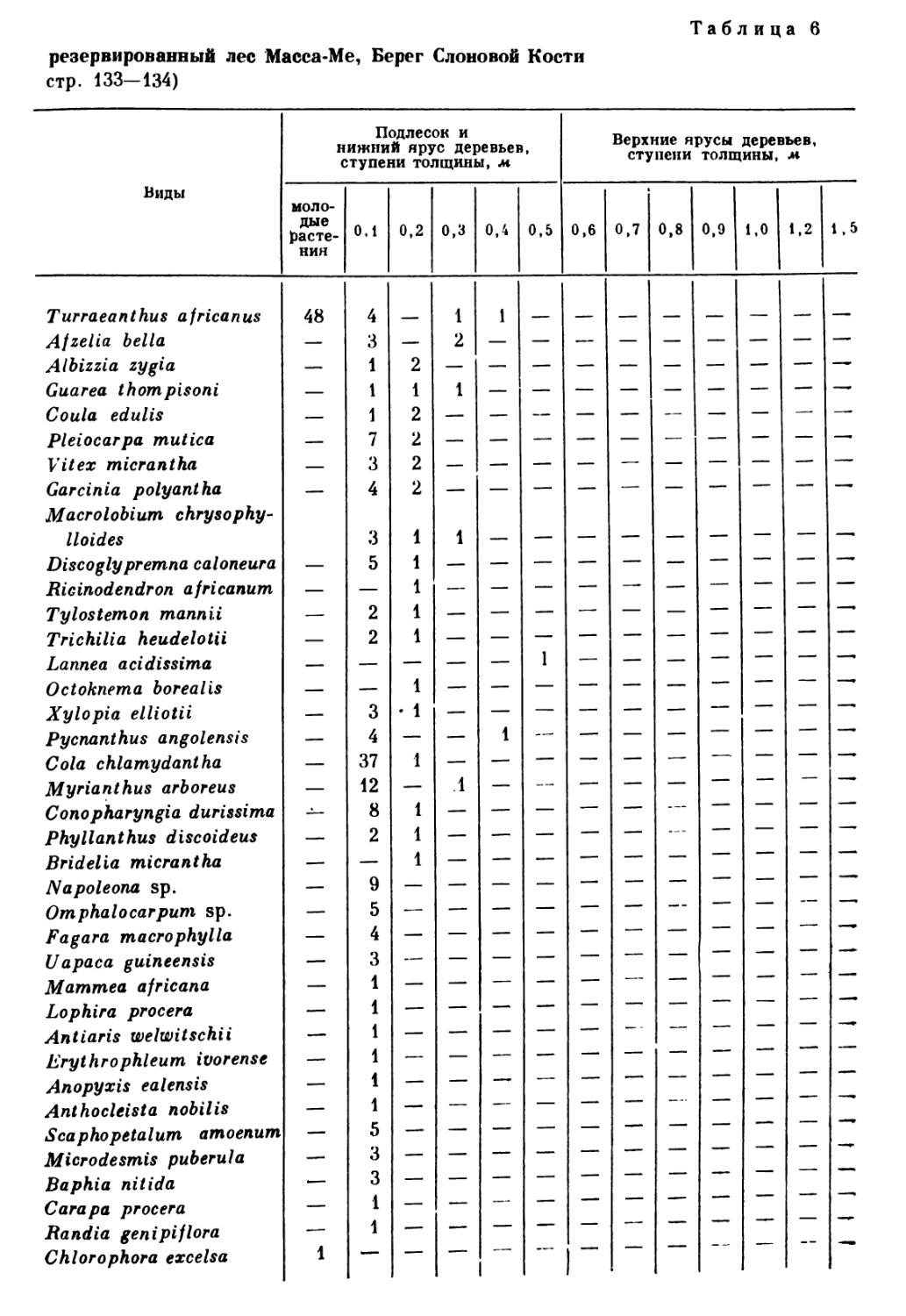
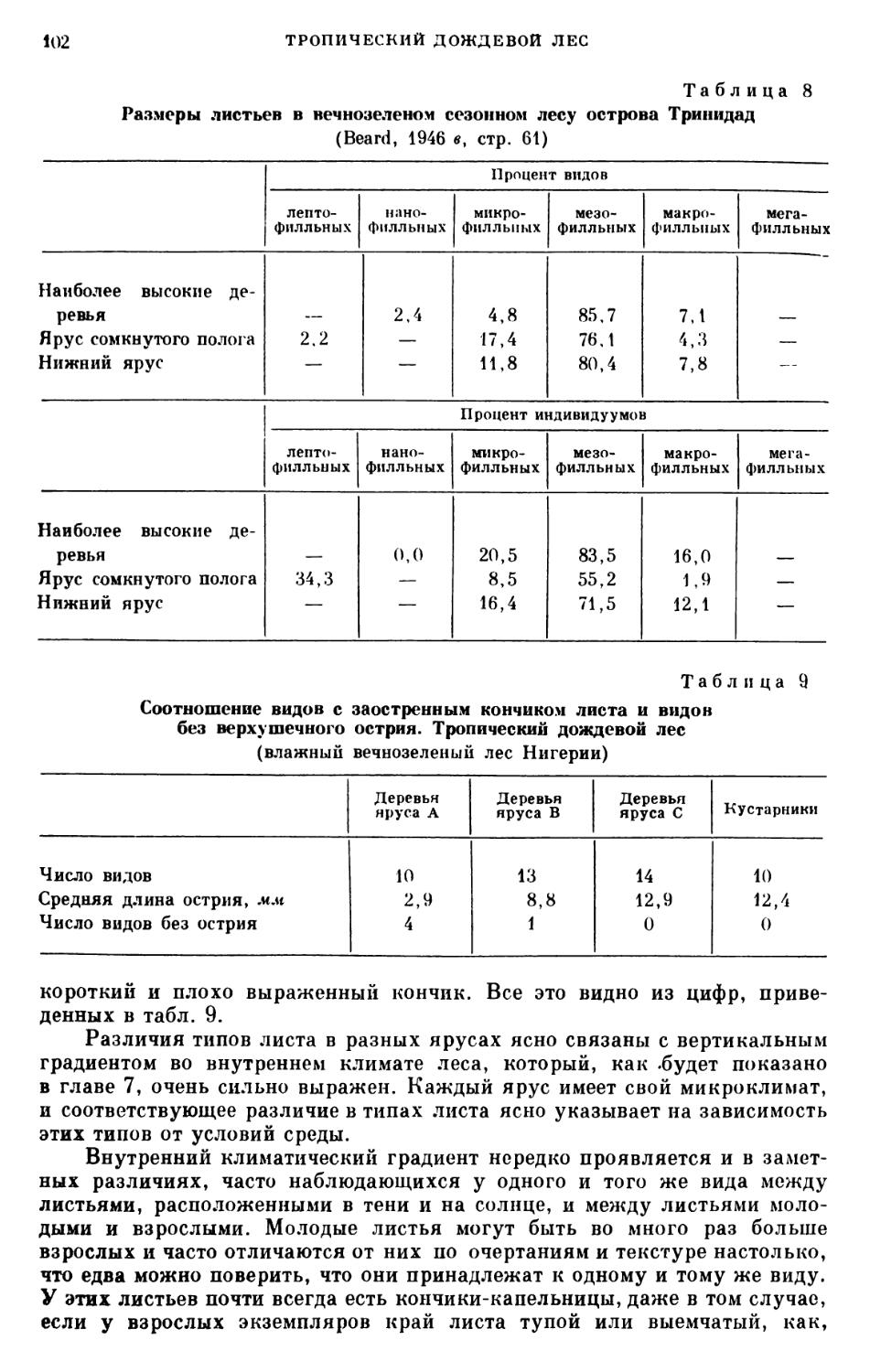
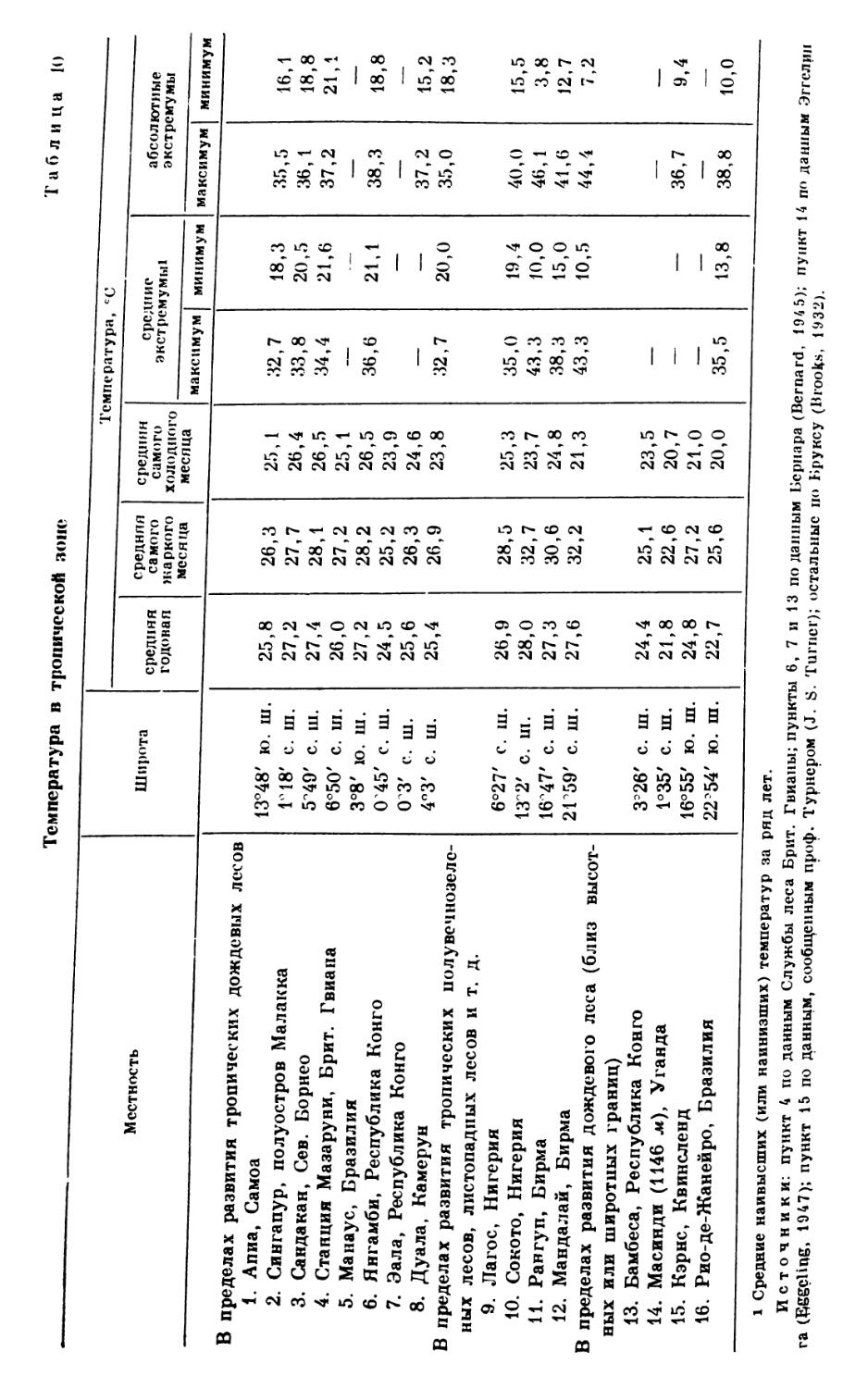
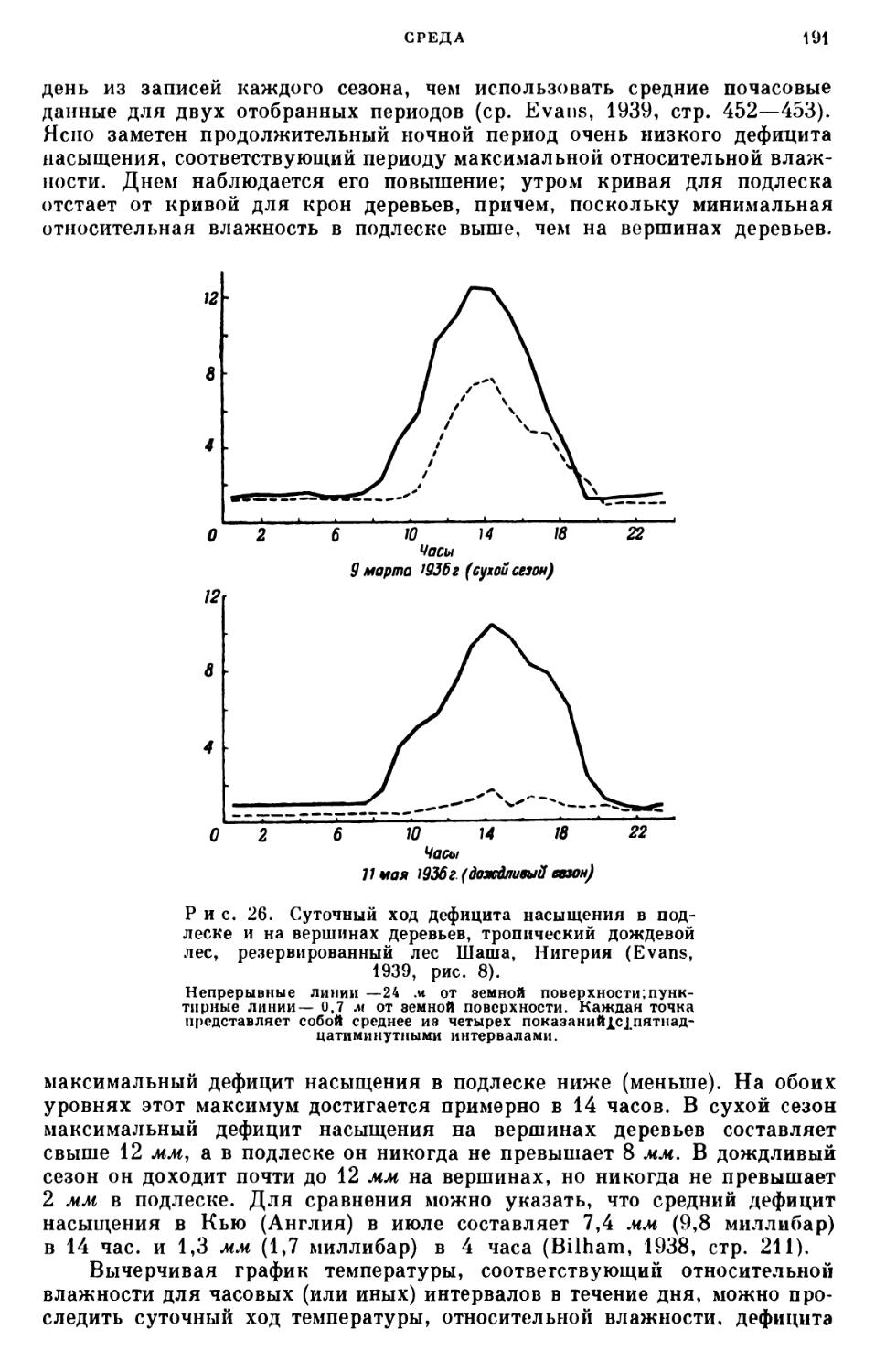
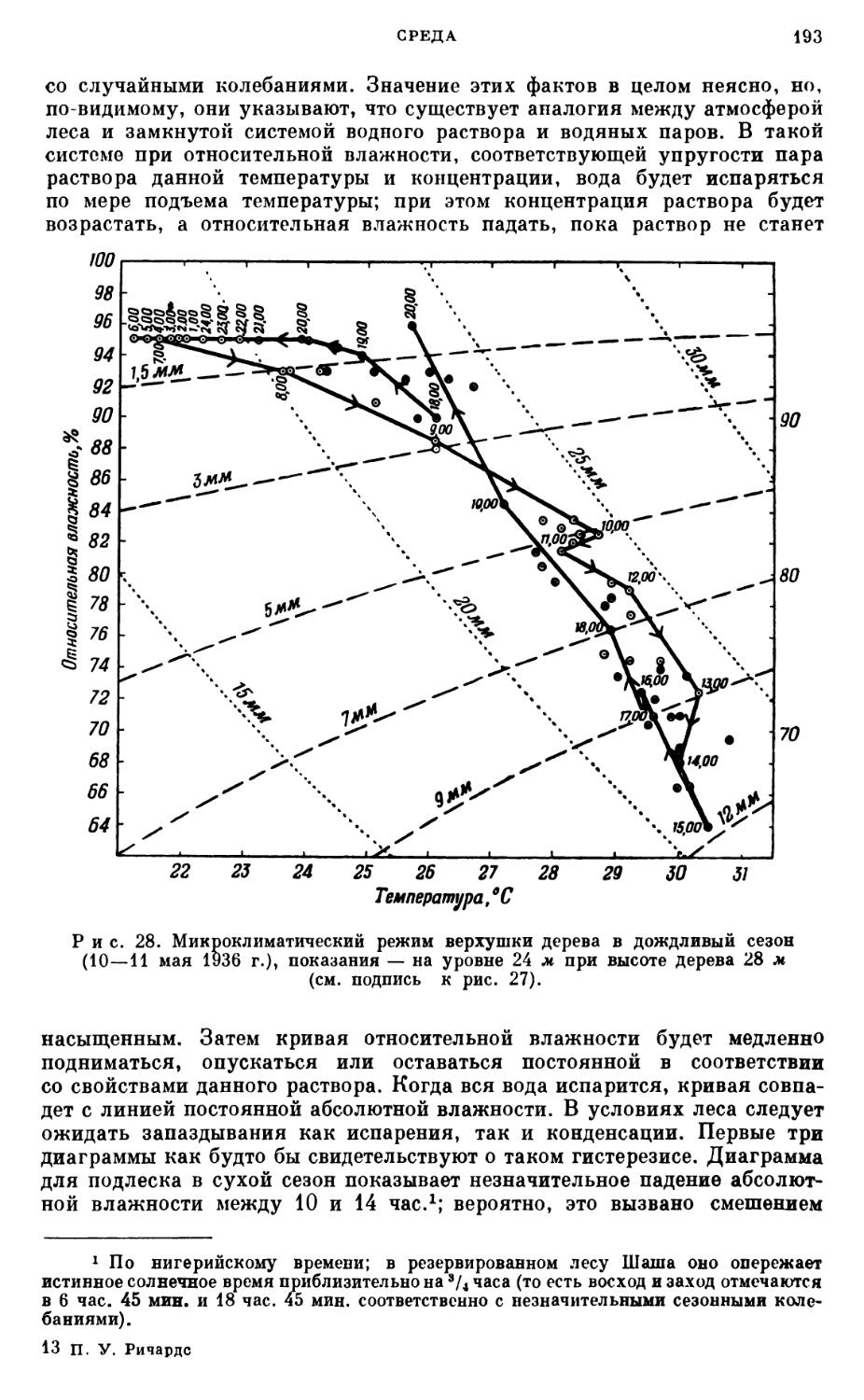
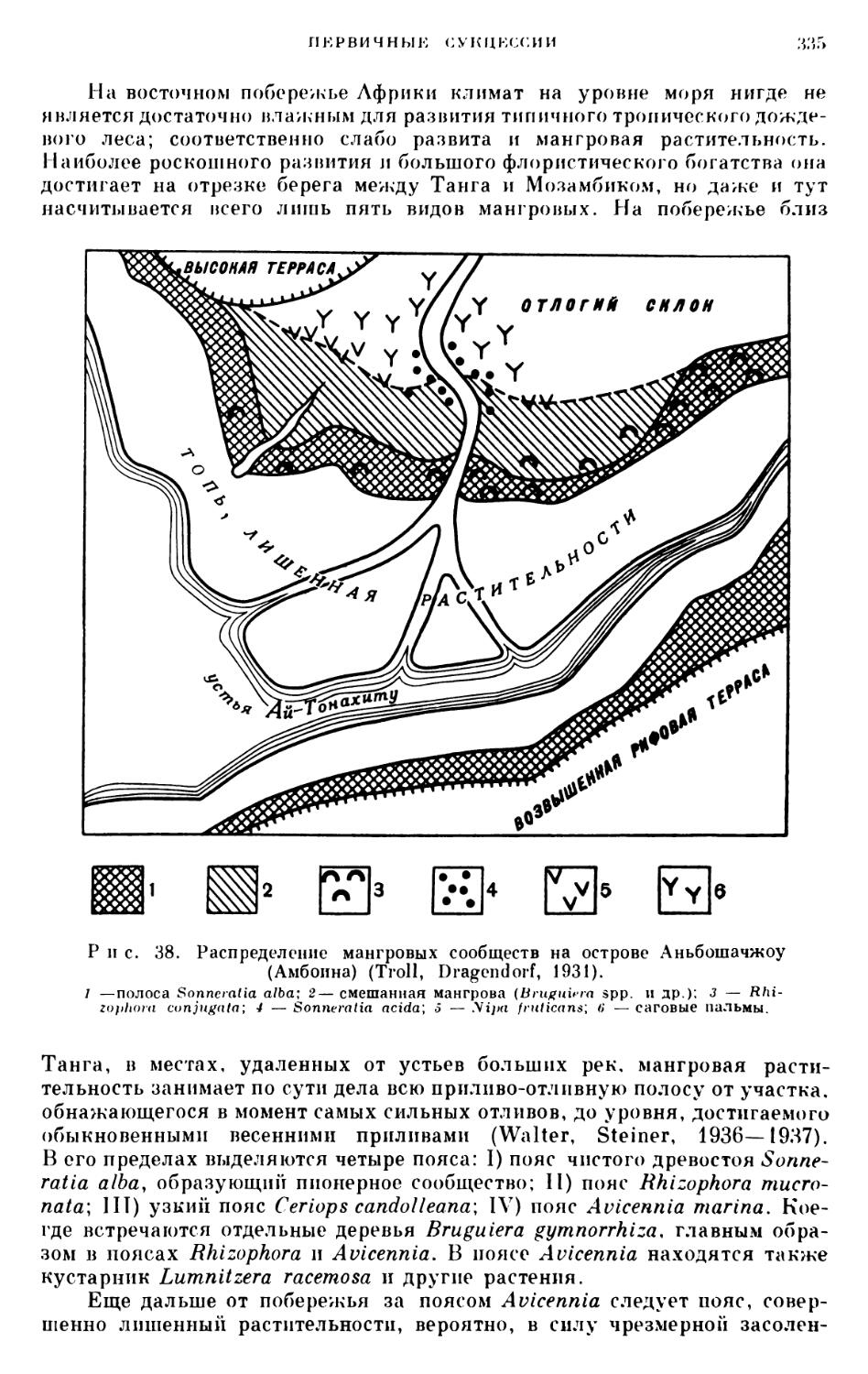
/





































































































































































































































































































































































































































































Текст
Р. П. RICH А R J> S
THE
TROPICAL RAIN FOREST
AN ECOLOGICAL STUDY
CAMBRIDGE 1957
п. У. РИЧАРДС
ТРОПИЧЕСКИЙ
ДОЖДЕВОЙ
ЛЕС
Перевод с английского Т. П. КОМОВА и т. И. ПОДОЛЬСКОЙ
Предисловие и редакция А и. А. ФЕДОРОВА
ИЗДАТЕЛЬСТВО ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва 1961
I
АННОТАЦИЯ
Книга профессора ботаники П. У. Ричардса — известного исследователя флоры влажных тропиков представляет собой фундаментальный труд о формациях влажнотропического (тропического дождевого) леса, их распространении, флористическом составе, структуре и взаимодействии с окружающей средой, который по праву может считаться классическим. Называя свою работу экологическим исследованием, автор уделяет максимум внимания этой стороне проблемы, подробно анализируя богатейший фактический материал и существующие теории. Труд базируется не только на личных, весьма разносторонних, исследованиях самого автора, он является систематизированной сводкой ценнейших и дотоле разрозненных сведений, накопленных многочисленным отрядом ученых и работников службы леса.
Учитывая все возрастающий интерес к изучению тропиков н большое научное значение книги Ричардса, эта монография, исключительно нужная для специалистов экологов и фнтоценологов, безусловно, завоюет себе широкое признание и среди преподавателей высших и средних учебных заведений.
* - 7 ’ .
„0 тркам г№графпх
Фото 1. Тропический дождевой лес па высоте 800 м над уровнем моря, горы Арфах, Новая Гвинея.
ПРЕДИСЛОВИЕ
ЫХ0Д в свет двух изданий (1952, 1957 гг.) книги английского ботаника профессора Пола Уэстмэкотта Ричардса «Тропический дождевой лес» явился крупным событием в развитии современных экологии, фитоценологии и ботанической географии. Особую роль играет она в ботаническом исследовании тропиков. В этой области значение книги Ричардса трудно переоценить. Тем читателям (в том числе и ботаникам), которые до спх пор знакомились с растительным миром тропиков по описаниям, сделанным путешественниками, и по компилятивным или слишком старым работам, книга Ричардса, если можно так сказать, открывает глаза на растительность тропиков и дает действительно правильное понятие о тропическом дождевом лесе взамен того фантастического и искаженного преувеличениями представления, которое, к сожалению, еще и теперь находит соответствующее отражение в литературе.
П. У. Ричардс является одним из наиболее выдающихся знатоков тропического леса и тропиков вообще наряду с такими специалистами, как А. Обревиль — во Франции, Ван-Стеенис — в Нидерландах, Ж. Леб-рэн — в Бельгии и некоторые другие ботаники в различных странах. Но даже и среди этих крупнейших ученых, занимающихся флорой, фитоценологией и фитогеографией тропических стран, П. У. Ричардс выделяется тем, что именно он впервые дал капитальную монографию по экологии и фитоценологии тропического дождевого леса в пределах всей области распространения этой группы формаций.
Ричардс выполнил эту задачу благодаря настойчивым и целеустремленным исследованиям, проведенным в течение ряда лет почти во всех главнейших районах, где имеется тропический дождевой лес. В этом отношении весьма любопытна биография профессора Ричардса1 — биография, довольно типичная для многих английских ботаников, которые нередко еще в молодости начинали свои самостоятельные исследования именно в тропической зоне.
П. У. Ричардс родился в 1908 г. в Уолтоне, графство Суррей, Англия, и получил высшее образование в Тринити-Колледже в Кембридже в период с 1933 по 1937 г. После этого до 1945 г. он состоял в Кембриджском университете в должности «демонстратора по ботанике» (что соответствует ассистенту в наших высших учебных заведениях). Уже в 1945 г. Ричардс — лектор по ботанике в Кембридже (то есть доцент по нашей терминологии), а в 1949 г._профессор в Университете Северного Уэльса в Бангоре, где
в настоящее время он заведует кафедрой ботаники.
Исследования П. У. Ричардса в тропиках начались с 1929 г., когда он выехал в Брит. Гвиану в составе научно-исследовательской экспедиции Оксфордского университета. Исследователю в это время было всего
1 s I е е n i7, C.G.G. J- van, Flora Malesiana, 1, Djakarta, 1950, p. 43!.
6
ПРЕДИСЛОВИЕ
20 лет, но появившаяся затем в печати работа по экологии и фитоценологии тропического дождевого леса Брит. Гвианы, выполненная Ричардсом совместно с ботаником Т. А. Дэвисом, свидетельствовала о его зрелости как ботаника и представляла собой весьма ценный труд, в котором впервые был применен и обоснован интересный метод исследования тропических лесов — метод «профильных диаграмм» (см. соответствующие главы книги), широко используемый ныне всеми фптоценологами, работающими в тропиках. Столь же плодотворные исследования тропического дождевого леса П. У. Ричардсу удалось вести в период с января 1932 г. по 1933 г. в Брит. Борнео (Саравак, гора Дмолит и пр.). Эти работы, как и исследование лесов в Гвиане, также проводились Оксфордской экспедицией. Попутно Ричардсу удалось побывать на Малаккском полуострове с его едва ли не самым роскошным в мире тропическим дождевым лесом. В последующие годы начиная с 1947 г. П. У. Ричардс несколько раз выезжал в Африку (Нигерия, Гана и пр.), где исследовал тропические дождевые леса и близкие к ним формации по тому же самому плану, что и во время более ранних экспедиций в тропической Америке и Азии. Свои выезды в тропики П. У. Ричардс продолжает и в настоящее время. Совсем недавно он побывал в Таиланде (Бангкок), где принял участие в работах очередного Тихоокеанского конгресса, затем, в составе комиссии ЮНЕСКО,—на Цейлоне. На Цейлоне после состоявшейся в Кэнди конференции по тропикам Ричардс вместе с другими ботаниками осматривал, между прочим, один из типичных участков тропического дождевого леса — «лес Кан-нелия». По-видимому, Ричардс, как почти никто до него, имел возможность видеть собственными глазами и подробно исследовать тропический дождевой лес всех частей света. Это наложило яркий отпечаток на все его творчество в области изучения растительности тропиков, в особенности же на характер его книги о тропическом дождевом лесе.
Эта книга, как и все предшествовавшие ей работы П. У. Ричардса, написана с большим вдохновением и представляет собой вполне оригинальную научную монографию, которую уже сейчас можно с полным основанием отнести к ряду классических исследований. Следует добавить, что П. У. Ричардс, кроме занятий экологией тропической растительности и фитогеографией, посвятил много труда систематике мхов — области, где он является общепризнанным специалистом1.
Конечно, нет особой надобности указывать в настоящем предисловии на основные научные достижения Ричардса, которые подытожены в его книге. Читатель сам увидит все это. Тем не менее, хотелось бы подчеркнуть некоторые моменты, характерные для Ричардса как исследователя. П. У. Ричардс на многих страницах своей книги показывает себя последовательным и строгим дарвинистом. Особое внимание уделяет он влиянию наследственных свойств растительного организма в их взаимодействии со средой. Ричардс настойчиво предостерегает от поспешных выводов, иногда содержащихся в различных «антропоморфных» гипотезах и отдельных взглядах различных исследователей на роль среды в видоизменении растительного организма и в выработке адаптивных свойств и признаков. Очень показателен в этом отношении раздел книги, где рассматривается проблема возникновения и жизненной роли так называемых досковидных
1 Гербарные коллекции, собранные Ричардсом в тропиках, хранятся в Англии в Королевском ботаническом саду в Кью, близ Лондона, а также в Оксфорде, Кембридже и в Бангоре. Часть дублетных коллекций находится на Борнео, в Музее Саравака, другая часть — в Сингапуре. Свои основные исследования II. У. Ричардс публиковал преимущественно в «Экологическом журнале» («The Journal of Ecology»).
ПРЕДИСЛОВИЕ
7
корней у тропических деревьев. Тщательно анализируя научную ценность пяти гипотез, П. У. Ричардс пока не склонен отдавать ни одной из них особого предпочтения, подчеркивая, что наиболее распространенный взгляд о прямой адаптивности досковидных корней как подпорок ствола не имеет для своего обоснования серьезных научных данных. С такой же осторожностью и критикой излагаются и другие проблемы биологии, экологии, фитоценологии и автэкологии тропического дождевого леса. Тем не менее важная роль среды во всем ее объеме в формировании растительного мира в труде Ричардса выступает очень ясно.
Большой заслугой П. У. Ричардса является выработка правильного научного представления о внешнем виде, экологическом строе и экосистеме (биогеоценозе по В. Н. Сукачеву) тропического дождевого леса. Ричардс развеял миф, создавшийся вокруг понятий об этом растительном сообществе как о каком-то феерическом парадизе, переполненном массами лиан, густейших зарослей эпифитов, пальм, где деревья якобы достигают в высоту чуть ли не 70 м и более и где органическая жизнь в разнообразнейших формах заполняет все экологические ниши и целиком все пространство (так называемый «horror vacui»), и что лес этот без применения топора будто бы совершенно неприступен и пр. и пр. Все это при трезвом анализе оказалось совершенно неверным. Между тем для большого числа книг, в особенности популярных, посвященных растительности тропиков, еще до сих пор довольно характерны неумеренно восторженные, фантастические описания тропических лесов и тропиков вообще.
Правдивое изображение тропического дождевого леса, даваемое Ричардсом, невольно напоминает замечательную книгу его великого соотечественника, соратника Ч. Дарвина,— А. Р. Уоллеса «Тропическая природа», не раз издававшуюся на русском языке. Из прежних авторов именно Уоллес дал замечательно верное, прекрасное описание тропического дождевого леса без всяких преувеличений и вымысла. Остается лишь пожалеть, что популяризаторы, писавшие о тропиках, видимо, часто забывали о книге Уоллеса, пользуясь сведениями из малодостоверных источников. Книга Ричардса, вне всякого сомнения, будет служить наилучшим руководством как для исследователей, так и для составителей учебников и популярных книг по ботанической географии тропиков.
Посетив тропические леса Китая (южная Юньань, остров Хайнань)1, а также Цейлона1 2, автор этих строк воочию убедился в том, что обычные представления о якобы присущих этим лесам оранжерейной роскоши, обилии красок и форм, непроходимости и сплошной путанице лиан совершенно не соответствуют действительности. После ознакомления с диптерокарповым тропическим дождевым лесом («лес Каннелия» и «лес Коттава», Цейлон) убеждаешься, что основное впечатление, оставляемое тропическим дождевым лесом, — это впечатление скорее довольно мрачного однообразия, а вовсе не роскоши и богатства. Лиан в этих лесах много, но они не мешают ходить по лесу. Эпифитов же много только в кронах деревьев, на стволах их почти нет. Листья большинства деревьев удивительно однообразны и приближаются по величине к типу «мезофилла». Все листья темно-зеленые, гладкие и блестящие. Поэтому общее впечатление монотонно темной зелени не нарушается даже в том случае,
1 Федоров Ан. Л., Влажнотропические леса Китая, «Ботанический журнал», 10, 43, 1958.
2 Федоров Ан. А., Диптерокарповый экваториальный влажнотропический лес Цейлона, Сборн., посвящ. 80-летию В. Н. Сукачева, 1960.
ПРЕДИСЛОВИЕ
8
если кое-где виднеются яркие цветы или молодые листья, которые обычно бывают красноватыми. Внутри леса с первого взгляда не замечаешь ничего кроме слоя мертвых опавших листьев, одиночных маленьких пальм (на Цейлоне обычно Calamus) и совсем негустого подлеска. К тому же и крупные досковидные корни в цейлонском лесу попадаются не часто. Поэтому сначала бросается в глаза не отличие тропического леса от наших голарктических лиственных лесов, а скорее их сходство. У тропического дождевого леса есть даже какое-то отдаленное сходство с сосновым бором, вызванное темной зеленью крон деревьев, прямизной и гладкостью стволов, а также тем, что некоторые деревья обладают моноподиальным ростом, придающим своеобразный облик кронам лиственных пород и делающим их похожими на кроны хвойных деревьев. Однако под внешним однообразием скрывается действительно поразительное богатство флористического состава, полностью унифицированного биологической конвергенцией. Деревья, принадлежащие к совершенно различным видам, родам, семействам, имеют удивительно сходный внешний облик. Тропический дождевой лес вовсе не есть беспредельное и хаотическое собрание чуть ли не всех жизненных форм растений. Наоборот, в этом сообществе явно господствуют фанерофиты. Другие жизненные формы подавлены, а некоторые (геофиты, криптофиты, терофиты) вообще отсутствуют. То, что называют кустарниковым подлеском (shrub stratum), в большинстве случаев является ярусом маленьких деревьев с невысокими, но прямыми и маловетвящимися стволами. Это сообщество состоит главным образом из энтомофильных растений, хотя часть растений опыляется при помощи птиц и летучих мышей. Наблюдается и самоопыление. Цветы в большинстве случаев не очень крупные и неяркие. Плоды и семена, как правило, приспособлены к распространению животными. Взаимосвязи растений и животных в биогеоценозе тропического дождевого леса, видимо, исключительно прочны.
В противоположность голарктическим сообществам, зачастую представляющим во флористическом отношении гетерогенный набор таксономически мало связанных видов из родов и семейств, также систематически разобщенных, флористический состав тропического дождевого леса именно тем и замечателен, что виды, роды и семейства, к нему относящиеся, часто образуют естественные группы сродства. Целые семейства в огромной массе своих представителей часто не выходят за пределы флористического состава тропического дождевого леса. В свою очередь многие тропические семейства, к которым относятся виды и роды растений этого леса, также обнаруживают в пределах названной формации свои первоначальные филогенетические связи. Особенно интересно в указанном отношении семейство Dipterocarpaceae, наиболее характерное для тропиков Азии. За исключением немногих листопадных пород, свойственных сухим муссонным лесам, подавляющее большинство родов и видов семейства Dipterocarpaceae сосредоточено в тропических дождевых лесах. Тут распространены обширные роды Dipterocarpus, Shorea, Нореа, Vatica, а на Цейлоне также род Duna, образующие в пределах одной и той же формации ряды генетически связанных между собой видов. Тут же в пределах тропического дождевого леса встречаются и почти все монотипные и олиго-типные роды этого семейства. По ряду признаков различных тропических родов семейство Dipterocarpaceae приближается к семействам Theaceae, Guttiferae, Malvaceae, Ochnaceae, также очень характерным для тропического дождевого леса. В равной степени и многие другие семейства в пределах тропического дождевого леса таксономически связаны между собой.
ПРЕДИСЛОВИЕ
»
Все это объясняется тем, что тропический дождевой лес, несомненно, развивался в течение миллионов лет как единое целое. Вероятно, современный тропический дождевой лес есть наследие и результат изменения других сообществ, но важно подчеркнуть, что весь путь его развития был строго автохтонным и явился фитоценотическим выражением, а также и фоном грандиозного процесса формирования основного ядра флоры покрытосеменных растений, родина которых, конечно, находится в тропиках. Голарктическая и антарктическая флоры по отношению к тропической представляют собой вторичное явление, возникшее в результате приспособления растений к менее благоприятным условиям в процессе вычленения и миграции из флоры тропиков1.
Касаясь вопроса о коренных или зрелых растительных сообществах (климакс) и процесса смены сообществ (сукцессии) при восстановлении растительного покрова после различных нарушений, Ричардс пользуется терминологией Клементса, создавшего учение о климаксе и сукцессии. Однако Ричардс прибегает к терминологии Клементса лишь в тех случаях, где эти обозначения действительно помогают объяснению динамики развития растительности, и самому понятию о климаксе, видимо, не придает того почти абсолютного значения, на котором настаивал Клементс. В таком относительном понимании климакса с учетом его изменяемости в ходе эволюции растительного покрова нет того метафизического оттенка, который чувствуется в некоторых построениях Клементса. Но полезность самой идеи о существовании коренных или зрелых сообществ и об определенных стадиях, которые претерпевает восстанавливающийся после нарушения растительный покров, очевидна. Заметим кстати, что к этой мысли давно приходили русские и советские ботаники. Например, понятие о коренной ассоциации в русской фитоценологии уже давно ввел в употребление акад. В. Н. Сукачев. Им же, как и другими фитоценоло-гами, широко применялось учение о сменах растительных сообществ, также созданное русскими учеными. Другое дело — терминология Клементса. Она трудно усвояема и очень громоздка. Терминов, предложенных Клементсом для обозначения характера климакса и фаз сукцессии, по-ви-димому, даже слишком много. Здесь мы сталкиваемся с тем случаем, когда обильная терминология уже не разъясняет природных явлений, к которым она относится, а в известной степени затемняет их понимание. Большое затруднение возникает и при переводе терминов Клементса на русский язык. Хотя некоторые из этих терминов были введены в русскую ботаническую литературу Н. Г. и В. В. Алехиными при переводе книги Мак-Дуголла1 2 3 * * * *, а затем В. Д. Александровой при изложении учения Клементса, все же русская транскрипция этих терминов не может считаться окончательно установившейся и нуждается в усовершенствовании.
Мы сочли возможным, вслед за Алехиными и Александровой, переводить термины Hydrosere, Xerosere и подобные им, как гидросерия, ксеросерия и т. п., хотя, возможно, лучше было бы слово «sere» передать по-русски как-нибудь иначе, чтобы избежать слишком для данного случая определенного смысла серии или ряда. У Клементса8 этим терминам соот
1 Федоров Ан. А., Флора Юго-Западного Китая и ее значение для позна-
ния растительного мира Евразии, «Комаровские чтения», 10, Изд. АН СССР, 1957.
3 М а к-Д у г о л л, Экология растений, М., 1935; Александрова В. Д.,
Учение Клемантса о климаксе, «Ботанический журнал», 5, 33, 1958.
’Clements F. Е., Plant Succession and Indicators, New York, 1928.
Clements F. E., Nature and Structure of the Climax, «J. Ecol.f,
24, 1936.
ПРЕДИСЛОВИЕ
10
ветствуют скорее не ряды сообществ, а различные временные фитоценозы, последовательно возникающие и исчезающие в ходе сукцессии. Неопределенное по смыслу слово «sere», взятое Клементсом из греческого языка, хотя и приравнивается им к латинскому слову «series», то есть ряд, имеет и другие значения. Клементс использует корень «ser», общий греческому и латинскому языкам, для удобства словообразования, а также для придания созданной таким способом терминологии благозвучности. Но то. что благозвучно в английском тексте, не всегда хорошо звучит по-русски. Во всяком случае, английское произношение всех клементсовских терминов для русского языка совсем не годится. Поэтому при переводе этих терминов на русский язык пришлось в большинстве случаев просто воспроизводить русскими буквами их первоначальную транскрипцию, ориентируясь не на английское, а в основном на латинское произношение.
Перевод некоторых других терминов, употребляемых Ричардсом, также нуждается в оговорках. Это прежде всего касается заглавия. Было решено сохранить его дословное написание «Тропический дождевой лес», несмотря на то, что в русской ботанической литературе утвердилось и стало до известной степени традиционным обозначение «влажнотропический лес», хотя оно не вполне соответствует впервые введенному Шимпером названию «tropischer Regenwald» и его английскому эквиваленту «tropical rain forest». Не останавливаясь на достоинствах и недостатках того и другого, скажем лишь, что это вызвано стремлением сохранить стройность авторской классификации и самобытность его терминологии.
Термин «hill forest», так же как и «submontane rain forest», переводится нами как «лес предгорий», которому они вполне морфологически соответствуют.
Очень трудным оказалось подыскать русский эквивалент обозначению «Heath forest», взятому Ричардсом из работы немецкого ботаника Губерта Винклера1 о лесах Борнео и являющимся дословным переводом на английский язык немецкого термина «Heidewald». Этим названием в его первоначальном смысле обозначался сосновый бор с покровом из вереска (Calluna vulgaris). Леса этого типа (Calluna Typus Каяндера) очень характерны для Германии (а также и для СССР, особенно его западной приморской части и средней России). Винклер обозначил термином «Heidewald» обедненный и полусухой (Subxerophile) — вариант тропического дождевого леса Борнео. Важно указать, что в этом лесу нередко в большом количестве встречается внешне весьма похожая на сосну Casuarina sumatrana. Поэтому естественно сравнение этого леса с сосновым бором. Имея все это в виду, мы переводим «heath forest» как «боровой» лес, приводя это выражение в кавычках.
От случая к случаю Ричардс использует французское обозначение тропического дождевого леса —«foret dense». Мы переводим этот термин словами «сомкнутый лес», а не «густой лес», как иногда делали переводчики ранее изданных книг. Дело в том, что французские ботаники ввели понятие «foret dense» для отграничения тропического дождевого или экваториального леса (foret equatoriale) от сообществ светлых лесов с несомкнутым древесным ярусом (foret claire), или лесистых саванн. Правда, следует помнить, что под словами «foret dense» обычно подразумевают не только типичный вечнозеленый тропический дождевой лес, но и полули-
1 Winkler Н., Die Pflanzendecke Sudosl-Borneos. «Bot. Jb.», 50, 1914, 188-208.
ПРЕДИСЛОВИЕ
И
стопадные его варианты. Это ясно видно хотя бы из работ А. Обревиля1 о тропических лесах Африки1 2.
Травяные сообщества, возникающие на местах, где некогда были развиты тропические леса, Ричардс называет термином «Grassland». Так как автор книги воздерживается по тем или иным соображениям называть эти фитоценозы саванной, а другие травяные сообщества, например луга и степи, в тропиках в сущности отсутствуют, то мы решили переводить термин «Grassland» словом «злаковник». Название это уже было предложено для употребления приблизительно в этом же смысле А. П. Г1льин-ским3. Сходный по неопределенности своего значения термин «Woodland» переводится нами как «редколесье» пли «редкостойный лес».
Некоторые геоботанические понятия у Ричардса толкуются иначе, чем принято в русской литературе. Например, под термином «подлесок» (undergrowth) Ричардс подразумевает не только маленькие деревья и кустарники, составляющие самый нижний древесный ярус леса, но также травяной покров и крупный подрост, а иногда только травяной покров (herbaceous undegrowth). В зависимости от значения термина «undergrowth», взятого в контексте, мы переводим это слово то как «подлесок», то как «травяной покров». Травяной покров со всходами и сеянцами древесных пород и лиан Ричардс называет «полевым ярусом» (field layer). Конечно, дословный перевод на русский язык термина «field layer» нельзя вводить в обиход. Поэтому мы выражение «field layer» переводим словами «травяной покров» или «травяной ярус», оговаривая, где это нужно, что в нем присутствуют также сеянцы деревьев.
Широко пользуясь различными узко местными названиями, употребляющимися в различных тропических областях для обозначения как климаксного тропического дождевого леса, так и различных стадий сукцессии лесной растительности, а также различных особых типов тропических лесов, Ричардс тем самым поставил переводчиков и редактора настоящего русского издания в весьма затруднительное положение. Все эти местные обозначения вроде «лес Морабуцеа», «лес Уаллаба», «лес Гринхарт», «паданг», «пинкадо», «индаинг» и пр., оставлены, конечно, без перевода и даны нами русскими буквами соответственно английской транскрипции, так как зачастую совершенно нельзя угадать правильное произношение этих терминов. Пытаться найти этим названиям какие-либо смысловые русские эквиваленты было бы такой же неблагодарной задачей, как переводить на английский язык всевозможные русские лесные термины, разные «груды», «гряды», «рамени», «субори», «бучины», «пихтачи», «листвя-ги» и т. п.
Монография П. У. Ричардса о тропическом дождевом лесе носит характер преимущественно экологического п фитоценологического, или, лучше сказать, пользуясь термином, предложенным В. Н. Сукачевым, биогеоценологического исследования. Может быть, поэтому в ней почти не затронута проблема происхождения и эволюции тропического дождевого леса. Конечно, вопрос этот очень труден и все еще не получил удовлетворительного объяснения. Приходится согласиться с Э. Д. X. Корнером4, что место и время возникновения этой формации представляет собой
1 Aubreville A., Climats, forets et desertification de L’Afrique tropicale, Paris, 1949.
2 О б p e в и л ь А., Тропическая Африка, сб. География лесных ресурсов земного шара, ИЛ, М., 1960.
3 Ильинский А. П., Растительность земного шара, М—Л., 1937.
‘Corner Е. J. Н., The Evolution of Tropical Forest, Evolution as a process. J. Haxley, A. Hardy, E. Ford (edit.), London, 1954.
12
ПРЕДИСЛОВИЕ
такую же тайну, как и родина покрытосеменных растении вообще. Известно лишь, что покрытосеменные появляются неожиданно, как «deus ex machine», начиная с мела, и уже господствуют на протяжении третичного времени. Где и когда возникла флора тропиков, которую можно было бы считать прародительницей современной тропической флоры, неизвестно. В этом отношении очень интересна попытка Корнера объяснить эту загадку. Обращаем внимание на цитированную выше статью Корнера об эволюции тропического леса, написанную с редким красноречием и убежденностью. В основу своих вглядов по указанному вопросу Корнер положил собственную так называемую «дурьян-теорию» происхождения покрытосеменных растений. По мысли Корнера, первичными покрытосеменными растениями были тропические пахикаульные (мясисто- или толстостебельные) и сравнительно невысокие примитивные деревья, по внешности похожие на современное дынное дерево Carica papaya1. Они обладали саговникообразной или пальмообразной формой роста и имели не-ветвящийся ствол, несущий на верхушке подобие «розетки» листьев. Эти пахикаульные прадеревья имели плоды и семена, снабженные сочными ариллусами наподобие плода видов современного тропического рода Durio (откуда п «дурьян-теория») из семейства Bombacaceae. Тип плода дурьяна Корнер считает наиболее примитивным среди всех остальных типов плода покрытосеменных растений. Развитие сочного ариллуса (у Durio к тому же с необыкновенно сильным своеобразным запахом) оказалось свойством, содействовавшим распространению первичных тропических растений различными животными, поедавшими сочные плоды, и сопряженной с животным миром эволюции этих растений. Из взглядов Корнера следует, что и сложность структуры тропического дождевого леса зависит не только от реакции растений, проявляющейся в их строении, на действие особо благоприятных условий окружающей среды, представляющей в данном случае «конгломерат местообитаний», но в особенности от взаимодействия растений и животных внутри лесной экосистемы. Поэтому эволюция тропического леса есть главным образом биологическая проблема.
Возникшие в ходе эволюции тропического дождевого леса настоящие деревья с высоким, полностью одревесневшим стволом составили уже близкое к современному сообщество тропического дождевого леса, пахикаульные же формы редуцировались и оттеснились в подлесок, а частично даже превратились в сапрофиты и паразиты (например, Rafflesia'). Загадка отсутствия ископаемых остатков каких-либо покрытосеменных растений, которых можно было бы признать примитивными (по сравнению с известными до сих пор), возможно тем и объясняется, что пахикаульные растения просто не оставили следов в пластах горных пород, так как, отмирая, быстро разрушались.
Процесс эволюции покрытосеменных растений, рисуемый Корнером (от пахикаульных примитивных деревьев к настоящим деревьям и далее в умеренных областях к травам; от неветвистого ствола к ветвистому; от каулифлории к обычному расположению цветков; от сложного листа через пальматный к простому; от мясистых крупносемянных плодов с ариллусом к сухим плодам без ариллусов; от крупных семян без периода покоя к мелким с продолжительным периодом покоя; от монокарпических форм к поликарпическим), и его концепция происхождения однодольных
1 К этому же типу относятся некоторые представители подлеска из семейства Araliaceae, например виды родов Schefjlera и Brassaiopsis.
ПРЕДИСЛОВИЕ
13
не от Ranales (или от Helobiae), а от анцестральных саговникообразных пахикаульных форм принципиально отличаются от той схемы (и системы) развития покрытосеменных, которая основана на ныне почти общепринятой «стробилярной» или «граналиевой» доктрине. Все основные принципы «стробилярной» филогении и систематики, как каждому ясно, противоречат концепции Корнера. Тем не менее эту точку зрения нельзя замалчивать, хотя бы потому, что для ее обоснования Корнер приводит много интересных доводов, и она в какой-то мере объясняет загадку отсутствия ископаемых остатков первоначальных покрытосеменных растений. Критика же этой концепции, как показала статья одного из творцов стробилярной теории Джона Паркина1, оказалась способной в качестве главных аргументов приводить лишь обычные доводы филогенетиков этого направления, которые в сущности почти в такой же степени декларативны, как и положения «дурьян-теории», но зато и прямо ей противоположны.
В конце своей книги П. У. Ричардс обращает внимание на необыкновенно быстрое, даже катастрофическое, истребление тропических дождевых лесов, происходящее в последнее время. Он справедливо считает, что эти леса могут совсем исчезнуть, если не принять экстренных мер к их сохранению, уже на протяжении жизни одного поколения людей. А между тем консервация возможно более крупных участков этих лесов и их быстрое исследование могли бы иметь очень большое значение как для науки, так и для практических надобностей. Значение тропического дождевого леса для развития ботаники, в частности филогенетической систематики, необычайно велико. Некоторые западные ботаники — специалисты по тропикам, например Ван-Стеенис, вообще считают, что ботаника, в том числе и фитоценология, ныне имела бы совсем иной и более полноценный характер, если бы она разрабатывалась с самого начала на основе тропической флоры и растительности тропических областей. Помимо этого, тропический дождевой лес таит в своих недрах громадные возможности отыскания новых видов растительного сырья, новых полезных веществ растительного происхождения, ценнейших и еще даже не изученных древесин, смол, камедей, красок, физиологически активных веществ и т. п. Уже настало время видеть в остатках тропического дождевого леса не строевой лес, подлежащий рубке, а рассадник, маточный питомник различных ценных древесных пород и других растений для последующего их размножения и разведения в культуре.
Ан. А. Федоров
1 Parkin John, The Durian theory — A Criticism, «Phytomorphology», 3, 1953, p. 1—2.
Глава 1
ВВЕДЕНИЕ
большинство исследований растительности проводилось и Европе, и я думаю, что благодаря скудности материала отц исследования с самого начала исходили из неправильной предпосылки.
Сравнивая и сопоставляя различные типы растительности, логически следовало бы начинать с наиболее богатого типа, лишь затем изучая менее сложные, обедненные, возникшие из первого путем отбора. Наиболее богатый тип растительности по количеству видов, объему и густоте находится в тропиках. Обедненная, антропогенная растительность Европы не должна была бы служить отправной точкой исследований.
ВАН-СТЕЕНИС
(С. G. G. J. van Sleenis, 1937 а)
«Земли здесь высокие, и на них множество высочайших гор. Даже остров Тенериф не может сравниться с Эспаньолой.
Все эти горы необыкновенно красивы, формы их бесконечно разнообразны, все они проходимы, все заросли деревьями бесчисленных пород и такой высоты, что кажется, будто они достигают неба. Мне уже говорили, что они никогда не теряют свою листву, и я сам мог в этом удостовериться, ибо я видел их такими же зелеными и прекрасными, какими они бывают в мае в Испании. Некоторые были в цвету, другие с плодами, прочие в ином состоянии, сообразно их природе»1.
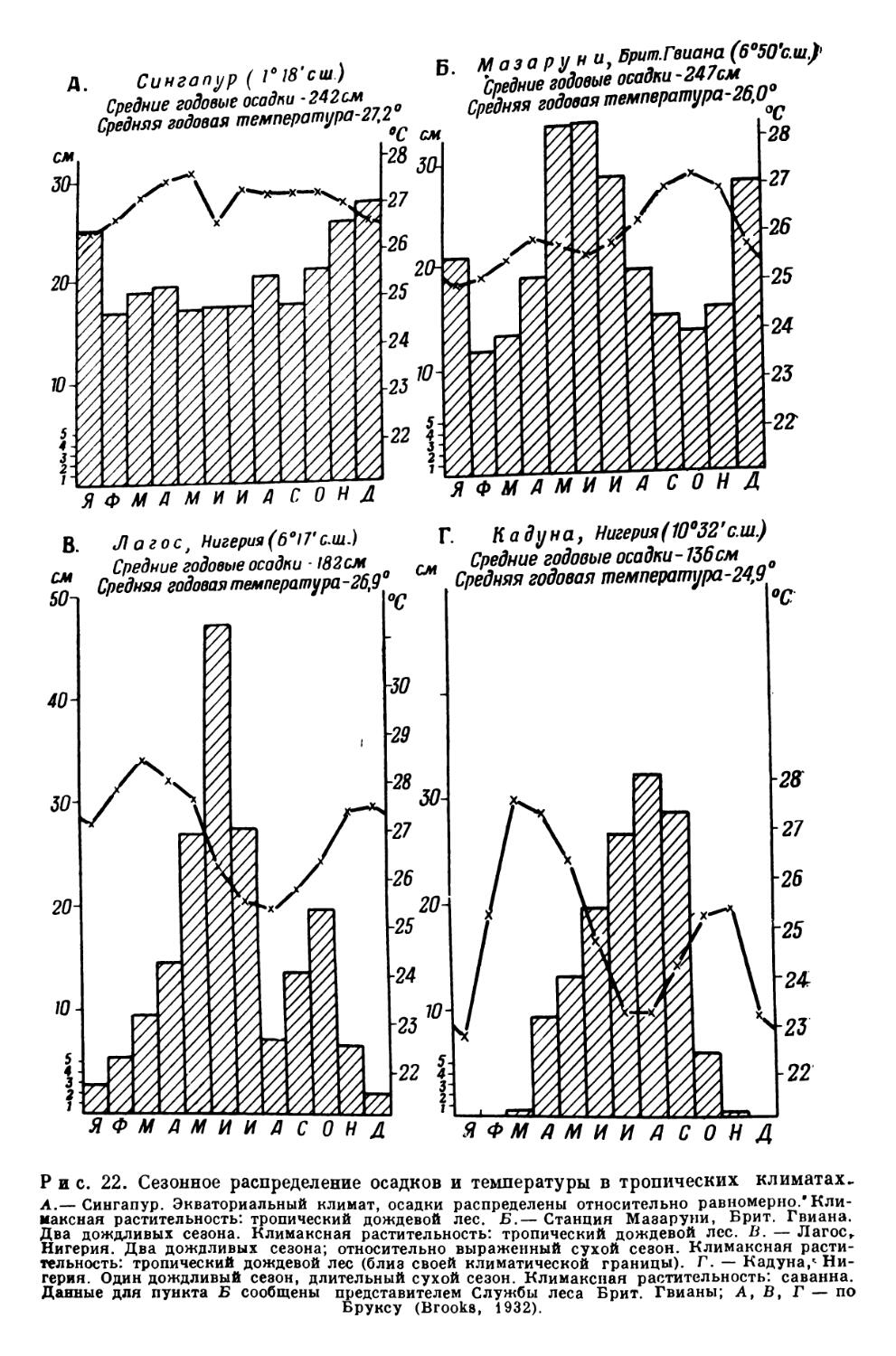
В приведенных строках, запечатлевших картину тропического ландшафта и написанных Христофором Колумбом в 1493 г. при знакомстве с островом Эспаньолой, содержится, возможно, самое раннее описание растительности, характеристике которой посвящена эта книга. Вечнозеленый тропический лес, произраставший во времена Колумба на большей части вест-индских островов, в настоящее время занимает обширные пространства приэкваториальных низменностей Южной и Центральной Америки, Африки и Юго-Восточной Азии. Эта характерная для влажных тропиков растительность покрывает или покрывала всю поверхность суши с достаточно жарким климатом и достаточно обильными равномерно распределенными осадками, за исключением небольших пространств, где почва слишком болотиста или где, как, например, на молодых вулканических лавах, она еще не успела развиться. В экологическом отношении вечнозеленый лес представляет собой климакс-ассоциацию в условиях экваториального климата.
1 «Путешествия Христофора Колумба» (Дневники, письма, документы), изд. II. Географгиз, М., 1952, етр. 64 (письмо Сантанхелю и Санчесу).
16
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
В научной литературе он чаще всего называется «тропическим дождевым лесом» (tropische Regenwald) — термин, предложенный Шимпером в его классической «Географии растений» (1898, 1903); иногда пользуются также латинским эквивалентом этого названия: «pluviisylva». Гумбольдт, один из первых научно описавший южноамериканский тропический лес, назвал его «гилеей» (от греческого vlq—лес). Немецкие авторы часто обозначают этим термином вечнозеленый тропический лес Африки. Часто пользуются они и словом «UrwaUb (первоначальный, первобытный лес). Последний термин может относиться к любому первобытному лесу, но по установившейся традиции применяется прежде всего именно к тропическому дождевому лесу. Англичане, живущие в тропиках, обыкновенно называют этот лес словом «bush», или «jungle», в Австралии он известен как «brush», или «scrub». [? Ред.]
Понятие «дождевой лес» обыкновенно относится не только к вечнозеленому лесу влажных тропических низменностей — растительной формации, или более точно, типу растительной формации, о которой идет речь в этой книге,— но также и к несколько менее роскошному вечнозеленому лесу, покрывающему нижние и средние участки склонов тропических гор, и к вечнозеленым лесам влажных субтропиков юго-западного Китая, южного Чили, южной Африки, Новой Зеландии и восточной, внетропи-ческой Австралии. В дальнейшем изложении эти типы формации будут называться горными дождевыми лесами и субтропическими дождевыми лесами; термин «дождевой лес» без специальной оговорки повсюду будет употребляться в смысле тропического дождевого леса.
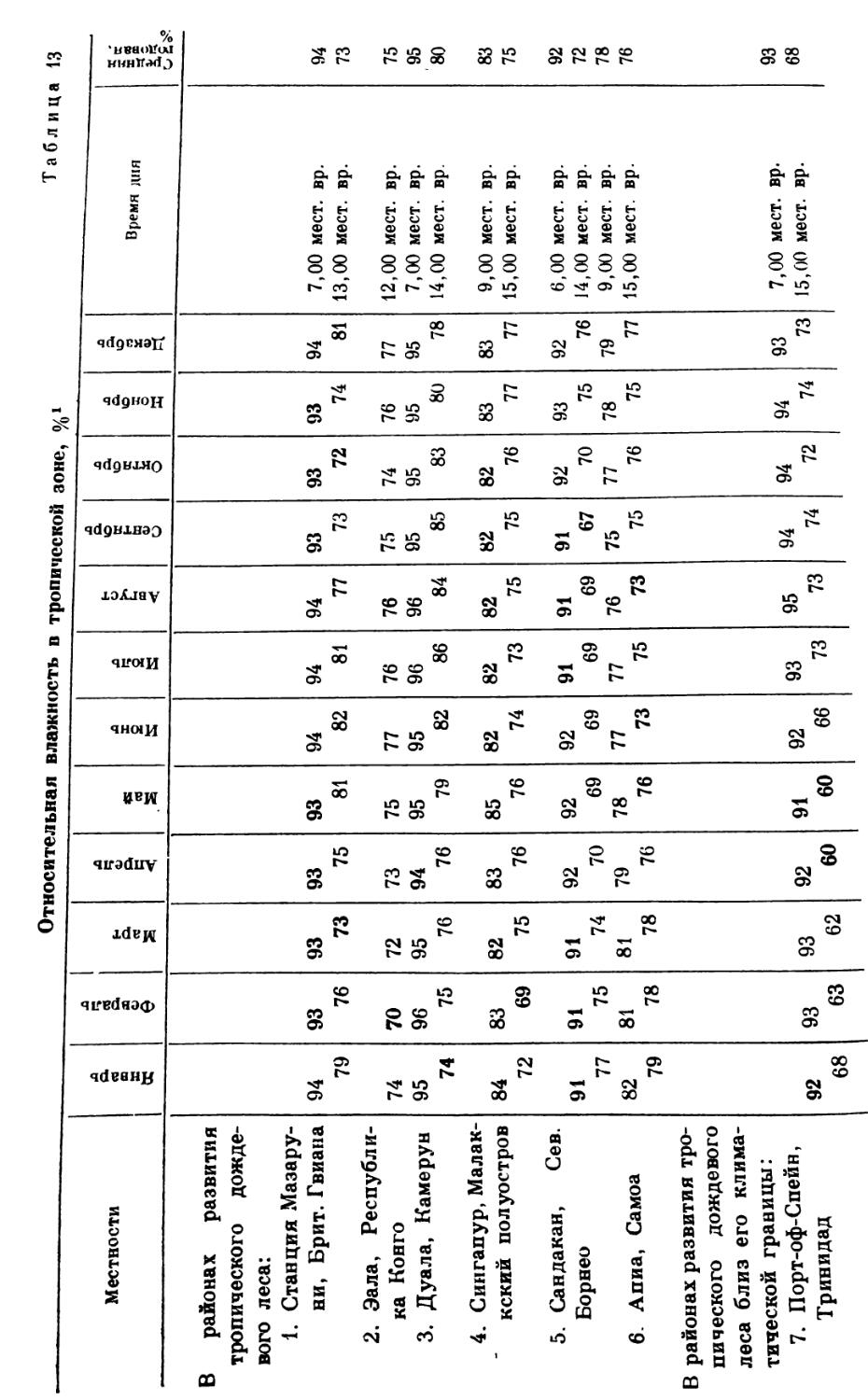
Что же представляет собой этот лес? Шпмпер (A. F. W. Schimper, 1903, стр. 260)1 характеризует его следующим образом: «вечнозеленый, гигрофильный по своему характеру, достигающий не меньше 30 м высоты, но обыкновенно гораздо более высокий, изобилующий толстоствольными лианами и как деревянистыми, так и травянистыми эпифитами». Указанное определение соответствует представлению о тропическом дождевом лесе, которое вкладывало в этот термин большинство авторов со времен Шимпера. Оно наиболее приемлемо и для целей данной книги, хотя ряд современных ученых предпочитает использовать его в менее широком смысле или же полностью отказывается от него. В более узком смысле понятие «тропический дождевой лес» должно было бы отвечать лесной растительности тропических климатов, развивающейся в условиях почти полного отсутствия сезонных колебаний метеорологических элементов и наиболее равномерного распределения осадков. В этом случае вечнозеленые леса, произрастающие в районе с заметно выраженным сухим сезоном, составляют обособленную группу. К этому вопросу мы еще вернемся в главах 15 и 16, где более подробно характеризуется различие между тропическим дождевым лесом, с одной стороны, и горным дождевым лесом, субтропическим дождевым лесом и другими, в той или иной степени листопадными тропическими лесными формациями, классифицированными Шимпером как леса муссонные, саванновые и колючие.
Но краткое, хотя и точное, определение дает лишь слабое представление о тропическом дождевом лесе. Это особенно относится к тем, кто знаком лишь с растительностью умеренных стран Европы и Северной Америки. Каковы же его характерные особенности? Какое впечатление производит он на наблюдателя?
1 Автор имеет в виду работу Шимпера «Plant-geography upon a physiological basis», перевод с немецкого W. R. Fischer, Oxford.
Фото 2. Полог тропического дождевого леса, Морабалли-Крик, Брит. Гвиана. Вид с высоты приблизительно 34 м над уровнем земли. На горизонте с левой стороны снимка ясно выделяется крона одного ив наиболее крупных деревьев, возвышающегося над окружающим лесом.
Фото 3. Подлесок леса Мора, Морабалли-Крик, Брит. Гвиана.
На снимке выделяется дерево Mora excelsa с досковидными корнями, принадлежащее к ярусу А. Маленькое дерево справа (Duguetia эр.) имеет облик, характерный деревьев яруса С.
ВВЕДЕНИЕ
17
Большинство людей, непосредственно не знакомых с тропической растительностью, составляют представление о ней по описаниям путешественников, к сожалению, часто предвзятым или преувеличенным, а то и совершенно неверным. Эти ошибки возникают либо вследствие слишком поспешных обобщений на основании осмотра небольшой территории, либо вследствие неверного предположения, что растительность внутри леса не отличается от растительности, окаймляющей берега рек, по которым, как правило, и пробирались путешественники. И все же основной виновницей неточности описаний является сама тропическая растительность, имеющая фатальную тенденцию склонять к риторическим преувеличениям всех, кто занимается ее изучением. Лишь немногим авторам удается устоять против искушения расцветить свою рукопись «блестящими пассажами», и большинство в потоке превосходных степеней теряет представление о реальности, описывая несуществующие вещи или неправильно истолковывая то, что действительно существует. При попытке нарисовать верную картину дождевого леса необходимо отметить и исправить некоторые из этих обычных ошибок.
Одной из замечательных особенностей тропического дождевого леса и, конечно, всей остальной растительности влажных тропиков является подавляющее преобладание в нем древесных растений. Древесные виды не ограничиваются деревьями, доминирующими в сообществе дождевого леса, к ним принадлежит и большинство лазящих растений, а также некоторые эпифиты. В значительной мере из деревянистых растений—сеянцев и подроста деревьев, а также из кустарников и молодых деревянистых лиан—состоит и подлесок. К травянистым растениям относятся лишь некоторые эпифиты и небольшое число видов подлеска. Растения, принадлежащие к семействам, которые в умеренной зоне представлены исключительно травянистыми, приобретают тут размеры и характер деревьев. Спрус (Spruce, 1908, I, стр. 256) так говорит о лесах Амазонии: «Здесь почти каждая естественная группа растений имеет серевья среди своих представителей. Травы (бамбуки) достигают 40, 60 и более футов высоты; их стебли стоят вертикально или переплетаются, образуя плотную стену колючих зарослей, сквозь которые не может пробраться даже слон. Вербеновые представляют собой раскидистые деревья с дланевидными листьями, подобными листьям конского каштана. Деревянистые лианы из семейства истодовых поднимаются к вершинам высочайших деревьев, увенчивая их гирляндами своих ароматных цветов. Обычный для вас барвинок заменяют красивые деревья, выделяющие млечный сок, у одних обладающий лечебными свойствами, у других смертельно ядовитый; те же качества присущи и их плодам. Фиалковые достигают размеров яблони. Маргаритки (или то, что могло бы показаться маргаритками) расцветают на деревьях, напоминающих ольху». В сообществе дождевого леса деревянистые растения образуют не только большую часть взрослой лесной растительности, но и играют главную роль во всех стадиях его развития с момента заселения обнаженных участков горных пород или почвы, а также при зарастании озер и болот.
Деревья дождевого леса крайне многочисленны по видовому составу и разнообразны по размеру. Размеры тропических деревьев иногда сильно преувеличивались. Средняя высота наиболее крупных деревьев дождевого леса редко превышает 150—180 футов (46—55 ле), хотя не являются исключением и деревья, превосходящие 200 футов (60 ле); кроме того, достоверно известно, что встречаются экземпляры, достигающие 300 футов (92 ле).
18
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Согласно Джонсу (Е. W. Jones, 1945), главнейшие древесные породы Европы и Северной Америки обычно не превышают 100 футов (30 м) высоты и лишь при исключительно благоприятных условиях вырастают до 150 футов (46 м). Следовательно, хотя деревья дождевого леса обыкновенно выше деревьев умеренных лесов, однако они никогда не достигают гигантских размеров калифорнийского красного дерева1 или австралийских эвкалиптов. По окружности и диаметру ствола самые крупные деревья тропического дождевого леса также далеко отстают от деревьев более высоких широт. Наиболее крупный из измеренных стволов имел в обхвате 56 футов (17 .и). Но деревья более 1 м в обхвате редки в сообществе дождевого леса, и он, пожалуй, более примечателен тонкостью стволов слагающих его древесных пород. Данные относительно размеров особенно крупных деревьев тропического дождевого леса и других деревьев приведены в табл. 1.
В лесах Европы и Северной Америки главные древесные породы принадлежат лишь к нескольким, а часто и только к одному виду; крайне редко лес состоит из 20—25 видов (Braun, 1941, стр. 236); в отдельных случаях число высоких деревьев достигает 25—30 видов на 0,5 га (лес «Атлантик» в Северной Америке, Schimper, 1935, стр. 831). Напротив, на 1 га тропического дождевого леса обычно насчитывается не менее 40 различных видов деревьев с диаметром ствола более 4 дюймов (10 см); иногда же на этой площади сосредоточивается более сотни видов. А. Р. Уоллес (А. В. Wallace, 1878, стр. 65) говорит: «Если путешественник заметит какой-нибудь вид и захочет найти несколько экземпляров его, самые тщательные поиски часто бывают напрасны. Его окружают деревья различнейшей формы, величины и окраски, но один и тот же вид повторяется редко. Сплошь да рядом он подходит к дереву, похожему на искомое, но тотчас же убеждается в их различии. Может быть, он и найдет второй экземпляр через полмили или же совсем его не найдет, но в другой раз может натолкнуться на него совершенно случайно»* 2.
Богатство древесной флоры является наиболее важной чертой дождевого леса, в непосредственной связи с которой находятся многие другие его особенности. Характерно для него и то, что деревья самых различных видов смешаны в довольно равномерной пропорции; гораздо реже один или два вида представлены более обильно, чем остальные.
Таким образом, тропический дождевой лес обыкновенно представляет собой сообщество с большим числом кодоминант, и лишь иногда в нем отчетливо преобладают только одна или две доминанты. Сообщества дождевого леса со многими доминантами мы будем называть смешанными лесами, а сообщества с одним господствующим видом — монодоминант-ными лесами (см. гл. 10, 11). Хотя деревья дождевого леса и относятся ко множеству видов, в целом их физиономичность и внешний вид отличаются исключительным однообразием. Их стволы, обычно прямые и тонкие, разветвляются лишь у самой вершины. У основания их обыкновенно имеются досковидные корни, плоские боковые выросты, в высшей степени характерные для деревьев дождевого леса. У представителей других растительных формаций такие опоры встречаются очень редко. Кора, как правило, тонкая и гладкая, редко имеет глубокие трещины или заметные чечевички. Огромное большинство зрелых деревьев, а также кустарников и молодняка характеризуется крупными кожистыми темно-зеле-
’ Sequoia tempervirens.— Прим. ред.
2 А. Р. Уоллес, Тропическая природа, М., Географгиз, 1956, стр. 76.
Id
Таблица I
Размеры наиболее крупных деревьев тропического дождевого леса1
Высота, м Виды Местонахождение Источник
84 Koompassia excelsa Саравак (остров Борнео) Fox worthy, 1927, стр. 84
81 К. excelsa Полуостров Малакка Foxworthy, 1927, стр. 84
71 Eucalyptus deglupta Новая Британия Lane-Poole, 1925а, стр. 214
70 Koompassia excelsa Саравак Beccari, 1904, стр. 330
70 Agathis alba Целебес Van der Koppel, 1926, стр. 529
59 Entandrophragma cylindri-cum Нигерия Kennedy, 1936, стр. 176
56 Desbordesia glaucescens Камерун Mildbraed, 1922, стр. 105
111 Sequoia sempervirens Калифорния Tiemann, 1935, стр. 903
107 Eucalyptus regnans Виктория (Австралия) Tiemann, 1935, стр. 904
75 Agathis australis Новая Зеландия Van der Koppel, 1926, стр. 530
46 Fagus sylvatica Франция Elwes and Henry, 1906, 1, стр. 13
40 Quercus robur Франция Elwes and Henry, 1907, 2, стр. 310
Ок-
ружность ствола- м Виды Местонахождение Источник
17 Entandrophragma cylindri-cum Нигерия Unwin (Kennedy, 1936, стр. 176)
14 Entandrophragma cf. ango-lense var. macrophyllum Нигерия Richards
13 Bertholletia sp> Бразилия Spruce, 1908, 1, стр. 18
123 Balanocarpus heimii Полуостров Малакка Foxworthy, 1927, стр. 53
~12э Dryobalanops aromatic a Полуостров Малакка Foxworthy, 1927, стр. 45
17 Tilia sp. Полуостров Малакка Kannegiesser (Biisgen and Munch, 1929, стр. 38)
15 Castanea sativa Полуостров Малакка Kannegiesser (Biisgen, MGnch, 1929, стр. 38)
23* Sequoiadendron giganteum (экземпляр «Генерал Шерман») Калифорния Tiemann, 1935, стр. 909
12 Quercus robur Уэльс Elwes, Henry, 1907, 2, стр. 309
23® Agathis australis Новая Зеландия Tiemann, 1935, стр. 911
1 Даны размеры самых крупных деревьев дождевого леса из указанных в литературе. Для сравнения приведены размеры крупнейших экземпляров нетропических деревьев.
2 «Это самое крупное дерево из всех виденных нами на полуострове» (Фоксуорси).
3 Окружность ствола вычислена по размерам диаметра.
4 Окружность ствола вычислена по размерам диаметра на высоте 10 футов над землей.
3 Окружность ствола вычислена по размерам диаметра.
2*
20
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ными, цельнокрайними или почти цельнокрайними листьями, напоминающими по размеру, форме и текстуре листья лавровишни (Primus lauroeerasus). Даже листочки, составляющие сложные листья деревьев из семейства бобовых, очень близки к ним по своему внешнему виду. Внешний облик листвы оказывается настолько однообразным, что наблюдатель неботаник легко может ошибиться, приняв самые различные деревья за виды лавра. Такая монотонность объясняется главным образом сходством формы и темной окраской большинства листьев. Крупные ярко окрашенные цветки встречаются редко, большинство деревьев и кустарников имеет не привлекающие внимания зеленоватые или беловатые цветки.
Однако на фоне значительного однообразия большинства деревянистых видов дождевого леса выделяется сравнительно небольшое число своеобразных растений, отличающихся по своему внешнему виду и строению от нормального типа. Это особенно относится к пальмам и видам Dracaena и Pandanus. Но так как эти растения редко встречаются в таком количестве, чтобы сильно повлиять на внешний вид сообщества в целом, и к тому же весьма часто вовсе отсутствуют, они гораздо менее бросаются в глаза в типичном дождевом лесу, чем об этом можно было бы судить на основании многих описаний.
Благодаря большим различиям высоты деревьев тропический дождевой лес обычно имеет многоярусное строение. Чаще всего наблюдается три яруса (кроме ярусов кустарников и трав), а иногда только два. Таким образом, в дождевом лесу, согласно часто цитируемой фразе Гумбольдта, располагается «лес над лесом». В главе 2 эти ярусы будут рассматриваться подробно.
Подлесок состоит из кустарников, травянистых растений и огромного количества подроста и всходов древесных пород. Книги, описывающие путешествия в тропиках, часто создают неправильное представление о густоте тропических лесов. Спелый (первичный) дождевой лес обыкновенно вполне проходим. Правда, по берегам рек или на прогалинах, где на почву падает большее количество света, образуются очень густые и часто совершенно непроходимые заросли, но внутри старых, ненарушенных лесов нетрудно двигаться во всех направлениях, лишь по временам раздвигая руками ветки и местами делая обходы, чтобы обогнуть упавший ствол или массу соскользнувших сверху лиан. Нож, или так называемый паранг, который принято брать с собой в лес, используется чаще всего для оставления ориентирных меток на деревьях, а не для того, чтобы прорубать себе путь. Продвижение в лесу замедляется не столько густотой растительности, сколько скользкой глинистой почвой и обилием упавших стволов и ветвей. Преувеличенное представление о густоте подлеска дают и фотографии; в лесу обыкновенно можно видеть друг друга на расстоянии по крайней мере 20 м. Внутри леса господствует сумрак, но когда сияет солнце, почва покрывается солнечными бликами. Гизенхаген (Giesenhagen, 1910, стр. 728) удачно сравнивает освещение дождевого леса с освещением европейского букового леса в его «весенней зелени». Наземные травянистые растения встречаются спорадически; количество видов трав на единицу площади обыкновенно меньше, чем в английском лесу; меньше и число индивидуумов. Почва покрыта только тонким слоем мертвых листьев; часто попадаются пятна оголенной земли. Тропические леса, в которых почва была бы «покрыта годами скоплявшейся гниющей растительностью», представляют собой редкое исключение или просто игру воображения. Кроме деревьев, кустарников и травянистого покрова, дождевой лес состоит еще из лазящих растений различного облика и размеров, а также эпифитов,
ВВЕДЕНИЕ
21
растущих на стволах, ветвях и даже на живых листьях деревьев и кустарников.
Обилие лазящих растений является одной из наиболее характерных его черт. Большинство их представлено деревянистыми видами лиан. Для них характерны очень длинные и толстые стволы. Нередко встречаются стволы толщиной в человеческое бедро. Некоторые лианы тесно прилегают к поддерживающим их деревьям, но большинство довольно свободно взбирается вверх к пологу крон, протягиваясь наподобие канатов, или свисает вниз петлями и фестонами. Количество видов лазящих растений огромно, причем по форме и строению они очень разнообразны.
Эпифитная растительность, кроме свойственных лесам умеренной зоны водорослей, мхов, печеночников и лишайников, состоит из большого количества орхидей и других цветковых растений, а также множества папоротников. Эпифиты, растущие на вершинах деревьев (часто невидимые с земли), представлены как кустарниковыми, так и травянистыми видами, а также полупаразитическими Loranthaceae и курьезными «удушителями» (главным образом видами Ficus), начинающими свое существование в качестве эпифитов, но впоследствии часто становящимися независимыми деревьями, укореняющимися в почве. Ни в каком ином растительном сообществе, за исключением некоторых типов горных и субтропических дождевых лесов, не наблюдается такого обилия и роскоши эпифитов.
Итак, тропический дождевой лес отличается крайне сложной структурой и состоит из растений главным образом деревянистых, но относящихся к самым разнообразным жизненным формам; причем некоторые из них совершенно не похожи на те, которые встречаются среди растительности умеренной зоны. Однако первое впечатление, которое производит это действительно наиболее своеобразное из всех растительных сообществ как по структуре, так и по богатству видового состава, есть впечатление мрачности и однообразия. Такое впечатление, обусловливаемое главным образом подавляющим преобладанием деревянистых растений и однообразием листвы почти у всех жизненных форм, усиливается отсутствием заметных сезонных изменений. Здесь нет ни зимы, ни весны, а только вечное лето в разгаре; растительность в основном сохраняет один и тот же вид в любое время года. Правда, имеются сезоны максимального цветения, когда в цвету находится большее количество видов, чем в другое время, а также сезоны максимального образования молодых листьев, но, как правило, рост и размножение растений происходят непрерывно и всегда можно встретить то или иное дерево в цвету. Это прямое следствие климата, отличающегося очень слабыми сезонными изменениями, а также высокой температурой и влажностью. Жизнедеятельность растений не прерывается ни холодом, ни засухой.
Считается, что тропический дождевой лес есть тип формации, или, согласно Клементсу (Clements), панклимакс. Этот тип формации объединяет три отдельные формации, занимающие основные области земного шара с экваториальным климатом: формацию дождевого леса Центральной и Южной Америки, формацию дождевого леса тропической Африки и формацию дождевого леса Юго-Восточной Азии, Австралии и островов Тихого океана. В каждой из этих формаций почти все виды, и многие из родов и семейств свойственны именно данной формации и отсутствуют в остальных двух. Однако их структура и последовательные стадии развития весьма сходны между собой. Каждая из них, как мы увидим, изменяется соответственно различиям климата и почвы, обнаруживая при этом удивительный параллелизм, несмотря на неоднородный состав флоры.
22
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
В любой области, Где тропический дождевой лес потенциально является климатическим климаксом, спелым лесом бывает покрыта только часть пространства. Наряду с ним существуют большие территории, занятые болотами, озерами, поймами рек и заболоченными эстуариями, растительность которых находится в различных стадиях развития в направлении дождевого леса. Подобные же условия создаются на обнаженных горных породах и молодых вулканических лавах, где еще только формируется почвенный слой. В ряде мест особенности почвы и рельефа по каким-либо причинам делают невозможным развитие типичного дождевого леса даже при наличии соответствующего климата. Здесь можно встретить сообщества, напоминающие верещатники или саванну. Постоянные сообщества подобного рода называются эдафическими климаксами.
Во всех частях зоны дождевого леса, кроме самых отдаленных и наименее обитаемых мест, имеются обширные пространства, где лес был уничтожен под пашню. Многие из когда-то возделывавшихся земель были затем заброшены и вновь покрылись естественной растительностью. Эта растительность вскоре принимает внешний вид первоначального леса, но все же в течение многих лет может быть легко распознана; это так называемый вторично выросший или вторичный лес в противоположность девственному, или первичному, лесу. Термины «первичный* и «климакс-ный» (или спелый) лес не являются синонимами, так как в неспелом лесу человек мог и отсутствовать, и поэтому он является первичным, хотя не климаксным. В I, II, III и V частях книги будет рассмотрен главным образом спелый, или климаксный, дождевой лес, а в IV и VI частях — первичные и вторичные сукцессионные сообщества.
БИОЛОГИЧЕСКИЙ СПЕКТР ДОЖДЕВОГО ЛЕСА
Чтобы точно определить диапазон жизненных форм в тропическом дождевом лесу и долю участия каждой из них в создании леса как целого, было бы желательно построить биологический спектр для небольшого количества типичных представителей первичного леса, пользуясь или хорошо известной «эфармонической» классификацией жизненных форм Раункиэра (Raunkiaer, 1934), или одной из недавно предложенных более чисто «физиономических» систем, вроде классификации «главных жизненных форм» Дю-Риэ (Du Rietz, 1931). К сожалению, в нашем распоряжении нет исчерпывающих сведений о жизненных формах ни одного из участков дождевого леса. Это особенно касается травянистых и более мелких деревянистых растений. В таблицах 2 и 3 представлены данные, на основании которых можно составить себе некоторое, правда весьма приблизительное, представление о биологическом спектре.
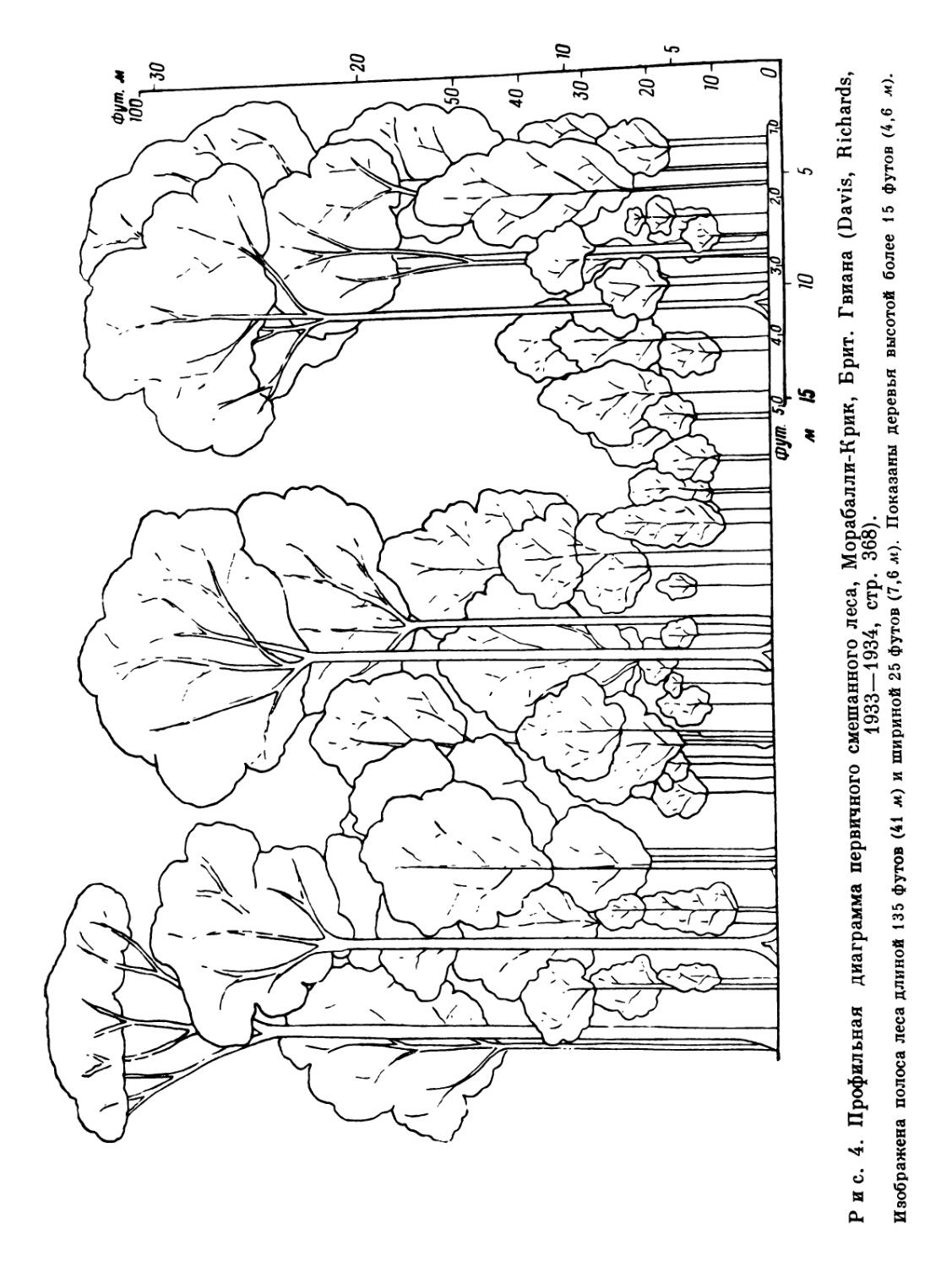
Таблица 2 основана на сборах цветковых растений, произведенных Сэндвизом в 1929 г. в Брит. Гвиане в местности Морабалли-Крик. Коллекция была получена с площади радиусом приблизительно в 5 миль (8 км), целиком покрытой лесом, за исключением очень маленьких прогалин (старых лагерей лесорубов и т. п.). Дождевой лес на этом участке местами является первичным (но не сплошь принадлежащим к одному типу, см. гл. 10), а отчасти вторичным. Большая часть сборов производилась в первичном лесу. Этот список, содержащий свыше 400 видов цветковых растений, конечно, не может считаться исчерпывающим, причем некоторые компоненты флоры (высокие деревья, лианы и эпифиты) представлены менее полно, чем другие («кустарники» и флора травянистого покрова). Данные, использованные для составления таблицы 3, относятся
ВВЕДЕНИЕ
23
Таблица 2
Жизненные формы цветковых растений в тропическом дождевом лесу, Морабалли-Крик, Брит. Гвиана1
Количество видов Жизненные формы по Раункиэру (1934) Жизненные формы по Дю-Риэ (1931)
Деревья:
Двудольные Высокие пальмы 163 3 Мега- и мезофанерофиты Очень высокие, высокие и низкие деревья
«Кустарники» и наземные травы:
Зеленые травы2 и «кустарники» 80 Микро- и нанофанерофиты, хамефиты Низкорослые и карликовые деревья, ктонофит-ные кустарники (?), травянистые растения
Сапрофиты 13 — —
Паразиты 1 — —
Адвентивные виды 8 Терофиты Травянистые растения
(на прогалинах)
Лианы и другие лазящие растения 83 Мезо- и мегафанерофиты Деревянистые и травянистые лианы
Эпифиты:
Кустарниковые (включая полу эпифиты) около 10 Эпифиты Эпифитоидные кустарники , эпифитоидные карликовые кустарники и т. д.
Травянистые около 81 Эпифиты Эпифитоидные травянистые растения
Полупаразиты (Loranthaceae) 7 Эпифиты Паразитические карликовые кустарники
1 Данные Сэндвиза (N. Y. Sandwith).
2 Зеленые в смысле не паразиты и не сапрофиты.—Прим. ред.
Данные, помещенные в этой таблице, не дают точных сведений об относительном количестве фанерофитов и хамефитов. Однако если принять, что число хамефитов равно половине количества «зеленых трав и кустарников» плюс паразиты и сапрофиты (безусловно, завышенная оценка), а вся флора, без учета адвентивных растений, составляет 441, то биологический спектр Раункиэра будет выглядеть следующим образом:
Стебле-суккуленты Эпифиты Фанерофиты Хамефиты Гемикриптофиты Геофиты Гело- и гидрофиты Терофиты
0 22 66 7 12 8 0 0 0 0
только к одному пробному участку величиной 400 х400 футов (122 х Х122 м), выбранному в дождевом лесу южной Нигерии.
При всех своих недостатках таблицы 2 и 3 хорошо иллюстрируют некоторые отличительные черты растительности дождевого леса. В них
24
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Таблица 3
Жизненные формы цветковых растений на опытном участке (400x400 футов) смешанного дождевого леса, Никрова, Нигерия1
Количество видов Жизненные формы по Раункиэру (1934) Жизненные формы по Дю-Риэ (1931)
Деревья более 4 дюймов (10 см) в диаметре:
Двудольные 70 Мега- и мезофанерофиты Очень высокие, высокие и низкие деревья
Высокие пальмы 0 — —
«Кустарники» 9 Нанофанерофиты Низкорослые и карликовые деревья, ктоно-фитные кустарники (?)
Наземные травы:
Зеленые травы 16 Нанофанерофиты, хамефиты Травянистые растения
Сапрофиты 0 — —
Паразиты 0 — —
Адвентивные виды 0 — —
Лианы и другие ла- Многочис- Мега- и мезофанеро- Деревянистые и травя-
зящие растения ленны фиты нистые лианы
Эпифиты Многочисленны Эпифиты Эпифитоидные карликовые кустарники, эпифитоидные травянистые растения
1 Данные Ричардса (Richards, 1939).
отчетливо проявляются отдельные важные различия между этим лесом и лесными сообществами умеренной зоны (см. также рис. 1). Хотя приведенные данные относятся только к двум небольшим участкам, у нас нет основания полагать, что выводы, извлеченные из них, не имеют общего значения.
Таблицы и рис. 1 прежде всего показывают огромное преобладание во флоре дождевого леса фанерофитов («деревьев», по классификации Дю-Риэ). Если бы сравнительные данные, которыми мы располагаем, относились не к видам, а к индивидуумам растений, то такое преобладание оказалось бы подавляющим. Во-вторых, «кустарниковые» и травянистые ярусы дождевого леса состоят главным образом из растений таких жизненных форм, которые отличаются от жизненных форм соответствующих ярусов лесов умеренной зоны. Дальнейшие исследования, вероятно, покажут, что большинство так называемых кустарников не есть настоящие кустарники, как их определяет Дю-Риэ («гелоксилы.., не дающие отдельного главного ствола, со стеблем, ветвящимся от базальной части, над или под поверхностью»), а миниатюрные деревья с одним главным стволом (см. стр. 95). В то время как в лесах умеренной зоны флора травяного покрова содержит большую пропорцию гемикриптофитов и геофитов, в тропическом дождевом лесу гемикриптофиты почти или пол
ВВЕДЕНИЕ
25
ностью отсутствуют, а геофиты встречаются редко (см. стр. 116); напочвенный покров леса состоит почти исключительно из фанерофитов и хамефитов.
Терофиты совершенно отсутствуют, за исключением прогалин, тропинок и т. п., где они иногда попадаются. Количество эпифитов (группы, в которую в дождевом лесу входят растения нескольких различных жизненных форм) значительно превосходит число наземных трав.
Таким образом, характерные черты биологического спектра тропического дождевого леса достаточно ясны; очевидно, что они находятся в тесной связи и являются отражением постоянно благоприятных для роста растений климатических условий, не знающих сезонных изменений Это видно из отсутствия жизненных форм, приспособленных к сезонным колебаниям климата, приносящим холод или засухи (гемикриптофиты, геофиты, терофиты) и из обилия вечнозеленых фанерофитов — жизненной форумы, наименее способной сопротивляться неблагоприятным условиям. Крайний случай полной неприспособленности к неблагоприятным климатическим условиям представляют пальмы. Крона пальмы является одиночно переннирующей почкой; в области кроны нет запасных почек, так что если вершина окажется поврежденной, то отмирает весь побег. «Подобно безрассудному игроку»,— говорит Шрётер (Schroeter) (цит. Рикли, Rikli, 1943, стр. 30), — «пальма все ставит на одну карту». Неветвящиеся одноствольные (Schopfbaumchen, см. стр. 57), представляющие собою своеобразный, весьма характерный для дождевого леса тип двудольных фанерофитов, по той же причине приспособлены только для роста в постоянно благоприятной среде. Преобладание фанерофитов в большинстве стадий сукцессий, ведущих к утверждению сообщества дождевого леса (гл. 12 и др.), служит убедительным доказательством их соответствия существующим условиям климата. Сумрак, создаваемый вечнозеленым пологом наряду с жестокой конкуренцией корней, вероятно,
39
Рис. 1. Сравнение биологического спектра флоры тропического дождевого леса М орабал л и-К рик, Б рит.
Гвиана (7), с биологическим спектром флоры листопадных лесов Германии (77).
Каждая колонка представляет число видов той или иной жизненной формы в процентах ко всему флористическому составу. Спектр леса Морабалли-Крик носит приближенный характер и составлен по способу, объясненному в примечании к табл. 2. Спектр листопадных лесов представляет собой среднее из спектров шести листопадных лесных ассоциаций северо-западной Германии (Тйхеп, 1929).
обусловливает бедность напочвенной флоры.
Однако постоянно влажная атмосфера и высокая температура делают
возможным развитие обильной, главным образом травянистой, эпифит-
ной растительности.
26
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
РАСПРОСТРАНЕНИЕ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА В ПРОШЛОМ И НАСТОЯЩЕМ
Сведения о распространении дождевого леса как. типа формации далеко не полны. На прилагаемой карте (рис. 2) показано пространство, в пределах которого дождевой лес представляет собою при современных климатических условиях клпмаксную растительность. Однако из-за малого масштаба эта карта не может претендовать на очень большую точность.
В ряде наиболее плотно населенных тропических районов первоначальная растительность в такой степени была заменена культурными и антропогенными сообществами, что представляется трудным, а то и совсем невозможным точно реконструировать первоначальный климатический климакс. В то же время некоторые отдаленные области так мало исследованы, что их растительность почти неизвестна. Многие из опубликованных карт растительности тропических стран не вызывают большого доверия, тем более что разные авторы весьма различно истолковывали понятие «тропический дождевой лес» (и другие наносимые на карту сообщества). Хотя дальше, в главах 15 и 16, будет идти речь о границах дождевого леса, некоторое представление о пределах его распространения небесполезно дать и здесь, конечно, в той мере, как это представляется возможным.
Тропический дождевой лес опоясывает земной шар, рассекаясь экватором на две несколько неравные половины, так что большая его часть оказывается расположенной в северном полушарии. Благодаря наличию горных цепей и плато, а также в силу неравномерного распределения основных климатических факторов (зависящих в свою очередь от неравного распределения суши и моря) пояс этот прерывается в нескольких местах. Северные и южные границы дождевого леса не совпадают с какими бы то ни было ясно определяемыми широтными пределами; в некоторых местах они не достигают географических тропиков, в иных они слегка выходят за их пределы.
Самый крупный монолитный массив тропического дождевого леса находится в бессейне реки Амазонки. Он простирается на запад до предгорий Анд и на восток до Гвианы, прерываясь лишь сравнительно небольшими участками саванн и листопадных лесов. Великая южноамериканская гилея, или дождевой лес, на юге вторгается в область Гран-Чако, а на севере по восточному краю Центральной Америки проникает в южную Мексику и на Антильские острова. На крайнем северо-западе Южной Америки (Эквадор, Колумбия) узкая полоса тропического дождевого леса тянется вдоль западных склонов Анд. Вторая прибрежная полоса дождевого леса располагается на восточном берегу Бразилии (от 6° ю. ш. до тропика Козерога, несколько выходя за его пределы), отделяясь от амазонского массива широким поясом листопадного леса. Однако, по-видимому, границы тропического дождевого леса в Америке известны хуже, чем в любой другой из главных тропических областей. Наиболее современная карта распространения леса этого типа (Smith, Johnston, 1945) не более подробна и, вероятно, не многим более точна, чем карта, помещенная в шимперовской «Географии растений» (1898, 1935).
В Африке дождевой лес в основном сосредоточен в бассейне Конго. Оттуда он простирается на запад в пределы бывшей Французской Экваториальной Африки, Габон и Камерун. Узкая полоса дождевого леса, тянущаяся параллельно берегу Гвинейского залива, проникает еще дальше
Рис. 2. Районы распространения тропического дождевого леса.
В Америке — по Смиту и Джонстону (Smith, Johnston. J945);» по Чемпиону (Champion, 1936) и Ван-Стеенису (van Steenie, 1935а); в Австралии — по Френсису (Francis, 1929); на островах Тихого океана ж в Африке — из различных
В Африке указанна площадь наряду с тропическим дождевым лесом включает и смешанный листопадный лес (сухой вечнозеленый лес), так как граница между ними не вполне исследована.
ВВЕДЕНИЕ
27
на запад в Нигерию, Гапу, Либерию и Гвинейскую республику. Эта полоса на участке между западной частью Нигерии и восточным районом Ганы прерывается областью с сухим климатом. В восточном направлении дождевой лес едва достигает области Великих озер, совершенно не встречаясь в своем типичном выражении к востоку от Великого африканского грабена. Южнее бассейна Конго дождевой лес представлен главным образом пост-климаксными галерейными лесами, уходящими в сторону Родезии.
Вопрос о границах центральноафриканского дождевого леса и о степени проникновения в него саванн был предметом больших споров. Более старые исследователи, вроде Стэнли (Stanley), полагали, что непрерывный лесной массив покрывал большую часть бассейна Конго. Позднее Вильдеман (De Wildeman Е., 1913, 1926) высказал предположение, что такой взгляд является неверным и объясняется тем, что большинство путешественников двигалось по рекам. По его мнению, предполагаемый центральноафриканский лесной массив состоит главным образом из узких полос галерейного леса, ограничивающихся приречными участками, а местности, лежащие в промежутках, как правило, заняты саванной. С этой точки зрения большая часть африканского дождевого леса не находится в соответствии с современными климатическими условиями, и есть скорее пост-климакс — результат благоприятных почв и топографии. Вильдемановская «Теория галерейного леса» сильно оспаривалась Милд-брэдом (Mildbraed, 1914, 1923). Вообще современные бельгийские исследователи стоят на старой точке зрения, считая, что африканский лес является целостным массивом, хотя в ряде мест и нарушается пятнами саванн, возделываемой почвы и вторичной растительности. Предмет спора был критически пересмотрен Лебреном (Lebrun, 1936 а). Составленная им карта дождевого леса бассейна Конго является, вероятно, наиболее верной изо всех до сих пор опубликованных1. Карту растительности Французской Западной Африки составил Шевалье (Chevalier, 1912, 1938 а и т. д.).
Тропический дождевой лес представляет собой естественный климакс по крайней мере для части Сейшельских островов, острова Реюньон и острова Маврикий, а также для восточного побережья Мадагаскара, но первичный лес на этих островах в настоящее время почти не сохранился.
В восточной Азии тропический дождевой лес произрастает на Цейлоне, в Западной Индии, Таиланде, остальной территории Индокитая и Филиппинских островах, а также на островах Индонезии и в Новой Гвинее. Самые обширные пространства его находятся на Малаккском полуострове и прилегающих островах — Суматре и Борнео, где индомалайский дождевой лес достигает исключительной роскоши и флористического богатства, и в Новой Гвинее. Площадь, занятая тропическим дождевым лесом, в Индии невелика; местами он встречается в Западных и Восточных Гатах, а также в значительной степени распространен в низкогорной зоне Восточных Гималаев, в горах Кхаси и в Ассаме. В Бирме, Таиланде и Индокитае дождевой лес встречается только местами. Здесь преобладает формация муссонного леса. В восточной части Зондских островов от западной Явы до Новой Гвинеи развитию тропического дождевого леса препятствует засушливый сезон, обусловленный сухим восточным муссоном из Австралии. Произрастание дождевого леса здесь возможно лишь
1 Бернар (Bernard, 1945) убедительно показал необоснованность взгляда о несоответствии леса Конго современным климатическим условиям.
28
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
в форме пост-климакса при благоприятных местных условиях. Хорошая карта растительности Индонезии с подробными границами распространения тропического дождевого леса была опубликована Ван-Стеенисом (van Steenis, 1935 а).
В Австралии индо-малайский тропический дождевой лес тянется к югу в виде узкой полосы вдоль восточного берега. Он произрастает и на островах западной части Тихого океана (Соломоновы, Новые Гебриды, Фиджи, Самоа и др.).
В указанных выше районах тропический дождевой лес, по-видимому, является естественным климаксом при климатических условиях, существующих в настоящее время. Известно несколько ограниченных областей, где площадь дождевого леса, по-видимому, в настоящее время увеличивается; упомянем, например, южную часть Конго (Robyns, 1936), Уганду (Eggeling, 1947), но в целом вследствие деятельности человека площадь, занимаемая первичным лесом, быстро уменьшается. Существуют обширные области, где этот лес совершенно исчез, а также области, вроде южной Нигерии, где лес, когда-то представлявший собою непрерывный массив, оказался разбитым на изолированные островки. Интенсивное истребление леса происходило главным образом в течение последних 50—100 лет, и, следовательно, еще совсем недавно девственный дождевой лес покрывал значительную часть земной поверхности. Судя по различного рода данным изменения климата, в не меньшей степени, чем разрушительная деятельность человека, сократили пространство, занятое ранее тропическим дождевым лесом; крупные территории, где в настоящее время развитие его уже невозможно, в более ранние периоды при иных климатических условиях были покрыты лесами, похожими на современный тропический дождевой лес.
В особенности многочисленны подобные указания для Африки. Много писалось относительно «высыхания» Африки (см. Stebbing, 1937 а, 1937 Ь; Lavauden, 1937) и о предполагаемом значительно большем распространении лесов в прошлом. Считают, что в западной части Африки постепенное иссушение климата и расширение Сахары в южном направлении продолжаются и в наши дни. Это вызвало соответствующее отступление вечнозеленого леса (вместе со смешанным листопадным лесом, саванной и колючим лесом, образующими экотон дождевого леса-пустыни), и местности, которые даже в историческое время были окружены лесом, теперь находятся в саванне или в пустыне. Эти гипотезы были критически рассмотрены Обревилем (Aubreville, 1938).
Дело в том, что вопрос этот, имеющий первостепенное значение, является вместе с тем в высшей степени спорным и может быть рассмотрен только вкратце. Перед нами встает двоякая проблема: во-первых, можно ли доказать, что изменения климата и перемены в распределении клима-ксных растительных формаций происходили на протяжении исторического периода и происходят еще и теперь, и, во-вторых, имели ли место такие изменения в недавнее геологическое время (четвертичный период)? Обре-виль говорит, что, хотя за тот короткий исторический период, относительно которого у нас имеются достаточные данные, в Западной Африке, возможно, и наблюдались колебания климата, однако по этим колебаниям еще нельзя судить о вековой направленности климатических изменений. Описание северной Нигерии, относящееся к 1822—1824 гг. и сделанное* путешественниками Денгамом (Denham) и Клапертоном (Clapperton), свидетельствует о наличии в то время климатических условий, совершенно тождественных современным. Это противоречит представлению
ВВЕДЕНИЕ
29
о быстром ухудшении и иссушении климата, на котором настаивали такие ученые, как, например, Стеббинг (см. об этом у Джонса, Jones, 1947). Если в действительности наблюдается постепенное передвижение на юг пустыни, саванны и лесов, то это может быть целиком отнесено за счет человеческой деятельности (подсечно-огневое земледелие, пастбищное хозяйство, пожары), ведущей к постпенному замещению климатически обусловленных формаций биотическими климаксами (см. гл. 15). Совершенно иным является вопрос об изменениях климата и растительности с плейстоцена до современности, и доказательства тут носят другой характер. Как геологические данные (остатки растений, ископаемые почвы, геоморфология и т. д.), так и биогеографические данные (реликтовые виды и т. д.) указывают на существование в свое время в тропической Африке влажных (дождливых), периодов, когда вечнозеленый лес простирался далеко за его теперешние границы. Хотя мысль о синхронности по крайней мере некоторых из этих влажных периодов с плейстоценовым оледенением Европы очень заманчива, определенных доказательств в пользу существования подобной взаимосвязи крайне мало (Zeuner, 1945).
Данные о распространении вечнозеленого тропического леса на востоке Африки во время дождливых периодов были рассмотрены Моро (Moreau, 1933). В горах Узамбара, Нгуру и Улугуру находятся небольшие (общая площадь их составляет меньше 300 кв. миль, или 777 кв. км) изолированные участки леса, похожие по своему характеру на тропический дождевой лес, но которые, пожалуй, лучше всего было бы назвать предгорным дождевым лесом (см. гл. 16). Как фауна, так и флора этих лесов, хотя она и состоит из эндемичных видов, обнаруживает поразительное сходство с фауной и флорой тропического дождевого леса в западной и центральной частях Африки. На основании изучения главным образом птиц Моро делает вывод, что эти восточноафриканские дождевые леса представляют собой реликты обширного лесного массива, который когда-то составлял одно целое с лесом Центральной Африки. Его развитие было бы возможным при увеличении среднего годового количества осадков, выпадающих в Восточной Африке на 15—20 дюймов (38—50 см). Чэпин (Chapin, 1932) также на основании изучения птиц высказал предположение, что тропический дождевой лес, занимая в прошлом значительно большую территорию, соединялся с лесом восточных тропических районов через Аравию и Индию. Составленная им карта изображает предполагаемое распространение леса в Африке во время влажного периода в плейстоцене. Фитопалеонтологические данные свидетельствуют (см. van Steenis, 1935 а) о существовании на низменностях Явы во время третичного периода дипте-рокарповых лесов, типичных сейчас лишь для ассоциаций тропического дождевого леса Малайи, Суматры и Борнео. Современные низинные леса Явы, почти целиком замененные сельскохозяйственными угодьями и тиковыми плантациями, представляют собой, за исключением крайнего запада, скорее муссонные, чем настоящие дождевые леса, и Dipterocar-расеае, встречаются в них только в незначительном количестве. Хотя многие диптерокарповые характерны и для лесов сезонного типа, их преобладание в прошлом может указывать на более влажный и более ровный климат того времени, а также на прежнее распространение тропического дождевого леса к востоку от его современной границы.
Еще раньше по свидетельству ископаемой флоры в меловом периоде, эоцене и олигоцене, даже в странах столь удаленных от тропиков, как Англия, существовала растительность, которая по флористическому составу, а возможно, и по внешнему облику близко напоминала современ
30
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ный тропический дождевой лес. Но обсуждение этого вопроса здесь завело бы нас слишком далеко.
Весьма вероятно, что по крайней мере в некоторых областях, где в настоящее время тропический дождевой лес является климатическим климаксом, подобная растительность неизменно существовала с очень отдаленного геологического времени. Согласно Постумусу (Posthumus, 1931, см. также vanSteenis, 1935 а), третичные леса Малезии только очень незначительно отличались по своему флористическому составу от современного дождевого леса.
ЗНАЧЕНИЕ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА ДЛЯ ЭКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Тропический дождевой лес оказался одним из последних в ряду основных типов формаций растительности, подвергнутых изучению экологами. Но весьма возможно, что по мере накопления данных по его экологии могут быть добыты новые важные сведения, которые дадут много нового относительно растительных сообществ умеренной зоны, до сих пор являвшихся основным объектом экологических исследований. Обилие видов в климаксных сообществах тропиков создает большие трудности для эколога, впрочем, компенсируемые важными преимуществами. Изучение в высшей степени сложных взаимоотношений видов друг с другом и со средой во флористически богатых сообществах тропиков не могут не обнаружить многого такого, что относится к законам, управляющим конкуренцией и доминированием видов в смешанных сообществах. Уже одно только изобилие видов в ассоциациях тропического дождевого леса представляет, как мы это увидим ниже, счастливую возможность разобраться в отношении форм растений к окружающей среде, в частности, например в отношении размера и формы листа к климату и микроклимату. Хотя форма растения в значительной степени определяется генетическими факторами, характерными для семейства, рода или вида, она отчасти регулируется также и средой, действующей или косвенно, через естественный отбор, или прямо, через развитие индивидуума. В сообществах, вроде дождевого леса, где имеется много видов из различных систематических групп, но сходной жизненной формы, конвергентные (или эфармонические) сходства между неродственными видами выступают гораздо более ясно, чем у сообществ, состоящих только из небольшого числа видов. Как указывал Ван-Стеенис, было бы логичнее начинать экологические исследования с неспециализированных, флористически богатых растительных сообществ экваториальной области и идти от них по направлению к специализированным, обедненным и сильно измененным сообществам умеренных областей, а не обратно. Но тропическая растительность еще настолько мало известна, что ее значение для экологии в общем еще слабо выявлено.
Ботаники пытались рассматривать тропическую растительность как атипичную, а растительность умеренных зон — как типичную, просто потому, что они были более знакомы с последней. Браун (W. Н. Brown, 1919) говорит: «Человек, выросший в стране с умеренным климатом, будет считать листопадный широколиственный лес обычным, а тропический лес —странным и причудливым. С другой стороны, обитатели влажных тропиков считают листопадные леса умеренных областей весьма странными. Конечно, было бы логично рассматривать широколиственные вечнозеленые леса тропиков обобщенным типом, а листопадный лес уме-
ВВЕДЕНИЕ
3U
ренных областей — как приспособленный к тому, чтобы противостоять зимним условиям, точно так же, как колючая растительность пустынных областей приспособлена к сухому климату. Нет также никаких оснований рассматривать наличие пальм, древовидных папоротников и эпифитов-чем-то более своеобразным, чем их отсутствие, в особенности, если это отсутствие связано с неблагоприятными условиями. Равным образом, густой лес из многих видов не должен рассматриваться чем-то более особенным по сравнению с более открытым, состоящим из небольшого числа< видов».
Тропический дождевой лес является приютом преимущественно для» широколиственных вечнозеленых деревьев, то есть формы, от которой, кажется, произошли все, или большая часть других цветковых растений. Различные данные указывают, что наши умеренные флоры прямо или» косвенно имеют тропическое происхождение. Следы этого до сих пор* сохранились у многих видов умеренной зоны или в их фенологии (Diels, 1918) или в их структуре. Бьюс (Bews, 1927), например, выдвинул сильный довод в пользу того, что влажнотропическая флора Африки филогенетически старше, чем флора более сухих и более холодных областей; его статистическое сравнение ископаемой и современной флор заставляет думать, что с начала мелового периода и до конца третичного большая часть земного шара имела «фанерофитный климат», и что растительность была, по крайней мере физиономически, похожа на растительность современного тропичесого дождевого леса. Вполне возможно, как полагает Бьюс, что самые ранние покрытосеменные были похожи экологически и по их жизненным формам на современную флору тропического дождевого леса,, рассматривающуюся как древний тип растительности, от которого сравнительно недавно возникла флора более сухих тропических и умеренных областей. Огромное флористическое богатство тропического дождевого-леса, несомненно, в значительной мере связано с его глубокой древностью; это был центр эволюции растений в течение крайне долгого времени.
Часть I
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Чтобы «понять» данное растительное сообщество, еще недостаточно просто знать, в каких условиях оно находится. Гораздо важнее проникнуть в суть того, как оно построено и какова его структура. Наука о растительности есть морфология растительных сообществ».
МОР 3 ЕЛЬ (Н. MEUSEL, 1935, стр. 269)
Глава 2
СТРУКТУРА: СИНУЗИИ И ЯРУСВОСТЬ
СИНУЗИИ
Строение растительного сообщества иногда сравнивают со структурой человеческого общества, хотя ни у кого не вызывает сомнения, что такое сравнение не может идти дальше чисто внешней, формальной параллели.
В более сложных растительных сообществах виды растений образуют экологические классы, или группы. В сообществе, как целом, размеры, облик и жизненные формы видов различны, но внутри одной экологической группы среди составляющих ее растений наблюдается сходство жизненных форм и отношения к окружающей среде. Эти экологические группы мы будем называть «синузиями», термином, введенным Гамсом в 1918 г. (Gams, 1918). Следовательно, синузия представляет собой группу растений весьма близкой жизненной формы, экологически однородных и играющих одинаковую роль в сообществе, часть которого они составляют. По словам Сэкстона (Saxton, 1924), применявшего этот термин в несколько более широком смысле, чем Гаме, синузия представляет собой совокупность видов (или особей), предъявляющих одинаковые требования к условиям местообитания. Виды одной синузии, часто весьма различающиеся по систематическому положению, являются в значительной мере эквивалентными экологически.
В таком растительном сообществе, как умеренный летнезеленый лес, составляющие его синузии легко различимы и имеют сравнительно простое пространственное расположение. Обыкновенно древесные растения образуют два яруса — один из высоких деревьев, а другой — из кустарников или более низкорослых деревьев. Ниже располагается один или большее число ярусов, состоящих из трав и полукустарников (наземные или покровные ярусы) и ярус мхов и печеночников. Кроме этих независимых растений, каждый ярус которых может рассматриваться как отдельная синузия, имеются еще лазящие растения и эпифиты (последние в данном случае, как правило, состоят целиком из бессосудистых растений) и синузии сапрофитов и паразитов, слагающиеся из небольшого числа цветковых растений, а также грибов и бактерий.
В тропическом дождевом лесу, где синузии более многочисленны, их пространственное расположение выражено гораздо менее отчетливо. Хотя тропические леса, подобно лесам умеренной зоны имеют определенную структуру, на первый взгляд они производят впечатление хаотического нагромождения растительности; согласно часто цитируемому выражению Юнгхуна (Junghuhn), природа здесь так лихорадочно стремится заполнить все стеблями и листьями растений, что кажется одержимой боязнью пространства. Тем не менее более пристальное изучение показывает, что в дождевом лесу, как и в других сложных растительных сообществах, растения образуют ограниченное число синузий с хотя и более сложным, но отчетливым расположением в пространстве. Такое строение
з*
36
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
с незначительными местными различиями свойственно всему типу формации тропического дождевого леса. Структура первичного тропического дождевого леса Америки, Африки и Азии по существу тождественна. Леса всех трех континентов образуют сходные синузии, которые, как будет показано в этой главе, имеют аналогичное пространственное распределение. Виды соответствующих синузий различных географических областей сходны как по своим жизненным формам, так в значительной мере и по своему внешнему облику. Таким образом основной тип структуры тропического дождевого леса повсеместно сохраняется.
Как уже говорилось, наши сведения о флористическом составе тропического дождевого леса все еще настолько неполны, что на основании их оказывалось невозможным построить полный биологический спектр даже для отдельного ограниченного его участка. Это одинаково справедливо как для эфармонической системы жизненных форм Раункиэра, так и для чисто физиономической, вроде предложенной Дю-Риэ. Но из этого следует, что, поскольку синузия представляет собой группу растений, относящихся к одной жизненной форме, исчерпывающая классификация синузий тропического дождевого леса при современном уровне знаний невозможна. Поэтому в практических целях приходится пользоваться приблизительной, крайне упрощенной схемой. Одна из таких схем, представляющая собой несколько видоизмененную классификацию, применявшуюся автором в предыдущих работах, приводится ниже. Ее достоинство заключается в удобстве и легкости применения.
Синузии тропического дождевого леса
А. Автотрофные растения (хлорофиллоносные)
1. Механически независимые растения
а) Деревья и «кустарники» ]
б) Травы ) °°РазУют *РУСЫ
2. Механически зависимые растения
а) Лазящие
б) У душители
в) Эпифиты (включая полупаразитические эпифиты) Б. Гетеротрофные растения (лишенные хлорофилла)
1. Сапрофиты
2. Паразиты
В основу этой схемы положен способ, посредством которого растение удовлетворяет потребность в углеводах. Каждая синузия использует собственный метод борьбы за пищу.
Автотрофные растения, сами вырабатывающие необходимые им углеводы, непосредственно зависят от света и по способу его достижения делятся на две группы. Механически самостоятельные или независимые растения достигают света без помощи других растений; механически зависимые растения (соответствующие товариществам, или Genossenschajten, Шимпера) этого делать не могут. Независимые растения могут быть подразделены на несколько ярусов, число и структура которых рассматриваются ниже. Зависимые растения обнаруживают менее ясную ярусность, определяемую главным образом ярусностью независимых растений, служащих им поддержкой. Одна часть их — синузия лазящих растений — состоит из растений, укореняющихся в почве, но нуждающихся в поддержке для своих слабых стеблей. Другая часть — эпифиты — состоит из растений с более или менее короткими стеблями. Они не могут
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 37
расти вблизи почвы и укореняются на стволах и ветвях других растений, Полупаразитические эпифиты, представленные единственным семейством Loranthaceae, получают воду и минеральные вещества от деревьев, на которых они развиваются (это не касается или почти не касается органической пищи). Однако большинство эпифитов использует поддерживающие их растения лишь в качестве механической опоры. Наконец, третья группа зависимых растений, называемая здесь удушителями. является переходной. Эти растения, начинающие жизнь в качестве эпифитов, впоследствии укореняются в почве; некоторые из них достигают значительных размеров и могут в конце концов заглушить и заместить своего «хозяина», как это делают, например, удушающие фикусы (Ficus spp.). Следовательно, удушители начинают жизнь как зависимые растения, но с течением времени становятся независимыми; все же их нельзя резко разграничить с полуэпифитами, вроде многих Агасеае, которые хотя и укореняются в почве, но никогда не становятся независимыми.
Класс гетеротрофных растений образуют сапрофиты и паразиты. Они не обязательно лишены способности реагировать на свет, но, получая органическую пищу прямо или косвенно от других растений, они не нуждаются в нем для своего питания и совершенно устраняются с арены борьбы за него.
Более подробная характеристика этих различных групп будет дана дальше, в 4 и 5 главах, где, в частности, рассматриваются пх физиономические признаки.
Насколько известно, нет такого климаксного сообщества, к которому может быть применено название тропического дождевого леса и в котором отсутствовала хотя бы одна из перечисленных экологических групп. Точно так же нет такого участка дождевого леса, где наблюдались бы синузии, отсутствующие в других районах. Различия в плане или схеме структуры касаются главным образом степени развития той или иной синузии. Так могут, например, встречаться сообщества, где эпифиты представлены большим числом видов и индивидуумов и обнаруживают большее разнообразие форм, чем в других; некоторые дождевые леса богаче лазящими растениями, чем другие, и т. п. Пространственное расположение синузий также изменяется в большей или меньшей степени.
ЯРУСНОСТЬ
Структура, общая всякому климаксному тропическому дождевому лесу, наиболее ясно обнаруживается в основной черте его архитектуры, а именно в ярусности деревьев, кустарников и покровных трав. Давно стало общим местом учебников, что в тропических лесах деревья образуют несколько расположенных друг над другом ярусов (применяются также и другие термины: слой, этаж, полог), тогда как в лесах умеренной зоны никогда не бывает более двух ярусов деревьев, в ряде случаев наблюдается лишь один. Впрочем, по этому поводу высказывались различные мнения. Так категорическому утверждению о наличии в тропическом дождевом лесу трех древесных ярусов противостоит поддерживаемое несколькими специалистами мнение, что их больше трех. Браун (Brown, 1919, стр. 31—32), например, следующим образом характеризует ярус-ность диптерокарпового леса на Филиппинах: «Деревья располагаются тремя довольно отчетливыми ярусами. Первый, или доминирующий, образует сомкнутый полог; под ним расположен следующий ярус из крупных
38
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
деревьев, также образующий сомкнутый полог. Еще ниже находится ярус, состоящий из небольших разбросанных деревьев». Описания некоторых авторов создают впечатление, что ярусы тропического дождевого леса также определенны и легко распознаваемы как в английской лесосеке с маячными деревьями, но Броун продолжает: «наличие трех ярусов, состоящих из различных деревьев, не обращает на себя внимания при беглом наблюдении, потому что состав всех ярусов весьма сложен, а деревья лишь в редком случае обладают какими-либо обращающими на себя внимание особенностями; к тому же более мелкие деревья верхнего яруса постоянно встречаются в ниже расположенном ярусе и между различными ярусами».
Ряд авторов утверждают (или высказывают предположение), что любая группировка деревьев по их высоте является произвольной и что ярусов в действительности не существует. Такой точки зрения придерживается Милдбрэд (Mildbraed, 1922, стр. 103—-104), который, конкретно ссылаясь на лес южного Камеруна, недвусмысленно говорит: «Часто утверждают, что тропический дождевой лес состоит из нескольких ярусов, или этажей. Это легко может создать ложное представление у всякого, кто сам не знаком с действительным положением вещей. На деле это означает лишь то, что деревянистые растения можно по желанию сгруппировать в 3, 4, 5, а возможно, и большее число классов по высоте. Мысленно вполне можно представить себе пространство, состоящее по высоте пз интервалов в 5, 10 или 20 м\ существуют и виды деревьев, достигающие при полном росте в нормальных условиях этих высотных интервалов. Но поскольку наряду с ними произрастают деревья, занимающие промежуточное положение по высоте, и примесь таких деревьев велика, то эти гипотетические высотные интервалы никогда в действительности не имеют вида обособленных ярусов. Вернее было бы сказать, что все пространство более или менее плотно заполнено растительностью». Подобного же мнения придерживается Шевалье (Chevalier, 1917, стр. 354—355) и другие.
В настоящей главе будет сделана попытка показать, что в большинстве сообществ нормального первичного тропического дождевого леса действительно, как правило, существуют три древесных яруса. Эти ярусы, хотя и всегда присутствующие, выражены, однако, плохо и редко распознаются при беглом наблюдении. Кроме древесных ярусов, имеется еще ярус, состоящий из «кустарников» и гигантских трав (таких, как высокие Scitamineae), и ярус низких трав и полукустарников.
Под ярусом разумеется строй деревьев, кроны которых варьируют по высоте около какой-то средней величины. В лесном сообществе с несколькими ярусами каждый ярус отличается особым флористическим составом, но так как лес непрерывно растет и возобновляется, то известная часть, а возможно, и большинство отдельных деревьев, находящихся в нижнем ярусе, принадлежит к видам, достигающим в зрелом состоянии более высокого яруса. Кроны молодых деревьев более высокого яруса и исключительно высокие деревья ниже расположенного яруса могут быть обнаружены в промежутке между ними. Если таких индивидуумов много, как это часто и бывает, то переход между смежными ярусами будет малозаметен и ярусность окажется сильно завуалированной. Деревья одного яруса могут образовывать как сомкнутый, так и разреженный полог. В первом случае кроны в большинстве своем будут находиться в боковом соприкосновении одна с другой, во втором — расположены с широкими промежутками. В ряде случаев в качестве синонима яруса применяется слово
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 39
«полог». Но этот последний термин обозначает более или менее сплошной слой древесных крон приблизительно одинаковой высоты. Иногда вся сомкнутая поверхность, или кровля, леса называется пологом. В тропическом дождевом лесу, как мы увидим, эта поверхностть может быть образована либо кронами деревьев самого высокого яруса, либо, что наблюдается чаще, кронами самого высокого и смежного с ним яруса.
Расхождение мнений относительно существования, природы и числа ярусов дождевого леса объясняется главным образом трудностью отчетливого обозрения леса в профиль. Находясь в лесу на земле, наблюдатель,
136-145
126-135
3 116-125 ^106-115 -g 96 -105 L
75 -
26- 35 -
100
150
16-25„ п
О Ю 20 30 40 50
65 -
55 -
36- 45 -
76-85
I 66-
I 56-
46—
Рис. 3. Общая высота древостоя и высота расположения самых нижних листьев деревьев на расчищенных участках близ Морабалли-Крик (Davis, Richards, 1933— 1934, стр. 367).
Сплошная линия характеризует распределение деревьев по высотным интервалам; прерывистая линия — высоту самых нижних листьев для всех деревьев выше 15 футов (4,6 м) на площади 400x50 футов (122X15,3 м).
200
как говорит Милдбрэд, является пленником. Видимость очень ограничена как в вертикальном, так и в горизонтальном направлениях. Из-за хаоса листьев, ветвей и стволов только иногда удается увидеть кроны более высоких деревьев. Правда, то тут, то там брешь, оставленная в пологе гибелью крупного дерева, приоткрывает завесу над верхними ярусами; а рассекающие заросли река или вырубка позволяют увидеть поперечное сечение леса сверху донизу. Однако такие поперечные разрезы могут ввести в сильное заблуждение. В местах, где свет получает возможность достигнуть более низких уровней леса, в подлеске происходит мощная вспышка роста, лазящие растения спускаются вниз с полога и приподнимаются с почвы, пока вся открытая поверхность не покроется плотной завесой листвы, которая совершенно скрывает от наблюдателя расположение крон деревьев. Структура леса на речном берегу всегда отличается от структуры внутри леса.
Делались попытки изучения ярусности тропического дождевого леса путем нанесения на диаграмму встречаемости деревьев, разбитых на произвольно установленные классы по высоте. Так, Буберг (Booberg,
40
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
1932) учел высоты всех занумерованных деревьев в резервированном лесу на Яве, пользуясь способом Курдерса (Koorders). Анализ полученной им непрерывной кривой не выявил какой-либо закономерности в характере распределения деревьев по высоте. Из этого он сделал вывод, что в лесах Явы нельзя различить «этажей» (ярусов), описанных Дильсом (Diels) и Рюбелем (Rubel). Сходную кривую (рис. 3), но обнаруживающую легкую тенденцию к максимальной встречаемости у известного класса высоты1, получили Дэвис и Ричардс (Davis, Richards, 1933—1934, стр. 367), пользуясь измерениями деревьев на сплошь вырубленных участках в смешанном дождевом лесу Брит. Гвианы. Воган и Впэ (Vaughan, Wiehe, 1941, стр. 131. 137) показали, что непрерывные кривые не опровергают наличия ярусности. Они нашли, что диаграмма в «нагорном климаксном лесу» острова Маврикия, отражающая частоту встречаемости деревьев в зависимости от размеров диаметра, представляет собой непрерывную кривую, хотя при визуальном наблюдении ясно выявляется заметная ярусность (изображено в их статье в виде точного перспективного рисунка).
Профильная диаграмма
Так как непосредственное наблюдение ярусности дождевого леса обыкновенно представляет непреодолимые трудности, то Дэвис и Ричардс (Davis, Richards, 1933—1934) предложили способ построения профильной диаграммы для передачи в масштабе на основании точных измерений положения, высоты, ширины, а также высоты кроны всех деревьев, находящихся на узких пробных полосах. Этот метод, впервые примененный в лесах Брит. Гвианы, впоследствии использовался многими исследователями в различных частях тропиков, причем он оказался ценным средством изучения и сравнения структуры сообществ тропического леса. Метод этот вкратце состоит в следующем.
Узкая прямоугольная полоса леса обносится веревкой, причем прямые углы строятся при помощи призматического компаса. В дождевом лесу длина полосы должна составлять не менее 200 футов (61 м) при ширине 25 футов (7,6 ле). Весь мелкий подлесок и деревья мельче произвольно выбранного нижнего предела вырубаются. Оставшиеся деревья наносятся на план с указанием размера диаметра. Затем измеряются общая высота каждого дерева, высота его до первой (крупной) ветви, нижний предел кроны и ее ширина. Часто эти измерения оказывается возможным получить, только срубив все деревья избранной полосы, причем они должны валиться в тщательно продуманном порядке с таким расчетом, чтобы более тяжелые деревья не сокрушили при падении более мелкие. Иногда необходимые измерения удается с достаточной точностью произвести с земли, используя уровень Абнея. Преимущество рубки состоит в возможности сбора гербария для определения видов.
В предыдущей главе указывалось, что, хотя тропический дождевой лес чаще всего имеет смешанный состав, где главные породы представлены очень большим числом видов, существуют и такие типы леса, в которых доминирует лишь один вид, представители которого безусловно преобладают или же полностью составляют самый высокий ярус. Структура смешанного тропического дождевого леса и монодоминантного леса различна в некоторых отношениях и будет рассмотрена отдельно.
1 Класс 76—85 футов (23—27 лс); это соответствует приблизительно ярусу В (см. ниже).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 41
Ярусность смешанного дождевого леса
Рис. 4—6 представляют собой профильные диаграммы типичных смешанных дождевых лесов. Рис. 4 характеризует смешанный лес Брит. Гвианы, рис. 5—смешанный диптерокарповый лес на острове Борнео, где доминируют представители семейства Dipterocarpaceae (см. гл. 10), рис.6— смешанный лес в южной Нигерии. Леса Брит. Гвианы и Борнео являются примерами типичного экваториального дождевого леса [«дождевой лес» в узком смысле Кия и Джонса (Keay, Jones), Бирда (Beard) и т. д., «экваториальный лес» бельгийцев (см. стр 361)], в то время как лес в Нигерии относится к слегка отличающемуся климаксному типу леса, свойственному климатам с несколько менее равномерным распределением осадков («влажный вечнозеленый лес», «тропический лес» бельгийцев). Тщательное изучение этих трех диаграмм даст, вероятно, более ясное представление об ярусности нормального дождевого леса, чем словесное описание, но все же желательно вкратце описать структуру каждого избранного примера, дополнив сведения, даваемые диаграммами, замечаниями, вытекающими из непосредственных наблюдений на месте.
Первичный смешанный лес, Морабалли-Крик, Брит. Гвиана (рис. 4). На диаграмме показаны три яруса деревьев (обозначим их буквами А, В и С, сверху вниз). Самый нижний из них (С) является сплошным и довольно хорошо выражен, верхние два — более или менее разрежены и неясно отделены по вертикали друг от друга. В первоначальном описании (Davis, Richards, 1933—1934, стр. 362—372) они рассматривались как один ярус неправильного профиля. Большинство окон в самом верхнем ярусе А закрыто деревьями яруса В; следовательно, вместе ярусы А п В образуют сомкнутый полог.
Средняя высота деревьев яруса А составляет на диаграмме около 35 м, но она может быть и выше (вплоть до 42 м). Ярус В имеет около 20 м высоты, а ярус С, включающий деревья между 20 м и произвольной нижней границей в 4,6 м (15 футов), достигает в среднем приблизительно 14 м высоты. На профильной полосе (длина 135 футов, или 41 м) находится 66 деревьев высотой более 15 футов; 7 из них можно отнести к первому ярусу, 12—ко второму, а остальные — к третьему.
Кроны яруса А только местами находятся в боковом соприкосновении, но следует помнить, что их полог более сомкнут, чем это кажется при взгляде на диаграмму, потому что на ней не показаны деревья, чьи кроны частично покрывают пробную полосу, а основания находятся вне ее. Деревья яруса А принадлежат ко многим видам и семействам (Lecythidaceae, Lauraceae, АгаПасеаеит.д.). Их кроны, обычно больше развитые в ширину, чем в высоту, приближаются к зонтичной форме.
Ярус В является более сомкнутым, но все же имеет кое-где окна. Подобно ярусу А он состоит из видов, принадлежащих к многочисленным семействам (большей частью другим, чем в ярусе А). Наряду с ними значительную группу составляют молодые особи, достигающие яруса А. Кроны больше развиты в высоту, чем в ширину.
В ярусе С очень мало окон, и плотность листвы и ветвей тут больше, чем на любом другом уровне леса, безразлично, более высоком или более низком. Более половины общего количества индивидуумов составляют молодые деревья видов, принадлежащих, по сути дела, более высоким ярусам; остальные представлены низкорослыми видами, свойственными собственно ярусу С; они относятся главным образом к семействам, едва представленным в ярусах А и В (в частности, Аппопасеае и Violaceae).
Рис. 4. Профильная диаграмма первичного смешанного леса, Морабалли-Крик, Брит. Гвиана (Davis, Richards, 1933-1934, стр. 368).
Изображена полоса леса длиной 135 футов (41 м) и шириной 25 футов (7,6 м). Показаны деревья высотой более 15 футов (4,6 м).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 43
Как молодые деревья ярусов А и В, так и виды, действительно относящиеся к ярусу С, обыкновенно имеют длинные, заостряющиеся конические кроны, гораздо более развитые в вышину, чем в ширину.
Ниже трех ярусов деревьев, представленных на профильной диаграмме, располагаются еще два яруса, состоящие преимущественно или частично из деревянистых растений; оба эти яруса плохо выражены. Верхний, в среднем достигающий приблизительно 1 м высоты, может быть назван ярусом кустарников (D), хотя в его состав входит много молодых деревьев, маленьких пальм, высоких трав (Marantaceae и др.) и крупных папоротников, а также небольших деревянистых растений, принадлежащих к жизненной форме кустарников или «карликовых деревьев» (см. стр. 95). Самый нижний наземный ярус (Е) состоит главным образом из проростков деревьев, где травянистые растения (двудольные, однодольные, папоротники и Selaginella) образуют лишь незначительную часть общего количества индивидуумов. Подобно кустарниковому этот ярус обычно отличается разреженностью, за исключением открытых мест и участков, где виды, растущие сообществами, образуют густые заросли.
На лесной почве мхи отсутствуют. За исключением пятен мхов, вроде Fissidens spp., на нарушенном почвенном покрове (вывороченные деревья, гнезда броненосцев и т. д.) мохообразные ограничиваются поверхностью живых или мертвых деревьев (см. стр. 144—145).
Для проверки профильной диаграммы ярусность леса исследовалась с помощью подъема на высокие деревья, расположенные в этом же лесу, но не примыкающие к опытной полосе. Здесь на высоте 110 футов (33,6 м) были сделаны следующие наблюдения: «Полог не представляет собой плоской поверхности, выделяются два более или менее ясно выраженных (верхних) яруса, но, поскольку оба они являются разреженными, создается впечатление очень неровной поверхности. Любые два высоких дерева могут быть отделены друг от друга одним или большим числом более низких деревьев. Более низкие деревья не растут непосредственно под более высокими и не сильно затеняются ими. Практически все более низкие деревья полога покрыты и переплетены между собой лианами. Последние обыкновенно не достигают деревьев самого высокого яруса. Кроме обычных деревьев верхнего полога (яруса А), встречаются одиночные деревья, заметно возвышающиеся над всеми другими. С нашей точки наблюдения были видны два таких дерева; их кроны ясно отделялись от крон окружающих деревьев».
На фото 2 показан вид, открывающийся с наблюдательного пункта; одно из упомянутых «маячных» деревьев видно на горизонте. На профильной диаграмме эти деревья отсутствуют. Они, вероятно, встречаются в среднем не чаще, чем по одному на квадратный километр, и достигают 40—45 м высоты. Обычно они принадлежат к видам, редко встречающимся в ярусе А (например, Нутепаеа courbaril, Peltogyne pubescens).
Результаты непосредственного наблюдения с высоты 110 футов подтверждают правильность профильной диаграммы. Оба метода убеждают в трехъярусном строении леса; редкие «маячные» деревья не образуют самостоятельного яруса.
Первичный смешанный диптерокарповый лес, гора Дьюлипг, Борнео (рис. 5). Первоначальное описание структуры леса дано Ричардсом (1936). Здесь также имеются три яруса, но вместо выраженного яруса С и плохо выраженных А и В, самый высокий ярус, хотя и разреженный, хорошо выражен и резко отделяется в вертикальном направлении от яруса В, ярус же В трудно отделить от яруса С. Средние высоты этих трех ярусов
44
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
составляют 35, 18 и 8 м соответственно. На пробной полосе было 7 деревьев в ярусе А, и 86—в В и С (около 48 в В и 38 в С); эти цифры нельзя непосредственно сравнивать с цифрами гвианского профиля, потому что здесь полоса была длиннее (200 футов, 61 м) и, кроме того, измерялись только деревья высотой в 25 футов (7,6 м) и более (вместо 15 футов и более).
Кроны деревьев яруса А большей частью довольно хорошо отделяются друг от друга и отчетливо возвышаются над очень плотным вторым ярусом. Поэтому при обозрении леса сверху (например, с гребня откоса) отдельные кроны деревьев яруса А видны поодиночке на большом расстоянии друг от друга. Большинство деревьев этого яруса принадлежит к Dip-terocarpaceae. Кроны длиной в среднем около 6 м обычно имеют большие' размеры в ширину, чем в высоту. Ярус В отличается почти полной сомкнутостью, каждая крона составляющих его деревьев находится обыкновенно в боковом соприкосновении с несколькими другими; полог почти так же плотен, как и полог яруса С. Видовой состав яруса очень разнообразен и образован многочисленными семействами. Имеется и небольшое количество молодых диптерокарповых. Кроны в 4, 5—Gm длины, как правило, больше развиты в высоту, чем в ширину.
Ярус С также сомкнутый. Он состоит как из молодняка видов, достигающих во взрослом состоянии ярусов А и В, так и из низкорослых деревьев, редко превышающих 45 футов высоты; причем первые преобладают. И те и другие деревья отличаются очень высокими и узкими кронами обычно конической формы. Низкорослые деревья, характерные для этого яруса, принадлежат ко многим семействам, но интересно отметить, что, как и в лесу Гвианы, здесь в изобилии встречаются представители семейства Аппопасеае. Пространство между ярусами В и С заполнено деревьями промежуточной высоты, причем наблюдения, произведенные в пределах всего района, показывают, что здесь имеются две совершенно отдельные группы видов — виды яруса В, достигающие во взрослом состоянии в среднем высоты около 18 м и не цветущие, пока они еще слишком маленькие, и виды яруса С со средней высотой во взрослом состоянии около 8 м, начинающие цвести задолго до достижения этой высоты. Таким образом, отсутствие разрыва по вертикали между средним и самым нижним ярусами деревьев объясняется отчасти обилием не вполне зрелых деревьев, нормально свойственных ярусу В, и отчасти наличием деревьев,, обычных в ярусе С, но превышающих их средний рост.
Ниже яруса С плотность растительного покрова заметно уменьшается. В подлеске уже не заметно ярусности, но его удобно подразделить на слой кустарников высотой в среднем до 4 м, состоящий из собственно кустарников, пальм и молодняка, и наземный слой, достигающий 1—2 м высоты и состоящий из сеянцев древесных пород и трав (включая папоротники и Selaginella). Здесь же изредка встречается один или два вида очень маленьких пальм. Число видов травянистых растений значительно, но сеянцы древесных пород превосходят их по количеству; на десяти опытных квадратах (размер каждого 1 кв. м) было найдено 184 сеянца древесных пород, кустарников и других деревянистых растений и 135 травянистых растений (побеги). Плотность наземного яруса неравномерна; на больших пространствах он представлен растениями, распространенными диффузно; местами же довольно густ, хотя редко достигает плотности, характерной для покрова в европейском листопадном лесу. В смешанных диптерокарповых лесах, растущих на низких местах, нет напочвенного яруса мхов, но последние иногда встречаются на очень крутых склонах.
I !
фут. 50 25 5
м 15 Ю 5
I' и с 5. Профильная диаграмма первичного смешанного диптерокарпового леса, гора Дьюлш, Р
Иаображена полоса леса длиной 200 футов («1 м) и шириной 25 футов (7,в лО. Нонаваны деревья высотой более
46
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Первичный1 смешанный лес («влажный вечнозеленый лес»), резервированный лес Шаша, Нигерия (рис. 6). Структура леса подробно описана Ричардсом (1939). Подобно лесам Гвианы и Борнео, лес Нигерии имеет три яруса. Сомкнутым является только самый нижний ярус (С), включающий деревья высотой до 15 м (в среднем около 10 м}. Над ним располагается беспорядочная масса деревьев различной высоты, самые высокие из которых достигают 46 м. Кроны этих высоких деревьев иногда соприкасаются своими краями, но выше яруса С не существует сомкнутого полога. Наблюдения над всем пространством, часть которого составляла пробная полоса, показали, что эта беспорядочная масса может быть разделена на два яруса: ярус А — из деревьев высотой 37—46 м (в среднем около 42 м) и ярус В — из деревьев высотой 15—37 м (в среднем около 27 м). Первый на профильной диаграмме представлен только одним деревом.
Таким образом, верхний ярус состоит из очень крупных одиночно разбросанных деревьев, но заметно более многочисленных, чем «маячные» деревья лесов Гвианы. Кроны этих деревьев имеют зонтичную форму и очень тяжелы; они достигают 25 м и более ширины, редко соприкасаются краями и отчетливо отделяются от крон яруса В. Ярус А насчитывает сравнительно небольшое число видов. На изученном пространстве это были главным образом Lophira procera и Erythrophleum ivorense.
Ярус В также открытый, и кроны деревьев в его пределах соприкасаются только иногда. На диаграмме следует обратить внимание на окно в этом слое под кроной крупного дерева яруса А. Деревья яруса В имеют маленькие, узкие кроны шириною обыкновенно менее 10 м. Виды многочисленны и принадлежат к самым различным семействам.
Ярус С отличается сомкнутостью и почти полным отсутствием окон. Кроны вплотную прилегают друг к другу и обыкновенно сильно переплетены лианами. Большинство деревьев никогда не достигает вышерасположенного яруса; они принадлежат к различным семействам, но наблюдается тенденция к местному преобладанию одного какого-нибудь вида (виды эти меняются в разных местах). Остальные деревья яруса представлены главным образом молодыми деревьями, нормально свойственными ярусу В. Молодые деревья, обычные в ярусе А, здесь, по-видимому, поразительно редки. Виды яруса С как в Гвиане, так и на острове Борнео имеют преимущественно маленькие конические кроны, но старые экземпляры в ряде случаев обладают замечательно широкими и тяжелыми кронами.
Ярус кустарников (D) состоит в значительной мере из молодых деревьев, обычно относящихся к ярусам В и С, и поэтому с трудом отделяется от яруса С. Видов, свойственных именно ярусу D (большая часть их представлена скорее карликовыми деревьями, чем настоящими кустарниками, см. стр. 95), немного. Плотность яруса кустарников очень изменчива; в девственных лесах он никогда не бывает настолько плотным, чтобы затруднить движение сквозь него, а в некоторых местах как кустарниковый, так и наземный ярусы почти отсутствуют.
1 Недавние исследования автора (Richards, 1948, не опубликованы) наводят на мысль, что этот лес подвергся в прошлом какому-то нарушению и является, таким образом, скорее древним вторичным, чем истинно первичным; однако он, вероятно, достаточно спелый и мало отличается по своей структуре от первичного (климаксного) леса области. То же самое можно сказать и почти о всех так называемых девственных, илн первичных, лесах Нигерии и, возможно, всей Западной Африки в целом. Лес, который никогда, ни в какое время не был использован для земледелия, существует в Западной Африке лишь в болотистых местах (лес пресноводного болота, см. гл. 13) или на крайне ограниченных участках крутых скалистых склонов и тому подобных малодоступных районов.
Рис. 6. Профильная диаграмма первичного смешанного леса. Р®®°Р®“Р°Ва"’^ 'А^'^Лра р,«-
Изображена полоса леса длиной 200 Футов 2 Акомбе (но определен); Ь'Ь ~ t «i’,',,>n,lZluJia JbD'U'dif'-Sliombosia pustu-
Ключ к обозначениям: Al> - I'awdnygalia или С о. 17mnllьр.. Ла,_АКО омое I j. ti lora„- Ip - SHon bo.ta ар . Л °” .sp .
cra'.Ep -Я.попа ep. (cp. dentata)' Er - l^c.ahma umb,:f .. V)*"''rWXwlopla «uinlaaii; Oa - £""«₽W,U’ in‘tu'p‘a>
lata-, 6d - Bcoll. llm kamerunensir, От - Rinorra up. (cp. obl0n8O°‘*°’j (op ы t,;(/ea).
48
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Самый нижний наземный ярус леса (Е) состоит из растений, высота которых колеблется от нескольких см до 1 м и выше. Компонентами его являются: сеянцы древесных пород (обычно преобладают), двудольные и однодольные травы, папоротники. Этот ярус развит еще более неравномерно, чем ярус D. Лесная почва на большом протяжении может оставаться почти совершенно оголенной, но местами, особенно на осветленных участках, она покрывается густым слоем травянистых растений и сеянцами древесных пород. Мхов в напочвенном покрове нет.
Снимок (фото 4), сделанный с высоты 78 футов (24 м) над уровнем почвы, изображает лес, подобный представленному на профильной диаграмме; на нем мы видим густую массу низкорослых деревьев с переплетенными кронами высотой около 9—12 м. Над этим компактным слоем на различную высоту поднимаются более высокие деревья, но они не образуют сомкнутого полога на каком бы то ни было уровне, так что с высоты более И .и на известном расстоянии можно ясно рассмотреть все вокруг. Если, взбираясь навысо кое дерево, миновать плотный ярусС, оказываешься среди яркого дневного освещения, и открывающаяся при этом картина ярусности весьма похожа на ту, которая изображена на профильной диаграмме.
Замечательную черту этого леса представляет большая изреженность двух верхних ярусов. Изреженность яруса А является, вероятно, следствием сравнительно сильной летней засухи и, может быть, представляет собой черту западноафриканского вечнозеленого леса. В самом деле, Обре-виль (Aubreville, 1933) описывает «сомкнутый лес» Берега Слоновой Кости как состоящий из плотной массы растительности высотой 20—30 м, над которой изолированно возвышаются более крупные деревья; он сравнивает его структуру с так называемыми taillis-sous-jutaie (лесосеки с маячными деревьями). Нужно также отметить, что в «вечнозеленом сезонном лесу» (ассоциация Carapa-Eschweilera) на Тринидаде (Beard, 1944 а, 1946 Ь; см. также рис. 6) самый высокий ярус деревьев очень разрежен. Но имея в виду, что леса заповедника Шаша сильно изменены выборочной рубкой и расчисткой, возможно, что открытость верхнего яруса образовалась либо из-за удаления части более крупных деревьев, либо эти леса представляют собой более высокую стадию вторичной сукцессии (см. гл. 17) и не являются настоящим первичным лесом.
Три только что приведенных примера, характеризующих смешанный дождевой лес разных районов, показывают сходство основных черт их ярусного строения, хотя они и обнаруживают существенные различия. На основании этих данных, а также сведений, почерпнутых из других источников, можно высказать следующие положения относительно структуры смешанного дождевого леса:
1. В смешанном дождевом лесу насчитывается пять ярусов самостоятельных растений (см. выше, схема на стр. 36); из них три яруса, состоящие исключительно из деревьев, называются соответственно ярусами А, В и С1; ярус D представлен главным образом деревянистыми растениями, но в нем часто обнаруживается и примесь травянистых видов. Этот ярус принято называть кустарниковым, хотя только немногие из его компонентов являются кустарниками в сколько-нибудь точном смысле слова. Последний наземный ярус (Е) состоит из трав и сеянцев деревьев.
1 Термины «доминантный» и «субдоминантный», обычно употребляемые приме-нительно к ярусам в лесоводственной литературе, не используются здесь в этом смысле во избежание путаницы с их экологическим толкованием.
Фото 4. Полог тропического дождевого леса, резервированный лес Шаша, Нигерия. Вид с высоты 24 м над землей.
На первом плане выделяются деревья яруса С. плотно переплетенные лианами, на втором плане— деревья ярусов А и В.
Ф о т о 5. Подлесок тропического дождевого леса, резервированный лес Окому, Нигерия.
Крупное дерево Entandrophragma angolense var. macrophyllum с характерными поверхностными корнями.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 't9
2. Высота каждого яруса меняется от места к месту, но в узких пределах. Так, в приведенных выше примерах ярус А достигает 30 м или больше в гвианском лесу, около 35 м — на острове Борнео и около 42 м — в Нигерии; высота яруса В соответственно составляет 20,18 и 27 м, а яруса С — 14, 8 и 10 м.
3. Ярус А обыкновенно отличается большей или меньшей разреженностью, причем степень ее, что, в частности, выявляется из сопоставления профилей, значительно варьирует. Возможно, что в самом высоком и наиболее роскошном дождевом лесу (например, в некоторых частях Малаккского полуострова, где средняя высота верхнего яруса деревьев, возмож-но, превосходит 200 футов—61 л<) этот ярус может быть практически сомкнутым. Имеются некоторые данные относительно того, что ярус А становится все более разреженным по мере приближения к климатическим границам тропических дождевых лесов (см. гл. 15). Ярус В может быть сомкнутым или более или менее разреженным, ярус С всегда более или менее сомкнутый и часто является самым плотным ярусом леса.
4. Смежные ярусы могут отчетливо отделиться один от другого или незаметно переходить друг в друга. Так, в гвианском лесу существует заметный разрыв между ярусами В и С и небольшой разрыв между А и В. В лесу острова Борнео наблюдается заметный разрыв между ярусами А и В, но его нет между В и С; в Нигерии, как и в Гвиане, большой разрыв замечается между ярусами В и С. Вертикальные границы кустарникового и наземного ярусов всегда выражены нечетко.
5. Каждый ярус имеет характерный для него флористический состав, но во всех ярусах, за исключением ярусов А и В, большую часть общего количества растений составляют молодые экземпляры видов, которые во взрослом состоянии достигают более высоких ярусов.
6. Деревья каждого яруса характеризуются особой формой кроны. Кроны яруса А развиваются предпочтительно в ширину и часто принимают зонтичную форму. В ярусе В их ширина равна длине или же длина превосходит ширину. В ярусе С кроны имеют коническую суживающуюся кверху форму, то есть их длина намного больше ширины.
В главе 4 будет показано, что деревьям каждого яруса присущи своп характерные физиономические черты, не зависящие от их принадлежности к тем или иным группам систематики.
Ярусность монодоминантных лесов
Климаксные сообщества, в древостое которых преобладает какой-либо один вид деревьев, в некоторых случаях составляющий более 80% самого высокого яруса, встречаются, как теперь известно, во всех главных географических областях пояса тропического дождевого леса. Флористический состав этих сообществ подробно характеризуется в гл. И, где показано, что некоторые из них покрывают сотни квадратных миль страны и могут быть названы консоциациями, в то время как другие представляют собою местные группы в смешанной ассоциации, и их лучше рассматривать как сообщества. Структура этих монодоминантных лесов, вероятно, всегда значительно отличается от структуры более широкораспространенных смешанных сообществ, причем их ярусность, как следует ожидать, должна быть лучше выраженной. Но пока по этому вопросу у нас еще мало точных сведений.
Недавно Бирд (Beard, 1946 а) описал ярусность консоциации острова Тринидад с доминантной Mora ezcelsa, деревом из семейства Leguminosae,
Ключ к обозначениям: A. -/’.hi mhor.a; R. - ('lalhrotropis brachyprlalu; speciosa'. II. m.— Pcntacb thra macioloba; II. p. - Suartzia pinnata; 11. t.___ H
t'b. —Guana glabra; Coe. -- Maxi mi liana rl<gan " - liroirnt'a lull fol la-
in.—Protium guiaru ns< ; L. s.— Ocotra ua-clunhcimii; La.— Cmdia sp.; M.— Mora excelsa; Ma.— Slrrculia cai ibaea', Me.— Euterpe langloisii; Mk.— Sapium aucupa-rium\ Mi.— Miconia sp.; Л.— ('alliandra guildingii; P. — Tabtbuia serrati folia;
Pd.—Inga sp.; S.— Rlitcdia sp.; W. — W'arsztuiczia coecinra; Ц', (J. —Touomi/a H’.ro. — Rubiaceae зр.;П . o.--7 vrminalia ainazonia; Y. o.— H uclu navia capital a
35-
30
25
В. ch.
Ma.
15
20
60
80
фут.
м 30
10
л* фут rl20
710
700
90
80
20-
70
-60
--50
-40
10-
5
30
-20
-10
0
Рис. 7. (а, 6). Профильные диаграммы климаксного вечнозеленого леса на острове Тринидад, В стт-II иди я. а) ,нч \р< Гуатекаре (ассоциация Сагара — E*chu-eilera), б) — лес Мора (консоциация Mora ejcelsa), округ Maiiapo (Heard, b • > а, рш-и<.шпМН пмагпамми изображает полосу леса длиной 200 фуги, (6t at) и шириной 2^ Футоу (7.6 ч )
52
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
могущим достигать высоты по крайней мере 58 м. На рис. 7 представлена профильная диаграмма этого сообщества; для сравнения там же показана одна из смешанных ассоциаций Carapa-Eschweilera (смешанный дождевой лес или вечнозеленый сезонный лес по терминологии Бирда), чередующаяся с лесом Мора на низменностях Тринидада. Как ясно видно на диаграмме, самый высокий ярус леса Мора имеет сомкнутый полог без окон с небольшим их числом на высоте 37—43 м над землей. Каждая крона «подлаживается своей формой к кронам соседних деревьев, причем они так плотно пригоняются одна к другой, что образуют своего рода мозаику... Сверху полог леса напоминает волнистую поверхность моря» (Beard 1946 а, стр. 173). В этом ярусе Mora excelsa составляет обыкновенно 85—95 % всех деревьев и 62% деревьев, имеющих диаметр 1 фут (30 см) и больше. Ниже яруса Mora excelsa располагаются два очень разреженных яруса деревьев высотою 12—25 и 3—9 м соответственно. В кустарниковом и наземном ярусах доминируют всходы и подрост Mora при почти полном отсутствии других растений.
Консоциация Мора, свойственная лесам Тринидада, состоит таким образом из трех ярусов деревьев подобно ассоциациям смешанного тропического дождевого леса, но ярус А здесь является гораздо более сомкнутым, а ярусность в целом — более отчетливо выраженной. Структура леса Мора в Брит. Гвиане (см. стр. 260) не изучалась методом профильных диаграмм, но она несомненно похожа на описанный выше тип ярусности.
Рис. 8, а воспроизводит неопубликованную ранее профильную диаграмму леса Уаллаба в Брит. Гвиане, построенную на основании данных, любезно предоставленных Дэвисом (Т. A. W. Davis). В этой консоциации доминирует Eperua falcata — дерево из семейства бобовых; обильно представлены также два других вида Eperua (Е. grandiflora и Е. jenmani). Состав и характер этого весьма своеобразного сообщества описаны на стр. 263 (см. также Davis, Richards, 1933—1934). На диаграмме видно, что, как и в лесу Мора, ярус А гораздо более выровнен, чем тот же ярус смешанного леса, и отличается почти полной сомкнутостью. Виды Eperua, безусловно, преобладают среди деревьев этого яруса. Ниже яруса А отчетливо выделяется ярус небольших деревьев высотой около 8—15 м, соответствующий ярусу С смешанного леса, но ярус В, занимающий промежуточную высоту, едва можно распознать. Таким образом, можно сказать, что деревья практически образуют лишь два яруса, и лес имеет структуру А С (вместо структуры АВС смешанного леса). Под древесными ярусами расположен довольно плотный ярус кустарников и разреженный наземный ярус из трав, всходов древесных пород и подроста. Подрост Eperua очень обилен.
К структуре А С приближается и лес Dacryodes—Sloanea Малых Антильских островов (рис. 8, б), весьма напоминающий по строению лес Уаллаба в Гвиане; в нем также заметно преобладание одного вида, правда, менее выраженное.
Хотя в консоциациях Мора и Уаллаба присутствие одного сильно доминирующего вида ведет к развитию более плотного яруса А и частичному или полному подавлению яруса В, в лесу из железного дерева Борнео и Суматры (вероятнее всего, климаксное сообщество с доминированием железного дерева EusideroxyIon zwageri, характерного для острова Борнео, см. стр. 282) ярус В, к которому принадлежит этот доминирующий вид, является крайне плотным, а ярус А почти отсутствует. Грессер (Gresser, 1919) пишет, что этот лес имеет плоскую компактную поверхность крон,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 53 поддерживаемую стволами высотой в 15—20 м\ она образуется главным образом кронами Eusideroxylon. Только изредка полог прорывается отдельными высокими Koompassia, Shorea \ или Intsia, заметно возвышающимися над общим уровнем. Разреженный подлесок характеризуется обилием молодых экземпляров доминирующего вида.
В дождевом лесу тропической Африки известны два широко распространенных монодоминантных климаксных сообщества — консоциация Cynometra alexandri и консоциация М acrolobium dewevrei. Они занимают обширные пространства восточной части Республики Конго. Консоциация Cynometra захватывает также и часть территории Уганды. Подробное описание структуры леса Cynometra, иллюстрируемое профильной диаграммой, было недавно опубликовано Эггелингом (Eggeling, 1947); в то же время проф. Ж. Луи (J. Louis) любезно разрешил мне просмотреть еще неопубликованные профильные диаграммы обеих консоциаций, составленные им совместно с д-ром Дони (С. Donis) для Конго. Консоциация Cynometra имеет весьма отчетливую трехъярусную структуру, которая отличается от структуры смешанного леса главным образом особенностями флористического состава; если в смешанном лесу ярусы А и В сложены разными видами, то в лесу из Cynometra подавляющее большинство деревьев обоих ярусов представлено видом Cynometra. Средняя высота яруса А, образующего почти сомкнутый полог, достигает 36 м, хотя и здесь иногда встречаются гораздо более высокие деревья. Ниже располагается средний ярус высотой 11—21 м, достаточно сомкнутый, за исключением участков, непосредственно затеняемых деревьями яруса А, и ярус С, состоящий из равномерно распределенных, но не образующих сомкнутого полога деревьев высотой до 11 м.
Консоциация М acrolobium отличается гораздо более своеобразной структурой. Доминирующий вид достигает высоты 35—40 м и может составлять свыше 90% древостоя. Macrolobium dewevrei сильно ветвится и имеет широко распростертую крону, под густой тенью которой угнетаются все другие растения. Поэтому остальные ярусы развиты очень плохо.
В связи с изучением структуры монодоминантных сообществ дождевого леса можно упомянуть о лесе из A Itingia excelsa на Яве и Суматре и лесе из Eucalyptus deglupta в Новой Британии, хотя они и не относятся к кли-максному тропическому дождевому лесу. Лес из Altingia представляет собой обособленное сообщество или локализованную консоциацию в смешанном вечнозеленом лесу, встречающуюся, согласно Ван-Стеенису (van Steenis, 1935 а), на высотах около 1000—1700л< над уровнем моря. Таким образом, это, вероятно, скорее тип среднегорного, а не тропического дождевого леса (см. гл. 16). Замечательной чертой его структуры является то, что кроны доминирующего вида отчетливо вздымаются над общей массой растительности, и, за исключением случайных индивидуумов Podocarpus imbricata, самый высокий ярус состоит только из Altingia, точно также в лесах из Eucalyptus deglupta (Lane-Poole, 1925 а, Ь) господствующая порода целиком слагает верхний ярус. Эвкалипты (Eucalyptus), достигающие приблизительно 70 м высоты, значительно возвышаются над компактным вторым ярусом, образованным смесью вечнозеленых видов, соответствующих ярусу А окружающего смешанного леса. Сообщество Е. deg-lupfy встречается только в Новой Британии, и существование его, видимо, завиЬит от периодически повторяющихся пожаров. Следовательно, его нельзя рассматривать как сообщество климатического климакса.
КЛЮЧ К обозначениям: (а). Ат — Ainaioua guianensis; А»—Aspidoeperma слсг/янпг, Н -- Uyrsonima sp.; (' — Cassia pteridophylla; (’а __ Cabmhmnut чр •
D — Duguetia neglect a't E -- Eperua /а/cahf (мягкая уаллаба); Eg — Eperua grandiflora (уллаба итури); Ес — Ecclinusa psilophyl la\ Em'-Euimohun fagi folium', Es — Eschueilera sp.; L — Licania hderomorpha’, M — Matayba inelegant, Ma — Marlierea shomb ui ghiana', Ms— Matayba' sp.; О -- Ocotra sp.; Or — Onnosia coutinhoi', P — Pouteria sp.; Sir — Swartzia sp.; 7’ — Toromita cepbalostigma', U — неопределенный вид; ЛЬ —Aniba bradi'ala-, Up-He-il9Chmiedia pendula', D — Dacryodcs excelsa', Fo — Faramea occidentalism Gm — Guarca macruphylla; И —Inga ingoides\ L — Lauraceae (вид не определен); pm — Pouteria multiflora', Ps — Pouteria seme carpi fol ia,—Quararibea lurbinata; Sa—Simaruba amara', Sc — Sterculia caribaea’, Ta — Tapura antillnna.
a
6
Р и с. 8 (а, б). Профильные диаграммы первичного дождевого леса: а — лес Уаллаба (консоциация Ерегиа), Барабара-h рик, река Мазаруни, Брит. Гвиана (Т. A. W. Davis, не опубликовано); б — ассоциация Dacryodes-Sloanea, остров Доминика, Вест-Индия (Beard, 1949, рис, 8).
Каждая диаграмма изображает полосу леса шириной 25 футов (7,6 .и).
56
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Отношение ярусности к флористическому составу
Солсбери (Salisbury, 1925) показал, что максимальные высоты деревьев и кустарников в Англии образуют почти непрерывный ряд от бука высотой в 36 м до полукустарников высотой меньше 1 м. Естественное английское редколесье тем не менее всегда содержит три довольно резко отграниченных яруса — деревьев, кустарников и трав. Такую простоту структуры Солсбери объясняет бедностью флоры и наличием только очень небольшого числа древесных видов в любом месте. Причиной этого является «не столько отсутствие видов, дающих градацию по высоте, сколько неспособность всех видов, за исключением небольшого числа деревьев, достичь достаточного обилия, чтобы существенно повлиять на структуру леса» (loc. cit., стр. 336). Это объяснение вполне устраивает, когда речь идет об английских или большинстве других лесов умеренной зоны1. Но оно становится явно неудовлетворительным при рассмотрении тропического дождевого леса с его огромным видовым богатством. Правда, в монодоминантных лесах, то есть таких лесах, где один вид «достигает достаточного обилия, чтобы существенно повлиять на структуру леса», ярусность выделяется более ясно, чем в смешанном лесу. Однако она все же совершенно очевидна и в последнем; причем структура большинства монодоминантных лесов не отклоняется столь далеко, как того можно было бы ожидать, от трехъярусного плана, общего смешанным лесам. По-видимому, приходится допустить что факторы, лежащие в основе возникновения ярусности, до сих пор еще полностью не раскрыты.
Структура климаксного сообщества, находящегося в равновесии с его средой, является выражением сложного баланса, в котором важную роль играет не только борьба за свет, но, вероятно, и другие факторы, в частности конкуренция корней (см. стр. 211—212), о чем нам еще очень мало известно. Природа процесса восстановления, о котором будет идти речь в следующей главе, несомненно также тесно связана с причиной, обусловливающей структуру леса.
1 Некоторые леса умеренной зоны имеют более одного яруса деревьев, см. Джонс (Е. W. Jones, 1945).
Глава 3
ВОЗОБНОВЛЕНИЕ ЛЕСА
Так как тропический дождевой лес представляет собой климатический климакс, то он, в соответствии с этим определением, должен находиться в состоянии равновесия. Отмершие деревья замещаются деревьями того же или других видов: флористический состав небольшого участка леса может с течением времени изменяться, но его средний состав на большой площади остается постоянным. Следовательно, несмотря на смену индивидуумов, сообщество в целом сохраняется, и надо думать, что по крайней мере на части занимаемого им пространства его структура и флористический состав остаются в значительной степени неизменными в течение тысяч, а возможно, и миллионов лет1.
Хотя дождевой лес, если не принимать во внимание вмешательства человека, рассматривается как стабильное сообщество, его равновесие является динамичным и лишь относительным. С точки зрения человеческого масштаба времени, состав леса кажется постоянным, но, подобно всем климаксным растительным сообществам, тропический дождевой лес подвержен очень медленному и постепенному изменению. Наряду с переменами, обусловливаемыми вековыми изменениями климата, в нем происходят и другие изменения, связанные с эволюцией растений и распространением видов. Одни виды формируются, другие исчезают или сокращают свой ареал. Такие изменения настолько медленны, что о них можно судить только по данным фитогеографии. Иногда какой-нибудь «агрессивный» вид приобретает возможность для сравнительно быстрого распространения на новых пространствах. Это, как думают Бирд (1946 а) и другие, по-видп-мому, произошло с Mora excelsa на острове Тринидад.
Консоциация с господством этого вида, по всей вероятности, сравнительно недавно появившегося на острове, в настоящее время захватывает и замещает окружающий смешанный лес. Но, насколько известно, подобные случаи встречаются редко: вообще же сообщества тропического дождевого леса, по-видимому, обладают способностью самовозобновляться, не изменяясь.
Процесс естественного возобновления в дождевых лесах, несомненно, отличается крайней сложностью, и хотя практическое значение его для лесоводства очевидно, существующие о нем сведения исключительно ограничены. Большая часть того, что было написано относительно так называемого «естественного возобновления» дождевого леса, относится к небольшому количеству наиболее важных в экономическом отношении видов, произрастающих в более или менее искусственных условиях, возникающих вследствие эксплуатации леса. Прежде чем понять или научиться научно управлять возобновлением при таких искусственных условиях,
1 На острове Тринидад, например, миоценовая флора, видимо, очень мало отличалась от современной флоры (Hollick, 1924; Berry, 1926).
58
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
необходимо знать, что происходит в ненарушенной природной среде, но в этой области нам почти ничего не известно.
Первостепенное значение имеют вопросы о том, какого среднего возраста достигают деревья в различных ярусах, какой возрастной класс преобладает среди главных доминант в зрелом ненарушенном дождевом лесу, каково должно быть относительное обилие видов в различных классах возраста, как изменяется скорость роста крупного дерева тропического дождевого леса во время последовательных стадий его развития, на какой стадии наступает наибольшее отмирание и, следовательно, наиболее интенсивный естественный отбор, и, наконец, остается ли флористический состав микроучастков постоянным или же смешанный лес должен рассматриваться как огромная мозаика, на отдельных участках которой происходит беспрерывная смена комбинаций видов? Разрешение этих и подобных вопросов является наиболее неотложной задачей, стоящей в настоящее время перед ботаниками, занимающимися изучением экологии дождевого леса. Из-за отсутствия достаточных сведений все выводы этой главы должны рассматриваться как предварительные.
ОТМИРАНИЕ СТАРЫХ ДЕРЕВЬЕВ И ОБРАЗОВАНИЕ ОКОН
Сведения о среднем и максимальном возрасте деревьев тропического дождевого леса очень скудны. В тропических лесах с заметным засушливым сезоном возраст деревьев в ряде случаев может быть приблизительно определен по годичным кольцам (то же можно сделать у небольшого количества видов в «вечнозеленом сезонном» типе тропического дождевого леса), но в типичном дождевом лесу с его относительно несезонным климатом деревья не обладают отчетливыми годичными кольцами (см. стр. 220). Поэтому приблизительный возраст деревьев в нем может быть определен лишь посредством экстраполяции ряда тщательных производимых в течение нескольких лет измерений скорости роста отдельных индивидуумов различного размера. Но и в лучшем случае этот прием является малоудовлетворительным. Наблюдения Брауна (Brown, 1919, стр. 156) за скоростью роста Parashorea malaanonan, важной доминанты филиппинского диптеро-карпового леса, показали, что средний индивидуум этого вида достигает за 197 лет диаметра 80 см\ так как деревьев толще 80 см не было обнаружено, то можно думать, что наибольший возраст представителей этого вида равняется приблизительно 200 лет. Уотсон (Watson, 1937), измерявший на полуострове Малакка скорость роста другого вида диптерокарповых, сравнительно быстро растущей Shorea leprosula (меранти тембага), определил ее средний максимальный возраст приблизительно в 250 лет. Какие-либо другие данные такого рода, по-видимому, отсутствуют. Согласно измерениям Брауна и Мэтьюза (Brown, Matthews, 1914), деревья второго яруса на Филиппинских островах в общем обнаруживают более медленный рост, чем доминирующие диптерокарповые, но определений их максимального возраста не делалось. Быстрорастущие, очень светолюбивые деревья, характерные для молодого вторичного леса, гораздо менее долговечны, чем виды первичного тропического дождевого леса (см. стр. 407).
На месте погибшего крупного дерева дождевого леса в древесном ярусе, к которому оно принадлежало, образуется окно. Эти окна имеют очень важное значение в процессе возобновления леса. Если дерево отмирает медленно, окно оказывается незначительным, но когда внезапный порыв ветра опрокидывает крупное и, по-видимому, еще относительно жизне
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
59
способное дерево, окно может быть значительно крупнее. В девственном лесу наряду с многочисленными, естественно образовавшимися окнами повсюду видны остатки упавших деревьев, хотя из-за большой скорости распада они разлагаются скорее (если только порода не обладает особо устойчивой древесиной), чем в лесах умеренной зоны.
Образование окна в одном из верхних ярусов ведет к развитию под ним густого подлеска вследствие повышенной освещенности и, возможно, местного ослабления корневой конкуренции. Из молодых деревьев этого подлеска более светолюбивые виды развиваются интенсивнее, чем теневыносливые, рост которых может быть временно или постоянно подавлен. При появлении более крупных окон, кроме видов, которые займут постоянное место в одном из трех ярусов, тут временно укореняются и те быстрорастущие виды вторичного леса, которые способны быстро расселяться. Именно таким образом в окнах первичного леса Южной Америки появляются виды Cecropia', в Африке подобную же роль играет Musanga сесго-pioides, а на полуострове Малакка и в Малайском архипелаге — виды Macaranga. В окнах климаксного леса эти виды вскоре подавляются медленнее растущими, но более теневыносливыми доминантами первичного леса. На более обширных прогалинах они становятся доминантами ранних стадий во вторичной сукцессии, но и здесь в конце концов эти деревья уступают место видам первичного леса (см. гл. 17). Естественное окно является, таким образом, местом, на котором в очень маленьком масштабе происходит развитие вторичной сукцессии.
Отношение между нормальным процессом возобновления в окнах и вторичной сукцессией иллюстрируется интересными наблюдениями Крамера (Kramer, 1933) в нагорном дождевом лесу, произрастающем на горе Гедех, остров Ява. На искусственно осветленных площадках размером меньше 10 акров возобновление доминант первичного леса происходило хорошо. Если же размеры открытых мест достигали 20—30 акров, то возобновление полностью подавлялось роскошным ростом видов вторичного леса. В свою очередь Блэнфорд (Blanford, 1929) заметил, что наилучшее восстановление доминант в вечнозеленом лесу Малаккского полуострова происходило в окнах не более 20 футов (6 м) в поперечнике; в более крупных окнах восстановления доминант в центре не наблюдалось.
Образование окон вследствие отмирания старых деревьев — закономерное явление в ходе естественного возобновления; оно, вероятно, необходимо для выживания более светолюбивых видов, входящих в состав климаксного леса, где наиболее теневыносливые виды способны восстанавливаться и под сомкнутым пологом. Так среди деревьев климаксного дождевого леса Брит. Гвианы экземпляры, высота которых часто превышает большинство других деревьев яруса А (стр. 44) и принадлежащих, например, к видам Нутепаеа, являются светолюбивыми и, по-видимому, целиком зависят от наличия окон для своего восстановления: подрост таких видов редко встречается под ненарушенным пологом леса. Другие виды, например Ocotea rodiaei, для того чтобы развиться, нуждаются по крайней мере в небольших прогалинах, третьи, например Licania venosa, безразличны к затенению и, вероятно, способны возобновляться как в окнах, так и под сомкнутым пологом. Наиболее теневыносливые растения не переносят слишком сильного света (и колебания влажности?) и не могут расти на обширных прогалинах. Подрост Ocotea rodiaei дает малый прирост на больших прогалинах и, как видно, страдает от недостатка влаги (Т. A. W. Davis).
60
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
СОСТАВ ВОЗРАСТНЫХ КЛАССОВ
Независимо от того, имеются окна или нет, возобновление деревьев в дождевом лесу возможно лишь при наличии достаточного количества всходов и молодняка первого и второго (жердняк) классов, чтобы обеспечить замещение. Хотя возрастной состав деревьев тропического дождевого леса не может быть изучен непосредственно (благодаря трудности определения их возраста), уже поверхностное изучение его состава по классам величин показывает, что среди обычных доминант существует огромное разнообразие в количестве молодняка, изменяющееся от вида к виду и иногда внутри одного и того же вида в зависимости от места. Какой-либо вид, составляющий значительную часть верхнего яруса, может иметь многочисленные всходы и молодые деревья, в то время как другой, тоже достаточно заметный по числу взрослых особей, в подросте представлен очень скудно. Иногда изобилующий сеянцами вид совершенно отсутствует как спелое дерево. В исключительных случаях обычная доминанта ярусов А и В кажется полностью лишенной возобновления. Например, в вечнозеленом лесу Берега Слоновой Кости молодые индивидуумы некоторых видов так редки, что местные жители говорят, что эти деревья «не имеют малышей» (Aubreville, 1938). ©возможных причинах такого загадочного явления, характерного не только исключительно для Африки, говорится ниже. В то же время в «вечнозеленом сезонном лесу» Тринидада самые распространенные виды древесных ярусов, по-видимому, являются наиболее обыкновенными среди подроста (Beard, 1946b, стр. 63).
Как указывалось ранее, в смешанном дождевом лесу в нижних ярусах (С, D, Е) молодые экземпляры видов, принадлежащих к ярусам А и В, превышают число видов, свойственных нижним ярусам, причем, по имеющимся данным, виды яруса А в общем представлены значительно хуже, чем виды яруса В. С другой стороны, в монодоминантных лесах сеянцы и молодняк доминирующих видов почти всегда очень обильны. Так, в консоциации Mora gonggrijpii Брит. Гвианы и консоциации М. excelsa Гвианы и Тринидада густые заросли сеянцев этих пород представляют их характерную черту. Таблица 4 иллюстрирует состав четырех типов монодоминантных лесов в Морабалли-Крик, Брит. Гвиана, по размеру деревьев, замерявшемуся на пробных площадках. Из таблицы видно, что доминирующие виды хорошо представлены во всех мелкоразмерных ступенях толщины, за исключением консоциации Ocotea, для которой, несмотря на «весьма обильные всходы», развившийся подрост (жердняк) мало характерен. Даже в этих монодоминантных сообществах не существует тесного соотношения между обилием всходов и обилием спелых деревьев. Большое количество всходов доминирующей породы свойственно и другим моно-доминантным лесам, например консоциации Eusideroxylon Борнео и Суматры и консоциации Cynometra alexandri Уганды1 (Eggeling, 1947). В сообществах Dryobalanops aromatica на Суматре (стр. 283) почва везде покрыта всходами деревьев (van Zon, 1915).
Различия в размерах и возрастном составе разных деревьев тропического дождевого леса есть, несомненно, результат видовой (наследственной) идиосинкразии. Эти различия зависят от способа размножения видов
1 Но в консоциации Parinari ezcelsa — монодоминантном сообществе равнинного типа тропического дождевого леса, встречающегося в Уганде на высоте между 4500 и 5000 футов (1400—1500 м) (см. стр. 282),—как всходы, так и молодые экземпляры доминирующих пород очень редки (Eggeling).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
61
Таблица 4
Размеры доминирующих видов на пробных участках площадью 400 x 400 футов (122 Х122 м) четырех консоциаций в Мо раба л ли-Крик, Брит. Гвпана
Davis, Richards, 1933—1934
Консоциации Среднее количество всходов до 2 .At высоты на 1 кв. м Количество молодых деревьев меньше 4 дюймов (10 см) в диаметре при высоте более 1 5 футов (4,6 л<) Количество деревьев с диаметром 4 дюйма (10 см) и более
4 — 8 дюймов (1 0—20 см) 8—12 дюймов (20 — 31 см) 1 2—16 дюймов (31 — 41 cat) 16—24 дюйма (И-61 с.и) 2't дюйма (61 с.ч и более)
Mora excelsa (лес Мора) 4,7 120 1 28 19 17 24 21
Mora gonggrijpH (лес Мора-буцеа) 11,7 318 39 17 9 38 16
Ocotea rodiaei (лес Грин-харт) (всходы исключи- тельно обильные) 16 1 2 5 10 38 18
Eperua falcata (лес Уаллаба) 1,8 80 1 47 43 37 64 3
1 На этих участках деревья данного размера подсчитывались на двух полосах, которые вместе составляли 1/в общей площади, поэтому полученные цифры были умножены на 8.
и от экологической выносливости (жизнеспособности) деревьев в молодых стадиях развития.
Различные виды тропического дождевого леса значительно отличаются друг от друга как по повторяемости семенных лет, так и по количеству образующихся семян. Некоторые виды цветут один раз в год, другие через более длинные или более короткие промежутки (см. гл. 8). Дэвис (Т. A. W. Davis) пишет относительно леса Брит. Гвианы следующее: «В пологе семена обыкновенно образуются в определенные периоды, но в подлеске небольшие количества цветков и плодов появляются почти непрерывно (см. гл. 8) или же через неправильные, но частые интервалы. Однако большинство деревьев, образующих главный полог (ярусы А и В), цветет один раз в год, или раз в два года, или даже трижды в год, хотя семена не всегда образуются, а если и образуются, то не всегда созревают. Эти деревья, вероятно, продуцируют большое количество полноценных семян по крайней мере один раз в три года. Как правило, часто дают семена наиболее распространенные виды; некоторые, например большинство видов Esch-weilera, Licania venosa и Ocotea rodiaei, цветут один или два раза в год, регулярно принося небольшое количество плодов, хотя хороший урожай наблюдается только раз в два или три года: другие виды, например Mora excelsa и Mora gonggrijpii, цветут примерно каждые 18 месяцев или два года и обыкновенно каждый раз дают хороший урожай семян... Цветение отнюдь не всегда сопровождается созреванием даже хотя бы небольшого количества семян. Иногда плод не завязывается, вероятно, ввиду неблагоприятной для оплодотворения погоды. По-видимому, это наиболее часто случается у видов с очень коротким периодом цветения. Урожай семян, когда они вызревают, как правило, способен без ущерба для возоб-
62
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
новления покрыть потери от таких причин, как неблагоприятная погода или насекомые. Однако целый ряд деревьев серьезно страдает от попугаев, которые кормятся главным образом незрелыми плодами и в особенности падки на семена бобовых. Мы наблюдали целую стаю маленьких длиннохвостых попугаев (Brotogeris chrysopterus), изо дня в день неделями кормившихся нежными и сочными незрелыми плодами Bombax surinamense. Вероятно, некоторые деревья таким путем нередко полностью лишаются урожая семян. Подобные явления несомненно встречаются и в других областях тропического дождевого леса. Годы хорошего и плохого урожая семян представляют собою обыкновенное явление и не обязательно зависят от ритма цветения; они наблюдаются, например, у некоторых малайских дпптерокарповых, у западноафриканского Triplochiton scleroxylon и, несомненно, у многих других деревьев дождевого леса.
Прорастание, по-видимому, редко является критической стадией в возобновлении. Семена деревьев дождевого леса обладают обыкновенно высокой всхожестью; по характеру прорастания они подразделяются па «одновременный» тип (Salisbury, 1929, стр. 219), когда все семена обыкновенно немедленно после опадания прорастают вместе [дружная всхожесть— Ре7.], например у Mora excelsa, и «постепенный» тип, при котором прорастание растягивается на продолжительный период, как у Ocotea rodiaei (Т. A. W. Davis). В любом случае лесная почва редко, да и то на короткое время, бывает слишком сухой для успешного прорастания и укоренения проростков. Чаще наблюдается обратная картина. Аяр (Aiyar, 1932) констатирует, что в Западных Гатах (Индия) семена деревьев, падая как раз перед муссоном или во время муссона, не прорастают или прорастают крайне медленно, пока не минует самый влажный период. У Mesua ferrea, например, семена осыпаются перед муссоном, в мае или июне, но обыкновенно не прорастают до сентября, когда начинается сухой сезон; Palaquium ellipticum роняет свои семена в период муссонных дождей, и хотя они прорастают сразу же, рост всходов идет медленно вплоть до прекращения слишком сырой погоды. Всходы часто бывают весьма обильны, но всегда большое количество их гибнет, и только очень небольшая часть выживает и укореняется. Эггелинг (Eggeling, 1947, стр. 59) говорит о консоциации Cynometra alexandri в Уганде следующее: «К концу дождливого сезона трудно пройти в пределах консоциации, не наступая на всходы Cynometra. Четыре-пять месяцев спустя, в конце сухого сезона, нужны тщательные поиски, чтобы вообще найти какое-нибудь молодое растение». В диптеро-карповых лесах Малайи в первый год от различных причин гибнет огромное количество всходов (Blanford, 1929). Согласно Уотсону (Watson, 1937, стр. 147), только очень небольшой процент всходов светолюбивых Shorea выживает в течение более двух лет. Так как семена этих деревьев вызревают нерегулярно (приблизительно раз в пять лет), «то можно представить себе картину смены последовательных волн недолговечных сеянцев, из которых каждый заранее является обреченным, если он случайно не окажется в пределах окна, образовавшегося в пологе леса. Но подобное совпадение происходит один раз за долгие годы. Ведь тут нет такого положения, как в правильно эксплуатируемом лесу, где такие окна регулярно образуются при последовательной вырубке деревьев, достигших товарной спелости». Более теневыносливые деревья для своего развития, возможно, меньше нуждаются в окнах, но степень выживаемости среди них, вероятно, не выше. Исключительно неблагоприятная погода может привести к гибели всего возобновления; так, в одной из местностей Брит. Гвианы засуха 1926 г. уничтожила целую заросль молодняка Mora
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 63
gonggrijpii (Т. A. W. Davis). Но такой случай является, вероятно, совершенно исключительным; обычная причина гибели лежит, по-видимому, в истощении растений вследствие конкуренции корней и недостатка света.
При рассмотрении относительного обилия различных возрастных классов деревьев (и лиан) тропического дождевого леса надо иметь в виду широкую амплитуду условий среды, действию которых подвергаются растения более высоких ярусов в течение различных стадий своего развития. Как мы увидим в главе 7, между верхними и нижними ярусами существуют сильные контрасты не только в отношении освещения, но также в атмосферной влажности и движениях воздуха. Вследствие этого часто случается, что высокоствольные виды деревьев оказываются исключенными из ярусов А и В не потому, что они не могли выжить, достигнув взрослого состояния, а из-за того, что условия в подлеске были неблагоприятны для их развития в молодом возрасте. В то же время возможно превосходное развитие молодых деревьев в обстановке, неподходящей для их взрослой стадии. В Брит. Гвиане всходы некоторых видов пальм, которые в виде вполне взрослых деревьев встречаются только на болотах, представляют собой совершенно обычное явление во всех типах лесов, куда птицы заносят их плоды. В Морабалли-Крик молодые экземпляры бесстебельной пальмы Jessenia oligocarpa (которая не является болотным видом) изобилуют в консоциации Уаллаба, где, однако, во взрослом состоянии эта пальма не встречается или встречается очень редко. Часто поразительное различие формы и структуры листьев (а также и их расположения) у молодых и взрослых индивидуумов деревьев и лиан ярусов А и В (см. гл. 4, 5) отражает именно это резкое изменение среды их обитания. Такие различия в наиболее сильной степени бывают выражены у теневыносливых видов.
СКОРОСТЬ РОСТА НА РАЗЛИЧНЫХ СТАДИЯХ РАЗВИТИЯ
Если не считать редких случаев, когда в пологе леса имеется подходящее окно, которое может оказать благоприятное влияние на развитие растений подроста, молодым деревьям дождевого леса приходится переживать длительный период угнетения, прежде чем они займут место в соответствующем ярусе. Хотя в течение этого времени количество новых листьев может несколько увеличиться, но рост в высоту практически отсутствует или, в лучшем случае, отличается крайней замедленностью. В результате молодое деревце лишь очень постепенно пробивается вверх через подлесок. Многие молодые деревья из числа уцелевших в период массовой гибели всходов, вероятно, погибнут в период угнетения, длящегося годы или даже десятки лет, но зато оставшийся подрост многих видов деревьев тропического дождевого леса, очевидно, сохраняет в течение удивительно долгого времени способность быстро развиваться при наступлении более благоприятных условий. Такая способность переживать долгий период угнетения роста является характерным признаком и, несомненно, представляет собой важную биологическую особенность деревьев дождевого леса. Обычные для высокоствольных видов (стр. 113) большие тяжелые семена, содержащие обильный запас пищи, обладают большими преимуществами, давая возможность всходам вырасти до размера, при достижении которого они уже имеют некоторые шансы пережить период угнетения. Герберт (Herbert, 1929) высказывает вполне правдоподобное предположение, что одна из причин слабого проникновения в тропический дождевой лес видов
64
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Eucalyptus заключается в отсутствии у них способности пережить период угнетения.
Наблюдения Брауна и Мэтьюза (Brown, Matthews, 1914) и Брауна (Brown, 1919) за ростом деревьев в диптерокарповых лесах на Филиппинах весьма ясно показали существование периода угнетения и большое изменение в скорости роста на различных стадиях развития. Хотя результаты этих исследований относятся только к одному пункту и лишь к очень небольшому количеству видов, их, вероятно, можно распространить на все деревья дождевого леса вообще.
В течение четырех лет производились тщательные измерения диаметров целого ряда деревьев трех видов в вечнозеленом лесу горы Маквилинг (высота над уровнем моря около 100 л<). Один из этих видов, типичное дип-терокарповое Parashorea malaanonan. доминировал в ярусе А; два других— Diplodiscus paniculatus и Dillenia philippinensis — были наиболее обычными в ярусе В. Одновременно производились измерения группы деревьев Parashorea. росших на открытом месте на краю леса. Диаметры измерялись раз в год с точностью до 0,5 мм. Экземпляры каждого вида группировались по ступеням толщины, и для каждой такой ступени определялся средний годичный прирост диаметра; из этого вычислялось среднее число лет пребывания на каждой ступени. Браун полагал, что индивидуальные различия в скорости роста деревьев, находящихся на ступени толщины 0—5 см. были так велики, что при определении возраста у деревьев этого класса получалась большая ошибка, которая, вероятно, сглаживалась при определении возраста деревьев, принадлежащих к более крупным ступеням толщины»
Таблица 5
Скорость роста трех видов деревьев в диптерокарповом лесу на высоте около 100 лс, гора Маквилинг, Филиппинские острова
(Brown, 1919 г.)
Виды Ступени толщины, слс Число измеренных экземпляров1 Средний годовой прирост диаметра, СЛ1 Среднее число лет, проведенных в данной ступени толщины
Parashorea malaanonan (в лесу) 0-5 4 0,07 71
5—10 10 0,27 19
10-20 21 0,38 26
20—30 19 0,49 20
30- 40 12 0,74 13,5
40—50 28 0,82 12
50-60 7 0,94 11
60—70 10 0,75 13
70—80 1 0,84 12
Parashorea malaanonan (на откры- 0-5 13 0,42 12
том месте) 5—10 27 0,55 9,1
10-15 7 0,73 6,8
Diplodiscus paniculatus 0—10 14 0,15 66,7
10-20 20 0,31 32,3
20—30 8 0,44 22,7
Dillenia philippinensis 0-10 1 0,46 22
10—20 1 0,55 18
20—30 6 0,39 26
30-40 1 0,24 42
1 Не все деревья измерялись каждый год в течение четырехлетнего периода.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
65
Результаты измерений приводятся в табл.З. Дополнением ей служит график (рис. 9), характеризующий скорость роста Parashorea и Diplodiscus. Верхние точки кривой для Parashorea, растущей на открытом месте, были вычислены, исходя из предположения, что деревья крупного диаметра росли одинаково как в лесу, так и на открытом месте.
Данные, относящиеся к Parashorea, весьма наглядно показывают влияние периода угнетения. Деревья, находящиеся в лесу, растут крайне медленно, пока не достигнут диаметра 5 ем. После этого скорость роста намного увеличивается. Для достижения этого диаметра они тратят почти
Рис. 9. Скорость роста Parashorea malaanonan и Diplodiscus pa-niculatus на высоте 300 м; гора Маквплинг, Филиппинские острова (Brown, 1919, рис. 2).
столько же времени, сколько на дальнейшее наращивание его от 5 до 40 см. У деревьев, произрастающих на открытом месте, скорость роста велика и в молодом возрасте, оставаясь почти постоянной на протяжении всего их развития. Медленный рост деревьев, развивающихся под пологом леса до достижения ими диаметра в 5 см, в значительной степени объясняющийся конкуренцией между корнями и борьбой за свет, ясно связан с неблагоприятными условиями среды. Деревья с диаметром 5—10 см являются уже «выдающимися», а деревья, чей диаметр превышает 20 см, находятся уже в самом высоком ярусе и в условиях, которые, по крайней мере в отношении освещения, приближаются к условиям открытого места. Данные о Diplodiscus свидетельствуют о несколько более медленном росте, чем у Parashorea, но и здесь также наблюдается заметный период угнетения. Вычисленные скорости роста для меньших ступеней толщины оказываются выше, чем для Parashorea, что объясняется отсутствием измерений деревьев Diplodiscus диаметром менее 5 ем; при наличии таких измерений, безусловно, был бы выявлен более медленный рост Diplodiscus по сравнению с Parashorea на всех стадиях развития. Данные о Dillenia philippinensis основываются на очень небольшом числе измерений, но они также указы
66
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
вают на более медленный рост этого вида, чем рост Par ashore а. Браун и Мэтьюз, как уже упоминалось выше, пришли к заключению, что деревья яруса В растут в общем медленнее, чем доминирующие дпптерокарповые.
То обстоятельство, что даже очень маленькие деревья Parashorea быстро растут на открытом месте, показывает, как и надо было ожидать, что медленный рост молодых деревьев этого вида в лесу объясняется не врожденными свойствами, а зависит от внешних условий. Скорость роста дерева на всех стадиях его развития определяется двумя группами факторов — факторами среды и факторами наследственности. Последние изменяются от вида к виду, и, как мы видели, светолюбивые растения при наличии достаточного освещения растут обыкновенно быстрее, чем теневыносливые.
Почти ничего не известно о количестве деревьев тропического дождевого леса, гибнущих во время периода угнетения, но, по всей вероятности, оно велико. Если это так, то в процессе развития этих деревьев существуют два критических момента, когда их гибель достигает высоких показателей; один приходится на первый или второй год жизни, когда они находятся в стадии всходов, а другой, возможно, падает на конец периода угнетения. По-видимому, молодое деревце, пережившее период угнетения и достигшее высоты 2 м пли около того, имеет высокую степень вероятности стать спелым, плодоносящим деревом. Солсбери (Salisbury, 1930) установил, что у многих британских травянистых растений почти все случаи гибели не достигших зрелости экземпляров приходятся на стадию всходов. Наряду с гибелью массы всходов существует высокая степень вероятности, что те растения, которые вышли за пределы некоторой очень ранней стадии, достигнут зрелого состояния. Например, у розеточных растений экземпляры, у которых уже заложились розетки, обыкновенно достигают фазы размножения. Филлипс (Phillips, 1931) также придерживается мнения, что в вечнозеленом лесу Книсна, Южная Африка, процент гибнущих растений гораздо больше среди молодых, чем среди старых деревьев возобновления. С другой стороны, деревья тропического дождевого леса, хотя они также более всего страдают на стадии всходов, находятся в неопределенном положении относительно достижения репродуктивной стадии до тех пор, пока они не переживут период угнетения.
МОЗАИЧНАЯ ТЕОРИЯ ВОЗОБНОВЛЕНИЯ
Как мы уже видели, молодые экземпляры доминант тропического дождевого леса могут быть представлены скудно или даже совершенно отсутствовать; флористический состав верхних ярусов часто весьма отличается от флористического состава популяции всходов и молодого подроста деревьев, подготовляющих смену для деревьев верхних ярусов. Эти факты побудили Обревиля (Aubreville, 1938) выдвинуть теорию, которую можно назвать мозаичной, или цикличной теорией возобновления.
Согласно его точке зрения, любая частная комбинация видов, доминирующих на данном небольшом пространстве смешанного тропического леса, не является постоянной ни в пространстве, ни во времени. Доминирующие ярусы изменяются по своему составу как в разных местах (насколько это верно обсуждается в гл. И), так, согласно Обревилю, и на одном и том же месте за длительный промежуток времени. За комбинацией видов, доминирующих в данном месте и в данное время, следует иная комбинация их. Ни одна комбинация видов не находится в состоянии постоянного равнове-
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 67
спя со средой. «Заимствуя сравнение из алгебры, можно сказать,что состояния равновесия между определяющими факторами местообитания и многочисленными независимыми переменными, создаваемыми характерными видами сообщества, образуют ряд уравнений, число которых гораздо меньше числа независимых переменных. Таким образом, возможны различные решения, удовлетворяющие уравнениям равновесия» (Aubreville, loc. cit., стр. 140 англ, перевода). Согласно этой теории, обширное пространство смешанного леса может рассматриваться своего рода мозаикой, каждый кусочек которой представляет собой различную комбинацию доминирующих видов. На любом небольшом участке различные комбинации будут следовать одна за другой более или менее циклично. Если эта точка зрения окажется достаточно доказанной, это приведет к значительным изменениям в обычных представлениях о климатическом климаксе как о стабильном растительном сообществе с неизменным флористическим составом.
Теория Обревиля, безусловно заслуживающая серьезного рассмотрения, основывается главным образом на изучении флористического состава так называемого сомкнутого леса (foret dense) — дождевого леса и смешанного листопадного леса — на Берегу Слоновой Кости. Обревиль приводит в качестве примера пробный участок размером 1,4 га внутри «ненарушенного первичного леса» (табл. 6). На этом участке находилось 74 вида деревьев; самым обычным видом среди группы, относящейся к наибольшей степени толщины, был Piptadenia africana. Было найдено 7 экземпляров этого вида диаметром 0,5 м и больше, и только два — диаметром 0,1 — 0,5 м\ молодых растений диаметром менее 0,1 м не было. Подобное же распределение по ступеням толщины обнаружилось у Canarium schweinfurthii, Parkia bicolor и Parinari tenuijolia. С другой стороны, Combretodendron africanum был представлен лишь четырьмя экземплярами диаметром 0,5 м и более и 10 диаметром 0,1—0,5 м\ К hay a ivorensis — двумя крупными деревьями (диаметром более 0,5 м), одним деревом диаметром 0,1 .и и 12 молодыми деревьями. Эти цифры позволяют заключить, что по мере гибели существующего в настоящий момент поколения доминирующих деревьев состав древостоя будет изменяться, и место прежних доминант займет новая комбинация господствующих деревьев.
Распределение, подобное только что описанному, по-видимому, является обычным для африканских лесов и было отмечено в других местах. Милдбрэд (Mildbraed, 1930а), ссылаясь главным образом на леса Камеруна, замечает, что по общему впечатлению более крупные лесные деревья восстанавливаются очень слабо; лес состоит из сравнительно небольшого числа крупных деревьев и из многочисленного подроста молодых деревьев, начиная с толщины тросточки до толщины в бедро, причем молодые экземпляры крупных деревьев составляют лишь незначительную часть всей массы. Милдбрэд объясняет такое скудное возобновление слабой освещенностью нижних слоев леса, которая препятствует прорастанию или, если всходы успели появиться, вскоре убивает их. На пробном участке смешанного леса в резервированном лесу Шаша в Нигерии (Richards, 1939) в самом высоком ярусе (А) наиболее обильно представленным деревом былаErythro-phleum ivorense, но все имевшиеся па участке 13 деревьев имели диаметр ствола 61 см или более, причем не было замечено ни всходов, ни подроста молодых деревьев или более развитого их молодняка (жердняк), хотя вряд ли при осмотре можно было их пропустить. Подобное же, но менее резко выраженное распределение по ступеням толщины характерно для большем части доминирующих видов во влажных вечнозеленых лесах провинции Бенин. Уотсон (Watson, 1937), говоря о лесах полуострова Малакки,
5*
Состав проэного участка ненарушенного первичного леса, (Aubr6ville, 1938,
Виды Подлесок и нижний ярус деревьев, ступени толщины, л» Верхние ярусы деревьев, ступени толщины, м
молодые растения 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1.0 1,2 1,5
Pipladenia africana 1 1 1 1 1 1 1 2
Combretodendron africa-пит — 4 3 2 1 1 2 — 1 — — — —
Khaya ivorensis 12 1 — — 1 1 — — — —
Canarium schweinfurthii 1 1 —
Klainedoxa gabonensis — — — — 1 — 1 — — — — — —
Parkia bicolor 1
Parinari tenuifolia 1
Pachylobus deliciosa — 46 37 И 2 — 1 —
Funtumia africana — 3 2 2 — 1 1 —
Diospyros sanza-minika — 45 27 2 —
Strombosia pustulata — 43 13 4 2 —
Scott cilia coriacea — 15 7 3 2 2 —
Mo nod ora myristica — 3 6 3 2 —
Parinari kerstingii — 2 4 2 1 2 —
Hannoa klaineana — 16 6 2 —
Allanblackia parviflora — 15 3 3 1 —
Scytopetalum tieghemii Calpocalyx brevibractea- — 3 3 1 4 —
tus — 16 5 2
Cola maclaudii Daniella sp. aff. thuri- — 3 6 1 — — — — — — — — —
fera Prot omegabar ia stapfi- — 3 2 1 1 2 — — — — — — —
ana — 5 3 1 2 — — — — — — — —
Trichoscypha arborea — 2 5
Panda oleosa — — 3 — 2 — — — — — — — —
Cola nitida — 6 5 — —
Irvingia gabonensis En tandrophragma ango- — 2 3 1 1
lense 3 3 2 2 —
Diospyros 'kamerunensis Guarea cedrata 7 22 7 4 2 1 — — — — — — —•
Tarr let ia utilis 53 6 2 1 — — — — — — — —
Amphimas pterocarpoides — 6 2 1 — —
Phialodiscus plurijugatus — 2 2 — 1 — — — — — — — —
Baphia pubescent — 10 3 — —
Enantia polycarpa — 3 3 — —
Iso Iona cam panu lata — 5 3 — —
Aporrhiza sp. Entandr ophragma cylin- — 3 3
drieum 1 — 2 —
резервированный лес Масса-Ме, Берег Слоновой Кости стр. 133—134)
Таблица 6
Виды Подлесок и нижний ярус деревьев, ступени толщины, м Верхние ярусы деревьев, ступени толщины, м
молодые растения 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,2 1,5
Turraeanthus africanus 48 4 1 1
Afzelia bella — 3 — 2 — — — — — — — — —
Albizzia zygia — 1 2 —
Guarea thorn pisoni — 1 1 1 —
Coula edulis — 1 2 — — — — — — — — —- —-
Pleiocarpa mutica — 7 2 —
Vitex micrantha — 3 2
Garcinia polyantha — 4 2
Macrolobium chrysophy-lloides 3 1 1
Discoglypremna caloneura — 5 1
Ricinodendron africanum 1
Tylostemon mannii — 2 1
Trichilia heudelotii — 2 1 — — — — — — — — — —
Lannea acidissima 1
Octoknema borealis — — 1 — — —
Xylopia elliotii — 3 • 1 — — — — — — — — — —
Pycnanthus angolensis — 4 — — 1 — — — — — — —•
Cola chlamydantha — 37 1 — — — — — — — — — —•
Myrianthus arboreus — 12 — 1 — — — — — — — — —*
Conopharyngia durissima — 8 1 — — — — — — — — — —•
Phyllanthus discoideus — 2 1 — — — — — — — — — —г
Bridelia micrantha — — 1
Napoleona sp. — 9 — — — ——
Omphalocarpum sp. — 5 — - — —
Fagara macrophyIla — 4 — — —-
Uapaca guineensis — 3 — — —
Mammea africana — 1 — — — — — — —
Lophira procera — 1
Ant laris welwitschii — 1 —
Erythrophleum ivorense — 1
Anopyxis ealensis — 1
Anthocleista nobilis — 1 — — — — — — —
Scaphopetalum amoenum — 5 — — — — — — — —
Microdesmis puberula Baphia nitida <— 3 3 — — — —
Сагара procera — 1 — — — — — — — — —
Randia genipiflora Chlorophora excelsa 1 1 — 1 1 — — — — — ; — —
70
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
пишет, что малочисленность в подлеске представителей видов, доминирующих вверху, является часто отличающей особенностью, исключительно свойственной девственным, особенно тропическим лесам.
Ченгапа (Chengapa, 1934). описывая восстановление диптерокарповых лесов (скорее полувечнозеленого, чем вечнозеленого типа) Андаманских островов, отметил преобладание спелых и переспелых деревьев над «совершенно неадекватным» количеством деревьев более молодого возраста и почти полное отсутствие сеянцев и молодняка подроста деревьев, за исключением полосы отчуждения вдоль железной дороги, обочин дорог, покинутых мест, где были расположены лагери, и недавно вырубленных участков.
Джонс (Е. W. Jones, 1945) в статье, посвященной восстановлению девственных лесов северной умеренной зоны, пришел к выводам, сходным с выводами Обревиля. В этих лесах, как и в лесу Берега Слоновой Кости,обильно восстанавливающиеся виды часто отличаются от видов, доминирующих в спелом древостое; часто совершенно отсутствуют всходы одного или большего количества видов спелых деревьев. Обилие всходов вовсе не обязательно указывает на то, какие виды действительно возобновляются; очень обильные всходы какой-либо породы могут погибнуть, если им не удается укорениться, и, с другой стороны, там, где средняя продолжительность жизни доминирующего вида составляет 300 лет, достаточно очень небольшого числа молодняка, чтобы обеспечить беспрерывное существование сообщества. Однако на основании имеющихся данных можно предполагать, что в девственном лесу северной умеренной зоны возобновление доминирующего вида, возможно, требует редкой комбинации благоприятных обстоятельств. Если такая комбинация наступает через достаточно долгие промежутки времени, то флористический состав сообщества будет непрерывно изменяться, и ни один вид (или группа видов) не будет постоянно доминирующим. Так, например, в климаксном буковом лесу через поколение в одной части занимаемого им пространства доминантой может стать серебристая ель, а в другой — канадская ель. Таким образом, климакс будет представлять собой мозаичное сообщество как во времени, так, возможно, и в пространстве.
Маловероятно, что калейдоскопическая концепция Обревиля может быть применена ко всем ассоциациям смешанного дождевого леса. В смешанном лесу Брит. Гвианы, например, все обильно распространенные виды более крупных ступеней толщины являются также обильными и в меньших ступенях, как это видно из табл. 7, хотя относительное обилие видов в меньших ступенях толщины отличается от относительного обилия в более высоких ярусах. Непосредственные исследования (Davis, Richards, 1930—1934, табл. IV) также показывают, что ни один вид, способный достичь крупных размеров, не бывает представлен в классах мелких размеров, что нельзя сказать о более крупных классах. Вероятно, флористический состав смешанной ассоциации, подобной лесу Брит. Гвианы, будет оставаться неизменным в течение неопределенного времени.
Сам Обревиль соглашается, что стабильные эдафические сообщества, вроде спелой мангровы или ассоциации Cynometra megalophylla, Ну те по-stegia spp., Pterocarpus santalinoides и т. п., характерные для берегов рек Берега Слоновой Кости, не могут быть изменяющимися по своему составу. В этих ассоциациях одна комбинация доминант, состоящая из видов с особыми требованиями или выносливостью в отношении к почве, видимо, способна сохраняться постоянно. То же можно сказать и о большинстве сообществ моно доминантно го климаксного тропического дождевого леса, например о консоциациях Mora excelsa и Ерегиа в Гвиане, а также о кон
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
71
социации Cynometra alexandri в Африке. В этих консоциациях, как уже упоминалось, более молодые возрастные классу доминирующих видов неизменно хорошо представлены, и нет оснований йолагать, что доминанта будет когда-либо замещена другим видом, пока среда остается без изменения.
Вопрос о том, дает ли мозаичная теория верное объяснение фактов или нет, и если верное, то насколько широко она может быть применена к ассоциациям смешанного дождевого леса, пока остается нерешенным.
Скудное возобновление доминирующих видов в африканских лесах указывает, по всей вероятности, на то, что состав сообщества меняется. Если лес по крайней мере в пределах участка, охарактеризованного в табл. 6, действительно является «нетронутым и первичным», как считает Обревиль, то, согласно мозаичной теории, изменения в нем должны носить циклический характер.
Таблица 7
Распределение по ступеням толщины восьми самых обычных видов деревьев на пробном участке величиной 400 у 400 футов (122x122 л<) смешанного дождевого леса, Морабалли-Крик, Брит. Гвиана
(Davis, Richards, 1933—1934)
Виды Среднее число молодых деревьев диаметром менее 4 дюймов (10 сл<) и более 15 футов (4,6 at) ВЫСОТЫ1 Число деревьев диаметром 4 дюйма (10 см) и больше
ступени толщины
4—8 дюймов (10—20 см) 8 — 12 дюймов । (20-31 см) 1 2—16 дюймов (31-4 1 см) 16—24 дюйма (4 1—64 tut) 24 дюйма (64 см) и более
Eschweilera decolorans2 Е. pallida 2 | 128 19 14 7 1 1
sagotiana 48 5 6 14 13 1
Licania laxiflora 24 18 13 5 6 —
L. heteromorpha var. perplexans 176 29 12 — — —
L, venosa 128 26 16 .9 10 —
Ocotea rodiaei 16 8 2 5 10 3
Pentaclethra macroloba 80 31 20 16 6 —
Данных относительно всходов нет
1 Деревья этой ступени толщины были сосчитаны на двух пробных полосах, составляющих 1/в часть участка, и полученные цифры умножены на 8.
2 Эти виды не различались в поле.
С другой стороны, если сообщество подверглось нарушениям в прошлом, то настоящая комбинация видов может быть одной из стадий восстановления, а изменения — частью нормального (не циклического) процесса развития в направлении к стабильному климаксу.
В некоторых случаях отсутствие возобновления может указывать на то, что вид находится в процессе исчезновения. Это почти наверняка правильно в отношении редкого эндемичного дерева Canarium mauri-tianum, произрастающего в «нагорном климаксном лесу» острова Маврикия (Vaughan, Wiehe, 1941, стр. 136), но здесь условия необычны в том отношении, что площадь леса была сокращена до предела, а естественные условия изменены вмешательством человека.
Глава J
ОБЛИК ДЕРЕВЬЕВ И КУСТАРНИКОВ
ЭКОЛОГИЧЕСКАЯ МОРФОЛОГИЯ ФЛОРЫ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Любой человек, впервые увидевший вечнозеленый тропический лес. неизбежно поражается некоторыми необыкновенными чертами этого сообщества, которые совершенно не свойственны или лишь изредка встречаются в других растительных формациях. Эти особенности замечаются повсюду как в дождевых лесах Старого света, так и Америки. Они действительно типичны для тропического дождевого леса, где бы он ни находился, и не приурочены к какой-либо одной географической области. Как указывалось во введении, это однообразие внешнего вида дождевого леса составляет резкую противоположность его таксономическому разнообразию.
В тропическом дождевом лесу бросаются в глаза крылообразные выросты, дополняющие основания стволов большей части крупных деревьев. Более близкое знакомство с таким лесом показывает, что деревья с крылообразными выростами или досковидными корнями не принадлежат к какому-либо одному семейству или группе семейств; они встречаются у видов самой разнообразной систематической принадлежности. У многих деревьев дождевого леса, в особенности у более мелких, цветки появляются прямо на стволе или крупных ветвях, причем почки пробиваются через кору. Такое необычайное явление, носящее название каулифлории, в высшей степени характерно для тропического дождевого леса и, как и досковидные корни, встречается у видов, принадлежащих ко многим совершенно неродственным семействам. Кроме таких бросающихся в глаза черт, как наличие досковидных корней и каулифлория, у деревьев тропического дождевого леса имеются и другие своеобразные морфологические особенности, которые в совокупности придают им легко распознаваемый облик, до известной степени схожий с деревьями таких родственных дождевому лесу типов формации, как горный дождевой и субтропический дождевой леса. Это справедливо не только в отношении деревьев, но в меньшей степени и в отношении других компонентов сообщества — кустарников, покровных трав, эпифитов и лиан, которые имеют свои собственные характерные черты, часто не менее замечательные.
Так как морфологические признаки, о которых идет речь, типичны для деревьев всех тропических дождевых лесов и не зависят от таксономической принадлежности видов, то они должны быть так или иначе связаны с окружающей средой. Из этого не следует, что они обязательно являются приспособительными в обычном смысле этого слова. Еще неизвестно, многие ли из этих признаков имеют большое значение в смысле выживания. По крайней мере некоторые из них, вероятно, «причинно» связаны со средой; в этом случае они не обязательно полезны для растения,но каким-то еще не совсем ясным для нас образом зависят от факторов местообитания. Морфологические признаки, о которых сейчас идет речь, могут быть названы эфармоническими в первоначальном значении этого термина (Ves-
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 73
que, 1882) без всякого предположения относительно происхождения «гармонии» между растением и средой.
Физиономичность, или экологическая морфология, флоры тропического дождевого леса была предметом многочисленных наблюдений. Действительно, внешний вид растительности тропического дождевого леса привлекал к себе до сих пор больше внимания, чем внешний облик какой-либо другой растительности. Бесчисленные авторы, как путешественники, так и профессиональные ботаники, отмечали наличие специальных признаков у растений дождевого леса, пытаясь каждый на свой лад «объяснить» их происхождение. Их догадки, иногда правдоподобные, но часто далекие от истины, обыкновенно основывались на предположении, что эти признаки нужны растениям, обладающим ими. В пределах одной главы невозможно дать обзор обширной литературы, посвященной этому вопросу. Более полный перечень ее находится в самом последнем издании шимперовской «Географии растений» (Schimper, 1935).
Надо отметить, что хотя все деревья тропического дождевого леса обладают многими общими чертами, однако каждый ярус имеет особый облик. Например, тип листвы, характерный для самого высокого (А) яруса деревьев, непохож на тип листвы, характерный для более низкого яруса деревьев и яруса кустарников (С и D); досковидные корни свойственны главным образом деревьям верхнего и среднего ярусов (А и В) и отсутствуют у деревьев нижнего яруса (С). С другой стороны, каулифлория характерна в основном для нижнего яруса деревьев и яруса кустарников (С и D). Таким образом, физиономичность в значительной степени является функцией яруса, к которому принадлежит растение, и описание внешнего облика каждого яруса служит дополнением к описанию самой ярусности, данному в главе 2.
ОБЛИК ДЕРЕВЬЕВ
При характеристике деревьев дождевого леса прежде всего следует обратить внимание на их внешний вид и форму. Это почти невозможно сделать в глубине леса, но вполне удается на прогалинах. Надо, однако, помнить, что изолированные деревья или деревья, находившиеся в течение некоторого времени на открытом месте, могут сильно видоизмениться и принять облик, весьма отличающийся от облика деревьев, выросших в их естественном окружении.
Во всех ярусах стволы у деревьев совершенно прямые или слегка извилистые. Трудно подыскать надлежащее сравнение, но можно сказать, что подавляющая масса деревьев тропического дождевого леса отличается большей тонкостью ствола по отношению к высоте, чем у деревьев умеренной зоны. Это особенно бросается в глаза в ярусе деревьев А и придает лесу в целом какой-то этиолированный вид. Прямизна и тонкость очень ясно выступают при сравнении отдельных видов тропического дождевого леса с родственными видами других растительных сообществ. Например, виды Eucalyptus, растущие в дождевом лесу, отличаются от остальных видов этого рода тем, что имеют более «колоннообразные» стволы (Herbert, 1929).
Впечатление устремленности ввысь, создаваемое крупными деревьями дождевого леса, частично зависит от значительной высоты неветвящейся части ствола. У спелых деревьев крона обыкновенно отличается меньшими размерами, чем у европейских деревьев, и первые ветви появляются срав
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
нительно близко к вершине. Отсутствие ветвей в нижней части ствола зависит. конечно, от конкуренции между соседними деревьями и низкой интенсивности освещения под пологом леса. Южноамериканская Eperua falcata, например, в естественных условиях имеет длинный неветвящийся ствол, характерный и для других высоких деревьев тропического дождевого леса, но в ботаническом саду Порт-оф-Спейн, на Тринидаде, ее ветви, несущие листья п цветки, начинались на высоте нескольких футов от земли, и внешний вид этого дерева напоминает облик деревьев английских парковых лесов. На берегах рек на стороне, обращенной к воде, деревья также ветвятся почти от основания ствола.
Как уже упоминалось в главе 2, кроны деревьев разных ярусов имеют различную форму, что зависит от высоты, на которой начинается ветвление, от длины ветвей и угла их расхождения. Это в свою очередь в значительной мере зависит от положения дерева в лесу. Вообще чем выше дерево, тем более широкой и более плоской бывает его крона. В ярусе А ширина крон обыкновенно превышает высоту (это заметно при взгляде на них в профиль), и они часто приближаются к зонтичной форме, как, например, у южноамериканской Peltogyne pubescens и западноафриканской Lophira procera. Однако зонтичная форма никогда не бывает так заметно выражена у деревьев тропического дождевого леса, как у деревьев более сухих типов тропической растительности.
В ярусе В кроны обыкновенно более удлиненные; в среднем их высота равна ширине. Наконец, в ярусе С, как правило, встречаются узкие кроны конической формы, чья высота значительно превышает ширину. В верхнем ярусе большинство ветвей первого порядка образует острый угол со стволом, в то время как в третьем ярусе они имеют тенденцию отходить от дерева под прямым углом.
На протяжении жизни дерево яруса А обычно последовательно принимает тип кроны, преобладающий в том ярусе, где оно в данный момент находится. Так, в юности оно имеет длинную, суживающуюся крону, характерную для яруса С, в почти спелом состоянии — более или менее закругленную, а по достижению полной спелости — крону уплощенной формы, характерную для яруса А. Поразительное различие формы крон у молодых и старых деревьев одного и того же вида в особенности хорошо заметно у некоторых малайских диптерокарповых, например у Dryobalanops аго-matica (Foxworthy, 1927, рис. к стр. 46).
У некоторых тропических деревьев, названных Корнером (Corner, 1940) «деревьями-пагодами», ветви образуют ряд ярусов, напоминающих крыши пагоды. Такой своеобразный облик получается главным образом вследствие перемежающегося роста побегов, который создает мутовчатое, или подобное мутовчатому, расположение главных ветвей. Пагодообразный облик крон исключительно ясно выражен у Terminalia catappa, а также у азиатской Alstonia scholaris, но не характерен для деревьев типичного спелого дождевого леса.
Деревья тропического дождевого леса значительно отличаются от деревьев умеренной зоны меньшей степенью разветвленности. Сложный узор веток и ветвей, характерный для большинства европейских деревьев, является редкостью в тропиках, и многие виды не имеют ветвей выше третьего порядка. Согласно Визнеру (Wiesner, 1895, стр. 676—678), обратившему на это особое внимание, ветвление до пятой или восьмой степени обычно для европейских деревьев, тогда как у деревьев дождевого леса наивысшей степенью ветвления является вторая, третья или в крайнем случае четвертая; ветвление выше пятой степени не встречается никогда. Однако,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
согласно Шимперу (Schimper, 1935), наблюдения Курдерса (Koorders) показали, что в исключительно редких случаях встречаются степени ветвления и выше пятой. Для деревьев тропического дождевого леса крайнее ограничение степени ветвления не является чем-то необычным. Во втором ярусе некоторых дождевых лесов растут высокие пальмы (например, виды Oenocarpus и многие другие, Южная Америка), совершенно лишенные ветвей. Кроме пальм в самом нижнем ярусе дождевого леса низменностей и кроме древовидных папоротников в горных дождевых лесах, нередко можно встретить небольшие деревья из двудольных, едва ли имеющие какие-либо ветви даже первого порядка — так называемые одноствольные — «Schopfbaumchen» Милдбрэда (Mildbraed, 1922, стр. 112). Такие деревья часто имеют исключительно крупные листья, сгруппированные у вершины. Это характерно для Clavija spp. из Южной Америки и Phyllo-botryum soyauxianum из Западной Африки. Ряд крупных деревьев дождевого леса, имеющих во взрослом состоянии нормально разветвленные кроны, остаются неразветвленными, с листьями, сосредоточенными на верхушке ствола, пока не достигнут высоты 6—7 м или более, как например южноамериканские Sterculia pruriens и S. rugosa.
Стремление листьев располагаться на концах побегов свойственно не только неветвящимся деревьям; оно достаточно хорошо проявляется и у большого числа видов с нормальным ветвлением, например у многих малайских Anacardiaceae (Campnosperma и Melanorrhoea spp.), а также у Terminalia superba — дерева, обычного для первого яруса западноафриканского тропического дождевого леса.
Деревья дождевого леса обыкновенно очень легко дают порослевые побеги, если главный ствол был повален или сгнил. Одним из наиболее замечательных примеров в этом отношении является Dicymbe corymbosa — дерево из семейства бобовых, доминирующее на обширных пространствах тропического дождевого леса во внутренней части Брит. Гвианы и известное также в районе Рио-Негро, бассейн Амазонки. У этого вида образование порослевых побегов наблюдается не от случая к случаю, а вполне закономерно. Первичный главный ствол, достигающий 30 и более метров высоты, с диаметром, превышающим 0,5 м, оказывается окруженным группой порослевых побегов, растущих от его основания. Когда с течением времени главный ствол отмирает, один или несколько порослевых побегов занимают его место и, разрастаясь, достигают размеров материнского ствола. Вокруг вторичного главного ствола растет новое поколение побегов; один из них также становится главным стволом, и так далее. В результате такого необычного способа роста каждый индивидуум оказывается состоящим из группы стволов различных размеров, чем и объясняется местное название этого дерева — «групповая Уаллаба». В Брит. Гвиане на пробной площадке размером в 800 кв. м было зарегистрировано 26 индивидуумов этого вида дерева и 61 ствол с диаметром в 2 дюйма (5 см) и более (Т. G. Tutin).
В болотистом дождевом лесу (Западная Африка), произрастающем на очень мягкой рыхлой почве, у дерева Grewia coriacea ствол нередко наклоняется до горизонтального положения, и на нем образуются порослевые побеги. Все они могут стать крупными деревьями, и когда породивший их ствол сгнивает, он оказывается замещенным рядом молодых независимых индивидуумов (Richards, 1939, стр. 45). Подобный тип побегообразования свойственен Dimorphandra conjugate, и Cyrilla antiliana, растущим в болотистом лесу Брит. Гвианы (Т. A. W. Davis).
76
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
КОРА
Деревья тропического дождевого леса имеют обыкновенно замечательно тонкую и, как правило, гладкую светлоокрашенную кору. Толстая кора, вроде коры европейского дуба пли сосны, в тропиках есть нечто необычное, и даже у крупных деревьев первого яруса она часто измеряется лишь несколькими миллиметрами. Фоксуорсп (Foxworthy, 1927) приводит измерения толщины коры у целого ряда деревьев Малайского архипелага, дающих деловую древесину (большей частью крупных видов яруса А). Среднее из этпх измерений составляет 0,4 дюйма (10 мм), максимальное — более 1 дюйма (25 мм) и минимальное — 0,15 дюйма (4 мм). В местности Морабалли-Крик в Брит. Гвиане была отмечена самая тонкая кора всего лишь в 3 мм толщиной. У деревьев с достаточно большим диаметром ствола развитие коры иногда бывает настолько слабым, что сквозь нее виден хлорофилл клеток, находящихся под корой. Это, например, наблюдается у Ficus cordifolia (Schimper, 1935, стр. 461). Деревья ярусов В и С, как и следовало ожидать, имеют более тонкую кору, чем деревья самого высокого яруса.
Гладкость, представляющая собою столь обычное свойство коры дождевого леса,— несомненное следствие ее тонкости. Слой феллогена здесь обыкновенно поверхностный и непрерывный, поэтому кора не трескается и сбрасывается обычно очень маленькими, едва заметными отдельностями.
Иногда, но значительно реже, встречается и шелушащаяся, растрескивающаяся кора. У Distemonanthus benthamianus и, согласно Милд-брэду (Mildbraed, 1922, стр. 111), у Ochna calodendron — двух западноафриканских видов — кора отделяется в виде крупных чешуек, как у платана. Кора Planchonella obovata, растущей в районе Малайского архипелага, лупится продольными широкими слоями, в то время как у видов Tri-stania, обычных в некоторых типах тропического дождевого леса Малайского архипелага, кора расщепляется на продольные слои, сохраняющиеся у основания дерева. Кора с бороздами или продольными трещинами гораздо более обыкновенна, чем кора, опадающая кусками; кора этого типа свойственна, например, многим малайским диптерокарповым и западноафриканской Mimusopsdjave. Можно было бы привести еще много других исключений из общего правила. Степень тонкости и гладкости коры деревьев тропического дождевого леса отчетливо выявляется при сравнении Lophira procera, высокого дерева, обыкновенного для западноафриканского дождевого леса, с близким его родственником (оба они часто рассматриваются как один вид), L. alata, встречающимся в парковых лесах и саваннах. Первый вид имеет тонкую, слегка отстающую отдельными кусками кору, а у второго кора толстая, покрытая глубокими бороздами и трещинами.
Благодаря тонкому покрову из беловатых корковых лишайников цвет коры иногда кажется светлее, чем он есть на самом деле. Но даже и там где нет лишайникового покрова, кора тропических деревьев обыкновенно окрашена светлее, чем у большинства деревьев умеренной зоны. Иногда кора верхних ветвей высокого дерева оказывается окрашенной светлее, чем кора ствола; это, по-видимому, является прямым следствием солнечного света, обесцвечивающего пигмент, который придает коре ее окраску. Габерландт (Haberlandt, 1926, стр. 96) считал, что светлая окраска коры тропических деревьев предупреждает излишний нагрев камбия, расположенного под корой. Хотя светлоокрашенная кора у тропических деревьев преобладает, некоторые деревья дождевого леса, например Lissocarpa guianensis, и многие виды Diospyros имеют темную, иногда почти черную, как уголь, кору.
СТРУКТУРА И ВНСШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 77
Изредка в дождевом лесу встречаются стволы с шипами, что более характерно для сухих тропических лесов (см. гл. 15); среди них Нуlode ndron gabunense и виды западноафриканской Fagara. У маленького африканского дерева Citropsis articulata и у Balanites wilsoniana молодые и вообще более мелкие ветви вооружены острыми шипами. Многие лесные пальмы, встречающиеся в тропическом дождевом лесу, также имеют шиповатые стволы.
ДОСКОВИДНЫЕ КОРНИ И КОРНЕВЫЕ СИСТЕМЫ
Почти все авторы, писавшие о тропическом дождевом лесе, высказывали свои соображения относительно происхождения досковидных корней, но до последнего времени это явление не было исследовано достаточно тщательно, и дело ограничивалось отрывочными указаниями на предполагаемое значение этих корней в качестве опор, или «якорей». Наиболее интересные статьи по этому предмету написаны Чипом (Chipp, 1922), Сенном (Senn, 1923), Фрэнсисом (Francis, 1924), Наве (Navez, 1930) и Петчем (Petch, 1930). Досковидные корни имеют большое экологическое значение и настолько важны в качестве характерного признака деревьев тропического дождевого леса, что необходимо несколько подробнее остановиться на их структуре и развитии.
Стволы деревьев дождевого леса часто на большой части своей длины являются желобчатыми или ребристыми. В крайних случаях ствол на поперечном сечении может быть отчетливо звездчатым, как например у южноамериканского вида Aspidosperma excelsum (Аросупасеае). Подобные желобки и ребра, однако, не включаются в понятие досковидных корней. Этот термин обычно применяется только к выростам, ограничивающимся нижней частью ствола. Местные расширения нижней части ствола (empattements французских авторов), вро^де тех, что имеются у деревьев умеренной зоны, часто встречаются и у тропических деревьев. Подобные выступы бывают более расплывчатыми и менее определенными по своим очертаниям, чем истинные досковидные корни, но составляют незаметный к ним переход. Выросты у основных стволов могут быть разделены на два главных класса.
1. Хо1ульные корни. Это крепкие, деревянистые, придаточные корни, отходящие от основного ствола на расстоянии приблизительно 1 м или больше от земли, часто прихотливо изогнутые вниз и проникающие в почву. Над почвой они ветвятся и иногда анастомизируют; в почве они дают начало боковым корням второго и третьего порядка. Они могут быть округлыми в сечении и более или менее концентрическими по строению илп же благодаря интенсивному вторичному росту в вертикальной плоскости оказываются уплощенными. Наиболее известными деревьями с округлым в сечении концентрическим типом ходульных корней являются обычные растения мангровы Rhizophora (Rhizophoraceae). Но подобные же корни встречаются также и у многих видов болотистого (с пресноводным питанием) и нормального хорошо дренированного тропического дождевого леса: например, у видов Clusia (включая неэпифитные виды) и Tovomita (Gutti-ferae) — в Южной Аамерике, Мacaranga barteri и Uapaca spp. (Euphorbia-ceae) — в Западной Африке, Elaeocarpus littoralls (Tiliaceae), Xylopia jerruginea (Annonaceae), виды Dillenia (Dilleniaceae); Eugenia (Myrtaceae) и т. д.— в Малайе.Подобные же, но более прямые ходульные корни встречаются у некоторых пальм, например у южноамериканской Iriartea
78 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и Pandanus (Pandanaceae). Уплощенные ходульные корни свойственны южноамериканской Virola surinamensis (Myristicaceae), африканским Bridelia micrantha (Euphorbiaceae), Musanga cecropioides (Moraceae), San-tiriopsis trimera (Burseraceae) и малайской Casuarina sumatrana. Некоторые травы тропического дождевого леса также имеют ходульные корни, например африканская Culcasia striolata (Агасеае), некоторые малайские Zingi-Ьегасеае и виды Mapania. Таким образом, ходульные корни наблюдаются у представителей самых разнообразных семейств и родов.
Большинство деревьев с ходульными корнями имеет небольшой или средний размер и относится к ярусу С; сравнительно немногие, например малайская Норса mengarawan и некоторые виды Dillenia, являются крупными деревьями ярусов А и В.
2. Собственно оосковисные корни. Это более или менее плоские треугольные пластины, расположенные углом между стволом и боковыми корнями, проходящими близ поверхности почвы. Они образуются крайним эпинастическим вторичным ростом верхней стороны боковых корней, причем первоначальный рост (как это видно на поперечном разрезе) всегда происходит вдоль основания. Деревья тропического дождевого леса с досковидными корнями настолько многочисленны, что нет надобности приводить их примеры. У нас нет точных цифр, характеризующих относительную частоту образования досковидных корней среди деревьев различных классов размера или различных ярусов, но наблюдения в Брит. Гвиане, на Борнео и в Нигерии показывают, что наибольшее количество деревьев с досковидными корнями свойственно самому высокому (А) ярусу — деревья выше 30 м. У деревьев яруса В досковидные корни все еще очень обычны, но число видов без досковидных корней тут выше, чем в ярусе А. В самом нижнем ярусе деревьев, не считая входящих в его состав молодых экземпляров, которые, став спелыми, будут принадлежать к более высоким ярусам, досковидные корни почти отсутствуют. Следовательно, имеется общее (но отнюдь не точное) сортношение между высотой дерева и образованием досковидных корней.
Ходульные и досковидные корни сходны тем, что и те и другие представляют видоизменение боковых корней. Различие между ними состоит в том, что ходульные корни образуются от придаточных корней, возникающих на главной оси над землей, в то время как досковидные корни развиваются на крупных боковых корнях, появляющихся у самой поверхности почвы или как раз над нею. Раз это так, то не удивительно, что встречаются и структуры промежуточного типа. Если ходульные корни подвергнутся большому утолщению в вертикальной плоскости, то они приобретают сходство с досковидными корнями. С другой стороны, хотя нижний край досковидного корня бывает обыкновенно погружен на несколько сантиметров в почву, может случиться, что между нижним краем корня и почвой на проксимальном конце окажется свободный просвет, тогда досковидный корень будет напоминать очень широкий ходульный корень. Иногда у одного и того же индивидуума могут быть как ходульные, так и досковидные корни; примером этого является африканская Grewia coriacea
Из способа образования досковидных корней ясно, что они могут возникать только у деревьев с поверхностной корневой системой и хорошо развитыми боковыми корнями, имеющими тенденцию проходить в горизонтальном направлении; ходульные корни предпочтительно встречаются у видов, где более крупные боковые корни сгибаются вертикально или косо книзу. Уилкинсон (Wilkinson, 1939) делит корневые системы деревьев малайского леса на четыре группы, у которых: 1) хорошо развиты боковые
Фото 6. Дерево с досковидными корнями (Mora excelsa), Морабалли-Крик, Брит. Гвиана.
Фото 7. Дерево с ходульными корнями (Tovomita sp.)> Морабалли-Крик, Брит. Гвиана.
СТРУКТУРА II ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
79
корни, стержневой же корень отсутствует; 2) боковые корни хороню раз виты, стержневой корень сохраняется; 3) боковые и стержневой корни одинаково хорошо развиты и сохраняются; 4) боковые корни слабые или
отсутствуют.
Подобную же классификацию для яванских деревьев предлагает Костер (Coster, 1932). При рассмотрении многочисленных представителей каждой группы Уилкинсона обращает на себя внимание то обстоятельство,
что все виды с явными досковидными корнями относятся к группам 1—3 и, следовательно, имеют мощные боковые корни, в то время как все виды с ходульными корнями принадлежат к 4 группе. Можно добавить, что существуют некоторые данные, в общем свидетельствующие о том, что более крупные деревья дождевого леса имеют плоско-полусферические пли блюдцеобразные поверхностные корневые системы и часто обходятся без стержневого корня. Деревья же меньшего размера отличаются более глубокими не очень раскидистыми корнями, так что их корневая система имеет форму узкого опрокинутого конуса. Если это является общим правилом, то преимущественное образование досковидных корней у деревьев верхнего яруса, а ходульных корней — у деревьев нижнего яруса становится понятным.
Досковидные корни встречаются гораздо чаще, чем ходульные, и имеют более важное экологическое значение. Все же, прежде чем перейти к более подробному описанию первых, следует коротко охарактеризовать последние.
Хотя ходульные корни являются вполне обычной принадлежностью деревьев многих семейств, однако у каждого данного вида существует известная пластичность в этом отношении; при некоторых
условиях ходульные корни развиваются,
а при других отсутствуют. Например, Ми- р 11 с- Продольный разрез до-sanga cecropioides, неизменно обладающая СК°“^"рМ<ЛГР(к7апс^7»24) ходульными корнями в Западной Африке,
в Уганде их часто лишена (Brasnett). В Малайе некоторые виды деревьев (Eugenia longijlora, Archytaea vahlii, Myristica elliptica, M. main-gayi, Macaranga spp.), которые, как правило, имеют ходульные корни, произрастая в болотистых лесах, обходятся без них на хорошо дренированной почве (Corner). Атапоа caribaea и О.гу there pallida, произрастающие на острове Доминика (Вест-Индия) на болотистых подзолистых почвах, образуют ходульные корни, но в низинном лесу с нормальным дренажем они развивают досковидные корни (Beard).
Встречаемость деревьев с ходульными корнями до известной степени связана с почвенными условиями. Как известно, деревья с ходульными корнями гораздо более обильны в заболоченной мангрове, чем в нормаль
8(1
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ном дождевом лесу. Они также обычны в лесах, растущих на пресноводных болотах. В южной Нигерии они определенно более свойственны лесу, произрастающему на пресноводном болоте с мягкой, зыбкой почвой, чем смешанному лесу с его плотными глинистыми или суглинистыми почвами. На пробном участке в болотистом лесу деревья с ходульными корнями составляли 7,3% всех деревьев с диаметром ствола 4 дюйма (10 сл*) и более; на участке смешанного леса, развитого на латеритной глине, соответствующая цифра составляла 1,3 ?о, а на участке смешанного леса на песчаном суглинке она равнялась 1,2°6 (Richards, 1939, стр. 45). В болотистых лесах Малайи в изобилии встречаются виды с ходульными корнями, а на острове Доминика описан «подтип» дождевого леса на слабо дренированной почве, имеющий (в хорошо развитом виде) почти чистый древостой из двух видов с ходульными корнями — Symphonia globulifera и Tovomita plumieri (Beard, 1944, с, стр. 144). В тропическом дождевом лесу на хорошо дренированных местообитаниях деревья с ходульными корнями очень редко встречаются в таком количестве, чтобы накладывать отпечаток на характер сообщества; в этих местообитаниях не наблюдается и приуроченности их к какому-либо одному типу почв. В тропическом дождевом лесу (Малайский архипелаг) Корнер (Е. J. Н. Corner) отметил на хорошо дренированной почве только три вида с ходульными корнями — Dillenia grandifolia, D. reticulata и Xylopia jerruginea, В местности Морабалли-Крик в Брит. Гвиане, где насчитывается пять типов первичного леса на различных почвах — от переувлажненной, подверженной затоплению, илистой до плотных глинистых и песчаных латеритов — не наблюдалось разницы в обилии деревьев с ходульными корнями во всех пяти типах леса.
У внетропической растительности ходульные корни встречаются, по-видимому, очень редко, но определенных сведений по этому вопросу мало.
Явные преимущества, доставляемые ходульными корнями дереву, растущему на зыбкой болотистой почве, могли бы с первого взгляда внушить мысль о том, что они представляют собой ясный случай приспособления, развившегося под действием естественного отбора. Однако при ближайшем рассмотрении соответствующих фактов возникают трудности, не позволяющие принять какое-либо простое «объяснение». В самом деле, если ходульные корни имеют важное значение в качестве опоры, то удивительно, что они чаще встречаются у небольших деревьев, растущих в подлеске под защитой густого полога, чем у высоких деревьев с тяжелой кроной, которым эти корни, казалось бы, должны быть особенно необходимы. Как будет упомянуто дальше, высокие даже вполне здоровые деревья дождевого леса часто валятся ветром, так что эффективная система укоренения, несомненно, была бы для них полезна.
Среди возможных причин, могущих объяснить сравнительно большой процент ходульных корней у деревьев тропического дождевого леса, следует указать на постоянно влажную атмосферу в более низких ярусах леса, что может способствовать образованию таких корней. На существование связи между влажностью и образованием ходульных корней указывает, между прочим, тот факт, что в болотистом дождевом лесу полуострова Малакка максимальная высота прикрепления ходульных корней совпадает с наивысшим уровнем паводка при наводнении в этой области; в некоторых местах эта высота может доходить до 30 футов (9 м) (Corner). Подобное же наблюдение было сделано Дэвисом (Т. A. W. Davis) в Брит. Гвиане. Возможно, что образование ходульных корней облегчается также тонкостью коры, обычной для деревьев тропического дождевого леса. Обратимся
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 81
теперь к рассмотрению собственно досковидных корней. В остальной части этой главы слово «опора» будет относиться только к ним.
Строение и развитие1 досковидных корней
Древесина, из которой состоят досковидные корни, обычно более тверда, чем древесина ствола того же дерева. Благодаря своей твердости досковидный корень при ударе по нему топором издает своеобразный «пустой» звенящий звук. Согласно Фрэнсису (Francis, 1924, стр. 31), исследовавшему досковидные корни Argyrodendron trifoliolatum из австралийского дождевого леса, ткани, составляющие досковидные корни, подобны тканям ствола. Кора досковидных корней, как правило, тоньше, чем кора ствола; у четырех изученных пород деревьев тропического дождевого леса Квинсленда кора ствола была в 1,6—2,5 раза толще коры досковидных корней (Francis, 1924, стр. 32).
Как отмечалось, внутреннее строение взрослого досковидного корня отчетливо эпинастическое с центром роста у основания (рис. 10). Когда в древесине досковидного корня ясно заметны годичные кольца, то видно, что ранние кольца являются правильно кольцеобразными; ненормальное утолщение внешней стороны начинается только спустя некоторое время. Следовательно, корни, от которых образуются досковидные выросты, вначале имеют обычное строение; специфические изменения наступают лишь позже. Это имеет известное значение в связи с возможным «причинным» объяснением образования досковидных корней. I
В целом стадия роста дерева, на которой начинается образование досковидных корней, различна у разных видов, но, по крайней мере в некоторых случаях, процесс этот начинается очень рано. У целого ряда квинслендских видов деревьев досковидные корни оказываются уже развитыми, когда ствол составляет х/17—х/9 своего максимального диаметра и 2/ь—х/4 максимальной высоты (Francis, 1924, стр. 25—26). Равным образом Петч (Petch, 1930, стр. 278), наблюдая (в ботаническом саду?) за деревьями Delonix regia, обнаружил у них досковидные корни в возрасте 2—3 лет, когда их высота не превышала 12 футов (3,7 м). Мак-Грегор (MacGregor, 1934, стр. 63—64) заметил, что на плантациях в Нигерии у Terminalia superba досковидные корни начинали образовываться раньше, чем дерево достигало пятилетнего возраста, когда его высота составляла менее 8 м. Следовательно, иногда высказываемое предположение, что досковидные корни развиваются только тогда, когда дерево становится высоким и приобретает мощную крону, неправильно.
Весьма примечательно, что у дерева с хорошо развитыми досковидны-мп корнями ствол, вместо того чтобы постепенно суживаться снизу вверх, всегда более или менее заметно суживается от верхнего уровня отхождения досковидных корней не только вверх, но и вниз, по направлению к земле. Фрэнсис (Francis, 1924, стр. 31) пашел, что у Echinocarpus woollsii в Квинсленде диаметр ствола, достигавший 60,4 см над досковидными корнями, равнялся только 22,6 см у поверхности почвы. Сужение ствола книзу отсутствует у молодых деревьев и становится выраженным только при достижении досковидными корнями крупных размеров. Причина этого явления но ясна; одно из возможных объяснений мы рассмотрим ниже.
У нескольких деревьев с досковидными корнями, например у Сапа-rium commune и Ceiba pentandra. в углах между ветвями и главным стволом, а иногда, видимо, и непосредственно на главном стволе имеются крылообразные расширения, похожие на досковидные корни.
82
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Число досковидных корней на одном стволе колеблется от одного до десяти; хотя точные данные о их среднем количестве отсутствуют, надо полагать, что обычно бывает от трех до пяти досковидных корней. Тот факт, что у взрослых деревьев число досковидных корней обычно бывает больше одного пли двух, охотно используется сторонниками мнения о их приспособительном значении, поскольку механически действенная опора создается минимум при трех досковидных корнях.
Размер, толщина и форма досковидных корней изменяются в весьма широких пределах. Хотя эти признаки заметно варьируют у различных индивидуумов одного и того же вида, однако они, как это впервые подчеркнул Чип (Chipp, 1922), являются до известной степени постоянными в пределах вида или более крупных систематических единиц. К сожалению, досковидные корни редко упоминаются в работах по систематике, и в нашем распоряжении находится мало сведений по этому предмету, но все, кто знаком с тропическими деревьями, знают, что досковидные корни представляют в определенных пределах значительную диагностическую ценность при полевой работе.
Размер досковидных корней у данного вида в значительной мере зависит от возраста дерева. У вполне взрослого дерева досковидные корни могут достигать размеров огромных досок, тянущихся вверх по стволу до 9 м и более и почтп на такое же расстояние в стороны. Широкие досковидные корни некоторых видов, например малайской Koompassia excelsa, употребляются, в частности, для изготовления больших обеденных столов. Размер досковидных корней имеет иногда диагностическое значение при различении близко родственных видов. Так, Mora gonggrijpii, обычное дерево гвианских лесов, всегда имеет досковидные корни меньшего размера, чем близкородственный, но более широко распространенный вид М. excelsa (см. рис. 11).
Толщина досковидных корней также сильно меняется и имеет порой диагностическое значение. У некоторых видов досковидные корни, даже когда они достигают больших размеров, имеют толщину всего в несколько сантиметров, в то время как у других видов они бывают гораздо толще. Между ними существуют разнообразные переходы от тонких, как лезвие ножа, пластинок до широких, бесформенных желобков или ребер, напоминающих выступы стволов у таких европейских деревьев, как Taxus и Саг-pinus.
Форма досковидных корней в профиль сильно меняется и часто весьма характерна для отдельных видов. Чип (Chipp, 1922) усиленно подчеркивал диагностическое значение отношения высоты досковидных корней к их основанию (или наклона наружной кромки). Но при этом он упустил из виду тот факт, что у многих, если не у всех, пород деревьев это отношение меняется: с увеличением возраста дерева досковидные корни становятся выше по сравнению с их шириной. Мы уже упоминали о наблюдениях Фрэнсиса (Francis) над изменением этого отношения у Argyrodendron tri-joliolatiun, подобное же изменение было отмечено автором у Mtora gonggrijpii в Брит. Гвиане и у Terminalia superba в Западной Африке. Тем не менее при сравнении деревьев в одинаковой стадии роста наклон наружного края досковидных корней имеет некоторое значение в качестве видового признака. Так Couratari palchra может быть легко отличена по своим высоким, очень круто наклоненным досковидным корням, от других Lecythi-daceae в лесах Морабалли-Крик, в Брит. Гвиане. Подобные же очень крутые досковидные корни характерны и для западноафриканской Alstonia congensis. Можно было бы привести и другие примеры.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
83
Досковидные корни встречаются у видов, принадлежащих к большому числу неродственных между собою семейств; в то же время ряд семейств, представленных в тропическом дождевом лесу, обнаруживает большую склонность к образованию досковидных корней, чем [другие. Dipterocarpaceae, Leguminosae и Sterculiaceae включают много видов, обыкновенно характеризующихся большими досковиднымп корнями, в то время как у тропических Aimonaceae, F'agaceae (Lithocarpus bennettii Малаккского полуострова, см. Corner, представляет собой исключение) и Lauraceae хорошо развитые досковидные корни обычно отсутствуют. Досковидные корни также отсутствуют у Myristicaceae по крайней мере у видов Малаккского полуострова (Corner). Между различными родами одного и того же семейства опять-таки существуют большие различия, например в составе семейства Bombacaceae, наряду с Bombax spp. и Ceiba pentandra с их огромными досковидными корнями, имеется также род Catostemma, многие виды которого, представленные в гвианском дождевом лесу, совсем не имеют досковидных корней. Аналогичным же образом, несмотря на то, что большинство деревьев тропического дождевого леса, принадлежащих к семейству бобовых, в большей или меньшей степени обладает развитыми досковидными корнями, у малайского рода Sindora (включающего большое количество видов с деловой древесиной) и у ряда других малайских родов досковидные корни всегда отсутствуют (Corner). У Dipterocarpaceae роды Dipterocarpus, Нореа и Shorea имеют виды как с досковидными корнями, так и без них. То же самое можно сказать и о видах крупных родов Diospy-ros (Ebenaceae) и Eugenia (Myrtaceae), Малаккский полуостров (Corner). В пределах вида наличие досковидных корней является, по-видимому, весьма постоянным признаком, но некоторые виды обнаруживают в этом отношении пластичность. Иногда эта пластичность настолько велика, что у одного и того же вида некоторые индивидуумы имеют большие доско-видные корни, а у других их нет совсем. До сих пор мы в этом отношении располагаем лишь небольшим количеством наблюдений, но у Гескье (Ghescuiere, 1925, стр. 2) имеются сведения о том, что африканские виды Combretodendron ajricanum и Tetrapleura tetraptera могут быть как с досковидными корнями, так и без них. Eschweilera sagotiana в Брит. Гвиане обычно почти лишена досковидных корней, но иногда у нее наблюдается один или большее количество крупных досковидных корней (Davis). Петч (Petch, 1930, стр. 278) указывает, что молодое дерево Delonix regia. выросшее из семени материнского дерева без досковидных корней, имело досковидные корни.
Из изложенного можно сделать .вывод, что стремление образовывать досковидные корни есть наследственный признак, зависящий от генетической конструкции дерева. Размеры, в которых это свойство реализуется, как мы увидим позже, в значительной степени зависят от окружающей среды.
Досковидные корни и среда
Хотя образование досковидных корней представляет собою в высшей степени характерную черту деревьев тропического дождевого леса, оно, однако, не ограничивается исключительно ими. Границы географического распространения деревьев с досковидными корнями показывают, что климатические условия, в сильной степени способствующие их развитию, заключаются в сочетании большого количества осадков (не обязательно в течение всего года) с высокой средней температурой. В пределах тропи
6*
84
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ков досковидные корни преобладают у деревьев низин и там, где выпадает большое количество осадков. Присутствие досковидных корней типично для дождевых лесов, но они встречаются также у деревьев некоторых типов муссонных лесов, например в Бирме. На Цейлоне деревья с досковидными корнями (например, Bombax malabaricum) встречаются как на открытых «патанах», так и в лесу (Petch, 1930, стр. 277). В западноафриканском смешанном листопадном лесу с осадками около 1200—1500 мм (см. стр. 361) деревья с досковидными корнями так же пли почти так же часты, как и в типичном дождевом лесу. В сухих тропических областях деревья с досковидными корнями находятся главным образом в галерейных лесах вдоль русел рек. В нагорных дождевых лесах тропиков досковидные корни или отсутствуют, или представляют собой редкое явление. Так в «моховом лесу» горы Дьюлит в Сараваке, покрывающем вершину горы и спускающемся до высоты 970—1100 м над уровнем моря, деревья с досковидными корнями совершенно отсутствуют. Шрив, изучавший нагорный дождевой лес голубых гор острова Ямайка (Shreve, 1914 а, стр. 24), также не обнаружил деревьев с досковидными корнями. В Австралии, где дождевой лес тянется без значительных перерывов от тропической до умеренной зоны, количество деревьев с досковидными корнями уменьшается с увеличением географической широты. На севере, в Квинсленде, деревья с досковидными корнями встречаются повсюду, где годовая сумма осадков превышает 60 дюймов (1524 мм) (Francis, 1924, стр. 33). В тропическом дождевом лесу северной части Нового Южного Уэльса известно сравнительно немного деревьев с досковидными корнями (Fraser, Vickery, 1938, стр. 155). Очевидно, даже у одного и того же вида способность к образованию досковидных корней уменьшается в южном направлении; например, у Laportea gigas, произрастающей в северной части Нового Южного Уэльса, имеются досковидные корни, а у ее южных экземпляров они отсутствуют (N. A. Burges).
Считается, что досковидные корни не встречаются у деревьев умеренной зоны, хотя они и образуются иногда даже у некоторых видов европейских деревьев, правда, никогда не достигая таких огромных размеров и яркой выраженности, как у тропических деревьев. Хорошо заметные досковидные корни имеются, в частности ,у Populus italica (сравн. Senn, 1923) и видов Ulmus (см. фото Warming, Graebner, 1933, рис. 399). Досковидные корни меньшего размера можно иногда наблюдать у Fagus sylvatica. и у других видов.
Досковидные корни и почва
Только недавно было обращено внимание на тесное соотношение, существующее между количеством видов с досковидными корнями, размером этих корней и свойствами почвы, хотя часто наблюдалось, что деревья, имеющие досковидные корни, как правило, в большем количестве встречаются в болотистом лесу, чем на хорошо дренированной почве. Вильдеман (De Wildeman, 1930, стр. 997) сделал общее предположение, что образование досковидных корней по крайней мере отчасти зависит от характера почвы. В дальнейшем Дэвису и Ричардсу (Davis, Richards, 1933—1934, стр. 126 и др.) удалось показать, что в тропическом дождевом лесу Брит. Гвианы существует тесное и определенное соотношение между размером досковидных корней у наиболее распространенных видов деревьев и типом почвы; эти авторы обнаружили также, что и число подчиненных видов, имеющих досковидные корни, также меняется параллельно изменениям
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 85
почвы. В местности Морабалли-Крик существует пять типов леса, где соответственно доминируют Eperua jalcata. Ocotea rodiaei, Eschweilera sagotiana. Mora gonggrijpii и M. excelsa. Лес из Eperua занимает участки, покрытые грубыми белыми песками на гребнях и плато, лес из Ocotea развивается на бурых песках с большей примесью мелкозема, лес из Eschweilera приурочен к суглинистым почвам, лес из Mora gonggrijpii — к тяжелым глинистым почвам, а лес из М. excelsa произрастает на затопляемых илистых почвах приречных равнин. В этом ряду, от типа Eperua и до Mora excelsa. образование досковидных корней у доминирующих или наиболее распространенных видов деревьев с диаметром 16 дюймов (41 см) и более идет постепенно, но заметно возрастая, как это можно видеть на схеме (рис. 11). В то же время возрастает и пропорция подчиненных видов с досковидными корнями. Eperua jalcata почти целиком лишена досковидных корней, и с ней ассоциируется сравнительно небольшое число видов с мощными досковидными корнями. Крайнее положение в упомянутом ряду занимает Mora excelsa. имеющая очень крупные досковидные корни; с пей ассоциируются другие виды с мощными досковидными корнями. Три остальных типа лесов являются в этом отношении промежуточными.
Подобное же соотношение между образованием досковидных корней и характером почвы было найдено при сравнении двух типов тропического дождевого леса на острове Борнео (Richards, 1936). На белых песчаных почвах («боровой» лес) пропорция видов с досковидными корнями была ниже, а сами корни в среднем были меньше, чем в лесу, растущем на суглинистой почве (смешанный лес). Однако в Нигерии при сравнении леса, растущего на пресноводном болоте, со смешанным лесом (Richards, 1939, стр. 45) было найдено, что деревья с досковидными корнями на болоте встречались не чаще, чем на сухих местах; скорее даже наблюдалось обратное соотношение. Причина такого отклонения от того, что можно считать общим правилом, неясна. Аналогичные наблюдения были сделаны Корнером (Е. J. Н. Corner) на Малаккском полуострове. Здесь у дерева Koompassia malaccensis из семейства бобовых, когда оно растет в болотистом лесу, досковидные корни развиваются в большей степени по сравнению с экземплярами, произрастающими на склонах холмов. Растущее на болотах дерево Pometia alnifolia имеет более крупные досковидные корни, чем Р. pinnate. встречающаяся на плотных дренированных почвах по берегам рек и на склонах холмов.
Фактором, с которым теснее всего связано образование досковпдных корней, является, вероятно, не структура почвы как таковая, но ее глубина. В европейских лесных почвах, по словам Солсбери (Salisbury, 1925, стр. 352), «...опускающийся корень встречает увеличивающееся сопротивление субстрата и нарастающее затруднение дыхания. С другой стороны, все более и более благоприятным становится снабжение водой и минеральными солями. Равновесие между благоприятными и неблагоприятными условиями и определяет глубину, на которую проникает корневая система». Поэтому можно ожидать, что чем более пористой и более дренированной является почва, тем толще будет хорошо аэрированный слой и тем глубже окажется уровень, на котором достигается упомянутое равновесие. Можно предположить, хотя это и не исследовалось, что у пяти упомянутых типов леса в местности Морабалли-Крик средняя глубина корневых систем является наибольшей в лесу из Eperua на пористом белом песке и наименьшей в лесу из Mora excelsa — на затопляемых илистых почвах. Приуроченность леса Мога к маломощным почвам видна также из того, что лес этот иногда встречается на крутых каменистых склонах холмов; на таких
О / 2 з 4 5м
Рис. И. Дисковидные корни взрослых экземпляров наиболее распространенных видов деревьев в пяти типах леса, Морабалли-Крик, Брит. Гвиана (Davis, Richards, 1933—1934, рис. 2, стр. 127).
А —'Mora excel на Benth (консоциации Mora excelsa); Б — Moia gonggrijpii (Kleinhoonte) Sandwith. (консоциация Mora gonggrijpii)’. li — Exchwrilrra mgotiana Miers (ассоциация смешанного леса); Г — Ocoha todiaei (Schomb.) Mez (консоциация Ocolra rodiari); Д — Epcrua falcata Aubl. (консоциация Eprrua falcata).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕС,\ 87
склонах почва скорее физически, чем «физиологически» маломощна. Нахождение деревьев с крупными досковидными корнями на крутых кряжах (где почва обыкновенно очень маломощна) было отмечено и в других местах. В Брит. Гвиане у Ocotea rodiaei досковидные корни встречаются чаще на плотных, глинистых почвах, чем на пористых песчаных, да и корневая система в целом располагается на них ближе к поверхности (Т. A. W. Davis).
Интересные наблюдения Костера (Coster, 1933, 1935 Ь) свидетельствуют о поверхностном расположении корневых систем тропических деревьев п зависимость этого от действительной глубины почвы. На Яве было установлено, что конкуренция корней представляет собою очень важный экологический фактор. В лесах низменностей благодаря высокой температуре и влажности почва обыкновенно слабо аэрирована, и конкуренция корней за кислород, видимо, преобладает над конкуренцией за воду и соли, которая, как показали недавние исследования, играет столь важную роль в экологии лесных сообществ умеренной зоны.
Поскольку, как указывалось раньше, досковидные корни могут развиваться только у деревьев с крупными поверхностными боковыми корнями, наличие связи между образованием досковидных корней и действительной глубиной и аэрацией почвы легко объяснимо. Не удивительно также, что у большинства, хотя не у всех, деревьев, имеющих досковидные корни, нет стержневого корня. Это уже давно было замечено Спрусом (Spruce, 1908, I, стр. 21) в Южной Америке и сравнительно недавно Фрэнсисом {Francis, 1924) в Квинсленде и Петчем (Petch, 1930) на Цейлоне. Возможность изучения корневых систем деревьев, обладающих досковиднымп корнями, представляется редко, но как на Борнео, так и в Нигерии автор всегда замечал, что у деревьев с досковидными корнями нет и признаков стержневого корня. Например, у огромного поваленного дерева Ceiba pentandra на берегу реки Они, в Нигерии, имевшего мощные досковидные корни, корневая система целиком состояла из поверхностных боковых корней с маленькими, опускающимися вниз ответвлениями; наибольшая глубина проникновения корней в почву не превышала, вероятно, и полуметра. Однако имеются и исключения. Фоксуорси (цит. Чиппом, Chipp, 1922) сообщает, например, о дереве Dracontomelon dao с Филиппинских островов, которое, несмотря на наличие досковидных корней, имело и стержневой корень, а Фрэнсис (Francis, 1924, стр. 32) видел экземпляр Argirodendron trifoliolatum в Квинсленде с шестью досковидными корнями и развитым стержневым корнем. Однако это дерево с диаметром ствола всего лишь в 9,6 см, возможно, лишится стержневого корня по достижении спелости. Eugenia grandis и Scaphium affine с полуострова Малакка также обладают стержневыми корнями при наличии умеренно развитых досковидных корней (Corner). Хотя деревья с досковидными корнями имеют обыкновенно хорошо развитые поверхностные корневые системы, наличие поверхностной корневой системы не обязательно влечет за собой образование досковидных выростов. Корни некоторых деревьев тропического дождевого леса тянутся в горизонтальном направлении над поверхностью почвы на огромные расстояния, тем не менее эти деревья не имеют доско-видных корней. У Entandrophragma angolense var. macrophyllum в резервированном лесу Окому, в Нигерии (фото 5), поверхностные корни отходят в стороны от ствола более чем на 20 м, но это дерево почти никогда не имеет досковидных корней. Такие же поверхностные корни характерны для южноамериканской Caryocar nuciferum, дерева, отличающегося лишь незначительными досковидными корнями.
88
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Теории образования досковидных корней
Для «объяснения» образования досковидных корней было выдвинуто четыре более или менее убедительные теории.
1. Теория приспособления. До совсем недавнего времени досковидные корни обычно рассматривались как специальное приспособление, сообщающее дереву устойчивость. Вначале сторонники этой теории полагали, что основная функция досковидных корней заключается в том, чтобы оказать сопротивление на сжатие, то есть действовать как опора. Однако современные авторы, касавшиеся этого вопроса, большее значение придают сопротивлению па растяжение (Ghesquiere, 1925). Там, где досковидные корни равномерно распределены вокруг ствола, как это часто и бывает, часть их, конечно, подвергается сжатию, а другая часть — растяжению; там, где ветры не отличаются постоянством направления, эти функции выполняются одними и теми же досковидными корнями попеременно. С точки зрения теории приспособления это различие не имеет большого значения.
Приняв логические следствия теории приспособления, нельзя не согласиться, что механическая устойчивость, придаваемая досковидными корнями, должна быть достаточно велика, чтобы сообщить видам деревьев, обладающим ими, преимущество в смысле действия отбора по сравнению с другими видами. Досковидные корни (или скорее потенциальная возможность образования их при соответствующих условиях окружающей среды} могут возникнуть одиночной мутацией или рядом мутаций, сохраненных и накопленных естественным отбором. Конечно, вряд ли можно отрицать, что досковидные корни имеют значение в качестве механических опор и что деревья с досковидными корнями обладают преимуществом перед другими. Однако целый ряд соображений заставляет сомневаться в том, что это преимущество достаточно велико с точки зрения выживания.
Наиболее важным из них является малое развитие досковидных корней у внетропических деревьев. Калифорнийская секвойя и гигантские австралийские эвкалипты, гораздо более высокие, чем любое дерево тропического леса (см. гл. 1), не имеют досковидных корней. Хотя в отдельных районах пояса тропических дождевых лесов и случаются торнадо, сильные ветры в местах их распространения, например в Брит. Гвиане, менее обычны, чем в умеренных областях (см. гл. 6). Надо иметь в виду и то, что деревья дождевого леса часто плотно переплетаются деревянистыми лианами, что существенно увеличивает их устойчивость. Всякому, наблюдавшему рубку деревьев дождевого леса, известно, что крупное дерево иногда оказывается настолько связанным лианами с соседними деревьями, что не падает, даже будучи срубленным у основания. Правда, некоторые наиболее высокие деревья часто не отягощаются лианами, но и досковидные корни у них не всегда отличаются мощным развитием. Милдбрэд (Mildbraed, 1922), являющийся сторонником этой теории, чувствовал указанное противоречие. Пытаясь объяснить его, он указывал, что, хотя большая высота тропических деревьев, сила тропических торнадо и, наконец, неровность полога, не оказывающая деревьям дождевого леса такой же надежной защиты, какая существует у деревьев европейских лесов, делают понятным присутствие досковидных корней, однако должны иметься и какие-то особые факторы, свойственные исключительно дождевому лесу и благоприятствующие образованию досковидных корней. Он заметил, что слабая интенсивность света и высокая влажность в подлеске тропического дождевого леса способствуют образованию воздушных при
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
даточных корней, и предполагал, что из них-то и развились досковидпые корни. В частности, плоские ходульные корни, вроде корней у Santi-riopsis trimera, он рассматривает как стадию эволюции досковидных корней из ходульных корней округлого типа.
Другим доводом, заставляющим сомневаться в том, что досковидные корни имеют важное приспособительное значение, является то, что эти корни вовсе не бывают наиболее мощными там, где они, казалось бы, больше всего нужны. Так, досковидные корни, должны были бы лучше развиваться у деревьев на мягкой, податливой почве, чем у деревьев па плотном субстрате. В то же время, как показано выше (см. стр. 84—85), хотя образование досковидных корней обыкновенно более выражено в лесу на пресноводном болоте, чем в дождевом лесу на хорошо дренированной почве, и в Брит. Гвиане и на острове Борнео у деревьев, произрастающих на рыхлых песчаных почвах, досковидные корни образуются реже, чем у развивающихся на плотных глинах и суглинках. Равным образом следовало ожидать, что образование досковидных корней будет особенно ясно выражено у деревьев открытых местообитаний, подверженных действию ветра. Однако в местности Морабалли-Крик, Брит. Гвиана, досковидные корни лучше всего развиты в лесу из Mora, растущем в укрытых речных долинах, и меньше всего в лесу из Ерегиа, расположенном на сравнительно открытых песчаных гребнях.
Уитфорд (Whitford, 1906), занимавшийся изучением горных лесов Филиппин, обнаружил, что досковидные корни совершенно не свойственны деревьям, растущим на открытых гребнях. Больше того, основываясь на обширных наблюдениях в Брит. Гвиане, Дэвис (Т. A. W. Davis) пришел к убеждению, что деревья, снабженные досковидными корнями, по сути дела, чаще валятся ветром, чем деревья без этих корней.
Несмотря на это, очевидно, что теория приспособления не может быть совершенно оставлена; существующие сомнения не являются непреодолимыми, хотя они и заставляют нас вместе с Милдбрэдом искать какое-то «причинное» объяснение для ее дополнения. Полезность досковидных корней, возможно, скорее случайная, чем первичная причина их наличия. Вполне вероятно, что отсутствие досковидных корней компенсируется чем-то другим, например корневой системой, хорошо приспособленной к слабо аэрированной почве. Такая корневая система может проникать на большую глубину и сохранять стержневой корень при таких условиях среды, которые заставляют корневые системы большинства растений оставаться поверхностными. Ниже приводятся три теории «причинного» объяснения наличия досковидных корней.
2. Теория отрицательного геотропизма. Фрэнсис (Francis, 1924, стр. 34) высказал предположение, что «верхней части главных поверхностных корней у видов, имеющих досковидные корни, могут сообщаться прямо или косвенно через ствол отрицательный геотропизм и фототропизм; таким путем может возникнуть перпендикулярное натяжение, образующее досковидные корни». Эта теория не вызывает доверия. Известно, что отрицательный геотропизм корней обнаруживают, например пневматофоры мангровых, но не существует таких примеров, где бы стимул тяготения вызвал вертикальное распространение горизонтального органа (нормальной реакцией в таком случае было бы, конечно, искривление). Большинство корней не чувствительно к свету, а те, которые к нему чувствительны, реагируют на свет обыкновенно отрицательно; во всяком случае трудно предположить, чтобы вполне развитый корень, покрытый корой, мог быть чувствительным к свету.
90 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
3. Теория проводящего тока. Автором этой теории является Петч (Petch, 1930). Она основывается главным образом на наблюдениях, проведенных в Пераденийском ботаническом саду и других местах Цейлона. Отправным пунктом этой теории является то, что у деревьев с досковидными корнями обыкновенно отсутствует стержневой корень. Петч высказал предположение, что у дерева, большая часть корневой системы которого состоит из поверхностных боковых корней, транспирационный ток, несущий из почвы растворенные питательные вещества, ограничивается в стволе определенными путями, расположенными на тех же радиусах, что и боковые корни. Эта теория предполагает, что подобное расположение проводящих путей в стволе сравнительно неэффективно. Это предположение подкрепляется как экспериментами Калдуэла (Caldwell, 1930), так и доказательствами, выдвинутыми самим Петчем. Токи воды и питательных веществ благоприятствуют росту камбия в секторах, расположенных на тех же радиусах, что и боковые корни, за счет промежуточных секторов, что ведет к образованию ребер, становящихся со временем досковидными корнями. Частота образования досковидных корней во влажных тропиках связана с насыщенностью почвы влагой, что препятствует развитию стержневых корней. Не все виды имеют досковидные корни, потому что у некоторых из них стержневой корень приспособлен к условиям переувлажнения.
Теория проводящего тока, как и первые две, не свободна от возражений. Уже приводились примеры (стр. 87) деревьев, одновременно с доско-впдными корнями имеющих и стержневой корень. Теория проводящего тока не объясняет, почему в умеренной зоне деревья с неглубокой корневой системой без стержневого корня так редко образуют досковидные корни. Хотя она и дает приемлемое объяснение неравномерного роста ствола, но остается неясным, почему в самом корне рост должен происходить главным образом по верхнему краю. Не понятно, какое участие принимает камбий ствола в образовании досковидных корней, и сам Петч (Petch, 1930, стр. 280) приводит примеры корней Canarium commune, у которого досковидные корни образовались на некотором расстоянии от ствола, а не на самом стволе.
Теория Петча имеет одно преимущество по сравнению с другими «причинными» теориями: она дает возможное объяснение сужению нижней части ствола — признаку, свойственному как деревьям с ходульными, так и с досковидными корнями. Представим себе, что, как и предполагает теория Петча, боковая проводимость тока веществ в стволе сравнительно недостаточна. Тогда секторы камбия, расположенные между ограниченными полосами, составляющими неразрывное целое с боковыми корнями, будут сравнительно скудно снабжены водой и солями; но выше по стволу восходящие токи могут постепенно распространиться, так что над определенным уровнем весь камбий окажется одинаково хорошо снабженным раствором питательных веществ и будет поэтому расти равномерно1 . Этот уровень расположен над местом прикрепления самых высоких ходульных или досковидных корней. Приблизительно таким путем можно было бы объяснить, почему, как говорит Фрэнсис, «недостаточно снабжаемый участок нижней части ствола перешел в досковидные корни».
1 Хармон (Harmon, 1942) нашел, что у американских деревьев общая сумма роста больше на радиусах, совпадающих с направлением боковых корней, чем между ними.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО Д ОЖД Е ВОГО Л ЕСА 91
4. Теория напряжения. Согласно Уитфорду (Whitford, 1906), Сенну (Senn, 1923), Наве (Navez, 1924, 1930) и другим, образование досковидных корней является прямой реакцией дерева на механическое напряжение, вызываемое ветром. Наиболее полное обоснование этой теории принадлежит Сенну (Senn, 1923), который после поездки на Яву, вызвавшей у него интерес к этой проблеме, занялся в Швейцарии подробным изучением досковидных корней у ломбардского тополя {Populus italica). Досковидные корни у этого вида меньше и тоньше, чем у большинства тропических деревьев, но вообще они схожи, и нет оснований предполагать, что причины их возникновения различны. Сенн тщательно изучил направление досковидных корней у большого числа деревьев в различных частях страны. Он не обнаружил соотношения между направлением досковидных корней и распределением воды или питательных веществ в почве; с другой стороны, удалось установить явную зависимость этих корней от одностороннего излучения тепла, причем образование досковидных корней стимулировалось близ поверхностей, отражающих тепло, например стен. Однако гораздо более поразительным оказалось соотношение, существующее между направлением досковидных корней п направлением ветров, замечательно постоянных во многих долинах Швейцарии.
Из 461 экземпляра исследованных деревьев 443 (96,1 %) имели более или менее отчетливые досковидные корни, причем у 407 (91,9?о) эти корни были развиты главным образом с наветренной стороны, противостоящей преобладающим ветрам. Сенн сделал из этого вывод, что образование досковидных корней объясняется создающимся под влиянием ветра натяжением, стимулирующим рост камбия на верхней стороне проксимального конца боковых корней и в прилегающей части ствола.
Исключение составляли восемь деревьев, располагавшихся в непосредственной близости от обрыва крутого берега или скалы, со стороны которых и развивались досковидные корни. Сенн (Senn) дает остроумное объяснение таких случаев. При постоянных ветрах одного направления нижняя часть ствола слегка наклоняется благодаря сопротивлению берега или скалы; при этом она действует как рычаг, силящийся вырвать корни на подветренной стороне и погрузить их в землю на наветренной. Возникающие таким образом напряжения противоположны тем, которые существуют в стволе дерева, растущего на открытой поверхности. Поэтому и направление досковидных корней оказывается противоположным обычному. Еще три случая, относительно которых нельзя сказать, что они обусловлены действием ветра, могут быть объяснены влиянием одностороннего теплового излучения. К тому же, как отмечал и сам Сенн, нельзя ожидать наличия математически точного соотношения с направлением ветра, так как досковидные корни образуются на уже существовавших раньше корнях, направление которых не было вначале обусловлено влиянием ветра.
Сенн также установил, что образование досковидных корней было особенно заметно в том случае, если нижняя часть ствола была затенена и, следовательно, находилась в окружении более влажного воздуха. Это обстоятельство делает понятным частоту нахождения досковидных корней в тропическом дождевом лесу.
Нельзя сказать, что взаимосвязь между направлением досковидных корней и преобладающими ветрами очень уж ясно выражена у тропических деревьев. Петч (Petch) решительно отвергает существование подобного соотношения у исследованных им в Перадении экземпляров Сапа-rium commune (1930, стр. 280); но вопрос этот может быть удовлетвори
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
тельно изучен лишь при использовании статистического метода. Сенн полагал, что фотографии тропических деревьев, снятые им и другими лицами, выявляют указанное соотношение, но единственным достоверным свидетельством являются тщательные наблюдения Наве (Navez, 1930) над большим числом деревьев Ceiba pentandra на Кубе. Эти деревья подобно швейцарским тополям, обнаружили определенную тенденцию к образованию досковидных корней на стороне, обращенной к преобладающим ветрам. Данные Наве носят статистический характер. На Малаккском полуострове и в Брит. Гвиане у деревьев, склоненных над водой, самые крупные досковидные корни, конечно, лишь у видов, для которых эти корни характерны, находились на стороне, противоположной реке, то есть на стороне, испытывающей наибольшее напряжение от тяжести склонившегося дерева (Corner, Т. A. W. Davis). Интересно также то, что у Canarium commune и Ceiba pentandra в пазухах более крупных ветвей имеются крылообразные расширения, которые, подобно досковидным корням, также могли возникнуть под влиянием натяжения.
Таким образом, имеются положительные данные, говорящие в пользу последней теории, но и в данном случае целый ряд затруднений мешает принять ее безоговорочно. Поскольку образование досковидных корней в какой-то степени, по-видимому, является генетически обусловленным признаком, встречающимся лишь у некоторых семейств, родов и видов, приходится предположить, что существуют какие-то наследственные факторы, управляющие чувствительностью растения к воздействию натяжения. В этом отношении некоторые растения отличаются, вероятно, большей чувствительностью, чем другие. Теория недостаточно объясняет, почему досковидные корни так мало развиты у деревьев умеренной зоны,, хотя наблюдения Сенна, свидетельствующие о том, что образованию досковидных корней у Populus italica способствуют высокая температура и влажность, могут служить ключом к объяснению этого обстоятельства. По-видимому, можно также предположить, что семейства и роды, наиболее чувствительные к упомянутому воздействию, оказались бессильными приспособиться к условиям умеренной зоны и поэтому в своем распространении ограничиваются только тропиками. Другое затруднение заключается в том, что теория натяжения не объясняет, почему досковидные корни развиваются у многих деревьев тропического дождевого леса, когда они еще молоды и находятся в подлеске, надежно защищенными от внешних влияний. Трудно поверить, чтобы стволы подобных деревьев испытывали натяжения, достаточные для образования досковидных корней.
Итак, в настоящее время не существует достаточно удовлетворительной теории, объясняющей развитие досковидных корней у деревьев тропического дождевого леса. Имеющиеся данные в целом говорят в пользу какой-либо «причинной» теории, причем они заставляют думать, что полезность досковидных корней для дерева является скорее случайной, чем первичной причиной их развития. Для окончательного решения этой проблемы необходимо произвести большое количество точных наблюдений и запастись экспериментальными данными. Надо не упускать из виду возможность того, что важную роль в образовании досковидных корней могли играть ростовые гормоны, и данные относительно образования и перемещения таких веществ у тропических деревьев могли бы содействовать решению этого вопроса.
Близкая связь, существующая, как выяснено, между образованием досковндных корней, корневой системой и характером почвы, делает
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
93
это явление особенно интересным для эколога, что и заставило нас довольно подробно остановиться на этой проблеме, с первого взгляда имеющей чисто морфологический интерес. Надеемся, что все сказанное здесь о досковидных корнях обратит внимание на этот предмет, находящийся в некотором пренебрежении. Мимоходом заметим, что досковидные корни имеют и экономическое значение, поскольку при валке ценных сортов деревьев, обладающих досковидными корнями, разрез по необходимости приходится делать выше их прикрепления, что приводит к большим потерям древесины. Поэтому установление факторов, влияющих на образование досковидных корней, должно представлять интерес для тропического лесоводства.
Пневматофоры
В тропических дождевых лесах болотистого типа (но, не в таких, где почва сравнительно хорошо дренирована) характерной чертой некоторых деревьев являются так называемые пневматофоры, или дыхательные корни. У некоторых видов они принимают форму «колен» или петлеобразных выростов, образованных небольшими отрезками поверхностных боковых корней и возвышающихся над почвой; на одном корне может образоваться несколько таких «колен». Pterocarpus officinalis (тропическая Америка) имеет пневматофоры в виде маленьких, полукруглых, крылоподобных выростов. Они появляются на боковых корнях на некотором расстоянии от места погружения в почву досковидных корней. У других видов пневматофоры представляют собою короткие, суживающиеся образования, направленные вертикально в воздух. Они возникают из боковых ответвлений поверхностных боковых корней, обладают отрицательным геотропизмом и имеют ограниченный рост. Как известно, у мангровых насчитывается несколько типов пневматофоров. Коленчатые корни встречаются у видов Bruguiera’. воздушные корни с отрицательным геотропизмом свойственны Avicennia и Sonneratia spp. и др. Менее известно, что пневматофоры часто встречаются у деревьев, произрастающих в лесах, разивающихся на пресноводных болотах.
Торенар (Thorenaar, 1927) обратил внимание на наличие пневматофор в лесах на заторфованных болотах (см. стр. 316) близ Палембанга на Суматре. Согласно Ван-Стеенису (van Steenis, 1935 а), пневматофоры чрезвычайно характерны для деревьев болотистых лесов Индонезии. Пневматофоры, похожие на пневматофоры Sonneratia и Avicennia. описанные Полаком (Е. Polak, 1933, стр. 27 и рис. 6, 7), встречаются у Ploiarium alternifolium. Cratoxylon и Tristania spp. в лесу, растущем на торфяном болоте на западе острова Борнео. Автор наблюдал оба типа пневматофор в аналогичном лесу в Сараваке у различных, точно не определенных видов деревьев, а коленчатые корни он видел у Mitragyna ciliata (Bubiaceae) в лесу на незаторфованном болоте в Западной Африке (рис. 12). У Symphonia globulifera. дерева, обычно произрастающего на засоленных и пресноводных болотах в Брит. Гвиане, петлеобразные корни нередко образуются в таком большом количестве, что затрудняют ходьбу (Т. A. W. Davis). Адамсон (Adamson, 1910) описал пневматофоры у Terminalia arjuna. растущего по урезам рек в южной Индии, а Лем (Lam, 1945, стр. 48) — у ротанговой пальмы (пальма-лиана), встречающейся в лесах пресноводных болот в Новой Гвинее. Следовательно, коленчатые корни и другие типы пневматофор представляют собой обычное явление в лесах, развивающихся на пресноводных болотах тропических районов. Коленчатые
9'<
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
корни Taxodium distichum, произрастающего на болотах умеренных и субтропических широт, конечно, хорошо известны.
Пневматофоры мангровых обыкновенно считаются вентилирующими органами, помогающими деревьям существовать на почве, почти лишенной кислорода. Их строенпе (многочисленные чечевички или постоянно слущивающаяся кора, обилие межклетников) подтверждает это предположение. К тому же выводу приводят и опыты Карстена (Karsten. 1891),
Р и с. 12. Петлеобразные воздушные корни (пневматофоры) Mitragyna ciliata, западноафриканского дерева, растущего на пресноводном болоте. Резервированный лес Шаша, Нигерия.
С фотографии (i/з натуральной величины).
продемонстрировавшего, что они действительно в большом количестве выделяют углекислый газ (см. также Troll W., Dragendorf, 1931; Chapman, 1944). Однако Троль (Troll W., 1930) недавно показал, что пневматофоры мангровых выполняют также и другую функцию , которая, вероятно, имеет такое же, если не более важное значение. В мангровых болотах уровень почвы непрерывно поднимается (в расположенной на Суматре местности, которую исследовал Троль, ежегодный прирост уровня почвы составлял не менее 15—35 мм), и пневматофоры играют при этом важную роль, давая возможность дереву приспособиться к такому подъему субстрата. У Sonneratia и Bruguiera (согласно Чапману, то же самое относится и к Avicennia) огромное количество тоненьких корешков располагается не на боковых корнях, а на подземной части пневматофор. В результате поглощающая часть корневой системы располагается близ поверхности почвы, а более глубокие корни, вероятно, выполняют функцию прикрепления дерева к субстрату. По мере поднятия уровня почвы пневматофоры мангровых растут вверх, образуя новые корешки соответственно на все более и более высоких уровнях; таким образом активная часть корневой системы сохраняется на постоянном уровне под поверхностью почвы. Хотя пневматофоры деревьев, растущих на пресноводных тропических болотах, никем не исследованы с этой точки зрения, все же можно предположить, что они выполняют аналогичные функции, так как по своей структуре они подобны пневматофорам мангровых; сомнительно лишь, что скорость прироста субстрата в условиях пресноводных
СТРУКТУРА II ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
95
болот, особенно в лесах, растущих на торфяных болотах, может быть равной скорости, которая наблюдается в зоне развития. Интересно отметить, что, как обнаружил автор, в заболоченных лесах Нигерии коленчатые корни Mitragyna cillata образовывались в большем количестве на более влажных местах. Это относится и к Symphonic globulijera, растущей в Брит. Гвиане. У этого дерева на более сухих участках коленчатые корни почти не образуются, за исключением отдельных случаев, когда оно произрастает в местных понижениях рельефа (Т. A. W. Davis).
ОБЛИК КУСТАРНИКОВ
Кроме молодых экземпляров деревьев, относящихся к более высоким ярусам, в ярусе кустарников тропического дождевого леса находятся растения самого разнообразного облика. Некоторые виды, например высокие Scitamineae, экологически принадлежащие к ярусу кустарников, морфологически относятся к травам. Деревянистые члены этого яруса также очень разнообразны по своему внешнему виду. Некоторые из них, например различные Rubiaceae, не имеют главного ствола и ветвятся у уровня почвы или невысоко над ним, причем растение имеет закругленный или обратноконический профиль. Это настоящие кустарники в физиономическом смысле, или, согласно определению Дю-Риэ (Du Rietz, 1931, стр. 46), «низкорослые кустарники»; их отличие от европейских кустарников состоит только в том, что они меньше ветвятся. Другие кустарники тропического дождевого леса, вероятно большинство, имеют отчетливо выраженный главный ствол и напоминают деревья в миниатюре. Эти виды (согласно классификации жизненных форм Дю-Риэ) относятся к «низкорослым деревьям» и «карликовым деревьям» (Du Rietz, 1931, стр. 45). У некоторых из них, относящихся к последней группе, ствол совершенно не ветвится, а листья группируются на верхушке растения, как, например, у западноафриканского Angylocalyx oligophyllus (Leguminosae) и Sphenocentrum jollyanum (Menispermaceae). Эти «кустарники» отличаются от уже упоминавшихся (стр. 75) Schopjbaum-chen только меньшими размерами. Близкое сходство общей формы этих видов и настоящих молодых деревьев создает соблазн рассматривать их с эволюционной точки зрения как преждевременно развившиеся деревья. Как бы то ни было, но присутствие этих миниатюрных деревьев составляет неотъемлемую особенность дождевого леса и резко отличает его от лесов умеренной зоны.
В пределах кустарникового яруса часто встречаются маленькие пальмы. В тропической Америке эта группа представлена многочисленными видами Badris^ Geonoma и других родов, а в районе Малайского архипелага обычны представители Eugeissonia, Licuala. Pinanga и др., местами доминирующие в этом ярусе. Так как эти маленькие пальмы не ветвятся, то, подобно карликовым видам Pandanus и Dracaena, встречающимся иногда в подлеске тропического дождевого леса, они по внешности очень похожи на только что упомянутые миниатюрные деревья.
ЛИСТЬЯ ДЕРЕВЬЕВ И КУСТАРНИКОВ
Деревья и кустарники тропического дождевого леса имеют весьма характерную листву не только в отношении формы, размера и структуры взрослых листьев, но и в отношении их расположения, способа распу
96
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
скания, а также степени защиты от внешних воздействий в почке. Многие из особенностей, присущих листьям тропического дождевого леса, несомненно, связаны адаптивно, или «причинно», со средой. Большинство листьев имеет важное диагностическое значение при определении тропического дождевого леса как растительного сообщества, отличного от других растительных формаций. Свойство быть вечнозелеными, представляющее собою наиболее важный признак листьев древесных растений дождевого леса, будет рассмотрено в главе 8 в связи с сезонными явлениями вообще.
Почки
Как и следовало ожидать, почки деревьев и кустарников тропического дождевого леса защищены гораздо меньше, чем почки деревьев в других климатах. Вследствие этого, исключая размер, они часто весьма похожи друг на друга как в состоянии покоя, так и в активно распускающемся состоянии. На связь, существующую между защитными структурами почки и климатом, обратил особое внимание Раункпэр (Raun-kiaer, 1934), деливший свои фанерофитные жизненные формы на три класса: 1) вечнозеленые фанерофиты без почечных чешуй, 2) вечнозеленые фанерофиты с почечными чешуями и 3) листопадные фанерофиты. Переходя от постоянно жаркого и влажного климата тропического дождевого леса к менее увлажненным тропической и субтропической областям, эти три класса фанерофитов по очереди занимают господствующее положение, но нигде один класс не становится преобладающим до полного исключения двух других. Так в лесах бассейна Конго число деревьев с защищенными почками увеличивается от влажного центра к периферии, где наблюдается отчетливая сезонная засуха (Lebrun, 1936 b, стр. 174).
Даже в наиболее влажном тропическом дождевом лесу, где отсутствует холодный и не резко выражен сухой сезон, все же, во всяком случае у более высоких деревьев, листья в почкосложении до известной степени страдают от высыхания. Надо сказать, что, хотя почки деревьев тропического дождевого леса, несомненно, защищены в гораздо меньшей степени, чем у деревьев других климатов, и здесь у значительного числа видов почки все-таки так или иначе защищаются. У многих деревьев имеются волосистые почки, у других они покрыты слизистыми или смолистыми выделениями. Многим видам свойственны более специализированные средства защиты, вроде крыльев на черешке, прилистников, образующих капюшон (например, у Musanga cecropioides и т. п.). Некоторые из этих образований были описаны Потером (Potter, 1891), показавшим, что если удалить прилиственный капюшон, защищающий почки Artocarpus incisa, то молодой лист оказывается недоразвитым и деформированным. У ряда тропических деревьев наблюдается даже наиболее специализированный тип покрова почки, а именно почечные чешуи (то есть листья, целиком приспособленные для защиты почек). Холтер-ман (Holtermann, 1907, стр. 183), например, описал почки, снабженные чешуями у чисто тропического рода Litsea (Lauraceae) и других деревьев влажной зоны Цейлона.
Покров почек у тропических деревьев (если он присутствует) имеет обыкновенно нежную и сочную консистенцию, а не жесткую и сухую, как у европейских деревьев. В то же время Резволл (Resvoll, 1925) обнаружил, что покоящиеся почки пяти видов Quercus в дождевом лесу Явы имели сухие почечные чешуи и весьма незначительно отличались по своему строению или внешнему виду от почек Q. robur умеренной зоны,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
97
за исключением того, что чешуи у них перекрывались менее плотно. Так как такие почки, видимо, не приспособлены к постоянно влажному климату Явы, то Резволл полагает, что их наличие обусловливается внутренними факторами, общими всему роду Quercus в целом.
Молодые листья
Распускающиеся молодые листья деревьев представляют собой одну из наиболее поразительных картин тропического дождевого леса. Цвет их обыкновенно сильно отличается от темно-зеленого цвета взрослых листьев, и даже после достижения ими окончательного размера они некоторое время часто оказываются лишенными хлорофилла и могут иметь ярко-красный или малиновый цвет. Реже встречается другая окраска листьев: так у африканской Pancovia harmsiana... молодые листья имеют аметистовый оттенок (Mildbraed, 1922, стр. 110), а у различных маленьких деревьев острова Борнео и Малаккского полуострова молодые листья отличаются своеобразным синевато-стальным цветом. Шталь (Stahl, 1893, стр. 144) сообщает о белых молодых листьях у Cynometra cauliflora и Humboldtia laurijolia, а автор видел такие лпстья у видов Duguetia в Гвиане и у других тропических деревьев. Виды Duguetia со свежей листвой выглядят как бы осыпанными обрывками белой бумаги. У некоторых видов листья имеют чрезвычайно бледный зеленый цвет с легким красноватым отливом.
Когда листья, как это часто бывает, образуются в один и тот же сезон, и у многих деревьев одновременно появляется новая листва, то красота расцветки дождевого леса не уступает северному лесу в его осеннем уборе. Яркость окраски листвы часто может обмануть даже опытного наблюдателя, заставив его думать, что деревья находятся в цвету.
Молодые листья, окрашенные в красный цвет, встречаются у представителей самых различных семейств, но, подобно досковидным корням, они до известной степени представляют собой видовой признак, встречаясь у некоторых видов или родов и отсутствуя у других, близко родственных. Такие листья свойственны не только двудольным, но встречаются, например, и у некоторых маленьких пальм. В травяном ярусе они не так часты, но автор наблюдал молодые красные листья у папоротника Tectaria maingayi на острове Борнео. Эта характерная окраска молодых листьев наблюдается в тропическом дождевом лесу всех географических областей. На Борнео она свойственна отдельным деревьям всех ярусов; в Гвиане она также замечается очень часто. В западноафриканском дождевом лесу ярко-красная окраска листьев, быть может, менее обычна, но она хорошо выражена, например, у Cynometra vogelii.
Для объяснения этой особенности тропического дождевого леса был предложен целый ряд умозрительных теорий, правда не особенно убедительных. Так, например, предполагали, что красный цвет, обусловленный наличием в клеточном соке пигмента антоциана, защищает хлорофилл от разрушительного действия слишком яркого света или что он, вызывая поглощение тепла, повышает температуру листьев, а это, как предполагают, благотворно сказывается на увеличении скорости транспирации. Указанные теории были критически рассмотрены Смитом (А. М. Smith, 1909), который показал, что температура молодых тонких листьев различных тропических деревьев может превышать или быть ниже температуры толстых, взрослых листьев одного и того же вида. Он нашел, что красные молодые листья действительно имеют несколько
98
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
более высокую температуру, чем зеленые листья такой же толщины и строения. Бюннинг (Banning, 1947, стр. 101—102) высказал мысль, что красная окраска может иметь важное значение в качестве защиты против ультрафиолетового излучения, так как оно сильно поглощается антоциановыми пигментами.
Другой особенностью, присущей молодой листве деревьев тропического дождевого леса, является то, что в течение нескольких дней после того, как они распустятся, и до достижения ими окончательного размера они часто выглядят слабыми и висят, как увядшие. Только позже, по мере приобретения листьями нормальной зеленой окраски, они становятся жесткими и принимают стоячее положение. Это свойство листьев, в той же степени как п красная окраска, характерно для деревьев дождевого леса всех тропических районов, ио оно, подобно красной окраске, менее распространено у деревьев Западной Африки. Лучше всего оно выражено у различных деревьев из семейства Leguminosae, но замечается также и у других семейств. У некоторых Caesalpinioideae, например у Amher-stia nobilis, у видов Brownea и Saraca не только листья, но и молодые ветви вначале бывают вялымп и свисающими. У этих деревьев почки распускаются очень быстро, часто в течение ночи, так что Тройб (ТгеиЬ) говорил, что у них листья как бы «высыпают» на дереве (цпт. Шимпером, 1935, стр. 484). Функе (Funke, 1929) сообщает интересные данные относительно времени, необходимого молодым листьям различных видов, чтобы стать зелеными и жесткими.
Вялость молодых побегов Amherstia объясняется отсутствием механических тканей, а не отсутствием тургора (Czapek, 1909), а у Brownea grandiceps Бюзген (Busgen, 1903, стр.438) установил в период распускания увеличение веса как в сухом, так и в свежем состоянии, что указывает как на настоящий рост, так и на увеличение уже образовавшихся клеток. Вероятно, у всех этих вялых повисших молодых листьев и побегов происходит задержка в дифференциации; распускание и дифференциация, вместо того чтобы протекать более или менее одновременно, как у большинства молодых листьев, происходят последовательно, будучи разделены значительным промежутком времени.
Эта своеобразная последовательность событий в развитии листа и побега наблюдается у деревьев всех ярусов. Она встречается и в других формациях, как, например, у видов Daniellia и Isoberlinia в западноафриканских саваннах, но редко и в гораздо менее выраженной степени. Различные авторы рассматривали это явление как приспособление к яркому свету, чрезмерному нагреву, сверхсильным осадкам или как свойство, помогающее лучшему использованию ограниченного пространства (КееЫе, 1895; Czapek, 1909). Все эти разнообразные мнения подтверждают мысль, что истинное объяснение должно быть причинным, учитывающим влияние климата; приспособительное значение, если оно вообще в данном случае существует, может быть просто случайным. На такую точку зрения встал Гёбель (Goebel, 1924, стр. 230—236), считавший, что вялость и обвислость молодых побегов (то же истолкование может быть применено и к молодым листьям) являются следствием каких-то внутренних наследственных свойств, которые в менее благоприятных местообитаниях могут быть быстро устранены естественным отбором. Побеги свисают потому, что они должны свисать, а не потому, что это для них выгодно и при условии высокой влажности и температуры в тропическом лесу задержка в приобретении тканями жесткости, которая могла бы стать вредоносной для растения в более суровой среде, здесь остается «безна
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 99
казанной». Если, говорит Гебель, на колючку ежа иногда попадает яблоко, то значит ли это, что колючка специально приспособлена для того, чтобы нести на себе яблоки? С этой точки зрения можно расценивать и многие другие признаки растений тропического дождевого леса, считающиеся приспособительными.
Размер и форма листьев
Хотя в тропическом дождевом лесу, как и в других растительных сообществах, имеются разнообразные типы листвы, среди них, безусловно, преобладают цельные склерофилльные листья, относящиеся к «мезофилль-ному» классу размера по классификации Раункиэра (Raunkiaer, 1934, стр. 368—378). Типом листа, характерным для огромного большинства
Р и с. 13. Листья деревьев первичного смешанного диптерокарпового леса (Ve натуральной величины), река Капах, гора Дмолит, Борнео. Каждый представляет собой типичный вполне развившийся лист различных пород деревьев. Яруоы, к которым они принадлежат, обозначены буквами.
деревьев и кустарников, независимо от их систематических связей, является «лавровый тип» листа, как его обозначает Варминг (Warming): его можно было бы, пожалуй, назвать мегасклерофпллом в отличие от микро-склерофилла («миртовый тип» Варминга), характерного для средиземноморского климата. В типичных случаях этот лист имеет интенсивно темнозеленый цвет и продолговато-ланцетную (до эллиптической) форму пластинки, край цельный или мелкозубчатый, часто имеется длинное, отчетливо выраженное острие, образующее кончик-капельницу. Лист отличается жесткой, часто более или менее кожистой текстурой; верхняя поверхность у него гладкая и часто сильно глянцевитая; войлочек и воло
100
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ски развиваются только на нижней поверхности. В случае сложного листа отдельные листочки стремятся приблизиться по величине и форме к преобладающему типу простого листа. Одно из наиболее заметных исключений составляют многие Mimosoideae, в изобилии встречающиеся в тропическом дождевом лесу, особенно в Африке и Америке; их листочки имеют нанофплльные или лептофилльные размеры.
Рис. 14. Листья растений дождевого леса двух местностей южной Нигерии (1/в натуральной величины).
Каждый представляет собой типичный вполне развившийся лист различных видов.
Принимались в расчет только листья деревьев, «кустарников» и наземных трав. Ярусы, к которым они принадлежат, обозначены буквами.
Все перечисленные факты наглядно выявляются при изучении образцов листьев деревьев и кустарников тропического дождевого леса. Бекер (Baker, 1938) опубликовал несколько интересных фотографий, демонстрирующих удивительное однообразие размера и формы листьев двудольных в лесу Синхарая па Цейлоне. Из 41 исследованного вида, принадлежащих к 20 семействам, 90 % имели листья с более или менее отчетливыми кончиками-капельницами. Таким же однообразием отличаются образцы листьев тропического дождевого леса острова Борнео и резервированного леса Шаша («влажного вечнозеленого леса») в Нигерии (см. рис. 13, 14).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА Ю1
Анализ размеров листьев показывает, что в нормальном дождевом лесу по крайней мере 80% видов (и индивидуумов) деревьев имеет, по классификации Раункиэра, листья мезофилльного класса величины (площадь листовой пластинки 2025—18 225 кв. мм). Макрофилльные и микрофилльные классы представлены скудно, а нанофилльнып и лепто-филльный классы, как правило, почти или совсем отсутствуют. Мега-филлы, часто считающиеся особенно характерными для влажных тропиков, сравнительно редки в первичном низинном тропическом дождевом лесу. Типичное распределение листьев по классам величин показано в таблице 8, составленной для вечнозеленого (сезонного леса) острова Тринидад с выраженным сухим сезоном, а также на рис. 15. Статистическое сравнение размеров листьев вечнозеленого сезонного тропического дождевого леса и более равномерно влажного истинного дождевого леса, вероятно, показало бы, что средний размер листа будет в последнем несколько больше, чем в первом.
Преобладание цельнокрайности так же обращает на себя внимание, как и однообразие размера листа. В большой коллекции листьев из двух местностей тропического дождевого леса (влажный вечнозеленый лес) Нигерии 80% видов имело цельные листья, а из видов с нецельнокрайними листьями лишь 4?6 обладало лопастными или надрезанными листовыми пластинками. В диптерокарповом лесу на высоте 450 м, гора Мак-вилинг, Филиппины, Браун (Brown, 1919, стр. 381) обнаружил, что 76 % видов и 80% индивидуумов имели цельнокрайние листья. Подобные же цифры могут быть получены для любого спелого тропического дождевого леса. Отклонения от нормального типа скорее подчеркивают, чем нарушают господствующее однообразие; не удивительно, что по утверждению Спруса (Spruce, 1908, I, стр. 37) в лесах Амазонки раздельный лист представляет собой «нечто редкостное».
Замечательно, что среда тропического дождевого леса, видимо, придает особую, своеобразную форму листве всех видов, подверженных ее влиянию. Это хорошо заметно при сравнении тропических видов с близкородственными видами, растущими в умеренном климате. Например, у Quercus листья сильно изменяются по размеру, форме и текстуре, но у близкородственного, в основном тропического, рода Lithocarpus почти все виды имеют цельные, кожистые, обыкновенно удлиненно-заостренные листья мезофилльного размера (Brenner, 1902). Листья некоторых видов Lithocarpus, например L. blancoi, совершенно точно соответствуют нормальному типу листа тропического дождевого леса. Другим ярким примером является Acer niveum, дерево дождевого леса с цельными, тонко заостренными листьями, весьма отличающимися от листьев большинства видов этого рода, растущих в умеренной зоне.
Хотя однообразие размера и формы листьев характерно для деревьев всех ярусов тропического дождевого леса, однако надо иметь в виду, что между разными ярусами имеются небольшие, но определенные различия. Этот факт редко был предметом обсуждения, но его легко обнаружить в любом сообществе дождевого леса. При переходе от небольших деревьев и подлеска (яруса С) к ярусу В и А средний тип листа становится постепенно меньше, толще, делаясь более кожистым по своей текстуре. Изменение размера сопровождается еще более заметным изменением типа кончика листа. В ярусе С почти все листья удлиненно-заостренные; в ярусе В кончик более короткий или его совсем нет, в то время как в ярусе А у большинства листьев кончика совсем не бывает или бывает только
102
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Таблица 8
Размеры листьев в вечнозеленом сезонном лесу острова Тринидад (Beard, 1946 <?, стр. 61)
Процент видов
лепто-филльных нано-филльных микро-филльных мезо-филльных макро-фИЛЛЬНЫХ мега-филльных
Наиболее высокие де-
ревья — 2,4 4,8 85,7 7,1 —
Ярус сомкнутого полога 2.2 — 17,4 76,1 4,3 —
Нижний ярус — — 11,8 80,4 7,8 —
Процент индивидуумов
лепто-филльыых нано-филльных микро-филльных мезо-филльных макро-филльных мега-филльных
Наиболее высокие де-
ревья — 0,0 20,5 83,5 16,0 —
Ярус сомкнутого полога 34,3 — 8,5 55,2 1,9 —
Нижний ярус — — 16,4 71,5 12,1 —
Таблица 9
Соотношение видов с заостренным кончиком листа и видов без верхушечного острия. Тропическим дождевой лес (влажный вечнозеленый лес Нигерии)
Деревья яруса А Деревья яруса В Деревья яруса С Кустарники
Число видов 10 13 14 10
Средняя длина острия, мм 2,9 8,8 12,9 12,4
Число видов без острия 4 1 0 0
короткий и плохо выраженный кончик. Все это видно из цифр, приведенных в табл. 9.
Различия типов листа в разных ярусах ясно связаны с вертикальным градиентом во внутреннем климате леса, который, как -будет показано в главе 7, очень сильно выражен. Каждый ярус имеет свой микроклимат, и соответствующее различие в типах листа ясно указывает на зависимость этих типов от условий среды.
Внутренний климатический градиент нередко проявляется и в заметных различиях, часто наблюдающихся у одного и того же вида между листьями, расположенными в тени и на солнце, и между листьями молодыми и взрослыми. Молодые листья могут быть во много раз больше взрослых и часто отличаются от них по очертаниям и текстуре настолько, что едва можно поверить, что они принадлежат к одному и тому же виду. У этих листьев почти всегда есть кончики-капельницы, даже в том случае, если у взрослых экземпляров край листа тупой или выемчатый, как,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 103
Рис. 15. Размер листьев деревьев и кустарников тропического дождевого леса по классификации Раункпэра.
Колонки иллюстрируют процентное соотношение видов каждого класса. Б и Ь* (Beard, 1946b); Г—Е (Brown, 1919).
А _ резервированный лес Шаша, Нигерия. Влажный, вечнозеленый лес: Б — Тринидад. Вечнозеленый сезонный лес (наиболее высокие деревья); В — Тринидад. Вечнозеленый сезонный лес (ярусом ниже); Г — гора Макви-линг, Филиппинские острова. Тропический дождевой диптерокарповый лес. 450 л<; Д — гора Маквилинг, Филиппинские острова. Предгорный дождевой (Quercus — Neolit sea) лес, 700 л; Е — гора Маквилинг. Филиппинские острова.
Горный (моховой) дождевой лес, 1000
например, у западноафриканской Lophira procera и Сatostemma jragrans в Брит. Гвиане (рис. 16). Нередко молодые листья бывают раздельными или сложными и нецельнокрайными, а взрослые — цельными и нераздельными. Так, у молодых деревьев малайского Artocarpus elastica листья глубоко надрезанные, а у спелого дерева — цельные. У €atostemma commune в Брит. Гвиане молодые листья состоят из трех-пяти листовых пластинок, а взрослый лист — только из одной. Такие поразительные изменения в форме листа встречаются главным образом у деревьев самого
104
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
высокого яруса; у деревьев третьего яруса форма листа совсем не меняется, а если и меняется, то очень мало.
Мы не располагаем количественными показателями относительно колебаний среднего размера или границ изменений других характерных признаков листьев в различных типах тропического дождевого пли же в различных местообитаниях внутри одного и того же типа листьев. Однако такие различия, вероятно, существуют, и их можно было бы обнаружить путем тщательного статистического исследования. Возможно, например, что у деревьев болотистого тропического дождевого леса листья крупных размеров встречаются чаще, чем в лесу, произрастающем на более дренированной почве. Так, листья деревьев «борового» леса на острове Борнео — сообщества, приуроченного к белым песчаным почвам (стр. 267), в среднем меньше, толще и жестче по своей текстуре, чем в смешанных лесах на глинах и суглинке (Richards, 1936, стр. 28). Интересно, что у видов, растущих по урезам рек, существует тенденция — она особенно ясно выявляется в лесах Малайского архипелага — иметь линейные или ланцетные лйстья, более узкие, чем у родственных видов, растущих в глубине леса. Это относится к Dipterocarpus oblongifolius, видам Fagraea (Loganiaceae), Garcinia (Guttiferae), Phoebe (Lauraceae), Syzygium и Eugenia (Myrtaceae) и т. д.; это же заметно и у листочков перистых листьев видов Dysoxylum. Явление не ограничивается только какими-нибудь отдельными семействами цветковых растений, но до известной степени заметно и у всех прибрежных папоротников. Беккари (Beccari, 1904, стр. 392), впервые наблюдавший эту особенность в Сараваке, назвал ее стенофиллией; затем ее отметил Лем (Lam, 1945, стр. 67—68) в Новой Гвинее и Страгнел и Мид (Strugnell, Mead, 1937, стр. 133) на Малаккском полуострове. Стенофилльная листва относится к общим признакам растений, произрастающих по берегам быстрых речек в Камеруне и других местах Западной Африки; автор недавно отметил много стенофилльных деревьев и кустарников в Ква-Фаллз, близ Калабара, среди которых были роды Deinbollia, Psychotria и другие. Согласно Беккари (Beccari), стенофиллия «возникла благодаря действию непрерывных токов воздуха, постоянно движущихся вдоль рек, а также благодаря периодическим затоплениям», но если она и имеет какое-либо приспособительное значение, то надо думать, что это последнее скорее связано с сопротивлением растения действию текущей воды, чем с воздушными токами.
При переходе от тропического дождевого леса к другим формациям, даже наиболее ему близким, вроде предгорного и горного тропического дождевого леса, преобладающий тип листа сразу меняется. Это прекрасно показал Браун (Brown, 1919) в своем весьма тщательном исследовании отношений типа листа к высоте в горах Филиппинских островов. Статистическому изучению были подвергнуты типы листьев в девственном диптерокарповом лесу, произрастающем на высоте 450 м (тропический дождевой лес), в лесу Quercus — Neolitsea, или предгорном лесу (среднегорный тропический дождевой лес), на высоте 700 м и в моховом лесу (горный тропический дождевой лес) на высоте 1000 м. По мере увеличения высоты наблюдается заметное увеличение процента мелких листьев и уменьшение процента цельнокрайних листьев.
Необычайное сходство листьев у деревьев тропического дождевого леса самой разнообразной систематической принадлежности не может быть случайным явлением. Тот факт, что даже отдельные листочки крупных сложных листьев, свойственные, например, деревьям из семейства Leguminosae, часто уподобляются простым листьям как по размеру,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА Ю5
так и по форме, показывает, что это однообразие объясняется влиянием среды. Подробное изучение вопроса о преобладании в данной среде цельных, мезофилльных, склерофилльных (мегасклерофилльных) листьев увело бы в область физиологии и анатомии. Достаточо сказать, что грубо телеологическое объяснение, выдвинутое в прошлом столетии (приспособление к высокой влажности, сильным дождям, интенсивному солнечному освещению и т. д.), уже не кажется удовлетворительным; в то же
того же вида; В — лист проростка Catostemma fragrant Г — лист взрослого дерева того же вида. А и Б относятся к резервированному лесу Шаша в Нигерии, В и Г — к лесам Брит. Гвианы (i/з натуральной величины).
время накопленные сведения не позволяют выдвинуть взамен какую-либо удовлетворительную теорию. Бьюс (Bews, 1927, стр. 15—17) считает тип листа тропического дождевого леса ксероморфным и связывает его признаки с низкой удельной проводимостью воды древесиной, что, по его мнению, характерно для тропических и субтропических вечнозеленых деревьев. Стокер (Stocker, 1935 Ь) провел интересные наблюдения над транспирацией у освещенных и затененных листьев в тропическом дождевом лесу острова Явы.
<06
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Чрезмерно заостренные кончики листьев многих деревьев пояса тропических дождевых лесов, или кончики-капельницы, издавна привлекают к себе внимание исследователей. Эта особенность встречается как у папоротников (рис. 17), так и у цветковых растений. В любой флоре при описании тропического дождевого леса пестрят выражения: folia longe acuminata, folia caudato-acuminata. Кончики-капельницы, подобные тем. что имеются у современных растений, встречаются и у ископаемых листьев некоторых третичных флор. За пределами тропического дождевого леса отчетливо выраженные кончики-капельницы редки, представляя собою спорадическое явление. Редко встречаются они и в «горном» дождевом лесу Ямайки1 (Shreve, 1914 а, Ь) и даже в своеобразном «нагорном климаксном лесу» острова Маврикия (Vaughan, Wiehe, 1941, стр. 152). Установлено, что кончики-капельницы чаще образуются и лучше развиваются в нижних ярусах леса и у молодых экземпляров высоких деревьев. Это заставляет сделать предположение о их тесной связи с определенной комбинацией влаги и тепла, характерной для климата тропического дождевого леса.
Юнгнер (Jungner, 1891) впервые высказал мнение, что кончики-капельницы являются приспособлением для быстрого осушения смоченной поверхности листа. В горных лесах Камеруна, куда он прибыл специально для изучения влияния тропических ливней на растительность, его поразило обилие кончиков-капельниц. Согласно его наблюдениям, листья с кончиками-капельницами скорее высыхают и реже покрываются слоем водорослей, грибов, лишайников и мхов, чем листья без капельниц. Присутствие же эпифилльных организмов, по его мнению, до такой степени влияет на ассимиляцию, что наносит серьезный ущерб растениям. Он считает, что кончики-капельницы, позволяя быстро стекать воде, тем самым очищают поверхность листа от спор эпифилльных организмов, от яиц и личинок насекомых, а также от растворимых веществ, которые могли бы благоприятствовать росту эпифилльных организмов. Шталь (Stahl, 1893) впоследствии экспериментально показал, что удаление кончиков-капельниц значительно увеличивало время, необходимое для высыхания листа, но он скептически относился к существованию какой-либо взаимосвязи между наличием кончиков-капельниц и развитием эпифилльных организмов. По его мнению, главное назначение кончиков-капельниц состоит в устранении с поверхности листа водяной пленки, которая, понижая температуру листа, могла понизить и скорость транспирации, что, как полагал Шталь, в свою очередь задерживает поглощение корнями минеральных солей и поэтому неблагоприятно для растения. Однако все это еще не доказано (см. стр. 210).
Впоследствии взгляды как Юнгнера, так и Шталя сильно критиковались, в особенности Шривом (Shreve, 1914 b). Изучая растения подлеска «горного» дождевого леса на Ямайке, Шрив не смог показать ,что удаление кончиков-капельниц сильно влияет на скорость высыхания. Он нашел, что эпифилльные организмы были обильны у листьев любого типа, включая и листья с кончиками-капельницами; встречаемость эпифилльных организмов, по его наблюдениям, зависит только от влажности атмосферы, окружающей растение (см. стр. 101 —102), а понижение температуры листьев благодаря наличию поверхностной водяной пленки слишком мало, чтобы повлиять на транспирацию. Однако водяная пленка уменьшает поглощение воды листьями из стебля, вероятно, отчасти благодаря остановке
1 См. стр. 390.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕС,\ 107
кутикулярной транспирации (которая «составляет немногим более половины потери воды листьями гигрофитов тропического дождевого леса») и отчасти благодаря фактическому поглощению воды слегка кутикуля-ризированным эпидермисом. Маклин (McLean, 1919, стр. 31) придерживается той же точки зрения.
К сожалению, листья, исследованные Шривом, как показывают фотографии, не были типичными мегасклерофиллами и не имели резко заостренной вершины, обычной длялистьев кустарников и мелких деревьев низинного тропического дождевого леса. У более жестких и более глянцевитых листьев кончики-капельницы могли заметнее способствовать скорости высыхания. Тут, очевидно, требуется больше экспериментальных данных.
Заключения Шрива о встречаемости эпифилльных организмов могут быть легко подтверждены любым наблюдателем. Эпифилльные организмы встречаются в большом количестве у всех долгоживущих листьев независимо от формы их вершины. В то же время сомнительно, чтобы они оказывали серьезное влияние на растение хозяина, так как покров из них обычно становится густым лишь на старых, увядающих листьях. Результаты, полученные Шривом относительно транспирации, применимы и fortiori к более склерофилльным листьям, у которых кутикулярная транспирация, вероятно, ничтожна. Поэтому вряд ли можно стать на точку зрения Шталя. С другой стороны, можно представить и другие возможные невыгоды продолжительного увлажнения. Согласно неопубликованной работе Накано (Nakano), цитированной Шимпером (Schimper, 1935, стр. 126), листья с удаленными кончиками-капельницами фотосинтезируют в увлажненном виде менее активно, чем неповрежденные; предполагается, что это вызвано некоторым поглощением света водяной пленкой, дольше сохраняющейся на листьях с отрезанными капельницами.
Тщательное изучение воздействия температуры и влажности на развитие молодых листьев могло бы, пожалуй, навести на мысль о возможной роли кончиков-капельниц. В связи с этим большой интерес представляет работа Холтермана (Holtermann, 1907). Этот автор установил, что на Цейлоне большинство растений тропического дождевого леса имеет длиннозаостренные листья, в то время как в сухой зоне и на вершинах гор растения этих капельниц не имеют. Часто листья в сухих местах обитания были даже выемчатыми. Такое различие в форме лпстьев отмечено не только у различных видов одного и того же рода, но даже в пределах одного и того же вида. Так, например, виды Ficus, растущие во влажной зоне, все без исключения имели более или менее длинные кончики-капельницы, в то время как виды сухой зоны имели тупые листья: у экземпляров Memecylon varians из дождливых местностей кончики-капельницы у листьев достигали длины 2—3 см, тогда как у экземпляров, растущих в сухих местообитаниях на вершинах гор они совершенно •отсутствовали. У многих видов кончики-капельницы были свойственны молодым листьям, а позже они исчезали (см. рис. 16). Кончик-капельница развивается гораздо раньше остальной части листовой пластинки. Если у молодого листа искусственно удалить кончик-капельницу, то во взрослом состоянии он становится выемчатым. Холтерманн также установил, что во всех исследованных случаях, где листья в зрелом •состоянии были тупыми или выемчатыми, это было следствием отсыхания острия у молодого листа. Отсыхание может быть объяснено сравнительно скудным водоснабжением острия, так как единственным путем подачи воды является главная жилка. Необходимы дальнейшие исследования «онтогении листьев с копчиками-капельницами. Троль (W. Troll, 1938,
108
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
стр. 1028) характеризует кончики-капельницы термином Nachldufber-spitze, рассматривая их как образования, появившиеся благодаря продолжению роста кончика листа в результате задержки дифференциации. Такой задержке дифференциации, вероятно, благоприятствует однообразно влажная атмосфера. Исходя из причинных оснований легко объяснить, почему листья «не образуют» кончики-капельницы в сухой среде.
Листовые подушечки. Функе (Funke, 1929, 1931) недавно обратил внимание на тот факт, что «сочленения» или листовые подушечки (на черешках или у сложных листьев у основания листочков), представляют собой типичный признак листьев тропических деревьев и лиан. Автор заметил эти образования и у различных папоротников в тропическом дождевом лесу. Сочленения сравнительно редки у растений умеренной зоны, но Фрейзер и Виккери (Fraser, Vickery, 1938, стр. 154) упоминают о них как об «имеющих почти всеобщее распространение» среди деревьев субтропического леса Нового Южного Уэльса. Согласно Функе (Funke), который работал на Яве, наличие одного, двух или даже трех сочленений гораздо более типично для тропических листьев, чем кожистая текстура, светоотражающая кутикула и тому подобные, часто упоминаемые признаки. Строение сочленений разнообразно, и Функе различает по крайней мере 15 различных типов. Несомненно, что эти структуры выполняют роль листовых подушечек и содействуют занятию пластинкой листа соответствующего положения по отношению к свету, что, как известно, осуществляется по-разному различными растениями. Нет никаких предположений относительно того, почему такой механизм, как листовые подушечки, особенно обычен у тропических растений, и не распространен среди растений умеренной зоны.
ЦВЕТКИ II ПЛОДЫ
В ботанике считается общепризнанным, что органы размножения растений более «консервативны», то есть менее подвержены изменяющим воздействиям среды, чем вегетативные органы, поэтому можно предполагать, что число общих признаков у цветов и плодов деревьев и кустарников дождевого леса будет невелико. В общем это верно, но все же у них существует несколько характерных особенностей, возможно, имеющих экологическое значение.
Самой обыкновенной и наиболее замечательной из таких особенностей является каулифлория, то есть образование цветков непосредственно на деревянистых ветвях и стволах. Для иллюстрации этого явления обычно упоминают Theobroma cacao — дерево яруса С, чья родина находится в тропическом дождевом лесу Америки. Жителя умеренной зоны каулифлория поражает как одна из самых своеобразных черт тропических деревьев. У большинства европейских деревьев цветки обыкновенно образуются только на побегах текущего, или в крайнем случае предшествующего года; они появляются исключительно на маленьких побегах, отсутствуя на толстых ветвях или на главном стволе. У многих деревьев тропического дождевого леса, наоборот, цветки или соцветия появляются непосредственно на стволах или же на очень коротких безлистных побегах, прикрепленных прямо к главному стволу или к крупным ветвям, тогда как более мелкие ветви, а иногда и вся крона, остаются чисто вегетативными.
Каулифлория может возникнуть двумя путями, но в любом случаецветущие побеги развиваются из почек, первоначально окруженных
Фото 8. Каулифлория у дерева Napoleona imperialism Иданре, Нигерия.
Фото 9. Эпифитный папоротник (Asplenium africanum резервированный лес Шаша, Нигерия.
СТРУКТУРАМ ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО .1Е(\ |09
листвой. Чаще каулифлория развивается при наличии на ветках пазушных почек, производящих цветки из года в год, вплоть до того времени, пока ветка не станет толстой ветвью или даже главным стволом. Таким образом дерево может иметь цветки повсюду от кроны до основания; у старого дерева более старые почки превращаются в короткие побеги с чешуйчатыми листьями и без видимых междоузлий. При развитии на них дополнительных пазушных почек могут образоваться толстые, похожие на галлы выросты. Второй способ заключается в том, что цветочные почки, формирующиеся в пазухах листьев, в течение долгого времени остаются спящими и начинают развиваться только после того, как несущий их побег достигнет значительного диаметра. В этом случае цветочные почки действительно пробиваются сквозь кору. Эволюционной проблеме каулифлории посвящены работы Томпсона (Thompson, 1943, 1946).
Термин «каулифлория» обыкновенно объединяет ряд несколько отличающихся одно от другого явлений. Милдбрэд (Mildbraed, 1922, стр. 115), подробно воспроизведено у Варминга и Гребнера (Warming and Graebner, 1933, стр. 741—748) в превосходной сводке, посвященной этому предмету, сделал попытку провести различие между ними и дать им определение. Простой каулифлории соответствует образование цветков в любом месте стебля, на стволе, на крупных и мелких ветвях, что объясняется непрекращающейся активностью почек в течение неопределенного периода. Простая каулифлория наблюдается у Theobroma, у видов Angylocalyx, Diospyros, Dry petes и многих других деревьев тропического дождевого леса.
При рамифлории цветки развиваются на более крупных ветках и сучьях, но отсутствуют на главном стволе; в этом случае почки остаются активными только в течение ограниченного периода. Милдбрэд в качестве примера приводит африканский вид Turraeanthus zenkeri (Meliaceae). Автор заметил такое же явление у Polyalthia insignis (Annonaceae) и у других видов, растущих на острове Борнео. Транкифлории соответствует образование цветков только на главном стволе. Она наблюдается у африканского вида Omphalocarpum procerum (Sapotaceae), у видов Diospyros (Ebenaceae), Cola (Sterculiaceae) и у многих других деревьев. Базифло-рией называется развитие цветков только у основания ствола, как, например, у африканского Uvariopsis sessilijlora (Annonaceae) ' (Mildbraed, 1922) и Polyalthia jlagellaris (Annonaceae) с острова Борнео. В другом случае, называемом идиоклаЗантией, цветки появляются на сравнительно длинных безлистных побегах или побегах, имеющих только листовые чешуйки, прикрепленных к стволу или к нижней части олиствленных ветвей, как, например, у Couroupita guianensis (Lecythidaceae) и Аппо-nidium mannii (Annonaceae). Виды, у которых соцветие представляет собой безлистную метелку, конечно, сюда не относятся, кроме тех, близкие родственники которых имеют одиночные пазушные цветы. Наконец, при флагеллифлории цветки образуются на длинных, похожих на хлыст ветвях с чешуйчатыми листьями и длинными междоузлиями, лежащих на поверхности почвы, например, как у Paraphyadanthe flagelliflora (Fla-courtiaceae) (Mildbraed, 1922).
Эти термины имеют определенную ценность, учитывая разнообразие существующих типов каулифлории, но надо помнить, что встречаются всевозможные переходные типы, и трудно отнести любой данный род каулифлории к какому-либо одному из указанных разделов классификации. Вероятно, чаще всего каулифлория проявляется в образовании цветков на стволе и более крупных сучьях, отсутствуя на мелких веточках, как,
но
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
например, у видов Polyalthia и многих других Аппопасеае. Более того» один и тот же вид может обнаруживать в этом отношении изменчивость: автор, например, обнаружил, что у Diospyros «confertiflora» (Ebenaceae) в Нигерии наблюдается как простая (в смысле Милдбрэда) каулифлория, так и рамифлория.
Милдбрэд предполагал, что рамифлория является наиболее примитивным видом каулифлории. В связи с этим представляется, конечно, очень заманчивым заключение, что рамифлория, простая каулифлория, транкифлория и базифлория представляют собой последовательные стадии эволюции в сторону возрастающей специализации. С этой точки зрения большой интерес представляет род Ficus, в пределах которого наблюдается большое разнообразие каулифлории. В подробном исследовании о малайских видах подрода Covellia Корнер (Corner, 1933) показал, что тут можно установить все типы каулифлории. У Ficus lepicarpa цветоложи располагаются парами в пазухах листьев; у F. fistulosa и F. scortechinii незначительная часть цветолож может находиться на олист-вленных побегах, но в большинстве своем они развиваются на более старых ветвях или на стволе, образуя компактные гроздья; у F. obpyra-midata nF. miquelii все соцветия находятся на более старых, не имеющих листьев ответвлениях ствола, причем главные плодущие ветви достигают гораздо большей длины, а гроздья соцветий напоминают огромные кисти винограда. У F. hispida соцветия находятся главным образом на лишенных листьев ответвлениях главного ствола; длина некоторых из них достигает 3 футов или около того, при этом они свободно висят, неся соцветия на всем своем протяжении. В конце рассматриваемого ряда находятся так называемые «геокарпические» виды, например/", beccarii и/", geocarpa. У них плодущие ветви возникают только в нижней части главного ствола не выше 3 футов от земли и представляют собой мало ветвящиеся хлыстообразные побеги, достигающие 20 футов (6 м) длины при толщине у основания приблизительно в один дюйм (2,5 см).Около точки роста у этих ветвей образуются опадаюшие прилистники, но листья на них отсутствуют. По мере роста они свисают книзу и затем ползут по земле; когда на их пути встречаются препятствия вроде земляной насыпи, они, подобно корням, зарываются в землю. Согласно Корнеру, описанные типы каулифлории могут представлять отдельные стадии эволюционного процесса.
Каулифлория в широком смысле этого слова представляет собой обыкновенное явление у деревьев и кустарников тропического дождевого леса, встречаясь у многих неродственных семейств. Милдбрэд (Mildbraed, 1922) только в тропической Африке обнаружил приблизительно 278 видов с каулифлорией и оценивает общее число их во всем мире более чем в тысячу. Поразительно то, что огромное большинство этих видов представляет собой деревья малого или среднего размера или же высокий кустарник, принадлежащие к ярусу С, или ярусу кустарников. Среди деревьев яруса А типичная каулифлория очень редка, хотя рамифлория отмечается у малайских видов Durio, Coelostegia, Myristica и Ficus, а внешний вид Polyalthia напоминает геокарпические фикусы. Некоторые виды Alexa Брит. Гвианы являются рамифлорными (Т. A. W. Davis). Каулифлория свойственна и некоторым лианам (см. стр. 126); аналогичный характер цветения встречается и у некоторых покровных трав дождевого леса (см. стр. 120). При всяких попытках объяснения каулифлории следует помнить об относительной частоте ее нахождения в самом нижнем ярусе.
С точки зрения эколога каулифлория представляет интерес главным образом благодаря тому, что она так характерна именно для тропиче
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА (11
ского дождевого леса, хотя в некоторых тропических областях, например в южной Нигерии, она встречается сравнительно редко. За пределами пояса тропических дождевых лесов каулифлория, по-видимому, крайне редка, хотя известное иудино дерево, Cercis siliquastrum, и небольшое число других деревьев умеренной зоны также обладает каулифлорией неспециализированного типа. Существует множество предположений о значении каулифлории, но большинство их совершенно не подтверждается экспериментальными данными. Милдбрэд (Mildbraed, 1922) сделал обзор ряда более вероятных теорий. Наиболее известной из них является теория Уоллеса (Wallace, 1878, стр. 34—35), предполагающая, что каулифлория представляет собой приспособление, посредством которого более мелкие деревья, лишенные возможности выбраться на свет и простор верхних ярусов, приобретают способность образовывать свои цветки в таких местах, где они лучше видны и более доступны для тенелюбивых бабочек.
Самый нижний ярус деревьев, как уже указывалось в главе 2, обычно отличается наибольшей густотой, в то время как подлесок сравнительно мало сомкнут, так что эта теория a priori обладает известной вероятностью. Точно установлено, что многие виды, обладающие каулифлорией,— некоторые из них являются двудомными,— нуждаются в перекрестном опылении насекомыми, хотя возможно, что эту роль выполняют муравьи, а не бабочки. С позиций теории Уоллеса получает объяснение и другое любопытное явление, названное Милдбрэдом (Mildbraed, 1922) пенду-лифлорией. В этом случае цветы располагаются на длинных шнуркообразных ножках, могущих достигать более 2 м длины и свисающих с ветвей; в результате цветы ясно выделяются среди листьев кроны. Это наблюдается у Eperua falcata (Leguminosae) — высокого дерева Южной Америки— и у африканской лианы Мисипа flagellipes (Legiminosae). Милдбрэд упоминает также об южноамериканском дереве Couepia longipendula (Rosa-сеае), африканском дереве Parkia pendula (Leguminosae) и об африканском дереве или кустарнике Uragoga peduneularis (Rubiaceae).
Другие теории, например теория Хаберландта (Haberlandt, 1926) и Клебса (Klebs, 1911, стр. 61—62), подходят к вопросу о каулифлории с физиологической точки зрения. Согласно Хаберландту, вечнозеленые деревья, у которых новые листья никогда не появляются сразу в большом количестве, а образуются постепенно, содержат в своих стволах и крупных ветвях только такое количество питательных веществ, которое необходимо для развития цветков и плодов. Если последние непосредственно развиваются на стволах и ветвях, то необходимость обратного перемещения пищи в ветки отпадает, п таким образом сберегаются время и энергия.
Самое очевидное возражение против этой теории состоит в том, что чем выше дерево, тем более выгодной должна была бы быть для него каулифлория, а она, как уже говорилось, встречается главным образом у небольших деревьев. Клебс считает, что образование цветков может происходить лишь при некотором избытке продуктов ассимиляции по отношению к количеству минеральных солей. У тропических деревьев, где ассимиляция в течение года происходит непрерывно, в стволе и ветвях накапливается большое количество ассимплятов. Так как в то же самое время посредством листьев осуществляется постоянный отвод минеральных солей, то в коре всегда должны иметься условия, благоприятные для образования цветков при наличии почек. Это одна из немногих теорий, свободных от налета телеологии.
112
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
То обстоятельство, что у тропических деревьев обыкновенно очень тонкая кора (см. стр. 76), может, конечно, до известной степени объяснить возможность пробуравливания ее цветочными почками, но это еще не объясняет наличие каулифлории. Во всяком случае вопрос о каулифлории в целом явно нуждается в экспериментальном исследовании.
Биология цветка деревьев и кустарников тропического дождевого леса мало исследована. Известно, что птицы (колибри, нектарницы и птицы из многих других семейств), а также насекомые тропической зоны играют значительную роль в опылении. Несомненно, что вследствие обилия птиц и насекомых, посещающих растения во всякое время года, анэмофпльные цветки встречаются редко. Растения, принадлежащие к семействам, чьи представители в умеренной зоне являются анэмофи-лами, в тропиках посещаются и, вероятно, опыляются насекомыми, хотя их цветки не имеют нектара. Например, мужские цветки малайских дубов (Quercus и Lithocarpus spp.), имеющие приятный запах, привлекают тучи маленьких пчел, мух и жуков; женские цветки почти наверняка опыляются ими, хотя эти цветки и не имеют запаха (Corner, 1940). Травянистое растение Pariana radiciflora, произрастающее в гвианском лесу, усиленно посещается, и, вероятно, опыляется насекомыми. Так как фауна птиц и насекомых неоднородна, а крайняя тишина воздуха в подлеске неблагоприятна для опыления ветром, то пропорции видов, опыляемых различными агентами, будут, вероятно, различными в разных ярусах, но определенные сведения об этом отсутствуют.
Семена и плоды деревьев и кустарников тропического дождевого леса и способы их распространения нуждаются в планомерном статистическом исследовании. Сомнительно, чтобы у приспособлений для распространения деревьев и кустарников тропического дождевого леса имелись какие бы то ни было общие признаки. В молодом вторичном лесу (см. гл. 17) почти все доминирующие виды имеют семена, снабженные ярко выраженными приспособлениями для распространения ветром или животными (главным образом птицами и летучими мышами); эти приспособления позволяют деревьям вторичного леса с необычайной быстротой расселяться на обнаженной почве или в окнах первичного леса. В первичном тропическом дождевом лесу распространение с помощью ветра и животных встречается реже, а многие деревья и кустарники, видимо, вообще не имеют никаких специальных приспособлений для расселения. По-видимому, характер приспособлений зависит от принадлежности к различным ярусам. Ветровое распространение, вероятно, почти исключительно ограничивается деревьями и лианами верхнего яруса; этого, собственно, и следовало ожидать, так как только этот ярус в той или иной степени подвержен значительному воздействию воздушных течений. В тропическом дождевом лесу Нигерии автор у большей части высоких деревьев обнаружил семена или плоды, видимо, приспособленные для распространения ветром, в то время как в нижних ярусах анэмофилия совершенно отсутствует; тут многие виды имеют тяжелые семена, падающие к основанию материнского дерева (впоследствии они могут быть распространены мелкими млекопитающими). Вероятно, то же самое наблюдается и в аналогичных лесах Малайского архипелага, где доминирующее семейство деревьев первого яруса Dipterocarpaceae имеет крылатые (распространяемые ветром) плоды. В южноамериканском лесу распространяемые ветром семена и плоды представляют собой редкость в любом ярусе; в Брит. Гвиане единственными обыкновенно распространяющимися с помощью ветра деревьями являются Aspidosperma
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА ЦЗ
excelsum и Bombax surinamense, но часто встречается распространение посредством птиц, летучих мышей и млекопитающих (Т. A. W. Davis). В Гвиане и, вероятно, в других частях тропиков многие деревья, в особенности принадлежащие к верхнему ярусу, имеют очень крупные семена, тяжело падающие на землю. Несомненно, тяжелые семена представляют некоторое преимущество для высоких деревьев в том отношении, что они не могут задержаться в листве ниже расположенных ярусов, кроме того, это позволяет им содержать большой запас пищи для ростка, что позволяет последнему быстро достичь достаточного размера, чтобы выдержать предстоящий ему продолжительный период угнетения (см. гл. 3). Но крупные, тяжелые семена могут воспрепятствовать растению занять подходящее местообитание. Так, Витками (Witkamp, 1925) считает, что Eusideroxylon zwageri (Lauraceae), дерево, являющееся единственной доминантой на больших пространствах лесов Суматры и Борнео (см. стр. 282), не растет на склонах холмов потому, что его тяжелые плоды скатываются по склону вниз. Медленное распространение Mora excelsa, расширяющей свой ареал на острове Тринидад, Маршалл (Marshall, 1934) считает зависящим от веса и размера ее семян.
ЗАКЛЮЧЕНИЕ
Из обзора физиономических признаков деревьев и кустарников тропического дождевого леса видно, что эти признаки так или иначе отчетливо связаны со специфическими условиями окружающей среды, в которой и развивается сообщество. Многие из этих условий значительно меняются по частоте встречаемости в разных ярусах, что и неудивительно, если иметь в виду существование больших микроклиматических различий в последних (см. гл. 7). Хотя физиономические черты, о которых идет речь, связаны с местом обитания, так как они встречаются у большого числа неродственных видов, однако в большинстве случаев эти черты не могут считаться результатом прямого «приспособления» к условиям среды. Некоторые из них могут иметь «причинное» происхождение, в настоящее время для нас непонятное. Другие особенности, наблюдаемые у растений тропического дождевого леса, вероятно, могут быть истолкованы примерно таким же путем, каким Гёбель (Goebel) объяснял «пони-клость» молодых побегов у Amherstia и других видов, предполагая, что они возникают вследствие действия каких-то неясных внутренних причин и существуют лишь благодаря благоприятным условиям тропического дождевого леса, неизбежно устраняясь отбором в более суровой среде. В связи с этим может считаться показательным то, что наибольшее разнообразие листвы в дождевом лесу наблюдается в подлеске, где температура и влажность крайне постоянны, а наибольшее однообразие — в верхних ярусах с переменчивым микроклиматом. Так или иначе для нас важнее не более или менее гипотетическое «приспособительное» или «причинное» происхождение структурных признаков деревьев и кустарников тропического дождевого леса, а диагностическое значение этих признаков для характеристики как сообщества в целом, так и образующих его ярусов. Уже достаточно сказано о том, что тропический дождевой лес, как тип формации, и различные ассоциации и консоциации, из которых он состоит, имеют множество в высшей степени характерных структурных черт, более или менее независимых от систематической принадлежности обладающих ими видов. Они-то и придают сообществу тропического дождевого леса его особый облик.
Глава 5
ТРАВЯНОЙ ПОКРОВ И ЗАВИСИМЫЕ СИНУЗИИ
Травяной покров тропического дождевого леса и синузии, зависящие от других растений в отношении опоры или питания (лазящие растения, эпифиты, сапрофиты и т. д.), находятся в специфических экологических условиях, и составляющие их виды в высшей степени специализированы по своей структуре и физиологии. Лазящие растения, или лианы, представляют собой характерную особенность почти всех дождевых лесов и активно конкурируют с деревьями за свет и пространство; в то же время эти весьма различные группы растений играют лишь незначительную роль в структуре и общем строе сообщества. Несмотря на это, они представляют большой интерес для эколога не только по причине действительного или предполагаемого богатства структурными приспособлениями (большей частью прекрасно известными и лишь вкратце рассматриваемыми здесь), но и по причине той точности, с которой распределение их отражает изменения условий микроклимата.
ТРАВЯНОЙ ПОКРОВ
Синузия травяного покрова не должна отождествляться с представлением о самом нижнем ярусе леса, поскольку, как указывалось в предыдущей главе, большинство растений этого яруса является подростом молодых деревьев, кустарников и деревянистых лиан. С другой стороны, не все растения, которые морфологически можно отнести к травам, принадлежат к описываемому ярусу. Часть их представляет собой эпифиты и не укореняется в почве, другие, например, некоторые из более крупных Scitamineae, хотя стебли их и не являются деревянистыми, достигают высоты в 6 м и более и с экологической точки зрения образуют часть кустарникового яруса или даже яруса самых низких деревьев. В настоящем разделе будут рассматриваться только травы, действительно составляющие часть самого нижнего яруса леса и, следовательно, не превосходящие по высоте приблизительно двух метров.
Уже указывалось, что в тропическом дождевом лесу редко наблюдается сомкнутый покров травянистой растительности, такой обычный в лесах умеренной зоны. В тропических дождевых лесах низменностей роскошная травянистая растительность встречается лишь на маленьких прогалинах, вдоль тропинок, по берегам рек и речек и в других местах, получающих больше солнечного света. Внутри леса находятся главным образом разрозненные экземпляры травянистых растений; иногда они почти совершенно отсутствуют, а виды, произрастающие куртинами и образующие сомкнутый полог, встречаются лишь от случая к случаю. На крутых склонах и в горных дождевых лесах, где по той или иной причине древесный полог оказывается менее густым, травянистая растительность, как правило, более обильна.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 115
Большинство травянистых видов, в изобилии встречающихся на открытых местах, может быть найдено и под пологом леса, за исключением самых темных его участков; в то же время ряд видов, приспособившихся к жизни в тени, например Trichomanes и некоторые другие папоротники, никогда не растут на открытых местах. Отсюда, по-видимому, следует, что среди трав тропического дождевого леса присутствуют два экологических элемента, возможно, заслуживающие того, чтобы их рассматри^ вать как отдельные синузии,— виды теневыносливые и виды тенелюбивые. Первая группа достигает максимального развития в условиях довольно сильного освещения, вторая, видимо, угнетается при наличии сильного света. Однако свет, как таковой, не всегда является главным фактором, определяющим разделение этих групп; важное значение может иметь также корневая конкуренция и влияние почти постоянно насыщенной влагой атмосферы (см. гл. 7). Многие теневыносливые растения склонны произрастать группами, в то время как тенелюбивыми являются преимущественно растения-одиночки. К группе теневыносливых принадлежит огромное большинство травянистых растений тропического дождевого леса, и их совокупность в известной степени аналогична видам травянистых растений, характерных для лесных опушек (Salisbury, 1925) европейских листопадных лесов.
Синузия травяного покрова, кроме двудольных и однодольных, включает многочисленные папоротники, а во многих местностях весьма постоянными ее компонентами являются виды Selaginella. Даже если сложить вместе теневыносливые и тенелюбивые травянистые растения дождевого леса, то и тогда число их семейств и видов оказывается ограниченным, хотя, конечно, оно сильно изменяется от района к району. В самой богатой области число это, по-видимому, никогда не превышает лишь некую часть от общего количества деревьев и кустарников, растущих на том же пространстве. Так, например, в самом нижнем (наземном) ярусе первичного леса (все типы) на площади в несколько квадратных километров, Морабалли-Крик, Брит. Гвиана, являющейся, вероятно, довольно типичной в этом отношении, было встречено менее тридцати обычных цветковых растений, от десяти до двадцати обыкновенных папоротников и один вид Selaginella при наличии нескольких сотен разнообразных деревьев и кустарников. На подобной же площади первичного леса в Нигерии, вероятно, наблюдается сходное соотношение, однако точные сведения по этому вопросу отсутствуют.
В то же время травяной покров тропического дождевого леса Малайского архипелага (если район горы Дьюлит на острове Борнео является типичным), значительно богаче, чем в лесах Америки или Африки. Флористический состав синузии здесь очень непостоянен, сильно меняясь от места к месту часто без явной связи со сменой почвенных и других условий. Многие виды, встречающиеся лишь на одном или двух соседних хребтах, видимо, нигде больше в пределах этой области не произрастают.
Даже там, где насчитывается сравнительно большое число видов травянистых растений, количество семейств цветковых растений, представленных в синузии, невелико. Во всех тропических лесах среди травяного покрова хорошо представлены Rubiaceae. Постоянно присутствует небольшое количество злаков (Gramineae), но в девственных лесах они немногочисленны как по видовому составу, так и по количеству индивидуумов, что является важным отличительным признаком, позволяю-
116
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
щим разграничить дождевой лес и некоторые более сухие типы тропического леса. В тропическом дождевом лесу Брит. Гвианы, кроме упомянутых двух семейств, обыкновенно представлены Сурегасеае, Gesneria-ceae, Marantaceae, Orchidaceae и Melastomaceae. В южной Нигерии самыми обычными семействами являются Acanthaceae, Агасеае, Commelinaceae и Liliaceae, хотя семейство Rubiaceae численно превосходит любое другое семейство двудольных. На острове Борнео представлено несколько большее число семейств, из которых наиболее обильны Агасеае, Begoniaceae, Сурегасеае, Gesneriaceae, Melastomaceae и Zingiberaceae.
Многие, хотя и не все, папоротники синузии травяного покрова являются тенелюбивыми, и количество их заметно увеличивается в особенно сырых и затененных местах — долинах водотоков и на крутых скалистых склонах. Большинство папоротников принадлежит к семейству Polypodiaceae, но часто встречаются и Trichomanes. хотя они редко представлены в большом количестве.
Ограниченное число видов и семейств, формирующих травяной покров, является, вероятно, следствием особых условий среды, настолько специализированной, что только сравнительно небольшое число семейств оказалось способным выделить виды, сумевшие к ней приспособиться.
Трудно дать общую характеристику и описать экологическую морфологию покровных трав ввиду большого разнообразия их форм. Их жизненные формы значительно отличаются от форм, встречающихся в лесах умеренной зоны. Растения компактного облика, вроде розеточных, редки и, как и следовало ожидать, учитывая однообразие климатических условий в течение года, очень мало растений с покоящимися почками на поверхности почвы или под ней. Большинство трав имеет сравнительно удлиненные надземные побеги, сохраняющиеся, вероятно, в течение нескольких лет и часто обнаруживающие слабо выраженную тенденцию становиться деревянистыми. Статистические данные относительно жизненных форм синузии травяного покрова тропического дождевого леса отсутствуют, но, вероятно, большинство трав, по терминологии Раункиэра, является травянистыми фанерофитами, а не гемикриптофитами или геофитами, как травы европейских листопадных лесов. Часто встречаются растения с подземными корневищами, которые приспособлены скорее для целей размножения и распространения, чем для переннирования. Эти корневищные виды склонны произрастать сообществами и образовывать густые заросли на площади в несколько квадратных метров и больше, вытесняя другие растения. К ним принадлежат Leandra divaricate, (Melastomaceae) в Брит. Гвиане, злак Leptaspis cochleata, виды Dorstenia и Geophila в Западной Африке, а также Susum malayanum (Flagellariaceae) и многие Zingiberaceae в пределах Малайского архипелага. Они, а вероятно, и большинство других растений, входящих в состав травяного покрова первичного леса, размножаются главным образом вегетативным путем.
Хотя растения с подземными корневищами, луковицами, клубнями или клубне-луковицами довольно обычны, однако истинные геофиты, у которых надземные побеги ежегодно отмирают на известный промежуток времени, редки и, по-видимому, встречаются в основном вблизи климатических границ тропического дождевого леса, где относительно выражен сухой сезон. Таким образом, они, по-видимому, полностью отсутствуют в низинных тропических дождевых лесах острова Борнео и Брат. Гвианы; в то же время в вечнозеленом лесу южной Нигерии, произрастающем в условиях более ясновыраженных сезонных изменений,
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА Ц7
встречаются различные Агасеае (Anchomanes и Amorphophallus spp.), которые являются клубне-луковичными геофитами, чьи наземные органы отмирают на несколько месяцев ежегодно; что касается соцветий, то у некоторых видов они образуются до, а у других после появления новых листьев. Cyanastrum cordijolium (Tecophilaceae) — обычная трава лесов Западной Африки — ведет себя как истинный геофит в резервированном лесу Шаша (Richards, 1939, стр. И), переннируя посредством своих клубнелуковиц, а в лесах Бенина и в Камеруне, возможно, вследствие более влажного климата листья этого растения остаются зелеными в течение
Рис. 17. Конвергенция формы листа растений тропического дождевого леса различной систематической принадлежности. А — перышки Pieris preussii — папоротника дождевого леса Нигерии; Б и В — листья двух Сурерасеае из подлеска диптерокарпо-вого леса, гора Дьюлит, Борнео; Г — лист Goniothalamus malayanus (Annonaceae) —кустарника из семейства двудольных, гора Дьюлит, Борнео (2/з натуральной величины).
всего сухого сезона. В тропическом дождевом лесу полуострова Малакка геофиты представлены видами Тасса и некоторыми Агасеае (Arisaema и Amorphophallus spp.) с клубне-луковицами или клубнями. В периодически затопляемых лесах Конго произрастают травянистые растения с луковицами или корневищами, дающими надземные побеги только во время сухого сезона (Lebrun, 1936 а, стр. 26); они могут рассматриваться как геофиты, или как гидрофиты.
Стебли трав тропического дождевого леса обычно сочны и хрупки, что, без сомнения, объясняется малым содержанием одревесневшей ткани, но иногда бывают и деревянистыми, как, например, многие малайские виды Didymocarpus и Sonerila, У некоторых видов стебель у основания поддерживается воздушными «опорными» корнями, как у некоторых малайских Zingiberaceae.
118
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Травы тропического дождевого леса имеют разнообразные листья, отличаясь в этом отношении от монотонных по листве деревьев и кустарников. Листья большинства трав отличаются тонкостью и мягкостью; среди них едва ли можно встретить крупный, блестящий, склерофилльный тип листа. Несомненно, листья трав менее долговечны, чем листья деревьев и кустарников, что подтверждается редким нахождением на них эпи-филльных организмов.
Большее разнообразие листьев среди растений травяного покрова по сравнению с листьями деревьев и кустарников в основном проявляется скорее в окраске листовых пластинок, в характере поверхности и форме краев, а не в размерах или общем очертании листьев. Хотя цифровые данные об относительном количестве листьев различных размеров по шкале Раункиэра (см. стр. 103) отсутствуют, заметно выявляется определенное преобладание мезофилльного класса. Листья меньших размеров, как и следовало ожидать, встречаются тут чаще, чем у деревьев и кустарников, а листья более крупные, чем мезофилльные, отсутствуют или очень редки. Так же, как и в более высоких ярусах, в травяном покрове преобладают листовые пластинки широко-эллиптической и продолговато-ланцетовидно-заостренной формы, однако нецельнокрайние листья встречаются здесь более часто. Хотя в большинстве случаев листовые пластинки травянистых растений в известной степени заострены, однако выраженные кончики-капельницы встречаются здесь реже, чем у кустарников и более мелких деревьев. Общая тенденция к образованию широких листьев в травяном покрове тропического дождевого леса выражена очень ярко у Gramineae и Сурегасеае; большинство видов этих семейств, обычно встречающихся в первичном дождевом лесу, имеют листья, резко отличающиеся по форме от листьев их родичей в других местообитаниях. Листья всех типичных злаков тропического дождевого леса (рис. 18) и, в частности, листья Ichnanthus panicoides и Pariana spp. из Южной Америки, Leptaspis cochleata — из Западной Африки, принадлежащих к так называемому типу «Commelinaceae» (АгЬег, 1934), совсем непохожи на узкие листья злаков тропической саванны или умеренной зоны; весь облик растений напоминает скорее Scitamineae или Commelinaceae, чем Gramineae. Равным образом, некоторые из Сурегасеае (рис. 17) обладают удивительно широкими заостренными листьями.
Создается впечатление, что листья злаков и осок тропического дождевого леса, пытаются как бы приноровиться к «модной» эллиптической форме, несмотря на унаследованный ими тип строения.
Листья трав дождевого леса, по-видимому, никогда не бывают так глянцевиты, как листья большинства деревьев и кустарников. В общем их поверхность нельзя назвать ни особенно глянцевитой, ни особенно матовой. Листья ряда трав, как это часто отмечалось, имеют бархатистую поверхность; это свойственно некоторым Агасеае и Orchidaceae, а также Neckia sp. (Ochnaceae) с Бррнео; однако такие виды не очень многочисленны. Бархатистая поверхность образуется клетками верхнего эпидермиса, которые, как и у лепестков с бархатистой поверхностью, выступают в форме сосочков. Согласно Шталю (Stahl, 1896), благодаря этим сосочкам вода, падающая на лист, сразу распространяется по его поверхности в виде тонкой пленки. Кроме того, они действуют и как «уловители», повышая поглощение листом тепла и света; Шталь рассматривал их как первичное приспособление для увеличения скорости транспирации.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА Ц9
Гораздо более общим признаком листьев травяного покрова тропического дождевого леса является пестрота их окраски. Они могут быть покрыты пятнами или полосами белого или бледно-зеленого цвета, как, например, у видов Begonia, некоторых южноамериканских видов Cephae-lis (Rubiaceae), африканской Dracaena phrynioides (Liliaceae); могут
В
Рис. 18. Листья трав тропического дождевого леса.
А — неизвестный вид смешанного диптерокарпового леса горы Дыолит, Борнео; Б — Leptaspis cochleata из резервированного леса Шаша, Нигерия; В — Pariana campestris из Суринама (из гербария Кембриджского университета) (i/з натуральной величины).
быть 'испещрены красными пятнами или точками, как у индонезийского вида Sonerila (Melastomaceae). Пестрая листва характерна для травяного покрова тропических дождевых лесов большей части тропиков и 'очень редка в других синузиях, хотя небольшое количество эпифитов, например южноамериканские Peperomia и Vriesia spp., также имеет пестроокрашенные листья. Шталь (Stahl, 1896) считал, что пестрота окраски так же, как и бархатистость листа, представляет собой приспособление для увеличения скорости транспирации. Другой заслуживающей упоминания особенностью окраски листьев травяного покрова дождевого леса является металлический синевато-зеленый блеск, наблюдающийся у некоторых видов Selaginella, обычно разводимых в оранжереях. Он встречается также и у видов Mapania (Сурегасеае), например у М. monostachya с Борнео, у папоротников рода Trichomanes и, вероятно,
120
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
у других родов. Синеватый блеск обусловливается присутствием зерен кутина в стенках клеток эпидермиса; Гентнер (Gentner, 1909) рассматривал это явление как приспособление к своеобразным условиям освещения под пологом тропического дождевого леса, однако экологическое значение его, если вообще о нем можно говорить, далеко не ясно.
Таким образом, листья травяного покрова тропического дождевого леса имеют целый ряд особых признаков, которые, как полагают, являются приспособлением к своеобразному микроклимату их обитания; некоторые из этих признаков еще раз будут рассмотрены в главе 7.
Цветки у растений травяного покрова, подобно цветкам древесных растений, редко бывают красивыми, но имеют несколько отличительных черт. Общим особенно для травянистых растений индонезийского тропического дождевого леса является то, что цветки их появляются у самой поверхности или непосредственно над поверхностью почвы на цветоножках, лишенных листьев или имеющих только чешуйчатые листья; эту особенность можно сравнить с каулифлорией у деревьев и кустарников. Во флоре Борнео виды Curculigo (Amaryllidaceae) и Forrestia (Gommeli-пасеае) наряду co многими другими имеют соцветия, развивающиеся от основания растения. У Cyrtandra pendulijlora гибкая ось соцветия располагается на поверхности почвы; у многих Zingiberaceae соцветие ответвляется от подземной части растения на некотором расстоянии от олиствленных побегов. Значение этой особенности совершенно неясно.
ЛАЗЯЩИЕ РАСТЕНИЯ
Распространение и облик
Благодаря своим размерам и обилию лазящие растения и особенно лианы, обладающие большими деревянистыми стеблями, представляют собой одну из самых выразительных черт тропического дождевого леса; их необычайные часто фантастические формы в сильной степени содействуют приданию этому лесу его своеобразного вида. Плющ, ломонос и другие вьющиеся растения лесов умеренной зоны дают лишь слабое представление о разнообразии подобных растений в тропиках. Лианы, иногда называемые ползучими или плетевидными кустарниками1, всегда присутствуют в типичном дождевом лесу; их стебли, часто имеющие вид веревок или проволоки толщиною иногда с руку или бедро, исчезают вверху в массе листвы и, свисая вниз, образуют здесь и там гигантские петли. Часто эти растения перекидываются с дерева на дерево и так прочно связывают их кроны, что дерево, даже срубленное у основания, продолжает стоять. Стволы тропических лиан могут достигать невероятной длины. Они могут, взобравшись да одно дерево, опуститься до земли и вновь взобраться на другое. Тройб (Treub, 1883, стр. 175) измерил лазящую пальму, ствол которой достигал 240 м длины. Возможно, что такая длина является редким исключением, большинство же тропических лазящих растений, вероятно, лишь изредка превышает 70 м. Лазящие растения образуют вполне определенную группу; это, однако, не мешает существованию переходных форм между ними и полностью независимыми растениями. К ним относятся деревья с неустойчи
1 Термин «лазящие кустарники», применяемый в некоторых работах по систематике растений, может ввести в заблуждение и едва ли является подходящим.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 121
выми стволами, вроде южноамериканской Annona haematantha (Annonaceae), и кустарники, вроде африканской Icacina trichantha (Icacinaceae). Равным образом нельзя провести резкую границу между лазящими растениями и эпифитами. Многие эпифиты имеют стволы как у лазящих растений, а некоторые лазящие растения, вроде различных Агасеае, американской Carludovica (Cyclanthaceae) и малайской орхидеи Dipodium pictum начинают свою жизнь, укоренившись в почве, а затем, утратив в конце концов связь с нею, становятся эпифитами. Некоторые полуэпифиты (см. стр. 144.), достигшие окончательных размеров, весьма похожи на крупные деревянистые лианы.
Лазящие растения встречаются в любом климате, где в растительном сообществе имеются деревья, могущие служить им поддержкой, но эти растения наиболее обильны в тропическом дождевом лесу. Шенк (Schenck, 1892—1893, стр. 57) считает, что 10/п всех видов лазящих растений находится в тропиках. В Вест-Индии, где тропический дождевой лес и подобные ему формации являются преобладающим типом растительности, к деревянистым лианам относится около 8% всех цветковых растений (Grisebach, цит. Шенком, Schenck 1892—1893, стр. 57). В Европе лазящие растения составляют менее 2% флоры. Согласно Шенку, наиболее богат лианами южноамериканский тропический дождевой лес, тогда как азиатский занимает второе место. Вряд ли можно удивляться обилию лиан в тропиках, имея в виду то, что подобная жизненная форма представляет большие преимущества в сомкнутом и очень высоком сообществе, каким является тропический дождевой лес. Лазящее растение быстро проникает в хорошо освещенные верхние ярусы леса при большой экономии материала, идущего на образование ствола, и, следовательно, способно очень быстро достичь зрелости.
Хотя на ранних стадиях своего развития лианы тропического дождевого леса, безусловно, подвергаются сильному затенению, однако огромное большинство их относится к явно светолюбивым растениям. Поэтому количество лиан всегда возрастает с увеличением освещенности. В девственном дождевом лесу нормальной сомкнутости лианы, хотя и присутствуют, встречаются не слишком часто; они становятся достаточно обильными, чтобы препятствовать продвижению лишь на хорошо освещенных местах, вроде вырубок или речных берегов. Вдоль многих тропических рек, особенно в Южной Америке, лианы образуют плотные завесы, совершенно скрывающие внутренность леса. При вырубке лесов лазящие растения всегда временно становятся и более обильными и более роскошными. Пространства, где были удалены крупные деревья, быстро заполняются непроницаемым переплетением лазящих растений, сквозь которые с трудом пробиваются молодые деревья; ненормальное обилие лазящих растений представляет собою один из признаков молодого вторичного леса. На плантациях лианы часто являются настоящим бичом.
В местности Морабалли-Крик, Брит. Гвиана, в лесу Ерегиа (Уал-лаба, стр. 263), произрастающем на грубых белых песках, лианы заметно малочисленнее, чем в лесах других типов, развивающихся на более тяжелых почвах в том же районе (Davis, Richards, 1933—1934, стр. 125). Такое же различие существует и между «боровым» лесом, растущим на белых песках (см. стр. 267), и смешанным лесом на глинистых и суглинистых почвах на острове Борнео (Richards, 1936, стр. 32). Сравнительная бедность лиан в лесах этих типов не связана с интенсивностью света, потому что в обоих случаях средняя освещенность была выше, чем в нормальном тропическом дождевом лесу, а, возможно, объясняется несколько
122
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
меньшей влажностью воздуха в подлеске. Так, в лесу Eperua. Морабалли-Крик, дефицит насыщения обнаруживает тенденцию быть больше, чем в других, произрастающих по соседству, типах леса (Davis, Richards, 1933—1934, табл. VI, стр. 376—377). Вероятно, на Борнео в «боровом» лесу влажность ниже, чем в смешанном. Возможно, что лианы в целом плохо противостоят сильному испарению в подлеске.
Лазящие растения обыкновенно классифицируются в зависимости от способа прикрепления их к поддерживающим растениям. С этой точки зрения они делятся на взбирающиеся, вьющиеся, лазящие при помощи придаточных корней и при посредстве прицепок. Взбирающиеся лианы не вьются, не имеют каких-либо специальных чувствительных органов прикрепления, хотя многие из них обладают приспособлениями, пассивно помогающими им взбираться — ветвями, расходящимися под прямым углом и загнутыми шипами. У вьющихся растений прицепки обыкновенно также отсутствуют, но верхушка молодых стеблей обладает вращательным движением в постоянном направлении, так что растение винтообразно закручивается вокруг своей опоры. Растения, лазящие при помощи воздушных придаточных корней, цепляются ими за поверхность, вблизи которой они произрастают. Лазящие растения, имеющие прицепки, наиболее специализированы, располагая органами, чувствительными к присутствию опоры; с их помощью растения активно прикрепляются к опоре, обыкновенно завиваясь вокруг нее. Прицепки имеют весьма разнообразную морфологическую природу. Некоторые лазящие растения могут быть отнесены одновременно к нескольким классам, так как они обладают сразу несколькими средствами прикрепления, например виды Hoy a (Asclepiadaceae), которые вьются и одновременно имеют на узлах цепкие придаточные корни.
С экологической точки зрения способ лазания менее важен, чем размер и максимальная высота, которой может достичь лазящее растение. Лазящие растения тропического дождевого леса распадаются на две не всегда резко отделяемые одна от другой группы: группу крупных, почти всегда деревянистых лиан, достигающих крон деревьев яруса В (реже яруса А) и, следовательно, во взрослом состоянии более или менее полно использующих солнечный свет, и группу гораздо более мелких, главным образом травянистых, лазящих растений, редко выходящих из тени подлеска. Эти две группы различаются не только по микроклимату, в котором они находятся, но и по их отношению к другим растением сообщества; их, возможно, следовало бы рассматривать как отдельные синузии. И в ту, и в другую группу входят лазящие растения всех четырех морфологических типов, но в различной пропорции. Растения, лазящие при помощи придаточных корней, например, сравнительно реже встречаются в синузии лиан, чем в синузии мелких лазящих растений.
Синузия крупных лиан содержит гораздо больше видов, чем синузия мелких лазящих растений. Большинство видов, входящих в первую синузию, принадлежит к различным семействам и родам двудольных. Здесь особенно хорошо представлены семейства Аппопасеае, Аросупасеае, Bignoniaceae, Combretaceae, Connaraceae, Convolvulaceae, Leguminosae, Menispermaceae, Sapindaceae. Среди однодольных лазящих растений наиболее важное место занимают пальмы. Ротанговые, или лазящие, пальмы встречаются во всех частях тропиков, но особенно обильны они в лесах Малайского архипелага. Здесь они отличаются исключительным богатством видового состава и играют большую роль в общем облике леса. Многочисленные кроны пальм, вкрапленные в общий полог леса,
Фото 10. Лес с многочисленными лианами, резервированный лес Шаша, Нигерия.
Видны корпи удушающего фикуса (Ficus sp.), прикрепленные к стволу большого дерева.
ото И. Каулифлория у лианы Pararistolochia flos-avis, резервированный лес Шаша. Нигерия
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 123
как правило, принадлежат не прямостоячим пальмам, а ротангам. В дождевом лесу Квинсленда лазящие пальмы, известные под названием «Lawyer vines», также представляют характерную черту леса. В лесах Южной Америки и Африки ротанги не столь обычны, а в глубине ненарушенного первичного леса Западной Африки они обыкновенно вообще отсутствуют, встречаясь главным образом по берегам рек и на болотах. Другими семействами, представленными среди лиан из числа однодольных, являются: Orchidaceae (Vanilla), Pandanaceae (Freycinetia)', следовало бы упомянуть и Агасеае, хотя большинство лазящих видов этого семейства принадлежит к синузии мелких лазящих растений. Некоторые виды Gnetum также относятся к лианам. Несколько видов лазящих папоротников, например виды из родов Stenochlaena, Teratophyllum, Lygodium, достигают достаточной высоты, чтобы быть включенными в эту же синузию.
Всходы и молодые растения крупных деревянистых лиан сначала бывают прямостоячими, и некоторое время их трудно отличить по внешнему виду от молодых деревьев или кустарников. Позже верхние междоузлия сильно удлиняются, и гибкий молодой ствол начинает искать опору. Молодые стволы нередко имеют крайние длинные междоузлия и очень маленькие листья; удлинение первых ускоряется, а последних задерживается; следовательно, лианы представляют собой растения с сильно этиолированными главными осями. Характер опоры, вероятно, не имеет большого значения; это может быть дерево, или кустарник, или ствол другого лазящего растения. Растения, лазящие с помощью придаточных корней, прикрепляются как к деревьям, так и к скалам. Фрейзе (Freise, 1939) отмечает, что в тропическом дождевом лесу южной Бразилии определенные виды лиан всегда ассоциируются с определенными видами поддерживающих их деревьев; это наблюдение нуждается в подтверждении. Высказывалось предположение относительно наличия подобной же связи между видами деревьев и живущими на них эпифитами (см. стр. 137). Вьющиеся растения могут взбираться на деревья лишь определенных диаметров, которые они способны обвить. Если поблизости нет подходящей опоры, то молодое растение склоняется книзу и ползет по земле, пока не найдет нечто, к чему оно может прикрепиться; в этом случае оно очень быстро начинает расти вверх, пока, если опора достаточно высока, оно не окажется на сильном свету в верхних ярусах леса. Малое количество лиан промежуточного возраста между стадией всходов и взрослыми растениями объясняется краткостью времени, необходимого для прохождения средних стадий роста. Некоторые крупные лианы, если им посчастливится прорасти на открытом, хорошо освещенном месте, где отсутствуют возможные опоры, остаются в виде низкого прямостоящего кустарника и могут цвести, достигнув в высоту немногим более 1 м. Это было замечено автором у Agelaea trifolia в Нигерии; внутри леса опа достигает 30 м и более; на заброшенном же поле, на краю фермы тот же самый вид встречается в виде прямостоящего кустарника без видимой тенденции к лазанию.
Главные стволы крупных деревянистых лазящих растений часто совершенно не ветвятся вплоть до достижения полога верхних ярусов, где сейчас же в изобилии появляются ветви и может образоваться большая крона. У многих видов наблюдается резкое различие между вегетативными и цветущими ветвями, с одной стороны, и сильно удлиненными лазящими стволами, с другой. Последние у взрослых лиан являются обыкновенно безлистными, ио у молодых они могут иметь или чешуйчатые листья, или нормальную листву. У многих лиан ветви кроны не обнару
124
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
живают стремления лазать; у них нет также прицепок или других специальных приспособлений для лазания, какие имеются у главного ствола. Последний по достижении взрослого состояния нередко также утрачивает свои специальные органы прикрепления. Несомненно, что такое изменение характера побега объясняется увеличением интенсивности света; лазящие стволы — это «теневые» побеги, а стволы кроны — «солнечные». Листья этих двух типов побегов иногда поразительно различаются (см. ниже стр. 125). Цветение обыкновенно начинается не раньше достижения лианами полога крон поддерживающих их деревьев.
Старые лианы нередко оказываются прикрепленными к дереву-опоре на значительной высоте, так что с вершин деревьев свисают вниз длинные, ничем не поддерживаемые стволы лиан. В таких случаях лиана первоначально должна была пользоваться опорой, которая затем сгнила и исчезла, или же она прикрепилась к стволу молодого дерева, которое своим последующим ростом подняло ее кверху.
Роль лазящих растений в структуре леса
Деревянистым лазящим растениям принадлежит значительное место в структуре леса. Кроны тропических деревьев, как уже указывалось (стр. 73), менее ветвисты и менее облиствлены, чем кроны соответствующего размера деревьев умеренной зоны. Лианы содействуют образованию сомкнутого полога и, следовательно, перекрывая разрывы между соседними деревьями, сильно увеличивают затененность. Они конкурируют с деревьями за пространство и свет, и нередко их присутствие обусловливает появление однобоких и уродливых крон. Вьющиеся растения иногда оказывают заметное сжимающее действие; если удалить такие растения со стволов упавших деревьев, то иногда на коре можно обнаружить глубокие спиральные борозды. Своей тяжестью лазящие растения могут ломать ветви поддерживающих их деревьев; тогда вся масса лианы соскальзывает вниз и может отмереть или начать вновь взбираться к пологу крон верхних ярусов.
Так как лазящие растения конкурируют с деревьями и влияют на симметрию их роста, то с лесоводственной точки зрения они считаются сорняками. В связи с этим подрезка стволов лиан является важной операцией в практике лесного хозяйства. Освещение полога в результате разреживания древостоя обыкновенно сильно стимулирует рост лазящих растений, и при таких условиях всегда имеется опасность полного заглушения подроста деревьев.
Синузия мелких лазящих растений, никогда не достигающих значительной высоты над почвой и совершающих свой жизненный цикл в тени дождевого леса, имеет гораздо меньшее значение; число видов таких растений невелико, и их участие в структуре леса ничтожно. Это по большей части вьющиеся растения, или растения, лазящие с помощью придаточных корней, или же сочетающие в себе оба этих типа приспособлений. Как правило, синузия мелких лиан в дождевом тропическом лесу представлена главным образом однодольными и папоротниками, но в некоторых хорошо освещенных типах леса вроде «борового» леса Борнео, довольно многочисленны двудольные лазящие растения. Здесь в синузию входит насекомоядное Nepenthes. Среди небольших однодольных лазящих растений наиболее важными являются Агасеае. В Южной Америке среди них иногда выделяются виды Carludovica (Cyclanthaceae); они взбираются при помощи воздушных корней на высоту 5—10 ле, а затем, утратив связь
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 125 с почвой, могут перейти в синузию теневыносливых эпифитов. Папоротники являются весьма постоянным членом синузии небольших лазящих растений; среди них наиболее важную роль играют роды Lomariopsis, Lygodium, Stenochlaena и Trichomanes.
Морфология и анатомия лиан
Стволы лазящих растений, в особенности крупных деревянистых лиан, обнаруживают большое разнообразие как по внешнему виду, так и по внутреннему строению. Они сильно изменяются по толщине и могут иметь круглое, угловатое или плоское поперечное сечение. Иногда они приобретают вид спирали или каната, иногда обладают крылообразными выступами или напоминают цепи. Некоторые из южноамериканских Bauhinia (Leguminosae), образно называемые гвианскими индейцами «следом черепахи», похожи на ленты с глубоко волнистыми краями. По своему внутреннему строению стволы лиан, как известно, обнаруживают необыкновенные аномалии вторичного роста. Свойственные им анатомические и морфологические особенности часто оказываются характерными для того или иного семейства или рода и облегчают их определение. Согласно Шенку (Schenck, 1892—1893)1, подробно описавшему анатомию лиан, структура ствола мало связана со способом лазания, и аномалии строения часто наблюдаются лишь у удлиненных лазящих стволов лианы, отсутствуя у коротких «солнечных побегов» ее кроны. У глав ного ствола ксилема обыкновенно состоит из ряда частично или полностью изолированных друг от друга сосудисто-волокнистых пучков, вкрапленных в более мягкую ткань. Таким образом ствол лианы как по внутреннему строению, так и снаружи часто напоминает веревку или канат, соединяя гибкость с большой прочностью на растяжение. Оба эти свойства необходимы растению, если иметь в виду его особый образ жизни.
Насколько известно, у подземных корней лазящих растений нет каких-либо специальных признаков, но воздушным корням лиан часто свойственны особые черты. Например, у Агасеае могут быть корни двух видов: прикрепляющие и питающие. Первые растут более или менее горизонтально в направлении от света и прикрепляют растение к его опоре; последние обладают ярковыраженным положительным геотропизмом. Оба вида корней резко отличаются по внутреннему строению, причем механические ткани лучше развиты у корней прикрепляющих, а проводящие — у питающих.
Листья лазящих растений тропического дождевого леса обыкновенно относятся к мезофилльным и по своему строению приближаются к листьям, преобладающим в том ярусе деревьев, которого они достигают. Но по форме листья лиан отличаются от листвы дерева-опоры. Простые эллиптические или продолговато-ланцетовидные листья довольно редки; тропические лианы так же, как и все вообще лазящие растения, склонны образовывать короткие листья с наибольшей шириной у почкообразного или сердцевидного основания. Некоторые лазящие растения имеют щитовидные листья. Главная жилка обыкновенно дланевидно расходится от места прикрепления черешка. Вершина может быть заостренной или же тупой, но не редки и листья с хорошо выраженными кончиками-капельницами. Пластинка листа обыкновенно располагается под •большим углом к черешку.
1 Хорошее описание внешней морфологии лиан дал В. Троль (W. Troll, 1938).
126
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Описанный тпп листа настолько преобладает среди принадлежащих к разнообразным семействам лазящих и в особенности вьющихся растений, что трудно поверить в случайный характер этого явления. Даже при наличии сложных листьев, как это наблюдается у бобовых и представителей семейства Соппагасеае, отдельные листочки стремятся приблизиться к преобладающей форме простого листа. Шенк не приводит никаких доводов для объяснения преобладания этого типа листа, но Линдманн (Lindmann, 1900) высказал предположение, что широкий лист, наиболее расширенный у основания, имеет преимущество перед другими, обладая большой площадью транспирации (что названный автор считает необходимым ввиду сопротивления, оказываемого транспирации длинным тонким стволом), а также позволяя листу легко включаться в уже имеющуюся мозаику листьев.
Среда, в которой живет лиана на ранней стадии своего существования в подлеске, сильно отличается от той, которая ее окружает во взрослом состоянии (см. гл. 7); к тому же растение, как уже упоминалось, очень быстро переходит из одних условий в другие. Поэтому не удивительно, что у многих лиан имеется весьма ясно выраженное различие между листьями теневыми, или «батифилльными», и листьями солнечными, или «акрофилльными» (термины предложены Холтумом, Holttum, 1938 а). Такой тип гетерофиллии в резко выраженной форме наблюдается у некоторых растений, лазящих с помощью придаточных корней, например у некоторых Агасеае (Goebel. 1913, стр. 388—391), у некоторых видов Ficus, подрод Synoecia (Corner, 1939), и у Marcgravia. Оба вида листьев у названных растений настолько отличаются друг от друга, что иногда эти растения ошибочно относили к различным родам. Молодое растение Marcgravia имеет маленькие, прямоугольные листья мягкой текстуры с бархатистой поверхностью; они всегда тесно прижаты краями к коре дерева-опоры. Позже, когда растение достигнет полога верхних ярусов и начнет ветвиться, появляются листья совершенно иной формы и текстуры — удлиненные, склерофилльные, глянцевитые, свободно располагающиеся в пространстве. Молодые листья, несомненно, выполняют охранительную функцию, защищая находящиеся под ними воздушные корни, и, в частности, помогают удерживать вокруг них воду. В Гвиане автору приходилось наблюдать, что, как и следовало ожидать, сорванные молодые листья вянут гораздо скорее, чем взрослые.
Замечательные примеры гетерофиллии у лазящих папоротников приводит Холтум (Holttum, 1938 а, стр. 427—429). У видов Lomariopsis, Lomagramma и Teratophyllum батифиллы отличаются от акрофиллов почти так же, как у Marcgravia, причем интересно отметить, что различия между батифиллами разных видов выражены в гораздо большей степени, чем между акрофиллами, подобно тому, как листья растений травяного покрова тропического дождевого леса более разнообразны, чем листья деревьев и кустарников. По словам Холтума, «эти лазящие папоротники наилучшим образом пользуются средой; корни их располагаются во влажной почве леса и дают систему вай, приспособленную к условиям сильного затенения и увлажнения; в то же время, пользуясь своими сильными лазящими стеблями, они выбираются на более светлое место на стволах деревьев, которые иначе остались бы незанятыми, и тут образуют более крупные и более устойчивые бесплодные и плодущие вайи» (loc. cit. стр. 428—429).
Такие крайние случаи гетерофиллии нельзя считать уж очень обычными, но менее резкие различия между молодыми и старыми листьями
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 127
представляют обыкновенное явление как у лиан, так и у деревьев. Например, у некоторых южноамериканских видов Bauhinia молодые листья расщеплены до основания на две доли, причем каждая из них вытягивается в длинный кончик-капельницу, тогда как взрослые листья имеют цельную листовую пластинку и оканчиваются просто острым кончиком.
Что касается цветков и плодов лазящих растений, то лишь немногие их признаки представляют интерес для эколога. Можно, например, упомянуть, что некоторые лианы, например различные Menispermaceae и западноафриканские виды Pararistolochia (Aristolochiaceae (фото И), обладают каулифлорией, так как их цветки развиваются только на неразвет-вленной, лишенной листвы части ствола. Виды с каулифлорией в количественном отношении, вероятно, составляют среди лиан приблизительно ту же часть, как и среди деревьев; по-видимому, и в том, и в другом случае может быть предложено одно и то же объяснение этого явления.
«РАСТЕНИЯ-УДУШИТЕЛП»
«Удушители» представляют собою одну из наиболее замечательных экологических групп тропического дождевого леса. Эта жизненная форма не имеет аналогов в европейских лесах. К ней принадлежат растения, начинающие свою жизнь как эпифиты, а позже опускающие вниз, в почву, корни и становящиеся независимыми или почти независимыми, нередко полностью заглушая дерево, служившее им опорой. Хотя и невозможно провести резкую границу между удушителями и полуэпифитамп (см. стр. 144), также посылающими в почву корни, но никогда не приобретающими независимости в смысле способности поддерживать себя, однако обе эти группы совершенно различны по своему значению в общей структуре леса. Удушители образуют синузию, располагающуюся на границе между зависимыми и независимыми растениями; большинство из них относится к родам Ficus (Могасеае), Schefflera (Araliaceae) и Clusia (Gut-tiferae). К подобному же типу относятся южноамериканские виды Cous-sapoa (Могасеае), Posoqueria (Rubiaceae) и некоторые виды Metrosideros (Myrtaceae).
В тропических дождевых лесах Африки, индо-малайской области и Австралии обильно представлены «фикусы-удушители» (Ficus spp.) как по количеству видов, так и индивидуумов, играя значительную роль в общем облике леса. Их семена прорастают на больших деревьях высоко над почвой, часто в развилине между стволом и крупной ветвью. Проросток кустится, посылает книзу воздушные корни, обладающие положительным геотропизмом, из которых некоторые держатся близ ствола дерева-хозяина, а другие свисают перпендикулярно вниз наподобие канатов; с течением времени эти корни достигают почвы и разветвляются в ней. Опускающиеся корни ветвятся и анастомозируют, пока ствол поддерживающего их дерева не окажется заключенным в сеть, состоящую из крайне крепких деревянистых переплетений; крона фикуса тем временем становится большой и тяжелой. Спустя известное время дерево-хозяин обыкновенно отмирает и сгнивает, оставляя на своехМ месте фикус, который хотя и не имеет главного ствола, но является совершенно независимым деревом. Непосредственная причина гибели «хозяина» неясна, но, по-видимому, она связана с затенением и может быть отчасти со сжиманием дерева фикусом, а также с конкуренцией со стороны его корней. Фикусы различаются по степени разрушительного воздействия
128
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
на своих «хозяев» (Corner, 1940, стр. 664—665). Иногда они становятся деревьями большого размера, относящимися к самому высокому ярусу леса.
По наблюдениям автора, в южноамериканском тропическом дождевом лесу удушающие виды фикуса немногочисленны, да и те представляют собой скорее полуэпифитные кустарники, постепенно укореняющиеся в почве, чем настоящие удушители. Наибольшее значение среди растений-удушителей этого района имеет род Clusia, представленный, по крайней мере в Гвиане и Вест-Индии, многочисленными видами, очень обильными по числу индивидуумов. Их всходы, подобные всходам удушающих фикусов, появляются высоко над землей на деревьях первого или второго ярусов. Воздушные корни опускаются вниз к почве и анастомозируют; дополнительные короткие корни-перемычки прочно прикрепляют растение к поддерживающему дереву. Клузии образуют большие кроны, но не создают такой плотной сети воздушных корней, как фикусы-удушители, и не всегда становятся самостоятельными растениями. Они редко полностью заглушают своих хозяев, хотя часто сильно деформируют и угнетают их крону. Интересное детальное описание вест-индской Clusia rosea дал Шимпер (Schimper, 1888, стр. 56—ВО).
ЭПИФИТЫ
Эпифиты тропического дождевого леса растут, прекрепляясь к стволам и ветвям деревьев, кустарникам и лианам, а некоторые — даже к поверхности живых листьев. В сомкнутом лесу эпифитный образ жизни является единственным выходом для растений, соединяющих в себе малые размеры со сравнительно высокими требованиями в отношении света; однако, получая преимущество хорошего освещения, они вынуждены мириться с ненадежным водоснабжением и отсутствием почвы. Небольшое количество эпифитов растет в подлеске дождевого леса, где условия освещения такие же или немного лучше, чем на уровне почвы. Это те эпифиты, которые не переносят конкуренции корней или опасности быть заглушенными непрерывным опадом мертвых листьев, или же те, корни которых нуждаются в лучшей аэрации, чем корни растений травяного покрова. Эпифиты имеют в высшей степени специализированный образ жизни. Поэтому эпифиты из числа цветковых растений и папоротников тропического дождевого леса сильно отличаются как по внешнему виду, так и по физиологии от растений травяного яруса, с которыми они и не имеют близкого таксономического родства. Шимпер (Schimper, 1888) считал, что в целом тропические эпифиты произошли от наземных растений, росших во влажных тенистых лесах. Однако резко выраженное отсутствие теневыносливости большинства эпифитов свидетельствует против такого взгляда, и Питтендрай (Pittendrigh, 1948) недавно представил сильные аргументы в пользу теории (первоначально предложенной Tietze), согласно которой представители Bromeliaceae, крупнейшего семейства американских тропических эпифитов, произошли от наземных растений, произраставших в полупустынных условиях. То же самое, по-видимому, верно и в отношении эпифитных Cactaceae, и, возможно, некоторых других групп цветковых эпифитов.
На подходящих открытых местах (голых скалах, морских берегах, в саваннах) многие эпифиты могут успешно расти на почве, но в лесу они целиком зависят от своих «хозяев», служащих им опорой. Типичные эпи
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 129
фиты не нуждаются в получении органической пищи от других растений, чем и отличаются от синузии паразитов. Только Loranthaceae (омелы), обильно представленные во всех тропических лесах, являются как полупаразитами, так и эпифитами.
Хотя эпифиты целиком зависят от других растений, сами они оказывают лишь незначительное воздействие на свою опору. Правда, дерево, тяжело нагруженное эпифитами, пожалуй, легче может быть опрокинуто сильным порывом ветра, чем дерево, их не имеющее; задерживая воду, эпифиты, возможно, содействуют нападению паразитических грибов на растение-хозяина, но в типичном тропическом дождевом лесу эпифиты редко бывают достаточно обильны, чтобы оказать серьезное влияние на деревья каким-либо из этих путей. Следовательно, хотя эпифиты и представляют характерный элемент в структуре леса, роль их в его жизни незначительна. Они интересны главным образом потому, что, с одной стороны, их распределение находится в тесной связи с экологическими факторами (микроклиматом, характером субстрата), а с другой стороны, благодаря своей необычайной структурной специализации, которая, возможно, больше, чем у любой другой группы растений, действительно заслуживает названия приспособления.
Эпифиты представляют интерес не только для ботаника; они заслуживают внимания и в том отношении, что являются основным, а в некоторых случаях и единственным местообитанием некоторых животных, играющих важную роль в экосистеме тропического дождевого леса. Корни эпифитных цветковых растений и папоротников являются одним из главных мест, где гнездятся древесные муравьи. Многие Bromeliaceae содержат большое количество перегноя и воды в своих розетках из перекрывающих друг друга листьев, служащих местообитанием разнообразной фауны. Сухопутные животные живут в перегное поверх уровня воды, а сама вода служит местом, где разводятся лягушки, большое количество личинок москитов и другие водные насекомые. В тропической Америке Bromeliaceae так обильны, что их «водоемы» образуют настоящие «воздушные болота» (Picado, 1912, 1913)1. Поскольку москиты, размножающиеся на Bromeliaceae, включают и переносчиков малярии, такие «болота» играют важную роль в распространении этой болезни.
Эпифиты всегда присутствуют в тропическом дождевом лесу, но количество их весьма изменчиво. Часто на каком-либо определенном участке наблюдается большее видовое разнообразие и большее число экземпляров эпифитов по сравнению с травянистыми напочвенными растениями. Обилие и разнообразие эпифитов составляет одну из наиболее ярких отличительных черт тропического дождевого леса от лесов умеренной зоны. Наряду с водорослями, грибами и мхами эпифитная флора дождевого леса содержит большое количество папоротникообразных и цветковых растений. Именно присутствие этих сосудистых эпифитов особенно отличает тропический дождевой лес от сообществ растений умеренной зоны. В Англии папоротник Polypodium представляет собой единственное высшее растение, являющееся настоящим эпифитом, хотя в некоторых случаях и здесь в качестве эпифитов могут успешно расти на деревьях и даже достигать зрелости различные цветковые растения. Почва — нормальное местообитание таких цветковых растений, и их присутствие на деревьях всегда объясняется чем-то более или менее слу
1 Имеются в виду скопления воды на самих растениях из семейства Bromeliaceae, например в пазухах листьев,— Прим, ред,
п. У. Ричардс
130
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
чайным. По терминологии Оливера (Oliver, 1930), такие растения называются «случайными», или «эфемерными», но не «типичными» эпифитами. Лишь Polypodium, хотя он часто растет па скалах или на почве, достигает взрослого состояния и плодоношения только на деревьях и, следовательно, может называться «типичным» эпифитом. Огромное большинство эпифитов тропического дождевого леса — «типичные»; «случайные», или «эфемерные», эпифиты встречаются здесь редко, что, может быть, объясняется бедностью и специализированной природой напочвенной флоры. Среди эпифитов тропического дождевого леса насчитывается несколько кустарников, достигающих значительных размеров (например, некоторые Ericaceae и Melastomaceae); присутствие этих кустарниковых эпифитов рассматривалось Шимпером (Schimper, 1935, стр. 403) как черта, отличающая тропический дождевой лес от других тропических лесных формаций.
Обилие эпифитов как сосудистых, так и бессосудистых характерно для горного и субтропического дождевого леса — двух наиболее близких к тропическому дождевому лесу растительных формаций. В таких более холодных и, возможно, обладающих более постоянной влажностью сообществах, например в горном лесу Восточных Гималаев, районе горного массива Кинабалу1 и в других горах Малайского архипелага, эпи-фитная растительность, хотя и менее богатая видами, достигает такого пышного развития, какое редко приходится наблюдать в тропическом дождевом лесу низменностей. Сведения о лесах, в которых деревья склоняются вниз под тяжестью почти скрывающих их эпифитов, относятся вообще к горным и субтропическим лесам, а не к настоящему тропическому дождевому лесу.
Эпифитные цветковые и папоротникообразные растения
Эпифитные цветковые растения тропического дождевого леса относятся к большому числу родов, принадлежащих, однако, к очень ограниченному числу семейств. Шимпер (Schimper, 1888) перечисляет около 33 семейств и 232 рода эпифитов, из которых большинство находится в тропиках1 2. Кроме этого, Шимпер перечисляет приблизительно 20 родов папоротникообразных. Число эпифитных родов значительно увеличилось со времени исследований Шимпера как последующим делением известных тогда родов, так и открытием новых, но составленный им список семейств до сих пор считается достаточно полным. Наибольшее количество родов и видов эпифитов тропического дождевого леса содержат следующие семейства цветковых растений: Araceae, Bromeliaceae и Orchidaceae — из однодольных, и Asclepiadaceae, Cactaceae, Ericaceae (включая Vacci-nioideae), Gesneriaceae, Melastomaceae и Rubiaceae — из двудольных. Известно также несколько эпифитных видов Pandanus (Pandanaceae) и Solanum (Solanaceae). Эпифитные папоротникообразные дождевого леса включают, кроме многочисленных папоротников, виды плаунов — Lycopodium и псилотов — Psilotum.
Большинство семейств эпифитов встречается как в Старом, так и в Новом свете, но Bromeliaceae — один из наиболее обильных и хорошо представленных семейств тропической Америки — совершенно отсутст
1 Хребет Крокер, северная часть острова Борнео.— Прим. ред.
2 Несколько родов, включенных в шимперовский список, является полуэлифи-тами, которые со временем становятся независимыми. Они рассматриваются здесь как удушители (см. выше стр. 127).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА Ш
вует на других континентах1. Эпифитпые Asclepiadaceae ограничиваются восточными тропиками. Cactaceae, представленные в тропической Америке несколькими родами и многими видами эпифитов, в Старом свете обладают единственным родом Rhipsalis, встречающимся в тропической Африке, на Маскаренских островах и Цейлоне.
Обилие и богатство эпифитной флоры значительно меняется в разных географических областях тропического дождевого леса, а также в различных местах одного и того же района. Надо отметить, что даже при благоприятных условиях только небольшая часть деревьев типичного тропического дождевого леса несет сосудистые эпифиты. Приводимые ниже данные обследования деревьев на вырубленных участках смешанного дождевого леса являются, вероятно, типичными: в местности Морабалли-Крик, Брит. Гвиана, на двух участках из 193 деревьев высотой 5 м и более 30 (16%) имели сосудистые эпифиты, а на другом участке из 55 деревьев высотой 14 м и более эпифиты были найдены на 21 дереве (38 % } (Davis, Richards, 1933—1934, стр. 378). На горе Дьюлит, Саравак, эпифиты располагались на 13% деревьев одного участка и 11% —другого» (Richards, 1936, стр. 16). Два участка в Нигерии дали соответственно 15 и 24% (Richards, 1939, стр. 27). Число различных видов эпифитов на одном дереве редко бывает большим даже в самых богатых ими районах. Наибольшие цифры, зарегистрированные автором, достигают 15 для Брит. Гвианы и около 13 для Нигерии. В лесу Будонго, Уганда, Эггелинг (Eggeling, 1947, стр. 55) насчитывал на одном дереве 40—45 видов цветковых растений и папоротников; Лонгман и Уайт (Longman, White, 1917) зарегистрировали на одном, не очень богатом эпифитами дереве тропического дождевого леса в Квинсленде1 2 16 видов сосудистых эпифитов и 34 бессосудистых. Мы не располагаем сведениями относительно числа индивидуумов эпифитов для тропиков, но в Новом Южном Уэльсе (цит. Longman, White, 1917, стр. 64) Турнер (Turner) насчитал более 200 экземпляров эпифитных орхидей на одном дереве.
Из основных областей распространения тропического дождевого леса Америка наиболее богата, а Африка наиболее бедна эпифитами. Хотя, конечно, было бы рискованно делать обобщения, основываясь на сведениях, касающихся какого-нибудь одного ограниченного района, однако следующие данные довольно ярко иллюстрируют упомянутое выше положение. В местности Морабалли-Крик, Брит. Гвиана, на территории, занимающей лишь несколько квадратных километров, Оксфордская экспедиция собрала 98 видов эпифитных цветковых растений п 34 эпифитных папоротника; считают, что действительное количество зна^ чительно больше. На несколько большей площади в южной Нигерии удалось обнаружить лишь 21 сосудистый эпифит (включая папоротники); предполагается, что в действительности их насчитывается не более 353. Оба эти района, вероятно, вполне типичны для низинного тропического дождевого леса соответствующих областей и вполне сравнимы во многих отношениях. Исследование областей распространения дождевого леса
1 Шевалье (Chevalier, 1938 Ъ) сообщает о нахождении сухопутных видов Pit-cairnia в Гвинейской республике.
2 В горных и предгорных дождевых лесах на каждом дереве присутствует гораздо большее число видов эпифитов. Бюннинг (Biinning, 1947) нашел более 50 видов только папоротникообразных эпифитов на одном дереве в лесу (по-видимому, предгорном)-в горах Суматры.
3 Эти цифры могут дать преувеличенное представление о бедности африканского леса эпифитами. В лесу Будонго (Уганда), занимающем площадь 352 кв, км, Эггелинг (Eggeling, 1947, стр. 55) нашел около 100 видов сосудистых эпифитов.
9*
132
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
в тропической Азии или Австралии даст, вероятно, промежуточные результаты.
Эти региональные различия в количестве видов эпифитной флоры не связаны с различиями процента деревьев, заселенных эпифитами, пли с различиями в числе эпифитов, находимых на дереве, что вполне подтверждается приведенными цифрами. Местные различия в богатстве и обилии эпифитной флоры явно связаны с климатическими факторами, но непосредственная причина этих региональных различий менее ясна. Вследствие зависимости водоснабжения эпифитов от непосредственного выпадения атмосферных осадков развитие богатой эпифитной растительности требует в первую очередь равномерного их распределения, сопровождаемого низким значением испарения. Такие условия наблюдаются во всей зоне развития тропических дождевых лесов, хотя известно, что большей части африканских дождевых лесов свойствен более продолжительный и более ясно выраженный сухой климатический сезон. Сравнительная бедность африканского леса эпифитами, возможно, и объясняется климатическими причинами, но вряд ли эти причины могут быть положены в основу различий, существующих между обилием эпифитов в тропиках Старого и Нового света вообще; тут, по-видимому, сказывается действие каких-то исторических причин. Большее богатство эпифитов в Новом свете в значительной мере обусловливается дополнительным присутствием многочисленных Bromeliaceae, развивающихся наряду с эпифитами из пантропических семейств. По каким-то причинам в Старом свете не возникло семейства, способного играть роль Bromeliaceae, которые часто являются пионерами и процветают в условиях, непригодных для других эпифитов.
Местообитание и физиология эпифитов
По характеру местообитания эпифиты во многих отношениях отличаются от растений, произрастающих на почве. Уже тот факт, что субстрат (большие ветви и стволы), на котором развивается растение, поднят над поверхностью почвы и оказывается наклоненным под разными углами или даже принимает вертикальное положение, ставит семена в особые условия как в смысле их распространения, так и в смысле укоренения. Кроме того, этот субстрат обыкновенно бывает более или менее гладким, что затрудняет прикрепление к нему; правда, встречающиеся тут и там выемки и трещины представляют собой удобные места для расселения эпифитов.
Другой важной чертой местообитания является недостаток почвы. Эпифиты неизбежно находятся в зависимости от небольшого количества органических остатков и минеральных частиц, скапливающихся в трещинах и углублениях на дереве, или почвы, которую они сами собирают. Как известно, многие эпифиты («скобочные эпифиты», «гнездовые эпифиты») в состоянии накапливать почву, которую они всегда очень интенсивно осваивают. Корневые системы почти всех эпифитов являются прибежищем муравьев. Муравьи собирают опавшие листья, семена и всякого рода остатки, играя, вероятно, значительную роль в обеспечении эпифитов почвой. Почти вся «почва», имеющаяся в распоряжении эпифитов, состоит из перегноя, образующегося из мертвых остатков других растений. Перегной, скапливающийся на деревьях, имеет важное значение для эпифитов и как источник минеральных питательных веществ, и в силу его способности удерживать воду. Недостаток свободных неорганических
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 133
ионов, доступных для эпифитов тропического дождевого леса, еще усиливается выщелачивающим действием частых сильных ливней. Потребность в перегное у различных видов эпифитов сильно варьирует. В своем исследовании эпифитной растительности среднегорного леса Тьюбодас на Яве Вент (Went, 1940) показал, что некоторые виды, как, например, Pholidota ventricosa, па открытых местах произрастают на больших скоплениях перегноя, а в затененных они могут развиваться при малом его количестве; для этих эпифитов перегной, по-видимому, является не столько источником питания, сколько резервуаром для воды.
Кроме минеральных веществ, высвобождающихся при разложении перегноя, эпифиты, как правило, правда, в очень небольших количествах могут использовать минеральные вещества, попадающие на них вместе с пылью. Согласно Диксону (Dixon, 1882), листья крупных эпифптных папоротников из Нового Южного Уэльса имеют исключительно высокое содержание золы (более 12%), а в почве, собираемой ими, может быть до 62% «песка». Для роскошных колоний эпифитов на расположенных вдоль дорог деревьях (например, на «дождливом дереве» Samanea saman) минеральные вещества, поступающие с пылью, несомненно, имеют важное значение (Pessin, 1925, стр. 32—33), но этот источник питания едва ли может играть большую роль внутри леса, где воздух исключительно чист. Лаусберг (Lausberg, 1935) показал, что листья многих растений выделяют значительные количества кальция, калия и других элементов. Если деревья тропического дождевого леса ведут себя подобным же образом, то выделяемые ими минеральные вещества могут составлять существенное дополнение к скудному питанию эпифитов. В таком случае эти выделения должны определять специфичность соотношения, иногда наблюдаемую между видами деревьев и составом их эпифитной флоры (см. стр 137—138). Миэ (Miehe, 1911), исследовавший гумус с корней яванских эпифитов, не обнаружил каких-либо отклонений в микробиологических процессах, протекающих в них, от процессов, свойственных нормальным почвам. Хотя существование Azotobacter не было определенно доказано, не подтвердился и вывод об отсутствии азотфиксирующей способности. Были обнаружены нитрифицирующие и разлагающие целлюлозу организмы, функционирующие так же, как и в других почвах. Согласно Спаннеру (Spanner, 1939), все истинные эпифиты, по-видимому, получают азот в виде ионов аммония, в водной культуре эпифит Myrmecodia лучше развивался в растворах солей аммония, чем в растворах той же концентрации, но содержащих нитраты.
Относительно малый объем почвы, которым приходится довольствоваться корням эпифитов, в соединении с чрезмерным вымыванием и часто наблюдающейся высокой интенсивностью испарения приводит к тому, что эпифиты нередко оказываются в условиях острого недостатка воды, в чем и заключается одно из основных различий между их местообитанием и местообитанием наземной флоры тропического дождевого леса. Даже в тех местах, где дождь идет почти ежедневно, эпифиты могут терять большое количество воды в промежутках между ливнями. Так как в большей части тропических дождевых лесов засухи, длящиеся несколько дней или недель, не представляют собой редкости, то эпифиты, не сумевшие сохранить свой запас воды и к тому же не обладающие засухоустойчивостью, не смогут выжить. Во время засухи 1923 г. в северном Квинсленде погибло большое количество эпифитов (Herbert, 1935, стр. 350). Эпифит должен обладать способностью быстро поглощать воду и не расходовать ее слишком быстро. Отсутствие сосудистых эпифитов в боль-
134
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
шпнстве областей умеренного климата, несомненно, обусловлено не низкими температурами, но длительным периодом зимней «физиологической» засухи. В таких климатах могут процветать только более мелкие эпифиты из числа споровых растений — мхи, печеночники и лишайники — с их малой потребностью в воде и замечательной способностью переносить длительную сухость воздуха.
Местообитание эпифитов до известной степени сходно с местообитанием скальных растений. В обоих случаях недостаток почвенного субстрата сочетается с нерегулярным водоснабжением. Нередко и там и тут наблюдаются одинаковые условия освещения и атмосферной влажности. Поэтому не удивительно, что флора подобных мест весьма близка по составу и что некоторые тропические эпифиты встречаются также и на скалах.
Камерлинг (Kamerling, 1912), исследовавший транспирацию эпифитных орхидей, установил, что ее интенсивность была очень низка; это поддерживает общепринятое мнение, что эпифитам, как группе растений, всегда свойственна замедленная транспирация. Однако Спаннер (Spanner, 1939) в своем интересном исследовании по физиологии Myrmecodia и Hydnophytum показал, что эти два эпифита, внешне очень похожие один на другой и часто растущие на одном и том же месте, значительно •отличаются поведением устьиц и интенсивностью транспирации. Реакция устьиц, очень медленная у Hydnophytum, отличается быстротой .у Myrmecodia. Транспирация всего растения Hydnophytum достигает приблизительно лишь х/3 максимальной интенсивности ее у Myrmecodia. Результаты исследования позволили Спаннеру сделать вывод, что представление о низкой интенсивности транспирации у большинства эпифитов неправильно и что среди эпифитов, как и среди большинства других экологических групп растений, имеются значительные физиологические различия, в связи с чем и скорость транспирации может быть низкой или высокой. Осмотическое давление у эпифитов {Myrmecodia и Hydnophytum) было исследовано‘Сенном (Senn, 1913), Гаррисом (Harris, 1918) и сравнительно недавно Спанне ром (Spanner, 1939). В общем величины осмотического давления оказались низкими. Блюм (Blum, 1933) нашел, что у эпифитов среднегорного леса в Тьюбодас (Ява) осмотическое давление клеток листа у эпифитов с хорошо развитой водоудерживающей тканью равнялось 8,4 атм, а у растений, без водоудерживающей ткани — 16,4 атм.
Факторы, определяющие распределение эпифитов в лесу
Местообитание эпифита почти всегда имеет лучшие условия освещения и более низкую и более изменчивую влажность, чем местообитание растения, живущего в том же климате на земле; в любом сомкнутом растительном сообществе существует микроклиматический градиент, направленный вверх от поверхности почвы. В тропическом дождевом лесу средняя освещенность и средняя интенсивность движения воздуха возрастает от напочвенного покрова вплоть до самого высокого яруса деревьев. В главе 7 будет показана параллельность хода градиентов температуры и дефицита насыщения. Условия, создающиеся в более низких ярусах леса, отличаются постоянством и резко отличаются от условий верхних ярусов, приближающихся к условиям открытого пространства.
Различные виды эпифитов предъявляют неодинаковые требования к микроклимату. Это было показано, например, Визнером (Wiesner, 1907, стр. 135—138) в его обширных исследованиях, посвященных пове
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 135
дению эпифитов в отношении света, и Вептом (Went, 1940) в работе по це-нологии эпифитов в Тьюбодас, на Яве (рис. 19). Распределение некоторых видов ограничивается очень хорошо освещенными или очень тенистыми местами, в то время как другие обнаруживают значительную пластичность в этом отношении; третьи избегают как сильного света, так и глубокой тени. Главным образом этими видовыми различиями в жизненных требованиях или степенью выносливости и объясняются весьма заметные изменения в составе эпифитной растительности как по высоте, так и на различных деревьях. Вертикальные микроклиматические градиенты не могут быть постоянными и изменяются от места к месту, вследствие неправильной ярусности леса и различиям в густоте крон у разных видов и экземпляров деревьев. Поэтому внутри леса эпифиты не обладают правильной ярусностью, но образуют ряды расположенных одно над другим сообществ с весьма изменчивым вертикальным распределением. Иногда важное значение для эпифитной растительности приобретают и горизонтальные микроклиматические различия; так, в лесу Будонго, Уганда, эпифитная флора болотистых лесов отличается от эпифитной флоры типичного дождевого леса, и некоторые виды, например Angraecum infundibulare, встречаются только вблизи воды (Eggeling, 1947, стр. 56).
Различия в степени наклона и внешнпх особенностях разных участков одного и того же дерева оказывают влияние на эпифитную флору не только через посредство большей или меньшей возможности для расселения семян и спор, но и благодаря равноценным условиям освещения и испарения. В результате эпифитная растительность на вертикальном стволе должна отличаться от такой же растительности на горизонтальных ветвях. Крупные эпифитные папоротники, вроде Asplenium nidus и Л, africanum склонны предпочитать стволы ветвям, но многие эпифиты обнаруживают противоположное стремление. Наклон поверхности влияет также на скорость накопления перегноя и отток воды с дерева. Уже давно известно, что некоторые виды эпифитов являются «перегнойными» и встречаются только там, где скапливаются сравнительно большие его количества. Вент (Went, 1940) показал, что среди эпифитов леса Тьюбодас, Ява, имеются выраженные гумифилы, с одной стороны, и гумифобы, с другой; последние предпочитают голую кору и, видимо, избегают мест с большим скоплением перегноя, где в свою очередь поселяются характерные ассоциации. Некоторые эпифиты оказывают явное предпочтение путям стока воды на дереве; в Брит. Гвиане маленькая Urticularia sc him-peri растет только на толстых подушках печеночников в «дренажных каналах» под развилинами ствола деревьев.
На изолированных деревьях в тропиках, как и в более высоких широтах, эпифиты неравномерно распределяются вокруг стволов, размещаясь в зависимости от расположения отдельных частей дерева по отношению к солнцу, ветру и преобладающим осадкам (van Оуе, 1921). Внутри леса, как и следовало ожидать, такое правило нарушается, и, по наблюдениям автора, экспозиция даже самых верхних и наиболее открыто расположенных ветвей оказывает незначительное влияние на эпифитную растительность; в результате у деревьев отсутствует «покрытая мхом сторона», помогающая путнику определять направление1.
Эпифитная растительность на дереве зависит также от вида дерева и его возраста. Вент (Went, 1940, стр. 22), исследуя растительность Явы,
1 Как известно, в лесах умеренной зоны и особенно субарктической, эпифитные мхи обычно наиболее часто и в большем количестве встречаются на северной стороне древесных стволов.— Прим. ред.
%
Рис. 19. Встречаемость эпифитов в зависимости от степени освещенности в предгорном дождевом лесу, Тыободас, Ява (VVenl, 1940).
Ордината- ьетвсй в процентах к освещенности вне леса-
Свешенное™Г дТ„™'о ’ ,Hccywi,!c Различные виды эпифитов в зависимости от степени ; ,„о = ndu-.ula ongu.di/.dia и Appindicula ,ап,ит. Е.
lalum-Elaphogl,p,dv,l„tum. / Ц,„ sli,ldii = A, v-l.ynanll.ut ho, *ЦМИ. Trihos .o. um
/tulclii uni—41 srhyna ntlms pulclua.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 137
установил, что «ассоциация» эпифитов на молодых деревьях Altingia excelsa, имеющих гладкую кору, отличается от эпифитов на чешуйчатой коре более старых деревьев. Изменение характера коры — лишь один из факторов, обусловливающих изменение в составе эпифитов по мере роста дерева. Некоторые эпифиты могут подготавливать место для других. Например, Bromeliaceae благодаря их замечательной водоудерживающей способности (стр. 143) часто создают условия, благоприятные для роста других, менее выносливых эпифитов. Шимпер (Schimper, 1888, стр. 58) отмечает, что на острове Доминика удушитель, или полуэпифит, Clusia rosea, почти всегда начинает свое развитие у основания крупных эпифитных бромелиевых. Первыми поселенцами на дереве обыкновенно бывают водоросли и лишайники, затем появляются листостебельные мхи и печеночники и, наконец, папоротники и цветковые растения. Таким образом создается сукцессия эпифитов, но она является неопределенной и не очень постоянной. Когда эпифиты достигают слишком пышного развития, то вся их масса может свалиться вниз, на почву, и процесс заселения повторяется снова.
Часто наблюдается поразительное различие в обилии и флористическом составе эпифитной растительности, развивающейся на деревьях разных видов. Так, на крупном бамбуке Dendrocalamus и на других деревьях с очень гладкой и твердой поверхностью сукцессия эпифитов редко переходит стадию лишайников. В Уганде деревья с лохматой корой, вроде Entandrophragma utile, заселены сильнее, чем виды с гладкой корой; правда, орхидеи Ancistrochilus rothschildianus и Eulophiopsis lurida предпочитают стволы деревьев с гладкой корой (Eggeling, 1947, стр. 56). Часто среди деревьев тропических городов заметно выделяется по обилию эпифитов Samanea saman. Шимпер (Schimper, 1888) заметил, что на юге Соединенных Штатов Америки эпифит Epidendrum conopseum предпочтительно, хотя не исключительно, встречается на Magnolia. Считается, что на Филиппинах папоротник Stenochlaena areolaris встречается в качестве эпифита исключительно на Pandanus utilissimus (Copeland, 1907).
До последнего времени обычно считали, что различие эпифитной растительности на деревьях разных видов объясняется действием чисто физических факторов: степенью грубости коры, ее водоудерживающей способности и способом слущивания. Однако Вент (Went, 1940) в своей замечательной работе об эпифитах среднегорного дождевого леса в Тыо-бодас привел убедительные доказательства в пользу того, что не менее важную роль играют и химические факторы. Эпифитная флора многочисленных деревьев была обследована им при помощи полевого бинокля и 42-х кратного телескопа. Оказалось, что таким способом можно определить почти все виды сосудистых эпифитов. Обилие эпифитных видов каждого дерева заносилось на произвольно выбранную шкалу, после чего состав эпифитных «ассоциаций» анализировался на диаграмме, где учитывались порода деревьев, грубость коры и другие факторы. В результате были обнаружены очень большие различия между эпифитами разных видов деревьев и установлено, что, несмотря на тенденцию эпифитной флоры каждого вида деревьев изменяться по мере их роста, различие в составе эпифитов разных древесных пород оказалось большим, чем между индивидуумами одного и того же вида. В некоторых случаях набор эпифитов был столь характерен, что давал возможность определить вид. В частности, таким путем можно установить виды рода С as tanop sis. Анализ эпифитной флоры на деревьях-хозяевах не оправдывает
138
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
предположения, что ее неоднородность связана только с физическими свойствами коры, густотой кроны и т. п. Вент пришел к выводу, что эти различия в значительной степени зависят от химического состава коры или характера стока воды по ее поверхности. Можно было бы легко продемонстрировать присутствие растворимых веществ в этой воде, например танина у Castanopsis argentea. Интересно отметить, что виды эпифитов, растущие на больших скоплениях перегноя, не обнаруживают явной приуроченности к какому-либо одному виду деревьев.
О существовании специфической приуроченности эпифитной растительности к строго определенному дереву-хозяину, на какую указал Вент, раньше и не подозревали, хотя она может иметь характер закономерности. Существуют данные, указывающие на то, что отношения между эпифитными мхами, печеночниками и лишайниками областей умеренной зоны и видами деревьев также являются до известной степени специфическими.
Спнузии эпифитов
Сложное взаимодействие микроклиматических условий с особенностями субстрата, о котором мы только что говорили, обусловливает очень сложное распределение эпифитов в пределах данной лесной территории, но в общем, как уже указывалось, они образуют ряд довольно определенных расположенных одно над другим сообществ. Эти сообщества были описаны Шимпером (Schimper, 1888), назвавшим их ступенчатое расположение «этажами» (ярусами) — термином, могущим повести к недоразумениям. Здесь они будут рассматриваться как синузии, причем с практическими целями вполне достаточно различать лишь три из них: а) теневые эпифиты, б) солнечные эпифиты и в) крайне ксерофильные эпифиты.
Теневые эпифиты располагаются главным образом в ярусе С; на стволах и ветвях деревьев яруса С, на молодых деревьях ярусов А и В и на стволах крупных лиан. Сосудистые эпифиты встречаются лишь изредка на нижней части стволов крупных деревьев. У очень крупных густо-облиствленных деревьев яруса А синузия теневых эпифитов иногда вновь появляется над первой развилиной в центре кроны. Это сообщество живет в условиях глубокого затенения в атмосфере с низким, мало изменяющимся дефицитом насыщения — то есть в микроклимате, мало отличающемся от микроклимата травяного яруса. Сообщество теневых эпифитов, как правило, и в основном состоит из папоротников, среди которых обычны папоротники с листьями, покрытыми проницаемой для воды кутикулой. Орхидеи и другие цветковые растения редко встречаются в изобилии, но в Брит. Гвиане компонентами этой синузии являются крошечная Peperomia emarginella и орхидея Cheiradenia cuspidata (которую всегда находят цепляющейся за крупные стволы на высоте приблизительно 1—2 м от земли). На Малаккском полуострове, в особенности на деревьях Saraca, растущих по берегам рек и ручьев, иногда в изобилии произрастают теневые эпифиты, включающие Hedychium longicor-nutum, Medinilla spp., Lycianthus parasitica, и многие маленькие орхидеи (Corner). *
Большинство теневых эпифитов не обнаруживает следов ксеромор-фности; напротив, они имеют в высшей степени гигроморфное строение. Их водный режим малоизучен. Шимпер (Schimper, 1935, стр. 24) утверждает, что у эпифитных папоротников с тонкими листьями, растущих
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 139
в тени, вроде папоротников с гигроскопической поверхностью листа, как потеря, так и поглощение воды незначительны, и суточный ход транспирации близок к ходу испарения. Осмотическое давление клеток корпя и листьев низко (обыкновенно не более 4—6,5 атм). Как известно, упомянутые выше папоротники имеют проницаемую для воды кутикулу; клетки могут пропускать сквозь свои стенки большое количество воды и также быстро утрачивать ее; поэтому вайи закручиваются в сухую погоду и распускаются вновь при возрастании влажности. Водный режим этих папоротников похож таким образом на водный режим мхов и печеночников.
Синузия солнечных эпифитов, как правило, представляет собой самое богатое как по количеству видов, так и индивидуумов сообщество среди эпифитной флоры. Она располагается преимущественно в центре крон и на более крупных ветвях деревьев ярусов А и В. На очень крупных деревьях солнечные эпифиты иногда опускаются вниз по стволу ниже первой развилины. Они живут в микроклимате, промежуточном между микроклиматом подлеска и открытого пространства, и затеняются в гораздо меньшей степени, чем теневые эпифиты. Они находятся в условиях сравнительно высокого и очень колеблющегося дефицита насыщения. Эта синузия включает много папоротников и очень большое число видов орхидей и других цветковых растений.
Солнечные эпифиты весьма различаются по образу жизни (см. стр. 128— 129) и структуре. Очень многие из них являются то в меньшей, то в большей степени ксероморфными. По своему водному режиму эти эпифиты, вероятно, почти так же изменчивы, как и в отношении структуры. Здесь трудно делать какие-либо обобщения, учитывая скудость существующих сведений (см. Schimper, 1935, стр. 24—29). Многие из них обнаруживают способность выдерживать большую потерю воды; некоторые виды могут терять без вреда более 50% своего сырого веса. Осмотическое давление в клетках листьев обычно значительно выше, чем у теневых эпифитов (Faber, 1927; Senn, 1913).
Крайне ксерофильные эпифиты живут на самых верхних сучьях и ветках высоких деревьев; на исключительно крупных по высоте деревьях они могут занимать большую часть кроны. Они почти полностью открыты для действия солнца и ветра и живут в условиях, близких к условиям открытого места. Так как едва ли какое-либо количество перегноя или растворенных солей может быть смыто сверху и достигнуть их, то им приходится существовать в условиях постоянного недостатка питательных веществ. Эта синузия содержит сравнительно мало папоротников и состоит главным образом из цветковых растений высоко специализированного типа. Например, в Морабалли-Крик, Брит. Гвиана, главными представителями этой синузии являются узколистные виды Tillandsia, вроде Т. bulbosa (Bromeliaceae), Aechmea spp. (Bromeliaceae), Codonanthe crassijolia (Gesneriaceae), Rhipsalis cassytha (Cactaceae) и отдельные орхидеи. Некоторые из этих видов являются суккулентами, и все они обнаруживают более или менее заметную ксероморфность. Водный режим компопентов этой синузии не был изучен, но, вероятно, в эту группу должны войти некоторые из солнечных эпифитов и «эпифитов, растущих в местах обитания с крайними условиями», о водном режиме которых говорит Шимпер (Schimper, 1935, стр. 24—29)1.
1 См. также работу Меца (Mez, 1904) о водном режиме «крайних атмосферных» тилландсий.
140
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Вертикальное распределение синузий эпифитов
Хотя в распределении эпифитов нет правильной ярусности и их вертикальное размещение меняется от дерева к дереву, однако средняя вертикальная амплитуда синузий имеет тенденцию быть постоянной в пределах одного типа леса, изменяясь с изменением последнего. В сообществе, где ярусы А и В образуют плотный полог, средняя вертикальная амплитуда расселения эпифитов всегда высока, а в более открытых типах леса она понижается. Это с поразительной отчетливостью выявляется в пяти типах первичного леса в Морабалли-Крик, Брит. Гвиана. Средняя освещенность подлеска (в процентах к полному дневному свету)1 в этих типах леса характеризуется следующими цифрами: лес Мора — 1,33; лес Морабуцея — 0,67; Смешанный лес — 0,67; лес Гринхарт — 0,81; лес Уаллаба — 1,43. В соответствии с ними находится и средняя высота расположения эпифитов. В трех слабоосвещенных типах леса (лес Морабуцея, Смешанный лес и лес Гринхарт) с земли видно лишь небольшое количество эпифитов; теневые синузии развиты мало, а типичные солнечные эпифиты, вроде Tillandsia spp., редко встречаются ниже приблизительно 18 м над уровнем почвы, за исключением отдельных экземпляров, которые не цветут (нижний предел высоты, где были встречены взрослые растения, составлял 9 лс). В более освещенных типах леса (Мора и Уаллаба) эпифиты спускаются гораздо ниже, и цветущие индивидуумы Tillandsia spp. могут быть встречены на высоте 3 ле от почвы. Было также замечено, что в затененных типах леса солнечный эпифит, случайно упавший с дерева на землю, почти всегда вскоре погибает, в та время как в более освещенных типах леса он продолжает процветать и на земле. В лесу Уаллаба иногда встречаются экземпляры Tillandsia, видимо, укоренившиеся в почве; при тщательном исследовании их на почве всегда обнаруживались остатки сгнившей ветви.
Подобно этому на острове Борнео эпифиты в хорошо освещенном «боровом» лесу (стр. 267) растут на более низких уровнях, чем в более темном смешанном лесу. В первом они иногда развиваются фактически на почве; так, в одном пункте был найден Asplenium nidus, укоренившийся в песчаной почве на дороге, проходящей через «боровой» лес (Richards, 1936, стр. 32). На Борнео на песчаном участке, поросшем кустарником, Б. Полак (В. Polak, 1933, стр. 26) обнаружил растущие в почве виды Bulbophyllum и другие эпифиты.
Во всех частях тропиков на деревьях вдоль рек и на отдельных деревьях, одиноко стоящих среди возделываемых полей, эпифиты располагаются на гораздо меньшей высоте над почвой, чем внутри леса.
Вертикальное распределение эпифитов в большей степени связано с интенсивностью освещения, чем со средней влажностью воздуха, которая изменяется в лесу более или менее обратно пропорционально яркости света (Davis, Richards, 1933—1934, стр. 382). Питтендрай (Pittendrigh, 1948), исследовавший вертикальное и географическое распространение Bromeliaceae в лесах и на плантациях какао острова Тринидад, также пришел к выводу, что основным фактором является свет.
1 На высоте груди по показаниям фотографического экспозиметра (типа актинометра).
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 141
Экологическая морфология
В отличие от других синузий тропического дождевого леса эпифиты обнаруживают мало единообразия как в форме листьев, так и в морфологии. Многим из них, однако, свойственна в высшей степени интересная и разнообразная специализация, очевидно, связанная с образом их жизни и определенно могущая рассматриваться как «приспособительная» в том смысле, что она полезна, а во многих случаях, вероятно, и необходима как часть растительного организма, развивающегося в специфических условиях местообитания. Однако эти приспособления развились не путем постепенного изменения организма в условиях среды, характерных для современного местонахождения эпифитов. Не вызывает сомнений, что по крайней мере некоторые из этих приспособлений, например водовместилища и гигроскопические волоски Bromeliaceae, были унаследованы от наземных растений, произраставших в полупустынных условиях. Следовательно, растения, от которых произошли эпифиты, были «заранее специализированы», что и дало им возможность развиться в эпифитную флору тропического дождевого леса (см. выше, стр. 128).
Структурные приспособления эпифитов впервые были подробно описаны Шимпером в его классической работе «Die epiphytische Vegetation Amerikas» (1888). Мы остановимся на них только вкратце, поскольку они описаны во многих учебниках; более подробная их характеристика дается Гебелем (Goebel, 1889) и Карстеном (Karsten, 1895, 1925). Можно также упомянуть работу по экологии тропических эпифитных папоротников Холтума (Holttum, 1938 а) и отослать желающих к списку литературы, имеющемуся в самом последнем издании шимперовской географии растений (1935).
Семена и всходы эпифитов обладают многими характерными признаками, связанными со способами их распространения и укоренения. Эпифиты, как впервые отметил Шимпер, почти все без исключения имеют переносимые ветром споры, плоды или семена, приспособленные для распространения животными или ветром. Мясистые плоды, часто с клейкой мякотью, облегчающей прикрепление семян к коре, свойственны Rhip-salis (Cactaceae) и многим Bromeliaceae; семена последних нередко в большом количестве откладываются птицами на телеграфных проводах, к которым они легко пристают и иногда успешно прорастают. Споры папоротников и очень маленькие семена Aeschynanthus, Orchidaceae и т. д. хорошо приспособлены для переноса их ветром. Семя эпифитной орхидеи Stanhopea oculata весит только 0,000003 г (Ulbrich, 1928, стр. 153), а семя Aeschynanthus — 0,00002 г (Beccari, цит. Гебелем, Goebel, 1889, стр. 153). У различных Gesneriaceae, Rubiaceae, Bromeliaceae (подсемейство Til-landsioideae) встречаются семена с летучками. Единственным в своем роде способом распространения обладает Tillandsia usneoides, так называемый «испанский мох» субропической и тропической Америки1, семена у него образуются редко, а корни имеются только у всходов; растение расселяется с помощью ветра, который отрывает и переносит похожие на бороду массы тилландсии или же при помощи птиц, использующих ее для постройки своих гнезд (Schimper, 1888).
Растения без специальных приспособлений для распространения семян, вероятно, были бы неспособны просуществовать как эпифиты; в то же время у эпифитов нет приспособлений, свойственных исключи
Он редко встречается в собственно тропическом дождевом лесу.
142
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
тельно им. Как указал Шимпер, наличие эпифптного образа жизни у растений различных семейств, на первый взгляд случайное, становится попятным, если принять во внимание, что растения всех семейств, включающих эпифиты, обладают подходящими приспособлениями для распространения семян. Семейства, вроде Leguminosae, чьи представители обладают тяжелыми семенами, не имеют эпифитов. С другой стороны, некоторые семейства, вроде злаков и сложноцветных (за очень редкими исключениями), не включают эпифитов, несмотря на наличие у их представителей соответствующих средств распространения; это, очевидно, объясняется цтсутствием каких-то других необходимых при эпифитном образе жизни свойств.
У Aeschynanthus и у полупаразитического Loranthus часть проростка образует сплющенный диск, снабженный многочисленными корневыми волосками, помогающими ему укорениться. Во взрослом состоянии большинство эпифитов укореняется при помощи очень развитой корневой системы, но у некоторых Bromeliaceae корни отсутствуют, и прикрепление осуществляется иным путем. У многих Агасеае, Orchidaceae и других корневая система подразделяется на корни питающие и прикрепляющие, резко различные как по характеру тропизма, внешнему виду и анатомическому строению, так и по выполняемым ими функциям. У более специализированных Bromeliaceae, впитывающих воду главным образом своеобразными многоклеточными волосками на листьях, корни, имеющие вид шнурков или проволоки, являются весьма совершенными, органами прикрепления, но, вероятно, поглощают очень мало воды; Шимпер (Schimper, 1888, стр. 68) показал, что растения, у которых были обрезаны корни, сохраняли свой водный баланс на том же уровне, что и неповрежденные растения.
Модификация структуры эпифитов наиболее ярко и значительно проявляется именно в отношении к воде и к накоплению почвы. Шимпер различал четыре класса эпифитов (вдобавок к полупаразитическим эпифитам): протоэпифиты, гнездовые и скобочные эпифиты, резервуарные эпифиты и полуэпифиты.
Протоэпифиты не имеют специальных приспособлений для накопления воды или почвы и не опускают в почву воздушных корней. Они представляют собою наименее специализированную группу и в очень малой степени защищены от влияния периодических засух и недостатка почвы. Некоторые из них, например виды Peperomia, и многие папоротники имеют ползучие стебли, которые позволяют им использовать большую площадь субстрата. Многие протоэпифиты отличаются ксероморфной структурой и снабжены водоудерживающими органами различного типа. Виды Peperomia, Dischidia и другие имеют суккулентные листья; у многих орхидей наблюдаются раздутые междоузлия, известные под названием ложных луковиц. Корневые клубни характерны для малайских видов Vaccinium и Pachycentria. У некоторых протоэпифитов быстрому поглощению воды содействует специальная омертвевшая ткань — веламен, расположенная с наружной стороны воздушных корней. Клетки веламена во время дождя наполняются водой, а в сухую погоду содержат воздух и выполняют роль изолирующей прослойки, предохраняющей корень от излишнего нагревания и потери воды. Веламен особенно характерен для корней эпифитных орхидей, но особенно интересно то, что он встречается также и у небольшого числа напочвенных травянистых растений (Schimper, 1888; Goebel, 1922). Возможно, что эти растения являются эпифитами, возратившимися к жизни на почве, но всего вероятнее, что
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 143
веламен представляет собой морфологический признак, развившийся в наземной среде, и что, подобно некоторым способам распространения семян, он представляет собой один из признаков, наличие которого позволило некоторым видам растений приспособиться к эпифитному образу жизни. У гнездовых и скобочных эпифитов растение построено таким образом, что оно накапливает перегной и растительные остатки, из которых корни добывают воду и минеральные вещества. У гнездовых эпифитов, примерами которых является крупный папоротник восточных тропиков Asplenium nidus и многие другие папоротники, ароидные и орхидные, корни образуют плотно переплетенную массу, напоминающую птичье гнездо; среди этих корней обитает масса муравьев и постепенно скапливается перегной. У скобочных эпифитов все листья или часть их похожи на скобки. Широко распространенный род папоротников — Platycerium имеет два типа листьев, причем листья одного из этих типов, а именно листья покровные, образуют скобки. У видов малайского рода Conchop-hyllum (Asclepiadaceae) все листья выпуклы и несколько отступают от коры. В промежутке между листьями и корой скапливаются растительные остатки. Своеобразная Dischidia rafflesiana также напоминает скобочные эпифиты. Часть ее листьев, обладающих мешковидной формой, имеет воронкообразное отверстие. В этих мешках, где муравьи устраивают гнезда, постепенно скапливается почва; корни врастают в эту почву и поглощают из нее воду и питательные вещества.
По своей экологии тождественны скобочным эпифитам очень своеобразные мирмекофиты Myrmecodia и Hydnophytum. принадлежащие к семейству Rubiaceae. Несколько видов этих родов являются обычными членами синузий солнечных и крайне ксерофильных эпифитов в тропических дождевых лесах Малайского архипелага. Стебель этих причудливых растений образует крупный клубень, испещренный отверстиями-ячеями, в которых неизменно поселяются муравьи. Ячейки образуются без всякого участия муравьев, что доказывается их возникновением при отсутствии последних. Клубни в значительной мере состоят из водянистой ткани, и вряд ли можно сомневаться, что первоначально они функционировали как водоудерживающие органы; можно, правда, предположить, что растение поглощает питательные вещества, извлекаемые из экскрементов муравьев и перегноя, скапливающихся в полостях клубней. Однако растение совершенно не зависит от этого способа питания, так как было доказано, что оно развивается и без участия муравьев. Эти курьезные эпифиты не раз были предметом исследования [см. литературу в работах Шимпера (Schimper, 1935, стр. 253—255) и Шпаннера (Spanner, 1939)]. У некоторых эпифитных папоротников родов Lecano-pteris и Polypodium корневище уплощено, сильно раздуто и изрешечено полостями, в которых живут муравьи, как у Myrmecodia и Hydnophytum.
Эпифиты, имеющие вместилища, или резервуары для воды, представлены лишь одним семейством Bromeliaceae, представители которого в изобилии произрастают в лесах тропической Америки, составляя одну из характернейших их черт. У этих эпифитов, за исключением Tillandsia usneoides и подобных ей видов, длинные, узкие и жесткие листья, образующие розетку, настолько плотно заходят один за другой, что создают водонепроницаемый резервуар, который у больших растений может содержать до 5 л воды. У Т. bulbosa внутренние листья смыкаются поверх резервуара, так что он может удерживать воду при любом положении розетки; вода проникает через щели между листьями, и растение способно расти, даже будучи перевернутым «вверх ногами». В резервуар бромелие-
144
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
вых наряду с частицами перегноя попадают насекомые; вместе с экскрементами и остатками мертвых животных, выводящихся внутри резервуаров, они пополняют количество питательных веществ, находящихся в воде. Чешуйкообразные волоски на листьях, у многих видов образующиеся только у основания листьев, выстилающих резервуар, поглощают воду и растворенные вещества, но не позволяют воде просачиваться сквозь листья; таким образом волоски действуют как односторонний клапан {Trichompompe, Mez, 1904). Корни, как указывалось выше, часто выполняют только механическую функцию, при этих условиях волоски оказываются единственными абсорбирующими органами. Эта в высшей степени удачная система собирания и поглощения воды и питательных веществ делает бромелиевые в значительной мере независимыми от субстрата, что является одной из причин их пышного развития даже там, где могут утвердиться лишь немногие другие эпифиты. Эффективность этой системы доказывается и большими размерами, которых достигают некоторые Bromeliaceae. Например, у Glomeropitcairnia erectiflorac, острова Тринидад соцветие может достигать 9 футов (3 м) высоты (Pittendrigh, 1948, стр. 67).
Полуэпифитам — последнему классу эпифитов из установленных Шимпером — свойственны длинные воздушные корни, достигающие почвы. Как только устанавливается связь растений с почвой, для них уже не возникает затруднений в снабжении водой и питательными веществами, с которыми приходится встречаться большинству других эпифитов. Некоторые из них, например различные Агасеае, являются травянистыми, а некоторые, подобно тропическим Coussapoa fagijolia. представляют собой крупные кустарники и, как и следовало ожидать, мало отличаются, если не считать их курьезных корней, от растений, живущих в почве. Все они связаны промежуточными типами, с одной стороны, с удушите-лями (стр. 127), иногда становящимися независимыми растениями, а, с другой стороны, с нормальными деревьями. Так, малайская Pyrus granulosa может в зависимости от условий быть независимым деревом или эпифитом со спускающимися корнями, находясь как бы на полпути между растением-удушителем и настоящим полуэпифитом (Corner, 1940, стр. 529).
Бессосудистые эпифиты
Лишайники, водоросли, мхи и печеночники характерны для эпифитной растительности тропического дождевого леса не в меньшей степени, чем сосудистые растения. Некоторые из этих бессосудистых растений встречаются в тесной ассоциации с папоротниками и цветковыми растениями и могут рассматриваться как составная часть одной и той же синузии; что касается размеров, то более крупные мхи и печеночники не уступают эпифитным гигрофильным папоротникам и конкурируют с ними на равных основаниях. Однако ради удобства в этом разделе будут охарактеризованы все виды бессосудистых эпифитов.
В тропическом дождевом лесу все лишайники и почти все мхи, печеночники и водоросли являются эпифитами. Как указывалось в главе 2, в лесу на почве обыкновенно отсутствует ярус мхов, хотя на очень крутых склонах и высоко в горах на поверхности почвы можно встретить некоторые мохообразные. Основная причина отсутствия напочвенных мхов, вероятно, заключается в угнетающем действии постоянного опада мертвых листьев; это подтверждается тем, что вообще мхи располагаются на стволах деревьев вплоть до уровня почвы, а также встречаются на кам
СТРУКТУР/Х II ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 145
нях и поваленных деревьях, с которых опадающие листья легко соскальзывают. На участках с недавно нарушенным почвенным покровом, где еще не успел накопиться толстый слой опавших листьев, например на гнездах термитов, на земле, выброшенной роющими животными, или на корнях поваленных деревьев также часто поселяются маленькие мхи (в особенности Fissidens spp.) и печеночники.
Кроме сообществ («ассоциул» Клементса, Clements, 1936), образуемых бессосудистыми растениями тропического дождевого леса на камнях, гнилых стволах и древесных пнях, они встречаются в качестве теневых и солнечных эпифитов, а также эпифилльных организмов, развивающихся на живых листьях. Сообщество бессосудистых теневых эпифитов располагается па стволах и ветвях древесно-кустарниковой растительности подлеска, а также, подобно сосудистым теневым эпифитам, расселяется в кронах крупных деревьев с густой листвой. Бессосудистые солнечные эпифиты встречаются на ветвях деревьев ярусов А п В, часто распространяясь на более мелкие ветки. Эпифилльные организмы ограничиваются главным образом тенистым подлеском и редко достигают более высоко расположенных ярусов деревьев. Все эти сообщества брио-фптов, лишайников и водорослей подвержены сукцессионному развитию, и их можно найти как на сериальных, так и на климаксных стадиях развития.
В теневом сообществе доминируют бриофиты многих семейств (мхи, Lejeuneaceae, Plagiochila spp. и т. д.); водоросли и лишайники, встречающиеся в небольшом количестве, не играют в этих сообществах почти никакой роли, за исключением ранних стадий сукцессии. В отличие от соответствующих сообществ цветковых растений и папоротников теневое сообщество бессосудистых растений гораздо богаче видами, чем солнечное. Сучья и ветви кустарников и деревьев яруса С нередко так густо покрыты мхами и печеночниками, что подними совершенно невидно коры; в то же время стволы крупных деревьев редко бывают покрыты сплошным покровом мохообразных. Кроме мохообразных, растущих пучками или образующих своего рода ковер, то есть с довольно обычными во флорах умеренной зоны формами роста1, характерную черту тропического теневого сообщества бессосудистых растений составляют свисающие мхи, принадлежащие главным образом к семейству Meteoriaceae; их первичный стебель, ползущий вдоль ветки, образует повисающие в воздухе ответвления, достигающие у некоторых видов метра и более в длину. Наличие свисающих мхов, встречающихся только в тропическом дождевом лесу и других формациях влажных лесов, несомненно, связано с постоянной высокой влажностью, так как висячее положение представляет максимальные возможности для испарения. Мохообразные теневого сообщества имеют типичную гигроморфную структуру; для них свойственны широкие листья и (у большинства видов) очень большие клетки с тонкими стенками.
Бессосудистые солнечные эпифиты составляют сильный контраст по сравнению с теневыми эпифитами, и только очень небольшое число видов является общим для обоих сообществ. Лишайники обильны (включая крупные листовые формы, вроде видов Parmelid) и сохраняются вплоть до более поздних стадий сукцессии. Мхи (например, виды Macro-mitrium) и печеночники (Frullania spp., Lejeuneaceae) также встречаются
1 Формы роста мхов тропического дождевого леса Явы были описаны Гизенха-геном (Giesenhagen, 1910).
10 II. у. Ричардс
146
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
в большом количестве, но число видов внутри каждой конкретной области обыкновенно ограничено. Большинство мохообразных имеет компактную ползучую или пучковатую жизненную форму; свисающие виды отсутствуют или редки. Растения отличаются ксероморфной структурой; для них характерны маленькие в сухом состоянии тесно прижатые чешуйчатые листья, клетки с толстыми стенками и маленькой клеточной полостью. Различия между теневыми и солнечными мохообразными в пределах тропического дождевого леса напоминают различия между солнечными и теневыми эпифитами европейских лесов (Olsen, 1917), но более сильно выражены.
Наличие эпифилльных организмов (Busse, 1905) — в высшей степени характерная черта тропического, горного и субтропического дождевых лесов; вне тропиков и субтропиков они известны в вечнозеленых лесах Японии и островов Макронезии (Richards, 1932; Allorge, 1939). Для существования эпифилльных организмов необходимо наличие вечнозеленых листьев с большой продолжительностью жизни, развивающихся в условиях постоянно высокой влажности и температуры. В сравнительно сухих вариантах тропических дождевых лесов, вроде влажного вечнозеленого леса Нигерии, эти организмы отсутствуют, встречаясь лишь по берегам водотоков и на заболоченных местах. Как уже указывалось, эпифилльные организмы поселяются главным образом в нижних (С, 1) и Е) ярусах леса. Главным их местообитанием являются листья кустарников и небольших деревьев.
Сообщество эпифилльных организмов состоит из водорослей (например, видов Trentepohlia. Phycopeltis и т. д.), лишайников (например, Strigula и многих других родов) и листовых печеночников (главным образом Lejeuneaceae и видов Raduld)\ эпифилльные мхи (например, Crossomitrium spp.— тропическая Америка, Taxithelium spp.— Малайский архипелаг) встречаются относительно редко. Эпифилльные виды не склонны располагаться на коре, но мхи, растущие на коре, иногда впоследствии распространяются на листья, где они не могут обосноваться, как на первичном субстрате. Лишайники и водоросли заселяют листья в первую очередь, и в сравнительно сухих местах сукцессия замирает на этой стадии. Однако при более благоприятных условиях появляются печеночники, которые в дальнейшем обыкновенно вытесняют лишайники и водоросли. Со временем лист может покрыться густым войлочком эпифилльных мохообразных, в котором иногда прорастают семена цветковых растений (например, орхидей, Bromeliaceae), никогда, разумеется, не достигающих взрослого состояния. Как только лист опадает, жизни маленького сообщества приходит конец.
Эпифилльные организмы встречаются только на вечнозеленых листьях с жесткой консистенцией, отличающихся большой продолжительностью жизни, и, за исключением случаев, когда листья располагаются в вертикальной плоскости, эпифилльные организмы поселяются только на верхней поверхности листа. Их количество очень непостоянно и меняется в зависимости от типа листьев. Оказывая определенное предпочтение некоторым типам листьев, они избегают другие по причинам, не сразу понятным; по-видимому, это зависит или от характера кутикулы или же от веществ, выделяемых листьями. Эпифилльные организмы поселяются и на листьях тонко расчлененных (например, листьях Mimosoideae) или опущенных, хотя не могут образовать на них сплошного слоя. Как отмечалось в главе 4, не существует зависимости между наличием эпифилльных организмов и отсутствием кончиков-капельниц. Эпифилльные мохо-
Ф о т о 12. Сапрофит Leiphaimos aphylla на древесном пне, Морабалли-Крик, Брит. Гвиана.
Ф о т о 13. Thonningia sanguinea, паразитирующая на корнях дерева, резервированный лес (Наша. Нигерия.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 147
образные, подобно эпифитным цветковым растениям, рассмотренным ранее, не являются паразитами, поскольку какие-либо указания на то, что они получают органическую пищу от растений, на которых живут, отсутствуют. Они оказывают лишь незначительное влияние на выполняемую листьями ассимиляцию и, предупреждая быстрое высыхание поверхности листа, по-видимому, содействуют нападению грибных паразитов; однако, как указывалось в предыдущей главе, любое неблагоприятное воздействие эпифилльных мохообразных имеет, вероятно, ничтожное значение. Совсем иную роль играют некоторые эпифилльные водоросли и лишайники. Водоросль Cephaleuros virescens, хотя и обладает зеленой окраской, является, как известно, частичным паразитом и вызывает сильные заболевания чая и других растений. Другие эпифилльные водоросли проникают в ткани растения-хозяина и, безусловно, являются частичными паразитами, хотя бы только «пространственными паразитами». Фиттинг (Fitting, 1910), исследуя отношения эпифилльных лишайников к листьям, на которых они поселяются, нашел, что они распадаются на три группы: более или менее глубоко проникающие в ткани (а), например Strigula, у которой водорослевым компонентом является Cephaleuros virescens; проникающие в кутикулу, но не в эпидермис (Ь), не проникающие в кутикулу (с). Только группа (с) является абсолютно не паразитической. Наибольшее количество представителей насчитывает группа (Ь).
Эпифилльные организмы обнаруживают много интересных структурных модификаций, по-видимому, помогающих прикрепляться к субстрату. У многих эпифилльных печеночников существуют приспособления, позволяющие им удерживать большие количества воды с помощью особых капилляров, или между листьями и субстратом, или в «водяных мешках». Подробное описание строения эпифилльных организмов было дано Гёбелем (Goebel, 1888, 1889), Карстеном (Karsten, 1895) и Массартом (Massart, 1898).
САПРОФИТЫ
В тропическом дождевом лесу, как и в лесах умеренной зоны, огромное большинство сапрофитов принадлежит к низшим организмам — бактериям и грибам. Однако в нем обыкновенно присутствует синузия небольших сапрофитных цветковых растений1. Эти цветковые растения, хотя и широко распространенные по наблюдениям автора, никогда не растут в изобилии, и иногда высказываемое утверждение, что в дождевом лесу цветковые растения замещают сапрофитные грибы, является, конечно, преувеличением; богатство тропического дождевого леса сапрофитными грибами обыкновенно сильно недооценивается благодаря сезонному появлению их плодовых тел (Corner, 1935). Хотя роль сапрофитных цветковых растений в жизни леса, несомненно, крайне незначительна, они все же представляют интерес с нескольких точек зрения.
Это сообщество образовано очень ограниченным числом семейств. Из однодольных здесь находятся Burmanniaceae (Gymnosiphon, Thismia и другие роды), Liliaceae (Petrosavia — только в пределах Малайского архипелага), Orchidaceae (различные роды) и Triuridaceae (Sciaphila, Triuris); из двудольных — Gentianaceae (Leiphaimos, фото 12, Voyria
1 В Брит. Гвиане небольшой папоротник (Schizaea jluminensis) часто ассоциируется с сапрофитными цветковыми растениями и, возможно, сам является частичным сапрофитом (Davis, Richards, 1933—1934, стр. 372).
10*
148
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и т. д.) и Polygalaceae (Epirrhizanthes). Некоторые из этих родов являются пантропическими. Многие из этих маленьких растений имеют причудливую внешность. Их морфологией, анатомией и циклом развития усиленно занимались многие ученые (Johow, 1885, 1889; Ernst, Bernard и др., 1910—1914). Большинство этих растений является холосапрофитами; они почти или полностью лишены хлорофилла, но по крайней мере некоторые орхидеи являются частичными сапрофитами (они содержат некоторое количество хлорофилла и, по-видимому, способны к фотосинтезу в ограниченных размерах. Эти растения бывают белого (Gymnosiphon), ярко-желтого (Leiphaimos aphylla), розового и синего цветов или же обладают пурпурным оттенком (Sciaphila).
Члены этой синузии развиваются на почве в лесу или иногда на мертвых стволах или пнях. Они предпочитают интенсивную тень или места, где опавшие листья собираются более толстым слоем, чем обыкновенно, например в небольших впадинах. Излюбленным местообитанием многих их них являются промежутки между досковидными корнями крупных деревьев. В Морабалли-Крик сапрофиты особенно обильны в лесу Мора-буцея (Mora gonggrijpii), самом темном из первичных лесов, встречающихся в этой области. Неизвестно, оказывает ли свет какое-либо непосредственное воздействие на распределение этих растений; их приуроченность к очень затененным местам может объясняться тем, что они не переносят, хотя бы и временно, ненасыщенной влагой атмосферы. Отсутствие или слабое развитие сапрофитов в тропических дождевых лесах с заметно выраженным сухим сезоном, например в юго-западной Нигерии, заставляет предполагать, что они не в состоянии пережить даже легкое высыхание почвы леса. Характер расселения сапрофитов в местах их развития отличается специфическими особенностями. Часто они совершенно отсутствуют на больших пространствах или бывают представлены только редкими, изолированными экземплярами наиболее обычных видов; однако то в одном, то в другом месте встречаются участки обычно величиной только в несколько квадратных метров, на которых группой произрастают более или менее многочисленные представители трех или четырех видов. Такой тип распространения был замечен на Борнео, в Брит. Гвиане и других местах. Видимо, это зависит не исключительно от густоты тени или толщины лесной подстилки; сапрофиты часто отсутствуют там, где условия для них, казалось бы, вполне благоприятны. Подземные органы всех этих сапрофитных цветковых растений имеют микоризы, которые могут оказывать на них свое действие. Было высказано предположение (van der Pijl, 1934), что распространение сапрофитов пятнами связано с факторами, оказывающими в свою очередь влияние на развитие микоризы. Заслуживают особого исследования отношения, существующие между распространением сапрофитов и определенных видов деревьев.
Следует мимоходом заметить, что у некоторых сапрофитов, о которых идет речь, корневая система располагается в поверхностном слое перегноя, в то время как у других она располагается в подстилающем минеральном слое почвы иногда на глубине 15—20 см (Johow, 1889, стр. 486—487).
Все упомянутые сапрофиты представляют собою невзрачные растения, которые неопытному человеку легко не заметить. За исключением некоторых орхидей, почти все они бывают ниже 20 см. Лишь сапрофитная орхидея, Galeola altissima, представляет единственное в своем роде исключение: это лазящее при помощи корней растение достигает высоты около 40 м.
СТРУКТУРА И ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА 149
ПАРАЗИТЫ
Кроме грибов и бактерий, в тропическом дождевом лесу имеются две синузии паразитов, а именно корневые паразиты, растущие на почве, и полупаразиты, располагающиеся, подобно эпифитам, на деревьях. Почвенные полупаразиты в тропическом дождевом лесу не наблюдались.
Синузия корневых паразитов не имеет большого значения, так как отличается малым видовым разнообразием и редко насчитывает большое количество индивидуумов. Корневые паразиты представлены лишь двумя семействами: Balanophoraceae и Rafflesiaceae. Первое насчитывает несколько родов: Helosis cayennensis широко распространен в тропической Америке, a Thonningia sanguinea (фото 13) часто встречается в вечнозеленом лесу Западной Африки. Из Rafflesiaceae в дождевом лесу встречается лишь малайская Rajjlesia, широкоизвестная своими гигантскими цветами, и Mycetanthe.
Некоторые из упомянутых корневых паразитов имеют ограниченный круг хозяев. Виды Rafflesia, например, встречаются главным образом на Vitaceae; другие корневые паразиты вполне индифферентны в этом отношении, как, например, Thonningia sanguinea. Обыкновенно в любой области встречается не более одного вида этих корневых паразитов. Бедность паразитических цветковых растений в тропическом дождевом лесу представляет собой удивительное и до сих пор необъясненное явление.
Эпифитные полупаразиты дождевого леса относятся только к одному семейству Loranthaceae, хотя оно представлено многочисленными родами и видами, число индивидуумов которых иногда бывает крайне велико. Loranthaceae встречаются во всей области распространения тропических дождевых лесов, и в любом ее округе часто можно найти большое число видов.
Большинство тропических Loranthaceae имеет очень большой круг хозяев и встречается на деревьях самых разнообразных родов и семейств; некоторые виды являются иногда гиперпаразитами, атакующими членов своего собственного семейства (Koernicke, 1910). Были даже отмечены примеры гиперпаразитизма второй степени (паразит на паразите паразита). Хотя тропические Loranthaceae обнаруживают лишь незначительные ограничения в выборе своих хозяев, они имеют весьма четкие пределы вертикального распределения. В тропическом дождевом лесу нормальной густоты и тенистости они всегда встречаются на сучьях и ветвях деревьев первого яруса и более высоких деревьях второго; они никогда не встречаются на более низких уровнях, за исключением открытых мест и отдельно стоящих деревьев. В более открытых типах леса они могут спускаться почти до уровня почвы. В хорошо освещенном «боровом» лесу на Борнео автор нашел Macrosolen beccarii, растущим на кустарниках и небольших деревьях на расстоянии 1—2 м от почвы, тогда как в более тенистом смешанном диптерокарповом лесу Loranthaceae повсеместно отсутствовали ниже 20 м, за исключением вырубок и мест, расположенных по урезам рек (Richards, 1936, стр. 32). В Нигерии Loranthus leptolobus обычен на высоте нескольких метров на отдельно стоящих деревьях какао и колы на возделываемых землях. Таким образом, вертикальное распределение эпифитных полупаразитов вполне совпадает с распределением аутотрофных солнечных эпифитов. Распределение тех и других определяется, по-видимому, главным образом отсутствием теневыносливости.
150 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Loranthaceae тропического дождевого леса представляет собой более или менее деревянистые растения, изменяющиеся по своему облику от стоячих до полустелющихся кустарников. Листья цельные, кожистые и. разумеется, вечнозеленые; они обыкновенно очень приближаются к типу, преобладающему у деревьев того яруса, где находятся Loranthaceae. Сходство их листьев с листьями хозяев нередко ведет к тому, что когда они не цветут, их существование вообще трудно заметить. Но в момент цветения их яркая окраска обращает на себя внимание. Дерево, покрытое Loranthaceae, — одно из самых прекрасных зрелищ в тропическом дождевом лесу Малайского архипелага.
Часть II
СРЕДА
Глава 6
КЛИМАТ1
Климат тропических дождевых лесов обладает двумя основными» особенностями: высокой постоянной температурой и обильными осадками, выпадающими в течение большей части года. Хотя в пределах этого типа климата существуют значительные вариации, главным образом в сезонном распределении осадков и температуры, ведущие его характеристики остаются более или менее постоянными во всей области развития дождевых лесов.
Распространение типа формаций тропического дождевого леса было показано в главе 1. Эти леса покрывают несимметричную площадь, не имеющую точных широтных границ. Иногда, как в южной Бразилии, они доходят до географических тропиков и даже пересекают их; в других местах границы дождевых лесов простираются только на несколько градусов к югу или к северу от экватора. Это и неудивительно, поскольку тропики не являются естественными климатическим границами, и в конечном счете связано с неравномерным распределением воды и суши. В климатологическом смысле тропическую зону определяют не широтные границы, а изотермы средней годовой температуры в 20°. В пределах тропиков в этом смысле наблюдаются значительные различия как в количестве, так и в сезонном распределении осадков и соответственно большое разнообразие растительного покрова,— от роскошных вечнозеленых лесов-до бесплодной пустыни.
Тропический дождевой лес может существовать только там, где годовое количество осадков превышает определенный минимум и где не бывает длительной засухи; там же, где засушливый сезон бывает долгим и сильно выраженным, вечнозеленый лес замещается растительной формацией, приспособленной к периодическому недостатку влаги. С возрастанием широты и удалением от моря влажный экваториальный климат сменяется тропическими климатами со все менее равномерным распределением и меньшим количеством осадков, что отражается в постепенном переходе от вечнозеленого леса относительно несезонного типа («истинный дождевой лес» Бирда, Beard, 1944 а) к несколько менее роскошному вечнозеленому сезонному лесу, затем к последовательно сменяющим друг друга листопадному саванновому и колючему лесу и, наконец, к пустыне (см. гл. 15). Часто тропический вечнозеленый лес внезапно переходит в открытую саванну, что, как увидим далее, обусловливается главным образом биотическими или эдафическими причинами, а не определяется исключительно климатом, как это обычно предполагают. По мере
1 Ценная работа Бернара (Bernard, 1945) об экоклимате центральной части бассейна Конго, к сожалению, слишком поздно попала к автору и не могла быть полностью использована при написании этой главы.
154
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
увеличения высоты над уровнем моря тропический дождевой лес низменности уступает место дождевому лесу предгорий и горному дождевому лесу (см. гл. 16); это вызвано скорее изменениями температуры, чем количества осадков. Таким образом в целом можно сказать, что северные и южные климатические границы типа формаций тропического дождевого леса почти повсюду определяются осадками, его высотные границы — преимущественно температурой.
Одна из выдающихся особенностей всех влажных тропических климатов состоит в том, что сезонные колебания температуры незначительны по сравненпю с сезонными изменениями количества осадков. Здесь нет ни лета, ни зимы, а лишь смена дождливых и сухих сезонов. Этим определяется не только сезонный ритм растительности, но и в значительной мере деятельность людей. Сравнительно маленькая амплитуда сезонных колебаний в климате тропических дождевых лесов очень наглядно показана на рис. 20, где средние месячные температуры сопоставлены со средними месячными осадками. В некоторых районах тропического дождевого леса, например в Сингапуре и на ряде тихоокеанских островов, ничтожны даже сезонные изменения в количестве осадков, в связи с чем здесь практически отсутствуют времена года.
Годовой цикл дождливых и сухих периодов в тропической зоне непосредственно связан с высоким положением солнца и обычно повторяется из года в год с величайшей регулярностью. Внезапные, неустойчивые и часто непредвиденные суточные изменения погоды, столь типичные для умеренных стран с морским климатом, вроде Британских островов, редко наблюдаются в тропиках. Хотя в экваториальных климатах годовые колебания выражены, вероятно, в гораздо меньшей степени, чем в умеренных, они возрастают по мере приближения к поясу тропических дождевых лесов, и ими ни в коем случае нельзя пренебрегать. Гедерт (Goedert, 1938) показал, что в Республике Конго, например, годовые колебания в количестве осадков возрастают от центральной части бассейна Конго к побережью. Хотя, как указывает Моро (Moreau, 1938), последствия необычно засушливых или исключительных в другом отношении лет могут меньше сказаться на естественных растительных сообществах, находящихся в равновесии со средой, чем на сельскохозяйственных культурах, все же они, вероятно, значительнее, чем обычно полагают. Засушливые годы хорошо известны даже в глубине экваториального лесного пояса, но их воздействие, по-видимому, наиболее сильно ощущается в переходной зоне между двумя различными растительными формациями. Так засушливые годы благоприятствуют вторжению африканской саванны (климакс пожарища, см. гл. 15) в вечнозеленый лес.
ТЕМПЕРАТУРА
На тропических низменностях средняя годовая температура колеблется между 20 и 28°. Вообще средние температуры только очень постепенно изменяются от одного места к другому; поэтому температурные градиенты сглажены, и изотермы далеко отстоят друг от друга. Близ экватора разница между средними температурами самых жарких и самых холодных месяцев составляет менее 5° даже в местностях в глубине материка, таких, как Манаус в долине Амазонки; на маленьких тропических островах она иногда не превышает 1° (на островных группах Гилберта и Эллис в Тихом океане, 0,2°). Наиболее низкие средние температуры
30
Рис. 20. Сезонные колебания температуры и осадков в умеренном океаническом климате (Кью), несезонном тропштеском (дождевой тропический лес) климате (Сингапур) и сезонном тропическом (листопадный лес) климате (Рангун) (вгоок ,
СРЕДА
15Л
обычно наблюдаются в дождливом сезоне, наиболее высокие — в сухом. По мере удаления от экватора сезонные колебания температуры возрастают особенно в областях с резко выраженным сухим и дождливым периодом, но даже в районах с самым неровным климатом они редко превышают 13°. Средняя суточная амплитуда колеблется от 3 до 16" в различных частях тропической зоны. Хотя минимальные температуры вблизи экватора высоки, максимальные оказываются ниже, чем в южной Европе, и редко превосходят 33—34°; ближе к тропикам средние годовые максимумы гораздо выше и могут превышать 50°.
В области, занимаемой тропическими дождевыми лесами, средняя температура составляет около 26° при средней для самых холодных месяцев обычно не ниже 25°, но там, где влажный лес более или менее тропического типа простирается за пределы собственно тропиков, годовая средняя может составлять около 20°. Кеппен (Кбрреп, см. стр. 170) считал среднюю температуру самого холодного месяца в 18° нижним температурным пределом для выделенного им «влажного тропического климата». Температурные условия в типичных низменных районах тропического дождевого леса показаны в табл. 10.
О распределении температур внутри вертикальных растительных поясов тропической зоны известно сравнительно мало. В тропиках средняя температура понижается примерно на 0,4—0,7° на каждые 100 м подъема. Так в Индонезии, согласно Мору (Mohr, 1944, стр. ИЗ), средняя температура на низменностях (0—200 м, тропический дождевой лес п муссонный лес) составляет примерно 25—27°, в поясе предгорий (200— 1000 м, тропический дождевой лес и дождевой лес предгорий) — 19—24°, в горном поясе (1000—1800 м, дождевой лес предгорий и горный дождевой лес) — 13—18°. Браун (Brown, 1919) дает максимумы и минимумы для различных поясов растительности на горе Маквилинг, на Филиппинах, приведенные в табл. И.
Абсолютные минимумы температуры в районе тропических дождевых лесов, вероятно, имеют небольшое значение для растительности. Хотя внутри тропического пояса в отдельных местностях и отмечались нулевые и отрицательные температуры на небольших высотах, например в континентальной части Южной Америки, но вряд ли они когда-либо наблюдались в пределах области тропического дождевого леса. Таким образом, собственно тропический дождевой лес в целом (исключая дождевой субтропический лес, например в восточной Австралии) лежит в пределах безморозной зоны. Тем более примечательно, что в то время как для ряда тропических растений положительная температура, близкая к нулю, считается губительной (Molisch, 1896), некоторые виды, характерные для тропических дождевых лесов, например Eucalyptus deglupta (Lane-Poole, 1938), проявляют известную степень морозоустойчивости.
Незначительность сезонных колебаний температуры, в большей или меньшей степени типичная для всей тропической зоны, частично зависит от незначительности годовых колебаний в продолжительности дня. Даже непосредственно на линиях тропиков наиболее короткий день длится примерно 101/2 часов, а самый длинный — только 131/* часов. Другими важными факторами являются термостатическое влияние океанов, занимающих почти три четверти тропической зоны, а также почвы. В Индонезии (Mohr, 1944, стр. 45) температура почвы в лесу на уровне моря равна 25—27°, а на глубине одного метра она постоянна в течение года, более того, она «в основном постоянна» даже на глубине 60 с.и. В Янгамби, Республика Конго, средняя температура почвы на глубине
т Местность емпература в - Широта 13°48' ю. ш. 1°18' с. ш. 5П49' с. ш. 6°50' с. ш. 3°8' ю. ш. 0 45' с. ш. 0 3' с. ш. 4°3' с. ш. 6°27' с. ш. 13 2' с. ш. 16 47' с. ш. 2Г59' с. ш. Зэ26' с. ш. 1°35' с. ш. 16°55' ю. ш. 22?54' ю. ш. тропической зоне 1 “блица 1о Температура, сс "
средняя годовая 25,8 27,2 27,4 26,0 27,2 24,5 25,6 25,4 26,9 28,0 27,3 27,6 24,4 21,8 24,8 22,7 средняя самого жаркого месяца 26,3 27,7 28,1 27,2 28,2 25,2 26,3 26,9 28,5 32,7 30,6 32,2 25,1 22,6 27,2 25,6 средняя самого холодного месяца 25,1 26,4 26,5 25,1 26,5 23,9 24,6 23,8 25,3 23,7 24,8 21,3 23,5 20,7 21,0 20,0 средние экстремумы! абсолютные экстремумы
в пределах развития тропических дождевых лесов 1. Апиа, Самоа 2. Сингапур, полуостров Малакка 3. Сандакан, Сев. Борнео 4. Станция Мазаруни, Брит. Гвиапа 5. Манаус, Бразилия 6. Янгамби, Республика Конго 7. Эала, Республика Конго 8. Дуала, Камерун В пределах развития тропических полувечнозеле-ных лесов, листопадных лесов и т. д. 9. Лагос, Нигерия 10. Сокото, Нигерия 11. Рангун, Бирма 12. Мандалай, Бирма В пределах развития дождевого леса (близ высотных или широтных границ) 13. Бамбеса, Республика Конго 14. Масинди (1146 м), Уганда 15. Кэрнс, Квинсленд 16. Рио-де-Жанейро, Бразилия максимум 32,7 33,8 34,4 36,6 32,7 35,0 43,3 38,3 43,3 35,5 минимум 18,3 20,5 21,6 21,1 20,0 19,4 10,0 15,0 10,5 13,8 максимум 35,5 36,1 37,2 38,3 37,2 35,0 40,0 46,1 41,6 44,4 36,7 38,8 | минимум 16,1 18,8 21,1 18,8 15,2 18,3 15,5 3,8 12,7 7,2 9,4 10,0
1 Средние наивысших (или наинизших) температур за ряд лет. Источники: пункт 4 по данным Службы леса Бпит Гвианы* птнкты к 7 и га (Eggeling, 1947); пункт 15 по данным, сообщенным проф. Турнером (J. S. Turner); остальные по Бруксу^г^,’ X ПУ"КТ ‘ 4 по данным Эггелии
СРЕДА
157
Таблица 11
•Средние максимальные н минимальные температуры у вершин высоких деревьев, произрастающих на различной высоте над уровнем моря на горе Маквилинг, Филиппинские острова
(октябрь 1912 г. январь 1915 г.) (Brown, 1919, стр. 227)
Высота, м Характер растительности Средний максимум, С Средний минимум, °C
80 Паранг (вторичные сообщества) 38,2 24,0
300 Диптерокарповый лес (тропический дожде-
вой лес) 32,4 20,0
450 Тот же тип леса 31,3 19,6
740 Среднегорный лес (дождевой лес предгорий) 30,4 18,1
1050 Моховой лес (горный дождевой лес) 28, 1 17,3
50 см составляет 26,2° при годовой амплитуде в 1,5Э, а в 50—75 см от поверхности исчезают все признаки суточных колебаний (Bernard, 1945).
ОСАДКИ
В тропической зоне осадки чрезвычайно широко варьируют в отношении количества и сезонного распределения. На значительной площади годовые суммы осадков составляют 250—400 см, но в отдельных местностях они иногда превышают 10 м (Черрапунджи, Ассам, — 1161 см, Дебунджа, у подножия вулкана Камерун, — 1017 см); в то же время в районах пустыни у северных границ зоны наблюдается очень малое количество осадков. Сезонность в распределении осадков не всегда связана с размерами их годовой суммы. Очень большие суммы осадков могут быть строго сезонными по распределению (в Черрапунджи четыре месяца подряд выпадает гораздо менее 10 см), с другой стороны, значительно меньшие суммы могут быть распределены относительно равномерно; но в общем в тропических областях с низкой суммой осадков к тому же наблюдается и неравномерное распределение их (сезонные или нерегулярные). Для растительности характер годового распределения осадков гораздо важнее, чем их годовая сумма.
В самых общих чертах сезонное распределение осадков в тропической области связано с широтой: на экваторе дожди выпадают во все времена года (хотя, вероятно, в тропиках нет мест, где осадки распределялись бы вполне регулярно); в зоне примерно от 3 до 10° к северу и югу от экватора наблюдаются два дождливых и два сухих сезона, а еще дальше от него в году выявляются только один дождливый и один сухой сезон (рис. 21). Эти широтные изменения сезонного распределения осадков зависят в основном от положения солнца в то пли иное время года; периоды, когда солнце находится в зените, обычно бывают периодами обильных осадков, а промежуточные периоды — сравнительно засушливыми или даже совершенно бездождными.
Широтная зона, где солнце стоит прямо над головой в любое время года, прогревается сильнее, чем зоны, лежащие к северу и к югу, и поэтому является ареной восходящих токов воздуха, место которых замещает воздух, приходящий извне; эта область сравнительно слабых ветров (экваториальная зона затишья) примыкает к двум поясам сильных
158
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и постоянных ветров (пассатные ветры). Над Индийским океаном эта система усложняется муссонами — периодическими ветрами, которые вызываются наличием летней барической депрессии в Центральноп Азии. В меньшей степени развитию муссонов способствуют барические условия Сахары и Австралийской пустыни.
Месяцы
ЯФМДМИИДСОНД
ЯФМАМИИДСОНД Месяцы
Рис. 21. Широтное распределение дождливых и сухих сезонов в тропиках.
Дождливые сезоны показаны штриховкой (по Мартонну).
Благодаря адиабатическому расширению восходящие воздушные токи приэкваториальной полосы максимального нагрева охлаждаются, что приводит к выпадению дождей. Таким образом, осадки тропической зоны, как правило, бывают конвекционными, вызываясь охлаждением воздуха, поднимающегося над нагретой поверхностью суши, хотя их могут породить и другие причины, например отклонение восходящих вертикальных токов воздуха в горных районах или же конвергенция воздушных потоков, слегка разнящихся по плотности, обычно вследствие небольшой разницы температур. В силу такого происхождения тропический дождь редко бывает слабым или длительным; он имеет общие черты с грозовыми ливнями умеренных стран и действительно часто сопровождается громом. Дожди во многих тропических районах, как и следовало ожидать, учитывая их происхождение, поразительно постоянны по времени выпадения и наблюдаются обычно после полудня или ранним вечером.
СРЕДА
159
Если на данной широте периоды, когда солнце находится в зените, отстоят далеко друг от друга, должно наблюдаться два сезона дождей, если они следуют друг за другом, то два дождливых сезона сливаются в один, хотя сезонная кривая осадков может показывать два заметных максимума. В этом и проявляется связь между сезонным распределением осадков и широтой, на которую уже было указано. Однако действительные связи не всегда так просты, как можно было бы ожидать, и сутцест* вуют местные аномалии в распределении осадков, которые не всегда легко объяснить. В Юго-Восточной Азии (включая Малайский архипелаг) в тех широтах, где должны были бы быть два дождливых сезона, благодаря муссонам наблюдается только один. В Брит. Гвиане (6е с. ш.) два максимума осадков совпадают с наименьшей, а не с наибольшей высотой солнца. Некоторые из этих аномалий, возможно, связаны с неравномерным распределением моря и суши и влиянием океанических течений. Подобными причинами можно объяснить наличие засушливых районов в северной Венесуэле, в части северо-восточной Бразилии, в Гане и т. д., где сумма осадков и время их выпадения не соответствуют широте. Благодаря местным факторам большие различия в количестве и ходе осадков могут быть свойственны двум рядом расположенным местностям и даже разным частям одного и того же города.
Поскольку, как уже сказано, тропический дождевой лес зависит от обильного и равномерного выпадения осадков в течение года, он является климатическим климаксом в экваториальной зоне сильных и равномерных осадков и в большей части зоны с двумя дождливыми сезонами; в зоне с одним сезоном дождей он развивается лишь в той мере, в какой орографические или другие местные условия увеличивают осадки в относительно сухом сезоне или насколько позволяют другие компенсирующие факторы.
Изучение цифр в табл. 12 показывает, что в типичных районах произрастания тропического дождевого леса годовая сумма осадков составляет не менее 200 см, хотя как в сумме, так и в сезонном распределении их существуют значительные колебания. В некоторых местностях, таких как Дебунджа, Камерун, благодаря особенностям рельефа сумма осадков намного превосходит 200 см. В других местах, например на станции Мазаруни, Брит. Гвиана, и в Сингапуре средняя месячная сумма осадков никогда не бывает ниже 10 см. С другой стороны, в Акилле, южная Нигерия, где годовая сумма осадков достигает значительной цифры в 208 см, пять месяцев подряд выпадает меньше 10 см осадков, и в течение трех из них — менее пяти. Вероятно, существование тропического дождевого леса в климате с такими сильными засухами возможно только при наличии какого-то компенсирующего фактора, в Акилле, может быть, благодаря относительно высокой влажности атмосферы в течение этих засушливых месяцев (см. стр. 340). Пример компенсации недостаточного количества осадков другими факторами дают также вечнозеленые приречные или галерейные леса (см. стр. 166—167), сходные с тропическим дождевым лесом по структуре и физиономии, но простирающиеся далеко за пределы его нормальных границ в виде узких полос вдоль рек в районах листопадного леса или саванны. В галерейном лесу недостаток осадков или неравномерность их распределения компенсируется избытком почвенной влаги. Подобные растительные сообщества следует рассматривать как постклимаксные.
В пределах зоны тропических дождевых лесов длительность и сила засух выражаются в отчетливых, хотя и не очень значительных разли-
* Ослики Н < Таблиц а 12
Местность Широта Средние месячные осадки, см ~
январь февраль март апрель май июнь июль август сентябрь октябрь ноябрь л ю я X ф 41ч <- я * 0-0 о реднее ко-нчество дне! осадками
i районах развития тропиче- Оно О ч о
ского дождевого леса:
1. Станция Мазаруни,
Брит. Гвиана 6°5О' с. ш. 21,0 11,5 13,1 18,4 32,2 33,3 27,5 19,6 14,4 12,8 15,0 27,2 246,9 417,5 212 194
2. Виктория, Камерун 3. Янгамби, Республика 4°Г с. ш. 2,7 6,7 15,2 18,8 34,5 70,6 106,9 80,0 44,6 22,8 10,3 4,3
Кон 1Х) 4. Зала, Республика Кон- 0°45' с. ш. 8,0 10,0 15,5 15,3 14,7 11,3 12,3 14,2 18,6 20,7 17,4 П,1 169,1 —
го 0°3' с. ш. 8,6 13,3 12,1 16,5 16,4 13,5 7,6 16,2 17,6 22,4 19,1 16,5 179,8 —
5. Сингапур, полуостров
Малакка 1°18' с. ш. 25,2 16,8 18,8 19,3 17,0 17,3 17,3 19,1 17,3 20,6 25,2 26,9 240,8 177
6. Понтманак, Борнео 0°6' ю. ш. 26,9 21,6 24,4 28,2 27,2 22,6 16,5 21,8 21,8 37,3 40,9 33,3 322,5 182
7. Сандакан, Борнео 5°49' с. ш. 46,7 24,4 20,3 10,4 15,0 18,5 16,5 20,6 23,9 25,4 37,3 45,0 304,0 181
8. Апиа, Самоа 13°48' ю. ш. 42,4 39,6 34,0 25,9 14,2 13,2 6,9 8,9 12,7 16,0 24,4 34,5 272,7 210
5 районах развития тропиче-
ского дождевого леса близ его климатической границы:
9. Порт-оф-Спейн, Тринидад 10°42' с. ш. 6,9 3,8 4,6 4,8 8,9 19,3 22,1 24,1 18,5 17,0 18,0 12,2 160,2 192
10. Остров Мартиника («мезофитный лес») 14°40' с. ш. 19,0 21,1 2,8 11,4 20,3 3,5 8,4 20,6 7,3 21,3 20,0 16,8 23,3 9,9 25,4 4,6 28,1 3,3 28,1 3,6 34,9 11,7 40,6 11,4 21,2 20,8 267,7 165,4 290 172
11. Манаос, Бразилия 12. Акилла, Нигерия 3°15' ю. ш. 6°40' с. ш. 9,6 18,0 19,6 40,7 41,9 13,4 25,2 24,2 8,0 1,1 208,0 125
Продолжение табл. 12
Местность Широта Средние месячные осадки, сл1 Средние годовые суммы осадков, см Среднее количество дней с осадками
январь февраль г март апрель май июнь июль август сентябрь октябрь ноябрь декабрь
13. Будонго, Уганда2 1°35'с. ш. 2,3 7,7 12,9 18,8 17,3 13,4 9,8 15,5 15,3 16,0 13,2 7,3 149,5 167
14. Джакарта, Ява 6°22' ю. ш. 30,2 34,0 20,3 14,2 10,4 9,4 6,6 4,1 7,1 11,4 14,5 19,3 181,5 138
15. Кэрнс, Квинсленд В районах развития тропического листопадного леса, саванн и т. п.*. 16. Остров Мартиника 16°55' ю. ш. 42,2 39,4 46,0 29,5 10,9 7,1 4,1 4,3 4,3 5,3 0,7 21,8 224,6
(«ксерофитный лес») 14°40' с. ш. 10,7 5,2 5,2 5,2 14,1 13,1 21,7 23,4 23,3 21,9 21,4 10,8 176,0 250
17. Кумаси, Гана 6°50' с. ш. 1,6 5,6 13,6 14,2 18,6 22,4 12,8 17,3 18,6 20,2 0,0 3,2 148,0 133
18. Сокото, Нигерия 13°2' с. ш. 0,0 0,0 0,3 0,5 5,2 10,4 16,3 20,3 10,7 1,0 0,0 0,0 64,6 55
19. Рангун, Бирма 6°47' с.ш. 0,5 0,5 0,8 3,6 30,7 46,7 54,6 50,0 39,1 18,5 7,1 0,8 252,9 122
20. Мандалай, Бирма 21°59' с. ш. 0,3 0,3 0,5 2,8 15,0 14,0 8,4 11,7 14,5 11,9 4,1 1,0 84,5 51
21. Пасириан, Ява 7°41' ю. ш. 24,4 30,0 20,3 13,2 8,9 5,8 2,8 0,5 0,8 2,0 6,4 16,0 130,9 94
1 Месячные суммы ниже 10 см (4 дюйма) даны курсивом; максимальные средние месячные суммы отмечены полужирным шрифтом.
2 1146 м выше уровня моря.
Источники. Пункт 1—по данным Службы леса Брит. Гвианы; пункт 2—цифры по данным Лесного управления Нигерии, пункты 3, 4 по дан ным Бернара (Bernard, 1945); пункты 6, 21-по данным Браака (Вгаак, 1936); пункт 9-по данным Бирда (Beard, 1946); пункт 12-по данным ичардса (Richards, 1939); пункт 13-по данным Эггелинга (Eggeling, 1947); пункт 15-по данным, сообщенным проф. Турмером (J. S. Turner); пункты iu. данным Стеле (Stehld, 1945, стр. 302, 369); пункт 17—по данным Фогги (Foggle, 1947); остальные—по данным Брукса (Brooks. ).
162
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
чиях в структуре и облике растительности. Лес в областях с долгими периодами (4—5 месяцев) незначительных осадков или засухи по составу растений мало отличается от (относительно) несезонных «истинно дождевых лесов» областей с более равномерно распределенными осадками; менее благоприятный климат засушливых областей проявляется в меньшей роскоши растительности и в наличии определенной пропорции таких растений, как листопадные деревья и клубневые геофиты, характеризующихся периодом покоя во время засушливого сезона. В районах с годовой суммой осадков, превышающей определенный минимум (для тропического дождевого леса может быть около 160 сл<), характер климаксной растительности больше зависит от сезонного распределения осадков, чем от их суммы. Даже в тех областях, где годовая сумма намного превышает минимум, необходимый для существования тропического дождевого леса, может развиваться такая засухоустойчивая растительность, как листопадный лес. Несомненно, что определяющим фактором всегда является длительность и сила засух, зависящие главным образом, но не всецело, от сезонного распределения осадков.
О характере изменения осадков в горах тропической зоны с увеличением высоты над уровнем моря известно мало, хотя установлено, что, как и в других частях света, в горах выпадает больше осадков, чем в прилегающих низменностях и что их количество увеличивается с возрастанием высоты. В некоторых тропических районах, например на восточной Яве и восточной части Зондских островов, более обильные осадки и более короткий сухой сезон создают условия для развития вечнозеленого тропического дождевого леса в горах, тогда как на низменностях произрастает листопадный лес. На Яве количество осадков увеличивается от подножия гор до высоты 610—1220 м, где наблюдаются максимальные величины (Junghuhn, цит. по Шимперу, Schimper, 1935, стр. 1261), а затем уменьшается по направлению к вершине; по более поздним данным Бра-ака (Braak, 1936, стр. 20) максимальное количество осадков выпадает не на какой-либо определенной высоте, а примерно в средней части каждой данной возвышенности. На горе Маквилинг, Филиппины, на высоте 740 м иногда выпадает больше осадков, чем на высоте 1000 м (Brown, 1919, стр. 256). Подобный ход осадков с достижением максимума на определенной высоте над уровнем моря и последующим уменьшением их но мере дальнейшего возрастания высоты, даже если он носит характер закономерности, вероятно, составляет особенность только наиболее крупных гор.
АТМОСФЕРНАЯ ВЛАЖНОСТЬ
Лучшим показателем влажности атмосферы для ботаника-эколога является дефицит насыщения воздуха — мерило его испаряющей способности. Однако данные о дефиците насыщения не всегда легко получить, и обычно приходится довольствоваться измерениями относительной влажности. Поскольку дефицит насыщения является производным температуры и относительной влажности воздуха, то одна и та же относительная влажность будет соответствовать совершенно различным дефицитам насыщения при различных температурах. Следовательно, если в изучаемом районе температуры непостоянны во времени и пространстве, то показатели относительной влажности не могут дать правильного представления об испаряющей способности воздуха — этого жизненно важного фактора с точки зрения физиологии растений. Однако в тропическом дождевом
СРЕДА
163
лесу температурные колебания (суточные, сезонные и территориальные) настолько незначительны, что обычно позволяют считать относительную влажность приблизительным критерием испаряющей способности. Таким образом, краткое рассмотрение данных по относительной влажности поможет воссоздать отчетливую картину природной среды, в которой развивается дождевой лес.
На открытых участках в пределах области распространения тропического дождевого леса относительная влажность обычно находится на высоком уровне в течение большей части дня даже в засушливый период. Она сильно колеблется в различные часы дня и имеет сезонный ритм, параллельный ходу осадков. Суточные и сезонные колебания подробно рассматриваются в следующей главе, посвященной микроклимату. Большинство опубликованных данных об относительной влажности в тропической зоне позволяет получить лишь весьма туманное общее представление о ее величинах; часто это всего лишь средние месячные показатели, которые с экологической точки зрения не очень убедительны, если только они не относятся к определенному часу в середине дня.
Таблица 13 дает средние месячные величины относительной влажности для нескольких избранных районов в области развития тропического дождевого леса; цифры даны для раннего утра (когда относительная влажность близка к суточному максимуму) и для середины дня (когда она обычно близка к суточному минимуму).
В целом в области развития тропического дождевого леса даже на открытых пространствах ночью относительная влажность всегда соответствует точке насыщения или близка к ней; днем в сухую погоду она падает до величин порядка 65%, кратковременно она может упасть до 55% и даже ниже. В такой сырой местности, как Богор, Ява (средняя годовая сумма осадков — 436,7 см; средняя месячная самого сухого месяца — 22,7 см), зарегистрирован абсолютный минимум относительной влажности в 28% (Вгаак, 1924). Средний годовой минимум относительной влажности в Сингапуре — 51%, а в Понтианаке, Борнео,— 35% (Вгаак, 1931, табл. 5). В Зала, в исключительно влажном районе центральной части бассейна Конго, средняя относительная влажность в самом сухом месяце (марте) составляет 56% (Bernard, 1945, стр. 114).
Характер растительности, постоянная сырость почвы и плесень, покрывающая книги, одежду, кожаные вещи и пр., если их не проветривать и не сушить на солнце, являются наглядными показателями высокой средней относительной влажности в области тропического дождевого леса. Поэтому трудно отчетливо представить себе, как велика амплитуда суточных колебаний и как незначительна бывает влажность на открытых местах в течение коротких промежутков дня в сухую погоду. Эти повторяющиеся периоды низкой относительной влажности, наблюдающиеся в большинстве даже наиболее сырых участков тропического дождевого леса и возможные в дождливый сезон, несомненно, должны, несмотря на свою кратковременность, сильно влиять на растительность, и, вероятно, в прошлом их значение недооценивали. Поскольку суточный минимум относительной влажности совпадает со временем дня, когда температура близка или достигает своего дневного максимума, соответствующий дефицит насыщения бывает значительным. Автор, проводивший наблюдения на большой вырубке в тропическом дождевом лесу Борнео, нашел, что в течение десяти дней в октябре (начало дождливого сезона) средняя минимальная влажность составляла 61 ?о, что при температурах, преобладающих в середине дня, соответствует дефициту насыщения примерно
Относительная влажность в тропической зоне, %* Таблица 13
Местности Январь Февраль Март Апрель Май Июнь Июль 1 Август Сентябрь Октябрь Ноябрь Декабрь Время 1 ДНЯ ’редням адовая, о
В районах развития
тропического дожде-
вого леса:
1. Станция Мазару-
ни, Брит. Гвиана 94 93 93 93 93 94 94 94 93 93 93 94 7,00 мест. вр. 94
79 76 73 75 81 82 81 77 73 72 74 81 13,00 мест. вр. 73
2. Зала, Республи-
ка Конго 74 70 72 73 75 77 76 76 75 74 76 77 12,00 мест. вр. 75
3. Дуала, Камерун 95 96 95 94 95 95 96 96 95 95 95 95 7,00 мест. вр. 95
74 75 76 76 79 82 86 84 85 83 80 78 14,00 мест. вр. 80
4. Сингапур, Малаккский полуостров 84 72 83 69 82 75 83 76 85 76 82 74 82 73 82 75 82 75 82 76 83 77 83 77 9,00 15,00 мест. мест. вр. вр. 83 75
5. Сандакан, Сев. Борнео 91 77 91 75 91 74 92 70 92 69 92 69 91 69 91 69 91 67 92 70 93 75 92 76 6,00 14,00 мест. мест. вр. вр. 92 72
6. Апиа, Самоа 82 79 81 78 81 78 79 76 78 76 77 73 77 75 76 73 75 75 77 76 78 75 79 77 9,00 15,00 мест. мест. вр. вр. 78 76
В районах развития тро-
пического дождевого леса близ его климатической границы: 7,00 15,00 мест. мест. вр. вр. 93
7. Порт-оф-Спейн, Тринидад 92 68 93 63 93 62 92 60 91 60 92 66 93 73 95 73 94 74 94 72 94 74 93 73 68
Продолжение табл. 13
Местности Январь Февраль Март Апрель Май Июнь Июль Август Сентябрь Октябрь Ноябрь Декабрь Время дня Средняя годовая, %
8. Лагос, Нигерия 84 83 82 81 83 87 87 85 86 86 85 86 9,00 мест. вр. 85
65 69 72 72 76 80 80 76 77 76 72 68 15,00 мест. вр. 74
9. Масинд и, Уганда2 70 66 77 80 82 82 88 85 84 81 80 77 8,30 мест. вр. 79
43 42 57 60 66 67 67 65 66 62 56 54 14,30 мест. вр. 59
10. Джакарта, Ява 90 92 90 89 89 89 87 84 82 81 84 87 8,00 мест. вр. 87
В районах развития тропического листопадного леса, саванн и пр: 11. Остров Мартиника («ксерофитный 75 76 73 71 69 68 65 62 62 65 69 73 14,00 мест. вр. 6
лес») 87 85 85 79 81 80 83 84 85 89 90 90 7,00 мест. вр. 85
74 70 70 72 73 72 75 78 78 81 83 79 17,00 мест. вр. 75
12. Кумаси, Гана 95 95 96 96 96 96 96 96 96 96 96 97 7,00 ср. вр. Гриив. 96
55 50 58 65 68 71 73 75 72 68 66 59 14,00 ср. вр. Гринв. 63
13. Сокото, Нигерия 30 29 24 40 59 67 79 85 79 63 38 31 7,00 мест. вр. 52
13 И 13 18 31 42 54 59 55 31 20 14 14,00 мест. вр. 30
14. Рангун, Бирма 77 83 85 83 83 89 92 91 90 84 83 78 8,00 мест. вр. 85
51 50 54 64 75 86 88 88 86 77 71 61 18,00 мест. вр. 71
15. Пасириан, Ява 84 86 85 81 78 77 73 69 66 67 71 79 8,00 мест. вр. 76
72 73 71 67 65 63 58 57 55 57 63 70 14,00 мест. вр. 64
1 Минимальные средние месячные значения отмечены полужирным шрифтом.
* 1146 м выше уровня моря.
Источники. Пункт 1—по данным Службы леса Брит. Гвианы; пункт 2—по данным Бернара (Bernard, 1945); ПУПКТ 1945?
1934); 9—по данным Эгге линга (Eggeling, 1947); пункты 10, 15—по данным |Ван-Стеениса (yan Steenls); пункт И—по Стэлс (Ыеще, )
пункт 12—по данным Фогги (Foggie, 1947); остальные данные были любевно предоставлены Лондонским метеорологическим управлением.
166
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
в 12 мм (Richards, 1936, стр. 8). При средней дневной температуре, равной 15°, в умеренном климате это соответствовало бы относительной влажности в 6%. Пока еще в нашем распоряжении мало данных о дефиците насыщения в тропических местностях. Бекер и Харрисон (Baker, Наг-risson, 1936, стр. 459—460) указывают средний месячный дефицит насыщения для 2 часов пополудни в местности на Новых Гебридах. Бернар (Bernard, 1945) недавно опубликовал данные о дефиците насыщения для ряда местностей в Республике Конго. Наблюдения Ивенса (Evans, 1939) над дефицитом насыщения в тропическом дождевом лесу Нигерии рассматриваются в главе 7.
В более сухих частях тропической зоны средняя относительная влажность в целом ниже, чем в области развития тропического дождевого леса; очень низкая влажность бывает чаще и держится дольше. Уменьшение атмосферной влажности от побережья в глубь материка является одной из отличительных особенностей климатического градиента в Западной Африке, что и подтверждают данные Джонса и Кия (А. Р. D. Jones, R. W. J. Кеау), приводимые ими в недавнем (неопубликованном) исследовании зон растительности и климата в Нигерии (табл. 14).
Таблица 14
Осадки и относительная влажность в различных растительных зонах Нигерии
По неопубликованным данным Джонса и Кия (А. Р. D. Jones, R. W. J. Кеау)
Зона Годовая сумма осадков, см Число месяцев с осадками менее 3 см Наименьшая средняя месячная относительная влажность, %
средняя за несколько лет наименьшая зарегистрированная
1 Свыше 230 0—1,3 78—85 70—80
2 140—230 1-2,3 71—84 60—70
3 110-230 1,9—4,3 58—81 40—65
4 110—150 3,8—5,0 44 63 30-55
5 95—145 4,2-5,8 26-35 22—30
В зоне 1, ближайшей к морю, развивается типичный тропический дождевой лес, в зоне 2 — влажный вечнозеленый лес (см. главу 15), а в зоне 3 — сухой вечнозеленый лес (смешанный листопадный лес, см. стр. 361). В остальных зонах (4—5) растительный покров представлен саванновыми лесами и т. п., в значительной степени видоизмененными в результате пожаров и воздействия биотических факторов. Имеются доказательства того, что наличие вечнозеленых и полувечнозеленых лесов в прибрежных зонах (1—3) зависит от высокой минимальной влажности в течение засушливого периода, противодействующей отрицательному влиянию длительной засухи. Наблюдения Бакстона и Льюиса (Buxton, Lewis, 1934, стр. 228—231) над относительной влажностью и дефицитом насыщения в Гадау, северная Нигерия, показывают, насколько различными могут быть условия среды в районе внутриматериковых саванн в засушливый сезон. Автор настоящей работы (Richards, 1939, стр. 9—10) уже показал, что распределение тропических дождевых лесов
СРЕДА
167
и смешанных листопадных лесов южной Нигерии не обнаруживает тесной связи с длительностью сухого сезона, и предположил, что определяющим фактором может быть степень проникновения в глубь материка влажного морского воздуха. Наблюдения в Сенегале, согласно Анго (Angot, 1928, стр. 148), наглядно демонстрируют ярко выраженное умеряющее влияние ветров, периодически дующих с моря, на тропический климат с сильными засухами; морской ветер в сухой сезон может вызвать после полудня повышение относительной влажности на 60% или более при одновременном значительном понижении температуры.
На горах в тропиках влажность повышается с увеличением высоты над уровнем моря, и высотный пояс начинается там, где постоянная облачность и туман не дают влажности упасть заметно ниже величины насыщения. Данные Брауна (Brown, 1919, стр. 272—275) для горы Маквилинг, Филиппины, иллюстрируют влияние высоты на влажность; у подножия горы (80 м) средняя относительная влажность под покровом порослевого леса составляла 82,2% при среднем суточном минимуме в 68,6%, в то время как на открытом месте на высоте 450 м относительная влажность выражается числом 87,9% при среднем суточном минимуме в 75,5%. В «моховом лесу» на высоте в 1000 м воздух месяцами остается на уровне насыщения или близко к нему, и самая низкая относительная влажность, зарегистрированная здесь когда-либо, составляет 91 ?6. Среднее суточное испарение соответственно понижается с увеличением высоты. Несомненно, подобные же результаты можно получить и на других горах тропической зоны.
х ВЕТЕР
Важная экологическая роль ветра выявляется с нескольких точек зрения. Движение воздуха влияет на величину и скорость испарения и поэтому является одним из основных факторов, регулирующих водный режим растений. В некоторых частях тропиков огромное влияние на растительность оказывают иссушающие ветры, например восточный муссон на Зондских островах и харматан в Западной Африке, которые ежегодно на короткое время значительно понижают влажность воздуха даже в области развития тропических дождевых лесов. Механическое воздействие сильных ветров также имеет экологическое значение и даже может препятствовать нормальному развитию климаксной растительности.
В общем для тропической зоны характерны более низкие средние скорости ветра, и очень сильные ветры в ее пределах наблюдаются реже, чем в умеренных областях, исключая лишь отдельные местности, вроде «пояса ураганов» Вест-Индии. Средние годовые скорости ветров в области распространения тропических дождевых лесов обычно не достигают 5 км/час, редко превосходя 12 км/час. Многим тропическим районам свойственны частые грозы, которым нередко предшествует шквальный ветер (торнадо), длящийся всего несколько минут, но развивающий чрезвычайно большую скорость. Группы деревьев и даже участки леса площадью в несколько гектаров, поваленные подобным шквалом,— нередкое зрелище в Нигерии и Гвиане. В зонах развития ураганов и тропических циклонов действие ветров выражено в еще большей степени. Воган и Вийе (Vaughan, Wiehe, 1937, стр. 301—302) противопоставляют губительное воздействие циклонических ветров на посевы и плантации тому ничтожному ущербу, который терпят коренные растительные сооб
168
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
щества, приспособившиеся к ветрам, благодаря длительному отбору. Бирд (Beard, 1945) назвал «ураганным лесом» субклимаксный лес, лишенный высоких деревьев, расположенный выше 500 м над уровнем моря на склонах вулкана Суфриер, остров Сент-Винсент (Вест-Индия), где частые ураганы наряду с крутизной склонов делают невозможным развитие типичного тропического дождевого леса. Как структура, так и состав растительности частично определяются ветром.
На открытом морском побережье сильные ветры могут деформировать кроны деревьев и способствовать развитию полосы характерного редколесья или кустарника. Бирд (Beard, 1946b, стр. 108—110) описывает полосу прибрежного леса шириной в 20—400 м на наветренном берегу Тринидада; своеобразие облика флористического состава этого сообщества он приписывает действию пропитанного солью пассата. Крайнее измельчение растительности на изолированных вершинах и хребтах тропических гор также отчасти может быть вызвано наличием сильных ветров, хотя, вероятно, есть и другие причины повсеместно наблюдаемых признаков угнетения деревьев по мере увеличения высоты над уровнем моря (см. стр. 173).
ИНСОЛЯЦИЯ И ОБЛАЧНОСТЬ
Продолжительность дня на экваторе составляет 12 часов лишь с незначительными сезонными колебаниями в длине сумерек; с широтой сезонные колебания продолжительности дня увеличиваются и на линиях тропиков составляют 2 часа. Хотя возможная длительность инсоляции никогда не составляет менее 10 часов, количество прямой радиации оказывается гораздо меньше предполагаемых величин благодаря сильной облачности. Вопреки обычному мнению для тропической зоны вовсе не характерно вечно безоблачное небо; это особенно относится к экваториальному поясу, который за свою облачность заслужил у французских матросов название «pot au noir». Приэкваториальная полоса в сущности является более или менее постоянным поясом облачности, где редко бывают ясные дни; средняя облачность над сушей составляет 5,2 балла в зоне от 0 до 10° с. ш. и 5,6 — в зоне от 0 до 10° ю. ш. Севернее и южнее облачность уменьшается, доходя до минимума у обоих тропиков [между 20 и 30° с. ш. над сушей — 3,4 балла; между 20 и 30° ю. ш.— 3,8 балла (Brooks, 1927а)]. Процент облачности в тропиках сильно колеблется в течение года; обычно он бывает наивысшим в дождливый сезон, хотя в некоторых районах наибольшая облачность совпадает с сухим сезоном.
Поскольку легкие облака лишь в незначительной степени уменьшают освещение, то число часов яркого солнечного сияния, зарегистрированное соответствующим прибором, имеет более важное значение с экологической точки зрения, чем процент облачности. К сожалению, такими данными располагают лишь сравнительно немногие станции в области развития тропических дождевых лесов. Следующие цифры (1920—1929 гг.) для станции Мазаруни, Брит. Гвиана, по-видимому, можно считать довольно типичными:
Среднее дневное солнечное сияние 5,5 часа
Среднее для наиболее солнечных месяцев (сентябрь, сухой сезон) 6,3 »
Среднее для наименее солнечных месяцев (июнь, дождливый сезон) 4,4 »
СРЕДА
169-
Данные Бекера и Харрисона (Baker, Harrisson, 1936, стр. 458) для Новых Гебридов показывают аналогичную дневную продолжительность солнечного сияния, но несколько большие колебания в средних месячных. В дождевом лесу Панамы, согласно данным Элли (Allee, 1926 а, стр. 276), продолжительность солнечного сияния значительно больше, чем на упомянутых станциях (70% возможного солнечного сияния в сухой сезон, 40% — в сезон дождей). Бернар (Bernard, 1945) считает, что в Янгамби, Республика Конго, среднее дневное солнечное сияние составляет около 5,1 часа, то есть 42,3% возможной инсоляции. Соответственно средняя величина для ряда тропических станций, расположенных на низменностях с морским климатом, составляет 58% [данные взяты Бернаром из книги Кёппена и Гейгера (Кбрреп, Geiger, Handbuch der Klimatologie)].
Поразительно малое количество яркого солнечного света особенно бросается в глаза при попытке фотографировать в тропическом дождевом лесу. Автор по собственному опыту знает, что облака непрерывно набегают на солнце даже в сухой сезон, и часто бывает трудно отыскать-периоды непрерывного освещения для достаточно долгой экспозиции.
Интенсивность освещения и общая величина солнечной радиации в тропических климатах изучена очень мало, но имеющиеся данные показывают, что, несмотря на высокое стояние тропического солнца, радиация на низменностях тропической зоны (на уровне земли), пожалуй, даже менее интенсивна, чем в умеренных областях. Борэма (Boerema, 1919) на основании пиргелиометрических данных установил, что в Богоре, на Яве (6°35' ю. ш.), общая радиация была значительно меньшей, чем в Вашингтоне, округ Колумбия (38°52z с. ш.), при близкой высоте солнца над горизонтом. Бюннинг (Bunning, 1948, стр. 96—98), сопоставив свои собственные и чужие измерения интенсивности освещения в тропиках, целиком принимает вывод Орта (Orth, 1939) о том, что в общем в Европе освещение ярче. Бернар (Bernard, 1945) провел углубленное изучение радиации в бассейне центрального Конго и пришел к выводу, что как средние, так и максимальные (кратковременные) величины общей радиации, вероятно, не выше, чем в умеренных областях в летние месяцы, причем здесь наблюдается явный дефицит синей и ультра-фиолетовой радиации. Полученные относительно низкие общие показатели радиации в этой части Африки Бернар относит за счет высокого содержания водяных паров и загрязненности (песок и вулканическая пыль, дым от многочисленных пожарищ и пр.) нижних слоев атмосферы. Вопрос о том, применимо ли подобное же объяснение ко всей тропической зоне в целом, остается открытым.
КЛИМАТИЧЕСКИЕ ГРАНИЦЫ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
В предыдущих разделах дано общее представление о климате тропического дождевого леса, основные особенности которого вполне понятны;, однако при попытках дать точное определение этого климата и провести границу между тропическим дождевым лесом, с одной стороны, и листопадным лесом, дождевым лесом предгорий и субтропическим дождевым лесом — с другой, приходится сталкиваться с большими затруднениями. В настоящее время эти трудности главным образом связаны с незнанием точного положения границы тропического дождевого леса и недостаточностью данных о климате местностей, расположенных близ этой предполагаемой границы.
170
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
В главе первой указывалось, что тропический дождевой лес мог бы произрастать на гораздо большей площади, чем та, которую он фактически занимает. Существуют большие безлесные площади, где климатические условия вполне подходящи для развития тропического дождевого леса: на некоторых из них он был уничтожен человеком и замещен посевами или вторичными сообществами; на других он отсутствует в силу других причин, таких, как неразвитость, или, наоборот, деградация почвы, или же неблагоприятные местные топографические условия. В таких местах он замещается саванной или зарослями вечнозеленого кустарника. Определяя климатические границы формации тропического дождевого леса, необходимо принимать в расчет всю площадь, на которой он потенциально может произрастать или действительно произрастает. В то же время нужно помнить, что само наличие большого лесного массива до некоторой степени изменяет климат, так что климат области потенциального развития тропического дождевого леса мог бы быть другим при существовании в ней лесного покрова.
Некоторые климатологи и ботаники в целях определения границ, внутри которых можно успешно выращивать определенные сельскохозяйственные культуры, пытались классифицировать климат посредством эмпирических «климатических индексов», включающих несколько климатических факторов. Подобные индексы применялись для определения климата тропического дождевого леса и других основных типов растительности. Наиболее известная и, вероятно, наиболее пригодная для экологических целей классификация климата была предложена Кёппеном (Корреи, 1918; 1936), который дает следующие параметры для влажного тропического климата, или климата дождевых лесов (А — климаты): средняя температура самых холодных месяцев (t) выше 18°, осадки (в мм) свыше 2^ + 14. Этот обширный класс климатов он подразделяет на Af (жаркие сырые лесные климаты) со средней суммой осадков в наиболее сухие месяцы не менее 60 мм\ Ат (лесные климаты, муссонный тип) и Aw (климаты саванн с периодическими засухами). Для климатов Ат и Aw обычна сезонная засуха, и средняя сумма осадков самого сухого месяца составляет менее 60 мм. Но в климатах Ат, занимающих промежуточное положение между Af и Aw, годовая сумма осадков способна «компенсировать» сезонные засухи (так, если в самый сухой месяц выпадает 40 мм, то годовая сумма должна превышать 1500 мм, если — 20 мм, то годовая сумма должна превышать 2000 мм и т. д.). В климатах Aw сухой сезон не компенсируется годовой суммой осадков. В общем, в местностях с климатами Af произрастают тропические дождевые леса; с климатами Ат — вечнозеленые сезонные, муссонные и т. п. лесные сообщества, а климатам Aw соответствуют саванны. Бернар (Bernard, 1945) детализировал эту схему применительно к растительному покрову центральной части бассейна Конго. Здесь климат Af свойствен узкому поясу, тянущемуся параллельно экватору и охватывающему территорию по обе стороны от него, которая простирается к югу примерно на 1° дальше, чем к северу. С юга и с севера к этому поясу примыкают области с климатами Ат, сменяющиеся в свою очередь климатами Aw. Область Af довольно точно соответствует площади настоящего омброфильного леса — («veritable foret ombrophile»), области Am — тропофильному лесу («foret tropophile») (вероятно, эквивалент смешанного листопадного леса Западной Африки) (см. гл. 15), а области Aw — саваннам.
Предлагалось и много других «климатических индексов», при помощи которых с той или иной степенью успеха старались установить связь
СРЕДА
171
климата с естественной растительностью; среди них — «фактор осадков» (осадки/средняя годовая температура) Ланга (Lang), «индекс аридности» С температура + 10) ^аРтонна (Martonne), коэффициент Майера (Меуег) {осадки/средний дефицит насыщения) и показатели «эффективности температуры» и «эффективности осадков» Торнтвэйта (Thornthwaite).
Моро (Moreau, 1938), сделавший чрезвычайно полезный обзор этих классификаций климата с биологической точки зрения, указал на целый ряд недочетов в них. Некоторые климатические индексы требуют таких данных, как средние дефициты насыщения, которые известны для очень немногих мест; другие не учитывают такие факторы, как «оккультные» (скрытые), или «горизонтальные» осадки, порождаемые туманами и влажными ветрами, которые могут иметь большое местное значение, изменяя воздействие осадков и температуры, а через них и самые биологические границы. Все эти индексы построены на «нормалях» с малым учетом аномальных или экстремальных метеорологических условий, которые могут оказаться решающими в экологии. Из опыта известно, что, хотя некоторые из этих климатических индексов в общих чертах довольно хорошо связывают предлагаемые климатические границы с распределением главных растительных сообществ, таких, как тропический дождевой лес, в деталях корреляция редко бывает удовлетворительна. Экологу, работающему в поле, все попытки установить тесную связь между биологическими границами и суммой климатических факторов представляются обреченными на неудачу, так как он знает, что распределение растительных сообществ (или, вернее, экосистем) зависит от многих как климатических, так и почвенных факторов. Взаимодействие экологических факторов так сложно, что нельзя надеяться на возможность простого количественного определения климатических границ или вообще какое-либо точное определение их при существующем, весьма ограниченном, знании климатических элементов, за исключением температуры и осадков.
На своих северных и южных границах тропический дождевой лес, как уже указывалось, обычно и, как правило, постепенно переходит в листопадный лес или более резко — в саванну. В главе 15 будет показано, что саванна, вероятно, никогда не бывает обусловлена климатическими факторами, и, следовательно, не существует и «климата саванн»; саванна возникает либо вследствие периодических пожаров, либо под влиянием определенных, но пока еще малопонятных особенностей почвенного покрова. Поэтому о климатической границе между тропическим дождевым лесом и саванной не приходится говорить. В то же время граница между тропическим дождевым лесом и листопадным лесом является действительным климатическим пределом. Существуют веские доказательства того, что эта граница определяется в основном сезонными засухами и что прямое воздействие температуры играет здесь незначительную роль. Иначе обстоит дело с высотными границами дождевого леса в горах и широтными границами в некоторых местностях, например восточной Австралии, где он очень постепенно переходит не в листопадный лес или саванну, а в субтропический вечнозеленый лес; эти границы почти наверняка частично обусловлены непосредственным влиянием температуры, хотя и не известно, какие именно ее показатели (средняя температура, месячный минимум, абсолютный минимум и т. д.) оказываются решающими.
Абсолютно все согласны, что климатическая граница тропического дождевого леса зависит от длительности сухого сезона, который соответствует числу «сухих месяцев», но в определении понятия «сухого месяца»
Б. М а з о ру » «, fW
’Средние идете осиЛш '^^.я,кпе гЛя^аа 9пИпаая температура'26,0
Рис. 22. Сезонное распределение осадков и температуры в тропических климатах-А.— Сингапур. Экваториальный климат, осадки распределены относительно равномерно.’Климаксная растительность: тропический дождевой лес. Б.— Станция Мазаруни, Брит. Гвиана. Два дождливых сезона. Климаксная растительность: тропический дождевой лес. В. — Лагос, Нигерия. Два дождливых сезона; относительно выраженный сухой сезон. Климаксная растительность: тропический дождевой лес (близ своей климатической границы). Г. — Кадуна,11 Нигерия. Один дождливый сезон, длительный сухой сезон. Климаксная растительность: саванна. Данные для пункта Б сообщены представителем Службы леса Брит. Гвианы; А, В, Г — по Бруксу (Brooks, 1932).
СРЕДА
173
мнения расходятся. Если только осадки в течение данного периода превосходят определенный минимум, достаточный, чтобы предотвратить снижение уровня почвенной влаги ниже критической величины, то дальнейшее увеличение осадков, вероятно, уже не существенно. Этот минимум будет зависеть от ряда факторов, включая испаряющую способность воздуха, транспирационную способность растительности, особенности почвы и топографии. Бирд (Beard, 1946 Ь) эмпирически установил, что месячную сумму осадков, равную 4 дюймам (10 см), в общем можно считать критической для Вест-Индии и что сухой период будет соответствовать числу следующих друг за другом месяцев с количеством осадков меньше упомянутого. Возможно, что эта цифра действительна и для других частей тропиков, хотя, пытаясь определить климатическую границу между вечнозеленым лесом и смешанным листопадным лесом (стр. 361) в южной Нигерии, автор (Richards, 1939, стр. 9) установил, что она зависела не •столько от числа последовательных месяцев с количеством осадков менее 10 см, сколько от суммы годовых осадков. Годовые суммы осадков сами по себе вряд ли могут оказаться решающим фактором. Более вероятно, что границу определяет средняя влажность в течение самых сухих месяцев, которая непосредственно не зависит от количества осадков, но находится в соотношении с ними. А. С. Томас (A. S. Thomas, 1932), изучая сухой сезон в Гане как фактор, лимитирующий рост какао (в диком состоянии произрастает в тропическом дождевом лесу), предложил определять сухой сезон по «степени влажности», используя формулу:
осадки в дюймах х число дней с дождем 100
Установленные этим способом и разными другими путями корреляции .между «сухими месяцами» и распределением тропических растительных формаций необходимо изучать в возможно большем числе районов.
Поскольку критическим фактором роста растений является отношение водоснабжающей способности почвы к расходу воды на транспирацию, топографические и эдафические факторы наравне с климатом должны иг рать большую роль в определении границ тропического дождевого леса. Так Бирд (Beard, 1946 Ь) обнаружил, что на острове Тринидад полувечнозеле-ный сезонный лес произрастает на известняках в тех климатических областях, где на менее водопроницаемых почвах развит вечнозеленый сезонный лес. Стэмп и Лорд (Stamp, Lord, 1923) и Стэмп (1925) подчеркивают важность как почвенных, так и климатических факторов в распределении полувечнозеленого и листопадного лесов в Бирме. Растительное сообщество, подобно отдельному виду, непременно будет более чувствительно к почвенным условиям на границе области своего распространения, нежели в глубине ее. Это настолько важно, что в Австралии, согласно Фрэнсису (Francis, 1929, стр. 6), влияние почв на развитие тропического дождевого леса все более и более возрастает по мере удаления от экватора. Таким образом, нельзя считать, что лимит засушливости тропического дождевого леса находится в точном соответствии с одними только климатическими факторами, будет ли это единичный фактор или их комбинация.
Факторы, лимитирующие вертикальное распространение дождевого леса в горах и его распространение до естественных широтных границ, пока мало изучены, если не считать весьма тщательного исследования Брауна (Brown, 1919) о размещении растительности на горе Маквилинг, Филиппины. На этой горе диптерокарповый лес (тропический дождевой лес) уступает место «среднегорному лесу» (влажному лесу предгорий)
174
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
на высоте около 600 м, а этот в свою очередь — «моховому лесу» на высоте около 900 м. Наиболее заметная особенность, бросающаяся в глаза при сравнении вертикальных поясов, состоит в сильном снижении высоты деревьев от диптерокарпового леса (средняя высота деревьев яруса А — 27,2 м) до карликового «мохового леса» (средняя высота полога — около 5,9 м). Тщательно изучив все климатические факторы и почвенные условия на различных высотах, Браун пришел к выводу, что с возрастанием высоты над уровнем моря влажность атмосферы увеличивается, сопровождаясь увеличением облачности с соответствующим уменьшением средней интенсивности освещения и понижением температуры. Возрастание влажности и падение температуры приводили к быстрому снижению величины испарения по мере подъема. Изучение роста деревьев наводит на мысль, что их измельчение нельзя приписать только действию температуры, а следует ставить в зависимость от комбинированного -воздействия температуры и интенсивности освещения. Высота деревьев тесно согласуется с индексами «освещение — температура» (результат умножения интенсивности освещения на «индексы температуры» для роста растений)* 1, за исключением меньших абсолютных высот, где более низкий по сравнению с ожидаемым рост деревьев, вероятно, находится в связи с влиянием сезонных засух. Другая поразительная особенность вертикальной поясности проявляется в увеличении количества эпифитов, а также более пышном развитии травяного покрова по мере увеличения высоты. Первое Браун объясняет снижением интенсивности испарения, а второе — увеличением почвенной влаги в сочетании с меньшим испарением.
Хотя не представляется возможным делать общие выводы по одним только данным Брауна, весьма вероятно, что температура — не единственный фактор, определяющий как высотные, так и широтные границы тропического дождевого леса. «Пояс туманов», или «пояс облаков», который существует на некоторых тропических горах, по-видимому, объясняющий (см. гл. 16) часто внезапный переход от леса предгорий к горному дождевому лесу, вряд ли может определить верхнюю границу тропического дождевого леса, которая обычно расположена ниже. Понижение поясов растительности, включая верхнюю границу тропического дождевого леса на обособленных горах, особенно вблизи моря, по сравнению с большими горными хребтами (стр. 370), несомненно, связано с соответствующим понижением климатических поясов.
Вопрос о климатических границах тропического дождевого леса представляет собой обширное поле для дальнейших исследований. Более точное, чем то, какое возможно сделать сейчас, определение границ будет чрезвычайно ценно не только для эколога, изучающего существующие в настоящее время растительные сообщества, но и для палеоботаника, пытающегося по ископаемой флоре определить климаты прошлого. В качестве примера можно упомянуть соображения Рида и Чендлера (Reid, Chandler. 1933) о климате, соответствующем флоре лондонской глины (эоцен). Тщательное изучение неизбежно приводит к выводу, что эта ископаемая флора, включающая виды рода Nipa (стр. 323) и много других
1 Согласно Б. Е. и Г. Дж. Ливингстон (В. Е. and G. J. Livingston, 1913) и Б. Е. Ливингстон (В. Е. Livingston, 1916), «экспоненциальныйтемпературный индекс» состав-2t—40 .
ляет ——— , где t — «нормальная средняя суточная температура», если брать за еди-
1 о
ницу скорость роста при 40е F (4,4° С) и принять, что скорость роста удваивается при каждом повышении на 10°. Был использован и «физиологический температурный индекс», установленный ранее при экспериментальной работе* с рассадой кукурузы.
СРЕДА
17Г>
растений, родственных теперешней Индо-Малайской флоре, была сходна с современной тропической (или по крайней мере субтропической) флорой зоны эстуариев и должна была существовать в климате гораздо более теплом, чем тот, который свойствен любой части нынешней Европы. Вряд ли можно оспаривать вывод Рида и Чендлера, что флора лондонской глины должна была существовать при обильных и равномерно распределенных осадках в условиях безморозной зимы и при средней годовой температуре не намного ниже 70° F (21° С). Однако более точные сведения об этом климате могут быть получены лишь после дальнейшего изучения современных климатических границ тропического дождевого леса и других родственных растительных сообществ1.
ЗАМЕЧАНИЯ О НЕКОТОРЫХ ФИЗИОЛОГИЧЕСКИХ ВОЗДЕЙСТВИЯХ КЛИМАТА ТРОПИЧЕСКИХ ДОЖДЕВЫХ ЛЕСОВ
Поскольку климат тропических дождевых лесов так сильно отличается от климата умеренных областей, то очевидно, что для эколога чрезвычайно важно иметь представление об общем воздействии тропических условий на основные физиологические процессы, протекающие в растениях. Здесь не представляется возможным заняться подробным рассмотрением физиологии тропических растений, но несколько кратких замечаний по этому вопросу помогут выяснить связь между климатом дождевых лесов и растительностью или по крайней мере указать на те проблемы, которые требуют разрешения.
Человека, впервые попавшего в экваториальную область, неизменно поражает буйная роскошь растительности — несомненный результат воздействия влажного теплого климата. В глаза особенно бросается необыкновенное изобилие разнообразнейших крупных и высоких растений. В какой-то мере впечатление пышности вызывается очень быстрым ростом некоторых элементов флоры, в частности видов, преобладающих на ранних стадиях вторичной сукцессии.
Пустыри, обочины дорог и заброшенные пашни не остаются практически незаросшими в течение многих месяцев или лет, как в умеренном поясе, а за несколько недель покрываются густой растительностью высотой в несколько метров. Некоторые виды растут поразительно быстро. У бамбука Dendrocalamus giganteus в течение примерно двух месяцев наблюдения ежедневный прирост в среднем составлял 22,9 см, а у одного экземпляра достигал 57 см в день (Kraus, 1895). Многие деревья вторичных лесов из семейства двудольных также растут очень быстро. Африканская Musanga cecropioides за 14 лет становится солидным деревом высотой в 16 м. На Яве наблюдали Albizzia falcata, которая за 6 лет достигла высоты в 25 м, а за 10 лет — 35 м (Koorders, Valeton, 1894; Haberlandt, 1926, стр. ill). Очень быстрый рост гораздо менее заметен в растительных сообществах, достигших полного развития. Помимо основных лесообразующих пород, которые, как отмечалось в главе 3, могут
1 Попутно следует отметить, что заключение о климате, сопутствовавшем развитию третичной флоры, выглядит основательнее, когда оно выводится из статистического изучения размеров листьев и тождественности внешнего облика (ср. Bews. 1927), чем когда оно базируется на анализе плодов и семян (Reid, Chandler), то есть на таксономическом родстве, а не на физиономических особенностях, которые, как но крайней мере свидетельствует современная растительность, являются исключительно чутким показателем условий среды.
176
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
годами существовать, не увеличиваясь сколько-нибудь заметно в размерах, известно много других видов, растущих весьма медленно. Клебс (Klebs, 1911) приводит множество подтверждающих это примеров. Браун (Brown, 1919) обнаружил, что на Филиппинах древесная порода из семейства диптерокарповых — Parashorea malaanonan — на ранней стадии развития росла медленнее, чем типичные североамериканские деревья — желтый тополь и белая сосна,— хотя в дальнейшем она обгоняла их в скорости роста. Крайним примером медленно растущего тропического растения является эпифитная орхидея Taenophyllum obtusum. у которой главными органами фотосинтеза служат уплощенные воздушные корни. В серии измерений роста этих корней при различной интенсивности освещения Визнер (Wiesner, 1897) установил максимальный прирост в 0,283мм в день, что составляет примерно 0,001 по сравнению с приростом вышеупомянутого бамбука. Согласно Лебрэну (Lebrun, 1936 b, стр. 176), диаметр деревьев в девственных экваториальных лесах Конго ежегодно увеличивается на 0,5—2 см. средний прирост в высоту составляет около 1,4 см.
Можно считать, что скорость роста большинства тропических растений — по крайней мере в первичных ненарушенных лесах — находится в пределах того, что обычно считается «нормой», хотя некоторым представителям флоры этих лесов свойствен исключительно быстрый рост; это выдвигает физиологические проблемы, которые в основном все еще не решены. Напрашиваются три возможных объяснения этого чрезвычайно высокого темпа роста: либо средняя величина ассимиляции (увеличение сухого веса всего растения на единицу площади поверхности листа в день) превосходит аналогичный показатель у любых растений умеренного пояса; .либо на увеличение площади листьев расходуется большая, чем у растений умеренного пояса, часть всех ассимилированных веществ (что приводит к более быстрому увеличению размеров растения в целом); либо же объяснение заключается просто в длительном непрерывном активном росте, который в некоторых случаях может и не прерываться периодами отдыха.
Данные, которыми можно было бы проверить эти три предположения, пока еще очень скудны. Из последних исследований — ни работа Гутенберга (Guttenberg, 1931), ни работа Стокера (Stocker, 1935 а) не дают величин уровня ассимиляции, превосходящих нормы, свойственные растениям умеренного пояса. В частности, у Стокера указываются совсем небольшие цифры, вероятно, благодаря специфике примененной им газометрической техники. Хотя метод «половины листа», примененный Гутенбергом, имеет то преимущество, что растения не находятся в искусственной внешней •среде, он оставляет сомнение относительно как количества, так и предназначения вещества, перемещенного из листьев. Кроме того, использованные Гутенбергом растения не принадлежали к наиболее быстрорастущим тропическим видам. В настоящее время не представляется возможным разграничить или исключить две первые из вышеупомянутых возможностей. Необходимы детальные исследования, которые дали бы картину не только ассимиляционной способности листьев, но также и схему роста всего растения у некоторых особенно быстрорастущих видов.
Вполне вероятно, что на ранних стадиях развития быстрорастущие тропические растения ведут себя примерно так же, как и однолетние растения умеренного пояса, такие, как Helianthus annuus. который при благоприятных условиях еженедельно удваивает свой сухой вес, пока не наступит пора цветения. После цветения и осыпания семян Helianthus погибает, достигнув за свою примерно трехмесячную жизнь размеров, сопоставимых •с размерами быстрорастущих тропических растений; в отличие от него
СРЕДА
177
последние обычно продолжают расти и после цветения. В связи с этим интересно вспомнить, что многие быстрорастущие тропические деревья (см. стр. 405) недолговечны и живут только несколько лет. Возможно, что интенсивно развивающиеся однолетние растения умеренных зон, завершающие свой жизненный цикл за несколько месяцев, представляют крайнюю ступень в ряду быстрорастущих недолговечных растений, преобладающих во вторичных сообществах тропиков.
Процесс дыхания листьев тропических деревьев был исследован Стокером (Stocker, 1935 а), который нашел, что средняя интенсивность дыхания освещенных солнцем листьев у трех видов, один из которых — Stele-choearpus burahol — характерен для тропического дождевого леса, составляла 1,3 мг СО2 на 1 кв. дм в час при 30° С. Эта величина весьма близка к полученным в Дании для Fagus silvatica (1,0 мг) и Fraxinus excelsior (1,2 мг). Однако Стокер считает доказанным, что в действительности интенсивность дыхания тропических деревьев ниже, чем европейских при той же температуре, и что тропическое растение при 30° дышит не более интенсивно, чем арктическое при 10°. Поскольку интенсивность дыхания, выраженная в этих показателях, так сильно колеблется у различных видов в одном и том же климате, теорию Стокера о «физиологической адаптации» дыхания к климату в настоящее время нельзя считать прочно обоснованной. Не удивительно, что при отсутствии достоверных сведений об интенсивности ассимиляции и дыхания у тропических растений сравнительные данные относительно момента компенсации (когда ассимиляция уравновешивает дыхание) у тропических и умеренных видов в настоящее время слишком скудны для окончательных выводов; такие данные были бы, однако, весьма ценны для понимания связи между тропическими тенелюбивыми растениями и окружающей их средой.
Некоторые ботаники-физиологи пытались установить общие закономерности, касающиеся интенсивности транспирации у тропических растений. Габерланд (Haberlandt, 1898, 1899), например, утверждал, что интенсивность транспирации у растений влажных тропиков в среднем в 2—3 раза ниже, чем в Центральной Европе. Однако видовые различия в этом отношении настолько значительны, что подобные утверждения почти не имеют смысла. У тропических растений, существующих в условиях сильного дефицита насыщения, таких, как деревья верхних ярусов дождевого леса или растения, произрастающие на обработанных участках и других открытых местах, можно наблюдать такую же и еще более высокую интенсивность транспирации, чем у европейских растений; с другой стороны, тенелюбивые растения подлеска тропического дождевого леса могут обладать более низкой интенсивностью транспирации, чем типичные европейские тенелюбивые растения. Большие колебания влажности воздуха в различных тропических микроклиматах (см. гл. 7) только отчасти объясняют столь несходное поведение различных видов. Интенсивность транспирации зависит не только от разности в величинах парциального давления водяного пара в воздухе, непосредственно окружающем мезофилл ьные клетки листа, и того же пара в атмосфере на определенном расстоянии от растения, но также и от сопротивления воздуха диффузии пара при движении его от листа в атмосферу. По-видимому, именно в этом сопротивлении диффузии и заключается постоянное различие в транспирации между растениями тропической и умеренной зоны. К сожалению, с этой точки зрения вопрос еще не был исследован, а это могло бы пролить некоторый свет на причину возникновения большой амплитуды интенсивности транспирации у тропических растений, населяющих различные микрокли-12 п. У. Ричардс
178
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
маты. Стокер (Stocker, 1935 b) также находит основания полагать, что, приспособляясь как с помощью «эволюции поверхности» (то есть отношения поверхности к объему), так и с помощью изменения соотношения содержания воды к площади поверхности, растения всех климатов способны удерживать расход влаги в определенных границах; другими словами, интенсивность транспирации по отношению к общему содержанию влаги показывает лишь незначительные изменения. Едва ли собственные данные Стокера подтверждают этот взгляд, так как у наблюдавшихся им тропических тенелюбивых растений в тени величина транспирации была гораздо ниже, чем у светолюбивых на солнце, хотя содержание влаги на единицу площади поверхности у них было гораздо выше.
В заключение можно упомянуть о косвенном влиянии тропического климата на рост растений, связанном со снабжением минеральными питательными веществами. Как будет показано в главе 9, сильные и постоянные ливни влажных тропиков приводят к непрерывному вымыванию растворимых солей из почвы. Поскольку сама почва обладает кислой реакцией п часто имеет высокую водопроницаемость, запасы минеральной пищи для растений всегда недостаточны. Физиологические последствия большой бедности запасов минерального питания для неокультуренных тропических растений (в отличие от возделываемых) все еще не изучены. Однако весьма вероятно, что борьба между корнями за скудные запасы питательных веществ у тропических растений должна быть еще ожесточенней, чем у растений умеренного пояса (ср. стр. 211) — это в свою очередь может серьезно влиять как на скорость роста растений, так, возможно, и на другие стороны их физиологии.
Г ла в а 7
МИКРОКЛИМАТЫ
Предыдущая глава была в основном посвящена рассмотрению климатических условий, характерных для сообществ тропического дождевого леса в целом, иными словами, речь шла о так называемом стандартном, или обобщенном климате, областей, занятых формациями этого леса. Такой стандартный климат — всего лишь один из многих климатов, сосуществующих в одной и той же местности в различных условиях; это климат, зарегистрированный примерно на уровне 1 м над поверхностью земли приборами, установленными, согласно правилам метеорологии, на более или менее обширных открытых территориях. На высоте 100 м над поверхностью земли или непосредственно на почве климат будет иным, чем на высоте 1 м, а в подлеске сомкнутого леса он окажется совсем другим, чем на больших открытых пространствах. В сущности стандартный климат выбран произвольно из множества отдельных климатов или микроклиматов главным образом потому, что он приближается к тем условиям, в каких находится человек на открытом воздухе. В пределах такого сложного растительного сообщества, как тропический лес, может существовать бок о бок много микроклиматов, и некоторые из них очень отличаются от стандартного климата той же местности.
Растения испытывают прямое воздействие только их непосредственного окружения. Поэтому, хотя листва зрелых деревьев, преобладающих в лесном сообществе, существует в условиях, близких к стандартному климату, у молодых деревьев, от которых зависит восстановление сообщества, и у многих второстепенных видов листва находится в совершенно других условиях. Таким образом, знание микроклиматов в пределах сообщества необходимо для истинного понимания связей между его компонентами и климатом. Важность этого факта осознали только сравнительно недавно, и изучение микроклиматов все еще находится на очень низкой ступени. Микроклиматы некоторых типов леса умеренных широт систематически исследованы лишь в некоторой степени. Все еще очень скудны и сведения о микроклиматах тропического дождевого леса, хотя зоологи много сделали в этом направлении в основном в связи с изучением насекомых, в частности москитов. Постепенно выясняется, что одна и та же площадь леса может давать приют двум совершенно различным фаунам — фауне верхушек деревьев и фауне подлеска (Allee, 1926 b; Hingston, 1932; Paulian, 1946, 1947) — с разнообразными экологическими связями. Основная цель этой главы — по возможности объединить разрозненные данные по указанному вопросу и попытаться дать связное представление о вертикальных и горизонтальных изменениях микроклимата в пределах сообщества тропического дождевого леса и таким образом обрисовать условия жизни разнообразных синузий, входящих в его состав.
180
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
При сопоставлении внутренних климатов леса со стандартным климатом выявляется, что в нескольких метрах над самым верхним ярусом древесных крон существуют практически те же условия, что и на высоте 1 м над землей на открытых вырубках. Полог крон образует барьер, задерживающий, но все же не предотвращающий конвекционные токи воздуха, возникающие между наружной атмосферой и пространством под кронами; он также оказывает сильное сопротивление горизонтальным движениям воздуха. Деревья перехватывают значительную долю осадков, частично стекающих по их стволам, а частично испаряющихся, не достигнув почвы. Кроме того, они поглощают часть солнечной радиации и земного излучения. Днем листья поглощают большую часть падающей световой и тепловой энергии. Хотя ночное излучение тепла в основном осуществляется кронами самых высоких деревьев, внутренность леса охлаждается за счет опускания относительно плотного холодного воздуха сверху. Росообра-зование, если оно наблюдается, происходит на вершинах деревьев, а не на уровне земли. В результате кроны деревьев создают в пространстве под ними климат более ровный, чем на открытых местах. Для внутренних участков леса характерны меньшие колебания температуры и относительной влажности (или дефицита насыщения), а все изменения, протекающие во внешней атмосфере, «приглушаются» и лишь спустя много времени в смягченном виде проявляются и внутри леса. В сущности микроклиматы пространства под пологом леса представляют промежуточное звено между климатом открытых участков и климатом почвы с его наименее изменчивыми условиями. Микроклимат изменяется с расстоянием от земли, что наглядно выявляется в существовании непрерывного или прерывистого вертикального климатического градиента между поверхностью земли и уровнем вершин самых высоких деревьев. Поскольку все ярусы деревьев различны по степени сомкнутости, микроклимат изменяется и в горизонтальном направлении.
Масштабы этих влияний и характер вертикальных климатических градиентов были довольно подробно исследованы Гейгером (Geiger, 1927)1 в европейских лесах с наипростейшей структурой, а именно в одновозрастных насаждениях, где деревья образуют единый ярус с сомкнутым ровным пологом. Установленные климатические градиенты оказались неодинаковыми; их величина менялась соответственно расстоянию от земли, причем характер градиентов был неодинаков для различных климатических факторов. Характер градиента зависит и от интенсивности того или иного фактора за пределами леса; так, градиенты скорости ветра и относительной влажности внутри леса изменяются в соответствии с внешними скоростями ветра или значениями относительной влажности.
Характер климатических градиентов внутри леса, конечно, зависит от его структуры. В тропическом дождевом лесу градиенты сложнее, чем в большей части умеренных лесов. Профильные диаграммы, приведенные в главе 2, показывают, что в тропическом дождевом лесу благодаря его большей высоте и многоярусности горизонтальный барьер крон гораздо толще и гуще, чем в лесах, изучавшихся Гейгером. Такой барьер, по-видимому, оказывает соответственно большее воздействие, препятствуя движениям воздуха и влияя на радиационный обмен. Поэтому следует (ролагать, что микроклиматические различия между нижними ярусами Тропического дождевого леса и свободной атмосферой будут значительнее,
1 Имеется русский перевод с третьего значительно переработанного издания: Р. Гейгер, Климат приземного слоя воздуха, ИЛ, М., 1960.
СРЕДА
181
чем соответственные различия для леса умеренной зоны. Характер градиентов, несомненно, меняется в зависимости от степени сомкнутости крон (верхних А и В) ярусов (ср. гл. 2).
ДВИЖЕНИЯ ВОЗДУХА
Одной из наиболее заметных особенностей нижних ярусов леса является исключительная неподвижность воздуха, очень усиливающая ощущение духоты. В тропическом дождевом лесу Брит. Гвианы, который показателен в этом отношении, слабый ветер почти непрерывно по крайней мере днем овевает верхушки деревьев, так что листва всегда колышется, в то же время в подлеске настолько тихо, что дым от папиросы поднимается совершенно вертикально, а обрывки бумаги остаются неподвижны. Но шквалы, которые часто внезапно налетают перед дождем, могут проникнуть достаточно глубоко, чтобы вызвать колыханье листьев и в подлеске. Так, в течение сухого солнечного периода в октябре 1929 г. наблюдался такой сильный ветер, что на протяжении нескольких дней по утрам листья вблизи земной поверхности шевелились по нескольку часов подряд. Значительный градиент скорости ветра должен быть чрезвычайно важен для растительности вследствие своего влияния на транспи^цию. Механическое воздействие движущегося воздуха, вероятно, также имеет некоторое значение; нежные тонкие листья, встречающиеся у некоторых видов подлеска, вскоре оказались бы разорванными в относительно ветреных верхних ярусах леса.
В Германии в одноярусном лесу с деревьями, достигающими 14 м высоты, Гейгер (Geiger, 1927) установил, что при небольшой скорости ветра над лесом ее величина под пологом крон не уменьшалась, оставаясь приблизительно одинаковой на всех уровнях примерно ниже 12 м; это показывает, что основное тормозящее действие производит верхняя (наружная) поверхность крон. При более высоких скоростях ветра на высоте около 6—8 м возникал незначительный вторичный максимум скорости, где ее величина была несколько большей, чем в кронах или у земли. В тропическом дождевом лесу с его более сложной структурой, вероятно,
Таблица 15
Скорости горизонтального перемещения воздуха в лесу, остров Барро-Колорадо, Панама
(Allee, 1926 а, стр. 279)
Положение анемометра Горизонтальное перемещение воздуха, к.и Средняя горизонтальная подвижка воздуха, к.н/24 часа Максимальная горизонтальная подвижка воздуха
6,5 футов (2 ле) над поверхностью земли (подлесок) 75 футов (23 м) над поверхностью земли («в середине крон») Открытая местность (соседний участок) 34,8 372,2 Около 1,6 Около 16,0 384 7,7 «ле/24 часа 28,6 км/24 часа 38,4 км/час
182
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
существует другое соотношение. В частности, можно ожидать, что при любых скоростях ветра вне лесного массива будет наблюдаться значительное различие между скоростями в кронах верхнего яруса и в подлеске. По-видимому, это подтверждается данными Элли (Allee, 1926 а) для тропического дождевого леса Панамы (табл. 15). Период наблюдения составлял 23 дня в течение сухого сезона, характеризующегося наивысшими скоростями ветра.
Для дождевого леса Бразилии (22°58' ю. ш.), возможно, субтропического по характеру флоры, но, вероятно, совершенно сходного по структуре с типичным тропическим дождевым лесом, Фрейзе (Freise, 1936, стр. 302) приводит следующие цифры средней годовой скорости ветра (см. табл. 16).
Таблица 16
Скорость ветра в двух районах вечнозеленого леса южной Бразилии (Freise, 1936, стр. 302)
Средняя скорость ветра на высоте 5 м над поверхностью земли, м/сек
Положение анемометра
округ Мамбукаба (1908-1935 гг.)
Серра-ду з-Айморес (1914-1935 гг.)
На открытой местности в 150 м от лесной опушки
В глубине леса:
100 м
1 100 м
2150 м
8500 м
12 000 м
2,3
0,5
Слишком мала для измерения
2,0
0,5
Слишком мала для измерения
Слишком мала для измерения
Слишком мала для измерения
Во время бури в местности Мамбукаба (там же, стр. 346) средняя скорость ветра, достигавшая 28 м/сек на высоте 5 м вне леса, снизилась при углублении в лес на 12 000 ле до 1,9 м/сек на той же высоте.
ПЕРЕХВАТ ОСАДКОВ
Любые деревянистые или травянистые растения улавливают известную долю осадков и уменьшают их количество, достигающее земли, причем степень перехвата зависит от числа, формы и расположения листьев и ветвей. Так как большинство растений поглощает воду только через корни, осадки, не достигшие земли, оказываются потерянными для них и имеют значение лишь в той степени, в какой они влияют на величину транспирации. Вопрос о перехвате осадков в умеренных лесах был тщательно изучен Хоппе (Geiger, 1927), Бургером (Burger, 1933) и другими. Исследуя 88-летний буковый лес в Германии, Хоппе установил, что количество осадков, проходящее через крону, и количество, стекающее по
СРЕДА
183
стволам, колебалось в зависимости от силы дождя. При очень легком дожде свыше 50% осадков проходит насквозь, при более сильном дожде свыше 60% проходит насквозь и свыше 10% стекает по стволам.
Фрейзе (Freise, 1936, стр. 303—304) изучал перехват осадков в вечнозеленом лесу1 Бразилии (см. табл. 17). Количество осадков, поступавшее
Таблица 17
Приблизительный средний баланс перехвата осадков в вечнозеленом лесу южной Бразилии, % (Freise, 1936, стр. 303)
Количество осадков: достигающих дождемера, расположенного на высоте 1,5 .и 33
испаряющихся непосредственно с кроны деревьев 20 сбегающих по стволам 46—испаряющихся
с поверхности 9,2 поглощающихся корой 9,2
достигающих основания деревьев 27,6—непосредственно
достигающих уровня грунтовых вод через трещины и т. п. 6,9 поглощающихся корнями 20,7
в дождемер, расположенный над пологом леса на высоте 1,5 м, значительно колебалось в соответствии с силой дождя. В тот месяц, когда дожди выпадали преимущественно в виде грозовых ливней, дождемер улавливал 24,7% осадков, в месяц с затяжными (обложными) дождями — 35,5%. Эти данные расходятся с наблюдениями Вогана и Вийе (Vaughan, Wiehe, 1947), которые установили, что в нагорном дождевом лесу острова Маврикия сравнительно большее количество воды достигало лесной подстилки (именно в сезон сильных ливней).
Конечно, осадки, достигшие почвы, не могут быть полностью использованы растениями, так как часть их теряется благодаря просачиванию и стеканию по поверхности. Мак-Лин (McLean, 1919, стр. 130—133) попытался вычислить величину так называемых «эффективных осадков» в дождевом лесу южной Бразилии. Он определяет их как процент суммы осадков, выпадающих на данной площади при учете не только механической задержки воды листьями, но и влияния дренажа в самой почве, который играет эффективную роль в реальном водоснабжении почвы. Для двух станций им были получены величины 16,6 и 27,5% соответственно при осадках в 13,8 мм.
1 Фрейзе называет этот лес «субтропическим», но, по-видимому, он не очень отличается по структуре от тропического дождевого леса.
184
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Как упоминалось в предыдущей главе, в тропических дождевых лесах может выпадать сильная роса. Она образуется только на кронах более высоких деревьев и никогда не наблюдается внутри леса. Вряд ли когда-нибудь роса бывает так обильна, чтобы стекать по стволам или капать с листьев на землю, но, вероятно, она имеет значение для солнечных эпифитов и, может быть, частично понижает величину транспирации с поверхности листьев в течение части суток.
ТЕМПЕРАТУРА
Среди многочисленных опубликованных наблюдений над температурой в глубине тропического дождевого леса наиболее существенны следующие: для дождевого леса низменности — наблюдения Мак-Лина (McLean, 1919, южная Бразилия), Элли (Allee, 1926 а, Панама), Дэвиса и Ричардса (Davis, Richards, 1933—1934, Брит. Гвиана), Бекера и Харрисона (Baker, Harrison, 1936, Новые Гебриды), Ричардса (Richards, 1936, Саравак),. Ивенса (Evans, 1939, юго-западная Нигерия), Хэддоу, Джиллета и Хайто-на (Haddow, Gillett, Highton, 1947, западная Уганда), Питтендрая (Pit-tendrigh, 1948, Тринидад). Для нагорного (включая горный лес и лес предгорий) дождевого леса существуют очень полные данные Брауна (Brown, 1919) для различных высот на Филиппинах и наблюдения Моро (Moreau, 1935 а) над «переходным лесом» Узамбара (Восточная Африка), а также Вогана и Вийе (Vaughan, Wiehe, 1947) над «нагорным климакс-ным лесом» острова Маврикий. Хотя большая часть упомянутых исследований относится только к коротким промежуткам времени, все же благодаря незначительным сезонным колебаниям температуры эти работы дают вполне правдивую картину температурных условий.
В табл. 18 приводится ряд типичных максимальных и минимальных температур на различной высоте над землей в разное время года. Как и следовало ожидать, в подлеске колебания температуры меньше, чем на более высоких уровнях леса; разницу создает главным образом более низкий максимум, минимум же в подлеске выше или лишь слегка ниже.
Днем форма и величина температурного градиента определяется в основном количеством солнечной радиации, перехваченной листьями и ветвями на различных уровнях. Следует ожидать устойчивого понижения температуры от первого яруса книзу; наблюдения показывают, что такой градиент действительно существует и среди дня температура в подлеске на 2—3° ниже, чем в кронах деревьев первого яруса. Ночью радиационный обмен усложняется. Гейгер (Geiger, 1927) показал, что в европейском лесу с одним ровным пологом крон деревьев ночью температура обыкновенно понижается от крон деревьев к земле; прежнее представление о существовании прямо противоположного градиента было результатом неточных измерений. Этот ночной градиент получается следующим образом. Земля и нижние ярусы леса находятся под прикрытием, и ночное излучение тепла в основном осуществляется кронами деревьев. Благодаря своей высокой плотности холодный воздух, окружающий кроны, опускается, и таким образом нижние ярусы леса становятся холоднее, чем верхние. Если пространство мржду кронами и землей несвободно и препятствует опусканию охлажденного воздуха, то возникает обратный градиент, поскольку в этом случае тепло с поверхности крон будет излучаться быстрее, чем охлажденный воздух оттекать к земле. Данные Ивенса (Evans) показывают, что такие обратные градиенты действительно суще-
Таблица 18
Температура па различных уровнях в пределах тропического дождевого леса, ° С
Местность Остров Барро-Колорадо, Панама (первичный дождевой лес) Близ Акиллы, южная Нигерия (первичный смешанный дождевой лесу
Период наблюдений 9/II—9/Ш, 1924 г. (сухой сезон) 17/11—22/11, 1924 г. (сухой сезон) 5/III—11/III, 1935 г, (сухой сезон) 9/V— 16/V, 1935 г. (дождливый сезону
Источник данных Элли (АПее, 1926 а) Ивенс (Evans, 1939)
Высота над землей, м 0,2 17 26 0,2 17 26 0,7 24 0,7 24
Подле-сок Второй ярус Верхняя часть полога Под-лесок Второй ярус Верхняя часть полога Под-лесок Ярус Bi Травяной покров Ярус В1
Средний максимум — — — 27,1 30,8 35,7 29,7 33,9 26,8 30,9
Средний минимум — — — 26,0 24,0 27,1 23,9 24,0 23,3 21,8
Средняя суточная амплитуда 2,9 7,3 11,8 1,1 6,8 8,6 5,8 9,9 5,5 9,2
Местность Гора Маквилинг, Филиппины (Первичный диптерокарповый лес, 300 м над уровнем моря)
Период наблюдений Октябрь 1912 г.— 27/П—27/Ш, январь 1915 г. 1914 г. 8 14/VIII—11/IX, 1914 г. 2
Источник данных | Браун (Brown, 1919)
Высота над землей, м — около 18 около 35-40 — около 18 около 35-40 — около 18 около 35—40
Под-лесок Де-рево второго яруса . Вершина высокого дерева од-лесок Дерево второго яруса Вершина высокого дерева Под-лесок Де-рево второго яруса Вершина высокого дерева
Средний максимум 26,3 27,0 32,4 27,5 27,5 32,5 26,9 27,3 31,3
Средний минимум 20,6 21,0 20,0 19,9 20,8 19,6 21,0 21,4 20,6
Средняя суточная амплитуда 5,7 6,0 12,4 7,6 6,7 12,9 5,9 5,9 10,7
1 Ср. профильную диаграмму (рис. 6) и фотографию этого дерева (фото 4).
2 Эти периоды ванты автором из данных Брауна.
186
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ствуют в некоторых тропических дождевых лесах. В лесу южной Нигерии (ср. стр. 46 и рис. 6) ярусы А (наиболее высокий) и В не имеют сомкнутых крон, но ярус С (самые низкие деревья) очень густой, и его плотная поверхность связана переплетающимися лианами. Можно предположить поэтому, что вертикальные движения воздуха происходят без помех лишь до поверхности яруса С, а между землей и кронами яруса С очень затруднены. Если это так, то вполне понятно, почему подлесок на высоте 0,7 м по ночам всегда теплее, чем ярус В на высоте 24 м (ср. Evans, 1939, стр. 450). Кривые суточных колебаний температуры в типичные дни (рис. 23) ясно показывают смену температурного градиента, происходящую
Рис. 23. Суточный ход температуры в подлеске и у вершин деревьев в первичном тропическом дождевом лесу. Резервированный лес Шаша, Нигерия (Evans, 1939, рис. 7).
Диаграмма показывает ход температуры с 18 час. одних суток до 20 час. следующих; А и А1—24 м над землей; Б и Б]—на высоте 0,7 м\ Д, Б — сухой сезон (8—9 марта 1936 г.); Д], Б1 — дождливый сезон (10 — 11 мая 1936 г.).
непосредственно перед заходом солнца и тотчас же после восхода. В сезон дождей эта смена происходит примерно на час позднее, вероятно из-за большей облачности.
Несколько труднее истолковать аналогичные данные Элли (Allee). Подобно данным Ивенса они неизменно показывают, что по ночам подлесок теплее, чем ярус В. В то же время оказывается более теплой, чем подлесок или ярус В, и верхняя часть полога (26 м над землей, ветка дерева, возвышающегося над окружающим лесом).
Если допустить, что минимальные температуры на различных уровнях дают вполне точную картину ночного градиента, то будет не лишним познакомиться с данными Брауна (Brown, рис. 24). В противовес данным Ивенса и Элли они характеризуют подлесок более прохладным по ночам, чем ярус В в любое время года, а ярус А оказывается еще более прохладным, чем ярус В и подлесок.
Очевидно, что в тропическом дождевом лесу существует несколько типов температурного градиента. Несомненно, различные типы его связаны с различиями в структуре леса, но для выявления этих связей нужны дополнительные данные.
СРЕДА
187
Цифры, полученные Ивенсом и Брауном, дают представление о величине сезонных амплитуд температуры на различных уровнях. Наблюдения
J//7 28/U 28/111 25/IV 25/V 20/VI 18/VII 15/VIII 12/IX IU/X //XI 5/XII 2/1 четырехнедельные периоды
V нс. 24. Средние недельные максимумы и минимумы температуры в ярусах А и В и в подлеске, первичный (диптерокарповый) лес на высоте 300 м, гора Маквилинг, Филиппинские острова (Brown, 1919, рис. 6).
Ивенса показывают, что в дождливый сезон максимальные температуры в подлеске ниже, чем в сухой сезон, тогда как минимальные температуры остаются примерно одинаковыми; с другой стороны, на высоте 24 м как
188
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
минимум, так и максимум в дождливый сезон примерно на 2—3° ниже, чем в сухой. Средняя из минимальной и максимальной температуры для обоих периодов наблюдения (которые, как уже говорилось, не дают представления о полной сезонной амплитуде) составляет в подлеске 26,8° в сухой сезон и 25,1°— в дождливый; на высоте 24 м — 29,0° в сухой сезон и 26,4е— в дождливый. Таким образом, на вершинах деревьев наблюдается чуть большая сезонная разница температур, чем в подлеске. Местности, изученные Брауном, лежат в климате с меньшими сезонными колебаниями. В табл. 18 представлены два типичных периода, один из которых совпадает с сезоном наивысших, а другой — с сезоном наиболее скудных осадков. Из этих данных видно, что сезонные различия здесь меньше, чем в местности, изученной Ивенсом. На всех трех уровнях максимум немного ниже, а минимум немного выше в дождливый сезон, чем в сухой. Сезонная разница средних температур очень незначительна и примерно одинакова на всех уровнях.
ОТНОСИТЕЛЬНАЯ ВЛАЖНОСТЬ, ДЕФИЦИТ НАСЫЩЕНИЯ, ИСПАРЕНИЕ
Тремя основными мерилами атмосферной влажности являются относительная влажность, обычно вычисляемая по показаниям смоченного и сухого термометров или измеряемая волосяным гигрометром, дефицит насыщения, вычисляемый по одновременным показаниям относительной влажности и температуры и величина испарения, измеряемая эвапорометром. Основные данные об относительной влажности в тропическом дождевом лесу можно найти для леса низменностей в работах Мак-Лина (McLean, 1919, южная Бразилия), Дэвиса и Ричардса (Davis, Richards, 1933— 1934, Брит. Гвиана), Ричардса (Richards, 1936, Саравак), Ивенса (Evans, 1939, южная Нигерия), Питтендрая (Pittendrigh, 1948, Тринидад); для нагорного леса — в работах Брауна (Brown, 1919, различные высоты на горе Маквилинг, Филиппины), Фабера (Schimper, 1935, Тьюбодас, Ява) и Стокера (Stocker, 1935b, Тьюбодас, Ява). Наблюдения над дефицитом насыщения в тропическом дождевом лесу были опубликованы Дэвисом и Ричардсом (Davis, Richards, 1933—-1934, Брит. Гвиана), Ричардсом (Richards, 1936, Саравак), Бекером и Харрисоном (Baker, Harrisson, 1936, Новые Гебриды), Ивенсом (Evans, 1939, южная Нигерия) и Хэддоу,. Джиллеттом и Хайтоном (Haddow, Gillett, Highton, 1947, Уганда) МакЛин (McLean, 1919, южная Бразилия) и Стокер (Stocker, 1935b, Тьюбодас, Ява) опубликовали ряд одновременных показаний температуры и относительной влажности, по которым можно вычислить дефицит насыщения. Браун (Brown, 1919, Филиппины) и Элли (Allee, 1926а, Панама) приводят данные наблюдений над испарением на различных высотах в тропическом дождевом лесу.
Атмосферная влажность интересует ботаников-экологов главным образом из-за своей связи с транспирацией, поэтому наибольшее значение будут иметь те ее показатели, которые можно сопоставить с величиной транспирации. Вероятно, непосредственно измеренные величины испарения имеют непревзойденную ценность для эколога, но на практике их значение весьма ограничивается трудностью или невозможностью сравнения результатов, полученных испарителями различных систем или даже одним и тем же инструментом, но в различных условиях. Поэтому здесь будут рассмотрены лишь данные об относительной влажности и дефиците насыщения.
СРЕДА
189
1' а б л и ц а 19
Относительная влажность в первичном тропическом дождевом лесу близ Акиллы, южная Нигерия
(Evans, 1939)
('ухой сезон Дождливый сезон
Высота над землей, м 0,7 24 0,7 24
Средний максимум, % 11 ро дс > л ж и те л ь ноет ь максимума, 94,7 97,6 96,2 94,9
часы 1 \, 1 11,0 17,8 11,1
Средний минимум, % Средний «получасовой мини- 68,7 61,7 87,4 64,5
мум» ], % 76,6 63,9 92,0 66,9
Средняя суточная амплитуда, % 18,1 33,7 4,1 28.0
1 См. стр. 1 Эи.
Наиболее целесообразно рассмотреть наблюдения Ивенса (Evans, 1939) над относительной влажностью и дефицитом насыщенпя в Нигерии, хотя и охватывающие только короткие периоды, но в некоторых отношениях представляющие самые полные из существующих серий наблюдений. Термографические и гигрографические исследования производились в первичном тропическом дождевом лесу в 24 м над землей, на дереве, и одновременно в подлеске, на высоте 0,7 м вблизи подножия того же самого дерева. Записи были тщательно проверены и исправлены по показаниям психрометра. Наблюдения проводились в течение определенного периода в конце сухого сезона и повторно в течение определенного периода в начале дождливого сезона. Так как эти периоды не приходились ни на самые сухие, ни на самые дождливые месяцы, то наблюдения не дают величины полной сезонной амплитуды.
Таблица 19 суммирует данные по относительной влажности; рис. 25 показывает суточные колебания относительной влажности в течение двух дней, один из которых типичен для сухого сезона, а другой — для дождливого.
Легко заметить, что в течение дня наблюдаются многочисленные колебания, четко выраженные в ходе кривых; в промежуток времени продолжительностью в несколько минут относительная влажность может возрастать или падать на целых 10% выше или ниже средней величины для часового периода. Эти колебания, вероятно, вызываются главным образом порывами ветра; их физиологическое воздействие должно быть ничтожно. Так как абсолютная суточная минимальная влажность обычно достигается в одно из этих кратких колебаний, то возможно, что «получасовой минимум», то есть минимальная относительная влажность, регистрировавшаяся непрерывно в течение получаса или же в несколько приемов общей продолжительностью в полчаса, важнее, чем абсолютный минимум; именно этот минимум используют для вычисления суточной амплитуды влажности. Наибольший интерес вызывает наличие длительного ночного периода, во время которого влажность воздуха близка к насыщению, что свойственно как вершинам деревьев, так и подлеску, хотя в последнем средняя продолжительность этого периода значительно больше. На вершинах
190
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
деревьев влажность начинает уменьшаться вскоре после восхода; в подлеске этот процесс начинается позднее и идет медленнее. Аналогичным образом вечерний максимум влажности устанавливается гораздо раньше в подлеске, чем на вершинах деревьев, где во время сухого сезона его продолжительность к тому же часа на три меньше. На вершинах деревьев
Часы
Часы
Часы
Рис. 25. Относительная влажность (записано гигрографом) в тропическом дождевом лесу близ Акиллы, южная Нигерия.
А1 и А2— на дереве, 24 м над поверхностью земли; Б1 и Б2—в подлеске, 0,7 м над поверхностью земли; Aj и Б1—9 марта 1935 г. (сухой сезон); А% и Б2 —11 мая 1935 г. (дождливый сезон).
продолжительность периода с максимальными значениями влажности примерно одинакова в сухой и в дождливый сезон, а в подлеске она возрастает с 14 до 18 час.
В сухой сезон минимум в подлеске примерно на 7% выше, чем на вершинах деревьев; в дождливый сезон разница здесь опять-таки больше (25 %). Таким образом, и длительность периода максимальной влажности и величина минимума в подлеске дают более значительные сезонные колебания, чем на вершинах деревьев, хотя суточная амплитуда влажности на вершинах больше в любом сезоне.
Суточный ход дефицита насыщения в подлеске и на вершинах деревьев показан на рис. 26. Для его характеристики удобнее выбрать типичный
СРЕДА
191
день из записей каждого сезона, чем использовать средние почасовые данные для двух отобранных периодов (ср. Evans, 1939, стр. 452—453). Ясно заметен продолжительный ночной период очень низкого дефицита насыщения, соответствующий периоду максимальной относительной влажности. Днем наблюдается его повышение; утром кривая для подлеска отстает от кривой для крон деревьев, причем, поскольку минимальная относительная влажность в подлеске выше, чем на вершинах деревьев.
Рис. 26. Суточный ход дефицита насыщения в подлеске и на вершинах деревьев, тропический дождевой лес, резервированный лес Шаша, Нигерия (Evans, 1939, рис. 8).
Непрерывные линии —24 м от земной поверхности;пунктирные линии— 0,7 м от земной поверхности. Каждая точка представляет собой среднее из четырех показанийхс].пятнад-цатиминутными интервалами.
максимальный дефицит насыщения в подлеске ниже (меньше). На обоих уровнях этот максимум достигается примерно в 14 часов. В сухой сезон максимальный дефицит насыщения на вершинах деревьев составляет свыше 12 мм, а в подлеске он никогда не превышает 8 мм. В дождливый сезон он доходит почти до 12 мм на вершинах, но никогда не превышает 2 мм в подлеске. Для сравнения можно указать, что средний дефицит насыщения в Кью (Англия) в июле составляет 7,4 мм (9,8 миллибар) в 14 час. и 1,3 мм (1,7 миллибар) в 4 часа (Bilham, 1938, стр. 211).
Вычерчивая график температуры, соответствующий относительной влажности для часовых (или иных) интервалов в течение дня, можно проследить суточный ход температуры, относительной влажности, дефицита
192
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
насыщения и абсолютной влажности (содержание водяных паров в воздухе) на одной диаграмме. Полученные таким образом многоугольники (полигоны) дают сведения относительно изменений влажности, вызванных транспирацией (перемешиванием различных слоев воздуха и т. д.). Семейства кривых, проведенных через многоугольники, показывают соответственно постоянный дефицит насыщения и постоянную абсолютную влажность.
Р и с. 27. Микроклиматический режим верхушки дерева в сухой сезон (8—9 марта 1936 г.), показания — на уровне 24 м при высоте дерева 28 м. Температура и относительная влажность в тропическом дождевом лесу, резервированный лес Шаша, Нигерия (рис. 27—30) (Evans, 1939, рис. 3—6).
Пунктирные линии показывают постоянный дефицит насыщения; точечные линии— постоянную упругость пара (мм рт. ст.). Показания до полудня обозначены значком^;
показания после полудня—®; 5,00, 20,00—часы.
В обоих полигонах для вершин деревьев (рис. 27 и 28) и в полигоне, характеризующем сухой сезон в подлеске (рис. 29), видно, что верхняя их часть располагается более или менее параллельно оси температуры, тогда как нижняя часть более или менее следует за константой абсолютной влажности; это говорит о том, что ночью, несмотря па изменение температуры, относительная влажность составляет примерно 100%, а абсолютная влажность изменяется; днем абсолютная влажность остается приблизительно одинаковой, а температура и относительная влажность изменяются. Это не заметно на рис. 30 (подлесок в дождливый сезон), поскольку суточная амплитуда относительной влажности очень незначительна в сравнении
СРЕДА
193
со случайными колебаниями. Значение этих фактов в целом неясно, но, по-видимому, они указывают, что существует аналогия между атмосферой леса и замкнутой системой водного раствора и водяных паров. В такой системе при относительной влажности, соответствующей упругости пара раствора данной температуры и концентрации, вода будет испаряться по мере подъема температуры; при этом концентрация раствора будет возрастать, а относительная влажность падать, пока раствор не станет
Рис. 28. Микроклиматический режим верхушки дерева в дождливый сезон (10—11 мая 1936 г.), показания — на уровне 24 м при высоте дерева 28 м (см. подпись к рис. 27).
насыщенным. Затем кривая относительной влажности будет медленно подниматься, опускаться или оставаться постоянной в соответствии со свойствами данного раствора. Когда вся вода испарится, кривая совпадет с линией постоянной абсолютной влажности. В условиях леса следует ожидать запаздывания как испарения, так и конденсации. Первые три диаграммы как будто бы свидетельствуют о таком гистерезисе. Диаграмма для подлеска в сухой сезон показывает незначительное падение абсолютной влажности между 10 и 14 час.1; вероятно, это вызвано смешением
1 По нигерийскому времени; в резервированном лесу Шаша оно опережает истинное солнечное время приблизительно на 3/4 часа (то есть восход и заход отмечаются в 6 час. 45 мин. и 18 час. 45 мин. соответственно с незначительными сезонными колебаниями).
13 п. У. Ричардс
194
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
воздуха внутренних участков леса с более сухим внешним воздухом благодаря усилению ветра и конвекции в течение утра.
В подлеске движения воздуха настолько слабы, а барьер между атмосферой внутри и вне леса так ощутим, что сравнение с замкнутой системой здесь вполне допустимо. Еще более поразительно то, что эта аналогия сохраняет свое значение также и для вершин деревьев как в дождливый,
так и в сухой сезон, что, может быть, объясняется присутствием влажного морского воздуха (ср. стр. 162), который постоянно поддерживает высокую абсолютную влажность.
Данные об относительной влажности и дефиците насыщения в других тропических дождевых лесах особенно для более высоких уровней столь
СРЕДА
195
отрывочны, что не дают возможности сделать сколько-нибудь детальное сравнение с условиями Нигерии, однако они все же позволяют сделать несколько общих замечаний. Для всех тропических дождевых лесов как в сухие, так и в дождливые сезоны характерен длительный ночной период высокой влажности, близкой к состоянию насыщения или равной ему.
Рис. 30. Микроклиматический режЩи подлеска в дождливый сезон (10—11 мая 1936 г.), показания — на уровне 0,7 м над землей (см. подпись к рис. 27).
Разумеется, продолжительность этого периода колеблется в соответствии с местными условиями, сезоном и высотой над уровнем земли. В дождливый сезон, а в сырых климатах в любое время года, в подлеске период максимальной влажности очень долог, и наблюдается лишь незначительное понижение влажности в течение нескольких часов среди дня. Суточная минимальная относительная влажность или максимальный дефицит насыщения наблюдается примерно в 14 час. как в подлеске, так и выше; его фактическая величина в подлеске обыкновенно не превышает 10 мм. В более высоких ярусах дефицит насыщения, конечно, больше, но даже в сухой сезон его средняя суточная величина редко превосходит 16 мм. Как в подлеске, так и выше относительная влажность и дефицит насыщения колеблются в течение всего дня.
Таким образом, в тропическом дождевом лесу ночной градиент влажности отсутствует, так как атмосфера на всех уровнях близка к насыщению. Утренний подъем температуры сопровождается увеличением дефи
13*
193
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
цита насыщения (или понижением относительной влажности), начиная с верхнего яруса леса. За утро прогревание нижних слоев атмосферы и смещение нижних и верхних слоев воздуха благодаря ветру и конвекционным течениям ведут к увеличению дефицита насыщения даже на низких уровнях, но испарение с поверхности почвы и процесс транспирации в сочетании с пониженной температурой в тени и меньшим движением воздуха препятствуют увеличению дефицита насыщения в подлеске в такой степени, как в верхних ярусах. Амплитуда влажности поэтому резко снижается от верха к низу, п устанавливается такой градиент влажности, крутизна и характер которого меняются в течение дня. Вечером в подлеске темнота наступает несколько раньше, чем у вершин деревьев, и поэтому величина дефицита насыщения снижается на нижних уровнях раньше, чем на верхних.
Суточная амплитуда колебаний как влажности, так и температуры в нижних ярусах меньше, чем в верхних. Сезонная амплитуда температуры больше в верхних ярусах, чем в нижних; в то же время, если результаты Ивенса имеют общее значение, то сезонная амплитуда влажности в подлеске должна быть больше, чем в верхних ярусах. Однако сезонная амплитуда как температуры, так и влажности на всех уровнях очень мала в сравнении с суточной амплитудой.
Наблюдения Хэддоу, Джиллетта и Хайтона (Haddow, Gillett, High-ton, 1947) над относительной влажностью, дефицитом насыщения и температурой в тропическом дождевом лесу на западной границе Уганды хорошо согласуются с данными Ивенса для Нигерии. Измерения проводились на уровне земли и на высотах 16 футов (5 л<), 31 фут (9 м) и 54 фута (16,5 м). Между уровнем земли и 16 футами высоты, так же как между 31 и 54 футами, наблюдались лишь незначительные различия в микроклимате, но между 16 и 31 футом эти различия были ярко выражены, хотя и здесь они заметно сглаживались в дождливую погоду. В этом лесу, таким образом, существовали два резко разграниченных микроклиматических яруса, или, иными словами, микроклиматический градиент носил черты прерывистости.
СВЕТ
Одной из особенностей тропического дождевого леса является контраст между сумраком подлеска и ослепительной яркостью света на вершинах деревьев и вырубках. Хотя прежде влияние освещения на структуру леса преувеличивали, не вызывает сомнения, что точное знание вертикальных и горизонтальных колебаний интенсивности освещения существенно для понимания экологии тропического дождевого леса в целом и распределения его отдельных компонентов.
К сожалению, измерение интенсивности освещения в лесу — очень сложная проблема. Прежде всего сами условия крайне сложны, так что полная картина освещенности может быть получена лишь в результате многочисленных разнообразных измерений. Свет, который не отражается непосредственно от наружной (верхней) поверхности леса, может проникать сквозь ткани листьев или же проходить между ними, образуя под пологом крон блики, величина и форма которых непрестанно меняются в течение дня. Внутри леса свет отражается листьями и ветвями — поверхностями с очень разнообразными оптическими свойствами. Вдобавок интенсивность и спектральный состав света, падающего на лес, конечно,
СРЕДА
197
меняются соответственно времени года, времени дня и степени облачности.
Помимо сложностей, связанных с самим характером освещения, существуют еще технические трудности, возникающие в процессе конкретных измерений. Нет необходимости обсуждать здесь преимущества и недостатки различных типов фотометров, применявшихся для экологических целей (ср. Shirley, 1935). Ясно, что идеальный прибор должен давать показания, которые легко можно превратить в абсолютные единицы, и обладать большой чувствительностью к лучам спектра, поглощаемым зернами хлорофилла. Еще не делалось измерений интенсивности освещения в лесах с помощью инструментов, вполне отвечающих этим требованиям. Визнер (Wiesner, 1895), Мак-Лин (McLean, 1919), Фабер (Faber, 1915), а также Дэвис и Ричардс (Davis, Richards, 1933— 1934) пользовались актинометрами, в которых потемнение светочувствительной бумаги сравнивалось со стандартным образчиком оттенка потемнения. Браун (Brown, 1919) применял актинометр Ливингстона с черным шариком, Элли (Allee, 1926а) и Моро (Moreau, 1935а) применяли люксометры, позволявшие производить визуальное определение степени освещенности, сравнивая ее с освещением от стандартного источника света. Единственный тип фотометра, свободный от субъективных погрешностей,— это фотоэлектрическая камера, или фотоэлемент. Этот прибор был использован Картером (Carter, 1934) в Брит. Гвиане и Ивенсом (Evans, 1939) в южной Нигерии.
Ниже дается лишь общий обзор полученных данных без каких-либо попыток критической оценки их.
В глубине тропического дождевого леса наблюдаются бесчисленные колебания интенсивности освещения. Поскольку разные типы леса часто очень разнятся по структуре, они также очень различны и по средней интенсивности освещения на уровне земли; эти колебания будут рассмотрены позднее. Однородному по структуре лесу также свойственна неравномерная освещенность благодаря прерывистости яруса А, а иногда и В и из-за неравномерного распределения деревьев. Рубка или естественная гибель деревьев создают просветы; иногда световые окна образуются в результате обнажения кроны листопадного дерева. Наряду с этими значительными колебаниями освещения повсюду видны узоры, создаваемые комбинацией солнечных бликов и тени.
Когда светит солнце, даже в самом тенистом лесу на земле бывают солнечные блики. Большое разнообразие их размеров и яркости затрудняет точное измерение их площади, но по оценке Картера (Carter, 1934) в Брит. Гвиане в зависимости от типа леса от 0,5 до 2,5% площади лесной подстилки оказывается освещенной солнечными бликами. Наблюдения Ивенса (Evans, 1939) над дневными колебаниями освещенности в подлеске дождевого леса Нигерии обнаружили, что солнечные блики с яркостью свыше 100% средней интенсивности освещения встречаются лишь на протяжении 4—5 часов в середине дня. Как указывалось во вступительной главе, интенсивность света в тени, как правило, больше предполагаемой. Возможно, что сравнительно сильное рассеянное освещение леса частично вызывается, как полагает Карстен (Karsten, 1924—1926), большим количеством света, отраженного глянцевитыми поверхностями склерофилльных листьев.
Прежние измерения интенсивности освещения в подлеске в местах отсутствия солнечных бликов дали результаты порядка 0,5—1 % полного дневного света. Мак-Лин (McLean, 1919) в южной Бразилии, пользуясь
198
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
актинометром с фильтром, «задерживающим все актиничные лучи за пределами X 5000», получил следующие результаты:
В «солнечных бликах», % В «глубокой тени», %
12 0,72
Визнер (Wiesner, 1895), также пользуясь актинометром, нашел, что в тропическом дождевом лесу «фанерогамный лимит» (самая низкая интенсивность освещения, при которой встречаются высшие растения напочвенного покрова) в общем составлял около 0,83—0,77?6. Элли (Allee, 1926 а, стр. 293) с помощью люксометра получил следующие результаты на острове Барро-Колорадо, Панама, (в фут-свечах):
Открытое место «Солнечный блик» в полдень «Глубокая тень»
4000—20 500 Среднее: 11 000 325—20 500 0,8—670 Среднее: 325 (2,95%)
Недавние наблюдения с фотоэлементом дали довольно сходные результаты. В лесах различного типа в Брит. Гвиане Картер (Carter, 1934) получил цифры, колеблющиеся от 10,4 до 72 % для солнечных бликов и 0,18—0,80°о — для затененных мест.
Р и с. 31. Суточный ход освещенности в подлеске тропического дождевого леса, резервированный лес Шаша, Нигерия.
Измерения производились па высоте 0,4 м над поверхностью земли 4 марта 1936 г. с интервалами в 10 сек. (от 7 час. 40 мин. до 14 час. 12 мин.) и с интервалами в 15 сек. (от 14 час. 12 мин. до 17 час. 41 мин.) посредством фотометра Уэстона с пластинкой из молочного стекла. Каждая горизонтальная линия на диаграмме представляет среднее из всех измерений за четверть часа (Evans, 1939, рис. 13) .
Ивенс (Evans, 1939) очень подробно изучил освещенность подлеска первичного леса в южной Нигерии. Измерения велись в продолжение полного 10-часового периода (рис. 31). Колебания интенсивности, очень регулярные в течение двух первых и двух последних часов наблюдения,
СРЕДА
199
были менее регулярны в середине дня, когда солнце поднялось выше и появились солнечные блики. Большую часть дня интенсивность освещенности остается на уровне примерно 0,5—1 % интенсивности ее на открытом пространстве. В отдельные короткие периоды, составляющие около 1% светлого времени суток, интенсивность возрастала более чем вдвое, а в еще более короткие периоды — более чем в пять раз. Результаты Ивенса для периодов, близких к полудню, более подробно характеризуются табл. 20.
Можно заключить, что и тогда, когда солнце закрыто облаками и когда оно ярко светит, освещение в затененных местах подлеска всегда меньше 1% наружного освещения, яркость солнечных бликов может быть по меньшей мере в 100 раз больше, чем интенсивность света в тени.
Рассеянный свет подлеска проходит сложный путь многократных отражений и фильтрации и, естественно, должен очень отличаться от наружного света по спектральному составу. Однако, сведения по этому вопросу очень неполны. Мак-Лин (McLean, 1919) посредством спектроскопа исследовал свет, отраженный от листьев, и нашел, что интенсивность красных лучей уменьшилась в меньшей степени, чем интенсивность лучей другой длины. Предположив, что отражение от листьев дает больше света, чем фильтрация сквозь них, он пришел к заключению, что свет в подлеске должен быть сравнительно богат красными лучами. Картер (Carter, 1934) в Брит. Гвиане непосредственно исследовал спектр освещения в подлеске, пользуясь фотоэлементом Бернхейма и различными фильтрами. Он установил, что свет был сравнительно беден красными лучами и заключил, что большая часть их, возможно, прошла через листья, не отразившись от их поверхности. Ивенс (Evans, 1939), используя фотоэлемент Уэстона и фильтры, сделал следующее вычисление состава света в подлеске дождевого леса в одном из пунктов в пределах Нигерии:
Минимальный %
Синий (3200—5000 А) 7,6
Зеленый (4700—5900 А) 22,4 У 75,3
Красный (за пределами 6000 А) 45,3 1
Остаток (24,7%) не определен, так как цифры подсчитывались для максимальной проницаемости фильтров; фактически он разделен между различными областями спектра таким образом, что его невозможно учесть. Когда часть этого остатка пытались распределить, то на синюю и зеленую области спектра досталась такая ничтожно малая доля, что большая часть этого остатка (то есть около 40%), видимо, должна лежать в области максимальной проницаемости красного фильтра выше 7000 А. Это указывает на увеличение проницаемости лесного полога непосредственно за пределами красного участка видимого спектра. Ивенс получил и другое подтверждение этого заключения. Незначительность пропорции красных лучей, установленная Картером, объяснялась вероятно тем, что примененный е им фотоэлемент был очень мало чувствителен к длинам волн в 7000 А и совсем нечувствителен к волнам с длиной около 7300 А. Фотоэлемент Уэстона, примененный Ивенсом, сохранял 11% своей максимальной чувствительности к волнам длиной 7000 А и 3—4% — к волнам 8000 А. Высокий процент световых волн, близких к инфракрасным, свойственный подлеску, вероятно, не играет никакой роли при фотосинтезе, поскольку эти лучи находятся вне предела поглощения хлорофиллом.
Бюнинг (Biinning, 1947), работая в диптерокарповых лесах северной Суматры, получил результаты, сходные с данными Ивенса; он противопо-
Таблица 20
Освещенность в подлеске тропического дождевого леса. Резервированный лес Шаша, Нигерия
Ивенс (Evans, 1939, табл. VII)
Пункт 1 2 3 4 5 6 7 8 Погода
I 12/IV 12,06—12,32 20,0 26,4 26,4 26 0 0 0 Сплошная облачность
II 16/IV 11,41—12,17 7.9 27,9 252 36 — 10,7 6,0 Продолжительность солнечно-
го сияния 31 мин.
III 16/IV 12,51—13,30 16,3 43,1 209 39 — 3,3 2,0 То же 20 ъ
22/IV 10,50—11,22 12,8 33,9 33,9 32 1,7 0 0 » » 27 »
9/V 10,00—10,36 3,7 31,1 44,0 34 1,5 1,0 0 » » 5 а
105 3,2 4,3 2,0
IV 22/IV 12,40—13,06 7,9 17,1 17 1 26 0 0 0 » » 23 а
9/V 11,54—12,48 6,0 12,1 14,8 54 2,5 0 0 » » 48 а
24/V 11,40—12,42 1,6 12,2 20,5 62 1,0 0 0 » » 3 »
142 3,5 0 0
V 22/IV 12,01—12,38 5,2 11,2 244 32 13,7 8,0 2,7 » » 25 »
9/V 11,00—11,52 5,1 12,5 13,8 52 4,7 0 0 » » 35 »
24/V 11,16—11,38 4,0 10,0 10,0 22 0 0 0 С плошная облачность
106 18,3 8,0 2,7
Итоги для пунктов II -V 389 25 23 10,7
Колонки: 1—дата и интервалы времени; 2—минимальные величины (цА); 3—максимальные величины, исключая цифры в колонках 6—8 (цА);
4—максимальные величины (цА); 5—длительность наблюдения (мин.); 6—длительность видимых солнечных бликов на поверхности фотоэлемента (мин.); 7—длительность наблюдения, в два раза большая средней продолжительности наблюдения в тени при безоблачном небе (мин.); 8—длительность наблюдения в пять раз большая средней продолжительности наблюдения в тени при безоблачном небе (мин.). Измерения производились фотометром Уэстона; данные для 5 пунктов. Поверхность фотометра находилась в 0,3 м над землей. Пункты II-V примыкают к участку, характеризуемому профильной диаграммой на рис. 6.____________________________________________________________________________________________________________
СРЕДА
201
ставляет «серый сумрак» тропического подлеска преобладающему зеленому освещению в европейских широколиственных лесах.
На более высоких уровнях леса средняя интенсивность освещения, конечно, гораздо выше, чем у земли; следовательно, в дождевом лесу существует резко выраженный градиент освещенности, изменяющейся от полного дневного света самого высокого яруса деревьев до низких интенсивностей в подлеске. Данных о характере этого градиента очень мало, но общие наблюдения заставляют думать, что в лесах одного типа может идти постепенное уменьшение средней интенсивности света сверху вниз, в то время как в лесах другого типа, возможно, существует какой-то уровень, на котором интенсивность падает сравнительно резко. Последнее, вероятно, имеет место в тропическом дождевом лесу Нигерии, где ярусы А и В открыты, а ярус С очень малопроницаем. Если влезть на дерево в этом лесу, минуя полог яруса С, то кажется, что находишься на ярком дневном свету (ср. стр. 48). Единственные опубликованные данные измерения интенсивностей освещения на разных уровнях тропического дождевого леса получены Элли (Allee, 1926а, стр. 293) на острове Барро-Колорадо (Панама) люксометром Макбета.
Ярусы Лесная подстилка Ярус «маленьких деревьев» «Вершины среднего яруса» «Верхний лесной полог»
Высота Освещенность в свечах на кв. фут (по отношению к освещенности лесной подстилки) 0 1 20—30 футов (6-9 м) 5 40—60 футов (12—18 м) 6 75—80 футов (23—25 м) 25
УГЛЕКИСЛОТА
Хотя концентрация углекислоты в воздухе не является климатическим фактором, она рассматривается наряду с элементами микроклимата, поскольку, как и интенсивность освещения, является лимитирующим условием фотосинтеза и, следовательно, роста растений. Кроме того, в пределах многоярусного растительного сообщества концентрация углекислоты колеблется, что, подобно другим микроклиматическим факторам, в значительной степени зависит от препятствий, создаваемых растениями для свободных движений воздуха.
В последнее время стало известно, что атмосфера в приземном слов леса как в умеренных областях, так и повсюду несколько больше насыщена углекислотой, чем наружный воздух. В нижних слоях европейских лесов наблюдались концентрации до 0,04% (по объему) (Feher, 1927, стр. 316), что примерно в 1,3 раза превышает нормальную концентрацию (0,03% по объему); более высокие цифры (примерно до 0,08% по объему) были зарегистрированы Лундегардом (Lundegardh, 1931, стр. 271). С другой стороны, есть свидетельства, что в верхних ярусах леса концентрации бывают ниже нормы, вероятно, оттого, что при энергичной ассимиляции листья забирают из воздуха большие количества углекислоты. Поскольку при большой интенсивности освещения скорость фотосинтеза оказывается
202
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
почти прямо пропорциональной концентрации углекислоты, подобные отклонения последней от нормы могут иметь большое экологическое значение. Даже в условиях очень слабой освещенности в тени тропического дождевого леса наличие высокой концентрации углекислоты должно было бы, вероятно, значительно повысить величину фотосинтеза.
Судя по общим данным, в нижних ярусах тропического дождевого леса следует ожидать высоких концентраций углекислоты. Отсутствие мощной лесной подстилки из мертвых листьев и незначительный процент
006
0,05
о °>04
со
О § ^0.03 t
X
X
G X X
X о х
X О 0
«Нормальная » концентрация
0,01
0 ------1----1-----1-----1----1----1-----1----1-----1----1-----1----I—
6 7 8 9 Ю 12 14 16 18
Время суток, часы
Рис. 32. Концентрация углекислоты в атмосфере подлеска, резервированный лес Шаша, Нигерия (Evans, 1939, рис. 1).
х — первичный лес; 0— вторичный лес.
гумуса в почве показывают, что разложение и выделение углекислоты микроорганизмами происходит очень энергично. Многоярусное расположение растительности, как уже отмечалось, оказывает сильное сопротивление движениям воздуха, так что перемешивание атмосферы верхних и нижних уровней очень затруднено. Прежние определения концентрации углекислоты в тропическом дождевом лесу давали в сущности необычайно высокие цифры. Так, производя наблюдение в тихую погоду в лесу около Рио-де-Жанейро (Бразилия), Мак-Лин (McLean, 1919) на высоте 1,5 ле обнаружил концентрацию углекислоты, равную 0,14 объемного % вечером и 0,34 объемного % на следующее утро, что было соответственно в 4,6 и 11,3 раза выше нормы. Фабер (Schimper, 1935, стр. 146) на высоте 0,5 м в среднегорном дождевом лесу в Тьюбодас, Ява, получил почти такие же высокие цифры.
СРЕДА
203
Последними работами Стокера (Stocker, 1935 b), Ивенса (Evans, 1939) и Бюнипга (Bunning, 1947) существование столь высоких концентраций не подтверждается. Наблюдения Стокера, проведенные как и Фабером в Тьюбодас, не обнаружили величин, превышающих нормальную концентрацию хотя бы вдвое ни на высоте 7 см, ни на высоте 1,7 м над землей. Более того, многие цифры оказались ниже нормы. Из пятнадцати наблюдений, проведенных на высоте 1,7 м, десять дали величины ниже нормы, и только три — выше нормы; на высоте 7 см все величины оказались выше нормы, но лишь одна превышала ее в полтора раза. Стокер делает вывод, что прежние взгляды были ошибочны и что для внутренних частей дождевого леса более типичны низкие, а не высокие концентрации. Результаты наблюдений Ивенса, проводившихся в южной Нигерии на высотах 0,6—1 м над землей, показаны на рис. 32. В среднем обнаруженные концентрации оказались несколько выше, чем у Стокера, но все они колеблются между нормой и двойной нормой (0,036—0,057 объемного % в первичном лесу, 0,033—0,058 — во вторичном). Незначительная разница, все же существующая между результатами Ивенса и Стокера, может объясняться тем, что последние были получены на высоте 1300 м, тогда как Ивенс проводил исследования в местности, расположенной близ уровня моря, где средняя температура была не менее чем на 9° выше; кроме того, лес отличался большей густотой. Ивенс указывает, что результаты МакЛина основаны только на двух разрозненных определениях и что примененная им методика была ненадежной. К сожалению, метод, которым были получены данные Фабера, не установлен. Наблюдения Бюнинга в целом согласуются с наблюдениями Стокера и Ивенса. В низменностях и на более низких частях горных склонов в очень влажной местности Суматры в лесу на высоте 0—30 см над землей он установил концентрацию примерно до 0,1 объемного % в утренние часы. В лесах несколько более сухого типа, подобных тем, какие изучал Ивенс, были найдены более низкие концентрации. У самой поверхности земли местами встречались концентрации до 0,2 объемного ?о.
Следовательно, какие-либо веские основания для утверждения об исключительно высоких концентрациях углекислоты в тропическом дождевом лесу отсутствуют; напротив, эти концентрации, по-видимому, почти не отличаются от концентраций на тех же уровнях в лесах умеренной зоны. Мнение Фабера (Schimper, 1935, стр. 147) о том, что в подлеске дождевого леса высокая концентрация углекислоты компенсирует низкую интенсивность освещения, вероятно, следует отвергнуть.
Данные Стокера, Ивенса и Бюнинга показывают наличие суточных колебаний концентраций углекислоты. Самые низкие концентрации отмечались Стокером утром; от 6 до 9 час. наблюдался подъем концентрации, за которым шло постепенное понижение, продолжавшееся до захода солнца. В то же время Ивенс обнаружил наиболее высокие концентрации между 6 и 7 час., затем концентрация равномерно падала до вечера. Понижение концентрации в течение дня наблюдал и Бюнинг. Понижение может быть объяснено процессами ассимиляции углерода растениями и, несомненно, также перемешиванием внутреннего и наружного воздуха конвекционными токами. Низкую концентрацию в ранние часы Стокер объясняет тем, что за ночь она падает благодаря переходу углекислоты в связанное состояние. По его мнению, это может происходить из-за растворения каплями росы небольшого количества щелочей, осаждающегося на листьях. Путем количественного подсчета Ивенс показал, что эта гипотеза несостоятельна, и предположил, что низкие величины, наблюдав
204
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
шиеся Стокером рано утром, возможно, вызваны интенсивным притоком воздуха в почву, охлажденную излучением, что должно замедлять наружную диффузию углекислоты.
Хотя наблюдения над концентрацией углекислоты в верхних ярусах тропического дождевого леса не производились, нет причин думать, что она должна быть иной, чем на высоких уровнях в умеренных лесах; возможно, что она оказалась бы еще ниже.
ВЛИЯНИЕ ТИПА ЛЕСА НА МИКРОКЛИМАТ
Следует ожидать, что в тропических дождевых лесах, разных по структуре и флористическому составу, микроклиматические условия на равной высоте над поверхностью земли будут неоднородными. Существование таких различий продемонстрировали Дэвис и Ричардс (Davis, Richards, 1933—1934, табл. VI и др.), сравнивая микроклиматические условия в подлеске первичного леса четырех типов в местности Морабалли-Крик, Брит. Гвиана. Наблюдения велись в приречном лесу Мора (консоциация Mora excelsa), лесу Уаллаба (консоциация Eperua jalcata), произрастающем на белых песках горных кряжей и плато, лесу Морабуцеа (консоциация Mora gonggrijpii) и смешанном лесном сообществе промежуточных местообитаний. Эти сообщества различаются как по флористическому составу (см. гл. 10), так и по структуре. Из имеющихся неполных данных был сделан вывод, что: 1) средняя температура, самая низкая в подлеске леса Мора, последовательно повышалась в лесу Морабуцеа и смешанном лесу, достигая максимума в лесу Уаллаба; 2) средний дефицит насыщения, наименьший в лесу Мора, достигает максимума в лесу Уаллаба; 3) колебания температуры и дефицита насыщения значительнее всего в лесу Уаллаба.
Помимо этих наблюдений, в подлеске четырех вышеназванных типон леса, а также в лесу Гринхарт (консоциация Ocotea rodiaei) была измерена интенсивность освещенности. Для того чтобы получить грубо приближенную величину средней освещенности, учитывая распределение тени и солнечных бликов, актинометр передвигался в пределах определенной дистанции. Этим методом удалось выявить следующую картину распределения; освещенности (в % от полного дневного света):
Лес Мора Лес Морабуцеа Смешанный лес Лес Гринхарт Лес Уаллаба
1,33 0,61 0,67 0,81 1,43
Таким образом, подлесок консоциаций Мора и Уаллаба — двух крайних типов ряда — освещен значительно лучше1.
На примере первичных лесных сообществ Морабалли-Крик видно, что колебания температуры и дефицита насыщения, с одной стороны, и степени освещенности подлеска — с другой, не идут параллельно.
1 Картер (Carter, 1934), который работал в другой части Брит. Гвианы и применял другую методику, определяя по отдельности интенсивность освещенности в затененных участках и на солнечных бликах в лесу Уаллаба, смешанном лесу, в трех подтипах леса Мора и в молодом вторичном лесу, пришел к несколько иным результатам.
СРЕДА
205
Температура и дефицит насыщения в нижних уровнях леса, вероятно, •отчасти зависят от структуры леса, а отчасти — от различий во влажности почвы, связанных с колебаниями уровня грунтовых вод и разнородностью ее водоудерживающих свойств. В то же время освещенность должна зависеть главным образом от числа и плотности ярусов деревьев. Лес Мора, характеризующийся сравнительно разреженными ярусами А и В, расположен на затопляемой равнине с постоянно высоким уровнем грунтовых вод и относительно водонепроницаемой почвой. Структура леса Мора-буцеа точно неизвестна, но, по-видимому, он обладает очень плотным пологом и, хотя также произрастает на водонепроницаемой почве, лучше дренируется. Смешанное сообщество (от которого лес Гринхарт лишь слегка отличается по структуре и составу) имеет три яруса деревьев, из которых А и В вместе образуют полог умеренной плотности (см. рис. 4); здесь почва и лучше дренирована и более проницаема для воды, чем в лесах типа Мора и Морабуцеа. Лес Уаллаба обладает только двумя хорошо развитыми ярусами деревьев, которые образуют примерно такой же разреженный полог, как в лесу Мора; почва отличается высокой водопроницаемостью, а расположение на горных хребтах ведет к исключительно хорошему дренажу. В лесу Уаллаба как почва, так и структура создают в подлеске менее постоянные климатические условия, более близкие к наружным, чем в лесах других типов. Можно считать, что степень различий между температурой и влажностью в подлеске и внешними климатическими условиями в большой степени зависит от плотности барьера, разделяющего атмосферу леса и наружную атмосферу. В лесу Уаллаба этот барьер менее выражен, чем в смешанном лесу и лесу Морабуцеа. В лесу Мора он тоже менее развит, но это с избытком компенсируется излишками почвенной влаги.
СВЯЗЬ ФИЗИОЛОГИИ РАСТЕНИЙ С МИКРОКЛИМАТОМ
Дальнейшее микроклиматическое изучение тропического дождевого .леса несомненно принесет многочисленные новые открытия, но некоторые важные факты установлены уже сейчас. Верхние ярусы леса находятся в сравнительно изменчивых условиях: они совершенно открыты воздействию солнца и ветра и испытывают резкие колебания температуры и влажности. С другой стороны, в нижних ярусах условия гораздо постояннее, здесь как суточная, так и сезонная амплитуда температуры и влажности поразительно малы. Зоолог-эколог (Allee, 1926 а, стр. 299) пишет: «Животным в глубине леса... нужно лишь избегать солнечных бликов, чтобы находиться в такой постоянной среде, которая должна возбудить зависть всякого опытного исследователя, пытающегося изучить влияние среды на наземных животных в лабораторных условиях».
Таким образом, в пределах единого растительного сообщества существуют два очень отчетливых, расположенных один над другим типа микроклимата, или, иными словами, здесь присутствует ярко выраженный вертикальный градиент среды. Кустарники и травы (включая эпифиты) должны приспосабливаться лишь к одному из вариантов среды, но более высокие члены сообщества, деревья ярусов А и В и большинство лазящих растений, вынуждены на различных стадиях своего развития быть выносливыми как к неизменному климату слабо освещенного подлеска, так и к более разнообразным условиям, в которые они попадают в зрелом возрасте. В лесах с густыми ярусами А и В деревья яруса С живут в уело-
203
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
виях, сходных с условиями жизни кустарника, подлеска и трав, но там, где (как, например, в смешанном лесу Нигерии) (стр. 46) верхние ярусы более открыты, их среда мало отличается от условий жизни деревьев ярусов А и В.
Вероятно, с физиологической точки зрения растения тропического дождевого леса, как правило, хорошо приспосабливаются к обычной среде своего обитания. На вырубках высокие деревья обыкновенно продолжают пышно расти, но многие из них, а может быть и большинство, для прорастания и прохождения ранних стадий роста требуют затенения или по крайней мере такой же влажной атмосферы, в которой развивается подрост в ярусе подлеска (гл. 3). Кустарники и травы ярусов D и Е (возможно, это относится также к большинству деревьев яруса С), по-види-мому, трудно приспосабливаются к микроклиматам, резко отличающимся от тех, в каких они развивались до вырубки леса. Интересно, что какао (Theobroma cacao), естественная среда которого — ярус С гвианского леса (Myers, 1930),— лучше всего культивируется в атмосферных условиях с наименьшими колебаниями влажности и поэтому нуждается в притенении и соответствующей защите от ветра (Hardy, 1935, стр. 175). Существование теневыносливых и тенелюбивых видов в подлеске тропического дождевого леса (гл. 5) показывает, что даже у растений одного и того же яруса могут быть различные способы приспособления к условиям микроклимата.
Характер и масштабы физиологических различий между растениями разных ярусов еще далеко должным образом не изучены, дальнейшее изложение ставит целью показать лишь существование известного диапазона физиологического поведения, отражающего вертикальный градиент микроклимата.
Водообмен
Уже говорилось о больших колебаниях величины средней скорости транспирации у различных растений тропического дождевого леса (гл. 6). Это, по-видимому, в известной степени связано с условиями влажности и температуры, свойственными тому ярусу, к которому принадлежат растения. Все новейшие работы по этому вопросу приводят к выводу, что у растений верхних ярусов дождевого леса, как и в умеренных лесах, транспирация происходит быстрее на единицу поверхности листа, чем у растений нижнего яруса, даже когда они находятся в одинаковых условиях среды. Подобным же образом транспирация происходит быстрее у солнечных листьев, чем у теневых. Критический обзор процесса транспирации у тропических растений был опубликован Стокером (Stocker, 1935 b), работавшим в Богоре (Ява). Три вида деревьев, растущих на открытом пространстве, он сравнил с тремя травами, растущими в тени. Деревья Calophyllum inophyllurn, обычное дерево прибрежных редколесий (стр. 321), Cassia fistula, обитающая (во вторичном произрастании) в злаковниках и тиковых лесах, и Elaeodendron glaucum, встречающийся в тиковых и листопадных смешанных лесах восточной Явы, к сожалению, не принадлежат к породам тропических дождевых лесов, но, весьма вероятно, по своим особенностям не очень отличаются от деревьев верхних ярусов последних. Травы — Begonia isoptera, Cyrtandra pendula и пока еще не определенное точно растение семейства Acanthaceae — были обычными представителями тенелюбивой флоры дождевого леса предгорий, характерного для окрестностей Тьюбодас (близ Богора).
СРЕДА
207
У деревьев в течение ночи транспирация была крайне низка, поднимаясь до больших величин ранним утром. Среди дня в солнечную погоду скорость транспирации (на единицу площади листовой поверхности) понижалась, хотя не было никаких признаков увядания. У тенелюбивых трав в их естественном местообитании суточный ход транспирации имел совершенно иной характер. Ночью, как и у деревьев, транспирация падала до нуля, но даже в течение дня она оставалась устойчивой и очень низкой, быстро повышаясь лишь в момент попадания на растение солнечного блика.
Таблица 21
Осмотическое давление клеток листвы растений в тропическом дождевом лесу Тьюбодас
(Blum, 1933)
Осмотическое давление эпидермальных и мезофилльных клеток листьев, ат
Среднее Минимальное Максимальное
Мелкие травы 5,4 2,6 7,4
Крупные травы 6,1 4,7 6,7
Деревянистые лианы 12,4 8,9 19,6
Маленькие деревья 13,0 2,6 25,5
Крупные деревья 15,1 8,1 30,9
Два из трех исследованных видов имели толстые склерофилльные листья; листья трав, также толстые, отличались большой мягкостью. Стокер показывает, что у склерофилльных листьев деревьев, как у большей части листьев подобного типа, наблюдается высокое содержание влаги по отношению к площади поверхности (примерно 1,1f г кв. дм), а содержание влаги у толстолистных трав оказалось еще выше (1,90 и 1,94 г, кв. см в двух случаях). По отношению к содержанию влаги величина транспирации у крайне склерофилльных Calophyllum и Elaeodendron ниже, чем у более мягколистной Cassia fistula, хотя по отношению к площади листовой поверхности наблюдается обратное соотношение. У всех трех деревьев транспирация по отношению к содержанию влаги в листьях была гораздо большей, чем у тенелюбивых трав. Наивысшие цифры для деревьев достигали 16—20 мг/г/мин, что эквивалентно 96—12О?6 влаги, проходящей через лист в течение 1 часа. Эти цифры не могут считаться исключительно высокими, если вспомнить о таких растениях умеренных стран, как, например, Helianthus tuberosus, у которого при меньшем отношении площади листа к сырому весу величина транспирации в солнечный день превосходит 150% от всего влагосодержания листа. У тенелюбивых растений отношение транспирации к содержанию влаги никогда не превышает весьма низкой цифры, равной 2 мг^/мин, или 1296 содержания влаги за час.
Осмотическое давление клеток листа было исследовано Блюмом (Blum, 1933) у ряда растений дождевого леса предгорий в Тьюбодас (Ява).
Осмотическое давление эпидермальных и мезофильных клеток закономерно изменяется соответственно ярусу, к которому принадлежит растение; чем выше растение, тем больше осмотическое давление. Табл. 21 подводит итоги некоторых наблюдений Блюма. Блюм считает, что градиент осмотического давления в основном зависит от градиента атмосферной влажности.
208
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Ассимиляция углерода и рост
Судя по резко выраженному вертикальному градиенту средней освещенности в тропическом дождевом лесу, можно было бы ожидать больших различий в ходе ассимиляции у светолюбивых и теневыносливых растений, но достаточные данные по этому вопросу отсутствуют. Фабер (Schimper, 1935, стр. 106, рис. 24) опубликовал «кривые ассимиляции» для Cyrtandra picta, типичной травы наземного яруса тропического дождевого леса на Яве, и для тропического светолюбивого растения Lantana сатага, которое хотя и не свойственно первичному тропическому дождевому лесу, но изобилует во многих частях тропиков на ранних стадиях сукцессии после его вырубки. Относительно методов, которыми были получены эти данные, подробных сведений нет, но для светолюбивых растений кривая ассимиляция углерода — интенсивность освещения круто идет вверх, достигая примерно половины суммарной освещенности (последняя определялась как «прямая плюс рассеянная освещенность примерно на 7° ю. ш. в ботаническом саду в Тьюбодас, Ява, в сухое время года»). Кривая, характерная для тенелюбивых растений, поднимается менее круто, и примерно после достижения суммарной освещенности дальнейшего подъема не наблюдается, то есть освещение перестает быть фактором, лимитирующим ассимиляцию.
По-видимому, более определенные данные по этому вопросу никем не публиковались, но работа Гутенберга (Guttenberg, 1931) об ассимиляции в листьях многих тропических растений ботанического сада в Богоре дает доказательства значительной неоднородности в процессах метаболизма среди представителей различных ярусов. Изученные растения можно разделить на четыре группы в соответствии с характером и количественной стороной ассимиляции в листьях: 1) те, у которых листья откладывают много крахмала в течение дождливого сезона (по-видимому, этот крахмал является резервом, используемым в сухие сезоны); крахмала в листьях больше, чем сахара; 2) те, которые накапливают и сахар и крахмал, но содержат больше сахара; 3) те, которые образуют крахмал в небольших количествах в течение дня (за ночь он исчезает); содержание сахара в этих листьях всегда высокое, а содержание крахмала периодически колеблется; 4) те, у которых листья никогда не образуют крахмала. Первая группа включает такие типичные растения тропического дождевого леса, как Palaquium philippense и Pausinystalia yohimbe, обладающие жесткими или кожистыми листьями и сходные по свойствам со склерофитами средиземноморского типа климата. Вторая группа включает ряд лазящих растений и эпифитов дождевого леса. Гутенберг считает, что их свойства могут быть связаны с почти постоянной ассимиляцией, прерывающейся лишь на короткое время. Третья группа преимущественно состоит из мяг-колистных видов, главным образом трав, а также кустарников и деревьев •средних ярусов тропического дождевого леса. По-видимому, углеводный метаболизм не отличается от многих европейских растений. Четвертая группа включает много ксерофитов, причем в нее входят как деревья, так и травы, а не только травы, как в Европе. Из представителей этой группы, по-видимому, ни один не является типичным видом дождевого тропического леса, за исключением, пожалуй, эпифитной орхидеи Coelogyne Pulverula.
Эти факты могут иметь экологическое значение. Хотя у членов одного и того же яруса или синузии наблюдается большое разнообразие свойств, очевидно, существует какое-то соотношение между углеводным обменом и склерофилльным или мягким строением листа. Поскольку, с другой сто
СРЕДА
209
роны, для каждого яруса свойственно характерное строение листа (гл, \ и 5), должно существовать соотношение между ярусом и типом углеводного обмена в листьях. Если такое соотношение существует, то оно. вероятно, зависит от различия в микроклиматах, в особенности от длительности периодов высоких величин испарения в различных ярусах.
Отсутствие данных не позволяет сравнивать точку компенсации (интенсивность освещения, при которой ассимиляция уравновешивает дыхание при данной температуре) у растений различных ярусов дождевого леса. Стокер (Stocker, 1935 а) установил точку компенсации Stelecho-carpus burahol, дерева тропического дождевого леса Явы. Оказалось, что она не выше, чем у деревьев Центральной Европы. Было бы чрезвычайно интересно знать, распространяется ли на дождевые леса закономерность, свойственная лесам умеренной зоны и выражающаяся в том, что точка компенсации выше у солнцелюбивых, чем у тенелюбивых растений, или, другими словами, в том, что при такой интенсивности освещения, когда листья светолюбивых растений испытывают безусловную потерю углеводов, листья тенелюбивых растений все еще могут находиться в условиях преобладания ассимиляции над дыханием.
Что касается скорости роста, наблюдения за ней, хотя и проведенные на множестве различных тропических растений, все еще недостаточны, чтобы сделать какие-либо общие выводы относительно средних ее величин в различных ярусах. Можно, правда, сослаться на данные Брауна (Brown, 1919), который изучал скорость роста тропических деревьев в разные периоды их жизни (см. гл. 3).
Физиологические особенности подлеска
Физиология подлеска тропического дождевого леса всегда вызывала большой интерес, и Мак-Лин (McLean, 1919) в некоторой степени рассмотрел этот вопрос. Благодаря постоянно высокой влажности, относительно неизменной температуре и низкой интенсивности освещения травы, кустарники и самый нижний ярус деревьев находятся в очень своеобразной среде. Ботаники-физиологи прошлого века, искавшие повсюду очевидных проявлений приспособления, ожидали найти в тенелюбивой флоре особенности строения, позволяющие увеличить транспирацию. Они считали, что интенсивность и размеры поступления пищи из почвы должны прямо или косвенно зависеть от величины транспирации, и поэтому предполагалось, что у подлеска тропического дождевого леса, находящегося в атмосфере с очень низкой испаряющей способностью, непременно будет проявляться минеральное голодание. Шталь (Stahl) и другие обратили внимание на то, что на листьях растений подлеска часто встречаются гидатоды, бугорчатые эпидермальные клетки, «капельные острия» и т. п., что принималось за приспособления для усиления транспирации. Некоторые из этих особенностей уже были рассмотрены (гл. 5). Мак-Лин резко критиковал эту точку зрения. Занимаясь исследованиями в южной Бразилии, он установил, что средняя величина транспирации у теневыносливых растений была весьма низкой, но тем не менее содержание золы в них и, следовательно, размеры минерального питания были не меньше или даже больше, чем у светолюбивых растений одного и того же сообщества. Выделение воды в жидком состоянии не наблюдалось, и листья «не обнаруживали каких-либо структурных особенностей, пригодных для увеличения транспирации до возможного ее предела. Более того, эпидермис листьев и другие
210
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
приспособления обычно защищают их от чрезмерной водопотери» (McLean, 1919, стр. 53).
Этот вывод нашел всеобщее признание; весьма правдоподобно, что теневыносливая флора тропического дождевого леса, несмотря на обычно очень низкую транспирацию, не терпит никаких лишений, поскольку речь идет о ее минеральном питании. Если у нее и есть какие-то особенности строения, способствующие усилению транспирации, то они, безусловно, не очень заметны. Можно было бы ожидать, что у листьев этих растений по крайней мере окажется большая площадь поверхности в сравнении с их весом или объемом (в гл. 5 было показано, что они обычно имеют больший размер), но Стокер (Stocker, 1935 Ь) установил, что у трех теневыносливых растений из леса в Тьюбодас (стр. 207) отношение площади поверхности к сырому весу было исключительно мало.
Теневыносливые растения тропического дождевого леса не только, как правило, обладают низкой величиной транспирации, они, очевидно, и не могут поддерживать высокую ее величину, не испытывая при этом постоянного внутреннего дефицита влаги, от которого растение уже не может оправиться. Примером служит поведение Icacina trichantha и вида Rinorea — двух кустарников, обычных в подлеске вечнозеленого леса южной Нигерии (Evans, 1939, стр. 479—480). Последнее из этих растений ограничивается в своем распространении лишь затененными участками леса, а первое иногда растет и на сухих открытых склонах на вырубках. В потометре, помещенном в лесной тени, срезанные побеги обоих кустарников в течение нескольких дней оставались живыми и почти не проявляли признаков увядания. Однако при перемещении в сухой воздух лаборатории, где величина транспирации у обоих видов возросла в 4—5 раз, Icacina еще просуществовала в потометре несколько дней, но у Rinorea стал развиваться все увеличивающийся дефицит влаги, и вскоре это растение засохло. Если поведение Rinorea типично для тенелюбивых видоч подлеска (отличных от теневыносливого типа, к которому принадлежи! Icacina)то, по-видимому, при исключительно сильных засухах или даже при длительном пребывании на солнечных бликах, тенелюбивым растениям может угрожать серьезное повреждение или гибель, так что «защитные» особенности, отмеченные Мак-Лином, менее парадоксальны, чем это могло показаться на первый взгляд.
Вредное воздействие быстрой транспирации на тенелюбивые растения может частично объясняться тем, что их листья бедны сосудами. Мак-Лин (McLean, 1919, стр. 42—43) установил, что у трех типичных тенелюбивых видов южнобразильского тропического дождевого леса отношение площади листа и площади поперечного сечения ксилемы в черешке в четыре с лишним раза больше, чем у трех типичных светолюбивых растений.
Поскольку транспирация у тенелюбивых растений в целом очень низка, не удивительно, что, согласно Блюму (Blum, 1933), клетки листвы тенелюбивых растений в Тьюбодас обладают самым низким осмотическим давлением из когда-либо измеренного у наземных растений.
Можно было бы также ожидать, что тенелюбивые растения тропического дождевого леса проявят специфические особенности в отношении ассимиляции углекислоты. При скудости освещения подлесок дождевого леса отличается неожиданной пышностью. Можно было бы предположить, что у тенелюбивых растений должны быть особые приспособления, помогающие им энергично ассимилировать углекислоту при низких интенсивностях освещения. Теория «углекислотной флоры» уже была рассмотрена и оказалась несостоятельной. Ясно, что листья тропических тенелюбивых
СРЕДА
211
растений, достигшие полного развития, часто обладают теми же анатомическими особенностями, что и листья тенелюбивых растений в других климатах, а именно тонким мезофиллом (паренхимой), чечевицеобразными палисадными клетками и т. п. Фабер (Schimper, 1935, стр. 114) констатирует, что некоторые типичные тропические тенелюбивые растения открывают свои устьица даже при таких низких интенсивностях освещения как 1/70—1/50 суммарной освещенности. Однако этих данных совершенно недостаточно для общей оценки их ассимиляционных свойств. Освещение в тропическом дождевом лесу еще слишком мало изучено, чтобы можно было решить, действительно ли оно менее благоприятно для тенелюбивых растений этого леса, чем в лесах других типов. Поскольку речь идет о функциональных приспособлениях в самом процессе фотосинтеза, надо иметь в виду, что у тропических тенелюбивых растений свет становится лимитирующим фактором ассимиляции при более низкой интенсивности освещения, чем у светолюбивых растений. Следует ожидать, хотя это еще не проверено, что точка компенсации здесь лежит ниже, чем у светолюбивых растений. Таким образом, мы пока не знаем, ограничивает ли освещение ассимиляцию углерода у тропических тенелюбивых растений, всегда ли это так, насколько простирается действие света как основного фактора, лимитирующего рост и распределение тенелюбивых растений.
Эта проблема настоятельно требует дальнейших исследований, поскольку начинают оспаривать важность освещения как основного фактора, определяющего структуру и состав подлеска, что прежде не вызывало сомнения. Так, например, обычно считали, что наличие определенных видов подлеска в глубокой тени, а других видов только на хорошо освещенных открытых местах определяется исключительно средней интенсивностью освещения. Действительно, при расчистке подлесок всегда становится гораздо гуще и пышнее и на первый взгляд может показаться, что это вызывается только лишь усиленным освещением. Однако последние исследования о сообществах умеренного леса заставляют полагать, что скудость подлеска некоторых умеренных лесов, вероятно, объясняется не недостаткохм света, а конкуренцией корней деревьев в почве. Если устранить воздействие корней круговым глубоким окапыванием по квадрату, то травянистая растительность при том же освещении разрастается буйно и пышно (Watt, Fraser, 1933). Работы Костера (Coster, 1935 а) и Уилкинсона (Wilkinson, 1939) в свою очередь подтверждают, что конкуренция корней может быть причиной различий в составе и густоте подлеска тропических лесов. Костер (Coster, 1935 Ь) сравнил густоту подлеска на маленьких опытных участках под деревьями на Яве и нашел, что различия больше зависели от вида дерева, чем от интенсивности освещения. В густых зарослях Lager str oemia speciosa, где освещение составляло 1,5—2 ?6 суммарной освещенности (методика измерения не указана), подлесок был густой п разнообразный, а под Swietenia macrophylla при 2—7% суммарной освещенности он полностью отсутствовал. Под большим деревом Ficus kurzii наблюдался пышный подлесок при 1,5—2% суммарной освещенности и даже под теми участками кроны, где интенсивность освещения составляла всего 0,4?о, были найдены кустарники и лазящие Агасеа. Костер считал, что эти различия были вызваны неоднородным характером корневой системы у различных видов деревьев. Эксперименты по окапыванию на тиковых плантациях дали результаты, сходные с результатами, полученными в европейских и американских лесах.
Если согласиться с тем. что влияние корней деревьев так велико, как это кажется, все же механизм их воздействия остается непонятным. Может
14*
212
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
происходить борьба корней за влагу, соли или другие дефицитные вещества; или же корни могут оказывать благоприятное или неблагоприятное влияние на подлесок посредством каких-нибудь растворимых выделений. Костер полагает, что основным фактором является борьба за кислород, которого не хватает для роста корней в условиях влажных тропиков.
Костер проводил наблюдения на сравнительно сухих низменностях Явы, и не известно, получились ли бы подобные результаты в типичном тропическом дождевом лесу более сырых местообитаний. Пока не будут проведены опыты, невозможно сказать, чем объясняется слабое развитие подлеска в тропическом дождевом лесу и встречаемость определенных видов только на открытых участках — борьбой ли корней или же недостаточностью освещения. Если дальнейшие исследования подтвердят результаты Костера, то, по-видимому, окажется, что тенелюбивые виды подлеска произрастают только в тени, потому что они не переносят периодов большого дефицита насыщения, а вовсе не потому, что они боятся высоких интенсивностей освещения; эта особенность должна сочетаться с выносливостью при интенсивной конкуренции корней. Теневыносливые виды, которые разрастаются на вырубках, вероятно, относительно легко выдерживают конкуренцию корней и более выносливы также к большим дефицитам насыщения.
Глава 8
СЕЗОННЫЕ ИЗМЕНЕНИЯ РАСТИТЕЛЬНОСТИ
Растительность умеренной зоны изменяется по временам года, и эти изменения более или менее совпадают с сезонными колебаниями климата. Тропический дождевой лес меняется гораздо меньше; как сообщество он не знает никаких ясно выраженных сезонных «аспектов» и никаких периодов «покоя». Весь год листва остается темно-зеленой, и каждый месяц цветут какие-нибудь растения; лишь время от времени отдельное дерево, стоящее без листьев или же покрытое яркой молодой листвой, вносит некоторое разнообразие в этот пейзаж. Все времена года — лето, зима, осень и весна — как бы совмещаются здесь и наблюдаются одновременно. Только вблизи климатических границ типа формаций тропического дождевого леса, где осадки имеют отчетливый сезонный ритм, периодические изменения становятся важным элементом природной среды. Но и здесь сезонные изменения растительности сравнительно малозаметны и проявляются главным образом в верхних ярусах леса; в подлеске же, где колебания температуры и влажности очень незначительны, внешний вид растительности остается почти неизменным круглый год.
Хотя в целом растительность как будто и не подчиняется сезонному ритму, жизнедеятельность отдельных растений более или менее периодична. Большинство видов являются вечнозелеными в том смысле, что у них никогда листья не опадают полностью, и многие из этих растений цветут почти непрерывно круглый год. Но все же очень многие пз них имеют периоды «покоя» и видимой активности. Периодичность проявляется в появлении новых листьев и побегов и, менее заметно, в цветении п других функциях растительного организма. Таким образом, несмотря на общий неизменный облик растительности, жизнедеятельность отдельных видов настолько регулярна, что может совершенно точно датироваться. На острове Эспириту-Санто (Новые Гебриды) сезонные колебания климата так мало заметны, что местные жители никогда не исчисляют свой возраст в годах, но проводят сев в регулярные сроки, определяемые цветением или плодоношением определенных деревьев. Ямс сажают, когда цветет Erythrina indica, а батат — когда плодоносит Alphitonia zizyphoides (Baker J. R., Baker I., 1936). Подобным же образом на Малайе цветение Sandoricum koetjape еще не так давно было сигналом к посеву риса (Corner).
Периодичностью физиологических функций обладают и грибы. Корнер (Corner, 1935) установил, что на юге Малаккского полуострова у шляпочных грибов каждый год обычно бывает два общих сезона «плодоношения». Каждый из ряда видов, развивающихся примерно в течение трех месяцев, имеет свой собственный период плодоношения, длящийся около недели.
Основное отличие периодчности физиологических функций у растений тропического дождевого леса и растений других климатов состоит
214
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
в том, что эти функции у различных видов, различных особей одного вида и даже различных частей одного и того же растения менее синхронны и часто, по-видимому, не зависят непосредственно от условий среды. Саймон (Simon, 1914) указывает, что в Богоре, Ява, вероятно, нет ни одного месяца в году, когда тот или иной вид не пробуждается от покоя к деятельности. В Перадении, Цейлон, не бывает месяца, когда бы каждый вид был полностью покрыт листьями (Wright, 1905). В любой данный момент деревья одного и того же вида и ветви одного и того же дерева могут находиться на разных стадиях развития, хотя в пределах одной и той же местности все особи одного вида часто ведут себя одинаково. Одну и ту же фазу развития в течение года проходит попеременно то одна, то другая часть флоры; в умеренных областях она подчиняется ритмическим сезонным колебаниям.
Чем отчетливее выражены сезонные изменения климата в зоне тропиков, тем синхроннее периодичность в жизни растений и тем больше она согласуется с сезонностью климата. В тех местах влажного вечнозеленого леса Нигерии (стр. 361), где в течение пяти месяцев подряд может выпадать меньше 10 см осадков, наблюдается отчетливо выраженное пробуждение растительности в начале дождливого сезона, хотя многие виды находятся в полном расцвете и в течение засушливых месяцев. В подлеске здесь встречаются даже геофиты, увядающие во время сухого периода (стр. 116). Это зона перехода к условиям тропического листопадного, или муссонного леса, где для огромного большинства видов сухой сезон является периодом покоя; периодичность влажного вечнозеленого леса Нигерии как целого тесно согласуется с периодичностью климата.
Обычно полагают, что основным внешним фактором, регулирующим периодичность физиологических процессов тропической растительности, является сезонная засуха, а не температура, как в умеренных климатах. Вероятно, однако, что фотопериодические реакции на продолжительность дня не менее существенный фактор, воздействующий на физиологический ритм растений как в сезонных, так и в несезонных тропических климатах. Бюнинг (Banning, 1947,1948) указывает, что у тропических растений, которые по необходимости являются растениями короткого дня, приспособленными к чередованию света и темноты примерно одинаковой длительности, фазы адаптации к свету и к темноте переходят одна в другую более резко, чем у растений умеренного климата; следовательно, тропические растения более чувствительны к незначительным изменениям продолжительности дня. Если это так, то изменения в продолжительности дня, возможно, являются причиной регулярного сезонного хода физиологических процессов некоторых растений в тех климатах, где нет заметных сезонных колебаний температуры или осадков (например, регулярное ежегодное цветение Erythrina indica на Новых Гебридах, о котором говорилось выше). Согласно Бюнингу (Banning, 1947, стр. 15—16) существуют некоторые данные о том, что местные или акклиматизировавшиеся виды, обычно свойственные умеренной зоне, но встречающиеся в горах тропической области, в большей или меньшей степени нечувствительны к продолжительности дня. Знаменательно также, что среди папоротников, включающих сравнительно большее, чем у цветковых растений, число видов, встречающихся и в высоких и в низких широтах, не наблюдается никаких фотопериодических реакций.
Уже давно установлено, что не все жизненные функции растений непременно замирают одновременно. Даже европейские листопадные деревья зимой не обязательно находятся в состоянии полного покоя. Это
СРЕДА
215
также относится и к тропическим растениям. Периодичность появления листьев и периодичность цветения, хотя зачастую связаны между собой, все же в большой степени проявляются независимо и поэтому должны быть рассмотрены порознь.
СМЕНА ЛИСТЬЕВ
Опубликовано много наблюдений над сменой листьев у растений как в ботанических садах, так и в естественных местообитаниях. В добавление к данным Шимпера (1903) для различных тропических районов имеются данные Клебса (Klebs, 1911), Фолькенса (Volkens, 1912), Саймона (Simon, 1914) и Габерландта (Haberlandt, 1926) для Явы; Райта (Wright, 1905), Хольтермана (Holtermann, 1907) и Смита (Smith, 1909) для Цейлона; Холтума (Holttum, 1931, 1938 b, 1940) для Малаккского полуострова; Дж. Р. и И. Бекер (Baker. J. R., Baker!., 1936) для Новых Гебридов; Шеф-лера (Scheffler, 1901), Милдбреда (Mildbraed, 1922) и Ричардса (Richards, 1939) для тропической Африки; Иэринга (von Ihering, 1923) для Бразилии; Дэвиса и Ричардса (Davis, Richards, 1933—1934) для Брит. Гвианы и Бирда (Beard, 1946 b) для Тринидада. Клебс (Klebs, 1912, 1915, 1926), Динглер (Dingier, 1911) и Швейцер (Schweizer, 1932) изучали вопрос экспериментально.
Очевидно, что среди тропических деревьев существует много весьма различных способов облиствления. Средняя продолжительность жизни листьев у тропических видов, согласно Вармингу и Гребнеру (Warming, Graebner, 1933), составляет 13—14 месяцев, но лишь у ничтожного количества видов, если они вообще существуют, возобновление листьев происходит равномерно в течение года. Фолькенс (Volkens, 1912) и Клебс (Klebs, 1911) утверждали, что в Богоре некоторые виды, например Albizzia fal-cata, являются «вечнорастущими», так как они совсем не обнаруживают какой-либо периодичности в смене листвы. Саймон (Simon, 1914) показал впоследствии, что Albizzia только в молодости ведет себя подобным образом; у старших особей появление листвы явно периодично. Согласно Фаберу (Schimper, 1935, стр. 383), то же самое оказалось и у других предполагаемых «вечнорастущих» видов. В действительности у большинства деревьев тропического дождевого леса новые лисГья появляются не непрерывно, а периодически, так что один побег несет одновременно несколько «генераций» листьев.
У некоторых видов все или большая часть старых листьев опадают задолго до того, как появляются новые, так что дерево остается обнаженным недели или месяцы, как, например Bombax flammeum, В. malabaricum, Cedrela toona var. australis, Hymenaea courbaril, Koompassia malaccensis, Piptadenia africana, Terminalia superba и многие другие. В то же время, и это, вероятно, наиболее характерно для видов тропического дождевого леса, старые листья могут опадать почти одновременно с появлением новых или же всего за несколько дней до этого. У некоторых деревьев старые листья опадают вскоре после полного развития новых, как, например, у Polyphragmon sericeum (Volkens, 1912, стр. 72).
Как и у европейских листопадных деревьев, старые листья тропических деревьев опадают после образования отделительного слоя. Как правило, они не опадают, пока еще совсем зеленые, хотя Ficus glabella, согласно Шимперу, является исключением (Volkens, 1912, стр. 69). Обычно при опадании листья желтеют или покрываются желтыми пятнами,
216
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
а иногда становятся ярко-красными (Volkens, 1912, стр. 69; Mildbraed, 1922, стр. 110). Корнер (Corner, 1938, стр. 119—121) заметил, что старые листья деревьев Малайского архипелага принимают цвета, чрезвычайно характерные для определенных видов и родов, чем способствуют их опознаванию. У яванских Pterocymbium javanicum и Sterculia foetida крона никогда полностью не обнажается; старые листья желтеют и, вероятно, теряют способность к фотосинтезу за несколько недель до опадания; другие, например Ehretia javanica, Planchonia valida, в течение длительного времени стоят почти без листвы (Simon, 1914).
Иногда старые листья опадают очень быстро. Фолькенс (Volkens, 1912, стр. 70) сообщает, что у Ficus variegata, Ява, казавшегося совсем свежим и зеленым, все листья облетели за 4 дня; однако большей частью этот процесс происходит гораздо медленнее.
Появление новых листьев, если оно периодично, может происходить через равные или неравные промежутки времени. Холтум (Holttum, 1931, 1938 Ь) установил, что в Сингапуре у Parkia roxburghii новые листья появлялись регулярно каждые 12 месяцев со стандартным отклонением, равным 0,24 за 10 лет; Cratoxylon poly anthum, Тamarindus indica и некоторым другим видам также свойствен годичный период облиствления. У Terminalia. catappa, Ficus variegata и Peltophorum jerrugineum новые листья появлялись с интервалами примерно 6 месяцев. У одного из деревьев Ficus variegata период облиствления составлял 6,6 месяца на протяжении 10 лет, а у других особей этого же вида — почти 6 месяцев. Изучая кустарник Breynia cernua в Богоре Смит (Smith, 1923) установил, что его старые листья и ветки опадали, а новые листья появлялись в среднем каждые 5,5 месяца, и за 5 лет растение проходило И полных циклов. Был также обнаружен ряд видов, чьи циклы не соответствовали году и не укладывались в году целым числом раз; у дерева Delonix regia в Сингапуре был регулярный период в 9 месяцев, а у Heritiera macrophylla наблюдались три последовательных периода в 2 года и 8 месяцев каждый (Holttum, 1931, 1938 Ь, 1940).
Каучуковое дерево {Hevea brasiliensis) и манго {Mangifera indica) представляют хорошо известный пример деревьев, у которых новые листья появляются через очень неравномерные промежутки времени. У различных особей каучукового дерева обычно бывают различные промежутки, зависящие также от их возраста (Holttum, 1931; Schweizer, 1932), хотя в климате южной Нигерии с его относительно выраженными сезонами, новые листья у деревьев этого вида ежегодно появляются более или менее одновременно к концу сухого времени года на всех особях независимо от возраста. У манго разные ветви одной и той же особи покрываются новой листвой с большими промежутками, так что смена листьев у всего дерева может тянуться по нескольку недель. Еще заметнее это выражено у Ceiba pentandra. В Западной Африке на этом дереве часто можно увидеть наряду с совершенно обнаженными ветвями и ветви, сплошь покрытые листвой. Разводимая в Сингапуре местная раса или подвид не обладает регулярностью облиствления, в то время как африкано-американский подвид меняет листья регулярно и равномерно (Corner). Произрастающая в Сингапуре Pterocarpus indicus сменяет листву одновременно на одной-двух ветвях, а в северной части Малаккского полуострова эта порода является полностью листопадной (Corner, 1940). Даже у тех тропических видов, которые в целом более или менее подчиняются одной и той же периодичности, распускание почек на различных ветвях, вероятно, редко происходит столь же одновременно, как у большинства деревьев
СРЕДА
217
умеренной зоны. Вероятно, возрастные изменения периодичности в смене листьев, как у Hevea, встречаются весьма часто.
Различия в облиствлении деревьев тропического дождевого леса настолько велики, что нельзя точно разграничить вечнозеленые и листопадные виды. Для удобства вечнозелеными можно считать те, у которых дерево покрыто большим количеством листьев в любое время года; листопадными — те, которые остаются обнаженными или почти обнаженными хотя бы на несколько дней. Фолькенс (Volkens, 1912) предложил другое определение, считая вечнозелеными только виды, одновременно покрытые листьями хотя бы двух «генераций» (Schiibe) в течение года. Согласно этому определению, деревья, которые сразу теряют все листья и тут же без перерыва покрываются новыми, могут быть отнесены к листопадным. Такая классификация может иметь преимущества с физиологической точки зрения, но менее пригодна для других целей.
Листопадные деревья в принятом здесь смысле изобилуют в тропических дождевых лесах всех тропических областей; они встречаются даже в климатах с почти равномерно распределенными осадками, как в Сингапуре и на Новых Гебридах. Корнер (Corner, 1946) полагает, что в вечнозеленых лесах тропиков встречается больше листопадных видов, чем в умеренных листопадных лесах. В типичном тропическом дождевом лесу почти все листопадные деревья принадлежат к ярусу А; ярусы В, С и подлесок почти полностью состоят из вечнозеленых пород. С приближением к климатическому пределу (определяемому продолжительностью сухого сезона, см. стр. 173) распространение тропического дождевого леса, количество листопадных видов постепенно возрастает, сначала в верхнем, а затем и в нижних ярусах. В Республике Конго, например, количество листопадных видов возрастает от центра лесного массива к окраинам, где засухи продолжительнее и сильнее (Lebrun, 1936а). Во влажном вечнозеленом лесу Нигерии, где сухой сезон весьма продолжителен, некоторые деревья яруса В, например Monodora tenuijolia и М. myristica являются листопадными, хотя большинство видов в его пределах относится к вечнозеленым; в ярусе А большая часть видов сбрасывает листву на несколько недель или месяцев в сухое время года. Бирд (Beard, 1942) установил пропорцию листопадных деревьев в четырех лесных формациях на острове Тринидад (см. табл. 22). Соотношение листопадных и вечно-
Таблица 22
Пропорция листопадных (включая полулистопадные) видов в лесах острова Тринидад
(Beard, 1942, стр. 14)
Тип леса % видов % отдельных экземпляров
«Верхний ярус» «Нижний(ие) ярус(ы)» «Верхний ярус» «Нижний(ие) ярус(ы)»
Вечнозеленый сезонный лес1 24 Ничтожный 6 Ничтожный
Лес Мора 12 0 Ничтожный 0
Полувечнозеленый сезонный лес 33 10 16,5 10
Листопадный сезонный лес 50 10 66 25
1 Названия формаций соответствуют последней классификации Бирда (Beard, 1946 Ь)
218
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
зеленых деревьев в различных ярусах является ценным диагностическим признаком для различения тропических лесных формаций, и можно лишь сожалеть о недостаточности цифровых данных подобного рода.
Много спорят об относительной важности внешней среды и внутренних пли автономных факторов, вызывающих и регулирующих ритм смены листьев у тропических деревьев. Тот взгляд, что внешние факторы почти полностью ответственны за это, особенно поддерживался Клебсом (Klebs, 1911, 1912, 1915, 1926), который пытался показать, что даже в наименее сезонных тропических климатах внешняя среда целиком контролирует периодичность физиологических процессов у растений. Согласно Клебсу, когда значение любого из факторов среды, обусловливающих жизнедеятельность растения, падает ниже минимальной величины, растение входит в фазу покоя. Это могут быть как эдафические, так и климатические факторы; когда листопад нельзя связать с каким-либо климатическим фактором, то его, вероятно, можно объяснить периодическим истощением питательных веществ, непосредственно получаемых растением из почвы.
Не вызывает сомнения, что внешние факторы в большой степени влпяют на периодичность физиологических процессов у растений даже в сравнительно несезонных климатах. Среди этих внешних факторов, вероятно, определенную роль играют и почвенные условия, хотя, пожалуй, главным образом в связи с водоснабжением. Так, в Бразилии деревья, растущие на известковых почвах, теряют листья в большей степени, чем растущие на других почвах (Warming, 1892). Однако Клебс, конечно, придерживается крайних взглядов, так как существуют очевидные доказательства того, что наряду с внешней средой большую роль в периодичности поведения растений играют и малоизученные внутренние факторы (Schimper, 1903; Volkens, 1912; Simon, 1914; Coster, 1923). Чередование фаз покоя и активности, по-видимому, составляет неотъемлемое свойство всех живых организмов. Среда может контролировать наступление и длительность этих периодов; чем яснее выражена сезонность климата, тем больше приходится наследственной периодичности организма приспосабливаться к ритму климата, но в климате тропического дождевого леса внутренние факторы как бы раскрепощаются, поскольку сезонные изменения столь незначительны, что нет такого времени года, когда внешние условия могли бы принудить растение перейти в фазу покоя, если оно побуждается к активности внутренними факторами, и наоборот.
Тот факт, что среди деревьев тропического дождевого леса в одних семействах встречается больше листопадных пород, чем в других, заставляет думать, что внутренние факторы генетически обусловлены и наследственны. Например, многие Sterculiaceae и Bombacaceae являются листопадными, в то время как Lauraceae — почти всегда вечнозеленые. Эксперименты Швейцера (Schweizer, 1932) также показывают, что у Hevea наследственные факторы играют очень важную роль в периодичности. В стадии сеянцев новые листья у отдельных экземпляров появляются разновременно, но поведение особей, размноженных черенками от одного и того же дерева, очень сходно, и новые листья появляются в этом случае почти одновременно.
Очевидно, что относительная значимость внутренних факторов в сравнении с факторами внешней среды различна у разных видов, следовательно, в указанном отношении некоторые виды более, некоторые менее пластичны. В ясно выраженных сезонных климатах тик (Т ectona grandis) и Bombax malabaricum длительное время в течение сухого сезона стоят без листьев, но в Сингапуре и на западной Яве, где осадки распределены
СРЕДА
219
исключительно равномерно, Bombax все же остается оголенным в течение примерно 3 месяцев, a Tectona становится вечнозеленым или почти вечнозеленым. Костер (Coster, 1923) экспериментально установил, что появление листьев на обнаженных ветвях Bombax можно вызвать только к концу периода покоя, a Tectona при подходящих условиях может воспроизвести листья в любое время, часто для этого достаточно поставить его ветку в воду. Бирд (Beard, 1946 Ь) подразделяет деревья Тринидада на деревья с обязательным и необязательным листопадом. У последних цикл опадания листьев колеблется год от года, соответственно выраженности сухого сезона. Обычно их листья облетают постепенно по мере обострения засухи вплоть до тех пор, пока они не окажутся совершенно оголенными, но в дождливый год явного листопада может и не быть. Виды с обязательным листопадом свойственны по большей части формации листопадного леса, виды с необязательным листопадом — преимущественно полувечнозеленому сезонному лесу (стр. 344).
Листопад у деревьев умеренной зоны является их устойчивым свойством, и внешняя среда сравнительно мало влияет на него. На Яве и в других тропических районах такие деревья, как бук (Fagus sylvaticd), сохраняют периодичность роста и появления листьев, хотя сроки и длительность периода покоя меняются, и разные ветви одного и того же экземпляра распускаются не одновременно, что, как мы видели, часто бывает у местных тропических видов (Schimper, 1935, стр. 387; Coster, 1926 Ь).
Вряд ли можно сомневаться в том, что у деревьев умеренных и других выраженных сезонных климатов листопад является адаптацией в том смысле, что он позволяет растению избегать чрезмерной транспирации в такие периоды, когда трудно возместить водопотерю. В тропических климатах, за исключением мест с очень сильными засухами, это преимущество в значительной мере теряет свой смысл. В вечнозеленых лесах Западной Африки у многих видов деревьев, которые некоторое время стоят без листьев, нежные молодые листья появляются задолго до конца сухого сезона, иногда даже в исключительно засушливый период харматана, когда опасность иссушения особенно велика. Даже в долине среднего течения Нигера с ее длительным сухим сезоном Acacia albida фактически теряет листья, когда начинаются дожди и снова покрывается листвой в начале сухого периода (Roberty, 1946, стр. 96). На острове Тринидад ни один вид не теряет листву в дождливый сезон, но некоторые из них обнажаются всего на несколько дней или недель в начале засухи, полностью покрываясь листьями в самом ее разгаре (Beard, 1946 b). На Цейлоне и в той части Явы, которая находится в сфере влияния «восточного муссона», многие листопадные деревья также развертывают свои нежные молодые листья задолго до конца сухого сезона, когда им еще грозит повреждение (Wright, 1905; Coster, 1923). Динглер (Dingier, 1911), экспериментируя на Цейлоне, сохранил на время сухого сезона листву у нескольких обычно листопадных видов и обнаружил, что она не испытала никакого повреждения. Из таких фактов можно сделать только один вывод — в относительно несезонных климатах, таких, как климат тропического дождевого леса, листопад одинаково возможен в любое время года, и деревья в сущности могли бы сохранить листву круглый год без всякого риска для себя.
На Тринидаде, согласно Бирду (Beard, 1946 b), нет листопадных видов, ограниченных в своем распространении только вечнозеленым лесом; наоборот, чаще всего они встречаются в листопадных и полулистопадных формациях. То же самое, вероятно, относится к листопадным видам и дру
220
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
гих тропических районов. Но если смена листьев в одинаковой степени зависит от внутренних и внешних факторов, то нельзя признать вполне обоснованной аргументацию Иэринга (Ihering, 1923), который, ссылаясь на данные по флоре южной Бразилии, утверждал, что листопадные виды проникли в вечнозеленый лес из областей с более сезонным климатом.
РОСТ И АКТИВНОСТЬ КАМБИЯ
Активность камбия у деревьев тропического дождевого леса была исследована Саймоном (Simon, 1914). У видов, полностью теряющих листву, вроде Tetrameles nudi flora камбий ветвей прекращает деление на все время, пока дерево обнажено, и вторичный рост возобновляется не раньше, чем распустится молодая листва. Подобным образом у Termi-nalia catappa, листья которой подолгу бывают обесцвеченными и, вероятно, физиологически неактивными, деятельность камбия на это время прекращается. Даже у Dillenia indica и Ficus variegata. чьи листья бывают совсем зелеными и, очевидно, активными большую часть года, опадая лишь в момент распускания почек пли незадолго до этого, деятельность камбия надолго, вплоть до появления новых листьев, прекращается. Единственным вполне вечнозеленым видом, который изучил Саймон, был Ficus annulata var. valida\ у этого дерева периодическому появлению новых листьев, по-видимому, всегда предшествует перерыв в деятельности камбия. Саймон сделал вывод, что по крайней мере у тех тропических деревьев, которые периодически обновляют листья, деятельность камбия также периодична. Более обширная работа Костера (Coster, 1927—1928) дополняет и подтверждает этот вывод. Она показывает, что, когда листья находятся в активном состоянии, камбий одинаково активен как у вечнозеленых, так и у листопадных видов.
Костер провел также обширное исследование слоев или колец роста у тропических деревьев. У деревьев в несезонных тропических климатах редко встречаются годичные кольца, какие наблюдаются у деревьев умеренной зоны. У некоторых кольца вообще отсутствуют; у других они не образуют замкнутых окружностей, у третьих отчетливые кольца не соответствуют годичному циклу. Костер установил, что в общем отчетливые кольца роста попадаются только у листопадных видов, то есть у тех, у которых бывает определенный перерыв в деятельности камбия. Однако обратного соотношения нет; у тех видов, рост камбия которых периодически прерывается, не всегда имеются отчетливые кольца роста. Анатомическое строение наблюдающихся колец очень разнообразно и внутри видов п между различными видами. Определение возраста по кольцам роста у тропических деревьев очень ненадежно, ибо даже в том случае, когда годичные слои или кольца хорошо очерчены, они иногда образуются только по достижении деревом определенного возраста.
У тропических деревьев рост побегов в длину сопутствует развертыванию листьев или следует за ним, и поэтому дальнейшее обсуждение этого вопроса излишне. Саймон (Simon, 1914) и Костер (Coster, 1925) изучили резервы питания тропических деревьев в связи со сменой листьев и периодическим ростом.
ЦВЕТЕНИЕ
Сведения о периодах цветения в тропическом дождевом лесу скудны и неопределенны. Некоторое внимание было уделено цветению отдельных
СРЕДА
221
видов, особенно тем случаям, когда оно носит массовый характер, но цветение в формациях и сообществах в целом мало изучено.
В вечнозеленом тропическом лесу цветение обычно происходит круглый год, и во всякое время года можно наблюдать цветение тех пли иных видов, причем некоторые цветут почти непрерывно; но даже в климатах с наименее выраженными сезонными различиями существует определенное время максимального цветения, которое, однако, не так бросается в глаза,
Месяцы
СОНДЯФМАМИИД
Corymborchis (_ © ________
veratri folia ф • V “ "чР
Ficus copiosa • • • • • • •-------• •---
Piper methysticum ® ® ф ф-----ф ф ф
Garuga fLori bund a ______ --_______________
Castanospermum / $ м ™ oust rale 5 ф ч? чя
Eugenia sp. • ф $
Barring t onia samoensis
Psychotria sp.
Geophila herbacea
Рис. 33. Периоды цветения и плодоношения у девяти наиболее распространенных растений в несезонном тропическом климате, Новые Гебриды (J. R. Baker, I. Baker, 1936, стр. 508).
Среди отобранных растений находятся деревья и травы. В феврале четыре вида растений выпали из наблюдений. Цветение обозначено розеткой, плодоношение — черным кружком.
чтобы обратить на себя внимание случайного наблюдателя. Как и следовало ожидать, эти максимумы заметнее всего там, где бывает ясно выраженный сухой сезон. В тропическом дождевом лесу на Новых Гебридах, где климат исключительно ровный, существует два основных периода цветения; первый длится с сентября по январь, в самое дождливое время года, совпадающее с небольшим повышением температуры после более прохладного сезона, а второй, более короткий и менее ясно выраженный, наблюдается в мае и июне, когда выпадает мало осадков и температура падает (Baker J. R., Baker 1., 1936, см. рис. 33). Эггелинг (Eggeling. 1947, стр. 64) рисует следующую картину цветения в консоциации Cynometra alexandri леса Будонго, Уганда (стр. 281), где бывает два сухих сезона, один из которых длится три месяца подряд и характеризуется осадками менее 10 см, второй — более короткий и менее засушливый: «лишь очень немногие виды цветут в промежутке между двумя сезонами дождей и в начале периода коротких дождей; к концу последнего зацветают полукустарники, и в течение почти двух месяцев повсюду виднеются длинные белые соцветия Whitjieldia elongata. В это же самое время цветут и многие Acanthaceae, но доминирующее травянистое растение Lanke-
222 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
steria elegans расцветает только после отцветания Whitfieldia. Яркие оранжевые цветки Lankesteria оживляют подлесок до самого конца сухого сезона. В феврале, когда Lankesteria еще в цвету, зацветают деревья нижнего яруса, и одним из первых Lasiodiscus mildbraedii, за которым следует Rinorea ardisiaeflora, чьи цветки распускаются с началом дождей. С наступлением дождливого сезона расцветает большая часть крупных деревьев, п вскоре весь полог белеет от массы соцветий Cynometra».
Хотя, как апл видели, основная пора цветения на Новых Гебридах приходится на более дождливое время года, в большинстве районов влажного тропического климата, включая районы с выраженным сухим сезоном, цветение происходит главным образом в сухое время года. Главный максимум цветения часто приходится на конец сухого сезона, захватывая п начало дождливого сезона, но иногда наблюдается и другой максимум в начале сухого сезона, как раз после прекращения дождей. Что является сигналом к началу цветения — влажность атмосферы или почвы или же легкие колебания средней температуры (которая обычно повышается в сухое время года и падает с наступлением дождей) — неизвестно.
Шимпер (Schimper, 1903, стр. 254) проанализировал данные Курдерса и Валетона о периодах цветения на Яве. Для острова в целом (на котором есть районы листопадного леса с очень длительным сухим сезоном) он установил, что 63% видов цветет в засушливое время, неполных 8% — только в сезон дождей и 29% — в разное время. «Флора Цейлона» Трай-мена (Trimen) дает сходные данные; в «влажных округах», где апрель, май и октябрь являются наиболее дождливыми месяцами, основной порой цветения древесных растений оказываются февраль — апрель и сентябрь. Максимальное цветение травянистых растений приходилось на февраль и сентябрь. По данным Шомбурга (Schomburgk), в Брит. Гвиане цветение доходит до максимума в начале и конце короткого сухого сезона (февраль и апрель) и в начале длительного сухого сезона (сентябрь). В поясе тропических дождевых лесов Нигерии пора максимального цветения наступает, вероятно, в начале сезона дождей и вторично — в конце его (Richards, 1939). В «сомкнутом лесу» на Берегу Слоновой Кости, где бывает два сезона дождей, цветение происходит главным образом в короткий сухой сезон (с июля по август) и с августа по декабрь, когда начинается продолжительный сухой сезон (Aubreville, 1938, стр. 151—152). Цветение заметно ослабляется как в разгар длительной засухи (январь, февраль), так и в самые дождливые месяцы года (май и июнь). Цветение деревьев, растущих по берегам рек, например Pterocarpus santalinoides, имеет меньшую связь с климатическими сезонами.
Есть данные, что цветение в верхних ярусах леса гораздо периодичнее, чем в нижних. Так в Брит. Гвиане в ярусах А и В цветение приурочено к двум основным периодам, в то время как деревья яруса С, кустарники и травы, по-видимому, цветут равномерно круглый год (Davis, Richards, 1933—1934, стр. 359). Если это так, то причина, несомненно, заключается в более ровных условиях температуры и влажности в пределах нижних ярусов.
Отдельные виды тропического дождевого леса чрезвычайно разнообразны по характеру цветения, но подавляющее большинство цветет, вероятно, через очень равномерные промежутки времени. У многих деревьев отдельные экземпляры цветут в самое различное время как в одной и той же местности, так и в разных местностях; ветви одного и того же дерева также могут цвести в разное время. Если говорится, что определенный вид цветет круглый год, это не означает, что каждый
СРЕДА
223
экземпляр цветет непрерывно, хотя для некоторых видов беспрерывное цветение является нормой. Так же как и при смене листьев, неопределенность периодов цветения у всего сообщества в целом в основном является следствием недостатка синхронизации этого процесса у видов и индивидуумов.
Корнер (Corner, 1940) подразделяет деревья Малайи на «вечноцветущие» виды с непрерывным воспроизводством, у которых цветки и плоды появляются на одном и том же экземпляре либо одновременно (Wormia suffruticosa, Adinandra dumosa), либо поочередно через очень короткие промежутки времени (Ficus spp.), и виды, цветущие периодически. У последних цветение может происходить раз в год, как у многих деревьев малайских лесов, дважды в год (Fagraea fragrans, Calophyllum inophyllum), чаще двух раз в год (Eugenia grandis, Rhodamnia} или же с интервалом больше года (различные Dipterocarpaceae). «Вечноцветущие» виды могут начать цветение в очень юном возрасте и цвести вплоть до отмирания; Adinandra dumosa, живущая, вероятно, 100 или больше лет, зацветает в возрасте 2—3 лет, Wormia sutfruticosa зацветает примерно в восемнадцатимесячном возрасте и непрерывно цветет по меньшей мере 40—50 лет (Corner). Многие небольшие деревянистые растения, вероятно, ведут себя подобным же образом, но более крупные деревья обычно цветут периодически и зацветают только совсем взрослыми. Периодически цветущие виды иногда имеют очень большой интервал между цветением; согласно Ридли (Schimper, 1935, стр. 394—395), некоторые малайские виды Нореа и Shorea (Dipterocarpaceae) цветут регулярно каждый шестой год. Отдельные экземпляры Homalium grandiflorum цветут только один раз в 10—15 лет (Corner). Некоторые периодически цветущие деревья цветут регулярно каждые 7—10 месяцев, но цветение у разных индивидуумов происходит не одновременно (Delonix regia, Lagerstroemia speciosa). Очевидно, что в подобных случаях оно обусловлено внутренним ритмом, не связанным непосредственно с периодичностью климата. На Берегу Слоновой Кости Обревиль (Aubreville, 1938, стр. 152) также установил, что одни впды цветут непрерывно, а другие — в точно установленные периоды: раз в год, дважды в год, или с большими интервалами.
Особый интерес представляют тропические растения, у которых наблюдается массовое цветение, когда все экземпляры цветут одновременно с большими или меньшими промежутками времени иногда на площади во много квадратных километров. Это довольно обычное явление среди растений тропического дождевого леса. Спрус (Spruce, 1908, I, стр. 257) следующим образом характеризует амазонские Myrtaceae: «Они замечательны своими одновременно распускающимися эфемерными цветками. В один день все миртовые определенного вида, разбросанные по лесу, покрываются снежно-белыми душистыми цветками; на следующий день от цветков сохраняются только увядшие лепестки. Поэтому, если ботаник не сумеет собрать миртовые в день их расцвета, ему нечего надеяться пожать среди них «лавры». В некоторых случаях была прослежена связь между массовым цветением и условиями погоды. Корнер (Corner, 1940, стр. 537) так описывает цветение трех культурных видов Coffea в Малайе: «В определенные дни года внезапно распустившиеся цветки превращают жалкий кофейный куст в благоухающую белизну. Все кусты кофе одного вида расцветают в одной и той же местности в одни и те же дни одновременно, причем массовое цветение у различных видов наступает в различные дни. Причина, по-видимому, заключается в необходимости стимула в виде определенных условий погоды для того, чтобы заставить раскрыться
224
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
цветочные бутоны. Бутоны развиваются до определенной величины, а затем как бы дремлют, пока погода не даст им нужного стимула; тогда они пробуждаются и через определенный период распускаются одновременно. Характер стимула или же интервал между стимулом и раскрытием бутона должны быть различными у разных видов, отсюда — различные дни цветения». Хорошо известным примером массового цветения могут служить различные бамбуки, которые цветут с большими интервалами и отмирают после созревания плодов, а также виды Sthrobilanthes. У S. сег~ nuus, Ява, массовое цветение происходит примерно через каждые 9 лет (van Steenis, 1942), у 5. sexennis, Цейлон, примерно через каждые 12 лет (Petch, 1924). У дерева Fagraea fragrans, Сингапур, массовое цветение наблюдается каждый год в мае с исключительной регулярностью; в октябре и ноябре бывает вторичное (не столь пышное) цветение. Холтум (Holttum, 1935) показал, что основной период цветения наступает примерно 4 месяца спустя после паузы в дождливом сезоне, которая обычно бывает в январе; в течение более чем 8 лет сдвиг в сроках этой паузы точно соответствовал сдвигу во времени цветения. На о. Маврикия Воган и Вийе (Vaughan, Wiehe, 1937, стр. 302) обнаружили, что массовое цветение Homalium paniculatum наступает после сильных циклонов.
Массовое цветение лучше всего изучено на примере орхидеи Dendro-bium crumenatum и некоторых других малайских эпифитных орхидей (Coster, 1926 а, а также указанная у этого автора литература). Массовое цветение этих орхидей охватывает большую площадь леса; у каждого вида цветки живут всего один день или еще меньше. Почти всегда цветение наблюдается после грозы, которой заканчивается короткий период засухи. Промежуток между грозой и цветением может составлять 8, 9, 10 или 11 дней в зависимости от вида орхидеи; для некоторых видов он бывает на 1—2 дня короче. Экспериментально доказано, что влияние гроз проявляется не в прямом воздействии дождя или атмосферного электричества, а во внезапном падении температуры. У двух видов бутоны вырастают до определенного размера и затем входят в фазу покоя, как это было показано выше на примере Coffea, и только понижение температуры стимулирует их к выходу из этого состояния. Чем дольше бутоны находятся в состоянии покоя, тем быстрее они потом распускаются, если же стимул слишком задерживается, бутоны могут раскрыться самопроизвольно.
Массовое цветение тропических деревьев и орхидей, видимо, имеет много общего с «пульсирующим» цветением рода Juncus, произрастающего в умеренном климате; у этого растения все экземпляры одного и того же вида в данной местности открывают свои цветочные бутоны с промежутками в несколько дней без всякой видимой связи с изменениями погоды.
Часто наблюдается явная связь между цветением и вегетативной деятельностью. У некоторых тропических листопадных деревьев созревание плодов регулярно следует за опаданием листьев. Многие виды цветут всегда во время листопада; иногда у деревьев, ветви которых теряют листву не одновременно, цветение происходит только на безлистных ветках; это часто наблюдается у Ceiba pentandra и иногда у Couratari pulchra, обычного дерева гвианского леса. Бирд (Beard, 1946 Ь, стр. 51) установил, что среди листопадных деревьев Тринидада примерно половина видов цветет в период полного облиствления, а плодоносит после листопада; около четвертой части цветет, когда нет листьев, и плодоносит в этот же или в следующий сухой сезон, вторая же четверть видов цветет в сухой сезон, находясь без листьев, и плодоносит в следующий дождливый сезон. У вечнозеленых дубов (Quercus и Lithocarpus) и диптерокарповых Малайи
СРЕДА
225
и у Mangifera indica в Сингапуре цветение не совпадает с появлением молодой листвы, но у других деревьев, таких, как виды Myristica и Saraca. цветки и новые листья появляются одновременно (Corner). В районах Явы, подверженных влиянию восточного муссона, где многие мелкие деревья и'‘кустарники являются «вечноцветущими», Костер (Coster, 1923) обнаружил, что в целом виды, у которых рост побегов идет непрерывно, тоже Являются «вечноцветущими», а когда появление побегов происходит периодично, то и цветение оказывается периодичным. Высказывалась мысль, что между цветением и листопадом существует причинная связь, объясняемая явным антагонизмом между этими явлениями, так что наступление генеративной фазы ведет к полному или частичному прекращению вегетативной деятельности. Однако более вероятно, что оба явления вызываются общей физиологической причиной.
Периодическое цветение у тропических растений, по-видимому, отчасти зависит от изменений в окружающей среде, но факты, приведенные выше, заставляют думать, что наряду с ними действуют также и внутренние причины. Имеющиеся данные показывают одновременное воздействие внешних и внутренних факторов как при периодическом наступлении генеральной фазы, так и при сбрасывании листьев. Возможно, что в действительности внутренние факторы — основной двигатель обоих явлений, но, как указывают Дж. Р. и И. Бекер (J. R. Baker, I. Baker, 1936), внутренний физиологический ритм не может быть настолько точен, чтобы определять сроки регулярной периодичности, которая часто наблюдается. Внешняя среда должна регулировать внутренний ритм и время от времени как бы «подводить часы». В умеренном или тропическом климате с резко выраженными сезонами влияние среды гораздо заметнее, чем в однообразном, относительно неизменном климате, в связи с чем действие внутренних, или эндогенных, ритмов менее проявляется. Некоторые подробности участия внутренних факторов в периодических явлениях у растений были проанализированы Бюнингом (Biinning, 1948), общие выводы которого совпадают с данными этой главы.
Как указывалось в главе3, периодическое и массовое цветение является одной из причин (но не единственной) хорошего или плохого урожая семян, поэтому оно заслуживает дальнейшего изучения благодаря своему значению для естественного и искусственного размножения. Поскольку количество времени, нужное для созревания плодов, различно у разных видов, период обсеменения не обязательно определяется периодом цветения. В сезонных климатах семена чаще всего осыпаются в сухую погоду, которая создает наиболее подходящие условия для их рассеивания, независимо от времени цветения. Прорастание может произойти тотчас же или позднее, при более благоприятных условиях.
Глава 9
ПОЧВЫ
Почвы влажных тропиков (даже если рассматривать только низменности) различны по структуре и по своим физическим и химическим свойствам не менее, чем почвы любой другой климатической зоны. Почва и естественная растительность всегда тесно связаны между собой, и тропический лес так же точно отражает почвенные условия, как и лес умеренного пояса. В сущности, обычное представление (может быть, вызванное внешним однообразием лесного покрова) о том, что и почва и растительность влажных тропиков более однородны, чем в других местах,— мало обосновано. Возможно, что почвенные факторы, наиболее явственно влияющие на растительность тропических дождевых лесов, в своем большинстве иные, чем те, с какими имеют дело экологи Европы или Северной Америки, но воздействие этих факторов тем не менее очевидно.
Несмотря на различие многих свойств, большая часть почв пояса тропических дождевых лесов обладает рядом важных общих особенностей. Они отличаются ярко-красным или желтым цветом; по механическому составу они обыкновенно глинисты или суглинисты, но в верхних слоях часто бывают песчаными; обычно они бедны основаниями и питательными веществами; они почти неизменно кислые; перегноя в них мало, причем он содержится главным образом в самых верхних горизонтах; глинистая фракция относительно богата глиноземом и бедна кремнеземом. Чтобы понять эти особенности почв дождевых лесов, необходимо познакомиться с их эволюцией и прежде всего рассмотреть действие почвообразующих факторов в климате влажных тропиков. Те многочисленные свойства почвы, которые хотя и представляют исключительный интерес для почвоведа, но, по-видимому, не существенны с экологической точки зрения, не будут рассматриваться в этой главе. Ее тема ограничивается разбором лишь тех свойств почвы, которые воздействуют на естественную растительность тропиков и имеют преимущественно экологическое значение как факторы, определяющие распределение группы формаций тропических дождевых лесов и составляющих их ассоциаций, консоциаций и отдельных видов. Вопрос о тропических почвах с более широкой точки зрения трактуется в работах Фагелера (Vageler, 1933), Мора (Mohr, 1944) и Робинсона (Robinson, 1949).
ПОЧВООБРАЗУЮЩИЕ ФАКТОРЫ
Все почвы постепенно развиваются из материнских пород, причем их свойства могут коренным образом меняться при переходе от юности к зрелости. Главными факторами, непосредственно влияющими на ход этой эволюции, являются материнская порода, из которой образовалась почва, растительность, произрастающая на ней, и климат (или, скорее, микроклимат), в котором развивается почва. Рельеф, особенно уклон
СРЕДА
227
поверхности, также имеет важное значение, создавая разнообразные условия дренажа и эрозии. Косвенное влияние оказывают на почву человек и животные, которые изменяют растительность. Существуют самые различные мнения о сравнительной важности этих разнообразных факторов в эволюции почвы. Некоторые отдают предпочтение климату, считая, что конечный результат развития почвы в данных климатических условиях всегда будет один и тот же безотносительно к материнской породе и растительному покрову. Другие приписывают большее значение влиянию материнской породы или растительности. Однако лучше рассматривать почву,, растительность, животный мир, климат и материнскую породу как компоненты единой «экосистемы» (Tansley, 1935), а эволюцию почвы — как результат совместного влияния всех компонентов этой экосистемы. В разное время и в разных условиях преобладает то один, то другой компонент. Если это верно, то было бы ошибочно особо подчеркивать роль материнской породы, растительности или климата, поскольку все они вместе приводят к конечному результату. Из этого следует, что в такой равновесной экосистеме, как климаксный тропический лес, изменение любого компонента неизбежно приводит к соответствующему изменению всех других компонентов. Так, вырубка или прореживание леса неизбежно приведет к изменению почвенного профиля. С этой точки зрения не вызывает удивления и тот факт, что даже под мощным влиянием тропического климата как зрелая почва, так и растительность могут быть разными на материнских породах различного литологического состава.
Выветривание в условиях влажных тропиков
Выветривание, приводящее, в частности, к образованию почвыг обычно протекает под совместным воздействием как физических, так и химических процессов, но во влажных тропиках выветривание — почти исключительно химический процесс. Такой фактор, как мороз, в других климатических условиях играющий важнейшую роль в физическом выветривании, отсутствует в условиях тропического дождевого леса. За исключением первичного разрушения обнаженной породы, которая подвергается попеременному нагреванию и охлаждению, основной процесс выветривания вызывается водой и растворенными в ней веществами, причем в породе, сложенной преимущественно из силикатов, выветривание завершается гидролизом. Это происходит в климате, отличительные особенности которого — высокая равномерная температура и постоянная повышенная влажность. Данные о температуре почвы тропического дождевого леса очень скудны, но можно считать, что повсюду в его пределах температура непосредственно ниже поверхности почвенного покрова редко намного, превышает 25°; в Индонезии на уровне моря температура лесной почвы на глубине 1 м составляет 25—27° (Mohr, 1944, стр. 45). При таких температурах и при избытке влаги гидролиз идет очень быстро, и на ровной местности кора выветривания может достигнуть 15 и более метров глубины даже при отсутствии физических агентов. В связи с этим следует помнить, что во многих тропических районах процесс выветривания может иметь чрезвычайно большую давность, тогда как в большей части Европы он прослеживается только лишь с последнего ледникового периода.
Не всю кору выветривания, покрывающую исходную горную породу, можно считать почвой или даже незрелой почвой. В тропиках почти всегда бывает трудно провести границу между собственно почвой и материнской породой; даже при знакомстве с наиболее глубоко выветренными!
15*
228
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
поверхностными слоями вызывает сомнение возможность проникновения собственно почвы (то есть слоя, населенного организмами и используемого корнями) глубже, чем на один, самое большее — 2 м. Массив частично выветренной породы (зона разрушения), который может быть во много раз толще самой почвы, лучше всего рассматривать как подпочву или почвообразующую породу.
В климате влажных тропиков эрозия, как и выветривание, протекает быстро, и на крутых склонах слой почвы, как правило, оказывается очень тонким даже под покровом леса. Например, в лесу на крутых склонах близ Рио-де-Жанейро (Бразилия) средняя глубина почвы составляет меньше 10 см (McLean, 1919, стр. 122). О большой интенсивности эрозии в тропиках свидетельствует мутность тропических рек и большое количество взвешенного материала, откладывающегося при разливах и наводнениях. Лем (Lam, 1945) приводит цифры, данные в табл. 23, иллюстрирующие, какое огромное количество твердого материала переносится тропическими реками. Фагелер (Vageler, 1933) подчеркивал, что в тропиках благодаря высокой интенсивности осадков эрозия может развиваться п на очень отлогих склонах. Если осадки превышают количество, которое может поглотить почва, то медленно движущаяся полоса воды широким фронтом распространяется по поверхности земли, постепенно разрушая ее. Этот тип эрозии называется плоскостной; воздействуя в течение длительного времени, она должна оказывать значительное влияние на почву.
Таблица 23
Перенос реками твердого материала (Lam, 1945, стр. 19)
Рейн Мамберамо (Новая Гвинея) Амазонка
Расход реки, куб.м!сек 2 400 10000 120 000
Твердый сток, тыс. т/год Снос твердого материала с 1 кв.км 2 000—5 000 15 000 1 000 000 —2 300 000
водосборной площади, кг)год Среднее понижение уровня по- 15 000 150 000 2 000 000
верхности земли за год, см 0,006 0,06 0,08
В результате химического выветривания пород, не являющихся подобно песчанику высоко кремнистыми, образуются продукты гидролиза сложных силикатов — оснований, кремнезема, глинозема, каолиновых минералов (гидраты алюмосиликатов и пр.),— а также окислы железа И вещества, подобные кварцу (если таковые имеются), относительно устойчивые к выветриванию. Эти первичные продукты скапливаются в подпочве, непосредственно над невыветренной исходной породой; их последующая судьба определяется главным образом свойствами почвенных вод и теми веществами, которые они несут в растворе.
В климате, где количество осадков превышает величину испарения, почвенные воды имеют в основном нисходящее движение, хотя возможность значительных передвижений воды, конечно, зависит от строения почвы1. Когда испарение преобладает над выпадением осадков, то нисхо-
1 Некоторые тропические почвы обладают твердой, малопроницаемой поверхностью, не допускающей сколько-нибудь значительного впитывания воды (Chenery, Hardy, 1945).
СРЕДА
229
дящее движение воды прекращается и могут даже возникнуть незначительные восходящие ее токи. В зоне тропических дождевых лесов количество осадков в целом превосходит испарение в течение большей части года, и только в те месяцы, когда количество осадков меньше 4 дюймов (10 см), можно считать, что испарение больше количества осадков. Таким образом, нисходящее движение воды в почве продолжается почти круглый год, и даже в зоне тропических дождевых лесов с явно выраженным сухим сезоном оно прерывается не более чем на 4 месяца в году. Поэтому все почвы тропических дождевых лесов, исключая наиболее неразвитые, сильно выщелочены и имеют элювиальный горизонт, или зону выщелачивания, залегающий над иллювиальным горизонтом, в который вода вмы-вает подвижные вещества.
Важно осознать, что состав почвенной воды не одинаков по всему разрезу почвы. В верхних слоях почвы нисходящий поток состоит из дождевой воды, несущей в основном двуокись углерода и органические вещества, извлеченные из гумуса на поверхности почвы. Ниже она пополняется растворимыми веществами, извлекаемыми из выветренной материнской породы, и в результате обогащается продуктами гидролиза силикатов.
Все первичные продукты гидролиза силикатов растворимы в почвенной воде при определенных условиях, но степень их растворимости весьма различна. Основания наиболее растворимы, в связи с чем они быстро уносятся дренажной водой, если только их не поглощают корни деревьев. Таким образом, содержание оснований в развитых почвах тропических дождевых лесов всегда очень низкое, и почвы здесь неизменно кислые, поскольку между реакцией и содержанием оснований существует прямая связь.
Кремнезем и полуторные окислы железа и алюминия (Fe2O3, А12О3) тоже растворимы и также уносятся нисходящим током почвенной воды. Все три вещества движутся в виде коллоидных золей и содержатся в растворе или осаждаются в зависимости от содержания электролитов и других коллоидов в почвенной воде. Кремнезем и силикаты алюминия относительно лучше растворимы в чистой дождевой воде, но имеют тенденцию осаждаться в виде необратимого геля в растворах, содержащих гуминовые кислоты или электролиты. С другой стороны, окислы алюминия и железа относительно мало растворимы в чистой дождевой воде, но могут переноситься водой, содержащей гумус, который действует как защитный коллоид, препятствующий их осаждению электролитами. Каолинитовые минералы, связанные с первичным выветриванием породы, не подвергаются, как часто думают, дальнейшему гидролизу до свободного кремнезема и глинозема, но являются устойчивыми конечными продуктами (F. Hardy).
Итак, в нормальных условиях тропического дождевого леса нисходящий ток почвенной воды вследствие постоянного выщелачивания содержит очень небольшие количества электролитов. По причинам, которые вскоре будут объяснены, он содержит также очень мало гумуса. У кремнезема и алюмосиликатов всегда существует тенденция к просачиванию вниз, а у полуторных окислов алюминия и железа — к осаждению в верхних слоях. Таким образом, в целом чем старше тропическая почва, тем она беднее кремнеземом, каолинитовыми минералами и основаниями и богаче полуторными окислами. Конечным результатом процесса является почва, состоящая почти исключительно из окислов алюминия и железа вместе с некоторым количеством кварца. Соотношение между количествами кремнезема и полуторными окислами (или одним глиноземом) является поэтому
230
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
важным показателем степени зрелости тропической почвы. Поскольку степень зрелости зависит от интенсивности различных почвообразующих процессов так же, как и от времени, в течение которого они действуют, модуль кремнезем/полуторные окислы не является непосредственным мерилом возраста почвы. В силикатах материнской породы модуль часто численно равен примерно 6, но в ходе эволюции почвы в условиях тропиков его значение может упасть ниже 2. Поскольку выщелачивание оснований и других питательных веществ происходит быстрее, чем выщелачивание кремнезема, более зрелая тропическая почва обладает, как правило, более низким плодородием и меньшей сельскохозяйственной ценностью.
Избирательное выщелачивание кремнезема из почвы носит название латеритизации. Противоположный процесс — вымывание полуторных окислов железа или алюминия с аккумуляцией кремнезема — называется оподзоливанием (podzolization). Это типичный процесс почвообразования в прохладных влажных климатах, хотя при известных условиях он может также происходить и на низменностях тропиков.
Тропические красноземы
Место не позволяет подробно остановиться на разборе различных значений, которые придавались терминам «латерит» и «латеритный». Это спорный вопрос, в котором все еще много путаницы (см., в частности, Shantz, Marbut, 1923: Martin, Doyne, 1927; Thorp, Baldwin, 1940; Pendleton, Sharasuvana, 1946). Латеритом можно назвать конечный продукт латеритизации — смесь глинозема и окисей железа с незначительными примесями. Однако под покровом тропического дождевого леса этот конечный продукт никогда не получается; действительно, в этом смысле латерит — скорее составная часть определенного горизонта в некоторых тропических почвах, чем почва сама по себе. Хотя истинный латерит, вероятно, не встречается в условиях тропического дождевого леса, многие (возможно, большинство) почвы влажных тропиков являются латеритными. Латеритную почву1 можно определить как одну из фракций глины, у которой модуль кремнезем/глинозем меньше 2,0 (Martin, Doyne, 1927).
Красный цвет, обычный у тропических почв, образовавшихся в условиях свободного дренажа, связан с избытком окислов железа. Оттенок цвета, вероятно, зависит от степени гидратации окислов; согласно Робинсону (Robinson, 1949, стр. 408), в тропических почвах окислы железа менее гидратированы, чем в почвах умеренного климата. Вся группа тропических почв красной окраски, проявляющая некоторую степень латеритизации, известна как тропические красноземы. Фагелер (Vageler, 1933) проводит разграничение между тропическими красными глинами и тро
1 Некоторые современные американские исследователи фонических почв (Byers et al., 1938; Roberts, 1936; Hardy, 1945) определяют почвы обширных площадей тропической Америки как подзолистые; в американском толковании (Byers et al., 1938) подзолизация — это процесс, связанный с: а) образованием торфяной подстилки из органического вещества на поверхности почвы и б) перемещением вниз (вымыванием) и часто раздроблением железистых соединений и глинозема. Это менее определенная концепция, чем данная выше, к тому же не учитывающая соотношения кремний/полу-торные окислы. Многие почвы тропического дождевого леса, несомненно, являются подзолистыми согласно этому определению, которое в основном подразумевает выщелачивание или удаление глинистого материала из почвы. Если принять американскую точку зрения, то латеритная почва также может быть названа подзолистой. В этом смысле термин Мора (Mohr, 1944, стр. 144) «ликсивиум», означающий «радикально выветренные и выщелоченные неорганические остатки» имеет бдлыпие достоинства.
СРЕДА
231
пическими красноземами. Первые менее развиты и поэтому менее латери-зованны, чем последние. Поскольку существует соотношение между степенью латеритизации и выщелачиванием питательных веществ, это разграничение имеет некоторый смысл с агротехнической точки зрения, но его значение для экологии естественной растительности еще не доказано. Некоторые почвы тропического дождевого леса носят скорее желтый оттенок и иногда именуются тропическими желтоземами. Этот желтый цвет обусловлен более высокой гидратацией окислов железа, чем в красноземах; по-видимому, между этими двумя группами почв нет более серьезных различий. Mop (Mohr, 1930, стр. 36) полагает, что желтоземы представляют собой стадию развития красноземов, так как желтые окислы железа неустойчивы, а красные — устойчивы.
Тропические почвы, подверженные попеременному увлажнению и высыханию, часто имеют характерную пятнистость.
Латеритные красноземы (в употребляемом здесь значении), может быть, являются самым распространенным типом почвы в районах развития тропического дождевого леса, но отнюдь не все тропические почвы латеритные. Многие виды материнской породы не могут подвергаться латеритному выветриванию. Например, в песчаниках и некоторых кислых изверженных и метаморфических породах содержание кремнезема так высоко, что характерная десиликация латеритных почв задерживается на неопределенное время. Из этих пород, как показано дальше, могут развиться почвы, не имеющие ничего общего с красноземами. Массивные изверженные породы, выветриваясь, создают в основном каолинитовую кору выветривания, чьи составные вещества, как сказано выше, устойчивы к процессу гидролиза. Не развиваются латеритные почвы и из глиносодержащих пород. Латеритное выветривание, которое создает типичные тропические красноземы, лучше всего проявляется на обломочных вулканических породах (F. Hardy).
Типы почв, определяемые материнской породой
На некоторых материнских породах развиваются почвы, совершенно отличные от тропических красноземов. В этих случаях, когда влияние почвообразующей породы преобладает над влиянием климата, формируются так называемые азональные почвы (в противоположность климатическим или зональным).
Харди и Фоллетт-Смит (Hardy, Follett-Smith, 1931) получили интересные данные по этому вопросу, исследуя почвенные профили в Брит. Гвиане. В условиях влажного тропического климата, где осадки превышали 100 дюймов (254 сл<), наблюдалось пять почвенных профилей, сформировавшихся на разнообразных изверженных породах от амфиболов (основные породы) до гранитов (кислые породы). Развитие почвы на основных и промежуточных породах, с одной стороны, и на гранитах — с другой, шло разными путями. На первых наблюдается быстрая латеритизация, создающая на поверхности породы корку «первичного гибситового (гидраргил-литового) латерита» (гибсит = А12О3-ЗН2О). При хорошем дренаже этот первичный латерит скапливается на значительной глубине. Там, где дренаж хуже (то есть частично затруднен), как это обычно бывает в низинах, происходит ресиликация латерита, очевидно, благодаря восходящим токам воды, поднимающим в засушливый период растворимый кремнезем или щелочные силикаты. Эта ресиликация ведет к образованию каолинитовых земель, окрашенных окислами железа в красный или желтый цвет.
232 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
I
В то же время на граните первоначальная потеря кремнезема или совсем отсутствует, или наблюдается лишь в очень незначительной степени, в связи с чем образование «первичного латерита» не происходит, и на материнской породе непосредственно возникают красные каолинитовые земли. Таким образом, несмотря на тождественный конечный результат почвообразования как на основных, так и на кислых породах, достигается он совершенно различными путями.
Наиболее наглядным примером тропической почвы, кардинально отличающейся от типичного краснозема не только процессом развития, но и особенностями зрелого профиля, является выщелоченный песок, который, как ныне известно, широко распространен в области тропического дождевого леса (Richards, 1941). В Брит. Гвиане такие почвы характерны для леса Уаллаба (Ерегиа) (см. стр. 263), в Сараваке (Борнео) — для «борового» леса (стр. 267). Как было указано Ричардсом (Richards, 1936), выщелоченный песок «борового» леса весьма близок к подзолам умеренного климата; прежде считали, что подзолы в тропиках встречаются только на больших высотах в прохладном климате. Несколько позднее Хардон (Hardon, 1936, 1937) дал подробное описание нескольких выщелоченных песчаных профилей, наблюдавшихся им невысоко над уровнем моря в восточном Борнео и на острове Банка (Малайский архипелаг), близ экватора, и убедительно доказал, что эти пески следует считать подзолами.
Почвы, изученные Хардоном, сформировались в климате со средней температурой, равной примерно 26°, и другими признаками, типичными для климата тропического дождевого леса. Средняя годовая сумма осадков в обеих местностях составляла около 300 см, причем ни один месяц не имел менее 10 см. Растительность представляла собой так называемый паданг1, подобие верещатника или редколесья. Паданг, хотя и очень отличается от тропического дождевого леса, типичного для этой области, все же, вероятно, является устойчивым эдафическим климаксом (см. стр. 289 и 348). В экологическом и флористическом отношении растительность паданга имеет много общего с «боровым» лесом. По структуре и внешнему облику он очень сходен с «мури» Брит. Гвианы — зарослями кустарников, где преобладает Humiria floribunda var. guianensis. Сообщество мури, по-видимому, также является эдафическим климаксом и находится в таком же отношении к лесуУаллаба (Ерегиа), произрастающему там же, как паданг к «боровому» лесу1 2.
Описание и аналитические данные по двум профилям паданга воспроизведены в табл. 24. Эти данные показывают, что пески имеют все типичные признаки подзола. В профилях выделяется сильно выщелоченный горизонт А (элювиальный), верхняя часть которого испещрена черноватыми пятнами гумуса; ниже расположен иллювиальный горизонт В, в котором наблюдается накопление гумуса и полуторных окислов. В одном из профилей горизонт В сцементирован в твердую плиту. Почвы имеют грубый механический состав и отличаются пористостью; процент илистой фракции в горизонте В выше, чем в А, но все же и там не превышает 10%.
1 «Паданг» по-малайски означает просто луг или открытое пространство без деревьев. Этот термин употребляется здесь за неимением лучшего; вместо него можно предложить термин «тропическая пустошь». Песчаные почвы, с которыми связаны «боровой» лес и растительность паданга, известны в Сараваке под названием «керан-гао.
2 «Мури» в отличие от паданга часто связывается как с саванной, так и семенным высокоствольным лесом (стр. 348).
СРЕДА
233
Модуль кремнезем/полуторные окислы в илистой фракции горизонта А высокий (3,62—8,64) , а горизонта В — низкий (0,31—0,42); это указывает на выщелачивание из горизонта А полуторных окислов, а не кремнезема: отложение полуторных окислов происходит в горизонте В. Это коренным образом отличается от процессов, протекающих в профиле латеритного краснозема. Как и следовало ожидать при наличии обильных осадков и пористой структуры, содержание оснований и питательных веществ в этих почвах крайне низкое даже по сравнению с другими тропическими почвами.
Поверхность почвы покрыта очень кислым гумусом, отличающимся высоким отношением углерода к азоту (57), что указывает на неполное разложение органических веществ. Очевидно, этот гумус скорее принадлежит к типу грубого гумуса. Процент содержания гумуса в более глубоких частях профиля не выше, чем в других почвах тропических низменностей, но гумус проникает глубже, и небольшие скопления его встречаются в горизонте В.
Паданговые почвы восточного Борнео и острова Банка развились на аллювиальных кварцевых песках, а подзолы «борового» леса в Сараваке — на миоценовых песчаниках. Согласно Хардону (Hardon), предпочтительное формирование на падангах подзолистых почв вместо тропических красноземов связано в основном с пористостью материнской породы и с изначальной бедностью основаниями. Это обеспечивает быстрое нисходящее движение воды и более энергичное, чем в большинстве других тропических почв, вымывание оснований из гумуса. Очевидно, что при этих условиях даже в жарком влажном климате в некоторой степени кислый гумус может аккумулироваться.
Видимо, тропические подзолы широко распространены в восточных тропиках, так как Хардон сообщает, что паданговая растительность встречается как в Таиланде, так и во многих частях Малайского архипелага. «Боровой» лес на подзолах широко распространен на Борнео.
Выщелоченные кварцевые пески, которые скорее всего являются тропическими подзолами, так же часто встречаются в области развития тропических дождевых лесов Америки. В Брит. Гвиане имеются большие площади «белых песков», поросших саванной, мури, лесом Уаллаба (Eperua) или другими специфическими разновидностями тропического дождевого леса (гл. 11). «Песок Валенсия» на Тринидаде (Hardy, 1940), вероятно, является почвой подобного же рода. Профили этих песков очень сходны с профилями паданговых почв, описанных Хардоном. Прежде считали, что «белые пески» Гвианы, которые достигают значительной глубины на гребнях и плато, являются непосредственным продуктом выветривания гранита in situ. Однако новейшие данные показывают, что эти продукты разрушения гранита находятся во вторичном залегании и скорее всего относятся к третичным аллювиальным отложениям, сформировавшимся в мелком море. В ходе своего формирования они, возможно, потеряли большую часть тонких фракций, вместе с которыми была вымыта и большая часть первоначально содержавшихся в них оснований.
На низменностях Брит. Гвианы в районах распространения выщелоченных песков произрастает особый тип леса, где преобладает Dimorphandra conjugata. Поверхность почвы здесь покрыта слоем торфянистого гумуса мощностью около 1 фута (30 см) (Т. A. W. Davis). Профили этих весьма интересных почв не были исследованы, но возможно, что они являются подзолами с необычайно развитым горизонтом А (сырой гумус). Поскольку эти почвы не затопляются и не находятся в условиях постоянного пере-
Профили тропических подзолов низменностей (Hardon, 1937)
Таблица 24
Местность Горизонт Глубина, см Описание % песка (0,05— 2 л<.м) 1 % илистой фракции (0,0005-0,005л1л<) 1 Органическое вещество, % Молекулярный показатель S1O2 А120з 4- Fe2O2 (для илистой фракции) pH СаО
Па дан г (Лоейей, восточное Борнео) Ао 0—10 Коричневато-черный полуразло-жившийся органический материал — — — — 2,8 —
Аа 10—120 Рыхлый белый кварцевый песок 97,3 0,5 0,1 — 6,1 0,008
в 120—ГО Желтовато-коричневый кварцевый песок 93,9 2,8 1,2 — 5,4 0,002
Остров Банка Ав 0-10 Полуразложившийся черный органический материал в смеси с крупным песком — — — 3,62 2,7 —
10-25 Рыхлый серовато-черный гумусовый кварцевый песок 95,6 1,6 — 7,17 3,9 0,022
а2 25-40 Рыхлый серовато-белый кварцевый песок 94,0 0,6 — 8,64 6,1 0,032
Вх 40—70 Сцементированный слой. Темно-коричневый очень плотный кварцевый песок 86,9 7,2 5,2 0,42 3,9 0,029
в» 70—100 Рыхлый светло-коричневый кварцевый песок 92,1 4,6 0,31 4,6 0,035
i Фракции >2 мм и мелкозема (0,005—0,05 мм) опущены.
СРЕДА
235
увлажнения, скопление таких больших количеств гумуса трудно объяснить (стр. 280). Возможно, причина заключается в высокой водопроницаемости (пористости) почвы, благодаря которой быстрое высыхание поверхности препятствует разложению перегноя. Отчасти это может быть связано с химическими и физическими особенностями листьев Dimorphandra.
Выщелоченные пески, аналогичные гвианским, широко распространены в бассейне Амазонки, особенно в районе Рио-Негро (Spruce, 1908, стр. 206-207, 303—304).
«Белый» песок, сходный с выщелоченными песками Южной Америки и восточных тропиков, встречается на острове Мафия у побережья Танганьики (Восточная Африка). Он покрыт верескоподобной растительностью, основной аспект которой создает Philippia mafiensis — кустарник из семейства вересковых (Greenway, 1938).
Поразительной особенностью выщелоченных песков всех частей тропиков является то, что реки, текущие по ним, отличаются черным цветом; их прозрачная вода кажется почти черной в отраженном свете, а при просвечивании оказывается оранжево-коричневой. «Черноводные» реки особенно хорошо известны в тропической Америке; именно благодаря черному цвету получила свое название Рио-Негро. В то же время в реках, пересекающих район распространения красноземных почв, вода, прозрачная или мутная, всегда бывает бесцветна. Коричневая окраска, очевидно, вызвана примесями гумуса в тонко дисперсном коллоидном состоянии: там, где «черная река» вливается в море, коричневое вещество осаждается электролитами морской воды. Еще не выяснено, почему гуминовые вещества этих песков легко растворяются в проточной воде. Предполагалось (Richards, 1936, стр. 26), что это связано с отсутствием в выщелоченных песках как электролитов, способных осадить их, так и илистой фракции и окислов железа, которые образовали бы с ними адсорбционные комплексы. Хардон (Hardon, 1937) относит коричневый цвет проточных вод зоны падан-гов на счет гуматов, содержащих железо, которые не осаждаются в горизонте В почвы благодаря его высокой кислотности. «Черные реки» иногда вытекают и из тропических болотных лесов (торфяные болота, см. стр. 316), но там, где такие болота, по всей видимости, отсутствуют, «черная вода» является достоверным показателем наличия выщелоченных песков.
Многочисленные данные подтверждают, что подзолы в противоположность прежнему представлению о их приуроченности лишь к прохладным климатам южнее и севернее 40 параллели или же к высокогорным районам близ экватора часто встречаются на тропических низменностях. Их присутствие в тропиках, где с точки зрения климата их существование кажется абсурдным, очевидно, больше связано с характером почвообразующей породы, чем с другими факторами почвообразования.
Пока еще не известно, существуют ли в области тропического дождевого леса и другие типы почв, определяемые характером материнской породы. Согласно Харди (Hardy, 1935, стр. 157), во влажных тропиках на известняках образуются почвы, очень сходные с красноземами и желтоземами, развившимися на породах с меньшим содержанием оснований. Выщелачивание вскоре удаляет почти весь карбонат кальция и другие основания, и почва становится кислой. Окончательные свойства почвы в значительной степени определяются примесями, содержащимися в исходном материале — известняке. Однако обнаженные известняковые породы в тропиках имеют особую флору (стр. 244), а почвы, покрывающие известняк благодаря высокой водопроницаемости последнего, часто несут более «сухой» тип растительности, чем другие почвы.
236
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Болотные лесные почвы
Поскольку водный режим играет в развитии почвы такую важную роль, тропические почвы, образовавшиеся в условиях затрудненного дренажа или преимущественно горизонтального оттока, существенно отличаются от почв с наличием беспрепятственного вертикального дренажа. Влияние затрудненного дренажа на ресиликацию «первичного латерита» уже рассматривалось; в этом разделе будет охарактеризован наиболее ярко выраженный тип болотных почв дождевого леса, в которых горизонт воды редко опускается больше чем на несколько дюймов ниже поверхности.
Эти болотные почвы мало исследованы, но они, по-видимому, делятся на две довольно резко обособленные группы — слабогумусные болотныо почвы и торфянистые болотные почвы (с поверхностным слоем торфа или гумуса). Последние развиваются в олиготрофных (крайний дефицит минеральных веществ) условиях; первые в эвтрофных (то есть там, где вода относительно богата основаниями и т. п.).
Слабогумусные болотные почвы большей частью являются тяжелыми почвами с преобладанием пылеватой или илистой фракций и отличаются от красноземов хорошо дренированных мест своим сероватым, беловатым или бледно-желтым цветом. Хотя преобладающая окраска их может быть-названа красной, они включают красновато-коричневые, оранжевые или желтые вкрапления. Согласно Мору (Mohr, 1930, стр. 23), эти почвы в основном содержат каолин и кремниевую кислоту, причем запасы питательных веществ, используемых растительностью, поступают главным образом с водой. Слой гумуса очень тонок, органическое вещество не может проникнуть сколько-нибудь глубоко. По-видимому, эти слабогумусные болотные почвы тропиков имеют много общего с глеевыми почвами других климатов. С экологической точки зрения они отличаются от лучше дренированных почв избытком воды, плохой аэрацией и, может быть, своими химическими свойствами.
Торфянистые болотные почвы встречаются гораздо реже слабогумусных, но тем не менее они лучше изучены. Пока Потонье и Курдерс (Poto-nie, Koorders, 1909) не обратили внимания на существование на Суматре торфяных болот значительной мощности, считалось, что при температурах^ свойственных тропическим низменностям, образование торфа невозможно (стр. 239—240)1. После опубликования названной статьи обширные торфяные болота были обнаружены во многих частях Малайского архипелага и Малаккского полуострова, в тропической Америке и тропической Африке. Эти торфяные болота имеют много общего с торфяниками умеренных областей, и, поскольку их растительный покров обычно состоит преимущественно из деревьев, их можно назвать тропическими болотными лесами (Waldmoore).
Полак (Е. Polak, 1933) дал превосходный обзор торфяных отложений и болотных лесов Индии. Он выделяет два основных типа болот — омбро-генный и топогенный. Первые встречаются на прибрежных равнинах Суматры, Явы и, вероятно, Новой Гвинеи; для их развития нужны обильные осадки — по-видимому, не менее 2000 мм в год. Вторые, менее распространенные, зависят главным образом от рельефа. Они встречаются по берегам озер как на низменностях, так и на возвышенностях. Расти-
1 Потонье считал, что европейские угли каменноугольного периода могли образоваться при тропических температурах в условиях, сходных с теми, какие существуют в современных тропических торфяных болотах.
СРЕДА
237
дельность омброгенных болот — высокий вечнозеленый лес; в сущности, это особый тип дождевого тропического леса (гл. 13); растительность топогенных болот часто травянистая. Sphagnum встречается только на топогенных болотах возвышенностей1.
Омброгенные болота часто располагаются непосредственно за прибрежной полосой мангровых зарослей, но грунтовые воды в них не имеют признаков засоления. Торфяные залежи занимают значительную площадь {наиболее обширные из них имеют, по-видимому, несколько километров в диаметре). Торфяная масса нарастает от краев по направлению к почти плоскому центру, так что болото в целом имеет форму двояковыпуклой линзы, нижняя поверхность которой лежит ниже уровня окружающей местности, а верхняя —выше. В средней ее части мощность торфа может превышать 7 м. Этот торф представляет собой красноватую или серовато-коричневую кашеобразную массу; только древесные корни создают здесь твердую опору. От центральной части болота по радиусам растекаются «черноводные» потоки, вода в которых отличается очень кислой реакцией (pH—3,0); водоснабжение главным образом определяется осадками. Эти болота крайне олиготрофны — при сгорании торф дает минимальный остаток минерального вещества.
Исследование профилей некоторых из этих болот указывает на то, что, по-видимому, в течение всего их развития на них существовала лесная растительность; остатки деревьев составляют основную массу торфа во всех горизонтах. Торф подстилает выщелоченная глина.
Как указывает Полак, форма, гидрография и химические особенности этих омброгенных болот показывают их сходство с верховыми болотами умеренной зоны. Топогенные болота, развивающиеся во впадинах и имеющие вдавленный, не линзоподобный профиль, богаты питательными веществами и являются тропическим эквивалентом болот Англии в Кембриджшире.
Аккумуляция торфа при тропических температурах, несомненно, частично обусловлена избытком воды и, следовательно, анаэробными процессами. Согласно Мору (Mohr, 1944, стр. 108), тропические почвы, образовавшиеся при избытке влаги, неблагоприятны для большинства грибов и для всех бактерий, за исключением факультативных или облигатных анаэробов. Таким образом, разложение органических остатков происходит медленно по сравнению со скоростью их образования (ср. стр. 239). Так как во многих тропических болотах аккумуляция гумуса отсутствует, очевидно, что, помимо избыточного увлажнения, для образования торфа необходим еще и какой-то эдафический фактор. По мнению Мора, этим фактором является вода, бедная питательными веществами и совершенно лишенная оснований. Правда, Полак не согласен с этим мнением, но близкое соседство болотных лесов и выщелоченных подзолов на Борнео заставляет думать, что это предположение близко к истине.
Известно, что в различных частях тропической Америки встречаются торфяные болота, вероятно, сходные с олиготрофными омброгенными болотами Индии. Уле (Ule, 1908 а) и Бульен (Bouillienne, 1930), описывают подобные болота бассейна Амазонки. «Пегассовые» болота Брит. Гвианы, очевидно, аналогичны им. Они занимают обширные площади в прибрежных «болотистых саваннах» (стр. 346) и встречаются внутри •страны в верховьях рек в области развития белых песков. «Пегасе»
1 Sphagnum, вероятно, встречается в болотных лесах Южной Америки, а также яа влажных участках некоторых типов саванн.
238
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
состоит из мягкого влажного торфа мощностью в несколько футов; грунтовые воды залегают иногда на глубине лишь одного фута от поверхности даже в сухую погоду.
ПОЧВЫ ГОРНЫХ РАЙОНОВ
По мере увеличения высоты над уровнем моря и почва и растительность меняются в соответствии с изменением климата. Тропический дождевой лес уступает место дождевому лесу предгорий и, наконец, горному или «умеренному» дождевому лесу (гл. 16). Основным фактором, определяющим в этом случае как растительность, так и почву, является понижение средней температуры. Из-за крутизны склонов горные почвы обычно подвержены интенсивной эрозии и часто отличаются незрелостью. О почвообразующих процессах и развитых почвах этого пояса известно мало. На Яве, согласно Мору (Mohr, 1930, стр. 124), с подъемом в горы красноземы сменяются желтоземами, а затем — выщелоченными почвами. С возрастанием высоты в почвах заметно повышается количество гумуса, а латеритизация, по всей видимости, уступает место подзолообразованию как основному процессу в развитии горных почв. Некоторые исследователи отмечали наличие подзолообразовательного процесса в высокогорных районах тропиков. Сенстиус (Senstius, 1930—1931) обнаружил, что на Яве и Филиппинах оподзоливание наблюдается на почвах, расположенных выше 2000 м над уровнем моря, хотя типичные подзолы не образовывались и там; в то же время Хардон (Hardon, 1936) в горах Новой Гвинеи отмечает типичные подзолы на высоте примерно 2000 м.
Почвы горного дождевого леса очень мало изучены. В горном «умеренно-теплом» дождевом лесу Узамбарских гор, Восточная Африка, Питт-Шенкель (Pitt-Schenkel, 1938, стр. 55) обнаружил как тропические лесные подзолы, так и почвы, напоминающие коричневые лесные почвы умеренной зоны.
БИОТИЧЕСКИЕ ФАКТОРЫ В РАЗВИТИИ ПОЧВЫ
Влияние растительности на почву сказывается прежде всего в изменении микроклимата. Условия развития почвы в первичном тропическом дождевом лесу на возделываемой земле или на участке, покрытом разреженным покровом вторичной растительности в одной и той же макроклиматической зоне, будут совершенно иными. Роль лесного покрова в перехватывании осадков и в изменении температуры и влажности на поверхности земли была показана в гл. 7. Мы еще вернемся к этому вопросу в связи с рассмотрением процессов разрушения дождевого леса и возникновения вторичных сукцессий. Кроме того, растительность влияет на почву, способствуя накоплению перегноя. Значение перегноя для развития почвы зависит отчасти от его коллоидных свойств, а частично от наличия оснований и других неорганических веществ, высвобождающихся в процессе минерализации.
Перегной образуется при разложении слоя отмерших листьев, лежащих на поверхности почвы. Обычное представление о якобы характерной для тропических лесов значительной мощности этого слоя совершенно ошибочно. Средняя толщина опада в тропическом дождевом лесу составляет всего 1—3 см. Обычно опавшая листва даже не покрывает всей
СРЕДА 239
поверхности земли, так что местами просвечивают участки обнаженной почвы. Только во впадинах накапливается более толстый слой. Разрушение подстилки происходит такими быстрыми темпами, что в значительной степени уравновешивает непрерывное опадание мертвых листьев. Разложение древесины, коры и отмерших листьев с превращением их в перегной и последующая минерализация с образованием углекислоты, воды и минеральных веществ являются в основном результатом деятельности почвенной флоры и фауны, но возможно, что происходит также чисто химическое окисление перегноя, в которое не вовлечены микроорганизмы (Mohr, 1944, стр. 103—105). Природа биологических и биохимических процессов, происходящих в условиях тропического дождевого леса, еще очень мало изучена. Биологическая сторона этой проблемы обсуждалась Корбетом (Corbet, 1935), а Фагелер (Vageler, 1933, стр. 131—139) кратко разбирает вопрос об органическом веществе почвы с биохимической точки зрения.
Флора тропических почв состоит из бактерий и грибов, а иногда также и водорослей. Фауна включает простейших, земляных червей и разнообразных насекомых, особенно муравьев и термитов, а также более крупных животных, таких, как землеройные млекопитающие и земноводные. Оценивая значение почвенной флоры, можно сказать, что как бактерии, так и грибы играют большую роль в разложении органического вещества. Почвенные грибы дождевого леса мало изучены, но считают, что при высоких температурах их роль обычно менее существенна, чем роль бактерий (Mohr, 1944, стр. 107). Насекомые, особенно термиты и муравьи, занимают почетное место среди разрушителей растительных остатков. Термиты очень многочисленны и проявляют активную деятельность, одинаково хорошо расселяясь как на стволах и ветвях деревьев, так и в самой почве. Разрушение мертвой древесины в основном зависит от них, причем их работа так эффективна, что отмершие древесные стволы, пни и ветви, упавшие на землю, обычно сохраняются очень недолго. Земляные черви не всегда встречаются в почве дождевого леса и никогда не отличаются изобилием, их деятельность, по-видимому, не имеет большого значения.
Хотя еще далеко не ясны все детали воздействия почвенной флоры и фауны, некоторые общие связи между их активностью в целом и условиями среды, особенно температурой, постепенно выявляются. С повышением температуры скорость накопления живого органического вещества растительными сообществами увеличивается до максимума, затем падает. То же самое можно сказать о процессах гумификации и минерализации этого органического вещества. Следовательно, существуют максимальные, минимальные и оптимальные температуры и для образования перегноя и для его разложения, но их количественное выражение неодинаково для этих двух процессов. Согласно Мору (Mohr, 1930, стр. 17—20)1, накопление перегноя в почве начинается при 0° и достигает своего оптимума примерно при 25, а максимума примерно при 40°. Разложение перегноя происходит при более высокой температурной амплитуде, так как почвенные микроорганизмы начинают активизироваться при температурах между 10 (или немного ниже) и 15°. Оптимум их активности в лучшем случае достигается только при 35°, а максимум — при температурах, превышающих обычную температуру почвы. В результате при 0—20° наблюдается все возрастающее накопление перегноя, выше 20° его
1 См. также Mop (Mohr, 1944, стр. 96—109).
240
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
прирост уменьшается, а при 25° кривые накопления и разложения пересекаются, что указывает на невозможность его накопления при более высоких температурах. Однако эти температурные соотношения действительны только при совокупности достаточного дренажа и аэрации; в анаэробных условиях, какие создаются в болотных почвах, рассмотренных выше, как уже говорилось, наблюдаются другие соотношения.
В тропическом дождевом лесу, там, где температура у поверхности земли составляет около 25° и влага содержится в избытке круглый год, не должно происходить аккумуляции перегноя при условии достаточной аэрации почвы. На практике перегноя бывает гораздо больше, чем можно предположить теоретически, но, за исключением поверхностного слоя почвы глубиной не свыше 10—20 см. содержание органического вещества редко превышает 1—2?6. Согласно Фагелеру (Vageler, 1933, стр. 7), тропические почвы часто содержат больше перегноя, чем можно предположить по их цвету. Это связано с тем, что цвет перегноя в тропических почвах часто бывает гораздо светлее, чем в почвах умеренной зоны, и легко маскируется светлой окраской окислов железа. Хотя в почвах тропического дождевого леса подавляющая часть перегноя обычно сосредоточена в 10-сантиметровом поверхностном слое, глубина проникновения колеблется в зависимости от строения почвы. В пористом песке перегной обычно вмывается гораздо глубже, чем в менее проницаемых почвах.
Возможно, что именно благодаря скорости разложения перегной красноземных или желтоземных почв тропического дождевого леса обычно дает не очень кислую реакцию. Пожалуй, он больше напоминает муль (мягкий гумус) почв умеренного леса.
Хотя в условиях тропиков количество образовавшегося перегноя обычно невелико, он оказывает значительное влияние на развитие почвы. Уже было отмечено, что присутствие перегноя замедляет выщелачивание кремнезема. Таким образом, перегной играет консервативную роль в развитии почвы, и под покровом тропического дождевого леса латеритизация не может дойти до крайних пределов. В то же время перегной способствует поддержанию плодородия почвы. В нем содержатся значительные количества оснований и питательных веществ, которые по мере минерализации перегноя поступают в верхние слои почвы. Затем эти вещества вмываются в нижние, более глубокие слои, где они поглощаются корнями растений, поднимаются вверх к стволам и листьям и снова возвращаются в почву в форме перегноя. Следовательно, растительность противодействует вымыванию растворимых веществ и возвращает их снова в поверхностные слои. Перегной благодаря своим адсорбционным свойствам также помогает замедлить вымывание, удерживая растворимые основания в верхней части почвы.
Возможно, что именно улавливание и осаждение оснований, осуществляемое перегноем, объясняет менее кислую реакцию верхних слоев почвы по сравнению с более глубокими участками почвенного профиля, хотя повышение кислотности с глубиной в тропических почвах может быть вызвано также и другими причинами (Doyne, 1935).
КРУГОВОРОТ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ
В климате влажных тропиков, как и во всех климатах, где вода в почве имеет преимущественно нисходящее движение, развитие почвы всегда идет в сторону ее истощения. Растворимые вещества непрерывно
СРЕДА
241
вымываются в более глубокие слои почвы и уносятся грунтовыми водами. Таким образом, самая важная общая особенность всех почв области развития тропического дождевого леса, как красноземного, так и подзолистого типа (за исключением, пожалуй, некоторых очень молодых почв),— это низкое содержание в них питательных веществ. В свете этого вывода кажется парадоксальным, что растительность тропического дождевого леса отличается таким пышным развитием. Эти выщелоченные и истощенные почвы влажных тропиков оказываются в состоянии нести на себе великолепный лес, в то время как гораздо более плодородные почвы менее влажных тропических областей покрыты саванной или значительно обедненным лесом. Эта проблема применительно к африканским лесам была рассмотрена Вальтером (Walter, 1936) и Мильном (Milne, 1937), применительно к лесам Вест-Индии — Харди (Hardy, 1936 а); эти авторы пришли к аналогичному выводу о том, что в нормальных условиях тропического дождевого леса сама растительность путем создания круговорота питательных веществ вызывает процессы, стремящиеся противодействовать истощению почвы. В результате почва под покровом естественной растительности достигает состояния равновесия, в котором ее истощение если не совсем приостанавливается, то протекает крайне замедленно.
При разрушении материнской породы непрерывно высвобождаются новые питательные вещества. Если корни деревьев в состоянии достичь этого горизонта, то растворимая часть питательных веществ поглощается растительностью. Некоторые из этих веществ идут на построение тканей растений и, в частности, на построение клеточных стенок, другие остаются растворенными в клеточном соке. В конечном счете все эти вещества вновь возвращаются в почву в результате отмирания и последующего разложения растений или их частей. Таким образом верхние слои почвы беспрерывно обогащаются питательными веществами, извлеченными первоначально из более глубоких слоев. Поскольку большинство корней, включая почти все «питающие» корни, располагается в верхних слоях почвы, подавляющая часть «свободных от гумуса» питательных веществ может быть почти сразу поглощена растительностью и использована для ее дальнейшего роста. Если потери и существуют, то они должны быть ничтожны; Мильн (Milne, loc. cit., стр. 10) показал, что в районе тропического дождевого леса Узамбары содержание электролитов в поверхностных водотоках очень низкое. Следовательно, при наличии вполне развитой почвы основная масса питательных веществ содержится в живой растительности и в слое перегноя, между которыми существует почти замкнутый круговорот. Ресурсы материнской породы нужны только для того, чтобы восполнить небольшие потери, вызванные дренажем.
Существование этого замкнутого цикла объясняет, почему почва, производящая роскошный тропический дождевой лес, после расчистки и культивации может оказаться далеко не плодородной. При вырубке леса, содержавшийся в нем запас питательных веществ уничтожается или, поступая в почву, вновь переходит в свободное состояние, в то же самое время слой перегноя часто подвергается разрушению в результате пожаров, губительного воздействия солнечных лучей. Как говорит Мильн (Milne, loc. cit., стр. 10), — «все движимое имущество ликвидируется и, как бывает на аукционах, спускается по дешевке, причем выручка совершенно не соответствует его стоимости». В результате на вырубках дождевого леса можно снять несколько хороших урожаев, используя временное плодородие почвы, но в дальнейшем, если не принять особых мер, эти земли окажутся непригодными для возделывания
242 ТРОПИЧЕСКИЙ ДОЖмг.и^,..
сельскохозяйственных культур. При обычной системе подсечно-огневого земледелия, практикуемой в областях развития тропического дождевого леса, редко можно снять больше двух-трех урожаев подряд без длительного промежуточного периода кустарниковой залежи. Даже в Гондурасе, где годовая сумма осадков не превышает примерно 180 см, урожай кукурузы на лесных вырубках падает примерно от 800—1000 фунтов (350—450 кг) в первом году примерно до 400—600 фунтов (180— 270 кг) на третьем году (Charter, 1941, стр. 16). На очень бедных пористых почвах тропического дождевого леса, таких, как подзол леса Уаллаба в Брит. Гвиане (стр. 263), возможно, не удастся получить и одного урожая. Изменения, вызываемые вырубкой или прореживанием леса, подчеркивают неустойчивость равновесия между почвой и растительностью в естественном тропическом дождевом лесу.
После того как было установлено, что почвы тропического дождевого леса бедны питательными веществами, многие авторы стали придавать большое значение наличию у многих входящих в его состав растений микоризы, которая часто встречается и у растений других сообществ. Янсе (Janse, 1897) исследовал 75 видов из первичного леса на западной Яве и установил наличие микоризы у 69 из них. Несмотря на отсутствие аналогичных данных по другим областям, не приходится сомневаться, что тщательное исследование корневой системы растений пояса тропических дождевых лесов показало бы, что очень большой процент из них обладает микоризой. Роль микориз изучена так мало, а вопрос в целом так сложен, что бесполезно обсуждать его здесь. Микоризы растений тропического дождевого леса могут иметь большое экологическое значение, но в настоящее время вряд ли можно сказать что-либо определенное.
В связи с этим следует вспомнить и об изобилии деревьев из семейства бобовых во многих тропических дождевых лесах. Поскольку почвы этих лесов бедны питательными веществами, наличие корневых клубеньков, содержащих связывающие азот организмы, должно быть большим преимуществом. В лесах пяти типов в местности Морабалли-Крик, Брит. Гвиана (стр. 258) Дэвис и Ричардс (Davis, Richards, 1933—1934) обнаружили большие различия в количестве представителей семейства бобовых. На пробном участке среди деревьев, имевших в диаметре 4 дюйма и больше, оказалось следующее количество бобовых:
Тип леса Мора Морабуцеа Смешанный Гринхарт Уаллаба
% бобовых (Leguminosae), отдельные экземпляры 59 33 15 14 53
Таким образом, больше всего представителей Leguminosae насчитывается в лесах Уаллаба и Мора. Сообщество Мора произрастает на болотистой почве, часто затопляемой водой; сообщество Уаллаба — на выщелоченном подзоле; леса других типов приурочены к красным или коричневым разновидностям тропического краснозема (см. гл. 10), где, возможно, нитрификация более активна, а выщелачивание менее интенсивно. Вероятно, эти факты имеют какое-то отношение и к общему распространению Leguminosae в тропическом дождевом лесу.
В консоциациях дождевого леса бобовые часто выступают в качестве единственной доминирующей породы.
243
ПОЧВА КАК ДИФФЕРЕНЦИАЛЬНЫЙ ЭКОЛОГИЧЕСКИЙ ФАКТОР
Ранее уже неоднократно говорилось или подразумевалось, что во влажных тропиках, как и везде, почвенные различия проявляются в различном составе растительных сообществ. Эдафические факторы не только изменяют флористический состав в пределах самого тропического дождевого леса, но, как о том свидетельствуют данные, могут быть причиной развития кустарниковых сообществ, например паданга, и сообществ типа «мури», упомянутых выше (стр. 232), или же некоторых типов саванны (гл. 15), которые существуют в качестве эдафических климаксов в климате тропического дождевого леса. Это противоречит общепринятому в течение длительного времени взгляду, что в тропиках почвенные различия не имеют большого экологического значения и что это значение, во всяком случае, несомненно меньше, чем в умеренной зоне. Лишь в последнее время сведения о составе тропических растительных сообществ увеличились настолько, чтобы рассеять это неправильное представление.
Этот вопрос будет еще рассмотрен в последующих главах; пока же-достаточно будет показать на двух примерах, насколько тесна связь между почвой и климаксной растительностью.
Среди пяти первичных лесных сообществ Морабалли-Крик, Брит. Гвиана, два проявляют особенно поразительную корреляцию с почвенными условиями. Лес Уаллаба (консоциация Eperua falcata) приурочен к выщелоченным «белым» пескам, покрывающим плато и кряжи; лес Гринхарт (консоциация Ocotea rodiaei) встречается на коричневом песке, который обычно оконтуривает кряжи. Эти два сообщества иногда соседствуют, и в этих случаях между ними проходит исключительно отчетливая граница, точно соответствующая переходу от выщелоченного песка к коричневому. В одном из профилей полный переход осуществлялся в пределах дистанции, равной 150 м (Davis, Richards, 1933—1934, рис. 1. стр. 121). В других местах Брит. Гвианы наблюдалась еще более резкая граница между этими типами тропического дождевого леса; Вуд (Wood, 1926, стр. 4—5) пишет следующее, имея в виду леса округа Бартика-Кабури:
«Как правило, можно было одним шагом перейти с белого песка, где лес Гринхарт никогда не встречается, на коричневый песок, где он произрастает, что случается обычно в тех местах склонов, где они спускаются от плоского водораздела к речкам. Наиболее разительный пример представлял собой муравейник akushi (муравьи-листорезы), разместившийся на площади около 30 кв. футов (9 кв. м). Верхний край муравейника лежал на белой почве, нижний — на коричневой, причем линия раздела была настолько резкой, что ее можно было указать рукой. Тут же в непосредственной близости один от другого, так что человек мог только боком протиснуться между ними, росли два дерева, одно из которых укоренилось на белой почве, а другое — на коричневой. Верхнее дерево было первым из сочленов сообщества Уаллаба, а нижнее — первым Гринхарт на этом склоне». Хотя границы между другими типами леса в местности Морабалли-Крик были обычно не такими резкими, как между консоциациями Уаллаба и Гринхарт, тесная связь между типом леса и почвой проявлялась повсюду.
Другим примером может служить распределение на Борнео и Суматре леса из железного дерева — особой консоциации с преобладанием Eusi-deroxylon zwageri, почти полностью вытеснившего все другие виды деревьев (стр. 282). Грессер (Gresser, 1919) заметил, что этот тип леса приуро-16*
242
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
сельскохозяйственных культур. При обычной системе подсечно-огневого земледелия, практикуемой в областях развития тропического дождевого леса, редко можно снять больше двух-трех урожаев подряд без длительного промежуточного периода кустарниковой залежи. Даже в Гондурасе, где годовая сумма осадков не превышает примерно 180 см, урожай кукурузы на лесных вырубках падает примерно от 800—1000 фунтов (350—450 кг) в первом году примерно до 400—600 фунтов (180— 270 кг) на третьем году (Charter, 1941, стр. 16). На очень бедных пористых почвах тропического дождевого леса, таких, как подзол леса Уаллаба в Брит. Гвиане (стр. 263), возможно, не удастся получить и одного урожая. Изменения, вызываемые вырубкой или прореживанием леса, подчеркивают неустойчивость равновесия между почвой и растительностью в естественном тропическом дождевом лесу.
После того как было установлено, что почвы тропического дождевого леса бедны питательными веществами, многие авторы стали придавать большое значение наличию у многих входящих в его состав растений микоризы, которая часто встречается и у растений других сообществ. Янсе (Janse, 1897) исследовал 75 видов из первичного леса на западной Яве и установил наличие микоризы у 69 из них. Несмотря на отсутствие аналогичных данных по другим областям, не приходится сомневаться, что тщательное исследование корневой системы растений пояса тропических дождевых лесов показало бы, что очень большой процент из них обладает микоризой. Роль микориз изучена так мало, а вопрос в целом так сложен, что бесполезно обсуждать его здесь. Микоризы растений тропического дождевого леса могут иметь большое экологическое значение, но в настоящее время вряд ли можно сказать что-либо определенное.
В связи с этим следует вспомнить и об изобилии деревьев из семейства бобовых во многих тропических дождевых лесах. Поскольку почвы этих лесов бедны питательными веществами, наличие корневых клубеньков, содержащих связывающие азот организмы, должно быть большим преимуществом. В лесах пяти типов в местности Морабалли-Крик, Брит. Гвиана (стр. 258) Дэвис и Ричардс (Davis, Richards, 1933—1934) обнаружили большие различия в количестве представителей семейства бобовых. На пробном участке среди деревьев, имевших в диаметре 4 дюйма и больше, оказалось следующее количество бобовых:
Тип леса Мора Мора-буцеа Смешанный Гринхарт Уаллаба
% бобовых (Leguminosae), отдельные экземпляры 59 33 15 14 53
Таким образом, больше всего представителей Leguminosae насчитывается в лесах Уаллаба и Мора. Сообщество Мора произрастает на болотистой почве, часто затопляемой водой; сообщество Уаллаба — на выщелоченном подзоле; леса других типов приурочены к красным или коричневым разновидностям тропического краснозема (см. гл. 10), где, возможно, нитрификация более активна, а выщелачивание менее интенсивно. Вероятно, эти факты имеют какое-то отношение и к общему распространению Leguminosae в тропическом дождевом лесу.
В консоциациях дождевого леса бобовые часто выступают в качестве единственной доминирующей породы.
СРЕДА
243
ПОЧВА КАК ДИФФЕРЕНЦИАЛЬНЫЙ ЭКОЛОГИЧЕСКИЙ ФАКТОР
Ранее уже неоднократно говорилось или подразумевалось, что во влажных тропиках, как и везде, почвенные различия проявляются в различном составе растительных сообществ. Эдафические факторы не только изменяют флористический состав в пределах самого тропического дождевого леса, но, как о том свидетельствуют данные, могут быть причиной развития кустарниковых сообществ, например паданга, и сообществ типа «мури», упомянутых выше (стр. 232), или же некоторых типов саванны (гл. 15), которые существуют в качестве эдафических климаксов в климате тропического дождевого леса. Это противоречит общепринятому в течение длительного времени взгляду, что в тропиках почвенные различия не имеют большого экологического значения и что это значение, во всяком случае, несомненно меньше, чем в умеренной зоне. Лишь в последнее время сведения о составе тропических растительных сообществ увеличились настолько, чтобы рассеять это неправильное представление.
Этот вопрос будет еще рассмотрен в последующих главах; пока же-достаточно будет показать на двух примерах, насколько тесна связь между почвой и климаксной растительностью.
Среди пяти первичных лесных сообществ Морабалли-Крик, Брит. Гвиана, два проявляют особенно поразительную корреляцию с почвенными условиями. Лес Уаллаба (консоциация Eperua falcata) приурочен к выщелоченным «белым» пескам, покрывающим плато и кряжи; лес Гринхарт (консоциация Ocotea rodiaei) встречается на коричневом песке, который обычно оконтуривает кряжи. Эти два сообщества иногда соседствуют, и в этих случаях между ними проходит исключительно отчетливая граница, точно соответствующая переходу от выщелоченного песка к коричневому. В одном из профилей полный переход осуществлялся в пределах дистанции, равной 150 м (Davis, Richards, 1933—1934, рис. 1, стр. 121). В других местах Брит. Гвианы наблюдалась еще более резкая граница между этими типами тропического дождевого леса; Вуд (Wood, 1926, стр. 4—5) пишет следующее, имея в виду леса округа Бартика-Кабури:
«Как правило, можно было одним шагом перейти с белого песка, где лес Гринхарт никогда не встречается, на коричневый песок, где он произрастает, что случается обычно в тех местах склонов, где они спускаются от плоского водораздела к речкам. Наиболее разительный пример представлял собой муравейник akushi (муравьи-листорезы), разместившийся на площади около 30 кв. футов (9 кв. м). Верхний край муравейника лежал на белой почве, нижний — на коричневой, причем линия раздела была настолько резкой, что ее можно было указать рукой. Тут же в непосредственной близости один от другого, так что человек мог только боком протиснуться между ними, росли два дерева, одно из которых укоренилось на белой почве, а другое — на коричневой. Верхнее дерево было первым из сочленов сообщества Уаллаба, а нпж-нее — первым Гринхарт на этом склоне». Хотя границы между другими типами леса в местности Морабалли-Крик были обычно не такими резкими, как между консоциациями Уаллаба и Гринхарт, тесная связь между типом леса и почвой проявлялась повсюду.
Другим примером может служить распределение на Борнео и Суматре леса из железного дерева — особой консоциации с преобладанием Eusi-deroxylon zwageri, почти полностью вытеснившего все другие виды деревьев (стр. 282). Грессер (Gresser, 1919) заметил, что этот тип леса приуро-
16*
244
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
чей к чистым пескам пли супесям. Последние исследования Виткампа (Witkamp, 1925) показали, что Eusideroxylon практически может быть использован в качестве геологического индикатора. В округе Коэтаи на Борнео породы представлены третичными песками, песчаниками, глинами и глинистыми сланцами с вкраплениями известняка и мергеля. С переходом от песка или песчаника к глине, сланцу, мергелю или известняку железное дерево исчезает или встречается все реже и реже; его исчезновение, если только переходы в характере самих пород не постепенны, всегда бывает внезапным. Большая часть местности представляет собой антиклиналь, где песчаник, глина и прочие породы располагаются параллельными полосами. В точном соответствии с этими длинными узкими полосами протянулись и сообщества железного дерева, отмечая выходы песка и песчаника.
Эти далеко не единственные примеры ярко иллюстрируют решающую роль, которую играет почва в распределении сообществ тропического дождевого леса. Влияние почвенных различии часто не замечают, так как типы тропической растительности могут быть внешне очень схожи, и нужен детальный флористический анализ, чтобы выявить их разнородность.
Несомненное значение почвенных различий для состава тропических растительных сообществ вовсе не означает, что наиболее действенные в условиях тропиков эдафические факторы обязательно должны быть тождественны характерным для умеренной зоны. Для последней (исключая приморскую растительность) важнейшими эдафическими факторами обычно считаются активная кислотность почвы (pH), содержание карбоната кальция и других оснований, структура почвы, ее водоудерживающая способность и аэрация, а в последнее время была высказана мысль о важном экологическом значении восстановительно-окислительного потенциала почвы. Сомнительно, чтобы во влажных тропиках кислотность или щелочность повсеместно имели бы большое экологическое значение, поскольку там почти все почвы отличаются кислой реакцией с более или менее выраженным дефицитом оснований. Дифференциальными эдафическими факторами здесь с точки зрения наших нынешних весьма несовершенных знаний, по-видимому, чаще всего являются водоудерживающая способность, аэрация и глубина почвы — следовательно, скорее ее физические чем химические свойства. Правда, Хардсон (Hard-son, 1937) доказал, что растения, произрастающие на характерных для низменностей крайне выщелоченных тропических подзолах падангов Индо-Малайской области (стр. 232), содержит значительно меньше оснований, чем те же виды, растущие на более плодородной почве; это внушает мысль, что различные степени дефицита оснований могут иногда оказаться экологически важными. Исключительная бедность тропических подзолов питательными веществами (они гораздо беднее, чем даже обычные почвы тропического дождевого леса) может быть одним из факторов, объясняющих специфичность их растительности. Наличие во многих, хотя и не во всех, тропических почвах больших количеств алюминия (см. ниже), возможно, является другим химическим эдафическим фактором, имеющим дифференциальное значение.
Вопрос о присутствии среди растений влажных тропиков кальцефи-лов и кальцефобов еще не разрешен окончательно. Некоторое внимание уделил ему в своей работе Ван-Стеенис (van Steenis, 1935а). Климаксные сообщества тропического дождевого леса не встречаются на известняковых почвах, так как во влажном тропическом климате зрелые и даже не вполне зрелые почвы никогда не содержат большое количество извести.
СРЕДА
245
В некоторых тропических областях, например на Тринидаде и в Гондурасе, лес, покрывающий известняковые хребты, значительно отличается от леса, развивающегося на окружающих низменностях, сложенных другими породами, но это различие вызвано главным образом чрезмерным дренажем, которому не препятствует тонкий почвенный покров, лежащий на пористом известняке, а не каким-либо химическим фактором. Так, в более влажных частях Тринидада на известняковых хребтах растет полувечнозеленый сезонный лес, отличающийся от вечнозеленого сезонного леса песчаных и глинистых почв, произрастающего в том же самом климате, но в местах с меньшим количеством осадков аналогичный вечнозеленый лес встречается на лишенной извести глине, залегающей не на известняках (Beard, 1946b). Решающим фактором здесь, несомненно, является водоудерживающая способность почвы. Чартер (Charter, 1941) также указывал, что виды, встречающиеся в восточном Гондурасе исключительно на известняковых почвах, в более сухом климате Тринидада произрастают на песках и глинах.
На Малаккском полуострове, на Яве и Борнео встречаются обрывистые известняковые утесы с чрезвычайно характерной флорой. Почва на них развивается лишь в углублениях, и условия для произрастания высокоствольного леса отсутствуют. Клейн (Klein, 1914) обратил внимание на ряд видов, вероятно, специфичных для этих утесов, которые он считал истинными кальцефилами, то есть зависящими в основном от химических, а не от физических свойств известняка. Ван-Стеенис (loc. cit.), однако, полагает, что большая часть этих растений — скорее крем-нефиты (растения скальных расщелин), чем кальцефилы; по его утверждению, некоторые из них встречаются в сходных местообитаниях на не известняковых породах. Наши собственные наблюдения над флорой известняковых утесов и песчаниковых уступов в Сараваке (Борнео) скорее подтверждают взгляды Клейна, чем Ван-Стеениса; известняковые утесы в Биди и Бау обладают чрезвычайно специфичной флорой, включающей знаменитую Moultonia singularis и папоротник Phanerosorus (Mata-nia) sarmentosus, которые свойственны выходам известняка, расположенным в значительном отдалении друг от друга, и, по-видимому, никогда не встречаются на неизвестняковых породах. Данная проблема нуждается в более критическом рассмотрении, чем это делалось до сих пор.
Если существование кальцефильных видов во влажных тропиках все еще спорно, то наличие в пределах тропического дождевого леса растений кальцефобов точно установлено. Уже отмечалось, что Eusi-deroxylon zwageri избегает известняк. Есть указания на то, что семейство Dipterocarpaceae в целом избегает известняковые породы (van Steenis, 1935, стр. 59). Правда, Саймингтон (Symington, 1943), подтверждая отсутствие диптерокарповых на известняковых возвышенностях юга Малаккского полуострова, связывает это лишь с неблагоприятными для засухоустойчивых видов диптерокарповых условиями увлажнения на них. На севере Малаккского полуострова, в непосредственной близости к полулистопадным и листопадным лесам Бирмы и Таиланда, на известняковых холмах встречается несколько видов из этого семейства, причем по крайней мере один из них — Нореа jerrea, по-видимому, оказывает явное предпочтение известняку.
В работах Ченери (Chenery, 1948 и неопубликованных) о растениях, аккумулирующих алюминий, поднимается вопрос о том, что избыток алюминия в тропических почвах, возможно, действует как дифференциальный экологический фактор. Установлено, что многие тропические
246
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
виды, как древесные, так и травянистые, принадлежащие в основном к обширным космополитическим или пантропическим семействам (Ru-biaceae, Melastomaceae), нескольким семействам папоротникообразных, аккумулируют большие количества алюминия в своих тканях. Такие растения изобилуют в первичных и вторичных сообществах тропического дождевого леса, и Ченери считает, что они играют существенную роль в сукцессиях и почвообразующих процессах в тропиках. Возможно, что именно различием в запасах алюминия отчасти обусловлено несходство между растительностью таких богатых алюминием тропических почв, как латеритные красноземы и кремнеземистые почвы, относительно бедные алюминием (например, подзолы).
Как уже говорилось, несмотря на очевидную значимость химических свойств почвы, представляется вероятным, что и физические ее свойства, особенно те, которые влияют на водообеспеченность растения и снабжение его корней кислородом, имеют большое значение в экологии тропических растений. Развитие одних характерных растительных ассоциаций по берегам рек и на затопляемых землях, других — на местах с беспрепятственным дренажем, тесная связь, установленная в Брит. Гвиане между структурой почвы и климаксными лесными сообществами (гл. 10),— все это экологические явления, вызванные хотя и не исключительно, но преимущественно физическими эдафическими факторами. Истинная природа этих факторов может быть выяснена лишь при дальнейших исследованиях.
Харди (Hardy, 1936b), ссылаясь на климаксные лесные сообщества Тринидада, высказал аналогичный взгляд на относительную важность химических и физических эдафических факторов. В южной части центрального района острова он изучил размещение двух разновидностей полувечнозеленого сезонного леса и трех—вечнозеленого сезонного леса. При сопоставлении карты почв с картой растительности стали очевидны некоторые совпадения. Отмечено, что отдельные виды встречаются только на песчаных, глинистых, иловатых, мергелистых почвах или на почве грязевых потоков. Харди считает, что основной фактор, регулирующий распределение видов и лесных сообществ в этом районе,— водоудерживающая способность почвы, которая в одинаковой степени зависит как от ее физических свойств, так и от рельефа и климата. Почва, климат и рельеф находятся в сложном взаимодействии, иногда стремясь противостоять друг другу, иногда действуя в одном направлении.
Значимость типа почвы выявляется главным образом в отношении тех физических свойств, которые определяют ее водный режим. Химические свойства и признаки почвы имеют значение лишь постольку, поскольку они воздействуют на режим почвенной воды. Из химических особенностей в этом отношении важны, очевидно, только содержание извести и окиси магния и количество перегноя. Другие химические факторы (запас азота, фосфатов, калия и т. д.) являются второстепенными в распространении типов растительности, хотя они приобретают важное значение при выращивании на бывших лесных землях сельскохозяйственных культур (Hardy, loc. cit. стр., 28).
В своей более поздней работе, где также решительно подчеркнута первостепенная важность физических свойств почвы, Бирд (Beard, 1946b) делает вывод, что распределение растительности в тропических странах, таких, как Тринидад, главным образом регулируется водным режимом, который является производным трех факторов — почвы, рельефа и климата.
СРЕДА
247
В Гондурасе известны две главные группы лесных сообществ: одна произрастает на почвах, развившихся на известняках, другая — на почвах, развившихся на гранодиоритах, сланцах, кварцитах и песчаниках. Хотя эти почвы разнятся по содержанию оснований и по другим химическим свойствам, все же Чартер (Charter, 1941) считает, что распределение растительности здесь регулируется водным режимом.
По некоторым данным аэрация не в меньшей степени, чем водоудерживающая способность тропических почв, является мощным экологическим фактором. Это должно быть особенно верным применительно к наиболее влажным климатам и к территориям, где топографические условия или структура почвы препятствуют дренажу. Ченери и Харди (Chenery, Hardy, 1945) в своем интересном описании «профилей влажности» некоторых почв Тринидада показали, что среди исследованных ими почв только три легко дренируемые песчаные почвы оказались адекватно аэрированы по всей глубине профиля во все времена года. Другие были пропитаны водой, по крайней мере в дождливый сезон, и неадекватно аэрированы, за исключением верхних слоев. Разные виды растений, несомненно, проявляют различную выносливость к плохо аэрированным почвам. Костер (Coster, 1935b) предположил, что деревья яванских низменностей испытывают меньшую потребность в кислороде почвы, чем деревья более пористых и лучше дренированных нагорных почв. Выше мы уже ссылались на его мнение о том, что конкуренция за кислород между корнями деревьев — очень существенна в экологии тропического леса (стр. 211—212).
Еще один фактор имеет прямое отношение к глубине аэрации почвы. Хотя, вообще говоря, тропическим деревьям свойственна неглубокая корневая система, глубина проникновения корней у них сильно варьирует. В гл. 4 было показано, что существует тесная связь между развитием досковидных корней и глубиной корневой системы, так как досковидные выросты развиваются на поверхностных боковых корнях и редко наблюдаются у деревьев с хорошо развитым главным корнем; также было показано, что в тропических дождевых лесах различного типа частота встречаемости досковидных корней находится в определенной связи с эффективной глубиной почвы. При маломощной почве или при высоком стоянии грунтовых вод, «фактически маломощная почва», корневая система поверхностна и досковидные корни обычны. У Сайкса (Sykes, 1930) имеется описание Gossweilerodendron — Cylicodiscus — типа смешанного тропического дождевого леса, приуроченного к глубоким песчаным почвам Нигерии. Он считает, что эта приуроченность объясняется наличием у господствующих видов этого сообщества длинных главных корней, нуждающихся в значительной мощности рыхлого слоя. Хотя и возможно, что глубина почвы иногда может влиять таким чисто механическим путем, но она, несомненно, должна взаимодействовать с аэрацией, ограничивающей толщину почвенного слоя, который может быть использован корнями деревьев.
Часть 1П
ФЛОРИСТИЧЕСКИЙ СОСТАВ
КЛИМАКСНЫХ СООБЩЕСТВ
Die uebergrosse Mannichfaltigkeit der bluehten-reichen Waldflora verbietet die Frage woraus diese Walder bestehen.
«Огромное разнообразие цветковых растений делает праздным вопрос о флористическом составе тропического леса»
ГУМБОЛЬДТ (ЦИТ. ПО КУРЦУ, KURZ, 1878)
Глава 10
СОСТАВ ПЕРВИЧНОГО ДОЖДЕВОГО ЛЕСА (I)
Важнейшей особенностью тропического дождевого леса является поразительное богатство его видового состава. Будем ли мы рассматривать огромную территорию вроде бассейна реки Амазонки п острова Борнео или небольшой пробный участок в один-два гектара, число видов, особенно деревьев, окажется значительно большим, чем в лесном сообществе любого другого типа. Такое огромное флористическое богатство в какой-то степени связано с условиями, способствующими интенсивному видообразованию, и, в частности, с климатическими условиями, благоприятными для роста и размножения растений в любое время года, но, вне всякого сомнения, в значительной мере оно обусловлено большой древностью располагающихся здесь крупных массивов суши, позволившей растительности сохраниться почти неизменной с какого-то неизвестного, но, во всяком случае, отдаленного геологического периода.
Точное количество видов, входящих в состав флоры тропического дождевого леса какой-либо отдельной страны или области, неизвестно, но косвенное представление об этом дают приводимые ниже цифры. В Индонезии, исключая Яву, преобладают тропические дождевые леса; здесь, согласно Эндерту (Endert, 1933), насчитывается около 3 тыс. видов деревьев со стволами диаметром более 40 см, принадлежащих приблизительно к 450 родам. На Малаккском полуострове, с его значительно меньшей территорией, сосредоточено около 2,5 тыс. видов деревьев (Foxworthy, 1927). Какие-либо новые данные относительно гилеи Амазонии отсутствуют; Губер (цит. Милдбрэдом, Mildbraed, 1930а) предполагает, что здесь имеется по крайней мере 2,5 тыс. видов «крупных деревьев» (в одном штате Пара их более 1 тыс.) Цифры эти, вероятно, занижены. В «сомкнутом лесу» («foret dense» — термин, объединяющий «смешанный листопадный лес» и настоящий вечнозеленый лес Берега Слоновой Кости) насчитывается 596 описанных видов деревьев с диаметром, достигающим 10 см и более, принадлежащих к 276 родам и 61 семейству (Aubre-ville, 1938). Число кустарников, трав, лиан и эпифитов в этих двух районах тропиков точно не известно, но, хотя оно и значительно, несомненно все же меньше числа деревьев. Ван-Стеенис (van Sleenis, 1938) считает, что вся цветковая флора Малайского архипелага (включая Малаккский полуостров и Новую Гвинею) включает приблизительно 20 тыс. видов. Только на одном Малаккском полуострове, большая часть которого была до недавнего времени равномерно покрыта девственным тропическим дождевым лесом, насчитывается не менее 9 тыс. видов (Corner). На Тринидаде, острове площадью 1754 кв. миль (4542 кв. км), известно 1289 видов цветковых растений, из которых 364 представляют собой деревья, в том числе и пальмы (Beard, 1946b).
252
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Производя обзор растительных сообществ умеренной зоны, не легко найти цифры, которые можно было бы сопоставить с этими, но наверняка можно сказать, что, возможно, лишь за исключением склерофильных скрабов Южной Африки и Австралии, ни одно растительное сообщество не может превзойти тропический дождевой лес по богатству видами.
Но внутри самой области распространения тропического дождевого леса наблюдаются заметные колебания в видовом разнообразии. Индомалайский тропический дождевой лес, занимающий меньшую площадь, чем такие же леса Америки или Африки, вероятно, богаче видами. Африканский лес флористически сравнительно беден и однообразен. Общее количество цветковых растений в пределах Республики Конго и Руанда-Урунди — обширной территории, включающей наряду с тропическим дождевым лесом огромные массивы листопадных лесов, саванн и горной растительности,— составляет только 9705 видов, принадлежащих к 1631 роду и 170 семействам (Robyns. 1946). В то же время флора Борнео, то есть флора гораздо меньшей и менее разнообразной по природным условиям области, оценивалась Меррилом (Merrill, 1915) не менее, чем в 10 тыс. видов. Сравнительная флористическая бедность Африки наиболее ярко проявляется в отношении некоторых семейств, которые обыкновенно рассматриваются как особенно характерные для областей тропического дождевого леса. Согласно Робэну, в Республике Конго имеется только 8 Lauraceae и 30 видов пальм. Orchidaceae, представленные в других частях тропиков в ошеломляющем разнообразии, здесь известны лишь в сравнительно скромном количестве, включая только 377 видов по сравнению с 5 тыс. видами Малайского архипелага (van Steenis, 1938). Эти региональные различия во флористическом богатстве не легко поддаются объяснению. Мильдбрэд (Mildbraed, 1922) приписывает бедность флоры африканского дождевого леса влиянию климатических изменений, происходивших в прошлом. Трудно сказать, насколько правильно такое объяснение, но, во всяком случае, указанное обстоятельство вряд ли можно отнести на счет какого-либо экологического фактора, проявляющегося в настоящее время. Тропический дождевой лес таких отдаленных островов, как остров Маврикия или группа островов Фиджи, также флористически беден ввиду долгой изоляции и трудности иммиграции.
Некоторые приблизительно установленные цифры числа видов для небольших пробных участков тропического дождевого леса даны в табл. 25. Речь о них будет идти в следующей главе.
Растительные формации обыкновенно складываются из отдельных синэкологических единиц, консоциаций, характеризуемых одним доминирующим видом, и более гомогенных группировок, ассоциаций, в которых ни одна доминанта не составляет выраженного большинства во всей популяции. Уже было показано, что тропический дождевой лес также состоит из консоциаций и ассоциаций, но благодаря большому количеству видов и трудности определения многих из них в поле часто бывает нелегко различить эти единицы. До самого последнего времени тропический дождевой лес рассматривался как какая-то хаотическая смесь, в которой нельзя или почти нельзя различить какой-либо порядок. Поскольку установить отдельные ассоциации или консоциации не удавалось, молчаливо предполагалось, что тропический дождевой лес, например южноамериканский, состоит из одной смешанной ассоциации, состав которой подвержен местным изменениям. Успехи систематики и применение количественных методов фитоценологии за последние 20 лет
Таблица 25
Число видов деревьев на небольших пробных участках первичного тропического дождевого леса
Местность Тип леса Число видов деревьев с диаметром ствола: Размер пробного участка, га Источник данных
4 дюйма £ (10 cjh) I и болше 1 дюймов м [20 см) и больше j 12 дюй-юв (около 30 см) и больше 16 дюймов (41 см) и больше
Южная Америка 1 Davis, Richards (1933—
Морабалли-Крик, Брит. Ассоциация смешанного дожде- 91 55 39 32 1,5
Гвиана вого леса 1934)
То же Лес Морабуцеа (консоциация Mora gonggrijpii) 71 45 29 21 1,5 Davis, Richards, (1933— 1934)
То же Пес Уаллаба (консоциация Ере- 74 49 30 16 1,5 Davis, Richards (1933— 1934)
rua falcata)
Африка Aubreville (1938)
Резервированный лес Ассоциация смешанного дожде- 74 58 35 около 23 около 1,4
Масса-Ме, Берег Слоновой Кости вого леса
Резервированный лес Око- Ассоциация смешанного дож- около 43 — 21 — 0,5 Richards (1939)
му, Нигерия девого леса около 60 70 51 32 40 31 1,0 1,5 Richards (1939) Richards (1939)
Фернандо-По Ассоциация смешанного дож- — — 15 — 1,0 Mildbraed (1933 b)
девого леса Jentsch (1911)
Камерун Ассоциация смешанного дож- — — 34 - 0,5
Остров Маврикий девого леса «Нагорный климаксный лес» 52 33 18 11 1,01 Vaughan, Wiehe (1941)
Азия
Гора Дьюлит, Саравак Смешанная диптерокарповая — 98 — 32 1,5 Richards (1936)
Резервированный лес Ма- ассоциация «Боровой» лес — 56 — 12 1,5 Richards (1936)
руди, Саравак Гора Маквилинг, Филип- Смешанная диптерокарповая ас- 43 27 15 11 0.25 Brown (1919)
пины социация (ассоциация Рага-shorea-Di plodiscus)
1 Сумма десяти маленьких участком.
254
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
позволили в настоящее время поставить на твердую почву дело распознавания ассоциаций и консоциаций в пределах тропического дождевого леса.
МЕТОДЫ ИЗУЧЕНИЯ
Ясно, что методы изучения флористического состава тропических дождевых лесов сильно отличаются от методов, применяемых при изучении растительных сообществ умеренной зоны. Они обыкновенно менее точны, но важно, чтобы они были, сколь это возможно, количественными, и включали бы подсчет индивидуумов на пробных единицах (площадках или полосах) известной площади. Огромное большинство видов сообщества тропического дождевого леса представляет собой высокие деревья, цветки и плоды которых расположены высоко над головой наблюдателя и не видны невооруженным глазом. При осмотре таких деревьев большую помощь оказывает сильный бинокль, но часто о цветении дерева можно догадываться только по приятному запаху да по жужжанию насекомых пли опавшим лепесткам. Единственными надежными способами получения материала для определения являются валка деревьев или залезание на них; иногда применяется отстрел ветвей из ружья или же используются специально дрессированные обезьяны. Благодаря исключительно ровному климату и относительной невыраженности сезонов цветение продолжается большую часть года. Как указывалось в гл. 8, некоторые деревья цветут реже одного раза в год, а небольшое число видов — не более одного раза в 10 лет. Таким образом, сбор гербарного материала всех видов даже на небольшом участке леса требует затраты большого количества времени, а подчас оказывается просто невозможным. Поэтому обыкновенно довольствуются точным определением лишь части имеющихся налицо видов, ограничиваясь относительно других лишь приблизительной количественной оценкой.
При повседневной работе по определению видов необходимо широко использовать такие заметные с земли дополнительные признаки, как особенность коры и ствола. Те, кому известна только немногочисленная флора деревьев умеренной зоны, возможно, будут удивлены, что эти признаки внешней морфологии достаточно разнообразны для того, чтобы ими можно было пользоваться, имея дело с древесной флорой порядка тысяч видов. Ствол и кора тропических деревьев дают большое количество разнообразных полезных диагностических признаков. Ствол, прямой или изогнутый, гладкий или с желобками, досковидные корни, когда они имеются, цвет и текстура коры, расположение чечевичек, цвет, запах и рисунок внешней поверхности заболони, макроскопический вид древесины — все это, вместе взятое, встречаясь в весьма характерных комбинациях, позволяет опытному глазу определять огромное большинство видов деревьев даже в богатой флоре. Пользуясь этими признаками, можно составить ключи для определения древесной флоры целых областей, как это сделал Фоксуорси (Foxworthy, 1927) в своей книге о деревьях Малаккского полуострова, дающих деловую древесину1. Правда, нужно отметить, что, несмотря на обычное совпадение отличительных
1 Можно также сослаться на работу Эндерта о деревьях Индонезии (Endert. 1928), в которой в качестве диагностических широко использованы указанные дополнительные признаки.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
255
особенностей ствола и коры с основными признаками, используемыми систематиками, существуют многочисленные исключения; например, иногда виды одного и того же рода сильно различаются по характеру ствола, коры и другим внешним признакам, а неродственные виды могут быть очень похожи по тем же признакам. Оценка всех этих незаметных, часто едва уловимых особенностей, требует острого глаза и значительного опыта. Всему, что при этом требуется от наблюдателя, нельзя научиться в течение нескольких недель или месяцев; в результате лишь немногие европейцы становятся в этом деле такими же знатоками, как местные жители, проводящие всю свою жизнь в лесу. Для последних различение сотен видов деревьев составляет часть их повседневной деятельности, и большинство видов поэтому имеет местные названия.
Например, у индейцев племени аравак в Брит. Гвиане в ходу несколько сот местных названий деревьев, причем большей частью каждое из них строго соответствует какому-либо одному виду. Ряд местных названий объединяет несколько родственных видов, например название «баромалли» относится к двум родственным видам Catostemma; «варакуза»— к видам Inga (за исключением небольшого числа очень легко различимых видов). Лишь небольшое число местных названий включает представителей различных родов или неродственных семейств; так, название «кокеритибалли» относится к нескольким родам Sapotaceae, «кулашири»— к различным Meliaceae, Flacourtiaceae и Sapindaceae. В ряде стран некоторые деревья не имеют местных названий или группируются под неопределенными сборными именами, например «меданг», в Малайе. Названия чаще всего присваиваются тем деревьям, которые употребляются в качестве строительного материала, как источники лекарственных и ядовитых веществ и т. д. или же в силу того, что с ними связаны какие-либо суеверия.
Надо подчеркнуть, что в то время, как любой местный житель лесов знает названия некоторых деревьев, только немногие знатоки помнят названия большого числа видов. В тропических странах с хорошо организованной лесной службой некоторые из таких местных экспертов получили подготовку для совместной работы с европейскими лесничими. Путем постоянной практики они приобрели необходимые сведения относительно определения видов деревьев и кустарников их округа, причем осведомленность этих местных знатоков поражает своим объемом и точностью. Они редко колеблются, называя то или иное дерево, сомневаясь лишь в том редком случае, когда им попадется совершенно неизвестный вид. Опытные «искатели деревьев» из местных жителей есть в Брит. Гвиане, Нигерии, Малайе и на Берегу Слоновой Кости и, несомненно, в большинстве других тропических стран.
После того как удастся установить местные названия деревьев в пределах пробного участка, необходимо сделать перевод этих названий для возможно большего числа видов на научный язык. Это — трудоемкий процесс, связанный с большой работой по проверке и сравнению. Использование каждого названия должно быть неоднократно проверено в поле, для того чтобы установить, какие виды или группы видов ему соответствуют. Должно быть собрано возможно большее количество гербарного материала для последующего иссследования специалистами. Надо помнить, что одно и то же местное название может в различных округах обозначать различные виды. Поэтому название, тщательно проверенное в пределах одной области, не может без дальнейшей проверки использоваться в другой. Когда местное название относится к группе сходных
254
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
позволили в настоящее время поставить на твердую почву дело распознавания ассоциаций и консоциаций в пределах тропического дождевого леса.
МЕТОДЫ ИЗУЧЕНИЯ
Ясно, что методы изучения флористического состава тропических дождевых лесов сильно отличаются от методов, применяемых при изучении растительных сообществ умеренной зоны. Они обыкновенно менее точны, но важно, чтобы они были, сколь это возможно, количественными, и включали бы подсчет индивидуумов на пробных единицах (площадках или полосах) известной площади. Огромное большинство видов сообщества тропического дождевого леса представляет собой высокие деревья, цветки п плоды которых расположены высоко над головой наблюдателя и не видны невооруженным глазом. При осмотре таких деревьев большую помощь оказывает сильный бинокль, но часто о цветении дерева можно догадываться только по приятному запаху да по жужжанию насекомых или опавшим лепесткам. Единственными надежными способами получения материала для определения являются валка деревьев или залезание на них; иногда применяется отстрел ветвей из ружья или же используются специально дрессированные обезьяны. Благодаря исключительно ровному климату и относительной невыраженности сезонов цветение продолжается большую часть года. Как указывалось в гл. 8, некоторые деревья цветут реже одного раза в год, а небольшое число видов — не более одного раза в 10 лет. Таким образом, сбор гербарного материала всех видов даже на небольшом участке леса требует затраты большого количества времени, а подчас оказывается просто невозможным. Поэтому обыкновенно довольствуются точным определением лишь части имеющихся налицо видов, ограничиваясь относительно других лишь приблизительной количественной оценкой.
При повседневной работе по определению видов необходимо широко использовать такие заметные с земли дополнительные признаки, как особенность коры и ствола. Те, кому известна только немногочисленная флора деревьев умеренной зоны, возможно, будут удивлены, что эти признаки внешней морфологии достаточно разнообразны для того, чтобы ими можно было пользоваться, имея дело с древесной флорой порядка тысяч видов. Ствол и кора тропических деревьев дают большое количество разнообразных полезных диагностических признаков. Ствол, прямой или изогнутый, гладкий или с желобками, досковидные корни, когда они имеются, цвет и текстура коры, расположение чечевичек, цвет, запах и рисунок внешней поверхности заболони, макроскопический вид древесины — все это, вместе взятое, встречаясь в весьма характерных комбинациях, позволяет опытному глазу определять огромное большинство видов деревьев даже в богатой флоре. Пользуясь этими признаками, можно составить ключи для определения древесной флоры целых областей, как это сделал Фоксуорси (Foxworthy, 1927) в своей книге о деревьях Малаккского полуострова, дающих деловую древесину1. Правда, нужно отметить, что, несмотря на обычное совпадение отличительных
1 Можно также сослаться на работу Эндерта о деревьях Индонезии (Endert. 1928), в которой в качестве диагностических широко использованы указанные дополнительные признаки.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
255
особенностей ствола и коры с основными признаками, используемыми систематиками, существуют многочисленные исключения; например, иногда виды одного и того же рода сильно различаются по характеру ствола, коры и другим внешним признакам, а неродственные виды могут быть очень похожи по тем же признакам. Оценка всех этих незаметных, часто едва уловимых особенностей, требует острого глаза и значительного опыта. Всему, что при этом требуется от наблюдателя, нельзя научиться в течение нескольких недель или месяцев; в результате лишь немногие европейцы становятся в этом деле такими же знатоками, как местные жители, проводящие всю свою жизнь в лесу. Для последних различение сотен видов деревьев составляет часть их повседневной деятельности, п большинство видов поэтому имеет местные названия.
Например, у индейцев племени аравак в Брит. Гвиане в ходу несколько сот местных названий деревьев, причем большей частью каждое из них строго соответствует какому-либо одному виду. Ряд местных названий объединяет несколько родственных видов, например название «баромалли» относится к двум родственным видам Catostemma\ «варакуза»— к видам Inga (за исключением небольшого числа очень легко различимых видов). Лишь небольшое число местных названий включает представителей различных родов или неродственных семейств; так, название «кокеритибалли» относится к нескольким родам Sapotaceae, «кулашири»— к различным Meliaceae, Flacourtiaceae и Sapindaceae. В ряде стран некоторые деревья не имеют местных названий или группируются под неопределенными сборными именами, например «меданг», в Малайе. Названия чаще всего присваиваются тем деревьям, которые употребляются в качестве строительного материала, как источники лекарственных и ядовитых веществ и т. д. или же в силу того, что с ними связаны какие-либо суеверия.
Надо подчеркнуть, что в то время, как любой местный житель лесов знает названия некоторых деревьев, только немногие знатоки помнят названия большого числа видов. В тропических странах с хорошо организованной лесной службой некоторые из таких местных экспертов получили подготовку для совместной работы с европейскими лесничими. Путем постоянной практики они приобрели необходимые сведения относительно определения видов деревьев и кустарников их округа, причем осведомленность этих местных знатоков поражает своим объемом и точностью. Они редко колеблются, называя то или иное дерево, сомневаясь лишь в том редком случае, когда им попадется совершенно неизвестный вид. Опытные «искатели деревьев» из местных жителей есть в Брит. Гвиане, Нигерии, Малайе и на Берегу Слоновой Кости и, несомненно, в большинстве других тропических стран.
После того как удастся установить местные названия деревьев в пределах пробного участка, необходимо сделать перевод этих названий для возможно большего числа видов на научный язык. Это — трудоемкий процесс, связанный с большой работой по проверке и сравнению. Использование каждого названия должно быть неоднократно проверено в поле, для того чтобы установить, какие виды или группы видов ему соответствуют. Должно быть собрано возможно большее количество гербарного материала для последующего иссследования специалистами. Надо помнить, что одно и то же местное название может в различных округах обозначать различные виды. Поэтому название, тщательно проверенное в пределах одной области, не может без дальнейшей проверки использоваться в другой. Когда местное название относится к группе сходных
256
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
видов, может случиться, что в данной области или на данном пробном участке имеется налицо только один из видов этой группы; в этом случае данное местное название может вполне безошибочно считаться присвоенным какому-либо одному виду. Не следует пренебрегать даже и приблизительными сведениями; часто бывает полезно выяснить присутствие того или иного рода, даже если невозможно решить вопрос о видовом составе и количественном распределении видов.
При составлении списка видов для какого-нибудь пробного участка или профиля должны приводиться как местные названия, так и научные, даже если эти научные названия были внесены позже. Доказательства правильности отождествления местных названий с научными будут, по всей видимости, различны в смысле их полноты. Таким образом, если имеется всего один гербарный экземпляр какого-либо вида, носящего местное название, то правильность присвоенного ему научного эквивалента будет менее очевидной, чем в случае наличия десяти экземпляров. Отождествление названий, основанных на материале, собранном фактически на том же участке или профиле, который в данном случае рассматривается, более надежно, чем если оно основывается на материале, собранном в другом пункте того же округа или в другом округе. Можно составить специальную схему для указания степени надежности применяемых названий (см., например, Richards, 1939, стр. 53 и табл. 6—9). Когда определение основывается на собранном гербарном экземпляре, надо приводить номер этого экземпляра и место хранения гербария.
Систематики, привыкшие работать с флорами умеренной зоны, возможно, усомнятся в надежности методов изучения флористического состава, предусматривающих услуги местных «искателей деревьев» и использование местных названий. Однако такие методы позволяют получать результаты, имеющие большое научное значение при условии соблюдения известных предосторожностей. При любом экологическом исследовании, предусматривающем составление списка видов, которые не могут быть определены непосредственно в поле, ошибки могут возникнуть на двух этапах работы: при определении собранных образцов и при решении вопроса о том, какие индивидуумы с пробной площади соответствуют собранным экземплярам. Хотя возможность ошибки на втором этапе работы зависит от осведомленности в систематике и опытности исследователя, совершенно устранить эти ошибки невозможно. Когда пользуются услугами кого-либо из местных «искателей деревьев», вероятность ошибки, конечно, несколько увеличивается, но, если проявить соответствующую осторожность, список видов, хотя, несомненно, менее полный и точный, чем такой же список для хорошо исследованной флоры умеренной зоны, все же может иметь высокую степень достоверности.
Методика выбора пробных площадей в тропических лесах аналогична применяемой в других растительных сообществах. Поэтому здесь она будет рассмотрена лишь вкратце. Лесные службы тропиков обыкновенно используют профильный метод. Но так как большинство первичных дождевых лесов чередуется с пространствами вторичных лесов (по краям старых заброшенных полей, на ветровалах и т. д.) и пересечены многочисленными речными долинами, где флористический состав отличается от флористического состава первичного леса, то для экологических исследований часто лучше предпочесть компактные пробные участки. В тропическом дождевом лесу, как и в других растительных сообществах, количество видов сначала возрастает параллельно увеличению площади, а затем падает, так что кривая всей площади имеет почти логарифмиче
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСПЫХ СООБЩЕСТВ
257
скую форму [рис. 34; см. также кривые Эггелинга (Eggoling, 1947) для лесных сообществ Уганды и Вогана, Виз (Vaughan, Wiehe, 1941) — для «нагорного климаксного леса» острова Маврикий!. Теоретически идеальный пробный участок должен соответствовать участку, на котором
Рис. 34. Число видов на единицу площади в ассоциациях тропического дождевого леса в Нигерии (Richards, 1939, рис. 7). Цифры 2 — 4 указывают номер пробного участка. Непрерывные линии даны для деревьев диаметром 4 дюйма (10 см) и больше, прерывистые линии —для деревьев диаметром 12 дюймов (30 с.и) и больше.
число видов впервые достигает своей максимальной величины. На практике избранный размер неизбежно представляет собой компромисс между теоретическими требованиями и имеющимися в распоряжении временем и возможностями. На основании опыта автора, квадратные участки 400x400 футов (около 1,5 га) дают удовлетворительные результаты для большинства типов тропического дождевого леса. В целях быстрой разведки удобно регистрировать все деревья по обеим сторонам тропинки на расстоянии 0,5—1 км (Aubreville, 1938, стр. 208).
При изучении флористического состава лесного сообщества желательно охватывать не только все древесные ярусы (видовой состав которых следует по возможности записывать в отдельные списки), но также ярусы
258
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
кустарников и трав, лиан и эпифитов. Для изучения ярусов кустарников и лиан не требуется каких-либо особых методов. В обоих этих ярусах большинство индивидуумов представляет собой молодые деревья; в связи с этим могут возникнуть значительные трудности при разграничении деревьев и кустарников, когда они не цветут. Во многих случаях достаточно составить списки видов только более низких ярусов и ограничиться сбором образцов тех видов, которые не могут быть определены в поле; частота их встречаемости устанавливается на глаз с применением обычных сокращенных обозначений (a., l.f., v.r. и т. д.). Более точные результаты могут быть получены методом малых квадратов, но в силу того, что и кустарники и травы не образуют сплошного покрова, эти квадраты должны быть во всяком случае больше 1 кв. м. Подсчет числа экземпляров, процента покрытия и т. д. для каждого вида может производиться теми же методами, которыми пользуются европейские экологи. При простом составлении списка флористического состава нижних ярусов, может использоваться весь пробный участок, выбранный для учета деревьев; при применении специальных квадратов они размещаются либо наугад, в границах крупного участка или профиля, либо же через произвольно выбранные правильные интервалы. Иногда более низкие ярусы лесного сообщества оказываются менее однообразными по составу, чем ярусы деревьев (светлые и тенистые участки, болотистые и более сухие пространства и т. д.); в таком случае различные типы подлеска должны изучаться отдельно.
Синузии лиан и эпифитов представляют особенные трудности для изучения. Лианы могут быть удовлетворительно изучены только в том случае, если участок сплошь вырублен, но даже и в этом случае они часто образуют переплетенную массу, в которой трудно определить количество экземпляров каждого вида. Для сплошь вырубленного участка можно также составить список эпифитов, а если необходимо, то и сосчитать их; обыкновенно в любой момент цветет только небольшая часть видов. В некоторых открытых типах леса иногда оказывается возможным распознать эпифиты и определить частоту их встречаемости непосредственно с земли при помощи бинокля (см. стр. 137).
СООБЩЕСТВА ПЕРВИЧНОГО ЛЕСА В БРИТ. ГВИАНЕ
Перед тем как обобщить данные об ассоциациях и консоциациях тропического дождевого леса (гл. И), полезно описать сообщества первичного леса двух ограниченных территорий, которые автор имел возможность непосредственно изучить, а именно Морабалли-Крик в Брит. Гвиане и бассейна реки Барам в Северном Сараваке (Борнео). Подробные описания этих районов и свойственных им леспых сообществ были опубликованы ранее (Davis, Richards, 1933—1934; Richards, 1936). Здесь мы ограничимся только кратким резюме.
Морабалли-Крик представляет собой небольшой приток, длиной каких-нибудь 24 км, впадающий в реку Эссеквибо, приблизительно в 80 км выше ее устья. Изучавшееся пространство расположено по радиусу в нескольких километрах от бывшей базы Оксфордской экспедиции (61Г с. ш.). По своей топографии это низкая, волнисто-холмистая равнина, большая часть которой, однако, не является плоской или болотистой. В обе стороны от реки местность повышается, примыкая к небольшим кряжам и плато с обрывистыми склонами; самые высокие из этих возвышенностей достигают около 100 м ад уровнем моря.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
259
Повсюду в пределах района подстилающими породами являются гранит и гнейс, но почвы заметно варьируют. В долинах ручья и его главных притоков почва представляет собой грязно-белый аллювиальный ил с тонкой структурой. Эта почва подвергается затоплению в любое время года, и даже в сухие периоды уровень грунтовых вод близок к ее поверхности. На более высоких местах располагаются почвы, в большинстве своем принадлежащие к типу тропических красноземов; среди них выявляется ряд переходов от очень тяжелой вязкой ярко-красной глины до гораздо более легкого, более рыхлого и пористого суглинка. Суглинок вообще имеет тенденцию встречаться на более высоком уровне и дальше от водотоков, чем глина. Остальные почвы представляют собой бурые и выщелоченные («белые») пески. Бурые пески, крупнозернистые и пористые, окрашены окисями железа. Их принадлежность к группе тропических красноземов считается спорной1. Бурый песок встречается главным образом на крутых склонах на незначительных пространствах. О выщелоченном песке, который относится к группе тропических подзолов низменностей, уже говорилось в гл. 9. Располагаясь на поверхности кряжей и плато, он часто достигает значительной глубины. При переходе от русла ручья к водоразделу почва обыкновенно, хотя и не всегда, изменяется в следующем порядке: ил, красная глина, красный суглинок, бурый песок, выщелоченный песок.
Насколько известно, климат всего округа является типично экваториальным. Годовая сумма осадков превышает 250 см. В годовом ходе наблюдаются два максимума осадков и два сухих сезона, однако средняя месячная сумма осадков никогда не опускается ниже 10 см1 2.
Дождевой лес, большая часть которого является первичным или только слегка тронутым рубкой, покрывает весь округ и тянется без перерыва по направлению к западу, где он смыкается с лесом бассейна Ориноко, и к востоку, через Суринам и Франц. Гвиану, чтобы соединиться с огромным лесом Амазонии. Северный край гвианского леса расположен приблизительно в 50 км к северу от Морабалли-Крик.
В первичном лесу можно различить пять основных сообществ. Хотя некоторые из них слегка варьируют по своему составу, образуя так называемые фации, в целом эти сообщества достаточно хорошо выражены. Переход от одного сообщества к другому в большинстве случаев осуществляется постепенно, но известны случаи п резкого разграничения их (см. стр. 243). Обычно при движении от русла к водоразделам последовательно встречаются: консоциация Мора (доминанта Mora excelsa), консоциация Морабуцеа (доминанта Mora gonggrijpii), ассоциация смешанного леса (полидоминантная), консоциация Гринхарт (доминанта Ocotea rodiaei) и консоциация Уаллаба (доминанта Eperua falcata). Некоторые из этих сообществ широко распространены в тропической Америке, другие приурочены лишь к ограниченным местообитаниям. Сообщество Мора, очень характерное для прибрежных районов Брит. Гвианы, проникает внутрь страны до гор Паракаима, где оно встречается в качестве галерейных лесов в саваннах (Myers, 1936), па восток в Суринам и на запад до дельты Орп-
1 Доктор Э. М. Чевери (Е. М. Chen cry) высказал предположение, что бурый песок мог произойти при вторичной пропитке выщелоченного песка водами, содержащими железистые соединения, вымытые из слоя песка, расположенного выше, то есть он представляет собой своего рода горизонт В тропических подзолов, покрывающих кряжи.
2 Метеорологические данные станции Мазаруни (удалена от места наблюдения на 27 км) (см. табл. 10—12 и рис. 22).
260
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ноко; оно вновь появляется в несколько измененной форме на Тринидаде (Beard, 1946а). Лес Морабуцеа встречается в различных частях Брит. Гвианы и в Суринаме. Гринхарт. как вид Ocotearodiaei, встречается только в Брит. Гвиане и в соседних частях Суринама и Венесуэлы; вероятно, он доминирует в консоциациях только близ центра их ареала. Лес Уаллаба, вероятно, широко распространен на северо-востоке Южной Америки. Леса, развивающиеся па выщелоченных песках в области Рио-Негро, бассейн Амазонки (Spruce, 1908, 2, стр. 206—207 и 303—304) и известные под названием каатинги, по-видимому, очень похожи на лес Уаллаба (каатингу из области Рио-Негро нельзя смешивать с каатингой южной Бразилии, которая представляет собой листопадный лес).
Количественные данные в последующих описаниях основываются на цифрах, полученных на типичных пробных участках — 400 х z<400 футов (около 1,5 га),— заложенных внутри каждого сообщества.
Лес Мора занимает самые низкие места, образуя широкие полосы на илистой почве затопляемой равнины по обеим сторонам речки и ее главных притоков. Иногда он также встречается на крутых скалистых склонах с очень неглубокой и малопроницаемой глинистой почвой. Общей чертой этих местообитаний является, по-видимому, маломощная почва, обусловленная в одном случае высоким уровнем грунтовых вод, а в другом — процессами эрозии.
Структура леса подробно не исследовалась, но она, вероятно, похожа на структуру леса Мора на Тринидаде, описанную Бирдом (см. рис. 7,6 и стр. 49), за исключением того, что верхние ярусы деревьев в лесу Брит. Гвианы менее сомкнуты и подлесок оказывается в лучших условиях освещения (средняя освещенность на высоте груди 1,33%; см. стр. 204).
Ярус А, чья высота превышает 30 м, состоит главным образом из доминирующих видов; ниже его располагаются нечетко разграниченные ярусы В и С, кустарниковый (D) ярус и травяной покров, который можно приблизительно подразделить на ярус высоких и ярус низких трав. Характерно роскошное развитие высоких трав вроде Carludovica sp. и различных Scitamineae, Mora excelsa образует свыше половины древостоя среди деревьев более крупных ступеней толщины и значительный процент деревьев среди меньших ступеней толщины, как это видно из следующих цифр:
Ступени толщины Mora excelsa, % Ступени толщины Mora excelsa, %
4-7 дюймов (10—19 см) 13,6 Все деревья от 4 дюймов
8—11 12—15 » (20—29 см) » (30—40 см) 13,5 35,4 (10 см) и более 23,4
16—23 дюйма (41—60 см) 54,6 Все деревья от 16 дюймов
24 лее » (61 см) и б о- 91,3 (41 см) и более 67,2
Фактически Mora excelsa является наиболее обильно представленным видом для любой ступени толщины, за исключением ступени 8— 11 дюймов. Всходы ее также обильны; в среднем на 1 кв. ярд участка на более сухих местах насчитывалось 2,7% сеянцев высотой ниже 3 футов, и 2% — высотой 3—6 футов; в местах более влажных их было меньше. Второе место после Mora занимают Pterocarpus officinalis и Pentacle-
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
261
thra macroloba; оба вида принадлежат к ярусу В. Каждый из них образует 10,2% древостоя среди деревьев диаметром от 4 дюймов и более. В целом на участке насчитывалось около 60 видов деревьев диаметром не менее 4 дюймов. Большинство их, включая Mora и Pentaclethra, как правило, встречается и в четырех других сообществах первичного леса. Pterocarpus и несколько других видов, видимо, свойственны исключительно консоциации леса Мора.
Лес Морабуцеа, Это сообщество, в котором доминирует другой вид Mora, имеет менее определенное местообитание. Лес Морабуцеа обычно расселяется на склонах более низких, плоских холмов, в отдельных случаях занимая и их вершины; он избегает затопляемые равнины и предпочитает тяжелые глинистые красноземы, хотя иногда, например на низкорасположенных местах у подножия склонов, он развивается на суглинистой или даже супесчаной почве.
Структура этого леса почти не изучена, но, по всей вероятности, он состоит из трех ярусов деревьев. Благодаря большой сомкнутости ярусов А и В подлесок освещен очень скудно (в среднем 0,61 %); это самый темный из всех пяти типов первичного леса. Характерными чертами его являются густой подрост доминирующих видов, сравнительно толстая подстилка из опавших листьев, обилие небольших сапрофитов и скудость всех других покровных трав.
Mora gonggrijpii является наиболее обильным видом для почти каждой ступени толщины, но среди деревьев больших диаметров его преобладание несколько менее выражено, чем преобладание М. excelsa в консоциации Мора. Для различных ступеней толщины наличие Mora gonggrijpii с соответствующим диаметром ствола выразится в следующих цифрах:
Ступени толщины Mora gonggrijpii, % Ступени толщины Мота gonggrijpii, %
4—7 дюймов (10—19 см) 17,6 Все деревья от 4 дюймов
8—11 » (20—29 см) 16,5 (10 см) и более 25,9
12—15 » (30—40 см) 19,6
16—23 дюйма (41—60 см) 61,3 Все деревья от 16 дюймов
24 » (61 см) и бо- (41 см) и более 60.7
лее 59,3
Второе место после доминанты занимают Eschweilera sagotiana (7,4% деревьев диаметром 4 дюйма и более) и Catostemma commune (5,0%). Оба вида принадлежат к ярусу А. Ни один другой вид не достигает 5% обилия. Общее число видов диаметром 4 дюйма и более равнялось 71. Среди них вряд ли был хотя бы один вид, свойственный исключительно лесу Морабуцеа.
В целом список видов очень близок к аналогичному списку для смешанного леса; главное различие заключается в меньшем видовом разнообразии и в высоком проценте Mora gonggrijpii.
Смешанный лес. Не имеющая определенной доминанты ассоциация смешанного леса характерна (у Морабалли-Крик) для довольно вязких суглинистых почв, промежуточных между тяжелыми глинами, которым свойственна консоциация Морабуцеа, и бурыми песками леса Гринхарт. Смешанный лес покрывает большую часть невысоко приподнятой ход-
262
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
мистой местности исследуемого района и встречается на вершинах холмов, там, где они не покрыты выщелоченными песками.
Структура этого леса уже была подробно описана (стр. 41, рис. 4). Примечательная черта его состоит в значительно большем количестве деревьев на единицу площади по сравнению с любым из двух охарактеризованных выше сообществ (в консоциации Мора 310 деревьев диаметром 4 дюйма и более на 1 га. в консоциации Морабуцеа — 309, в смешанной ассоциации 432). Подлесок освещен скудно (0,67%), но несколько лучше, чем в лесу Морабуцеа; возможно, вследствие этого травяной покров гораздо более обилен.
Самой характерной особенностью флористического состава смешанного леса является большое число доминирующих видов. Среди деревьев диаметром 16 дюймов (41 см) и более на первом месте по числу особей стоит Eschweilera sagotiana. составляющая 15,6% древостоя. Но среди деревьев диаметром 4 дюйма и более этот вид образует только 6,1% древостоя, в то время как Pentaclethra macroloba (ярус В) — 11,3%. Пять видов составляют здесь свыше 5?6. Вероятно, состав смешанной ассоциации колеблется в различных частях исследуемого района, и Eschweilera и Pentaclethra. возможно, не всегда являются наиболее обильно представленными видами. Общее количество видов деревьев на пробном участке равнялось 91, то есть являлось наивысшим по сравнению с любым из сообществ, за исключением консоциации Гринхарт. В это число входит большинство видов, характерных и для других типов леса, за исключением некоторых видов, свойственных лесам Мора и Уаллаба, и, возможно, очень немногих, ограничивающихся в своем распространении лесами Морабуцеа и Гринхарт.
Лес Гринхарт (Ocotea rodiaei). Как уже было сказано, это сообщество в пределах исследуемого района встречается лишь на одном типе почв, а именно на легких красновато-бурых песках, залегающих главным образом на верхних частях склонов плато и кряжей, вершины которых покрыты выщелоченным песком — субстратом, на котором произрастает лес Уаллаба. Граница между лесом Гринхарт и Уаллаба может быть очень резкой (см. стр. 243).
Структура леса Гринхарт специально не изучалась, но она, вероятно, очень сходна со структурой смешанного леса, хотя в лесу Гринхарт деревья большого размера встречаются несколько чаще. Число деревьев с диаметром ствола 4 дюйма и более составляет 519 на 1 га. что значительно больше, чем в смешанном лесу, но гораздо меньше, чем в лесу Уаллаба. Травяной покров хорошо развит, причем его видовой состав отличается от видового состава покровных трав других типов леса.
Ocotea rodiaei является наиболее обильным видом по числу особей среди более крупных деревьев, но среди деревьев с небольшим диаметром ствола Ocotea составляет лишь небольшую часть древостоя.
Ступени толщины Ocotea rodiaei, % Ступени толщины Ocotea rodiaei, %
4—7 дюймов (10—19 см) 0,5 Все деревья от 4 дюймов
3—11 * (20—29 см) 3,3 (10 см) и более 9,4
12—15 » (30—40 см) 1,1
16—23 дюйма (41—60 см) 38,4 Все деревья от 16 дюймов
24 » (61 см) и более 60,0 (41 см) и более 43,4
Ф.ото 14. Лес Морабуцеа (консоциация Mora ganggrijpii), Морабалли-Крик, Брит. Гвиана. г
Подлесок в основном состоит из всходов ч подроста доминирующей породы.
Фото 15. Лес Уаллаба (консоциация Eperua /ale ata), Морабалли-Крик, Брит. Гвиана.
В середине рисунка видны два экземпляра Eperua sp., бросается в глаза отсутствие досковидных корней. Злакоподобное растение Til-landsia sp. слева от ближайшей Ерегиа является эпифитом, упавшим с вершин к дерева и укоренившимся в почве: другая Tillandsia растет в нескольких футах от поверхности земли на маленьком дереве слева
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
263
Среди подчиненных видов наиболее обильными по числу экземпляров являются Pentaclethra macroloba (9,1 % деревьев диаметром 4 дюйма п более), Eschweilera sagotiana (7,6%) и Licania venosa (5,6%). Все эти виды наиболее обильно представлены и в смешанном лесу. Вообще лес Гринхарт по составу так близко напоминает ассоциацию смешанного леса, что он может рассматриваться скорее как специальная фация или «локация», чем как отдельная консоциация. Общее количество видов деревьев на участке равняется 95, что превышает соответствующие цифры для всех других рассмотренных сообществ. Правда, это лишь ненамного больше, чем в смешанном лесу. При тщательном изучении этой консоциации можно было бы выявить несколько видов, свойственных исключительно лесу Гринхарт.
Лес Уаллаба — последний из пяти изученных типов леса — представляет собой консоциацию Eperua falcata — дерева, которое, подобно двум видам Mora, принадлежит к семейству Leguminosae. По своей структуре, флористическому составу и условиям местообитания лес Уаллаба очень своеобразен и отличается от других сообществ в большей степени, чем они отличаются друг от друга. Этот лес строго ограничивается в своем распространении выщелоченными песчаными почвами (подзолами), исчезая вместе с их исчезновением. В местности Морабалли-Крик лес Уаллаба был единственным типом первичного леса, произраставшего на выщелоченных песках, но в других районах тропического дождевого леса Гвианы на подобных почвах встречаются другие сообщества (см. стр. 279—280).
Структура леса Уаллаба, для которой характерно почти полное отсутствие яруса В, была уже описана в гл. 2 (рис. 8, б). Особой чертой, свойственной исключительно этому лесу, является крайне большое число деревьев на единицу площади (617 экземпляров диаметром 4 дюйма и больше на 1 га), что почти вдвое больше, чем в лесу Мора. Ярус кустарников исключительно густ. Травянистых растений немного как по числу видов, так и индивидуумов, но они в высшей степени характерны. Часть этих видов свойственна только лесу Уаллаба. Скудное развитие травяного покрова зависит не от недостатка света; освещенность здесь выше, чем в любом другом типе леса, и составляет 1,43%. Другой отличительной чертой леса Уаллаба является редкость деревьев с досковидными корнями (рис. 11 и стр. 85) и малочисленность крупных лиан. Некоторые виды лиан, по-видимому, характерны для этой консоциации.
Ступени толщины Eperua falcata, % Ступени толщины Eperua falcata,
4—7 дюймов (10—19 см) 8—11 » (20—29 см) 12—15 » (30—40 см) 16—23 дюйма (41—60 см) 24 » (61 см) и более 10,0 17,3 32,7 66,7 75,0 Все деревья от 4 дюймов (10 см) и более Все деревья от 16 дюймов (41 см) и более 21,1 67,0
Eperua falcata преобладает среди деревьев всех диаметров, исключая лишь самые тонкомерные, причем среди деревьев более крупного диаметра этот вид составляет подавляющее большинство. Eperua falcata хорошо возобновляется; шесть выборочных подсчетов на пробном участке дали в среднем 1,8% сеянцев ниже 2 м высотой на 1 кв, м. Второе место по чис
264
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ленности занимают Catostemmafragrans nLicania buxifolia. Среди деревьев диаметром 16 дюймов и более они составляют менее 5%, зато среди деревьев с диаметром ствола 4 дюйма и больше им принадлежит 15,1 и 12,2%. Следовательно, они образуют значительную часть всего древостоя. Среди видов, формирующих этот тип леса, проявляется склонность к так называемому «групповому расположению» — сосредоточенность на единице площади большого числа экземпляров, принадлежащих к незначительному числу видов, выражающаяся в изобилии особей некоторых второстепенных видов, а также в наличии одной доминанты (табл. 29). На участке насчитывалось 74 вида деревьев диаметром 4 дюйма и более, то есть гораздо меньше, чем в смешанном лесу и в лесу Гринхарт, но несколько больше, чем в лесу Мора и Морабуцеа. Около 3/4 видов, в том числе и доминанта Eperua jalcata, свойственны и другим типам леса. Остальные, включая Catostemma fragrans и Licania buxifolia, видимо, встречаются только в лесу Уаллаба, по крайней мере в пределах изученного района.
Таким образом, консоциация Eperua отличается специфическим флористическим составом. Не говоря о видах, которые в своем распространении ограничиваются только, или почти только ею, даже те виды, которые встречаются в других консоциациях, располагаются здесь в весьма своеобразном количественном соотношении. Интересно, что существуют кустарники, травы и, вероятно, лианы, свойственные исключительно лесу Уаллаба.
Основные черты пяти первичных лесных сообществ района Морабалли-Крик суммируются в табл. 26. Сравнение их флористического состава заставляет сделать целый ряд выводов. Большое количество видов является общим для всех пяти сообществ (см. Davis, Richards, 1933—1934, табл. II—VI), но их относительная пропорция неодинакова в каждом из них. Некоторые виды, вероятно, ограничиваются в своем распространении только каким-либо одним из этих пяти сообществ, но так как списки составлялись только для одного участка каждого сообщества, то было бы ошибочным пытаться перечислить эти виды. Различия в видовом составе различных сообществ не одинаково велики; леса Морабуцеа, смешанный и Гринхарт гораздо более сходны по видовому составу, чем леса Мора и Уаллаба.
Все пять сообществ произрастают в одинаковых климатических условиях и, вероятно, мало изменены деятельностью человека. Очевидно, что состав каждого из них определяется свойствами почвы или же комбинацией почвы и рельефа. Границы почв и границы сообществ часто удивительно точно совпадают. Степень сходства или различия между почвами очень хорошо отражается на степени различия во флористическом составе. Почвы лесов Морабуцеа, смешанного и Гринхарт имеют много общего, но почвы лесов Мора и Уаллаба весьма отличаются как друг от друга, так и от всех остальных.
Примечателен также тот факт, что все пять типов леса образуют правильный ряд с закономерным изменением их признаков в последовательности: Мора, Морабуцеа, смешанный, Гринхарт, Уаллаба. Некоторые из этих признаков более или менее равномерно изменяются, то усиливаясь, то сглаживаясь по всему ряду; например, число деревьев на единицу площади равномерно возрастает от минимума в лесу Мора до максимума в лесу Уаллаба. Другие признаки достигают своего максимума или минимума к середине ряда и ослабляются или усиливаются по направлению к обоим его концам. Таким образом, процент дающих наибольшее
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
265
Таблица 26
Типы леса в районе Морабалли-Крик, Брит. Гвиана
Тип леса Мора Морабуцеа Смешанный Гринхарт Уаллаба
Характеристика местообитания: структура почвы Легкий Тяже- Легкий Песок Рыхлый
показатель структуры (Hardy) ил 40 лый ил 44 суглинок 18 5 песок 0
освещенность на высоте груди (%) 1,33 0,61 0,67 0,81 1,43
Флористический состав: Mora Mora Ocotea Е региа
доминанта или наиболее обиль- excelsa gonggrij- «ч к. <3 2 -с л rodiaei falcata
ный вид % деревьев диаметром 4 дюйма (10 см) и более 23,4 pii 26,6 о* Eschwei sagotiai 2 Penlaclel mac role 9,4 21,1
% деревьев диаметром 16 дюймов (41 см) и более 67,2 60,7 15,6 6,7 43,4 67,0
число видов деревьев диаметром 4 дюйма (10 см) и более (приблизительно) 60 71 91 95 74
число видов деревьев диаметром 16 дюймов (41 см) и более (приблизительно) И 21 32 33 16
Другие особенности: число деревьев диаметром 4 дюйма (10 см) и более на 1 га 310 309 432 519 617
число деревьев диаметром 16 дюймов (41 см) и более па 1 га 45 60 60 87 67
обилие видов деревьев достигает максимума на концах ряда; он неуклонно падает в направлении к середине и достигает минимума в ассоциации смешанного леса, где не существует какой-либо одной доминанты. В свою очередь количество видов на единицу площади оказывается наибольшим в середине ряда, постепенно снижаясь к его концам. Степень доминирования одиночного вида и общее количество видов связаны и изменяются обратно пропорционально (см. стр. 284).
В конечном счете обе эти характеристики (см. табл. 26) зависят, по-видимому, от свойств почвы. Есть основания предполагать, что почвы лесов Мора и Уаллаба сравнительно мало благоприятны для роста растений: первая в силу своей маломощности и слабой аэрации, вторая — в силу излишней пористости и, вероятно, ввиду того, что она, подобно другим подзолам, сильно выщелочена и страдает недостатком необходимых для растений питательных веществ. В то же время почва смешанного леса является, вероятно, наиболее благоприятной в ряду рассмотренных лесных сообществ. Об этом, в частности, свидетельствует тот факт, что
266
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
индейцы при выборе места для подсечно-огневого земледелия отдают предпочтение смешанному лесу. Попытки сельскохозяйственного использования почв леса Уаллаба потерпели полную неудачу (Milne, 1940). Почвы Морабуцеа и Гринхарт занимают, по-видимому, промежуточное положение. Таким образом, оказывается, что на лучших почвах развивается наиболее «смешанный» тип леса, а неблагоприятные свойства почвы обусловливают монодоминантность других типов. Эта закономерность, видимо, распространяется на все сообщества тропического дождевого леса. Получается, что на наиболее благоприятной почве успешно развиваются весьма разнообразные по степени выносливости виды, а на менее благоприятных почвах действие отбора создает преимущество видам относительно выносливым.
СООБЩЕСТВА ПЕРВИЧНОГО ЛЕСА В САРАВАКЕ
Сравнение двух типов первичного низинного тропического дождевого леса, изученных автором у Дьюлит и Маруди в бассейне Барама в Северном Сараваке, с пятью первичными типами леса Брит. Гвианы является очень поучительным. Климат этих мест имеет много общего < климатом Морабалли-Крик, но флористический состав здешней растительности, конечно, совершенно иной; нет ни одного вида, свойственного двум этим областям.
На склонах и в предгорьях Дьюлит — куэсты, достигающей приблизительно 1100—1400 м высоты и расположенной к юго-западу от реки Тинджар, притока Барама, — были изучены два типа леса, известные под названием смешанного и «борового». «Боровой» лес изучался также в окрестностях Маруди (Клодтаун) на реке Барам, приблизительно в 80 км от Дьюлит в низкорасположенной местности.
За исключением мест, где естественная растительность была уничтожена земледельческой культурой, почти все низменности острова Борнео покрыты дождевым лесом. В местности Дьюлит подсечно-огневой способ земледелия уничтожил большую часть первичного леса, расположенного на равнине, но на более крутых склонах холмов девственный лес сохранился до сих пор. Среди первичных лесов Борнео выделяются: 1) низинный смешанный дождевой лес, 2) «боровой» лес, 3) предгорный дождевой лес и 4) горный или «моховой» лес. Леса типов 3 и 4 встречаются лишь выше 450 м над уровнем моря. Их подробная характеристика дается в гл. 16. Смешанный и «боровой» леса располагаются на низменностях в сходных климатических условиях, дифференцируясь по признаку почв. Смешанный лес произрастает на вязких красных или желтых суглинках типа тропических красноземов или желтоземов, занимая преобладающее положение на низменностях, тогда как «боровой» лес встречается на грубых выщелоченных песках, похожих по своим свойствам на почву леса Уаллаба и также принадлежащих к типу низинного тропического подзола. В местах резкого разграничения двух этих типов почв наблюдается и внезапный переход от одного типа леса к другому. «Боровой» лес был встречен только в двух местах: на крутом склоне горы Дьюлит на высотах 750—1100 м и у Маруди, приблизительно на уровне моря.
Как вязкий желтый суглинок, так и выщелоченный песок образовались из миоценового песчаника, и не ясно, почему в одном случае развитие почвы шло в сторону латеритизации, а в другом — в сторону оподзоливания. Возможно, что это было вызвано незначительными различиями в механическом составе отдельных слоев песчаника.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
267
Смешанный и «боровой» леса заметно отличаются по своему внешнему облику и структуре, а также по флористическому составу, но благодаря разнообразию флоры и недостаточной ее обработке сравнение может быть проведено только в весьма общих чертах.
Сметанный лес. Этот преобладающий в районе горы Дьюлит тип леса представляет собой характерный для всей области Малайского архипелага первичный лес. Структура его была подробно описана в гл. 2, Наиболее яркая особенность этого леса состоит в исключительном изобилии видов деревьев и малой пропорции каждого из них в составе древостоя. В нем, следовательно, воплотился крайний пример смешанного леса или ассоциации тропического дождевого леса. На пробной площадке было найдено более 98 видов деревьев с диаметром ствола 8 дюймов (20 см) и более1 и по крайней мере 32 вида диаметром 16 дюймов (41 см) и более. Судя по этим данным, видовое разнообразие деревьев более крупных диаметров смешанного леса Борнео измеряется примерно теми же цифрами, что и в смешанном лесу Гвианы, но видовое разнообразие более мелких деревьев намного больше. Из 261 дерева диаметром около В дюймов не более 12 (4,5%) принадлежало к одному и тому же виду. Большинство деревьев пробного участка принадлежало к ярусу В и имело меньше 16 дюймов в диаметре. Из общего числа деревьев диаметром 16 дюймов и более, равного 62 (9,7%), два наиболее обильных особями вида (оба не были определены) были представлены каждый шестью индивидуумами, но они составляли соответственно только 3,4 и 2,7% общего древостоя с диаметром 8 дюймов и больше. Следовательно, смешанный лес горы Дьюлит еще дальше отстоит от монодоминантности, чем смешанный лес Гвианы. Примечательно, что, несмотря на очень незначительную пропорцию каждого вида в составе древостоя, по крайней мере 17°о деревьев диаметром 8 дюймов и более и 44 % деревьев диаметром 16 дюймов и более принадлежат к семейству Dipterocarpaceae. Этим обеспечивается «доминирование» диптерокарповых в верхнем ярусе, что позволяет назвать его, как и большинство низинных индо-малайских лесов, смешанным диптерокарповым лесом. По-видимому, «доминирование семейств» является довольно обыкновенной чертой как монодоминантных, так и смешанных тропических лесов.
«Боровой лес». Название «боровой лес» (Heidewald) присвоено Винклером (Winkler, 1914) характерному типу «субксерофильного первичного леса», свойственному песчаным почвам на юго-востоке острова Борнео. Климаксное сообщество Саравака, произрастающее на выщелоченных песках, соответствует винклеровскому описанию как по видовому составу, так и по многим физиономическим признакам. Термин «боровой лес» был предложен потому, что это сообщество, подобно европейским борам, приурочено к песчаной почве и многие из его компонентов имеют ксеро-морфный облик (маленькая ксерофильная листва и т. п.). Характер климата, по-видимому, исключает предположение о том, что ксероморфность «борового» леса, целиком являющегося вечнозеленым, имеет отношение к сезонной засухе. Скорее всего в ее основе лежат те же явления, которые вызывают и недостаточно выясненную «физиологическую сухость» боровых и болотных растений умеренной зоны, растущих на очень кислой почве, бедной минеральными солями.
1 Из-за трудностей исследования оказалось невозможным подсчитать деревья с диаметром ствола меньше 4 дюймов, как это было сделано для леса в местности Af орабалли-Крик.
268
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
«Боровой» лес районов Дьюлит и Маруди резко отличается от смешанного леса своим более густым, низкорослым подлеском (ярусы D—Е), лучшей освещенностью более низких ярусов, меньшим количеством деревьев с досковидными корнями и большим числом деревьев на единицу площади. Ярусность не изучалась, но весьма вероятно, что здесь существуют только два хорошо развитых яруса деревьев вместо трех, характерных для смешанного леса. Лиан мало, но и те, которые есть, отличаются небольшими размерами.
Травяной покров развит очень слабо, причем ему, так же как и ярусу кустарников, свойственны виды, которые являются редкими или отсутствуют в смешанном лесу.
Для двух пробных участков — в долине Койан (гора Дьюлит) и в Маруди были составлены списки видов. Эти участки оказались несколько различными по составу, что и можно было ожидать, учитывая разницу высоты и общих условий местообитания, но в основных чертах было выявлено большое сходство. Оба они менее богаты видами, чем смешанный лес; на участке Койан имелось около 55 видов деревьев диаметром 8 дюймов и более, на участке Маруди — 56 по сравнению с 98 на участке смешанного леса. Более важным является то обстоятельство, что оба участка обнаруживали тенденцию к монодоминантности, что выражалось в значительно большей пропорции какого-либо вида в составе древостоя, чем любого вида в смешанном лесу; правда, эта пропорция была меньше, чем пропорция доминанты гвианских консоциаций. На участке Койан хвойное Agathis borneensis составляло 15,2% деревьев диаметром 8 дюймов и более и 35,2% деревьев диаметром 16 дюймов и более. На участке Маруди деревьев Agathis было тоже много, но наиболее обильно представленным видом оказался не определенный вид семейства диптерокар-повых, к которому относилось 12% деревьев диаметром 8 дюймов и более и 35,9% деревьев диаметром 16 дюймов и более. Таким образом, хотя оба участка являлись примером скорее смешанных ассоциаций, чем консоциаций, все же тот или другой вид в них образует существенную часть древостоя. Как и в гвианских консоциациях тенденцией к «групповому расположению» обладало несколько видов. Так, например, второй наиболее обильно представленный вид на участке Койан составляет 12,4% деревьев диаметром 8 дюймов и более и 30,8% деревьев диаметром 16 дюймов и более (табл. 29). Аналогичным образом в лесу Маруди второй по обилию вид (из семейства диптерокарповых) составлял 11,4% деревьев диаметром 8 дюймов и более. На участке Маруди наблюдается заметное преобладание семейства диптерокарповых (40,5% деревьев диаметром 8 дюймов и более и 51,3% деревьев диаметром 16 дюймов и более). На участке Койан диптерокарповых значительно меньше, что, несомненно, объясняется большей высотой его над уровнем моря (Dipterocarpaceae представляют собой характерное семейство низменностей).
На обоих участках «борового» леса было определено достаточно видов, чтобы убедиться, что его флора в общем сильно отличается от флоры смешанного леса и, вероятно, содержит много видов, свойственных исключительно ей, как из числа лиан, так и из числа деревьев, кустарников и трав. Например, Nepenthes, характерные для «борового» леса, никогда не наблюдались в смешанном лесу горы Дьюлит. Agathis и несколько других видов, типичных для саравакского «борового» леса, в изобилии встречаются и в «боровых» лесах южной и юго-восточной частей острова Борнео (Winkler, 1914; Diels, Hackenberg, 1926), удаленных не меньше, чем на 700 км.
Таблица 27
Сравнение эдафической дифференциации типов леса Брит. Гвианы и Саравака (остров Борнео)
Брит. Гвиана Саравак-
Тип леса смешанный Уаллаба смешанный Койан «боровой» Маруди «боровой»
Почва Структура Легкий суглинок Показатель текстуры (Hardy) 18 Освещенность на высоте груди 0,67% Структура и физиономия (древесный ярус): Число деревьев диаметром 8 дюймов (20 см) и более на 1 га 232 Число деревьев диаметром 24 дюйма (61 см) и более на 1 га 6 Подлесок Редкий Травяной покров Мало видов, много индивидуумов Лианы Крупные, много- численные Деревья с досковидны- Рыхлый песок 0 1,43% 325 2 Плотный Очень мало видов, мало индивидуумов Небольшие, редко встречаются Суглинок 18-27 Умеренная 184 29 Редкий Много видов и индивидуумов Крупные, многочисленные 1!е< 5—13 сок 0
Хорошая
232 29 246 7
Плотный
Небольшие, редко встречаются
ми корнями Много деревьев Мало деревьев Много деревьев Мало деревьев
с Флористический состав: Число деревьев диаметром 8 дюймов (20 см) и более (приблизительно) 55 Число деревьев диаметром 16 дюймов (41 см) и более (приблизительно) 32 мощными д 49 16 осковидными 98 32 корнями 55 18 56 12
270
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Продолжение табл. 27
Тип леса Брит. Гвиана Саравак
смешанный Уаллаба смешанный Койа и «боровой» Маруди «боровой»
% наиболее обильных Pentaclethra Eperua «Меданг- Agathis «Те кам»
видов среди деревьев macroloba falcata лит» borne-
диаметром 8 дюймов (20 см) и более 13 32 5 ens i s 15 12
% наиболее обильных Eschweilera Eperua «Мераитп Agathis «Текам»
видов среди деревьев sagotiana falcata дагинг» и borne-
диаметром 18 дюймов (41 см) и более 16 67 «Ма раках бату» по отдельности 10 ensis 35 36
Сравнение «борового» и смешанного леса показывает, что как на острове Борнео, так и в Гвиане лес на выщелоченных песках (подзолах) более беден видами и отличается меньшей смешанностью по составу, чем соответствующий смешанный тип на тропическом красноземе. Как на Борнео, так и в Гвиане лес на сравнительно неблагоприятных почвах обнаруживает тенденцию к монодоминантности, но на Борнео, вероятно благодаря большему флористическому богатству области в целом, эта тенденция выражена гораздо слабее. На Борнео, как и в Гвиане, выщелоченные пески обладают избирательностью в отношении растений: на них вместе с видами, никогда не встречающимися в смешанном лесу, могут произрастать и более выносливые виды смешанного леса.
Весьма показательно, что, кроме тенденции к монодоминантности, «боровой» лес Борнео обнаруживает очень далеко идущее сходство с лесом Уаллаба Брит. Гвианы и в других отношениях. Некоторые из этих общих черт показаны в табл. 27. Эти два сообщества настолько похожи по внешнему виду, что «боровой» лес выглядит так, как будто каждое отдельное растение леса Уаллаба заменено здесь растением сходного облика лишь иной таксономической принадлежности. «Боровой» лес и лес Уаллаба не только весьма схожи друг с другом, они в одинаковой степени отличаются и от соответствующих типов смешанных лесов. «Боровой» лес и лес Уаллаба, расположенные в различных полушариях, не имеют общих видов и содержат лишь очень небольшое количество общих родов, так что их сходство целиком обязано сходству климатических и почвенных условий. Случаи сходства между растительностью отдаленных областей с различной флорой, но со сходным климатом хорошо известны и прекрасно иллюстрируются единообразием структуры и внешнего вида средиземноморской склерофильной растительности в различных частях света или сходством ассоциаций смешанного тропического дождевого леса тропической Америки, Африки и индо-малайской области. На примере «борового» леса и леса Уаллаба мы встречаемся с явлением, имеющим не меньшее значение, но менее известным и состоящим в параллельной эдафической дифференциации растительных сообществ при сходных климатических условиях.
Глава 11
СОСТАВ ПЕРВИЧНОГО ДОЖДЕВОГО ЛЕСА (II)
Как уже упоминалось, большинство тропических дождевых лесов имеет смешанный состав, хотя вопреки предположениям более ранних исследователей они не всегда состоят из полидоминантных ассоциаций.
Несмотря на то что первичные монодоминантные сообщества встречаются, как мы увидим, во всех основных географических областях, где произрастает тропический дождевой лес, Брит. Гвиана, видимо, представляет собой нечто исключительное как в отношении числа монодоминантных сообществ, так и по большим размерам площади, которую они занимают. Но даже у Морабалли-Крик смешанный лес является наиболее широко распространенным типом и занимает, вероятно, большую часть территории по сравнению с любым другим сообществом. В некоторых тропических областях, например во всей Западной Африке, первичные монодоминантные сообщества, по-видпмому, отсутствуют и весь лесной массив на хорошо дренированных почвах образует одно очень обширное сообщество смешанного леса, изменяющееся по своему составу в зависимости от места, но почти не поддающееся разделению на отчетливые ассоциации. Следовательно, смешанные леса Брит. Гвианы и Борнео представляют собой типичный тропический дождевой лес в том смысле, что они вряд ли обнаруживают какую-либо тенденцию к монодоминантности.
ФЛОРИСТИЧЕСКИЙ СОСТАВ СМЕШАННОГО ДОЖДЕВОГО ЛЕСА
Видовой состав малых пробных участков
Флористический состав двух пробных участков типичного смешанного леса был довольно подробно описан в предыдущей главе. В табл. 28 данные, относящиеся к этим участкам, сравниваются с данными, полученными на некоторых других пробных участках смешанных сообществ; табл. 25 (гл. 10) дает количественную характеристику видов на пробных участках смешанного леса различных местностей. Из этих таблиц можно извлечь несколько интересных показателей, но значение их в настоящее время не легко оценить. Табл. 28 показывает, что даже в далеко отстоящих областях смешанный дождевой лес обнаруживает значительное однообразие в общих чертах своего состава. Факты заставляют также предполагать, что смешанный лес различных географических областей обнаруживает мало местных особенностей в отношении своего строения, хотя, пользуясь данными, собранными с небольшого числа пробных участков, было бы неразумно делать уж очень определенные выводы.
Число видов на единицу площади колеблется в значительных пределах, особенно среди деревьев небольшого диаметра. В каждом отдель
"272 ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ном округе число это довольно постоянно, а различия между разными округами, вероятно, зависят главным образом от флористического богатства области в целом. Последнее обстоятельство, как мы говорили раньше, по-видимому, объясняется скорее историческими факторами, чем условиями окружающей среды, существующей в данное время. Только для западноафриканского леса собрано значительное число цифр, характеризующих количество видов на единицу площади; помимо данных, приведенных в таблице, известны цифры, опубликованные Йенчем (Jentsch, 1911), Милдбрэдом (Mildbraed, 1930а, 1930b, 1933b) и Ричардсом (Richards, 1939). Нельзя утверждать, что сведения, содержащиеся в таблице, охватывают все разнообразие флористического состава. Участки, исследованные Иенчем в Камеруне (полученные им данные нельзя непосредственно сравнивать с данными автора), возможно, даже богаче по видовому составу смешанного леса горы Дьюлит. С другой стороны, не исключено, что некоторые небольшие территории тропического дождевого леса могут содержать меньше видов на единицу площади, чем сравнительно бедный флористически остров Маврикий (бедность последнего, вероятно, обусловлена его изолированным положением).
Число видов, конечно, частично зависит от числа индивидуумов на единицу площади. Это соотношение может изменяться в широких пределах и, судя по сообществам Морабалли-Крик, находится в зависимости от почвы и других условий среды. В смешанном вечнозеленом сезонном лесу Тринидада число деревьев на 1 акр в более влажных местах почти вдвое больше, чем в более сухих (Beard, 1946b, стр. 63).
Отношение числа индивидуумов к числу видов Милдбрэд (Mildbraed, 1930а) назвал коэффициентом смешения (Л/ischungsquotient), считая этот показатель весьма важным для характеристики различных типов сообществ тропического дождевого леса. Однако он упустил из виду тот очевидный факт, что в то время, как число индивидуумов изменяется в зависимости от размеров пробного участка, число видов остается неизменным (см. кривые числа видов на единицу площади, рис. 34). Поэтому коэффициентом смешения можно пользоваться только при сравнении участков одинакового размера. В примерах, фигурирующих в табл. 28, если исключить участок леса на острове Маврикий с его ненормально большим количеством индивидуумов на единицу площади при небольшом числе видов, величины вариаций не очень значительны. На всех пробных участках среднее число индивидуумов, приходящееся на тот или иной вид, больше для деревьев меньших диаметров, чем для деревьев крупного диаметра.
Процент наиболее обильных по числу особей видов обнаруживает ряд черт, вероятно, имеющих определенное значение. На всех участках наиболее обильно представленные виды ступени толщины в 4 дюйма и более отличаются от наиболее обильных видов для ступени в 16 дюймов и более. Таким образом, виды, близкие к доминированию среди более крупных деревьев, не составляют такую же часть среди более Мелких деревьев. Каждый ярус, вероятно, имеет свой, преобладающий по числу особей вид. Среди деревьев диаметром 16 дюймов и более ни один вид не превышает по количеству особей (приблизительно) 1/6 древостоя. Исключение в этом отношении составляет лес на острове Маврикий, где один из видов образует около х/4 всех деревьев. На трех участках из показанных в таблице, наиболее обильно представленные виды ступеней толщины «4 дюйма и более» и «8 дюймов и более» составляют меньшую часть в этих ступенях, чем наиболее обильные виды ступени «16 дюймов и более»;
Состав смешанного тропического дождевого леса
Таблица 28
Азия Южная Америк; 1 Африка
Гора Дьюлит, Саравак 1 Морабалли-Крик, Брит. Гвиана 2 Резервированный лес Окому, Нигерия 3 Резервированны! лес Масса-Ме, Берег Слоновой Кости 4 । Кроун-Лэнд Макабе, остров Маврикий (550 над уровнем моря)5
Размер участка, га (приблизительно) Число деревьев на 1 га: 1,5 1,5 1,5 Около 1,4 530 1,0 1710
диаметром 4 дюйма (10 см) и более — 432 390
диаметром 8 дюймов (20 см) и более 184 232 223 214 331
диаметром 16 дюймов (41 см) и более Число видов на 1 га: 44 60 47 39е 60 52
диаметром 4 дюйма (10 см) и более — 91 70 74
диаметром 8 дюймов (20 см) и более 98 55 51 58 33
диаметром 16 дюймов (41 см) и более Отношение виды/индивидуумы: 32 32 7,1 31 6,4 Около 23 10,0 5,2 2.4 Pachylobus deliciosus и 32,9 10,0 5,5 Eugenia glornerata
виды диаметром 4 дюйма (10 см) и более —
виды диаметром 8 дюймов (20 см) и более 2,7 6,0 5,6
виды диаметром 16 дюймов (41 см) и более Процент наиболее обильных видов: 1,9 2,8 Pentaclelhra macroloba 2,3 Slrombosia relivenia
диаметром 4 дюйма (10 см) и более «Меданг лит» И Pentaclethra 30 Strombosia 15 pachylobus 18 Eugenia
macroloba retivenia deli с iosus glornerata
диаметром 8 дюймов (20 см) и более 5 «Ма раках бату» То же 13 Eschweilera sagotiana 35 Pausinystalia sp. (или spp.) 17 Pi p/a denia africana 19 M im usops maxima
диаметром 16 дюймов (41 см) и более 1 10 1 10 16 14 (или меньше) 13 25
1 Richards, 1936.
* Davis, R i с h a rd s, 1933—1934.
8 Richards, 1939.
4 Aubrdville, 1938.
b Vaughan, Wiehe, 1941.
• Для деревьев диаметром 40 см и более.
Количество видов, представленных 1, 2, 3. . . индивидуумами в смешанных и монодоминаптных дождевых^™"4“
- - — - Число индивидууме в более 100
1 1—5 1 —10 1 1-20 21-30 31-40 41-50 51-60 61—70 71—80 81-90 91-100
Смешанные леса 1. Морабалли-Крик, Брит. Гвиана 2. Резервированный лес Окому, Нигерия 21 23 68 48 79 58 4 7 4 2 1 1 2 — 1 1 1 — — —
3. Гора Дьюлит, Борнео1 41 85 97 1 — 1
Монодоминантные леса (или ассоциации с тенденцией к монодоминантности) 4. Лес Мора (Mora excelsa), Морабалли-Крик, Брит. Гвиана 22 41 47 9 1 2 1
5. Лес Морабуцеа (Mora gonggrijpii), Морабалли-Крик, Брит. Гвиана 25 49 61 3 3 1 1
6. Лес Гринхарт (Ocotea rodiaei), Морабалли-Крик, Брит. Гвиана 31 63 78 6 — 3 2 2 2
7. Лес Уаллаба (Eperua jalcata), Морабалли-Крик, Брит. Гвиана 14 45 55 И 4 — 1 — — 3
8. Заболоченный лес на пресноводном болоте, резервированный лес Шаша, Нигерия И 19 24 6 2 3 1 1
9. «Боровой» лес, долина Койан, Борнео 1 18 39 47 7 — — 2 — — — — — —
10. «Боровой» лес, Маруди, Борнео1 13 42 49 3 1 2 1 — — — — — —
11. Лес из железного дерева (Cynometra ale-xandri), Буданго, Уганда 3 3 5 2 — — 1 1 — — — 2
I Для деревьев диаметром 8 дюймов (20 см) и более (другие данные для деревьев диаметром 4 дюйма (10 см) и болей. Данные дли пунктов 1 — 10 — из Дэвиса и Ричардса (Davis, Richards, 1933—1 934), Ричардса (Richards, 1936, 1939); для 11—из 3j гелии га (Fggcling, 1947).
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
275
другими словами, среди нижних ярусов леса замечается меньшая тенденция к монодоминантности, чем среди верхних. На двух западноафриканских участках, где было обнаружено обратное соотношение, верхние ярусы могли быть искусственно нарушены вырубкой.
«Смешанный» характер лесов может быть выражен еще и другим путем, а именно определением числа видов на каждом участке, представленных 1, 2, 3... индивидуумами. Результаты такого подсчета для нескольких участков смешанного леса (и для сравнения нескольких монодоминантных участков) приведены в табл. 29. Надо заметить, чт<г в тропических дождевых лесах всех типов огромное большинство впдов-представлено очень малым числом индивидуумов. Как в смешанном, так и в монодоминантном лесах по крайней мере половина видов на участке размером 400 х400 футов представлена лишь пятью или даже меньшим числом экземпляров (что составляет менее 1—2% древостоя в зависимости от числа индивидуумов на единицу площади) и значительно больше половины — десятью индивидуумами и меньше. В смешанном лесу горы Дьюлит 85 видов из общего количества, равного 98, диаметром 8 дюймов и более, представлены пятью индивидуумами или меньшим их числом. В смешанном лесу редко встречаются виды, насчитывающие более чем 100 индивидуумов (участок смешанного леса в резервированном лесу ©кому в Нигерии составляет в этом отношении исключение), но в монодоминантных лесах такого обилия могут достичь несколько видов (три в гвианском лесу).
Изменения флористического состава смешанного леса на больших площадях
До сих пор предметом нашего обсуждения были лишь небольшие однородные пробные площадки. Однако не менее важно познакомиться с флористическим составом леса на обширных территориях. Надо установить, подвергается ли флористический состав смешанного леса закономерным изменениям на обширных территориях или в пределах целой географической области или же он испытывает лишь незначительные случайные колебания от места к месту. Если закономерные изменения существуют, то можно ли в пределах какой-либо области выделить смешанные ассоциации, зависящие от различий в почвенном покрове или климатических условий? Связаны ли изменения в составе ярусов деревьев с изменениями состава подлеска и травяного покрова? Ответы на эти вопросы различаются в зависимости от взглядов авторов на природу ассоциации.
В сущности время для определенного ответа на эти вопросы еще не наступило, но некоторые данные все же заслуживают рассмотрения.
Мейер Дрийс (Meijer Drees, 1938) произвел статистическое сравнение состава небольших пробных участков «семенного леса» (смешанный дождевой лес в Индонезии) и пришел к выводу, что он состоит только из одной ассоциации. В низинном дождевом лесу Тринидада (вечнозеленый сезонный лес Бирда) Маршал (Marshall, 1934) установил три «ассоциации»: ассоциацию Сагара guianensis — Eschweilera subglandulosa, ассоциацию Mora excelsa и ассоциацию Сагара guianensis — Licania biglandulosa. В первой из них он выделяет четыре различающиеся по составу подгруппы, каждая из которых отражает незначительные изменения в почвах и рельефе. Маршаловская «ассоциация» Mora представляет собой монодоминантное сообщество и фигурирует здесь как консоциация. Бирд 18*
276
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
(Beard, 1946b), рассматривающий весь вечнозеленый сезонный лес как одну ассоциацию Сагара — Eschweilera с четырьмя «фациациями», составил новую классификацию лесных сообществ Тринидада, в которой все «ассоциации» и «подгруппы» Маршала, включая монодоминантное сообщество Mora, относятся к фациациям.
«Сомкнутый лес» Ганы, согласно Чипу (Chipp, 1927), образует несколько сообществ: ассоциацию Cynometra — Lophira и «пре-климаксы»— Lophira—Entandrophragma, Entandrophragma — Khaya и Tri-plochiton — Piptadenia, Последний вполне соответствует типу леса, который обычно называется смешанным листопадным лесом (см. гл. 15), и применение к нему термина «пре-климакс» может вызвать критические замечания. Другие сообщества соответствуют смешанному тропическому дождевому лесу Нигерии и Берега Слоновой Кости, но надо заметить, что виды, давшие названия этим сообществам, не являются единственными пли даже главными доминантами; это просто более или менее произвольно выбранные, обильно встречающиеся виды яруса А.
Предложенная Чипом схема была критически разобрана Обреви-лем (Aubreville, 1938), считающим, что в смешанном дождевом лесу Западной Африки нельзя выделить истинных ассоциаций. Перед тем как перейти к рассмотрению взглядов Обревиля, полезно познакомиться с его собственным описанием «сомкнутого леса», (foret dense) Берега Слоновой Кости, сливающегося с лесами Ганы. Это — наиболее полная из имеющихся характеристик различий в составе смешанных лесов, раскинутых на огромном пространстве.
Обревиль показывает, во-первых, что отдельные виды деревьев, составляющих дождевой лес, имеют различные типы распространения. Многие, например Lophira procera, распределяются более или менее равномерно по всей площади леса; распространение других видов носит более локальный характер. Некоторые компактно располагаются на какой-либо территории, иногда сравнительно большой, как, например, у Chid-lovia sanguinea, или же исчисляемой лишь несколькими квадратными километрами, как в случае с Stemonocoleus micranthus. Причины такого загадочного различия в распределении видов, внезапно возникающего среди, казалось бы, однообразного и однородного лесного пространства, неясны и, видимо, носят исторический характер [теория, объясняющая приблизительно такое же явление в распределении гвианского тропического дождевого леса, была выдвинута Дэвисом (Davis, 1941)].
Ввиду таких различий в распространенности видов флористический состав леса неизбежно должен изменяться от места к месту. Хотя в целом в пределах исследованной части леса Берега Слоновой Кости насчитывалось приблизительно 596 видов крупных деревьев, их число в каждом данном месте значительно меньше. На 1 га обычно размещается около 40 видов (размеры диаметров не устанавливались), а на несколько большей однородной территории их было около 701. Каждая местность имеет характерный набор обильно представленных видов, число которых, однако, не превосходит 20. Часто популяция содержит 1—4 «доминирующих» вида (термин «доминанта» употребляется Обревилем в ином смысле, чем принято в этой книге) и 1—6 обильно представленных видов; огромное число других видов представлено одним или очень немногими
1 Число видов на исследованном автором пробном участке (площадью 1,5 га) смешанного леса, резервированный лес Окому в Нигерии, было равно 70 (диаметром 4 дюйма и более).
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
277
индивидуумами. «Доминирующие» и «обильные» виды меняются от места к месту; вид, «обильный» в одной местности, может почти отсутствовать в соседней. На обширной территории встречаются всевозможные комбинации видов, но «доминанты» всегда выделяются из группы, включающей, вероятно, не более 20 видов. Таким образом, смешанный дождевой лес состоит из большого числа видов со сходными экологическими требованиями, причем эти виды встречаются в разнообразных сочетаниях, многократно меняющихся на территории лесного массива. Эти сочетания варьируют не только в пространстве, но, согласно Обревилю, и во времени; поэтому любой участок лесного сообщества непрерывно изменяется по своему составу. Теория регенерации леса Обревиля, которую мы назвали мозаичной теорией, была уже рассмотрена в гл. 3.
Если изложенное верно в отношении всей обширной территории смешанного леса, то в нем нельзя выделить истинных ассоциаций. В таком случае «ассоциации» и «пре-климаксы» Чипа представляют собой лишь незначительное число произвольно выбранных комбинаций видов, непостоянных как во времени, так и в пространстве, и смешанный лес в целом должен рассматриваться как одна ассоциация колеблющегося состава.
Обревиль допускает возможность существования истинных ассоциаций в местах развития специфического почвенного покрова. В них популяция видов оказывается более гомогенной, чем в смешанном лесу, и иногда можно встретить почти чистые древостои, состоящие из одного вида. Примером таких «эдафических ассоциаций» являются приречные заросли в пределах Берега Слоновой Кости, включающие Cynometra megalophylla. Hymenostegia emarginata и т. д. Составляющая их комбинация видов должна, вероятно, рассматриваться скорее как фрагмент прибрежного сообщества, аналогичного прибрежной части леса Мора в Гвиане и леса пресноводного болота Западной Африки, описанного в гл. 13, чем как ассоциация смешанного леса1.
Описание флористического состава леса Берега Слоновой Кости прекрасно согласуется с характеристикой леса Ликомба в Камеруне (Mildbraed, 1930 а, 1930 b), занимающего гораздо меньшую площадь. Хотя Милдбрэд нашел в отдельных местах заметное преобладание некоторых видов, он все же пришел к выводу, что этот лес представляет собой скорее смесь различных видов в разной пропорции, чем ряд истинных ассоциаций.
Чтобы сообщество смешанного леса могло быть признано отдельной ассоциацией, оно должно в пределах обширной территории сохранять постоянный состав, в котором, в частности, не происходила бы смена наиболее обильно представленных видов. Нужно также, чтобы характерный флористический состав оставался постоянным во всех ярусах, включая кустарники и травяной покров. Ясно, что действительность не соответствует этой схеме, но следует признать также, что пока еще в смешанном тропическом дождевом лесу не установлено и наличие нескольких ассоциаций в пределах одной географической области.
1 Группы характерных видов, окаймляющих берега рек п более мелких водотоков, встречаются, конечно, во всех типах тропического дождевого леса, подобно тому как Alnus, Salix и т. д. свойственны рекам умеренной зоны. Примером могут служить сараковые водотоки, окаймленные Saraca spp., и «реки Мерам», чьи берега заросли Dipterocarpus oblongifolius, в смешанном лесу Малаккского полуострова (Corner, 1940, стр. 42). Некоторые из этих видов, вероятно, ограничиваются собственно урезом реки и не встречаются в более широкой полосе прибрежного леса.
278
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Доминирование семейств в смешанном лесу
Явление «доминирования семейств» в некоторых типах смешанных лесов, проявляющееся в численном преобладании видов одного и того же семейства или группы родственных родов, уже упоминалось. Наиболее ярким примером его служит доминирование семейства Dipterocarpaceae в низинных тропических дождевых лесах индо-малайской области,к западу от линии Уоллеса. Доминирование этого семейства резко выражено в некоторых тропических дождевых лесах (главным образом смешанных по составу) от Цейлона до Индокитая и Борнео; для любого участка этих лесов характерно присутствие целого ряда видов, относящихся к одному пли нескольким родам семейства диптерокарповых, среди наиболее обильно представленных видов.
Можно говорить и о доминировании семейства Leguminosae в некоторых южноамериканских лесах или Meliaceae в некоторых лесах Западной Африки. Причина этого, видимо, широко распространенного явления еще не установлена. Обыкновенно полагают, что конкуренция наиболее сильна между близко родственными видами растений или животных. В этом случае не понятно, почему несколько видов одного и того же рода или семейства встречается в качестве кодоминант в одном и том же местообитании. Доминирование семейств, вероятно, заслуживает дальнейшего исследования, особенно с точки зрения характера конкуренции в сообществах смешанного леса. Вопрос этот до сих пор остается еще очень неясным.
СОСТАВ МОНОДОМИНАНТНОГО ДОЖДЕВОГО ЛЕСА
Среди сообществ первичного леса Брит. Гвианы и острова Борнео, описанных в предыдущей главе, четыре типа, а именно Мора, Морабуцеа, Гринхарт и Уаллаба из Морабалли-Крик, характеризовались наличием определенного вида деревьев, который образует существенную часть всего древостоя и в силу этого может рассматриваться как единственный доминирующий вид; в другом типе — «боровом» лесу острова Борнео, несмотря на то, что ни один вид не может быть назван действительно доминирующим, существует «тенденция» в сторону монодоминантности.
По-видимому, эти пять типов леса, представляют собой климаксные сообщества, находящиеся в устойчивом равновесии со средой обитания. Нет никаких указаний на то, что какое-либо из них представляет собой стадию первичной или вторичной сукцессии. Сообщество Mora excelsa, развивающееся на затопляемой равнине (не смешивать с сообществом, произрастающим на каменистых склонах холмов), представляет собой тропический эквивалент европейского пойменного леса или «спелого болотного редколесья» и является конечной стадией гидросерии (см. гл. 13). Американские экологи рассматривали бы такое сообщество как «субклимакс», но так как оно не обнаруживает тенденции развиться в ассоциацию смешанного леса (или во что-либо другое), то, пожалуй, лучше рассматривать его как «физико-географический» или «эдафиче-ский» климакс (см. стр. 315). Аналогичные сообщества встречаются и на затопляемых равнинах в других частях тропической Америки, но являются ли они монодоминантными или обнаруживают тенденцию к монодоминантности — не известно.
Помимо четырех известных монодоминантных сообществ района Морабалли-Крик, имеются сведения о целом ряде типов тропического
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
279
дождевого леса с одной или в небольшом числе случаев с двумя доминантами как в Брит. Гвиане, так и других частях тропиков, но об их состоянии, требованиях к местообитанию, площади распространения и флористическом составе большинства из них почти ничего не известно. Некоторые из них, вероятно, представляют собой стадии вторичных сукцессий. В сукцессиях, следующих за вырубкой или выжиганием дождевого леса (см. гл. 17), временное доминирование одного вида представляет собой обычное явление. На ранней стадии таких сукцессий какой-либо вид иногда образует почти чистый древостой, примером чего служат сообщества Musanga cecropioides, Trema guineense и т. д. в тропической Африке. Некоторые типы монодоминантного леса, производящие поначалу впечатление первичных, безусловно, относятся к вторичным. Так, например, весьма интересный дождевой лес из Eucalyptus deglupta (Е. naudi-niana) острова Новая Британия, видимо, встречается главным образом на выжженных пространствах, а почти чистые, часто равновозрастные древостои Octomeles sumatranus, характерные для некоторых частей Новой Гвинеи, встречаются на недавно отложенном аллювии (Lane-Poole, 1925а, Ь). Однако для части монодоминантных типов отсутствуют достаточно убедительные данные, позволяющие предполагать их вторичную природу. Теперь те скудные сведения по этому предмету, которые имеются в нашем распоряжении, могут быть подытожены.
Среди еще не упоминавшихся монодоминантных типов Брит. Гвианы лучше всего известен тип, образованный Dicymbe corymbosa (групповая Уаллаба),— деревом из семейства бобовых. Этот вид отличается замечательным свойством активно размножаться вегетативным путем, так что большая часть взрослых деревьев представляет собой группу, объединяющую до 12 крупных стволов, отходящих от общего основания (см. гл. 4). Dicymbe доминирует в Брит. Гвиане на очень больших пространствах; она не встречается ни вблизи морского побережья, ни в местности Морабалли-Крик, но чрезвычайно характерна для района Бартика-Потаро. Майерс (Myers, 1936) говорит о нахождении этого вида далеко внутри страны близ гор Серра-Пакараима. Он также встречается и, по-видимому, местами доминирует в долине реки Ваупес, бассейн Амазонки. Степень его доминирования очень высока. Тютин (Т. G. Tutin) установил, что в пределах Брит. Гвианы Dicymbe составляла на двух пробных участках размером 40 х20 м каждый, 57 и 52% всех стволов диаметром 2 дюйма (5 см) и более1. На обоих участках субдоминантной была Eperua jalcata (Уаллаба), причем ассоциированные с ней виды были аналогичны характерным для консоциации Уаллаба (без Dicymbe). Экологические требования сообщества Dicymbe неясны; в некоторых местах оно встречается на выщелоченных песках бок о бок с консоциацией Eperua (которая является агрессивной); в других местах оно произрастает на тропических красноземах, обычном субстрате консоциации Mora gonggrijpii} иногда Dicymbe и М. gonggrijpii образуют смешанное сообщество.
Близ прибрежных саванн Брит. Гвианы, от реки Корантейн на востоке и до Эссекибо на западе (стр. 347), большие площади заняты консоциациями Dimorphandra conjugata семейство Leguminosae. Этот же тип леса вновь появляется на реке Померун и внутри страны, в области Эссекибо-Кабури, но, очевидно, отсутствует на реке Рупунуни или в других еще более удаленных местах. Доминанта представляет собой дерево
1 37 и 29% соответственно, если рассматривать каждую «группу» как одно дерево.
280
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
высотой около 38 м, обильно размножающееся, главным образом вегетативным путем, а именно корневыми отпрысками и побегами, возникающими из спящих почек на упавших стволах. Она образует почти чистые древостои или может смешиваться с родственной Dimorphandra hohen-kerkii\ другими деревьями, ассоциирующимися с доминантой, являются бания (Swartzia sp.?), какаруа (Sapotaceae) и Catostemma fragrans — одна из субдоминант леса Уаллаба. Иногда Dimorphandra conjugata образует только скудный кустарник высотой 4—5 м. Консоциация в своем распространении ограничивается выщелоченными песками и примечательна наличием толстого слоя грубого перегноя или торфа, обычно образующего своего рода оправу вокруг оснований стволов. В этой консоциации, как и в консоциации Уаллаба, часто встречаются группы характерного кустарника, [так называемый «мури-буш», «muri bush» (Т. A. W. Davis, стр. 265 и 324)].
В низовьях реки Корантейн и в районе Махаикони в нижних частях склонов холмов местами встречаются леса с преобладанием Aspidosperma excelsum (Apocynaceae) (Т. A. W. Davis). Близ саванн Гвианского нагорья Майерс (Myers, 1936) наряду со смешанным лесом и консоциациями Mora excelsa и Dicymbe corymbosa обнаружил типы леса с преобладанием порознь в каждом из них Peltogyne sp. и дерево, известное под местным названием ашероа. Первая порода образует почти чистые древостои на песчаных затопляемых равнинах и представляет собой, вероятно, вторичную сукцессию; вторая же, покрывающая обширные плоские пространства на высоте 1300—3000 футов (400—900 м) и избегающая склонов, возможно, образует климаксный тип леса. Надо заметить, что все доминанты чистых консоциаций в Брит. Гвиане, за исключением Ocotea (Гринхарт), Aspidosperma excelsum и, возможно, ашероа, принадлежат к семейству Leguminosae.
По-видимому, какие-либо определенные сведения относительно монодоминантных лесов в материковой части тропической Америки, за исключением Гвианы, отсутствуют, но на острове Тобаго известен свойственный исключительно ему, вероятно, климаксный тип тропического дождевого леса, развивающийся на маломощных вулканических почвах открытых мест, расположенных выше 800 футов (240 м) над уровнем моря, названный Бирдом (Beard, 1944b) «ксерофитным дождевым лесом». В нем почти половину древостоя среди деревьев в 4 фута (120 см) и более в обхват составляет Manilkara bidentata. Ассоциация Dacryodes—Sloanea Малых Антильских островов также имеет тенденцию образовывать монодоминант-ное сообщество у границы ее распространения (см. стр. 285).
В Западной Африке и Республике Конго крупные пространства занимает лес, который можно сравнить с лесом Мора, произрастающим на затопляемой равнине в местности Морабалли-Крик; этот лес тоже подвергается затоплению и также, вероятно, представляет собой конечную (климаксную) стадию гидросерии. Флористический состав этих лесов, о которых будет идти речь в гл. 13, весьма отличается от флористического состава смешанных лесов на хорошо дренированных почвах. Интересно, что в некоторых из этих лесов замечается тенденция к монодоминантности или даже к образованию почти чистых древостоев из одного вида. В резервированном лесу Шаша, Нигерия, на пробном участке леса, подверженного затоплению, общее число видов по сравнению с участками смешанного леса того же района (за исключением одного, который, вероятно, не представлял собой климаксного сообщества) было низким: 38 деревьев диаметром 4 дюйма и более; И деревьев 16 дюймов и более.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
281
На хорошо дренированных землях африканского дождевого леса монодоминантные консоциации обнаруживают интересную локализацию в распространении. В поясе вечнозеленого леса, окаймляющего Гвинейский залив, практически не наблюдалось ни одного монодоминантного сообщества климаксного характера, но в некоторых районах огромного экваториального лесного массива Центральной Африки существуют обширные площади, по-видимому, монодоминантного климаксного леса. Лебрэн (Lebrun, 1936а) делит экваториальный лес бассейна Конго на три «подобласти»: 1) центральный бассейн; 2) северо-восточный бассейн и 3) восточный бассейн (переходный к горным лесам Восточной Африки). В подобласти 1, флористически самой богатой, лес носит смешанный характер и монодоминантность практически отсутствует, но в подобласти 2 по крайней мере 2/3 площади занято лесами, в которых доминирует Macrolobium dewevrei или Cynometra alexandri.
Консоциации Macrolobium dewevrei были впервые описаны Милд-брэдом (Mildbraed, 1914, 1922), сообщившим об их наличии к западу от Рувензори, в лесу Итури и в ряде местностей южного Камеруна. Дальнейшие сведения о них дали Вермозен (Vermoesen, 1931) и Лебрэн (Lebrun, 1936а). М. dewevrei часто составляет подавляющую часть древостоя; на одном пробном гектаре в районе Уеле-Итимбири Лебрэн нашел, что этот вид включает 94% деревьев диаметром более 20 см. М. dewevrei представляет собой высокое дерево, ветвящееся почти от основания, что придает лесу своеобразный вид. Травяной покров хорошо развит; в изобилии встречаются всходы М. dewevrei. Консоциация произрастает на мощных песчаных почвах (Vermoesen, Lebrun) или, согласно Милдбрэду, на желтой или желтовато-серой суглинистой глине и, несомненно, теснейшим образом связана с эдафическими условиями. Согласно карте Лебрэна, она покрывает площадь во много сотен квадратных километров, протянувшись в западном направлении широкой полосой вдоль восточной и северной окраин леса Конго. Иногда доминанта так же, как и в чистой консоциации, небольшими группами или разбросанно, встречается в смешанном лесу. Как составная часть смешанного леса консоциация Macrolobium dewevrei тянется на запад до Нигерии, где ее местонахождение, однако, ограничивается берегами рек и заболоченными пространствами.
Подобные же консоциации, но несколько меньшего размера и менее чистые, образует Cynometra alexandri. Они встречаются на крайнем северо-востоке лесного массива Конго, к северу и северо-западу от Рувензори, на высоте 900—1000 м над уровнем моря (Lebrun, 1936а), а также распространяются в западпую Уганду. Эггелинг (Eggeling, 1947) дал подробное описание состава и строения консоциации Cynometra (железное дерево) в лесу Будонго (Уганда). Здесь доминанта образует 20—35% всех деревьев диаметром 4 дюйма (20 см) и более и 75—90% деревьев диаметром 16 дюймов и более. На исследованном Лебрэном участке в пределах Республики Конго эта консоциация составляла около 50% деревьев диаметром более 20 см. Число других видов деревьев в лесу из Cynometra в Будонго невелико; на одном пробном участке (400 х400 футов) Эггелинг нашел только 11 видов, а на другом — 25 по сравнению с 58 и 53 видами, встреченными на двух таких же участках в смешанном лесу. Сообщество имеет три яруса (стр. 49), но ярус А состоит исключительно из наиболее крупных деревьев Cynometra, выступающих из яруса В. Из других видов в пределах яруса В наиболее обильно представленными являются Strychnos sp. и три вида Celtis. В ярусе С обыкновенно доминирует Las iodise us mildbraedii, но иногда он замещается Lepidoturus laxiflorus. Этот тип
282
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
леса отличается специфическим внешним видом. Лианы в нем встречаются редко, разреженность яруса С и яруса кустарников позволяет просматривать лес на далекое расстояние; травяной покров развит плохо, причем во время сухого сезона напочвенные травы сохнут и поникают, как в лесах гораздо более сухого типа. Почва принадлежит к типу тропических красноземов, причем более глубокая ее часть вблизи верхней границы горизонта В2 (30 см) суглинистая и имеет пылеватую структуру.
Cynometra alexandri встречается также в лесу Будонго в качестве составной части смешанного леса; интересно отметить, что она достигает тут больших размеров, чем в монодоминантной консоциации1. Важное значение имеет доказанный Эггелингом факт, что в Будонго, где лес в целом распространяется за счет злаковников, смешанный лес представляет собой сериальное сообщество, а консоциация Cynometra — климакс. Последняя медленно внедряется и замещает первую.
Групповое расположение, подобное групповому расположению Macrolobium dewevrei и Cynometra alexandri. свойственно также видам африканского дождевого леса—Berlinia ledermannii, В. polyphylla, Bra-chystegia sp. и Tessmannia parvifolia. Из замечаний Милдбрэда (Mild-braed, 1914, 1922) можно сделать вывод, что ни один из этих видов не образует больших сообществ, которые можно было бы сравнить с двумя только что описанными. В Уганде сообщество дождевого леса встречается на высоте 4500—5000 футов (1400—1500 м над уровнем моря). В этом сообществе «около 80% полога» (Eggeling) составляет Parinari excelsa (Rosaceae). Пока не ясно, есть ли основания относить это сообщество к тропическому дождевому лесу; возможно, что это местный вариант переходного леса в смысле Лебрэна (стр. 381).
Интересен и. возможно, показателен тот факт, что все виды, играющие роль монодоминант на хорошо дренированных местообитаниях в африканском тропическом дождевом лесу, принадлежат (за исключением Parinari).как и большинство гвианских монодоминант,к семейству бобовых.
В тропическом дождевом лесу индо-малайской области монодоминант-ные сообщества не сериальных стадий редки и встречаются только на сравнительно малых площадях. Лесные сообщества, отличающиеся от преобладающей смешанной диптерокарповой ассоциации, распространены в местообитаниях, близких по условиям к затопляемой равнине в Гвиане, на которой произрастает лес Мора, но относительно их состава трудно сказать что-либо определенное. О заболоченных лесах тропических верховых торфяников будет сказано ниже (стр. 316).
Среди малайских типов леса, развивающихся на хорошо дренированных почвах, особый интерес представляют леса с преобладанием железного дерева острова Борнео, Eusideroxylon zwageri, так как эта доминанта дает деловую древесину большой экономической ценности. Этот вид широко распространен на Суматре и Борнео, встречаясь в виде отдельных экземпляров в ярусе В смешанного диптерокарпового леса. Однако местами железное дерево приобретает значительное преобладание над другими породами и может образовать почти чистые леса (Gresser, 1919; Witkamp, 1925; Koopman, Verhoef, 1938). Своеобразная структура лесов из железного дерева Борнео была описана в гл. 2. Лишь время от времени над общим уровнем этого леса высоко вздымаются одиночные кроны Koompassia, Shorea
1 В местности Морабалли-Крик, Брит. Гвиана, Mora excelsa также достигает больших размеров, когда произрастает в качестве обособленных индивидуумов в смешанном лесу, чем когда она является членом образованной ей консоциации (Davis, Richards, 1933-1934, стр. 111).
Фото 16. Тропический дождевой лес на высоте 350—550 м над уровнем моря полуостров Малакка.
Предгорный диптерокарповый лес с группами Shorea curtieei (светлые кроны) в центре.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
283
и Intsia. Согласно Ван-дер-Лаану (van der Laan, 1925, 1926а, 1926b, 1927), опубликовавшему некоторые данные о составе этих лесов, на 1 га может приходиться 33 дерева из числа доминант диаметром свыше 20 см, которые образуют до 30% запаса деловой древесины. Грессер говорит, что в районе Джамби, Суматра, рощи Eusiderorylon обычно занимали участки размером менее 100 га, но одна среди них раскинулась на площади 373, а другая — 1068 га. Купман и Верхоуф указывают, что в районах Джамби и Палембанг имеются «комплексы» Eusideroxylon до 10 тыс. га. Доминанта дает обильный подрост, другие виды в подлеске развиваются слабо.
Леса из борнеосского железного дерева обыкновенно занимают плоские приречные равнины и непосредственно прилегающие к ним пологие склоны. Грессер (Gresser) ставит явную неспособпость этого дерева расселяться на крутых склонах в зависимость от его тяжелых яйцеобразных плодов, скатывающихся вниз (стр. ИЗ). Как упоминалось в гл. 9 (стр.244), консоциации Eusiderorylon ограничиваются в своем распространении песчаными почвами и обнаруживают тесную связь с распределением песчаников и песков.
Другой монодоминантный тип леса в районе Малайского архипелага образует Dryobalanops arornaticatK8myp,vjWL борнеосское камфарное дерево,величественная древесная порода, принадлежащая к семейству Dipterocarpaceae. Подобно Eusideroxylon,она также встречается в виде отдельных экземпляров в смешанном лесу, но в качестве монодоминанты свойственна лишь мелким, резко локализованным группам. Рощи Dryobalanops обнаружены в различных районах полуострова Малакка, главным образом в восточной его части и на Суматре. Наиболее известный участок леса из Dryobalanops в настоящее время сильно измененный окультуриванием, находится у Кэнчинга, близ Куала-Лампур. Ван Зон (Van Zon, 1915) описал леса Dryobalanops для района Бенкалис, Суматра. Наиболее крупный участок их не превышал здесь 20 га, а каждый из семи других занимал меньше 5 га. Все они были расположены среди типичного смешанного диптеро-карпового леса. На пробном участке площадью 0,25 га из 31 дерева диаметром более 20 см 10 оказались Dryobalanops. Возобновление было обильным. Группы Dryobalanops, видимо, не приурочены к каким-либо особым почвенным условиям. Подобное же, но менее ясно выраженное групповое расположение свойственно и другим диптерокарповым дождевого леса, например Shorea curtisii (фото 16), которая доминирует в небольших обособленных сообществах, развивающихся среди массива дождевых лесов Малаккского полуострова, на крутых склонах холмов. Деревья Shorea curtisii выделяются своей бледной синевато-зеленой листвой (Foxworthy, 1927, стр. 39).
У Стеупа (Steup, 1930) имеется описание найденного им в районе Позо, остров Целебес, монодоминантного, по-видимому, климаксного типа леса; правда, не исключена возможность, что его лучше было бы определить как составную часть листопадной или полулистопадной формации, чем как тип настоящего тропического дождевого леса. В качестве доминант выступали Diospyrus sp. или Diospyros в ассоциации с пальмой Livistona rotundifolia при почти полном исключении других видов. Этот лес встречался на крутых каменистых склонах и гребнях, иногда занимая площадь в несколько гектаров. Стеуп (Steup, 1932) описал также тип леса (климаксный?)для Целебеса с «тенденцией» к монодоминантности,подобной той, что наблюдается у «борового» леса Саравака (гл. 10). В Гунонг-Клабат на равнине, сложенной рыхлыми песками вулканического происхождения, имеется лес, в котором на пространстве около 1570 га наиболее обильно
284
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
представленным видом является Elmerrillia ovalis. На пробных трансептах, охвативших площадь 25,3 га, к этому виду принадлежало 11 % деревьев диаметром более 20 см.
В Папуа, Новая Гвинея, у подножия гряды Гойдро существует тип леса, в котором Intsia bijuga и Anisoptera polyandra являются кодоминанта-ми (Lane-Poole, 1925а,Ь). Первая образует 51% древостоя (диаметр не установлен), а последняя — 43%; прочие виды — только 6%.
Ясно, что необходимы более подробные сведения относительно состава монодоминантных лесов; точная информация имеется только относительно четырех консоциаций Морабалли-Крик в Брит. Гвиане и леса Cynometra alexandri в Восточной Африке. Основываясь на этом материале, было бы преждевременным делать очень широкие обобщения.
В монодоминантных типах леса Морабалли-Крик пропорция доминирующего вида среди более крупных деревьев (диаметром 16 дюймов и более) составляет от 43 до 67 %, а среди всего древостоя (деревья диаметром 4 дюйма и более) — от 9 до 27%; в сообществе Cynometra, согласно данным Эггелинга, процент доминирующего вида несколько выше, чем в любом из упомянутых сообществ Морабалли-Крик; возможно, то же самое наблюдается в консоциации Macrolobium dewevrei тропической Африки и в консоциации Eusideroxylon Борнео и Суматры. Между типичными смешанными лесами и сообществами вроде саравакского «борового» леса, у которых имеется только «тенденция» к монодоминантности, с одной стороны, и сообществами, приближающимися к чистым древостоям, состоящим из одного вида,— с другой, существуют многочисленные переходы. Отношение видов к индивидуумам (коэффициент смешения) для гвианского монодоминантного типа изменяется на пробном участке площадью 400 х 400 футов от 3,9 до 6,3 для деревьев диаметром 16 дюймов и больше; на двух пробных участках леса Cynometra оно соответственно равно 18,2 и 7,5. Это, конечно, более высокие показатели, чем в сообществах смешанного леса (табл. 28); в то же время в монодоминантных сообществах Морабалли-Крик среди деревьев диаметром 4 дюйма и больше это соотношение изменяется от 6,5 до 12,4, что не очень отличается от соответствующих цифр для некоторых участков смешанного леса. На одном из исследованных Эггелингом участков леса Cynometra коэффициент смешения оказался равным 53,9.
На единицу площади в консоциации Cynometra и во всех монодоминантных консоциациях Морабалли-Крик приходится меньшее число видов деревьев, чем в соответствующих типах смешанного леса. Оно также будет меньше в саравакском «боровом» лесу, чем в смешанном лесу того же района. Этого следовало ожидать, так как чем большая часть древостоя образуется одним каким-либо видом, тем меньше места должно оставаться для других.Этот момент наглядно иллюстрируется ассоциацией Dacryodes— Sloanea, климаксным сообществом низинного тропического дождевого леса Малых Антильских островов. В табл. 30 показано соотношение числа видов на участке в 10 акров с процентом числа особей Dacryodes excelsa, являющимся наиболее обильно представленным видом во всей ассоциации. Как видно из таблицы, общее число видов в сообществе приблизительно обратно пропорционально проценту особей наиболее обильно представленного вида. Примечательно, что ассоциация Dacryodes — Sloanea отличается наиболее богатым флористическим составом на острове Доминика, являющемся центром ее ареала; с увеличением расстояния от центра доминанта начинает занимать все большее место в сообществе, значительно сокращая число подчиненных видов.
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
285
Таблица 30
Изменение состава ассоциации дождевого леса Ttacryodes—Sloanea (по данным J. S. Beard)
Острова % Dacryodes— Sloanea Число видов и а 10 акров (4,05 га)
Доминика 20 60
Сент-Люсия И 41
Сент-Винсент 25 39
Гренада 40 23
Сент-Киттс 40 18
В некоторых (но не всех) монодоминантных типах леса виды — субдоминанты также стремятся образовывать значительную часть древостоя. Эта тенденция составляет заметную черту лесов Мора и Уаллаба, а также саравакского «борового» леса, но отсутствует в лесу, образованном Mora gonggrijpii (Морабуцеа) или ассоциации Cynometra', в этих последних типах леса доминанта почти не имеет конкурентов. «Доминирование семейств», которое, как уже говорилось, является характерной чертой смешанных лесов, свойственно также и некоторым монодоминантным лесам. Например, в гвианском лесу Уаллаба с доминантой Eperua falcata, где другой вид Eperua, Е. jenmani, также представлен весьма обильно, а несколько часто встречающихся видов принадлежат к тому же семейству, что и доминанта, Leguminosae образуют 53% всего древостоя. В лесу Mora доминанта, три субдоминанты и несколько подчиненных видов, образующих 59% древостоя, принадлежат также к семейству Leguminosae.
Монодоминантные сообщества, как и следовало ожидать, отличаются от смешанных лесов как по структуре, так и по флористическому составу; об этих различиях уже говорилось в гл. 2.
ЗАВИСИМОСТЬ МЕЖДУ СМЕШАННЫМИ И МОНОДОМИНАНТНЫМИ ПЕРВИЧНЫМИ СООБЩЕСТВАМИ
На основании данных, сообщенных в этой и предыдущей главах, можно попробовать, правда очень предположительно, дать общую картину флористического состава тропического дождевого леса в целом. В каждом из его основных географических подразделений (или формаций) — американском, африканском, индо-малайско—австралийском — большая часть первичного леса состоит из смешанных ассоциаций, представленных очень большим числом видов, ни один из которых не является доминантой; на небольшом пробном участке такого леса огромное большинство этих видов представлено очень небольшим числом индивидуумов. Неспособность любого из видов получить преобладание в древостое, как полагает Обревиль, возможно, объясняется тем, что все они отличаются весьма сходными экологическими требованиями и почти одинаково реагируют на незначительные изменения среды. При таких условиях на основании теории вероятности можно, собственно, ожидать, что состав этих смешанных сообществ будет колебаться как в пространстве, так, по-видимому, и во
286
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
времени. Эти колебания не имеют экологического значения и просто указывают, что смесь находится в состоянии динамического равновесия. В пределах любого географического ареала смешанный лес образует одну ассоциацию.
Однообразие этих исключительно интенсивно смешанных ассоциаций нарушается по речным долинам, болотам и на территориях с постоянно высоким уровнем грунтовых вод, где им на смену приходят типы леса различного состава, часть которых является консоциациями. Помимо них, на определенных, не очень обширных площадях с хорошим или нормальным дренажем развиваются монодоминантные типы леса или леса с тенденцией к монодоминантности; они очень неоднородны по протяженности, индивидуальности и «степени доминирования». С одной стороны, это — сообщества вроде леса Уаллаба в Брит. Гвиане, леса с господством Cynometra alexandri в Африке и «борового» леса в Сараваке, имеющие в высшей степени своеобразный флористический состав и занимающие большие территории; такие сообщества можно, пожалуй, сравнить с консоциациями лесов умеренной зоны. Другую крайность представляют леса, подобные лесу из Dryobalanops Суматры и полуострова Малакка, вряд ли большее, чем местные уклонения с доминированием одного вида при групповом его распределении в лесу.
Имеющиеся данные, по-видимому, указывают на то, что монодоминантные сообщества обыкновенно бывают приурочены к строго определенным почвам или же встречаются при особой комбинации почв и рельефа; дальнейшие исследования, возможно, покажут, что это верно и для всех сообществ. В предыдущей главе была выдвинута гипотеза, что в сообществах тропического дождевого леса монодоминантность обусловлена неблагоприятными почвенными условиями, но эта гипотеза еще не может считаться окончательно доказанной. Правда, для монодоминантных сообществ Брит. Гвианы она выглядит очень убедительно; что же касается других частей тропиков, то нужны еще дополнительные данные. Если принять эту идею, по крайней мере в виде рабочей гипотезы, то можно считать, что местообитание обладает своего рода избирательным, или селективным, действием в отношении весьма многочисленных потенциальных доминант формаций тропического дождевого леса. В смешанном лесу условия среды почти одинаково благоприятны или неблагоприятны для большинства этих доминант, но при особых условиях, ведущих к монодоминантности, некоторые виды целиком устраняются, тогда как другие приобретают относительное преимущество.
Если нарисованная нами картина верна, то должна существовать близкая аналогия между тропическим дождевым лесом с его обширными смешанными ассоциациями и относительно локализованными консоциациями, образованными из родственного типа смешанного леса, и «смешанным мезофитным лесом» Северной Америки, как его описала Браун (Е. Braun, 1935). По флористическому богатству этот лес местами приближается к тропическому дождевому лесу. Большая часть его состоит из смешанной ассоциации со значительным числом доминирующих видов. В южном Огайо, например, кодоминантами в более или менее равной степени является шесть видов. По направлению к границам ареала смешанного мезофит-ного климаксного леса (или формации) начинают появляться варианты ассоциации; это происходит не в силу какой-либо определенной перегруппировки доминант, но благодаря исчезновению некоторых из них. С другой стороны, на пространстве максимального развития мезофитного леса «разнообразие во внешнем виде, возникающее благодаря изоляции
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
287
и перегруппировке доминант, создает множество на первый взгляд совершенно непохожих друг на друга сообществ с разными доминантами» (Braun, стр. 514). Между «выпадением доминант» близ границ распространения мезофитного леса и обеднением, сопровождаемым тенденцией к монодоминированию, на периферии ассоциации Dacryodes — Sloanea тропического дождевого леса на Малых Антильских островах (стр. 285) имеется поразительное сходство.
В то же время «разнообразие во внешнем виде» в местах максимального развития североамериканского мезофитного леса может быть сравнено с параллельным смешанной ассоциации развитием нескольких консоциаций при различных почвенных условиях в тропическом дождевом лесу Брит. Гвианы. Более мезофитные варианты североамериканского леса Браун считает климатическими климаксными сообществами; другие, менее влаголюбивые и в силу этого ограничивающиеся сухими склонами и гребнями, рассматриваются ею как «субклимаксы», или «физико-географические климаксы». Эти варианты Браун называет «сегрегатными ассоциациями»; она считает, что они постепенно выделились со времени третичного периода из однообразно смешанного и недифференцированного климаксного сообщества. Применительно к Европе концепция недифференцированного смешанного климаксного леса бессмысленна, так как плейстоценовое оледенение целиком уничтожило третичный смешанный лес, остатки которого, возможно, сохранились лишь в некоторых районах вроде западного Кавказа.
В тропическом дождевом лесу, возможно, также происходил подобный процесс дифференциации, и монодоминантные типы, включая типы только с легкой тенденцией к монодоминантности, вроде «борового» леса острова Борнео могут рассматриваться как «сегрегатные ассоциации». Смешанные ассоциации, конечно, представляют собой недифференцированный климакс. Масштабы процесса сегрегации были неодинаковы в различных географических областях. В Гвиане этот процесс развивался успешно и привел к созданию нескольких, в высшей степени дифференцированных сегрегатных ассоциаций. О причине этого можно только догадываться; такой причиной могла быть древность участка суши, где происходил процесс развития растительных сообществ; это могло также зависеть от местных условий, ведущих к резко выраженной дифференциации почвенных типов. В тропической Африке и Азии сегрегация проявляется гораздо слабее. Типы лесов вроде леса с преобладанием Dryobalanops, возможно, являются зародышем сегрегатных ассоциаций.
ТЕОРИЯ КЛИМАКСА
Предположения, выдвинутые в предыдущем разделе, возбуждают следующие вопросы; какие типы первичного дождевого леса представляют собой климатический климакс? Какое отношение к климатическому климаксу имеют типы, подобные гвианскому лесу Уаллаба с одной доминантой, и типы, подобные «боровому» лесу острова Борнео с тенденцией к монодоминантиости? Куда надо их отнести и как следует их рассматривать с точки зрения теории климакса?
Под климатическим климаксом вообще понимается состояние стабильности в развитии растительности. Стабильность рассматривается применительно к человеческому масштабу времени: с точки зрения геологических масштабов все растительные сообщества испытывают постепен
288
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ные изменения. Существуют два разных воззрения относительно развития климаксов в данном климате. Согласно так называемой теории моноклимакса, вся растительность данной климатической области развивается в направлении одного и того же климатического климакса, за исключением тех случаев, где специфические почвенные условия, например сильно выщелоченная песчаная почва, или такие особенности рельефа, как крутизна склонов, постоянное заболачивание и т. п., препятствуют нормальному развитию и ведут к установлению субклимаксов. С точки зрения так называемой поликлимаксной теории считается, что развитие растительности идет в направлении ряда климаксных типов, одинаково стабильных, но приспособленных к различным почвенным условиям, характеру рельефа, особенностям дренажа и т. д.
Теория климатического климакса, как и все вообще экологические концепции и теории, была построена на основании изучения растительности умеренной зоны, главным образом в Северной Америке и Евразии. Изучение тропических растительных сообществ, несомненно, внесет значительные изменения в эти концепции и теории, но существующие сведения еще слишком недостаточны, чтобы, основываясь на них, можно было как следует заняться вопросом теории климакса применительно к формациям дождевого леса.
Во всяком случае, то, что известно о флористическом составе сообществ тропического дождевого леса, не увязывается с моноклимаксной теорией. Так, все пять типов первичного леса Морабалли-Крик находятся в одних и тех же климатических условиях, и нет основания считать какой-либо из них менее стабильным, чем другие. Невозможно представить какой-либо процесс развития или изменения почвенных условий, который мог бы, например, превратить лес Уаллаба на выщелоченном песке в смешанный лес на красном суглинке, или наоборот. Лес Мора, подверженный затоплению, не обнаруживает тенденции развиться в смешанный лес, а почва леса Мора не может превратиться в почву смешанного леса. То же самое можно сказать и об отношении «борового» леса острова Борнео к смешанному лесу. Видимо, все первичные типы лесов Морабалли-Крик должны рассматриваться как эквивалентные, что и требует поликлимакс-ная теория. Каждый из этих типов зависит от различных, но в равной степени постоянных комбинаций почвы и рельефа, развивающихся при той же самой комбинации климатических условий. На примере лесов Морабалли-Крик можно познакомиться со своего рода цепью климаксов.
Существует почти полная аналогия между развитием почвы и развитием климакса. Раньше предполагали, что при одном и том же климате может развиться один и только один тип почвы; эта точка зрения дополняет моноклимаксную теорию. В настоящее время общепризнано, что, помимо климата, такие факторы, как почвообразующая порода, рельеф и т. п., могут играть преобладающую роль в развитии почвы, так что в пределах одной и той же климатической области рядом могут развиваться различные типы зрелой почвы. Примером этого является тропические красноземы и низинные тропические подзолы, описанные в гл. 9. Английская школа экологии растений приняла компромисс между моноклимаксной и поликлимаксной теориями развития растительности, рассматривая ту климаксную формацию в каждой климатической области, которая развивается при более широкораспространенной или «нормальной» комбинации почвы и рельефа, как климатический климакс, а все другие формации той же области — эдафическими или физико-географическими климаксами. С этой точки зрения смешанный лес Морабалли-Крик (и другие ассоциации
ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
289
смешанного тропического дождевого леса) можно рассматривать как климатический климакс, а консоциации Мора и Уаллаба — как климаксы эдафические. Сообщества Морабуцеа и Гринхарт следует также рассматривать как эдафические климаксы или, поскольку они сравнительно мало отличаются от смешанного леса, их можно назвать«лоциациями» (Clements, 1936) климатического климакса. В связи с этим уместно вспомнить общепринятое теперь различие между «зональными» и «азональными» почвами. Зональные почвы являются «нормальным» климатическим типом каждой климатической области, в развитии которых главную роль играют климат и растительность; азональные почвы вроде низинных тропических подзолов находятся под преобладающим воздействием материнской породы, возраста или рельефа. Аналогичным образом ассоциации смешанного тропического дождевого леса можно рассматривать как зональную климак-сную формацию, а другие стабильные типы первичного дождевого леса, зависящие от менее распространенных комбинаций почвы и рельефа, как формации азональные.
Позже (гл. 13, 15) будет показано, что и другие типы растительности, кроме типов первичного леса, рассмотренных в двух предыдущих главах, например некоторые типы саванны торфяных болот, мури, падангов и лесов, о которых уже говорилось в гл. 9, по-видимому, представляют собой стабильные типы растительности, развивающиеся в тех же климатических условиях, что и дождевой лес, но при специфических условиях почвы и рельефа. В этом случае они не в меньшей степени заслуживают названия климаксных и могут рассматриваться как азональные климаксы.
Часть IV
ПЕРВИЧНЫЕ СУКЦЕССИИ
Глава 12
ПЕРВИЧНАЯ КСЕРОСЕРИЯ И ВОССТАНОВЛЕНИЕ РАСТИТЕЛЬНОСТИ НА ОСТРОВЕ КРАКАТАУ
Сукцессии, или серии, ведущие к установлению стабильного климаксного дождевого леса, делятся, возможно несколько произвольно, на первичные сукцессии, или присерии, впервые появляющиеся на почве, до тех пор не заселенной растениями, например на почве, вновь образовавшейся на обнаженной породе или возникшей из илистого субстрата при обмелении озер и морей, и на вторичные сукцессии, или субсерии, развивающиеся в местах, где ранее существовавшая растительность была уничтожена или повреждена вырубкой, выжиганием, затоплением и т. д. Субсерии дождевого леса, обязанные своим появлением главным образом вмешательству человека, будут описаны в гл. 17; эта и две следующие главы посвящены информации о присериях.
ВОССТАНОВЛЕНИЕ РАСТИТЕЛЬНОСТИ НА ОСТРОВЕ КРАКАТАУ
Наиболее ярким примером когда-либо описанного развития первичной сукцессии растений является восстановление растительности на вулканическом острове Кракатау после полного уничтожения его первоначальной растительности грандиозной катастрофой 1883 г.1
Хотя многие факты, сюда относящиеся, уже хорошо известны, мы все же полагаем, что краткий очерк развития растительных сообществ после извержения послужит хорошим введением к рассмотрению присерий, ведущих к развитию тропического дождевого леса.
История острова после извержения
Восстановлению растительности на Кракатау посвящена обширная литература, отчасти благодаря тому, что предмет этот, к сожалению, подал повод к ожесточенному спору (Backer, 1929, Ernst, 1934, Doctors van Leeuwen, 1936; см. также Turrill, 1935). Предметом спора был вопрос о том, пережила ли извержение хоть какая-либо часть первоначальной растительности острова. Все более ранние посетители острова полагали, что она была совершенно уничтожена, но Баккер (Backer, 1929) выдвинул соображения, в которых он выразил сомнение в правильности такого заключения; он предполагал, что в лощинах на наиболее высоких частях острова, которые были впервые исследованы только много лет спустя после извержения, могла сохраниться по крайней мере часть первоначальной флоры в виде семян, спор, подземных частей растений или их фрагментов.
1 Хотя остров до извержения был покрыт растительностью, восстановление ее на нем после катастрофы рассматривается как присерия, так как поверхность острова оказалась покрытой новыми почвообразующыми породами.
294
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Сомнения в полном уничтожении фауны были высказаны ранее и совершенно независимо Р. Ф. Шарффом (R. F. Scharff). Эти аргументы, в особенности выдвинутые Баккером, не легко опровергнуть. С точки зрения самого характера данных строгое доказательство «гипотезы стерилизации» привести невозможно; как и в большинстве вопросов географии растений мы должны довольствоваться лишь высокой степенью вероятности без полной уверенности. Однако в свете контраргументов, выдвинутых Эрнстом (Ernst, 1934) и Доктерс-ван-Лееувеном (Docters van Leeuwen, 1936), мы вынуждены признать, что вероятность полной стерилизации и в самом деле весьма велика. Тому, кто хочет проследить за аргументами, выдвигаемыми той и другой стороной, следует обратиться к перечисленным выше работам, здесь же будет дан лишь краткий и непредубежденный отчет о развитии растительности на острове. Хотя для изучения вопросов распространения растений и животных при наличии морских барьеров спор о том, принимали ли участие в восстановлении растительности острова Кракатау немногие, пережившие катастрофу остатки старой флоры или нет, является решающим, с интересующей нас точки зрения это имеет второстепенное значение. Впрочем, каков бы ни был исход спора, ученый, изучающий растительную сукцессию, не согласится с Баккером (Backer, 1929, стр. 286), что «...проблема Кракатау вообще не имеет никакого значения для ботаники», но определенно выразит сожаление, что более ранние исследования острова не были более частыми и более подробными.
Кракатау принадлежит к группе мелких вулканических островов, расположенных в Зондском проливе, между Явой и Суматрой, приблизительно в 40 км от них. В начале 1883 г. он имел 9 км в длину и 5 км в ширину, возвышаясь на 2728 футов (822 м) над уровнем моря. В то время весь остров был покрыт роскошной растительностью. О природе и составе ее почти ничего не известно, но есть все основания предполагать, что в основном это был тропический дождевой лес того же характера, что и произрастающий сейчас на Суматре. Близ берега, несомненно, существовали различные прибрежные растительные сообщества; дождевой лес, вероятно, дифференцировался по направлению к вершине на предгорный и горный типы (см. гл. 16). В мае 1883 г. вулкан, уже долгое время считавшийся потухшим, начал действовать; деятельность его постепенно возрастала, пока не достигла своего максимума 26 и 27 августа. В эти дни произошло знаменитое извержение, грохот которого был слышен на Цейлоне и в Австралии. Более половины острова погрузилось в море, пик его раскололся надвое, хотя наивысшая точка горы сохранилась; территория южного побережья увеличилась на 4,6 кв. км; оставшиеся части Кракатау оказались покрыты слоем пемзы и пепла мощностью в среднем около 30 м. В течение вулканической деятельности подавляющая масса растительности была, конечно, уничтожена. Как уже указывалось, до недавнего времени все исследователи единодушно считали, что остров был полностью стерилизован и что все организмы, найденные там после извержения, должны были мигрировать через море. Растительность двух соседних островков— острова Верлатен и острова Ланг — была почти наверняка уничтожена в то же самое время. Растительность острова Себеси, расположенного в 19 км от Кракатау, сильно пострадала, но не погибла полностью.
После катастрофы 1883 г. вулканическая деятельность прекратилась, и сильные дожди (свыше 260 см осадков в год) вскоре прорыли глубокие овраги в новом покрове из пепла и пемзы. Некоторое время остров все еще оставался без какой бы то ни было растительности, и Е. Котто (Е. Cot-teau), побывавший на Кракатау в мае 1884 г., увидел перед собой лишь
ПЕРВИЧНЫЕ СУКЦЕССИИ
295
пустыню; единственным живым существом, попавшимся ему на глаза, был паук.
Однако когда в июне 1886 г. на Кракатау прибыл ботаник Тройб (Treub), то там уже было значительное количество растительности. В своем отчете (1888) Тройб подчеркнул уже существовавшее резкое различие между флорой отмелей и флорой внутренней части острова. На побережье было зарегистрировано девять следующих видов цветковых растений: Calophyllum inophyllum1, Cerbera manghas, Erythrina sp., Hernandia peltata, Ipomoea pes-carpae.Scaevola jrutescens,z лак Pennisetum macros tacky am и два неопределенных вида Сурегасеае. Кроме того, он нашел семена и плоды нескольких других видов. За исключением Pennisetum, все виды, как отметил Тройб, обычны для недавно возникших коралловых островов и являются широко распространенными пантропическими или индо-малайскими видами, произрастающими на морском побережье. Во внутренних частях острова бросилось в глаза обилие папоротников; Тройб обнаружил среди них 11 видов, включая Acrostichum aureum, Pteridium aquilinum, Pityrogramma calomelanas, Dryopteris и Pteris spp., из цветковых растений были найдены Blumea humifusa, два вида Conyza, Scaevola frutescens, Tournefortia argentea, Wedelia biflora и злаки Neyraudia madagascariensis var zollingeri и Pennisetum macrostachyum. Эти растения встречались спорадически и были представлены гораздо меньшим числом экземпляров, чем папоротники. Некоторые цветковые растения и папоротники принадлежали к видам, обыкновенно растущим близ моря. Доктерс-ван-Лееувен высказал предположение, что нахождение их вдали от берега может отчасти объясняться отсутствием конкуренции, но, конечно, при условии экологического сходства их местообитаний с песчаным побережьем. Кроме сосудистых растений, Тройб нашел также два неизвестных вида мха и обнаружил, что поверхность пепла и пемзы всюду была покрыта пленкой сине-зеленых водорослей (Cyanophyceae): было собрано шесть видов, принадлежащих к родам Anabaena, Нуpleothrix, Lyngbya, Symploca и Tolypothrix. Студенистый, гигроскопический слой, образованный этими водорослями, имел, вероятно, важное значение, являясь благоприятным субстратом для прорастания семян и спор. Следовательно, в этот момент растительность состояла из двуслойной ассоциации: верхний слой состоял главным образом из папоротников, а нижний — из Cyanophyceae. Общее число зарегистрированных видов сосудистых растений было равно 26. Из-за непроходимости местности Тройбу не удалось проникнуть далеко в глубь острова.
Дальнейшее изучение растительности Кракатау было продолжено только в 1897 г., то есть 14 лет спустя после катастрофы. Среди лиц, посетивших остров в этом году, был снова Тройб; результаты работ в виде небольшой статьи были опубликованы Пенцигом (Penzig, 1902). В промежуток времени, прошедший с 1886 г., растительность сильно развилась, на острове начали появляться сомкнутые растительные сообщества.
На отмелях хорошо развилась «формация Pes-caprae» (ассоция с доминированием Ipomoea pes-caprae) — характерное сообщество песчаных побережий тропиков (см. стр. 320). В ее составе были найдены некоторые виды, включая Terminalia catappa и Barringtonia asiatica, принадлежащие к «формации Barringtonia» (литторальное лесное сообщество, см. стр.321). Calophyllum inophillum — дерево, характерное для этого сообщества,
1 Эти и следующие определения выправлены Доктерс-ван-Лееувеном (Docters van Leeuwen, 1936).
296
ТРОПИЧЕСКИЙ дождевой лес
отсутствовало, хотя в 1886 г. Тройб встречал на побережье его молодые экземпляры. Уже укоренилась Casuarina equisetijoliа — другие весьма характерное дерево прибрежных лесов; на острове Верлатен оно в это время уже начало образовывать сомкнутый древостой.
Внутренняя часть острова теперь имела совершенно иной вид по сравнению с тем, который предстал перед Тройбом десять с половиной лет назад. Покров папоротников уступил место густому злаковому травостою, местами превышавшему человеческий рост. Эта саванна, или «злаковая степь», на нижних частях склонов состояла главным образом из Saccharum spontaneum, Neyraudia madagascariensis var. zollingeri и Pennisetum macro-stachyum, а на более высоких местах из Imperata cylindrica var. major (хорошо известный злак аланг-аланг) и Pogonatherum paniceum. В предгорной зоне вперемешку со злаками росли различные двудольные, в том числе лианы. Среди двудольных было несколько растений, типичных для побережий, например Scaevola frutescens и Vigna marina. Выше, на холмах, вперемешку со злаками росли изолированные кустарники, различные двудольные травы и папоротники. На обрывистых туфовых стенах растительность мало изменилась. Она состояла из папоротников и пленки водорослей. Среди них был найден целый ряд Cyanophyceae, которых не было в сборах Тройба, а также пять видов диатомовых водорослей. Хотя последние не были зарегистрированы в 1886 г., можно думать, что их попросту не заметили. Кроме водорослей и приблизительно 64 видов сосудистых растений, экспедицией 1897 г. было обнаружено два вида мхов, один вид печеночников и один вид пластинчатых грибов. Как и в 1886 г., оказалось невозможным проникнуть достаточно далеко в глубь острова.
Следующее подробное исследование растительности острова было предпринято в 1906 г., когда Кракатау посетила группа ботаников. Результаты этой экспедиции были изложены в хорошо известной книге Эрнста (Ernst, 1908).
Как в прибрежной растительности, так и в растительности внутренней части острова за это время произошло много дальнейших изменений.
Растительные сообщества, свойственные побережью, теперь занимали более широкий пояс, вероятно потому, что и сам берег расширился благодаря процессам аккумуляции. Теперь растительность отмели состояла из двух обособленных зон. Внешняя представляла собой ассоцпю Pes-саргае, теперь лучше развитую и более богатую видами, чем в 1897 г. За ней располагалась «формация Barringtonia», образующая прерывистый, но хорошо развитый пояс редколесья, не везде сомкнутый, но в существенных чертах весьма сходный с соответствующим сообществом побережья острова. Большая часть сомкнутого древостоя этого редколесья, видимо, состояла из более или менее чистого насаждения Casuarina equisetijolia', в других местах преобладающими деревьями были Barringtonia asiatica, Calophyllum inophyllum, Hibiscus tiliaceus и Terminalia catappa. Наряду с деревьями произрастали лианы, кустарники, злаки и другие травы. Интересно отметить наличие кокосовых пальм, хотя они могли быть и посажены.
Описание растительности внутренней части острова выглядит у Эрнста намного схематичнее, чем описание прибрежных ассоциаций, но оно дает представление о том, что до значительной абсолютной высоты здесь по-прежнему простиралась саванна, состоявшая из тех же видов злаков, что и отмеченные Пенцигом. К ним присоединялись некоторые Сурегасеае, вьющиеся растения вроде Vigna, Canavalia и Cassytha, кустарники
ПЕРВИЧНЫЕ СУКЦЕССИИ
297
Tournefortia и Scaevola и различные папоротники. «За пределами прибрежного леса полого поднимающаяся поверхность юго-восточной стороны острова была покрыта описанной нами степеобразной растительностью, густые заросли которой заполняли расселины и по крутым склонам поднимались по направлению к вершине. Их однородная масса, состоящая из живых и отмерших стеблей злаков и тростника, лишь изредка нарушалась упавшим деревом или кустом» (Ernst, 1908, стр. 31). Глубокая лощина, расположенная приблизительно на полпути до вершины, привлекала особое внимание пышными зарослями деревьев и кустарников, но достичь ее оказалось невозможным и, подобно предшествовавшим экспедициям, группа Эрнста не смогла приблизиться к пику.
Дальнейшие сведения о растительности внутренней части острова были сообщены Баккером (Backer, 1909, 1929), вновь посетившим Кракатау два года спустя после экспедиции Эрнста, участником которой он был. На юго-востоке острова за прибрежной растительностью он обнаружил местами узкие полосы смешанного редколесья, состоявшего из Ficus fistulosa, F. fulva, Macaranga tanarius, Melochia umbellata и Pipturus incanus, высотой от 5 до 15 м. Все эти деревья обычно характеризуют вторичные дождевые леса Явы и всей области Малайского архипелага вообще (стр. 414). Выше этого смешанного леса простиралась злаковая саванна с господством Saccharum spontaneum и с отдельными экземплярами упомянутых пяти видов деревьев смешанного леса.
Баккеру удалось исследовать ту заросшую лесом лощину, в которую не удалось проникнуть Эрнсту, и подняться до высоты приблизительно 400 м. Лес, растущий в лощинах, был лучше всего развит на высоте между 300 и 400 м над уровнем моря; он состоял из очень небольшого числа видов деревьев и имел травяной покров, подобный уже описанному. Среди десяти видов, не обнаруженных предыдущими экспедициями, был найден кустарник Cyrtandra sulcata, относящийся к семейству Gesneriaceae, который, как мы увидим в дальнейшем, сильно размножился. Согласно данным одного из спутников Баккера по экспедиции, растительность, расположенная выше 400 м, состояла из деревьев и папоротников. Следовательно, весьма вероятно, что сообщество папоротников, покрывавшее в 1886 г. большую часть острова, еще сохранилось на значительных высотах, куда оно было вытеснено злаковой саванной.
С 1908 по 1919 г. ботанических исследований Кракатау не производилось. Растительность острова подверглась за это время лишь незначительному культурному воздействию. В 1916 г. один немец, получив концессию на разработку строительного камня, поселился на острове и оставался на нем в течение нескольких лет. Он развел сад и, вероятно, интродуцировал несколько новых растений; вообще же его пребывание на острове оставило, кажется, мало следов. В 1919 г. случился пожар, сжегший значительную часть сухой злаковой саванны, но, согласно Доктерс-ван-Лееувену, деревья и кустарники были только опалены, и нанесенный им вред был ничтожен. Вероятно, естественный ход развития растительных сукцессий на Кракатау не был серьезно нарушен человеком. В 1919 г. группа, в которую входил Доктерс-ван-Лееувен, произвела более подробное изучение растительности острова, чем это делалось ранее. Исследованию подверглись все районы острова, включая верхнюю часть склонов. Была достигнута, наконец, и сама вершина Кракатау. Начиная с 1919 г. Кракатау стал очень часто посещаться ботаниками, тщательно изучавшими развитие его растительности. Это было весьма кстати, потому что в течение этого периода на острове произошли очень важные изменения. В 1936 г.
298
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Доктерс-ван-Лееувен (Docters van Leeuwen, 1936) опубликовал подробный отчет о состоянии острова в 1919 г. и о последующих изменениях вплоть до 1932 г.
Прибрежные сообщества 1919 г. мало изменились по сравнению с 1906 г., но в связи со значительной абразией побережья ареалы формаций Pe.s-caprac и Barringtonia сильно сократились; первая в 1919 г. занимала только узкую полосу, вторая во многих местах была смыта; состав обоих сообществ в 1919 г. был вполне типичным, хотя даже в 1932 г. они не содержали всех видов, свойственных подобной растительности на Яве и Суматре. Доктерс-ван-Лееувен считает сообщества Pes-caprae и Barringtonia перманентными, стабильными и не способными к дальнейшему преобразованию (см. ниже, стр. 321—322). В то же время консоция Casuarina, безусловно, является сериальной, так как, несмотря на то что она еще существовала в 1919 г., во многих местах ей на смену пришел смешанный лес М acaranga— Ficus, который впервые был замечен Вакером в 1908 г. Высокие старые деревья Casuarina, заглушенные лианами, частично уже упали, тогда как другие явно находились в состоянии отмирания. Под ними, вероятно из-за густой тени, подрост молодых деревьев этого вида отсутствовал; вместо них здесь развивались Мacaranga, Ficus spp., Pipturus incanus, Premna integrifoliav.1. д. C 1919 г. замещение консоций казуарины продолжалось, хотя в некоторых местах появились новые леса из этого дерева. Согласно Доктерс-ван-Лееувену (loc. cit., стр. 254), за консоцией Casuarina следует иногда не ассоция Macaranga—Ficus, а сообщество Barringtonia.
Растительность нижней части склонов, смыкавшаяся с полосой прибрежных сообществ, которая к 1919 г. мало изменилась по сравнению с 1898 г., в 1932 г. совершенно преобразилась. Лес Мacaranga — Ficus, впервые замеченный в 1908 г., развился и распространился. Он стал настолько тенистым, что в нем почти исчез травяной покров, хотя молодые деревья имелись в изобилии. Средняя часть острова в 1919 г. в основном все еще представляла собой злаковую саванну с доминированием Saccharum spontaneum в ассоциации с Imperata и различными двудольными травами; однако везде среди злаков были разбросаны экземпляры Мacaranga, Ficus и т. д., растущие поодиночке или группами. Они служили предвестниками превращения саванны в смешанное редколесье. В некоторых местах тень под деревьями была достаточной, чтобы подавить злаки и позволить развиваться теневыносливым видам вроде орхидеи Nervilia aragoana. Эта тенденция в сторону развития леса была гораздо более заметна в лощинах, где буйный рост деревьев и кустарников заглушал злаки и где замечалось даже некоторое скопление гумуса. В лощинах, расположенных невысоко над уровнем моря, Macaranga была самым обычным деревом; выше состав становился более смешанным. В то же время на отвесных туфовых склонах развитие сукцессий отставало; в местах с достаточно стабильной поверхностью доминировал злак Pogonatherum.
После 1919 г. группы деревьев неуклонно продолжали разрастаться, пока, наконец, почти вся саванна не оказалась превращенной в смешанный лес Macaranga — Ficus. В 1928 г. вся юго-восточная сторона острова была покрыта лесом приблизительно до высоты 400 м над уровнем моря. Лес простирался как вверх, так и в стороны со дна лощин. При переходе от саванны к лесу местами по временам доминировал папоротник Nephro-lepis. Кое-где на гребнях все еще сохранялась консоция Saccharum, но к 1931 г. она почти исчезла, появляясь лишь как стадия субсерии в местах развития оползней. Смешанный лес, по-видимому, в большинстве своих признаков был тождествен вторичному лесу (см. гл. 17), который
ПЕРВИЧНЫЕ СУКЦЕССИИ
299
в Малайской области появляется везде после вырубки первичного дождевого леса. Несмотря на обилие лиан, смешанный лес все же проходим. Во многих местах главным растением подлеска был Nephrolepis, один или вместе с Selaginella plana. Здесь и там встречался Costus sericeus. Хотя этот смешанный лес богаче видами, чем злаковая саванна, его видовое разнообразие значительно уступает первичному тропическому дождевому лесу.
Одним из наиболее ценных результатов экспедиции Доктерса-ван-Лееувена в 1919 г. было исследование самой высокой части острова. Почти вся эта часть оказалась покрытой густой растительностью, состоящей из кустарника Cyrtandra sulcata с очень небольшой примесью других видов. В оврагах, расположенных выше 400 м, это растение становилось все более и более обычным, а на еще больших высотах оно доминировало всюду, за исключением гребней, где находились Saccharum, папоротники и небольшое количество других трав. Ветви Cyrtandra были отягощены эпифитными папоротниками и мохообразными, придававшими сообществу внешность «мохового» горного леса (см. гл. 16). С 1919 г. консоция Cyrtandra подверглась такому же поразительному изменению, как злаковое сообщество нижней части склонов. Дерево Nauclea purpurascens в 1919 г. лишь изредка, преимущественно в лощинах, попадавшееся среди зарослей Cyrtandra, начало становиться все более и более обычным. В 1929 г. оно уже доминировало в зоне Cyrtandra до высоты 750 м, причем старые деревья достигали 25—30 м высоты. В 1932 г. Nauclea распространилась еще более, и ясно, что в недалеком будущем значительная часть вершины зарастет лесом Nauclea с подлеском из Cyrtandra.
Общий обзор сукцессий
Из приведенного выше описания видно, что, несмотря на наличие невосполнимых пробелов, можно создать достаточно полную картину основных черт развития сукцессий после извержения Кракатау. Правда, так и осталось неизвестным, когда и при каких обстоятельствах консоция Cyrtandra заместила сообщество папоротников, которое, как это можно предположить, предшествовало ей. Ряд сведений, пожалуй, можно было бы получить, изучая становление растительности на Анак-Кракатау, вулканическом кратере, возникшем из моря близ Кракатау в 1927 г. во время возобновления вулканической активности. Общий ход событий на Кракатау показан на диаграмме, изображенной на рис. 35.
Сукцессии на соседних островах — Верлатен и Ланг,— насколько известно, не отличались от сукцессий Кракатау.
Развитие растительности на Кракатау еще не достигло стабильной климаксной стадии, однако с известной уверенностью можно предсказать общий ход будущих изменений, по крайней мере для средней и верхней части острова. Можно предполагать, что в первой лес М acaranga — Ficus путем ряда изменений разовьется в стабильный климаксный тропический дождевой лес, до известной степени аналогичный смешанному первичному дождевому лесу соседних частей Суматры и Явы.
Трудно сказать, сколько времени займет этот процесс, но, основываясь на изучении вторичных сукцессий (см. гл. 17), можно предположить, что он потребует гораздо больше времени, чем то, которое уже прошло с момента катастрофы. Имея в виду изолированное положение острова, нельзя ожидать, что видовое разнообразие покрывающего его леса когда-либо
300
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
будет сравнимо с видовым богатством дождевого леса крупных островов, в прошлом связанных с азиатским материком. Растительность на высотах свыше 400 м над уровнем моря, очевидно, развивается в направлении нескольких климаксов и, всего вероятнее, будет похожа на индо-малайский горный дождевой лес (см. гл. 16); на крупных горных массивах рас-стительность этого типа появляется только на сравнительно больших высотах, но на отдельных пиках, на небольших островах, вроде Кракатау, она может быть встречена и в пределах немногих сотен метров над уровнем моря (см. стр. 370).
Сукцессия средней зоны Кракатау служит превосходным примером ксеросерии, или первичной сукцессии, развивающейся на обнаженной
Эдафические климаксы ?
Спелая Слепая формация формация Pescaprae Bornngtonia
Г^лимаксный низинный дождевой лес]
Гклимаксный предгорный дождевой лес]
1952г.
1919г.
1906г,
1897г.
1886г.
1885г.
Пионерные злаковые ассоции (НассРатит +Neyraudia и т.п.)
формация Pes-caprae
В ысотные Отлоги й берег
зоны
Консоции Nauclea
Консоции Cyrtandra
Ассоции папоротников /Ассоции папоротников
* Cyanophyceae и +Cyanophyceae?]
Полное уничтожение растительности—
Средняя зона (до 400м )
Верхняя зона (выше 400м)
Рис. 35. Диаграмма развития сукцессий Кракатау после катастрофического извержения 1883 г.
породе, но, за исключением самых ранних стадий, она мало отличается (см. гл. 17) от вторичной сукцессии на заброшенных пашнях и т. д. Отличительной чертой этой сукцессии является последовательное доминирование сначала споровых — в данном случае главным образом водорослей и папоротников, позже травянистых цветковых растений и, наконец, деревьев. Это типично почти для всех ксеросерий, развивающихся в районах, где естественным климаксом является лес; но во вторичной сукцессии, возникающей на уже существующем почвенном покрове, стадия споровых выпадает. На обнаженной породе лишайники и мохообразные обыкновенно развиваются совместно с водорослями и участвуют как пионеры в почвообразовании; мохообразные наблюдались на Кракатау уже в 1886 г., но кажется, ни они, ни лишайники не имели большого значения на ранних фазах сукцессии. Напротив, водоросли играли при этом важную роль, и, хотя Тройб в 1886 г. наряду с водорослями обнаружил папоротники и цветковые
ПЕРВИЧНЫЕ СУКЦЕССИИ
301
растения, все же, вероятно, самыми ранними поселенцами были сине-зеленые или диатомовые водоросли с бактериями; зеленые водоросли не встречались. Последнее не удивительно, так как, согласно Фритчу (Fritsch, 1907, 1936), во влажных тропиках на обнаженной породе или почве обыкновенно доминируют Cyanophyceae.
Временное доминирование папоротников встречается редко. Оно характерно для некоторых, но не для всех вторичных сукцессий области тропического дождевого леса. На Кракатау оно отчасти могло быть связано с ранним появлением спор, легко распространяемых ветром, а отчасти объясняться приживаемостью папоротников на почвах с недостатком минеральных питательных веществ (см. стр. 420).
Хотя в развитии растительности средней зоны Кракатау выделяются отчетливые фазы поочередного доминирования споровых трав и деревьев, интересно отметить тот факт, что раункиэровский спектр жизненных форм (для всех трех зон острова, взятых вместе) оставался приблизительно постоянным в этих трех фазах. Доктерс-ван-Лееувен приводит цифры (табл. 31), характеризующие соотношение различных жизненных форм.
Таблица 31
Жизненные формы цветковых растении на Кракатау (Docters van Leeuwen, 1936, стр. 191)
Год Число видов цветковых растений Фанерофиты Хамефиты Гемикриптофиты Криптофиты Терофиты
%
1886 15 59 7 20 7 7
1897 49 70 6 14 6 4
1906 73 78 6 8 3 5
1934 219 65 6 12 2 15
Из этих цифр видно, что фанерофиты составляли большинство видов флоры Кракатау с самых ранних стадий сукцессий. Из статистических данных, приведенных в гл. 1, ясно видно подавляющее преобладание фанерофитов в климаксном дождевом лесу; существенно, что такое преобладание, хотя и в меньшей степени, обнаруживается и в первичной ксеро-серии. В следующих главах будет показано, что преобладание фанерофитов характерно для большей части стадий первичных и вторичных сукцессий, которые развиваются в направлении становления тропического дождевого леса. Это согласуется с мнением Раункиэра (Raunkiaer, 1934, см. стр. 197—198), считавшего, что биологический спектр первичной и вторичной сериальной растительности (в силу того, что он определяется прежде всего климатом) подобен биологическому спектру всей флоры той же области. Однако надо заметить, что гемикриптофиты и терофиты, отсутствующие во флоре климаксного тропического дождевого леса, играют значительную роль в сериальной растительности Кракатау.
В сукцессиях Кракатау, как и вообще во всех сукцессиях, развивающихся на обнаженной почве в весьма влажных климатах, происходил процесс постепенной замены открытых сообществ сомкнутыми. Уже ко времени второго посещения острова Тройбом (14 лет спустя после извержения) во многих местах существовали сомкнутые растительные сообщества. Впоследствии все большая и большая часть растительного покрова становилась сомкнутой, хотя и в настоящее время в местах постоянных
302
ТРОППЧЕСКПЯ ДОЖДЕВОЙ ЛЕС
подвижек почвы и на отвесных склонах, где сукцессия отстает в своем развитии, можно наблюдать открытые сообщества.
Следует отметить одно существенное отличие сукцессий Кракатау от большинства, а возможно, даже и от всех первичных сукцессий умеренных областей, которое проявляется в непрерывном росте числа видов. Поскольку точные данные о количестве видов, составляющих каждое сообщество по отдельности, отсутствуют, можно воспользоваться цифрами Доктерс-ван-Лееувена (Docters van Leeuwen, цит. работа, стр. 209) (по покрытосеменным и голосеменным) для всего Кракатау и двух соседних островков Ланг и Верлатен.
1883 г. 1886 г. 1897 г. 1908 г. 1920 г. 1928 г. 1934 г.
0 26 64 115 184 214 271
В умеренных областях при появлении растительности на обнаженной почве незначительное вначале число видов постепенно возрастает до максимума, но затем, при приближении к климаксу, снова снижается (Warming, 1909, стр. 356). Это выпадение видов происходит обыкновенно одновременно с изменением от открытого к сомкнутому сообществу. На Кракатау, по всей вероятности, это одинаково верно как для побережья и вершины, так и для средней зоны, не заметно каких-либо следов выпадения видов, наоборот, здесь происходит неуклонное возрастание их числа. Консоция Saccharum по числу видов богаче, чем предшествовавшая ей ассоция папоротников, а лес М acaranga — Ficus в свою очередь богаче консоции Saccharum. Если, как предполагается, лес Мacaranga — Ficus будет замещен смешанным дождевым лесом, то видовое разнообразие, несомненно, увеличится еще больше. Непрерывное увеличение числа видов в ходе сукцессии вплоть до стадии климакса, без последующего их выпадения, не представляет собою особенности, свойственной исключительно Кракатау, но характерно для тропических сукцессий вообще.
На Кракатау развитие открытых сообществ в сомкнутые сопровождается заменой смешанных ассоций монодоминантными; так, смешанная ассоция папоротников уступила место злаковому сообществу с единственной доминантой Saccharum spontaneum. Однако позже снова появляются смешанные сообщества; лес Мacaranga — Ficus, подобно климаксному дождевому лесу, который может прийти ему на смену, есть сообщество полидоминантное. Выпадение видов на последней стадии в большинстве сукцессий умереиной зоны обыкновенно приписывалось увеличению интенсивности конкуренции, но так как интенсивность конкуренции возрастает также и по мере развития тропических сукцессий без какого бы то ни было выпадения видов, то это не может быть единственной причиной описанного явления. По-видимому, меньшее число видов на конечной стадии по сравнению со средней у сукцессий умеренной зоны на самом деле связано с тем, что умеренные климаксы являются обыкновенно монодоминантными или имеют лишь небольшое число доминант, что является следствием бедности флоры видами древесных растений.
Анализируя причины, скрытые в основе развития любой растительной сукцессии, нужно считаться с тремя группами факторов: во-первых, с количеством семян, плодов и иных средств размножения, во-вторых, с влиянием растений друг на друга в смысле конкуренции или коопера-
ПЕРВИЧНЫЕ СУКЦЕССИИ
303
цпи и, наконец, с изменениями местообитания, происходящими вследствие изменения климата и реакций со стороны самой растительности. Взаимодействие этих трех групп факторов, конечно, крайне сложно. Виды появляются и исчезают, и в любом данном случае обыкновенно невозможно обнаружить абсолютно все факторы, обусловливающие их появление или исчезновение.
При восстановлении растительности на Кракатау количество зачатков, очевидно, должно было иметь очень важное значение. Если предположить, что первоначальная флора была полностью уничтожена, то можно лишь с известной степенью вероятности установить наиболее возможный путь проникновения на остров зачатков растений — безразлично в виде ли семян, плодов или спор — в зависимости от способа их распространения с помощью ветра, воды, животных или человека. На этом основании и составлена табл. 32. Из приведенных в ней цифр видно, что количество
Таблица 32
Способы распространения семенных растений (покрытосеменных п голосеменных), произрастающих на Кракатау и островах Ланг и Верлатен
(Docters van Leeuwen, 1936, стр. 240)
Способы распространения % ВИДОВ
1886 г. 1897 г. 1908 г. 1920 г. 1928 г. 1934 г.
Ветер 62 44 28 35 42 41
Море 38 47 52 36 33 28
Животные 0 9 20 19 23 25
Человек 0 0 0 9 2 6
Общее число видов 26 64 115 184 214 271
видов, распространяемых ветром, было в 1886 г., вероятно, значительно больше, чем позже. Виды, распространяемые животными, сначала вообще отсутствовали, а виды, распространяемые человеком, появились только после прибытия на остров в 1916 г. немецкого поселенца. На основании этих данных, безусловно, можно сделать вывод, что трудность распространения имела значение ограничительного фактора на скорость развития сукцессии; другими словами, некоторые виды появились поздно не потому, что для них было неподходящим местообитание, а потому, что на остров просто не проникли их зачатки. Отсутствие на Кракатау Loranthaceae — семейства паразитов, обильно распространенных как на Яве, так и на Суматре,— наилучшим образом доказывает невозможность преодолеть морской барьер для многих видов, только поэтому не входящих в состав флоры Кракатау. Многие виды, для которых остров является подходящим местообитанием, несомненно, никогда на него не попадут.
Изменения среды
Хотя способы распространения видов, конечно, оказали большое влияние на сукцессии растений Кракатау, нельзя недооценивать и роль, которую сыграло при этом изменение среды. В 1883 г., непосредственно
304
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
вслед за катастрофой, Кракатау не был особенно привлекательным местообитанием для растении. Вся поверхность острова на глубину многих метров была покрыта рыхлым, подвижным, крайне пористым материалом — продуктом извержения вулкана; материал этот состоял из кусков пемзы разной величины и вулканического пепла, или пемзового песка. Непривлекательность этого субстрата усугублялась безраздельным господством солнца и ветра. Его химический состав был вовсе не так неблагоприятен, как это можно было бы предполагать, так как анализ (Ernst, 1908, стр. 49) показал, что в нем содержалось большинство обычных питательных веществ, за исключением связанного азота и фосфора, причем многие из этих веществ находились в растворимой форме. Со временем пемзовый песок, первоначально представлявший собой лишь почвообразующий материал, развился в настоящую почву. Проведенное в 1928 г. исследование почвенного профиля (van Baren, 1931) обнаружило весьма заметное выщелачивание натрия и калия. Отношение кремния к алюминию (стр. 229—230) оказалось меньше, чем в материнской породе, что указывало на несомненное развитие процесса латеритного выветривания, столь характерного для вулканических пород во влажном тропическом климате.
В 1886 г. под действием денудации из первоначально однообразного покрова уже образовалось четыре различных типа субстрата, а именно пемзовый песок, все еще покрывавший большую часть площади, морской песок, смешанный с пемзой на побережье, почва лощин, частично состоящая из старой невыветренной вулканической почвы, а частично из эродированного пемзового песка, и, наконец, обнаженная невыветрившаяся лава. Под действием климата, бактерий и вновь появившейся растительности пемзовый песок постепенно превратился в среду, более или менее благоприятную для роста растений. Азотсодержащие соединения, сначала поступавшие в почву исключительно с дождем,позже стали появляться и благодаря деятельности азотфиксаторов. Время появления первых почвенных бактерий неизвестно, но Эрнст (Ernst, 1908, стр. 51), анализировавший в 1906 г. образцы почвы Кракатау, нашел, что бактерии в них были почти так же многочисленны, как и в почвах Явы; среди этих бактерий были обнаружены микроорганизмы нескольких различных биологических типов, включая азотфиксаторы. Обилие Cyanophyceae на ранних стадиях восстановления растительности можно считать показательным, так как существует мнение, что некоторые роды этих водорослей обладают способностью фиксировать атмосферный азот. Происхождение фосфора так и осталось невыясненным; возможно, он попал с морскими брызгами или с атмосферной пылью.
С появлением довольно плотного растительного покрова среда быстро становится благоприятной для растений. Затенение почвы растениями полностью изменяет микроклимат на ее поверхности, увеличивая среднюю влажность и понижая амплитуду колебаний температуры. Корни растений связывают песок, делая его менее рыхлым и подвижным. Остатки отмерших растений снабжают почву перегноем, изменяя тем самым как физические, так и химические ее свойства. По мере того как растительность становилась выше, в результате смены травянистых растений деревьями воздействие ее на почву все более усиливалось.
Следовательно, как и во всех первичных сукцессиях, растения изменили среду, сделав ее более благоприятной для себя. Растения, менее требовательные, уступили место более требовательным, с которыми они уже не могли больше конкурировать при условиях, созданию которых
ПЕРВИЧНЫЕ СУКЦЕСС ИИ
305
они сами содействовали. Устанавливалась общая тенденция в направлении от среды ксерофитной к более мезофитной, другими словами, от условий более крайних к условиям менее крайним.
ДРУГИЕ КСЕРОСЕРИИ
Большая часть настоящей главы была посвящена сукцессиям Кракатау, поскольку они представляют собой наиболее ярко выраженные и лучше всего документированные примеры сукцессий, которые удалось проследить с момента их появления на обнаженной породе в климате, где тропический дождевой лес является климаксным типом растительности. Однако возможность наблюдения ксеросерий в тропиках представляется довольно часто, в особенности в областях вулканической деятельности, хотя материалов по такого рода сукцессиям собрано мало. Весьма благоприятны для этих исследований вулканические острова группы Самоа в Тихом океане, где имеются потоки застывшей лавы различной давности, причем относительно некоторых известна дата образования. Острова находятся в условиях влажного тропического климата, и в их пределах можно наблюдать различные стадии развития дождевого леса на потоках лавы. Согласно Фаупелю (Vaupel, 1910), потоки лавы на северной стороне острова Савайи, образовавшиеся во время извержения 1905 г., при исследовании их в 1906—1907 гг. были совершенно лишены растительности. Другие потоки лавы на том же острове, имевшие возраст около 100—150 лет, были уже покрыты на благоприятных для растительности местах затеняющими почву деревьями, укоренившимися в трещинах лавы, на почве, скопившейся на ее поверхности или же, возможно, проникшими прямо в погребенную почву, находящуюся под лавой. Старшие поколения поселенцев на острове помнят, что 50 лет тому назад на этих пространствах не было ничего, кроме папоротников и мелких кустарников. О молодых лавовых полях острова Самоа упоминает также Рехин-гер (Rechinger, 1910, стр. 225—227), который говорит, что такие поля местные жители называют «му» (жгучими) из-за высокой температуры, приобретаемой обнаженной черной лавой на солнце. Растительность (явно на весьма ранней стадии сукцессии) в своем описании он изображает ксерофитной, состоящей главным образом из низких кустарников с жесткими кожистыми листьями вроде Morinda citrifolia, видов Gardenia, Fagraea и Loranthus, произрастающих вместе с ксерофильными папоротникообразными вроде Nephrolepis nrjescens и Lycopodium сегпиит, а также небольшого количества злаков и паразитической Cassytha jiliformis. Другие потоки лавы несут на себе хорошо развитую сомкнутую растительность, состоящую из густого подлеска, над которым возвышаются гигантские деревья Eugenia spp., Rhus simarubifolia, Ficus spp., Calophyllum speciosum и т. д. В этих лесах на почве обыкновенно встречаются мхи, орхидеи и злаки, а па стволах деревьев — папоротники и виды Freycinetia.
Бирд (Beard, 1945) описал сукцессии на вулкане Суфриер (остров Сент-Винсент) в Вест-Индии, развившиеся после извержения 1902 г. Сукцессии этой горы обнаруживают большое сходство с сукцессиями на •соответствующих высотах Кракатау. На Суфриере, в высотном поясе, где климаксом является тропический дождевой лес, первичная сукцессия близко напоминала вторичную сукцессию той же области. Самыми ранними деревьями-поселенцами являются светолюбивые виды, обладающие эффективными средствами распространения семян, например Ochroma 20 П. У. Ричардс
306
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и Cecropia spp.; те же виды часто встречаются в качестве доминант в молодом вторичном лесу на некогда возделывавшихся, но потом покинутых землях (стр. 418—419). В более высоких поясах гор сериальные сообщества приближаются по своему облику к таким формациям, которые в других местах встречаются в качестве климаксов на гораздо больших высотах; так, здесь наблюдаются ассоции, подобные «лесу эльфов» (см. гл. 16), парамо или альпийским тропическим лугам и альпийской «тундре». Все эти типы растительности похожи на встречаемые обычно гораздо выше. Бирд приписывает это явление действию менее благоприятных, более ксерофильных условий, существовавших на ранних стадиях сукцессии. Харди (Hardy, 1939) исследовал почвы Суфриера в 1933 г. и нашел, что по содержанию азота и органических веществ их можно сравнить с возделываемыми почвами острова Сент-Винсент. Следовательно, в течение каких-нибудь 30 лет развилась почва, видимо, способная поддерживать климаксную растительность.
Прочие сведения относительно ксеросерий во влажных тропиках очень скудны. Доктерс-ван-Лееувен (Docters van Leeuwen, 1936) дает обзор ряда примеров восстановления растительности в местах, опустошенных деятельностью вулканов, но так как в большинстве случаев первоначальная растительность подвергалась лишь частичному уничтожению, то последующие сукцессии являются скорее вторичными или смешанными, чем истинно первичными. Воган и Виэ (Vaughan, Wiehe, 1937, 1941) описали различные растительные сообщества острова Маврикий, которые они сочли стадиями ксеросерии, ведущей к нагорному климаксному лесу (типу тропического дождевого леса, см. гл. 2). Они подчеркивают, что это сукцессия, где доминируют древесные растения; травы и злаки участвуют в ней мало. Шевалье (Chevalier, 1909) дал краткое описание интересной сукцессии на обнаженной гранитной породе в горах Фута-Джаллон в Западной Африке. Здесь, вероятно, климаксом является скорее смешанный листопадный лес (см. гл. 13), чем истинный вечнозеленый тропический дождевой лес. Первым обосновавшимся здесь растением была Trilepispilosa. В период успешного развития этого растения сукцессия почти не изменялась, но стоило только засухе или лесным пожарам уничтожить его, как начинали прорастать семена различных других растений. В результате возникло сообщество травянистых растений, позже вытесненное древесными растениями. Подобную же сукцессию можно наблюдать в южной Нигерии, например в горах Иданре.
Глава 13
ГИДРОСЕРИИ
В пресноводных озерах, прудах, на плесах рек с медленным течением суша ведет постоянное наступление на воду. В этом процессе превращения в сушу (Verlandung) или образования новых земель за счет водной площади существенное участие принимает растительность. Водные растения, оказывая сопротивление движению воды, замедляют его и таким образом увеличивают интенсивность отложения взвешенных в воде частиц. Позже растения-амфибии помогают консолидации осадков и, добавляя к ним свои мертвые остатки, изменяют структуру этих осадков и увеличивают их массу. Это ведет к общему подъему уровня почвы относительно уровня воды, что сопровождается появлением гидросерии, или растительной сукцессии, начинающейся укоренением различных типов водной растительности и обыкновенно заканчивающейся в области господства лесного климата типом леса, сходным, но не вполне тождественным с климатическим климаксом. В процессе развития гидросерии происходит постепенное изменение растительности от стадии открытой воды до стадии сравнительно сухих условий, и, как и в ксеросерии, растительность стремится создать мезофитную среду.
Для тропиков гидросерии, иногда грандиозных масштабов, чрезвычайно характерны. На маленьких речках, в болотах и лагунах, занимающих большие пространства на многих тропических низменностях, нередко можно наблюдать все стадии сукцессии: от открытой воды до сомкнутого леса. На берегах больших тропических рек также иногда видны хорошо развитые гидросерии, но так как эти берега в большинстве случаев плотно заселены, то естественная растительность часто бывает совершенно уничтожена или сильно изменена.
Как правило обстоятельства не всегда позволяют производить непрерывное наблюдение за изменением растительности в каком-нибудь одном месте, но о последовательных стадиях развития гидросерии можно судить с достаточной степенью определенности по поясному распределению растительных сообществ.
Полная история развития гидросерии едва ли была исследована где-либо в пределах распространения тропического дождевого леса, но имеющиеся в нашем распоряжении многочисленные более или менее отрывочные наблюдения этого процесса могут быть сведены воедино. Это до известной степени облегчается хорошо известным фактом, что водные и болотные растения обыкновенно распространены гораздо более широко, чем растения сухопутные. Например, в Новом свете многие виды, входящие в состав гидросерий, распространяются по всей тропической зоне; то же самое относится к тропической Африке и Индо-Малайской области. Некоторые виды водных растений являются общими для умеренной и тропической зон, а некоторые главнейшие роды тропической гидросерии пред-20*
308
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ставлены в Старом и Новом свете близко родственными видами, например Raphia. Сагара. Symphonia. Следовательно, у гидросерий наблюдается тенденция быть однородными даже в весьма отдаленных областях не только в общем развитии, но и в отношении структуры иоблика их последовательных фаз. На более ранних стадиях различия в климате мало отражаются на их развитии.
Встречающиеся местные изменения, вероятно, зависят от различий в уровне воды (то есть от сезона и высоты паводка), характере отлагаемых осадков и химическом составе растворенных в воде веществ. Влияние этих факторов изучено мало.
ГИДРОСЕРИИ ТРОПИЧЕСКОЙ АМЕРИКИ
Типичная гидросерия была описана Кенойером (Кепоуег, 1929) для озера Гатун в Панамском канале. Было установлено пять стадий:
1) Плавающая водная ассоция. Salvinia auriculata. Jussieua natans. Utricularia mixta. Pistia stratiotes. Eichornia azurea.
2) Ассоция водяной лилии. Nymphaea ampla вместе с предыдущими видами.
3) Выступающая из воды водная ассоция. Наиболее обильные виды: Typha angustifolia и папоротник Acrostichum danaeifolium в сочетании с Crinum erubescens. Hibiscus sororius и Sagittaria lancifolia.
4) Ассоция тростникового болота. Состоит из Cyperus giganteus, Scirpus cubensis и других Сурегасеае вместе с крупными злаками вроде Phragmites communis и Gynerium sagittatum. травы Jussieua suffruticosa из семейства двудольных и папоротников.
5) Ассоция «кустарников на маршах». Dalbergia ecastophylla и высокое ароидное Montrichardia arborescens.
Ассоция «кустарников на маршах», вероятно, со временем уступит место болотистому лесу, но последнее сообщество еще не развилось.
Эта исключительно простая сукцессия очень похожа на многие гидросерии умеренной зоны, причем в ней участвует несколько родов и даже видов, свойственных умеренным широтам. Она также очень похожа на гидросерии, которые развивались в сторону теперь совершенно уничтоженного «мезофитного леса» в Пуэрто-Рико (Gleason, Cook, 1926). На Тринидаде встречается последовательный ряд сукцессий, развивающихся от травяного болота до заболоченного леса пресноводных болот. Они были кратко описаны Маршалом (Marshall, 1934). Бирд (Beard, 1946b) описывает также болотные и «маршевые» (сезонные болота) сообщества Тринидада, но не входит в обсуждение их сукцессий. «Болотистые саванны» или «водные саванны», занимающие большие пространства в прибрежной области Суринама (Lanjouw, 1936), в Брит. Гвиане и Венесуэле, представляют собой безлесные болота и являются стадиями гидросерий. Но сукцессии и здесь не были изучены.
Гидросерии, расположенные по берегам больших рек, несколько отличаются от гидросерий озер и болот, так как условия здесь осложняются размывающим действием потока, сезонными колебаниями уровня воды и, близ моря, приливами. Возможно, наиболее ярким примером сукцессии подобного рода являются furos. или протоки в дельтовой зоне между устьем собственно Амазонки и Рио-Пара. Вода тут подвергается влиянию приливов и отливов, но остается более или менее пресной. Здесь происходит непрерывное образование все новых и новых аллювиальных остров
ПЕРВИЧНЫЕ СУКЦЕССИИ
ЗОЭ
ков, которые постепенно укрепляются поселяющейся на них растительностью. Растения-пионеры, первыми освоившие эти островки, с течением времени вытесняются другими, которые могут укорениться только под их защитой. По мере развития сукцессии площадь островов увеличивается и ранее поселившиеся растения вытесняются к воде, образуя пояс, расположенный с краю. Следовательно, вокруг более крупных островов происходит образование поясов растительности, представляющих последовательные стадии гидросерий. Подобную же сукцессию с поясным расположением растительности можно наблюдать на отлогих берегах меандр. Превосходное описание этих сукцессий дал Губер (Huber, 1902), интересные подробности сообщает также Буйен (Bouillienne, 1930).
Обычно первыми поселенцами на илистых отложениях оказываются Montrichardia arborescens, мощное ароидное, достигающее 2—3 м высоты, и Drepanocarpus (Machaerium) lunatus, кустарник из семейства бобовых. Эти два вида образуют практически чистые сообщества, причем одно, по-видимому, исключает другое; часто эти консоции чередуются через каждые 10 м или около того. Оба вида имеют плавающие семена, в изобилии встречающиеся в речном потоке. Прорастание происходит необыкновенно быстро, и всходы могут укорениться в промежуток времени между двумя приливами; в этом отношении эти растения похожи на мангровые. Под прикрытием, создаваемым Montrichardietum и Drepanocarpe-tum, развивается растительность совершенно иного рода, а именно масса плавающих растений, или «плавающий газон», состоящий из скопления злака Hymenachne amplexicaulis, произрастающего совместно с Eichomia azurea и Е. crassipes. Это последнее сообщество особенно широко развито в тех частях Амазонки, где уже не сказывается влияние приливов. Здесь оно может укорениться и играть определенную роль в сукцессии, тогда как в области furos это сообщество не имеет сколько-нибудь важного значения.
Вторая стадия сукцессии знаменуется вторжением Rhizophora race-mosa, которая постепенно вытесняет сообщества Montrichardia и Drepanocarpus. Rhizophora racemosa образует консоцпю, простирающуюся вверх по реке до пределов распространения приливной волны; она не ограничивается, как большинство мангровых, солеными и солоноватыми водами (см. стр.328). Местами Rhizophora замещается другими мангровыми, Avicen-nia tomentosa, A. nitida и Laguncularia; последние два вида также растут в воде, которая, по-видимому, никогда не является соленой или даже солоноватой. Мангровые вскоре заглушают сообщества Montrichardia и Drepanocarpus, превращая их в весьма угнетенный подлесок.
Спустя некоторое время под мангровыми укореняются всходы пальм Euterpe oleracea и Mauritia jlexuosa и двудольных деревьев Cecropia palmata и С. paraensis. Cecropia является одним из родов, играющих очень важную роль во всех вторичных сукцессиях дождевого леса Америки (см. стр. 419). Пальмы и цекропии очень быстро перерастают и подавляют мангрову; они могут непосредственно привести к установлению Варзеа (varzed) — леса пресноводных болот, подверженного затоплениям. Такой лес состоит из гигантских деревьев и похож по общему облику на климаксный тропический дождевой лес. В других случаях (причины этого не совсем ясны) развитие сукцессии, по-видимому, идет иным путем и за сообществом Rhizophora следует сообщество из пальм Raphia vinifera var. taedigera. Позади пояса Rhizophora или Raphia может находиться или пояс с доминированием высокой пальмы Mauritia jlexuosa, образующий чистое сообщество, или же растение в смеси с другими пальмами вроде Euterpe
310
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
oleracea, Manicaria saccijera и Maximiliana regia, или же пояс из сообщества Cecropietum с доминированием двух упомянутых выше видов. Для пояса пальм и Cecropia характерно присутствие деревьев Virola surinamensis и Сагара guianensis. Хотя причины этих изменении в развитии промежуточных стадий сукцессии неизвестны, ясно, что любые из них в конце концов приводят к становлению леса Варзеа. Характер леса Варзеа неоднороден в разных местах. Его средняя высота колеблется от 15 до 30 м. Весьма характерным для него является неправильный профиль, на котором выделяются огромные, куполообразные кроны Ceiba pentandra. Другими обращающими на себя внимание деревьями этого болотистого леса являются Сагара guianensis, Hevea brasiliensis и Н. guyanensis, Symphonia globulijera, Virola surinamensis и многие Leguminosae. Часто встречаются пальмы, а там, где лес достигает уреза воды, нередко наблюдается кайма из Pachira aquatica.
Лес Варзеа области /иго$на Амазонке, описанный Губером и Буйен-ном, очень похож по структуре и флористическому составу на лес Мора затопляемых равнин Брит. Гвианы, о которых шла речь в гл. 10. Однако в лесу Варзеа сама Mora совершенно отсутствует и вообще не известно, имеется ли тут какая-либо тенденция к монодоминантности. Как лес Варзеа, так и лес Мора могут считаться эдафическими климаксами.
Гораздо меньше изучены гидросерии нижней Амазонки вне области дельты, хотя кое-что по этому поводу можно почерпнуть из статей Буйена (Bouillienne, 1930) и Уле (Ule, 1908 а). Здесь приливы ослаблены, но сезонные паводки вызывают сильные колебания уровня воды. Климат здесь более континентальный, чем в области дельты. Вдоль реки тянется restinga, или «дамба», образованная отложением наиболее грубых осадков; она возвышается над лежащей ниже уреза реки заболоченной местностью. Для этого района были описаны следующие сериальные сообщества: 1) различные типы плавающей водной растительности, включая сообщество, где доминирует гигантская водяная лилия Victoria amazonica и «плавающий газон» из Echinochloa polystachya и Eichornia\ 2) «campos de varzea»— злаковые болота, в которых, так же как и в «плавающем газоне», доминируют главным образом Echinochloa polystachya, Hymenachne amplexicaulis и Paspalum repens\ 3) зона кустарников Salix humboldtiana и Alchornea castaneifolia, оконтуривающая берега реки. Сообщество «campos de varzea» непосредственно обрамляется лесом. Окраину этого леса составляет зона 4) cecropietum, образованная несколькими видами, включая Cecropia palmata и С. paraensis\ эта окраина состоит из групп деревьев, не образующих непрерывного пояса. Каждая группа деревьев обыкновенно имеет одинаковый возраст, и Cecropietum существует в продолжении только одного поколения (см. стр. 420). Следующее сообщество (5) представляет собой лес Варзеа, растущий главным образом на restingas или дамбах, вдоль реки, и, следовательно, располагающийся узкими полосами. Различают две последовательные стадии в развитии этого леса: стадию молодого леса, когда главными деревьями являются быстрорастущие виды вроде Calyco-phyllum spruceanum, Inga spp. Triplaris surinamensis, и стадию спелого леса, подобного по своему составу лесу области дельты, за исключением того, что наиболее обильной пальмой является здесь A strocaryum jauari, a Mauritia почти отсутствует. В восточной части нижней Амазонки эти леса значительно менее роскошны, чем леса дельты, вероятно, благодаря более сухому климату, но далее вверх по реке, где климат становится более влажным, они снова становятся более высокоствольными.
ПЕРВИЧНЫЕ СУКЦЕССИИ
311
Уле (Ule, 1908а) кратко описал сукцессию, характерную для песчаных берегов Журуа, притока верхней Амазонки. Первой стадией здесь является травянистая растительность маршей, состоящая из злаков и различных других трав. Там, где берег повышается, появляются Salix hum-boldtiana и Alchornea castaneifolia', за ними следуют виды Cecropia. На «черноводных» притоках Амазонки вроде Рио-Негро прибрежная растительность, согласно Уле (loc. cit.), сильно отличается от прибрежной растительности главного русла реки и «светловодных притоков»; здесь совершенно отсутствуют сообщества Cecropia и некоторые самые обыкновенные виды, характерные для других рек.
Данные относительно сукцессий на речных берегах в Брит. Гвиане имеются у Родвея (Rodway, 1894), Дэвиса (Davis, 1929) и Майерса (Myers, 1935). На участках гвианских рек с более медленным течением ранние стадии заселения аллювия похожи на те же стадии в дельте Амазонки. Сначала появляется Montrichardia arborescens, за которой следует скраб, состоящий из Drepanocarpus lunatus и одного или двух видов бобовых. Как и на Амазонке, существует масса плавающих растений (плавающий газон), прикрепляющаяся к Montrichardia или к кустарникам. Среди плавающих растений доминирует Panicum elephantipes. Более поздние стадии сукцессии исследованы не были, но Дэвис описал целый ряд сообществ северо-западного района Брит. Гвианы, которые, вероятно, представляют собой поздние стадии гидросерии. В местах с солоноватой водой берега рек окаймляются узкой полосой мангровы из Rhizophora mangle. За ней в более или менее солоноватых болотах развивается лес «truli bush», состоящий из двудольных деревьев с подлеском, в основном образованным пальмой Manicaria saccifera. Там, где обширные болота сочетаются с довольно низким уровнем воды, «truli bush» замещается «kiri-kowa bush», где доминирует Iryanthera paraensis. К другим типам болотистого леса этого района принадлежат «саванна Ите (Ite)» с доминирующей пальмой Mauritia flexuosa и подлеском из Clusia cuneata и т. д. и лес Мора с доминантой Mora excelsa. Этот лес напоминает лес Мора местности Морабалли-Крик, описанный в гл. 10, за исключением того, что на Амазонке больше пальм.
Майерс (Myers, 1935) описал последовательную смену типов растительности на берегах гвианских рек при продвижении вверх по течению. Вблизи их эстуариев расположен Rhizophoretum, иногда ассоциирующийся с плавающими растениями (sudd). Затем следует кайма из Pachira aquatica и Pterocarpus officinalis, перемежающихся зарослями Montrichardia и Drepanocarpus lunatus. Еще выше по течению располагается пояс из смешанного болотистого леса с недифференцированной окраиной и, наконец, в значительном отдалении от моря, появляется высокоствольный тропический дождевой лес, спускающийся непосредственно к урезу реки. Эти различные типы растительности, конечно, не отражают прямой последовательности в стадиях гидросерии, но, как указал Майерс, они отчасти зависят от таких факторов, как уменьшающаяся ширина потока, понижающаяся соленость и изменяющийся характер отложений. Изменение растительности по берегам гвианских рек совершенно аналогично изменениям, наблюдаемым, судя по данным Корнера (Corner, 1940, стр. 42) и Саймингтона (Symington, 1943, стр. XVIII), при движении вверх по малайским рекам, где мангрова уступает место прибрежному лесу из Dipterocarpus oblongifolius (нерам) и т. д., и, наконец, появляется сообщество, в котором обильно представлены виды Saraca («потоки сарака»).
312
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ГИДРОСЕРИИ ТРОПИЧЕСКОЙ АФРИКИ
Весьма удобными для наблюдений за стадиями гидросерий являются озера и медленно текущие реки Африки, в особенности на западном побережье, где на сотни миль тянутся цепи мелких пресноводных лагун,, в которые впадают реки. Однако имеющиеся здесь сукцессии мало изучены.
В юго-западной Нигерии (Richards, 1939) сукцессия в лагунах и непосредственно в реках начинается с развития погруженных и свободноплавающих водных сообществ. Первые состоят из таких растений, как Ceratophyllum demersum. Utricularia spp., а вторые — из хорошо развитого «плавающего газона», представленного злаками Pistia и т. д. Плавающий газон образует обширные поля и может стать серьезным препятствием для навигации. Когда уровень воды снижается в достаточной степени, появляется укореняющееся сообщество с плавающими листьями, где обыкновенно доминирует Nymphaea lotus. За этими чисто водными сообществами следует травяное болото, состоящее из злаков или из Cyperus papyrus или же сообщество из Pandanus sp. Иногда за ним может развиться сообщество с доминированием кустарников типа Alchornea cordijolia, но обычно следующую стадию составляет пальмовое болото с доминированием одного (или нескольких?) видов Raphia. Эта стадия подготовляет развитие высокоствольного леса пресноводных болот, состоящего главным образом из двудольных деревьев.
Этот заболоченный лес имеет общее сходство с климаксным тропическим дождевым лесом, но обыкновенно бывает более разреженным и непостоянен по структуре; флористически он беднее, чем первичный лес на сухой почве, но оба они имеют много общих видов. Лес, растущий на пресноводных болотах Западной Африки, вероятно, подвержен сильным изменениям в зависимости от местных условий; Ричардс (Richards, 1939,. стр. 42) описал, по-видимому, широко распространенный тип его, представленный, например, в резервированном лесу Шаша, Нигерия. В этом лесу в изобилии имеются высокие деревья, однако они распределяются неравномерно, так что группы, дающие густую тень и, следовательно, имеющие скудный подлесок, перемежаются в нем с окнами, поросшими густыми зарослями кустарника и лиан. Доминирующим растением этих зарослей является Leea guineensis, раскидистый кустарник, поддерживаемый опорными корнями. Самым обыкновенным деревом является Mitra-gyna sp. (вероятно, М. ciliata), которое, вряд ли являясь доминантой, все же составляет36% древостоя среди деревьев с диаметром ствола 16 дюймов и более. Обилие экземпляров этого вида или видов, ему близко родственных, характерно для обширных пресноводных болот тропической Африки. На островах и по низким, затопляемым речным берегам в Бас-Касаи и в районе озера Леопольда II, Республика Конго, Mitragyna sti-pulosa (или, скорее, недавно выделенный замещающий вид М. ciliata} образует чистый древостой «вроде искусственных насаждений» ^Ver-moesen, 1931, стр. 181, цит. Е. Laurent, М. Laurent). Некоторые из соподчиненных видов болотистого леса Шаша ограничиваются болотами, другие встречаются также и на сухих местах. К последней группе относится целый ряд видов, которые на сухой почве более обычны для вторичного, чем для первичного спелого леса. Частота нахождения их в болотистом лесу, вероятно, объясняется сравнительно хорошим освещением.
Все стадии сукцессии, развивающейся от сообществ открытой воды до высокоствольного леса пресноводного болота, могут в сильно умень-
Фото 17. Лес, развивающийся на пресноводном болоте, Акилла, Нигерия.
На прогалине, которая видна па переднем плане, доминирующим растением является Cyrloaperma unrgah-nw, широколиственное дерево — МП rag у па cillata, а пальма — Haphia зр.
Фото 18. Лес, развивающийся ыа пресноводном болоте, резервированный лес Шаша, Нигерия.
Видны обладающие ходульными корнями экземпляры Uapaca
9laudlii.
ПЕРВИЧНЫЕ СУКЦЕССИИ 31,‘>
шенном виде наблюдаться на берегах крупных рек Нигерии, где растительность располагается соответствующими поясами. Например, на Оссе эти пояса последовательно включают: 1) погруженные водные растения и плавающий газон, состоящий из Pistia и т. п.; 2) Pandanetum; 3) Raphie-tum; 4) лес пресноводного болота.
Чипп (Chipp, 1927) в своем описании растительности Ганы упоминает целый ряд сериальных стадий, очень похожих на те, которые наблюдаются в юго-западной Нигерии. Среди поздних стадий, которые он перечисляет, отметим: 1) «ассоцию лагун на маршах», состоящую главным образом из злаков, причем одной из доминант является Anadelphia', 2) ассоцию Calamus — Ancistrocladus — Raphia или же чистую консоцию Raphia hooker С которая, по-видимому, очень похожа на пальмовое болото Нигерии; 3) ассоцию Tarrietia — Anopyxis, соответствующую лесу пресноводных болот Нигерии.
Госсвейлер и Мендонса (Gossweiler, Mendonca, 1939) описали целый ряд сообществ Анголы, очевидно похожих на стадии гидросерий Нигерии и Ганы. Эггелинг (Eggeling, 1935) довольно подробно описал гидросерию болота Наманве на озере Виктория в Восточной Африке. Хотя абсолютная высота достигала 3730 футов (1140 м), однако общий ход первичной сукцессии оказался весьма похожим на развитие западноафриканских гидросерий. Здесь установлены следующие гидросерии:
1) Водные растения с плавающими листьями и погруженная водная растительность. Виды Nymphaea, Ceratophyllum, Тгара и т. д.
2) «Сообщество папоротников и осоковых». Смесь различных папоротников, осок, злаков и других травянистых растений.
3) Болото Limnophyton. Ассоция с доминированием Cyperus papyrus и злака Miscanthidium violaceum с участием Limnophyton obtusijolium в качестве субдоминанты.
4) Папирусное болото. Доминирует Cyperus papyrus, имеется много второстепенных видов.
5) Болото Phoenix. Деревья высотой 20—30 футов (6—Эле). Из двудольных присутствует Mitragyna stipulosa, из однодольных — пальма Phoenix reclinata.
6) Дождевой лес.
Папирусное болото может непосредственно переходить в болото Phoenix, но иногда развивается промежуточная стадия с доминированием Miscanthidium. Болото Phoenix образует узкий пояс между сообществом Papyrus и Miscanthidium и окружающим лесом. По направлению к опушке леса встречается другая пальма — Raphia monbuttorum.
В крупном массиве дождевого леса бассейна Конго стадии гидросерий должны быть сильно развиты, но сведения о них очень скудны. Лебрэн (Lebrun, 1936а), однако, различает два главных типа болотистого леса в области Конго: а) лес на почве с сезонным затоплением, в остальное время года твердой и сухой; б) заболоченный лес. Первый тип, по-видимому похожий на описанный выше болотистый лес Нигерии, имеет менее густой полог деревьев, чем лес, встречающийся на хорошо дренированных плотных землях (terra firma). Он относительно беден видами; из основных родов деревьев следует отметить ChrysophyHum, Cynometra, Maba. Cuban-guia, Uapaca. Травяной покров отсутствует, если не считать нескольких луковичных и корневищных, которые развивают надземные побеги во время сезона низкого стояния воды. Второй тип, более богатый видами, имеет большее количество деревьев на единицу площади, чем лес, растущий на сухой почве, хотя ни одно из этих деревьев не становится очень
314
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
крупным. Большинство стволов имеет досковидные пли ходульные корни. «Доминантами» являются Chrysophyllum laurentii, Uvariastrum pynaertii и роды Parinari Mitragyna, Copaifera и Uapaca. Виды этих трех последних родов обнаруживают тенденцию образовывать более или менее однородные древостои. Подлесок богат видами, но травяной покров не является сплошным. В упомянутой статье Лебрэна описывается также ряд водных сообществ и сообществ, растущих по берегам рек.
Тротпэн (Trochain, 1940) дал полное описание гидросерий и водных растительных сообществ Сенегала. Хотя климаксом здесь является «климатическая саванна», однако более ранние сериальные стадии тождественны аналогичным стадиям лесных областей.
ГИДРОСЕРИИ ВОСТОЧНОГО РАЙОНА ТРОПИКОВ
В восточных районах тропиков экологические и фитогеографические исследования сильно затруднялись исключительным разнообразием флоры. Поэтому не удивительно, что мы обладаем крайне скудными сведениями относительно гидросерий и пределах распространения дождевых лесов Индо-Малайской области. По-видимому, не существует ни одной опубликованной сводки о гидросерии этих районов, хотя многочисленные сообщества растений на маршах и болотах описывались неоднократно.
Ван-Стеенис и Рутнер (van Steenis, Ruttner, 1932) изучали различные погруженные в плавающие сообщества озер Явы и Суматры, однако большинство этих озер расположено довольно высоко над уровнем моря и нельзя с уверенностью сказать, похожа ли их растительность на растительность низинных водоемов, где сукцессия может привести к становлению того или иного типа тропического дождевого леса. На медленно текущих реках Борнео развивается плавающая растительность, которая вместе с различными типами лесных и болотных сообществ упоминается Ван-Стеенисом (van Steenis, 1935а) в его хорошо документированном перечислении типов растительности Малайского архипелага; правда, в этой работе он не анализирует их сукцессионные взаимосвязи.
Различные типы болотистого леса, большинство которых должно рассматриваться как поздние стадии гидросерий, были описаны Эндертом (Endert, 1920) в его работе о лесных типах Палембанга (Суматра) и Чемпионом (Champion, 1936) в его работе о классификации лесов Индии и Бирмы. Бирманские болотистые леса были описаны Курцем (Kurz, 1875), Стэмпом и Лордом (Stamp, Lord, 1923). Корнер (Corner, 1940, стр. 42) сообщает сведения, относящиеся к некоторым интересным типам болотистых лесов Малаккского полуострова. Так как о сукцессионных и экологических отношениях этих типов болотистого леса почти ничего не известно, то было бы бесполезно подробно на них останавливаться. Однако можно отметить, что во многих из этих лесов число видов деревьев сравнительно ограничено, причем наблюдается тенденция к монодоминантности. Во многих районах Малайского архипелага Melaleuca leucadendron образует характерную консоцию болотистой почвы; это сообщество часто является стадией субсерии или сукцессии, развивающейся после выжигания растительности (Endert, 1932), и, по видимому, не принадлежит к первичной гидросерии.
На низменностях Новой Гвинеи обширные пространства заняты болотной растительностью, включающей леса из саговой пальмы (Met-
ПЕРВИЧНЫЕ СУКЦЕССИИ
315
roxylori), дающей основную пищу местным племенам. Кроме заметок Лема (Lam, 1945, гл. 4) и Лейн-Пула (Lane-Poole, 1925b), у нас почти нет данных относительно этого интересного типа растительности.
КЛИМАКСЫ БОЛОТИСТОГО ЛЕСА
В главе 9 указывалось, что большинство тропических болотных почв не является торфяными, содержа в лучшем случае лишь немного больше перегноя, чем почвы с нормальным дренажем; но там, где дренажные воды являются очень олиготрофными (бедными растворенными минеральными веществами), может происходить образование торфа, которое приводит к развитию омброгенных болотистых лесов, представляющих собой тропический эквивалент верховых болот умеренной зоны. Эти леса развиваются на линзообразных массах торфа в несколько футов толщиной. По-видимому, в соответствии с двумя типами болотной почвы нормальной (не торфянистой) с относительно эвтрофным обводнением и торфянистой с крайне олиготрофным обводнением — находятся два различных типа гидросерий, ведущих к различным типам климаксного леса.
В ходе развития любой гидросерии уровень почвы постепенно повышается, пока не достигнет или даже несколько не превысит уровня нормального стояния воды. В эвтрофных водах это повышение уровня происходит главным образом за счет скопления неорганических отложений, в олиготрофных оно почти целиком обязано скоплению растительных остатков. Однако и в том и в другом случае уровень почвы не может превысить максимального горизонта воды, так как при этом исчезли бы условия, необходимые для накопления или образования торфа. Следовательно, гидросерия заканчивается образованием «сухой» почвы, в которой, однако, грунтовые воды располагаются близко к поверхности по крайней мере в течение части года. В тропиках такая почва способна нести на себе высокоствольный лес, более или менее похожий по структуре и внешнему виду на климаксный, но отличающийся от него по флористическому составу, так как по крайней мере большинство произрастающих здесь видов должно обладать специальными качествами, позволяющими им развиваться в условиях постоянно пересыщенной влагой почвы или же при наличии застойных вод на небольшой глубине. Такой лес должен рассматриваться скорее как эдафпческий, чем как климатический климакс. Конечная стадия гидросерии не может, следовательно, представлять собой типичный тропический дождевой лес (то есть климатический климакс) при отсутствии каких-либо существенных изменений в среде (например, ведущих к общему понижению уровня грунтовых вод).
Болотистый лес неторфяных болот
Примеры заболоченных климаксных лесов, произрастающих на неторфяных болотах, уже указывались. К ним принадлежат, например, лес Мора Брит. Гвианы (гл. 10), лес Варзеа бассейна Амазонки, нигерийский пресноводно-болотистый лес и т. д. Все эти сообщества, по-видимому, являются конечными стадиями (эдафическим климаксом) гидросерий в сравнительно эвтрофных водах, то есть в условиях, неподходящих для образования торфа.
316
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Болотистый лес верховых болот (торфяной болотистый лес)
Леса, растущие на верховых болотах и, следовательно, развивающиеся при наличии олиготрофных вод (см. гл. 9), широко распространены в зоне тропического дождевого леса, в особенности на Малаккском полуострове, на островах Малайского архипелага и в Новой Гвинее. Ввиду трудностей, сопряженных с исследованием этих лесов, они до сих пор мало изучены. Полак (Е. Polak, 1933), давший подробное описание стратиграфических и гидрографических условий, в которых находятся леса верховых болот Суматры и Борнео, только коротко касается самой растительности; некоторые сведения о ее флористическом составе можно почерпнуть у Эндерта (Endert, 1920), Севандоно (Sewandono, 1937) и Сай-мингтона (Symington, 1943) (другие ссылки см. van Steenis, 1935а, стр. 89-90).
Это вечнозеленые леса, в которых доминируют двудольные деревья. Их высота у края болота, иногда достигающая 30 ж, постепенно понижается по направлению к центру, где часто развиваются лишь карликовые формы деревьев (Kriippelholz), заросли которых тут и там прерываются окнами открытой воды (Polak, loc. cit., стр. 16). Хотя в среднем деревья этих лесов имеют меньший диаметр и высоту, чем деревья типичного дождевого леса, они дают значительное количество деловой древесины; на островах Рио и восточном побережье Суматры эти леса широко эксплуатировались, главным образом китайскими лесорубами, применявшими так называемую систему «панглонг». Благодаря обильному подросту основных видов деревьев, а также вследствие отсутствия злаков вроде Imperata, заполняющих лесные вырубки на сухой почве (гл. 17), болотистый лес очень быстро возобновляется (van der Koppel, 1945). Многочисленные коленчатые корни и пневматофоры (стр. 93) деревьев делают лес труднопроходимым.
В состав флоры входят пальмы, виды Pandanus и Podocarpus, а также представители большинства главных семейств, включая Dipterocarpaceaer обычных для индо-малайского тропического дождевого леса; но, вероятно, большинство видов свойственно исключительно этому типу растительности; наблюдается сходство с флорой «борового» леса (стр. 267). Общее число видов ограничено. Севандоно считает, что на Суматре в болотистом лесу рассматриваемого типа насчитывается меньше ста видов. Ярко выражена тенденция к групповому расположению видов, что приводит к развитию на больших пространствах почти чистых сообществ. Флористический состав постепенно изменяется от края болота по направлению к центру. В соответствии с этим на верховом болоте полуострова Панех, Суматра, располагаются следующие зоны: 1) «плавающий лес», 2) лес на торфяном слое толщиной менее 0,5 м\ 3) лес на более мощном слое торфа, состоящий из следующих подзон: а) лес с густым подлеском из пальм Licuala и Zalacca', б) сомкнутый лес; в) высокоствольный лес из деревьев с тонкими стволами, смешанных с карликовыми деревьями; г) карликовый лес с доминантой Tristania (Polak, loc. cit., стр. 17). Эндерт указывает, что Campnosperma macrophy Пит образует почти чистые леса в дельте Моэзи на Суматре; в центре болот, где слой торфа обладает наибольшей мощностью, доминируют Tristania obovata и Ploiarium alternifolium. До самых вершин деревьев взбирается лиана Nepenthes ampullaria.
Концентрические зоны растительности, надо полагать, представляют собой различные стадии гидросерии, но детали в развитии сукцессии не были исследованы. Изучение стратиграфии торфа, проведенное Полаком,
ПЕРВИЧНЫЕ СУКЦЕССИИ
317
показало, что древесные растения доминируют с ранних стадий развития этих верховых болот.
Известно, что в тропической Америке существуют обширные торфяные болота, однако сведений о них почти нет. Буйен (BouilKenne, 1930) описывает лес Игапо (igapo) (торфяное болото) в дельте Амазонки. Деревья тут достигают высоты 30 м, имеют маловетвящиеся кроны и отличаются наличием пневматофор. В изобилии встречаются пальмы, эпифиты и обычные травянистые болотные растения вроде Montrichardia arbores-cens.
Растительность так называемых «пегасовых болот» (pegass swamp) Брит. Гвианы (стр. 237) и своеобразный лес Карапите «carapite» острова Доминика в Вест-Индии (Beard, 1944с), возможно, близки к растительности тропических верховых болот. Лес Карапите произрастает в насыщенном водой грунте, подпираемом подпочвенным пластом железняка; для него характерен «исключительно толстый поверхностный слой сырого перегноя». Деревья достигают в высоту приблизительно 90 футов (27 ,н), причем наиболее обильно представленными видами являются Атапоа caribaea (carapite) и Tapura antillana.
ОБЩИЕ ЧЕРТЫ ТРОПИЧЕСКИХ ГИДРОСЕРИЙ
Ввиду отсутствия данных о гидросериях олиготрофных вод речь пойдет только о сукцессиях в сравнительно эвтрофных водах. Из тех очень неполных сведений, которые имеются в нашем распоряжении, можно заключить, что гидросерии тропиков повсюду весьма сходны по происхождению и в основном следуют той же линии развития, что и в умеренных областях.
Первая стадия состоит из сообществ свободноплавающих и полу-погруженных водных растений. В большинстве случаев их структура и флористический состав очень близки к аналогичным типам растительности умеренной зоны; для них характерны многие одинаковые или близко родственные роды и даже виды. Своеобразной чертой тропических гидросерий являются так называемые плавающие газоны, в которых обыкновенно доминируют злаки. Полное описание этого типа растительности в его классическом выражении на Ниле дано Дойерлингом (Deuerling, 1909). Единственными сообществами вод умеренной зоны, напомнающими сообщества плавающих растений тропиков, являются так называемые «hover» английских топей и речных разливов и плавни дельты Дуная (Pallis, 1916).
В местах интенсивного заиления с низким уровнем вод появляется укореняющаяся растительность с плавающими листьями, состоящая из водяных лилий и т. п.; она подготовляет путь для сообществ растений, которые, хотя и укореняются под поверхностью воды, частично выступают из нее. Там, где свободноплавающая и погруженная растительность, как и растительность с плавающими листьями, не может развиться, например в дельте Амазонки, сукцессия может начаться со стадии водных растений, выступающих из воды.
Эта последняя стадия сменяется кустарником пли сообществами низкорослого леса, похожими на аналогичные сообщества умеренной зоны. С течением времени они уступают место конечной стадии гидросерии.
Таким образом, в ранних фазах гидросерии обыкновенно доминируют травянистые растения; позже они замещаются деревянистыми. В гидро
318
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
сериях как умеренной, так и тропической зоны на промежуточных стадиях важную роль играют однодольные. В умеренных областях они доминируют в стадии тростникового болота; в тропиках, где также часто наблюдается стадия тростникового болота, более обычной является стадия с доминированием пальм. Местами на этих средних стадиях важное значение имеют виды Pandanus. Это временное доминирование однодольных может объясняться, по-видимому, свойственной многим из них исключительной выносливостью к плохо аэрированной среде, окружающей корни. Именно это в соединении с их ограниченным ростом в высоту исключает однодольные из более поздних стадий сукцессии.
Последней фазой сукцессии является высокоствольный лес с доминированием двудольных деревьев. На этой фазе уровень воды находится все еще у поверхности почвы или совсем близко от нее даже во время сухого сезона. Спелый болотистый лес безусловно является тропическим эквивалентом пойменного леса или ивовых зарослей, занимающих аналогичные местообитания в Европе при доминировании видов Salix, Alnus и т. д. С первого взгляда такой лес кажется весьма похожим как по структуре, так и по общему виду на первичный смешанный дождевой лес, растущий на местах с хорошим дренажем; действительно, многие виды оказываются общими для этих типов тропического леса. Однако при более подробном исследовании между ними выявляетя целый ряд важных различий. В болотистом лесу на единицу площади приходится большее число деревьев, но, несмотря на это, он имеет более открытый и менее правильный по очертаниям полог, чем лес, растущий на сухой почве. Вследствие хорошего освещения подлесок, как правило, оказывается более густым; он включает светолюбивые виды, среди которых встречаются виды, характерные для молодого вторичного леса, растущего на местах с нормальным дренажем. Флористически болотистые леса отличаются от первичного смешанного леса сухих местообитаний не только присутствием во всех ярусах многочисленных видов, характерных исключительно для болот, но и тенденцией к монодоминантности. Общее число видов на единицу площади тут меньше, чем в лесах, растущих на сухой почве, но больше, чем на более ранних стадиях гидросерии, потому что в тропической гидросерии, как и в ксе-росерии, наблюдается прогрессивное увеличение видового разнообразия.
Эта конечная стадия гидросерии с доминированием высокоствольных деревьев, отличающаяся наибольшим флористическим богатством, представляет собой, как мы видели, стабильный эдафический климакс. При отсутствии значительных изменений среды, ведущих к устойчивому понижению уровня грунтовых вод, дальнейшее развитие в климатический климакс тут невозможно, так как высокий уровень воды, сопровождающийся плохой аэрацией почвы, исключает возможность появления большинства характерных доминант смешанного тропического дождевого леса.
Глава 14
ПРИБРЕЖНЫЕ СУКЦЕССИИ
Отправной точкой развития прибрежных сукцессий являются морские отмели, сложенные илом, песком или кораллами и располагающиеся ниже или выше максимального уровня приливов. Как и в других первичных сукцессиях, в их развитии отражается постепенный переход от условий, которые могут переносить только немногие специализированные виды, к условиям, пригодным для успешного существования гораздо большего числа видов. На ранних стадиях этих сукцессий такие факторы среды, как влияние приливов, воздействие волн, морских ветров и соленой воды, а также характер субстрата, играют гораздо большую роль в определении типа растительности, чем климат. В дальнейшем значение этих неклиматических факторов по сравнению с климатическими уменьшается, и растительность постепенно становится похожей на климатический климакс района. Это объясняет, почему более ранние стадии прибрежных сукцессий проявляют сходство не только в местах развития тропических дождевых лесов, но и далеко за их пределами. Так, «формация Pes-caprae*— растительное сообщество, характерное для песчаных берегов экваториальной зоны,— выходит далеко за пределы тропиков, а мангровая растительность, правда в сильно обедненном виде, простирается на север вплоть до Бермудских островов, залива Акабы в Красном море и южной Японии; ее южная граница пересекает Наталь и северную часть Новой Зеландии (Schimper, 1935, стр. 585—587). Таким образом, эти прибрежные сукцессии оказываются в областях с разнообразными типами климата, в пределах которых климаксная растительность изменяется от тропического дождевого леса до пустынных ассоциаций, в связи с чем их поздние стадии, естественно, будут различными.
Основы сведений о прибрежной растительности тропиков были заложены Шимпером (Schimper, 1891) в его классической работе об индомалайской береговой флоре. В соответствии с его классификацией прибрежные сообщества подразделяются на формации Pes-caprae и Barringtonia. характерные главным образом для открытых песчаных берегов, и на формации мангровых и пальмы Nipa, встречающиеся преимущественно на укрытых илистых берегах. Первые два из этих сообществ могут укорениться лишь в местах, расположенных выше среднего уровня высоких приливов; другие могут начать свое развитие в полосе отлива. В промежуточных типах местообитания развиваются смешанные или переходные сообщества; возможно, что некоторые типы береговой тропической растительности нелегко поместить в рамки этой классификации. Сукцессии скалистых морских берегов родственны скорее ксеросерням, чем другим береговым сукцессиям, и не могут быть рассмотрены здесь.
320 ТРОПИЧЕСКИЙ дождевой лес
РАСТИТЕЛЬНОСТЬ ПЕСЧАНЫХ БЕРЕГОВ
На песчаных берегах Индо-Малайской области формация Pes-caprae образует зону низкорослых травянистых растений, среди которых наиболее постоянным (и часто доминирующим) видом является Ipomoea pes-caprae. За ней, в местах, где естественная растительность не была уничтожена, часто располагается узкий пояс прибрежного редколесья, для которого характерны Barringtonia speciosa и В. racemosa. Оба эти сообщества широко распространены по берегам тропической части Индийского и западной части Тихого океана. В Вест-Индии формация Pes-caprae появляется вновь в окаймлении древесной растительности, близко соответствующей формации Barringtonia. Обе эти формации могут быть подразделены и далее, причем переход от одной к другой бывает как постепенным, так и резким. Поясное расположение растительности лучше всего проявляется на широких отлогих берегах, где часто развивается система песчаных дюн. В местах сужения отлогого берега ширина полос сильно уменьшается, а иногда одна из полос выклинивается. После Шимпера тропические растительные сообщества на песчаных берегах неоднократно описывались (Ulte, 1901; Cheeseman, 1903; Schenck, 1904; Tansley, Fritsch, 1905; Guppy, 1906; Diels. 1906; Rechinger. 1908, 1910, стр. 207— 210; Borgesen, 1909; Gleason, Cook, 1926; Christophersen, 1927; Chipp, 1927; Daniker, 1929; Raunkiaer, 1934; Stehle, 1935; Chapman, 1944 и т. д.); ссылки на остальную литературу о прибрежной растительности Индо-Малайской области можно найти у Ван-Стеениса (van Steenis, 1935а).
Формацию Pes-caprae правильнее было бы назвать ассоцией Ipomoea pes-caprae — Canavalia, поскольку как в восточных, так и в западных тропиках в ее составе почти также постоянно, как и Ipomoea, присутствуют виды рода Canavalia из семейства бобовых. Это сообщество является тропическим аналогом прибрежной растительности Европы и Северной Америки, в которую входят такие виды, как Agropyron junceum и Arenaria peploides; по крайней мере один вид (Salsola kali) одинаково характерен для обоих типов растительности. Формация Pes-caprae представляет собой смешанное сообщество растений, из которых многие, включая Ipomoea и Canavalia, являются стелющимися растениями с длинными побегами, распростертыми на поверхности песка. Некоторые виды, в частности Ipomoea, иногда встречаются и внутри страны, в то время как другие строго ограничиваются лишь морским побережьем. Большинство видов отно сится, вероятно, к галофитам, которым не вредит временное погружение в морскую воду и высокое содержание соли в почве. Подобно другим галофитам, часть из них является суккулентами. Семена и плоды большинства этих растений хорошо плавают в морской воде и распространяются именно этим способом; они прорастают главным образом в наносах с запасами перегноя и влаги.
Кроме нескольких видов Ipomoea и Canavalia, здесь обыкновенно находятся различные другие двудольные, например Sesuvium portulacastrum, и злаки, включая Sporobolus virginicus\ в восточных тропиках обычны Spinifex spp., Thuarea involuta, Zoisia matrella и др. Поодиночке или группами встречаются такие кустарники, как Suriana maritima,Scaevola frutes-cens и Tournefortiag naphalodes. Вокруг более высоких растений этого сообщества может скапливаться песок, образуя зачатки дюн; сливаясь, последние превращаются в настоящие дюны.
Заметки о внешнем облике и биологии этих береговых растений можно найти в нескольких упомянутых выше статьях, а также у Шимпера (Schim-
ПЕРВИЧНЫЕ СУКЦЕССИИ
321
per, 1903, 1935). Их жизненными формами занимался Раункиэр (Raun-kiaer, 1934, гл. 5).
Формация Pes-caprae может образовать сплошной покров на почве, но вблизиморя она обычно является более или менее открытым сообществом растений. На песчаных побережьях Цейлона (Tansley. Fritsch, 1905) можно различить внутреннюю и внешнюю зоны формации Pes-caprae, причем первая является открытой и целиком состоит из видов, ограничивающихся в своем распространении морскими берегами, а последняя — сомкнутой и наряду с типичными прибрежными видами содержит различные сорняки, свойственные внутренним районам острова. На береговых песчаных дюнах Барбадоса (Gooding, 1947) также наблюдаются две зоны, сравнимые с цейлонскими, а именно: а) внешняя, без Ipomoea, состоящая из одиночных низкорослых растений, среди которых преобладают Sporo-bolus virginicus, Philoxerus vermicularis и Euphorbia buxifolia, и б) внутренняя, где доминирует Ipomoea, сопровождаемый различными подчиненными видами, включая в некоторых местах Canavalia maritima1, а на стороне, обращенной к морю, нередко образуется пояс из Tournejortia gnaphalodes\ растительный покров здесь сомкнут или становится сомкнутым по мере увеличения расстояния от моря. Еще дальше от берега располагается полоса скраба с доминированием Coccoloba uvifera (см. ниже, стр. 322). Раункиэр (Raunkiaer, 1934, гл. 5) различает несколько «фаций» формации Pes-caprae.
В сухих климатах, например на многих коралловых островах Тихого океана (Christophersen, 1927), сообщества низкорослых галофптов часто тянутся далеко в глубь суши, покрывая обширные пространства. Большинство видов этих сообществ встречается также в типичном узком поясе формации Pes-caprae.
Часто на тропических побережьях по мере удаления от берега кустарники идеревья становятся все более и более многочисленными, в результате чего возникает плавный переход от преимущественно травянистого сообщества Pes-caprae к преимущественно древесному типу растительности. Однако в ряде мест переход этот является внезапным, так что прибрежный лес, или формация Barringtonia, образует сплошную стену позади низкорослой растительности, расположенной в зоне приливо-отливной полосы.
Формация Barringtonia в своем типичном выражении развивается на песчаных побережьях более влажных тропических районов Индийского и Тихого океанов в местах с редким населением; там, где население значительно, деревья обыкновенно вырубаются на топливо или при расчистке земли для плантаций кокосовых пальм. Она представляет собой сомкнутый лесной пояс или же редколесье, где группы деревьев перемежаются открытыми пространствами, примерно как в саванне. Иногда, например на Соломоновых островах и по берегу Новой Гвинеи, формация Barringtonia образует узкую полосу деревьев, приближающихся по высоте к деревьям тропического дождевого леса внутренних районов страны. Наиболее постоянными и характерными деревьями этого сообщества являются: Barringtonia speciosa и В. racemosa, Calophyllum inophyllum, Terminalia catappa, Thespesia populnea, Hibiscus tiliaceus, Tournejortia argentea, Pan-danus tectorius и Casuarina equisetijolia. Последний вид часто образует исключительно чистые древостои в особенности на быстро нарастающих
1 На Барбадосе этот вид встречается редко, тогда как на других островах Вест-Индии он доминирует в этой зоне.
322
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
береговых отмелях вроде песчаных кос, расположенных в устьях рек. Большая часть этих видов никогда или почти никогда не встречается вдали от побережья, но некоторые из них не выносят затопления морской водой. Баркилл (Burkill, 1928) заметил, что в Паганге (п-в Малакка) после затопления исключительно высоким приливом листья многих деревьев в полосе Barringtonia пожелтели и опали. Многие стволы искривились, изогнувшись или наклонившись в сторону моря. Обилие лиан и густота подлеска делают лес труднопроходимым; в областях с большим количеством осадков эпифиты на ветвях представляют обычное явление. Биологией видов формации Barringtonia подробно занимались Шимпер (Schimper, 1891), Тэнсли и Фритч (Tansley, Fritsch, 1905) и др. Подобно формации Pes-caprae. формация Barringtonia может быть подразделена на зоны; более мелкие деревья и кустарники, например Scaevola frutescens, Tournefortia и Panda-nusspp., обыкновенно растут по окраине, обращенной к морю, а более высокие виды располагаются дальше от берега. В некоторых местах наблюдается постепенный переход от леса Barringtonia к характерному для данной местности лесу внутренних районов.
На берегах Вест-Индии также существует полоса древесной растительности, во многом соответствующая формации Barringtonia восточных тропиков. Иногда она состоит из деревьев, достигающих 20 м высоты и перевитых лианами, но часто, в особенности на очень открытых берегах, эта растительность отличается никзорослостью, образуя скраб или заросли. Обыкновенно здесь, по крайней мере на обращенной к морю стороне, доминирует маленькое дерево, или кустарник, Coccoloba uvifera, хотя не менее характерны и другие виды, например Hippomane mancinella и Chryso-balanus icaco.
На обращенной к морю стороне по границе с формацией Pes-caprae Coccoloba часто образует почти чистое сообщество с ничтожной примесью других видов, но по мере удаления от моря ее доминирование становится менее выраженным; в изобилии появляются другие деревья, кустарники и травы. Некоторые из соподчиненных видов, например Terminalia catappa и Thespesia populnea, являются пантропическими, встречаясь и в формации Barringtonia восточных тропиков, а несколько родов представлено сходными видами как в сообществах Coccoloba. так и Barringtonia. На Тринидаде Бирд (Beard, 1946b) различает два литторальных сообщества песчаных отмелей — ассоциацию Coccoloba—Hippomane в кустарниковой или порослевой форме и более высокую ассоциацию Manilkara bidentata с пальмой Roystonea oleracea. Последняя ассоциация обыкновенно располагается дальше от моря и смыкается с лесом внутренних районов страны. Ширина полосы Coccoloba изменяется в зависимости от степени открытости побережья и, по-видимому, контролируется влиянием морских ветров.
Хотя о растительности песчаных берегов в тропиках писалось много, на ее сукцессионные взаимосвязи почти не обращалось внимания. Ясно, что полосы растительности между обнаженной приливо-отливной полосой и лесом или иным сообществом внутренних районов представляют собой, по крайней мере потенциально, стадии сукцессии. Поскольку характер различных прибрежных сообществ, видимо, определяется содержанием соли в почве, степенью и частотой затопления морской водой и сильными ветрами, несущими соленые брызги, то надо думать, что сукцессия не будет прогрессировать, пока береговая линия остается постоянной и не продвигается в море. Когда под действием ветров, волн и течений, а также дюнообразующей деятельности самой растительности новые накопления песка расширят береговую зону, растительные сообщества смогут подверг
ПЕРВИЧНЫЕ СУКЦЕССИИ
323
нуться быстрому изменению и станут тогда действительными стадиями сукцессии.
В связи с этим очень интересно познакомиться с наблюдениями, проведенными Гудингом (Gooding, 1947), на песчаных дюнах острова Барбадос. Здесь, как и на прибрежных песчаных дюнах других мест, содержание соли в почве уменьшается с увеличением расстояния от моря; в то же время благодаря поступлению в почву растительных остатков, содержание воды в ней увеличивается, как это видно из следующих цифр (для одного пункта):
Среднее содержание соли в почвенной воде, % Средняя обводненность песка, %
Полоса пионерной растительности (50 м от берега моря) 5,3 0,26
Полоса Ipomoea pes-caprae (100 м от берега) 1,8 0,48
Полоса Coccoloba (150 м от берега) 0,4 1,65
Характер почвы совместно с действием ветра (что проявляется в карликовых, изломанных ветром формах кустарника Coccolobd), вероятно, препятствует соответствующим полосам растительности распространяться в направлении приливо-отливной полосы, но на небольших ограниченных пространствах иногда можно видеть настоящую сукцессию, которая показывает, что три упомянутые полосы растительности действительно представляют собой стадии псаммосерии или сукцессии, похожей на ту, которая наблюдается на песчаных побережьях Европы и Северной Америки. В этой сукцессии важную роль играют сами растения; около них скапливается песок, а к почве присоединяются их отмершие остатки. Там, где не возникает настоящей сукцессии, каждая полоса находится в равновесии со средой и может рассматриваться как эдафический климакс, не вторгающийся в соседние зоны.
Подобная же картина, несомненно, наблюдается и в других областях. На Кракатау, как мы видели, на морском берегу наблюдались активные сукцессии. В общем, вероятно, полосы растительности на песчаных берегах тропической зоны можно рассматривать как стадии сукцессии, которая при определенных условиях могла бы стать активной.
МАНГРОВАЯ РАСТИТЕЛЬНОСТЬ Н «ФОРМАЦИЯ NIP А»
Слово мангрова применяется как для обозначения экологической группы видов, обитающих в приливо-отливной полосе, так и для обозначения растительных сообществ, состоящих из таких видов. В первом смысле мангрова — это вечнозеленые деревья и кустарники, принадлежащие к нескольким таксономически далеко отстоящим семействам. Их роднит предпочтение, оказываемое одинаковому местообитанию и схожий облик; они, следовательно, сходны по физиологическим признакам и структурным приспособлениям; у большинства из них имеются пневматофоры, или «дыхательные корни», и более или менее выраженная тенденция к живо
324
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
рождению. Мангрова является, может быть, наиболее замечательным примером растений с эфармонической конвергенцией1, или сходством между неродственными видами, произрастающими в одинаковых условиях среды. С настоящими мангровыми ассоциируются виды со сходными, но менее сильно выраженными признаками; для таких видов Тэнсли и Фритч (Tansley, Fritsch, 1905) предложили название полумангров.
Мангровая растительность, или «мангровая формация» Шимпера, Представляет собой комплекс растительных сообществ, покрывающих большие пространства и окаймляющих полосу мелководья у тропических берегов. Мангрова изменяется от леса, достигающего 30 и более метров высоты, до хилого кустарника (скраба), едва лишь поднимающегося до 2 м. Монотонно темная, блестящая зелень полосы мангровы и, по-видимому, совершенно непроницаемая путаница воздушных корней представляют одну из наиболее характерных черт ландшафта многих тропических берегов. Термин «приливный лес», который иногда присваивается мангровой растительности, является не совсем подходящим, потому что, укореняясь временами ниже приливо-отливной полосы, она может встречаться и выше зоны приливов, а также и на берегах, где вообще приливы отсутствуют. По эстуариям мангровая растительность проникает далеко в глубь страны. Типичные мангровы находятся на укрытых илистых берегах, где суша часто очень интенсивно наступает на море, но встречаются также на коралловых рифах и на песчаных берегах, где нарастание суши идет очень медленно (Chapman, 1944). Там, где происходит увеличение площади суши, мангровы принимают в этом активное участие. Преграждая течение, они увеличивают интенсивность отложения осадков; связывая почву корнями и добавляя перегной, они содействуют ее укреплению; может даже образоваться нечто вроде морского торфа. По мере поднятия уровня почвы мангровы все далее проникают в сторону моря и, следовательно, играют в соленой и солоноватой воде роль, сходную с топ, которую в пресной воде выполняют тростники, пресноводные гпдросериальные сообщества.
В солоноватоводных эстуариях обширные пространства занимают полумангровы, образованные, например, пальмой Nipa fruticans, они часто присоединяются к сообществам настоящих мангров. Эти полумангровые сообщества называют, согласно Шимперу, «формацией Nipa», хотя настоящая консоция Nipa представляет собой только одно, свойственное восточным тропикам сообщество этого рода. В Америке и Африке имеются аналогичные сообщества, например болота Manicaria saccifera северо-восточной части Южной Америки и сообщество Pandanus западного побережья Африки (стр. 330).
Как уже указывалось, мангровая растительность распространяется приблизительно до 32° с. ш., а в южном полушарии она проникает в области, еще более удаленные от экватора, но наибольшей роскоши и флористического богатства она достигает во влажных тропиках, где дождевой лес представляет собой климатический климакс; в своем наивысшем выражении она представлена на берегах Малаккского полуострова и соседних островах. Можно различать две мангровые формации — восточную на берегах Индийского и западной части Тихого океана и западную на берегах Америки, Вест-Индии и Западной Африки. Австралийские мангровы в некоторых отношениях отличаются от остальной части восточной формации и, возможно, заслуживают того, чтобы их считали отдельной «субфор
1 В русской литературе это явление чаще обозначается термином «биологи еская конвергенция».— Прим. ред.
ПЕРВИЧНЫЕ СУКИ ЕСС И И
325
мацией». Восточные и западные формации в основном схожи по их внешнему виду и экологическим отношениям, но восточные мангровы гораздо богаче видами. Хотя все роды растений западных мангров находятся и в восточных, их видовой состав оказывается совершенно различным. Тихоокеанские острова Фиджи и Тонга представляют собой нечто исключительное в том отношении, что в их флоре наряду с восточным видом рода Rhizophora (R. mucronata) существует и западный R. mangle. Мангровы благодаря их многочисленным структурным и физиологическим особенностям представляют выдающийся научный интерес; они имеют также большое хозяйственное значение, не только косвенное, проявляющееся в том, что они «отвоевывают» сушу у моря, но и прямое, так как они дают топливо, деловую древесину и дубильное корье. Вследствие этого лишь немногие типы тропической растительности были изучены больше, чем мангровые сообщества. Здесь нет места подробному рассмотрению этого предмета, который затрагивает множество вопросов, не имеющих прямого отношения к основной теме книги. Среди наиболее ценных современных работ в обширной литературе по мангровым следует назвать работы Уатсона (Watson, 1928), Дэвиса (Davis, 1940) и Чапмэна (Chapman, 1944). Весьма интересны также сводки Ван-Стеениса (van Steenis, 1935а) по мангровам Индонезии и общий обзор Фабера (Schimper, 1935). Гашги (Guppy, 1906) подробно изучал вопросы географического распространения мангров, а Фабер (Faber, 1923), Тролль и Драгендорф (Troll, Dragendorf, 1931), Вальтер и Штейнер (Walter, Steiner, 1936—1937), Чапмэн (Chapman, 1944) занимались различными вопросами, связанными с их физиологией и аутэкологией. Нас сообщества мангровых интересуют главным образом с точки зрения их сукцессий.
Одна из наиболее характерных черт мангровых болот проявляется в расположении доминирующих видов более или менее параллельно береговой линии. У внутреннего края мангровы обыкновенно наблюдается постепенный переход к лесу пресноводных болот или к какому-либо другому типу неморской растительности. Это по аналогии с песчаными побережьями заставляет предполагать существование здесь по крайней мере потенциальной сукцессии, а в местах быстрого приращения берегов легко наблюдать и настоящие сукцессии. Во время развития этих сукцессий многие факторы среды, и в особенности относительный уровень суши и воды, изменяются. Эти факторы находятся в весьма сложном взаимодействии, вследствие чего и мангровые сукцессии не отличаются простотой. Природу этих сукцессий легче всего познать, если сосредоточить внимание на двух наиболее интенсивно исследованных областях — мангровых лесах Малаккского полуострова (Watson, 1928) и Флориды (Davis, 1940). Удобнее сначала рассмотреть мангровые леса Флориды, так как небольшое число составляющих их видов упрощает задачу.
Мангровая растительность Флориды проникает на север вплоть до южной границы «губительных» заморозков. В ее составе присутствуют все виды западной мангровой формации, располагающиеся, как и в других местах, поясами, которые могут быть поставлены в связь с частотой воздействия приливной волны, природой субстрата, активностью процессов приращения суши и эрозии и засоленностью почвенных вод. Развитие растительности в одном поясе подготовляет условия для ее роста в другом. Следовательно, одно сообщество следует за другим, пока не установится окончательный тип растительности, который свойствен внутренней части страны и не переносит затопления морской водой. Можно различить следующие главные типы мангровых сообществ.
326
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
1) «Исходное (пионерное) семейство Rhizophora». Оно состоит главным образом из молодых растений Rhizophora mangle, растущих на почти постоянно затопленной почве. С Rhizophora ассоциируются полупогруженные морские покрытосеменные Thalassia testudinum и Cymodocea manatorum, в некоторых местах злак Spartina alterniflora. Часто встречаются всходы мангровых Avicennia и Laguncularia. Живородящие проростки Rhizophora плавают в море и укореняются на мелях, песчаных банках и т. п., а также по внешнему краю спелого мангрового леса при условии достаточно тихой и не слишком глубокой воды. Молодые растения прочно прикрепляются к рыхлому субстрату с помощью многочисленных опорных корней. Обыкновенно на 100 кв. футов приходится от20до 30молодых растений Rhizophora.
Осаждение и накопление растительных и животных остатков поднимает уровень почвы и укрепляет ее, так что с укоренением растения уже могут выдерживать сильный напор волн.
2) «Консоция спелой Rhizophora». Постепенно пионерное семейство Rhizophora развивается в спелое сообщество, то есть превращается в лес высотой до 30 футов (9 м). Уровень почвы к этому времени становится значительно выше, и ходульные корни деревьев так переплетаются, что оказываются способными улавливать все без исключения остатки. Теперь деревья укоренились уже настолько основательно, что вырвать их может только сильный ураган.
Морские водные покрытосеменные здесь менее обильны. Сообщество Rhizophora, которое может долгое время сохраняться в виде почти чистой консоции, может и быстро измениться под влиянием инвазии других видов, способных произрастать на твердой почве в укрытии, создаваемом Rhizophora. При изменении сообщества может развиться внутренний пояс из мангровой Avicennia в ассоциации с низкорослыми растениями, обычно растущими на засоленных маршах.
Во многих частях побережья вне берега не существуют какие-либо пионерные стадии, помимо спелой консоции Rhizophora. В таких местах, хотя мангровое сообщество и не распространяется, как обычно, при помощи развивающихся уже на дереве проростков, деревья могут посылать корни в более глубокие воды, где среди этих корней будут скапливаться отложения, и таким образом мангровое болото станет медленно распространяться в море.
3) «Ассоция Avicennia засоленных маршей». За внешней консоцией Rhizophora на суше, затопляемой через правильные промежутки времени или лишь от случая к случаю, обычно располагается пояс с доминированием Avicennia nitida. Это растение образует открытый лес, составляющий яркий контраст при сравнении с плотно переплетенной консоцией Rhizophora, с подлеском из суккулентных кустарников вроде Batis maritima, Salicornia perennis и злаков, характерных для засоленного марша. В местах с низким уровнем почвы подлесок может быть скудным, но очень характерной является густая заросль, располагающаяся среди пневматофор Avicennia и состоящая из растений засоленных маршей. Ассоция Avicennia лучше всего развивается на участках, не подверженных регулярному затоплению приливом. Естественное возобновление Avicennia идет медленно, и, по мере того как старые деревья отмирают, лес становится все более и более открытым, давая место для внедрения растительности засоленных маршей и для инвазии видов из ассоции Conocarpus, которая часто граничит с внутренним краем пояса Avicennia. При полном исчезновении деревьев место мангрового болота может занять луговая растительность или растительность засоленного марша.
ПЕРВИЧНЫЕ СУКЦЕССИИ
327
4) «Переходная ассоция Conocarpus». Полумангр Conocarpus erectus, хотя и не всегда бывает обильно представлен, является наиболее характерным видом этого сообщества; он произрастает в местах, редко затопляемых приливами, располагаясь непосредственно за ассоцией Avicennia. Подобно этой последней, ассоция Conocarpus представляет собой открытое сообщество, состоящее из деревьев и кустарников с подлеском из низкорослых растений засоленного марша. В этом сообществе растет большое количество видов деревьев, кустарников, злаков и покровных трав, а также папоротников и эпифитов. Некоторые из этих растений свойственны также и «лесам хэммок»1 (вечнозеленые леса внутренних районов); многие деревья слабо развиты и представляют собой просто случайно занесенные экземпляры из сообществ, существующих внутри страны.
Во многих местах, хотя и не везде, можно найти доказательства того, что почва ассоции Conocarpus была создана мангровой, и поэтому названная ассоция является частью мангровой сукцессии. По внутреннему краю пояс Conocarpus переходит в негалофитные сообщества: «леса хэммок» (стр. 390), сосновые леса, ассоциации пальм и пород с твердой древесиной или дюнную ассоцию. В некоторых «лесах хэммок» есть признаки, свидетельствующие о том, что почва этих лесов подстилается почвой мангров, но в других местах не наблюдается каких-либо очевидных сукцессионных связей между мангровой растительностью и формациями внутренней части страны. Иногда мангровая растительность непосредственно переходит в климаксный лес внутренней части страны, минуя ассоцию Conocarpus.
5) «Ассоциация спелого мангрового леса». В некоторых местах на побережье Флориды растет очень высокий, роскошный мангровый лес, в котором 60% высоких деревьев образует Rhizophora и около 30 ?о — Avicennia. Кроме них, в виде незначительной примеси в нем также встречаются мангровые Laguncularia racemosa, Conocarpus и различные пальмы, а также другие растения, характерные для внутренних районов. В пределах сообщества не наблюдается признаков поясности в распределении растений. Почва регулярно и на большую глубину затапливается высокими весенними приливами, а во время влажного летнего сезона обильно пропитывается пресными водами, но обычные приливы не достигают этих мест, и вода редко бывает очень соленой. Характерной чертой сообщества является торфянистая почва толщиной 10—14 футов (3—4 м), образованная остатками мангров. Это сообщество, видимо, очень стабильно, и нет основания думать, что оно будет развиваться дальше.
Кроме упомянутых пяти главных сообществ флоридской мангровы, есть и другие, имеющие менее важное значение. Так, например, местами, под влиянием не вполне определенных факторов развивается чистый древостой Laguncularia. Известны также сообщества чахлой, редкостойной мангровы, но они неясно отличаются от нормальных высоких мангровых сообществ.
Сукцессионные отношения между различными мангровыми сообществами носят сложный характер и еще не вполне выяснены, но в них различают несколько гипотетических серий. Предположения Дэвиса о соотношениях между основной мангровой и ассоциированными с нею сообществами графически переданы на рис. 36. Основная линия сукцессии, видимо, идет от «исходного (пионерного) семейства Rhizophora» к «консоции спелой Rhizophora» и от нее через «ассоцию засоленных маршей Avicennia» и пере-
1 Хэммок (hammock) — так называются в южных штатах Северной Америки, особенно во Флориде, древостои, состоящие из пород с твердой древесиной. — Прим. ред.
328
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ходную ассоцию Conocarpus к «лесу хэммок» — климатическому климаксу района (стр. 390). Положение «ассоциации спелого мангрового леса» неясно. Дэвис рассматривает ее как стабильный «субклимакснып» тип, не имеющий тенденции превращаться в какой-либо иной тип растительности и поэтому не участвующий в главной сукцессии, развивающейся от сообществ, открытой морской воды до леса внутренних районов. Мы
Ассоции отлого-
Дюнные ассоции
Тропический лес Климаксная ассоциация
Ассоции спелого мангрового ' леса
CONOCARPUS Переходные ассоции
Эдафическая субклимаксная ассоциация сосны
Ассоции п/ясноводных маршей
Ассоции открытой (соленой) воды----------- •
Консоиии Lagunadana
Ассоции прерии Марль
AVICENNIA Ассоции засоренных маршей
Rhizophora лоции пресноводных маршей
Rhizophore лшцш солоноващых маршей
7Jpe3nutT уровень высоких приливов
/ мангрового скраба
Консоции спелой RHIZOPHORR
Средний уровень низких приливов
Пионерное семейства RHIZ0PH0RR
I
Морские ассоции
Рис. 36. Сукцессионные соотношения мангровых и некоторых ассоциированных с ними растительных сообществ Флориды (J. Н. Davis, 1940).
можем, пожалуй, рассматривать этот субклимакс как эдафический или физико-географический климакс, аналогичный лесу торфяных болот или тропическому верховому болоту, о котором говорилось в предыдущей главе, но развивающийся в условиях солоноватых или соленых вод.
Если допустить, что четыре сообщества — исходное сообщество Rhizophora, консоция Rhizophora, ассоция Avicennia и ассоция Conocarpus — представляют собой последовательные стадии сукцессии, то становятся совершенно очевидными сильные изменения среды в ходе развития этой сукцессии. Благодаря взаимодействию растительности и окружающей ее среды уровень почвы повышается. На исходной стадии растительность почти непрерывно находится под водой, но затем затопляемость уменьшается, пока в стадиях Avicennia и Conocarpus приливы вообще становятся редкостью. Эти изменения относительного уровня суши и воды
ПЕРВИЧНЫЕ СУКЦЕССИИ
329^
сопровождаются изменениями солености почвы, как это можно видеть из табл. 33. Средняя соленость, достигающая максимума на стадии Avicennia, в дальнейшем снижается приблизительно до величины, характерной для почвы внутренней части страны. Ни в одном иЗ мангровых сообществ соленость не является постоянной. Она больше всего изменяется у консоций Avicennia, где грунтовые и поверхностные воды только иногда восполняются приливом. В сухую погоду соленость может подняться здесь до очень большой величины, а после дождя упасть очень низко. Кроме замечательной способности всходов укореняться в условиях приливо-отливной полосы и выносливости растений к продолжительному затоплению на всех стадиях развития, главным физиологическим свойством мангровых является их способность расти в среде с высокой, часто изменчивой соленостью. Но так как Rhizophora и даже Avicennia могут в естественных условиях развиваться в местах с совершенно пресными почвенными водами (см. стр. 308), можно сделать вывод, что по крайней мере американские^ мангровые являются солевыносливыми, а не солелюбивыми растениями, или, иными словами, они скорее факультативные, чем облигатные галофиты. Такая точка зрения подтверждается некоторыми экспериментальными данными1.
Таблица 33
Условия местообитания мангровых сообществ во Флориде (J. Н. Davis, 1940, табл. 6, стр. 349)
Сообщество Число образцов Соленость, %
Почвенный раствор Поверхностные воды
Исходное (пионерное) семейство Rhizophora 7 2,91 3,185
Консоция спелой Rhizophora 14 3,56 3,488
Ассоция Avicennia засоленных маршей 13 5,11 3,684
Переходная ассоция Conocarpus 8 1,80 0,144
Ассоциация «леса хэммок» 8 0,35 Поверхностные воды отсутствуют
За пределами Флориды западные мангровые сообщества нигде не исследовались достаточно подробно, но, судя по имеющимся данным, поясное расположение растительности в них, как и общий ход сукцессии, везде в основном одинаковы. Низкорослая растительность засоленных маршей Флориды, ассоциирующаяся главным образом с поясом Avicennia, отсутствует вблизи экватора. Глизон и Кук (Gleason, Cook, 1926) на острове Пуэрто-Рико обнаружили на обращенной к морю стороне мангровы пояс, состоящий из Rhizophora mangle, который в направлении от берега последовательно сменялся поясом Avicennia и поясом Conocarpus. Папоротник Acrostichum аигеит, изредка встречающийся в сообществах Avicennia
1 Фабэр (von Faber, 1923), а также Шимпер (Schimper, 1935) не согласны сданной точкой зрения; возможно, что восточные мангровы, которые эти авторы имеют в виду, в этом отношении действительно отличаются от западных.
330
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и Conocarpus на полуострове Флорида, здесь доминирует, образуя обширные куртины или полосы по внутреннему краю мангровы. Хотя с первого взгляда заросли папоротника могут показаться стадией первичной сукцессии, следующей за сообществом Conocarpus, Глизон и Кук пришли к выводу о их более вероятном вторичном происхождении на местах вырубки мангровы. Аналогичные пояса, но образованные Acrostichum, характерны для внутреннего края мангровы и в других частях земного шара. Когда вмешательство человека отсутствует, то на Пуэрто-Рико, по-видимому, за сообществом Conocarpus должен следовать лес, растущий на солоноватом болоте с доминантой Pterocarpus officinalis и, наконец, «мезофитный» лес, то есть климатический климакс. В сравнительно сухом климате Гваделупы (Stehle, 1935) за мангровыми сообществами следует Acrostichum, в свою очередь уступающий место сообществу влажного луга с Paspalum spp., Kyllinga и т. д.; не ясно, представляет ли этот луг стадию первичной сукцессии.
На Тринидаде и в Брит. Гвиане, как, несомненно, и на низменных побережьях многих других районов тропической Америки, часто в направлении внутренних частей страны наблюдается постепенный переход от мангровы к травянистой или лесной растительности пресноводного болота. Вероятнее всего, как и в Пуэрто-Рико, этот переход представляет собой настоящую сукцессию. Согласно Чапмэну (Chapman, 1944), на Ямайке мангровые сообщества могут быть в конце концов заменены «прибрежнодюнной растительностью», колючим кустарником (скраб) или пресноводным тростниковым болотом с Typha domingensis.
Обширные мангровые леса Западной Африки состоят из тех же видов, что и тропические американские мангровы, и поэтому, как уже было сказано, они могут рассматриваться как часть той же формации. Сведений относительно поясного расположения растительности и экологии западноафриканской мангровы очень мало. Вальтер и Штейнер (Walter, Steiner, 1936—1937) опубликовали наблюдения о мангровой растительности Камеруна и некоторые заметки Шимпера о мангрове устья Конго. Короткое сообщение о мангровых Конго сделал и Пинаэр (Pynaert, 1933). В Камеруне Rhizophora mangle образует пионерное сообщество непосредственно у кромки воды, за ним располагается Avicennia. Еще дальше от берега мангрова уступает место солоноватоводному сообществу с доминированием Pandanus, наряду с которым обычно произрастают и пальмы (Raphia sp., Calamus sp., Phoenix reclinata). Это сообщество напоминает некоторые из тех сообществ, о которых уже говорилось (стр. 312) как о составной части пресноводной гидросерии Западной Африки. Сходные сообщества наблюдаются и в устье Конго, причем следует отметить, что здесь, как и в Америке, Rhizophora иногда растет в совершенно пресной воде.
Обратимся теперь к более сложной восточной формации мангров и рассмотрим мангровые сообщества Малаккского полуострова, описанные Уатсоном (Watson, 1928).
На берегах Малаккского полуострова растут исключительно высокие, роскошные мангровые леса, содержащие гораздо больше видов, чем леса Флориды; Уатсон перечисляет 17 «главных» и 23 «второстепенных» вида. Эти сообщества обыкновенно образуют отчетливые пояса, показанные на рис. 37, хотя иногда встречается и иной порядок расположения. Здесь можно выделить пять главных типов сообществ:
1) Тип Avicennia—Sonneratia griffithii. На полуострове Малакка исходные (пионерные) сообщества образуют не виды Rhizophora, a Avicennia alba и A. intermedia или иногда на мощных илистых отложениях, богатых
ПЕРВИЧНЫЕ СУКЦЕССИИ
331
органическим веществом, Sonneratia grijjithii. Эти леса укореняются на мелях или песчаных банках, обнажающихся в мелководье, или же по внешнему, обращенному к морю краю уже имеющихся лесов. Avicennia intermedia растет на сравнительно твердом глинистом субстрате, по которому свободно можно ходить, A. alba и Sonneratia — на более мягком и более темном иле. На глинистой почве за Avicennia обычно следует Bruguiera caryophylloides, но там, где пионером является Sonneratia, за ней обыкновенно следует Rhizophora mucronata.
2) Тип Bruguiera caryophylloides. Встречается на более высоком абсолютном уровне, чем предыдущий, и образует почти непрерывную полосу па западном побережье Малаккского полуострова позади леса Avicennia, отсутствуя лишь на небольших участках, где лес Avicennia непосредственно переходит в лес Rhizophora. Он произрастает на твердой глинистой почве, в местах, не затопляемых обычными приливами; вода держится здесь лишь один или два дня до и после весенних паводков. Растительность этого типа обыкновенно отсутствует как на мелях, так и в лесах, растущих по берегам рек.
3) Тип Rhizophora. В нем доминируют Rhizophora conjugata п R. mucronata. Этот тип встречается на землях, затопляемых обычными высокими приливами; следовательно, он располагается на несколько более низких абсолютных отметках, чем тип Bruguiera caryoplylloides. Свойственные этой местности многочисленные небольшие поверхностные водотоки содействуют распространению проростков. В пределах полуострова Малакка лес Rhizophora распространен шире, чем какой-либо другой тип мангрового леса. Обычно он растет на темноцветных богатых перегноем почвах с примесью тонкого песка; этот лес плохо развивается на твердых глинистых почвах, но сухой субстрат не является для него помехой, вследствие чего он способен также далеко проникать внутрь страны, как и любое другое из мангровых сообществ, за исключением типа Bruguiera gymnorrhiza. Вероятно, Rhizophora conjugata занимает больший ареал, чем R. mucronata, предпочитающая более влажные места вроде берегов протоков и ручьев. Среди ассоциирующихся с Rhizophora видов можно назвать Bruguiera parvijlora и Xylocarpus granatum. Более старые древостои Rhizophora обыкновенно подвергаются инвазии со стороны папоротника Acrostichum аигеит, который стремится обосноваться на холмиках, сооружаемых креветками при рытье норок, и растет на местах, доступных только самым высоким приливам.
4) Тип Bruguiera parvijlora. Может образовывать чистые древостои в более влажных местах. Некоторые полагают, что он предшествует типу Rhizophora (3), а другие думают, что он за ним следует. Тип Bruguiera parvijlora можно считать «оппортунистическим» сообществом, занимающим то более высокое, то более низкое положение, чем отличающееся большей продолжительностью жизни и более теневыносливое сообщество ризофоры, которая способна вытеснять сообщество Bruguiera (4), там, где условия благоприятствуют распространению ее всходов. Сообщество Bruguiera parvijlora часто растет на местах, прежде занятых Rhizophora, но оно могло заселить их после вырубки
5) Тип Bruguiera gymnorrhiza. Этот тип, несомненно, представляет собой последнюю стадию развития мангровых лесов и знаменует начало перехода к тропическому дождевому лесу внутренних районов. Доминанта является самым крупным и, вероятно, наиболее долгоживущим видом семейства Rhizophoraceae; на ранних стадиях сукцессии она встречается только спорадически. Этот тип мангровы очень теневынослив и может
Разрез по E-F
Объяснения обозначении
0 rhizophora J MUCRONA та Ф RHI20PH0RA £ CONJUGАТА Y BRUGUIERA 1 PARVIFLORA 1 * RRUGUIERA L caryophyuoides BRUGUIERA ERIOPETALA BRUGUIERA . GYMNORHIZA Ф CARA PA A OBOVATA ♦ CARAPA к MOLUCCENSIS fl INTSIA A BIJUGA ? GERIOPS J- CANDOLLEANA °P SONNCRATIA 1 GRimrun 9 AVICENNIA 1 INTERMEDIA t AVICENNIA 1 ALBA ACROSTICRUM Aim AUREUM
Рис. 37. Диаграмма типичного распределения наиболее важных видов мангровых лесов Малаккского полуострова (VValson, 1928).
1 — всегда затопленная полоса; 2 -отлив; з — наинизший уровень прилива; 4 —средний уровень прилива; .5 — максимальный (в дни весеннего равноденствия) уровень прилива.
Фото 19, Пионер мангровы, остров Лейден, Батавский залив, Ява.
Виден варослый вкаемпляр и всходы Rhizophora etyloea.
Фото 20. Спелая мангрова (консоциация Rhizophora mucronata) при низком приливе, залив Ароэ, восточная Суматра.
Деревья достигают 40 м высоты.
обосноваться в сообществах Rhizophora, где затенение настолько интенсивно, что препятствует возобновлению самой Rhizophora. Однако всходы Bruguiera gymnorrhiza чувствуют себя не совсем хорошо в тени материнского дерева. Подлесок состоит главным образом из Acrostichum аигеит. Почва здесь настолько приподнята отложением осадков и скоплением перегноя, а также благодаря действию креветок, что находится выше уровня максимальных приливов. Переход к лесу внутренних районов заметен по инвазии таких видов, как Xylocarpus moluccensis, Intsia bijuga, Ficus retusa, Daemonorops leptopus, Pandanus spp. и многих других. Сукцессия часто прекращает свое существование в результате расчистки леса под сельскохозяйственные культуры.
Из сказанного следует, что отношения, существующие в сукцессиях этих пяти мангровых сообществ, никоим образом не являются простыми, но основная линия первичной сукцессии, вероятно, выглядит следующим образом:
Тип
Avicennia-Sonneralia
I____________________
Тип
Bruguiera caryophylloides
Смешанный тропический дождевой
гт. Гип
Тип
—> Bruguiera
Rhizophora gymnorrhiza
Положение типа Bruguiera parvijlora, как уже указывалось, является несколько неопределенным. В сукцессии наблюдаются многочисленные местные вариации, и было бы неправильным думать, что все без исключения мангровые болота последовательно проходят через все стадии развития или что в сукцессии обязательно присутствуют все типы мангровых сообществ даже в обширных лесах.
Наиболее существенное изменение среды, происходящее входе развития сукцессии, состоит в поднятии уровня почвы, влекущем за собой уменьшение частоты и интенсивности затопления. В отношении реакции к затоплению приливами виды растений мангровы могут быть классифицированы следующим образом: 1) виды, произрастающие на землях, затопляемых всеми приливами; при таких условиях нормально не может существовать ни один вид, выживает лишь одна Rhizophora mucronata; 2) виды, произрастающие на землях, затопляемых «приливами средней высоты». Сюда относятся: Avicennia alba, A. intermedia, Sonneratia grijjithii и растущая по берегам рек Rhizophora mucronata; 3) виды, растущие в местах, затопляемых только «обыкновенными высокими приливами»; в этих условиях растет большинство видов мангровы, а ризофоры обнаруживают тенденцию быть доминирующими. Sonneratia grijjithii хотя и произрастает в этих условиях, но не может размножаться; 4) виды, растущие на землях, затопляемых только весенними приливами. Такие места, видимо, слишком сухи для ризофор и больше подходят для Bruguiera gymnorrhiza и В. caryophylloides; 5) виды, произрастающие на землях, затопляемых только равноденственными или другими исключительными приливами. Здесь господствует Bruguiera gymnorrhiza, но выживают также Rhizophora conjugata и Xylocarpus moluccensis.
Нужно заметить, что стадия, на которой доминирует Bruguiera caryophylloides и которая считается (что может быть доказано) более ранней по времени, чем стадия ризофоры, развивается на более высоком абсолютном уровне, более редко подвергающемся затоплению. Объяснение этого обстоятельства не вполне ясно, но Уатсон предполагает, что строение корневой системы Bruguiera caryophylloides способствует большому скопле
334
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
нию перегноя и, следовательно, поднятию уровня почвы. Затем система протоков создает дренаж, направленный в противоположную от моря сторону. Эрозия размывает скопленные отложения ила, что ведет к вторичному понижению уровня почвы. Когда это произойдет, образуется почва, богатая перегноем и более подходящая для укрепления на ней Rhizophora, чем до появления Bruguiera,
В обширных мангровых болотах побережий Суматры и Борнео произрастают оба рассматриваемых сообщества, причем их соотношения в ходе сукцессии оказываются почти такими же, как и на Малаккском полуострове. По восточному берегу Суматры, согласно Троллю и Драгендорфу (Troll, Dragendorf, 1931), тянется почти непрерывный пояс мангровых, местами достигающий нескольких километров в ширину. Доминирующим видом в пионерном сообществе является Avicennia alba, часто ассоциирующаяся с Sonneratia alba. По направлению в глубь страны Avicennia уступает место видам Rhizophora, Bruguiera, Xylocarpus и т. д., а по внутренней окраине болот обыкновенно располагается пояс пальмы Nipa fruticans. с которой часто ассоциируется Sonneratia. Здесь грунтовые воды солоноватые, а не соленые. Зона распространения приливной волны и пределы проникновения соленых и солоноватых вод не совпадают, и, как подчеркивают Тролль и Драгендорф, некоторые мангровы, в особенности Bruguiera gymnorrhiza, Sonneratia acida и полумангрова Nipa, подобно западным мангровам, могут расти в совсем пресной воде. Еще далее в глубь страны, где естественная растительность не была уничтожена, за поясом Nipa следует широкий пояс лесного верхового болота (стр. 316). На Амбоине и Моллук-ских островах (Troll, Dragendorf, 1931) (рис. 38) мангровые сообщества в основном похожи на такие же сообщества Суматры, причем пионерным видом является Sonneratia alba. На полуострове Малакка и Больших Зондских островах восточная мангрова достигает своего наивысшего и наиболее сложного развития. В других местах, на берегах Индийского и Тихого океанов, особенно за пределами ареала тропического дождевого леса, условия среды обычно оказываются менее благоприятными, почему здесь присутствует меньшее количество видов мангров. Флористическое богатство восточной мангровы, достигающее максимума в Малайской области, уменьшается как в западном направлении, к Африке, так и в восточном, северном и южном направлениях в Тихом океане, пока, наконец, у самых отдаленных границ ее распространения, например на побережье Красного моря и в южной Японии, оно не сведется всего к нескольким видам. В центральной части Тихого океана формация мангров, как таковая, исчезает совершенно, и на острове Таити, например, места, подходящие для мангровы, занимаются зарослями Hibiscus tiliaceus — дерева, обыкновенно входящего в состав прибрежного леса и сообщества полумангровы. С обеднением флоры поясность и сукцессии мангровых сообществ соответственно упрощаются.
Часто в восточной формации мангров пионерами являются Avicennia alba и виды Sonneratia, но надо заметить, что в некоторых местах, например на островах Фиджи (Mead, 1928) и в Новой Каледонии (Daniker, 1929), эту роль, как и в западной мангрове, играет Rhizophora (R. mucronata). По мнению Шимпера (Schimper, 1891), основанному главным образом на его собственных наблюдениях в лагунах Тжилатжап (Ява) и Негомбо (Цейлон), Rhizophora всегда является пионером в образовании восточной мангровы, между тем как Тролль и Драгендорф (Troll, Dragendorf, 1931) полагают, что она всегда следует за каким-либо другим видом; однако ни одна из этих точек зрения пока не доказана.
П Е Р В И Ч Н Ы Е СУК Ц ЕСС И И
На восточном побережье Африки климат на уровне моря нигде не является достаточно влажным для развития типичного тропического дождевого леса; соответственно слабо развита и мангровая растительность. Наиболее роскошного развития и большого флористического богатства она достигает на отрезке берега между Танга и Мозамбиком, но даже и тут насчитывается всего лишь пять видов мангровых. На побережье близ
9
Р н с. 38. Распределение мангровых сообществ на острове Аньбошачжоу (Амбоина) (Troll, Dragendorf, 1931).
1 —полоса Sonneratia alba; 2— смешанная мангрова (Bruguiera spp. и др.); 3 — Rhizophora conjugata; 4 — Sonneratia acida; 5 — Nipa f rut icons; 6 — саговые пальмы.
Танга, в местах, удаленных от устьев больших рек, мангровая растительность занимает по сути дела всю приливо-отливную полосу от участка, обнажающегося в момент самых сильных отливов, до уровня, достигаемого обыкновенными весенними приливами (Walter, Steiner, 1936—1937). В его пределах выделяются четыре пояса: I) пояс чистого древостоя Sonne-ratio, alba, образующий пионерное сообщество; II) пояс Rhizophora mucronata* III) узкий пояс Ceriops candolleana\ IV) пояс Avicennia marina. Кое-где встречаются отдельные деревья Bruguiera gymnorrhiza. главным образом в поясах Rhizophora и Avicennia, В поясе Avicennia находятся также кустарник Lumnitzera racemosa и другие растения.
Еще дальше от побережья за поясом Avicennia следует пояс, совершенно лишенный растительности, вероятно, в силу чрезмерной засолен
336
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ности грунта, которую неспособно перенести какое бы то ни было растение» В местах, где естественная растительность сохранилась, этот пояс сменяется зарослямп негалофильного вечнозеленого кустарника (скраб), в составе которого характерным деревом является баобаб Adansonia digitata. Близ устьев рек поясность несколько иная. Здесь пионером оказывается Rhizophora. а по внутреннему краю мангровы располагается пояс из Acrostichum аигеипг, мангровая растительность незаметно переходит впресновод-ные болотные сообщества. Весьма вероятно, что в сухом климате побережья Восточной Африки за мангровой растительностью в ходе сукцессии следуют исключительно негалофпльные типы растительности, развитие которых на влажных местообитаниях вблизи устьев рек обусловлено эда-фическими факторами; в других местах конечной стадией сукцессии является сообщество Avicennia, которое должно рассматриваться как эда-ч|)ический (или физико-географический) климакс.
Сказанного о мангровой растительности западных и восточных тропиков вполне достаточно, чтобы показать общий ход сукцессии. Прй известных условиях, которые обыкновенно имеются на укрытых от действия прибоя илистых берегах, в районах развития тропических дождевых лесов за мангровой растительностью следует лес пресноводного болота или в некоторых случаях непосредственно климаксный тропический дождевой лес. При других условиях сами мангровые сообщества образуют стабильный климакс, связанный с местными условиями рельефа. В первом случае конечные стадии как мангровой сукцессии, так и первичной гидросерии могут быть тождественны. Вообще же мангровая сукцессия наряду с интересными чертами сходства обнаруживает и важные различия с пресноводной гидросерией эвтрофных вод. В обоих типах сукцессии наблюдается постепенный переход от крайне неблагоприятных к более приемлемым условиям среды; в обоих случаях первоначальные доминанты, чью роль выполняют один или два вида пионеров, постепенно уступают место более смешанному сообществу древесных растений. Важное отличие мангровой сукцессии от гидросерии состоит в том, что травянистые растения и пальмы играют в ней лишь незначительную роль. Пионерами, а также доминирующими видами, следующими за ними, являются двудольные деревья, что служит дальнейшим доказательством подавляющего преобладания древесных растений и фанерофитной жизненной формы в тропической растительности.
Ч а с т ь V
ТРОПИЧЕСКИЙ
ДОЖДЕВОЙ ЛЕС
ВБЛИЗИ
КЛИМАТИЧЕСКИХ ГРАНИЦ ЕГО СУЩЕСТВОВАНИЯ
Глава 15
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС, ЛИСТОПАДНЫЙ ЛЕС И САВАННА
Шимпер (Schimper, 1903, стр. 260; 1935) различает четыре тропические «климатические формации» (низменностей), которые он определяет следующим образом:
«Дождевой лес» — вечнозеленый, гигрофильный по характеру, не менее 30 м высоты, а обычно и выше, изобилующий толстоствольными лианами, а также древесными и травянистыми эпифитами.
«Муссонный лес» — более или менее безлистный в сухое время года (особенно ближе к его концу), по характеру тропофильный, обычно не столь высокий, как дождевой лес, но тоже изобилующий деревянистыми лианами, богатый травянистыми эпифитами, хотя и бедный древесными.
«Саванновый лес»—более или менее безлистный во время сухого периода, изредка вечнозеленый, обычно не ниже 20 л, часто гораздо выше, на поминающий парк, очень бедный подлеском, лианами и эпифитами, но отличающийся пышным травяным покровом, в составе которого выделяются злаки.
«Колючий лес» характером листвы и средней высотой напоминает саванновый лес, но более ксерофилен, с хорошо развитым подлеском, тонкоствольными лианами, скудным травяным покровом (особенно мало злаков) п обыкновенно малым количеством эпифитов. Колючие растения всегда в изобилии.
В добавление к этим четырем формациям низменностей, по структуре более или менее близким к лесу или редколесьям, Шимпер различает еще две другие, где деревья не составляют основного фона: тропический злаковник и тропическую пустыню; первый встречается либо в виде «саванны» (злаковое сообщество с равномерно или неравномерно распределенными деревьями), либо в виде «степи» (злаковник без деревьев).
Эти шесть формаций в сходной форме встречаются в тропиках Америки, Африки и Азии, различаясь, однако, по флористическому составу, и Шимпер считает, что каждая из них является вполне определенной и легко распознаваемой по своей физиономии и структуре. «При осадках не менее 180 см безраздельно господствует высокий лес. В отношении местности с осадками 150—180 см не существует никаких данных. При осадках от 90 до 150 см идет борьба между ксерофильным редколесьем и злаковни-ком. Когда в течение вегетационного периода преобладают сильная жара и длительная засуха, победа остается за ксерофильным редколесьем, а при более умеренных температурах и более равномерном распределении осадков в период вегетации, а также при господстве ветреных, сухих или холодных периодов побеждает злаковник» (Schimper, 1903, стр. 282). Эти шесть формаций, по мнению Шимпера, определяются отчетливо выраженным типом климата; так, в пределах тропической зоны различаются климат дождевого леса, климат муссонного леса, климат злаковников и т. д. Последующие исследования выяснили, что в отдельных случаях эдафические факторы могут преодолевать или изменять воздействие климата. Напри
22*
340
ТРОПИЧЕСКИЙ дождевой лес
мер, полосы вечнозеленого леса [галерейные леса в смысле Швайнфурта (Schweinfurth)] часто вдоль речных долин и русел проникают далеко в область саванн; это обусловливается постоянной влажностью почвы, которая компенсирует сезонную засушливость климата.
Достоинство изложенной классификации Шимпера заключается как в ее простоте, так и в том, что она значительно продвинула вперед описание тропических лесных сообществ; но в настоящее время она устарела. С экологической точки зрения самый большой ее недостаток состоит в том, что одна из этих «климатических формаций», а именно тропический злаков-ник, скорее всего не имеет непосредственного отношения к климату. Тропический злаковник (который, согласно Шпмперу, включает и саванну и безлесную «степь») может непосредственно соприкасаться как с роскошным дождевым лесом, так и с лесом гораздо более «сухого» типа, встречаясь в самых разнообразных климатических условиях. Фактически, как показал недавно Бирд (Beard, 1946b), «тропический лесостепной климат» вообще не существует. Накапливаются данные в пользу того взгляда, что все тропические злаковники низменностей и «открытые» саванны представляют собой либо биотические климаксы, обычно возникающие на месте пожарищ, либо эдафические климаксы, развивающиеся на почвах, почему-либо неблагоприятных для произрастания деревьев, либо стадии гидросерии. Крайне сомнительно, чтобы какое-нибудь из этих злаковых сообществ действительно являлось истинным климатическим климаксом.
За последнее время было сделано несколько попыток создания новой классификации и соответствующей терминологии для тропической растительности; среди них наибольшего внимания заслуживают схемы Чемпиона (Champion, 1936) и Барт Дэви (Burtt Davy, 1938). Однако ни одна из них еще не получила всеобщего признания.
Различные формации, составляющие переход от тропического дождевого леса к пустыне, образуют последовательный ряд, почти сходный во всех областях тропиков, но, как подчеркивали Ричардс, Тэнсли и Уатт (Richards, Tansley and Watt, 1940), для составления классификации, применимой и к Старому и к Новому свету, нужны многочисленные дополнительные сведения об этих растительных сообществах. Наиболее важны данные о структуре (которую можно точно зафиксировать посредством профильной диаграммы) и о внешнем облике составляющих сообщества видов (что также поддается точному измерению или описанию). В конечном итоге, вероятно, можно свести все точные характеристики тропических растительных формаций к количеству и высоте свойственных им ярусов, спектру жизненных форм, процентному соотношению вечнозеленых и листопадных деревьев, мезофилльных и микрофилльных видов и т. п.
До тех пор пока все эти подробности неизвестны, было бы бесполезно обсуждать в деталях сложные проблемы классификации и номенклатуры. Цель этой главы состоит в том, чтобы применительно к ряду областей нарисовать картину связей тропического дождевого леса с более сухими типами леса (вне его климатических границ) и с саванной.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС, ЛИСТОПАДНЫЙ ЛЕС И САВАННА В СЕВЕРО-ВОСТОЧНОЙ ЧАСТИ ЮЖНОЙ АМЕРИКИ
Типы леса на острове Тринидад
Остров Тринидад, отделенный от материка Южной Америки проливом шириной 11 км, особенно благоприятен для изучения тропического дождевого леса вблизи его климатических границ. Хотя площадь острова
Фото 21. Аэрофотоснимок галерейного леса и саванн, район Сиба
Уганда. ’
В саванне доминирует Pennisetum purpureum.
Ф о т о 22. Южноамериканская саванна южнее реки Ориноко, дорога из Упаты к Эуасипати, Венесуэла*
Среди деревьев доминируют С urate На amcricana с примесью Pyrsonima crassifolia и Howdichia wgilioides. Напочвенный покров состоит из Trachypogon низкорослых дер нови иных злаков, верхний слой почвы — белый песок—покрывает водонепроницаемую красную глину, количество осадков составляет около 130 cat в год.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМ/\ТИЧЕСКИХ ГРАНИЦ
341
составляет всего 4543 кв, км (1754 кв. миль), его рельеф, почвенные и климатические условия поразительно разнообразны. Годовая сумма осадков колеблется от 250 см до 120 см, и, что еще существеннее для растительности, в их сезонном распределении наблюдается большая неравномерность; в одних частях острова каждый месяц выпадает свыше 4 дюймов (10 см) осадков, в других — ежегодно бывают длительные засухи. Примерно 23% площади острова составляет резервированный лес, который благодаря сохранившейся до недавнего времени низкой плотности населения в большей своей части является первичным, сравнительно мало измененным человеком.
Леса Тринидада впервые были описаны Маршалом (Marshall, 1934), а позднее Бирдом (Beard, 1944а, 1946b), пополнившим множеством точных данных сведения об их структуре, внешнем облике и флористическом составе. На низменностях и в «нижнем горном поясе» [предгорья высотой примерно до 2500 футов (760 л)] Бирд различает четыре климатические формации, каждая из которых является, по-видимому, самостоятельным климаксом и представлена одной или несколькими ассоциациями или консоциациями, например: 1) предгорный дождевой лес, 2) вечнозеленый сезонный лес, 3) полу вечнозеленый сезонный лес и 4) листопадный сезонный лес. Эти формации образуют ряд, в котором первая занимает районы с особенно благоприятным климатом, последняя — районы с наиболее сильными сезонными засухами, а две остальные — районы с промежуточными условиями. На соседних островах Галф и на материке, в Венесуэле, существуют еще две ксерофильные формации того же самого ряда, а именно — колючее редколесье и кактусовый кустарник, отсутствующие на Тринидаде. Помимо перечисленных главных климатических формаций, на Тринидаде известно также прибрежное редколесье, обусловленное местным климатом морского побережья, и ряд эдафических формаций, которые не будут здесь рассматриваться.
Хотя точные сведения о климатических границах этих формаций отсутствуют, некоторые условия, необходимые для их существования, можно определить следующим образом:
Предгорный дождевой лес Вечнозеленый сезонный лес Полу вечнозеленый сезонный лес Листопадный сезонный лес
Годовая сумма осадков, см Длительность сухого периода Свыше 180 Не бывает Свыше 180 3 месяца; в каждом месяце менее 10, но более 5 см осадков 130—80 5 месяцев; в каждом месяце менее 10, но более 2,5 см осадков 80—130 5 месяцев: в каждом месяце менее 10, в двух менее 2,5 см осадков
О климатических условиях колючего редкостойного леса и кактусового кустарника известно мало. Эффективность осадков в пределах этих формаций в значительной степени определяется местными почвенными условиями (см. стр. 244—245), причем различия в осадках отчасти компенсируются различиями во влажности и водоудерживающей способности почвы.
м фут.
стпоиныи лес скраи
Рис. 39. Профильные диаграммы климатически обусловленных растительных формаций низменностей Тринидада, Вест-Индия, и прилегающей части Венесуэлы (Beard, 1944а).
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 343
1) Предгорный дождевой лес. Это весьма типичный дождевой лес, хотя измельчавший и несколько измененный воздействием ветра. Почти все виды, а число их очень велико, вечнозеленые; лиан и эпифитов мало. В Тринидаде эта формация представлена только одной ассоциацией — Byrsonima spicata—Licania.
2) Вечнозеленый сезонный лес. Эта формация живо напоминает типичный дождевой лес и, подобно ему, состоит из трех ярусов деревьев (рис. 7, а, стр. 49), соответственно расположенных на высотах 30 м и выше, 12—27 и 3—9 л; это сообщество несколько ниже и не столь роскошно, как вполне развитый дождевой лес.
Но основное отличие от «истинного» дождевого леса заключается в том, что самый высокий ярус деревьев здесь еще более разрежен. Хотя диаметр некоторых деревьев может достигать 3 и более метров, большие экземпляры немногочисленны. «Общее впечатление таково, что отдельные гиганты окружены более мелкой порослью, и представление о тесном ряде колонн, как в дождевом лесу, здесь отсутствует» (Beard, 1944b, стр. 138).
Ветви у более крупных деревьев расположены заметно ниже, и стволы без сучьев на высоте свыше 20 м встречаются редко. Эта формация преимущественно вечнозеленая, но у некоторых видов больших деревьев бывает короткий безлистный период, длящийся от нескольких дней до нескольких недель, почему их можно считать полулистопадными. В самом высоком ярусе примерно 17% видов являются листопадными, 10% —полулистопадными, но из отдельных особей только 3 и 0,4% — соответственно листопадные и полу листопадные. В нижних ярусах преобладание вечнозеленых видов еще заметнее.
Так же, как и в дождевом лесу, досковидные корни являются характерной чертой многих деревьев первого яруса, и листья подавляющего большинства деревьев принадлежат по размерам к «мезофилльной» группе Раункиэра (Raunkiaer, стр. 84). Лианы и эпифиты весьма многочисленны. Различают четыре «фациации» вечнозеленого сезонного леса, в которых встречаются более или менее одинаковые виды, но в различных пропорциях. В одной, рассматривающейся здесь как несомненная консоциация, доминирует Mora excelsa1 (см. стр. 49 и 259).
Из вышеприведенного описания видно, что различие между вечнозеленым сезонным лесом и типичным дождевым лесом низменностей очень незначительно. Многие лесные сообщества Америки и других стран (включая, вероятно, дождевой лес Нигерии, см. стр. 46 и др.), которые обычно считаются тропическими дождевыми лесами, сходны с вечнозеленым сезонным лесом Тринидада и должны причисляться к аналогичной формации. В то же время не следует забывать, что, несмотря на несомненную пользу выявления различий между вечнозеленым сезонным лесом и более роскошным тропическим дождевым лесом, встречающимся в климатах с менее выраженным сухим сезоном, было бы весьма нежелательным настолько сужать термин «дождевой лес», чтобы исключать из него сообщества, подобные вечнозеленому сезонному лесу. Гораздо предпочтительнее сохранить твердо установленное широкое понятие о тропическом дождевом лесе и рассматривать вечнозеленый сезонный лес как «субформацию» или, может быть, используя терминологию Клементса, как фациацию формации тропического дождевого леса.
1 В статье Бирда от 1944 г. «фациация Могол вечнозеленого сезонного леса рассматривалась как консоциация «истинного дождевого леса».
344
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
3) Полувечнозеленый сезонный лес. Эта и следующая формация, видимо, приблизительно соответствует муссонному лесу в смысле Шимпера; однако, как указывает Бирд, представляется желательным не применять термин «муссонный лес» к лесным сообществам за пределами Юго-Восточной Азии.
В полувечнозеленом сезонном лесу существует только два яруса деревьев: ярус высотой 20—26 м, образующий более или менее плотный полог, и нижний ярус высотой 6—14 м. Деревья значительной толщины редки, средний диаметр спелых деревьев составляет около 0,5 м. Ствол ветвится на незначительном расстоянии от земли, кроны имеют зонтиковидную форму. Верхний ярус состоит как пз вечнозеленых, таки из листопадных видов; в соответствии с типом ассоциации от 26 до 42 °о видов и от 21 до 30 % особей являются листопадными. В первом ярусе большинство деревьев факультативно листопадные и лишь некоторые могут быть названы истинно вечнозелеными. Интенсивность листопада меняется из года в год; в дождливый год большинство деревьев почти не теряет листву, а в сильную засуху полог в целом производит впечатление безлистного. Нижний ярус, в большинстве своем вечнозеленый, также включает значительное количество листопадных видов. Листья верхнего яруса преимущественно «мезофиллы», а в нижнем ярусе заметна примесь «микрофиллов».
В описываемом сообществе изредка встречаются деревья с большими досковидными корнями; колючие растения не очень характерны, но некоторые виды обоих ярусов имеют колючие стволы. Лианы более обильны, чем в других формациях Тринидада; все большие деревья густо увешаны ими. Эпифитов сравнительно мало.
Существует шесть ассоциаций, принадлежащих к формации полувеч-нозеленого сезонного леса. Вероятно, все они свойственны местообитаниям^ подверженным более сильной сезонной засухе, чем местообитания вечнозеленого леса, причем недостаток влаги здесь вызывается не исключительно климатическими, но сочетанием топографических, эдафических и климатических факторов.
। 4) Листопадный сезонный лес. В самом Тринидаде формация листопадного сезонного леса представлена двумя сообществами, которые, видимо, являются вторичными ассоциями, значительно измененными деятельностью человека. Но на соседнем острове Малый Тобаго эта формация, вероятно, является климаксной (Beard, 1944b), и описание Бирда частично основано на исследовании названного района.
Листопадный сезонный лес невысок и имеет два яруса деревьев — верхний, открытый, состоящий из отдельно расположенных деревьев высотой до 20 л, и нижний высотой в 3—10 м. Стволы раздваиваются или ветвятся низко над землей и часто бывают искривлены; многие деревья нижнего яруса растут группами. Тут мало очень больших деревьев; максимальный диаметр примерно 0,5 м.
Свыше 2/3 деревьев верхнего яруса листопадные; листопад происходит с неизменной регулярностью в каждый сухой сезон. Нижний ярус почти целиком вечнозеленый. Примерно половина листьев верхнего яруса — «мезофиллы», половина — «микрофиллы»; большинство листьев нижнего яруса — «мезофиллы», хотя и велика примесь «микрофиллов».
Деревья с досковидными корнями отсутствуют; стволы некоторых видов покрыты шипами или колючками. Лианы довольно редки, а эпифитов нет или мало.
5) Колючее редколесье. Эта формация, как уже было сказано, на Тринидаде отсутствует, встречаясь в близлежащих частях Венесуэлы в виде
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 345
весьма открытого низкорослого сообщества, состоящего в основном из жестколистных, микрофилльиых, вечнозеленых деревьев высотой в 3— 10 At, принадлежащих главным образом к семействам Leguminosae (Mimo-soideae, Caesalpineae); большинство деревьев колючие. Злаки, обычно характерные для подобных типов растительности других частей света, здесь отсутствуют, и напочвенный покров состоит лишь из некоторых Вго-metiaceae и суккулентных растений.
6) Кактусовый кустарник. Эта формация встречается на острове Патос, между Тринидадом и Венесуэлой, ина материке. Она представляет собой растительность открытого типа с преобладанием колонковидных кактусов и опунций, смешанных с разнообразными микро- или лепто-филльными кустарниками и напочвенными Bromeliaceae. Здесь злаки отсутствуют, и почва часто оказывается обнаженной.
Шесть климатических формаций, описанных Бирдом, видимо, образуют непрерывный ряд, где один тип постепенно переходит в другой. Хотя границы между ними проводятся довольно произвольно, все же деление, предложенное Бирдом, в общем представляется удобным как с практической, так и с теоретической точек зрения.
В целом этот ряд выявляет отчетливую взаимосвязь между структурой и внешним обликом растительности, с одной стороны, и климатом — с другой (рис. 39). По мере усиления сезонной засухи (безразлично, только ли за счет климата или же в результате специфического сочетания климата с другими факторами среды) растительность мельчает и ее структура упрощается одновременно с уменьшением числа ярусов. Сначала в верхних ярусах, а затем и в нижних появляется все больше листопадных деревьев. Тот факт, что нижние ярусы еще остаются вечнозелеными, когда в верхнем ярусе начинают преобладать листопадные виды, легко объясняется их более влажным микроклиматом. Наблюдается общая тенденция к измельчанию листьев; «микрофиллы», редкие в дождевом лесу, встречаются все чаще по мере продвижения к сухому варианту ряда. Досковидных корней становится все меньше и меньше; возможно, что это является всего лишь следствием уменьшения среднего размера деревьев, а шиповатость становится все более и более обычным явлением.
Растительные формации Тринидада, несомненно, являются аналогами формаций, получивших большее развитие на материке Южной Америки. Сезонные леса, более или менее сходные с лесами Тринидада, широко распространены во внутренних районах Брит. Гвианы (Davis, 1933; Myers, 1936); в бассейне Амазонки (Ducke, 1938, цит. Beard, 1944а), в Венесуэле (Myers, 1933; Pittier, 1939, цит. Beard, 1944а), в Колумбии (Dugand, 1934), южной Бразилии (Warming, 1892) и в Центральной Америке, а также на Кубе (Bennett, Allison, 1928) и других Антильских островах. Колючее редколесье и кактусовый кустарник наблюдаются в различных частях Вест-Индии, особенно на Больших Антильских островах, например Ямайке (Brockmann-Jerosch, 1925), а также в Венесуэле и других местах материка Южной Америки. Сходны с этими сообществами хорошо известные каатинги южной Бразилии (Liitzelburg, 1925—1926; Ule, 1908 b, 1908 с) (не смешивать с каатинговыми лесами области Рио-Негро, см. стр. 260). Таким образом, можно допустить, что вдоль границ американского тропического дождевого леса повсюду, где естественная климаксная растительность сохранилась или не подверглась радикальному изменению под воздействием человека, располагается ряд формаций, очень сходных с формациями Тринидада. Будущее покажет, насколько предложенная Бирдом классификация применима ко всей тропической области Нового света.
346
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Саванна в Вест-Индии и северо-восточной части Южной Америки
До сих пор еще ничего не было сказано об отношении саванны к тропическому дождевому лесу на Тринидаде и северо-восточной части Южной Америки. Это — сложная проблема, которую здесь невозможно исчерпать.
Следует заметить, что термин «саванна», так же как и ряд других обозначений, является словом местного наречия (вероятно, карибского происхождения), и очень трудно или почти невозможно подыскать ему точный научный эквивалент. В Вест-Индии «саванной» фактически обозначают всякую низкорослую растительность с преобладанием злаков или других травянистых растений; это слово бытует даже применительно к городским паркам и газонам. Пользуются им и для наименования сообществ с преобладанием злаков или Сурегасае, растущих в постоянно заболоченных местностях. Сюда же относятся «водные саванны» дельты Ориноко и прибрежной области Брит. Гвианы (Myers, 1933) и «болотные саванны» на побережье Суринама (Lanjouw, 1936), так же как «savanes basses» и «savanes tremblantes» Франц. Гвианы (Benoist, 1925). Вероятно, сюда же тяготеет и «campos de varzea» нижней Амазонки (Bouillienne, 1924, 1930). Все эти «саванны» представляют собой стадии гидросерии и имеют мало общего с теми саваннами, которые будут рассмотрены здесь.
В типичных саваннах преобладают злаки, иногда осоки (Rhynchospora spp. и т. п.) обычно со значительной примесью двудольных трав и мелких кустарников. Деревья и кустарники, обычно более или менее ксероморф-ные, редко совершенно отсутствуют на обширных площадях. Характерно их произрастание разбросанными группами или полосами вдоль рек и потоков (окаймляющий, или галерейный лес), что придает ландшафту облик парка (фото 22). В «садовых саваннах» низкорослые, часто искривленные деревья равномерно разбросаны по злаковому покрову, так что возникает сходство с запущенным яблоневым садом. Среди деревьев южноамериканской саванны обращают на себя внимание пальмы Mauritia и Copernicia. Определение саванны, предложенное Ланжоу (Lanjouw, 1936),— «саванны — это равнины на островах Вест-Индии и северной части Южной Америки, покрытые более или менее ксероморфными травами и мелким кустарником, с немногочисленными деревьями и более крупным кустарником» — удобно, хотя вряд ли имеет смысл ограничивать употребление термина лишь одним географическим районом.
Но даже в рамках этого определения саванны очень разнообразны по внешнему облику, флористическому составу и экологии. Хотя за последние годы было опубликовано несколько превосходных описаний саванн Южной Америки, все же они изучены еще слишком мало, чтобы можно было составить их удовлетворительную классификацию.
На Тринидаде существуют два типа настоящей саванны (Beard, 1946b; см. также Myers, 1933 и Marshall, 1934) — низинная и горная. В низинных саваннах Пьярко, Эрина и некоторых других растительность состоит из злаков (виды Paspalum, Axonopus, Trachypogon ligularis и т. д.) и других травянистых растений, среди которых попадаются корявые деревья типичных для саванны видов (преимущественно Curatella americana и Byrsonima crassifolid). В хорошо известных саваннах Арипо почва покрыта Сурегасеае и злаками вперемешку с видами Polygala, Utricularia, Dro-sera, пятнами сфагнума и многими другими растениями. Попадаются случайные купы кустарников и разрозненные рощи высоких пальм Mauritia, но Curatella отсутствует. Горная саванна в Сент-Жозеф представляет собой злаковник из Trachypogon ligularis с обособленно] растущими кустами
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 347
и деревьями; часть этой площади, как Пьярко и т. д., покрыта главным образом Curatella и Byrsonima, но в других местах преобладают Myrcia ste-посагра и Roupala montana, которые не встречаются в низинных саваннах.
Саванны Тринидада невелики и встречаются в виде прогалин среди обширных лесных массивов (преимущественно вечнозеленые сезонные леса); очень трудно приписать их присутствие климатическим факторам. По-видимому, лес не стремится проникать в саванны, и хотя некоторые из них часто выгорают, но не все они подвержены пожарам; Бирд констатирует, что саванны Арипо никогда не горели. Известно, что злаковники, встречающиеся в некоторых частях Тринидада, появились в результате уничтожения леса при подсечно-огневом земледелии и последующих повторных выжиганиях. В этих злаковниках, представляющих собой стадии отклонившейся сукцессии (см. гл. 17) и называемых вторичными саваннами, доминирующей травой является Imperata brasiliensis, никогда не встречающаяся в изобилии в настоящих саваннах (близко родственный вид играет очень заметную роль во вторичных и отклонившихся сукцессиях в тропиках Старого света, см. гл. 17). Как травяная, так и древесная флора здесь совершенно иная, чем в тех первичных саваннах, о которых идет речь. Богатая флора первичных саванн, включающая ряд эндемичных видов, или же видов с прерывистым распространением, является веским доказательством их естественного происхождения и большой древности. В низинных саваннах дренаж затруднен и почва часть года оказывается заболоченной (хотя и не настолько, как в вышеупомянутых «водных саваннах»), а в остальное время сильно высыхает; типична ее «покрытая рябью» кочковатая поверхность. Горная саванна у Сент-Джозефа произрастает на лучше дренированной, но очень неглубокой песчаной почве, подстилаемой слоем водонепроницаемого кварцита, которая очень иссушается во время засухи. Вероятно, именно чередование заболоченности и чрезмерной сухости почвы препятствует произрастанию леса в местах развития саванны1. Бирд считает, что саванны Тринидада занимают древние почвы, которые не были покрыты лесом со времен плейстоцена1 2.
В Брит. Гвиане известны две основные области распространения саванн {помимо «водных саванн»), которые занимают обширные пространства: а) саванны Бербисе к западу от реки Бербисе, протянувшиеся примерно на 60—90 миль в глубь страны; б) внутренние саванны, расположенные в бассейне Рупунуни и в окрестностях гор Пакараима и Параима; через границу Бразилии они проникают в бассейн Рио-Бланко. Саванны Бербисе были описаны Мартином (Martyn, 1931) и Фолетт-Смитом (Follett-Smith, 1930); они занимают волнистые равнины на высоте 80—90 футов (24—28 м) над уровнем моря и состоят главным образом из злаков и Сурегасеае. Местами попадаются «островки» кустарников и деревьев, а в долинах рек встречаются полосы галерейного леса. Несколько иная растительность располагается в более влажных понижениях, называемых «блюдцами», и на крупных муравейниках, где поселяются кустарники и деревья. Равнина сложена в основном бурыми песками; там, где им на смену являются грубые выщелоченные пески (сходные с выщелоченными песками
1 Аналогичным же образом объясняется отсутствие деревьев на «dambos» или в долинных злаковниках северной Родезии (Michelmore, 1939, стр. 283—284).
2 Чартер (Charter, 1941) выразил сходный с Бирдом взгляд на происхождение так называемой «сосновой гряды» — типа саванны в Брит. Гондурасе (Центральная Америка), где главенствует Pinus caribaea. Он считает почвы сосновой гряды перезрелыми (старческими) и полагает, что на более ранней стадии своего развития они несли вечнозеленый лес.
348
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
леса Уаллаба, см. стр. 263), типичная саванновая флора замещается кустарником «мури» — угнетенной порослью кустарников и маленьких деревьев (главным образом Huniiria floribunda var. guianensis, Clusia nemorosa), чередующихся с участками очень скудной травянистой растительности, включающей травы, папоротник Schizaea incurvata и лишайники. Кустарник «мури», как указано в гл. 9, внешне очень напоминает так называемую паданговую растительность Малезии, которая развивается в весьма сходных почвенных и климатических условиях. Саванна окружена вечнозеленым дождевым лесом, и, как во многих случаях, переход от саванны или кустарника «мури» к лесу почти всегда совершается внезапно. Участки саванны или «мури», иногда величиной всего в несколько гектаров, могут встречаться в виде прогалин среди девственного леса, вдалеке от основных саванновых областей. Согласно Дэвису (не опубликовано), в непосредственной близости к саваннам Бербисе обычно располагается лес Дакама с преобладаниемDimorphandra conjugate, (стр. 279). Несколько далее обычно встречается консоциация типа Уаллаба {Eperua). Здесь наблюдается поясное расположение растительных сообществ, причем типична следующая их последовательность при переходе от леса к саваннам: 1) консоциация Eperua; 2) ассоциация Eperua—Dimorphandra; 3) консоциация Dimorphandra; 4) переходная зона, в которой преобладает «bania» (Swartzia sp.), Catostemma fragrans. Licania sp. и «какаруа» (Sapotaceae); 5) кустарник «мури»; 6) саванна. Саванны Бербисе не граничат ни с листопадным, ни даже с полулистопадным типом леса, и какие-либо признаки перехода по типу вечнозеленый лес — листопадный лес — саванна здесь отсутствуют.
Внутренние саванны гораздо обширнее. В области Рупунуни—Рио-Бланко (Myers, 1936) они представляют собой пространства с сине-зеленым покровом преобладающего тут злака Trachypogon plumosus, произрастающего в сообществе с другими злаками и многими видами двудольных травянистых растений. Среди редко разбросанных деревьев чаще всего встречается Curatella americana и виды Byrsonima. Злаковник перемежается полосами галерейного леса и «островками» низкого редколесья, поперечник которых колеблется от 20—30 м до нескольких километров. В более влажных понижениях (baixas) господствующая ассоциация представлена в основном видами семейства Сурегасеае; рощицы двудольных деревьев тут замещают пальмы Mauritia flexuosa. Галерейные леса и другие лесные массивы, примыкающие к саваннам Рупунуни—Рио-Бланко, не являются дождевым лесом, включая большой процент листопадных видов (Myers, 1933; Davis, 1933), и их, вероятно, следовало бы отнести к типу полувечно-зеленого сезонного леса; флора древесцых «островков» несколько иная, там меньше листопадных видов. Нагорные саванны гор Пакараима сходны с саваннами Рапунуни, но растительность их более разнообразна.
Саванны Суринама (Lanjouw, 1936, см. ниже, стр. 349) и Франц. Гвианы (Benoist, 1925), очень сходные с саваннами Бербисе, частично состоят из открытых злаковников, а частично из кустарника «мури». В суринамских саваннах преобладают не злаки, а Сурегасеае; прилегающий лес вечнозеленый. Подобные же участки саванны вновь появляются в районе нижнего течения Амазонки (Bouillienne, 1926).
Самые обширные саванны северной части Америки — это льяносы Венесуэлы, знакомые многим по классическому описанию Гумбольдта (Humboldt, 1852). Современный обзор их растительности сделан Майерсом (Myers, 1933), который различает два типа: а) саванны из рыхлокустовых злаков, где в основном преобладают злаки Hyparrhenia rufa и Andropogon
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 349
condensatus-, деревья, особенно Curatella americana, Bowdichia virgilioides и пальма Copernicia tectorum, произрастают в галерейных лесах, а также и в других местах в виде изолированных групп различных размеров: б) высокотравные саванны; здесь безраздельно господствует высокий злак Paspalum fasciculatum. Леса, граничащие с льяносами, относятся преимущественно к сухому листопадному типу (Schimper, 1935).
Экологические особенности саванны в северо-восточной части Южной Америки
Саванны попеременно считали то климатической климаксной формацией, то биотическим климаксом, возникающим главным образом под воздействием пастьбы и периодических пожаров, то, наконец, эдафическим климаксом, зависящим от специфических почвенных условий. Шимпер (Schimper, 1935, стр. 424) так определяет климатические условия саванны: годовая сумма осадков равна 90—150 см, выраженный сухой сезон длится 4—5 месяцев, а средняя месячная амплитуда температуры может достигать 14°. Согласно Шимперу, при этих условиях между саванной и лесной растительностью идет борьба, исход которой решается отчасти такими климатическими факторами, как количество осадков в вегетационный период, а отчасти эдафическими факторами. С этой точки зрения, климат является важнейшим фактором, определяющим наличие саванны. Весьма вероятно, а может быть и определенно, что для рассматриваемых сейчас южноамериканских саванн определяющий фактор их формирования не всегда один и тот же, причем в пользу главенствующей роли климата очень мало данных. Существование саванны в чрезвычайно разнообразных климатических условиях, ее частое соседство с типичным дождевым лесом, внезапность перехода от саванны к лесу и многочисленность мелких вкраплений саванны в больших массивах высокоствольного леса — все это факты, свидетельствующие против климатической теории.
Как мы уже видели, экологические условия саванн Тринидада трудно примирить с взглядами Шимпера, к такому же заключению приводит и весьма удачный обзор саванн Суринама, данный Ланжоу (Lanjouw, 1936); годовая сумма осадков в саваннах Суринама колеблется от 2056 до 2219 мм (по данным трех станций), а в засушливый период 2—3 месяца подряд выпадает меньше 100 мм. Если сопоставить эти цифры с цифрами, приведенными в табл. 12 (стр. 160—161), то станет очевидно, что саванна здесь находится внутри климатической границы вечнозеленого тропического дождевого леса, который часто произрастает и в местах с гораздо более длительной сезонной засухой. С другой стороны, есть много свидетельств того, что как эдафические условия, так и пожары в значительной степени определяют характер растительности. Суринамские саванны расположены на низких плато с хорошо дренированными, более или менее пористыми почвами, представленными крупнозернистыми выщелоченными песками или же суглинками. Благодаря обильным осадкам эти почвы сильно выщелочены и, вероятно, очень бедны питательными веществами. По-видимому, они принадлежат к типу подзолистых почв и напоминают почвы леса Уаллаба Брит. Гвианы, «борового» леса Борнео и паданга (см. гл. 9), но. может быть, благодаря плоскому рельефу саванны на этих почвах подвержены сезонному заболачиванию. Растительность здесь часто выжигается пожарами, и отчетливая граница между саванной и окружающим лесом вряд ли может быть обусловлена чем-либо, кроме огня. Ланжоу полагает, что прежде на месте саванн произрастал дождевой тропический лес, подоб
350
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ный тому, который до сих пор существует по соседству. Выщелачивание почвы вызвало замещение этого леса «саванновым лесом» (возможно, что этот тип растительности был ближе к кустарнику «мури» или лесу Дакама, упомянутым выше (стр. 348), а не к климатическому периодически листопадному лесу в смысле Шимпера). Саванновый лес более дождевого леса, подверженный действию огня, был превращен повторными пожарами в саванну, которая является, таким образом, климаксной растительностью пожарища, хотя первоначально ее локализация определялась эдафически-ми причинами.
Становление и существование саванн Бербисе в Брит. Гвиане, очень схожих с саваннами Суринама, вероятно, можно истолковать таким же образом. Природные условия и режим осадков в них аналогичны саваннам Суринама, хотя сезонная засуха обычно выражена несколько сильнее. Фоллетт-Смит (Follett-Smith, 1930) довольно детально изучил почвы этих саванн и установил три основных типа: кислые бурые пески, серые пески и крупные белые пески (как уже говорилось, последний тип связан с распространением кустарника «мури»). В этих саваннах регулярно пасут скот; они подвергаются как преднамеренному (с целью вырастить молодую траву), так и случайному выжиганию. По-видимому, и здесь саванновая растительность обусловлена не климатом, а частично характером почвы, частично пожарами и использованием площади под пастбища.
Саванны Рупунуни — Рио-Бранко, в Гвиане, как и льяносы Венесуэлы, встречаются в областях с меньшей годовой суммой осадков (вероятно, около 1150—1500 мм), чем в Суринаме и саваннах Бербисе, а сезонная засуха выражена сильнее. Саванны Рупунуни — Рио-Бранко не так интенсивно используются под выпас, но в течение длительного времени они часто и регулярно выжигались. Майерс (Myers, 1936) несомненно прав, считая нынешнюю растительность этих саван «климаксом пожарищ», хотя и почвенные условия, которые, кстати, мало изучены, так же могли иметь значение. Согласно Майерсу (Myers, 1933), периодические пожары сильно воздействуют и на льяносы, где к тому же пасется скот, а в отдельных местах в результате перевыпаса растительный покров оказывается полностью разрушенным. «Высокотравные» саванны периодически затопляются, и весьма вероятно, что все льяносы, как и саванны низменностей Тринидада, в определенное время года подвергаются заболачиванию. По немногочисленным имеющимся данным можно заключить, что растительность здесь, как и в других рассмотренных нами саваннах, первоначально определялась эдафическими факторами, но была в значительной степени изменена пожарами. И в области Рупунуни — Рио-Бранко и в льяносах Венесуэлы климатическим климаксом, очевидно, является листопадный или полулистопадный лес.
Сведения о климате, почвах, частоте пожаров и интенсивности выпаса в саваннах северо-восточной части Южной Америки и соседних островов все еще чрезвычайно скудны, но из всего сказанного ясно видно, что ни одну из саванн этой области нельзя считать климатическим климаксом. На большую часть их сильно воздействовали пожары. Майерс (Myers, 1936, стр. 176) говорит: «В Южной Америке мне никогда не приходилось встречать саванны, которая не носила бы следов более или менее частых пожаров, как бы она ни была мала, изолирована или отдалена от населенных пунктов». Возможно, это утверждение преувеличивает действительное положение дел, так как некоторые саванны, например саванпа Арипо на Тринидаде, очевидно, не пострадали от огня; учитывая обширность многих саванн, трудно предположить, что распространению лесной расти
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 351
тельности в их пределах предшествуют одни только пожары. Частые палы создали резко очерченные границы саванн и, возможно, благоприятствовали разрастанию злаков и другой травянистой растительности за счет древесных растений, но в целом саванны северо-восточной части Южной .Америки скорее всего первоначально сложились как эдафический климакс, который впоследствии под воздействием многократных выжиганий превратился в климакс пожарищ. Саванны типа Арипо могут дать представление о характере первоначальных сообществ эдафического климакса, от которого произошли другие типы саванн.
ДОЖДЕВОЙ ЛЕС, ЛИСТОПАДНЫЙ ЛЕС И САВАННА В ЮГО-ВОСТОЧНОЙ АЗИИ
В Азии наиболее типичный тропический дождевой лес произрастает на Малаккском полуострове и островах Суматра и Борнео. В пределах этой обширной области низменности находятся в условиях экваториального климата и характеризуются обильными и равномерными осадками. К северу и западу от нее, в Таиланде, Бирме и в тропической и субтропической Индии, несмотря на наличие значительных массивов вечнозеленого леса, климат в целом более сезонный, для которого свойствен ежегодный сухой период со средней месячной суммой осадков ниже 4 дюймов (10 см). Вследствие этого растительность здесь преимущественно более или менее тропофильная (в смысле Шимпера), и климаксные типы колеблются от полулистопадного и листопадного муссонного леса до колючего редколесья. Местами, например в центре «засушливой зоны» Бирмы и в наиболее сухих местах Индии, преобладает ясно ксерофитная полупустынная растительность.
На Малаккском полуострове первые признаки перехода от вечнозеленого тропического дождевого леса к листопадному муссонному лесу проявляются в северных княжествах Малайской федерации (Перлис, Кедах, Калантан — в северной части), где все чаще и чаще попадаются типичные «бирманские» деревья, такие, как Albizzia lebbek иLagerstroemia floribunda\ некоторые из этих видов встречаются и южнее. На южных и восточных островах Малайского архипелага экваториальный климат также сменяется климатом с сухим сезоном, и вместо вечнозеленого леса климаксом становится листопадный лес. На Зондских островах граница между несезонным тропическим дождевым лесом и сезонным муссонным лесом пересекает Яву, западная часть которой с ее влажным климатом дождевого леса (хотя в ее пределах дождевой лес низменностей почти полностью уничтожен) контрастирует с муссонным климатом восточной части.
Громадный массив вечнозеленого леса, центром которого условно может быть назван Сингапур, делит, таким образом, сезонный муссонный лес Юго-Восточной Азии на две далеко отстоящие друг от друга области; поэтому поразительно, что между листопадными лесами Таиланда и Бирмы, с одной стороны, и аналогичными лесами восточных Зондских островов, с другой (см. van Steenis, 1935, стр. 83—85), наблюдается исключительное флористическое сходство. Например, такое типичное дерево муссонного леса, как тик (Tectona grandis), в изобилии встречается в обеих областях, между тем как в промежуточной области дождевого леса он даже не поддается разведению.
Сезонные (или мусонные) леса восточных Зондских островов недостаточно изучены, и в большинстве случаев они так изменены пожарами и другими биотическими факторами, что трудно восстановить их перво
352
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
начальную структуру. По этой причине здесь будет подробно рассмотрена лишь бирманская переходная зона от дождевого леса к муссонному, который наблюдается в Бирме. Даже в Бирме так широко проявляется влияние пожаров и подсечно-огневого земледелия, что распознать климатические клпмаксные типы чрезвычайно трудно.
Типы лесов Бирмы хорошо известны благодаря работам Курца (Kurz, 1875). Стэмпа (Stamp, 1925) и подробному исследованию части «приречной полосы» долины Иравади Стэмпом и Лордом (Stamp, Lord, 1923). Чемпион (Champion. 1936) также опубликовал обширную и тщательно разработанную классификацию, в которой предлагает единую систему номенклатуры для лесных сообществ всей Индии и Бирмы.
В Бирме существует определенная градация климата от влажной области Араканского побережья на западе и от Тенассерима и дельты Иравади на юге до «сухого пояса», занимающего центральную часть бассейна Иравади. На западе и на юге — обильные осадки и короткий сухой период, тогда как в «сухом поясе» осадки местами составляют всего 20 дюймов (51 см), выпадая преимущественно проливными дождями в некоторые дни дождливого сезона, а остальная часть года почти совершенно бездождная. Согласно Стэмпу (Stamp, 1925), Бирму можно разделить на три части, используя изогиеты в 40 (102 см) и 80 (203 см) дюймов.
I. Области с осадками выше 80 дюймов. Здесь климаксной растительностью является вечнозеленый лес, обычно обозначаемый как тропический дождевой лес. Климат неблагоприятен для произрастания типичных деревьев муссонного леса — «на плантациях тик вырастает быстро, но во многих случаях деревья вскоре становятся дуплистыми и корявыми» (Stamp, 1925, стр. 7).
II. Области с осадками 40—80 дюймов. Здесь климаксные типы соответствуют определению муссонного леса в понимании Шимпера (стр. 339), где преобладающие древесные виды теряют листья в засушливый сезон (старые листья опадают в январе или феврале, а новые появляются примерно за месяц до начала дождей в июне). В пределах этой области находятся основные тиковые леса.
III. «Сухой пояс» с осадками ниже 40 дюймов. В этой области отсутствуют высокоствольные леса; климаксная растительность меняется от низкого редколесья, соответствующего шимперовской категории саваннового леса, до колючего леса и кустарниковой полупустыни.
В пределах этих трех областей можно выявить ряд климатических климаксов или формаций. Главные из них, согласно схеме Стэмпа, показаны в табл. 34. Следует отметить, что при построении этой схемы Стэмп за основу взял четыре основных подразделения Шимпера — дождевой лес, муссонный лес, саванновый лес и колючий лес (см. Stamp, Lord, 1923). Во всех установленных климаксных формациях доминируют древесные растения. Хотя в Бирме встречаются саванны или подобные им сообщества, например каинговые злаковники или злаковники из слоновой травы, они, всего вероятнее, являются сериальными стадиями и составляют фазы биотических и отклонившихся сукцессий. То же самое, хотя и с меньшей уверенностью, можно сказать о различных типах бамбуковых зарослей и зарослей из видов пальмы Calamus. Основные флористические и физиономические особенности формаций, данных в табл. 34 и описанных Курцем (Kurz, 1875) и Стэмпом (Stamp, 1925), могут быть сведены к следующему:
1. Вечнозеленый диптерокарповый лес. Высокоствольный смешанный лес с преобладанием Dipterocarpus spp. и других Dipterocarpaceae. Флори-
Основные климатические климаксы низменностей Бипмы (Stamp, 1925) 1
Таблица 34
Осадки в дюймах t
Климаксный тип
1. Вечнозеленый диптерокарповый лес [южный тропический влажный вечнозеленый] (Различные Dipterocarpaceae и т. п.)
2. Пайнкадо, или полувечнозеленый лес [южный тропический полувечнозеленый] (Xylia xylocarpa)
Климатическая формация (Schimper)
s
—30-
5.
6.
7.
8.
9.
3- Влажный тиковый лес
[южный тропический влажный листопадный] (Tectona grandis, Xylia xylocarpa)
4. Сухой тиковый лес
[южный тропический сухой листопадный] (Tectona grandis, Terminalia tomentose)
Полуиндаинг
[южный тропический сухой листопадный] (Dipterocarpus tuberculatus, Pentacme suavis, Shorea obtusa)
„ I
Индаинг [южный тропический сухой листопадный] (Dipterocarpus tuberculatus)
Лес из Diospyros, или лес тэ
[южный тропический сухой листопадный] (Dios руг os burmanica) к - 1
Колючий лес ша-дахат [южный тропический колючий] (Acacia catechu, Tectona hamiltoniana)
10. Сухой листопадный лес без тика [южный тропический сухой листопадный]
(Vitex spp., Hetero-phragma adenophyl-turn)
И. Тхан-дахат
[южный тропический сухой листопадный] (Terminalia oliveri, Tectona hamiltoniana)
8.
9.
12.
у
33
Колючий лес ша-дахат
[южный тропический колючий] (Acacia catechu, Tectona hamiltoniana)
Колючий кустарник ша [южный тропический колючий] (Acacia catechu)
si
Колючий кустарник ша (южный тропический колючий] (Acacia catechu)
Песок
Суглинок
Молочайниковая полупустыня [южный тропический колючий] (Euphorbia antiquorum и др.)
Глина
1 Названия, соответствующие терминологии Чемпиона (Champion, 1 936), даны в квадратных скобках. Главные (доминанты или наиболее рас п рос. пане ные) виды даны в круглых скобках.
354
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
стическп эта формация значительно богаче всех других; по словам Курца, он обнаружил 300—350 видов деревьев на площади в каких-нибудь 8—9 кв. миль (20—30 кв. км), тогда как на подобной же площади самого богатого листопадного леса едва ли найдется 70—80 видов. Здесь три яруса деревьев, не считая кустарников и трав различной высоты. Древостой верхнего яруса редко бывает ниже 150—200 футов (45—60 л») и состоит из смеси вечнозеленых и листопадных видов; некоторые виды, например Dipte-rocarpus spp., ежегодно сбрасывают листья, но новые листья распускаются прежде, чем опадают старые. Средние и нижние ярусы целиком состоят из вечнозеленых деревьев: «высокие деревья образуют как бы листопадный открытый лес над нижним ярусом вечнозеленых деревьев» (Kurz, 1875, стр. 31). Пальмы, включая виды Calamus и бамбуки, составляют часть подлеска. Травяной покров скуден и состоит главным образом из папоротников и двудольных травянистых растений при полном отсутствии злаков; множество эпифитов и лиан.
Из этого описания ясно, что вечнозеленая диптерокарповая формация Бирмы очень похожа на типичный дождевой тропический лес, но отличается большим количеством листопадных деревьев; очевидно, она очень сходна с вечнозеленым сезонным лесом Тринидада и тропической Америки.
Эта формация занимает часть южной Бирмы, встречаясь как в Пегу-Йома с годовой суммой осадков 80 дюймов (203 еле) и более, так и в Тенас-сериме, где осадки составляют 150—200 дюймов (380—500 см).
2. Полувечнозеленый лес или пайнкадо (pyinkado). Типичным деревом этой формации является Xylia xylocarpa.. Это листопадное дерево из семейства бобовых, распространенное в Бирме так же широко, как и тик (Tectona grandis), но более влаголюбивое. Этот вид часто господствует. Среди других деревьев обычны Lagerstroemia, Dipterocarpus alatus и D. turbinatus. Встречаются бамбуки, изобилуют лианы и эпифиты. Структура этого леса детально не изучена, но во многих отношениях он, по-видимому, сходен с предыдущим, за исключением еще большего количества листопадных деревьев.
Обычно полувечнозеленый лес развивается при годовой сумме осадков 80—95 дюймов (203—242 см), но на неглубоких пористых почвах он может встречаться и при значительно большем количестве осадков.
3. Влажный тиковый лес. Листопадная порода Tectona grandis — наиболее характерный вид в этой и следующей формациях, но она лишь в исключительных случаях является доминантной и редко составляет более 10% древостоя. Во влажном тиковом лесу Ху На xylocarpa — самый распространенный сопутствующий вид, а численно он даже превосходит тиковое дерево. Тик, произрастающий на песчаниках в Пегу-Йома, достигает 120 футов (37 лс) высоты, а в менее благоприятных условиях— всего лишь 80—90 футов (24—27 м). Лес в целом может не терять листьев до конца марта, хотя иногда он обнажается уже в феврале, но безлистный период не продолжается более 3—4 месяцев. Кустарниковый ярус не очень развит, но бамбуки играют заметную роль. Травяной покров обычно отсутствует, мало и папоротников. Лианы довольно многочисленны, хотя видовое разнообразие их невелико, и они не изменяют внешний вид леса. Орхидеи и эпифиты мало заметны.
Влажный тиковый лес по структуре и физиономии, пожалуй, можно сопоставить с пол увечнозеленым сезонным лесом Тринидада. Он широко распространен в тех районах Бирмы, где годовая сумма осадков колеблется между 60—90 дюймами (152—229 см) [в северном Тенассериме до 100 дюймов (254 еле)].
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ З55
4. Сухой тиковый лес. Хотя и в этой формации характерным видом является Tectona grandis, большинство сопутствующих видов отличается от тех, что произрастают во влажном тиковом лесу, где подавляющая часть видов обычно входит в состав и вечнозеленого диптерокарпового и полувечнозеленого леса; здесь же это преимущественно виды леса индаинг (формация 6) и других лесов сухого типа. Бамбук (Dendrocalamus strictus) является обязательным, типичным компонентом сухого тикового леса, тогда как Bambusa polymorpha и Cephalostachyum pergracile типичны для влажного тикового леса. Почва оказывает здесь более сильное влияние, чем в случае влажных формаций, и сухой тиковый лес третичных песчаников очень ясно отличается от таких же лесов на древних гнейсах, сланцах и известняке.
Сухой тиковый лес широко распространен; нижний предел осадков, при которых он может развиваться, составляет около 40 дюймов (102 см); верхний непосредственно зависит от характера почвы; на малоплодородных песчаных почвах сухой тиковый лес можно встретить даже при годовой сумме осадков 75 дюймов (191 см), а на суглинистых и вообще более плодородных почвах он вытесняется влажным тиковым лесом уже при 50—55 дюймах (127—142 см) осадков.
5— 6. Полуиндаинг и индаинг. Первая из этих формаций является переходной к формации 4 и вряд ли может считаться самостоятельной. Самым распространенным деревом в формациях 5 и 6 обычно бывает «ин» (Dipterocarpus tuberculatus; индаинг = лес из «ина»), часто составляющее консоциации, порою почти чистые. В дополнение к D. tuberculatus характерными видами в лесу типа полуиндаинг являются Pentacme suavis, Shorea obtusa, Terminalia tomentosa, а в индаинг—Pentacme suavis, Melanorrhoea usitata и Terminalia tomentosa. Индаинг — открытый и солнечный лес; деревья стоят редко, лианы и подлесок не препятствуют видимости. Средняя высота деревьев колеблется от 30 до 80 футов (8—24 м), за исключением самого «ин» и нескольких других видов, стволы у деревьев искривленные и невысокие, а ветви непропорционально толсты. В сухое время года индаинг остается совсем без листьев, хотя у многих деревьев молодые листья распускаются задолго до начала дождей; большая часть деревьев цветет после листопада. Нижний ярус часто состоит из зарослей злаков, а также включает виды Cycas и иногда пальму Phoenix paludosa. Попадаются обширные площади, на которых совсем нет бамбуков. Флора покрова бедна, и почва во многих местах остается обнаженной. Обычно много эпифитов, особенно орхидей.
Полуиндаинг и индаинг распространены в областях с осадками от 25 дюймов (64 см), как в некоторых местах «сухого пояса» до 60—80 дюймов (152—203 см), как, например, в Пегу-Йома, но почвенный покров влияет на их распределение не меньше, чем климат. Чаще всего они встречаются на рыхлых песках с твердой, содержащей соединения железа подпочвенной прослойкой, но могут произрастать также на латеритах, известняках, на безводных гнейсовых кряжах и на твердых песчаниках с тонким почвенным слоем.
7. Лес из Diospyros, или лес тэ. Это формация смешанного состава, для которой особенно типична Diospyros burmanica, или тэ; Pentacme suavis и Terminalia tomentosa также многочисленны. По структуре и физиономии лес Diospyros мог бы сойти за типичный саванновый лес (в смысле Шимпера), так как деревья тут низкорослые, а подлесок почти полностью состоит из злаков (преимущественно виды Andropogon). Прямоствольный Calamus и бесстебельный Phoenix являются теми немногими пальмами,
356
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
которые растут здесь. Встречаются тут бамбуки и разнообразные лианы, а также некоторые эпифиты. Папоротники, напочвенные и эпифитные, распространены широко, но по числу особей не обильны.
Амплитуда осадков в пределах этой формации составляет примерно 34—38 дюймов (86—97 см); почва песчаная, сходная с почвой, типичной для формаций 5 и 6.
8. Колючий лес ша-дахат. Этот лес обладает характерной структурой колючего леса, хотя иногда бывает открытым наподобие саванны. Основные породы — Acacia catechu, ша, которая обычна также для формаций 7 и И и встречается здесь как в древесной, так и в кустарниковой форме, и Tectona hamiltoniana (дахат) (которая здесь представляет собой маленькое дерево). Распространение Acacia leucophloea ограничено восточной стороной «сухого пояса». Почти все деревья, за исключением Tectona hamiltoniana,— колючие, и ни одно не превышает в высоту 30 футов (9 м). Много деревянистых лиан; травяной покров представлен главным образом злаками.
В «сухом поясе» этот тип растительности хорошо развит на глинистых и суглинистых почвах при меньшем количестве осадков, чем требуется для развития формации И, а на песке эта растительность встречается при еще меньшем количестве осадков (меньшем, чем в случае формации 7).
9. Колючий кустарник ша. Это низкий кустарник, в котором самым характерным растением является Acacia catechu', в вышину она не превышает 1,5—2 м, а в центре «сухого пояса» оказывается еще мельче и образует менее густые заросли. В восточной части «сухого пояса» A. catechu замещается A. leucophloea. Tectona hamiltoniana встречается в виде кустарника ниже 1 м высотой. Чуть ли не весь нижний ярус состоит из покрова злаков, чередующегося с пятнами обнаженной почвы. Встречаются некоторые розеточные растения.
Верхний предел осадков для этой формации составляет около 40 дюймов (102 см). На засоленных почвах «куатти-мие» («kyattimye»), образовавшихся из пегуанских глин в условиях недостаточного увлажнения, она развивается при 35 дюймах (89 см) осадков. На песчаной почве предел осадков выше, и здесь леса индаинг и леса из Diospyros существуют при таких же количествах осадков, каких в случае глинистых почв хватило бы лишь для развития кустарника.
10. Сухой листопадный лес без тика. Эта формация по структуре и внешности сходна с сухим тиковым лесом; она встречается в тех же климатических условиях, что и леса полуиндаинг и индаинг, но на склонах холмов с бедными каменистыми суглинистыми почвами. Наиболее обычны виды Vitex и Heterophragma adenophyllum, но трудно назвать какие-либо особенно характерные виды. Бамбуки часто бывают так обильны, что подавляют все другие растения; типичный вид — Dendrocalamus strictus.
11. Лес тхан-дахат. Это — саванновый лес, имеющий некоторое сходство с английскими порослевыми редколесьями на глинистой почве. Главные деревья — Tectona hamiltoniana (дахат) и Terminalia oliveri (тхан); весьма обычна также Acacia catechu. Tectona hamiltoniana ветвится невысоко над землей, и его средняя высота не превышает 30 футов (9 м); Terminalia oliveri представляет здесь неправильно растущее и корявое дерево. Dendrocalamus strictus — обычный, но и единственный здесь бамбук. Нижний ярус почти полностью состоит из злаков, преимущественно Andropogon apricus. Лианы часто бывают многочисленны.
Лес тхан-дахат покрывает обширные площади на плотных глинах и развивается при осадках 35—39 дюймов (89—99 см). Этот лес иногда
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 357
растет бок о бок с гораздо менее ксерофильным индаингом, правда развитым на песчаных почвах.
12. Молочайниковая полупустыня. Для этого сообщества типичны колючие молочаи вроде Euphorbia antiquorum-, встречаются также различные кустарниковые виды и иногда дерево Bombax insigne. Растения часто отстоят далеко друг от друга, а почва между ними обнажена или частично покрыта злаками. Эта формация встречается в центре «сухого пояса», но только лишь на засоленных (особенно если в подпочве имеется гипс) и глинистых почвах.
Таким образом, в Бирме существует постепенная смена растительности от вечнозеленого диптерокарпового леса (близкого к типичному дождевому лесу Малайской области) до кустарниковой полупустыни. Характер климаксной растительности в данной области указывает на отчетливую связь ее с годовой суммой осадков, хотя фактически он, несомненно, больше зависит от длительности и силы периодических засух, чем непосредственно от количества осадков; количество осадков, выпадающее в течение дождливого сезона, по-видимому, не имеет особого значения.
Стэмп и Лорд (Stamp, Lord, 1923) в своей работе отчетливо показали, что распределение формаций в значительной степени зависит не только от климата, но и от почв. Свойства почвы, особенно ее глубина и пористость (которая, по-видимому, обусловливает ее влагоемкость во время сухого сезона), существенно изменяют воздействие осадков, так что влияние климата и почвы в значительной мере взаимно компенсируется. В одном и том же климате, но на различных почвах можно встретить различные формации; например, саванновый лес тхан-дахат, колючий лес ша и колючий кустарник ша могут существовать бок о бок в одной и той же местности, находясь фактически в одинаковых климатических условиях. Подобным же образом одно и то же сообщество может встречаться на сходных почвах в разных климатах. Чем дальше отойти от условий тропического дождевого леса и приблизиться к более сухому почвенно-климатическому комплексу, тем сильнее сказывается влияние почв: так, вечнозеленый диптерокарповый, полувечнозеленый (папнкадо) и влажный тиковый леса, очевидно, относительно независимы от характера почвы, но сухой тиковый лес (при осадках около 100—90 см) уже испытывает значительные изменения в зависимости от почвенных разностей, а при еще меньших осадках эта формация совершенно различна на разных почвах.
При сравнении типов растительности этого ряда с растительностью Тринидада выявляется большое сходство. В обоих рядах древесные растения господствуют во всех климаксных типах; в Бирме с уменьшением осадков травяной ярус становится все значительнее, а на Тринидаде травы вообще не являются важной составной частью какой-либо климаксной формации. Как в Бирме, так и на Тринидаде с увеличением сухости среды растительность мельчает и становится более разреженной, появляется все больше листопадных древесных видов сначала в верхних, а затем в нижних ярусах сообщества. При тех ничтожных сведениях, какими мы располагаем относительно структуры и физиономических особенностей бирманских формаций, невозможно что-либо добавить к этому, и при нынешнем состоянии наших знаний было бы явно неблагоразумно пытаться применить обычную терминологию к климаксным формациям Бирмы и Тринидада.
Ряд климатических климаксных формаций, близко напоминающих формации Бирмы, встречается в Индии (см. Champion, 1936) и, несом
358
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ненно, также в Таиланде и т. д. Связь климаксной растительности с климатом и почвой в Индии не так четко выражена благодаря значительным изменениям, какие претерпел лес в результате человеческой деятельности. Названия для индийских эквивалентов различных формаций по номенклатуре Чемпиона (Champion, 1936), встречаемых в Бирме, включены в табл. 34.
Саванна в Юго-Восточной Азии
Лесные формации Бирмы были описаны довольно подробно, так как они очень отчетливо показывают экотон или постепенный переход климаксной растительности от вечнозеленого леса на одном конце ряда до полупустыни на другом. Изменения в растительности вызваны возрастанием длительности и интенсивности ежегодного засушливого периода, причем в Бирме этот процесс не осложняется наличием злаковников, не считая вторичных сообществ, которые, несомненно, являются следствием отклонившихся сукцессий. Относительная бедность злаковниками и саванными по сравнению со сходными в климатическом отношении тропическими районами Африки и Америки характерна для большей части Индо-Малайской области, хотя местами, например на нагорьях Цейлона (патаны), на Малых Зондских островах и в юго-восточной части Новой Гвинеи, существуют весьма обширные площади злаковников. Нет необходимости подробно обсуждать происхождение и характер этих пространств злаковников. Многие из них, включая, вероятно, площади с преобладанием злака аланг-аланг (Imp erat a cylindrical, явно представляют собой вторичные сообщества, появляющиеся после истребления тропического дождевого или листопадного леса в результате подсечно-огневого земледелия и выжигания. Так, небольшие участки с напоминающей саванны растительностью на юго-восточном Борнео (Winkler, 1914) и злаковники долин Раму и Маркхам на Новой Гвинее, несомненно, произошли именно этим образом. Патаны, или нагорные злаковники Цейлона (Pearson, 1899), по-видимому, ближе к саваннам северной части Южной Америки и, может быть, в одинаковой степени обязаны своим существованием как эдафическим причинам, так и огню. Отсутствуют какие-либо признаки того, что злаковники Юго-Восточной Азии являются климатически обусловленными климаксными сообществами.
КЛИМАТИЧЕСКИЙ ЭКОТОН В АФРИКЕ
Анализ взаимосвязи между тропическим дождевым лесом, листопадным лесом и саванной в Африке был оставлен напоследок отчасти из-за огромной площади, занимаемой двумя последними формациями, а отчасти потому, что человек до такой степени изменил растительность Африки, что здесь гораздо труднее распутать сложное взаимодействие биотических факторов, климата и почвы, чем в тропической Америке или Юго-Восточной Азии.
В Африке, как показано в гл. 1, существует центральный массив «сомкнутого леса», занимающий обширную часть бассейна Конго, бывшей Французской Экваториальной Африки, Габона и Камеруна и тянущийся узкой полосой к западу вдоль Гвинейского залива до 12° з. д. и 8° с. ш. Даже в самом центре «сомкнутый лес» почти повсеместно нарушается отдельными участками саванны различных размеров. По краям этот лес
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 359
зачастую внезапно переходит в широкий пояс саванны или «паркового леса».
Термин «сомкнутый лес» (который удобен для использования в данной работе) аналогичен так называемому foret dense современных французских авторов и Hylaea — немецких; он включает все типы низинного леса, в котором деревья образуют сомкнутый полог [в развитых сообществах высота верхнего яруса редко бывает ниже 120 футов (37 м)], а травяной покров отсутствует. В своей центральной части сомкнутый лес состоит главным образом из вечнозеленых деревьев, но по направлению к периферии пропорция листопадных видов постепенно возрастает, и флористический состав изменяется; вследствие этого обычно проводят разграничение между двумя типами сомкнутого леса — вечнозеленым лесом, или истинным тропическим дождевым лесом, и смешанным листопадным лесом (subxerophiler Tropenwald, Mildbraed, 1922).
В тропической Африке к западу от Камеруна наблюдается постепенный переход от климата западной прибрежной зоны с относительно коротким сухим сезоном, в течение которого влажность воздуха остается сравнительно высокой, к сухому с отчетливо выраженными сезонами климату внутренней части страны, достигающему своего полного развития в пустыне Сахара. Вследствие этого создается чередование хорошо очерченных зон растительности, располагающихся примерно параллельно берегу (и экватору). Здесь растительность изучена лучше, чем в Восточной (где климатическая зональность осложняется рельефом) и в Центральной Африке.
Это обстоятельство и правильная зональность побуждают нас рассмотреть преимущественно область Западной Африки. Мичельмор (Michel-more, 1939) дал ценный обзор злаковников южных, восточных и центральных районов тропической Африки.
Зональность в Западной Африке
Последовательно расположенные зоны растительности классифицировались и именовались по-разному. Приведенная схема, следующая общеизвестной классификации Шевалье (Chevalier, 1900, 1938), представляется лучшей из ныне существующих (данные о горных областях опущены):
I. Зона сомкнутого леса. Она охватывает площадь сомкнутого леса (в данном ранее определении), включая как высокий вечнозеленый лес на побережье, так и смешанный листопадный лес в глубине страны. Эта зона образует пояс различной ширины. От точки, лежащей чуть восточнее Лагоса, в Нигерии, до западной точки, к северу от Аккры, в Гане, ее прерывает длинная полоса саванны, доходящей здесь до самого моря. Большая часть сомкнутого леса возделывается или возделывалась, и оставшийся лес фактически весь является вторичным. Исключительно интенсивное подсечно-огневое земледелие местами привело к возникновению отклонившейся сукцессии (см. стр. 412). Стадии вторичных сукцессий характеризовались вторжением злаков, а воздействие неоднократных пожаров привело к образованию типа растительности, который Джонс (А. Р. D. Jones, 1945) именует производным саванновым редколесьем. Физиономически и отчасти флористически это сообщество сходно с саванной последующей зоны, хотя его происхождение выдает присутствие немногочисленных экземпляров масличной пальмы (Elaeis) и других деревьев, характерных для зоны сомкнутого леса.
360
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
II. Гвинейская зона. Здесь, в условиях хорошего дренажа, растительность на невозделанных землях варьирует от открытой саванны с разбросанными, преимущественно листопадными, устойчивыми к пожарам деревьями до саваннового редколесья с злаковым покровом. Там, где влагоемкость почвы выше средней, встречаются островки сомкнутого’ леса (Кеау, 1947). Многие из них приурочены к руслам рек, и поэтому часть этих лесов правильнее называть галерейными, или окаймляющими, лесами. Большая часть этой зоны, исключая лишь незначительные площади, подвержена частым пожарам, и ее растительность следует скорее считать климаксом пожарищ, чем климатическим климаксом. Гвинейскую зону удобно разделить на две части, хотя они и не всегда четка разграничиваются:
а) Южногвинейская подзона. Обычные для саванны деревья представлены здесь видами: Daniella oliveri, Lophira alata, Terminalia glauce-csens и т. п. Большая их часть таксономически близка к видам сомкнутого леса. Остатки высокоствольного леса в пределах этой подзоны напоминают по флористическому составу смешанный листопадный тип сомкнутого леса (включают Khaya grandifoliola, Triplochiton scleroxylon и т. д.),. но обычно не так высоки и чаще состоят всего из двух (а не трех) ярусов. Южногвинейские саванны обычно открыты, но в малонаселенных районах сохранились участки саваннового редколесья, в которых не все деревья устойчивы к пожарам; такие участки, вероятно, приближаются к климатическому климаксу.
б) Северогвинейская подзона. Здесь преобладают деревья Isober-linia doka, I. tomentosa, Monotes kerstingii, Uapaca somon и т. д. Они идентичны или родственны видам редкостойного леса «миомбо»1 Brachyste-gia — Isoberlinia южных и восточных районов тропической Африки; виды, близкие к видам сомкнутого леса, встречаются гораздо реже, чем в южногвинейской подзоне. Но как и в этой последней, почти вся растительность здесь сильно изменена огнем; лишь небольшие площади саваннового редколесья, вероятно, не очень отличаются от климатического климакса. Островки леса встречаются главным образом вдоль рек (галерейный лес); их подлесок состоит преимущественно из видов сомкнутого леса, а верхний древесный ярус — из видов, преобладающих в южногвинейских саваннах (Terminalia glaucescens, Anogeissus schimperi, Malacantha alni-folia и т. д.) вместе с несколькими видами, встречающимися как в сомкнутом лесу, так и в саванне, например Khaya senegalensis, Afzelia africana, Diospyros mespiliformis.
III. Суданская зона. Островки вечнозеленого леса почти совсем отсутствуют. Растительность, значительно измененная человеком, представляет собой открытую саванну. Часть деревьев относится к узколистным колючим видам, преобладающим в зоне Сахел, часть — к широколистным видам без колючек, которые встречаются и в саваннах северной Гвинеи. Наиболее распространены баобаб (Adansonia digitata), Lan-пеа microcarpa, Balanites aegyptiaca, Combretum glutinosum, Sclerocarya birrea и др.
IV. Зона Сахел. Климаксной формацией этой зоны, вероятно, является колючее редколесье с преобладанием колючих деревьев с тонкорассе-ченными перистыми листьями, как у видов Acacia. Этот лес обычно настолько разрежен, что между деревьями свободно может проехать авто
1 Милдбрэд (Mildbraed, 1933а) применяет термин «миомбовальд» (Miombowald) к аналогичному саванновому редколесью в северном Камеруне.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 361
мобиль (Aubreville, 1938); напочвенный покров — из злаков или других травянистых растений — не полностью покрывает землю или и вовсе отсутствует. Растительность, как и в предыдущих зонах, очень сильно изменена человеком и пасущимися животными, но благодаря разреженности растительного покрова пожары не могут оказать существенного влияния на его развитие. Небогатая по видовому составу флора резко отличается от флоры Суданской зоны; правда, у воды встречаются такие суданские деревья, как Adansonia, Diospyros mesptliformis и Anogeissus schimperi, но они редко образуют галерейные леса.
V. Зона Сахары. Здесь существует только скудная и чрезвычайно специфическая флора пустыни.
В настоящей работе внимание будет сосредоточено на зоне сомкнутого леса и границе между ним и саванной («производной» саванной и гвинейскими саваннами).
Экотон внутри сомкнутого леса
£ Внутри сомкнутого леса очевиден переход, или экотон, от роскошных, почти полностью вечнозеленых лесов самых влажных областей к менее пышным окаймляющим саванны Гвинейской зоны листопадным лесам, которые принято называть «смешанным листопадным лесом> (Triplochiton — Piptadenia, предклимакс Чиппа, Chipp, 1927). Климатический градиент, создающий экотон, уже рассматривался (гл. 6). Изменение в растительности касается преимущественно ее флористического состава: структура и внешний облик сомкнутого леса повсеместно остаются очень сходными. Термин «смешанный листопадный лес» применялся к различным сообществам, существующим в разнообразнейших климатических условиях, одна часть которых, вероятно, близка к клпмаксному состоянию, другая — к сериальному; поскольку термин «смешанный листопадный лес» является, таким образом, неопределенным, многие предпочитают избегать его. Джонс и Кий (неопубликовано) в пределах сомкнутого леса Нигерии выделяют дождевой, влажный (вечнозеленый) и сухой (вечнозеленый) лес. Эта терминология, пожалуй, лучше всякой другой и может, хотя бы условно, быть принята здесь, но надо иметь в виду, что влажный вечнозеленый лес и, вероятно, сухой вечнозеленый, так же как и «истинный дождевой лес» Джонса и Кия, включаются в принятое в этой книге понятие о тропическом дождевом лесе.
Большая часть того, что рассматривалось как тропический дождевой лес Западной Африки, включая вышеописанный лес Нигерии (гл. 2), в этом смысле является влажным лесом. Он был подробно описан Чипом (Chipp, 1927), Обревилем (Aubreville, 1938) и Ричардсом (Richards, 1939)1. Первичный лес1 2 по флористическому составу, очевидно, всегда смешанный, и, пожалуй, его можно рассматривать как единую ассоциацию. В то время как большинством черт своей структуры и внешности он напоминает дождевой лес тропической Азии и Америки, ряд его особенностей, наверное, связан с относительно сильной сезонной засухой, свойственной данному климату (до пяти месяцев кряду с осадками менее 10 см). Растительность здесь не столь высока и роскошна, как в лучших американских и азиатских дождевых лесах, а ярусы А и В менее сомкнуты
1 Лес Ганы, описанный Фогги (Foggie, 1947), вероятно, лучше всего классифицировать как влажный лес, хотя по характеру он может быть промежуточным между влажным и сухим лесом.
2 См. примечание на стр. 46.
362
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
(хотя нет уверенности, что это не было вызвано вмешательством человека в прошлом). Деревья, ежегодно сбрасывающие листья на несколько недель, многочисленны, хотя и не встречаются в таком изобилии, как в молодом вторичном лесу и на вырубках. Многие виды покрываются молодой листвой одновременно с опаданием старых листьев. Следовательно, сезонная активность или периодичность жизненных процессов растительности в целом выражена сильнее, чем в типичном дождевом лесу. О присутствии геофитов, переживающих сухой период в форме многолетних подземных корневых утолщений, или клубней, уже упоминалось ранее (стр. 116). Лес бассейна Конго, вероятно, имеет много общего с западноафриканским влажным лесом, который в свою очередь очень близок к вечнозеленому сезонному лесу Бирда на Тринидаде.
В Нигерии «истинный дождевой лес», в понимании Джонса и Кия, занимает лишь небольшую площадь к западу от Камеруна и детально не изучался. Некоторые леса округа Обан-Калабар, вероятно, также следует отнести сюда. Такого же типа лес простирается (или недавно простирался) до дельты Нигера и южной границы провинции Бенин; весьма вероятно, что аналогичный характер имеют часть лесов Либерии и примыкающих районов Сьерра-Леоне. К востоку от Камеруна «истинный дождевой лес», должно быть, встречается в Гвиане, экваториальной Африке и центральной части бассейна Конго. Истинный дождевой лес обладает более богатым видовым составом, чем влажный лес, и содержит ряд характерных видов, которые не встречаются во влажном или сухом вечнозеленых лесах. За исключением флористического состава, он очень мало отличается от влажного леса, хотя ярусы А и В, возможно, более сомкнуты, а примесь настоящих листопадных деревьев еще меньше; что касается таких «сезонных» растений, как геофиты, они редки или отсутствуют.
Сухой вечнозеленый лес Западной Африки располагается, главным образом, на северной окраине сомкнутого леса, хотя он ни в каком отношении не составляет перехода к саваннам Суданской зоны. Полное описание его структуры и состава еще никем не производилось, но в работах Чипа (Chipp, 1927), Маккея (Mackay, 1936), Мак-Грегора (MacGregor, 1934, 1937), Обревиля (Aubreville, 1938) и Ричардса (Richards, 1939) содержится много данных о лесе этого типа. Трудно говорить определенно о его характерных особенностях, так как он настолько истреблен или изменен подсечно-огневым земледелием, что редко встречается в первичном или просто в спелом состоянии.
Деревья, вероятно, образуют три яруса, но верхние ярусы менее густы, и их средняя высота, возможно, меньше, чем во влажном лесу. Основное отличие от влажного леса, как указывает само название «смешанный листопадный лес», заключается в большей пропорции деревьев, безлистных в сухое время года, но вечнозеленые виды также многочисленны, а безлистные периоды у листопадных видов не совпадают, так что лес никогда не бывает столь обнаженным, как умеренный листопадный лес. Кустарники и нижние ярусы деревьев почти полностью вечнозеленые. Лиан много, но эпифиты, как и следовало ожидать, немногочисленны как по составу видов, так и по числу индивидуумов. Многие виды деревьев встречаются и в сухом и во влажном лесу, но флора сухого леса беднее, в ней нет многих видов, типичных для истинных дождевых лесов и влажных лесов. В частности, в них отсутствует Lophira procera. Некоторые виды встречаются только в сухом лесу или же бывают там многочисленнее, чем в других типах леса; среди них важно упомянуть Triplochiton
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИН 363 scleroxylon, который очень здесь распространен и местами может составлять 20% древостоя; также характерны Chlorophora excelsa, Manso-nia altissima, Sterculia rhinopetala и другие Sterculiaceae. Некоторые деревья, обычные для сухого леса, встречаются и во влажном лесу, главным образом на вырубках и во вторичных сообществах. Вдоль водотоков п в других эдафически благоприятных местообитаниях некоторые виды влажного леса, распространяясь к северу, проникают в сухой лес точно так же, как виды сомкнутого леса образуют изолированные островки в благоприятных местообитаниях внутри гвинейских саванн. Травяной покров сухого леса, в дождливый сезон столь же пышный, как и во влажном лесу, в сухой сезон обычно увядает, причем большинство видов, входящих в его состав, отмирает полностью.
Как говорит Обревиль (Aubreville, 1938, стр. 123—124), различие между сухим и влажным сомкнутым лесом в основном флористическое, которое не отражается существенно на внешнем облике. Переход от одного сообщества к другому обычно бывает постепенным. В лесном массиве Шаша, Нигерия, одна формация неощутимо переходит в другую, но там, где смена почвы и климата совпадает, переход может быть более резким.
Об экологии так называемого сухого вечнозеленого леса существовали различные мнения. Вряд ли примесь листопадных деревьев в нем достаточно велика, чтобы причислить его к муссонному лесу, как это делает, например, Мак-Грегор (MacGregor, 1937); и действительно, согласно Милдбрэду (Mildbraed, 1922), в Африке нет высокоствольных сомкнутых листопадных лесов, которые можно непосредственно сопоставить с тропическими листопадными лесами Индии и Бирмы. Они ближе к полувечнозеленым сезонным лесам Тринидада. Поскольку структурные различия между сухим и влажным лесом и между влажным и истинным дождевым лесом ничтожны, а флористические различия носят в основном количественный характер, то вряд ли получится правильная картина, если трактовать все эти три типа сомкнутого леса как отдельные самостоятельные формации. А так как различия между климатическими условиями сухого и влажного леса вполне реальны, хотя и незначительны, то сухой лес логически нельзя считать преклимаксом влажного, как это делает Чип (Chipp, 1927). Пользуясь терминологией Клементса (Clements, 1936, стр. 274—275), все три типа сомкнутого леса можно рассматривать как фациации одного климакса, или формации, или, пожалуй, в этом случае можно было бы воспользоваться термином «субформация».
Граница между сомкнутым лесом и саванной
В большинстве мест сухой вечнозеленый лес составляет северный край сомкнутого леса, и граница между сомкнутым лесом и саванной — все равно, производной ли (см. выше стр. 359) или истинной гвинейской саванной — обычно бывает очень резкая, зачастую столь же отчетливая, как граница между покрытой лесом и возделываемой территориями в Европе. В непосредственном соседстве с лесом саванна представляет собой злаковник с разбросанными купами деревьев или, еще чаще, очень открытое низкорослое редколесье с покровом из высоких трав.
Флористический состав, структура и внешний облик сомкнутого леса и саванны резко контрастируют. Громадное большинство доминирующих и подчиненных видов в этих сообществах различны, но несколько родов представлено в сомкнутом лесу и в саванне «замещающими» видами,
364
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
очень резко отличными по физиономии, но близких таксономически. Так, Khaya представлена видами К. ivorensis и К. anthotheca во влажном вечнозеленом лесу, видом К. grandifoliola — в сухом лесу и видом К. senegalensis—в саваннах. Небольшое огнестойкое дерево Lophira alata, столь характерное для производных и южногвинейских саванн, так похоже по своим таксономическим особенностям на высокую L. procera влажного леса, что многие авторитеты считали оба растения сборным видом |связь между ними была рассмотрена Обревилем (Aubreville, 1936)]. Последний пример характерен тем, что обычно ни тот, ни другой вид не встречается в промежуточном сухом лесу. Эти «замещающие» виды, может быть, следует считать экотипами.
Большие различия между растительностью сомкнутого леса и саванны, конечно, зависят от больших различий среды обитания, причем одним из важнейших ее факторов безусловно является огонь1. В сомкнутом лесу нелегко вызвать пожар, и практически этот лес не поддается огню; напротив, все западноафриканские саванны подвержены ежегодным или даже еще более частым пожарам. Эти пожары, соразмерно своей интенсивности, разрушают наземные части травянистой растительности и большую часть подлеска, включая всходы деревьев. Сами деревья, хотя обычно и не погибают, сильно повреждаются; именно этими повреждениями объясняется их искривленный, изуродованный вид. Не вызывает сомнения, что флора саванны должна выдерживать жестокую борьбу за существование; выжить могут только виды, устойчивые к огню. Практически вся растительность африканской саванны, за исключением тех участков, где она представляет собой стадии вторичных сукцессий, является биотическим климаксом, определяемым пожарами (климакс пожарищ). Важный вопрос о пожарах в кустарниковых зарослях здесь не может быть подробно обсужден. Интересующимся следует ознакомиться с докладом Лебрена (Lebrun, 1947), в котором детально описано влияние огня на главные растительные формации тропической Африки и на различные физиономические категории растений.
Какова же связь между климаксами пожарищ и естественным климатическим климаксом этой области и в какой степени естественна резкая граница между сомкнутым лесом и саванной? Выжигание травянистой растительности саванны настолько широко распространено и так давно практикуется в Западной Африке (это отметил еще Ганнон во время своего путешествия около 470 г. до н. э.), что очень трудно ответить на оба эти вопроса. Вероятно, при наличии только тех данных, какими мы теперь располагаем, невозможно точное решение ни одной из этих проблем.
Следует ясно представить, что многочисленные, более или менее изолированные островки вечнозеленого или полувечнозеленого сомкнутого леса внутри саванн нельзя рассматривать как реликты климатического климакса, за исключением производных саванн, которые топографически и климатически находятся внутри пояса сомкнутого леса (в некоторых случаях — в областях влажного леса). Участки сомкнутого леса в гвинейских саваннах встречаются в виде галерейных лесов вдоль рек и в виде рощ поодаль от воды. Последние, если они не посажены искусственно местными жителями, обычно указывают на благоприятные
1 Здесь следует сослаться на важную работу Морисона, Хойля и Хоуп-Симп-сона (Morison, Hoyle, Hope-Simpson, 1948), которые очень подробно рассматривают экологические факторы, включая огонь, влияющие на растительность саванны в юго-западном Судане.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИН 365 эдафические или топографические факторы, некоторые из них не так давно соединялись с галерейными лесами перемычками. Все эти островки леса связаны со специфическими условиями местообитания.
Мак-Грегор (W. D. MacGregor, 1937) экспериментально подошел к проблеме климакса в западноафриканских саваннах. В Олокемеджи, Нигерия, близ границы сомкнутого леса в саванне были заложены три опытных участка. Участок А начисто выжигали ежегодно в марте, В — подвергался частичному ежегодному выжиганию в ноябре, а С — был полностью огражден от огня. Через шесть лет А мало изменился, зато в С значительно увеличилась высота растений и появились многочисленные виды сомкнутого леса, хотя в начале эксперимента их не было и в помине. На участок В также проникли виды сомкнутого леса, но не в таком количестве, как в С. Из результатов этого эксперимента и из наблюдений над окружающей местностью Мак-Грегор сделал вывод, что полная защита растительности от огня вызовет превращение саванны в сомкнутый лес; возникнет лесное сообщество, а травяной покров исчезнет. Поэтому сомкнутый лес (сухой вечнозеленый лес) следует считать климатическим климаксом данного района.
Поскольку саванны Олокемеджи, несомненно,— производные и лежат в климатических границах сомкнутого леса, приведенные данные никак не разрешают проблему климатического климакса в области саванн Гвинеи, которые по флоре и климатическим условиям отличаются от производных саванн. Следует признать очень желательным проведение подобных же экспериментов в южно- и северогвинейских подзонах, хотя за их результаты и нельзя поручиться, так как естественная растительность в этих областях нарушена в такой степени, что свободные от огня участки вряд ли смогли бы быть обсеменены растениями, совокупность которых составляет климакс. Следует также помнить, что длительное непрерывное выжигание, вероятно, очень сильно изменило свойства почвы, которая стала резко отличаться от почвенных разностей климатических климаксных сообществ.
На основании имеющихся данных можно, по крайней мере в отношении южногвинейской подзоны, согласиться с мнением Обревиля (Aubreville, 1936, 1937, стр. 96—98), что если бы каким-то чудом повторные пожары прекратились, а почва в то же время вернулась бы к своему первоначальному состоянию, то открытая саванна и саванновое редколесье гвинейской зоны были бы вытеснены листопадным лесом, относительно открытым, с преимущественно древесным подлеском и скорее лишь с отдельными куртинами злаков, чем со сплошным злаковым покровом («foret claire tropicale» — светлый тропический лес Обревиля; сомкнутый саванновый лес Джонса и Кия, 1946). Преобладание трав в современной растительности — бесспорно, результат выжигания. В последней работе Кия (Кеау, 1947) подчеркиваются флористические и экологические различия между южной и северной подзонами гвинейской зоны, и поэтому кажется вероятным, что в пределах этого гипотетического климакса должен быть постепенный переход от юга к северу. В южной под' зоне естественным климаксом вполне может быть «светлый лес» Обревиля; севернее, в более засушливом климате, климаксом, возможно, становится саванновое редколесье, более похожее на редкостойные леса миомбо южных и восточных районов тропической Африки и напоминающее наименее нарушенные редкостойные леса этой области.
При допущении полной защиты от огня можно предположить, что в этом случае наблюдался бы очень постепенный переход от высокостволь
366
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ного вечнозеленого и полувечнозеленого леса к более низкорослому, преимущественно листопадному «светлому лесу» и сообществу «миомбо»; в этих условиях открытая злаковая саванна должна отсутствовать. Огонь создал резкую границу, часто наблюдающуюся между лесом и открытой саванной; этот внезапный переход в сильной степени обусловлен весьма различными биологическими свойствами и экологическими требованиями растений сомкнутого леса и флоры саванны, состоящей в основном из сформировавшихся путем отбора огнестойких видов.
Многочисленные данные свидетельствуют о том, что в большей части Западной Африки, как и в других районах континента, площадь саванн за последнее время расширилась за счет отступления сомкнутого леса. И процесс этот все еще продолжается; Скаэтта (Scaetta, цит. De Wilde-man, 1934) указывает, что в Руанда-Урунди лес отступает примерно на 1 км в год. В нескольких Местах, наоборот, отмечено внедрение леса в саванну, например в южной части Республики Конго (Lebrun, 1936) и в Уганде (Eggeling, 1947). Как указывалось в гл. 1, не вызывает сомнений, что быстрое сокращение площади африканского леса вызвано главным образом подсечно-огневым земледелием, выжиганием саванны и другими проявлениями человеческой деятельности; Данных, свидетельствующих в пользу того мнения, что причина заключается в «усыхании Африки» или «наступлении Сахары», ничтожно мало. Если даже вековое изменение климата все еще продолжается, его влияние должно сказываться гораздо медленнее, и им нельзя объяснить тот быстрый процесс изменения границ леса, какой наблюдается сейчас;
Саванны лесного массива Конго
Хотя здесь в основном рассматриваются взаимоотношения леса и саванны в западной части тропической Африки, следует кратко упомянуть о вкраплениях саванны в центральной части бассейна Конго, интересный обзор которых дал Робэн (Robyns, 1936). Климатическим климаксом в большей части центрального лесного массива Республики Конго, безусловно, является сомкнутый лес (дождевой лес, влажный вечнозеленый или сухой вечнозеленый лес), хотя внутри него существуют площади злаковников различного характера. Робэн классифицирует их следующим образом:
I. Наземные формации:
А. Эдафические саванны
В. Климатические саванны
С. Вторичные саванны
II. Водные формации.
Водные формации являются стадиями сукцессии, ведущей к становлению вечнозеленого болотного леса как эдафического климакса, поэтому их можно сопоставить с южноамериканскими «водными саваннами» и т. п. Из наземных формаций вторичные саванны — как показывает название — также являются сериальными стадиями, создающимися в основном под воздействием человека; иногда они возникают после потравы слонами. Предоставленные самим себе, эти формации снова превращаются в лес. Климатические саванны находятся на северных и южных границах леса и встречаются только в западной части центральной части бассейна Конго. Их экология, по-видимому, сходна с экологией саванн гвинейской подзоны в Западной Африке.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 36"
Эдафические саванны представляют особый интерес, так как они, видимо, являются ближайшей параллелью южноафриканских саванн, которые были ранее рассмотрены в этой главе. Различают два типа эда-фических саванн: саванны на «латерите» и саванны эзобес. О первых почти ничего не известно; они встречаются только в районе Убанги, и в них преобладает злак Ctenium newtonii. Саванны эзобес занимают небольшие участки. Типичный образец такой саванны близ Кокийавиля занимает площадь около 3 га, располагаясь в наиболее низинной части центрального бассейна, обычно вблизи рек, они никогда не встречаются на плато или водоразделах. Флора эзобес очень однообразна; преобладающее растение — злак Hyparrhenia diplandra, с которым ассоциируются другие травянистые растения, кустарники и низкорослые чахлые деревья. Обычно эзобес занимают песчаные отмели высохших стариц реки; полоса галерейного леса обычно располагается между эзобес и берегом реки. Механический состав почвы колеблется от песка до глины; в некоторых случаях это «чистый белый песок» (напоминает почву саванны Бербисе в Гвиане). Эзобес не выжигают, и Робэн считает некоторые из них «естественными» саваннами, в которых росту деревьев препятствуют специфические почвенные условия. Однако иногда в эзобес внедряется окружающий их лес. Может быть, их следует рассматривать как стадии сукцессии, при которой неблагоприятные почвенные условия не препятствуют внедрению деревьев, а лишь замедляют его. Какова бы ни была истинная экология эзобес, в некоторых отношениях они проявляют поразительное сходство с прогалинами саванн и сообществами мури в гвианском тропическом дождевом лесу (стр. 348).
ВЫВОДЫ
Этот весьма неполный обзор взаимоотношений вечнозеленого леса с листопадным лесом и саванной в различных частях тропиков обнаружил исключительную сложность проблемы. Можно критиковать широкие обобщения Шимпера, но существующие знания не позволяют заменить их какими-либо новыми ясными или определенными выводами.
В целом кажется вероятным, что там, где почвенные условия не препятствуют росту деревьев и где естественная растительность не подвергается частым выжиганиям, дождевой тропический лес у своих климатических границ уступает место листопадному лесу, листопадный лес — саванновому редколесью, а с дальнейшим увеличением сухости климата саванновое редколесье в свою очередь замещается колючим редкостойным лесом и растительностью пустыни. Переход от тропического дождевого леса к листопадному обычно бывает очень постепенным и характеризуется не каким-либо резким или внезапным изменением внешнего облика, а изменением флористического состава, при котором уменьшается количество видов и вместо вечнозеленых начинают преобладать деревья с более или менее регулярным листопадом; постепенно становится все более и более открытым и в конце концов исчезает и ярус А.
Саваннами называют растительные сообщества различной физиономии и экологии, встречающиеся в разнообразных климатических условиях; некоторые из них представляют собой сериальные стадии, другие, несомненно, являются устойчивыми климаксами. Саванны, в которых преобладают деревья (при сплошном злаковом покрове или без него), могут быть климатическим климаксом, но многие типы саванн следует
368
считать климаксами пожарищ; они часто возникают из полулистопадного или даже вечнозеленого леса ка\; результат ot:v<5\ovs.\\\x-шихся (измененных) сукцессий, вызванных земледельческой л повторным выжиганием. Открытые саванны с разбросаннимк или купами деревьев и злаковники без деревьев метут возникнуть даря деградации леса или саваннового редколесья, вслеткствххе
иоп сельскохозяйственной обработки или выжигания. Ни 'в случаях они, вероятно, являются эдафическими климаксамуг, местными почвенными условиями, неблагоприятными, для роътя
Природа других факторов не выяснена. Известно лишь, -что т\нотт<а. -е. ’эгх’йлк направлении действует сезонное заболачивание, сменнют^есн.
нием в остальное время гора.
Тот взгляд, что низинные тропические злаковники всетд^а. 'svkh.vchsvvx тиматическим климаксом, находящимся в равновесии с каиновым климатом», не имеет основательного подтверждав».^-, лххуъ’хххлк ледует считать, что злаковник занимает определенное %
ном климатическом экотоне от тропического
мне.
Глава 16
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ СВОИХ ВЫСОТНЫХ И ШИРОТНЫХ ГРАНИЦ
Изменения в растительности с увеличением высоты во влажных тропиках не менее поразительны, чем в умеренных областях. По мере подъема на лесистую тропическую возвышенность меняется и внешний облик преобладающих видов и структура растительности. Высокоствольный пышный тропический дождевой лес низменностей уступает место другим сообществам, тоже вечнозеленым, но не столь высоким, более простым по структуре и более бедным флористически. Чисто тропическая флора замещается горной флорой, многие роды и даже виды которой обычно характерны для умеренной зоны. Тропический дождевой лес сменяется формацией, которую часто называют предгорным дождевым лесом; он в свою очередь замещается горным дождевым лесом. Во многих местах, особенно на гребнях хребтов и одиночных вершинах, горный дождевой лес состоит из карликовых искривленных деревьев, густо покрытых несметным количеством эпифитов, особенно печеночников и листостебельных мхов; этот очень характерный тип растительности часто называют моховым лесом, или карликовым редколесьем. Предгорный и горный дождевые леса не всегда четко разграничиваются, и иногда оба именуются умеренным дождевым лесом, но было бы предпочтительнее сохранить это название для формаций за пределами тропиков.
На наиболее высоких горных хребтах выше горного дождевого леса может располагаться горный лес другого типа, который достигает климатической границы древесной растительности; выше этого предела встречаются лишь сообщества низкорослых альпийских растений, голые скалы и вечные снега. Таким образом, от тропических низменностей до линии вечных снегов размещается пять основных вертикальных поясов растительности.
Эта вертикальная поясность, которая в общих чертах, видимо, одинакова в Старом и Новом свете, зависит от изменении климата, но связь между ходом климатических элементов и высотой над уровнем моря отнюдь не простая. Колебания некоторых климатических показателей с высотой были рассмотрены в гл. 6; можно напомнить, что в тропиках средняя температура понижается примерно на 0.4—0,7° С на каждые 100 м подъема. Однако важно осознать, что, помимо температуры, почти все другие климатические факторы, влияющие на рост растений, включая осадки, атмосферную влажность, скорость ветра и интенсивность солнеч-ной радиации, также изменяются с увеличением высоты, причем их изменения носят значительно более сложный характер. Так, в районе высоких горных хребтов тропической зоны количество осадков, вначале возрастая, достигает максимальных величин на определенной высоте над уровнем моря, а затем вновь уменьшается. Поскольку растительность, фауна, климат и почва являются частями единой взаимосвязанной эко-24 п. У. Ричардс
3'0
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
системы, то характер почвы также изменяется с увеличением высоты; об этом вкратце упоминалось в гл. 9.
Так как величина изменения климатических элементов с высотой варьирует от места к месту, то фактически границы одного и того же вертикального пояса растительности располагаются на неодинаковой высоте. На небольших одиночных вершинах и гребнях пояса расположены ниже, чем на крупных хребтах; на прибрежных горах они также
Р и с. 40. Продольный (с востока на запад) профиль Северного хребта, остров Тринидад, Вест-Индия, показывающий колебание вертикальных границ поясов растительности (Beard, 1946b).
Вертикальный масштаб: 2000 футов в 1 дюйме; горизонтальный масштаб: 4 мили в 1 дюйме
ниже, чем на горах, удаленных от моря, где проявляется так называемый эффект Massenerhebung1, хорошо известный в Альпах и на других европейских горных хребтах. Понижение верхней границы тропического дождевого леса и горного пояса на горах тропиков является точной параллелью снижения границы древесной растительности в Предальпах по сравнению с центральными хребтами Альп. Изменение границ вертикальных поясов раетительности на горных хребтах тропической зоны, по крайней мере частично, можно объяснить их расположением относительно преобладающих ветров. Бирд (Beard, 1946b, стр. 41) показал, что на восточной оконечности северного хребта Тринидада, обращенной к господствующим пассатным ветрам, пояса растительности понижаются.
ПОЯСНОСТЬ НА ГОРНЫХ ХРЕБТАХ МАЛАЙСКОГО АРХИПЕЛАГА
Значительная часть площади индо-малайского и новогвинейского дождевого леса гористая, и вертикальная поясность растительности изучена здесь лучше, чем в каком-либо другом месте тропиков. Лишь немногие вершины этого района превышают 3500 м, и большинство из них не достигает верхней климатической границы древесной растительности, лишь одиночная вершина Кинабалу, на северном Борнео, и высокие горные цепи Новой Гвинеи превосходят 4000м, причем только последние покрыты вечным снегом. Частично или полностью лишены деревьев вершины многих вулканов Зондских островов, превышающих 3000 м,
1 То есть увеличение абсолютной высоты границ вертикальных поясов растительности в результате возрастания континентальности климата при удалении от моря.— Прим. ред.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 371
но отсутствие древесной растительности, вероятно, вызвано пожарами или местными почвенными условиями; сомнительно, чтобы горные вершины какого-нибудь из Зондских островов достигали климатической границы деревьев.
Ван-Стеенис (van Steenis, 1935b) опубликовал детальный обзор высотной поясности на горах Малайского архипелага, преимущественно с флористической точки зрения. Пользуясь методом, применявшимся Сендтнером (Sendtner) при изучении европейских Альп, он попытался объективно установить естественное разграничение поясов. Была составлена таблица верхних и нижних высотных границ примерно для 800 горных растений, и обнаружилось, что большая часть их стремилась достигнуть своих верхних или нижних границ на определенных критических уровнях или в пределах узких поясов, а не в промежуточных высотных интервалах. Эти критические уровни можно считать естественными границами между поясами.
Из таблицы можно заключить, что в пределах Малайского архипелага существует пять вертикальных поясов растительности (причем два нижних подразделяются) и что критическими высотными границами являются 1000, 2400, 4000 и 4500 м. Хотя эти пояса установлены скорее флористическим, чем фитоценологическим или экологическим методами, они вполне согласуются с теми, которые, как уже было указано, очевидно, существуют во всей области влажных тропиков; это показано в табл. 35.
Таблица 35
Вертикальные пояса растительности в пределах Малайского архипелага
Высота, Флористические пояса Ван-Стеениса Вертикальные пояса растительности (типы формаций)
0—1000 0— 500 Тропический пояс Район низменностей Тропический дождевой лес
500—1000 Район предгорий Предгорный дождевой лес (средне-горный лес Брауна)
1000—2400 Горный пояс Горный дождевой лес (включая моховой лес или карликовое редколесье)
1000—1500 2400—4000 Среднегорный район Субальпийский пояс Тропический субальпийский лес
Климатическая граница распространения древесной растительности
4000—4500 4500 и выше I Альпийский пояс Климатическая граница ] Нивальный пояс I Тропический альпийский кустар-1 ник, хомофитные формации и т. д. вечных снегов Нивальная хомофитная растительность
Критические высоты, вычисленные Ван-Стеенисом, безусловно, лишь очень приблизительны; следует помнить, что вертикальные границы поясов растительности сильно колеблются в различных частях Малезим. Эти колебания зависят не от широты, а преимущественно от упомянутого
24*
372
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
эффекта Massenerhebung (van Steenis, 1932, стр. 176, и 1935b, стр. 298— 300; Richards, 1935, стр. 345). Например, на маленькой изолированной возвышенности Генонг Сантубонг, на побережье Борнео, переход к моховому горному лесу наблюдается на высоте 750 м, на Генонг Белумут, в Джохоре.— на высоте 840 л (Holttum, 1924), но на главной горной цепи западной Явы подобный тип леса не встречается до высоты 1650 м (Sei-friz, 1923), а в восточной части Новой Гвинеи — до высоты 1500—2400 м (Lane-Poole, 1925а, Ь). Границы других зон испытывают подобные же колебания.
Характерной чертой горной растительности Малайского архипелага является резкий контраст между флорой низменностей и флорой гор в отношении их географического сродства. В низинном дождевом лесу вся флора в целом принадлежит к индо-малайским родам, и это в большой степени верно также и для предгорного пояса, но в горном лесу наряду с индо-малайскимп и северными умеренными элементами появляется значительный процент таких видов, как Leptospermum, Tristania, Phyllocladis и др. Следовательно, австралийский элемент, вопреки ожиданиям, увеличивается внезапно по достижении границы горного леса. В пределах Малайского архипелага индо-малайская и австралийская флоры так же сильно обособлены в экологическом отношении, как и в субтропической Австралии (см. ниже, стр. 396 и Richards, 1943). Происхождение и история горной флоры Малайского архипелага исчерпывающе освещены Ван-Стеенисом (van Steenis, 1934, 1935, 1936).
Поясность на высоких хребтах Новой Гвинеи
Горы Новой Гвинеи наиболее благоприятны для изучения вертикальной поясности растительности в восточных тропиках. Они настолько высоки, что достигают границы вечных снегов, и, следовательно, здесь можно наблюдать полную последовательность поясов; отсутствие вулканов способствует развитию ненарушенной климаксной растительности, росту которой не препятствуют извержения или неблагоприятные почвенные условия. Новая Гвинея имеет еще и то преимущество, что ее растительность испытала сравнительно незначительное воздействие человека, а местами сохранилась в девственном виде.
Хотя Новая Гвинея еще очень плохо исследована как с географической, так и с ботанической точек зрения, все же о растительности крупных горных цепей накопилось весьма много данных главным образом благодаря работе Гиббса (Gibbs, 1917) о горах Арфак, Лема (Lam, 1945) и Ван-Стеениса (van Steenis, 1937b) о Центральном хребте и Лейн-Пула (Lane-Poole, 1925а, Ь) о Папуа и восточной части Новой Гвинеи.
В восточных горах Лейн-Пул различает следующие пояса:
I. Низинный лес 0—1000 футов (300 м). Это — первичный тропический дождевой лес, покрывающий большую часть поверхности острова, за исключением «сухих поясов» с осадками ниже 180 см. Смешанный по составу, очень высокий и пышный, он обладает обычными признаками тропического дождевого леса.
II. Лес предгорий 1000—5500 футов (300—1650 м). Этот лес менее высок и состоит главным образом из деревьев небольшого диаметра. Немногочисленные деревья с досковидными корнями так же, как виды Ficus и Alstonia, следует считать случайно попавшими сюда из пояса низменностей. Преобладают двудольные деревья. Один из самых распространенных и характерных видов — дуб, Quercus junghuhnii, кото-
Фото 23. Горный дождевой лес на высоте около 2000 м с хвойными деревьями, горы Арфак, Новая Гвинея.
рый иногда образует чистые древостои площадью до 1 акра (0,4 га); другие наиболее распространенные деревья: Cedrela toona, Vatica рариапа, виды Albizzia, Elaeocarpus, Eugenia, Archidendron и др. Хвойные появляются только у верхней границы пояса. Напочвенный покров состоит преимущественно из папоротников и видов Elatostemma\ лиан и эпифитов не так много, как в низинном лесу.
III. Среднегорный лес — от 5500 до приблизительно 7500 футов (1650—2250 м). Нижняя граница этого пояса совпадает с облачной зоной. Лес представляет собой хвойно-дубовую ассоциацию. Виды Quercus (включая Lithocarpus^) являются одними из распространеннейших двудольных деревьев; характерно и хвойное дерево Araucaria cunninghamii, особенно на гребнях; среди других хвойных встречаются виды Podocarpus и Phyllocladus. Средний размер листьев меньше, чем у деревьев в нижних поясах, и сложные листья встречаются реже; деревья с досковидными корнями отсутствуют. Наземный покров состоит из Elatostemma, пленчатых папоротников и мхов, включая очень крупную Polytrichum-подобную Dawsonia. Лианы мелки и немногочисленны. Эпифитный лишайник (Usnea sp.?) часто встречается в кронах деревьев.
IV. Моховой лес. Его нижняя граница находится на высоте 5000— 8000 футов (1500—2500 м) (в среднем около 7500 футов, 2250 м); верхняя граница 10 000—11 000 футов (3000—3350 м). Этот горный дождевой лес характерен для гор всей Малезии, Он состоит из одного яруса суковатых искривленных деревьев высотой не более 6 м; стволы и ветви скрыты под очень толстым покровом, состоящим в основном из печеночников и мхов, слой которых удваивает их диаметр. Хвойное Podocarpus thevetiifolia местами составляет 80% древостоя; часто встречаются и два вида Eugenia. Попадаются бамбуки и древесные папоротники, но ротанговых пальм нет, а другие лианы встречаются редко. Средняя температура этого пояса около 10° С; основная особенность климата проявляется в постоянной сырости и упорных туманах.
V. Высокогорный лес. Нижняя граница проходит на высоте 10 000— И 000 футов (3000—3350 м). Этот лес более высокостволен, чем моховой; он состоит из крупных деревьев, большая часть которых принадлежит к хвойным из родов Dacrydium, Libocedrus, Phyllocladus и Podocarpus, Встречаются некоторые двудольные деревья, как, например, виды Eugenia. Высокогорный лес обычно не составляет непрерывного пояса, а образует массивы или участки, разделенные злаковниками с разбросанными по ним кустарниками и древовидными папоротниками. Он расположен выше пояса туманов и находится в более солнечном и, вероятно, более сухом климате, чем моховой лес.
Выше 12 000 футов (3600 м) деревья отсутствуют, и растительность состоит из злаковников, болот и низкорослого кустарника на скалистых участках. Обширные высокогорные злаковники, расположенные выше границы древесной растительности в восточной части Новой Гвинеи, на первый взгляд кажутся настоящими альпийскими лугами, то есть климатическим климаксом, но Лейн-Пул приводит веские доводы в пользу того, что в действительности эти злаковники являются биотическим климаксом, созданным пожарами, которые вызывали туземцы, охотясь на уаллоби (мелкие кенгуру). Некоторые наблюдения заставляют думать, что в этих горах деревья могут подниматься по меньшей мере до 4050 м, за исключением очень крутых склонов и болотистых мест. Термин «горная саванна», употребленный Лейн-Пулом для обозначения этих высокогорных злаковников, оказывается, таким образом, вполне уместным.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 373
рый иногда образует чистые древостои площадью до 1 акра (0,4 га)\ другие наиболее распространенные деревья: Cedrela toona, Vatica рариапа, виды Albizzia, Elaeocarpus, Eugenia, Archidendron и др. Хвойные появляются только у верхней границы пояса. Напочвенный покров состоит преимущественно из папоротников и видов Elatostemma\ лиан и эпифитов не так много, как в низинном лесу.
III. Среднегорный лес — от 5500 до приблизительно 7500 футов (1650—2250 м). Нижняя граница этого пояса совпадает с облачной зоной. Лес представляет собой хвойно-дубовую ассоциацию. Виды Quercus (включая Lithocarpus?) являются одними из распространеннейших двудольных деревьев; характерно и хвойное дерево Araucaria cunninghamii, особенно на гребнях; среди других хвойных встречаются виды Podocarpus и Phyllocladus. Средний размер листьев меньше, чем у деревьев в нижних поясах, и сложные листья встречаются реже; деревья с досковидными корнями отсутствуют. Наземный покров состоит из Elatostemma, пленчатых папоротников и мхов, включая очень крупную Polytrichum-подобную Dawsonia. Лианы мелки и немногочисленны. Эпифитный лишайник (Usnea sp.?) часто встречается в кронах деревьев.
IV. Моховой лес. Его нижняя граница находится на высоте 5000— 8000 футов (1500—2500 м) (в среднем около 7500 футов, 2250 л); верхняя граница 10 000—11 000 футов (3000—3350 м). Этот горный дождевой лес характерен для гор всей Малезии. Он состоит из одного яруса суковатых искривленных деревьев высотой не более 6 м; стволы и ветви скрыты под очень толстым покровом, состоящим в основном из печеночников и мхов, слой которых удваивает их диаметр. Хвойное Podocarpus thevetiifolia местами составляет 80% древостоя; часто встречаются и два вида Eugenia. Попадаются бамбуки и древесные папоротники, но ротанговых пальм нет, а другие лианы встречаются редко. Средняя температура этого пояса около 10° С; основная особенность климата проявляется в постоянной сырости и упорных туманах.
V. Высокогорный лес. Нижняя граница проходит на высоте 10 000— 11 000 футов (3000—3350 м). Этот лес более высокостволен, чем моховой; он состоит из крупных деревьев, большая часть которых принадлежит к хвойным из родов Dacrydium, Libocedrus, Phyllocladus и Podocarpus. Встречаются некоторые двудольные деревья, как, например, виды Eugenia. Высокогорный лес обычно не составляет непрерывного пояса, а образует массивы или участки, разделенные злаковниками с разбросанными по ним кустарниками и древовидными папоротниками. Он расположен выше пояса туманов и находится в более солнечном и, вероятно, более сухом климате, чем моховой лес.
Выше 12 000 футов (3600 м) деревья отсутствуют, и растительность состоит из злаковников, болот и низкорослого кустарника на скалистых участках. Обширные высокогорные злаковники, расположенные выше границы древесной растительности в восточной части Новой Гвинеи, на первый взгляд кажутся настоящими альпийскими лугами, то есть климатическим климаксом, но Лейн-Пул приводит веские доводы в пользу того, что в действительности эти злаковники являются биотическим климаксом, созданным пожарами, которые вызывали туземцы, охотясь на уаллоби (мелкие кенгуру). Некоторые наблюдения заставляют думать, что в этих горах деревья могут подниматься по меньшей мере до 4050 за исключением очень крутых склонов и болотистых мест. Термин «горная саванна», употребленный Лейн-Пулом для обозначения этих высокогорных злаковников, оказывается, таким образом, вполне уместным.
374
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Наблюдения Лема (Lam, 1945), произведенные в горах Дормантоп (3580 .и) в западной части Новой Гвинеи, выявили наличие поясности, тождественной установленной Лейн-Пулом для восточных хребтов. Переход от высокоствольного дождевого леса к карликовому моховому лесу происходит здесь на высоте примерно 1420 м. Лесной пояс, непосред-
Рис. 41. Распределение хвойных пород в горных лесах Новой Гвинеи (Lam, 1945).
Ag — Agathis alba", Ar. — Araucaria; D. — IJacrydium; L.—Libocedru8;Ph.— Phyllocladus; Po.— Podocarpus.
ственно примыкающий к верхней границе древесной растительности, здесь также состоит в основном из хвойных пород; по мере подъема наблюдается постепенное увеличение количества хвойных (рис. 41). У высоты 2600 м многие виды достигают своей верхней границы; тут же начинают появляться различные травянистые растения и кустарники, типичные для безлесного пояса. Выше границы леса, которая очень четко очерчена, все еще попадаются от дельные мелкие деревья Libocedrus, а в укромных местах — виды Casua-rina, но большая часть растительности состоит из пустошных и болотоподобных сообществ, а также отдельных карликовых растений в расщелинах голых скал. В горах Дормантоп нет горной саванны и отсутствуют какие-либо свидетельства искусственного возникновения границы древесной растительности; правда, безлесность вершины может быть обусловлена скорее каменистым характером субстрата, чем климатом. Лем высказывает мнение, что климатическая граница леса (но не отдельных деревьев) в этой части Новой Гвинеи лежит на высоте около 3000 м.
Наблюдения над растительностью Карстен-Топпена (5030 л<),снеговой вершины в западной Новой Гвинее, произведенные Ван-Стеени-сом(уап Steenis, 1937), подтверждают
тот взгляд, что высокогорные злаковники не являются климатической формацией, и показывают, что климатическая граница древесной растительности располагается здесь не ниже 4200 я. Говорят, что на этой горе местные жители никогда не поднимались выше 3200 м, так что на растительность больших высот не мог повлиять огонь. Лес у границы деревьев носит смешанный характер и отличается редкостойкостью; в отличие от других высоких гор Новой Гвинеи он состоит не из хвойных, а из двудольных деревьев.
Несмотря на неполноту приведенных данных, они все же позволяют
составить довольно ясное представление о поясности на высоких горных хребтах Новой Гвинеи. Тропический дождевой лес простирается по ниж
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 375
ней части горных склонов до 1000 м, где он сменяется лесом предгорий. Те скудные сведения, которые о нем имеются, заставляют думать, что его следует считать самостоятельной формацией, может быть, сопоставимой со среднегорным лесом Филиппин (см. ниже, стр. 376); к этому сообществу условно можно применить термин «дождевой лес предгорий». На больших высотах, как и во всех влажных горах Малайского архипелага, в горах Новой Гвинеи наблюдается переход к низкорослому одноярусному моховому лесу, но поскольку огромное количество эпифитных мохообразных не является универсальным признаком этой формации, то лучше называть этот лес горным дождевым лесом. Для этого пояса Новой Гвинеи характерно обилие хвойных деревьев, хотя они и не преобладают в древостое. Еще выше расположен пояс с более сухим климатом, менее подверженный туманам; обычно, хотя и не всегда, здесь господствуют хвойные деревья. Поскольку этот пояс простирается до климатической границы леса, его можно назвать субальпийским. Климатическая граница древесной растительности проходит на высоте 4200 м или выше; над ней располагается безлесный альпийский пояс, где растительность состоит главным образом из кустарников и двудольных травянистых растений.
Поясность на более низких горах Малайского архипелага
Более низкие вертикальные пояса, представляющие для нас наибольший интерес, удобно изучать на примере более низких горных хребтов Малайского архипелага, растительность которых известна гораздо лучше, чем новогвинейская. Без сомнения, наиболее полный обзор поясности, приложимый к горной растительности любого района влажных тропиков, дал Браун (Brown, 1919) в своем очерке о растительности горы Маквилинг, на Филиппинах. Эта работа выделяется не только обилием точных данных о составе и структуре растительности на различных высотах, но также полнотой анализа климатических условий и попыткой установить физиологические воздействия климата и зависимость облика растительности от этого фактора. Мы лишь кратко коснемся его выводов.
Он выделил четыре пояса: 1) паранг — 0—200 м\ 2) диптерокарпо-вый лес — 200—600 м; 3) среднегорный лес — 600—900 м', 4) моховой лес — 900—1140 м.
Территория паранга первоначально была занята дождевым лесом, аналогичным диптерокарповому лесу второго пояса, но впоследствии он был вырублен, и, за исключением обрабатываемых участков, эта площадь покрылась смешанными вторичными сообществами (сериальные стадии) от открытого злаковника с преобладанием Jmperata (аланг-аланг) и Saccharum spontaneum до сомкнутого вторичного дождевого леса.
Диптерокарповый лес, занимающий следующий вертикальный пояс, представляет собой смешанную вечнозеленую лесную ассоциацию, где три вида Dipterocarpaceae составляют значительную часть деревьев первого яруса: это — тот тип смешанного диптерокарпового леса (см. гл. 10 и 11), который является климатическим климаксом в большей части Малайского архипелага. По структуре это обычный тропический дождевой лес, состоящий из трех ярусов деревьев, средняя высота которых достигает соответственно 27, 16 и 10 м\ высота наиболее крупных деревьев примерно 36 м. Нижний ярус состоит в значительной степени из молодых деревьев подроста и ротанговых пальм, но выше 500 м и в ущельях наблюдается покров из папоротников и других травянистых растений. Очень
376
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
много лиан, особенно ротангов, но эпифиты встречаются сравнительно редко и главным образом на больших ветвях самых высоких деревьев. Мхов и печеночников на древесных стволах не так много. Хорошо выражены такие физиономические особенности дождевого тропического леса, как досковидные корни и каулифлория. Огромное большинство растений, превышающих 2 м, имеет листья, относящиеся ио своим размерам к «мезо-филльному» классу Раункиэра (см. рис. 15, стр. 103); но встречается и некоторое количество «макрофилльных» или «микрофилльных» растений.
Флористически диптерокарповый лес горы Маквилпнг принадлежит к единой ассоциации с многочисленными доминантами; это — типичный смешанный дождевой лес. На опытном участке (0,25 га) было найдено 92 древесных вида высотой свыше2.и. Поскольку самым распространенным видом в первом ярусе является Parashorea malaanonan (Dipterocarpaceae), а во втором ярусе — Diplodiscus paniculatus (Tiliaceae), Браун назвал эту ассоциацию Par ashore a-Diplodiscus.
Переход от диптерокарпового леса к среднегорному происходит постепенно. Среднегорный лес также представляет собой смешанное вечнозеленое сообщество, в пределах которого различаются две ассоциации; Quercus-Neolitsea и Astronia rolfei, отличающиеся от ассоциации Parashorea-Diplodiscus своей двухъярусной структурой.
В ассоциации Quercus-Neolitsea средняя высота деревьев уменьшается с подъемом; на высоте 700 м первый ярус достигает приблизительно 17 л, а второй —4 м\ длина самого большого из измеренных деревьев составляла 21,8 м. Таким образом, высота верхнего яруса примерно равна высоте второго (В) яруса диптерокарпового леса; в сущности, ассоциация Quercus-Neolitsea очень похожа на ассоциацию Parashorea-Diplodiscus, за исключением того, что в ней отсутствует ярус с преобладанием диптерокарповых. Верхний полог все же более открыт, чем в дип-терокарповом лесу. Подлесок гораздо менее густой, но травяной покров и папоротники развиты лучше и часто образуют на земле толстый ковер; лианы многочисленны. Одна из наиболее достопримечательных особенностей ассоциации Quercus-Neolitsea состоит в обилии эпифитов, число которых возрастает с подъемом, но все же остается меньшим, чем в моховом лесу; их значительно больше, чем в диптерокарповом лесу, причем в своем распространении они не ограничиваются кронами деревьев. Как и в диптерокарповом лесу, здесь преобладают «мезофилльные» листья; незначительный процент образуют «микрофиллы» и «макрофиллы». В ассоциации Quercus-Neolitsea виды Quercus (особенно Q. soleriana) и Neolitsea villosa являются самыми распространенными деревьями верхнего яруса. На опытном участке (0,25 м) было насчитано 70 древесных видов высотой более 2 м. Многие из них характерны и для диптерокарпового леса.
На высоте от 750 до 850 м среднегорный лес состоит из ассоциации Astronia rolfei. Она ниже, чем ассоциация Quercus-Neolitsea, и верхний ярус у нее менее развит; ее, вероятно, следует рассматривать как вариант той же самой формации («локации» по терминологии Клементса), свойственный крутым склонам. Вершина Маквилинг, превышающая 900 м, покрыта моховым лесом — очень своеобразным сообществом, которое больше отличается от диптерокарпового и среднегорного леса, чем эти две формации отличаются друг от друга. Моховой лес образует лишь один ярус деревьев со средней высотой 6—10 м. Толстый слой эпифитов (преимущественно мхов на ветвях и печеночников и мелких папоротников на стволах) и сильно развитые воздушные корни придают карликовому
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 377
лесу фантастический вид. Средняя величина листьев гораздо меньше, чем в среднегорном и диптерокарповом лесах; количество «микрофиллов» и «мезофиллов» примерно одинаково, а «макрофиллы» отсутствуют. Для этой формации свойствен довольно густой почвенный покров из папоротников и цветковых травянистых растений, причем здесь, так же как в других моховых лесах Малайского архипелага, отсутствует отчетливая грань между наземной флорой и эпифитами, и виды, обычно являющиеся наземными, иногда растут как эпифиты, и наоборот. Многочисленные лазящие растения, как эпифиты, так и укореняющиеся в земле, принадлежат к незначительному числу видов.
Моховой лес образует единую ассоциацию Cyathea-Astronia\ в ней нет явно господствующих видов, но более распространенным является древовидный папоротник Cyathea caudata и виды Astronia (Melastomaceae). Из 21 вида деревьев, встречающихся в моховом лесу, 8 можно найти также и в диптерокарповом лесу (входит в состав подлеска); в целом же флора сильно отличается от флоры всех других формаций.
Основные различия в структуре и составе трех вертикальных поясов растительности на горе Маквилинг показаны в табл. 36. Из нее видно, что главное изменение структуры растительности по мере подъема заключается в уменьшении высоты сообщества, которому сопутствует уменьшение числа ярусов. Интересно отметить, что переход от трехъярусной структуры к одноярусной происходит не путем сближения ярусов, а благодаря постепенному исчезновению сначала верхнего, затем второго яруса. Рост количества стволов древесных растений на единицу площади (в моховом лесу их число почти вдвое больше, чем в диптерокарповом), безусловно, зависит от уменьшения размера отдельных деревьев. О количественном увеличении эпифитной растительности по мере подъема какие-либо данные отсутствуют.
Таблица 36
Сравнение пробных площадок (размер 0,25 га) на горе Маквилинг, Филиппинские острова (Brown, 1919)
Диптерокар-повый лес (450 м) Среднегорный лес (700 м) Моховой лес (1020 м)
Количество ярусов деревьев 3 2 1
Высота самого крупного дерева на участке, м 36 22 13
Средняя высота ярусов, м 27, 16, 10 17,4 6
Количество особей древесных растений высотой более 2 м 353 539 610
Количество видов древесных растений высотой более 2 м 92 70 21
Помимо изменений в структуре, происходят сдвиги во внешнем облике и флористическом составе. Наиболее любопытное явление, наблюдающееся с увеличением высоты, состоит в уменьшении среднего размера листа (рис. 15, стр. 103) и исчезновении таких особенностей, как досковидные корни и каулифлория. Серьезное флористическое изменение проявляется в значительном уменьшении числа видов, однако при этом не проявляется никакой тенденции к монодоминантности (ср. гл. 11).
378
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Связь между вертикальной поясностью растительности на горе Маквилинг и климатическими факторами вкратце обсуждалась в гл. 6. Измельчание деревьев по мере подъема, по мнению Брауна, вызвано не столько падением средней температуры, сколько комбинированным воздействием пониженной температуры и ослабленного освещения (благодаря возросшей облачности). По данным, приведенным Брауном, можно заключить, что почва в диптерокарповом лесу (и наверное, также и в среднегорном) принадлежит к типу тропического краснозема или желтозема, а в моховом лесу — к типу подзола. Вероятно, почва мохового леса отличается высокой кислотностью и крайне бедна питательными веществами. Браун не придает большого значения почвенным различиям как факторам, вызывающим изменения в растительности по мере подъема, но все же следует отметить, что моховой лес горы Маквилинг, как и горный лес всех других тропических районов (см. Richards, 1936, стр. 354— 355), обладает особенностями, обычно связанными с «физиологической сухостью» (измельчание, многочисленность видов с мелкими кожистыми листьями и пр.). Эти признаки аналогичны наблюдающимся в умеренном климате у растительности кислых, бедных минеральными веществами почв; невольно хотелось бы искать объяснения этого в каких-либо химических особенностях субстрата, таких, как крайняя бедность минеральными веществами или слабая нитрификация, а не исключительно в климатических факторах, как это делает Браун. Подобные же взгляды были недавно выдвинуты для объяснения «физиологической ксероморф-ности» европейских болотных растений (см. Steemann-Nielsen, 1940).
Общие черты вертикальной поясности растительности горы Маквилинг типичны и для других средневысотных гор Филиппин, Борнео, полуострова Малакки и др. На горе Дьюлит, Борнео (Richards, 1936), у подножия и на нижних склонах горы развит такой же смешанный дипте-рокарповый лес, как и на Маквилинг, но, поскольку здесь он не подвергся интенсивному разрушению, пояс паранга отсутствует. Структура и состав леса до высоты 450 м почти не изменяются, но в промежутке между ней и нижней границей мохового леса, располагающейся на высотах 970— 1100 .и, находится лесной пояс, который отличается от смешанного дипте-рокарпового по составу и, вероятно, по структуре; он не был подробно изучен, но, по-видимому, подобен среднегорному лесу горы Маквилинг (Браун). Горный лес отделен здесь чрезвычайно резкой границей и по общему виду очень сильно отличается как от низинного, так и от среднегорного леса. Деревья, высота которых тут редко превышала 15 м, а на открытых местах была еще меньше, составляли скорее два не очень отчетливых яруса, а не один, как на горе Маквилинг. Досковидные корни и другие признаки, характерные для тропического дождевого леса, отсутствовали, а листья были мелкими и кожистыми, иногда даже эрикоид-ными. Основной чертой, как и в других моховых лесах, оставалось исключительно пышное развитие эпифитной растительности; большую часть эпифитов составляли печеночники, хотя листостебельных мхов, папоротников и цветковых растений также было множество.
Вертикальную поясность на Малаккском полуострове описали Хол-тум (Holttum, 1924), Саймингтон (Symington, 1936, 1943) и другие. Dipterocarpaceae, преобладающее семейство в дождевых лесах низменностей, согласно Саймингтону, достигает своей верхней границы на высоте около 4000 футов (1200 м) на главных хребтах и примерно на половине этой высоты — на изолированных вершинах. Верхнюю границу диптерокар-пов можно использовать для приблизительного подразделения тропи
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 379
ческого дождевого леса низменностей и «горного леса». Саймингтон различает три вертикальных подпояса диптерокарпового леса: 1) диптеро-карповые леса низменностей, характеризующиеся преобладанием в верхнем ярусе типично «низинных» видов диптерокарпов (главным образом видов Shorea из группы «красных меранти»); 2) диптеро-карповые леса низких предгорий, в которых господствуют «холмовые» меранти вроде Shorea curtisii, этот пояс редко достигает 1000 футов (300 м) высоты; 3) диптерокарповые леса высоких предгорий. Здесь Shorea curtisii становится редкой и вместо нее появляются такие специфические диптерокарпы, как Shorea platyclados и S. ciliata. Выше верхней границы Dipterocarpaceae располагаются два пояса «горного леса»: горные дубняки и горные эрикоидные леса. Диптерокарпы не всегда присутствуют в их составе. Горные дубовые леса простираются до высоты 5000—6000 футов (1500—1800 м) на более высоких горах; они высоко-ствольны, состоят из двух ярусов и характеризуются обилием дубов (виды Quercus и Lithocarpus'). Горные эрикоидные леса, встречающиеся на самых высоких горах, выше дубовых, низкорослы и отличаются обилием мхов, дубы встречаются в них редко, преобладают деревья семейства вересковых — виды Pieris, Vaccinium и Rhododendron. Три подпояса диптерокарпового леса Саймингтона явно принадлежат к формации тропического дождевого леса; горные дубовые леса, сходные с лесами предгорий на Новой Гвинее и со среднегорным лесом горы Маквилинг, представляют формацию, именуемую в книге дождевым лесом предгорий. Горный эрикоидный лес подобен горному дождевому лесу других гор Малайского архипелага. На обширных горных хребтах полуострова Малакка благодаря эффекту Massenerhebung границы соответствующих поясов обычно располагаются гораздо выше, чем на таких обособленных горах, как Маквилинг и Дьюлит.
Растительность высоких вулканов Явы была неоднократно описана (см. Schimper, 1935; Docters van Leeuwen, 1933; Seifriz, 1923 и 1924). На западной Яве обычно различают три лесных пояса: тропический дождевой лес, который на высоте примерно 1500—2200 м переходит в умеренный дождевой лес; этот лес беднее видами и отличается от тропического дождевого леса отсутствием деревьев с досковидными корнями, меньшими размерами листьев, большей толщиной стволов и большим количеством эпифитов, а также и флористически; здесь преобладают дубы, а также виды Castanopsis, Litsea javanica и Podocarpus imbricatus. Выше него располагается карликовый редкостойный или облачный лес, покрывающий наиболее высокие вершины и являющийся типичным карликовым моховым лесом. На днищах кратеров лес сменяется злаков-никами или кустарником с господством Anaphalis javanica] эти сообщества, вероятно, обязаны своим существованием повторным пожарам или особым эдафическим условиям.
В более сухом климате восточной Явы поясность несколько иная Тропический дождевой лес, более сухого и не столь роскошного типа, как на западной Яве, встречается в самом нижнем поясе, а вместо умеренного дождевого леса западной Явы здесь произрастает сообщество^ отличающееся присутствием Casuarina junghuhniana', по внешнему виду оно похоже на саванновое редколесье. Самые высокие вершины покрыты злаковниками и карликовым кустарником. Имеются доказательства того, что редкостойный лес из Casuarina, по крайней мере частично, является климаксом, возможно, что злаковники и кустарник самого верхнего пояса также не представляют истинного климатического климакса.
380
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ СВОЕЙ ВЫСОТНОЙ ГРАНИЦЫ В АФРИКЕ
Высокогорная растительность Африки с ее эффективными древовидными Senecio, лобелиями и Ericaceae достаточно хорошо известна, что нельзя сказать о вертикальной поясности многих африканских горных хребтов в тропических районах.
Для изучения изменений растительности у верхнего предела распространения тропического дождевого леса Африка не является благоприятной областью. Большинство высоких африканских гор расположено в районах с относительно сухим климатом, и не тропический лес, а скорее саванна и аналогичные более или менее ксерофитные сообщества покрывают их нижние склоны. Даже там, где дождевой лес достигает подножия гор, как у восточного края бассейна Конго, пояса сухого и влажного климата так сложно переплетаются, что последовательность растительных формаций, связанную с высотными градациями климата, нелегко отграничить от последовательности, вызванной местными колебаниями в количестве осадков. Далее, поскольку значительная часть горной растительности в Африке произрастает на молодых вулканических почвах, на которых она еще не достигла своего полного развития, а значительная часть изменена огнем, пасущимися животными и подсечно-огневым земледелием, то часто бывает трудно даже распознать климакс-ные типы.
Условия у верхней границы африканского сомкнутого леса (тропического дождевого леса, ср. гл. 15) лучше всего изучены, особенно благодаря работе Лебрена (Lebrun, 1935, 1936а), на западных склонах высоких вулканов, отделяющих бассейн Конго от Великого африканского грабена.
Здесь обширный низинный тропический дождевой лес Центральной Африки во многих пунктах близко подступает к горам, хотя значительная часть его уже истреблена, и восточный край поэтому смещен к западу. На средних высотах гор существует зона вечнозеленого нетропического дождевого леса, где заметная часть флоры по своим родственным связям скорее субтропическая или умеренная, чем тропическая; аналогичные леса, принадлежащие к той же самой формации и именуемые то субтропическими, то теплыми умеренными, то умеренными или горными дождевыми лесами (полная синонимика дана у Питт-Шенкеля, Pitt-Schenkel, 1938), разбросаны повсеместно в восточно-африканских горах; условно все они могут быть отнесены к формации того же самого типа, что и горные дождевые леса Малайского архипелага.
Согласно карте, данной Лебреном (Lebrun, 1935, рис. 1), в нескольких местах замечается непрерывный переход между горными лесами и низинным тропическим дождевым лесом Конго, что отчетливо видно к северо-западу от Рувензори и к западу от озера Киву. Чаще, однако, низинный и горный леса разделяются узкой полосой ксерофильной саванны и злаковников. Согласно Лебрену, ее происхождение связано с фенами, которые дуют с гор и создают местный пояс сухого климата. Там, где нормальный высотный климатический градиент не нарушается, средняя температура падает примерно на 0,6° на 100 м подъема, а годовая сумма осадков у высоты 2400 м достигает максимума, равного 225 см, и вновь уменьшается на больших высотах. Вследствие увеличения облачности величина инсоляции по мере подъема снижается и доходит до минимума в зоне максимального выпадения осадков.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 381
Там, где низинный и нагорный дождевые леса смыкаются, можно выделить следующие пояса:
I. Пояс тропического дождевого леса (Foret equatoriale Лебрена), верхняя граница 1100—1300 м\
II. Переходный лесной пояс (Foret de transition), нижняя граница 1100—1300 м, верхняя граница 1650—1750 м\
III. Пояс горного дождевого леса (Foret mesophile de montagne), нижняя граница 1650—1750 м. верхняя граница 2300—3400 м.
Верхняя граница горного дождевого леса, которая совпадает с уровнем максимального выпадения осадков, отмечена очень характерной полосой бамбуковых зарослей (нижняя граница 2200—2400 м, верхняя — 2600 ле), образуемых очень плотной консоциацией Arundinaria alpina, обычно встречающейся на более высоких горах Восточной Африки. Выше 2600 м бамбуковый лес сменяется поясом Ericaceae (нижняя граница 2600—3100 л, верхняя — 3700—3800 л), где преобладают древовидные виды Erica и Philippia, или же (особенно в восточной группе гор Вирунга) поясом Hagenia (2600—3100 м) — типом низкорослого открытого редколесья; еще выше располагается альпийский пояс — главное местообитание древовидных Senecio и лобелий (нижняя граница 3700— 3800 м, верхняя — 4600 м), и пояс вечных снегов1.
Переходный лес впервые был выделен как определенное сообщество Лебреном (Lebrun, 1934), но до сих пор является почти не изученным. Это сообщество не так роскошно, как низинный тропический дождевой лес и отличается от последнего меньшей высотой и меньшим диаметром деревьев; запас древесины на единицу площади в нем также меньше (см. ниже). Флористически переходный лес представляет собой смешанную ассоциацию, очень богатую видами, многие из которых эндемичны; некоторые виды часто встречаются и в низинном лесу (например, Сагара grandiflora. Cynometra alexandri. Symphonia gabonensis, Uapaca guineensis и пр.); другие проникают сюда из верхней горной зоны.
Если у верхней границы леса переходного типа сухой фёновый пояс отсутствует, он постепенно уступает место горному дождевому лесу, высокоствольному сомкнутому сообществу, состоящему из двух ярусов деревьев (strate dominante и strate dominee). одного или двух ярусов кустарников и яруса травянистых растений. Средняя высота верхнего яруса деревьев составляет примерно 25 м\ кроны деревьев компактны и разобщены друг от друга; в отличие от тропического дождевого леса их не переплетают лианы. Следующие цифры дают представление о некоторых особенно показательных различиях между горным лесом переходного типа и тропическим дождевым лесом.
В горном дождевом лесу подавляющее большинство деревьев является вечнозелеными, и очень немногие имеют досковидные корни. Он явственно отличается от тропического дождевого леса отсутствием или малочисленностью лиан. Обилие эпифитов (мхов, лишайников, папоротников, орхидей, Loranthaceae) увеличивается с высотой местности, но разрастание мхов на стволах деревьев не доходит до такой степени, как в поясе Ericaceae, развитом на значительно больших высотах. Кустарниковый ярус очень густой, а травяной покров, исключая участки вырубок, очень разрежен и включает много полукустарников. Как и в тропическом
1 На горах Кения и Абердаре (Fries R. Е., Fries Т. С. Е., 1948) наблюдается аналогичная поясность, но, поскольку эти горы возвышаются над «базальтовым плато» с сухим климатом, зоны I и II замещаются злаковником («степь тхемеда»).
382
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Таблица 37
Сопоставление формаций вечнозеленых лесов на горах восточной части Республики Конго
(Lebrun, 1936а, стр. 151)
Тропический дождевой лес Лес переходного типа Горный дожде ной лес
Число стволов (более 20 см в диаметре) на 1 га 115 180 220
Средняя высота ветвления стволов, м 13 12 10
Средняя высота крон, м 30 25 20
Средний диаметр стволов, м 60 40 35
Запас древесины на 1 га, куб. м 400—600 300 200
дождевом лесу, на земле отсутствует слой мхов, хотя они и встречаются у подножия деревьев и на валунах.
Флористически горный лес сильно отличается от тропического дождевого леса тем, что в его составе наряду с растениями, одинаково характерными и для тропических лесов низменности, встречаются виды, родственные растениям умеренной зоны. Этот лес является смешанной ассоциацией с многочисленными доминирующими видами.
Лебрен выделяет три подпояса (горизонта) первичного горного дождевого леса: нижний (1600—1900 м). средний (1900—2100 м) и верхний (2100—2400 м), Нижний близко напоминает лес переходного типа. Деревья здесь выше и подлесок гуще, чем в двух других, 30% деревьев обладает сложными листьями; присутствует некоторое количество листопадных видов. Среди главных доминант первого яруса отметим: Entandrophragma speciosum. Ficalhoa laurijolia. Lebrunia bushaie. Ocotea usambarensis и Strom-bosia grandijolia, Много древовидных папоротников. Среди травянистых растений попадаются растения умеренной зоны, например Dryopteris jilix-mas и Sanicula europea. Для этого подпояса характерны некоторые лианы. Хотя эпифитов много, они не так роскошны, как на больших высотах, а лишайники из рода Usnea незаметны. В среднем подпоясе деревья более высокие, чем в верхнем, но ниже, чем в нижнем (цифры для горного леса, приведенные в табл. 37, относятся к этому подпоясу). Здесь процент деревьев со сложными листьями падает до 25%. Основные доминанты первого яруса: Entandrophragma speciosum. Ficalhoa lauri-folia. Ocotea viridis и О, usambarensis. Parinari mildbraedii. Podocarpus milanjianus и P, usambarensis. Strombosia grandifolia и Symphonia globu-lijera, В ущельях распространены древовидные папоротники, часто в комбинации с Ensete edule, Травяной покров очень разрежен и включает представителей родов умеренной зоны. Крупных лиан очень мало. Эпифитов множество, но виды рода Usnea незаметны. Лес верхнего подпояса имеет некоторое сходство с карликовым редкостойным лесом. Высота деревьев не превышает 10—15 м. их стволы и ветви искривлены. Листья деревьев, как правило, мелкие, кожистые и цельнокрайние; только 15% видов имеют сложные листья. Типичными деревьями первого яруса являются: Ekebergia ruppeliana. Ficalhoa laurijolia. Olea hochstetteri. Parinari mildbraedii. Podocarpus milanjianus. Sideroxylon adolji-jriederici и Symphonia gabonensis. Кустарниковый подлесок хорошо развит, но напочвенный травяной покров скуден.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 38?,
Поучительно сопоставить вертикальную поясность восточной части Республики Конго с поясностью гор Узамбара, Танганьика, описанной Моро (Moreau, 1935 b1). Эти горы представляют собой два небольших самостоятельных массива — восточный, достигающий почти 4900 футов высоты (1500 л€), и западный, превышающий 7000 футов (2000 м)\ оба массива являются островками влажного климата в сухой области, для которой характерна растительность типа «листопадного паркового леса» (саванны). Хотя значительная часть первичного леса уничтожена сельскохозяйственной обработкой, роскошный тропический дождевой лес еще сохраняется на восточных склонах гор. Во влажных частях Узамбар Моро выделил три лесных пояса: низинный — до 2500 футов (760 промежуточный — 2500—4500 футов (760—1370 м) и нагорный — более 4500 футов (1370 м). В древостое низинного леса преобладают Antiaris usambarensis, Ficus spp., Sterculia appendiculata, Trema guianensis, Chlo-rophora excelsa и Albizzia spp. Такой видовой состав наводит на мысль о вторичном происхождении этого сообщества; первичный лес, возможно, скорее походил на западноафриканский сухой вечнозеленый (смешанный листопадный) лес, чем на «истинный тропический дождевой лес». Подлесок низинного леса включает колючие виды и состоит главным образом из Acacia pennata и Harrisonia abyssinica\ Olyra lalifolia образует густые заросли в несколько футов вышиной, других травянистых растений мало. Лиан много, но эпифиты редки и только местами попадаются Platycerium angolense. Лес промежуточного типа наиболее роскошный из всех трех типов. Моро даже считает, что это самый пышный лес Восточной Африки. Виды деревьев здесь очень многочисленны, и многие из них являются единственными представителями соответствующих родов и эндемичны; среди доминант следует отметить Macaranga usambarica, Allan-blackia stuhlmannii, Piptadenia buchananii, Isoberlinia scheffleri и Pari-nari spp., много древовидных папоротников. На более открытых местах напочвенный покров разнообразен и богат. Обращают на себя внимание лианы и фикусы-удушители; эпифитов много как по обилию особей, так и по числу видов. Общий вид нагорного леса, который достигает полного развития только в западных Узамбарах, несколько напоминает лес промежуточного типа, хотя и отличается от него флористическим составом. Среди основных деревьев, доминирующих совместно или порознь, находятся Podocarpus spp. и Ocotea usambarensis. В этой же зоне встречаются леса из Juniperus procera («кедр»). Пит-Шенкель (Pitt-Schenkel, 1938) подробно описал эти нагорные леса, которые он называет «теплыми умеренными дождевыми лесами».
Выявление других деталей в структуре и составе лесов Узамбара, безусловно, очень желательно, пока же, чисто условно, низинный лес, пожалуй, может быть уравнен с «экваториальными лесами» (foret equa-toriale) восточной части Республики Конго, промежуточный лес — с лесом переходного типа Лебрена и нагорный лес — с горным дождевым лесом.
На более высоких горах Западной Африки непосредственный переход от тропического дождевого леса к нагорным формациям наблюдается очень редко благодаря замене естественной растительности возделываемыми землями на большей части предгорий.
1 Дополнительные сведения, любезно предоставленные II. Дж. Тринвеем (Р. J. Greenway), используются в дальнейшем.
384
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
На острове Сан-Томе в Гвинейском заливе тропический дождевой лес, вероятно, первоначально покрывал всю поверхность вплоть до наивысшей точки (2024 .«). В пределах острова выделяются три пояса (Exell, 1944): пояс низинного тропического дождевого леса (0—800 м), пояс горного дождевого леса (800—1400 м почти повсеместно) и пояс облачного леса (1400—2024 лг). Низинный дождевой лес в настоящее время почти полностью истреблен. По-видимому, это была скорее смешанная ассоциация типа влажного вечнозеленого леса (стр. 361), чем «истинный тропический дождевой лес», и по составу она, вероятно, походила на низинный лес острова Фернандо-По и соседних частей континента. Это сообщество, вероятно, постепенно переходило в горный лес, причем переход, видимо, «характеризовался главным образом изменениями в составе леса, чем в его общем облике» (Exell, 1944, стр. 18). В горном лесу деревья достигают большой высоты и образуют густой полог. Подлесок в значительной мере состоит из Chasalia doniana высотой до 1 м. Напочвенный покров изобилует наземными орхидеями и папоротниками, причем последние как по числу видов, так и особей, по-видимому, достигают здесь максимального развития по сравнению с другими местами в Африке. Встречаются и некоторые древовидные папоротники. Много крупных лиан, а эпифитные мхи, лишайники и цветковые растения так многочисленны, что почти полностью покрывают стволы. Флористический горный лес очень богат; особенно много Bubiaceae и Euphorbiaceae, тогда как Leguminosae отсутствуют или же редки. Облачный лес находится в зоне максимальных осадков, где туман почти -никогда не рассеивается. Деревья здесь мельче, чем в нижних зонах, а эпифиты еще более многочисленны, чем в горном лесу. Флора этой зоны мало изучена, однако известно, что для нее характерны многие виды, которые, скорее всего, отсутствуют на меньших высотах, причем большая часть их эндемична. Встречаются некоторые виды умеренной зоны.
На горах острова Фернандо-По (3291 м) и на горе Камерун (4067 м) на. африканском континенте наблюдается аналогичная поясность, но эти горы гораздо выше, чем горы на острове Сан-Томе, и верхние склоны их лишены деревьев. На горе Камерун лес поднимается выше 2000 м. Над границей деревьев находится открытый злаковник с редкими кустарниками и низкорослыми деревьями (Maitland, 1932), который ежегодно выжигается местным населением. Самая высокая часть горы представляет собой «альпийскую пустыню» из обнаженной лавы с редкими колониями злаков и других растений. Лес на нижних склонах, который до высоты примерно 1000 м, вероятно, был аналогичен лесу на прилегающих низменностях, большей частью сведен с целью расчистки места для плантаций и полей местных жителей. Примерно от уровня 1000 м по направлению вверх еще сохранился непрерывный лесной пояс, но большая часть его покрывает лаву недавнего или сравнительно недавнего происхождения и, вероятно, не представляет собой климакса. Нижние участки этого леса высокоствольны, но и там его флористический состав значительно отличается от низинного тропического дождевого леса; в подлеске встречаются такие, свойственные умеренной зоне виды, как Sanicula еигораеа и Viola abyssinica', лиан мало, но мхи многочисленны. Обращают на себя внимание группировки древовидного папоротника Cyathea manniana, развивающиеся на участках с нарушенным лесным покровом и в ущельях. Выше 1500 м количество древесных видов сильно уменьшается, и на обширных площадях встречаются почти чистые древостои Syzygium, часто с густым подлеском из Мimulopsis violacea и других Acanthaceae. Близ
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 385
границы леса деревья становятся карликовыми и искривленными; среди них преобладают Hypericum leucop tychodes, Lasiosiphon glaucus, Rapanea neurophylla, Pittosporum mannii и Schejflera hookeriana. Этот лес сильно напоминает моховой лес Малайского архипелага, за исключением того, что бриофиты, правда, многочисленные, здесь далеко не так роскошны; очевидно, это — один из типов горного дождевого леса. Граница древесной растительности в этих местах, безусловно, не является климатической и, вероятно, частично определяется наличием молодых лавовых потоков, а частично — ежегодными пожарами, препятствующими внедрению деревьев в злаковник.
Приведенные данные показывают, что в тех районах Африки, где тропический дождевой лес непосредственно соприкасается с нагорным вечнозеленым лесом, то есть там, где его верхняя граница действительно определяется высотой над уровнем моря, а не какими-либо привходящими климатическими или биотическими условиями, общие черты перехода оказываются сходными с теми, какие наблюдаются в пределах Малайского архипелага. По-видимому, высокоствольный лес переходного типа в среднегорной зоне Восточной Африки, горный лес на Сан-Томе и лес на высоте примерно 1000—1500 м на горе Камерун имеют много общего как во внешнем облике, так и в структуре с дождевым лесом предгорий Малайского архипелага. Сходны между собой и горные леса Африки и Малайского архипелага.
Карликовый горный лес Восточной Африки (верхний подпояс Лебрена), видимо, во многих отношениях сопоставим с низкорослым моховым типом горного дождевого леса Малайского архипелага. Однако пока еще, может быть, преждевременно применять одинаковые термины к аналогичным формациям этих географически удаленных областей.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ СВОЕЙ ВЫСОТНОЙ ГРАНИЦЫ В ЮЖНОЙ АМЕРИКЕ И ВЕСТ-ИНДИИ
Леса Амазонии простираются к западу до подножия Анд, где характер и состав растительности почти сразу же изменяются. С увеличением высоты над уровнем моря тропический дождевой лес уступает место другим вечнозеленым лесным формациям, включающим и нетропическую флору. На влажных восточных склонах Анд в Колумбии, Перу и Боливии обычно различают следующие области: I — область тропического дождевого леса подножий гор (montana), только слегка отличающаяся по виду и флоре от тропического дождевого леса Амазонии (montana real, гилея Гумбольдта); эта область простирается вверх примерно до 1200—1500 м. Над ней лежат две области вечнозеленого леса, в которых и флора и общий вид растительности совершенно иные, чем в нижней; II — это переходная область (medio yungas) и III — область вершины (ceja de la montana). Ceja начинается в Перу на высоте 1800—2000 м и достигает 3400—4б00 м (Weberbauer, 1911); в Боливии ее границы лежат на высотах 2200—2800 и 3400 м (Cardenas, 1945).
Еще выше располагается альпийская область тропических Анд, состоящая преимущественно из открытых парамо (paramo), или степей, для которых характерны древовидные Compositae (главным образом виды Espeletia и Culcitium), поразительно сходные по внешнему облику с древовидными соответствующей области африканских гор.
Дождевой тропический лес подножий Анд отличается от леса низменностей тем, что в нем отсутствуют некоторые виды и роды. Верхняя гра-
386
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
яйца многих типично тропических элементов, таких, как пальмы Iriartea, Phytelephas и Bactris, совпадает с верхней границей тропического дождевого леса подножий, но некоторые проникают и выше. Таким образом, область medio yungas обладает смешанной флорой, в которой присутствуют и тропические и умеренные элементы. Именно здесь находятся знаменитые леса хинного дерева; обращают на себя внимание также бамбуки (виды Chusquea) и древовидные папоротники. По-видимому, леса medio yungas имеют значительное сходство с высокоствольными горными (умеренными) лесами Восточной Африки. В лесах ceja de la montana (JBuschwald, Gebiisch и Gestrauch, Weberbauer, 1911) преобладают низкорослые деревья с суковатыми и искривленными стволами, обильно поросшими эпифитными мхами; листья деревьев преимущественно мелкие, кожистые и цельные. Фактически они представляют собой типичный «облачный лес», или формацию карликового редколесья; этот лес, очевидно, близко сходен с моховым лесом Малайского архипелага, на который он действительно очень похож по флористическому составу.
Эти вечнозеленые лесные формации были неоднократно описаны (Weberbauer, 1911, 1945; Herzog, 1923; Cuatrecasas, 1934 и др.), но, поскольку имеющиеся данные касаются главным образом флористического состава, дальнейшее обсуждение их здесь было бы излишним.
Горные леса Вест-Индии, особенно Тринидада и Малых Антильских островов, изучены лучше, чем леса Анд. Лесные формации и их вертикальная поясность исследовались Глизоном и Куком (Gleason, Cook, 1926), Стэле (Stehle, 1935), Бирдом (Beard, 1944а, 1946b) и другими. Хотя эти растительные формации весьма сходны по внешнему облику с формациями, развитыми в более широких масштабах в Андах, они располагаются в Вест-Индии с ее сравнительно изолированными прибрежными горами, где отсутствует эффект М assenerhebung на гораздо меньших высотах.
На Тринидаде Бирд (Beard, 1946b) различает три пояса растительности: «нижнегорный дождевой лес» — от 800 до 2500 футов (250—760 лс), «горный дождевой лес» — от 2500 до 2900 футов (760—880 м) и карликовое редколесье (только на вершине Арипо на высоте более 2900 футов). Вертикальные границы этих формаций и ширина переходной полосы от одной к другой несколько колеблются; как упоминалось ранее (ср. стр. 370, рис. 40), все пояса снижаются в направлении восточного наветренного края Северного хребта.
Нижнегорный дождевой лес Бирда следовало бы включить в формацию тропического дождевого леса в нашем понимании. Он отличается от типичного низинного дождевого леса лишь несколько меньшей высотой деревьев. Средняя высота спелых деревьев доминантных видов составляет примерно 90 футов (27 м), исключая затененные места; самый высокий ярус образует плотный полог на высоте 70—100 футов (22—30 м); над ним находится прерывистый нижний ярус деревьев (рис. 42). Физиономические черты, типичные для деревьев тропического дождевого леса, плохо развиты; так, досковидные корни мало заметны, а каулифлория встречается редко. Лиан и эпифитов не очень много. Пальмы и древовидные папоротники мало характерны. Здесь развита только одна ассоциация: Byrsonima spicata —Licania ternatensis. На 100 акрах (40 га) этого леса было зарегистрировано 87 видов, что несколько меньше обычного количества видов в низинном «вечнозеленом сезонном лесу» Тринидада (стр. 343). Примерно 28% древостоя составляет Licania ternatensis.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 387
«Горный дождевой лес» занимает ограниченную площадь и встречается только в пределах массива Арипо. Этот лес состоит из двух ярусов (рис. 42, б), и его средняя высота еще меньше, чем высота нижнегорного леса. Верхний древесный ярус образует сомкнутый полог на уровне 50— 65 футов (15—20 лг), причем ни одно дерево в его составе не превышает 75 футов (23 м). Досковидные корни и другие типичные черты тропического дождевого леса в большинстве случаев отсутствуют, но лианы и эпифиты очень многочисленны и роскошны, причем среди эпифитов много мхов. Листья по величине колеблются от «мезофиллов» до «микрофиллов», листовые пластинки преимущественно кожистые. Под вторым древесным ярусом располагается ярус, состоящий преимущественно из пальм и древовидных папоротников. Наиболее многочисленны в этой ассоциации два вида — Richeria grandis и Eschweilera trinitensis.
Оба вида типичны именно для этого пояса. Флора здесь не отличается богатством и включает всего 28 видов древесных растений.
Формация карликового редколесья представлена консоциацией Clusia intertexta — дерева с ходульными корнями,— которая приурочена непосредственно к вершине Арипо. Господствующее дерево образует сомкнутый верхний ярус высотой 20—25 футов (6—7,5 ле); ниже развиваются густые заросли древовидных папоротников и маленьких пальм, достигающие в среднем 10 футов (3 м) высоты. Эпифиты, представленные мхами и лишайниками, исключительно пышны и целиком покрывают стволы и ветви. Помимо господствующего вида, здесь насчитывается всего шесть других двудольных деревьев; вся древесная флора (включая древовидные папоротники и пальмы) ограничивается 11 видами.
Три климаксные формации гор Тринидада, различаемые Бирдом, по-видимому, составляют весьма близкую аналогию с соответствующими поясами восточных тропиков. Нижнегорный дождевой лес — явный эквивалент несколько измененного варианта смешанного диптерокарпо-вого леса типа «диптерокарповых лесов низких и высоких предгории» Саймингтона, развитого на нижних склонах гор малайского архипелага. Формация Тринидада, которую Бирд называет «горным дождевым лесом», по структуре и флористическим связям аналогична среднегорному лесу, горному дубовому лесу и т. п. в пределах Малайского архипелага, для которых мы предлагаем термин «дождевой лес предгорий». Сходство в структуре и физиономии между карликовым редколесьем Тринидада и «моховым» типом горного дождевого леса Малайского архипелага очевидно, хотя, как и следовало ожидать, с флористической точки зрения между ними мало общего.
Поясность Тринидада и других островов Вест-Индии сходна в своих основных чертах. Между ними существуют лишь некоторые флористические различия. Так, на Малых Антильских островах ассоциация Вуг-sonima spicata—Licania ternatensis, относящаяся к поясу нижнегорного дождевого леса, замещается ассоциацией Dacryodes excelsa—Sloanea. но внешний облик и структура сообщества испытывают лишь незначительные изменения. Подобным же образом на Малых Антильских островах формация карликового редколесья часто представлена ассоциацией Didymopanax—Charianthus вместо ассоциации Clusietum, характерной для Тринидада (Beard, 1945 и др.). На более высоких островах благодаря «эффекту Мassenerhebung», пояса шире, и на больших высотах присутствуют безлесные формации. В Гваделупе (Stehle, 1935), например, «гигрофитный лес» простирается до высоты примерно 1000 .и. В своей нижней части это сообщество сходно с нижнегорным лесом Тринидада;
м фут
а
а _ тропический дождевой лес (нижнегорный дождевой лес Бирда);
лл фут
г™ 50 ~
25 --80
Р и с. 42(я, б). Профильные* диаграммы тропического дождевого и предгорного дождевого лесов, Северный хребет, Тринидад. Вест-Индия (Beam, 1946b).
б предгорный дождевой лес (горный дождевой лес Вирда)
390
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
выше оно, видимо, напоминает дождевой лес предгорий («горный дождевой лес» Бирда). Примерно от 1000 до 1120 м располагается карликовое редколесье с преобладанием видов Clusia (С. venosa), как и на Тринидаде. У своей верхней границы карликовое редколесье сменяется саванноподобным сообществом Lobelietum guadeloupense, которое физиономически, видимо, сходно с парамо Анд. Еще выше располагается Pitcairnietum, состоящий из Bromeliaceae, папоротников и пр. при наличии в напочвенном покрове Sphagnum, Breutelia и других мхов. Он уступает место растительности вулканических вершин и плато, где господствует Sphagnum и другие мхи и печеночники с очень незначительной примесью цветковых растений.
На некоторых островах сообщества с преобладанием пальм располагаются выше дождевого леса предгорий, например леса из Euterpe glo-hosa на Пуэрто-Рико, развитые на уровне 650—750 м (Gleason, Cook, 1926). Некоторые из этих пальмовых сообществ, вероятно, сериальные, но их происхождение и экология требует дальнейших исследований. Происхождение и экология лесов из Pinus caribaea на Больших Антильских островах также пока неясны. На некоторых островах Вест-Индии, например на Сент-Винсенте (Beard, 1945), ураганы и вулканические извержения истребили значительную часть климаксной растительности, сменившейся сериальными сообществами (см. стр. 305—306).
Шрив (Shreve, 1914а) детально изучил физиологическую экологию леса на высоте 4500—7420 футов (1373—2262 м) на Голубых горах Ямайки. Исследованное сообщество было названо им горным дождевым лесом; согласно употребляемой нами терминологии часть этого леса, вероятно, относится к предгорному дождевому лесу, а часть является истинным горным дождевым лесом.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ СВОИХ ШИРОТНЫХ ГРАНИЦ
В восточной Австралии, Юго-Восточной Азии (южный Китай, северная Бирма и т. п.) и на восточном побережье Южной Америки вечнозеленый дождевой лес достигает и пересекает географические тропики, глубоко вклиниваясь в северном и южном направлениях в субтропические районы. Эти внетропические участки тропического вечнозеленого леса в некоторых отношениях отличаются от истинного тропического дождевого леса и, вероятно, должны рассматриваться как самостоятельный тип формации — субтропический дождевой лес. Название «умеренный» следует сохранить только для таких формаций, как субантарктические леса юго-восточной Австралии и Новой Зеландии, южного Чили и т. д., в которых, хотя они и принадлежат к вечнозеленым, преобладают такие деревья умеренной зоны, как виды Nothofagus, и для которых характерна флора нетропическая (каково бы ни было ее первоначальное происхождение). Помимо этих обширных внетропических пространств вечнозеленого леса, которые более или менее непосредственно продолжают главный пояс тропического дождевого леса, тут и там в благоприятных местах внутри субтропической зоны встречаются небольшие островки широколиственного вечнозеленого леса, которые, по крайней мере флористически, напоминают тропический дождевой лес. Примерами этого являются «висячие леса», или леса хэммок, Флориды (Davis, 1943) и участки интересного реликтового галерейного леса на берегах Ла-Платы (34° 47' ю. ш.), описанные Кабрера и Даусоном (Cabrera, Dawson. 1944).
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 391
Сведения о субтропических дождевых лесах преимущественно касаются их флористического состава, и подробное обсуждение их здесь было бы неуместным. Однако субтропический дождевой лес Австралии, связанный прямым переходом с истинным тропическим дождевым лесом, расположенным севернее, представляет большой интерес.
Хотя значительная часть австралийского тропического дождевого леса уничтожена, его все еще можно проследить в виде узкой прерывистой полосы, идущей параллельно восточному побережью от Квинсленда до Нового Южного Уэльса и Виктории (подробнее см. у Фрэнсиса, Francis, 1929). В тропическом Квинсленде этот лес сильно напоминает индомалайский тропический дождевой лес не только общим обликом, но, если не считать полного отсутствия диптерокарповых, и флористическим составом. С приближением к югу растительность меняется: наряду с уменьшением количества видов (особенно деревьев) к индо-малайской флоре примешивается все больше субтропических и умеренных элементов, а такие характерные признаки тропического дождевого леса, как досковидные корни, каулифлория и пр., исчезают или становятся менее заметны. Согласно Фрэнсису (Francis, 1929, стр. 4), тенденция к монодоминантности сопровождается уменьшением числа видов; так, на нагорье Дорриго (30—31° ю. ш.) Ceratopelatum apetalum «иногда почти монополизирует древостой». Эти изменения в характере дождевого леса происходят постепенно и проявляются сначала примерно на широте реки Ричмонд, в северной части Нового Южного Уэльса (29° ю. ш.) (Francis, loc. cit.) Преобладающий тип листвы остается вечнозеленым и «мезофил льным» по всему поясу дождевого леса; лианы и эпифиты многочисленны. Таким образом, даже у своих южных границ субтропический дождевой лес еще в значительной степени сохраняет внешний облик тропического леса, хотя и несколько обедненного.
Флора и внешний облик субтропического дождевого леса резко контрастируют с примыкающими к нему формациями — склерофилль-ным лесом, где доминируют виды Eucalyptus, и субантарктическим влажным лесом с преобладанием Nothofagus. Даже в местах обоюдного внедрения этих формаций нетрудно проследить границу между ними. По направлению к своей южной границе субтропический дождевой лес оказывается все в большей и большей зависимости от благоприятных почвенных и климатических условий. Как уже говорилось в гл. 6, почвы на базальтах благоприятствуют развитию субтропического дождевого леса, а почвы с меньшей влагоемкостью, вроде почв, развивающихся на песчаниках, способствуют становлению склерофилльных сообществ (Fraser, Vickeri, 1938). В некоторых районах такую же или еще большую роль играет микроклимат (Osborn, Robertson, 1939). Субантарктический влажный лес приспособлен к более прохладным условиям, чем субтропический дождевой лес, и встречается на больших высотах и дальше к югу. Таким образом, в субтропической Австралии существует отчетливое экологическое отчленение индо-малайской флоры дождевого леса от преимущественно склерофилльной эндемической австралийской флоры; это совершенно аналогично тому, что наблюдается в пределах малайского архипелага, где, как уже говорилось, австралийская флора ограничена преимущественно горным лесом и некоторыми характерными местообитаниями на низменности.
Превосходный обзор лесов верхнего течения реки Вильямс и окрестностей горы Баррингтон в Новом Южном Уэльсе (32° ю. ш.), сделанный Фрезером и Викери (Fraser, Vickeri, 1937, 1938, 1939), дает возможность
392
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
провести детальное сравнение субтропического дождевого леса Австралии и тропических дождевых лесов. Изученная территория гориста и местами достигает примерно 5000 футов (1525 м). Средняя годовая сумма осадков в округе составляет около 150 см. На более низких высотах средняя минимальная температура самого холодного месяца (июль) немного ниже 40° F (4° С), а средний годовой максимум колеблется между 80 и 90° F (26—32° С). В наиболее возвышенных районах этой области наблюдаются гораздо более низкие температуры и зимой выпадает снег.
Здесь различаются три климаксные формации: смешанный субтропический дождевой лес, субантарктический дождевой лес и эвкалиптовый (склерофилльный) лес. Субтропический дождевой лес покрывает дно и склоны долин, выходящих на юго-восток, восток и северо-восток, начиная примерно с 300 и до 1000—3000 м над уровнем моря. Субантарктический лес, консоциация Nothojagus moorei, располагается выше субтропического; распространению его книзу, вероятно, препятствуют высокие летние температуры, а распространению субтропического леса кверху — низкие зимние температуры. Эвкалиптовый лес встречается в долинах ниже субтропического дождевого леса, а также на гребнях и в верхней части склонов горных кряжей и плато.
Эти формации повсюду четко разграничены, хотя многие виды являются общими для нескольких формаций. Большие различия в физиономии и составе формаций хорошо видны по биологическим спектрам (рис. 43). На всей этой территории субтропический дождевой лес наступает на эвкалиптовый лес, исключая лишь его верхние пределы, и в некоторых местах быстро вторгается в него. Фрезер и Викери полагают, что эта экспансия является следствием процессса восстановления субтропического леса после его отступления в конце третичного периода, вызванного орогеническими движениями. Некоторые склерофилльные элементы дождевого леса, как, например, два вида Eucalyptus, являются, вероятно, реликтами в этом поступательном движении.
Субтропический дождевой лес района верхнего течения реки Вильямс представляет собой высокий, густой вечнозеленый лес, состоящий из деревьев многих семейств, родов и видов. Основной древесный ярус образует сомкнутый полог и располагается на высоте 60—80 футов (18—24 м)\ поверхность полога очень неровная; отдельные деревья возвышаются над ним, достигая примерно 100 футов (30 м). Этот ярус состоит из смеси видов Dysoxylum, Ficus, Cedrela и других родов с типичной для дождевого леса лавроподобной листвой; за исключением трех, все эти виды вечнозеленые. Над основным (В) пологом намечается очень прерывистый ярус А, состоящий из двух несравненно более высоких видов Eucalyptus sa-ligna и Syncarpia laurifolia,— первый из которых достигает высоты 130— 170 футов (40—52 лс); оба эти дерева в изобилии встречаются в соседнем эвкалиптовом лесу и физиономически находятся ближе к склерофилльной флоре, чем к флоре дождевого леса. Благодаря этому ярусу, состоящему из деревьев другого физиономического типа, чем основной полог, лес напоминает «лес камерере» Новой Англии (стр. 53), у которого аналогичный ярус А из Eucalyptus deglupta перекрывает густой смешанный полог из типичных видов тропического дождевого леса. Ниже яруса В находятся ярус С с деревьями высотой 15—20 футов (5—6 м) (включает виды Diospyros, Drimys, Eupomatia и др., а также древовидный папоротник Alsophila leichhardtiana) и кустарниковый ярус. Напочвенный покров состоит из папоротников, многолетних травянистых растений и всходов деревьев.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 393
Доминирующие породы (ярус В) обладают многочисленными физиономическими признаками деревьев тропического дождевого леса. Они отличаются прямыми и тонкими стволами, наличием досковидных корней (правда, эти образования вполне развиты только у деревьев, произрастающих в лощинах с очень влажной почвой). Листья большинства
Рис. 43. Биологические спектры Раункиэра для субтропического, субантарктического и тропического дождевых лесов.
А —субтропический дождевой лес, район верхнего течения реки Вильямс, Новый Южный Уэльс; Б — тропический дождевой лес, Морабалли-Крик, Брит. Гвиана. В — субантарктический дождевой лес, район верхнего течения реки Вильямс, Новый Южный Уэльс; А и В — по данным Фрезера и Викери (Fraser, Vickery, 1938); Б — дано как в табл. 2 (гл. 1). Проценты исчислялись из всей флоры в целом; папоротники не включены.
видов принадлежат к «мезофилльной» группе, многие из них имеют капельные острия и листовые подушки (см. стр. 108). Однако ряд особенностей, например каулифлория, отсутствует. Лианы, порой очень крупные, представленные многими видами, редко достигают яруса А. Много эпифитных растений — орхидей, папоротников, мхов и др.
Субтропический дождевой лес является смешанным. На пробных площадках (общая площадь не указана) насчитывалось 181 дерево высо-
394
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
той более 30 футов (10 м), эти деревья принадлежали к 34 видам; отношение числа видов к числу индивидуумов составляло, таким образом, 5 : 3. Можно сопоставить это с показателями для опытного участка тропического дождевого леса на горе Дьюлит, Борнео (стр. 267); где 261 дерево диаметром 8 дюймов (20 см) относится к 98 видам (коэффициент — 2,7). Наиболее обильный особями вид Schizomeria ovata составляет около 10% древостоя; двум другим принадлежит гораздо меньшая доля. Отсутствие всякой тенденции к монодоминантности составляет резкий контраст по отношению к субантарктическому дождевому лесу той же области, в котором Nothofagus moorei составляет 76% деревьев, достигающих более 30 футов высоты. Хотя состав субтропического леса на реке Вильямс неодинаков в разных местах, его рассматривают как единую ассоциацию.
Биологический спектр (рис. 43) субтропического дождевого леса очень сходен со спектром дождевого тропического леса. В обеих формациях наблюдается ярко выраженное преобладание фанерофитов и эпифитов над всеми другими жизненными формами. Несмотря на наличие в субтропическом лесу значительного количества гемикриптофитов — жизненной формы, характерной скорее для умеренной, чем для тропической зоны,— черты, отличающие его от типичного экваториального тропического дождевого леса, проявляются намного слабее, чем черты сходства.
СРАВНИТЕЛЬНЫЙ ОБЗОР ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА ВБЛИЗИ ЕГО ВЫСОТНЫХ И ШИРОТНЫХ ГРАНИЦ
Гумбольдт (Humboldt, 1817) впервые высказал мысль о том, что вертикальные пояса растительности соответствуют широтным ее зонам, меняющимся от экватора к полюсам. Он полагал, что увеличение высоты над уровнем моря примерно на 1000 м в тропиках эквивалентно разнице примерно в 9° 30' по широте, или же 100 м подъема эквивалентны примерно 1°; таким образом, на экваторе высота около 8800 м должна фитогеографически соответствовать полюсам. Действительно, между внешним обликом горной растительности и растительности равнин в других широтах, но в условиях одинаковых средних температур существует заметное сходство, но это соответствие отнюдь не точно, и, как указал Лем (Lam, 1945, стр. 82), концепция Гумбольдта слишком упрощает дело. Климат эквивалентных вертикальных поясов и широтных зон никогда не бывает одинаков, поскольку в данном высотном поясе на экваторе долгота дня и сезонные колебания — если взять только эти два фактора — иные, чем в высоких широтах. К. Троль (С. Troll, 1948) также подчеркнул различие между климатом тропических гор и низменностей умеренной зоны; первый (71ageszeitenklima) характеризуется небольшими сезонными и относительно большими суточными колебаниями температуры, последний (Jahreszeitenklima) — большими сезонными и относительно малыми суточными колебаниями. Троль, однако, указывает, что в некоторых отношениях, а именно по повторяемости дней с морозом и незначительности колебаний средней месячной температуры, климаты тропических гор гораздо более похожи на климаты холодных субантарктических областей, чем на северный умеренный климат. Он предполагает, что этим можно объяснить тот факт, что южные умеренные (включая субантарктические) элементы гораздо шире представлены в тропической горной флоре, чем северные умеренные элементы. Даже растительные
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ 395
сообщества тропических гор напоминают сообщества южного полушария; например, горный дождевой лес тропиков проявляет много сходства с холодным умеренным дождевым лесом Новой Зеландии, южного Чили и т. д., но, как показывают факты, он очень далек от любого из северных умеренных лесных сообществ. Согласно Тролю, существует асимметрия между растительными зонами северного и южного полушарий.
Данные, использованные в этой главе, слишком неполны, чтобы дать соответствующую оценку теории Гумбольдта или ее видоизмененному варианту, предложенному Тролем, но беглое сравнение сообществ, примыкающих к дождевому тропическому лесу у его высотных и широтных границ, будет весьма интересным.
С подъемом, по мере перехода от низменности к горному дождевому лесу, структура и флористический состав климаксной растительности подвергаются изменению. Средняя высота деревьев уменьшается, причем степень уменьшения сильно зависит от экспозиции по отношению к преобладающим ветрам. Вместо трех ярусов низменного леса вначале сохраняется лишь два, так как ярус А постепенно редеет и наконец исчезает. Средний размер листьев становится все меньше, и в горном дождевом лесу обычны микрофиллы и даже еще более мелкие листья. Капельные острия, досковидные корни и каулифлория быстро исчезают. Крупные лианы встречаются все реже и, наконец, пропадают. Количество эпифитов возрастает (по крайней мере до подножия горного леса), но цветковые растения в их сообществах играют все меньшую и меньшую роль, а папоротники и бриофиты выступают на первое место. Основные флористические изменения заключаются в увеличении количества умеренных видов и уменьшении общего числа видов, особенно деревьев. Оскудению флоры обычно не сопутствует выраженная тенденция к монодоминантности, как это бывает в эдафически неблагоприятных местообитаниях на низменности.
Между австралийским субтропическим лесом и тропическим лесом существует много подобных различий. Высота субтропического и тропического лесов почти одинакова; этим подтверждается тот взгляд, что измельчание горного леса в первую очередь зависит от низкой интенсивности освещения, связанной с облачностью (как полагает Браун), и от воздействия ветров, а не от одного только понижения температуры. Субтропический лес на реке Вильямс, как и тропический лес, имеет три яруса деревьев, но верхний ярус очень разрежен; вероятно, большинство субтропических лесов двухъярусны. Досковидные корни и каулифлория уменьшаются с увеличением широты так же, как и с подъемом. Мы не располагаем точными сведениями о размерах листьев в субтропическом лесу, но, поскольку здесь преобладают мезофиллы, вероятно, в этом отношении не существует резких различий между субтропическим и тропическим лесом. Капельные острия у листьев не исчезают, как это бывает при увеличении высоты. Флористические различия между субтропическим лесом Австралии и тропическим лесом аналогичны различиям, связанным с высотой. В обоих случаях уменьшается количество видов и тропическая флора смешивается с умеренной. Однако в субтропическом лесу иногда наблюдается тенденция к монодоминантности вида.
Часть VI
ЧЕЛОВЕК
И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Глава 17
ВТОРИЧНЫЕ И ОТКЛОНИВШИЕСЯ СУКЦЕССИИ
Повсюду, где тропический дождевой лес является климатическим климаксом, и особенно в густонаселенных областях, обширные площади «девственного», или первичного, леса истреблены и замещены культурной растительностью или же вторичными сообществами, видоизменяющимися от зарослей злаков и других травянистых растений до сомкнутого леса, напоминающего первоначальную первобытную растительность. Эти сообщества, за исключением тех, где преобладают злаки (как правило, результат отклонившихся сукцессий), обычно называют «вторичной порослью», или «вторичным лесом». Высказывались соображения, что многие так называемые вторичные сообщества, возникающие в результате неоднократного повторного истребления первичной естественной растительности, фактически являются третичными, четвертичными и т. д. Но термин «вторичный лес» удобен и к тому же слишком давно введен, чтобы стоило заменять его каким-либо новым, например понятием «замещающий лес», предложенным Лебреном. Во многих странах для вторичных сообществ употребляют местные названия; так, в Брит. Гвиане вторичный лес называют «низким кустарником» или майнапом (mainap), в восточной Бразилии — капоэйрой (capoeira), на Филиппинах — парангом (parang), а в пределах Малайского архипелага — белукаром (belukar), или блука (Ыика) [иногда также утан-муда (utan muda)].
Вторичные сообщества, происшедшие от тропического дождевого леса — имеют ли они вид леса, кустарника, саванны или же (как это часто бывает) хаотической чащи деревьев, трав и лиан,— всегда более или менее неустойчивы и, таким образом, являются стадиями вторичных сукцессий. Предоставленные самим себе и защищенные от вырубания, выжигания и поедания скотом, они постепенно заселяются видами первичного леса, и нет сомнения, что в конце концов климатический климакс вновь утвердился бы, но, вероятно, лишь спустя очень большой промежуток времени. Там, где растительность подвергается воздействию скота или периодических пожаров, появляются отклонившиеся сукцессии, ведущие, очевидно, к устойчивым биотическим климаксам.
Хотя многие типы вторичной растительности, происшедшей от тропического дождевого леса, описаны, они редко подвергались тщательному изучению; очень мало систематических наблюдений проводилось и над теми сукцессиями, к которым они принадлежат. Это достойно сожаления, так как ни один другой аспект экологии тропического дождевого леса не имеет большей практической ценности и не сулит более важных теоретических результатов. Из существующих отрывочных наблюдений очевидно, что эти вторичные и отклонившиеся сукцессии сложны п очень изменяются от места к месту в зависимости от условий среды и истории е развития. Только тщательные количественные наблюдения в течение
400
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
длительного периода могут полностью разрешить эти вопросы. Такие наблюдения помогли бы заполнить наиболее серьезный пробел в современном познании тропического леса.
Истребление первичного леса, порождающее вторичные сукцессии, может происходить различными путями п по разным причинам. Главнейшей причиной уничтожения леса вплоть до настоящего времени была система подсечно-огневого земледелия, которая практикуется почти во всех тропических странах, за исключением нескольких районов, в частности, тропической Индии и низменностей Явы, где развилась интенсивная система оседлого земледелия. Отдельные приемы подсечного земледелия имеют местные особенности, но общий принцип повсюду одинаков. Определенный участок леса вырубается, за исключением наиболее крупных деревьев, которые обычно щадят, то ли оттого, что валка их берет много труда, то ли из-за суеверия. Срубленные деревья сжигают и затем сажают такие культуры, как маниок, маис, ямс, бананы или суходольный рис. Поскольку перегной в таких случаях быстро разрушается под действием солнца, а земля обычно возделывается примитивно и совсем не удобряется, почва вскоре истощается и становится бесплодной. После снятия одного или нескольких урожаев (их число зависит от природного плодородия почвы и интенсивности процессов выщелачивания и эрозии) участок забрасывают, и начинается развитие вторичной поросли. В процессе роста сукцессии плодородие почвы частично восстанавливается, и через несколько лет вторичный лес снова может быть вырублен.
Выбор нового участка в местах прежнего посева или же непосредственно в девственном лесу диктуется, конечно, плотностью населения и количеством пригодных для посевов земель. Лейн-Пул (Lane-Poole, 1925а) говорит, что в Новой Гвинее наблюдается сокращение земледельческого цикла, что приводит к преждевременному вырубанию вторичного леса, когда плодородие почвы еще недостаточно восстановилось. Где бы ни распахивался новый участок, площадь залежей (или вторичной растительности) оказывается непропорционально велика в сравнении с фактически обрабатываемой землей, поэтому с точки зрения охраны лесов и землепользования эта система чрезвычайно опустошительна.
Сельскохозяйственные плантации в тропиках устраивают или стремятся устраивать на постоянных местах, но в действительности они зачастую оказываются такими же временными, как и участки при подсечно-огневом земледелии, что ведет к безрассудному разрушению плодородия почвы. Когда плантация перестает приносить доход ввиду истощения почвы, эрозии, болезней растений или по экономическим причинам, ее забрасывают и на ее месте возникает вторичная сукцессия. Хотя такие многолетние культуры, как кофе или каучук, в течение некоторого времени конкурируют с растительностью вторичной заросли, развитие сукцессии идет по существу так же, как при подсечно-огневом земледелии. В большинстве тропических стран существуют обширные площади заброшенных плантаций, наглядно показывающие различные стадии вторичных сукцессий.
Гибель леса может вызываться не только освоением земель под культурные растения. Вопрос о том, может ли гореть нетронутый дождевой лес в отличие от сухих тропических лесов, был предметом горячих споров. Возможно, что пожары в его пределах всегда имеют лишь местное значение и могут случаться только при исключительных условиях погоды; пожары не как постоянное явление, ведущее к полному уничтожению
Ф ото 24. Кукурузное поле на расчищенном участке дождевого леса (подсечно-огневое земледелие) близ Акиллы, Нигерия.
Фото 25. Вторичная сукцессия, развившаяся на залежи пятилетней давности близ Акиллы, Нигерия.
На переднем плане справа видны Vernonia conferta (крупные листья) и молодой экземпляр Fagara sp. (перистые листья).
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС 401
деревьев, безусловно, не так редки в тропическом дождевом лесу Брит. Гвианы и, несомненно, бывают также и в других частях тропиков. Иногда участки леса валятся сильными ветрами — торнадо и ураганами, что расчищает путь для возникновения вторичных сукцессий, это, в частности, характерно для западной Индии.
Таким образом, площадь, где появилась вторичная сукцессия, могл? до этого испытать воздействие самых различных факторов. Последующий ход событий зависит от размеров расчищенной площади, степени ее удаленности от нерасчищенных площадей, местных климатических условий, рельефа и типа почвы, а также от степени разложения перегноя, интенсивности эрозии и истощенности почвы. В свою очередь истощение почвы зависит от нескольких факторов, особенно от того, сколько времени и каким способом возделывалась площадь (если это имело место), была ли она выжжена и какое количество щепы и «обрубков» остается на земле после расчистки. На эрозию почвы влияют уклон поверхности, количество и интенсивность осадков и многие другие факторы. Не удивительно, что развитие вторичной сукцессии, состав и физиономия сообществ, которые могут возникнуть, также чрезвычайно различны.
Площади первичного дождевого леса, используемые для выборочной рубки, можно назвать обедненным лесом. Прогалины, оставшиеся после вывоза древесины, заселяются видами вторичного леса и таким образом образуется сообщество весьма неправильной структуры, состоящее из перемежающихся участков первичного и вторичного леса. Обедненный лес подобного рода занимает обширные площади, например в округах Нигерии, производящих красное дерево, махагони (Richards, 1939, стр. 12), и в районах развития леса Гринхарт Брит. Гвианы; подобное же сообщество (gelichteter Urwald) для Борнео описано Винклером (Winkler, 1914, стр. 205). Иногда в результате обеднения наступает бурное разрастание уже существующего подлеска без внедрения посторонних видов.
ОБЩИЕ ПРИЗНАКИ ВТОРИЧНОГО ДОЖДЕВОГО ЛЕСА
Прежде чем перейти к характеристике самих сукцессий, необходимо кратко перечислить некоторые общие признаки «типичного» вторичного леса, то есть ранних сериальных стадий, встречающихся на площадях, которые обрабатывались или использовались для получения древесины, но после этого не выжигались и не служили пастбищами. Хотя, как уже подчеркивалось, вторичные сообщества на участках тропического дождевого леса чрезвычайно разнообразны, вторичный лес имеет свои особые, резко выраженные черты и, как правило, его легко распознать.
Прежде всего вторичный лес ниже и в среднем состоит из деревьев меньших размеров, чем деревья первичного леса (ср. термин «низкая поросль»). Было бы бесполезно приводить точные цифры, но всегда можно подозревать, что лес, состоящий преимущественно из более мелких деревьев, чем соседние леса, расположенные в аналогичных местообитаниях того же района, является вторичным. Поскольку участки начисто вырубленного или полностью уничтоженного огнем леса относительно редки, в пределах вторичного леса обычно встречаются отдельные высокие деревья, намного превосходящие средние размеры. При взгляде на вторичный лес сверху бросается в глаза более ровная поверхность его полога. Например, молодой вторичный лес на окультуренном участке долины
402
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Тинджар острова Борнео — если смотреть на него с горы Дьюлит (1200 .w)— выглядит гладкой лужайкой; в более темноокрашенном первичном лесу кроны отдельных деревьев можно различить на гораздо большем расстоянии.
Очень молодой вторичный лес часто поразительно правилен и однороден по структуре, хотя обилие мелких лиан и молодого подроста делает его густым и непролазным в отличие от нетронутого первичного леса. Для более поздних стадий сукцессии типична чрезвычайно беспорядочная структура; сплетение лиан, заросли осок с острыми, как*ножи, листьями и стеблями (виды Scleria) и другие подобные растения разбросаны среди высоких куп деревьев, отбрасывающих густую тень и характеризующихся скудным подлеском. Неправильная структура, как уже говорилось, является одним из ведущих признаков обедненного леса; можно было бы ожидать, что там, где вторичная сукцессия появляется на участке, более пли менее основательно очищенном от первоначальной растительности, она должна быть одновозрастной и обладать правильной структурой. И действительно, на самых ранних стадиях сукцессии очень обычны одновозрастные древостои, состоящие из одной или нескольких древесных пород. Так, на Южной Суматре Ван-Стеенис (van Steenis, 1933) встретил вторичный лес с правильной двухъярусной структурой; верхний ярус составляли деревья вида М acaranga (bancana?) высотой около 10 м, а нижний — густые заросли Lantana сатага 2—3 м высотой. Это сообщество, по-видимому, представляло собой одновозрастное лесонасаждение, выросшее на площади, полностью очищенной от первоначальной ра тительности.
С течением времени эти одновозрастные древостои пионерных видов стареют и часто оказываются неспособными восстанавливаться в новых экологических условиях, которые они же создали. В результате большинство деревьев на значительной площади может одновременно быть повалено ветром или погибнуть по какой-либо другой причине, и тогда обширные прогалины временно заселяются лианами, впоследствии постепенно заполняясь медленно растущими деревьями, преобладающими в следующей стадии сукцессии. Это — одна из причин обилия лиан, характерного для вторичного леса. Там, где растения-пионеры принадлежат к видам с различной продолжительностью жизни, прогалины появляются неодновременно, что и приводит к возникновению лоскутной, неправильной структуры, характерной для более поздних стадий сукцессии. Равномерная смесь возрастных групп, встречаемая в спелом лесу, достигается только после длительного периода развития.
Еще легче распознать вторичный лес по флористическому составу. Иногда указывают на то, что господствующие виды, типичные для вторичного леса, отсутствуют в первичном лесу. Это неверно; если такие виды и существуют, их очень мало, зато многие виды первичного леса отсутствуют в молодом вторичном лесу. Небольшое число вторичных видов встречается в естественных окнах в первичном лесу, например вследствие гибели больших деревьев; однако во вторичном лесу эти виды гораздо многочисленнее, чем в первичном, так что общая «флористическая картина» получается другая. Состав вторичного леса будет подробно рассмотрен при обсуждении самой сукцессии, пока же можно отметить, что в Америке указанием на вторичный лес служит обилие таких деревьев, как Cecropia, Vismia guianensis, Miconia spp., некоторые виды Inga и Byrsonima и многие другие; в Западной Африке — Musanga cecropioi-des, Trema guineense, Fagara macrophylla, Harungana madagascariensis.
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
403
Macaranga barteri, Pycnanthus angolensis, Stere ulia, tragacantha и пр., а в пределах Малайского архипелага — Мacaranga, Mallotus, Glochidion, Trema, Rhodamnia, Elaeocarpus и др. Анализ состава вторичного леса примерно тридцатилетнего возраста в Сингапуре, данный Баркиллом (Burkill, 1919), показывает, что в сравнении с первичным лесом там представлены совсем другие семейства. Во вторичном лесу наибольшим количеством видов были представлены семейства Euphorbiaceae и Lrticaceae, наибольшим количеством особей — Myrtaceae, Rhizophoraceae, Laura-сеае и Urticaceae (в указанной последовательности). Ни одно из этих семейств, кроме Myrtaceae и Lauraceae, не числится среди наиболее часто встречающихся семейств в первичном лесу Малайи. Dipterocarpaceae, столь типичные для малайского климаксного леса, абсолютно отсутствовали в этом вторичном лесу, a Rubiaceae, богато представленные в первичном лесу, занимали только десятое место по числу индивидуумов.
Несмотря на отсутствие каких-либо цифровых материалов, пригодных для сравнения количества видов, приходящихся на единицу площади во вторичном и первичном лесах, несомненно, что вторичный лес в среднем гораздо беднее видами, чем первичный.
По приблизительному подсчету, сделанному Шевалье (Chevalier, 1909), вторичный лес Берега Слоновой Кости состоит примерно из 30 видов деревьев, а «девственный» лес — из 250—300 (последняя цифра явно преуменьшена, а первая, вероятно, недалека от истины). Иногда, хотя далеко не всегда, вторичный лес бывает монодоминантным или состоит лишь из нескольких доминирующих пород. Как указывалось в гл. 11, многие из описанных монодоминантных типов тропического дождевого леса могут быть стадиями во вторичных или иных сукцессиях или же климаксами пожарищ.
Таким образом, вторичный лес (в том ограниченном понимании, в каком мы его сейчас рассматриваем), обладает резко выраженными признаками, по которым он может быть легко распознан. Старый вторичный лес трудно или невозможно отличить от ненарушенного девственного леса, но, поскольку вторичный лес представляет собой стадии сукцессии, ведущей в конечном счете к восстановлению климатического климакса, именно этого и следует ожидать. Биотические климаксы, являющиеся результатом отклонившихся сукцессий, могут близко походить на другие климатические климаксы, но только не на тот тропический дождевой лес, от которого они произошли. Иногда принадлежность определенных растительных группировок к вторичным сообществам наряду с указанными признаками может быть подтверждена наличием остатков строений и укреплений, некогда построенных местными племенами, осколков глиняной посуды и другими свидетельствами человеческой деятельности. В районе Мазаруни, Брит. Гвиана, Вуд (Wood, 1926) обнаружил, что «низкая поросль» иногда встречается в тех местах, которые, по словам индейцев, никогда не возделывались. Установлено, что речки, направляющиеся к этим участкам, были в свое время расчищены для судоходства; можно предположить, что «низкая поросль» занимала места, где первичный лес был вырублен или испытал вмешательство человека. Обширные пространства, покрытые пустошной растительностью (см. ниже, стр. 418.) близ озера Тоба на Суматре и на плато Дижёнг на Яве на первый взгляд казались устойчивым климатическим климаксом, но присутствие пней выдавало их вторичное происхождение на месте тропического дождевого леса, уничтоженного в результате возделывания и выжигания (van Steenis, Ruttner, 1932, стр. 245).
4 04
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ОСОБЕННОСТИ ДЕРЕВЬЕВ ВТОРИЧНОГО ЛЕСА
Доминирующие деревья ранних стадий вторичной сукцессии во многих отношениях резко отличаются от доминант первичного леса. Деревья вторичного леса в сущности обладают рядом общих признаков, которые не связаны с их таксономической принадлежностью. Таким образом, подобно деревьям первичного леса (см. гл. 4), они образуют естественную биологическую группу. Некоторые из этих особенностей явно адаптивные в том смысле, что обладающие ими виды не могли бы без них играть соответствующую роль в растительном сообществе. Существуют, вероятно, и другие общие особенности, значение которых не вполне ясно.
Прежде всего почти все деревья вторичного леса являются светолюбивыми и не переносят затенения. Они хорошо, растут на просеках пли вырубках достаточного размера, но в конце концов их вытесняют теневыносливые виды первичного леса или поздних стадий вторичной сукцессии. Многие вторичные виды, как будет видно из дальнейшего, не могут возобновляться в тени материнских растений, и жизнь сообщества в этом случае ограничивается лишь одним поколением.
Еще две особенности деревьев вторичного леса составляют существенную часть их биологического оснащения, а именно эффективное рассеивание семян и быстрый рост. Эффективное рассеивание позволяет им немедленно заселять все вновь появившиеся вырубки, прогалины и заброшенные пашни. До того как большие площади первичного леса были уничтожены человеком (что произошло совсем недавно), единственными, в основном небольшими и временными местообитаниями, доступными для вторичных видов, были естественные прогалины, обнаженные участки на берегах рек и т. п., приживаемость на которых обеспечивалась лишь быстрой колонизацией. Быстрый рост позволяет видам ранних стадий сукцессии утвердиться прежде, чем их покроют своей тенью медленно растущие виды вторичного леса.
К сожалению, мы не располагаем достаточными данными, чтобы сравнить распределение различных способов рассеивания семян среди видов первичного и вторичного леса. Тщательное статистическое изучение этого вопроса было бы чрезвычайно интересно и могло бы значительно помочь в определении возраста поздних стадий во вторичных сукцессиях. Однако каждый может наблюдать, что у большинства деревьев, характерных для молодого вторичного леса, семена или плоды хорошо приспособлены для переноса ветром или животными. Так, среди обычных (тропических) американских вторичных видов семена Vismia guianensis разносятся птицами и плодоядными летучими мышами, Didymopanax morototoni, Gua-zuma ulmifolia, Miconia spp., Byrsonima spp. и др.— птицами, Ochroma spp.— ветром. Из распространенных в Западной Африке деревьев вторичных сообществ семена Musanga cecropioides (подобно своему американскому двойнику — Cecropia) очень эффективно разносится птицами и плодоядными летучими мышами, Trema обладает очень мелкими легкими семенами, Pycnanthus angolensis и, вероятно, Мacaranga barteri разносятся птицами. Из обычных азиатских видов Alstonia и Anthocephalus рассеиваются ветром; Melastoma malabathricum и большинство видов Мacaranga. Mallotus. Trema. Rhodamnia и Rhodomyrtus разносятся птицами. В то же время, хотя господствующие виды первичного леса часто обладают крылатыми или легкими семенами (см. гл. 4), они редко разносятся животными (кроме, разве, обезьян) и часто имеют тяжелые плоды или семена без всякого специального приспособления для рассеивания; как уже
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
405
было отмечено, их средства рассеивания так несовершенны, что фактически препятствуют распространению семян в подходящие местообитания.
Хотя эффективность рассеивания плодов и семян является основной причиной быстрого расселения видов вторичного леса на любой прогалине, несмотря на расстояние от материнских деревьев, это не уменьшает интереса к сообщению Саймингтона (Symington, 1933) о том, что в верхних слоях почвы первичного леса, который, насколько известно, никогда прежде не расчищали и не повреждали, им были обнаружены покоящиеся семена видов белукара (вторичный лес). В этой связи может иметь значение большой выход семян у видов вторичного леса и то, что они, видимо, плодоносят круглый год. Солсбери (Salisbury, 1942, стр. 209) показал, что у видов британской флоры «периодически доступных местообитаний» продукция семян значительно выше, чем у видов постоянных местообитаний, особенно теневыносливых. Аналогичное соотношение почти наверняка можно было бы обнаружить между видами первичных и вторичных тропических лесов. Вторичные виды всегда размножаются посредством семян и плодов, но иногда их быстрое распространение после первого появления в лесу вызывается вегетативным воспроизводством; например, Musanga размножается отводками (Ghipp, 1913).
Быстрый рост вторичного леса неоднократно подчеркивался. Существует много отрывочных наблюдений над исключительно быстрым ростом некоторых видов. Так, Доктерс-ван-Лееувен (Docters van Leeuwen, 1936, стр. 72) отметил, что М acaranga tanarius, одно из распространенней-ших деревьев вторичных зарослей Малайского архипелага, в Богоре достигло высоты 8 м за 2 года. Саймингтон (Symington, 1933) нашел, что восемь особей того же вида на полуострове Малакка за 3 года выросли в среднем до 11 м\ Trema «amboinense» за тот же срок достигла высоты 7 м. Чип (Ghipp, 1927, стр. 58) видел в Гане деревья Ceiba pentandra высотой 12 м на участке, заброшенном только за 3 года до этого. В южной Нигерии Росс (неопубликованные данные) обнаружил, что Musanga cecropioides за 15—20 лет достигает 24 м высоты и 2—3 м в обхвате. Американская бальза (виды Ochromd) растет с такой же быстротой (Сох, 1939). Средняя высота вторичного леса 30-летнего возраста в Сингапуре, изученного Баркиллом (Burkill, 1919), составляла 14—15 м. Точное сравнение коэффициентов роста деревьев вторичного и первичного леса возможно благодаря исключительно тщательным измерениям, проведенным Брауном и Мэтьюзом (Brown and Matthews, 1914) и Брауном (Brown, 1919) в климаксном диптерокарповом лесу и паранге (вторичный лес) на Филиппинах. Результаты подытожены в табл. 38. Эти данные показывают, что коэффициент роста у видов вторичного леса, особенно у тех, диаметр которых в зрелом возрасте превышает 20 см, оказывается неизменно выше (в 2—3 раза), чем у видов диптерокарпового леса. Согласно Брауну, среднее дерево вторичного леса на исследованной площади достигает 30 см в диаметре за 14,5 лет; это примерно в четыре раза больше средней нормы для обычных североамериканских деревьев строевого леса. Быстрый рост деревьев вторичного леса, вероятно, возможен потому, что на ранних стадиях своего развития они не затенены, хотя деревья типичного первичного леса не растут так быстро даже в благоприятных условиях.
Небольшая максимальная величина и диаметр, достигаемые деревьями вторичного леса, никоим образом не связаны с коэффициентом их роста, а являются следствием непродолжительности их жизни. Средние деревья
406
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Таблица 38
Средний ежегодный прирост диаметра у деревьев вторичного и первичного тропического дождевого леса на горе Маквилинг, Филиппинские острова
(Brown, 1919)
Диаметр, с.н
0-5 5-10 10—20 20-30 30-40 40-50 50-60 60—70 70-80
Виды паранга (вторичный лес): а) диаметром меньше 20 см
Dysoxylum decandrum — — — — — — — — —
Ficus hauili 1,76 2,01 2,28 — — — — — —
Ficus ulmifolia 0,36 1,89 — — — — — — —
Ficus sp. 0,28 — — — — — — — —
Leea manillensis 0,58 0,44 — — — — — — —
Leucaena glauca — 1,29 — — — — — — —
Melanolepis multiglan- — 0,82 — — — — — — —
dulosa
Melochia umbellata — 3,27 — — — — — — —
Mussaenda philippica 0,77 — — — — — — —
Pipturus arborescens — 1,81 — — — — — — —
Premna cumingiana 0,50 0,92 — — — — — — —
Psidium guajava 0,22 0,77 0,92 — — — — — .—
Voacanga globosa 0,66 1,60 — — — — — — —
В среднем 0,64 1,40 1,60 — — — — — —
б) диаметром более 20 см
Alstonia sholaris — 3,38 — — — — — —
Antidesma ghaesembilla — — 1,97 — — — — —
Artocarpus cumingiana 1,00 1,55 1,77 — — — — — —
Bauhinia malabarica — 2,25 2,40 2,81i — — — — —
Canarium villosum — 1,93 — — — — — — —
Columbia serratifolia 2,09 1,91 2,05 2,80i — — — — —
Cordia myxa — — 1,76 — — — — — —
Ficus nota — — 2,37 — — — — — —
Glochidion philippicum — 1,96 — — — — — — —
Lit sea gluminosa 0,76 1,86 — — — — — — —
Litsea perrottetii — — 2,15 — — — — — —
Mai lotus philippensis — — 1,43 — — — — — —
Mallot us ricinoides 1,70 1,89 1,79 3,59i — — — — —
Polyscias nodosa — 1,70 — — — — — — —
Premna odorata — — 2,12 — — — — — —
Sapindus saponaria — 1,68 — — — — — — —
Trema orient alls — — 4,03 1,7P — — — — —
В среднем 1,39 2,08 2,19 2,59 — — — — —
Виды смешанного диптерокарпового (первичного) леса
Dillenia philippinensis 0,46 0,55 0,39 0,24 — — — —
Diplodiscus paniculatys 0Д5 0,31 0,44 — — — — —
Parashorea malaanonan 0,07 0,27 0,38 0,49 0,74 0,82 0,94 0,75 0,84
Parashorea malaanonan 0,42 0,55 0,73 — — — — — —
1 Эти намерения относятся к деревьям диаметром 20—30 или 35 см.
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
407
вторичного леса на Филиппинах, согласно Брауну (Brown, 1919, стр. 138), достигают своего полного развития при диаметре ствола 30—35 см и в дальнейшем начинают терять жизнеспособность. Исключительно быстрорастущая Melochia umbellata — одно из более мелких деревьев вторичного леса, достигает наибольших размеров, вероятно, в течение 3—4 лет, а потом отмирает (там же, стр. 148). Желательно было бы иметь больше сведений по этому вопросу, но и сейчас очевидно, что в общем деревья вторичного леса недолговечны, рано созревая и рано размножаясь. Во вторичном тропическом дождевом лесу Нигерии Росс (неопубликованные данные, см. стр. 409) обнаружил, что на опытных участках экземпляров Musanga cecropioides в возрасте 17,5 лет было гораздо меньше, чем в возрасте 14,5 лет; что связано с массовым отмиранием этого дерева без какой-либо видимой причины в возрасте 15—20 лет и неспособностью его возобновляться в тени материнского полога.
Мягкость и низкая плотность древесины, составляющие общеизвестные особенности большинства деревьев вторичного леса, несомненно, являются следствием их быстрого роста. У многих деревьев вторичного леса древесина чрезвычайно мягкая и легкая; особенно легка древесина бальзы (виды Ochroma), распространенной во вторичном лесу тропической Америки. Часто думают, что лесоматериалы из вторичного леса благодаря своей мягкости не имеют большой экономической ценности; однако существует много исключений, среди которых, помимо бальзы, можно назвать окоуме (Аисоитеа klaineana), дерево, образующее обширные чистые древостои на участках заброшенных плантаций в Габоне и обладающее ценной древесиной, которая широко используется в фанерном производстве. В области сухого вечнозеленого леса юго-восточной Нигерии деловую древесину дают три самых распространенных вида, заселяющие заброшенные плантации,— Chlorophora excelsa, Treminalia superba и Triploch iton scleroxylon.
Помимо вышеупомянутых особенностей — светолюбивости, эффективного рассеивания семян и быстрого роста, — деревья вторичного леса имеют и другие общие свойства, которые не носят, однако, столь явных адаптивных черт. Им уделяли мало внимания, но общие наблюдения заставляют думать, например, что существуют различия в характере листвы в первичном и вторичном лесу. Как мы видели (см. гл. 4), листья деревьев первичного леса отличаются удивительной однородностью размера, формы и строения. Листья вторичного леса более разнообразны; они бледнее по окраске — факт, давно отмеченный Спрусом (Spruce, 1908, I, стр. 355), а статистическое исследование, наверное, показало бы, что во вторичном лесу рассеченные и нецельнокрайние листья более обычны, чем в первичном лесу. Сравнение размеров листьев в первичном и вторичном лесу также могло бы дать интересные результаты. Исследования Брауна на Филиппинах (Brown, 1919, стр. 378—390) показали, что в паранге (вторичный лес) процент растений с крупными листьями больше, чем в девственном диптерокарповом лесу. С другой стороны, измерения листьев 20 древесных видов высотой 2 фута (61 см) и выше, произведенные Баркиллом (Burkill, 1919, стр. 152) во вторичном лесу 30-летнего возраста в Сингапуре, показали, что листва большинства видов принадлежит к «микрофилльному» классу Раункиэра, 2—4 вида имели «мезо-филльные», а один вид — «нанофилльные» листья. В первичном лесу подавляющая масса видов обладает «мезофилльными» листьями (стр. 99—100). Эти наблюдения не позволяют сделать каких-либо определенных выводов, хотя вполне возможно, что среди деревьев вторичного леса «макрофилль-
408
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ные» и «микрофилльные» листья встречаются чаще, чем в первичном лесу, и, следовательно, являются более разнообразными по размерам.
Интересно, что виды, типичные для «более сухих» (то есть более сезонных) типов леса иногда встречаются в сериальных стадиях сукцессии, развивающейся в районах дождевого леса. Так, в Западной Африке на вырубках и во вторичных сообществах в пределах массива дождевого леса (влажный вечнозеленый лес) часто можно видеть Chlorophora excelsa и некоторые другие обычные доминанты сухого вечнозеленого леса (см. стр. 362—363). В южной Индии на вырубках во влажном вечнозеленом лесу распространено Kydia calycina — дерево, обычно встречающееся в сухом листопадном лесу.
ПРОИСХОЖДЕНИЕ ВТОРИЧНОЙ ЛЕСНОЙ ФЛОРЫ
Весьма важны подробные исследования родственных связей и происхождения видов флоры дождевого леса в географическом отношении. В общем виды вторичного леса распространены шире, чем виды первичного леса. Некоторые из них являются пантропическими или распространены в пределах всей неотропической либо палеотропической области, и лишь очень немногие эндемичны или имеют узкоместное распространение. Напротив, виды первичного леса, как правило, оказываются приуроченными к четко ограниченным территориям; хотя в порядке исключения, ряд их встречается и в Старом и Новом свете, но практически среди них отсутствуют пантропические виды. Во многих тропических районах часть ведущих видов на ранних стадиях вторичных сукцессий представлена одичавшими культурными растениями или пришельцами, случайно занесенными из других областей. Так, в молодых вторичных сообществах Малайского архипелага три наиболее многочисленных растения — травянистые Eupatorium pallescens и Ageratum conyzoides и кустарник Lantana camara — лишь недавно интродуцированы из Америки. На многих тихоокеанских островах первоначальный низинный девственный тропический дождевой лес исчез или же сильно сократил свою площадь, невозделанные участки на его территории широко заселены ввезенными извне растениями, многие из которых имеют американское происхождение. На Фиджи, например, большие площади на месте былого леса покрыты кустарником Psidium guajava, Lantana, Metastoma malabathricum и другими чужеземными растениями.
Широкое распространение видов вторичного леса, безусловно, связано с хорошим рассеиванием их плодов и семян и успешным заселением земель с нарушенным покровом первоначальной растительности. Те же самые биологические особенности их, которые дают им возможность использовать любую прогалину или любое доступное временное местообитание еще до интенсивного вырубания дождевого леса, позволяют им и в новых условиях осваивать огромные площади и зачастую проникать во все уголки мира. Ван-Стеенис (van Steenis, 1937) считает флору всех «антропогенных» растительных сообществ Малезии (включая такие доминанты климакса пожарищ, как Melaleuca leucadendron, Pinus merkusii и Tectona grandis, а также флору белукара) результатом определяемого средой отбора из общей массы видов, населяющих смешанный лес. Согласно этому взгляду, все виды вторичного леса в небольших количествах изначально существовали в первичном тропическом дождевом лесу; вырубка и выжигание больших участков леса создают лишь возможность отбора тех видов,
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС 40$>
которые способны использовать создавшиеся условия. Подобная трактовка, вероятно, вполне правильна применительно к большинству видов вторичного леса, но некоторые из них, особенно чужеземные и пантропические виды, скорее всего являются выходцами из других растительных формаций и первоначально не встречались в тропическом дождевом лесу.
ЗАМЕЧАНИЯ О ВТОРИЧНЫХ СУКЦЕССИЯХ
Уже отмечалось, что было опубликовано несколько детальных исследований вторичных сукцессий, ведущих к становлению тропического дождевого леса. Кроме того, существуют некоторые разрозненные наблюдения, из которых faute de mieux следует воссоздать картину общего хода событий. По-видимому, наиболее интересными из них являются данные Р. Росса, полученные им в Нигерии в 1935 г.1
Вторичные сукцессии Нигерии
Росс производил наблюдения в резервированном лесу Шаша, расположенном на низменности и составляющем часть некогда сплошного пояса тропического дождевого леса (в этом месте около 60 км шириной)г тянущегося параллельно побережью океана непосредственно за окаймляющими его снаружи лагунами, мангровыми зарослями и пресноводно-болотными лесами. Рельеф, климат, почвы и первичная растительность этого пояса были достаточно полно описаны в другом месте (Richards, 1939)г а некоторые детали структуры спелого леса уже обсуждались в этой книге (см. стр. 46 и след.). Это район с относительно сильной сезонной засухой (годовая сумма осадков составляет 208 см при пяти месяцах кряду с осадками меньше 10 см). и первичный лес здесь развивается скорее по типу влажного вечнозеленого, чем «истинного дождевого леса» (см. стр. 361); однако нет никаких оснований думать, что вторичные сукцессии в его пределах сколько-нибудь серьезно отличаются от тех, которые имели бы место в менее резко сезонном климате, где климатическим климаксом является «истинный дождевой лес».
В лесу Шаша разбросанные села и деревушки существовали еще до того, как он был резервирован. Каждая деревня окружена кольцом обрабатываемых земель. За исключением находящихся в непосредственном соседстве с домами постоянных плантаций колы и фруктовых деревьев^ эти земли возделываются по типичной подсечно-огневой системе.
Поскольку земельные наделы, в пределах которых разрешена сельскохозяйственная деятельность, невелики в сравнении с количеством населения, повторной обработке подвергаются участки, оставленные еще совсем недавно; фактически все необрабатываемые наделы покрыты вторичным лесом различного возраста. Когда землю очищают для посева, то несколько крупных деревьев, особенно видов с твердой древесиной, как. например, Lophira procera. оставляют несрубленными; впоследствии они могут погибнуть от огня. Все обрубки, кроме стволов больших деревьев, сжигаются. Основными культурами являются: кукуруза, ямс, маниок, бананы, пизанги и колоказия. В небольших количествах выращиваются
1 Вскоре после первого издания этой книги в 1952 г. работа Росса была опубликована (Ross, 1954).
410
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
и другие растения. Спустя несколько сезонов, когда земля становится менее производительной, обрабатываемый участок оставляется и на нем начинает постепенно развиваться вторичная сукцессия.
Исследуя ряд опытных участков на сопоставимых площадях, для которых известна дата прекращения обработки, можно с некоторой степенью точности восстановить ранние стадии сукцессии. В своей работе Росс проводит сравнение исследованных им участков площадью 200X \ 100 футов (60x30 м), не возделывавшихся в течение 5,5, 14,5 и 17,5лет, и одного участка неизвестного, но, вероятно, еще большего возраста. Нижеследующее описание сукцессии основывается главным образом на этих результатах и на менее подробных наблюдениях, проводившихся в других районах этой же области.
Оставленная пашня вначале, если не считать нескольких выживших деревьев и многолетних культурных растений, подобных бананам, которые держатся еще некоторое время, оказывается обнаженной, но уже через несколько месяцев земля покрывается густой массой низкорослой растительности; через несколько лет она оказывается покрытой вторичным лесом высотой примерно 16 м. Пока земля обнажена, любой вид в пределах радиуса рассеивания его семян может занять эту площадь при условии, что его всходы смогут приняться. Следует помнить, что условия для прорастания семян здесь совсем иные, чем в первичном лесу. Конкуренция со стороны других растений минимальная, но земля не защищена от солнца, ветра и дождя; следовательно, освещение и суточные колебания температуры и влажности увеличиваются. Кроме того, в процессе обработки почва прямо и косвенно оказывается измененной в различных отношениях. Эти различия в среде будут далее рассмотрены более полно.
Первичные растения-поселенцы делятся на три группы: а) травянистые сорняки, характерные для обработанной земли, например Phyl-lanthus и виды Solanum; б) виды, преимущественно древесные, типичные для вторичных местообитаний, но встречающиеся также в высокоствольном лесу на небольших прогалинах, созданных падением и гибелью крупных деревьев, например Musanga cecropioides, Trema guineense, Vernonia conferta и V. frondosa, Fagara macrophylla (это — «типичные деревья вторичного леса», описанные на стр. 404—408); в) светолюбивые виды высокоствольного леса, семена которых могут приняться на открытых местах, например Erythrophleum ivorense, Khaya ivorensis, Lophira procera.
Группа травянистых сорняков может очень быстро образовать сомкнутое сообщество, но доминирование этих растений — явление преходящее, и в сукцессии они играют незначительную роль. Виды группы «б», если не становятся господствующими сразу, приобретают доминирование очень скоро и сохраняют его в течение 15—20 лет. Это происходит потому, что они достаточно высоки для того, чтобы подавить травянистые сорняки, и растут быстрее, чем виды высокоствольного леса (группа «в»), к этим последним относятся Vernonia conferta и V. frondosa — деревья из семейства сложноцветных, отличающиеся крупными листьями; обычно они первыми становятся доминантами, причем Vernonia conferta особенно часто образует беспримесные лесонасаждения. Иногда среди первых доминант встречается Trema guineense, а порой и другие виды. Один маленький участок, совершенно очищенный от растительности в марте 1935 г., к маю уже покрылся густой порослью всходов Trema высотой около 1 м с некоторой примесью более мелких всходов Albizzia gummifera, Carica papaya и Sola-num sp. Доминантность тех или других видов в фазе вторжения, вероятно,
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
411
частично зависит от случая, например от того, какие виды обильнее плодоносят в то время, когда участок окажется доступным для поселения на нем растений (правда, имея в виду вышеупомянутую работу Саймингтона, можно допустить, что немалое количество растений развивается из покоившихся в почве семян). С момента образования сомкнутого сообщества фактор случайности, по-видимому, уменьшается, и тогда флористический состав определяется главным образом конкуренцией.
Примерно через три года доминантой на каждом исследованном участке становилась Musanga cecropioides. Этот наиболее многочисленный и характерный вид вторичных сообществ тропической Африки, очевидно, неспособен поселяться на голой земле, но как только образуется растительный покров, его исключительно быстрый рост дает ему большое преимущество, несмотря на сравнительно открытый полог. При господстве Musanga большую роль начинает играть Macaranga barteri — другой вид группы «б», — который растет не так быстро и не бывает так высок, как Musanga. Интересно, что как Musanga. так и Macaranga обладают ходульными корнями. Под сенью древесного яруса располагается густой второй ярус из кустарников и молодых деревьев, включающий Conopharyngia penduliflora, Discoglypremna caloneura (на желтоземах) и Rinorea sp. и Rau-ivolfia womitoria. Через 5 лет второй ярус достигает уже 4—5 м высоты. В первые годы в фазе Musanga мелкие травы (виды Geophila, Leea guine-ensis, Gommelinaceae и др.) гораздо многочисленнее, чем в первичном лесу, и образуют сомкнутый покров. Позднее, с увеличением затененности (и может быть, благодаря усилению конкуренции между корнями) травянистый покров становится все более редким.
Через 15—20 лет деревья Musanga погибают; так, на участке 17,5-летней давности количество экземпляров этого вида было уже значительно меньше, чем на участке 14,5 лет.
Причиной смерти, видимо, является только старость; перед самой гибелью деревьев кора на ходульных корнях начинает шелушиться, но каких-либо других симптомов заболевания не наблюдается. Обыкновенно во время сезона торнадо (с марта по май) отмершие и отмирающие деревья валятся ветром, и во вторичном лесу примерно пятнадцатилетнего возраста повсюду можно видеть упавшие деревья. Погибшие деревья не оставляют потомства, поскольку Musanga. очевидно, неспособна возобновляться в тени собственного полога. Таким образом, консоция Musanga является сообществом, дающим только одно поколение.
К этому времени другие виды достигают высоты 20—25 м. среди них: Albizzia gummifera, Anthocleista vogelii, Diospyros «con ferti flora», Disco-glypremna caloneura. Funtumia elastica и Sarcocephalus diderrichii. Некоторые из них относятся к группе «б» и представляют собой обычные элементы высокоствольного леса. Появляются также и молодые особи Diospyros insculpta, Erythrophleum ivorense. Lophira procera и виды Scottellia. На 17,5-летнем участке было зарегистрировано 35 видов группы «в» и 32 вида группы «б», но среди последних наблюдалось несколько большее число экземпляров диаметром свыше 2 дюймов (5 см), чем среди первых.
Таким образом, вторичная сукцессия на вырубках лесного массива Шаша явственно стремится к восстановлению климатического климакса. Дальнейший ход развития после первых 20 лет не был прослежен. В последующей фазе, по-видимому, должно произойти уменьшение видов группы «б» и соответственный рост группы «в», ведущий к развитию высокоствольного леса. Молодой высокоствольный лес все еще будет неустойчив, поскольку он включает ненормально большее количество светолюбивых
412
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
видов, которые поселились еще в начале сукцессии; в сомкнутом сообществе у этих видов возобновление возможно только на прогалинах. Недавние наблюдения автора в другой части Нигерии заставляют думать, что прежде чем водворится устойчивое равновесие, должен пройти еще целый ряд изменений в течение долгих лет.
Вторичная сукцессия, таким образом, состоит в Нигерии из трех стадий:
I. Краткий период колонизации (invasion), во время которого устанавливается сомкнутый растительный покров.
II. Стадия господства Musanga, когда один вид становится, безусловно, преобладающим в течение одного поколения.
III. Стадия медленного замещения деревьев вторичного леса деревьями первичного. Третья стадия тянется, несомненно, гораздо дольше, чем две другие, но ее фактическая длительность неизвестна. Следует отметить, что в сериальных сообществах древесные растения начинают преобладать почти немедленно; если травянистая фаза и наблюдается, то она бывает очень короткой. Травы (например, такие теневыносливые виды, как Lep-taspis cochleata, которые встречаются и в первичном лесу) могут участвовать в сукцессии, но большого значения не имеют; никогда не бывает даже временного доминирования высоких дерновинных злаков вроде Impera-ta, присутствие которой составляет важную особенность вторичных и отклонившихся сукцессий в некоторых частях тропиков. Каждая стадия сукцессии прокладывает дорогу для следующей. Таким образом, сомкнутый растительный покров, развившийся в стадии I, создает необходимые условия среды для появления Musanga. Глубокая тень консоций Musanga создает подходящий микроклимат для видов, которые становятся преобладающими в стадии III. Стадии I и II характеризуются быстрорастущими светолюбивыми видами, типичными для вторичного леса. В стадии III происходит постепенное возвращение к господству медленнее растущих видов первичного леса.
Вторичные сукцессии в Конго
Отрывочные наблюдения во многих районах вечнозеленого леса тропической Африки показывают, что если не нарушать ход развития вторичной сукцессии, возникающей на месте заброшенной пашни, то в своих основных чертах она будет очень походить на вышеописанную сукцессию в Нигерии. Если же она подвергается периодическим выжиганиям, используется в качестве пастбища или же вновь вовлекается в сельскохозяйственное производство спустя всего несколько лет после первой культивации, то развивается отклонившаяся сукцессия, где господствует злаковое высокотравье, для которого особенно характерна Imperata cylindrica var. africana (разновидность травы лаланг Малайского архипелага). Отклонившиеся сукцессии могут привести к вытеснению леса открытым злаковником с разбросанными по нему деревьями и кустарниками или же без них. Подобный злаковник, заслуживающий название вторичной саванны1, может существовать неограниченно долго в качестве биотического климакса, если соответствующий участок используют под пастбище или периодически выжигают.
1 Эквивалент производной саванны А. П. Д. Джонса (см. стр. 359).
Фото 26. Прогалина во вторичном лесу, резервированный лес Окому, Нигерия.
Прогалина образовалась в результате вырубки или бурелома. Впоследствии пасущиеся слоны, повреждая деревья, способствовали вторжению трав
Фото 27. Старый вторичный лес, резервированный лес Окому, Нигерия.
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС 413
Отчетливая картина развития этих сукцессий в Конго дана Вермё-сеном (цит. Лебреном, Lebrun, 1936а), который различает среди сукцессий два расходящихся «ряда» — «прогрессивный» и «регрессивный». Флористический состав различных стадий этих «рядов» очень разнообразен, изменяясь под влиянием местных почвенных различий и других причин. «Прогрессивный» ряд встречается там, где между последовательными периодами окультуривания земли наблюдается длительный перерыв, а «регрессивный» — в местах с коротким перерывом. Первый соответствует вторичной сукцессии, ведущей к восстановлению климаксной лесной растительности; второй — не что иное, как отклонившаяся сукцессия, приводящая к более или менее полному господству злаков или же папоротника Pteridium.
При развитии «прогрессивной» сукцессии заброшенная пашня прежде всего заселяется быстрорастущими травянистыми растениями; они завершают свой жизненный цикл в несколько недель. За ними следует стадия многолетних и «полукустарничковых» трав, изобилующих травянистыми и древесными лазящими растениями. Появляются и молодые деревца, выросшие из семян или возникшие от пней, сохранившихся на участке; остаются и некоторые недолговечные травянистые растения первой стадии. Через несколько месяцев кусты и молодые деревья начинают явно преобладать и вскоре образуют выраженный полог; начинается третья, или древесная, стадия сукцессии. В древесной стадии Вермёсен выделяет три фазы: 1 — с преобладанием Musanga cecropioides, Trema guineensis, llarungana madagascariensis и Pycnanthus angolensis-, 2— с преобладанием видов Bosqueia, Conopharyngia, Alstonia, Funtumia, Albizzia, Pentacle-thra, Sterculia, Ricinodendron, Fagara, Ficus и др.; 3 — характеризующуюся постепенным возвратом к преобладанию типичных для первичного леса видов. Вермёсен устанавливает следующий оптимальный период и продолжительность этих фаз: фаза 1: оптимум 10—20 лет, общая продолжительность 20—30 лет; фаза 2: оптимум 20—30 лет, продолжительность 50 лет; деревья первичного леса становятся преобладающими (фаза 3) через 60—100 лет. Лебрен считает эти цифры завышенными. Шевалье (Chevalier, 1917) нарисовал аналогичную картину хода сукцессии в Габоне.
На реке Убанги «регрессивная» сукцессия развивается следующим образом: участок леса начисто вырубают и выжигают: собирают несколько урожаев кукурузы, но когда урожайность падает, землю забрасывают, и начинает утверждаться только что описанная «прогрессивная» сукцессия. Однако когда она доходит до первой фазы древесной стадии (что здесь обычно сопровождается преобладанием Trema), то землю опять расчищают и выращивают на ней другую культуру. В период травянистой стадии сукцессии после первой обработки земли злак Imperata встречается отдельными дернинками и не цветет; в последующих кустарниковой и древесной стадиях этот злак глохнет, но не погибает, сохраняясь вплоть до второго периода обработки почвы. Как только землю забрасывают вторично, Imperata становится доминантой и может образовать почти беспримесные консоции; это сообщество держится более или менее неограниченное время в качестве биотического климакса, оказываясь вполне устойчивым, хотя оно возникает и поддерживается искусственно. На сырых глинистых почвах Imperata замещается саванной из Pennisetum, а на песках — вторичным злаковником, в свою очередь уступающим место обширным зарослям Pteridium.
414
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Как увидим ниже, происхождение вторичных сообществ, образованных злаками или папоротником Pteridium в Конго, вряд ли отличается от пути происхождения некоторых почти идентичных сообществ Малай» ского архипелага, Новой Гвинеи и других тропических районов1.
Вторичные п отклонившиеся сукцессии Малайского архипелага
В пределах Малайского архипелага существуют огромные площади вторичного леса «белукара», а в ряде мест на территории бывшего дождевого леса развиваются кустарники и злаковники; но сукцессии, стадиями которых являются эти вторичные сообщества, мало исследованы. Однако почти не приходится сомневаться, что в этой части Азии, как и в тропической Африке, после истребления первичного леса сукцессия ведет к восстановлению климатического климакса, безразлично, следует за этим период культивации или нет. Когда вторичные сообщества используют под пастбища или повторно выжигают или когда периоды повторной культивации следуют друг за другом с очень короткими промежутками, то сукцессия отклоняется и, по-видимому, возникают устойчивые биотические климаксы, обычно с преобладанием трав. Часто местные жители выжигают вторичные злаковники через правильные промежутки времени, так как вырастающие после этого молодые побеги трав особенно хороши для откорма скота.
Йохеме (Jochems, 1928) в своей работе о залежной растительности на месте бывшей табачной плантации у Дели, Суматра, дал очень полное описание ранних стадий вторичной сукцессии. Район Дели славится отличным качеством и высокой урожайностью табака, и здесь практикуется тщательно разработанная система земледелия. Каждый участок земли дает сбор табака не чаще, чем раз в 8 лет. За сбором табака обычно следует единичный посев богарного (неполивного) риса или кукурузы, после чего земле дается полный отдых до следующей посадки табака; таким образом, вторичная сукцессия беспрепятственно развивается в течение 7 лет. На некоторых плантациях делались попытки заменить бесполезный белу кар (вторичный лес) самопроизвольно развивающимся в период отдыха плантации «искусственным белукаром» из древонасаждений, ценных своей древесиной или способностью улучшать почву (Legumi-noseae). Сезон посадки табака начинается приблизительно в январе, и за несколько месяцев до посадки участок очищают от белукара, а деревья складывают штабелями и сжигают. Затем почву обрабатывают до глубины примерно 50 см, что провоцирует густую поросль сорняков, или же производят посев покровной культуры Mimosa invisa. Непосредственно перед посадкой табака сорняки или защитный посев удаляют и почву тщательно обрабатывают еще раз. Табак остается на корню примерно до июля; в начале его развития землю тщательно выпалывают, но поближе ко времени сбора дают взойти сорнякам. Если после табака собираются сеять рис или маис, то землю расчищают, оставляя только отдельные разбросанные деревья вторичного леса. Урожай собирается в декабре. В течение последующего продолжительного периода отдыха земли возникает
1 Можно добавить, что на окраинах дождевого леса Будонго в Уганде (Eggeling, 1947), там, где лес расширяется за счет окружающего злаковника, сукцессия, развивающаяся от злаковника к лесу, резко отличается от вышеописанных африканских сукцессий. Правда, нет уверенности, что эта сукцессия действительно является вторичной, поскольку неизвестно, возвращается ли тропический дождевой лес па площади, где он недавно был вырублен, или же климатические изменения или какая-либо другая причина вызвали его вторжение на неосвоенную им ранее территорию.
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
415
сукцессия, в которой можно различить три стадии: I — травянистая стадия, флористически богатое сообщество, состоящее преимущественно из недолговечных травянистых растений и молодых деревьев; II — стадия Trema — Blumea, где главными растениями являются мелкие деревья Trema sp., Blamea balsamijera и Abroma angustum*, III — древесная стадия, являющаяся типичным белукаром. Когда продовольственные культуры не выращивают, то стадия не получает такого полного развития и, минуя II стадию, непосредственно переходит в III. В III стадии господствует один или несколько видов; по флористическому составу различают следующие пять типов белукара: Trema {Trema sp.), Мacaranga {М. tana-rius или M. denticulata), Callicarpa (C. tomentosa), Melochia {M. umbel-lata) и смешанный (без доминанты). Наиболее характерными сопутствующими видами деревьев во всех пяти типах являются: Commersonia bertra-mia, Ficus jistulosa, F. toxicaria, Millettia atropurpurea, Pithecellobium lobatum. Из кустарников преобладают Desmodium polycarpum и Lantana camara*, среди травянистых видов господствует папоротник Dryopteris arida и лаланг Imperata cylindrica var. major. Травянистые лианы Argy-reia capitata, Lygodium scandens, Merremia vitifolia, M. umbellata и Peri-campylus glaucus играют большую роль в этих сообществах. Йохеме дает детальную характеристику состава ряда опытных площадок.
На отдельных участках табачных плантаций встречается множество сеянцев крупного дерева Koompassia excelsa, одного из компонентов окружающего первичного леса. Их появление связано с близостью плантации маточных деревьев, но Йохеме считает это свидетельством того, что вторичная сукцессия привела бы в конечном счете к восстановлению первичного леса, если бы этому не препятствовал последующий период обработки почвы. Заслуживает внимания, что злак Imperata, хотя и обычен на табачных плантациях в районе Дели, редко становится доминантой; согласно Йохемсу, это случается только при выжигании вторичной растительности. Подземные стебли Imperata устойчивы к огню, и, таким образом, после пожара он получает преимущество перед конкурентами.
Описываемая сукцессия, в общем достаточно типичная для вторичных сукцессий Малайского архипелага, несомненно, в некоторой степени определяется своеобразной и сложной сельскохозяйственной системой, часть которой она составляет.
Ценное, но неполное исследование вторичной растительности в Кепон-ге, полуостров Малакка, было опубликовано Саймингтоном (Symington, 1933). В этой местности вторичные сообщества в различных стадиях развития занимают заброшенные огороды на тех участках, которые за несколько лет до расчистки были покрыты дождевым тропическим лесом, но впоследствии подвергались выжиганию, скашиванию злаков и т. п. Подразумевается, что первая стадия сукцессии знаменовалась развитием травянистых растений, включая злаки и ползучие растения. Затем начинали господствовать кустарники, в частности Metastoma polyanthum, натурализовавшаяся в Азии, и Lantana camara, которые сменялись вторичным лесом из видов Мacaranga и аналогичных деревьев; последний, постепенно преобразовываясь в течение многих лет, приближался к высокоствольному климакспому лесу. Хотя общий ход сукцессии, таким образом, довольно легко восстанавливается, несколько важных проблем все еще остаются нерешенными; среди них особенно трудным является вопрос, почему Imperata в одних местах становится доминантой, задерживая развитие сукцессии на длительный период, а в других — нет. Наблюдения, производимые в течение трех лет на ряде опытных участков
416
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ лес
площадью 1 чейн (20 w. .и) с целью выяснить эти вопросы, не дали сколько-нибудь определенных результатов.
Два участка были расположены на обрабатывавшейся в течение нескольких лет территории и заброшенной незадолго до начала наблюдений. При первом обследовании эти участки представляли собой смешанное сообщество, состоявшее примерно из 52 видов, включая Trema «amboinensis», Macaranga tanarius, Bridelia tomentosa и другие молодые деревья, Musa malaccensis, кустарники, некоторые уцелевшие культурные растения и разнообразные травянистые растения, включая папоротники, осоки и злаки. Было встречено несколько небольших пятен лаланга {Imperata) и один-два клочка земли со злаком Brachiaria distachya. В течение первых нескольких месяцев наблюдений поколения травянистых растений быстро следовали друг за другом. Через год преобладание травянистых сорняков стало уменьшаться отчасти благодаря росту деревьев, кустарников и Musa, отчасти благодаря разрастанию Imper ata, которая размножается посредством корневищ. Через 3 года, когда участки освидетельствовались в последний раз, деревья, иногда с нижним ярусом, состоящим из Lantana, образовали на части площади плотный полог, под которым травянистый покров фактически отсутствовал; на остальной площади почти безраздельно господствовала Jmperata, хотя в одном месте колония Brachiaria долго сопротивлялась ее вторжению. От первоначального смешанного травянистого сообщества остались только следы. Таким образом, в пределах опытных участков Imperata утвердилась как стадия сукцессии лишь на части площади.
Соседний участок располагался в совершенно чистом сообществе Jmperata, история становления которого осталась неизвестной. Хотя на первый взгляд казалось, что Imperata вытеснила все остальные растения, среди ее покрова все же присутствовали изолированные кустарники, травянистые и ползучие растения. Основное изменение за 3 года наблюдений заключалось в том, что Imperata стала заметно менее активной и на части площади была вытеснена папоротником Nephrolepis biserrata. Кусты Metastoma достигли высоты 3 м и теперь давали достаточно тени, чтобы умерить энергию растущей под ними Imperata. Ничто не свидетельствовало, что на этом участке могут обосноваться какие-либо древесные виды после образования сомкнутого покрова Imperata’, отмеченный переход от господства Imperata к господству древесных видов был вызван разрастанием растений, обосновавшихся на ранней стадии сукцессии.
Заслуживает упоминания еще один опытный участок, исследованный Саймингтоном; он был заложен на окраине первичного леса на небольшой вырубке, почва которой никогда не подвергалась обработке. Было важно выяснить, по какому пути пойдет развитие сукцессии на ненарушенной лесной почве и будет ли она отличаться от сукцессии на истощенной почве заброшенной пашни. До расчистки растительность состояла из видов первичного леса с небольшой примесью видов белукара, вторгшихся из соседних сообществ вторичной растительности. Когда 3 года спустя участок был обследован, там утвердился типичный вторичный лес с преобладанием М acaranga tanarius. В первой стадии сукцессии господствовали травянистые растения Physalis minima и Amaranthus zlridis, под покровом которых наблюдались массовые всходы Trema, Musa и других видов белукара. Позднее Trema стала доминантой, но в конечном счете ее подавила Macaranga. Примечательно, что Imperata совсем не появилась на этом участке.
Вторичные сукцессии на Филиппинах были описаны Уитфордом (Whitford, 1906) и Брауном (Brown, 1919). Значительная часть земель
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
417
ной площади этих островов (около 40%) покрыта злаковниками — так называемыми когоналес (cogonales) — или вторичным злаковником с преобладанием Imperata (называемой здесь когон) или Saccharum spontaneum, дерновинного злака сходного типа. Большая часть низменностей покрыта парангом — смесью злаковников и вторичного леса, подобного малайскому белукару. Как когоналес, так и паранг, вероятно, занимают место бывшего вечнозеленого диптерокарпового леса; при отсутствии дальнейшего вмешательства человека и пожаров обработанная земля и злаков-ник вновь быстро превращаются в лес. Несколько следующих один за другим пожаров отдают землю во владение злакам, и в паранге деревья стремятся распространиться в промежутках между пожарами; таким образом, паранг представляет собой «пояс напряжения», или буферную зону, между злаковником и лесом, сохраняющийся годами, причем деревья то побеждают, то отступают. Среди злаковника встречаются отдельные экземпляры устойчивых к огню деревьев (Antidesma ghaesembilla, Bauhinia malabarica и Acacia farnesiana), стремящихся распространиться и сгруппироваться. При отсутствии пожаров мелкие деревья, кустарники и лианы наводняют злаковник, но прежде чем образуется сомкнутый древесный полог, Imperata и Saccharum замещаются злаками, образующими более высокие, но не столь густые травостои.
Вторичный лес на Филиппинах — это «густая разнородная чаща», состоящая из очень разнообразных деревьев, кустарников и лиан. Среди деревьев многочисленны Ficus, Litsea glutinosa, Macaranga tanarius, Melochia umbellata, Trema orientalis и многие другие. Большинство из них — типичные небольшие, быстрорастущие деревья вторичного леса с мягкой древесиной. Исключение составляют очень высокая Bischofia javanica и Psidium guajava, дерево с весьма твердой древесиной. Состав вторичного леса отличается резко выраженной изменчивостью, причина которой не ясна. Предполагается, что он разовьется в климаксный вечнозеленый лес, но ход развития этой сукцессии не прослежен. На расчищенных, но не обработанных площадях наблюдается гораздо менее сложная сукцессия, чем в злаковниках.
Другие данные о вторичных сукцессиях рассматриваемого района где климаксом является тропический дождевой лес (или какой-либо сходный тип вечнозеленого леса), вполне согласуются с общей картиной, выявляющейся при суммировании накопленных сведений. На востоке Новой Гвинеи (Lane-Poole, 1925a,b) местные жители обычно практикуют систему подсечно-огневого земледелия с циклом не короче 8—10 лет. Между периодами повторной обработки почвы вырастает густой вторичный лес из видов Trema, Macaranga, Geunsia, Pipturus и пр. Если этот цикл сокращается, наблюдается вторжение Imperata и других злаков. При их появлении местные жители стараются ежегодно выжигать траву, чтобы облегчить охоту за дичью. Таким образом, вскоре создается вторичный злаковник, являющийся стабильным биотическим климаксом. Обширные злаковники долин Маркхам (Магг, 1938) и Раму, несомненно, возникли подобным образом.
Заброшенные культурные земли в Южном Бантаме (Ява) вначале обычно заселяет Imperata, а позднее кустарники Metastoma malabathricum и Lantana (Backer, 1913). За кустарниковой стадией следует развитие типичного вторичного леса с видами Vitex, Grewia, Dillenia и др.
Фрей-Уисслинг (Frey-Wyssling, 1931) дает интересное описание пустошной растительности, или маквисоподобного кустарника, состоящего из
418
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
видов: Leptospermum flavescens. Metastoma decemfidum. Rhodomyrtus tomentosa и др.; встречается на плато Хабинсаран (от 800 м над уровнем моря) в округе Батак, Суматра. Подобные же сообщества покрывают большие площади в районе озера Тоба. Фрей-Уисслинг, сопоставляет этот кустарник со злаковникамп, встречающимися в очень сходных экологических условиях на плато Каро, и делает вывод, что оба сообщества развиваются на месте вечнозеленого дождевого леса, причем кустарник появляется там, где антропогенное влияние (огонь п пасущийся скот) незначительно, а злаковник — там, где благодаря большей плотности населения оно более интенсивно.
Не менее ценные сведения о вторичных сообществах и сукцессиях дали Винклер (Winkler, 1913, 1914), Баркилл (Burkill, 1919), Эндерт (Еп-dert, 1920), ван Розендель я Торенар (Roosendael, Thorenaar, 1924), ваи дер Лаан (van der Laan, 1925) и Ван-Стеенис (van Steenis, 1932, 1933, 1937а).
Из всего сказанного становится ясно, что в настоящее время было бы бесполезно делать обобщения относительно вторичных сукцессий Малайского архипелага, но очевидно, что между сукцессиями здесь и в тропической Африке есть много общего. Намного большее видовое разнообразие вторичных лесов Малайи по сравнению с африканскими, несомненно, зависит от того, что малайская флора в целом гораздо богаче. По-видимому, более существенную роль играет различие между продолжительностью стадии злаковника с господством видов Imperata. Для вторичных сукцессий Малайского архипелага характерно иногда очень длительное преобладание лаланга (imperata cylindrica var. major}, что, очевидно, является нормальной особенностью неотклонившейся вторичной сукцессии, тогда как в тропической Африке I. cylindrica var. africana. видимо, участвует только в отклонившихся сукцессиях, возникающих после усиленной обработки почвы под культурные растения и т. п. Роль, которую играет в сукцессии злак Imperata. не приносящий никакой пользы и являющийся сущим бичом, составляет одну из важнейших экологических проблем в тропиках. Дальнейшие исследования покажут состоятельность любопытного предположения Саймингтона (Symington, 1933) о том, что нежелательную фазу господства Imperata во вторичной сукцессии можно было бы обойти, засеяв обнаженную почву семенами подходящих деревьев вторичного леса. За отсутствием места не представляется возможным детально обсудить экологию Imperata. тем более, что недавно вышла ценная работа, посвященная этому вопросу (Gray, 1944).
Вышеописанные сукцессии развиваются на «нормальных» почвах Малайского архипелага, то есть тропических красноземах или желтоземах. На тропических подзолах, где климаксом является «боровой» лес (см. гл. 9, стр. 231—236, и гл. 10 и И), встречаются вторичные сообщества различной внешности и очень разного состава. Они никем не описывались с достаточной полнотой, хотя широко распространены в некоторых частях Борнео, например вокруг Мири и Клодтауна, в Сараваке. Вторичный лес ксерофитных Myrtaceae, растущий на белых песках восточного Борнео и упоминаемый Эндертом (Endert, 1925), очевидно, принадлежит к сообществам этого рода; Tristania obovata и Eugenia bankensis — самые распространенные здесь деревья. Пустошная паданговая растительность на песчаных почвах Борнео, Бангка и др., упомянутая выше (стр. 231), иногда, пожалуй, может быть биотическим климаксом, возникшим из отклонившейся сукцессии.
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
419
Вторичные сукцессии тропической Америки
На вырубках и заброшенных пашнях в американских тропиках развивается вторичный тропический дождевой лес, очень сходный по структуре с вторичным дождевым лесом Старого света, хотя, конечно, совсем иной по своему флористическому составу. Сукцессии здесь мало изучены, несомненно лишь, что они очень различаются в зависимости от почвы.
Повсюду в более влажных частях тропической Америки характерной особенностью молодого вторичного леса являются быстрорастущие деревья с мягкой древесиной из рода Cecropia. Эти деревья могут образовать беспримесные древостои, которые, подобно сообществам Musanga cecropioi-des тропической Африки, вероятно, являются сообществами одного поколения (Bouillienne, 1926, стр. 82). Цекропии очень напоминают Musanga (с которой они близки таксономически) не только формой листьев и общим обликом, но и своими экологическими особенностями. Злак Jmperata cylindrica, играющий такую важную роль во вторичных и отклонившихся сукцессиях Старого света, в Южной Америке встречается только в Чили: в остальных районах произрастает I. brasiliensis и другие аналогичные лалангу злаки. Ползучая осока из рода Scleria, известная как трава-бритва из-за острых краев своих листьев, часто образует непроницаемые заросли во вторичном лесу Гвианы.
Кенойер (Кепоуег, 1929) проследил путь развития’сукцессии на старых вырубках на острове Барро-Колорадо, Панама. В первой стадии заселения основными растениями были злаки (около 20 видов) и осоки (виды Суper us и Scleria) с примесью многочисленных двудольных трав (Amarant-haceae, Euphorbiaceae, Compositae, Solanaceae, Mimosoideae и др.) Большинство их недолговечно. Через год появляются крупнолистные однодольные, среди которых выделяются яркие гелпконии и панамская пальма Carludovica palmata; характерно обилие всходов деревьев, преобладающих в следующей стадии (виды Trema, Cecropia, Apeiba, Ochroma, Cordia и пр.) и разнообразных ^травянистых растений, отсутствовавших в первой стадии. При участии лиан эта растительность, уже почти достигающая высоты человеческого роста, превращается в непроходимую чащу.
Два года спустя начинают преобладать деревья, среди которых наиболее выделяются Cecropia mexicana. и С. longipes. Образуется молодой вторичный лес, где наряду с доминантами развиваются Ochroma Нтопеп-se (бальза) и пальма Attalea gomphococca. Через 15 лет это сообщество становится гораздо богаче видами. Появляются Ficus spp., 8—10 видов Inga. многие мелкие деревья и кустарники из семейства меластомовых. Protium spp. и др. Изобилуют лианы, включая примечательную лентоподобную Bauchinia excisa. Кустарниковый подлесок и травяной покров очень богаты. К этому времени уже приживается ряд видов первичного леса, но пока сообщество остается все еще более густым и непроходимым, чем первичный лес. Многое указывает на то, что эта сукцессия в конечном счете приведет к развитию климаксного леса.
Вторичные сукцессии на лесных вырубках Тринидада (Marshall, 1934, стр. 38—39) и в Гвиане очень напоминают сукцессию Панамы; Бенуа (Ве-noist, 1925) дал беглое описание аналогичной сукцессии во Франц. Гвиане.
На островах Вест-Индии очень распространены вторичные сообщества всех возрастов, но сукцессии мало изучены. Стеле (Stehle, 1935) говорит, что в Гваделупе растениями-пионерами, заселяющими открытые пространства в лесах на небольших и средних высотах (соответствует место
420
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
положению нижнегорного дождевого леса Бирда, см. стр.- 386), являются сорняки Ageratum conyzoides, Ipomoea nil, Hyptis spp., Lantana aculeata и др. В «средней области» различают два типа более старых вторичных сообществ: а) вторичный лес, состоящий главным образом из Cecropia peltata, Hibiscus tulipiflorus, Ochroma pyramidale и Oreopanax dussii, встречающихся там, где сохранился первоначальный слой перегноя; б) кустарник с преобладанием Croton corylifolius, С. flavens и видов Miconia— на нпзкорасположенных местах прежней расчистки, где слой перегноя был нарушен. В сообществе присутствуют древовидные папоротники Cyathea spp., а также Gleichenia sp.; последняя распространена во вторичных сообществах многих влажных тропических районов. Значение папоротников во вторичной растительности на истощенных почвах во влажных климатах подчеркивал Уордло (Wardlaw, 1931), который установил, что на заброшенных банановых плантациях в Санта-Лючия спустя 2 года преобладающими деревьями были древовидный папоротник Cyathea sp.1, а также Cecropia palmata и Scheffler a sp. вместе с кустарниками из родов Miconia и Peperomi .
Фрейзе (Freise, 1938) подробно описал капоэйра (capoeira), или вторичный лес, развитый на заброшенных пахотных угодьях прибрежной Бразилии (от Пернамбуко до Санта-Катарина). По-видимому, вторичная сукцессия здесь в некоторых отношениях отличается от сукцессии в области тропического дождевого леса Центральной Америки и северной части Южной Америки. Первая фаза знаменуется заселением почвы злаками (Andropogon spp., Chloris bahiensis, Brachiaria reptans) и другими травянистыми растениями вроде Thalia и Alternanthera spp., к растениям-пионерам вскоре присоединяются исключительно быстрорастущие лазящие Canavalia, Pachyrrhizus и Phaseolus spp. Некоторые кустарники, пережившие последнюю расчистку леса, вместе с вторгающимися сюда светолюбивыми деревьями образуют островки, которые стремятся к разрастанию и затеняют траву. В развивающемся вторичном молодом лесу встречается несколько видов Cecropia, каждый из которых характерен для определенных почвенных условий: они живут 8—12 лет. Из остальных деревьев вторичного леса некоторые (например, Euphorbiaceae, Sterculiaceae и др.) достигают возраста 25—30 лет, другие живут дольше; большинство их характеризуется мягкой древесиной.
Говорят, что некоторые виды вторичного леса никогда не встречаются в первичном лесу, так же как во вторичном лесу отсутствуют некоторые виды первичного. По мнению Фрейзе, эта сукцессия ведет к становлению несколько иного климакса, чем первоначальный девственный лес. Это могло бы объясняться чрезвычайно экстенсивным уничтожением первичного леса, при котором остается слишком мало подходящих материнских деревьев. Максимальный возраст вторичного леса оценивается в 150— 200 лет; за это время в нем может произойти смена 5—8 поколений недолговечных и 3—5 поколений более долговечных древесных видов.
Вторичный лес этой области часто подвергается рубке, выжиганию и т. п. В результате зарождаются отклонившиеся сукцессии, ведущие к развитию злаковников, сообщества из Pteridium, или же к обнажению материнской породы.
1 Обилие древовидных папоротников во вторичных сообществах на относительно небольших высотах характерно для Малых Антильских островов (Beard).
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
421
ОБЩИЕ ЧЕРТЫ ВТОРИЧНЫХ СУКЦЕССИЙ В ОБЛАСТИ РАЗВИТИЯ ДОЖДЕВОГО ЛЕСА
Вышеописанные детальные наблюдения над вторичными и отклонившимися сукцессиями при своей неполноте все же позволяют сделать некоторые общие выводы.
Во всех областях развития тропического дождевого леса вторичные сукцессии стремятся к восстановлению климатического климакса, хотя выжигание, пастьба и истощение почвы, вызванное чрезмерной сельскохозяйственной обработкой, могут стимулировать появление отклонившейся сукцессии и привести к стабилизации биотических климаксов.
В первой фазе сукцессии господствуют сорняки, включая обычно недолговечные и скорее эфемерные, чем однолетние злаки, поскольку их жизненный цикл часто гораздо короче года. В следующей фазе могут преобладать кустарники, но нередко господство непосредственно переходит от травянистых растений к деревьям. Деревья могут образовать полог путем массового вторжения или же постепенно, утверждением вначале единичных пионеров (последнее, видимо, случается при наличии густого травяного покрова, созданного видами вроде Imperatd). В любом случае в конце концов вырастает лес с деревьями, преимущественно недолговечными, быстрорастущими, рассеивающими семена при помощи ветра или животных; этот лес, медленно изменяясь, развивается в сообщество, аналогичное климатическому климаксу данной области. Молодой вторичный лес имеет тенденцию быть одновозрастным и монодоминантным; первые доминанты могут образовать сообщество одного поколения, которое исчезает, не дав потомства, и замещается сообществами с другими доминантами. Постепенно вторичный лес становится более смешанным по возрастной структуре и флористическому составу, пока не приблизится к состоянию спелого леса.
Таким образом, вторичные сукцессии воспроизводят в большем масштабе те перемены, которые совершаются при нормальной регенерации первичного леса, где прогалины, образованные гибелью крупных старых деревьев, временно заселяются теми же быстрорастущими, легко размножающимися видами, какие преобладают во вторичном лесу. В «обедненном лесу», где большое количество деревьев подвергается выборочной рубке, создаются промежуточные условия и образуется неполный вторичный сукцессионный ряд.
Развитие вторичной сукцессии отчасти зависит от внутренних свойств почвы и протекает различно, например, на тропических красноземах и тропических подзолах. Чрезвычайную роль играет история участка в период между уничтожением первоначального леса и началом вторичной сукцессии; если промежуточный период расчистки был настолько длительным, что привел к изменению структуры почвы и уничтожению в ней запасов перегноя, или же если наблюдалась ускоренная эрозия, последующая сукцессия будет отличаться от той, которая зарождается непосредственно после вырубки леса.
Сопоставление вторичных сукцессий с первичными ксеросериями, описанными в гл. 12, показывает их однородность. Чем сильнее вырождение почвы в тот период, когда земля после уничтожения первичного леса пустует или обрабатывается, тем больше будет сходство между вторичной сукцессией и первичной ксеросерией.
Превходящие факторы изменяют развитие сукцессии, сообщая одним видам преимущество перед другими. Это часто приводит к замещению
422
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
леса злаковником, очень похожим на эдафический или климатический климакс. Частичное уничтожение леса огнем также способствует развитию некоторых видов за счет других и, таким образом, приводит к большим изменениям во флористическом составе сообщества.
Конечно, между вторичными и отклонившимися сукцессиями во всех областях, где лес является климатическим климаксом, существует значительное сходство. В Англии и в тропиках последовательность стадий травянистых растений, кустарников и деревьев оказывается аналогичной. Доминанты сериальных лесных сообществ в умеренных странах обладают многими особенностями доминант вторичного дождевого леса, а именно светолюбивостью, быстрым ростом, непродолжительностью жизни и эффективными средствами рассеивания семян. Так, березы (Betula spp.) в этом отношении имеют много общего с деревьями тропического вторичного леса. Возникновение в ходе отклонившейся сукцессии злаковников и некоторых типов сосновых лесов в умеренных областях совершенно аналогично соответствующим процессам в дождевом тропическом лесу.
Сведений относительно возраста вторичных сукцессий тропиков очень мало. Шевалье (Chevalier, 1948) указывает, что лес на месте древнего города Анкор-Ват, Камбоджа, разрушенного, вероятно, пять или шесть веков тому назад, теперь напоминает девственный тропический лес этой же местности, хотя и обнаруживает некоторые отличия. В общем ясно, что чем длительнее период между уничтожением первичного леса и появлением вторичной сукцессии и чем больше изменились почва и среда в целом в течение этого периода, тем больше времени требуется для восстановления климаксной растительности.
ИЗМЕНЕНИЯ СРЕДЫ В ХОДЕ РАЗВИТИЯ ВТОРИЧНЫХ И ОТКЛОНИВШИХСЯ СУКЦЕССИЙ
Как уже неоднократно подчеркивалось, тропический дождевой лес представляет собой великолепный пример экосистемы, в которой климат, почвы, растительность и фауна являются компонентами исключительно сложного комплекса, находящегося в динамическом равновесии. Следовательно, когда один из компонентов — дождевой лес — частично или полностью уничтожают рубкой или выжиганием, то другие компоненты экосистемы претерпевают столь же радикальные изменения. При последующих вторичных сукцессиях микроклимат, почва и фауна развиваются параллельно растительности, пока вновь не восстанавливается равновесие.
Уничтожение лесного покрова сразу изменяет освещенность лесной почвы, измерявшуюся небольшими величинами до полного дневного света. Колебания температуры резко возрастают, а средняя и минимальная влажность воздуха сильно понижаются. Вместо сложной системы микроклиматов, характерной для высокоствольного леса (см. гл. 7), создаются условия, очень близкие к стандартному климату данной местности. Обнаженная почва, открытая воздействию солнца и дождей, быстро меняет свои свойства. На более крутых склонах начинается эрозия, разрушающая верхние слои почвы или вымывающая более тонкие ее частицы. Повышение температуры почвы приводит к быстрому разложению перегноя. Согласно Корбе (Corbet, 1935), разложение перегноя в обнаженной почве скорее химический, чем микробиологический процесс; количество бактерий в почве почти одинаково до и после вырубки леса (см. стр. 238).
ЧЕЛОВЕК И ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
423
С потерей перегноя связана потеря азота почвы, удаляющегося преимущественно в виде газа (Corbet, 1935).
Уменьшение в почве связанного азота, происходящее вслед за расчисткой леса, составляет лишь часть общего процесса истощения почвы. Как видно из предыдущего, в первичном лесу почва и растительность находятся в динамическом равновесии, причем питательные вещества циркулируют по почти замкнутому кругу, так как небольшие количества растворимых питательных веществ, потерянные вследствие выщелачивания, восполняются минеральными веществами из невыветренных слоев. Когда лес уничтожают, этот круг разрывается; если растительность остается гнить па поверхности почвы, то значительные количества минеральных веществ, высвобождающихся при этом, вероятно, уносятся дренажными водами.
Если вторичная сукцессия возникает сразу же после вырубки леса, то опасность эрозии невелика и сомкнутый растительный покров образуется в течение нескольких недель; процесс истощения почвы приостанавливается прежде, чем он зайдет слишком далеко, и почва и микроклимат сразу начинают возвращаться к своему первоначальному состоянию. Если, с другой стороны, после расчистки следует длительный период обработки почвы, особенно если посевы не создают адекватного напочвенного покрова, то продолжительная обнаженность почвы вместе с потерей питательных веществ в собранном урожае ведет к таким большим изменениям в структуре, содержании перегноя в составе и количестве питательных веществ в почве, что время, потребное для восстановления равновесия между почвой и растительностью (даже при отсутствии сильной эрозии), очень удлиняется. Это делает понятным большое значение длительности периода обработки почвы, размеров и степени разрушения перегноя для хода развития и продолжительности последующей сукцессии.
Общий характер вышеописанных изменений среды хорошо известен, но подробных сведений о его особенностях очень мало. С экологической точки зрения было бы особенно важно получить более полное представление об очень сложных процессах, происходящих в почве непосредственно после уничтожения леса и при развитии последующих сукцессий. Чуть ли не единственные данные, имеющиеся по этому вопросу, содержатся в отчете об исследовании почв лесного массива Арена, Тринидад, сделанном Дути, Харди и Родригесом (Duthie, Hardy, Rodriguez, 1936).
Климаксной растительностью здесь является смешанный лес Сагара guianensis — Eschweilera subglandulosa— Maximiliana caribaea (фация вечнозеленого сезонного леса в смысле Бирда, см. стр. 343). Песчаная почва (тип «песка кароим») отличается рыхлой структурой. Было проведено сравнение почв на следующих четырех опытных участках за двухлетний период: а) естественный лес; б) расчищенный лес, не подвергавшийся выжигу и распашке; в) расчищенный лес, подвергавшийся выжигу, но не распахивавшийся; г) расчищенный лес, выжженный и засеянный «продовольственными культурами». При исследовании изучались следующие основные свойства почвы: содержание органического вещества, общее содержание азота, отношение углерода к азоту и количество имеющихся питательных веществ (определяемое измерением электропроводимости экстрактов почвы в определенных условиях).
Было установлено, что расчистка, видимо, вызывает немедленное уменьшение общего количества органического углерода и азота и соответ-
424
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Таблица 39
Изменения почвы на расчищенных, выжженных и обработанных участках в лесном массиве Арена, Тринидад (Duthie, Hardy, Rodriguez, 1936)1
Органическое вещество, % Общее содержание азота, %
0—6 дюймов (0— 1 5 см) 6-12 дюймов (15—30 см) 12—24 дюй-ма (30-60 см) 0—6 дюймов (0—15 см) 6—12 дюймов (1 5—30 см) 12—24 дюй- ма (30-60 см)
Участок 1. Естественный лес 26 июня 1933 г. 18 июля 1935 г. Участок 2. Выжженный 26 июня 1933 г. 18 июля 1935 г. Участок 3. Невыжженный 26 июня 1933 г. 18 июля 1935 г. Участок 4. Выжженный и вспаханный 26 июня 1933 г. 18 июля 1935 г. 2,37 2,19 1,41 0,93 1,31 1,64 1,57 1,15 0,97 1,07 1,35 0,93 0,73 0,99 0,81 0,83 0,64 0,79 0,87 0,77 0,52 0,52 0,53 0,59 0,13 0,09 0,05 0,05 0,05 0,08 0,03 0,05 0,07 0,03 0,05 0,04 0,03 0,05 0,03 0,04 0,05 0,05 0,04 0,03 0,02 0,03 0,02 0,03
Отношение углерод/азот Доступные питательные вещества (величины э л ектроп ро во димости)
0—6 дюймов (0—15 см) 6—12 дюймов (15—30 см) 12—24 дюйма (30—60 см) 0—6 дюймов (0—15 см) 6—12 дюймов (15—30 см) 12—24 дюй- ма (30—60 см)
Участок 1. Естественный лес 26 июня 1933 г. 18 июля 1935 г. Участок 2. Выжженный 26 июня 1933 г. 18 июля 1935 г. Участок 3. Невыжженный 26 июня 1933 г. 18 июля 1935 г. Участок 4. Выжженный и вспаханный 26 июня 1933 г. 18 июля 1935 г. 11,2 13,7 14,5 13,7 15,1 11,7 15,6 13,1 9,1 11,2 14,8 14,0 14,0 11,6 17,6 11,6 7,7 9,7 12,7 14,3 13,1 11,1 18,3 11,7 41 52 42 59 35 51 31 25 28 35 43 56 30 43 27 35 25 37 36 31 56 34 30 39
1 Участки были расчищены 28 мая 1933 г.; участки 2 и 4 выжжены 23 августа 1933 г.
ственное понижение отношения углерод/азот. В данной почве эти изменения наблюдались до глубины 2 футов (62 см). Так как валка леса проводилась в начале дождливого периода, растительность вскоре восстановилась путем естественного возобновления, после чего содержание углерода и азота и отношение углерод/азот стали увеличиваться. Возможно, это
ПОСТСКРИПТУМ
425
объясняется присоединением органического вещества из листьев, опавших с поваленных деревьев. В последующий засушливый сезон этот процесс ускорился, и ко второму периоду дождей биохимические процессы происходили преимущественно в нижних слоях почвы, где отношение углерод/азот продолжало уменьшаться. К началу третьего периода дождей (дата последнего эксперимента) увеличение содержания органического углерода и азота было значительным, но отношение углерод/азот не достигло своего первоначального уровня. Установлено, что минеральные питательные вещества, обильно выделяемые в периоды быстрого органического распада, утрачивались на протяжении влажного сезона из-за выщелачивания.
На расчищенном и выжженном, но не вспаханном участке количество органического углерода и азота и отношение углерод/азот оставались на более низком уровне, чем на участке, расчищенном без выжигания. Это наблюдалось до глубины по меньшей мере 1 фута (30 см). Выжигание высвободило соли со щелочной реакцией и значительно уменьшило кислотность почвы, которая, впрочем, восстановилась спустя 2 года; выжигание не повлияло на нормальный процесс освобождения минеральных питательных веществ при гниении поваленных деревьев.
Изменения почвы на расчищенном, выженном и обработанном участке были аналогичны изменениям на двух расчищенных, но необработанных участках, за исключением того, что восстановление органического вещества в почве заметно приостановилось из-за меньшего количества перегноя, внесенного в почву при посеве, по сравнению с количеством, доставляемым естественным растительным покровом.
Из этой работы явствует, что в условиях исследованной площади всякое ухудшение, вызванное одной только расчисткой леса, очень быстро ликвидируется. Последствия продолжительной обработки гораздо более стойки.
Постск риптум
БУДУЩЕЕ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Тропический дождевой лес состоит из ряда растительных формаций, различных по флористическому составу, но очень сходных по структуре и физиономии; по этой причине их объединяют в один тип формаций, или панклимакс. Спелые или первичные тропические дождевые леса представляют собой чрезвычайно сложные растительные сообщества и находятся в состоянии очень подвижного и тем не менее весьма устойчивого динамического равновесия с другими компонентами экосистемы — климатом, почвой и животным миром.
О прошлом формаций тропического дождевого леса известно очень мало, но есть основания думать, что они существовали в неизменном виде или же, изменяясь чрезвычайно медленно, с отдаленнейших времен. Вековые изменения климата иногда расширяли, а иногда суживали границы тропического дождевого леса, но, вероятно, этим формациям редко приходилось приспосабливаться к внезапным или очень резким изменениям среды. До самого последнего времени человек почти не оказывал влияния на дождевой лес, обширные площади его были совершенно безлюдны или обитаемы племенами, собирающими лесные плоды, чье воздействие на растительность было не большим, чем воздействие представителей животного мира.
426
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Просуществовав таким образом миллионы лет, экосистема дождевого леса совсем недавно — в большей части мест только в последние 100 лет или около того — была грубо потревожена вторжением цивилизации в тропики. Массовое истребление леса и замещение его плантациями каучука, кофе, какао и аналогичных культур, возделываемых на экспорт, сопровождалось, пожалуй, еще большим, экстенсивным вырубанием его в процессе подсечно-огневого земледелия, осуществляемом местным населением. Так за очень небольшой промежуток времени сообщества первобытного климаксного леса на огромных пространствах были замещены плантациями, рудеральными сообществами и сериальными стадиями. До этого тропический дождевой лес находился примерно в той стадии развития, которую переживали леса Европы в мезолитический период, когда были заселены только окраины леса и ограниченные участки по берегам рек. Последующее повсеместное истребление леса в тропиках можно сравнить с расчисткой европейского леса земледельческими племенами, появившимися в период неолита, за исключением того, что первый процесс был завершен в несколько десятилетий, тогда как второй длился тысячи лет.
Хотя подсчитано (Heske, 1938—1939), что тропический дождевой лес все еще составляет около половины мировой площади леса, уничтожение сохранившегося леса идет со все возрастающей быстротой. Если не будут приняты решительные меры, чтобы остановить это истребление, то весь тропический лес в целом может исчезнуть уже при нынешнем поколении, за исключением некоторых недоступных мест и маленьких «резервированных лесных массивов», искусственно поддерживаемых главным образом для производства древесины. В лучшем случае сохранится только небольшая часть первоначального леса, а основная масса лесной формации, покрывавшей огромную часть земной поверхности в течение миллионов лет, обречена на гибель. Каких последствий следует при этом ожидать?
Наиболее опасным может оказаться влияние на ход эволюции растительности всего земного шара. Флора тропического дождевого леса с ее неимоверным богатством видами, принадлежащими к тысячам родов и множеству семейств, в прошлом составляла резервуар генетического разнообразия и потенциальной изменчивости. В течение по крайней мере позднейших эпох истории земли она была тем средоточием эволюционной активности, откуда пополнялась вся остальная флора мира. Это не означает, что не было других источников эволюции растений, но, как мы видели в гл. 1, есть точные доказательства того, что значительная часть флоры умеренных областей прямо или косвенно произошла из тропиков; другие центры, такие, как Южная Африка и Австралия, оказывали гораздо меньшее влияние за пределами своих границ. Таким образом, тропический лес играл особую роль в сравнении с другими главными растительными формациями, быть может оттого, что в нем эволюция не прерывалась сезонными остановками в жизнедеятельности растений или такими вековыми климатическими изменениями, как ледниковые периоды. Возможно также, что высокие постоянные температуры в дождевом лесу особенно благоприятны для выживания мутаций.
Эту свою роль — роль источника снабжения генетическим материалом и новыми формами растительной жизни — тропический дождевой лес в недалеком будущем уже не сможет более играть или же она станет гораздо менее значительной. Площадь тропического дождевого леса настолько уменьшилась, а соседствующие формации, способствовавшие миграции растений из влажных тропиков в другие области настолько изме-
ПОСТСКРИПТУМ
427
иены или истреблены, что проникновение в субтропические и умеренные области растений тропического происхождения, теряет свое значение, если еще не полностью прекратилось. Поэтому, возможно, что истребление тропического дождевого леса, происходившее за последние 100 лет. существенно изменит будущий ход эволюции растений и закроет многие пути эволюционного развития.
Другое следствие разрушения дождевого леса, быть может очень важное, связано с его влиянием на климат; это спорный вопрос, и здесь он обсуждаться не будет. Влияние лесов на климат в общих чертах разбирали Зон (Zon, 1912), Брукс (Brooks, 1927b) и Никольсон (Nicholson, 1936). Вопросу о влиянии тропического дождевого леса на осадки особое внимание уделил Бернар (Bernard, 1945).
Более очевидные и прямые результаты истребления дождевого тропического леса относятся к его непосредственной экономической ценности. Во-первых, он является источником многочисленных материалов, особенно древесины, топлива, целлюлозы и «второстепенных древесных продуктов», таких, как смолы, каучук, камфора, раттан и пр., и во-вторых, он защищает почву, особенно в гористых районах, от ускоренной плоскостной и линейной эрозии.
Тропический дождевой лес является единственным источником такой ценной древесины, как гринхарт, многих сортов красного дерева (маха-гони) и пр., но все же его значение как поставщика сырья меньше, чем можно было бы ожидать. Частично это происходит оттого, что в мировом спросе на древесину основное место принадлежит мягкой древесине хвойных пород, которые обычно не встречаются в лесах тропических низменностей. Хеске (Heske, 1938—1939) подсчитал, что в 1937 г. хвойная древесина составляла 92,5% мирового экспорта древесины и 88% импорта частично оттого, что, как мы видели, дождевые тропические леса обычно представляют собой смешанные ассоциации с очень большим количеством доминант, ни одна из которых не составляет значительной части всего древостоя. Эти доминанты различны по своей товарной ценности, к тому же в настоящее время только малая часть их находит применение в экономике. Аппель (Appel, 1938—1939) указывает, что из 500 видов деревьев в тропическом дождевом лесу Камеруна только какие-нибудь 30% имеют товарную ценность. В будущем, вероятно, найдет применение гораздо большее число видов дождевого леса; уже сейчас развитие фанерного производства в тропиках делает возможным использовать гораздо больший процент древесных пород, чем раньше. Можно предполагать, однако, что будущее развитие производства древесины в тропиках пойдет в сторону создания искусственных сообществ, состоящих из сравнительно немногих ценных видов, выращиваемых путем посадок или путем лесотехнической обработки существующих первичных, обедненных или вторичных смешанных лесов. В формировании и поддержании таких сообществ определенную помощь может оказать экологическое изучение естественных монодоминантных дождевых лесов.
Ботанику политика концентрации внимания на немногих «экономически ценных» видах внушает некоторые опасения. Среди многих тысяч видов деревьев тропического дождевого леса существует лишь незначительное количество таких, чьи свойства — механические, технические, биохимические и др. — хоть приблизительно известны. Среди большинства быстроисчезающих «бесполезных» видов, вероятно, многие могут оказаться неожиданно ценными — одни как источники древесины для специальных целей, другие — как сырье для извлечения побочных хими
428
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ческих продуктов и, наконец, некоторые — в качестве создателей новых типов деревьев путем гибридизации. Флоре тропического дождевого леса угрожает опасность исчезнуть прежде, чем скрытые в ней ценности будут окончательно выявлены.
Экономическая ценность тропического дождевого леса, выявленная и потенциальная, сама по себе достаточна, чтобы стремиться сохранить оставшиеся площади первичного леса с гораздо большим рвением, чем это делалось до сих пор, но, кроме того, существуют еще эстетические и научные основания для этого. Тропический лес — незаменимое поле для биологических исследований, в нем скрыт тот ключ к научному познанию, которого больше нигде не найти. Корнер (Corner, 1946) красноречиво убеждал в необходимости серьезного расширения исследовательской работы по ботанике в тропиках. Несомненно, что существующие ботанические теории в слишком большой степени основывались на ограниченной и обедненной флоре умеренных областей, и что все еще полностью не осознано, какие богатейшие материалы для наблюдения и эксперимента сосредоточены в вечнозеленых лесах тропиков. «Я боюсь,— говорит Корнер,— чтобы весь девственный низинный лес тропиков не был уничтожен прежде, чем ботаника проснется». Современная ситуация требует создания системы естественных заповедников под национальным или международным контролем, в которых естественная растительность была бы полностью ограждена от посягательств со стороны лесопромышленника или местного земледельца. Подобные заповедники должны занимать обширную площадь, и в них должны быть созданы благоприятные условия для ботанических и зоологических исследований. Национальный парк Альберт в Республике Конго представляет собой отличный пример природного заповедника; если образцы древних растительных сообществ тропиков должны быть сохранены, чтобы грядущие поколения изучали их и восхищались ими, совершенно недостаточно ограничиться лишь относительно небольшими лесными резервациями, эксплуатация которых осуществляется под контролем.
БИБЛИОГРАФИЯ
Adamson R. S. (1910), Note on the roots of Terrninalia arjuna Bedd., «New Phytol.», 9, 150—156.
A i у a r T.V.V. (1932), The Sholas of the Palghat Division, Parts I, II, «Indian For.», 58, 414—432, 473—486.
Allee W. C. (1926a), Measurement of environmental factors in the tropical rain— forest of Panama, «Ecology», 7, 273—302.
Allee W. C. (1926b), Distribution of animals in a tropical rain-forest with relation to environmental factors, «Ecology», 7, 445—468.
A 1 1 о r g e V., Aliorge P. (1939), Sur la repartition et 1’ecologie des hepatiques epiphylles aux Azores, «Bol. Soc. Broter» (ser. 2) 13, 211—231.
A n g о t A. (1928), Traite elementaire de meteorologic (4th ed, revised by С. E. Brazier), Paris.
Appel E., (1938—1939), Waldnutzung im westafrikanischen Urwald, «Z. d. Welt-forstwirtsch.», 6, 95—101.
A r b e r A. (1934), The Gramineae, a study of cereal, bamboo and grass, Cambridge.
A ubr e ville A. (1933), La foret de la Cote d’Ivoire, «Bull. Com. Afr. occid. frang.», 15, 205—261.
Aubreville A. (1936), La flora forestiere de la Cote d’Ivoire, Paris.
Aubreville A. (1938), La foret coloniale: les forets de 1’Afrique occidental franchise, «Ann. Acad. Sci. colon.», Paris, 9, 1—245.
Backer C. A. (1909), De flora van het eiland Krakatau, «Jversl. topogr. Dienst Ned.- Ind.», Batavia (Java).
Backer C. A. (1913), Een lastige vreemdeling, «Trop. Natuur», 2, 27—31, 33—36.
Backer C. A. (1929), The problem of Krakatao as seen by a botanist, Weltevreden (Java) and The Hague.
Bain F. M. (1934), The Rainfall of Trinidad, Dept. Agric. Trinidad and Tobago.
Baker J. R. (1938), Rain-forest in Ceylon, «Kew Bui.» (1938), 9—16.
Baker J. R., Baker I. (1936), The seasons in a tropical rain-forest (New Hebrides), Part 2, Botany, «J. Linn. Soc. (Zool.)», 39, 507—519.
Baker J. R., Harrisson T. H. (1936), The seasons in a tropical rain-forest, Part I, Meteorology, «J. Linn. Soc. (Zool.)», 39, 443—463.
Baren J. van (1931), Properties and constitution of a volcanic soil, built in 50 years in the East-Indian Archipelago, «Med. Landbouwhoogesch. Wageningen», 35 (Verb. 6), 1—29.
Beard J. S. (1942), The use of the term «deciduous» as applied to forest types in Trinidad, B.W.I., «Emp. For. J.», 21, 12—17.
Beard J. S. (1944a), Climax vegetation in tropical America, «Ecology», 25,127— 158.
Beard J. S. (1944b), The natural vegetation of the island of Tobago, British West Indies, «Ecol. Monogr.», 14, 135—163.
Beard J. S. (1944c), Forestry in the Windward Islands, «Development and welfare in the West Indies», Bull, No. 11.
430
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Beard J. S. (1945), The progress of plant succession on the Soufriere of St. Vincent, «J. Ecob, 33, 1—9.
Beard J. S. (1946a), The Mora forests of Trinidad, British West Indies, «J. Ecol.», 33, 173—192.
Beard J. S. (1946b), The natural vegetation of Trinidad, «Oxf. For. Mem.», No. 20.
Beard J. S. (1949), The natural vegetation of the Windward and Leeward Islands, «Oxf. For. Mem.», No. 21.
В ecca ri 0. (1904), Wanderings in the great of Borneo, Ed. and transl. by F.H.H. Guillemard, London.
Bennett H. H., Bennett R. V. (1928), The soils of Cuba, Trop. Res. Foundn., Washington.
В en oist R. (1925), La vegetation de la Guyane frangaise, «Bull. Soc. bot. Fr.», 72, 1066—1078.
Bernard E. (1945), Le climate ecologique de la Cuvette Centrale Congolaise, Publ. Inst. Nat. Agron. Congo Beige, Brussels.
В e г г у E. W. (1925), Tertiary flora of Trinidad, «Studies in Geology», 6, 71 — 161.
В ews J. W. (1927), Studies in the ecological evolution of the Angiosperms, «New Phytol.», 26, 1—21 и след.
В i 1 h a m E. G. (1938), The Climate of the British Isles, London.
Blanford H. R. (1929), Regeneration of evergreen forests in Malaya, «Indian For.», 55, 333—339, 383—395.
Blum G. (1933), Osmotische Untersuchungen in Java, I, «Вег. schweiz. bot. Ges..», 42, 550—680.
Boerema J. (1919), Intensiteit der zonnestraling, «Hand, eerste ned.-ind. natuurw. Congr.», 99—101.
Booberg G. (1932), Grondvormen, etages, en phytocoenosen van Java’s vegetatie, «Hand. 6de ned.- ind. natuurw. Congr.», 1931, 329—346.
Bergesen F. (1909), Notes on the shore vegetation of the Danish West Indies, «Bot. Tidsskr.», 29, 201—217.
Bouillienne R. (1926), Savanes equatoriales en Amerique du Sud, «Bull. Soc. Belg.», 58, 217-223.
Bouillienne R. (1930), Un voyage botanique dans le Bas-Amazone, «Arch. Inst, bot. Univ. Liege», 8, 1—185.
В raak C. (1924), Het klimaat van Nederlandsch Indie, Deel I, «Verh. magn. met. Obs. Batavia», 8.
В г а а к C. (1931), Klimakunde von Hinterindien und Insulinde, в книге W. К oppen, R. Geiger, Handbuch der Klimatologie, 4, Berlin.
Braun E. L. (1935), The undifferentiated deciduous forest climax and the association-segregate, «Ecology», 16, 514—519.
Braun E. L. (1941), The differentiation of the Deciduous Forest of the eastern United States, «Ohio J. Sci.», 41, 235—241.
В r e n n e г W. (1902), Klima und Blatt beider Gattung Quercus, «Flora», 90, 114—160.
Brockman n-J erosch H., Brockman n-Jerosch M. (1925), Jamaika, «Vegetationsbilder», Reihe, 16, Hft. 5—6.
Brooks C.E.P. (1927a), The mean cloudiness over the earth, «Mem. Roy. Met. Soc.», 1, 127 — 138.
Brooks C.E.P. (1927b), The influence of forests on rainfall and run-off, «Quart. J. R. Met. Soc.», 54, 1—17.
Brooks C.E.P. (1932), Climate; A Handbook for Business Men, Students and Travellers, 2nd ed., London.
Brown W'. H. (1919), Vegetation of Philippine mountains, Manila.
Brown W. H., M a t t h e w s D. M. (1914), Philippine Depterocarp forests, «Philipp. J. Sci.», 9, (sect. A.), 413—561.
БИБЛИОГРАФИЯ
431
В ii n n i n g E. (1947), In den Waldern Nord-Sum a Iras, Bonn.
В ii n n i n g E. (1948), Entwicklungs- und Bewegungsphysiologie der Pflanze, Lehr-buch der Pflanzenphysiologic, 2, 3, Berlin, Gottingen, Heidelberg.
Burger H. (1933), Waldklimafragen, II, HI, «Mitt, schweiz. Zent.-Anst. forstl. Versuchsw.», 18, 7—54, 153—192.
Burkill I. H. (1919), The composition of a piece of well-drained Singapore secondary jungle thirty years old, «Gdns’ Bull. Singapore», 2, 145—157.
Burkill I. H. (1928), The main features of the vegetation of Pahang, «Malay. Nat.», 2, 11—21.
Burtt Davy J. (1938), The classification of tropical woody vegetation types, «Imp. For. Inst. Paper», No. 13.
В ii s g e n M. (1903), Einige Wachstumsbeobachtungen aus den Tropen, «Вег., dtsch. bot. Ges.», 21, 435—440.
В ii s g e n M., M ii n c h E., (1929), The structure and life of forest trees, Transl. by T. Thomson, London.
Busse W. (1905), Ueber das Auftreten epiphyllischer Kryptogamen im Regenwald-gebiet von Kamerun, «Вег. dtsch. bot. Ges.», 23, 164—172.
Buxton P. A., Lewis D. J. (1934), Climate and tsetse flies; laboratory studies upon Glossina submorsitans and tachinoides, «Philos. Trans.», Ser. B, 224, 175-240.
Byers H. G., Kellogg С. E., Anderson M. S., T h о r p J. (1938), Soil formation, «Soils and Men.», Yearb. U.S. Dep. Apric., 948—978.
Cabrera A., Dawson G. (1944), La selva marginal de Punta Lara, «Rev. Mus. La Plata», (N. S.), Sec. Bot., 5, 267—382.
Caldwell J. (1930), The movements of food materials in plants, «New Phytol.», 29, 27—43.
Cardenas M. (1945), Aspecto general de la vegetacion de Bolivia, in F. Verdoorn, «Plants and plant science in Latin America», Waltham, Mass., 312—313.
Carter G. S. (1934), Reports of the Cambridge Expedition to British Guiana, 1933. Illumination in the rain forest at ground level, «J. Linn. Soc. (Zool.)», 38, 579— 589.
Champion H. G. (1936), A preliminary survey of the forest types of India and Burma, «Indian For. Rec.» (N. S.), 1, 1—286.
Chapin J. P. (1932), Birds of the Belgian Congo, Part I, «Bull. Amer. Mus. Nat. Hist.», 65.
Chapman V. J. (1944), Cambridge University Expedition to Jamaica, 1939; Part I: A study of the botanical processes concerned in the development of the Jamaican shoreline; Part ILA study of the environment of Avicennia nitida Jacq. in Jamaica; Part III: The morphology of Avicennia nitida Jacq. and function of the pneumatophores, «J. Linn. Soc. (Bot.)», 52, 487—533.
Charter C. F. (1941), A reconnaissance survey of the soils of British Honduras, Government of British Honduras, Trinidad.
Cheeseman T. F. (1903), The flora of Rarotonga, «Trans. Linn. Soc. Lond. (Bot.)» 6, 261-313.
Ch en er у E. M. (1948), Aluminium in plants and its relation to plant pigments, «Ann. Bot. Oxford», N. S., 12, 121 —136.
Chenery E.M., Hardy F. (1945), The moisture profile in some Trinidad forest and cacao soils, «Trop. Agriculture, Trin.», 22, 100—115.
Chengapa B. S. (1934), Andaman forests and their reproduction, «Indian For.», 60, 54—64, 117—129, 185—198.
Chevalier A. (1900), Les zones et les provinces botaniques de 1’Afrique Occidentale fran^aise, «С. R. Acad. Sci.», Paris, 130, 1205—1208.
432
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Chevalier А. (1909), L’extension et la regression de la foret vierge de 1'Afrique tropicale, «С. R. Acad. Sci.», Paris, 149, 458—461.
Chevalier A. (1912), Carte botanique, forestiere et pastorale, «Geographic», 26, pl. I (пояснения, стр. 276—277).
Chevalier A. (1917), La foret et les bois du Gabon, «Les vegetaux utiles d’Afri-que tropicale fran^aise», Fasc. 9, Paris.
Chevalier A. (1924), Sur la foret primitive tropicale et la foret secondaire, «С. R. Soc. Biogeogr.», 1, 39—40.
Chevalier A. (1938a), Flore vivante de 1’Afrique Occidentale fran^aise, 1, Paris.
Chevalier A. (1938b), Sur la presence d’une Bromeliacee spontan£e en Guinee fran^aise, «Bull. Soc. bot. fr.», 85, 489—490.
Chevalier A. (1948), Biogeographic et ecologie de la foret dense ombrophile de la Cote d’Ivoire, «Rev. Bot. appl.», 28, 101—115.
Chipp T. F. (1913), The reproduction of Musanga smithii, «Kew Bull.», (1913), 96.
Chipp T. F. (1922), Buttresses as an assistance to identification, «Kew Bull.» (1922), 265-268.
Chipp T. F. (1927), The Gold Coast Forest, A study in synecology, «Oxf. For. Mem.», 7.
Christophersen E. (1927), Vegetation of Pacific Equatorial Islands, «Bishop Mus. Bull.», 44.
Clements F. E. (1936), Nature and structure of the climax, «J. Ecol.», 24, 252—284.
Copeland E. B. (1907), Comparative ecology of the San Ramon Polypodiaceae, «Philipp. J. Sci.» (sect. C), 2, 1—76.
Corbet A. S. (1935), Biological processes in tropical soils, with special reference to Malaya, Cambridge.
Corner E. J.H. (1933), A revision of the Malayan species of Ficus: Covellia and Neo-morphe, «J. Malay Br. Asiat. Soc.», 11, 1—65.
Corner E.J.H. (1935), The seasonal fruiting of agarics in Malaya, «Gdns’ Bull. Singapore», 9, 79—88.
Corner E.J.H. (1938), The systematic value of the colour of withering leaves, «Chronica Bot.», 4, 119—121.
Corner E.J.H. (1939), A revision of Ficus, subgenus Synoecia, «Gdns* Bull. Singapore», 10, 82—161.
Corner E.J.H. (1940), Wayside trees of Malaya, 2 vols., Singapore.
Corner E.J.H. (1946), Suggestions for botanical progress, «New Phytol.», 45, 185 — 192.
Coster C. (1923), Lauberneuerung und andere periodische Lebensprozesse in dem trockenen Monsun-gebiet Ost-Javas, «Ann. Jard. bot. Buitenz.», 33, 117—189.
Coster C. (1925), Die Fettum wand lung in Baumkorper in den Tropen, «Ann. Jard. bot. Buitenz.», 35, 71—104.
Coster C. (1926a), Periodische Bliiteerscheinungen in den Tropen, «Ann. Jard. bot. Buitenz.», 35, 125—162.
Coster C. (1926b), Die Buche auf dem Gipfel des Pangerango, «Ann. Jard. bot. Buitenz.», 35, 105—119.
Coster C. (1927—1928), Zur Anatomic und Physiologic der Zuwachszonen und Jah-resringbildung in den Tropen, «Ann. Jard. bot. Buitenz.», 37, 49—160; 38, 1-114.
Coster C. (1932), Wortelstudien in de Tropen. I. De jeugdontwikkeling van het wortelstelsel van een zeventigtal boomen en groenbemesters, «Tectona», 25, 828— 872.
Coster C. (1933), Wortelstudiёn in de Tropen. III. De zuurstofbehoefte van het wortelstelsel, «Tectona», 26, 450—497.
Coster C. (1935a), Licht, ondergroei en wortelconcurrentie, «Tctona», 28, 612—619.
БИБЛИОГРАФИЯ
433
Coster С. (1935b), Wortelstudien in de Tropen. IV. Wortelconcurrentie, «Tectona», 28, 861—878.
Cox H. A. (1939), A handbook of Empire timbers, Dep. Sci. and Industr. Res., London.
C ua t recasas J. (1934), Observaciones geobotanicas en Colombia, «Trab. Mus. Cienc. nat., Madr.», Ser. Bot., 27.
Czapek F. (1909), Ueber die Blattentfaltung der Amherstien, «S.-В. Akad. Wiss. Wien», 118, 201—230.
1) an iker A. U. (1929), Neu-Caledonien, Land und Vegetation, «Vjschr. naturf. Ges. Zurich.», 74, 170—197.
Davis J. H. (1940), The ecology and geologic role of mangroves in Florida, «Publ. Carneg. Instn.», No. 517, 303—412.
Davis J. H. (1943), The natural features of southern Florida, especially the vegetation and the everglades, «State of Florida, Dep. of Conservation, Geol. Bull.», No. 25.
Davis T.A.W. (1929), Some observations on the forests of the North West District, «Agric. J. Brit. Guiana», 2, 157—166.
Davis T.A.W. (1933), Report on the Balata industry in British Guiana with special reference to the Rupununi District, «British Guiana, Second Legislative Council, Third Sess.», 1932.
Davis T.A.W. (1941), On the island origin of the endemic trees of the British Guiana peneplain, «J. Ecol.», 29, 1 — 13.
Davis T.A.W., Richards P. W. (1933—1934), The vegetation of Moraballi Creek, British Guiana; an ecological study of a limited area of Tropical Rain Forest. Parts I, II, «J. Ecol.», 21, 350—384; 22, 106—155.
Deuerling O. (1909), Die Pflanzenbarren der Afrikanischen Fliissemit Berucksich-tigung der Wichtigsten pflanzlichen Verlandungserscheinungen, «Munch, geogr. Studien», No. 24 («Abstr. Bot. Zbl.», 1910, 114, 91—94).
Diels b. (1906), Die Pflanzenwelt von West-Australien, «Veget. d. Erde», 7.
Diels L. (1918), Das Verhaltnis von Rhythmik und Verbreitung bei den Perennen des europaischen Sommerwaldes, «Вег. dtsch. bot. Ges.», 36, 337—351. (См. обзорную работу Schimper, 1935, 626—629.)
Diels L., Hackenberg G. (1926), Beitrage zur Vegetationskunde und Floristik von Slid—Borneo, «Bot. Jb.», 60, 293—316.
Dingier H. (1911), Versuche uber die Periodizitat einiger Holzgewachse in den Tropen, «S.-В. bayer. Akad. Wiss. math.-naturw. KI.», Munchen, 127—143.
Dixon W. A. (1882), On the inorganic constituents of some epiphytic ferns, «Proc. Roy. Soc. N. S. Wales», 15, 175 — 183.
Doctors van L e e u w e n W. M. (1933), Biology of plants and animals occurring in the higher parts of Mount Pangrango—Gedeh in West-Java, «Verh. Akad. Wet.», Amst. (sect. 2), 31, 1—270.
Doctors van L e e u w e n W. M. (1936), Krakatau, 1883 to 1933. A. Botahy, «Ann.
Jard. bot. Buitenz.», 46—47, 1—506.
Doyne H. C. (1935), Studies in tropical soils. Increase of acidity with depth, «J. Agric. Sci.», 25, 192—197.
D u g a n d A. (1934), The Transition Forests of Atlantico, Colombia, «Trop. Woods.», 40, 1-14.
Du Rietz G. E. (1931), Life-forms of terrestrial flowering plants, I, «Acta phytogeogr. suec.», 3 (1).
D u t h i e D. W., Hardy F., Rodriguez G. (1936), Soil investigations in the Arena Forest Reserve, Trinidad (Typescript. Imp. Coll. Trop. Agric. Trinidad, Summary by R. L. Brooks). «Imp. For. Inst. Paper», No. 6, 1937.
434
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Е g g е 1 i n g W. J. (1935), The vegetation of Namanve Swamp, Uganda, «J. Ecol.», 23, 422-435.
E g g e 1 i n g W. J. (1947), Observations on the ecology of Budongo rain forest, Uganda, «J. Ecol.», 34, 20—87.
E 1 w e s H. J., H e n г у A. (1906—1907), The trees of Great Britain and Ireland, 1 (1906); 2 (1907), Edinburgh.
Endert F. H. (1920), De woudboomflora van Palembang, «Tectona», 13, 113—160.
Endert F. H. (1925), Verslag van de Midden—Oost—Borneo—Expeditie, 1925, Batavia.
Endert F. H. (1928), Geschlachtstabellen voor Nederlandsch-Indische Boomsoorten naar vegetatiev kenmerken, «Meded. Proefst. Boschw.», Batavia, 20, 1—242.
Endert F. H. (1932), De proefbaanmetingen in de panglonggebieden van Bengkalis (Sumatra’s Oostkust) en Riouw, «Tectona», 25, 713—785.
Endert F. H. (1933), Eenige resultaten van de boschverkening in de Buitengewesten, «Tectona», 26, 391—421.
Ernst A. (1908), The new flora of the volcanic island of Krakatau, Transl. by A. C. Seward, Cambridge.
Ernst A. (1934), Das biologische Krakatau Problem, «Vjschr. naturf. Ges, Ziirich», 79 (прилож.), 1—187.
Ernst A.,Bernard С. и др. (1910—1914), Beitrage zur Kenntnis der Saprophyten Javas, «Ann. Jard. bot. Buitenz», 23, 20—61; 24, 55—97; 25, 161 —188; 26, 219-257; 28, 99-124.
Evans G. C. (1939), Ecological studies on the rain forest of Southern Nigeria. II. The atmospheric environmental conditions, «J. Ecol.», 27, 436—482.
Exell A. W. (1944), Catalogue of the vascular plants of S. Tome (with Principe and Annobon), London.
Faber F. C. (1915), Physiologische Fragmente aus einem tropischen Urwald, «Jb. wiss. Bot.», 56, 197—220.
Faber F. C. (1923), Zur Physiologic der Mangroven, «Вег. dtsch. bot. Ges.», 41, 227— 234.
Faber F. C. (1927), Die Kraterpflanzen Javas in physiologisch-dkologischer Beziehung, «Arbeit. Treub-Lab.», Buitenzorg, 1.
F eh er D. (1927), Untersuchungen fiber die Kohlenstoffernahrung des Waldes, «Flora», (N. F.), 21, 316-333.
Fitting H. (1910), Ueber die Beziehungen zwischen den Epiphyllen Flechten und den von ihnen bewohnten Blattern, «Ann, Jard. bot. Buitenz.» (приложение 3), 505-518.
F о g g i e A. (1947), Some ecological observations on a tropical forest type in the Gold Coast., «J. Ecol.», 34, 88—106.
Follett-Smith R. R. (1930), Report of an investigation of the soils and of the mineral content of pasture grasses occurring at Waranama Rahch, Berbice River, «Agric. J. Brit. Guiana», 3, 142—159.
Foxworthy F. W. (192J), Commercial timber trees of the Malay Peninsula, «Malay. For. Rec.», 3.
Francis W. D. (1924), The development of buttresses in Queensland trees, «Proc. Roy. Soc. Qd.», 36, 21—37.
Francis W. D. (1929), Australian rain-forest trees, Brisbane.
Fraser L., Vickery J.W, (1937), The ecology of the Upper Williams River and Barrington Tops Districts, I. Introduction. «Proc. Linn. Soc. N. S. W.», 62, 269— 283.
БИБЛИОГРАФИЯ
435
Fraser L. Vickery J, W. (1938), The ecology of the Upper Williams River and Barrington Tops Districts. II. The Rain-Forest Formations, «Proc. Linn. Soc. N.S. W.», 63, 139—184.
Fraser L. Vickery J, W. (1939), The ecology of the Upper Williams and Barrington Tops Districts. III. The Eucalypt Forests and general discussion, «Proc. Linn. Soc. N. S. W.», 64, 1—33.
Freise F. (1936), Das Binnenklima von Urwalder in subtropischen Brasilien, «Petermanns Mitt.», 82, 301—307.
Freise F. (1938), Beobachtungen in Zweitwuchsbestanden a us dem Kiistenwaldge-biet Brasiliens, «Z. Weltforstwirtsch.», 6, 281—299.
Freise F. (1939), Einige Bemerkungen uber soziologische Verhaltnisse im Urwald, «Z. d. Weltforstwirtsch.», 6, 603—621.
F r e y-W у s s 1 i n g A. (1931), Over de struikwildernis van Habinsaran, «Trop. Natuur», 20, 194—198.
Fries R. E., Fries T. С. E. (1948), Phytogeographical researches on Mt Kenya and Mt Aberdare, British East Africa, «К. svenska Vetensk-Akad. Handl.», ser. 3, 25, 1—83.
Fritsch F. E. (1907), A general consideration of the эиЬаёпа! and fresh-water algal flora of Ceylon, Part I, ЗиЬаёпа! algae and algae of the inland fresh-waters, «Proc. Roy. Soc.», B, 79, 197—254.
Fritsch F. E. (1936), The role of the terrestrial alga in nature, «Essays in geobotany in honor of William Albert Setchell», Berkeley (California), 195—217.
F и n к e G. L. (1929), On the biology and anatomy of some tropical leaf joints, I, «Ann. Jard. bot. Buitenz.», 40, 45—74.
Funke G. L. (1931), On the biology and anatomy of some tropical leaf joints, II. «Ann. Jard. bot. Buitenz.», 41, 33—64.
Gams H. (1918), Prinzipienfragen der Vegetationsforschung, «Vjschr., naturf. Ges. Zurich», 63, 293—493.
Geiger R. (1927), Das Klima der bodenahern Luftschicht, Braunschweig.
Gentner G. (1909), Ueber den Blauglanz auf Blattern und Friichten, «Flora», 99, 337—354.
Ghesquiere (1925), Note sur les racines tabulaires ou accotements ailes de quelques arbres congolais, «Rev. zool. afr.»; 13 (приложение 2), 1—2.
Gibbs L. S. (1917), A contribution to the phytogeography and flora of the Arfak Mountains, etc., London.
Giesenhagen K. (1910), Die Moostypen der Regenwalder, «Ann. Jard. bot. Buitenz.» (приложение 3, фото 2), 711—790.
Gleason H. A., Cook M. T. (1926), Plant ecology of Porto Rico, «Scientific Survey of Porto Rico and the Virgin Islands», 7, Part I, N. Y. Acad. Sci., New York.
Goebel K. (1888), Morphologische und biologische Studien, I, Ueber epiphytische Fame und Muscineen, «Ann. Jard. bot. Buitenz.», 7, 1—73.
Goebel K. (1889), Pflanzenbiologische Schild erungen, III, Epiphyten, Marburg.
Goebel K. (1913), Organographie der Pflanzen. Ite Teil. Allgemeine Organo-graphie, Jena.
Goebel K. (1922), Erdwurzeln mit Velamen, «Flora», (N. F.), 15, 1—26.
Goebel K. (1924), Die Entfaltungsbewegungen der Pflanzen, Jena.
Goedert P. (1938), Lessols de 1’Afrique Centrale, sp6cialment du Congo Beige, Introduction — Le regime pluviale au Congo Beige, «Publ. Inst. Nat. pour L’Et. agron. Congo Beige, Hors serie.».
Gooding E.G.B. (1947), Observations on the sand dunes of Barbados, British West Indies, «J. Ecol.», 34, 111 — 125.
436
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
G osswei 1 er J., Mendonca F. А. (1939), Carta fitogeografica de Angola, Lisbon.
Gray A. P. (1944), Ecology, Chapter IV in Imperata cylindrica, taxonomy, distribution, economic significance and control, «Imp. Agric. Bur. Joint Publ.», No. 7, Oxford and Aberystwyth.
Greenway P. J. (1938), Mafia Island, its natural history, etc. Unpubl. Memoir, E. Afr. Res. Sta. Amani.
Gresser E. (1919), Resumeerend rapport over het voorkomen van ijzerhout op de olieterreinen Djambi. I, «Tectona», 12, 283—304.
Guppy H. B. (1906), Observations of a naturalist in the Pacific between 1896 and 1899. 2. Plantdispersal, London.
Guttenberg H. (1931), Beitrage zur Kenntnis der Laubblattassimilation in den Tropen, «Ann. Jard. bot. Buitenz.», 41, 105—184.
Haberlandt G. (1898), Ueber die Grosse der Transpiration in feuchten Tropenklima, «Jb. wiss. Bot.», 31, 273—288.
Haberlandt G. (1899), Erwiderung, «Jb. wiss. Bot.», 33, 166—170.
Haberlandt G. (1926), Eine botanische Tropenreise, Leipzig.
H a d d о w A. J., G i 1 1 e t t J. D., H i g h t о n R. B. (1947), The mosquitoes of Bwamba County, Uganda. V. The vertical distribution and biting-cycle of mosquitoes in rain-firest, with further observations on microclimate, «Bull. Ent. Res.», 37, 301—330.
Hard on H. J. (1936), Podsol profiles in the tropics, «Natuurk. Tijdschr. Ned.-Ind.», 96, 25—41.
Hardon H. J. (1937), Padang soil, an example of podsol in the Tropical Lowlands, «Verb. Akad. Wet. Amst.», 40, 530—538.
Hardy F. (1935), The chemical and ecological researches on cacao, «Trop. Agriculture», Trinidad, 12, 175—178.
Hardy F. (1936a), Some aspects of cacao soil fertility in Trinidad, «Trop. Agriculture», Trinidad, 13, 315—317.
Hardy F. (1936b), Some aspects of tropical soils, «Trans. 3rd Int. Congr. Soil Sci.», Oxford, 1935, 2, 150-163.
Hardy F. (1939), Soil erosion in St Vincent, B.W.I., «Trop. Agriculture», Trinidad, 16, 58-65.
Hardy F. (1940), A provisional classification of the soils of Trinidad, «Trop. Agriculture», Trinidad, 17, 153—158.
Hardy F. (1945), The soils of South America, «Plants and plant science in Latin America», (ed. by F. Verdoorn), Waltham, Mass.
Hardy F., F о 11 e t t-S m i t h R. R. (1931), Studies in tropical soils. II. Some characteristic igneous rock soil profiles in British Guiana, South America, «J. Agric. Sci.», 21, 739—761.
Harmon M. H. (1942), A study of some growth factors affecting symmetrical growth in trees, «Butler Univ. Stud.», 5, 134—144. (Abstr. «Biol. Abstr.», 17, No. 684, 1943.)
Harris J. A. (1918), On the osmotic concentration of the tissue fluids of phanerogamic epiphytes, «Amer. J. Bot.», 5, 490—506.
Herbert D. A. (1929), The major factors in the present distribution of the genus Eucalyptus, «Proc. Roy. Soc. Qd.», 40, 165—193.
Herbert D. A. (1935), The climatic sifting of Australian vegetation, «Rep. Melb. Meeting, Aust, and N. Z. Ass. Adv. Sci.», 349—370.
Herzog T. (1923), Die Pflanzenwelt der bolivischen Anden und ihres ostlichen Vorlan-des, «Veget. d. Erde», 15.
БИБЛИОГРАФИЯ
437
Н esk е F. (1938—1939), Dertropische Wald als Rohstoffquelle, «Z.d. Weltforstwirtsch.», 6, 413-485.
Kingston R.W.G. (1932), A naturalist in the Guiana Forest, London.
H о 1 1 i с к A. (1924), A review of the fossil flora of the West Indies, «Bull. N.Y. Bot. Gdn.», 12, 259—323.
Holtermann C. (1907), Der Einfluss des Klimas auf den Bau der Pflanzengewebe. Anatomisch-physiologische Untersuchungen in den Tropen, Leipzig.
Holttum R. E. (1924), The vegetation of Gunong Belumut in Johore, «Gdns’ Bull. Singapore», 3, 245—257.
Holttum R. E. (1931), On periodic leaf-change and flowering of trees at Singapore, «Gdns’ Bull. Singapore», 5, 173—206.
Holttum R. E. (1935), The flowering of Tembusu trees (Fagraea fragrans Roxh. in Singapore 1928—1935. «Gdns’ Bull. Singapore», 9, 73—78.
Holttum R. E. (1938a), The ecology of tropical Pteridophytes, in F. Verdoorn, «Manual of Pteridology», 420—450, The Hague.
Holttum R. E. (1938b), Leaf-fail in a non-seasonal climate (Singapore), «Proc. Linn. Soc. Lond.», 150th session 1937—1938, 78—81.
Holttum R. E. (1940), On periodic leaf-change and flowering of trees at Singapore. II, «Gdns’ Bull. Singapore», II, 119—175.
Huber J. (1902), Contribugao a geographia physica dos Furos de Breves e da parte occidental de Marajo, «Bol. Mus. Paraense Hist. Nat.», 3, 447. (Резюме в «Bot. Zbl.», 93, 1903, 235—237).
Humboldt A. von (1817), De distributione geographica plantarum secundum coeli temperiem et altitudinem montium prolegomena, Paris.
Humboldt A. von (1852), Personal narrative of travels to the equinoctial regions of America, 2. Transl. T. Ross, London.
Ihering H. (1923), Das periodische Blattwechsel der Baume in tropischen und subtropischen Siidamerica, «Bot. Jb.», 58, 524—598.
J a n s e J. M. (1897), Les endophytes radicaux de quelques plantes javanaises, «Ann. Jard. bot. Buitenz.», 14, 53—201.
J en tsch (1911), Der Urwald Kameruns, Folgerungen aus den auf der Expedition 1908—1909 gewonnenen Erfahrungen in Bezug auf den Zustand und die Nutz-barmachung des Waldes, «Beih. Tropenpfl.», 12, 1—199.
J och em s S.C.J. (1928), De begroeiing der tabakslanden in Deli en hare beteeknis voor de tabakscultuur, «Med. Deli. Proefstat.» (2 de ser.), No. 59.
J ohow F. (1885), Die chlorophyllfreien Humusbewohner Wrest-Indiens, «Jb. wiss. Bot.», 16, 415—449.
J oh ow F. (1889), Die chlorophyllfreien Humuspflanzen nach ihren biologischen und anatomisch-entwicklunsgeschichtlichen Verhaltnissen, «Jb. wiss. Bot.», 20, 475— 525.
Jones A.P.D. (1945), Notes on terms for use in vegetation description in southern Nigeria, «Farm, and Forest (Nigeria)», 6, 130—136.
Jones A.P.D. (1947), Botanica nigerica, «Farm and Forest», 8, 10—16.
Jones A.P.D., К ea у R.W.J. (1946), Descriptive terms for the vegetation of the drier parts of Nigeria, «Farm and Forest», 7, 34—40.
Jones E. W. (1945), The structure and reproduction of the virgin forest of the North Temperate Zone, «New Phytol.», 44, 130—148.
Jungner J. R. (1891), Anpassungen der Pflanzen an das Klima in den Gegenden der regenreichon Кamerungebirge, «Bot. Zbl.», 47, 353—360.
438
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ лес
К а ш е г 1 i n g Z. (1912), De verdamping van epiphyte Orchideen, «Natuurk. Tijdschr. Ned.-Ind.», 71, 54—72.
Karsten G. (1891), Ueber die Mangrove-Vegetation im malayischen Archipel, «Bib-liogr. bot.», Heft 22.
Karsten G. (1895), Morphologische und biologische Untersuchungen fiber einige Epiphytenformen der Molukken, «Ann. Jard. bot. Buitenz.», 12, 117—195.
Karsten G. (1924—1926), Das Licht im tropischen Regenwalde, «Vegetationsbilder», Reihe 16, Hft. 3.
Karsten G. (1925), Ueber manlelformige Organe bei Epiphyten und Wurzeiklettern, ♦Flora», (N. F).. 18—19 («Goebel Festschr.»), 300—311.
Keay R.W.J. (1947), Forest vegetation in the savannah regions of Nigeria, «Fifth Empire Forestry Conference» (1947).
К eeble F. W. (1895), The hanging foliage of certain tropical trees, «Ann. Bot.», Oxford, 9, 59—94.
Kennedy J. D. (1936), Forest flora of Southern Nigeria, Lagos.
Kenoyer L. A. (1929), General and successional ecology of the Lower Tropical Rain-Forest at Barro Colorado island, Panama, «Ecology», 10, 201—222.
Klebs G. (1911), Ueber die Rhythmik in der Entwicklung der Pflanzen, «S.-В. hei-delberg. Akad. Wiss., math.— nat. KL», 23.
Klebs G. (1912), Ueber die periodischen Erscheinungen tropischer Pflanzen, «Biol. Zbl.», 32, 257—285.
Klebs G. (1915), Ueber Wachstum und Ruhe tropischer Baumarten, «Jb. wiss. Bot.», 56, 734—792.
Klebs G. (1926), Ueber periodisch wachsende tropische Baumarten, «S.-В. Heidelberg. Akad. Wiss., math.-nat. KL», 2.
Klein W. C. (1914), Kalkplanten in Nederlandsch-Indie, «Trop. Natuur.», 3, 133—135.
Koernicke M. (1910), Biologische Studien an Loranthaceen, «Ann. Jard. bot. Buitenz.», приложение 3, 665—697.
Koopman M.J.F., Verhoef L. (1938), Eusideroxylon zwageri T. and B., het ijzerhout van Borneoen Sumatra, «Tectona», 31, 381—399.
Koorders S. H., Valeton T. (1894), Bijdrage tot de kennis der boomsoorten van Java, No. 1, «Medd. s’Lands Plantentuin», 2, Buitenzorg.
Koppel C. van der (1926), Winning van copal in het Gouvernement Celebes en onderhoorigheden, de uitvoer uit Makassar, en eenige details over het gebruik van copal., «Tectona», 19, 525—574.
Koppel C. van der (1945), Forestry in the Outer Provinces of the Netherlands Indies, in F. Verdoorn, «Science and scientists in the Netherlands Indies», New York.
Koppen W. (1918), Klassifikation der Klimate nach Temperatur, Niederschlag und Jahreslauf, «Petermanns Mitt.», 64, 193.
Koppen W. (1936), Das geographische System der Klimate. В книге W. Koppen, R. Geiger, «Handbuch der Klimatologie», 1, Teil C., Berlin.
Koppen W., Geiger R. (1928), Klimakarte der Erde, Gotha.
Kramer F. (1933), De natuurlijke verjonging in het Goenoeng-Gedehcomplex, «Tectona», 26, 156—185.
Kraus G. (1895), Physiologisches aus den Tropen, I, Das Langenwachstum der Bam-busrohre, «Ann. Jard. bot. Buitenz.», 12, 196—210.
Kurz S. (1875), Preliminary report on the forest and other vegetation of Pegu, Calcutta.
L a a n E. van der (1925), De bosschen van de Zuider- en Ossterafdeeling van Borneo, «Tectona», 18, 925—952.
L a a n E. van der (1926 a), Analyse der bosschen in de onderafdeeling Pleihari van de afdeeling Bandjermasin der Zuider- en Oosterafdeeling van Borneo, «Tectona». 19. 103—123.
БИБЛИОГРАФИЯ
439
L а а n Е. van der (1926 b), De bosschen van de onderafdeeling Tanah Boemboe van be afdeeling Zuidoostkust der residentie Zuider- en Ossterafdeeling van Borneo, «Tectona», 19, 215—238.
L a n n E. van der (1927), De analyse des bosschen van de eilanden PoeloeLaoet en Seboe-koe der Residentie Zuider- en Oosterafdeeling van Borneo, «Tectona», 20, 19—36.
Lam H. J. (1945), Fragmenta Papuana (Observations of a naturalist in Netherlands New Guinea), Transl. L. M. Perry, «Sargentia», 5.
Lan e-P о о 1 e С. E. (1925 a), The forests of Papua and New Guinea, «Emp. For. J.», 4, 206—234.
Lan e-P о о 1 e С. E. (1925 b), The forest resources of the Territories of Papua and New Guinea, Parliament of the Commonwealth of Australia, Canberra.
Lan e-P о о 1 e С. E. (1938), Kamerere, «New Guinea Agric. Gaz.», 4, 15—17.
Lanjouw J. (1936), Studies of the vegetation of the Surinam savannahs and swamps, «Ned. kruidk. Arch.», 46, 823—851.
Laus berg T. (1935), Quantitative Untersuchungen fiber die kutikulare Exkretion des Laubblattes, «Jb. wiss. Bot.», 81, 769—806.
Lavauden L. (1937), The equatorial forest of Africa, its past, present and future, «J. Roy. Afric. Soc.», 36 (extra Suppl.).
Lebrun J. (1935), Les essences forestieres des regions montagneuses du Congo oriental, «Publ. Inst. Agron. Congo Beige», Ser. sci. I.
Lebrun J. (1936a), Repartition de la foret equatoriale et des formations vegetales limitrophes, Brussels.
Lebrun J. (1936b), La foret equatoriale congolaise, «Bull, agric. Congo beige», 27, 163—192.
Lebrun J. (1947), La vegetation de la plaine alluviale au sud du Lac fidouard, «Exploration du Parc National Albert, Mission J. Lebrun (1937—1938)», Fasc. I, Brussels.
L i n d m a n n C.A.M. (1900), Vegetationen i Rio Grande do Sul (Sydbrasilien), Stockholm.
Livingston В. E. (1916), Physiological temperature indices for the study of plant growth in relation to climatic conditions, «Physiol. Res.», I, 399—420.
Livingston В. E., Livingston G. J. (1913), Temperature coefficients in plant geography and climatology, «Bot. Gaz.», 56, 349—375.
Longman H. A., W h i t e С. T. (1917), The flora of a single tree, «Proc. Roy. Soc. Qd.», 29, 64—69.
Lundegardh H. (1931), Environment and plant development, Trans. E. Ashby. L ii t z e 1 b u r g P. von (1925—1926), Estudo botanico do Nordeste, «Minist. Viagao e obr. publ.», Publ., 57, Ser. 1 A, 1—3. (Abstr. in «Bot. Jb.», 61, 1927, Literatur-ber, 21-31.)
MacGregor W. D. (1934), Silviculture of the Mixed Deciduous Forests of Nigeria, «Oxf. For. Mem.», 18.
MacGregor W. D. (1937), Forest type and succession in Nigeria, «Emp. For. J.», 16, 234—242.
Mackay J. H. (1936), Problems of ecology in Nigeria, «Emp. For. J.», 15, 190—200.
McLean R. C. (1919), Studies in the ecology of Tropical Rain-forest: with special reference to the forests of South Brazil, «J. Ecol.», 7, 15—54; 121—172.
Maitland T. D. (1932), The grassland vegetation of the Cameroons Mountain, «Kew Bull.» (1932), 417-425.
Marr С. C. (1938), Agricultural survey of the Markham valley, «New Guinea Agric Gaz.», 4, 2—12.
Marshall R. C. (1934), The physiography and regetation of Trinidad and Tobago, «Oxf. For. Mem.», 17.
440
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Martin F. J., D о у п е Н. С. (1927), Laterite and lateritic soils in Sierra Leone» «J. Agric. Sci.», 17, 530—547.
Martyn E. B. (1931), A botanical survey of the Rupununi Development Company’s ranch at Waranama, Berbice River, «Agric. J. Brit. Guiana.», 4, 18—25.
Massart J. (1898), Les vegetaux epiphylles, «Ann. Jard. bot. Buitenz.», приложение 1, 103-108.
Mead J. P. (1928), The forests of the Fiji islands «Emp. For. J.», 7, 47—54.
Meijer Drees E. (1938), Plantensociologie, boschbouw en hdutteelt, «Tectona», 31 166-205.
Merrill E. D. (1915), A contribution to the bibliography of the botany of Borneo,. «Sarawak Mus. J.», 2, 99—136.
Meusel H. (1935), Wuchsformen und Wuchstypen der europaischen Laubmoose, «Nova Acta Leopoldina», (N. F.), 3 (12), 123—277.
M e z C. (1904), Physiologische Bromeliaceen-Studien, 1, Die Wasser-Oekonomie der extreme atmospharischen Tillandsien, «Jb. wiss. Bot.», 40, 157—229.
M ich elm or e A.P.G. (1939), Observations on tropical African grasslands, «J. Ecol.»» 27, 282—312.
M i eh e H. (1911), Javanische Studien — Zur Frage der mikrobiologische Vorgange-im Humus einiger humussammelnder Epiphyten, «Abh. sachs. Ges. (Akad.) Wiss., math.-phys. KI.», 32, 376—398.
Mildbraed J. (1914), Die Vegetationsverhaltnisse im Sammelgebiet der Expedition, «Wiss.' Ergeb. d. deutsch. Zentral-Afrika-Expedition 1907—1908», 2» 603—691, Leipzig.
Mildbraed J. (1922), Wissenschaftliche Ergebnisse der zweiten deutschen Zentral-Afrika-Expedition 1910—1911 unter Fuhrung Adolf Friedrichs, Leipzig* Mildbraed J. (1923), Das Regenwaldgebiet im aquatorialen Afrika, «Notizbl.
bot. Gart», 8, Berlin, 574—599.
Mildbraed J. (1930a), Zusammensetzung der Bestande und Verjiingung im tropi-schen Regenwald, «Вег. dtsch. bot. Ges.», 48 (Generalversammlungs-Heft), 50—57.
Mildbraed J. (1930b), Sample plot surveys in the Cameroons rain-forest. (TransL by H. M. Heyder.), «Emp. For. J.», 9, 242—266.
Mildbraed J. (1933a), Zur Kenntnis der Vegetationsverhaltnisse Nord-Kameruns, «Bot. Jb.», 65, 1—52.
Mildbraed J. (1933b), Ein Hektar Regenwald auf Fernando Po, «Notizbl. bot. Gart.», 11, Berlin, 946—950.
Milne G. (1937), Essays in applied pedology. I. Soil type and soil management in-relation to plantation agriculture in East Usambara, «East Afr. Agric. J.» (July 1937).
Milne G. (1940), Report on a journey to parts of the West Indies and the United States for the study of soils. Afric Res. Station, Amani.
Mohr E.C.J. (1930), Tropical soil-forming processes and the development of tropical soils, with special reference to Java and Sumatra. TransL by R. L. Pendleton» «College of’Agric., Univ. Philippines, Exp. Sta. Contrib.», No. 655 (mimeographed).
Mohr E.C.J. (1944), The soils of equatorial regions with special reference to the Netherlands East Indies. TransL by R. L. Pendleton, Ann. Arbor, Michigan.
Molisch H. (1896), Das Erfrieren von Pflanzen fiber dem Eispunkt, «S. - B. Akad. Wiss. Wien», 105, 82—95.
Moreau R. E. (1933), Pleistocene climatic changes and the distribution of life ir> East Africa, «J. Ecol.», 21, 415—435.
Moreau R. E. (1935a), Some eco-climatic data for closed Everggreen Forest in Tropical Africa, «J. Linn. Soc. (Zool.)», 39, 285—293.
БИБЛИОГРАФИЯ
44f
Moreau R. E. (1935b), A synecological study of Usambara, Tanganyika Territory» with particular reference to birds, «J. Ecol.», 23, 1—43.
Moreau R. E. (1938), Climatic classification from the standpoint of East African biology, «J. Ecol.», 26, 467—496.
Morison C.G.T., Hoyle А. С., H о p e-S i m p s о n J. F. (1948), Tropical soilvegetation catenas and mosaics. A study of the south-western part of the Anglo-Egyptian Sudan, «J. Ecol.», 36, 1—84.
Myers J. G. (1930), Notes on wild cacao in Surinam and in British Guiana, «Kew. Bull.» (1930), 1-10.
Myers J. G. (1933), Notes on the vegetation of the Venezuelan Llanos, «J. Ecol.»,. 21, 335—349.
Myers J. G. (1935), Zonation of vegetation along river courses, «J. Ecol.», 23, 356—360.
Myers J. G. (1936), Savannah and forest vegetation of the interior Guiana Plateau, «J. Ecol.», 24, 162-184.
N a v e z A. (1924), La foret equatoriale bresilienne, I, «Bull. Soc. Bpt. Belg.», 57r 7—17.
N a v e z A. (1930), On the distribution of tabular roots in Ceiba (Bombacaceae), «Proc. Nat. Acad. Sci. Wash.», 16, 339—344.
Nicholson J. W. (1936), The influence of forests on climate and water supply in Kenya, «Kenya For. Dep. Pamphlet», No. 2.
Oliver W.R.B. (1930), New Zealand epiphytes, «J. Ecol.», 8, 1—50.
Olsen C. (1917), Studies on the succession and ecology of epiphytic bryophytes on the bark of common trees in Denmark, «Bot. Tidsskr.», 34, 313—342.
Orth R. (1939), Zur Kenntnis des Lichtklimas der Tropen und Subtropen sowie des tropischen Urwaldes, «Beitr. Geophys.», 55, 52.
Osborn T.G.B., Robertson R. N. (1939), A reconnaissance survey of the vegetation of the Myall Lakes, «Proc. Linn. Soc. N. S. W.», 64, 279—296.
О у e P. van (1921), Influence des facteurs climatiques sur la repartition des epiphytes a la surface des troncs d'arbres a Java, «Rev. gen. Bot.», 33, 161—176.
P a 11 i s M. (1916), The structure and history of plain the floating fen of the deka of the Danube, «J. Linn. Soc. (Bot.)», 43, 233—290.
Paulian R. (1946), Preliminary survey of the West African rain-forest canopy, «Nature», London, 157, 877.
Paulian R. (1947), Observations ecologiques en foret de Basse Cote d’Ivoire, Paris.
Pearson H.H.W. (1899), The botany of the Ceylon patanas, «J. Linn. Soc. (Bot.)», 34, 300—365.
Pendleton R. L., Sh arasu v an a S. (1946), Analyses of some Siamese laterites, «Soil Sci.», 62, 423—448.
P e n z i g O. (1902), Die Fortschritte der Flora des Krakatau, «Ann. Jard. bot. Buitenz»., 18, 92-113.
P e s s i n L. J. (1925), An ecological study of the polypody fern Poly podium polypodioi-des as an epiphyte in Mississippi, «Ecology», 6, 17—38.
P e t c h T. (1924), Gregarious flowering, «Anp. R. Bot. Gdns», Peradeniya, 9, 101 —117 P e t c h T. (1930), Buttress roots, «Ann. R. Bot. Gdns», Peradeniya, 11, 277—285.
Phillips J. (1931); Mortality among plants and its bearing on natural selection, «Nature», London, 127, 851—852.
P i c a d о C. (1912), Les maires aeriennes de la foret vierge am er i caine: les Bromeli-acees, «Biologica», 11, 110—115.
P i c a d о C. (1913), Les Bromeliacees epiphytes, considerees comme milieu biologi-que, «Bull. sci. Fr. Belg.», Ser. 7, 47, 215—360.
442
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
р i j 1 L. van der (1934), Die Mycorrhiza von Burmannia und E pirrhizanthes und die Fortpflanzung ihres Endophyten, «Rec. Trav. bot. neerland.», 31, 761—779.
pi t t end r i gh C. S. (1948), The Bromeliad-Anopheles-Malaria Complex in Trinidad. I. The Bromeliad flora, «Evolution», 2, 58—89.
p i t t-S c h e n к e 1 C.J.W. (1938), Some important communities of Warm Temperate Rain Forest at Magamba, West Usambara, Tanganyika Territory, «J. Ecol.», 26, 50—81.
Polak B. (1933), Een tocht in het zandsteen gebied bij Mandor (West Borneo), «Trop. Natuur.», 22, 23—28.
Polak E. (1933), Ueber Torf und Moor in Niederlandisch Indien, «Verh. Akad. Wet.», Amsterdam, 30, 1—85.
P о s t h u m u s 0. (1931), Plantae in «Festbundel K. Martin, Leidsche Geol. Meded.», 5, 485-508.
Potonie H., Koorders S. H. (1909), Sumpfflachmoornatur der Moore des Pro-duktiven Karbons, «Jb. preuss. geol. Landesanst.», 30, 389—443.
Potter M. C. (1891), Observations on the protection of buds in the tropics, «J. Linn. Soc. (Bot.)», 28, 343—352.
P у n a er t L. (1933), La mangrove congolaise, «Bull. Agric. Congo Beige», 23, 184— 207.
Raunkiaer C. (1934), The life-forms of plants and statistical plant geography, Oxford.
Rechinger K. (1908), Vegetationsbilder aus dem Neu-Guinea-Archipel, «Vegeta-tionsbilder», Reihe 6, Hft. 2.
Rechinger K. (1910), Botanische und zoologische Ergebnisse einer Forschungs-reise nach den Samoainseln, dem Neuguinea-archipel und den Salomoninseln, III. Siphonogamen der Samoa-inseln, «Denkschr. Akad. Wiss., Wien», 85, 202—388.
Reid M. E., Chandler M.E.J. (1933), The London Clay flora, London.
R e s v о 11 T. R. (1925), Beschuppte Laubknospen in den immerfeuchten Tropenwal-dern, «Flora», (N. F.), 18—19 («Goebel Festschr.»), 409—420.
Richards P. W. (1932), Ecology, In F. Verdoorn, «Manual of bryology», 367—395, The Hague.
Richards P. W. (1936), Ecological observations on the rain forest of Mount Dulit, Sarawak, Parts I, II, «J. Ecol.», 24, 1—37; 340—360.
Richards P. W. (1939), Ecological studies on the rain forest of Southern Nigeria. 1. The structure and floristic composition of the primary forest, «J. Ecol.», 27, 1-61.
Richards P. W. (1941), Lowland Tropical Podsols and their vegetation, «Nature», London, 148, 129—131.
Richards P. W. (1943), The biogeographic division of the Indo-Australian Archipelago. 6. The ecological segregation of the Indo-Malayan and Australian elements in the vegetation of Borneo, «Proc. Linn. Soc. Lond.», (Sess. 154, 1941 —1942), 154-156.
Richards P. W. (1945), The floristic composition of primary tropical rain forest, «Biol. Rev.», 20, 1—13.
Richards P. W., T a n s 1 e у A. G., Watt A. S. (1940), The recording of structure, life form and flora of tropical forest communities as a basis for their classification, «J. Ecol.», 28, 224—239 (Also published as «Imp. For. Inst. Paper», No. 19, 1939).
Rikli M. (1943), Das Pflanzenkleid der Mittelmeerlander, 1, Bern.
Roberts R. C. (1936), Soil survey of Porto Rico, «U. S. Dep. Agric.» No. 8.
Roberty G. (1946), Les associations vdgetales de la valke moyenne du Niger, «Ve-roff. geobot. Inst. Rubel.», 22.
БИБЛИОГРАФИЯ
443
Hobinson G. W. (1949), Soils, their origin, constitution and classification (3 rd ed.), London.
ft о b у n s W. (1936), Contribution a 1’etude des formations herbeuses du District Forestier Central du Congo Beige, «Mem. Inst. Roy. Colon. Beige.», 5.
Robyns W. (1946), Statistiques de nos connaissances sur les spermatophytes du Congo Beige et du Ruanda-Urundi, «Bull. Jard. bot. Brux.», 18, 133—144.
R о d w a у J. (1894), In the Guiana forest, London.
Roosendael J. van, Thorenaar A. (1924), De natuurlijke verjonging van Ngerawan (Hopea mengarawan Miq.) in Zuid Sumatra, «Tectona», 17, 519—567.
Ross R. (1954). Ecological studies on the rain forest of Southern Nigeria, III. Secondary succession in the Shasha Forest Reserve, «J. Ecol.», 42, 259—282.
Salisbury E. J. (1925), The structure of woodlands. Festschr. C. Schroeter, «Ve-roff. geobot. Inst. Rubel.», Hft. 3, 334—354.
Salisbury E. J. (1929), The biological equipment of species in relation to competition, «J. Ecol.», 17, 197—222.
Salisbury E. J. (1930), Mortality amongst plants and its bearing on natural selection, «Nature», London, 125, 817.
Salisbury E. J. (1942), The reproductive capacity of plants, London.
Saxton W. T. (1924), Phases of vegetation under monsoon conditions, «J. Ecol.», 12, 1-38.
Scheffler G. (1901), Ueber die Beschaffenheit des Usambara-Urwaldes und uber den Laubwechsel an Baumen desselben, «Notizbl. bot. Gart.» 3, Berlin, 139—166.
Schenck H. (1892—1893), Beitrage zur Biologic und Anatomic der Lianen, I, Beit-trage zur Biologic der Lianen, «Bot. Mitt. Trop.», Hft. 4 (1892); II, Beitrage zur Anatomic der Lianen, «Bot. Mitt. Trop.», Hft. 5 (1893).
Schenck H. (1904), Strand vegetation Brasiliens, «Vegetationsbilder», Reihe 1, Hft. 7.
Schimper A.F.W. (1888), Die epiphytische Vegetation Amerikas, «Bot. Mitt. Trop», 2.
Schimper A.F.W. (1891), Die indo-malayische Strandflora, «Bot. Mitt. Trop.», Hft. 3.
Schimper A.F.W. (1898), Pflanzengeographie auf physiologischer Grundlage, Jena.
Schimper A.F.W. (1903), Plant-geography upon a physiological basis. Transl. by W.R. Fisher. Edited by P. Groom, I. B. Balfour, Oxford.
Schimper A.F.W. (1935), Pflanzengeographie auf physiologischer Grundlage, Jena.
Schweizer J. (1932), Ueber die Periodizitat des Blattwechsels bei tropischen Baumen, «Mitt, naturf. Ges. Bern», 1932, 41—46.
S e i f r i z W. (1923), The altitudinal distribution of plants on Mt. Gedeh, Java, «Bull. Torrey Bot. CL», 50, 283—305.
S e i f r i z W. (1924), The altitudinal distribution of lichens and mosses on Mt.Gedeh, Java, «J. Ecol.», 12, 307—313.
Senn G. (1913), Der osmotische Druck einiger Epiphyten und Parasiten, «Verh. naturf. Ges. Basel», 42, 179—183.
Senn G. (1923), Ueber die Ursachen der Brettwurzelbildung bei der Pyramiden-Pap-pel, «Verh. naturf. Ges. Basel», 35, («Festbd. H. Christ»), 405—435.
Senstius M. W. (1930—1931), Agro-geological studies in the tropics, «Soil Res.», 2, 10-56.
Sewandano M. (1937), Inventarisatie en inrichting van de veenmoerasbosschen in het panglonggebied van Sumatra’s Oostkust, «Tectona», 30, 660—675.
S h a n t z H. L., M a r b u t C. F. (1923), The vegetation and soils of Africa, «Amer. Geogr. Soc. Res. Series.», 13.
Shirley H. L. (1935), Light as an ecological factor and its measurement, «Bot. Rev.», 1, 355—381.
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Shreve F. (1914а), A montane rain-forest. A contribution to the physilogical plant geography of Jamaica, «Publ. Carneg. Instn.», No. 199.
Shreve F. (1914b), The direct effects of rainfall on hygrophilous vegetation, «J. Ecol.»,. 2, 82—98.
Simon S. V. (1914), Studien uber die Periodizitat der Lebensprozesse der in dauernd feuchten Tropengebieten heimischen Baume, «Jb. wiss. Bot.», 54, 71 — 187.
S m i t h A. C., J о h n s t о n I. M. (1945), A phytogeographical sketch of Latin America, in F. Verdoorn, «Plants and plant science in Latin America», Waltham, Mass.
Smith A. M. (1909), On the internal temperature of leaves in tropical insolation, with special reference to the effect of their colour on the temperature; also observations on the periodicity of the appearance of young coloured leaves of trees growing in Peradeniya Gardens, «Ann. R. Bot. Gdns. Peradeniya», 4, 229—298.
Smith J. J. (1923), Periodischer Laubfall bei Breynia cernua Muell. Arg., «Ann. Jard. bot. Buitenz.», 32, 97—102.J
Spanner L. (1939), Untersuchungen fiber den Warme- und Wasserhaushalt von Myr-mecodia und Hydnophytum, «Jb. wiss. Bot.», 88, 243—283.
Spruce R. (1908), Notes of a botanist on the Amazon and Andes (2 vols.), Ed. by A. R. Wallace, London.
Stahl E. (1893), Regenfall und Blattgestalt, «Ann. Jard. bot. Buitenz.», 11, 98—182.
Stahl E. (1896), Ueber bunte Laubblatter, Ein Beitrag zur Pflanzenbiologie, «Ann. Jard. bot. Buitenz.», 13, 137—216.
Stamp L. D. (1925), The vegetation of Burma, Calcutta.
Stamp L. D., Lord L. (1923), The ecology of part of the riverine tract of Burma,. «J. Ecol.», 11, 129-159.
Stebbing E. P. (1937a), The forests of West Africa and the Sahara, London and Edinburgh.
StebbingE. P. (1937b), The threat of the Sahara, «J. R. Afr. Soc.», 36 (extra SuppL).
S t e e m a n n-N i e 1 s e n E. (1940), Ueber die Bedeutung der sogenannten xeromor-phen Struktur im Blattbau der Pflanzen auf nahrstoffarmen Boden, «Dansk bot.. Ark.», 10, No. 2.
Steenis C.G.G.J. van (1932), Вotanical results of a trip to the Anambas and Natoe-na Islands, «Bull. Jard. bot. Buitenz.» (set. 3), 12, 151—211.
Steenis C.G.G.J. van (1933), Report of a botanical trip to the Ranau region, south Sumatra, «Bull. Jard. bot. Buitenz.», (Ser. 3), 13, 1—56.
Steenis C.G.G.J. van (1934), On the origin of the Malaysian mountain flora, Part I,. «Bull. Jard. bot. Buitenz.» (Ser. 3), 13, 135—262.
Steenis C.G.G.J. van (1935a) MaleischeVegetatieschetsen, «Tijdschr, ned. aardri-jksk. Genoot», Reeks 2, 52, 25—67,171—203, 363—398. (Карта напечатана также в «Atlas van tropisch Nederland, 1938», Weltevreden, Java.)
Steenis C.G.G.J. van (1935b), On the origin of the Malaysian mountain flora, Part 2, Altitudinal zones, general considerations and renewed statement of the problem, «Bull. Jard. bot. Buitenz.» (Ser. 3), 13, 289—417.
Steenis C.G.G.J. van (1936), On the origin of the Malaysian mountain flora, Part 3, Analysis of floristical relationships (1st instalment), «Bull. Jard. bot. Buitenz.» (Ser. 3), 14, 56-72.
Steenis C.G.G.J. van (1937a), De invloed van den mensch op het bosch, «Tectona»,. 30, 634—652.
Steenis C.G.G.J. van (1937b), Over de flora van den Carstensztop, in A.H. Colijnr «Naar de eeuwige sneeuw van tropische Nederland», Amsterdam.
Steenis C.G.G.J. van (1938), Recent progress and prospects in the study of the Malaysian flora, «Chron. Bot.», 4, 392—397.
библиография
445
8 teen is C.G.G.J. van (1942), Gregarious flowering of S trobilanthes (Acanthaceae) in Malaysia, «Ann. R. Bot. Gdn.», Calcutta (150 th Anniv. vol.), 91—97.
S t eenis C.G.G.J. van, Ruttner F. (1932), Die Pteridophyten und Phanerogamen der deutschen limnologischen Sunda-Expedition, «Arch. Hydrobiol.», XI, Tro-pische Binnegewasser, 3, 231—387.
Stehl e H. (1935), Essai d’ecologie et geographic botanique, «Flore de la Guadeloupe et dependances», I, Basse Terre (Guadeloupe).
Stehle H. (1945), Forest types of the Caribbean Islands, Part I, «Carib. For.», 6 (suppl.), 273—408.
S t eu p F.K.M. (1930), Bijdragen tot de kennis der bosschen vanJNorrd- en Midden-Celebes, 1, «Tectona», 23, 857—873.
S t e u p F.K.M. (1932), Bijdragen tot de kennis der bosschen van Noord- en Midden-Celebes, III, Het zoogenaande tjempaka-hoetan complex in de Minhasa, «Tectona», 25, 119—147.
Stocker O. (1935a), Assimilation und Atmung westjavanischer Tropenbaume, «Planta», 24, 402—445.
Stocker O. (1935b), Transpiration und Wasserhaushalt in verschiedenen Klimazonen, III, Ein Beitrag zur Transpirationsgrosse im javanischen Urwald, «Jb. wiss. Bot.», 81, 464-496.
St rael en V. van (1933), Resultats scientifiques du voyage aux Indes Neerlandaises de LL.AA.RR. la Prince et le Princesse Leopold de Belgique, 1, Introduction, «Mem. Mus. Hist. nat. Belgique», hors serie, Brussels.
Strugnell E. J.,Mead J.P. (1937), An ascent of Gunong Tahan, «Malay. Forester», 6, 131—140.
Swynnerton C.F.M. (1917), Some factors in the replacement of the ancient East African forest by wooded pasture land, «S. Afr. J. Sci.», 14, 493—518.
Sykes R. A. (1930), Some notes on the Benin forests of Southern Nigeria, «Emp. For. J.», 9, 101 — 106.
Symington C. F. (1933), The study of secondary growth on rain forest sites, «Malay. Forester», 2, 107—117.
Symington C. F. (1936), The flora of Gunong Tapis in Pahang, «J. Malay. Br. Asiat. Soc.», 14, 333—364.
Symington C. F. (1943), Foresters’ manual of Dipterocarps, «Malay. For. Rec.», 16.
T a n s 1 e у A. G. (1935), The use and abuse of vegetational concepts and terms, «Ecology», 16, 284—307.
Tansley A.G.,Fritsch F. E. (1905), Sketches of vegetation at home and abroad, I. The flora of the Ceylon littoral, «New Phytol.», 4, 1—17, 27—55.
Thomas A. S. (1932), The dry season in the Gold Coast and its relation to the cultivation of cacao, «J. Ecol.», 20, 263—269.
Thompson J. McL. (1943), A modern study of cauliflory, «Nature», London, 151, 481—482.
Thompson J. McL. (1946), The study of plant behaviourism; a common meeting ground for those engaged in botanical enquiry, «Proc. Linn. Soc. Lond. Session», 157 (1944—1945), 72—91.
Thorenaar A. (1927), Eigenaardige wortelvormigen in de moerasbosschen van Palembang, «Trop. Natuur.», 16, 73—82.
Thorp J., Baldwin M. (1940), Laterite in relation to soils of the tropics, «Ann. Ass. Amer. Geogr.», 30, 163—194.
T i e m a n n H. D. (1935), What are the largest trees in the world? «J. For.», 33, 903 — 915.
446
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
Т г е u b М. (1883), Observations sur les plantes grimpantes du Jardin Botanique de Buitenzorg, «Ann. Jard. bot. Buitenz.», 3, 160—183.
T г e u b M. (1888), Notice sur la nouvelle flore de Krakatau, «Ann. Jard. bot. Buitenz.», 7, 213—223.
Trochain J. (1940), Contribution a 1’etude de la vegetation du Senegal, «Mem. Inst. d’Afrique Noire», 2, Paris.
Troll C. (1948), Der asymmetrische Aufbau der Vegetationszonen und Vegetations-stufen auf der Nord- und Siidhalbkugel. «Вег. geobot. Inst. Rubel.», 1947 (1948), 46-83.
Troll W. (1930), Ueber die sogenannten Atemwurzeln der Mangroven, «Вег. dtsch. bot. Ges.», 48 (Generalversammlungs-Heft), 81—99.
Troll W. (1938), Vergleichende Morphologic der hoheren Pflanzen, 1, Berlin.
Troll W., Dragendorf 0. (1931), Ueber die Luftwurzeln von Sonneratia Linn, f. und ihre biologische Bedeutung, «Planta», 13, 311—473.
T u г г i 11 W. B. (1935), Krakatau and its problems, «New Phytol.», 34, 442—444.
Tux en R. (1929), Ueber einige nordwestdeutsche Waldassoziationen von regionaler Verbreitung, «Jb. geogr. Ges. Hannover», 1929, 55—116.
U 1 b r i c h E. (1928), Biologic der Friichte und Samen (Karpobiologie), Berlin.
U 1 e E. (1901), Die Vegetation von Cabo Frio an der Kiiste von Brasilien, «Bot. Jb.»r 28, 511-528.
U 1 e E. (1908a), Die Pflanzenformationen des Amazonas-Gebietes, «Bot. Jb.», 40, 114— 172; 398-443.
U 1 e E. (1908b), Catinga und Felsenformationen in Bahia, «Bot. Jb.», 40, Beibl. 93, 39-48.
U 1 e E. (1908c), Das Innere von Nordost-Brasilien, «Vegetationsbilder», Beihe 6, Hft. 3,
V a g e 1 e г P. (1933), An introduction to tropical soils, TransL by E. Greene, London.
Vaughan R. E., W i eh e P. O. (1937), Studies on the vegetation of Mauritius. I. A. preliminary survey of the plant communities, «J. Ecol.», 25, 289—342.
Vaughan R. E.,Wiehe P. 0. (1941), Studies on the vegetation of Mauritius. HI. The structure and development of the Upland Climax Forest, «J. Ecol.», 29, 127— 160.
Vaughan R. E., W i e h e P. 0. (1947), Studies on the vegetation of Mauritius IV. Some notes on the internal climate of the Upland Climax Forest, «J. Ecol.», 34, 126—136.
Vaupel F. (1910), Die Vegetation der Samoa-Inseln, «Bot. Jb.», 44, Beibl. 102, 47—58.
Vermoesen C. (1931), Les essences forestieres du Congo Beige. I. Manuel des essences de la region equatoriale et du Mayombe, Brussels.
V e s q u e J. (1882), L’espece vegetale consideree au point de vue de 1’anatomic compa-ree, «Ann. Sci. nat. (Bot.)», Ser. 6, 13, 5—46.
Volkens G. (1912), Laubfall und Lauberneuerung in den Tropen, Berlin.
V о t h P. D. (1939), Conduction of rainfall by plant stems in a tropical rain forest, «Bot. Gaz.», 101, 328—340.
Wallace A. R. (1878), Tropical nature and other essays, London.
Walter H. (1936), Nahrstoffgehalt des Bodes und natiirliche Waldbestande, «Forstl.
Wschr. Silva», 24, 201-205, 209-213.
Walter H., Steiner M. (1936—1937), Die Oekologie der Ost-Afrikanischen Mangroven, «Z. Bot.», 30, 65—193.
Wardlaw C. W. (1931), Observations on the dominance of pteridophytes on some St Lucia soils, «J. Ecol.», 19, 60—63.
БИБЛИОГРАФИЯ
447
Warming Е. (1892), Lagoa Santa, «К. danske vidensk. Selsk. Skr.», 6.
Warming E. (1909), Oecology of Plants, Transl. P. Groom., I. B. Balfour, Oxford.
Warming E., Gra ebn er P. (1933), Lehrbuch der okologischen Pfl an zen geographic, Berlin.
Watson J. G. (1928), The mangrove swamps of the Malay Peninsula, «Malay. For. Rec.», 6.
Watson J. G. (1937), Age-class representation in virgin forest, «Malay. Forester» 6, 146—147.
Watt A. S., F r a s e r G. K. (1933), Tree roots and the field layer, «J. Ecol.», 21, 404—414.
Weberbauer A. (1911), Die Pflanzenwelt der peruanischen Anden., «Veget. d. Erde», 12.
Weberbauer A. (1945), El mundo vegetal de los Andes Peruanos, Lima.
Went F. W. (1940), Soziologie der Epiphyten eines tropischen Urwaldes, «Ann. Jard. bot. Buitenz.», 50, 1—98.
Whitford H. N. (1906), The vegetation of the Lamao Forest Reserve, «Philipp. J. Sci.», 1, 373—431, 637—682.
Wiesner J. (1895), Untersuchungen fiber den Lichtgenuss der Pflanzen mit Riick-sicht auf die Vegetation von Wien, Kairo und Buitenzorg (Java), «S.-В. Akad. Wiss. Wien», Math.-naturw. Cl.», 104, 605—711.
Wiesner J. (1897), Pflanzenphysiologische Mittheilungen aus Buitenzorg., VII, Zur Physiologic von Taeniophyllum zollingeri, «S.-В. Akad. Wiss., Wien, Math.-naturw. CL», 106, Abt. 1, 77—98.
Wiesner J. (1907), Der Lichtgenuss der Pflanzen, Leipzig.
Wildeman E. de (1913), Documents pour Petude de la geobotanique congolaise, «Bull. Soc. Bot. Belg.», 51 (vol. jubilaire), fasc. 3.
Wildeman E. de (1926), Les forets congolaises et leurs principales essences econo-miques, Brussels.
Wildeman E. de (1930), Empattements, contreforts, racines-ech asses, «Bull. Acad. Belg.», Cl. Sci., ser., 5, 16, 989—995.
Wildeman E. de (1934), Remarques a propos de la foret equatoriale congolaise «Mem. Inst. Roy. Colon. Beige.», 2, 1 — 120.
Wilkinson G. (1939), Root competition and sylviculture, «Malay. Forester», 8, 11—15.
Winkler H. (1913), Die Pflanzenwelt der Tropen. В книге R. H. F r a n с e, Das Le-ben der Pflanzen., Stuttgart.
Winkler H. (1914), Die Pflanzendecke Siidost—Borneos, «Bot. Jb.», 50, 188—208.
W i t k a m p H. (1925), De ijzerhout als geologische indicator, «Trop. Natuur», 14, 97—103.
Wood B. R. (1926), The valuation of the forests of the Bartica-Kaburi area, «Report by the Conservator of Forests to the British Guiana Combined Court, Second Special Session», 1926, Georgetown, British Guiana.
Wright H. (1905), Foliar periodicity of endemic and indigenous tress in Ceylon, «Ann. Bot. Gdns, Peradeniya», 2, 415—517.
Z euner F. E. (1945), The Pleistocene Period, its climate, chronology and faunal successions, Ray Soc., London.
Z on R. (1912), Forests and water in the light of scientific investigation (Appendix 5 Final Rep. Nat. Waterways Commission), «U. S. Senate Document», 469, 62nd, Cong., Sess. 2, Reprinted 1927, U. S. Dep. Agric. Forest Service.
Z on P. van (1915), Mededeelingen omtrent den kamferboom (Dryobalanops aroma lieu', «Tectona», 8, 220—224.
ОГЛАВЛЕНИЕ
Предисловие......................................................... 5
Глава 1. Введение................................................
Часть I. СТРУКТУРА II ВНЕШНИЙ ОБЛИК ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Глава 2. Структура: синузии и ярусность............................ 35
Глава 3. Возобновление леса........................................ 57
Г лава 4. Облик деревьев и кустарников............................. 72
Глава 5. Травяной покров и зависимые синузии ..................... 114
Часть II. СРЕДА Глава 6. Климат................................................... 153
Г лава 7. Микроклиматы............................................ 179
Г лава 8. Сезонные изменения растительности................. . . 213
Г лава 9. Почвы................................................... 226
Часть III. ФЛОРИСТИЧЕСКИЙ СОСТАВ КЛИМАКСНЫХ СООБЩЕСТВ
Глава 10. Состав первичного дождевого леса (I).................... 251
Глава 11. Состав первичного дождевого леса (II) .................. 271
Часть IV. ПЕРВИЧНЫЕ СУКЦЕССИИ
Глава 12. Первичная ксеросерия и восстановление растительности на острове Кракатау .................................................... 293
Глава 13. Гидросерии............................................. 307
Глава 14. Прибрежные сукцессии.................................... 319
Часть V. ТРОПИЧЕСКИЙ ДОЖДЕВОЙ-ЛЕС
ВБЛИЗИ КЛИМАТИЧЕСКИХ ГРАНИЦ ЕГО СУЩЕСТВОВАНИЯ Глава 15. Тропический дождевой лес, листопадный лес и саванна..... 339
Глава 16. Тропический дождевой лес вблизи своих высотных и широтных границ....................................................... 369
Часть VI. ЧЕЛОВЕК II ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС Глава 17. Вторичные и отклонившиеся сукцессии..................... 399
Постскриптум. Будущее тропического дождевого леса................. 425
Библиография...................................................... 429
Ричарде ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС Редактор Л. И. КУДРЯШЕВА Художник Г. И. Мануйлов, Художественный редактор В. И. Шаповалов Технический редактор Н. А. Иовлрва. Сдано в производство 29/ХП—1960 г. Подписано к печати 15/VI—1961 г. Бумага 07X1081/16=15,1 бум. л., 41,4 печ. л. в т/ч. 17 вкл. Уч.-изд. л. 37,5. Изд. Л? 11/5065. Цена 2 р. 89 к. Зак. 858
ИЗДАТЕЛЬСТВО ИНОСТРАННОЙ ЛИТЕРАТУРЫ Москва 1-й Рижский пер., 2
Московская типография № 5 Мосгорсовнархоза, Москва, Трехпрудный пер., дом 9.
ОПЕЧАТКИ
Страница Строка Напечатано Следует читать
45 Рис. 5 1 снизу 5 футов 25 футов
165 Табл. 13 (продолж.), последний столбец 8 сверху 6 69
273 Табл. 28, второй столбец 20 сверху То же «Меранти дагинг»
Зак. 858