/





















































































































































































































































































Текст
.Я ^Л'_ .. '. iir»»ifci¥aMr>-wrgb«agyi,w.iii <мь^^^6^'зчря^и^и*симйийамй
/1зменчивость
|. ОРГАНИЗМОВ
I В ГЕОЛРГЙНЕЖЭМ '
I ПРОШЛОМ .'
Шд60)ЗЭС?СПЬ ЬЬ6 ЭЗОБОЗбОг-дОд d<5d<S330d
АКАДЕМИЯ НАУК ГРУЗИНСКОЙ ССР
гдс?от&отс?(пг>ооь обьйстло
ИНСТИТУТ ПАЛЕОБИОЛОГИИ
1970
е?. «?лзоо)л,эзое?о
обвббоъасоб
аэое?Од07)б
„30(36006026“
соьосзоьо
1970
Л. Ш. ДАВИТАШВИЛИ
ИЗМЕНЧИВОСТЬ
ОРГАНИЗМОВ
В ГЕОЛОГИЧЕСКОМ
ПРОШЛОМ
«МЕЦНИЕРЕБА»
ТБИЛИСИ
1970
56
56.57
Д 13
Палеобиология изучает историческое разви-
тие органического мира в течение геологическо-
го времени- Естественный отбор происходил на
основе изменчивости органических форм- Поэто-
му для выяснения хода эволюции и ее законо-
мерностей необходимо познание изменчивости
организмов в геологическом прошлом. После
краткого очерка истории этой проблемы рас-
сматриваются общие вопросы изменчивости.
Далее следуют главы, посвященные изменчи-
вости ископаемых организмов различных
групп (фораминифер, двустворчатых, брюхоно-
гих и головоногих моллюсков, рыб, пресмыкаю-
щихся, млекопитающих и других). Особенно
яркие примеры внутрипопуляционной изменчи-
вости приводятся из области палеобиологии
моллюсков, населявших морские .бассейны Чер-
номорско-Каспийской зоны в последовательные
века миоцена и плиоцена. Автор дока бывает воз-
можность устанавливать изменчивости ископа-
емых форм и зависимость этого явления от воз
действия факторов наружной среды — морской
пресноводной, наземной и подземной.
Изучение изменчивости и факторов, ее вы
зывающих, имеет большое значение для стра-
тиграфических исследований, а также для выяс-
нения условий осадконакопления в соответству*
ющие геологические века.
Изложение иллюстрируется многочисленны
ми рисунками.
Книга рассчитана на самый широкий крут
читателей, интересующихся эволюцией органичес
кого мира — биологов различных специальное
тей (включая палеонтологов), геологов, геогра
фов, философов и др.
2—10—1
4Q—71 мрстн.
ПРЕДИСЛОВИЕ
Эта книга посвящена внутривидовым наследственным изменениям
организмов прошлых геологических времен. Такие изменения представ-
ляют собой предмет генетики, и многим .покажется, по всей вероятнос-
ти, слишком дерзкой попытка рассмотреть это явление на основе иско-
паемого матерала. Тем не менее, нам представляется совершенно не-
обходимым сведение воедино палеобиологических данных об изменчи-
вости органических форм и научный анализ этих данных. Такая мысль
обосновывается двумя серьезными доводами.
Во-первых, реальные факты исторического развития органического
мира изучаются не необиологией, а палеобиологией. Палеобиология
— не паука о диковинных чудовищах и курьезных окаменелостях. Она
по своему содержанию близка к науке об эволюции живой природы и
неуклонно стремится к дальнейшему сближению с этой наукой. Она
повседневно сталкивается с фактами эволюции и изменчивости органи-
ческих форм. Факты изменчивости ископаемых видов имеют, конечно,
определенное значение для науки и заслуживают серьезного изучения:
это необходимо для всестороннего познания изменчивости и ее проис-
хождения, а следовательно и для освещения вопросов эволюции.
Во-вторых, палеобиологическое исследование явлений изменчивос-
ти имеет немаловажное значение для освещения вопросов исторической
геологии и в частности относительной геохронологии, для которой, ко-
нечно, важно отличать виды от форм индивидуальной изменчивости:
особи одного вида, даже одной популяции, могут сильно отличаться од-
на от другой, и для геохронологических выводов существенно знать,
что такая изменчивость является внутривидовой, не выходящей за рам-
ки данного вида. Далее, изменчивость некогда живших организмов мо-
жет указывать на факторы, которыми она была вызвана, что позволя-
ет подойти к выяснению условий накопления осадков, где захороня-
лись остатки организмов. Это тоже очень важно для стратиграфии и
для воссоздания хода геологического развития соответствующих участ-
ков земной коры.
Итак, изучение внутривидовой изменчивости имеет существенное
значение и для науки об эволюции (следовательно, и для палеобиоло-
гии, как эволюционной науки), и для геологии.
Я уверен в том, что в этой книге окажется много недостатков,
тем более, что она — первый опыт сводки и анализа данных об изменчи-
вости вымерших органических форм. Прошу читателей сообщать мне
о замеченных промахах и слабых сторонах моей работы.
Г. А. .Квалиашвилм прочел рукопись «книги и сделал много ценных
замечаний. Он же, К. Г. Багдасарян и Р. В. Ратиани читали .корректу-
ры. Я глубоко признателен этим товарищам за оказанную мне помощь.
При подготовке^ этой книги мне помогли многие лица. Т. А. Ло-
минадзе, И. Г. Тактакишвили, Е. В. Рухина и Н. Г. Химшиашвили да-
ли мне ценные советы, которыми я воспользовался. А. А. Али-Заде,
Е. Г. Ахвледиани, Ц. И. Бадзошвили, Н. Р. Джикия, Е. М. Жгенти,
Т. Г. Китовани и Л. В. Мусхелишвили предоставили мне фотографии ра-
ковин моллюсков, воспроизведенные в моей книге. Т. Т. Мелик-Шахна-
зарова и Ц. И. Экизашвили оказали мно помощь библиографической
консультацией. Всем этим товарищам приношу сердечную благодарность.
Буду искренне признателен всем читателям:, которые, познакомив-
шие с книгой, сообщат мне свои замечания о ее содержании.
Автор
ВВЕДЕНИЕ
Наследственность и 1из1менчивость «можно рассматривать с разных
точек зрения, под тем или иным углом зрения, и каждый такой способ
представляется нам вполне законным. Тот анализ явлений изменчи-
вости, который дается в этой книге, не подтверждает и не опроверга-
ет ни одной из существующих генетических теорий и совсем не каса-
ется биохимических процессов, ведущих к изменению того, что теперь
обычно называется «наследственной информацией» и что прежде с не-
меньшим успехом именовалось просто наследственностью. Не имеем ни
возможности, ни желания вникать в существо молекулярных процес-
сов, приводящих к изменению наследственности.
В настоящее время мы находимся лишь в преддверии нового, глу-
бокого и всестороннего, учения о наследственности и изменчивости. В
современной генетике многое изменяется очень быстро и глубоко. В ней
много гипотетичного, что вполне понятно, но лишает возможности го-
ворить о твердо установленных законах.
Палеобиолог (так же как и необиолог, по роду работы своей да-
лекий от генетики, но считающий возможным воздерживаться от изуче-
ния и толкования явлений изменчивости) может пойти по одному из
двух путей.
Один путь — некритическое и слепое принятие одной из генетичес-
ких теорий со всеми ее понятиями и идеями. По этому пути пошли не-
которые сравнительные анатомы, экологи, ботаники, а также разные
необиологи других специальностей. Этот путь чреват грозными опас-
ностями. Генетика в течение почти всего периода своего существования
многократно испытывала ломку своих принципиальных тезисов, сегод-
ня сжигая то, чему поклонялась вчера, с тем, чтобы завтра отбросить
тб, йто сейчас котируется как незыблемые истины. Биолог или палео-
биолог, доверчиво перенимающий фундаментальные законы генетики,
которые могут быть завтра презрительно отброшены специалистами-
генетиками как ненужная ветошь, нередко повторяет зады генетичес-
кой науки для построения эволюционных обобщений. И действительно,
даже самые осведомленные, просвещенные биологи-эволюционисты, не
работающие в области генетики, обычно оперируют устаревшими гене-
тическими понятиями, ныне вызывающими у компетентных генетиков
линщ снисходительную улыбку. И все-таки мы не можем утверждать,
что' -биологи-теоретики должны уклоняться от этого пути. Мы скорее
склоййы думать, что и его можно испробовать, но с надлежащими ме-
рами осторожности. Мы хорошо знаем, что некоторые биологи-теорети-
ки в недавнем прошлом совершенно добросовестно пытались пользо-
ваться, как основой своих рассуждений об изменчивости, главными
принципами мичуринизма, а теперь те же исследователи отрекаются
от своих собственных обобщений и выводов, сделанных в духе идей
мичуринизма.
6
Итак, в этом деле необходимы сугубая осторожность, тщательное
изучение идей генетики и, по возможности, их глубокий анализ.
Но существует и другой путь изучения явлений изменчивости. Это
— путь независимого от тех или иных генетических теорий изучения
изменчивости морфологических, физиологических и этологических осо-
бенностей органических форм, переходов между ними, а также связей
между этими явлениями и изменениями среды обитания. Такое иссле-
дование пользуется лишь более или менее достоверно и прочно уста-
новленными положениями генетики, каких в современной науке, к со-
жалению, пока довольно мало. При этом биологи избегают употреб-
ления терминов сомнительного или сугубо гипотетического значения,
которыми так богата и столь быстро продолжает обогащаться генети-
ка. Тем самым биолог, наблюдающий и описывающий явления изменчи-
вости, признает себя вынужденным отказаться от глубокого изучения
тонкостей молекулярно-биологических, биохимических и биофизичес-
ких процессов, с которыми сопряжены явления изменчивости. Такой
подход представляется нам вполне оправданным и законным, хотя бы-
ло бы нелепо требовать, чтобы по этому пути вели свои исследования
все биологи, сталкивающиеся с явлениями изменчивости. В этой кни-
ге мы придерживаемся именно этого пути. Читатель, надеемся, мо-
жет удостовериться в том, что и таким путём могут быть добыты и ос-
мыслены некоторые очень важные данные об изменчивости организ-
мов, особенно же организмов прошлых геологических времен.
Это, конечно, не исключает полезности сопоставления выводов,
полученных путем таких, существенно организмальных, исследований
с данными современной генетики.
Таким образом, в настоящее время существуют два аспекта проб-
лемы изменчивости. Один — аспект преимущественно эксперименталь-
ных исследований, представляющих собой предмет генетики; другой
— аспект преимущественно морфо-физиолого-этологических исследова-
ний, наблюдений изменчивости того, что называется фенотипами ор-
ганизмов. Только исследования этого второго направления могут в из-
вестной мере осуществляться в палеобиологии.
Здесь может возникнуть вопрос, насколько целесообразны попыт-
ки изучения изменчивости организмов прошлых геологических эпох.
Могут ли такие попытки дать ощутимые научные результаты? Пос-
кольку такие сомнения могут возникать не только у генетиков и специ-
алистов по молекулярной биологии, работающих на основе точного экс-
периментального метода, но и у необислогов вообще, считаем нужным
коснуться этого принципиального вопроса уже здесь, в начале нашей
книги. Да, такие исследования вполне возможны, они могут вести к
хорошо обоснованным выводам не только о наличии изменчивости у
вымерших организмов, но и о ее причинах.
Убедительно показать изменения организмов под воздействием
среды на палеобиологическом материале, конечно, — трудное и слож-
ное дело. Палеобиолог имеет перед собой только остатки тела орга-
низма, обычно — частично сохранившийся скелет, а чаще лишь остат-
ки небольшой доли твердых частей тела, способных к сохранению в
иског'аемом состоянии, или даже только отпечатки тех или иных час-
тей тела. По такому материалу, разумеется, нелегко восстанавливать
строение тела и его главнейших органов, а тем более — их функции.
Но для выяснения изменений некогда существовавших организмов под
воздействием среды требуется, кроме того, установление хода филоге-
нетического развития организмов, а также и хода развития среды на
7
данном участке земной коры. Отрывочный характер филогенетических
линий и отсутствие более или менее точных данных о ходе развития
среды на соответствующих участках очень часто делают задачу выяс-
нения изменяющего действия среды на «живые тела» прошлых времен
слишком трудной при нынешнем уровне знаний. Однако задача эта
принципиально вполне разрешима; по крайней мере в некоторых слу-
чаях работникам палеобиологии удавалось найти подход к изучению
этого вопроса, намечать пути его разработки и с достаточной убеди-
тельностью показывать воздействие условий среды на организмы гео-
логического прошлого. Такого рода примеры дает, в частности, палео-
биология населения морских бассейнов, существовавших в течение поз-
днетретичного и четвертичного времени на том обширном участке зем-
ной коры, который можно называть Черноморско-Каспийской облас-
тью, или провинцией.
В восточной части древнего великого Средиземного моря, более
или менее обособившейся от остального пространства этого моря в
связи с значительными тектоническими движениями земной коры, на
протяжении позднетретичной эпохи и четвертичного периода соверша-
лись глубокие физико-географические преобразования, протекавшие с
необычайно значительной скоростью и в разных направлениях. Резко
изменялись очертания морей, связь этой части великого Средиземного
моря с остальной, более западной, частью бассейна то становилась зат-
рудненной, то совершенно прекращаясь, то вновь восстанавливаясь, и
соответственно этому здесь резко и быстро изменялись гидрологические
условия, в частности соленость и pH. Морские полносоленые бассейны
сменялись полуморскими, с заметно пониженной соленостью, в некото-
рой мере сходными с современным Черным морем, эти последние —
сильно опресненными озерами — морями, или «каспиями», — по тер-
минологии Н. А. Андрусова, т. е. бассейнами до известной степени близ-
кими по солености к нынешнему Каспийскому морю, или даже пресно-
водными бассейнами. В другие моменты тут возникали вновь бассейны
полносоленые или с соленостью, близкой к нормальной морской, и т.
д. В то же время довольно значительные изменения испытывал и кли-
мат, особенно сильные колебания которого отмечаются в поздней
части верхнетретичной эпохи (в плиоцене). Все эти, необычайно резкие
и быстрые изменения условий среды должны были, естественно, вызы-
вать значительные и быстрые изменения в организмах, населявших эти
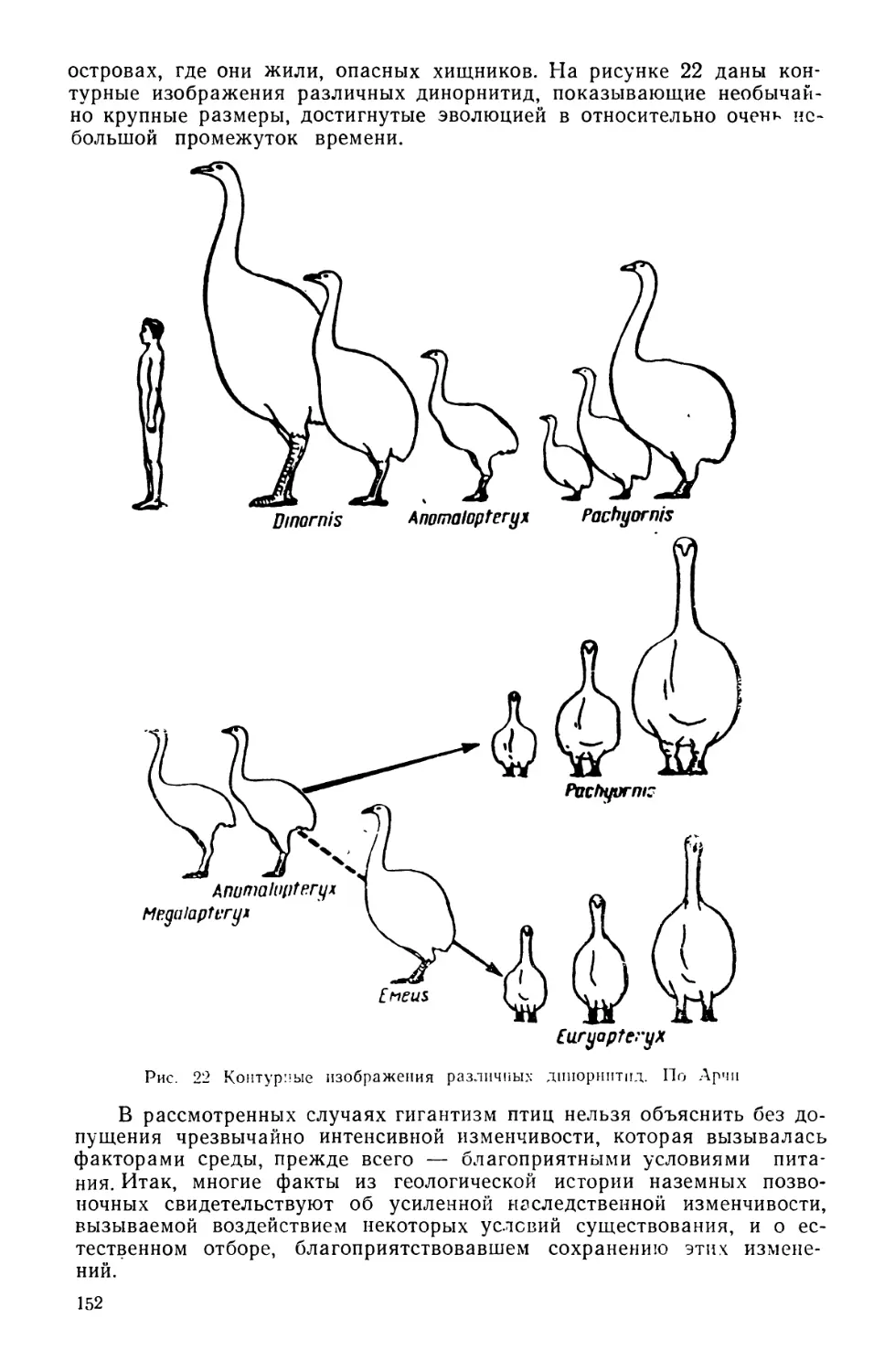
водоемы. И в самом деле, мы наблюдаем здесь исключительно быст-
рый процесс развития форм, связанный, как в этом легко убедиться, с
интенсивной, вызываемой изменениями среды, изменчивостью организ-
мов. Во многих случаях изменчивость имеет бесспорно направленный
характер и охватывает сразу множество особей. Иногда это явление
просто бросается в глаза. В одной из своих статей, написанной десятки
лет назад, мы (1936), описывая такие явления, пользуясь терминоло-
гией классической генетики, усматривали в подобных случаях направ-
ленное мутирование, связанное с массовым единообразным изменением
наследственности, вызываемым прямым воздействием условий сущест-
вования. Как в упомянутой статье, так и в предшествовавших своих
высказываниях мы склонялись к мысли, что это «направленное му-
тирование» вызывалось и поддерживалось в течение более или менее
длительного, в геологическом смысле, времени, благоприятными изме-
нениями климата, пищевых ресурсов и других важных для жизни изу-
чаемых форм условий существования. Речь идет о некоторых группах
моллюсков - - пластинчатожаберных и брюхоногих. Эти животные по-
8
падали, в результате геологических преобразований, в условия, значи-
тельно отличавшиеся от условий, для них обычных, причем процесс
изменения условий в определенном направлении неоедко тянулся в те-
чение довольно продолжительного времени,. При подобных обстоятель-
ствах многие формы более или менее быстро вымирали, другие же,
приспособляясь к новым условиям, необычайно быстро эволюировалп,
давая начало не только новым видам, но и новым родам и даже более
высоким таксномическим категориям, — подсемействам и семействам.
Поразительна, как это будет пояснено в одной из последующих глав,
изменчивость, например, брюхоногих группы палюдин (вивипарусов)
так называемых дуабских отложений киммерийского яруса среднего
плиоцена (Абхазия). Палюдины попали в область осаждения дуаб-
ских отложений, в обширное слабо соленое киммерийское озеро-море,
из пресноводных водоемов. Выходцы из пресной воды могли бы погиб-
нуть в условиях киммерийского «каспия», но его соленость была, оче-
видно, более или менее терпимой по крайней мере для некоторых из
палюдин, как и для некоторых других форм пресноводного происхож-
дения, попавших в киммерийское озеро-море и не погибших там пол-
ностью. Новое местообитание отличалось от исходного также и други-
ми существенными гидрологическими условиями; прежде всего, эго
был очень обширный, морского типа водоем, размерами не меньше
современного Черного моря. И вот палюдины проявляют необычайную
но интенсивную изменчивость. Стенки раковин утолщаются, резко и
существенно изменяется контур оборотов, и если взять наудачу деся-
ток раковин, то едва ли не каждая из них окажется отличающейся
от любой другой больше, чем раковины двух заведомо различных ви-
дов «нормальных» палюдин из обычных для этих моллюсков пресно-
водных местообитаний. Без огромного, совершенно исключительного
изменяющего действия среды на целые популяции никак нельзя понять
быстрого исторического развития палюдин и других брюхоногих ким-
мерийского и следующего за ним, куяльницкого, ярусов. Возникают,
совершенно новые роды. Для того, чтобы оценить необычайную быст-
роту исторического развития палюдин в нашем плиоцене достаточно
вспомнить, что именно пресноводные брюхоногие, включая сюда и
палюдин, развивались несравненно медленнее (если они оставались
обитателями («нормальных» биотопов материковых пресных вод), чем,
например, морские моллюски того же класса.
Однако такую повышенную изменчивость обнаруживают не толь-
ко галюдины, но и многие другие моллюски-выходцы из пресноводных
небольших водоемов, преимущественно из текучих вод, попавшие в
киммерийский бассейн в его юго-восточной части и выдержавшие но-
вый для них гидрологический режим. Такую бурную изменчивость про-
являют многие формы, например, неритинид, что подтверждается появ-
лением буквально множества новых видов и нескольких новых родов.
Поразительна также изменчивость тех форм морского происхож-
дения, которые выдержали значительное изменение привычного для их
непосредственных предков гидрологического режима при резком опрес-
нении и других изменениях, сопровождавших обычно отделение бассей-
нов Черноморско-Каспийской провинции от мирового океана или су-
жение тех проливов, через которые осуществлялась связь этих бассей-
нов с нормально-соленым морем. Таковы многие представители семейс-
тва Cardiidae.
Таким образом, «в новых, необычных для данного вида условиях
жизни, его изменчивость может в огромной степени повышаться, при-
9
чем характер изменчивости и ее направление зависят от характера и
силы воздействия, которыми вызывается эта изменчивость» (Дави-
ташвили, 1949, стр. 148).
В различных главах этой книги читатель найдет довольно подроб-
ное рассмотрение таких явлений в палеобиологической истории не
только неогеновых моллюсков Черноморско-Каспийской области, но и
других групп животных.
Итак, нет никакого сомнения в том, что изучение изменчивости
на палеобиологическом материале — реальная, серьезная научная за-
дача, «важная и с эволюционно-биологической, и с геологической точки
зрения. Вполне понятно, что такие исследования по своему характе-
ру далеки от экспериментальной работы генетика. Методологически
они близки к дисциплинам биологии, которые подходят к освещению
проблемы изменчивости путем наблюдений над морфофизиологически-
ми и этологическими особенностями организмов. Но, говоря так, мы
очень далеки от умаления значения экспериментального изучения нас-
ледственности и изменчивости, ныне успешно осуществляемого на ос-
нове новейших методов. Кроме того мы решительно отвергаем то пони-
мание изменчивости и ее причин, к которому нередко приходили ис-
следователи, шедшие путем наблюдений и их описания, и которое
сводится к утверждению абсолютно слитного характера наследственнос-
ти и к отрицанию ее дискретности.
Основная задача этой книги — рассмотрение наследственной внут-
ривидовой изменчивости ископаемых организмов, которая является
одним из обязательных факторов эволюции. Ненаследственная «моди-
фикационная» изменчивость не может подвергаться естественному от-
бору и служить основой для видообразования. Это, конечно, отнюдь
не значит, что модификации не играют никакой роли в эволюции ор-
ганизмов; по этому вопросу могут быть и действительно существуют
различные мнения. Нельзя также забывать, что способность давать
те или иные модификации есть наследственное свойство организмов
вида, претерпевающего, в тех или иных условиях среды, определенные
модификации. Однако несомненно и то, что прямое отношение к эво-
люции путём естественного отбора имеют именно наследственные изме-
нения особей данной популяции. Поэтому нас в первую очередь ин-
тересуют вариации, наследственный характер которых можно так или
иначе доказать.
Относительно нынеживущих форм это устанавливается с доста-
точной достоверностью соответствующими экспериментами. Если то
или иное изменение, морфологическое или физиологическое, замечен-
ное в данном поколении, повторяется в последующих поколениях не-
зависимо от условий среды, в которой они развиваются, то мы можем
сделать вывод о наследственном характере этой вариации. Она может
быть определена экспериментатором как длительная модификация, но
это не меняет сути дела: длительная модификация — тоже наслед-
ственное изменение и может быть подхвачена отбором.
Однако подобные эксперименты производились преимущественно
над формами, не имеющими большого значения для решения вопросов
геологической истории и её хронологии. Эти исследования, тем не ме-
нее, имеют прямое отношение к основной теме нашей книги и пото-
му затрагиваются нами. Однако встает «вопрос, нет ли возможнос-
ти устанавливать факт наследственной изменчивости внутри вида без
соответствующих экспериментов, которые совершенно невозможны в
палеобиологических исследованиях. Такие эксперименты почти никог-
10
да, к сожалению, не ставились и над теми современными организмами,
которые принадлежа^ к группам, имеющим существенное значение для
палеобиологии и стратиграфии, например, над моллюсками и форами-
ниферами. Заметим, что такие экспериментальные исследования были
бы весьма желательны, и надо надеяться, что недалеко то время,
когда они будут начаты в интересах геологической науки. Это было
бы одним из направлений исследований, которые немецкие ученые
иногда называют актуопалеонтологическими. Актуопалеонтология, по
мысли этих ученых [см., например, В. Шэфер (W. Schafer, 1962), стр.
2], призвана изучать ныне совершающиеся процессы для освещения
вопросов палеонтологии, исторического прошлого органического мира.
Но ведь всё это касается возможностей будущих исследований
развития жизни. Не можем ли мы, однако, в ожидании лучших для
нашей науки времен, уже теперь как-то подойти к изучению явлений
наследственной изменчивости без эксперимента? По нашему мнению,
такая возможность существует. Поясним нашу мысль. Когда перед
нами — компоненты популяции вида А, попавшие в несколько сво-
еобразные условия жизни и представляющие некоторое отклонение от
нормы, мы не имеем оснований для решения вопроса, наследственны
ли эти отклонения или являются лишь модификациями. Такой случай
часто представляют популяции фораминифер, отличающиеся от обыч-
ных форм данного вида по величине раковинки и ее скульптуре (орна-
ментации), о чем будет речь в одном из последующих разделов кни-
ги. Но если мы имеем хронологическое чередование популяций вида,
отличающихся одна от другой по тому или иному признаку, то тут
возникает вопрос, не являются ли такие последовательные из-
менения наследственными.
Представим себе, что в отложениях одного и того же бассейна
мы имеем стратиграфическую последовательность видов: А — Б —
В — Г — Ди что каждый вид связан незаметными переходами с
предыдущим и последующим. Если самостоятельность видов А, Б, В,
Г и Д установлена с достаточной достоверностью, а изменчивость
внутри каждого из них имеет общее для всех этих видов направление
— «общую тенденцию», то едва ли можно сомневаться в наследствен-
ном характере этой изменчивости. В дальнейшем мы рассмотрим мно-
го таких случаев в филогенетических ветвях различных беспозвоноч-
ных. Подчеркнём, что наличие внутривидовой изменчивости и наслед-
ственный характер вариаций выясняется несмотря на то, что точные
границы между видами могут вызывать серьезные сомнения.
Нетрудно видеть, что такой способ установления наследственной
внутривидовой изменчивости доступен преимущественно палеобиологии,
которая в определенных довольно многочисленных случаях вскрывает
непрерывные ряды хронологически чередующихся форм. В необиологии
он может применяться лишь в исключительно редких случаях, так как
она имеет дело почти исключительно с одновременно живущими орга-
ническими формами.
Глава первая
НЕКОТОРЫЕ ФАКТЫ ИЗ ИСТОРИИ ПРОБЛЕМЫ
ИЗМЕНЧИВОСТИ, ВАЖНЫЕ С ПАЛЕОБИОЛОГИЧЕСКОЙ
ТОЧКИ ЗРЕНИЯ
«Без критики ошибочных идей дальнейшие, исследо-
вания философских проблем биологии были бы очень
затруднительны. Поэтому неправы те, кто считает изли-
шним возвращаться к анализу ошибок прошлого. Без
проведения такой, далеко не увлекательной, работы груз
старых ошибок долго еще будет затруднять движение
вперед».
К. М. Завадский и А. С. Мамзин «Некоторые вопро-
сы развития биологии в СССР и диалектический мате-
риализм». — Научные доклады высшей школы. Философ-
ские науки. 1965, стр. 106.
First bandit: Let us first see peace in Athens; there
is no time so miserable but a man may be true.—W. Sha-
kespeare. Timon of Athens, Act IV, Scene III.
Первый разбойник: Давайте прежде дождемся мира в
Афинах. Человек может быть честным в любое, самое
скверное, время. — У. Шекспир «Тимон Афинский». Акт
IV, сцена III, стр. 500. Полное собрание сочинений. Мос-
ква, 1960 г.
ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
В этой главе мы рассмотрим некоторые крупнейшие факты из
истории изучения наследственности и изменчивости нынеживущих ор-
ганизмов — факты, о которых мы часто забываем и которые, тем не
менее, имеют важное значение для понимания нынешнего состояния
теоретических основ этих исследований. Говоря о забвении этих фак-
тов, мы понимаем, что это звучит невероятно: как это виднейшие учё-
ные-специалисты могут забывать о том, что важно для понимания
сути дела! Мы имеем в виду прежде всего те факты, которые в нас-
тоящее время считаются зачастую вымыслом, — дошедшим до нас в
качестве пережитка тех времен, когда какая-то внешняя для науки
сила навязывала биологам нелепые представления. Впрочем, присту-
пим сразу к констатации и оценке этих фактов.
12
ИДЕАЛИСТИЧЕСКАЯ «ВОЛНА» В БИОЛОГИИ НА ГРАНИЦЕ ДВУХ ВЕКОВ
Остановимся прежде всего на разительном факте бурного подъема
и распространения на Западе антидарвинизма разных оттенков как
реакции против материалистического учения Дарвина и классиков дар-
винизма; так развивались события примерно на границе 19-го и 20-го
веков. Подъем этой антидарвинистской волны мы считаем незыблемо
установленным фактом. Это иногда отрицается людьми, не дающими
себе труда познакомиться с литературой по эволюционизму и теоре-
тической биологии вообще. Некоторые ученые ныне рассуждают при-
мерно по такой схеме: «По-моему такой факт невозможен, а следо-
вательно он есть плод вымысла, а даже злонамеренного вымысла. Тео-
ретическая биология всегда развивалась и развивается совершенно
беспрепятственно в капиталистическом мире. Некоторые задержки мог-
ли иметь место и действительно происходили лишь у нас, в связи с
засильем безграмотных «диктаторов в биологии», действовавших под
флагом мичуринизма. А вне социалистических стран развитие биологии
могло итти и в действительности шло совершенно гладко: идеи совер-
шенствовались, дело шло неуклонно ко все более глубокому познанию
законов жизни и ее исторического развития». Такое понимание разви-
тия теоретической биологии и особенно эволюционного учения мы
(1968, стр. 189) назвали «теорией благорастворения воздухов».
Однако всё дело в том, что сформулированное нами положение
об идеалистической волне, направленной против материалистического
дарвинистского понимания органического мира, есть просто факт, —
факт точно установленный на основании тщательного анализа огромной
мировой литературы. В течение многих лет мы старательно изучали
труды биологов, опубликованные на разных языках — французском,
немецком, английском, испанском, итальянском и других, особенно тру-
ды палеонтологов, которые высказывались по важнейшим теоретичес-
ким вопросам биологии, но также и труды биологов других специаль-
ностей. И чем глубже мы изучали эти произведения, тем больше и
твёрже убеждались мы в широкой волне реакции против материалисти-
ческого учения Дарвина и его единомышленников именно в упомяну-
тую нами эпоху. Это, впрочем, многократно отмечалось и некоторыми
крупными представителями антидарвинистского лагеря, — конечно, с
большим удовлетворением. Фактами такого рода переполнена наша
объемистая книга «История эволюционной палеонтологии от Дарвина
до наших дней» (1948). Многие другие факты такого же рода были
приведены и в других наших работах (1966). Этот крупный факт прив-
лекал наше внимание и занимал нас на протяжении всей нашей рабо-
ты в области эволюционной палеонтологии, или палеобиологии; но мы
могли убедиться в том, что и в других областях биологии в широком
смысле господствовала та же тенденция. Мы ставили перед собой воп-
рос: если подавляющее большинство биологов-теоретиков на грани-
це двух столетий оказалось на стороне идеалистического понимания
процесса эволюции, то неужели не было в Северной Америке и Запад-
ной Европе тогда среди деятелей биологии борцов за дарвинистские
материалистические идеи? Упорные поиски убедили нас, что таких би-
ологов среди признанных корифеев науки было тогда мало. Это были
прежде всего, престарелый Эрнст Геккель в Германии, выдающийся
зоолог, который с 60-х гг. прошлого века открыто выступал за мате-
риалистическое понимание развития органического мира, и бельгийс-
кий палеонтолог, замечательный ученик Владимира Ковалевского, Луи
13
Ч. Дарвин
14
Долло. Из биологов Запада нам было бы трудно назвать еще каких-
либо борцов за дарвинизм, за материалистические принципы, из чис-
ла крупных деятелей биологической науки. Мы думаем, что многие
из видных биологов, работавших в западных странах, держались дар-
винистского, материалистического понимания эволюции и её факторов,
ко по каким-то причинам не высказывали этих своих взглядов, не бо-
ролись за них. Ученые, писавшие тогда по этим вопросам, в подавля-
ющем большинстве случаев придерживались тех или иных вариантов
идеи зависимости процесса эволюции от внутренней тенденции, дейст-
вующей якобы помимо влияния среды. Эти учёные не могли, конечно,
объяснить, какие же силы направляли эволюцию; одни из них верили
в управление эволюцией сверхъестественными факторами, другие умал-
чивали об этом, третьи даже думали, что такое ортогенетическое разви-
тие происходит от естественных причин, то есть пытались объяснить
его материалистически. Но как бы то йи было, все эти естествоиспы-
татели отказывались от попыток объяснения «механизма» эволюции
как естественного процесса. Всё это — факт, бесспорный, ясный и за-
мечательный факт. Он прямо показывает, что после некоторого пери-
ода господства идеи эволюции, как естественного, а не чудодейст-
венного, процесса наступила полоса реакции против материалистическо-
го понимания развития органического мира. В подтверждение этого
положения мы сослались на свои многолетние работы. Но подчеркнём,
что в наших книгах мы проанализировали лишь незначительную часть
литературы. Дальнейшее ознакомление с первоисточниками убедило
нас, что отступление от материалистического объяснения эволюции бы-
ло ещё более разительным. Это была своего рода пиррова победа над
дарвинизмом. Подчёркивая этот факт, мы воздержимся здесь от по-
пытки его причинного объяснения, путь к которому был указан В. И.
Лениным в его «Материализме и эмпириокритицизме». Воздержимся,
потому что, во-первых, мы неоднократно затрагивали этот вопрос в
прежних работах, во-вторых, здесь для нас важен самый факт, а его
объяснение может быть предметом специального изучения, заслужива-
ющим внимания историков науки.
Правда, иногда наши учёные отказываются верить в этот факт.
Иной биолог не хочет верить на слово тем, кто пишет об этом факте; а
самому проверить его недосуг. Это действительно, нелегкое дело—соб-
рать и прочесть массу писанного по данному вопросу на разных язы-
ках в разных странах. А утверждение, будто бы подавляющее большин-
ство учёных, писавших об эволюции и её факторах в течение значитель-
ного отрезка времени, держалось неправильного, по сути ненаучного,
идеалистического понимания этого явления, может, действительно, по-
казаться очень странным. Оно даже похоже на клевету — поклёп на
честных и талантливых работников науки. А раз так, то, пожалуй,
лучше просто отвергнуть существование этого, впрочем, бесспорного,
факта! Но те, кто знает этот факт, никак не могут отбросить его.
Итак, единственный выход из создавшегося положения—вновь и вновь
изучать теоретические работы эволюционистов периода от рубежа двух
столетий до настоящего времени. Проверить, например, соответствуют
ли действительности сведения, сообщенные нами в наших книгах и ста-
тьях, не присочинили ли мы чего-либо. Мы твёрдо убеждены, что такая
проверка установит, что на самом деле картина отхода от дарвинизма
как материалистического учения об эволюции, имеет гораздо более
внушительные размеры, чем об этом говорилось в наших работах .
15
Значит, скептикам из ведущих биологов надо только организовать
ознакомление с данными литературы по эволюционизму. Это, вероятно,
не потребует от них особенно большой затраты работы, поскольку к
их услугам — целые институты с весьма внушительными средствами,
г. они могут привлечь к этой работе многочисленных сотрудников. Мы
убеждены, что полученный нами результат будет подтвержден и ока-
жется только, что мы слишком сдержанно и скромно оценили факт
отступления крупнейших авторитетов от материалистического понима-
ния эволюции к идеалистическому.
Нам известно, что некоторые из наиболее упрямых скептиков
ставят вопрос так: «Если Вы утверждаете, что в биологии западных
стран примерно на рубеже двух веков решительно победило антидар-
винистское идеалистическое понимание эволюции, то будьте любезны
объяснить нам причину этого странного процесса попятного дви-
жения методологических основ науки. Если >вы дадите удовлетворяющее
нас объяснение этого явления, то мы готовы поверить, что оно сущест-
вует; если же такого объяснения вы нам не преподнесете, то нечего
говорить и о факте, подлежащем объяснению — такой факт лишен
всякого значения». Едва ли надо доказывать несостоятельность такого
рассуждения. Разве мыслимо отрицать факт, если вы считаете неудач-
ным существующее объяснение причин этого факта?
Существует еще один подход к тому же самому вопросу.
Известно, что не столь давно некоторые товарищи проявляли склон-
ность резко преувеличивать роль учёных нашей страны в развитии на-
уки и столь же резко преуменьшать значение исследований, которые
велись за рубежом. Это была, конечно, недостойная тенденция, роняв-
шая честь отечественной науки. Это не был патриотизм в высоком
смысле слова, а было то, что М. Е. Салтыков-Щедрин называл пат-
риотистикой. Когда у нас было довольно широко принято говорить о
преимуществах отечественной биологии перед зарубежной, биологи
нередко присоединяли к этому декларации о реакционности основных
направлений биологической мысли в Западной Европе и США. Иног-
да кричали о реакционности самого Ч. Дарвина, даже о его «мрако-
бесии». Это было в высшей степени нелепо. Но это вовсе не означает
того, что русская научная мысль не могла опережать в определенных
направлениях науку самых передовых стран мира. В частности обще-
известно, что палеобиология, или эволюционная палеонтология, была
основана русским учёным В. О. Ковалевским. И всё-таки стремление
некоторых деятелей нашей науки утверждать то, что мы назвали «те-
орией благорастворения воздухов», согласно которой биологическая
мысль развивалась всегда плавно, одинаково успешно во все времена
всюду и везде, родилась отчасти как реакция против нелепого возве-
личения деятелей отечественной науки с принижением роли западных
биологов.
Итак, начало текущего века и первые его десятилетия характеризу-
ются широким распространением в западных странах различных
форм реакции против дарвинизма, как материалистического учения о
развитии живой природы. Это — факт, который нельзя игнорировать.
Следовательно, возникла обстановка, неблагоприятная для восходяще-
го развития подлинно-научной методологии эволюционизма, — своеоб-
разная «волна» реакционных идей.
Здесь будет уместно поставить вопрос: как влияла эта общая обс-
тановка на зарождение и первые шаги развития новой науки — гене-
тики, которая возникла как раз в эти годы? Ведь отход подавляющего
16
большинства эволюционистов от материалистического понимания эво-
люции мог охватить, частично или полностью, также и ленетиков уже
в самом начале существования этой отрасли биологии, — отрасли,
теснейшим образом связанной с учением об эволюции. Причина, выз-
вавшая антидарвинистскую реакцию в эволюционном учении, вполне
могла оказать такое же влияние и на генетику, на основные представ-
ления первых генетиков. Если мы обратимся к литературе по генетике
самого раннего этапа развития этой науки, то нам будет нетрудно убе-
диться, что такое влияние действительно осуществлялось и действовало
в совершенно определенном направлении.
Приятно, когда этот недостаток ранней генетики признают откры-
то и честно сами поборники этого направления биологических исследо-
ваний. В не столь давно вышедшей книге «Эволюция популяций и ра-
диация» (1966) крупный советский генетик Н. П. Дубинин так (наз-
вал один из разделов главы пятой этой книги: «Антидарвинизм и
метафизическая теория гена ,в генетике первых двух десятилетий XX в.»,
(там же, стр. 194).
В другой своей работе Дубинин (1968, стр. 8) писал: «Генетика пе-
реживала свой первый механистический этап развития- Новые открытия
трактовали метафизически. Иоганнсен, дав исчерпывающее доказа-
тельство ненаследуемости фенотипических уклонений, на базе которого
было создано представление о различиях между фенотипом и геноти-
пом, заявил, что его опыты по отбору в чистых линиях эксперименталь-
но разрушили основу дарвиновского учения. Отрицая за отбором ка-
кую-либо созидательную роль, Иоганнсен писал, что генетика вполне
устранила основу дарвиновской теории отбора и что таким образом
проблема эволюции в действительности является совершенно открытым
вопросом. Иоганнсен утверждает, что генетика, являясь точной наукой,
должна быть свободна от всякой эволюционной гипотезы. Эти идеи
Иоганнсена широко распространяются в генетике». Далее, сославшись
на величайшее значение достижений генетики уже в первых двух де-
сятилетиях XX века, Дубинин (там же) продолжал: «Однако ведущие
лидеры генетики того времени пытались истолковать эти новые данные
с идеалистических, во многом с махистских позиций, вырывая наслед-
ственность из общей связи явлений жизни. Процессы эволюции наслед-
ственности они видят только в комбинаторике генов, которые призна-
ются неизменными изначальными сущностями. Мутации рассматрива-
ют только как дезинтеграцию наследственности». Несколько дальше, в
связи с рассмотрением огромных достижений молекулярной биологии,
Дубинин (там же, стр. 13) писал: «Казалось бы, генетика вернулась к
исходным метафизическим и механистическим принципам понимания
живого и его эволюции. Идеалист Лотси мечтал свести эволюцию к ко-
мбинаторике вечных, неизменных, ограниченных в числе элементов —
генов».
Дубинин достаточно решительно и смело раскрыл картину убо-
жества теоретической мысли основоположников генетики, которым,
однако принадлежали крупные открытия в этой области. Он писал
(«Правда» от 11 июля 1968 г.): «История развития генетики в фило-
софском плане была исполнена противоречий. После открытия зако-
нов наследственности, обоснования теории гена, теории мутаций и
хромосомной теории долгое время на авансцену выходили метафизика
и идеализм» (подчеркнуто нами.—Л. Д.). Этот советской ученый пра-
вильно оценил старое учение о гене: «Основной методологической сла-
бостью старого учения о гене был механицизм и автогенез. Механи-
2- Л. Ш. Давиташвили 17
иизм пытался утвердиться в генетике, исходя из идеи о неделимости ге-
на, из представления, что генотип — это мозаика генов, а организм
(фенотип) — это мозаика признаков. Метафизика и автогенез в ста-
ром учении о гене более всего выражались в отрыве генов от об-
менных процессов в организме и от действия факторов внешней сре-
ды, что дало почву для автогенеза, игнорирования диалектических
связей внутреннего и внешнего. В старых представлениях каждый ген
изображался как вечно равный себе, неизменный элемент». Эта бес-
пощадная характеристика теоретической основы старой генетики не-
сомненно соответствует действительности, но не может быть признана
слишком суровой. Добавим, что, когда крупнейшие палеонтологи За-
пада приняли основные постулаты классической генетики, некоторые из
них заботливо перенесли черты идеализма и метафизики в свои ис-
следования. Об этом свидетельствуют, например, работы Г. Ф. Осборна
(Н. F. Osborn, 1917), который утверждал, что организм есть собрание
независимых, хотя и взаимосвязанных признаков, и Г. Г. Суиннертона
(Н. Н. Swinnertori, 1930, стр. 77; 1932), по которому «организм постро-
ен из многочисленных единичных признаков», в значительной мере не-
зависимых, хотя и сопряженных между собой.
Это коренные недостатки в дальнейшем, один за другим, преодоле-
вались, по меретого, как обнаруживалась, в процессе изучения фактов
науки, несостоятельность тех или иных исходных положений классичес-
кой генетики. Этот процесс, как мы думаем, еще далеко не завершен.
Отмеченные Дубининым исконные недостатки генетики раннего пе-
риода оказались очень живучими. Основоположникам этой науки нра-
вилась идея эмансипации генетики от эволюционизма. У. Бэтсон, как
известно, высказывался в пользу концепции Лотси. «Постоянство фак-
торов, с которыми все время имеет дело генетик в своих работах, —
писал Г. А. Левитский (1925, стр. 4), — сводит, как будто, всю проб-
лему изменчивости и эволюции к комбинированию данных изначала
генетических элементов. На этом, как известно, строит свою теорию
Лотси (Lotsy). Бэтсон не скрывает своей симпатии к этой попытке».
Соответственно этому своему отношению к эволюции, Бэтсон пренебре-
жительно писал об «эволюционной вере» (W. Bateson, 1922).
Нам могут сказать, что это было ведь лишь на заре новой науки
— генетики. При историческом подходе к пониманию развития идей
вполне понятны также слова великого советского генетика Н. И. Ва-
вилова (1927, стр. 419), который в своей замечательной работе о геогра-
фических закономерностях в распределении генов культурных расте-
ний писал: «Так мы приходим с иной стороны к мысли, выдвинутой
нашим учителем Bateson’oM о том, что процесс эволюции надо рассмат-
ривать, как процесс упрощения, развертывания сложного клубка пер-
воначальных генов». Но ведь ту же мысль еще довольно недавно раз-
вивал крупный французский ученый А. Львов (A. Lwoff, 1944), писав-
ший, что эволюция есть ретроградация в направлении к упадку и смер-
ти. Наконец современный выдающийся ученый Н. Пири (1959, стр.
84) проводит совершенно ту же идею в своем докладе, читанном на
Международном симпозиуме 19—24 августа 1957 г. Вот подлинные его
слова: «Доказательство того, что большая часть эволюции сопровож-
дается скорее избирательной потерей, чем приобретением свойств,
было приведено Львовым (13); из этого можно заключить, что совре-
менные организмы происходят от одного или нескольких сверхор-
ганизмов (подчеркнуто нами. — Л. Д.) с более широкими возмож-
ностями, чем те, которые встречаются теперь. Необходимо сделать вы-
18 • >
вод, что эти сверхорганизмы были далеки от эобионтов и, вероятно,
создавались в результате симбиоза между некоторыми из них»;
Таким образом, эта идея «сверхорганизма», заключавшего в себе
все гены позднее возникавших форм, — решительно всех видов, кото-
рые появлялись на всем протяжении исторического развития органи-
ческого мира, обнаруживает удивительную живучесть. Если еще мож-
но понять, что Лотси, Бэтсон и их единомышленники могли склонять-
ся к принятию этой фантастической идеи в тот период развития гене-
тики, когда абсолютная неизменяемость гена была одним из незыбле-
мых принципов этой науки, то упорное стремление удерживать эту
идею теперь, когда ее несостоятельность доказана всем ходом разви-
тия биологии, заставляет нас задумываться о причинах такого упорст-
ва. Невольно возникает подозрение, что живет еще та неприязни к ма-
териалистическому пониманию эволюции, которая так резко усилилась
на грани прошлого и текущего веков.
Итак, в этой главе мы отметили некоторые факты, очень важные
для понимания нынешнего положения генетики и перспектив её даль-
нейшего участия в разработке вопросов эволюционного учения. Эти
факты могут быть, вкратце, сведены к следующему.
1. Основатели генетики мало заботились о применении результатов
их исследований к учению об эволюции органического мира. О Г.
^Менделе известно, что в его личной библиотеке была книга Ч. Дар-
вина «Происхождение видов», но ни в одном из произведений талан-
тливого учёного-монаха ни единым словом не упоминается эволюция,
нигде не говорится о развитии органических форм в природе. В опуб-
ликованных материалах переписки Менделя, например, с известным
эволюционистом-антидарвипистом К. Негели тоже ни в какой степени
не затрагиваются вопросы эволюции и видообразования. Далее, Мен-
дель думал, что каждому «фактору» (этот термин у него, по-видимо-
му, соответствует позднее появившемуся термину «ген») соответствует
один признак развивавшегося организма (фенотипа). Ранние генети-
ки, как Бэтсон, Лотси и другие, допускали мысль, что какие-то пер-
воначальные организмы, на заре истории живой природы, обладали
ьсем богатством генов и что эволюция живой природы, ими прямо не
отрицавшаяся, сводилась просто к сокращению этого первоначального
запаса и к гибридизации, ведшей к колоссальному разнообразию
форм, которые появлялись и исчезали на протяжении всего геологи-
ческого времени и которые произвели ныне существующий органичес-
кий мир — протистов, растений и животных. Эта идея была факти-
чески отрицанием эволюции. Дубинин справедливо утверждает, что
в течение первых двух десятилетий в генетике царил антидарвинизм.
2. Многие исследователи прямо указывали ранним менделистам
на их серьезные ошибки, подвергали их заблуждения научной критике,
но до крайности увлеченные своими идеями, генетики оставляли без
внимания эту критику, хотя она исходила от таких выдающихся дарви-
нмстов как А. Уоллес, К. А. Тимирязев и многих других биологов.
Правда, позже даже ревностные менделисты поняли наивность своих
первоначальных воззрений и постепенно отходили от них. При этом
они нередко даже возмущались, что им «приписывают» взгляды, от
которых они уже отказались, хотя они избавились бы от своих заб-
луждений гораздо раньше, если бы прислушивались к критике.
3. В классической школе генетиков неукоснительно проводился
тот принцип, что хотя они выдвигали наивные несостоятельные основ-
ные положения, но критика этих ошибок была совершенно »ненуж1ной.
19
Вместо того, чтобы критиковать заблуждения классической генетики,
биологи, не согласные с их главными тезисами, должны были ждать
годами, десятками лет — в надежде, что генетики сами поймут свои
ошибки и устранят их. Классическая генетика, действительно, осво-
бождалась от некоторых своих исконных ошибок принципиального ха-
рактера, причем освобождалась от них постепенно, медленно, хотя на
ложность этих положений им указывалось неоднократно другими би-
ологами. Так из классической генетики выбрасывались представления:
(I) о точном соответствии одного гена одному признаку (с введением
1гсонятий тюлите-н.ии и плейотро1пии), (2) о неизменяемости генов, (3)
о независимости мутирования от воздействия условий внешней среды
Г. Мендель
(с введением понятия мутагенов, число которых весьма значительно
росло из года в год) и некоторые другие. Однако всякая ссылка на
то, что ведь критики принципов классической генетики указывали на
несостоятельность всех таких представлений задолго до того, как они
были отброшены самими поборниками этого направления, всегда вы-
зывала гневные выпады со стороны этих последних. Многие генетики
считали и считают себя вправе без всякого постороннего вмешательства
пересматривать основы своей науки, и, если нужно, отбрасывать их.
Этим товарищам, к сожалению, кажется, что свою школу надо во что
бы то ни стало оберегать от критики со -стороны инакомыслящих. Они
20
думают, что, если не защищать всячески свою школу, инакомыслящие
её разорят, и погибнет хороший очаг науки. Положим, в основе док-
трины данной школы лежат в корне неверные мысли, которые были
замечены инакомыслящими. Если приверженцы этой школы согла-
сятся признать правильность критики, то это будет уступка «врагу>,
которая может вести даже к капитуляции. Поэтому надо упорно защи-
щаться, отстаивать все основные положения доктрины своей школы,
пусть даже неправильные. Мы думаем, что такая тактика опасна для
науки и её прогресса, ложна, несостоятельна. Если крупная ошибка
обнаружена сторонниками другого направления, надо принимать кри-
тику немедленно и только с благодарностью. Это избавит нас от оши-
бок гораздо скорее, чем постепенное обнаружение и устранение заб-
луждений. В данном частном случае исправления в основные принципы
классической генетики можно было бы внести сразу же после убеди-
тельных критических замечаний К. А. Тимирязева, а также И. В. Ми-
чурина и некоторых его последователей, — десятками лет раньше, чем
это было частично сделано в действительности приверженцами клас-
сической генетики.
ЗАКОН ГОМОЛОГИЧЕСКИХ РЯДОВ Н И. ВАВИЛОВА И ЗАКОН
АНАЛОГИЧЕСКОЙ ИЗМЕНЧИВОСТИ Ч. ДАРВИНА
За последние годы значительно оживился интерес к закону гомо-
логических рядов, выдвинутому выдающимся советским генетиком
Н. И. Вавиловым. В советской литературе очень много статей, где рас-
сматривался этот закон. Часто о нем писалось, как об открытии, име-
ющем для биологии такое же значение, как периодический закон Мен-
делеева для химии.
Чтобы показать, как оценивается это обобщение Н. И. Вавилова в
нашей литературе последних лет, приведем лишь очень немногие выс-
казывания биологов. В книге —Современиные проблемы эволюционной
теории», недавно изданной под редакцией В. И. Полянского и Ю. И.
Полянского (1967), сказано: «На материалах, собранных преимущест-
венно в центрах многообразия, Вавиловым был открыт закон гомоло-
гических рядов в наследственной изменчивости...» (стр. 156). «Закон
этот, кроме глубокого теоретического значения, важен для практичес-
кой селекции, так как позволяет предвидеть нахождение различных
наследственных форм» (там же, стр. 454). В книге, где популярно из-
лагаются основы классической генетики, вышедшей с предисловием
академика Б. Л. Астаурова, В. М. Полынин сравнивает закон Вавилова
с законом Менделеева: «Когда Менделеев построил свою знаменитую
периодическую систему элементов, ее внутреннее совершенство требо-
вало, чтобы для некоторых элементов, которые людям не были извест-
ны, оставались свободные ячейки в таблице. Шли голы, и «вакантные
места заполнялись предсказанными элементами, причем, предсказанны-
ми не вообще, а с их индивидуальными признаками. Та же судьба ожи-
дала и вавиловскую таблицу» (стр. 237). Далее автор говорит, что «би-
ологи часто сопоставляют законы Менделеева и Вавилова». А затем
высказывает философскую мысль: «Не свидетельствует ли параллелизм
менделеевской и вавиловской систем об общности органической и неор-
ганической природы? О том, что обе они живут по одним законам бы-
тия» (там же, стр. 238).
А. Я. Ильин (1966, стр. 88) утверждает, что «закон Вавилова явил-
ся новым важнейшим вкладом в эволюционное учение» и что «откры-
21
тие закона гомологических рядов означало шаг вперед по пути прев-
ращения биологии из описательной науки в точную» (там же, стр.
92). Впрочем, этот автор, по-видимому, был склонен думать, что упо-
добление закона гомологических рядов периодическому закону хими-
ческих элементов «нельзя понимать абсолютно, поскольку не каждый
существующий вид непременно должен выявлять полный гомологичес-
кий ряд в рамках другого вида» (там же, стр. 92). Л. Д. Дергачёв
(1964, стр. 106) писал, что «закон гомологических рядов, открытый Н. И.
Вавиловым, был по существу дальнейшим развитием дарвинизма».
Н. Н. Воронцов (1966, стр. 221) утверждает, что «выдвинутый в
1922 году Н. И. Вавиловым закон гомологических рядов наследствен-
ной изменчивости, справедливо рассматривается многими биологами
как аналог периодической системы химических элементов Д. И. Мен-
делеева». Далее он пишет (стр. 222): «В своей исходной форме закон
гомологических рядов наследственной изменчивости Н. И. Вавилова
играет исключительную роль. Биологи подобно химикам конца XIX —
начала XX века, смогли предсказать существование еще не открытых
экотипов, рас, видов и родов и предсказывать их свойства».
В таком же духе писали о законе гомологических рядов в нас-
ледственной изменчивости Н. А. Майсурян (1967), Н. А. Базилевская
(1968)- и многие другие. При желании можно было бы привести еще
десятки подобных оценок закона гомологических рядов.
' Академик Н. П. Дубинин и В. С. Губарев (1966, стр. 25) писали
по поводу гомологических рядов Н. И. Вавилова: «Уже в 1920 году
Николай Иванович Вавилов сделал выдающееся открытие, значение
которого один из делегатов съезда селекционеров, на котором Николай
Иванович сделал доклад, определил очень лаконично: «Съезд стал ис-
торическим. Биология будет приветствовать своего Менделеева». Речь
идет о законе гомологических рядов. Н. И. Вавилов установил, что
наследственная изменчивость близких видов и родов растений во мно-
гом повторяют друг друга. Существует aaiKOMOieipiH спеть «в формообра-
зовании организмов. Среди мутаций, возникающих в растительном ми-
ре, нет хаоса. Есть определенные, четко установленные направления,
по которым идут процессы появления новых наследственных изменений.
Генетическое значение закона гомологических рядов огромно. Н. 11.
Вавилов создал «периодическую систему» наследственной изменчивости
растении».
Отмечая .важное значение этого закона, Дубинин (1970) возражал
против ложных его толкований.
Н. И. Вавилов формулировал «закон гомологических рядов» в 1920
году следующим образам: «Генетически близкие л.иннеевакие виды ха-
рактеризуются сходными и параллельными рядами наследственных
форм, и, как правило, наблюдается положение: чем ближе генетически
виды, тем резче и точнее проявляется сходство рядов морфологичес-
ких и физиологических признаков. Ближайшие генетически линнеоны
характеризуются, следовательно, одинаковыми рядами наследственной
внутривидовой из1мон'4*ивости» (1935, стр- 16). Далее сказано: «Вторая
закономерность в полиморфизме, вытекающая по существу из первой,
состоит в том, что не только генетически близкие виды, но и роды
проявляют сходство в рядах наследственной изменчивости» (там же,
стр. 21). Затем, на странице 34, читаем: «Подводя итоги рассмотрен-
ным закономерностям, мы приходим к следующим положения:.:: 1 Виды
и роды, генетически близкие, характеризуются сходными рядами
наследственной изменчивости с такой правильностью, что, зная ряд
22
форм в пределах одного вида, можно предвидеть нахождение парал-
лельных форм у других видов и родов. Чем ближе генетически распо-
ложены в общей системе роды и линнеоны, тем полнее сходство в ря-
дах их изменчивости. 2. Целые семейства в общем характеризуются
определенным циклом изменчивости, проходящей через все роды и ви-
ды, составляющие семейство» (см. также Н. И. Вавилов, 1967, стр. 35).
Поясняя сущность закона гомологических рядов Н. И. Вавилова,
А. В. Ильин (1966, стр. 86) писал: «Было установлено, что генети-
чески близкие виды характеризуются сходными и параллельными ряда-
ми наследственных форм и чем ближе друг к другу стоят виды в ге-
нетическом отношении, тем резче проявляется сходство между рядами
морфологических и физиологических признаков. Так, например, были
обнаружены сходные ряды наследственных форм пшеницы и ржи
(длиноостные, полуостистые, гладкочешуйные, длинноколосные, корот-
коколоюные и т. д., всего примерно 30 морфологически и физиологи-
чески различных форм)». Н. И. Вавилов «пришел к выводу, что гене-
тически близкие виды характеризуются сходными рядами форм в пре-
делах одного вида, можно предвидеть параллельные формы у других
сидов и родов» (там же).
Таков смысл закона гомологических рядов, как он был сформули-
рован Н. И. Вавиловым.
Рассмотрим теперь закон аналогичной изменчивости Ч. Дарвина,
как он излагается в трудах основателя эволюционного учения.
В дневнике Дарвин писал: «Поскольку все виды одного и того же
рода предполагаются происходящими от общего предка, можно было
бы ожидать, что они иногда будут варьировать аналогичным спосо-
бом, так что разновидности двух или нескольких видов будут похо-
дить друг на друга».
Этот вопрос Дарвин рассмотрел также в своем знаменитом «Про-
исхождении видов».
В этом своем произведении Дарвин (1939, стр. 384; 1882, стр. 125)
писал. «Различные виды обнаруживают аналогичные изменения, так
что разновидность одного вида часто приобретает признак, свойствен-
ный родственному виду, или возвращается к некоторым признаками
древнего предка». Самые различные породы голубей в странах, далеко
отстоящих одна от другой, как говорил Дарвин, образуют подпороды
с взъерошенными перьями на голове и перьями на ногах. Это Дарвин
считает аналогичными изменениями у двух или многих пород. Подоб-
ные аналогичные изменения, по Дарвину (там же), обязаны своим про-
исхождением унаследованной различными породами от общего предка
одинаковой конституции и наклонности к изменению при воздействии
одинаковых влияний, нам не известных. Такие же явления наблюда-
ются и в растительном царстве. Дарвин ссылается на то, что Нодэн
наблюдал много подобных случаев аналогичных изменений в семейст-
ве тыквенных, а различные авторы — у злаков. Подобные же случаи
отмечались, по Дарвину, и у насекомых.
Однако вполне естественно, что наибольшее внимание он уделил
этому вопросу в книге «Изменчивость животных и растений в состоя-
нии одомашнения». Для того, чтобы иметь возможность сопоставить
мысли Дарвина с законом Н. И. Вавилова, надо добросовестно поз-
накомиться с тем, что писал об «аналогической изменчивости» Ч. Дар-
вин.
Во втором томе «Изменчивости животных и растений при домести-
кации» этой теме посвящен специально довольно большой раздел гла-
23
вы 26-ой. Этот раздел носит соответствующее заглавие: «Аналогичная,
или (параллельная, изменчивость» (1921,стр. 340 и 1951, стр. 715). «Под
этим термином я подразумеваю, что сходные признаки иногда появля-
ются у нескольких разновидностей или рас, происходящих от одного
и того же вида, и в более редких случаях — у потомков значительно
различных видов (...). Случаи аналогичной изменчивости, поскольку
речь идет об их происхождении, можно объединить, оставляя в стороне
более мелкие подразделения, в две главные группы: во-первых, случаи,
вызванные неизвестными причинами, действующими на организмы,
сходно построенные и потому изменявшиеся сходным образом; во-вто-
рых, случаи, происшедшие от повторного появления признаков, кото-
рыми обладал более или менее отдаленный предок. Однако разграни-
чение этих двух главных категорий часто может проводиться лишь
предположительно, и как мы сейчас увидим, они постепенно переходят
одна в другую». Дарвин приводит случаи первой категории аналогич-
ных вариаций, не зависящих от реверсии, возврата к состоянию пред-
ка. Таковы многие случаи деревьев, принадлежащих к совершенно раз-
личным порядкам, но образовавших плакучие и пирамидальные разно-
видности. Бук, лещина и барбарис дали начало разновидностям с фио-
летовыми листьями. Далее Дарвин ссылается на сообщение, соглас-
но которому множество чрезвычайно различных растений дали разно-
видности с глубоко разрезными или рассеченными листьями. У разно-
видностей, происшедших от трех самостоятельных видов Brassica,
стебли, или так называемые корни, расширены в шаровидные массы.
Гладкий персик (нектарин) — потомок бархатистого персика, а разно-
видности бархатистых персиков и гладких персиков представляют заме-
чательный параллелизм: у плодов белая, красная или желтая мякоть,
косточки то не отделяются, то отделяются; цветки то крупны, то мел-
ки; листья то пильчаты, то зубчаты; снабжены то шаровидными, то поч-
кообразными железками, или совсем лишены железок. А ведь надо за-
метить, что каждая разновидность нектарина (гладкого персика) от-
нюдь не получила свой признак от соответствующей разновидности
бархатистого персика. Отдельные разновидности близкородственного
рода, а именно абрикоса, отличаются одна от другой приблизительно
такими же параллельными чертами. Нет основания полагать, что ка-
кие-нибудь из этих разновидностей просто вновь приобрели давно ут-
раченные признаки: в большинстве случаев этого наверняка не было.
«Три вида Cucurbita (тыкв, — Л. Д.) дали множество рас, приз-
наки которых так близко соответствуют друг другу, что, по настойчи-
вому утверждению Нодена, эти расы могут быть расположены почти
строго параллельными рядами. Отдельные разновидности дыни ин-
тересны тем, что они похожи важными признаками на другие виды то-
го же рода или родственных родов. Так, например, у одной разновид-
ности плод так похож и снаружи и внутри на плод совершенно иного ви-
да, а именно огурца, что их почти нельзя различить; у другой разно-
видности имеется длинный, цилиндрический плод, закрученный напо-
добие змеи; у третьей семена прикреплены к частям мякоти; у чет-
вертой плод, когда созреет, внезапно лопается и распадается на куски.
Все эти в высшей степени замечательные особенности характерны для
видов, принадлежащих к родственным родам. Едва ли можно объяс-
нить появление столь многих необычных признаков реверсией к одной
древней форме; но мы должны думать, что все члены семейства унас-
ледовали приблизительно сходную конституцию от какого-то раннего
24
предка. Подобные , же случаи наблюдаются у наших злаков и у мно-
I их других растений» (1921, стр. 341).
Мы привели те места из книги Дарвина, где кратко резюмируют-
ся случаи аналогичной изменчивости, подробно рассмотренные им в
других главах той же работы. Так, об аналогичной изменчивости у
злаков говорится в главе 9-ой. Почти все сорта или виды пшеницы из-
меняются совершенно параллельно (1921, т. 1, стр. 331; 1951, стр. 345):
«семена бывают покрыты пушком или гладки, различного цвета; цве-
точные же чешуи то несут ости, то лишены .их и т. д. Тот, кто пред-
полагает, что все сорта пшеницы произошли от одного и того же дико-
го вида, может объяснить эту параллельную изменчивость наследова-
нием сходной конструкции и вытекающей отсюда склонностью изме-
няться одинаковым образом; а тот, кто придерживается общей тео-
рии происхождения с видоизменением (то есть теории эволюции. —
Л. Д.), может распространить этот взгляд на различные виды пше-
ницы, если таковые когда-либо существовали в природном состоянии».
В первом томе упомянутого сочинения Дарвин подробно рассматрива-
ет явления и закономерности аналогичной изменчивости в разных груп-
пах растений.
Однако Дарвин не ограничивался изучением аналогичной изменчи-
вости у растений. Он рассматривал ее и у животных. И тут он тоже
выделяет случаи аналогичной изменчивости, не зависящие от прямой
реверсии. Такие случаи он отмечает у кур, голубей, канареек, лоша-
дей, рогатого скота, собак.
Затем Дарвин рассматривает случаи аналогичной изменчивости,
которые можно считать зависящими от реверсии. Так, у самых разно-
образных пород голубей иногда появляются разновидности, окрашен-
ные совершенно как прародительский дикий голубь (1951).
Дарвин говорил о «законе аналогичной изменчивости» (1921, стр.
442). Он придавал ему, как мы видели, большое значение и все же не
считал его всеобщим законом развития. Он писал (1882, стр. 126,
1939, стр. 286): «Признаки, обязанные своим происхождением исключи-
тельно аналогичной изменчивости, вероятно, не будут важными, ибо
сохранение всех функционально важных признаков будет определять-
ся естественным отбором в соответствии с различными формами образа
жизни видов». Таким образом, Дарвин указал на важнейшую причину,
ограничивающую роль аналогичной, или аналогической, изменчивости
в развитии органических форм.
Заметим, наконец, что термин — «гомологические ряды» задолго до
Н. И. Вавилова употреблял, почти в том же смысле, знаменитый аме-
риканский палеонтолог Э. Д. Коп. В отряде бесхвостых амфибий раз-
личаются два отряда: Arcifera и Fermisternia. Эги подотряды обнаружи-
вают, по Копу (Е. Д. Соре, 1904, стр. 70), интересные параллельные из-
менения в развитии различных скелетных частей. Такие параллельные
ряды Коп (Соре, 1868) называет гомологическими (1887, стр. 26,95 и
96; 1904, стр. 71).
Мы сравнительно подробно остановились на «законе аналогической
изменчивости» Дарвина для того, чтобы иметь возможность сравнить
этот закон с «законом гомологических рядов» Н. И. Вавилова. Впро-
чем, еще раз отметим, что взгляды Дарвина и его аргументацию мы
привели очень кратко. Интересующиеся могут обратиться к обширно-
му исследованию Дарвина «Изменчивость животных и растений при
доместикации». Там можно найти много поучительного. Под пером
великого естествоиспытателя-мыслителя оживали самые, казалось бы,
25
скучные материалы, как например, «аналогическая», или «гомологи-
ческая», изменчивость гладких и бархатистых персиков.
Дарвин писал о той закономерности, которую впоследствии Н. И.
Вавилов назвал «законом гомологических рядов изменчивости». Н. П.
Дубинин (1968, стр. 18) справедливо отметил, что, по Н. И. Вавилову,
«за параллелизмом изменчивости в основном кроется явление мутации
гомологических генов». У Дарвина этого, конечно, не было и не мог-
ло быть. Но едва ли в этом можно усмотреть коренное различие
между законом гомологических рядов изменчивости и законом анало-
гической изменчивости, ил.и какой-то недостаток в концепции Дарвина.
Свой «закон аналогической изменчивости» Дарвин обосновал
обильным и разнообразным материалом: многочисленными группами
растений и, вдобавок, явлениями, наблюдаемыми в животном мире.
Далее, Дарвин сразу же поставил вопрос о причинах возникнове-
ния аналогичных рядов изменчивости и постарался выяснить эти при-
чины; самые явления аналогичной изменчивости он различал по их
происхождению, — по причинам, ее вызывающим.
Наконец, Дарвин показал, что закон аналогической изменчивости
никак нельзя считать всеобщим законом развития живой природы и
установил, что он бесспорно имеет ограниченное значение, хотя заслу-
живает пристального внимания исследователей: если бы он думал
иначе, он не уделил бы этому закону так много места в своей книге
«Изменчивость животных и растений при доместикации».
Встает вопрос: почему многочисленные ученые, считающие «закон
гомологических рядов» крупнейшим, даже «величайшим открытием в
биологии, не сопоставляют этот закон с «законом аналогической из-
менчивости» Ч. Дарвина? Частично тут, быть может, играет роль обыч-
ное забвение трудов Дарвина. Но, так или иначе, в подавляющем
большинстве случаев совершенно исключается всякая тень недобросо-
вестности. Возможно, что некоторые биологи, имеющие желание пи-
сать о законе эволюции, просто не удосужились прочесть кажущий-
ся устаревшим огромный, двухтомный труд Дарвина. Такая работа
могла казаться им неоправданной потерей времени: ведь Дарвин не
знал генетики, не читал даже статей Менделя, писал в духе устаревшей
биологии XIX века. А. Л. Зеликман (1966) опубликовал статью «Незас-
луженно забытые идеи Ч. Дарвина в области изменчивости организ-
мов». Там он во многом совершенно справедливо отмечает «забвение»
многих высказываний Дарвина по части изменчивости организмов. Но
странно, что Зеликман имеет в виду лишь одну книгу Дарвина — его
«Происхождение видов». Раз автор задался целью «восстановит»
Дарвина» в правах по части изменчивости организмов, то как же можно
было полностью игнорировать замечательный огромный труд — рабо-
ту Дарвина, посвященную полностью именно этой проблеме? Зеликман
забыл об аналогической изменчивости Дарвина и о других идеях Дар»
вина в области изменчивости.
Устанавливая такие факты и забвение того, что думал и обосновы-
вал Дарвин, что он писал в широко известных своих истинно класси-
ческих произведениях, мы воздержимся тут от поисков причин этого при-
мечательного явления. Нам кажется, что здесь мы можем ограничить-
ся точным и бесспорным установлением фактов. Причинами же их мо-
гут заняться историки науки и особенно науковеды. В самом деле, как
это происходит, что честные работники науки совершенно забывают
концепции, которые развивались, обосновывались и громогласно про-
возглашались в трудах Дарвина, переведенных на многие языки и не-
26
однократно изданные, например, в русском переводе? Чем объясняет-
ся, что открытия и достижения величайшего гения биологии, Ч. Дарви-
на уподобились тому, что в римском праве называлось res nullius, —
ничья вещь, которую можно открывать вновь без ссылки на автора,
впервые сделавшего открытие и осмыслившего эту «вещь» гораздо все-
стороннее и глубже, чем позднейшие ее открыватели?
К ИСТОРИИ ВОПРОСА О НЕХРОМОСОМНОЙ НАСЛЕДСТВЕННОСТИ
В недавние еще времена считалось, что наследственность сосредо-
точены лишь в хромосомах ядра клетки. Представители классической
генетики 'непоколебимо верили в этот тезис. Однако наука строится на
фактах, и раз всплывают факты, нарушающие благолепие первоначаль-
ной хромосомной теории, то некоторое изменение этой теории в самом
существе ее становится досадной необходимостью. Можно, конечно, тя-
нуть и откладывать признание такой необходимости, но запретить всем
ученым заниматься изучением нехромосомной наследственности невоз-
можно: даже среди самых ревностных поборников классической генети-
ки всегда найдутся люди, желающие вести подобные исследования. По-
этому благоразумие требовало от сторонников классической генетики
взять в свои руки проблему нехромосомной наследственности: установ-
ленные факты настоятельно требуют внесения существенных принципи-
альных поправок в хромосомную теорию или замены ее какой-то новой
теорией, которая вобрала бы в себя, конечно, все положительные момен-
ты хромосомной теории.
При таких обстоятельствах странной и в то же время заслуживаю-
щей пристального внимания представляется нам позиция крупного ис-
следователя В. Л. Рыжкова.
Сборник «Актуальные вопросы современной генетики», под редак-
цией С. И. Алиханяна, содержит статью В. Л. Рыжкова «Цитоплазма-
тическая наследственность». Там этот ученый (1966а, стр. 358), справед-
ливо считаемый ведущим по цитоплазматической наследственности сре-
ди сторонников классической генетики, писал: «Ни на чем не основан
предрассудок, будто бы современная научная генетика отрицает роль
цитоплазмы или что она считает, что в наследственности главную роль
играет ядро. Такие представления не свойственны генетике. Более того,
знание цитоплазматической наследственности стало возможным только
благодаря открытиям Менделя. И сами основатели генетики открыли
роль цитоплазмы в наследственности». На следующей странице автор
пишет: «... все доказательства в' пользу того, что цитоплазма в том или
другом случае не играет роли в наследственной передаче признаков (...)
ничего не говорят в пользу монополии ядра в наследственности».
В предисловии к русскому переводу книги Дж. Джинкса «Нехро-
моссмная наследственность» В. Л. Рыжков (1966в, стр. 6) повторяет по
существу ту же мысль: «Существует ошибочное мнение, что классиче-
ская генетика рассматривает ядро в качестве единственного носителя
наследственности. На самом деле такие представления чужды генетике,
хотя некоторые биологи в свое время их и высказывали (...). Открытие
нехромосомной наследственности является заслугой основателей науч-
ной генетики». Итак, по Рыжкову, классическая генетика никогда не го-
ворила о монопольной^ роли ядра в наследственности, более того —
именно основоположники классической генетики открыли роль цито-
плазмы в наследственности, цитоплазматическая наследственность мог-
ла стать объектом изучения только благодаря открытиям Менделя.
27
Один из основоположников генетики Т. Морган (Т. Н. Morgan, 1926..
стр. 491) безоговорочно рекомендовал своим сторонникам игнориро-
вать цитоплазму в своих генетических исследованиях.
Откройте любой учебник генетики, принадлежащий перу одного из
представителей классической школы, и вы найдете там категорическое
утверждение, что именно хромосомы ядра являются носителями наслед-
ственности. Почти в каждом крупном труде, созданном в духе класси-
ческой генетики, 1вы встретите такое же утверждение. Чтобы далеко не
ходить, возьмем хотя бы как раз ту книгу Джинкса, которая была изда-
на на русском языке под редакцией того же В. Л. Рыжкова. Там, в рус-
ском переводе, предисловие автора Джинкса (1966, стр. 17) начинается
следующими словами: «Широко распространенное представление о не-
хромосомной наследственности как о явлении, настолько редко встре-
чающемся, что ему можно не придавать большого значения, теперь по-
колеблено». А в первой главе, говоря о фактических данных, которые
не подчиняются законам Менделя, Джинкс пишет: «Но хотя эти данные
правильно истолковывались как примеры нехромосомной наследствен-
ности, их наряду со многими другими наблюдениями, описанными
в последующие годы, долгое время рассматривали просто как своего ро-
да курьезы, никак между собой не связанные. В сущности, эти от-
дельные отклонения от законов Менделя почти не привлекали внима-
ния к проблеме нехромосомной наследственности (...). В настоящее
время клетку рассматривают как некое целое со свойствами более
сложными, нежели те, которые могли бы определяться простым сум-
мированием хромосомных и нехромосомных детерминантов» (Джинкс,
1966, стр. 20. Подчеркнуто нами. — Л. Д.).
Таким образом, опровержение успокоительных разъяснений предис-
ловия Рыжкова можно найти через несколько страниц в книге, издан-
ной под редакцией этого авторитетного ученого.
Сопоставьте прежнее мнение классических генетиков о ничтожном,
побочном значении нехромосомной наследственности, как о «курьезе»
с заявлением известного французского генетика Эритье (1964, стр. 251),
что «изучение нехромосомной наследственности — далеко не акцессор-
ная (побочная) область биологии, наоборот, в действительности оно
занимает центральное место. Оно находится в тесной связи с наиболее
фундаментальными проблемами». Это сопоставление указывает на ог-
ромный сдвиг. Появилось множество работ по нехромосомной наследст-
венности, основанных на точном эксперименте: С. Грэник (S. Granick,
1965); Р. Саиджер (R. Sager, 1964); Г. Бил, (1964 и 1966); К. Бан (К- S.
Bhan, 1964); Э. Щ. Гоукинс и Р. Коул (Sh. Е. Hawkins, R. Cole, 1965),
Р. Хагеманн (R. Hagemann, 1965 и 1968); Л. Бунур (L. Bounoure, 1964);
Дж. Даниелли (J. F. Danielli, 1960); Уилки Д. (D. Wilkie, 1964;
Ф. Шотц (F. Schotz, 1967); А. Мирский, (А. Е. Mirsky, 1963); М. Ренио
(М. Regnault, 1963); Г. Маккардт (Н. Marquardt, 1964) и др. За последние
годы усиленно разрабатывались вопросы так называемой кортикальной
наследственности, которая, как теперь установлено, является одной из
форм неядерной наследственности (ом., например, Кёртис А. С. Г.
(A. S. G. Curtis, 1965); X. Равен, 1964; А. Б. Калинина, 1968). Все эти
новейшие достижения ведут к признанию, что свойством наследствен-
ности обладает вся клетка в целом: ядро, цитоплазма и оболочка.
Таким образом, важность нехромосомной наследственности означа-
ет, что носителем свойства наследственности является отнюдь не толь-
28
ко ядро клетки, но й вся клетка. Только в самое последнее время эта
проблема начала привлекать внимание ученых школы классической ге-
нетики.
Возникает вопрос, почему же эти ученые в недавнем прошлом пре-
небрегали явлениями, которые со всей ясностью указывают на то, что
способностью передачи по наследству обладает, в той или иной форме
и в той или иной мере, вся клетка? С этим вопросом естественно свя-
зан и другой: все ли биологи игнорировали эту категорию явлений и
не было ли таких групп исследователей, которые смотрели на дело ина-
че? На эти вопросы ответить нетрудно. Представители мичуринского
направления всегда настаивали на том, что наследственностью облада-
ет вся клетка — и ядро, <и цитоплазма, что наследственность есть свой-
ство всей клетки ,в целом, всего живого тела. В этом утверждении не-
сомненно есть значительная доля истины, хотя несомненно и то, что
мно1ие представители мичуринского направления игнорировали веду-
щую роль хромосом ядра как носителя того, что теперь часто назы-
вают генетической информацией. И представителям классической ге-
нетики надо было бы прислушаться к голосу мичуринцев: это помог-
ло бы генетикам-неоменделистам быстрее и лучше разобраться в дан-
ном вопросе. Но вот тут-то и сказывается аномалия в состоянии науки
генетики: классическая и мичуринская школа находятся в положении
«воюющих сторон»: ни одна из этих школ не желает отнестись терпимо
к своим оппонентам, не прислушивается к голосу критики, если тако-
вая исходит от противного «лагеря». Некоторые генетики классичес-
кой школы считают недопустимым усматривать что-то разумное в выс-
казываниях представителей мичуринской школы, особенно же в выс-
казываниях Т. Д. Лысенко: все, что он писал должно быть обяза-
тельно сплошным и беспросветным заблуждением. Между тем, как
раз в вопросе о «монополии» ядра и его хромосом Лысенко был по
крайней мере отчасти прав, а генетики неоменделисты были неправы
до самого недавнего времени, когда пренебрегали явлениями «внея-
дерной» наследственности.
Дело доходило, как мы могли убедиться, до того, что даже авто-
ритетные исследователи отрицали самоочевидные факты: ярким приме-
ром такого явления может служить, как мы видели, позиция В. Л.
Рыжкова, который утверждал, что классическая генетика нисколько
не пренебрегала проблемой наследственности всей клетки в целом.
Необходимо, чтобы работники всех школ внимательно изучали
результаты исследований «противной» школы не только для того, что-
бы «отражать» ее атаки, но и для того, чтобы извлекать из этих ре-
зультатов все ценное, все нужное и полезное для науки. И когда мы
слышим, будто бы во всей продолжительной научной деятельности то-
го или иного крупного исследователя одной из «антагонистических»
школ нет ни единого светлого пятнышка, ни единого достижения, что
вся его продукция есть сплошной брак, никуда не годный мусор, у нас
всегда возникает сомнение в справедливости таких заявлений. В свое
время, к примеру, было принято изображать Вейсмана как совершенно
порочную фигуру в науке. А ведь в действительности А. Вейсман был
одним из крупнейших зоологов-эволюционистов, и произведения его
содержат много ценного для биологической науки.
Сделаем общие выводы из только что сказанного.
Вопрос о цитоплазматической наследственности имеет довольно
длинную историю. Однако в нашу задачу не входит изложение хода
развития этой идеи. Отметим только, что она долго признавалась ге-
29
нетиками классической школы в общем не заслуживающей, внимании.
Теперь, когда цитоплазматическая наследственность стала предметом
серьезного изучения и этой школой генетиков, можно, конечно, гово-
рить, что она всегда признавала явления цитоплазматической наслед-
ственности и уделяла им должное внимание. Но для того, чтобы убе-
диться в ошибочности такого представления, достаточно взглянуть
на курсы генетики, издававшиеся до 50-х годов текущего века.
Там либо не найдете никакого упоминания об этой проблеме, либо она
рассматривается как совершенно второстепенная, или даже третьесте-
пенная.
Один из основоположников классической генетики, выдающийся
американский ученый Т. Г. Морган (Morgan, 1926, стр. 491) писал:
«...мо^кет не казаться слишком экстравагантным предположение, что са-
ма протоплазма (исключая ее продукты секреции) состоит из единиц,
которые растут и делятся и наследуются. Думаю, что никто не отрицает
такой возможности, но удивительно, что те, кто отстаивает это воз-
зрение (или бросает вожделенные взоры в этом направлении) упуска-
ют из виду прекрасно установленные результаты генетического иссле-
дования. Именно так всегда, кроме редких случаев пластидного насле-
дования, наследование всех известных признаков может быть достаточ-
но объяснено присутствием генов в хромосомах. Одним словом, в ге-
нетике можно игнорировать цитоплазму» (подчерк-
нуто нами. — Л. Д.).
ПРОБЛЕМА ВЕГЕТАТИВНОЙ ГИБРИДИЗАЦИИ
В этой книге, посвященной вопросу об изменчивости организмов
прошлых геологических времен, нет особой надобности долго задержи-
ваться на проблеме вегетативной гибридизации, которая интересует
нас лишь постольку, поскольку она проливает свет на существо внут-
ривидовой изменчивости. Другие аспекты этой проблемы должны прив-
лекать к себе генетиков и селекционеров, которые ищут путей макси-
мального использования животных и растений для целей народного
хозяйства и здравоохранения.
Нам кажется несколько странным положение этой проблемы в сов-
ременной науке. С одной Стороны, имеются категорические заверения о
наличии определенных успехов в экспериментальном изучении этой
проблемы. С другой стороны, в солидных обобщающих трудах новей-
шего времени обычно она попросту замалчивается, а по каким сообра-
жениям замалчивается, об этом трудно судить неспециалисту. Неволь-
но возникает подозрение, что у некоторых исследователей имеется тен-
денция «снять» эту проблему, которая может вызвать сомнение в не-
которых принципах классической генетики.
Так, в изданной под редакцией С. И. Алиханяна (1966) книге, пос-
вященной актуальным вопросам современной генетики, — в коллек-
тивном труде, содержащем более 600 страниц большого формата, —
нет -ни -одной строчки о вегетативной гибридизации.
Несколько иначе подошел к этой проблеме автор капитального
курса генетики М. Е. Лобашев (1967). Он (там же, стр. 585) отмеча-
ет, что растения, измененные в результате прививки и взаимного вли-
яния подвоя и привоя, И. В. Мичурин, вслед за Ч. Дарвином и К. А.
Тимирязевым, называл прививочными, или «вегетативными, гибрида-
ми». Мичурин, действительно, критиковал некоторых ученых, которые,,
по его словам, слишком односторонне исследовали возможность яв-
30
ления вегетативной гибридизации у растений вообще и в особенности
у садовых плодовых деревьев. «Упомянутые ученые, — писал Мичурин
(Соч., т. 1, стр. 388), — или сомневаются, или совершенно отвергают
возможность явления вегетативных гибридов, но такой взгляд свиде-
тельствует об их слишком поверхностном знании жизни растений.
Напротив, на основании своих многолетних наблюдений, я категори-
чески утверждаю, что при соединении прививкой частей растений двух
различных форм и в особенности двух различных видов или родов рас-
тений, за редкими исключениями, почти постоянно наблюдаются явле-
ния изменения в строении соединенных частей с доминирующим укло-
нением в сторону одной из соединенных форм...». Далее Мичурин пи-"
сал, что изменение здесь имеется, за редкими исключениями, хотя оно
может оказаться очень слабым и заметным «лишь глубокому и опытно-
му в этом деле наблюдателю». Лобашев находит, что «при прививках,
возможно, происходят цитоплазматические изменения, которые могут
наследоваться при семенном размножении по типу длительных моди-
фикаций» (Лобашев, 1967, стр. 587). Он приходит к заключению, что
«основной вопрос: могут ли изменяться направленно гены привоя под
влиянием метаболитов подвоя — остается открытым». Итак» этот уче-
ный согласен признать возможность возникновения вегетативных гиб-
ридов на уровне длительных модификаций, но считает невыясненным
вопрос, могут ли таким путем изменяться гены. Таким образом, он
нахЪдит, чт<\ проблема вегетативной гибридизации заслуживает даль-
нейшего тщательного изучения. Это можно только приветствовать. Мы
ье можем- ^Даваться в рассмотрение вопроса о связи между вегета-
тивной гибридизацией и длительной модификацией, но считаем нужным
подчеркнуть, что само явление длительной модификации изучено пока
еще весьма слабо и занимает довольно неясное положение в системе
неоменделистского понимания наследственности и изменчивости. Следо-
вательно, определение сути вегетативной гибридизации через понятие
длительной модификации очень похоже на рассуждение того типа, ко-
торое известно в элементарной логике как ignotum per ignotius («не-
ведомое через еще более (неведомое»).
Между тем существование такого явления, как вегетативная гиб-
ридизация растений подтверждается исследованиями многих ученых:
И. Е. Глущенко (1948), Р. Георгиевой (1948), Р. Г. Главинич (1959),
Д. Поспишиловой (D. PospiSilova, 1957), И. Клотза (J. Klotz, 1957),
Д. Пастора (G. Pasztor, 1957), Н. Л. Удольской (1960), Иагишита Но-
бору (Yagishita Noboru, 1961), Ф. Д. Мампория (1969) и многих других.
В работах только что упомянутых авторов имеются ссылки на бо-
лее ранние исследования по вегетативной гибридизации. В общем не-
сомненно, что такие работы велись не без успеха в разных странах.
Мы, конечно, не можем уделить здесь много места этой проблеме, но
должны отметить наличие целого ряда работ по вегетативной гибриди-
зации и у животных. Явления этой категории изучались как советски-
ми специалистами — в частности А. М. Громовым (1963), Б. Г. Но-
виковым и его соавторами (1961), А. А. Чилингаряном и его соавто-
рами (1962), — а также многими компетентными специалистами и в
западных странах [см., например, Г. Катрфаж и Ж. Сабатье (Н. Qua-
trefages, G. Sabatier, 1961), П. Леруа (Р. Leroy, 1963), а также П. Лоу,
Дж. Карсон и С. Кинг (Р. Lowe, J. Carson, S. King, 1963)]. Уод-
дингтон, создатель теории генетической -ассимиляции, о которой мы по-
31
ворим и в другом месте этой книги, счел нужным уделить серьезное
внимание работам И. Е. Глущенко по вегетативной гибридизации расте-
ний и X. Ф. Кушнера по вегетативной гибридизации животных. Упомя-
нутый крупный английский генетик (С. Н. Waddington, 1963, стр. 145)
писал, что результаты этих экспериментов, которые он считает важными,
необъяснимы с позиций «конвенциональной» (т. е. традиционной) гене-
тики.
Опыты на курах проводились введением крови от одних пород пти-
цам других пород. Поскольку, однако, эритроциты птиц имеют ядра,
а следовательно и специфическую ДНК, то полученные у реципиентов
изменения иногда рассматривались как результат трансформирующего
влияния ДНК донора на воспроизводительные клетки реципиентов.
Однако К. Ф. Кушнер (1964, стр. 479) ссылается на ранее им опи-
санные опыты (Кушнер, 1960), в которых курам белый леггорн вместо
цельной крови иньецировали отцентрифугированную плазму, получен-
ную из крови кур породы нью-гемпшир: «Контрольные анализы пока-
зали, что в такой плазме практически отсутствует ДНК. И несмотря на
это, в потомстве реципиентов, начиная со второго поколения (хотя и
несколько реже, чем в опытах по переливанию цельной крови), стали
появляться особи с такими же отклонениями в окраске оперения. Сле-
довательно, преобразующее или трансформирующее влияние на воспро-
изводительные клетки реципиента оказывает не только цельная кровь
с ее форменными элементами, но и ее жидкая часть — плазма. Мож-
но с полным основанием допустить, что переливаемая плазма крови
в этом отношении во многом напоминает циркулирующие в растениях
жидкие пластические вещества, которые, как показывают многочис-
ленные опьцы по вегетативной гибридизации растений, также могут
влиять на наследственность прививаемых компонентов». Аналогичные
результаты быкли получены В. Павличенко (1963) у свиней, у которых,
как и млекопитающих вообще, эритроциты не имеют ядра.
Таким образом, представляется немыслимым полностью отрицать
явление вегётатив1ной гибридизации у растений и животных. Мы видели,
что М. Е. Лобашев (1967) довольствуется утверждением, что вегета-
тивная гибридизация есть, возможно, результат длительной модифика-
ции, а не изменения в составе хромосом ядра. Другие представители
той же школы Н. П. Дубинин и Я. Л. Глембоикий (1967, стр. 243)
применяют несколько иной способ для дискредитации самой идеи веге-
тативной гибридизации. Они вносят ее в группу «спорных методов»
селекции. Тем самым они воздерживаются от категорического отрица-
ния этого явления. Но они утверждают, что этот метод имел, в луч-
шем случае, лишь ничтожное практическое значение. «Таким образом,
— говорят они (воспроизводим это место их книги совершенно точно.
— Л. Д.), 30-летнее господство догм Лысенко смогло быть реализовано
в селекции самоопылителей всего лишь 4% сортов, происхождение ко-
торых не обошлось без отбора». Таким образом, Дубинин и Глембоц-
кий критикуют метод вегетативной гибридизации с очень важной, хо-
зяйственной, стороны, но не решаются отрицать это явление. Тем не
менее оно доставляет немало хлопот нашим генетикам.
Нельзя отрицать большое научное значение открытия Г. Менделя,
но нельзя и думать, что это открытие решает абсолютно все вопросы
учения о наследственности и изменчивости. Вегетативная гибриди-
зация — одно из явлений, сильно осложняющих решение задач этого
учения. Она очевидно относится к области внехромосомной наследст-
венности и напоминает генетикам о необходимости изучать отношения
32
между хромосомными и внехромосомными факторами наследственнос-
ти и изменчивости. Это осложнение задач генетики огорчает некоторых
ученых, тем более, что исследования в области вегетативной гибриди-
зации велись вне школы классической генетики и не могут быть запи-
саны в ее актив. Существует мнение, что не мешало бы предать заб-
вению вегетативную гибридизацию, «обратно закрыть» ее. Такую цель
преследовал, по-видимому, С. Я. Краевой, недавно опубликовав книж-
ку «Возможна ли вегетативная гибридизация растений посредством
прививок» (1967). Автор думает (стр. 44), что «вегетативные гибриды
не получаются потому, что нет такого биологического механизма, пос-
редством которого прививаемые компоненты могли бы обмениваться
между собой генами». Но как же быть с теми многочисленными опы-
тами, при которых наследственные изменения все-таки вызывались
посредством вегетативной гибридизации? И правильно ли думать, что
пзхменения могут совершаться только путем обмена генами между ком-
понентами? Автор твердо настаивает на том, что для прививочного (ве-
гетативного) гибрида «является обязательным слияние двух ядер веге-
тативных клеток, которые происходят соответственно из подвоя и
привоя» (там же, стр. ПО). Однако непонятно, почему автор хочет
вычеркнуть цитоплазматическую наследственность, ныне принимаемую
всеми генетиками. В общем автор проявляет сильное желание ликвиди-
ровать вегетативную гибридизацию: он не хочет ни на шаг отступить
ст ортодоксальных принципов хромосомной наследственности. А су-
ществование вегетативной гибридизации, действительно, не объяснимо
с таких теоретических позиций.
«НОВОЕ В УЧЕНИИ О БИОЛОГИЧЕСКОМ ВИДЕ» И ЗНАЧЕНИЕ ЭТОЙ
КОНЦЕПЦИИ В НАУКЕ О НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
Как уже было сказано, представители классической школы генети-
ков поступили бы хорошо, если бы в свое время обратили бы должное
внимание на критику, которой мичуринцы, и ib частности Т. Д. Лысен-
ко, подвергали допускавшуюся ими абсолютизацию «внутренних» при-
чин изменчивости или, во всяком случае, резкое преувеличение значения
этих причин. Лысенко и его единомышленники, в этом отношении
продолжая линию И. В. Мичурина, решительно подчеркивали тезис о
ведущем значении условий среды и их изменений, развивали ту мысль,
что живые тела и их наследственность изменяются под влиянием этих
экологических факторов. Мы видели также, что ортодоксальные неомен-
делисты пренебрегали этой критикой, отвергали ее без серьезного ана-
лиза критических положений, которые выдвигались Мичуриным, Лы-
сенко и другими представителями той же школы. Это пренебрежение
критикой на том лишь основании, что она исходит от представителей
«враждебного» лагеря, не способствовало успехам данной отрасли би-
ологии.
Мичуринцы развивали и другие тезисы, которые содержали значи-
тельную долю истины и представляли поэтому большой научный инте-
рес. Этих идей нам придется коснуться в других главах нашей книги.
Здесь мы остановимся на одном элементе воззрений Лысенко и
его соратников, который, на наш взгляд, отнюдь не может считаться
ценной чертой учения этой школы биологов.
Дело в том, что с 1950 года Лысенко стал усиленно пропаганди-
ровать свое, якобы новое, понимание видообразования и эволюции, ко-
торое он противопоставлял дарвиновскому. Надо прямо сказать, что
3. Л- Ш- Давиташвили 33
именно тогда и как раз этим он и оттолкнул от себя многих дарви-
нистов, которых безусловно подкупала защищаемая им идея зависи-
мости изменчивости организмов от условий их существования, — от
изменений этих условий.
Ныне всем довольно хорошо знакома концепция видообразования
и вообще формообразования, которую столь энергично защищали Лы-
сенко и его сторонники. Идея внезапного возникновения, вследствие
более или менее резкого изменения условий существования, совершен-
но новых видов и даже новых более высоких таксонов, возвращает
нас к додарвиновским временам. Здесь, в книге, посвященной вопро-
сам изменчивости, было бы неуместно даже упоминать о выдвигаемой
Лысенко теории видообразования, если бы внезапное возникновение
нового таксона не означало в сущности какой-то макромутации,
«грсссмутации», чего-то похожего на сальтации и онтомутации, т. е.
явления, безусловно относящегося к области учения о наследственнос-
ти и изменчивости.
Мы уже многократно отмечали несостоятельность этой странной
концепции (1956, 1959, 1966) и считаем излишним возвращаться к ее
критике, тем более, что она решительно отвергается подавляющим
большинством биологов. Ее не может принять, по нашему мнению,
последовательный эволюционист. Мы не можем понять, как биологи,
считающие себя материалистами, да еще диалектическими материа-
листами, могут допускать, например, превращение — от одного поко-
ления к непосредственно следующему, граба в лещину или сосны в
ель. Если особь одного определенного вида может породить особь
другого вида, даже другого рода, другого семейства, другого отряда,
то почему бы не допустить «порождение» сразу нового надотряда и
даже нового класса? Такое понимание возникновения видов несомнен-
но приближается к креационизму. Заметим, что аналогичные воззре-
ния существовали и существуют у многих противников дарвинизма
как материалистического учения. Кстати, сторонникам «нового в уче-
нии о виде» следовало бы иметь в виду, что их явно неудачная кон-
цепция может быть свободно использована против дарвиновского уче-
ния о происхождении человека. Если форма одного отряда может
внезапно породить форму другого отряда, то ведь человек мог произой-
ти не от менее высоко организованных приматов, а от других каких-
либо животных, например, от кошек.
Как, однако, могло случиться, что многие ученые определенной би-
ологической школы поддерживали в течение долгого времени явно
ошибочное, антидарвинистское понимание видообразования? Стоит
задуматься о причинах этого печального факта. Нам кажется, что ^то
могло произойти лишь в условиях, когда школа боится «выносить сор
из избы», опасается, как бы откровенная критика ошибки главы
школы не ослабила «линию обороны» против другой школы, школы
классической генетики. И в таких случаях, опять-таки, дело науки стра-
дает.
ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ И ОБЩИЕ СООБРАЖЕНИЯ
В этой главе мы рассмотрели лишь немногие вопросы истории на-
учной мысли в области изменчивости организмов и притом из числа
тех, которые, по нашему мнению, имеют существенное значение для по*
нимания изменчивости организмов в геологическом времени. Некото*
34
рые другие вопросы затронем в связи с вопросами о причинах и ха-(
рактере изменчивости организмов, преимущественна ископаемых.. ., .
Постараемся сделать выводы из сказанного в предшествующих
частях этой главы.
Для понимания хода развития проблемы изменчивости и ее совре-
менного положения очень важно ясно представить себе тот несомнен-
ный факт, что на границе текущего и предыдущего веков в развитии
биологической мысли имел место резкий перелом, суть которого заклю-
чается и пирровой победе идеалистического понимания эволюции наД|
материалистическим. Некоторые ученые ныне считают непозволитель-
ным называть те или иные биологические концепции идеалистически-'
ми. Это иногда .называют «приклеив заднем ярлычков», равносильным
преступлению в науке. Между тем, нет ничего оскорбительного в выра-
жениях «идеалистическая теория эволюции» «или идеалистическое по-.
нимание исторического развития органического мира». Как надо назы-
вать то понимание развития живой природы, которое исходит из тезиса
о вмешательстве божественных, сверхъестеоственных сил в эти пронес- ,
сы, об управлении эволюции божественным разумом? Биолог, верящий
в сверхъестественные силы, в божественное управление эволюцией ор-г
ганизмов, не усмотрит ничего оскорбительного в том, что мы называем'
его воззрения идеалистическими. Он, пожалуй, скорее обидится, если,
мы назовем его материалистом. Тут в признании идеалистических взгля-.
дов идеалистическими нет никакого «приклеивания ярлычка».
Многие ведущие биологи и историки биологической мысли засви- \
детельствовали тот факт, что приблизительно на рубеже 19-го и 20-го'
веков влияние материалистического эволюционного учения в общем (ни
не везде!) ослабело, а влияние идеализма в биологии резко усили-
лось. Об этом писали П. Жаннель (Р. Jeannel, 1950) и Фезсергилл.
(Ph. Fothergill, 1953) Дж. Монталенти (Р- Montalenti, 1965) и другие.
В этом убеждает всякого изучение соответствующей литературы этой
эпохи (см. по этому вопросу Л. Давиташвили, 1948 и 1966.)
И как раз в этот переломный исторический момент родилась на-
ука, получившая название генетики. Ее название, кстати сказать, ка-
жется нам подобранным не совсем удачно, ибо производимое от него
прилагательное «генетический» имеет гораздо более, широкое значение,
чем «генетика»: его широко употребляют в смысле «относящийся к.
происхождению» и во всех естественных науках, и в языкознании, и
в прочих гуманитарных науках. Более удобным был бы термин «гене-,
зиология», означающий «изучение явлений наследственности», •— тер-.
мин, предложенный еще раньше американским ученым А. Гайэттом
(A. Hyatt, 1893). Наука эта, © момент своего возникновения, оказалась
страдающей некоторыми врожденными пороками. Первоначально ге-,
нетики считали, что каждый ген соответствовал одному определенному
признаку. Утверждали, что организм состоит из определенного числа
таких признаков. Например, Г. Ф. Осборн, перешедший из лагеря
неоламаркистов в лагерь менделистов, думал, что организм есть соб-
рание независимых, хотя и сопряженных между собой единичных
признаков, или биопризнаков. Этим последним термином он решил за-
менить термин «единичный признак», употребляемый генетиками .—
менделистами (Осборн (Osborn, 1917), стр. 4191 «Биопризнаками яв-_
ляются, — писал он, те признаки, крупные и малые, которые, на осно-
вании данных онтогении, филогении или наследственности (менделиз-
ма), оказываются отделимыми или независимыми друг от друга, как
единицы в процессах наследования, эволюции и индивидуального раз-
* 35
витая» [Осборн (Osborn, 1917), стр. 450]. У Осборна, как и у многих
современных ему биологов, особенно же у генетиков, каждый единич-
ный признак живет своей самостоятельной жизнью. Такое понимание
признаков и генов теперь не пользуется популярностью и среди мен-
делистов; всякому непредубежденному биологу должна была бросать-
ся в глаза его резко выраженная, вопиющая механистичность. Но в
эпоху раннего менделизма оно было одной из основ классической ге-
нетики. Крупные деятели этого направления склонялись к идее неиз-
меняемости генов и тем самым тяготели к креационизму, к вере в
постоянство видов. Некоторые из корифеев этой науки были готовы
признать, что когда-то, в самом* начале истории органического мира,
существовавшие древние организмы обладали колоссальным богатст-
вом неведомо как возникших генов, а изменения, которые потом пре-
терпевал органический мир, сводились к утрате отдельными формами
многих генов и, следовательно, признаков, а также, быть может, к
перекомбинировке этих генов. Тем самым отрицалась подлинное разви-
тие, возникновение нового из недр старого. Это означало уже явно ме-
тафизическое понимание живой природы, резко противоречащее дар-
винизму. Такие, явно отрицательные, черты раннего менделизма, как
мы уже говорили, признавались и признаются авторитетными представи-
телями современной генетики, например, Н. П. Дубининым. Но ведь их
несостоятельность была ясна всем дарвинистам уже в первые годы
существования классической генетики. И не было никакой надобности
отмахиваться от критики таких принципиальных заблуждений даже в
те годы.
Возникает вопрос, не были ли эти недостатки как-то связаны с
общим падением теоретического уровня эволюционной биологии того
времени, точнее — не вызвано ли и это общее снижение уровня, и по-
роки ранней генетики какой-то общей причиной. По нашему мнению,
на этот вопрос надо дать утвердительный ответ. Эта общая причина
была посторонней для данной отрасли науки и состояла в давлении со
стороны социальных и политических условий, в которых развивалась
наука. Более детальное исследование этих причин — дело истории на-
уки и науковедения. Однако признание того факта, что генетика роди-
лась в условиях кризисного состояния эволюционной биологии, имеет
большое значение для общей объективной оценки классической генети-
ки: эта наука с самого же начала ее существования была отмечена
некоторыми крупными недостатками, она несла на себе печать метафи-
зики. В дальнейшем те, кто заботился о ее восходящем развитии, дол-
жны были принимать меры к ее исцелению от врожденных изъянов.
Считаем нужным отметить, что развитие науки в области позна-
ния наследственной изменчивости сильно страдает из-за нетерпимости
отдельных школ. Это наблюдается, к сожалению, не только за рубе-
жом, но и у нас. Борьба против этого общего зла и его преодоление
значительно ускорят прогресс научных исследований. Примером иде-
ологического давления на совесть ученого может служить смена гос-
подствующих направлений в биологии США. До смерти Г. Ф. Осборна
(1857—1935), выдвинувшего теорию аристогенеза, мы не встречали в
американской литературе, при самом усидчивом ее изучении, серьез-
ных возражений против этой теории, привлекавшей чудо к объяснению
эволюции органического мира. А ведь еще при Осборне, очевидно, у
многих американских ученых сформировались материалистические
постнеодарвинистские представления, явно противоречащие аристогене-
зу (но по существу механистические). Затем, довольно скоро, постнеодар-
36
винизм поднял голову и восторжествовал. В лице Дж. Г. Симпсона он
не только отверг аристогенез и ему подобные теоретические построе-
ния сонма других биологов-идеалистов, но и занял по отношению к
подобным концепциям резко отрицательную, мы сказали бы, даже
порой вызывающую, позицию. Стали приписывать этим концепциям
стультификацию, открыто заговорили об их ненаучности. Почему же,
однако, в США не было несколько раньше таких критических оценок
различных откровенно идеалистических, автогенетических воззрений?
Мы можем объяснить это лишь тем, что агрессивный антидарвинизм
поборников ортогенетических идей был еще в силе, хотя, быть может,
сила их начала уже убывать. Авторитет Г. Ф. Осборна, однако, едва ли
был единственным или хотя бы главным фактором, сдерживавшим
подъем механистического материализма «синтетической теории эволю-
ции». И тем не менее смерть этого «властителя дум», этого «Дарвина
новых времен» (как его при жизни называли) знаменательным образом
совпала с началом восхождения постнеодарвинизма.
Справедливость требует признания того, что в нашей стране раз-
витие эволюционной мысли протекало в иных условиях. Приблизитель-
но на границе двух веков, правда, раздавались голоса некоторых иде-
алистически мыслящих биологов, и даже несколько позже, уже после
первой мировой войны и Октябрьской революции, отдельными учены-
ми изредка высказывались мысли, созвучные идеям Осборна и других
западных биологов — идеалистов. Но обстановка, возникшая после
Октябрьской революции, способствовала не развитию идеализма в био-
логии, а укреплению у нас сильной материалистической традиции, но-
сителями которой были выдающиеся деятели науки; среди них
особенно видное место занимал К. А. Тимирязев.
И у нас были сменявшие одна другую доминантные школы гене-
тиков. Было время — мы имеем в виду преимущественно двадцатые
годы,—когда генетики были искренне убеждены в роковой отсталос-
ти классического дарвинизма. Однако им не удавалось подавить «ста-
ромодный» эволюционизм: в частности А. Н. Северцов и его ученики
вели успешно исследования в области сравнительной анатомии, кото-
рую А. Н. Северцов развивал как эволюционную науку, руководя шко-
лой эволюционной морфологии животных. Однако господство класси-
ческой генетики было довольно кратковременным: росла и крепла ми-
чуринская школа. Т. Д. Лысенко объединил вокруг себя агробиологов,
которые чувствовали недостатки классической генетики. Однако, к со-
жалению, дело не ограничилось борьбой или столкновением мнений,
творческим спором, научной дискуссией вокруг главных принципов ге-
нетики, а развернулась борьба за господствующее, командное положе-
ние. Прекратилось господство классической генетики. Хорошо извест-
но, что в конце 40-х годов, вопрос «кто кого?» решился — на время!
— в пользу школы Т. Д. Лысенко, и она стала чинить расправу над
своими идейными противниками.
Мы видели, что победа, лысенковцев, приведшая их к временному
безраздельному господству в биологии, не пошла на пользу этой шко-
ле. В условиях полного отсутствия критики внутри школы и суровых
оборонительно-наступательных мероприятий по отношению к инакомыс-
лящим биологам оказался возможным возврат к додарвиновской кон-
цепции внезапного, чудодейственного возникновения таких таксонов,
как вид, род, семейство и даже отряд без всякого участия отбора. Это
«новое в учении о виде» явилось подготовкой к отказу от идеи прои-
схождения человека от (приматов. Учение об антропогенезе было
37
бтодвй^уто на задворки науки. Настали трудные дни для эволюцион-
ноЙИмйсли. Такая деградация -наушной мысли стала возможной имеино
вследствие неограниченного господства олигархии в науке, с полным
исключением критики и самокритики даже внутри данной школы.
Но такое привилегированное положение одной школы не могло
существовать долго. Семья советских биологов вновь обрела право
свбббДно развивать исследования, — свободу научного -слова. И вот
Скоро другая, постнеодарвинистская школа начала претендовать на
привилегированное положение. Однако в настоящее время общие ус-
ловия'не благоприятны для какой бы-то ни было диктатуры в биоло-
гической науке.
! Многие генетики классической школы склонны замалчивать доста-
точно неприглядное прошлое менделистской генетики. При этом они
обычно строго придерживаются «теории благорастворения воздухов.»
Они замалчивают обстоятельную обоснованную научную критику «ран-
него менделизма», содержащуюся в трудах многих отечественных и
зарубежных биологов.
Генетики классической школы иногда изображают явные недостат-
ки этого направления как его достоинства. Так было, например, с ци-
топлазматической наследственностью. До недавнего времени в генетике
классической школы это явление игнорировалось как ничтожное и не
заслуживающее внимание, или вовсе отрицалось. Когда, однако,, на-
копилось много результатов специальных исследований, доказываю-
щих, что факты наследственности не укладываются полностью в хро-
мосомную теорию, пришлось заговорить о нехромосомной наследствен-
ности. Тут, казалось бы, надо было признать, что ученые, критиковав-
шие хромосомную теорию, были хоть отчасти правы, когда они утвер-
ждали, что «наследственность есть свойство живого тела» (Лысенко,
1965, стр. 633). Это, как писал Лысенко (там же), «в мичуринской
биологии известно уже несколько десятилетий».
Некоторые наши постчеодарвинисты начали прибегать к совершен-
но не нужному преувеличению значения научной деятельности отдель-
ных деятелей классической генетики и к умалению заслуг других био-
логов. Здесь мы имеем в виду, прежде всего, забвение достижений Ч.
Дарвина, игнорирование его научного наследства.
Нездоровым симптомом мы считаем замалчивание заслуг Ч. Дар-
вина в области учения о наследственности, в изменчивости и в частнос-
ти, — замалчивание его закона аналогической изменчивости. Может
быть, некоторые товарищи просто не удосужились познакомиться с
трудами Дарвина и особенно с его «Изменчивостью животных и расте-
ний» — не просмотрели эту книгу даже тогда, когда было желание по-
казать «незаслуженное забвение» идей Дарвина.
Попав в положение господствующего научного направления, наши
биологи •— постнеодарвинисты порой поддаются соблазну «командо-
вать» в науке, повелевать, как в свое время повелевали генетики ми-
чуринской школы. Это — очень опасно прежде всего для самой класси-
ческой генетики: отсутствие критики способствует созданию условий
застоя и укоренению неправильных воззрений. Мы видели, к чему эго
привело внутри школы Т. Д. Лысенко. Совершенно то же может прои-
зойти и со школой классической генетики.
Быстрое восходящее развитие биологической мысли в области поз-
нания изменчивости и ее законов протекает в условях свободы иссле-
дований, когда нет никаких групп и школ, претендующих на привиле-
гированное положение.
*38
Глава вторая
НЕКОТОРЫЕ ДАННЫЕ ОБ ИЗМЕНЧИВОСТИ ОРГАНИЗМОВ,
ВАЖНЫЕ В ПАЛЕОБИОЛОГИЧЕСКОМ ОТНОШЕНИИ
ВОПРОС О ВОЗНИКНОВЕНИИ РЕЗИСТЕНТНОСТИ МИКРОБОВ ПО
ОТНОШЕНИЮ К АНТИБИОТИКАМ
Развитие резистентности у бактерий и некоторых насекомых по
отношению к веществам, которые применяются для их истребления,
изучалось преимущественно в последние два десятилетия, в связи с ис-
пользованием антибиотиков, с одной стороны, и инсектицидов, с дру-
юй. Правда, наблюдения над подобными фактами производились и
раньше, но они не были предметом систематического изучения. И как
раз эти факты имеют особенно важное значение для исследования фак-
торов внутрипопуляционной изменчивости органических форм.
Изучение жизни организмов не выше одноклеточных приводит к
веским доводахМ в пользу идеи наследования особенностей, вызванных
влиянием среды на данную популяцию. Такие факты далеко не опровер-
гнуты приверженцами классической неоменделистской генетики. Неко-
торые из этих ученых справедливо настаивают на том, что закономер-
ности, установленные в развитии микробов, нельзя механически рас-
пространять на более высоко организованные формы жизни. Тем не
менее, данные по наследованию резистентности у микробов могут су-
щественно ограничить область применения неоменделистской концеп-
ции. «Явления комбинаторики, — пишет крупный английский ученый
С. Хиншел1вуд (С. Hinshelwood, 1958, стр. 316—317),—не могут играть
большую роль у микроорганизмов, размножающихся простым делени-
ем. Как ни плодотворна связь между свойствами клетки и строением
генов, всё же суть доктрины, покоящейся на этом допущении, в неко-
тором отношении несовершенна, как, правда, и несовершенен сам по
себе каждый раздел науки. Получается картина, в основном, статичес-
кая. Однако явления роста, приспособления и воспроизведения требу-
ют динамического объяснения». «Важное явление выработки сопротив-
ления к лекарственным веществам, — пишет этот автор в другом месте
(там же, стр. 317), — часто считается следствием отбора случайных
мутантов, хотя легко показать, на основании общеизвестных физико-
химических положений, что это явление должно возникать как авто-
матическая реакция вследствие того, что клетки перестроили свою ре-
активность». По Хиншелвуду, возражение против этого вывода «в силь-
ной степени основано на относительной стабильности таких изменений и
на доктрине, что наследственность будто бы изменяется путём спонтан-
ных генны?< мутаций». Под воздействием угнетающих веществ проис-
ходит изменение свойств микроорганизмов, их приспособление к дейст-
39
бию этих веществ. Объяснение такого приспособления сложным рядом
множественных мутащий потребовало бы, по Хи1ншел;вуду, принятия
весьма маловероятных допущений. Он утверждает, что отрицание всех
механизмов кроме отбора случайных мутантов и благоприятных комби-
наций генов есть злоупотребление принципами генетики (там же,
стр. 319).
Бактерии быстро развивают резистентность по отношению к анти-
биотикам, например, к стрептомицину, и это вызывает необходимость
применять разные антибиотики в лечении одного и того же больного.
Но нельзя отбрасывать с порога идею возникновения новых, резистен-
тных к данному антибиотику штаммов путём отбора уже существовав-
ших в исходной популяции малочисленных резистентных особей. Прав-
да, поначалу может показаться, что выживание штамма объясняется
прямым физиологическим результатом воздействия на данную попу-
ляцию соответствующего антибиотика, но вопрос о возможной роли
отбора безусловно заслуживает изучения.
Вопрос о причинах возникновения резистентности у болезнетвор-
ных бактерий, вполне естественно, вызвал животрепещущий интерес не
только у микробиологов, но и у биологов вообще. Возникла дискуссия,
один из важнейших моментов которой был отражен на лондонском
симпозиуме 1957 года. Материалы симпозиума содержат много данных,
которые свидетельствуют об адаптивном изменении микробов под вли-
янием антибиотиков и других ядовитых препаратов, вызывающих, по-
видимому, развитие сопротивляемости к этим веществам у бактерий.
В этом отношении весьма интересны сведения, сообщаемые А. Ч.
Дином и С. Хиншелвудом (А. С. R. Dean, Hinshelwood, 1957, стр. 4—
24). Эти авторы неоднократно развивали мысль о том, что вредные для
бактерий препараты могут непосредственно вызывать резистентность у
этих микроорганизмов. В то же время они не отрицают возможность
отбора спонтанных мутаций. «Мы никогда не отрицали, — писали они
(там же, стр. 4), — что могут возникать, и действительно возникают,
структурные мутации, ведущие к повышению резистентности или к улуч-
шенной утилизации питательных веществ; очевидным результатом тако-
го процесса является то, что возникающие так мутанты быстро подвер-
гаются отбору в соответствующей среде». И в то же время авторы ос-
паривали допущение, что беспорядочное «рандомное» мутирование и
отбор — единственный механизм (или хотя бы даже только главный
механизм) адаптации к новой среде или развития сопротивляемости к
ядам. Они указывали более непосредственно действующие механизмы и
приводили хорошие, по их мнению, доказательства в пользу существо-
вания таких механизмов.
Авторы проводят весьма поучительную и интересную параллель
между спором о возникновении резистентности к антибиотикам у мик-
робов и историей теории флогистона в химии. «Некоторые аспекты спо-
ра между мутационизмом и адаптационизмом, — писали Дин и Хин-
шельвуд (там же, стр. 7), — представляют интересную аналогию с ис-
торией теории флогистона. Сторонники этого учения, насколько нам из-
вестно, так и не заявили открыто об отказе от него. Однако под давле-
нием фактов они изменяли его, делали добавления, подпирали его вспо-
могательными гипотезами до тех пор, пока оно не стало неотличимо
от соперничающего с ним учения во всем, кроме излишней номенклату-
ры, которая вскоре была позабыта». Авторы продолжают: «Не проис-
ходит ли нечто похожее на это с некомпрометированной версией мута-
ционной теории? Вначале мутации были катастрофическими, разовыми
40
событиями, происходящими совершенно шальным образом в чрезвычай-
но ничтожмой, крошечной, части популяции и преимущественно во вре-
мя случайностей деления ядра. Очертания картины постепенно смяг-
чались. Мутации могут совершаться в отсутствии деления ядра и могут
охватывать всю популяцию (...): они могут индуцироваться медикамен-
том, к которому позже мутант обнаруживает резистентность, что пред-
ставляет заметное отклонение от шального характера (равдомности)»
(там же).
Постепенное отступление поборников теории мутации и отбора от
основных принципов этой теории было отмечено Дином и Хиншелву-
дом чрезвычайно метко. Сопоставление с падением теории флогистона
тоже весьма удачно. Это касается не только ортодоксально-генетичес-
ких идей в области изучения резистентности микробов по отношению к
антибиотикам и другим ядам: совершенно то же самое мы видим в раз-
витии всей классической генетики и так называемой «синтетической
теории эволюции» в целом.
Отнюдь не отрицая менделевских принципов в отношении расщеп-
ления при гибридизации, ни в какой мере не умаляя значения хромо-
сом, Дин и Хиншелвуд приводят серьезные доказательства в пользу
прямого влияния антибиотиков на природу микробов, выражающееся и
в возникновении у этих органзмов резистентности к соответствующим
препаратам. Кратко упомянем лишь немногие из доводов, приводимых
авторами, потому что полное изложение их аргументации, несомненно
представляющей большой интерес, завело бы нас слишком далеко в
сторону от основной задачи нашей книги.
По наблюдениям авторов, продолжительное воздействие медика-
ментов. в частности антибиотиков, на бактерии вызывает стабильное
приспособление, резистентность против этих веществ. Но эта стабиль-
ность имеет различные степени в зависимости от длительности воздей-
ствия. Если, например, бактериальные клетки только что приобрели
способность расти в присутствии препарата, то они легко утрачивают ее
в среде, свободной от такового. По мере возрастания длительности воз-
действия возврат к исходному состоянию становится все менее легким,
и в конце концов адаптация представляется относительно стойкой,
стабильной. Такой ход развития был выяснен при адаптации АегоЬас-
ter aerogenes к профлавину и к сульфаниламиду. Когда реверсия име-
ет место, она не обязательно оказывается полной, но может устанавли-
ваться более низкий уровень иммунитета, «состояние равновесия», ко-
торое сохраняется в течение значительного времени (там же, стр. 9.).
Дин и Хиншелвуд «приучали» тот же вид бактерий к умеренно высо-
ким концентрациям профлавина, пропамидина и хлорамфеникола, а
затем держали эти «приученные» (тренированные) штаммы в течение
очень долгого времени (около тысячи поколений) в среде, содержащей
препарат. Приобретенные адаптации, хотя они имели очень значи-
тельную стабильность, утрачивались после продолжительного пребыва-
ния в среде, свободной от препарата. Отсюда авторы заключили, что
стабильная наследственность еще не может указывать на неправиль-
ность теории ответа на воздействие среды. Более того, вся последова-
тельность явлений в подобных экспериментах, по словам авторов, лег-
че объясняется гипотезой адаптаций, чем теорией мутаций и возврат-
ных мутации. Такой вывод представляется нам достаточно убедитель-
ным и весьма осторожным.
Авторы (стр. 10) ссылаются на случаи, когда в связи с адаптаци-
ей к новым 'источникам углеводного питания масса возрастала пример-
41
но на 40% без увеличения числа бактериальных клеток. Принимая во
внимание, что клетка едва ли возрастает значительно более, чем вдвое
против своей первоначальной величины, можно заключить, что по мень-
шей мере значительная доля популяции участвовала в процессе адап-
тации.
Дин и Хиншелвуд привели и другие веские соображения в поль-
зу теории непосредственной адаптации бактерий к антибиотикам и дру-
гим вредным для них веществам.
Остальные участники симпозиума высказывали различные мнения.
Некоторые из них просто уклонились от рассмотрения каких-ли-
бо гипотез происхождения резистентности, кроме выдержанной в духе
кл ассичеокой генетики.
Так, Л. Л. Кавалли-Сфорца (L. L. Cavalli-Sforza, 1957, стр. 40)
находит, что некоторые случаи могут быть объяснены и на основе клас-
сической генетики, и как «негенетические адаптации». Но в исследо-
ванных им случаях возникновения резистентности к стрептомицину и
хлорамфениколу «гипотеза генетической адаптации» (выдержанная в
духе классической генетики) вполне приемлема, а потому, по словам
Кавалли-Сфорца, «не возникала надобность в каких-либо дополнитель-
ных альтернативных объяснениях». М. Демерец (1957, стр. 57) рассудил
дело примерно так же: «Никакие из этих данных, — писал он, — не ис-
ключают возможности участия внегенных механизмов в дополнение к
генным механизмам в развитии резистентности к препаратам». И все-
таки он не останавливается на рассмотрении роли «внегенных» факто-
ров, которая, по его мнению, должна быть второстепенной. По В. Д.
Дэвису (там же, стр. 60) генетики не станут возражать против возмож-
ности, что среда может вызвать направленное наследственное измене-
ние, но он утверждал, что «тяжесть доказательства все же лежит на
тех, кто утверждает, будто бы сопротивляемость против препаратов
или какое-либо иное наследственное изменение у бактерий может быть
вызвано направляющим влиянием среды». Однако совершенно непонят-
но, почему в данном случае Дэвис допускает привилегированное по-
ложение неоменделистов, почему-то свободных от «тяжести доказатель-
ства» их воззрений, возлагая эту тяжесть всецело на инакомыслящих-
Воо-бще же говоря, сторонники классически-генетическото толкова-
ния причин резистентности к антибиотикам считали необязательным
доказывать свою правоту и ложность мнения инакомыслящих. Нам ка-
жется, что основная причина такой линии поведения заключается в
признанной гегемонии одной школы над всеми другими.
Однако другие участники симпозиума занимали в дискуссии более
объективные позиции. Так, Дж. Ледерберг (J. Lederberg, 1957, стр.
62), правда довольно неохотно, признал, что картина, изображенная
Хиншелвудом, встречается в действительности, а потому нет никакой
надобности доказывать существование механизмов, о которых толко-
вал Хиншелвуд. Маргарет Дж. Торнлей, Дж. Синай и Дж. Юдкин
(J. М. Thornley, J. Sinai, J. Yudkin, 1957, стр. 152) рассмотрели различ-
ные способы возникновения резистентности у бактерий и посвятили осо-
бый раздел «Ламарковской наследственности», где они соглашаются с
исследователями, признающими возможность приобретения наследст-
венной резистентности путем «прямой индукции».
Эксперименты и доводы Дина и Хиншелвуда, а также некоторых
других непредубежденных исследователей представляются нам имею-
щими значительную силу убеждения. Но эти исследования должны
42
быть продолжены. Нам думается, что постепенность в развитии резис-
тентности, многочисленность «шагов» (мутаций), необходимость допус-
тить множество ступеней-мутаций реверсии при постепенной утрате ре-
зистентности — 1все это совершенно исключает возможность «обойтись»
худосочной формулой «мутация — отбор».
Упомянутый нами видный исследователь Дж. Ледерберг (Leder -
berg, 1967, стр. 624) утверждал, что «направленное мутирование у выс-
ших организмов уже давно дискредитировано, и установлена «коренная
неопределенность» мутирования, присущая как спонтанным, так и ин-
дуцированным рентгеном мутациям. Однако возникновение резистент-
ности, несомненно индуцированной лекарствами, оживило иллюзии, что
гены бактерий можно направленно изменять. Этот вывод должен был
подорвать концепцию генов для этих организмов». Ледерберг призна-
вал, что «мы еще мало знаем о непосредственных причинах возникно-
вения как спонтанных мутаций, так и мутаций, индуцированных радиа-
цией или химическими веществами» (там же, стр. 625). Но все же он,
как ему показалось, окончательно решил загадку механизма резистент-
ности к лекарствам у бактерий. Ледерберг подсчитал, что в большой
популяции чувствительных к стрептомицину бактерий на 10 млн. бак-
терий одна оказалась резистентной уже в исходной популяции, не под-
вергнутой еще действию антибиотика; он извлёк эту бактерию и заста-
вил её делиться, что привело к образованию весьма многочисленной
популяции, причем вся популяция оказалась резистентной к стрептоми-
цину.
Ледерберг (Lederberg, 1967, стр. 625) писал: «Каждый хорошо изу-
ченный случай наследуемой резистентности можно объяснить спонтан-
ным возникновением резистентных мутаций и их последующим отбором
под действием лекарства. Кроме того, при благоприятных условиях,
если ухитриться выделить резистентные мутанты без того, чтобы они
когда-либо входили в соприкосновение с лекарством, можно четко до-
казать их спонтанное происхождение».
Нам кажется, что для окончательного решения этого вопроса нуж-
ны основательные дополнительные исследования. Многое, впрочем, го-
ворит за правильность мнения Хиншелвуда. Например, — общеизвест-
ный факт, что приспособление бактерий к антибиотику может привес-
ти к потребности этих бактерий в присутствии его. Едва ли эта пот-
ребность могла существовать уже у исходных форм, предки которых
развивались всегда, конечно, в отсутствии этого вещества. Это — один
из фактов, говорящих против «преадаптации», которую везде ищут
постнеодарвинисты: довольно странно думать, что в геноме микроба
хранились неприкосновенно не только ген резистентности против опре-
делённого антибиотика, но и ген определяющий необходимость этого
антибиотика для развития данного вида микроба.
По поводу спора между Хиншелвудом и его сотрудниками, с одной
стороны, и сторонниками «синтетической теории эволюции», с другой,
К. Гесслер (1967, стр. 167) справедливо писал: «Дискуссия, развернув-
шаяся на основе полученных данных, показала, что результаты экспе-
риментов нельзя объяснить с помощью мутационной теории посредст-
вом отбора предсуществовавших мутантов».
После провозглашенной Ледербергом окончательной победы не-
оменделистской концепции генезиса резистентности Дин и Хиншелвуд
{Dean, Hinshelwood, 1963 и 1964) продолжали энергично защищать
43
свое объяснение этого процесса. Не может быть и речи о победе идеи
«спонтанная мутация плюс отбор» в том разделе биологии, которая
занимается изучением сопротивляемости по отношению к антибиотикам
и другим медикаментам у микробов.
«ДЛИТЕЛЬНЫЕ МОДИФИКАЦИИ» У ГЕТЕРОТРОФНЫХ ПРОТИСТОВ
На третьем международном конгрессе протозоологов довольно'
видное место было уделено обсуждению проблемы длительных модифи-
каций. Это явление получило широкую известность после исследований
Иоллоса (V. Jollos, 1921 и 1932), которому принадлежит и самый тер-
мин «длительная модификация» (см. Давиташвили, 1936). Т. Криппа-
Франчески (1969, стр. 112) вполне справедливо подтверждает выска-
зывание Соннеборна о том, что длительные модификации «оказали очень
незначительное влияние на генетику». Более того, было принято утвер-
ждать, что исследования Иоллоса были проведены недостаточно тща-
тельно, а потому их результаты не заслуживают внимания. Это дава-
ло по старинке мыслящим генетикам некоторый покой и поддерживало
в них уверенность в незыблемости основных принципов их науки. Но
этому покою пришел конец. Криппа-Франчески (1969, там же) пишет:
«Работа Иоллоса, тщательно выполненная, насыщенная
экспериментальными данными (подчеркнуто нами. — Л.
Д.) и даже сегодня считающаяся в целом приемлемой, содержит выво-
ды, довольно революционные для того времени и иногда не очень убе-
дительные». Далее Криппа-Франчески находит, что большинству авто-
ров, выступавших с подтверждением теории длительных модификаций,
«не удалось улучшить положение, потому что они скорее пытались най-
ти новые примеры этого явления, чем стремились лучше понять основ-
ные, уже описанные случаи. Мы, однако, думаем, что собирание при-
меров» длительной модификации» было делом бесспорно законным и
полезным для науки. Было нужно показать, насколько распространено
в природе это явление, и накопление фактов такого рода никак нельзя
признать вредным или бесполезным, если только не считать основной
задачей генетики обеспечение от критики и пересмотра однажды приня-
тых принципов этой науки.
«Различные роды, виды и штаммы, — пишет далее Криппа-Фран-
чески (там же), — вырабатывают устойчивость к условиям среды и
сохраняют эту устойчивость на протяжении сотен вегетативных поко-
лений после удаления индуцирующего фактора, а согласно некоторым
авторам, — и после полового процесса. В настоящее время трудно ска-
зать, не сохраняется ли эта устойчивость в некоторых случаях навсег-
да». Подчеркнем, что тут речь идет о том, что автор считает длитель-
ными модификациями, которые, по его словам (там же), «возникают и
в условиях ежедневной изоляции (без отбора), если животные нахо-
дятся в постепенно ухудшающихся условиях». Все это достаточно
красноречиво свидетельствует о том, что «длительная модификация»
есть явление, с которым генетик обязан считаться, если он ставит своей
задачей познание действительности, а не сохранение догм, их защиту
от фактического материала.
Далее автор говорит, что существуют две гипотезы, пытающиеся
объяснить такие факты: макронуклеарная и цитоплазматическая. Ж-
Женермон, придерживающийся макронуклеарной гипотезы, «обнару-
жил, что его чистые линии, проанализированные на устойчивость к
хлористому кальцию перед адаптацией, обнаруживают генетическую
44
гетерогенность на этапе формирования новых карионид». По Криппа-
Франчески, принимаемая Женермоном макронуклеарная гипотеза» мо-
жет объяснит многое, но не все факты, касающиеся длительных моди-
фикаций» (там же, стр. 113); она не объясняет некоторых фактов, опи-
санных самим Женермоном. «Помимо этого, — продолжает Криппа-
Франчески (там же), макронуклеарная гипотеза никак не объясняет
модификации, возникающие без отбора». Для объяснения таких случа-
ев и прежде всего многих случаев индуцированной устойчивости к
температуре, наблюдавшихся Франчески, она выдвигает цитоплазмати-
ческую (гипотезу». Точное экспериментальное исследование дало ей «до-
казательства решающей роли цитоплазмы в детерминации изменения
устойчивости» (там же). «Сравнивая эту цитоплазматическую теорию
с макронуклеарной, автор (там же, стр. 113) не видит необходимости
привлекать дифференцировку макронуклеуса для объяснения возникно-
вения специфических длительно сохраняющихся изменений у простей-
ших».
Ж. Женермон (1969) изучал явления длительной изменчивости у
простейших, наблюдаемые при адаптациях к повышенной температуре
и к хлористому кальцию. «Потеря приобретенной резистентности, —
говорит он (там же, стр. 111), — происходит постепенно во время
вегетативного размножения, но ускоряется при половых процессах
(конъюгации или автогамия)». Это, по его мнению, указывает, что
макронуклеус «играет важную роль в существовании длительных моди-
фикаций». Тем не менее он признает, что; если раса культивируется
достаточное время в присутствии яда, а затем снова переводится на
обычную среду, то уровень ее резистентности часто остается выше нор-
мальной. «Лишь через многие десятки поколений после дезадаптации
резистентность вновь становится нормальной» (там же).
Таким образом, не остается сомнений в том, что «макронуклеарная
гипотеза», отмечающая значение ядра клетки для явлений длительной
модификации, отнюдь не может претендовать на полное объяснение
этого явления. Явления длительной модификации представляют собой
бесспорное наследственное изменение простейших, индуцированное фак-
торами среды (повышение температуры, введение хлористого кальция).
Связь между явлениями, происходящими в ядре (макронуклеусе),
и явлениями «длительных модификаций» с дарвинистской точки зре-
ния в высшей степени вероятна, но это не спасает «классической» точки
зрения, устаревших догм. Даже при стремлении удержать прежнюю
схему, исследователи признают существование этих «длительных моди-
фикаций». Так, К. М. Суханова (1969, стр. 190) отмечает, что, хотя «из-
менения тепло- и холодоустойчивости свободноживущих и паразитиче-
ских простейших, происходящие быстро под влиянием окружающей тем-
пературы, носят ярко выраженный модификационный характер», моди-
фикационная изменчивость по этим признакам «может усложняться
появлением длительных модификаций».
ГИПОТЕЗА ГЕНЕТИЧЕСКОЙ АССИМИЛЯЦИИ УОДДИНГТОНА
Нам кажется здесь уместным объективный анализ той идеи «гене-
тической ассимиляции» и «канализации», которую долго и упорно раз-
вивает Ч. Уоддингтон в целом ряде своих работ. Общее впечатление, к
которому приводит многих читателей ознакомление с этой концепцией,
сводится к тому, что она представляет собой стыдливое прикрытие вы-
нужденного признания наследования приобретенных признаков. Однако
45
Уоддингтон убежден, что она дает вполне удовлетворительное, с точки
зрения ортодоксальной генетики- толкование явлений, которые могут
лишь казаться объяснимыми с позиций «ламаркизма». Свою гипотезу
генетической ассимиляции автор излагает в целом ряде работ. Она
встречает некоторое сочувствие со стороны отдельных эволюционистов
постнеодарвинистской школы, но в большинстве случаев обнаруживает-
ся довольно холодное к ней отношение со стороны крупных представи-
телей этого направления. Ей уделяет видное место Дж. С. Уильямс
(G. С. Williams, 1966), однако во многих обобщающих работах крупных
теоретиков постнеодарвинизма она просто не упоминается (напр., Сим гр
сон (G. G. Simpson, 1953, 1964); Олсон (Е. С. Olson, 1965); Росс
(Н. Н. Ross, 1962).
Рассмотрение гипотезы генетической ассимиляции начнем с того
несколько упрощенного объяснения этого предполагаемого явления, ко-
торое содержится в книге Эрлиха и Холма «Процесс эволюции» (1963;
русский перевод 1966).
Вот отрывок, где в упомянутой книге (1966, стр. 169) дается это
объяснение:
«Как можно объяснить формирование высокогорных растений, се-
верных рас бабочек и другие сходные явления? Вызванные средой из-
менения не могут быть непосредственно переданы последующим поко-
лениям; ламаркистское представление о наследовании приобретенных
признаков давно отвергнуто. Уоддингтон постулировал процесс, в ре-
зультате которого такие приобретенные признаки могли бы п р и пос-
редстве отбора ассимилироваться в генотипе, и это предположе-
ние подкрепляется рядом экспериментов самого Уоддингтона и других
исследователей. Сущность этой генетической ассимиляции
лучше всего можно пояснить на следующем примере. Лабораторную по-
пуляцию одной линии Drosophila melanogaster дикого типа подвергали
тепловому шоку на стадии куколки. У отдельных мух, выводившихся иэ
таких куколок, обнаруживался аномальный разрыв жилок на крыльях
(фенотип crossvoinless). В качестве родителей для получения следующей
генерации использовали только особей, обладавших приобретенным фе-
нотипом «кроссвейнлесс». После второй дюжины поколений частота осо-
бей «кроссвейнлесс» выводившихся из подвергнутых тепловому шоку
куколок, превышала 90%, но, кроме того, отдельные мухи «кроссвейн-
лесс» начали появляться из необработанных куколок линии, .в которой
шел отбор. Скрещивания между такими мухами дали линии с высокой
частотой фенотипов «кроссвейнлесс» при отсутствии теплового шока.
Могло бы показаться, что приобретенный признак стал наследственным.
На самом деле в этом эксперименте отбор, по-видимому, благоприят-
ствовал тем генотипам, которые обусловливали низкий порог индукции,
желательного (с точки зрения отбора) фенотипа. В конце концов этот
порог снизился настолько, что уже не требовалось никакого теплового»
шока для того, чтобы развитие шло в направлении фенотипа «кроссвей^н-
лесс». Аналогичные результаты были получены при изучении других
признаков дрозофилы».
Приведенные строки, действительно, вызывают сомнения относи-
тельно возможности объяснения описываемого явления на основе прин-
ципов ортодоксальной генетики по формуле «спонтанные наследствен-
ные изменения плюс отбор». Нелегко понять, -как порог, соответствую*
щий определенным генотипам, мог снизиться настолько, что развитие
шло и без теплового шока в направлении фенотипа «кроссвейнлесс».
Иное дело, если -мы допустим индуцированное шоком мутирование в.
46
направлении указанного фенотипа, то есть мутагенное действие теплово-
10 шока: такое объяснение было бы, по меныпей мере, понятным. В про-
тивном случае надо было ,бы думать, что у какой-то части исходной «попу-
ляции уже был генотип, обусловливавший фенотип crossveinless, но
он ничем не проявлял себя до применения теплового шока.
Между тем сам Уоддингтон (Waddington, 1960, стр. 389) утверж-
дает, что приобретенные признаки «согласно неоменделистской теории,
действуют лишь как генетический «шум» в том смысле этого термина,
какой принят в теории информации». Обратимся теперь к тому, что пи-
сал сам Уоддингтон по поводу свой концепции генетической ассимиля-
ции. «Недавно, — говорит он (I960, стр. 390), •— были произведены экс-
перименты, в которых популяции дрозофилы были испытаны на присут-
ствие генетической изменчивости способности отвечать онтогенетически-
ми изменениями на стимулы, исходящие от различных абнормальных
типов среды, причем была изучена эффективность действия отбора на
эту генетическую изменчивость». Во всех случаях, несмотря на разнооб-
разие средовых стимулов, была обнаружена генетическая изменчивость
способности организма регулировать, поддающейся действию отбора. В
одном из своих экспериментов Уоддингтон попытался вызвать адаптацию
популяции дрозофилы к высокой концентрации хлористого натрия в
среде, где живут личинки. Выяснилось, между прочим, что с повыше-
нием концентрации соли в среде возрастают величины анальных па-
пилл. Когда подвергшиеся отбору штаммы выращивались в нормальной
среде, они уже не возвращались полностью к состоянию штаммов, не
подвергавшихся отбору. Такое отсутствие гибкости в системе онтоге-
нетического развития Уоддингтон называет канализацией.
Тепловой шок, которому подвергались личинки, вызывал появле-
ние изменений в жилках крыльев. Путем отбора можно было создавать
штаммы, реагирующие высокой частотой определенного онтогенети-
ческого отклонения. Удавалось продуцирование штаммов с определен-
ным видоизменением даже в отсутствие стресса.
«Мы можем спросить себя,—рассуждает Уоддингтон (Wadding-
ton, 1960. стр. 397), — отчего происходит эта генетическая изменчи-
вость. Не возникла ли она в ходе эксперимента?» По его мнению, име-
ются достаточно веские основания думать, что это не так в случае с
возникновением фенокопий «кроссвейнлесс». Типы «кроссвейнлес» спон-
танно возникают в некоторых диких популяциях. Эксперименты, по сло-
вам Уоддингтона (Waddington, 1960, стр. 397), говорят против «ламар-
ковского наследования приобретенного признака».
Однако примечательно, что сам Уоддингтон говорит о случаях, где
«ген, действующий в направлении отбора, появляется в ходе эксперимен-
тов» (там же, стр. 398). «Поскольку, — рассуждает он, — эксперимен-
ты производились над сотнями тысяч мух, появление таких мутаций не
столь неожиданно, чтобы мы приписывали это самому оредовому стрес-
су». Мутации, как он думает, происходили нормальным образом, то есть
беспорядочно, как попало. Таким образом, и в случаях, когда опыты на-
водят на мысль о возникновении новых мутаций, Уоддингтон считает
возможным оставаться в рамках неоменделистского понимания причин
изменчивости и ее характера. Он убедился в том, что воздействием раз-
личных факторов среды (поваренной солью, теплом, парами эфира)
можно вызвать развитие определенных фенотипов, которые передаются
по наследству и после устранения соответствующего стресса. Хорошо^
с его точки зрения, когда в подобных случаях так или иначе1 можно до-
пустить отсутствие мутирования. Но на худой конец возможны же и му-
47
тации, совершающиеся хаотически, «шальным» способом, который нео-
менделисты до сих пор считают единственно нормальным.
Пусть взятые сами по себе опыты Уоддингтона укладываются в
прокрустово ложе неоменделистской концепции. Но ведь это возможно
лишь до тех пор, пока речь идет об одном изменении в данном направ-
лении. Если же мы представим себе целый ряд подобных изменений, их
восходящую серию, то тут уже рассуждения Уоддингтона окажутся
бессильными спасти основную идею ортодоксальной генетики. Палео-
биологическая история многих групп животных, в частности — моллюс-
ков, живших в замкнутых или полузамкнутых бассейнах Черноморско-
Каспийской области и всего Паратетиса, изобилует такими примерами.
Здесь гипотеза Уоддингтона совершенно неприменима. Совместно с
такими случаями,—о них идет речь в других местах этой книги,—
опыты по генетической ассимиляции недвумысленно свидетельствуют
о последовательно направленном развитии изменчивости под воздейст-
вием факторов среды.
НАПРАВЛЕННЫЕ НАСЛЕДСТВЕННЫЕ ИЗМЕНЕНИЯ, ИНДУЦИРУЕМЫЕ У
НЕМАТОД РОДА CAENORHABDITIS
Самого пристального внимания заслуживают исследования фран-
цузского ученого Ж. Л. Брена над нематодами [Брен (J. L. Brun, 1966,
1967]; В. Ни гон, Брен, (V. Nigon, Brun, 1967). Эти наследования имеют
большое значение для понимания закономерностей и факторов наслед-
ственной изменчивости, а следовательно и эволюции организмов вооб-
ще.
Брен (Brun, 1966а, стр. 128) считает совершенно изумительным
тот факт, что имеется лишь мало исследований, касающихся возник-
новения приспособления животных к повышенным температурам. Этот
факт, действительно, примечателен, но его нетрудно объяснить, мы ду-
маем, господством идей постнеодарвинизма в современной эксперимен-
тальной биологии. Поскольку эволюция понимается как процесс, в ос-
новном зависящий от отбора случайных «ошибок» в наследовании ге-
нома, едва ли можно удивляться тому, что подавляющее большинство
представителей экспериментальной биологии решительно воздержива-
ется изучать проблему мутагенного влияния «обычных» факторов сре-
ды на наследственную природу организмов: такая возможность мыслит-
ся этими учеными как априори исключенная.
Брен утверждает, что в литературе упоминается один единствен-
ный случай «генетического приспособления» к повышенным температу-
рам среди многоклеточных. Он имеет в виду работу А. М. Банта (1939)
о копеподах вида D phnia longispina: в половом клоне, происходя-
щем от партеногенетического предка, содержавшемся на протяжении
14 лет в лабораторных условиях, было замечено появление, после лет-
него перерыва, терморезистентных особей, которые в работе рассмат-
риваются как мутанты в виду их стабильности.
При таких обстоятельствах, как справедливо замечает Брен (Brun,
1966а, стр. 128), понятно, что авторы часто сомневаются в возмож-
ности получать у многоклеточных животных экспериментально выз-
ванную адаптацию к повышенным температурам.
Однако скудость экспериментальных успехов в этой области, как
думает Брен, зависит преимущественно от трудностей, связанных с над-
лежащей постановкой экспериментального исследования на большом
числе поколений. Caenorhabditis elegans — исключительно подходя-
48
щий объект исследования в этом направлении. Во-первых, изучение
многочисленных поколений облегчается благодаря краткости репро-
дукционного цикла у этих нематод (четыре дня при 23,5°С). Во-вторых,
поведение этих организмов можно наблюдать как на уровне особи,
так и на уровне популяции, благодаря их высокой плодовитости. В-тре-
тьих, гермафродитное самооплодотворение этого вида позволяет экспе-
риментатору избегать затруднения, о которое спотыкаются многие ис-
следователи: тут Брен имеет в виду различную термическую чувстви-
тельность у самцов и самок рездельнополых организмов. Брен ставит
вопрос, является ли внешний фактор первопричиной привыкания или
он играет роль лишь отбирающего фактора, выявляя предсуществую-
щие адаптивные возможности?
Брен думает, что в случае С. elegans эта проблема может быть ре-
шена наилучшим образом благодаря особенностям репродукции (раз-
множения) этой формы. В самом деле, это животное можно содержать
в культуре в виде самооплодотворяющихся гермафродитов.
До опытов, предпринятых Бреном, штамм, который был впослед-
ствии использован, содержался в лаборатории более, чем 12 лет (око-
ло 700 поколений). Следовательно, должна была возникнуть полная
генетическая однородность животных. При таких условиях эту немато-
ду вполне можно уподобить чистой линии. Между тем, неэффективность
отбора чистой линии, по Брену, вполне доказана.
Поэтому приспособление этой формы нематод вследствие отбора
через изменение температуры, по-видимому, исключается. Возникнове-
ние адаптивных механизмов должно в данном случае зависеть от тер-
мического воздействия. Это и побудило Брена заняться изучением дан-
ной проблемы на нематодах вида С. elegans.
С. elegans — свободноживущая (в почве) форма, в состоянии по-
ловой зрелости она достигает в длину около 1 мм. Это — протандриче-
ский автогамный гермафродит, у которого лишь очень редко встреча-
ются самцы (приблизительно один самец на 1000 гермафродитных осо-
бей).
Автор описывает постановку экспериментов, исключающую, надо
думать, возможность ошибки. В каплю агар-агара вводится один-
единственный гермафродитный производитель-
Культура никогда не содержит более сотни потомков, происходя-
щих от одного прародителя и принадлежащих к одному и тому же по-
колению.
Исходный штамм поддерживался при постоянной температуре 18,J.
От этого штамма автор вывел линии, «приспособленные» к различным
более высоким температурам: 22°—23°—23,5°—24°—24,5° (там же, сто.
136).
Линия С. elegans, приспособившаяся к некоторой повышенной
температуре, всегда характеризуется плодовитостью более мощной, чем
плодовитость штамма, содержимого в обычных условиях, то есть при
18°С. Число поколений, в течение которых линия выводится при повы-
шенной температуре, определяет: (1) степень приспособленности этой
линии к дайной температуре, а также (2) «возможность привыкания к
еще более высоким температурам.
Поскольку повторение экспериментов в тождественных и различ-
ных условиях температуры показывает, что репродукционная способ-
ность очень велика, отношение между причиной и следствием устанав-
ливается с весьма высокой степенью вероятности.
4- Л. Ш- Давиташвили 49
Если зародыш, полученный при температуре в 18°, переносится в,
среду с температурой в 24,5°С, он развивается до взрослого состоя-
ния, но оказывается совершенно бесплодным, стерильным. Если такой
же зародыш переносится в среду с 23°С, он развивается во взрослое
животное, но с очень пониженнной плодовитостью. Если животные пе-
реносятся из среды с 18°С в среду с 22°С, то сначала констатируется
прогрессивное падение плодовитости в ходе последовательных поко-
лений. Потом, однако, плодовитость постепенно повышается. Таким
путем получается линия, которая, по-видимому, вполне приспособлена
к температуре в 22°С. Если делается попытка получить от этой линии
приспособление к 23°, то результаты оказываются отрицательными с
животными, привыкшими к температуре в 22° в течение менее 90 поко-
лений. Но если животные взяты после 95 поколений, живших при 22°г
то нематоды, доведенные до 23°, выдерживают фазу пониженной пло-
довитости, отмечаемой в первых поколениях после перевода; достигши
минимума, плодовитость прогрессивно поднимается, и возникает, в кон-
це концов, штамм, стабильный при 23°.
Начиная от этого штамма, приспособленного к 23°, те же резуль-
таты наблюдаются при переводе к 23,5°С: приспособление возникает
лишь в том случае, если попытка делается начиная с 252-го поколения,
воспитанного при 23°.
Путем такой процедуры можно в конце концов получить линию,
беспредельно плодовитую при 24,5°.
Итак, если С. elegans происходит непосредственно от штамма, со-
держимого на протяжении нескольких лет в лаборатории при темпера-
туре 18°С, он не может размножаться долго при температуре выше 22°.
Но прогрессивно, очень медленно повышая температуру культуры, мы
можем получить линии, постоянно размножающиеся, сохраняющие
плодовитость до 24,5° [Брен, Brun, 1966 б), стр. 267]. Стабильность-
созданных таким путем адаптивных механизмов возрастает вместе с
продолжительностью культуры при повышенной температуре. Брен
(Brun, 1966в, стр. 439) утверждает, что строго автограмное размноже-
ние этого гермафродитного вида нематод делает невозможным искание
причин генетической изменчивости в высоко гетерозиготном состояния.
При этом питательная среда играет лишь ничтожную роль. Стабиль-
ность акклиматизации зависит, по-видимому, от длительности пребы-
вания штамма при определенной температуре. Акклиматизация возни-
кает в результате кумулятивного эффекта действия температуры. Это
действие — очень медленное. Например, при 23,5°С требуется не менее
100 поколений для того, чтобы плодовитость животных была в четыре
раза больше, чем в первых поколениях после перехода из среды с 23°С.
Нематоды, выведенные от штамма, существовавшего при 18°С, могут
выдерживать 245°С только после 1081 поколения.
Справедливо сознавая, что эги исследования подсказывают очень
ответственные выводы, Брен обстоятельно рассматривает возможности
объяснения процессов приспособления ценорабдитов к повышенной тем-
пературе. Выводы эти представляются неожиданными, и они явно про-
тиворечат господствующим в генетике представлениями о наел еде г-
венней изменчивости организмов.
В то же время чрезвычайно интересно сопоставить всю цепь экспе-
риментов Брена над нематодой С. elegans с последовательностью исто-
рического развития многих групп моллюсков, обитавших в водоемах
Паратетиса на протяжении неогена и антропогена. В обоих случаях
наблюдается постепенное приспособление к определенным условиям;
50
среды: в опытах Брена к последовательному повышению температуры,
а в истории моллюсков Паратетиса к определенно направленному из-
менению солености (обычно к ее понижению). Изученные еще.Неймай1-
ром палюдины очевидно тоже постепенно приспособлялись к химизму
вод, где они обитали. j ,
ПЕРЕХОД ФИТОФАГИЧЕСКИХ НАСЕКОМЫХ К НОВЫМ КОРМОВЫМ
РАСТЕНИЯМ
Переход насекомых-стенофагов от одной растительной пиши к
другой наблюдался специалистами обычно у вредных насекомых, кото-
рые иногда изменяют вид растительной пищи и тем самым . могут
наносить вред культурам, какие до того были для них недоступными.
В редких случаях агробиолог может быть заинтересован в переводе
полезного насекомого от одного вида растительной пищи, недостаточно
представленного в данной местности, к другой, там более обильной. В
любом из этих случаев наблюдения могут способствовать изучению* воп-
роса об изменчивости насекомых и значении этой изменчивости для
эволюционного учения.
Профессор физиологии животных Ч. Лэдд Проссер (С. L. Prosser,
1960, стр. 586) писал: «Насёкомые имеют склонность откладывать яй-
ца на растении того же самого вида, на котором они питались, когда
были личинками; если им предлагается непривычная пища на протя-
жении нескольких поколений, то личинки могут принимать ее с возрас-
тающей готовностью». Эта чрезвычайно скупо выраженная мысль под-
тверждается многими исследованиями. В своей президентской i речи,
прочитанной перед Лондонским королевским энтомологическим обще-
ством, Б. П. Уваров (В. Р. Uvarov, 196b стр. 52) сказал: «Несмотря на
ряд случаев, когда было доказано, что вызванные факторами среды
изменения оказываются, по крайней мере временно, наследственными,
существует нежелание признать факты, которые могут показаться бла-
гоприятствующими сильно оклеветанному ламарковскому принципу».
Далее этот ученый призывает энтомологов «изучать свои проблемы без
опасения нарушить какие-то устаревшие каноны генетики учебных ру-
ководств» (там же).
Г. С. Андреварта и Л. С. Берч (Н. G. Andrewarta, L. С. Birch, 1954,
стр. 664) пишут, что некоторые насекомые, если они воспитывались на
несвойственной им пище, обнаруживали предпочтение к этому виду
пиши, когда они начинали искать места для кладки яиц.
Литература по прикладной энтомологии, как говорят эти авторы,
содержит указания на то, что определенные виды насекомых быстро,
за период в несколько лет, расширяли набор растений, которыми они
питаются, включив сюда и те виды или разновидности, которыми они
до того пренебрегали. Так, яблонная плодожорка, Cydia pOmonella, до
1918 г. в США не указывалась на грецком орехе. ОДнако немного поз-
же, в 1926 г. Г. Куэйл (Н. Quoyle, 1926) сообщил, что грецкий орех
постоянно подвергается нападению этого вредителя, причем некоторые
обстоятельства указывали на то, что это могло происходить вследствие
появления новой расы Cydia, — расы, предпочитающей орехи другой
пище. Но, надо заметить, ни у личинок, ни у взрослых он не мог найти
какие-либо морфологические различия между особями, собранными с
яблонь, и особями, собранными с орехов. Когда он предоставлял этим
насекомым, воспитанным на орехе, выбирать между яблонями и ореха-
51
ми, он не обнаруживал у них особой склонности откладывать на орехе,
а не на яблоне. Клоп Lygus pabulinus — обыкновенное насекомое в
Восточной Англии. До 1917 г. его иногда находили на смородине, но
никогда не встречали на яблонях. К 1926 г. этот вид стал вредителем
смородины, а также вполне обычным и на яблонях (У. Г Торп. W. Н.
Thorpe, 1930). Английский ивовый клоп Plesiocoris rugicollis по-
видимому, впервые обратился к яблоне около 1918 г., после чего стал
настоящим вредителем насаждений этого плодового дерева (Э. Майр,
Е. Мауг. 1942, 1947, стр. 322).
В западных штатах США бабочки вида Colias philodice питались
преимущественно разными клеверами и другими бобовыми. До 1948 г.
в верхо1вьях р. Арканзас (начали замечать личинки этого вида © качест-
ве серьезного вредителя люцерны (У. Гованитц, W. Howanitz, 1949).
Можно было бы указать так же и на многие другие примеры. Торп
(Thorpe, 1930 и 1931), Г. Г. Андреварта и Л. С. Берч (Andrewartha,
Birch, 1954, стр. 691) вполне резонно замечают, что первое проявление
новой повадки у некоторых особей популяции, быть может, не опреде-
ляется генетически, хотя оно и может иногда вести к дифференциации
рас и даже видов.
Ихнеумонид (наездник) Nemeritis canescens в природном состоя-
нии есть хищник^ откладывающий яйпа в тело гусеницы —огневки Ep-
hestia kiihniella. У. Г. Торп и Ф. Г. У. Джонс (1937) воспитывали
несколько особей Nemeritis на гусеницах бабочки Meliphora grisella,
а затем сравнивали их поведение с поведением нормальных взрослых,
воспитанных на Ephestia. Обе группы были привлекаемы запахом гу-
сениц Meliphora, однако экспериментальные особи привлекались также
запахом Ephestia. Эта последняя способность совершенно отсутствова-
ла у контрольных особей.
Торп (Thorpe, 1939) воспитывал Drosophila melanogaster в среде,
содержащей перечную мяту; те, которые были воспитаны в этой спе-
циальной среде привлекались ею, когда испытывались в ольфактометре.
Гусеница горностаевой моли Hyponomeuta padella может быть
найдена питающейся на яблоне, терне или боярышнике. Торп (Thorpe,
1930 и 1931) предлагал гусеницам, собранным с боярышника, листья
яблони. Гусеницы не ели листьев яблони при наличии боярышника, но
в отсутствии предпочтительной пищи их можно было воспитывать до
взрослого состояния на альтерна™в1ной.
Берч (Andrewartha, Birch, 1954, стр. 509 и 691) воспитывал боль-
шой «штамм» рисового долгоносика Calandra oryzae на кукурузе, а
другой штамм — на пшенице в течение примерно 15 поколений. Когда
насекомым было предоставлено выбирать между пшеницей и рисом
для откладки яиц, долгоносики, воспитанные на кукурузе, отложили в
ьукурузе 28% яиц, а воспитанные на пшенице отложили на кукурузе
15% яиц.
Приведя подобные данные, Андреварта и Берч (Andrewatha, Birch,
1954, стр. 692) заключают, что это явление освоения насекомыми
нового вида пищи не является всеобщим; но оно, по их словам, «все же
дает возможное объяснение быстрого распространения новой повадки —
черты, которая являлась общей во всех приведенных выше случаях —
особенно потому, что факты, по-видимому, иногда не указывают на ге-
нетические различия, по крайней мере в начальных стадиях».
Переведя молодых гусениц коконопряда L^siocampa quercus — ви-
да, нормально питающегося листьями дуба, на сосновую хвою, А. Пик-
те (A. Pictet, 1912) вывел расу, питающуюся сосновой хвоей. Сначала
52
большинство гусениц погибло; позже, когда они начали поедать хвою
с верхушки, они оказались способными обосноваться на сосне. Уже
второе поколение обнаруживало предпочтение сосне» и принадлежащие
к нему особи лишь с трудом удалось заставить вернуться к дубу.
Т. Армстронг (Т. Armstrong, 1945) сообщил о новой расе коконо-
прядов из одного грушевого сада в Онтарио. Бабочки в этом саду дос-
тигают взрослого состояния тремя неделями ранее, чем живущие на
яблонях в других местах той же области.
Ф. Г. Крейгхэд (F. G. Craigheard 1921) разводил жуков-дрово-
секов семейства Cerambycidae на определенных видах древесины. Он
нашел, что через 4—6 лет формировались расы, которые обнаружива-
ли возрастающее предпочтение к тому сорту лесоматериалов, на кото-
ром их разводили.
Эти довольно старые сведения представляют, конечно, большой ин-
терес, и они едва ли поддадутся объяснению по схеме: сначала «спон-
танные» мутации, потом отбор. Откуда взялся у особей, перенесенных
с дуба на сосну, ген «тяготения» к сосне и «отвращения» к дубу?
Андреварта и Берч (Andrewartha, Birch. 1954, стр. 716) прихо-
дят к выводу, что некоторые насекомые, если они воспитывались на не-
свойственной им пище, могут обнаружить предпочтение к этому виду
пищи, когда они начинают искать места для кладки яиц.
«Поскольку изменение в поведении может быть запечатлено у не-
которых особей в ходе развития одного поколения, то такое изменение,
-по Андреварта и Берчу (Andrevartha, Birch 1954, стр. 693), — не
следует объяснять отбором или каким-либо иным генетическим меха-
низмом». Подобные явления хорошо известны и отмечаются и у других
насекомых.
Такие соображения привели Торпа (Thorpe, 1945) и других иссле-
дователей к допущению, что новые расы и, в конечном счете, новые ви-
ды могут возникать в отсутствии географического (пространственного)
барьера в тех случаях, когда часть популяции в силу какого-то необы-
чайного обстоятельства оказывается вынужденной жить и воспитывать-
ся в несвойственном данной форме месте или на несвойственной ей пи-
ще. Затем предпочтение к данному типу местообитания или к данному
виду пищи передается потомству. В течение ряда поколений это пред-
почтение усиливается не только вследствие повторения первоначаль-
ного процесса, но и в силу отбора. Со временем предпочтение, по сло-
гам этих авторов, становится настолько сильным, что оно образует со-
ответствующий изолирующий механизм.
Торп (Thorpe, 1945, стр. 68) дает следующий гипотетический при-
мер того, как, в его понимании, может возникать изоляция среди птиц.
«Представьте себе, — говорит он (там же), — участок- где имеются два
типа биотопов а и б (например, два типа растительности) в мозаичном
распределении, и что.один вид ограничен в своем распространении био-
топом а в пределах указанного участка. В некоторых исключительных
обстоятельствах кризиса или в результате какого-то слабого зароды-
шевого изменения (изменения генотипа) отдельные особи вида проник-
нут fB биотоп б, и воспитанная там молодь испытывает на себе влияние
среды или вообще специализируется в соответствии с новой нишей. Ес-
ли в этой нише достаточно пространства для экспансии, птицы нового
биотопа быстро заполняют его; в течение этого времени они будут в
значительной мере репродуктивно изолированы от птиц биотопа а.
Приводя эти слова Торпа, Андреварта и Берч (Thorpe, Andrewartha,
Birch, 1954, стр. 717) отмечают, что эта идея не совсем (нова, ибо она
53
была намечена выдвинутой Дж. М. Болдвином (1902) теорией органи-
ческого отбора; она была выкристаллизована экспериментами Торпа,
которые дали ей эмпирическое обоснование. Но до сих пор она еще не
подтверждена убедительными наблюдениями над организмами в естест-
венных условиях. Нам, однако, кажется несколько странным такое мне-
ние: наследственное изменение, вызванное воздействием пищи или био-
топа, по* нашему мнению, не соответствует основной идее «эффекта Бол-
дуина»: тут всюду шла речь о прямом изменяющем воздействии на ор-
ганизм со стороны некоторых факторов среды.
Таким образом, в экспериментах Торпа мы видим отнюдь не «сов-
падающий отбор», а бесспорное наследование приобретенных свойств.
Ведь сам Торп прямо утверждал, что в некоторых случаях образование
новой формы происходило без участия отбора. Тут «органический», или
«совпадающий» отбор (эффект Болдуина) может служить для того, что-
бы автор не был обвинен или заподозрен в склонности к «ламаркизму».
Особенно же знаменательно то, что во всех рассмотренных случаях
наследовалась адаптация, — полезное организму свойство. Несомненно
заслуживает внимания эта категория фактов, показывающая, что в оп-
ределенных специальных случаях адаптация сохраняется и закреп-
ляется в штамме без отбора.
Некоторые «синтетисты», встречая затруднения в применении своей
теории к объяснению явлений эволюции, прибегали и прибегают к тому
или иному варианту идеи «органического», или «совпадающего» отбора
(эффекту Болдуина). В этом мы не видим ничего удивительного, пос-
кольку в очень многих случаях недостаточность постнеодарвинистского
толкования процессов эволюции выступает наружу. Однако идея воз-
никновения «генокопий», там, где имеются «фенотипические» приспосо-
бительные изменения, слишком наивна. Она вполне удовлетворяет да-
леко не .всех поборников «синтетической теории эволюции». Дж. Г. Симп-
сон (Simpson, 1953в) писал, что с точки зрения со1временных теорий
эволюции «эффект Болдуина» вполне вероятен, но не известен ни один
случай, где «эффект Болдуина» несомненно действовал, и нет никаких
наблюдений, объяснимых только этим путем. Он, по Симпсону, вероят-
но, имел место, но удивительно мало конкретных фактов, которые ука-
зывали бы на то, что его можно считать часто действующим и важны л
элементом механизма эволюции (Симпсон, Simpson, 19536, сгр. 115).
Как альтернатива неоламаркизма или мичуринизма, «эффект Болдуи-
на», по словам Симпсона, предполагает, что аккомодация (адаптивная
сомация) параллельна генетическим изменениям со сходными результа-
тами. В действительности же, как вполне справедливо замечает Симп-
сон, это — совсем не альтернатива, и основной вопрос остается откры-
тым (там же).
Однако независимо от оценки «эффекта Болдуина» в целом, от чего
мы тут воздержимся, привлечение это^о принципа для объяснения слу-
чаев очень быстрого (в течение времени развития нескольких поколе-
ний) возникновения новых наследственных свойств нельзя считать оп-
равданным. II совершенно ясно, что попытки думать об «эффекте Бол-
дуина» в тех случаях, когда, как утверждают Андреварта и Берч, нас-
ледственное изменение фиксируется в течение жизни одного поколе-
ния, совершенно безнадежны.
Явления изменения фигофагического питания и некоторые других
признаков наблюдались и у других насекомых. Немалый интерес пред-
ставляют в этом отношении эксперименты с тлями — насекомыми,
очень тесно связанными с определенными видами растений, и живуши-
54
’ми на кормовых растениях, где у них происходит смена поколений, пре-
имущественно партеногенетических.
Здесь прежде всего упомянем тщательные и хорошо документиро-
ванные исследования Е. С. Смирнова (1961), а также Г. В. Самохвало-
вой (1954). В опытах Г. В. Самохваловой (1951 и 1954) у тлей Neomy-
sis circumflexus после перенесения их с гороха на перец или на гречи-
ху, уже в первом поколении началось изменение рисунка склеротиза-
кии кутикулы спинной поверхности тела, а к десятому поколению все
особи имели рисунок иной, чем на горохе. После перемещения тлей в
условия, одинаковые с контролем, на горох й на вику, новый характер
рисунка сохранялся не менее, чем у 15 последовательных поколений.
Такое изменение рисунка сопряжено с размерами тела, плодовитостью
и скоростью развития. По данным Смирнова (1961, стр. 177), его опы-
ты с тлями показывают, что «обратимая наследственная реакция пере-
ходит в необратимую лишь после некоторого переломного момента в
ходе изменений».
К числу различных вариантов изменений, получаемых в результа-
те внешних воздействий, Смирнов относит следующую категорию изме-
нений: «Изменения, возникающие при таких внешних воздействиях, ко-
торые ставят организм на грань его регулятивной способности. Наблю-
даются при расшатывании наследственной основы и имеют необрати-
мый характер. Служат материалом для естественного отбора, сохраняю-
щего наиболее удачные варианты» (Смирнов, 1961, стр. 178). Тли —
очень быстро размножающиеся насекомые, теснейшим образом связан-
ные с определенными формами кормовых растений, — в общем весьма
малоподвижные организмы, в жизненном цикле которых решающую
роль играет партеногенез. Все это говорит за то, что тли — особенно
благоприятный объект для изучения воздействия внешних условий на
наследственную основу.
Исследования Смирнова и его сотрудников не вызывают у беспри-
страстного читателя ни малейших сомнений, но хорошо, что эти работы
были продолжены ученым, которого никак нельзя заподозрить в особью
симпатиях к «ламаркизму» и который в своих последних статьях стоит
всецело на позициях «синтетической теории эволюции». Мы имеем в ви-
ду ценные и интересные работы Г. X. Шапошникова (1965, 1966 и 1967).
В одной из статей он (1965, стр. 22) ссылается на то, что некоторы-
ми исследователями и прежде всего Смирновым и его сотрудниками эк-
спериментально были получены изменения в морфологии тлей, возни-
кавшие в результате питания тлей, без участия естественного отбора.
Однако при возвращении тлей в прежние условия эти изменения ока-
зывались обратимыми (?), и, как выражается Шапошников, «трудно
сказать», возможно ли образование новых форм таким путем.
В опытах Шапошникова Dysaphis anthrisci maikopica в течение
50 поколений воспитывались на своем хозяине, зонтичном Anthriscus
nemorosa и параллельно на малопригодном кормовом растении Chaero-
phyllum bulbosum, а затем, адаптировавшись к последнему, — на быв-
шем ранее совершенно непригодным Chaerophyllum maculatum. Эти из-
менения в пище вызвали возникновение новой формы. Они были, по
Шапошникову, по-видимому необратимыми. Во всяком случае к вось-
мому поколению взрослые тли полностью утратили способность питать-
ся на своем старом хозяине. Возникли морфологические различия. Все
эти изменения сохранились на протяжении всего эксперимента, до 46-
го поколения, и Шапошников полагает, что они могли существовать в
течение неопределенно долгого времени (1965, стр. 17). Указанное чрез-
55
вычайно быстрое возникновение новой формы, неспособной жить на
прежнем кормовом растении, никак не могло произойти без индуциро-
вания наследственных свойств воздействием внешней среды, прежде
всего, — новой пищи, к которой тли были вынуждаемы, в процессе эк-
сперимента, приспособляться. Надо помнить, что у этих подопытных на-
секомых смена поколений происходит чрезвычайно быстро. В опыте с
Dysaphis anthrisci maikopica в результате жизни в течение 8 поколе-
ний на малопригодном растении (Chaerophyllum bulbsum), тли адап-
тировались к нему и одновременно потеряли способность жить на своем
старом хозяине (Anthriscus) и приобрели способность жить на новом,
ранее совершенно непригодном растении (Ch. maculatum).
Таким образом, в науке накоплен богатый материал, недвусмыс-
ленно свидетельствующий о прямом индуцирующем влиянии пищи на
наследственную основу организма. Но совершенно ясно, что это ни в
какой 'мере нельзя толковать в ламаркистском смысле — как возни-
кновение видов без участия отбора. Отбор 'несомненно действует как
ведущий фактор формообразования в естественных условиях.
РАЗВИТИЕ УСТОЙЧИВОСТИ К ЯДАМ У ВРЕДНЫХ НАСЕКОМЫХ
Практика применения антибиотиков оказалась весьма ценной для
изучения теоретического вопроса о возможности наследования адапта-
ций, приобретаемых организмами. Правда, речь идет лишь о микробах,
а законы развития этих форм могут оказаться существенно отличаю-
щимися от законов развития более высоко организованных живот-
ных.
Общеизвестно применение ядов против вредных насекомых. Но эти
ядьь инсектициды, вызывают у насекомых устойчивость, аналогично
тому, что наблюдается у микробов при воздействии на -.них антибиоти-
ками. Встает вопрос о происхождении устойчивости. Возникает ли она
вследствие отбора предсуществовавших генов или вызывается прямым
воздействием факторов среды — в данном случае изменением ее хи-
мизма? Очень часто специалисты отвечают на этот вопрос категориче-
ски отрицанием возможности возникновения резистентности вследствие
воздействия химического фактора. Другие придают значение лишь это-
му второму фактору. Однако заслуживает внимательного рассмотрения
также и третья возможность — естественный отбор резистентных осо
бей данной популяции, устойчивость которых возникла как прямой ре-
зультат воздействия * ядов.
Появление первых устойчивых против инсектицидов форм насеко-
мых вызвало сенсацию среди ученых.
В этом отношении интересна статья профессора И. А. Рубцова
(1948, стр. 62—63), который имел возможность в 1947 г. познакомиться
с работой Института санитарии (Istituto superiore di sanita) в Риме.
В изолированной ферме, на расстоянии около 30 км ст Рима, решили изба-
виться от мух. После первых и притом ошеломляющих успехов приме-
нения ДДТ было решено воспользоваться этим инсектицидом. Однозре-
•менно преследовалась и другая цель: уничтожение комаров. Через неско-
лько дней после обработок цель, казалось, была достигнута: нигде не об-
наружили ни одной мухи, ни одного москита, ни одного комара. Это было
в 1944 г. Сотрудники Института санитарии, как они сами рассказыва-
ли И. А. Рубцову, праздновали победу сразу над малярийным комаром
и домашней мухой. В следующем 1945 г. сплошной очистке подверглись
смежные районы. Однако в 1945 г. в местах прошлогодней обработки
56
ядом были замечены единичные мухи. Снова была произведена сплош-
ная повсеместная обработка тем же ДДТ площадей, где появлялась
муха. Через несколько дней мухи опять стали редкостью, но на этот
раз, к удивлению сотрудников, исчезновение мух не было полным, как
это казалось в 1944 г. В L946 г. мух появилось уже гораздо больше, чем
в предыдущем. В третий раз по той же методике была произведена сплош-
ная обработка ядом снова всей площади. Однако на этот раз ре-
зультаты были явно неудовлетворительными. Гибла лишь часть мух,
значительная же часть их здравствовала. Возникла устойчивость мух:
они не погибали от обычных дозировок. Пробыв же на чистом яде в
течение нескольких минут, они частью пибли, частью оправлялись, как
только уходили от непосредственного контакта с ядом. В 1947 г. Руб-
цов там же наблюдал, что после пульверизации обычным, смертельным
для обыкновенной мухи, составом яда (керосин пятипроцентный и
ДДТ), мухи были в изобилии и не обнаруживали никаких признаков
отравления.
Механизм появления этой устойчивости, по словам Рубцова (там
же, стр. 63), еще не ясен. Рубцов думает, что в каждом случае он имеет
особое объяснение. Но мыслимая схема появления устойчивости пред-
ставляется. по Рубцову, в следующем виде.
В обычной популяции существуют отдельные, стойкие к яду особи
или отличающиеся поведением, позволящим избегать отравления. Они
менее конкурентноспособны и в нормальных условиях очень редки. При
массовом уничтожении обычных, по стойкости к яду, особей, редкие ус-
тойчивые особи размножаются более интенсивно без конкурентов на
освободившейся жизненной арене.
Такое объяснение, казалось бы, довольно логично; но возникнове-
ние устойчивости к яду все-таки вызывает смущение у специалистов.
«Удивительной представляется, — пишет Рубцов (там же, стр. 63),
скорость, с которой возможна дифференцировка новой формы из сме-
шанной популяции». «Существование высокостойких к ДДТ форм внут-
ри гида — продолжает Рубцов, — весьма удивительно, принимая во
внимание чрезвычайно высокую токсичность яда для целого ряда отря-
дов насекомых: мух, жуков, перепончатокрылых и др. Механизм стой-
кости, равно как и механизм токсичности ДДТ, остаются до сих пор
неясными». Далее этот ученый справедливо заявляет: «Остается неяс-
ным, как могли возникнуть в популяции стойкие к яду формы. Эту устой-
чивость нельзя себе представить как неожиданно появившиеся мута-
ции». Видимо автор чувствует, что если исключить возможность возник-
новения новых мутаций, то появление устойчивости к яду становится
загадочным. «Явление устойчивости к яду, — продолжает он далее —
представляется слишком сложным механизмом для того, чтобы его мож-
но было представить в виде спонтанно и сразу возникшей мутации». С
этим, по нашему мнению, вполне можно согласиться: подобная «спон-
танная мутация» была бы странным событием. «Гораздо естественнее и
правдоподобнее допустить, — заключает Рубцов, — что явление устой-
чивости к яду у части популяции выработалось постепенно, исторически,
как попутное в известных условиях приспособление. Но в таком случае
мы должны принять и существование соответствующих внешних причин
п условий, т. е. хотя бы периодического и локального существования в
среде обитания мухи (например, навозе) органических соединений, ана-
логичных ДДТ по своему действию на организм насекомого». Из этого
отрывка первое предположение представляется нам в общем довольно
приемлемым: устойчивость вырабатывалась очевидно исторически, шаг
57
за шагом. Но ведь этот тезис едва ли совместим с чисто постнеодарви-
нистским пониманием эволюции. Что же касается второго предложения,
то оно содержит мысль, кажущуюся нам сомнительной. Резистентность
могла возникать не до воздействия на насекомых инсектицидом, а в
процессе применения этого яда, постепенно накопляясь и усиливаясь.
Ныне известно уже множество инсектицидов, и многие насекомые
быстро приобретают устойчивость к этим инсектицидам. Можно ли до-
пустить мысль, что в каждой популяции данного вада (насекомых, име-
ется гол1ный комплект генов резистентности ко всем существующим ин-
сектицидам и ко всем инсектицидам, которые имеют быть изобретен-
ными в будущем? Такая исчерпывающая преадаптация представляет-
ся нам маловероятной.
Явления резистентности против некоторых ядов наблюдались у на-
секомых еще в 1916 г., когда было установлено уменьшение чувствитель-
ности по отношению к синильной кислоте у популяций оранжерейной
щитоносной вошки Aonidiella aurantii и Saissetia oleae. Отмечалось
уменьшение чувствительности по отношению к некоторым инсектицид-
ным средствам также у других насекомых. Но это не имело большого
практического значения, и явлению резистентности специалисты уделя-
ли мало внимания. Положение существенно изменилось за последние со-
рок лет. когда было синтезировано, один за другим', множество сильно
действующих инсектицидов. Агрономы и медики рассчитывали, что эти
инсектициды позволят нам разрешить многие вопросы сельского хозяй-
ства и здравоохранения (С. Сърбова и Светла Аврамова, 1965, стр. 36).
Однако через два года после того, как началось широкое применение
ДДТ в борьбе против домашней мухи, в 1946 г. в Швеции были отмече-
ны первые мухи, резистентные к этому яду. После этого число резистент-
ных видов, — точнее, видов, в которых отмечались штаммы, резистент-
ные к определенным' инсектицидам, быстро росло. По данным Дж. Ф.
Крау (J. F. Crow, 1966, стр. 31), к 1948 г. было известно всего 12 видов,
резистентных по отношению к инсектицидам; в 1951 г. (таковых было
уже 16, в 1954 г. их число достигло 25, в 1957 г. — 76, в 1960 уже 137, в
1963 г. их было 157. Число таких видов неуклонно растет. В 1962 г. ре-
зистентностью обладал 81 вид насекомых медицинского значения (Сър-
бова и Аврамова, 1965 г., стр. 36). Эти цифры не совсем точны. Но так
или иначе, число резистентных видов быстро растет.
Количество разнообразных инсектицидов, применяемых в борьбе с
сельскохозяйственными вредителями и распространителями болезней,
тоже постоянно возрастает. И соответственно развивается резистент-
ность против этих веществ. Резистентность может быть по отношению к
одному или немногим инсектицидам и не быть сопряженной с резистент-
ностью к другим.
У насекомых, в частности у Musca domestica, Culex и Anopheles,
развивается резистентность к любым разнообразным инсектицидам’ ко-
торые применяются для их уничтожения. Естественно встает вопрос:
происходит ли тут всего лишь отбор — дарвиновский отбор через вы-
живание особей, почему-то заранее наделенных геном или генами рези-
стентности, которые до того находились в резерве и получили биологи-
ческое значение только из-за действия инсектицида, или здесь соверша-
ется приспособление, похожее на привыкание организма к ядам, по ана-
логии с так называемым митридатизмом (привыканием организма к
сильно действующим токсическим веществам при постоянном последо-
вательном увеличении доз)? Нынешние исследователи иногда формули-
руют этот вопрос очень кратко, хотя и неточно: постадаптация (прио-
.58
сретение наследственной адаптации в жизни особи) или преадаптация
(наличие соответствующего гена, или соответствующих генов еще до на-
чала применения инсектицида)?
Исследователи, стоящие на позициях постнеодарвинизма, обычно
самым решительным образом высказываются за «преадаптацию». И это
неудивительно: принятие идеи «постадаптации» означало бы признание
не только мутагенного действия инсектицидов, но и наследуемости при-
обретенной адаптации, — признание того, что по крайней мере в неко-
торых случаях полезная для организма адаптация оказывается наслед-
ственной. Правда, тут можно и нужно оговориться, что речь идет об
узко ограниченной категории изменений. Но и это долж-
но казаться постнеодарвинистам непростительной уступкой «ламаркиз-
му» или мичуринскому учению. В действительности же тут нет отступ-
ления от того, что часто называют ортодоксальным дарвинизмом.
Специалисты различают по крайней мере три основных вида ре-
зистентности насекомых к инсектицидам. Поведенческая резис-
тентность [см., напр., Д. Бюсвин (J. R. Busvine 1959), стр. 4999]
заключается в способности организма избегать» не допускать внутрь
клеток летальные дозы ядовитых веществ. Это связано с изменением
не только повадок насекомых, но и их реакций. Однако, по Бюсвину
(там же), это — очень редкое явление. Другой тип резистентности —
вигорная толерантность, или жизнеспособная стойкость —
состояние низкой чувствительности ко многим, практически даже ко всем
существующим инсектицидам. Такая резистентность, по-видимому, не
бывает резко выражена, и ее, быть может, надо было бы называть как-
то иначе. Третий, и можно сказать, основной вид резистентности может
быть назван физиологической резистентностью, реаль-
ность которой — вне сомнения и которая включает в себя высокую спе-
цифическую стойкость по отношению к одному определенному яду или
к одной группе ядов. «Самые разительные случаи очень высокой ре-
зистентности — по словам Крау (Crow, 1966, стр. 31), — зависят ог
некоего детоксифицирующего механизма». Так, мухи, резистентные к
ДДТ, обладают особым энзимом, который удаляет НС1 из молекулы,
причем остается нетоксическое соединение. Существует также энзим,
который устраняет токсичность некоторых органических фосфатов.
Если данный штамм под действием одного инсектицида повышает ре-
зистентность по отношению к различным инсектицидам, то говорят о
перекрестной резистентности, или устойчивости (Сърбова и Авра-
мова, 1965, стр. 37).
Многие исследователи не только убеждены в зависимости резистент-
ности от определенных генов, *цо даже ставят и решают вопрос о том,
подчинена ли резистентность одному фактору или многим факторам. В
первом случае они говорят обычно о монофакториальности, во втором—
о полифакториальности (позволим себе заметить, что более правиль-
ными мы считаем термины унифакториальность и мультифакториаль-
ность). [См.,. например, Дэвидсон (Davidson, 1959), стр. 5051.
Нередко исследователи совершенно категорически утверждают, что
резистентность к инсектицидам устанавливается в результате отбора—
элиминации чувствительных к инсектициду особей и выживания резис-
тентных. В результате сохраняются и размножаются особи, обладаю-
щие соответствующими генами (см., например, A. A. Brown, Браун,
1959, стр. 498 и Крау, Crow, 1966, стр. 32).
Но, как ни существенна защита этого мнения для сохранения ус-
тоев постнеодарвинизма, сторонники этого течения испытывают беспо-
59
койство, опасаясь, что такое объяснение может оказаться недостаточ-
ным или даже несостоятельным.
Майр (Мауг, 1963, стр. 193) писал, что быстрота развития резис-
тентности — один из факторов, терзающих сторонников теории естест-
венного отбора. Для того, чтобы преодолевать трудности, связанные, с
проблемой резистентности, в полеводстве и садоводстве необходимо
постоянно вводить все новые и новые инсектициды (там же). По сло-
вам Крау (Crow, 1966, стр. 32), «короткая полезная служба инсектици-
да означает постоянную потребность в новых ядах».
Говоря о быстроте развития резистентности, мы прежде всего
должны подчеркнуть всеобщий характер этого процесса: очевидно нет
такого инсектицида, против которого у насекомых не может развивать-
ся резистентность, как и нет такого вредителя—насекомого, который не
был бы способен приобрести резистентность против любого инсектици-
да. Musca domestica приобретает резистентность ко всем применяемым
в настоящее время инсектицидам (Дербенева-Ухова, 1959, стр. 738).
Быстрота возникновения резистентных штаммов под воздейст-
вием инсектицидов, действительно поразительна. В. П. Дербенева-Ухо-
ва и В. П. Морозова (1950) описали случай обнаружения резистентнос-
ти комнатной мухи в конце первого же сезона широкого применения
ДДТ. В обзорной статье о появлении у насекомых устойчивости к хло-
рированым углеводородам В. И. Вашков (1957, стр. 157) отмечает, что
в некоторых местностях Северной Америки резистентность к диэлдри-
ну появилась очень быстро — через два месяца. Африканский носитель
малярии, Anopheles gambiae по словам Дж. Рейда (J. A. Reid, 1960,
стр. 218), уже через 18 месяцев после начала обрызгивания диэлдри-
ном в некоторых местах дал резистентные популяции. А через два года
свыше 90% комаров этого вида были уже резистентны, почти все в го-
мозиготном состоянии. У нормальных комаров Anoph. gambiae, как
утверждает Рейд (там же), ген резистентности к диэлдрину присутст-
вовал лишь у 0.04—6.0% особей. За 25—30 поколений (полтора-два
года) отбор, по словам этого исследователя, поднял распространение
этого полудоминантного гена резистентности от менее одного на тыся-
чу комаров в гетерозиготном состоянии почти до 100% при 90% гомо-
зиготных особей в популяции. Диэлдрин стал бесполезным для борьбы
с малярией.
Реид сообщает также, что в Малайе, в Пенанге, вредный комар
Culex fatigans за два года применения одного из инсектицидов удеся-
терил резистентность к этому инсектициду. Примечательно, однако,
чю всего лишь через десять поколений воспитания этих комаров бе°
данного инсектицида резистентность пала, и теперь она оказывается
нормальной. Как понять это с точки зрения идеи отбора уже существо-
вавших в исходной популяции генов резистентности к данному инсекти-
циду? Как можно объяснить «нормализацию» резистентности, точнее —
восстановление чувствительности за срок, по-видимому, менее года?
Рейд полагает, что гены резистентности имеют другие, «слегка вред-
ные» действия. Эта плейотропия, очевидно, вытесняет носителей таких
генов, дает решающее преимущество комарам, свободным от этого
двусмысленного гена, а такие комары с необычайной быстротой совер
шенно вытесняют предыдущих победителей. Таким образом, скажем мы
в дополнение к рассуждениям Рейда, в этом случае применения инсек-
тицида комары-носители гена резистентности в конечном счете спаса-
ют всю популяцию от окончательного вымирания и обеспечивают вос-
становление нормального населения без резистентных генов.
60
Быстрота развития резистентности отмечается многими исследова-
телями, смущая, по-видимому, постнеодарвинистов.
Майр (Мауг, 1963, стр. 259) писал, что если даже резистентность
зависит от одного гена, развитие резистентности к разным инсектици-
дам у Musca, Anopheles и Culex в два года или менее того означает
чрезвычайно быструю эволюцию в дикой популяции. Ф. П. В. Уинтеп-
ингэм (F. Р. W- Winterigham. 1959, стр. 506) отмечал как знаменатель-
ное явление быстроту и степень резистентности, которая у насекомых
возникала при соответствующих экспериментах. Тысячекратная резис-
тентность к ДДТ, или гаммаизомеру гексахлороциклогексана, была
достигнута за 20 поколений или еще быстрее.
Эта бросающаяся в глаза быстрота развития резистент-
ности серьезно озадачивала даже тех исследователей, которые не
хотели сомневаться в правильности постнеодарвинистского объясне-
ния такого процесса. По Уинтерингэму (там же, стр. 507), уровень дос-
тигнутой резистентности столь высок, а первоначальная популяция
иногда столь малочисленна, что возникает вопрос, могла ли единичная
особь первоначальной популяции выдерживать больше, чем какую-то
долю той дозы яда, какую успешно выдерживали последующие поколе-
ния. Поэтому, по словам Уинтерингэма, возникает подозрение, что ин-
сектицид, быть может, обладает мутагенными свойствами, то есть, ина-
че говоря, вызывает какое-то наследственное изменение в направлении
устойчивости. Этот автор думает, что никакие эксперименты с действи-
ем инсектицидов на насекомых не исключают «мутагенного действия»
при высоком давлении отбора, ведущем к высоким уровням резистент-
ности.
Уинтерингэм ссылается тут на Уоддингтона (Winterigham, 1956,
стр. 1210), по которому можно предположить, что способность насеко-
мого отвечать на инсектицидное давление детоксифицирующей энзим-
ной адаптацией может находиться под генетическим контролем, и край-
няя энзимная адаптация может стать генетически «фиксированной». Та-
кой механизм, по Уинтерингэму (там же, стр. 507—508), может объ-
яснить развитие очень высоких уровней резистентности без постулиро-
вания существования соответствующих высоко резистентных мутантов
в данной популяции. Заметим, что такое объяснение едва ли придется
по вкусу стойким последователям постнеодарвинизма: они, действи-
тельно, не присоединяются к Уинтерингэму и Уоддингтону, хотя эти два
исследователя стараются искать объяснения факторов, не выходящего
за рамки «синтетической теории эволюции».
Не особенно легко объяснить, с позиций этой теории также заме-
чательно высокий уровень резистентности, достигаемый некоторыми
штаммами насекомых. Так, у комнатной мухи можно вызвать факти-
чески полный иммунитет к диэлдрину (эпоксигексахлоротетрацикло-
децену), — универсальному яду, который при сравнимых тканевых
концентрациях столь же токсичен для млекопитающих, как и для насе-
комых.
Быстрота, сила возникновения резистентности, высокий уровень,
которого она может достигнуть и способность организмов вырабатывать
устойчивость к самым разнообразным (практически ко всем существую-
щим) ядам, — все это плохо укладывается в посгнеодарвинистскую ги-
потезу и требует какого-то дополнительного объяснения.
Видимо, в отличие от постнеодарвинистов, занимающихся други-
ми вопросами эволюции, сторонники «синтетической теории», работаю-
щие в области изучения резистентности, ощущают необходимость как-
61
то парировать альтернативное, подлинно дарвинистское, объяснение
развития этого свойства.
Так, они говорят, что резистентность не достигается, если в попу-
ляции отсутствуют особи с соответствующими резистентными генами
[Дж. Рейд, (Reid,) стр. 217], что якобы наблюдается в восточноафри-
канских популяциях Anopheles gambiae. Некоторые ученые, напри-
мер, Крау (Crow, 1966, стр. 31), ссылаются на эксперименты, при кото-
ных рез1И1СтентнО|Сть не разв.ивается, несмотря на -при1ме1нен!ие нелеталь-
ных доз в течение нескольких поколений. Это, как будто прямо говорит
против прямого физиологического эффекта инсектицида. Г. Люерс
(Н. Liiers, 1959, стр. 502) писал, что обработка Drosophila melanogaster
сублеталыными дозами ДДТ в течение 50 поколений не привела ни к
каким изменениям чувствительности. Позже эксперимент был продол-
жен и доведен до 150 поколений. Этот экспериментатор, очевидно,
склонен видеть здесь подтверждение тезиса о преадаптации. Но сели
даже приобретение резистентности ^через воздействие инсектицидов и
вполне возможно, то для .успешного осуществления такого эксперимен-
та необходимо, очевидно, строгое соблюдение определенных условий—
количества доз, времени их подачи и т. д. Однако имеются основания
полагать, что такие эксперименты не проводились с должным упорст-
вом и тщательностью на протяжении достаточно длинного ряда поко-
лений. Так, Уинтерингэм (Winterigham 1959, стр. 506) писал, что
«подвергая непрерывно или через промежутки относительно малые по-
пуляции насекомых сублетальным или почти летальным концентраци-
ям токсических веществ в лаборатории, исследователи смогли получить,
«резистентные» популяции, фактически иммунные в отношении универ-
сальных животных ядов». Из этих слов явствует, что в таких экспери-
ментах действовали не «убийственными» дозами инсектицидов. Поло-
жительные результаты получались, а от чего они зависели — от «пря-
мого воздействия» на насекомых или от выживания исконно резистент-
ных при элиминации чувствительных, — таким путем установить едва
ли возможно. Указывалось также, что отбор оказывался безрезультат-
ным в инбредных линиях, лишенных изменчивости, и это тоже считалось
доводом против прямого влияния яда на организм. Однако и этот довод
не убедителен: по-видимому, отсутствие у потомства резистентности
служило главным доводом в пользу инбредного характера линии. Нам
кажется, что свидетельство в пользу «преадаптивного» присутствия ге-
нов резистентности у насекомых представляется сомнительным даже
ортодоксальным сторонникам постнеодарвинизма. Так, Браун (Brown,
1959- стр. 497), утверждающий, что все факты говорят в пользу разви-
тия резистентных популяций насекомых через «дарвиновский отбор» и
отрицающий наличие каких-либо данных в пользу «постадаптивного*
привыкания», мутагенного действия .инсектицидов на насекомых, тем не
менее писал о необходимости проверить, «не может ли чрезвычайно бы-
строе развитие резистентности к диэлдрину у комнатной мухи зависеть
от постадаптивного элемента в дополнение к преадаптивному» (там же,
стр. 498). Действительно, слишком уж быстро «привыкают» мухи к диэл-
дрину.
Мы уже упомянули о том, что Уинтерингэм (Winterigham, 1959,
стр. 507) высказывал такие же сомнения и полагал необходимым про-
верить, не обладают ли инсектициды мутагенными свойствами.
Болгарские исследовательницы С* Сърбова и С. Аврамова (1965,
стр. 40) тоже не решаются полностью примкнуть к «преадаптационной»
гипотезе. Они цитируют следующее место из упомянутой нами статью
62
Брауна: «Мы принуждены принять парадоксальный на первый взгляд
выбсд, что адаптация, обусловливающая устойчивость, не вызывается
инсектицидом». А далее авторы резонно ставят такой вопрос: «Если
резистентность обусловлена детоксифицирующим энзимом и если осо-
би, которые вырабатывают этот фермент, обладают специальным, ге-
ном, то откуда же взялся этот ген!» (подчеркнуто нами. —
Л. Д.).
На семинаре по борьбе с насекомыми медицинского значения, сос-
тоявшемся в 1962 г. в Женеве советский специалист профессор В. П. Дер-
бенева-Ухова сообщила следующее: «В результате применения субле-
тальных доз ДДТ к взрослым особям Musca domestica мухи станови-
лись, хотя и медленно, в продолжение нескольких поколений, резистент-
ными». По Дербеневой-Уховой (1959), резистентность возникает не толь-
ко вследствие отбора: адаптация и модификации родителей тоже дей-
ствуют на потомство.
Объяснение возникновения резистентности к инсектицидам в духе
постнеодарвинизма представляется, таким образом, довольно искусст-
венным и малсхвороятным.
Таким образом, против этого объяснения говорят следующие сооб-
ражения.
Резистентность возникает почти всюду и всегда при употреблении
инсектицидов для уничтожения насекомых.
Она развивается чрезвычайно быстро — в полтора или два года
после начала применения соответствующего инсектицида, а то и гораз-
до быстрее. Такая быстрота возникновения резистентности в популя-
ции насекомых, размножающихся половым путем, делает довольно сом-
нительным мнение об установлении резистентности путем отбора пред-
существовавших мутантов. Тут может быть применена математическая
методика, которая, по всей вероятности, даст более точный и реши-
тельный ответ на вопрос о происхождении резистентности.
Трудно думать также, что в хромосомах насекомых всегда может
найтись нужный ген резистентности против соответствующего инсек-
тицида: ведь инсектицидов теперь множество, и они достаточно разно-
образны, и ко всем этим инсектицидам насекомые могут приобретать
устойчивость.
Не менее трудно понять, почему после прекращения обрызгивания
данным инсектицидом резистентность быстро утрачивается и геном воз-
вращается в нормальное состояние. Мы, видели, что существует попыт-
ка объяснить такое восстановление исходного положения слабым вред-
ным действием гена резистентности, в силу чего сохранившиеся где-то
чувствительные к инсектициду особи получают небольшое преимущест-
во над резистентными и вытесняют их. Однако чрезвычайно трудно до-
пустить, что такой механизм, связанный с плейотропией гена резистент-
ности, является обязательным для каждого гена резистентности, что
такой ген обязательно должен быть в то же время немного (именно—
немного ) вредным, чтобы быстро самоустраниться по миновании в нем
надобности.
Мы видели, что слабость такого якобы дарвинистского толкование
резистентности сознают многие специалисты, отнюдь не склонные ни
к ламаркизму, ни, тем более, к мичуринизму.
Гораздо более вероятным представляется нам допущение, что
адаптация к данному инсектициду способна передаваться по наслед-
ству следующим поколениям. В этом смысле приобретает особый инте-
рес сообщение Дербеневой-Уховой (см. Сърбова и Аврамова, 1965) о
63
том, что сублетальные дозы ДДТ вызывают резистентность именно у
женских особей. Не исключена возможность, что такое воздействие вы-
зывает изменение именно в плазме, а не в ядре. Возможно также, что
возникшая таким путем наследственная изменчивость остается недол-
говечной и сама собой отпадает, если прекращается действие вещества,
вызывавшего это свойство.
Вопрос о происхождении резистентности насекомых к инсектици-
дам могут успешно изучать лишь специалисты, работающие в этой об-
ласти прикладной энтомологии. Мы имели возможность познакомить,-
ся только с небольшой частью литературы по этому вопросу. При этом
мы не нашли в литературе указаний на работы по изучению возникно-
вения у насекомых стойкости по отношению к репеллентам, т. е. ве-
ществам, «отпугивающим», «отталкивающим» насекомых, но не убиваю-
щим, не отравляющим их. Нам кажется, что, поскольку в таких случаях
яды не попадают внутрь тела насекомых, едва ли репелленты могут
вызывать, даже при долгом их применении, резистентность, которая, в
нашем представлении, может возникать лишь в результате сложных
биохимических процессов.
Мы, конечно, нисколько не претендуем на решение вопроса о проис-
хождении резистентности у насекомых при употреблении инсектицидов.
Нам хотелось только показать недостаточность позиции ортодоксальной
«синтетической теории» в этом вопросе и необходимость искать какого-
то иного объяснения этого явления.
В то же время следует подчеркнуть, что мутагенное возникновение
нового свойства, сразу же являющегося адаптацией, т. е. свойством
обязательно полезным, приспособительным, никак не укладывается в
«синтетическую теорию» и находится в непримиримом противоречии с
нею. Как мы постараемся показать это в последующих разделах кни-
ги, возникновение подобных свойств в порядке постадаптации нисколь-
ко не противоречит основам дарвиновского учения об эволюции, хотя
подобные изменения могут возникать, по-видимому, лишь в случаях
одной, специфической категории. Широчайшее распро-
странение способности насекомых становиться резистентными против
разнообразнейших синтетических ядов, старательно изучается работ-
никами сельскохозяйственной и медицинской энтомологии. Развитие
устойчивости к ядам внушает тревогу этим исследователям. Она на-
столько велика, что раздаются даже призывы не поддаваться панике
(Р. Висманн, 1957, стр. 3). Однако эта проблема представляет огром-
ный интерес также для науки об эволюции организмов и особенно для
проблемы изменчивости.
НАСЛЕДСТВЕННЫЕ ИЗМЕНЕНИЯ НЕКОТОРЫХ КУЛЬТУРНЫХ РАСТЕНИЙ,
ВЫЗЫВАЕМЫЕ ФАКТОРАМИ СРЕДЫ
Очень многие исследователи сталкивались с фактами наследствен-
ной изменчивости растений, вызываемой факторами среды. А. Деррент
(A. Durrant, 1958а, 19586, 1960) утверждал, что растения льна испы-
тывают наследственные изменения в зависимости от характера удоб-
рений, вносимых в почву. Он выращивал растения льна на искусствен-
но удобренных почвах и установил, что растения передавали изменен-
ный тип не только потомству, порожденному через самооплодотворе-
ние, но также и потомству, полученному путем скрещивания, причем
передача происходила в равной мере и через пыльцу и через яйцеклет-
64
ки. Такой измененный тип удерживался в последовательных поколе-
ниях, как это показали Деррент и Тайсон (A. Durrant and Н. Tyson,
1964), которые применяли всевозможные комбинации количеств азота,
фосфора и калия. Г. Р. Хайкин (Н. R. HighKin, 1958) писал, что наслед-
ственные свойства гороха могут изменяться под влиянием температур-
ного режима. Эти изменения удерживались>в неослабленном состоянии
в инбредных линиях гороха. Р. А. Бринк (R. А. Вгтк, 1958 и 1960) разли-
чал два типа изменения пигментации у кукурузы, зависящие, как он
думал, от некоторых аллелей локуса Р. Одно из них, по его словам, пред-
ставляет собой обычное явление мутации: оно спорадично, ступеньчато
(то есть не распадается на ряд постепенных преобразований) и носит
ненаправленный характер; оно приводит к появлению новых аллелей,
которые столь же стабильны, как и те, из которых они возникли. Дру-
гой тип изменения отличается от первого во всех этих отношениях: оно
является определенным, при известных условиях возникая в 100% слу-
чаев; оно может дать начало постепенному ряду преобразованных про-
дуктов; оно является направленным в том смысле, что данная аллель
всегда изменяется одним и тем же способом при одних и тех же усло-
виях; измененные аллели, которым дает начало такое изменение, оказы-
ваются скорее метастабильными, чем стабильными, так как наблюдга-
лось ослабление эффекта. Этот второй тип изменения Бринк назвал
Апарамутацией». Он думал, что существует некий ортохроматин, изме-
нения которого дают начало настоящим мутациям, и парахроматинг ко-
торый, правда, не представляет существенный элемент структуры гена,
но влияет на выражение деятельности гена и который, претерпевая из-
менения, дает начало парамутациям. К. Мазсер (К. Mather, 1961, стр.
405) находит, что парамутационные изменения в хромосомах могут
играть важную роль в развитии, но сами по себе они не позволяют нам
понять, почему две клетки, происшедшие от деления одной материнской
клетки» следуют двум различным путям развития. Поэтому мы вынуж-
дены, по словам Мазсера, в поисках объяснения такой дифференциации,
обратиться опять-таки к цитоплазме. Мы должны, говорит он, изучить
вопрос, могут ли изменения в цитоплазме вызывать изменения в ядре.
Он думает, что «парамутация, как и любое иное изменение в развитии,
должно зависеть непосредственно от действия цитоплазмы (или, по
меньшей мере, от внехромосомной плазмы) на ядерные материалы и
лишь косвенным образом на гены, которые первоначально вызывали из-
менение и преобразование этой цитоплазмы». Если окажется, что цито-
плазматическое изменение может обусловить парамутационные преоб-
разования хромосом то, как говорит Мазсер (там же, стр. 406), «воз-
можно, что внешнее вмешательство может, воздействуя на цитоплазму,
тоже вызывать прямые изменения в наследственных материалах».
Отсюда, нам кажется, с необходимостью вытекает заключение, что
Мазсер, как и другие авторы, не могут уклониться от необходимости
признать, что некоторые наследственные изменения (пусть они называ-
ются парамутациями или как-либо иначе) могут охватывать значитель-
ную долю популяции (по словам Мазсера, даже 100% испытуемых осо-
бей) и что такие изменения являются определенно направленными.
На такую же мысль наводит интересная статья Дж. Хилла (J. HiH,
1965) посвященная индуцированию средой наследственных изменений
у Nicotiana rustica.
Этот исследователь исходит из того положения, что' нормальное
воздействие среды не приводит к постоянному изменению фенотипа, но
существуют, по его словам, два исключения из этого общего правила:
5. Л. Ш- Давиташвили 65
индуцированная мутация и кондиционирование. Оба эти явления пред-
ставляют собой фенотипические преобразования, вызванные специфи-
ческим набором условий среды и сохраняющиеся после удаления этих
условий. Индуцированные хромосомные мутации возникают врассып-
ную, вразброс, беспорядочно, а кондиционирование, как и большинство
индуцированных внехромосных мутаций, есть процесс не хаотический
(non random), который, как мы видели,сравнивался Мазсером с пара-
мутацией, обнаруженной Бринком у кукурузы. Хилл главное место в
своей статье уделяет кондиционированию. Такой называет «направлен-
ное изменение в развитии фенотипа, вызываемое действием внешних
агентов» (там же, стр. 733). Такое изменение может быть временным,
то есть фенокопией, длящейся лишь в течение жизни индивидуума, или
оно может оказаться более перманентным, если эффект передается от
родителя к потомству. Однако такой относительно перманентный эф-
фект, по Хиллу, может постепенно ослабляться на протяжении последо-
вательных поколений, как это происходит, найример, с длительными мо-
дификациями. Но он может оставаться неослабленным даже после
нескольких циклов полового размножения. Был поставлен эксперимент
с целью выяснить, нельзя ли индуцировать наследственные изменения в
инбредных расах Nicotiana rustica посредством различных комби-
наций обработки удобрениями. Эти расы были инбредными еще в нача-
ле 40-х гг. и с того времени поддерживались таковыми на протяжении
приблизительно двадцати поколений. «Таким образом, — по словам
Хилла (там же, стр. 733), — не может оставаться никакого сомнения
относительно инбредного характера использованного материала». Автор
очень подробно описывает этот эксперимент. Результаты показали, что
изменения, вызванные различными способами удобрения у одной расы
Nic. rustica, должны быть признаны наследственными, потому что эти
изменения передавались двум поколениям без применения удобрений.
Причины кондиционирования в настоящее время могут быть, по Хиллу,
лишь предметом догадок. Но он находит, что когда индуцированные
изменения такого рода возникли, они вполне могут оказаться необрати-
мыми при нормальном диапазоне условий среды.
Итак, исследователи, которых мы не можем заподозрить в склон-
ности к «ламаркизму» и неверности в отношении принципов ортодок-
сальной генетики, устанавливают явления, которые нельзя понимать
инаие, как наследственные индуцированные изменения растительных
организмов. Упомянутые в предыдущих строках ученые добросовестно
стараются объяснить эти случаи в духе неоменделизма и убедить себя,
что «длительные модификации», «парамутации», кондиционирование и
подобные им термины вполне могут обеспечить незыблемость генети-
ческих догм. Однако мы видим, что подобные наследственные изменения
возникают во множестве (а не в виде единичных мутантов) как реак-
ция на воздействие определенных факторов наружной среды. Они да-
леко не всегда затухают, а наоборот — во многих случаях неуклонно
сохраняются на протяжении неопределенно долгого времени в серии по-
колений даже при половом размножении. Нет никаких доказательств
того, что эти наследственные изменения не закрепляются в хромосомах
ядра, а в некоторых случаях имеются основания предполагать такое:
закрепление.
66
ЯВЛЕНИЕ ГИБРИДНОЙ МОЩИ, ЖИЗНЕННОСТИ, И ИЗМЕНЧИВОСТЬ
Проблема так называемого гетерозиса, имеющая громадное тео-
ретическое и практическое значение, не может быть тут предметом спе-
циального рассмотрения. Коснемся лишь того аспекта этой великой
проблемы, который имеет прямое отношение к учению об изменчивости
внутри видов.
Гетерозис— «гибридная мощность», повышение жизнеспособности
и мощности гибридных организмов, порождаемых в результате скре-
щивания неродственных особей, которые могут принадлежать к разным
расам, разным породам животных или разным сортам растений. Полу-
ченные таким путем гибриды превосходят родителей по некоторым
признакам. Однако дальнейшее скрещивание гибридов между собой
ведет к затуханию этой мощности в последующих поколениях.
Гибридизация может служить одним из способов изменчивости ор-
ганических форм и вести, через естественный отбор, к формированию
новых видов как растений, так и животных. Однако рассмотрение воп-
роса о значении гибридизации для эволюции организмов выходит за
рамки задачи, которой посвящена эта книга. Здесь мы коснемся лишь
одного аспекта проблемы гибридной мощности, или гетерозиса, — ас-
пекта, который редко затрагивается в трудах представителей ортодок-
сальной постнеодарвинистской школы. Мы имеем в виду изменение по-
томства организмов одного и того же вида, одного и того же подвида
или одной и той же расы, даже особей, близко родственных, но вырос-
ших до состояния половой зрелости в несколько различных обстанов-
ках, в неодинаковых условиях.
В знаменитой книге Ч. Дарвина «Изменения животных и растений
в состоящим доместикации» глава восемнадцатая «О преимуществах и
невыгодах изменения условий существования. Бесплодие от разных
причин» начинается небольшим разделом, носящим заглавие: «О поль-
зе, приносимой небольшими изменениями в условиях существования».
Там Дарвин [Ch. Darwin, 1921, стр. 127: 1951, стр. 556)] говорит, что, в
связи с обоснованием положения о той выгоде, которая получается от
скрещивания, он задумывался о пользе, получаемой организмами «ог
небольших изменений в условиях их существования». Развивая эту
мысль, он писал/ «Нет двух индивидуумов, а тем более двух разновид-
ностей, абсолютно тождественных по своей конституции и строению, и
когда зародыш (the germ) одного из них оплодотворяется мужским
элементом другого, можно думать, что он подвергается действию, до
некоторой степени сходному с тем, какому подвергается индивидуум в
слегка измененных условиях (...). Есть веские доказательства, что пе-
ренесение семян, клубней, луковиц и черенков растений из одной почвы
или местности в другую, насколько возможно менее сходную, приносит
большую выгоду».
По Дарвину (1951, стр. 581), «скрещивание ведет к увеличению
размеров тела, мощности и плодовитости потомства», в то время как
«продолжительное тесное родственное разведение (инбридинг) умень-
шает крепость конституции потомков, их размеры и плодовитость.
Т. Д. Лысенко уделял очень большое внимание этим соображениям
Дарвина, которые он постарался положить в основу своего учения о
жизненности организмов. Он писал (1949, стр. 518): «Перекрест-
ное опыление, как это было неопровержимо доказано Дарвином,
как правило, полезно для организма. Потомство из семян, по^
лучаемых путем перекрестного опыления, более жизненно. Сог-
67
ласно разъяснению Дарвина, различные организмы, развиваясь в отно-
сительно различных условиях, по-разному формируют себя из окру-
жающей пищи. Получаются относительно различные организмы, а от-
сюда и разные половые клетки. Объединение таких несколько различ-
ных по своей наследственности половых клеток дает более жизнеспо-
собные организмы» (подчеркнуто автором. — Л. Д.).
Лысенко (1952, стр. 640) утверждал, что степень жизненности жи-
вого тела обусловливается его противоречивостью. «Живое тело, — по
словам этого ученого (там же), — только потому и обладает жизнен-
ным импульсом, что ему свойственны внутренние противоречия»; «опло-
дотворение создает жизненность, жизненный импульс». То положение,
что «при узкородственном разведении, в особенности длительном, все
без исключения животные, также растения перекрестноопылители, обя-
зательно превращаются в маложизненные, малоподвижные, вырожден-
ные формы, есть — по Лысенко (1952, стр. 709), — важнейший закон
живой природы, вскрытый Дарвином».
По Кушнеру (1964, стр. 343), «мичуринское учение о жизненности,
как его формулирует Т. Д. Лысенко, исходит из того, что источником
развития живого тела является его внутренняя противоречивость, ко-
торая в большинстве случаев создается в процессе оплодотворения —
объединения в зиготе различающихся половых клеток самца и самки.
Создаваемая вследствие этого разнокачественность внутри зиготы и
является импульсом для дальнейшего ее развития. При близкородст-
венном разведении животных объединяются сходные, мало различаю-
щиеся в биологическом отношении половые элементы. Поэтому и внут-
ренняя противоречивость развивающейся зиготы недостаточна — жиз-
ненный импульс мал, организм может оказаться нежизненным или бу-
дет развиваться очень вяло. При неродственном внутривидовом скре-
щивании, когда объединяются относительно различные половые клет-
ки, .в зиготе создается значительная внутренняя прот.и1во|речи1вость, со-
ответственно повышается жизненный импульс, в организме энергично
осуществляются процессы ассимиляции-диссимиляции, рост и развитие
происходят более интенсивно. Довольно часто в этих случаях потомст-
во по своим размерам, продуктивности, жизнеспособности и плодовито-
сти превосходит исходные родительские формы».
Далее Кушнер ссылается на опыты Лысенко с черенкованием ржи,
когда наследственность расчеренкованных растений оставалась прак-
тически одинаковой: в таких случаях, говорит Кушнер, речь идет о фи-
зиологической дифференциации гамет в результате изменений условий
жизни. «В других случаях, например при скрещивании животных раз-
ных пород или растений разных сортов, речь идет о различиях гамет
также и по их наследственности» (там же).
Немного выше (стр. 342) приводится опыт Лысенко, безусловно
очень интересный:
В опытах Лысенко (1948) и его учеников на многих видах пере-
крестноопыляющихся растений (рожь, свекла, морковь и др.); «эти рас-
тения при вынужденном самоопылении (инцухте) полностью или почти
совсем не дают семян, а если семена и образуются, то из них выраста-
ют очень хилые маложизненные растения. Однако достаточно было
расчеренковать взятое для опыта материнское растение, вырастить че-
ренки в различных условиях и ко времени цветения дать этим частям
возможность перепылить друг друга, как было получено довольно боль-
шое количество нормальных по жизнеспособности семян.
68 *
Аналогичные опыты были затем проведены на кроликах и курах
(Лебедев, 1940; Аракелян, 1950; Кушнер и Китаева, 1953). Стало очевид-
ным, что природу вырождения потомства при инбридинге нельзя свя-
зывать с теорией летальных генов».
В свое время Дарвин приводил достаточно убедительные доказа-
тельства вреда инбридинга и пользы скрещивания особей, воспитан-
ных в разных условиях.
Мы должны, однако, заметить, что «учение о жизненности», в том
виде как оно преподносится в произведениях Лысенко, не вполне при-
емлемо для нас. Несколько лет назад (1961, стр. 470) мы писали, >ito
«в проблеме жизненности многое остается пока неясным и требующим
дальнейших упорных исследований». Мы ставили вопрос о том, «в ка-
кой мере жизненность можно противопоставлять наследственности»,
поскольку сама жизненность все-таки наследственна. Лысенко ду-
мает, что образование жизненности связано с внесением разнокачест-
венное.™ в живое тело, что создает «внутреннюю противоречивость»
организма; при этом Лысенко ссылается на закон борьбы противоре-
чий, имеющий общее значение в материалистической диалектике. Та-
кой подход нам кажется неверным. Нельзя же думать, что в каком-то
состоянии, на какой-то стадии развития или существования живое
тело утрачивает диалектические свойства, перестает подчиняться самым
общим диалектическим законам и переходит или стремится перейти в
некое метафизическое состояние. И вот, слияние гамет спасает живое
тело от метафизики, возвращает его в лоно диалектики. Эта часть уче-
ния о жизненности была недостаточно продумана его автором. Никог-
да действие закона диалектики не ослабевает, и в природе метафизика
не одолевает диалектику.
«В растениеводстве мичуринское учение о жизненности организмов
явилось предпосылкой разработки таких важных и эффективных прие-
мов повышения урожайности, как внутрисортовое скрещивание само-
опыляющихся растений, искусственное дополнительное опыление пере-
крестноопыляющихся, периодическая смена семенного материала, со-
ответствующий подбор родительских компонентов для гибридизации и
др. В животноводстве учение о жизненности оказалось наиболее пло-
дотворным при разработке системы межпородных скрещиваний для
товарных ферм, приемов закрепления гетерозиса в следующих после
первого поколениях, для целей совершенствования чистопородных жи-
вотных племенных ферм путем «освежения крови», для создания мето-
дов борьбы с отрицательными последствиями инбридинга в тех слу-
чаях, когда к этой системе спаривания приходится прибегать и т. д.»
(Кушнер, 1964, стр. 344).
Б. Л. Астауров (1967а, 19676, 1968а, 19686) в последнее время уде-
лял большое внимание вопросу о так называемом «физиологическом ге-
терозисе», который соответствует «жизненности» в понимании Т. Д. Лы-
сенко. Астауров, по наше1му мнению, проявил некоторую степень бес-
пристрастия в поисках правильного решения этого вопроса. Он пришел
к заключению, что существование этого явления нельзя считать дока-
занным. Он пишет: «требуются весьма строгие эксперименты для до-
казательства того, что при скрещивании географически разобщенных
отродий наблюдающийся положительный эффект идет за счет негене-
тического влияния различающихся внешних условий (физиологчеоко-
го гетерозиса), а не за счет обычной генетической дифференцировки.
Подобных опытов, к сожалению, не известно». (Астауров, 1967а, стр.
89). В другом месте той же статьи он пишет: «В пользу существования
69
у тутового шелкопряда так называемого физиологического гетерозиса,
т. е. гетерозиса, не сводимого к генетическим различиям родителей и
их гамет и обусловленного какой-то физиологической «разнокачествен-
ностью» половых клеток, вызванной различиями в жизненных условиях
родителей, безупречных доказательств нет» (подчеркнуто нами. —
Л. Д.). На этом основании он делает вывод, что «реальность так назы-
ваемого физиологического гетерозиса не доказана» (там же, стр. 94).
Буквально то же самое автор повторяет в другой своей статье (19686,
стр. 235), где он критикует опыты над рожью, сахарной свеклой (стр.
222), и «ад некото|ры)М1и домаш1Н1И1М!и жи1вотны1М'и (там же, сгр. 222 и
223). Однако эти эксперименты проводились приверженцами мичури-
низма. Почему же Астауров или его сторонники не могут поставить та-
кие же исследования, в условиях, не вызывающих сомнений у этих уче-
ных? Меж тем и в другой своей работе Астауров говорит то же самое:
«Для того, чтобы концепция физиологического гетерозиса получила
право на безусловное признание, мало одних псевдофило-
софских рассуждений, необходимы строгие и прямые, эксперименталь-
ные доказательства» (подчеркнуто нами. — Л. Д., 1968а, сгр. 9'5). Если
так, то почему же он и его сторонники не поставят такие эксперимен-
ты?
Приходится думать, что он исходит из того же принципа, что и Дэ-
вис, высказываний которого мы коснулись в связи с вопросом о насле-
довании резистентности к антибиотикам у микробов: «тяжесть до-
казательства» должна — мол лечь на мичуринистов, которые за-
щищают концепцию «жизненности». А правоверным генетикам в это
дело вмешиваться нечего: во-первых, они уже занимают господствую-
щее положение в науке, а во-вторых, соответствующие эксперименты,
если они будут проведены с надлежащей тщательностью, чего доброго,
«приведут к «нежелательным!» результатам-
Следует отметить, что мы считаем неправильным противопостав-
ление «жизненности» понятию наследственности. Когда по тем или
иным причинам (чаще всего вследствие скрещивания представителей
двух популяций с существенно неоднородной наследственностью или
живущих -в неод<и.на1ко:вых условиях среды), поднимаются мощь потом-
ства и его плодовитость, такое явление нельзя не признать формой из-
менения наследственности. Это изменение не ограничивается одним по-
колением — оно передается по наследству, в той или иной мере, и бли-
жайшим последующим поколениям. Именно этот аспект «гибридной
мощи» и представляет наибольший интерес с точки зрения проблемы
изменчивости организмов, имеющей первостепенное значение для нау-
ки об эволюции.
Проблема «гибридного вигора», или «жизненности», по нашему
мнению, должна занять видное место в разработке вопросов этой нау-
ки. Нам кажется, что с традиционной, наиболее распространенной, точ-
ки зрения, нельзя удовлетворительно объяснить столь обычные явления,
как смена источников питания, например, у тлей, их обязательный пе-
реход с одного кормового растения на другое, принадлежащее к дру-
гому виду или даже к другому крупному таксону, а также многие слу-
чаи перехода эндопаразитов от одного хозяина к другому в обязатель-
ном цикле развития этих паразитов. С дарвиновской же точки зрения
эти явления вполне объяснимы.
Работники школы Лысенко, как уже сказано, производили опыты,
которые, по их убеждению, доказывают наличие изменений организмов
в зависимости от условий их индивидуального развития и передачи этих
70
изменений следующему поколению. Если эти опыты были поставлены
вполне правильно, то полученные таким путем выводы имеют огромное
научное и практическое значение. Они, в частности, явились бы новым
прекрасным доказательством того, что факторы среды могут вызывать
массовую наследственную изменчивость организмов — растений и жи-
вотных. Такие выводы оказались бы в полном соответствии с результа-
тами исследований по различным другим направлениям. Но мы должны
считаться с тем, что такой авторитетный исследователь как Астауров
находит эксперименты, произведенные мичур'и1ни1стами, не вполне без> -
пречными. Необходимо, следовательно, повторить эти опыты в условия::,
не вызывающих никаких сомнений у специалистов. Нам кажется, что
этим должны заняться и представители ортодоксальной генетики.
Если они получат положительные результаты, им, конечно, придется
несколько отступить от одной из своих догм, но зато выиграет научная
истина, а следовательно — и практика, которая базируется на теорети-
ческих тезисах,
А пока мы вынуждены сказать, что, хотя обобщения мичуринистов
подвергаются сомнению со стороны компетентных исследователей друго-
го направления, эти обобщения в целом гармонируют с основными прин-
ципами дарвинистского учения об эволюции и ее факторах.
ОБЩИЕ ВЫВОДЫ
В этой главе были рассмотрены некоторые результаты преимущест-
венно экспериментальных исследований изменчивости организмов. Мы
подчеркнули, что явления изменчивости не могут быть правильно поня-
ты без учета достижений генетики, — науки о наследственности и из-
менчивости. Эта наука возникла на основе открытий Г. Менделя. Еще
Дарвин писал о том, что теперь понимается под дискретностью наслед-
ственности. Он говорил, что, по его мнению, «элементы обоих родитель-
ских видов находятся в каждом гибриде в двояком состоянии, а именно
и слитно, и совершенно раздельно» (Дарвин, 1951, стр. 476). Мы дума-
ем, что правильное понимание наследственности и изменчивости пред-
полагает признание единства дискретности и слитности.
Полное игнорирование успехов ортодоксальной генетики вело к
крупным ошибкам в понимании изменчивости и ее происхождения.
По нашему мнению, английский биолог Д. Мичи близок к правиль-
ной оценке современного состояния биологической мысли в области изу-
чения наследственности и изменчивости организмов. В сборнике, посвя-
щенном столетию учения Дарвина, Мичи опубликовал статью под загла-
вием «Третья стадия в развитии генетики» (D. Michie, 1958).
Мичи счел нужным, прежде всего, рассмотреть то понимание нас-
ледственности и изменчивости, которого придерживался Ч. Дарвин. Он
отмечает, что эти идеи Дарвина остаются почти не известными боль-
шинству современных исследователей. О них ничего не говорят профес-
сора своим студентам, если не считать шуточек, — попыток высмеять
якобы примитивные взгляды Дарвина. Мичи утверждает, что идеи
Дарвина были в пренебрежении просто потому, что
они опередили свое время на 110 л ет. Концепция Дарви-
на о способе возникновения наследственных изменений организма, в ко-
торой сильно подчеркнута роль среды, оказывалась в тени сначала из-за
«бессмертной зародышевой плазмы» Вейсмана, потом — «незапятнан-
ных гамет», далее «чистых линий» и, наконец, «хромосомной теории».
Свои соображения Мичи поясняет графическими схемами, которые
он заимствует из работы известного генетика Ч. Д. Дарлингтона, преду-
71
преждая, однако, читателя, что он собирается воспользоваться этими
схемами для пояснения идей, диаметрально противоположных идеям
Дарлингтона.
Схема (рис. 1) поясняет сущность двух теорий наследственности.
Верхняя половина рисунка иллюстрирует ту теорию наследственности,
по которой факторы среды, влияющие на живые тела, могут действо-
вать и на генетический материал, согласно гипотезам Ламарка и Дар-
вина. Эта схема допускает «наследование приобретенных свойств» и
обещает экспериментатору возможность изменения срироды растений и
животных путем сознательного контроля над условиями их воспитания.
Рис. 1. Схема, поясняющая две теории наследственности (по Д. Мичи): Наверху:
система, согласно которой влияния, действующие на тело, могут действовать на
генетический материал (по гипотезам Ламарка и Дарвина). Внизу: тип
системы, постулируемый Вейсманом и большинством современных
биологов (по Д. Мичи).
Связь между зародышевой плазмой и телом (сомой) тут взаимная, и
потому Мичи назвал эту теорию «двухпутной теорией» наследственнос-
ти. Это была концепция, из которой исходил Дарвин, хотя многие, как
говорит Мичи, удивятся, что Дарвин был до такой степени ламаркистом.
Еще в очерке 1844г.Дарвин писал, что в некоторых условиях существо-
вания организмы в течение своей индивидуальной жизни несколько от-
клоняются от своих обычных признаков, и многие из особенностей, при-
обретенных таким путем, передаются потомкам. Этот взгляд он разви-
вал в «Происхождении видов» и в «Изменчивости растений и животных
в условиях доместикации». Когда в 1876 году он опубликовал ряд опы-
тов над 54 видами растений, доказывающий благотворное действие пе-
рекрестного оплодотворения, в противоположность самооплодотворению,
то это явление он приписал скорее различиям в истории среды, чем в
генетической истории родителей, участвующих в скрещивании: высокая
жизнеспособность потомков зависит, по Дарвину, прежде всего и глав-
ным образом от того, что родители воспитывались не в одинаковых, а не-
сколько различных условиях.
Нижняя половина рисунка иллюстрирует ту теорию, согласно кото-
рой зародышевая плазма порождает зародышевую плазму, отпочковы-
вая в каждом поколении «смертное» тело, сому, в качестве временного
носителя зародышевой плазмы. Тут наследственность определяет при-
72
роду тела, но преобразования сомы не оставляют никакого следа в г/о-
томстве, не влияют на потомство. Новые изменения могут возникнуть
лишь как внутренние «ошибки» в том процессе, через который зароды-
шевая плазма копирует зародышевую плазму в нисходящем ряду поко-
лений.
Ткани тела состоят из мириадов микроскопических клеток. В каж-
дой клетке содержатся цитоплазма и ядро, в котором находятся ни-
тевидные хромосомы. Ядра зародышевых клеток содержат гаплоидное,,
половинное, число хромосом, имеющихся в соматических клетках, ко-
торым свойствен диплоидный набор хромосом.
Блестящие экспериментальные исследования Т. Моргана и его пос-
ледователей расшифровали сложные маневры и явления обмена, че-
рез которые хромосомы репродуктивных клеток осуществляют распре-
деление и рекомбинацию наследственных признаков. Так создалась»
«хромосомная теория наследственности». Эта теория, по словам Мичи
(там же, стр. 59), характерна для той стадии развития генетики, ко-
торую он называет «классической стадией» (приблизительно от начала
столетия до второй мировой войны). Схема, изображенная на рисунке
2, показывает, что «хромосомная теория» есть усложненный вариант
Рис. 2. Схема поясняющая хромососную теорию наследственности, —
«однопутную теорию» (по Д- Мпчи).
теории Вейсмана. Генный материал передается неизменным от оплодо-
творенного яйца потомкам. Ядерный аппарат не испытывает существен-
ных превратностей развития и дифференциации, за исключением ред-
ких и «рандомных» (шальных, беспорядочных) мутаций. Это — «одно-
путная» теория наследственности. Область исследования эксперимен-
тальной генетики была разработана, по словам Мичи, почти до ис-
черпания.
Вторая стадия развития генетики, по Мичи, охватывает приблизи-
тельно послевоенный период. Она характеризуется тем, что центр
внимания переносится на те явления наследственности, которые не ук-
ладываются в классическую картину, — на передачу внехромосом-
ных эффектов в половом -воспроизведении, на «изучение роли цитоплаз-
мы в наследственности, то есть на явления, которые обнаруживаются в
73
эмбриональной дифференциации или в пропагации клонов одноклеточ-
ных организмов (простейших, бактерий, грибов). Эта стадия, которая,
по Мичи, завершается только теперь (вспомним, что работа Мичл
была опубликована в 1958 году, а с того времени в генетике произош-
ло много нового), была «царством дуалистической философии в би-
ологии» (рис. 3). Генетик-дуалист, думает, что существуют две системы
наследственности: хромосомная, соответствующая «однопутной» теории
Вейсмана, и цитоплазматическая система, соответствующая «двухпут-
ной?? (обоюдосторонней) теории Ламарка и Дарвина. Мичи приводит
случаи наследования, которые не соответствуют представлениям «клас-
сической» стадии, в частности, изученные Хиншелвудом факты «трени-
ровки» бактерий, которые «научаются» справляться с новой пищей пли
ядами и передают свои вновь приобретенные биохимические свойства
своим отдаленным потомкам, факты «ламаркистского» поведения колов-
раток (Rotifera) и другие.
Заключительное состояние дуалистической концепции в генетике
Мичи иллюстрирует схемой, которая показывает, что наследственность
контролируется как хромосомным аппаратом ядра, так и цитоплазмой.
Via с л е Э с т в е м>-«о с. т о линии через яЗро и хромозО!
— *Клеточная наслеЗс’’депност0 мере* - цитоплазма
Рис. 3. Схема, поясняющая дуалистическую концепцию наследственности. Сплошными
линиями показана наследственность, идущая от ядра и хромосом. Прерывистой —
клеточная наследственность, идущая через цитоплазму (носит весьма случайный
характер в передаче свойств). По Д. Мичи-
Мичи находит, что выдвинутая Дарвином гипотеза пангенезиса, в
ю же время является гипотезой, приковывающей к себе внимание и
пророческой. Поскольку она может служить стимулом для новых по-
пыток найти унитарную теорию наследственности, Мичи считает нуж-
ным остановиться на рассмотрении основных черт этой дарвиновской
концепции.
В процессе формирования различных тканей тела их клетки, как
выражается Дарвин, выбрасываются крошечные крупинки, или атомы,
которые свободно циркулируют по всей системе и, когда получают
74
подходящее питание, размножаются самоделением. Эти крупинки, гра-
нулы или «геммулы», передаются от родителей потомству.
Эта модель дифференциации и наследственности допускает насле-
дование приобретенных свойств. Мичи находит, что дарвиновская ги-
потеза пангенезиса была для своего времени замечательным достиже-
нием. Во-первых, она постулировала са1мовоспроиз1водящиеся частицы,
которые соответствуют хромосомным генам, а существование этих
последних было установлено лишь через 50 лет после того, как Дарвин
создал эту гипотезу. И прошло еще 25 лет до того, как в науке нача-
лось серьезное обсуждение возможности того, что подобные
«геммулы» (плазмагены) могут играть роль в эмбриональном разви-
тии. Во-вторых, гипотеза пангенезиса есть, по словам Мичи, «герои-
ческая» попытка связать наследственность зародышевого пути и раз-
витие тела организма в единую систему, где зародышевый путь не рас-
полагается в стороне, как нечто нерушимое, а подвергается силам раз-
вития и распада, которые управляют всем остальным телом. На протя-
жении целого столетия подобные представления решительно исключа-
лись из биологической науки. Но наступил день, как утверждает Мичи,
когда они начали восстанавливаться.
Дуалистический принцип, ныне господствующий в генетике, не мо-
жет быть сохранен: генетики должны, по словам Мичи, стремиться к
созданию единой, монистической теории двухпутного типа. Такая те-
ория должна охватывать и онтогенез и наследственность. Она должна
признать, что зародышевая плазма не отгорожена непроницаемой сте-
ной от сомы и поддается влияниям, исходящим от «смертного тела».
Новая генетическая теория должна пересмотреть прежние взгляды на
зародышевую плазму и сому, должна отвергнуть дуализм, должна вы-
яснить пути воздействия цитоплазмы на хромосомные гены. Совре-
менная биология уже идет к открытию этих путей.
Итак, заключает Мичи, началась третья стадия в развитии генети-
ки. Нам кажется, что Мичи учитывает новейшие достижения биологии
и особенно генетики и приходит к правильным выводам. Мичи за мо-
нистическое понимание наследственности. Сому нельзя считать, подоб-
но Вейсману и представителям классической генетики, каким-то побоч-
ным образованием, не влияющим на зародышевую плазму: влияние
тут обоюдное, и в ту ив другую сторону.
Мичи не считает возможным отмахиваться от гипотезы пангенези-
са, которую выдвинул Дарвин и в которой геммулы подобны генам
современной генетики. В общем, третья стадия генетики есть, в извест-
кой мере, возврат к идеям Дарвина.
Мы рассмотрели некоторые категории фактов, которые в своей
совокупности говорят именно за необходимость перехода к принципам
лого, что Мичи называет «третьей стадией» развития генетики. Экспе-
риментальное изучение изменчивости микробов, нематод, насекомых и
других организмов приводит специалистов к выводу о том, что наслед-
ственные изменения вызываются факторами среды и могут носить опре-
деленно направленный характер, причем они могут возникать не у
единичных, а у многочисленных особей данной популяции.
Анализ таких явлений, наблюдаемых в каждой отдельной группе
органических форм, показывает, что наиболее вероятным оказывается
объяснение изменчивости воздействием внешних условий. Но посколь-
ку такое толкование представляется единственно вероятным в отноше-
нии всех более или менее тщательно изученных видов, а иные гипоте-
75
зы при этом отпадают, вероятность все более и более приближается к
полной достоверности.
Мичи, по-видимому, был прав, когда он писал, что Дарвин опере-
дил науку на сто десять лет. Это» впрочем не значит, что все факты,
противоречащие принципам классической генетики, нельзя описывать
и толковать путем введения новых понятий и дополнительных допу-
щений, с помощью которых можно подпирать не особейно прочное
здание этой науки. Но это не устранит необходимости перестройки хо-
дячих представлений на основе диалектического понимания изменчи-
вости и ее причин.
Мы уже отметили, что английские ученые Дин и Хиншелвуд (Dean,
Hinshelwood, 1957) очень удачно и в высшей степени остроумно срав-
нивали современное состояние классической генетики с закатом теории
флогистона. Обзор современного состояния научной мысли в этой об-
ласти — по необходимости очень краткий — дает полное подтвержде-
ние этой оценки.
Мы должны со всей прямотой заявить, что нам не под силу вы-
яснять ход биохимических процессов, молекулярных изменений, кото-
рые происходят в процессе влияния внешних условий на организмы.
Впрочем мы думаем, что такие тонкости в основном пока еще недос-
тупны и специалистам по молекулярной генетике. Эти исследователи
просто описывают наблюдаемые факты, пользуясь терминологией
классической генетики. Можно думать, что любые факты изменения
организмов вследствие воздействия на них факторов среды можно опи-
сывать на языке классической генетики (в широком смысле этого сло-
ва,, с (включением сюда и всей современной генетики), который в колос-
сальной степени усложнялся за время существования этой .науки, т. е.
с начала текущего столетия, особенно же за последние два десяти-
летия.
Образцы такого изощрения языка у работников генетики и их пок-
лонников можно указать в работах М. Г. Агаева. Он, например, пишет:
«при длительном воздействии на растения хлебных злаков необычными
температурными условиями (путем коренного изменения сроков посева)
в течение ряда поколений гены типа развития нередко переходят в
высокомутабильное состояние, что иногда и обеспечивает высокий
выход мутантных форм с (измененным типом развития» (Агаев, .1969»
стр. 1370). А непосредственно дальше этот автор делает заявление,
показывающее, что кроется за его флогистонизированным способом
изложения: «Причины и механизм специфического температурного ре-
жима на хлебные злаки (индуцирование мутаций генов, детерминирую-
щих яровой и озимой типы развития), пока еще не изучены»
(подчеркнуто нами.—Л. Д.). Позволительно спросить: почему в таком
случае автор, как и все приверженцы данной генетической школы, счи-
тают для всех обязательным выражать приведенное положение-
на языке неоменделистской генетики? Что изменится от того, если, вы-
ражая абсолютно ту же самую мысль, исследователи будут употреб-
лять более нейтральные, простые выражения, не обязывающие их при-
нять неоменделистскую схему? Почему вместо «гены типа развития пе-
реходят в высокомутабильное состояние», нельзя сказать, что организ-
мы испытывают повышенную наследственную изменчивость? Раз при-
чины и механизмы мутагенного действия «пока еще не изучены», то
есть попросту неизвестны, то не будет никакого греха, если исследова-
тели не прибегнут к ортодоксальной генетической терминологии, а бу-
дут выражаться без таких претенциозных терминов как «высокомута-
76
Сильное состояние» и «гены типа развития». Но нам кажется, что в то
же время нельзя возражать против этих выражений, если они достав-
ляют удовольствие некоторым авторам, поскольку это не ведет к пута-
нице. То же самое можно сказать и о «мутагенной роли изменений
некоторых обычных экологических факторов» (там же, стр. 1369). Надо
протестовать не столько против флогистонизации научного языка,
сколько против требования считать эту флогистонизацию для всех обя-
зательной.
Сторонники догм генетики и соответствующей терминологии не
приводят существенных возражений против неортодоксального понима-
ния явлений индуцированной изменчивости, или их возражения лишены
убедительной силы.
Примером могут служить опят-таки работы Агаева. Так, он наста-
ивает на том, что «в ходе взаимопревращения яровых и озимых рас-
тений строго сохраняется основная Характеристика мутационного про-
цесса — ненаправленность (объективная случайность) мутаций» (там
же,стр. 1372). Он утверждает, что эта закономерность мутационного
процесса не нарушается даже в тех случаях, когда мутации искус-
ственно индуцируются в строго контролируемых условиях. При этом он
ссылается на С. А. Алиханяна (1967, стр. 69), который писал: «Све-
дения о мутагенезе, добытые уже сегодня, не дают основания надеяться
на получение под действием какого-нибудь мутагена 100% мутантов
одного определенного фенотипа, хотя и можно считать
реальным получение преимущественно одних типов изменений по срав-
нению с другими» (подчеркнуто нами. — Л. Д.).
Нам кажется очень странным, что Алиханян говорит тут о «100%
мутантов одного определенного фенотипа», а другой серьезный иссле-
дователь, Агаев, считает такую аргументацию справедливой. Неужели
для признания направленного действия «мутагена» необходимо, чтобы
все сто процентов возникших под его влиянием особей пред-
ставляли один определенный фенотип? Сомневаемся, чтобы какой-либо
сторонник идеи направленного изменения наследственной природы ор-
ганизмов под влиянием того или иного фактора среды утверждал, что
сто процентов их потомства представляют один определенный фено-
тип. Если противники этой идеи ссылаются на то, что не все сто про-
центов потомков одинаково и единообразно указывают на соответст-
вующее изменение популяции, то это уже, по нашему мнению, — не
более как соломинка, за которую хватаются эти люди науки. Приз-
нание того, что «можно считать реальным преимущество одного типа
изменений по сравнению с другими», — это как раз то, против чего до
самого недавнего времени восставали ортодоксальные генетики «синте-
тической» школы и как раз то, что отстаивали их оппоненты из числа
дарвинистов.
Разбирая работу Ш. Райки (1967), где автор описывает свои весь-
ма тщательно поставленные эксперименты, доказывающие превраще-
ние, в определенных условиях, яровой пшеницы в озимую, Агаев пи-
шет: «Представленный в книге Райки обширный и разнородный фак-
тический материал показывает, что под влиянием осеннего посева в
генетически чистых (по генам типа развития) линиях типично яровой
пшеницы сорта Лютесценс 62 действительно возникают озимые формы»
(там же, стр. 1374). Далее, по словам Агаева, «подробный анализ ма-
териалов Райки показывает, что в проведенных им опытах превращение
яровых пшениц в озимые нередко характеризовалось большой часто-
той» (там же, стр. 1375). Райки, по Агаеву (там же), обнаружил
77
«отдельные варианты и многочисленные подлипни (представленные
сотнями и тысячами особей), в которых изменение типа развития про-
изошло почти у всех растений» (там же, стр. 1375; подчерк-
нуто нами. — Л. Д.). Наконец, примечательно и то, что, по Агаеву
(там же), «общие итоги исследований Райки по «озимизации» заслу-
живают высокой оценки, так как эти данные получены в опытах, стро-
гих в методических отношениях». Далее о результатах исследований
Райки Агаев (там же, стр. 1376) говорит: «Очевидно, это открытие поз-
волит глубже анализировать форомообразовагельные закономерности
процесса «озимизации», чем до сих пор. Вместе с тем оно является од-
ним из наиболее убедительных доказательств того, что в исследованиям
по экспериментальной «озимизации» действительно может происхо-
дить наследственное преобразование яровых растений в озимые».
Таким образом, Агаев дает блестящую оценку экспериментам Рай-
ки, который, действительно, поднял на более высокий уровень методи-
ку изучения явлений преобразования злаков. И тем не менее Агаев
заключает свою статью громовыми обвинениями в адрес Райки. Первое
обвинение гласит, что «Райки бездоказательно считает, что озимизации
является экспериментальным подтверждением «феномена Ламарка»,
т. е. принципа наследования приобретаемых признаков». «Яровые рас-
тения, — по Агаеву (там же), — приобретают озимость как новый
фенотипический признак, только в результате соответствующих мута-
ционных изменений их наследственности». Нетрудно, однако, видеть,
что тут вносится лишь словесная поправка (и тритом совершенно из-
лишняя, не меняющая ни на йоту сути дела) в положение, доказыва-
емое Райки. Суть же заключается в том, что в результате воздействия
фактором среды вызывается определенно направленное изменение ос-
новной массы посеянных растений, — изменение, обеспечивающее их
произрастание в новых условиях развития. Очевидно Агаев снял бы
обвинение Райки в «бездоказательности» только в том случае, если бы
было доказано, что определенным образом изменились не большинство
особей, даже не почти все особи, а обязательно все сто процентов осо-
бей. Раз этого нет, то можно, по Агаеву, настаивать на применении в
данном случае классической «синтетической» схемы.
Тем самым становится понятным второе обвинение, предъявляемое
Райки, который якобы «бездоказательно утверждает, что к анализу про-
цесса превращения яровых растений в озимые не применимы гене-
тические концепции гена и мутации». Это громко звучащее обвинение,
конечно, тоже может служить лишь словесной поправкой, притом не-
нужной, к формулировкам Райки: суть процесса Райки описал совер-
шенно точно, но, по Агаеву, он обязан говорить не о превращении яро-
вых растений в озимые, а о мутации генов развития организмов.
Третье торжественно провозглашаемое Агаевым обвинение, гласит,
будто Райки бездоказательно заявляет, что результаты экспериментов
по «озимизации» обусловлены не естественным отбором, а приспособи-
тельной («адекватной») изменчивостью». Между тем сам Агаев подчер-
кивает, что исходный материал, над которым работал Райки был и с-
ключительно генетически яровой пшеницей: только это и имелось
в виду, когда Райки утверждал, что озимизация не была вызвана естест-
венным отбором: раз в посевном материале не было озимых элементов,
то в данном случае отбор не мог привести к их приумножению и к эли-
минации яровых элементов.
Окончательное заключение Агаева гласит, что предлагаемая Рай-
ки «интерпретация по экспериментальной «озимизации», названной авто-
78
ром «генетической», по-существу, является «антигенетической», а следо-
вательно несостоятельной» (там же, стр. 1377). Мы, однако, видели, чго
эта разгромная критика исследований Райки по существу является тре-
бованием излагать идеи на «синтетическом »языке, с применением клас-
сической генетической терминологии, хотя Райки достаточно ясно изло-
жил и обосновал свои положения об «озимизации» яровой пшеницы.
Здесь перед нами — один из ярких примеров флогистонизации
мышления в области классической генетики. Современная литература
неуклонно обогащается такими примерами, которые в недалеком буду-
щем должны будут полностью овладеть всем содержанием этого учения.
Эта флогистонизированная доктрина будет, очевидно, существовать до
тех пор, пока она не будет предана забвению. Нам кажется, что об этом
процессе надо помнить, но едва ли следует бороться с такой тенденци-
ей излагать научные материалы флогистонизированным языком: надо
ей дать возможность тихо отойти в область небытия.
Глава третья
АКТУОПАЛЕОНТОЛОГИЧЕСКИЙ АНАЛИЗ БИОТОПОВ,
БЛАГОПРИЯТСТВУЮЩИХ ИНТЕНСИВНОЙ МАССОВОЙ
ИЗМЕНЧИВОСТИ ОРГАНИЗМОВ
ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Р. Рихтер (R. Richter, 1929) ввел уже давно довольно громоздкий
термин «актуопалеонтология», что должно означать изучение тех ны-
нешних явлений в жизни организмов, которые помогают понять жизнь
прошлых времен. Такое направление исследований представляется нам
очень нужным для понимания жизни и развития органического мира в
геологическом времени. Актуопалеонтологические работы, освещающие
современнную жизнь с палеобиологической точки зрения, так ска-
зать, — по заданию палеобиологии, призваны заниматься тем.и вопроса-
ми биологии, которые особенно важны для истории биоса и для био-
стратипрафии. Нагример, изучение современных наутилусов (.корабли-
ков) представляет огромный интерес для познания не только наутило-
идей кайнозоя и мезозоя, но и более древних родичей этих моллюсков,
а также и аммоноидей мезозоя и палеозоя.
Ставя перед собой задачу выяснения явлений изменчивости у ис-
копаемых животных, мы, естественно, должны коснуться вопроса об
условиях, способствующих изменчивости современных организмов и в
частности — о тех типах местообитаний, которые особенно благоприят-
ны для интенсификации подобных процессов. Существует мнение, будто
бы условия некоторых типов среды настолько постоянны, константны,
что там не может быть изменчивости, стимулируемой экологическими
факторами. Тем самым ставится под сомнение значение среды для из-
менчивости, что может толкать малоискушенных людей к идее «абсо-
лютно внутренних» факторов эволюции, если не к полному отрицанию
развития органических форм.
В этой главе мы постараемся показать, что различные типы биото-
пов могут способствовать и в действительности способствуют повыше-
нию изменчивости нынеживущих форм, а следовательно должны были
иметь такое же значение и в геологическом прошлом.
ИЗМЕНЧИВОСТЬ ЖИВОТНЫХ АБЕРРАНТНЫХ НЕМОРСКИХ БИОТОПОВ
Существуют типы среды, сильно отличающиеся от основных эколо-
гических областей земного шара, — участки, где по крайней мере не-
которые существенные для организмов условия жизни оказываются
сильно отклоняющимися от обычных норм, — от условий, господствую-
щих на смежных пространствах, — и все же годные для обитания по
крайней мере некоторых организмов.
80
Живые существа, попавшие на такие участки и там прижившиеся,
иногда прочно колонизируют эти участки, не только выживают там, но
и приспособляются к их своеобразным условиям. В ходе экогенеза
такие бцртопы являются скорее вторичными: туда организмы, претерпе-
вая эволюцию, попадают обычно лишь после того, что их предки дол-
го эволюировали на других участках, где существует комплекс более
или менее нормальных условий питания, освещения, тепла, газового ре-
жима.
Изучение населения таких участков показывает, что многие формы
испытывали там повышенную изменчивость. Интенсивность такой измен-
чивости у некоторых форм иногда бывает несравненно более высокой,
чем изменчивость ближайших родичей этих форм, живущих и эволюи-
рующих в условиях, которые мы можем назвать нормальными.
Что это за аберрантные биотопы?
Это, в воде, — солоноватоводные бассейны различной величины:
некоторые заливы океанов и морей, полузамкнутые моря типа Черного
моря, обширные реликтовые замкнутые моря, которые И. И. Андрусов
называл чаще всего озерами-морями (такое наименование кажется
нам удачным, по крайней мере в применении к замкнутым обширным
морям прошлого), а иногда «каспиями»: нынешним бассейном такого
типа является Каспийское море. Затем, к числу аберрантных немор-
ских биотопов можно отнести более или менее обширные озера, вроде
Байкала и больших озер Восточной Африки, далее — сравнительно не-
большие озера с более или менее богатым населением.
Вполне возможно, что к категории аберрантных биотопов прибли-
жаются некоторые заливы и другие морские акватории, имеющие до-
вольно широкую связь с океаном, но все же хотя бы периодически час-
тично от него обособляющиеся, более или менее отделяемые от него
барьерами, нередко временными, существующими недолго, но
достаточно продолжительно для обособления биотопов местных рас
различных, преимущественно бентонных, организмов.
Что же касается пространств суши, то там более или менее обособ-
ленные участки, где в условиях полной или относительной изоляции
развивались местные формы организмов, чаще возникали на территори-
ях, окруженных водой, чем на отдельных участках материков, отделен-
ных от остальной сушн орографическими барьерами — высокими горны-
ми цепями, которые иногда действуют как фильтр, пропускающий одни
формы, и препятствующий продвижению других.
Поскольку нас интересуют места, где должна была осуществляться
интенсивная изменчивость вследствие воздействия необычных, аберрант-
ных условий среды, наше внимание должно быть привлечено, в первую
очередь, не только к таким обособленным материкам, как Австралия и
Южная Америка, но и к островам, где, как показывают биогеография
и палеонтология, нередко возникали очаги эволюции своеобразнейших
групп организмов, что могло зависеть от воздействия особенных, абер-
рантных, абиотических и биотических условий.
На материках иногда возникают водоемы, где некоторые организмы
эволюируют с необыкновенной быстротой в зависимости от усиления
изменчивости. Это — почти всегда озера, существующие относительно
длительное время. В таких водоемах развивается эндемичная фауна
моллюсков, членистоногих, рыб и других органических форм. Таковы
юзера Северо-Восточной Африки, Байкал, Охридское озеро и другие.
Высокие, иногда чрезвычайно высокие темпы видообразования в таких
Я Л. Ш. Давиташвили
водоемах возможны только при условии изменчивости гораздо более
интенсивной, чем в других, обычных материковых водоемах. Мы не бу-
дем останавливаться на вопросе о формировании широко известных
своеобразных фаун в упомянутых современных озерных водоемах, огра-
ничимся лишь ссылкой на некоторые труды, где освещаются эти вопро-
сы. (См., напр., Кожов, 1962, Станкович, 1962). Поскольку основным
предметом этой книги является изменчивость организмов геологичес-
кого прошлого, мы рассмотрим особо в последующих главах отдельные,,
наиболее яркие примеры ископаемых фаун древних озер.
Однако на суше существуют и другие весьма своеобразные, абер-
рантные биотопы, обособленность которых не зависит от изоляции этих
биотопов горными цепями. Существуют организмы, живущие не на по-
верхности суши, а под этой поверхностью, ведущие подземный образ
жизни — в расселинах скал, трещинах поверхности, в почве, и наконец,
— в пещерах. К обитателям этой категории аберрантных биотопов при-
мыкают организмы, живущие внутри подстилки лесов. Такие биотопы
возникали в самых разнообразных местах всех материков, где сущест-
вовали условия для образования больших и малых углублений, годных
для жизни в них хотя бы немногих органических форм.
Здесь мы коснемся вопроса о некоторых относительно мало извест-
ных биотопов наземной среды, имеющих значение для понимания измен-
чивости организмов и факторов, ее вызывающих.
В подстилке и гумусе мощных лесов тропической Африки, на вы-
соте 3000 м, существует гумикольная фауна. Особенно примечательны
данные о Pselaphidae [Жаннель и Лелеп (Jeannel, Leleup 1952); Жаннель
(Jeannel, 1952); Лелеп (Leleup, 1956); Вандель (Vandel, 1964, стр. 515[.
У некоторых живущих в гумусе микропселафид наблюдается четко вы-
раженный половой диморфизм: самки баскрылы и лишены глаз, а у сам-
цов—и крылья и глаза. У других видов имеется поэци ландри я: в
пределах одного и того же вида у одних самцов имеются крылья и гла-
за, а у других представителей того же пола нет ни глаз, ни крыльев. У
третьих самцы с крыльями и глазами исчезают, и тогда оба пола аноф-
тальмны и бескрылы.
Примечательно, что эта регрессивная эволюция протекала не под
землей, а в гумусе тропических лесов.
Фреатобиология изучает обширную область явлений — фауну
(правда, довольно однообразную), которая населяет обширные прост-
ранства, характеризующиеся весьма своеобразными, специфическими
условиями, лишь до некоторой степени сближающими эту среду со сре-
дой пещер.
Систематическое исследование фреатических фаун было начато
сравнительно недавно.
Румынский биолог К. Моташ (С. Motas, 1957, 1958, 1962) назвал
фреатобиологией новый раздел лимнологии, который изучает с точек
зрения систематики, экологии и географического распределения фреа-
тикольных животных, то есть животных, обитающих во фреатических
горизонтах. Фреатический водный горизонт — наиболее близкий к по-
верхности почвы водоносный горизонт, — питающий обычные колодцы*
которые не пересекают непроницаемые слои. Фреатический горизонт
встречается повсюду в долинах на глубине от нескольких дециметров до
100 м и более. Фреатических рыб описывает Ж. Тинес (9. Thines, 1969).
Животные из низших ракообразных при переходе к фреатической,
жизни испытали постепенную депигментацию вплоть до полного обес-
82
цвечения кожи, а также редукцию органов зрения вплоть до полного
их исчезновения [К. Моташ (Motas, 1962), стр. 150].
Насколько интенсивно происходили процессы изменчивости на уча-
стках подобных аберрантных типов?
Жизнь ив особенности эволюция большинства таких фаун наземной
среды изучены пока весьма недостаточно. Поэтому этими скудными
данными трудно воспользоваться, для освещения вопросов изменчивос-
ти. Много лучше обстоит дело с биоспеологией, хотя и тут можно
было бы сделать еще очень много для .выяснения факторов, обусловли-
вающих возникновение наследственных, генетических изменений орга-
нических форм. В дальнейшем мы постараемся рассмотреть некоторые
факты биоспеологии с этой точки зрения.
БИОСПЕОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ, ПРОЛИВАЮЩИЕ СВЕТ НА
ПРИЧИНЫ ПОВЫШЕННОЙ ИЗМЕНЧИВОСТИ ПЕЩЕРНЫХ ОРГАНИЗМОВ
Биоспеологические исследования имеют огромное значение для ос-
вещения наследственной изменчивости, ее закономерностей и причин.
Биоспеология имеет одно существенное преимущество перед палеобио-
логическим изучением филогенетической истории морей с соленостью,
уклоняющейся от нормальной; науке о жизни в пещерах доступны жи-
вые организмы: она может изучать эти организмы, их строение, их он-
тогенез, может экспериментировать над ними и выяснять особенности
их генетики. Поэтому биоспеология в состоянии внести неоценимый
вклад в познание изменчивости и эволюции органического мира.
Французский биолог Вандель, автор недавно вышедшего обобщаю-
щего труда «Биоспеология» (Vandel, 1964), как нам кажется, сумел наг-
лядно показать общебиологическое значение биоспеологии для раскры-
тия законов органического мира. Действительно, биоспеология вскрыва-
ет факты изменчивости организмов и параллельно с этим выясняет те
особенности среды, от которых могут зависеть эти превращения живот-
ных пещер.
Вандель пришел к заключению, что эволюцию пещерных животных,
нельзя понимать как процесс, в основном зависящий от изменений гено-
типов и естественного отбора. Впрочем, Вандель, как известно, придер-
живается антидарвинистского понимания эволюции, с нашей точки зре-
ния неправильного,
Недавно по вопросу о значений биоспеологии для эволюционного,
учения выступил известный генетик-биоспеолог, немецкий ученый К.
Коссвиг* профессор Гамбургского университета, опубликовавший статью
«Генетика и регрессивная эволюция» (С. Kosswig, 1965). В этой статье
автор выдвигает новое понимание так называемой регрессивной эволю-
ции пещерных животных, которое он обосновывает многими фактами
биоспеологической науки. Нам кажется, что статья эта во многом ис-
ключительно поучительна. Интересны рассматриваемые в ней фактичес-
кие данные, собранные, в значительной мере, самим автором, и то лк о-,
вание этих материалов, к которому пришел Коссвиг.
Животные, обитающие в пещерах, изменяются в определенных на-
правлениях (обесцвечивание покровов тела, редукция органов зрения,
определенные физиологические изменения, развитие рецепторов, более;
приспособленных к пещерной среде, чем глаза, и т. д.). Изменчивость
в этих направлениях осуществляется очень быстро, с совершенно нео-
бычайной скоростью, и уже это вынуждает исследователей задумывать-
ся о причинах столь повышенной изменчивости. Все пещерные животные
происходят от форм, живших в эпигейных условиях, то есть не внут-г
83
ри пещер, а на дневной поверхности — на суше или в воде; попав в ус-
ловия мрака, высокой влажности, относительно постоянной температуры
и скудости пищевых ресурсов [Л. Фаж (Fage, 1955, стр. 79—84], живот-
ные, если они не гибнут, эволюируют в сторону приспособления к этим
условиям. Депигментация покровов тела и редукция глаз имеют столь
широкое распространение и происходят так быстро, что первое объясне-
ние, которое проходило в голову некоторым биоспеологам, заключается
в гипотезе, по которой такие изменения представляют собой прямой ре-
зультат воздействия резко измененных факторов среды.
Такая мысль как бы напрашивается сама собой, и некоторые уче-
ные принимали это объяснение без строгого критического анализа. Так,
Жаннель (Jeannel, 1930, стр. 178) писал: «Среди большинства групп пе-
щерных организмов наследственный эффект неупотребления достаточен
для объяснения того, что после многочисленных поколений, живших в
полной темноте, глаз атрофировался или исчез». Правда, тут же этот
автор продолжает: «У других форм, в особенности у насекомых, эта
эволюция, по-видимому, зависела, во всяком случае, от более сложных
причин». Ж. де Латтен (Lattin, 1940, стр. 13) писал, что для объяснения
происхождения явлений приспособления и особенно — происхождения
типа пещерного животного исследователи прежде всего думали о воз-
действии факторов наружной среды, поскольку наблюдается непреры-
вное нарастание пещерных признаков от животных, обитающих у входа
в пещеру к обитателям ее внутренних частей. В соответствии с этим
становление подобных признаков /можно было бы представить себе так,
что у надземных видов, которые случайно попадали в темную среду, в
ходе поколений редуцировались, как бесполезный балласт, пигмента-
ция тела и органы зрения, причем нередко развивались компенсирую-
щие Органы, которые хотя бы частично восполняли потерю такого важ-
ного органа чувств как глаз. Такое развитие, по Латтену, шло тем быс-
трее и полнее, чем резче выражены условия соответствующего пещер-
ного биотопа- «Поскольку .мы теперь знаем, — продолжает де Лат ген
(там же), — что многие из этих признаков пещернего животного нас-
ледственны, то эти признаки, первоначально появившиеся как модифи-
кации, должны были постепенно становиться наследственными».
Однако такое понимание признаков троглобионтов, как признает
де Латтен (там же), находится в резком контрасте со всем тем, что мы
знаем сегодня относительно видо- и расообразования надземных форм.
Поэтому Коссвиг, немецкий генетик, вместе с некоторыми своими уче-
никами, занявшийся проблемой троглобионтов, поставил ряд сравни-
тельных и экспериментальных исследований, и его исследования, как ут-
верждает де Латтен, с большой убедительной силой показали, что ла-
маркистское объяснение во многих отношениях несостоятельно и потому
от него нужно отказаться. Итак, де Латтен был уверен в том, что гене-
тик Коссвиг встал на правильный путь изучения проблемы происхожде-
ния и эволюции троглобионтов и окончательно разбил так называемое
ламаркистское объяснение этого процесса. Исследования Коссвига, дей-
ствительно заслуживают особого внимания. Правда и то, что он упор-
но искал объяснения генезиса пещерных животных на основе ортодок-
сальной классической генетики. И в высшей степени показательно, чго
сам Коссвиг в конце концов убедился в полной несостоятельности орто-
доксального объяснения и отверг его. Но эволюции воззрений Коссвига
мы коснемся несколько далее, потому что она имеет большое значение
для правильного толкования этой биоспеологической проблемы.
Посмотрим, насколько применима к эволюции обитателей пещер
'-классическая постнеодарвмнистская схема «спонтанные изменения и
84
естественный отбор». В принципе не следует отрицать возможность рег-
рессивной эволюции, выражающейся в редукции глаз, депигментации и
других процессах, вследствие естественного отбора спонтанных мута-
ций. Можем вполне согласиться, например, с Ж. де Латтеном (Lattin,
1940, стр. 26) в том, что в надземных популяциях животных мутации,
тызвающие дегенерацию глаз и крыльев, будут иметь почти при всех
обстоятельствах определенно отрицательное селективное значение: в
подавляющем большинстве случаев носители таких отклонений будут
элиминироваться, устраняться. В подземном же жизненном пространст-
ве такая мутация, если только она одновременно не вызывает падения
жизнеспособности, отнюдь не будет иметь отрицательного селективного
значения для своего носителя, потому что глаза и пигментация тут со-
вершенно бесполезны. И, по-нашему, де Латтен опять-таки прав, когда
он высказывает мнение, что утрата таких образований в изменившихся
условиях среды, то есть при переходе от надземной жизни к подземной,
имеет даже положительное значение, потому что таким путем организм
освобождается от затрат на бесполезные признаки. Оставляя в сторо-
не другие, .крайне апорные(постнеодарвини1стские толкования эволюцион-
ного процесса в пещерной среде, мы думаем, что такое объяснениетене-
зиса троглобионтов было бы вполне совместимо с дарвинистским пони-
манием эволюции, если только у нас было бы более или менее вероят-
ное представление об источниках изменчивости. Сказать просто: «.мута-
ции возникают случайно», значит не дать никакого объяснения явлени-
ям изменчивости.
. Между тем многие пещерные животные, особенно, по-видимому,
сравнительно недавно поселившиеся в этой среде, обнаруживают нео-
бычайно высокую степень индивидуальной изменчивости. Например, де
Латтен (Lattin, 1940, стр. 14 и 29) писал об удивительной изменчивости
в степенях и форме редукции глаз и в пигментации покровов у несом-
ненно очень недавно поселившегося в пещерной среде Asellus aquaticus
cavernicol js, а также о высокой изменчивости других изэпод и некоторых
рыб. Однако такую же изменчивость, судя по литературным данным, об-
наруживают и многие другие обитатели пещер: турбеллярии, копеподы,
псевдоскорпионы, опилиониды, ложнопауки, хилоподы, жуки и, нако-
нец, рыбы.
Заслуживает внимания чрезвычайно высокая изменчивость популя-
ций Asellus aquaticus cavernicolus, занимающих ареалы, близкие один к
другому. Иногда это объяснялось тем, что более светлые формы селятся
преимущественно внутри пещер, а более темные — в слабо освещенной
зоне входа в пещеру.
Еще более примечательные данные об измечивости троглобионтов
приводит Коссвиг (Kosswig, 1965). Деятельность этого ученого пред-
ставляет собой поучительный пример отказа крупного специалиста от
постнеодарвинистского понимания эволюции троглобионтов после дол-
гих лет верности «синтетической» теории эволюции. Оказалось, что фак-
ты, которые тщательно изучал этот исследователь, нельзя примирить с
таким пониманием эволюции. И в конце концов Коссвиг счел себя вы-
нужденным отказаться от попыток добиться этого. Мы довольно под-
робно рассмотрим эту эволюцияю воззрений Коссвига и его новейшее
толкование развития пещерных животнык. Одновременно нам придется
затронуть вопрос об изменчивости троглобионтов и изложить некотр-
рые данные, приводимые Коссвигом, поскольку они, на нащ взгляд,
очень важны для понимания происхождения изменчивости обитателей
пещер
85
В своих болёе ранних работах этот авторитетный специалист, гене-
гик-биоспеолог, как мы уже сказали, пытался объяснить явления пещер-
ной эволюции на основе неодарвинистского понимания эволюции: спон-
танная, во всяком случае случайная изменчивость, то есть мутации,
плюс отбор. Но позже это толкование показалось ему крайне невероят-
ным. Особенно смущала его гомозиготность регрессивных ’ мутаций. И
вот, не желая примкнуть ни к идеалистам-автогенетикам типа Ванделя,
ни «к ламаркистам, к которым он, конечно, причислял и дарвинистов,
Коссвиг стал толковать эволюцию троглобионтов с позиций «мутацио-
низма» (который отличается от мутационизма де Фриза и других «клас-
сиков» этого бесплодного направления) — накоплением индифферент-
ных мутаций в одном и том же направлении без всякого естественного
отбора, который, в крайнем случае, мог лишь элиминировать, устрани г о
неприспособленные, негодные мутанты.
Мутации, по Коссвигу (там же, стр. 229), — внезапные, редкие и
случайные изменения в генах. Однако он должен же был видеть, изу-
чая собственный материал, что в пещерных условиях «мутации» не ред-
ки, наоборот — изменения (наследственные) иногда чрезвычайно часты
и между край|ни»м1и вариантами в той же популяции встречаются все-
возможные переходы. Кроме того он сам признается, что чрезвычайную
генетическую изменчивость пещерных организмов нельзя объяснять
только гибридизацией: гибридизация, вполне возможно, участвовала, но
лишь дополнительно, как второстепенный источник изменчивости в соот-
ветствии с принципами менделизма.
Почему же генотип троглобионтов с течением времени прогрессив-
но изменяется в определенном направлении (там же, стр. 228)? Это, по
словам Коссвига, происходит благодаря мутациям.
Прибегая к своеобразному «мутационизму», Коссвиг ищет выхода
в полигенической интерпретации мутаций.
Коссвиг различает два тит а троглобионтов: рецентных, то есть ста-
новящихся, и «старых», уже определившихся. Только вторые отделены
от соответствующей предковой эпигейной (надземной) формы репродук-
ционным барьером.
Переход к подземной жизни совершается на основе «преадапта-
ций», под которыми Коссвиг понимает черты строения и функций над-
земных форм, могущие пригодиться в подземных условиях, по-видимо-
му, при относительно неглубокой перестройке. Такими '«преадаптация-
ми» могут быть: фотофобия, гигрофилня, стенотермия, преобладание
сенсорных органов, действующих без света, над органами зрения. Ре-
дукция органов, становящихся рудиментарными, вестигиальными, вы-
зывается по мнению Коссвига, вследствие случайных мутаций.
Согласно другой, «неодарвинистской», теории пищевые резервы, до-
ступные обитателям пещер, настолько скудны, что в такой среде могут
Выживать лишь формы со значительным замедлением метаболизма. Од-
нако, возражает Коссвиг, метаболические скорости троглобионтов могут
быть не только нормальными, но и повышенными — по крайней мере у
некоторых рецентных каверникол, например, у рыбы Anoptichthys.
«Неодарвинистскую» концепцию Коссвиг называет несколько упро-
щенной (стр. 236).
' Объяснение, которое дает теперь Коссвиг, оказывается достаточно
простым. Он думает, что надземный вид, проникший в подземный био-
топ, и‘благодаря своим «преадаптациям» там укоренившийся, испытыва-
ет’в этом биотопе процесс изменения, дающий терпимые, не вредные и
не полезные мутации. Сама по себе мутация не имеет позитивного се-
лективного значения, она может, самое большое, воспроизводиться, раз-
86
множатьюя, сохраняя .свой биологически нейтральный характер. Тут да-
вление отбора заменяется давлением мутационным, то есть -появлением
новых мутаций, аналогичных'мутациям, существующим и тоже терпимым
в своеобразных условиях подземной среды, ибо они не имеют селекцион-
ного значения. Мутационное давление позволяет повышенному числу
неаллельных генов способствовать увеличению числа нейтральных генов
подземной популяции. Мы, по словам Коссвига, стоим перед несомнен-
ным фактом: гомозиготным характером троглобионтов в отношении их
регрессивных признаков (Заметим, что это нисколько не удивительно:
если новые, регрессивные признаки возникли не путем скрещивания,
формы, выражаясь терминами генетики, могут быть лишь гом озигот-
ньими, а не гетерозиготными, ибо гетерозиготное состояние предполага-
ет совмещение в одном диплоидном наборе аллелей различного проис-
хождения). Тут Коссвиг делает замечание, заслуживающее нашего при-
стального внимайия: «Итак, хотя неодарвинистские концепции завели
нас в тупик, мы. считаем нужным, вопреки всему этому, искать генети-
ческие объяснения этих явлений» (стр. 236).
Коссвиг ставит вопрос: существует ли средство, которое дало 6oi
нам возможность открыть другие генетические механизмы и на их ос-
нове объяснить гомозиготность регрессивных признаков? Несколько
лет назад, как признается сам автор, он попытался дать такое объяс-
нение. Тогда он предложил теорию, согласно которой надо принять во
внимание прежде всего плейотропию генов. Плейотропия — явле-
ние хорошо известное генетикам. Оно означает свойство гена влиять
не на один только признак, а на многие признаки, которые могут даже
быть независимыми один ит другого. Итак, тогда Коссвиг предполагал,
что тот же ген, который редуцирует in игм е н т а ц.и ю, равным об-
разом влияет и на плодовитость, но только в положительном смысле.
Соответственно этому носители регрессивного гена уже обладали бы
положительным признаком в отборе, и они размножались бы с более
значительным успехом, чем остальные особи популяции, чис-
ленность которых в ходе поколений постепенно
сокращалась бы. Следовательно, эволюция в сторону депигмен-
тации вела бы к росту численности депигментированных особей за счет
численности пигментированных.
Коссвиг подчеркивает, что структуры, подвергающиеся регрессии,
представляют огромнейшее разнообразие: глазки планарий, сложные
глаза насекомых и глаза ры§ резко различаются одни от других как
морфологически, так и функционально. Поскольку регрессия проявляет-
ся у животных форм до такой степени различных, пришлось бы предпо-
ложить. что гены, ответственные за редукцию этих органов, обладают в
каждом отдельном случае позитивным селекционным
значением в силу плейотропии. Но ведь такая гипотеза, как
признается Коссвиг, чрезвычайно мало вероятна (там же, стр. 237).
Различия, о которых мы только что говорили, обнаруживаются в .составе
пигментов троглобионтов разных таксономических групп. Меланины—
производные тирозина; с другой стороны, темные пигменты декапод и
амфипод суть каротиноиды. В высшей степени невероятно, что
потеря этих пигментов в каждом отдельном случае
вызывалась системой генов плейотромического эф-
фекта, прямо или косвенно влияющих на скорость
размножения в положительном смысле. Заметим, что и
без этих действительно убедительных соображений Коссвига, надуман-
ность его прежней теории представляется нам ясной. В самом деле, его
87
теория плейотропии кажется какой-то соломинкой, за которую ортодок-
сальный генетик ухватился только для того, чтобы обойтись без уступок
толкованиям, не совместимым с постнеодар1винизмом. Почему регрессив-
ные признаки, раз1вивающиеся в пещерных условиях, должны обязатель-
но плейотропически сопровождаться повышением плодовитости? И Кос-
свиг совершенно справедливо заявляет (стр. 237), чго мы должны также
отбросить эту инспирированную «неодарвинизмом» гипотезу, которая ос-
нована на идее гетерозиса. Ведь гетерозис может играть важную роль
лишь в случае гетерозиготности, троглобионты же обнаруживают высо-
кую степень гомозиготности в своих регрессивных признаках.
Изучение полученных в 1933 году экземпляров слепой |рыбы Sty-
gicola dentatus (Brotulidae) из пещер о. Кубы, с чрезвычайно из-
менчивой окраской от явно пигментированной до белесой поверхности
тела, побудило Коссвига выдвинуть гипотезу, что такал изменчивость
была терпимой в подземных условиях и что существование всех пере-
ходов от совершенной пигментации до полной депигментации зависит
от влияния многих генов, действующих в одном и том же направлении.
Весьма значительная изменчивость установлена у Asellus aquaticus тз
ьекоторых пещерах Югославии. Изучая популяции этого членистоно-
гого, Коссвиг наблюдал все переходы от нормальной окраски надземной
формы до совершенной прозрачности в разных местах надзем-
ной реки. Существует некоторая корреляция между степенью депиг-
ментации и степенью редукции глаза (стр. 251). Совершенно слепы
лишь особи сильно депигментированные (рис. 4). Коссвиг приводит
замечательные случаи асимметричной редукции глаз: у одной и той же
особи правый и левый глаз дегенерированы не в одинаковой степени:
один больше, другой меньше. Это, по-нашему мнению, один из нагляд-
ных примеров очень резкой, бурной изменчивости пе-
щерной популяции. Среди представителей Asellus с нормально
развитыми глазами встречается довольно много особей с ослабленной
пигментацией, до практически совершенно обесцвеченных; окраска же
глаз варьириет от красной до фиолетовой и черной.
Еще выше по течению той же подземной реки, всего в нескольких
километрах от предыдущего местообитания, живет другая подземная
популяция: поверхность тела у особей этой популяции совершенно де-
пигментирована, а редукция глаз представляет картину колоссальной
изменчивости.
Коссвиг упоминает еще одну популяцию того же самого вида. Тут
г.се особи депигментированы, а строение глаз в большинстве случаев
чрезвычайно редуцировано.
Таким образом, разные популяции Asellus aquaticus, живущие в од-
ной и той же области, совсем неподалеку одна от другой, обнаруживают
разнообразнейшие степени изменчивости регрессивных признаков. Это,
очевидно, дало некоторым биологам повод говорить об ортогенети-
ческой дегенерации видов, ведущей к их вырождению и выми-
ранию. Такое объяснение, как мы видели, для Коссвига неприемлемо, —
сн ищет действительно научного, материалистического толкования этих
процессов.
Не менее примечательны результаты изучения рыб из пещер Куева
Чика в центральной Мексике. Они тесно связаны с надземной рыбой из
той же области — Astyanax mexicanus. Эти пещерные рыбы выделялись
обычно в особый род Anoptichthys и описывались как Anoptichthys ant-
robius (из пещеры Пачон) и Anoptichthys jordani (пещера Чика). На ос-
нове чисто морфологических признаков можно было бы описать еще
88
много видов этих пещерных рыб. Популяция, обитающая в пещере Чи-
ка, представляет колоссальное разнообразие по своим регрессивным
признакам. Некоторые особи оказались пигментированными и имеющи-
ми нормально развитые глаза; другие, наоборот, представляют класси-
ческий пещерный тип. Между этими двумя крайностями наблюдаются
переходные формы, у которых глаза редуцированы то более, то ме-
нее, а пигментация обнаруживает весьма разнообразные стадии. Две
особи, представляющие один и тот же фенотип, редки. Некоторые, тож-
дественные по пигментации, очень сильно отличаются друг от друга по
строению глаз. Вог/реки мнению некоторых систематикой, «Anoptichthys»
нельзя» по Коссвигу, считать видо-м, отличающимся от «Astyanax»:
Гранд-Дом (слева) и Черной Пещеры (справа), в Югославии. Величины выражены в
процентах. А — депигментированные особи; В—очень слабо пигментированные особи;
С—слабо пигментированные особи; D — хорошо пигментированные особи; Е — особи
очень сильно пигментированные- Для каждой степени пигментации показано
пропорциональное распределение случаев слепоты и различных окрасок глаз.
По Коссвигу
это скорее происшедшая от последнего экологическая раса, что подтве-
рждается и интерфертильностью обеих форм. Таким образом между эти-
ми двумя зоологическими типами, по словам Коссвига, еще не возник
сексуальный барьер, несмотря на большие морфологические и физиоло-
гические различия. После обнаружения популяции, населяющей пещеру
89»
Чика, были найдены и другие популяции, что дало систематикам повод
к описанию двух новых видов.
Все эти пещерные харациды живут на относительно небольшой
площади, представляющей, вероятно, остаток прежней гидрографичес-
кой сети. Наибольшую изменчивость обнаруживает популяция пеще-
ры Чика, самой близкой к надземному течению системы. Другие попу-
ляции (Сабинос, Тинайа, Пачон) состоят из настоящих, типичных тро-
глобионтов, депигментированных и слепых. Произведенное Коссвигом
скрещивание рыб из пещер Сабинос и Пачон, с одной стороны, и над-
земной Astyanax mexicanus, с другой, дало поколение жизнеспособных
гибридов с нормальной плодовитостью. Как и следовало ожидать, эго
поколение представляло однообразный фенотип. Зато следующее по-
коление, F2, содержало разнообразнейшие фенотипы и обнаружило из-
менчивость, еще более высокую, чем популяция пещеры Чика. Коли-
чественные признаки (депигментация, диаметр глаза и др.) обнаружи-
вали все стадии перехода и все формы рекомбинации признаков.
Коссвиг полагает, что период инстабильности (быть может, этот
период соответствует расшатанности наследственности — явлению, ко-
торое отмечалось генетиками и селекционерами как у нас, так и за ру-
бежом) совпадает с началом регрессивной эволюции (стр. 254). Он
думает, что популяция в ряде последовательных поколений прогрессив-
но достигает высокой степени гомозиготности в одной и той же филети-
ческой линии. Так понимает теперь Коссвиг эволюцию регрессивных
форм и развитие у них гомозиготности; по регрессивным признакам.
Коссвиг вполне допускает, что гибридизация может способствовать
усилению изменчивости в процессе эволюции пещерных форм. Но в то
же время он резонно подчеркивает, что усиленную изменчивость, столь
широко распространенную у каверникол, нельзя толковать исключи-
тельно как результат гибридизации. Но это рассуждение вплотную
подводит нас к вопросу: чем же в таком случае можно объяснить столь
необыкновенно бурную генетическую изменчивость при переходе жи-
вотных к пещерному образу жизни, точнее — откуда берутся возника-
ющие тут в массовом количестве мутации? Ведь такого возникновения
«случайностных» мутаций и такого усиленного их накопления никто
нигде не замечал у непосредственных эпигейных предков пещерных жи-
вотных. Неужели тут можно уклониться от вывода, что мутирование
вызывалось особенными условиями, непривычными, необычными для
надземных анцестральных видов?
В своей последней работе Коссвиг (Kosswig, 1965) считает возмож-
ным толковать так называемую регрессивную эволюцию как процесс,
развертывающийся на основе «широкой полигении», зависящий, глав-
ным образом, от того, что в данных экологических условиях лишь не-
которые мутации являются нейтральными, индифферентными в отно-
шении выживаемости своих носителей. Но слабость этого толкования,
которое Коссвиг называет мутационистским, противополагая его «не-
одарвинистскому» (постнеодарвинистскому), заключается в том, что та-
кой «мутационизм», резко подчеркивающий мутационное давление, туг
занимающее место давления отбора, оставляет нетронутой загадку
причины усиленного мутирования. Нам кажется, что Коссвиг близок к
пониманию этого слабого места его новой теории. Об этом свидетель-
ствует одна фраза в заключительной части его интересной статьи (стр.
255): «Прежде чем принять интерпретацию, которая учитывает какой-
либо селективный фактор или более или менее ясно выраженную ла-
маркистскую тенденцию, следует, кажется, доказать, что эти сообра-
90
жения одновременно экологического и этологического порядка ошибоч-
ны» (под «этими соображениями» автор подразумевает свою новую
гипотезу регрессивной эволюции троглобионтов). Но недостаточность
упомянутых соображений Коссвига едва ли подлежит сомнению. И
дальнейшее усовершенствование воззрений этого исследователя, мы ду-
маем, должно заключаться в признании того бесспорного факта, чго
усиленное мутирование рецентных троглобионтов зависит от новых для
них экологических условий.
Специалистов-биоспеологов озадачивает неимоверно интенсивная
изменчивость многих животных, обитающих в пещерах. С этой повы-
шенной изменчивостью как-то связана весьма быстрая эволюция таких
организмов. Обычно даваемые объяснения этого процесса представля-
ются нам недостаточными.
Прежде всего мы должны были отвергнуть гипотезу, развиваемую
крупным биоспеологом А. Ванделем (Vandel, 1964). Суть этой гипотезы
сводится к тому, что возникновение пещерных животных есть результат
процесса общей деградации всех филогенетических линий органическо-
го мира, ведущей к редукции и исчезновению различных органов. Пер-
вая фаза филогенеза выражается прогрессивной эволюцией, а
последняя — регрессивной, или геронтократической, эволюцией, кото-
рая завершается смертью данного органического типа (там же, стр.
547). Регрессивные формы должны были искать приюта в подземной
среде, особенно в пещерах. Только там еще могли продолжать свое су-
ществование «филетически старческие» группы.
Сторонники «синтетической» теории эволюции решительно отверга-
ют такие объяснения, и в этом с ними нельзя не согласиться. Свое мне-
ние о гипотезе А. Ванделя мы высказали в книге о состоянии эволюци-
онного учения (1966, стр. 72), и едва ли следует возвращаться здесь к
этому вопросу. Идея Ванделя связана с концепцией «филетического
старения», или броккизма, о котором мы писали в целом ряде наших
работ. Это — чисто идеалистическая теория, совершенно неприемлемая
для естествоиспытателей, ищущих рационального объяснения явлений
живой природы. Тем не менее многие биоспеологи склонны считать
чуть ли не главной причиной возникновения троглобионтов «бегство»
некоторых форм из освещенной эпигейной среды в темную подземную.
Так, по В. Людвигу (Ludvig, 1942, стр. 453), вполне вероятно, что не-
пигментированные животные, чувствуют себя в темноте несколько луч-
ше, чем при свете. При прочих равных условиях светобоязнь у них нес-
колько сильнее, чем у пигментированных. Поэтому пигментированные
должны были вселяться в пещеры несколько реже, а кроме того, должны
были (выходить обратно на свободу быстрее .и чаще, чем лишенные пиг-
ментации наружных покровов. Если же с потерей пигмента и глаз со-
пряжена более сильная светобоязнь, то такие признаки пещерных жи-
вотных, как обесцвечивание кожи и утрата зрения, должны накоплять-
ся в пещерных популяциях за очень короткое время (стр. 454). Это объ-
яснение, как нетрудно видеть, довольно близко к концепции Ванделя.
По Людвигу, дело тоже сводится к «уходу» неудачников в подземелье.
Он не объясняет также, почему у очень многих, весьма разнообразных
беспозвоночных и позвоночных животных признаки регрессивного раз-
вития, как депигментация и анофтальмия (отсутствие глаз), должны
быть интимно сопряжены со светобоязнью. Да и вообще трудно пред-
ставить себе, как целый комплекс пещерных признаков, включая пол-
ную слепоту и полное обесцвечивание покровов, возникал © надзем-
ных условиях в непосредственной близости к пещерам. Довольно сход-
91
но с этим объяснение, которое дает О. Штребель (Strebel, 1964, стр.
342 происхождению пещерных признаков у некоторых членистоногих
— коллембол (хвостоножек, из подкласса бескрылых насекомых). Он
допускает, что, в связи с потеплением после ледникового времени, эта
бескрылые насекомые стали подыскивать пещеры, где они на-
ходили условия, соответствующие их потребностям. По Штре-
белю (стр. 343), еще эпигейные предки пещерных животных приобрели
мутации, которые делали их способными жить в мелких углублениях и
ь пещерах. Этот вариант гипотезы «подыскивания биотопа» едва ли
многим лучше других. Во-первых, пещеры, населенные троглобионта-
ми, встречаются и за пределами области четвертичного оледенения, во-
вторых, случайные мутации никак не могли дать то разнообразие форм
и приспособлений, которые имеются у пещерных обитателей. Впроче^м,
гипотеза, поддерживаемая Штребелем, не носит финалистского харак-
тера и стоит на уровне постнеодарвинизма.
Самое слабое место всех постнеодарвинистских гипотез происхожде-
ния троглобионтов, по нашему мнению, — их неспособность объяснить
чрезвычайно бурную и интенсивную изменчивость этих обитателей пе-
щер, особенно в начальный период их эволюции (по проникновении их
предков в подземную среду), когда они дают нередко вспышку измен-
чивости.
Этим мы отнюдь не хотим умалить значение естественного отбора в
эволюции пещерных животных. Даже в развитии «регрессивных» изме-
нений, как депигментация покровов тела и анофтальмия, конечно, дол-
жен был постоянно участвовать отбор. Но он мог действовать там, где
была достаточная для этого изменчивость. Без естественного отбора не
могли развиться многие приспособительные особенности пещерных жи-
телей, — такие как удлинение конечностей и антенн и дополнительные
органы чувств {де Латтен, (Lattin, 1940), стр. 13; Штребел, (Strebel,
1964), стр. 340], у различных членистоногих или сильное развитие ман-
тии и возникновение на ее поверхности многочисленных папилл у слиз-
ней Troglolestes sokolovi (С. И. Левушкин и П. В. Матёкин, 1965, стр. 42).
Однако Коссвиг убедительно показал, что эволюцию пещерных оби-
тателей нельзя объяснить на основе «неодарвинизма», то есть постнео-
дарвинизма. Особенно важно, что к такому выводу пришел ученый,
вполне владеющий методом классической генетики и успешно его при-
меняющий к изучению биоспеологических материалов. Он установил,
что е данном случае большое разнообразие форм и высокую изменчи-
вость никак нельзя объяснить гибридизацией, которая в этом процессе
могла принимать лишь второстепенное участие. Типичные троглобион-
ты — гомозиготны по своим регрессивным признакам, и потому тут
нельзя усматривать последствия гетерозиса: ведь гетерозис может иг-
рать существенную роль лишь при гетерозиготном состоянии популя-
ций. Коссвиг производил гибридизацию пещерных животных и показал,
что в некоторых случаях во втором поколении происходит расщепление
ь соответствии с законами генетики. Но при всем желании остаться на
позициях-постнеодарвинизма и классической генетики он был вынуж-
ден, как мы видели, признать, что основным фактором эволюции трог-
лобионтов является «мутационное давление», а не давление отбора.
Однако Коссвиг не объясняет, почему это давление мутаций должно
быть столь мощным. Ведь, судя по его статье 1965 г., он понимает ге-
незис мутаций в духе классической генетики и постнеодарвинизма —
это спонтанные случайные изменения. В пещерах они должны были бы
происходить с такой же скоростью, как и вне пещер. Но если дело обг
92
стояло именно так, то откуда могло взяться мощное мутационное дав-
ление, которое, якобы, вызывало весьма быстрое формообразование?
Изменчивость многих пещерных животных поистине колоссальна!
Она иллюстрируется, между прочим, материалами, изученными Кос-
свигом, например, — разнообразнейшими формами редукции глаз у
представителей Asellus некоторых пещер Югославии, — из популяций,
живущих на небольших расстояниях одна от другой. В нормальных ус-
ловиях, то есть в обычной эпигейной обстановке столь разнообразные
формы изменчивости были бы, конечно, невозможны. Такая изменчи-
вость, действительно, могла вести развитие популяций в определенных
направлениях. Сторонники классической школы генетиков могут ска-
зать, что, имея столь богатое разнообразие форм в популяциях одной
пещеры или нескольких пещер, отстоящих на небольшом расстоянии од-
на gi другой, мы не можем уверенно различать в таком материале чи-
сто индивидуальную изменчивость и таксоны. Допуская, что тут при-
сутствует не один подвид или даже, быть может, не один вид, мы дол-
жны все же признать, что и в таком случае перед нами случай весьма
интенсивной изменчивости. Можно также подозревать, что здесь перец
нами не генетическая изменчивость, а только модификационная, ненас-
ледственная. Но, воздерживаясь от решительного ответа на такое заме-
чание, мы скажем определенно, что изменения, ведущие от зрячих форм
к совершенно слепым, нельзя считать сплошь, без исключения, моди-
фикациями в понимании классической генетики: в сериях таких изме-
нений должны обязательно присутствовать и наследственные.
Итак, в материале Коссвига и других биоспеологов нельзя не ви-
деть исключительно бурной генетической изменчивости. Такая измен-
чивость могла быть основой для столь быстрого формирования новых
1 аксонов — подвидов и видов.
Коссвигу принадлежит, по нашему мнению, большая заслуга: он
прекрасно показал несостоятельность постнеодарвинистского объясне-
ния причин эволюции пещерных форм. Он понял большое значение по-
вышенной изменчивости в образовании пещерных форм. Но он, нам ка-
ж.ется, снижает значение отбора, который действует в видообразова-
нии и троглобионтов, и, заговорив о повышенной изменчивости обита-
телей пещер, воздерживается от объяснения причин этой изменчивости.
При таком подходе к изучению биоспеологического материала он ос-
тавляет повышенную изменчивость троглобионтов неразрешенной за-
гадкой. Из его работы вытекает, что здесь, в пещерах, не дейсд1ВО1вал|И
никакие факторы изменчивости, кроме «случайности», которая, по воз-
зрениям постаеодарв!И1Н1исто1в, действует везде и всюду. Можно думать,
что Коссвиг просто не решился сделать неизбежный вывод о том, чго
особенные условия жизни в подземной среде резко ускорили процесс из-
менения особей и вели этот процесс в определенном направлении. Он
опасается дарвиновского «ламаркизма», но, как мы видели, уже не от-
крещивается от него.
Те животные, которые, попадая в подземную среду, там приживают-
ся, подвергаются действию совершенно необычных условий этой абер-
рантной среды и потому испытывают бурные изменения.
К числу самых примечательных из этих изменений принадлежат
обесцвечивание поверхности тела и редукция органов зрения. Такие из-
менения испытывают представители самых разнообразных групп живот-
ных — беспозвоночных и позвоночных. Эти изменения вызываются усло-
виями среды в пещерах. Нужно, пожалуй, большое мужество, чтобы
после десятилетий служения постнеодар1в<И1НИ,зму признать его несостоя-
93
тельность; это по существу и было сделано Коссвигом. Но, по-видимому,
необходимо еще большее мужество для того, чтобы признать факт нас-
ледственного изменения организмов под влиянием вновь возникших ус-
ловий среды.
Изменения, которые происходили у троглобионтов под влиянием
факторов среды и их изменений, были массовыми и направленными.
Они были более или менее постепенными: нельзя же думать, что исчезно-
вение глаз у рыб Astyanax совершилось внезапно от одного поколения к
следующему, — факты говорят против такой внезапности; нельзя ду-
мать, что нормально пигментированное животное столь же внезапно ста-
новилось совершенно обесцвеченным.
Нам приходится признать, что в данном случае вызванные экологи-
ческими условиями изменения организмов становились наследственны-
ми. Мы не знаем, каков интимный механизм такого наследственного из-
менения. Изучение этого вопроса слишком далеко от доступной нам лич-
но сферы научных исследований. Этим должны заниматься, по-нашему,
прежде всего сами генетики и работники молекулярной биологии.
Итак, биоспеологические исследования проливают обильный и яркий
свет на явления изменчивости организмов, тем самым оказывая огром-
ную услугу научному, дарвинистскому объяснению живой природы и ее
развития. Но рассмотренные в этой главе данные имеют и более близкое,
специальное, отношение к проблеме изменчивости ископаемых организ-
мов. Находки пещерных беспозвоночных прошлых геологических перио-
дов, правда, очень редки. Но в относительно недавнем геологическом
прошлом жило много пещерных позвоночных; особенно большой инте-
рес представляют для нас млекопитающие, которые проводили в пеще-
рах значительную часть своей жизни. Этот пещерный, или полупещер-
ный, образ жизни таких зверей наложил на них определенный отпеча-
ток. Сделанный н предыдущих страницах анализ фактов изменчиности
и развития нынеживущих беспозвоночных обитателей пещер поможет
нам осмыслить значение изменчивости живших там же млекопитающих,
что мы попытаемся сделать в одной из последующих глав.
МОРСКИЕ БИОТИПЫ, БЛАГОПРИЯТНЫЕ ДЛЯ ПОВЫШЕННОЙ
ИНДИВИДУАЛЬНОЙ ИЗМЕНЧИВОСТИ И СООТВЕТСТВЕННО
УСКОРЕННОЙ ЭВОЛЮЦИИ ОРГАНИЗМА
Общеизвестно, что в некоторых морских биотопах, вследствие отно-
сительного или полного обособления бассейнов от мирового океана и
возникновения в этих бассейнах своеобразных гидрологических режи-
мов, возникали условия, особенно благоприятные для изменчивости и
быстрой эволюции органических форм. Яркие примеры таких процес-
сов дает история жизни в некоторых морских бассейнах позднего па-
леозоя и неогена как в Евразии, так и в Южной Америке. Эти явления
были освещены в ряде классических трудов Н. И. Андрусова (1961,.
1963, 1964, 1965). О них же писали выдающийся советский геолог-па-
леонтолог В. П. Колесников (1939, 1941, 1949) и другие советские уче-
ные. Их общий обзор был дан в недавно опубликованной нашей книге
(1969).
В последующих главах этой книги мы довольно подробно рассмот-
рим характерные примеры усиленной изменчивости многих пластин-
чатожаберных и б|рюхонопих моллюсков, живших в неогеновых бассей-
нах Паратетиса. Эти процессы вызывались преимущественно отклоне-
нием солености от нормальной морской, что вызывало значительные из-
94
менения в биоценозах. Анализ подобных случаев, надеемся, даст нам
возможность в немалой степени разобраться в их причинах; а здесь мы
ограничимся лишь замечанием, что относительно быстрое изменение
гидрологического режима вызывало вымирание многих форм морских
организмов, усиление изменчивости и ускоренную эволюцию морских
форм, выдержавших это изменение, и усиленную изменчивость и эво-
люцию форм пресноводного происхождения, для которых эта среда ста-
ла доступной вследствие значительного понижения солености вод.
Нам могут сказать, что только что упомянутые нами случаи явля-
ются исключительными и что в «нормальных» условиях полносоленых
морей, пресноводных водоемов и на суше, нет предпосылок для массо-
вой направленной изменчивости. Такое утверждение было бы, однако,
неверным. В полносоленом море изменчивость донных животных, прав-
да, значительно менее интенсивна, чем в более или менее замкнутых
водоемах с соленостью, существенно отклоняющейся от нормальной. Но
ьсе-таки на отдельных более или менее обширных участках подобного
моря могут возникать условия, особо благоприятные для усиления мас-
совой направленной изменчивости, а такая изменчивость дает материал
для видообразования путем отбора. Подобные участки более или менее
обособлены от окружающего пространства, и поскольку условия жизни
на них несколько отклоняются от обычных, господствующих в других
частях данного моря или океана, на таких участках может иметь место
относительно интенсивная и направленная изменчивость, охватываю-
щая заметную часть популяции того или иного вида. Вопрос о том, как
возникает изоляция отдельных участков, моря, изучен довольно слабо.
В некоторых случаях для обособления достаточно наличия барьера в
виде подводной возвышенности. Даже в открытом заливе моря солен-
ность может заметно отклоняться от нормальной, а это в свою очередь
обусловливает направленную изменчивость обитателей дна. Если вдоль
берега идет холодное течение, которое следует прямо, не заворачивая
в существующий здесь залив, то можно представить себе, что и в от-
сутствии морфологического барьера популяции некоторых донных форм,
обитающих в заливе, будут отрезаны от прилежащего моря. Средняя
температура в заливе может оказаться более высокой, чем на соседних
участках того же океана. Впадающая в относительно небольшой за-
лив большая река может заметно влиять на соленость его вод и на ха-
рактер грунта, что с течением времени окажет влияние на изменчи-
вость населяющих дно этого залива животных.
От воображаемых случаев перейдем к конкретным примерам. Об-
ратимся к интересным данным, которые были сообщены И. И. Пуза-
новым (1954) относительно изменений организмов, попавших в лиман-
ные условия. 1931 год был годом низкого стояния уровня лиманов и
сильного осолонения Черного моря; после началось опреснение, дос-
тигшее максимума в 1941 году, когда уровень Хаджибейского лимана
был более чем на 2 м выше уровня моря, а соленость соответствовала
приблизительно солености Одесского залива. Взрыв дамбы в 1941 го-
ду привел к установлению на целых два года связи лимана с морем. За
это время в лиман проникли морские организмы, и некоторые из них
прекрасно прижились там и обильно размножились. Из моллюсков та-
кими оказались Cardium, Mytilus и Mytilaster. И. И. Пузанов и его
сотрудники занялись изучением вопроса, насколько успели измениться
морские животные, попавшие в лиман, после 1941 года все более и бо-
лее осолонявшийся и в настоящее время имеющий соленость втрое
большую солености Одесского залива Черного моря. Оказалось, что не-
95
которые морские животные, проникшие в лиман, и там прижившиеся,
испытали за совершенно ничтожный промежуток времени заметные
морфологические и физиологические изменения. Так, камбала — Pleu-
ronectes flesus luscus Pall. несколько измельчала (особенно —
самцы), у нее изменилась окраска верхней стороны тела, рыба стала
заметно более высокотелой и короткоперой (все плавники укороти-
лись), брюшные плавники сильно подались вперед, увеличилась длина
рыла, увеличилось число плавниковых лучей в грудных плавниках, и
т. д. Кроме того, рыба достигает поло1воз(релост1И уже в начале (второго
года жизни, а не в трехлетием возрасте, как ib море. Наконец, весьма
примечательна отмеченная автором «огромная мутабильность» — «весь-
ма характерный признак для глоссы (камбалы), попавшей в необычные
для нее условия», увеличение размаха индивидуальных колебаний раз-
личных морфологических признаков. Как бы ни понимал это явление
сам автор, — а он склонен строго придерживаться норм классической
генетики, от которых отошли или отходят теперь самые правоверные се
представители, тут перед нами яркий пример расшатанной наследствен-
ности и бурной изменчивости вследствие изменения некоторых условий
существования. Некоторые изменения претерпели и другие животные,
прижившиеся в лимане, — бычки, мидии, крабы. И. И. Пузанов (там же,
стр. 30) признает несомненным, что «ответы организма на измененную
среду» усиливаются из поколения в поколение, хотя п думает, что такие
«ответы» безусловно не наследственны (там же).
По поводу изменений форм, попавших в Хаджибеевский лиман и за
немного лет сильно изменившихся, мы должны сделать еще одно заме-
иание. Нельзя валить в одну кучу все такие водоемы как лиманы, ла-
гуны и пр. Бывают лиманы и лиманы; бывают лагуны и лагуны. Если
речь идет об относительно крупных лагунах, или об озерах-морях, су-
ществовавших в течение геологически ощутимых отрезков времени
(очень долго с точки зрения масштабов истории человеческого общест-
ва — сотни тысяч лет, даже миллионы лет), то тогда развившиеся *
них формы — потомки морских животных — особенно бентонные мол-
люски, окажутся настолько сильно пригнанными к специфическим ус-
ловиям данного озера-моря, данного солоноватоводного или пере-
осоленного водоема, или данной лагуны, что этим формам будет очень
трудно прижиться в водах какого-либо морского бассейна. В таком слу-
чае переход лагунной формы в море мало вероятен, он возможен лишь
как исключительное явление. Более того, нелегко представить себе воз-
можность того, что формы, развившиеся в одном замкнутом бассейне,
свободно приживутся в другом замкнутом бассейне, ибо гидрологические
условия каждого такого бассейна строго специфичны для него и обычно
существенно отличаются от гидрологических условий других таких во-
доемов, хотя бы даже они были расположены на небольшом расстоя-
нии один от другого. Иное дело — небольшие, прибрежные участки мо-
ря, иногда более или менее обособляющиеся от него, но только нена-
долго, то есть не отделенные от открытого моря относительно прочны-
ми^ и долговечными барьерами. Условия, господствующие на таких
участках (которые иногда представляют собой заливы или даже лима-
ны, лагуны и пр.), могут не столь резко отличаться от гидробиологи-
ческих условий моря (или, лучше, по своим гидрологическим условиям
такие участки заливы, лагуны, даже лиманы и пр. могут не столь силь-
но отличаться от моря). Достаточно относительно небольшого откло-
нения от обычных в данном море условий, например, солености, или
температуры, и несколько повышенной кормности, для того, чтобы тут
96
произошел отсев многих форм, живущих на соседних участках моря, и
чтобы в то же время существенно усилилась изменчивость прижив-
шихся форм, имеющая массовый и определенно направленный харак-
тер.
При очередном воссоединении с морем формы, начавшие обособ-
ляться или даже обособившиеся от родительских видов, могут выйти за
пределы залива или лимана, попасть в открытое море и там прижиться.
Аналогичные факты можно привести в огромном количестве из ис-
тории самых разнообразных групп ископаемых беспозвоночных кайно-
зоя, мезозоя и палеозоя. Так, была отмечена повышенная изменчивость
некоторых форм рода остракод, — раковинчатых ракообразных из ниж-
некаменноугольных отложений Волго-Уральской области, причем ав-
тор, изучавший это явление, связывает таковое с расшатанной наследст-
венностью в условиях начавшегося опреснения бассейна Камско-Ки-
нельокой В1пади:ны и сменой карбонатного осадконакопления терриге-
новым обломочным (М. М. Грачевский, 1958).
Таким образом, палеобиология может и должна изучать явления
изменчивости органических форм и причины явлений, в этом отноше-
нии существенно дополняя исследования генетиков и селекционеров.
Палеобиологический материал показывает, что значительная массовая
направленная изменчивость организмов на более или менее обширных
участках, вызванная изменением условий существования, — довольно
часто наблюдаемое явление. Наследуемость же таких изменений при
нынешнем состоянии палеобиологических знаний особенно убедительно
и наглядно иллюстрируется историей моллюсков замкнутых и полузам-
кнутых бассейнов, о которой мы уже говорили.
Относительно медленное изменение абиотических условий водной
среды морей, связанных с мировым океаном и имеющих соленость
нормальную или близкую к нормальной, вполне согласуется с относи-
тельно медленной эволюцией морских животных, в частности беспозво-
ночных, особенно —бентонных моллюсков. Однако даже в огромных
морских водоемах могут возникать и в действительности возникают
участки, где абиотические условия несколько отклоняются, хотя бы на
короткое время, от нормальных морских бассейнов. Такие изменения
стимулируют изменчивость. Вообще же условия, вызывающие и нап-
равляющие изменчивость у обитателей полносоленых морей, изучены
еще очень слабо.
ОБЩИЕ ВЫВОДЫ ОБ УСЛОВИЯХ СРЕДЫ, ВЫЗЫВАЮЩИХ ПОВЫШЕННУЮ
ИЗМЕНЧИВОСТЬ
Мы видели, что обстановка, благоприятствующая усиленной из-
менчивости организмов, может возникать в биотопах различных типов.
Изменения физико-географические и биоценотические могут создавать
аберрантные экологические условия на различных участках суши — в
надземвых и подземных биотопах. Мы видели, что в таких случаях
многие нынеживущие формы испытывают процесс необычайно буркой
изменчивости, которую нельзя толковать как только модификационную.
Такая интенсификация изменчивости охватывает более или менее зна-
чительные части соответствующей популяции и не может быть названа
спонтанной, хотя в то же время нельзя отрицать и ее случайный харак-
тер. В других случаях (например, в морях нормальной солености) no-
т. Л. Ш. Давиташвили 97
вышение изменчивости, во многом зависящее не только от физико-гео-
графических преобразований, но и от изменения биотических условии
среды, может быть, не столь резким, но все же достаточным для одно-
временного появления однозначных отклонений не только у разрознен-
ных единичных особей, а у относительно большой доли популяции дан-
ного вида.
Глава четвертая ,
ЯВЛЕНИЯ ИЗМЕНЧИВОСТИ У ИСКОПАЕМЫХ ФОРАМИНИФЕР
Исследования фораминифер, ископаемых и современных, обнару-
живают много случаев расшатывания наследственности и морфологи-
ческих отклонений, причем такие изменения нередко оказываются тес-
но связанными с некоторыми изменениями в абиотических и даже био-
тических условиях среды.
Э. Болтовской изучал комплексы фораминифер, населяющих бухту
Сан-Блас и залив Сан-Хорхе (Атлантический океан, южное побережье
Аргентины). Фораминиферы залива Сан-Хорхе и в особенности бухты
Сан-Блас обнаруживают явную тенденцию к малорослости и в некото-
рых случаях их раковинки совершенно теряют ребристость наружной
поверхности. У отдельных форм наблюдаются значительные отклонения
от нормального строения. Эти изменения носят характер индивидуаль-
ной изменчивости, зависящей от условий существования, — по-видимо-
от недостаточного питания (Болтовской, Е. Boltovskoy, 1954). То же
мнение Болтовской высказывает и в своих более поздних работах (нап-
ример, Boltovskoy, 1963, стр. 43).
Аналогичные явления отмечались и на ископаемом материале. Так,
Л. Оньибен, (L. Ogniben, 1958), изучавший фораминифер из третичных
(галеогено1вых и .миоценовых) отложений Казерты, в Италии, (наблюдал
малорослость и аномалии в строении раковинок этих корненожек. При-
чиной этих явлений он склонен считать интенсивность поступления обло-
мочного материала с суши в море. По Майерсу, (В. Н. Myers, 1936),
уменьшение величины организма у фо)ра1М'И1нифер зависит прежде всего
от ухудшения условий питания. В сублиторальной зоне условия питания
гораздо лучше, чем .в литоральной и соответственно раковинки Elphi-
dium crispum в первой их этих зон значительно крупнее, а число камер у
них больше, чем во второй зоне. Близ Плимута (Майерс, Myers, 1943)
весной, когда пищи много, образуются длинные и узкие камеры. Позд-
ней осенью, когда количество пищи менее обильно, камеры короткие и
широкие, <С. Г. Лаликер (С. G. ЬаИскег, 1948) рассмотрел вопрос о кар-
ликовости у фораминифер. Он высказал предположение, что возникно-
вение карликов в этой группе организмов зависит от воздействия внеш-
них условий: количества пищи, температуры воды, химического состава
и физических условий воды, концентрации ионов водорода. Он про-
анализировал данные о влиянии колебаний этих факторов на размеры
и форму организма у ныне живущих простейших. Лаликер считает
возможным, что некоторые «карликовые фауны» простейших возникали
вследствие колебания концентрации иона водорода в воде. В глинистом
сланце одной толщи олигоцена прибрежной Равнины Мексиканского
залива Heterostegina texana тонкостенны и малорослы — около 1,5
9?
мм в ширину, в то время как та же форма, встречающаяся в известко-
вистом песчанике, имеет в ширину около 4,3 мм.
По Р. Саид (R. Said, 1953) карликовость у этих организмов объяс-
няется замедлением роста, которое вызывается низким метаболизмом.
Понижение же метаболизма зависит от неблагоприятных условий, в
частности — недостатка растворенного кислорода. Г. Колом (G. Co-
lom, 1959) сообщил о том, что в провинции Кастельон (Испания) в
пресноводных отложениях был обнаружен современный вид форами-
нифер Discorinopsis aguayoi. Род этот близок к Discorbis. При этом
большинство особей — малорослые, обычно со слабо кальцини-
рованными раковинками. Нормальной средой обитания для этого
вида является морская среда. Автор думает, что эта форма есть
реликт, /приспособившийся к пресноводному биотопу. В. Покорный
(V. Рокоту, 1958, стр. 125) находит, что форам1ин1иферы не очень чувст-
вительны к недостатку кислорода, но при значительном недостатке
кислорода может понижаться метаболизм, что вызывает появление кар-
ликовых особей.
Ф. Беттенштедт (F. Bettenstedt, 1962), думает, что неблагоприят-
ные условия, например, недостаток кислорода, вызывает карликовость
фораминифер.
Многие другие исследователи также считают основной причиной
уменьшения размеров раковинок ухудшение экологических условий
[В. А. Крашенинников, 1948; Р. Тодд (R. Todd, 1957); А. В. Фурсенко
1959; Курт (G. Kurg, 1961 и другие].
Иногда, однако, высказывается и противоположное мнение, соглас-
но которому неблагоприятные условия жизни вызывают появление осо-
бей более крупных размеров. В этом смысле высказывались Дж. С.
БрэДшоу (j. S. Bradshow, 1957), Дж. Гантер ( G. Gunter, 1957), Р. Р.
Лэнкфорд (R. R. Lannford, 1959), Д. Никол (D. Nicol, 1944), Ф. Б.
Фледжер (F. В. Phleger, 1960) и другие. Такие суждения так же, как
им противоположные, подкрепляются ссылками на фактический мате-
риал. Э. Болтовской (BoltovsKoy, 1965, стр. 93) полагает, что эти проти-
воположные мнения можно согласовать между собой следующим обра-
зом: «Более крупные экземпляры возникают в несколько неблагопри-
ятных условиях, когда они достигают зрелости немного позже нормы;
средние экземпляры, так же, как и несколько менее крупные, соответ-
ствуют оптимальным условиям; карликовые экземпляры с признаками
угнетенной фауны образуются тогда, когда одно или несколько усло-
вий значительно хуже, чем нормальные. В настоящее время трудно ре-
шить, до какой степени правильно это предположение, поскольку эта
проблема в высшей степени сложна и требует дальнейших детальных
исследований». С этим последним выводом нужно, очевидно, согласить-
ся.
В общем исследования современных и ископаемых фораминифер
показывают значительную индивидуальную изменчивость фораминифер
и к тому же зависимость этой изменчивости от условий среды. Имеют-
стя прямые наблюдения /над изменчивостью. Г. Фризе (Н. Friese, 1951)
изучал изменчивость ископаемых форм родов Elphidium, Robulus,
Nonion и некоторых других. Расположив переходные экземпляры в
непрерывные ряды, он смог проследить изменения многих морфологи-
ческих .признаков. Сведения об изменчивости фораминифер сообщают-
ся многими другими учёными, в частности В. А. Крашенинниковым
(19586) и X. М. Саидовой (1960а). Очень значительные изменения
100
возникали в культурах Cibicides lobatplus и Gipsina, которые
изучал К. Г. Нихольм (К. G. Nyholm, 19Q1 и 1962) в условиях, близких
к нормальным ‘ (табл. I).
Болтовской (BoltovsKoy, 1965) отмечал, что необычайно высокая
изменчивость раковинки у фораминифер ставит едва ли преодолимые
препятствия к их использованию для стратиграфических целей. С эти-
ми фактами следует считаться всякому специалисту, изучающему вопро-
сы геохронологии на основе фораминифер. Тут мы очевидцу встре-
чаемся с исключительно интенсивной внутрипопул$щионной изменчи-
востью.
Другое дело, насколько вариации в строении раковинок форамм-
нифер я)вляются наследственными. Различение наследственной, мута-
ционной, и ненаследственной, -модификационной, изменчивости у фора-
минифер — нелегкое дело. Попытки такого различения делались неко-
торыми специалистами. Но нам кажется? что методологический подход
к таким исследованиям иногда оказывался ненадежным. Например,
Беттенштедт (F. Bettenstedt, 1962), исследования которого более под-
робно мы рассмотрим несколько дальше, склонен думать, что изменен-
ные признаки, в начале замечаемые лишь у единичных экземпляров, а
затем становящиеся все более и более распространенными, частыми,
должны представлять собой мутации. И наоборот, признаки, сразу ох-
ватывавшие значительную часть данной «популяции, являются, по его
мнению, заведомо модификационными, ненаследственными. Нам же ка-
жется, что о и ©наследственном характере новых признаков можно пред-
полагать лишь в том случае, если в последующем, в немного более мо-
лодых отложениях, эти признаки исчезают. Беттенштедт различает да-
же длительные модификации у ископаемых форм, если измененный
признак удерживается в более поздних популяциях. Однако мы не зна-
ем, что дает автору право сделать такой вывод.
Если признак, несколько измененный в популяции данного геологи-
ческого уровня, последовательно усиливается в более молодых слоях,
то мы не видим возможности решить, что данное изменение является
модификалрьнным. Вообще надо признать, что применение генетичес-
кой терминологии требует от палеобиолога большой осторожности. В то
же время мы не можем понять, почему постепенно нарастающий и
постепенно распространяющийся признак является обязательно му-
тацией в отличие от модификации. Тут исследователем, видимо, руко-
водит убеждение, будто всякое «по-настоящему» наследственное изме-
нение должно быть мутацией в прямом смысле слова, и что такая мута-
ция является спонтанной и не должна зависеть от воздействия условий
среды.
О. И. Джанелидзе (1953, стр. 138) приводит многочисленные слу-
чаи аномального строения и цланктонны^ и бентрнных форамицифер.
Эти отклонения от нормального строения автор объясняет из-ме-
нениями гидрологического режима. Hq материале из аредтдего
миоцена О. И. Джанелидзе (1968) и из сармата Л. С. Майсура'дзе
(1968) показывают, что повышенная изменчивость многих форамини-
фер зависела от их жизни в условиях своеобразных, почти обособлен-
ных или замкнутых бассейнов. Это касается, например, тарханс-кой
Quinqueloculina boueapa d'Orb., Sigmojlina tenujs Gzizek, мцргцх чркрнн-
ских форм и сарматских Quinqueloculina ц Porospnonjop.
Интересные сведения о повышенной изменчивости некоторых фр-
рами1Н1ифер из третичных отложений территории СССР были рпубди-
кованы О. В. Окропилидзе (1956, стр. 338—341). Он отмечает заметные
морфОлори^ескме отклонения в строении раковинок глобигерин, раз-
витие дополнительных аномальных камер, зависящие от каких-то фак-
торов среды, возможно, — от условий питания или температуры.
Не менее интересны наблюдения М. Рушелли (М. Ruschelli, 1966)
над фораминиферами аквитанско-гельветских отложений (нижний ми-
оцен и нижняя часть среднего миоцена) Асти, в Италии. У некоторых
видов автор отмечает аномальное нарастание раковинки, повышенную
изменчивость в форме и размерах, нарушение порядка в расположении
камер и т. д., и все эти отклонения от -нормального строения он припи-
сывает действию необычных и неблагоприятных для фораминифер ус-
ловий как на дне, так и на поверхности воды. Малорослость некоторых
фораминифер из плиоцена одной местности близ Александрии, в Север-
ной Италии, П. Асколи (Р. Ascoli, 1957) объясняет некоторым похоло-
данием и, возможно небольшим понижением солености.
, . ф. Беттенщтедт (F. Bettenstedt, 1962, стр. 389) считает возможным
говорить о мутациях на основании изучения ископаемого материала,
раковин, скелетов, зубов. Это, правда, может показаться малообосно-
ванным генетику, который имеет возможность подвергать исследова-
нию живые организмы и производить над ними эксперименты. Однако
в некоторых случаях палеонтологические данные, по мнению этого ис-
следователя, .более или менее соответствуют понятиям генетики популя-
ций, и едва ли можно сомневаться в том, что многие изменения ископа-
емых apiraiHiH3(MoiB имеют значение результатов мутаций. Сюда относят-
ся прежде всего, очень редкие случаи спонтанного появления у малочис-
ленных особей .нового признака, встречаемость которого постепенно за-
тем расширяется и. в конце концов становится господствующей во всей
популяции ископаемого вида. Следующие критерии, по Беттенштедту,
помогают устанавливать наличие мутаций в ископаемом материале:
1) .непосредственное появление нового признака сначала у единичной
особи популяции; 2) повторное появление одного и того же изменения
признака у представителей геологически более молодых популяций то-
го же самого вида, даже в разнофациальных осадках, что позволяет
установить сохранение признака при различных условиях среды и,
следовательно, его наследственный характер; 3) распространение ново-
го признака на все особи вида (что указывает на селекционную цен-
ность мутировавшего аллеля организма). Автор (1962, стр. 391) исходит
из того положения, что «все филогенетические преобразования сводятся
исключительно к мутациям».
Популяция ископаемого вида фораминифер, по Беттенштедту, всег-
да охватывает смесь многочисленных поколений со всеми вариантами и
локально возможными модификациями: она представляет собой усред-
ненную морфологическую картину вида за много тысячилетий. Мало
вероятно, .чтобы за такой промежуток времени постоянно одинаковые,
но ддя; вида крайние воздействия среды давали бы односторонне ис-
каженную картину вида, иными словами, чтобы существовали долго-
вечные длительные модификации. Тут, по мнению автора, не может быть
модификаций.
Кодь скоро эволюция достигает межвидового масштаба, объяснение
ее модификацией отпадает. Ненаследственные модификации не могут
складываться, а потому ход филогенеза не может быть ими объяснен.
Автор придает большое значение указаниям на то, что новый признак,
вёдуЩий’к новому роду, первоначально появляется у одного единствен-
ного* йли?у немногих особей популяции. Этот процесс невозможно обь-
102
яснить как модификативные приспособления, поскольку можно было
бы ожидать, что влияние биотопа действовало на всю фауну и соответ-
ственно на все остальные варианты того же самого вида, а эти послед-
ние не обнаруживают никакого изменения признаков. Следовательно,
категорически утверждает автор (там же, стр. 392), «перед нами здесь
несомненно спонтанные мутации».
Беттенштедт (там же, стр. 396) находит, что филогенетические пре-
образования, вопреки повторным изменениям условий среды и жизни,
продвигаются прямолинейно (ортогенетически) и неудержимо, «как бы
в силу внутреннего стимула» в однажды взятом направлении развития.
Хотя автор не считает этот вывод неизбежным, но это все-таки, по его
словам, доказывает, что филогенетические процессы нельзя считать
процессами модификативного приспособления к неорганической среде,
морфологические преобразования в филогенетических рядах сводятся
к многочисленным мутациям.
Однако богатый материал микропалеонтологических наблюдений,
как говорит автор, позволяет нам говорить о некоторых изменениях
раковинок как о модификациях. Таковы случаи возрастания величины
»раковинки при благоприятных условиях жизни, например, при относи-
тельно высокой температуре вод. Это наблюдается, например, в верхах
верхнего готерива Северо-Западной Германии, которые очень богаты
микроскопическими ископаемыми по сравнению с подстилающими и пе-
рекрывающими отложениями и содержат много видов с крупными рако-
винками. К числу таких форм принадлежит Hechtina antiqua (Reuss),
которая к тому же является стратиграфически важной формой. Позже
эта форма встречается в среднем барреме, но лишь редко и лекально.
К тому же в среднем барреме этот вид обычно представлен малорос-
лыми особями. Малорослые нуммулиты, в позднем эоцене проникшие
благодаря климатическому оптимуму в море Северной Германии, дале-
ко не достигают величины, свойственной им в южногерманских и се-
вероафриканских эоценовых отложениях. «В обоих случаях, — заклю-
чает Беттенштед (1962, стр. 396), — высокая температура воды была,
во-видимому, прямо или косвенно действовавшим фактором среды».
Беттенштедт приводит и другие случаи, в которых некоторые факторы
(мелководье, более сильное движение вод, предположительно высокая
температура) приводят к увеличению размеров бентонных форамини-
фер мелководных отложений верхнего сантона Ганновера-Лерте. В
пользу модификативного увеличения размеров раковинок говорит и то,
что этим изменением были охвачены многие виды — не менее восьми
из двадцати. Таким образом Беттенштедт находит, что во многих слу-
чаях изменения ископаемых раковинок были модификационными, то
есть не наследственными. Возможно даже, в некоторых случаях, приб-
лизительное определение продолжительности существования длитель-
ных модификаций. В одном случае, по его данным, увеличение рако-
винок, вызванное внешними факторами, затухает после отложения тол-
щи осадков мощностью в 5 м. Накопление этой толщи продолжалось не
менее 10 000, но скорее более 50 000 лет, может быть 100 000 лет. И
все-таки автор думает, что вызываемые столь продолжительным дейст-
вием факторов среды изменения фораминифер не становились наслед-
ственными. Это он считает палеонтологическим вкладом в вопрос, ко-
торый Вейсман пытался решить своим знаменитым экспериментом по
отрубанию хвостов у мышей.
Попытка Беттенштедта применить основные положения генетики к
изучению ископаемых фораминифер, представляет, по нашему мнению,
103
большой интерес. Таких попыток в палеонтологических исследованиях
было довольно мало, хотя не было недостатка в употреблении генети-
ческих терминов при изучении ископаемых животных — беспозвоноч-
ных и позвоночных. Мы видели, что Беттенштедт различает такие поня-
тия как модификации, длительные модификации и мутации. Он усмат-
ривает эти явления в истории изучаемых им фораминифер. Как следует
оценить этот опыт применения генетики в палеонтологических иссле-
дованиях фораммн|Ифе|р?
Отметим сначала то, что мы считаем безусловно положительным
ненным вкладом в изучение ископаемых корненожек.
Беттенштедт, подтверждая и развивая результаты исследования
Б. Граберт (В. Grabert, 1959), убедительно показывает постепенность
филогенетического развития фораминифер. Здесь мы воспроизводим
некоторые из его иллюстраций, показывающих постепенное филогене-
тическое развитие в разных группах фораминифер (рис. 5, 6, 7, 8).
ю 11
Рис. 5. Ряд форм, иллюстрирующих развитие от Gaudryina dividens к Spiroplectinata
lata. Видно последовательнее увеличение числа двурядно расположенных камер. Фиг.
0—10. Gaudryina dividens Grabert. фиг. 11. Spiroplectinata lata Grabert. Верхний апт
(фиг. 0—9), нижний, альб (фиг. 10) и средний альб (фиг. 11) северо-западной Германии.
По Беттенштедту и Грабер
Материал, изученный Б5аттенштедт0|М, привел его к филогенети*
ческим построениям, исключающим внезапное резкое изменение стро-
ения в процессе эволюции фораминифер.
104
Рис. 6. Вариационные ряды Frondicularia simplicissima Dam. Четыре рчда (А, В, С, D),
в восходящем псрядке, характеризуют соответственна зоны верхнего готерива
северо-западной Германии. Из Цедлера и Беттенштедта
Мы, однако, не можем согласиться с его толкованием тех случаев
развития форм, которые он понимает как модификации, длительные мо-
дификации и мутации.
Прежде всего, длительные мутации всегда понимаются как измене-
ния, которые передаются потомству, по их возникновении, в течение не-
которого числа поколений. Иного понимания длительных модификаций
Dauermodifikationen) в генетике не существует, и это, конечно, извест-
но Беттенштедту. В таком случае, если длительные модификации, опи-
сываемые этим автором, существовали, как он думает от десяти тысяч
лет до ста тысяч лет, то это были модификации, однажды, в начале это-
го периода их существования, возникшие вследствие воздействия каких-
то факторов среды, а после передававшиеся по наследству, возможно,
на протяжении десятков и сотен тысяч поколений. Следовательно,
«длительные модификации» трудно отличить, по их поведению, от му-
таций, которые ведь тоже не вечны. Коротко говоря, отпадает всякая
возможность противопоставления длительных модификаций самым нас-
тоящим мутациям.
1'2 15 2,0 2,3 2,9 3,5
Рис. 7. Ряд, иллюстрирующий развитие Lagena hauteriviana Bartenstem et Brand.
Видно постепенное возрастание длины; цифры показывают отношение длины к
ширине. Верхний готерив северо-западной Германии. По Беттенштедту
Мы не можем согласиться также с тем, что мутационные изменении
фораминифер являются спонтанными, то есть фактически независимы-
ми от среды и ее факторов, а Беттенштедт на этом настаивает.
По Беттенштедту, мутации сначала возникают в единственном
числе или весьма малочисленными (а потом численность мутан-
тов популяции возрастает в силу естественного отбора). Это он и счи-
тает доказательством мутантного характера изменений. Естественный
отбор — основной фактор эволюции и видообразования, в этом мы с
Беттенштедтом вполне согласны. Некоторое сомнение вызывает его ука-
зание на численность первоначальных мутантов: один или несколько. Об
одном, конечно, не может быть и речи: можно говорить лишь об отно-
сительной малочисленности нового варианта. Но вот немаловажный
вопрос: как возникают такие варианты? Всего прбще было бы объ-
яснить это явление в духе Шиндевольфа и Г. Ф. Осборна — как та-
инственную эволюцию зародышевого вещества, но Беттенштедт, по-
видимому, не хочет возвращаться к этим неовиталистическим концеп-
циям. А современная наука ищет толчка к таким изменениям в сре-
де, где живут организмы. Если это объяснение правильно, то тем более
106
затруднительно усматривать возможность различать по ископаемому-
материалу длительные модификации и мутации.
Тут мы должны отметить еще один положительный момент в ис-
следованиях автора: на целом ряде примеров из истории меловых фо-
раминифер, представленных в Северо-Западной Германии, сщ устанав-
ливает явление массового направленного и к тому же в сущности нас-
ледственного изменения раковинок. Решение вопроса о том, какую
роль играли эти изменения в эволюции соответствующих групп фора-
минифер, зависит от установления филогенетических цепей видов. Нам
кажется, что и этот вопрос, судя по работам Беттенштедта., близок к.
своему окончательному решению.
Изложенное приводит нас к следующим выводам относительно
изменчивости фораминифер.
У современных и ископаемых видов фораминифер наблюдается
большая внутривидовая изменчивость, которая в некоторых случаях
очень сильно повышается. Однако далеко не всегда эта изменчивость
оказывается бесспорно наследственной. Наблюдения над ныне живу-
щими фораминиферами доказывают наличие изменчивости, на не ус-
танавливают ее наследственного характера, для чего требовались бы
специальные экспериментальные исследования.
Изучение популяций отдельных видов ископаемых фораминифегг
позволяет устанавливать вариации в величине раковинок и в морфоло-
гических особенностях, но не приводит к выяснению вопроса о наслед-
ственности этих изменений, если исследованию не подвергаются одно-
временно предшествующее и последующее звенья филогенетической
цепи. Если же изучаются хотя бы три звена филогенетической цепи,,
то могут быть установлены, во-первых, факт необратимого фи-
логенетического развития видов, а во-вторых, определенная направлен-
ность внутривидовых изменений, что доказывает их наследственный
характер.
Длительные модификации, о которых говорит Беттенштедт, вполне
п:огут оказываться определенно направленными индивидуальными из-
менениями; для того, чтобы удостовериться в этом, необходимо иссле-
довать те же явления в смежных звеньях филогенетической цепи. Ес-
ли при этом выясняется, что в ряду звеньев происходит наращивание
одних и тех же признаков, их последовательное усиление, то трудно
сомневаться в наследственном характере наблюдаемых индивидуаль-
ных изменений.
Стратиграф-геоисторик должен учитывать в своей работе не толь-
ко наследственные изменения и изменения, наследуемость которых
представляется вероятной, но и те изменения, которые, согласно тер-
минологии современной генетики, являются чисто модификационными,
ненаследственными. Чтобы пользоваться данным видом для целей ге-
охронологии, нужно знать по возможности все многообразие его внут-
ривидовой изменчивости; это необходимо для того, чтобы правильно*
определять вид, несмотря на множество колебаний в морфологических:
признаках его раковинок.
Глава пятая
НЕКОТОРЫЕ УКАЗАНИЯ НА «БУЙНУЮ» ИЗМЕНЧИВОСТЬ
(«РАСШАТАННУЮ НАСЛЕДСТВЕННОСТЬ») У ИСКОПАЕМЫХ
БЕСПОЗВОНОЧНЫХ
Необиологам и в особенности генетикам не следует относиться с
пренебрежением к палеобиологическому методу изучения явлений из-
менчивости. Мы видели, что этот метод оказывается во многих отно-
шениях весьма действенным. Он может подкреплять, а в иных случаях
опровергать мнения, возникающие на основе исследования лишь ныне-
живущих организмов. Он может оказываться полезным и там, где
возможность его применения на первый взгляд представляется исклю-
ченной. Возьмем один пример. Известно, что современная генетика в
лице представителей различных направлений, признает явление так на-
зываемой расшатанной наследствнности. О ней говорят и мичуринис-
ты, и представители ортодоксальной генетики. Нам кажется, что палео-
биология располагает материалом, изучение которого может пролить
свет на этот интересный вопрос.
В научной литературе имеется немало сведений об изменчивости
тетракораллов.
Так, Т. А. Добролюбова (1948) отмечала изменчивость кораллов
ряда Dibunophyllum bipartitum — Caninia okensis. Эгот исследователь
различает «революционные» этапы в процессе филогенеза ка-
менноугольных кораллов и утверждает (там, же, стр. 162), что
«все кораллы «революционного» этапа филогенеза являются переход-
ными формами с «расшатанной», еще не закреплённой наследствен-
ностью и не могут относиться к видам, родам и семействам, как они по-
нимаются в палеонтологической и зоологической систематике. Эти ко-
раллы являются не только межвидовыми, но одновременно и межродо-
выми и межсемейственными формами». Изменчивость возрастает до
необычайных размеров в течение сравнительно короткого времени
(стешевское время раннего карбона), охватывая признаки видовые,
родовые и даже семейственные. Эта изменчивость, по Добролюбовой
(стр. 167), «вызывает все большие и большие изменения структуры
и быстро приводит к полному трансформизму вида, рода и семейства».
В более поздней статье Добролюбова рассматривает формо-
образование у нижнекаменноугольных колониальных кораллов Litho-
strotion и Lonsdaleia. В северо-западном крыле Подмосковной кот-
ловины существовала неустойчивая прибрежная обстановка, где ус-
ловия обитания кораллов были иными, чем в открытом море, и корал-
лы, населявшие различные участки бассейна, вели себя по-разному в
смысле изменчивости, формообразования, а следовательно и видооб-
разования (Добролюбова, 1952, стр. 97).
109
Буйная изменчивость каменноугольных одиночных и колониаль-
ных кораллов зависела, по мнению Добролюбовой, от местных условий
среды, но в её работах мы не находим объяснения причин этой усилен-
ной изменчивости.
Н. П. Герасимов (1953, стр. 117) тоже писал о совершенно необык-
новенной изменчивости верхнепалеозойских (сакмарских) тетракорал-
лов. Он утверждал, что проведенными им совместно с сотрудниками
исследованиями «установлено совместное присутствие в одной ко-
лонии двух форм Diphystrotion, четырех выдов Thysanophyllum,.
пяти видов Orionastraea».
Не вдаваясь в критику данных Добролюбовой и Герасимова, мы
вынуждены признать, что в их выводы относительно таксономического
определения кораллитов вкрались какие-то ошибки. С чем связаны,
сшибки такого опытного исследователя как Т. А. Добролюбова, ска-
зать трудно. Не будем также строить предположения относительно про-
исхождения ошибок Герасимова. Однако принадлежность отдельных
частей одной и той колонии колониальных кораллов к разным видам,
даже к разным родам совершенно исключается всем опытом мировой
науки и теоретическими соображениями. То же самое надо сказать
и о такой изменчивости популяций, которая охватывает возникновение
форм не только разных видов, но и разных родов и даже разных се-
мейств.
Тем не менее вполне вероятно, что описываемые этими двумя авто-
рами случаи действительно указывают на повышенную внутривидовую
изменчивость, а потому возможно, что эти сообщения содержат зерно
истины. Оба автора пытаются объяснить замеченные ими явления из-
менчивости разнообразием среды.
Интересны также сообщения Герасимова (там же, стр. 117) отно-
сительно особенностей брюхоногих артинского и сакмарского ярусов.
По этому поводу он пишет: «В пестроте условий среды артинского
моря, документированных песчано-глинистыми отложениями урминс-
кого горизонта артинского яруса, однотипные раковины отдельных
«видов» обнаруживают устройство мягкого тела по нескольким пла-
нам, а в относительно устойчивых условиях сакмарского моря, доку-
ментированных известняками этого яруса, подобные же «виды» раз-
личий в устройстве мягкого тела не обнаруживают».
Таким образом, исследования Добролюбовой и Герасимова дают
нам не более как намеки на внутривидовую изменчивость и её обуслов-
ленность специфическими условиями тех частей водоемов, где они
обитали.
Весьма интересна повышенная изменчивость позднемеловых мор-
ских ежей ряда Micraster leskei—М. cortesudinarium—М. coranguinum
Изменчивость этих форм была описана в капитальной работе А. У. Роу
(A. W. Rowe, 1899), который отмечал наличие переходных форм, что
затрудняет проведение границ между видами. К- А. Кермак (К- А. Кег-
mack, 1954) пришел к выводу, что эволюция этих ежей шла в направлении,
совершенствования приспособления к роюшему образу жизни. Г. Л. Гоукинс
(Н. L. Hawkins, 1943) относил их к числу первых ежей, приспособив-
шихся к жизни внутри ила, и отмечал их необычайную изменчивость.
Глава шестая
КЛАССИЧЕСКИЕ ИССЛЕДОВАНИЯ ИЗМЕНЧИВОСТИ У
НЕОГЕНОВЫХ ПЛАНОРБИД И ВИВИПАРИД НЕМОРСКИХ
ВОДОЕМОВ
ПЛАНОРБИСЫ ШТЕЙНГЕЙМА
В Вюртемберге, в Западной Германии, имеется небольшой участок^
в поперечнике едва ли более 3—4 км, где есть -серия отложений, счи-
тающихся миоценовыми, точнее — верхнемиоценовым1и. Это — осадки
небольшого озера; среди них различаются горизонты, каждый из кото-
рых содержит характерный для него комплекс ископаемых моллюсков
пресноводного происхождения, преимущественно планорбисов. Так как
давно -была замечена интенсивная изменчивость этих моллюсков, они
привлекали внимание многих исследователей: Ф. Гильгендорфа (НИ-
gendorf, 1867 и 1877), А. Гайэтта (Hyatt, 1880 и 1881), Ф. Готтшика
(1919—1920), Л: Плате (Plate, 1919—1920), В. Венца (W. Wenz, 1922),
Г. Клэна (Klahn, 1923) и других. Исходные, древнейшие формы пла-
норбисов Штенгейма имели обычную для этого семейства плоскую ра-
ковину, но в слоях, лежащих выше, появляются формы, у которых ра-
ковина становится башенковидной (рис. 9). Все исследователи едино-
душно высказываются за то, что такие изменения были вызываемы воз-
действием внешних условий. И действительно, едва ли можно понимать
это явление иначе. Но какие конкретно изменения среды вызывали по-
вышенную изменчивость штейнгеймск1их планорбисов? По этому вопро-
су среди исследователей существуют значительные разногласия. Одни
ставят этот процесс под непосредственную зависимость от высокой тем-
пературы, объясняемой притоком вод горячих источников. Другие при-
дают большее значение изменению химизма вод, которое в свою оче-
редь могло зависеть от горячих источников. Мы не будем вдаваться в
рассмотрение этого вопроса. Для нас достаточно то, что необычайная
изменчивость штейнгеймских планорбисов вызывалась влиянием абио-
тических факторов внешней среды.
Тут, однако, встает другой вопрос: были ли те изменения, которым
подвергались штейнгеймские планорбисы, наследственными, или пред-
ставляют собой результаты реакции, последствия которой должны бы-
ли бы сходить на нет, если бы эти моллюски попали снова в обычные
для их непосредственных предков условия химизма и температуры? Мы
полагаем, что, поскольку для отдельных горизонтов штейнгеймской
толщи характерны определенные формы планорбисов, отсутствующие и
в нижележащих и вышележащих слоях, по крайней мере некоторые ба-
шенковидные формы представляют собой самостоятельные таксоны хо-
тя 'бы подвидового, а скорее — видового ранга (рис. 10).Но ученые,
занимавшиеся исследованием этого материала, высказывали различные
111
№ Зола д л аЗлЛлряд j -ый ao&WMtefiрл/ c ' ай яооочмЛ&ряд
10 Supre/ruzs supre/nuj | x depresses cr'wce/zs cuJfafrzj 1 /марл*
О Я ever/ел sr revert e/zj сгш'г&лз соЛсЛьл шсуол
д Oxystorza ojcestorza . i ,егела^ C/Wce/ZJ СШЛЛ/ЛЛ Мар/Г 1 1 ^dL'/z^fZa Sas
7 Ли#.ТЛ0С/Л - /углгсз croc‘/r/o/7/z/.s c/wee/zs cowatas 1 x t0r3t/ztf0/v/uj I jf/wasst
6 dejMeTrocki - /от л frvc/zpzrwzzz crtf0e(f/z0 cos fa faj Jtraassi
£ Р/а/гагЗ/- /отлш р?алол&рл7ШЗ Л/6/гсл1££и//га /traassi
Pufcatas jufeato'7 su6/ic//za>rto//za tfj'a/zssi
3 7'e/iuz^ {еаии 1 Su3/ze//zzsfo//z a /zrazzssi
а JteZfc/tel/A! - 1 /zez//ze/z- 1 sfc/oAez/zze/zs^ sfeut/zez/xe/MM
1 i e/zszs /г Зелл .sis (мал <pip.wq/ l/wofyfaj
Рис. 9. Изменчивость штейнгеймских планорбпссв. Схема стратиграфического распределения ф)рм Giraulus trochiformis. По Венцу
7
Рис. 10. Предпологаемые филогенетические ряды штейнгемских планорбисов
рода Gyraulus. По Гайэтту
8. Л. Ш. Давиташвили
113
мнения. Гильгендорф и Гайэтт придерживались того мнения, что тут
мы умеем перед собой настоящую эволюцию планорбисов. Готтшик
(1919—1920) находил, что правильность воззрений Гильгендорфа под-
тверждается .материалами филетического музея в Иене: там различ-
ные формы Planorbis IGyraulus] multiformis оказываются приурочен-
ными точно к отдельным .пачкам, где эти формы встречаются. Венц (Wenz„
1922, стр. 156) склонен думать, что здесь не происходило видообразова-
ние. Вполне возможно, по его мнению, что весь процесс 'Представляет
собой лишь далеко идущую реакцию на абнормальные внешние усло-
вия» и что это — «реактивные формы», которые при нормальных усло-
виях возвращаются к исходной форме. Г. Клэн (Klahn, 1923, стр. 59)
считает существенным то, что в штейнгеймских отложениях определен-
ные стадии развития планорбисов связаны с определенными зонами
осадков и что тем самым можно говорить об эволюции этих моллюсков
(рис. 10, табл. II). Тем самым, по его словам, возникает положение, ко-
торое кажется парадоксальным: этапообразный филогенез, зависящий
от внешних факторов. Нас не должно удивлять, что Клэн заговорил о
парадоксальности эволюции под влиянием факторов среды. Дело в том,
что этот автор с похвальной откровенностью писал (там же, стр. 58),
что он придерживается «виталистического воззрения» относительно при-
чин эволюции.
Так или иначе, нельзя считать вполне доказанным, что штейнгей-
мский материал представляет собой уполне достоверный случай эво-
люции моллюсков» хотя мы и считаем это весьма вероятным.. Следова-
тельно, он имеет для нас немалый интерес как установленный факт вы-
сокой изменчивости планорбисов под влиянием изменившихся условий
существования. Однако из осторожности оставляем пока открытым во-
прос о наследственном характере отдельных вариаций планорбистов
миоцена Штейнгейма.
ИЗМЕНЧИВОСТЬ ФОРМ РОДА ViviparUS из ОЗЕРНЫХ ОТЛОЖЕНИЙ
НЕОГЕНА
По этому вопросу можно было бы написать огромную монографию.
Мы, однако, полагаем достаточным здесь отметить лишь некоторые наи-
более важные моменты, чтобы показать, как далеко может идти измен-
чивость представителей этого рода (палюдин), в нормальных условиях
пресноводных биотипов обычно сохраняющих гладкие и выпуклые обо-
роты даже у своих крайних разновидностей.
Моллюски рода Viviparus являют один из ярких и общеизвестных
примеров изменчивости организмов под влиянием изменившихся усло-
вий существования. Ознакомление с этой главой, полагаем, убедит чи-
тателя в том, что данному вопросу можно было бы посвятить по мень-
шей мере один объемистый том.
Мы должны признаться, что возможно полное изучение палеобио-
логической истории и необиологии этого рода представляется нам очень
заманчивой работой для эволюциониста. Но здесь мы ограничимся тем,
что совершенно необходимо для освещения изменчивости вивипарид в
соответствии с основной темой нашей книги.
Рассмотрим наиболее выпуклые факты изменчивости вивипарид в
неогеновое время в Славонии, а также .на о. о. Кос и Родос. Отдельно,
(в другой гла1ве этой книга, постараемся проанализировать данные по из-
менчивости вивипарид из плиоценовых солоноватоводных отложений
Паратетиса. Это — факты, которые совершенно недвусмысленно свиде-
114
Vukolinovici
('f Pauli)
( °ovulum)
(” Viquesneh)
arthritira
Zplebori
(:* rudis)
trPilari)
CStrossnwiyeri)
avellana
Hbrnesi
ornate
SluriJ
Herbichi
iMojsisGvicsi
alia
Forbesl ,
. ahecannata
ч oncopliora
Deimanniana
notha
ambigua
Hippocratis
melanthopsis
grandi
Wolfi
Lenzi
Bru sinai
ryrtomaphora
| spun
obumea
la'tosiraca
Suessi
Fuchsl
Sadleri
Coa
striclurafa
bifarcinata
lignttaruni
pannomca
Neumayri
•rhailnoidri
Рис. 11. Филогенез форм рода Viviparus из палюдиновых слоев
Славонии. По Неймайру
115
тельствуют об изумительно высокой изменчивости представителей рода,
[(опадавших в непривычные для него условия существования обширных
озерных бассейнов к почему-то там прижившихся.
Палюдины (вивипарусы) — типичные моллюски текучих вод, глав-
ным образом рек.
Широчайшую известность получили классические исследования
М. Неймайра (Neumayr, 1875) по филогенезу вивипарусов из палюдино-
вых отложений Славонии (которая расположена в восточной части Хор-
ватии, в Югославии). Рисунки, показывающие филогенез славонских
палюдин, даются в учебниках палеонтологии, в общих руководствах по
эволюционному учению. Другое дело — насколько соответствуют дей-
ствительности предлагаемые Неймайром филогенетические построения
(рис. 11). Вполне возможно, что формы, выделенные Неймайром в ка-
честве видов, не заслуживают признания даже подвидами. Сам Ней-
майр приводит изображения двух разновидностей современной палю-
дины Margarya inargeriana из большого озера Талифу, в южнокитай-
ской провинции Юннань, причем одна из этих разновидностей имеет
почти совершенно гладкую раковину, с негл боким спиральным же-
лобком в самой верхней части оборота, а другая — с довольно резко
выраженной скульптурой. Возможно, что эти две формы одного и того
юмического значения (рис. 12). Тем
не менее несомненно, что ряды Ней-
майра показывают и бурную измен-
чивость славонских вивипарусов и
быстрое их видообразование. Ней-
майр вполне справедливо утверж-
дает (1889, стр. 59), что в рядах
славонских вивипарусов «расстоя-
ние между начальным и конечными
членами гораздо значительнее, чем
наивысшая степень изохронной (од-
новременной) изменчивости, кото-
рая встречается у животных той же
группы».
Неймайр (1875, стр. 100) под-
черкивает, что во многих случаях
мы можем наблюдать постепенное
изменение штамма через многие
«мутации» в одном и том же разре-
зе. Он указывает на замечательный
параллелизм в этих постепенных изменениях в пределах рода Viviparus,
а также, в И31вестной мере, между Viviparus и Melanopsis. Этот парал-
лелизм выражается прежде всего в утолщении раковины и появлении
мощной скульптуры (Неймайр, 1875).
Классические исследования замечательного палеонтолога-дарви-
ниста до сих пор не утратили своего значения. Они дали убедительней-
шее и яркое доказательство направленной изменчивости форм под вли-
янием факторов среды, в (которой организмы живут и развиваются.
Рис. 13 и табл. III, воспроизводимые нами из работы Неймайра,
показывают большое разнообразие вивипарусов из плиоцена Славонии.
Тут безусловно не может быть речи об однОхМ ряде форм. Во многих
случаях", по словам Неймайра (1875, стр. 100), с полной несомненностью
наблюдается постепенное изменение филогенетической ветви через мно-
гочисленные вариации. Во всех хорошо изученных случаях устаназли-
же вида не имеют никакого
Рис. 12. Viviparus margerianus из боль-
шого сзера Талифу в южнокитайской
провинции Юннань. 1—килеватая раз-
новидность; 2—гладкая разновидность.
По Неймайру ,
116
Рис. 13. Некоторые гладкие и килеватые формы Viviparus из палюдинсвых слоев
Славонии. 1 и 2. Viv. lignitarum. Зи 4—форма, промежуточная между Viv. lignitarumu
и Viv. spurius. 5 и 6 Viv. spurius, близкая к Viv. lignitarum 7 и 8—типичная Viv.
spurius. 9 и 10—форма Viv. ambiguus, отклоняющая ст типа. И и 12—типичная Viv.
ambiguus. 13 и 14—форма, промежуточная между Viv. nothus и Viv.Jsturi. 15 и 16-
форма Viv. sturi, еще на достигщая типичного состояния. 17 и 18—типичная Viv. sturi.
По Неймайру
117.
вается самостоятельное параллельное развитие форм. Так, в западном
бассейне Западной Славонии Viviparus nothus развивается в Viv. sturi,
а в восточном бассеийе, она, по Неймайру, переходит в Viv. ornatus; в
одном районе Viv. sadleri развивается в Viv. wolfi, в другом в Viv. len-
zi,.;B третьем в Viv. cyrtomaphorus, в четвертом в Viv. altus и Viv.
gfandis;
Неймайр уделял внимание и (вопросу о причинах этих изменений.
Прежде западнославонский бассейн был солоноватоводным, а после, в
период отложения палюдиновых слоев, он постепенно опреснялся, что
следует из исчезновения солоноватоводных форм, место которых зани-
мали чисто пресноводные обитатели. Он утверждал (стр. 102), что мощ-
ная скульптура и утолщение раковины не наблюдаются там, где нет
опреснения. Этот вывод, как будет видно из дальнейшего, нельзя счи-
тать доказанным — в действительности картина гораздо сложнее, чем
это представлялось Неймайру на основании тех сравнительно скудных
данных, которым он располагал.
Однако более общее положение, которое отстаивает Неймайр, мож-
но считать твердо установленным: .«Поэтому, — говорит он (1875, стр.
1-02), — в рассмотренных случаях мы можем признать прямое воздей-
ствие внешних условий важнейшей причиной изменения... мы видим, что
долго продолжающееся воздействие внешних условий жизни и их мед-
ленное изменение вызывает одни и те же преобразования у различных
родов моллюсков...».
Дальнейшие рассуждения Неймайра сугубо гипотетичны, хотя и
небезынтересны. Он ссылается на сведения о высокой степени постоян-
ства в удержании направления изменчивости без воздействия внешних
условий и думает, что это не позволяет нам приписывать сохранение
направления изменчивости лишь прямому воздействию условий су-
ществования (там же, стр. 103). По его словам, мы должны считаться
с возможностью того, что и без такого воздействия, вызванное в
начале направление изменчивости, иногда продолжается далее. Хотя
эти соображения заслуживают внимательного изучения, к ним надо от-
нестись с осторожностью, чтобы не впасть в ортоламаркизм, безуслов-
но чуждый Неймайру, как стойкому дарвинисту. К этому вопросу нам
придется вернуться в другом месте.
Нам представляется очень существенным тот факт, что в различ-
ных более или менее обособленных пресноводных водоемах Западной
Славонии одна и та же форма, один и тот же вид, давал начало раз-
личным потомковым формам. Тем самым подтверждается, что разви-
тие отдельных форм шло на основе изменчивости, индуцируемой мест-
ными условиями, а таковые не могли быть абсолютно тождественными
в различных, хотя и близких друг к другу, водоемах.
Установленные Неймайром филогенетические отношения между
формами палюдин вызывали впоследствии некоторые возра?кения.
Э. Дакке (Dacque, 1921) отметил, что выпады неймайра иногда учены-
ми воспринимались упрощенно: они были склонны думать, < что тут
речь шла о едином филогенетическом ряде. Дакке правильно указывает
на то, что изученные Неймайром формы были найдены в различных
местах, и они жили в различных водоемах. Нельзя утверждать, чго в
нижних горизонтах встречаются везде гладкие формы, а в верхних
только сильно окульптированные. «Филогенетическое древо» тех же
славонских палюдин, даваемое В. Францом (Franz, 1932), мало отли-
чается от ’построения Неймайра.
Нетрудно, однако, видеть, что сложная картина стратиграфическо-
го распределения различных форм нисколько не опорачивает основного
118
вывода о чрезвычайно высокой изменчивости палюдин плиоцена Сла-
вонии и о зависимости этой изменчивости от условий среды. Нельзя,
наконец, сомневаться в наследственном характере выделяемых Ней-
майром «мутаций», потому что наиболее сильно скульптированные фор-
мы не могли возникнуть непосредственно и сразу от совершенно глад-
ких: тут, конечно, необходимо признать ряд этапов эволюции. Последо-
вательное развитие в сторону все более усиливавшейся скульптирован-
ности и накопления сопряженных с нею признаков было бы совершен-
но невозможно, если бы отдельные ^мутации» были лишены наслед-
ственного значения. ?
Неймайр (iNeumayr, 1880) изучал также палюдин из неогеновых от-
ложений о. Кос, в Эгейском море.
Палюдины озерного бассейна о. Кос, как думает А. П. Павлов
(1925), эволюировали в ином направлении, чем в Славонии. «Viviparus
tournoueri, Viv. forbesi и Viv. munieri из отложений острова Кос, — по
словам В. В. Богачева (1961, стр. 69), — проходит стадии structuratus и
nothus сла|во.нюких палюдин, a Viviparus gonceixi из более высокого го-
ризонта отвечает стадии верхнепалюдиновых пластов».
Различия в .вивипарусах с. Коса и Славонии вполне естественны
при разъединенности бассейнов. Прежде, говорит Богачев (1,961, стр.
69), он «думал о возможности оттока вод из Черноморской котловины
в Средиземное море» и предполагал возможность миграции палюдин.
Но даже в самых нижних горизонтах о. Кос виды с гладкими ракови-
нами обнаруживают лишь общее сходство с формами нижних слоев
Славонии, «не достигая полного видового сходства, так что вполне ес-
тественно и различие в формах последующих стадий» (там же, стр. 69).
Заметим, что идея миграции вивипарусов из Славонии в область
эгейского о. Кос совершенно невероятна. А теперь, когда изучение гео-
логического строения области Паратетиса весьма значительно продви-
нулось вперед со времен Неймайра и других исследователей конца
XIX века, нетрудно понять, почему вивипарусы славонского типа не
встречены в плиоцене причерноморских стран. Очевидно тут мы имеем
дело с чисто автохтонным развитием фауны вивипарусов.
Это подтверждается данными о вивипарусовых фаунах более юж-
ного острова Эгейского моря — о. Родоса.
По Гейза Буковскому (1893, стр. 62), на о. Родосе существовало,
по-видимомс, два самостоятельных, отдельных бассейна, в которых от-
лагались левантинские осадки и жили вивипарусы. В одном, северном,
бассейне обитала более многочисленная фауна 'моллюсков, из которых
автор упоминает один вид Viviparus с четырьмя разновидностями, пять
форм Melania, три вида Melanopsis, один вид Limnaea, два вида Neritina,
два вида Hydrobia, один вид Pyrgula, три формы Unio и одни вид Dre-
issena.
Другой палюдиновый (вивипарусовый) бассейн был расположен в
южной части Родоса; он был совершенно изолирован от северного
весьма значительными горными сооружениями и, как и первый, обры-
вался к морю. Буковский нашел единственное местонахождение иско-
паемых, откуда он собрал: три вида Viviparus четыре формы мёланий,
два видя меланопсисов, две формы неритин, один вид гидробий и один
вид Unio.
В первом бассейне жил один вид — Viviparus clathratus Desh. —
чрезвычайно своеобразная скульптированная форма, поверхность обо-
ротов покрыта рядами бугорков или спиральных ребер. В пределах
этого вида наблюдается сильная изменчивость (рис. 14). Помимо ти-
119
Рис. 14. Некоторые фермы рода Viviparus из левантинских отложений о Родоса. 1 и 2—Viv. clathratus. 3—Viv.
clathratus var. langonianus. 4 и 5—-Viv. clathratus, типичная. 6—9—Viv. clathratus var. camirensis. 10—Viviparus
rhodiensis (край устья сохранился неполностью). По Буковскому
личной формы Буковский выделяет пять разновидностей, которые, од-
нако, едва ли имеют значение подвидовых таксономических единиц, хо-
тя дальнейшее развитие (если бы озеро продолжало существовать
дольше), привело бы, быть может, к образованию таковых.
Во втором бассейне жили, по-видимому, совершенно иные предста-
вители рода. Во всяком случае из отложений этого бассейна Буковский
собрал раковины трех видов: Viv. rhodiensis Buk., Viv. acramiticus Buk.
и Viv. (Tulotoma) forbesi Tourn.
Судя по описаниям, мы допускаем, что, во-первых, выделяемые
Буковским разновидности Viv. clathratus не выходят за рамки внутри-
популяционной изменчивости этого изменчивого вида, a Viv. forbesi все
же очень близок к Viv. clathratus. Это наводит нас на мысль, что оба
вивипарусовых бассейна на о. Родос не были столь резко обособлены,
как думает автор, хотя и представляли два самостоятельных местооби-
тания. Ведь Буковский имел возможность лишь совсем бегло ознако-
миться с местонахождениями и довольно поспешно собрал там фауни-
стические остатки. Более тщательное исследование могло бы привести
к несколько иным результатам относительно взаимоотношений этих
бассейнов и населяющих их вивипар сов. Мы знаем, что, например, да-
же в пределах Керченского полуострова наблюдаются существенные
зоогеографические различия в киммерии, а моллюсковая фауна куяль-
ника Гурии заметно отличается от таковой Абхазии. Рассмотрение этих
подробностей не входит в нашу задачу.
Для нас достаточно констатировать, что вивипарусовый комплекс
северного и южного местонахождений о. Родоса заметно отличаются
один от другого, а роскошно представленный вид Viv. clathratus отли-
чается от видов всех других известных озерных местонахождений поло-
сы Паратетиса .и является бесспорно эндемиком.
В понтических отложениях окрестностей Балатонского озера —
озера Балатон в Венгрии — встречаются вивипарусы, сильно откло-
няющиеся от исходных пресноводных форм (Halavats, Г. Галавач,
1903). Не вдаваясь в описание этих форм, воспроизводим некоторые
изображения (из монографии Галавача) Viviparus sadleri Partsch, Viv.
cyrtomaphorus, Viv. kurdensis Lorent. и некоторые другие (рис. 15).
У нас отнюдь нет уверенности в том, что все эти формы действи-
тельно имеют таксономическое значение: дальнейшая их ревизия, быть
может, установит принадлежность их к’гораздо меньшему числу энде-
мичных видов. При описании Viv. sadleri Partsch Галавач делает ин-
тересные замечания: «Часто встречающиеся в местонахождениях об-
ласти Балатонского озера, в Тихани, Кенесе, гладкие вивипарусы столь
сильно разнообразны, что могут быть установлены целые крупные ряды
постепенных переходов. Один крайний пункт такого ряда образуют
стройные конические формы, а противоположный — коренастые, ци-
линдрические формы, и между этими двумя крайностями стоят формы,
склоняющиеся либо к одной конечной форме, либо к другой конечной
форме. Коренастые, цилиндрические конечные формы с лестницевид-
ным завитком Брусина назвал Viv. cyrtomaphora, соответственно более
стройные конические формы должны обозначаться именем Viv. sadleri,
которое было введено Парчем. Это — не первый случай столь сильного
варьирования солоноватоводных моллюсков в отдельных местонахож-
дениях, что конечные члены рядов форм, получают особые названия;
это надо считать тем более оправданным, что обе крайние формы
встречаются часто только в значительно удаленных местах без каких-
121
либо переходных форм». В данном случае, по словам Галавача, Viv.
•sadleri встречается в Радманесте, a Viv. cyrtomaphorus у подножия
’Фонио-хеди (Halavats, Галавач, 1903, стр. 40). При любой интерпре-
тации несомненным остается факт повышенной изменчивости, которую
автор правильно связывает с обитанием этих форм в солоноватых во-
дах.
Здесь-мы рассмотрели данные тех исследований изменчивости иско:
паемых пресноводных моллюсков, которые можно назвать классически-
ми. Эти исследования были выполнены преимущественно еще в прош-
лом веке и в начале текущего века известными палеонтологами — Гиль-
гендорфом, Гайэттом, Неймайром и другими. Все эти ученые с полной
уверенностью говорили о том, что органические формы изменялись
постепенно под влиянием факторов среды — температуры, солености,
питания и других.
Рис. 15. Некоторые формы рода Viviparus из понтических отложений
окрестностей о. Балатон. 1, 2, 5, и 6—Viv. kurdensis. 3 и 4—Viv.
sadleri. 7 и 8—Viv. gracilis. 9, 10, 11, и 12—Viv. cyrtomaphorus. По
Г алавачу
В дальнейшем мы рассмотрим аналогичные факты, которые были
изучены исследователями последующих поколений. Мы имеем в виду
прежде всего материалы по изменчивости неогеновых и четвертичных
моллюсков, живших в морях Паратетиса, где соленость иногда сильно
.122
понижалась, приближаясь в этом отношении к пресноводным бассей-
нам. Там мы коснемся изменений, которые испытали некоторые соло-
новатоводные моллюски пресноводного происхождения, в частности —
вивипарусы, что даст нам возможность сопоставить эти данные с теми
изменениями ископаемых форм, которые были рассмотрены в этой
главе.
Глава седьмая
ТЕМПЫ ИЗМЕНЧИВОСТИ И ПРОДОЛЖИТЕЛЬНОСТЬ
ЭВОЛЮЦИИ ОРГАНИЧЕСКОГО МИРА В ТЕЧЕНИЕ
ГЕОЛОГИЧЕСКОГО ВРЕМЕНИ
Здесь мы подходим к рассмотрению вопроса, очень важного для
понимания общего процесса эволюции органического мира на Земле:
соответствуют ли те .или иные представления об изменчивости органи-
ческих форм результатам, которые были достигнуты развитием всех ор-
ганизмов — протистов, растений и животных — в течение геологичес-
кого времени? Естественный отбор мог действовать лишь на основе
наследственных изменений организмов. Для того, чтобы отбор привел к
определенным результатам, нужно, чтобы изменчивость создавала дос-
таточную базу для его действия. Некоторые биологи д мают, что при
этом не достаточны очень мелкие, незначительные изменения и что для
достижения эволюцией существенных результатов в смысле образова-
ния новых видов нужны глубокие коренные изменения, большие скачки
которые получали различные наименования: Р. Гольдшмидт (Gold-
schmidt, 1961, стр. 486) писал о системной мутации, имеющей якобы
место тогда, когда мы имеем особые мутации, а именно макромутации,
вызывающие очень большое отклонение от нормального поведения. Та-
кие макромутации в своем ограниченном действии на часть тела или на
весь организм, по Гольдшмидту, ясно показывают, что возможны «чрез-
вычайно сильные отклонения», внезапные скачки сразу, одним шагом
прочь от данного вида к другому, новому. Так возникают, как выража-
ется Гольдшмидт, «многообещающие чудовища», которые могут вести
к новой эволюционной линии. А. М. Дальк (А. М. Dalcq, 1949 и 1955)
допускал, что скачкообразное возникновение нового типа строения про-
исходит через онтомутацию. Л. Убиш (L. Ubisch, 1953) подчеркивал
значение «гроссмутаций» (маифомутаций) для эволюции и находил, что
«гроссмутации широчайшего масштаба» могут создавать жизнеспособ-
ные организмы без поддержки естественного отбора (там же, стр. 434).
Этот автор, как и другие сторонники внезапного появления совершенно
новых таксонов, ссылаются на тщетность поисков «недостающих звень-
ев» в филогенетических рядах: таких звеньев, по его мнению, и .не было.
Подобные сальтационистские воззрения до сих пор имеют довольно
широкое распространение. Против такой идеи справедливо возражали
многие эволюционисты, в частности Э. Майр (Мауг, 1960, стр. 357) и
Симпсон [см., например, (Simipson, 1960), стр. 149].
Сторонники постнеодарвинизма находят, что эволюция идет, в ос-
новном,' путем естественного отбора мелких, в фенотипе малозаметных
или даже незаметных мутаций, незначительных «ошибок» в воспроизве-
дении «генетического кода». И нам кажется, что эти ученые несравнек-
124
но ближе к истине, чем сторонники Сальтационизма, согласно которым
филогенез происходит (может быть, даже без существенного участил
отбора) на основе внезапных ма1К1ромутаций, сразу создающих новый
тип строения организма. Однако и у сальтационистов есть аргумент
против постепенной эволюции путем отбора мелких и мельчайших му-
таций: не будет ли такой ход эволюции слишком уж медленным для
объяснения общего филогенеза органического мира на протяжении ге-
ологического времени? Ведь «многообещающие чудовища» Гольдшмид-
та приводят почти мгновенно к образованию филогенетической новинки
— совершенно нового типа строения и функций организма. Тем самым
сальтационизм как будто приближает нас к объяснению хода филогене-
за больших таксонов. Но сальтационисты забывают, что разительное из-
менение организма в любой его части без столь же существенных изме-
нений других систем органов данного существа неизбежно создает не-
жизнеспособные существа, которые мгновенно гибнут или, в лучшем
случае» быстро отметаются естественным отбором. Если мы допустим,
что коренные глубокие изменения возникают сразу во многих или во
всех системах органов животного, то такое представление окажется в
высшей степени невероятным и близким к признанию чуда в развитии
жизни на Земле. Попытки сальтационистов ссылаться на отсутствие, в
палеонтологической истории органической жизни, переходных форм,
незаметных переходов от одного типа строения к другому основаны на
недоразумении. Недаром к такому аргументу прибегают чаще всего
и усерднее всего сальтационисты из числа ученых, не занимающихся
изучением палеонтологических объектов и геологической истории. Им
легче отрицать неполноту геологической летописи.
Между тем геологи и палеонтологи на каждом шагу встречаются с
фактами, показывающими неполноту наших знаний о группах ископа-
емых организмов. Ограничимся здесь лишь одним примеро;м изъянов в
наших познаниях об организмах прошлого. В недавно опубликованной
книге, посвященной причинам вымирания организмов (1969), мы рас-
смотрели, между прочим, проблему вымирания рудистов. Мы постара-
лись показать, что оно не было внезапным и геологически строго одно-
временным. Но в момент писания этой главы мы не знали, что совсем не-
давно были обнаружены рудисты верхнего палеоцена (И. А. Коробков,
1967). Между тем все до того опубликованные данные о последних
рудистах гласили о том, что в нижнетретичных отложениях эти формы
нигде не известны, а многие авторы утверждали, что рудисты вымерли
сразу в конце мелового периода. Отсутствие между очень многими так-
сонами бесспорных промежуточных звеньев никоим образом нельзя
считать подтверждением сальтационизма: такие зленья существовали!
Если, однако, мы исключим сальтационизм, то ведь и обычные тол-
кования процесса видообразования, даваемые постнеодарвинистами
(«синтетистами»), встречают большие затруднения. По признанию этих
ученых, единичные мутации всегда вредны, а потому их носители не
могут сохраняться при действии естественного отбора. Даже полезное
изменение может сохраниться лишь в том случае, если параллельно с
ним соответственно изменяются другие, сопряженные с ними признаки
и их генетическая основа. Малая вероятность такого совпадения по-
буждала видных генетиков и эволюционистов неоменделистского нап-
равления выдвигать какие-то дополнительные теоретические построе-
ния, которые делают эволюционный процесс зависящим от действия
различно направленных сил. Если к этому прибавить еще невероятность
достаточно большого количества одновременно возникающих, хотя
125
бы и мелких, спонтанных мутаций, нужного для обеспечения полового»
размножения соответствующих мутантов, то такое понимание эволюции
окажется связанным с представлением об очень и очень медленном
ходе эволюции. Работа стабилизирующего фактора оказывается более
успешной, чем работа естественного отбора как фактора изменения, раз-
вития таксонов.
Мы не решаемся здесь категор1ичесюи ответить на вопрос, возможно,
ли вообще видообразование на основе факторов, признаваемых постне-
одарвинистами, но думаем, что, по только что приведенным соображе-
ниям, вероятность такого видообразования очень невелика. Однако при
нынешнем уровне знаний нужно поставить вопрос несколько иначе; мож-
но ли на основе постнеодарвинистской теории объяснить весь процесс
формообразования на протяжении всего геологического времени от поз-
днего докембрия доныне, — процесс, приведший к нынешнему органи-
ческому миру? Если на этот вопрос придется дать отрицательный от-
вет, то «синтетистское» понимание наследственной внутривидовой из-
менчивости надо будет признать несостоятельным.
Можно считать точно установленным, что эволюция происходит на
основе очень мелких изменений, но эти незначительные изменения ох-
ватывают, очевидно, сразу разные стороны строения и деятельности
живого тела. Мы принимаем ту концепцию эволюционного развития
которая некоторыми нашими учеными называлась презрительно «плос-
кой эволюцией». Наше понимание «плоской эволюции» существенно от-
личается от неоменделистского тем, что основным материалом, подпа-
дающим под действие отбора, мы считаем не обособленные изменения
отдельных признаков, а изменения целых комплексов признаков, прав-
да, чрезвычайно незначительные. Неоменделистская концепция, кото-
рая некогда начала с формулы «один ген — один признак», неуклонно,
хотя и медленно, приближается к только-что высказанному нашему по-
ниманию эволюционного процесса. Возьмем, например, то понимание
наследственной изменчивости, подвергаемой отбору, которое содержит-
ся в работах Э. Майра (Мауг 1959, 1960) и других эволюционистов, пол-
ностью принимающих принципы классической генетики. Эти исследова-
тели учитывают то, что для сохранения в процессе естественного отбора
особи должны обладать не одним изолированным полезным признаком,
а комплексом признаков. Любое, ничтожнейшее полезное изменение
глаза, как органа зрения, может дать своему обладателю определен-
ное преимущество лишь в том случае, если оно в некоторой мере охва-
тывает все основные части этого органа и сопряжено с соответствую-
щим изменением в нервной системе, а также и в других органах, де-
ятельность которых непосредственно связана с функцией зрения. Любое,
хотя бы самое ничтожное полезное изменение любой кости запястья
может быть удержано в процессе естественного отбора лишь в том
случае, если одновременно соответственно изменятся не только соседние
кости пясти и предплечья, но и некоторые мускулы, нервы, кровенос-
ные сосуды. Тут можно призвать на помощь явление плейотропии генов,
то есть множественности действия каждого гена, хотя обычно плейпро-
пия понимается как свойство, охватывающее различные действия»
физиологически между собой не связанные. Тем не менее введение по-
нятия плейотропии мы рассоматриваем как попытку приблизиться к дей-
ствительности. Так или иначе отбор может подхватывать и закреплять
те или иные признаки лишь при условии их слаженности, гармоничном
действии с другими признаками организма. Суть дела заключается, по
нашему мнению, в единстве дискретности наследственных факторов, ко-
126
торая была признана, как мы видели, еще Дарвином и которая была
подчеркнута открытиями Г. Менделя, с целостностью организма.
Тут, однако, мы попытаемся подойти к проблеме наследования из-
менений со стороны геоисторического развития органического мира и от-
ьетить на вопрос, в какой мере возможно объяснить историческое раз-
витие организмов в течение геологического времени теми данными об
изменчивости и наследственности, которыми располагает современная
генетика, то есть на основе идеи мелких, совершенно незначительных
наследственных изменений, а не на основе фантастической идеи саль-
таций. В поисках ответа на этот вопрос нам надо прежде всего попы-
таться выяснить, какова скорость видообразования, понимаемого с по-
зиций классической генетики. С этой точки зрения основной путь обра-
зования новых наследственных изменений — спонтанные мутации, не
зависящие от характера среды. Но геология третичной системы показы-
вает, что там, где среда изменяется относительно медленно, животные
изменяются тоже медленно и наоборот, чем глубже геологические изме-
нения характера среды, тем глубже изменения организмов. Тут вспоми-
нается опять-таки поучительное сопоставление моллюсков полносолено-
го моря Тетиса с моллюсками. Паратетиса в течение неогена. Многие
формы, жившие в бассейне Северной Италии в раннем миоцене, почти
не изменились до настоящего времени и относятся к тем же самым ви-
дам. Это значит, что, согласно определениям специалистов-палеонто-
логов, виды оставались практически неизменными в течение проме-
жутка времени около 30 млн лет. Впрочем, не может быть и речи об аб-
солютной неизменности видов за этот период времени: некоторые из-
менения могли очень мало отразиться на раковине. На протяжении вре-
мени от раннего миоцена доныне в среде очевидно происходили изме-
нения, которые могли вызывать изменчивость органических форм. Если
бы не было никаких изменений среды, в данном случае — гидрологи-
ческих условий и биоса в морском бассейне, — то и изменчивость орга-
низмов отсутствовала бы, но такая абсолютная неподвижность совер-
шенно исключается. В некоторых экологических областях условия сре-
ды гораздо более разнообразны и более изменчивы, чем они были в
полносоленом море Тетиса и соответственно эволюция, зависящая от из-
менчивости и отбора, была гораздо более интенсивной.
Древнейшие остатки жизни известны в породах, возраст которых
определяется в 2 700 млн. лет [Р. Фюрон, (R. Furon, 1967), стр. 971].
Длительность фанерозоя составляет приблизительно 500—600 млн. лет.
Встает вопрос: могли ли бы за такой промежуток времени возникнуть
и развиться до нынешнего состояния все филогенетические линии про-
тистов, растений и животных, если бы эволюция всегда протекала в
силу отбора в основном спонтанных, случайных и «шальных» наслед-
ственных изменений («ошибок»)? Мы затрудняемся ответить на этот
вопрос, но вообще говоря, такой ответ возможен — нужно только под-
готовить условия для математической обработки фактических дан-
ных. Прежде всего необходимо с максимальной возможной точно-
стью найти вероятные варианты ответа на вопрос, сколь длительна
должна быть эволюция от одного вида к другому, ближайшему, при
условии, что она будет происходить на основе лишь спонтанных мута-
ций, не зависящих от воздействия условий среды. Затем нужно будет
хотя бы ориентировочно наметить, сколько видовых звеньев будут
находиться между соседними родами, а затем — через сколько родов
прошла эволюция данного отряда о г его возникновения до появления
ближайшего, более позднего отряда. Восстанавливая хронологию раз-
127
вития того или иного крупного таксона, мы должны еще учитывать
длительность цикла индивидуального развития от одного поколения до
следующего, ближайшего; а эти величины, как известно, сильно варьи-
руют. При нынешнем уровне знаний исторического развития трех
царств организмов — протистов, растений и животных — данные, хотя
бы грубо ориентировочные, могут быть получены лишь для немногих
филогенетических линий, например,—для лошадиных, хоботных и
некоторых других групп млекопитающих, а также для аммоноидей из
головоногих моллюсков. К каким результатам приведут эти, действи-
тельно очень полезные, математические выкладки» заранее сказать
трудно, но мы имеем все основания опасаться, что результаты, к ко-
торым придут таким путем математики, будут во много раз превышать
то число лет, которое в действительности потребовалось для эволюции
от одной группы к другой.
С точки зрения этой проблемы поучительны данные симпозиума
Уистарского института в США (1987), в частности доклад математи-
ка С. М. Юлэма «Как формулировать математически проблемы тем-
пов эволюции» и дискуссия по этому докладу, в которой принимали
живейшее участие некоторые самые видные представители «синтети-
ческой теории эволюции». Докладчик должен был ответить на вопрос,
достаточно ли одного миллиарда лет (фанерозой плюс часть позднего
протерозоя) для того, чтобы, в соответствии с принципами неодарви-
низма, совершилась та эволюция органического мира, которая свиде-
тельствуется палеобиологической историей жизни, а также нынешними
фаунами и флорой. Юлэм справедливо подчеркнул стоящие перед ним
трудности, но все же сделал некоторые предварительные, заведомо
сверхупрощенные, допущения и попытался подойти к математическому
решению вопроса о том, сколько времени требовалось бы факторам»
принимаемыми «синтетической теорией эволюции» для того, чтобы бы-
ли осуществлены наличествующие результаты всеобщего филогенети-
ческого процесса развития жизни на Земле. Он предположил, что
жизнь единичной особи т равна одному дню. Если весь тот промежу-
ток времени, в течение которого на нашей планете существовала
жизнь, обозначить буквой Т, то общее число -поколений от самой древ-
ней, добактериальной, формы жизни до нашего времени выразится
формулой Т/т. Дифференциальное преимущество, даваемое одной
«благоприятной» мутацией, Юлэм обозначает через у- Если гамма рав-
на 10-6 то для того, чтобы большинство популяции обладало этим
крошечным преимуществом, потребуется 106 поколений. Через 107 по-
колений почти все особи будут обладать этим преимуществом. Но для
развития такого органа, как скажем, глаз, нам требуется не одно улуч-
шение, а, скажем, 106 улучшений, и число нужных поколений соответ-
ственно возрастает. Добавим от себя: жизненный цикл поколения мо-
жет оказаться и очень часто оказывается у хордовых, обла-
дающих парными глазами, равным многим годам, а не одному дню,
как условно принимает Юлэм. «Надо помнить,—сказал Юлэм (там же,
стр. 24), — что нам нужно не одно улучшение, а, скажем, 106 в после-
довательном порядке. Так что нам требуется, скажем, около 1013 поко-
лений, а не гораздо больше».
Остановимся на этой величине — 1013 поколений, то есть десять
тысяч миллиардов поколений. Не будем упускать из виду, что Юлэм
на всю восходящую эволюцию органического мира уделяет один мил-
лиард лет (там же, стр. 22), что, конечно, является грубым упроще-
нием — даже ископаемая летопись, которая начинается отнюдь не с
первого появления жизни на земле, охватывает, как мы видели, не
128
менее 2.7 миллиардов лет. Но можно сказать, что миллиард лет на-
зад на земле были лишь весьма низко организованные, преимуще-
ственно одноклеточные, организмы, а первые достоверно установлен-
ные находки многоклеточных животных относятся ко времени менее
700 млн лет. Так или иначе, за последний миллиард лет, по словам
Юлэма, должна была осуществиться последовательность приблизи-
тельно в десять тысяч миллиардов поколений. Сам Юлэм считает воз-
можным принять среднюю длительность времени от одного поколения
к другому равной приблизительно одному дню. Но и в таком случае
развитие столь большого числа поколений Потребовало бы около 25
миллиардов лет. Можно, к тому же, решительно оспаривать среднюю
продолжительность жизни поколения животных, скажем, силура, да-
же ордовика или кембрия (не говоря уже о ‘более поздних животных)
в один день. Уже в ордовике существовали панцирные бесчелюстные
первичноводные позвоночные — продолжительность их жизни не мог-
ла быть менее года. А ордовик начался почти полмиллиарда лет на-
зад (около 490 млн. лет назад). На кораллах девона различаются
«кольца», соответствующие годам и даже суткам. Уже в среднем де-
воне существовали древесные растения, очевидно многолетние. Многие
животные мезозоя и кайнозоя жили десятки лет, если не более. Для
развития, например, лошадеобразных путем «шальных», беспорядоч-
ных мутаций или «рекомбинаций» требовались бы поистине астроно-
мические сроки эволюции. Не сомневаемся, что тут перед нами об-
ласть будущих интереснейших математических и кибернетических ис-
следований. Однако уже при нынешнем уровне знаний постнеодарви-
нистское объяснение эволюции должно быть признано невероятным.
Один из подходов к освещению этого вопроса связан с выяснением
времени достижения половой зрелости различными представителями
современной фауны. Такие сведения недавно были собраны Г. И. Шпе-
том (1968). Воспользуемся его данными. Крысы и мыши дают в год
до 4—5 и более поколений, причем молодые самки серой крысы уже в
возрасте трех месяцев могут приносить детенышей. Морские свинки
становятся половозрелыми через два месяца после рождения. Но для
полного созревания землеройки, крота и ежа требуется целый год. У
крупного рогатого скота молодые становятся способными к размноже-
нию уже в 1,5—2 года, так же как и у обыкновенного оленя и север-
ного оленя. Дикая свинья созревает на 19-ом месяце. Двугорбый вер-
блюд для своего развития требует пяти лет, а дромадер даже 6—7 лет.
Азиатский слон достигает половозрелости лишь в 16—20 лет и прино-
сит по одному детенышу раз в три года. Многие домашние животные,
(свинья, коза) созревают быстрее своих диких родичей.
Среди пресмыкающихся даже более мелкие виды становятся спо-
собными к размножению лишь через несколько лет. У гаттерии мо-
лодь выходит из яйца более чем через год после откладки. У некото-
рых змей половая зрелость наступает лишь на четвертый год. У евро-
пейской черепахи — на седьмой-девятый год жизни или позже. У ниль-
ского крокодила — на 20-м году. У амфибий, при относительно не-
больших размерах тела, половая зрелось достигается в 3—5 лет, реже
в 2 года.
Эти данные говорят в пользу того, что пойкилотермные четвероно-
гие в общем требуют довольно длительного времени для достижения
зрелости. А древние формы, особенно из числа крупных стегоцефалов
и пресмыкающихся, достигавших большой величины тела, по всей ве-
роятности, соэревли еще медленнее.
Из современных рыб медленно растут, по-видимому, акулообраз-
9. Л. Ш. Давиташвили 129
ные. Самка небольшой акулы Squalus acanthias L., длиной до 120 см,
достигает зрелости на 12—14 году жизни. Имеется ряд косвенных дан-
ных о медленном росте двоякодышащих, кистеперых, каймановых,
амий, химер. Из костных рыб хондростоидеи созревают обычно в 12—
18 лет (самки). Белуга и калуга созревают в возрасте 15—20 лет
(Г. В. Никольский, 1944, стр. 118; И. С. Строганов, 1962, стр. 388).
Костистые растут в общем быстро. Ио различные виды лососей созре-
вают на З.м—6 м году; байкальский омуль, нельма, муксун соответст-
венно в 6—7, 13—14 и 6—12 лет.
У круглоротых рост происходит очень медленно. Личинка миноги,
пескоройка, растет 3—4 и даже 5 лет.
Очень медленно растут многие членистоногие, хотя другие из них
достигают зрелости быстро. Но анализ таких данных о различных
группах членистоногих и других групп беспозвоночных требует спе-
циального рассмотрения, от которого мы тут воздержимся.
Приведенные нами сведения о продолжительности созревания
различных позвоночных подсказывают мысль, что, оставляя в сторо-
не особые случаи приспособления к быстрой смене поколений, средняя
продолжительность развития от рождения до половой зрелости у боль-
шинства ископаемых форм едва ли была ниже одного года. При этом
надо учесть, что менее высоко стоящие группы в общем развиваются,
при прочих разных условиях, медление, чем выше организованны.?.
Первые позвоночные появились, по-видимому, не позднее начала
ордовика, а, может быть, много раньше. Задолго до них должны бы-
ли начать свою эволюцию их предки, примитивные хордовые, которые,
может быть, существовали уже миллиард лет назад. Если даже мы
примем, что средняя продолжительность индивидуального развития
хордовых составляла 1/100 года (что надо признать очень заниженной
оценкой), то и тогда для эволюции всех хордовых от примитивнейших
до наиболее высоко организованных, потребовала бы, согласно пред-
положенному Юлэмом суммарному числу поколений в 1013, многие
миллиарды лет! Если же мы, допустим, что половая зрелость то-
стигалась в среднем невЗ—4 дня, а в 1/10 года, то общая длительность
такой эволюции достигла бы фантастической величины в тысячу милли-
ардов лет. Таким образом, суммарное число поколений органических
форм, существовавших на протяжении геологического времени, ска-
зочно возрастает. Миллиард лет, о которых говорил докладчик, оказы-
вается смехотворно малым. Не удивительно, что эти выкладки, кото-
рыми представители постнеодарвинизма надеялись подтвердить свою
теорию, их сильно смутили. Когда Юлэм упомянул астрономические
числа поколений, нужных для освоения серии улучшений, которые
могли вести к образованию такого сложного объекта, как глаз, изве-
стный зоолог-эволюционист Э. Майр перебил его вопросом: «А не могли
они (эти улучшения. — Л. Д.) происходить все одновременно?» На
это Юлэм ответил: «Это чрезвычайно невероятно» (Юлэм, 1967, стр.
24). И нам кажется, что он ответил справедливо. Вполне естественно,
что доклад Юлэма вызвал протесты со стороны присутствовавших на
совещании сторонников постнеодарвинизма. Ведь, если бы они согла-
сились с докладчиком, то им оставалось бы лишь отказаться от «син-
тетической теории эволюции». Уоддингтон заявил, что докладчик по-
ставил вопрос верх дном (там же, стр. 28). Некоторые участники сим-
позиума отмечали, что трудности, о которых говорил Юлэм, возника-
ют лишь при бесполом размножении; а там, где существует процесс
оплодотворения, трудности отпадают. Председатель Мидоуэр заявил,
что математики, по-видимому, неправильно понимают процесс есте-
130
ственного отбора. Они думают, что объектом отбора является мутант,
а в действительности отбору подвергается рекомбинант (там же, стр.
29).
Доклад Юлэма, конечно, не решил судьбу постнеодарвинизма.
Этот математик, по нашему мнению, нащупал слабую сторону «синте-
тической теории эволюции», но не смог доказать ее несостоятельность.
Дискуссия по докладу Юлэма показала необходимость математи-
ческой ревизии того понимания наследственности и изменчивости ор-
ганизмов, которого придерживаются многие современные ученые. Эта
дискуссия продемонстрировала ненадежность идеи эволюции на ос-
нове отбора спонтанных («шальных») наследственных изменений, но
не опровергла эту идею.
Нам кажется, что ревизию существующих представлений о фак-
торах изменчивости надо начинать с обследования филогенетического
развития высокоорганизованных организмов преимущественно кайно-
зойской эры. Особенно полезным для этой цели материалом могут
послужить ископаемые лошадиные и хоботные, а также некоторые
группы парнопалых копытных, китообразных, приматов и других зве-
рей, филогенез которых протекал очень быстро.
Вся совокупность имеющихся у нас данных по истории животно-
го мира делает крайне невероятной эволюцию на основе случайных,
беспорядочных «ошибок» в воспроизведении генетической информа-
ции. Однако окончательное решение вопроса возможно лишь путем
тщательных исследований филогенеза всего биоса на протяжении гео-
логического времени при серьезном использовании математического
метода.
Глава восьмая
О ВНУТРИПОПУЛЯЦИОННЫХ ИЗМЕНЕНИЯХ ВЕЛИЧИНЫ ТЕЛА
ЖИВОТНЫХ ПРОШЛЫХ ГЕОЛОГИЧЕСКИХ ВРЕМЕН
НЕКОТОРЫЕ ОБЩИЕ СООБРАЖЕНИЯ
Возрастание и уменьшение средней величины тела, характерной
для популяции животных данного вида, очень часто наводило иссле-
дователей, палеонтологов, геологов и зоологов, на мысль, что эти из-
менения размеров зависели от условий существования организмов:
обычно думают, что возрастание вызывалось действием каких-то бла-
гоприятных факторов, а уменьшение — ухудшением условий жизни.
Мы считаем необходимым рассмотреть этот (Вопрос, поскольку его изу-
чение может пролить свет на изменчивость организмов и ее причины.
В геологической литературе при описании тех или иных толщ и
содержащихся в них фаунистических остатков нередки упоминания о
так называемых «угнетенных» фаунах, причем важнейшим признаком
угнетенности считается обычно уменьшение величины особей против
размеров, обычных для данного вида. Встречаются также указания на
случаи возрастания величины тела ископаемых организмов за пределы
свойственных данному виду колебаний размеров. Иноща говорят да-
же о гигантизме животных данного вида на том или ином участке
ареала его распространения.
Отклонения в сторону карликовости могут быть, как обычно вы-
ражаются, чисто модификациоными, то есть не наследственными или,
наоборот, наследственными. Их изучение представляется существен-
ным для исторической геологии. Даже модификационные изменения
нормальной величины тела заслуживают пристального внимания гео-
логов-стратиграфов и биостратиграфов. С одной стороны, исследова-
ние таких изменений может указывать на условия осадконакопления,
их индуцировавшие, и тем самым способствовать решению геохроно-
логических задач, потому что выяснение относительного геологическо-
го возраста зависит от познания развития обстановки, в которой жи-
ли соответствующие виды организмов. С другой стороны, изменения,
наследственный характер которых не доказан и которые могут ока-
заться простыми модификациями, имеют также прямое и непосред-
ственное значение для решения стратиграфических вопросов. Так,
если в двух точках выходов отложений данный вид представлен двумя
модификациями, заметно отличающимися одна от другой по величине
тела и если это .различие зависит от воздействий на организмы мест-
ных условий, то оно не должно препятствовать стратиграфической
идентификации этих двух точек. Несмотря на это различие данный
вид вполне может быть привлечен для целей параллелизации пластов
пород. Если бы перед нами были две формы, принадлежащие к очень
близко родственным, но различным таксонам, то их ценность для'
132
стратиграфической шараллелизации была бы, конечно, существен-
ной яной)- То же самое, но .в еще большей мере касается случаев, когда
перед нами не одна карликовая форма, а целый комплекс карлико-
вых форм. Если их карликовые размеры имеют общее происхождение
и зависят от одной и той же экологической причины, действовавшей в
данной точке, то мы все же можем сопоставлять эту толщу с другими,
где соответствующий комплекс ископаемых представлен особями нор-
мальных размеров. В то же время возможны случаи, когда целый
комплекс ископаемых форм уменьшенных, против нормы, размеров,
характерен для всего ареала распространения этих форм даже в том
•случае, когда имеются основания пред|пола1гать ненаследственный ха-
рактер этой «карликовости».
Но изучение внутривидовой изменчивости величины ископаемых
организмов имеет не меньшее значение для освещения процессов эво-
люции. Такие исследования, как это будет показано в дальнейшем,
могут вскрывать факторы изменчивости, ее зависимость от внешних
условий, и ее роль в ходе филогенеза.
Установление наследственного характера изменений, индуцирован-
ных факторами среды, представляет, конечно, большие трудности для
палеобиолога. При этом одним из положительных указаний служит
широкое распространение популяций измененных форм и исторически
длительное их существование. Если в популяции, занимающей более
или менее обширный ареал, все или почти все особи характеризуются
измененной, — например, уменьшенной, — величиной тела, то уже это
в известной мере говорит за то, что данные пределы колебаний яв-
ляются характерными для всей тю1пул.Я1ЦИ1и, а потому наследственны-
ми. Если к тому же соответствующий диапазон изменчивости величи-
ны тела выдерживается в течение определенного отрезка геологиче-
ского времени, то наследственный характер этого свойства становится
еще более вероятным. Однако окончательно установленным это считать
нельзя, так как все же возможно, что те или иные измененные
условия существования вызывали обратимое уменьшение (или увели-
чение) размеров во всей популяции, занимавшей данный ареал рас-
пространения. Иное дело, если уменьшение или увеличение тела на-
блюдается не в одном, а в двух или нескольких последовательных го-
ризонтах: тогда сомневаться в индуцированной наследственной измен-
чивости величины организма уже трудно. Если отклонение от разме-
ров, нормальных- для данной филогенетической ветви, очень значи-
тельно, то и такой факт говорит в пользу наслед!СТ1ве1Н1Н1О1го характера
такого отклонения: чем резче отклонение, тем вероятнее его наслед-
ственный характер.
Таковы некоторые предварительные общие соображения о внутри-
популяционных изменениях величины тела особей ископаемых живот-
ных. Изучение этих явлений имеет немалое значение для познания
изменчивости организмов в геологическом прошлом.
Все это побуждает нас посвятить особую главу вопросу о внутри-*
популяционных изменениях величины тела организмов и о условиях
среды, их индуцирующих.
О ЯВЛЕНИЯХ КАРЛИКОВОСТИ У ИСКОПАЕМЫХ БЕСПОЗВОНОЧНЫХ
Исследователи, геологи и палеонтологи, нередко указывали на
конкретные случаи заметного уменьшения размеров органических
форм. При этом карликовость, то есть необычайно малые размеры
особей, обычно объяснялась неблагоприятными условиями среды, в
которых происходило индивидуальное развитие особей.
азз
В нашу задачу не входит рассмотрение развития малой величины
тела, как результата естественного отбора. Малая величина может
представлять адаптивное преимущество и становиться постоянным ви-
довым признаком.
В статье, посвященной карликовым фауанам, Г. В. Шаймер (Н. V.
Shimer, 1908) указывал, что неблагоприятными условиями, вызываю-
щими карликовость, могут быть отклонения от нормального гидроло-
гического режима, изменение в химическом составе воды, вызван-
ное опреснением, увеличение солености, содержания железа или
других ионов, а также возрастание количества сероводорода или
других газов; колебания температуры и др. По Ф. Б. Лумису
(F. В. Loomis, 1903, стр. 920), фауна пиритового слоя горизонта Тул-
ли (низы верхнего девона) в западной части штата Нью-Йорк, содер-
жащая более 45 видов, состоит из особей величиной в среднем около
1/15 величины средней величины особей того же вида из подстилаю-
щих слоев толщи Гамильтон (верхи среднего девона). Эта карликовая
фауна содержит 1 форму из бластоидей, 15 плеченогих, 12 двустворок,
8 брюхоногих, 7 головоногих, 1 трилобита и две формы из остракод.
Все экземпляры — крошечных размеров. В качестве причины карли-
ковости указывают на присутствие большого количества растворенно-
го железа и газов, происходивших от разложения органических ве-
ществ. Аналогичный случай представляет, по Лумису (там же), фауна,
остатки которой сохранились в оолитовой железной руде слоев Клин-
тон (силур) в Рочестере (штат Нью-Йорк). Встреченный здесь ком-
плекс ископаемых беспозвоночных состоит из особей, диаметр кото-
рых в среднем равен одной трети диаметра представителей того же
вида из непосредственно перекрывающих и подстилающих пластов
(Лумис, стр. 865).
Вполне допуская измельчание беспозвоночных из только что наз-
ванных толщ вследствие неблагоприятных условий жизни, мы не уве-
рены в том, что указываемая исследователями причина этого явления
соответствует действительности. Г. А. Купер (Cooper, 1957, стр. 804)
сообщает, что, согласно исследованиям минералогов, пирит Тулли,
быть может, есть продукт замещения известняковых линз пиритом.
Это делает сомнительным значение железа как причины карликово-
сти в данном случае. Нет, однако, никаких сомнений в том, что из-
мельчание вызывалось в этих случаях каким-то изменением условий
существования морских беспозвоночных.
Исследователи нередко указывают на подобные случаи уменьше-
ния тела компонентов тех или иных комплексов денных моллюсков.
Такие комплексы обычно именуются геологами и палеонтологами «уг-
нетенными фаунами» (см., например, М. Э. Ноинский, 1932). Мы отме-
чали (Давиташвили, 1936, стр. 181) малую величину некоторых мол-
люсков из слоя бурого железистого песчаника дуабских слоев у слия-
ния Дуаба с Мокви, в Абхазии. «Здесь карликовыми размерами отли-
чаются в частности Didacna crassatellata Desh. catephes Dav., Prionopleura
priniopleura (Andrus.) acetae (Dav.), Prosodacna longiuscula Sen. var“.
Ранее мы (1933, стр. 374) писали: «Возможно, что карликовые раз-
меры некоторых кардиид из слоя железистого песчаника дуабского раз-
реза объясняются неблагоприятными для развития условиями, возмож-
но — вредным действием солей железа».
Этим соображениям в известной мере соответствуют данные о со-
временных морских бассейнах. При рассмотрении фауны Балтийского
моря Л. А. Зенкевич (1947, стр. 229) отмечает «правило уменьшения
13k
размеров при уменьшении солености». Так, Mytilus edulis, имеющий у
берегов Англии до 150 мм в длину, в Кильской бухте имеет не более
ПО мм, а у берегов Финляндии не больше 40 мм, в дистальных же
частях Ботнического и Финского заливов только до 21—27 мм. Одна-
ко, по словам Зенкевича (там же), на некоторых формах это правило
не проявляется. На это указывают размеры Macoma baltica в Север-
ном море и в различных частях Балтийского моря. «Видимо, — за-
ключает Зенкевич (там же, стр. 229 и 230),—в противоположность
упомянутым выше Mytilus, Муа и Cardium, Macoma подчиняется воз-
действию изменения солености лишь в слабой степени». Нелегко ска-
зать, какие изменения испытывали донные моллюски в Черном море
при происходивших там изменениях гидрологических условий. Этот во-
прос требует специального изучения тем более, что имеются указания
на некую «перечеканку» средиземноморского -облика исходных форм
моллюсков на черноморский (Зенкевич, 1947, стр. 271). Но никто из
исследователей современных морских фаун моря не обнаруживает ка-
кого-либо сомнения в обусловленности изменения величины тела бен-
тонных моллюсков изменениями условий среды.
Мы не будем детально рассматривать вопрос о тех факторах сре-
ды, которые могут вызывать уменьшение или увеличение тела живот-
ных. Этот вопрос был детально изучен П. Ташем (Tasch, 1953 и
1957). Он нашел, что факторами, влияющими на рост животных (преи-
мущественно водных), могут быть: физиологическое действие три-
гидрола воды (ЗН2О), воздействие сероводорода, катионов железа и
других металлов, солености воды, pH, а также скученность населения
па данном участке и т. д. Таш (Tasch, 1953 и 1957) тщательно иссле-
довал «лимонитовую» фауну из пенсильванских отложений штата
Канзас. Эта фауна считалась карликовой, но в такую общую харак-
теристику Таш внес существенные поправки. Если относительно мел-
кие экземпляры окажутся просто юными, не достигшими взрослого
состояния, то карликовость их, конечно, — мнимая. Только некоторые
формы из этой фауны оказались действительно карликовыми. Из 21
элемента здесь, по-видимому, только один, Imitoceras grahamense, как
кажется, содержит карликовые экземпляры. Остальные элементы ли-
бо не поддаются изучению с этой стороны, либо представлены не
карликовыми взрослыми, а лишь юными экземплярами. Единообразие
в размерах тела ископаемых животных из данного местонахождения
может зависеть от седиментационных процессов, — сортировки остат-
ков организмов [Таш (Tasch, 1953), стр. 402]. Тем не менее Таш (там
же, стр. 409) признает, что у морских беспозвоночных наблюдается
прямое отношение между средней величиной тела и соленостью вод:
эти животные обычно мельче в воде с относительно низкой соленостью,
чем те же виды в воде с более высокой соленостью (Гунтер, (Gjn-
ter), 1945, стр. 55 .и 56].
Таш утверждает, что многие случаи карликовости ископаемых фаун
документированы недостаточно, а потому не могут быть приняты в рас-
чет при изучении явления уменьшения величины тела у животных прош-
лых геологических времен. Общий вывод Таша, который попытался соб-
рать все примеры ископаемых карликовых форм, сводится к тому, что
даже «классические», чаще всего упоминаемые случаи карликов-ости жи-
вотных генолог-ического прошлого, оказываются плохо документирован-
ными, что якобы явно недостаточно обоснована их «карликовость».
Это пессимистическое заключение Таша, основанное, правда, на
изучении лишь морских фаун прошлого, показывает, что вопрос о кар-
135
ликовом характере отдельных ископаемых форм и целых комплексов
таковых заслуживает тщательного критического пересмотра. Изучение
этого вопроса на материале древнее неогена, по-видимому, представ-
ляет большие трудности. Это относится, в частности, к так называемым
карликовым ам'мовитовым фаунам из среднего и верхнего триаса [А. А.
Бони (A. A. Boni), 1940 и 1942; Б. Куммель (В. Kummel), 1948].
Тем не менее в палеобиологии известны совершенно бесспорные
случаи значительно измельчавших беспозвоночных. Это — двустворки
и брюхоногие из некоторых среднемиоценовых и мэотических отложе-
ний Паратетиса. Нам неоднократно приходилось касаться этого вопро-
са. В одной из наших работ мы (1936, стр. 192) писали: «Наиболее
яркий пример этого явления представляет по моему мнению морская
моллюсковая фауна мэотического яруса. Здесь почти все родовые
группы (имеются в виду группы морского происхождения. — Л. Д.)
представлены весьма мелкими, совершенно карликовыми формами:
Area, Modiola, Tapes (Paphia.—Л. Д.) Dosinia, Mactra, Trochus, Nassa
и т. д.». Еще раньше мы писали (1933, стр. 160), что в значительно
опресненном море не было условий, благоприятных для широкого рас-
селения и распространения морских форм» проникших в бассейн из
какого-то связанного с океаном моря: «О том же свидетельствует и
«малорослость» мэотической фауны: морские виды представлены обыч-
но маленькими экземплярами. Некоторые формы являются совершен-
но карликовыми; таковы, например, известные доныне представители
Trochus и Nassa. В общем представители морской фауны в мэотисе
мельче, чем в чокракских и конкских отложениях (средний миоцен.—
Л. Д.), где фауна также известна малоросл остью».
Нам и позже приходилось возвращаться к этому интересному во-
просу, особенно в статье о воздействии среды на природу организмов
(1949) и в работе о некоторых вопросах изменчивости и видообразо-
вания моллюсков Паратетиса (1965). Моллюски, попавшие в конкский
бассейн (конкский век непосредственно предшествовал сарматскому
или даже, по мнению некоторых исследователей, был начальной
частью сарматского времени), не имели здесь времени, достаточного
для сколько-нибудь интенсивного видообразования (Давиташвили,
1949, стр. 143); вследствие изменения среды одни виды измельчали,
другие быстро вымерли.
В мэотисе измельчание выразилось гораздо сильнее. Оно охвати-
ло всю группу моллюсков морского происхождения: Ostrea sp., Area
miopliocenica Dav., Modiolus minor Andrus., Loripes, pseudoniveus (Andrus.),
Cardium (Cerastoderma) mithridatis Andrus., Card, mithridatis ukrainicum
Ossaul., Card, maeoticum Dav., Dosinia maeotica Andrus., Paphia curta
(Andrus.), Paphia (Venerupis?) abichi Andrus., Abra tellinoides (Sinz.),
Mactra superstes Dav., Ervillia minuta Sinz., Sphenia cimmeria Andrus.,
Gibbula maeotica (Andrus.), Potamides (Pirenella) disjunctum (Sow.), Pot.
(Pir) disjunctoides (Sinz.), Pot. (Pir.) novorossicum (Sinz.), Bittium reticu-
latum (Costa), Alvania pseudalvania (Andrus.), Alvania maeotica Badz.,.
Nassa retowskii Andrus., Nassa andrussovi Dav., Nassa krokosi Ossaul.,
Tornatina minima Ossaul. и др.
Мы писали, что «формы морского происхождения, за исключени-
ем, может быть, двух форм рода Cardium, очень близких одна к дру-
гой, не обнаруживают никаких признаков видообразования или обра-
зования хотя бы подвидов, представляя собой каждая дериват одного
136
определенного морского вида» (Давиташвили, 1965, стр. 34). Впрочем,
Ц. И. Бадзошвили, специально изучавшая морскую фауну мэотиса
Грузии считает возможным проследить видообразование в группе пота-
мидов: от средиземноморского вида Potamides disjunctus, по ее мнению,
произошли Pot. disjunctoides (Sinz.), Pot. novorossicum и Pot. maeoticum
Badz-Она также думает, что Cerithium maeoticum Badz. филогенетически
связан с Cer. rubiginosum Eichw. Однако в обоих случаях новый вид от-
личается от предполагаемого предка упрощенной, ослабленной скульп-
турой. Кроме того относительно интенсивное формообразование этих
моллюсков может зависеть и от способности более успешно выдержи-
вать понижение солености по сравнению с настоящими морскими мол-
люсками. Ц. И. Бадзошвили (1966), посвятившая вопросу о карлико-
вости мэотических моллюсков особую статью, попыталась даже ко-
личественно выразить степень уменьшения раковин отдельных видов
по сравнению со средиземноморскими родичами этих видов, ныне жи-
вущими в морской среде. Отношение длины экземпляров мэотического
вида к длине раковины соответствующего современного вида она назва-
ла «коэффициентом карликовости» (там же, стр. 633). Ее данные по-
казывают, что, во-первых, разные формы сильно различаются между
'собой (по «коэффициенту карликовости», а, во-вторых, б пределах от-
дельных видов этот коэффициент подвержен весьма значительным гео-
графическим колебаниям. Все это показывает, что морские моллюски,,
попавшие в мэотичеокое сильно опресненное море и там не вымершие,
испытывали на себе неблагоприятные условия новой среды не в одина-
ковой степени и отнюдь не совсем одинаково на них реагировали. Но
все они измельчали. К этому надо прибавить, что то более, то менее
ограниченное распространение отдельных видов нельзя обьяснять толь-
ко неполнотой палеонтологической летописи. Этот последний фактор
имеет, конечно, определенное значение, но в то же время едва .ли мож-
но думать, что только в силу неполноты этой летописи, при относи-
тельно равномерном распределении видов во всех аналогичных биото-
пах мэотического моря, отдельные формы были бы представлены —
то обильно, то скудно — на отдельных небольших участках и полно-
стью отсутствовали бы на всех остальных. А между тем, как известно,
одни виды морских моллюсков мэотиса имеют широкое распростране-
ние на всей площади развития мэотических отложений, а другие ха-
рактерны лишь для отдельных «пятен», как Западная Грузия, Таман-
ский полуостров, бассейн Ингульца и т. д. По-видимому, некоторые
виды морских мэотических моллюсков были местными формами, энде-
миками, жившими только в отдельных’ местах морского дна. Такая
картина (показывает, что морские моллюски, проникшие в мэотичеокое
море, где они нашли много свободного пространства, незанятого близ-
кими к ним аборигенами (ибо таковые все вымерли к концу позднего
сармата), испытывали на себе «угнетение» в различной степени. Одни
из них, хотя в общем измельчали, но все-таки создали обильные по-
пуляции и получили более или менее широкое распространение (на-
пример, Dosinia rriaeotica и Modiolus minor), а другие оказались в по-
ложении локальных реликтов.
В такой обстановке понятно измельчание и его причина — оно бы-
ло вызвано прямым воздействием внешней среды. Очень трудно до-
пустить, что изменения были лишь модификациями, — онтогенетичес-
кими реакциями на раздражения со стороны гидрологических факто-
ров, не передававшимися по наследству от поколения к поколению.
Ведь речь идет о геологическом веке, длительность которого выража-
137
лась либо миллионами, либо, в крайнем случае — сотнями тысяч лет.
Даже чисто эндемичные формы занимали немалые участки дна мэо-
тического моря, а другие были распространены почти на всей площади
распространения осадков мэстического яруса. Мы не говорим уже о том,
что ученые выделяли и выделяют различные виды мэотических мор-
ских моллюсков, то есть не сомневались в таксономическом значении
измельчавших форм. Очень трудно представить себе возникновение
тождественных модификаций в течение огромного отрезка времени на
обширной площади, занимаемой мэотическими пластами. Мы не ис-
ключаем возникновения модификаций, то есть ненаследственных изме-
нений, но наряду с таковыми должны были происходить более стойкие
преобразования моллюсков, — называйте их, если угодно, длительны-
ми модификациями, или мутациями. Иначе трудно представить себе
продолжительное, <в геологическом смысле, существование форм, ко-
торые многими исследователями, начиная от И. Ф. Синцова и
Н. И. Андрусова, принимались за особые виды. Но дополнительные со-
ображения, подтверждающие наследственный характер изменений, выз-
ванных у моллюсков мэотиса воздействием среды, будут приведены
после рассмотрения явлений возрастания величины тела у ископаемых
беспозвоночных.
Мы сочли нужным уделить особое внимание данным о карлико-
вости мэотических моллюсков морского происхождения: эти данные
показывают, что, несмотря на довольно пессимистический вывод, к ко-
торому пришел Таш в результате своих тщательных исследований, па-
леобиология располагает бесспорными доказательствами значитель-
ного уменьшения размеров тела морских беспозвоночных, вызванного
определенным воздействием внешней среды.
Существуют, конечно, и многие другие свидетельства в пользу
изменения тела моллюсков в сторону карликовости под влиянием сре-
ды. Здесь мы сошлемся еще лишь на работу Д. М. Раузер-Черноусовой
(1928), которая предположила, что уменьшение и некоторые другие
изменения абсолютных размеров раковины форм у Cardium edule из
Соленого озера, близ Севастополя, зависели от повышения солености
и перехода от песчаного субстрата к илистому.
На табл. IV изображены особи Strombus bubonius из района остро-
вов Зеленого мыса и из тирренских отложений близ Аликанте (Испа-
ния), сильно различающиеся по размерам. Популяции, где преобла-
дают экземпляры малой величины, могут навести на мысль, о наличии
особей карликовой формы этого вида. Но, конечно, вопрос о том, име-
ют ли это колебания какое-либо таксономическое значение, требует в
каждом отдельном случае специального изА/чения.
ОБ УВЕЛИЧЕНИИ ТЕЛА У ДВУСТВОРЧАТЫХ И БРЮХОНОГИХ МОЛЛЮСКОВ
Существует довольно обширная литература о так называемом ги-
гантизме двустворок и брюхоногих. Однако, когда речь идет о гиган-
тизме, нередко подразумеваются изменения величины тела моллюсков
в процессе филогенеза, вызываемые естественным отбором. Такие при-
способительные изменения могли происходит, строго говоря, на основе
отклонений, происхождение которых путем воздействия внешних усло-
вий остаются невыясненными. Нас же в данном случае интересуют лишь
изменения, зависящие от влияния факторов внешней среды. Поясним
это одним примером. П. Ю. Родда и У. Л. Фишер, (Р. U. Rodda, 19G4)
в интересной статье о роде брюхоногих Athleta, из эоценовых отложе-
ний Прибрежной Равнины Мексиканского залива на обширном ма-
138
териале (1600 экземпляров) установили, что филогенез шел в направ-
лении возрастания размеров раковины, которое сопровождалось после-
довательными изменениями различных морфологических элементов ра-
ковины. Однако мы не знаем ничего определенного о происхождении
индивидуальных изменений, на основе которых в данном случае про-
текала эволюция. Поэтому мы не можем воспользоваться этим мате-
риалом для выяснения генезиса внутривидовой изменчивости и ее ха-
рактера.
Нередко, однако, исследователи более или менее убедительно
обосновывают положение о том, что увеличение размеров моллюсков
зависело от факторов внешней среды. Это касается и наземных, и прес-
новодных, и солоноватоводных, и мороких моллюсков.
Изучение этого материала показывает, что в одних случаях уве-
личение тела и раковины происходило без коренных- изменений в мор-
фологическом строении животного, а в других вместе с общим увели-
чением тела происходило и глубокое изменение в строении, функциях
и образе жизни моллюсков.
Рассмотрим сначала вопрос о возрастании величины тела без ко-
ренных морфологических, физиологических и этологических изменений
у наземных моллюсков. В этом отношении большой интерес представ-
ляют исследования испанского ученого Г. Колома (Colom, 1955, 1957 и
1961), который изучал явления гигантизма у наземных моллюсков
острова Майорки из Балеарских островов. Полученные им результаты
и выводы из них сводятся к следующему. По крайней мере некоторые
из гигантских форм связаны с гористыми биотопами возвышенности
Сиер'ра Норте, а также соседних массивов высотой более 1000 м.
Гига1нтиз1м четко выражен у двух видов, относимых к семейству
Helicidae — Archelix punctata (Miiller) и Asteromphalus aspersus. У
этих видов гигантизм зависит, по Колому, от высоты биотопов — он
тесно связан с особой средой гористых местностей. Климат Сиерра
Норте, несмотря на высоту, скорее сухой. Однако на горах постоянные
в этой зоне дожди и туманы, вызываемые относительно низкими, по
сравнению с низинными участками, температурами, поддерживают
достаточно обильную растительность до наступления лета. Поэтому тут
гелициды имеют постоянный источник питания около 10 месяцев в го-
ду. На низменностях же растительность обычно исчезает к середине
мая. Соответственно гелициды низменных участков подвержены про-
должительной летней диапаузе, которая может длиться четыре или
даже пять месяцев, от середины мая до конца октября, когда начи-
нают выпадать первые осенние дожди. Живущие же в гористой зоне
популяции остаются вне влияния столь длительной засухи, и для них
летняя диапауза длится не более одного-полутора месяцев. На эти
благоприятные условия популяции гористых биотопов отвечают, по
Колому, увеличением тела и соответственно раковины.
Таким образом, как думает Колом, в данном случае существенное
увеличение размеров моллюсков зависит от обилия пищи и сокра-
щенной диапаузы, и этому не препятствуют относительно низкие тем-
пературы, даже волны холода и снегопад.
Однако Колом (Colom, 1961, стр. 296) говорит об этом явлении
как об «аномальном развитии этих гигантских форм» и не затрагивает
.-вопросы о его значении для эволюции. На самом деле, в данном случае
нет доказательств наследственного характера гигантизма. Возможно,
что тут налицо лишь следствие улучшения условия питания и сокра-
щения весенне-осенней диапаузы. Но в то же время никак нельзя
считать доказанным чисто модификационное значение гигантизма ге-
139
лицид острова Майорки. Этот вопрос безусловно заслуживает дальней-
шего углубленного изучения.
Рассматривая вопрос об изменчивости наземных моллюсков и ее
происхождении, мы не можем умолчать о редкой находке двух форм,
встреченных нами в дуабских отложениях. Это были две раковины,
которые были переданы нами для изучения покойному В. А. Линд-
гольму незадолго до его смерти и позднее утраченные. Одну из них
можно было без колебаний отнести к роду Pomatias, о другой же В. А.
Линдгольм сообщил мне, что она напоминает по облику раковины ро-
да Elona и принадлежит либо к Helicidae, либо к Zonitidae. «Во вся-
ком же случае,—писали мы (1933, стр. 375),—обе формы обладают по-
разительно крупными, гигантскими размерами по сравнению с подоб-
ными им ныне живущими моллюсками. Исключительная величина
этих раковин, быть может, говорит (. . .) за теплый и влажный климат
суши, Прилегающей к этой части киммерийского бассейна». Мы, к со-
жалению, не знаем ни о каких новых находках этих гигантских на-
земных форм в дуабских слоях или в каких-либо иных местонахожде-
ниях отложений киммерийского возраста. Между тем, изучение их
могло бы пролить свет на физико-географические условия этой при-
брежной полосы суши в киммерийский век. Нам представляется весь-
ма вероятным, что несомненно огромная величина раковин представи-
телей рода Pomatias указывает на существовавшие там очень благо-
приятные условия климата и питания. Найденная нами раковина была
так велика, что, исходя из данных о самых крупных помациасах (ци-
клостом), ископаемых и современных, мы не можем представить
себе внезапный переход от величины «нормальной» к величине поисти-
не исполинской. Должны были существовать какие-то промежуточ-
ные, по величине, звенья. А это наводит на мысль о наследственном
характере изменчивости дуабских помациасов. То же самое, пожалуй,
можно сказать и о другой раковине, с гладкой наружной поверхностью
оборотов.
Палеобиология располагает обильными материалами, свидетель-
ствующими об изменении размеров моллюсков в сторону увеличения,
гигантизма.
Однако эти филогенетические изменения обычно сопровождаются
значительными изменениями морфологическою строения и адаптаций.
Возрастание величины раковин наблюдается во многих группах дву-
створок и брюхоногих позднего кайнозоя Паратетиса. Поскольку из-
менчивость неогеновых и четвертичных форм этих двух классов рассма-
тривается особо в другом разделе нашей книги, мы здесь приведем
лишь некоторые наиболее наглядные случаи и общие сведения о раз-
витии этих организмов к сторону гигантизма.
Спаниодонтеллы. Происхождение древнейших спаниодонтелл от
лютеций, известных с эоцена, считается установленным.
Рисунок 16 дает сильно упрощенное и схематизированное изобра-
жение развития от древнейших спаниодонтелл миоцена, которые были
почти столь же крошечными двустворками, как лютеции, в сторону бо-
лее крупных чокракских форм, которые позже дают начало разнооб-
разным, но в общем более крупным караганским формам. Эти послед-
ние, после некоторой адаптивной радиации, вымирают в конце кара-
ганского времени. Многие караганские спаниодонтеллы были подлин-
ными гигантами в этой группе двустворок. Увеличение размеров бы-
ло, однако, связано с экогенетической экспансией и дивергенцией в фи-
логенезе спаниодонтелл, и можно было бы думать, что здесь совер-
шался процесс эволюции на основе совершенно случайных, шальных,.
140
мутаций. Но для сильно убыстренной эволюции путем естественного
отбора были необходимы многочисленные наследственные изменения в
сторону увеличения тела, м такие изменения должны были идти непре-
рывным потоком на протяжении всего времени от раннего миоцена до
позднего карагана.
Мелкие спаниодонтеллы вновь появляются в конкское время, но
это уже не потомки форм предыдущего, караганского времени, а но-
вые вселенцы из полносоленого моря, откуда пришла вся конкская
фауна морских моллюсков.
Эрвилии миоцена. Мелкие фор-
мы морского миоцена, жившие в
полносоленом море, дали начало
более крупным — сначала в чок-
раке, где развивалась относитель-
но крупная Ervilia prapodolica, вы-
мершая в чокраке, не оставив по-
томства, а затем в нижнем сарма-
те, где существовала Erv. dissita.
Из этой последней развилась Erv.
concinna, которая жила на относи-
тельно ограниченном участке средне
сарматского моря. Увеличение
размеров имело очевидно адаптив-
ное значение, но и здесь оно могло
совершаться лишь на основе доста-
точно обильной внутривидовой из-
менчивости (рис. 17).
Мактры сармата. В течение сар-
матского века мактры эволюиро-
вали необычайно быстро. Одни
формы сохраняли небольшие раз-
меры, другие быстро увеличива-
вались, но особенно характерен для
сарматских мактр процесс чрезвы-
чайно интенсивной дивергентной
эволюции. Очень схематизирован-
ный рисунок 18 дает некоторое
представление лишь о филогенети-
ческом увеличении тела, характер-
ном для отдельных линий сармат-
ских мактр. Так или иначе
Рис. 16. Схема, поясняющая уве-
личение размеров спаниодонтелл в
процессе филогенеза. 1—Spaniodon-
tella nitida, предкева i ферма.
2—формы из чокрака. 3. формы
из карагана. 4—фермы из
конки. X 1/2
что многие мактры в
примечательно,
среднем сармате, имевшие относительно малорослых предков в ран-
нем сармате, оказываются очень крупными представителями рода. Со-
вершенно несомненна интенсивная внутривидовая изменчивость, кото-
рая вела к гигантизму или по крайней мере заметному увеличению
многих мактр в среднем сармате (хотя довольно крупные формы из-
вестны уже в верхах нижнего сармата и попадаются также местами в
верхнем сармате).
Пафии сармата. Аналогичную картину представляют филогенети-
ческие ветви, ведущие от Paphia modesta к среднесарматским пафи-
ям — Paphia vitaliana d'Orb. и Р. gregalia Partsch. И в этой груп-
пе происходила довольно значительная радиация, но вполне заметно
общее увеличение размеров форм, особенно форм, обитавших на песча-
нистых грунтах.
141
Кардииды сармата. Эта группа достшла в раннем и особенно
среднем сармате исключительно пышного расцвета. Исследователю
бросается в глаза разнообразие адаптаций сарматских кардиид, с чем
связано было возникновение множества видов, большое число которых
все еще ждет монографического описания. Мало изучены формы, оби-
тавшие на тонкозернистых грунтах. И все-таки во многих филогенети-
ческих ветвях наблюдается увеличение тела, а противоположного яв-
ления мы почти не знаем, по крайней мере при переходе от раннего
сармата к среднему. В среднем сармате некоторые филогенетические
ветви приводят к очень крупным формам (рис. 19). Все это, по нашему
мнению, показывает, что кардииды нижнего и особенно среднего сар-
мата жили и эволюировали в благоприятных для их существования
условиях, что и вызывало, надо думать, множество индивидуальных из-
менений тела в сторону увеличения. Закреплявший эти изменения ес-
тественный отбор вел к образованию разнообразных видов, которые в
общем были более крупны, чем
раннесарматские.
Рис. 18. Сильно упрощенная схема,
поясняющая возрастание величины в
некоторых f филогенетических линиях
рода Mactra в течение сармата. 1—Mac-
tra basteroti 2—М. eichwaldi Lask. bug-
lovensis Lask. 3—M. eichwaldi einchwal-
di Lask. 4—M. vitaliana d’Orb. I—Пол-
носоленый морской средний миоцен.
II—конка. Ill—нижний сармат.
IV—средний сармат, х 1/2
Рис. 17. Сильно упрощенная схема фи-
логенеза рода Ervilia в миоценовых бас-
сейнах Паратетиса. Показана лишь ли-
ния, ведущая к крупной среднесармат-
ской форме Erv. concinna. 1—Erv. pu-
silia Phil. 2—Erv. trigonula Sokol.
3—Erv. dissita Eichw. 4—Erv. concinna
Eichw. I.—Полносоленый средний мио-
цен. II—конка. Ш—нижнии сармат.
IV—средний сармат
Моллюски солоноватоводного плиоцена. Известно, что некоторые
виды, встречающиеся в понтических и вышележащих киммерийских
отложениях, достигают в киммерии более крупной величины, чем в
142
понте. Примерами могут служить Plagiodacna carinata Desh. и Dreisse-
nomya aperta Desh. Примечательно, что в таких случаях ни один
автор, насколько нам известно, не предлагал относить понтические и
киммерийские экземпляры к различным таксонам, хотя бы к различ-
ным подвидам. По всей вероятности, такие случаи представляют собой
явление увеличения роста, которое здесь едва ли может быть объясне-
но без допущения, что оно зависело от воздействия каких-то благо-
приятных внешних условий. В дальнейшем, оно могло быть закреплено
естественным отбором. Почти все
киммерийские формы двустворок за-
метно крупнее их понтических роди-
чей и предшественников, от которых
они могли произойти. Происхожде-
ние всех основных элементов ким-
мерийской моллюсковой фауны от
понтических ни у кого не вызывает
сомнений. Довольно искусственным
было бы обьяснение преобладания
в киммерии более крупных форм от-
бором «шальных» мутаций. Прави-
льнее, нам кажется, предположе-
ние, что возрастание величины те-
ла тут происходило вследствие воз-
действия каких-то, в общем благо-
приятных, условий существования,
среди которых видное место зани-
мало, надо полагать, обилие пищи,
зависевшее от длительного господ-
ства теплого и влажного климата.
Однако следует обратить вни-
мание на то обстоятельство, что
более или менее значительное уве-
личение тела у моллюсков понта и
киммерия в большинстве случаев
было сопряжено с заметными изме-
нениями морфофизиологического
характера. Понтические формы за-
частую существенно отличаются от
своих киммерийских преемников по
своим морфологическим чертам.
Особенности, приобретенные в
результате воздействия факторов
внешней среды, могут оказаться по-
лезными и потому закрепляться че-
Рис.' 19. Сильно упрощенная схема фи-
логенетических отношений некоторых
сарматских форм рода Cardium, показы-
вающая увеличение раковин. 1 и 2—С.
ruthenicum Hilb. 3.—С. lithopodolicum
Dub., 4 и 5—С. sarmaticum Barb- 6—С.
repartitum koles. 7—C. laevigato-loweni
koles. 8—C. loveni. xl/2
рез естественный отбор.
Еще Н. И. Андрусов (1897, стр. 648; 1964, стр. 369) писал об эво-
люции конгерий: «Параллельно с выработкой синупаллиатного, под-
вижного, типа, замечается стремление противоположного рода, а имен-
но к образованию крупных неподвижных форм... Сильный биссус
Congeriae mytiliformes представлял уже достаточную гарантию против
механического действия волн». Далее Андрусов говорит, что увеличе-
ние размеров раковин конгерий группы triangulares «было лишь след-
ствием благоприятных условий питания и вместе с тем прекрасным
приспособлением против врагов, какими несомненно являлись рыбы.
143
Увеличение толщины раковины и образование острых или выдающихся
килей делало, конечно, затруднительным разгрызание и проглатывание
раковин». У некоторых ископаемых конгерий массивность, толстостен-
ность крупных раковин делали, по Андрусову, излишним биссус, кото-
рый испытывал редукцию. У Congeriae rhomboideae следов биссуса не
замечается, «и лишь крупная толстая раковина служила у них одно-
временно защитой от врагов и волн» (там же).
В этих словах замечательный геолог-палеобиолог дает правильное,
по нашему мнению, объяснение филогенетического изменения конге-
рий в сторону увеличения тела и раковины: изменчивость вызывалась
благоприятными условиями питания, а поскольку это было выгодой,
отбор вел эволюцию <в стосрану возрастания размеров и утолщения ра-
ковин.
Наблюдаемые у многих двустворок понта-киммерия-куяльника яв-
ления увеличения тела происходили параллельно с морфофизиологи-
ческими и экологическими изменениями, то менее, то более глубоки-
ми. Это можно иллюстрировать рассмотренными нами примерами фи-
логенеза группы ,,Didacna“ multistriata—«Did.» caeleno, а также групп
Digressodacna digressa и Tchaudia. Дальнейшие примеры можно было бы
взять из истории групп ,,Didacna“ (Crassadacna) crassatellata, Limnocar-
dium moquicum, Prosodacna macrodon, Congeria caucasica и т. д.
Таким образом, у двустворок солоноватоводного плиоцена Черно-
морско-Каспийской области филогенетическое увеличение общих раз-
меров раковины наблюдается преимущественно v форм тех ветвей, ко-
торые испытывали более или менее интенсивное видообразование или
даже филогенетическую радиацию (но, конечно, возрастание тела не
всегда сопровождало интенсивный процесс видообразования).
Сопоставим теперь то, что было сказано о развитии карликовости
(нанизма) с тем, что мы видим у форм, испытывавших развитие в на-
правлении гигантизма. У первых мы не замечали сколько-нибудь ин-
тенсивного видообразования, и могла бы даже возникнуть мысль, не
являются ли карликовые формы простыми модификациями, ненаслед-
ственными изменениями, возникавшими в результате реакции орга-
низма на воздействие тех или иных факторов среды (преимущественно
абиотических гидрологических факторов). У вторых размеры тела из-
менялись, в противоположную сторону, но параллельно существенно
изменялись морфофизиологические и этологические особенности орга-
низма. Если развитие в сторону гигантизма было всегда связано с из-
менением наследственных черт животных, с формированием новых так-
сонов, то естественно предположить, что и нанизм развивался как на-
следственное свойство и становился характерным для таксонов мало-
рослых моллюсков, эволюировавших от форм моллюсков с нормальной
величиной тела.
Итак, разбор данных'об ископаемых карликовых и гигантских
моллюсках и о причинах изменений величины их тела приводит нас
к следующим общим выводам:
1. Изменения величины тела как в сторону карликовости (наниз-
ма), так и в сторону гигантизма зависят от условий жизни, более или
менее отклоняющихся от обычной нормы. В первом случае эти условия
становились неблагоприятными, а во втором, наоборот, более благо-
приятными, чем в норме.
2. Вызванные факторами среды изменения размеров тела по край-
ней мере иногда становились или оказывались наследственными. Че-
рез естественный отбор устанавливалась новая норма величины тела.
144
Нам остается только добавить, что в некоторых случаях суще-
ствует возможность привлечения экспериментального метода к реше-
нию вопроса о том, является ли отклонение величины тела от обыч-
ной нормы наследственной или ненаследственной, модификационной.
В принципе для этого достаточно поставить некоторое количество осо-
бей гигантской или карликовой популяции вида в гидрологические, ус-
ловия, обычные для предков данной формы, обладавших «нормальны-
ми» размерами. Возврат этой популяции изучаемого вида к обычной
величине непосредственных предков был бы доказательством модифи-
кационного характера уменьшенных или увеличенных размеров. Про-
тивоположный результат говорил бы в пользу признания того, что
воздействие некоторых факторов на ранней стадии онтогенеза вызы-
вает иногда наследственные изменения. Однако мы вполне отдаем се-
бе отчет в том, что постановка и проведение таких экспериментов (на-
пример, над некоторыми моллюсками Черного моря) сопряжено с
большими трудностями.
Изложенное в этой главе, по нашему мнению, подтверждает тезис
о возможности наследственного изменения величины тела у значитель-
ной части популяции данного вида вследствие воздействия некоторых
факторов среды на развивающихся особей моллюсков.
Этот тезис не опорачивается тем фактом, что изменения величи-
ны тела в сторону увеличения или в сторону уменьшения могут быть
полезными.
ВНУТРИВИДОВЫЕ ИЗМЕНЕНИЯ ВЕЛИЧИНЫ ТЕЛА У ИСКОПАЕМЫХ
ТЕТРАПОД
Многие исследователи указывали на изменения средней величи-
ны — карликовость и гигантизм ископаемых четвероногих позвоноч-
ных — млекопитающих, птиц и рептилий. Тут мы имеем в виду не очень
мелких или очень крупных животных вообще, а только тех, которые,
судя по их филогенетической истории, являются либо сильно измель-
чавшими, карликовыми, либо сильно увеличившимися, гигантскими, в
сравнении с обычными представителями тех же групп, близкими роди-
чами этих карликов или гигантов. Наше внимание в первую очередь
привлекают животные сравнительно поздних геологических времен —
антропогена и неогена. И само собой разумеется что, в соответствии с
основной темой этой книги, мы будем рассматривать эти материалы с
определенной точки зрения — с точки зрения индивидуальной из-
менчивости некогда существовавших популяций. Это, конечно, не зна-
чит, что нельзя заметить нанизм у представителей совершенно вымер-
ших групп животных.
Палеонтологи и геологи, изучающие историю органического мира,
зачастую исходят из убеждения в том, что и нанизм и гигантизм не-
редко развиваются как прямой результат воздействия факторов сре-
ды. При этом эти ученые обычно считают излишним доказывать пра-
вильность такого понимания происхождения карликовости и гигантиз-
ма: оно представляется им совершенно бесспорным. Так, Г. Ф. Осборн
(Osborn, 1929, т. 1, стр. 29), который писал о карликовости у титано-
териев. (бронтотериоидей), утверждал, что нанизм, так же как и
гигантизм возникает вследствие воздействия факторов среды. «Изве-
стно, — писал он (там же), что гармоническое возрастание или умень-
шение величины всего животного сопряжено с некоторыми изменения-
ми в местообитании; часто это явно зависит от того, что благоприят-
ная среда способствует развитию более крупных рас, а неблагоприят-
ная ведет к карликовости».
10. Л. Ш. Давиташвили 145
Внимание исследователей нередко привлекали карликовые гиппо-
потамы и карликовые слоны, жившие на Средиземноморских островах.
Как будет видно из дальнейшего, причины резкого уменьшения их ве-
личины мы считаем до сих пор недостаточно изученными. Абель (Abel,
1912, стр. 24) писал: «Скопление остатков карликовых слонов в пеще-
рах отдельных средиземноморских островов можно понять только в
том случае, если мы примем, что эти пещеры были местообитаниями
тех рас слонов, которые при их изоляции на средиземноморских остро-
вах не только сделались карликовыми расами, но и перешли к пещер-
ному образу жизни подобно ставшим карликовыми гиппопотамам этих
островом». По словам Абеля (Abel, 1912, стр. 559), изучение многочис-
ленных местонахождений ^ископаемых костей карликовых гиппопота-
мов и карликовых слонов средиземноморских островов позволило ему
сделать заключение о пещерном образе жизни этих животных. Если
кости животных этих двух групп встречаются в речных отложениях, то,
по некоторым наблюдениям, как утверждал Абель, они были вынесе-
ны потоками из пещер и теперь находятся во вторичном залегании.
Нам кажется, что при изучении этого вопроса заслуживают внимания
интересные соображения И. Г. Пидопличко (1951, стр. 27) об инфан-
тилизме некоторых слонов. «Есть указания, — пишет он, — что в преж-
нее время бивни африканских слонов попадались весом 120—130 кг. В
возможности такой величины бивней современных слонов принято
сомневаться. Однако, если учесть явление инфантилизма у современ-
ных слонов, как и у многих других животных, возникшее в результате
преследования их человеком, то подобное сомнение не оправдывает-
ся». Инфантилизм в данном случае понимается как сохранение у
взрослых особей бивней, по величине (и по форме) соответствующих
бивням юных особей более древних популяций, от которых произошли
нынеживущие слоны. По Пидопличко, это явление зависит от антропи-
геского фактора. Мы думаем, что не исключено действие этого фак-
тора также и по отношению к расам, которые дали начало карликовым
расам слонов и гиппопотамов средиземноморских островов. Но вопрос о
том, мог ли влиять человек на популяции четвертичных животных, нап-
равляя их развитие в сторону «инфантилизма» или карликовости, тре-
бует дополнительного тщательного изучения.
Остатки карликовых слонов изучены в общем подробнее и тща-
тельнее, чем остатки гиппопотамов. Эти слоны жили на островах Си-
цилии, Мальте, Сардинии, Кипре и Крите. Расселение слонов не могло
здесь зависеть от факторов переноса особей подобно многим мелким
беспозвоночным и позвоночным суши — всякие мысли о заносе слонов
какими-нибудь пловучими деревьями или ветром были бы совершенно
нелепы. Они могли попасть на острова лишь в том случае, если бы
таковые имели связь, хотя бы кратковременную, с материком. Но не
обязательно допустить, что территории всех перечисленных островов
образовали один сплошной массив суши, примыкавшей к территории
нынешнего материка Евразии. Возможно, например, что между Кип-
ром и более западными островами, которые были заселены слонами,
не было непрерывной связи и что популяции слонов, жившие на этих
островах, проникли туда независимо одна от другой с соседних мате-
риков. Этот вопрос требует особого изучения. Высказывалось мнение,
что родственные между собой, но самостоятельные виды пигмеев-сло-
нов и пигмеев-гиппопотамов на далеких друг от друга островах пред-
ставляют собой пример независимой, параллельной, эволюции [см. Ос-
борн, (Osborn, 1910), стр. 410] с некоторой дивергенцией от общих:
предков.
146
На средиземноморских островах было- отмечено несколько форм
карликовых слонов. Palaeoloxodon [Elephas] mnaindriae был описан
впервые с острова Мальты, а его указывали на острове Сицилии и да-
же на Крите. Palaeoloxodon meitensis Falconer первоначально был описан
тоже с острова Мальты, а затем этот вид указывали также из Сицилии.
Кроме того, некоторые исследователи отождествляли с ним Paleol.
lamarmorae, найденного на острове Сардинии, и Palaeol. creticus с
Крита. Palaeoloxodon falconeri Busk, первоначально' был описан по
остаткам, собранным на острове Мальты, а потом' был определен по
остаткам, найденным на острове Сицилии. К этому же виду относят
Palaeol. Cypriotes Bate, описанный по материалу с острова Кипра. Са-
мым малорослым из описанных карликовых островных слонов был
Palaeol. falconeri, средняя высота которого не превосходила 0,9 м. Не-
которое представление о величине и общем виде трех карликовых сло-
нов дает рисунок 20, где дается реставрация Palaeoloxodon mnaindriae,
Palaol. melitensis и Palaeol. falconeri. На рисункже 21 даются схемати-
ческие изображения бивней трех карликовых слонов (Palaeol. falconeri,
Palaeol. melitensis и Palaeol. mnaindriae и одного слона Palaeol. antiquus,
обладавшего бивнями нормальных, не уменьшенных размеров. Этот ри-
сунок показывает, что бивни карликовых слонов испытали значительное
уменьшение в различной степени.
Все три вида жили в плейстоцене, а остатки их сохранились пре-
имущественно в пещерных отложениях.
Заметим, что, если американские авторы (Г. Ф. Осборн и Дж. Г.
Симпсон) относят рассматриваемую тут форму к роду Palaeoloxodon,
то многие е®'ро1пей|сиие исследователи [например, Р. Вофрей (Vaufrey,
1958)] рассматривают их как представителей рода Elephas. Этих кар-
ликовых слонов обычно рассматривают как подвиды, или расы Elep-
has [Palaeoloxodon] antiquus. Мы, конечно, не будем тут входить в так-
сономический анализ этих форм и подчеркнем только, что наличие
взаимных переходов между отдельными формами, очевидно, затрудня-
ет проведение границ между таксонами видового и родового порядков.
Быстрое, в геологическом смысле, уменьшение размеров у карли-
ковых слонов — вне сомнения. Но мы затрудняемся указать на веро-
ятную основную причину этого филогенетического процесса. Почти все
исследователи, рассматривавшие этот вопрос, ссылаются на неблаго-
приятные условия среды. Одни из авторов имеют в виду прямое воз-
действие факторов, например, недостаток пищи, другие пытаются объ-
яснить это явление естественным отбором. Так, Ч. Ф. Сакки (Sacchi,
1961), ссылаясь на уменьшение тела у средиземноморских островных
форм — слонов, гиппопотамов, лисиц, оленей и других,—высказал
мысль, что, когда этот нанизм был наследственным, мы в праве толко-
вать его как адаптацию вида к неблагоприятной среде: у более мел-
ких особей меньшие метаболические потребности и потому. малорос-
лые виды могли иметь более многочисленные популяции.
Известное значение, быть может, имел тот факт, что слоны и не-
которые другие млекопитающие приспособлялись к пребыванию в пе-
щерах, а входы в пещеры относительно редко бывают достаточно ши-
рокими и высокими для очень крупных зверей.
Полагая, что этот вопрос требует новых исследований, мы все же
думаем, что обилие карликовых форм крупных зверей и быстрота про-
цесса, ведшего к карликовости, недвусмысленно указывают на повы-
шенную изменчивость в направлении уменьшения тела. Иначе нелегко
представить себе развитие, например, такого маленького слона как
147
Рис. 20. Карликовые слоны: 1— Palaeoloxodon antiquus mnaindriae. 3—Palaeob
antiquus melitensis. 2—Palaeol. antiquus falconeri. По Осберну.
Palaeoloxodon falconeri за столь короткий промежуток времени как не-
которая часть плейстоцена.
Если по существующим данным о возникновении карликовости у
млекопитающих затруднительно с полной уверенностью утверждать,
что е этом процессе существенную роль играло повышение изменчи-
вости внутри вида, то примеры филогенетического увеличения тела у
некоторых наземных четвероногих позвоночных являют убедительные
доказательства изменчивости в направлении гигантизма. Любопытно,
что такие примеры дают нам изучение опять-таки островных фаун.
Рис. 21. Схематические изображения бивней кар.:иксвых с ленов. 1—Palaeoloxodon
antiquus falconeri. 2—Palaeol. antiquus melitensis. 3—Palaloi. antiquus mnaindriae.
4—Palaeol. antiquus „нормальней величины". По Вофрей. х 1/20
Сошлемся прежде всего на гигантоких черепах островов Галапа-
гос вида Testudo elephantopus Harlan, в котором различают много
подвидов. Трудно представить себе, чтобы гигантская величина этих
черепах могла возникнуть без И1нтенси1вной индивидуальной изменчиво-
сти в этом направлении [Г. Вермут и Р. Мертенс (Н. Wermuth,
R. Mertens, 1961) У. Test elephantopus elephantopus (северная окончен-
ность острова Альбемарле) карапакс достигает 110см, в то время как Tes-
tudo elegans Schoepfer (Передняя Индия и Цейлон) имеет карапакс длиной
всего 25 см. У Testudo chilensis—около 22 см. Некоторые другие виды
могут быть гораздо крупнее, но все же далеко не достигают размеров
только что упомянутого подвида Test, elephantopus. Test, elephantopus
abingdonii Gunther, с острова Абингдон имеет карапакс в 100 см, Test,
elephantopus ephippium Giinther, с острова Дункан—НО см, Test, elephan-
topus galapagoensis Baur—110 см и т. д. Иззестна крупная черепаха
Сейшельских островоз Testudo gigantea Schweigger. Testudo gigantea
daudinii Dumeril et Birbon, живущая на южных острозах группы Альдаб-
ра, имеет карапакс длиной около 100 см, a Test, gigantea elephatina
Dumeril et Birton, на островах Мозамбикского пролива, достигает еще
более крупных размеров—ее карапакс имеет в длину около 125 см.
Столь крупных и даже более крупных размеров достигают некото-
рые морские черепахи рода Chelonia Latreille, но мы не будем здесь
касаться этой группы черепах. Здесь нас интересуют лишь гигантские
островные формы, которые, как мы видим, встречаются на различных
островах тропического пояса. Все они происходят от материковых
предков, имевших более умеренные размеры. Значительная, даже ги-
гантская, величина могла представлять для этих животных некоторые
преимущества; она, казалось бы, должна была бы быть столь же вы-
годной и для черепах соседних материков (Южной Америки и Афри-
149
ки). Потому естественно возникает вопрос, почему гигантской величи-
ны достигали островные формы, а родственные им материковые сохра-
няли Свои умеренные размеры. Можно предположить, что в островной
обстановке преимущество в борьбе за существование имели гиганты, а
р материковой — формы умеренных размеров. Но это предположение
приобретает некоторое значение только в том случае, если есть основа-
ние считать материковую среду относительно неблагоприятной для со-
ответствующих форм, если в ней действуют факторы, препятствующие
широкому процветанию этих черепах.
Наиболее простым и вероятным представляется то мнение, что на
островах экологические условия способствовали изменчивости в сто-
рону возрастания величины наземных черепах в большей мере, чем на
материках. Можно думать, что на островах эти животные не только
находили обильную пищу, но и встречали там гораздо меньше врагов
и.конкурентов, чем их родичи, жившйе на соседнем обширном масси-
ву суши. Можно полагать, что относительная легкость добывания пи-
щи была одним из факторов, вызвавших необычайные размеры остров-
ных черепах.. Другим фактором было очевидно уменьшение давления
со стороны врагов, которые могли нападать на черепах на разных ста-
диях их онтогенеза. Объяснить же гигантскую величину только есте-
ственным отбором обычных случайных изменений без повышения ин-
дивидуальной изменчивости представляется нам неправ ильным. К то-
му же филогенетическое увеличение тела происходило в таких случаях
очень быстро.
Обратимся теперь к рассмотрению вопроса об изменчивости пред-
ков некоторых островных птиц.
На Маскаренских островах еще недавно, примерно до конца 17-го
века, жили своеобразные нелетающие птицы, принадлежавшие к раз-
личным видам, но известные -под общим название^м дронтов- На
острове Маврикия жил Raphus cucullatus ( = Didus ineptus), на острове
Родригес Pezophaps solitarius, а на острове Реюньон Raphus solitarius
( = Didus borbonicus). Это были птицы с сильно редуцированными кры-
льями, безусловно лишенные способности летать. Их чаще всего относят
к отряду голубей (Дементьев, 1964), хотя они несравненно крупнее лю-
бых форм этого отряда — крупные дронты достигали в высоту 75 см.
Лютшвагер (Lilttschwager, 1961) оспаривает их отнесение к отряду
голубей, думая, что они скорее принадлежат к отряду пастушковых. Он
даже не исключает возможности, что для дронтов надо установить от-
дельный отряд.
Таким образом, дронты чрезвычайно сильно отклонились от сво-
их предков из летающих птиц. За какой же отрезок геологического
времени протекала эволюция этой исключительно своеобразной остров-
ной группы птиц? Эволюция в направлении утраты способности поле-
та могла начаться лишь после того, как предки этих птиц поселились
на территории островов Маскаренской группы. Если эта группа некогда
представляла какую-то часть единого крупного массива суши, то там
едва ли началась эволюция дронтов, как нелетающих птиц — они
слишком ясно несут на себе признаки форм, которые могли жить и
развиваться только в условиях изоляции, условиях отсутствия угрозы
со стороны активных соперников и хищников. Существует мнение, что
Мадагаскар, Сейшельские и Маскаренский острова представляют со-
бой остатки некогда существовавшего массива суши, который распал-
ся в послеэоценовое время.
Дату вселения летающих предков в этот островной район устано-
вить трудно. Но нет сомнения в том, что их эволюция в направлении
150
нелетающего образа жизни и увеличении размеров тела протекала
чрезвычайно, почти неимоверно быстро. В отсутствии сильных врагов
могли выживать относительно малоподвижные формы, эволюция кото-
рых привела к возникновению неуклюжих жирных птиц, которые потом
были быстро истреблены человеком, —дронтов не только отстреливали
мушкетами, ло,в1или в сетях, но и били просто палками [Ерииуэй, (J. С.
Greenway, 1958)]. Такой процесс эволюции мог происходить только при
исключительно усиленной индивидуальной изменчивости, зависевшей
очевидно от обильного питания при отсутствии отбора на быстроту
движений. История дронтов указывает на важное значение условий
существования, индуцировавших изменчивость.
Из сказанного явствует, что палеобилогическое изучение дронтов
может иметь огромное значение для решения вопроса о причинах из-
менчивости животных и о значении изменчивости, индуцированной
факторами среды, для эволюции органичеокго мира. Однако в истории
дронтов очень многое останется неясным до тех пор, пока не будет
лучше изучена геология Маскаренских островов. При нынешнем же
уровне знаний по этому вопросу мы можем строить лишь гипотезы,
которые, впрочем, не лишены интереса. Два из этих островов—о-ва
Реюньон и Маврикия находятся на общем цоколе, а третий — о-в Род-
ригес лежит в 638 км к востоку от о-ва Маврикия. Все три острова
представляют собой в основном вулканические сооружения. На о-ве
Реюньон вулканическая деятельность, по Э. Кренкелю (Е. Кгепке!,
1957, стр. 221) еще продолжается. Извержения — смешанного, гавай-
ско-стромболианского, типа. У подножия одной из гор указывалось
присутствие метаморфизированных известняков, включенных в базальт.
О-в Маврикия — тоже вулканический, но признаков недавней деятель-
ности не заметно. Упоминалось присутствие невулканических пород в
двух местах: глинистого сланца и смятого в складки хлоритового
сланца. О-в Родригес образован лавами и туфами. Существует предпо-
ложение, что этот остров расположен на гранитном фундаменте.
Когда возникли эти маленькие острова? На этот вопрос геология
не дает ответа. Вполне возможно, что они появились разновременно
и, что, например, о-в Родригес является наиболее древним из троих.
Другой вопрос, когда поселились на них предки дронтов. Учитывая
вулканическую деятельность, которая на о-ве Реюньон была интенсив-
ной еще в четвертичном периоде, мы считаем вполне возможным, что
первые поселения предков дронтов появились там сравнительно не-
давно, — может быть, в конце плейстоцена или даже в начале голоце-
на. Но доказанным считать это никак нельзя. Решение этого вопроса —
дело будущих исследований. Если подтвердится, что предки дронтов
прилетели на острова в позднем плейстоцене или еще позже, то можно
представить себе, какое огромное значение будет иметь такой палео-
биологический вывод для проблемы изменчивости в целом. Но если
даже эволюция дронтов тянулась миллион или два миллиона лет, то и
в таком случае темпы этого процесса надо признать исключительно
высокими и необъяснимыми без допущения направленной изменчиво-
сти, зависевшей от определенного воздействия факторов среды.
Нелетающие птицы Мадагаскара и особенно Новой Зеландии, суще-
ствовавшие еще в недавнем прошлом, испытывали чрезвычайно бы-
струю эволюцию в направлении увеличения роста, что тоже нельзя
представить себе без усиленной индивидуальной изменчивости в на-
правлении увеличения тела. Эта изменчивость была, по-видимому, вы-
зываема обилием легко доступной пищи. Но необходимым условием
эволюции гигантских эпиорнисов и динорнисов было отсутствие на
151
островах, где они жили, опасных хищников. На рисунке 22 даны кон-
турные изображения различных динорнитид, показывающие необычай-
но крупные размеры, достигнутые эволюцией в относительно очень не-
большой промежуток времени.
Curyaptery*
Рис. 22 Контурные изображения различных динорнитид. По Арчи
В рассмотренных случаях гигантизм птиц нельзя объяснить без до-
пущения чрезвычайно интенсивной изменчивости, которая вызывалась
факторами среды, прежде всего — благоприятными условиями пита-
ния. Итак, многие факты из геологической истории наземных позво-
ночных свидетельствуют об усиленной наследственной изменчивости,
вызываемой воздействием некоторых условий существования, и о ес-
тественном отборе, благоприятствовавшем сохранению этих измене-
ний.
152
Глава девятая
ИЗМЕНЧИВОСТЬ МОЛЛЮСКОВ ПАРАТЕТИСА В МИОЦЕНЕ
ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Моллюски, жившие разновременно в водах бассейнов Паратетиса^
обычно обнаруживают высокую внутривидовую изменчивость. Это яв-
ление ярко выражено у представителей многих родов, существовавших в
миоцене и плиоцене. Здесь мы рассмотрим лишь очень немногие приме-
ры внутривидовой изменчивости некоторых моллюсков карагана, сар-
мата, понта, киммерия, куяльника, гурия и чауды. Предварительно, од-
нако считаем нужным сделать некоторые общие замечания.
Очевидно еще до рассмотрения отдельных примеров надо сказать
следующее.
Об изменчивости видов моллюсков Паратетиса можно судить на
основании двух методов.
Во-первых, на основании установления рядов форм, из которых од-
на естественно и с необходимостью выводится из другой, менее продви-
нутой в филогенезе, и ведет к третьей, непосредственно с ней связанной
и из нее выводимой. Такие филогенетические ветви форм данного вида
могут иметь сложное, даже очень сложное, строение и все же представ-
лять собой внутривидовую картину изменчивости.
"Но вернее было бы начать с того, что популяции одного вида иног-
да являют картину интенсивной изменчивости, которая, однако, по всем
признакам, не выходит за пределы данного вида. Тут могут быть такие
варианты, которые не укладываются в один чертеж, в одну филогенети-
ческую линию, ибо изменчивость имеет многосторонний характер и идет
в разнообразных направлениях. И в таких случаях надо постараться
построить схему, но не всегда это удается, или получается явно упро-
щенная схема, не отражающая действительных отношений между фор-
мами. Такие отношения наиболее просто строятся для современных
популяций видов, когда особи живут совместно и можно предпола-
гать, что они входят в одну популяцию совместно размножающихся
особей. Для ископаемого же материала этот вопрос осложняется: даже
в случае большого скопления особей в одной прослойке или в одном
сласте трудно утверждать, что особи принадлежат к одной на месте раз-
вивавшейся популяции или хотя бы к популяциям, непрерывно сменяв-
шим одна другую, развивавшихся одна йз другой. Все же такое пред-
положение представляется более или менее вероятным, если в одной
прослойке находим множество совместно захороненных раковин.
Однако изучение внутривидовой изменчивости возможно и тогда,
когда нет возможности установить прямую преемственность между по-
пуляциями, но все же изучение материала приводит исследователей к
их отнесению к одному и тому же виду.
153.
Заметим, что для ископаемых форм по самоочевидным причинам
трудно дать более или менее полную картину изменчивости, которая
может быть чрезвычайно богатой и разнообразной.
Во-вторых, возможно установление изменчивости, как внутривидо-
вого явления и в тех случаях, когда в потоках эволюции, в филогенети-
ческих линиях нет возможности проводить границы между видами, но
все же можно утверждать, что перед нами не один вид, а два, несколь-
ко или много видов, приндлежащих к одной филогенетической цепи. С
этим случаем мы очень часто встречаемся при изучении ископаемого ма-
териала. В таких случаях может, конечно, возникнуть мысль, что перед
нами лишь одна полиморфная форма, и нечего тут искать разных так-
сонов даже подвидового ранга. Однако некоторые факты иногда прямо
указывают, что мы имеем дело со многими или, по крайней мере, с дву-
мя подвидовыми или видовыми единицами. Примерами могут служить
спаниодонтеллиды родов Spaniodontella и Savanella из караганского
горизонта среднего миоцена, Sinzowia, Kishinewia и Barbotella из сред-
него сармата, Provalensiennesia и Valensiennus из солоноватоводных отло-
жений верхнего миоцена и нижнего и среднего плиоцена, а также рас-
сматриваемые в этой книге кардииды понтического и киммерийского
яруса. Хотя во многих случаях очень трудно установить границы вида с
полной точностью, как это часто возможно при изучении рецентных
форм, не может быть никаких сомнений, что материал содержит ракови-
ны особей, различающихся между собой как представители одной и той
же популяции, являющие лишь внутривидовую изменчивость.
ЛЮТЕЦИИ, СПАНИОДОНТЕЛЛЫ, САВАНЕЛЛЫ
В морских палеогеновых, нижне- и среднемиоценовых отложениях
встречаются представители группы мелких, в общем, двустворок, исход-
ные, эоценовые, формы которых, принадлежащие к роду Lutetria (рис.
23), отличались крошечными размерами — около 1 мм в диаметре. Поя-
вившись в нашем миоцене, формы этой группы по началу оставались
очень мелкими. Может быть, именно миниатюрная величина этих дву-
створок, живших в полносоленых морях, и была одной из причин того,
что они до сих пор изучены мало. В морях нашего времени эта груг-па,
по-видимому, не представлена, а предполагаемые связи ее с некоторыми
современными мелкими двустворками довольно сомнительны. Вполне
возможно, что эту группу придется выделить в отдельное семейство лю-
тециид, тем более, что уже теперь известны не менее, чем четыре рода,
входящие в ее состав: Lutetia Deshayes, Alveinus Conrad, Spaniodon-
tella Andrussow и Savanella Zhghenti. Вопрос о том, вымерла ли эта
группа полностью (в миоцене?), или какие-то ее представители продол-
жают существовать, оставаясь до сих пор незамеченными, из-за своей
малой величины, специалистами по нынеживущим моллюскам полносо-
леных морей, остается пока не решенным.
Мы уже упоминали спаниодонтелл как пример филогенетического
увеличения размеров моллюсков, связанного, как мы утверждали, с уси-
ленной изменчивостью этих двустворок в водах среднемиоценовых бас-
сейнов Черноморско-Каспийской области. Здесь мы разберем вопрос
об их индивидуальной изменчивости более подробно. После небольшой
нашей статьи о спаниодонтеллах (1934) эту группу специально изучала
154
Е: М. Жгенти, которая пока лишь частично опубликовала результаты
своих исследований (1968). Она любезно предоставила нам .некоторые
свои еще не опубликованные данные и изображения, воспроизводимые
Рис. 23. Lutetia umbonata Desh. из лютетского яруса Парижского бассейна. Фиг. 1,2—•
левые створки. Фиг. 3—экземпляр, приближающийся к строению спаниодонтелл.
Сильно увеличено. По Жгенти.
в этой книге. На рис. 24 изображены формы спаниодонтелл из самых
низов чокрака, среднего и верхнего горизонтов чокрака и самых верхов
этого горизонта, а также из вышележащих караганских (в широком
Рис. 24. Ряд форм Spaniodontella intermedia—Span- pulchella. 1,2, 3,4 Span, interme-
dia. Самые маленькие спаниодонтеллы из самых низов чокрака Ставропольского края.
5, 6—Span, intermedia. Экземпляры обычной величины, находимые в среднем и верхнем
чокраке Ставропольского края в массе вых количествах. 7 и 8—Span- minima. Встреча-
ются в самых верхах чокрака. 9, 10—Span- pulchella. Встречаются в карагане (вклю-
чая варненские слои). Увел. 5 раз. По Жгенти
смысле) отложений. Этот ряд показывает постепенное увеличение раз-
меров раковин в водах с несколько пониженной, против нормальной
морской, соленостью. Возрастание величины наблюдается не только от
горизонта к горизонту, но и на различных уровнях одного и того же
чокракского, горизонта.
На рис. 23 изображены левые створки Lutetia umbonata Desh. из
отложений лютетского яруса Парижского бассейна. Одна из этих ство-
рок, согласно замечанию Жгенти, показывает уже некоторое отклонение
в сторону спаниодонтелл. Очевидно уже внутривидовая изменчивость
этого эоценового вида представляла собой предварительное условие,
благоприятное для развития, в подходящей экологической обстановке,
в сторону спаниодонтелл. Древнейшей в нашем Черномороко-Кас1пий-
ском миоцене формой этой группы была Spaniodontella intermedia Andrus,
(рис. 24). Здесь, в новых для этих 'моллюсков условиях, усилива-
ется, по словам Жгенти, внутривидовая изменчивость, направленная в
155
сторону увеличения размеров тела. В самых верхах чокрака наряду с
только что названным видом, встречаются и довольно крупные формы
(до 3 мм в поперечнике), то есть спаниодонтеллы вполне караганскога
облика. В караганском, спаниодонтелловом, веке появляются новые ви-
ды этого рода. Если у лютеций поперечник раковины едва превышал
1 мм, у типичной Span, intermedia он достигал 2мм, а у караганских
форм он иногда превышает 10 мм. Замковый аппарат становится более-
мощным, массивным, изменяются очертания раковины, она вытяги-
вается то в переднезаднем направлении, то в дорзовентральном,
иногда образуются массивные выпуклые макушки. В средней части тол-
щи KaparaiHCKHx отложений изменчивость спаииодонтелл, по мнению*
Жгенти, испытывает дальнейшую интенсификацию: там она, по словам
этого автора (1968, стр. 101), «в большинстве случаев имела вполне
определенную направленность, — в сторону появления грубой кон-
центрической ребристости, утолщения стенок раковины» и других черт,,
закрепляющихся в новом роде Savanella, типом которого является
Spaniodontella andrussovi Toala. Саванелл ы, по Жгенти, возникли в
прибрежной зоне моря, где преобладали грубозернистые грунты, а за-
тем быстро расселились по другим участкам моря. Нн основании стра-
тиграфического распространения саванелл, в результате изучения экоге-
неза рода Spaniodontella Жгенти (1961) выделила отдельную геохроно-
логическую единицу под названием варненских слоев, или варненского
горизонта (1967). Не вдаваясыв морфологические детали, здесь мы вос-
производим некоторые фотографии Жгенти, показывающие очень интен-
сивную внутривидовую изменчивость спаниодонтелл и саванелл (рис.
25, 26, табл. V).
Уже те немногие данные, которые мы могли рассмотреть здесь, в этом
разделе^ посвященном изменчивости среднемиоценовых спаниодонтеллг
показывают, что эволюция моллюсков этой группы протекала постепен-
но, без внезапных скачков, хотя и скачкообразно. Относительно хорошо
изученный материал по двустворкам-эндемикам одной, восточной, части
Паратетиса при довольно хорошей сохранности этих, в общем обычно
не тонкостенных, хотя и мелких моллюсков, позволил исследователям
установить филогенетические связи между отдельными таксонами, не
только внутривидовыми, но и надвидовыми. Трудно сомневаться в са-
мостоятельности видов этой группы, но совершенно плавные, незамет-
ные переходы между ними порой ставили исследователей в тупик, и они
иногда отказывались признавать эти виды. Когда нет резкой, бросаю-
щейся в глаза разницы между двумя смежными видами, которые свя-
заны промежуточными формами, границу между видами провести, дей-
ствительно очень трудно. Это и побудило столь опытного специалиста
как Эмилия Коюмджиева (1965), отрицать возможность пользоваться
формами видов караганских спаниодонтелл с гладкой наружной поверх-
ностью и ребристыми формами саванелл для различения горизонтов в
широко понимаемом карагане. Но не надо забывать, что такие плавные
переходы между видами существовали во всех решительно случаях фи-
логенеза, и если бы геологическая летопись сохраняла нам все такие
промежуточные формы, то, приняв точку зрения упомянутого исследо-
вателя, мы должны были бы всюду и везде отказываться от проведения
геохронологических границ.
Изложенное показывает, что в группе спаниодонтелл и родственных
им форм мы встречаем несомненные доказательства широкого распрост-
ранения изменчивости, которая у них достигла высокой степени интен-
сивности. Это особенно бросается в глаза на фоне медленной эволюции.
156
Рис. 25. Спаниодонтеллы карагана. 1—10—Span, pulcuella. Фиг. 2—форма, несколько вытянутая дорзовентра-
льно. Фиг. 5—высота раковины заметно превосходит ее ширину- От фиг. 6 (типичная форма) тянется цепочка
форм, изменчивость которых направлена в сторону вытягивания брюшного края кпереди и увеличения толщины
раковины. Караган, включая варненский горизонт. Фиг. 1], 12 и 13—нормальные представители Span, pulchella.
Фиг. 14—16—типичные представители Span, tapesoides. Этот ряд показывает изменчивссть очертаний раковины
в сторону уподобления роду Paphia (Tapes)- Сильно увеличено. По Жгенти.
предков спаниодонтелл — лютеций, которые, хотя они и обнаруживают
способность к внутривидовой изменчивости, эволюировали очень мед-
ленно от эоцена до миоцена, так как жили и развивались в мало изме-
Рис. 26. Savanella andrussovi. Сомкнутые створки со спинной стороны.
Заметна изменчивость раковины от яйцевидной (фиг. 1) до почти ша-
ровидной (фи. 4). Варненские слои Западной Грузии.
По Жгенти.
нявшихся условиях морей с соленостью нормальной или близкой к нор-
мальной.
ИЗМЕНЧИВОСТЬ СРЕДИ САРМАТСКИХ МОЛЛЮСКОВ
Мы не имеем возможности подвергнуть здесь систематическому
разбору весь колоссальный материал по изменчивости моллюсков верх-
него кайнозоя Паратетиса.
Остановимся на некоторых группах моллюсков, живших в Сармат-
ском море.
Это море, несмотря на его колоссальные размеры, было по-видимо-
му совершенно замкнутым. Во всяком случае в нем жило множество
эндемичных беспозвоночных. Поскольку моллюски, двустворки и брю-
хоногие, обладают раковинами, способными особенно хорошо сохра-
няться в ископаемом состоянии, из всех беспозвоночных сармата лучше
всего и в наибольшем изобилии сохранились ископаемые именно этих
двух классов. Когда это море обособилось от мирового океана, оно не
могло оставаться полносоленым. Соленость вод Сарматского моря бы-
ла заметно ниже нормальной морской и очевидно испытывала сущест-
венные колебания на протяжении сарматского века. Но все же она бы-
ла терпимой для многих морских моллюсков по крайней мере в ранней
части сармата. Поэтому многие моллюски морского происхождения
продолжали существовать в Сарматском море и испытывали там, в но-
вых для них и притом довольно изменчивых условиях, быстрое эволюци-
онное развитие. Все это делает сарматских двустворок и брюхоногих
интереснейши1м объектом для изучения изменчивости и факторов, ее
стим у ли р ую щи х.
При рассмотрении явлений филогенетического увеличения общих
размеров тела животных мы коснулись, с этой стороны, некоторых фак-
тов исторического развития сарматких двустворок: эрвилий, мактрид,
пафий и кардиид. Тут, за невозможностью уделить больше места сар-
матским представителям этого класса, мы обратимся к некоторым при-
мерам изменчивости форм класса брюхоногих, живших в сарматском
море. Но и в этом случае, ограничимся, по тем же соображениям, рас-
смотрением лишь немногих групп.
Заслуживают внимания эндемичные роды брюхоногих, известные
из среднего сармата, особенно Barbotella, Kishinewia и Sinzowia.
158
Barbotella представляет собой необычайное изменение в направле-
нии турбообразной раковины, и крайние формы этой группы напомина-
ют, по облику, раковины Oxystele orientalis из так называемых тортон-
ских отложений западной части Украины, а еще больше похожи на Tur-
bo mammi laris, Bolma meinardi и В. perangulata оттуда же. Между тем не-
посредственная филогенетическая связь барботелл с «Trochus» [Callistoma]
podolicus не подлежит сомнению. Фридберг (Friedberg, 1928, стр. 515, табл.
33, рис. 1) описал и изобразил под названием Callistoma podolicum Dub-
var. А несомненную барботеллу, похожую на Barbotella grossocostata Rad.
et Pavb/He считая, очевидно, возможным выделить эту форму из рода Cal-
Рис. 27. Некоторые формы рода Barbotella. 1 и 2—Barbotella omaliusii d’Orb. 3—5—
Barb, hornesi Barb- 6—8—Barb- grossocostata Rad. et Pavl. 9 и 10—Barb- laevigato-
omaliusii Koles. 11 и 12—Barb, intermedia Rad. et Pavl. Средний сармат Молдавии.
По Колесникову.
listoma и семейства Trochidae. Колесников (1928) первоначально всю
эту группу относил к роду Trochus под названием Тг. omaliusii. Он
находил, что нет надобности выделять эту группу даже в особый под-
род Barbotella Cossmann. В нашей работе (1932) она фигурирует под
названием Trochus (Barbotella) omaliusii. Позже (1935) Колесников счел
нужным рассматривать эту группу как самостоятельный род Barbotella
Cossmann, и поступил, по нашему мнению, совершенно правильно. Однако-
159
связь этой своеобразной группы с Callistorna podolicum не подлежит ни
малейшему сомнению, а те формы, которые ранее выделялись лишь как
подвид, или разновидности вида Trochus omaliusii, были правильно приз-
наны Колесниковым самостоятельными видами. Не входя в рассмотрение
особенностей этих форм, даем рис. 27, где изображены Barbotella oma-
liusii, Barb, laevigatoomaliusii, Barb, hornesi, Barb, grossocostata, Barb, in-
termedia. Колесников дает следующую филогенетическую схему барботелл
(рис. 28):
BarfiotePPa i/iter/neotia Pad et PavP
BarPotePPa grossocostata Pad et ParP
BarPotePPa Pdsvp&rt BarP.
'asPotePPa Pa eta -
'gatoo/naP/as/l PoPes.
PasPotePPa o/77aPtastt ct’PrP
Рис. 28. Филогенетическая схема барботелл. По Колесникову.
Заметим, что безусловно возможны и несколько иные толкования
филогенетических отношений этих форм. Несомненно, однако, то, что,
как это признавали Фридберг (Friedberg, 1928). Колесников (1928 и
1935) и Давиташвили (1935), барботеллы, через наиболее примитивный
вид Barb, omaliusii, происходят от каллистом группы Call, podolicum.
Переход от этой последней формы к ранним барботеллам — достаточно
постепенный. Барботеллы жили на огромном пространстве Сарматского
моря (от Балканского полуострова до Казахстана), в его мелководной
полосе. Они обнаруживают необычайную индивидуальную изменчи-
вость, заметную на образцах из одного и того же .местонахождения, как
это видно на рис. 29, где изображены три экземпляра Bar. intermedia,
заметно отличающиеся один от другого по числу, расположению и фор-
ме ребер.
Во всяком случае нет сомнения в том, что в сарматском бассейне
развитие одной группы трохид пошло по линии уподобления раковины
некоторым представителям семейства Turbinidae, а именно подсемей-
ству Turbininae и его родам Turbo и Astrea, к которому относят вид
Astrea (Bolma) rugosa (Linne), внешне похожий на некоторых барботелл.
По самоочевидным причинам мы не можем согласиться с И. А. Короб-
ковым, который относит род Barbotella к сем. Turbinidae и даже к под-
семейству Turbininae (Коробков, 1955, стр. 112—114). Сходство есть, но
существуют и несомненные отличия, а главное — барботеллы филогене-
тически неразрывно связаны с трохидами. В частности барботеллы, как
дериваты трохид, лишены известковой крышечки, характерной для тур-
бинид.
160
В. Венц (Wenz, 1938, стр. 353, рис. 834) считает Barboletta ошибоч-
но подродом рода Turbo. Тип подрода — Turbo (Barbotella) hoernesi
Barbot. To же самое повторяется в «Основах палеонтологии», — томе
«Моллюски — брюхоногие» (1960, стр. 105).
Совершенно ясно, что такая невероятно быстрая эволюция могла
происходить только при наличии необычайно повышенной изменчивости,
которая не имеет никакого подобия, по своей интенсивности, с измен-
чивостью моллюсков полносоленых морей.
Рис. 29. Barbotella intermedia Rad- et Pavl. Средний сармат Молдавии, с. Петрикана.
По Л. Мусхелишвили.
Предков барботелл надо искать несомненно в огромной и разнооб-
разной группе преимущественно среднесарматских форм, к которой
принадлежит Call, podolicum (Dub.) и многочисленные другие виды,
связанные между собой постепенными переходами. Систематика их
представляет неимоверные трудности. Крупнейший знаток сарматских
моллюсков В. П. Колесников это учитывал, но не успел предпринять
новую коренную ревизию этих трохид. Такая ревизия, при условии прив-
лечения богатого материала из всех местонахождений, где только изве-
стны формы, похожие на Call, podolicum, обещает дать результаты,
очень ценные и сратиграфически и эволюционно-палеобиологически. На
рис. 30 изображена одна из этих форм, опреде-
ленная как Call, sulcatopodolica (Koles.).
Итак, группа «Trochus» podolicus представляет
огромное разнообразие форм, сильно затрудняющее
построение их филогенетической систематики. Это
отразилось на работах В. П. Колесникова. В своей
более ранней работе (1928)этот азтор различал
Trochus podolicus (Dub.) Pusch, Tr. podolicus var. lae-
vigatopodolicus Koles., Tr. podolicus var. sulcatopodoli-
cus Koles., Tr. hommairei d'Orb., Tr. caucasicus Eichw.
Согласно схеме, содержащейся в этой работе Ко-
лесникова, от Trochus podolicus происходят непос-
редственно Tr. podolicus var. sulcatopodolicus, Tr.
podolicus var. hommarei, Tr. podolicus var. caucasica,
Рис. 30. callistoma
sulcatopodolica (ko-
les.). Средний сар-
мат Молдавии, с.
Петрикана. По Л.
Мусхелишвили.
. Tr. omaliusii и Tr. podolicus var. laevigatopodolicus.
В монографии, посвященной моллюскам сармата, Колесников (1935,
стр. 138 и 139) усматривает уже две совершенно самостоятельные груп-
пы «подоликообразных» форм и выводит эти две группы от различных
предковых форм.
И. Л. Ш- Давиташвили 161
Первую (рис. 31) группу Колесников производит от вида Trochus
sarmates Eichw.:
ТУ'ос/z/zs sasT/Mt te# fzc/z w.:
Тгаг/яайге/zxzs
Trto/rwztfzsrz
Tjzcaacazzza^ffic^w. ^Tae^a^o/zocroeicasTbres.
Tr /wctoftcaj/'jfaty
Tr cojrfierta/zus tf’OrS.
Tr/za/zifrzz z5zc^
Tr szz r'/zzates #zc/?vz-----1
Рис- 31. Происхождение первой группы «подоликообразных» форм. Но
Колесникову.
Вторую группу (рис. 32) автор ведет, как от общего предка, от
Tr. podolicoformis:
Тг. teaurw/rfzz.
Tr frargi/zatorz/zzvwz raTTs
Tr pseudo/zo/rrzazrrz jfofes.
Tr Utyoetatus Mfes.
*—----------Tr. роУгТаиуагши JTrfes
Рис. 32- Происхождение второй группы «подоликообразных форм- По
Колесникову.
Несмотря на свои тщательные филогенетические изыскания, Колес-
ников не был уверен в правильности построенной им схемы. После рас-
смотрения связей между нижнесарматским ,,Trochus“ podolicoformis Ко-
162
les. и пятью среднесарматскими видами—Тг. beaumontii, Tr. sulcatopo-
dolicus, Тг. insperatus, Тг. pseudohommairei и Тг. marginatosinzowi—этот
автор писал: „Родственная связь между этими среднесарматскими видами
и Trochus podolicoformis Koles., несомненна, но схема их соотношений мне
не ясна. Вполне возможно, что все 5 среднесарматских видов являются
тем же Trochus podolicoformis Koles., но несколько изменившимся в сред-
несарматских условиях" (1935, стр. 167).
При описании чрезвычайно характерного среднесарматского вида
Trochus podolicus (Dub.) Pusch В. П. Колесников (1935, стр. 181) пи-
сал: «Изменчивость раковин описываемого вида очень велика. При
уменьшении выпуклости оборотов намечается постепенный переход к
Trochus hommaierei d'Orb. и Trochus caucasicus Eichw., которые отличают-
ся друг от друга, главным образом, неодинаковой удлиненностью. При
сглаживании бугорков намечается не менее постепенный переход к Тг.
laevigatopodolicus Koles. И, наконец, при округлении килевой (линии.—•
Л. Д.]раковины приобретают настолько сходный вид с Barbotella oma-
liusii, что невольно закрадывается мысль об их род-
ственности/4
Рис. 33. Схема’строе-
ния раковины Sinzo-
Таким образом, тща-
тельное и глубокое, про-
должительное изучение
отношений между фор-
мами «подоликообраз-
ных» трохид и барботел-
лами сармата заставляло
Колесникова по-разному
строить схемы их фило-
генетических взаимоотно-
шений и даже допускать,
что многие формы этой
группы могут оказаться
принадлежащими к одно-
му и тому же виду. Все
это во всяком случае
свидетельствует о колос-
сальной, резко выражен-
Рис. 34. Sinzowia striata-tuber-
culata (Sinz-). Средний сармат.
ной внутривидовой из-
менчивости этих моллюс-
wia. По Колесникову. КОВ В сарматском море, хЗ. По Колесникову,
создававшей возмож-
ность их изумительно быстрой эволюции, которая пела к возникновению
Не ТОЛЬКО .НОВЫХ (ВИДОВ, но м новых (родов.
Остановимся теперь на выделенных Колесниковым родах Sinzowia
(рис. 33 и 34) и Kishinewia (рис. 35 и 36), — формах, которые до иссле-
дований этого ученого были по традиции относимы к роду Phasian£lla,
хотя и не имеют ничего общего с этим родом. Коробков (1955, стр. 106)
объединяет эти формы в один род Sinzowia, низведя Kishinewia до сте-
пени подрода этого рода. Воздержимся от рассмотрения этого таксоно-
мического вопроса. Колесников и Коробков вполне правильно включают
эти формы в семейство Trochidae. Тут перед нами случай резкого удли-
нения раковины, которая приняла облик, напоминающий, в крайних
163
случаях удлинения, род Turritella, что видно из сравнения рис. 33 и 34,
изображающих представителей рода Sinzowia (Давиташвили, 1932, тб.
10, рис. 20—23 и 27) и рис. 37, где представлена Turritella terebralis.
Отмечаем это наружное сходство, связанное, вероятно, со сходством
в образе жизни этих башенковидных форм. Вполне возможно, что изу-
чение материала покажет наличие малозаметных , переходов от более
или менее «нормальных» трохид к сильно вытянутым в длину на подо-
бие туррителлы. Уже в пределах родов Sinzowia и Kishinewia наблюда-
ются различные степени вытянутости в высоту башенковидных раковин.
Но существуют и такие формы, как Trochus woronzowii, которые, не
представляя перехода от обычных трохид к
туррителлообраз1НЫ1М, в то же время показыва-
ют значительное изменение в сторону башен-
ковидного удлинения раковины (Колесников,
табл. 23, рис. 21—23).
Рис. 35. Схема строения
раковины Kishinewia. х 3.
По Колесникову,
Рис. 36. Kishinewia bessarabica (d’Orb).
Средний сармат. Кишинев. По
Колесникову.
Подобные преобразования могли совершиться лишь па основе ис-
ключительно высокой индивидуальной изменчивости сарматских трохид.
Рассмотренные нами примеры показывают, что трохиды, сохранив-
шиеся в сарматском море после вымирания на границе между конкой и
сарматом, стали родоначальниками некоторых групп чрезвычайно ин-
тенсивно эволюировавших форм, причем наблюдается любопытнейшее
уподобление этих явно эндемичных форм великого замкнутого Сарматс-
кого моря разным родовым группам моллюсков морей и нормальной
соленостью, в частности, родам турбинид, фазианеллид и туррителлид.
Появление за такой короткий промежуток геологического времени
(преимущественно средний сармат) столь аберрантных форм приходит-
ся признать несомненным свидетельством резко повышенной внутри-
видовой изменчивости. Эта изменчивость, по-видимому, вызывалась су-
щественными изменениями гидрологических условий; ей благоприятст-
вовали отсутствие некоторых обычных обитателей полносоленых морей и
наличие свободных экологических ниш, способствовавшее быстрому рос-
ту численности популяций, а следовательно и появлению многочислен-
ных мутантов.
164
Внутривидовая изменчивость не менее сильно выражена и в других
группах моллюсков сармата, особенно — среднего. Однако мы должны
воздержаться от попыток проаналиаировать эти явления в других груп-
пах сарматских форм. Этот вопрос затрагивается некоторыми специа-
Рис. 37. Turritella terebralis Lanak. Мио-
цен Аквитании, х 1. По Ксробкову.
листами, например, — Л. В. Мусхелишвили (1968), которая рассмотре-
ла вопрос об изменчивости у сарматских брюхоногих семейства
Nassidae.
Глава десятая
РЕЗКО ПОВЫШЕННАЯ МАССОВАЯ ИНДИВИДУАЛЬНАЯ
ИЗМЕНЧИВОСТЬ В НЕКОТОРЫХ ГРУППАХ ДВУСТВОРОК
ПЛИОЦЕНА РИОНСКОГО ЗАЛИВА
Основные стратиграфические единицы морского, точнее — солоно-
ватоводного, плиоцена, прослеживаются в Черноморском бассейне до-
вольно широко: понт, киммерий, куяльник, и, вероятно, гурий представ-
лены как в южных, так и северных частях этого бассейна. То же самое
с бесспорностью относится и к чауде, которую очень часто включают
в плейстоцен, хотя мы не уверены в правильности отнесения, к различ-
ным основным хронологическим единицам гурия и чауды. Несмотря на
это, комплексы одновременно существовавших форм моллюсков, извест-
ные в разных районах, заметно отличаются один от другого. Более
того, многие факты указывают на то, что в различные века плиоцена
в Черноморском бассейне существовали отдельные очаги видообразова-
ния двустворчатых моллюсков. Наличие некоторых форм лишь в одном
или немногих местонахождениях не доказывает, конечно, эндемичности
этих форм, так как вполне возможно, что они жили и на других участ-
ках и только пока не найдены нами нигде, за пределами определенных
местонахождений. Ведь известны случаи, когда формы, в изобилии
встречающиеся в одном месте выхода данного горизонта, попадаются в
других (Местах очень редко и лишыв немногих точках. Примерами могут
служить хотя бы Moquicardium moquicum (Sen.) и Oraphocardium огар-
bense Dav. Однако в некоторых случаях спорадическое распростране-
ние ископаемых форм плиоцена Черноморского бассейна следует приз-
нать свидетельством их эндемичного характера.
Даже беглое ознакомление с фаунами моллюсков, например, пон-
та и киммерия, свойственными разным участкам, где представлены со-
ответствующие толщи, убеждает всякого исследователя в наличии зоо-
географического своеобразия каждого из этих участков несмотря на то,
что имеются также и формы, общие для всех их. И совершенно особ-
няком стоит солоноватоводная толща Румынии, известная под названи-
ем дакийского яруса: моллюсковая фауна этих отложений настолько
своеобразна, что точная параллелизация их с толщами, развитыми на
территории Юга нашей страны, представляет пока еще не преодолен-
ные трудности.
Вопрос о существовании в нашем солоноватоводном плиоцене отно-
сительно обособленных очагов эволюции двустворок мы -рассмотрим на
примере дрейссенид и кардиид киммерийского и к у ял ьницко го ярусов.
Для того, чтобы подойти к изучению об изменчивости форм, в
данном случае прежде всего необходимо установить факт их эндемич-
ности. Л на. ископаемом материале это всегда нелегко, так как невоз-
166
можно с точностью установить детали явлений прохореза от «очага»
возникновения данного вида к различным другим местам и дальнейших
перемещений популяции. Это особенно затруднительно, в частности, по-
тому, что нам ничего, или почти ничего не известно о распространении
плиоценовых солоноватоводных отложений к западу от юго-западной
границы Грузинской ССР. Особая осторожность в решении вопроса об
эндемичности тех или иных форм нужна еще и потому, что многие соло-
новатоводные плиоценовые формы, встречающиеся на территории СССР
лишь в Западной Грузии, включая сюда и Абхазскую АССР, известны
также в Румынии, а иногда и в Югославии. Так, в понте Рионского за-
лива встречаются Congeria digitifera Andrus., Cong, rhomboidea M.
Hoern., Cong, rumana Sabba. Эти виды еще недавно считались характер-
ными для понта Румынии и Северной Югославии. «Мы полагаем, — пи-
сал И. Г. Тактакишвили (1968, стр. 731), — что при более тщательном
изучении понтической фауны Западной Грузии в ней можно будет най-
ти гораздо больше «румынских» элементов».
И тем не менее нет никаких оснований отказываться от попыток
установления эндемичности форм из солоноватоводного плиоцена Чер-
номорского бассейна, выяснения очагов их возникновения, и путей их
расселения. Результаты таких исследований, если они оказываются
успешными, создают необходимые предпосылки для изучения изменчи-
вости форм моллюсков.
В пользу эндемичности форм в данном случае могут свидетельст-
вовать следующие факты: 1) многочисленность форм, встречающихся
только в данном районе и не известных в других местах; 2) наличие
на определенном участке не только изолированных форм, но и многих
видов данного таксона, филогенетически близких друг к другу и не
встреченных за пределами этого района; 3) физико-географическое
своеобразие изучаемого района, заставляющее предполагать возникно-
вение там условий, благоприятствовавших повышенной изменчивости
и ускоренному видообразованию; 4) факт повышенной изменчивости
исследуемых форм — заметно более интенсивной, чем их внутривидо-
вая изменчивость за пределами данного участка.
Разберем сначала, возможно полно, примеры из истории соло-
новатоводных кардиид киммерия и куяльника Западной Грузии (см.
стратиграфическую схему).
Для киммерия Рионского залива характерны виды группы Moqui-
cardium moquicum Sen. (верхний горизонт слоев Дуаба). Помимо это-
го вида, здесь встречаются: М. submoquicum Ebers. (нижний горизонт
киммерия, Гудаутский район Абхазии), М. praemoquicum Dav. (нижний
горизонт слоев Дуаба, Абхазия), М. privum Ebers. (дуабские слои),М.
ozurgeticum Dav. (киммерий близ г. Махарадзе в Гурии), М. guriense
Dav. (Гурия, река Oipano). Вне Грузии из всего этого разнообразия
форм встречена только одна, определяемая А. Г. Эберзиным как М.
moquicum и притом лишь в одном месте южного побережья Керченско-
го полуострова — Кызаулский рудник (два экземпляра).
В киммерии Западной Грузии известны по меньшей мере три вида
рода Prosodacna, не встреченные нигде за пределами этого района нес-
мотря на большое обилие представителей этого рода, в более северных
частях области распространения киммерийских пластов: Prosodacna lon-
giuscula Sen., P. leptopsamatha Dav. и P. callopistes Dav. Заметим, что у
этих форм наблюдается определенная внутривидовая изменчивость, осо-
бенно по величине тела.
167
Обращает на себя внимание обилие представителей рода Pachy-
dacna в киммерии Западной Грузии; но некоторые формы этого рода
известны и из других, более северных местонахождений.
Исключительно разнообразно представлен вид Crassadacna crassatel-
lata (Desh.) в дуабских отложениях киммерия Абхазии. Тут дела-
лись попытки выделять много таксонов ранга подвидов. Вполне возмож-
но, что некоторые из подвидов установлены обоснованно, но относитель-
но других сохраняются сомнения, не представляют ли они лишь индиви-
дуальную изменчивость, наследственную или ненаследственную. Так или
иначе, интересно, что повышенная изменчивость, ведшая, быть может,
к образованию новых подвидов, отмечается именно в юго-восточной час-
ти киммерийского бассейна.
Таким образом, Рионскому заливу киммерийского озера-моря свой-
ственны многие локальные формы кардиид. Поскольку они отсутствуют
в более северных частях Кавказа и на Керченском полуострове, можно
думать, что это были местные, эндемичные формы, характерные для
данного участка.
Нельзя умолчать о том, что в киммерии того же района встречает-
ся много дреиссенид, не попадающихся в других местонахождениях это-
го яруса, даже богатых ископаемой моллюсковой фауной. Загадкой пред-
ставлялось нахождение здесь очень крупной Congeria caucasica, о кото-
рой иногда думали, что она переселилась сюда с запада. В более север-
ных районах распространения киммерийских отложений конгерии совер-
шенно отсутствуют. Между тем теперь в понте Рионского залива изве-
стны конгерии, родственные Congeria subcarinata Desh., но в то же вре-
мя гораздо более крупные. Эти понтические конгерии представляют со-
бой, очевидно, переход к верхнедуабским конгериям группы Congeria
caucasica, которые, кстати сказать, обнаруживают очень высокую измен-
чивость, побуждавшую К. Сенинского к установлению не одного, а двух
видов: С. caucasica и С. abchasica. Однако в нашей коллекции мы зат-
рудняемся обособить один от другого эти предполагаемые виды, хотя
вполне возможно, что тут мы имеем случай, представляющий началь-
ный этап такого эволюционного процесса. Нигде за пределами очень не-
большой часта Рионского залива эти крупные конгерии не были встре-
чены, хотя в киммерии прилегающих участков Западной Грузии изве-
стно много дреиссенид. В исключительно высокой изменчивости кон-
герий этой группы убеждает нас выделение Г. Ф. Челидзе (1952) но-
вых форм: С. caucasica, С. caucasica Sen. var. perpatula Tchel., C. post-
subcarinata Tchel., C. acuta Tchel., C. gabuniai Tchel. Когда индивидуаль-
ная изменчивость очень сильна, почти каждый экземпляр кажется пред-
ставляющим особый вид или, по крайней мере, подвид. Быть может, неко-
торые из выделенных Челидзе таксонов намечают начало новых филоге-
нетических линий, которые, однако, не успели осуществиться. Но весь ма-
териал в совокупности несомненно свидетельствует о бурной изменчивос-
ти, по крайней мере отчасти наследственной (рис. 38, 39). Все дает нам
основание думать, что крупные киммерийские конгерии дуабскихслоев —
локальные эндемичные формы, развивавшиеся из эндемичных же конге-
рий понта, к которым принадлежит вид И. Г. Тактакишвили (1963).
В Рионском заливе обитали также и другие эндемичные формы дрейс-,
сен и конгерий, представляющие значительное разнообразие: С. mira-'
bilis Sen., С.» turgidopsis Andrus., C. ebersini Tchel. и другие. Известно,
что вне Рионского залива конгерии в киммерии всюду отсутствуют или,
по крайней мере, нигде не встречены.
168
Среди киммерийских дрейосен Рионюкого залива также имеются фор-
мы, которые приходится считать эндемиками. Это Dreissena cyclorhamp-
Рис. 38. Крупные конгерии из верхней части дуабских слоев
киммерия. 1 и 2—„Congeria caucasica“ Sen. 3 и 4 „Congeria
abchasica" Sen. Оба экземпляра принадлежат, по-видимому,
к одному и тому же виду Con. caucasica Sen. По Сени некому.
ha Andrus., D. semilunaris Sen., D. obliqua Sen., D. dilatata Andrus., D.
pseudomirabilis Tchel. и др. Особенно интересна эта последняя форма,
169
напоминающая Congeria mirabilis, но отличающаяся от нее отсутствием
апофизы,—структуры, характерной для рода Congeria.
Таким образом, трудно сомневаться в эндемичности некоторых ха-
рактерных киммерийских дрейссенид Рионского залива,
дуабских слоев.
особенно же —
Рис. 39. Схематизированные (контурные) изображения конгерий группы Congeria cauca-
sica Sen. из верхних слоев дуабской толщи киммерия (Мокви, Абхазия). 1 и 2—„Cong,
caucasica Sen. var. perpatula“ Tchelidze, 3 и 4—„Cong caucasica Sen. var. subcaucasica"
Tcb^l., 5 и 6—„Cong. postsubcarinata“ Tchel., 7 и 8—„Cong. acuta“ Tchel.,
10— „ Cong. gabuniani“ Tchel. Все изображенные образцы принадлежат, по-видимому,
к одному виду Cong, caucasica Sen., давая некоторое представление о его внутривидо-
вой изменчивости. Уменьшено. По Челидзе.
9 и
Такую же картину представляет фауна моллюсков из куяльницких
отложений Западной Грузии. С этой стороны наибольшего внимания
заслуживают Chartoconcha postcimmeria Dav., вся группа “Didacna" me-
deae Dav., ,,D.“ caeleno Dav. и Didacnomya phasiaca Dav. Эти формы
170
-обильно представлены в куяльнике Западной Грузии и не найдены ниг-
де за ее пределами. Их отсутствие в одновозрастных отложениях, пред-
ставленных к северу и северо-западу от Рионского залива, служит дос-
таточным указанием на их эндемичный характер. Одна из них, Did.
caeleno, была очевидно эндемиком особенно ограниченного распростра-
нения, так как она встречается лишь в Гурии и отсутствует в более се-
верных местонахождениях куяльника. Примечательно, что исходная,
наиболее древняя форма Did. medeae была встречена в киммерии то-
же Рионского залива.
Ограничиваясь только что приведенными фактами — их число мож-
но было бы значительно приумножить даже на основе уже имеющихся
данных, — сделаем некоторые выводы. Все имеющиеся материалы го-
ворят о том, что в киммерии и куяльнике Рионский залив был одним
из важнейших очагов возникновения и центром ареалов новых двуство-
рок, откуда они иногда распространялись в смежные области Черномор-
ского бассейна. Однако очень многие формы по тем .или иным причинам
оставались эндемиками Рионского залива как в киммерии, так и в ку-
яльнике.
Аналогичную картину мы наблюдаем в истории более молодых го-
ризонтов — гурия и чауды. Отметим, что, хотя моллюсковая фауна этих
двух веков была, по-видимому, гораздо менее разнообразной, чем в ким-
мерии и куяльнике, все-таки характерными чертами ее исторического
развития была очень интенсивная изменчивость и, соответственно, высо-
кие темпы эволюции. За относительно короткое время возникли новые
роды ( Digressodacna Dav. et Kit. и Tschaudia Dav.), виды которых ока-
зывались чрезвычайно изменчивыми. Тот факт, что большинство мес-
тонахождений гурия и чауды находятся в Гурии, где, к тому же, наб-
людаются постепенные переходы между формами последовательных
горизонтов и наибольшее морфологическое разнообразие раковин, го-
ворит в пользу того, что именно в Гурии был основной очаг развития
турийских и чаудинских моллюсков.
Таким образом, на всем протяжении времени от понта до чауды
Рионский залив был участком, где особенно интенсивно протекала эво-
люция двустворок. Там возникали энедемичные виды и даже роды.
Иногда, впрочем, отпрыски этих форм, путем прохореза, проникали и в
другие области Черноморско-Каспийского бассейна. Необычайно быст-
рая эволюция могла совершаться только на основе очень высокой ин-
дивидуальной изменчивости. И мы, действительно, можем наблюдать, та-
кую изменчивость на материале из кардиид и дрейссенид, рассматри-
ваемом в этой книге. Какие факторы внешней среды вызывали усилен-
ную изменчивость двустворок плиоцена именно в Рионском заливе, при
нынешнем состоянии знаний мы сказать не можем. То, что известно о
растительности прибрежной полосы этой области, говорит о теплом кли-
мате и обилии растительного вещества, поступавшего с суши. Связь
между повышением изменчивости и ускорением эволюции совершенно
несомненна.
Рассмотрим теперь, более подробно, формы некоторых отдельных
групп кардиид киммерия, куяльника, а также гурия и чауды. На этих
примерах можно наглядно показать явления повышенной изменчивости
и зависящую от нее эволюцию путем естественного отбора.
Мы уже видели, что, при наличии очень интенсивной внутривидовой
изменчивости у представителей филогенетических линий, более или ме-
нее полно представленных в палеобиологической летописи, оказывается
трудным проведение границ между смежными, непосредственно родст-
171
венными, видами. В таких случаях, при постепенности переходов между
видами, некоторые исследователи, обладающие огромным опытом, от-
казываются от попыток найти границу между видами и, как следствие,
от различения двух видов.
Займемся сначала так называемыми дидакнами группы „Didacna"
multistriata Rouss. (киммерий—куяльник).
«Didacna» multistriata, как известно, характерна для киммерия, а
«Didacna» medeae — для куяльницких отложений Западной Грузии.
На табл. VI, VII, VIII изображены различные представители группы
«Did.» medeae. На этих рисунках наглядно видны изменчивость и фило-
генетическое развитие в определенном направлении — в сторону увели-
чения размеров раковины, ее массивности (толстостенности) и мощности
ее кардинальных зубов.
В своей капитальной монографии, посвященной солоноватоводным
кардиидам СССР, А. Г. Эберзин (1962, стр. 92) объединяет в один вид
„Did." medeae Dav." „Did." gelaenoDav.,отрицая какое-либо таксономичес-
кое значение этих двух форм. Он, по-видимому, думает, что эти две
формы представляют собой фенотипические реакции, зависящие от нес-
колько различных условий существования: если молодь попадала в зону
«волноприбоя или заметных течений» на грубозернистый песчаный или
галечный грунт, то развивались формы «celaeno», в условиях же мень-
шего мелководья на песчанисто-глинистом грунте сублиторали онтогенез,
приводил к формированию «medeae». Молодые же особи «celaeno», по
Эберзину, менее отличались от «medeae». «Чем моложе особи, — гово-
рит Эберзин (там же), — тем большее сходство обнаруживается у них
по отношению друг к другу». Эти соображения Эберзина представляют
интерес ,и должны быть приняты во внимание при изучении эволюции в
пределах данной группы. Но последовательно проводя эту идею, мы
должны были бы обратить внимание на примитивные формы «Did.»
medeae, которые очень близки к «Did.» multistriata и могут быть с таким
же успехом отнесены к этому виду, особенно в том случае, если перед
нами — молодые экземпляры «medeae». Однако надо заметить, что да-
же у солоноватоводных кардиид никто еще не замечал столь резко
разнящихся норм реакции в пределах одного вида. Если бы Эберзин
был прав, то это говорило бы об огромной изменчивости одного из ви-
дав солоноватоводных кардиид нашего плиоцна. В действительности
же как ни велика эта изменчивость, она едва ли доходит до той степе-
ни, которую предполагает Эберзин.
Если, однако, мы и не можем вполне согласиться с Эберзиным в по-
нимании данной группы, то все же надо признать, что, проводя границы
между видом medeae и celaeno, а также между multistriata и medeae,
мы все же должны признать не только значительную меру постепеннос-
ти в эволюции этих форм, но и чрезвычайно высокие темпы внутривидо-
вой изменчивости.
Аналогичную картину являют многие другие группы кардиид
нашего плиоцена. Так, Эберзин (1962, стр. 88) отмечает «переходные
особенности» нижнекиммерийских представителей вида «Didacna gurief-
fi с азовского побережья Керченского полуострова, сближающие этот
вид с верхнепонтическими „Did." sulcatina sibsulcatina Andrus. Рако-
вины этого вида, по Эберзину (там же, стр. 85), очень сходны с «Did.»
«gurieffi». Последний вид, по словам Эберзина, — представляет дальней-
ший этап в развитии форм sulcatina—subsulcatina—gurieffi как в отно-
шении увеличения размеров створок, так и строения замка и ребер. Тут
172
перед нами опять-таки наглядный пример определенно направленной ин-
тенсивной изменчивости и видообразования.
Очень высокую изменчивость обнаруживают представители вида
Plagiodacna modiolaris Rouss., встречающиеся в киммерии Запад-
ной Грузии. В популяциях, живших севернее Абхазии, изменчивость это-
го вида, довольно значительная, была все же не так высока, как на уча-
стках киммерийского моря, соответствующих территориям Абхазии и
Гурии. Этот вогрос был освещен в статье Е. Г. Ахвледиани (1958). Не-
которое, но далеко неполное, представление об изменчивости этого вида
в популяциях, живших в указанной южной части бассейна, можно по-
лучить по табл. IX. Заметно варьирует величина раковины. Изменчивы
ее очертания, часто очень неправильные. Так, верхне-задний угол ее при-
поднят в различной мере, иногда лишь образуя крыловидное расшире-
ние (см. табл. IX, фиг. 8). Варьирует высота раковины (ср. на той же
табл. фиг. 3 и 13). Сильно изменчива величина субапикальной пластин-
ки (там же ср. фиг. 8 и 10). Варьирует длина кардинального зуба. Из-
менчива массивность раковины.
Приведем еще один пример интенсивной изменчивости у кардиид
киммерия, — опять-таки преимущественнно в южной части области рас-
пространения отложений этого яруса. Stenodacna angusticostata Rouss.
обнаруживает особенно сильную изменчивость в степени мощности зам-
ка, в величине кардинальных зубов и в толщине передней части створки
(рис. 40). О соотношении между Sten. praeangusticostata Fbers., из ниж-
него киммерия Кубанской области, и Sten. angusticostata Rouss.
А. Г. Эберзин (1951. стр. 100) писал: «между обоими видами существуют
такие переходные формы, которые, с одной стороны еще более подкреп-
ляют уверенность в тесной родственной связи этих видов, а с другой —
стирают явственность отличительных признаков между ними». Тот же
автор (там же), пишет: «Геологически наиболее молодые представители
Sten. angusticostata Rouss. из верхнекиммерийских отложений обнару-
живают, по сравнению с типичными, среднекиммерийскими, формами,
крайнюю степень развития некоторых признаков. Например, макушка у
них еще сильнее приближается кпереди, а неравносторонность при этом
еще более возрастает».
Очень высокая изменчивость наблюдается и в других киммерий-
ских и куяльницких родах кардиид, живших в юго-восточной части
бассейна, на территории нынешней Западной Грузии. Это вовсе не зна-
чит, что повышенной изменчивости не было в популяциях, живших к се-
веро-западу от названного участка. Мы хотим только сказать, что в
южных популяциях изменчивость была особенно интенсивной.
Палеобиология располагает данными, свидетельствующими о весь-
ма интенсивной изменчивости двустворок, в частности, — кардиид, в
Черноморском бассейне также и в более поздние времена. Здесь мы ог-
раничимся указанием лишь на некоторые, стратиграфически и эволюци-
онно-палеобиологически особенно важные, таксоны кардиид гурия и
чауды. Чаудинские кардииды филогенетически очень тесно связаны с
более древними, гурийскими. Тем не менее очень часто гурийский гори-
зонт относится стратиграфами к верхнему плиоцену, а чаудинский — к
плейстоцену. Неразрывная связь между моллюсками гурия и моллюска-
ми чауды, существование плавных переходов между теми и другими го-
ворят за то, что гурийское время не могло быть отделено от чаудинско-
го сколько-нибудь значительным пробелом. Однако тесные филогенети-
ческие взаимосвязи между моллюсковыми фаунами гурия, с одной сто-
роны, и чауды, с другой, вовсе не исключают возможности отнесения
173
гурия к плиоцену, а чауды — к плейстоцену, если это будет достаточно
обосновано геологически и палеобиологически: нельзя же думать, что в
истории Земли каждая новая эпоха открывалась мгновенными катаст-
рофическими событиями широкого распространения. Итак, этот геохро-
нологический вопрос мы оставим здесь открытым, и займемся теми дан-
ными, которые свидетельствуют о силе и характере индивидуальной из-
менчивости в гурии и чауде.
Рис. 40. Внутривидовая изменчивость Stenodacna
Западная Грузия (Гурия, сел. Джиханджири).
angusticostata Rouss. Киммерий.
Фиг. 1—9. По Е. Ахвледиани.
В статье Л. Ш. Давиташвили и Т. Г. Китовани (1964) были кратко-
рассмотрены взаимоотношения форм двух родов кардиид, один из ко-
торых, Digressodacna Dav. et Kit., очень характерен для гурия, а другой,.
Tschaudia Dav.,—для чауды. В этой статье мы утверждали, что род
Tschaudia происходит от рода Digressodacna.
Мы писали (там же, стр. 387), что огромная изменчивость пред-
ставителей Digressodacna digressa Liv., хорошо показанная А. Г.
Эберзиным (1962), не носит совершенно беспорядочного характера
«шальных» вариантов: намечается господствующая направленность из-
менений в сторону увеличения выпуклости раковины, возрастания ее
размеров в редукции синуса мантийной линии. Формы рода Tschaudia
тоже обнаруживают интенсивную индивидуальную изменчвость. Т. Г.
Китовани, которая продолжает детально изучать представителей обоих
174
родов, склонна выделить в составе каждого из них немало форм видо-
вого значения. Воздерживаясь здесь от рассмотрения таксономических
вопросов, мы, с любезного разрешения Т. Г. Китовани, приведем здесь
изображения изучаемых ею форм; здесь видно, что крайние формы,
резко различающиеся одна от другой, связаны между собой постепен-
ными переходами. На воспроизводимых здесь рисунках представлены
лишь формы рода Tschaudia и их ближайшие предки из дигрессодакя
(табл. X).
Глава одиннадцатая
РЕЗКО ПОВЫШЕННАЯ МАССОВАЯ ИНДИВИДУАЛЬНАЯ
ИЗМЕНЧИВОСТЬ В НЕКОТОРЫХ ГРУППАХ БРЮХОНОГИХ
ПЛИОЦЕНА РИОНСКОГО ЗАЛИВА
ЯВЛЕНИЯ ИЗМЕНЧИВОСТИ У НЕКОТОРЫХ ПЛИОЦЕНОВЫХ НЕРИТИНИД
Неритиниды Черноморско-Каспийского плиоцена, понимаемого в
андрусовском объеме, привлекают к себе внимание палеонтологов и гео-
логов прежде всего по наличию среди них иногда очень крупных рако-
вин и форм, сильно отклоняющихся по своему строению от обычных не-
ритинид пресноводных, и даже солоноватоводных. Некоторые из таких
«аберрантных» групп неритинид были описаны еще Н. И. Андрусовым
(N. Andrussoff, 1912), другие были им замечены, но он не успел их опи-
сать, третьи были обнаружены после Н. И. Андрусова.
Эти формы сильно отличаются от «нормальных» неритинид, и чуть
ли не каждая из них стоит как-то особняком среди неогеновых, четвер-
тичных и современных неритинид.
Остановимся на некоторых из этих форм и рассмотрим вопрос о
том, какую роль играла индивидуальная изменчивость в их эволюции.
Прежде всего, постараемся ответить на вопрос: имели ли эти фор-
мы автохтонное происхождение, то есть были ли они эндемиками, или
они филогенетически связаны с какими-нибудь внешне сходными с ни-
ми формами из других стран, — быть может, стран весьма отдаленных.
Действительно, некоторые из брюхоногих нашего плиоцена похожи на
современные формы Восточной и Юго-Восточной Азии и Полинезии.
Много лет назад мы рассмотрели вопрос о возможности кровного родст-
ва между этими формами и пришли к отрицательному заключению.
Н. И. Андрусов, как и К. Сенинский (1905), был определенно склонен ус-
матривать генетическую связь закавказской фауны брюхоногих с «экзо-
тическими» формами Дальнего Востока, Юго-Восточной Азии и Поли-
незии.
Андрусов (Andrussoff, 1912, стр. 17) различал подрод Neritaea в
пределах рода Neritina и отнес к этому подроду вид Neritina (Neri-
taea) colchica. Это—необычайно крупная, для неогена Европы и Запад-
ной Азии, неритина. «Если родство Neritaea colchica с Clypeolum
подтвердится, — писал он (1912, стр. 17; 1964, стр. 595), — то в нашем
распоряжении будет еще один пример родственных связей гастроподо-
вой фауны кавказских плиоценовых отложений с фаунами островов Ти-
хого и Индийского океанов». Позже Андрусов совершенно уве-
ренно высказывался о родстве Ner. colchica с клипеолями. „Neritaea
colchica, — писал он (1928; 1962, стр. 596), — родственна с видами
рода Clipeolum Reel., распространенного в Японии и на островах Тихого
176
океана». Однако некоторые брюхоногие из дуабских отложений, по
Андрусову, «конечно, автохтонного происхождения, в особенности это
касается своеобразных неритинид (Neritonyx, Brusinaella), которые, по-
видимому, представляют дальнейшую крайнюю форму развития рода
Ninnia» (1962, стр. 596).
Нам еще придется коснуться вопроса о воображаемых связях брю-
хоногих живших в озерах-морях нашего плиоцена, с сходными «экзо-
тическими» формами. Но уже тут отметим, что Андрусов, склонный к
признанию таких связей, все же отрицал возможность таковых в отно-
шении некоторых форм, а кроме того, писал (там же): «В таких бас-
сейнах, как понтический, киммерийский и им подобные, они (пресно-
водные элементы. — Л. Д.) являются иммигрантами из впадающих
рек, и поэтому присутствие одинаковых форм не является доказатель-
ством прямого сообщения одновременных водных бассейнов».
Neritonyx и Brusinaella так своеобразны и настолько сильно отли-
чаются от всех ископаемых и современных неритинид, что выделение
этих форм в самостоятельные роды является, действительно, вполне оп-
равданным (рис. 41 и 42). О неритониксе Андрусов писал, что этот
род характеризуется совершенно плоской ракови-
ной, очертаниями напоминающей человеческий но-
готь. Нижний край раковины несет килевидный ва-
лик, начинающийся вблизи завитка. Брузинелла,
как правильно утверждает Андрусов (1964, стр.
588), «стоит также совершенно изолированно среди
Рис. 41. Neritonyx um-
guiculata Andrus. Из
верхней части слоев р.
Дуаба, киммерии (с.
Мокви, Абхазия). По
Андрусову.
Рис. 42. Brusinaella petasata Andruss. Из
верхней части слоев р. Дуаба, киммерий
(с. Мокви, Абхазия). По Андрусову.
неритинид». Раковина у нее тоже очень плоская. Достаточно взглянуть
на изображение раковины, чтобы оценить оригинальность этой необык-
новенной формы, самой плоской из всех доныне описанных неритинид.
Неритеа похожа на «нормальных» цинний, но сильно отличается от всех
известных видов этого рода очень крупной раковиной.
Чрезвычайно необыкновенные неритиниды известны также из дру-
гих ярусов нашего солоноватоводного плиоцена. Очень своеобразны
формы, для которых нами был выделен особый род Meganinnia: Meg. shio
(рис. 43) и Meg. corrugata. Так как эти формы известны в весьма нич-
тожном числе экземпляров —один образец Meg. corrugata был найден
12- Л- Ш. Давиташвили 177
в чаудинских отложениях, другой же — в гурийских отложениях Гурии,,
a Meg. shio — в одном экземпляре в чауде Гурии — у нас нет уверен-
ности в том, что обе эти формы» заметно отличающиеся одна от другой,
должны считаться принадлежащими к одному и тому же роду. Meg.
corrugata внешне несколько похожа на ныне живущую Neritina ves-
pertina, форму, описанную из ручьев Сандвичевых островов (рис. 44)
и определенно отличается от всех описанных ископаемых форм нерити-
нид наличием двух уховидных отростков, придающих меганинниям сим-
метричный облик.
Очень трудно допус-
тить общность происхож-
дения чаудинских форм
Черноморского бассейна
и внешне сходных с ними
дальневосточных нерити-
нид. То же самое при-
ходится сказать и о род-
ственных связях дуабской
Neritaea colchica, с одной
стороны, и похожих на
нее форм, которые, по
словам Андрусова (1964,
Рис. 44. Конвергентное сходство двух форм нерити-
нид. Фнг. 1 и 2—Meganinnia corrugata Dav. из чаудин-
ских отложений Гурии. 3—4—Neritina vespertina Nut-
Рис. 43. Meganinnia shio
Dav. из чаудинских отло-
жений Гурии, х 2.
tal—Сандвичевы острова (в ручьях), х 2.
Тихого и Индийского океанов, с другой, стр. 595), живут на островах
Зная способность организмов к развитию, в процессе филогенеза, на ос-
нове аналогической изменчивости сходных форм, гораздо естественнее
предположить, что чисто внешнее сходство раковин есть результат па-
раллельного развития. Тут мы не -можем даже сослаться на сходство
экологических условий: «экзотические» крупные неритиниды живут в
.небольших озерах, ручьях и даже во влажных местах суши.
Из киммерийских отложений Мегрелии Л. К. Габуния (1949) опи-
сал род Manana — исключительно своеобразную форму с резко выра-
женными спиральными ребрами, которые несут острые тонкие шипы и
бугорки. В куяльницких слоях Гурии тот же исследователь (1949) на-
шел раковины, принадлежащие к роду Mananopsis, который характерн-
ее
зуется спиральными ребрами и верхним килевым ребром, усаженным
острыми шипами.
Итак, мы должны признать, что упомянутые нами плиоценовые не-
ритиниды Черноморско-Каспийского бассейна — настоящие эндемики.
К вопросу об эндемизме этих форм мы вернемся по рассмотрении неко-
торых других брюхоногих пресноводного происхождения, остатки кото-
рых встречаются в нашем солоноватоводном плиоцене. А здесь мы толь-
ко подчеркнем, что формирование в течение относительно небольшого
отрезка геологического времени, не только новых видов неритинид, но
и более крупных таксонов — подродов и даже родов — могло произойти
лишь при условии усиленной внутривидовой изменчивости: без этого ес-
тественный отбор не мог бы дать такие результаты. Усиленная измен-
чивость должна была зависеть от каких-то экологических условий, при-
чин которых мы попытаемся коснуться несколько дальше в этой главе.
ОБ ИЗМЕНЧИВОСТИ У ВИВИПАРИД ЧЕРНОМОРСКО-КАСПИЙСКОГО
СОЛОНОВАТОВОДНОГО ПЛИОЦЕНА
Рис. 45. Euxinomar-
garya mandarinica
(Sen.) из куяльниц-
ких слоев Гурии. х 2.
наших плиоценовых
Виьипариды довольно хорошо представлены в отложениях Черно-
морско-Каспийского солоноватоводного плиоцена, начиная от понта. В
понте и киммерии более северных местонахождений этих ярусов формы
рода Viviparus имеют раковины более или менее нормального строения,
гладкие с выпуклыми оборотами. Только в южной части области рас-
пространения этих отложений на территории СССР встречаются формы,
сильно отклонившиеся от обычных вивипарид Европы и Западной Азии,
с резко изменившимся очертанием поперечного сечения оборотов и зна-
чительно скульптированные. Мы остановимся на таких формах, потому
что именно их эволюция должна была зависеть от существенно изменяв-
шихся внешних условий.
В дуабских отложениях киммерийского яруса встречаются формы,
схожие с различными плиоценовыми вивипарусами Румынии и Славонии
и обычно определяемые как Viv. dezmanianus Brus., Viv. sadleri Partsch,
Viv. neumayri Brus. Это в большинстве некрупные формы, но стенки ра-
ковины у них толще» чем у обычных пресноводных
вивипарусов, а боковая поверхность оборотов бо-
лее или менее уплощена. Можно думать, что такие
раковины лучше приспособлены к жизни в условиях
сзера-моря с более или менее сильным прибоем.
Наше внимание, однако, больше привлекают за-
метно рёзЧё аберрантные формы, которые выделя-
ются в особые эндемичные роды Euxinomargarya
Akhvl. и Suchumica Sen. (рис. 45 и 46), характер-
ные для куяльницких отложений Рионского зали-
ва. Сходство эуксиномаргарий с маргариями из озе-
ра Талифу (провинция Юннань, Южный Китай) и
из озера Ланао (остров Минданао, Филиппины)
бросается в глаза при сравнении рис. 45 и рис. 47.
Любопытно также сравнить Suchumica gracilis Sen.
с Viviparus mearnsi Bartsch из озера Ланао (рис. 48).
Возможны более или менее близкие филогене-
тические связи форм из плиоценовых отложений
Рионского залива со сходными формами
плиоцена Румынии, но общность происхождения
форм и форм, живущих на одно-м из Филиппинских островов или в
Южном Китае, конечно, совершенно исключается (Давиташвили, 1932).
179
Примечательно, что, согласно новейшим исследованиям, эвксино-
маргарии начинают появляться уже ib киммерии Рионского залива (Е. Г.
/Ахвледиани), а расцвета достигают в куяльнике того же района. Суху-
мики же известны пока только из куяльника Рионского залива.
Рис. 46. Suchumica gracilis Sen. из куяльника Абхазии, х 3.
Рис.* 47, Margarya melanoides Neviil из оз. Талифу. Китай. Несколько уменьшено.
180
Так или иначе трудно усомниться в том, что подобные формы, резко
отличающиеся от своих родичей, живущих и живших в нормальных для
вивипарид условиях текучих пресных вод, могли возникнуть лищь на
основе весьма интенсивной внутривидовой изменчивости. Эволюция в
данном случае чрезвычайно быстро привела к возникновению новых
родов.
Рис. 48. Viviparus mr.arnsi Bartsc'i. Из
озера Лана- , на о-ве Минданао (Филип-
пины). Увеличено.
НЕКОТОРЫЕ ОБЩИЕ СООБРАЖЕНИЯ О ВНУТРИВИДОВОЙ ИЗМЕНЧИВОСТИ
ПЛИОЦЕНОВЫХ БРЮХОНОГИХ, ХАРАКТЕРНЫХ ДЛЯ
РИОНСКОГО ЗАЛИВА
В солоноватоводных плиоценовых отложениях западной части Гру-
зии, кроме уже перечисленных форм, встречаются еще многие другие
виды брюхоногих, возникшие в результате очень быстрой эволюции и
сильно отличающиеся от своих родичей, живших и живущих в Европе
и Западной Азии. Сюда относятся формы, обычно определяемые как
Melanopsis nobilis Sen., Mel. acuminata, Melania andrussowi Sen., Amphi-
melania aff. Gajii Brus. и другие. Некоторые из этих форм похожи
на отдельных представителей плиоцена более западных частей Па/ра-
тетиса и, возможно, имеют с ними общее происхождение.
Здесь мы должны подчеркнуть тот факт, что многие из форм плиоце-
на Рионского залива известны до сих пор только из этой области, где
они встречаются иногда в большом изобилии. Их мы не находим в более
северных местонахождениях соответствующих горизонтов. Такие специ-
фически южные формы брюхоногих известны уже в верхнем понте Гу-
рии, но особенно характерны они для киммерийских, куяльницких, гу-
рийских и чаудинских отложений. Едва ли можно думать, что они жи-
ли как в Рионском заливе, так и к северу от него. Против такого допу-
щения* говорит поразительное отсутствие «южных» форм брюхоногих в
киммерии и куяльнике Краснодарского края, Таманского и Керченско-
го полуостровов (при наличии там «нормальных» форм тех же групп),
181
в то время как многие кардииды известны и в южных, и в северных
частях области распространения наших солоноватоводных плиоценовых
осадков.
Поэтому наиболее вероятно предположение, что формы «южных»
брюхоногих возникали на юге и по тем или иным причинам не могли ус-
пешно распространяться на север. Их чрезвычайно усиленная внутриви-
довая изменчивость — необходимое условие быстрого видообразова-
ния — индуцировалась какими-то экологическими условиями, которые
существовали на юге и отсутствовали в более северных частях Черно-
морского бассейна в понте, киммерии, куяльнике, гурии и чауде. Воп-
рос о том, какие же это были условия, вызывавшие повышенную из-
менчивость, заслуживает специального изучения. В настоящее время мы
не можем дать определенный ответ ина этот вопрос. Вполне возможно,
что именно в южной части солоноватовод.ного. Черноморского бассейна
изменчивости брюхоногих'пресноводного происхождения благоприятство-
вали относительно высокие температуры вод, которые оставались дос-
таточно теплыми в течение всего года.
Твердо установленным можно считать тот факт, что на протяже-
нии довольно длительного времени от позднего понта до чауды в облас-
ти Рионского залива существовали условия, благоприятствовавшие
очень интенсивной массовой, направленной индивидуальной изменчивос-
ти моллюсков, на основе которой и происходила эволюция, приводившая
к возникновению новых видов, новых подродов и родов. За время, соот-
ветствующее одному геологическому веку, формировались даже роды.
Повышенную изменчивость брюхоногих плиоцена Рионского зали-
ва интересно сопоставить с аналогичным явлением, наблюдаемым у
моллюсков из плиоценовых пластов некоторых районов, лежащих к за-
паду от Черного моря.
Резко повышенная изменчивость наблюдается также у различных
брюхоногих, в частности у вивипарусов из плиоценовых солоноватовод-
ных слоев Румынии. Ограничимся здесь ссылкой на вивипарусов дакий-
ских отложений бассейна Брашова. Эти формы были описаны Э. Иеке-
лиусом (Е. JeKelius, 1932). Особенно интересно существование здесь об-
щей конической формы с уплощенными боковыми поверхностями обо-
ротов и резким нижнем килем, — Viv. pseudovukotinovici Lorentey.
Она напоминает Viv. vukotinovici, но существенно отличается от нее не-
которыми особенностями (более плоское основание, более мощное раз-
витие нижнего киля, наличие второго киля, не нарушающего однако об-
щего конического очертания раковины и т. д.). Большое сходство заме-
чается также между описываемой Иекелиусом формой из бассейна Бра-
шова и уже упомянутой нами Suchumica из куяльницких отложений
Западной Грузии.
Примечательно, что и у Иекелиуса нет сомнений относительно роли
условий среды в возникновении резкой изменчивости румынских виви-
парид и других форм пресноводного происхождения из дакийских от-
ложений Румынии.
«Моллюсковые фауны многочисленных позднеплиоценовых морских
бассейнов Юго-восточной Европы содержат, наряду с видами с простым,
стабильным строением, многочисленные бросающиеся в глаза формы,
обнаруживающие большую изменчивость, с килями и бугорками, с раз-
ворачивающимися раковинами и т. д.» (Иекелиус, JeKelius, 1932, стр.
42). «Когда мы ищем причины этого поразительного развития специаль-
но в отношении юго-восточно-европейских неогеновых пресноводных
фаун, то мы находим различные широко распространенные изменения
182
в физико-химическом характере их жизненных округов» (там же, стр<
43—44). Предпосылкой развития этих плиоценовых пресноводных фаун
Иекелиус считает крупные события, которые в первой половине неогена
коренным образом изменили химические и физические отношения среды
моллюсков внутренних бассейнов. «Вследствие этого, — говорит автор
(стр. 44), — консервативные формы автохтонного европейского фауни-
стического фонда были вынуждаемы известным образом реагировать на
различную минерализацию воды, особенно высокое содержание изве-
сти, сильный волноприбой в водах крупных внутренних озер и т. д.
и т. д.».
Тут Иекелиус, — как почти все другие исследователи моллюсковых
фаун Паратетиса и в частности обширных озерных водоемов, представ-
лявших собой остатки крупного морского бассейна, — без всяких коле-
баний усматривает причину изменчивости в изменениях физических и
химических свойств среды.
Некоторые группы моллюсков нашего неогена сильно отличаются
от всех моллюсков, представленных в тех же горизонтах, а также в под-
стилающих и перекрывающих отложениях. Тогда может возникнуть
мысль о том, что они произошли от форм, не встречающихся в сериях
осадков Паратетиса. Например, Valencienniidae .нашего неогена настоль-
ко сильно отличаются от всех брюхоногих этой полосы, что высказыва-
лась мысль об их происхождении от те^х или иных аллопатрических
предков, каким-то образом иммигрировавших сюда из полносоленого
моря. Между тем, тщательный анализ этого материала приводит иссле-
дователей к выводу (И. Г. Тактакишвили, 1967), что это семейство
могло возникнуть лишь в Паратетисе и представляет собой пример глу-
бокого изменения некоторых представителей семейства Lymnaeidae,
очень мало похожих на валенсиенниид. Эти последние не могут иметь
иного происхождения. Значит, совершалась чрезвычайно быстрая эволю-
ция на основе исключительно богатой и более или менее направленной
внутривидовой изменчивости.
В процессе эволюции пресноводные брюхоногие семейства лимне-
ид должны были перейти от дыхания кислородом воздуха к дыханию
кислородом, растворенным в воде, перейти к донному образу жизни,
причем некоторые из этих форм селились на довольно глубоких участ-
ках дна огромных «каспиев», а также претерпеть многие другие глубо-
кие морфологические и физиологические изменения. Все такие преобра-
зования могли осуществляться лищь на основе бурной индивидуальной
изменчивости определенно направленного и массового характера. Толь-
ко при такой изменчивости естественный отбор мог привести к образо-
ванию нового семейства Valencienniidae в составе нескольких родов,
заметно отличающихся между собой по многим существенным призна-
кам.
Глава двенадцатая
ИЗМЕНЧИВОСТЬ МАКТРИД АКЧАГЫЛА И ЗНАЧЕНИЕ
АНАЛОГИЧЕСКОЙ ИЗМЕНЧИВОСТИ В ЭВОЛЮЦИИ
МОЛЛЮСКОВ ПАРАТЕТИСА
В предыдущих главах мы неоднократно отмечали интенсивную из-
менчивость морских моллюсков, попадавших в необычайные для них ус-
ловия существования, если изменение среды не вызывало вымирания
этих моллюсков.
Мы встречались такж£ с явлением аналогической изменчивости,
которую испытывали формы» более или менее близкие одна и другой, ког-
да они оказывались в аберрантных условиях жизни. Такое явление наб-
людается, например, в эволюции некоторых брюхоногих пресноводного
происхождения, попадавших в солоноватоводные бассейны, в своеобраз-
ные «каспии» киммерийского, куяльницксго, гурийского и чаудинского
веков. Вспомним хотя бы то, что было сказано в предыдущей главе о
неритинидах киммерия и куяльника Рионского залива, о своеобразных
вивипаридах куяльницких отложений того же района, а также о причуд-
ливых неритинидах гурия и чауды Гурии.
Такие явления обнаруживаются при сопоставлении основных групп
моллюсков, живших в огромном море акчагыльского века в позднем
плиоцене, с одной стороны, с теми же группами, обитавшими в великом
сарматском море позднего миоцена, с другой. Примечательный парал-
лелизм в филогенетическом развитии этих моллюсков сармата и акчагы-
ла привел некоторых опытных исследователей даже к неправильному
выводу о непосредственных филогенетических связях сарматских и ак-
чагыльских членов этих групп.
В действительности же, как в этом нетрудно будет убедиться, в дан-
ном случае, перед нами яркий пример той аналогической изменчивости,
•о которой писал еще Дарвин и которую имел в виду крупнейший совет-
ский генетик Н. И. Вавилов, когда он говорил о законе гомологической
изменчивости.
Чрезвычайно интересные данные об акчагыльском комплексе мол-
люсков приводит А. А. Али-Заде (1967) в своей крупной монографии об
акчагыле Туркменистана. Этот исследователь описал множество форм
из акчагыльских отложений Туркменистана, фауну которых он произво-
дит от сарматской. Основным доводом для такого вывода служит сход-
ство многих акчагыльских форм с сарматскими моллюсками. В нашу за-
дачу, однако, не входит разбор монографии А. А. Али-Заде. Анализ всех
его выводов потребовал бы слишком много места в этой книге. Отметим
только, что ни в одном случае сходство между изученными этим автором
акчагыльскими и сарматскими формами .не является настолько глубо-
ким, чтобы его нельзя было считать результатом параллелизма в разви-
тии.
А. А. Али-Заде из акчагыла Туркменистана описал 117 видов рода
Cardium и 9 видов кардиид рода Replidacna. Мы не уверены в том,
что все эти виды выделены на вполне достаточном основании; болыпин-
184
ство их соответствует, вероятно, лишь вариантам индивидуальной из-
менчивости или разновидностям. Однако нельзя не видеть, что сообщае-
мые этим исследователем данные говорят об очень интенсивной инди-
видуальной изменчивости акчагыльских кардиид. Далее, при наличии
столь высокой степени изменчивости и таких темпов формообразования
вполне естественно, что эволюция акчагыльских кардиид могла привести
к формам, внешне похожим на те или иные виды сарматских кардиид.
Ведь и сарматские, и акчагыльские кардииды произошли от настоящих
морских форм и испытали ускоренную радиацию. Таким образом в
туркменистанском акчагыле оказались формы, обнаруживающие сходст-
во, в общем довольно поверхностное, с сарматскими Card, ingratum Ko-
les., Card, vindobonense (Partsch), Card, plicatofittoni Sinz., Card, prae-
plicatum Hilb., Card, plicaturn Eichw., Card, laevigatoloweni Koles., Card,
inflatum Sinz., Card, aviculare Sinz., Card, desperatum Koles. и Card, bar-
boti Hoern. Ведь число возможных вариантов изменения формы и величи-
ны раковины у кардиид, в конце концов, далеко не бесконечно: возмож-
ны различные степени увеличения раковины, ее толстостенно1сти, ее вы-
пуклости (вздутости), «скульптура» может быть представлена то более
частыми, то более редкими ребрами (вплоть до полного их исчезнове-
ния), ребра могут быть чешуйчатыми, шиповатыми и гладкими, верхне-
задний угол может быть либо не расширен, либо более или менее рас-
ширен. Все подобные варианты многочисленны, но число их все-таки не
бесконечно, ограничено. Аналогичные варианты могли возникать в раз-
личных морях с пониженной соленостью, если исходные формы были
обычными видами морских кардиид. Но доходило ли дело до полной ви-
довой тождественности или хотя бы до такого сходства, которое давало
бы нам право говорить о близких филогенетических отношениях соответ-
ствующих сарматских и акчагыльских форм? На этот вопрос надо отве-
тить отрицательно, хотя мы можем понять, что А. А. Ал и-Заде должно
было поразить сходство между соответствующими парами сарматских и
•акчагыльских форм.
На табл. XI мы воспроизводим изображения Avimactra acutecarina-
ta Andrus., по фотографиям, которые любезно передал нам А. А. Али-
Заде. Этот автор относит только что упомянутый вид к роду Cryptomac-
tra Andrus. с чем мы не можем согласиться, так как киль, наблюдае-
мый у обеих форм, вполне мог возникнуть у каждой из них совершен-
но самостоятельно. Среди описанных А. А. Али-Заде мактрид имеется
«Lutraria» andrussovi, которая, действительно, напоминает собой по
очертанию раковины формы рода Lutraria, но никак не может быть от-
несена к этому роду по другим признакам (в частности—по совершенно
иному характеру мантийной линии), которые, наряду с другими особен-
ностями, заставляют нас признать эту форму состоящей в тесном родст-
ве с остальными акчагыльскими мактридами.
Таким образом, происходящие от морских форм моллюски акчагы-
ла развились в этом море в результате очень быстрого филогенетичес-
кого развития от немногих предков, попавших в этот бассейн не позд-
нее начала акчагыльского века. Эволюция мактрид сармата и акчагы-
-ла— один из ярких примеров параллелизма в филогенетическом разви-
тии, которое осуществлялось на основе аналогической изменчивости, —
явления, о котором мы говорили в первой главе нашей книги.
При изучении различных форм акчагыльских мактрид нельзя не за-
метить, что близкие виды незаметно переходят один в другой. Это явле-
ние мы наблюдали на материале из так называемых таманских слоев,
185
который был собран на южном побережье Таманского полуострова при
нашем участии И. М. Губкиным и М. И. Варенцовым; и оно произвело
на нас глубокое, неотразимое впечатление. Подобные плавные перехо-
ды могут заставить исследователя усомниться в возможности различать
такие виды, как Avimacra subcaspia Andrus., A. karabugasica Andrus, и
A. ossoskovi Andrus. Эти очень тесные связи между упомянутыми видами
отмечали исследователи акчагыльских мактрид. Так, В. П. Колесников
(1950, стр. 46) писал: „Вполне возможно, что A. ossoskovi является свя-
зующей формой между A. subcaspia и A. veniukovi. Необходимо отметить,
что наиболее удлиненные экземпляры по форме почти не отличимы от
A. subcaspia". А. А. Али-Заде тоже признает сходство между A. ossos-
kovi и A. subcaspia, а также между A. ossoskovi и A. veniukovi.
Считаем уместным более детально рассмотреть взаимоотношения
между A. subcaspia, A. karabugasica и A. ossoskovi по материалам из
среднего акчагыла Южной Кахетии, пользуясь набором изображений,
любезно предоставленными в наше распоряжение Н. Р. Джикия (табл.
XII, XIII, XIV). На таблице XII изображены правые створки A. subcas-
pia и A. karabugasica. Фигуры 1 —10 представляют 'более или менее типич“
ные особи A. subcaspia, довольно короткие и слабо неравносторонние. Фи“
гуры 11,12 и 13 показывают постепенный переход к более удлиненным и
более неравносторонним раковинам, которые более плоски, чем типич-
ные представители вида. Это — формы, промежуточные между А.
subcaspia и A. karabugasica. На фигурах 14—20 изображены плоские,
удлиненные и сильно неравносторонние раковины—типичные формы А.
karabugasica.
Таблица XIII дает некоторое представление об изменчивости A. os-
soskovi. Раковины отличаются между собой по степени выпуклости ство-
рок и по очертанию. На фигурах 1, 2 и 3 показаны короткие раковины,
а на фигуре 15, наоборот, удлиненная форма. На таблице XIV изображе-
ны удлиненные формы с довольно резко выраженным килем и оттяну-
тым задним концом. Обращает на себя внимание утолщенность, мас-
сивность нижних боковых зубов (фиг. 7—15).
Плавные, почти незаметные переходы между формами, выделяемы-
ми в качестве самостоятельных видов, наблюдаются в разных (иногда
значительно отдаленных друг от друга) районах распространения ак-
чагыльских отложений. Это обстоятельство безусловно заслуживает
внимания. Оно заставляет нас поставить вопрос: являются ли, напри-
мер, A. subcaspia и A. karabugasica самостоятельными видами и не пра-
вильнее ли считать их принадлежащими к одному полиморфному виду?
Колесников писал (1950, стр. 31): «А. subcaspia обладает настолько из-
менчивыми раковинами, что Н. И. Андрусов, устанавливая этот вид и
изображая его, дал большее количество фотографий переходных форм к
A. karabugasica, чем типичных раковин, 'относящихся собственно к опи-
санному им виду. Этим Андрусов затемнил понятие о самом виде A. sub-
caspia». Так или иначе, колоссальная изменчивость видов акчагыльских
мактрид служила основой для быстрого и широкого видообразования.
Типичные формы разных, но сходных между собой, видов так сильно
•отличаются между собой, что нет возможности отрицать наличие мно-
гочисленных видов.
Акчагыльские мактриды, так же как и другие группы моллюсков
этого замечательного и до сих пор во многом загадочного морского бас-
186
сейна, представляют собой чрезвычайно заманчивый объект палеобио-
логического исследования, которое может привести к освещению некото-
рых важнейших проблем эволюционного учения, в частности— пробле-
мы изменчивости и ее генезиса. Но уже теперь можно сказать, что изу-
чение мактрид акчагыльского века приводит к данным, доказывающим
зависимость массовой изменчивости от своеобразия экологических усло-
вий и закон аналогической изменчивости Ч. Дарвина.
Глава тринадцатая
ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ У АММОНОИДЕП
ЮРЫ И МЕЛА
Известно, что аммоноидеи — особенно ценная для стратиграфичес-
кой параллелизации группа ископаемых животных. По различным
признакам их раковин устранавливается относительно очень быстрый
ход их эволюционного развития. Такая интенсивная эволюция могла
совершаться лишь на основе весьма высокой внутривидовой изменчи-
вости. В отношении темпов филогенеза аммоноидеи стоят выше таких
групп беспозвоночных моря, как, например, большинство пластинча-
тожаберных и плеченогих, хотя и эти группы всегда широко привле-
каются для определения геологического возраста и геохронологических
сопоставлений. Нам могут сказать, что относительно очень высокие
темпы эволюции аммоноидей — явление кажущееся и зависящее от то-
го, что раковина у них несет больше признаков, характерных для от-
дельных таксонов, чем ископаемые остатки других моллюсков и плече-
ногих. Правда, кроме внешней формы раковины аммонита и ее орнамен-
тации для определения таксонов аммоноидей очень важны также пере-
городочные линии. Кроме того при достаточно удовлетворительной сох-
ранности по раковине аммонита можно судить о его онтогенетических
стадиях, что также очень важно для изучения систематики этих форм.
На способных сохраняться в ископаемом состоянии раковинах других
моллюсков признаков, отражающих строение тела и онтогенетическое
развитие особи, замечается гораздо меньше. Но аммоноидеи, как и дру-
гие головоногие, эволюировали действительно быстро. Довольно быстро
сменялись во времени формы белемноидей, которые в большинстве слу-
чаев сохранялись в виде ростков, мало, казалось бы, говорящих о строе-
нии тела и о процессе филогенетического развития. Аммоноидеи принад-
лежали к числу наиболее прогрессивных групп беспозвоночных в мезо-
зойских морях. Организация их была сложной, как и жизнедеятель-
ность: обладая способностью ползать по дну, подобно большинству брю-
хоногих моллюсков, они могли кроме того совершать значительные пе-
редвижения в водной массе морей — вертикальные и горизонтальные.
Высокая степень подвижности позволяла им попадать в самые разно-
образные обстановки, какие только существовали в мезозойских морях
и океанах, во многие морские биотопы, с которыми были связаны все-
возможные биоценозы. Это открывало возможность воздействия на ам-
монитов, в различных стадиях онтогенеза, тех или иных факторов сре-
ды. Таким путем вызывались наследственные изменения, которые вслед-
ствие естественного отбора становились признаками новых таксонов.
Без высокой изменчивости внутри популяций невозможно объяснить
высокие темпы филогенеза аммоноидей. Существует, конечно, мнение,
будто бы эволюция этих животных была управляема какой-то вну-
188
тренней тенденцией, не зависящей от условий среды. Но подавляющее
большинство палеобиологов и необиологов теперь вполне сознает, что
идея такого ортогенеза выходит за рамки научного исследования раз-
вития жизни и уводит сторонников этой идеи в область мистицизма.
Изучение аммоноидей показало исследователям, что эти моллюски
действительно обладали высокой индивидуальной изменчивостью.
Примечательно, что исследователи аммоноидей отмечали постепен-
ность перехода от одного вида к другому, — наличие промежуточных
форм.
Большое внимание было уделено многими исследователями вопросу
об изменчивости среднеюрских аммонитов семейства Cosmoceratidae.
Р. Бринкманн (R. Вппкгпапп, 1929), на основе тщательного изучения
зональных послойных сборов в юрских отложениях Англии, установил
переход от одного вида рода Cosmoceras к другому. Оскар Кун
(О. Kuhn, 1939. стр. 524) отметил существование «всех переходов»
между тремя видами Cosmoceras: Cosm. jason, Cosm. castor и Cosm.
pollux. Дж. К. Калломон (J. H. Callomon, 1955) подтвердил данные
Бринкманна. Большое место изменчивости внутри видов космоцератид
нижнего и среднего келловея Западной Европы уделил А. Тентам
(И. Tintant, 1963) в своей капитальной монографии, посвященной это-
му семейству. Изучив представителей рода Cosmoceras из последова-
тельных горизонтов он подчеркивает изменчивость этих форм в геологи-
ческом времени. «Если географическую изменчивость космоцератид ус-
тановить затруднительно, — пишет он (там же, стр. 427), — то их из-
менчивость во времени, наоборот, является быстрой и непрерывной, как
это превосходно доказал Бринкманн в своем мастерском труде о кос-
моцерасах Петербороу в Англии». Тентам отмечает обилие промежуточ-
ных форм, которые зачастую трудно отнести к тому или иному таксону.
Такие плавные переходы он указывает между Cosmoceras (Zugocosmo-
ceras) medea Callomon и Kosm. (Zugocosm.) jason (Rein.). Этот факт
наличия особей, составляющих подлинное промежуточное звено между
популяциями из смежных горизонтов, привел Тентана к выводу, что
«только определение средних форм из каждого позволяет уточнить их
таксономическое положение» (стр. 428). Исходя из такого понимания
вида, автор считает необходимым отказаться от присвоения видового
названия каждому экземпляру: точное определение, по его словам, воз-
можно только для совокупности форм из данного горизонта.
Б. Циглер (1959, стр. 229—235), изучавший аммонитов из верхней
юры Южной Германии, отмечает, что «нет никакой прерывистости» в
эволюции Creniceras dentatum (Reinecke), очень характерного для ниж-
него киммериджа этой области. От самых низов до верхов нижнего ким-
мериджа наблюдается совершенно постепенное возрастание величины
взрослых экземпляров и уменьшение ширины пупка.
О. Гаас (О. Haas, 1946) посвятил работу внутривидовой изменчи-
вости и онтогении Prionotropis woolgari Meek, Prionocyclus wyomingen-
sis Meek из туронских отложений. Этот ученый подчеркивает «ши-
рокий диапазон внутривидовой изменчивости» (там же, стр. 147) обо-
их этих видов, зависящей, в значительной мере, от различий в ско-
рости развития: определенные фазы орнаментации могут появляться
раньше или удерживаться дольше у одних особей, чем у других. Таким
образом возникает «почти умопомрачительное», по выражению авто-
ра, обилие форм, иногда сильно отличающихся одна от другой даже
в одной и той же стадии роста. Гаасу, как он заявляет, хорошо изве-
189
стно, что некоторые, а может быть и все формы, которые он включает,
в вид Prionotropis woolgari, особенно если бы они были найдены в раз-
ных местах, многими специалистами по аммонитам могли быть отнесе-
ны к различным видам или даже к различным подродам.
Мы привели лишь небольшое число случаев внутривидовой измен-
чивости юрских и меловых аммоноидей. Но такие данные достаточно
ясно показывают, что изменчивость этих форм весьма велика и, буду-
чи направлена в определенную сторону, может вести от одного подвида
к другому, от одного вида к другому. Она зачастую затрудняет прове-
дение границ между видами, делая переходы между смежными вида-
ми — членами одной и той же филогенетической цепи — совершенно не-
заметными, плавными. Б. Циглер (1959, стр. 235) отмечал, что эволю-
ция аммонитов протекала в большинстве случаев плавно, без перерывов.
Однако он же утверждал, что в очень немногих случаях эволюция была,
по всей вероятности, прерывистой: прерывистая эволюция, по его мне-
нию, должна была зависеть от ускоренного развития. И он тут же при-
бегает к допущению, что оно могло происходить в малочисленных по-
пуляциях, где новые мутации имеют высокие шансы распро-
странения и укрепления. Здесь мы не будем обсуждать вопрос о воз-
можности особой, прерывистой формы эволюции. А теперь лишь заме-
тим, что быстрая, даже чрезвычайно быстрая эволюция отнюдь не дока-
зывает прерывистости эволюционного процесса в понимании Циглера и
сторонников так называемой синтетической теории эволюции: самая
быстрая эволюция может в то же время быть плавной, сплошной, неп-
рерывной в том смысле, что эволюционный процесс всегда состоит из
скачков, но только скачки эти — малые и обычно почти незаметные.
Решаясь высказаться относительно отсутствия сальтаций в разви-
тии аммонитов, мы должны признать, что дальнейшие исследования не
должны ограничиваться небольшим числом наудачу выхваченных при-
меров. Необходима капитальная сводка по вопросу о внутривидовой из-
менчивости аммонитов. Такую сводку могут дать специалисты — «ам-
монитологи».
Наши примеры были взяты из истории юрских и меловых аммони-
тов. Однако сделанные нами выводы, во всяком случае, не расходятся,
с тем, что известно и о более древних аммоноидеях. Так, А. К. Налив-
кина (1936) приводит случаи незаметного, по сути, как она выражается^
постепенного перехода одного вида в другой среди девонских гониати-
тов и климерий Мугоджар.
Такой же характер имела, по-видимому, эволюция других голово?
ногих. Д. П. Найдин (1957) утверждает, что в некоторых группах позд.-
немеловых белемнитов наблюдается постепенное развитие, и когда воз-
никает много промежуточных форм, нельзя указать точную границу
между двумя видами. Впрочем, по мнению этого автора, видообразова-
ние иногда происходило внезапно, и тогда граница между видами уста-
навливается очень легко. Нам, однако, кажется, что Найдин не дает
убедительного доказательства полной внезапности видообразования в.
случаях этой второй категории. По всей вероятности, мы имеем тут са-
мый обычный случай неполноты наших сведений об ископаемых живот-
ных. Если в данном местонахождении мы имеем два близких друг к
другу вида А и Б, стратиграфически приуроченных к двум последова-
тельным горизонтам, то чаще всего вид А не является непосредствен-
ным предком вида Б: форма, давшая начало виду Б, могла жить дале-
ко от данного местонахождения. Возникнув где-то в более или менее от-
даленном районе, вид Б мог затем испытать прохорез, и одна из его ко-
190
лоний могла попасть в район данного местонахождения. Поэтому чет-
кая граница между видами А и Б, встречающимися в одном и том же
разрезе, есть, скорее всего, результат перемещения популяций, а не
внезапного возникновения вида Б.
Сделаем общие выводы из того, что было тут изложено об измен-
чивости аммоноидей мезозоя.
У аммонитов юры и мела очень часто наблюдается весьма высокая
изменчивость внутри видов. Нередко значительно отличаются одна от
другой коллекции особей одного и того же вида, собранные в двух пос-
ледовательных пластах, налегающих один на другой. При такой внутри-
видовой изменчивости в вертикальном направлении еще нет оснований
различать два таксона, если отсутствуют другие данные, которые позво-
лили бы провести такое различение. При наличии двух самостоятельных
видов, связанных между собой постепенными переходами, трудно ука-
зать точную границу между видами — таковая проводится более или
менее условно и приблизительно. Таким образом, эволюция аммонитов
в хорошо изученных случаях оказывается непрерывной, оплошной, плав-
ной. Когда сторонники взрывного видообразования говорят презритель-
но о «плоской эволюции», то они осуждают саму природу явления, а не
сторонников идеи поступательного развития органических форм.
Аммониты были высоко активными животными, способными быст-
ро передвигаться в водной массе и в горизонтальном, и в вертикальном
направлении и таким образом попадать в различные по гидрологичес-
ким условиям места моря. Соответственно этому их связь с определен-
ными биоценозами была относительной и непостоянной. Неудивитель-
но, что при таком образе жизни аммониты, особенно в ранних стадиях
своего онтогенеза, подвергались прямому воздействию факторов внеш-
ней среды, которые и вызывали у этих моллюсков те или иные измене-
ния. Эти изменения служили материалом, на основе которого естест-
венный отбор вел к образованию новых низших таксонов — подвидов
и видов.
Однако при нынешнем уровне наших знаний слишком трудно отве-
тить на вопрос, какие именно факторы внешней среды вызывали внутри-
видовую изменчивость аммонитов. Это не удивительно: аммониты жи-
ли преимущественно в морской воде нормальной солености, и если бас-
сейн оставался полносоленым морем, нелегко установить, по ископае-
мой фауне и по литологическим и геохимическим признакам, какие
именно экологические изменения могли вызывать усиленную изменчи-
вость аммонитов в том или ином направлении. Это — дело будущих ис-
следований.
Глава четырнадцатая
ЯВЛЕНИЯ ИНДИВИДУАЛЬНОЙ ИЗМЕНЧИВОСТИ У
ИСКОПАЕМЫХ ПОЗВОНОЧНЫХ
ПРЕДВАРИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
В этой главе мы рассмотрим некоторые примеры повышенной из-
менчивости у ископаемых, позвоночных, по всем признакам зависевшей
от услоовий существования. Хорошо известно, что попав в новые, часто
относительно ограниченные, биотопы, например на острова или в озера,
животные обычно претерпевали интенсивную эволюцию, выходившую
далеко за пределы того, что многие называют, довольно неудачно, ми-
кроэволюцией. Напомним читателю отдельные, хорошо известные слу-
чаи такого развития, вследствие которого возникли многие ценозы,
ныне существующие в разных странах света.
Хорошо известно широкое распространение явлений эндемизма сре-
ди животных различных озер, например Байкала, фауна которого опи-
сывалась многими исследователями в частности М. М. Кожовым (1962).
Д. Н. Талиев (1948), изучивший байкальских Cottoidei подчеркивал,
что «батифильные (живущие на глубинах более 200 м — Л. Д.) виды
байкальских бычков обладают весьма высокой внутривидовой изменчи-
востью» (там же, стр. 152), как, впрочем и многие другие животные,
обитающие в этом озере. О многочисленных эндемичных формах,
живущих в Охридском озере, в Югославии, писал С. £танкович (1959).
Очень яркие примеры эндемизма и изменчивости видов представляют
фауны озер Восточной Африки, о чем писали многие исследователи, из
которых упомянем Дж. Фрайера (D. Fryer, 1960) и Л. Бидла (L. С.
Beadle, 1962).
Не меньший интерес представляют, с рассматриваемой здесь точки
зрения, фауны островов, рассматриваемые многочисленными исследова-
телями, например, Ч. Сакки (С. F. Sacchi, 1961), который писал о явле-
ниях «мик1роэ>волюции» животных мелких Средиземноморских островов,
М. Радовановичем (М. Radovanovic, 1961), рассматревшим фауны остро-
вов Адриатического моря с точки зрения эволюционизма, Р. Маргале-
фом (R. Margalef, 1961), изучавшим биоценозы островов Балеарских и
Колумбретес (близ восточного побережья Испании), М. У. Холдгейтом
(М. W. Holdgate, 1960), исследовавшим фауну океанических островов
Атлантического океана, расположенных в южной холодно-умеренной
зоне (Тристан-да-Кунья, Инасессибл, Найтингейл и Гоф), Ж. Дорстом
(J. Dorst, 1959 а и 1959), давшим интересные работы о значении фауны
знаменитых Галапагосских для освещения проблем эволюционного уче-
ния и другими.
’Y&
Во всех подобных случаях наблюдается широкое распространение
эндемизма: существуют местные подвиды, виды, даже роды и более вы-
сокие таксоны. Едва ли можно думать, что они возникли без всякого
участия мутирования, только в силу перераспределения ранее сущест-
вовавших генов. Не говоря уже о том, что исследования последних деся-
тилетий привели большинство исследователей к резкому ограничению
или к полному отрицанию значения «генетико-автоматических» процес-
сов или «.генетического дрифта» в эволюции органического мира — это
отмечает, между прочим Сакки (Sacchi, 1961, стр. 331) в упомянутой на-
ми статье о микроэволюции мелких средиземноморских островов, —
очень трудно представить себе направленную эволюцию без отбора, при-
ведшую к возникновению таксонов выше вида. Отсюда естественно до-
пустить, что одним из существенных факторов эволюции эндемиков яв-
ляется повышенная изменчивость, стимулируемая необычными для дан-
ных форм условиями существования. К тому же некоторые биотопы,
где живут эндемики, возникли геологически недавно; например, мелкие
островки Адриатического моря, где обитают эндемичные подвиды яще-
риц, изученные Радов ан ови чем (Radovanovi£, 1961)—Lacerta sicula ragu-
sae, L. melisellensis lissana и другие—возникли, по геологическим данным
20 000—30 000 лет назад. Заслуживает внимания также то, что эти яще-
рицы и теперь обнаруживают очень высокую степень изменчивости. На-
пример, популяция Lacerta melisellensis, населяющей островок Горня
Дейница, характеризуется очень сильной изменчивостью окраски. Мож-
но ли связать это явление с «эффектом Сьюэлла Райта», когда речь идет
(> популяции, населяющей один крошечный островок? Подобные факты
говорят, по-нашему, скорее за изменчивость, индуцируемую несколько
измененными условиями жизни.
Палеобиологические данные, к рассмотрению которых мы здесь
приступаем, подтверждают то положение, что изменчивость позвоночных
существенно повышается при изменении условий обитания, если соот-
ветствующие популяции выдерживают это изменение.
В этой главе мы коснемся проблемы изменчивости ископаемых
представителей классов рыб, пресмыкающихся и млекопитающих. Неко-
торые случаи изменчивости у вымерших птиц были приведены в главе
VI, посвященной внутривидовым изменениям величины тела у живот-
ных, и здесь мы не вернемся к этому вопросу, хотя он имеет, по наше-
му мнению, существенное значение для понимания факторов, вызываю-
щих наследственную изменчивость организмов.
УСИЛЕННАЯ ИЗМЕНЧИВОСТЬ У РЫБ НЕКОТОРЫХ РЕЧНЫХ СИСТЕМ В
ЧЕТВЕРТИЧНОМ ПЕРИОДЕ
Очень интересные данные по этому вопросу содержатся в работах
советского ихтиолога Г. У. Линдберга (1955), который изучал истори-
ческое развитие рыб Восточной Азии, Сахалина и Японии на основе раз-
работанного им биогеографического метода. Считаем нужным кратко
изложить те из важнейших результатов его исследований, которые име-
ют прямое отношение к интенсивной изменчивости рыб, живших в бас-
сейнах четвертичного периода.
Этот ученый исходит из того положения, что дно океанов в тече-
ние четвертичного периода подвергалось существенным тектоническим
процессам, с которыми были связаны крупные морские регрессии и
трансгрессии. При регрессиях материковая отмель оголялась до глу-
бины в 200—300 м. Затем затоплялись равнинные участки суши до вы-
13- Л. Ш- Давиташвили 193
соты в 180 и 80 м над современным уровнем океана. При таких событиях
происходили значительные изменения гидрологических условий (колеба-
ния температуры и другие), не говоря уже о резких изменениях берего-
вой линии.
В ту длительную фазу регрессии, которая имела место в конце
плиоцена, на Дальнем Востоке реки материка и реки островов сообща-
лись между собой, образуя единую речную систему, в которой обитала
единая по своему составу фауна типично пресноводных рыб. В резуль-
тате трансгрессии» охватившей рав(нину до высоты 180 м над уровнем мо-
ря, значительные участки рек оказались затопленным морем. Вследствие
этого часть типично пресноводной ихтиофауны погибла; но другая часть
выжила и отступила к верховьям за пределы воздействия трансгрессии.
Фауны, обособленные одна от другой (восточная и западная), эволюи-
ровали в изменившихся условиях существования. Возникли существенно
изменившиеся формы. Позже, в фазу регрессии, эти новые формы име-
ли возможность расселяться по восстановленной речной системе. Затем-
в фазу новой трансгрессии (до высоты в 80 м над уровнем моря), ихтио-
фауна вновь распалась на две части, изолированные одна о^ другой в
верховьях рек. Все это привело к тому, что в реках Японии из общего
числа 18 родов рыб пять являются эндемичными для Японии; там от-
мечается 4 эндемичных подвида и 29 эндемичных видов (Линдберг,
1955, стр. 37—43).
Такое же бурное видообразование происходило и в материковых
реках Дальнего Востока.
Интенсивная эволюция рыб-эндемиков, с образованием не только
новых подвидов и видов, но и родов за столь геологически короткий
отрезок времени, как четвертичный период, не может быть объяснена
генетико-автоматическими процессами или «эффектом Сьюэлла Райта»:
таксоны выше вида могли возникать лишь при условии последователь-
ного появления, одной за другой, многих форм, составляющих филоге-
нетическую цепь; ограниченного исходного генфонда для этого было яв-
но недостаточно. Следовательно, неизбежно допущение одновременного
массового появления в популяциях «мутантов», индуцируемых «давле-
нием» определенных факторов среды, — направленного «мутирования»
особей, которые могли служить материалом для интенсивного видооб-
разования путем естественного отбора.
Аналогичный процесс мог совершаться и в развитии морских ихтио-
фаун, когда морской бассейн оказывался на некоторое время обособлен-
ным от океана.
НЕКОТОРЫЕ ДАННЫЕ ОБ ИНДИВИДУАЛЬНОЙ ИЗМЕНЧИВОСТИ
ИСКОПАЕМЫХ ДИНОЗАВРОВ
Наблюдения над индивидуальной изменчивостью мезозойских реп-
тилий затруднительны прежде всего потому, что находки остатков мно-
гочисленных особей, о которых можно было бы предполагать, что они
жили в составе одной и той же популяции, представляют большую ред-
кость. И все же некоторые выводы о внутривидовой изменчивости иног-
да, по нашему мнению, вполне возможны. Это касается, например, нахо-
док черепов верхнемеловых динозавров. Приведем фактические данные,
а потом попытаемся, на основе морфологического анализа, сделать не-
которые выводы.
Начнем с утконосых динозавров, или гад р оз аиров. В книге, посвя-
щенной теории полового отбора (1961, стр. 260—296), мы постарались
доказать, что многие черты аспекта черепа у этих древних животных
194
развивались под влиянием аллестэтического фактора, то есть вч зависи?
мости от восприятия этих особенностей другими животными того же
Рис. 49. Череп Lambeosaurus lambei Parks, х 1/8. По Лоллу и Рай,
самого и иных видов. Это касается особенно гребней черепа. Рисунки
49, 50, 51 дают некоторое представление о большом разнообразии этих
Рис. 50. Череп Lambeosaurus clavinitalis Sternberg, х 1/8. по ЛоллужРайт.
структур в пределах рода Lambeosaurus Parks. В коллекции черепов
Lamb, lambei, изученной американскими исследователями, нельзя най-
ти такие два экземпляра, у которых очертания предчелюстной и носовых
195
костей были бы хоть приблизительно одинаковыми. Вообще греб-
ли ламбеозавро/в необычайно сильно изменчивы. У представите-
лей рода Corythosaurus гребень на черепе был высокий, узкий, сжатый
с боков, шлемообразный. Cor. casuarius Brown и близкие к нему фор-
мы обнаруживают огромную изменчивость в структурах, связанных
с гребнем черепа. Гребень может быть большим и высоким, но иногда
Рис. 51. Череп Lambeosaurus magnicristatus Sternberg, х 1/8. По Лоллу
и Райт.
он оказывается определенно более низким. Варьирует также наклон пе-
реднего края черепа. Колеблются и другие показатели гребня (рис. 52).
Что касается Coryth. excavatus Gilmore (рис. 53), то у голотипа
верхняя часть предчелюстной кости не достигает верхушки гребня, а у
другого экземпляра, плезиотипа,, достигает. Cor. intermedins Parks
был описан по одному почти полному черепу (голотип) и по другому,
менее полному, а также по частям посткраниального скелета (рис. 54 и
55). Эта форма отличается от Cor. casuarius преимущественно меньшей
переднезадней длиной гребня и присутствием шипа на его задней сто-
роне, а также некоторыми другими чертами. Паратип, изображенный
на рисунке 55 , достаточно отличается от голотипа (см. рис. 54) доволь-
196
но неправильным очертанием гребня, на задней стороне которого имеет-
ся остроугольный выступ, хорошо видный у Cor. excavatus (рис. 53).
Другой экземпляр Cor. intermedins отличается от голотипа большей
крутизной переднего края гребня, а также, вероятно, и другими чертя-
ми. Своеобразна задняя сторона гребня этого экземпляра: здесь есть
его выпуклая нижняя часть, четко выделяющаяся в очертании задней
части гребня и напоминающая этим только что упомянутый паратип, но,
в отличие от него, совершенно лишенная шпоровидного шипа [Лолл и
Райт, (Lull, Wright, 1942), стр. 202]. Рассмотрение этих и других форм
рода Corythosaurus привело нас (1961, стр. 282) к выводу, что в пределах
Рис. 52. Череп Corythosaurus casu; rius Brown, x 1/8. По Лоллу и Райт.
этого, наиболее богатого изученными видами, рода шлемоносных гад-
розаврид, «хотя в черепе разных форм рода имеется немало отличи-
тельных черт, самые существенные и крупнейшие черты различия свя-
заны со шлемом». Чрезвычайная изменчивость гребней весьма примеча-
тельна. Почти каждый из видов был описан по одному или по двум
экземплярам. В тех случаях, когда кроме голотипа в. распоряжении ис-
следователей имелся также плезиотип или паратип (Cor. casuarius, Сог.
excavatius, Cor. intermedins), дополнительный экземпляр оказывался за-
метно отличающимся от голотипа. Это заставляет нас признать, с одной
стороны, усиленную индивидуальную изменчивость коритозавров (по
признакам гребня и связанных с ними структур), а с другой, — убыст-
ренную эволюцию, которая выражалась прежде всего в резких измене-
ниях тех же самых структур.
197
И йовышенная индивидуальная изменчивость, и необычайно быст-
рая эволюция гребня свидетельствуют о том, что он представлял собой
образование, исторически развивавшееся в силу своего аллэстетическо-
гр значения и притом, скорее всего, как «украшение», возбуждающее и
привлекающее особей противоположного пола, а не как элемент предо-
стере.гающего облика, который мог действовать на представителей дру-
гих* таксонов животных.
'г 1 * :
Рис. 53. Череп Corythosauris excavatus Gilmore, х 1/8. По Лоллу и Райт.
Коснемся теперь вопроса о повышенной изменчивости некоторых
структур аллэстетического значения у представителей другой группы
позднемеловых динозавров — рогатых динозавров, или цератопсий. У
них необычайно высокую индивидуальную изменчивость мы наблюдаем
на так называемом воротнике и отчасти — на рогах. За сравнительно
короткое время эволюция этой группы привела к изумительному разно-
образию в развитии этих структур.
Чрезвычайно высокая индивидуальная изменчивость в морфологи-
ческом строении рогов и особенно воротника наблюдается, например, у
Chasmosaurus из толщи Белли Ривер, в Альберте. Касаясь цератопсий
вообще, Лолл (Lull, 1933, стр. 103) писал, что вариации черепов, «за-
висящие от возраста, пола или чисто индивидуальные (подчеркнуто на-
ми — д Д.), таковы, что, по-видимому, нельзя найти два одинаковых
черепа». Тот в сущности скудный материал, которым до настоящего
времени располагают исследователи цератопсий, свидетельствует о
чрезвычайно интенсивной изменчивости именно воротника у этих живот-
ных, превосходящей изменчивость их носовых рогов. Поскольку носо-
вые рога не могли быть структурами чисто аллестетического значения
и должны были приносить пользу не только в боях между самцами, но
и в качестве оружия против хищников, более высокая индивидуальная
198
изменчивость такой преимущественно аллэстетической структуры, как
воротник, вполне понятна с точки зрения дарвиновского понимания
эволюции.
Аллэстетические структуры усиленно варьировали во многих видах
теплокровных животных. У птиц яркое оперение многих представителей
родов, другие формы которых могли оставаться лишенными импозант-
ного оперения, эволюировало, очевидно, на основе очень высокой инди-
видуальной изменчивости. Такая эволюция во многих случаях вела к
большому разнообразию перьевого наряда в группах близко-родствен-
ных видов. У млекопитающих повышенную изменчивость проявляли, на-
пример, ветвистые рога многих оленей, утратившие или почти утратив-
шие свое первоначальное боевое значение. В качестве примеров бурной
индивидуальной изменчивости можно было бы привести также формы,
у которых бивни служили тоже не столько в качестве оружия, сколько
«украшениями». Такими, возможно, были бивни некоторых хоботных
(см. Давиташвили, 1961, стр. 428 и 429).
Рис. 54. Череп Corythosaurus intermedins Parks. Голотип, х 1/8. По Лоллу
и Райт.
Таким образом, в случаях, когда эволюция происходила путем от-
бора аллэстетических структур и особенностей поведения животных, в
частности путем полового отбора, наблюдается усиленная внутривидо-
вая изменчивость именно этих структур. Это, как мы видели, вполне
убедительно подтверждается и палеобиологическими наблюдениями, так
как нельзя же допустить, что почти каждый экземпляр ископаемого че-
репа утконосого динозавра или цератопсии принадлежал к особому ви-
ду. Это значит, что существует какая-то связь между повышением измен-
чивости и формой естественного отбора, действующей в данном случае.
199
Дарвин (1953, стр. 325) писал, что «вторичные половые признаки
исключительно изменчивы». «Это различие бивней у различных рас или
видов слонов, большая изменчивость рогов у оленей, особенно у север-
ного оленя, редкое наличие рогов у самки Antilope bezoartica и их частое
отсутствие у самок Anticocarpa americana, присутствие двух бивней у
некоторых самцов нарвала, полное отсутствие клыков у некоторых самок
моржа—все эти случаи представляют собой примеры крайней изменчи-
вости вторичных половых признаков и их склонности различаться у
самых близких форм» (там же, стр. 562).
Рис. 55. Череп Corythosaurus intermedins Parks. Паратип. х 1/8. По
Лоллу и Райт.
Тут весьма высокая степень изменчивости сопряжена с ускоренным
эволюционным развитием. Особенно быстрые и значительные филогене-
тические преобразования испытывают аллэстетические перигамические
структуры.
Чем же можно объяснить повышенную индивидуальную изменчи-
вость перигамических аллэстетических особенностей животных?
Дарвин объяснял быстрое филогенетическое их развитие тем, что
половой отбор зависит «от элемента, подверженного изменчивости»
(там же, стр. 531), «от столь колеблющегося элемента, как вкус» (там
же, стр. 551), то есть от восприятия этих особенностей самками.
Однако в только что приведенных словах Дарвина речь идет об от-
боре, как факторе филогенетического развития; вопрос же о возникно-
вении изменчивости, ее характере и темпах тут не затрагивается. Но
200
нам кажется, что Дарвин приблизился и к пониманию причин повышен-
ной изменчивости в тех случаях, когда действует половой отбор на ал-
л эст ети ч ес ки е особ ен ности ж ив от н ы х.
Дарвин писал (1953, стр. 531), что измененные условия жизни вида
вызывают «в нем в известном размере колеблющуюся изменчивость (а
certain amount of fluctuating variability)44.
Думаем, что этот вопрос заслуживает пристального внимания гене-
тиков. Мы же не можем решить его. Выскажем лишь чисто гипотетичес-
кое предположение. Возникновение или существенное усиление отбора
на определенную структуру, или «паттерн» поведения, должны влечь
за собой выживание и распространение многих обладателей этих осо-
бенностей, которые могут существовать не в одном, а во многих, хотя и
близких между собой, вариантах. Эти варианты не элиминируются от-
бором, что может способствовать разнообразию дальнейшей изменчи-
вости. Кроме того, поскольку половой отбор на аллэстетические особен-
ности зависит от колеблющегося фактора — «вкусов» самок, а не от
жестких условий питания, размножения, сохранения особей, этот отбор
может благоприятствовать не только определенным вариациям, но и
повышению изменчивости в данном направлении.
Это — наши гипотетические соображения, а непреложным фактом
является чрезвычайно высокая внутривидовая изменчивость в популя-
циях, где интенсивно действует половой отбор на аллэстетические осо-
бенности животных.
Надо полагать, что то же в известной мере имеет место и в других
случаях, когда в отборе действует аллэстетический фактор, например,
в эволюции апосематической окраски и вообще сематического или апо-
сематического облика, кри1птических признаков и вообще тех особеннос-
тей организ/Мов, которые зависят от восприятия облика зрительными ре-
цепторами других животных.
ИЗМЕНЧИВОСТЬ У ИСКОПАЕМЫХ МЛЕКОПИТАЮЩИХ
Эта книга, конечно, не претендует на равномерное освещение явле-
ний индивидуальной изменчивости во всех важнейших группах органи-
ческого мира. Мы обсуждаем здесь основные вопросы изменчивости на
палеонтологическом материале, привлекая к анализу лишь отдельные
примеры. Систематическое исследование изменчивости во всех основ-
ных таксонах животных и растений — дело будущих исследований. В
этой главе мы рассматриваем некоторые характерные примеры изменчи-
вости среди наземных четвероногих прошлых геологических эпох. Это,
как мы думаем, позволит нам существенно расширить наши представле-
ния об изменчивости организмов и ее связи с действием факторов сре-
ды.
Вопрос о повышенной изменчивости у млекопитающих представля-
ет, конечно, немалый интерес. Более или менее говышенная изменчи-
вость наблюдается в некоторых биотопах, которые отклоняются от не
изолированных или слабо изолированных участков материковой суши.
Такими аберрантными биотопами можно считать некоторые острова
(куда когда-то попали некоторые анцестральные формы млекопитаю-
щих, там прижившиеся и давшие начало местным формам), относитель-
но резко обособленные участки высокогорий или вообще возвышенных
горных сооружений, некоторые впадины, окруженные горами и т. д.
Большой интерес представляют также пещеры, которые, правда, за
•очень редкими исключениями, не могут служить удобным местообита-
120
нием для каких-либо зверей. А. Вандель (Vandel, 1964, стр. 285) назы-
вает всего лишь один род, по-видимому, подземных крыс, Neopoma, с
двумя видами Neop. magister и Neop. pennsylvanica, которые лишь изред-
ка .встречаются на поверхности. Глаза у них имеют нормальное строение,
хотя способность зрения у них пониженная, как это наблюдается, впро-
чем у многих грызунов.
Что же касается летучих мышей, то пещеры служат для многих
из них убежищами, особенно в зимнее время. В некоторых тропических
пещерах накопляются значительные массы их гуано.
Но если среди млекопитающих очень мало настоящих троглобион-
тов, .постоянно живущих в пещерах, то звери, подолгу пребывавшие в
пещерах, нередко испытывали значительные изменения в строении, фи-
зиологических свойствах и повадках.
В этом отношении выдающийся интерес представляют прежде всего
пещерные медведи четвертичного периода.
Однако необходим, по нашему мнению, более широкий подход к
этому вопросу.
Прежде всего отметим, что повышенная изменчивость наблюдается
не только у пещерных медведей, но и в других группах медведей. Так,
исследователи сообщают данные об изменчивости медведей в Северной
Америке. Прежде всего бросается в глаза очень большое число видов
и подвидов медведей, которые указываются некоторыми авторами. Тут
невольно возникает мысль, что по крайней мере часть этих таксонов
выделена без достаточных оснований и являет собой картину внутри-
видовой изменчивости. В монографии о серых и крупных бурых медве-
дях Северной Америки Мерриам (С. Н. Merriam, 1918) различает не
менее 86 видов и подвидов. Однако, по В. Маринелли (W. Marinelli,
1931, стр. 368), который познакомился с материалом Мерриама, в На-
циональном музеев Вашингтоне, не может быть сомнений в высокой из-
менчивости этих животных, причем как самый материал, так и данные
о местах находок не дают доказательств видовых различий отклоняю-
щихся типов. Произведенное Мерриамом выделение таксонов, основанное
лишь на чисто морфологических критериях, могло, по Маринелли, при-
вести к тому, что к разным таксонам оказались отнесенными очень близ-
ко родственные особи, быть может, даже животные одного и того же
помета. Отнесение таких экземпляров хотя бы даже к разным подви-
дам, а не видам, означало бы, как выражается Маринелли, сведение
систематики к абсурду. Нам кажется, что даже то понимание система-
тики североамериканских медведей, какого придерживался Мерриам,
делает неизбежным признание очень высокой изменчивости этих зверей.
Пещерный же медведь вообще и прежде всего те его особи, остатки
которых были изучены авторами коллективной монографии о Микснитц-
кой пещере [(О. Абель и Г. Кирле (О. Abeb G. Kyrle, 1931], — едва
ли не более яркий пример внутрипопуляционной изменчивости медве-
дей. Встает вопрос, нет ли какой-то общей причины повышения индиви-
дуальной изменчивости у современных североамериканских и ископае-
мых пещерных медведей Евразии? Невольно возникает мысль, что та-
кой общей причиной мог быть постепенный отход от чисто плотоядного
питания к потреблению в пищу растений, вплоть до трав (Abel, Абель,
1935, стр. 592).
Маринелли пришел к выводу, что большое морфологическое разно-
образие особей, остатки, которых сохранились в Микснитцкой пещере,
отнюдь не исключает возможности кровного родства между этими осо-
бями. Иначе говоря, возможно, что тут перед нами картина подлинно
202
внутрипопуляционной изменчивости, и г/ритом чрезвычайно высокой
даже для медведей (Marinelli, Маринелли, 1931, стр. 425). В то же вре-
мя заметны общие тенденции изменчивости, которые выражаются, в
частности, в развитии выпуклости лобной кости с формированием так
называемой глабеллы и в возрастании общих размеров тела. К. Эрен-
бург (К. Е. Ehrenberg, 1931, стр. 571) находит, что пещерные медведи
шревратились в преимущественно фитофагичеоких зверей и в связи с
этим у них возросла жевательная поверхность задних зубов. Подобно
Маринелли, Эренбург подчеркивает изменчивость пещерных медведей,
называя ее их характерной особенностью (Ehrenberg, 1931, стр. 537).
Изменчивость микснитцких медведей Эренбург изучил только на
задних зубах. Он отмечает чрезвычайную изменчивость этих зубов. Во-
обще же этот пещерный медведь по своей чрезвычайно высокой индиви-
дуальной изменчивости представляет собой, как думает Эренбург
(Ehrenberg, 1931, стр. 570), совершенно исключительное явление среди
диких животных. Примечательно, что этот исследователь отмечает и
направление изменчивости задних зубов пещерного медведя, — его по-
пуляций, известных г.о остаткам в Микснитцкой пещере. Так, на пос-
леднем верхнем моляре и последнем нижнем развиваются многочислен-
ные боковые бугорки. Передние премоляры у непосредственных предков
этого вида были настолько рудиментарны, что они не могли усилиться
для привлечения к жевательной функции. М3 был тоже утрачен уже у
предков.
Безусловно заслуживают внимания соображения О. Абеля о «деге-
нерации» пещерных медведей Микснитца. Надо только помнить, что
этот палеонтолог отвергал участие естественного отбора в эволюции
новых форм организмов и думал, что роль этого фактора сводится толь-
ко к сохранению уже существующего вида, стабильности вида. Дегене-
рацию пещерных медведей Абель объясняет чрезвычайно благоприят-
ными, оптимальными, условиями их существования. Они жили, по Абелю
(Abel, 1931, стр. 733—734), в теплое межледниковье, рисс-вюрмское, мно-
го времени проводя в пещерах, в условиях достаточного обилия корма и
отсутствия очень опасных врагов. Серьезным противником был только
человек раннего палеолита, но едва ли человек отваживался выходить
против медведя в открытом бою. Абель отмечает огромную изменчи-
вость всех скелетных элементов, обилие патологических явлений и, на-
конец, появление карликовых форм в самых молодых слоях.
Нам кажется, что мы еще далеки от правильной и хорошо обосно-
ванной оценки основных факторов развития биоценотических отношений
времени существования пещерных медведей в Европе. В этой области
надо еще очень и очень много поработать. При этом нужно стараться
определить количественное значение каждого фактора, с привлечением,
по возможности, кибернетического метода. А пока мы можем сказать
лишь немногое, и то, в значительной мере, лишь предположительно.
Вполне вероятно, что эпоха расцвета популяций пещерного* медве-
дя была временем очень хороших, оптимальных условий их существо-
вания. К такому состоянию мог привести ход развития биотопов и их
населения. Некоторое ослабление естественного отбора могло обусло-
вить выживание особей с теми или иными недостатками и заболевания-
ми, патологическим развитием зубов, скелета и разных внутренних ор-
ганов.
Многочисленность популяций уже сама по себе способствовала про-
явлениям изменчивости в различных направлениях. Однако в еще боль-
шей мере повышение изменчивости должно было зависеть от изменения
203
отношений между организмом и средой. Животные проводили значи-
тельную часть вемени внутри пещер, где они были малоподвижны. На
организм влиял и влажный воздух пещер, их слабая освещенность, от-
носительно ограниченный диапазон температурных колебаний и пр.
Очень существенное значение для индуцирования изменчивости имеет
пища: в даном случае — переход к фитофагии как основному способу
гитания. Мы сказали, что усиленная изменчивость наблюдается у сов-
ременных медведей, изученных Мерриамом; еще более интенсивная
изменчивость отмечена у пещерного медведя. Надо полагать, что в обо-
их случаях должна была действовать, по крайней мере частично, одна
и та же группа факторов; мы склонны думать, что усиленная изменчи-
вость пещерных медведей зависела прежде всего от процесса перехода
от чисто плотоядного к фитофагическому, в основном, питанию.
Может возникнуть вопрос, не являются ли изменения, наблюдаемые
у медведей и в частности у пещерного медведя, чисто модификационны-
ми, ненаследственными. Мы думаем, что значительные изменения, про-
исходившие при переходе от Ursus ceningeri к Ursus spelaeus, а
также приуроченность этих видов к двум различным моментам геологи-
ческого времени достаточно убедительно говорят о том, что этот процесс
мог совершаться лишь на основе наследственной изменчивости.
Индивидуальная изменчивость отмечается в самых разнообразных
группах ископаемых млекопитающих, — практически всюду, где толь-
ко имеется достаточное количество материала, костей или зубов, при-
надлежавших более или менее многочисленным особям млекопитаю-
щих. Еще О. Абель (Abel, 1929, стр. 140) писал о мастодонтах, что три
«вида» „Bunolopphodon,,—В. angustidens Cuvier, В. longirostris Каир,
В. arvernensis Croizet et Jobert—„теснейшим образом связаны между
собой полной серией переходных форм“.
Л. К. Габуния обратил наше внимание на то, что коренные зубы
поздних южных слонов Archidiskodon meridionalis Nesti, известных из
раннего плейстоцена, связаны постепенными переходами с коренными
Elephas trogontherii Pohlig, наиболее поздние представители которого,
ь свою очередь, почти не отличимы, по строению зубов, от ранних форм
Elephas primigenius Blumenbach из среднего плейстоцена (у них почти
одинаковы форма зубов, число и частота пластин).
Постепенные переходы часто отмечаются и в историческом разви-
тии очень древних млекопитающих, хотя их остатки сравнительно редка
встречаются в массовом количестве, делающим возможным биометри-
ческое их изучение. Примерами таких переходов между таксонами, без
резких скачков и пробелов в эволюции, могут служить Arctocyonidae
(семейство отряда Condylarthra, представленое преимущественно впалео-
цене), которых изучал И. Вейгельт (I. Weigelt, 1947 и 1950),
Таким образом, тщательный анализ данных по ископаемым млеко-
питающим подтверждает то положение, что эволюция этих животных
протекала на основе незначительных, почти незаметных изменений, пос-
тепенно, что, конечно, нисколько не свидетельствует об отсутствии скач-
ков в филогенетическом развитии животных.
Маргалеф (Margalef, 1961) указывает на очень большую изменчи-
вость копытного Myotragus balearicus, жившего в четвертичном перио-
204
де на о-вах Майорке и Минорке, о чем свидетельствует ископаемый
костный материал. Этот автор думает, что, поскольку многие остатки
обнаруживают деформации, вызванные, очевидно, заболеваниями, по-
вышенная изменчивость могла зависеть от отсутствия «контроля со сто-
роны хищников». Впрочем мелкие хищники Балеарских островов,
Martes martes latinorum и Genetta genetta balearica, тоже оказываются
очень изменчивыми. По Маргалефу, повышенная изменчивость мелких
хищников зависела от упрощения биоценотических отношений, в связи
с которым эти животные «заняли высший трофический уровень». Так или
иначе, попав в условия, отличные от тех. которые существовали в исход-
ных материковых биотопах этих млекопитающих, они испытывали (а не-
которые из них и теперь испытывают) повышенную индивидуальную
внутривидовую изменчивость, от которой, по-видимому, и зависело обра-
зование, за короткое время, новых подвидов.
Интродукция млекопитающих может вызвать чрезвычайно интен-
сивную их изменчивость в новой жизненной обстановке. Можно было
бы привести много примеров этого явления, но мы ограничимся одним.
Зеленая мартышка Cercopithecus aethiops saoacus населяет опре-
деленную территорию Западного побережья Африки. Э. Г. Аштон (1957)
обритил внимание на изолированную колонию этих обезьян, живущую
на о-ве Сент-Киттс (Малые Антильские острова), куда предки этих жи-
вотных были завезены человеком, вероятно, в XVII веке. Часть этих
мартышек одичала. Лет 15 назад около сотни черепов мыртышек о-ва
Сент-Киттс были доставлены в музей Лондонского медицинского кол-
леджа. Эти черепа сравнивались с черепами зеленых мартышек из За-
падной Африки. Промеры черепов и зубов показали, что мартышки
о-ва Сент-Киттс заметно отличаются от популяций той же формы,
обитающей в Африке. Аштон считает вполне возможным, что процесс
изменения размеров черепа приведет к обособлению самостоятельного
подвида.
Заметим, что в последние годы многие исследователи уделяли осо-
бое внимание вопросам изменчивости позвоночных, особенно — млеко-
питающих. С этой стороны нас очень заинтересовала книжка А. В. Яб-
локова (1966), специально посвященная изменчивости у млекопитаю-
щих. Мы, однако, не нашли в ней данных об изменчивости, от которой
зависит процесс эволюции путем естественного отбора. Зато большой
интерес представляют многие статьи сборника «Внутривидовая измен-
чивость наземных позвоночных животных и микроэволюция», возникше-
го в результате работы Всесоюзного совещания по этой проблеме, сос-
тоявшегося в 1964 г. в Свердловске. С точки зрения палеобиологии
млекопитающих особого внимания заслуживает статья К. Л. Паавера
(1965), где рассматриваются случаи «вековой» изменчивости млекопи-
тающих Прибалтики в голоцене. Паавер установил достоверные остео-
метрические различия между субфоссильными млекопитающими При-
балтики и совершенными представителями тех же видов. Так, прибал-
тийский лось раннего голоцена (VII—VI тысячелетия до н э.), отличал-
ся крупными размерами. Аналогичные данные сообщаются автором и
относительно бобра, кабана, косули. У барсука же размеры тела от
раннего голоцена до настоящего времени увеличивались. Автор укло-
няется от суждения о генетическом характере таких изменений, но ссы-
лается на существующее мнение, что «установленное в голоцене измене-
ние остеометрических признаков в западноприбалтийских популяциях
некоторых видов млекопитающих (. . .) в основном наследственно обус-
ловлено» (там же, стр. 79).
ЗАКЛЮЧЕНИЕ
ЗНАЧЕНИЕ ИЗМЕНЧИВОСТИ ИСКОПАЕМЫХ ОРГАНИЗМОВ ДЛЯ РЕШЕНИЯ
ЗАДАЧ СТРАТИГРАФИИ И ГЕОХРОНОЛОГИИ. ОБЩИЕ ВЫВОДЫ
Читатель не мог не заметить, что красной нитью в этой книге про-
ходит идея зависимости наследственных изменений организмов от фак-
торов окружающей среды. Надо сказать, что эта идея далеко не нова
для исследователей, работающих в области исторической геологии; бо-
лее того, она доминирует в сознании большинства этих ученых. Но вся-
кий, кто внимательно познакомился с содержанием этой книги, должен
был убедиться в том, что мы очень далеки от умаления значения генети-
ческого изучения организмов и от отрицания основных положений, до-
казанных генетическими исследованиями. Не может быть ни малейшего
сомнения в том, что то, что теперь принято не совсем удачно называть
«генетической информацией», содержится преимущественно в ядре клет-
ки и его хромосомах. Не может быть никакого сомнения и в том, что на-
зывается дискретностью наследственности, хотя в то же время нас-
ледственность представляет собой единство дискретности и слитности.
Бессмысленно отрицать значение открытий Г. Менделя и его продол-
жателей, хотя столь же бессмысленно преувеличивать значение этих
открытий. Надо признать прискорбным тот факт, что в настоящее вре-
мя некоторые крупные ученые проявляют склонность игнорировать
открытия в области изучения наследственности и изменчивости.
В связи с только что сказанным считаем нелишним отметить не-
обходимость устранения одного из распространенных ныне методоло-
гических заблуждений. В науке об эволюции существует, как извест-
но, немало различных школ. Эти школы различаются одна от другой
основными принципами, на которых построено учение каждой из них„
но иногда также и по предмету изучения и по применяемой ими эк-
спериментальной методике. Например, изучая явления наследствен-
ности и изменчивости, работники одних школ уделяют много внимания
экспериментам по так называемой вегетативной гибридизации, совер-
шенствуют методику эксперимента, в то время как другие школы, на-
ходя, очевидно, эти опыты бесперспективными, считают излишним серь-
езно заняться их проверкой. Почти то же самое надо сказать об экспе-
риментах, направленных к выяснению возможности приобретения на-
секомыми резистентности по отношению к различным инсектицидам.
Работники тех школ, которые отрицают возможность возникновения
резистентности путем «воспитания», уклоняются от затраты средств и
времени на производство соответствующих экспериментов с достаточ-
ной -тщательностью и, по-видимому, ограничиваются лишь довольно
«беглой» экспериментальной проверкой экспериментов своих оппонен-
тов. Это естественно; раз нет надежды добиться желаемого результата,.
206
то стоит ли тратить силы и время на такую работу? А между тем эк-
сперименты, которые были бы достаточно тщательно поставлены про-
тивниками идей вегетативной гибридизации и при соблюдении всех тон-
костей соответствующих опытов, могли бы дать решающие результаты.
Нечего бояться и того, что эти результаты могут оказаться несоответ-
ствующими основным установкам школы: истина дороже всего!
Эта своеобразная косность принимает особенно угрожающий ха-
рактер, когда школа занимает господствующее положение в науке и
пользуется неограниченным авторитетом. В таких случаях у предста-
вителей данной школы иной раз возникает уверенность в том, что не они
должны экспериментально проверять выводы своих оппонентов, а эти
последние несут «тяжесть доказательства» принимаемого ими тезиса.
Изложенный в предыдущих главах материал показывает, что ана-
лиз явлений изменчивости органических форм геологического прошло-
го невозможен без учета данных геохронологии, стратиграфии и ус-
ловий накопления осадков в тех биотопах, где жили и эволюировали
организмы. Изучение изменчивости ископаемых организмов требует
знания хода исторического развития соответствующих участков земной
коры в течение геологического времени. В то же время и геология,
включая стратиграфию, нуждается в познании изменчивости ископае-
мых организмов.
Внутрипопуляционная изменчивость ископаемых организмов, если
даже она оказывается значительной и бросается в глаза, не может
иметь такого стратиграфического значения, как возникновение новых
таксонов — не только новых видов, но и подвидов, хотя бы они отлича-
лись от других подвидов лишь незначительными, малозаметными приз-
наками. Впрочем геолог-палеонтолог может устанавливать не момент
возникновения нового низшего таксона, а появление его на данном
участке. Если в соответствующем слое мы наблюдаем появление форм,
которых нет в непосредственно подстилающих отложениях, то это, ко-
нечно, еще не дает нам права проводить стратиграфическую границу
непосредственно ниже этого слоя. Для этого необходимо, прежде всего,
проследить и установить те же палеонтологические отношения на доста-
точно обширной территории. Но все-таки наличие вида, присущего дан-
ному слою и отсутствующего в смежных пластах, надо считать фактом,
который, совместно с другими данными, может послужить для проведе-
ния стратиграфической границы. Если же мы имеем основание думать,
или даже только подозревать, что перед нами не отдельные таксоны, под-
виды или виды, а только внутрипопуляционная изменчивость, то исполь-
зование такого материала для стратиграфических и геохронологических
целей представляется, вообще говоря, значительно более затруднитель-
ным. Мы видели, что точное решение вопроса о том, наследственны ли
те или иные явления изменчивости, на ископаемом материале обычно не-
возможно. В лучшем случае, имея перед собой хорошо подобранные
почти незаметные (или практически незаметные) переходы между двумя
бесспорно самостоятельными видами, мы можем сказать, что тут несом-
ненно имела место наследственная изменчивость форм. Но едва ли воз-
можно сказать определенно, где кончается ряд вариации одного, предко-
вого, вида и где начинаются вариации другого, дочернего, вида. Надо
прямо сказать, что в палеобиологии нет возможности устанавливать, что
те или иные особи были мутантами или вообще отличались от других
особей генетически, — генотипически. Такая точность палеобиологичес-
кому исследованию недоступна. Поэтому во многих случаях мы вынужде-
ны заключать, что некая изучаемая нами особь Н является промежуточной
207
между подвидом (или видом) М и подвидом (или видом) О, хотя генети-
ческое изучение этой особи могло бы установить, что несмотря на фено-
типические ее особенности, она определенно принадлежит лишь к од-
ному из смежных таксонов, — анцестральному или потомковому. Иначе
говоря, несмотря на довольно широкие возможности изучения эволюции
и изменчивости, палеобиология не может входить в точное изучение не-
которых интимных подробностей изменения генотипа ископаемых орга-
низмов.
Мы можем утверждать, что вид А дал начало виду Б в том случае,
если особи второго вида бесспорно отличаются от первого по своему от-
ношению к среде хотя бы незначительными, но характерными деталя-
ми, и притом постоянно.
Изучение изменчивости ископаемых форм приводит нас к еще од-
ному выводу, существенному для решения стратиграфических вопросов.
Мы видели, что при наличии достаточно обильного палеобиологического
материала переход от одного вида к другому происходит не внезапно,
а мелкими скачками, которые не нарушают впечатления постепенности
развития. Было бы, конечно, удобно для биостратиграфов, если бы меж-
ду видами существовали легко уловимые или резкие границы — это в
заметной степени облегчило бы определение геологического возраста и
сопоставление толщ. Но нет никакой пользы в сохранении иллюзии
таких резких границ, раз их нет и быть не может. В предыдущих главах
мы привели немалое число таких случаев. Ф. Беттенштедт (F. Betten-
stadt, 1958) писал, что во всех случаях, когда мы «имеем сплошное, без
пропусков, филогенетическое развитие от одного вида к другому, перед
нами оказываются непрерывные переходы, мельчайшие мутационные
шаги и постепенные перемещения диапазонов вариации. Это, по словам
Беттенштедта (там же, стр. i33), устанавливается не только в палеон-
тологии фораминифер и остракод, которыми занимаются микропалеон-
тологи — это выясняется при изучении решительно всех групп иско-
паемых организмов, когда биометрически обрабатывается большое чис-
ло особей. Г. Геберер (G. НеЬегег, 1957, стр. 907—908), также, как и
Симпсон (Simpson, 1944), отвергает значение сальтаций и системных
мутаций в эволюционном процессе, и мы вполне согласны с ними в
этом, хотя и для нас неприемлемо даваемое этими авторами объясне-
ние того, что переходные формы встречаются сравнительно редко (они,
по нашему мнению, преувеличивают значение быстроты эволюции в ма-
лых популяциях).
Таким образом, изучение внутривидовой изменчивости необходимо
для стратиграфических исследований и в то же время она не может
быть освещена без выяснения стратиграфических отношений слоев, где
встречаются соответствующие органические остатки да и без более ши-
рокого изучения геологического строения и геологической истории соот-
ветствующих территорий.
Вместе с тем изучение изменчивости ископаемых форм необходимо
и для решения некоторых основных во*гросов науки об эволюции и эво-
люционной генетики. Без таких палеобиологических данных эволюцион-
ная генетика не может дать всестороннее освещение вопросов наследст-
венности и изменчивости организмов.
Только при помощи со стороны палеобиологии, опирающейся на
геологию, биология может получить полную картину воздействия биото-
пов на организмы и их популяции, вызывающего массовые направлен-
ные изменения наследственных особенностей живых существ. Мы виде-
ли, что такие «аберрантные» биотопы, разновременно возникавшие и на
208
суше и в водной среде, были ареной усиленной направленной изменчи-
вости, которая, подхватываемая естественным отбором, вела к образо-
ванию новых форм.
Мы убедились в том, что общий итог эволюции органического мира
от древнейших докембрийских времен до настоящей эпохи не мог быть
достигнут без фактора массовой направленной изменчивости: для того,
чтобы такой результат был осуществлен на основе естественого отбора
«шальных», беспорядочных мутаций, нужны, были бы, по-видимому, по
меньшей мере целые десятки, если не сотни или тысячи, миллиардов
лет. Тут ортодоксальная постнеодарвинистская теория, «синтетическая
теория эволюции», встречает непреодолимое препятствие со стороны так
называемой абсолютной геологической хронологии. Палеобиология по-
казывает основательность этого грозного предостережения. Однако пол-
ное выяснение «меры несоответствия между итогом всеобщей филогении
органического мира, с одной стороны, и постнеодарвинистским ее объ-
яснением, с другой, — дело будущих исследователей, которые смогут
количественно изучать ход эволюции живой природы на всем протяже-
нии геологического времени.
Палеобиология, поддерживаемая геологией, с одной стороны, и
необиологией, с другой, дает убедительнейшие доказательства зависи-
мости изменчивости организмов от факторов среды. Она показывает,
как изменения, индуцируемые средой, поставляли материал для есте-
ственного отбора — ведущего фактора эволюции.
Палеобиология показывает также, что роль естественного отбора
не ограничивается фиксацией и закреплением того материала, который
в готовом виде дается ему изменчивостью. Сама изменчивость зависит
от естественного отбора и его форм. Когда естественный отбор элими-
нирует неблагоприятные в данной жизненной обстановке отклонения и
фиксирует благоприятные варианты, он тем самым способствует умно-
жению и усилению последних и сокращению возможности появления
первых. При изучении ископаемого материала, проводимом совместно
с. исследованием условий среды, существовавших на данном участке в
течение соответствующего отрезка геологического времени, может быго
выяснена связь между характером и направленностью индивидуальных
изменений и формой действовавшего тут отбора. В частности удается
установить, что чрезвычайно высокая изменчивость некоторых аллэсте-
тических структур, многих ископаемых рептилий (например, динозав-
ров верхнего мела) и многих вымерших млекопитающих (например, хо-
ботных плиоцена) была связана с действием полового отбора.
Трудно представить себе серьезное изучение степени изменчивости
организмов без использования палеобиологического материала.
Итак, палеобиология должна принимать существенное участие в
исследовании явлений внутривидовой изменчивости. Однако при этом
надо всегда иметь в виду специфику палеобиологического метода, ко-
торый изучает лишь прошлое органического мира, пользуясь при этом
мертвым ископаемым материалом. В этом — определенная ограничен-
ность палеобиологических исследований индивидуальной изменчивости,
этим подчеркивается ведущее значение необиологического изучения яв-
лений наследственности и изменчивости методами современной генети-
ки, которая все глубже и глубже проникает в суть этих явлений благо-
даря применению новейших методов молекулярной биологии.
Различные методы изучения одного и того же предмета взаимно
дополняют и укрепляют друг друга.
Мы можем утверждать, что, прослеживая изменчивость и формо-
образование ископаемых животных, можно, по крайней мере в некото-
14- Л. Ш. Давиташвили 209
рых случаях, устанавливать, что тут перед нами не затухающие дли-
тельные модификации, а бесспорно наследственные изменения, которые
дают начало новым видам: изменения в тех же самых направлениях,,
возникая и у последующих, дочерних видов, обычно подхватываемые
естественным отбором, вели к образованию целых филогенетических це-
пей, развивавшихся в течение более или менее продолжительного, в
геологическом смысле, времени.
Итак, в подобных случаях, изучая палеобиологический материал,
мы можем настаивать на том, что здесь речь должна идти не о том,
что в генетике называют длительными модификациями, а изменениями
генотипа.
В то же время палеобиолог, устанавливающий явления наслед-
ственной изменчивости, не имеет никакой возможности входить в оцен-
ку тех понятий, которые развиваются в генетике на основе экспери-
ментов: он не может толковать о природе и составе генов и о кариоло-
гических процессах, связанных с наследственностью, о тонких биохими-
ческих явлениях, от которых по данным молекулярной генетики зави-
сит передача «наследственной информации». Он не может высказывать-
ся ни за ни против того или иного понимания этих интимных процес-
сов, стоящих всецело вне области палеобиологических и стратиграфи-
ческих исследований.
Мы думаем, что развиваемые нами основные положения вполне
можно излагать языком современной генетики. В этой книге мы говорим
о массовой направленной изменчивости, зависящей от факторов среды.
Такое понимание причин изменчивости можно выразить с помощью по-
нятия о генах-мутаторах, действие которых по Н. П. Дубинину (1970,
стр. 271), «в состоянии резко изменить <мута1бильность организмов».
По мнению этого автора, «действие, гена-мутатора, по-видимому, связа-
но с тем, что он изменяет свойства ДНК-полимеразы, что ведет к массо-
вости мутаций». Это, вероятно, не меняет существа дела. Мы утвержда-
ем, что воздействие факторов среды ведет к массовой направленной из-
менчивости. Молекулярная генетика» может и должна изучать интимные
биохимические механизмы таких процессов.
В заключение подчеркнем, что для биостратиграфии и палеобио-
логии имеют определенное научное значение и такие наблюдения над
изменчивостью вымерших организмов и их популяций, которые не мо-
гут дать ответа на вопрос, является ли эта изменчивость наследствен-
ной. Если даже замечаемые изменения, отклонения от типа вида, бу-
дучи массовыми, все же окажутся ненаследственными, они тем не ме-
нее дают возможность судить о факторах среды, их вызывающих, и та-
ким образом позволяют судить об условиях осадконакопления на дан-
ном участке (например, об условиях солености, температуры бассейна),
а это безусловно существенно для решения определенных вопросов гео-
логической истории и стратиграфии.
210
ЛИТЕРАТУРА
Агаев М. Г. 1969. К проблеме экспериментального взаимопревращения яровых и
озимых растений. — Бот. журн., 54, № 9, 1364—1378.
Али-3 а д е А. А. 1967. Акчагыл Туркменистана. Том 2. М.
А л и х а н я н С. И. 1967. Современная генетика. М.
Андрусов Н- И. 1897- Ископаемые и живущие Dreissensidae Евразии. — Тр-
СПб- о-ва естес|в-, отд. геол, и минер., т- 25, стр. 1—683.
А н д р у с о в Н. И. 1961. Избранные труды, т. 1. М.
Андрусов Н. И. 1963. Избранные труды, т. 2. М-
Андрусов Н. И. 1964- Избранные труды, т. 3- М- \
Андрусов Н- И- 1965- Избранные труды, т. 4. М-
Аракелян М. М. 1950- О биологической роли семени как ментора у животных. —
Советская зоотехния, № 9-
Астауров Б. Л. 1967 а. О негенетических концепциях гибридной мощности в связи с
вопросом о реальности так называемого физиологического гетерозиса. — Бюлл-
Моск- о-ва испыт. природы, отд- биол., т- 74(5), стр- 77—96.
Астауров Б. Л- 1967 б- О так называемом физиологическом гетерозисе в свете дан-
ных современной генетики- — Вести, с -х. науки, № 3, стр. 24—30-
Астауров Б. Л- 1968 а. Физиологический гетерозис и генетика. — Вопросы фило-
соф-, № 3, стр. 85—96-
Астауров Б- Л- 1968 б- О так называемом физиологическом гетерозисе. — В сб-:
«Гетерозис: теория и практика», стр. 215—238, Л-
Ахвледиани Е. Г- 1957. О филогенетических связях между некоторыми брюхоно-
гими киммерийского и куяльннцкого ярусов. — Сообщ. Акад, наук Груз. ССР,
т- 19, № 4, стр. 451—458-
Ахвледиани Е. Г. 1968. Явления индивидуальной изменчивости у некоторых Саг-
diidae Черноморского плиоцена. Общие вопросы эволюционной палеобиологии,
IV, стр- 41—52. Тбилиси.
БадзошвилиЦ. И- 1966- К вопросу о карликовости некоторых групп моллюсков- —
Сообщ. АН Груз. ССР, т. 41, № 3, стр. 631—638-
Базилевская Н. А- 1968- Научное наследие Н- И- Вавилова и его значение для
современной биологии- — Научн докл. высш, школы. Биол. науки, № 2, стр. 7—
17.
Берман 3- И, Завадский К- М., Зеликман А. Л., Пара манов А. А. и
Полянский Ю- И. 1967. Современные проблемы эволюционной теории. Л.
Богачев В. В. 1961- Материалы к истории пресноводной фауны Евразии. Киев-
В г ви лов Н. И. 1920. Закон гомологических рядов в наследственной изменчивости- Л.
Вавилов Н И- 1927. Географические закономерности в распределении генов куль-
турных растений. — Тр- по пр. бот., генетике и селекции, 17, № 3, 411—428-
Вавилов Н. И. 1935- Научные основы селекции пшеницы. М-Л.
Вавилов Н. И- 1967. Закон гомологических рядов в наследственной изменчивости.
Линнеевский вид как система. Л.
Башков В- И. 1957- О появлении у насекомых устойчивости к хлорированным угле-
водородам. — Тр. Центр, науч.-исслед. дезинфекционного ин-та, вып. 10, стр.
155—163.
Воронов Н. Н. О гомологической изменчивости. — В сб.: Проблемы кибернетики,
вып. 16, стр. 221—229. М. ,
211
Габуния Л. К- 1949. О представителях семейства Neritidae из среднеплиоценовых
отложений Западной Грузии. — Бюлл. /Лоск- s-ва исп. прир-, отд. геол-, т- 24
(6), стр< 40—48.
Г а б у н и я Л. К 1950. Об окраске моллюсков среднеплиоценовых отложений Запад-
ной Грузии. — Бюлл. Моск, о-ва исп. прир., отд. геол-, т. 25(5), стр. 71—76-
Гсоргиева Р. 1948- Гибридная изменчивость при трансплантации некоторых пасле-
новых- М- (Пер. с болгарского).
Герасимов Н. П- 1953. О некоторых факторах видообразования и о значении их
для стратиграфии. — Материалы Палеонт. совещания по палеозою 14—17 мая
1951 г. М
Гесслер К. 1967. О сущности жизни. Пер. с нем-, М.
Г л а в и н и ч Р- Г. 1959. Изменение наследственности растений путем прививки. М-
(Пер. с сербского).
Глущенко И- Е- 1948- Вегетативная гибридизация растений. М-
Грачевский М- М. 1958. Новый вид остракод из Малиновских слоев Куйбышевско-
ю Заволжья. — Докл. АН СССР, т. 120, № 6, стр. 1322.
Громов А. М. 1969. Изменение организма птицы переливанием крови. — Птицевод-
ство, № 3, стр. 25—26.
I р э н и к С. 1965. Цитоплазматические единицы наследственности. — Агробиология,
№ 4, стр. 630—633.
Давиташвили Л- Ш- 1932. Руководящие ископаемые нефтеносных районов Крым-
ско-Кавказской области. Сарматский ярус- М -Л.
Давиташвили Л- Ш- 1933. О стратиграфическом подразделении киммерийского
яруса. — Бюлл. Моск, о-ва испыт. прир., отд. геол., т- 11 (4), стр. 368—384:
Давиташвили Л- Ш- 1934- О происхождении рода Spaniodontella Andrussow.
Тр. Зак- геол.-гидрогеодез. треста- Тбилиси-
Давиташвили Л. Ш. 1936. К изучению закономерностей изменения величины те-
ла в филогенетических ветвях. Проблемы палеонтологии, т. 1, стр. 179—198. М.
Давиташвили Л. Ш- 1940. Некоторые проблемы дарвинизма. — Под знаменем
марксизма, № 2, стр. 114—164.
Давиташвили Л. Ш- 1948- История эволюционной палеонтологии от Дарвина до
наших дней- М-
Давиташвили Л- Ш- 1949- Палеобиология и проблема воспитывающего действия
среды на природу организма. — Вестник Моск, ун-та, № 10, стр. 139—148-
Давиташвили Л- Ш. 1961- Теория полового отбора. М-
Давиташвили Л- Ш. 1965- Некоторые вопросы изменчивости и видообразования
в свете палеобиологической истории населения Паратетиса. — Бюлл. Моск, о-ва
испыт- прир-, отд- геол-, т. 40 (1), стр. 28—45.
Давиташвили Л- Ш. 1966. Современное состояние эволюционного учения на За-
паде. М-
Давиташвили Л- Ш- 1968. Вопросы методологии в учении об эволюции органи-
ческого мира. Тбилиси.
Давиташвили Л. Ш- 1969. Причины вымирания организмов- М-
Давиташвили Л. Ш. и Китовани Т- Г. 1964. О филогенезе некоторых Cardi-
idae из гурийских и чаудинских отложений. — Сообщ. АН Груз. ССР, т. 26.
№ 2, стр. 385—392.
Дарвин Ч- 1939. Сочинения- Т. 3. Происхождение видов путем естественного отбора.
Дарвин Ч- 1951- Сочинения. Т. 4. Изменения домашних животных и культурных
растений. М.
Дарвин Ч. 1953. Сочинения. Т- 5. Происхождение человека и половой отбор. М-
Дементьев Г- П- 1964. Класс Aves.Птицы. — Основы палеонтологии. Земноводные,
пресмыкающиеся, птицы- М.
Дербенев а-У х о в а В- Н. 1959. Резистентность переносчиков к инсектицидам до
212
материалам Всемирной организации здравоохранения. — Медиц. паразитология
и паразит, болезни, т. 28, № 6, стр. 737—741-
Дербенева-Ухова В. П. и Л и н е в а В. А- 1951. Приобретение устойчивости к
ДДТ у комнатной мухи Musca domistica Предварительное сообщение- — Медиц.
паразитология и паразит, болезни, т. 20, вып. 4, стр. 335—339-
Дербенева-Ухова В. П. и Морозова В. П. 1950. Случай появления ДДТ ус-
тойчивости у комнатной мухи. — Мед. паразитология и паразит, болезни, т. .19,
стр. 464—467-
Дергачев Л- П- 1964. Диалектико-материалистические черты мировоззрения Нико-
лая Ивановича Вавилова. — Некоторые вопросы филос. Межвуз. филос., Kg V,
стр. 103—114-
Джанелидзе О. И- 1953. Фораминиферы тарханского горизонта Грузии — Тр. Ин-
та палеобиологии АН Груз- ССР, т. 4, стр. 117—155.
Джинкс Дж. Л- 1966- Нехромосомная наследственность. М.
Добролюбова Т- А- 1948. Изменчивость кораллов филогенетического ряда
Dibunophyllum bipartitum (McCoy)—Caninia okensis Stuck.—Изв. аАН СССР, cep.
биол-, № 2, стр. 149—167.
Добролюбова Т- А- 1952. Формообразование у нпжнекаменноугольных кораллов
Lithostrotion и Lonsdaeleita в свете мичуринского учения. — Изв. АН СССР,сер.
биол., № 6, стр- 95—НО.
Дубинин Н. П- 1966. Эволюция популяций и радиация- М-
Дубинин Н. П- 1968. Некоторые методологические проблемы генетики. М-
Дубинин Н. П. 1968. Н. И. Вавилов как генетик. — Генетика, № 3, стр. 18—27.
Дубинин Н- П- 1970 Общ? я генетика. М.
Дубин ин Н. П. и Глембоцкий Я. Л. 1967. Генетика популяций и селекция. М.
Дубинин Н. и Губарев В. 1966. Нить жизни. Очерки о генетике. М.
Жгенти Е. М- 1961- Новый род Savanella Zhgh. и его стратиграфическое значе-
ние. — Сообщ. АН Груз. ССР, т- 27, № 1-
Жгенти Е. М. 1967. Экогенез рода Spaniodontella. Тезисы докладов Всесоюзно-
го совещания. Ин-т палеобиологии АН Груз. ССР. Тбилиси.
Жгенти Е- М- 1968. Значение изучения экогенеза и изменчивости для расчленения
среднемиоценовых отложений Черноморско-Каспийской области. — Общ. вог-
росы эвол. палеобиологии, т- IV, стр. 97—116- Тбилиси-
Женермон Ж. 1969. Проблема длительных модификаций у простейших. —Успехи
протозоологии- Л., НО—112-
Зеликман А. Л- 1966- Незаслуженно забытые идеи Ч- Дарвина в области измен-
чивости организмов. — Генетика, № 3, стр- 142—153.
Зенкевич Л. А- 1947. Фауна и биологическая продуктивность моря. Советская
наука-
Ильин А- Я- 1966- О теоретическом значении закона гомологических рядов Н. И- Ва-
вилова. — Вопросы философии, № 5, стр- 85—92.
Калинина Л- Б. 1968- О кортикальной наследственности у простейших. — Цито-
логия, т. 10, № 1, с гр. 3—19-
Кож о в М- М. 1962- Биология озера Байкал. М-
Колесников В- П. 1930- О сарматских представителях Trochidae — Тр. Геол-
музея АН СССР, т. 6, стр. 37—67.
Колесников В. П. 1935. Сарматские моллюски. — Палеонтология СССР т. 10,
2- Л.
Колесников В. П. 1939. О закономерностях развития замкнутых бассейнов. —
Докл- АН СССР, т- 23, № 8, стр. 808—810. I
Колесников В. П- 1941. О «цикличности» развития третичных фаун. — Докл,
АН СССР, т 31, № 1, стр. 36—38.
Колесников В- П- 1949. О некоторых проблемах палеонтологии. — Бюлл. Моск,
о-ва испыт. природы, отд. геол., т. 24, стр- 3—45.
213
Коробков И А. 1955- Справочник и методическое руководство по третичным мол-
люскам- Брюхоногие- Л.
Коробков И- А. и Макаренко Д. Е- 1967. Paramonopleura gen. nov. (Rudistae)
из верхнего палеоцена Украины. Палент. журн-, № 4, стр. 135—138-
Коюмджиева Ем. 1965. Върху никои особенности на стратиграфията на кара-
гана във Варненско. — Известия на научно-изслед. геологич. ин-т, т. 2, стр-
49—60. София-
Краевой С. Я- 1967- Возможна ли вегетативная гибридизация растений посредст-
вом прививок? М-
Крашенников В. А- 1958. Стенофациальные и эврифациальные виды форамини-
фер. — Труды ВНИГРИ, вып. 9, Палеснт- сб- № 2.
К р и п п а-Ф ранчески. 1969. Длительные модификации. — Успехи протозоологии-
Л-, 112—113.
Кушнер X- Ф- 1964. Наследственность-сельскохозяйственных животных. М-
Кушнер X. Ф- и Китаева О. Н. 1953. Новые экспериментальные данные по воп-
росу о родственном и неродственном разведении кур- — Тр. Ин-та генетики АН
СССР, № 20-
Кушнер X. Ф-, Толоконникова Е- В. и Моисеева И. Г. I960. Влияние внедрения <у-'
рам различных компонентов чужеродной крови на тип оперения их потомства. —
Тр. Ин-та генетики АН СССР, т- 27, стр- 145.
Лебедев М. М. 1940. Влияние условий развития на результат родственного спари-
вания животных- — Яровизация, № 5-
Левитскпй Г. А. 1925. Карно — и генотипические изменения в процессе эволюции —
Тр. по пр. бот. и селекции, 25, № 5, 3—28.
Левушкин С- И- и Матекин П- В- 1965 Biospeologica Sovietica. XXV. Troglolestes
sokolovi gen. n., sp. n.—Бюлл. Моск, о-ва испыт. прир., отд. биол., т. 70(3), стр.
35—46.
Линдберг Г. У- 1955. Четвертичный период в свете биогеографических данных. Л.
Лысенко Т. Д. 1965. О цитоплазматической наследственности. № 4, стр. 633—636.
Майсурян Н. А. 1967. Выдающийся ученый нашего времени. — Природа,
№ 9, стр. 48—53-
М а м п о р и я Ф. Д. 1969. Проблема вегетативной гибридизации растении.
Сухуми-
Мусхелишвили Л. В. 1968. К вопросу об изменчивости сарматских нас-
сид. Общие вопросы эволюц- падеобиол., т- IV, с гр- 21—29. Тбилиси.
Н а л и в к и н а А. К- 1936. Верхнедевонские гониатиты и климении Мугоджар. —
Тр. ВНИГРИ, нов. сер., вып. 72, стр. 60—125.
Новиков Б. Г.. Чепинога О. П. и Любарская М. А. 1961. Эффект инъек-
ции чужепородной ДНК у уток.—Журн. общей биол., т. 22, № 4, стр. 317—320.
Н о и и с к и й М. Э. 1932. Краткий очерк истории изучения недр Татарскойй
республики.
Окропиридзе О. В. 1956. К вопросу о дополнительных камерах у глобиге-
рин. — Докл. АН СССР, т. 106, № 2, сгр. 338—341.
Основы палеонтологии. 1960. Брюхоногие. М-
Паавер К. Л. 1965. О вековой изменчивости как форме группой изменчивости жи-
вотных во времени. — В сб.: Шварц С. С. (отв. ред.), Внутривидовая измен-
чивость наземных позвоночных животных и микроэволюция, 77—82. Свердловск.
Павличенко В. 1963. Соматические гибриды свиней. — Международный с.-х. жур-
нал, № 3.
Павлов А. П. 1925. Неогеновые и послетретичные отложения южной и восточной
Европы. — Мемуары геол. отд. о-ва люб. естеств. антроп. и этногр., вып. 5.
Пидопличк^о И. Г. 1951. О ледниковом периоде. Вып. 2. Киев.
Пири Н. 1959. Химическое многообразие и преблема происхождения жизни, возник-
новение жизни на Земле — Тр. Междунар. симпоз. 19—24 авг 1957, М., 79—87.
214
П о л ы ни н В. М. 1967. Мама, папа и я. М.
Пузанов И. И. 1954. О некоторых изменениях морских организмов, попавших в
соленые лиманы — Бюлл. Моск, о-ва испыт. прир., отд. биол., том 59, вып. 4,
стр. 23—31.
Раевская О. Г. 1953. Влияние близкородственного размножения и скрещивания на
жизненность некоторых дрожжей. — Автореферат диссертации. Ин-т генетики
АН СССР.
Раузе р-Ч е р н о у с о в а Д. М. 1928. Об одном ряде мутаций. Изв. Асе. науч.-исслед.
ин-т физ.-мат., I МГУ.
Рубцов И. А. 1948. Появление устойчивой к ДДТ формы домашней мухи и ре-
зультате трехлетнего применения инсектицида. — Природа, № 4, стр. 62—63.
Рыжков В. Л. 1966 а. Предисловие к русскому изданию книги Дж. Джинкса
«Нехромосомная наследственность», стр. 5—15- М-
Рыжков В. Л. 19666. Цитоплазматическая наследственность. В сб. Акту-
альные вопросы современной генетики, стр. 358—391- М.
Саидова X. М. 1960. Распределение фораминифер в донных отложениях
Охотского моря. — Труды Ин-та океанов. АН СССР, т. 32.
Самохвалова Г. В. 1951. Получение наследственных изменений у тлей при пере-
мене кормовых растений. — Журн. общей биол., т. 12, № 3, стр. 176—191.
Самохвалова Г. В. 1954. Получение направленных наследственных изме-
нений у тлей при перемене кормовых растений. — Сообщ. второе. — Зээл
жури., т. 33, № 5, стр. 1032—1040.
Сенинский К. 1905. Новые данные о неогеновых пластах Юго-западного Закав-
казья. — Тр. О,ва естествоисп. При Юрьевском ун-те, т. 16.
С; им пеон Дж. Г- 1948- Темпы и формы эволюции. М-
Смирнов Е. С. 1961. Наследование приобретенной ритмики размножения у
Neomysis corcuflaxus Bunt. (Aphidae). Журн. общей биол., т. 22, № 3, стр.
164—178.
Станкович С. 1959. Охридското озеро и иеговист жив свет. Скопле.
Строганов Н. С- 1962. Экологическая физиология рыб. Т. 1. М-
Суханова К. М. 1969. Температурные адаптации у простейших. — Успехи
протозоологии. Тезисы докладов и сообщен, на III Междунар. конгрессе про-
тозоологов. Л., 189—190.
Сърбова С. и Аврамова С. 1965. Сървменнии насоки на изеледване проблемата
за инсектипидината резистентност. — Природа (болг.), кн. 5, стр. 36—42.
Такта киш вили И. Г- 1967- Историческое развитие семейства валенсиенниид. —
Тр. Ин-та палеобиологии АН Груз. СССР, Тбилиси.
Тактакишвили И. Г. 1968. «Румынские» элементы в понтической фауне
Западной Грузии. — Сообщ. АН Груз. ССР, т. 50, № 3, стр. 729—733.
Т а л и е в Д. Н. 1948. К вопросу о темпах и причинах дивергентной эволюции
байкальских Cottoidei. — Тр. Байкальской лимнолстанции. М.—Л., 12,
106—158.
У дольская Н. Л. 1960. Основные положения учения Дарвина о гибридизации
растений. — Вести, с.-х. науки, № стр. 29—33.
Фурсенко А. В. 1959. Подкласс Subclasses Foraminifera, общая часть. — В:
«Основы палеонтол.», том «Общая часть. Простейшие.
Хиншелвуд К. 1958. Некоторые взаимоотношения физических и биологи-
ческих наук. — Журн. общей биол., т. 19, № 5, стр. 315—319.
Челидзе Г. Ф. 1952. Некоторые двустворчатые дуабских слоев. Тр. Геол,
ин-та АН Груз. ССР, сер. геол., т. 6 (II).
'Чилингарян А. А., Павлов Е. Ф. и Мкртчян Л. П. 1962. Наследственная
трансформация окраски и изменение количества ДНК у пекинских уток,
вызванное инъекциями изолированных ядер эритроцитов уток другого вида. —
Изв. АН Арм. ССР, т. 15, № 12, стр. 25—31.
215
Шапошников Г. X. 1965. Морфологическая дивергенция и конвергенция в.
эксперименте с тлями. — Энтомол. обозрение. Том 44, вып. I, стр. 3—35.
Шапошников Г. X. 1966. Возникновение и утрата репродуктивной изоля-
ции и критерий вида. — Энтомол. обозр., т- 45, вып. 1, стр. 3—33-
Шапошников Г. X. 1968. Эволюция тлей в связи со специализацией и сменой хозя-
ев. — Автореферат диссертации на соискание ученой степени доктора биологи-
ческих наук. Л.
Шпет Г. И. 1968. Биологическая продуктивнссть рыб и других животных. Киев.
Шпет Г. И. 1968. Об ускорении роста в эволюции некоторых групп животных.—
В сб.: Темп индивидуального развития животных и его изменения в ходе
эволюции. Ред. С. В. Емельянов, стр. 266—275. М.
Эберзин А Г. 1947 Род Limnccardium Stoliczka в плиоцене Понто-Каспийского-
бассейна. — Труды Палеонт. ин-та АН СССР, т. 13, № 4, М.
Эберзин А. Г. 1951. Солоноватоводные кардииды плиоцена СССР. Часть 2. — Тру-
ды Палеонт. ин-та АН СССР, т. 31. М.-Л.
Эберзин А. Г. Солоноватоводные кардииды плиоцена СССР. Часть 3. Proso-
dacna, prionopleura, Pachydacna. — Труды Палеонт. ин-та АН СССР, т. 74. М.
Эберзин А. Г. 1962. Солоноватовидные кардииды плиоцена СССР. Часть 4. Род
Didacna Eichwald, подроды Pontalmyra и Crassadacna — Труды Палеонт. ин-та
АН СССР, т. 91. М.
Эберзин А. Г. 1965. О находке Ecericardium в плиоцене Румынии. — Палеонт.
сборник Львовского гос. ун-та, № 2 , вып. 1, стр. 14—17.
Эберзин А. Г. 1967. Солоноватовидные кардиды плиоцена СССР. Часть 5.
Pseudocatillus, Didacnomya, Macrodacna. — Труды Палеонт. ин-та АН СССРг
т. 112. М.
Эрлих П. иХолм Р. 1966. Процесс эволюции. Пер. с англ. М.
Яблоков А. В. 1966. Изменчивость млекопитающих. М.
Abel О. 1912. Grundzuge der Palaejiologie der Wirbeltiere. Stuttgart.
Abel O. 1929. Palaobiologie und Stammesgeschichte. jena.
Abel O. 1931. Die Degeneration des Hohlenbaren von Mixnitz und deren wahrschein-
liche Ursache. In: Abel O. u. Kyrle G. Die Drachenhohle bei Mixnitz. Spelaol.
MonOgr., Bd. 7 u. 8. pp. 719—744. Wien.
Abel O. 1935. Vorzeitliche Lebensspuren. Jena.
Abel O. und Kyrle G. 1931. Die Drachenhohle bei Mixnitz.—Spelaologische Mo-
nogr., Bd. 7 u. 8. Wien.
Andrewart h a H.G. and Birch L. C. 1954. The distribution and abundance
of animals. Chicago.
Andrussoff N. 1912. Ober einige Neritiniden aus neogenen Ablangerungen des po
to-caspischen Gebietes. Зап. СПб. мин. об-ва, т. 41, стр. 1—17.
An t on i us О. 1922. Uber die Variability des Hohlanbarenshadels. Palaontol. Zt.,
Bd. 4, pp. 104—106.
Arkell W. J., Kummel B. and Wright C. W. 1957. Treatise on invertebrate
paleontology, part L, Mollusca 4, Cephalopoda. Ammonoidea, pp. L 129—L465.
Armstrong T. 1945. Differences in the life history of codling moth Carpocaps
pomonella (L.) attacking pear and apple. Canad. Ent., 77: 231—33, p. 692.
A s с о 1 i P. 1957. Microfaune del tortoniano di Mombisaggio e della serie plioceni-
ca di Volpegeroni. — Rev. ital. paleont. e stratigr., 63, No. I, pp. 3—28. Toronto-
Alessandria.
Barnett S. A. (ed. by.) 1958. A century of Darwin. London.
Bateson W. 1922. Evolutionary faith and modern doubts. — Sci., № 1412.
Beadle L. C. 1962. The evolution of species in the lakes of East Africa. — Uganda
Journ., 26, № 1, 44—54.
Beale G. H. 1964. Inheritance without genes? — Discovery, 25, №. 11, pp. 27—28,.
Beale G. H. 1966. The roile of the cytoplasma in heredity.—Proc. Rov. Soc., В 164
№ 995, pp. 209—218.
216
Bettenstaedt F. 1958. Phylogenetische Beobachtungen in der Micropalaontologie.
Palaontol. Zeitschr., Bd. 32, Nr. 3—4, pp. 115—140.
Bettenstaedt F. 1962. Evolutionsvorgange bei fossilen Foraminiferen. — Mitt.
Geol. Staatsinst. Hamburg. № 31, pp. 385—460.
В h a n K. S. 1964. Cytoplasmic inheritance. — Bot. Rev., v. 30, No. 2, pp. 312—332.
Boltovskoy E. 1954. Foraminiferos del golfo San Jorge. — Rev.Inst. nac. invest,
cienc. natur. Gienc. geol., 3, № 3, pp. 79—246.
Boltovskoy E. 1963. Foraminiferos у sus relationes con el medio. — Revista Mus
Argentine de ciencias naturales «Bern. Rivadavia». Hidrobiologia, tome I, № 2,
Buenos Aires.
Boltovskoy E. 1965. Los foraminiferos recientes. Buenos Aires.
Boltovskoy E. 1965. Twilight of foraminiferilogy. — Journ. Paleont., 39, № 3,
383—390.
Boni A. A. 1940. Fauna anisica pigmea scoperta nelle Prealpi Byesciane. —Soc. geol.
Italiana, B, v. 58, pp. 321—428.
Boni A. A. 1942. Faune pigmee triassiche: contribute alia paleobiologia del nanismo
di faune marine. — Soc. geol. Italiana, B. v. 61, pp. 228—272.
Bonner D. M. 1961. Heredity. Englewood Cliffs, New Jersey.
Bounoure L. 1964. L’heredite cellulaire. — Sci. et enseignem. Sci., v. 5, № 33,
pp. 43—47.
Bradshaw J.S. 1957. Laboratory studies of the rate of growth of the Foraminifera
Streblus beccarii (Linne), var. tepida.—Cushman Journ. Paleont., 31, 6.
Brink R. A. 1958. Paramutation at the R. locus in maize plants. Cold. Spring Harb.
Symp. Quant. Biol., 23, 379.
Brink R. A. 1960. Paramutation and chromosome organization. — Quart. Rev. Biol.
35, 120.
В r i n к m a n h R. 1929. Statistisch-biostratigraphische Untersuchungen an mittel-
jurassischen Ammoniten. Uber Artbegriff und Stammesentwicklung. — Abh. Ges.
Wiss. Gottingen. Math. -Phys. KL, N. F., Bd. 13, Nr. 4.
Brown A. W. A. 1958. Physiological mechanisms in evolution of resistance.—Proc.
15-th Internat. Congr. of Zoology. London. 16—23 July, pp. 497—499. London.
Brun J. L. 1965. Genetic adaptation of Caenrhabditis elegans (Nematoda) to high
temperatures. — Science, v. 150, № 3702, p. 1467.
Brun J. L. 1966. L’adaptation aux temperatures elevees chez un nematode: Caenorh-
habditis elegans Maupas 1900. — Annales de Biol, anim., bioch., biophys., 6 (2),
pp. 127—158; 6 (3), pp. 267—300; 6(4), pp. 439—466. Paris.
Brun J. L. 1967. Influence des conditions de milieu sur la fecondite de Caenorhabitis
elegans a differentes temperatures.—Nematologica, 12 (1966), pp. 539—556.
Bukowski Geiza v. 1893. Die Levantinische Molluskenfauna der Insel Rhodus. —
Denkschr. math.-naturw. Classe d. Keiserl. Akademie der Wissensch., Bd. 60.
Wien.
Busvine J.R. 1959. The detection and measurement of insecticide resistance. XV
the Internat. Congr. of Zool., pp. 499—502. London.
Callomon J.H. 1955. The Ammonite succession in the Lower Oxford clay and Kel-
laways Beds at Kidlington, Oxfordshire, and the zones of the Callovian stage. —
Phill. Trans. Roy. Soc. London, ser. B, Biol. Sci., № 664, vol. 239, pp.
215—264.
C a v a 1 1 i-S f о r z a L. L. 1957. Indirect selection and origin of resistance. In: Drug
resistance in micro-organisms. Mechanism of development (For Ciba Foundation),
pp. 30—40.
Colom G. 1955. Sobre la existencia de dos razas gigantes de Helicidos en las zonas
de maximas alturas de la Sierra Norte de Mallorca. — Bol. Soc. Hist. Nat. de las
Baleares. Palma. Fasc. 1—4, pp. 11—20.
Colom G. 1957. Biogeografia de las Baleares. La formacion de las islas у el origen de
su flora у su fauna. Palma de Mallorca. Estudio general Luliano.
217
Colom G. 1959. Notas micropaleontologicas у ecologicas sobre algunas formaciones
continentaes espaDolas. — Estud.. geol, v. 15, № 41—44, pp. 93—106.
Colom G. 1961. Les populations geantes d’helicides de la haute montagne de Majorquc
(Baleares). Colloques internat. C. N. R. S. 94. Le peuplement des ties Mediterra-
neennes et le probleme de I’insularite, pp. 295—298. Paris.
Cooper G. A. 1957. Brachiopods (Annotated bibliograpy). In: Ladd H. S. Treatise
on marine ecology and paleoecology. Vol. 2. Paleoecology. — Geol. Soc. Amer.,
memoir 67, pp. 801—804.
Cope E. D. 1887. The origin of the fittest. Essays on evolution. London and
New Jork.
Cope E. D. 1904. The primary factors of organic evolution. Chicago.
Craigheard F. G. 1921. Hopkin’s host-selection princuple as related to certain
Cerambycid beetles. — Journ. Arg. Research, 22: 189—220. p.. 692.
Crow J. F. 1966. The evolution of resistance in hosts and pests. — Agric. Chemisry,
v. 21, № 4, pp. 31—32.
Curtis A. S. G. 1965. Cortical inheritance in the amphibian Xenopus laevis: prelim-
nary results. — Arch, biol., v. 76, № 2—4, pp. 523—546.
DacqueE. 1921. Vergleichende biologische Formenkunde der fossilen niededen Tiere.
Berlin.
D a 1 c q A. M. 1949. L’apport de 1’err b’yologie causale au probleme de devolution.
Port. Acta biol., Vol. Jub. Pr. Goldschmidt, pp. 367—400. Coimbra.
D a 1 c q A. M. 1955. Les ontomutations a 1’origine des Mammiferes.—Bull, de la Soc.
zoologique de France, t. 79, № 4, pp. 240—255.
D a n i e 1 1 i J. F. 1960. Some new approaches to inheritance and evolution. — Proc.
Roy Inst. Gr. Brit., v. 38, № 3, pp. 271—276.
Darwin Ch. 1882. Origin of species. London.
Darwin Ch. 1888. The descent of man and selection in relation to sex. London.
Darwin Ch. 1921. The variation of plants and animals under domestication. 10-th
impression of the second edition. In two volumes. London.
Davidson G. 1959. The inheritance of resistance to insecticides in anopheline mos
quitoes. XVth Internat. Congr. zool., pp. 504—506. London.
Davis B. D. 1957. In: Wolstenholme О. В. E. and O’Connor (editors for Ciba Foun-
dation). Drug resistance in micro-organisms. London (Discussion), p. 60.
Dean A. C. R., Hinshelwood C. 1957. Aspects of the problem of drug resistan-
ce in Bacteria. In: Drug resistance in micro-organisms. Mechanism of development
(for Ciba Foundation), pp. 4—24. London.
Dean A. C. R. and Hinshelwood C. 1963. Integration of cell reactions. — Na-
ture, v. 199, pp. 7—11.
Dean A. C. R., Hinshelwood C. 1964. What is heredity? — Nature, v. 202,
pp. 1046—1012.
D о r s t J. 1959a. La signification des Galapagos dans 1’etude de 1’evolution. — Actes
Soc. helv. sci. natur., 139-e session, Lausanne. Aaran, 98—106.
D о r s t J. 195£b. Quelques affinites de la faune des lies Galapagos. — Compte rend.
Soc. hiogeogr., № 2, 57—60.
Durrant A. 1958a. Environmental conditioning of flax. — Nature, 181, № 4613,
928—929.
Durrant A. 1958b. Environmentally induced inherited changes in flax.—Proc. X-th
internat. congress of genetics. Toronto, 2.
^Ehrenberg K. 1931. Die Variability der Backenzahne beim Hohler.bahren. In:
Abel O. und Kyrle G. Die Drachenhohle bei Mixnit—Speleolog. Monogr., Bd. 7 u.
8, pp. 537—573. Wien.
Ehrlich P. R. and Holm R. W. 1963. The process of evolution. New York.
F a g e L. 1955. A postscript on subterranean evolution. — Bull. Nat. Inst. Sci. India,
№ 7, pp. 79—84.
Fothergill Ph. 1953. Historical aspects of organic evolution. New York.
218
Friedberg W. 1928. Mieczaki mioceriskie ziem polskich. Zeszyt piaty. Poznan.
F r i e s e H. 1951. Zur Foraminiferen-Fauna der Meeresmolasse des unteren Inngebietes.
—Geol. Dienst Berlin, Abh., N. F., 227.
Fryer G. 1960. Some controversial aspects of speciation of African Cichlid fishes. —
Proc. Zool. Soc. London, 135, № 4, 569—578.
F u г о n R. 1967. La geologie et la biogenese, pp. 91 —101, Paris.
GoldschmidtR.B. 1961. Theoretische Genetik. Bearbeitet von F. Schwanitz.
Berlin.
Grabert B. 1959. Phylogenetische Untersuchungen an Gandryina und Spiroplectinata
(Foram.) — Abhandl. Senckenberg. Naturforsch. Gesellschaft., 498, Frankfurt am
Main.
G r a n i с к S. Cytoplasmatic units of inheritance. — Science, v. 147, pp. 911—913.
Greenway J.C. 1959. Extinct and vanishing birds of the world. New York.
Gunter G. 1945. Summary of ecological studies of invertebrates of the Texas coast.
Nat. Research Council, Rept. Comm, on Mar. Ecology as related to paleontology,
pp. 55—56.
Gunter G. 1957. Temperature. In: Treatise Marine Ecol. Paleoecol., v. I. Geol. Soc.
Amer. Mem., 67.
Haas O. 1946. Intraspecific variation and ontogeny of Prionotropis woollgari and
Prionocyclus wyomingensis. —Bull. Amer. Mus. Nat. History, v. 86, art. 4 New
York.
Hagemann S. 1964. Plasmatische Vereebung. Genetic, Grundlagen, Ergebnisse und
Probleme in Einzeldarstellungen. Jena.
Hagemann R. 1967—1968. Das Plasma als Trager genetischer Information. — Biol.
Rundschau., Bd. 5, pp. 97—112; Bd. 6, pp. 1 —14.
Halavats, GyulaV. 1903. Die Fauna der Pontischen Schichten in der Umgebung
des Balatonsees. — Resultate der wissensch. Erforschung des Balatonsees, I. Bd.,
I Teil, pp. 1—80, Budapest.
Hawkins Sh. E., Cole R. J. 1965. Studies on the basis of cytoplasmic inheritance
in amoebae. — Experimental cell research^ 37, pp. 26—38.
Heberer G. 1959. Theorie de additiven Typogenese. In: Die Evolution der Organis-
men. 2. erweiterte Aufl. (Herausgegeben von G. Heberer.. Stuttgart), pp. 856—914.
L’H er i t i er P. 1964. Le probleme de Theredite non chromosomique. Eiicarpia. Pa-
ris. 1962, pp. 237—251.
H i g h к i n H. R. 1958. Temperature-induced variation in peas. — Amer. Journ. of
Botany, 45, № 8, 626—631.
H i 1 g e n d о г f Dr. 1867. Cber Planorbis multiformis in Steinheimer Siiwasserkalk. —
Monatsber. d. kdnigl. Preussischen Akad. d. Wissensch. zu Berlin. Um dem Jahre,
1866, pp. 474—504.
Hilgendorf F. 1877. Noch einmal Planorbis multiformis. — Zeitschrift d. Deutsch
geol. Ges., 29. Bd., pp. 50—62. Berlin.
Hill J. 1965. Environmental induction of heritable changes in Nicotiana rustica- —
Nature, 207, 732—734.
H о 1 d g a t e M. W. 1960. The fauna of the mid-Atlantic islands, Proc. Roy. Soc., B-
152, 550—567.
H о v a n i t z W. 1949. Change of host preference in Colias philodice. — Journ. Econ.
Ent., 41: 980—81, pp. 691, 692.
Holder H. 1955. Die Gattung Taramelliceras in suwestdeutschen Unter—und Mittel-
malm. — Palaeontographica (Stuttgart). —Abt. A, Bd. 106, pp. 17—153.
Hyatt A. 1880. The genesis of the Tertiary species of Planorbis at Steinheim.—Anni-
vers. memoirs Boston soc. nat. hist. 1830—1880, pp. 1—114.
Hyatt A. 1881. Transformations of Planorbis at Steinheim, with remarks on the effect
of gravity upon the from of shells and animals.—Proc. Amer. Assoc, for the advan-
cement of science. 29-th meeting held at Boston, Mass., aug. 1880. Salem, pp.
527—549.
219
Hyatt A. 1894. The phylogeny of an acquired characteristic.—Proc. Amer. Philosopfr.
Soc., v. 32, № 143, pp. 349—647.
Jeannel R. 1950. La marche de revolution. — Publ. Museum nat. histoire natur.,
№ 15. Paris.
Jeannel R. 1952. Pselaphides recueillis par N. Leleup au Congo beige IV. Faune de
» I’Itomvwe et. du Rugege. — Ann. Mus. Congo beige. Tervuren, zool. II, pp.
1—295.
Jeannel R.et Leleup N. L’evolution souterraine dans la region mediterraneene'
et sur les montagnes du Kivu.—Notes biospeologiques, VII, pp. 7—13.
J e к e 1 i u s E. 1932. Fauna Neogena a Romanei. Die Molluskenfauna der Dazischen
Stufe des Bekens von Brasov. Bucuresti.
J i с к e 1 i C. F. 1902. Die Unvolkommenheit des Stoffwechsels als Grundprinzip fur
Werden und Vergehen im Kampf uns Dasein.
J i с к e 1 i C. F. 1910. Die Unvollkommenheit des Stoffwechels als Grundprinzip im
Werden und Vergehen der Schneckenschalen. —Abhandl. d. Senckenberg. Natur -
forsch. Gesellsch., Bd. 32.
J о 1 1 о s V. 1932. Uber den gegenwartigen. Stand des Evolutions-problems. — Forsch.
und Forrsch., №11.
Klahn H. 1923. Palaontologische Methoden und ihre Anwendung auf die palaontolo-
gischen Verhaltnisse des Steinheimer Beekens. Berlin.
Klotz J. 1957. К problemu vegetativni hybridisace rostlin. — Vesmir, 36, № 6, pp..
182—184. I
К r e n к e 1 E. 1957. Geologie und Bodenschatze Afrikas. Leipzig.
Kuhn O. 1939. Die Ammoniten des frankischen Calloviums. — Nova Acta Leopold.
N. F., Bd. 6, Nr. 43. Halle (Saale).
К u m m e 1 B. 1948. Environmental significance of dwarfed Cephalopods. — Journ. Se-
diment. Petrology, v. 18, № 2, pp. 61—64.
К u r g G. 1961. Foraminiferes et Ostracodes de 1’etang de Thau.—Inst. Peches Marit.
Rev. Trav., 25.
L a 1 i с к e r C. G. 1948. Dwarfed Protozoan faunas. — Journ. Sedim. petrology, v.
№ 2, pp. 51—55.
Lankford R.R. 1959. Distribution and ecology of Foraminifera from east Missis-
sippi delta margin. — Amer. Assoc. Petr. Geol. Bull., 43, 9.
Lattin G. de. 1940. Uber die Evolution der Hohlentiercharactere. Sitzungsber.
Geselschaft Naturf. Freunde Berlin, Jahrg. 1939, pp. 11—41.
Lederberg J. 1957. In: Wolstenholme О. В. E. and O’Connor (editors for Ciba
Foundation). Drug resistance in micro-organisms. London (Discussion), p. 61.
Leroy P. 1963. L’hybridation vegetative chez les oiseaux. — Annee biol., v. 2,
№ 7—8, pp. 337—370.
Loomis F. B. 1903. The dwarf fauna of the pyrites layer at the horizon of the Tully
limestone in western New York. — New York State Mus. Bull., 69, pp. 892—920.
Lowe P. C., Carson J. R. and King S. C. 1963. Effect of blood tranfusions о
plumage color in chicken. — Journ. Heredity, v. 54 № 1, pp. 17—22.
Ludwig W. 1942. Zur evolutorischen Erklarung der Hohlentiermerkmale durch Al-
lelelimitation. — Biol. Zblatt, Bd. 62, pp. 447—455.
L u e r s H. 1959. Genetics in relation to inherited resistance. XV th Internat. Congr.
zool., Proceed, pp. 502—503. London.
Lull R. S. and Wright N. E. 1942. Hadrosaurian dinosaurs of North America.
Geol. Soc. America spec, paper, № 40.
Liittesschwager J. 1961. Die Drontenvogel. — Neue Brhem-Biicherei, Nr. 276.
L w о f f A. 1944. L’evolution physiologique. — Etude des pertes de functions chez les
microorganismes. Paris.
Margalef R. 1961. Modalites de devolution en rapport avec la simplification des
biocenoses insulaires. — Colloq. intern. Centre nat. rech. scient. № 94, 313—317.
220
М^а г i n е 1 1 ь W. 1931. Der Schade! des Hohlenbaren von Mixnitz und deren wahrschein-
liche Ursache. In: Abel O. u. Kyrle G. Die Drachenhohle bei Mixnitz.—Speleo-
log. Monogr., Bd. 7 u. 8., pp. 537—573. Wien.
M^a rquardt H. 1964. Die Deutung eines Failes plasmatischer Vererbung bei Stre-
ptocarpus als Kern-Plasma-Operonsystem. — Biol. Zblt., Bd. 83, H. 1, pp,
1 — 18.
Mather K. 1961. Nuclear materials and nuclear change in differentiation.—Natu-
re, 190,404—406.
M а у г Ё. 1942. Systematics and the origin of species. New York. pp. 665, 691, 694,
695, 708, 709, 710, 711.
M а у r E. 1959. Isolation as an evolutionary factor. — Proc. Amer. Philos. Soc., v.
103, № 2, pp. 221—230.
M а у r E. 1960. The emergence of evolutionary novelties. In: Evolution after Darwin.
Vol. I. The Evolution of life, pp. 349—380. Chicago.
Merriam С. H. 1918. Review of the Grizzly and big bears of North America. —
U. S. Dep. of Agricult., N. am. Fauna, № 41.
Michie D. 1958. The third stage in genetics. In: Barnett (editor), A century of Dar-
win,' pp. 56—84. London.
M i r s к у A. E. 1963. Austauschvorgange zwischen Zellkern und Zellplasma.—Natur-
wissenschaften, Bd. 50, Nr. 7, pp. 277—282.
Morgan T. H. 1926. Genetics apd the physiology of development.—The Amer. Na
tural., vol. 60, № 571, pp. 489—515.
M о t a § C. 1958. Freatobiologia, a noua ramura a Limnologici (La Phreatobiologie,
une nouvelle branche de la Limnologie). — Natura, t. 10, f. 3 (Comunic. 18
octob. 1957).
M о t a ? C. 1962a. Procede des sondages phreatiques. Division du domaine sonterrain
—Classification ecologique des animaux sonterrains—Le Psammon.—Acta Musei
Macedonici scientiarum naturalium. Tom VIII, № 7 (75), pp. 135—173.
M о t a § C. 1962b. Sur les Acariniens phreatiques, leur distributio geographique, leur
origine. — Zool. Anzeiger, Bd. 168, 71, 7—10, pp. 325—350.
M о t a § C. 1963. La notion de nappe phreatique, la phreatobiologie,les debuts, son ob
ject. — Analele stiintifice ale Universitatii «А1. I. Cuza»» din Iasi (Serie Nova)
Sectiunea II (Stiinte naturale). Biologie. Tom IX. Anul 1963. Fasc. I, pp.
57—61.
Myers E. H. 1936. The life of Spirillina vivipera Ehrenberg, with notes on morpho-
genesis, systematics and distribution of the Foraminifera.—Roy. Mier. Soc. J 56.
Myers E. H. 1943. Life activities of Foraminifera in relation to marine ecology.
—Amer. Phil. Soc. Proc., 86, 3.
N e u m а у r M. 1880. Uber den geologischen Bau der Insel Kos.—Denkschr. d. Kais
Akad. d. Wissensch., math.-naturw. Classe, Bd. 40. Wien.
Neumayr M. 1889. Die Stamme des Thierrei ches. Bd. I. Wien und Prag.
Neumayr M. und Hornes M. 1880. Uber den geologischen Bau der Insel Kos
und uber die Gliederung der jungtertiaren Ablagerungen des Archipels. — Denk-
schr. Akad. Wissensch., Bd. 40. Wien.
Neumayr M. und P a u 1 С. M. 1875. Die Congerien—und Paludienenschichten Sla-
voniens und deren Faunen. Ein Beitrag zur Deszendenz-Theorie.—Abhandl. der
К- K. geologischen Reichsanstalt, Bd. VII, Heft Nr. 3.
N i с о 1 D. 1944. New west American species of the foraminiferal genus Elphidium.—
Journ. Paleont., 18.
N igonV. etBrunJ. 1967. Cenetique et evolution des nematodes libres. Perspec-
tives tirees de Fetude de Caenorhabditis elegans. — Experientia, v. 23, № 161,
pp. 1 —10.
N у h о 1 m K. G. 1961. Morphogenesis and biology of the Foraminifer Cibicides loba-
tulus. —Zooll. Bidr. Uppsala, 33.
221
N у h о 1 m К. G. 1962. A study of the foraminifer Gypsina. —Zool. Bidr. Uppsala,
33.
О g n i b e n L. 1958. Stratigrafia e microfauna del Terziario della zona di Coiazzo
Caseota). — Rev. ital. paleont. e stratigr 64., № 3, pp. 199—286.
Olson E. C. 1965. The evolution of life. London.
Osborn H. F. 1910. The age of Mammals in Europe, Asia and North America. New
York.
Osborn H. F. 1917. Biocharacters as separable units of organic structure. — Amer
Natural., v. 51, № 608, pp. 449—456.
Osborn H. F. 1929. The Titanotheres of ancient Wyoming, Dakota and Nebraska.
Vol. I. — Geol. Survey Monograph. 55. Washington.
Osborn H. F. 1942. Proboscidea. Vol. 2. New York.
P h 1 e g e r F. B. 1960. Ecology and distribution of recent Foraminifera. Baltimore.
Pictet A. 1912. Un nouvel exemple de I’heredite des caracteres acquis. — Compt.
rend Soc. phys. nat. Geneve, 28: 36—38, p. 692.
Plate L. 1919—1920. Bemerkungen fiber die deszendenztheoretische Bewertung der
Umwandlungen von Planorbis multiformis.—Jenaische Zeitschr. F. Naturwissen-
schaft. 56. Bd., Neue Folge, 49. Bd., pp. 217—224. Jena.
Pokorny VI. 1958. Grundziige der zoologischen Mikropalaontologie. Bd. I. Berlin.
P о s p i s i 1 о v a D. 1957. Predbezna zprava о vysledkoch posobenia mentora na ml-
ade hybridne semenaca. vinica — Pol’nohospodarstvo, 4, № 3, pp. 565—582.
Prosser C. Ladd. 1960. Comparative physiology in relation to evolutionary the-
ory.— Evolution after Darwin, vol. I, pp. 569—594. Chicago.
QuatrefagesH.et Sabatier G. 1961a. Contribution a 1’etude des modifica-
tion de certains caracteres hereditaires des volailes par injections de sang. —
Cour Avic., 188.
QuatrefagesH.et Sabatier G. 1961b. Quelques observations interessantes
faites sur les volailles par injection de sang. — Cour. Avicole, 188.
Q u a у 1 e H. J .1926. The codling moth in walnut. — Bull. Calif. Agr. Exper. Sta.,
№ 402, pp. 690, 191.
RadovanovicM. 1961. Resultats des recherches faites dans les lies Adriatiques
sous le jour de I’evolutionnisme. — Bull. Acad, serbe des Sciences et des Arts,.
Classe Sci. math, et natur. 26, № 8, 59—90.
Regnault M. 1963. Une heredite sans chromosomes. — Sci. et avenier, № 202,
pp. 816—819.
Reid J. A. 1960. Mosquitoes, insecticides and evolution.—Proc. Centenary and Bio-
centenary Congr. of Biology. Singapore, Dec. 2—9, 1958, pp. 217—219. Sin-
gapore.
Richter R. 1929. Grfidung und Aufgaben der Forschungsstelle fiir Meeresgeologie
«Senckenberg» in Wilhelmhaven.—Natur. u. Museum, Bd. 59, pp. 1—0. Frank-
furt a. M.
Robertson F. W. 1966. The ecological genetics of growth in Drosophila. 8. Abap-
tation to a new diet... — Genetical Research, vol. 8, № 2, Oct., pp. 165—179
Rodda P. U. and Fischer W. L. 1964. Evolutionary features of Athleta (Eoce-
ne, Gastropoda) from the Gulf Coastal Plain. — Evolution, v. 18, № 2, pp..
235—244.
Ross H. H. 1962. A synthesis of evolutionary theory. Englewood Cliffs, N. J.
R u s c h e 1 1 i 1956. La serie aquitaniano — elveziana del rio Mainia, Asti, Parte I„
Stratigrafia. — Rev. ital. paleont. e stratigr., 62, № I, pp. 11—51.
S a с c h i C. F. 1961. Considerations sur les phenomens microevolutifs des animaux des.
petites lies mediterranneennes. Colloques internat. C. N. R. S. 44. Le peuplem
des iles Mediterranneennes et le probleme de I’insularite. Banyuls-Sur-Mer, pp.
321—335. Paris.
Sager Ruth. 1964. Nonchromosomal heredity — New England J. Med. v. 271, № 7
pp. 352—357.
222
Said R. 1953. Foraminifera of Great Pond East Falmouth, Massachussets.—Chshman
Lab. for. Foram. Res., Spec. Publ., 41.
Schafer W. 1962. Aktuo-Palaontologie nach Studien in der Nordsee, Frankfurt am
Main.
S c h о t z F. 1967. Extrachromosomale Vererbung. — Ber. Dtsch. Bot. Ges., Bd. 80,
H. 8, pp. 523—538.
S h i m e г H. V. 1908. Dwarf faunas. — Amer. Naturalist, v. 42, № 499, pp. 472—
490.
Simpson G. G. 1944. Tempo and mode in evolution. New York.
Simpson G. G. 1953a. The Baldwin effect.—Evolution, v. 7, № 2, pp. 110—117.
Simpson G. G. 1953b. The major features of evolution. New York.
Simpson G. G. 1960. The history of life. In: Evolution flfter Darwin. Vol. I.
The evolution of life, pp. 117—180. Chicago.
Simpson G. G. 1964. This view of life. New York.
Strebel O. 1964. Hohlentiermerkmale an Springswanzen. — Mikrokosmos, Bd. 53,
Nr. II, pp. 339—343.
Swinnerton H.H. 1930. Outlines of paleontology. 2-nd ed. London.
Swinnerton H.H. 1932. Unit characters in fossils. — Biol. Reviews, v. 7, № 4
Tasch P. 1953. Causes and palececological significance of dwarfed fossil marine
invertebrates. — Journ. Paleontology, v. 27, № 3, pp. 356—444.
Thornley J. M., Sinai J. and Y u d к i n J. 1957. Multiple mechanisms of
aquired drug resistance. In: Woltenholme О. В. E. and O. Connor (editors for
Ciba Foundation). Drug resistance in micro-organisms, p. 152. London.
Thorpe W. H. 1930. Biological races in inspctes and allied groups.-— Biol Rev.,
5: 177—212, pp. 691, 693.
Thorpe W. H. 1931. Biological races in insects and their significance in evolution.
—Ann. Appl. Biol., 18: 406—14, pp. 691.
Thorpe W. H. 1939. Further studies on pre-imaginal olfactory conditioning in in-
sects. — Proc. Roy. Soc. London, B, 127: 24—33, p. 691.
Thorpe W. 1945. The evolutionary significance of habitat selection. — Journ.
Anim. Ecol., 14: 67—70, pp. 693, 716.
Thorpe W. H. and Jones F. G. W. 1937. Olfactory conditioning in a parasitic
insect and its relation to problem of host selection. — Proc. Roy. Soc. London,
B. 124: 56—81, p. 691.
T i n t a n t H. 1963. Les Kosmoceratides du Callovien inferieur et moyen d’Europe Oc-
cidentale. — Publ. de I’universite Dijon. 29.
Todd R. 1957. Foraminifera from Carter Creek, northeastern Alaska. — U. S. Geol.
Surv. Prof. Paper 294-F.
О b i sc h L. 1953. Die Bedeutung von Metamorphose, Generationswechsel und Poly-
morphismus fur die Abstammungslehre.—Zool. Anz. 17. Supplementbd. Verhandl.
Dtsch. zool. Ges. 2—8, pp. 430—434.
U 1 a m S. M. 1967. How to formulate mathematically problems of rate of evolution?
Wistar Inst. Sympos. Monogr., № 5, pp. 21—31.
Uvarov В. P. 1961. Presidential address. — Proc. Roy. Entomol. Soc. London./v.
25, p. 52.
V a n d e 1 A. 1964. Biospeologie. La biologie des animaux cavernicoles. Paris.
Vaufrey R. 1958. Proboscidiens, etude systematique. In: Traite de paleontologie.
Tome VI, deuxieme volume, pp. 203—295. Paris.
Waddington С. H. 1960. Evolutionary adaptation. Evolution after Darwin. Vol.
I. The evolution of life. pp. 381—402. Chicago.
W e i g e 1 t J. 1947. Biometrische Studien an palaozenen Saugetiere und ihre Bedeu-
tung fiir die Beurteilung des Evolutionsgeschehens. — Fortsch. und Fortschr.
21/23. Berlin.
223
W e i g е 1 t J. 1950. Die tektonische Pradestination des Lebensraumes alttertiaren Wir-
beltierfaunen Deutschlands. —Zeitschr. Deutsch. Geol. Ges., 100. Stuttgart.
W e n z W. 1922. Die Entwicklungsgeschichte der Steinheimer Planorben und ihre Be-
deutung fur die Deszendenzlehre.—Aus Natur. und Museum. Senckenb. Natur-
forsch. Ges. 1817—1922. 52- Bericht, H. 3—4, pp. 135—158. Frankfurt am Main.
W e n z W. 1938—1944. Gastropoda. — Handbuch der Palaozoologie. Bd. 6, Teile 1 —
7. Berlin.
Wermuth H. und Mertens R. 1961. Schildkroten. Krokodile. Briickenechsen.
Jena. ,
Wilkie D. 1964. The cytoplasm in heredity. London.
Williams G. C. 1966. Adaptation and natural selection. A critique of some curren
evolutionary thought. Princeton, New Jersey.
Winterigham F. P. W. 1959. Physiological-chemical implications of insect resi-
tance. XV th Internat. Congr. zool. London. 16—23. July 1958, pp. 506—510.
London.
Y a g i s h i t a Noboru. 1961. Studies on graft hybrids of Capsicum annuum L. I. Va-
riation in fruit shape caused by grafting and the effects in the first and second
generations. — Bot. Mag., Tokyo, 74, № 873, pp. 122—130.
Ziegler B. 1959. Evolution in Upper Jurassic Ammonoides. — Evolution, vol. 13,
№. 2, pp. 229—235.
ГЛАВНЕЙШИЕ СТРАТИГРАФИЧЕСКИЕ ПОДРАЗДЕЛЕНИЯ
Группы Системы Отделы Ярусы (и другие подразделения) Абсол. возраст рубежей (мил. лет)
Четвер- тичная Голо- цен Современный 1,5—2 10 26 36 57 65 - 105 135
Плей стоцен Верхний
Средний
Нижний
Третич- ная Нео- ген Плиоцен Верхний Средний Нижний
Миоцен Верхний Средний Нижний
Пале- оген Олигоцен Верхний Средний Нижний
Эоцен Верхний Средний Нижний
Палеоцен Верхний Нижний
Мезозой- ская Меловая Верхний Датский
Сен он Верхний Нижний Маастрихтский Кампанский Сантонский Коньякский Туронский Сеноманский
Нижний Альбский Аптский
Неоком Барремский Готеривский Валанжинский
15. Л. Ш. Давиташвили
225
Группы Системы Отделы Ярусы (и другие подразделения) 1 Абсол. возраст рубежей (мил. лет)
Верхний, Титон Верхний волжский Нижний волжский
или мальм Кимериджский Оксфордский Келловейский Лузитанский
Мезозойская Юрская Средний, или доггер Батский Байосский Ааленский
Верхний Тоарский
Нижний, или Средний Домерский Плинсбахский
лейас Нижний Лотарингский Синемюрский Геттангский 195
Верхний Рэтский Норийский Карнийский
Триасовая Средний Ладинекий Анизийский
Нижний, или скифский Оленекский Индский ОЛА
Палеозой- Верхний Татарский Казанский
ская Пермская Нижний Кунгурский Артинский
Сакмарский 1 Сакмарский 1 Ассельский 285.
Оренбургский
Каменно- угольная Верхний Жигулевский Гжельский Касимовский
Средний Московский Башкирский, или каяльский
Нижний Намюрский Визейский Турнейский 350-
226
Группы Системы Отделы Ярусы (и другие подразделения) Абсол. возраст рубеже (мил. лет)
Девонская Верхний Фаменский Франский 400 430 ' 490 . 600(?) 1600 , 2600 более 3500
Средний Живетский Эйфельский
Нижний' » .Кобленцский ЖединСкий j
Силурий- ская Верхний Лудловский
Нижний Венлокский Ландоверский
Ордовик- ская Верхний Ашгильский Карадокский Ландейльский
Средний
Нижний Аре-нигский Тремадокский
Кембрий- ская Верхний Средний Не выделены
Нижний Ленский Алданский
Протерозой- ская Верхняя подгруппа
Нижняя подгруппа
Архейская
Примечание: Схема стратиграфических подразделений, утвержденная Межведэм-;
ственным стратиграфическим комитетом СССР для „Основ палеон-
тологии", здесь дополнена указаниями на абсолютный возраст ру-
бежей (в млн. лет), проверенными М. М. Рубинштейном, которому
мы приносим за это глубокую благодарность.
227
СТРАТИГРАФИЧЕСКАЯ СХЕМА НЕОГЕНА
ЧЕРНОМОРСКОЙ ЗОНЫ
X
W
Д
о
н
с
X
PJ
д
о
£
Верхний
Чаудинский горизонт
Гурийский горизонт
Таманские слои
Куяльницкий ярус
Средний [ Киммерийский ярус
Нижний
Верхний
Средний
] Понтический ярус
Г Мэотический ярус
[ Сарматский ярус
“Конкский горизонт
Караганский горизонт
Чокракский горизонт
Тарханский горизонт
Горийский горизонт
_Коцахурский горизонт
Нижний [ Сакараульский горизонт
Примечание 1. Некоторые исследователи рассматривают «горий-
кзкий» горизонт как нижнюю часть тархана. 2. Верхняя часть караган-
ского горизонта выделяется некоторыми исследователями в самостоя-
тельный «варненский» горизонт. 3. Нижняя часть конкского горизонта
иногда выделяется в самостоятельный картвельский горизонт. 4. Мэоти-
ческий ярус, возможно, принадлежит уже к плиоцену. 5. Таманские
слои представляют собой эквивалент акчагыльского яруса Каспийской
зоны (или его части). 6. Тесная связь между фауной гурийского и чау-
динского горизонтов делает сомнительным проведение границы между
плиоценом и плейстоценом по подошве чауды.
(ПбайБоъаслд aam^mtao'06 ^абь^’во
А Э % о з Э о
^0360 808503605700 саЛ^сб^^г <дсо68д2>о6 030573 2)050оо2юЬо£оЭ о ^066^57 зд<>>-
57соз°£)|6 5060080.
I 0003*80 30660577)2700 *6003036000 00305760*6606000
860*836357003060 <3’\j(5° 030573605000606 36006573806 065^006005006. 05060*86'535700
ообзоБоЪЭсло 0305736050006060 500 33^27 с) 6°°^ ^6800503360000 3063000063ЗоЭо
„050305706(^7)60 £05770“ XIX 5ОО XX 60*5337)63000 Эо^БоЪд.
II 0003*80 ^068005033605700 об 80060038360 000Б0Э3760033 0063060*683606
03057360700606 *8360636, 60083570000 066360000 860*8363570060 ojgoo Бо-
ЭобБ 0063060*68000 030573605000606 3033606000306.
III 00030 80000036 оЗ бооофооЗдбоб о^ф’дЗо^зсобфсо^^Зо'дб 060570*66, боо-
8573600 635760363570 обооб оо6зо6о*ЬЭсг-о 3085703.6367)570 63°27Э6070060600030b
(ЭооЭофдбт^о 06703077)0277)60 0305736070060600030b 306603^300633000
Ьд5760363570 6835730006, 80^06^33*80 060*6737)60 2°° *6737)60 бософообдбо)
IV 00030 80873605700 боЗоббо *36007)^6370*060 <063060*683606 (тробофдЬ-
020 7^6080607363606) 030573607006060780.
V 0003*80 зо2рЗсо(ззЗ’352?°о 3^°6оз^)6 0033522330000 0320333&0, 63003360b
ф&£)6о 60273^3^00 БоЗобЬо 3^(332220003360060 300522^)6332>оЬ Planorbidae 500
Viviparidae-b (ззо222з&о5ооо&оЬ ^зЬоЬз^.
VI 0003*80 3066027^327°° 030573^05000^06 йзЭЗзЬоЬ 3600^27380 500 ЭоЬо £оо-
80030503'&£)273&O 006306^3570 ЬоЭдобооЬ Э003270 33<*>£7£)(joob Ьо6з6327оЗ'00&оЬооо6
3300270030^360 506000b 306303570030*80. 2о°2ооЬ(®)£)6зЗ^з2700, 600Э 3b 33С°^?'Э6П°
96 *830827360 8008620060300 66^3270)020 Зоооф£)6о2о, „£5^3660300, 300306(^^570 Q30-
57зЗо2оооЗоЬ“ Ьоюз£) 83357*153.
VII 0003*80 30060570*63Зэд2700 ^066*727 3300270030^36 50600000 ообзобоЪЭдЗоЬ
Ь6з£)27°6 605005006 оЬфоо6о£)27° (330573Зо50ооЗоЬ «до^фзЗо ЗооЗ^)57053006 <3063-
273<?>*8o 500 5ооЬо&^з003^3)27°° °С?Зо оЭ 5330573ао5ооо&оооо 50080030503573<5obo 306g-
800 Зо6оо?>з&оооБ, 6050053 3000065035)00500 оЭ 0063060*68000 065003050’53’5383^0.
VIII 00030 808503605700 ЗобофзоообоЬ 80057^363000 065003050^3057^360 (33^-
5732)0500060605080 8000533606 30680357002)0*80.
IX 0003*80 3050800533Эд2700 500 30060570*1532)^32700 6000Б06 З570005336-
об оо66оз50£)57ооБ 80057^363000 Эо6оо2)6озо 065003050^3057^360 (ззо£7з2>0200о<5оЬ
030^360.
X 0003*80 3066057*35700 03033 8003573632)0 *838663*3570 8^303570003316006 8со-
2^'^363000 66305006630 £(УЗ<зз2>*Эо 08033 ^2>о6 357осо0зб*Эо.
XI 0003*80 0Э033 °°3dC?6o‘lf)6o6ooo 3066057^32700 0^603057’5360 о’^дЪоЬ Mactri-
dae 500 30605^3000606 8го57*з6зз2)о6 060570030’5360 0зо57д&о5осо2)о6 60300060.
229
XII ooog7)o £ОоБо^оЪд&£)£рос> о£)&£)^»о ^оо (_p6(j£)£?o ЗдЛооо^одЬоЬ Ъсо£о-
дбооо оЭсоБофоЬ об£оозо£0£)0£р£)Ло (ggo^pgcbo^ocobob ЬодоооЬо.
XIII 00030 ЭоЗ^одБо^оо ЬЭд^доооЬ ЬдЛЬдЭ^рооБооо oE^oogo^o'Qo^'Q^o (330-
£РЗ?>02ооо5)оЬо£оЭо.
£О0 &СО£Р<пЬ, £о6Ьо£г>£)£рОО БоЭоЛЬо СоЛ^дБсЛЭд 2юЬ оБ£003020£РОР’£)Ло (J3OCP0-
?>о(ооо&оЬ Эсод^дБоооо ЭБо^дЭдоусо^о oScoqoEoooo 30200^339^)0-
Ьоо>зоЬ (оо 9oaQg9^)Qpoo ^о&1)£)£^оЬ соЛ^оБоЪЭооо (ЗЗоорд&о^осо&оЬ зоБЬосз’зоЬ 'Эд-
С?33°С? Эо(пд2>£)£ро Log Лоо со ^oobggEg&o.' БоБддБд&оо, ЛгоЭ Зо^рд ooiocn £роо £О£) Л о
ЭоЬо^роЬ 'ЭдЬ^одсрооо з^роБ^д&д ЭоСсойЛозо (330^3^o^ocobob оооЭоадо^од&^д^д&о
оБсоодооо’зЭооо £0630000632>Ьо ^оо (зЬоодбд&оЬ ЗобсоЬд&Ъд i6cn£co6(j °^3
ЬЗд^рдоооЬ £о6дЗсоЗо.
VARIATION OF ORGANISMS IN THE PAST GEOLOGICAL TIMES
ABSTRACT
The book deals with the problem of the variation of the organic forms
of the geologic time.
Chapt. I discusses some fundamental facts pertaining to the history
of the problem of variation inasmuch as they are important^ from the pa-
leobiological standpoint. There was a „surge" of idealistic conceptions on
the boundary of XIX and XX centuries.
Chapt. II contains a brief discription of the data of the variation of
living forms most essential for the explanation of causes of variation of
the fossil organisms.
Chapt. Ill is devoted to the actuopaleontological analysis of the bio-
topes favourable for the intensive variation of organisms (terrestrial and
subterranean extramarine biotopes as well as marine biotopes with conditions
favouring the variation).
Chapt. IV discusses material relating to the variation of fossil’Protis-
ta (predominantly foraminifers).
Chapt. V is giving an account of the classical investigations of varia-
tion of fossil gastropods of the families Planorbidae’and Viviparidae from the
Neogene lacustrine deposits.
Chapt. VI discusses the problem of tempo of variation with reference
to the total duration of the evolution of the organic world during the geo-
logic time. The outcome of the discussion is that this process can not be
explained on the basis of utterly random variation.
In chapt. VII an analysis is made of the data relative to the histori-
cal changes of body size of organisms within populations of the past times;
such changes are shown to have depended on the environmental conditions
in which individuals were being developed.
Chapt. VIII deals with the individual variation of mollusks that in-
habited Parathetis throughout the Miocene.
Chapt. IX is setting forth the facts of mass individual variation of the
bivalve mollusks during the Pliocene in the Rion bay.
Chapt. X is considering similar phenomena that occured in various
groups of the Pliocene gastropods in the same bay.
In chapt. XI forms of the family Mactridae that inhabited the Akcha-
ghilian basin are discussed from the same point of view.
231
Chapt. XII analyses data on the individual variation in some ammo-
nites of the Jurassic and the Cretaceos.
Chapt. XIII is devoted to the individual variation of the vertebrates.
In the last chapter of the book the bearing of the individual varia-
tion of fossil organisms in the solution of the problems of the geochrono-
logy is discussed. General conclusions are formulated as to the importance
of the variation of ancient organisms for the theory of evolution. It is
stressed that the thorough study of the paleobiological material shows the
dependence of the mass individual variation of the conditions of develop-
ment and life of the individual organisms in both aqueous and terrestrial
environments.’
УКАЗАТЕЛЬ АВТОРОВ
Абель О. 146, 202—204
Аврамова С. 58, 59, 62, 63
Агаев М. Г. 76—78, 219
Али-Заде 5, 184, 185, 219
Али-Заде А. А. 5, 184, 185, 219
Андреварта Г. С. 51—54
Андрусов Н- И. 8, 81, 94, 138, 143,
176—178, 186, 219
Аракелян М. М. 69, 219
Армстронг Т- 53
Арчи 152
Асколи П. 102
Астауров Б. Л. 21, 69, 70, 71, 219
Аштон Э. Г. 205
Ахвледиани Е. Г. 5, 173, 174, 180, 219
Бадзошвили Ц. И. 5, 137, 219
Базилевская Н. А. 22, 219
Бан К. С- 28
Бант А. М. 48
Берч Л- С. 51—54
Беттенштедт Ф. 100—108, 208
Бил Г. Г. 28
Богачев В. В. 119, 219
Болдвин Дж. М. 54
Болтовской Э. 99—101
Бони А- А. 136
Браун А. А. 59, 62
Брен Ж. Л. 48—51
Бринк Р- А. 65, 66
Бринкманн Р. 189
Брусина С. 121
Брэдшоу Дж. С. 100
Буковский Гейза 119—121
Бунур Л. 28
Бэтсон В. 18, 19
Бюсвин Д. 59
Вавилов Н- И. 21—23, 25, 26, 184, 219
Вандель А. 82, 83, 86, 91, 202
Варенцов М. И. 186
Вашков В. И. 60, 219
Вейсман А- 29, 73, 74
Вейгельт И. 204
Венц В. 111, 112, 114, 161
Вермут Г. 149
Висманн Р. 64
Воронцов Н. Н. 22, 220
Вофрей Р. 147
144,
Гаас О- 189
Габуния Л. К. 178, 204, 220
Гайэтт А. 35, 111, 113, 114, 122
Галавач Г. 121, 122
Гантер Дж. 100
Геберер Г- 208
Геккель Э. 13
Георгиева Р. 31, 220
Герасимов Н. П. ПО, 220
Гесслер К. 43, 220
Гильгендорф Ф. 111, 114, 122
Главинич Р. Г. 31, 220
Глембоцкий Я- Л. 32
Глущенко И. Е. 31, 32, 220
Гованитц У. 52
Гольдшмидт Р. 124, 125
Готтшик Ф- 111, 114
Гоукинс Э. Ш. 28
Граберт Б. 104
Грачевский М. М. 97, 220
Гринуэй Дж. 151
Громов А. М. 31, 220
Грэник С. 28, 220
Губарев В- С. 22
Губкин И. М. 186
Давиташвили Л. Ш. 10, 35, 44, 134, 136,
137, 160, 163, 164, 174, 179, 199, 220
Дакке Э. 118
Дальк А. М. 124
Даниелли Дж. 28
Дарвин Ч. 13, 16, 19, 23—27, 30, 38, 67—69,
71, 72, 74—76, 127, 184, 187, 200, 201,
220
Дарлингтон Ч. Д. 71, 72
Дементьев Г. П. 150, 220
Демерец М. 42
233
Дербенева-УхОва В- П. 60, 63, 221
Дергачев Л. Д- 22, 221
Деррент А. 64, 65
Джанелидзе О. И. 101, 221
Джикия Н. Р. 5, 186
Джинкс Дж. 27, 28, 221
Джонс Ф. Г. У. 52
Криппа-Франчески Т. 44, 45, 222
Куммель Б. 136
Кун О. 189
Купер Г. А. 134
Кург Г. 100
Кушнер X. Ф. 32, 68, 69, 222
Куэйл Г. 51
Дин А. Ч. 40, 41, 43, 76
Добролюбова Т. А- 109, ПО, 221 Лаликер С- Г. 99
Долло Л- 15 Ламарк Ж-—Б. 72, 74
Дорст Ж- 192 Латтен Ж. де 84, 85 92
Дубинин Н. П- 17—19, 22, 26, 32 36, 221 Лебедев М. М. 69, 222
Дзвидсон Г. 59
Дэвис В. Д. 42, 70
Жаннель П. 35, 82, 84
Жгенти Е. М. 5, 155—158, 221
Женермон Ж. 44, 45, 221
Завадский К. М. 12
Зеликман А. Л. 26, 221
Зенкевич Л- А. 134, 135, 221
Иагишита Но-бору 31
Иекелиус Э- 182, 183
Ильин А. Я- 21, 23, 221
Иоганнсен В. 17
Иоллос Ф. 44
Кавалли-Сфорца Л. Л. 42
Калинина А. Б. 28, 221
Калломон Дж. К- 189
Карсон Дж. 31
Катрфаж Г. 31
Квалиашвили Г. А. 5
Кертис А. С- Г- 28
Кинг С. 31
Кгрле Г. 202
Китаева О. Н. 69
Китовани Т. Г. 5, 174, 175
Клстз И. 31
Клэн Г- 111, 114
Ковалевский В. О. 13, 16
Кожов М. М. 192, 221
Колесников В. П. 94, 159—164, 186, 221
Колом Г. 1005 139
Коп Э. Д. 25
Коробков И. А. 125, 160, 163, 165, 222
Коссвиг К. 83—93
Коул Р. 28
Коюмджиева Э. 156, 222
Краевой С- Я. 33, 222
Крау Дж. Ф. 58—60
Крашенников В. А. 100, 222
Крейгхед Ф. Г. 53
Кренкель Э. 151 К
Левитский Г. А. 18, 222
Левушкин С. И. 92, 222
Ледерберг Дж. 42, 43
Лелеп Н. 82
Ленин В. И. 15
Леруа П. 31
Линдберг Г. У. 193, 194, 222
Линдгольм В- А. 140
Лобашев М. Е. 30—32
Лолл Р. С. 195—200
Ломинадзе Т. А. 5
Лотси И. П. 17—19
Лоу П. С. 31
Лумис Ф. Б. 134
Людвиг В. 91
Люерс Г. 62
Лютшвагер И. 150
Львов А. 18
Лысенко Т- Д. 29, 32, 33, 37, 38, 67—70,
222
Лэнкфорд Р.. Р. 100
Мазсер К. 65, 66
Майерс Б Г. 99
Майр Э. 52, 60, 61, 124, 126, 130
Майсурадзе Л. С. 101
Майсурян Н. А. 22, 222
Маккардт Г. 28
Мамзин А-'С. 12
Мампория Ф. Д. 31, 222
Маргалеф Р. 192, 204, 205
Маринелли В. 202, 203
Матекин П- В. 92, 222
Мелик-Шахназарова Т. Т. 5
Менделеев Д. И. 21, 22
Мендель Г. 19, 26—28, 32, 71, 127, 206
Мерриам С. Г. 202, 204
Мертенс Р. 149
Мидоуэр 130
Мирский, А. 28
Мичи Д. 71—76
Мичурин И. В. 21, 30, 31, 33
Морган Т. Г. 28, 30, 73
234
-Морозова В. П- 60
Моташ К- 82, 83
Мусхелишвили Л. В. 5, 161, 165, 222
Найдин Д. П. 190
Наливкина А. К. 190, 222
Негели К. 19
Неймайр М. 115—119, 122
Никол Д. 100
Никольм К. Г. 101
Никольский Г. В- 130
Новиков Б- Г. 31, 222
Ноден Ш. 23
Ноинский М. Э. .134, 222
Оцропиридзе О. В. 102, 222
Олсон Е. С. 46
Оньибен Л. 99
Осборн Г- Ф. 18, 35—37, 106, 145—148
Паавер К. Л. 205, 222
Павлов А. П. 119, 222
Павличенко В- 32, 222
Пастор Д- 31
Пидопличко И. Г. 146, 222
Пикте А. 52
Пири Н. 18, 222
Плате Л. 111
Покорний В. 100
Полынин В. М. 21, 223
Полянский В- И. 21
Поспишилова Д. 31
Проссер Ч- Л. 51
Пузанов И. И- 95, 96, 223
Равен X. 28
Радованович М. 192. 193
Райки Ш. 77—79
Райт Сьюэлл 193—200
Ратиани Р. В. 5
Раузер-Черноусова Д. М. 138, 223
Рейд Дж. 60, 62
Реньо М. 28
Рихтер Р. 80
Родда П- Ю. 138
Росс Г. 46
Рубинштейн М- М- 212
Рубцов И. А. 56, 57, 223
Рухина Е. В. 5
Рушелли М. 102
Рыжков В. Л. 27—29, 223
Сабатье Ж. 31
Саид Р. 100
'Саидова X. М. 100, 223
Сакки Ч. Ф- 147, 192, 193
Салтыков-Щедрин М. Е- 16
Самохвалова Г- В. 54, 223
Северцов А. Н. 37
Сейджер Р. 28
Сенинский К. 168, 169, 176, 223
Сербова С. 58, 59, 62, 63, 223
Симпсон Дж. Г. 37, 46, 54, 124, 208, 223
Синай Дж. 42
Синцов И. Ф. 138
Смирнов Е. С. 55, 223
Соннеборн Т- М. 44
Станкович С. 192, 223
Строганов Н. С. 130, 223
Суиннертон Г. Г. 18
Суханова К. М. 45, 223
Тайсон Г. 65
Тактакишвили И. Г. 167, 168, 183, 223
Талиев Д. Н. 192, 223
Таш П. 135
Тентан А- 189
Тимирязев К. А- 19, 21, 30, 37
Тодд Р. 100
Торн лей М. Дж- 42
Торп У. Г. 52—54
Убчш Л. 124
Уваров Б. П. 51
Удольская Н. Л. 31, 223
Уилки Д. 28
Уильямс Дж. С. 46
Уинтерингэм Ф. П- В. 61, 62
Уоддингтон Ч. Г. 31, 45—48, 61, 130
Уоллес А. 19
Фаж Л. 84
Фишер У. Л. 138
Фледжер Ф- Б- 100 f
Фрайер Дж- 192
Фридберг В. 159, 160
Фризе Г. 86, 100
Фурсенко А. В. 100, 223
Фюрон Р. 127
Хагеманн Р. 28
Хайкин Г. Р. 65
Хилл Дж. 65, 66
Химшиашвили Н. Г. 5
Хиншелвуд К. 39—43, 74, 76, 223
Холдгейт М. У. 192
Холм Р- У. 46
Цедлер 105
Циглер Б. 189, 190
235
Челидзе Г. Ф. 168, 170, 223
Чилингарян А. А. 31, 223
Шаймер Г. В. 134
Шапошников Г. X. 55, 224
Шекспир У. 12
Шефер В. 11
Шиндевольф О. 106
Шотц Ф. 28
Шпет Г. И- 129, 224
Штребель О- 92
Эберзин А. Г. 167, 172—174, 224
Экизашвили Ц. И. 5
Эренбург К. Е. 203
Эритье П. 28
Эрлих П. Р. 46, 224
Юдкин Дж. 42
Юлэм С. М. 128—131
Яблоков А. В- 205, 224
УКАЗАТЕЛЬ ТЕРМИНОВ
Ааленский ярус 211 Баррем 1СЗ, 210
Аберрантные биотопы 81—83, 184, 201, 208 Ьат 107, 2 Л
Абиотическая среда 81, 97, 111 Башк.\ скин яр\с 211
Абсолютная геологическая хронология 209 Бентос 103
Абсолютно внутренние факторы эволю-Биология 10, 12
ции 80
Автогамия 45
Автогамный гермафродит 49
Автогенез 17, 18
Агробиология 37
Адаптационизм 40
Адаптация 41, 42, 44, 54, 55, 63, 142
Адаптивная радиация 140
Адаптивная сомация 54
«Адекватная изменчивость» 78
Ажгильский ярус 212
Акклиматизация 50
Аккомодация 54
Актуопалеонтология 11, 80
Акчагыл 184—186
Алданский ярус 212
Аллель 65, 102
Аллестэтический фактор 195, 199—201
Аллопатрические предки 183
Альб 104, 210
Аналогическая (аналогичная) изменчи-
вость 21, 23—26, 178, 184, 187
Аналогичные ряды изменчивости 26
Анофтальмия 91, 92
«Аь тагонистические» школы 29
Антибиотики 39, 41—43
Антигенетическая интерпретация 79
Антидарвинизм 13, 19, 83
Аьтропический фактор 146
Антропоген 50
Антропогенез 37
Апосематическая окраска 201
Апт 104, 210
Аренигский ярус 212
Ар исто генез 36
Артинский ярус 211
Ассельский ярус 211
Байосский ярус 211
Биометрия 204
Биопризнак 35
Биос 131
Биоспеология (биоспелеология) 83, 92, 94
Биостратиграфия 132
Биотоп 81, 92, 94, 97, 139, 203, 205, 208
Биофизика 7
Биохимические процессы 64
Биохимия 7
Биоценоз 192
Биоценотические отношения 97, 203, 205
Ботаника 6
Валанжинский ярус 210
Варненский горизонт 155—157, 214
Вегетативная гибридизация 30—33
Венлокский ярус 212
Взрывное видообразование 191
Вигорная толерантность 59
Вид 23, 24, 31, 34, 44, 52, 90, 95, 102, 103,
108, 111, 186, 190, 191, 193, 194, 207
Видообразование 10, 19, 34, 84, 95, 106,
114, 116, 126, 136, 144, 167, 173, 190
Визейский ярус 211
Внутренняя тенденция развития 15
Внутривидовая изменчивость ПО, 153—
—155, 165, 189, 209
Внутривидовое скрещивание 68
Внутривидовой таксон 156
Внутривидовые наследственные измене-
ния 5
Внутрипопуляционная изменчивость 4, 101
Гамета 71
«Геммулы» 75
Ген 17—19, 33, 35, 36, 40, 43, 56. 58-60,
65, 75, 76, 78, 193, 212*
Ген полудоминантный 60
Ген резистентности 58, 60
«Ген типа развития* 77
237
Генезиология 35 Дивергенция 140, 146
Генетика 6, 8, 10, 17, 20, 22, 27—30, 35—Дискретная наследственность 71, 126, 206
38, 42, 44, 71, 76, 77, 83, 84, 90, 92—91, Диэлдрин 60, 61, 62
96, 101 —103,’ 106, 126, 205—207 “ '
Генетика популяций 102
Генетико-автоматические процессы
194 '
«Генетическая ассимиляция» 31, 45, 46,
Генетическая изменчивость 93
«Генетическая информация» 131, 206
Генетическая теория 6
«Генетический дрифт» 193
«Генетический код» 124
«Генетическое приспособление» 48
Геном 43
Генотип 18, 32, 46, 53, 207
Генофонд 194
Геологическое время 130, 209
Геология 145, 207
Геохронология 101, 108, 132, 156, 206, 207
Гермафродит 49
Геттангский ярус 211
Гетерозиготность 60
Гетерозис 67, 69, 92
Гетеротрофный протист 44
Гжельский ярус 211
Гибридизация 19, 30
«Гибридная мощность» 67, 70
«Гибридный вигор» 70
Гигантизм 132, 139, 140, 144, 145,
150, 152
Гидрологический режим 9
Голоцен 205, 210
Гомологическая изменчивость
Гомологические ряды 22, 23,
Горийский горизонт 214
Готерив 103, 105, 210
«Гроссмутация» 34
Гурий (гурийский
171, 173, 174, 181,
184
25, 26
горизонт)
182, 184, 214
153,
19, 22, 33, 34,
Длительная модификация 10, 31, 44, 45,
66, 102, 104, 106, 212
193 ДНК 32
Доггер 211
48 Домерский ярус 211
Доместикация 23, 25, 67, 72
Донор 32
Дуабская толща 140, 170
Дуалистическая философия
74
«Единичный признак» 35
Естественный отбор 10, 17
53, 59, 60, 85, 86, 93, 95^
131, 144,
194, 199,
151, 183,
201 203, 205, 209
> ’ ’
Жединский
Живетский
Жигулевский ярус 211
Жизненность' 67—70
Жизнеспособность 68
ярус 212
ярус 212
40,
106,
188,
44, 46, 47,
125,
191,
126,
193>
аналогической изменчивости 23,
Вавилова 21
гомологических рядов 21
наследственности 17
Закон
Закон
Закон
149 3аКОН
' «Зародышевая плазма» 71, 72, 75
Зародышевый путь 75
Зигота 68
26.
вулканические 151
гавайско-стромболианского*
63, 64,
Дарвинизм 13, 15,
118
«Дарвиновский отбор» 62
Датский ярус 210
«Двухпутная теория наследственности»
72, 74, 75
Девон 134, 190, 212
Дегенерация глаз 85
Дегенерация крыльев 85
Дезадаптация 45
Депигментация 84, 91
Детерминанты 28
Детоксифицирующий механизм 59, 63
134, 136, 138
6, 8, 12,
133, 144, 149,
171—174,
188—193, 197—200,
22, 93—95,
151, 153,
176, 179, 181,
202—209,
расы 66
Идеализм 13, .16—18, 35
Извержения
Извержения
типа 151
166, Измельчание
Изменчивость
119, 132,
167, 168,
185, 186,
212
Инбредные
Инбридинг 62, 69
Индский ярус 211
Индуцированные изменения 66
Инсектицид 58—60, 62, 63, 206
Интенсификация изменчивости 97
Инфантилизм 146
Инцухт 68
Историческая геология 5, 206
Ихтиология 193
Ихтиофауна 194
114,
16U
18а,
238
Казанский ярус 211
Кайнозой 129, 131, 140, 210
Каменноугольная система 211
Кампан 210
«Канализация» 45
Караган 141, 153—157, 214
Карадокский ярус 212
Кариологические процессы 212
Карликовость 100, 133—138, 144—147
Карликовые слоны 148 *
«Карликовые фауны» 99, 134
Каротиноид 87
Картвельский горизонт 214
Касимовский ярус 211
«Каспий» 8, 183, 184
Каяльский ярус 211
Келловейский ярус 211
Кембрий 129, 212
Кибернетика 203
Кимериджский ярус 211
Киммерий 9, 140, 142, 143, 153, 166—163,
170, 171, 173, 177—182, 184, 214
Класс 34
Кобленцкий ярус 212
Кондиционирование 66
Конкский горизонт 136, 214
Коньякский ярус 210
Конъюгация 45
Кормность 96
Кортикальная наследственность 28
Коцахурский горизонт 214
Коэффициент карликовости 137
Кризис в эволюции 36
Криптический признак 201
«Кроссвейнлесс» 46
Кунгурский ярус 211
Куяльник 9, 153, 171, 172, 178, 179, 181,
182, 184, 214
Лагуна 96
«Ламаркизм» 54—56, 63, 66, 84, 93
Ламарковская наследственность 42, 47
Ландейльский ярус 212
Ландоверский ярус 212
Лейас 211
Ленский ярус 212
Лиман 95, 96
Линнеон 23
Локальный реликт 137
Локус 65
Лотарингский ярус 211
Лудловский ярус 212
Лютетский ярус ’ 155
Маастрихский ярус 210
Макромутация 34, 124
Макронуклеарная гипотеза 44, 45
Макронуклеус 45
Малорослость 99
Мальм 211
Массовая индивидуальная изменчивость*
166, 187
Массовая направленная изменчивость 97
Материализм 13, 15, 16, 19, 34, 36
«Материализм и эмпириокритицизм» 15
«Махистские позиции» 17
Медицинская энтомология 64
Межледниковые эпохи 203
Межпородное скрещивание 69
Межродовая форма 109
Межсемейственная форма 109
Мезозой 129, 210, 211
Мел 188, 190, 191, 210
Меланины 87
Менделизм 55, 36, 86
Менделизм ранний 36, 38
Местонахождение 190
Метаболизм 86, 100
Метастабильные изменения 65
Метафизика 18, 69
Метафизические идеи 17
Методология эволюционизма 16, 206
Механизм токсичности 57
Механицизм 17
Миграция 119
Микропалеонтология 208
Микроэволюция 192, 193
Миоцен 101, 102, 136, 142, 154, 165, 184,.
210, 214
Митридатизм 58
Мичуринизм 6, 13, 29, 33, 63, 68, 71
«Многообещающее чудовище» 125
Модификация 10, 11, 45, 63, 84, 93, 101 —
103, 106, 108, 132, 137, 138, 145
Молекулярная биология 7, 17, 209
Молекулярная генетика 76
Монофакториальность 59
Морфология 7, 10, 22, 51, 55, 139
Морфо-физиолого-этология 7, 10
Московский отдел 211
Мультифакториальность 59
«Мутагенное действие» 61, 76
Мутагены 20, 61, 77
Мутант 43, 61, 66, 194, 207
Мутационизм 40, 86, 90
Мутационная теория 43
Мутационное давление 87, 92
239”
Мутационный процесс 77 Палеозой 211
Мутация 40, 41, 43, 47, 57, 78, 85, 86, 101, Палеолит 203
102, 104, 108, 116, 124, 125, 128, 141, Палеонтология 145
143, 193, 208
Мэотический ярус 137, 214
Надвидовой таксон 156
Намюрский ярус 211
Нанизм 144, 147
Направленная эволюция 193
Направленное мутирование 8, 194
Наследование приобретенных свойств
Палеоцен 210
Пангенезиса гипотеза 74
Параллелизм в филогенетическом разви-
тии 184, 185
Параллелизм изменчивости 26
Параллельная эволюция 146
Параллельные изменения 25
Парамутация 65, 66
2 Паратип 196
Наследственная внутривидовая измени и* Партеногенез 55
' вость 10 11 30 Перекрестная резистентность 59
Наследственная изменчивость 22, 71, 193 Перекрестное оплодотворение 72
«Наследственная информация» 6
Наследственность 12, 48, 75, 206, 208
Наследственные изменения 11
«Негенетическая адаптация» 42
Необиология 6, 109, 209
Неоген 50, 111, 122, 176, 210
Неодарвинизм 86, 87, 90, 92
Неоламаркизм 35
Неоменделизм 29, 39, 43, 47, 76, 126
Нехромосомная наследственность 27 1
32, 33
Перекрестное опыление 67
Перигамическая структура 200
Периодическая система элементов 21
Пермская система 211
Пещерная популяция 88
Пигмей-гиппопотам 146
Пигмей-слон 146
Пигментация 87
Плазмаген 75
?8, Плейотропия 20, 87, 88, 126
Плейстоцен 147. 149, 151, 174, 204, 210
Ниша экологическая 53
Плинсбахский ярус 211
«Нормальные» биотопы 9 Плиоцен 8, 102, 114, 144, 154, 166, 167,
174, 176, 177, 179, 181—183, 209, 210, 214
Обратимая наследственная реакция 55 Плодовитость 50
Объективная случайность 77 «Плоская эволюция» 126, 191
«Однопутная теория наследственности» Поведенческая резистентность 59
73. 74 Подвид 111, 194, 205, 207
«Озимизация» 78, 79
Оксфордский ярус 211
Оленекский ярус 211
Олигоцен 99, 210
Подрод 190, 191
Позднетретичное время 8
Пойкилотермные животные 129
Полигения 20
Ольфактометр 52
Онтогенетическая реакция 137
Онтогения 35
Организмальная биология 7
«Органический отбор» 54
Ордовик 129, 130, 212
Оренбургский ярус 211
Ортогенез 37, 188
Ортогенетическая дегенерация видов
Ортоламаркизм 118
Осадконакопление 4, 132, 212
Остеометрия 205
Относительная геохронология 5
Отряд 34
«Ошибка» в наследственности 48,
127, 131
Палеобиология 5, 7, 8, 10—13, 16,
133, 144, 151 161, 207, 208
’ ’ , ’
Полиморфизм 22
Полиморфный вид 186
Полифакториальность 59
Полносоленые бассейны 8
Полносоленые моря 154, 161, 183, 191
Половой отбор 200, 201, 209
Понт 143, 153, 172, 177, 182, 214
Популяция 10, 11, 65, 75, 89, 98, 102, 106,
88 НО, 132, 133, 167, 191, 203,’ 205
Порода 23
Постадаптация 58, 59, 62
Постнеодарвинизм 36—38, 46, 48, 54, 59,
61, 85, 90, 92, 93, 124, 126, 129, 131, 209
12| «Преадаптация» 43, 58, 62, 86
Прививочный (вегетативный) гибрид 33
Прогрессивная эволюция 91
101, Пропамидин 41
Протерозой 128
240
Стрептомицин 43
Сублетальная доза 62
Сублиторальная зона 99
Субфоссильные млекопитающие 205
Сульфаниламид 41
Таксономический анализ 147
Таманские слои 185, 214
Тарханский горизонт 214
Татарский ярус 211
Темпы видообразования 81
«Теория благорастворения воздухов» 13,
16
Теория информации 47
Теория флогистона 40
Терморезистентность 48
Тоарский ярус 211
Трансгрессия 194
Трансформизм 109
Тремадокский ярус 212
Третичная система 101, 210
Триас 136, 211
Троглобионт .84—86, 91—94, 202
Трофический уровень 205
Турнейский ярус 211 . .. и:
Туронский ярус 210
«Угнетенная фауна» 132
15^ Унифакториальность 59
Устойчивость 56, 57, 59, 60
Фаменский ярус 212
Фанерозой 128
Фенокопня 66
«Феномен Ламарка» 78
Фенотип 7, 18, 19, 46, 65, 77, 78, 90, 208
Фенотип «кроссвейнлесс» 46
Фенотипические уклонения 17
Физиологическая резистентность 59
43, физиологические особенности 7
Физиологический гетерозис 69, 70
Физиология 10, 22, 139
Филогенез 109, 114, 115, 125, 131, 133,
140, 188
Филогенетика 8
Филогенетическая ветвь 11, 116, 142
Филогенетическая радиация 144
Филогенетическая цепь 108, 190
Филогенетические построения 116
103, Филогенетическое развитие 7, 104, 204
Филогенетическое увеличение тела 149
Профлавин 41
Прохорез 171, 190
Радиация филогенетическая 185
Раса 52, 66
Расообразование 84
Расшатанная наследственность 97, 109
Регрессивная форма 91
Регрессивная эволюция 83, 91
Регрессивный признак 88—90
Регрессия моря 193, 194
Редукция глаза 88
Резистентность 39—43, 57, 59, 60—64, 206
Репродукционный барьер 86
Репродукционный цикл 49
Ретроградация 18
Рецептор 83
Реципиенты 32
Рисс-вюрм 203
Род 24, 31, 34, 44, 177, 189, 194
Сакараульский горизонт 214
Сакмарский ярус 110, 211
Сальтационизм 124, 125, 190, 208
Самооплодотворение 49, 72
Самоопылители 32
Сантон 103, 210
Сармат 101, 136, 137, 141 — 143, 153,
159, 161 — 165, 184, 185, 214
Сверхорганизмы 18, 19
Сельскохозяйственная энтомология 64
Сематический облик 201
Семейство 23
Сеноманский ярус 210
Сенон 210
Силур 129, 134, 212
Синемюрский ярус 211
«Синтетическая теория эволюции» 37,
54, 61, 64, 79, 85, 91, 128, 130, 190
Систематика 89, 161, 202
Системная мутация 124
Скачок в развитии 204, 208
Скифский ярус 211
Слитность 71, 206
«Совпадающий отбор» 54
Солоноватоводный бассейн 182
Сома 72, 75
«Спонтанная мутация» 57, 85, 101,
127
«Спонтанные наследственные
46, 84, 92, 102, 131
Сравнительная анатомия 6
Стратиграфия 5, И, 118,. 132,
206—208, 212 ’
16. Л. Ш. Давиташвили
изменения» Филогения 35, 113, 118, 131, 144, 147, 150,
154, 158, 162, 173, 179, 188, 200, 211
Финализм 92
161, 190, Фитофагическое насекомое 51, 54
Фитофаги я 203, 204
241
Флогистонизация 79
Флогистонизированный язык 79
Франский ярус 212
Фреатический водный горизонт 82
Фреатобиологця 82
Функция 7
Хлорамфеникол 41
Хромосома 28, 29, 33, 63, 65, 66, 75, 206
Хромосомная наследственность 33, 74
«Хромосомная теория» 71, 73
Целостность организма 127
Ценоз 192
Цитоплазма 27, 28, 30, 45, 65
Цитоплазматическая наследственность
29, 30, 44, 45
Эволюционная палеонтология' 13, 16
Эволюционное учение 19, 21, 51
Эволюция 5, 6, 10, 15, 17—19, 64, 67, 84,
85, 91, 95, 97, 104, 106, 128, 129, 161, 168,
179, 181, 188—190, 192, 203, 204, 208, 209
Эйфельский ярус 212
«Экзотические» формы 176, 178
Экогенетическая экспансия 140.
Экологическая ниша 164
Экологические условия 150
Экология 6
Эндемизм 181, 121, 164, 166—168. 171,
192, 193
Энзимная адаптация 61
Эобионты 19
27, Эоцен 103, 210
Эпигейные формы 83, 86, 93
Эпоксигексахлоротетрациклодецен 61
Эритроциты 32
Этология 139, 144
«Эффект Болдуина» 54
«Эффект Сьюэлла Райта» 193
Юра 188—191, 211
Ядерный материал 65
Ядро 28, 32, 45, 65, 66
Чауда 153, 166, 171, 173, 174, 181, 182. 184,
214
Четвертичный период 8, 122, 176, 193, 204, Этологические особенности 7
210
Чокрак 136, 141, 155, 214
Чувствительность к ядам 60, 61
Штамм 40, 41, 44 60, 116
Эволюционная биология 36
Эволюционная гипотеза 17
указатель латинских названий
Abra tellinoides 136
Aerobacter aerogenes 41
Alvania maeotica 136
A. pseudalvania 136
Alveinus 154
Amphimelania aff. gajii 181
Anomalopteryx 152
Anopheles 58, 61
A. gambiae 60, 62
Anoptichthys 86, 89
A. antrobius 88
A. iordani 88
Anthriscus nemorosa 55, 56
Antilocarpa americana 200
Anti lope bezoartica 200
Aonidiella aurantii 58
Area 136
A. miopliocenica 136
Archelix punctata 139
Archidiskodon meridional is 204
Arcifera 25
Arctocyoniade 204
Asellus 88, 93
A. aquaticus 88, 89
A. aquaticus cavernicolus 85
Aster omphalus aspersus 139
Astrea 160
A. (Bolma) rugosa 160
Astyanax 89, 94
A. mexicanus 88, 90
Athleta 138
Avimactra acutecarinata 185
A. karabugasica 186
A. ossoskovi 186
A. subcaspia 186
A. veniukovi 186
Barbotella 154, 158—161
B. grossocostata 159, 160
B. hornesi 159, 160
B. intermedia 159—161
B. laevigatoomaliusii 159, 160
B. omaliusii 159, 160
Bittium reticulatum 136
Bolma meinardi 159
B. perangulata 159
Brassica 24
Brusinaella 177
B. petasata 177
Bunolophodon 204
B. arvernensis 204
B. angustiadens 204
B. longirostris 204
Caenorhabditis 48
C. elegans 48—50
Calandra opyzae 52
Callistoma 159
C. podolicum 161
C. podolicum var. 159, 160
C. sulcatopodolica 161
Caninia okensis 109
Cardiidae 9
Cardium 95, 135, 136, 143, 184
C. aviculare 185
C. barboti 185
C. desperatum 185
C. edule 138
C. ingratum 185
C. inf latum 185
C. laevigatoloweni 143, 135
C. lithopodolicum 143
C. loveni 143
C. maeoticum 136
243
C. (Cerastoderma) mithridatis 136
C. mithridatis ukrainicum 136
C. plicatofittoni 185
C. plicatum 185
C. praeplicatum 185
C. repartition 143
C. ruthenicum 143
C. sarmaticum 143
C. vindobonense 185
Cercopithecus althiops satacus 205
Cerambycidae 53
Cerithium maeoticum 137
C. rubiginosum 136
Chaerophyllum bultosum 55, 56
C. maculatum 55, 56
Cartoconcha postcimmeria 170
Chasmosaurus 198
Chelonia 149
Cibicides lobatulus Ю1
Clypeolum 176
Colias philodice 52
Congeria 170
C. abchasica 168, 169
C. acuta 168, 170
C. caucasica 144, 168—170
C. caucasica var. perpatula 168, 170
C. caucasica var. subcaucasica 170
C. digitifera 167
C. ebersini 168
C. gabuniai 168, 170
C. mirabilis 168, 169
C. mytiliformes 143
C. postsubcarinata 168, 170
C. rhomboidea 144, 167
C. rumana 167
C. subcarinata 168
C. triangulares 143
C. turgidopsis 168
Condylarthra 204
Cory thesaurus 196
C. casuarius 196, 197
C. excavatus 196—198
C. intermedins 196, 197, 199, 200
Cosmoceras 189
C. castor 139
C. jason 189
244
C. pollux 189
C. (Zugocosmoceras) medea 189
C. (Zugocosmoceras jason 189
Cosmoceratidae 189
Cottoidei 192
Crassadacna crassatellata 168
Creniceras dentatum 189
Cryptomactra 185
Cuciirbita 24
Culex 58, 61
C. fatigans 60
Cydia pomonella 51
Daphnia longispina 48.
Dibunopyllum bipartitum 109
,,Didacna“ caeleno 144, 170—172
D. (Crassadacna) crassatellata 144
D. crassatellata catephes 134
D. gurieffi 172
D. medeae 170, 172
D. multistriata 144, 172
D. sulcatina sibsulcatina 172
Didacnomya phasiaca 170
Digressodacna 171, 174
D. digressa 144, 174
Dinornis 152
Diphystrotion 110
Discrobis 100
Discorinopsis aguayoi 100
Dosinia 136
D maeotica 136, 137
Dreissena 119
D. cyclorhampha 169
D. dilatata 169
D. pseudomirabilis 169
D. semilunaris 169
D. obliqua 169
Dreissenomya aperta 143
Drosophila melanogaster 46, 52, 62
Dysaphis anthrisci maikopica 55, 56
Elephas 147
E. (Palaeoloxodon) antiquus 147
E. primigenius 204
E. trogontherii 204
Elphidium 100
E. crispum 99
Elona 140
Emeus 152
Ephestia kiihniella 52
Ervilia 142
E. concinna 141, 142
E. dissita 141, 142
E. minuta 136
E. praepodolica 141
E. pusilia 142
E. trigonula 142
Euryapteryx 152
Euxinomargarya 179
E. mandarinica 179
Formisternia 25
Frondicularia simplicissima 105
Gaudryina dividens 104
Genetta genetta balearica 205
Gibbula maeotica 136
Gipsina 101
Gyraulus 113
G. crescens 112
G. costatus П2
G. denudatus 112
G. depressus 112
G. elegans 112
G. kleini 112
G. kraussi 112
G. major 112
G. oxy stoma 112
G. planorbiformis 112
G. revertens 112
G. sulcatus 112
G. steinheimensis 112
G. steinheimensis involutes 112
G. subhemistoma 112
G. supremus 112
G. tenuis 112
G. trochiformis 112
G. turbiniformts 112
Hechtina antiqua 103
Helicidae 140
Heterostegina texana 99
Hydrobia 119
Hyponomeuta padella 52
Imitoceras grahamense 135
Kishinewia 154, 158, 163, 164
K. bessarabica 164
Lacerta melisellensis lissana 193
L. sicula ragusae 193
Lagena hauteriviana 106
Lambeosaurus clavinitalis 195
L. lambei 195
L. magnicristatus 196
Lasiocampa quercus 52
Lenticulina (Planularia) beieriana 107
Limnaea 119
Lithostrotion 109
Lonsdaleia 109
Loripes pseudoniveus 136
Lutetia 154
L. umbonata 155
Lutraria 185
L. andrussovi 185
Lygus pabulinus 52
Lymnaeidae 183
Macoma 135
M. baltica 135
Mactra 136, 142
M. basteroti 142
M. eichwaldi eichwaldi 142
M. eichwaldi buglovensis 142
M. superstes 136
M. vitaliana 142
Mananopsis 178
Margarya margeriana 116
M. melanoides 180
Martes martes latinorum 205
Magalapteryx 152
Meganinnia 177
M. corrugata 177, 178
M. shio 177, 178
Melania 119
M. andrussowi 181
Melanopsis 116, 119
M. acuminata 181
M. nobilis 181
Meliphora grisella 52
Modiola 136
Modiolus minor 136, 1-37
Moquicardium guriense 167
M. moquicum 166, 167
245
M. ozurgeticum 167
М. praemoquicum 167
М. privum 167
М. submoquicum 167
Musca 61
M. domestica 58, 60, 63
Mya 135
Myotragus baleoricus 204
Mytilaster 95
Mytilus 95
M. edulis 135
Nassa 136
N. andrussovi 136
N, krokosi 136
N, retowskii 136
Nassidae 165
Nemeritis canescens 52
Neomysis circumflexus 55
Neopoma 202
N. magister 202
N. pensylvanica 202
Neritaea 176
Neritina 119, 176
N. (Naritaea) colchica 176, 178
N. vespertina 178
Neritonyx 177
Nicotiana rustica 65, 66
Ninnia 177
Nonion 100
Oraphocardium oraphense 166
Orionastraea 110
Ostrea sp. 136
Oxystele orientalis 159
Pachydacna 168
Pachyornis 152
Palaeoloxodon 147
P. antiquus 147, 149
P. antiquus falconeri 148, 149
P. antiquus melitensis 148, 149
P. antiquus mnaindriae 148, 149
P. creticus 147
P. Cypriotes 147
P. falconeri 147, 149
P. lamarmorae 147
P. melitensis 147
P. (Elephas) mnaindriae 147
Paphia (Tapes) 136, 157
P. (Venerupis?) abichi 136
P. curta 136
P. gregaria 141
P. modesta 141
P. vitaliana 141
Phasianella 163
Rerophaps solitarius 150
Plagiodacna carinata 143
P. modiolaris 173
Planorbis (Gyraulus) multiformis 114:
Plesiocoris rugicollis 52
Pleuronectes flesus luscus 96
Pomatias 140
Porosononion 101
Potamides djsjunctus 137
P. (Pir.) disjunctoides 136
P. (Pirenella) disjuncum 136, 137
P. maeoticum 136.
P. novorossicum 136, 137
Prionocyclus wyomingensis 189
Prionopleura priniopleura aeetae 134
Prionotropis woolgari 189
Prosodacna 167
P. callopistes 167
P. leptopsamatha 167
P. longiuscula 167
P. longiuscula var. 134
P. macrodon 144
Provalensiennesia 154
Pselaphidae 82
Pyrgula 119
Quinqueloculina 101
Q. boueana 101
Raphus cucullatus ( = Didus ineptus)
150
R. solitarius ( = Didus borbonicus) 150
Replidacna 184
Robulus 100
Rotifera 74
Saisetia oleae 58
Savanella 154, 156
S. andrussovi 158
Sigmoilina tenuis 101
Sinzowia 154, 158, 163, 164
246
5. striatotuberculata 163
Spaniodontella 154, 156
S. andrussovi 154, 156
S. intermedia 155, 156
S. minima 155
S. nitida 141
S. pulchella 155, 157
S. tapesoides 157
Sphenia cimmeria 136
Spiroplectinata lata 104
Squalus acanthias 130
Stenodacna angusticostata 173, 174
Strombus butonius 138
Stygicola dentatus (Brotulidae) 88
Suchumica 179, 182
S. gracilis 179, 180
Tapes 136
Testudo chilensis 149
T. elegans 149
T. elephantopus 149
T. elephantopus abingdonii 149
T. elephantopus elephantopus 149
T. elephantopus ephippium 149
T. elephantopus galapagoensis 149
T. gigantea 149
T. gigantea daudinii 149
T. gigantea elephatina 149
Tornatina minima 136
Trochidae 159, 163
Trochus 136, 159
T. armavirensis 162
T. beaumontii 162, 163
T. caucasicus 161 —163
T. cordierianus 162
T. hommairei 161 — 163
T. insperatus 162, 163
T. marginatosinzowi 162, 163
T. omaliusii 159—161, 163
T. podolicoformis 162, 163
T. (Callistoma) podolicus 159, 161 — 163
T. podolicus var. laevigatopodolicus
161, 163
T. podolicus var, sulcatopodolicus 161
T. praearmavirensis 162
T. pseudohommairei 162, 163
T. sarmates 162
T. sulcatopodolicus 162, 163
T. woronzowii 164
Troglolestes sokolovi 92
Tschaudia 144, 171, 174, 175
Turbinidae 160
Turbo 160, 161
T. (Barbotella) hoernesi 161
T. mammilaris 159
Turritella 164
T. terebralis 164, 165
Thysanophyllum 110
Unio 119
Ursus deningeri 204
U. spelaeus 204
Valencienniidae 183
Valenciennius 154
Viviparus 114—117, 119, 120, 122,
179
V. achatinoides 115
V. acramiticus 121
V. altecarinatus 115
V. altus П5, 118
V. ambiguus 115, 117
V. arthriticus 115
V. avelianus 115
V. bifarcinatus 115
V. brusinai 115
V. clathratus 119—121
V. clathratus var.
V. clathratus var. camirensis 120
V. coa 115
V. cyrtomaphorus 118, 121, 122
V. dezmanianus 115, 179
V. eburneus 115
V. (Tulotoma) forbesi 115, 119, 121
V. fuchsi П5
V. gorceiti 119
V. gracilis 122
V. grandis 115, 118
V. herbichi 115
V. hippocratis 115
Viviparus hornesi 115
V. kurdensis 121
247
V. lenzi 115, 118
V. lignitarum 115, 117
V. leiostracus 115
V. margerianus 116
V. mearnsi 179, 181
V. melanthopsis 115
V. mojsisovicsi 115
V. munieri П9
V. neumayri 115, 179
V. nothus 115, 117—119
V. oncophorus 115
V. ornatus 115, 118
V. ovulum П5
V. pannonicus 115
V. ,,pauli“ 115
V. ,,pilari“ П5
V. pseudovukotinovici 182
V. rhodiensis 120, 121
V. ,,rudis“ П5
V. sadleri 115, 118, 121, 122, 179
V. sturi 115, 117, 118
V. spurius 115, 117
V. stricturatus 115, 119
V. ,,strossmayeri“ 115
V. suessi 115
V. tournoueri П9
V. ,,viquesneli“ 115
Viviparus vukotinovici 115, 182
V. wolfi И8
V. zelebori 115
Zonitidae 140
ТАБЛИЦЫ
ОБЪЯСНЕНИЕ ТАБЛИЦ
Табл. 1. Морфологические варианты Cibicides lobatus. По Нигольну из Болтовского.
Табл. II. Некоторые формы Gyraulus trochiformis из Штейнгейма. По Венцу.
Табл. III. Некоторые килеватые формы Viviparus из палюдиновых слоев Славонии.
1 и 2 — Viv. dezmannianus, 3 и 4—форма, переходная от V. defmannianus к Viv.
zelebori, 5 и 6 — переход к Viv. zelebori, 7 и 8 —форма очень близкая к Viv.
zelebori, 9 и 10 — форма Viv. zelebori с нижним килем, еще лишенным бугор-
ков, Ни 12—типичный Viv. zelebori, 13—18—Viv. dezmannianus, 19 и 20 — пе,
реход от Viv. dezmannianus к Viv. altecarinata, 21 и 22 —Viv. altecarinata,
23—24—Viv. arthriticus. По Неймайру.
Табл. IV. Изменчивость форм группы Strombus bubonius. 1 — Strombus aff. bubonius
Lamk. А1иоцен Испании, 2—Str. coronatus Depr. Плиоцен, 3 и 4—Str. bubonius
Lamk., форма из тирренского яруса II; 3 — крупная форма, 4 —малая форма.
5 —Str. bubonius из тирренского яруса II—карликовая форма, 6 — Str. bubo-
nius Lamk., современная, малая форма. Острова Зеленого Мыса. 7 — Str. bub-
nius Lamk., современная. Острова Зеленого Мыса. Крупная форма. Все изоб-
ражения — в 2/3 натуральной величины. По Л. Императори.
Табл. V. Изменчивость и развитие в направлении Savanella. 1, 2, 3 — Span, pulchella
opisthodon. Поверхность покрыта довольно грубыми концентрическими ребрами.
В самых низах варненского горизонта попадаются особи, гладкие в начальной
стадии онтогенеза раковины. Встречаются в верхней части собственно карагана
(непосредственно ниже варненских слоев) и в варненских слЪях.
4—19 — Savanella andrussovi. Поверхность покрыта типичными для этого рода
грубыми высокими ребрами с широкими межреберными желобками. Варненские
слои Западной Грузии. По Жгенти.
Табл. VI. «Didacna» medeae Dav. Из куяльницких отложений Гурии. На фиг. 1—14—
различные образцы, показывающие изменчивость по величине тонкостенности ра-
ковины и по мощности замка. Экземпляры, изображенные на фиг. 1 и 2, довол-
ноМолодые, более похожи на предковый вид «Did», multistriata Rouss., чем бо-
лее крупные. X 1. По Е. Ахвледиани.
Табл. VII. Крупные экземпляры «Did», medeae Dav. жиг. 1—8 показывают изменчи-
вость в направлении вида «Did», celaeno Dav., к которому близки экземпляры,
изображенные на фиг. 5, 6, 7 и 8. X I. По Е. Ахвледиани.
Табл. VIII. «Didacna» celaeno Dav. Фиг. 1—6 показывают изменчивость и различные
степени мощности замка, который наиболее толст у экземпляра, изображенного
на рис. 1 и 2. X I. По Е. Ахвледиани.
Табл. IX. Внутривидовая изменчивость Plagiodacna modiolaris Rouss. Киммерий. За-
падная Грузия (1 —12 — Гурия; фиг. 13 и 14 — Абхазия).
Табл. X. Изменчивость форм рода Tschaudia. Западная Грузия (Гурия). Фиг. 1—4 —
ближайшие предки чаудий, принадлежащие еще к роду Digressodacna (1, 2,
3—из гурийского горизонта, 4 — из слоев, переходных от гурия к чауде).
Фиг. 5—34 — различные формы рода Tschaudia. По Т. Г. Китовани.
Табл. XI. Avimactra acutecarinata (Andrus.). Акчагыл. Малый Балхан, Даната, Крас-
новодск, северный склон Копет-Дага. 1а, 16, 2, 3, 4, 7 — правые створки, 5, 6
и 8 — левые створки. X 2. По А. А. Али-Заде.
251
Табл. XII. Раковины представителей рода Avimactra, правые створки. Средний акчагыл
Восточной Грузии (южная Кахетия, Чатма-Байда). 1 —10 — A. subcaspia And-
rus., 11—13—формы, переходные от A. subcaspia Andrus, к A. karabugasica And-
rus, 14—20 — A. karabugasica Andrus. X 2. По Н. Р. Джикия.
Табл. XIII. Avimactra ossoskovi Andrus. Средний акчагыл Восточной Грузии (южная
Кахетия, Чатма-Байда). 1—7 — относительно короткие раковины, 8—12 — нес-
колько более удлиненные, 13—15 — удлиненные, с более резко выраженным
килем. X 2. По Н. Р. Джикия.
Табл. XIV. Avimactra ossoskovi Andius. Средний акчагыл Восточной Грузии (южная
Кахетия, Чатма-Байда). 1—6 — вид снаружи, удлиненные сильно неравностен-
ные раковины с четко выраженным округлым килем и оттянутым задним концом,
7—15 — вид изнутри. Нижние боковые зубы толстые, массивные. X 2. По
Н. Р. Джикия.
Таблица 1
Таблица II
Таблица Ill
со
о
Таблица IV
I
2
Таблица IV (продолжение)
3
4
Таблица IV (продолжение)
5
б.
Таблица'ТУ (продолжение)
7
Таблица V
Таблица VI
Таблица VII
Таблица VIII
6
Таблица IX
Таблица X
Таблица X (продолжение)
22
28
34
Таблица XI
6
Таблица XII
Таблица XIII
Таблица XIV
ОГЛАВЛЕНИЕ
Предисловие ....................................................... 5
Введение .......................................................... g
Глава первая
НЕКОТОРЫЕ ФАКТЫ ИЗ ИСТОРИИ ПРОБЛЕМЫ ИЗМЕНЧИВОСТИ,
ВАЖНЫЕ С ПАЛЕОБИОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ.....................12
Предварительные замечания ...............................12
Идеалистическая «волна» в биологии на границе двух веков......13
Закон гомологических рядов Н. И. Вавилова и закон аналогической
изменчивости Ч. Дарвина.......................................21
К истории вопроса о нехромосомной наследственности ...........27
Проблема вегетативной гибридизации ...........................30
«Новое в учении о биологическом виде» и значение этой концепции в
науке о наследственности и изменчивости.......................33
Заключительные замечания и общие соображения .................34
Глава вторая
НЕКОТОРЫЕ ДАННЫЕ ОБ ИЗМЕНЧИВОСТИ ОРГАНИЗМОВ, ВАЖНЫЕ В
ПАЛЕОБИОЛОГИЧЕСКОМ ОТНОШЕНИИ.................................39
Вопрос о возникновении резистентности микробов по отношению к анти-
биотикам ...........•.........................................39
«Длительные модификации» у гетеротрофных протистов.......... 44
Гипотеза генетической ассимиляции Уоддингтона ................45
Направленные наследственные изменения, индуцируемые у нематод рода
Caenorhabditis ...............................................48
Переход фитофагических насекомых к новым кормовым растениям ... 51
Развитие устойчивости к ядам у вредных насекомых .............56
Наследственные изменения некоторых культурных растений, вызываемые
факторами среды ..............................................54
Явление гибридной мощи, жизненности и изменчивость ...........67
Общие выводы . ...............................................71
Глава третья
АКТУОПАЛЕОНТОЛОГИЧЕСКИЙ АНАЛИЗ БИОТОПОВ, БЛАГОПРИЯТС-
ТВУЮЩИХ ИНТЕНСИВНОЙ МАССОВОЙ ИЗМЕНЧИВОСТИ ОРГАНИЗ
МОВ......................................................... 80
Изменчивость животных аберрантных неморских биотопов..........80
Биоспеологические исследования, проливающие свет на причины повыше-
нной изменчивости пещерных организмов.....................• • 83
Морские биотопы, благоприятные для повышенной индивидуальной измен-
чивости и соответственно ускоренной эволюции организмов ..... . . 94
Общие выводы об условиях среды, вызывающих повышенную изменчивость 97
253
Глава четвертая
ЯВЛЕНИЯ ИЗМЕНЧИВОСТИ У ИСКОПАЕМЫХ ФОРАМИНИФЕР .... 99
Глава пятая
НЕКОТОРЫЕ УКАЗАНИЯ НА «БУЙНУЮ» ИЗМЕНЧИВОСТЬ («РАСШАТАН-
НУЮ НАСЛЕДСТВЕННОСТЬ») У ИСКОПАЕМЫХ БЕСПОЗВОНОЧНЫХ.. 109
Глава шестая
КЛАССИЧЕСКИЕ ИССЛЕДОВАНИЯ ИЗМЕНЧИВОСТИ У НЕОГЕНОВЫХ
ПЛАНОРБИДИ ВИВИПАРИД НЕМОРСКИХ ВОДОЕМОВ................111
Планорбисы Штейнгейма ..................................111
Изменчивость форм рода Viviparus из озерных отложений неогена . . .114
Глава седьмая
ТЕМПЫ ИЗМЕНЧИВОСТИ И ПРОДОЛЖИТЕЛЬНОСТЬ ЭВОЛЮЦИИ
ОРГАНИЧЕСКОГО МИРА И ТЕЧЕНИЕ ГЕОЛОГИЧЕСКОГО ВРЕМЕНИ.. 124
Глававосьмая J
О ВНУТРИПОПУЛЯЦИОННЫХ ИЗМЕНЕНИЯХ ВЕЛИЧИНЫ ТЕЛА ЖИВОТ-
НЫХ ПРОШЛЫХ ГЕОЛОГИЧЕСКИХ ВРЕМЕН ...................132
Некоторые общие соображения ............................132
О явлениях карликовости у ископаемых беспозвоночных.....133
Об увеличении тела двустворчатых и брюхоногих моллюсков.138
^Внутривидовые изменения величины тела у ископаемых тетрапод . . 145
Глава девятая
ИЗМЕНЧИВОСТЬ МОЛЛЮСКОВ ПАРАТЕТИСА В МИОЦЕНЕ ................153
Предварительные замечания .................................153
Лютеции, спаниодонтеллы, саванеллы .....................154
Изменчивость среди сарматских моллюсков ................158
Глава десятая
РЕЗКО ПОВЫШЕННАЯ МАССОВАЯ ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИ-
ВОСТЬ В НЕКОТОРЫХ ГРУППАХ ДВУСТВОРОК ПЛИОЦЕНА РИОН-
СКОГО ЗАЛИВА .......................................166
Главаодиннадцатая [ J j !
РЕЗКО ПОВЫШЕННАЯ МАССОВАЯ ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИ-
ВОСТЬ В НЕКОТОРЫХ ГРУППАХ БРЮХОНОГИХ ПЛИОЦЕНА РИОН-
СКОГО ЗАЛИВА..................................... 176
Явления изменчивости у некоторых плиоценовых неритинид .176
Об изменчивости у вивипарид Черноморско-Каспийского солоноватовод-
ного плиоцена ..........................................179
Некоторые общие соображения о внутривидовой изменчивости плиоцено-
вых брюхоногих, характерных для Рионского залива .......181
Глава двенадцатая
ИЗМЕНЧИВОСТЬ МАКТРИД АКЧАГЫЛА И ЗНАЧЕНИЕ АНАЛОГИЧЕН________
КОЙ ИЗМЕНЧИВОСТИ В ЭВОЛЮЦИИ МОЛЛЮСКОВ ПАРАТЕТИСА. . 184
254
Глава тринадцатая
ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ У АММОНОИДЕЙ ЮРЫ И МЕЛА. . 188
Глава четырнадцатая
ЯВЛЕНИЯ ИНДИВИДУАЛЬНОЙ ИЗМЕНЧИВОСТИ У ИСКОПАЕМЫХ
ПОЗВОНОЧНЫХ ................................... 192
Предварительные замечания .......................192
Усиленная изменчивость у рыб некоторых речных систем в четвертич-
ном периоде.....................................193
Некоторые данные об индивидуальной изменчивости ископаемых динозав-
ров . ......................................... 194
Изменчивость у ископаемых млекопитающих.........201
Заключение .... ................................206
ЗНАЧЕНИЕ ИЗМЕНЧИВОСТИ ИСКОПАЕМЫХ ОРГАНИЗМОВ ДЛЯ
РЕШЕНИЯ ЗАДАЧ СТРАТИГРАФИИ И ГЕОХРОНОЛОГИИ. ОБЩИЕ
ВЫВОДЫ..........................................206
Литература 211
ГЛАВНЕЙШИЕ СТРАТИГРАФИЧЕСКИЕ ПОДРАЗДЕЛЕНИЯ .........225
СТРАТИГРАФИЧЕСКАЯ СХЕМА НЕОГЕНА ЧЕРНОМОРСКОЙ ОБЛАСТИ . 228
СоЛ^оБоЪЭсЛО
(6 3 Ъ О 2 Э з) ................................229
Variation of organisms inthe past geological ti
mes............................................ 231
Указатель авторов . ................................233
Указательтерминов ..................................237
Указатель латинских названий .......................243
Объяснение таблиц ..................................251
Таблицы I—XIV .......................................
Лео Шиович Давиташвили
ИЗМЕНЧИВОСТЬ ОРГАНИЗМОВ В ГЕОЛОГИЧЕСКОМ ПРОШЛОМ
Напечатано по постановлению Редакционно-Издательского Совета
Академии наук Грузинской ССР
*
Редактор издательства Л. К. К о б и д з е
Художник Г. А. Надирадзе
Техредактор Н. А. Эбралидзе
Корректор Л. М. Ча га н а ва
Сдано в набор 10.6.1970; Подписано к печати 10.11.1970;
Формат бумаги 70Х1081/32; Печатных л. 22.26; Уч.-Издат. л. 21.19;
УЭ 11457; Тираж 1500; Заказ 1522;
Цена 1 руб. 88 коп.
„SggGoQingdo”, ooboQ^obo, 60, ^ф^'ЬсодоЬ j., 15
Издательства «Мецниереба», Тбилиси, 60, ул. Кутузова, 15
boj. ЬЬ6. ojo£>g3oob ЬфоЭ&д cndoQ^obo 60, J£)C)E)<b<r>3ob 15
Типография Академии наук ГССР, Тбилиси, 60, ул. Кутузова, 15