуются высоким отношением активности фосфогликолатфосфа-
тазы к активности ФГК-фосфатазы. И наоборот, в экстрактах,
полученных из листьев С4-растений, отмечено высокое отноше-
ние активности ФГК-фосфатазы к активности фосфогликолат-
фосфатазы. У Сграстений ФГК-фосфатаза локализована в ос-
новном в клетках мезофилла, тогда как фосфогликолатфосфа-
таза почти вся находится в клетках обкладки проводящих пуч-
ков. Роль ФГК-фосфатазы у С4-растений не выяснена. Согласно
одной из гипотез, этот фермент облегчает челночный перенос
ФГК из клеток обкладки проводящих пучков в клетки мезофил-
ла (разд. 11.6). По другой гипотезе ФГК-фосфатаза вместе с
глиоксилатредуктазой участвует в синтезе серина у С4-расте-
ний. Основная часть метки, которая появляется в составе сери-
на после фиксации ИСО2, находится в карбоксильной группе.
Исходно, в процессе фиксации 14СО2 по карбоксильной группе
должна метиться главным образом ФГК. Дальнейшие метабо-
лические превращения ФГК в серии через глицерат и гидрокси-
пируват должны привести к тому, что метка появится в составе
карбоксильной группы серина. У Сз-растений радиоактивная
метка после фиксации ИСО2 практически одинаково' распреде-
ляется между различными положениями углеродных атомов в
серине и глицине. Это согласуется с тем, что они синтезируются
при участии гликолатиого пути. В процессе фиксации ИСО2
в ВПФ-цикле равномерное включение метки наблюдается у
РуБФ. После взаимодействия РуБФ с О2 метка должна вклю-
чаться в оба положения гликолата. Очень важно отметить, что
значительное включение метки в карбоксильную группу серина,
которое наблюдается у С4-р астений, вовсе не означает, что глав-
ным путем синтеза серина является его образование из ФГК.
По мере фиксации 41СО2 удельная активность ФГК намного
превысит удельную активность гликолата, так что включение
метки из этих двух предшественников в серин не будет отра-
жать действительное соотношение потоков углерода, включаю-
щегося в серин по тому или иному метаболическому пути.
Поскольку квантовый выход фотосинтеза у С4-растений оди-
наков как при 21% О2, так и при 2% О2 (разд. 13.12), можно
вполне аргументированно говорить о том, что при характерном
для нормальных условий уровне СО2 в атмосфере гликолатный
путь практически не функционирует. Если С4-р астения подверг-
4S4
ГЛАВА 13
нуть стрессовому воздействию, такому, как недостаток воды
(при этом увеличивается сопротивление устьиц), или такому,
как повышенная температура, внутриклеточная концентрация
СО2 может резко понизиться. При высоких температурах [CO21]
в водной фазе будет понижаться из-за уменьшения ее раствори-
мости в воде. И в том и в другом случае скорость фотосинтеза
будет уменьшаться вследствие ограниченного поступления СО2
из атмосферы в клетки обкладки проводящих пучков. Это, веро-
ятно, приведет к тому, что в [Этих клетках увеличится отноше-
ние О2/СО2, возрастет относительный поток углерода к гликола-
ту и еще в большей степени усилится повторное циклическое
использование СО2 (повторная фиксация при участии ФЕП- или
РуБФ-карбоксилазы).
В этих условиях, когда доступ СОг лимитируется, квантовый
выход фотосинтеза у С4-растений должен обязательно понизить-
ся. Даже если не наблюдается явной утечки СО2 из листа
Сграстений, при возросшем циклическом использовании СОг
необходима дополнительная энергия, чтобы фиксировать эту
СО2. При этом наблюдалось бы заметное снижение квантового
выхода.
Источники данных: [1, 2, 31, 36, 50, 57, 64].
13.10. Ингибирование фотосинтеза кислородом
и две его составляющие
В 1920 г. Варбург обнаружил, что у Chlorella кислород инги-
бирует дыхание. Исследования, проведенные в шестидесятые
годы Бьеркманом, Даунесом и Хескетом и Кротковым и др., по-
казали, что при нормальных атмосферных условиях О2 ингиби-
Рис. 13.7. Зависимость углекис-
лотного компенсационного пункта
(Г) у Сз-растения от концентра-
ции Ог.
рует фотосинтез у одних высших
растений (позднее стало извест-
но, что у Сз-растений) и совер-
шенно не действует на другие
высшие растения (известные ны-
не под названием Сграстений).
В 1966 г. Форрестер и др. и Тре-
гунна и др. предположили, что
кислородное ингибирование фо-
тосинтеза складывается из двух
составляющих. Одну из этих со-
ставляющих обозначили как
кислородное ингибирование за
счет фотодыхания. Ее существо-
вание вытекало из того факта,
что увеличение [О2] приводит к
увеличению переходного скачка
ФОТОДЫХАНИЕ
425
Рис. 13.8. Эффективность карбоксилирования (ЭК) у С3-растения.
после выключения света и к прямо пропорциональному смеще-
нию углекислотного компенсационного пункта (Г) в область
более высоких концентраций СО2 (рис. 13.7). Вторую составля-
ющую обозначили как кислородное ингибирование эффектив-
ной фиксации СО2 (эффективности карбоксилирования, ЭК).
ЭК определяют по начальному наклону кривой на графике за-
висимости наблюдаемого фотосинтеза (НФС) от концентрации
СО2 (рис. 13.8):
ЭК = [СО„] —Г (13.29)
Таким образом, эта зависимость описывается вогнутой кривой,
и ЭК убывает с увеличением концентрации О2 (рис. 13.9).Если
построить график зависимости 'НФС от внутриклеточной [СО2]
и затем определить ЭК, то величина, обратная ЭК (1/ЭК), бу-
дет равна гт (сопротивление мезофилла):
НФС = 1-^2.]”путр~ [c°iJxn (13.30)
r m
где [СО2]впутр — внутриклеточная концентрация СО2 во внут-
реннем воздушном пространстве листа, [СО2]хп—концентра-
ция СО2 в хлоропластах, гт — сопротивление мезофилла (скла-
дывается из сопротивления переносу через водную фазу .и сопро-
тивления карбоксилированию).
426
ГЛАВА 13
Если принять, что [СО2]хп примерно равна Г, то
гт
НФС = г
Г m
[СО2]Вцутр'— Г
НФС
= 1/ЭК
(13.31)
(13.32)
Следовательно, увеличение концентрации О2 ведет к уменьше-
нию ЭК или к увеличению сопротивления мезофилла. Обратная
зависимость между ЭК и гт сохраняется только' в том случае,
когда ЭК рассчитывают, исходя из внутриклеточной ’[СОг] и
когда интенсивность фотосинтеза измеряют при сравнительно
низкой [CO2J (начальный участок графика зависимости интен-
сивности фотосинтеза от 1[СО2] представляет собой прямую ли-
нию) .
Идея о наличии двух составляющих кислородного ингибиро-
вания фотосинтеза получила широкую поддержку еще за не-
сколько лет до получения доказательств биохимического харак-
тера. Одна из составляющих отражает прямое ингибирование
карбоксилазы кислородом, а вторая обусловлена потерей СО2
в процессе гликолатного пути (разд. 13.3).
Одно время считали, что химическое подавление гликолат-
ного пути можно было бы использовать для подавления фото-
дыхания и, таким образом, для стимуляции фотосинтеза. Нет,
однако, никакой информации о том, что, подавив гликолатный
путь, можно уменьшить конкурентное ингибирование фотосин-
теза кислородом. Хотя и было обнаружено, что некоторые инги-
биторы блокируют определенные этапы гликолатного' пути
(разд. 13.4), нет никаких данных, которые позволили бы сде-
лать вывод о том, что фотосинтез возрастает после подавления
Рис. 13.9. Зависимость эффектив-
ности карбоксилирования (ЭК) у
С3-растения от концентрации О2.
обмена гликолата. Так, напри-
мер, бутил-2-гидрокси-З-бутино-
ат, эффективный ингибитор гли-
колатоксидазы, вызывает накоп-
ление гликолата и ингибирует
фотосинтез (разд. 13.4). Очень
важно то, что если гликолат об-
разуется, то он в основном ме-
таболизируется с образованием
трнозофосфатов, и углерод та-
ким образом возвращается и
ВПФ-цикл. Некоторые исследо-
ватели считают, что химически
модифицированная РуБФ-карб-
оксилаза-оксигеназа, даже если
у нее худшие кинетичс'жчо ’'-па-
метр ы (Хм и Vшах) ДЛЯ СВЯЗЬ!-
ФОТОДЫХАНИЕ
427
вания О2, может обеспечивать повышенные скорости фотосинте-
за у Сз-растений. Другие по-прежнему думают, что фотодыха-
ние может приносить даже некоторую выгоду растению в су-
ществующих условиях окружающей среды. Эти аспекты будут
рассмотрены в разд. 13.13.
Сейчас, когда мы лучше понимаем причины кислородного
ингибирования фотосинтеза, можно проанализировать, с каки-
ми биохимическими механизмами связаны изменения интенсив-
ности фотосинтеза при изменении [СО2] и фО2]. Фотосинтез в
листе можно описать уравнениями, которые включают в себя
кинетические константы для карбоксилазы и две составляющие
кислородного ингибирования фотосинтеза. Эти уравнения можно
сравнить с экспериментальными данными, характеризующими
фотосинтез в листе (разд. 14.3). Так, например, в отсутстгиеО2
и при условии, что зависимость интенсивности фотосинтеза в ли-
сте от [СО2] описывается кинетической кривой Михаэлиса —
Ментен,
ИФС0»/о О2 = .. (13.33)
-|-/<М(СО2)
где НФС — истинная скорость фотосинтеза, Vmax — максим аль-
пая скорость карбоксилазной реакции, [СО2] —концентрация
СО2 и лм(со2) — константа Михаэлиса карбоксилазы для СО2.
В обратных величинах уравнение (13.33) можно выразить так:
1/ИФС0% 0!!- 1 /^ах+йЩ- (13.34)
LVAJ2J итах
На графике двойных обратных величии (зависимость ИФС0о/о0
от [СО2] в координатах Лайнуивера — Берка) точка пересечения
с осью абсцисс, в которой 1/ИФСо%о2 = 0, соответствует величи-
не —1/Ам- Результаты исследований фотосинтеза в листьях
Сз-растений при низкой концентрации О2 и у С4-растений гово-
рят о том, что этот подход можно использовать для определе-
ния величины /<м(со2) (гл. 14, рис. 14.5).
Для тех случаев, когда в атмосфере присутствует О2, кото-
рый подавляет фотосинтез у Сз-растений, можно вывести сле-
дующее уравнение:
НФС = Vmax‘ ТСО2] + ^М(со2)] (1 + [О2]/Л> (о2)) (13’35)
Это уравнение можно вывести, зная зависимость между НФС
(наблюдаемая скорость фотосинтеза), скоростью карбоксилаз-
ной реакции (Ук), скоростью оксигеназной реакции (Vo) и Г
.[26], когда
V______________[|-О2]____________/1 о ос\
К - [СО2] + ЛМ(Со2) • (1 + £О2]/Лл (о2) 1 • °-'
428
ГЛАВА 13
Hnax'IQPgJ
К . "* Л'Мка:к (С02)
(13.37)
^Мкаж—это-кажущаяся Ли для СОз при различных концент-
рациях О2.
^Мкаж (С02) = ^ЖС02г(1ЧНО^Ж1(02)) (13.38)
Точно таким же образом
у ____________________________Ур max' [Og]_______
° W +/<М(02)-(1 +ICO2]/Kj (СОг))
(13.39)
где Vo — скорость оксигеназной реакции, VOmax—скорость ок-
сигеназной реакции при насыщающей концентрации субстрата,
Лм(о2>—константа Михаэлиса для 02 и /С1(со2) — константа ин-
гибирования для С02 (разд. 13.3). Отсюда следует, что
НФС = УК—0,5Vo
(13.40)
В точке, соответствующей Г, VK=0,5Vo, так как на каждую мо-
лекулу СО2, связавшуюся в результате карбоксилазной реак-
ции, приходится две молекулы гликолата, образовавшегося в
ходе оксигеназной реакции. Последующие метаболические пре-
вращения этих двух молекул гликолата в гликолатном пути
приводят к тому, что высвобождается 1 молекула СО2.
Отсюда также следует, что
г_^1СОД И
Тогда
/
Если вместо Ук подставить ее значение из уравнения (13.37),
то
, J-_____[' -Х']______
НФС - Умах-[СОа]____________[СО2] +ХМкаж (сОа) ... .
[СОЛ-ЛМкаЖ(с02) [СО2]
После преобразований получаем
НФС=У
Г max
[COJ-Г
[СО2] Км„аж (со2)
(13.41)
В этом уравнении учтены обе составляющие кислородного ин-
гибирования: одну из них, ту, что связана с. фотодыханием, от-
ражает вычитание Г в числителе дроби, а другую, т. е. непосред-
ФОТОДЫХАНИЕ
423
Рис. 13.10. Модель, описывающая фотосинтез в листе С3-растения [по урав-
нению (13.42), когда 1/НФС = 1/1/].
ствениое конкурентное ингибирование карбоксилазы, вводят,
внося в знаменатель значение кажущейся /<м(со2)- Данное урав-
нение справедливо для случаев, погда концентрация РуБФ яв-
ляется насыщающей или остается постоянной.
В обратных величинах уравнение (13.41) выглядит так:
1 /НфС = --уу j у • .Г + Л^мсо2) ' (1 з.42)
Kmax LlAJ2J 1 Kmax
Тогда на графике двойных обратных, величин ./[зависимость
1/НФС от 1/([СО2]—Г)] точка пересечения с осью абсцисс,
в которой 1/НФС=0, будет Соответствовать величине
-1/(/<мкаж(Со2)+Г) (рис. 13,10).
Суммарное ингибирование фотосинтеза кислородом, конку-
рентное ингибирование кислородом и ингибирование, обуслов-
ленное фотодыханием, можно выразить в следующем виде:
ИФСо% о2—НФСз1% о2 =
= Суммарное ингибирование кислородом (21%Оа).
ИФС0% Оз — ИФС21 % о2 =
= Конкурентное ингибирование кислородом (21% О2)
ИФС21%. о2—НФС21 %. о2 = Ингибирование,. обусловлен-
ное фотодыханием в присутствии 21 % О2
где ИФС0 %о2 —истинная скорость фотосинтеза в присутствии
0% О2, ИФС21%о2 —истинная скорость фотосинтеза в присут-
ствии 21 % О2, НФСа! %о2 — скорость .наблюдаемого фотосинте-
430
ГЛАВА 13
за при 21 % О2. Эти величины можно выразить в форме урав-
нений
ИФС„о, = [со^З„ (13.33>
Иф^°'=жг?С^ <13'43’
НФС21% оа = VmaxWjg^------ (13.41)
Данные уравнения могут быть применены для получения кри-
вых, отражающих зависимость НФС и ИФС от [СО2]. Величи-
ны Vmax и Км можно определить либо в прямых опытах с кар-
боксилазой, либо рассчитать по графику зависимости 1/НФС
от 1/(|[СО2]—Г), а Г определить экспериментальным путем.
Графики зависимости 1/НФС от 1/([СО2]—Г) (которые анало-
гичны графикам зависимости 1/7 от 1/S, применяемым в фер-
ментативной кинетике), полученные при разных концентрациях
О2, свидетельствуют о том, что О2 является конкурентным инги-
битором (рис. 14.5, В) и что величина кажущейся Кмкаж(со2>
возрастает с увеличением [О2], как и следует ожидать для кон-
курентного ингибитора (рис. 14.5, Б и 14.6). При высоких кон-
центрациях СО2 в фотосинтезе целых листьев могут происхо-
дить такие изменения, что фотосинтез никогда не будет дости-
гать скорости, соответствующей Утах карбоксилазы. Это связано^
с ограниченным притоком энергии фотохимических процессов
(рис. 14.6).
Экспериментально измерить НФС21 %Оа и ориентировочно
оценить ИФСоо/оо2 очень просто. Разность между этими двумя
величинами выражает суммарное ингибирование фотосинтеза
кислородом. ИФСо%о2 обычно измеряют в присутствии 1 или
2% О2, так как у некоторых видов растений устьица могут не
открыться, если в атмосфере совсем не будет О21}. И наоборот,
непосредственно измерить ИФС21%о2 очень трудно. Если бы
можно было измерить фотодыхание, то сумма НФС21%02 и ско-
рости фотодыхания составила бы ИФС21%02.
Большинство методов, применяемых для измерения фотоды-
хания, позволяет провести лишь качественную оценку и не от-
*> Следует отметить, что концентрация О2 в хлоропластах очень близка к
его концентрации в атмосфере, несмотря на то что в процессе фотосинтеза
происходит выделение О2. Скорости оттока О2 из листа и притока СО2 в лист
равны. Максимальная разница между концентрацией СО2 в атмосфере и внут-
ри хлоропластов составляет примерно 0,03%. Если принять, что сопротивле-
ния для диффузии СО2 и О2 одинаковы, то [О2] в листе должна в лучшем
случае превышать [OJ в атмосфере не более чем на 0,03%.
ФОТОДЫХАНИЕ
431
ражает истинную величину скорости фотодыхания в стационар-
ных условиях при нормальном содержании СО2 в атмосфере.
Одной из проблем является то, что часть СО2, высвобождаемой
в ходе фотодыхания, снова связывается в процессе фотосинтеза.
Людвиг и Канвии разработали метод с двойной меткой. Исполь-
зуя 12СО2 и иСО2, они определяли этим методом наблюдаемую
скорость фотодыхания при стационарном состоянии фотосинте-
за. Для этого в растение вводили 12СО2 в равновесных услови-
ях, измеряли ее поглощение с помощью инфракрасного газоана-
лизатора и определяли НФС21 %Оа. Затем вводили ИСО2 и од-
новременно в течение 30 с измеряли скорость поглощения мет-
ки, что позволяло определить ИФС21%Оа . Полагали, что за та-
кой короткий период продукты фиксации ИСО2 не успевают ис-
пользоваться в процессе фотодыхания. Разность между ско-
ростью фиксации ИСО2 и 12СО2 принимали за скорость фотоды-
хания. Эти эксперименты подтвердили представление о том, что
существуют две составляющие кислородного ингибирования фо-
тосинтеза— обусловленная прямым ингибированием и связан-
ная с фотодыханием, — которые вносят почти одинаковый вклад
в суммарное подавление фотосинтеза кислородом при нормаль-
ных атмосферных условиях. Остается неясным, однако, в какой
степени 12СО2, выделяющаяся в ходе фотодыхания, использует-
ся повторно во внутреннем цикле, и какова доля ассимилиро-
ванной 14СО2, которая вовлекается в процесс фотодыхания в
различных условиях эксперимента. Поэтому предложенный вы-
ше способ, вероятно, имеет ограниченное применение, особенно
при определении относительной интенсивности фотодыхания при
различных концентрациях СО2 и О2 или при различных темпе-
ратурах. В ряде работ, где применяли эту методику, авторы
выдвигали предположение о том, что скорость фотодыхания в
атмосфере с 21 % О2 остается неизменной при увеличении кон-
центрации СО2 вплоть до нормальной. Непосредственная оцен-
ка скачкообразного увеличения СО2 после выключения освеще-
ния и расчетные значения скорости этого процесса в неравновес-
ной системе свидетельствуют, однако, о том, что интенсивность
фотодыхания постепенно убывает с увеличением [СО2] выше Г.
ИФС21 %о2 можно определить, прибавив скорость скачкообраз-
ного прироста СО2 после выключения освещения при той или
иной [СО2] к скорости НФС21о/0о2. При нормальном содержа-
нии СО2 в атмосфере на долю фотодыхания приходится около
1/3 суммарного ингибирования фотосинтеза кислородом. Опыты
по определению скорости поглощения 18О2 также свидетельству-
ют о том, что фотодыхание ослабевает с увеличением i[СО2].
Поскольку СО2 конкурирует с О2 в реакции, катализируемой
РуБФ-карбоксилазой-оксигеназой, вполне логично, что при вы-
432
ГЛАВА 13
соких концентрациях СО2 фотодыхание подавляется. В процес-
се фотодыхайия в присутствии один из атомов О2 включает-
ся в состав карбоксильной группы фосфогликолата. [уравнение
(13.1) ], а затем и в карбоксильные-группы других метаболитов
гликолатного пути. Следует отметить, что метка из 18О2 может
включаться и в продукты фотосинтеза в результате реакции Ме-
лера (разд. 5.8). Поэтому нельзя говорить о том, что поглоще-
ние 13О2 на свету обеспечивается исключительно за счет фото-
дыхания.
Необходимо еще раз возвратиться к тому, как же связано
значение эффективности карбоксилирования (ЭК) с уравнения-
ми (13.33) и (13.41). По определению величина ЭК соответст-
вует наклону начального отрезка кривой на графике зависимо-
сти НФС от [СОа], и, как уже отмечалось выше, ЭК убывает с
увеличением [Ог]. В отсутствие О2 из. уравнения (13.33) выте-
кает, что при низкой ;[СОг] концентрация субстрата становится
ничтожно малой по сравнению с величиной Км(со2)> так что
ИФСо%о2 ~ Утах- [СОг] /Км(со2у По- этой причине начальный
отрезок кривой, отражающей интенсивность фотосинтеза при
малых значениях [СОг], оказывается прямолинейным, а ЭК =
= ИФС0%о2/[СО2] = Ктах/Км(со 2). По-видимому, уменьшение ЭК
при увеличении [Ог] обусловлено непосредственным конкурент-
ным ингибированием фотосинтеза кислородом. По определению
йри конкурентном ингибировании наклон начального участка
кривой на графике зависимости скорости реакции от концентра-
ции субстрата уменьшается благодаря увеличению Км, так что
Рис. 13.11. Изменение эффективности карбоксилирования НФС/([СО2]—Г)
по мере того, как концентрация СО2 снижается до Г [из уравнения (13.41),
см. текст].
ФОТОДЫХАНИЕ
433
Рис. 13.12. Определение Кцо-i фотосинтеза in vivo у С3- растения.
ЭК= Утах/Дмкаж, а кажущаяся Км возрастает с увеличением
концентрации ингибитора. Б присутствии Ог из уравнения
(13.41) следует, что наклон кривой на графике зависимости
НФС21%оа от [СО2]—Г равен Ктах/([СО2]+Кмкаж(сог)). По
мере того как [СОг] приближается к Г, наклон кривой прибли-
жается к величине Vmax/(Г+Кмкг1ж <со2) ) Поскольку величи-
на Г относительно мала по сравнению с величиной Кмкаж(со2ц
наклон начального отрезка кривой отражает величину ЭК [т. е.
Утах/Кмкагк(СО2)] (рис. 13,11). Графики двойных обратных вели-
чин, описывающие зависимость интенсивности фотосинтеза от
'[СОг]—Г, свидетельствуют о том, что углекислотная кривая
подчиняется кинетике Михаэлиса — Ментен до тех пор, пока не
достигается очень высокая [СОг] (рис. 14.5). В исследованиях
фотосинтеза в целых листьях было показано, что с увеличением
[О2] ЭК уменьшается (или увеличивается гт) таким образом,
как если бы О2 являлся конкурентным ингибитором. ЭК (или
иначе УшагУКмкаж) ПРИ различных [О2] можно оценить по гра-
фику зависимости 1/V от 1/([СОг]—Г), изображенному на
рис. 13.10. Зависимость 1/ЭК от [О2] линейна, что позволяет
определить величину Кдо2) (рис. 13.12).
1 /эк = /СМк'-1,к (с°2) = Л'М(СО^ -]_ [о I.
Кпах Кпах 2 Клах' Kl (О,)
Связь фотосинтеза в целом листе с карбоксилированием и фото-
химическим потенциалом у Сз- и С^-растений более детально
рассматривается в гл. 14.
Источники данных: [4, 13, 14, 15, 23, 26, 27, 29, 37, 46, 62]..
28-234
-434
ГЛАВА 13
13.11. Доля ингибирования фотосинтеза кислородом
НфС2% О2 НФС21% О2
НФС2% Оа
(100) =
= % ингибирования фотосинтеза кислородом
Это уравнение хорошо отражает величину кислородного ин-
гибирования фотосинтеза в различных условиях. С увеличением
[СОа] доля вызываемого О2 ингибирования фотосинтеза снижа-
ется от 100% при Г до величины, близкой к нулю, при концен-
трации СО2, в 2—3 раза превышающей нормальный уровень
СО2 в атмосфере (т. е. в присутствии 0,1% СО2). При повыше-
нии температуры кислородное ингибирование постоянно нарас-
тает (от 20—30% при 20 “"С до 40—60% при 35°C). Это свиде-
тельствует о том, что фотосинтез у Сз-растений более чувстви-
телен к Ог при более высоких температурах. Причины данного
явления неясны, однако, возможно, это связано с увеличением
отношения растворимостей О2/СО2 или же с изменением кине-
тических констант у РуБФ-карбоксилазы-оксигеназы, более бла-
гоприятным для оксигеназной реакции (гл. 14). Фотосинтез у
С4-растений, обладающих механизмом концентрирования СОг,
позволяет преодолеть такую чувствительность к О2 при высо-
ких температурах, свойственную фотосинтезу С3-растений
(разд. 14.5). С увеличением интенсивности освещения абсолют-
ная величина кислородного ингибирования возрастает так же,
как и скорость фотосинтеза. В то же время доля ингибирования
фотосинтеза кислородом при этом практически не меняется или
лишь слегка убывает.
Источники данных £23, 27, 37, 39].
13.12. Влияние кислорода на квантовый выход
Обычно кислородное ингибирование фотосинтеза у Сз-рас-
тений оценивают при высокой интенсивности освещения. Вместе
с тем исследуют и влияние О2 на квантовый выход (КВ) фото-
синтеза (т. е. число’ молей фиксированной СО2 на моль погло-
щенных квантов, см. разд. 3.9). Но в последнем случае измере-
ния проводят при низкой интенсивности освещения, не превы-
шающей 5% прямой инсоляции. При любом заданном значении
[CO2J и температуры доля ингибирования фотосинтеза кислоро-
дом при сильном освещении равна доле ингибирования КВ. От-
сюда видно, что доля кислородного ингибирования при высоких
интенсивностях освещения не столь уж велика, как этого следо-
вало бы ожидать, если бы его функция заключалась только в
диссипации избытка энергии. Даже при очень слабом освеще-
нии Оя понижает эффективность использования поглощенной
ФОТОДЫХАНИЕ
435
Ингибирование квантового выхода, %
Рис. 13.13. Связь между ингибиро-
ванием квантового выхода и ингиби-
рованием фотосинтеза иа сильном
свету при увеличении [Os] (Bjorkman,
1966).
Рис. 13.14. Высокая концентрация
СО2 снимает кислородное ингибиро-
вание квантового выхода (Ehlerin-
ger, Bjorkman, 1977).
энергии. Кислородное ингибирование КВ снимается при увели-
чении [СОг] выше ее нормального уровня в атмосфере, причем
одновременно снимается и кислородное ингибирование фотосин-
теза (на сильном свету). Это свидетельствует о том, что дейст-
вие Ог во многом зависит от относительного содержания О2 и
СО2 (рис. 13.13 и 13.14). Снижение КВ под действием О2 может
быть связано с потерями энергии на зависимые от О2 флуорес-
ценцию и фотодыхание. Повышенное излучение флуоресценции
в присутствии О2 может быть обусловлено кислородным инги-
бированием ферментов ВПФ-цикла, в результате которого
уменьшается использование энергии фотохимических процессов.
У Сз-растений доля ингибирования КВ под влиянием О2 воз-
растает с увеличением температуры. Это очень напоминает
увеличение доли ингибирования фотосинтеза кислородом, ко-
торое наблюдается на ярком свету при повышении температу-
ры. Сходным образом и повышенное ингибирование КВ при бо-
лее высоких температурах может быть связано с увеличением
отношения растворимостей О2/СО2 при повышении температуры
или же с изменением кинетических констант для О2 и СО2, бла-
гоприятным для взаимодействия с О2 (рис. 13,15; гл. 14).
У Сграстений КВ остается одинаковым как при низком (2%),
так и при высоком (21%) содержании О2 в атмосфере, а темпе-
ратура в интервале 15—35 °C никак не влияет на КВ. Это впол-
не согласуется с данными о том, что у С4-растений отсутствует
кислородное ингибирование фотосинтеза.
28*
436
ГЛАВА 13
U Температура писта
?Рис. 13.15. Зависимость квантового выхода от температуры у Сз-растелий
(Ku, Edwards, 1978).
При 30 °C в присутствии 21% О2 и нормальном содержании
СО2 в атмосфере величины КВ у С3- и С4-рчстений очень близ-
ки. В присутствии 2% О2 КВ у С3-растений возрастает, а у
Сграстений он одинаков при 2 и 21% О2 (табл. 13.2). Поэтому
при 30 °C, низкой интенсивности освещения и в присутствии
.2% О2 Сз-растения более эффективно (36%; табл. 13.2) исполь-
зуют поглощенную ими энергию, чем Ct-pастения. В общих чер-
тах эти различия можно объяснить следующим образом. В при-
сутствии 2% б2 фотосинтез не ингибируется кислородом ни у
С3-, ни у С^-растений. В этом случае энергетические затраты со-
ставляют теоретически 3ATP/2NADPH на каждую молекулу
фиксируемой СО2, которая усваивается в ВПФ-цикле у Сз-рас-
тений, и 5ATP/2NADPH на одну молекулу СО2 у С4-растений.
Поэтому квантовая эффективность у С4-растений ниже, чем у
Ca-растений; Однако в присутствии 21 % О2 квантовая эффек-
тивность у Сз-растений снижается из-за кислородного ингибиро-
вания фотосинтеза. Рассмотрим в качестве примера 30%-ное
ингибирование фотосинтеза кислородом. Относительная ско-
рость фотосинтеза в присутствии 2% О2 = Ю0, а в присутствии
21% О2 = 70. В этом случае ингибирование фотосинтеза кисло-
родом в процентах составляет:
100—70
—ТОО (700)^30% . .
ФОТОДЫХАНИЕ
437
Таблица 13,2. Усредненные величины квантового выхода у ряда С3- .
и С|-растениЙ при 30 °C, нормальном содержании СО2 в атмосфере ..
и в присутствии 2 или 21% Оз (Ehleringer, BjSrkman, 1977)
Квантовый выход Число квантов, необходи- мых для фиксаций одной молекулы СО2
21% О2 2% О2 21% О2 ‘ 2% О2
Сз-Растения 0,0524 0,0733 19,1 13,6
СгРастения 0,0534 0,0538 18,7 18,6
При постоянном поглощении энергии в присутствии 2 или
21% Ог энергетические затраты на фиксацию СОз и последую-
щие превращения до уровня триозофосфатов должны возрасти
ют 3ATP/2NADPIH иа одну молекулу СОз, фиксируемой при
2% Ог, до 4,3ATP/2,9NADPH (3ATP/2NADPH иа 0,7 молекулы
фиксируемой СОг) иа одну молекулу СО2, фиксируемой при
21% Ог. При атмосферном уровне О2 для фиксации 10 молекул
СО2 у Сз-растений потребуется 43 молекулы АТР и 29 молекул
NADPH, а у С4-растений — 50 молекул АТР и 20 молекул
NADPH (разд. 11.6). Следовательно, в присутствии 21 % О2
суммарное потребление энергии при фиксации одной молекулы
СОг у Сз- и у С4-растений становится одинаковым.
Измеряемые квантовые затраты у С3- и С4-растений намного
выше, чем величины, рассчитанные теоретически. Это может
быть связано с расходованием энергии па другие процессы, вы-
ходящие за рамки фиксации СОг до триозофосфатов (такие, как
ассимиляция азота, синтез макромолекул, дыхание), а также с
неспецифическим поглощением энергии (например, клеточными
стенками) в листьях. Рассмотрим следующий пример: на одну
молекулу NADPH приходится 4 кванта и Р/2е-=1,33 (когда
Н|/АТР = 3, см. разд. 5.14). В этом случае теоретически расход
энергии в присутствии 2% О2 у Сз-растений составит 9, а у
Сграстений —15 квантов. В присутствии 2% О2 для С3-растений
(3ATP/2NADPH):
2NADPH, 2.66АТР = 8 квантов
ЗАТР— 2.66АТР = (0,34АТР) (ЗВ+/АТР) = 1Н+ = 1 квант
Итого: 9 квантов (ср. с экспериментальной величиной 13,6,.
: . приведенной в табл. 13,2)
Аналогичный расчет для С3-растения, находящегося в атмо-
сфере с 21% О2, с учетом того, что на одну молекулу связан-
ной СО2 приходится 4,3ATP/2,9NADPH, дает величину — 13
438
ГЛАВА 13
квантов (ср. с экспериментальной величиной 19,1, приведенной
в табл. 13.2).
Для С4-растений в присутствии 2 или 21% О2 (5АТР/'
/2NADPH):
2NADPH, 2,66АТР = 8 квантов
5АТР— 2.66АТР = (2.34АТР) (ЗН+/АТР) = 7Н+= 7 квантов
Итого: 15 квантов (ср. с экспериментальной величиной 18,6,
приведенной в табл. 13.2)
В нормальных атмосферных условиях при снижении темпе-
ратуры ниже 30 “С у Сз-растений наблюдается снижение кисло-
родного ингибирования КВ. Это приводит к тому, что КВ у этих
растений становится выше, чем у С4-растений. Поэтому при.
ограниченном освещении и при низкой температуре С3-растения
более эффективно, чем С4-растения, используют энергию света.
И наоборот, при высоких температурах и при слабом освещении
С4-растения усваивают энергию света более эффективно, чем.
Сз-растения (из-за того, что при повышении температуры КВ у
Сз-растений снижается).
Хотя степень кислородного ингибирования фотосинтеза оди-
накова при разных интенсивностях освещения, лимитирующее
действие СО2 У Cj-растений проявляется сильнее при ярком ос-
вещении. При высокой интенсивности освещения атмосферного,
уровня СО2 может не хватать для насыщения фотосинтеза у
Сз-растений даже тогда, когда нет кислородного ингибирования:
фотосинтеза (разд. 14.5). По этой причине иа сильном свету
С4-растения, обладающие механизмом концентрирования СО2,.
более эффективно используют имеющуюся в наличии энергию,
чем это делают Сз-растения. При оценке относительной эффек-
тивности использования энергии этими двумя группами фотосин-
тезирующих организмов следует учитывать все имеющиеся фак-
торы: температуру, освещение, содержание СО2 и О2.
Источники данных: [25, 38].
13.13. Предполагаемая роль фотодыхания
Вопрос о том, почему одни растения способны к фотодыха-
нию, а другие нет, дебатируется с давних пор. Одно из объяс-
нений заключается в том, что фотодыхание устраняет избыточ-
ную энергию, возникающую за счет фотохимических процессов,
и таким образом предотвращает фотоингибирование фиксации
СО2, которое может вызываться фотоокислением и разрушением
фотохимического аппарата. Освещение фотосинтезирующих кле-
ток в атмосфере азота в отсутствие СО2 приводит к необратимо-
му нарушению их способности фиксировать СО2. Кислород мо-
жет защищать клетки от такого фотохимического повреждения.
ФОТОДЫХАНИЕ
439
Конечно, при низкой освещенности фотоингибирование невели-
ко, и в этих условиях, когда скорость фотосинтеза лимитирует-
ся не доступом СО2, а поступлением энергии, кислородное ингиби-
рование фотосинтеза, по-видимому, незначительно. Однако от-
носительное ингибирование фотосинтеза кислородом на слабом
свету и при сильном освещении одинаково. В связи с выдвину-
тым выше объяснением возникла идея о том, что фотодыхание
защищает растение от фотодеструкции в тех случаях, когда
ограничение доступа СОг выражено особенно сильно — во время
недостатка воды и (или) при высокой температуре. Так, напри-
мер, при очень низкой [СО2], т. е. в углекислотном компенсаци-
онном пункте, энергия рассеивается без реальной фиксации СО2
(см. рис. 6.11).
Возможно, что гликолатный путь играет какую-то роль в
синтезе сахарозы и таких аминокислот, как глицин и серин. Од-
нако скорость поступления углерода в реакции гликолатного
пути [которая в среднем составляет 30—40 мкмоль выделяю-
щейся СОг- (мгХл) -1 -ч-гораздо выше, чем это требуется для
-синтеза аминокислот. Значение гликолатного пути для синтеза
сахарозы пока не ясно. На первый взгляд очевидно, что путь
синтеза сахарозы из триозофосфатов гораздо проще и короче.
Кроме того, у С4-растеиий гликолатный путь функционирует с
гораздо меньшей скоростью, и это свидетельствует о том, что
этот путь не очень важен для поддержания высокой скорости
синтеза сахарозы н аминокислот.
Некоторые гипотезы основаны на том, что фотодыхание в
паше время ие дает растению никаких преимуществ. По- одной
из гипотез предполагают, что РуБФ-карбоксилаза-оксигеиаза
возникла в ходе эволюции миллионы лет назад, когда отноше-
ние СО2/О2 в атмосфере было намного выше и поэтому более
благоприятствовало фотосинтезу, а не фотодыханию. ВПФ-
цикл представляет собой единственный известный карбоксили-
рующнй механизм, который может функционировать автоката-
литически (разд. 6.3). Неизбежным следствием этого могла
быть чувствительность РуБФ к окислению. С изменением усло-
вий окружающей среды (уменьшение отношения СО2/Ог) обмен
веществ у растений мог идти даже лучше без такого окисления,
как это видно на примере С4-растений. У С4- и САМ-растений
в ходе эволюции возникла более эффективно работающая, ус-
тойчивая к О2 карбоксилаза (ФЕП-карбоксилаза), но этот
фермент не может функционировать независимо от РуБФ-кар-
боксилазы, так как у этих растений отсутствует независимый
автокаталитический цикл.
Голдсуорси считал, что гликолатный путь возник в резуль-
тате адаптации у тех водорослей, которые находились в состоя-
нии симбиоза с бактериями. В ограниченном пространстве с низ-
440
ГЛАВА 13
кой [СОг] и высокой [Ог] синтез гликолата водорослями мог
оказаться взаимно выгодным обоим симбионтам. Гликолат мог
быть полезным для бактерий и служил для них источником уг-
лерода, тогда как бактерии могли снабжать примитивные водо-
росли необходимым метаболитом, например какой-нибудь ами-
нокислотой. Когда фотосинтезирующие организмы стали само-
стоятельными, им уже не нужно было синтезировать гликолат,
но они сохранили эту способность благодаря наличию у них
РуБФ-карбоксилазы, одного из основных ферментов ВПФ-
цикла.
Источники данных: [1, 2, 4—8, 30, 35, 44, 49, 59].
13.14. Стимуляция роста Сз-растений в средах
с пониженным содержанием О2
или обогащенных СО2
Кислородное ингибирование фотосинтеза можно устранить,
понизив [Ог] до 2% либо увеличив в 3 раза [СОг] в атмосфере.
Б этих условиях подавляется синтез глицина и стимулируется
Таблица 13.3. Влияние концентрации 02 и СО2 на включение метки
в некоторые продукты (глицин и сахарозу) ассимиляции 14СО2
Л. В присутствии 21% при разных концентрациях СОз1) (Lee, Whittingham, 1974) Доля суммарного включения метки, %
COs. м. д. относительная скорость фото- синтеза глицин <нхароза
100 0,08 36,5 3,6
300 0,20 15,6 12,5
950 0,63 4,9 32,4
В. При разных концентрациях СО2 п О22) (Osmond. BJdrkman, 1972) Доля суммарного включения, %
COs, м. д, 02, % фотосинтез, мкмоль иСО2’(дм2'Б мин)-1 (ГЛИЦИП сахароза
302 21 28,2 8,0 21,6
302 2 42,4 2,1 30,9
1068 20 86,0 2,1 36,9
1038 2 82,0 0,3 42,3
9 Продукты определяли после 3 мин фиксации иСОа на свету у Сз-растения Lyco*
persicon esculeniutn.
2) Продукты определяли после 5 мин фиксации НСО2 на свету у Сз-растения Atrlplex
patula.
ФОТОДЫХАНИЕ 441
"синтез сахарозы (табл, 13.3), Все эти биохимические изменения
фотосинтеза в целом листе хорошо согласуются с предположе-
нием о подавлении обмена веществ, связанного с фотодыханием.
Если фотодыхание приводит лишь к потере только что фиксиро-
ванной СОа и не дает растению никаких преимуществ (можно
предположить, например, что в результате нормального фото-
синтеза и «темнового» дыхания удовлетворяются потребности
растения в АТР и других метаболитах), то, вероятно, растение
будет лучше расти в атмосфере с повышенным содержанием СО2
при высоком отношении СО2/О2. В течение многих лет при вы-
ращивании растений в теплицах практиковалось выдерживать
их в атмосфере, обогащенной СОг, что в несколько раз увели-
чивало урожай. Все это были или садовые растения, или расте-
ния, применяемые в цветоводстве, о которых сейчас известно,
что это Сз-растения. За исключением кукурузы, сорго, сахарного
тростника, просяных культур и некоторых пастбищных злаков,
которые относятся к числу С4-растений, все основные сельско-
хозяйственные культуры принадлежат к С3-растениям, например
соя культурная, картофель и такие зерновые культуры, как рис,
ячмень и пшеница (разд. 11.8). После того как были открыты
Сз- и С4-пути фотосинтеза и были выявлены явные различия в
их ответной реакции иа содержащиеся в атмосфере Ог и СОг,
исследователи сконцентрировали внимание на сравнительном
изучении поведения С3- и С4-растений в различных средах. Так,
например, Бьеркман и его сотрудники в Институте Карнеги об-
наружили, что вес сухого вещества у различных видов Phaseo-
lus и Mimulus (Сз-растения) возрастает в 1,5—2 раза, если эти
растения выращивают при пониженной концентрации О2 в ат-
мосфере. В то же время у Zea mays (С4) такого эффекта не
наблюдалось. Точно так же Акита и Танака в Японии, проводя
интенсивные сравнительные исследования Сз- и С4-растений,
обнаружили, что масса сухого вещества у Сз-растений возрас-
тает в 2—5 раз после обогащения атмосферы СО2 (до 1000 или
2500 м.д.), а масса сухого вещества у С4-растений при этом
практически не изменяется.
При выращивании сои культурной (С3-растеиие) в атмосфе-
ре, обогащенной СОг, наблюдается резкая стимуляция вегета-
тивного и репродуктивного роста и ассимиляции N2 (табл. 13.4).
Усиленная ассимиляция N2 при обогащении атмосферы СО2 или
при ее обеднении Ог, очевидно, обусловлена стимуляцией фото-
синтеза и большей доступностью фотосинтетических продуктов
для азотфиксирующих симбиотических бактерий в корневых
клубеньках.
Снижение [О2] в атмосфере также стимулирует вегетативный
рост и фиксацию N2 у сои культурной. Совершенно' неожидан-
ным эффектом оказалось подавление репродуктивного роста и у
42
ГЛАВА 13
Таблица 13.4. Влияние С02 и 02 на вегетативное и репродуктивное развитие
Sorghum и Glycine max и на симбиотическую фиксацию N2 У Glycine max
(Quebedeaux, Hardy, 1973; 1975; Quebedeaux et al., 1975)
О». % СОг, М. Д. Вегетативный рост (листья, стебли, корпи), 1 г сухого вещест ва/растеиие Репродуктивный рост (семена), 1 г сухого вещеотва/ра- стение
A. Sorghum (С4-растение) Вегетативный и репродуктивный рост”
5 300 95,7 0,0
10 300 88,7 1,6
21 300 109,9 7,7
Б. Glycine max (С3-растение) Вегетативный и репродуктивный рост2’
5 300 55,3 0,0
21 300 10,0 12,9
5 2000 117,2 0,0
21 2000 102,0 20,1
В. Glycine max (Са-растение) Ассимиляция N23)
О2, % СО2, м. д. 1мг фиксированного N2/
/растение через 83 дня
21 300 155
5 300 350
21 1200 700
J) Оценивали после выращивания в течение 168 дней. Корпи выдерживали в обыч-
ной атмосфере, а надземную часть растений помещали, как указано, в различные усло-
вия, Дневная и ночная температуры составляли соответственно 30 и 21 °C.
2) Оценивали через 83 дня, при этом надземную часть растений помещали па 14—
83 дня в различные условия, а корни выдерживали просто па воздухе. Дневная и ночная
температуры составляли соответственно 30 и 21 °C.
3 Авторы суммировали данные, представленные в мг фиксированного N2, приходя-
щегося па I растение за 1 день, чтобы получить общее количество мг фиксированного
Na, приходящегося на одно растение. Растения выращивали соответственно днем при
29 °C, а ночью — при 18 °C.
сои, и у сорго (С4-растение) при снижении [О2]. Причины вы-
сокой потребности в О2 для репродуктивного роста непонятны,
но, по-видимому, это не связано с обменом веществ в процессе
фотодыхания.
Любое подавление роста, связанное с кислородным ингиби-
рованием фотосинтеза, в значительной степени снимается при
повышении концентрации СО2 в атмосфере до величины, в 2 ра-
за превышающей нормальную. Естественно, что степень стиму-
ляции роста при подавлении фотодыхания зависит и от других
факторов окружающей среды, таких, как свет, температура и
условия питания. За период с 1860 по 1960 г. средний уровень
ФОТОДЫХАНИЕ
443
СО2 в атмосфере постепенно вырос от 285 до 325 м. д. и продол-
жает расти со скоростью около 0,7 м.д. в год, в основном из-за
сжигания ископаемых запасов топлива. Небольшое увеличение
урожайности сельскохозяйственных культур частично может
объясняться увеличением фСО2] в атмосфере. Буферная емкость
океана, поглощающего СО2, и стимуляция фотосинтеза при по-
вышенных концентрациях СО2 — вот те факторы, которые будут
определять скорость возможного увеличения уровня СО2 в ат-
мосфере в обозримом будущем.
ОБЩАЯ ЛИТЕРАТУРА
1. Chollet R., Ogren IF. L. Regulation of photorespiration in Сз and C< species,
Bot. Rev,, 41, 137—179 (1975).
2. Jackson W. A., Volk R. J. Photorespiration, Ann. Rev. Plant Physiol., 21,
385—432 1970.
3. Lorimer G. IL, Woo К. C., Berni/ J. A., Osmond C. 13. The C2 photorespiratory
carbon oxidation cycle in leaves of higher plants: pathway and consequences.
In: Fourth International Congress of Photosynthesis (eds. D. O. Hall,
J. Coombs and T. W. Goodwin), pp. 311—322, London, The Biochemical
Society, 1978.
4. Schnarrenberger C., Fock H. Interactions among organelles involved in pho-
torespiration, Encyclopedia of Plant Physiology, Vol. 111, Transport in Plants
(eds, C. R. Stocking and U. Heber), pp. 185—234, New York, Springer-Ver-
lag, 1976.
5. Tolbert N. E. Microbodies — peroxisomes and glyoxysomes, Ann. Rev. Plant
Physiol., 22, 45—74 (1971).
6. Whittingham С. P. Photosynthesis, London, Oxford University Press, 1971.
7. Wolf F. T. Photorespiration, the C4 pathway of photosynthesis and related
phenomena, Adv. Frontiers Plant Set, 26, 161—231 (1970).
8. Zelitch I. Photosynthesis, Photorespiration and Plant Productivity, New York,
Academic Press, 1971.
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
9. Akita S., Tanaka I. Studies on the mechanisms of differences in photosynthe-
sis among species. IV. The differential response in dry matter production
between C3 and C4 species to atmospheric carbon dioxide enrichment, Proc.
Crop Sci. Soc. Japan., 42, 288—295 (1973).
10. Arron G. P., Spalding M H„ Edwards G. E. Stoichiometry of carbon dioxide
release and oxygen uptake during glycine oxidation in mitochondria isolated
from spinach (Spinacia oleracea) leaves, Biochem. J., 184, 457—460 (1979).
11. Badger M. R., Andrews T. J. Effects of CO2, O2 and temperature on a high-
affinity form of ribulose diphosphate carboxylase-oxygenase from spinach,
Biochem. Biophys. Res. Commun., 60, 204—210 (1974).
12. Badger M. R., Lorimer G. H. Activation of ribulose-l,5-bisphosphate oxyge-
nase. The role of Mg2-!-, CO2 and pH, Arch. Biochem. Biophys., 175, 723—
729 (1976).
13. Berry J. A., Badger M. R. Direct measurement of photorespiration as a func-
tion of CO2 concentration, Carnegie Inst. Wash. Year Book, 1978, pp. 175—
178, 1979.
14. Berry J. A., Osmond С. B., Lorimer G. H. Fixation of I8O2 during photorespi-
ration, Kinetic and steady-state studies of the photorespiratory carbon oxida-
444
ГЛАВА 13
tion cycle with intact leaves and isolated chloroplasts of C3 plants, Plant
. Physiol., 62, 954—967 (1978).
15. Bjorkman 0. .The effect of oxygen concentration on photosynthesis in higher
plants, Physiol. Plant., 19, 618—633 (1966). .
16. ' BjMitnan O„ Gauhl E„ Hiesey W. M., Nicholson F., Nobs M. A. Growth of
. Mimulus, Marchantia and Zea under different oxygen and carbon dioxide
levels, Carnegie Inst. Wash. Year book, 67, 477—479 (1969).
17. Bolin B., Keeling C. D. Large scale atmospheric mining as deduced from the
seasonal and meridianal variations of carbon dioxide, J. Geophys. Res., 68,
' 3899—3920, 1963. . . . •
18. Bowes G., Ogren W. L., Hageman R. H. Phosphoglycolate production catar
lyzed by ribulose diphosphate carboxylase, Biochem, Biophys. Res. Commun,
45, 716—722 (1971).
19. Bowes G., Ogren IP. L., Hageman R. H. pH dependence of the Km(C02) of
ribulose 1,5-dlphosphate carboxylase, Plant Physiol., 56, 630—633 (1975).
20. Canvin D. T., Fock H., jdoyd N. D. H. Environmental treatment and carbon
flow in the glycolate pathway, Fourth International Congress on Photosyn-
thesis (eds, D. O. Hall, J. Coombs and T. W. Goodwin), pp. 323—334, The
Biochemical Society, London, 1978.
21. Decker I. P. A rapid, post-illumination deceleration of respiration in green
leaves, Plant Physiol., 30, 82—84 (1955).
22. Delaney M. E., Walker D. A. A comparison of the kinetic properties of ribu-
lose bisphosphate carboxylase in chloroplast extracts of spinach, sunflower
and four other reductive pentose phosphate pathway species, Biochem. J., 171,
477—482 (1978).
23. Doehlert D. C„ Ku M. S. B., Edwards G. E. Dependence of the post-illumina-
tion burst of CO2 on temperature, light, CO2, and O2 concentration in wheat
(Triticum aestivum), Physiol. Plant., 46, 299—306 (1979).
24. Douce R„ Moore A. L., Neuburger Л1. Isolation and oxidative properties of
intact mitochondria isolated from spinach, Plant Physiol., 60, 625—628
(1977).
25. Ehleringer J., Bjorkman D. Quantum yields for CO2 uptake in C:t and C«
plants. Dependence on temperature, CO2, and O2 concentration, Plant Phy-
siol., 56, 86—90 (1977).
26. Farquahctr G. D., von Caemmerer S., Berry J. A. A biochemical model of
photosynthetic CO2 assimilation in leaves of C3 species, Planta, 149, 78—90
(1980).
27. Forrester M. L., Krotkov G., Nelson C. D. Effect of oxygen on photosynthesis,
photorespiration and respiration in detached leaves, I. Soybean, Plant. Phy-
siol., 41, 422—427 (1966).
28. ' Frederick S. E., Newcomb E. H. Microbody-like organelles in leaf cells,
Science, 163, 1353—1355 (1969).
29. Gerbaud A., Andre M. Photosynthesis and photorespiration in whole plants
of wheat, Plant Physiol,, 64, 735—738 (1979),
30. Goldsworthy A. Riddle of photorespiration, Nature, 224, 501—502 (1969).
31. Huang A. H. C,, Beevers H, Microbody enzymes and carboxylases in sequen-
tial extracts from C4 and C3 leaves, Plant Physiol., 50, 242—248 (1972).
32. Jensen R, G., Bahr J. T, Ribulose 1,5-bisphosphate carboxylasc-oxygenase,
Ann. Rev. Plant Physiol., 28, 379—400 (1977).
33. Kerr M. IF., Groves D. Purification and properties of glycolate oxidase from
Pisum sativum leaves, Phytochemistry, 14, 359—362 (1975),
34. Keys A. J., Bird J, F., Cornelius M. J., Lea P. J., Wallsgrove R. M„ Mif-
lln B. J. Photorespiratory nitrogen cycle, Nature, 275, 741—743 (1978),
35. Krause G. H„ Lorimer Q. H., Heber U„ Kirk M. R. Photorespiratory energy
dissipation in leaves and chloroplasts. In: Fourth International Congress on
Photosynthesis (eds. D. O. Hall, Jl Coombs' and T. W. Goodwin), London*
’ Biochemical Society, pp. 299—310, 1978.
ФОТОДЫХАНИЕ
445-
36. Ku S. В., Edwards G. E. Photosynthesis in mesophyll protoplasts and bundle
sheath cells of various types of C4 plants. IV. Enzymes of respiratory meta-
bolism and energy utilizing enzymes of carbon assimilation, Z. Pflanzen-
pbysiol., 11, 16—32 (1975).
37. Ku S. B., Edwards G. E. Oxygen inhibition of photosynthesis. II. Kinetic
characteristics as affected by temperature, Plant Physiol., 59, 991—999'
(1977). •
38. Ku S. B., Edwards G. E. Oxygen inhibition of photosynthesis. III. Tempera-
ture dependence of quantum yields and its relation to O2/CO2 solubility
ratio, Planta', 140, 1—6 (1978).
39. Ku S. B., Edwards G. E., Tanner С. B. Effect of light, carbon dioxide, and
temperature' on photosynthesis, oxygen inhibition of photosynthesis, and’
transpiration in Solatium tuberosum, Plant Physiol., 59, 868—-872 (1977).
40. Laing IF. A., Ogren W. L., Hageman R. H. Regulation of soybean net pho-
tosynthetic CO2 fixation by the interaction of CO2, Os and ribulose-l,5-dipnos-
phatc carboxylase, Plant Physiol., 54, 678—685 (1974).
41. Lea P. I., Miflin B. J. Photosynthetic ammonia assimilation. In: Encyclopedia
of Plant Physiology, New series. Vol. 6, Photosynthesis II. Photosynthetic
carbon metabolism and related processes (eds. M. Gibbs and E. Latzko),.
pp..445—455, New York, Springer-Verlag, 1979.
42. Lee R. B., Whittingham С. P. The influence of partial pressure of carbon
dioxide upon carbon metabolism in the tomato leaf, J. Exp. Bot., 24, 277—
287 (1974).
43. Leech R. M The isolation of structurally intact chloroplasts. Biochim. Bio-
phys. Acta, 79, 637—639 (1964).
44. Lorimer G. II., Andrews T. Plant photorespiration—an inevitable conse-
quence of the existence of an oxygen atmosphere, Nature, 243, 359—360
(1973).
45. Lorimer G. EL, Andrews T. J., Tolbert N. E. Ribulose diphosphate oxygenase.
II. Further proof of reaction products and mechanism of action, Biochemistry,
12, 18—23 (1973). .
46. Ludwig L. I., Catwin D. T. An open gas-exchange system for the simulta-
neous measurement of the CO2 and MCO2 fluxes from leaves, Can. J, Bot.,
49, 1299—1313 (1971).
47. Miflin В. I., Beevers H. Isolation of intact plastids from a range of plant
tissues, Plant Physiol., 53, 870—874 (1974),
48. Ogren IF. L., Bowes G. Ribulose diphosphate carboxylase, regulates soybean
photorespiration, Nature, New Biol,, 230, 159—160 (1971).
49. Osmond С. B., Bjlirkman O. Simultaneous measurements of oxygen effects
on net photosynthesis and glycolate metabolism in C3 and C4 species of
Atriplex, Carnegie Inst. Wash. Year book, 1971, 141—148, 1972.
50. Osmond C. B„ Harris B. Photorespiration during C4 photosynthesis, Biochim.
Biophys. Acta, 234, 270—272 (1971).
51, Plass G. N. Carbon dioxide and climate, Sci. Amer., 201, 41—45 (1959).
52. Pritchard G. G.. WhUtinghum С. P., Griffin W. J. Effect of isonicotinyl hydra-
zide on the path of carbon in photosynthesis, Nature, 190, 553—554 (1961).
53. Quebedeaux B., Hardt/ R. IF. F, Oxygen as a new factor controlling repro-
ductive growth, Nature, 243, 477—479 (1973).
54. Quebedeaux B., Hardt/ R. IF. F. Reproductive growth and dry matter produc-
tion of Glycine max (L.), Merr. in response to oxygen concentrations, Plant
Physiol., 44, 102—107 (1975).
55. Quebedeaux B„ Havelka U. D„ Livak K. L., Hardy R. W. F. Effect of altered
pO2 in the aerial part of soybean on symbiotic N2 fixation, Plant Physiol,,
56, 761—764 (1975).
56. Radmer R. I., Kok B. Photoreduction of O2 primes and replaces CO2 assimila-
tion, Plant Physiol,, 58, 336—340 (1976).
446
ГЛАВА 13
57. Randall D. D., Tolbert N. Б. Two phosphatases associated with photosynthesis
and the glycolate pathway. In: Photosynthesis and Photorespiration, (eds.
M. D. Hatch, С. B. Osmond and R. O. Slatyer), pp. 259—266, New York,
Wiley-Interscience, 1971.
58. Servaites J. C., Ogren W. L. Chemical inhibition of the glycolate pathway
in soybean leaf cells. Plant Physiol., 60, 461—466 (1977).
59. Smith B. N. Evolution of C4 photosynthesis in response to changes in car-
bon and oxygen concentrations in the atmosphere through time, Biosystems,
8, 24—32 (1976).
<60 . Ting 1. P., Rocha V., Mukerji S. R., Curry R. On the localization of plant
cell organelles, In: Photosynthesis and Photorespiration (eds. M. D. Hatch,
С. B. Osmond and R. O. Slatyer), pp. 534—540, New York, Wiley-Interscience,
1971.
<61 . Tolbert N. E. Isolation of leaf peroxisomes, Methods In Enzymol., 23, 665—
682 (1971).
62 . Tregunna E. B., Rroikov G., Nelson C. D. Effect of oxygen on the rate of
photorespiration in detached tobacco leaves, Plant Physiol,, 19, 723—733
(1966).
63. Usuda H., Arron G. P., Edwards G. E. Inhibition of glycine decarboxylation
by aminoacetonitrile and its effect on photosynthesis in wheat, J. Exp. Bot,,
31, 1477—1483 (1981).
64, Usuda 11., Edwards G. E. Localization of glycerate kinase and some enzymes
for sucrose synthesis in C3 and C4 plants, Plant Physiol,, 65, 1017—1022
(1980).
• 65, Wallsgrove R. M., Lea P. L, Miflin В. I. Distribution of the enzyme of nitro-
gen assimilation within the pea leaf cell, Plant Physiol,, 63, 232—236
(1979).
66. Whittingham С. P., Hiller R. G., Bermingham M. The production of glycolate
during photosynthesis, In: Photosynthetic Mechanisms of Green Plants,
pp. 675—683, Publ. 1145, Natl. Acad. Sci.— Natl. Res. Council, Washington,
D.C., 1963.
• 67. Wittwer S. H., Robb W. Carbon dioxide enrichment of greenhouse atmosphe-
res for food crop production, Econ. Bot., 18, 34—56 (1964).
68, Woo R. C., Osmond С. B. Glycine decarboxylation in mitochondria isolated
from spinach leaves, Aust. J. Plant Physiol,, 3, 771—785 (1976).
<69. Zelitch I. The relationship of glycolic acid to respiration and photosynthesis
in tobacco leaves, J, Biol. Chem., 234, 3077—3081 (1959),
ГЛАВА 14
Первичные карбоксилазы
и регуляция фотосинтеза
и транспирации факторами внешней среды
Краткое содержание
Биологию С3- и С4-фотосинтеза, как и влияние О2 на фото-
синтез у Сз- и С4-растений, следует рассматривать в связи с
уровнем СО2 и О2 в атмосфере. Зная характеристики ФЕП- и
РуБФ-карбоксилазы, можно создать модели, описывающие про-
текание фотосинтеза в зависимости от уровня СО2 и О2.
РуБФ-карбоксилаза использует СО2 (Лм = 5—15 мкМ), а ФЕП-
карбоксилаза связывает бикарбонат (Ам = 0,3—0,4 мМ или даже
ниже). При гидратировании СО2 образуется бикарбонат, осо-
бенно при высоких значениях pH. При низкой концентрации О2
(например, при 2%) и pH 8,0 относительное сродство этих двух
карбоксилаз к атмосферной СО2 может быть одинаково. При
атмосферной концентрации О2 (21%) РуБФ-карбоксилаза на-
ходится в невыгодном положении, так как с увеличением кон-
центрации О2 возрастает /<м(со2)*
Количество СО2, которое доступно для первичных карбокси-
лаз в листе С3- или С4-растений, зависит от ее концентрации
в атмосфере, сопротивления диффузии СО2 через устьица, тем-
пературы листа и сопротивления диффузии СО2 через водную-
фазу. У С4-растеиий ФЕП-карбоксилаза локализована, как и
карбоангидраза, в цитозоле клеток, а у С3-растений карбоангид-
раза находится в хлоропластах, и возможно, в цитоплазме.
И у С3- и у С4-растений сопротивление диффузии СО2 может
быть сведено к минимуму под действием карбоангидразы, кото-
рая способствует быстрому приведению СО2 в равновесие с
НСО3“, а, как известно, высокая концентрация НСО3~ облегчает
диффузию.
В том случае, когда температура листа и внутриклеточная
концентрация СО2 известны, можно рассчитать концентрацию-
этого газа в растворе, омывающем поверхность клеток мезо-
филла. А после этого можно рассмотреть и модели фотосинтеза
в листьях С3- и С4-растений, отражающие зависимость интенсив-
ности этого процесса от концентрации доступной для клеток
СО2 и от содержания в них карбоксилаз.
При обычном содержании СО2 в атмосфере интенсивность,
фотосинтеза у С3-растений явно меньше, чем их карбоксилирую-
448 ГЛАВА 14
(СО.I
Рис. 14.1. Изменения наблюдаемого фотосинтеза (НФС) у Cs- и Сд-растений
в ответ на изменение концентрации СО2. Cs- и Ci-растения совершенно по-
разному отвечают на изменение [СО2] в атмосфере — это видно из кривых за-
висимости скорости фотосинтеза от [СО2]. Имеющийся у С4-растеиий механизм
концентрирования СО2 позволяет им вести фотосинтез с максимальной ско-
ростью при относительно низкой [СО2] в окружающей среде. Для насыщения
фотосинтеза у Сз-растений необходима высокая [СО2], тогда как при малой
[СО2] фотосинтез сильно подавляется за счет фотодыхания (гл. 13). Факто-
рами, контролирующими [СО2], поступающей в фотосинтезирующие ткани
Cs- и Ci-растений, являются водный дефицит, который ведет к закрыванию
устьиц, и температура.
щий потенциал. Это связано с тем, что О2 ингибирует РуБФ-
карбоксилазу и способствует окислению гликолата в ходе фото-
дыхания. При насыщающем уровне СО2 интенсивность фотосин-
теза возрастает по мере того, как снимается ингибирование,
вызванное О2. Но даже при насыщающей концентрации СО2
максимально возможный карбоксилирующий потенциал не до-
стигается из-за ограниченного поступления ассимиляционной
силы, необходимой для того, чтобы регенерировать РуБФ.
При естественной концентрации СО2 и О2 в атмосфере
РуБФ-карбоксилаза С4-растений функционирует почти с макси-
мальной активностью благодаря наличию механизма концентри-
рования СО2 в С4-цикле. Нечувствительность ФЕП-карбоксила-
зы к ингибированию кислородом и наличие механизма, концен-
трирующего СО2, позволяют растению фотосинтезировать с от-
носительно высокой скоростью даже при таких низких концент-
рациях СО2, которые наблюдаются при повышенных температу-
рах или при вызванном водным дефицитом увеличении сопротив-
ления устьиц. Относительно невысокая скорость фотосинтеза у
некоторых С4-растений при пониженных температурах может
быть обусловлена недостаточным содержанием РуБФ-карбокси-
.лазы, повышенной лабильностью, пируват,, ортофосфат—дикина-
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И. РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
449
зы на холоду, или же какими-то еграничеииями транспорта мета-
болитов.
Повышение температуры оказывает отрицательное действие
па фотосинтез у Сз-растений, так как при этом уменьшается
растворимость СОг, увеличивается отношение растворимостей
Ог/СОг и, возможно, меняются кинетические константы для
РуБФ-карбоксилазы-оксигеназы. Точно так же, при водном де-
фиците лимитирующим фактором становится концентрация СО2
в межклеточном пространстве; при этом создаются условия,
явно невыгодные для фотосинтеза, особенно у Сз-растений.
Сопротивление мезофилла, которое включает в себя карбо-
ксилирующий компонент фиксации СО2, сказывается только на
фиксации СО2, но ие на транспирации. Поэтому при прочих
равных условиях низкое сопротивление мезофилла отражается
в невысоком значении соотношения транспирация/фотосинтез,
как это показано для С4-растений.
14.1. Введение
Какие преимущества дает растению С4-механизм фотосинте-
за и дает ли он их вообще? Интенсивность фотосинтеза лимити-
руется концентрацией СО2 во внешней среде. Этот фактор при-
обретает особое значение при высоких температурах, когда
уменьшается растворимость СО2, и в условиях стресса (водный
дефицит или солевой шок), когда возрастает сопротивление
устьиц и атмосферная СО2 становится менее доступной. С точки
зрения фотосинтеза С4-растения более эффективно функциони-
руют при относительно высоких температурах, а С3-растения
лучше функционируют при сравнительно невысоких температу-
рах. О2 ингибирует фотосинтез у Сз-растений, однако такое ин-
гибирование снимается, когда концентрация СО2 в атмосфере
превышает нормальный уровень (гл. 13). В С4-цикле влияние
пониженной концентрации СО2 компенсируется тем, что внутри
клеток обкладки проводящих пучков создается микросреда,
обогащенная СО2.
14.2. Состав атмосферы и растворимость газов
Различия между С3 и С4-фотосинтезом гораздо проще понять,
если соотнести их с некоторыми физическими характеристиками
окружающей среды и, в частности, связать эти процессы с кон-
центрацией СО2 и О2 в атмосфере и с температурой. Концентра-
ция СО2 в атмосфере очень невелика и в среднем составляет
примерно 0,0325% (325 мкл/л, 325 мкл/100 мкл, 325 м.д.'), хотя
и наблюдаются некоторые небольшие, отклонения, связанные с,
29—234
450
ГЛАВА 14
географическим положением той или иной местности.
Состав атмосферы по объему (%)
N2 78,084
О2 20,946
СО2 0,033
Аг 0,934
99,997
Растворимость газа в воде зависит от его коэффициента абсорб-
ции (а, или коэффициент Бунзена, который равен объему газа,
поглощенного одним объемом воды при давлении 1 атм). Как
было замечено Стрэнгом (Strang, 1981), единственным коррект-
ным способом определения сродства того или иного фермента
к газообразному субстрату является выражение этой величины
через истинную концентрацию (в молях) данного газа, находя-
щегося в контакте с ферментом, а не через общую концентра-
цию газа в газовой фазе. Растворимость газов убывает по мере
увеличения температуры (снижается коэффициент Бунзена).
Например, при 15°C и концентрации СО2, равной 325 м.д., кон-
центрация свободной СО2 в воде составляет 14,8 мкМ., а при
35 °C она снижается до 8,59 мкМ.
Количество СО2 (в мкмолях), содержащееся в 1 л воды при
15 °C и давлении 1 атм, рассчитывают по формуле
. -Рсо2 («) (10е)
СО2 (мкмоль/л) =------------
где 22,4 — количество литров, приходящихся на 1 моль СО2 при
обычной температуре и давлении, Рсо2— парциальное давление
СО2, равное 0,0325% .....° =0,000325. Если температура
воды составляет 15 °C и а—1,019 мл газа/мл воды, то
СО2 (мкмоль/л) == <1Q6L = 14,78 мкМ
При любой заданной температуре количество газа, раство-
ренного в воде, пропорционально парциальному давлению этого
газа над раствором (закон Генри). Отклонения от этого закона
наблюдаются только при повышенном давлении, превышающем
нормальное атмосферное давление, или при высокой концентра-
ции растворенных веществ в растворе. Растворимость О2 или
СО2 на поверхности клеток мезофилла при заданном парциаль-
ном давлении в межклеточном пространстве должна не очень
отличаться от растворимости этих газов в воде, так как соли,
присутствующие в растительном соке ( — 0,1 М), снижают рас-
творимость этих двух газов всего лишь на 5—10% (Rabino-
witch, 1945). Поскольку коэффициент абсорбции уменьшается с
увеличением температуры, должно наблюдаться и пропорцио-
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
451
Рис. 14.2. Влияние температуры на растворимость атмосферной СОг
(0,0325%) и О2 (21%) и отношение растворимостей О2/СО2 в воде. Данные
рассчитаны по величине коэффициента Бунзена для О2 и СО2 при разных
температурах (Ku, Edward, 1977а).
нальиое уменьшение химической активности (или иначе актив-
ной массы) данного газа, и соответственное снижение его кон-
центрации в растворе. Для практических целей закон Генри
вполне применим, и это позволяет рассчитывать концентрацию
СО2 на поверхности клеток мезофилла при условии, что извест-
ны концентрация СО2 в воздушном пространстве листа и тем-
пература листа.
Отношение концентраций О2/СО2 в атмосфере в присутствии
21% О2 и 0,0325% СО2 составляет 646/1, а в воде при 25°C оно
равно 24/1 (264 мкМ О2/11 мкМ СО2). Это различие связано с
тем, что СО2 лучше растворяется в воде, чем О2 (т. е. коэффи-
циент абсорбции для СО2 гораздо выше). Обратите внимание
на то, что при увеличении температуры растворимость СО2 па-
дает быстрее, чем растворимость О2 (рис. 14.2). Это приводит к
тому, что отношение растворимостей О2/СО2 с ростом темпера-
туры увеличивается. Концентрация иона НСО3~, находящегося
в равновесии с растворенной СО2, зависит от pH, при этом pKi
~6,33, а р Кг—10,38.
Н2О Ц-СО2 i==5 Н+4-НСО,- H++CCV-
Так же как pH по определению — отрицательный логарифм кон-
центрации ионов водорода (разд. 1.7), так и рК по определе-
29*
Л52
ГЛАВА 14
нию — отрицательный логарифм К, где К —кажущаяся констан-
та равновесия (разд, 2,6) для диссоциации кислоты (НА) в со-
ответствии с уравнением НА Н+4-А". Поэтому
/< = [Н[йА)'1 > а рК == — 1g X, или 1g (1 /К)
Термины рК и pH можно связать через уравнение Гендерсона —
Хассельбаха
рН= pK-|-lg-~y-
Из этого уравнения видно, что рК кислоты — это то значение
pH, при котором диссоциирует половина всех молекул кислоты,
т. е. в том случае, когда [А~] = [НА], уравнение Гендерсона —
Хассельбаха упрощается до. уравнения рН=рК.
Некоторые различия в величинах рКь встречающиеся для
бикарбоната в тех или иных работах, отчасти могут быть свя-
заны с изменением pKi при изменении температуры. Умбрейт и
др. (Umbreit et al., 1972) .считают, что в области температур от
20, до 40 °C pK,i изменяется со скоростью 0,0005 единица/градус.
Эта поправка будет использована в последующих примерах.
Температура, °C а СО2 РКХ
20 0,878 6,392
25 0,759 6,365
' ... 30 0,665 6,348
. 35 0,592 6,328
40 0,530 6,312
(По Umbreit et al., 1972.)
Такие различия в значении рК лишь незначительно влияют на
расчетную величину Хм(соа) Для РуБФ-карбоксилазы in vitrio,
так как' этот фермент использует в качестве субстрата свобод-
ную СО2. При pH 8,0, 35°C, общей концентрации неорганиче-
ского углерода (СО2 + НСО3_) в среде 0,4 мМ и pKi = 6,328 рас-
четная концентрация растворенной СО2 равна 8,33 мкМ. Просле-
дим на этом примере, как ее рассчитывают. . :: т.
; ' ' . ' Пример •
PH-pK+lg JS^ .
8,0 = 6,328+lg (°’4 Л' . -А
где X—концентрация СО2
1 ’ : Д;672=.1е ^°’У^;-. г)\ : ...
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
453
1,672 = 1g 47
0,4 мМ .
X = 0,00833 мМ = 8,33 мкМ
Если pH равен 8 или ниже, рКя можно игнорировать при рас-
четах, так как концентрация СОз2- очень мала. In vivo кон-
центрация СО2, доступная для клеток мезофилла, зависит от
парциального давления СО2 в межклеточном воздушном про-
странстве листа и не зависит от величины pH. В то же время pH
сильно влияет на выравнивание концентраций СО2 и бикарбо-
ната. В атмосфере, содержащей 0,0325% СО2, при 30 °C кон-
центрация растворенной СО2 равна 9,65 мкМ (рис. 14.3).
ГНСО.-1
Тогда при pKi=6,348 pH^6,348-|-lg 3 J
При pH 6,348 концентрация НСО3~=9,65 мкМ
При pH 7,348 концентрация НСО3- = 96,5 мкМ
При pH 8,0 концентрация НСО3~=432 мкМ . .
Значение этих физических характеристик газов станет более
явным, если мы рассмотрим энзимологию карбоксилаз, участ-
вующих в С3- и С4-фотосинтезе.
Источники данных: [29, 47, 50].
Рис. 14.3. Концентрация СО2 и бикарбоната в воде, при'разных’значениях pH.
Температура воды 30°C. ;
454
ГЛАВА 14
14.3. Сз-Растения: РуБФ-карбоксилаза
РуБФ-карбоксилаза связывает свободную СОг, а не ион
НСОз~ (разд. 8.12). Что касается карбоангидразы, то не ясно,
участвует ли она в катализе переноса СО2 от поверхности кле-
ток мезофилла к месту фиксации СО2 в хлоропластах Сз-расте-
ний или же выполняет другие функции. Этот фермент, который
катализирует гидратацию СО2, локализуется как в хлороплас-
тах, так и в цитоплазме клеток мезофилла у С3-растений:
СО£ 4- Н2О ^=±: Н+ + НСО3- (14.1)
СО2 находится в равновесии с ионами Н+ и НСО3_, при этом
равновесная концентрация НСО3_ зависит от pH раствора.
Карбоангидраза, действуя как катализатор, значительно уско-
ряет процесс достижения равновесия.
Рассмотрим на примере, как же функционирует карбоангид-
раза в клетках мезофилла у С3-растений и как она связана с
РуБФ-карбоксилазой. Пусть температура равна 30°C, межкле-
точная [СО2] 250 м.д., а pH в цитозоле и в строме хлороплас-
тов 8,0. По мере того как СО2 будет растворяться на поверхно-
сти клеток мезофилла, [СО2] растет и становится равной
7,4 мкМ. Карбоангидраза обеспечивает быстрое приведение
гсо21 внутр
250 м.д.
pH 8,0
Рис. 14.4. Предполагаемый основной путь переноса СО2 (жирные стрелки)
в Сз-листе при участии карбоангидразы, сдвигающей равновесие реакций.
Реакция СО2ч=ьНСОз ’ представляет собой краткую форму записи катализи-
руемой карбоангидразой реакции СО2-ЬН2О=т±НСОз~-)-Н';'.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
455
концентраций СО2 и НСО3~ в равновесие, в результате чего ко-
нечная концентрация НСО3~ достигает 331 мкМ. Диффузия
СО2 или НСОз- в хлоропласты и внутри них происходит по гра-
диенту, от более высокой концентрации к более низкой (нет ни-
каких данных об активном транспорте). Диффузия зависит от
градиента концентрации и от коэффициента диффузии для этих
молекул. Поскольку концентрация НСО3~ примерно в 45 раз
выше, чем концентрация СО2, для диффузии НСО3~ создается
большой потенциал. Однако, поскольку молекула НСО3~ имеет
заряд, сопротивление диффузии для ее проникновения через
оболочку хлоропласта гораздо выше, чем для свободной СО2.
Быстрое приведение СО2 в равновесие с НСО3~ в присутствии
карбоангидразы в цитозоле или внутри хлоропласта может во
многом облегчать диффузию этих соединений в соответствую-
щие компартменты клетки. Кроме того, это предотвращает
также локальное истощение СО2 в хлоропласте в процессе фото-
синтеза (разд. 8.12, рис. 14.4). С улучшением методов опреде-
ления для Дм(С02) РуБФ-карбоксилазы получали все меньшие
значения (разд. 6.5). Последние цифры близки к 10 мкМ (в ат-
мосфере N2); они приблизительно соответствуют величине рас-
творимости 0,0325% СО2 при ЗО^С. В настоящее время пробле-
мы, касающиеся РуБФ-карбоксилазы, связаны уже не с ее срод-
ством к СО2 или максимальной активностью (Ушах), а скорее
всего с тем, что О2, как это было показано, является конкурент-
ным ингибитором фермента (разд. 13.3).
Точные значения /<м(со > и /(цог) (соответственно концентра-
ция СО2, необходимая для достижения полумаксималы-юй ак-
тивности, и кинетическая константа, характеризующая ингиби-
рующее действие О2) для РуБФ-карбоксилазы неизвестны. Ми-
нимальные значения /<м(СОа) варьируют от 6 до 16 мкМ, а мини-
мальные величины /Слог) лежат в пределах от 200 до 400 мкМ.
Обе эти константы близки по величине к растворимости данных
газов в воде при нормальном их содержании в атмосфере. Как
уже обсуждалось в гл. 13, О2 является и конкурентным ингиби-
тором РуБФ-карбоксилазы, и субстратом этого фермента, если
учесть, что РуБФ-карбоксилаза может выполнять функции и
РуБФ-оксигеназы. Для того чтобы связать кинетические кон-
станты карбоксилазы с интенсивностью фотосинтеза в листьях,
следует проанализировать кинетику фотосинтеза в листе в зави-
симости от [СО2]. При таком анализе следует учитывать ряд
факторов, которые определяют интенсивность фотосинтеза в
листе: сопротивление пограничного слоя; сопротивление устьиц
(так, например, сопротивление устьиц часто возрастает при
увеличении концентрации СО2 в атмосфере); фотодыхание, в ре-
зультате которого углекислотная кривая фотосинтеза пересека-
ется с осью абсцисс не в нулевой точке, а в углекислотном ком-
456
ГЛАВА 14
пенсационном пункте (Г); сопротивление переносу СО2 через
водную фазу фотосинтезирующей ткани и возможное лимитиро-
вание фотохимических процессов при высокой концентрации СО2.
Ни один из этих факторов не действует на карбоксилазу при ее
определении in vitro. Для того чтобы оценить величину /(м(соа)
для РуБФ-карбоксилазы in vivo можно использовать следующую
процедуру.
а) Получите углекислотную кривую в атмосфере, содержа-
щей 2% О2. В этих условиях подавляется фотодыхание и Г сме-
щается почти к нулю.
б) Постройте график зависимости скорости фотосинтеза от
рассчитанной концентрации СО2, растворенной на поверхности
клеток мезофилла. Для этого нужно определить внутриклеточ-
ную [СО2] и измерить температуру листа. При любом значении
[СО2] в окружающей среде [СО2] в межклеточном пространст-
ве листа, в непосредственной близости к клеткам мезофилла,
можно рассчитать по методу Гаастры (Gaastra, 1959):
[СО2]ЕНуТр = [СО2]иаруЖН НФС (ra -j- rs) (14.2)
где [СО2] IiapyH<n= [СО2] в окружающем воздухе, НФС —ско-
рость наблюдаемого фотосинтеза, га— сопротивление погранич-
ного слоя для диффузии СО2 и rs — сопротивление устьиц для
диффузии СО2.
Эти четыре параметра можно измерить при любом значении
[СО2] наружи (разд. 14.6) и затем рассчитать [СО2]виутр. Из
уравнения
НФС — 1СО^наРУжн— [COJxn f 14 3)
можно вывести уравнение
НФС = ^СОгкэружи [СО2]пнутр [СОа]внутр~~~ [CO2]xn (j z]
ra + rs rm
где [СО2]хп= [CO2] в хлоропласте, a rm— сопротивление мезо-
филла.
в) Постройте график обратных величин (зависимость 1/НФС
от 1/([СО2]—Г)) для определения величины /См(соа) (см. так-
же разд. 13.10).
Эксперимент подобного рода, проведенный при 30 °C с рас-
тениями пшеницы, иллюстрирует рис. 14.5, А, Б. В этом случае
Км(со2)^5 мкМ, что соответствует концентрации СО2 в межкле-
точном пространстве приблизительно 190 м. д. Нарушения ли-
нейности на графике двойных обратных величин наблюдаются
только при высокой [СО2], в результате чего эксперименталь-
ные значения Утах (рис. 14.5, А) становятся ниже, чем расчет-
ная величина Утах (рис. 14.5,5.). Это может быть связано с ка-
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И.РЕГУЛЯЦИЯ ФОТОСИНТЕЗА 457
кими-то фотохимическими ограничениями, о которых уже гово-
рилось, когда мы сходным образом анализировали процессы,
происходящие в хлоропластах. Во всяком случае, любое сопро-
тивление переносу СОг (сопротивление диффузии СО2 через
водную фазу), по-видимому, не является существенным'факто-
ром, иначе Д'м(С<>2) была бы еще меньше, чем считают в .настоя-
щее время. Если такие эксперименты провести при различных
концентрациях О2 и построить график зависимости 1/НФС от
1/([СО2]— Г) [где в разности ([СО2]—Г) учитывается вклад
фотодыхания (разд. 13.10], то можно определить величину
Дцог) и оценить увеличение кажущейся /<м«|Ж(СО2) иод влияни-
ем О2. Из такого рода опытов вытекает, что /<1(О2) близка к
200 мкМ (разд. 13.10). В присутствии 21% О2 кажущаяся
Дм(сог) возрастает почти до 15 мкМ (рис. 14.5, Б).
Результаты анализа процесса фиксации СО2 в целых'.листьях
н кинетические свойства РуБФ-карбоксйлазы-оксигеназы мож-
но использовать для создания обобщенной модели фотосинтеза
в листьях Сз-растеппп (рис. 14.6). Вопрос о том, каковы точные
значения Д'м(со-), Д’ко2) и Для РуБФ-карбоксилазы у раз-
ных растений iv vivo и in vitro, все еще остается открытым.
Кривая I на рис. 14.6 отражает теоретически возможную про-
изводительность фотосинтеза в отсутствие О2 (если исходить из
величины карбоксилирующего потенциала). Кривая II отражает
возможную производительность фотосинтеза в присутствии
21% О2. Кривая III описывает скорость наблюдаемого фотосин-
теза в присутствии 21% О2, когда нет никаких фотохимических
ограничений. Кривая IV отражает скорость фотосинтеза in vivo
при различных концентрациях СО2. В этом примере раствори-
мость СО2 на поверхности клеток мезофилла при нормальном
содержании СО2 в атмосфере составляет 7,4 мкМ, что соответ-
ствует скорости фотосинтеза в листе —250 мкмоль СО2-
• (мгХл)-1-ч-1. Ингибирование фотосинтеза кислородом при
нормальном содержании СО2 в атмосфере (т. е. в этом случае
7,4 мкМ) обусловлено одновременно и кислородным ингибиро-
ванием РуБФ-карбоксилазы (кривые I и II), и потерей СО2 за
счет фотодыхания (кривые II и III). Эти две составляющих ин-
гибирования взаимосвязаны, так как на каждые две молекулы
О2, взаимодействующие с РуБФ (разд. 13.10), приходится одна
молекула СО2, высвобождающейся в ходе фотодыхания. Отно-
сительный вклад этих двух составляющих в ингибирование фо-
тосинтеза меняется с изменением [СО2]. Это подтверждается
тем, что число молекул СО2, дополнительно взаимодействующих
с РуБФ в отсутствие О2, вовсе не соответствует активности ок-
сигеназы в присутствии О2. Последнее было бы возможно толь-
ко в том случае, если бы кинетические константы [7<м, A’i И
Утах!'уравнения (13.36), (13.39)] для О2 и СО2 были бы одни
458
ГЛАВА 14
Рис. 14.5. Фотосинтез в листьях пшеницы (С3) и сорго (С4) при 30 °C и раз-
личных [СО2]. Изменяли содержание СО2 в газовой фазе. [СО2] иа поверхно-
сти клеток мезофилла рассчитывали по [СО2] в межклетниках с учетом тем-
пературы листа. А. Фотосинтез в листьях пшеницы в присутствии 2 и 21% О2.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА 459
Б. График двойных обратных величин для зависимости интенсивности фото-'
синтеза у пшеницы от [СО2] (разд. 13.10). В. Фотосинтез в листьях сорго в
присутствии 21 % О2. Г. График двойных обратных величин для зависимости
фотосинтеза в листьях сорго от [СО2] в межклетниках. Плотность квантового
потока 145 нмоль квантов-см-2-с-1 (400—700 нм). Содержание Хл на еди-
ницу поверхности листа 0,3 г-см-2. Стрелкой на оси абсцисс на графиках А
и Б отмечена [СО2] внутри листа, которая создается при обычном содержа-
нии СОа в атмосфере. (Данные любезно предоставлены S. В. Ku; см. также
Ku, Edwards, 1977b, 1978.)
( CO 2 | ,MK M
Рис. 14.6. Модель фотосинтеза в листьях С3-растения, в которой рассматри-
вается связь этого процесса с потенциальной активностью РуБФ-карбокси-
лазы. Кривая I — активность РуБФ-карбоксилазы в отсутствие О2, когда
_____Утах [СОд]
v ~ [СОВ] -( -/(мссо,)
Кривая II—активность РуБФ-карбоксилазы в присутствии 21% Од, когда
_______________Утах [СОа]__________
v- [СОа] + /<М(СО2) (1 + [02]//<г (О2)) '
Кривая III — фотосинтез в целых листьях в присутствии 21% О2 при задан-
ной [СО2] на поверхности клеток мезофилла, без какого бы то ни было огра-
ничения доступа ассимиляционной силы, когда
_ [СО,,]—Г
НФС21% 02 - 7шах- [СО2] + ЛМ(Со2) (1 + [О2]/Кг (о2))
[см. уравнение (13.35)]
Кривая IV — фотосинтез в целых листьях в присутствии 21% О2 при огра-
ниченном поступлении ассимиляционной силы, образование которой лимити-
руется при высокой [СОг]. Такое ограничение должно лимитировать скорость
регенерации РуБФ — субстрата карбоксилазы. В результате этого при высо-
кой [СО2] кривая IV отклоняется от кривой III.
Жирная точка соответствует скорости фотосинтеза в целых листьях в при-
сутствии 2% О2 при обычной концентрации СО2 в атмосфере. Стрелка на осн
абсцисс указывает [СО2] 0,0325%. Детально уравнения для карбоксилазы и
для фотосинтеза в листьях рассматриваются в разд. 13.10, где обсуждается
также связь этих процессов с кислородным ингибированием фотосинтеза. Ис-
ходные данные: [СОг] в воздухе=0,0325%; [СО2] в межклеточном простран-
стве^,025%; температура листа=30°С; растворимость СО2 на поверхности
клеток мезофилла=7,4 мкМ; растворимость 21% О2 в листе—245 мкМ; Г —
углекислотный: компенсационный пункт — соответствует [СО2]=60 м. д.=
=2 мкМ; кинетические константы для РуБФ-карбоксилазы: Км(со >=5мкМ
(величина, полученная при кинетическом изучении целых листьев, типичная
величина для .изолированного фермента составляет —10 мкМ, разд. 13.3),
Л1(о1!)=2001 мкМ, Утах Для РуБФ-карбоксилазы=800 мкмоль СО2-(мгХл)-1-
•ч-1. Это значение. принимают за величину Утазс для РуБФ-карбоксилазы в
условиях in vivo при насыщающей [СО2]. Реальная величина Утах для экст-
рагированного фермента может быть выше, чем в условиях in vivo, если эти
условия далеки от оптимальных (например, концентрация РуБФ или
Mg2+, pH).
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
461
и те же. Количество молекул.'..СО2,, дополнительно1 . вааимоДей:-
ствующих с карбоксилазой в отсутствие. О2,. зависит от ’[СО2]
и от кинетических констант для карбоксилазы. При высокой
[СО2] составляющая, обусловленная конкуренцией СО2 и О2 за
РуБФ-карбоксилазу-оксигеназу, убывает гораздо медленнее, чем
составляющая, обусловленная фотодыханием. Это свидетельст-
вует о том, что при высокой [СО2] активность оксигеназы от-
носительно невелика по сравнению с карбоксилирующей способ-
ностью, которая возрастает в отсутствие О2. Как было показано
в опытах с Сз-растениями, максимальной скорости фотосинтеза,
в условиях in vivo можно достичь, либо повысив [СО2] в атмо-
сфере выше нормального уровня, либо понизив [О2] (рис. 14.5).
Совершенно очевидно, что отношение СО2/О2 в листе . имеет
большое значение для регуляции скорости фотосинтеза.
Подлинная кривая фотосинтеза для целых листьев (кри-
вая IV на рис. 14.6) не совпадает с кривой, характеризующей
потенциальную производительность РуБФ-карбоксилазы в при-
сутствии 21% О2 (кривая II на рис. 14.6), по двум причинам.
При низкой [СО2] скорость фотосинтеза уменьшается за счет
фотодыхания, и это явление наиболее выражено при приближе-
нии [СО2] к углекислотному компенсационному пункту. При
[СО2], превышающей ее содержание в атмосфере, фотосинтез
достигает насыщения прежде, чем карбоксилаза. Это связано1 с
недостатком ассимиляционной силы. При атмосферной [СО2]
количественное соотношение между фотохимическим аппаратом
и потенциалом ассимиляции углерода может быть таково, что
ассимиляционная сила будет производиться в достаточном ко-
личестве только при относительно высокой интенсивности света.
При насыщающей [СО2], однако, ассимиляционная сила может
быть лимитирующим фактором даже при максимуме прямой
инсоляции. Если в этих условиях создать еще и дополнительное
освещение, то это ограничение доступа энергии можно снять,
если только оно не связано с насыщением какого-либо перенос-
чика в цепи транспорта электронов пли со световым насыщени-
ем процесса при максимальной инсоляции. Хотя влияние раз-
личных концентраций СО2 очень интенсивно исследовалось,
обычно при этом принимали, что интенсивность света должна
быть эквивалентна полному солнечному освещению или даже
несколько ниже. На самом же деле недостаток ассимиляцион-
ной силы при высокой [СО2] должен сказываться на [РуБФ],
так что этот субстрат становится лимитирующим фактором для
карбоксилирования.
1 В нескольких работах, в которых было измерено содержание
РуБФ в клетках или в листьях, было сделано предположение
о том, что [РуБФ] может снижаться настолько, что этого суб-
страта не хватит для заполнения всех активных центров. РуБФ-
462
ГЛАВА 14
карбоксилазы в хлоропластах. Само по себе это не означает,
что РуБФ становится лимитирующим фактором, так как по
меньшей мере часть участков связывания субстрата, по-види-
мому, занята шестиуглеродным интермедиатом этой реакции.
Полагают, что концентрация участков связывания субстрата на
РуБФ-карбоксилазе в хлоропластах необычайно высока, при-
близительно 3 мМ (если считать, что на 1 мг Хл приходится
около 5 мг белка фракции I, мол. масса равна 550 000, объем
стромы составляет — 25 мкл/мгХл, а на каждую молекулу фер-
мента приходится 8 участков связывания, т. е, по одному
участку на каждой большой субъединице). Км(со2)> равная
примерно 10 мкМ, меньше, чем концентрация участков связыва-
ния субстрата на РуБФ-карбоксилазе в хлоропластах. Если до-
бавить 10 мкМ СО2 в пробу, в которой концентрация РуБФ-
карбоксилазы эквивалентна ее содержанию в строме хлороплас-
тов, то, очевидно, будет занята лишь незначительная часть
участков связывания. Однако, поскольку этот фермент обладает
сравнительно высоким сродством к СО2, в стационарных усло-
виях фотосинтеза может быть занято гораздо большее число
участков, особенно если скорость притока СО2 из атмосферы
намного превышает скорость обращения фермента. Таким об-
разом, в хлоропластах может находиться всего несколько мик-
ромолей свободной СО2, а с ферментом может быть связано в
1000 раз больше.
Степень активации РуБФ-карбоксилазы в различных услови-
ях in vivo неизвестна. Вполне естественно, что если степень ак-
тивации этого или других ферментов изменяется с изменением
I [СО2] или [О2], то это должно влиять и на кинетику фотосин-
теза.
Существуют как эмпирические, так и механические модели,
описывающие зависимость фотосинтеза в целом листе от внеш-
них факторов. Нетрудно вывести одно или несколько математи-
ческих уравнений, которые бы согласовывались с эксперимен-
тальными данными, но это отнюдь не означает, что они соответ-
ствуют и биохимии данного процесса. Так, например, модель
Тенхунена и др. описывает возможную ответную реакцию Сз-рас-
тения (пшеница) на изменение [СО2] и [О2]. Эти авторы, од-
нако, сделали ряд допущений, облегчивших расчет величины
Лмссоа) для карбоксилазы, и получили —0,1 мкМ, что примерно
на два порядка меньше, чем минимальное из ее значений, опре-
деленных экспериментальным путем. Было сделано допущение
о том, что экспериментально измеряемая величина Vmas фото-
синтеза эквивалентна Ушах карбоксилазы (такое же допущение
делали Джонс и Слетьер). Сейчас, однако, стало известно, что
потенциальная активность карбоксилазы намного выше, чем
измеряемая величина Ушах- Это, вероятно, связано с фотохими-
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
463.
ческими ограничениями при высокой [СО2]. Кроме того, пред-
полагали, что Утах карбоксилазы (скорость при насыщающей
концентрации СО2) равна Утах оксигеназы (скорость при насы-
щающей концентрации О2); однако Ушах карбоксилазы оказа-
лась в несколько раз выше, чем Уюах оксигеназы. Считали так-
же, что сопротивление переносу СО2 через водную фазу настоль-
ко значительно, что [СО2] в хлоропласте должна быть намного
ниже, чем на поверхности клетки. Хотя для диффузии действи-
тельно необходим некоторый градиент концентрации, нет ника-
ких данных о том, что сопротивление переносу СО2 очень вели-
ко. Если функция карбоангидразы заключается в том, чтобы об-
легчать диффузию СО2, то [СО2] на участке карбоксилирова-
ния должна быть очень близка к ее концентрации на поверхно-
сти клетки (рис. 14.4).
Поскольку при построении любой модели исследователи ис-
ходят из определенных предположений и допущений, а иногда
строят ее чисто эмпирическим путем, всем моделям присущи
определенные ограничения, которые затрудняют их интерпрета-
цию в связи с биохимией фотосинтеза. Выяснение механизма
фотосинтеза и ответной реакции разных растений на изменяю-
щиеся условия внешней среды могут дать много полезной ин-
формации для моделирования фотосинтеза.
Источники данных: [3, 6, 20, 22, 26, 29, 30, 31, 36, 48, 51].
14.4. С4-Растения: ФЕП- и РуБФ-карбоксилазы
РуБФ-карбоксилаза С4-растений по своим свойствам, по-ви-
димому, ие отличается от аналогичного фермента Сз-растений в
том смысле, что она тоже ингибируется О2 и имеет относительно
низкую Км(соз). Активность этого фермента, однако, и количест-
во белка (белка фракции I) у С4-растений могут быть значи-
тельно меньшими, чем у С3-растений (при расчете на единицу
Хл, единицу поверхности листа или в процентах общего содер-
жания растворимых белков). Так, например, кукуруза и Pani-
cum miliaceum (С4-растения) имеют — 1 мг белка фракции
I/мг Хл листьев, а пшеница, табак и картофель (Сз-растения) —
от 3 до 6 мг белка фракции 1/единица Хл листьев. Остается не-
ясным, насколько эти различия отражают действительные раз-
личия в карбоксидирующем потенциале in vivo. В моделях фо-
тосинтеза для Сз-растений обычно использовали величину кар-
боксилирующего потенциала (в расчете на единицу Хл), кото-
рая в 2 раза превосходила величину карбоксилирующего потен-
циала для С4-растений (ср. рис. 14.6 и 14.8).
Одно время считали, что способность С4-растеиий фиксиро-
вать СО2 с очень высокой скоростью (особенно при ярком осве-
щении и при повышенных температурах) вполне можно объяс-
464-
глава 14
нить -тем, что ФЕП-карбоксилаза обладает высоким сродством
к COs. В настоящее время, однако, из литературы известны та-
кйё значения КмСсОг/нсозЩДЛ-я -этих, двух.карбоксилаз (ср. разд.
6:5й 13.3), которые никак не соответствуют , этому объясне-
нию' (см. ниже).
ФЕП-карбоксилаза из. С4-растений имеет следующие свойст-,
вй: а) о'на использует в качестве субстрата ;НСО3~, а не свобод-
ную СOi; б)'она, по-видимому, имеет /(м(нсо3“)^0,2—0,4 мМ;
в) Vinax ФЕП-карбоксилазной реакции при насыщающих кон-
центрациях ФЕП и НСО3~ составляет 800—1200 мкмоль-
• (мгХл)-1-ч”-1 при 30 “С и т) в отличие от РуБФ-карбоксилазы,
она пе ингибируется О2. Поэтому преимущество- ФЕП-карбок-.
сил азы над РуБФ-царбоксилазой с точки.зрения эффективности
карбоксилирования может состоять в том,'что первая не обла-
дает оксигеназной активностью и, возможно, имеет более высо-
кую Умах; правда, величина Ртах для РуБФ-карбоксилазы еще
Не определена: Безусловно, что Утах ФЕП-карбоксилазной реак-
ции при насыщающей .концентраций ФЕП и НСО3~ значитель-
но'выше, чем скорость фотосинтеза в листьях.
Как можно интерпретировать значение /(м(нсоз“) для ФЕП-
карбоксилазы, равное 0,2—0,4 мМ, о чем свидетельствует оно,
о низком или высоком сродстве к СО2, присутствующей в атмо-
сфере?. На концентрацию иона НСО3~, доступного для ФЕП-
карбоксилазы in vivo, влияет несколько факторов: концентрация
СО2 в межклеточном воздушном пространстве листа, темпера-
тура листа, pH цитозоля и содержание карбоангидразы.
У С4-растений свыше 90% карбоангидразы находится в клет-
ках мезофилла, где Она локализуется в цитоплазме. Следова-
тельно, и ФЕП-карбоксилаза, и карбоангидраза локализованы в'
ФЕП р.
Рис. 14.7. Предполагаемый основной путь переноса СО2 (жирные стрелки)
в листе Ci-растений при участии карбоангидразы, сдвигающей равновесие
реакции. Реакция СО2 ** НСОа~ представляет собой краткую форму записи
катализируемой карбоангидразой реакции СО2-|-Н2Оч^НСОз~-|-Н+.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
465-
одном и том же компартменте клетки. Как только СО2 входит
в цитозоль клетки, она тотчас же гидратируется до НСО3п-|-Н+
И СО2, и НСО3~ диффундируют по градиенту концентрации во
всем цитозоле. Так как потенциальный градиент для НСО3~ на-
много больше, чем для СО2, бикарбонат как субстрат гораздо
более доступен для ФЕП-карбоксилазы (рис. 14.7). Если ката-
литическая активность карбоангидразы намного превышает ак-
тивность ФЕП-карбоксилазы, то конечная концентрация НСОз“
в цитозоле будет определяться концентрацией СО2 в воздуш-
ном пространстве между клетками мезофилла, а также темпера-
турой и pH цитозоля. Так, например, при концентрации СО2 в
воздушном пространстве листа — 250 м.д. и при-темпер ату ре
листа 30°C растворимость свободной СО2 равна 7,4 мкМ. При
pH 8,0 такая свободная СО2 в цитозоле находилась бы в рав-
новесии с 331 мкМ НСО3~, что очень близко по величине к
Л'м(нсо~8) для ФЕП-карбоксилазы. При насыщающей концентра-
ции ФЕП это обеспечивало бы скорость ~500 мкмоль СО2-
• (мгХл)-1 •ч~1. Следовательно, ФЕП-карбоксилаза может обла-
дать довольно высоким сродством к атмосферной СО2. Вполне
естественно, что [НСО3'_] в клетках мезофилла варьирует в за-
висимости. от сопротивления устьиц, температуры листа и pH
цитозоля. pH цитозоля еще не определен, известно лишь, что,
активность ФЕП-карбоксилазы из С4-растений в условиях in
vivo максимальна при pH около 8,0 и очень мала при pH 7,0.
В отличие от Сз-растений фотосинтез у С4-р астений близок
к насыщению при нормальной концентрации СО2 в атмосфере..
Рассмотрим, как скорость фотосинтеза у сорго (С4-растеппя)
связана с межклеточной i [СО2], и проанализируем кинетику
этого процесса, построив график двойных обратных величин
[зависимость НФС от ([СО2]—Г)]. В этом случае Дм(со2) со-,
ставит ~2 мкМ при 30°C, что соответствует 68 м.д. СО2 в меж-
клеточном пространстве (рис. 14.5,В, Г). Следовательно, лист.
С4-растения может обладать чуть большим сродством к СО2,
чем лист Сз-растения, даже если его сравнивать с листом Сз-рас-
тения при низкой концентрации О2. При 30 °C 2 мкМ СО2
должна находиться в равновесии с 0,09 мМ НСОз- (при усло-
вии, что pH цитозоля в клетках мезофилла равен 8,0). Это
намного меньше, чем величина /<м(нсо"5)> которую обычно при-
водят в работах с экстрагированной ФЕП-карбоксилазой.
При обычном содержании СО2 в воздухе и температуре 30 °C
скорость фотосинтеза у С4-растения будет выше, чем у Сз-рас-
тения (рис. 14.5). Это можно объяснить кислородным ингибиро-
ванием фотосинтеза у Сз-растений. В то же время скорость фо-
тосинтеза в листе Сз-растения при насыщающей концентрации
СО2 выше, чем в листе С4-растения. Это, по-видимому, связано-
30—234
466
ГЛАВА 14
Рис. 14.8. Модель фотосинтеза в листе С4-растения, в которой рассматрива-
ется связь этого процесса с потенциальной активностью ФЕП- и РуБФ-кар-
боксилазы (Hatch, Osmond, 1976). Кривая I— потенциальная скорость ФЕП-
карбоксилазной реакции в соответствии с кинетикой Михаэлиса—-Ментен при
насыщающей концентрации ФЕП и pH цитоплазмы, равном 8,0, когда
Кмгнао- ) для ФЕП-карбоксилазы составляет 0,3 мМ, что эквивалентно
6,6 мкМ’СОа (эта величина может снижаться до 2 мкМ в соответствии с
результатами кинетического анализа фотосинтеза в целых листьях,
рис. 14.5, Г). Кривая II— реальная скорость фотосинтеза в листьях при из-
менении [СО2] в клетках мезофилла (график, построенный по эксперимен-
тальным данным, например данным рис. 14.5,5). Кривая III— скорость фик-
сации COs за счет РуБФ-карбоксилазы в присутствии 21% О2 и ее изменение
в ответ на изменение [СО2] в клетках обкладки проводящих пучков, когда
v___ ___________Утах [СО,]___________
— [СО3] -ф- Ам.(СО2) (1 + 102]/7<i (02j)
[гл. 13, уравнение (13.36)]. Стрелкой на оси абсцисс показана концентрация
СОа, которая доступна для клеток мезофилла при обычном содержании СОа
в воздухе. При любом заданном значении скорости фотосинтеза соответст-
вующая [СОа] па кривых II и III будет отражать [СО2] в клетках мезофилла
(кривая II) или в клетках обкладки проводящих пучков (кривая Ш).
Исходные данные. ФЕП-карбокснлаза: Км(нсо“а)= 0,3 м-М, Ушах=1000 мкмоль-
• (мгХл)-'-ч-1; РуБФ-карбоксилаза: /(м(аоа) = 5 мкМ, Kt(Oa)=200 мкМ,
Ушах—400 мкмоль-(мг Хл)-’-ч-1; [C02]B„YTp = 0,0250/o; температура листа
30 °C; растворимость СО2 в мезофилле=7,4 мкМ СО2, 331 мкМ НСО3_; рас-
творимость О2=245 мкМ.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
467
с более высоким уровнем РуБФ-карбоксилазы в листьях Сз-рас-
тений. Установив определенные значения кинетическим констан-
там для ФЕП- и РуБФ-карбоксилаз у С4-растений, можно со-
здать общую модель С4-фотосинтеза (рис. 14.8). Как видно из
рис. 14.8, при низких [СОа] ФЕП-карбоксилаза имеет боль-
шой карбоксилирующий потенциал. Этот фермент связыва-
ет СОг атмосферы с более высокой скоростью, чем РуБФ-кар-
боксилаза у Сз-растенпй, карбоксилирующая активность кото-
рой подавляется Ог. Принято считать, что благодаря С4-циклу
создается высокая [СО2] в клетках обкладки проводящих пуч-
ков. Из исследований по фиксации ИСО2 следует, что суммар-
ный пул СО2 в листьях С4-растеиий (при расчете на вес сырого
вещества) приблизительно в 10 раз больше, чем у С3-р астений,
и доходит до 60 мкМ СО2 в клетках обкладки проводящих пуч-
ков при стационарном состоянии фотосинтеза. Как показано на
рис. 14.8, при обычном содержании СОг в воздухе (указано
стрелкой под осью абсцисс) фотосинтез в листе достигает насы-
щения при скорости 310 мкмоль• (мгХл)"1-ч-1, что соответству-
ет [СО2] в клетках обкладки проводящих пучков —40 мкМ.
Реальная скорость С4-фотосинтеза ниже, чем скорость ФЕП-
карбоксилазной реакции, особенно при более высоких [СО2].
In vivo ФЕП, по-видимому, не насыщает фермент, так как реге-
нерация ФЕП лимитируется скоростью ВПФ-цикла. При дале-
ком от насыщающего уровне субстрата этот фермент находится
под аллостерическим контролем (разд. 12.8).
При 30 °C и одинаковом сопротивлении устьиц скорость фото-
синтеза у С4-растений выше, чем у С3-растений (ср., например,
скорость процесса в присутствии 6 мкМ СО2, рис. 14.5, 14.6,
14.8). Если [СО2] в листе уменьшается из-за увеличения сопро-
тивления устьиц или из-за повышения температуры, то скорость
фотосинтеза у С4-растений может возрасти в 2-—3 раза по срав-
нению со скоростью фотосинтеза у Сз-растений.
Остается неясным, существует ли строгая «один-к-одному»
зависимость между поступлением углерода в С4- и в ВПФ-цик-
лы. Если вся атмосферная СО2, которая связывается в ВПФ-
цикле в листьях С4-растеиий, поступает в С4-цикл, то скорость
последнего должна быть по меньшей мере равна скорости функ-
ционирования ВПФ-цикла, Даже при самой минимальной диф-
фузии СО2 из клеток обкладки проводящих пучков скорость
функционирования С4-цикла должна быть выше, чем скорость
ВПФ-цикла. Поскольку вполне вероятно, что сопротивление
диффузии СО2 из клеток обкладки проводящих пучков доста-
точно высоко, убыль СО2 и дополнительное ее использование в
С4-цикле могут быть очень незначительны. После освещения
листьев С4-растений С4-цикл должен функционировать с более
высокой скоростью, чем ВПФ-цикл. Это необходимо для того,
30*
468
ГЛАВА 14
чтобы [СОг] в клетках обкладки проводящих. пучков могла
стать выше, чем в клетках мезофилла. Иногда ,С4-цикд называ-
ют «СОг-насосом», причем, как.и в случае других известных на-
сосов, для.этого процесса необходима энергия.
Источники данных: [4—6,28—31,34].
14.5. Влияние температуры, света и водного дефицита
на карбоксилирование
С4-растения особенно хорошо растут при температуре 25—
35 °C, тогда как Сз-растения— при температуре 15—25ЬС. Это
хорошо иллюстрируют данные Каванабе, который сравнил отно-
сительную скорость роста растений умеренного и тропического
пояса при трех разных температурных режимах (табл. 14.1).
Одно из наиболее ранних и наиболее исчерпывающих исследо-
ваний зависимости фотосинтеза в целых листьях от температу-
ры, света и [СО2] было проведено Гаастрой в пятидесятых го-
дах. В своих работах он применял инфракрасный газовый ана-
лизатор.- Эксперименты этого и других авторов, проведенные
на таких растениях, как томаты, шпинат и огурцы (которые
сейчас относят к Сз-растениям), показали, что фотосинтез лими-
тируется [СО2], особенно при повышенных температурах
(рис. 14.9 и 14.10).
Гщах РуБФ-карбоксилазы у Сз-растений должна быть выше
при относительно более высоких температурах (в норме Vmax
Рис. 14.9. Типичные температурные кривые фотосинтеза у С3- и Сч-растений.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
469.
Таблица 14.1. Относительная скорость роста проростков ряда теплолюбивых
и холодостойких трав при трех разных температурных
(дневная/ночная температура) режимах11
Подсемейство и вид расгепля Температурный режим, "С2)
1П/10 27,/22 36/31
Сз-травы
Festucoideae
Agropyron trichophorum (Link) Richt. 71 100 62
Bromus inermis Lcyss. 77 100 66
Festuca arundinacea Schreb. 77 100 56
Poa pratensls L. 69 100 63
Trisetum splcatum (L.) Richt. 58 100 64
Phalaris arundinacea L. 71 100 66
Stipa hyalina Nees. С^тровы Eragrostoideae 54 100 71
Chlorls gayana Kuntli. Panicoideae 22 91 100
Seiaria sphacelata Stapf. and Hub- bard 5 88 100
Cenchrus clltaris L. 11 £86 100
Paspalum dllatalum Poir. 23 90 100
Panicum coloratum Walt. 35 82 100
Panicum maximum Jacq. 23 89 100
Digitaria argyrogranta (Nees) Stapf. 32 86 100
Sorghum almurn Parodi 26 ') Il упрощенном индо из работы Kawanabc, 1968. 77 100
а)Днсвпая/ночпая температура; для всех Са-растеиий приняты за 100 данные, полу-
чепные при 27/22 °C, а для всех С^-растений — данные, полученные при 36/31 °C.
ферментативной реакции возрастает примерно в 2 раза при по-
вышении температуры на 10°С, т. е. Qio=2). Однако скорость
фотосинтеза при высоких температурах лимитируется из-за ог-
раниченной растворимости СО2 и из-за увеличения отношения
растворимостей О2/СО2. Более высокая чувствительность фото-
синтеза к О2 у Сз-растений при повышении температуры может
быть отчасти связана с физическими явлениями, обусловленны-
ми увеличением отношения растворимостей О2/СО2 (рис., 14.2).
470
ГЛАВА 14
Рис. 14.10. Типичные температурные кривые фотосинтеза у Сз-растеннй при
обычном и при насыщающем уровнях СО2 в воздухе.
Было высказано также предположение о том, что происходит
какое-то биохимическое изменение кинетических констант
РуБФ-карбоксилазы-оксигеназы, которое может благоприятство-
вать при более высоких температурах протеканию оксигеназной
реакции. Однако сообщения о такой возможности противоречат
друг другу, возможно, в связи с тем, что технически очень труд-
но одновременно определить активность оксигеназы-карбоксила-
зы при заданных концентрациях О2 и СО2 при той или иной
температуре. Так, например, было показано, что температура
влияет либо на энергию активации, которой определяется отно-
сительное изменение для оксигеназы по сравнению с Vmax
для карбоксилазы, либо на величину кинетических констант
Хм(со2) для карбоксилазы или /<м(02) для оксигеназы. В другой
работе температура сходным образом влияла на Дм(со2)И Лм(Оа),
если их значение вычисляли, исходя из концентрации субстра-
тов в водной фазе (что указывает на отсутствие дифференци-
ального влияния температуры на биохимическом уровне). Если,
однако, для расчетов использовали парциальное давление О2
ИЛИ СО2 В газовой фазе, ТО 7<М(СО2 в газовой фазе) при увеличении
температуры возрастала в большей степени, чем Дм(02 в газовой
Фазе) (что говорит о влиянии температуры на растворимость этих
газов) [10,11,29,35].
Темновое дыхание с увеличением температуры усиливается,
и это может в значительной степени снижать реальную скорость
фотосинтеза на свету. Однако поскольку темновое дыхание до-
стигает насыщения при низкой [О2] (разд. 13.6), повышенная
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
471
чувствительность фотосинтеза в присутствии 21% Ог (по срав-
нению с 2% О2) скорее всего не связана со стимуляцией темно-
вого дыхания.
Очевидно, фотосинтез реагирует иа концентрацию газов (СОа
и О2) в атмосфере как единый и целостный процесс. Биохими-
ческие ответные реакции фотосинтеза на эти газы можно пра-
вильно проанализировать только в том случае, если известны те
концентрации газов, которые доступны для фотосинтезирующей
ткани в водной фазе при различных температурах.
При низкой температуре эффективность фотосинтеза у
Сз-растеиий возрастает; это связано с увеличением растворимо-
сти СОг и уменьшением доли кислородного ингибирования фо-
тосинтеза. По-видимому, у Сз-растений уровень РуБФ-карбок-
силазы, рассчитанный на единицу Хл в листе, выше, чем у С4-
растений. При низких температурах, когда лимитирующее дей-
ствие СОг выражено в меньшей степени, повышенное содержа-
ние РуБФ-карбоксилазы у С3-растений может обеспечивать им
равную или даже более высокую, чем у Сграстепий, фотосин-
тетическую активность. При высокой температуре и насыщаю-
щей концентрации СОа ферментативная активность, выражен-
ная на единицу белка РуБФ-карбоксилазы, должна быть доста-
точно высока, так что для поддержания высокой скорости фото-
синтеза требуется меньшее количество фермента. При высоких
температурах механизм, концентрирующий СОг, способствует
тому, что уровень СОг в воздухе ие лимитирует скорость про-
цесса.
Qio для фотосинтеза (т. е. показатель того, насколько ско-
рость фотосинтеза возрастает при увеличении температуры на
10°) может изменяться даже при насыщающей концентрации
СОо. Результаты исследований, проведенных па водорослях и
иа изолированных хлоропластах С3-растений, свидетельствуют
о том, что величина Q10 при температурах ниже 15 °C варьиру-
ет от 9 до 14, в интервале температур 15—25 °C она приближа-
ется к 2, а при более высоких температурах становится еще
меньше. Подобные цифры свойственны автокаталитическим про-
цессам, к которым принадлежат реакции ВПФ-цикла (разд. 6.3
и 6.14). Таким образом, при повышении температуры иа 10°C
скорость всех этапов ВПФ-цикла должна удваиваться, если Qio
для всех ферментов равен 2, ио при этом одновременно должна
возрастать и скорость реакций обратной связи, в результате
чего при более высокой температуре каждому из ферментов
требуется больше субстрата. При еще более высокой темпера-
туре эта тенденция будет ограничиваться скоростью поступле-
ния ассимиляционной силы (гл. 5/. Конечным результатом всех
этих эффектов является то, что фотосинтетическая ассимиляция
углерода в хлоропластах Сз-растений перестает подчиняться за-
472
ГЛАВА 14
Рис. 14.12. Типичная световая кривая-
фотосинтеза для отдельного листа
Сз-растения при нормальном содер-
жании Ог и СОз в атмосфере. Стрел-
кой на оси абсцисс Отмечен макси-
мум прямой инсоляции (Akita et al.,
1969; Bohlar-Nordenkampf, 1976; Ga-
astra, 1959; Ku et al., 1977; Singh
et al„ 1974).
Рис. 14.11. Типичные световые кривые
фотосинтеза для отдельного листа
Ci-растения при высокой (25—35 °C)
и низкой (10—15 °C) температуре и
при нормальной концентрации О2 и
СО- в атмосфере. Стрелкой на оси
абсцисс отмечен максимум прямой
инсоляции (Akita et al,, 1969; Singh
et al„ 1974).
кону Аррениуса, согласно которому существует прямо пропор-
циональная зависимость между логарифмом максимальной ско-
рости фотосинтеза и обратной величиной температуры, выра-
женной в градусах Кельвина.
У Сграстений основным лимитирующим фактором при оп-
тимальной температуре является свет, так как обычно фотосин-
тез не достигает насыщения даже при самом ярком солнечном
освещении (рис. 14.11). Свет очень важен для активации ряда
фотосинтетических ферментов, для открывания устьиц и для об-
разования ассимиляционной силы. Так, например, степень ак-
тивации светом таких ферментов С4-пути, как пируват, ортофос-
фат— дикиназа и NADP-малатдегидрогеназа, зависит от интен-
сивности освещения. Относительно яркое освещение требуется и
для максимального открывания устьиц у таких С4-растений, как
кукуруза; у С3-растений, например у пшеницы, устьица откры-
ваются при гораздо меньшей освещенности. По мере увеличе-
ния интенсивности освещения должно возрастать производство
энергии и обеспечение ею С4- (СО2-насоса) и ВПФ-циклов. Та-
ким образом, хотя СО2 уже перестает лимитировать скорость
С4-фотосинтеза, зависимость этого процесса от освещения оста-
ется.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
473
Рис. 14.13. Предполагаемые
световые кривые фотосинтеза
для отдельного листа С3-рас-
тения при оптимальной темпе-
ратуре и изменяющемся уров-
не О2 и СО2. Стрелкой на оси
абсцисс отмечен максимум
прямой инсоляции (Bohlar-
Nordenkampf, 1976; Gaastra,
1959; Ku et al., 1977).
Интенсивность падающего излучения
При низких температурах фотосинтез у С4-растений обычно
достигает насыщения задолго до того, как достигается макси-
мум прямой инсоляции (рис. 14.11). При низких температурах
максимальная скорость фотосинтеза в листьях С4-растений мо-
жет становиться даже меньше, чем у Сз-растений. Это связано
не с лимитирующим действием света или [СО2], а скорее всего
с ограниченным транспортом метаболитов, инактивацией неко-
торых ферментов на холоду и снижением активности фермен-
тов. Так, например, транспорт пирувата в хлоропластах мезо-
филла Сграстения, росички кровяной, очень чувствителен к по-
нижению температуры. У многих С4-растений пируват,ортофос-
фат— дикиназа очень лабильна на холоду и при низкой темпе-
ратуре диссоциирует, превращаясь из тетрамера в димер или в
мономер. Содержание белка фракции I (РуБФ-карбоксилаза)
у некоторых С4-растений составляет всего 1/4 его содержания
у Сз-растений (разд. 14.4). Поэтому при низких температурах
каталитическая активность этого фермента у С4-растений может
быть намного ниже, чем у С3-растений.
При нормальной концентрации СО2 и О2 в атмосфере фото-
синтез у Сз-растений достигает насыщения или приближается
к нему при сравнительно слабом освещении в широком диапа-
зоне температур (рис. 14.12).
Выше точки светового насыщения фотосинтез у С3-растений
лимитируется [СО2] и вследствие кислородного ингибирования
,'этого процесса. При нормальной концентрации СО2 в атмосфе-
ре доля кислородного ингибирования фотосинтеза (в присутст-
вии 21% О2 по сравнению с 2% О2) одинакова или слегка убы-
вает при увеличении интенсивности освещения. Поэтому кривые
светового насыщения одинаковы как в присутствии 21% О2, так
и 2% О2 (рис. 14.13).
4 74
ГЛАВА Н
При низкой концентрации Ог и высокой интенсивности осве-
щения СО2 может все еще лимитировать скорость фотосинтеза у
Сз-растений. Это особенно справедливо в случае относительно
высоких температур (когда лимитируется растворимость СО2)
или сравнительно большого сопротивления устьиц (когда огра-
ничивается диффузия СО2 в лист). Поэтому Сз-растения долж-
ны извлекать несомненную выгоду из обогащения атмосферы
СО2 при относительно высокой температуре. Когда [СО2] яв-
ляется насыщающей, С3-растения способны вести себя подобно
С4-растениям, точно так же реагируя на увеличение интенсивно-
сти освещения вплоть до максимума прямой инсоляции
(рис. 14.13). Механизм, концентрирующий СО2 у С4-растений,
имеет двоякий эффект: во-первых, он устраняет кислородное ин-
гибирование фотосинтеза и, во-вторых, он способствует поддер-
жанию насыщающей концентрации СО2.
Очень часто бывают интересны и исключения из общих пра-
вил. Такие Сз-растения, как подсолнечник, по неизвестным для
нас причинам при обычном содержании О2 и СО2 в воздухе
дают световые кривые, достигающие насыщения при отно-
сительно сильном освещении (возможно, это связано с толщи-
ной листа или с содержанием Хл в расчете на площадь поверх-
ности листа). Вместе с тем у С4-растения Panicum capillare све-
товое насыщение наступает при сравнительно слабом освеще-
нии. И у Сз-, и у С4-растений имеется большое число адаптив-
ных приспособлений к различным условиям окружающей среды,
особенно к температуре. Так, C4-pacTei:ne Tidestroma oblongtfo-
lia, которое растет в Долине Смерти в Калифорнии, выдержи-
вает очень высокие температуры (до 50°C). Многие из исследо-
ванных С4-растений чувствительны к понижению температуры
до 10—15 °C. В то же время некоторые растения, такие, как
Atriplex sabulosa, A. confertifolia, Panicum virgatum и Spartina
townsendii, приспособились к низким температурам (5—20°C).
По этой причине очень важны сравнительные исследования ре-
гуляции фотосинтеза у С3- и С4-растений, принадлежащих к
различным систематическим группам и произрастающих в са-
мых разных местообитаниях.
Источники данных: '[1, 2, 7, 8, 10—14, 17, 19, 23, 24, 27, 30,
32, 33—35, 37, 39, 42, 43, 45, 46, 49, 52, 53].
14.6. Эффективность использования воды
Транспирацию и фотосинтез можно выразить в терминах за-
кона Ома:
Скорость =
Разность потенциалов
Сопротивление
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
475
Транспирацию можно выразить формулой
^L—Га
r'a + r's
(14.5)
где Т — скорость транспирации, выраженная через количество
воды, потерянной листом, например в мг воды-см-2 поверхности
листа-с-1 (рис. 14.14). Ее определяют по увеличению относи-
тельной влажности после продувания воздуха, влажность кото-
рого известна, через камеру, в которой находится лист.
IVl— концентрация водяных паров в устьичной щели листа,
например в мг воды-см-3. Считают, что эта величина отражает
насыщающую концентрацию водяных паров в устьичной щели,
которая зависит от температуры листа.
IFa — концентрация водяных паров в воздухе, например в мг
воды см-3.
г'&— сопротивление пограничного слоя для диффузии воды.
Эта величина связана с наличием воздушного слоя (не имеюще-
го возмущений), который примыкает к листу. Она зависит от
формы листа и от скорости ветра. Сопротивление пограничного
слоя минимально, когда листья очень узкие и тонкие и когда
скорость ветра достаточно высока. Чтобы максимально понизить
величину г'й, при изучении газообмена в дыхательной камере
очень важно поддерживать достаточно высокую скорость воз-
душного потока. При высоком значении г'й температура листа
резко возрастает, что может пагубно отразиться на результатах
измерения интенсивности фотосинтеза. При подходящей скоро-
сти воздушного потока вклад сопротивления пограничного слоя
(например, 0,5 с-см-1 и меньше) в общее сопротивление диффу-
зии газов в нормальных условиях минимален. г'а можно опре-
делить, сделав из промокательной бумаги реплику листа и за-
тем насытив ее водой. В этом случае сопротивление реплики к
потере воды (испарению) в дыхательной камере будет отражать
величину г'л. Кроме того, величину г'а можно рассчитать по сле-
дующему уравнению:
,__,,, 7 / Ширина листа (см)
а — ' V Скорость ветра (см-с-1)
(14.6)
где К' — константа, величина которой определяется тем, какие
единицы используют при измерении; например, для диффузии
воды она равна 0,96 с1/2см-1. Поэтому сопротивление погранич-
ного слоя для испарения воды из листа шириной в 2 см при
скорости воздушного потока 100 см-с-1 будет составлять 0,13 с-
-см-1.
r's — сопротивление устьиц для диффузии воды. Оно зависит
от числа устьиц и от степени их раскрытия. /в для любого лис-
476
ГЛАВА 14
Рис. 14.14. Графическое изображе-
ние составаляющих, из которых
складывается общее сопротивле-
ние для потери воды в ' ходе
транспирации.
(со21 в атмосфере
Рис. 14.15. Графическое изображе-
ние составляющих, из которых
складывается общее сопротивле-
ние для диффузии СО2.
10О2] в хлоропласта
та можно рассчитать (в с-см-1) после измерения всех остальных
величин, которые входят в уравнение (14.5). Можно также ис-
пользовать диффузионные порометры, датчики которых при-
крепляют к листьям и таким образом измеряют убыль воды в
зависимости от степени раскрытия устьиц. Градиент потенциала
’[уравнение (14.5)] можно выразить и не через разность парци-
альных давлений. Обычно для этой цели вполне достаточно
разности концентраций, особенно в тех случаях, когда темпера-
тура листа и воздуха одинакова или когда налицо значительный
градиент, концентраций. Строго говоря, величина этого градиен-
та определяется разностью парциальных давлений водяных па-
ров, которая зависит от абсолютной температуры и концентра-
ции паров.
Для фотосинтеза: Р — ^0^Ь°-]дух 19°^х.п (14.7)
raTrSTrm
где Р — скорость фотосинтеза, выраженная, например, в нг фик-
сированной СО2-см-2 поверхности листа-с-1; [СО2] воздух — кон-
центрация СО2 в воздухе, выраженная, Например, в, нг-см~3;
[СО2] хп — концентрация СО2 в хлоропласте, выраженная, на-
пример, в нг-см-3; rs — сопротивление устьиц для диффузии СО2,
выраженное, например, в с-см-1. Его можно рассчитать, зная
величину r's, по уравнению 1,56/8=4. [Для воды, молекулярная
масса которой меньше, чем у СО2, коэффициент диффузии D
несколько выше, так как
_ дн2о Г 1/мол. масса НВО ~|1/2 , Г 1/18 11/2 .
5 ^СО2 L 1/мол. масса СО2 J ~~ [ 1/44 J ~~ ’
Поэтому при прочих равных условиях вода диффундирует в
1,56 раза быстрее, чем СО2.]
< 4—сопротивлений пограничного слоя листа для диффузии
-СО2, выраженное, например, ,в с-см"1. Его можно рассчитать,
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
477
Таблица 14.2. Данные по сопротивлению листьев потоку СОг
у некоторых С3- и С4-растений (Gifford, 1974, S. В. Ku, личное сообщение)
Вид растения Минимальное из известных значений, С •CM*’1
г 1) m ra + rs
Сз-растепия Glycine max 2—3 . 1,3 .
Atriplex hastata 2,6 1,1
Phaseolus spp. 2,6 1,1
Tritlcum aestivum 2,8 1,1
Solatium tuberosum 5,4 2,5
Medicago saliva 2,8 1,0
Сграстеиия Amaranthus spp. 0,3
Atriplex spongiosa 0,4—0,6 1,9
Zea mays 0,7—0,9 1,5
Saccharum of ficinarum 0,3 1,5
Sorghum bicolor 1,8 1,8
Panicum vlrgatum 2,0 2,9
'11 Эти величины rm ие эквивалентны 1/ЭК, так как измерения проводили при обыч-
иой концентрации СО2 в воздухе, а при этом остаточное сопротивление включает в себя’
любые фотохимические ограничения (см. разд. 13.10, где описаны условия, при которых
гт = 1/ЭК).
зная величину г'а, по уравнению га=1,35г'а- [Отношение Dmo/
IDaoz возводят в степень 2/3. (Онао/Осо,)0'66 == (1.56)°>66= 1,35,
поскольку необходимо учесть и ту составляющую переноса че-
рез пограничный слой, которая связана ие с диффузией, а с
турбулентностью. Эта поправка носит несколько произвольный
характер, так как степень турбулентности в пограничном слое
определяется и геометрией листа, и скоростью потока воздуха
через камеру.]
rm — сопротивление мезофилла для диффузии СО2, выражен-
ное, например, в с-см~‘. Это остаточное сопротивление (иногда
называемое неведомым сопротивлением, так. как очень трудно
измерить те величины, из которых оно складывается), которое
включает в себя сопротивление диффузии СО2 через водную
. фдзу клеток мезофилла (сопротивление переносу.)сопротивле-
ние карбокецлированию (сопротивление, обусловленное ограни-
чёйным. потеидиалом •карбоксилирования) и фотохимическое
•478
ГЛАВА 14
сопротивление. При лимитирующей [СОг] и высокой интенсив-
ности освещения фотохимическое сопротивление устраняется,
и тогда гт=1/ЭК, где ЭК — эффективность карбоксилирования
(разд. 13.10). Величину rm можно определить из уравнения
(14.7). Определению поддаются все величины, входящие в это
уравнение, за исключением [СО2]хп, которую обычно принима-
ют равной величине Г, т. е. углекислотному компенсационному
пункту. Ясно, что, если очень занизить величину, приписывае-
мую [СОа] хп> реальная величина rm будет завышена.
Для Сграстений характерны более низкие значения rm, чем
для С3-растений (табл. 14.2). Это может быть связано с тем,
что ФЕП-карбоксилаза С4-растений обладает высокой карбок-
• силирующей способностью, имеет высокое сродство к СОг и не
чувствительна к ингибированию кислородом (разд. 14.4 и 14.5).
По мере уменьшения уровня Ог в атмосфере величина rm у
Сз-растений уменьшается и приближается к величинам, харак-
терным для С4-растений. Поскольку О2 ингибирует РуБФ-карбо-
ксилазу, при увеличении [О2] будет возрастать и сопротивление
.мезофилла для поглощения СО2.
Таблица 14.3. Эффективность использования
воды разными растениями
(Shantz, Piemeisel, 1927, Black et al., 1969)
Вид растения и группа, к которой оно было отнесено впоследствии Н20/Образованиое сухое вещество, г/г
С3-р астения
Tritlcum aesilvum 557
Oryza sativa 682
Chenopodium album 658
Citrullus vulgaris 577
Cucumis sativ-us 686
Phaseolus vulgaris 700
Medicago sativa 844
С4-растения
Set aria italica 285
Zea mays 349
Amaranthus graecizans 260
Portulaca oleracea 281
Растения с более низким rm имеют то преимущество, что они
«более эффективно используют воду (число граммов транепирируе-
мой воды на грамм фиксированной СОг), поскольку величина rm
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
479'
существенна для фотосинтеза, а не для транспирации. Умень-
шение величины rm при заданной величине ra+ri ведет к сти-
муляции фотосинтеза, тогда как транспирация при этом остает-
ся постоянной. Для С4-растений характерны более высокие зна-
чения сопротивления мезофилла и более высокая эффективность
использования воды, чем у С3-растений. Данные, собранные
Шантцем и Пимейзелем более чем полвека назад, хорошо ил-
люстрируют тот факт, что Сграстения более эффективно ис-
пользуют воду, чем Сз-растения (табл. 14.3).
При сравнении минимальной величины сопротивления у ряда
Сз- и С4-растений видно, что rm у С4-растений ниже, чем у Сз-рас-
тений, a г&+гв, наоборот, чуть выше у С4-растений (табл. 14,2).
Не ясно, с чем связано более высокое сопротивление диффузии
у С4-растений, то ли с меньшим числом устьиц на единицу по-
верхности листа (разд. 15.14), то ли с разной величиной устьич-
ных щелей. При низком гт и относительно высоком сопротивле-
нии диффузии у С4-растений создаются условия, способствую-
щие более эффективному использованию воды (транспирация/
/единица фотосинтеза). При сравнении разных видов растений
более высокое отношение общего сопротивления листа испаре-
нию к общему сопротивлению для поглощения СО2 указывает
на более эффективное использование воды тем или иным рас-
тением:
Поскольку испарение воды в ходе транспирации способствует
удалению избыточного тепла, при увеличении сопротивления
устыщ температура листа может повышаться. А это в свою оче-
редь приводит к тому, что возрастает градиент концентрации
водяных паров между листьями и окружающим их воздухом,
поскольку увеличивается значение ТГц, и, кроме того, это ока-
зывает положительное влияние на транспирацию, которое не
учитывается в уравнении (14.8). Поэтому при увеличении со-
противления устьиц конечная скорость транспирации будет за-
висеть и от величины r's, и от величины Ж- Изменения темпера-
туры листа, обусловленные изменением га, тоже влияют иа ско-
рость фотосинтеза. Описанные здесь взаимодействия рассмат-
ривались в самых разных моделях. Действительная эффектив-
ность использования воды (транспирация в расчете на единицу
образуемого сухого вещества) зависит и от других факторов.
Здесь можно упомянуть потери воды, связанные с дыханием,
и относительную влажность среды. В климатических местностях
с низкой относительной влажностью градиент водяных паров-
между листьями и атмосферой должен быть выше, а эффектив-
480
ГЛАВА 14
ность транспирации ниже по сравнению с обычными усло-
виями.
Хотя минимальное сопротивление диффузии (ra-|-rs) у С4-рас-
тений может быть чуть выше, чем у Сз-растений, при довольно
высоких температурах общее сопротивление притоку СОг У пер-
вых становится ниже и, следовательно, скорость их фотосинтеза
должна увеличиваться.
Рассмотрим два примера, которые помогут нам понять, поче-
му эффективность использования воды у С4-растений может
•быть выше, чем у Са-растений.
Пример I
Группа растений Сопротивление, с* см-1
га rs 1 rm | общее
Сз-растепие 1 1 3 5
С4-растение 1 3 1 5
В этом случае общее сопротивление поглощению СО2 одинако-
во у обоих растений, а ото значит, что и скорости фотосинтеза
у них должны быть одинаковы. Но сопротивление для потери
воды у С4-растения в два раза больше, чем у С3-растения; по-
этому эффективность использования воды у первого должна
•быть в 2 раза выше, чем у второго, если все другие условия
одинаковы [т. е. при одной и той же величине градиента концен-
трации водяных паров (IFl—№а)]. Из этого примера видно, что
С4-растение в состоянии создавать повышенное сопротивление
устьиц (например, при различного рода водном дефиците) и
даже поддерживать такую скорость фотосинтеза, которая наб-
людается у Сз-растения в отсутствие каких бы то ни было
стрессовых воздействий.
Для второго примера возьмем случай, когда сопротивление
пограничного слоя и сопротивление устьиц у обоих растений
совершенно одинаковы, а это значит, что при заданной величи-
не градиента концентрации водяных паров скорость транспира-
ции у этих растений должна быть одинакова.
Пример II
Группа растений Сопротивление, с«см--1
''а ] rs | rm 'общее
Сз-растение I 1- 3 S
С4-растение 1 ' 1 1 3
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
48/.
Однако общее сопротивление у С4-растений ниже. Это связано
с тем, что у них меньше гш, благодаря чему лист С4-растений
осуществляет фотосинтез со скоростью в 2 раза большей, чем
у Сз-растений. В этом примере эффективность использования
воды в листе С4-р астения должна быть выше, так как у него
выше скорость фотосинтеза при заданной скорости транспира-
ции. В естественных условиях при довольно высоких темпера-
турах (25—35 °C) может создаваться промежуточная ситуация,
если сопротивление устьиц у С4-растения чуть выше.
Температура влияет на фотосинтез как С3-, так и ^-расте-
ний, поскольку с изменением температуры меняются сопротив-
ление мезофилла и сопротивление устьиц. При повышении тем-
пературы могут увеличиваться значения rm и у Сз-растений. От-
части это обусловлено уменьшением карбоксилирующего потен-
циала, связанным с уменьшением растворимости СО2. В ка-
честве других причин можно назвать также увеличение отноше-
ния растворимостей О2/СО2 (при этом возрастает степень кис-
лородного ингибирования) и изменение кинетических свойств
РуБФ-карбоксилазы-оксигеназы (разд. 14.4). Такой эффект
температуры может частично компенсироваться за счет приро-
ста производительности карбоксилирования при повышении тем-
пературы (т. е. за счет увеличения Утах).
У С4-растений величины rm сравнительно малы при повы-
шенных температурах. А при сравнительно низкой температуре
(10—20 °C) rm у С4-растений может достигать значительной ве-
личины из-за уменьшения Утах некоторых реакций (возможно,
РуБФ-карбоксилазной реакции) или же инактивации ряда фер-
ментов при понижении температуры. С повышением температу-
ры уменьшается величина отношения rm(C4)/rm(C3); это указы-
вает на то, что высокая температура более благоприятна для
С4-фотосинтеза. Зависимые от температуры изменения rm, не-
сомненно, влияют и па фотосинтез, и на транспирацию у С3- и
С4-р астений.
Источники данных: [9, 16, 25, 41, 44, 54].
14.7. С3-, С4-фотосинтез и урожайность
сельскохозяйственных культур
Очень большой интерес привлекает возможность использо-
вать С4-фотосиитез для получения высоких урожаев (или же
для создания агрономических методов борьбы со злостными
сорняками, относящимися к С4-растениям). При селекции сель-
скохозяйственных культур очень важной агрономической харак-
теристикой является высокая производительность карбоксили-
рования (отражением чего является низкая величина rm).
В этом случае растение более эффективно использует воду и,
31-234
482
ГЛАВА 14
если все остальные условия благоприятны, осуществляет фото-
синтез с высокой скоростью.
При оптимальной температуре скорость фотосинтеза у С4-рас-
тений выше, чем у Сз-растений, при любом способе выражения
этой величины (на единицу поверхности листа, на единицу Хл
листа или на единицу азота в листе). В неоптимальных услови-
ях С4-растения способны поддерживать повышенное сопротив-
ление в устьицах, сохраняя таким образом воду. Благодаря это-
му фотосинтез у них происходит со скоростью, близкой к скоро-
сти фотосинтеза у Сз-растений (разд 14.6). В частности, более
высокая скорость фотосинтеза у С4-растений, если ее рассчиты-
вать на единицу азота, может быть связана с тем, что содержа-
ние белка фракции I у этих растений ниже, чем у С3-растеиий.
Сз- и С4-сельскохозяйственные культуры пробовали выращи-
вать в самых различных условиях, однако интерпретация полу-
ченных данных затруднительна, поскольку скорость роста и
рост растений контролируются слишком большим числом фак-
торов. Температура, интенсивность освещения, ярусная струк-
тура посева, потери, связанные с дыханием, и показатель уро-
жайности (экономический урожай/биологический урожай) —
вот некоторые из тех факторов, которые следует учитывать при
сравнении продуктивности разных видов растений. При этом
необходимо отличать кратковременное выращивание от роста
растения в течение всей его жизни.
С4-растения особенно удобны для выращивания в некоторых
естественных условиях окружающей среды. В оптимальных ус-
ловиях, т. е. при высоких температурах и при очень ярком осве-
щении, такие растения, как кукуруза и сахарный тростник, рас-
тут с очень высокой скоростью. Вместе с тем некоторые Сз-рас-
тения, такие, как картофель и сахарная свекла, наиболее про-
дуктивны в странах с умеренным климатом (см. также
разд. 3.11).
Источник данных: [15—18, 21, 38].
ОБЩАЯ ЛИТЕРАТУРА
1. Bjorkman О. Comparative studies on photosynthesis in higher plants, Pho-
tophysiology. (ed. A. Giese), Vol. 8, pp. 1—63, New York, Academic Press,
1973.
2. Bjorkman 0., Berry J. High efficiency photosynthesis, Sci. Amer., 229, 80—
93 (1973).
3. Black С, C, Photosynthetic carbon fixation in relation to net COa uptake,
Ann. Rev. Plant Physiol., 24, 253—286 (1973).
4, Hatch M. D., Osmond С. B. Compartmentation and transport in C4 photosyn-
thesis. In: Encyclopedia of Plant Physiology, Vol. 3. Transport in Plants.
New series (eds. C. R. Stocking and U. Heber), pp. 144—184. New York,
Springer-Verlag, 1976.
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
483
5. Jensen R. G., Bahr J. Т. Ribulose 1,5-bisphosphate carboxylase-oxygenase,
Ann. Rev. Plant Physiol., 28, 379—400 (1977).
6. Jolivet E. Les differentes modalitfes de carboxylation photosynth&tique chez
les especes de type C3 et de type C4, Physiol. Veg., 14, 563—594 (1976).
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
7. Akita S., Miyasaka A., Muraia Y. Studies on the differences of photosynthesis
among species. I. Differences in the response of photosynthesis among spe-
cies in normal oxygen concentration as influenced by some environmental
factors, Proc. Crop Sci. Soc., Japan, 38, 509—523 (1969).
в. Akita S., Moss D. N. Differential stomatai response between Сз and C4
species to atmospheric CO2 concentration and light, Crop Sci., 12, 789—
793 (1972).
9. Akita S., Tanaka 1. Studies on the mechanisms of differences in photosynthe-
sis among species. IV. The differential response in dry matter production
between Сз and C4 species to atmospheric carbon dioxide enrichment, Proc.
Crop. Sci. Soc., 42, 288—295 (1973).
30. Badger M. R-, Andrews T. J. Effects of CO2, O2, and temperature on a high
affinity form of ribulose diphosphate carboxylase-oxygenase from spinach.
Biochem. Biophys. Res. Commim., 60, 204—210 (1974).
11. Badger M. R., Gollatz G. J. Studies on the kinetic mechanism of ribulose
1,5-bisphosphate carboxylase and oxygenase reactions, with particular refe-
rence to the effect of temperature on kinetic parameters, Carnegie Institute
Yearbook, 1976, pp. 355—361, 1977.
12. Baldry C. W., Bucks C., Walker D. A. Temperature and Photosynthesis.
I. Some effects of temperature on carbon dioxide fixation by isolated chloro-
plasts, Biochim. Biophys. Acta, 126, 207—213, 1966.
13. Bird I. F., Cornelius M. J„ Keys A. J. Effects of temperature on photosynthe-
sis by maize and wheat, J. Exp. Bot., 28, 519—524 (1977).
14. Bjorkman O., Pearcy R, IV., Harrison A. T., Mooney H. Photosynthetic adap-
tation to high temperatures. A field study in Death Valley, California, Scien-
ce, 175, 786—789 (1972).
15. Black С. C. Ecological Implications of dividing plants into groups with
distinct photosynthetic production capacities, Advan, Ecol. Res., 7, 87—114
(1971).
16. Black С. C„ Chen T. M., Brown R. H. Biochemical basis for plant competi-
tion, Weed Sci., 17, 338—344 (1969).
17. Bolhar-hJordenkampf H. R. Measurements and mathematical models for the
CO2 gas exchange of Phaseolus vulgaris var. nanus L. with special conside-
ration of photorespiration and atrazine effect, Biochem. Physiol. PElanzen,
169, 121—161 (1976).
18. Brown R. H. A difference in N use efficiency in Сз and C4 plants and its
implications in adaptation and evolution, Crop Sci., 18, 93—98 (1978).
19. Caldwell M. M., Osmond С. B., Hott D. L. C4 pathway photosynthesis at
low temperatures in cold-tolerant Atriplex species, Plant Physiol., 60, 157—
164, 1977.
"20 . Gaastra P. Photosynthesis and crop plants as influenced by light, carbon
dioxide, temperature, and stomatai diffusive resistance, Meded Land Hogesch
Wageningen, 59, 1—68 (1959).
'21 . Gifford R. M. A comparison of potential photosynthesis, productivity and
yield of plant species with differing photosynthetic metabolism, Aust. J.
Plant Physiol., 1, 107—117 (1974).
"22 . Hogetsu D., Miyachi S. Role of carbonic anhydrase in photosynthetic CO2
fixation in Chlorella, Plant Cell Physiol., 20, 747—756, 1979.
31
484
ГЛАВА 14
23. Huber S. С., Edwards G. E. Transport in C4 mesophyll chloroplasts. Charac-
terization of the pyruvate carrier, Biochim. Biophys. Acta, 462, 583—602
(1977).
24. Johnson H„ Hatch M. D. Properties and regulation of leaf NADP-nialate
dehydrogenase and malic enzyme in plants with the C4-dicarboxylic acid
pathway of photosynthesis, Biochein. J., 119, 273—280, 1970.
25. Jones H. G. Crop characteristics and the ratio between assimilation and
transpiration, J. Appl. Ecol., 13, 605—622 (1976).
26. Jones H., Slatyer R. Estimation of the transport and carboxylation compo-
nents of the intracellular limitation to leaf photosynthesis, Plant Physiol.,
50, 283—288 (1972).
27. Kawanabe S. Temperature responses and systematics of the Gramineae,
Proc. Japan. Soc. Plant Taxonoin., 2, 17—20 (1968).
28. Ku S. B., Edwards G. E. Photosynthesis in mesophyll protoplasts and
bundle sheath cells of various types of C4 plants. IV. Enzymes of respira-
tory metabolism and energy utilizing enzymes of carbon assimilation, Z.
Pflanzenphysiol., 77, 16—32 (1975).
29. S. B., Edwards G. E. Oxygen inhibition of photosynthesis. I. Tempera-
ture dependence and relation to O2/CO2 solubility ratio, Plant Physiol., 59,
986—990 (4977a).
30. Ku S. B.. Edwards G. E. Oxygen inhibition of photosynthesis. II. Kinetic
characteristics as affected by temperature, Plant Physiol., 59, 991—999
(1977b).
31. Ku S. B., Edwards G. E. Photosynthetic efficiency of Panicutn hlans and
Panicutn tnilioides in relation to C3 and C4 plants, Plant, and Cell Physiol.,
19, 665—675 (1978).
32. Ku S. B., Edwards G. E„ Smith D. Photosynthesis and non-structural
.carbohydrate concentration in leaf blades of Pantoum virgatum L. as affec-
ted by night temperature; Can. J. Bot., 56, 63—68 (1978).
33. Ku S. B., Edwards G. E., Tanner С. B. Effects of light, carbon dioxide, and
temperature on psotosynthesis, oxygen inhibition of photosynthesis, and
transpiration in Solanuni tuberosum, Plant Physiol., 59, 868—872 (1977).
34. Ku S. B., Schmitt M. R., Edwards G. E. Quantitative determination of
RuBP carboxylase-oxygenase protein in leaves of several C3 and C4 plants,
J. Exp. Bot., 30, 89—98 (1979).
35. Laing U'z. A., Ogren W. L., Hageman R. H. Regulation of soybean net pho-
tosynthetic CO2 fixation by the interaction of CO2, O2, and ribulosc-1,5-
diphosphate carboxylase, Plant Physiol., 54, 678—685 (1974).
36. Lilley R. McC., Walker D. A. Assimilation by leaves, isolated chloroplasts
and ribulose bisphosphate carboxylase from spinach, Plant Physiol., 55,
1087—1092 (1975).
37. Long S. P., Incoll L. D., Woolhouse II. W. C4 photosynthesis in plants from
cool temperature regions, with particular reference to Spartlna townsciidli.
Nature, 257, 622—624 (1975).
381 Monteith J. L. Reassessment of maximum growth rates for C3 and C4
crops, Expl. Agric., 14, 1—5 (1978).
39. Murata У., lyama J. Studies on the photosynthesis of forage crops. II. In-
fluence of air-temperature upon the photosynthesis of some forage and
grain crops, Proc. Crop Sci. Soc., Japan, 31, 315—322 (1963).
40. Rahinowitch E. I. Photosynthesis and Related Processes, Vol. 1, pp. 326—
351, New York, Wiley-Interscience, 1945.
41. Rawson H. M., Begg J. E. The effect of atmospheric humidity on photo-
synthesis, transpiration and water use efficiency of leaves of several plant
species, Planta, 134, 5—10 (1977).
42. Selwyn M. J. Temperature and photosynthesis. II. A mechanism for the
effects of temperature on carbon dioxide fixation, Biochim. Biophys. Acta,
126, 214—224 (1966).
ПЕРВИЧНЫЕ КАРБОКСИЛАЗЫ И РЕГУЛЯЦИЯ ФОТОСИНТЕЗА
485
43. Servaites J. С., Ogren W. L. Oxygen inhibition of photosynthesis and stimu-
lation of photorespiration in soybean leaf cells, Plant Physiol., 61, 62—67
(1977).
44. Shantz II. L., Piemeisel L. N. The water requirements of plants at Akron,
Colorado, J. Agric. Res, 34, 1093—1189 (1927).
45. Shirahashi K., Hayakawa S, Sugiyatna T. Cold lability of pyruvate, ortho-
phosphate dikinase in the maize leaf, Plant Physiol., 62, 826—830 (1978).
46. Singh M., Ogren IP. Wildholm J. M. Photosynthetic characteristics of
several C3 and C4 plant species grown under different light intensities,
Crop. Sei, 14, 563—566 (1974).
47. Strang R. H. C. Estimation of Km values of enzymes requiring molecular
O2 as a substrate, Biochem. J. Letters, 193, 1033—1034 (1981).
48. Tenhunen J. D., Weber J. A., Yocum C. S, Gates D. Af. Solubility of gases
and the temperature dependency of whole leaf affinities for carbon dioxide
and oxygen. An alternative perspective, Plant Physiol, 63, 916—923
(1979).
49. Terri J. A,, Stowe L. G. Climatic patterns and the distribution of C4 gras-
ses in North America, Oecologia, 23, 1—12 (1976).
50. Umbreit W. W., Burris R. H., Stauffer I. F. Manometric techniques and
related methods for the study of tissue metabolism, Minneapolis, Minnesota,
Burgess Publishing Co, 1972.
51. Walker D. A., Utley R. McM. Ribulose bisphosphate carboxylase—an enig-
ma resolved? In: Perspectives in Experimental Biology, Vol. 2 (cd. N. Sun-
derland), pp. 189—198, Oxford, Pergamon Press, 1975.
52. Williams G. I. Photosynthetic adaptation to temperature in C3 and C4
grasses, Plant Physiol, 04, 709—711 (1974).
53. Yamamoto E., Sugiyatna T„ Miyachi S. Action spectrum for light activation
of pyruvate phosphate dikinase in maize leaves, Plant Cell Physiol, 15,
987—992 (1974).
54. Zelitch I. Photosynthesis, Photorespiration, and Plant Productivity, pp. 347,
New York, Academic Press, 1971,
ГЛАВА 15
Метаболизм кислот
у семейства толстянковых
(САМ-метаболизм)
Краткое содержание
История изучения растений, имеющих САМ-метаболизм, на-
чалась задолго до того, как были открыты фотодыхание и
С4-фотосинтез. Информация о накоплении органических кислот
в темноте 'Содержалась уже в самых первых из известных ра-
бот. Приводимые в них же значения дыхательного коэффициен-
та свидетельствовали о том, что в темноте поглощаются и СОг,
и Oj. Все это было' известно за много лет до того, как была ус-
тановлена непосредственная связь между темновой фиксацией
СОг и накоплением органических кислот в темноте.
Как теперь известно, накопление органических кислот в
темноте сопровождается деградацией крахмала, образованием
фосфоенолпирувата (ФЕП) в ходе реакций гликолиза и карбо-
ксилированием этого предшественника в результате катализи-
руемой ФЕП-карбоксилазой реакции с СО2, находящейся в воз-
духе. Образующийся при этом оксалоацетат восстанавливается
до малата и затем накапливается в вакуоли, где он и хранится.
На свету малат декарбоксилируется под действием малатдегид-
рогеназы или же превращается в оксалоацетат и затем декар-
боксилируется под действием ФЕП-карбоксикиназы. После это-
го освободившаяся СОг поступает в ВПФ-цикл.
В определенных условиях окружающей среды некоторые
CAM-растения могут переключать свой обмен веществ и фото-
синтезировать почти как Сз-растения. В «САМ-состоянии»
устьица открываются ночью и закрываются днем, а в «С3-со-
стоянии» все происходит наоборот. Очень часто газообмен у
CAM-растений происходит таким образом, что СОг поглощается
и днем, и ночью. Переходу в CAM-состояние способствуют ус-
ловия водного дефицита и резкие перепады температур днем и
ночью. Чтобы различить эти два способа ассимиляции СОг,
очень удобно воспользоваться данными об изотопном составе
(13С/12С) растения. Дело в том, что ФЕП-карбоксилаза почти не
различает 12СОг и 13СОг и фиксирует оба соединения одинаково',
тогда как у РуБФ-карбоксилазы степень дискриминации выра-
жена гораздо' сильнее.
С экологической точки зрения САМ-метаболизм лучше все-
го рассматривать как адаптивное приспособление к условиям
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
487
Рис. 15.1. Метаболизм кислот у семейства толстянковых. Растения с САМ-
метаболизмом (Crassulacean Acid Metabolism, названным так по Crassula-
сеае, у которых он был впервые обнаружен) очень экономно расходуют воду,
что обусловлено метаболической адаптацией процесса фотосинтеза. Ночью
СОа связывается в составе малата. Днем малат декарбоксилируется, а выде-
ляющаяся при этом СО2 ассимилируется в ВПФ-цикле. Это позволяет расте-
нию днем держать устьица в закрытом состоянии, т. е. тогда, когда потен-
циальная потеря воды возрастает из-за повышения температуры. Фотосин-
тез у САМ- и у Сь-растений имеет общую особенность: у тех и у других
углерод С4-кислот передается в ВПФ-цикл.
водного дефицита, которое способствует запасанию воды в су-
хих местообитаниях. Для CAM-растений характерна более вы-
сокая, чем у С3- или Сграстений, эффективность использования
воды, однако, их продуктивность намного ниже. Вода в расте-
нии запасается ночью, так как градиент концентрации водяных
паров между листьями и окружающим их воздухом в это время
меньше всего. Относительно высокое сопротивление устьиц и
низкое сопротивление мезофилла (последнее связано с участием
ФЕП-карбоксилазы в фиксации СОг) способствуют созданию
условий, благоприятных для поддержания минимального отно-
шения транопирация/фотосинтез.
15.1. Введение
Мы все привыкли к мысли о том, что зеленые растения днем
(в результате фотосинтеза) поглощают СОг и выделяют Ог,
а ночью (в результате дыхания) поглощают О2 и выделяют СО2.
Одно время было даже принято на ночь выносить цветы из
488
ГЛАВА 15
больничных палат, так как считалось, что растения ночью ухуд-
шают состав воздуха. При этом совершенно забывали о том,
что незеленые ткани растения постоянно выделяют СОг, и о том,
что количество СОг, выделяемой букетом цветов, скорее всего
намного меньше того количества СО2, которое выдыхает один
человек. На самом деле, и зеленые листья, вероятно, дышат по-
стоянно, но в обычных условиях, при ярком освещении, фикса-
ция СО2 намного превышает, а поэтому и маскирует выделение
СОа. До сих пор еще не пришли к единому мнению о том, может
или ие может «темновое» дыхание продолжаться и на свету.
Этот вопрос слишком сложен, и мы, к сожалению, здесь не мо-
жем на нем подробно остановиться. Пожалуй, следует лишь
сказать, что скорее всего в ткани растения, одновременно осу-
ществляющего фотосинтез (а возможно, и фотодыхание, гл. 14),
«темновое дыхание» изменяется, но нет никаких данных, что при
этом прекращает функционировать цикл трикарбоновых кислот,
также, впрочем, как и данных, подтверждающих, что он продол-
жает работать по-прежнему. К тому же абсолютно бесспорно,
что большинство растений способны расти при постоянном ос-
вещении; для этого необходимо, чтобы они снабжались проме-
жуточными продуктами ЦТК (например, сукцинатом, который
служит предшественником 6-аминолевулиновой кислоты, ис-
пользуемой для синтеза хлорофилла).
Практически нет никаких сомнении в том, что многие зеле-
ные ткани растений фиксируют некоторую часть СО2 в темноте.
И здесь можно обнаружить включение ИСО2 в органические
кислоты. Обычно, однако, на долю этого процесса приходится
не более 1 % общей фиксации СО2 при фотосинтезе, а выделение
СО2 в ходе дыхания целиком его маскирует. У некоторых рас-
тений, однако, ночью наблюдается значительная фиксация СО2
в составе малата (и некоторых других кислот), а днем происхо-
дит своеобразное «раскисление», в результате которого СО2,
высвобождающаяся в процессе декарбоксилирования, вновь
связывается за счет процесса фотосинтеза. Такие суточные ко-
лебания очень хорошо выражены и лучше всего исследованы у
растений, принадлежащих к сем. толстянковых (Crassulaceae).
Поэтому в статьях Мейрион Томас термин «метаболизм кис-
лот», который был ранее предложен Беннет-Кларком, превра-
тился в термин «метаболизм кислот у толстянковых» (Crassu-
lacean Acid Metabolism), который (или его английское сокра-
щение «САМ») сейчас и применяют чаще всего для обозначения
обмена веществ у тех растений, которые ведут себя сходным
образом. Следует отметить, однако, что такой вариант обмена
веществ не ограничивается только семейством толстянковых.
В норме этот тип обмена веществ связан с суккулентностыо,
однако известны многие суккуленты (например, галофиты),
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
489
Рис. 15.2. Микрофотографии поперечных срезов листа САМ-растеиия {Ananas
comosus, ананас), полученные при помощи светового микроскопа; Х288.
А. Срез, сделанный по направлению к эпидермису верхней поверхности листа.
Б. Срез, сделанный по направлению к эпидермису нижней поверхности листа.
Диаметр клеток мезофилла (/) составляет приблизительно 30 мкм (с любез-
ного разрешения S. В. Ku).
у которых не проявляется САМ-метаболизм, и есть много таких
CAM-растений, которые, в самом лучшем случае, можно назвать
лишь полусуккулентами.
У CAM-растений очень часто встречаются очень крупные
(-—100—200 мкм в диаметре) клетки мезофилла, размер кото-
рых в несколько раз превышает размеры клеток мезофилла
у С3- или Сграстений. Вакуоли в таких клетках увеличены (так
как в них запасается малат) настолько, что иа долю цитоплаз-
мы остается сравнительно мало места. Анатомическое строение
листа у CAM-растений очень разнообразно. Сами по себе ана-
томические исследования мало что юворят о различиях между
Сз- и САМ-растениями (в то время как растения семейства зла-
ковых можно вполне определенно отнести либо к С3-> либо к
Сграстениям только на основании изучения особенностей ана-
томического строения; здесь задача облегчается тем, что среди
490
ГЛАВА 15
злаков не найдены САМ-растения). Так, например, ананас,
очень прибыльное для торговли растение с САМ-метаболизмом,
относится к суккулентам, но имеет сравнительно мелкие клетки
мезофилла (рис. 15.2), которые не имеют никаких характерных
анатомических особенностей, что не позволяет легко отличить
его от Сз-растений (рис. 10.8, 10.10).
CAM-метаболизм обнаружен у представителей следующих
семейств: Agavaceae, Aizoaceae, Asclepiadaceae, Bromeliaceae,
Cactaceae, Compositae, Crassulaceae, Cucurbitaceae, Dideraceae,
Euphorbiaceae, Liliaceae, Orchidaceae, Polypodiaceae, Portulaca-
ceae, Vitaceae и Welwitschiaceae. Предполагают, что CAM-мета-
болизм свойствен ряду растений, относящихся к семействам
Geraniaceae, Labiatae, Oxalidaceae и Piperaceae. Без каких бы
то ни было доказательств к CAM-растениям относят и некоторые
(или даже всего один) виды семейств Butaceae, Caryophylla-
ceae, Chenopodiaceae, Convolvulaceae и Plantaginaceae. Иногда
CAM-растениями можно назвать лишь очень немногие виды
того или иного семейства. В зависимости от возраста растения
или от условий окружающей среды очень многие САМ-растения
либо фотосинтезируют так же, как С3-растения, либо осуществ-
ляют специфический CAM-метаболизм (так, например, САМ-
метаболизм может быть не облигатным, а факультативным, см.
разд. 15.11). Очевидно, такая «гибкость» обмена веществ от-
нюдь не облегчает выявление растений, способных к САМ-мета-
болизму, которые гораздо труднее идентифицировать, чем С3-
или Сграстения. Считают, что некоторые растения осуществля-
ют САМ-метаболизм всегда (т. е. они облигатные САМ-расте-
ния). В этом случае, как, например, у некоторых видов Cacta-
ceae и Crassulaceae, изменение условий внешней среды сказы-
вается только на интенсивности САМ-метаболизма.
Источники данных: [7, 14, 15, 16, 54, 59].
15.2. Открытие САМ-метаболизма
Разговор о CAM-метаболизме имеет очень долгую историю.
Это замечательный образчик того, как целый ряд исследовате-
лей оказались неспособными или не пожелали уложить извест-
ные факты в рамки вполне приемлемой гипотезы. Нам могут
возразить, что легко рассуждать после того, как все уже сдела-
но, но, к сожалению, по горькому опыту большинства научных
исследователей, мы видим, что очень часто они не способны
увидеть то, что маячит перед ними, до тех пор, пока не ткнутся
В ЭТО1 носом.
О поглощении двуокиси углерода в темноте впервые стало
известно еще в 1804 г., когда де Соссюр, основываясь на более
ранней работе Лавуазье с животными, провел очень большое
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
491
исследование газообмена в органах растений. После этого в те-
чение долгих лет для оценки объемного отношения СО2/О2 в
процессе дыхания животных и растений использовали термин
«дыхательный коэффициент» (ДК). В настоящее время показа-
тель ДК очень часто применяют только для оценки измеряемо-
го газообмена, связанного с такими, например, процессами на
молекулярном уровне, как окисление сахаров или липидов,
а для оценки общего газообмена в целых органах или у целого
организма, когда суммируется несколько одновременно иду-
щих процессов, обычно пользуются терминами «кажущийся
или наблюдаемый ДК», или «коэффициент газообмена» (КГО).
Де Соссюру удалось совершенно правильно объяснить поглоще-
ние О2 растением и сопутствующее ему выделение СО2 в темно-
те, рассмотрев их с точки зрения того процесса, который мы
теперь называем «дыханием» или «темновым дыханием (т. е.
сопровождающееся окислением потребление органических запас-
ных веществ, в результате которого рост растений и другие
процессы обеспечиваются энергией и метаболитами). Он не су-
мел, однако, объяснить причины одновременного поглощения
СО2 и О2 в темноте (так называемый отрицательный ДК), кото-
рое он наблюдал, когда помещал кактус-опунцию в камеру с на-
сыщенной СО2 газовой фазой. Точно так же и Хейн, который в
1813 г. весьма своеобразно открыл закисление/раскисление при
переходе темнота/свет, прибегнув к такому довольно эксцент-
ричному средству, как дегустация листьев Bryophyllum, рас-
сматривал в качестве возможного объяснения ие включение
СО2, а поглощение и выделение О2. Ниже мы приводим письмо,
которое Хейн послал А. Б. Ламберту, эсквайру, вице-президен-
ту Линнеевского общества, и которое было оглашено секрета-
рем общества на заседании 20 апреля 1813 г.
Милостивый государь!
Не так давно я имел возможность поговорить с Вами о-
замечательном раскислении листьев растений при дневном
освещении. Поскольку само по себе это обстоятельство
очень любопытно и проливает свет на взгляды тех прослав-
ленных философов, которые писали на эту тему, я хотел бы
вкратце изложить его в этом письме, которое Вы, если Вам
будет угодно, можете в виде выдержек или любым другим
способом, который Вы сочтете наиболее подходящим, пред-
ставить на суд Общества.
Листья у котиледона чашечкового, того растения, кото-
рое мистер Солсбери называет Bryophyllum calycinum. и ко-
торое в целом имеет травянистый вкус, утром такие же кис-
лые, как у щавеля, если даже не более. По мере того как
наступает день, они теряют свою едкость и к середине дня
492
ГЛАВА 15
совсем безвкусны, а к вечеру становятся почти горькова-
тыми. Так обстоит дело в Индии, где это растение довольно'
широко выращивается в наших садах, и остается лишь вы-
яснить, имеет ли место то же самое в оранжереях Англии,
куда оно’ было вывезено1 позднее.
Я видел это растение и в нашей стране, но всего один
раз; это было у мистера Лоддигеса, в Хакни, примерно в две-
надцать часов дня, когда я обнаружил, что оно почти без-
вкусно. Отдаленность этого места от моего жилища до сих
пор не позволяет мне посетить его в более ранний час ут-
ром, но я почти не сомневаюсь, что это растение будет та-
ким же кислым, каким, как я писал, оно бывает в Индии.
Я вряд ли должен добавить что-нибудь к тому, что ту
едкость, которой обладают эти листья утром, нельзя при-
писать чему-либо другому, кроме кислорода, который расте-
ние поглотило во мраке ночи или который был перенесен
за это время из других составных элементов растения.
Я думаю, что кислород поглощается, ио при этом он соеди-
няется со своим основанием настолько непрочно, что даже
свет дня оказывает на него немедленное действие и снова
его высвобождает.
И Пристли, и Ингенхоуз после многочисленных опытов
пришли к заключению, что все растения днем выделяют
животворящий воздух, а иочыо связывают воздух или газ
угольной кислоты; но эти выводы некоторыми подвергают-
ся сомнению из-за изменчивости результатов опытов, проде-
ланных с тех пор по этому поводу. То, о чем я сейчас рас-
сказываю, не лишено интереса, так как оно, по-видимому,
неопровержимо доказывает правоту теории этих прослав-
ленных философов.
Я уповал узнать что-нибудь новое или относящееся к
существу этого интересного вопроса в работе Шпренгеля
«Строение и природа растений», но, к вящему моему разо-
чарованию, не нашел в ней ничего, кроме того, что уже
было высказано двумя вышеупомянутыми философами, а в
более поздние времена — Соссюром и Сенебье. Шпренгель
много разглагольствует о выделении и поглощении угольно-
го газа и только однажды упоминает о кислороде, когда он
останавливается на наблюдениях Сенебье, согласно кото-
рым ночью растениями в замкнутых сосудах выдыхается
больше угольного газа, чем кислорода, который высвобож-
дается на солнце.
Я прошу позволения заметить далее, что растение, ко-
торое было рассмотрено1 выше, представляет по моему мне-
нию один из истинных видов котиледона, с которым оно
совершенно сходно и по габитусу, и по родовым признакам;
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
493
при этом единственное различие касается числа органов
плодоношения, которое у котиледона чашечкового на одну
пятую меньше, чем у других видов этого рода, различие,
которое, однако, в соответствии с принципами линнеевской
системы, пе дает достаточных оснований для его отделения.
Имею честь быть и пр.
20-е апреля 1813 г.
Бенджамин Хейн
Письму Хейна не придали особого значения, и оно игнорирова-
лось почти полвека, а наблюдения де Соссюра объясняли по-
ступлением двуокиси углерода в (кислый!) клеточный сок как-
туса. Тем не менее в конце девяностых годов прошлого столе-
тия Обер и одновременно Краус обнаружили корреляцию меж-
ду степенью закисления в темноте и степенью ассимиляции на
свету в течение предшествующего дня. Оба этих автора показа-
Рис. 15.3. График, с помощью которого Репсов показал, что накопление ма-
лата в листьях Bryophyllum можно объяснить фиксацией СО2 (переделанный
рис. 19.3 из книги Thomas et al., 1973). Измерения проводили в темноте при
12 °C. / — суммарное поглощение фиксированной СО2, которое равно иистин-
ному поглощению СО2 (кривая /г) плюс убыль СО2 в ходе дыхания (рас-
считано по кривой ///). ZZ—иакоплеиие малата, измеренное по титруемой
кислотности, /// — поглощение 02 во время дыхания (которое рассматрива-
ют как нормальное окисление углеводов). IV — истинное поглощение С02|
начиная с 5% СО2 в воздухе. Проведя большое число опытов с растениями,
принадлежавшими к нескольким родам, исследователи ньюкаслской группы
рассчитали, что иа 1 мг связанной СО2 приходится около 2,5 мг образовав-
шегося малата (если посчитать молярное соотношение, то оно будет при-
ближаться к 1:1). На основании этих данных был сделан вывод о том, что
.накопление малата может быть объяснено'фиксацией СО2. .
494
ГЛАВА 15
ли, что растения подобного рода накапливают крахмал, если их
освещать в камере с воздухом, лишенным СО2, и оба они пред-
положили, что раскисление включает в себя окисление органи-
ческих кислот и высвобождение СО2, которая затем снова свя-
зывается в процессе фотосинтеза. Несмотря на то что Варбург
(Warburg, 1886) тоже показал, что закисление коррелирует с
поглощением СО2, а раскисление — с ее высвобождением, пи
один из этих ученых, так же, как и Беннет-Кларк, Викери и
Вольф (которые работали в этой области в тридцатые и соро-
ковые годы), не смог установить той окончательной связи меж-
ду этими явлениями, которая сейчас вполне очевидна. Томас и
его сотрудники в сороковые годы вновь открыли «эффект де
Соссюра», когда обнаружили, что Bryophyllum в темноте одно-
временно поглощает и СО2, и О2. Уже было известно, что это
растение накапливает кислоты, и это привело Томаса к выводу
о том, что малат образуется из внутренней и наружной СО2 в.
результате карбоксилирования некоего акцептора, который об-
разуется при расщеплении крахмала и сахара (рис. 15.3). С по-
мощью только что открытой Вудом и Беркманом реакции
[уравнение (15.3)] исследователям предстояло продемонстри-
ровать биохимическую осуществимость подобного рода карбо-
ксилирования.
Источники данных: [I, 8, 14, 41, 59—62, 65].
15.3. Реакция Вуда и Беркмана
В 1936 г. Вуд и Беркман обнаружили, что при сбраживании
глицерола пропионовокислыми бактериями используется СО2.
Последующие исследования с применением тяжелого изотопа
углерода 13С показали, что СО2 включается в пропионовую кис-
лоту, пропиловый спирт и сукцинат (в последнем в карбоксиль-
ные группы). СО2 связывалась и при аэробной диссимиляции
пирувата в грудной мышце голубя. Эти наблюдения привели к
предположению о том, что первоначальная реакция включает
в себя карбоксилирование пирувата или производного пирува-
та. Кроме того, Калницкий и Беркман предположили, что необ-
ходимая для такой реакции энергия может поступать от фосфо-
енолпирувата. Когда из бактерий и ряда других организмов бы-
ли выделены белки, катализирующие реакцию
Оксалоацетат > Пируват -|- СО2 (15.1)
некоторые исследователи пришли к неправильному выводу
о том, что указанные выше данные можно объяснить
обращением этой реакции. При этом они совершенно иг-
норировали тот факт, что равновесие реакции смещено в дру-
гую сторону и не благоприятствует карбоксилированию пирува-
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
49 5
та, так как АГ7~-г6 ккал. Теоретически эту трудность можно
преодолеть, если связать данную реакцию с другой, очень благо-
приятной для нее реакцией, например катализируемой малатде-
гидрогеназой:
Пируват+ СО2 --->- Оксалоацетат (15.1)
Оксалоацетат NADH + Н+----> Малат -|- NAD+ (15.2)
И на самом деле Герберту удалось продемонстрировать вклю-
чение некоторого количества меченой СО2 в присутствии подхо-
дящей смеси ферментов. Однако скорость включения была не-
соразмерно мала, и стало ясно: для того чтобы такая сопряжен-
ная система хорошо функционировала, она должна полностью
удовлетворять всем кинетическим, а также термодинамическим
критериям.
Если равновесие неблагоприятной реакции Ач==^В нужно
сместить вправо за счет ее сопряжения с реакцией Вт—-С,
а суммарная последовательность реакций должна катализиро-
ваться двумя ферментами, то существенно, чтобы второй из них
имел очень высокое сродство к конечному продукту (В) первой
реакции. Если дело обстоит иначе, катализ второй стадии будет
осуществляться настолько медленно (в присутствии ничтожно
малого количества субстрата, образуемого на первой стадии),
что окончательное равновесие в суммарной системе не будет
достигнуто в разумный промежуток времени. Поскольку снача-
ла исследователи недооценили эти аспекты проблемы и поэтому
потерпели неудачу, карбоксилирование пирувата заслужило
сомнительную славу называться не реакцией Вуда и Беркмана
(the Wood and Werkmann reaction), а «реакцией человека, ко-
торый не желал бы работать» («the wouldn’t work man» reac-
tion; игра слов, основанная на созвучии этой фразы с фамилия-
ми людей, описавших реакцию). Эта шутка подорвала-таки ре-
путацию первооткрывателей данной реакции, которые никогда
не сомневались в том, что настоящий механизм еще предстоит
охарактеризовать. У высших растений эту проблему решили
Баидурски и Грейнер, выделившие ФЕП-карбоксилазу из шпи-
ната. Они открыли реакцию, для которой впервые выполнялись
благоприятное положение равновесия и катализ ферментом, об-
ладающим высоким сродством к СО2. И только тогда все поня-
ли, что реакция Вуда и Беркмана является реальностью.
Источники данных: [8, 11, 27, 30, 78].
15.4. Путь углерода у САМ-растений
По современным представлениям, деградация крахмала и
образующихся при этом сахаров и их производных (которые,
в конечном итоге, превращаются в триозофосфаты) у САМ-рас-
496
ГЛАВА 15
тений идет точно так же, как и у всех остальных растений, т. е.
по пути Эмбдена — Мейергофа — Парнаса (или, иначе, глико-
лиза) и окислительного пентозофосфатного цикла. Последний
из этих двух путей может вносить свой вклад в окисление уг-
леводов и без двойного карбоксилирования, существование ко-
торого предполагали вначале (разд. 15.8). Значительная часть
образующегося при этом фосфоенолпирувата превращается да-
лее в малат в результате совместного действия ФЕП-карбокси-
лазы и малатдегидрогеназы:
ФЕП СО2 Н2О -------> ОксалоацетатР( (15.3)
ОксалоацетатNADH-|-Н+----->- Малат-|-NAD+ (15.2)
Все эти события происходят в темноте, и именно ими объясня-
ется фиксация СО2, закисление и последующее изменение вкуса
листьев, которое наблюдал Хейн в начале прошлого века. На
следующий день малат декарбоксилируется и высвобождаемая
при этом СО2 вновь используется в реакциях фотосинтеза. В де-
карбоксилировании могут участвовать:
а) NADP-малатдегидрогеназа (декарбоксилирующая), описанная Очоа:
Малат NADP+ -------->- Пируват + NADPH -|- Н+ + СО2 (1 5.4)
б) NAD-малатдегидрогеназа (декарбоксилирующая):
Малат 4- NAD+ -----> Пируват + NADH + Н+ + СО2 (1 5.5)
в) малатдегидрогеназа и ФЕП-карбоксикиназа, объединенные вместе:
Малат -|- NAD+ ---> Оксалоацетат -|- NADH -|- Н+ (15.2)
Оксалоацетат-|-АТР ----> ФЕП -|- ADP -|- СО2 (15.6)
В последнем случае продуктом декарбоксилирования является
ФЕП, который может либо снова ассимилироваться и превра-
титься в углеводы в результате обращения реакций гликолиза,
либо вступить в дальнейшие превращения в цикле трикарбоно-
вых кислот. Если продуктом декарбоксилирования является пи-
руват, как в случае реакции, катализируемой малатдегидроге-
иазой (декарбоксилирующей), то возникает еще один дополни-
тельный барьер для глюконеогенеза, но его можно преодолеть
за счет следующей реакции:
Пируват -|- АТР + Р( -> ФЕП -|- AMP -|- PPj (15.7)
Причем пируват,фосфат—дикиназа, необходимая для протека-
ния этой реакции, у ряда САМ-растений МДГ-типа обнаружена.
Если образуется ФЕП любым из вышеупомянутых способов, то
вполне очевидно возникает необходимость как-то компартмен-
тализов^тЬ' или как-то контролировать поступление ФЕП, чтобы
избежать его немедленного потребления в процессе ФЕП-кар-
боксилазной реакции, иначе цикл был бы бесполезным. Счита-
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
497
ют, что трехуглеродный продукт декарбоксилирования в основ-
ном превращается в крахмал. Степень возможной реассимиля-
ции СО2 определяется тем, в какой мере этот трехуглеродный
продукт окисляется в ЦТК-
Источники данных: [4, 28, 41].
15.5. Двуокись углерода как метаболит
темновых реакций
В недавнем прошлом радиоактивные изотопы настолько ши-
роко попользовали для выяснения последовательности различ-
ных метаболических реакций, что временами даже не верится,
что гликолитический путь и цикл трикарбоновых кислот сфор-
мулированы практически без применения радиоактивных ме-
ток. То же самое можно сказать и о темновой фиксации СО2 у
CAM-растений. И в этом случае все основополагающие резуль-
таты были получены до применения изотопов. Так, например,
если малат образуется за счет темновой фиксации, то, очевид-
но, количество фиксированной СО2 должно соответствовать ко-
личеству образующегося малата. Это и было показано в 1945—
1947 гг. Биверсом и Рэнсомом в лаборатории Томаса. Хотя То-
мас и упомянул об этих данных (и о сделанном им выводе о
том, что СО2 является метаболитом) в третьем издании своего,
учебника, который вышел в свет в 1947 г., в связи с,‘тем, что
ему было совершенно несвойственно спешить с публикацией
своих статей, работы, написанные им вместе с этими соавтора-
ми, появились соответственно лишь в 1949 и 1954 гг. В это же
время совершенно независимо к аналогичному выводу пришли
Боннер и Боннер, работавшие в США. В 1948 г. они показали,
что накопление малата меняется в зависимости от концентра-
ции СО2 (0—0,1%) в окружающем воздухе. В конце концов ра-
диоактивные метки заняли подобающее им место в исследова-
ниях, и было показано, что Bryophyllutn в темноте поглощает
ИСО2 и включает ее в малат. Позднее Солтмаи и его сотрудни-
ки, использовав те же самые методы, которые ранее применя-
лись для прослеживания пути углерода при фотосинтезе, обна-
ружили, что после инкубации с меткой всего лишь в течение
6 с радиоактивность почти целиком оказывается в малате и ас-
партате, и заключили, что эти соединения возникают в резуль-
тате восстановления и переаминироваиия общего предшествен-
ника,— вывод, который совсем не противоречит тому, что сна-
чала в результате карбоксилирования ФЕП образуется оксало-
ацетат.
Источники данных: [17, 46, 59—64, 69]..
32—234
498
ГЛАВА 15
15.6. Обратная пропорциональность между количеством
субстрата и количеством продукта
Считают, что малат у САМ-растений образуется в резуль-
тате темнового карбоксилирования некоего субстрата, появля-
ющегося в процессе катаболизма углеводов. Огромная работа,
проведенная Вольфом, Беннет-Кларком, Викери и др., позволи-
ла обнаружить, что в темноте при закислении происходит рас-
пад крахмала и что, когда закисление достигает максимума,
высвобождение СО2 сводится к минимуму. Напрашивался вы-
вод о том, что СО2 используется для образования кислот (рис.
15.4).
Источники данных: [9, 14, 45, 65].
15.7. ферменты, участвующие в накоплении
органических кислот и в их последующем
использовании
Как уже говорилось выше, есть данные о том, что один из
ферментов, обеспечивающих синтез малата у САМ-растений,
должен карбоксилировать некий продукт расщепления углево-
дов и давать другой продукт, превращающийся в аспартат так
же легко, как и малат. В 1956 г. Уокер предположил, что эту
роль играет ФЕП-карбоксилаза, которая к тому времени была
выделена из Kalanchoe и Sedutn. (Впервые этот фермент был
найден в экстрактах шпината Бандурски и Грейнером в 1953 г.)
В 1960 г. Уокер предсказал также участие в процессе декарбок-
силирования малатдегидрогеназы (декарбоксилирующей).
Из работ в Ньюкасле следовало, что карбоксилаза, уча-
ствующая в этом процессе, должна обладать высоким срод-
ством к СО2 (поскольку в темноте CAM-растения способ-
ны поглотить всю СО2, находящуюся в небольшом замкнутом
пространстве) и что одна или несколько реакций этого пути
должны быть чувствительны к концентрации СО2 и подавляться
при ее повышении (поскольку темновое закисление обладает
сходной чувствительностью, рис. 15.5). Оба этих свойства при-
сущи ФЕП-карбоксилазе.
Хотя Бандурски и Грейнер так же, как Чен и Венне-
сленд, много раз высказывались по поводу высокого сродства
ФЕП-карбоксилазы к СО2 (выявленной ими соответственно в
препаратах из шпината и проростков пшеницы), первые ре-
альные измерения этой величины были проведены Уокером и
Брауном (Walker, Brown, 1957), которые определили величину
Кмкаж (для бикарбоната) в экстрактах Kalanchoe, равную
0,22 мМ (pH 7,4; см. также разд. 14.4). Позднее был получен
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
499
Рис. 15.4. Обратно пропорциональ-
ные дневные колебания у Bryophyl-
lutn. Содержание крахмала и pH воз-
растают днем и снижаются ночью.
Суммарное содержание органических
кислот (особенно малата) снижается
днем и увеличивается ночью (Tho-
mas, 1947).
целый ряд значений этой величины, вплоть до такого низкого,
как 0,02 мМ. Создается впечатление, что этот фермент может
существовать в двух формах, сродство которых к ФЕП совер-
шенно несоизмеримо. Не исключено, что эти различия отража-
ют свойства активной и неактивной форм данного фермента.
Ингибирование малатом может само по себе приводить к сни-
жению активности фермента днем. Это должно уменьшать ве-
роятность работы совершенно бесполезного цикла, в котором
за счет немедленного рекарбоксилирования при участии ФЕП-
карбокоилазы нарушалось бы декарбоксилирование малата, яв-
ляющееся неотъемлемым свойством САМ-метаболизма. Вместе
с тем вероятность бесполезного цикла уменьшалась бы и при
одновременном увеличении величины Км и для бикарбоната, и
для ФЕП. В настоящий момент, однако, вопрос об активации
ФЕП-карбоксилазы, сравнимой с активацией РуБФ-карбокси-
лазы (разд. 6.11), вообще не ставится, вероятно, потому, что
максимальные значения Kni, которые были зарегистрированы
до сих пор, по-прежнему, свидетельствуют об очень высоком
32“
500
ГЛАВА 15
Рис. 15.5. Темновое закисление в листьях (кривая I) стимулируется СОг при
повышении ее концентрации примерно до 3% и затем постепенно подавляет-
ся. ФЕП-карбоксилаза (кривая II) тоже ингибируется при высокой [СОг],
в то время как карбоксилирование пирувата, которое катализирует фермент
Очоа (кривая III) очень незначительно при низкой [СОг] и достигает макси-
мума по мере того, как темповое закисление сводится к нулю. Все. данные
получены для Kalanchoe (пли экстрактов из Kalanchoe) в лаборатории То-
маса в Ньюкасле (Wolf, 1960).
сродстве этого фермента к бикарбонату. Выше уже упомина-
лось о том, что в декарбоксилировании могут участвовать ма-
латдегидрогеназа или ФЕП-карбоксикиназа (разд. 15.4). Дан-
ные Вольфа, который обнаружил, что поглощение кислорода
возрастает, а КГО (.коэффициент газообмена) снижается после
введения пирувата (либо во время закисления, либо во время
раскисления) в кусочки листьев Bryophyllum calycinum, можно
в некотором смысле объяснить наличием пируват,фосфат — ди-
киназы. Поскольку при окислении пирувата образование СО2
должно стимулироваться в большей степени, чем поглощение О2,
некоторые исследователи утверждают, что пируват влияет иа
какой-то другой процесс и что наиболее вероятной из имею-
щихся возможностей является его использование для образо-
вания малата. Первым этапом глюконеогенеза считается во-
влечение пирувата в пируват,фосфат —дикпназную реакцию,
однако вполне возможно, что образующийся из пирувата ФЕП
тотчас же потребляется для синтеза малата в том случае, ког-
да нарушена нормальная компартментация процессов обмена
веществ или когда в результате поступления веществ снаружи
через поврежденную поверхность среза устраняется нормаль-
ный регуляторный контроль. Следует, однако, иметь в виду,
что дикиназа относится к числу тех ферментов, которые акти-
вируются светом, и поэтому кажется маловероятным, чтобы она
эффективно функционировала в темноте.
Источники данных: (7, 9, 11, 55, 62, 66—70].
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
501
15.8. Гипотеза двойного карбоксилирования
Включение МСО2 в ходе ФЕП-карбоксилазной реакции при-
водит к тому, что образуются малат и аспартат, специфически
меченные в С-4-положении (p-карбоксильная группа). ФЕП-
карбоксилазу 'иногда называют p-карбокоилазой, потому что
СО2, входящая в состав продукта, появляется в р-положении,
т. е. у С-З-атома, если нумерацию атомов углерода вести от
уже имеющейся карбоксильной группы субстрата; обозначение
атомов углерода греческими буквами начинают с С-2 и, таким
образом, указывают на положение того или иного атома угле-
рода по отношению к С-1-атому:
HOOC-CO-CIL-COOH
12 3 4
Установлено, однако, что у растений и малат, и аспартат ме-
тятся по обеим карбоксильным группам. При этом независи-
мо от условий включения и продолжительности инкубации с
14С метка распределяется между С-1 и С-4 примерно в соот-
ношении 1/3 : 2/3. Существует множество путей, по которым уг-
лерод, сначала включившийся в С-4-положейие, затем может
быть перенесен в положение С-1. Так, например, фумарат яв-
ляется симметричной молекулой, и взаимопревращение малата
и фумарата, которому способствует фумараза, может приво-
дить к рандомизации метки.
О такой возможности обычно говорили, но практически ею
пренебрегали и отбрасывали из-за того, что соотношение С1/С4
было постоянно. В более поздних работах было установлено,
что использование для определения внутримолекулярного рас-
пределения радиоактивности Lactobacillus plantarum ненадеж-
но. Во многих опытах реально наблюдали предпочтительное
включение метки в С-4-положение, как это и следует из пред-
положения о р-карбокенлнроваиги. Однако постоянство соотно-
шения метки необходимо было приемлемо объяснить, в связи
с чем Рейсом, Бредбир и Стиллер выдвинули гипотезу о двой-
ном карбоксилировании. Согласно той схеме, которую они
предложили, несколько видоизмененный пентозофосфатный цикл
функционировал таким образом, что рибулозобпсфосфат обра-
зовывался и карбоксилировался в темноте. В присутствии 14СО2
по этой схеме образовывались две молекулы ФГК, одна из них
была меченой, а вторая — нет ’[уравнение (6.15)]. После того
как эти молекулы превращались в ФЕП, они снова карбокси-
лиро’вались, так что в конечном итоге в одной из них атом 14С
•.входил в состав обеих карбоксильных групп, а в другой — толь-
ко в одну из них (p-карбоксил). Одним словом, наблюдаемое
распределение метки должно было быть результатом появле-
502
ГЛАВА 15
ния молекул малата, меченных по обеим карбоксильным груп-
пам, а не следствием того, что образуется смесь молекул, ме-
ченных только по С-1- или только по С-4-атому углерода.
Это предположение не получило никакой поддержки от
Сэттона и Осмонда, равно как и от Клюге и др., которые сти-
мулировали включение 14С в малат, вводя ФЕП и ФГК в сре-
ды для листьев Bryophyllum, но не обнаружили никаких изме-
нений в распределении метки внутри молекулы малата. В кон-
це концов Кокберн и Мак-Оли провели очень изящный и стро-
гий эксперимент, использовав для этой цели масс-спектрограф'
и 13С. Этот метод позволил им взвесить интересующие их мо-
лекулы и вполне определенно показать, что ни у одной из них
нет лишней массы, которая должна была бы появиться, если
бы в одной и той же молекуле малата были два атома 13С.
Поэтому гипотеза двойного карбоксилирования была отбро-
шена и забыта, хотя до сих пор остается загадкой, почему
-—2/3 метки постоянно обнаруживают в С-4-положеиии малата.
Источники данных: [18, 20, 29, 31, 53, 76].
15.9. Накопление органических кислот на свету
В обычных условиях яблочная кислота (малат) потребля-
ется на свету. Пытаясь понять, как же регулируется закисле-
ние— раскисление, Томас и Биверс обнаружили, что процесс
раскисления замедляется, если увеличить содержание СО2 в
воздухе. Поскольку недавно стало известно (разд. 15.14), что
в процессе раскисления [СО2] в листе возрастает, вполне воз-
можно, что СО2 как-то контролирует скорость раскисления in
vivo.
К тому же Ренсон показал, что если ткань листа после ее
раскисления поместить в камеру, насыщенную СО2, то после
освещения произойдет закисление этой ткани. Максимальное
закисление наблюдается в присутствии 5% СО2. Цифры, близ-
кие к максимальной, получаются даже в присутствии 10—20%
СО2, и это несмотря на то, что в такой атмосфере в значитель-
ной степени подавлено нормальное фотосинтетическое погло-
щение СО2 (и связанное с ним выделение О2), а интенсивность
темновой фиксации не превышает 18—38% максимальной ве-
личины. Томас и его сотрудники предположили, что подавле-
ние фотосинтеза при высоких [,СО2] можно объяснить тем, что
при этом ингибируется образование АТР и NADPH. Такое ин-
гибирование скорее всего благоприятствует не восстановлению'
ФГК до триозофосфатов, а ее превращению в ФЕП. Почему
СО2 подавляет возникновение ассимиляционной силы, авторы
вовсе не обсуждают, они лишь сообщают о том, что при высо-
кой концентрации СО2 понижается pH и поэтому ингибируется
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
503
фотосинтетическая ассимиляция углерода. Если же учесть ре-
зультаты работ, проведенных на реконструированных системах
хлоропластов, то можно прийти к выводу о том, что к повы-
шению концентрации СО2 чувствительна РуБФ-карбоксилаза
(как и ФЕП-карбоксилаза). Представление о том, что ФЕП на
свету может карбоксилироваться в какой угодно степени, очень
трудно увязать с предложенной для него ролью в глюконеогене-
зе, берущем свое начало от малата. Вполне возможно другое
объяснение, а именно что при высоком уровне СО2 может сме-
щаться равновесие реакций декарбоксилирования. Так, напри-
мер, было показано, что в присутствии 10% СО2 интенсивность
образования малата из пирувата, катализируемого малатдегид-
рогеназой (декарбоксилирующей), может доходить до (и даже
превышать) 80% ее максимальной величины.
Источники данных: [;7, 61, 62, 68].
15.10. Энергетические затраты при САМ-метаболизме
Энергетические затраты, приходящиеся иа фиксацию СО2
у САМ-растении, можно рассматривать как затраты иа: 1) тем-
новое закисление; 2) декарбоксилирование С4-кислот; 3) фик-
сацию СО2 в ВПФ-цикле и 4) превращение С3-прсдшественни-
ков, образующихся после декарбоксилирования, в крахмал.
а) Темновой закисление
При расщеплении крахмала и при образовании малата
ночью (рис. 15.6) ие происходит никаких изменений восстано-
вительной силы. Для восстановления оксалоацетата в малат
используется NADH, который образуется в процессе гликолиза
при участии NAD-трнозофосфатдегидрогсназы. Если в рас-
щеплении крахмала участвует фосфорилаза, на каждую моле-
кулу образующегося ФЕП в конечном итоге приходится по 0,5
молекулы синтезируемой АТР. Если же крахмал гидролизует-
ся различными амилазами, то тогда не должно синтезироваться
никакого АТР. И в том и в другом случае в ходе темнового за-
кисления не происходит практически никаких изменений в со-
держании ассимиляционной силы. Механизм, обеспечивающий
накопление малата в вакуоли в ходе закисления, остается неяс-
ным; для него, возможно, и нужна энергия.
б) Декарбоксилирование (Д-кислот
Если в декарбоксилировании участвует NADP-малатдегид-
рогеиаза, то па каждую молекулу выделившейся СО2 приходит-
' ся по одной молекуле образовавшегося NADPH [уравнение
(15.4)]. Для превращения малата в ФЕП под действием NAD-
504
ГЛАВА 15
Pj (Фосфорилаза)
АТР '
ADP
ФБФ
ТОМНоеое закисление
малатдегидрогеназы и ФЕП-
карбоксикиназы требуется од-
на молекула АТР, и при этом
образуется одна молекула
NADH {уравнения (15.2) и
(15.6)].
в) ВПФ-цикл
При фиксации СО2 в ВПФ-
цикл е на каждую молекулу
СО2 приходятся 3 молекулы
АТР и 2 молекулы NADPH?
(гл. 6). Если при этом проис-
ходит кислородное ингибиро-
вание фотосинтеза, то затра-
ты энергии будут еще выше
(разд. 13.6). Но САМ-расте-
ния, как и Сгр астения, могут
в процессе декарбоксилирова-
ния С4-кислот выделять доста-
точно СО2, чтобы подавить
кислородное ингибирование
фотосинтеза (разд. 15.14).
г) Превращение С^-продукта
декарбоксилирования
в крахмал
Рис. 15.6. Темновое закисление и свя- в том случае КОГда декар-
занные с ним энергетические затраты. боксилазой "является NADP-
малатдегидрогеназа, на каж-
дую молекулу пирувата для его превращения в крахмал должно
расходоваться по 3,5 молекулы АТР и по одной молекуле
NADH. При этом три молекулы АТР из этого числа необходи-
мы для того, чтобы превратить пируват в триозофосфат [сум-
ма уравнений (15.7—15.13)]:
Пируват -|- АТР + Р( ->-
---> ФЕП 4-АМР-р РРг (пируват, ортофосфат—цикиназа) (15.7)
РР; ----->- 2Р[ (пирофосфатаза) (15.8)
АМР 4-АТР --------> 2ADP (адснилаткиназа) (15.9)
ФЕП -----> 2-ФГК (енолаза) (15.10)
2-ФГК ------>- 3-ФГК (фосфоглицеромутаза) (15.11)
3-ФГК + АТР -------> ДФГК 4-ADP (ФГК-киназа) (15.12)
МЕТАБОЛИЗМ. КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
505
ДФГК + NADH + Н+------>-
----> ТФ + NAD+ -|- Pj (NAD-триозофосфатдегвдрогеназа) (1 5.13)
В итоге: Пируват-|-ЗАТР + NADH + Н+--------------->
---> ТФ + NAD+ + 3ADP 2Р, (15.14)
Оставшиеся 0,5 молекулы АТР необходимы для того, чтобы
превратить Сз-эквпвалеит триозофосфата в крахмал, так как
для синтеза ADP-глюкозы, которая является предшественни-
ком в синтезе крахмала, требуется потратить 1 молекулу АТР
иа каждую молекулу глюкозофосфата.
Глюкозо-1 -фосфат АТР --->
---> ADP-глюкоза -f- РР, (глюкозо-1-фосфат—аденилилтрансфераза)
В том случае, когда декарбоксилазой служит ФЕП-карбокспки-
иаза, иа каждую молекулу ФЕП для его превращения в крах-
мал необходимо потратить 1,5 молекулы АТР и одну молекулу
NADH. В конечном итоге энергетические затраты иа фиксацию
одной молекулы СО2 у САМ-р астений, использующих NADP-
малатдегидрогеназу (декарбоксилирующую), должны состав-
лять 6,5 АТР, 2 NADPH, а у CAM-растений, использующих
ФЕП-карбоксикиназу, 5,5 АТР, 2 NADPH. Энергетические за-
траты на фиксацию СО2 должны увеличиваться при значитель-
ном повышении температуры, когда возрастают потери СО2 в
ходе дыхания, и в условиях жестокой засухи, когда устьица
остаются закрытыми, а углерод, который высвобождается в хо-
де дыхания, циклически •используется для САМ-метаболнзма.
Среди изученных САМ-растений можно найти виды, у кото-
рых главной декарбоксилазой служит NADP-малатдегидроге-
наза (МДГ), и виды, у которых эту роль играет ФЕП-карбокси-
кииаза. Поэтому САМ-растеиия можно, как и С^-растеиия, от-
носить либо к растениям NADP-МДГ-, либо к растениям ФЕП-
КК-типа (гл. 11). Есть данные и о том, что у ряда САМ-расте-
ний имеется NAD-малатдегидрогеназа. Неясно, однако, како-
ва доля участия этого фермента в суммарном процессе декарб-
оксилирования. В настоящее время ие обнаружено ни одного
САМ-растеиия, которое можно было бы сравнить с NAD-МДГ-
растеииями, относящимися к С«-растениям ( т. е, ие показано,
что NAD-МДГ является главной декарбоксилазой). Энергети-
ческие затраты при катализируемом NAD-малатдегидрогеназой
декарбоксилировании, если оно действительно происходит в хо-
де CAM-метаболизма, должны быть сравнимы с расходом энер-
гии иа декарбоксилирование, идущее при участии NADP-малат-
дегидрогеиазы.
Внутриклеточная локализация ферментов фотосинтеза у
CAM-растений изучена еще. недостаточно. А ведь именно лока-
506
ГЛАВА 15
ВАКУОЛЬ —► 2NADPH
2NADPH
Рис. 15.7. Модельная схема внутриклеточного транспорта и фотосинтетиче-
ских путей у САМ-растений. (Штриховыми стрелками показан темновой ме-
таболизм, сплошными — световой метаболизм.) Заметьте, что в сумме энерге-
тические затраты иа фиксацию одной молекулы СО2 сводятся к тому, чтобы
фотохимически генерировать энергию, необходимую для запуска системы ас-
симиляции углерода. Как изображено на рисунке, С3-продукт, который обра-
зуется в результате декарбоксилирования малата, прямо превращается в
крахмал. Триозофосфат, образовавшийся из С3-продукта декарбоксилирова-
ния, скорее всего входит в общий пул триозофосфатов в хлоропласте. Об-
суждение схемы см. в тексте. [С некоторыми модификациями по данным для
Sedum praealtum (NADP-МДГ-типа) и нескольких видов ФЕП-КК-типа
(Spalding et al., 1979а; Ku et al., 1980).]
лизация ФЕП-карбоксилазы и декарбоксилаз определяет затра-
ты энергии на транспорт метаболитов между хлоропластами и
цитозолем.
В процессе темнового закисления в вакуолях клеток запа-
сается малат. Результаты последних исследований внутрикле-
точной локализации ферментов свидетельствуют о том, что у
САМ-растений ФЕП-карбоксилаза, NADP-МДГ и ФЕП-карбок-
сикиназа находятся в цитоплазме, а NAD-МДГ — в митохонд-
риях. Поэтому завершающие реакции закисления и исходная
реакция раскисления происходят вне хлоропластов (рис. 15.7).
Энергетические затраты на фиксацию одной молекулы СОг
у САМ-растений, по-видимому, сравнимы или чуть выше, чем
затраты у Сграстений (разд. 11.6). А энергетические затраты
на фиксацию одной молекулы СО2 у С3- и С4-растений при
нормальных атмосферных условиях почти одинаковы, так как
кислород уменьшает квантовый выход фотосинтеза у Сз-расте-
ний (разд. 13.12). Поэтому все эти три группы растений при
нормальных атмосферных условиях расходуют на фотосинтез
почти одинаковое количество энергии.
Несмотря на то что уже многое известно об обмене веществ
у САМ-растений, регуляция этого процесса практически не изу-
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
507
Рис. 15.8. Изменения чувствительности ФЕП-карбоксилазы к подавлению ма-
латом у растения Mesembryanihemum crysiallinum, у которого можно инду-
цировать CAM-метаболизм, Фермент экстрагировали из полностью распра-
вившихся листьев, выращенных при фотопериоде 12 ч темноты/12 ч света. На
рисунках представлены содержание малата (А), активность растворимой
ФЕП-карбоксилазы при pH 8,0 и 2,0 мМ ФЕП в отсутствие (5) и в присут-
ствии 2 мМ малата (В). [Эти неопубликованные данные любезно предоста-
вил д-р Клаус Винтер, см. также (Winter, 1980).]
чена. Эту регуляцию составляют регуляция ферментативной
активности в темноте и на свету, механизм запасания и высво-
бождения малата из вакуоли, транспорт метаболитов в хлоро-
пластах, а также переключение синтеза сахарозы на синтез
крахмала и обратно. Так, например, ФЕП-карбоксилаза и де-
карбоксилазы (т. е. NADP-малатдегидрогеназа и ФЕП-карбок-
сикиназа) действуют в цитоплазме клетки. Катализируемое эти-
508
ГЛАВА 15
ми ферментами карбоксилирование и декарбоксилирование ре-
гулируется таким образом, что эти ферменты обычно функцио-
нируют в разное время; в противном случае все время работал
бы совершенно бесполезный цикл карбоксилирования — декар-
боксилирования. Последние работы Винтера показали, что свой-
ства ФЕП-карбоксилазы у САМ-растений меняются при пере-
ходе от света к темноте, причем днем этот фермент становится
особенно чувствительным к малату и ингибируется им. Эти
данные очень важны для понимания регуляции САМ-метабо-
лизма (рис. 15.8).
Источники данных: [15, 24, 25, 32, 50, 52, 74].
15.11. Переключение обмена веществ от САМ-метаболизма
на Сз-фотосинтез
Некоторые из САМ-растений проявляют такую гибкость фо-
тосинтеза, которая ие наблюдается ни у С3-, ни у С4-растеиий.
В зависимости от условий окружающей среды такие САМ-рас-
тения могут менять соотношение между темновой и дневной
фиксацией СО2 из воздуха. Считают, что при дневной фиксации
атмосферная С02 связывается у САМ-растений так же, как у
Сз-растений, т. е. при непосредственном участии РуБФ-карбок-
силазы.
Показано, что такие факторы внешней среды, как темпера-
тура, водный дефицит и продолжительность фотопериода, влия-
ют иа направленность процессов фотосинтеза у некоторых
САМ-растений. Высокая дневная температура, низкая ночная
температура и водный дефицит более благоприятны для тем-
новой ассимиляции СО2. У некоторых видов из рода Kalanchoe
короткий день и длинная ночь способствуют САМ-метаболпзму
и ускоряют цветение. У некоторых видов Sedan, наоборот, цве-
тению, а возможно, и переходу к САМ-метаболпзму способст-
вуют длинный день и короткая ночь.
Темновую фиксацию СО2 у некоторых САМ-растений мож-
но вызвать, повысив соленость среды, что имитирует водный
дефицит. Так, например, и водный дефицит, и полив растений
очень соленой водой могут вызывать у растений Mesembryan-
themutn crystallinum и Portulacaria afra переход от дневной
фиксации атмосферной СО2 к CAM-метаболизму. При такого
рода воздействии наблюдались и дневные колебания кислотно-
сти листьев, и ночная фиксация СО2.
Влияние температуры на фотосинтез у САМ-растений мож-
но продемонстрировать на примере работ Нилеса, проведенных
им на ананасе. Наиболее интенсивная фиксация СО2 воздуха в
темноте наблюдалась при пониженных ночных температурах
(рис. 15.9). Уменьшение скорости реальной фиксации СО2 в
темноте при более высокой температуре может быть связано
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
509
Рис. 15.9. Фиксация СОг у ананаса. Дневная температура 25 °C, продолжи-
тельность дня: I—14 ч темноты/10 ч света; II—8 ч темноты/16 ч света
(с модификациями по Neales, 1973b).
ие с уменьшением скорости САМ-метаболизма, а с увеличени-
ем интенсивности дыхания. В среднем, минимальная ночная
температура во время вегетационного периода ананаса на
Гавайях составляет 10 °C, в Австралии 12 °C, а в Южной Аф-
рике 5°C. Это говорит о том, что ананасы в этих широтах рас-
тут в основном за счет САМ-метаболизма.
Осмонд (Osmond, 1978) заметил, что у многих' САМ-расте-
иий, когда они функционируют по САМ-путн, в течение 24-часо-
вого цикла можно наблюдать четыре фазы метаболизма. Фа-
за 1 заключается в фиксации СО2 и синтезе органических кис-
лот в темноте. Фаза II происходит в самом начале светового
периода и представляет собой начальное скачкообразное увели-
чение фиксации СО2 из воздуха. В это время могут активно
функционировать как ФЕП-, так и РуБФ-карбоксилаза. Фа-
за III (фаза раскисления) идет на свету и заключается в том,
что при закрытых устьицах происходят декарбоксилирование
Сгкислот и фиксация высвободившейся при этом СО2. Фаза
IV осуществляется на последних этапах светового периода, ког-
да раскисление уже завершено; эта фаза состоит в том, что
по мере увеличения проводимости устьиц фиксация СО2 из воз-
духа происходит преимущественно за счет РуБФ-карбоксилазы..
Источники данных: [2—5, 19, 22, 26, 35, 38—40, 43, 44, 56,.
72].
15.12. Фракционирование изотопов углерода у САМ-,
Сз- и С4-растений
В атмосферной СО2 содержится небольшое количество тя-
желого изотопа углерода 13С (1% содержания 12С). Измерения
содержания 13С/12С в различных материалах и в атмосфере ос-
510
ГЛАВА 15
нованы на сравнении полученных данных с содержанием этих
изотопов в известняках мелового периода в Южной Каролине
(ископаемые остатки карбонатного скелета Belemnitella ате-
ricana). На самом деле в воздухе содержится меньше 13СО2,
чем в ископаемом стандарте. Поскольку эти различия очень
малы, относительное содержание 13С/12С в пробе принято обо-
значать как б13С и выражать в %о (в промилле, т. е. в частях
на тысячу). По отношению к стандарту б13С для СО2 возду-
ха ~—7%о. Величина 13С/12С в воздухе слегка меняется, но в
среднем это означает, что в наше время в воздухе содержится
на 0,007 13СО2 меньше, чем во время мелового периода. Хотя
эта разница очень мала, содержание 13С в пробе (по отноше-
нию к содержанию 12С) можно очень точно определить при по-
мощи масс-спектрометра.
А13Г ( (18С/1аС)в пробе_1 1 1 ЛЗ
U ( /13С/12Г\ 1 IV
I W М в стандарте )
Если в стандарте содержится 1,116% 13С, а в воздухе в сред-
нем находится 1,108% 13С, то
§1зг _ Г 11.08/1000 । 1 j 0з у о/
° 11,16/1000 1 J ‘ /о°
[т. е. содержание 13СО2 в воздухе на 0,007 меньше, чем в пробе
7/1000-(1,116%) =0,0078%
1,116%—0,008% = 1,108% 13С в воздухе].
В то время как величина б13С отражает изотопный состав по
•отношению к данному стандарту, фракционирование изотопов
(или их дискриминация) выражается через разность величин 6
для исходного источника и конечного продукта какой-либо ре-
акции или того или иного процесса:
» 6 (источник) — 6 (продукт)
Фракционирование = (ис^чник) 1 обб~1 '
Если величина 6 для источника составляет —7%о, а для про-
дукта она равна —27°/о0, то фракционирование равно + 2О%о.
’Отсюда следует, что величина б для продукта зависит от сте-
пени фракционирования и от количества минорного изотопа в
источнике; для этого и вносят поправку в знаменатель дроби.
Эта поправка становится очень значительной в том случае, ког-
да в источнике содержится относительно большое количество
минорного изотопа.
В процессе ассимиляции углерода все растения в той или
иной степени различают 13СО2, причем у Сз-растений такая
.дискриминация выражена сильнее, чем у Сг или САМ-расте-
2ний. Средние значения для тканей Сз-растений находятся в
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
511
Таблица 15.1. Факторы, действующие на начальной стадии поглощения СО2,
которые могут влиять на изотопный состав углерода у растений
(см. также O’Leary, 1981)
Фактор Величина фракцио- нирования, %о
Диффузия СО2 из воздуха (СО2 вВешн/СО2 внутр) +4
Достижение равновесия между СОг и НСОз" -8
(СО2+Н2ОЧ*НСО8-+Н+)
ФЕП-карбоксилазаа> 4-2
РуБ Ф -кар боксилаз а8> 4-34
') Болес тяжелый изотоп обычно концентрируется там, где связи внутри соединения^
сильнее; в этом случае в НСОз~» а не в COs.
2) Величина фракционирования Н-2%0 справедлива в том случае, если в качестве
стандарта используется НСО3-«
3) Величина, приведенная для РуБФ-карбоксилазы, представляет собой среднее зна-
чение, полученное из рядя работ, в которых величины фракционирования варьировали
в пределах 28—38%0, Нет строгих данных о том, что температура влияет па степень
фракционирования для этой карбоксилазы.
пределах от —25 до —35%q, для Сграстений — в пределах от
—10 до —17%о, а для САМ-растений (во время активного за-
кисления) они приближаются к цифрам, получаемым для
(^-растений. В табл. 15.1 рассматриваются те этапы между
поглощением СО2 из воздуха и ассимиляцией углерода, на ко-
торых может происходить фракционирование. Поскольку фрак-
ционирование изотопов у РуБФ-карбоксилазы выражено в.
большей степени (4~34%о), чем у ФЕП-карбоксилазы (4~2%о),.
полагают, что различия в изотопном составе углерода у Сз-,.
С4- и САМ-растений отражают именно различия в дискримини-
рующей способности этих главных карбоксиалаз. Но разницу в
изотопном составе фотосинтезирующей ткани нельзя объяснить-
только за счет этих различий. В том случае, когда существует
определенное сопротивление поглощению СО2 листом (т. е.
ограничения для диффузии), фракционирование может дости-
гать 4%о. У Сз-растений субстратом для РуБФ-,карбоксилазы
служит ие НСОз~, а СО2. Если ассимиляция углерода целиком
и полностью определяется активностью РуБФ-карбоксилазы, а’
не скоростью диффузии, то следует ожидать, что величина
613С будет составлять —41%о {I—7%0 (в атмосфере) —34%о (для’
РуБФ-карбоксилазы)]. Эту величину можно получить вычита-
нием фракционирования из б13С для воздуха.
613С(для продукта) = 61SC (в воздухе)—Фракционирование
Если же ассимиляция углерода в процессе Сз-фотосинтеза це-
ликом зависит только от скорости диффузии, то следует ожш
512
ГЛАВА 16
дать, что величина 6 будет не более, чем —11%о (—7%0 —4°/оо).
Поэтому величина б может быть и выше, чем —41%о, все за-
висит от того, насколько процесс фотосинтеза лимитируется за
счет диффузии. У некоторых морских трав, у которых СО2 свя-
зывается в процессе С8-фотосинтеза, фотосинтезирующие ткани
практически не отличаются по изотопному составу (—11%о) от
свободной СО2, растворенной в морской воде (разд. 10.7). Счи-
тают, что в этом случае самым главным этапом, который ли-
митирует ассимиляцию углерода, является проникновение СО2
в ткань за счет диффузии, так что почти вся поглощенная СО2
(13С+12С) связывается РуБф-карбоксилазой.
У С4- и САМ-растеиий фракционирования изотопов углеро-
да в ходе ФЕП-карбоксилазной реакции почти не наблюдается.
Получаемые величины б13С в этом случае варьируют в преде-
лах от —10 до —17°/оо, что может быть связано с фракциониро-
ванием изотопов в процессе диффузии или во время достиже-
ния равновесия между СО2 и НСОз-. ФЕП-карбоксилаза ис-
пользует НСОз", а превращение СО2 в НСО3 " ведет к обогаще-
нию 13С. При этом величина фракционирования приближается
к —8°/оо. Минимальная величина, которую следовало бы ожи-
дать, если вся система лимитируется скоростью диффузии, со-
ставляет — 11%о (—7%о—4°/оо); если же процесс лимитируется
скоростью карбоксилирования, то максимальная из теоретиче-
ски возможных величин будет составлять —1%о:
[—7%0 —( 8%о (ДЛЯ превращения СО» в НСО~3) 2%о (для ФЕП-карбоксилазы)]
Тяжелый изотоп 13С диффундирует в воздухе гораздо медлен-
нее, чем 12С, поэтому фракционирование, связанное с диффу-
зией, происходит до тех пор, пока не выравнивается концент-
рация СО2 снаружи и внутри листа (в межклетниках). Фрак-
ционирование, обусловленное диффузией, у С4-растений может
быть несколько выше, чем у Сз-растений. Это связано с тем,
что при нормальных атмосферных условиях концентрация СО2
в межклетниках у С4-растений (~100 м. д.) намного ниже,
чем у Сз-растений (~220—250 м. д.).
Для строгой оценки фракционирования изотопов вплоть до
пункта карбоксилирования необходимо, чтобы изотопный со-
став атомов углерода, входящих в СО2„ определяли после того,
как пройдут первые этапы ассимиляции. Как правило, однако,
в статьях сообщается только об изотопном составе всей ткани
или отдельных ее компонентов. Так, в одной из работ обнару-
жено, что величина б13С для С-4-атома малата у Kalanchoe
daigremontiana (CAM-растение) составляет приблизительно
—8%о, что вполне попадает в доверительный интервал ожидае-
мых величин (М. O’Leary, личное сообщение). По мере того
как углерод включается дальше в биомассу, происходит после-
МЕ1АВОЛИЗМ кислот У семейства толстянковых
51
Таблица 15.2. Изменения уровня малата (Д Малат) ночью и величины б13С
в разные дни и месяцы 1977 г. у полностью развитых листьев однолетнего
растения Mesembryanthemum crystallinum, растущего на берегу
Средиземного моря в окрестностях Цезареи, Израиль (в измененном виде
по Winter et al.. 1978, Winter, 1979)
Сезон Дата Л Малат
мкэкв-(г сырого вещества)-1 й1аС, %о
Период дождей 26/27 января -3 -26,1
Период дождей 24/25 февраля -4 -26,7
Период дождей 17/18 марта —1 -25,4
Период засухи 26/27 апреля +113 -22,7
Период засухи 25/26 мая +113 -19,1
Период засухи 26 июня Не определяли -15,7
дующее фракционирование изотопов, в результате чего величи-
на б13С может приближаться к —13%о.
У Сз-растений продукты РуБФ-карбоксилазной реакции
обогащены 12С (если сравнивать их с СО2 воздуха), потому
что этот фермент связывает 12С быстрее, чем 13С. У С4-растений,
однако, в замкнутом пространстве обкладки проводящих пуч-
ков связывается практически вся СО2, которая высвобождает-
ся в результате декарбоксилирования. По этой причине изо-
топный эффект фракционирования сглаживается и степень обо-
гащения продуктов 13С приближается к той, которая характерна
для оксалоацетата, образующегося в результате ФЕП-карбокси-
лазной реакции в клетках мезофилла.
Показано, что у многих растений САМ-метаболизм являет-
ся факультативным, т. е. дневные колебания содержания кис-
лот у них сильно выражены лишь при недостаточном водо-
снабжении. Сходным образом варьируют и величины 613С, так
что (как у С4нрастений) значение б наименее отрицательно
тогда, когда дневные колебания выражены в наибольшей сте-
пени (табл. 15.2). Благодаря различиям в величинах б можно
отличить сахарозу, синтезируемую Сз-растением (например,
сахарной свеклой), от сахарозы, синтезируемой С4-растением
(например, сахарным тростником). Такие различия во фрак-
ционировании изотопов у разных видов в какой-то мере можно
использовать при классификации растений и их делении на фо-
тосинтетические группы. Одним из преимуществ, связанных с
применением данного метода для классификации, является то,
что для подобных исследований ие нужны живые ткани расте-
ния и поэтому можно работать на обширном материале гер-
33-234
514
ГЛАВА Io
CO; атмосферы <>IJC = -7
C.i Растение;
C.i -Растение'
САМ-Растенио
САМ-Состоянии С, -Состояние
(ночная фиксация) (дневная фиксация)
CO?
▼
РуБФ-
карбоксилаза
▼
Продукты
6' •’ C -30
—НСО3
ФЕП-
карбоксилаза
РуБФ-
карбоксилаза
I
Продукты
6, JC -- -13
,^-нсо
*
V
ФЕП-
карбоксилаза
I
РуБФ-
карбоксилаза
Продукты
б' -’С^ -13
СОг
▼
РуБФ-
карбоксилаза
V
Продукты
6 ’ ’ С =4 - 30
Рис. 15.10. Фиксация растениями СОа, участие в этом процессе карбоксилаз
и происходящие при этом изменения изотопного состава углерода в расти-
тельной ткани.
барных образцов. Так, например, Смит и Браун отнесли мно-
гие виды семейства злаковых либо к Сз-, либо к С4-растениям
исключительно на основании величин б13С (дело облегчается
тем, что ни один из видов Gramineae не относится к САМ-рас-
тениям). Этот метод хороню коррелирует с другими методами,
которые используют для того, чтобы отличить Сз- и Сграсте-
ния, например, методами определения углекислотного компен-
сационного пункта, различий в анатомическом строении и т. д.
У САМ-растений 'величины б13С попадают в интервал меж-
ду типичными значениями б для Сз- и ф-растений. Все зави-
сит от того, какая доля СО2 фиксируется ночью (по типу
CAM-метаболизма) по сравнению с прямой фиксацией СО2
днем при участии РуБФ-карбоксилазы (С3-фотосиитез). Поэто-
му величину б13С для CAM-метаболизма используют как пока-
затель участия ночной фиксации СО2 за счет ФЕП-карбокси-
лазы п дневной фиксации СО2 воздуха за счет РуБФ-карбокси-
лазы в суммарном связывании двуокиси углерода. Сначала
считали, что наиболее высокая величина б13С у Сграстеиий
свидетельствует в пользу механизма транскарбоксилироваиия
(разд. 10.2). При декарбоксилировании С4-кислот и последую-
щей реф'ик’сации СО2 РуБФ-карбоксилаза, которая в этом слу-
чае служит вторичной карбоксилазой, еще способна дискрими-
нировать изотопы углерода и связывать преимущественно |2СО2.
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
515
В настоящее время принято считать, что и у С4-, и у САМ-рас-
тений декарбоксилирование С4-кислот происходит в строго ог-
раниченном пространстве (у С4-растений в клетках обкладки
проводящих пучков, а у САМ-растений в листе при закрытых
устьицах), В результате вся высвобождающаяся СО2 (13СО2+
+ В * * * 12СО2) может связываться почти полностью, даже несмотря
на то что РуБФ-карбоксилаза способна к некоторой дискрими-
нации 13СО2 (рис. 15.10). У CAM-растений часто наблюдаются
промежуточные (между значениями 6 для С3- и С4-растений)
значения б13С. Изящное доказательство того, что ночная фик-
сация приводит к увеличению, а дневная фиксация СО2 из воз-
духа, наоборот, к уменьшению величины б у САМ-растений,
было получено Нальборчиком, Ла Круа и Хиллом. Они постави-
ли очень простой опыт. Растения Kalanchoe daigremontia ин-
кубировали с СО2 на свету или в темноте или же и на свету,
п в темноте. После этого анализировали изотопный состав уг-
лерода в листьях. Полученные данные представлены ниже.
Условия инкубации с СО2 61SC
В темноте —10,6
На свету —25,9
На свету+в темноте —15,0
Источники данных: [12, 13, 23, 34, 35, 37, 42, 44, 48, 49, 71].
15.13. Связь САМ-метаболизма с С4-циклом
Наиболее очевидной особенностью С4- и САМ-растений яв-
ляется то, что обе эти группы растений сначала включают СО2
в оксалоацетат с помощью ФЕП-карбоксилазы, а затем снова
фиксируют ее (после декарбоксилирования в той или иной фор-
ме), ио уже в реакциях ВПФ-цикла. У С4-растений все эти
процессы — первоначальная фиксация СО2, декарбоксилирова-
ние органических кислот и повторная фиксация высвободив-
шейся СО2 — происходят одновременно, но первый этап фик-
сации СО2 пространственно отделен от двух последних.
У САМ-растений все эти три процесса происходят в одной и
той же клетке, но исходная фиксация СО2 идет в темноте, а
два других процесса протекают на свету. У С4-растений про-
дукт декарбоксилирования С4-кислот превращается в ФЕП, ко-
торый затем прямо используется ФЕП-карбоксилазой. А у
САМ-растений ФЕП образуется в результате расщепления
крахмала в процессе гликолиза. ФЕП, образующийся после
декарбоксилирования С4-кислот на свету, превращается в крах-
мал и в таком виде хранится, пока не наступит темновой пе-
риод. У С4-р астений пул С4-кислот составляет 1—2 эквивален-
33
516
ГЛАВА 15
та на грамм сырого вещества, а период их полуобновления ti/3
равен 5 с, У CAM-растений пул С4-кислот приближается к
100—200 эквивалентам на грамм сырого вещества, a t равен
10 000 с.
Источники данных: [5, 33].
15.14. Экологическое значение САМ-метаболизма
CAM-растения особенно хорошо приспособлены к полуза-
сушливым условиям окружающей среды. Характер обмена ве-
ществ у этих растений таков, что днем в жару они запасают ту
СО2, которая высвобождается в процессе дыхания, а ночью,
когда потери воды сведены к минимуму, они ассимилируют
СО2 из воздуха. Некоторые суккулентные растения могут да-
же переходить к Cg-фотосинтезу и использовать этот процесс
как дополнительное средство для роста в условиях водного изо-
билия. Отношение транспир ация/фотосинтез (граммы траиспи-
рируемой воды/граммы фиксируемой СО2) для САМ-растений
(когда идет САМ-метаболизм) составляет примерно 50—100,
для С4-растений 250—300 и для С3-растений 400—500. Мы при-
водим типичные величины, которые встречаются в литературе.
Следует отметить, что эти цифры ие отражают и не учитывают
те различия, которые могут быть связаны с изменениями тем-
пературы листа и концентрации водяных паров в воздухе в
момент измерения. Более строгим было бы сравнение величин
rs(H2o)/ra(co2)+/'m, где г8(н3О) отражает устьичное сопротивление
для диффузии воды, rs(co2) — устьичное сопротивление для
диффузии СО2, а Гт — сопротивление мезофилла для фиксации
СО2. Более высокое устьичное сопротивление и более низкая
величина гт должны способствовать снижению соотношения
транспирация/фотосинтез (разд. 14.6).
При любом заданном значении устьичного сопротивления
САМ-растение сможет запасти больше воды, если оно будет
фиксировать СО2 ночью, а не днем. Потенциальная потеря во-
ды листом определяется тем, какова разность между концент-
рацией водяных паров в листе и воздухе. Поэтому в жаркой
засушливой зоне, где температура листа высокая, а влажность
воздуха минимальна, опасность потери воды очень велика. При
высокой освещенности и минимальной транспирации темпера-
тура листа может быть даже выше, чем температура воздуха.
[Транспирация уменьшает опасность перегрева, так как избы-
точное тепло тратится на превращение воды из жидкости в
пар.]
Рассмотрим, например, кактус, у которого температу-
тура ткани листа днем составляет 40 °C, а ночью 20 °C. При
40 °C концентрация водяных паров в межклеточном простраист-
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
517
ве листа в момент насыщения равна 51 г/м3, а при 20 °C она
снижается до 17 г/м3. Если, например, концентрация водяных
паров в воздухе 10 г/м3, градиент для потери воды растением
составит 51—10 = 41 г/м3 днем и 17—10 = 7 г/м3 ночью. В этом
случае потенциальная потеря воды днем в шесть раз выше, чем
ночью.
Реальную скорость транспирации можно выразить следую-
щим уравнением (разд, 14.6):
Г'а 4- r's
где Т — транспирация, Wl — концентрация водяных паров в
листе, НА— концентрация водяных паров в воздухе, га'— соп-
ротивление пограничного слоя для диффузии воды, rs' — усть-
ичное сопротивление для диффузии воды,
Как следует из этого уравнения, чем выше сопротивление
пограничного слоя 'И сопротивление устьиц, тем -ниже транс-
пирация. Число устьиц у САМ-растений гораздо меньше, чем
у С3- или Сграстений (табл, 15.3). Для САМ-метаболизма в
условиях водного дефицита наиболее характерны минималь-
ные значения сопротивления листа, приближающиеся к
100 с/см. Правда, в оптимальных условиях зарегистрированы
еще меньшие значения, вплоть до 2 с/см,
САМ-растения приспособлены не только к сбережению во-
ды, они также запасают углерод, и, кроме того, у них сведены
Таблица 16.3. Частота и локализация устьиц у некоторых САМ-,
Са- и С4-растений (Meidner, Mansfield, 1968; Ting' et al., 1972;
см. также Kluge, Ting, 1978)
Вид растения Число устьиц/мм2
эпидермис на верхней поверх- ности листа эпидермис на нижней поверх- ности листа
САМ-растения Agave amerlcana 21 21
Crassula argenta 24 33
Portulacaria afra 45 33
С4-растения Zea mays 98 108
Ca-p астения Melianthus annuus 150 230
Quercus robur 0 340
Vida faba 153 170
518
ГЛАВА 15
Рис. 15.11. Типичная картина газообмена у Сз- и С4-растений (А) и у САМ-
растений с облигатным CAM-метаболизмом (Б).
к минимуму потери той СО2, которая высвобождается в про-
цессе дыхания. Когда эти растения фотосинтезируют по типу
CAM-метаболизма, они могут проходить полный суточный
(24 ч) цикл без явного оттока СО2 из растения (рис. 15.11).
Ночью происходит истинное поглощение СО2, а СО2, выделяю-
щаяся в процессе дыхания, может снова связываться ФЕП-
карбоксилазой. Днем устьица закрыты, а СО2, высвобождаю-
щаяся при декарбоксилировании Сгкислот или в процессе ды-
хания, ассимилируется ш ВПФ-цикле.
Следует отметить, что в результате декарбоксилирования
С4-кислот, которое происходит при закрытых устьицах, во вре-
мя CAM-метаболизма может создаваться такая концентрация
СО21 которая будет насыщать РуБФ-карбоксилазу. Здесь мы
имеем механизм, аналогичный механизму концентрирования
СО2 у Сграстений (разд. 12,4, 14.4), который сводит к мини-
муму кислородное ингибирование фотосинтеза. Недавно Кок-
берн и др., отобрав пробы газов у ряда САМ-растений, расту-
щих в природных условиях, проанализировали их с помощью
газовой хроматографии и показали, что [СО2] в листьях САМ-
растений днем колеблется в пределах 0,5—2%, а ночью пада-
ет ниже ее уровня в атмосфере. Повышенная [СО2] в листьях,
которая наблюдалась днем, была связана с повышенным усть-
ичным сопротивлением и обнаруживалась лишь тогда, когда
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ i
была повышена и кислотность (содержание малата). Ограни-
чение фиксации СО2 ночью приводило к понижению кислотно-
сти ткани листа и к снижению [СО2] в листе на следующий
день. Спалдииг и др. определили и [,О2], и [СО2] в пробах,
полученных из листьев ряда САМ-растений во время раскис-
ления, и установили, что уровень О2 доходит до 42%, а содер-
жание СО2 достигает 0,35%. В среднем отношение СО2/О2 в
листе было примерно в 4 раза выше, чем в окружающем воз-
духе. Увеличение [СО2] внутри листа и повышение отношения
СО2/О2 в листьях САМ-растений, которое наблюдается во вре-
мя декарбоксилирования малата, способствуют созданию ус-
ловий, благоприятствующих фотосинтезу и препятствующих
кислородному ингибированию этого процесса (разд. 13.3, 14.3),
В том случае, когда фотосинтез идет по CAM-пути, сопротивле-
ние устьиц может непосредственно контролироваться той кон-
центрацией СО2, которая создается в листе в результате де-
карбоксилирования малата. Увеличение [СО2] в листьях САМ-
растений в течение дня коррелирует с увеличением устьичного
сопротивления. Если поместить Opuntia ficus-indica в камеру с
2% СО2 в воздухе и выдержать там растение в течение ночи,
то устьица закроются. Об этом свидетельствует резкое увели-
чение сопротивления устьиц от -~8 до 150 с/см. При угрожаю-
щем водном дефиците некоторые САМ-растеиия поддержива-
ют высокое сопротивление устьиц в течение всего суточного
цикла, но при этом у них наблюдаются колебания кислотности
ткани, очень похожие на САМ-метаболизм при переходе свет —
темнота. Возможно, что СО2,. выделяющаяся ночью во время
дыхания, улавливается в процессе образования малата, а за-
тем ее углерод днем передается в ВПФ-цикл. Было обнаруже-
но, что колючие подушечки Opuntia basilaris днем содержат
много СО2, а ночью концентрация СО2 в них заметно снижает-
ся. В этом случае функция устьиц может регулироваться ка-
ким-либо другим дополнительным фактором (или факторами),
поскольку известно, что такие устьица даже ночью остаются
закрытыми, несмотря на то что концентрация СО2 внутри рас-
тения понижается.
Источники данных: [2—4, 21, 22, 26, 38—40, 43, 51, 56—58].
ОБЩАЯ ЛИТЕРАТУРА
1. Brtiinsma 7.'Studies ди-the Crass ulacean Acid Metabolism, Acta Botanica
Neerlandica 7, 531—590, а также и в North Holland Pub. Co., Amsterdam.
[В этом перечне приводится много ссылок на литературу XIX века
(Aubert, Kraus, de Saussure, de Vries, Warburg и т. д.)], 1958.
2. Burris R. H., Black С. C. CO2 Metabolism and Plant Productivity, Baltimore,
Maryland, University Park Press, 1976.
3. Kluge M., Ting I. P„ Crassulacean Acid Metabolism. Analysis of an ecolo-
gical adaptation, pp. 209. Berlin, Springer-Verlag, 1978.
520
ГЛАВА IF
4. Marcelle R. Environmental and Biological Control of Photosynthesis, The
Hague, Junk, 1975.
5. Osmond G. B. Crassulacean Acid Metabolism; A curiosity in context, Ann
Rev. Plant Physiol., 29, 379—414 (1978).
6. Ranson S. L„ Thomas M. Crassulacean Acid Metabolism, Ann. Rev. Plant
Physiol., 11, 81-110 (1960).
7. Thomas M., Ranson S. L., Richardson J. A. Plant Physiology, Fifth Edition,
London, Longman, 1973.
8. Walker D. A. Pyruvate carboxylation and plant metabolism, Biol. Rev., 37,
215—256 (1962).
9. Wolf J. Der Diurnale Saurerhythmus, Encyclopedia of Plant Physiology
(ed. von W. Ruhland Vol. XII/2), pp. 809—889, Berlin, Springer-Verlag.
[В этом обзоре тоже приводится множество ссылок на ранние работы,
как и у Бртоинсма.], 1962.
10. Wood Н. G., Utter М. F. The role of СО2 fixation in metabolism, Essays in
Biochem. 1, 1—27. London, Academic Press, 1965.
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
11. Bandurski R. S., Greiner С. M. The enzymatic synthesis of oxalacetate
from phosphorylenolpyruvate and carbon dioxide, J. Biol. Chem., 204, 781 —
786 (1953).
12. Bender M, M, Variations in the 13C/12C ratios in relation to the pathway of
photosynthetic carbon dioxide fixation, Phytochemistry, 10, 1239—1244
(1971).
13. Bender M. M., Rouhani I., Vines H. M., Black С. C. 13C/12C ratio changes
in Crassulacean acid metabolism plants, Plant Physiol., 52, 427—430
(1973).
14. Bennet-Clark T. A. The role of organic acids in plant metabolism, New
Phytol., 32, 37—71, 128—161, 197—230 (1933).
15. Black С. C. Photosynthetic carbon fixation in relation to net CO2 uptake,
Ann. Rev. Plant Physiol., 24, 253—286 (1973).
16. Black С. C., Williams S. Plants exhibiting characteristics common to Cras-
sulacean acid metabolism. In: CO2 Metabolism and Plant Productivity
(eds. R. H. Burris and С. C. Black), University Park Press, Baltimore, 407—
424, 1976.
17. Bonner IE, Bonner J. The role of carbon dioxide in acid formation by suc-
culent plants, Amer. J. Bot., 35, 113—117 (1948).
18. Bradbeer J. W., Ranson S. L,, Stiller M. Malate synthesis in Crassulacean
leaves. I. The distribution of 14C in the malate of leaves exposed to 14CO2
in the dark, Plant Physiol., 33, 66—70 (1958).
19. Brulfert J., Guerrier D„ Queiroz O. Photoperiodism and enzyme rhythms:
kinetic characteristics of the photoperiodic induction of Crassulacean acid
metabolism, Planta, 125, 33—44 (1975).
20. Cockburn W., McAuley A. The pathway of carbon dioxide fixation in Crassu-
lacean plants, Plant Physiol., 35, 67—89 (1975).
21. Cockburn W., Ting I. P., Sternberg L. O, Relationships between stomatai
behaviour and internal carbon dioxide concentration in Crassulacean acid
metabolism plants, Plant Physiol., 63, 1029—1032 (1979).
22. Crews С. E„ Williams S. L., Vines H. M„ Black С. C. Changes in the meta-
bolism and physiology of Crassulacean acid metabolism plants grown in
controlled environments. In CO2 Metabolism and Plant Productivity (eds.
R. H. Burris and С. C. Black), pp. 235—250, Baltimore, Maryland, Univer-
sity Park Press, '1976.
23. Deleens E. La discrimination du 13C et les trois types de metabolisme des
plantes, Physiol. Veg., 14, 641—655 (1976).
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
521
24. Dittrich Р. Nicotinamide adenine dinucleotide-specific “malic enzyme” in
Kalanchoe daigremontiana and other plants exhibiting Crassulacean Acid
Metabolism, Plant Physiol., 57, 310—314 (1976).
25. Dittrich P., Campbell IF. H., Black С. C. Phosphoenolpyruvate carboxyki-
nase in plants exhibiting Crassulacean acid metabolism, Plant Physiol., 52,
357—361 (1973).
26. Hartsock T. L., Nobel P. S. Watering converts a CAM plant to daytime
CO2 uptake, Nature, 262, 574—576 (1976).
27. Herbert D. Oxalacetic decarboxylase and carbon dioxide assimilation in
bacteria, Symp. Soc. exp. Biol., 5, 52—71 (1951).
28. Heyne B. On the deoxidation of the leaves of Cotyledon calycina, Trans.
Linn. Soc. London, 213, 815—816 (1815).
29. Jolchine G. Sur la distribution du 14C dans les molecules d’acide malique
synthetisees par fixation de UCO2 dans les feuilles de Bryophyllum daigre-
montianum Berger. Bull, Soc. Chim. Biol., 41, 227—234 (1959).
30. Kalnitsky G„ Werkman С. H. Enzymatic decarboxylation of oxaloacetate
and carboxylation of pyruvate, Arch. Biochem., 4, 25—40 (1944).
31. Kluge M., Kriebitzsch C. H„ Willert D. J. V. Dark fixation of CO2 on Cras-
sulacean acid metabolism: are two carboxylation steps involved?, Z. Pflan-
zenphysiol., 72, 460—465 (1974).
32. Ku M. S. B., Spalding M. FL, Edwards G. E. Intracellular localization of
phosphoeiiolpyruvate carboxykinase in leaves of C4 and CAM plants, Plant
Sci. Letts., 19, 1—8 (1980).
33. Laeisch IF. M. Chloroplast structural relationships in leaves of C4 plants.
In: Photosynthesis and Photorespiration (eds. M. D. Hatch, С. B. Osmond
and R. C. Slatyer), pp. 323—349, New York, Wiley-Interscience, 1971.
34. Lerman I. C. How to interpret variations in the carbon isotope ratio of
plants: biologic and environmental effects, In: Environmental and Biolo-
gical Control of Photosynthesis (ed. R. Marcelle), pp. 323—335, The Ha-
gue, Junk, 1975.
35. Lerman I. C., Queiroz O. Carbon fixation and isotope discrimination by a
crassulacean plant: dependence on the photoperiod. Science, 183, 1207—
1209 (1974).
36. Meidner H„ Manfield T. A. Physiology of Stomata, p. 478, London, McGraw-
Hill, 1968.
37. Nalborczyk E., LaCroix L. I., Hill R. D. Environmental influences on light
and dark CO2 fixation by Kalanchoe daigremontiana, Canad. J. Bot., 53,
1132—1138 (1975).
38. Neales T. F. The effect of night temperature on CO2 assimilation, transpi-
ration and water use efficiency in Agave americana, L. Aust. J. Biol. Sci.,
26, 705—714 (1973a).
39. Neales T. F. Effect of night temperature on the assimilation of carbon dio-
xide by mature pineapple plants, Ananas comosus (L) Merr, Aust. J. Biol.
Sci., 26, 539—546 (1973b).
40. Neales T. F., Patterson A. A., Hartney V. J. Physiological adaption to
drought in the carbon assimilation and water loss of xerophytes, Nature,
219, 469—472 (1968).
41. Ochoa S„ Mehler A., Kornberg A. Reversible oxidative decarboxylation of
malic acid, J. Biol. Chem., 167, 871—872 (1947).
42. O’Leary M. H. Carbon isotope fractionation in plants, Phytochemistry, 20,
553—567 (1981).
43. Osmond С. B. CO2 assimilation and dissimilation in the light and dark in
CAM plants, In: CO2 Metabolism and Plant Productivity, pp. 217—234,
Baltimore, Maryland, University Park Press, 1976.
44. Osmond С. B., Bender M. M„ Burris R. H. Pathways of CO2 fixation in the
CAM plant Kalanchoe daigremontiana. III. Correlation with 18C value
522'
ГЛАВА 15
" during growth and waler stress, Aust, J. Plant Physiol., 3, 787.—799
(1976).
45. Pucher G. IF., Leavenworth С. H. S., Ginter W. D., Vickery H, B. The diur-
nal variation in organic acid and starch content of Bryophyllum calycinum,
Plant Physiol., 22, 360—376 (1947).
46. Saltman P., Kunitake G., Spolter H., Stitte C. The dark fixation of CO2
by succulent leaves: the first products, Plant Physiol., 31, 464—468 (1956).
47. de Saussure T. H. Recherches chlmiques sur la vegetation, Paris, Nyon,
1804.
48. Smith B. N. Natural abundance of the stable isotopes of carbon in biolo-
gical systems, Bioseience, 22, 226—231 (1972).
49. Smith B. N„ Brown \V. V. The Kranz syndrome in the Gramineae as indi-
’ cated by carbon isotopic ratios, Amer. J. Bot., 60, 505—513 (1973).
50. Spalding M. H., Schmitt M. R., Ku S. B., Edwards G. E. Intracellular loca-
lization of some key enzymes of Crassulacean Acid Metabolism in Sedum
praealtum, Plant Physiol., 63, 738—743 (1979a).
51. Spalding M. H., Schmitt M. R., Ku S. B., Burris R. H., Edwards G. E.
• Grassulacean acid metabolism and diurnal variations of internal CO2 and
O2 concentrations in Sedum praealtum DC, Aust. J. Plant Physiol., 6, 557—
567 (1979b).
52. Sutton B. G. Glycolysis in CAM plants, Aust. J. Plant Physiol., 2, 489—
492 (>1975).
53, Sutton B. G., Osmond С. B. Dark fixation of CO2 by Crassulacean plants —
evidence for a simple carboxylation step, Plant Physiol., 50, 360—365
' (1972).
54. Szarek S. R., Ting I. P. The occurence of Crassulacean acid metabolism
among plants, Photosynthetica, 11, 330—342 (1977).
55. Tchen T. T., Vennesland B. Enzymatic carbon dioxide fixation into oxaloace-
tate in wheat germ, J. Biol. Chem., 213, 533—546 (1955).
56. Ting I. P., Hanscom Z. Induction of acid metabolism in Portulacaria afra.
Plant Physiol., 59, 511—514 (1977).
57. Ting I. P., Johnson H. B., Szarek S. R. Net CO2 fixation in Crassulacean
acid metabolism plants, In: Net Carbon Dioxide Assimilation in Higher
Plants (ed. С. C. Black), pp. 26—53, Raleigh, North Carolina, Cotton Inc,
1972.
58. Ting I. P„ Thompson M„ Dugger IF. Leaf resistance to water vapor trans-
fer in succulent plants: Effect of thermoperiod, Amer. J. Bot., 54, 245—251
(1967).
59. Thomas M. Plant Physiology, 3e, pp. 320—340, London, J. and A. Churchill,
. 1947.
60. Thomas M. Carbon dioxide fixation and acid synthesis in Crassulacean acid
metabolism, Symp. Soc. Exp. Biol., 5, 72—93 (1951).
61. Thomas M., Beevers H. Physiological studies on acid metabolism in green
plants. II. Evidence of CO2 fixation in Bryophyllum and the study of diur-
.. паГfluctuation of acidity in this genus, New Phytol., 48, 421—427 (1949).
62. Thomas AL, Ranson S. L. Physiological studies on acid metabolism in
green plants. III. Further evidence of CO2 fixation during dark acidification
of plants showing Crassulacean acid metabolism, New Phytol., 53, 1—30
(1954).
63: Thurlow J., Bonner /. Fixation of atmospheric CO2 in the dark by leaves of
Bryophyllum, Arch. Biochem., 19, 500—511 (1948).
64. Varner J. E., Burrell R. C. Use of C14 in the study of the acid metabolism
si-Bryophyllum calycinum, Arch. Biochem., 25, 280—287 (1950).
65. Vickery H. B. The behaviour of isocitric acid in excised leaves of Bryophyl-
lum calycinum during‘culture in alternating light and darkness, Plant Phy-
-< Si&l„ 27, 9—11 (1952)." . . .
МЕТАБОЛИЗМ КИСЛОТ У СЕМЕЙСТВА ТОЛСТЯНКОВЫХ
623
66. Walker D. A. Malate synthesis in a cell-free extract from a Crassulacean
plant, Nature, 178, 593—594 (1956).
67. Walker D. A. Physiological studies on acid metabolism. 4. Phosphoenolpy-
ruvate carboxylase activity in extracts of Crassulacean plants, Biochem. J.,
67,73—79 (1957).
68. Walker D. A. Physiological studies on acid metabolism. 7. Malic enzyme
from Kalanchoe crenata: effects of carbon dioxide concentration, Biochem.
J., 74, 216—223 (1960).
69. Walker D. A., Brown J. M. A. Physiological studies on acid metabolism.
5. Effects of carbon dioxide concentration on pliosphoenolpyruvic carbo-
xylase activity, Biochem. J., 67, 79—83 (1957).
70. Walker D. A., Ranson S. L. Physiological studies in acid metabolism in
green plants. Transaminases in cell —free extracts from Kalanchoe leaves,
Plant Physiol., 33, 226—230 (1958).
71. Whelan T., Sackett UK M„ Benedict C. R. Enzymatic fractionation of car-
bon isotopes by phosphoenolpyruvate carboxylase from C4 plants, Plant
Physiol., 51, 1051—1054 (1973).
72. Winter K. C02-fixation metabolism in the halophytic species Mesembryan-
themum crystallinum grown under different environmental conditions, Plan-
ta, 114, 75—85 (1973).
73. Winter /(. Photosynthetic and water relationships of higher plants in a
saline environment. In: Ecological Processes in Coastal Environments (eds.
R. L. Jefferies, A. J. Dary), pp. 297—320, Oxford, Blackwell Scientific
Publications, 1979.
74. Winter K. Malate sensitivity of PEP carboxylase during CAM, Plant Phy-
siol., 65, 792—796 (4980).
75. Winter R., Foster L G., Edwards G. E., Holturn J. A. M. Intracellular, loca-
lization of enzymes of carbon metabolism in Mesembryanthemum crystal-
linum exhibiting C3 photosynthetic characteristics or performing crassula-
cean acid metabolism, Plant Physiol., 69, 300—307 (1982).
76. Winter R., Randler O. Misleading data on isotope distribution in malate-^C
from CAM plants caused by fumarase activity of Lactobacillus plantarum,
Z. Pflanzenphysiol., 78, 103—112 (1976).
77. Winter R., Luitge U., Winter E., Troughton J. M. Seasonal shift from C3
photosynthesis to Crassulacean acid metabolism in Mesembryanthemum
crystallinum growing in its natural environment, Oecologia, 34, 225—237
(1978).
78. Wood FL G., Werkman G. H. The utilization of CO2 in the dissimilation of
glycerol by the propionic acid bacteria, Biochem. J., 30, 48—53 (1936).
ГЛАВА 16
Сравнительное изучение С3 - и
Ct-метаболизма в других тканях
(помимо листа)
Краткое содержание
Участвуя в фотосинтетической ассимиляции СО2, рибулозо-
бисфосфат (РуБФ) и РуБФ-карбоксилаза выполняют у рас-
тений уникальную функцию. Этого нельзя сказать о фосфое-
нолпирувате (ФЕП), ФЕП-карбоксилазе и Сгкислотах, роль
которых в процессах обмена веществ у растений гораздо более
многогранна и, как это видно на ряде примеров, далеко выхо-
дит за пределы их участия в САМ,- или Сгфотосинтезе.
В процессе открывания устьиц у некоторых видов растений
синтезируется малат при участии ФЕП-карбоксилазы. В ре-
зультате этого синтеза на свету концентрация малата в замы-
кающих клетках устьиц резко возрастает. Благодаря такому
увеличению концентрации малат наряду с фондом ионов К+
вносит свой вклад в создание повышенного осмотического по-
тенциала в замыкающих клетках, что совершенно необходимо
для того, чтобы устьица открылись.
Плоды, например перикарпий мелких семян злаков, зеле-
ные томаты и стручки бобовых, обладают значительной актив-
ностью ФЕП-карбоксилазы, а главным продуктом ассимиляции
14СО2 у них является малат. Основная функция такого 04-ме-
таболизма может заключаться в том, чтобы сводить к миниму-
му потери углерода, в ходе дыхания.
Синтез малата играет очень важную роль в питании расте-
ний. Обычно содержание неорганических катионов у растений
несколько выше, чем содержание неорганических анионов, а
избыточный положительный заряд при этом компенсируется
органическими анионами (чаще всего малатом). К тому же при
превращении нитрата в аминный азот происходит защелачива-
ние клетки, которое также может нейтрализоваться за счет
синтеза яблочной кислоты.
Когда в проростках начинает функционировать глиоксилат-
ный цикл, в процессе деградации липидов образуется малат,
который служит промежуточным продуктом для синтеза саха-
розы. При таком метаболическом превращении из оксалоаце-
тата образуется ФЕП. В этой реакции участвует ФЕП-карбокси-
киназа, которая обеспечивает поступление Сз-предшественника
для глюконеогенеза. ФЕП-карбоксилазу и соответственно мета-
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И СГМЕТАБОЛИЗМА
525
Рис. 16.1. У высших растений метаболические превращения двуокиси углеро-
да происходят ие только в листьях. Многие ткани растений, в которых содер-
жатся хлоропласты, могут ассимилировать СОз при участии ВПФ-цикла. СОг
может связываться также в составе С4-кислот. После декарбоксилирования
таких кислот и их метаболических превращений в ЦТ К она может и выде-
ляться.
болизм малата можно обнаружить в любой растительной тка-
ни. РуБФ-карбоксилаза также встречается не только в листь-
ях, но и в других тканях, однако она распространена ие столь
повсеместно, как ФЕП-карбоксилаза. В дополнение к той уни-
кальной роли, которую дикарбоновые кислоты играют в фото-
синтезе у Сграстений (разд. 10.7) и в CAM-метаболизме (разд.
15.4), С4-кислоты могут выполнять в различных тканях совер-
шенно особые функции.
Источник данных: [43].
16.1. Устьица
Малат синтезируется в замыкающих клетках эпидермаль-
ной ткани. Он вносит свой вклад в создание тургора, который
необходим для того, чтобы открыть усты-ща. В замыкающих
клетках обычно имеются хлоропласты, ио, по-видимому, эти
клетки в основном гетеротрофны, так как восстановленный уг-
лерод поступает в них главным образом за счет идущего в ли-
сте фотосинтеза. Поэтому в отличие от Сгфотосиитеза или
526
ГЛАВА 16
CAM-метаболизма обмен Сгкислот в замыкающих клетках
устьгщ не связан непосредственно с ВПФ-циклом.
Есть очень веские доказательства, что увеличение содержа-
ния растворенных веществ в замыкающих клетках, которое на-
блюдается во время открывания устьиц, связано с увеличением
в них концентрации калия, хлорида и малата. Относительная
доля яблочной кислоты и хлорида может варьировать от вида
к 'Виду, так, например, у Vicia faba анионы представлены глав-
ным образом малатом, а у кукурузы противоионом для калия
служит хлорид, который накапливается в довольно больших ко-
личествах.
Для того чтобы устьица открылись, нужно, чтобы в замыка-
ющих клетках произошли существенные осмотические измене-
ния. Увеличение концентрации растворенных веществ внутри
замыкающих клеток приводит к поглощению воды из примы-
кающих к ним клеток. Поэтому поглощение воды, которое обыч-
но сопутствует повышению концентрации растворенных ве-
ществ, приводит к увеличению тургора в замыкающих клетках
(гидростатическое давление в замыкающих клетках возраста-
ет по сравнению с давлением в примыкающих к ним клетках)
и к открыванию устьиц. У двудольных растений наружные кле-
точные стенки замыкающих клеток (которых обычно пара)
тоньше, чем их внутренние стенки (рис. 16.2). Изменения фор-
мы замыкающих клеток у двудольных, которые происходят
при открывании устьиц, напоминают изменения формы воздуш-
ного шарика, если его надувать, прижав одной стороной к твер-
дой поверхности. У злаков стенки замыкающих клеток в сере-
дине толще, а на концах тоньше. При увеличении осмотическо-'
Рис. 16.2. Типичное строение устыщ у двудольных растений.
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Ci- И СгМЕТЛБОЛИЗМА
527
Замыкающею
клетка
Рис. 16.3. Типичное строение устьиц
у злаков.
го давления концы кл,еток
растягиваются, и устьица от-
крываются (рис. 16.3).
В хлоропластах замыкаю-
щих клеток обычно содер-
жится крахмал, который у Ca-
ll у Сграстений синтезирует-
ся ночью и частично разруша-
ется днем. В этом случае мы
имеем картину, обратную той,
которая наблюдается в про-
цессе метаболизма крахмала
в клетках мезофилла (С3, С<
и САМ) и в клетках обкладки
проводящих пучков листа.
Откуда поступает малат во
время открывания устьиц, неясно. Предполагают, что при этом
в замыкающих клетках разрушается крахмал, в результате че-
го образуется ФЕП, необходимый для образования малата под
действием ФЕП-карбоксилазы и малатдегидрогеназы. По-види-
мому, это происходит аналогично образованию малата во вре-
мя темновой фазы САМ-метаболизма (разд. 15.5, 15.6). Есть
данные о том, что в замыкающих клетках эпидермальной тка-
ни локализована ФЕП-карбоксилаза. Яблочная кислота, кото-
рая синтезируется в замыкающих клетках, должна поставлять
те протоны, которые необходимы для обмена на ионы К+, по-
ступающие из соседних побочных клеток (рис. 16.4).
Ночью Сгкислота декарбоксилируется, а Сз-предшественни-
ки снова превращаются в крахмал. Выделяющаяся при этом
СО2 скорее всего не ассимилируется вновь в ВПФ-цикле, как
у САМ- пли С4-растений, а просто теряется. Для превращения
пирувата в крахмал необходима энергия, которую может обес-
печить дыхание митохондрий.
Замыкающие клетки (например, у Tulipa spp. и Commelina
spp.), по-видимому, почти не способны к истинной ассимиляции
СО2 в ВПФ-цикле. Основными первичными продуктами фикса-
ции МСО2 в иеотделеиной или отделенной от листа эпидермаль-
ной ткани являются малат и аспартат. Скорость фиксации
ИСО2 в полосках эпидермиса, отделенных от остальной ткани'
листа, намного ниже, а на свету она всего лишь в два раза вы-
ше, чем в темноте. Вместе с тем скорость появления ассимили-
рованной 14СО2 в эпидермальной ткани, не отделенной от ос-
тальных тканей листа, на свету в 100—200 раз выше, чем в
Темноте. Если полученные данные выразить, в расчете на 1 г.
азота, то скорость появления меченых метаболитов в иеотдё-
леипой эпидермальной ткани вполне сравнима со скоростью их
528
ГЛАВА ie
Яблочная Малат" ’ -f- Н* —---► Н1
кислота
к'«-----к+
Замыкающая клетка
Соседняя клетка
Рис. 16.4. Синтез малата и обмен Н+/К+ во время открывания устьиц.
появления в ткани мезофилла (у Commelina. sp.; Thorpe, Milt-
horp е) .
Очевидно, in vivo определенная часть углерода, ассимили-
руемого в ткани мезофилла листа, быстро транспортируется в
эпидермальную ткань. Аутло и др. показали, что в хлороплас-
тах замыкающих клеток Vicia faba практически нет ни РуБФ-
карбоксилазы, ни фосфорибулокиназы, ни NADP-триозофос-
фатдегидрогеназы. Возможно, что хлоропласты в замыкающих
клетках нужны днем главным образом для обеспечения этих
клеток энергией, а их каталитическая роль в истинной ассими-
ляции углерода в ВПФ-цикле не существенна. Высказывалось
и такое чисто спекулятивное предположение, что в плазматиче-
ской мембране, которая разделяет замыкающие клетки устьиц
и побочные клетки, имеется система, осуществляющая АТР-за-
висимый обмен ионов Н+—К+.
Есть несколько работ, в которых описана физиология усть-
иц у САМ-растений. При CAM-метаболизме устьица открыва-
ются ночью и закрываются днем. Поэтому крахмал в замыка-
ющих клетках распадается только ночью, в результате чего
образуется тот предшественник, который используется для
синтеза малата. Днем крахмал вновь синтезируется. В этом
случае можно выявить определенный параллелизм между на-
блюдаемой картиной обмена крахмала и тем, что происходит в
ткани мезофилла в листьях САМ-растений. Поэтому очень ин-
тересно выяснить, способны ли замыкающие клетки устьиц у
таких растений осуществлять CAM-метаболизм. Следует отме-
тить тот факт, что очень часто устьица у Сз- и Сграстений за-
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И СгМЕТАБОЛИЗМА
52»
крываются иа свету тогда, когда повышается [СО2] в воздухе,
и, наоборот, открываются в темноте, когда [СО2] становится
незначительной. Вполне понятно поэтому, что именно [СО2]
в подустьичном пространстве является фактором, контролирую-
щим деятельность устьиц у САМ-растений, и что поведение
устьиц подчинено тем изменениям i[CO2] внутри листа, которые
происходят в процессе темнового закисления и светового рас-
кисления (разд. 15.14).
В регуляции работы устьиц участвуют самые разнообразные
факторы; например, закрывание усты-щ контролируется содер-
жанием СО2 в воздухе; иа открывание устьиц влияет длина
волны света, синий свет более эффективен в этом отношении,,
чем красный, а такие регуляторы роста, как абсцизовая кисло-
та, влияют на степень закрывания устьиц при водном дефици-
те. Механизмы, которые лежат в основе того или иного эффек-
та, равно как и роль метаболизма углерода в физиологии усть-
иц, изучены далеко недостаточно. Не исключено, что у разных
видов растений деятельность устьиц контролируется различны-
ми факторами.
У разных растений хлоропласты присутствуют в замыкаю-
щих клетках в разных количествах; например, у Vicia faba их
намного больше, чем у кукурузы. Поэтому вовсе необязатель-
но, чтобы фотосинтетический метаболизм и энергетический об-
мен в пластидах замыкающих клеток устьиц был одинаков у всех
видов.
Источники данных: ;[1, 11, 17, 27, 28, 30—32, 39, 46, 47].
16.2. Сз- и Ci-фотосинтез в тканях
репродуктивных органов
а) Фотосинтез в колоске злаков
В исследованиях фотосинтеза в соцветии (колосе или ме-
телке) таких мелкозерных злаков, как ячмень, пшеница и рис,,
достигнуты значительные успехи. Колосок у этих злаков со-
стоит из верхней цветковой чешуи, плода, нижней цветковой
чешуи, остей и колосковых чешуй; вероятно, главный вклад в-
ассимиляцию СО2 в колоске вносят три последних компонента
цветка (рис. 16.5).
Фотосинтез в метелке злаков имеет ряд преимуществ. Во-
первых, продукт фотосинтеза образуется вблизи того места,
куда он поступает (т. е. вблизи плода), и не нужно перено-
сить его на далекое расстояние; во-вторых, колосок выступает
над пологом трав и максимально освещается (обычно он и ста-
реет позже, чем листья). У некоторых сортов хлебных злаков
почти половина всех продуктов фотосинтеза, которые поступа-
34—234
530.
ГЛАВА ID
Верхняя
цветковая чешуя
Плод
Ость
Нижняя
цветковая чешуя
Колосковая
чешуя
Колосок
Рис. 16.5. Общее строение колоска в метелке зерновых культур.
Ют в зерно, образуется за счет фотосинтеза, происходящего в
колоске. Для остистых сортов характерна более высокая ак-
тивность фотосинтеза в колоске.
Те ткани колоска, в которых есть хлоропласты, по-видимо-
му, способны фиксировать СО2 в ВПФ-цикле. Уирс и др. об-
наружили РуБФ-карбоксилазу и те продукты, которые образу-
ются в результате фиксации ИСО2 в ВПФ-цикле, в перикарпии
(т. е. в наружном слое плода, который образуется из стенки за-
вязи), верхней цветковой чешуе, нижней цветковой чешуе и в
колосковых чешуях овса и пшеницы. Активность ФЕП-карбо-
ксилазы в этих тканях колоска была гораздо выше, чем в тка-
ни листа. То же самое можно сказать и об отношении актив-
ностей ФЕП-карбоксилазы и РуБФ-карбоксилазы. Уровень де-
карбоксилаз С4-кислот при этом был очень невысок. Верхняя
и нижняя цветковые чешуи и перикарпий могут заново усваи-
вать некоторую часть той СО2, которая выдыхается во время
развития репродуктивных органов. В отличие от других фото-
.синтезирующих тканей цветка злаков в перикарпии большая
часть СО2 связывается в составе яблочной кислоты (см. ниже).
Источники данных: [12, 18, 24, 38, 44, 48].
б) Фотосинтез в плодах
Развивающиеся плоды преимущественно гетеротрофны;
обычно более 95% всего углерода поступает в них из листьев.
Окисление такого привнесенного углерода в ходе дыхания спо-
собствует образованию предшественников, необходимых для
синтеза органических кислот, в том числе малата (который от-
личается по происхождению от малата, образуемого у С4- или
у, САМ-растений). Потери углерода, связанные с дыханием, от-
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ С,- И СгМЕТАБОЛИЗМА
531
части снижаются за счет фиксации некоторого количества СО2,
выделившейся в ВПФ-цикле.
Исследуя развитие плода у ячменя, Натбим и Дюффус при-
шли к заключению, что обмен веществ в ткани перикарпия по
некоторым признакам напоминает С4-фотосинтез. Если ткань
перикарпия выделяли из соцветий через 30 дней после цвете-
ния (зерно обычно созревает через 60 дней после цветения)
п инкубировали ее иа свету с ИСО2 в течение 1 мин, то в малат
включалось 84% метки. Введение |4СО2 при большей продол-
жительности включения приводило к тому, что метка появля-
лась п в сахарозе. Поскольку в наружных слоях развивающе-
гося плода нет устьиц, часть выдыхаемой СО2 могла улавли-
ваться ФЕПжарбоксилазой, что должно сводить к минимуму
связанные с дыханием потери углерода. Чтобы установить по-
следовательность реакций, входящих в этот путь, необходимо
проследить, как включается метка в промежуточные продукты
п как она распределяется в них. Если углерод из Соположе-
ния малата поступает в С-1-положение ФГК, то, по-видимому,
передача углерода происходит так же, как при С4-фотосинтезе
(разд. 10.2, 12.3). Однако перенос метки из ИСО2 в малат и да-
лее в сахарозу может происходить н без передачи углерода
малата в ВПФ-цикл (например, как при метаболизме ИСО2 в
эндосперме клещевины обыкновенной, см. разд. 16.6).
У томатов, которые относятся к Сз-растеииям, скорость тем-
нового дыхания в зеленых плодах обычно очень высока; ско-
рость дыхания на свету у них гораздо меньше, так как при
этом происходит фиксация СО2. В отделенном от остальной
ткани наружном слое зеленого томата скорость фиксации 14СО2
на свету в 3 раза выше, чем в темноте. Причем в обоих слу-
чаях основная часть метки включается в малат. При этом на-
блюдается достаточно интенсивное включение метки в метабо-
литы ЦТ1< (цитрат) и ВПФ-цикла. Относительное содержание
ФЕП- и РуБФ-карбоксилаз и NADP-малатдегидрогеназы (де-
карбоксилирующей) в наружном слое зеленого плода почти
такое же, как в листьях С4- и САМ-растений. Если внутрь пло-
да ввести радиоактивный малат, то он метаболизируется fl-
ЦТК, а возможно, и декарбоксилируется при участии NADP-
малатдегидрогеназы. При этом часть выделившейся СО2 снова
усваивается в ВПФ-цикле. Поскольку у плодов томатов нет
устьиц, в данном случае фиксация СО2 может служить тем
механизмом, с помощью которого улавливается выдыхаемая
СО2 п сводятся к минимуму потери углерода. Малат, образу-
ющийся в результате такой фиксации, может либо запасаться
внутри вакуоли, либо декарбоксилироваться с последующей
передачей СО2 в ВПФ-цикл, либо включаться в ЦТК, способ-
ствуя сохранению постоянного пула его метаболитов.
34*
532
ГЛАВА 16
Плоды таких бобовых растений, как фасоль и горох, фикси-
руют ИСО2 и па свету, и в темноте. На ранних стадиях раз-
вития плода на свету идет поглощение СО2 из воздуха, а на бо-
лее поздних стадиях свет может подавлять процессы, которые
ведут к потере выдыхаемой СО2 (об этом свидетельствуют дан-
ные, полученные при помощи инфракрасного газового анализа-
тора). Подобно плодам томатов или зернам ячменя, бобы спо-
собны повторно усваивать часть выдыхаемой СО2. Для стенки
боба, которую называют тестой (или, иначе, семенной кожу-
рой), как и для зародыша, характерно значительное содержа-
ние ФЕП- и РуБФ-карбоксилаз; при этом активность ФЕП-
карбоксилазы несколько выше. Вполне возможно, что семенная
кожура наиболее эффективно ассимилирует СО2, причем в се-
менах сои этот процесс идет с большей скоростью, чем в семе-
нах гороха. Флинн и др, предположили, что при выращивании
гороха в условиях искусственного освещения -~20% всего уг-
лерода его плода поставляется за счет фотосинтеза в самом
бобе, В семенной кожуре боба есть два фотосинтетически ак-
тивных слоя: наружный слой, состоящий из хлоренхимы мезо-
карпия, и внутренний слой, состоящий из внутреннего эпидер-
миса эндокарпия (внутриплодника). Хлоропласты мезокарпия
ассимилируют СО2 воздуха, а хлоропласты внутреннего эпи-
дермиса усваивают СО2, выдыхаемую семенем. Самбо ш др.
установили, что количество СО2, фиксируемой в бобах сои, со-
ставляет всего ~4% того углерода, который поступает из
листьев; в то же время в бобах может усваиваться до 50—70%
выдыхаемой СО2. Устьичное сопротивление и сопротивление по-
граничного слоя (разд. 14.6) в бобах примерно в два раза вы-
ше, чем в листьях сои.
В ягодах винограда органические кислоты (особенно ма-
лат) накапливаются в самом начале роста. Затем по мере со-
зревания ягод органические кислоты распадаются и синтези-
руются сахара. В ягодах обнаруживается активность ФЕП-
карбоксилазы, ФЕП-карбоксикиназы и NADP-малатдегпдроге-
назы (декарбоксилирующей). Накопление органических кислот
в этом случае может быть связано с фиксацией СО2 при уча-
стии ФЕП-карбоксилазы (не исключено и участие ФЕП-карб-
оксикиназы) и малатдегидрогеназы. В процессе раскисления
С4-кислоты могут декарбоксилироваться под действием ФЕП-
карбоксикиназы и (или) NADP-малатдегидрогеназы, а образу-
ющиеся при этом Сз-продукты, т. е. соответственно ФЕП и пи-
руват, превращаются в сахара за счет глюконеогенеза. Для
того чтобы превратить пируват—-продукт декарбоксилирова-
ния под действием NADP-малатдегидрогеназы — в сахар, нуж-
но, чтобы под действием .пируват,ортофосфат — дикиназы об-
разовался ФЕП (разд. 15.4, 15.10). Не ясно, присутствует ли
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И С4-МЕТАБОЛИЗМА
533
Сахароза
Рис. 16.6. Некоторые из возможных путей синтеза и метаболизма малата в
плодах.
этот фермент в ягодах винограда, но он был обнаружен в пе-
рикарпии зерен ячменя. Вполне возможно, что закисление и
раскисление в ягодах идет так же, как у С4- или САМ-расте-
ний; в этих процессах принимает участие один и тот же набор
ферментов. По-видимому, эффективность фиксации СО2 в ВПФ-
цикле, которая должна следовать за декарбоксилированием
С4-кислот, в ягодах гораздо ниже, чем уСг или САМ-растений.
Для многих плодов характерна относительно высокая ско-
рость дыхания. Поступивший в них углерод в составе сахаров
и аминокислот может затем ассимилироваться в основном
в форме липидов, белков и олигосахаридов, которые образу-
ются с использованием энергии, выделяющейся в процессе ды-
хания. Для функционирования ЦТК необходим непрерывный
синтез оксалоацетата, который взаимодействует с ацетил-СоА.
Если нет потерь метаболитов ЦТД в каждом обороте этого
цикла, то оксалоацетат будет постоянно регенерироваться:
Ацетил-СоАД-Оксалоацетат—-> 2СО2Оксалоацетат (16.1)
534
ГЛАВА 16
Вместе с тем метаболиты этого цикла используются и в про-
цессах биосинтеза, поэтому для постоянного функционирования
ЦТК необходим источник С4-кислот. Таким источником может
служить малат, который синтезируется из С3-предшественника.
Некоторые плоды кислы по самой своей природе, так что на
какой-то из стадий их развития, по-видимому, синтезируются и
запасаются малат и другие органические кислоты. У плодов,
в которых запасается большое количество лимонной кислоты
(например, в лимонах, томатах), можно ожидать интенсивный
синтез таких С4-кислот, как оксалоацетат или малат. Для син-
теза цитрата в ЦТК необходимо поступление в цикл эквивалент-
ного количества пирувата и С4-кислоты. Во время гликолиза
ФЕП может превращаться в пируват за счет синтеза С4-кис-
лот и их декарбоксилирования, что, по-видимому, необходимо
для превращения NADH и NADPH (разд. 16.4). В зеленых
плодах фотосинтез является вторичным процессом, в ходе ко-
торого может реасснмилироваться некоторая часть выдыхае-
мой СО2 или СО2, образующейся после декарбоксилирования
малата. В такой реассимиляции СО2 участвует ВПФ-цикл
(рис. 16.6).
Источники данных: [2, 3, 9, 13—16, 20, 21, 29, 35, 36, 45].
16.3. Зеленеющие листья
Зеленые листья в темноте, а этиолированные листья и в
темноте, и на свету фиксируют 14СО2 главным образом в виде
органических кислот; при этом значительная часть метки вклю-
чается в малат. Тамас и др. обнаружили, что главными про-
дуктами ассимиляции 14СО2 в выращенных в темноте листьях
ячменя (Сз-растение) являются малат, аспартат, глутамат и
цитрат. Такое включение метки наблюдалось и в темноте, и на
свету. После освещения ткани листа в течение 2—4 ч синтез
малата, аспартата и глутамата в листе резко возрастал, а пер-
вичным и основным продуктом ассимиляции был малат. Син-
тез всех этих органических кислот может быть очень важен
для дыхательного метаболизма и для образования белков и
липидов в процессе развития хлоропластов. После предвари-
тельной экспозиции на свету в течение 6 ч главным продуктом
фиксации 14СО2 в ткани листьев, выращенных в темноте, ста-
новится сахароза, точно так же, как и в зрелых листьях Сз-
растений.
16.4. Корни
В корнях малат является таким же метаболитом ЦТК, как
и в других тканях. После образования из Сз-предшественников
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И СгМЕТАБОЛИЗМА
536
малат может входить в ЦТК дополнительно к тому пирувату,
который поглощается митохондриями,
Тинг и Даггер предположили, что синтез малата и его де-
карбоксилирование в ткани корня приводят к тем же резуль-
татам, что и трансгидрогеназные реакции, обеспечивающие об-
разование NADPH, когда он необходим для биосинтетических
процессов, NADH, образовавшийся во время гликолиза, может
превратиться в NADPH в результате сопряжения реакций, ка-
тализируемых NAD-малатдегидрогеназой и NADP-малатде-
гидрогеназой (декарбоксилирующей). Для этого требуется од-
на молекула АТР, которая образуется в процессе гликолиза
при участии пируваткииазы (рис. 16.7).
Одна из гипотез, объясняющих устойчивость растений к за-
топлению (к умеренному анаэробиозу), связывает это яв-
ление с наличием ФЕП-карбоксилазы и с синтезом малата. Во
время гликолиза в анаэробных условиях синтезируется этанол,
который, накапливаясь в больших количествах, становится ток-
сичным для ткани корня. Синтез этанола, который катализиру-
ет алкогольдегидрогеназа, идет за счет избыточной восстанови-
тельной силы, в результате чего гликолиз ие прекращается и
•образуется АТР. Было высказано предположение, что у рас-
Рис. 16.7. Синтез малата и его де-
карбоксилирование в процессе глико-
лиза приводит к такому же резуль-
тату, что и реакция, катализируемая
трансгидрогеназой, которая превра-
щает NADH в NADPH (Ting, Dug-
ger, 1965;. 1967). :.r'
Рис. 16.8. Одна из гипотез, согласно
которой устойчивость растений к за-
топлению связана с синтезом малата,-
536
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И С,-МЕТАБОЛИЗМА
тений, устойчивых к затоплению, ФЕП, который образуется в
ходе гликолиза, используется ФЕП-карбоксилазой, а образую-
щийся оксалоацетат превращается в малат под действием ма-
латдегидрогеназы. Это должно влиять и на циклические пре-
вращения пиридиновых нуклеотидов, и на синтез этанола (рис.
16.8), в результате чего может происходить переключение об-
мена веществ, обеспечивающее накопление коферментов. Пони-
жение уровня малатдегидрогеназы (декарбоксилирующей) у
устойчивых к затоплению видов растений способствует накоп-
лению малата.
Было высказано предположение, что длительное затопление
приводит к некоторому увеличению концентрации малата в
корнях устойчивых к затоплению растений. Но результаты опы-
тов Авадхани и др., исследовавших устойчивые к затоплению
проростки риса, ие подтверждают это предположение. Экспо-
зиция таких проростков в среде с низкой '[О2] приводила лишь
к незначительному увеличению содержания малата, а концент-
рация этанола в ткани после однодневной инкубации при пони-
женной концентрации О2 становилась в 20—30 раз выше кон-
центрации малата.
Источники данных: [4, 5, 23, 25, 40, 41].
16.5. Ионный баланс и поддержание постоянного pH
Содержание неорганических катионов в растениях обычно
намного превышает уровень неорганических анионов. Электро-
химическое равновесие достигается за счет органических анио-
нов и во многих случаях за счет аниона малата. Такую зависи-
мость можно описать уравнением С—A=Y, где С — содержа-
ние катионов, выраженное в миллиэквивалеитах (например, К+,
Na+, Са2+, M.g2+), А — содержание неорганических анионов,
выраженное также в миллиэквивалентах (например, ЫОз~,
НРО42~, SO42- и С1~), Y — число миллиэквивалентов органи-
ческих анионов. Для каждого вида растения Y — довольно по-
стоянная величина. Типичная величина Y, выраженная в
мэкв/кг сухого вещества, для трав составляет 1000, для бо-
бовых, не имеющих клубеньков, она равна 2000, а для сахарной
свеклы 4000. Лумис и Геракис обратили внимание на то, что
благодаря большой потребности в синтезе органических анио-
нов растения имеют возможность удалять вместе с ними боль-
шое количество собственных отходов жизнедеятельности. Так,
например, поглощение калия корнями, вызванное накоплением
избыточного количества неорганических анионов, может по-
влечь за собой обмен К+—Н+. В процессе синтеза яблочной
кислоты могут возникать протоны, которые обмениваются на
калий и выходят из корней, поэтому в ткани образуется малат
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сз- И Сз-МЕТАБОЛИЗМА
53/
калия. В таких случаях стехио-
метрический синтез малата мо-
жет компенсировать недостаточ-
ное поглощение анионов.
Было высказано предположе-
ние о том, что и в корнях, и в
листьях поддержание строгого
баланса между синтезом и рас-
падом малата необходимо для
того, чтобы контролировать ве-
личину pH в клетках. Метаболи-
ческие превращения или погло-
щение метаболитов, в результа-
те которых защелачивается со-
держимое клеток, могут компен-
сироваться за счет синтеза ма-
лата, в котором участвуют ФЕП-
карбоксилаза и NAD-малатде-
гидрогеназа. Закисление содер-
жимого клеток может компенси-
ФЕП
NADH
NAD
▼
Яблочная ------у,
кислота
Малат
N AD(P)^
NAD(P)H
Пируват + СО;
Рис. 16.9. Потенциальная спо-
собность к синтезу малата и
к его декарбоксилированию
обеспечивает регуляцию pH в
клетках.
роваться с помощью декарбоксилирования малата под дейст-
вием NAD- или NADP-малатдегидрогеназы (декарбоксилиру-
ющих) (рис. 16.9). При расссмотреиии этой гипотезы следует
учитывать компартментацию ферментов в клетке и то, в какой
степени pH может контролировать ферментативные реакции,
лежащие в основе закисления или раскисления.
Синтез органических кислот в клетках растений большей
частью связан с восстановлением нитрата. Когда нитрат пре-
вращается в конечном итоге в белковый азот, возникает ион
гидроксила, который может нейтрализоваться за счет образо-
вания малата.
NO.,’' - NADH -[- Н+ ----------->- NO2" -[- NAD+ -J- НаО
NO2" + 6Н+ бе" ---->• NHS + Н2О + ОН"
(16-2)
Если корни поглощают KNO3, который затем транспортируется
в листья, где нитрат превращается в аминный азот, то при
этом образуется КОН. Синтез яблочной кислоты приводит к
тому, что образуется малат калия, который может храниться в
вакуолях. В этом случае количество синтезируемого малата
должно быть эквивалентно количеству нитрата, которое в ходе
ассимиляции превратилось в азот аминогруппы. У некоторых
растений главную роль в компенсации защелачивания, проис-
ходящего в процессе восстановления нитрата (рис. 16.10), мо-
гут играть другие органические анионы (например, оксалат).
Вместо того чтобы сохраниться в тканях листа, малат ка-
лия может транспортироваться в корни растения и там де-
638
ГЛАВА 16
Рис. 16.10. Защелачивание, связанное с восстановлением нитрата, может
быть нейтрализовано за счет синтеза яблочной кислоты.
карбоксилироваться при участии малатдегидрогеназы. При этом
образуется НСО3~, который может выходить из корней наружу
в обмен на NO3~, поглощаемый корнями. Из НСО3~ образуется
свободная СО2, а это в свою очередь ведет к появлению ионов
гидроксила. Суммарный эффект всех этих превращений заклю-
чается в том, что ионы гидроксила переносятся из листьев в-
почву (рис. 16.11).
Источники данных: [8, 10, 19, 22, 26, 33, 34, 42] .
16.6. Метаболизм малата в глиоксилатном цикле
проростков
Малат является промежуточным продуктом глиоксилатного
цикла, который связан с метаболическими превращениями ли-
пидов в сахарозу в ткани эндосперма (например, в эндоспер-
ме клещевины) или в семядолях проростков. Он синтезируется
при участии малатсинтетазы глиоксилатного цикла:
Ацетил-СоА Ч-Тлиоксилат->• Малат-|-СоА (малатсинтетаза) (16.3}
Затем малат используется в качестве предшественника для
синтеза сахарозы. В этом процессе участвуют малатдегидроге-
наза и ФЕП-карбоксикиназа, а в конечном итоге образуется
ФЕП.
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ С.г И СгМЕТАБОЛИЗМА
539
Рис. 16.11. Связанный с поглощением нитрата С4-ЦНКЛ, который функциони-
рует между листьями и корнями.
В ткани эндосперма клещевины обнаружены активности
ФЕП-карбоксикиназы, ФЕП-карбоксилазы и РуБФ-карбокси-
лазы. Это единственная не содержащая хлорофилла ткань рас-
тения, в которой обнаружена активность РуБФ-карбоксилазы.
Активность этого фермента возрастает в процессе прорастания
и достигает максимальной величины через четыре дня. Точно
так же возрастает активность малатсинтетазы и изоцитратлиа-
зы глиоксилатиого цикла. Вместе с тем ВПФ-цикл, по-видимо-
му, ие имеет особого значения для ткани эндосперма.
Если в ткань эндосперма вводили В * * * * * 14СОг, то метка включа-
лась в малат, а затем и в сахарозу. Малат может включать
метку либо за счет ФЕП-карбоксикиназной реакции (возможна
или реакция карбоксилирования, или обмен ФЕП+14СО2+
4-АПР=₽=14С-Оксалоацетат4-АТР), либо за счет сопряженных
реакций, катализируемых ФЕП-карбоксилазой и малатдегид-
рогеназой. Большая часть метки в гексозном остатке сахаро-
зы находится в С-3- и в С-4-положеииях. Это можно объяснить
тем, что метка, включившаяся в малат, рандомизуется и рас-
пределяется между С-1- и С-4-атомами в результате фумараз-
ной реакции (разд. 15.8). Впоследствии декарбоксилирование,
катализируемое карбоксикиназой, приводит к появлению мет-
540
ГЛАВА 16
ки в углероде карбоксильной группы ФЕП, а затем в резуль-
тате глюконеогенеза в С-3- и С-4-положении гексозы. Если бы
значительная часть СО2 фиксировалась в ВПФ-цикле либо сра-
зу же, либо после декарбоксилирования Сгкислот, то должны
были бы равномерно метиться все метаболиты этого цикла, что-
в конечном итоге приводило бы к рандомизации метки, вклю-
чившейся в сахарозу. Этого, однако, ие наблюдается, и поэто-
му можно предположить, что сахароза синтезируется скорее
всего не за счет ВПФ-цикла, а из Сз-продукта декарбоксилиро-
вания малата.
Источники данных: [5, 6, 7, 26].
СПЕЦИАЛЬНАЯ ЛИТЕРАТУРА
1. Allaway №. G. Accumulation of malate in guard cells of Vicia faba during,
stomatai opening, Planta, 110, 63—70 (1973).
2. Andrews A. K., Svec L. V. Photosynthetic activity of soybean pods at dif-
ferent growth stages compared to leaves, Canad. J, Plant Sci., 55, 501—
505 (1975).
3. Atkins C. A., Kuo I., Pate I. S., Flinn A. M„ Steele T. №. The photosynthe-
tic pod wall of peas (Pisum sativum L.): Distribution of CO2 fixing enzy-
mes in relation to pod structure, Plant Physiol., 60, 779—786 (1977).
4. Avadhani P. N., Greenway H., Lefroy R., Prior L. Alcoholic fermentation
and malate metabolism in rice germination at low oxygen concentrations,.
Aust. J. Plant Physiol., 5, 15—25 (1978).
5. Beevers H. Respiratory Metabolism in Plants. 232 pp. White Plains, New
York, Row, Peterson and Co., 1961.
6. Benedict C. R. The presence of ribulose 1,5-bisphosphate carboxylase in
the non-photosynthetic endosperm of germinating castor bean, Plant Phy-
siol., 51, 755—759 (1973).
7. Benedict C. R., Beevers H. Formation of sucrose from malate to phospho-
enolpyruvate, Plant Physiol., 36, 540—544 (1961).
8. Ben-Zioni A., Lips S. H., Vaadia У. Correlations between nitrate reduction,
protein synthesis and malate accumulation, Plant Physiol., 23, 1039—1047
(1970).
9. Crookston R. K„ O'Toole J., Ozbun J. L. Characterization of the bean pod
as a photosynthetic organ, Crop. Sci., 14, 708—712 (1974).
10. Davies D. D. Control of and by pH, In: Symp. Soc. Exp. Biol., 27, 513—
529 (11973).
11. Dittrich P., Raschke K. Malate metabolism in isolated epidermis of Com-
melina communis L. in relation to stomatai functioning, Planta, 134, 77—81
(1977).
12. Enyl В. A. C. The contribution of different organs to grain weight in
upland swamp rice, Ann. Bot., 26 , 529—531 (1962).
13. Farineau J., Laval-Martin D. Light versus dark carbon metabolism in cherry
tomato fruits. II. Relationship between malate metabolism and photosynthe-
tic activity, Plant Physiol., 60, 877—880 (1977).
14. Flinn A. M., Atkins C. A., Pate J. S. The significance of photosynthetic and
respiratory exchanges in the carbon economy of the developing pea fruit.
Plant Physiol., 60, 412—418 (1977).
15. Harvey D. M., Hedley C. L., Keely R. J. Photosynthetic and respiratory
studies during pod and seed development in Pisum sativum L, Ann. Bot.,
40, 993—1001 (1976).
СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ Сг И СгМЕТАБОЛИЗМА
541
16. Hedley С. L., Harvey D. М., Reely R. J. Role of PEP carboxylase during
seed development in Pisum sativum, Nature, 258, 352—354 (1975).
17. Humble G. D., Raschke /(. Stomatai opening quantitatively related to potas-
sium transport. Evidence from electron probe analysis, Plant Physiol., 48,
447—453 (1971).
18. Jennings V. M„ Shibles R. M. Genotypic differences in photosynthetic con-
tributions of plant parts in grain yield in oats, Crop Set, 8, 173—175
(1968).
19. Rirkby E. A., Mongol K. Ionic balance in different tissues of tomato plant
in relation to nitrate, urea, or ammonium nutrition, Plant Physiol., 42,
6-14 (1967).
20. Lakso A. N., Kliewar IF. AL The influence of temperature on malic acid
metabolism in grape berries, Plant Physiol., 56, 370—372 (1975).
21. Laval-Martin D., Farineau J., Diamond J. Light versus dark carbon metabo-
lism in cherry tomato fruit. I. Occurence of photosynthesis. Study of the
intermediates, Plant Physiol., 60, 872—876 (1977).
22. Loomis R. S., Gerakis P. A Productivity of agricultural ecosystems. In:
Photosynthesis and Productivity in Different Environments (ed. J. P. Coo-
per), pp. 145—172, Cambridge, Cambridge University Press, 1975.
23. Me Manmon M., Crawford R. M. M. A metabolic theory of flooding tole-
rance: the significance of enzyme distribution and behaviour, New Phytol.,
70, 299—306 (1971)..
24. Nutbeam A. R„ Duffus С. M. Evidence for C4 photosynthesis in barley peri-
carp tissue, Biochem. Biophys. Res, Cominun., 70, 1198—1203 (1976).
25. Osmond С. B. Ion absorption and carbon metabolism. In: Encyclopedia
of Plant Physiology, Vol. II. New series. Transport in Plants (eds. U. Luttge
and M. G. Pitman), pp. 347—372, New York, Springer-Verlag, 1976.
26. Osmond С. B., Akazawa T., Beevers H. Localization and properties of ribu-
lose diphosphate carboxylase from castor bean endosperm, Plant Physiol.,
55, 226—230 (1975).
27. Outlaw IF. H., Manchester J„ DiCamellia C. A., Randall D. D., Rapp B„
and Vcilh G. M. Photosynthetic carbon reduction pathway is absent in chlo-
roplasts of Vicia faba guard cells, Proc. Natl. Acad. Sci. USA, 76, 6371—
6375 (1979).
28. Pearson C. J. Daily changes in stomatai aperture and in carbohydrates and
malate within epidermis and mesophyll of leaves of Commellna cyanea and'
Vicia faba, Aust. J. Biol. Sci., 26, 1035—1044 (1973).
29. Quebedeaux B., Chollet R. Growth and development of soybean (Glycine-
max (L.) Merr.) pods. CO2 exchange and enzyme studies, Plant Physiol.,,
55, 745—748 (1975).
30. Raghavendra Л. S„ Das V. S. R. Control of stomatai opening by cyclic pho-
tophosphorylation, Curr. Sci., 41, 150—151 (1972).
31. Raschke /(. Stomatai action, Ann. Rev. Plant Physiol., 26, 309—340 (1975).
32. Raschke R„ Dittrich R. 14C carbon-dioxide fixation by isolated leaf epider-
mis with stomata closed or open, Planta, 134, 69—75 (1977).
33. Raven J. A., Smith F. A. Nitrogen assimilation and transport in vascular-
land plants in relation to intracellular pH regulation, New Phytol., 76,.
415—431 (1976).
34. Raven J. A, Smith F. A Cytoplasmic pH regulation and electrogenic H+
extrusion, Curr. Adv. Plant Sci., 24, 649—660 (1976).
35. Ruffner H. P., Kliewer W. M. Phosphoenolpyruvate carboxykinase activity-
in grape berries, Plant Physiol., 56, 67—71 (1975).
36. Sambo E. Y., Moorby J., Milthorpe F. L. Photosynthesis and respiration-
of developing soybean pods, Aust. J. Plant Physiol., 44, 713—721 (1977).
37. Tamas I. A, Yemm E. W„ Bidwell R. G. S. The development of photosyn-
thesis in dark-grown barley leaves upon illumination, Canad. J. Bot., 48,.
2313—2317 (1970).
S42
ГЛАВА lb
38. Thorne G. N. Photosynthesis in ear and flag leaf in wheat and barley, Ann.
Bot, 29, 317—329 (1965).
39. Thorpe N., Milthorpe F. L. Stomatai metabolism. CO2 fixation and respira-
tion, Aust. J. Plant Physiol., 4, 611—621 (1977).
40. Ting !. P., Dugger W. M. Trans hydrogenation in root tissue: mediation
by carbon dioxide, Science, 150, 1727—1728 (1965).
41. Ting !. P., Dugger W. M., Jr. CO2 metabolism in corn roots. I. Kinetics of
carboxylation and decarboxylation, Plant Physiol., 42, 712—718 (1967).
42. Ulrich A. Metabolism of non-volatile organic acids in excised barley roots
as related to cation-anion balance during salt accumulation, Amer. J. Bot.,
28, 526—537 (1941).
43. Walker D. A. Pyruvate carboxylation and plant metabolism, Biol. Rev., 37,
215—256 (1962).
44. Walpole P. R., Morgan D. G. Physiology of grain filling in barley, Nature,
240,416—417 (1972).
45. Willmer С. M., Johnston IF. R. Carbon dioxide assimilation in some aerial
plant organs and tissues, Planta, 130, 33—37 (1976).
46. Willmer С. M., Kanai R., Pallas J. E. Jr., Black С. C. Detection of high levels
of phosphoenolpyruvate carboxylase in leaf epidermal tissue and its signi-
ficance in stomatai movements, Life Sci., 12, 151—155 (1973).
47. Willmer С. M., Pallas J. E., Black С. C. Carbon dioxide metabolism in leaf
epidermal tissue, Plant Physiol., 52, 448—452 (1973).
48. Wirth E„ Kelly G. J., Fischbeck G., Latzko E. Enzyme activities and pro-
ducts of CO2 fixation in various photosynthetic organs of wheat and oat,
Z. Pflanzenphysiol., 82, 78—87 (1977).
ПРИЛОЖЕНИЕ А
Выделение хлоропластов
и критерии их интактности
АЛ. Введение
Для того чтобы исследовать метаболизм углерода, транс-
порт метаболитов в хлоропластах и компартмеитацию фермен-
тов в клетке (чему посвящена большая часть этой книги), не-
обходимо прежде всего получить изолированные хлоропласты,
которые при этом должны быть интактными, в достаточной
степени очищенными от других субклеточных структур и био-
химически активными. В некоторых случаях требуется очень
высокая степень очистки хлоропластов (например, для уста-
новления локализации тех ферментов, которые присутствуют
только в очень небольших количествах), поэтому может ока-
заться, что гораздо важнее освободиться от каких бы то ни
было примесей и загрязнений, чем получить хлоропласты, спо-
собные осуществлять фотосинтез с очень высокой скоростью.
При других обстоятельствах можно поступиться степенью очи-
стки ради сохранения максимального уровня активности, но'
наличие примеси цитоплазматических ферментов в препарате
может очень усложнить интерпретацию полученных результа-
тов (гл. 8). К тому же важно ие забывать два обстоятельства,
которые часто упускают из виду, несмотря на то что это само'
собой разумеется. Во-первых, абсолютно невозможно получить
«хорошие» хлоропласты из «плохого» исходного материала
(см. разд. А.З). Во-вторых, «хорошие» хлоропласты будут ра-
ботать хорошо только тогда, когда с ними будут правильно об-
ращаться. Если оглянуться назад, то станет совершенно ясно,
что после того, как в начале 60-х годов в практику были снова
введены приемы Хилла, использовавшего в качестве осмотиче-
ски активного вещества сахара, в плане улучшения методики
выделения хлоропластов было сделано очень мало, если, ко-
нечно, не считать предварительного получения протопластов
(см. разд. А.5). Все последующие улучшения методики, спо-
собствующие получению более активных хлоропластов, каса-
лись главным образом методики определения активности, и в
частности были связаны с введением в среду неорганического
пирофосфата (разд. 8.14).
Методика выделения хлоропластов имеет решающее значе-
ние для получения достоверных результатов. В настоящее вре-
544
ПРИЛОЖЕНИЕ А
мя исследователи разработали относительно эффективные ме-
тоды только для некоторых Сз- и Сграстений. Выделить же
функционально активные хлоропласты из САМ-растений прак-
тически не удается. Это может быть связано с повышенной
кислотностью фотосинтезирующей ткани у этих растений и с
наличием в ней таннинов или других веществ, находящихся
внутри вакуоли, которые могут пагубно влиять на хлоропласты
с самых первых этапов их выделения.
Сейчас принято считать, что хлоропласта являются тем ме-
стом, где СО2 фиксируется при участии ВПФ-цикла (см.
гл. 8). Это относится ко всем хлоропластам Сз- и САМ-расте-
ний и к хлоропластам клеток обкладки проводящих пучков у
Сграстений. В хлоропластах мезофилла Сграстений ВПФ-цикл
отсутствует. В присутствии пирувата и ФЕП-карбоксилазы ци-
тозоля на свету в этих хлоропластах протекает карбоксилиру-
ющая стадия Ci-цикла (гл. 12).
Для оценки эффективности фотосинтеза в изолированных
хлоропластах разумнее всего сравнить ее с эффективностью
фотосинтеза в той ткани, откуда были выделены эти органел-
лы. Эти данные приобретут наибольший смысл, когда все из-
мерения будут проведены на исходной ткани растения (опре-
деление обмена СО2 при помощи инфракрасного газоанализа-
тора) и на препарате хлоропластов, выделенных из той же са-
мой ткани (определение фиксации ИСО2 или измерение выде-
ления О2 при помощи кислородного электрода), при одинако-
вых освещении, температуре и концентрации СО2. Совершенно
•непрактично делать такие сравнения при абсолютно строгом
режиме или ежедневно. Для общей оценки эффективности ра-
боты изолированных хлоропластов можно воспользоваться ус-
редненными значениями интенсивности фотосинтеза в ткани
.листа различных растений (табл. А.1).
Таблица А.1. Типичная скорость фотосинтеза у трех групп
фотосинтезирующих растений при оптимальных условиях
Группа фотосинтезирую- щих растений Количество фиксированной СО2. мкмоль«(мг Хл)-’’Ч-Н Температура, °C
Сз-растения 100-200 (200—400)1> 15-25
С4-растения 200—400 20-35
САМ-растения 30-80 15-35
') Скорость фотосинтеза в атмосфере, насыщенной COs, у Сз’растений будет выше,
чем при нормальной концентрации СО2 в воздухе. Поэтому интенсивность фотосинтеза
в хлоропластах при насыщающей концентрации СО2 следует сравнивать с интенсивностью
фотосинтеза в исходной ткани в условиях насыщения СО2.
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
545
Отсюда сразу видно, что приводившиеся в прошлом в неко-
торых работах цифры, согласно которым скорость фиксации
СОз в изолированных хлоропластах САМ-растений была равна
1—5, а в хлоропластах Сз- и (Д-растений 10—20 мкмоль-
- (мгХл)-1-ч-1, явно не соответствуют истине. Пока ие получе-
ны хлоропласты с достаточно высокой интенсивностью фотосин-
теза, их метаболизм в условиях in vitro никак нельзя сравни-
вать с тем, что на самом деле происходит в исходной ткани.
Это наглядно видно иа примере потребности в Pi (гл. 8). Ско-
рость фотосинтеза в хлоропластах Сз-растений в отсутствие Pi
очень невелика. Обнаружение того факта, что Pi необходим
для хлоропластов, внесло существенный вклад в наши пред-
ставления о том, каков же механизм фотосинтеза in vivo.
А.2. Методы
В общем все методы, которые применяют для выделения
хлоропластов, основаны либо иа механическом разрушении
самой ткани листа, либо на разрушении в щадящих условиях
тех протопластов, которые были предварительно из нее полу-
чены. Такой прием был разработан лишь недавно, и в этом
случае для получения протопластов используют ферменты
(целлюлаза + пектиназа), которые растворяют клеточную стей-
ку клеток мезофилла. Для того чтобы получить интактные ор-
ганеллы, протопласты осторожно разрушают в щадящих усло-
виях.
А.З. Выращивание растений
Это, пожалуй, первый и чрезвычайно важный фактор, ко-
торый определяет возможность получения функционально ак-
тивных хлоропластов. Вполне естественно, что условия, необ-
ходимые для оптимального роста различных растений, неоди-
наковы. В большинстве случаев различная интенсивность фото-
синтеза (и противоречивость сделанных выводов) у одного и
того же растения может быть связана с различиями исходно-
го растительного материала у разных исследователей. Так, на-
пример, для того чтобы получить хорошие хлоропласты шпина-
та, нужно обратить особое внимание на продолжительность
фотопериода и на условия питания растения во время его вы-
ращивания. Когда .хлоропласты выделяют из протопластов, воз-
никает еще одно обстоятельство. При определенных условиях
выращивания ткань листа может становиться очень устойчивой
к действию ферментов, которые переваривают клеточную стен-
ку. При получении протопластов из листьев табака или томатов
следует обращать особое внимание на такие факторы, как ос-
35—234
546
ПРИЛОЖЕНИЕ А
вещенность, влажность и условия питания. А для злаков наи-
больших успехов в получении протопластов удалось достичь
путем использования относительно молодой ткани листа
(в возрасте от одной до трех недель в зависимости от вида).
Исходный материал
Для многих целей до сих пор предпочитают использовать
шпинат. Это связано, во-первых, с. тем, что в этом растении
практически отсутствуют фенольные соединения, которые обыч-
но загрязняют получаемые препараты, и, во-вторых, с тем, что
исследованию хлоропластов именно этого растения посвящено
громадное число работ. Еще совсем недавно шпинат был един-
ственным растением (за исключением, пожалуй, молодых
листьев гороха), из которого удавалось получать действитель-
но интактные и активные хлоропласты. Для некоторых целей
очень удобными оказались растения гороха, хотя с каждым го-
дом становится все яснее, что хлоропласты, выделенные из мо-
лодых побегов гороха, отличаются по проницаемости от хлоро-
пластов, полученных из взрослых растений шпината. Аврон и
др. очень широко использовали в своих исследованиях салат,
ио главным образом для изучения транспорта электронов и фо-
тофосфорилирования. Приемы получения протопластов (разд.
А.5) позволили выделить «хорошие» хлоропласты и из многих
других растений.
Культивирование
Хотя огородный шпинат (Spinacia oleracea) используется
почти повсеместно, его очень трудно вырастить. В некоторых
местах Северной Америки, в Австралии, Индии и Южной Евг
ропе это растение можно вполне успешно выращивать в откры-
том грунте даже в самой середине лета, но в более северных
широтах (например, в Англии), где продолжительность дня
летом гораздо больше, шпинат практически не образует листь-
ев до самого цветения. Поэтому для экспериментальных целей
его обычно выращивают в таких условиях, когда продолжи-
тельность дня не превышает 8,5 ч. Если нет никаких особых
устройств, позволяющих ограничивать длину дня, продолжи-
тельность сезона выращивания можно чуть-чуть увеличить за
счет использования длиннодневных сортов шпината. Вряд ли
можно рекомендовать какой-то один сорт, который бы хорошо
рос в самых разных условиях окружающей среды. Во многих
случаях вполне годится сорт U. S. Hybrid 424 (марка компа-
нии Ferry-Morse Seed Со. P.O.Box 100, Mountain View, Cali-
fornia 94042), который продается также под названием НуЬ-
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
547
Таблица А,2. Питательный раствор для водной культуры шпината
Номер sPacTBop Концептрацдя исход- ного раствора Объем на 20 л пита- тельного раствора, приготовленного на ди- стиллированной воде, мл
1 KNO3 1М 120
2 Са (NOa)a 1М 80
3 MgSO4 1М 40
4 KH2PO4 1М 20
5 MgCl2 1М 80
6 Микроэлементы (В, Мп, Z11, Си, Мо) 20
7 Na.Fe-ЭДТА (3,86 г на 250 мл) 20
Микроэлементы
Количество в 2S0 мл Н2О, мг
Н3ВО3
М.пС12'4Н2О
ZnSO4-7H2O
CuSO4-5H2O
NaMoO4-2H2O
715
452
55
20
7,25
rid 102 Yates (244-254 Horsley Road, Milperra, N.S.W. 2214,
Australia или The Seed Centre, Withyfold Drive, Macclesfield,
Cheshire, United Kingdom). Так, например, Ульриху Хеберу
удалось получить максимальное из когда-либо зарегистриро-
ванных значение интенсивности ассимиляции 1 углерода
(^350 мкмоль-(мгХл^'-ч-1 при 20°С), взяв для опыта рас-
тения шпината Yates 102, которые были выращены, в Канберре
в условиях водной культуры. Значительного улучшения резуль-
татов добились и в Шеффилде (Англия), когда тот же самый
сорт шпината вырастили в той же самой среде. Обычно ско-
рость фотосинтеза при этом превышала 100 мкмоль-(мгХл)-1-
•ч-1, а иногда и 200 мкмоль- (мгХл)-1-ч-1. Такую интенсивность
фотосинтеза в принципе можно получить и в опытах со шпи-
натом, выращенным в открытом грунте, однако нет никаких
сомнений в том, что во многих случаях не стоит жалеть тех
затрат, которые связаны с использованием водной культуры’,
Стандартная питательная смесь, которую сейчас применяют в
Шеффилде (табл. А.2), разработана иа основе той смеси, ко-
торую использовали в Канберре. Для подачи питательного рас-
твора обычно применяют одни из двух возможных способов.
35*
548
ПРИЛОЖЕНИЕ. А
Либо корни растений погружают в плоские неглубокие поддо-
ны, через которые медленно протекает аэрированный питатель-
ный раствор, либо растения выращивают в пластмассовых
горшках, заполненных вермикулитом (вспененная в результате
нагрева слюда) или смесью вермикулита и торфа, в которые
снизу периодически поступает питательный' раствор. Семена
проращивают на вермикулите, смоченном питательным раство-
ром, а когда семядоли полностью расправятся, проростки пере-
носят на. пробки из вспененного полиуретана. Пробки затем
вставляют в отверстия (диаметром 3,5 см), прорезанные в
крышке, которая накрывает продуваемые воздухом сосуды ак-
вариумного типа. В каждом из таких резервуаров можно раз-
местить от 12 до 20 растений. После того как на растениях об-
разуются листья размером 2—3 см, растения переносят в бо-
лее крупные (хотя и менее глубокие) резервуары или в обыч-
ные горшки. В летние месяцы продолжительность дня сокра-
щают до 8,5 ч. Для этого применяют автоматически открываю-
щиеся и.закрывающиеся жалюзи, которые затемняют растения
на рассвете и на закате дня. Зимой продолжительность дня
увеличивают, освещая растения лампами Wotan (на основе га-
логенида металла с диспрозием) мощностью 400 Вт, помещен-
ными на расстоянии 1 м, что дает приблизительно 50 Вт-м~2
фотосинтетически активного освещения в центре освещаемой
поверхности. |[До сих пор нам не удавалось вырастить шпинат,
из которого можно было бы получить действительно активные
хлоропласты типа А (разд. А.7), если использовать для осве-
щения только люминесцентные лампы.].
При максимуме прямой инсоляции высокая температура,
по-видимому, не подавляет рост растений, и • действительно,
иногда самый лучший материал удается получить при выра-
щивании растений под стеклом, когда температура на солнце
(летом) доходит до 50 °C. При скудном зимнем освещении тем-,
пература, при которой выращивают растения, не должна пре-,
вышать 25 °C.
Горох
Семена замачивают на ночь в воде, через которую продува-
ют воздух, и затем проращивают их в вермикулите в плоских
поддонах при плотности засева 125 г (сырого вещества семян)
на один поддон (размером 38X23X8 см). И в этом случае,
когда освещенность недостаточно высока, следует избегать по-
вышения температуры, но при относительно сильном освеще-
нии (свыше 50 Вт-м-2), температуре 20—25°C и фотоперио-
де 9—12 ч хороший материал можно получить уже через 9—
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
549
11 сут, т. е. тогда, когда высота побегов такого сорта, как Про-
гресс № 9 (Suttons Ltd., London Road, Earley, Reading, Uni-
ted Kingdom), достигнет 3—4 см.
A.4. Приемы механического разрушения
К таким приемам относятся прежде всего разрушение ис-
ходной ткани (обычно в ножевом блендере), когда всячески
стараются не повредить хлоропласты. Поэтому гомогенизацию
проводят очень недолго (5 с или даже меньше), заведомо зная,
что разрушение клеток будет недостаточно полным н выход
хлоропластов не превысит 1%. Несомненный выигрыш, по-вп-
днмому, может дать быстрое выделение органелл из гомогена-
та, поэтому хлоропласты обычно быстро отделяют (путем цент-
рифугирования) от той среды, в которой разрушали клетки, в
течение 2—3 мин после гомогенизации.
Сред, которые применяют для разрушения, очень много;
они различаются по составу, ио общим для всех сред является
то, что все они содержат осмотически активное вещество (разд.
8.10). Обычно это сахар (глюкоза или сахароза) или много-
атомный спирт (типа сорбитола или маннитола) в концентра-
ции 0,33 М. Кроме того, в среде обычно присутствует буфер,
который необходим для поддержания определенной концентра-
ции. ионов водорода (очень часто, хотя ц ие всегда, в среде
поддерживают слегка кислый pH), и антиоксидант типа аскор-
бата. Во многих случаях, по-видимому, необходим также и хе-
латный агент. Когда в качестве буфера используется неоргани-
ческий пирофосфат, то выполняется и это требование. Можно
внести в среду и другие защитные агенты, например бычий сы-
вороточный альбумин, но если у нас есть «хороший» шпинат,
то превосходные хлоропласты можно получить, взяв .раствор,
содержащий только 0,33 М сорбитол и 10 мМ IS^PaCK,. дове-
денный с помощью НС1 до pH 6,5.
• Механическое разрушение ткани не совсем подходит для
получения интактных хлоропластов из С^растсний. Процент-
ное содержание интактных хлоропластов в препарате, получен-
ном из клеток мезофилла, обычно очень невелико (судя по со-
держанию маркерных ферментов и по .пробе на интактность с
феррицианидом, разд. А.7). Клетки обкладки проводящих .пуч-
ков имеют очень толстую клеточную стенку и поэтому более
устойчивы к разрушению, чем клетки мезофилла, а следова-
тельно, и выход хлоропластов из таких клеток ничтожен.' При-
механическом разрушении клеток С4щастений возникает ..еще
одна трудность, связанная с перекрестным загрязнением полу-
чаемых препаратов хлоропластов . мезофилла й обкладки про-
водящих пучков (гл. 10). < , . . • 1.
36—234
650
приложение а
А.5. Получение хлоропластов из протопластов
Этот метод получения функционально активных хлоропла-
стов до сих пор еще не вышел из стадии становления. Он ока-
зался особенно удобным для работы с С4-растениями, так как
позволил получить в чистом виде протопласты мезофилла, из
которых затем можно выделить интактные хлоропласты. Этот
метод был успешно применен и в опытах с однодольными
Сз-растениями (пшеницей и ячменем). Его можно использовать
(правда, довольно ограниченно) и при работе с двудольными
Сз-растениями (шпинат, подсолнечник, табак и горох), и с
CAM-растениями, например с Sedum praealtum и Mesembry-
anthemum crystatlinum.
Получение ткани листа
Метод, которым получают ткани листа для дальнейшего1
ферментативного переваривания, зависит от вида растения.
Листья однодольных растений просто разрезают на кусочки
обычным лезвием бритвы (делая вручную поперечные срезы
через каждые 0,7—0,8 мм). Можно использовать и механичес-
кий резак, позволяющий получить совершенно однородные сег-
менты, что намного уменьшает возможные отличия получаемых
препаратов друг от друга. Эта задача кажется очень неслож-
ной, но каждый человек по-своему режет лист. У двудольных
растений эпидермальный слой можно иногда отделить от ниж-
ней поверхности листа, просто ухватив его пинцетом (напри-
мер, у табака и Kalanchoe daigremontiana). Можно потереть
поверхность листа наждачной бумагой или щеткой (например,
зубной), но очень осторожно, чтобы не повредить те ткани, ко-
торые лежат под эпидермисом. После этого ткань листа инку-
бируют не более 2,5—4 ч при 25—30 °C и слабом освещении.
Большая продолжительность инкубации нежелательна, так как
это может привести к утрате фотосинтетических функций. Для
некоторых видов растений проводят такую обработку, как вве-
дение среды, содержащей ферменты, непосредственно в сегмен-
ты листа путем вакуум-инфильтрации, или же встряхивание на
качалках во время инкубации. Ни один из этих методов, по-
видимому, не дает преимуществ тогда, когда берут мелко из-
мельченные листья однодольных растений.
В качестве примера приведем состав одной из возможных
сред выделения:
2%-ная целлюлаза (из Trichoderma viride)
Пектиназа (из Rhizopus sp.), 0,1%-ная для С4- и 0,3%-ная
для Сз- и САМ-растений
0,5 М сорбитол
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
551
1 мМ СаС12
20 мМ MES-буфер, pH 5,5
0,05%-ный бычий сывороточный альбумин (обезжиренный)
Ферменты указанных выше грибов есть в продаже. Оказа-
лось, что они удобны для работы со многими растениями,
включая многие однодольные. В некоторых случаях можно
увеличить стабильность протопластов и сделать более или ме-
нее постоянным выход этих структур, если предварительно
обессолить или слегка очистить используемый препарат фер-
ментов. Для видов, устойчивых к перевариванию данными
ферментами, иногда бывает полезно либо заменить одни фер-
менты на другие, либо изменить условия выращивания.
Выделение и очистка
После того как ткань проинкубируют с переваривающими
ферментами, среду выделения можно осторожно отделить и от-
бросить. Затем после нескольких промывок раствором осмоти-
чески активного вещества (например, 0,5 М сорбитолом, со-
держащим 1 мМ СаС12) получают смесь, состоящую из прото-
пластов, хлоропластов и проводящей ткани. Непереваренный
материал, в который входят эпидермальная и проводящая тка-
ни листа, удаляют, профильтровав полученную суспензию через
нейлоновое сито (материал, получаемый из Сз- и САМ-расте-
ний, фильтруют через сита с отверстиями размером 1 мм и
200 мкм, а материал, получаемый из Сграстений, — через си-
та с отверстиями размером 1 мм, 500 и 80 мкм). Тяжи об-
кладки проводящих пучков у С4-растений обычно устойчивы к
перевариванию, и их можно собрать на нейлоновом оите с от-
верстиями 80 мкм. Низкоскоростное центрифугирование получен-
ного экстракта в течение 5 мин при 100g позволяет отделить
осадок, состоящий из протопластов и хлоропластов. Есть два
способа очистки протопластов. Протопласты мезофилла неко-
торых растений всплывают, когда их ресуспендируют в раство-
ре с достаточно высокой плотностью. Так, например, если про-
топласты мезофилла пшеницы, ячменя или подсолнечника сус-
пендировать в растворе, содержащем 0,5 М сахарозу, и отцент-
рифугировать при низкой скорости (5 мин при 200 g), то они
всплывут и соберутся в верхней части пробирки. Если необхо-
димо, плотность среды можно увеличить, добавив в нее, напри-
мер, декстран (Т2о или Тдо) до концентрации 2—10% (вес/объем)
или фиколл. Если же перед центрифугированием на среду с
сахарозой наслоить раствор (например, сорбитола) с меньшей
плотностью, то после центрифугирования протопласты попадут
на границу этих двух фаз. После этого их можно легко со-
брать пастеровской пипеткой.
36*
552
ПРИЛОЖЕНИЕ А
Протопласты, которые всплывают хуже, можно очистить в
водной двухфазной системе, приготовленной после смешивания
двух полимеров: декстрана и полиэтиленгликоля. Такая смесь со-
стоит из.5,5%-кого (вес/объем) полиэтиленгликоля6000, 10%-но-
го (вес/объем) декстрана Т2о, 10мМ натрий-фосфатного буфера
(.pH 7,5) и 0,46 мМ сорбитола. Если тщательно смешать
0,6 мл препарата неочищенных протопластов и 5,4 мл двухфаз-
ной смеси и затем отцентрифугировать в течение 5 мин при
300 g и 4 °C, протопласты перейдут на границу раздела фаз, а
хлоропласты соберутся в нижней фазе. Отобранные из проме-
жуточной фазы протопласты можно снова ресуспеидировать и
затем промыть соответствующей средой.
Хранение
Протопласты мезофилла наиболее устойчивы, когда их хра-
нят при сравнительно низких pH и в присутствии некоторых
двухвалентных катионов (например, в 0,4 М сорбитоле, содер-
жащем 1 мМ CaCls и 20 мМ. MES, pH 6,0). Результаты иссле-
дований, проведенных на пшенице, ячмене и табаке, говорят о
том, что сорбитол вполне подходит для того, чтобы поддержи-
вать осмотические свойства среды по меньшей мере в течение
8—10 ч при условии, что протопласты хранят во льду. Если
препарат держат при 25 °C, то и протопласты, и выделенные из
них хлоропласты теряют фотосинтетическую активность, что
указывает иа непригодность сорбитола в этом случае. В то же
время протопласты не теряют своих фотосинтетических свойств
и при 25 °C по меньшей мере в течение 10—20 ч, если осмоти-
чески активным веществом среды является сахароза.
Выделение хлоропластов из протопластов
Средний диаметр протопластов мезофилла у Сз- и Сд-рас-
тений составляет примерно 30—40 мкм, а для протопластов
САМ-растений . эта величина равна по меньшей мере 60—
LOO мкм. Для выделения хлоропластов можно использовать
быстрый и эффективный прием, который заключается в том,
что суспензию протопластов несколько раз пропускают через
нейлоновое сито (с отверстиями размером 20 мкм, если рабо-
тают с Сз- и- Сграстениями, или размером 44 мкм, если рабо-
тают с САМ-растениями). Для этого можно натянуть нейлоно-
вую сетку на пластмассовый шприц одноразового применения
(объем шприца 1 мл), у которого кончик отрезан так, что раз-
мер отверстия приблизительно равен 3 мм. Часть протопластов
можно разрушить, если суспензию два-три -раза пропустить
через шприц. В этом случае в среду, выходят интактные хлр-
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
55»
ропласты (диаметром 3—5 мкм). Этот метод дает хорошие ре-
зультаты и. очень прост, ио наряду с ним можно использовать,
и другие методы. Так, например, в основном интактные хлоро-
пласты получают при пропускании суспензии протопластов че-
рез Yeda Press под давлением 75 фунтов на кв. дюйм
(5,2725 кг/см2). Протопласты САМ-растений можно разрушить,
пропустив их через шприц с иглой № 25. Для того чтобы полу-
чить осадок хлоропластов, практически освобожденный от
фракции цитозоля, экстракт протопластов можно отцентрифу-
гировать при низкой скорости, т. е. в течение 1 мни при 250 g.
Этого вполне достаточно для большинства исследований фото-
синтеза, в которых используются хлоропласты, выделенные из
разных растений. Для изучения внутриклеточной локализации
ферментов суммарный экстракт протопластов можно наслоить
на градиент концентрации сахарозы, а затем отцентрифугиро-
вать его и 'расфракциоиировать при помощи стандартных приё-
мов (разд. 12.1).
Исходный материал
Клеточные стенки состоят главным образом из целлюлозы,
гемицеллюлозы и белка, а пектин и пектат кальция обычно
рассматривают как цементирующий материал, расположенный
между клетками.
Ниже указаны фирмы, где можно приобрести те особые ма-
териалы, которые используют для выделения отдельных кле-
ток или протопластов. Коммерческие препараты перевариваю-
щих ферментов, которые используют для этой цели, обычно
представляют собой сложную смесь из нескольких ферментов.
В деполимеризации пектата и пектина могут участвовать де-
полимеразы нескольких типов, включая такие ферменты, как
полигалактуроназа, полиметилгалактуроназа, пектатлиаза и
пектиилиаза. Вполне очевидно, что разные виды растений су-
щественно различаются по составу пектии-пектата, который
как бы цементирует клетки, связывая их воедино, поэтому не
удивительно, что одни деполимеразы более эффективно дейст-
вуют иа клетки одних растений, а другие — на клетки других
растений. К сожалению до сих пор не удалось добиться мало-
мальски значительного успеха в ферментативном выделении
клеток из злаков.
Для того чтобы разрушить целлюлозу и различные геми-
целлюлозы, требуется целый ряд ферментов. Можно назвать
некоторые ферменты, участвующие в деградации целлюлозы:
эндо-1,4-^-глюканаза, целлобиогидролаза, целлобиаза и некий
фермент, который, возможно, превращает кристаллическую
целлюлозу в ее аморфную форму еще до того, как начнут дей-
554
ПРИЛОЖЕНИЕ А
1. Ферменты
Целлюлаза из Trichoderma viride (содержит ферменты, разрушающие целлю-
лозу и гемицеллюлозу).
Под названием “онозука RIOh RS” (Onozuka R10 и RS) Yakult Biochemical Co. Ltd. Enzyme Products 8-21, Shingikancho Nishinomiya Japan
Под названием “целлюлизин” (Cel- lulysin) Calbiochem-Behring Calbiochem-Behring Corp. Corp. Hoechst U. K. Ltd. P. 0. Box 12087 Hoechst House San Diego, CA Salisbury Road 92112 Hounslow, Middx. USA TW4 6JH England
Под названием “онозука R10” (Onozuka R10) Unwin and Co. Ltd. Prospect Place Welwyn, Bedfordshire, England
Под названием “мейселаза” (Mei- celase) Meiji Seika Kaisha Ltd. 8-2 Chomc Kyobashi, ChutbKu Tokyo, Japan
Пектиназа из Rhizopus sp. (содержит полигалактуроназу)
Под названием “мацерозим RIO” (Macerozyme RIO) Yakult Biochemical Unwin and Co., Ltd. Co., Ltd. (см. выше) (см. выше)
Под названием “мацераза” (Mace- rase) Calbiochem-Behring Corp. (см. выше)
Пектиназа из Aspergillus sp. Под названием "экстрактаза PC” (Extractase PC) Fermco Biochemics, Inc. 2638 Delta Lane Elk Grove Village, IL 60007 USA
Под названием “рохамент Р” (Ro- hament Р) Rohm G.m.b.H D-6100 Darmstadt Kirschenallee Postfach 4242 West Germany
Под названием “пектолиаза Y23- пектиназа” (Pectolyase Y23-Pecti- nase) из Aspergillus japonicus (ак- тивные компоненты —полигалак- Seishin Pharmaceutical Co., Ltd. Noda, Chiba Japan
туроназа, пектинлиаза и неиден-
тифицированный белковый фактор)
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
555
2. Нейлоновые сита 3. Декстран Ыспгу Simon, Ltd. P.O.Box 31 Stockport, Cheshire, SK3 ORT England Tetko, Inc. Precision Woven Screening Media 420 Saw Mill River Road, Elmsford NY 10523 USA
(Т20 — средняя мол. масса 20 000) (Т411 —- средняя мол. масса 40 000) US Biochemicals Corp. 21000 Miles Parkway Cleveland, OH 44128 USA (Декстран T20 и T4D) Pharmacia Fine Chemicals. Uopsala, Sweden (Декстран T4-:,)
ствовать другие ферменты. К гемицеллюлозам относится боль-
шая, очень сложная по составу гетерогенная группа полисаха-
ридов, в которую входят ксилан, арабпиаиы, галактаны, ман-
наны п глюкуроновая и галактуроновая кислоты. Различия в
составе гемицеллюлозы у разных видов могут очень сильно
влиять на эффективность разрушения клеточной стенки под
действием тех или иных имеющихся в продаже препаратов
ферментов.
А.6. Достоинства и недостатки методов механического
и ферментативного разрушения клеток
В настоящее время эффективность получения фотосинтети-
чески активных хлоропластов с применением любого из этих
двух способов во многом определяется тем, какое растение мы
берем. При помощи механического разрушения клеток можно
получить хлоропласты с очень высокой интенсивностью фото-
синтеза, особенно если взять такие Сз-растения, как шпинат и
горох.
В более поздних работах говорится также о том, что мож-
но получить активно функционирующие хлоропласта и из про-
топластов мезофилла этих двух видов растений. В рутинных
опытах использование протопластов для изучения ассимиляции
СО2 у шпината и гороха не дает никаких преимуществ. В то же
время этот прием очень удобен, когда нужно определить внут-
риклеточную локализацию каких-либо ферментов, поскольку
он позволяет получить большое (в процентном отношении) ко-
личество интактных органелл. Использование шпината в науч-
ных исследованиях, по-видимому, связано с теми же трудно-
стями, что и применение томатов и табака, а именно с боль-
шой вариабельностью получаемого растительного материала,
обусловленной колебаниями условий выращивания, которые
могут даже приводить к тому, что некоторые ткани становятся
устойчивыми к ферментативной обработке.
556
ПРИЛОЖЕНИЕ А
Относительно интактные и активные хлоропласты были вы-
делены из протопластов мезофилла таких Сз-растений, как
пшеница, ячмень, табак и подсолнечник. К тому же интактные
хлоропласты, выделенные из протопластов мезофилла таких
Сграстеиий, как Digitaria sanguinalis (росичка кровяная),
Eleusine indica (елеузнна индийская) и Urochloa panicoides,
эффективно катализируют зависимые от света реакции, свя-
занные с фотосинтетическим метаболизмом в С^цикле. Пригод-
ность протопластов для изучения С^фотосинтеза достаточно
полно обсуждается в гл. 10 и 12. Ни механическое, ни фермен-
тативное разрушение клеток не позволяют обычно получить та-
кие хлоропласты, из САМ-растений, которые были бы способны
ассимилировать СОг со скоростью, превышающей несколько
мкмоль-(мгХл)_1-ч-1. „Средняя скорость фиксации СО2 в хло-
ропластах, изолированных из протопластов САМ-растеиия Se-
dum praealtum, составляет около 20—30 мкмоль-(мгХл)"1 ч’г.
При исследовании внутриклеточной локализации ферментов
в клетках мезофилла ферментативное разрушение клеток да-
ет ряд преимуществ. Во-первых, устраняются митохондрии,
микротельца и такие потенциальные источники посторонних
ферментов, как проводящие и эпидермальная ткани. Во-вто-
рых, можно получить высокое процентное содержание интакт-
ных органелл. А это несомненное преимущество, когда надо
выяснить внутриклеточную компартментацшб ферментов. При
механическом разрушении более вероятны такие явления, как
агрегация органелл, неспецифическое связывание раствори-
мых ферментов с органеллами и формирование комплексов
между белками и таннинами. Вероятность такого рода собы-
тий намного уменьшается, если используют готовые протопла-
сты.
Протопласты могут служить очень удобной системой для
изучения и реконструкции обмена веществ между цитоплазмой
и органеллами. У С4-растеп.ий экстракты протопластов мезо-
филла способны осуществлять карбоксплирующую стадию
С4-цикла; для этого нужны и хлоропласты, и цитозоль (разд.
12.3). У Сз-растений экстракты протопластов или объединен-
ные вместе субклеточные фракции можно использовать для ис-
следования и реконструкции фотосинтетического метаболизма,
для которого необходимо взаимодействие между фракцией ци-
тозоля, митохондриями п пероксисомами (например, синтез са-
харозы и фотодыхание).
Если уж удалось получить препарат протопластов, то та-
кой гомогенный исходный материал можно использовать для
неоднократного выделения хлоропластов с более или менее
одинаковой активностью. Это позволяет свести к минимуму
возможные отклонения, которые обычно наблюдаются в пре-
ВЫДЕЛЕНИЕ ХЛОРОПЛЛСТОВ 55?
паратах, полученных после механического разрушения различ-
них тканей листа, и максимально увеличить вероятность, что-
наблюдаемые различия действительно обусловлены действием
того или иного фактора (условия выделения, ресуспендирова-
ния и определения). В ряде случаев результаты, полученные
в опытах с протопластами, можно использовать для разработ-
ки эффективных приемов механического разрушения ткани то-
го или иного растения.
К числу недостатков ферментативного метода получения
протопластов можно отнести: 1) невозможность применения
этого метода для некоторых видов; 2) необходимость затрачи-
вать дополнительное время На выделение протопластов (что
отнюдь не ограничивает возможность применения этого метода
в тех случаях, когда гарантированные результаты оправдыва-
ют затраченные усилия) и 3) ограниченный выход органелл
(1—5 мгХл на 10 г ткани листа). Сюда же относится и на-
стоятельная необходимость проводить переваривание в тече-
ние минимального времени н использовать для очистки прото-
пластов наиболее подходящий способ. Следует помнить о том,
что переваривающие, ферменты могут подавляться различного
рода ингибиторами, а сама обработка ферментами может да-
вать ряд побочных эффектов.
Это обсуждение касалось только тех методов, которые мож-
но использовать для-выделения хлоропластов. Однако из тех же
принципов следует исходить и при получении функционально-
активных митохондрий и пероксисом из листьев растений.
А.7. Критерии, по которым определяют степень
интактности хлоропластов
Изолированные хлоропласты могут находиться в самых раз-
личных состояниях как с точки зрения их морфологии, так и
с точки зрения их функции. Дэвид Холл предложил очень удоб-
ную терминологию для обозначения состояния хлоропластов
Сз-растений, которая связывает их вид под микроскопом или
их строение с их функцией. «Полные хлоропласты» типа А по-
лучают в растворах, содержащих сахара; у них сохранены ин-
тактные оболочки, и они фиксируют СОг со скоростью около
50—250 мкмоль- (мгХл)_1-ч-1. В такие хлоропласты не прони-
кают экзогенный NADP+ и феррицианид (окислитель в реак-
ции Хилла). «Неразрушенные хл-оропласты» типа В получают
в растворах, содержащих сахара или соли; скорость фиксации
СОг У них гораздо ниже. В хлоропластах сохраняются раство-
римые белки, но и феррицианид, и ADP проникают в них,
указывая на то, что оболочка таких хлоропластов утратила не-
которые из своих свойств, п в частности свою полупроницае-
-•558
ПРИЛОЖЕНИЕ А
мость. Хлоропластами типа С, D, Е и F называют разрушен-
ные хлоропласты различного типа (у всех у них утрачена на-
ружная оболочка), полученные разными способами.
Есть очень удобный способ, который позволяет отличить
.хлоропласты типа А от хлоропластов другого типа. Поскольку
'феррицианид не проникает через оболочку интактных хлоро-
пластов, у хлоропластов типа А не наблюдается зависимого
от феррицианида выделения О2. Если препарат хлоропластов
подвергнуть осмотическому шоку (быстро понизив концент-
рацию осмотически активного вещества и создав таким обра-
зом экзоосмос), то наружная оболочка хлоропластов повреж-
дается и можно наблюдать максимальное выделение Oj в при-
сутствии феррицианида. Измерив скорость зависимого от фер-
рицианида выделения О2 до (В) и после (А) осмотического
шока, можно рассчитать степень интактности хлоропластов
(В %):
д g
Степень интактности (%)==—5---100
А
Для того чтобы не происходило зависимого от СО2 выделения
О2, в среду добавляют глицеральдегид, а для разобщения пе-
реноса электронов и фосфорилирования и получения макси-
мальной скорости наблюдаемого процесса вносят NH4CI. На-
глядно этот метод определения степени интактности изображен
на рис. А.1, где приводятся данные для препарата хлороплас-
тов, полученных из протопластов пшеницы.
Если степень интактности определить феррицианидным ме-
тодом, то эта величина будет говорить лишь о том, что обо-
лочка хлоропласта не утратила своей полупроницаемости и ни-
как не будет коррелировать со способностью хлоропласта к
фиксации СО2. Есть данные о том, что в процессе выделения
часть хлоропластов повреждается, а затем вновь восстанавли-
вает свою полупроницаемую оболочку, но уже после того, как
•будет утеряна часть ферментов стромы. Поэтому такие хлоро-
пласты будут иметь высокую степень интактности, если судить
по феррицианидной пробе, но очень низкую активность, если
судить по зависимому от СО2 выделению О2. Это говорит с
том, что феррицианидная проба может давать завышенную ве-
личину процента интактных, функционально активных хлоро-
пластов. Поэтому в графики зависимости между содержанием
растворимых белков и степенью интактности (определенной по
феррицианидной пробе) следует вносить поправку, учитываю-
щую завышение последней величины.
Степень интактности, которую дает феррицианидная проба,
не всегда является показателем способности хлоропластов к
•фотосинтетической ассимиляции СО2 и по ряду других причин.
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
559
Рис. А.1. Определение степени интактиости хлоропластов с использованием
феррицианида в качестве окислителя в реакции Хилла. Хлоропласты были
выделены из протопластов пшеницы. I— экстракт протопластов после осмо-
тического шока, II—интактные протопласты, III — хлоропласты после осмо-
тического шока, IV — интактные хлоропласты. Расчетная степень интактио-
сти для протопластов равна 94%, для хлоропластов — 95%. Числа в скоб-
ках — количество выделившегося О2 в мкмоль-(мгХл)-1-ч~1 с учетом по-
правки па поглощение О2 кислородным электродом.
Так, например, хлоропласты подсолнечника, полученные после
механического разрушения ткани листа, имеют очень высокую
степень интактиости, но скорость фиксации СОг у них очень
мала. Вполне возможно, что у таких хлоропластов нарушены
какие-то другие свойства, например, неправильно функциониру-
ет система переносчиков в оболочке хлоропласта (гл. 8), кото-
рую можно в некоторых случаях повредить в процессе выделе-
ния хлоропластов. В группе так называемых хлоропластов ти-
па А могут существовать различные подгруппы. (Аналогичный
метод был использован Доусом и др. для определения степени
интактиости изолированных митохондрий. Для этого измеряли
активность сукцинат-цитохром-с — оксидоредуктазы до и после
осмотического шока, о которой судили по восстановлению экзо-
генного цитохрома с. В случае интактных митохондрий цито-
хром с не проникает через наружную мембрану. Для того что-
560
ПРИЛОЖЕНИЕ А
бы цитохром восстановился, нужно, чтобы он каким-то образом
получил доступ к внутренней мембране митохондрий.)
Когда хлоропласты выделяют из протопластов, для опреде-
ления степени интактности можно воспользоваться и другим
способом. Экстракт протопластов центрифугируют при низкой
скорости, чтобы отделить хлоропласты от тех ферментов, кото-
рые находятся вне этих органелл. Определяют активность та-
кого фермента, который, как считают, целиком и полностью ло-
кализован в хлоропластах in vivo, например NADP-глицераль-
дегидфосфатдегидрогеназы в экстракте, полученном из осад-
ка хлоропластов (ХП), и в надосадочной жидкости (НЖ):
хп
Степень интактности(%) = 100-
Точность этого метода определяется тем, насколько полно вы-
является активность исследуемого фермента во фракции хлоро-
пластов и в надосадочной жидкости. Некоторые ферменты ти-
па РуБФ-карбоксилазы при попадании в бесклеточный экст-
ракт становятся лабильными или ингибируются. Поэтому очень
важно, чтобы активность фермента в суммарном бесклеточном
экстракте (полученном без применения осмотически активных
веществ) была равна сумме активностей этого фермента во
фракции хлоропластов и в надосадочной жидкости.
Среды, применяемые для исследования фотосинтетического
метаболизма в изолированных хлоропластах, сильно различа-
ются по составу. Наиболее важные условия определения актив-
ности Сз- (гл. 8) и С^хлоропластов (гл. 12) были рассмотрены
нами в предыдущих главах.
А.8. Очистка
Независимо от того, как были выделены хлоропласты, т. е.
после механического разрушения ткани или из протопластов,
их следует отделить центрифугированием. Иногда этого бывает
вполне достаточно для получения интактных хлоропластов. Та-
ким образом можно почти полностью избавиться от тех фрак-
ций, которые загрязняют препарат хлоропластов (цитоплазма,
пероксисомы и митохондрии), поскольку они почти полностью
остаются в надосадочной жидкости, а интактные хлоропласты
седиментируют после центрифугирования в течение 1—2 мин
(при 300—2000g в зависимости от состава среды). Большая
часть разрушенных хлоропластов тоже остается в надосадоч-
ной жидкости, а в осадке после центрифугирования, в самых
оптимальных условиях содержится от 70 до 100% интактных
хлоропластов.
При необходимости можно удалить и тилакоиды. Для это-
го следует отцентрифугировать препарат в среде, не содержа-
ВЫДЕЛЕНИЕ ХЛОРОПЛАСТОВ
.561
щей никаких солей (0,33 М сорбитол, доведенный трисом до
pH 7,5); это помогает удержать поврежденные хлоропласты в
надосадочной жидкости. Но чтобы избежать значительной по-
тери активности при храпении, полученный в конце концов пре-
парат интактных хлоропластов все-таки следует хранить в
среде, содержащей соли.
Для очистки хлоропластов и других органелл применяют
центрифугирование в градиенте концентрации сахарозы. Одна-
ко при такой обработке хлоропласты подвергаются воздейст-
вию слишком высоких концентраций осмотически активного ве-
щества, что может привести к снижению фотосинтетической ак-
тивности. Для того чтобы создать высокую плотность при срав-
нительно низкой осмотической концентрации вещества в гра-
диенте, можно воспользоваться высокомолекулярными полиме-
рами. Так, например, очищенные и функционально активные
хлоропласты гороха и шпината можно получить после очистки
в перколле (коллоидном растворе частиц кремниевого золя, по-
крытых поливинилпирролндоном). Неочищенный препарат хло-
ропластов можно наслоить иа 40%-пый (по объему) перколл,
содержащий 0,33 М сорбитол и какой-нибудь буфер. После
центрифугирования при 2500 g в течение 1 мин материал, не
входящий в хлоропласты, будет находиться в надосадочной
жидкости, а в осадке соберутся интактные хлоропласты. Мож-
но провести центрифугирование и в линейном градиенте пер-
колла (приблизительно от 10 до 90%). Неочищенный препарат
хлоропластов наслаивают на такой градиент и затем центри-
фугируют в течение 15 мин приблизительно при 10 000 g. Ин-
тактные хлоропласты, сконцентрировавшиеся на участке гра-
диента, отвечающем их плавучей плотности, можно будет со-
брать, раскапав градиент по фракциям, и затем промыть их
для удаления перколла. Одним из самых последних достиже-
ний в области выделения органелл является метод отделения
хлоропластов от балластного материала в течение нескольких
секунд после лизиса протопластов (разд. 8.3). Этот прием осо-
бенно удобен для изучения компартмеитации метаболитов и
продуктов фотосинтеза в клетке во время ассимиляции угле-
рода.
ЛИТЕРАТУРА
1. Douce R. Е., Christensen L., Bonner 117. D. Preparation of intact plant mito-
chondria, Biochem. Biophys. Acta, 275, 148—160 (1972).
2. Edwards Q. E., Huber S. C., Gutierrez M. Photosynthetic properties of plant
protoplasts. In: Microbial and Plant Protoplasts (eds. J. F. Peberdy,
A. H. Rose, H. J. Rogers and E. C. Cocking), pp. 299—322, New York, Aca-
demic Press, 1976.
3. Edwards Q. E., Robinson S. P., Tyler N. J. C., Walker D. A. Photosynthesis
by isolated protoplasts, protoplast extracts and chloroplasts of wheat.
562
ПРИЛОЖЕНИЕ А
Influence of orthophosphate, pyrophosphate and adenylates, Plant Physiol.,
62, 313—317 (1978).
4. Edwards G. E., Lilley R. McC., Hatch M. D. Isolation of intact and functio-
nal chloroplasts from mesophyll and bundle sheath protoplasts of the Ct
plant Panicum miliaceum, Plant Physiol., 63, 821—827 (1979).
5. Hall D. 0. Nomenclature for isolated chloroplasts, Nature New Biol., 235,
125—126 (1972).
6. Huber S. C., Edwards 0. E. An evaluation of some parameters required for
the enzymatic isolation of cells and protoplasts with CO2 fixation capacity
from C3 and C4 grasses, Physiol. Planta, 35, 203—209 (1975),
7. Huber S. C„ Hall T. C., Edwards G. E. Light dependent incorporation of
UCO2 into protein by mesophyll protoplasts and chloroplasts isolated from
Pisum sativum, Z. Pflanzenphysiol., 85, 153—163 (1977),
8. Ishii S. Enzymatic maceration of plant tissues by endopectin lyase and
endo-polygalactuironase from Aspergillus japonicus, Phytopathology, 66,
281—289 (1975).
9. Kanal R„ Edwards G. Purification of enzymatically isolated mesophyll
protoplasts from Сз, C4 and Crassulacean Acid Metabolism plants using an
aqueous dextran polyethylene glycol two-phase system, Plant Physiol., 52,
484—490 (1973).
10. Lilley R. McC., Fitzgerald M. P., Rlenits K- G., Walker D. A. Criteria of
intactness and the photosynthetic activity of spinach chloroplast prepara-
tions, New Phytol., 75, 1—10 (1975).
11. Mills IV. R„ Joy K. W. A rapid method for isolation of purified, physiolo-
gically active chloroplasts, used to study the intracellular distribution of
amino acids in pea leaves, Planta, 148, 75—83 (1980).
12. Nagata 1., Ishii S. A rapid method for isolation of mesophyll protoplasts,
Can. J. Bot., 75, 1820—1823 (1979).
13. Nakatanl H. Y„ Barber J. An improved method for isolating chloroplasts
retaining their outer membranes, Biochim. Biophys. Acta, 461, 510—512
(1977).
14. Slabas A. R., Powell A. J., Lloyd C. 117. An improved procedure for the iso-
lation and purification of protoplasts from carrot suspension culture, Planta,
147, 283—286 (1980).
15. Spalding M. H., Edwards G. E. Photosynthesis in isolated chloroplasts of
the Crassulacean acid metabolism plant Sedurn praealtum, Plant Physiol.,
65, 1044—1048 (1980).
16. Spalding M. H., Scmltt M. R„ Ku S. B., Edwards G. E. Intracellular loca-
lization of some key enzymes of Crassulacean Acid Metabolism in Sedum
praealtum. Plant Physiol., 63, 738—743 (1979).
17. Takabe T., Nishimura M., Akazawa T. Isolation of intact chloroplasts from
spinach leaf by centrifugation in gradients of the modified silica '‘Percoll’’,
Agric. Biol. Chem., 43, 2137—2142 (1979).
18. Walker D. A. Preparation of higher plant chloroplasts. In: Methods in
Enzymology, Vol. 69, pp. 94—104, New York, Academic Press, 1980.
ПРИЛОЖЕНИЕ Б
Номенклатура ферментов
Из рекомендаций Международного биохимического союза-
по номенклатуре ферментов (1978), Academic Press, New York..
Порядок перечисления: рекомендуемое название, номер соглас-
но классификации ферментов, (другие названия), системати-
ческое название. [Реакция]
Б.1. Восстановительный пентозофосфатный путь
Рибулозобисфосфат-карбоксилаза, КФ 4.1.1,39 (карбок-
сидисмутаза), З-фосфо-Э-глицераткарбокси-лиаза (диме-
ризующая) .
[В-рибулозо-1,5-бисфосфат+СО2 =
= 2 З-фосфо-Э-глицерат. Фермент катализирует также
реакцию О-рибулозо-1,5-бисфосфат+О2=
= 3-фосфо-Г)-глицерат-[- фосфогликолат]
Фосфоглицераткиназа, КФ 2.7.2.3 (1,3-дифосфоглице-
рат—>-ADP — трансфосфатаза), АТР:3-фосфо-П-глицерат
1-фосфотрансфераза.
[АТР+3-фосфо-Ь-глицерат=
= АЭР-|-3-фосфо-П-глицероил фосфат]
Глицеральдегидфосфатдегидрогеназа (NADP+) (фосфо-
рилирующая), КФ 1.2,1.13 {триозофосфатдегидрогеназа
(NADP+)], D-глицеральдегид-З-фосфат : NADP+ оксидо-
редуктаза (фосфорилирующая).
[3-фосфо-П-глицероилфосфат-]-ЫАПРН-]-Н+=
= Е)-глицеральдегид-3-фосфат-|-ортофосфат-|-МАОР+]
Триозофосфатизомераза, КФ 5.3.1.1 (фосфотриозоизоме-
раза), D-глицеральдегид-З-фосфат кетол-изомераза,
[О-глицеральдегид-3-фосфат=дигидроксиацетонфосфат]
Фруктозобисфосфат-альдолаза, КФ 4.1.2.13 (зимогексаза),
Э-фруктозо-1,6-бисфосфат D-глицеральдегид-З-фосфат-
лиаза.
564
ПРИЛОЖЕНИЕ Б
[3-фосфр-В-глицероилфосфат+ЫАОРН-]-Н+=
=глицёральдегид»3-фосфат+ортофдсфат+ЫАВР+]. Фер-
мент катализирует еще одну реакцию ВПФ-пути: D-эрит-
розо-4-фосфат-(-дигидроксиацетонфосфат=
=седогептулозо-1,7-бисфосфат
Фруктозо-бисфосфатаза, КФ 3.1.3.11 (гексозодифосфата-
за), D-фруктозо-! ,6-бисфосфат 1-фосфогидролаза.
ID-фруктозо-1,6-бисфосфат4-Н2О—
= О-фруктозо-6-фосфат-|-ортофосфат]
Транскетолаза, КФ 2.2.1.1 (гликольальдегидтраисфераза),
седогептулозо-7-фосфат: D-глицеральдегид-З-фосфат гли-
кольальдегидтраисфераза.
[Седогептулозо-7-фосфат+О-глицеральдегид-3-фосфат =
= О-рибозо-5-фосфат-]-О-ксилулозо-5-фосфат]. Фермент
катализирует еще одну реакцию ВПФ-пути: D-фруктозо-
6-фосфат+В-глицеральдегид-3-фосфат=
= О-ксилулозо-5-фосфат+О-эритрозо-4-фосфат
Седогептулозобисфосфатаза, КФ 3.1.3.37, седогептулозо-
1,7-бисфосфат—1-фосфогидролаза.
[Седогептулозо-1,7-бисфосфат+Н2О=
=седогептулозо-7-фосфат+ортофосфат]
Рибозофосфат-изомераза, КФ 5.3.1.6 (фосфопентозоизо-
мераза), D-рибозо-б-фосфат кетол-изомераза.
[О-рибозо-5-фосфат=О-рибулозо-5-фосфат]
Рибулозофосфат—3-эпимераза, КФ 5.1.3.1 (фосфорибу-
лозоэпимераза), D-рибулозо-б-фосфат— 3-эпимераза.
[0-рибулозо-5-фосфат=0-ксилулозо-5-фосфат]
Фосфорибулокиназа, КФ 2.7.1.19 (фосфопентокиназа),
АТР—^D-рибулозо-б-фосфат—1-фосфотрансфер аза.
{АТР+О-рибулозо-5-фосфат=
=АОР+0-рибулозо-1,5-бисфосфат]
Б.2. Гликолатный путь
Фосфогликолатфосфатаза, КФ 3.1.3.18, 2-фосфоглико-
лат — фосфогидролаза.
[2-фосфогликолат+Н2О=гликолат+ортофосфат]
Гликолатоксидаз'а, КФ Г.1.3.1 (гликолат—>-О2 трансгид-
. рогеназа) , гликолат : кислород оксидоредуктаза.
[гликолат-|-О2=глиоксилат+Н2О2]
НОМЕНКЛАТУРА ФЕРМЕНТОВ
565-
Каталаза, КФ 1.11.1.6, перекись водорода : перекись водо-.
рода оксидоредуктаза.
ОаН-ЗНгО]
Глицинаминотрансфераза, КФ 2.6.1.4, глицин : 2-оксоглу-
тарат аминотрансфераза.
[глиоксилат+Б-глутамат=глйцин-|-2-оксоглутарат]
Серин-глиоксилат— аминотрансфераза, КФ 2.6.1.45, L-ce-
рин: глиоксилат.аминотрансфераза.
[Ь-серин+глиоксилат=3-гидроксипируват+глицин]
Глицин-синтаза, КФ 2.1.2.10, 5,10-метилентетрагидрофо-
лат: аммиак гидроксиметилтрансфераза (карбоксили-
рующая, восстанавливающая).
[тетрагидрофолат+глицинН-окйсленный . белок—перенос-
чик водорода=5,10-метилентетрагидрофолат+СО2+‘
4-ЫНз+"восстановленный белок—переносчик водорода]
Серин-гидроксиметилтрансфераза, КФ 2.1.2.1 (серин-
альдолаза), 5,10-метилентетрагидрофолат : глицин гид-
роксиметилтрансфераза.
[5,Ю-метилентетрагидрофолат+глицин4-Н20=
=тетрагидрофолат-]-Ь-серин]
Глиоксилатредуктаза, КФ 1.1.1.26 (NADH—э-глиокси-
лат—трансгидрогеназа), гликолат: NAD+ оксидоредук-
таза.
[гидроксипируват+NADH+H+ss
= D-rni-mepaT-|-NAD+]
[глиоксилат-|-ЫАОН-|-Н+=гликолат-|-.НАО+].
Примечание: восстанавливает гидроксипируват до D-гли-
церата или глиоксилат до гликолата.
Глицераткиназа, КФ 2.7.1.31, ATPiD-глицерат 3-фосфо-
трансфераза.
[АТР-|-15-глицерат=АОР+3-фосфо-О-глицерат]
Ферменты, катализирующие реакции, которые связаны
с гликолатным путем и необходимы
для челночной передачи восстановительной силы
Малатдегидрогеназа, КФ 1.1.1.37 (L-малат—>NAD—
трансгидрогеназа), L-малат: -NAD+ оксидоредуктаза.
[Ь-малат+ЫАО+=оксалоацетат-]-КАОН4-Н+]
Ферменты, необходимые для реассимиляции аммиака,
представлены в разд. Б.7.
566
ПРИЛОЖЕНИЕ Б
Б.З. Образование сахарозы из триозофосфатов
Триозофосфат-изомераза, КФ 5.3.1.1 (фосфотриозоизоме-
раза), D-глицеральдегид-З-фосфат кетол-изомераза.
[дигидроксиацетонфосфат=В-глицеральдегид-3-фосфат]
Фруктозобисфосфат-альдолаза, КФ 4.1.2.13 (зимогексаза),
О-фруктозо-1,6-бисфосфат D-глицеральдегид-З-фосфат-
лиаза.
[дигидроксиацетонфосфат+О-глицеральдегид-3-фосфат=
=0-фруктозо-1,6-бисфосфат]
Фруктозо-бисфосфатаза, КФ 3.1.3.11 (гексозодифосфата-
за), О-фруктозо-1,6-бисфосфат 1-фосфогидролаза.
[D-фруктозо-1,6-бисфосфат-|-Н2О =
= В-фруктозо-6-фосфат+ортофосф.ат]
Глюкозофосфат-изомераза, КФ 5.3.1.9 (фосфогексозо—
изомераза), D-глюкозо-б-фосфат кетол-изомераза.
[О-фруктозо-6-фосфат=О-глюкозо-6-фосфат]
Фосфоглюкомутаза, КФ 2.7.5.1 [глюкозо—(1->6)-фосфо-
мутаза], a-D-глюкозо-1,6-бисфосфат: глюкозо- 1-фосфат
фосфотрансфераза.
[а-В-глюкозо-6-фосфат-|-а-О-глюкозо-1,6-бисфосфат=
— a-D-глюкозо-1,6-бисфосфат-(-а-О-глюкозо-1-фосфат]
Глюкозо-1-фосфат—уридилилтрансфераза, КФ 2.7.7.9
(иОРглюкоза-пирофосфорилаза), UTP : a-D-глюкозо-1-
фосфат уридилилтрансфераза.
[иОР+а-О-глюкозо-1-фосфат=
=пирофосфат+иВР-глюкоза]
Нуклеозид-дифосфат—киназа, КФ 2.7.4.6 (АТР—>-нуклео-
зид-дифосфат.—трансфосфатаза), АТР : нуклеозид-дифос-
фат фосфотрансфераза.
[АТР + нуклеозид-дифосфат=ADP + нуклеозид-трифосфат]
Неорганическая пирофосфатаза, КФ 3.6.1.1, пирофос-
фат— фосфогидролаза.
[пирофосфат+Н2О=2 ортофосфат]
Сахарозофосфат-синтаза, КФ 2.4.1.14 (UDP-глюкоза-
фруктозофосфат глюкозилтрансфераза), UDP глюкоза:
D-фруктозо'-б-фосфат 2-а-Э-глюкозилтрансфераза
[ТЮР-глюкоза-|-В-фруктозо-6-фосфат=
=иВР+сахарозо-6-фосфат]
НОМЕНКЛАТУРА ФЕРМЕНТОВ
567
Сахарозофосфатаза, КФ 3.1.3,24, сахарозо-61?-фосфат фос-
фогидролаза.
[сахарозо-6в’-фосфат-]-Н2О=сахароза-]-ортофосфат]
Или иначе:
Сахарофосфатаза, КФ 3.1,3,23, сахарофосфат—фосфогид-
ролаза.
[сахарофосфат+Н2О=сахар-|-ортофосфат]
Примечание', в данном случае катализирует превращение
фруктозо-6-фосфата во фруктозу-]-Pi.
Сахарозосинтаза, КФ 2.4.1.13 (UDP-глюкоза-фруктоза—•'
глюкозилтраисфераза).
[UDP-глюкоза-]-0-фруктоза=иОР-]-сахароза]
Примечание: в листьях этот фермент может участвовать
' в разложении сахарозы.
Б.4. Синтез крахмала из триозофосфатов
Триозофосфат-изомераза, КФ 5.3.1.1 (фосфотриозоизоме-
раза), D-глицеральдегид-З-фосфат кетол-изомераза.
[О-глицеральдегид-3-фосфат=дигидроксиацетонфосфат]
Фруктозобисфосфат-альдолаза, КФ 4,1.2.13 (зиморексаза),
D-фруктозо-1,6-бисфосфат Е)-глицеральдегид-3,-фосфат-
лиаза.
[дигидроксиацетоифосфат+О-глицеральдегид-3-фосфат=
= D-фруктозо-1,6-бисфосфат]
Фруктозо-бисфосфатаза, КФ 3.1,3,11 (гексозодифосфата-
за), D-фруктозо-1,6-бисфосфат 1-фосфогидролаза,
[О-фруктозо-1,6-бисфосфат+Н2О= , ...
О-фруктозо-б-фосфат-фортофосфат]
Глюкозофосфат-изомераза, КФ 5,3,1.9 (фосфогексозоизо-
мераза), D-глюкозо-б-фосфат кетол-изомераза.
[О-фруктозо-6-фосфат=О-глюкозо-6-фосфат] '
Фосфоглюкомутаза, КФ 2.7.5.1 [глюкозо,—(1-^-6)-фосфо-
мутаза], а-О-глюкозо-1,6-бисфосфат: а-О-глюкозо-1-фос-
фат фосфотрапсфераза.
[а-0-глюкозо-6-фосфат-]-а-0-глюкозо-1,6-бисфосфат=
= а-О-глюкозо-1,6-бисфосфат-]-а-О-глюкозо-1-фосфат]
Глюкозо-1-фосфат—аденилилтрансфераза, КФ 2.7.7.27
(ADP-глюкозопирофосфорилаза), АТР : a-D-глюкозо-!-
фосфат аденилилтрансфераза.
1[АТР+«-О-глюкозо-1-фосфат =
= пирофосфат-]-ADP-глюкоза]
568
ПРИЛОЖЕНИЕ Б
Неорганическая пирофосфатаза, КФ 3.6.1.1, пирофосфат—
фосфогидролаза.
[пирофосфат-]-Н2О=2 ортофосфат]
Крахмал (бактериальный гликоген) — сиитаза, КФ 2.4.1.21
(АГ)Рглюкоза-крахмал—глюкозилтрансфераза), АВРглю-
коза: 1,4-а-0-глюкан 4-а-глюкозилтрансфераза.
[ADP-глюкоза-]- (1,4-а-В-глюкозил) п =
ADP+ (1,4-а-В-глюкозил)п+1]
Ферменты, участвующие в превращении триозофосфатов в гек-
„созофосфаты, катализируют одни и те же реакции как при
синтезе крахмала, так и при образовании сахарозы. Различают-
ся они только по их локализации в клетке: ферменты, участ-
вующие в синтезе крахмала, находятся в хлоропластах, а фер-
менты, нужные для образования сахарозы, — в цитозоле. Сле-
довательно, регуляторные свойства изозимов могут сильно раз-
личаться.
.Б.5. С4-путь
Фаза карбоксилирования
Пируват, ортофосфат—дикиназа, КФ 2.7.9.1, АТР : пиру-
ват, ортофосфат фосфотрансфераза.
[АТР-|-пируват+ортофосфат=
AMP-j-фосфоенолпируват+пирофосфат]
Неорганическая пирофосфатаза, КФ 3.6.1.1, пирофосфат—
фосфогидролаза.
• [пирофосфат-]-Н2О=2 ортофосфат]
Аденилаткиназа, КФ 2.7.4.3 (миокиназа), АТР: АМР фос-
фотрансфераза. ;
[ATP+AMP=ADP+ADP]
Фосфоенолпируват-карбоксилаза, КФ 4.1.1.31, ортофос-
фат: оксалоацетат карбокси-лиаза (фосфорилирующая).
[фосфбенолпируват-рСО2-]-Н2О=
=оксалоацетат-]-ортофосфат]
Малатдегидрогеназа (NADP+), КФ 1.1.1.82, L-малат:
: NADP+ оксидоредуктаза
[оксалоацетат-|-,Ь1АЕ)РН-]-Н+=Ь-малат-|-ЫАВР+]
Аланинаминотрансфераза, КФ 2.6.1.2 (глутамико-пирови-
ноградная трансаминаза), L-аланин : 2-оксоглутарат ами-
нотрансфераза.
[Ь-аланин-]-2-оксоглутарат= пируват-]- L-глутамат]-
НОМЕНКЛАТУРА ФЕРМЕНТОВ
569
Аспартатаминотрансфераза, КФ 2.6.1.1 (глутамикощаве-
левоуксусная трансаминаза), L-аспартат: 2-оксоглутарат
аминотрансфераза.
[оксалоацетат-)-Ь-глутамат=Ь-аспартат-|-2-оксоглутарат]
Фаза декарбоксилирования (NADP-малатдегидрогеназа)
Малатдегидрогеназа (декарбоксилирующая) (NADP+),
КФ 1.1.1.40 (яблочный фермент), L-малат: NADP+ окси-
доредуктаза (декарбоксилирующая оксалоацетат).
[L-мал ат+'NAD Р+—пируват-)- CO2-)-N ADPH+Н+]
Фаза декарбоксилирования (NAD-малатдегидрогеназа)
Аспартатаминотрансфераза, КФ 2.6.1.1 (глутамикощаве-
левоуксусиая трансаминаза), L-аспартат: 2-оксоглутарат
аминотрансфераза.
[Ь-аспартат-|-2-оксоглутарат=оксалоацетат-)-Е-глутамат]
Малатдегидрогеназа, КФ 1.1.1.37 (L-малат—>-NAD—
трансгидрогеназа), L-малат: NAD+ оксидоредуктаза.
[оксалоацетат-pNADH+H-1'—L-мaлaт-|-NAD+]
Малатдегидрогеназа (декарбоксилирующая), КФ 1.1.1,39
(яблочный фермент), L-малат:NAD+ оксидоредуктаза
(декарбоксилирующая).
[L-мал aT-)-iNAD+=пируват-)- CO2-)-NADH-)-H+]
Аланинаминотрансфераза, КФ 2.6.1.2 (глутамико-пирови-
иоградиая трансаминаза), L-аланин : 2-оксоглутарат ами-
нотрансфераза.
[ni'ipyBaT+L-rayTaMaT=L^aHHi-i+2-оксоглутарат]
Фаза декарбоксилирования (ФЕП-карбоксикиназа)
Аспартатаминотрансфераза, КФ 2.6,1.1 (глутамикощаве-
левоуксусная трансаминаза), L-аспартат: 2-оксоглутарат
аминотрансфераза.
П^-аспартат-|-2-оксоглутарат=оксалоацетат4-Ъ-глутамат]
Фосфоенолпируват-карбоксикиназа (АТР), КФ 4.1.1.49
(фосфопируват-карбоксилаза АТР), АТР : оксалоацетат
карбокси-лиаза (трансфосфорилирующая).
[АТР+оксалоацетат=АВР+фосфоенолпируват+СО2]
37-234
570
ПРИЛОЖЕНИЕ Б
Б.6. Метаболизм кислот у толстянковых
Накопление органических кислот
а-Амилаза, КФ 3.2.1.1 (диастаза), 1,4-а-О-глюкан глюка-
ногидролаза.
[Катализирует эндогидролиз 1,4-а-гликозидных связей в
полисахаридах, содержащих три или более остатков D-
глюкозы, соединенных 1,4-а-связями]
^-Амилаза, КФ 3.2.1.2 (диастаза), 1,4-а-глюкаи мальто-
гидролаза.
[Катализирует гидролиз 1,4-а-гликозидиых связей в поли-
сахаридах, последовательно отщепляя остатки мальтозы
от передуцирующих концов цепей]
a-D-гликозидаза, КФ 3.2.1.20 (мальтаза), a-D-глюкозид—
глюкогидролаза.
[Гидролизует 1,4-связи концевого нередуцирующего ос-
татка a-D-глюкозы с высвобождением а-глюкозы]
Гексокиназа, КФ 2.7.1.1 (АТР—s-глюкоза—трансфосфата-
за), АТР : D-гексоза 6-фосфотрансфераза.
[АТР-|-О-гексоза=АОР-|-О-гексозо-6-фосфат]
Фосфорилаза, КФ 2.4.1.1 [Р-фермеит (только для расти-
тельного фермента)], 1,4-a-D-глюкан : ортофосфат a-D-
глюкозилтрансфераза.
[(1,4-a-D-глюкозил )п+ортофосфат=
= (1,4-а-О-глюкозил) „-j+а-Э-глюкозо-1-фосфат]
Фосфоглюкомутаза, КФ 2.7.5.1 (глюкозофосфомутаза),
а-О-глюкозо-1,6-бисфосфат: а-О-глюкозо-1-фосфат фос-
фотрансфераза.
[а-О-глюкозо-1,6-бисфосфат-|-а-О-глюкозо-1-фосфат=
а-О-глюкозо-6-фосфат-|-а-О-глюкозо-1,6-бисфосфат]
Глюкозофосфат-изомераза, КФ 5.3.1.9 (фосфогексозо-
изомераза), D-глюкозо-б-фосфат кетол-изомераза.
[О-глюкозо-6-фосфат=О-фруктозо-6-фосфат]
6-Фосфофруктокиназа, КФ 2.7.1.11 (фосфогексокиназа),
АТР : D-фруктозо-б-фосфат 1-фосфотрансфераза.
[АТР-|-0-фруктозо-6-фосфат=
= ADP+D-фруктозо- 1,6-бисфосфат]
Фруктозобисфосфат-альдолаза, КФ 4.1.2.13 (зимогексаза),
номенклатура ферментов
571
D-фруктозо-!,6-бисфосфат D-глицеральдегид-З-фосфат-
лиаза.
[О-фруктозо-1,6-бисфосфат=
=дигидроксиацетонфосфат+В-глицеральдегид-3-фосфат]
Триозофосфат-изомераза, КФ 5.3.1.1 (фосфотриозоизоме-
раза), D-глицеральдегид-З-фосфат кетол-изомераза.
[б-глицеральдегид-3-фосфат=дигидроксиацетонфосфат]
Глицеральдегидфосфатдегидрогеназа, КФ 1.2.1.12 (три-
озофосфатдегидрогепаза), D-глицеральдегид-З-фосфат:
: NAD+ оксидоредуктаза (фосфорилирующая).
[О-глицеральдегид-3-фосфат+ортофосфат-|-КАО+=
—3-фосфо-О-глицероилфосфат4-ЫАОН+Н+]
Фосфоглицераткиназа, КФ 2.7.2.3 (1,3-дифосфоглице-
рат—>-ADP—траисфосфатаза), АТР : З-фосфо-Ь-глице-
рат 1-фосфотраисфераза.
[3-фосфо-0-глицероилфосфат+АОР=
АТР+З-фосфо-О-глицерат]
Фосфоглицеромутаза, КФ 2.7.5.3 [глицерат (3->-2)—фос-
фомутаза], 2,3-бисфосфо-О-глицерат: 2-фосфо-О-глицерат
фосфотраисфераза.
[2,3-бисфосфо-О-глицерат+3-фосфо-О-глицерат=
= 2-фосфо-О-глицерат-|-2,3-бисфосфо-О-глицерат]
Енолаза, КФ 4.2.1.11 (фосфопируватгидратаза), 2-фосфо-
D-глицерат гидро-лиаза.
[2-фосфо-О-глицерат=фосфоенолпируват4-Н2О]
Фосфоенолпируват-карбоксилаза, КФ 4.1.1.31, ортофос-
фат : оксалоацетат карбокси-лиаза (фосфорилирующая).
[фосфоенолпируват-]-СО2+Н2О = оксалоацетат4-орто-
фосфат]
Малатдегидрогеназа, КФ 1.1.1.37 (L-малат—э-NAD—
траисгидрогеиаза), L-малат : NAD+ оксидоредуктаза.
[OKcanoa4eTarr-]-NADH-]-H+=L-ManaT-|-NAD+]
Фаза декарбоксилирования (NADP-малатдегидрогеназа)
Малатдегидрогеназа (декарбоксилирующая) (NADP+),
КФ 1.1.1.40 (яблочный фермент), L-малат: NADP+ ок-
сидоредуктаза (декарбоксилирующая оксалоацетат).
^-малат+ЫАОР+==пируватН-СО2-[-^АОРН-|-Н+]
37*
572
ПРИЛОЖЕНИЕ Б
Фаза декарбоксилирования (NAD-малатдегидрогеназа)
Малатдегидрогеназа (декарбоксилирующая), КФ 1.1.1.39
(яблочный фермент), L-малат: NAD+ оксидоредуктаза
(декарбоксилирующая).
[Ь-малат+ЫАО+—пируват+СО2+КАОН+Н+]
Фаза декарбоксилирования (ФЕП-карбоксикиназа)
Аспартатаминотрансфераза, КФ 2.6.1.1 (глутамико-щаве-
левоуксусная трансаминаза), L-аспартат : 2-оксоглута-
рат аминотрансфераза.
[Е-аспартат-|-2-оксоглутарат=оксалоацетат-|-Е-глутамат]
Фосфоенолпируват-карбоксикиназа (АТР), КФ 4.1.1.49
(фосфопируват-карбоксилаза АТР), АТР : оксалоацетат
карбокси-лиаза (трансфосфорилирующая).
[АТР+оксалоацетат=АВР+фосфоенолиируват+С02]
Превращение С3-продукта декарбоксилирования
в крахмал
Пиру ват,ортофосфат—дикиназа, КФ 2.7.9.1, АТР : пиру-
ват,ортофосфат фосфотрансфераза.
[АТР+пируват+ортофосфат=
=АМР+фосфоенолпируват+пирофосфат]
Примечание-, фермент, предположительно участвующий
в превращении пирувата в ФЕП после декарбоксилиро-
вания, катализируемого NADP-или NAD-малатдегидроге-
назой.
Енолаза, КФ 4.2.1.11 (фосфопируват-гидратаза), 2-фос-
фо-В-глицерат гидро-лиаза.
[фосфоенолпируват+Н20=2-фосфо-В-глицерат]
Фосфоглицеромутаза, КФ 2.7.5.3 [глицерат (3->2)—фос-
фомутаза], 2,3-бисфосфо-В-глицерат: 2-фосфо-В-глицерат
фосфотрансфераза.
'['2,3-бисфосфо-В-глицерат-[-2-фосфо-В-глицерат=
= 3-фосфо-В-глицерат+2,3-бисфосфо-В-глицерат]
Фосфоглицерат-киназа, КФ 2.7.2.3 (1,3-дифосфоглице-
рат—>ADP—трансфосфатаза), АТР : З-фосфо-В-глицерат
1-фосфотрансфераза.
[АТР-|-3-фосфо-Ь-глицерат=
= ADP+3-фосфо-В-глицероил фосфат]
номенклатура ферментов
57-
Глицеральдегидфосфатдегидрогеназа, КФ 1.2.1.12 (трио-
зофосфатдегидрогеназа), ' D-глицеральдегид-З-фосфат:
: NAD+ оксидоредуктаза (фосфорилирующая).
[О-глицеральдегид-3-фосфат-]-ортофосфат-|-НАО+=
3-фосфо-0-глицероилфосфат-]-КАОН-|-Н+]
Дальнейшие превращения триозофосфатов в крахмал
представлены в разд. Б.4.
Б.7. Обмен нитратов
Нитратредуктаза (NADH), КФ 1.6.6.1 (ассимилирующая
нитратредуктаза), NADH : нитрат оксидоредуктаза.
[КАВН-|-нитрат+Н+=ЫАО+-|-нитрит-|-Н2О]
Нитратредуктаза [NAD(P)H], КФ 1-6.6.4 (NADH—>-нит-
рит траисгидрогеназа), NAD (Р)Н : нитрит оксидоредук-
таза.
[3NAD (P)H+3H++hhtPht=3NAD (P)++NH4OH+H2O]
Глутаминсинтетаза, КФ 6.3.1.2, L-глутамат: аммиак лига-
за (образующая ADP).
[АТР-|-Ь-глутамат-|-КНз=
=АБР-|-ортофосфат-]-Ь-глутамии]
Глутаматсинтаза (ферредоксин), КФ1.4.7.1, L-глута-
мат: ферредоксин оксидоредуктаза (переаминирующая).
[Ь-глутамин-|-2-оксоглутарат-|-2 восстановленный ферре-
доксин=2 глутамат+2 окисленный ферредоксин]
Глутаматдегидрогеназа [NAD(P)+, КФ 1.4.1.3 (дегидро-
геназа глутаминовой кислоты), L-глутамат: NAD (Р) +
оксидоредуктаза (дезаминирующая).
[L-^yTaMaT-]-HzO-]-NAD (Р)+=
= 2-оксоглутарат+НН3-|-МАО (Р) Н-]-Н+]
Список сокращений
ADP AMP НФС ИФС АТР Стпуть Ct-растения Аденозиндифосфат Аденозинмонофосфат Наблюдаемый фотосинтез Истинный фотосинтез Аденозинтрифосфат Путь превращений С.;-дикарбоновых кислот Растения, имеющие С4-путь превращения дикар- боновых кислот
Сз-растения Растения, ассимилирующие СО2 непосредственно в ВПФ-цикле
САМ Метаболизм кислот у растений сем. толстянковых (Crassulaccae)
ЭК CF0 Эффективность карбоксилирования Компонент сопрягающего комплекса, обеспечиваю-
CFi щий прохождение протонов через мембрану Компонент сопрягающего фактора, катализирую- щий синтез АТР
Хл ДБМИБ Хлорофилл Дибромтимохинон-2,5-дибром-3-метил-6-изопро- пил-п-бензохинон
ДХММ ДХФИФ (ДФИФ) ДГАФ ДФГК дтт Фд или Fd ФБФ ФГА или ГЗФ Г6Ф NAD+ 3-(3',4'-Дихлорфенил)-1,1-диметилмочевина Дихлорфенолиндофенол Дигидроксиацетонфосфат 1,3-Дифосфоглицерат Дитиотрейтол Ферредоксин Фруктозо-1,6-бисфосфат Г лицеральдегид-3 -фосфат Глюкозо-6-фосфат Никотинамидадениндинуклеотид (окисленная фор- ЖХО 1
NADH ИЛИ NADH21 Растения NAD-МДГ-типа Md ) Никотинамидадениндинуклеотид (восстановленная форма) Растения, у которых основной декарбоксилазой является NAD-малатдегидрогеназа (декарбоксили- рующая)
1 В восстановленной форме NAD и NADP содержат по два электрона и одно-
му протону (разд. 5.15). Наиболее точно эту форму отражает запись:
NADPH+H+. Однако чаще используют обозначения NADH2 и NADPH2.
СПИСОК СОКРАЩЕНИИ
575
NADP+ Никотинамидадениндинуклеотидфосфат (окислен- ная форма)
NADPH или NADPHs Растения NADP-МДГ-типа Никотинамидадениндинуклеотидфосфат (восста- новленная форма) Растения, у которых основной декарбоксилазой является NADP-малатдегидрогеназа (декарбок-
ОА Р680 Р700 Р/2е ФЕП Растения ФЕП-КК-типа 2-ФГК ФГК, 3-ФГК Pi пдс МФС ФС I, ФС II РР1 Q кв Га силирующая) Оксалоацетат Реакционный центр хлорофилла фотосистемы II Реакционный центр хлорофилла фотосистемы I Отношение АТР к двум электронам Фосфоенолпируиат Растения, у которых основной декарбоксилазой является ФЕП-карбоксикнназа 2-Фосфоглицерииовая кислота З-Фосфоглицериновая кислота Неорганический фосфат (ортофосфат) Протондвижущая сила N-Метилфеназоиийметосульфат Фотосистемы I и II Неорганический пирофосфат Первичный акцептор электронов фотосистемы II Квантовый выход Сопротивление пограничного слоя листа для диф-
Га' фузии СО2 Сопротивление пограничного слоя листа для диф-
РуБФ Гт ВПФ-путь гв Гв' ЦТК X 613С фузии воды Рибулозобисфосфат Сопротивление мезофилла для фиксации СО2 Восстановительный пентозофосфатный путь Сопротивление устьиц для диффузии СО2 Сопротивление устьиц для диффузии воды Цикл трикарбоиовых кислот Первичный акцептор электронов фотосистемы I Величина, характеризующая изотопный состав уг-
г лерода (13С/12С). Углекислотный компенсационный пункт, или СО2-
ПОФ компеисациониая точка Переносчик ортофосфата
Указатель латинских названий
Acanthaceae 343
Agavaceae 490
Agave americana 517
Agropyron repens 344
— trichophorum 469
Aizoaceae 343, 490
Amaranthaceae 343
Amaranthus 344, 477
— edulis 386
— graecizans 478
— retroflexus 314, 323
Ananas comosus 489
Andropogon geradii 345
— ischemum 282
— scoparius 345
Andropogoneae 345, 346
Aristida 346 ' ’
Aristoideae 345, 346, 436
Asclepiadaceae 490
Aspergillus japotilcus 554
Asteraceae 343
Atriplex has tat a 282, 477
— hortensis 282
— nitens 282
— rosea 282
— sabulosa 474
— spongiosa 308, 386, 477
Avena sativa 306, 338
Belemnitella americana 510
Beta vulgaris 281, 282
Boraginaceae 343
Bouteloua 346
— curtipendula 345
— gracilis 345
Brachiaria 346
Brassica oleracea 282
Bromeliaceae 490
Bromus inermis 469
Bryophyllum 494, 497, 499, 502
— calycinum 500
Buchloe 346
— dactyloides 345
Butaceae 490
Cactaceae 490
Capparidaceac 343
Caryophyllaceae 343, 490
Cenchrus 346
— ciliaris 469
Chenopodiaceae 305, 343, 490
Chenopodium album 344, 478
— bonus-henrlcus 282
Chlorella 43, 62, 75, 76, 294, 42-
Chlorideae 346
Chloris 346
— gayana 308, 323, 469
Citrullus vulgaris 478
Ccminelina 527, 528
Convolvulaccae 490
Compositae 490
Crassula argenta 517
Crassulaceae 488
Cucumis sativus 478
Cucurbitaceae 490
Cyndon dactylon 344
Cyperaceae 305, 343
С у per us 305, 344
— rotundas 344
Dideraceae 490
Digitaria 312, 346
— argyrogranta 469
— sanguinalis 362—364
Echinochloa 346
— colonuin 344
— crusgalli 344
Egeria densa 315
Eichorina crassipes 344
Eleusine 346
— coracana 344
— indica 363, 556
Eragrosteae 346
Eragrostis 346
Eragrostoideae 305, 345, 346, 469
Erlochloa 346
Euchlaena 346
Euphorbiaceae 343, 490
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
677
Festuca arundinacea 469
Festucoideae 345, 469
Fimbristylis 305
Froelichia gracilis 314
Fucus 169
Galanthus nivalis 49, 152
Geraniaceae 490
Glycine max 282, 442, 477
Gomphrena celosoidea 307, 309
— globosa 282, 359
Gramineae 305, 343, 344
Halophila 315
Heliant hus antiuus 282, 517
Hordeum vulgare 282, 338, 364
Hydrilla verticillata 315
Imperala cylindrica 344
Ipomoea 344
Kalanchoe 498, 508
— daigremontla 515, 550
Labiatae 490
Lactobacillus plantarum 501
Lantana camara 344
Leptochloa 346
Liliaceae 490
Lycoperslcon esculentum 282
Maydeae 345, 346
Medicago sativa 477, 478
Mesembryanthemum cryslallinum 507,
508, 513, 550
Mimulus 441
Muhlenbergia 346
Nicotiana tabacum 282
Nyctaginaceae 343
Opuntia basilaris 519
— ficus-indica 519
Oryza sativa 478
Orchidaceae 490
Oxalidaceae 490
Paniceae 346
Panicoideae 345, 469
Panicutn 346
— bulbosum 282
— capillare 342, 474
— coloraium 469
— maximum 323, 342, 344, 469l
— tnillaceum 282, 323, 339, 342, 344,
346, 463
— texanum 343
— virgatum 345, 474, 477
Paspalum 346
— • dilatatum 469
Pelargonium 218
Pennisetum .346
— typhoides 45, 46, 344
Plwlaris arundinacea 469
Phaseolus 441, 477
— mango 282
— vulgaris 478
Piperaceae 490
Pisum sativum 282
Planlaginaceae 490.
Poa pratensis 469
Polygonaceae 305, 343
Polypodiaceae 490
Potamogeton pectinatus 316
Porlulaca oleraceae 282, 314, 323, 337,
342, 478
Portulacaceae 343, 490
Portulacaria afra 508, 517
Quercus robus 517
Khizopus 550
Ricinus 169, 171
Saccharum 346
— officinarum 323, 335, 338, 477
Scenedesmus 123, 294, 295
Scrophulariacea 343
Sedum 498, 508
— praealtum 550, 556
Setaria 344, 346
— ilalica 478
— lutescens 338
— sphacelata 469
Solatium tuberosum 477
Sorghastrum 346
— natans 345
Sorghum 346, 442
— almum 469
— bicolor 323, 335, 338, 341, 477
— dochna 282
— halepense 344
— sudanense 337
578 УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
Spartina townsendii 474
Splnaceae oleracea 546
Sporoboleae 346
Sporobolus 346
Stipa hyalina 469
Syringa vulgaris 307
Thalassia testudinum 315
Tidestroma oblongifolia 474
Trichoderma viride 550
Triodia irritans 377
Triseium spicatum 4691
Trlticum aestivum 338, 363, 477, 478
Tulipa 527
Uloa 169
Urochloa 346
— panicoides 363, 556
Vicia faba 517, 526, 528, 529
Vitaceae 490
Welwitschiaceae 490
Xantium 344
Zea mays 306, 323, 335, 338, 441, 478,
517
Zoysia 346
Zoysieae 346
Zygophyllaceae 343
Предметный указатель
Автокатализ 166
— «производительная» реакция 113,
115, 214
— роль в индукции 180
Аденилатциклаза 322
— активация светом 381
Аденилаты, переносчик 254
Аденозиндифосфат (ADP) 31
Аденозинмоиофосфат (АМР) 31
Аденозинтрифосфат, ассимиляционная
сила 31
— богатый энергией 29
— гидролиз 30
— использование в фотосинтезе 87
— образование 84
— роль в индукции 184
— свободная энергия гидролиза 29,
30, 106
— синтез 80
----квантовый расход 108, 109
---- протонный градиент 101
----термодинамика 106
•---эксперимент Ягендорфа 101
— стабильность с точки зрения резо-
нанса 29
— транспорт в хлоропластах 243, 244
— фосфорилирование фосфоглицери-
новой кислоты 146
Аденозинтрифосфат/аденозиидифос-
фат (ATP/ADP), отношение, влия-
ние на ФГК-зависимое выделение
кислорода 199, 200, 209
— регуляция фотосинтеза 286, 289
Аденозинтрифосфатаза (АТРаза) 108
Аланин, влияние на проницаемость
хлоропластов 230
— образование аспартата 322
Аланин — аминотрансфераза 322
Аллостерический контроль 384
Альдолаза (фруктозобисфосфат-аль-
долаза) 240, 393
— катализируемые реакции 135, 136,
138, 143
Амилаза 156
Амилоза 156. См. также Крахмал
Амилопектин 156. См. также Крахмал
Аминокислоты, транспорт в хлоропла-
сты 243
а-Аминолевулиновая кислота 53, 54
Аммиак, выделение при фотодыхании
148, 149
— как разобщитель транспорта элек-
тронов 227
Аммония соли, поглощение 373
Анаболизм 20
— роль хлоропластов 214
Антимицин А 92, 380
Арсенат 104
Аспартат, декарбоксилирование 358
----у ФЕП-КК-растений 361
— как первичный продукт С4-фото-
синтеза 296, 297, 354, 356
Аспартат — аминотрансфераза 322,
572
Ассимиляция азота С3- и (^-растения-
ми 387
Ассимиляционная сила 31, 44, 146, 214
----восстановление фосфоглицери-
новой кислоты 146
-------СО2 86
----генерация 84—110
----использование у С4-растеиий
367
----квантовый расход 108
Атмосфера, газовый состав 449—451
Бактерии фотосинтезирующие 86
— хемосинтезирующие 86
Белок, содержащий железо и серу 58
— фракции I 473. См. также Рибуло-
зобисфосфат-карбокснлаза
Бензольное кольцо, распределение
электронов 27
----способы изображения 27, 28
Бикарбонаты, осмотическое сжатие
хлоропластов 233
— растворимость, влияние pH 453 :
Бунзена коэффициент 450
580
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Валиномицин
Виолаксантии 54
Вода, окислительно-восстановитель-
ный потенциал 33
— энергия образования и диссоциа-
ции 26
---- переноса водорода 27
— эффективность использования у
Сз- и С4-растений 478—480
Водный стресс и гликолатный путь в
С4-р астениях 424
Водород — кислород, соединение, вы-
свобождающаяся энергия 23
Возбужденное состояние 67—70
Восстановительный ' пентозофосфат-
ный цикл (ВПФ-цикл, цикл
Кальвина — Бенсона) 32, 113—
162
:------автокатализ 113, 115, 214
—------деградация крахмала 156
— •----затрата энергии 83, 413,
504
--------изменения в содержании
ФГК и РуБФ 126
-----------свободной энергии Г42—
144
— ------ингибирование пентозомопо-
фосфатом 239
--------карбоксилирование 113
--------кинетические исследования
125
----— локализация 328, 329
•-------открытие и употребление
терминов 122
—-------- повторное включение трп-
- озофосфата 147
--------промежуточные продукты
128
гу- — — путь углерода 127, 128
— — — расход АТР ‘32, 108, 109
-------- регенерация . метаболитов
.113, 4'15 . 7 ’. '
— — — "регуляция фосфоглицёрат-
киназой 208
--------связь с фотодыхательным
циклом .окисления углерода 147'
— ------синтез крахмала 152 >
е—------1 сахарозы 158
— — — схема 114 .
--------участие углерода 114, 128,
147, .152 ...
--------фазы 114 \ я
--------ферменты 128^.153,: .563
-—>-----фиксация углерода 115
--------оункционированйе J45''
— энергетика ЦО '
Вуда и -Беркмана реакция 494 t
)'у нкци о пиров анйе ,1451
Гексозомоцофосфаты 239, 394
Гексозофосфат-изомераза 154
Гендерсона — Хассельбаха уравнение
233, 452
Гибридизация С3- и С4-видов 348
Гибридные орбитали 29
Гликоальдегидфосфат 302
Гликолат 405
Гликолатный путь у С3-растеиий 407,
409
—-- -----глюкопеогепетическая
функция 407
— — ингибирование 426
— .-------локализация ферментов
415
---------митохондрии 417, 421
---------связь с обменом в хлоро-
пластах 420
—-------- — челночный перенос мета-
болитов 418
пероксисомы 421
— — -----роль 439
---------синтез сахарозы 419
-------С4-растений 422
----------при водном стрессе 424
----------ферменты 409—413, 564
Гликолатоксидаза 409, 564
Глиоксилатпый цикл 538
Глиоксилатредуктаза 410, 565
Глиоксисомы 416
Глицеральдегид как ингибитор асси-
миляции углерода 229, 241
Глицеральдегидфосфат 134, 135
Глицеральдегидфосфатдегидрогеназа
563, 573
— NADP-зависимая, активация све-
том 187, 268
Глицерат-1,3-бисфосфат 133
Глицераткиназа 410, 565
Глицериновая кислота, транспорт в
хлоропласты 243
Глицин 408, 409, 417
— транспорт в хлоропластах 243
Глицинамииотрансфераза 409, 565
Глицнп-сиитаза 409, 565
Глутамат 354, 358—361, 409, 410
Глутаматдегидрогеназа 388, 573
NAD-глутаматдегидрогеназа 392
Глутаматсиптаза 573
Глутамиисинтетаза 149, 388, 573
Глутатион, инактивация ферментов
264, 265
ч— темновое окисление 271
Глюкозр, выделяющееся тепло 25
'— транспорт в хлоропласты 240, 241
ADP-глюкоза 154, 155 . •
АО^-гуирцоза-крахмалсянтетаза 393
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
581
UPP-глюкоза 158
Глюкозо-1-фосфат — адеиилнлтраис-
фераза 154, 157, 234, 393
Глюкозо-1-фосфат — уридилплтраис-
фераза 393, 394
Глюконеогенез 118, 407
Горох, водная культура 548
Грамицидин D 104
Двойное карбоксилирование 501
Двуокись углерода (СО2), акцептор R
115
---- и накопление органических кис-
лот 502
-------рост Сз-растений 440
-------фотосинтез Сз- и С4-растеиий
447
---- перенос карбоангидразой 454,
465
----поступление к месту ассимиля-
ции углерода 231
----растворимость 452, 453
----резонансные структуры 116
---- сопротивление листьев потоку
СО2 477
----фиксация в реконструирован-
ных хлоропластах 204, 205, 209
—-------- зависимость от пирувата
363, 364
------- ингибиторы 368
------- индукция 201
-------при фотосинтезе 115
Действие масс, регуляция 273—278
Декарбоксилирование в клетках об-
кладки проводящих пучков 303
— С4-КИСЛОТ 293
— перепое углерода 325, 326
— у злаковых 345
Декарбоксилирующие ферменты, ре-
гуляция 385
Декстран 555
Детергенты 104
Дибромметилизопр оп ил беиз охипои
(ДБМИБ) 66, 92, 99
Дигидроксиацетонфосфат 135
— ингибирование транспорта фосфа-
та 252
— снятие ингибирования фотосинте-
за, вызванного ортофосфатом 192
— сокращение, индукции 183
— структура 135
— участие в челночном механизме
253, 255, 331
Дигитонин 66
Дикарбоновые кислоты, кинетические
' ” константы 254 ' ,
---- метаболизм 301 .
----переносчик 243, 253, 374, 376
----.— и челночный механизм 254
Диссипациоппые потери света 45
1,3-дифосфоглицериповая кислота
(ДФГК) 133
---- образование 167
Дифференциальное центрифугирова-
ние 416
Дихлорфевнлдиметилмоч евина
(ДХММ) 66, 91, 93, 380
— ингибирование фиксации СО2 368
— место действия 98
Дихлорфеполиндофепол (ДХФИФ)
78, 83, 93
Додецилсульфат 64
Донор электронов для фотосистемы 11
(Z) 56
Дыхание, образование Н — О-связей
13
— плодов 533
— роль транскетолазы 137
— темповое 43, 470
Дыхательный коэффициент 491
Енолаза 296, 322, 571, 572
Железо, окислительно-восстановитель-
ный потенциал 33
Закон действующих масс 22
-------и равновесие .22 i
Замыкающие клетки, раскрывание
устьиц 526
---- ферменты 528
Изоипкотипилгидразпп 408
Изопрен 52
Иммунофлуоресценция 312
Импульсная метка 296
Инактивация ферментов 185, 187, 270
Индукция в реконструированных хло-
ропластах 197, 205—207
----фотохимических процессах 173
— влияние изменения рГ1 171, 178
----интенсивности света 169, 170,
185, 188
----температуры 169, 170, 188, 189
— восстановление лаг-фазы 210
— выделения кислорода 19,5
— вызванная ортофосфатом 1,91
— кратковременная и длительная 168
— методы изучения 168, 169 ....
— молекулярные механизмы .17.8 —
582
ПРЕДМЕТНЫЙ указатель
— накопление экспортируемого мета-
болита 178—180
— последовательность событии 208
— при насыщении СОг 172
----удалении эпидермиса 172, 175
— простая 167, 168
— роль автокатализа 180
----ортофосфатов 189, 190, 234
----рпбулозобисфосфата 183, 185,
240
---- устьиц 170—172
— содержание АТР 184
— сокращение индукционного перио-
да 182—183, 225, 284
— фиксации СОг 201—204
— in vitro 177
Интенсивность света 38
Инфракрасная спектроскопия 168, 170,
172
Ионы, транспорт 245, 246
Ископаемое топливо, использование
человеком 14
Кальвина цикл 122, См, также Вос-
становительный пеитозофосфат-
иый цикл
Карбоангидраза как «защищенный»
фермент 233
— перепое СО2 454
— С4-растеиий, локализация 464
— роль в хлоропластах 233
Карбоксидисмутаза см. Рибулозобис-
фосфат-карбоксилаза (оксигеназ а)
Карбоксилирование в С4-пути 321
— восстановительное 116
— двойное 295
— константа равновесия 119
— фотосинтетическое, механизм 117,
122
— энергетика 116
p-Каротин, максимум поглощения 54
Каротиноиды 54
— и спектр действия фотосистемы 11
77
Катаболизм 20
Каталаза 95, 96, 409
Катализ, регуляция 261, 262, 269
Катализаторы, активация магнием
266—269
— инактивация глутатионом 264, 265
— медиаторы светового эффекта
266
Квантовый выход фотосинтеза 43
— расход 43, 44
----для Z-схемы 83
Кислород, влияние на квантовый вы-
ход фотосинтеза 434
-------фотосинтез 461
— выделение 70
----в отсутствие СОг 232
— —влияние аденнндипуклеотидов
287, 288
-------ATP/ADP 199
-------ФГК 199, 200, 234
— — индукция 195
----реконструированными хлоро-
пластами 195, 196, 198, 275—
277
----феррицианидзависимое 227
— и рост Сз-растений 440
— ингибирование фотосинтеза 424
Клетки обкладки проводящих пучков,
глнколатоксидаза 422
----------глиоксилатредуктаза 422
---- ----- декарбоксилирование 269,
370
---------- кукурузы 309
----------С4-растениЙ, ферменты
357
----------РуБФ-карбоксилаза 303
----------фотосинтетическая дея-
тельность 367
Клеточная стенка, состав 553
а-Коричные кислоты 374
Кории, метаболизм малата 534
Кранц-анатомия 304, 310
Кранц-клетки 305, 309, 314
Красный спад 75, 77
Крахмал 49
— визуальное обнаружение 217, 218,
220
— деградация 156
----в процессе САМ-метаболизма
498
---- регуляция 158
— изображение картин 218
— место образования 231
— накопление у кукурузы 309, 310
— образование в устьицах 220
— синтез в ВПФ-цикле 152
-------С4-растениях 393, 394
------ влияние маннозы 280—283
-------фосфатов 278—280, 284
----регуляция 153, 157, 158, 279
----ферменты 567, 568
---- энергетические затраты 504
Крахмал—синтаза 154, 393
Крахмал-фосфорилаза 156, 157, 393
Ксилоза, поглощение хлоропластами
240
Ксилулозо-5-фосфат (Ку5Ф) 138, 140,
141
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
583
Листья зеленеющие 534
— химический состав 14
Лютеин 54
Магний, активация катализаторов
266—269
-----фиксации СО2 в темноте 201
— ингибирование фотосинтеза 262
Малат, декарбоксилирование 358, 361
-----механизм 369, 371
— как продукт С4-фотосинтеза 295
— обмен в корнях 534, 535
— образование при САМ-метаболизме
497
— оксалоацетат, окислительно-восста-
новительный потенциал 33
— роль в устьицах 525
— синтез в С4-растещ1ях 353, 356
— транспорт, челночные механизмы
255
— фосфоеиолпируват, челночный об-
мен 359
Малатдегидрогеназа 565, 569, 571,
572
NAD-малатдегидрогеназа 324, 504,
572
— перенос углерода 326
— фотохимические затраты 329
NADP-малатдегидрогеназа (малик-
энзим, яблочный фермент) 296,
301, 303, 304, 324, 337, 338, 571
— активация светом 381, 382
— аллостерические свойства 386
— в САМ-метаболизме 496
— в плодах 531
— перенос углерода 326
— С4-растений 304
— фотохимические затраты 329
Малик-энзим см, NADP-малатдегид-
рогеназа
Малатсинтетаза в прорастающих се-
менах 539
Малонат, ингибирование фотосинтеза
295
Мальтоза, участие в деградации крах-
мала 157, 158
Манноза, влияние на С3- и С4-расте-
ния 282
— поглощение хлоропластами 240
— стимуляция синтеза крахмала
280—283
Марганец, недостаточность 56
— состояние в адаптированных к
темноте водорослях 72
Марганец-содержащий белок 55, 56,
71
NAD-МДГ-растения, замедленная
флуоресценция 342
— метаболиты в клетках обкладки
проводящих пучков 359
------С4-ццкла 355
— обмен между клетками 320
— фиксация СО2 363
— фотосистема 11 342
— хлорофилл/Р 700, отношение 342
NADP-МДГ-растения, декарбоксили-
рование малата и аспартата 358
— метаболиты клеток обкладки про-
водящих пучков 357
— — С4-цикла 354
— обмен между клетками 320
— перенос электронов 335
— распространение среди злаковых
347
— синтез аспартата 354, 355
— фиксация СО2 363
— ФС1/ФС11, отношение 340
— F08s/F73o, отношение 338
Медиаторы светового эффекта 266
Мезофилл С4-растеиий, использование
ассимиляционной силы 367
------ферменты 351—356
— фотосинтетическая деятельность
362
Мелера реакция 93, 95, 380
Мембрана, проницаемость для прото-
нов 103
— структура 49, 50
— тилакоидов, препараты 49
— — состав 49
-схема 50
— хлоропластов, проницаемость
247
Мембранный потенциал и транспорт
протонов 107
Менадион 90
Мерсалнл 373
САМ-метаболиэм в замыкающих
клетках 528
— влияние условий окружающей сре-
ды 486, 508, 509
— газообмен 518
— двойное карбоксилирование 501
— декарбоксилазы 505
— декарбоксилирование С4-кислот
503
— образование крахмала, энергетиче-
ские затраты 504
— — органических кислот 502
— открытие 490
— путь углерода 495
— распространение у растений 490
— регуляция 506, 507
584
предметный указатель
— связь с Ct-циклом 515
— фазы 509
— ферменты 498, 570
— экологическое значение 516—519
— энергетические затраты 503
Метаболиты, роль в индукции 178—
180, 193
— транспорт из стромы хлоропла-
стов 252
Метгемоглобин, восстанавливающий
фактор 59
Метилвиологеи (паракват) 90, 95
Мстиоиинсульфоксимин 388
Минеральные соли в растениях 14
Митохондрии, поглощение малата 371
— транспорт метаболитов 371
Михаэлиса — Ментен уравнение 119
С02-насос 294, 321, 465
Неоксантин 54
Нигериции 104
Никотниамидаденинди нуклеотид
(NAD) 109
—‘ восстановленный (NADH), роль в
дыхании и фотосинтезе 85
Никотинамидадениндинуклеотидфос-
фат (NADP), восстановление 109,
НО
— восстановленный 61
------ ассимиляционная сила 31
— — окислительно-восстановитель-
ный потенциал 33
----роль в дыхании и фотосинтезе
85
---- свободная энергия при окисле-
' ' Нии’27' А '
— как окислитель в реакции Хилла
65, 243
— свободная энергия при восстанов-
лении 27
— структура 60
— • транспорт 243 '
Нитрат, обмен, схема 391
•Ннтратредуктаза 387, 388, 573
— ассимиляция аммиака, энергетиче-
ские затраты 413
Обмен у толстянковых 162, 486—519.
Cjw. также САМ-метаболизм
Окисление, перенос электрона 10, 11
—' — Н2 10, 11
Окислительно-восстановительные ре-
акции 10, 11
Окислительно-восстановительный по-
тенциал 32, 33 .
Окислительное фосфорилирование 85
Окислительный пентозофосфатный
цикл 285
Оксалоацетат 300, 355, 368
— как первичный продукт фотосин-
теза 295
— малат, челночный обмен 255, 390
Оксоглутарат 322, 324—326, 356, 358,
360
— глутамат, челночный обмен 326
Органические кислоты, накопление
486
Ортофосфаты, ингибирование фото-
синтеза 189, 190, 214—238
--------защита пирофосфатом 234—
238
----------промежуточными продук-
тами 190—193
— переносчик 251
— роль в индукции 193
Осмотики 229, 240
Осмотическое давление 47, 48
Осмотическое набухание и поглоще-
ние 230
Остерхаута — Хасса гипотеза индук-
ции 166, 179
Паракват (метилвиологеи) 90, 95
Пектиназа 550, 554
Переаминироваиие 300
Перепое электрона при окислении 11
---- — электролизе 15, 16
----солнечные (фотоэлектрические)
батареи 16
— энергии путем резонанса 73
Пероксисомы,' ферменты гликолатного
пути 415,. 417 '
Пигментная система, перенос энергии
73
------ разделение 66
----содержание хлорофилла 63
— — состав 66 ... . .
Пигмент (ы) фотосинтетического ап-
парата 56
— Р 680 (Р 683, Р 690), возбужде-
ние и локализация 50
— как реакционный центр ФСП 57
— Р 700 55, 66 .
------ возбуждение и локализация "50
----п выделение О2 65 .
---- окислительно-восстановитель-
ный' потенциал 33, 65
— —свойства 59 ..
—---у-Сграстеннй 335 '
Пноцианин 80, 91
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
585
Пиридоксаль-5-фосфат, . ингибирова-
ние переносчика ортофосфата
251-253
Пирофосфатаза 236, 322
Пирофосфаты, влияние на хлоропла-
сты 238
— защита от ингибирующего дейст-
вия ортофосфата 234
— подавление фотосинтеза 238
Пируват — аланин, челночный обмен
326
— перенос, ингибитор 373
— переносчик 372
Пируваткниаза в клетках обкладки
проводящих пучков 325
Пируват, фосфат — дикиназа 572
— активация светом 381
— в САМ-метаболизме 496
Питательный раствор для водной
культуры 547
Плазмодесмы, транспорт метаболитов
376, 377
Плазмолиз 47
Плазмоцианнн 50, 59
Пластогидрохипоп — пластохинон,
окисление — восстановление 98
Пластохинон 50, 57, 58, 79, 94, 97
Пластоцианин, окислительно-восста-
новительный потенциал 33
Плоды, фотосинтез 530
Плотность потока квантов 41
— светового потока 37
Поглощение света атомами 67
--- молекулами 68
Подвижное сопряжение 82
Подсолнечник, индукция 174
Постоянная Планка 36
Поток излучения 41
Пропионовокислые бактерии, исполь-
зование СОг 494
Протондвижущая сила 80, 96—101
Протонный градиент 101
— насос 208
Протопласты, время хранения 552
Протохлорофиллид 53, 54
Пшеница, индукция 170, 173
— протопласты 177
pH, активность ферментов 186
— изменение под действием света
178
— индукция 208, 209 .
— синтез малата 537
— феррициаиид-зависимый сдвиг 179
Радиоактивных изотопов метод 122
Радиометрические измерения 41
Разобщители фотофосфорилирования
104
Сз-растения, газообмен 518
— поглощение изотопов углерода 510,.
511
— рост в средах, обогащенных СО2.
440—442
С4-растепия, газообмен 518
— систематика 343
CAM-растения, газообмен 518
— изотопы углерода 509—514
— темновая фиксация 400
— физиология устьиц 528
Реакционный центр фотосистемы I.
65, 334
-------11 65
Регуляция активации спетом 263—
266, 382
---катализаторов ионами магния
266—269
— действие масс 274—278
— значение 285
— катализа 261, 262, 269
— принципы 262
— темновой инактивации 271, 272
— транспортом метаболитов 278
— ферментов 381
— фотосинтеза 447
— фотосинтетической ассимиляции
углерода 262—289
Резонанс, бензольное кольцо 27, 28
— распределение электронов 27
Рибоза, влияние па проницаемость.
хлоропластов 230
— транспорт в хлоропласты 240
Рибозо-5-фосфат 140, 239
Рибозофосфат-нзомераза 564
— катализируемые реакции и свойст-
ва 140
Рнбулозобисфосфат, карбоксилирова-
ние 203
— роль в индукции 184, 185, 240
— снятие подавления фотосинтеза,
вызванного ортофосфатом 191,
192
— содержание при изменении кон-
центрации СО2 и освещения 126,
127
Рибулозо-5-фосфат (РубФ) 141, 275,.
276,278
Рибулозобисфосфат-карбоксилаза
(РуБФ-карбоксилаза). 53, 563
— активация магнием 266
— — светом 186, 187, 268 • 1
— в плодах 531
.------Сграстениях 302—304,. 463,
471
38-234
586
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
----хлоропластах 267, 268
— влияние Оз 455
— ингибирование ортофосфатом 193
— листьев 121
— локализация 312
— отношение к изотопам углерода
513, 514
— перенос СО2 454
— свойства 405
— сродство к СО2 113, 266
— участки связывания 462
Рибулозофосфат — 3-эпимераза, свой-
ства и катализируемые реакции
141
Рост растений, опыты Вт Гельмонта
13, 14
Сахара, транспорт в хлоропласты
240
•Сахароза, синтез в цитоплазме 158
----У С8- и Ci-растений 393, 394
---- ферменты 566
Сахарозофосфат-синтетаза 158, 163,
566
•Сахарозофосфатфосфатаза 393, 567
Световая энергия 9
----активация ферментов 167, 186.
263, 265, 266, 272, 273, 381, 382
----единицы 37
----гидролиз АТР 29, 30
----изменения в биохимических ре-
акциях 142
----------ВПФ-цикле 144
----и равновесие 22—24
----использование растениями 34
----максимальная эффективность
45
----расщепление связей 9, 12
----фотосинтез 12, 13
----фотосинтетически активная ра-
диация 37, 40, 41
Седогептулоз о-1,7-бисфосфат 139
Седогептулозобисфосфатаза (СБФ-
аза) 139, 209, 564
Седогептулозо-7-фосфат 139
Сельскохозяйственные культуры, го-
дичная потребность человека 42
Серин, образование при фотодыхании
148, 149
— транспорт в хлоропластах 243
Серин-гидрокснметилтрансфераза
565
Серин-глиоксилат — аминотрансфе-
раза 409, 565
Солнечные батареи в космических ис-
следованиях 16
Солнечный свет, максимальная интен-
сивность 39, 40
----плотность потока у поверхно-
сти моря 39
---- фотосинтетическая активность
45
Сопротивление для диффузии СО2
476, 477
--------воды 475, 476
Сопрягающий фактор 102, 103, 105
----роль в фотофосфорилировании
105, 106
---- электростатический комплекс
104
Сорбитол, влияние на проницаемость
хлоропластов 230
Сорные травы, распространение Ci-ра-
стений 344
Спектр действия 74—77
----выделение кислорода 75
----красный спад 75, 77
---- хлорофилла 74
----эффекта усиления 78
— испускания хлоропластов 66
— поглощения и температура 68
---- хлоропластов 76
Средняя годовая облученность для
Англин 46
Стоксов сдвиг 70
Сукцииил-СоА и синтез хлорофилла
54
Сульфаты, переносчик 246
Супероксидный радикал 96
Z-схема 55, 78—81. См. также Фото-
система I, Фотосистема II
— аналогия 81
— влияние пиоциаиина 91
— запасание энергии 82
— квантовый расход 83
— образование АТР 81, 82
— пути транспорта электронов 90
— энергетическая диаграмма 79
— эффект усиления 77
Темновая инактивация ферментов
387
Температура, сопротивление устьиц
и мезофилла 581
Термодинамика, законы 19
— необратимая 20
— открытых систем 20
н-Тетраметил-п-фенилендиамин 95
Тилакоиды, ассимиляционная сила
31
— изменения pH 99
предметный указатель
587
— мембрана 49
— образование АТР 216
— осмотическое равновесие 48
— переносчик электронов 50
— происхождение слова 47
— эффект освещения 97, 98
Тиоредокснн 208, 266
— активация ферментов 264, 265
Тпофосфат 104
Толстянковые, обмел кислот, схема
487. Сж, также САМ-метаболизм
Транскарбоксилирование в С4-растс-
ниях 302, 303
Трансгидрогеназа 535
Транскетолаза 564
— катализируемые реакции 137, 140
— свойства 137
— субстраты 154
Транскрипция у САМ-растений 517
Транспорт внутриклеточный у САМ-
растений 506
— метаболитов 230
---- в хлоропласты 239
-------ходе С4-цикла 326
----изучение in vitro 221
----межклеточный 376
----методы изучения 216—221,
225
----регуляция ассимиляции СО2
278
----у С4-растений 371—378
----функция хлоропластов 214—
216
— протонов и транспорт электронов,
связь 107
— электронов в хлоропластах 90
---- нециклический 335
----псевдоциклический 84
----ускорение с помощью разобщи-
телей 102
----фосфорилирование, сопряжение
104
— — циклический 84
Трииитробеизолсульфоиат 251
Трнозофосфатдегидрогеиаза, свойст-
ва и катализируемые реакции
134
NADP-триозофосфатдегидрогеиаза,
активация светом 382
— локализация 329
— челночный механизм 39
Триозофосфат-изомераза 563, 566, 567,
571
— свойства и катализируемые реак-
ции 135
Триозофосфаты 153, 154, 156, 157, 240
— роль в индукции 193—195
— транспорт через оболочку хлоро-
пластов 216
Тритон Х-100 66
Тушитель флуоресценции (Q) 57, 72
Углекислотный компенсационный
пункт 149, 403, 404
Устойчивость к затоплению и синтез,
малата 535
Устьица злаков 526, 527
— образование крахмала 220
— открывание 526
— поступление малата 527
— роль в индукции 170—172
— силиконовые отпечатки 174, 175
— синтез малата в замыкающих,
клетках 525
— строение 526, 527
— хлоропласты 528, 529
— частота 517
Факторы внешней среды, регуляция
фотосинтеза и транспирации
477—482
Фепазиимоиосульфат (ФМС) 91
ФЕП-КК-растения, декарбоксилирова-
ние аспартата и малата 361
— распространение 346
— синтез аспартата 355
----- малата 356
— транспорт продуктов метаболизма
320
— фотосистема II 342
D-фермент 156, 157
R-фсрмеит 156, 157
Ферментативная реакция, влияние-
концентрации субстрата 119, 121
-----Ужо 119
Ферменты азотного обмена 387
— активация 186, 263 , 268—270
— в клетках мезофилла 323
— гликолатного пути 409—413, 422
— замыкающих клеток 528
— «защита» от окружающей среды
228
— инактивация 185, 187, 270
— - САМ-метаболизма 498
— номенклатура 563
— прорастающих семян 539
— разрушающие целлюлозу, препара-
ты 554
— регуляция 261, 262, 269, 270
— сродство к субстрату, /См 120
— у С4-растений 301, 351—356
Ферредоксин 50, 59
38’
588
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
— восстановление NADP 109
— катализ циклического фотофосфо-
рилирования 92
— окислительно-восстановительный
потенциал 33, 59, 60
ферредоксин-NADP — редуктаза 50,
60, НО
Феррицианид и сдвиг pH 179
— как окислитель в реакции Хилла
72
Фильтрация центрифугированием 222,
226
Флуоресценция в хлоропластах 338,
374
.— хлорофилла 70
Фосфаты и ассимиляция углерода
278—280, 283
— переносчик 193, 200, 215, 247
•— транспорт в хлоропласты 248, 252
----кинетические константы 248
.---стехиометрия 249
Фосфогликолатфосфатаза 564
,2-фосфогликолевая кислота 131, 405
Фосфоглнцераткиназа 133, 563, 571,
572
— активация светом 187, 268
— катализируемые реакции 133
— локализация 329
— регуляция ВПФ-цикла 208
— роль в индукции 208
Фосфоглицериновая кислота (ФГК.)
296
---- в процессе индукции 185
---- влияние на выделение О2 199
---- восстановление в хлоропластах
197
-------регуляция 200, 274, 275
----дигидроксиацетонфосфат, чел-
ночный обмен 321
----и выделение О2 232, 288, 289
----— фиксация СО2 364, 365
---- ингибирование транспорта фос-
фата 258
----как окислитель в реакции Хил-
ла 201, 234
----образование в ВПФ-цикле 113,
114, 125, 126, 295, 297
---- при изменении концентрации
СО2 и освещения 126, 127
----снятие ингибирования фотосин-
теза, вызванного ортофосфатом
191—192
---сокращение индукционного пе-
риода 228
----транспорт в хлоропласты 249
Фосфоглицератфосфатаза (ФГК-фос-
фатаза) в Сз- и С4-растепиях
423
Фосфоглицеромутаза 296, 322, 571,
572
Фосфоглюкомутаза 154
Фосфоеиолпируват (ФЕП) 296
— гидролиз, высвобождаемая энер-
гия 117
— транспорт в хлоропласты 374
— фотосинтетическое карбоксилиро-
вание 117
Фосфоенолпируват-карбоксикпназа
(ФЕП-КК) 118, 321, 572
— декарбоксилирование 361, 370,
387
— ингибитор 369
— обогащение изотопов углерода
512—514
— перенос углерода 326
— С4-растенпй 304, 320
— флуоресценция 342
— фотохимические затраты 329
— хлорофилл/Р 700, отношение 342
Фосфоеиолпнруваткарбоксилаза
(ФЕП-карбоксилаза) 120, 121,
233, 295, 301, 571
— в плодах 531
--- устьицах 527
— ингибирование малатом 507
- - локализация 312
— регуляция 381, 383
— уровень в Сз-растенпях 304, 385
С4-раетеинях 304, 325, 385,
463—465
— участие в САМ-мстаболпзме 496—
499
— формы 499
— эволюция 439
Фосфолипиды 51
Фосфофруктокпназа 158
Фосфорпбулокнназа 146, 564
— активация светом 187, 268, 382
— локализация 329
— свойства и катализируемые реак-
ции 141
Фосфорилирование 80. См. также
Окислительное фосфорилирова-
ние
— протоидвнжущая’сила 101
— разобщение 104
— циклическое 88
Фотодыхание 399—441
— выделение СО2 402
---NH3 148, 149
— гликолатный путь 409—41’4
— зависимость от температуры 43 -
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
589
— и ВПФ-ЦНКЛ, взаимодействие 147
— методы измерения 430, 431
— повторное использование аммиака
412
— поглощение О2 403
— роль 438—440
— синтез серина 148, 149
— увеличение концентрации СО2 401
— участие углерода 152
Фотолиз воды 9, 12, 87
----в небиологической системе 16
Фотометрические измерения 41
Фотон 41, 42
Фотоиасыщепие 62
Фотосинтез в колоске злаков 529
---- плодах 530
— и урожайность сельскохозяйствен-
ных культур 581
— ингибирование кислородом 400,
424, 429, 434
----обусловленное фотодыханием
429
— как источник органического угле-
рода 13, 14
----окислительно-восстановитель-
ный процесс 12
— в листьях в процессе позеленения
534
— насыщение светом 474
— разделение на две фазы 31, 47
— разрыв Н — О-связи 13
— регуляция факторами внешней
среды 468—474
— стационарная фаза 210
— скорость при оптимальных усло-
виях 544
----У С3-растепий 46, 465
----— С4-растепий 465
— уравнение 9, 11, 86
— фотонасыщение 62
- Qio 471
Cj-фотосинтез, квантовый выход
434—437
— модель 429, 457—460
— синтез крахмала 282
Сгфотосинтез, модель 458—460, 466
— открытие 293
— фотохимические затраты 329
САМ-фотосинтез, внутриклеточный
транспорт, модель 506
Фотосинтетическая единица и пиг-
ментная система 61, 63
----содержание хлорофилла 62
— эффективность 45
---- иа молекулярном уровне 44
т- — при синем свете 44
Фотосинтетически активная радиация
(ФАР) 37, 40, 41
Фотосинтетические ферменты, актива-
ция светом 166, 167, 263, 265, 266,
272, 273, 381, 382
Фотосинтетический аппарат, транс-
формация энергии 54
— контроль 104, 105
Фотосистема I (ФС1) 50, 57, 58, 63—
65
---- акцептор электронов 59
----квантовый выход флуоресцен-
ции 67
— — спектры действия для нее 76
----—испускания 66
Фотосистема I/фотосистема II, отно-
шение 334
Фотосистема II 50, 56—58, 63—65
------- ингибиторы 91
----марганецсодержащий белок 55
---- расположение 98
----спектр действия для нее 76
-------- испускания 66
Фотофосфорилирование 82
— нециклическое, доноры и акцепто-
ры 88, 93
— образование АТР 379
— при С4-фотоснитезе 379
— псевдоциклическое, определение и
схема 92
— сопряжение 84
— циклическое 82, 89—92
---- катализ ферредоксином 92
----кофакторы 90
------- in vitro 92
Фотохимический аппарат, структура
---- функции 55, 67
Фотоэлектролиз 16, 17
Фракционирование тканей 216—221
Фруктоза, поглощение хлоропластами
240
Фруктозо-1,6-бисфосфат 135
— аккумуляция хлоропластами и pH
262, 263
— гидролиз 136
— и выделение О2 275—277
Фруктозо-бисфосфатаза (ФБФ-аза)
566
— активация светом 137, 158, 187,
209, 268, 271, 272, 382
— катализируемые реакции 136
— свойства 137
— темповая инактивация 209, 271,
272
— холостой цикл функционирования
286
590
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Фруктозобисфосфат-альдолаза 563
Фруктозо-6-фосфат 136, 137, 153, 154,
156—158
Фруктозо-6-фосфаткиназа, холостой
цикл функционирования 286
Фут-свеча 37, 39
Хемиосмотическая гипотеза 89, 97,
101, 106
----градиент pH 102
Хемоосмос и протондвижущая сила
96—100
Хилла реакция, акцептор электронов
71, 72
---- в клетках обкладки проводя-
щих пучков 339
---- индукция 179
----окислители 61, 66, 284
---- скорость 173
---- уравнение 93
----фотохимия 71
Химические связи 9, 25, 26
Хлор-ион и фотосистема II 56
Хлорелла, спектр действия для выде-
ления Ог 75
Хлоропласты, влияние валиномицина
245
— возникновение 53
— восстановление NAD и NADP 109
— выделение 545, 552
---- из протопластов 551, 552
---- среда 550
— гликолатный обмен 420
— граны 47, 49
— деградация крахмала 157
— изолированные, образование кис-
лорода 87
— индукция 169, 176, 185, 197
— как самостоятельные фотосинтези-
рующие органеллы 231
•---транспортирующие органеллы
214—216
— межмембранное пространство 226,
247
— мембрана 47
— методы механического и фермента-
тивного разрушения клеток 555
•---отделения от цитоплазмы 221
— осмотические изменения объема
229
— очистка 560
— поглощение фосфорилированных
метаболитов 239
— получение из протопластов 550
— реконструированные, выделение
О2 195
— — индукция 205—207
— разрушение ткани 549
— Сграстений, декарбоксилирование
347
— — замедленная флуоресценция
339
----мезофилла 322, 337
•---обкладки проводящих пучков
337
— — свойства 338
---- спектры низкотемпературной
флуоресценции 341
•---типы 301
----фиксация СО2 338, 363, 364
— реакция на экзогенный фосфат
235
— сжатие, вызванное бикарбонатом
233
— спектр поглощения 76
— степень интактности 557, 558
— строма 47, 49
— структура 47
— транспорт АТР и NADP 243
— — ионов 245, 246
— — сахаров 240
----фосфатов, кинетические кон-
станты 248
---- электронов 90
— феррицнанидзависимое выделение
О2 227
— фиксация СО2 при осмотическом
шоке 201
— фотохимические различия 333
— хроматография 222, 223
— челночные механизмы 254—256
— электронная микроскопия 224
Хлорофилл, возбужденное состояние
10, 68—70, 72, 74
— локализация 47
— максимум поглощения 56
— обесцвечивание 70
— перенос энергии 73
— разделение на колонках 52
— светособирающие антенны 78, 79
— синтез 53, 54
— снятие возбуждения 70
— спектр действия 74
----поглощения 53
— структура 52
— флуоресценция 70
— число молекул на фотосинтетиче-
скую единицу 61, 62
— экстракция 218
— а 52
----спектр поглощения 53
— а/b, отношение и активность фо-
тосистем 334, 338
предметный указатель
591
— b 51, 52
— — спектр поглощения 53
Хлорофилл/Р 700, отношение 338, 340
гг-Хлорфепилртутьсульфонат 251
Холостой цикл функционирования 286
Хроматографическая карта 124
Хроматография 123, 222, 223
— водорослей 222
Целлюлоза 550, 554
Центрифугирование в силиконовом
масле 371, 372
ЦИТОХРОМ Й559 57
— bs 50
— / 33, 50, 58
Челночные механизмы 254—256
Шпинат, водная культура 547
— индукция 174
ЭДТА 102
Экзергонические реакции 24
Экзоосмос 229
Электрогеиный транспорт 373
Электролиз 15, 16
Электрои-транспортиая цепь 55, 84.
См, также Z-схема
Эндергонические реакции 24
Эидоосмос 229
Эндоплазматический ретикулум, уча-
стие в транспорте метаболитов
377
Энергетические затраты при ассими-
ляции нитратов 413
--------САМ-метаболизме 503
--------фотосинтезе 409
Энергия, единицы измерения 24
— запасание 82, 85
— и свет 34—46
— перенос в разных пигментных сио
темах 73
— потребность человека 42
Энтропия 19—21
Эриохром синий 267
Эритрозо-4-фосфат 138, 139
Эффект де Соссюра 494
Оглавление
Предисловие редактора перевода
Предисловие......................
ЧАСТЬ А
Глава 1. Введение......................................................Ю
1.1. Что такое фотосинтез............................ 10
1.2. Разрыв химических связей......................... Ю
1.3. Окисление и восстановление.......................10
1.4. Фотосинтез как окислительно-восстановительный процесс 12
1.5. Как заводится главная биологическая пружина . . 12
1.6. Фотосинтез как источник органического углерода ... 13
1.7. Некоторые аналогии и заключение..................15
Глава 2. Энергия и законы физики и химии..............................10
2.1. Законы термодинамики.............................19
2.2. Что такое система.....................................20
2.3. Энтропия (S)..........................................20
2.4. Свободная энергия.....................................21
2.5. Свободная энергия и равновесие........................22
2.6. Единицы энергии.................................24
2.7. Энергия образования углеводов....................25
2.8. Энергия связи...................................25
2.9. Расщепление воды................................26
2.10. Резонанс .............................................27
2.11. Свободная энергия гидролиза аденозинтрифосфата
(АТР)...............................................29
2.12. АТР как компонент ассимиляционной силы ... 31
2.13. Окислительно-восстановительный потенциал ... 32
Глава 3. Энергия и свет...............................................34
3.1. С чего все начинается..................................34
3.2. Свет — это волна........................................35
3.3. Свет — это поток частиц.................................36
3.4. Кванты...................................................36 '
3.5. Энергия света...........................................36
3.6. Интенсивность света....................................38
3.7. Солнечный свет и свеча.................................39
3.8. «Зеленый человек».......................................42
3.9. Квантовая эффективность.................................42
3.10. Эффективность фотосинтеза иа молекулярном уровне 44
3.11. Максимальная эффективность фотосинтеза на уровне
растения в целом...........................................45
3.12. Максимальная эффективность...........................45
ОГЛАВЛЕНИЕ
593
Глава 4. Фотохимический аппарат и его функции.....................47
Фотохимический аппарат.............................
4.1. Структура хлоропласта.........................
4.2. Тилакоидный компартмент ......................
4.3. Стромальный компартмент' .....................
4.4. Структура мембран.............................
4.5. Хлорофиллы . .............................
4.6. Синтез хлорофилла.............................
4.7. " Дополнительные пигменты ....................
4.8. Курица или яйцо...............................
4.9. Компоненты электрон-транспортной цепи
4.10. Фотосинтетическая единица....................
4.11. Пигментные системы '.........................
4.12. Реакционные центры...........................
4.13. Фотосистемы (ФС I и ФС II)...................
4.14. Поглощение света атомами.....................
4.15. Поглощение спета молекулами..................
4.16. Переход хлорофилла в возбужденное состояние
4.17. Снятие возбуждения...........................
4.18. Фотохимические реакции и выделение Ог
4.19. Перенос энергии в пигментных системах
4.20. Спектр действия..............................
4.21. Красный спад.................................
4.22. Эффект усиления..............................
4.23. Z-Схема......................................
4.24, Отношение Р/2е_..............................
4.25. Запасание энергии в Z-схеме..................
4.26. Квантовый расход для Z-схемы.................
47
47
49
49
49
51
54
55
61
63
65
65
67
68
68
70
70
73
74
77
77
78
81
82
83
Глава
5. Образование АТР; генерация ассимиляционной силы
84
Краткое содержание ....................................
5.1. Окислительное фосфорилирование....................
5.2. Зачем нужна ассимиляционная сила..................
5.3. Циклическое фотофосфорилирование..................
5.4. Циклическое фотофосфорилирование, катализируемое
ферредоксином..........................................
5.5. Циклическое фотофосфорилирование, in vivo
5.6. Псевдоциклическое фотофосфорилирование
5.7. Нециклическое фотофосфорилирование . . . .
5.8. Реакция Мел’ера' . . . .
5.9. Хемоосмос и установление электрохимической разности
потенциалов, или протондвижущей силы
5,10. Механизм образования АТР.........................
5.11. Сопряжение, разобщение и фотосинтетический контроль
5.12. Сопрягающий фактор...............................
5.13. Термодинамика синтеза АТР........................
5.14. Связь между транспортом протонов, транспортом элек-
тронов и энергией .....................................
5.15. Восстановление никотинамидадениндинуклеотидфосфа-
та (NADP)..............................................
Общая литература........................................
Специальная литература..................................
84
85
86
89
91
92
92
93
95
96
101
104
105
106
107
109
594
ОГЛАВЛЕНИЕ
ЧАСТЬ Б
Глава 6. Восстановительный пентозофосфатный цикл и связанные с ним
реакции ......................................................
Краткое содержание
6.1. Фотосинтетическая фиксация углерода................
6.2. Регенерация........................................
6.3. Автокатализ........................................
6.4. Энергетика.........................................
6.5. Сродство к СО2.....................................
6.6. Основные характеристики карбоксилирования
6.7. Изучение восстановительного пентозофосфатного цикла
6.8. Кинетические исследования.........................
6.9. Изменения в содержании ФГК и РуБФ . . . .
6.10, Включение метки в молекулы.......................
6.11. Ферменты.........................................
6.12. Свободные энергии................................
6.13, Функционирование ВПФ-цикла.......................
6.14, Использование ассимиляционной энергии . . . .
6.15. Повторное включение в цикл.......................
6,16. Связь между ВПФ-циклом и фотодыхательным циклом
окисления углерода . .......................
6.17. Участие углерода в ВПФ-цикле и в фотодыхательном
цикле ..................................................
6.18. Синтез крахмала...................................
6.19. Деградация крахмала...............................
6.20. Регуляция синтеза и деградации крахмала
6.21. Синтез сахарозы...................................
6.22. На пути ассимиляции углерода......................
Общая литература............................... . . .
Специальная литература .................................
113
115
115
115
116
119
122
122
125
126
127
128
142
145
146
147
147
152
152
156
158
158
159
163
164
Глава 7. Индукция ..................................................166
Краткое содержание........................................166
7.1. Индукция в целых растениях и листьях . . . . 167
7.2. Исторические аспекты.................................169
7.3. Роль устьиц........................................ 170
7.4. Отсутствие индукции в фотохимических процессах 173
7.5. Индукция in vitro....................................177
7.6. Молекулярные механизмы индукции...........178
7.7. Вклад автокатализа...................................180
7.8. Активация ферментов под действием света . . . 186
7.9. Влияние интенсивности света и температуры . . . 188
7.10. Индукция и ортофосфат..........................189
7.11. Снятие подавления, вызванного ортофосфатом, проме-
жуточными продуктами ВПФ-цикла.......................191
7.12. Природа подавления под действием ортофосфата . 193
7.13. Индукция в выделении кислорода и роль ФГК . . 195
7.14. Индукционные явления в фиксации СО2 .... 201
7.15, Индукция в реконструированной системе хлоропластов 205
1.16. Последовательность событий во время индукции . 208
7.17. Восстановление лаг-фазы..............................210
Общая литература ..........................................211
Специальная литература ................................... 211
ОГЛАВЛЕНИЕ
59^
Глава
8. Пластиды и внутриклеточный транспорт.....................214
Краткое содержание.....................................
8.1. Хлоропласт как транспортирующая органелла
8.2. Экспериментальная основа исследований транспорта —
фракционирование целых тканей .........................
8.3. Экспериментальная основа исследований транспорта
in vitro...............................................
8.4. Хроматографический анализ.........................
8.5. Фильтрация центрифугированием.....................
8.6. Косвенные методы изучения транспорта метаболитов .
8.7 Сокращение индукционного периода и снятие ингиби-
рующего действия ортофосфата...........................
8.8. Добавление ингибиторов.........................
8.9. Катализ интактными и разрушенными хлоропластами .
8.10. Осмотические изменения объема....................
8.11. Транспорт метаболитов............................
8.12. Двуокись углерода................................
8.13. Триозофосфаты и 3-фосфоглпцерат (ФГК) ....
8.14. Ортофосфат и неорганический пирофосфат
8.15. Пентозомонофосфаты и гексозомопофосфаты
8.16. Бисфосфаты сахаров...............................
8.17. Свободные сахара.................................
8.18. Карбоновые кислоты...............................
8.19. Аминокислоты.....................................
8.20. АТР и NADP.......................................
8,21. Транспорт ионов..................................
8.22. Специфическая проницаемость внутренней мембраны
8.23. Переносчик ортофосфата...........................
8.24. Переносчик дикарбоновых кислот...................
8.25, Переносчик аденилатов............................
8.26. Челночные механизмы..............................
Общая литература.......................................
Специальная литература ................................
214
214
216
221
222
222
225
225
227
228
229
230
231
234
234
239
240
240
242
243
243
245
247
247
253
254
255
256
257
Глава 9. Регуляция фотосинтетической ассимиляции углерода . . 261
Краткое содержание........................................261
9.1. Основные принципы....................................261
9.2. Регуляция катализа...................................262
9.3. Активация катализаторов..............................263
9.4, Различие между регуляцией катализа и активацией ка-
тализаторов ..............................................269
9.5. Темновая инактивация.................................270
9.6. Действие масс.......................................273
9.7. Регуляция транспортом...............................278
9.8. Значение регуляции...................................285
9.9. Роль адениновых нуклеотидов в регуляции . . . 286
Общая литература.........................................289
Специальная литература ................................. 290
Глава 10. Открытие С4-пути..................................... . 293
Краткое содержание................................... . 293
10.1. С4-днкарбоновые кислоты — ранние продукты фотосин-
теза .....................................................293
10.2. Вклад Хэтча и Слэка в проблему С4-фотосинтеза . . 296
10.3. Метаболизм С4-дикарбоиовых кислот.............301
10.4. Анатомия фотосинтезирующей ткани у С4-растений 304
396
ОГЛАВЛЕНИЕ
10.5. Выделение хлоропластов, протопластов и клеток из
С4-растений..........................................310
10.6. Современная упрощенная схема С4-фотосннтеза . . 313
10.7. Какие растения можно считать С4-растениями? . . 314
Общая литература......................................317
Специальная литература ................................... 317
Глава 11. Три подгруппы С4-растений: их биохимия, фотохимия и систе-
матика ........................................................... 3201
Краткое содержание....................................320
11.1. Стадия карбоксилирования в С4-пути..............321
11.2. Стадия декарбоксилирования в С4-пути .... 323
11.3. Краткое изложение основных предполагаемых путей пе-
реноса углерода с участием трех декарбоксилирующих
механизмов............................................... 325-
11.4. Сбалансированность зарядов при межклеточном транс-
порте метаболитов в ходе С4-цикла.........................326
11.5. Межклеточная локализация ВПФ-цикла у С4-растеиий 328
11.6. Фотохимические затраты при С4-фотосинтезе . . . 328
11.7. Фотохимические различия хлоропластов разных типов 333
11.8. Систематика.....................................343
11.9. Растения с промежуточными (С3/С4) характеристиками 347
Специальная литература ................................... 349
Глава 12. Интеграция функций при С4-фотосинтезе.................351
Краткое содержание....................................351
12.1. Внутриклеточная локализация ферментов С4-цикла в
клетках мезофилла...................................351
12.2. Локализация ферментов и внутриклеточный транспорт
метаболитов С4-цикла в клетках обкладки проводящих
пучков...............................................357
12.3. Доказательства фотосинтетической деятельности кле-
ток мезофилла........................................362
12.4. Доказательства фотосинтетической деятельности кле-
ток обкладки проводящих пучков.......................367
12.5. Механизмы внутриклеточного транспорта метаболитов 371
12.6. Механизм межклеточного транспорта метаболитов 376
12.7. С4-метаболизм, сопряженный с циклическим, псевдо-
циклическим и нециклическим фотофосфорилированием 379
12.8. Регуляция ферментов С4-цикла....................381
12.9. Ассимиляция азота: сравнение С4- и Сз-растений 387
12.10. Синтез крахмала и сахарозы у С4-растений . . . 393
Специальная литература ................................... 395
Глава 13. Фотодыхание...........................................399
Краткое содержание....................................399
13.1. Введение........................................400
13.2. Распознавание фотодыхания.......................401
13.3. Происхождение гликолата.........................405
13.4. Гликолатный путь у С3-растений..................407
.13.5. Упрощенная схема пути углерода в глнколатном цикле 408
13.6. Реакции гликолатного пути и энергетические затраты 409
13.7. Внутриклеточная локализация ферментов гликолатно-
го пути 415
13.8. Метаболизм в изолированных органеллах и его связь
с гликолатным путем.............................420
13.9. Гликолатный путь у С4-растений..................422
ОГЛАВЛЕНИЕ .597
13.10. Ингибирование фотосинтеза кислородом и две его со-
ставляющие ...................................... • • 424
13.11. Доля ингибирования фотосинтеза кислородом . . 434
13.12. Влияние кислорода на квантовый выход .... 434
13.13. Предполагаемая роль фотодыхания . ..... 438
13.14. Стимуляция роста С3-растеиип в средах с понижен-
ным содержанием Оа или обогащенных СО2 . . . 440
Общая литература .......................................443
Специальная литература . 443
Глава 14. Первичные карбоксилазы и регуляция фотосинтеза и транспи-
рации факторами внешней среды............................447
Краткое содержание......................................447
14.1. Введение . . . . . . 449
14.2. Состав атмосферы н растворимость газов . . . 449'
14.3. Сз-Растепия: РуБФ-карбоксилаза.............454
14.4. С4-Растснпя: ФЕП- и РуБФ-карбоксилазы .... 463.
14.5. Влияние температуры, света и водного дефицита па •
карбоксилирование ................................ 468
14.6. Эффективность использования воды...........474
14.7. Сз-, Сгфотоспитез п урожайность сельскохозяйствен-
ных культур........................................481
Общая литература................................ 482'
Специальная литература ................................ 483
Глава 15. Метаболизм кислот у семейства толстянковых (САМ-метабо-
лизм) . . . . ........... 486
Краткое содержание . . ' ............................486
15.1. Введение......................................... 487
15.2. Открытие САМ-метаболизма...................490
15.3. Реакция Вуда и Веркмаиа....................494
15.4. Путь углерода у САМ-растений...............495
15.5. Двуокись углерода как метаболит темновых реакций 497
15.6. Обратная пропорциональность между количеством суб-
страта и количеством продукта....................498
15.7. Ферменты, участвующие в накоплении органических
кислот и в их последующем использовании . . . 498
15.8. Гипотеза двойного карбоксилирования .... 501
15.9. Накопление органических кислот на свету . . . 502'
15.10. Энергетические затраты при САМ-метаболизме . . 503
15.11. Переключение обмена веществ от САМ-метаболизма
на Сз-фотоснитез......................................508
15.12. Фракционирование изотопов углерода у САМ-, С3- и
С4-растеинй.................................. 509'
15.13. Связь САМ-метаболизма с С4-циклом .... 515'
15.14. Экологическое значение САМ-метаболизма . . . 516
Общая литература...................................519
Специальная литература ............................... 520
Глава 16. Сравнительное изучение С3- и С4-метаболизма в других тка-
нях (помимо листа).........................................524
Краткое содержание.................................524
16.1. Устьица..........................................525
16.2. С3- и С4-фотосинтез в тканях репродуктивных органов 529
16.3. Зеленеющие листья................................534
16.4. Корпи............................................534
598
ОГЛАВЛЕНИЕ
16,5. Ионный баланс и поддержание постоянного pH . . 536
16.6. Метаболизм малата в глиоксилатном цикле проростков 538
Специальная литература ................................. 540
Приложение А. Выделение хлоропластов и критерии их интактиости 543
А.1. Введение..................................543
А.2. Методы....................................545
А.З. Выращивание растений......................545
А.4. Приемы механического разрушения .... 549
А.5. Получение хлоропластов из протопластов . . . 550
А.6. Достоинства и недостатки методов механического
и ферментативного разрушения клеток . . . 555
А.7. Критерии, по которым определяют степень интакт-
ности хлоропластов.............................557
А.8. Очистка...................................560
Литература..........................................561
Приложение Б. Номенклатура ферментов............................. 563
Б.1. Восстановительный пентозофосфатный путь . . 563
Б.2. Гликолатный путь..........................564
Б.З. Образование сахарозы из триозофосфатов . . 566
Б.4. Синтез крахмала из триозофосфатов .... 567
Б.5. С4-путь...................................568
Б.6. Метаболизм кислот у толстянковых .... 570
Б.7. Обмен нитратов.................................573
'Список сокращений.................................................574
Указатель латинских названий.......................................576
Предметный указатель............................................. 579
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформ-
лении, качестве перевода и другие просим присылать
по адресу:
129820, Москва, И-110, ГСП
1-й Рижский пер., д. 2,
издательство «Мир»
'Монография
Джерри Эдвардс, Дэвид Уокер
ФОТОСИНТЕЗ С3- И СгРАСТЕНИЙ:
МЕХАНИЗМЫ И 'РЕГУЛЯЦИЯ
Научные редакторы’ Т. И. Пономарева, Н. Н. Шафрановская
Мл. редактор О. В. Шагании
Художник А. В. Захаров
Художественный редактор А. Я Мусин
Технический редактор Е. С. Потапеикова
Корректор В. С. Соколов
ИВ № 5731
Сдано в набор 27.03.86. Подписано к печати 04.11.86. Формат
«60X90710. Бумага типографская Na 1. Печать высокая. Гар-
нитура литературная. Объем 18,75 бум. л. Усл. печ. л. 37.50.
Уел. кр.-отт. 37,50. Уч.-изд. л. 38.40. Изд. № 4/4534. Тираж
1700 экз. Зак. 234. Цена 6 р. 10 к.
ИЗДАТЕЛЬСТВО «МИР». 129820, ГСП. Москва. И-110.
1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при Го-
сударственном комитете СССР по делам издательств, поли-
графии и книжной торговли. 113105. Москва. Нагатинская
ул., д. 1.