/












































































































































































































































































Текст
RM, Миркин, Л.Г. Наумоиа, А_Л_ Мудлашсв
Высшие растения
Учебник
Б.М. Миркин, Л.Г. Наумова, А.А. Мулдашев
ВЫСШИЕ РАСТЕНИЯ:
КРАТКИЙ КУРС СИСТЕМАТИКИ
С ОСНОВАМИ НАУКИ
О РАСТИТЕЛЬНОСТИ
Рекомендовано Министерством образования Российской
Федерации в качестве учебника для студентов высших учебных
заведений, обучающихся по направлениям подготовки бакалавров
«Экология и природопользование» и «Сельскохозяйстввнныенауки»,
а также по специальностям подготовки дипломированных
специалистов «Биология», «Ботаника», «Экология»,
«Природопользование»
Москва • «Логос» • 2001
УДК 58
ББК28.5
М63
Рецензенты:
Доктора биологических наук
Л.Б. Заугольнова, В.Г Онипченко
Миркин Б.М., Наумова Л.Г., Мулдашев А.А.
М63 Высшие растения: краткий курс систематики с основа-
ми науки о растительности: Учебник.-М.: Логос, 2001.
264 с.: ил.
ISBN 5-94010-041-4
Рассматриваются основные положения систематики высших расте-
ний и науки о растительности. Дается обзор таксонов моховидных, сосу-
дистых споровых, голосеменных и цветковых. Особое внимание уделяет-
ся анализу адаптаций растений различных таксонов к неблагоприятным
условиям среды. Приводятся данные о практической значимости таксо-
нов. Кратко охарактеризованы основные положения современной науки
о растительности (экология видов, популяций, фитоценозов, флор).
Для студентов высших учебных заведений, обучающихся по направ-
лениям и специальностям «Биология» и «Сельское хозяйство». Представ-
ляет интерес для научных работников в области биологии, экологии, лес-
ного и сельского хозяйства.
ББК 28.5
ISBN 5-94010-041-4
© Центр «Интеграция», 2001
ПРЕДИСЛОВИЕ
В подготовке биологов в университетах исключительно
важны традиционные дисциплины - ботаника и зоология, кото-
рые изучают все студенты независимо от того, какую специали-
зацию они изберут - генетику, молекулярную биологию, фи-
зиологию человека и животных или один из разделов ботаники.
Эти дисциплины играют определяющую роль в формирова-
нии представлений о биоразнообразии и служат фундаментом ос-
воения принципов его охраны и рационального использования
как важнейшего невозобновимого ресурса биосферы.
К сожалению, в последние годы учебники по систематике
высших растений для вузов издавались небольшими тиражами
(А. Г. Еленевский и др., 2000), а те, которые были написаны и
опубликованы четверть века назад, чрезмерно перегружены
фактическим материалом, знание которого необязательно для
студента-биолога - «неботаника». По этой причине потребность
в кратком курсе систематики высших растений с обычно вклю-
чаемым в него дополнением по основам геоботаники сегодня
очень велика. Авторы попытались выполнить этот «социальный
заказ» биологических факультетов университетов. Впрочем, та-
ким компактным учебником могут пользоваться и студенты пе-
дагогических и сельскохозяйственных вузов.
В предлагаемом курсе авторы сознательно придерживаются
экологического и прагматического взглядов на описываемые
таксоны. Вопросы филогении либо вовсе не рассматриваются,
либо излагаются конспективно как дополняющие основное со-
держание. Вопросы происхождения разных групп высших рас-
тений, включая и цветковые, на сегодняшний день столь не яс-
ны, что при начальном ознакомлении с систематикой вряд ли
есть смысл предлагать читателю умозрительные и противоречи-
вые филогенетические гипотезы. Тем не менее, для общего раз-
вития в учебник включены как антиномии некоторые гипотезы
происхождения цветковых и однодольных.
В соответствии с этой целевой установкой особенностями
настоящего учебника являются:
1. Концептуальный характер изложения материала. Авторы
дают читателю возможность освоить «скелет» систематики рас-
тений и познакомиться с весьма ограниченным числом таксонов,
которые представляют разные варианты строения растений. По
3
сравнению с традиционными учебниками резко сужен список
рассматриваемых семейств и таксонов более высокого ранга
(при описании споровых растений). Сокращен и список понятий,
используемых для характеристики таксонов. Авторы полагают,
что, освоив этот «скелет» знания, все остальное читатель смо-
жет почерпнуть из дополнительной литературы.
2. Выбор большинства таксонов проведен с учетом их прак-
тической значимости, и поскольку не все читатели знакомы со
специальными курсами растениеводства или ботанического ре-
сурсоведения, особое внимание уделяется краткой характери-
стике ресурсных и культурных растений.
3. Значительное внимание уделено экологии таксонов, так
как именно условиями среды во многом объясняются особенно-
сти растений. При описании отделов специально анализируются
важнейшие адаптации растений, позволяющие им преодолевать
сопротивление среды (действие неблагоприятных факторов) на
разных стадиях жизненного цикла. Вопросы экологии рассмат-
риваются при характеристике опыления и распространения пло-
дов цветковых, при объяснении полупаразитизма и паразитизма
и особенностей таких семейств, как Nymphaeaceae или Cheno-
podiaceae.
4. Авторы сознают, что систематика растений не самый
увлекательный предмет, особенно если перегружать его деталя-
ми. Поэтому в текст вкраплены любопытные сведения под руб-
рикой «Это интересно».
В заключение отметим, что предлагаемый курс содержит
самые первые знания о высших растениях. Тем, кто проявит ин-
терес к ботанике, рекомендуем обратиться к более полному
учебнику А. Г. Еленевского и др. (2000) или к многотомному из-
данию «Жизнь растений». Список рекомендуемой литературы
приведен в конце учебника.
ЧАСТЬ 1
ВВЕДЕНИЕ В СИСТЕМАТИКУ ВЫСШИХ
РАСТЕНИЙ
Высшие растения это наиболее разнообразная
группа растений, и потому система, позволяющая ориентироваться в
этом разнообразии, особо важна для всего комплекса наук, связанных
с их изучением, использованием и охраной.
Дадим общую характеристику систематики растений как науки, а
высших растений как основных автотрофных организмов, адаптирован-
ных к жизни на суше. Именно наземная среда была фактором, под
действием которого сформировался комплекс признаков, присущих
высшим растениям.
Глава 1
ДЛЯ ЧЕГО НУЖНА СИСТЕМАТИКА ВООБЩЕ
И ВЫСШИХ РАСТЕНИЙ В ЧАСТНОСТИ
Систематика - это наука о разнообразии организмов, опре-
деляющая их место в системе органического мира. Существует
систематика животных, микроорганизмов, грибов, растений. В
ботанике различают систематику низших растений и высших
растений.
1.1. Понятие о таксономии
Наиболее распространенная система, которую сегодня ис-
пользуют ботаники, иерархическая строится по принципу
«коробка в коробке». Любая ступень иерархии системы называ-
ется таксономическим рангом.
Главным таксономическим рангом является вид. Над ним
располагаются род, семейство, порядок, подкласс, класс (табл. 1).
Ниже - подвид и разновидность. Кроме этих основных рангов,
в некоторых случаях используются дополнительные - секция,
объединяющая группу сходных видов в пределах рода, а также
подсемейство и триба - объединения сходных родов в пределах
семейства.
Любая система таксонов называется таксономией. Таксо-
номия должна быть в первую очередь удобной для хранения и
поиска информации о разнообразных растениях и позволять
5
быстро определить место в системе образца растения неизвест-
ного вида. Система эта - открытая, в нее могут включаться но-
вые ранее неизвестные науке виды растений или (после накоп-
ления значительного нового материала) новые роды.
Таблица 1
Основные таксономические ранги систематики
высших растений и примеры таксонов
Таксономический ранг Окончание в латинском названии Пример таксона
Отдел -ophyta Magnoliophyta
Класс —opsida Magnoliopsida
Подкласс -idae Magnoliidae
Порядок -ales Magnoliales
Семейство -aceae Magnoliaceae
Род Magnolia
Вид Magnolia grandiflora
1.2. Задачи, для решения которых
нужна систематика растений
1.2.1. Определение областей экстраполяции данных,
получаемых другими науками
Современное знание опирается на достижения многих наук,
исследующих природу растений: генетики, анатомии, физиоло-
гии, биохимии и др. Изучить кариотип (определить число хро-
мосом и их форму) или сделать анализ биохимического состава
каждого растения, входящего в растительный покров даже не-
большой территории, невозможно.
По этой причине исследования проводят выборочным мето-
дом: изучают отдельные модели - образцы растений, которые
выбирают из генеральной совокупности (растений одного так-
сона). Используя полученные данные, характеризуют все расте-
ния этого таксона, т. е. полученный на основе выборочной со-
вокупности результат экстраполируют на всю генеральную со-
вокупность.
Если границы генеральной совокупности определены невер-
но, то исследователь попадает в ситуацию, напоминающую по-
ложение отца Федора из «Двенадцати стульев» И.Ильфа и
Е.Петрова. Он искал бриллианты в стульях, которые были по-
хожи на гарнитур тещи Воробьянинова, но не входили в сово-
купность этого гарнитура. Менее анекдотичные, но постоянные
6
ошибки такого рода допускают ботаники-ресурсоведы, не дос-
таточно хорошо владеющие систематикой растений. Они могут
рекомендовать сбор растительного сырья из группы близких
видов, среди которых нужное химическое соединение содержат
растения лишь одного из видов.
Таким образом, именно знание систематики растений по-
зволяет проводить границы между генеральными совокупностя-
ми таксонов разного уровня - видов, родов, семейств или внут-
ривидового уровня - подвидов, разновидностей и т.д.
Систематика 'растений, называемая некоторыми представи-
телями молекулярной биологии и генетики «старой» и «описа-
тельной» наукой, на самом деле является важнейшей состав-
ляющей современного ботанического знания. Без ее развития
«пробуксовывают» самые современные науки, в которых иссле-
дования проводятся на молекулярном уровне. Без точного так-
сономического адреса изучаемых объектов все достижения этих
наук будут иметь невысокую ценность.
1.2.2. Изучение флор
Флора - это совокупность таксонов определенной террито-
рии. Размер этой территории может быть различным. Напри-
мер, флора Европы, флора России, флора Башкортостана или
Татарстана, флора Башкирского Предуралья, флора города
Уфы, флора городского лесопарка и т.д. Изучение флор необ-
ходимо для решения множества научных задач.
Во-первых, при этом человек получает информацию о нали-
чии в составе флоры ценных ресурсных растений, которые за-
тем использует как лекарственные, пищевые, кормовые, техни-
ческие и др.
Во-вторых, на основе знания флоры изучается раститель-
ность, т. е. совокупность растительных сообществ (фитоце-
нозов), которым посвящена ч. 3 учебника.
В-третьих, в настоящее время изучение флор, отражающих
характер воздействия человека на окружающую среду, является
элементом биологического мониторинга за состоянием экосистем.
За изменениями флоры необходимо вести постоянное на-
блюдение, так как они не только отражают изменения в окру-
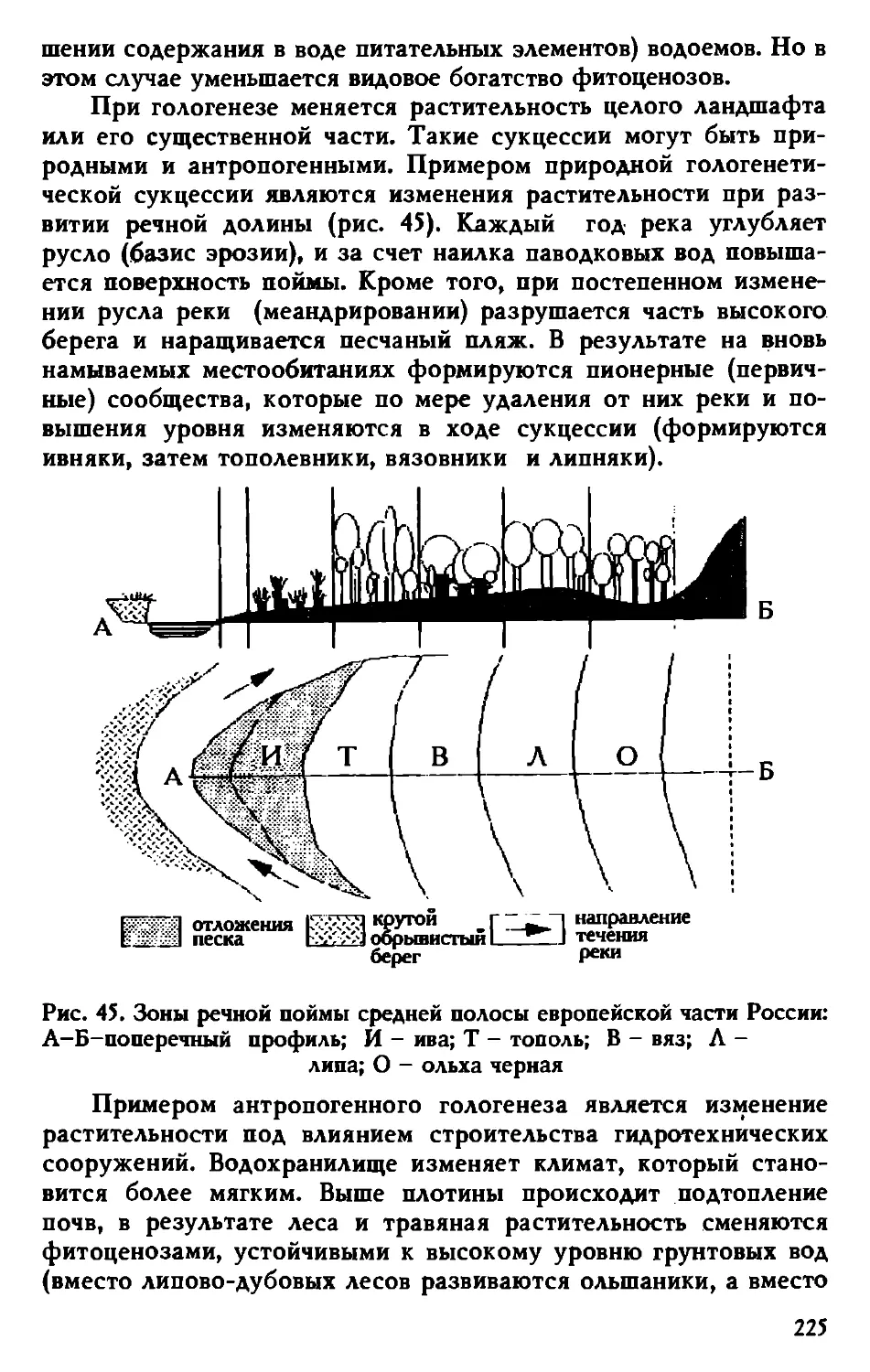
жающей среде, но нежелательны сами по себе. Исчезающие ви-
ды, которые сегодня считаются бесполезными, после дополни-
тельного изучения могут оказаться ценными ресурсными расте-
ниями. Виды-пришельцы, напротив, могут оказаться вредными
для экосистемы и для человека.
7
1.2.3. Прогностическая роль систематики растений
Систематика позволяет предсказывать существование неиз-
вестных науке таксонов или наличие у известных таксонов неиз-
вестных признаков. Великий генетик и растениевод Н.И.Вавилов
открыл закон гомологических рядов наследственной изменчиво-
сти. Суть этого закона - повторяемость одних и тех же призна-
ков (остистость колоса, опушение листа и цвет зерновки у злаков
и т.д.) у близких видов и даже родов. Знание того, какие виды
или роды являются близкими, позволило предсказать наличие
еще не описанных подвидов и разновидностей. После открытия
целебных свойств женьшеня (Panax ginseng) начался поиск целеб-
ных свойств у других видов семейства аралиевых (Araliaceae).
Оказалось, что в этом семействе есть другие виды, влияющие на
организм подобно женьшеню, например, заманиха (Oplopanax
elatus), элеутерококк колючий (Eleutherococcus senticosus) и дру-
гие. Примеров того, как на основе таксономической близости ви-
дов удалось предположить и обнаружить у них нужные химиче-
ские соединения, можно привести много.
1.3. История систематики растений
Систематика растений уходит корнями в доисторические времена.
Московский профессор А.А.Уранов считал, что начало классификации
растений надо отнести к «дочеловеческому» периоду. Способность от-
личать среди прочих нужные или, наоборот, вредные растения была
присуща предкам человека в виде инстинкта собирания. Этот инстинкт
хорошо развит у животных (насекомые собирают пыльцу и нектар с
определенных растений; белки собирают семена сосны сибирской и
плодовые тела шляпочных грибов; хомяки прячут в норах корни раз-
ных растений, а также плоды и семена злаков и бобовых; муравьи не
только собирают, но и выращивают растения и т.д.). Экзаменатором в
такой «школе систематики» был голод, и потому «хвостатые и крыла-
тые ученики» должны были твердо усваивать уроки того, что, где и ко-
гда можно есть.
Пещерный человек, который занимался собирательством, хорошо
различал растения. Чтобы найти съедобные корни и плоды, нужно бы-
ло знать ресурсные растения «в лицо» и иметь представления об их
местообитании.
Сознательный подход к классификации растений связан с появле-
нием их наименований (символический знак, рисунок, иероглиф, сло-
во), а зачатки систематики уже были у народов, которые культивиро-
вали растения и имели письменность (Египет, 5-4 тыс. лет до н. э.; Ки-
тай, 4-3 тыс. лет до н. э.; Индия, 2 тыс. лет до н.э.).
Первые крупные ботанические сочинения, дошедшие до нас и
имеющие историческое значение, принадлежат Теофрасту (Феофрасту)
из Эреса (370-280 гг. до н.э.). Он описал более 450 видов хозяйствен-
ных и лекарственных растений и считается одним из первых система-
8
тиков, названным Карлом Линнеем «отцом ботаники». (Имя Теофраста
увековечено в ботанике родом Theophrasta и семейством Theophra-
staceae.)
Историографы систематики различают четыре основных периода
ее развития:
описательных, или практических (утилитарных), классификаций -
от древних времен до XVI в.;
искусственных систем - с XVI по XVIII в.;
естественных систем - с конца XVIII до середины XIX в.;
эволюционной филогенетической систематики (последарвиновский
период).
Если рассматривать эти периоды как отражение общих законо-
мерностей развития науки, то их можно свести к трем этапам: прагма-
тическому, классическому и неопрагматическому.
На первом этапе ставилась практическая задача - научиться раз-
личать растения, чтобы с большей пользой использовать их в пищу и
как природные лекарства. В это время основное внимание уделялось
именно тому, как создать систему, которой удобно пользоваться, какие
признаки растений наиболее приемлемы для надежного различения
разных видов.
Этот этап завершил великий Карл Линней, создавший до сих пор
используемую бинарную номенклатуру растений (как, впрочем, и жи-
вотных) и предложивший формальную систему группирования видов в
24 класса, четко различающихся по основным признакам строения
цветка (тычинкам и пестикам):
классы с 1 по 13 — по количеству одиночных тычинок;
классы с 14 по 20 - по длине тычиночных нитей, по срастанию ни-
тей или пыльников и гинецея;
классы с 21 по 23 - по наличию цветков разных полов на растении;
класс 24 - тайнобрачные (водоросли, грибы, мхи, папоротники).
Однако в целом система Линнея, несмотря на ее простоту и удобст-
во (особенно при ограниченном числе видов, которых во времена Линнея
было описано всего 6 тысяч), была по современным оценкам довольно
грубой. Если некоторые классы достаточно естественны (класс 6 объеди-
нял крестоцветные, класс 10 — гвоздичные), то большинство классов бы-
ли пестрыми. Так, класс 3 включал ирисы и большинство злаков, класс
5 - примулу и незабудку, класс 21 - рогоз, тую и дуб.
В послелиннеевский период объем знаний быстро возрастал. В это
время наука становится классической: она ставит задачу понять и пол-
ностью описать картину окружающего мира. При таком подходе для
исследователя не может быть интересных и неинтересных объектов,
изучать нужно все. От ответа на вопросы «что?», «какой?», «сколько?»
наука перешла к более сложной задаче - выяснению «почему?».
В это время ботаники пытались создать естественную систему (в
которой растения объединяются в таксоны по группе признаков), а
затем и филогенетическую, все таксоны которой связаны отношениями
родства в единое филогенетическое древо.
В XX в. классическая наука породила множество вариантов фило-
генетических схем, противоречащих друг другу. Разные авторы исходи-
9
ли из моно- и полифилетической гипотез происхождения цветковых
(см. ч. 2); из разных гипотез происхождения однодольных и двудоль-
ных; по-разному интерпретировали одни и те же признаки цветка и ве-
гетативных органов (например, водопроводящие элементы у кукушкина
льна рассматривались как зачатки будущих сосудов и как результат их
редукции). В этом «лесу из филогенетических деревьев», которые с
трудом воспринимались всеми, кроме их авторов, было несложно за-
блудиться.
Видный методолог современной биологии С.В.Мейен с юмором пи-
сал о том, что накопленные систематиками знания не позволяют изо-
бражать никаких филогенетических деревьев. Ситуация такова, что в
лучшем случае можно соорудить кустарник, причем значительная часть
ветвей разбросана вокруг него, и систематики не знают, куда их при-
ставить.
1.4. Особенности современного этапа развития
Сложности создания филогенетической системы наиболее
ярко проявились в 60-70-е годы нашего столетия, когда объем
знания возрос настолько, что стал несоизмерим с ботаникой
времен Линнея. Количество описанных видов сосудистых расте-
ний приблизилось к 300 тысячам, т. е. возросло в 50 раз. Если
во времена Линнея в силу неполноты знания все известные ему
растения было несложно разбить на четко различающиеся виды
(их называют линнеонами), то теперь стало очевидно, что меж-
ду большинством «линнеонов» есть переходы.
Палеоботаническая летопись, на которую возлагались
большие надежды как на источник фактического материала для
воссоздания картины истории растений, оказалась слишком
неполной. Ее можно сравнить с библиотекой, которая вначале
подверглась нападению мышей, потом пожару и наводнению.
Все другие данные, которые пытались использовать для созда-
ния естественной филогенетической системы о современном
географическом распространении растений, об их биохимиче-
ском составе и генетическом сходстве - мало помогали, так как
разные ученые интерпретировали одни и те же данные совер-
шенно по-разному.
По словам крупного современного философа И.Пригожина,
пришлось признать, что наши знания находятся на самом на-
чальном этапе объяснения природы. Все это привело к измене-
нию общего настроя в науке, она приобрела «неклассический»
(неопрагматический) характер. Мир оказался слишком слож-
ным, чтобы его можно было познать до конца. И потому в пер-
вую очередь приходится изучать то, что связано с практически-
ми интересами человека.
Идеи филогенетической классификации теряют популяр-
ность. Профессор В.Н.Тихомиров, долгие годы возглавлявший
10
кафедру систематики высших растений МГУ, предложил систе-
му цветковых на основе морфологического и экологического
сходства, без попыток связать таксоны выше порядка филоге-
нетическими отношениями.
Неклассический характер современной систематики про-
явился и при понимании вида у растений. Долголетняя дискус-
сия о критериях «биологических видов», т. е. естественных ви-
дов, сотворенных самой природой и генетически изолированных
друг от друга, завершилась признанием того, что эта категория
неуниверсальна. Оказалось, что на «биологические виды» рас-
падается не все разнообразие растений. У самоопыляющихся
существуют микровиды-жорданоны, есть гибридизирующие со-
вокупности «полувидов» (сингамеоны), имеются «виды-двой-
ники» (морфологически сходные, но различающиеся генетиче-
ски) и «виды-близнецы» (морфологически различные, но с оди-
наковым генотипом).
В систематику пришло понятие «таксономический вид», ко-
торым обозначаются совокупности растений примерно одинако-
вого объема. Если существует «биологический вид», то «таксо-
номический вид» совпадает с ним по объему. Если совокупность
растений не разбивается на такие «хорошие виды», то виды вы-
деляются условно. Разумеется, в последнем случае определить
их трудно даже опытному ботанику.
Для одной и той же совокупности растений разные ученые
сегодня выделяют разное число видов (в более крупном и более
мелком объеме) и группируют эти виды в разное число родов,
семейств и порядков. Это вызывает частые дискуссии и ревизии
таксономии. Спастись от потока ревизий можно только «кол-
лективным договором» о едином понимании системы. Примером
такого «договора» являются два издания (в 1981 и 1985 годах):
«Сосудистые растения СССР» и «Сосудистые растения России
и сопредельных государств (в пределах бывшего СССР)» - спи-
ски видов всей территории гигантского распавшегося государ-
ства. Автором этих списков является С.К.Черепанов.
Глава 2
ОБЩАЯ ХАРАКТЕРИСТИКА ВЫСШИХ РАСТЕНИЙ
2.1. Отличия высших растений от низших
Высшие растения отличаются от низших многими призна-
ками, которые связаны с тем, что они по преимуществу - оби-
татели суши. И именно поэтому они несравненно сложнее и
разнообразнее обитающих в водной среде низших растений.
11
Соотношение количества видов наземных и водных организмов
(с учетом всего биологического разнообразия, включая живот-
ных, грибы, прокариоты) составляет 5:1. У растений оно еще
более значительно - 10:1 (300 тысяч высших и 30 тысяч низ-
ших). Еще более показательны различия вклада высших и низ-
ших растений в продуктивность и биомассу биосферы. Низшие
растения производят примерно треть первичной биологической
продукции, но поскольку длительность жизни большинства из
них невелика, биомасса низших растений составляет менее 1%
общей массы продуцентов биосферы.
Впрочем, разделение на высшие и низшие только по среде
обитания будет не совсем верным. Часть высших растений, на-
зываемых гидрофитами, ведет водный образ жизни. Среди них
много обитателей пресноводных водоемов (Nymphaeaceae,
Lemnaceae, Potamogetonaceae и др.). В то же время многие во-
доросли ведут наземный образ жизни и заселяют поверхность
почвы и даже скальных пород. Однако в обоих случаях водная
и наземная среды вторичны, т. е. гидрофиты - это высшие рас-
тения, которые перешли от сухопутного образа жизни к водно-
му, а наземные водоросли - водные растения, которые посели-
лись на суше. Все водоросли могут нормально функционировать
только тогда, когда субстрат их обитания обильно увлажнен. В
другое время они находятся в диапаузе.
2.2. Приспособления к наземному образу жизни
Наземная среда несравненно разнообразнее водной (вода в
любом случае «мокрая», хотя может различаться по насыщенно-
сти солями, в том числе элементами питания, по температуре, ос-
вещенности, скорости перемещения). В условиях суши в очень
широких пределах изменяются все экологические факторы, глав-
ным из которых для растений является обеспеченность водой.
Для обеспечения водой у высших растений выработалось
множество приспособлений. Основные из них - проводящие и
покровные ткани. Проводящие ткани развивались в процессе
эволюции - от трахеид к сосудам и от ситовидных клеток к си-
товидным трубкам с клетками-спутницами. Покровные ткани
растений тем совершеннее, чем засушливее условия, в которых
они обитают. У растений сухих местообитаний (ксерофитов)
формируется не только толстая кутикула, но и восковой налет,
опушение. Они выделяют в атмосферу ароматические вещества,
которые окутывают растения, тем самым уменьшая транспира-
цию. У ксерофитов совершенствуется устьичный аппарат.
Решив задачу «не засохнуть», растение на суше должно
еще и не упасть под влиянием силы тяжести и ветра. В водной
12
среде такой проблемы не возникает, поскольку даже гигантские
ламинарии, прикрепленные к морскому дну, поддерживаются
плотной водной средой. У высших растений сформировались
механические ткани (склеренхимные волокна), которые объеди-
нены вместе с проводящими тканями в систему сосудисто-
волокнистых пучков - стелу.
Разные представители высших растений обладают стелами
разных типов, причем при повышении уровня развития возрас-
тает площадь соприкосновения проводящих элементов с прочи-
ми тканями. Так, в ходе эволюции из примитивной протостелы
возникли сифоностела деревьев, эустела травянистых двудоль-
ных и атактостела однодольных растений.
У высших растений тело дифференцировано на органы: оно
имеет огромную фотосинтезирующую поверхность листьев, ко-
торые располагаются в несколько слоев-этажей (до пяти), и по-
верхность корней с корневыми волосками, поглощающими воду
с элементами почвенного питания.
Большую роль в жизни высших растений играют мутуали-
стические взаимоотношения с грибами. У некоторых видов,
кроме того, сложились аналогичные взаимополезные отношения
с азотфиксирующими бактериями, которые позволяют повысить
снабжение растений азотом.
Наконец, атмосфера как среда жизни стала причиной фор-
мирования многоклеточных органов спороношения - спорангиев
и полового размножения - гаметангиев (антеридиев и архего-
ниев, рис. 1а).
2.3. Жизненный цикл
Воздушно-почвенная среда обитания привела к формирова-
нию у всех высших растений единой схемы жизненного цикла
(цикла воспроизведения). Напомним, что у водорослей сущест-
вуют самые разные варианты жизненных циклов: гаплонтный,
при этом диплоидна только зигота, первое деление которой -
мейоз, и вся жизнь проходит в гаплоидной фазе (хламидомона-
да, спирогира, улотрикс); диплонтный гаплоидны только га-
меты, и вся жизнь проходит в диплоидной фазе (фукус); изо-
морфный диплогаплонтный - все особи внешне одинаковы, но
одни из них диплоидны и образуют споры, другие гаплоидны и
образуют гаметы (ульва, диктиота).
У всех высших растений один и тот же гетероморфный (га-
метофит и спорофит внешне различаются) диплогаплонтный (с
чередованием гаплоидного и диплоидного поколения) жизнен-
ный цикл (рис. 16). При этом, у всех них, кроме моховидных,
преобладает спорофит и редуцирован (тем больше, чем выше
13
уровень организации) гаметофит. У споровых растений (папо-
ротники, хвощи, плауны) гаметофит - автономный организм, а
у семенных он развивается («паразитирует») на спорофите, что
позволяет наилучшим образом обеспечить вероятность оплодо-
творения и развития зародыша дочернего спорофита. Семя -
высшее достижение приспособления растений к наземному
образу жизни.
Только у моховидных преобладающим поколением является
гаметофит, которому подчинен уменьшенный в размерах спо-
рофит, «паразитирующий» примерно так же, как гаметофит на
спорофите у семенных растений.
Теч
Меноз
Материнские
клетки спор (2п)
Спора
Гаметофит
(п) '
Антеридии со
сперматоэоидами( п )
Архегоиии с\ \
яйцеклетками (пд
Оплодотвортие
Зигота. (2п)
млн
лет
Мел 135
Юра 195
Триас 225
Пермь 280
Карбон350
Девон 400
Силур 440
Спорангии Зародыш
^Ч^Спорофит^х^
(2п)
Рис. 1. Общая характеристика высших растений: а - гаметангии: слева -
архегоний (1 - стенка, 2 - шейковые канальцевые клетки, 3 - брюшная
канальцевая клетка, 4 - яйцеклетка), справа - незрелый антеридий (1 -
стенка, 2 - сперматогенные клетки); б - схема жизненного цикла; в -
геологическая история современных отделов
2.4. Основные отделы
В составе высших растений выделяются несколько отделов,
которые объединены в три группы: бессосудистых споровых -
14
отдел моховидные (Bryophyta), сосудистых споровых - плауно-
видные (Lycopodiophyta), хвощевидные (Equisetophyta) и папо-
ротниковидные (Polypodiophyta), семенных - голосеменные
(Pinophyta) и покрытосеменные (Magnoliophyta). Кроме того, к
высшим растениям относятся вымерший отдел проптеридофитов
(Propteridophyta) и небольшой своеобразный отдел псилотовид-
ные (Psilotophyta), которые мы не рассматриваем.
Из рис. 1в, на котором показана геологическая история со-
временных отделов высших растений, очевидно, что пять пер-
вых отделов появились в разное время одного геологического
периода - девона, «золотого века» высших растений. Однако,
если до наших дней мхи дошли примерно в том же составе, что
были в палеозое, то разнообразие хвощей, плаунов и папорот-
ников уменьшилось: почти исчезли древовидные формы. Значи-
мость голосеменных растений сохранилась, но изменился их со-
став (произошли изменения на уровне порядков). Цветковые
появились сравнительно недавно (в меловом периоде) и быстро
заняли господствующее положение в биосфере.
2.5. Заключение: сравнение высших и низших растений
Таким образом, объединение растений в категорию высших и про-
тивопоставление их низшим вполне оправдано: высшие растения уст-
роены более сложно, чем низшие В табл. 2 показаны основные отличия
высших и низших растений.
Таблица 2
Сравнение высших и низших растений
Характеристика Низшие растения Высшие растения
Время появле- ния на Земле Одноклеточные - 1,5 млрд лет назад, много- клеточные - 700 млн лет назад Споровые - более 400 млн лет назад, покры- тосеменные - 200 млн лет назад
Высшие таксоны Algae (Chlorophyta, Phaeophyta, Bacillario- phyta, Rhodophyta и ДР-) Bryophyta, Propteri- dophyta, Psilotophyta, Equisetophyta, Lycopo- diophyta, Polypodiophy- ta, Pinophyta, Mag- noliophyta
Количество видов Около 30 тыс. Около 300 тыс.
Среда обитания и роль в экоси- стемах Преимущественно водная, основные про- дуценты пресноводных и морских экосистем Преимущественно на- земная, основные про- дуценты наземных эко- систем
Вклад в биоло- гическую продук- цию биосферы 32% 68%
Доля в фито- массе биосферы 0,2% 99,8%
15
Окончание табл. 2
Характеристика Низшие растения Высшие растения
Жизненный цикл Разнообразный - гап- лонтный, диплонтный, диплогаплонтный (изо- морфный и гетеро- морфный) Один - гетероморф- ный диплогаплонтный, с преобладанием спо- рофита (у моховидных - гаметофита)
Морфологиче- ская дифферен- циация тела и дифференциация тканей Слабая, строение слоевищное Сильная, у большин- ства тело дифференци- ровано на побег (сте- бель с листьями) и ко- рень, состоящие из тка- ней: покровных, прово- дящих, механических, ассимиляционных, за- пасающих и др.
Гаметангии У большинства одно- клеточные; если много- клеточные, то без сте- рильных клеток Многоклеточные ан- теридии и архегонии со стерильной стенкой
Спорангии Одноклеточные: если многоклеточные, то без стерильных клеток Многоклеточные со стерильной стенкой
Перенос муж- ских гамет (или гаметофитов) для оплодотворения Водой Ветром, насекомыми и другими животными, реже - водой
Оплодотворение Наружное, в водной среде Внутреннее, в архего- нии
Двудомность и однодомность Преобладают одно- домные организмы Преобладают одно- домные организмы, но много и двудомных
Распростране- ние диаспор Водой, ветром, ред- ко - насекомыми Ветром, насекомыми и другими животными, водой
Фотосинтези- рующие органы Слоевище Листья, часто много- ярусные, реже - стебли
Обеспечение минерального пи- тания Поглощение всей по- верхностью слоевища Поглощение развитой корневой системой
Роль симбио- трофии Некоторые водоросли в симбиозе с грибами образуют лишайники У большинства пред- ставителей развита ми- кориза, у некоторых групп есть симбиоз с аз отфиксирующими бактериями
В заключение отметим, что появление высших растений позволило
сформироваться огромному разнообразию животных и других гетеро-
трофных организмов, которые питаются их фитомассой - непосредст-
венно или опосредованной в биомассе фитофагов и зоофагов низших
порядков.
16
ЧАСТЬ 2
СПОРОВЫЕ И ГОЛОСЕМЕННЫЕ
Споровые (моховидные, плауновидные, папоротнико-
видные) и голосеменные растения - древние обитатели суши, появив-
шиеся еще в девоне. Они отличаются большим разнообразием строения
вегетативных и генеративных органов и особенностями жизненных
циклов. Интерес представляют те приспособления (адаптации), кото-
рые позволили представителям этих «ветеранов» растительного мира
дожить до наших дней, и поэтому в ч. 2 много внимания уделяется
способам их выживания.
Глава 3
МОХОВИДНЫЕ (BRYOPHYTA)
3.1. Общая характеристика
Эти древнейшие высшие растения появились около 400 млн
лет назад (остатки, достоверно относимые к моховидным, име-
ют возраст 370 млн лет). Они намного старше динозавров, их
разнообразие сформировалось до того, как произошло разъе-
динение материков. По этой причине бриофлоры (флоры мхов)
разных континентов имеют гораздо больше общего, чем флоры
сосудистых, и в особенности цветковых, растений.
Моховидные достаточно разнообразны (около 25 тыс. ви-
дов), распространены в увлажненных местообитаниях, но встре-
чаются и в пустыне, и на бесплодной скальной поверхности.
3.1.1. Гаметофит
В системе высших растений моховидные занимают особое
место. С одной стороны, они — типичные обитатели суши с при-
способленными к воздушной среде гаметангиями и спорангиями
и с четким чередованием поколений. С другой стороны, в жиз-
ненном цикле у моховидных по сравнению с остальными выс-
шими растениями все наоборот - преобладает не спорофит, а
гаметофит (рис. 2а). И стебель, и листья характерны не для
спорофитов, а для гаметофитов. Поскольку в одном случае ли-
стья образуются на диплоидном растении, а в другом - на гап-
лоидном, то очевидно, что сходство этих листьев внешнее, а не
17
основанное на общем происхождении. Поэтому даже было
предложено называть лист моховидных филлоидом, а стебель -
каулоидом, однако это не прижилось.
в) г)
Рис. 2. Моховидные: а- схема жизненного цикла (1 - протонема, 2 -
гаметофит, 3 - спорофит); б - Marchantia; в - Polytrichum commune
(1 - гаметофит, 2 - колпачок, 3 - спорогон - общий вид и разрез); г -
Sphagnum (1 - гаметофит, 2 - поперечный срез листа)
18
Кроме того, в отличие от всех остальных высших, моховид-
ные - это бессосудистые растения: у них нет развитой прово-
дящей системы (трахеид, сосудов, ситовидных трубок).
Моховидные - небольшие многолетние (за редким исключе-
нием) вечнозеленые растения, у которых отсутствуют корни. К
субстрату они прикрепляются ризоидами, представляющими со-
бой сильно вытянутые в длину одиночные клетки у примитив-
ных представителей (печеночников) и нити из одного ряда кле-
ток, разделенных косыми перегородками, у более совершенных.
Поглощение и испарение воды у моховидных осуществляет-
ся всей поверхностью гаметофита. Правда, у некоторых в стеб-
ле есть проводящий пучок из узких и длинных клеток - гидро-
идов и лептоидов, выполняющих функцию сосудов и ситовид-
ных трубок. Встречаются как одно-, так и двудомные мхи. На
гаметофитах образуются антеридии и архегонии обычного для
высших растений строения.
Благодаря сравнительно небольшой тканевой дифференциа-
ции, а также способности моховидных оживать после полного
высыхания, любая часть гаметофита может дать начало новрму
растению. Большинства мхов вегетативно размножаются с по-
мощью специальных органов (выводковых почек, листьев, час-
тей листьев, веточек), которые образуются на гаметофите. Эти
органы могут длительное время находиться в состоянии покоя и
переноситься на большие расстояния и водой, и ветром, тем са-
мым напоминая семена (хотя, конечно, не имея с ними ничего
общего!).
Как этап вегетативного размножения, можно рассматривать
прорастание многих почек на протонеме - нитевидной или пла-
стинчатой стадии гаметофита, развивающегося из споры. В резуль-
тате из одной проросшей споры образуется не один, а много гаме-
тофитов.
3.1.2. Оплодотворение и образование спорофита
Оплодотворение возможно только при наличии воды, по-
скольку только в ней могут двигаться сперматозоиды. У при-
митивных моховидных сперматозоиды плывут по сплошной
водной пленке и привлекаются в шейку архегония выделяемыми
им химическими веществами. У многих более совершенных мхов
группа антеридиев на верхушке стебля окружена «чашей» из
листьев, в которой скапливается дождевая вода. Сперматозоиды
выходят в воду и рассеиваются брызгами от попавших в «чашу»
дождевых капель. Капельки воды с большим количеством спер-
матозоидов могут переносить с растения на растение и насеко-
мые (это похоже на насекомоопыление цветковых растений).
Развивающийся из зиготы спорофит называется спорогоном
и представляет собой коробочку с ножкой, расширенной в ниж-
19
ней части в стопу-гаусторию, с помощью которой он, присасы-
ваясь к гаметофиту, живет за его счет. Однако в период созре-
вания он может иметь зеленую окраску и фотосинтезировать,
т.е. «паразитизм» спорофитного поколения неполный.
Связь спорофита и гаметофита очень органична. Гаметофит
не только питает, но и защищает спорофитное поколение, и
помогает в рассеивании спор (образование так называемой
«ложной ножки», которая поднимает коробочку над растени-
ем). Это заставляет вспомнить отношения гаметофита со спо-
рофитом у семенных растений.
Вместе со спорогоном у моховидных разрастается архего-
ний, который, разрываясь, своим брюшком в виде колпачка
прикрывает коробочку. Таким образом, колпачок формально к
спорогону не относится, поскольку представляет собой отмер-
шую часть гаметофита.
Процесс оплодотворения мхов, особенно двудомных, ос-
ложнен обязательностью наличия капельно-жидкой среды. Если
ее нет, то половой процесс не происходит. По этой причине в
жизни мхов большую роль играет вегетативное размножение.
Размножаться вегетативно способен и спорофит. В экспе-
рименте из рассеченной на 133 части ножки спорогона 113 ку-
сочков проросли и дали новые растения.
3.2. Обзор основных таксонов
Отдел Bryophyta имеет следующую систематику:
Класс Bryopsida (настоящие мхи)
Подкласс Bryidae
Подкласс Sphagnidae
Подкласс Andreaeidae
Класс Hepaticopsida (печеночники)
Класс Anthocerotopsida (антоцеротовые мхи).
Не рассматривая последний класс, отметим лишь, что это -
тропические и субтропические мхи теломного строения, близ-
кие к печеночникам, но отличающиеся тем, что полости их тела
заполнены не воздухом, а слизью, в которой развивается сим-
биотически связанная со мхом азотфиксирующая цианобакте-
рия носток.
3.2.1. Класс Hepaticopsida (печеночники)
Название классу дано в средние века, когда считалось, что
сходство формы растения и какого-то органа человека - «под-
сказка» всевышнего о лечебном предназначении растения. Теми
печеночниками, которые по своей форме напоминают печень
человека, пытались лечить заболевания этого органа.
20
В этот класс входит около 6 (10) тыс. видов (300 родов и 50
семейств). Главная особенность печеночников - дорзовентраль-
ное (т.е. с различающимися верхней и нижней сторонами)
строение тела. В клетках присутствуют особые образования -
масляные тельца. Печеночники разделяются на слоевищные
(подкласс Marchantiidae) и листостебельные (подкласс Junger-
manniidae), причем последние преобладают по числу видов и
распространены преимущественно в тропиках и субтропиках.
В средней полосе чаще встречаются слоевищные печеночни-
ки, например маршанция многообразная (Marchantia polymo-
rpha, рис. 26), которая обитает в сырых местах: на древесине,
почве у родников и колодцев. В городах маршанцию можно
найти в щелях между каменными плитами старых лестниц. На
ее ремневидном слоевище расположены подставки - гаметофо-
ры, напоминающие миниатюрные грибы со шляпками, рассечен-
ными на лопасти. Гаметофоры, несущие антеридии, называются
антеридиофорами, а несущие архегонии архегониофорами.
Архегониофоры более рассечены, их шляпки похожи на много-
лучевые звездочки, шляпки антеридиофоров похожи на зонти-
ки. Антеридии расположены на верхней стороне шляпок, а ар-
хегонии - на нижней.
Спорогон состоит из короткой ножки и спорангия, в кото-
ром, кроме спор, образуются стерильные клетки со спиральными
утолщениями - элатеры. Элатеры играют роль пружинок, кото-
рые во влажную погоду скручены, а в сухую раскручиваются и,
разрыхляя споровую массу, способствуют ее рассеиванию.
Вегетативное размножение у маршанции происходит либо
при разделении слоевища на части, либо с помощью специаль-
ных выводковых почек, которые вызревают на верхней части
слоевища в специальных выводковых корзиночках и разносятся
дождевой водой.
522. Класс Bryopsida (настоящие мхи)
В этом классе, насчитывающем около 15 тыс. видов (800 ро-
дов, относимых к 80 семействам), различаются два главных под-
класса: сфагновые (Sphagnidae) и бриевые (зеленые, Bryidae).
Кроме того, в этот класс входят андреевые (Andreaeidae) черные
мхи (около 100 видов) очень маленькие растения с длиной
стебельков в несколько миллиметров, которые в настоящем
курсе не рассматриваются.
Для всех представителей класса характерна особая «юно-
шеская» стадия развития гаметофита - протонема.
Подкласс Bryidae (бриевые мхи). Бриевые мхи наиболее раз-
нообразны (число видов превышает 14 тыс. и приспособлены к
21
самым разным условиям жизни - от холодных скал в Арктике до
жарких пустынь. Однако в неблагоприятных условиях мхи актив-
но растут только в то время года, когда достаточно тепла и вла-
ги. А длительные неблагоприятные периоды переносят в состоя-
нии криптобиоза (анабиоза), которое может длиться годами.
Строение зеленых мхов традиционно рассматривают на
примере кукушкина льна (Polytrichum commune, рис. 2в). Его
гаметофит, достигающий высоты 20-40 см, представляет собой
неветвящийся стебель, густо усеянный жесткими узкими листь-
ями. Архегонии и антеридии расположены группами на вер-
хушках разных стеблей.
В состав спорогона входит стопа, которой он прикреплен к
гаметофиту, ножка и сложно устроенная коробочка, прикрытая
золотисто-бурым колпачком. В коробочке можно выделить
шейку, урночку и крышечку. Внутри урночки проходит колонка,
верхняя часть которой образует пленку {эпифрагму), прикры-
вающую устье урночки; кольцевидный спорангий прикреплен
нитями к колонке и стенкам урночки.
По краю урночки располагаются зубцы, составляющие пе-
ристом. Зубцы перистома совершают гигроскопические движе-
ния (во влажную погоду прижаты, в сухую - отгибаются) и тем
самым способствуют рассеиванию спор, которые разносятся
ветром (этому помогает и длинная упругая ножка, раскачиваю-
щаяся при движении воздуха). Количество спор в спорангии
очень велико (порядка 50 млн штук).
Развитие нового гаметофита проходит в две стадии. Вначале
из споры образуется разветвленная нитчатая протонема, напо-
минающая зеленую водоросль. Затем на протонеме закладыва-
ются почки, из которых вырастают листостебельные побеги с
антеридиями и архегониями.
Подкласс Sphagnidae (сфагновые мхи). В этом подклассе
всего один род сфагн (Sphagnum, рис. 2г), или сфангум, насчи-
тывающий 300 видов растений бледно-зеленого цвета, за что их
называют белыми мхами. На пластинчатой протонеме сфагна
закладывается почка, развивающаяся впоследствии в листосте-
бельный побег.
Для взрослых гаметофитов характерны большие морфоло-
гические, анатомические и физиологические отличия от брие-
вых мхов. Все представители подкласса - влаголюбивые расте-
ния, обладающие специальными приспособлениями для накоп-
ления и сохранения влаги.
Стебель - прямостоячий с многочисленными густо облист-
венными боковыми ветвями. На верхушке ветви скучены в плот-
ную головку, ниже собраны в пучки. Одна часть ветвей каждого
22
пучка оттопырена в сторону, благодаря чему побеги соединяют-
ся в дерновинки, другая свисает вдоль стебля, образуя своеоб-
разный фитиль, по которому передвигается вода.
Однослойные листья образованы из чередующихся клеток
двух типов: узких живых фотосинтезирующих и широких мерт-
вых водоносных (гиалиновых). Из гиалиновых клеток состоит
и прозрачная кожица, покрывающая стебельки и веточки. На-
личием гиалиновых клеток объясняется замечательная способ-
ность сфагна быстро и в большом количестве впитывать воду
(во влажном состоянии вес сфагна может в 20-40 раз превы-
шать вес в сухом состоянии). В сухое время года гиалиновые
клетки обезвоживаются и рост мха прекращается.
Ризоидов у взрослого растения нет. Побег неограниченно
долго нарастает в верхней части и постепенно отмирает снизу.
При этом вещества, содержащиеся в отмирающей части, ис-
пользуются растущей живой частью растения (т.е. сфагн как бы
постоянно «ест собственные ноги»).
Антеридии и архегонии образуются на особых ветвях, при
этом ветви с антеридиями часто бывают ярко окрашенными в
различные цвета (красный, фиолетовый, зеленый).
У коробочки спорогона, имеющей колонку, отсутствует пе-
ристом. Поскольку ножка спорогона короткая, он выносится
вверх сильно вытягивающейся верхушкой гаметофита - ложной
ножкой. При созревании коробочка ссыхается, давление возду-
ха внутри нее повышается и крышечка резко, с хлопком отбра-
сывается. При этом из коробочки вылетает облачко спор на
расстояние до 10 см.
Как самое нетребовательное к питательным элементам рас-
тение, сфагн может обходиться той Пылью, которая попадает
на его листья. При этом, как правило, мох растет в экологиче-
ски чистых местах, в удалении от городов и дорог, там, где ко-
личество пыли бывает ничтожно малым. В то же время боль-
шинство видов сфагна не выдерживает присутствия в воде со-
лей кальция и за счет выделяемых всей поверхностью расте-
ния органических кислот подкисляет окружающую среду до
pH = 3,5-4,5.
Сфагн формирует самые мощные (толщиной в несколько
метров) торфяные залежи верховых болот, отличающиеся край-
не низкой зольностью (5-10%) и потому бедные элементами пи-
тания. На сфагновых болотах произрастают специфические
растения, в первую очередь из семейства вересковых (Erica-
ceae), в том числе клюква (Oxycoccus palustris) и багульник
(Ledum palustre). Кроме того, на этих болотах обитает особая
карликовая форма сосны.
23
Общеизвестны целебные качества сфагна как сильного ан-
тисептика и прекрасного адсорбента (во время Отечественной
войны повязки со сфагном заменяли вату).
Заканчивая характеристику сфагновых мхов, приведем их
отличия от бриевых мхов (табл. 3).
Таблица 3
Различия бриевых и сфагновых мхов
Характери- стика Подкласс
Bryidae Sphagnidae
Особенно- сти эколо- гии Разнообразные место- обитания - от пустынь и скал до водоемов, однако наиболее массово в та- ежных лесах Специфические болотные местообитания с мягкой водой. Формируют самые мощные залежи торфа на верховых болотах
Число ви- дов Больше 14 тыс. Около 300
Основные таксоны Bryales, Andreaeales Sphagnales
Гаметофит Протонема нитчатая вет- вистая образует много по- чек. Взрослый гаметофит имеет ризоиды, рост вверх ограничен, гиалиновых клеток нет Протонема пластинча- тая образует одну почку. У взрослого гаметофита ри- зоидов нет, рост неограни- чен, в листьях и стеблях имеются гиалиновые (во- доносные) клетки
Спорофит Коробочки с перистомом Коробочки без перис- тома
Значение Формируют напочвенный покров тайги, используют- ся для биоиндикации Основные торфообразо- ватели верховых болот. Используются как тепло- изолирующий материал и заменитель ваты
3.3. Экологическое разнообразие
и механизмы устойчивости моховидных
Как уже отмечалось, моховидные обитают в очень широком
диапазоне условий среды. В их составе встречаются водные, бо-
лотные, боровые и луговые мхи, эпифитные мхи (растущие на
деревьях), а также мхи, растущие на камнях и на гниющей дре-
весине. Кроме того, существуют и другие экологические группы
мхов. Так, в пустынях есть бессубстратные мхи, которые растут
на песчинках, переносимых ветром. Есть мхи, которые поселя-
ются на навозе.
Специально рассмотрим приспособления, которые позволи-
ли мхам, несмотря на сложный жизненный цикл с уязвимостью
24
звеньев, сохранить разнообразие и роль в природе с палеозой-
ских времен. Для этого приведем табл. 4, в которой показаны
факторы, способные на разных стадиях жизненного цикла пре-
рывать его, и адаптации, помогающие мхам противостоять со-
противлению среды.
Таблица 4
Факторы риска и стратегия повышения устойчивости
к ним на разных стадиях жизненного цикла моховидных
Этап жизненного цикла Фактор риска Элементы стратегии устойчивости
Взрослый гаметофит Засуха Малые размеры, позволяющие подни- мать по растению воду без помощи спе- циализированных тканей Всасывание воды всей поверхностью Способность к криптобиозу Наличие водозапасающих клеток
Оплодотво- рение Отсутствие условий для пере- носа спер- матозоидов к архего- нию Небольшие размеры гаметофитов, повы- шающие шансы формирования капельно- жидкой среды Рост дернинками Выделение архегониями веществ, при- влекающих сперматозоиды Образование «чаш» из листьев вокруг антеридиев Использование насекомых для переноса сперматозоидов
Развитие спорофита Засуха, дефицит элементов питания Питание спорофита за счет гаметофита Защита спорофита гаметофитом: погру- жение его в такань гаметофита (печеноч- ники), разрастание архегония одновремен- но с ростом гаметофита (настоящие мхи)
Распростра- нение спор Недорас- сеивание Образование ложной ножки на гамето- фите Упругость ножки спорогона Специальные механизмы спорометания: элатеры, перистом, стреляющие коробочки
Развитие молодого гаметофита Попадание спор в ус- ловия, не- благопри- ятные для прораста- ния Огромное количество спор Репродукция гаметофитов на стадии протонемы Вегетативное размножение гаметофитов
25
3.4. Значение моховидных
Моховидные - активные участники экосистемных процессов
в лесах таежной зоны, а также на верховых и переходных бо-
лотах. Мхи входят в состав пионеров, которые заселяют скалы.
Как уже отмечалось, сфагновые мхи используются в медицине
и, кроме того, применяются населением для теплоизоляции при
строительстве деревянных домов.
В последние годы все большее значение приобретает спо-
собность мхов индицировать загрязнение среды фактом своего
присутствия или отсутствия и состоянием. В ФРГ используются
специальные живые приборы - бриометры - коробочки с рас-
тущими в них мхами, выставляемые на оживленных транспорт-
ных магистралях. По состоянию мхов оценивается интенсив-
ность загрязнения атмосферы выхлопами автотранспорта.
3.5. Заключение: Сравнение основных таксонов
Bryophyta - высшие растения небольших размеров с преобладанием в
жизненном цикле гаметофита, без корней и специализированных проводя-
щих тканей. Спорофит развивается («паразитирует») на гаметофите.
Большую роль играет вегетативное размножение. В табл. 5 приведены наи-
более общие признаки двух основных классов этого отдела.
Это интересно
Мхи и мухи. В большинстве своем споры мхов распространяются
ветром, и такие мхи - анемохорные растения. Но встречаются «зоо-
хорные» мхи, в распространении спор которых участвуют мухи. При-
мером служат представители рода сплахн (Splachnum), которые раз-
растаются на навозе (обычно, если навоз попал на сырой луг или боло-
то, куда забрела корова) в виде подушечки до 25 см в поперечнике. На
спорогоне сплахна ниже коробочки развивается яркое пурпурное,
оранжевое или желтое - кольцо, которое называется апофизой. Дости-
гая 2 см в диаметре, апофиза сильным запахом привлекает мух, к но-
гам которых приклеиваются липкие споры. Перелетая с одной «ле-
пешки» на другую, насекомые расселяют мох.
Светящийся мох. У некоторых видов мхов сформировались при-
способления для жизни в условиях крайне слабого освещения. Так,
мох схистостега перистая (Schistostega pennata), растущий в темных
пещерах, имеет долгоживущую пластинчатую протонему, состоящую из
линзовидных клеток и ориентированную перпендикулярно лучам света.
Клетки фокусируют лучи на хлоропласты, которые концентрируются у
задней стенки. Часть света при этом отражается, и вся протонема по-
блескивает нежным золотисто-зеленым цветом.
Далекая двудомность. Многие мхи двудомны. Однако вид Tortula
laevipila уникален: мужские особи одной из его разновидностей извест-
ны только в Европе, а женские - только в Северной Америке! По этой
причине он размножается только вегетативно.
26
Таблица 5
Характеристика двух классов отдела Bryophyta
Характе- ристика Класс Hepaticopsida Класс Bryopsida
Экологи- ческие ус- ловия оби- тания Влажные затененные ме- стообитания на почве, гни- лой древесине или на кам- нях Всюду, на разных суб- стратах - в воде, на кам- нях, на почве, на гнилой древесине, на навозе и т.д. Как правило, в хорошо ув- лажненных местообитани- ях, но растут даже в пус- тыне
Важней- шие таксо- ны Marchantiidae, Jungermaniidae* Bryidae, Sphagnidae, Andreaeidae*
Количе- ство видов 10 тысяч 15 тысяч
Особен- ности строения гаметофита Дорзовентральное рас- тение слоевищной или «облиственной» (у боль- шинства) формы из «стебля», вдоль которого расположены три ряда «листьев» (два спинных и один брюшной). В клетках присутствуют особые об- разования масляные тельца. Ризоиды однокле- точные. Антеридии и архе- гонии у некоторых распо- ложены на специальных подставках Радиально-симметричное растение, как правило, с вертикальным стеблем и многорядными (спирально расположенными) листья- ми. Ризоиды многоклеточ- ные. Антеридии и архего- нии расположены на вер- хушках стеблей
Особен- ности строения спорофита Коробочка без колонки и крышечки, раскрывается створками, рассеиванию спор помогают специаль- ные пружинки - элатеры Спорофит устроен более сложно: прикрыт колпач- ком (образован из архего- ния), имеется крышечка, колонка и перистом, регу- лирующий распростране- ние спор. Элатер нет
Примечание: звездочкой отмечены таксоны, которые в курсе не рассматриваются.
П
Глава 4
СОСУДИСТЫЕ СПОРОВЫЕ РАСТЕНИЯ
4.1. Общая характеристика
К этой группе относятся растения с проводящей системой,
которые в отличие от семенных растений расселяются спорами.
В жизненном цикле таких растений чередуются крупный хоро-
шо развитый спорофит и маленький просто устроенный гаме-
тофит - заросток.
В рамках сосудистых споровых растений четко дифферен-
цируются две основные группы таксонов по особенностям жиз-
ненного цикла: с одинаковыми спорами и с разными.
4.1.1. Гаметофиты
При равноспоровости огромное количество спор, как и у
моховидных, «бросается на ветер» (в прямом и переносном
смысле). И лишь незначительная часть этого богатого наследия
попадает в условия, где спора может прорасти и сформировать
заросток, как правило обоеполый. Однако у некоторых расте-
ний из числа хвощей возможно появление недоразвитых архе-
гониев или антеридиев, а также формирование однополых заро-
стков.
Равноспоровость достигает наивысшего развития у папо-
ротников, у которых многочисленные гаметофиты растут груп-
пами и развиваются быстро. За счет неодновременного созрева-
ния антеридиев и архегониев и растянутости этих процессов
происходит перекрестное оплодотворение.
У разноспоровых растений, которые есть среди папоротников и
плаунов, образуется два вида спор - крупные макроспоры, из ко-
торых развивается женский гаметофит, и мелкие микроспоры,
дающие начало мужскому гаметофиту. При этом спорофит в пер-
вую очередь «заботится» о женском гаметофите, который развива-
ется, не покидая оболочки макроспоры (эндоспорически), за счет
содержащегося в ней большого запаса питательных веществ (жи-
ров, белков, углеводов). Мужской гаметофит у разноспоровых рас-
тений тоже эндоспорический, но энергетические затраты спорофи-
та на него не сопоставимы с затратами на макроспоры, в которых
развиваются более крупные женские заростки. Количество микро-
спор и макроспор также несопоставимо: на одну макроспору при-
ходится много микроспор.
Разноспоровость в силу большей автономности женского за-
ростка от условий среды обеспечивает лучшие возможности для
выживания потомства, чем равноспоровость. В последнем случае
28
успех сохранения достигается только за счет очень большого ко-
личества спор, из которых лишь незначительная часть попадает в
условия, благоприятные для развития гаметофита.
4.1.2. Строение спорофита
У большинства сосудистых споровых растений имеются хо-
рошо развитые вегетативные органы: стебли, листья, корни, и,
как очевидно из самого названия этой группы, проводящие тка-
ни (ксилема и флоэма). Происхождение листьев двоякое: телом-
ное и энационное. Энационные листья (микрофиллы), имеющие
одну жилку, сформировались в результате поверхностного вы-
роста на теломе (такие выросты называются энациями). Телом-
ные листья (макрофиллы), образованные в результате слияния
системы теломов, имеют разветвленную систему жилок.
Макрофиллы характерны для папоротников, а микрофил-
лы - для плаунов. Природа листьев хвощей дискутируется (есть
мнение, что это редуцированные макрофиллы).
Единственной группой, у которой нет расчленения тела на
стебли, листья и корни, являются самые древние из высших
растений - проптеридофиты (Propteridophyta). Они появились
примерно 450 млн лет назад и исчезли через 100 млн лет.
4.2. Отдел Lycopodiophyta (плауновидные)
Плауновидные - это отдел споровых растений с «великим»
прошлым и сравнительно небольшим значением в современной
растительности. Во влажных мезозойских лесах, особенно в
карбоне, леса которого дали каменный уголь, плауновидные
были важной группой, представленной крупными древовидными
организмами. До наших дней дошло сравнительно немного тра-
вянистых представителей, вклад которых в биологическую про-
дукцию биосферы незначителен. К.Линней относил все плау-
ны ... к мхам!
Таксономия этого отдела выглядит следующим образом:
Класс Lycopodiopsida (равноспоровые)
Порядок Asteroxylales*
Порядок Lycopodiales (плауновые)
Порядок Protolepidodendrales*
Класс Isoetopsida (разноспоровые)
Порядок Isoetales (полушниковые)
Порядок Selaginellales (селагинелловые)
Порядок Lepidodendrales*
(Звездочкой отмечены ископаемые порядки, которые в учебнике
на рассматриваются.)
29
4.2.1. Порядок Lycopodiales (плауновые)
Этот центральный порядок отдела объединяет равноспоро-
вые растения. Главный род - плаун (Lycopodium, рис. За). В его
составе насчитывается около 200 видов (в России 7 видов),
связанных преимущественно с поясом тайги в Азии и Северной
Америке.
6)
в)
Рис. 3. Плауновидные: a Lycopodium clavatum (1 - спорофит, 2
спорофилл со спорангием, 3 - гаметофит, 4 - молодой спорофит на
гаметофите); б - Isoetes lacustris; в - Selaginella (1 - спорофит, 2 - ко-
лосок с микроспорангиями и макроспорангиями, 3 - макроспора с
женским гаметофитом, 4 - микроспора с мужским гаметофитом)
30
Для спорофита характерно дихотомическое ветвление стеб-
ля, который густо усеян листьями-мирофиллами (они узкие,
жесткие, имеют одну жилку и на зиму остаются зелеными; про-
водящая система стебля - плектостела). Особенность плаунов -
длинные надземные побеги с придаточными корнями. Растение
как бы плывет по земле, отчего пошло его русское название
плаун (в старых книгах его называли плывун). Научное назва-
ние плауна означает «волчья лапа».
Длина ветвящихся побегов может достигать десятков мет-
ров, а возраст формируемого ими клона - 300 лет. Иногда та-
кие клоны образуют «ведьмины кольца»: при центробежном
разрастании клона старые побеги в центральной части посте-
пенно отмирают.
На верхушках приподнимающихся ветвей образуются спо-
роносные колоски из более широких и светлых, чем вегетирую-
щие листья, спорофиллов. В пазухе каждого спорофилла созре-
вает спорангий почковидной формы с крупными желтыми спо-
рами. И только у некоторых видов спорангии формируются в
пазухах вегетирующих листьев.
Заростки плаунов слоевищные и бывают двух типов.
Первый тип - подземные заростки, которые образуются из
спор, попавших в почву на глубину 5 - 8 см с током воды или
благодаря дождевым червям (через их пищеварительную систе-
му споры проходят невредимыми). Подземные заростки - аб-
солютные сапротрофы, которые могут развиваться только в
симбиозе с грибом. Созревают такие заростки крайне медленно
(6-15 лет). На одном заростке может образоваться несколько
спорофитов.
Второй тип заростка - надземный. Такие заростки бледно-
зеленого цвета имеют листовидные выросты. Они фотосинтези-
руют, однако симбиоз с грибом сохраняет свою роль. Сущест-
вуют также переходные полуподземные формы заростков. Для
северных видов плаунов характерны только подземные заро-
стки.
Антеридии и архегонии развиваются неодновременно: сна-
чала заросток функционирует как мужской, затем - как жен-
ский. За счет этого происходит перекрестное оплодотворение.
Имеются факты, говорящие даже о межвидовом скрещивании
плаунов. Сперматозоид двужгутиковый.
Замедленность жизненного цикла плаунов - одна из причин
резкого сокращения их численности (некоторые виды занесены
в «Красные книги»).
До того как плауны попали на страницы «Красной книги»,
они имели практическое значение. Спорофиты были ритуаль-
ным элементом оформления гробов, а богатые маслом легкие и
сыпучие споры применялись в медицине как детская присыпка и
31
для пересыпания пилюль. Споры использовались также в пиро-
технике (под действием огня взвешенные в воздухе споры взры-
ваются ярким пламенем, причем достаточно безопасным, так
как при мгновенном сгорании спор воздух не успевает нагреть-
ся) и в фасонном литье металлических изделий (слои газов, об-
разующиеся при сгорании масла, не позволяют изделию прили-
пать к форме и придают металлу гладкую поверхность), на-
пример на заводе художественного литья в г. Касли.
422. Порядок Isoetales (полушниковые)
Два других порядка плауновидных Isoetales и Selaginel-
lales - включают разноспоровые растения, у которых имеются
макро- и микроспорагнии. Из спор, образующихся в этих спо-
рангиях, развиваются женские и мужские заростки.
Род шильник, или полушник (Isoetes, рис. 36), включает
около 70 видов (в России - 4 вида) водных или земноводных
некрупных (до 20 см, некоторые до 90 см) растений. Шильники
распространены в олиготрофных водоемах от Гренландии до
тропиков и могут образовывать сплошные заросли на глубине
до 3 м. При временном пересыхании водоема они способны
впадать в состояние криптобиоза.
Стебель шильника, называемый протокормом, представляет
собой веретеновидное тело, от верхней части которого отходят
многочисленные (до 400) спирально расположенные листья, а от
нижней - корни.
Различают стерильные и фертильные листья - микроспоро-
филлы с микроспорангиями и макроспорофиллы с макроспо-
рангиями. Спорангий расположен в нижней расширенной части
листа в овальном углублении, над ним имеется треугольный
язычок. Количество макроспор в макроспорангии достигает де-
сятков (у некоторых до 2000), а количество микроспор в микро-
спорангии - нескольких тысяч (у некоторых до миллиона). Спе-
циального механизма для вскрывания спорангия нет, споры ос-
вобождаются после разрушения его стенки и распространяются
водой. У некоторых Шильников возможна автоспория: в основа-
нии листа вместо спорангия вырастает молодое растеньице спо-
рофита.
Выше спорангия лист резко сужается и становится сильно
вытянутым и заостренным на конце. Форма листьев отражена в
русском названии - шильник. Латинское название означает
«всегда одинаковый» и отражает вечнозеленость растения.
Практическое значение этих своеобразных древних расте-
ний невелико. Их разводят в аквариумах, в оранжереях. На се-
вере шильники служат кормом оленям.
Все четыре вида шильника, встречающиеся в России, вклю-
чены в ее Красную книгу.
32
4.2.3. Порядок Selaginellales (селагинелловые)
Род селагинелла (Selaginella, рис. Зв) включает около 1000
(700) видов, распространенных преимущественно в тропических
и субтропических областях, хотя некоторые его представители
заходят на территорию России (Selaginella selaginoides и др.,
всего 8 видов). Селагинелла отличается редкой теневыносливо-
стью, некоторые виды могут расти там, где не выживают другие
растения - при освещенности 0,2% полного солнечного света.
Большинство видов растет на влажных почвах, хотя имеются и
устойчивые к засухе виды, произрастающие на гранитных ска-
лах (S. sanguinolenta в горах Азии). Все селагинеллы, как мхи
или шильники, обладают способностью к криптобиозу.
По внешнему облику селагинеллы очень похожи на миниатюр-
ные плауны, чем объясняется их название плаунок (умень-
шительное от плаун - Selago). Но есть и представители со стебля-
ми толщиной до 0,5-0,8 см, возвышающимися над землей на 1 м.
Однако у большинства селагинелл, в отличие от плаунов,
стебли имеют дорзовентальное строение и несут более крупные
боковые и более мелкие спинные листья. От стеблей отходят
длинные корнеподобные выросты ризофоры, которые дают
многочисленные придаточные корни.
Спорносные колоски образуются на верхушках боковых ве-
точек. В колосках в пазухах спорофиллов располагаются круг-
лые макроспорангии с четырьмя макроспорами и овальные мик-
роспорангии с многочисленными микроспорами. К моменту со-
зревания спор спорангий трескается, и споры выпадают или
даже «выстреливаются»: микроспоры на 2 см, а макроспо-
ры - на 10 см. Заростки, как у всех разноспоровых растений,
эндоспорические: развиваются внутри оболочки споры.
У некоторых видов селагинеллы в макроспорангии развива-
ется только одна макроспора, и ее прорастание начинается еще
внутри макроспорангия (колосок в это время может быть обло-
ман и лежать на земле). Это важное явление создает предпо-
сылки для образования семязачатка уже у споровых растений.
Практическое значение селагинеллы невелико. Некоторые
тропические виды используются в фитодизайне.
4.3. Отдел Equisetophyta (хвощевидные)
Хвощевидные - древняя группа растений, расцвет которой
пришелся на каменноугольный период (400-250 млн лет назад).
В это время древовидные хвощи были доминантами болотных
лесов, а их питательные стробилы - лакомством для динозав-
ров. У хвощевидных очень характерная внешность. Поскольку у
них членистый, легко разламываемый в узлах стебель, раньше
33
отдел назывался «членистостебельные». В зеленом фотосинте-
зирующем стебле хвощей имеются многочисленные полости, а
чешуевидные, сросшиеся в «воротничок», охватывающий узел,
листья утеряли способность к фотосинтезу.
Ископаемые хвощи достаточно многочисленны, из них наи-
более изучены роды каламит (Calamites) и клинолист (Spheno-
pyllum). (Относимый ранее к хвощам род гиения - Hyenia - по-
сле тщательного изучения оказался папоротником.) Сегодня
хвощевидные представлены всего одним родом хвощ (Equise-
tum, рис. 4), в составе которого около 20 видов, причем, среди
них уже нет крупных форм.
Рис. 4. Equisetum arvense (1 - вегетативный побег, 2 - спороносный
побег с колоском, 3 - спорангиофор, 4 - спора, 5 - группа спор)
Хвощи растения Северного полушария, в Южном полу-
шарии они встречаются редко. В Африке и Австралии хвощи
существуют только как заносные растения, а в Новой Зеландии
их нет совсем. В России насчитывается 10 видов, а также ряд
гибридов между ними.
Научное название роду - «лошадиный хвост» - дал Плиний
Старший, который увидел сходство морфлогии хвоща с конским
хвостом. От корня «хвост» происходит и русское название рода.
34
Среди хвощей преобладают невысокие растения (до 1 м),
однако у одного вида хвоща многощитинкового (Е. myrio-
chaetum) - длина стебля достигает 9 м. Эти стебли слабые и не
в состоянии самостоятельно удерживаться в вертикальном по-
ложении. Они опираются на соседние растения.
У разных видов хвоща по-разному дифференцированы спо-
роносные и вегетативные побеги. У хвоща приречного (Е. flu-
viatile) они не различаются, и спорангии образуются поздно на
верхушках побегов. До этого невозможно определить, на каком
побеге будут закладываться спороносные колоски. У хвоща по-
левого (Е. arvense), напротив, спороносные и вегетативные побе-
ги резко различаются. Розовато-бурые неветвящиеся споронос-
ные побеги появляются рано весной и развиваются за счет за-
пасов питательных веществ в клубеньках корневища. Вегетатив-
ные побеги появляются позже, после созревания спор и усыха-
ния спороносных побегов. У хвоща лесного (Е. sylvaticum) од-
новременно появляются вегетативные и спороносные побеги, и
после рассеивания спор спороносные побеги функционируют
как вегетативные.
У хвощей причудливо сочетаются ксероморфные и гигро-
морфные признаки. Как и для гигрофитов, для них характерны
полости внутри стебля, причем кроме главной центральной по-
лости внутри ребристого стебля имеются многочисленные более
мелкие полости (под ребрами и под ложбинками). Как у ксеро-
фитов (растений сухих местообитаний), у хвощей редуцированы
листья - они мелкие и незеленые. Кроме того, устойчивость к
засухе обеспечивается сложно устроенными устьицами, регули-
рующими испарение, и развитой механической тканью. Эпидер-
ма стеблей хвощей содержит кремнезем, что делает хвощи (в
особенности хвощ зимующий Е. hyemale) хорошим средством
для чистки кастрюль.
Все хвощи - многолетние растения и хорошо размножаются
вегетативно корневищами, которые бывают двух типов: гори-
зонтальные и вертикальные.
Спороносный колосок состоит из спорангиофоров, не по-
хожих на листья. Спорангиофор - это щиток с ножкой, на ко-
тором с нижней стороны сидят спорангии. В молодом колоске
щитки сомкнуты, а при созревании спор ось колоска удлиняет-
ся и щитки раздвигаются. Спорангии вскрываются на верхуш-
ках, и споры высыпаются. Кроме обычных двух оболочек, спо-
ры имеют еще дополнительную из двух лент (элатер), закру-
ченных вокруг споры. Ленты гигроскопичны и при высыхании
раскручиваются, при этом споры сцепляются и высыпаются
группами. (Движение элатер можно наблюдать под микроско-
35
пом: вытрясти споры из колоска на предметное стекло, поды-
шать на них и наблюдать при малом увеличении.)
Все споры у хвоща внешне одинаковы, и образующиеся из
них гаметофиты чаще всего обоеполые. Однако при неблаго-
приятных условиях архегонии могут быть недоразвиты или во-
обще не развиваются. Так появляются мужские заростки: они
очень маленькие (несколько миллиметров) и представляют со-
бой зеленую пластинку с небольшими лопастями. Иногда разви-
ваются женские заростки, они крупнее и сильнее рассечены.
Длительное время в учебной литературе образование одно-
полых заростков из одинаковых спор трактовалось как «физио-
логическая разноспоровость» и рассматривалось как ступень на
пути возникновения настоящей морфологической разноспоро-
вости. После опубликования работ литовского ботаника Й.Нау-
ялиса связь между условиями среды и полом заростков оспари-
вается. Причины появления однополых заростков не выяснены.
Заростки развиваются в течение нескольких недель, и после
перекрестного оплодотворения (у обоеполых заростков за
счет неодновременного созревания антеридиев и архегониев)
образуют молодые спорофиты.
Большинство хвощей ядовиты для домашних животных и
при массовом содержании в сене могут вызывать отравление
скота. Человек без риска навредить здоровью употребляет в
пищу спороносные побеги хвоща полевого. Несколько видов
хвощей, в особенности хвощ полевой, используются в народной
медицине. Жесткими стеблями хвощей иногда полируют рога и
древесину, очищают металлические поверхности. Некоторые
хвощи - злостные сорняки.
4.4. Отдел Polypodiophyta (папоротниковидные)
4.4.1. Общая характеристика
Этот обширный отдел сосудистых споровых растений вклю-
чает 300 родов и более 10 тысяч видов. Подобно мхам, папо-
ротники распространены в широком спектре экологических ус-
ловий - от пустынь, болот, рисовых полей и озер с солоноватой
водой до тропических лесов, причем именно с тропическим поя-
сом связано основное разнообразие папоротников.
Строение спорофита. Папоротники сильно различаются по
размерам. Среди них есть крупные растения высотой до 25 м и
диаметром ствола до 50 см и крошечные растеньица всего в не-
сколько миллиметров.
То, что обычно называется папоротником, это спорофит-
ное поколение. Как правило, спорофиты папоротников - много-
36
летние растения с подземным корневищем, однако есть неболь-
шое число однолетних видов. Жизненные формы папоротников
разнообразны, это «деревья» (ткани у них первичны по проис-
хождению, камбия нет), травы, лианы и многочисленные эпифи-
ты, без которых нельзя представить себе тропический лес.
В отличие от других сосудистых споровых растений у папо-
ротников по массе преобладают листья, которые и составляют
наиболее заметную часть спорофита. Листья крупные, в почке
сложены улиткообразно, нарастают верхушкой и несут споран-
гии. Такие листья, называемые плосковетками - вайями, явля-
ются макрофиллами. Молодые вайи покрыты волосками и че-
шуйками.
Большинство лесных папоротников умеренных областей
имеют мясистые, сифоностелические корневища, на которых
каждый год образуются новые пучки листьев. Листья чаще все-
го перистые, хотя есть папоротники с цельными листьями, на-
пример листовик («тещин язык» - Phyllitis scolopendrium).
Спорангии. Папоротники, за исключением нескольких ро-
дов, - равноспоровые растения. Спорангии у них располагаются
на листьях и различаются по положению и строению. Наиболее
древние формы имели верхушечное и краевое расположение
одиночных спорангиев, у большинства современных папоротни-
ков спорангии находятся на нижней стороне листа. При этом
обычно спорангии собраны в группы - сорусы. У древних папо-
ротников спорангии созревали одновременно, для современных
характерно последовательное созревание, начинающееся от
центра группы.
Групповое положение спорангиев позволяет обеспечить их
лучшее снабжение элементами питания за счет крупного прово-
дящего пучка, подходящего к сорусу. У многих родов сорусы
защищены особыми выростами листа - покрывальцами, или ин-
дузиями. Кроме того, в группе молодые спорангии, которые
только еще начинают развиваться, находятся под защитой уже
созревших спорангиев.
Защитную роль играет и спиральное свертывание молодых
листьев: внутри такой «улитки» уже начинают вызревать сору-
сы. У некоторых видов (мараттиевые папоротники) защита спо-
рангиев обеспечена еще более надежно - они срастаются в си-
нангии.
У некоторых папоротников (ужовниковых) спорангии обра-
зуются из группы клеток и имеют многослойную стенку, а у
большинства - из одной клетки и имеют стенку из одного слоя
клеток.
Для разных групп папоротников характерно свое приспо-
собление для разбрасывания спор. У наиболее просто организо-
37
ванных форм (ужовниковых) спорангии просто разрываются
поперечной щелью. У наиболее развитых папоротников (под-
класс полиподииды), например у щитовника (Dryopteris filix-
mas) или орляка (Pteridium aquilinum), спорангий вскрывается
с помощью специального механического кольца. Это кольцо из
клеток с неравномерно утолщенными оболочками опирается на
ножку спорангия, охватывая 2/3 его периметра. При высыха-
нии в клетках создается высокое давление (до 300 атм), при
этом кольцо сжимается и спорангий разрывается поперечной
трещиной. Кольцо внезапно распрямляется и играет роль ката-
пульты, разбрасывающей споры.
У папоротников количество спор несравненно меньше, чем у
моховидных. При этом чем сложнее строение папоротника и
лучше его приспособления к условиям среды, тем меньше спор
он образует. У ужовниковых в одном спорангии может быть до
15 тысяч спор, но у большинства папоротников число спор из-
меряется сотнями и десятками. Вероятность того, что спора
прорастет в гаметофит и в результате оплодотворения возник-
нет новый спорофит, у папоротников в тысячи раз выше, чем у
моховидных, где счет спор идет на миллионы.
Споры папоротников имеют многослойную оболочку и хо-
рошо переживают неблагоприятные условия, длительное время
сохраняя способность к прорастанию.
Гаметофиты. Споры равноспоровых папоротников дают на-
чало свободноживущим обоеполым гаметофитам. Они могут
быть автотрофными зелеными (как у большинства папоротни-
ков средней полосы) или гетеротрофными, как у ужовниковых.
Гетеротрофные гаметофиты, как и гаметофиты плаунов, разви-
ваются в симбиозе с грибом.
Автотрофные гаметофиты-заростки большинства папорот-
ников пластинчатые с ризоидами и имеют сердцевидную форму,
размером не более 5 мм. Несмотря на обоеполость таких гаме-
тофитов, самооплодотворение случается редко, так как антери-
дии появляются раньше и в области ризоидов, а архегонии
позже и вблизи выемки на переднем конце гаметофита. Разница
во времени формирования двух типов гаметангиев способствует
перекрестному оплодотворению, в итоге чего появляется более
жизнеспособный спорофит.
У разноспоровых папоротников, как и у разноспоровых
плаунов, однополые гаметофиты развиваются под облочками
микроспор и макроспор и сильно упрощены.
Кроме типичного жизненного цикла со стадиями «споро-
фит - спора гаметофит спорофит», многие папоротники
имеют дополнительные варианты размножения путем образова-
38
ния гаметофитов из вегетативных клеток спорофита (апоспо-
рия) и спорофитов из соматических клеток гаметофита (апога-
мия). Отмечаются и случаи партеногенеза — образования ново-
го спорофита из яйцеклетки гаметофита без оплодотворения.
Весь этот разнообразный арсенал приспособлений к небла-
гоприятным условиям при спороношении, развитии гаметофита
и спорофита объясняет высокую устойчивость папоротников к
влиянию разнообразных неблагоприятных факторов, включая
воздействие человека.
Основные таксоны. Рассматриваемые таксоны отдела Poly-
podiophyta соподчинены следующим образом:
Класс Ophioglossopsida (ужовниковые)
Класс Marattiopsida (мараттиевые)
Класс Polypodiopsida (полиподиопсиды)
Подкласс Polypodiidae (полиподииды)
Подкласс Marsileidae (марсилеевые)
Подкласс Salviniidae (сальвиниевые)
Ужовниковые это древние травянистые папоротники с
примитивным строением, мараттиевые - тоже древние, но дре-
вовидные папоротники, широко представленные в тропическом
поясе. Наконец, полиподиопсиды - самый большой и разнооб-
разный класс папоротников: в него входят не менее 270 родов и
до 10 тыс. видов. В этом классе, кроме подкласса Polypodiidae,
объединяющего наземные папоротники, есть особая группа вод-
ных разноспоровых папоротников, которые выделяются в два
подкласса - сальвиниевых и марсилеевых. Иногда их объединя-
ют в один порядок водных папоротников - Hydropteridales, од-
нако такое объединение неудачно, так как эти папоротники су-
щественно различаются строением споронесущих органов.
4.4.2. Класс Ophioglossopsida (ужовниковые)
Этот небольшой класс безусловно примитивных папоротни-
ков представлен одним порядком Ophioglossales, двумя семейст-
вами - Ophioglossaceae и Botrychiaceaea, соответственно с дву-
мя главными родами ужовник (Ophioglossum) и гроздовник
(Botrychium).
Ужовник (рис. 5а) включает около 50 видов, распространен-
ных и в умеренной, и в тропической зоне (в России - 4 вида).
Название Ophioglossum в переводе с латинского означает
«змеиный язык» (русское название рода ужовник тоже
имеет «змеиный» корень). Для флоры умеренной зоны обычен
О. vulgatum, растущий в укромных местах по лесным опушкам,
вырубкам, сырым заросшим мхами лугам.
Ужовники - многолетние растения с небольшим вертикаль-
ным корневищем (с микоризными придаточными корнями) и од-
39
ним двураздельным листом, одна часть которого фотосинтези-
рует, а другая спороносит. В молодом состоянии лист улитко-
образно не свернут. Спорангии, образуемые из нескольких
клеток спороносной части листа, имеют многослойную стенку и
лишены приспособлений для вскрывания: они просто разрыва-
ются щелью. Корневища живут долго, сохраняясь в почве в хо-
лодное время года. Время их жизни сравнимо с возрастом де-
ревьев и может превышать сто лет.
6)
в) г)
Рие. 5. Папоротниковидные: а - Ophioglossum; б - Dryopteris filix-mas
(1 — спорофит, 2 - нижняя сторона листа, 3 — сорус, 4 - спорангий,
5 гаметофит)^ в - Marsilea quadrifoliay г - Salvinia natans (1 спо-
рофит, 2 - женский гаметофит, 3 — сорусы микроспорангиев и макро-
спорангиев )
40
Шаровидные споры ужовников долго сохраняют всхожесть
и прорастают в темноте.
Червеобразный гаметофит живет в поверхностных слоях
почвы, лишен зеленой окраски и на ранних этапах онтогенеза
входит в симбиоз с грибом. Гаметофит обоеполый, но первыми
созревают антеридии, и фактически он функционирует как од-
нополый, вначале как мужской, а потом - как женский.
Гроздовник включает около 40 видов, они характерны для
тропического пояса, но большинство этих видов в небольшом
количестве встречаются и в умеренной зоне (в России - 8 ви-
дов). Редкий для России гроздовник простой (В. simplex) зане-
сен в Красную книгу.
Ужовниковые - «медлительные» растения: гетеротрофный
гаметофит формируется 10 - 20 лет (т.е. дольше, чем у плау-
нов); развивающйся из зиготы спорофит долго «вскармливает-
ся» на гаметофите и только через несколько лет образует пер-
вый, еще подземный лист.
Это интересно
«Ключ-трава». С ужовниковыми связано много разных небылиц.
Так, в США их считают «кладоуказателями», которые помогают нахо-
дить закопанные драгоценности. Там же полагают, что присутствие
гроздовника облегчает поиск одного из видов женьшеня.
4.4.3. Класс Marattiopsida (мараттиевые)
Это древняя группа тропических папоротников, история ко-
торых, в отличие от ужовниковых, прослеживается по ископае-
мым формам. Их расцвет связан с каменноугольным периодом,
когда они в большом количестве были распространены по всем
континентам. До нас мараттиевые дошли в обедненном составе
(хотя и не столь бедном, как ужовниковые). Современные по-
томки мараттиевых почти не отличаются от своих мезозойских
предков, что делает их изучение особенно интересным.
До наших дней дожили 7 родов, виды которых обитают
только в тропической зоне. Эти красивые величественные рас-
тения достигают крупных размеров за счет огромных листьев.
Их небольшие, часто клубневидные стебли частично погружены
в почву, но листья (которые в почке улиткообразно свернуты)
вырастают до 6 м в длину и имеют мощный черешок толщиной
в человеческую руку.
Корни мараттиевых образуют микоризу, но в отличие от
ужовниковых эта микориза факультативная, поскольку расте-
ние может нормально развиваться и без нее.
Спорангии у мараттиевых образуются, как и у ужовнико-
вых, из группы клеток и имеют многослойную стенку. Они рас-
41
положены с нижней стороны листа тесными рядами или сра-
стаются в «соплодия» - синангии. В одном синангии может
быть до 100 спорангиев.
Гаметофиты наземные, зеленые, обоеполые, но в отличие от
рассматриваемых далее современных папоротников полиподиид
живут долго (несколько лет), а иногда не отмирают и после
развития спорофита.
Наиболее известны роды ангиоптерис (Angiopteris - около
100 видов) и мараттия (Marattia - 60 видов), обитающие в тени-
стых ущельях и на влажных склонах дождевых тропических ле-
сов. В наших северных областях мараттиевые папоротники
можно увидеть только в оранжереях, где чаще всего разводят
ангиоптерис вознесенный (A. evecta).
4.4.4. Класс Polypodiopsida
Подкласс Polypodiidae (полиподииды). Полиподииды очень
разнообразны и разделяются на много порядков и семейств.
Только в России распространены представители 19 семейств, 46
родов и 120 видов. Наиболее крупными встречающимися у нас
родами являются Dryopteris (14 видов), Athyrium (13 видов),
Polystichum (8 видов), Asplenium (8 видов), а подавляющее
большинство родов представлено всего 1-2 видами.
Для всех полиподиид характерно строение, с которым у нас
и ассоциируется понятие «папоротник» (рис. 56). Они имеют
подземное корневище и надземные листья-вайи, на нижней сто-
роне которых на бугорках расположены сорусы, прикрытые
индузием. Спорангии снабжены уже упомянутым кольцом для
вскрывания. У небольшого числа родов, в том числе и у обыч-
ного для наших лесов Matteuccia (М. struthiopteris - страусиное
перо), есть специальные фертильные вайи (коричневого цвета,
поэтому они кажутся усохшими).
В экологическом отношении среди полиподиид различают
две группы: лесные папоротники (роды Dryopteris, Pteridium,
Matteuccia и др.) и наскальные (роды Cystopteris, Asplenium,
Polypodium, Woodsia и др.). Наскальные папоротники отличают-
ся высокой теневыносливостью и могут жить в темных расще-
линах скал и старых каменных стен. Это качество делает их
привлекательными для фитодизайна (Nephrolepis, Adianthum и
др.): они могут расти в темных частях помещений (под лестни-
цами и др.).
Некоторые наскальные папоротники (Asplenium septentrio-
nale, Polypodium vulgare и др.) могут переносить значительное
обезвоживание в период летних засух. Они усыхают до состоя-
ния, когда листья от прикосновения крошатся, но при этом бы-
стро восстанавливаются после дождей.
42
Наибольший ареал имеет орляк (Pteridium aquilinum), пред-
ставляющий редкий пример «естественного» космополитизма
(как правило, космополиты - это виды, распространение кото-
рых связано с человеком). Он встречается на всех материках,
кроме Антарктиды.
Практическое значение полиподиид невелико. Кроме фито-
дизайна, некоторые виды (например, орляк) употребляют в пи-
щу. Для этого заготавливают молодые листья-«улитки», кото-
рые после вымачивания (при этом удаляются ядовитые вещест-
ва) засаливают, получая продукт, по вкусу напоминающий гри-
бы. Корневища щитовника мужского (Dryopteris filix-mas) ис-
пользуют в медицине в качестве глистогонного средства.
Два дальневосточных вида - пиррозия язычковая (Pyrrosia
lingua) из семейства многоножковых (Polypodiaceae) и краекуч-
ник Куна (Cheilanthes kuhnii) из семейства птерисовых (Pteri-
daceae) - включены в Красную книгу России. Много других ви-
дов включено в ряд региональных Красных книг.
Подкласс Marsileidae (марсилеевые). Эта небольшая группа
разноспоровых папоротников обитает в воде или на побережьях
и названа К.Линнеем в честь итальянского ботаника Л.Марсиля.
В подклассе всего три рода, самый известный из которых мар-
силея (Marsilea, содержит около 70 видов, три из них встреча-
ются в России).
Для марсилеевых характерны спорокарпии, которых нет у
других папоротников. Спорокарпии — это твердые образова-
ния, в которых формируются сорусы микро- и макроспоранги-
ев. Оболочка спорокарпиев имеет листовое происхождение.
Благодаря очень твердой оболочке спорокарпии выдержи-
вают сильные механические воздействия и пересыхание, пре-
красно защищая спорангии, стенки которых тонки и прозрачны.
С помощью кольца в оболочке зрелый спорокарпий раскрыва-
ется на две створки, но только в воде за счет набухания осо-
бой ткани, поэтому прорастание спор происходит в воде. Сухие
спорокарпии не раскрываются, а споры в них сохраняют жиз-
неспособность десятки лет.
В южной части России на прибрежных сырых песках обыч-
на марсилея четырехлистная (Marsilea quadrifolia, рис. 5в). Ее
своеобразные листья отходят из стелящегося по поверхности
сырой почвы корневища. Каждый лист состоит из длинного че-
решка и четырех листочков, напоминающих листочки клевера
(ночью они «спят» складываются и повисают вниз), за что
марсилею называют водяным клевером.
Спорокарпии, сидящие на ножках в нижней части черешка,
по форме и размерам похожи на мелкие семена фасоли. В мик-
роспорангиях образуется по 64 микроспоры, в макроспоранги-
43
ях - по одной макроспоре, наполненной питательными вещест-
вами. Спорокарпии распространяются водными потоками и во-
доплавающими птицами, через пищеварительную систему кото-
рых они проходят неповрежденными (т. е. марсилеи являются
одновременно гидро- и зоохорными растениями).
Женский гаметофит представлен практически одним архего-
нием, его развитие начинается внутри макроспоры, находящейся
еще в макроспорангии (этот процесс напоминает развитие гаме-
тофита семенных растений), и происходит очень быстро.
Мужской гаметофит состоит из двух антеридиев с неболь-
шим (16) числом сперматозоидов, его вегетативная часть сведе-
на всего к одной клетке, которая называется проталлиальной.
Весь процесс развития этого крохотного заростка, включая дос-
тижение сперматозоидом яйцеклетки, завершается за 24 часа.
Сперматозоиды адаптированы к быстрому перемещению в вод-
ной среде (они закручены спиралью в 13-14 оборотов и имеют
множество жгутиков).
Особенности гаметофитов разноспоровых папоротников
хорошо иллюстрируют биологические и экологические преиму-
щества разноспоровости: чрезвычайно маленькие и очень про-
сто устроенные гаметофиты, «проскакивая» эту опасную в ус-
ловиях наземного существования стадию жизненного цикла,
быстро образуют зародыш дочернего спорофита. Шансы выжи-
вания зародыша повышаются тем, что на первое время для него
имеется достаточный запас питательных веществ, накопленных
материнским спорофитом и не израсходованных женским гаме-
тофитом.
Значение марсилей невелико. В дельте Волги они входят в
состав сообществ, которые затапливаются в период паводка и
служат нерестилищем для рыбы. Нередко они развиваются на
рисовых полях как сорняки. Используются марсилеи как аква-
риумные и оранжерейные растения. Некоторые виды их съе-
добны.
Это интересно
Хлеб из папоротника. В спорокарпиях австралийского вида Marsi-
lea drummondii накапливается так много крахмала, что местное населе-
ние готовит из него «нарду», подобное хлебу. О «нарду» упоминает
даже Жюль Верн.
Подкласс Salviniidae (сальвиниевые). Свое название эти па-
поротники получили в честь итальянского ботаника XVII в.
А.Сальвини. Как и марсилеевые, это водные разноспоровые рас-
тения с раздельнополыми сильно редуцированными гаметофи-
тами, развивающимися внутри оболочек микро- и макроспор.
44
Однако спорокарпиев у них нет, а микро- и макроспорангии
образуются в разных сорусах.
В подклассе всего два рода - сальвиния (Salvinia, 8 видов) и
азолла (Azolla, 6 видов), выделенные в отдельные семейства
Salviniaceae и Azollaceae.
Сальвиния распространена в основном в Африке и Амери-
ке, но один вид - сальвиния плавающая (S. natans) обитает в
России. Это небольшое растение имеет тонкий ветвящийся сте-
бель с мутовками из трех листьев, два из которых ярко-зеленые
и плавают на поверхности, а третий - корневидный, рассечен на
нитевидные доли и погружен в воду (рис. 5г).
Сорусы, образующиеся у основания корневидных листьев,
закрыты шаровидным двуслойным индузием и внешне напоми-
нают спорокарпии марсилеевых. После созревания они опуска-
ются на дно и там разрушаются. Освобожденные спорангии
поднимаются к поверхности, где происходит прорастание спор.
Мужской гаметофит, развивающийся из микроспоры, как и
у марсилеи, очень сильно редуцирован, хотя имеет не одну, а
две проталлиальные клетки. Антеридиев у него тоже два.
Женский гаметофит, как и у марсилеи, начинает разви-
ваться в макроспоре, которая не покидает макроспорангия. Од-
нако, разрастаясь, он разрывает оболочку макроспоры и ма-
краспорангия, высовывается в месте разрыва и зеленеет. При
этом гаметофит значительно увеличивается, превосходя разме-
рами макроспору. Затруднений в оплодотворении не бывает, так
как споры развиваются в воде, поэтому сперматозоид легко
достигает архегониев (их 3 - 5, и находятся они на открытой
части женского гаметофита).
Важную роль в хозяйстве южных стран играет азолла, ко-
торая ведет такой же образ жизни, что и сальвиния, но отлича-
ется тем, что в ее листьях живут азотфиксирующие цианобак-
терии из рода анабена (семейство Nostocaceae). По способности
накапливать азот азолла — бесспорный чемпион: ковер из па-
поротника, покрывающий воду, может накопить в год до
1000 кг/га азота (для сравнения: поле клевера не более
200 кг/га). Эта особенность азоллы широко используется в аг-
ротехнике рисосеющих стран: в Корее, Вьетнаме, Индии, Китае
и др. Азолла, растущая между стеблей риса, является хорошим
азотным удобрением, которое повышает его урожай.
Это интересно
Агрессивная сальвиния. В благоприятных условиях сальвинии бы-
стро разрастаются за счет вегетативного размножения. Завезенная из
Южной Америки в Африку сальвиния ушковидная (Salvinia auriculata)
45
настолько акклиматизировалась, что стала злом. Она так заполонила
водоемы, что препятствует судоходству, образуя на поверхности воды
слой толщиной до 25 см. Сальвиния забивает сети рыбаков, мешает ра-
боте турбин гидроэлектростанций, образует запруды в речных рукавах.
4.5. Экологическое разнообразие и механизмы
устойчивости сосудистых споровых растений
По экологическому разнообразию сосудистые споровые
растения значительно уступают моховидным. Среди них нет на-
стоящих ксерофитов. Это растения хорошо увлажненных ме-
стообитаний (или растущие в воде), часто теневыносливые.
Неустойчивость этих растений к засухе является основной
причиной ограниченности их распространения. И тем не менее,
эти «дети палеозоя», которые процветали в мезозое, особенно в
каменноугольный период (именно на продукции их фотосинтеза
сегодня работает большая часть тепловых электростанций ми-
ра!), дожили до наших дней.
Рассмотрим механизмы, которые позволили этой группе
растений сохраниться (табл. 6).
Таблица 6
Факторы риска и стратегия повышения устойчивости к ним
споровых сосудистых растений на разных стадиях жизненного цикла
Фаза Фактор риска Адаптации для преодоления
Спорофит Засуха, влияние человека Развитие корневой системы Переживание неблагоприятных условий в стадии корневищ Криптобиоз Редукция листьев (хвощи), совершенст- вование проводящей системы и покровов (папоротники)
Спороно- шение Засуха Защита спорангиев сомкнутыми споран- гиофорами (хвощи), индузием (папоротни- ки), формирование синангиев и спорокар- пиев (папоротники)
Распро- странение спор Неполное рассеива- ние, попа- дание спор в неблаго- приятные условия Возникновение различных вариантов «баллистических» аппаратов (папоротни- ки) Формирование покровов спор, позво- ляющих им длительное время сохранять способность к прорастанию и дожидаться благоприятного для прорастания стечения обстоятельств Вегетативное размножение, апогамия и апоспория, партеногенез (папоротники)
46
Окончание табл. 6
Фаза Фактор риска Адаптации для преодоления
Формиро- вание га- метофита Засуха, дефицит элементов питания Возникновение разноспоровости (плау- ны, папоротники) с повышением автоном- ности женского заростка. Он и форми- рующийся на нем дочерний спорофит обеспечиваются питанием за счет запасов в макроспоре Ускоренное развитие автотрофных обое- полых заростков Замедленное развитие гетеротрофных заростков в симбиозе с грибом
Успех оп- лодотворе- ния Отсутствие капельно- жидкой среды, са- мооплодо- творение Скученное произрастание заростков, рассеивание спор «хлопьями» за счет эла- тер (хвощи) «Прижимание» заростков к поверхности почвы или развитие их в почве «Протандрия »
4.6. Заключение: сравнение отделов
Мы завершили рассмотрение трех отделов сосудистых растений, в
которых наблюдается большое разнообразие строения спорофита, осо-
бенностей спороношения, развития и организации гаметофитов. При
этом общим для всех сосудистых споровых растений (как, впрочем, и
моховидных) является то, что оплодотворение возможно только при
наличии воды, в которой перемещаются сперматозоиды. Снять это ог-
раничение смогли только рассматриваемые в гл. 5 семенные растения,
что и позволило им стать господствующими на Земле.
Кроме того, сосудистые растения объединяет то, что в их жизнен-
ном цикле преобладает спорофит с развитыми проводящими тканями и
корневой системой. Автономный гаметофит (заросток) имеет малые
размеры. Обязательным условием полового процесса является наличие
капельно-жидкой воды. Различия отделов показаны в табл. 7.
Во всех трех отделах сосудистых споровых растений Eguise-
tophyta, Lycopodiophyta, Polypodiophyta наблюдается параллелизм
вариантов жизненного цикла. Во всех группах появляется разноспоро-
вость с образованием раздельнополых гаметофитов. При этом намеча-
ются предпосылки для перехода к семенному размножению:
гаметофиты сильно редуцированы, они быстро развиваются
внутри оболочек микро- и макроспор. Особенно упрощается мужской
гаметофит, который состоит из одной или нескольких проталлиальных
клеток и одного-двух антеридиев;
у женского гаметофита появляется и усиливается тенденция к
«выкармливанию» развивающегося зародыша. Женский гаметофит бо-
лее крупный, наполнен питательными веществами, накопленными мак-
роспорой за счет спорофита, и лишь незначительная часть этих запасов
расходуется на формирование архегониев, а большая часть идет на пи-
тание развивающегося зародыша;
47
- число макроспор в макроспорангии сокращается до одной (на-
пример, у рода шильник развивается много макроспор, у большинства
селагинелл - от 16 до 4, а у некоторых 2 и даже 1, у всех водных па-
поротников - 1);
- развитие женского гаметофита начинается еще тогда, когда мак-
роспора находится внутри макроспорангия. Так происходит у некото-
рых селагинелл и у марсилеи. Это можно рассматривать как прелюдию
к формированию семязачатка.
Таблица 7
Сравнительная характеристика сосудистых споровых растений
Признак Отделы
Equisetophyta Lycopodiophyta Polypodiophyta
Число видов Около 20 Около 1200 Более 10 тысяч
Географиче- ское распрост- ранение Преимущест- венно умерен- ные широты се- верного полу- шария Субтропики и тропики (Isoeto- psida) и умерен- ные широты (Lycopodiopsida) Преимущест- венно в тропи- ческом и суб- тропическом поясе, встреча- ются и в уме- ренных широтах
Листья Редуцирован- ные макрофил- лы, незеленые Микрофиллы Макрофиллы, у наземных вайи, у водных - про- стые или слож- ные листья
Тип спороно- шения Равноспоро- вость Равно- и разноспоровость
Тип заростка Автотрофный Гетерр- и ав- тотрофный Авто- и гете- ротрофный
Экологические условия Прибрежные местообитания, луга, леса, поля Леса Леса (от тро- пических до та- ежных), водо- емы, расщелины скал
Глава 5
ОТДЕЛ PINOPHYTA (ГОЛОСЕМЕННЫЕ)
5.1. Общая характеристика
Голосеменные, как и рассматриваемые далее покрытосемен-
ные (Magnoliophyta), - главные продуценты наземных экосистем
планеты, отличающиеся от споровых растений тем, что основ-
ным средством расселения у них являются не споры, а семена.
48
Семя - стадия жизни спорофита, особое образование, в ко-
тором в компактном и защищенном от неблагоприятных усло-
вий состоянии находится будущий взрослый спорофит - заро-
дыш, а также запас питательных веществ «на первый случай»
(т. е. до того, как молодой спорофит сможет обеспечивать себя
за счет фотосинтеза) - эндосперм.
Академик А.Л.Тахтаджян в «Жизни растений» пишет о се-
мени следующее: «Внутреннее оплодотворение, развитие заро-
дыша внутри семязачатка и появление новой, чрезвычайно эф-
фективной единицы расселения - семени - являются главными
биологическими преимуществами семенных растений, давшими
им возможность полнее приспособиться к наземным условиям и
достигнуть более высокого развития, чем папоротники и другие
бессемейные высшие растения. Если при размножении спорами
каждый раз образуется огромное их число, обычно миллионы,
то при размножении семенами число последних во много раз
меньше. Это вполне понятно, так как семя - несравненно более
надежная единица расселения, чем спора. В семени уже содер-
жится, причем в очень хорошей, надежной упаковке, зародыш -
крошечный спорофит с корешком, почечкой и зародышевыми
листьями (семядолями). Стоит ему освободиться из сковываю-
щей его семенной кожуры и просунуть наружу корешок, как он
легко укореняется и начинает самостоятельную жизнь. Но для
того, чтобы зародыш мог развиться настолько, чтобы разорвать
семенную кожуру, высунуться наружу и укорениться, ему ну-
жен некоторый первоначальный запас питательных веществ и
необходимый ферментативный аппарат. Этот запас питательных
веществ вместе с маленькой биохимической лабораторией заро-
дыш находит готовым в самом семени. В этом проявляется не-
что, отдаленно напоминающее заботу о потомстве в животном
мире, но «заботу», конечно, основанную не на инстинкте, а
чисто автоматическую, слепую, хотя также выработанную в
процессе эволюции. Семя - это поистине маленький шедевр
эволюции» (т. 4. 1978. С. 258).
Представители обширного отдела голосеменных составляют
до трети основных лесообразующих пород мира, их роль осо-
бенно велика в умеренных широтах, где они формируют биом
тайги.
Все голосеменные - это деревья или кустарники, как прави-
ло, автотрофные, хотя в тропиках есть два рода паразитических
голосеменных, а в Южном полушарии - очень мелкие кустар-
нички (из семейства Podocarpaceae), которые по размерам едва
превышают мхи. Исключение составляют и саговниковые, с тол-
стым неветвящимся колонно- или клубневидным, частично по-
груженным в почву стволом.
49
Древесина, за исключением представителей класса Gneto-
psida, состоит только из трахеид. Листья, как правило, узкие
(игольчатые) или чешуевидные, хотя есть роды с широкими ли-
стьями, напоминающими листья однодольных растений.
«Золотой век» голосеменных - это мезозой, и до нашего
времени они дошли в ограниченном разнообразии. При этом
современные голосеменные четко разделяются на две группы.
Первая, включающая саговниковые (Cycadopsida) и гинкговые
(Ginkgoopsida), - это «живые ископаемые». Как будет показано
далее, у них много особенностей, отсутствующих у представи-
телей второй группы - хвойных (Pinidae), которые на современ-
ной планете являются основными голосеменными.
Особняком в отделе стоит класс Gnetopsida (гнетовые), ко-
торый отнесен к голосеменным с немалой долей условности.
5.1.1. Особенности жизненного цикла
«Патронажная» роль спорофита по отношению к гамето-
фиту усилена уже у споровых растений (появление макроспор
со значительным запасом питательных веществ, развитие жен-
ских гаметофитов не только в оболочке макроспоры, но и в
макроспорангиях селагинелла, марсилея). У голосеменных
растений эта тенденция достигает еще более высокого уровня:
не только женский гаметофит не покидает оболочки макроспо-
ры, но и макроспора остается в макроспорангии. Таким обра-
зом, женский гаметофит не выходит (даже частично) наружу,
не соприкасается с окружающей средой и сохраняет постоян-
ную связь со спорофитом, что обеспечивает лучшие условия для
развития нового спорофитного поколения и, по существу, пре-
вращает женский гаметофит в стадию жизни спорофита.
Значительные изменения у голосеменных претерпевает и
мужской гаметофит, который, как и у разноспоровых растений,
развивается в оболочке микроспоры, но еще более редуцирован.
На смену многоклеточным антеридиям приходит новое образо-
вание из вспомогательных вегетативных клеток, которые об-
служивают гаметогенные клетки, дающие очень небольшое (как
правило, 2) количество мужских гамет.
Процесс оплодотворения у семенных растений избавляется
от «привычек прошлого», т. е. от связи с водной средой, кото-
рая теперь не нужна для переноса мужских гамет к женским.
Мужской гаметофит, называемый пыльцой, целиком переносит-
ся ветром к женскому гаметофиту, где и прорастает, используя
(в той или иной мере) питательные вещества женского гамето-
фита (которые образовались в свою очередь за счет спорофит-
ного поколения).
50
5.1.2. Репродуктивные органы
Семязачаток. У голосеменных, как и у рассматриваемых да-
лее цветковых (покрытосеменных), появляется новая структу-
ра - семязачаток. Его строение удобно рассмотреть на примере
семязачатка сосны (Pinidae).
Семязачаток состоит из макроспорангия нуцеллуса, за-
щищенного дополнительным покровом интегументом. На
верхушке семязачатка интегумент не замкнут, его края образу-
ют отверстие - микропиле. Внутри нуцеллуса развивается жен-
ский гаметофит, представляющий собой бесцветное многокле-
точное тело, клетки которого накапливают значительное коли-
чество запасных веществ, главным образом масел. На обращен-
ном к микропиле конце гаметофита образуются два погружен-
ных в его ткань архегония, в брюшке каждого из которых на-
ходится крупная яйцеклетка. У других более примитивных
хвойных могут быть десятки архегониев (у араукариевых - до
25, а у кипариса - до 200!).
В отличие от семязачатка хвойных семязачатки древних го-
лосеменных, которые в наше время представлены «живыми ис-
копаемыми» - саговниковыми (Cycadopsida) и гинкговыми
(Ginkgoopsida) (рис. 6а), имеют архегониальную камеру, в кото-
рую попадают сперматозоиды перед оплодотворением.
Семя. В результате оплодотворения яйцеклетки из семяза-
чатка формируется семя. В процессе его созревания из зиготы
развивается зародыш нового спорофита, ткань женского гаме-
тофита разрастается, обогащается запасными питательными
веществами и становится эндоспермом семени. В ходе развития
зародыша и эндосперма нуцеллус постепенно разрушается, и в
зрелом семени от него остается только сухая пленка. Покровы
семязачатка превращаются в плотную и кожистую семенную
кожуру.
Все эти части семени легко рассмотреть на примере кедро-
вого орешка - семени сосны сибирской (Pinus sibirica). Твердая
скорлупа - это преобразованный интегумент, коричневая пле-
ночка вокруг белого ядрышка - остатки нуцеллуса, съедобная
часть орешка — эндосперм. Зародыш обычно съедается вместе с
ним, но при желании его нетрудно выделить и рассмотреть.
У «живых ископаемых» семена опадают до полного созрева-
ния и даже до оплодотворения (это так называемые «яйцекладу-
щие» растения). У хвойных семя покидает материнский спорофит
в состоянии полной готовности к развитию дочернего спорофит-
ного растения (эти растения называют «живородящими»).
При этом, если для семян «яйцекладущих» растений харак-
терно отсутствие периода покоя после созревания, то у «живо-
51
родящих» растений семена обладают способностью длительное
время пребывать в состоянии покоя. Они долго не теряют
всхожести и могут дождаться наиболее благоприятного периода
для прорастания и развития нового растения. Это повышает
эффективность семенного размножения.
а) 6)
Рис. 6. Голосеменные: а - строение семязачатка саговниковых (слева) и
хвойных (справа) (1 - микропиле, 1 - интегумент, 3 - архегониальная
камера, 4 - нуцеллус, 5 - женский гаметофит); б - обоеполый стробил
беннеттитовых; в - сосна (1 - побег со стробилами^ 2 - часть женского
стробила, 3 - чешуя с семязачатками, 4 - семязачаток, 5 - пыльца, 6 -
чешуя с пыльником, 7 - часть мужского стробила, 8 - семя)
Пыльца. Как уже отмечалось, мужской гаметофит голосе-
менных чрезвычайно сильно редуцирован и развивается в мик-
роспоре еще до рассеивания. Вегетативной части у него нет, он
состоит из двух клеток - генеративной, которая функционально
заменяет целый антеридий, и клетки трубки. У «живых иско-
паемых» клетка трубки превращается в гаусторий, у хвойных -
в пыльцевую трубку. Роль гаустории и пыльцевой трубки раз-
лична. Если первый способствует питанию мужского гаметофи-
та (высасывая вещества из нуцеллуса), то вторая доставля-
52
ет мужские гаметы к яйцеклетке. Из генеративной клетки обра-
зуются сперматозоиды (у «живых ископаемых») или спермин (у
хвойных).
Пыльца некоторых хвойных (сосновые, подокарповые) име-
ет воздушные мешки, которые служат не только для ее распро-
странения по воздуху, но и для прорастания на семязачатках.
Семязачаток перед опылением выделяет капельку опылительной
жидкости и благодаря воздушным мешкам, действующим как
поплавки, пыльца принимает положение, наиболее удобное для
прорастания.
Пыльцы у голосеменных, особенно у хвойных, образуется
очень много, нередко она летит сплошным желто-зеленоватым
облаком, оседая пленкой на поверхности водоемов.
Стробилы (шишки). У голосеменных микро- и макроспоро-
филлы могут развиваться на одной (однодомные растения) или
на разных (двудомные растения) особях. Несмотря на разнооб-
разие их строения, просматривается общая закономерность: чем
древнее таксон, тем больше размеры макро- и особенно микро-
спорофиллов, которые могут быть даже перистыми, напоми-
нающими листья папоротников (как у вымерших беннеттитов).
У более развитых голосеменных спорофиллы становятся
чешуевидными и объединяются в стробилы (шишки), удобные
для созревания, защиты и рассеивания семян. Возникают при-
способления для распространения семян за счет покровов само-
го семени или частей семенных шишек. У всех ныне живущих
голосеменных стробилы однополые, мужские называются мик-
ростробилами, а женские - макростробилами.
5.1.3. Основные таксоны
Систематика голосеменных достаточно сложная. В отделе
выделяются несколько классов и много порядков. Приведем
список таксонов отдела Pinophyta, которые изучаются в на-
стоящем курсе:
Класс Pteridospermae* (семенные папоротники)
Класс Bennettitopsida* (беннеттитовые)
Класс Cycadopsida*** (саговниковые)
Класс Ginkgoopsida* (гинкговые)
Класс Pinopsida
Подкласс Cordaitidae* (кордаитовые)
Подкласс Pinidae (хвойные)
Порядок Araucariales* (араукариевые)
Семейство Araucariaceae
Порядок Pinales*** (сосновые)
Семейство Pinaceae
Порядок Cupressales** (кипарисовые)
Семейство Taxodiaceae
Семейство Cupressaceae
53
Порядок Taxales* (тисовые)
Семейство Тахасеае
Порядок Podocarpales* (подокарповые)
Семейство Podocarpaceae
Класс Gnetopsida* (гнетовые)
Порядок Welwitschiales (вельвичиевые)
Семейство Welwitschiaceae
Порядок Ephedrales (эфедровые)
Семейство Ephedraceae
Порядок Gnetales (гнетовые)
Семейство Gnetaceae
Примечание: *** - таксон рассматривается подробно, ** - таксону
дается общая характеристика,* таксону дается краткая характери-
стика.
5.2. Класс Pteridospermae (семенные папоротники)
Семенные папоротники - древняя вымершая группа голосе-
менных, обнаруженная в палеозойских и раннемезозойских от-
ложениях (их возраст составляет около 350 млн лет). По внеш-
нему облику эти растения были близки к папоротникам, но
имели семязачатки, которые располагались прямо на листьях,
что и дало повод назвать эту группу семенными папоротниками.
После оплодотворения семязачаток отделялся от растения.
Далее без периода покоя, характерного для настоящих семян,
из него уже на земле развивался спорофит. Поэтому семенные
папоротники нередко называют семязачатковыми растениями, а
не семенными. Наиболее изучены роды медуллоза (Medullosa) и
калимматотека (Calymmatotheca).
5.3. Класс Cycadopsida (саговниковые)
5.3.1. Общая характеристика
Саговниковые - одна из самых удивительных групп в соста-
ве ныне живущих голосеменных, истинные «динозавры» расти-
тельного мира, которые сохранились лишь в тропических и суб-
тропических районах всех пяти континентов земного шара. Эти
причудливые растения появились не позднее 285 млн лет назад
и были настолько многочисленными в мезозое, что его называ-
ют «временем саговников и динозавров».
Название класса происходит от греческого слова kykas
пальма и связано с внешним сходством этих растений с пальма-
ми. Не только Теофраст, но даже К.Линней считал эти растения
пальмами, т.е. древовидными покрытосеменными однодольными
растениями.
По палеоботаническим данным в мезозое саговниковые за-
селяли всю территорию России, вплоть до тундровой зоны, но
54
начиная с третичного периода из-за изменения климата они по-
теряли свои позиции и были вытеснены цветковыми растениями.
До наших дней дошло 10 родов саговниковых, в составе ко-
торых 120-130 видов. Любопытно, что на разных континен-
тах — свой набор родов. Это объясняется тем, что семена сагов-
никовых не могут сохранять жизнеспособность во время дли-
тельных морских путешествий (в отличие от такого растения-
путешественника, как кокосовая пальма, плоды которой пере-
носятся морскими течениями на тысячи километров).
Саговниковые, как правило, встречаются рассеянно, пооди-
ночке или небольшими группами в достаточно сухих местах -
саваннах и в составе растительности «средиземноморского ти-
па», т.е. сообществ с преобладанием жестколистных засухоус-
тойчивых кустарников. Жители средней полосы могут увидеть
саговниковые только в ботанических садах. В садах и парках
Черноморского побережья Кавказа (в меньшей степени Крыма)
распространен в культуре саговник поникающий (Cycas revo-
luta) родом из Японии. В условиях Батуми он дает зрелые
всхожие семена.
Другой вид саговника саговник завитой (С. circinalis)
также широко культивируется в ботанических садах и тропи-
ческих и субтропических парках. Он настолько популярен, на-
пример в штате Флорида (США), что его здесь, далеко от роди-
ны, называют «флоридской саговой пальмой». При этом выра-
щивают только женские экземпляры, размножая их вегетатив-
ным путем и не получая семян, поскольку необходимые для
опыления мужские особи имеют дурно пахнущие шишки.
5.3.2. Жизненная форма
Саговниковые имеют неповторимый облик и представляют
жизненную форму розеточных деревьев. Далеко не все роды
имеют высокий ствол, и у многих представителей имеется про-
токорм - клубневидное тело, от которого, как и у шильника,
отходят и корни, и листья. Возраст саговниковых достигает 100
лет (есть данные о старых деревьях возрастом до 2000 лет).
Из ныне живущих голосеменных только у саговниковых,
как и у семенных папоротников, листья папоротниковидные.
Количество листьев может достигать 150. Если высота саговни-
ков варьируется от менее 1 м до 10-12 м и выше, то размер ли-
стьев - от 5-6 см до 5-6 м. Для листьев характерны толстая ку-
тикула и глубоко погруженные устьица, что отражает приспо-
собление к условиям сухого климата.
Стволы саговниковых различаются по форме и высоте, од-
нако преобладают два основных типа: надземный колонновид-
ный (наиболее высокие из них достигают 20 м), обычно одетый
сплошным шершавым панцирем из остатков черешков листьев, и
подземный или полуподземный клубневидный.
55
Мощная кора состоит в основном из паренхимных клеток,
накапливающих крахмал, из таких же клеток состоит и зани-
мающая треть объема стебля сердцевина. Кроме того, в коре и
сердцевине содержится большое число слизевых каналов с во-
дянистой слизью. Древесина в стеблях развита слабо, однако
отличается большой длиной трахеид - до 10 мм, в этом с ними
могут сравниться только некоторые ископаемые беннеттитовые.
Корневая система представлена хорошо развитым ветвя-
щимся главным корнем, и только у некоторых саговниковых
главный корень рано отмирает и заменяется придаточными
корнями, возникающими из стебля. Примечательной особенно-
стью всех саговниковых являются особые растущие вверх и вы-
ступающие из почвы корни, внешне напоминающие кораллы,
кораллоиды, в которых живут микоризный гриб и азотфикси-
рующие микроорганизмы. Это уникальный в растительном мире
случай сожительства четырех и даже пяти различных организ-
мов: растения-хозяина, гриба, двух видов бактерий (Bacterium
radicicola, Azotobacter sp.) и цианобактерии (Nostoc punctiforme,
Anabaena cycadae и другие виды).
5.3.3. Генеративные органы
Все саговниковые двудомные растения. Находящиеся на
разных экземплярах микро- и макростробилы формируются на
верхушке ствола среди листьев. Исключение составляет род са-
говник (Cycas), у которого макроспорангии развиваются на лис-
товидных макроспорофиллах прямо на стволе.
Макростробилы обычно крупные, у некоторых видов просто
огромные (длиной до 1 м), а семязачатки могут достигать 5-6
см. Это самые крупные и тяжелые «шишки» в растительном ми-
ре: их масса может достигать 50 кг. Цвет стробилов, кроме зе-
леного или коричневатого, может быть ярким - желтым, оран-
жевым. Особенно нарядно выглядят стробилы, покрытые мел-
кими сосочками, придающими поверхности этих «шишек» се-
ребристый оттенок (например, у Encephalartos inopinus).
Хотя женский гаметофит у саговниковых, как у всех голо-
семенных, утерял возможность существовать самостоятельно,
именно у этих растений хорошо просматривается связь с «раз-
носпоровым прошлым»: у макроспоры формируется двуслойная
оболочка, обычная для спор бессемейных растений. А если хи-
рургическим путем извлечь эту макроспору, то на свету в ней
может развиться зеленый гаметофит!
По объему и количеству клеток женский гаметофит саговни-
ковых не уступает даже самым крупным свободноживущим гаме-
тофитам папоротников. На нем закладывается много материнских
клеток архегониев, это тоже архаическая черта. Правда, полного
развития достигает только около десятка архегониев, и они силь-
но редуцированы: в зрелом состоянии имеют только две шейковые
56
клетки. Яйцеклетка достигает огромных размеров - до 6 мм, ядро
в ней видно невооруженным глазом, как точка!
Микростробилы обычно мельче макростробилов и отлича-
ются от них по форме: как правило, они более узкие, вытяну-
тые в длину. Микроспорангии развиваются на нижней поверх-
ности чешуек (микроспорофиллов) небольшими группами (сору-
сами), как у папоротников. Их количество у разных представи-
телей различно: виды саговника имеют до 1000 микроспоран-
гиев на одной чешуе, а некоторые виды замии - всего по два.
Семена у большинства саговниковых крупные 3-4 см в
длину и 2-3 см в ширину и имеют яркую окраску - красную,
желтую или оранжевую, что, по-видимому, может рассматри-
ваться как приспособление к распространению животными.
Мощная семенная кожура состоит из двух слоев. Наружный
мясистый слой поедается животными, а внутренний твердый,
как косточка, предохраняет само семя от повреждений.
Семенная кожура, как и обильный эндосперм, формируется
не только до процесса оплодотворения, но даже до опыления, и
потому между семязачатком и семенем нет внешней разницы.
Как уже отмечалось, зародыш дозревает в опавшем семени в
течение длительного времени.
5.3.4. Значение саговниковых
В районах естественного произрастания человек издавна
разносторонне использовал саговниковые - в пищу, в лекарст-
венных целях, для различных поделок, в садоводстве, как риту-
альное растение. У большинства видов саговниковых наиболее
ценная пищевая часть крахмалоносная сердцевина ствола
(кафрский хлеб). Крахмал добывали, распиливая ствол на коль-
ца, просушивая их на солнце и затем растирая в муку, которую
многократно промывали. Конечный продукт этого производст-
ва - саго, которое представляет зерновидные белые комочки. (В
нашей стране «саго» изготовляют из картофельного крахмала
по особой технологии).
Кроме того, в пищу используются семена многих видов са-
говниковых (эндосперм содержит до 70% крахмала, а мясистая
часть кожуры - до 30% масла), а также молодые листья (осо-
бенно в Шри-Ланка).
Листья некоторых видов саговниковых - непревзойденный
материал для цветочных композиций. В Японии заготовка и пе-
реработка листьев саговника поникающего превратилась в ин-
дустрию. Листья срезают, некоторое время кипятят в воде, а
затем несколько дней вымачивают в специальном консервирую-
щем растворе и два месяца высушивают в тени. Ежегодно Япо-
ния экспортирует более 1000 огромных тюков, в каждом из ко-
торых десятки тысяч листьев.
57
Все саговниковые содержат ядовитые вещества, местные
жители знают об этом с давних пор и используют специальные
обезвреживающие способы приготовления из них пищи, однако
и в настоящее время случаются отравления даже со смертель-
ным исходом.
Отравляются и животные. Так, в начале нашего века в Ав-
стралии, массово погибал скот, который поедал листья этих
растений. По этой причине австралийские фермеры едва полно-
стью не уничтожили макрозамию (Macrozamia), которая была
обычным растением сухих пастбищ-малли с эвкалиптами.
В целом популяции саговников в большинстве районов мира
находятся в удручающем состоянии. Они являются уже не
столько ресурсными растениями, сколько объектами самой же-
сткой охраны.
Это интересно
Саговники и леопарды. Семенами саговниковых питаются многие
животные, в том числе обезьяны. В Африке после резкого сокращения
численности естественного врага - леопарда - обезьяны-бабуины очень
размножились, что привело к истощению популяции саговниковых.
Защита саговниковых в этом случае переросла в восстановление попу-
ляции леопардов.
5.4. Класс Bennettitopsida (беннеттиты)
Беннеттиты вымерли примерно 70 млн лет тому назад в пе-
риод расселения цветковых, а появились в начале мезозоя (око-
ло 200 млн лет назад). По своему облику они напоминали са-
говниковые, и длительное время их ископаемые остатки относи-
лись к этому классу. Однако со временем был накоплен матери-
ал, который показал значительное отличие беннеттитов от са-
говниковых.
Наряду с однополыми, как у саговниковых, стробилами
(род вильямсония Williamsonia), у беннеттитов имелись и
обоеполые (роды вильямсониелла - Williamsoniella, цикадеои-
дея - Cycadeoidea). Это единственная группа голосеменных, в
которой есть представители со стробилом, объединяющим мик-
ро- и макроспорофиллы (рис. 66).
В стробиле цикадеоидеи на мясистом семяложе располага-
лись многочисленные семеязачатки, разделенные стерильными
межсеменными чешуями; к основанию семяложа прикреплялись
перистые микроспорофиллы со многими микроспорангиями; сна-
ружи стробилы были защищены многочисленными густо опушен-
ными покроволистиками. Такой стробил отдаленно напоминал
цветок покрытосеменных растений, в котором макроспорофиллы
(пестики) и микроспорофиллы (тычинки) расположены примерно
так же. Поэтому многие ботаники рассматривали беннеттиты как
58
возможных предков цветковых растений. Последний вариант ги-
потезы беннеттитового происхождения цветковых был предло-
жен С.В.Мейеном и будет обсуждаться в ч. 3.
5.5. Класс Ginkgoopsida (гинкговые)
Эта своеобразная группа голосеменных представлена всего
одним видом - гинкго двулопастный (Ginkgo biloba). Научное
название виду дал К.Линней. В переводе с японского Ginkgo оз-
начает серебряный плод, так издавна назывались съедобные се-
мена этого дерева. Оно широко известно на Востоке как свя-
щенное и растет в парках, окружающих старинные храмы в
Японии, Китае и Корее. В этих странах гинкго известен с неза-
памятных времен, он упоминается в трудах китайских и япон-
ских ученых VII-VIII вв.
Гинкго - высокое (до 30 м) листопадное дерево с характер-
ными вееровидными двулопастными кожистыми листьями с ди-
хотомическим жилкованием, которые приобретают осенью пе-
ред сбрасыванием красивый золотистый оттенок.
Это растение - двудомное, хотя современные садоводы легко
исправляют «ошибку» природы и прививают побеги с мужских
растений на женские и наоборот. Впрочем, закладка генератив-
ных органов у дерева происходит поздно - на 25-30 году жизни,
и до этого времени определить пол растения невозможно.
Сережковидные микростробилы состоят из оси, на которой
спирально сидят микроспорофиллы, представляющие собой
тонкую ножку, на конце которой висят 2 (реже 3-4) микроспо-
рангия. Макростробилы состоят из длинной ножки, заканчи-
вающейся двумя семязачатками, из которых обычно только
один развивается в семя с мясистым покровом.
К осени семена окрашиваются в красивый янтарно-желтый
цвет, но приобретают неприятный запах из-за присутствия в мя-
систых покровах масляной кислоты, поэтому в садах и парках
разводят только мужские деревья, которые размножают вегета-
тивно. Семена с эндоспермом по вкусу напоминают сырой горох
и употребляются на Востоке в пищу в вареном и жареном виде.
Естественный ареал гинкго очень мал, это эндемик, сохра-
нившийся с мезозойских времен всего в одном районе Китая (в
горах Динь-Му-Шань). Однако благодаря усилиям человека
ареал гинкго расширился. Деревья гинкго живописны и привле-
кательны с ранней весны до поздней осени, и уже более 150 лет
он выращивается как декоративная культура не только в суб-
тропиках, но и в умеренных областях мира. Он долговечен, на
родине известны экземпляры возрастом 1000 лет и больше.
В Крыму в Никитском ботаническом саду гинкго выращи-
вают с 1818 г. В европейской части СНГ он хорошо растет и да-
ет всхожие семена до широты Киева. Взрослые экземпляры
гинкго есть и севернее, например в Риге у здания университета.
59
Гинкго перспективен для озеленения южных городов, так
как устойчив к пыли и газам, а также к грибковым и вирусным
заболеваниям, и, кроме того, практически не повреждается на-
секомыми. Он является сырьем для получения некоторых ле-
карственных препаратов.
Как «живое ископаемое», гинкго пользуется особым уваже-
нием у ботаников. Неоднократно международные ботанические
конгрессы использовали его лист в качестве эмблемы.
5.6. Класс Pinopsida, подкласс Pinidae (хвойные)
5.6.1. Общая характеристика
В этом классе, наряду с современным подклассом Pinidae,
имеется подкласс ископаемых голосеменных - Cordaitidae (кор-
даитовые), которые мы не рассматриваем. Отметим лишь, что
кордаитовые были представлены широким спектром жизненных
форм - от кустарников до деревьев высотой 30 м и были рас-
пространены во влажных приморских лесах. Расцвет кордаито-
вых связан с каменноугольным периодом, в изобилии они росли
и на протяжении следующего более сухого и холодного перм-
ского периода.
Хвойные - самый большой подкласс голосеменных с наи-
большим разнообразием таксонов (7 семейств, 55 родов, 560 ви-
дов), вклад которых в биологическую продукцию и биомассу
биосферы уступает только вкладу цветковых.
На обширных пространствах севера Евразии и Америки
хвойные формируют тайгу - леса, древостой которых чаще все-
го представлен одним видом. В южном полушарии хвойные
наиболее богато представлены в умеренных областях Новой Зе-
ландии, Австралии, Южной Америки, причем именно в бассейне
Тихого океана (Китай, Тайвань, Япония, Новая Каледония, Но-
вая Зеландия, Чили и др.) сосредоточено их основное видовое
разнообразие. А.Л.Тахтаджян объясняет это стабилизирующим
влиянием океана: в этой части планеты не было таких резких
колебаний климата, как в континентальных районах.
Таким образом, если основная биомасса хвойных сосредо-
точена в Северном полушарии (за счет небольшого числа видов
из родов ель, пихта, лиственница, сосна), то основное разнооб-
разие - в Южном, хотя здесь хвойные не образуют таких ог-
ромных массивов лесов, как сибирская тайга.
Хвойные - древесные растения размером от нескольких де-
сятков сантиметров до 120 м. Возраст гигантов может достигать
нескольких тысяч лет.
Жизненная форма. Стебель хвойных имеет тонкую кору и
массивную древесину, которая на 90—95% состоит из трахеид и
в отличие от покрытосеменных содержит очень мало паренхи-
мы. У многих хвойных и в коре, и в сердцевине имеются смоля-
60
ные каналы, наполненные эфирными маслами, смолой и бальза-
мами. Постоянные (не травматические) смоляные каналы в дре-
весине характерны только для семейства сосновых. У большин-
ства других хвойных они образуются только в ответ на поране-
ния ствола.
У большинства хвойных развивается мощный стержневой
корень, от которого отходят длинные боковые корни. Кроме
длинных корней, имеются короткие, мелкие, сильно ветвистые,
которые в основном и выполняют функцию всасывания и часто
бывают микоризными. А на корнях подокарповых есть клубень-
ки с бактериями, напоминающие клубеньки бобовых.
Листья большинства хвойных узколинейные или чешуевид-
ные и называются хвоей. Однако у тропических и субтропиче-
ских хвойных они совсем не похожи на хвою сосен, елей или
пихт. Так, у видов рода агатис (Agathis, Araucariaceae) они на-
поминают листья однодольных и достигают 18 см в длину и 6
см в ширину, а у рода подокарп (Podocarpus, Podocarpaceae) -
соответственно 35 и 9 см. Но рекорд по длине принадлежит все
же игловидным листьям североамериканской сосны болотной
(Pinus palustris) - 45 см.
У большинства хвойных листья вечнозеленые, плотные, бо-
лее или менее жесткие, кожистые и содержат крупные смоля-
ные каналы. В их строении ясно выражены ксероморфные при-
знаки - листья жесткие с толстой кутикулой, устьица погруже-
ны в углубления, которые заполнены воском, что уменьшает ис-
парение.
Генеративные органы. Микростробилы, часто называемые
«мужскими колосками», состоят из укороченной оси, на кото-
рой расположены сильно редуцированные микроспорофиллы.
На каждом микроспорофилле образуются от 2 микроспоранги-
ев (у сосновых) до 7 (и даже до 15 - у араукарии и агатиса). У
большинства хвойных мужские колоски мелкие (1-3 см), но у
араукарий они достигают 25 см в длину и 4-5 см в диаметре,
напоминая стробилы саговниковых. Строение пыльцы было
описано ранее.
Макростробилы у подавляющего большинства хвойных ком-
пактные, как хорошо известная всем шишка сосны. На ее оси
расположены кроющие чешуи, в пазухе которых находится «се-
менная чешуя», несущая на верхней стороне два семязачатка. К
моменту созревания шишек у большинства хвойных именно се-
менные чешуи деревенеют и служат защитой для семян. У тисо-
вых и подокарповых семенная чешуя становится сочной, мяси-
стой и ярко окрашенной, имитируя плод цветковых. Сходное яв-
ление наблюдается у можжевельника, у него семенные чешуи
становятся мясистыми, сладкими и окрашиваются в синий цвет.
Семена сильно различаются по величине и форме. У неко-
торых родов они снабжены одним большим крыловидным при-
61
датком, у других - двумя-тремя небольшими крыльями. Сочный
окрашенный покров на семенах некоторых хвойных способству-
ет их распространению животными.
5.6.2. Порядок Araucariales (араукариевые)
В порядке всего одно, очень своеобразное семейство Юж-
ного полушария - Araucariaceae, которое, как и саговниковые и
гинкговые, относится к «живым ископаемым» (хотя семязача-
ток у них типичный для хвойных). В составе семейства - два
рода: агатис (Agathis) и араукария (Araucaria).
Название араукарии дано по провинции Арауко в Южном
Чили (эту провинцию населяли индейские племена, называемые
арауканами). Араукарии - долгоживущие (до 2000 лет) высокие
(до 75 м) деревья с диаметром ствола 2-3 м, их ареал охватыва-
ет Южную Америку и тихоокеанские острова близ Австралии
(Новая Гвинея, Новая Каледония и др.).
Род агатис также распространен на этих островах, но пред-
ставлен и на материке Австралия. Примечательная особенность
видов этого рода - широкие ланцетные листья, напоминающие
листья однодольных растений.
Это интересно
«Загадка обезьян». Араукария чилийская (Araucaria araucana) име-
ет столь жесткие и колючие листья, что, по словам очевидцев, на нее
не садятся птицы. За это дерево получило название «загадка обезьян»
(имеется ввиду загадка «Как забраться на дерево?»), которое вошло в
ботанические словари. Крупные семена этой араукарии, тем не менее,
питательные и вкусные.
Каури. В Новой Зеландии бережно охраняют остатки лесов каури
(Agathis australis). Наиболее знаменитые экземпляры носят личные
имена и к ним, как и к мамонтовым деревьям в Калифорнии, прикреп-
лены дощечки с этими именами. Местные маори считают одно из де-
ревьев каури лесным богом. Окаменевшая смола каури подобна бал-
тийскому янтарю.
5.6.3. Порядок Pinales
В составе порядка - одно обширное семейство Pinaceae, ко-
торое содержит 11 родов и 250 видов, причем в подавляющем
большинстве сосновые распространены в Северном полушарии.
Они составляют основу наших таежных лесов, в которых пред-
ставлены роды ель (Picea), пихта (Abies), лиственница (Larix),
сосна (Pinus). Помимо лиственницы, остальные роды - вечнозе-
леные растения, активно участвующие в биосферных кругово-
ротах диоксида углерода и кислорода: их вклад в эти кругово-
роты сопоставим с вкладом вечнозеленых тропических лесов.
Все хвойные - микотрофы, т.е. живут в симбиозе с мико-
ризными грибами, которые относятся к базидиомицетам - шля-
62
почным грибам. Наиболее ценные среди них как объекты заго-
товки - масленок, рыжик, боровик.
Кратко охарактеризуем наиболее важные для России роды
и виды.
Сосна (Pinus). В составе этого самого обширного рода семей-
ства свыше 100 видов, на территории России распространена
«главная сосна» - сосна обыкновенная (Р. sylvestris). В Сибири
большую ценность представляют леса из сосны сибирской (Р. si-
birica), называемой в народе кедром (хотя настоящий кедр есте-
ственно распространен только за нашими пределами - в Сирии и
Ливане или на Гималаях). Корейским кедром называют сосну ко-
рейскую (Р. koraiensis), которая распространена на Дальнем Вос-
токе. Всего в России встречается около 10 видов сосны.
Пихта (Abies). На территории России встречаются 5 видов
пихты. Пихта сибирская (A. sibirica) отличается сравнительно мяг-
кими и широкими - до 3 мм - иголками, что делает это дерево
особо любимым в качестве новогодней «елки». Вид имеет огром-
ный ареал, включающий восток европейской части России, Сибирь,
север Монголии, северо-западный и северо-восточный Китай. Наи-
большая площадь пихтовых лесов - в Западной Сибири.
У пихты ценность представляет не только древесина, но и
хвоя, из которой выжимают пихтовое масло.
Ель (Picea). В России растет 7 видов ели, из которых наи-
большим ареалом обладает ель сибирская (Р. obovata). В куль-
туре распространены «серебристые ели» - формы двух амери-
канских видов (Р. pungens и Р. engelmannii) и «голубые ели» -
формы ели канадской (Р. canadensis).
Латинское название ели восходит к греческому «писса»
смола. Ель обыкновенная (Р. abies) - основной доминант евро-
пейской тайги. Ее ареал охватывает практически всю Европу
вплоть до Пиренеев. На севере и северо-востоке европейской
части России этот вид замещается елью сибирской. Промежу-
точные формы между этими видами - ель финская.
Еловая древесина - это не только прекрасный строительный
материал, но и уникальное сырье для производства музыкаль-
ных инструментов - скрипок, пианино, контрабасов и др. Ис-
пользуется специальная «резонансная» древесина с годичными
кольцами одинаковой ширины.
Лиственница (Larix). В России встречается несколько видов
лиственницы. Основной вид лиственница сибирская (L. si-
birica), которая на востоке замещается лиственницей Гмелина
(L. gmelinii). Ареал этого листопадного дерева связан с наибо-
лее суровыми климатическими условиями резко континенталь-
ных районов Азии. За счет сбрасывания листьев лиственница
легче переносит трескучие морозы, которых не выдерживают
вечнозеленые хвойные, гибнущие от иссушения за счет зимней
транспирации.
63
Лиственница ценится за прочную древесину, которая не
гниет в воде (ею были облицованы стенки каналов Мариинской
системы).
Человек в значительной степени расширил ареал лиственни-
цы на запад, где она хорошо растет в культуре. Посаженная в
XVIII в. Линдуловская роща под Санкт-Петербургом в настоя-
щее время объявлена заповедником. В этих созданных челове-
ком лесах деревья достигают высоты 50 м и диаметра ствола -
60-90 см.
Кедр (Cedrus). Этот род связан со странами Средиземномо-
рья (3 вида) и Гималаями (1 вид), но интродуцирован в ряде
районов России, особенно в Крыму и в Закавказье. Наиболее
знаменит кедр ливанский (С. libani), который был интродуциро-
ван в районы Южной Европы (Италия, Франция) еще в XVII
столетии, в 1826 г. первые кедры были высажены в Крыму.
Это интересно
Долгожитель. Среди сосен обнаружен старейший экземпляр де-
ревьев Калифорнии - самая старая сосна долговечная (Pinus longaeva)
имела возраст 4844 года!
Пиния. Видимо, самой древней культурой хвойных является пиния
(Р. pinea), которая в Италии стала называться «pino domestico» — пи-
ния домашняя. Ее начали высаживать в начале первого тысячелетия до
нашей эры. Пинии выращивались ради «орешков», хотя эти красивые
деревья с тонким стволом и раскидистой кроной были и остаются ук-
рашением итальянского ландшафта.
«Вечная» древесина. Старые деревья кедра ливанского могут до-
стигать в окружности 7 м, именно из их древесины делались деревян-
ные детали саркофагов египетских фараонов. Древесина столь долго-
вечна, что спустя 2,5 тыс. лет после захоронения она полностью сохра-
нилась.
Легенды о сосне. По греческой мифологии в сосну была превра-
щена нимфа Питие, которую бог ветра Борей приревновал к лесному
Пану. В Китае и Индокитае сосны считаются священными деревьями,
приносящими счастье и отводящими беды. В Древнем Вьетнаме вера в
могущество сосны заставляла сажать их вокруг дворца Императора как
символ долголетия и величественности царствующей династии.
5.6.4. Порядок Cupressales (кипарисовые)
В этот порядок, кроме семейства кипарисовых (Cupressa-
сеае), давшего ему название, входит семейство таксодиевых
(Taxodiaceae), представленное остатками голосеменных, кото-
рые вплоть до мелового периода были доминантами лесов Евра-
зии и Америки.
Семейство Taxodiaceae (таксодиевые). В семействе - 10 ро-
дов и 14 видов. Это в основном американские растения (роды
секвойя - Sequoia, метасеквойя - Metasequoia, секвойяденд-
рон - Sequoiadendron), но некоторые виды встречаются в Юго-
Восточной Азии.
64
Именно в этом семействе имеются виды, вошедшие в Книгу
рекордов Гиннеса. Секвойядендрон (мамонтово дерево) - самое
толстое и долгоживущее дерево мира (диаметр ствола до 10 м,
возраст до 4500 лет). Секвойи - самые высокие деревья мира,
их высота достигает 112 м. Некогда знаменитые североамери-
канские леса из этого вида (американцы называют секвойю
красной сосной) почти полностью уничтожены.
Секвойя и секвойядендрон интродуцированы в Европу, они
выращиваются в Никитском ботаническом саду (г. Ялта). Сегодня
посетители этого сада могут видеть деревья высотой до 30 м и
диаметром ствола до 1,5 м. Между тем, возраст их - всего 60 лет.
Это интересно
Гиганты. В естественных условиях сохранилось всего не более 500
экземпляров секвойядендрона. Все деревья взяты под охрану, а самые
старые из них получили собственные имена - «Отец лесов», «Генерал
Шерман», «Генерал Грант» и др. «Отец лесов» уже уничтожен, но, по
описанию, его диаметр достигал 12 м, а периметр — 132 м. Такое дере-
во содержало (и содержит у сохранившихся гигантов) до 1500 куб.м
древесины, для перевозки которой потребовалось бы 20—25 железно-
дорожных вагонов. На спиле одного из деревьев умещался негритян-
ский джаз-диксиленд, под музыку которого танцевали одновременно 30
пар. В Йосемитском парке в 1881 г. через ствол секвойядендрона был
проложен тоннель, по которому и сегодня свободно проезжают авто-
мобили.
Семейство Cupressaceae (кипарисовые). Это семейство не-
сравненно больше и разнообразнее таксодиевых. В него входят
19 родов и 130 видов, широко распространенных по обоим по-
лушариям.
Все кипарисовые - вечнозеленые деревья средних размеров
(но некоторые - очень высокие, до 40 и даже 70 м) или кустар-
ники. Самые крупные и важные роды - можжевельник (Juni-
perus) и кипарис (Cupressus).
В роде кипарис - 15-20 видов, обитающих на всех матери-
ках, кроме Австралии. Пирамидальный кипарис вечнозеленый
(С. sempervirens var. sempervirens), украшающий сады и парки
Кавказа и Крыма, это выведенная человеком культурная
форма. Дикая разновидность этого кипариса - С. sempervivus
var. gorisontalis. В Крыму кипарис рассадили греческие пересе-
ленцы еще до новой эры, хотя наибольшее распространение он
получил после второй интродукции, 200 лет назад. Родина этого
кипариса - Северный Ирак.
На Кавказе чаще всего встречается кипарис мексиканский
(С. lusitanica), введенный в культуру в Крыму в середине XIX в.
В Россию он попал через Испанский полуостров, куда был за-
везен из Америки.
В средней полосе кипарис не выдерживает зимы, но выра-
щивается другой представитель этого семейства — туя западная
(Thuja occiaentalis). Этот вид давно введен в культуру и сегодня
65
насчитывает не менее 120 садовых форм. Кроме того, выращи-
вается туя восточная (Т. orientals), родом из Средней Азии, ко-
торая в естественных условиях, видимо, не сохранилась.
Наиболее замечательным родом этого семейства является
можжевельник (Juniperus), среди 70 видов которого есть и высо-
кие деревья (до 30 м высоты) и низкие кустарники. В России
распространены около 14 кустарниковых (J. communis, J. sabina,
J. sibirica и др.) и древовидных (J. excelsa, J. oxycedrus и др.) ви-
дов. Особенностью можжевельников являются шишки, напоми-
нающие ягоды. Благодаря этим «ягодам» они расселяются пти-
цами и млекопитающими.
Древовидные можжевельники образуют светлые арчовые
(от «арча» - местного названия можжевельника) леса в Сред-
ней Азии. Можжевеловые леса средиземноморского типа со-
хранились в Крыму и охраняются в небольшом заповеднике
«Мыс Мартьян», который расположен рядом с Никитским бо-
таническим садом. Есть сосново-можжевеловые леса и в Се-
верной Америке.
На большой площади в горах Сихотэ-Алиня (Дальний Вос-
ток) встречается микробиота перекрестнопарная (Microbiota
decussata) - единственный эндемичный род хвойных в России.
Микробиота и оба вида кипариса включены в Красную книгу.
Это интересно
Чистый воздух. Можжевеловые леса отличаются высокой концен-
трацией в атмосфере эфирных масел, убивающих микробное население.
Подсчитано, что продукции паров масел с одного гектара такого леса
достаточно, чтобы очистить от бактерий атмосферу среднего по вели-
чине города. Однако, к сожалению, в городах можжевельники не рас-
тут, так как не выдерживают атмосферного загрязнения.
Запах можжевельника. Древесина можжевельника отличается
прочностью, красивым рисунком и приятным смолистым запахом, ко-
торый сохраняется в изделиях из этой древесины в течение десятков
лет. В Эстонии кружка из можжевельника, из которой пьют пиво в
сауне, обязательный атрибут оборудования этого традиционного
места отдыха эстонцев.
В Китае и Монголии в ритуалах ламаистских храмов используется
порошок из листьев одного из видов можжевельника (J. pseudosabina).
Этот порошок поджигают, он тлеет и испускает ароматы, которые на-
страивают верующих на общение с Буддой.
5.6.5. Порядок Podocarpales (подокарповые)
В этом порядке всего одно крупное семейство подокарпо-
вых (Podocarpaceae), включающее 140 видов. Подокарповые
растут во влажных тропических лесах австрало-азиатского ре-
гиона. Основной род - подокарп (Podocarpus). Подокарпы - де-
ревья (некоторые виды достигают высоты 80 м и диаметра ство-
66
ла 2 м) или кустарники. Особенность подокарпа - линейные или
ланцетные листья, напоминающие листья однодольных.
На территории СНГ некоторые виды подокарпа интродуци-
рованы на Черноморском побережье Кавказа в ботанических
садах. Есть подокарпы и в Никитском ботаническом саду.
5.6.6. Порядок Taxales (тисовые)
Главное семейство этого порядка - Тахасеае (тисовые),
распространенное в северном полушарии. Род тис (Taxus), дав-
ший название семейству, содержит 10 видов. На территории
СНГ распространено два вида. Тис ягодный (Т. baccata) обычен
в лесах Западной Европы и заходит в наиболее теплые районы
Украины, Белоруссии и Кавказа, где площадь лесов с участием
тиса превышает 1500 га. Другой вид - тис остроконечный
(Т. cuspidata) - растет на Дальнем Востоке.
В прошлом тис был более распространенным видом, ныне
значительная часть его популяций либо уничтожена, либо по-
дорвана интенсивными рубками. Причина этого - высокая
стоимость древесины тиса, которая имеет красивый рисунок и
используется для изготовления самого дорого шпона.
5.7. Класс Gnetopsida (гнетовые)
Как уже отмечалось, гнетовые - это группа растений, кото-
рая столь своеобразна, что помещается в отдел голосеменных ус-
ловно. Входящие в нее виды объединяет ряд черт, не свойствен-
ных другим голосеменным и сближающих их с покрытосеменны-
ми: вокруг стробилов есть покров, похожий на околоцветник; в
ксилеме присутствуют почти такие же сосуды, как у покрытосе-
менных; у гнетума и вельвичии отсутствуют архегонии.
Гнетовые представлены тремя сильно различающимися ро-
дами гнетум (Gnetum), эфедра (Ephedra) и вельвичия (Wel-
witschia).
Гнетум представлен 30 видами, встречающимися во всех
влажных тропических областях. Это деревья и лианы, крупные
кожистые листья которых напоминают листья двудольных.
Род эфедра содержит 35 видов (в России встречается 5 ви-
дов), обитающих в аридных областях. Они представляют собой
сильно разветвленные кустарники со слабозаметными мелкими
чешуевидными листьями и внешне напоминают хвощ.
Велъвичию удивительную (W. mirabilis), единственный вид
рода, ботаники называют «отщепенцем растительного мира».
Вельвичия растет только в очень сухих бесплодных каменистых
пустынях юго-западной Африки (Ангола, Намибия, ЮАР). Это,
наверное, самое причудливое из сосудистых растений. Большая
часть толстого (диаметром до 1,2 м) ствола растения скрыта в
земле, а на поверхность выступает его верхняя похожая на пень
67
часть, от которой отходят только два гигантских (до 6 м) кожи-
стых листа с параллельным жилкованием. Эти листья служат
растению всю жизнь (до 200 лет), с возрастом разрываясь на
ленты, отмирающие с концов.
Это интересно
Встреча с «невозможным». Португальский ботаник Ф.Вельвич, пер-
вым обнаруживший названное в его честь растение, говорил, что сна-
чала даже боялся дотронуться до него, опасаясь, что оно исчезнет.
Привезенное в Англию растение описал в 1863 г. ботаник Дж.Хукер.
Посетивший в 1971 г. пустыни юго-западной Африки английский бота-
ник Г.Раули назвал свою статью «Путешествие в невозможное я
встречаю вельвичию».
5.8. Заключение: сравнение саговниковых и хвойных
Сравнение «живых ископаемых» и современных голосеменных
ключ к пониманию особенностей и разнообразия этого отдела (табл. 8).
Таблица 8
Сравнительная характеристика Cycadopsida и Pinopsida
Признак Cycadopsida Pinopsida
Роль в биосфе- ре Незначительная Большая, формируют биом тайги и значи- тельные площади лесов в Южном полушарии
Число видов 130 560
Особенности экологии Растения сухих ме- стообитаний - саванны, «средиземноморская » растительность Растения холодных северных территорий или умеренного клима- та, реже - в тропиках
Жизненная форма «Розеточные дере- вья», часто с клубне- видным стеблем, от крупных (10-12 м) до мелких (несколько сан- тиметров) Деревья (до 120 м вы- соты) и кустарники
Древесина Преобладает паренхима Преобладают трахеиды
Семязачаток Имеется архегониаль- ная камера Архегониальной ка- меры нет
Пыльца Прорастает с образо- ванием гаустории Прорастает с образо- ванием пыльцевой труб- ки
Мужские гаметы Сперматозоиды Спермин
Семена Зародыш дозревает после опадения семени («яйцекладущие ») Зародыш развивается до опадения семени («живородящие »)
68
ЧАСТЬ 3
ЦВЕТКОВЫЕ
Прежде чем рассматривать семейства цветковых, да-
дим их общую характеристику, приведем гипотезы происхождения и
сравним однодольные и двудольные.
Задача отбора семейств для рассмотрения оказывается достаточ-
но сложной. Ограничивая список всего 27 семействами, расположен-
ными по системе А.Л. Тахтаджяна (1987), при их выборе авторы руко-
водствовались двумя критериями:
• представить основные варианты разнообразия цветка как органа,
имеющего первостепенное значение для диагноза таксонов цветко-
вых (цветок наиболее «консервативен», т.е. в меньшей степени, чем
другие органы, меняется под влиянием условий среды);
• из числа «претендентов» на иллюстрацию разнообразия цветковых
отобрать по преимуществу таксоны флоры средней полосы, наибо-
лее значимые для человека (включающие культурные растения, ре-
сурсные, сорные и др.).
Каждое семейство характеризуется «паспортом» в виде таблицы, в
которой указаны его основные признаки: число родов и видов (в скоб-
ках - число в России), обобщенные формулы цветков, наиболее типич-
ные для семейств, географический ареал, жизненная форма, особенно-
сти плодов и способов их распространения. Кроме того, в «паспорте»
приведены важнейшие (как правило, для естественной флоры нашей
страны) роды, а также роды культурных растений.
В формуле цветка использованы следующие условные обозначе-
ния: Р (от perigonium) - простой околоцветник, Са (от calyx) - чашеч-
ка, Со (от corolla) - венчик, А (от androeceum) - андроцей, G (от gynoe-
ceum) - гинецей. Цифры указывают количество элементов, знак оо
неопределенное число их, скобки - срастание элементов, плюс - рас-
положение в разных кругах. Черточка снизу - верхняя завязь, свер-
ху - нижняя завязь, справа - полунижняя. Звездочка - актиноморф-
ный цветок, стрелка - зигоморфный.
В текстовом описании семейств указываются порядки, к которым
они относятся, и общее число семейств в этих порядках. Наиболее
важные для человека семейства, такие как злаки, охарактеризованы
наиболее полно.
После описания видов основного семейства упоминаются близкие
к ним практически значимые виды из других семейств, как правило,
относящихся к одному порядку. Список наиболее распространенных из
нерассмотренных культурных растений с указанием семейств, к кото-
рым они относятся, приведен в конце книги.
69
Глава 6
ОБЩАЯ ХАРАКТЕРИСТИКА ЦВЕТКОВЫХ РАСТЕНИЙ
И ОБЗОР НЕКОТОРЫХ ГИПОТЕЗ
ИХ ПРОИСХОЖДЕНИЯ
6.1. Общая характеристика цветковых
Цветковые - основные вкладчики в биологическую продук-
цию и биомассу суши. Общее число описанных видов их превы-
шает 250 тыс. (при том, что число видов голосеменных составляет
около 700, всех папоротниковидных - около 10 тыс., моховид-
ных - 25 тыс., а водорослей - 30 тыс.). В действительности число
видов цветковых на планете еще больше, но не все они описаны,
поскольку не полностью изучен самый богатый видами регион
влажных тропических лесов. Более того, о многих из этих видов
мы так и не узнаем, так как они исчезают в процессе истребле-
ния лесов под натиском плантаций кофе, гевеи и бананов.
Цветковые поражают не только своим морфологическим
разнообразием (в «Системе магнолиофитов» А.Л.Тахтаджяна
указывается 166 порядков и 533 семейства), но и разнообразием
жизненных форм и широтой спектра условий среды, в которых
они могут обитать. Цветковые растут в самых экстремальных ус-
ловиях: в высокогорьях, в арктических и аридных пустынях, на
солончаках, в соленой воде на морских прибрежьях (Zostera
marina). У них наиболее развита стратегия патиентности (вынос-
ливости к экстремальным условиям, см. ч. 3) и эксплерентности
(способности к быстрому заселению незанятых местообитаний,
что позволяет им играть роль «ремонтной бригады» природы,
восстанавливая внезапно нарушенную растительность).
Размеры цветковых меняются от гигантов, высотой более
100 м (австралийские эвкалипты уступают только секвойям) до
крохотных растений (ряска малая - Lemna minor).
Роль цветковых как главых продуцентов биосферы объяс-
няется совершенством организации, причем не только уже от-
меченной способностью к переживанию стрессов и быстрому
расселению, но и особенностями жизненного цикла и еще
большей, чем у голосеменных, «заботой» спорофита о гамето-
фите. Все преимущества высших растений, о которых мы гово-
рили в ч. 1, достигают у цветковых наивысшего развития.
Кратко рассмотрим важнейшие черты организации этих
удивительных растений.
Совершенство системы обеспечения водой. За исключением
некоторых родов из Magnoliales и близких к нему порядков
(Drimys, Degeneria, Schisandra, Trochodendron и др.), все цветко-
70
вые имеют развитую проводящую систему с настоящими сосу-
дами, что позволяет обеспечивать растения влагой даже в усло-
виях ее недостатка в почве. Более того, проводящая система
дополняется эффективными покровными тканями (эпидермой и
перидермой) и устьичным аппаратом.
Высокая эффективность фотосинтеза. Развитый листовой ап-
парат цветковых имеет большую поверхность и при наличии дос-
таточного количества влаги и элементов минерального питания
располагается многоярусно, образуя 4-5 слоев над поверхностью
почвы, причем нижние слои адаптированы к использованию све-
та, который ослаблен и отфильтрован верхними слоями.
Совершенна и внутренняя структура листа. Мезофилл,
дифференцированный на богатую хлоропластами столбчатую
хлоренхиму и рыхлую губчатую хлоренхиму, пронизан густой
сетью проводящих пучков, обеспечивающих снабжение водой и
отток синтезированных веществ.
Наконец, в зависимости от обеспеченности влагой различа-
ются типы фотосинтеза: Сз у растений, хорошо обеспеченных
влагой, и С< у растений сухих местообитаний. Эти типы фото-
синтеза рассматриваются в курсе «Физиология растений», здесь
отметим лишь наиболее экстравагантный вариант его у пустын-
ных растений с сочными водозапасающими листьями и стеблями
(суккулентов). Эти растения фотосинтезируют при закрытых
устьицах за счет углерода, накопленного в ночное время, когда
прохладно и открывание устьиц для поступления в лист углеки-
слого газа не ведет к иссушению растения.
Минеральное питание. Корневая система цветковых раз-
ветвленная и может проникать в почву на большую глубину (до
10 м и более). Все цветковые — симбиотрофы и связаны с мико-
ризными грибами, бактериями ризосферы и, реже, с клубенько-
выми азотфиксирующими бактериями (представители семейства
бобовых).
Генеративные органы. Совершенство вегетативных органов
спорофита сочетается с новым специализированным органом -
цветком, обеспечивающим бесполое и половое размножение и
образование семян. Гаметофиты, развивающиеся в цветке, силь-
но редуцированы: мужской - до состояния двухклеточной
пыльцы, а женский - до уникального восьмиядерного образова-
ния, называемого зародышевым мешком. У цветковых развита
система переноса мужских гаметофитов к женским (перенос
пыльцы на рыльце пестика).
В строении цветка четко прослеживаются две линии спе-
циализации: энтомофильная (насекомоопыление) и анемофиль-
ная (ветроопыление). В свою очередь, у энтомофильных расте-
71
ний также есть две линии: на узкую специализацию к опреде-
ленным видам-опылителям (как у орхидных или бобовых) и на
широкую специализацию с доступностью пыльцы для разных
опылителей (сложноцветные). В первом случае цветок устроен
так, что доступен только определенным насекомым, во вто-
ром он достаточно открыт для посещения разными насеко-
мыми, и мелкие цветки объединены в соцветия, функционирую-
щие как единая «опылительная система» (насекомое ползает по
всему соцветию).
У ветроопыляемых растений увеличивается продукция
пыльцы (пыльники крупные) и поверхность рыльца (например, у
злаков или осоковых). Околоцветники у них невзрачные и ре-
дуцированные.
У цветковых существуют разнообразные приспособления к
перекрестному опылению и ограничению самоопыления: дву-
домность, самонесовместимость (ингибирование прорастания
собственной пыльцы), дихогамия - протандрия и протогиния
(неодновременное созревание тычинок и пестиков), херкогамия
(пространственное разобщение пыльцы и рыльца, как, напри-
мер, у губоцветных) и др. При этом у многих видов самоопыле-
ние возможно как «запасной вариант», который срабатывает в
конце цветения, если перекрестного опыления не произошло.
Более того, у представителей некоторых семейств (в осо-
бенности у сложноцветных) распространено явление апомикси-
са - девственного развития семян без оплодотворения.
Наконец, расцвету цветковых во многом содействовало со-
вершенство семян, способных к длительному покою и имеющих
разнообразные приспособления для распространения (ветром,
животными, водой, человеком). Способность к расселению рас-
тений с помощью семян дополняется способностью многих ви-
дов к вегетативному размножению за счет корневищ, клубней,
луковиц и др.
Все вместе эти качества цветковых и объясняют их широкое
распространение, хотя в некоторых районах (например в биоме
тайги) они уступают пальму первенства голосеменным. Впрочем,
последние сохраняют преимущества лишь при сравнительно
слабом влиянии человека. При интенсивном влиянии голосемен-
ные не выдерживают натиска человека, и им на смену приходят
более пластичные цветковые (еловые леса замещаются ольша-
никами и березняками).
6.2. Гипотезы происхождения цветковых
По меткому выражению А.Л.Тахтаджяна, гипотезы это
строительные леса для здания теоретического знания. Хотя, к
сожалению, далеко не в любых «лесах» это «здание» может
72
быть построено. И, видимо, происхождению цветковых так и
суждено остаться «отвратительной тайной» (abominable mistery,
выражение Ч.Дарвина из его письма, написанного в 1879 г.
Дж.Гукеру).
Во времена Ч.Дарвина о происхождении цветковых было
известно очень мало. Изучение палеоботанической летописи
только начиналось. На основании этих скудных сведений Дар-
вин считал, что цветковые появились внезапно - в меловом пе-
риоде и сразу на всей планете почти во всем современном раз-
нообразии.
К концу 40-х годов нашего столетия, когда сформулировал
свою гипотезу происхождения цветковых А.Л.Тахтаджян, па-
леоботаническая летопись также была все еще бедной. Толчок к
разработке гипотезы дали экспедиции Тахтаджяна в юго-
восточные районы Китая, где он обнаружил уже перечисленные
роды магнолиевых с гомоксильной («трахеидной») древесиной,
свойственной голосеменным. На палеогеографической карте ме-
зозоя территория юго-восточного Китая называется Катазией, и
гипотеза Тахтаджяна была названа катазионной. Все положе-
ния своей гипотезы Тахтаджян вывел дедуктивно как плод «иг-
ры воображения».
Спустя 30 лет, когда в 70-е годы родилась гипотеза
С.В.Мейена, палеоботаническая летопись уже была пополнена
множеством новых данных. Были, наконец, обнаружены пер-
вые ископаемые цветковые, которые оказались непохожими на
магнолиевые с гомоксильной древесиной. Эти первые цветковые
были распространены по всему тропическому поясу, откуда они
расселились в высокие широты к северу и к югу. Гипотеза Мей-
ена получила название гипотезы «тропической помпы» (фито-
спрединга). Таким образом, если Тахтаджян основывался на
данных о современных закономерностях распределения цветко-
вых-«живых ископаемых», то Мейен - на достаточно обильной
палеоботанической информации.
Кроме того, во времена формулирования гипотезы Тахтад-
жяна дарвинизм и синтетическая теория эволюции (СТЭ) пере-
живали (особенно в СССР) апогей развития, так как они в зна-
чительной мере укрепились благодаря борьбе с «теориями»
Т.Лысенко. К началу 70-х годов популярность дарвинизма резко
упала. Стало очевидным, что СТЭ не в состоянии объяснить
сформировавшееся разнообразие видов растений и животных,
так как модель «случайные ненаправленные мутации и отбор
полезных изменений условиями среды» описывает слишком
медленный процесс. В это время получают развитие ранее от-
вергавшиеся недарвинистские варианты объяснения эволюции -
номогенез (развитие по программе, заложенной в самой приро-
73
де живого вещества) и сальтационизм (допущение крупных
скачкообразных изменений формы). Этот новый уровень разви-
тия теории эволюции отражен в гипотезе Мейена.
В табл. 9 сформулированы в виде антиномий основные по-
ложения гипотез Тахтаджяна и Мейена.
Таблица 9
Основные антиномии гипотез происхождения цветковых
Вопрос Гипотеза
катазионная фитоспрединга
Время происхож- дения Нижний и средний мел Нижний - верхний мел
Время расселения Верхний мел Нижний - верхний мел
Место происхо- ждения Катазия (Ю-В Азия) Тропический пояс
Экологические условия Горы Различные, преиму- щественно равнинные
Предки Семенные папоротни- ки Беннеттиты
Древние формы Вечнозеленые деревья с одиночными крупны- ми цветками типа маг- нолиевых Различные древесные растения с невзрачными цветками, собранными в соцветия
Способ опыления древних форм Энтомофилия и осо- бенно кантарофилия (опыление жуками) Факультативное: эн- томофилия, анемофи- лия, самоопыление
Основной эволю- ционный меха- низм возникно- вения разнообра- зия СТЭ. Адаптации к различным горным ме- стообитаниям (по высо- те над уровнем моря, экспозиции). Сочетание мутаций и направлен- ного отбора. Популяци- онные волны. Генети- ческий дрейф. Чередо- вание периодов изоля- ции локальных популя- ций (демов) и гибриди- зации Сальтации: выжива- ние организмов с круп- ными изменениями фор- мы неадаптивного ха- рактера, откачивание их «тропической помпой» в высокие широты и адаптационные измене- ния в соответствии с СТЭ
Уязвимыми моментами обеих гипотез являются представле-
ния о предках цветковых. Тахтаджян считает таковыми семен-
ные папоротники, но выводит это благодаря жестким дедукци-
ям, путем последовательного исключения всех прочих групп го-
лосеменных. Еестественно, что невозможно исключить или при-
влечь те группы, которые еще неизвестны, а такое вполне веро-
74
ятно. Неполнота геологической летописи уменьшается, но весь-
ма медленно.
Мейен считает, что предками цветковых могли быть беннет-
титовые, стробил которых был обоеполым. При этом несоответ-
ствие строения макроспорофиллов беннеттитовых и цветковых
он пытается преодолеть гипотезой о «переносе фруктифика-
ций», т.е. о достаточно сложных миграциях генеративных орга-
нов внутри структуры «процветка». В.Н.Тихомиров (1987) на-
звал этот элемент гипотезы «экстравагантным».
В целом, безусловно, гипотеза Мейена на сегодняшний день
может оцениваться выше (именно на сегодняшний день, так как
появление новых фактов может качнуть чаши весов и дать пре-
имущества взглядам Тахтаджяна), как лучше подтверждаемая
фактами в вопросах о месте и времени возникновения цветковых.
6.3. Однодольные и двудольные
Подразделение цветковых на два класса: двудольные
(Magnoliopsida) и однодольные (Liliopsida), краеугольное по-
ложение их таксономии. Классы различаются столь резко
(табл. 10), что общего у них оказывается меньше, чем различий.
Объединение их в один отдел во многом формально.
Таблица 10
Сравнительная характеристика однодольных и двудольных
Признак Двудольные Однодольные
Количество семядо- лей в зародыше Две Одна
Цветок Тенденция к пяти- членности Тенденция к трех- членности
Стела Есть камбий; си- фоностела и эустела Камбия нет; атак- тостела
Жилкование листа Перистое или пальчатое, незамк- нутое Параллельное или дуговидное, замкну- тое
Корневая система Стержневая или кистекорневая Мочковатая
Жизненная форма От деревьев до однолетних трав Многолетние и од- нолетние травы, ред- ко древовидные
Число семейств 429 104
Число родов 10 тысяч 3 тысячи
Число видов 190 тысяч 63 тысячи
75
Происхождение однодольных столь же «отвратительная
тайна», что и происхождение цветковых. Чисто исторический
интерес представляют гипотезы гетерокотилии (недоразвитие
одной семядоли) и синкотилии (срастание семядолей). Сегодня
можно говорить о двух гипотезах происхождения однодольных
(табл. 11): А.Л.Тахтаджяна (от гидрофитов, близких к нимфей-
ным) и ХЖак-Феликса (независимое от двудольных происхож-
дение однодольных от разноспоровых плауновидных типа
шильника). Достоверность обеих гипотез невелика, хотя гипоте-
за Жак-Феликса привлекательнее своей простотой.
Таблица 11
Основные антиномии гипотез происхождения
однодольных растений
Вопрос Гипотеза
А .Л .Тахтаджяна ХЖак-Феликса
Природа семядо- ли Гомолог семядолей двудольных, сформиро- вавшийся в результате мутации Гомолог протокорма разноспоровых плауно- видных
Предок Двудольные, подоб- ные нимфейным Разноспоровые плау- ны, подобные шильнику
Происхождение От ювенильных ста- дий двудольных От ювенильных ста- дий плауновидных
Направление эволюции Гигрофильная с по- следующей вторичной мезофитизацией Адаптация к ухуд- шающимся условиям
Лист Теломный Энационный
Уровень органи- зации Выше двудольных Ниже двудольных
6.4. Заключение: причины успеха цветковых
Как уже отмечалось, цветковые на сегодняшний день являются
экологически и таксономически самой разнообразной группой высших
растений. В табл. 12 приведены признаки, которые позволили цветко-
вым занять ведущее положение в биосфере.
Таблица 12
Признаки, обеспечившие успех цветковых
Группа признаков Цель Способы ее реализации
Половой процесс Защита жен- ского гаметофита Образование завязи
Повышение ве- роятности опло- дотворения Адаптации к опылению насекомы- ми и ветром, самоопыление (вклю- чая клейстогамию), апомиксис
76
Окончание табл. 12
Группа признаков Цель Способы ее реализации
Плоды и семена Повышение ин- тенсивности рас- селения Адаптации к распространению се- мян животными, ветром, водой, че- ловеком
Переживание неблагоприятных условий Наличие в семенах гибридного эн- досперма, позволяющего зародышу первое время развиваться за счет этого запаса
Вегетатив- ные органы Повышение эффективности фотосинтеза Высокий листовой индекс, разви- тая хлоренхима, адаптации к высо- кой интенсивности солнечного излу- чения
Улучшение почвенного пита- ния Развитая проводящая система и средства экономии воды (кутикула, устьица, опушение) Развитая корневая система Симбиоз с грибами и бактериями
Расширение спектра условий местообитаний Разнообразие форм приспособле- ний к разным экологическим усло- виям (включая разные виды стресса и нарушения) — разные жизненные формы и стратегии
Активизация вегетативного размножения Образование корневищ, луковиц, корневых отпрысков, порослевое во- зобновление у деревьев, парцелля- ция надземных частей у трав и др.
Глава 7
КЛАСС MAGNOLIOPSIDA - ДВУДОЛЬНЫЕ
7.1. Семейство Magnoliaceae - Магнолиевые
Паспорт семейства
Родов - 14 (1), видов - 230 (1)
Цветок - *Саз С03+3 А№ Gs
Распространение - субтропики и тропики, центр разнообра-
зия - Юго-Восточная Азия
Жизненная форма деревья и кустарники, вечнозеленые,
реже листопадные
Опыление - перекрестное жуками (кантарофилия) и само-
опыление
Плоды - шишковидные, апокарпные. Семена распространя-
ются птицами (орешковидные плодики - ветром)
Важнейшие роды - Liriodendron, Magnolia
77
Семейство (рис. 7) представляет порядок Magnoliales, объе-
диняющий 3 семейства. По А.Л. Тахтаджяну, этот порядок
включает самые древние цветковые растения. Признаки древно-
сти он усматривает в особенностях цветка: длинная ось, неоп-
ределенное число элементов, спиральное расположение тычинок
и плодолистиков, опыление жуками. Как древний признак рас-
сматриваются размер и положение цветков - они крупные и
обычно сидят одиночно на концах ветвей. Околоцветник круго-
вой, 3-6-членный. Доли околоцветника одинаковые или наруж-
ные из них - чашечковидные. Их число в пределах семейства
варьирует от 12-18 (редко больше) до 6-7.
Рис. 7. Magnolia: 1 - цветок, 2 - андроцей и гинецей, 3 -тычинка,
4 - диаграмма цветка, 5 - плод
Цветки приспособлены к опылению жуками. Так, у многих
магнолий (Magnolia) рыльца готовы к восприятию пыльцы в бу-
тоне, когда только жуки могут проникнуть внутрь цветка. В
раскрытом цветке рыльца уже не функционируют. Питаются
жуки пыльцой, нектаром, выделениями лепестков, рыльцами.
В России в диком виде растет только один вид этого семей-
ства - магнолия обратнояйцевидная (Magnolia hypoleuca). В ви-
де небольшого дерева (до 10-12 м высотой) она встречается в
хвойно-широколиственных лесах о. Кунашир (Курилы). Как
редкий вид, включена в Красную книгу.
Ряд видов семейства с декоративными целями интродуциро-
ван на территории России, на юге страны они выдерживают ус-
ловия открытого грунта. Магнолии (М. grandiflora - магнолия
крупноцветковая и др.) - обязательный элемент садов и парков
курортных зон Кавказа и Крыма.
78
Тюльпанное дерево (Liriodendron tulipifera) - североамери-
канский вид, на родине может достигать 75 м высоты, а окруж-
ность колоннообразного ствола - 10 м.
Из других семейств, близких к Магнолиевым, отметим Лав-
ровые (Lauraceae, порядок Laurales), к которому относятся лавр
благородный (Laurus nobilis, листья лавра - обычная приправа,
используемая в кухнях многих стран), авокадо (Persea
americana, богатые витаминами, жиром и белком несладкие
плоды авокадо очень вкусны) и коричник (Cinnamomum), раз-
личные виды которого дают корицу, камфору, эфирные и жир-
ные масла, лекарственные вещества, ценную древесину и явля-
ются великолепными декоративными растениями.
Это интересно
Лауреат. Согласно древнегреческому мифу, лесная нимфа Дафна,
не желая стать женой бога поэзии и искусства Апполона, превратилась
в вечнозеленый лавр, в котором продолжало биться ее сердце. В па-
мять о прекрасной нимфе Аполлон сплел венок из лавра и предназна-
чил его людям для наград. В античной Греции лавровыми венками на-
граждали победителей олимпийских игр и называли их дафнофорами
(дафна по гречески - лавр), и только позднее римляне стали называть
победителей лауреатами, т.е. увенчанными лаврами. С лавром связано и
известное выражение «почивать на лаврах».
Бакалавр. Слово «бакалавр», которым обозначают первую ступень
высшего образования, переводится с латинского как «ягода лавра».
7.2. Семейство Nymphaeaceae - Нимфейные
Паспорт семейства
Родов - 6(3), видов - 50(10)
Цветок - *Са4.12Соа, G^j _
Распространение - пресноводные местообитания всех мате-
риков до зоны тундр
Жизненная форма - гидрофиты с плавающими на поверхно-
сти воды листьями и мощными корневищами
Опыление - кантарофилия, у некоторых - клейстогамия
Плоды - синкарпная ягодообразная много листовка. Семена
распространяются птицами, водой
Важнейшие роды - Nuphar, Nymphaea, Victoria
Семейство (рис. 8) входит в порядок Nymphaeales, в составе
которого еще два семейства. Нимфейные представляют особый
экологический тип растений, ведущих водный образ жизни
экологическую группу гидрофитов. У всех представителей от-
сутствуют сосуды в корневищах и черешках листьев, но развита
79
воздухоносная ткань - аэренхима. Поверхность листьев не сма-
чивается благодаря восковому налету и образованию бугорча-
тых выростов.
Нимфейные широко распространены по всему земному ша-
ру, и основным фактором, который ограничивает ареалы от-
дельных видов, является температура воды. Для тропического
рода виктория (Victoria) необходима вода с температурой не
ниже 28°, некоторые виды рода нимфея (Nymphaea alba), дости-
гающие 68°с.ш., могут расти в промерзающих водоемах. Период
пересыхания водоема нимфейные переживают в стадии покоя в
виде глубоко погруженных в грунт побегов-корневищ, хотя не-
которые виды — земноводные и могут образовывать листья да-
же при пересыхании водоема (если грунт остается хорошо ув-
лажненным).
Рис. 8. Нимфейные: а - Nymphaea (1 цветок, 2 переход от
лепестков к тычинкам, 3 - плод); 6 - Nuphar (цветок и диаграмма
цветка)
Размеры плавающих листьев у виктории достигают 2 м в
диаметре. Такой лист с загнутыми вверх краями (наподобие
сковородки) может выдерживать вес 35 кг.
Цветки нимфейных разнообразны. В пределах семейства
встречаются разные типы завязи: верхняя, полунижняя и ниж-
няя. Околоцветник у большинства нимфейных двойной и отли-
чается постепенностью перехода от лепестков к тычинкам. Вен-
чик и андроцей содержат неопределенное число элементов.
Цветки обладают способностью закрываться в определенные
80
часы (утром или вечером), становясь ловушками для жуков-
опылителей: они остаются до очередного открытия цветка, по-
едая тычинки и обсыпаясь пыльцой.
Плоды в форме урны крупные (у виктории могут достигать
размера головы ребенка) и содержат от 400 до 2000 семян. Со-
зревшие под водой семена, окруженные слизью, всплывают на
поверхность. Напоминая икру рыб, они привлекают основных
распространителей - водоплавающих птиц. Семена не погибают
даже в замерзшем иле и сохраняют всхожесть более 5 лет.
В России встречаются три вида нимфеи, или кувшинки
(N. alba, N. Candida, N. tetragona), четыре вида кубышки (Nuphar
lutea, N. pumila и др.) и эвриала устрашающая (Euryale ferox).
Эти растения в некоторых регионах взяты под охрану, так как
уничтожаются населением из-за красивых цветков (особенно
кувшинки, называемые в народе водяными лилиями). Два вида
включены в Красную книгу. Гибнут нимфейные и при загрязне-
нии водоемов.
К порядку Nymphaeales близок монотипный порядок Nelum-
bonales - Лотосовые с одним родом лотос (Nelumbo). Эти кра-
сивейшие растения представлены в России одним видом - ло-
тос орехоносный (N. nucifera), который охраняется в Астрахан-
ском заповеднике и в двух малых охраняемых территориях на
Дальнем Востоке. В районах массового произрастания лотос -
это пищевое растение. Так, в Китае, Японии, Индии семена и
корневища лотоса используют для изготовления муки, крахма-
ла, сахара и масла. Из корневищ варят суп, применяют их и как
гарнир.
Это интересно
Цветок-легенда. По числу легенд о цветке лотос, видимо, не имеет
себе равных. В переводе с греческого лотос означает «умытый» - та-
кова свежесть его красок. И если розу называют царицей цветов, то
лотос - это настоящий царь.
В Древнем Египте лотос считали цветком богов. Скипетр - знак
власти фараонов - делался в виде цветка лотоса на длинном стебле,
цветок и бутоны лотоса чеканились на монетах. На гербе Египта и сей-
час пять цветков лотоса.
В Индии богиню плодородия Лакшми, супругу Вишну, называли
рожденной лотосом, стоящей на лотосе, окрашенной лотосом, и изо-
бражали с цветком лотоса в волосах. Восхвалению лотоса посвящены
индийские ритуальные танцы: сложенные руки пальцами вверх на
уровне лица - это спящий лотос, если пальца слегка раздвинуты - ло-
тос просыпается.
Гомер в «Иллиаде», рассказывая о странствиях Одиссея, привел
его в страну лотофагов - людей, питавшихся лотосом. Отведав изы-
сканного кушанья, спутники Одиссея позабыли о родине и захотели
81
навсегда остаться на этой земле. В романе Г.Флобера «Саламбо» опи-
сан пир легионеров, на котором подавали вино из лотоса.
Пища из лотоса. Вот что в начале века писал об использовании ло-
тоса Н.Ф.Золотницкий: «Главными его потребителями являются япон-
цы и китайцы, у которых он разводится как овощ. В пищу идут корне-
вища, зерна, листья. Корневище едят сырым и вареным. Летом его едят
со льдом, как прохладительное, а на зиму маринуют. Содержа много
крахмала, оно чрезвычайно питательно и удобоваримо. На рынках в
Японии, Китае и Индокитае корневища эти лежат целыми грудами и
носят название «хазун». По вкусу он, когда сварен, несколько напо-
минает сельдерей или брюкву. Едят его также и печеным на угольях
или поджаренным как сладкие коренья, а кроме того, из него делают
еще муку, которой засыпают суп, как манной крупой или саго. Зерна
едят засахаренными как лакомство или приготовляют из него пирож-
ное. Что касается листьев, то осенью, когда прекращается цветение,
молодые, выходящие в виде побегов листья нарезают, связывают в пуч-
ки и продают на рынках. Едят их в вареном виде как спаржу. Кроме
того, китайцы едят его тычинки, стебель, корневище, считая, что еда
эта возвращает старикам красоту и молодость. Китаянки же, как неко-
гда и древние египтянки и финикиянки, украшают себя его цветами».
7.3. Семейство Ranunculaceae — Лютиковые
Паспорт семейства
Родов - 50(40), видов - 2000(400)
Цветок - от *Ра>Аа> Go, до *Са5 Со^ Gs (есть Т)
Распространение - прохладные области в основном Северно-
го полушария (лесная зона), часто по заболоченным местам
Жизненная форма - большей частью многолетние травы,
реже - лианы, редко одно-двухлетние травы и полукустар-
ники
Опыление - насекомыми, редко ветром (Thalictrum)
Плоды - многоорешек, многолистовка, сочная однолистов-
ка. Распространяются ветром, водой, животными (у лес-
ных видов - муравьями)
Важнейшие роды - Aconitum, Adonis, Anemone,
Anemonoides, Caltha, Clematis, Delphinium, Pulsatilla,
Ranunculus, Thalictrum
Культурные растения - Anemone, Clematis, Aquilegia
Семейство (рис. 9) входит в порядок Ranunculales, объеди-
няющий 8 семейств. Лютиковые растения умеренного и хо-
лодного климата северного полушария, хотя отдельные пред-
ставители встречаются и в тропиках. Преобладающая часть
лютиковых - многолетние травы, но есть однолетники и даже
полукустарники. Это чаще луговые, луговоболотные и лесные
виды.
82
д)
е)
Рис. 9. Лютиковые: а - Adonis (1 цветок, 2 - гинецей и часть андро-
цея); б - Trollius (1 - цветок, 2 - нектарник и тычинка); в - Ranunculus
(1 - цветок, 2 - лепесток, 3 диаграмма цветка, 4 плод); г
Aconitum (цветок и тычинка); д - Delphinium (1 - цветок, 2 - диаграм-
ма цветка, 3 - плод); е - Thalictrum (1 - цветущий побег, 2 - бутон и
раскрывшийся цветок, 3 - плодик)
Главный признак, на основании которого виды объединены
в семейство, - неопределенное число тычинок и пестиков в
цветке. Околоцветник разнообразный (причем у многих окра-
шенной бывает чашечка, а лепестки превращены в нектарники):
простой из многочисленных элементов (купальница - Trollius)
или небольшого их числа (5-6, сон-трава - Pulsatilla), двойной
обычного для двудольных пятерного типа (лютик — Ranun-
83
cuius), зигоморфный (аконит Aconitum, живокость
Delphinium, консолида - Consolida).
Почти все лютиковые - насекомоопыляемые растения, при-
чем виды с зигоморфным цветком могут опыляться только на-
секомыми с длинными хоботками (шмелями), однако есть и вет-
роопыляемые (например, васияистник малый - Thalictrum
minus), а некоторые тропические виды опыляются колибри. На-
секомых привлекает пыльца или нектар.
Среди лютиковых есть весенние первоцветы - эфемероиды
(т.е. многолетники, большую часть года находящиеся в состоя-
нии покоя в виде корневищ, клубней или луковиц), которые в
народе называются «подснежниками» (настоящий род подснеж-
ник - Galanthus - из порядка Amaryllidales). Наиболее распро-
странены эфемероиды широколиственных лесов ветреничка
лютиковидная (Anemonoides ranunculoides), в. дубравная (А. пе-
morosa) и в. алтайская (A. altaica). На сухих лугах обычна вет-
реница лесная (Anemone sylvestris), а в луговых степях - про-
стрел раскрытый (Pulsatilla patens).
Большинство лютиковых накапливают ядовитые алкадоиды
и потому имеют жгучий и горький вкус и обычно не поедаются
скотом (в весенний период, тем не менее, возможно отравление
телят). Алкалоиды лютиковых широко применяются в медицине,
а аконит по сей день считается «королем тибетской медицины».
Адонис весенний (Adonis vernalis) содержит гликозиды, исполь-
зуемые для лечения сердечно-сосудистых заболеваний.
Благодаря красивым цветкам многие лютиковые использу-
ются как декоративные растения, особенно ломонос (Clematis),
водосбор (Aquilegia), дельфиниум (Delphinium), консолида (Con-
solida), купальница азиатская (Trollius asiaticus), называемая
огоньком или жарком.
К семейству лютиковых близки семейства Барбарисовые
(Berberidaceae) и Маковые (Papaveraceae). Первое семейство
интересно прежде всего потому, что в его состав входит важ-
ный для озеленения населенных пунктов род барбарис
(Berberis). Его плоды используются в кондитерской промышлен-
ности (карамель «Барбарис») и как приправа в узбекской кухне.
К семейству маковых относится мак снотворный (Papaver
somniferum), семена которого также применяются в кондитер-
ском производстве, но более важен мак как источник наркоти-
ческих веществ, используемых в медицине (на их основе готовят
препараты папаверин, пантопон, морфин). Последнее заставило
поставить под жесткий контроль возделывание мака не только
84
на специализированных предприятиях лекарственной промыш-
ленности, но даже на индивидуальных садовых участках. К ма-
ковым относится и популярное лекарственное растение чисто-
тел большой (Chelidonium majus) рудеральный вид, распро-
страненный в нарушенных лесах и редколесьях, а также в насе-
ленных пунктах в тенистых местах у заборов и строений.
Это интересно
Ядовитый аконит. Ядовитые свойства аконита использовались в
военном искусстве в Древней Греции и Китае - из него получали яд
для стрел. А в Непале аконитом отравляли мясные приманки для по-
имки крупных хищников и питьевую воду для защиты от нападения
врагов. Ядовитые алкалоиды некоторых видов используются для про-
изводства лекарств, в частности, для лечения некоторых форм аритмии
сердца.
Лютиковые в фольклоре. Купальница европейская (Т. europaeus) -
национальное растение удмуртов, его имя носит популярный фольк-
лорный ансамбль - «Италмас». «Подснежникам» из лютиковых (сон-
трава, ветренички) посвящена башкирская песня «Умырзая».
Цветок грез. Снотворное действие мака было известно с глубокой
древности. Древние греки считали его атрибутом бога сна Гипноса и
его родного брата - бога смерти Танатоса. Древние римляне верили,
что бог сновидений Морфей навевает приятные грезы и усыпляет лю-
дей, прикасаясь к ним маковым цветком.
Опиум у мусульман. Кроме целительного действия, опиум облада-
ет еще одним свойством - он утишает голод, и это используется му-
сульманами во время строгого поста «рамадан». Обычай курить опиум
также возник прежде всего в мусульманских странах, где он заменил
запрещенное по закону Магомета употребление вина. Однако опиум,
прозванный магометанами «маш Алла» - даром Господним, оказался
«даром Дьявола»: по своим гибельным последствиям он во много раз
хуже всякого вина.
Разноликий символ. Древние народы обратили внимание и на чрез-
вычайную плодовитость мака: он служил символом плодородия и супру-
жества. Отголоском этого является обычай сыпать в обувь новобрачной
маковые зерна как пожелание не быть бездетной. Иногда количество се-
мян в маковой головке было олицетворением процветающего города, а в
форме маковой коробочки с вырезами у верхушки видели сходство с
зубчатыми стенами древних городов. Кроме того, маковое зерно до сих
пор символизирует все мельчайшее, незначительное или невыполнимое.
Желая показать степень голода, говорят: «У меня с утра маковой росин-
ки во рту не было». Желая выразить что-либо невыполнимое, что трудно
даже сосчитать, говорят: «Как маком усыпано».
85
7.4. Семейство Caryophyllaceae - Гвоздичные
Паспорт семейства
Родов - 80(40), видов -2000 (300)
Цветок - *Са, Со, ияи А,+, G(,,2)
Распространение - по всему миру, но главным образом уме-
ренные области Северного полушария
Жизненная форма - одно- и многолетние травы, реже кус-
тарники
Опыление - насекомыми, реже самоопыление
Плоды - коробочки. Семена распространяются в основном
ветром и муравьями
Важнейшие роды - Cerastium, Coronaria, Dianthus, Lychnis,
Melandrium, Saponaria, Silene, Stellaria, Steris
Культурные растения - Dianthus
Вместе с рассматриваемым далее семейством Маревые,
Гвоздичные (рис. 10) входят в одноименный порядок Caryophy-
llales, в котором объединено 17 семейств.
Среди гвоздичных много лесных и луговых трав. Встреча-
ются они и в засушливых районах - в степях, полупустынях и
пустынях, а также в тундрах и высокогорьях, где представлены
своеобразной подушечной формой. Растут эти растения-
подушки очень медленно: побеги удлиняются всего на несколь-
ко миллиметров в год, хотя возраст отдельных экземпляров
исчисляется столетиями.
Листья у подавляющего большинства гвоздичных супротив-
ные, простые, цельные, часто узкие, а цветки обычно собраны в
дихазиальные соцветия.
У гвоздичных четко выражен присущий двудольным пятерной
тип цветка. Завязь верхняя, обычно с многочисленными семяза-
чатками. Опыляются они насекомыми. Одни имеют открытые
цветки (звездчатка - Stellaria, ясколка - Cerastium и др.) и широ-
кий спектр опылителей, другие - цветки со сросшейся чашечкой
(смолевка - Silene, смолка - Steris и др.), а опылителями могут
быть насекомые только с длинным хоботком, преимущественно
бабочки. Цветки многих гвоздичных окрашены в различные от-
тенки красного цвета, который воспринимают эти насекомые;
цветки, опыляющиеся ночными бабочками, имеют белый цвет и в
ночное время источают сильный запах, привлекая опылителей.
Чтобы избавиться от непроизводительных затрат пыльцы и
нектара на прокорм ползающих насекомых, которые не опыля-
ют эти виды (в первую очередь муравьев), растения имеют
клейкий стебель, причем выделение клейкого вещества прекра-
щается как только цветение закончится.
86
a)
б) в) r)
Рис. 10. Гвоздичные: a Stellaria nemorum (1 цветущий побег, 2 -
чашечка, 3 - лепесток, 4 - андроцей и гинецей, 5 плод, 6 - диаграм-
ма цветка); б - Silene nutans (1 - цветущий побег, 2 - лепесток, 3
цветок без венчика); в - Dianthus (1 - обоеполый цветок, 2 - женский
цветок в разрезе); г - семена (1 - с присемянником, Moehringia; 2 -
с крылом, Spergula)
Опыление в основном перекрестное, но как запасной вари-
ант возможно и самоопыление (а у некоторых видов встречают-
ся закрытые - клейстогамные - цветки, в которых всегда про-
исходит самоопыление). У представтелей этого семейства, как и
некоторых других, распространена гинодиэция - есть два типа
цветков: крупные - обоеполые и мелкие - женские.
Семена распространяются ветром при раскачивании коро-
бочек, причем высыпаются они не сразу, а частями, и у некото-
рых видов коробочки в ненастье закрываются зубчиками, так
что вода не может повредить семенам. Есть среди гвоздичных и
жизненная форма «перекати-поле»: осенью сильные порывы
ветра отрывают качим метельчатый (Gypsophila paniculata) от
корня, и его рыхлые шаровидные кусты разносятся ветром на
большие расстояния. Семена некоторых видов разносятся му-
равьями (мерингия - Moehringia).
87
Некоторые представители рода гвоздика (Dianthus chinensis,
D. caryophyllus, D. barbatus) введены в культуру, в результате се-
лекции получены очень красивые формы с крупными цветками.
Это интересно
Гвоздичные-рекордсмены. Колобантус кито (Colobanthus quitensis)
может быть включен в Книгу рекордов Гиннеса: это растение входит во
флору цветковых Антарктиды, состоящую всего из двух видов! Stellaria
decumbens - второй рекордсмен, обнаруженный в Гималаях на высоте
6 тыс. м над уровнем моря.
Дом для гусениц. Опыление некоторых гвоздичных осуществляют
бабочки, которые откладывают яички в завязь. Появляющиеся вскоре
гусеницы питаются незрелыми семенами, а затем уходят из цветка, но
неповрежденных семян у растений остается достаточно много.
«Дрова» из гвоздичных. В горах (до 2000 м над уровнем моря) на
скалах растет качим аретиевидный (Gypsophila aretioides). Он образует
очень твердые подушки, похожие на камень, одетый лишайником. Диа-
метр подушки достигает 2 м, а масса 150 кг. Местные жители упот-
ребляют эти подушки как топливо.
Красная гвоздика. Крестовый поход французского короля Людо-
вика IX (1720 г.) оказался неудачным: во время осады Туниса в вой-
сках разразилась эпидемия чумы, спасения от которой никому не было.
И тогда на помощь пришла дикая гвоздика. По подсказке одного лека-
ря король распорядился поить отваром из гвоздики всех заболевших, и
чума была побеждена. Но больному королю отвары не помогли... С
вернувшимися на родину крестоносцами во Францию попал невидан-
ный прежде спасительный цветок, который стал там предметом покло-
нения, символом борьбы и победы. Впоследствии парижские коммуна-
ры умирали на баррикадах с эмблемой на груди - красной гвоздикой.
7.5. Семейство Chenopodiaceae - Маревые
Паспорт семейства
Родов -115 (35), видов - 1500 (190)
Цветок - *РЛ5 А2., G(2-^
Распространение по всему миру, но преимущественно
субтропики с засушливым климатом и засоленными почвами
Жизненная форма - от однолетних трав до небольших де-
ревьев
Опыление - ветром (но есть и случаи насекомоопыления)
Плоды - орешки или семянки, окруженные чашечкой
Важнейшие роды Atriplex, Chenopodium, Haloxylon,
Salicomia, Salsola, Suae da
Культурные растения - Beta vulgaris, Spinacia oleracea
88
Это семейство (рис. 11) объединяет ветроопыляемые расте-
ния (хотя у некоторых видов в цветках имеются нектарные дис-
ки и в опылении принимают участие насекомые). Цветки обое-
полые или однополые. Околоцветник чашечковидный, невзрач-
ный, со свободными или сросшимися долями. Число тычинок
обычно совпадает с числом долей околоцветника. Гинецей чаще
из двух плодолистиков.
в) г)
Рис. 11. Маревые: а - Beta vulgaris (1 - часть соцветия, 2 - цветок в
разрезе, 3 - соплодие, 4 - плод, 5 - семя); 6 - Chenopodium album (1 -
цветущий побег, 2 - обоеполый цветок); в Atriplex nitens (1 - муж-
ской цветок, 2 - женский цветок, 3 - женский цветок без околоцвет-
ника, 4 - побег с плодами, 5 - плод); г - Salicomia europaea (общий
вид и цветок)
Маревые распространены повсюду, но наиболее обильны в
пустынях и полупустынях, где их список возглавляет удиви-
тельное дерево саксаул (Haloxylon), имеющее важное значение
для закрепления песков. Маревые обладают разнообразными
приспособлениями к произрастанию в условиях почвенной и
атмосферной сухости пустынь и степей или физиологической
89
сухости засоленных почв и солончаков озерных и морских по-
бережий.
В их клетках создается высокое осмотическое давление,
причем клеточный сок содержит так много растворенных солей,
что зола многих маревых издавна служит источником соды и
поташа для населения (и даже для промышленного произ-
водства).
У большого числа маревых листья редуцированы, и ассими-
ляционную функцию выполняют членистые зеленые ветви (что
особенно ярко выражено у саксаула). При этом многие расте-
ния являются ярко выраженными суккулентами (например, со-
лерос - Salicornia, сведа - Suaeda, поташник - Kalidium).
Ослаблению испарения и защите от солнечных лучей слу-
жит и хорошо развитый покров из различных волосков (напри-
мер, у кокпека — Atriplex сапа, бассии - Bassia), а также пора-
зительная способность даже однолетних пустынных маревых
быстро одревесневать и образовывать перидерму (например,
разросшийся экземпляр солероса обыкновенного легко принять
за многолетнее растение).
Как и у других ветроопыляемых растений, цветки у маре-
вых невзрачные, одиночные или собранные в клубочки. Зачас-
тую чашечка не опадает, а остается при плоде, причем может
разрастаться, образуя крылья, шипы, рожки, и даже становить-
ся сочной и принимать яркую окраску (например, у жминды -
Chenopodium foliosum).
Плоды распространяются в основном ветром, чему способст-
вуют разнообразные крыловидные придатки, а также характер-
ная для многих маревых жизненная форма «перекати поле». Но
могут распространяться и животными как эндозоохорно (пти-
цами), так и эпизоохорно (млекопитающими). В первом случае
плоды съедобны, а во втором - имеют разнообразные прицепки.
Маревые - основные доминанты пустынь Азии и Африки,
главные кормовые растения пастбищ этой зоны, излюбленный
корм верблюдов. В настоящее время многие виды пустынных
маревых улучшены селекцией, введены в культуру и использу-
ются для создания «искусственных пустынь» и закрепления
песков (работы Н.Т.Нечаевой). Особенно на больших площадях
высевается саксаул (с использованием авиации).
К маревым относятся крупные сорные и рудеральные ро-
ды - марь (Chenopodium) и лебеда (Atriplex), в каждом из кото-
рых примерно по 250 видов. Они представляют стратегию руде-
ралов. Как сорняк особо опасна марь белая (Ch. album), кото-
рая образует огромное количество семян, формирующих семен-
ной банк в почве. Ее семена, как и семена многих других сор-
90
ных растений, обладают свойством гетероспермии (т.е. разносе-
мянности). Благодаря разной толщине кожуры они имеют раз-
ный период покоя, поэтому после каждого рыхления почвы
прорастает только определенная часть семян.
В голодные годы население использовало марь и лебеду как
суррогат хлеба (это отражено в поговорке «Не то беда, что не
хлеб, а лебеда - нет хуже беды, когда ни хлеба, ни лебеды»).
К маревым относятся важнейшие культурные растения из
рода свекла (Beta) - сорта столовой, кормовой и сахарной свек-
лы. Предком этих культур является ныне сохранившаяся в ди-
кой природе Средиземноморья свекла морская (В. vulgaris subsp.
maritima). Еще в VIII в. до н.э. в культуру была введена салат-
ная свекла мангольд. Лишь в XVI в. появилась столовая свекла
и в XVIII в. - сахарная, а производство сахара из нее было на-
чато в 1800 г. (хотя первый сахар получил немецкий химик
А.С.Маркграф в 1747 г.). К концу 1810 г. в России работало уже
8 сахарных заводов. В настоящее время из свеклы, в корнях
лучших сортов которой содержание сахара достигает 30%, по-
лучают треть мировой продукции сахара.
Ценной огородной культурой является шпинат (Spinacia
oleracea). Он содержит много белка, витаминов и микроэлемен-
тов (особенно железа) и является не только пищевым растением
и диетическим продуктом, но и укрепляющим средством при
истощении организма и малокровии.
К маревым близко семейство Амарантовые (Amaranthaceae),
которое знаменито родом амарант, или щирица (Amaranthus).
Большинство видов этого рода - однолетние сорняки (щирицы).
Несколько видов в тропических странах выращиваются как овощ-
ные культуры. Разные виды амаранта также изучаются как пер-
спективные силосные культуры, способные давать до 1000 ц/га
зеленой массы. Некоторые считают амарант культурой XXI в.
Это сомнительно, так как при высоких урожаях из почвы выно-
сится такое количество элементов питания, что для компенсации
этого необходимо вносить 3-5 т/га минеральных удобрений, что
недопустимо с экологической точки зрения.
У цветоводов популярен красивый петуший гребень (Celosia
argentea), а также некоторые декоративные виды амаранта. В
прошлом семена нескольких видов амаранта широко использо-
вались для производства муки и выпечки хлеба в Китае, Мекси-
ке и некоторых странах Европы.
Это интересно
Царь пустыни. Саксаул единственное дерево, встречающееся в
пустыне. Растет он медленно и к 200 годам достигает высоты 5 м. Дре-
весина саксаула такая твердая, что оставляет зазубрины на топоре, а
ее теплотворная способность выше, чем у бурого угля.
91
7.6. Семейство Polygonaceae - Гречишные
Паспорт семейства
Родов - 50 (20), видов - 800 (190)
Цветок - * Са6.3 Aj+j
Распространение - по всему миру, но основное разнообразие
- в умеренных областях
Жизненная форма - одно- и многолетние травы, реже дере-
вья, кустарники и лианы
Опыление - насекомыми, ветром
Плоды - орешки, часто крылатые. Распространяются вет-
ром, реже водой и животными (прилипают к ногам)
Важнейшие роды - Bistorta, Polygonum, Rumex
Культурные растения - Fagopyrum, Rheum
Семейство (рис. 12) входит в одноименный монотипный по-
рядок. Его отличает: трехдольный тип цветка, отсутствие венчи-
ка и наличие трубчатого влагалища раструба, окружающего
основание междоузлия. Большинство видов растет на лугах и во
влажных местообитаниях, однако некоторые гречишные «захо-
дят» в пустыню и в этом случае переходят на ветроопыление.
б)
Рис. 12. Гречишные: а - Rumex obtusifolius (1 - часть соцветия, 2 - цве-
ток, 3 - плод, 4 - раструб у основания черешка листа); б -Fagopyrum
(1 - цветок, 2 - внешний вид плода и его поперечный срез)
92
Гречишные обычно обильно цветут и плодоносят, их ореш-
ки имеют приспособления для распространения ветром (напри-
мер, у видов джузгуна - Calligonum), а плоды щавелей (Rumex)
прекрасно плавают, так как часто имеют «желвачки» - вздутия
из губчатой ткани. В высокогорьях гречишные (например, горец
живородящий - Bistorta vivipara) чаще всего размножаются ве-
гетативно: в нижней части соцветия образуются выводковые
почки, дающие начало новым растениям.
Из всех видов гречишных наиболее распространен спорыш
(Polygonum arenastrum) - чемпион по устойчивости к вытапты-
ванию. Эта его способность используется при озеленении: ковер
из спорыша - хорошее покрытие для игровых площадок (вклю-
чая и футбольные поля).
В состав семейства входит важная продовольственная куль-
тура - гречиха (Fagopyrum esculentum), ее начали возделывать
4000 лет назад в Китае и Индии. В России гречиха стала попу-
лярной в XI-XII вв. Кроме того, гречиха - ценный медонос,
причем приближение ульев к ее посевам повышает урожай.
Все части растений гречишных (особенно подземные) со-
держат танины, и некоторые из их видов (ревень - Rheum, го-
рец, щавель) используются как хорошие дубители, в этом отно-
шении перспективен широко распространенный щавель пирами-
дальный - R.thyrsiflorus.
Это интересно
Достоинства гречневой каши. Русский агроном А.Н.Энгельгардт
оценил энергетическую эффективность гречневой каши при работе
землекопов: она на 25% выше, чем ячневой, и в два раза выше, чем кар-
тофеля. Высоко оценивал роль гречневой каши в питании солдат
А.В.Суворов, поэтому иногда ее называют суворовской.
Самое ботаническое название гречневой каши. Особенность плодов
гречишных, в том числе и гречихи, - трехгранность, соответствующая
числу плодолистиков. В монгольском языке гречневая каша называется
«гурвульчен буда», что переводится как «треугольная каша».
7.7. Семейство Fagaceae — Буковые
Паспорт семейства
Родов - 8 (2), видов - 900 (15)
Цветок - *Р(з+з) А12-6 Go , *Р(з+з) Ao G(j)
Распространение - умеренная, тропическая и субтропиче-
ская области
Жизненная форма - деревья, реже кустарники
Опыление - ветром, иногда насекомыми
Плоды - орехи, распространяются животными
Важнейшие роды - Castanea, Fagus, Nothofagus, Quercus
Культурные растения - Castanea
93
Семейство (рис. 13) входит в одноименный монотипный по-
рядок. Наиболее близко к буковым семейство Березовые (поря-
док Betulales), которое рассматривается далее.
Представители буковых - это преимущественно деревья,
листопадные или вечнозеленые, основные лесообразующие по-
роды широколиственных лесов Евразии, Америки (как Север-
ной, так и Южной), Африки.
Рис. 13. Quercus robur: 1 - побег с мужскими сережками, 2 - часть
мужской сережки, 3 - мужской цветок, 4 — женский дихазий с при-
цветниками (общий вид и в разрезе), $ - плоды
Буковые представляют большую группу порядков ветроопы-
ляемых, преимущественно древесных, растений с обоеполыми или
редуцированными однополыми цветками. Цветки мелкие (безле-
пестные невзрачные или вообще без околоцветника), собранные в
соцветия разного типа (сережковидные, колосовидные, головча-
тые и др.). Женские соцветия у основания окружены чашевидной
плюской. В цветках часто бывают рудименты органов другого
пола. Околоцветник более или менее сросшийся.
Происхождению этой группы порядков посвященно много
публикаций. Есть мнение, что они являются древними цветко-
выми, берущими начало от гнетовых. По противоположной точ-
ке зрения (ее придерживается А.Л.Тахтаджян), эта группа вос-
ходит вероятнее всего к Magnoliales и ветроопыление этих де-
ревьев вторично. Обе позиции не обоснованы.
Плоды орехи, заключенные в древеснеющую плюску,
распространяются животными, которые, создавая запасы для
пропитания, либо забывают про часть хранилищ, либо теряют
плоды в процессе доставки к месту хранения.
Для умеренной полосы Восточной Европы наиболее важен
род дуб (Quercus), представленный здесь одним видом - дубом
черешчатым (Q. robur). Общее число видов дуба превышает 450,
основное видовое разнообразие - в Северной Америке и Юго-
94
Восточной Азии. В Сибири и Центральной Азии дубов нет (по
Уралу проходит восточная граница дуба черешчатого).
Среди дубов преобладают листопадные, но много и вечно-
зеленых видов (в особенности важен Q. ilex, который домини-
рует на обширных территориях Средиземноморья). Этот кус-
тарниковый дуб очень хорошо приспособлен к хозяйственному
влиянию человека и быстро отрастает (восстанавливается по-
рослью) после нарушения выпасом и пожаром. В Западном
Средиземноморье распространен широко известный дуб пробко-
вый (Q. suber), который для получения пробки выращивают за
пределами естественного ареала.
На территории ряда районов России в значительной мере
вырублены дубравы из дуба черешчатого, поскольку его древе-
сина ценится как топливо и материал для поделок. В благопри-
ятных условиях (на богатых почвах и при достаточном количе-
стве осадков) этот дуб достигает высоты 25-30 м. Особенно-
стью этого вида является высокая экологическая пластичность,
на южной границе ареала он представлен кустарниковыми
формами.
Название семейству дал род бук (Fagus), представленный в
Европе двумя видами: бук европейский (F. sylvatica), бук вос-
точный (F. orientalis). Эти теплолюбивые и требовательные к
почвенному плодородию деревья достигают 50 м в высоту и жи-
вут до 500 лет. Под густым пологом его развивается мало тра-
вянистых растений (поэтому буковые леса называют «мертво-
покровными»). Высоко ценится и древесина бука, имеющая кра-
сивый рисунок.
Третий важный для европейской части России и стран СНГ
род - каштан (Castanea). Наиболее известен каштан настоя-
щий, или посевной (С. sativa), плоды которого съедобны. Осо-
бенность этого вида - сочетание ветроопыления с опылением
насекомыми, которые привлекаются обильной пыльцой. Амери-
канский родственник европейского каштана каштан зубча-
тый (С. dentata) печально известное растение, с которым
произошла «самая крупная ботаническая катастрофа». Эта одна
из главных лесообразующих пород США была практически
полностью уничтожена в начале нашего века патогенным гри-
бом (Endothia parasitica), занесенным с интродуцированным из
Китая другим видом каштана.
От настоящего каштана следует отличать конский каш-
тан - Aesculus hippocastanum, который относится к семейству
Конскокаштановые (Hippocastanaceae). Это декоративное дере-
во, широко культивируемое в Западной Европе и на Украине,
весной покрывается крупными конусовидными белыми соцве-
95
тиями, красиво выделяющимися на фоне 5-7-пальчатосложных
листьев. Дерево достаточно холодостойко и плодоносит даже
на широте Москвы, однако его семена несъедобны.
Это интересно
Желуди. Желуди дубов играли важную роль в питании людей в
доисторические эпохи, а в более поздние времена они использовались
для выпечки хлеба, особенно в неурожайные годы. У некоторых среди-
земноморских дубов желуди не только съедобны, но и приятны на
вкус, и в ряде стран широко используются в пищу местным населением
(Испания, Португалия, Италия, Греция, Турция, Алжир). Едят желуди
«своих» дубов индейцы Америки, японцы, а китайцы приготовляют из
них национальное блюдо «до-фун» (кстати, листья дубов используются
для выкармливания шелковичного червя).
7.8. Семейство Betulaceae — Березовые
Паспорт семейства
Родов - 7 (6), видов - 150 (80)
Цветок - *Р2 A2.i Go , *Р0 Ао
Распространение - умеренные области Северного полушария,
отчасти тундра и высокогорья
Жизненная форма - деревья, кустарники и кустарнички
Опыление - ветром
Плоды - орешки и орехи, распространяются ветром и жи-
вотными
Важнейшие роды - Alnus, Be tula, Carpinus, Corylus
Культурные растения - Corylus
Это семейство (рис. 14) монотипного порядка Betulales объе-
диняет деревья, некоторое количество кустарников и даже кус-
тарничков {береза карликовая - Betula папа в тундровой зоне).
Все они однодомные, ветроопыляемые, листопадные, с простыми
очередными листьями и с крупными рано опадающими при-
листниками. Распространены березовые в умеренных областях
Северного полушария, являясь важными составляющими лесов.
Цветки у березовых однополые, мелкие, невзрачные, собра-
ны в соцветия. Йо внешнему виду мужские и женские соцветия
хорошо различаются: мужские - длинные, сережковидные, во
время цветения висячие; женские короткие, прямостоячие,
шишковидные или висячие. Цветут березовые одновременно с
распусканием листьев (береза) или даже немного раньше (ольха,
лещина). Оси мужских сережек сильно удлиняются, и они легко
раскачиваются от малейшего дуновения ветерка, высеивая лег-
кую ярко-желтую пыльцу, которая далеко разносится ветром.
96
Женские соцветия остаются упругими, только рыльца удлиня-
ются во много раз. Высовываясь из-под чешуи, липкие ярко-
красные рыльца прекрасно улавливают пыльцу. Не исключено,
что в переносе пыльцы участвуют и мелкие насекомые (пол-
зающая и летучая тля, мелкие жучки и дрД мириадами пере-
ползающие и перепархивающие с соцветия на соцветие.
Рис. 14. Березовые: а - Betula pendula (1 — мужские и женские соцве-
тия, 2 - мужской дихазий, 3 - женский дихазий, 4 - плод); б - Corylus
avellana (1 - мужские и женские соцветия, 2 - соплодие, 3 - плод);
в - Alnus incana (соплодие и корень)
Плоды (исключая род лещину - Corylus) мелкие и легкие,
производятся в огромном количестве и разлетаются на большие
расстояния. Парусность орешков увеличивается разнообразными
крыльями, образованными из частей цветка или прицветников.
97
У синзоохорного рода лещина плоды - крупные, тяжелые,
толстостенные орехи, окруженные плюской из сросшихся при-
цветных чешуи. При созревании они выпадают из плюски.
Наиболее распространенные в Европе и встречающиеся на
территории России виды - береза повислая, или бородавчатая
(Betula pendula) и береза пушистая, или белая (В. pubescens).
Первый вид распространен на сухих почвах, второй - по пере-
ходным болотам. При повреждении береза легко отрастает по-
рослью от основания ствола. Этим объясняется массовое разви-
тие березняков на месте вырубленных сосновых лесов (что ши-
роко распространено в России).
Белый цвет стволов березы связан с наличием в клетках
пробки бетулина. Береза ценится за древесину, которая в моло-
дом возрасте используется для производства досок и строи-
тельного бруса. Березовые дрова считаются самыми лучшими.
Весной заготавливается березовый сок, который консервируется
или перерабатывается в квас. Из бересты готовятся разнооб-
разные поделки. В медицине используются почки березы. Бере-
зовые веники являются обычным атрибутом бани.
Кроме березы, в состав семейства входят ольха (наиболее
широким ареалом обладают два вида: ольха черная Alnus
glutinosa и о. серая - A. incana) и уже упомянутая лещина (на
территории России - лещина обыкновенная - Corylus avellana).
Ольха черная - доминант низинных болот с богатым торфом,
ольха серая распространена в таежной зоне и относится к чис-
лу вторичных мелколистных пород, которые замещают ель по-
сле вырубания и способствуют ее возобновлению, выступая в
роли растений-нянь. Важной особенностью ольхи является сим-
биоз с азотфиксирующими микроорганизмами (в первую оче-
редь с актиномицетами). Клубеньки на корнях ольхи достигают
5 см в диаметре.
Лещина отличается от остальных видов семейства особен-
ностями плода (поэтому иногда выделяется в отдельное семей-
ство Corylaceae - лещиновые). Это важное ресурсное растение,
которое дает орехи. Культурное растение, являющееся естест-
венным гибридом между тремя видами лещины (С. avellana,
С. maxima, С. pontica) и называемое фундуком, имеет крупные
орехи. Основными поставщиками их являются южные районы
России, Крым, Кавказ, где имеются плантации фундуков. Орехи
используются в кондитерском производстве, как лакомство и
как сырье для производства ценнейшего орехового масла.
Значимость видов рода ольха как источника древесины
меньше, чем березы, тем не менее ольховые топи очень важны
для сохранения гидрологических характеристик ландшафтов.
98
Кроме того, эти топи являются прекрасными фильтрами, обез-
зараживающими и очищающими загрязненную воду (даже са-
мые токсичные стоки свиноферм).
Граб европейский (Carpinus betulus) распространен по всей
Западной и Восточной Европе, он очень теневынослив и образу-
ет второй ярус в широколиственных буковых и дубовых лесах.
7.9. Семейство Salicaceae - Ивовые
Паспорт семейства
Родов - 3 (3), видов - 550 (150)
Цветок - *Р0 А2.х Go , *Ро Ао G^
Распространение - главным образом умеренные и холодные
области Северного полушария
Жизненная форма - деревья, кустарники и кустарнички
Опыление - ветром и насекомыми
Плоды - коробочки, семена распространяются ветром
Важнейшие роды - Chosenia, Populus, Salix,
Культурные растения - Populus
Ивовые (рис. 15) выделяются в монотипный порядок Sali-
cales. Все они свето- и влаголюбивы и имеют цельные очередные
листья обычно с прилистниками.
Однополые цветки ивовых располагаются на разных особях
(двудомность) и собраны в соцветия, называемые сережками, и
представляющие собой кисть с мягкой поникающей осью и
очень короткими цветоножками. Сережки опадают целиком, у
мужских экземпляров - после цветения, а у женских - после
рассеивания семян.
Околоцветник у ив редуцирован полностью, вместо него
есть 1-3 маленьких нектарника, у чозении нет ни околоцветни-
ка, ни нектарников, а у тополя есть бокальчатый околоцветник.
Плоды-коробочки раскрываются двумя створками, мелкие
семена, имеющие при основании пучок волосков, почти не со-
держат запасов питательных веществ и быстро, в течение одних
суток прорастают при попадании на влажный и свободный от
растений субстрат (обычно - это свежий суглинистый или супес-
чаный наилок вдоль берегов рек). Также быстро растут и сеянцы,
которые за первый год жизни могут достичь высоты 30-60 см.
Если тополя (Populus) и чозения (Chosenia) опыляются вет-
ром, то ивы (Salix) - насекомыми, что делает их ранними медо-
носами: ивы цветут раньше, чем другие деревья и кустарники, и
представляют ценную кормовую базу для пчел.
99
Рис. 15. Ивовые: а - Salix саргеа (1 - мужское сецветие, 2 - мужской
цветок, 3 - женское соцветие, 4 - женский цветок, 5 - завязь в разре-
зе, 6 - плод, 7 - семя); б - Populus nigra (1 - мужское соцветие, 2 -
мужской цветок, 3 - женский цветок, 4 - раскрывшийся плод, 5 - семя)
В роде тополь центральное положение занимает тополь
дрожащий, или осина (Р. tremula). Пластинки листьев осины
широкие, а черешки длинные, отчего листья трепещут при лег-
ком дуновении ветра (откуда и пошло латинское название). Это
широко распространенная вторичная порода, которая образует
леса на месте вырубленных дубняков и липняков. Осина дает
обильную поросль от корней, и часто целые группы или рощи
осин представляют собой один клон. Древесина осины ценится
как строительный материал.
Второе место занимает тополь черный, или осокорь (Р. nig-
ra) - древесная порода прирусловой поймы, реже вместе с ним
встречается тополь белый (Р. alba).
Самый многочисленный род - ива, в его составе 350-370
видов. Важнейшие виды для средней полосы России ива
трехтычинковая (S. triandra), ива пятитычинковая (S. pentan-
dra), ива белая (S. alba), ива пепельная (S. cinerea), ива корзи-
ночная (S. viminalis), ива козья (S. саргеа).
100
Ивовые имеют огромное практическое значение. Тополь,
благодаря быстрому росту, считается само» перспективной дре-
весной породой для биологического варианта солнечной энерге-
тики. Гибриды тополя обладают рекордной скоростью роста,
тополевые плантации срезают в возрасте 7-8 лет, когда стволи-
ки имеют диаметр 6-10 см и высоту до 8 м. Тополем сегодня
занимаются несколько научно-исследовательских институтов
Европы и США. Как источники древесины, перспективны и не-
которые быстрорастущие гибриды ив.
Ивы выращивают как сырье для производства дубителей,
корзин и легкой мебели. В естественных экосистемах они слу-
жат кормом для лосей, оленей и зайцев. Ивовый веточный корм
неплохо едят и домашние животные. Тополя и ивы широко ис-
пользуются в городском озеленении. Они обладают высокой ус-
тойчивостью к загрязнению атмосферы и потому выращиваются
на территории промышленных предприятий.
Это интересно
Верба. Верба (Salix acutifolia), которая рано весной украшается
пушистыми сережками, почитается с языческих времен. Славяне упо-
минали ее в связи с именем своего главного божества Перуна. «Перу-
нова лоза» была непременным участником весенних обрядов. Позднее
праздник почитания вербы слился с христианским праздником, кото-
рый отмечается в шестую неделю Великого поста. По библейской ле-
генде в это время иудейский народ встречал Христа в Иерусалиме вет-
вями финиковой пальмы. В память об этом событии воскресный день
шестой недели Великого поста назван церковью Вербным (у католиков
он называется праздником пальмы).
7.10. Семейство Cucurbitaceae - Тыквенные
Паспорт семейства
Родов - 120 (8), видов - 700 (9)
Цветок - *Са^) Co(j) A(j.j) Go , * Са^) Со^) Ao Gf^J)
Распространение - тропические и субтропические области
Жизненная форма - однолетние и многолетние травы, лианы
Опыление - насекомыми
Плоды - ягода или тыквина, реже коробочки, семена рас-
пространяются животными
В естественной флоре умеренной полосы отсутствует
Культурные растения - Citrullus, Cucumis, Cucurbita, Luffa,
Melo
101
Семейство (рис. 16) относится к монотипному порядку Cu-
curbitales. Для его представителей характерны однополые цвет-
ки со спайнолепестным венчиком и нижней завязью (некоторые
виды двудомные). Крупные хорошо развитые нектарники жен-
ских цветков наполнены очень сладким нектаром и доступны
всем желающим, поэтому цветки тыквенных посещают около
150 видов насекомых. В мужских цветках насекомые кормятся
очень питательной пыльцой.
Как правило, тыквенные - это быстро растущие растения,
стебли у них лазающие с усиками (метаморфизированными по-
бегами) и крупными листьями. Плоды - тыквины - иногда дос-
тигают колоссальных размеров и массы более 100 кг.
Рис. 16. Cucumis: 1 - мужской цветок, 2 - женский цветок,
3 - плод (поперечный разрез)
Преимущественно в рудеральной флоре южной части Рос-
сии спорадически можно встретить около десяти одичавших или
заносных видов тыквенных. Среди них наиболее натурализо-
вавшимися являются переступень белый (Bryonia alba) и занос-
ный североамериканский вид пузыреплодник лопастнолистный
(Echinocystis lobata). Пузыреплодник, или эхиноцистис, очень
быстро растет и может за лето достичь высоты 10 м, опыляется
он не только насекомыми, но и ветром.
Однако семейство интересно в первую очередь культурными
пищевыми, декоративными и техническими растениями, такими
как тыква и кабачки (Cucurbita pepo, родом из Америки), огу-
рец (Cucumis sativus), дыня (Melo sativa), люффа (Luffa cylind-
rica, эти три культуры родом из Индии), арбуз (Citrullus lanatus,
родом из Африки).
102
Это интересно
Плюющие плоды. В южных степных районах России растет не-
обычное растение из семейства тыквенных «бешеный огурец»
(Ecballium elaterium), у которого семена, окруженные слизью, с силой
выбрасываются из плода под действием тургорного давления.
Самые крупные плоды. Во многих странах до настоящего времени
проводят конкурсы крупноплодных тыкв. В Канаде были получены
плоды массой 284 и 287 кг, а в США рекордный плод массой 302 кг
был выращен в 1986 г.
7.11. Семейство Brassicaceae (Cruciferae)- Капустные
(Крестоцветные)
Паспорт семейства
Родов - 380 (100), видов - 3200 (350)
Цветок - *Сй2+2 Coi А2+1 G(2)
Распространение - по всему миру, но главным образом в
умеренных и холодных зонах
Жизненная форма - одно- и многолетние травы, реже кус-
тарники и полукустарники
Опыление - насекомыми
Плоды - стручки и стручочки. Семена распространяются
ветром (в том числе, формы «перекати-поле»), животными
(в том числе муравьями), авто- и гидрохорно
Важнейшие роды Alyssum, Barbarea, Berteroa, Bunias,
Lepidium, Rorippa, Sisymbrium, Thlaspi
Культурные растения - Brassica, Crambe, Raphanus, Sinapis
Семейство (рис. 17) крестоцветных входит в порядок Cappa-
rales, включающий 4 семейства, и отличается редким однообра-
зием строения цветка. Примерно в современном объеме их как
«класс » выделил К.Линней.
От типичного пятичленого цветка двудольных цветок пред-
ставителей этого семейства отличается четырехдольностью око-
лоцветника, что и дало повод назвать их крестоцветными. Ты-
чинки двусильные: в наружном круге - две короткие и во внут-
реннем - четыре длинные. Цветки собраны в кистевидные или
щитковидные обычно безлистные соцветия.
Листья у крестоцветных очередные, но у большинства пред-
ставителей нижние листья образуют прикорневую розетку. Ши-
рокому расселению крестоцветных способствуют разнообраз-
ные приспособления: крылатые или пузыревидно вздутые пло-
ды, мелкие и легкие разносимые ветром семена, форма «пере-
кати-поле». Важной особенностью многих крестоцветных явля-
ется гетерокарпия, причем различаться могут как целые плоды
103
1
в)
Рис. 17. Крестоцветные: а - Brassica oleraceae (1 - часть соцветия, 2 -
цветок (в разрезе и без околоцветника), 3 диаграмма цветка, 4
плод); б - разновидности капусты огородной (1 - кочанная, 2 - савой-
ская, 3 - цветная, 4 - брюссельская, 5 кольраби); в - плоды (1
Raphanus raphanistrum, 2 - Capsella bursa-pastoris, 3 - Thlaspi arvense,
4 - Lunaria rediviva, 5 - Crambe maritime, 6 - Camelina, 7 - Bunias)
104
(например, у видов вайды - Isatis, сердечника - Cardamine), так
и их части (например, у морской горчицы — Cakile maritima).
Семейство играет важную роль в жизни человека - включа-
ет значительное количество культурных растений и несколько
злостных и трудноискоренимых сорняков.
Самым главным родом из культурных крестоцветных явля-
ется капуста (Brassica). Форма капусты огородной (В. oleracea),
родиной которой является Средиземноморье, в результате се-
лекции дала множество культурных форм с резкими морфоло-
гическими отличиями - капусту кочанную (var. capitata), брюс-
сельскую (var. gemmifera), цветную (var. botrytis), кольраби (var.
gongyloides). Другие культуры этого рода - горчица сарептская
(В. juncea) и черная (В. nigra), рапс (В. napus var. napus), турнепс
(В. гара).
На сегодняшний день наиболее «модной» культурой являет-
ся рапс. Его семена содержат до 50% масла, кроме того, рапс
зимостоек и быстро развивается, что позволяет использовать
его как кормовую и сидеральную (т.е. запахиваемую в почву в
качестве удобрения) культуру. Кроме рапса, выращивают и
другие быстро развивающиеся крестоцветные, способные нако-
пить значительную зеленую массу за короткий срок (например,
в год весенне-летней засухи с поздним началом дождей): горчи-
цу сарептскую, виды катрана (Crambe).
В Средней Азии включение крестоцветных в хлопчатнико-
вые севообороты имеет большое фитосанитарное значение,
уменьшая риск нападения на хлопчатник насекомых-фитофагов.
В хозяйственных целях используются естественные заросли
некоторых крестоцветных, например хрена дикого (Armoracia
rusticana). В декоративных целях издавна выращивают ночную
фиалку (Hesperis matronalis), левкой однолетний (Matthiola
annua) и др.
К полевым сорнякам относятся редька дикая (Raphanus
raphanistrum), дескурайния Софьи (Descurainia sophia), гулявник
Аезеля (Sisymbrium loeselii), гулявник лекарственный (S. offici-
nale), неслия метельчатая (Neslia paniculata), ярутка полевая
(Thlaspi arvense) и др.
Это интересно
Пищевые дикорастущие крестоцветные. Среди дикорастущих кре-
стоцветных есть пищевые овощные и витаминоносные растения. Свер-
бига восточная (Bunias orientalis) в начале мая образует нежные стебли,
которые едят в свежем виде, из листьев готовят салаты, а также заго-
тавливают их впрок (сушат, маринуют, солят). Из зеленых частей су-
репки обыкновенной (Barbarea vulgaris) готовят суп, пюре, гарниры.
Надземная часть пастушьей сумки обыкновенной (Capsella bursa-
105
pastoris) богата фитонцидами, микроэлементами и витаминами. Ее мо-
лодые листья используют в свежем виде, в супах и салатах и как на-
чинку для пирогов, а высушенные и измельченные — для заправки пер-
вых блюд (в Китае пастушью сумку уже более 100 лет разводят как
овощ). Используют в пищу также ярутку полевую (Thlaspi arvense),
клоповник сорный (Lepidium ruderale), сердечник луговой (Cardamine
pratense).
Горчица-путешественница. На морских побережьях весьма обычна
морская горчица (Cakile maritima), что связано с ее приспособленностью
к засоленным грунтам и разнообразием форм расселения: гидро-, анемо-
и антропохорно. В верхних члениках ее плодов формируется губчатая
ткань, поэтому они хорошо держатся на воде и разносятся морскими те-
чениями. Нижние членики плодов остаются на стеблях, которые после
усыхания отламываются и перекатываются ветром. Поскольку морская
горчица часто растет вблизи портов, верхние части плодов вместе с гру-
зом попадают на судна и разносятся на дальние расстояния. Таким пу-
тем эта «уроженка» Средиземноморья успешно натурализовалась в Аме-
рике и Австралии, куда прибыла с первыми колонистами. Даже на вул-
каническом острове, образовавшемся в 1963 г. в Атлантическом океане (в
20 милях южнее Исландии), первым сосудистым растением оказалась
морская горчица, обнаруженная там уже в 1965 г.!
7.12. Семейство СаппаЬасеае - Коноплевые
Паспорт семейства
Родов - 3 (3), видов - 3(3)
Цветок — *Pj A; Go , *P(sj Ao G(2)
Распространение - умеренные области Северного полушария
Жизненная форма - прямостоящие или вьющиеся травы
Опыление - ветром
Плоды - орешки, распространяются животными
Важнейшие роды - Cannabis, Humulus
Культурные растения - Cannabis, Humulus
Семейство (рис. 18) входит в порядок Urticales, в составе
которого еще четыре семейства, в том числе Вязовые - Ulma-
сеаеа (к которому в России относятся два рода: вяз - Ulmus и
дзельква Zelkova), Тутовые Могасеае (в состав которого
входит шелковица - Morns) и Крапивные (Urticaceae).
Семейство Коноплевые состоит всего из трех родов: коноп-
ля (представлена одним видом Cannabis sativa), хмель (в средней
полосе встречается только один вид Humulus lupulus) и недавно
установленного рода хумулопсис (Humulopsis).
Конопля - одна из первых технических культур, которую
выращивали на территории России с конца первого тысячеле-
тия. Это двудомное однолетнее растение (сейчас выведены од-
106
подомные сорта) за один сезон в хороших условиях вырастает
до 4 м и более (при этом скорость ее роста как у бамбука).
Лубяные волокна женских растений - матерки - грубые, из них
получаются крепкие канаты. Особое внимание возделыванию
конопли уделил Петр I, когда ему потребовались канаты для
первых русских военных кораблей. Ее культура распространи-
лась по всей России. Площадь посевов конопли настолько рас-
ширилась (до 1 млн га), что появилась возможность экспорти-
ровать конопляное волокно - пеньку. Пенька устойчива к гние-
нию и незаменима для изготовления вещей и материалов, по-
стоянно пребывающих в водной среде (рыболовные снасти, по-
жарные рукава, палатки и т.п.).
Рис. 18. Коноплевые: a -Cannabis sativa (1 - цветущий побег мужского
растения (поскони), 2 - мужской цветок, 3 -цветущий побег женского
растения (матерки), 4 — женский цветок, 5 - плод); б - Humulus lupulus
(1 — мужское соцветие, 2 - мужской цветок, 3 — женское соцветие, 4 -
женские цветки с кроющим листом, 5 - соплодие «шишка», 6 - плод)
107
Из более тонких волокон мужских экземпляров (поскони)
население пряло ткани для повседневной одежды, а конопляное
масло использовалось в пищу (сейчас оно идет на изготовление
олифы, лака, красок, мыла). В XVIII-XIX вв. улицы крупных
русских городов, в том числе Москвы, освещали светильники,
заправленные конопляным маслом. Остающийся после отжима
масла жмых - ценный концентрированный корм для скота. Ко-
нопляное семя - излюбленный корм певчих и декоративных
птиц. Отходы (пакля) широко использовались как теплоизоля-
ционный и обтирочный материал.
Конопля может дичать и засорять посевы разных культур
(эту форму иногда выделяют в самостоятельный вид С. ru-
der alis).
Печальную славу снискала себе конопля как источник нар-
котических веществ. Молодые побеги и женские соцветия коно-
пли - основа для изготовления анаши (марихуаны). Застывший
сок растения - гашиш - один из самых опасных наркотиков. С
возделыванием конопли как источника наркотических веществ в
странах Центральной Азии ведется широкая (и низко результа-
тивная, из-за высоких цен на наркотики) борьба. В условиях
умеренного и холодного климата конопля практически теряет
наркотические свойства.
Хмель применяется в производстве пива (в него добавляют
настой высушенных женских соцветий - «шишек» хмеля и имен-
но он придает напитку специфический пивной вкус) и культи-
вируется на всех континентах. Хмель является также лекарствен-
ным растением, идущим на изготовление препаратов с успокои-
тельным, мочегонным и противовоспалительным действием.
Это интересно
Древнейший напиток. Пиво - очень древний напиток. В Талмуде
указывается, что в Вавилоне плененные евреи спаслись от проказы
только потому, что пили пиво. Римляне, употреблявшие виноградное
вино, считали пиво «варварским напитком», недостойным цивилизо-
ванного человека, однако их потомки сегодня, как и весь мир, потреб-
ляют пиво.
«Скифская баня». Применение конопли как наркотического сред-
ства имеет тысячелетнюю историю. Геродот в своей «Истории» так
описывает обычай скифов одурманивать себя дымом конопли: «Взяв
конопляное семя, скифы подлезают под войлочную юрту и затем бро-
сают его на раскаленные камни. От этого поднимается такой сильный
дым и пар, что никакая эллинская баня не сравнится с такой баней.
Наслаждаясь ею, скифы громко вопят от удовольствия. Это парение
служит им вместо бани, так как водой они вовсе не моются».
108
7.13. Семейство Urticaceae - Крапивные
Паспорт семейства
Родов - 60 (7), видов - 1000 (25)
Цветок - *Р4 А4 Gg > Ад G(2)
Распространение - преимущественно тропические и суб-
тропические области, но часть видов встречается в умерен-
ной зоне (в основном по рудеральным местообитаниям)
Жизненная форма - травы, древесные растения
Опыление - ветром, насекомыми, возможен апомиксис
Плоды - ореховидные, распространяются животными
Важнейший род - Urtica
Культурные растения - Boehmeria, Pilea
Это большое тропическое семейство (рис. 19) во флоре уме-
ренной полосы представлено семью родами. Наиболее распро-
странены в роде крапива (Urtica) - к. двудомная (U. dioica) и к.
жгучая (U. urens). В азиатской части России к ним добавляется
крапива коноплевидная (U. cannabina) - наиболее «сердитое»
растение, оставляющее долго незаживающие ожоги. Европей-
ские виды крапивы космополитны, они связаны с нарушенными
местообитаниями на богатых и хорошо увлажненных почвах.
Рис. 19. Urtica dioica: 1 - побег с мужскими соцветиями,
2 - мужской цветок, 3 - женский цветок (общий вид и в разрезе)
Как и все представители группы порядков, которые объеди-
няются А.Л.Тахтаджяном в подклас Hammameiididae (во многих
учебниках они называются «однопокровными»), крапивные
это в подавляющем большинстве ветроопыляемые растения с
сильно редуцированными однополыми цветками. Соцветия - не
только сережки, но также и головчатые, и метельчатые. В
большинстве случаев крапивные опыляются перекрестно, но
возможно самоопыление и даже апомиксис развитие семян
109
без оплодотворения. У апомиктичных растений микропиле за-
растает и самязачаток оказывается неспособным принять пыль-
цевую трубку мужского гаметофита.
Распространяемые эндозоохорно плоды крапивных орехо-
видные, мелкие и сухие, хотя у некоторых тропических видов -
плоды сочные. Очень часто крапивные размножаются вегета-
тивно при помощи укоренения стеблей, корневых отпрысков,
подземных столонов и т.д.
Для листьев крапивных характерно наличие цистолитов -
беловатых образований, пропитанных карбонатом кальция. Ли-
стья и стебли покрыты жгучими волосками со стрекательными
клетками (количество их огромно до 1000 стрекательных
клеток на 1 мг массы листа). Жгучие волоски также имеют
дальневосточные растения лапортея (Laportea) и жерардиния
(Gerardinia).
Среди крапивных много прядильных растений, наиболее
ценное из них рами (Boehmeria nivea), из шелковистого во-
локна которого (длиной до 500 мм) изготавливают разнообраз-
ные ткани. Родом рами из Китая, но издавна культивируется во
многих странах (в том числе на территории бывшего СССР - в
Закавказье и Средней Азии).
Род пилея (Pilea) включает широко культивируемые изящ-
ные декоративные растения, среди которых особенно привлека-
тельны пестролистные формы.
Это интересно
Страшная лапортея. Дурной славой пользуется лапортея (Lapor-
tea) - самое жгучее тропическое крапивное. Ожоги, причиняемые раз-
ными видами этого древесного растения, настолько болезненны, что
приводят к обмороку, они не заживают в течение нескольких месяцев
и даже могут вызвать смерть ребенка. Жгучие волоски защищают эти
растения от поедания животными, но не от всех. Так, листья австра-
лийских лапортей без вреда для себя поедает крупный рогатый скот.
Доступное богатство. Крапива двудомная - синейшее пищевое и
лекарственное растение. В народной медицине ее используют для лече-
ния самых разных заболеваний.
Листья крапивы - природный витаминный концентрат, используе-
мый для профилактики и лечения авитаминоза. В них также много
хлорофилла, белка, крахмала, органических кислот, солей железа, ка-
лия, меди, марганца, титана, никеля. По питательной ценности листья и
молодые побеги близки к бобовым. При варке часть витаминов разру-
шается, поэтому листья лучше есть свежими в салатах (ошпаренные
кипятком они теряют жгучесть). Верхушки молодых побегов готовят
впрок засушивая, листья засаливают.
Из листьев крапивы добывают хлорофилл, который употребляется
как лекарственное средство, улучшающее обмен веществ и стимули-
110
рующее ранозаживление. Растение нашло широкий спрос в косметике.
Наконец, используются и своеобразные консервирующие свойства кра-
пивы - рыба, переложенная ее побегами, долго остается свежей.
В стеблях крапивы много прочного волокна, из которого можно
изготовлять веревки и грубые ткани (из крапивы должна была сплести
рубашки для своих заколодованных братьев героиня сказки Г.Х.Ан-
дерсена «Дикие лебеди»). И кто знает, если бы не появились синтети-
ческие волокна, то крапива могла бы стать промышленной прядильной
культурой.
7.14. Семейство Euphorbiaceae — Молочайные
Паспорт семейства
Родов - 330 (3), видов - 7000 (70)
Цветок — *Р0., Gq , *Po-s Ao Gq)
Распространение почти повсеместно, за исключением
Арктики, но особенно многочисленны в тропических и суб-
тропических областях
Жизненная форма - деревья, древовидные стеблевые сукку-
ленты, кустарники, многолетние и однолетние травы
Опыление - насекомыми (мухи, маленькие пчелы, тарака-
ны), редко ветром
Плоды коробочки. Семена распространяются птицами,
насекомыми (муравьями), летучими мышами
Важнейшие роды - Euphorbia
Культурные растения - Aleurites, Hevea, Manihot, Ricinus
Молочайные (рис. 20) входят в одноименный порядок, в ко-
тором еще три семейства. Для молочайных характерно чрезвы-
чайное богатство жизненных форм. Распространенные в Афри-
ке колонновидные суккулентные молочаи похожи на кактусы
(в этом проявляется один из законов экологии в сходных
экологических условиях у далеких в систематическом отноше-
нии видов может формироваться одна жизненная форма).
У большинства видов имеется млечный сок, содержащий
различные вещества, в том числе каучук. У гевеи бразильской
(Hevea brasiliensis) его содержание достигает 50%, это каучуко-
вое дерево из бассейна Амазонки широко культивируется в
тропической Азии и в некоторых африканских странах. Добы-
ваемый из гевеи каучук составляет 90% мирового производства
натурального каучука. Есть молочайные и без млечного сока,
например виды ветроопыляемого рода пролесник (Mercurialis).
В Краснодарском крае возделывается ценная лекарственная
и техническая культура - клещевина (Ricinus communis), масло
которой известно под названием касторового, или рицинового.
111
Рис. 20. Малочайные
Euphorbia virgata: 1
цветущее растение, 2 -
циатий, 3 циатий с
удаленными прицвет-
никами
Ценны в хозяйственном отношении виды рода тунг
(Aleurites), называемого масляным деревом. Из его семян добы-
вают очень ценное быстро высыхающее техническое масло. Два
интродуцированных вида тунга выращивают на Черноморском
побережье Кавказа. Пищевое растение маниок съедобный
(Manihot esculenta) широко культивируется в тропических рай-
онах, этот кустарник очень быстро растет и образует огромные
(массой до 15 кг) клубневидные корни, из которых получают
муку и крахмальную крупу - маниоковое саго.
Самый большой род семейства - молочай (Euphorbia, около
1500 видов) - широко распространен по всей Земле, кроме Арк-
тики. Представляет интерес соцветие молочая - циатий. Он со-
стоит из одного женского цветка и нескольких мужских цвет-
ков с редуцированными околоцветниками. Роль околоцветника
выполняют прицветники, которые чередуются с нектарными
железками.
Все виды молочая содержат в млечном соке ядовитое веще-
ство эуфорбин и могут вызывать очень тяжелые отравления.
Для скота молочаи опасны не на пастбищах, где животные их
избегают, а в сене. В России встречается около 70 видов этого
рода. Наиболее известен из них молочай лозный (Е. virgata) -
злостный сорняк полевых культур.
112
Это интересно
Молочайное масло. Из семян молуккского дерева (Aleurites
moluccana) получают очень вкусное масло. Семена этого растения на-
столько богаты маслом, что местное население Малайского архипелага
использует их для освещения: нанизанные на лучинку семена горят как
свечи.
7.15. Семейство Rosaceae - Розовые
Паспорт семейства
Родов- 100(45,), видов - 3000(560)
Цветок - * Са<1) Со^0 A?.» G^i
Распространение - по всему миру, но особенно - умеренная
и субтропическая области Северного полушария
Жизненная форма преобладают листопадные деревья и
кустарники, но много одно- и многолетних трав
Опыление - насекомыми (спектр опылителей широкий)
Плоды -листовка, костянка, яблоко и очень разнообразные
“сборные” (многоорешек, многокостянка и др.). Распрост-
раняются животными (чаще эндозоохорно)
Важнейшие роды Alchemilla, Cerasus, Crataegus,
Filipendula, Fragaria, Geum, Padus, Potentilla, Rosa, Rubus,
Sanguisorba, Sorbus, Spiraea
Культурные растения Amygdalus, Armeniaca, Cerasus,
Cydonia, Fragaria, Malus, Persica, Prunus, Pyrus
Семейство (рис. 21) входит в одноименный порядок Rosales, в
составе которого еще два семейства. Эти растения широко пред-
ставлены в естественной растительности (лесной, степной, луговой)
умеренной и субтропической зоны северного полушария.
Околоцветник - обычно пятичленный, тычинки - чаще мно-
гочисленные. Строение и количество плодолистиков в семействе
весьма разнообразно, что определяет и огромное разнообразие
плодов: встречаются как апокарпные, так и синкарпные. Завязи
бывают верхние и нижние.
Цветки у розовых открытые и опыляются самыми разными
насекомыми. Имеются и ветроопыляемые виды, а для некото-
рых родов характерен апомиксис (рубус - Rybus, лапчатка -
Potentilla, манжетка - Alchemilla, шиповник - Rosa).
В пределах семейства отмечается большое разнообразие
плодов. На этом основании выделяются подсемейства: спирей-
ные (листовка), шиповниковые (многокостянки и многоорешки,
в образовании которых часто принимает участие гипантий), яб-
лоневые (яблоко), сливовые (костянка) и др.
113
I
б)
Рис. 21. Розовые: а - Spiraea (1 - цветок, 2 - диаграмма цветка, 3 - ги-
нецей, 4 плодик); б - Rosa (1 - цветок, 2 - диаграмма цветка, 3 -
плод); в - плоды (1 - Geum rivale, 2 - Rubus idaeus, 3 — Fragaria vesca);
г - Prunus (1 - цветок, 2 - диаграмма цветка, 3 - плод); д - Malus (1 -
цветок, 2 - плод ), Pyrus (3 - диаграмма цветка)
Розовые - основа садоводства умеренной зоны и субтропи-
ков. Основные культурные виды:
абрикос - Armeniaca vulgaris,
айва обыкновенная — Cydonia oblonga,
алыча культурная - Prunus cerasifera
вишня садовая - Cerasus vulgaris,
114
груша домашняя - Pyrus communis,
земляника садовая - Fragaria magna (F. ananassa),
малина обыкновенная - Rubus idaeus,
миндаль обыкновенный - Amygdalus communis,
мушмула японская - Eriobotrya japonica,
персик - Persica vulgaris,
рябина черноплодная - Aronia melanocarpa,
слива домашняя — Prunus domestica,
черешня - Cerasus avium,
яблоня домашняя — Malus domestica.
Имеется несколько межродовых гибридов: вишня+черемуха
Cerapadus, вишня+слива Ceraprunus и др.
Среди декоративных культур особое место занимает роза. В
настоящее время существует не менее 25 тыс. культиваров (сор-
тов и форм) розы (даже сортов яблони - менее 10 тыс.).
Это интересно
Спасительница Карла Линнея. Великий Линней тяжело заболел
подагрой и от неминуемой смерти был спасен растением земляника
лесная (Fragaria vesca). Он ел ее плоды в течение всего периода плодо-
ношения и в таком количестве, в каком мог их добыть и съесть.
«Вкусная случайность». Садовая земляника (клубника, Fragaria
ananassa) - это гибрид, который спонтанно возник в садах Франции в
середине XVIII в. из двух видов американской земляники (F. chiloensis
и F. virginiana).
Дороже золота. Розовое масло - важнейший продукт для парфю-
мерии - получают из лепестков розы дамасской (Rosa damascena). Наи-
более знаменита культура этой розы в Казанлыкской долине (Болга-
рия). Для получения 1 кг масла необходимо 3 т лепестков. Не удиви-
тельно, что 1 г масла стоит дороже 1 г золота.
Культура древняя и прекрасная. Как декоративное растение, роза
является одной из самых древних культур. В Средней Азии ее начали
возделывать 4 тыс. лет назад (по сей день Персию называют Гюлиста-
ном - страной роз, от розы происходит и древнее название Сирии -
Суристон). Роза была культовым растением в Древнем Риме. Римляне
верили, что роза вселяет в сердца мужество, и потому вместо шлемов
на головы воинов надевали венки из роз, а на щитах выбивали изобра-
жение розы.
Тайна под розой. В Греции и Риме белая роза считалась цветком
тайны. Если во время пиршества на столе красовался этот цветок, все
знали, что произнесенные здесь слова должны сохраниться в тайне:
«sub rosa dictum - под розой сказано».
Падение Рима и роза. Во времена падения Рима из царственного
цветка роза сделалась цветком пьяных оргий, выразительницей низ-
менных чувств. Злоупотребление ею как украшением достигло крайних
пределов. В знаменитом обеденном зале императора Нерона на гостей
сыпались миллиарды свежих розовых лепестков. Вообще на всех обе-
115
денных пиршествах римлян столы и пол были усыпаны лепестками роз,
а гости и даже прислуживающие им рабы были украшены венками из
роз. Но всех превзошел император Гелиогобал: на одном из его засто-
лий знатные гости были забросаны таким множеством падавших с по-
толка розовых лепестков, что некоторые задохнулись под ними. Рим-
ские патриции усыпали лепестками розы даже поверхность моря, когда
отправлялись на прогулку на галерах. Обширные римские сады были не
в состоянии обеспечить такую потребность в розах, и приходилось
привозить их целыми кораблями из Александрии и Карфагена, где для
этого возникла даже «садовая индустрия».
Роза и христианство. С падением Рима культура розы пришла в
упадок, а первые христиане смотрели на нее как на цветок разврата и
гибели. Но благодаря дивной красоте и приятному запаху роза завое-
вала и их расположение. Через несколько столетий святые отцы объя-
вили розу райским цветком и посвятили ее Пресвятой Богородице. В
Священном писании сообщается, что архангел Гавриил сделал для нее
три венка. Венок из белых роз означает радость Богородицы, из крас-
ных - ее страдания, из желтых - ее славу.
В одном из сказаний о кресте Спасителя говорится о моховой ро-
зе, которая возникла из капель крови Христовой. Кровь, струившаяся
по кресту, собиралась ангелами в золотые чаши, но несколько капель
случайно упали на мох, который жадно впитал их в себя. Из этих бо-
жественных капель выросла моховая роза, красный цвет которой дол-
жен служить вечным напоминанием о пролитой за наши грехи крови.
Самая ботаническая война. В XIV в. тридцать лет воевали за анг-
лийский престол Ланкастеры и Йорки. Символами враждующих дина-
стий были белая и алая розы. Как символ примирения, английские са-
доводы вывели розу с белыми и алыми лепестками и назвали этот сорт
ланкастерйоркским.
7.16. Семейство Fabaceae - Бобовые
Паспорт семейства
Родов - 450 (552, видов - 12000 (750)
Цветок - ТСо]+2+(2) Gj
Распространение - по всему миру
Жизненная форма многолетние травы (редко однолет-
ние), кустарники и деревья
Опыление - насекомыми
Плоды - бобы. Распространяются животными, ветром, во-
дой
Важнейшие роды - Astragalus, Caragana, Coronilla, Genista,
Latbyrus, Medicago, Meli lotus, Onobrycbis, Oxytropis,
Trifolium, Vicia
Культурные растения Aracbis, Cicer, Galega, Glycine,
Indigofera, Lupinus, Medicago, Pbaseolus, Pisum, Robinia,
Trifolium, Vicia
116
Семейство бобовых (рис. 22) относится к одноименному
порядку Fabales, в котором три семейства (иногда эти семейства
рассматриваются как подсемейства в рамках более широкого
семейства бобовых: цезалъпиниевые — Caesalpinioideae, мимозо-
вые - Mimosoideae и бобовые - Faboideae).
Рис. 22. Бобовые: a — Pisum (1 - цветок, 2 - венчик, 3 - андроцей и ги-
нецей, 4 - гинецей, 5 - плод, 6 - диаграмма цветка); б - Trifolium (со-
цветие, цветок, плод); в - плоды (1 - Vicia, 2 - Medicago, 3 - Melilotus,
4 - Onobrychis); г - Lupinus (клубеньки на корнях)
Для бобовых характерны сложные листья с прилистниками
и устойчиво повторяющийся у всех видов тип цветка со срос-
шейся пятизубчатой чашечкой, «мотыльковым» венчиком (из
«паруса», двух «весел» и «лодочки» из двух сросшихся лепест-
ков), 10 тычинок (у большинства видов 9 из них срослись в
трубку и 1 свободная) и пестика, столбик которого выдается из
тычиночной трубки. Опыляются бобовые насекомыми с длин-
ным хоботком, в первую очередь шмелями. (Еще Ч. Дарвин знал
о связи урожая семян клевера с численностью мышей, которые
уничтожают гнезда шмелей.)
По числу видов бобовые уступают только сложноцветным и
орхидным и играют огромную роль как в естественных экосисте-
мах, так и в сельском хозяйстве, так как являются уникальными
117
посредниками между азотом атмосферы и живыми организмами,
не способными к усвоению молекулярного азота. Благодаря
азотфиксации симбиотическими клубеньковыми бактериями, жи-
вущими в клубеньках на корнях, бобовые обогащают почву азо-
том, что позволяет отказаться от энергетически дорогих и эколо-
гически опасных азотных минеральных удобрений.
Поле клевера способно в течение года накопить 200 кг/га
действующего вещества азота, а поле люцерны в южных рай-
онах (с использованием полива) - до 700 кг/га. Высокое содер-
жание белка в семенах и в вегетативной массе бобовых делает
их ценнейшим кормом для сельскохозяйственных животных.
Растительный белок бобовых (сои, фасоли арахиса, гороха и
др.) составляет основу белковой части рациона населения Ин-
дии и ряда других густонаселенных и бедных стран Азии и
Южной Америки.
Характеристика основных культурных растений семейства
приведена в табл. 13.
Из числа возделываемых бобовых самым знаменитым рас-
тением является соя. Семена сои, кроме белка, содержат много
жира, на основе которого производят маргарин, по качеству не
уступающий сливочному маслу. Из сои готовят заменители мя-
сопродуктов (сосиски) и молока. Ее семена в раздробленном
состоянии используются также для откорма скота. За период с
1950 по 1995 гг. площадь под соей в мире возросла в 6 раз. В
России возделывание сои, кроме районов Дальнего Востока и
Краснодарского края, еще не получило широкого распростра-
нения.
Фасоль - также важнейшая зернобобовая культура, осо-
бенно для развивающихся стран. Многочисленные сорта и сор-
тотипы ее относятся к двум родам: фасоль (Phaseolus) и вигна
(Vigna), по происхождению принадлежащие к двум очагам
американскому и азиатскому. В культуре наиболее распростра-
нена фасоль обыкновенная (Ph. vulgaris).
Основной бобовой культурой в России является горох, ко-
торый начал возделываться 1000 лет назад. Большие площади у
нас заняты бобовыми многолетними травами: клевером (Trifo-
lium), люцерной (Medicago), донником (Melilotus), эспарцетом
(Onobrychis), викой (Vicia), чиной (Lathyrus), козлятником
(Galega).
В сельском хозяйстве южных стран (включая и страны
Средней Азии) весьма значимы арахис (Arachis hypogaea), фа-
соль (Phaseolus) и нут (Cicer arietinum).
118
Таблица 13
Характеристика основных культурных растений
семейства бобовых
Культура Происхождение История введения в культуру Хозяйственное значение
Соя (Glycine max) Китай, где со- средоточено боль- шое количество примитивных форм и сортов Известна с пя- того тысячелетия до н.э. В Запад- ной Европе вы- ращивается с конца XVIII в., в России - с конца XIX в. Зерно содер- жит 35-55% бел- ка, из него полу- чают муку и мас- ло. Ценный корм для животных. Может выращи- ваться как сиде- рат. Первое ме- сто по производ- ству занимают США и Китай
Фасоль обыкно- венная (Phaseolus vulgaris) Америка (Г ва- темала, Аргенти- на), где до на- стоящего време- ни близкие виды встречаются в диком состоянии Еще ацтеки вы- ращивали ее вме- сте с кукурузой. В Европе стала распространяться после путешест- вий Колумба. В Россию проникла через Грузию и выращивается с XVII в. Используется в пищу как в спе- лом состоянии (сухие семена), так и в виде не- дозревших бобов. По распростра- нению в мире - вторая после сои бобовая культура
Бобы кормовые (Vicia faba) Первичный центр - Юго-Западная Азия и Индия, вторичный - Сре- диземноморье. Близкие дикорас- тущие виды име- ются в Алжире Известны с глубокой древно- сти в странах Азии. Первые находки в Европе относятся к эпо- хе неолита. Куль- тивировались в Древнем Египте и Древней Г реции. Издавна выращи- ваются в Грузии и на Русской равнине Продовольст- венная и кормо- вая культура. Семена содержат до 35% белка. В СНГ выращива- ется главным об- разом на Украине и в Белоруссии
Горох посевной (Pisum sativum) Имеет гибрид- ное происхожде- ние. Первичный центр - Средняя Азия, вторичный - Передняя Азия С глубокой древности выра- щивается в За- кавказье Культивируется для получения зерна, которое содержит 22-24% белка
119
В современном сельском хозяйстве бобовые играют все
большую роль как сидераты, зеленая масса которых запахива-
ется в почву в качестве удобрения. Для Нечерноземья лучший
сидерат - люпин (Lupinus), для Черноземья - виды донника.
Это интересно
Аллейные посевы с бобовыми. В тропических странах широко рас-
пространены аллейные посевы, в которых чередуются кулисы (высокие
рядки) из бобового дерева Leucaena leucocephala и посевы однолетних
культур. При этом деревья, восстанавливающие плодородие почвы, пе-
риодически срезают, что дает дополнительную древесину.
Геокарпия у арахиса. На побегах арахиса образуются многочис-
ленные недолговечные цветки, которые утром раскрываются, а после
обеда увядают. Большинство цветков оказываются стерильными, и
только самые нижние самоопыляются и завязывают плоды. При этом
основание завязи разрастается в стержень, который изгибается по на-
правлению к почве и, удлиняясь, проникает в нее на глубину 8-10 см.
Там и созревают бобы, содержащие по 2-4 семени. В отличие от бобов
гороха они не раскрываются, а их поверхность имеет характерную
ячеистую структуру.
«Мимоза» и «акация». Мимозу стыдливую (Mimosa pudica), скла-
дывающую листочки при малейшем прикосновении к ним, жители Рос-
сии могут увидеть только в оранжереях. А те букеты, которые под на-
званием «мимоза» привозят с Кавказа к 8 марта, принадлежат другому
растению из семейства мимозовых акации серебристой (Acacia
dealbata). «Акацией» население средней полосы называет другое бобо-
вое - карагану древовидную (Caragana arborescens), которая разводит-
ся около домов, на лесополосах, в садах и легко дичает.
Рекорд размера плода и семени. Этот рекорд принадлежит мимо-
зовым. У энтады лазящей (Entada scandens) - самый крупный плод в
мире, ее боб достигает в длину 1,5 м. Самые крупные в мире семена,
длина которых составляет 15-17 см, у моры маслоносной (Мога
oleifera).
Мясистые бобы. Мясистые плоды некоторых цезальпиниевых
сладкие или кислые. Так, рожковое дерево (Ceratonia siliqua) с притор-
но-сладкими на вкус бобами широко культивировалось в Среди-
земноморье, где «цареградские рожки» были излюбленным лакомством
населения. Ради кисловато-сладких бобов, богатых сахарами (до 40%) и
витамином С, во всех тропических странах выращивают тамаринд
(Tamarindus indica). Только Индия ежегодно собирает более 250 000 т
плодов. Родина тамаринда - сухие саванны Западной Африки, его ме-
стное название «dakar», и в честь этого растения даже названа столи-
ца Сенегала - Дакар.
120
7.17. Семейство Apiaceae (Umbelliferae) - Сельдерейные
(Зонтичные)
Паспорт семейства
Родов - 430 (93), видов - 3000 (230)
Цветок - * Ca(f^) Cof A; G р)
Распространение - по всему миру, но главным образом уме-
ренные области Северного полушария
Жизненная форма - одно- и многолетние травы, редко де-
ревья
Опыление - насекомыми
Плоды вислоплодник, распространяются животными,
ветром, водой
Важнейшие роды - Aegopodium, Antbriscus, Carum, Conium,
Eryngium, Heracleum, Peucedanum, Pimpinella, Seseli, Sium
Культурные растения Anetbum, Apium, Carum, Corian-
drum, Daucus, Foeniculum, Pastinaca, Petroselinum
Зонтичные (рис. 23) входят в порядок Apiales (Araliales), в
котором есть еще два семейства, в том числе Аралиевые
(Araliaceae). Это одно из самых естественных семейств с устой-
чивым пятерным цветком и, как правило, с соцветием, пред-
ставляющим собой сложный зонтик (хотя имеются виды с со-
цветием простой зонтик и даже с головчатым соцветием, на-
пример синеголовник - Eryngium planum). Для многих видов ха-
рактерно сочетание актиноморфных цветков в центре зонтика с
зигоморфными цветками по его краям, причем больший размер
имеет внешний лепесток (это можно рассматривать как прелю-
дию к формированию корзинки сложноцветных с разделением
цветков на центральные трубчатые и краевые ложноязычковые).
Цветки пятичленные, однообразные: лепестки обычно белые
или желтые, зубцы чашечки слабо развиты и мало заметны, ты-
чинки прикреплены к нектарному диску, находящемуся на вер-
хушке нижней двугнёздной завязи. От диска отходят два (по
числу плодолистиков) столбика. Нектар широко раскрытых
цветков доступен разным насекомым-опылителям, особенно ко-
роткохоботковым мухам, средством привлечения которых явля-
ется и специфический запах многих зонтичных.
Сухой плод - вислоплодник - при созревании распадается
на две половинки - мерикарпии, которые некоторое время ви-
сят на вильчатом карпофоре.
Стебель у зонтичных дудчатый с заметно утолщенными уз-
лами и ребристыми междоузлиями, может достигать 4 м в вы-
121
соту. Листья очередные, разделены на сильно расчлененную
пластинку, черешок и влагалище, охватывающее стебель.
Большинство видов зонтичных России - растения средней
полосы. К ним относится основной доминант травяного яруса
широколиственных лесов - сныть обыкновенная (Aegopodium
podagraria), в лесах распространены и крупные зонтичные
дудник лесной (Angelica sylvestris), дягиль (A. archangelica), а
также реброплодник уральский (Pleurospermum uralense, из его
цветочных стрелок делают «башкирскую флейту» - курай).
а) б)
в)
Рис.23. Зонтичные: а - Daucus carota (1 - соцветие, 2 - цветок (общий
вид и в разрезе), 3 - диаграмма цветка, 4 - плод (общий вид и в разре-
зе); б - Eryngium planum (соцветие и цветок); в - плоды (1 - Conium,
2 - Coriandrum, 3 - Carum, 4 - Heracleum, 5 - Cicuta)
На лугах обычны бедренец камнеломка (Pimpinella saxifraga),
тмин обыкновенный (Carum carvi), борщевик сибирский
(Heracleum sibiricum), в степях - жабрица порезниковая (Seseli
libanotis), в рудеральных местообитаниях болиголов крапча-
тый (Conium maculatum, сильно ядовитое растение), в при-
брежно-водных - поручейник широколистный (Sium latifolium)
и самое ядовитое зонтичное - вех ядовитый (Cicuta virosa).
Однако наиболее знаменито семейство зонтичных много-
численными культурными растениями, которые, кроме моркови
122
(Daucus carota), являются пряными и ароматическими. Специ-
фический аромат присущ всем зонтичным, что связано с содер-
жащимися в них эфирными маслами (особенно богаты этими
маслами плоды).
Список культурных растений (кроме уже названной морко-
ви) включает:
анис обыкновенный - Pimpinella anisum,
кориандр посевной - Coriandrum sativum,
пастернак посевной - Pastinaca sativa,
петрушка кудрявая - Petroselinum crispum,
сельдерей пахучий - Apium graveolens,
тмин обыкновенный - Carum carvi,
укроп огородный - Anethum graveolens,
фенхель обыкновенный - Foeniculum vulgare.
Семейство Зонтичных иногда объединяют с семейством
Аралиевых (80 родов, 850 видов), широко известное благодаря
лекарственным и декоративным представителям: женьшеню
(Panax ginseng), элеутерококку (Eleutherococcus senticosus), за-
манихе (Oplopanax elatus), аралии (Aralia), плющу (Hedera).
Это интересно
Корень жизни. В одной из многочисленных восточных легенд, по-
священных женьшеню, говорится, что он рождается от молнии. На
протяжении многих веков восточная медицина приписывала корню
женьшеня способность исцелять все болезни. Поэтому К.Линней и дал
этому растению научное название Рапах - от греческих слов «пан» -
все и «акос» - исцеляющий. Женьшень в переводе с китайского озна-
чает «человек-корень», и действительно, его корень напоминает чело-
веческую фигуру. Сейчас женьшень введен в культуру и выращивается
во многих странах, в том числе и у нас.
7.18. Семейство Solanaceae — Пасленовые
Паспорт семейства
Родов - 95 (10), видов - 2500 (30)
Цветок - *Ca(f) Co(f> A; G^i (редко слабо ?)
Распространение — по всему миру, но наибольшее разнообра-
зие в Южной Америке
Жизненная форма - одно- и многолетние травы, кустарни-
ки, лианы
Опыление - насекомыми
Плоды - ягоды или коробочки, распространяются живот-
ными
Важнейшие роды Atropa, Capsicum, Datura, Hyoscyamus,
Nicotiana, Solanum
Культурные растения Capsicum, Lycopersicon, Nicotiana,
Physalis, Solanum
123
Пасленовые (рис. 24) входят в одноименный порядок. Это
преимущественно «американское» семейство: большая часть ви-
дов его распространена в Центральной и Южной Америке.
Пятичленные актиноморфные с двойным околоцветником
цветки пасленовых различаются степенью срастания чашелисти-
ков и лепестков у разных родов. Опыляются цветки различными
насекомыми, а в тропических областях - птицами и даже мле-
копитающими.
4
а)
б)
Рис. 24. Пасленовые: а - Solanum tuberosum (1 - соцветие, 2 - цветок в
разрезе, 3 - андроцей, 4 - диаграмма цветка, 5 - плод); б - плоды (1 -
Physalis, 2 - Hyoscyamus, 3 - Nicotiana, 4 - Solanum melongena, 5 -
Datura, 6 - Atropa belladonna)
Семейство играет очень большую роль в жизни человека. В
его составе есть важные пищевые растения:
баклажан - Solanum melongena,
картофель - Solanum tuberosum,
помидор - Lycopersicon esculentum,
сладкий перец - Capsicum annuum,
физалис овощной - Physalis peruviana.
Кроме того, к этому семейству относится табак - Nicotiana
tabacum, N. rustica.
124
Все эти культурные растения (исключая баклажан, в диком
виде произрастающий в Индии) - выходцы из Америки.
Кроме того, в составе семейства есть три известных дико-
растущих вида красавка обыкновенная, или белладонна
(Atropa belladonna), белена черная (Hyoscyamus niger) и дурман
вонючий (Datura stramonium). Все они содержат ядовитые алка-
лоиды, которые в невысоких концентрациях являются ценными
лекарственными веществами (сильное действие алкалоидов бе-
лены на человека отражено в бытовых русских сказках, где
муж спрашивает неразумно поступающую жену: «Ты что, баба,
белены объелась?»).
В садах часто выращивают физалис Франше (Physalis fran-
chettii), у этого «японца» при плодах разрастается крупная пу-
зыревидно вздутая желтая или оранжевая чашечка, благодаря
чему растение используется в зимних сухих букетах.
Это интересно
История картофеля в Европе. Картофель - одно из первых куль-
турных растений - выращивался индейцами раньше, чем зерновые
культуры, так как его «агротехника» была предельно простой: нужно
было лишь бросить клубень в ямку и засыпать землей, а использовать
клубни выросшего растения можно было по мере необходимости. Кро-
ме привычного для нас картофеля, индейцы использовали еще около 20
видов рода паслен. У некоторых из них клубни были горькими и тре-
бовали специального приготовления (их промораживали, провяливали,
промывали в проточной воде, толкли и получали «чуньо» - однообраз-
ную серую массу, которая могла долго храниться).
В Европу картофель попал благодаря экспедиции Х.Колумба. Пер-
вое упоминание о нем датируется 1535 г. у Юлиана де Кастельяноса,
затем о нем пишет Сиеса де Лионе в «Хронике Перу». Однако в Испа-
нии картофель появился лишь спустя 30 лет: в 1565 г. клубни были по-
дарены испанским королем больному римскому папе Пию IV. Из Ис-
пании картофель распространился в Италию, Бельгию, Голландию,
Германию, Польшу. Англичане завезли себе картофель независимо от
испанцев.
Активное внедрение картофеля в культуру датируется XVIII в.,
причем непривычное растение завоевывало авторитет у населения с
трудом. Прусский король Фридрих Вильгельм I провозгласил выращи-
вание картофеля национальной задачей. Сопротивлявшимся грозили
отрезанием носов и ушей. В России внедрение картофеля привело к
«картофельным бунтам» (основная роль в этом принадлежала старооб-
рядческой церкви).
Видимо, наиболее безболезненно и даже по юмористическому сце-
нарию картофель был распространен во Франции благодаря фармацев-
ту-химику Антуану Пармантье. Во время семилетней войны аптекарь
попал в плен к немцам, отведал картофеля и решил во чтобы то ни
стало распространить его на своей родине.
125
Он начал со званого обеда, на который пригласил химика
А.Лавуазье и политика-демократа Б.Франклина. После того, как его
высокие гости положительно оценили блюда из картофеля, Пармантье,
опираясь на их авторитет, обратился к Людовику XVI за поддержкой.
Король выделил ему в пригороде Парижа участок земли, а на одном из
балов появился с цветком картофеля на груди. Прическа королевы бы-
ла украшена таким же цветком (это был один из первых и удачнейших
вариантов рекламной кампании!).
Далее хитрый аптекарь очень удачно использовал психологические
особенности французов (как, впрочем, и других наций, поскольку вы-
ражение «запретный плод сладок» - общечеловеческое). Он попросил
короля выделить охрану: днем «свирепые» солдаты зорко следили за
тем, чтобы никто не приближался к плантации экзотического растения,
а по ночам иногда «случайно» исчезали. Смельчаки стали красть кар-
тофель и высаживать его в своих садах.
Вклад Пармантье в распространение картофеля во Франции отме-
чен двумя памятниками - на месте первого картофельного огорода и
на родине аптекаря в городе Мондидье. На памятнике сделана подпись
«Благодетелю человечества» и высечены слова Людовика XVI: «По-
верьте мне, настанет время, когда Франция поблагодарит Вас за то, что
Вы дали хлеб голодающему человечеству».
Появление картофеля в Англии связано с именем легендарного
вице-адмирала (и пирата!) Френсиса Дрейка.
Картофель в России. Первый картофель в России был выращен в
Санкт-Петербурге на Аптекарском огороде (ныне Ботанический инсти-
тут РАН) в 1736 г. В это время картофель слыл изысканным деликате-
сом: сохранились документы дворцовой канцелярии об отпуске на бан-
кеты высокопоставленных лиц по ... фунту картофеля!
В распространении «второго хлеба» в России сыграли роль Петр I
и Екатерина II. Управляющий имениями Екатерины А.Т.Болотов, вы-
дающийся русский агроном, много сделал для пропаганды новой про-
довольственной культуры и опубликовал рекомендации по ее возделы-
ванию. Интерес правителей России к картофелю был столь велик, что
Правительственный Сенат только с 1765 по 1767 гг. рассматривал во-
прос о картофеле ... 23 раза!
Красивая и коварная. Карл Линней назвал белладонну именем
жестокой богини Атропы, перерезающей нить жизни, - род Atropa, а
видовой эпитет в названии вида - «belladonna» - он посвятил древне-
римским модницам, которые закапывали сок этого растения в глаза и
натирали им щеки. Щеки розовели, а глаза с расширенными зрачками
приобретали особую выразительность. Сейчас белладонну специально
выращивают, чтобы получать болеутоляющие средства. Изготовляемое
из белладонны лекарство - атропин - в руках врачей не обрывает нить
жизни, а укрепляет ее.
126
7.19. Семейство Boraginaceae - Бурачниковые
Паспорт семейства
Родов - 130 (43), видов - 2300 (170)
Цветок - *Ca(j)Co(f) A; Ggt (реже Т)
Распространение - тропики, субтропики и умеренные об-
ласти
Жизненная форма - деревья, кустарники, в умеренной поло-
се -одно- и многолетние травы
Опыление - насекомыми
Плоды - чаще ценобий, распадается на четыре орешка (эре-
ма), распространяются животными (эпизоохорно, мирме-
кохорно), ветром, водой
Важнейшие роды - Cynoglossum, Echium, Lappula, Myosotis,
Nonea, Pulmonaria, Symphytum
Культурные растения - Borago
Семейство (рис. 25) входит в одноименный порядок, в кото-
ром всего семь семейств.
2 _
Рис. 25. Бурачниковые: Symphytum (1 - соцветие, 2 - цветок);
Pulmonaria (3- цветок, 4 - гинецей); Cynoglossum (5 - плод)
Характерными особенностями бурачниковых являются же-
сткое щетинистое опушение растений и особый тип соцветия -
завиток, который до цветения обычно улиткообразно свернут.
Это энтомофильные растения с актиноморфными (реже зиго-
морфными, например синяк - Echium) цветками. У многих бу-
рачниковых, например у медуницы (Pulmonaria), окраска венчи-
ка меняется на протяжении срока цветения (розовый становит-
ся голубым, желтый или белый - красноватым). Значение этого
признака остается неясным, хотя известно, что окраска особен-
но заметно меняется после посещения цветков опылителями.
127
Завязь, образованная двумя плодолистиками, двугнездная,
но, как и у губоцветных, каждое гнездо разделяется ложной
перегородкой на две камеры. В результате образуется дробный
плод, распадающийся на 4 орешковидные доли. У большинства
растений при плодах остается и разрастается чашечка. Распро-
страняются плоды самыми различными способами. Большинство
бурачниковых - эпизоохоры (у них плодики покрыты крючко-
ватыми образованиями, с помощью которых они прикрепляются
к шерсти животных, за это один из родов назван липучкой —
Lappula), но много мирмекохоров, гидрохоров, анемохоров, есть
и баллисты (в умеренной полосе они не встречаются). Редко
встречаются костянки и коробочки.
Большинство бурачниковых умеренной полосы - рудераль-
ные растения, занимающие нарушенные местообитания. Единст-
венное культурное пищевое растение - огуречная трава, или бу-
рачник лекарственный (Borago officinalis). Йесной до появления
плодов огурца листья бурачника используются как их замени-
тель в салатах и окрошках. Вид легко дичает и подобно другим
видам семейства становится сорно-рудеральным растением. Бу-
рачник - прекрасный медонос, который выращивается на пасе-
ках и в садах для привлечения насекомых. Есть культурные
формы незабудок (Myosotis), которые используются в декора-
тивном садоводстве.
К Boraginales близок порядок Convolvulales, включающий
два семейства. Семейство Вьюнковые (Convolvulaceae) в умерен-
ной зоне представлено преимущественно лианами из родов
вьюнок (вьюнок полевой - Convolvulus arvensis - злостный сор-
няк), повой (Calystegia) и др. Ипомея пурпурная (Ipomoea
purpurea) - популярный декоративный вьющийся однолетник.
Другое семейство этого порядка Повиликовые (Cuscu-
taceae) - представлено одним родом повилика - Cuscuta. Эти
растения-паразиты, как и представители семейства заразихо-
вых, приспособлены к жизни на определенных видах растений-
хозяев.
Это интересно
Цветок памяти. Согласно греческому мифу, богиня Флора спусти-
лась на землю, чтобы одарить цветы именами. Когда она закончила ра-
боту и хотела удалиться, услышала слабый голосок: «Ты забыла меня,
Флора. Дай и мне имя!». С трудом нашла богиня в буйном разнотравье
обиженный маленький голубой цветок. Она назвала его незабудкой и
наделила чудесной силой возвращать память тем людям, которые нач-
нут забывать своих близких или свою родину.
128
Мала незабудка, но почитается веками. Во многих странах в ее
честь устраивались праздники. В Англии в этом празднике участвовали
король и королева, в Германии в день незабудки школьники и студенты
занимались неполный день. Во Франции незабудку дарят на память, и
этот подарок сохраняется как реликвия. А древние кузнецы в соке не-
забудки закаливали дамасские и толедские клинки, которые были уди-
вительно прочными и одновременно легкими и гибкими.
7.20. Семейство Scrophulariaceae - Норичниковые
Паспорт семейства
Родов - 300 (33), видов - 3000 (280)
Цветок - ^Ca(f-4) CO(f.4) А4 G(2) (реже почти *)
Распространение - по всему миру, но наиболее многочислен-
ны в умеренных областях Северного полушария и в горах
тропиков
Жизненная форма - одно- и многолетние травы, кустар-
нички
Опыление - насекомыми
Плоды - двугнездные коробочки. Семена распространяются
ветром и муравьями
Важнейшие роды Euphrasia, Linaria, Melampyrum,
Pedicularis, Rhinanthus, Scrophularia, Verbascum, Veronica
Культурные растения - Antirrhinum, Digitalis
Семейство норичниковых (рис. 26) входит в одноименный
порядок из 17 семейств и является самым крупным и разнород-
ным из них.
Норичниковые отличаются от губоцветных большим разно-
образием формы цветка. Чашечка варьирует от раздельнолист-
ной (наперстянка - Digitalis) до длиннотрубчатой с четырьмя
или пятью зубцами, венчик может иметь короткую трубку и
быть блюдцевидным (коровяк Verbascum), но чаще трубка
длинная, а верхняя часть венчика двугубая. В таком венчике
есть дополнительные выросты или выпуклости, которые закры-
вают доступ к нектару мелким насекомым, и только шмели и
крупные пчелы легко открывают цветки и опыляют их (напри-
мер, у мытника - Pedicularis, льнянки - Linaria, львиного зева -
Antirrhinum).
Тычинок - редко 5 (коровяк), чаще - 4, а иногда даже 2
(вероника - Veronica). Гинецей всегда из двух плодолистиков,
плоды у большинства - коробочки (хотя у некоторых африкан-
ских и американских представителей есть ягодообразные и да-
же костянкообразные плоды).
129
Рис. 26. Норичниковые: а - цветущий побег, цветок и диаграмма цветка
(1 Verbascum nigrum, 2 - Linaria vulgaris, 3 - Veronica chamaedrys);
б - цветки (1 - Scrophularia, 2 - Digitalis, 3 - Antirrhinum, 4 - Pedicu-
laris, 5 - Odontites)
Чтобы отличить двугубые норичниковые от губоцветных, нуж-
но знать что у них нет характерного для губоцветных аромата, ли-
стья, как правило, очередные, а стебли в сечении круглые.
Норичниковые - растения умеренных широт с прохладным
климатом и потому особенно разнообразны на альпийских гор-
ных лугах.
В составе семейства много видов-полупаразитов (погрем-
ки - род Rhinanthus, марьянник - Melampyrum, мытник, очан-
ка - Odontites). Полупаразиты имеют зеленые листья и фото-
синтезируют, но кроме того, питаются за счет растения-
хозяина, к корням которого присасываются специальными при-
сосоками-гаусториями. Их связь с растением-хозяином факуль-
тативная, и у каждого полупаразитного вида может быть до
сотни видов-хозяев.
Есть в составе семейства и полный корневой паразит
петров крест (Lathraea squamaria), который живет на корнях
лещины или ольхи. Это удивительное растение в течение 10-
130
25 лет скрыто в земле, где его корневища сильно разрастаются,
ветвятся, часто крестообразно, и образуют все новые и новые
гаустории. Только после этого растение выносит на поверх-
ность красноватые соцветия на толстых ножках, цветет и дает
семена. Время его цветения приходится на весну, когда у расте-
ния-хозяина сок особенно богат органическими веществами.
По строению цветка к норичниковым, особенно к роду мыт-
ник, близки паразитические виды из семейства Заразиховые (Ого-
banchaceae), в состав которого входят злостные сорняки полевых
культур. Заразиховые иногда рассматривают даже как подсемей-
ство норичниковых. В отличие от полупаразитических норични-
ковых заразихи - полные паразиты и не имеют хлорофилла.
Полупаразиты из числа норичниковых (особенно погремки)
могут наносить ущерб луговым травостоям, резко снижая их
урожайность, и потому с ними приходится бороться. Поскольку
это двулетние растения, то обычно достаточно в течение двух лет
рано скашивать травостой, чтобы не дать погремкам образовы-
вать семена. В то же время в естественных экосистемах полупа-
разиты повышают видовое богатство сообществ, так как сдержи-
вают конкурентную способность видов-доминантов (см. ч. 3).
Это интересно
Славящий Геракла. Согласно греческому мифу, цветок с необыч-
ным названием «львиный зев» создала Флора по воле богов в память о
первом подвиге Геракла - победе над Немейским львом. Под жарким
солнцем цветок открывает свой зев с ярким красным пятном и стано-
вится похожим на окровавленную пасть льва.
7.21. Семейство Lamiaceae (Labiatae) - Яснотковые
(Губоцветные)
Паспорт семейства
Родов - 210 (50), видов - J500 (260)
Цветок — Т Co(j) Co(j) А.4,2 Gf?)
Распространение - по всему миру, но наибольшее разнообра-
зие - в Средиземноморье и Центральной Азии
Жизненная форма - травы, полукустарники, кустарники
Опыление - насекомыми
Плоды - ценобий, распадается на четыре орешка (эрема),
распространяются животными, ветром, водой
Важнейшие роды Galeobdolon, Glechoma, Lamium,
Leonurus, Lycopus, Mentha, Origanum, Phlomis, Salvia,
Scutellaria, Stachys, Thymus
Культурные растения - Lavandula, Mentha, Ocimum, Thymus
131
Семейство (рис. 27) входит в одноименный порядок, в кото-
ром, кроме него, еще два семейства. Большинство губоцветных
встречаются в теплых сухих областях, наиболее богато они
представлены в Средиземноморье, включая и Крым, воздух ко-
торого буквально напоен ароматами губоцветных. В Крыму
массово разрастается на нарушенных местообитаниях розмарин
и возделывается лаванда.
Часть видов губоцветных распространена и в средней поло-
се, причем не только в степной, но и в лесной зоне. В широко-
лиственных лесах обычны будра плющевидная (Glechoma hede-
гасеа), яснотка белая (Lamium album), чистец лесной (Stachys
sylvatica), живучка ползучая (Ajuga reptans). На влажных лугах
обитают мята полевая (Mentha arvensis) и чистец болотный
(Stachys palustris). В степной зоне повсеместно встречаются ви-
ды тимьяна (Thymus), зопник клубненосный (Phlomoides tube-
rosa), шалфей (Salvia) и др.
Семейство отличается замечательной узнаваемостью входя-
щих в его состав видов. Цветок двугубый, стебель четырехгран-
ный, листья супротивные, преимущественно цельные. И, кроме
того, все губоцветные имеют специфический запах из-за содер-
жащихся в них эфирных масел. Сильный запах губоцветных
уменьшает их поедаемость фитофагами, а в условиях засушли-
вого климата пары эфирных масел, кроме того, играют роль
«одеяла», которое окутывая растение, уменьшает испарение во-
ды листьями.
Рис. 27. Губоцветные: а - Lamium album (1 - цветущий побег, 2 - цве-
ток, 3 - пестик, 4 - диаграмма цветка); б - цветки Salvia (1 - разрез
цветка с насекомым опылителем, 2 - обоеполый цветок, 3 - женский
цветок), Melissa (4), Ajuga (5), Thymus (6 - цветок и завязь)
б)
132
С эфирными маслами связано практическое использование
губоцветных как технических, лекарственных и ароматических
растений.
У кустарниковых шалфеев Мексики (шалфей - самый боль-
шой род в семействе, в его составе около 700 видов) эфирные
масла выделяются столь интенсивно, что подавляют другие ви-
ды растений (это явление называется аллелопатией). В резуль-
тате вокруг куртины кустарника в растительном покрове обра-
зуются «пролысины».
Венчик цветка губоцветных в нижней части трубчатый, вы-
ше - разделен на две губы, верхняя из которых образована
двумя, а нижняя тремя лепестками. Нижняя губа обычно
крупная и служит посадочной площадкой для опылителей. Ты-
чинки прикреплены к трубке венчика. Гинецей образован из
двух плодолистиков, но делится ложной перегородкой пополам,
поэтому завязь становится четырехлопастной с одним семяза-
чатком в каждой лопасти. Плод состоит из 4 орешковидных до-
лей, причем чашечка всегда остается при плодах, а у некоторых
губоцветных даже разрастается.
Насекомые-опылители привлекаются нектаром, который на-
ходится в нижней части венчика. У некоторых губоцветных есть
замечательные приспособления типа шлагбаума, которые от-
крывают доступ к нектару лишь после попадания пыльцы на
спинку насекомого. Эти приспособления наиболее совершенны
у шалфея, специализированные тычинки которого имеют особое
рычажное устройство, срабатывающее, когда насекомое просо-
вывает голову в трубку венчика. При этом гнездо пыльника
ударяет по спинке насекомого, высыпая на нее пыльцу.
Плоды многих губоцветных распространяются животными
как эндозоохорно, так и эпизоохорно (часто на ногах животных
и человека). Есть мирмекохоры, плоды которых имеют различ-
ные придатки, служащие пищей муравьям (живучки, яснотки).
У анемохорных губоцветных единицами распространения
могут быть отдельные орешки дробного плода, в таком случае
их аэродинамические свойства усиливаются волосистыми, кры-
ловидными или хохолковидными выростами. У этих видов стеб-
ли долго сохраняются в сухом состоянии, постепенно рассеивая
плоды даже в зимнее время. У других видов, наоборот, расто-
пыренно разветвленные стебли с плодоносящими соцветиями
легко обламываются у основания и перекатываются ветром по
133
степи, постепенно разбрасывая плоды. Такими «перекати-поле»
являются некоторые виды шалфея, зопника, котовника
(Nepeta). Чем дольше плоды не выпадут из чашечки, тем на
большее расстояние они перенесутся, поэтому в чашечках есть
приспособления для удерживания плодов - кольцо волосков
или загнутые внутрь зубцы.
В культуре возделываются пряные губоцветные мята
(Mentha piperita), мелисса (Melissa officinalis), душица (Origanum
vulgare), майоран (Majorana hortensis), тимьян (Thymus vulgaris),
базилик (Ocimum basilicum) и эфиромасличное растение - ла-
ванда (Lavandula angustifolia, L. latifolia).
Среди губоцветных есть опасные полевые сорняки - чистец
однолетний (Stachys annua), пикулъники ладанниковый (Ga-
leopsis ladanum) и двунадрезанный (G. bifida).
Это интересно
Продлевающий жизнь. В старинных легендах о шалфее говорится,
как о продлевающем жизнь. В Англии расказывают о человеке, кото-
рый питался только хлебом, маслом и шалфеем и прожил полтораста
лет. Шалфеем врачевали еще в Древнем Египте. А Плинию Старшему
приписывают слова: «Как может умереть человек, если в саду у него
растет шалфей?». Сейчас многие знают, что полоскание рта настоем
шалфея помогает от болезней зубов. Но даже в наше время открыва-
ются новые тайны этого растения.
В Молдове, где выращивается мускатный шалфей (Salvia sclarea,
кстати именно он придает неповторимый запах одеколону «Шипр»),
произошел такой случай. Две заболевшие туберкулезом лошади, на ко-
торых ветеринарные врачи уже махнули рукой, без присмотра паслись
на поле, куда вывозились остатки после переработки шалфея. Даже
лужи на этом поле были темно-зелеными от его настоя. И лошадям
полегчало, а когда ветеринар осмотрел их - туберкулеза как не быва-
ло... Так было открыто еще одно целебное свойство шалфея.
Пахнет мятой... Аромат мяты ценился в Древней Греции и Древнем
Риме. Перед приемом гостей столы натирали свежими листьями мяты,
а залы опрыскивали настоянной на ней водой. Считалось, что мятный
аромат создает доброе и жизнерадостное настроение. Плиний Старший
советовал своим ученикам даже носить венки из мяты: они возбуждают
деятельность мозга. Поверье это сохранилось до Средневековья, когда
студенты, готовясь к экзаменам и диспутам, тоже украшали свои го-
ловы мятными венками.
134
7.22. Семейство Asteraceae (Compositae) - Астровые
(Сложноцветные)
Паспорт семейства
Родов - 1510 (150), видов - 20000 (2000)
Цветок - * Са(о, хохолок) Со<>) A(f) G(2)
Распространение - по всему миру, но наибольшее разнообра-
зие - в умеренных и субтропических областях
Жизненная форма - одно- и многолетние травы, реже кус-
тарники и деревья
Опыление - насекомыми, редко ветром
Плоды семянки, распространяются животными (в том
числе муравьями), ветром, водой
Важнейшие роды Achillea, Arctium, Artemisia, Bidens,
Carduus, Centaurea, Cirsium, Leucanthemum, Scorzonera,
Senecio, Solidago, Sonchus, Tanacetum, Taraxacum, Tussilago
Культурные растения - Aster, Bellis, Chrisanthemum, Cynara,
Dahlia, Helianthus, Rudbeckia, Tagetes и dp.
Это семейство (рис. 28) входит в монотипный порядок Aste-
rales и является самым большим из двудольных (по числу видов
его можно сравнить только с семейством орхидных из одно-
дольных). Семейство может быть названо «всюдным», так как
его представители распространены не только на всех континен-
тах, но и в самых разных местообитаниях (от жарких пустынь
до холодной Арктики и Антарктики, в высокогорьях и солоно-
ватых водах).
Характерной особенностью сложноцветных является при-
сущее им «коллективное» соцветие - корзинка. Предвестниками
этого «выгодного» соцветия, позволяющего сократить расходы
на рекламную функцию цветка, можно считать соцветия моло-
чайных и даже зонтичных. В сложных зонтиках, как мы отме-
чали, краевые цветки имеют асимметричный венчик с более
крупными лепестками, обращенными наружу.
У корзинки имеется общее ложе и обертка, образованная
верхушечными листьями. Количество цветков в корзинке варьи-
рует в широких пределах - от тысячи (подсолнечник - Heliant-
hus) до двух (амброзия - Ambrosia) и даже до одного (шарого-
ловник - Sphaerocephalum). Размеры корзинки меняются от 30-
60 см (культурные сорта подсолнечника) до нескольких милли-
метров (полыни - Artemisia).
«Коллективная реклама» позволяет уменьшить размеры
околоцветника без риска снизить эффективность опыления.
Чашечка у многих сложноцветных превращена в хохолок, кото-
рый сохраняется на плодах и способствует их расселению вет-
135
г)
Рис. 28. Сложноцветные: а - Helianthus annuus (1 - часть корзинки, 2 -
трубчатый цветок, 3 - плод); б - цветки (1 - трубчатые, 2 - язычковые,
3 - воронковидные, 4 - ложноязычковые, 5 - андроцей); в - располо-
жение цветков в соцветиях (1 - Taraxacum, 2 - Achillea, 3 - Tussilago,
4 - Centaurea); г - плоды (1 - Taraxacum, 2 - Cirsium, 3 - Solidago,
4 - Bidens, 5 - Xanthium)
ром. Венчик пятичленный, но может иметь разную форму, что
позволяет различать трубчатые, язычковые, воронковидные,
ложновбронковидные и ложноязычковые цветки (два послед-
них типа - стерильные и чисто рекламные цветки, генеративные
органы которых редуцированы).
136
Тычинок пять, их нити свободные, но пыльники обычно
спаяны в трубку, внутри которой проходит столбик с рыльцем.
Завязь нижняя, образована двумя плодолистиками.
Существуют два разных вида корзинок из одинаковых
цветков (например, у одуванчика, Taraxacum, - из язычковых
цветков, у пижмы, Tanacetum, - из трубчатых) и из разных
цветков. Например, из трубчатых и ложноязычковых (подсол-
нечник, нивяник - Leucanthemum, ромашка - Matricaria и др.),
воронковидных и ложноворонковидных (василек - Centaurea,
серпуха - Serratula).
Большинство сложноцветных опыляются насекомыми, при-
чем спектр опылителей очень широк, так как корзинки доступ-
ны для посещения разными видами насекомых. Если перекрест-
ное опыление насекомыми не состоялось, возможно самоопы-
ление или даже апомиксис - образование семян без оплодотво-
рения.
В семействе есть ветроопыляемые роды, например полынь.
Корзинки полыни мелкие, невзрачные, кроме того, значительная
часть видов произрастает в пустынях и полупустынях, где насе-
комоопыление затруднено из-за крайней сухости воздуха,
обедняющей состав насекомых.
Плод семянка, распространяющаяся ветром, водой (у
прибрежно-водных растений, таких как нардосмия - Nardosmia
и череда - Bidens), животными. Часто распространяются целые
корзинки с плодами (лопух - Arctium, дурнишник - Xanthium).
Сложноцветные входят в состав самых разных раститель-
ных сообществ: луговых, степных, лесных, пустынных, тундро-
вых, солончаковых.
Особое значение для человека имеют виды сорных и руде-
ральных сложноцветных. В числе полевых сорняков - осот ого-
родный (Sonchus oleraceus), бодяк полевой (Cirsium arvense), мо-
локан татарский (Lactuca tatarica). К рудералам относятся ви-
ды родов чертополох (Carduus), татарник (Onopordum), латук
(Lactuca), полыни горькая и обыкновенная (Artemisia absinthium,
A. vulgaris).
Среди сложноцветных есть важные культурные растения:
подсолнечник (Helianthus annuus), топинамбур (Н. tuberosus).
Искусственно создан гибрид этих видов - тописолнечник, ис-
пользуемый как силосная культура с большой зеленой массой.
Среди сложноцветных много прекрасных декоративных
растений, которые занимают важное место в садово-парковом
хозяйстве (роды Cosmos, Aster, Calendula, Bellis, Chrisanthemum,
Tagetes и др.).
137
Это интересно
История подсолнечника. Как и картофель, этот «американец» по-
пал в Испанию как декоративное растение. С XVII в. подсолнечник
стали использовать как грызовую культуру, из семян которой приго-
товляли суррогат кофе.
В России подсолнечник появился и как декоративная, и как грызо-
вая культура, только во второй половине XVIII в. И именно в России
в его историю были вписаны самые замечательные страницы. На рубе-
же XVIII и XIX столетий в результате народной селекции появились
современные сорта с крупными корзинками и одним стеблем (декора-
тивные сорта имели много стеблей и мелкие корзинки). Хотя первое
масло в 1817 г. получил англичанин А.Бенуа, именно в России в это
время было начато его промышленное производство, поскольку в юж-
ных губерниях России и на Украине подсолнечник выращивался повсе-
местно.
Трижды над подсолнечником нависала угроза полной гибели, и
трижды он восстанавливался благодаря все той же народной селекции.
В ЗО-60-е годы прошлого столетия его посевы едва не погибли от
ржавчины (грибной болезни). Однако крестьяне отобрали сохранив-
шиеся растения и так были получены культивары подсолнечника, ус-
тойчивые к этому заболеванию. Площади посевов восстановились.
Спустя 10 лет на подсолнечник напал паразитный сорняк - зара-
зиха. И вновь «народные селекционеры» вывели линии, устойчивые к
заразихе.
Наконец, уже на рубеже нашего столетия посевы подсолнечника ед-
ва не погибли от подсолнечниковой моли. Однако в этом случае крестья-
нам помогли ученые: был выведен гибрид подсолнечника с его диким ка-
лифорнийским предком, который не повреждался этим вредителем.
В результате селекции всего за столетие период созревания под-
солнечника был сокращен с пяти месяцев до 70-80 дней, а содержание
масла в его семенах достигло 50%.
Наиболее замечательно то, что лишь в 20-е годы нашего столетия
подсолнечник в таком окультуренном состоянии вернулся на свою ро-
дину - в Америку и ныне на больших площадях возделывается в США
и Канаде.
Символ Японии и Китая. С хризантемой связано предание о про-
исхождении Японии. В давние времена узнал жестокий китайский им-
ператор, что на одном из островов растет хризантема, из которой
можно приготовить эликсир жизни. Но сорвать цветок под силу только
человеку с чистым сердцем и добрыми намерениями. Послал император
за цветком триста самых прекрасных молодых юношей и девушек. Они
нашли хризантему, но не вернулись. Очарованные благодатной приро-
дой острова они основали новое государство - Японию.
Хризантема изображена на государственном флаге Японии, на пе-
чатях и денежных знаках. Высшая награда в стране - Орден Хризанте-
мы. До недавнего времени даже одежду из тканей с изображением им-
ператорской хризантемы с 16 краевыми цветками имели право носить
лишь члены императорского дома. Октябрь в Японии по старому лун-
ному календарю - месяц хризантем.
138
Любят хризантемы и в Китае: ее именем назван девятый месяц го-
да, и в девятый день этого месяца проводится Праздник цветов. По по-
верью, именно в этот день хризантема приобретает магическую силу.
Есть храм хранительницы хризантем, где она изображена молодой де-
вушкой. По легенде эта девушка выпила вина с цветком хризантемы и
стала бессмертной.
Однако обожествление цветка не помешало сделать его овощем. В
Китае и Японии из него делают салаты и пирожные. Для этого выра-
щивают специальные сорта съедобных хризантем с мясистыми сочными
лепестками. Лепестки окунают в смесь взбитых яиц и муки, обмакива-
ют в горячее масло и раскладывают на бумаге, чтобы масло впиталось
в лепестки. Посыпают сахаром и подают на стол.
Глава 8
КЛАСС LILIOPSIDA - ОДНОДОЛЬНЫЕ
8.1. Семейство Liliaceae - Лилейные
Паспорт семейства
Родов - 10(8), видов - 750 (75)
Цветок - * Р3+3 А3+3
Распространение - умеренные и субтропические области Се-
верного полушария
Жизненная форма - многолетние травянистые растения с
луковицами
Опыление - насекомыми
Плоды - коробочки. Большинство - баллисты, есть мирме-
кохоры
Важнейшие роды - Fritillaria, Gagea, Lilium, Tulipa
Культурные растения - Erythronium, Lilium, Tulipa
Лилейные (рис. 29) входят в порядок Liliales, который у
разных авторов включает разное число семейств, близких по
строению цветка и потому ранее объединявшихся в одно семей-
ство Liliaceae.
У лилейных наиболее четко проявляются все признаки
класса однодольных: трехдольный цветок, линейные листья,
проводящая система из разбросанных закрытых проводящих
пучков, мочковатая корневая система и др.
Лилейные - геофиты, которые зимуют в стадии подземного
органа - луковицы. Луковицы могут быть многолетними и од-
нолетними. У однолетних луковиц к концу сезона все чешуи
отмирают и в старых оболочках остается новая замещающая
луковица.
139
a)
6)
Рис. 29. Лилейные: а - Lilium martagon (1 - цветок, 2 - диаграмма
цветка, 3 - стебель с плодами (канделябр), 4 - плод (общий вид и по-
речный разрез), 5 - луковица); б - Fritillaria ruthenica (1 - соцветие,
2 - плод)
Большинство луковиц имеют особые втягивающие (контрак-
тильные) толстые и сочные корни, которые при высыхании
укорачиваются в вертикальном направлении и втягивают луко-
вицу, часто на значительную глубину. У тюльпанов (Tulipa) за-
мещающая луковица втягивается в почву с помощью столона
заглубления, внутри которого она находится.
Среди лилейных есть растения-эфемероиды, которые быст-
ро проходят фазы вегетации, цветения и плодоношения в пери-
од, когда в почве еще достаточно весенней влаги (степные и
пустынные тюльпаны) или еще не распустились листья на де-
ревьях (гусиный лук - Gagea).
В составе семейства много прекрасных декоративных расте-
ний, многие из которых введены в культуру: лилия саранка
(L. martagon), тюльпаны, рябчики (Fritillaria) и др.
Отметим некоторые другие наиболее важные семейства по-
рядка Liliales.
Луковые (Alliaceae, 30 родов, 750 видов) распространены на
всех континентах, но большинство произрастает в умеренных
областях Северного полушария. Давший название семейству род
лук (Allium) включает много видов культурных и ресурсных
растений, у которых в пищу используются луковицы и зеленые
листья: лук репчатый (А. сера), чеснок (A. sativum), лук-порей
140
(A. porrum), лук-батун (A. flstulosum), лук косой (A. obliquum),
шнитт-лук (A. schoenoprasum), черемша (A. victorialis) и мн. др.
Характерный признак луковых - присутствие в зеленых ли-
стьях и в чешуях луковиц членистых млечников, заполненных
млечным соком - латексом. Кроме того, во всех тканях содер-
жится летучее чесночное (и несколько близких к нему) масло,
которое определяет специфический луковый и чесночный вкус
и запах.
Гиацинтовые (Hyacinthaceae) имеют важное значение в де-
коративном садоводстве благодаря таким красивоцветущим рас-
тениям, как гиацинт (Hyacinthus), мускари, или мышиный гиа-
цинт (Muscari), птицемлечник (Ornithogalum).
Ирисовые (Iridaceae) это большей частью корневищные
многолетники. Наибольшее значение имеют роды ирис (Iris) и
гладиолус (Gladiolus), представленные в культуре многочислен-
ными сортами.
Ландышевые (Convallariaceae) известны в нашей флоре та-
кими родами, как ландыш (Convallaria), майник (Maianthemum)
и купена (Polygonatum). Ландыш майский (С. majalis) - ценное
лекарственное растение. Он также введен в культуру как тене-
выносливый декоративный многолетник.
Спаржевые (Asparagaceae) отличаются от ландышевых
обычно древеснеющими стеблями, редуцированными и более
или менее чешуевидными листьями. Во многих странах Европы
и Северной Америки широко возделывается как овощная куль-
тура спаржа (Asparagus officinalis). В пищу употребляются ее
отбеленные (этиолированные) побеги. Ряд видов спаржи рас-
пространены в культуре как декоративные растения.
Это интересно
Страна тюльпанов. Тюльпан попал в Вену в 1554 г. из сада турец-
кого султана и менее чем за 100 лет распространился по Европе. Осо-
бенно благодатными для тюльпанов оказались условия Голландии, ко-
торую охватила настоящая «тюльпаномания». Тюльпаны разводились
повсюду, луковицы продавались по баснословным ценам, новые сорта
их можно было обменять на дома или даже целые имения. Негры Га-
арлема захотели иметь черный тюльпан, и он был выведен. Гонорар за
этот сорт составил 100 тысяч гульденов золотом. Голландские тюльпа-
ны потрясли красотой любителей цветов всего мира. Считают, что за
историю их культивирования около 12 тысяч сортов сменили друг дру-
га и примерно 3 тысячи существуют сейчас.
Целебный овощ. Лук - древнейшее культурное растение. Египтяне
считали лук и чеснок божественными растениями и приносили их лу-
ковицы на алтарь богам. Не менее почитаем был лук и в Древней Гре-
ции, где его не только приносили в дар богам, но и дарили новобрач-
141
ным. В Древнем Риме лук входил в обязательный рацион питания сол-
дат. В средние века лук распространился по всем странам Европы, но
особого расцвета «луководство» достигло в Х-ХП вв. в Испании, где и
по сей день есть сорта с луковицами, превосходящими все прочие по
величине и сладости.
Разные сорта лука репчатого отличаются формой луковиц и окра-
ской чешуй. Кроме желтых, есть сорта с белыми и фиолетовыми луко-
вицами. Размер их тоже различен. Луковицы валенсийского сорта (Ис-
пания) достигают массы 1 кг, а один немецкий огородник вырастил лу-
ковицу массой 2850 г. Сообщалось, что в Мексике получили луковицы
массой около 4 кг!
Целебные свойства лука хорошо отражены в народной поговорке
«Кто ест лук, тот избавлен от мук».
Самый питательный лук. В пустынных степях Центральной Азии,
где количество осадков составляет менее 200 мм/год, часто встречают-
ся так называемые луковые степи с доминированием лука многокорне-
вого (Allium polyrrhizum). Этот лук - бесспорный чемпион по содержа-
нию белка в листьях: он может достигать 26% сухой массы. Столь вы-
сокопитательное растение служит основой кормовой базы на пастби-
щах. Лук придает приятный запах мясу овец, которые паслись в луко-
вых степях, но ухудшает вкус популярного напитка из кобыльего мо-
лока - кумыса.
8.2. Семейство Orchidaceae — Орхидные
Паспорт семейства
Родов - 750 (40), видов - 25 000 (120)
Цветок - Т Coj Coj А3-1 G(j)
Распространение - по всему миру, главным образом в тропи-
ческих областях
Жизненная форма многолетние наземные травы, эпифи-
ты, лианы
Опыление - насекомыми
Плоды - коробочки. Семена очень мелкие и многочисленные,
распространяются ветром
Важнейшие роды - Cephalanthera, Cypripedium, Dactylorhiza,
Epipactis, Goodyera, Gymnadenia, Listera, Orchis, Platanthera,
Культурные растения - Vanilla
Семейство (рис. 30) входит в монотипный порядок Orchi-
dales. Это самое большое и космополитное семейство, хотя по-
давляющая часть видов растет в тропиках (к примеру, в Европе,
как и в России, число орхидных не превышает 120 видов).
Орхидные встречаются в самых разнообразных условиях.
Например, в Колумбии есть виды, обитающие на нетающих сне-
гах гор, в Австралии - под землей.
142
Рис. 30. Орхидные: а - Dactylorhiza (1 -растение, 2 - цветок, 3 - пестик
и тычинка, 4 - ножки поллиниев с липкими подушечками); б - Cypri-
pedium (1 - верхушка побега, 2 - опыление цветка (насекомое выползает
из «окна» с комком пыльцы на спинке); в - Ophrys insectifera (1 - расте-
ние, 2 - цветок, 3 - оса в процессе псевдокопуляции, 4 - головка осы с
прикрепившимися поллинариями); г — Epipactis palustris (1 - верхушка
побега, 2 — цветок, 3 - соответствие формы губы форме тела опылителя)
Орхидные умеренной полосы - это, как правило, растения с
неброскими цветками, хотя среди них есть и очень красивые,
например венерин башмачок (Cypripedium). Самые красивые
представители орхидных связаны с тропическим поясом, где
большая часть из них - эпифиты.
143
Эпифитные орхидеи сильно различаются по размеру: среди
них есть крохотные, едва различимые на ветвях деревьев рас-
теньица, и многометровые лианы, и лазающие растения. Так, у
видов американского рода онцидиум (Oncidium) свисающие со-
цветия имеют длину в несколько метров.
Эпифитные орхидеи имеют особые воздушные корни с тол-
стым слоем гигроскопической ткани, которая впитывает воду и
может дать приют азотфиксирующим микроорганизмам. Воз-
душные корни могут достигать земли и становиться обычными
питающими растение почвенными корнями. Однако основными
запасниками воды у эпифитных орхидей являются особые
утолщения стебля - «псевдобульбы».
Некоторые эпифитные виды связаны мутуалистическими
взаимоотношениями с древесными муравьями, которые защи-
щают их от фитофагов, а сами получают приют в утолщенных
стеблях растений и пищу.
Есть среди эпифитных орхидных и ксерофиты-суккуленты,
у которых фотосинтезируют зеленые стебли, воздушные корни
и даже цветки. У представителей с суккулентными листьями -
тот же тип фотосинтеза, что и у кактусов (так называемый
кислый метаболизм фотосинтез при закрытых устьицах за
счет углекислого газа, запасенного в ночное время).
Некоторые орхидные сапрофиты, т.е. бесхорофилльные
растения (никогда не выходящие на дневную поверхность), ко-
торые поселяются на гниющих растительных остатках и усваи-
вают из них необходимые питательные вещества за счет сим-
биоза с грибом. Впрочем, симбиоз с грибами характерен для
всех орхидных на ранних стадиях развития. В их необычайно
мелких семенах с недифференцированным зародышем нет ника-
ких пищевых запасов, поэтому они не способны прорастать без
дополнительного источника питания. До тех пор, пока эта обли-
гатность симбиоза с грибом не была выяснена, удавалось раз-
множать орхидеи только вегетативно. Первый опыт выращива-
ния орхидей из семян на питательной смеси, искусственно ин-
фицированной грибом, датируется лишь началом нашего столе-
тия. Ботаники говорят, что орхидея состоит из двух организмов
и «носит прекрасные одежды наверху, но зависит от невидимо-
го слуги внизу».
Наземные орхидеи умеренных широт имеют обычно два
подземных клубня с запасом питательных веществ, один из ко-
торых, прошлогодний и начавший сморщиваться, питает разви-
вающийся стебель и к концу сезона отмирает, а второй разрас-
тается и будет питать растение на следующий год.
144
Цветки орхидных редко бывают одиночными, обычно они со-
браны в соцветия (кистевидные, колосовидные, зонтиковидные) и
необыкновенно разнообразны. Своим видом цветки могут имити-
ровать различных насекомых - пчел, ос, бабочек, а также других
животных (ящериц, пауков, медуз, лягушек, лебедей).
Цветок состоит из трех чашелистиков, обычно лепестковид-
ных, трех лепестков, один из которых, так называемая губа,
сильно отличается от других и служит местом посадки для на-
секомых. Только у немногих орхидных сохранилось 3 или 2
тычинки, у большинства - тычинка одна, и она объединена со
столбиком и рыльцем в единую структуру - колонку.
Пыльца тоже своеобразная - она соединена в более или
менее плотные массы - поллинии (их бывает от 2 до 8, каждый
содержит сотни тысяч пыльцевых зерен, а консистенция полли-
ниев бывает от мучнистой до совершенно твердой).
Завязь нижняя, в процессе развития она скручивается и
цветок к концу бутонизации поворачивается на 180 градусов,
так что губа оказывается снаружи в удобном для насекомого
положении.
Изучение опыления орхидных - одна из самых увлекатель-
ных задач ботаника-натуралиста, которой занимался еще
Ч.Дарвин. В настоящее время накопилось уже много данных об
опылении разных орхидных, и очевидно, что среди всех насеко-
моопыляемых растений именно виды этого семейства достигли
наибольшего успеха в коадаптации с опылителями (этим они
отличаются от сложноцветных, где эволюция пошла по пути
развития неспециализированного насекомоопыления).
Самый замечательный способ опыления наблюдается у оф-
рисов (Ophris), цветки которых напоминают мух, пчел, шмелей,
пауков. У цветка офриса насекомоносной (О. insectifera) губа
похожа на тело насекомого со сложенными крыльями, боковые
леспестки имеют вид антенн, а ложные нектарники в основании
губы напоминают глаза насекомого. Цветки не «предлагают»
насекомым ни нектара, ни пыльцы: они выделяют летучие веще-
ства, подобные половым феромонам, а внешний облик губы
действует как визуальный стимул. Самцы жалящих перепонча-
токрылых, появляющиеся после перезимовки раньше самок, по-
буждаемые половым инстинктом летят на цветки офриса и ве-
дут себя так, как при копуляции с самками. В результате их
движений поллинии прилипают к головке насекомого. Украшен-
ный «рогами» самец для удовлетворения полового инстинкта
посещает другой цветок, производя перекрестное опыление.
145
У других орхидных, например у венерина башмачака, цвет-
ки представляют собой хитроумную ловушку, которая заставля-
ет насекомое произвести опыление, причем привлекаются насе-
комые с помощью обмана - яркий стаминодий служит для них
указателем несуществующего нектара.
Цветки тех орхидных, которые «расплачиваются» с опыли-
телями за работу нектаром, также поражают совершенством
приспособлений, обеспечивающих вынос опылителем поллиниев
и доставку их на рыльце в другой цветок {тайник - Listera,
дремлик - Epipactis, гнездовка - Neottia).
Среди множества приспособлений для перекрестного опы-
ления есть одно, универсальное для всего семейства, - длитель-
ность цветения. У большинства орхидных цветки в ожидании
опыления не увядают неделями и месяцами, а за счет того, что
цветки распускаются неодновременно, все соцветие имеет све-
жий вид еще дольше. Кроме того, запахи цветков чрезвычайно
разнообразны - от запаха гниющего мяса до тончайшего аро-
мата духов.
Несмотря на такое совершенство механизма опыления у ор-
хидных, его эффективность невысока: плоды завязываются у
одного из 300 (или даже из 1000) цветков. Особо опасным этот
вариант опыления оказался в условиях нарастающего антропо-
генного влияния, когда в результате загрязнения среды гибнут
насекомые-опылители.
Плоды орхидных - коробочки, разнообразные по величине:
от мелких сухих до сочных длинных, напоминающих бобы фа-
соли (как, например, у ванили - Vanilla planifolia). Микроско-
пически малые семена орхидных распространяются ветром и
даже, подобно пыльце, восходящими (конвекционными) токами
теплого воздуха в тропическом лесу.
Орхидные - украшение Земли, «аристократы» среди расте-
ний. Уже в XVI и XVII вв. завоз тропических орхидей в Евро-
пу стал доходным делом. Как отмечалось, вплоть до начала
XX в. орхидеи размножали только вегетативно. В настоящее
время выращивание орхидей из семян с использованием микро-
биологической техники и искусственным инфицированием гри-
бом - один из самых доходных видов бизнеса (особенно он
развит в Калифорнии).
Тем не менее, несмотря на индустрию производства орхид-
ных, давление на их популяции в природе сохраняется очень
высоким. В Красной книге нашей страны - 44 вида орхидных (в
том числе Cypripedium calceolus, Calypso bulbosa и др.).
Раздел ботаники об орхидных столь богат содержанием,
что выделен в самостоятельную науку орхидологию.
146
8.3. Семейство Cyperaceae - Осоковые
Паспорт семейства
Родов - 100 (20), видов - 4000 (370)
Цветок Carex - * Pq Ay Go , * Ро Ao G(j)
Распространение - по всему миру, но наиболее многочислен-
ны в умеренных и холодных областях
Жизненная форма - многолетние, реже однолетние травы
Опыление - ветром
Плоды - орешки, распространяются водой и животными
Важнейшие роды - Bolboschoenus, Carex, Cladium, Cyperus,
Eleocharis, Eriophorum, Kobresia, Rhynchospora, Scirpus
Культурные растения - Cyperus esculentus, Eleocharis dulcis
Семейство (рис. 31) входит в монотипный порядок Cyperales
и объединяет ветроопыляемые растения, которые по морфоло-
гическим признакам близки к злакам, за что их часто относят к
морфологической группе граминоидов (как и представителей
семейства Ситниковые - Juncaceae). Однако в отличие от зла-
ков с полыми соломинами и вздутыми междоузлиями, у осоко-
вых стебли преимущественно трехгранные, без вздутых узлов,
неполые, листья трехрядные (а не двухрядные), образующиеся в
основном в нижней части стебля. Кроме того, у осок влагалища
всегда замкнутые.
Большинство осоковых - гигрофиты, обитающие на боло-
тах, сырых лугах и в прибрежно-водных местообитаниях. Одна-
ко среди них есть виды, которые произрастают в степных усло-
виях, - осоки ранняя (Carex ргаесох), приземистая (С. supina),
стоповидная (С. pediformis), твердоватая (С. duriuscula). Целый
ряд видов осок входит в состав лесных сообществ (С. muricata,
С. pilosa, С. rhizina, С. sylvatica и др.). Имеются также горно-
тундровые, солончаковатые и даже пустынные виды.
Большинство осоковых - многолетние травы, образующие
клоны. Клоны могут состоять из побегов, тесно сближенных в
кочку (как у осоки дернистой — С. cespitosa), или из пятна по-
бегов, соединенных друг с другом достаточно длинными корне-
вищами, осоки ранняя, пузырчатая (С. vesicaria), острая
(С. acuta) и др.
Невзрачные цветки осоковых собраны в колоски, образую-
щие сложные соцветия: колосовидные, метельчатые, зонтико-
видные, головчатые. Цветки бывают обоеполыми (с околоцвет-
ником в виде щетинок, волосков или совсем без околоцветни-
ка) и однополыми (при этом растения могут быть одно- или
двудомными).
147
У самого большого рода семейства (и одного из самых
крупных родов цветковых) - осока (больше 2000 видов) - одно-
полые цветки сильно редуцированы. Мужской цветок состоит
из трех тычинок, сидящих в пазухе кроющей чешуи. Женский
цветок, также сидящий в пазухе кроющей чешуи, представлен
пестиком, заключенным в особое образование - мешочек, напо-
минающий кувшинчик с узким горлышком. Из горлышка вы-
ставляется часть столбика с рыльцами. Природа мешочка не ус-
тановлена, возможно, что это видоизмененный прицветный
лист. Как правило, осоки однодомны.
Рис. 31. Осоковые: а — цветки (1 Eriophorum, 2 Scirpus, 3
Cyperus, 4 - Carex); б - Carex vesicaria (1 - цветущий побег, 2 - жен-
ский цветок (общий вид и разрез))
Возможно, именно благодаря мешочкам род осока стал са-
мым многочисленным и широко распространенным из осоковых.
Мешочки защищают завязь и развивающийся плод, а у многих
видов они, кроме того, являются приспособлением к распро-
странению плодов.
В умеренной зоне широко распространен камыш озерный
(Scirpus lacustris) - высокое (до 2,5 м) растение с почти безли-
стными стеблями, образующее обширные заросли в реках, озе-
рах и прудах на глубине 1 м и более. Ассимилирующими орга-
нами у камыша являются стебли, а размножается он преимуще-
ственно вегетативно с помощью длинных корневищ. Вместе с
148
другими прибрежно-водными растениями камыш выполняет
важную функцию биологической очистки воды. Он является
одним из основных торфообразователей. Стебли камыша ис-
пользуются для изготовления плетеных изделий, а также в ка-
честве упаковочного материала. (В народе часто ошибочно ка-
мышом называют другое растение - рогоз, Typha. Этот род с
характерными удлиненными коричневыми «шишками» относит-
ся совсем к другому семейству - Typhaceae.)
Виды пушицы (Eriophorum) - обычные растения болот
преимущественно распространены в тундре, на севере лесной
зоны и альпийском поясе гор. Околоцветник пушицы состоит из
многочисленных шелковистых волосков, которые после
цветения удлиняются и колоски приобретают вид пушистых
головок снежно-белого или рыжеватого цвета.
Распространенная по всему земному шару болотница
(Eleocharis) обитает по мелководьям, отмелям, берегам водоемов,
травяным болотам. Один из ее видов - болотницу сладкую, или
водяной каштан (Е. dulcis) - возделывают в Юго-Восточной Азии
как пищевое растение, используя богатые крахмалом сладкие
клубневидные образования на ее корневищах.
Другое культурное пищевое растение из семейства - сыть
съедобная, или чуфа (Cyperus esculentus), возделывается в стра-
нах Средиземноморья. Ее «клубни» богаты не только сахаром и
крахмалом, но и маслом.
Сыть бурая (Cyperus fuscus) типичное растение-пионер,
заселяющее прибрежные отмели и галечники. Сыть очередно-
листная (С. alternifolius) известна как популярная комнатная и
оранжерейная культура (в быту ее нередко ошибочно называют
папирусом).
Клубнекамыш морской (Bolboschoenus maritimus) - растение
засоленных почв, широко распространенное в южной части
России. На его корнях имеются крупные запасающие клубни, с
этим связано название растения.
Доминантами высокогорных лугов Кавказа, Карпат и Цен-
тральной Азии являются виды рода кобрезия (Kobresia).
Это интересно
Самая древняя «бумага». Первая бумага изготовлялась в Египте
уже в начале III тысячелетия до н.э из стеблей папируса (Cyperus
papyrus) гигантского (высотой до 4-5 м) травянистого растения с
почти безлистными стеблями диаметром до 7 см. Технология была
проста: сердцевину стеблей разрезали на полоски шириной 10 мм и
раскладывали в два слоя - вдоль и поперек. Затем слои сдавливали, и
полоски склеивались клейким веществом, содержащимся в сердцевине
папируса. Полученные тонкие эластичные листы высушивали на солн-
149
це, полировали кусочками слоновой кости и склеивали в свитки ши-
риной 20—30 см и длиной 10-40 м.
Название «папирус» происходит от латинского слова «рари»
царский. От этого же корня происходит и английское слово - paper
(бумага), а также французское и немецкое - papier.
Папирус в древнем Египте служил не только для производства бу-
маги, его корневища использовались в пищу. 30 веков папирус служил
источником получения писчего материала. Его закат произошел в
VIII в., когда он был вытеснен изобретенной в Китае бумагой. В дель-
те Нила сохранилась всего одна небольшая заросль папируса. Сейчас
его выращивают только как декоративное растение в садах и парках
Египта, Бразилии и других стран с теплым климатом.
Лодки Тура Хейердала. Из папируса были сделаны лодки знамени-
того путешественника Т.Хейердала - «Ра-1», Ра-2», «Тигрис», которые
были точными копиями древнеегипетских судов. На этих лодках Хей-
ердал совершил дальние переходы через Атлантический (от Марокко
до берегов Америки) и Индийский океаны.
8.4. Семейство Poaceae (Gramineae)- Мятликовые (Злаки)
Паспорт семейства
Родов - 650(130), видов - 10 000 (810)
Распространение - по всему миру
Жизненная форма - одно- и многолетние травы
Опыление - ветром
Плоды - зерновки, распространяются животными, ветром
Важнейшие роды Agrostis, Alopecurus, Bromopsis,
Calamagrostis, Deschampsia, Elytrigia, Festuca, Koeleria,
Phragmites, Poa, Puccinellia, Setaria, Stipa
Культурные растения Avena, Hordeum, Oryza, Panicum,
Saccharum, Secale, Sorghum, Triticum, Zea
Злаки (рис. 32) относятся к монотипному порядку Poales.
Они занимают особое положение и в системе цветковых, и в
растительном покрове Земли. Это семейство объединяет типич-
ные ветроопыляемые растения с сильно редуцированным цвет-
ком. Он состоит из двух цветковых чешуй, двух пленочек - ла-
дикул (их рассматривают как результат редукции внутреннего
круга околоцветника), обычно трех (реже больше до 120 у
бамбуков, и меньше - 2 или 1) тычинок и пестика с двулопаст-
ным рыльцем (количество плодолистиков дискутируется - один
или два).
Цветки собраны в колоски, имеющие колосковые чешуи,
похожие на цветковые, но обычно крупнее. В одном колоске
может быть от одного до двадцати цветков. Колоски в свою
очередь образуют различные соцветия: колос, початок, метелку
150
и др. Иногда колоски бывают клейстогамными (Leersia,
Cleistogenes и др.). У некоторых злаков (Роа, Festuca и др.)
встречаются вивипарные (живородящие) колоски, образующие
выводковые почки, которые высыпаются на землю и вырастают
в новые растения.
б)
в)
Рис. 32. Злаки: а - колосок (1), цветок (2) и диаграмма цветка (3); б -
соцветия и колоски (1 - Triticum aestivum, 2 - Elytrigia repens, 3 -
Phleurn pratense, 4 - Avena saliva, 5 - Poa annua, 6 - Festuca pratensis;
в - Zea mays (1 - часть стебля с женским соцветием, 2 - женский ко-
лосок, 3 - часть стебля с мужским соцветием, 4 - мужской колосок)
151
Строение вегетативных органов также своеобразно. У
большинства злаков стебель полый - соломина (но есть расте-
ния и с неполым стеблем: кукуруза — Zea, сорго - Sorghum). Ли-
стья линейные, иногда сложенные пополам, иногда свертываю-
щиеся в трубочку и с опушением (у степных овсяниц).
Злаки - однолетние, а чаще многолетние травы. Последние
активно ветвятся (кустятся) и образуют клоны. Клоны могут
быть представлены плотной дерновиной диаметром 5-40 см у
так называемых плотнокустовых злаков (к ним относятся щучка
дернистая - Deschampsia cespitosa, типчак - Festuca valesiaca,
ковыли - Stipa и др.) или пятном размером в несколько десят-
ков сантиметров и даже метров у корневищных злаков, таких
как пырей ползучий (Elytrigia repens) или кострец безостый
(Bromopsis inermis). Клоны могут разделяться на самостоятель-
ные группы побегов, таким путем происходит вегетативное раз-
множение растений.
Различают два подсемейства злаков: настоящие злаки
(Pooideae) и бамбуковые (Bambusoideae). У бамбуковых побеги
более или менее одревесневают и могут достигать высоты 40 м.
Кроме того, побеги у них ветвятся, что для злаков малохарак-
терно. На Сахалине и Курилах встречается 5 видов дикорасту-
щего рода саза (Sasa), или курильский чай, образующих густые
заросли по склонам гор.
Учитывая, что злаки играют огромную роль в жизни био-
сферы (формируют биом злаковников, занимающий обширную
площадь) и человека (к злаковым относятся наиболее важные
культурные растения), мы рассмотрим это семейства более
подробно.
Злаки как доминанты злаковников. Злаковники биом,
объединяющий растительные сообщества с доминированием
злаков, в прошлом занимал до 30% площади суши планеты с
количеством осадков от 200 до 1700 мм/год. В настоящее время
их площадь сократилась до 20%, так как значительная часть
злаковников замещена сельскохозяйственными землями.
Большая часть злаковников первична, т.е. сформировалась
естественным путем. Это - степи (в Северной Америке их назы-
вают прериями), альпийские луга, саванны. В последнем случае
к травянистому покрову с доминированием злаков добавляется
разреженный полог деревьев и кустарников. Кроме того, суще-
ствуют и вторичные злаковники - луга, которые сформирова-
лись на месте лесов, вырубленных человеком. Такие злаковники
сохраняются до тех пор, пока их использует человек: косит
траву, пасет скот. В противном случае луга зарастают лесом.
Столь большая роль злаков в растительном покрове Земли
объясняется тем, что они хорошо приспособлены к отчуждению
152
(скусыванию или скашиванию) растительной массы и быстро
отрастают после этого (т.е. обладают высокой отавностью). По
этой причине формирование евразийских степей и американ-
ских прерий произошло при участии копытных животных-
фитофагов (в Америке - бизонов и вилорогов, в Евразии - тар-
панов, туров и сайгаков), без которых и сегодня эти сообщества
существовать не могут. Впрочем, теперь диких фитофагов сме-
нил домашний скот, который особенно при высоком поголовье
отрицательно влияет на травостои, вызывая обеднение видового
состава.
Степи России сформировались в основном за счет видов
злаков из родов ковыль (Stipa), овсяница (Festuca, из более ксе-
рофитных видов - типчаков’. F. valesiaca, F. rupicola, F. pseu-
dovina и др.), тонконог (Koeleria), житняк (Agropyron), змеевка
(Cleistogenes, в азиатской части).
Кроме высокой отавности, злаки отличаются устойчивостью
к действию огня, а в условиях степей и особенно саванн пожа-
ры явление достаточно обычное из-за регулярных гроз, а в
последнее столетие и из-за неаккуратных действий людей.
Наконец, для злаков характерна очень «выгодная» морфо-
логия. Листья у них узкие и расположены под острым углом к
солнечным лучам, что позволяет увеличивать общую фотосин-
тезирующую поверхность. Злаки активнее других трав могут
усваивать элементы минерального питания. Это позволяет им
доминировать в самых разных условиях среды - от мелководий
в долинах южных рек (плавни из тростника Phragmites
australis) и богатых почв прирусловой поймы, где формируются
урожайные луга из костреца безостого и двукисточника тро-
стниковидного (Phalaroides arundinacea), до самых сухих пус-
тынных степей Центральной Азии, где выпадает меньше
200 мм/год осадков.
Особо высоким фотосинтезом отличаются злаки естествен-
ных степей и саванн. В среднем биологическая продуктивность
таких сообществ оказывается в два раза выше, чем у посевов в
тех же условиях, и при этом без всяких затрат на обработку
почвы, посев и удобрение. Продуктивность злаковников в 2
раза выше среднемировой продуктивности суши (соответствен-
но 800 и 300 г/кв.м в год), хотя значительная ее часть занята
высокопродуктивными лесами (тропическими и умеренных ши-
рот). Иными словами, естественные злаковники - это экосисте-
мы с самым высоким «коэффициентом полезного действия».
Хлебные злаки. Хлеб и используемое для его выпечки зерно
(или заменители хлеба, также приготовляемые из зерна злаков,
пример которых рисовая или перловая каша) - основа питания
153
человечества. Сегодня на каждого землянина производится бо-
лее 300 кг зерна в год, хотя в разных странах этот показатель
сильно различается (в США 800 кг, в бедных африканских
странах - менее 200 кг).
Зерноводство одна из первых отраслей земледелия (но
все же, видимо, не самая первая, так как возделывание куль-
турных растений началось с выращивания клубнекорневых
культур, подобных картофелю или свекле). Появление зерна
злаков как длительно сохраняемого продукта питания, который
концентрирует в себе намного большее число калорий, чем
клубни или корни, было одним из самых мощных стимулов раз-
вития цивилизации. Французский ботаник Ж.Бросс пишет по
этому поводу: «Культура злаков, позволившая год от года на-
капливать запасы зерна, не только создала обеспеченность, ко-
торой дотоле не было, но и образовала новый резерв потенци-
альной энергии - накопленное зерно представляет собой самую
древнюю форму капитала. Эти остатки в свою очередь обеспе-
чивали человека энергией, которую можно было использовать
на другие цели, нежели производство пищи: отсюда зарождает-
ся разнообразие профессий, специализация и, в конечном итоге,
социальные классы... Понадобилось создать армию и военона-
чальника для управления армией... Богатство должно находить-
ся в обороте, а значит, возникает торговля; требовалось вести
учет, отсюда система условных знаков и книга записей, в ко-
торой его регистрировали. Понадобились писцы.» (1996, с. 121-
122).
Основные продовольственные злаковые культуры и их ха-
рактеристики перечислены в табл. 14.
За длительную историю выращивания злаки во многом из-
менились. Вплоть до начала нашего столетия велся искусствен-
ный отбор на повышение урожайности и устойчивости злаков,
т.е. их способности выдерживать засуху, давать урожай на поч-
вах, обедненных элементами питания, отрастать при нападении
насекомых-вредителей и грибных болезней. Земледельцы были
заинтересованы в получении сортов, которые отличались друж-
ным развитием и способностью подавлять сорные растения.
Однако, после того, как человек получил возможность обра-
батывать большие площади полей тракторами, вносить удобрения
и использовать пестициды, которые позволяли подавлять сорня-
ки, уничтожать насекомых-вредителей и защищать культуру от
болезней, направление селекции изменилось. Выводимые сорта
отличались все более высокой урожайностью, но становились все
более требовательными к обеспечению питанием и влагой (их
было необходимо поливать и удобрять) и неустойчивыми к болез-
154
Таблица 14
Характеристика основных пищевых злаковых культур
Культура Происхождение История введения в культуру Хозяйственное значение
Пшеница мягкая (Triticum aestivum) Первичный центр Абисси- ния, Передняя Азия, Средизем- номорье, вторич- ный - Китай, Япония Известна за 5 тыс. лет до н.э. в Египте и за 3 тыс. лет до н.э. в Китае. В Европе возделывается с Каменного века Главная продо- вольственная культура, рас- пространенная во всем мире. Ози- мые и яровые формы
Пшеница твердая (Triticum durum) Первичный центр Перед- няя Азия, вто- ричный Эфио- пия Как и мягкая пшеница, одна из древнейших куль- тур Второе место по распростране- нию после мяг- кой пшеницы. Озимые и яровые формы. Высокое содержание бел- ка и клейковины
Рожь по- севная (Secale cereale) Первичный центр Малая Азия и Закавка- зье, вторичный - Иран и Афгани- стан. Культурные сорта произошли от сорнополевой ржи. В настоящее время существует только в культуре Одно из наибо- лее окультурен- ных растений. В Европе известна с Бронзового ве- ка. На террито- рии нынешнего СНГ впервые от- мечена на Украи- не за 2-3 тыс. лет до н.э. Озимые и яро- вые формы, при- чем озимые со- ставляют 99,8% всех посевов ржи. Самая зи- мостойкая куль- тура
Ячмень культур- ный (Hordeum vulgare) Очаги форми- рования - Абис- синия и Китай- ско-Японский центр В культуре из- вестен около 6 тыс. лет. В Евро- пе одна из древнейших зер- новых культур Самая скоро- спелая культура из всех зерновых. Выращивается как крупяная и кормовая. В Ев- ропе преоблада- ют пивоваренные ячмени
Овес по- севной (Avena sativa) Родина Ки- тай. Сформиро- вался из сорно- полевого овса и сейчас известен только в культуре Первые упоми- нания датируют- ся IV в. до Н.Э. Культивируется для получения зерна, крупы, му- ки, а также как кормовая культу- ра
155
Окончание табл. 14
Культура Происхождение История введения в культуру Хозяйственное значение
Кукуруза (Zea majus) Первичный центр южная Мексика, вторич- ный западный Китай. Сейчас известна только в культуре Одна из самых древних культур. Первые посевы известны с 3400 г. до н.э. В Евро- пу завезена после второго путеше- ствия Колумба. На территории нынешнего СНГ впервые попала в Грузию Выращивается как пищевая и кормовая культу- ра. Более 80% зерна кукурузы производится в «кукурузном поясе» США
Просо обыкно- венное (Panicum miliaceum) Центральный и западный Китай В Китае из- вестно с 2700 г. до н.э. С глубо- кой древности выращивается в Закавказье Культивируется для получения зерна, которое дает крупу и му- ку, а также идет на корм. Засухо- устойчивый ксе- рофит
Рис посев- ной (Oryza sativa) Китай и Индия. Сейчас известен только в культу- ре В Юго-Восточ- ной Азии возде- лывается не ме- нее 4-5 тыс. лет, в Египте и Испа- нии - с VIII в., в США с 1865 г. В бывшем СССР - с 20-х годов текущего столе- тия Основная пи- щевая культура для более 2 млрд человек
ням, а также к насекомым-фитофагам. Они обладали низкой
конкурентной способностью и легко заглушались сорняками. По
этой причине для возделывания этих сортов было необходимо
применять в высоких дозах различные пестициды.
Новые сорта рекордсмены по урожайности - давали до
100 ц/га зерна, но это зерно обходилось очень дорого, так как
для его получения было необходимо вкладывать много антропо-
генной энергии. Кроме того, удобрения и пестициды вымыва-
лись с полей, окружающая среда загрязнялась. Под влиянием
высоких доз удобрений разрушались почвы. Новые сорта ока-
зались экологически опасными и экономически невыгодными,
что стало особенно очевидно после того, как в 60-70-е годы в
156
тропическом поясе прошла Зеленая революция. «Отец Зеленой
революции» - мексиканский генетик-селекционер Норман Бер-
лоуг - вывел сорт пшеницы «Мексикале», урожайность которо-
го оказалась в 3 раза выше, чем у других сортов, выращиваемых
в этом климате, благодаря тому, что в выведенном им сорте на
формирование зерна затрачивалось около 60% продуктов фото-
синтеза, а это в 1,5-2 раза выше, чем у традиционных сортов
пшеницы.
В настоящее время постепенно происходит переориентация
селекционеров с выведения «сортов-рекордсменов» на выведе-
ние «сортов-тружеников» (эти термины предложил итальянский
агроэколог Дж.Ацци). Современные сорта-труженики, как и
«народные сорта» прошлого столетия, дают умеренно высокий
урожай, но не требуют высоких затрат на защиту растений и
могут выращиваться без полива при невысоких дозах удобре-
ний. Такие сорта называются адаптивными, т.е. приспособлен-
ными к местным природным условиям и потому дающими на
каждую единицу затраченной антропогенной энергии макси-
мальную отдачу урожаем.
Пример адаптивного сорта рожь «Чулпан», выведенная
башкирским селекционером С.Кунакбаевым. Этот сорт засухо-
устойчив, способен заглушать сорняки и быстро отрастать при
нападении насекомых-вредителей.
Кроме хлебных злаков, в хозяйстве человека очень важны
кормовые злаки, которые высеваются на травяных полях сево-
оборотов или при коренном улучшении лугов. Важнейшими
кормовыми культурами являются кострец безостый (Bromopsis
inermis), овсяница луговая (Festuca pratensis), тимофеевка луго-
вая (Phleum pratense), ежа сборная (Dactylis glomerata), лисо-
хвост луговой (Alopecurus pratensis).
Отметим также сахарный тростник (Saccharum officina-
rum), плантации которого дают около 2/3 мировой продукции
сахара.
Это интересно
Первые земледельческие цивилизации возникли у шумеров в меж-
дуречье Тигра и Евфрата, у египтян в долине реки Нил, у индейцев
племени майя в Южной Америке. До наших дней дошли удивительные
легенды о том, как человек научился выращивать культурные растения.
О двух из них мы расскажем, использовав книгу С.М.Скорнякова «Зе-
леная родословная».
Победа богини Ашнан. Предки шумеров - духи земли апунаки - не
знали, что такое хлеб. Боги решили улучшить их жизнь и слепили из
глины пастуха Лахара и его сестру богиню зерна Ашнан. Брат с сестрой
стали ссориться, и чтобы помирить их, собрался совет богов. Его вер-
157
дикт был следующим: только хлебом можно насытиться, ибо он укрепля-
ет тело, а вино веселит сердце. Хлеб и вино важнее для жизни, чем мо-
локо и шерсть. Ашнан должна занимать первое место, а Лахар должен
чтить ее и признавать ее старшинство. Эта легенда очень современна: до-
ля вегетарианцев в будущем будет возрастать, так как к середине сле-
дующего столетия народонаселение увеличится настолько, что исполь-
зование зерна на корм скоту будет практически прекращено.
Откуда на земле рис. У жителей острова Калимантан есть забавная
легенда о появлении риса. В древности он был пищей богов и отведать
его могли лишь немногие счастливцы, которые побывали в гостях на
небе. Но боги зорко следили, чтобы гости не унесли с собой его семе-
на. Похитить рис у богов удалось хитрому юноше: он вынес несколько
бесценных зерен в трещинах своих пяток.
8.5. Семейство Агесасеае (Palmaceae) — Арековые
(Пальмовые)
Паспорт семейства
Родов - 210 , видов - более 3 000
Цветок - * Caj С03 А3+3 G(3>
Распространение - тропики, немногие виды - в субтропиче-
ских и умеренно теплых областях
Жизненная форма «одревесневшие травы» (высотой от
нескольких сантиметров до 70 м), есть лианы
Опыление - насекомыми и ветром
Плоды - разнообразные (сочные и ярко окрашенные). Рас-
пространяются животными, водой, человеком
Важнейшие роды - Cocos, Phoenix
Культурные растения - Arenga, Cocos, Phoenix
Карл Линней назвал пальмы (рис. 33) принцами раститель-
ного мира. Действительно, это удивительно красивые царствен-
ные растения, обычные в тропическом и субтропическом поясах
в составе самых различных сообществ, включая болота-мангры
в эстуариях рек. Они входят также в состав тропических лесов,
причем лазающие лианы из рода каламус (Calamus) имеют дли-
ну до 180 м.
Характерный облик позволяет почти безошибочно отличить
большинство пальм от других растений: прямой неветвящийся
одревесневший стебель с кроной крупных веерных или пери-
стых листьев на верхушке. Листья ясно расчленены на черешок
и пластинку. Как и все другие однодольные, пальмы не имеют
камбия, и в их стебле нет настоящей древесины. Стебель дости-
гает значительной толщины за счет первичного роста, он состо-
158
ит из корового слоя и многочисленных проводящих пучков и
волокон, рассеянных в основной паренхиме. Кроме того, утол-
щение ствола может происходить за счет деления и растяжения
клеток паренхимы.
На вершине пальмы находится верхушечная почка - так на-
зываемая пальмовая капуста, хорошо защищенная черешками
взрослых листьев. Размер листьев с черешком может дости-
гать 25 м.
Цветки раздельнополые, но большинство пальм - однодом-
ные растения. Многочисленные цветки собраны в бокоцветные
соцветия - это метелки с колосовидными, сережковидными или
початковидными ветвями. Несмотря на то, что цветки невзрач-
ные, как правило, они опыляются насекомыми, причем круг
опылителей достаточно широк.
Рис. 33. Пальмы: а - Phoenix dactylifera (1 - общий вид растения, 2 -
плод, 3 - семя); б - Lodoicea maldivica (1 - общий вид растения, 2 -
общий вид плода и эндокарп с семенем); в - Cocos nucifera (1 - общий
вид растения, 2 - плод в разрезе)
Среди пальм есть настоящие монокарпики, которые всю
жизнь готовятся к единственному цветению и после этого пол-
ностью отмирают. Так, корифа высокая (Corypha elata) зацвета-
ет в возрасте 30-40 лет, образуя соцветие до 4,5 м длины из
10 млн цветков. Общая сухая масса плодов, созревающих в те-
чение 18 месяцев, составляет 600 кг. У саговой пальмы (Metro-
xylon sagu) вегетативный рост продолжается 10-15 лет, а цве-
тение - около 2 лет, причем верхушечное метельчатое соцветие
достигает в длину 5-7 м.
159
Плоды пальм сочные и ярко окрашенные, распространяются
птицами, грызунами, летучими мышами и даже слонами. У неко-
торых пальм, в том числе кокосовой (Cocos nucifera), плоды рас-
пространяются морскими течениями. Без потери всхожести они
могут путешествовать до 4 месяцев и проплывать до 5 000 км.
Рассмотрим самые знаменитые пальмы.
Финиковая пальма (Phoenix dactylifera) - важнейшая плодо-
вая культура аридных районов Ближнего Востока и Африки.
Это «благословенное дерево», «королева пустыни». Произра-
стает в оазисах и, по арабской поговорке, эта красавица «купа-
ет ноги в воде, а прекрасную голову держит в огне солнца».
Финиковая пальма - хороший индикатор близкого расположе-
ния уровня грунтовых вод. В диком состоянии неизвестна. В
России широко культивируется в оранжереях и в больших по-
мещениях как декоративное растение.
Кокосовая пальма (Cocos nucifera) входит в число важней-
ших растений Океании. Ее стройные (до 30 м высотой) стволы
увенчаны кронами листьев и составляют обязательный элемент
ландшафтов тихоокеанских побережий.
Из незрелых кокосовых «орехов» получают напиток (коко-
совое молоко), свежий эндосперм используется в пищу (из него
изготовляют различные блюда и лакомства), из высушенного
эндосперма - копры - получают кокосовое масло. Волокна из
мезокарпа плодов - материал для изготовления прочных вере-
вок и канатов, циновок, ковров и щеток, а из твердого эндокар-
па делают посуду, пуговицы, различные поделки.
Из сладкого сока молодых соцветий получают сахар, вино,
спирт, уксус. Стволы используют для постройки жилищ, изго-
товления мебели, как топливо. Листья - материал для покрытия
крыш, плетения корзин, шляп, вееров.
Масличная пальма (Elaeis guineensis) в естественных услови-
ях обитает по берегам рек и озер, во влажных и болотистых
долинах, на опушках леса. Земледельцы, вырубая и расчищая
лес, способствовали ее расселению. И сейчас на больших пло-
щадях Африки она стала «пальмой покинутых стоянок челове-
ка». Масличная пальма особенно ценна тем, что дает два сорта
масел: из семян - пищевое масло, сходное с кокосовым, из мя-
коти плода - масло, богатое каротиноидами, оно используется
для его получения и в производстве мыла, свечей, а также как
смазочное. Из сладкого сока соцветий пальмы изготавливают
вина (за сезон одно растение может дать 240 л сока).
Винная пальма (Caryota urens) растет в Индии, Бирме, Не-
пале. Экономически важное растение, из сока соцветий которо-
160
го получают вино и сахар, а из ствола добывают саго. Эпитет
«urens» - жгучая - связан с обжигающей мякотью плодов, в
которой присутствуют многочисленные игловидные кристаллы
оксалата кальция. Эта пальма монокарпическая, она зацветает в
возрасте 12-15 лет и непрерывно цветет в течение 5-7 лет, по-
сле чего погибает.
Сахарная пальма (Arenga pinnata) издавна культивируется
по всей тропической Азии ради сока, получаемого из мужских
соцветий. В течение 2,5 месяцев с одного соцветия собирают в
сутки до 7 л сока, содержащего около 20% сахара, который до-
бывают выпариванием. Ферментацией сока получают вино,
спирт, уксус. Прочное и не гниющее в воде волокно из листо-
вых влагалищ - другой важный промышленный продукт сахар-
ной пальмы.
Сейшельская пальма (Lodoicea maldivica) - реликт Сейшель-
ских островов. Основание ствола пальмы имеет форму лукови-
цы, которая сидит в чаше диаметром около 80 см и глубиной до
0,5 м. Чаша имеет множество отверстий с трубчатыми отрост-
ками наружу, через которые корни пальмы проникают в землю.
Такие гнезда прочны и сохраняются столетиями. Уникальны и
созревающие 7-10 лет крупные плоды (до 18 кг, а иногда и 45
кг) с двулопастным эндокарпом. Семя такого плода прорастает
1-1,5 года, а молодое растение получает питательные вещества
из эндосперма в течение 3-4 лет.
По значимости для человека в тропическом поясе пальмы
уступают только злакам.
На Кавказе и в Крыму для декоративных целей использу-
ют более 20 видов пальм. Наиболее выносливы из них и хорошо
зимуют в открытом грунте трахикарпус (Trachycarpus fortunei)
и хамеропс (Chamaerops humilis). В комнатном цветоводстве вы-
ращивают ливистону (Livistona), рапис (Rhapis), сабаль (Sabal),
финиковую пальму, хамеропс и др.
Это интересно
Самые древние консервы. Малайцы и палинезийцы, отправляясь в
длительные морские путешествия, брали с собой кокосовые орехи. Это
был надежный запас пищи и питья в прекрасной естественной упаков-
ке. Туземцы не только питались кокосовыми орехами, но и высаживали
их на всех островах, которые посещали.
Крабы на деревьях. Самым удивительным фитофагом можно счи-
тать краба «пальмовый вор» (Birgus latro), который питается мякотью
незрелых плодов кокосовой пальмы. Краб разрывает волокна мощными
клешнями, пробивает отверстие и через него вытаскивает мякоть. Ино-
гда он разбивает эндокарп ударами о камни. Он даже залезает на
161
пальму, сбивая кокосовые орехи. Химическое исследование его жира
показало, что он напоминает кокосовое масло и имеет мало общего с
животным жиром.
Древнейшая жвачка. Бетелевая пальма (Агеса catechu) одно из
самых важных экономических растений Старого света. Ее семена со-
держат танины и алкалоиды, из которых арекоидин имеет токсические
свойства. Тонко нарезанные ломтики семян входят в состав бетелевой
жвачки вместе с бетелем (Piper betle) и гашеной известью. Эту жвачку
используют миллионы людей в тропической Азии и Африке. При же-
вании обильно выделяется слюна и вся полость рта окрашивается в
кроваво-красный цвет. Ради семян бетелевую пальму широко культи-
вировали с древних времен. В диком состоянии она неизвестна.
ЧАСТЬ 4
ОСНОВЫ НАУКИ О РАСТИТЕЛЬНОСТИ
В природе растения сообитают в сложных ансамб-
лях - популяциях, растительных сообществах и флорах, объединенных
в первую очередь условиями среды. Эта часть знакомит читателей с
основными положениями широкого междисциплинарного комплекса
наук об отношениях растений к условиям среды, называемого экологи-
ческой ботаникой, геоботаникой, или наукой о растительности (НОР)
(табл. 15).
Формы жизни на уровне ниже организма (тканевом, клеточном,
субклеточном) наукой о растительности не изучаются.
Таблица 15
Структура междисциплинарного комплекса
«наука о растительности»
Уровень организации объекта ис- следований Наука Объекты изучения
Организ- менный Экологичес- кая физиоло- гия растений Физиологические механизмы при- способления растений к экологиче- ским условиям
Популяци- онный Популяци- онная биоло- гия растений Популяции растений: их размер, генетическое и фенетическое разно- образие, пространственная структу- ра и механизмы регулирования плотности
Видовой Экология растений, бо- таническая география Жизненные формы, закономерно- сти распределения видов по различ- ным местообитаниям, географиче- ские ареалы, типы поведения (стра- тегий) видов
Фитоцено- тический Фитоцено- логия Природа растительных сообществ, их видовой состав, структура (мор- фология, фенология), продуктив- ность, экология, динамика, геогра- фия. Классификация растительных сообществ
Инфрацено- тический Флористика Флоры
163
Наука о растительности очень обширна и при ее изучении в соста-
ве курса «Высшие растения» студенты знакомятся лишь с некоторыми
ее разделами-науками: основными положениями экологии видов, попу-
ляций, сообществ, флор. Вопросы экологической физиологии растений
не рассматриваются.
Глава 9
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
9.1. Классификация экологических факторов
Экологические факторы - это условия внешней среды, су-
щественно влияющие на живые организмы.
Они делятся на абиотические, т.е. факторы неорганиче-
ской, или неживой, природы, и биотические - порожденные
жизнедеятельностью организмов. Совокупность абиотических
факторов в пределах однородного участка называется экото-
ном, вся совокупность факторов, включая биотические, - био-
топом.
К абиотическим факторам относятся:
климатические - свет, тепло, воздух, вода (включая
осадки в различных формах и влажность воздуха);
эдафические, или почвенно-грунтовые, - механический и
химический состав почвы, ее водный и температурный режим;
топографические - условия рельефа.
Климатические и эдафические факторы во многом опреде-
ляются географическим положением экотопа его удаленно-
стью от экватора и от океана и высотой над уровнем моря.
Прямые экологические факторы непосредственно влияют на
растения (увлажнение, температура, богатство почвы питатель-
ными элементами и др.).
Косвенные экологические факторы действуют на растения
опосредствованно - через прямые факторы среды, влияя на их
значения. Примеры косвенных факторов: географическая широта
и удаленность от океана, рельеф (высота над уровнем моря и
экспозиция склона), гранулометрический состав почвы. С подъе-
мом в горы изменяется климат (количество осадков и темпера-
турный режим); экспозиция и крутизна склона влияют на ин-
тенсивность прогревания поверхности почвы и режим ее увлаж-
нения, а гранулометрический состав почвы - на растения через
режим увлажнения и динамику питательных элементов и т.д.
164
9.2. Комплексные градиенты
Группа экологических факторов, которые изменяются со-
пряженно, называется комплексным градиентом. Р.Уиттекер пи-
сал, что нет экологических факторов, которые не объединялись
бы в комплексные градиенты.
Комплексные градиенты, как правило, формируются кос-
венными факторами, подобными высоте над уровнем моря или
географической широте. В них могут объединяться и прямодей-
ствующие экологические факторы. Например, повышение ин-
тенсивности выпаса вызывает уплотнение почвы, а на влажных
почвах в степных районах - и их засоление за счет усиления
капиллярного подъема воды, несущей соли к поверхности поч-
вы. Изменение увлажнения влияет на биохимические процессы,
протекающие в почве, и активность различных групп микроор-
ганизмов, которые осуществляют гумификацию или минерали-
зацию органического вещества. Изменение температуры почвы
также может вызвать цепную реакцию процессов изменения
режима увлажнения и физико-химических преобразований.
9.3. Ведущие экологические факторы
Комплексные градиенты, которые влияют на состав и
структуру фитоценозов в большей степени, чем другие, назы-
ваются ведущими. Ведущие комплексные градиенты всегда
включают в свой состав лимитирующие факторы среды, т.е. те
факторы, которые находятся в максимуме или минимуме и по-
тому сильнее прочих воздействуют на распределение и развитие
растений.
Так, в тундровой зоне основу ведущего комплексного гра-
диента составляет количество тепла, поскольку влаги там дос-
таточно, а обеспеченность элементами питания зависит от теп-
ла. Чем теплее субстраты, тем активнее идет в них процесс об-
разования гумуса и меньше накапливается неразложившихся
остатков растений.
В зоне тайги основу ведущего комплексного градиента
составляет обеспеченность почв питательными элементами.
Почвы, образующиеся на карбонатных породах, богатых каль-
цием и другими минеральными элементами, позволяют фор-
мироваться сообществам с высокой продуктивностью. Однако в
южной части зоны тайги возрастает роль второго ведущего
комплексного градиента, связанного с увлажнением.
В лесостепной и степной зоне ведущий комплексный гра-
диент объединяет экологические факторы, связанные с увлаж-
нением, так как здесь почвы (черноземы) богаты питательными
165
элементами. На этот комплексный градиент может наклады-
ваться влияние выпаса (как отмечалось, выпас - это также ком-
плексный градиент), а в южной части степной зоны - ком-
плексного градиента засоления почвы.
9.4. Биотические факторы
Биотические факторы - следствие взаимоотношений орга-
низмов, для растений - это конкуренция, влияние животных
(фитофаги, паразиты, опылители, распространители плодов и
семян), грибов (микоризные, паразитические) и микроорганиз-
мов (азотфиксирующие и болезнетворные бактерии, вирусы).
Более подробно влияние биотических факторов рассматривает-
ся в п. 12.1.2 и 12.1.3.
9.5. Антропогенные факторы
Факторы, связанные с влиянием человека, выделяются в от-
дельную группу антропогенных факторов. В настоящее время
роль антропогенных факторов резко возросла, и потому изуче-
ние последствий их влияния и разработка способов регулирова-
ния отношений человека и природы являются важнейшими
проблемами НОР. Более подробно этот вопрос рассматривает-
ся в п. 12.3.3.
Глава 10
ЭКОЛОГИЯ ВИДОВ
10.1. Основные подходы при изучении экологии видов
Экология видов - широкое понятие, которое включает мно-
го признаков, характеризующих отношение растений к услови-
ям среды. Главные из них:
жизненная форма - отражение внешних условий в морфо-
логии видов;
распределение видов по осям (градиентам) экологических
факторов - увлажнения, богатства почвы, засоления, освещен-
ности, реакции почвенного раствора, климатических факторов
(в первую очередь температурного режима);
география видов, т.е. ареалы, которые они занимают;
типы стратегии (поведения) видов, обеспечивающие их ус-
тойчивость в разных экологических условиях.
Экология каждого вида индивидуальна, нет двух видов, ко-
торые были бы одинаково распределены на градиентах экологи-
ческих факторов, имели бы идентичную жизненную форму и
полностью совпадающие географические ареалы. Нет двух ви-
166
дов с абсолютно одинаковой стратегией жизни. Открытие этого
феномена в начале текущего столетия связано с именами рос-
сийского ученого Л.Г.Раменского и американского ботаника
Г.Глизона.
Однако из соображений удобства виды со сходной эколо-
гией (жизненной формой, ареалом, стратегией) объединяют в
типы. Эта операция в науке называется идеализацией, или ре-
дукцией (сведением сложного к простому). Полезность ее мож-
но проиллюстрировать бытовым примером: несмотря на инди-
видуальность формы тела человека, для массового пошива раз-
работана система размеров одежды и обуви.
В дальнейшем мы будем оперировать именно такими упро-
щенными системами типов по каждому из признаков, характе-
ризующих экологию видов. Однако следует помнить об услов-
ности этих типов и неизбежности существования переходов
между ними.
10.2. Жизненные формы
Жизненная форма это внешний облик организма, ком-
плекс морфологических, анатомических, физиологических и по-
веденческих признаков, в котором отражается его приспособ-
ленность к условиям внешней среды. В сходных условиях среды
организмы даже из систематически далеких групп могут иметь
сходную жизненную форму (например, суккулентные кактусы и
молочаи, растения-подушки из бобовых, сложноцветных, гвоз-
дичных, зонтичных и др.).
Наиболее популярная система жизненных форм, предло-
женная К.Раункиером еще в начале столетия, сыграла в эколо-
гии не меньшую роль, чем работы К.Линнея в области система-
тики растений. Датский исследователь свел все разнообразие
жизненных форм к небольшому числу их макротипов, выде-
ленных всего по одному критерию - положению и способу за-
щиты почек возобновления в течение неблагоприятного периода
(холодного или сухого).
Типы жизненных форм Раункиера хрестоматийны и вошли
не только в экологическую и фитоценологическую литературу,
но и практически во все энциклопедические справочники и сло-
вари по биологии (рис. 34). Кратко прокомментируем их.
Фанерофиты (деревья, кустарники, эпифиты). Почки возоб-
новления расположены высоко над землей и защищены специ-
альными чешуями. Эта жизненная форма по высоте разделяет-
ся на ряд вариантов (мега-, мезо-, микро- и нанофанерофиты).
Хамефиты (кустарнички, полукустарнички, стелющиеся
растения, например клюква, брусника). Почки возобновления
167
Рис. 34. Жизненные формр растений по Раункиеру: 1 - фанерофит; 2 - хаме-
фиты; 3 - гемикриптофит; 4 - геофиты; 5 - терофит; 6,7 - гидрофиты (чер-
ным выделены зимующие части)
Таблица 16
Спектры жизненных форм флор основных биомов мира
(в %, по Уиттекеру, 1980)
Биомы Фане- рофиты Хаме- фиты Гемик- рипто- фиты Геофиты Терофи- ты
Тропический до- ждевой лес 96 2 0 2 0
Субтропический лес 66 17 2 5 10
Лес умеренно те- плой зоны 54 9 24 9 4
Лес умеренно холодной зоны 10 17 54 12 7
Тундра 1 22 60 15 2
Дубовое редколе- сье 30 23 36 5 6
Степь 1 12 63 10 14
Полупустыня 0 56 14 0 30
Пустыня 0 4 17 6 73
Глобальный, или нормальный, спектр 46 9 26 6 13
168
расположены не выше 20-30 см от почвы, так что зимой поми-
мо чешуи они защищены снегом.
Гемикриптофиты (большинство луговых и степных трав).
Почки возобновления расположены на уровне почвы и допол-
нительно защищены мертвым растительным опадом.
Криптофиты. Разделяются на геофиты, у которых почки
возобновления находятся в земле (луковичные, корневищные,
клубневые растения), и гидрофиты - водные растения.
Терофиты. Однолетники, которые зимуют в стадии семян.
Спектры жизненных форм отражают разнообразие эколо-
гических условий, в которых сформировалась растительность.
В экологии используется понятие типовых спектров жиз-
ненных форм, которые отражают особенности растительности
крупных территориальных единиц (биомов, см. п. 12.4.2). При-
меры таких спектров приведены в табл. 16.
Существуют и другие классификации жизненных форм рас-
тений. Так, по форме роста они разделяются на травы, кус-
тарнички, кустарники, деревья, а травы в свою очередь - на
корневищные (короткокорневищные - Роа pratensis и длинно-
корневищные - Agropyron repens), рыхлокустовые (Festuca
pratensis), плотнокустовые (Nardus stricta, Festuca valesiaca),
стержнекорневые (Rumex confertus), кистекорневые (все одно-
дольные) и т.д.
10.3. География видов
Географические ареалы видов индивидуальны. Практически
нет двух видов, которые имели бы полностью совпадающие
ареалы. Ареалы могут варьироваться по размеру (от космопо-
литных - у всюду встречающихся видов, до небольших - у эн-
демиков, распространенных на ограниченной территории, часто
имеющей площадь в несколько квадратных километров), быть
сплошными или разорванными (дизъюнктивными). Всеобщим
правилом является низкая устойчивость видов к воздействию
человека у границ ареалов. В этих условиях виды становятся
угрожаемыми и включаются в региональные Красные книги.
Универсальной системы ареалов нет. Автор одной из
лучших книг об ареалах растений А.И.Толмачев писал: «Опыт
создания некоторых основ формальной «системы ареалов» и ее
«наглядного» отображения при помощи формул свидетельству-
ет, по-видимому, о практической невозможности систематиза-
ции данных об ареалах, не подчиненной конкретным ботани-
ко-географическим задачам. Видимо, для сопоставления основ-
169
ных черт распространения растений в общеземном плане
нужны одни основы классификации, для рассмотрения ареалов
в деталях, в региональном плане - другие» (1974, с. 108).
По этой причине анализ ареалов видов - очень сложная за-
дача, для изучения географических закономерностей распреде-
ления видов на евразийском материке выявляют их отношение к
широтной зональности и географическим секторам, на которые
меридионально разделяется этот материк.
Для решения первой задачи используется система широт-
ных поясов, с разделением видов на:
арктоальпийские (в эту группу включаются виды, распро-
страненные в Арктике, а также в горных тундрах);
бореальные (например, Oxalis acetosella, Trientalis europaea,
Vaccinium vitis-idaea, V. myrtillus);
неморальные (например, Asarum europaeum, Stachys offici-
nalis, Cacalia hastata);
лесостепные (например, Centaurea jacea, Coronilla varia,
Fragaria vesca);
степные (например, Festuca valesiaca, Falcaria vulgaris, Hor-
deum brevisubulatum);
пустынные (например, виды рода Haloxylon);
плюризональные (т.е. распространенные в нескольких ши-
ротах; к этой группе относятся виды специфических местооби-
таний - водных, сегетальных и рудеральных; например, Alisma
plantago-aquatica, Avena fatua, Arctium lappa).
Во втором случае используется система долготных поясов с
разделением видов на западно-европейские, центрально-евро-
пейские, восточно-европейские, сибирские, дальневосточные,
северо-американские и голарктические (космополитные) виды.
К последним относятся виды, распространенные на евразийском
и североамериканском континентах (например, Hippuris vulgaris,
Inula britannica, Equisetum fluviatile).
Распространение видов в долготном направлении охватыва-
ет обычно несколько долготных секторов (такие виды называ-
ются плюрирегиональными). Так, виды Asarum europaeum и As-
paragus officinalis распространены во всей Европе; Stachys offici-
nalis и Bolboschoenus maritimus в Европе и Азии; Galium
aparine и Glechoma hederacea - в Европе и Сибири и т.д.
Кроме того, используется система высотных поясов с под-
разделением видов на равнинные, субмонтанные, монтанные
(т.е. горные, с разделением их на субальпийские, альпийские
и нивальные).
170
10.4. Экологические группы видов
В экологические группы объединяются виды со сходным
отношением к условиям среды. Поскольку, как мы уже говори-
ли, виды на градиентах факторов среды распределены индиви-
дуально, любая система экологических групп условна, но доста-
точна, чтобы ботанику было удобно характеризовать отношения
видов к разным факторам среды.
Так, по отношению к увлажнению все виды разделяются на
группы:
гидрофиты - водные растения (Nymphaea alba, Nuphar lu-
tea, Elodea, Lemna и др.);
гигрофиты (или гелофиты) - группа «земноводных» расте-
ний, обитающих по береговым и прибрежным местообитаниям с
избыточным или переменным увлажнением (Sagittaria
sagittifolia, Alisma plantago-aquatica, Butomus umbellatus, Caltha
palustris, Scirpus sylvaticus);
мезофиты - растения нормально увлажненных почв (это
луговые и лесные травы, а также кустарники и деревья (Bro-
mopsis inermis, Trifolium pratense, Geranium pratense, Aegopo-
dium podagraria, Convallaria majalis, Tilia cordata, Corylus avel-
lana);
ксерофиты - растения сухих почв (Stipa pennata, Festuca
valesiaca, Veronica incana, Salvia stepposa).
Система экологических групп по фактору увлажнения мо-
жет быть и более детальной, к пяти перечисленным группам до-
бавляются дополнительные группы, представляющие переходы
между ними: гигрогидрофиты и гидрогигрофиты, мезогигрофи-
ты и гигромезофиты, мезоксерофиты и ксеромезофиты. Для
отнесения видов к экологическим группам могут быть исполь-
зованы как прямые наблюдения в природе, так и достаточно
сложные приемы обработки полевых учетов или специальные
справочники.
Между системами экологических групп и жизненных форм
нет полного соответствия, поскольку к одним и тем же услови-
ям растения могут приспосабливаться по-разному. Так, среди
ксерофитов есть кустарники с жесткими листьями, листовые и
стеблевые суккуленты, плотнокустовые злаки, эфемеры и гео-
фиты-эфемероиды.
Нет прямого соответствия и между экологическими группа-
ми и распределением видов по широтным зонам. Так, в степной
зоне во влажных логах наряду с ксерофитами растут мезо- и
гигрофиты, а в водоемах - гидрофиты. В лесной зоне по юж-
ным хорошо прогреваемым меловым склонам могут произра-
стать ксерофиты и т.д.
171
Аналогично тому, как виды разделяются на группы по от-
ношению к фактору увлажнения, они могут быть подразделены
на группы и по отношению к другим факторам.
Так, по отношению к богатству почвы питательными эле-
ментами выделяют группы видов:
эутрофы - растения богатых почв (Festuca pratensis, Dacty-
lis glomerata, Trifolium pratense);
мезотрофы - растения умеренно богатых почв (Festuca ru-
bra, Trifolium hybridum, Alchemilla vulgaris);
олиготрофы - растения бедных почв (Nardus stricta, Anto-
xanthum odoratum).
По отношению к засолению почвы выделяются экологиче-
ские группы видов:
галофиты - растения засоленных почв (Salicornia herbacea,
Suaeda corniculata);
мезогалофиты - растения умеренно засоленных почв (Tri-
folium fragiferum, Cirsium esculentum, Triglochin maritimum);
гликофиты - растения незасоленных почв (см. эутрофы).
По отношению к освещенности различаются сцио- и гелио-
фиты. Растения кислых почв выделяются в группу ацидофилоб,
растения известковых почв калъцефилов, а каменистых
петрофитоб. Растения почв, богатых азотом, называются нит-
рофилами и т.д.
10.5. Стратегии видов растений
Тип стратегии (тип поведения) - одна из самых важных ха-
рактеристик вида. В стратегиях отражаются отношения видов к
абиотическим условиям среды, к биотическим факторам и к на-
рушениям.
Понятие экологической стратегии появилось в литературе в
50-е годы минувшего столетия и вначале применялось к живот-
ным. И только в 60-е годы начали говорить о стратегиях расте-
ний, хотя это явление, без использования термина «стратегия»,
изучалось еще в конце XIX в.
Прежде чем познакомиться с системами стратегий, которые
используются в НОР, необходимо усвоить содержание двух важ-
ных экологических понятий - конкуренция и экологическая ниша.
10.5.1. Конкуренция
Конкуренция — важный биотический фактор, оказывающий
влияние на растения. Это соревнование организмов за потреб-
ление ресурса, находящегося в дефиците. Между растениями
озера (Potamogeton crispus, Stratiotes aloides или Nymphaea alba)
172
нет конкуренции за воду, но степные растения (Stipa pennata,
Festuca valesiaca) борются за этот ресурс. В разреженных со-
обществах пустынных растений нет конкуренции за свет, но
злаки в густом травостое пойменного луга находятся в состоя-
нии острой конкуренции за него. Роль конкуренции как основ-
ного механизма естественного отбора отмечал Ч.Дарвин.
Различают несколько вариантов конкуренции.
Внутривидовая и межвидовая конкуренция. Принципиаль-
но эти два варианта конкуренции не различаются, хотя конку-
ренция между растениями одного вида в большинстве случаев
более мягкая, чем между растениями разных видов.
Симметричная и асимметричная конкуренция. Симметрич-
ная конкуренция возникает между растениями с примерно оди-
наковыми конкурентными способностями (например, между
Tilia cordata и Quercus robur в смешанном лесу), а асиммет-
ричная - между растениями с разными конкурентными способ-
ностями (например, между процветающими и угнетенными эк-
земплярами деревьев одного или разных видов). Обычно со
временем асимметричность внутривидовой конкуренции усили-
вается, и более слабый конкурент в конце концов погибает.
Диффузная конкуренция. Это борьба растений нескольких
видов одновременно за несколько ресурсов, основной тип кон-
куренции в природе. Ее можно сравнить с взаимоотношениями
пассажиров в переполненном автобусе, которые одновременно
толкают друг друга.
4
Конкуренции
нет
Увеличение богатства почвы
Увеличение освещенности
Рис. 35. Основной комплексный градиент конкуренции
(по Tilman, 1988)
173
Парциальная и интегральная конкуренция. Парциальной на-
зывается конкуренция за один фактор среды, а интегральной -
за несколько. На богатых почвах формируются сомкнутые тра-
востои, и растения конкурируют главным образом за свет,
причем, чем богаче почвы, тем острее конкуренция. На бедных
почвах (сухих или засоленных, на которых поступление воды и
питательных элементов в растения затруднено) формируются
разомкнутые травостои, и растения конкурируют не за свет, а
за почвенные ресурсы. В условиях среднего богатства почвы
растения одновременно конкурируют за элементы почвенного
питания и свет (рис. 35).
10.52. Экологическая ниша
Конкуренция - основной тип взаимоотношений растений при
их совместном произрастании. Уровень ее может быть снижен,
если у сообитающих видов будут разные экологические ниши.
До сих пор нет общепринятого определения понятия
«экологическая ниша». Ю.Одум (E.Odum) назвал ее «профес-
сией вида в экосистеме». Если рассматривать экосистему как
совокупность организмов и условий среды, в результате дея-
тельности которых производится органическое вещество, то
«профессия» каждого вида будет определяться тем, из какого
материала, в какое время, на какой территории и какое орга-
ническое вещество он производит.
Понятие ниши пришло в НОР из зоологии, благодаря ра-
ботам Дж.Гринелла (J.Grinell), Ч.Элтона (C.Elton) и Г.Хатчин-
сона (G.Hutchinson). Понимание ниши применительно к живот-
ным достаточно простое, так как различия положения их по-
пуляций в экосистеме очевидны: разные животные питаются
разной пищей и занимают разные территории. У растений
один тип пищи (водный раствор минеральных веществ, угле-
кислый газ и солнечный свет). Поэтому до начала 60-х годов
понятие экологической ниши применительно к растениям не
использовалось. Однако постепенно оно стало обычным и для
НОР, так как одинаковые ресурсы разные виды растений ис-
пользуют по-разному. Д.Тильман (D.Tilman) даже говорит о
том, что каждый вид растений имеет «свой тип питания».
В понятие «тип питания» включаются следующее признаки:
разный тип распределения корневых систем. В пустыне
Гоби, к примеру, есть сообщества из тростника (Phragmites
australis) и саксаула (Haloxylon ammodendron). Первый вид
гигрофит, который связан с грунтовыми водами на глубине 8-
10 м, второй - типичное пустынное дерево с приповерхност-
ным укоренением в сухих горизонтах почвы. В.Г.Онипченко
174
описал сообщества пустошей в высокогорьях Кавказа, где са-
мый верхний слой почвы занимают лишайники, использующие
как источник воды и минеральных элементов атмосферные
осадки, а в более глубоком горизонте расположена сплошная
сетка из корней злаков;
разное время вегетации. В период цветения и плодоноше-
ния растениям требуется больше ресурсов, и цветение в раз-
ное время снижает конкуренцию;
разная требовательность к свету. В лесу одни виды при-
урочены к затененным местам (Asarum europaeum, Paris
quadrifolia), а другие - к светлым прогалинам (Dactylis glome-
rata). Виды подлеска (Sorbus aucuparia, Corylus avellana, Euony-
mus verrucosa) довольствуются меньшим количеством света, чем
виды высокого древесного яруса;
разная требовательность к элементам минерального пита-
ния. Например, бобовые могут практически не получать азота из
почвы, так как в их корнях живут азотфиксирующие бактерии,
снабжающие их азотом, но азот необходим злакам.
Растения различаются также по взаимоотношениям с на-
секомыми-опылителями, фитофагами и по отношению к другим
факторам. Таким образом, экологическая ниша вида - это со-
вокупность занимаемого им пространства, потребностей в ре-
сурсах и временного ритма «работы по профессии».
Различают три вида ниши - фундаментальную, реализо-
ванную и регенерационную.
Фундаментальная - это та ниша, которую может занять
вид при отсутствии конкуренции. Фундаментальные ниши очень
широкие, поэтому в ботанических садах успешно выращивают
растения из разных районов земного шара: в этих условиях они
защищены от конкуренции с видами местной флоры.
Реализованная ниша - это часть фундаментальной, которую
вид занимает при наличии конкуренции с прочими видами.
Классическими примерами, показывающими различия
фундаментальной и реализованной ниш, являются опыты по
удобрению естественных лугов и искусственных чистых посевов
тех видов, которые слагают эти луга. При внесении минераль-
ных удобрений на луга лесной зоны (они сформировались после
расчистки леса на бедных подзолистых почвах) из травостоя
выпадают типичные для таких лугов олиготрофные виды - по-
левица тонкая (Agrostis tenuis) и душистый колосок (Antho-
xanthum odoratum) и разрастаются эутрофы, бывшие ранее
второстепенными видами с небольшой долей количественного
участия в травостое - ежа сборная (Dactylis glomerata) и пырей
ползучий (Elytrigia repens).
175
Однако и полевица тонкая, и душистый колосок в искус-
ственном чистом посеве положительно реагируют на внесенные
удобрения - их урожайность увеличивается в 2-4 раза. Объяс-
нить такое, на первый взгляд удивительное, явление можно,
зная различия фундаментальной и реализованной ниш этих ви-
дов. При отсутствии конкуренции виды бедных лесных почв
хорошо растут в условиях богатого минерального питания, а
при наличии конкуренции они вытесняются злаками богатых
почв. Иными словами, их фундаментальная ниша охватывает и
бедные, и богатые почвы, а реализованная - только бедные
почвы, на которых они, благодаря биологическим и экологи-
ческим особенностям, являются сильными конкурентами.
Аналогично, степные злаки-ксерофиты - ковыли (р. Stipa) и
типчак (Festuca valesiaca) - в чистом посеве при поливе дают
урожай выше, чем на сухом степном черноземе. Их реализован-
ная ниша охватывает условия засушливой степной зоны, где
выжить им помогает мощная корневая система, узкие листья,
опушение и другие приспособления для экономного расходо-
вания воды. При отсутствии конкуренции степные злаки хоро-
шо растут при обильном увлажнении, а в природе в таких ус-
ловиях они не встречаются, так как не выдерживают конкурен-
ции с приспособленными к увлажнению луговыми видами.
Рис. 36. Формирование бимодального распределения вида
на градиенте среды при появлении сильного конкурента
На рис. 36 показано формирование реализованной ниши за
счет вытеснения ценотически сильным видом более слабого ви-
да из оптимальных условий местообитания.
Регенерационная ниша - это набор условий, позволяющий
развиться из семени новому растению. Например, регенераци-
онная ниша тополей и ив включает свежие влажные отложения
аллювия вдоль берега реки. Если на аллювии сформируется
травяной покров, то легкие семена этих растений не смогут
достигнуть почвы и прорасти.
176
10.5 .3. Типы стратегий растений Маклиода — Пианки
В 1884 г. Дж. Маклиод (J.MacLeod) разделил все растения
на «пролетариев» и «капиталистов». К «пролетариям» он отнес
растения-малолетники, которые зимуют в виде семян, а к «ка-
питалистам» - растения, зимующие с капиталом органического
вещества - клубнями, толстыми стеблями, корневищами и т.д.
Спустя 70 лет Р.Макартур (R.McArthur) и Е.Уилсон (E.Wil-
son) независимо открыли типы стратегий Маклиода, описав
их как два типа отбора у видов - r-отбор и К-отбор. Позднее
представления об этих типах отбора подробно разработал
Э.Пианка (1981):
r-отбор - это эволюция организмов в направлении увели-
чения затрат на размножение;
К-отбор — это эволюция организмов в направлении увели-
чения затрат на поддержание жизни взрослой особи.
Несмотря на то, что стратегии г- и К-отбора были разра-
ботаны для животных, они в целом соответствовали стратегиям
растений - «пролетариев» и «капиталистов». Таким образом,
система стратегий растений Маклиода - Пианки основывается
на учете одного фактора - роли репродуктивного усилия в вы-
живании. «Пролетарии» образуют большое количество семян и
выживают за счет того, что всегда какая-то часть семян попа-
дает в благоприятные условия. Это растения-однолетники. «Ка-
питалисты» - это деревья и кустарники, они имеют зимующие
органы с запасом органических веществ, с помощью которых
переживают неблагоприятные условия и, являясь многолетни-
ками, многократно дают потомство. Растения с переходным ти-
пом стратегии, например многолетние луговые травы, характе-
ризуются достаточно высокой плодовитостью и умеренной до-
лей многолетних органов.
В целом эта система используется в современной НОР
сравнительно редко.
10.5 .4. Типы стратегий растений Раменского — Грайма
А. Г. Раменский в 1935 г. разделил все виды растений на
три «ценобиотических типа», которые назвал виолентами
(«львами», «силовиками»), патиентами («верблюдами», «вы-
носливцами») и эксплерентами («шакалами»). Его работа оста-
лась незамеченной даже в России, поэтому не удивительно, что
40 лет спустя английский эколог Дж. Грайм практически один к
одному повторил концепцию Раменского.
В отличие от системы Маклиода - Пианки, система Ра-
менского - Грайма двумерна, и предложенные ими типы стра-
тегий отражают отношение растений к двум факторам: к бла-
177
гоприятности условий местообитания и к нарушениям. На
рис. 37 показана эта система типов стратегий в виде так назы-
ваемого «треугольника Грайма». Буквы в углах треугольника
обозначают три первичных типа стратегии, сочетания из двух
и трех букв - переходные, вторичные типы.
Усиление нарушений
Ухудшение условий
винные типы стратегий;
CR, CS, RS, CRS - пере-
ходные (вторичные) типы
Содержание первичных типов стратегий сводится к сле-
дующему:
С (от англ, competitor - конкурент) - виолент, конку-
рент, «лев». Это мощные растения, чаще деревья, кустарники
или высокие травы (например, Phragmites australis в дельтах
рек), которые произрастают в благоприятных условиях (полное
обеспечение водой, элементами питания, теплый климат) при
отсутствии нарушений. Они имеют распростертую крону (или
корневища, как у тростника), за счет которой держат под кон-
тролем условия среды и полностью (или почти полностью)
используют обильные ресурсы таких местообитаний.
Виоленты всегда абсолютно доминируют в сообществах, и
примесь других видов незначительна. Пример типичного вио-
лента - бук (р. Fagus). В буковых лесах под пологом деревьев
сумрачно и почти нет трав и кустарников (такие леса называют
мертвопокровными). В зарослях тростника в дельте Волги био-
масса доминанта составляет 99%, другие виды встречаются
единично. У виолентов реализованная и фундаментальная ниши
совпадают, так как эти растения настолько сильные, что нахо-
дятся вне конкуренции с другими претендентами на эту нишу.
При ухудшении условий (просыхание почвы, засоление и
т.д.) или при нарушениях (рубка леса, высокие рекреационные
нагрузки, пожары, воздействие техники и т.д.) виоленты поги-
178
бают, так как не приспособлены для выживания под воздейст-
вием этих факторов.
Чистый виолент - редкость, в природе чаще распростра-
нены вторичные типы, переходные от виолента к другим типам
стратегий.
S (от англ, stress-tolerant - устойчивый к стрессу) - патиент,
«выносливец», «верблюд». Эти разнообразные растения, оби-
тающие в неблагоприятных условиях (засуха, засоление, дефицит
ресурсов минерального питания, холодный климат и т.д.), обла-
дают специальными морфологическими и физиолого-биохими-
ческими механизмами для преодоления стресса и экономного
расходования ресурсов. Например, кактусы, чтобы избежать вы-
сыхания, фотосинтезируют при закрытых устьицах (углекислый
газ они запасают в более прохладное ночное время).
Тонкие листья сциофитов содержат много хлорофилла.
Галофиты также имеют разные приспособления для выжи-
вания. Растениям солончаков сложно поглощать воду, и потому
для них характерны многие признаки ксерофитов (суккулент-
ность, толстая кутикула, опушение, кроме того - выделение из-
бытка солей на поверхность листьев и др.).
Олиготрофы в тундре имеют многолетние листья, из кото-
рых перед опадением питательные вещества переходят в сте-
бель. Подобным образом и сфагновый мох при неограниченном
росте вверх перекачивает питательные вещества из отмирающей
нижней части в живую.
Для переживания стресса перевыпаса у растений сформи-
ровались приземистые формы, делающие их недоступными па-
сущимся животным, многие из них имеют горький вкус, сни-
жающий поедаемость этих растений.
Фундаментальные и реализованные ниши у патиентов, как
и у виолентов, совпадают, но по иной причине. Если у вио-
лентов это связано с их большой конкурентной способностью,
то у патиентов - с отсутствием других претендентов на такие
сложные условия жизни. Патиенты формируют сообщества
пустынь, солончаков, тундр, высокогорий.
R (от лат. ruderalis - сорный) - эксплерент, рудерал, «ша-
кал». Эти растения замещают виоленты при сильных наруше-
ниях местообитаний. Большинство из них - однолетники (ре-
же - малолетники) с высоким репродуктивным усилием (т.е.
виды-«пролетарии» по терминологии Маклиода). Они способ-
ны формировать банк семян в почве, например, виды полыни
(р. Artemisia), мари (р. Chenopodium), лебеды (р. Atriplex), или
имеют приспособления для распространения плодов и семян,
например, летучки у одуванчика (р. Taraxacum), бодяка
179
(p. Cirsium) или прицепки у липучки (р. Lappula) и лопуха
(р. Arctium), плоды которых разносят животные и человек. Се-
мена некоторых растений распространяются водой. К интенсив-
ному семенному размножению нередко добавляется вегетатив-
ное, например корневищами и корневыми отпрысками у осота
(р. Sonchus).
Таким образом, рудералы первыми начинают восстанав-
ливать растительность при нарушениях: семена одних видов
уже имеются в почвенном банке, семена других быстро достав-
ляются на место нарушения ветром или другими агентами. Эта
важная для природы группа растений, подобно живице на по-
раненном стволе сосны, залечивает нанесенные природе раны.
Кроме типичных эксплерентов, существуют виды с та-
кой же стратегией, которые дают вспышки обилия в ненару-
шенных сообществах в периоды, когда ослаблено конкурентное
влияние со стороны постоянно обитающих в них видов. Пример
таких видов - однолетники-эфемеры, которые после дождей
покрывают зеленым ковром пустыню и за короткий срок закан-
чивают вегетацию. Влияния конкуренции со стороны пустын-
ных кустарников с разреженным пологом и глубокими корне-
выми системами эфемеры практически не испытывают.
Эксплерентами являются и весенние эфемероиды в лесах.
Они проходят основную часть цикла развития до того, как
распустятся листья на деревьях, и таким образом уходят от
конкурентного влияния виолентов.
Реализованная ниша эксплерентов приближается к нулю,
так как они имеют очень слабую конкурентную способность и
исчезают после того, как на нарушенное местообитание попадут
семена и разовьются растения с более высокой конкурентной
способностью.
К эксплерентам относятся все культурные растения (см.
п. 10.5.5) и полевые сорняки. Под влиянием человека, который
стремился очистить от. них поля, сорняки приобрели свойства,
дающие им преимущества в конкуренции с культурными расте-
ниями. Они быстрее отрастают с весны и, имея более разви-
тые и глубокие корневые системы, активнее усваивают воду с
питательными элементами. Другими словами, если за 10 тысяч
лет земледелия человек сделал культурные растения «нежен-
ками», то сорные растения, наоборот, «закалились», что по-
зволяет им сохраняться в посевах. Основной механизм приспо-
собления это огромные банки семян, достигающие сотен
миллионов на одном гектаре поля. Считается, что на одно высе-
янное зерно пшеницы в почве содержится в среднем около 200
семян сорняков.
180
Велика роль эксплерентов в городских и сельских экосисте-
мах, где они расселяются на многочисленных нарушенных и
нарушаемых местообитаниях, тем самым препятствуя развеива-
нию пыли с незакрепленной растительностью поверхности земли
и концентрируя атмосферные загрязнители в своих листьях. Ру-
деральный «бордюр» по краю поля служит пристанищем многим
полезным насекомым из числа «врагов наших врагов».
В табл. 17 приведена сравнительная характеристика трех
типов первичных стратегий растений системы Раменского
Грайма.
Как мы уже говорили, три первичных типа стратегий свя-
заны множеством переходных типов, которые называются вто-
ричными стратегиями. Видов с вторичными стратегиями боль-
ше, чем видов «чистых» первичных стратегий, особенно вио-
лентов. Примером вида, имеющего стратегию виолент-патиент
(CS), является сосна обыкновенная (Pinus sylvestris), которая
может формировать высокие древостои на бедных песчаных
почвах, а также все виды ели (р. Picea), которые произраста-
ют в холодном климате на бедных кислых (но хорошо ув-
лажненных) почвах.
Стратегию виолент-рудерал (CR) имеют такие виды, как
ольха серая (Alnus incana), которая разрастается на вырубках, и
крапива двудомная (Urtica dioica) - обычный доминант почв,
богатых азотом. Виды со стратегией рудерал-патиент (RS)
можно наблюдать на вытоптанных площадках вокруг колодцев
в пустынной зоне.
Наконец, смешанный тип стратегии - CRS - представляют
все луговые растения, которые сочетают в своем поведении чер-
ты виолентности, патиентности и эксплерентности, хотя эти ка-
чества у разных видов представлены в разном соотношении.
Например, у ячменя короткоостого (Hordeum brevisubulatum)
или бескильницы расставленной (Puccinellia distans) больше при-
знаков патиентности, а у пырея ползучего (Elytrigia repens)
эксплерентности и виолентности.
Некоторые виды обладают пластичностью стратегии. На-
пример, дуб (Quercus robur) в местообитаниях с оптимальными
условиями - типичный виолент, а у южной границы ареала
патиент в кустарниковой форме. Патиентом на засоленных поч-
вах является тростник (Phragmites australis), который в дельте
рек имеет стратегию настоящего виолента. Японское искусство
выращивания карликовых деревьев (бонсай) основано на пре-
вращении виолентов в патиенты. Природный бонсай создает-
ся из сосны (Pinus sylvestris) на болотах. На сфагновых кочках
растут сосенки, которые в возрасте 90-100 лет имеют высоту
несколько десятков сантиметров.
181
Таблица 17
Сравнительная характеристика типов стратегий растений
Признак Тип стратегии
виолент (С) патиент (S) эксплерент (R)
Абиотические условия среды Благоприят- ные Неблагопри- ятные Благоприят- ные
Наличие на- рушений Нет Нет Есть
Уровень взаи- моотношений растений Высокий Низкий Высокий
Жизненная форма Деревья, кус- тарники, реже травы мезо- морфного обли- ка с широким простиранием в пространстве, мощной корне- вой системой и большой листо- вой поверхно- стью Небольшие растения, кус- тарнички, дере- вья, многолет- ние травы ксе- роморфного об- лика, однолет- ние и многолет- ние суккуленты, лишайники, мхи Однолетние травы, реже многолетние травы с интен- сивным вегета- тивным размно- жением
Способ регу- ляции плотно- сти популяции Зависимостью от смертности Зависимостью от абиотическо- го стресса Пластично- стью популяции
Тип возрас- тного состава популяции Нормальный (реже инвази- онный или рег- рессивный) Нормальный (реже инвази- онный или рег- рессивный) Инвазионный
Наличие бан- ков семян Нет Нет Есть
Наличие бан- ков проростков Есть Нет Нет
Тип реагиро- вания на стресс Морфологиче- ский Физиолого- биохимический Морфологиче- ский
Экологическая ниша Широкая; по объему реали- зованная ниша близка к фун- даментальной, дифференциа- ция ниш выра- жена хорошо Узкая; по объему реали- зованная ниша приближается к фундаменталь- ной, дифферен- циация ниш не выражена Широкая; по объему реали- зованная ниша много меньше фундаменталь- ной, дифферен- циация ниш слабая
182
10.5 .5. Стратегии культурных растений
Стратегии культурных растений заслуживают специального
рассмотрения, так как в настоящее время их разнообразие
достигло примерно двух тысяч видов и потенциально может
быть увеличено примерно в пять раз за счет введения в культу-
ру новых видов лекарственных, технических, пищевых, декора-
тивных растений.
До конца прошлого столетия, пока энерговооруженность
человека была невысокой, при селекции учитывалась не только
продуктивность растений, но и их устойчивость к болезням,
вредителям и сорнякам. Мы уже говорили (ч. 2), что селекцией
удалось вывести сорта подсолнечника, устойчивые к заразихе
и грибковым заболеваниям.
Однако, когда человек обрел возможность использовать
тракторные плуги, минеральные удобрения, пестициды и про-
водить орошение на больших площадях, направление селекции
культурных растений изменилось. Отбор стал проводиться в
основном в направлении увеличения продукционного потенциа-
ла, т.е. способности растений давать высокий урожай зерна,
зеленой массы или клубней при максимальном обеспечении их
ресурсами минерального питания, влаги, защите от сорняков,
вредителей и болезней пестицидами.
Таким образом, человек развивал только свойство экспле-
рентности культурных растений, заменяя их способность к
защите от неблагоприятных факторов (т.е. к преодолению со-
противления среды) искусственными приемами удобрением,
орошением и химической обработкой посевов.
В последние десятилетия селекция направлена на повыше-
ние патиентности, т.е. на способность культурных растений
преодолевать сопротивление среды - быть устойчивыми к засу-
хе, кислой реакции почвенного раствора, невысокому уровню
обеспечения почвы элементами минерального питания, к вреди-
телям, болезням. За счет повышения виолентности (мощности
растений, дружности их развития) они становятся более устой-
чивыми к сорнякам.
Таким образом, современная селекция культурных растений
проводится в направлении развития у них свойств CRS.
183
Глава 11
ПОПУЛЯЦИИ РАСТЕНИЙ
11.1. Локальные и менделевские популяции
Популяция - это совокупность особей одного вида. В био-
логии как популяция чаще всего рассматриваются все особи од-
ного вида, которые связаны процессами обмена генами (для
растений — перекрестным опылением). Однако такое определе-
ние популяции (ее называют менделевской) неудобно для эколо-
гов, так как установление генетической общности совокупности
растений - дело слишком сложное и трудоемкое. В экологии
распространено более прагматическое понятие - локальная по-
пуляция, т.е. группа особей одного вида на определенной эко-
логически однородной территории. В дальнейшем термином
«популяция» будем обозначать именно локальную.
Соотношение размеров локальных и менделевских популяций
может быть различным. Если экотоп занимает обширные про-
странства (например, равнина, занятая еловыми лесами или ко-
выльными степями), то размер менделевских популяций может
быть меньше локальных. И, наоборот, если экотоп по размеру
невелик, то локальная популяция может быть меньше менделев-
ской. При этом одна менделевская популяция видов с широкой
экологической амплитудой может включать несколько локальных
популяций из расположенных рядом сходных экотопов.
11.2. Сравнение популяций растений и животных
Главное различие популяций растений и животных
(табл. 18) заключается в том, что обладающие подвижностью
животные могут более активно реагировать на складывающиеся
условия внешней среды, избегая неблагоприятных стечений об-
стоятельств или рассредотачиваясь по территории для компен-
сации снижения запаса ресурса на единицу площади. Подвиж-
ность облегчает им и защиту от хищников. Растения же при-
креплены к почве и должны реагировать на меняющиеся биоти-
ческие (хищники, паразиты, конкуренция с более сильным сосе-
дом) или абиотические (засуха, засоление и т.д.) условия за
счет морфологических или физиолого-биохимических адаптив-
ных реакций.
184
Таблица 18
Сравнительная характеристика популяций растений и животных
Признак Растения Животные
Границы осо- бей Не всегда четко опре- делены, и в качестве структурных элементов популяций выступают как генеты, так и раме- ты Четко определены
Продолжитель- ность периода роста Как правило, неогра- ничен за счет мери- стем, сохраняющихся в течение всей жизни, что ведет к нарастанию биомассы популяции в течение жизни особей Чаще всего ограни- чена (млекопитающие, насекомые). Рост за- вершается к фазе ре- продуктивной зрелости или раньше, что обес- печивает сравнительную стабильность биомассы популяции половозре- лых особей
Модульная структура Выражена. Модулями выступают побеги Отсутствует
Вариация раз- меров особей од- ного возраста Значительная Практически отсут- ствует
Факторы регу- лирования плот- ности Рождаемость и смерт- ность Плотность популя- ции, кроме того, меня- ется во время миграции особей
Отношение к стрессу (неблаго- приятным усло- виям среды) Повышение устойчи- вости за счет онтогене- тических адаптаций Адаптации дополня- ются способностью к миграциям
Резервные группы Имеются почвенные банки диаспор Как правило, отсутст- вуют
Забота о по- томстве Отсутствует Имеются многочис- ленные механизмы для этого
11.3. Плотность популяции
Плотность популяции это количество особей, приходя-
щееся на единицу площади. Определение плотности популяций
растений имеет свои особенности, так как у многих видов так
называемых клональных растений, кроме семенного, сущест-
вует вегетативное размножение (из почек на под- или надзем-
ных органах). Новые побеги, образующиеся вегетативным пу-
тем, называются раметами (в отличие от генетов - растений
семенного происхождения).
185
Раметы могут быть связаны с материнским растением или
со временем отделяются от него и становятся самостоятельны-
ми особями. В результате бывает трудно определить, какие по-
беги самостоятельны, а какие являются частями одного расте-
ния. С этим исследователь сталкивается, например, при изуче-
нии корневищных злаков костреца безостого (Bromopsis
inermis) или пырея ползучего (Elytrigia repens). В таких случаях
счетной единицей служит не особь, а побег.
Для определения плотности популяции используются проб-
ные площадки разного размера, который тем больше, чем
крупнее растение. Учет плотности популяций деревьев прово-
дится на площадках не менее 400 м2, подсчет кустарников
может быть проведен на площадке размером в 100 м2, а тра-
вянистых растений - от 10 до 0,1 м2. Плотность популяций
мхов определяют на площадках размером от 10 до 25 см2.
Существуют три основных способа регуляции плотности
популяций.
Зависимость от смертности. Этот способ регуляции плот-
ности популяции характерен для животных, но встречается и
у многих видов растений-виолентов, например у дуба (Quercus
robur). На 1 м2 леса может прорасти несколько десятков желу-
дей. По мере взросления растений происходит самоизрежива-
ние: часть особей гибнет в результате конкуренции. Поэтому
взрослые древостои дубового леса в сходных условиях (одина-
ковый климат и тип почвы) имеют примерно одинаковую плот-
ность. Человек способствует регуляции смертностью плотности
популяции дуба, вырубая слабеющие растения и таким обра-
зом улучшая условия для роста остальных деревьев.
Пластичность. У видов-эксплерентов плотность популя-
ций взрослых растений может быть очень разной, причем чем
она выше, тем меньше размеры особей. При высокой плотно-
сти происходит так называемая «миниатюризация». Например,
особи мари белой (Chenopodium album) из популяций с низкой
и очень высокой плотностью могут отличаться по весу в 20 ты-
сяч раз. Такой пластичностью обладают популяции однолетних
растений, которые экологи называют «оппортунистическими».
Экотопическая регуляция. У патиентов в условиях крайней
неблагоприятности, которые сдерживают рост и развитие рас-
тений, т.е. вызывают стресс (при сильном засолении, крайней
сухости, сильном затенении и т.д.), фактором регулирования
плотности популяций может быть и сам абиотический стресс. В
этом случае плотность популяции будет зависеть именно от
приспособленности вида к этим условиям (т.е. патиентности) и
степени неблагоприятности (сопротивления) среды.
186
Однако у большинства растений плотность популяций ре-
гулируется одновременно двумя или даже тремя способами:
зависимостью от смертности, пластичностью и сопротивлением
среды. Поэтому при более высокой плотности и при выражен-
ном стрессе особи бывают мельче, хотя и не настолько, как у
мари белой.
11.4. Масса популяции
Масса популяции, как и ее плотность, определяется на еди-
ницу площади. Популяции разных видов могут существенно
различаться по массе. Виды, популяции которых преобладают
по массе, называются доминантами, виды с малой массой по-
пуляции - второстепенными, а виды, занимающие промежу-
точное положение, - содоминантами.
Выраженность доминантов в фитоценозах различна. На-
пример, в дубовом лесу очевидный доминант - дуб (Quercus
robjir), а содоминант - сныть (Aegopodium podagraria), в приру-
словой пойме реки явным доминантом может быть кострец
безостый (Bromopsis inermis). В некоторых случаях популяции
многих видов имеют примерно одинаковую массу и доминанты
не выражены.
Существует несколько способов определения массы попу-
ляции. В травянистой растительности (луговой, степной и др.)
применяют следующий простой способ. На нескольких учетных
площадках размером 0,1-1 м2 скашивают травостой, полученные
укосы разбирают по видам, взвешивают растения каждого вида,
вычисляют их среднюю массу на учетную площадку и выража-
ют абсолютную массу популяции в г/м2 или относительную
массу в процентах от общей массы травостоя.
В лесах массу популяций древесных растений определяют
по специально выбранным модельным деревьям, у которых из-
меряют и взвешивают (обычно по фракциям) листья, ветви,
ствол. У ствола чаще определяют только объем, а массу вычис-
ляют с использованием показателя удельного веса. Наконец,
часто количественное участие популяции в составе травостоя
определяется глазомерно, используя показатель проективного
покрытия, т.е. той части поверхности почвы, которую занима-
ют проекции особей популяции.
Проективное покрытие может быть выражено в абсолют-
ных значениях - в процентах, но на практике чаще используют-
ся специальные шкалы, в которых баллами обозначены опреде-
ленные интервалы значений проективного покрытия. Шкалы со-
ставлены с учетом возможностей человека, который легко от-
187
личает на глаз 5 и 10% покрытия, но затрудняется в различе-
нии 70 и 80% покрытия. Наиболее распространена следующая
шкала:
-I— незначительное участие популяции вида,
1 балл - проективное покрытие до 5%,
2 балла - от 6 до 15%,
3 балла - от 16 до 25%,
4 балла - от 26 до 50%,
5 баллов - от 51 до 100%.
11.5. Распределение популяций в пространстве
Особи популяций разных видов распределены в пространст-
ве по-разному. Различают три основных типа распределения
(рис. 38).
При регулярном распределении особи популяции располо-
жены примерно на одинаковом расстоянии друг от друга. В
природе такое распределение встречается редко, но часто на-
блюдается в растительных сообществах, созданных человеком
(например, в посевах пропашных культур - кукурузы или са-
харной свеклы, когда расстояние между растениями в рядках
и между рядками одинаково; в сквере, где регулярно высажены
деревья).
Рис. 38. Типы распределений осо-
бей популяций в пространстве: а —
регулярное, б случайное, в
групповое (пятнистое)
При случайном распределении расстояния между особями
подчиняются закону случайного распределения и положение
каждой особи не зависит от положения других особей.
При групповом распределении (пятнистом, контагиозном)
особи распределены в пространстве группами (куртинами).
Случайное и групповое распределения популяций широко
распространены в природе, причем тип распределения зависит
от особенностей биологии вида, экологических условий и попу-
ляций других видов, с которыми данная популяция сообитает.
Например, биологические особенности (осыпание семян
вблизи материнского растения, разрастание корневищ) опреде-
ляют пятнистое произрастание бобовых - клевера (р. Trifolium),
люцерны (р. Medicado) и папоротника орляка (Pteridium
aquilinum). Особи популяций других видов имеют тенденцию к
случайному распределению, но при неблагоприятных условиях
а) б) в)
188
они могут образовывать группы в тех участках фитоценоза, где
слабее конкуренция и легче выжить.
Наиболее простым показателем пространственного распре-
деления вида является его встречаемость, метод изучения ко-
торой разработан К.Раункиером. Для оценки встречаемости ви-
да регулярно (т.е. через равные интервалы) или в случайном
порядке закладывается серия пробных площадок (обычно 50)
размером 0,1 м2. Затем рассчитывают отношение числа площа-
док, на которых встречен вид, к их общему числу и выражают
его в долях единицы или в процентах. Полученный показа-
тель - количественный, и его достоверность определяется по
формулам оценок эмпирических вероятностей, которые приво-
дятся в любом учебнике статистики.
11.6. Гетерогенность популяций
Естественные популяции (включая и те, которые сформи-
ровались под влиянием человека: заросли рудеральных расте-
ний, сорняки на полях) состоят из неодинаковых особей. Эта
неоднородность популяций определяется гено- и фенотипиче-
скими причинами. От генотипических причин зависят следую-
щие два типа неоднородности.
Наличие в популяции нескольких экотипов. Экотипы - это
генетически обусловленные экологические варианты вида, ко-
торые могут гибридизировать. Популяции луговых злаков, на-
прймер, полевицы гигантской (Agrostis gigantea), пырея ползуче-
го (Elytrigia repens), овсяницы луговой (Festuca pratensis), обыч-
но состоят из 3-7 экотипов, которые по-разному развиваются
в разные по климатическим условиям годы. Это генетическое
разнообразие растений повышает устойчивость популяций к
неблагоприятным факторам внешней среды. Влияние человека
(сильный выпас скота, удобрение, загрязнение) может привести
к выпадению некоторых экотипов, поэтому охрана генетиче-
ского состава популяций - одна из важных задач рационально-
го природопользования.
Из генетически пестрых природных популяций видов,
ставших сорняками полей, под влиянием гербицидов отобра-
лись экотипы, устойчивые к их действию, и это снизило эф-
фективность химических прополок. Говорить о том, что под
влиянием гербицидов возникают устойчивые к ним формы сор-
няков, неверно. Эволюция растений - процесс очень медлен-
ный. Эти формы существовали в природе до применения гер-
бицидов и были преадаптированы, т.е. как бы заранее подго-
товлены к новому фактору.
189
Половая дифференциация растений в популяции. У двудом-
ных растений соотношение мужских и женских особей в сред-
нем составляет 0,5:0,5. Однако в тех случаях, когда для попу-
ляции ухудшаются экологические условия, увеличивается доля
мужских особей, более устойчивых к действию неблагоприят-
ных факторов (т.е. регуляция плотности зависимостью от
смертности у растений разных полов происходит по-разному).
Такая закономерность изучена у можжевельника (р. Juniperus) и
тополя (р. Populus).
Два других типа неоднородности популяций зависят от
фенотипических причин.
Дифференциация особей по возрастному составу. Наличие
в популяции особей разного возраста - молодых (ювенильных),
половозрелых (генеративных), стареющих (сенильных) - связа-
но с процессом ее возобновления и поддержания устойчивого
состояния. Может быть выделено и большее число возрастных
состояний - до 7-9.
Наиболее просто определяется возраст у деревьев. Из
ствола специальным буром берут пробу древесины и по ней
подсчитывают число годичных колец. Определять абсолютный
возраст травянистых растений сложно, и для них часто уста-
навливают относительный возраст, т.е. тот, на который расте-
ние «выглядит». В угнетенном состоянии старым (сенильным)
может выглядеть растение, которое при благоприятных услови-
ях было бы отнесено к половозрелому и процветающему.
По соотношению особей разного возраста различают три
типа популяций (Работнов, 1992): инвазионные (внедряющиеся),
в которых преобладают молодые растения; нормальные, в кото-
рых одинаково представлены растения разного возраста; рег-
рессивные (отмирающие), которые состоят из старых особей.
Кроме трех основных типов популяций, Т.А.Работнов пред-
ложил выделять четверый - ложноинвазионные популяции. Они
образуются у тех видов, которые имеют в почве банк (запас) се-
мян или вегетативных зачатков (корневища, клубни, луковицы).
Обычно эти растения находятся в состоянии покоя и отсутствуют
в составе травостоя. Если на банк подействует фактор, способ-
ный вывести его из состояния покоя, то происходит массовое
развитие растений, и этот вид может стать доминантой.
Стимулирующим прорастание фактором может быть нару-
шение целостности верхнего слоя почвы. При этом в ней уве-
личивается количество воздуха, и аэробные микроорганизмы
переводят аммонийные формы азота в нитратные.
Дифференциация особей по виталитету. Показателем вита-
литета (жизненного состояния) особи в основном является ее
190
масса, поскольку чем лучше развито растение, тем оно круп-
нее. Виталитет выражается и в интенсивности цветения и обра-
зования семян (называемой репродуктивным усилием). Методы
изучения виталитета разработаны Ю.А.Злобиным. По весу все
особи популяции разделяются на три группы: крупные, средние
и мелкие. По соотношению этих групп популяции делятся на
процветающие (преобладают крупные и средние особи), рав-
новесные (особи трех весовых групп представлены одинаково) и
депрессивные (преобладают мелкие особи). На рис. 39 показа-
ны гистограммы популяций с разным виталитетным спектром.
Рис. 39. Примеры виталитетных спектров популяций: а - процветаю-
щая, б — равновесная, в - депрессивная; М - мелкие особи, С - сред-
ние особи, К - крупные особи; по вертикали - долевое участие
Виталитетный спектр популяции важная оценка ее со-
стояния. Например, если редкий или исчезающий вид представ-
лен депрессивной популяцией, то возможно его полное исчез-
новение. Необходимо выявить и исключить влияние фактора,
угнетающего эту популяцию (выпас, загрязнение, вытаптывание
туристами и т.д.).
Фенотипическая и особенно генотипическая неоднородно-
сти популяций повышают их устойчивость, так как они пол-
нее используют ресурсы среды и легче переносят неблагоприят-
ные условия. Поэтому в последние годы в практике сельского
хозяйства начали создавать неоднородные популяции культур-
ных растений. Например, выращивают генотипически неодно-
родные сортосмеси кукурузы (высокий и низкий сорт), пшени-
цы (сорта с разной засухоустойчивостью), которые дают устой-
чивые урожаи в разные по климату годы. Фенотипическую не-
однородность создают, высаживая вместе непророщенные и
пророщенные клубни картофеля, проводя дополнительную под-
191
садку семян огурцов в междурядья, высевая смесь из сухих и
замоченных семян моркови и т.д. Такие посевы полнее исполь-
зуют ресурсы почвы и света за счет снижения конкуренции.
11.7. Особенности популяций споровых растений
У папоротниковидных (папоротников, плаунов и хвощей)
гаметофит и спорофит представляют независимые организмы,
что делает изучение их популяций более сложным по сравне-
нию с семенными растениями. В этом случае организмы гаме-
тофитов и спорофитов представляют собой как бы «популяции
второго порядка», называемые в экологии когортами.
Литовский ботаник И.Науялис (J.Naujaiis) установил, что
заростки хвощей и папоротников имеют стратегию эксплерен-
тов, и в определении их численности главенствует сопротивле-
ние среды, преодолеть которое заросткам с крайне низким
биотическим потенциалом удается далеко не всегда. Их страте-
гия - ожидание, а поскольку длительность жизни спор, когда
они способны прорастать, ограничена (банки жизнеспособных
спор хвощей и папоротников возможны только в том случае,
если они постоянно пополняются), то и плотность когорт гаме-
тофитов невелика. Она не идет в сравнение с астрономическим
количеством спор, которое продуцируют спорофиты.
У плаунов заростки являются эксплерентами только на
первых стадиях развития, а затем они становятся патиентами.
Однако плата за патиентную стадию очень велика: если для
развития заростков хвощей достаточно месяцев, папоротни-
ков - двух-четырех лет, то время жизни заростка плауна может
достигать двух десятилетий.
Когорты заростков имеют достаточно сложную структуру,
особенно у папоротников. В их составе различаются ювениль-
ные, мужские, женские, двуполые заростки, старые стерильные
заростки и еще три состояния, отражающие процесс развития
спорофита из оплодотворенной яйцеклетки. При этом в разных
условиях структура популяций может меняться. Очевидно, что
КПД такой когорты ограничен, так как шансов породить новый
спорофит у женских заростков немного, не говоря о том, что
их совсем нет у мужских или стерильных. Реальными продуцен-
тами спорофитов оказываются лишь обоеполые заростки. При
такой структуре когорт устойчивость популяции гаметофитов
поддерживается только за счет большого количества спор и их
интенсивного прорастания.
В когортах заростков плаунов различается меньшее число
возрастных состояний - гаметофит до оплодотворения яйце-
клетки и три стадии после оплодотворения (с подземным побе-
192
гом, с надземным побегом, молодой спорофит). Когорты гаме-
тофитов хвощей наиболее сложны для изучения в природе и в
основном исследуются в лаборатории. Их развитие, как отме-
чалось, наиболее стремительно, и заростки, не успевшие за 4-6
месяцев дать потомство, гибнут от морозов. Когорты спорофи-
тов представляют типичные клональные растения, размножаю-
щиеся надземными и подземными побегами. Для папоротников
характерны пятна, для плаунов - «ведьмины кольца» (особенно
в лишайниковых борах), для хвощей - кусты, формирующиеся
на корневищах.
Глава 12
РАСТИТЕЛЬНЫЕ СООБЩЕСТВА (ФИТОЦЕНОЗЫ)
Растительные сообщества, или фитоценозы, - ансамбли ви-
дов в пределах однородного экотопа. В растительном сообщест-
ве виды могут сосуществовать в результате влияния разных
факторов:
экотопа;
взаимоотношений растений друг с другом;
гетеротрофных организмов (животных, грибов, бактерий);
различных нарушений.
На состав растительных сообществ влияет также возмож-
ность дифференциации экологических ниш видов, входящих в
их состав.
В разных сообществах роль этих факторов неодинакова. По
этой причине вначале мы рассмотрим отдельно каждый из фак-
торов организации фитоценозов, а затем познакомимся с раз-
ными моделями их организации.
В любом случае деление растительного покрова на сообще-
ства условно. В силу уже отмеченной индивидуальности эколо-
гии видов, при изменении условий среды видовой состав расти-
тельности изменяется постепенно, в результате чего формиру-
ется растительный континуум, или непрерывность раститель-
ности. Разбиение континуума на растительные сообщества пре-
следует практические цели.
12.1. Факторы и модели организации фитоценозов
12.1.1. Экотоп
Экотоп - это ниша фитоценоза в ландшафте, его местооби-
тание. Экотоп объединяет занимаемое фитоценозом простран-
ство и ресурсы, используемые для производства биологической
продукции. Экотоп формируется многими экологическими
факторами, которые рассмотрены в § 1.1.
193
В любом случае экотоп - главный фактор организации фи-
тоценоза, так как именно от него, т.е. абиотических факторов
среды, зависит, будет ли на участке лес или степь, будут
преобладать в лесном сообществе ель или липа, а в степном -
ковыли или разнотравье. Только при очень сильных нарушени-
ях местообитаний роль исходного экотопического комплекса
несколько уменьшается.
12.1.2. Взаимоотношения растений
Основным вариантом взаимоотношений растений является
конкуренция (см. п. 10.5.1). В процессе конкуренции растения
соревнуются в потреблении ресурсов и тем самым изменяют
среду друг для друга, оставляя конкуренту меньше света, воды,
питательных элементов, опылителей, чем бы он мог получить
при отсутствии конкуренции. Рассмотрим некоторые другие ва-
рианты взаимоотношений растений, которые также влияют на
фитоценозы.
Взаимное и одностороннее средообразование. Этот вариант
относится к неконкурентным взаимоотношениям растений. Как
и при конкуренции, при этом возможны асимметричные отно-
шения (растения изменяют среду друг для друга неодинаково^) и
симметричные (эти изменения среды примерно одинаковые).
Изменения условий среды {биотическая трансформация) могут
происходить в направлении их улучшения или ухудшения, при-
чем второй вариант распространен чаще.
Классическим примером взаимного отрицательного средо-
образования являются отношения сфагновых мхов и сосуди-
стых растений на сфагновом болоте. Между сфагновыми мхами,
которые отличаются неограниченным ростом вверх, и сосуди-
стыми растениями - вересковыми (Ledum paiustre, Chamaeda-
phne caiicuiata, Andromeda angustifoiia, Oxycoccus paiustre), со-
сной (Pinus syivestis f. pumiia), а также некоторыми осоками, -
складываются отношения антагонизма. Сфагновые мхи, повы-
шая уровень поверхности болота, ухудшают условия корневого
питания цветковых растений, хотя они и обладают специальной
защитной системой перемещения корневой системы как вверх,
так и в направлении участков болота, где нарастание его по-
верхности происходит не столь быстро.
Цветковые, в свою очередь, подавляют сфагновые мхи, ис-
сушая верхний слоя торфа и, особенно, затеняя мхи опадом ли-
стьев. Тем не менее, в ходе эволюции на сфагновом болоте
сформировалось равновесие между антагонистами: количество
опада цветковых не бывает большим, так как их рост сдер-
живается сфагновыми мхами, и продуктивность снижается (мы
194
уже говорили об особой патиентной карликовой форме сосны
на болоте.)
Рост цветковых также сдерживается очень медленной мине-
рализацией сфагнового торфа, который и без того, как торф
верховых болот, отличается крайней бедностью минеральными
элементами и кислой реакцией. Кроме того, за счет теплоизо-
ляционных свойств верхнего слоя торфа и густого стеблестоя
мха для горизонта расположения основной массы корней ха-
рактерна низкая температура. Таким образом, угнетая цветко-
вые растения, но не настолько, чтобы вытеснить их с болота,
сфагновые мхи тем самым защищают себя от их пагубного
влияния.
Примером одностороннего отрицательного средообразова-
ния является влияние деревьев-доминантов на виды мохового и
травяного яруса. Под пологом деревьев уменьшается освещен-
ность, повышается влажность воздуха. При разложении опада
деревьев почвы обедняются вследствие образования кислот,
способствующих вымыванию питательных элементов в глубь
почвенного слоя. Этот процесс особенно активен в таежном ле-
су под пологом густых елей.
Пример одностороннего положительного средообразования -
обогащение почвы азотом бобовыми, которые симбиотически
связаны с клубеньковыми бактериями-азотфиксаторами.
Паразитизм и полупаразитизм. В фитоценозах возможны
вертикальные (т.е. между организмами разных трофических
уровней) взаимоотношения между автотрофными и паразитны-
ми растениями. Из паразитных растений наиболее распростра-
нены полупаразиты (их много в семействах Lorantaceae,
Scrophulariaceae, Santalaceae), которые осуществляют фотосин-
тез, а у растений-хозяев заимствуют в основном воду и эле-
менты минерального питания и, возможно, некоторое количе-
ство органических веществ. Полные паразиты не имеют хлоро-
филла и являются гетеротрофами, полностью питающимися за
счет растений-хозяев. Паразитами представлены семейства Ого-
banchaceae и Cuscutaceae.
Полупаразиты могут обитать на разных видах растений-
хозяев, а у паразитов выражена специфичность в их выборе. Все
паразитные растения действуют на растение-хозяина угнетающе.
В естественных фитоценозах паразитные растения большой
роли не играют, хотя в некоторых случаях они регулируют
плотность популяций автотрофных видов (например, в пусты-
нях такую роль для саксаула - р. Haloxylon играют предста-
вители рода Cistanche). Опасны паразитные растения для посе-
вов: виды родов повилика (Cuscuta) и заразиха (Orobanche)
195
являются злостными сорняками многих культур. От заразихи
страдают плантации арбузов в Астраханской области, где
плантации приходится переносить на новые места, перемещая и
оросительную систему.
Снизить вред от растений-полупаразитов можно путем
раннего скашивания травостоя до созревания их семян (боль-
шинство полупаразитов - двулетники, и потому при отсутствии
поступлений новых партий семян в почву их плотность быстро
снижается).
Для очищения полей от растений-паразитов используют се-
вообороты: к моменту возвращения на поле культуры растения-
хозяина семена паразита теряют всхожесть. В США для борьбы
с паразитами из рода Striga применяют биологический метод.
Семена паразита прорастают только тогда, когда рядом в поч-
ве есть корень растения-хозяина, о чем они «узнают» по его
корневым выделениям. На поле вносится химическое вещество,
имитирующее корневые выделения растения-хозяина, семена
паразита прорастают, и проростки, не способные к автотроф-
ному питанию, погибают.
В целом паразиты опаснее для растений-хозяев, чем полу-
паразиты. Однако в естественных фитоценозах они встречаются
в небольшом количестве и обычно не приводят к выпадению из
фитоценоза популяций растений-хозяев, поскольку в результате
сопряженной эволюции последние адаптированы к паразитам.
Они лишь снижают продуктивность трав и сокращают срок
жизни деревьев, на которых паразитируют.
Взаимопомощь растений. Этот тип взаимоотношений расте-
ний в фитоценозе играет несравненно меньшую роль, чем кон-
куренция, но он достаточно распространен в природе. Как ре-
зультат взаимопомощи рассматривается известный растениево-
дам эффект группы - растения, высеянные группой, развива-
ются лучше, чем растения, выращиваемые порознь. Причины
такого эффекта могут быть разными. У группы растений легче
формируется симбиоз с бактериями и грибами, образующими
микоризу и ризосферу.
Однако по достижении растениями определенного возраста
эффект группы сменяется конкуренцией, растения в центре
группы развиваются хуже, чем расположенные по краю (в ре-
зультате возникает так называемый эффект «корыта»).
Произрастающие группой растения полнее опыляются насе-
комыми, так как повышается вероятность переноса пыльцы с
цветков одного растения на другое, а яркое цветовое пятно из
нескольких цветущих и выделяющих ароматические вещества
растений привлекает насекомых.
196
Очень интересен вариант односторонней помощи растения-
«няни» другому растению. Береза или ольха могут быть няней
для ели: няня защищает молодую елочку от прямых солнечных
лучей, без чего на открытом месте ель вырасти не сможет. Кроме
того, няни защищают всходы молодых елочек от выжимания их
из почвы морозом. В таких же отношениях состоят кустарники
(из семейств Lamiaceae и Asteraceae) и южно-американские как-
тусы. Молодые кактусы страдают от прямого солнечного света.
Обладая особым типом фотосинтеза, который происходит днем
при закрытых устьицах, они сильно перегреваются. Поэтому мо-
лодые кактусы могут развиваться только под защитой засухоус-
тойчивых кустарников-«нянь».
Роль растений-посредников. Отношения растений через по-
средников (в п. 12.1.3 будет описана роль посредников-живот-
ных) изучены недостаточно, однако, видимо, играют в природе
большую роль. В. Г. Карпов выяснил, что ель влияет на мхи
(Pleurozium schreberi и др.) через посредника - кислицу (Oxalis
acetosella). Когда в эксперименте обрубались корни ели, и кис-
лица получала возможность без конкуренции захватить большее
количество почвенных ресурсов, она подавляла мхи, которые
практически выпадали из фитоценоза. В то же время, в еловом
лесу без обрезки корней (на контроле) разрастание кислицы
сдерживалось конкуренцией с елью, корни которой расположе-
ны в том же приповерхностном слое почвы, что и у кислицы.
Аналогично дуб влияет через сныть на эфемероиды (виды ро-
дов Anemonoides, Corydalis, Scilla): в период вегетации эфемероидов
листья на деревьях еще не распустились, но летом они через ре-
жим освещенности влияют на распределение сныти. Сныть начина-
ет вегетировать рано, и потому на осветленных участках леса, где
ее меньше затенял дуб, она подавляет эфемероиды.
Аллелопатия. Этот термин обозначает взаимное влияние
растений друг на друга через изменение среды в результате вы-
деления в нее продуктов жизнедеятельности (летучих или рас-
творенных в воде, стекающей с листьев или омывающей корни).
Аллелопатические эффекты наглядно проявляются в экспе-
риментах, которые выполняют в замкнутых экологических сре-
дах - чашках Петри, под стеклянными колпаками, в вегетаци-
онных сосудах с использованием концентрированных вытяжек
из растений или смывов с них. Однако в природных фитоцено-
зах аллелопатические влияния существенной роли не играют,
так как там нет условий для накопления аллелопатических ве-
ществ (большинство из них являются летучими терпенами или
фенольными соединениями). Их концентрация постоянно пони-
жается ветром, водой, микроорганизмами. Кроме того, в есте-
197
ственных условиях практически невозможно отделить аллело-
патические взаимовлияния растений от конкуренции.
Дифференциация (упаковка) ниш в сообществе. Нами были
рассмотрены представления о нише у растений. Именно объеди-
нение в сообществе видов с разными нишами (т.е. «упаковка» ви-
дов) и раздел ими ресурсов (дифференциация ниш) - основной
механизм, который в ходе формирования сообщества способен
снизить уровень конкуренции. Это «выгодно» сообществу, так
как расходы на противодействие конкуренту требуют дополни-
тельной энергии, а следовательно, ведут к снижению продуктив-
ности отдельных компонентов и всего сообщества в целом.
Принцип дифференциации ниш используется в растение-
водстве и лесоводстве при создании поликультур (посевов и
посадок из нескольких видов растений). Дифференциация ниш в
искусственных фитоценозах, создаваемых человеком, обеспечи-
вает более полное использование пространства и солнечного
света или почвенных ресурсов. Это достигается за счет разной
глубины корневых систем (например, корни люцерны проника-
ют на глубину 5-7 м, а у злаков они расположены в слое до 30
см от поверхности) и разной требовательности к обеспечению
элементами минерального питания (например, злакам в пер-
вую очередь нужен азот, а бобовым фосфор и калий), за
счет создания «экологической очереди» фенологического раз-
вития растений.
В состав поликультуры многолетних трав обычно включают
2—5 видов растений с разными нишами, но иногда травосмесь
составляют из 10-15 видов. Распространены и однолетние по-
ликультуры, например посевы кукурузы с подсолнечником, го-
рохом, фасолью, соей, люпином. В ФРГ широко используются
сидеральные поликультуры (т.е. смеси, урожай которых запахи-
вается в почву как зеленое удобрение) из 5-10 видов трав, ко-
торые быстро развиваются и образуют большую биомассу за
два месяца вегетации.
Исследователи пастбищ Средней Азии (Н.Т.Нечаева и др.)
использовали принцип дифференциации ниш при создании в
пустыне искусственных фитоценозов («агропустынь») из видов
местной флоры. Естественные пустынные фитоценозы состоят
из 5-7 видов обычно одной жизненной формы: деревьев, кус-
тарников или трав. В «агропустынях» вместе выращиваются ви-
ды разных жизненных форм, дифференцированные по экологи-
ческим нишам, что позволяет без удобрения и полива полу-
чать урожай зеленой массы в 4-5 раз больше, чем с природных
фитоценозов.
198
Подсевные сидеральные культуры (зеленые удобрения) мед-
ленно развиваются под пологом основного возделываемого
растения, но после его уборки быстро «наверстывают упущен-
ное» и накапливают растительную массу.
В пастбищные травосмеси включают виды, которые в тече-
ние вегетационного сезона имеют разную скорость развития и
тем самым обеспечивают постоянную «зеленую волну» биоло-
гической продукции.
12.1.3. Роль гетеротрофных организмов
Несмотря на то, что гетеротрофные виды не входят в со-
став растительных сообществ, их вклад в организацию фитоце-
нозов может быть очень велик. В большинстве случаев гетеро-
трофы выступают как посредники, усиливая или ослабляя по-
зицию одного или нескольких конкурирующих видов.
Животные. На исход конкуренции между растениями суще-
ственно влияют фитофаги. Так, распространение ковылей (р.
Stipa) связано с их устойчивостью к выпасу крупных фитофа-
гов. В настоящее время из-за перевыпаса скота в степных паст-
бищах в конкуренции побеждают малопоедаемые мелкие полы-
ни (р. Artemisia), а в луговых травостоях - пастбищное низко-
травье (Plantago, Taraxacum и др.).
Однако в силу коадаптации организмов разных трофиче-
ских уровней далеко не всегда хорошо поедаемое растение про-
игрывает в конкуренции малопоедаемому. В саваннах, где оби-
тают крупные фитофаги (антилопы, жирафы, зебры и др.), хо-
рошо поедаемые растения имеют высокую скорость отрастания
и быстро восстанавливаются. Растения с медленным ростом за-
щищены от животных колючками, жестким опушением, горьким
вкусом и т.д. и поедаются плохо.
У некоторых растений (в том числе и у картофеля) после
поедания их частей фитофагами повышается концентрация ве-
ществ, защищающих их от поедания. И.В.Стебаев, подчеркивая
глубокую связь между растениями и фитофагами, формирую-
щими экологическое равновесие, пишет, что «...виды растений и
животных двигаются в эволюции подобно флотам, технически
соревнующимся в мощности брони и артиллерии, избегая при
этом губительных генеральных сражений» (Стебаев и др., 1993,
с. 172). Кроме того, влияние фитофагов на особо «вкусные»
виды уменьшается тем, что они могут быстро переключаться с
питания одним видом на другой, что способствует устойчи-
вости состава сообществ.
199
Большое значение для фитоценоза имеют насекомые-
опылители и агенты распространения плодов и семян. Как уже
отмечалось (ч. 2), узкоспециализированные к опылителям виды
при их исчезновении выпадают из состава сообществ.
Достаточно многочисленны факты влияния растений друг
на друга через посредство насекомых. Так, сорные растения,
которые привлекают хищных насекомых, могут положительно
воздействовать на культурное растение, защищая его таким об-
разом от нападения фитофагов и улучшая жизненное состояние
и продуктивность (разумеется, если плотность сорных растений
не высока и этот положительный эффект не перекрывается
конкуренцией за ресурсы). На этой основе за счет повышения
биологического разнообразия в посевах формируется система
обратных связей, которая может регулировать отношения
культурных растений, сорняков, фитофагов и зоофагов. По-
скольку возможности повышения биологического разнообразия
в самом посеве ограничены, большая роль отводится окраинам
полей. При уменьшении размера полей рудеральные, луговые
растения, кустарники и даже деревья могут через посредни-
ков насекомых-фитофагов и птиц - благотворно влиять на
культурные растения, повышая их продуктивность и виолент-
ность, что важно для установления баланса между культурой
и популяциями сорных растений.
Микоризные грибы. Большинство сосудистых растений ми-
котрофны. Однако это не всеобщее правило. Однолетники из
семейств Brassicaceae, Chenopodiaceae, Caryophyllaceae, Polygo-
naceae, которые дают вспышки обилия в условиях нарушений и
богатых нитратами почв, не имеют микориз. При их смене мно-
голетниками в процессе экологической сукцессии выделения
микоризных грибов многолетников подавляют однолетники. В
то же время эти выделения благоприятно влияют на виды мно-
голетников.
Для растений более бедных почв грибы-посредники, улуч-
шающие обеспечение минеральными элементами (в первую оче-
редь фосфором), уже необходимы. При этом они по-разному
влияют на конкурентную способность разных видов. Так, в
смешанном посеве из Lolium регеппе и Holcus lanatus при ино-
куляции грибами, формирующими микоризу, конкурентная
способность первого вида уменьшалась, а второго - увеличива-
лась. Аналогично инокуляция уменьшала конкурентную способ-
ность Koeieria pyramidata по сравнению с Andropogon gerardii.
В целом у большинства многолетних трав инокуляция по-
вышает семенную продуктивность и содержание в семенах пи-
200
тательных элементов, особенно фосфора, и наиболее эффек-
тивно у доминантов сообществ.
Весьма интересно, что при улучшении условий минерально-
го питания (внесение удобрений на лугах) типичные микотрофы
«отказываются» от микоризы и переходят к тому же типу пря-
мого минерального питания без гриба-посредника, как и расте-
ния-рудералы. Это не удивительно, так как затраты энергии на
микоризу у растений могут достигать трети продуктов фото-
синтеза, который осуществляет растение-хозяин.
Наконец, микоризный гриб может быть посредником между
растением-хозяином и растением-паразитом. Так, паразитное
растение подъельник (р. Monotropa) использует сосну как хо-
зяина не непосредственно, а через общий для него и сосны ми-
коризный гриб.
Бактерии-симбиотрофы. Бактерии-симбиотрофы из числа
азотфиксаторов менее значимы, чем микоризные грибы, и все-
таки иногда достаточно важны. При зарастании бедных азотом
субстратов - поверхностей отвалов пустой породы при горных
разработках, отложений аллювия вдоль речного русла - первы-
ми поселяются бобовые, симбиотически связанные с бактерия-
ми-азотфиксаторами и потому более конкурентно-мощные в
этих условиях, чем граминоиды.
Однако бобовые растения теплолюбивы и потому в услови-
ях Крайнего Севера имеют низкую конкурентную способность.
Здесь обогащение субстрата азотом достигается за счет жизне-
деятельности цианобактерий. Пионерами зарастания в таких
условиях являются осоки, злаки и разнотравье.
Паразитические микроорганизмы. Паразитические грибы
могут быть важными посредниками, зачастую определяющими
исход конкуренции сосудистых растений. Они могут настолько
ослабить растение, что под влиянием здоровых конкурентов
оно погибнет. Так, из пойменных лесов Средней России из-за
голландской болезни, которую вызывает патогенный гриб
Ceratocystis ulmi, выпал вяз. Опенок, поселившийся на корнях
липы или дуба, может ослабить их конкурентную способность.
В начале века в США вместе с интродуцированным кашта-
ном из Китая был завезен паразитический гриб Endonthia para-
sitica, который в течение 50 лет практически уничтожил амери-
канский зубчатый каштан широко распространенный доми-
нант широколиственных лесов США. Гибель каштана ускоря-
лась тем, что ослабленные паразитом растения проигрывали в
конкуренции другим доминантам лесов: дубам, кариям и др. В
201
результате этой самой крупной ботанической катастрофы по-
гибло 3,5 млрд деревьев.
Фактором, который коренным образом меняет характер от-
ношений между растениями, могут быть вирусы. Характерный
пример - распространение вируса «коровьей чумы» в саван-
нах Африки. Под влиянием этого вируса, который первона-
чально вызвал болезни у домашнего скота, а затем поразил
многие виды диких крупных копытных животных, резко снизи-
лась нагрузка фитофагов на растительность саванны, и в пер-
вую очередь - на ее древесно-кустарниковый компонент. Это
изменило его отношения с травяным покровом саванны. Кус-
тарники и деревья стали бурно разрастаться, подавляя травы,
что усилило влияние пожаров, которые при таком сомкнутом
древесном пологе стали более частыми. После пожаров дере-
вья отрастали плохо и замещались кустарниками, корневища
которых позволяли им сохраняться во время пожара.
Только в 70-е годы, когда удалось снять влияние на экоси-
стемы вируса «коровьей чумы», вылечив от болезни домашний
скот, поставлявший паразита популяциям диких животных че-
рез выделения слюны на пастбищах, процесс был остановлен.
Восстановилась плотность популяций веткоядных животных, а
вместе с ней и баланс между древесно-кустарниковым и травя-
ным компонентами саванны.
Этот пример представляет наиболее сложный вариант
трансбиотического регулирования взаимоотношений между рас-
тениями при нескольких взаимодействующих друг с другом по-
средниках (фитофаги, вирус, пожар).
12.1.4. Роль режима нарушений
Большинство фитоценозов подвергается различным наруше-
ниям. В лесах происходят пожары (от молний, а чаще - из-за
человека), ветровалы. Самые разнообразные нарушения естест-
венных фитоценозов связаны с деятельностью человека (рубка
леса, атмосферные загрязнения, воздействие транспорта и др.).
Не всякое нарушение и не для любого фитоценоза оказыва-
ется губительным. Выпадение отдельных деревьев в лесу спо-
собствует обогащению видового состава напочвенного покрова
за счет видов-рудералов.
Во многих случаях растительность легко восстанавливается
после периодически повторяющихся пожаров. В Южной и Се-
верной Америке, в Средиземноморье и Австралии есть особые
типы фитоценозов, для которых пожар - обязательное условие
202
существования. Составляющие их растения приспособлены к
воздействию огня - переживают пожар в виде семян или под-
земных органов. Такие растения называются пирофитами. По-
могают устоять против пожаров и муравьи, которые затаскива-
ют семена растений в свои подземные ходы.
Умеренный выпас является нормальным фактором для степей
и саванн: он не нарушает конкурентных отношений между расте-
ниями и не вызывает обеднения состава фитоценозов. В то же
время этот фактор серьезно нарушает лесные сообщества.
Существуют и особые типы фитоценозов, сформированные
эксплерентами на месте крупных нарушений растительности.
Они называются рудеральными и формируются на пустырях,
обочинах дорог, на буртах земли и навозных кучах.
Таким образом, нарушения — важный фактор организации
многих фитоценозов, и его влияние не всегда отрицательно.
Нарушения ослабляют более конкурентно мощные виды и по-
вышают видовое богатство фитоценоза.
12.1.5. Множественность моделей
организации фитоценозов
Мы рассмотрели факторы, которые организуют фитоценоз:
условия среды, предопределяющие состав входящих в фитоце-
ноз видов, горизонтальные отношения между видами растений,
влияние на эти взаимоотношения вертикальных отношений рас-
тений и животных, режим нарушений. Дифференциация эколо-
гических ниш смягчает конкуренцию и помогает сформировать-
ся фитоценозу с высоким видовым богатством.
В разных фитоценозах роль различных факторов организа-
ции неодинакова. Разнообразие моделей организации фитоце-
нозов показано в табл. 19, где тремя плюсами обозначены слу-
чаи, когда фактор является основным, двумя - дополнительным
и одним - когда слабым. Из таблицы видно, что условия среды
являются главным фактором организации любого фитоценоза,
так как в первую очередь от климата и почвы зависит, что
сформируется на данной территории - тайга, лес или степь.
В экстремальных условиях пустынь или в водной среде
(абиотическая S-модель) взаимоотношения между растениями
настолько слабые, что фитоценоз представляет собой группу
популяций, которые связаны только общим отношением к усло-
виям среды. Аналогично слабы взаимоотношения между видами
в сообществах сфагновых болот (биотическая S-модель), где
экстремальные условия созданы за счет биотической трансфор-
203
Таблица 19
Модели организации фитоценозов
Модель Вклад факторов организации
Экотоп Биоти- ческая транс- форма- ция Горизон тальные взаимо- отно- шения Верти- кальные взаимо- отно- шения Интен- сив- ность нару- шении Диффе- ренциа- ция ниш Пример
Абио- тиче- ская S +++ + + + + + Пусты- ни, вод- ные со- общест- ва
Биоти- ческая S +++ +++ + + + ++ Верхо- вые бо- лота
C-S +++ ++ +++ + + +++ Немо- ральные и боре- альные леса
G-B +++ ++ ++ +++ ++ ++ Степи, саванны
C-R-S +++ + + + ++ ++ После- лесные луга
R ++ + + + +++ + Руде- ральные и сеге- тальные сообще- ства
мации экотопа (формирование бедного, кислого и холодного
сфагнового торфа).
Роль горизонтальных взаимоотношений в организации раз-
ных фитоценозов неодинакова. Она наиболее важна в лесных
фитоценозах (C-S-модель), где деревья-доминанты обладают
сильным средообразующим эффектом, и в измененных влияни-
204
ем деревьев условиях выживают те виды трав, которые способ-
ны переносить затенение и подкисление почвы. Горизонтальные
взаимоотношения важны и для развития древостоя: конкури-
руют деревья как одного вида, так и разных видов.
Для организации степных фитоценозов (G-B-модель, от
grazer - травоядное и brouser - веткоядное) важен фактор вер-
тикальных взаимоотношений. Степи формировались под влия-
нием диких копытных, которых сегодня сменил домашний скот.
Большинство луговых фитоценозов формируются в результате
деятельности человека на месте лесов, которые он уничтожил, и
для них нарушение является важным фактором организации.
Исключение составляют альпийские луга и луга на засоленных
почвах, где лес расти не может.
Наконец, рудеральные группировки (R-модель) могут суще-
ствовать только при действии нарушений.
Условия среды в организации агрофитоценозов (также R-
модель) менее значимы, чем в организации естественной расти-
тельности: человек преобразует исходный комплекс условий
среды - пашет почву, вносит удобрения, орошает ее. Также
менее значимы исходные условия среды в организации руде-
ральных фитоценозов, в которых преобладают виды с широки-
ми экологическими амплитудами, позволяющими им произра-
стать в нарушенных местообитаниях с существенными разли-
чиями увлажнения или богатства почвы.
Что касается дифференциации экологических ниш, то этот
процесс наиболее развит в лесных сообществах (C-S-модель) и
наименее - в пустынных и водных сообществах (абиотическая
S-модель), а также в агрофитоценозах и рудеральных фитоце-
нозах (R-модель). Причина - низкий уровень диверсификации
видов и однотипность их стратегий выживания.
12.2. Основные признаки фитоценозов
12.2.1. Видовое богатство
Видовое богатство фитоценоза - это число видов, приходя-
щееся на единицу площади. Оно зависит от многих факторов,
первым из которых является размер учетной единицы.
Если выявлять видовое богатство с помощью вписанных
друг в друга квадратных или круглых площадок увеличиваю-
щегося размера, то с ростом площади учетной единицы будет
увеличиваться число выявленных в фитоценозе видов. Постро-
енная из полученных значений кривая называется кривой число
видов/площадь. Фитоценологами накоплен большой фактиче-
205
ский материал о форме этих кривых. В большинстве случаев
при заложении в фитоценозах «матрешек» площадок разме-
ром 0,1; 0,25; 1, 4, 9, 16, 25, 100 м2 и т.д. полученные кривые
вначале резко поднимаются вверх, а потом постепенно перехо-
дят на плато. Начало перехода на плато показывает, что на,
площадке этого размера в основном выявлены все виды фито-
ценоза. Чем фитоценоз богаче видами, тем меньше размер
площадки, при котором кривая уходит на плато (рис. 40).
В мелком масштабе наибольшим видовым разнообразием ха-
рактеризуются альварные луга, которые распространены в Эсто-
нии и Швеции. Эти луга связаны с неглубокими почвами на кар-
бонатных породах и образованы маленькими по размеру расте-
ниями. На учетной площадке размером 1 дм2 умещается до 40 ви-
дов. В курских степях В.В.Алехин (1931) на 1 м2 насчитывал до
100 видов. В крупном масштабе (сотни квадратных метров) наи-
более богаты видами тропические леса, где на 400 м2 может про-
израстать до двух тысяч видов деревьев, лиан и эпифитов.
Размер площадки, при котором происходит «перелом» кри-
вой (хотя не всегда он выражен четко), называется минималь-
ным. ареалом. Пробную площадку этого размера фитоценологи
используют для выполнения геоботанических описаний (основ-
ного фактического материала для характеристики растительно-
сти). Для фитоценолога геоботаническое описание подобно гер-
барному листу для ботаника-флориста.
Рис. 40. Кривые «число видов/площадь» для пустыни (а), пустынной (б)
и луговой (в) степи (★ - точка кривой, соответствующая минималь-
ному ареалу)
206
Размер площадки - это субъективный фактор, влияющий на
оценку видового богатства изучаемого фитоценоза, поскольку
ее размер исследователь выбирает сам. Существуют несколько
объективных факторов, определяющих число видов в фитоцено-
зе. Эти факторы взаимодействуют друг с другом, и потому
очень трудно построить математическую модель для прогноза
видового богатства. Рассмотрим эти факторы.
Флора. Потенциальный запас видов в данном районе, из ко-
торого могут отбираться виды для формирования того или
иного сообщества.
Экотоп. Экологические условия, в которых формируется
фитоценоз. При благоприятном экотопе - почвы достаточно
влажные, богаты питательными элементами и имеют близкую к
нормальной реакцию среды, а климат мягкий, - в одном фито-
ценозе потенциально могут произрастать многие виды. В экс-
тремальных условиях (пустыня, солончак, арктическая пустыня
и т.д.) потенциально может расти небольшое число видов, они
будут представлять группу патиентов. На рис. 41 показана зави-
Высота над уровнем моря, м
Волгоградское ]QQ
водохранилище
Число I--------1 I-----Л
ВИДОВ 1-------- 1------“-1
1-5 8-10
200 300 400 Каспий
Расстояние, км
W///////A ШЛ
11-15 16-20 21-25 26-30
Рис. 41. Изменение видового богатства сообществ (число видов на
100 м2) поймы в нижнем течении Волги (по данным Голуба, 1986)
207
симость видового богатства от условий среды - увлажнения и
засоления почвы - в низовьях реки Волги. Видовое богатство
снижается при увеличении засоления почв и при их переувлаж-
нении. Поэтому оно минимально в водных и прибрежно-водных
сообществах (нижняя часть рисунка) и максимальна в луговых
фитоценозах. Низким видовым богатством характеризуются и
окружающие пойму пустынно-степные сообщества пастбищ.
Стратегический спектр видов. В суровых услових экотопа в
фитоценозе будут преобладать патиенты. Но в благоприятных
условиях в составе фитоценоза может быть (или не быть) вио-
лент. При появлении виолента видовое богатство резко снижа-
ется. Так, в буковых лесах в напочвенном покрове крайне мало
видов, поэтому эти леса называют «мертвопокровными»; бедны
видами фитоценозы плавней с доминированием тростника в
дельтах рек.
Если виолент отсутствует, то экологические ресурсы мо-
гут быть разделены между разными видами за счет дифферен-
циации экологических ниш (как, например, в луговых степях), и
видовое богатство будет очень высоким.
Режим нарушений. Умеренный режим нарушений препятст-
вует усилению роли виолентов, и видовое разнообразие фито-
ценозов повышается. В травяном покрове широколиственного
леса, который умеренно посещается отдыхающими, видов боль-
ше, чем в заповедном лесу, где большую часть пространства
ниш захватывает сныть. Повышается видовое разнообразие лу-
гов и степей при умеренном выпасе. Если нагрузка нарушающе-
го фактора будет высокой, то видовое богатство фитоценоза
снизится, причем при периодическом действии фактора
(вспашка, проход техники с разрушением растительного покро-
ва и др.) будут преобладать эксплеренты, а при постоянном
(интенсивный выпас) - патиенты.
Возраст фитоценоза. Фитоценоз не развивается как единое
целое, поскольку виды в нем не приспосабливаются друг к дру-
гу. Они совмещены в сообществе в результате дифференциации
экологических ниш. Тем не менее, для того чтобы в фитоценоз
попали зачатки всех видов растений, которые потенциально мо-
гут обитать в нем, и эти виды дифференцировали свои эколо-
гические ниши, нужно время. Влияние возраста фитоценоза на
его видовое богатство может носить сложный (нелинейный) ха-
рактер. Несмотря на то, что в целом с возрастом число видов
увеличивается, в фитоценозе может появиться и усилиться
виолент, тогда видовое богатство на молодых стадиях развития
фитоценоза до появления виолента будет выше.
208
Видовое богатство на площадках, соответствующих мини-
мальному ареалу, изменяется в очень широких пределах: от 1-2
видов (в прибрежно-водной или водной растительности, в
пустынях или на злостном содовом солончаке) до 1000 видов (в
тропическом лесу).
Основные типы фитоценозов средней полосы России имее-
ют следующее видовое богатство:
Тип Число видов
Сенокосные луга.................... 40-60
Пастбищные луга.................... 20-30
Сенокосные степи................... 70-90
Пастбищные степи................... 20-40
Сегетальные пашенные сообщества..... 10-20
Рудеральные сообщества.............. 5-15
Широколиственный лес............... 25—40
Хвойный лес.......................... 15—25
Низинные травяные болота............. 10-20
Сфагновые болота..................... 10-20
В целом в современной растительности наблюдается тен-
денция снижения видового богатства фитоценозов из-за усили-
вающегося влияния на них человека (т.е. влияния антропогенно-
го пресса). Поэтому многие виды растений находятся в угро-
жаемом положении.
1222. Структура фитоценоза
Под структурой фитоценоза (синморфологией) понимается
количественное соотношение слагающих его видов, их взаимо-
расположение в пространстве и изменение этих параметров во
времени в результате обратимой (циклической) сезонной и раз-
ногодичной изменчивости. Соответственно, различают про-
странственную и временную структуру фитоценоза.
Временная структура связана со способностью фитоценоза
перестраиваться под действием сезонных изменений климата,
его колебаний от года к году и других периодически действую-
щих факторов без изменения флористического состава. Вре-
менную структуру нужно отличать от рассматриваемой далее
динамики фитоценозов, когда их изменения имеют направлен-
ность (векторизованы) и выражаются в изменении флористи-
ческого состава или в устойчивом изменении количественного
соотношения видов-доминантов.
209
В пространственной структуре различают вертикальную и
горизонтальную структуру, а во временной - сезонные и разно-
годичные изменения.
Вертикальная структура фитоценоза. Разновысотность рас-
тений и различия в распределении корневых систем по почвен-
ному профилю у разных видов - одна из главных осей диф-
ференциации экологических ниш в фитоценозе. В сомкнутом
фитоценозе в зависимости от высоты различается режим осве-
щения, а от глубины - режим увлажнения, засоления, содер-
жание элементов минерального питания в почвенном растворе.
Существует два основных варианта разновысотности:
ярусность, когда на глаз видно расчленение фитоценоза по
вертикали на достаточно четко отграниченные слои-ярусы, и
вертикальный континуум, когда такие слои в фитоценозе не
различаются (рис. 42). В подземной части фитоценоза ярусов в
распределении подземных органов, за очень редким исключе-
нием, не наблюдается.
Рис. 42. Ярусность в лесу (а) и вертикальный континуум на лугу (6)
Ярусность характерна для лесов умеренной полосы, где
четко различаются ярусы деревьев, подлеска и подроста, травя-
ной или травяно-кустарничковый, моховой или лишайниково-
моховой. Ярусность в природе встречается реже, чем верти-
кальный континуум, классическим примером которого является
вертикальная структура тропического леса. Континуальна вер-
тикальная структура луговых и степных травостоев, реже в
этих фитоценозах отмечается нечеткая размытая ярусность.
В фитоценозах, где ярусности нет или она выражена не-
достаточно четко, вертикальный континуум подразделяют на
210
условные слои - фитоценотические горизонты, число которых
зависит от задач исследования. При детальном изучении рас-
пределения по вертикали фитомассы в травяных фитоценозах
(например, для определения того, какая часть травостоя
окажется выше линии скашивания или может быть использова-
на определенным видом животного) растения последовательно
срезают с шагом всего 5 см, а в тропических лесах для изуче-
ния количества листьев, распределения видов фауны и т.д. бы-
вает достаточно выделить 4-5 слоев.
Важной универсальной синтетической характеристикой вер-
тикальной структуры фитоценоза, которая может использо-
ваться в фитоценозах как с ярусами, так и с вертикальным кон-
тинуумом, является индекс листовой поверхности (ИЛП) - от-
ношение площади поверхности листьев к площади поверхности
почвы, над которой они находятся. У разомкнутых сообществ
пустынь ИЛП составляет доли единицы, у большинства луговых
фитоценозов 4-6. В еловом лесу ИЛП может достигать 12, т.е.
на 1 га леса приходится 12 га листовой поверхности.
Разноглубинность проникновения корневых систем и раз-
личия в распределении максимальной массы корней характерны
для всех фитоценозов, хотя в фитоценозах умеренной полосы
зона корневого максимума многих видов находится на глубине
0-30 см. В то же время даже в луговом фитоценозе корни
Medicago falcata или Rumex confertus могут уходить в почву на
глубину нескольких метров. В растительности аридных терри-
торий существует особая группа фреатофитов
которые имеют глубокую корневую систему
грунтовых вод (например, верблюжья колючка
dalhagi, тростник обыкновенный - Phragmites australis и др.).
Эти виды используются для индикации уровня грунтовых вод
при бурении скважин.
Разноглубинность проникновения корней - яркий пример
дифференциации экологических ниш. Так, в солонцеватых сте-
пях в верхнем надсолонцовом слое расположены корни расте-
ний-гликофитов (типчака, ковылей, степного разнотравья и бо-
бовых), а в солонцовом горизонте корни галофитов, таких
как представители рода кермек (Limonium) или полынь Лерха
(Artemisia lerchiana).
Дифференциация экологических ниш по осям разновысот-
ности и разноглубинности используется в агроэкологии. В част-
ности, сидеральные культуры (бобовые, крестоцветные), кото-
рые имеют более глубокие корневые системы, чем зерновые
злаки, используются для возвращения в горизонт распростра-
растений,
и достигают
Alhagi pseu-
211
нения корней пшеницы или кукурузы питательных элементов
удобрений, вмытых в глубь почвы.
Такую же функцию в посевах выполняют сорные растения,
поэтому, если количество сорняков не превышает порога вре-
доносности (т.е. того количества, начиная с которого происхо-
дит существенное снижение урожаев), их наличие в агрофито-
ценозе полезно. Они не только возвращают в пахотный гори-
зонт вмытые в глубь почвы удобрения, но и накапливают в
своих запасающих органах вносимые удобрения. При перегни-
вании запасающих органов сорных растений питательные эле-
менты возвращаются в почвенный раствор.
Мы уже упоминали о работах Н.Т.Нечаевой по созданию
«агропустынь» - искусственных многовидовых продуктивных
фитоценозов из представителей местной флоры (иногда улуч-
шенных селекцией). Основной принцип конструирования этих
фитоценозов — подбор видов с разной высотой (деревья, кус-
тарники, кустарнички, травы) и разной глубиной корневых сис-
тем, чтобы обеспечить более полное использование ресурсов
солнечного света и почвы. Благодаря дифференциации экологи-
ческих ниш, продуктивность «агропустынь» в 4-5 раз выше,
чем естественных пустынь, видовое богатство которых значи-
тельно меньше.
В лесоводстве используются многоярусные поликультуры -
посадки из 2-3 видов древесных растений. Они не только пол-
нее используют ресурсы света и почвы, но и более устойчивы к
вредителям и болезням, а следовательно, быстрее накапливают
древесную массу и лучше «оздоравливают» атмосферу (выде-
ляют кислород, усваивают углекислый газ, а в пригородных зо-
нах поглощают пыль и токсичные примеси из воздуха).
Горизонтальная структура фитоценоза. Мы уже подчеркива-
ли, что регулярное и случайное распределение популяций в фи-
тоценозах встречается довольно редко. В большинстве фитоцено-
зов популяции распределены неслучайно и образуют группы. Но
чаще всего в итоге неслучайного распределения популяций скла-
дывается случайная горизонтальная структура фитоценоза.
Если в горизонтальной структуре фитоценоза выявляются
закономерно повторяющиеся пятна с различиями в составе
видов или их количественном соотношении, связанные с фито-
ценотическими причинами, то говорят о мозаичности фитоце-
ноза, а пятна называют микрог-руппировками.
Мозаичность следует отличать от пестротности фитоце-
ноза, когда микрогруппировки не выражены и неоднородность
носит случайный характер, и от комплексности растительно-
212
сти. При комплексности пятна, аналогичные микрогруппиров-
кам, возникают вследствие исходной неоднородности условий
среды. Примером могут служить, пятна солончаков в Прикас-
пийской пустыне, где на фоне полыни черной (Artemisia
diversifolia) формируются пятна полыни белой (Artemisia
lerchiana), или неоднородность растительности на южных скло-
нах гор, где чередуются небольшие участки почв разной мощ-
ности с разным содержанием хряща-скелета.
Различаются следующие варианты мозаичности фитоценозов.
Регенерационные мозаики — неоднородность фитоценоза,
связанная с возобновительным процессом — «моделью карусе-
ли». Эта модель описывает поочередное пребывание в одной
нише нескольких видов, причем разные части фитоценоза нахо-
дятся на разных фазах «карусели», и поэтому его общий видо-
вой состав остается постоянным. Масштаб этого процесса в
травянистых фитоценозах измеряется сантиметрами и деци-
метрами, а в лесах он достигает десятков и сотен метров.
При выпадении деревьев в лесу образуются «окна», зарастаю-
щие видами, специально приспособленными к такому «ремонту»
древостоев. Это наиболее характерно для тропических лесов, в ко-
торых существуют две группы видов: дриады - основные виды и
номады - виды, которые временно поселяются в «окнах» и под
пологом которых постепенно восстанавливаются дриады. Номады
образуют либо банк семян, либо банк проростков, т.е. всходов,
которые подолгу не идут в рост, дожидаясь образования «окна».
Если «окно» не образуется, то через 3-7 лет они гибнут и заме-
щаются новыми всходами. Такой банк имеется и у деревьев уме-
ренных широт, например у дуба (эти проростки у него называются
«торчками»). В тайге «окна» зарастают березой, ольхой и ивами,
играющими роль растений-«нянь» для ели.
Клановые мозаики - неоднородность фитоценоза, связанная
с вегетативным размножением растений. Полное описание
клоновых мозаик для папоротника орляка (Pteridium aquilinum)
сделал английский ботаник А.Уотт (A.Watt). Папоротник раз-
растается круглым пятном; постепенно, вследствие старения,
его продуктивность в центральной части пятна снижается, и
уменьшается мощность подстилки, защищающей корневища от
мороза. В результате в центре пятна папоротник вытесняется
злаками, и микрогруппировка имеет вид кольца из папоротника,
внутри которого растут злаки. Постепенно кольцо распадается
на отдельные части, каждая из которых разрастается в новый
круглый клон со стареющей центральной частью и т.д.
213
Разрастающиеся куртины степных кустарников также
формируют микрогруппировки. Так, в Монголии кустарник ка-
рагана мелколистная (Caragana microphylla) разрастается круг-
лыми куртинами, диаметр которых увеличивается с возрас-
том. Внутри куртины кустарника скапливаются эоловые нано-
сы и образуется холмик, на котором поселяются рудеральные
растения. Лучше, чем между куртинами, в этой микрогруппи-
ровке растет и ковыль Крылова (Stipa krylovii), так как там
он находит защиту от высоких нагрузок пасущихся животных.
Чешские исследователи (T.Herben, F.Krahulec) назвали виды,
способные разрастаться и формировать клоновые мозаики, «пар-
тизанами», противопоставив их видам-«сидням», которые дли-
тельное время произрастают в одном и том же месте в фитоце-
нозе (его называют микросайтом или микроместообитанием).
Фитоэнбайроментальные мозаики связаны с изменением
среды одним из видов и реагированием на Это изменение других
видов. Как правило, основным фактором формирования моза-
ичности выступает бобовое растение, так как симбиотически
связанные с ним азотфиксирующие бактерии повышают содер-
жание азота в почве.
В Башкортостане в пойме реки Белой изредка наблюдаются
микрогруппировки, образованные клевером горным (Trifolium
montanum). Внутри круглых пятен клевера горного диаметром
5-7 м в результате конкуренции за свет подавлены и злаки, и
разнотравье. Вокруг пятен клевера («центра») густым «ободом»
разрастаются злаки мятлик узколистный (Роа angustifolia),
типчак (Festuca valesiaca) и тонконог Делявиня (Koeleria
delavignei), которые здесь пользуются азотом, не испытывая
сильной конкуренции за свет и влагу. Между пятнами из «цен-
тра» и «обода» злаки, бобовые и разнотравье растут пример-
но в одинаковом соотношении.
Аллелопатические мозаики встречаются редко, поскольку,
как мы уже отмечали, аллелопатические эффекты редко могут
стать главным фактором организации фитоценоза. Опублико-
ваны описания нескольких вариантов аллелопатических мозаик,
из которых наиболее интересны мозаики шалфейного скраба в
Северной Америке.
Скрабы - это особый тип фитоценозов, где среди пышной
травяной растительности разбросаны куртины колючих кустар-
ников из родов шалфей (Salvia) или полынь (Artemisia). В шал-
фейном скрабе, изученном Г.Муллером (H.Muller), вокруг кур-
тин шалфея, который выделяет сильно пахнущие летучие аро-
матические вещества, образуются не занятые растительностью
214
«пролысины». Периодически в скрабе происходят пожары, по-
сле которых формируется равномерный покров травяной расти-
тельности, так как шалфей относится к пирофитам. Через 5-7
лет из сохранившихся в почве корневищ восстанавливается
шалфей, и вокруг его разрастающихся куртин образуются но-
вые «пролысины».
Феномен мозаичности используется, хотя пока еще сравни-
тельно редко, в луговодстве. Обычно при сплошном посеве
травосмеси из злаков и бобовых в первые два года разрастают-
ся бобовые, а злаки подавляются, поэтому приходится увели-
чивать количество семян злаков в высеваемой смеси. Однако
позднее раскустившиеся злаки начинают вытеснять бобовые из
травостоя, и снижается содержание белка в получаемом сене.
Опыт создания мозаичных посевов из чередующихся полос зла-
ков и бобовых или расположенных в шахматном порядке кле-
ток злаков и бобовых размером 4 м2 показывает, что эти посе-
вы устойчивее и продуктивнее сплошных.
Временная структура фитоценоза. Фитоценоз постоянно
изменяется во времени, подстраивая свою структуру и функ-
цию под изменения условий среды - суточные, сезонные, раз-
ногодичные. Кроме того, на временную структуру фитоценоза
влияют ритмы развития отдельных видов (периодичность цве-
тения, плодоношения и т.д.).
На наземные фитоценозы суточные ритмы влияют сравни-
тельно слабо. Так, у некоторых растений (латук татарский
Lactuca tatarica, клевер - р. Trifolium) в течение суток изменяет-
ся угол наклона листьев; у туранговых тополей (Populus diver-
sifolia, Р. pruinosa), растущих в поймах среднеазиатских рек, из-
меняется положение листовых пластинок, которые во время
солнцепека повернуты к солнечным лучам ребром, а утром и ве-
чером - широкой стороной. Происходят суточные изменения ха-
рактеристик сообществ речного планктона: днем они располага-
ются на глубине 1-2 м, а при снижении освещенности (вечером и
утром) концентрируются в приповерхностном слое.
Более значимы для наземных фитоценозов сезонная и раз-
ногодичная изменчивость окружающей среды.
В составе фитоценозов есть группы видов, имеющих разные
циклы сезонного развития, поэтому внешний облик фитоценоза
в течение сезона многократно изменяется (рис. 43). Например, в
широколиственных лесах весной в напочвенном покрове пред-
ставлены эфемероиды («подснежники») - ветренички (Anemo-
noides), хохлатка (Corydalis solida), гусиный лук (Gagea lutea),
чистяк (Ficaria verna). Они отцветают до распускания листьев
на деревьях.
215
май июнь июль август
31 10 20 30 10 20 31 10 20
Pukatilla flavescens
Myosotis suaveolens
Kobresia filifolia
Anemone sylvestris
Oxytropis strobilacea
Plantago canescens
Potentilla stipularis
Taraxacum ceratophonim
Poa pratensia
Rumex thyreiflonis
Bromus ircutensis
Vicia cracca
Lychinia sibirica
Seneck) jacobaea
Onobrychis sibirica
Pedicular» venusta
Aster alpimis
Achillea millefolaim
Hordeum brevisubulatum
Galium verum
Allium splendena
Geranium pratense
Dianthus versicolor
Leontopodium palibmianum
Silene repens
Campanula glomerate
31 10 20 30 10 20 31 10 20
1-H^n 2-H^n 5-К4И4Й 6-ESEZ3
a)
Число
июнь июль август
б)
Рис. 43. Фенологический спектор (а) и кривая цветения (б) остепненно-
го луга поймы Лены (по Кононову, 1982): 1 - вегетация, 2 - бутониза-
ция, 3 - цветение, 4 - созревание полодов и семян, 5 - осыпание пло-
дов и семян, 6 - отмирание
216
Неодновременно зацветают виды лугов и степей. В запо-
веднике «Стрелецкая степь», где охраняются участки северных
луговых степей на черноземах, поочередно зацветают сон-
трава (Pulsatilla patens) с фиолетовыми цветками, горицвет ве-
сенний (Adonis vernalis) - с желтыми, шалфей луговой (Salvia
pratensis) - с синими и т.д. Облик фитоценозов изменяется, и
тогда говорят о смене аспектов. Разновременное цветение явля-
ется одним из проявлений дифференциации экологических
ниш, снижающей конкуренцию между популяциями. В феноло-
гических изменениях фитоценозов отражаются их адаптации к
климату и почвенным условиям.
Для обозначения разногодичных изменений фитоценозов
используется термин «флюктуация». Флюктуации вызываются
различными факторами, внешними и внутренними по отноше-
нию к фитоценозу, продолжительность их цикла не превышает
10 лет (более длительные циклические изменения рассматрива-
ются как циклические сукцессии, представляющие переход от
флюктуаций к векторизованным изменениям). Т.А.Работнов
предлагает различать следующие типы флюктуаций.
Наиболее обычны экотопические (климатогенные) флюк-
туации. Облик луговых фитоценозов в результате экотопиче-
ских флюктуаций может изменяться до неузнаваемости.
Урожайность пойменного луга на гриве в долине реки Бе-
лой в разные годы изменяется в 10 раз. Во влажный год (высо-
кий паводок, обильные осадки) в послепаводковый период уро-
жайность луга достигала 40 ц/га сена, и в травостое преоблада-
ло разнотравье: виды лабазника (р. Filipendula), герань (Gerani-
um pratense), кровохлебка (Sanguisorba officinalis), василистники
(р. Thalictrum) и др. В сухой год урожайность снижалась до
4 ц/га, и травостой был представлен типчаком (Festuca valesiaca)
и мятликом узколистным (Роа angustifolia). Большая часть рас-
тений луга оставалась в покоящемся состоянии.
Экотопические флюктуации фитоценозов у северной гра-
ницы степей приводят к тому, что (при однократном учете)
граница во влажный год может сместиться на юг, а в сухой -
на север (луга и степи различаются по соотношению в фитоце-
нозах мезофильного разнотравья и дерновинных злаков: типча-
ка, ковылей - р. Stipa, тонконога стройного - Koeleria cristata).
В экстремальные годы граница может «смещаться» на 100 км.
Такое изменение облика фитоценозов в разные годы было од-
ной из причин отказа от классификации травяных фитоценозов
по составу обильных видов. В разные годы в одном и том же
фитоценозе обильными бывают разные виды, хотя при этом
217
фитоценоз остается все тем же, он лишь находится в разных
флюктуационных состояниях.
Экотопические флюктуации выражены и в пустынных фи-
тоценозах. В них периодически в благоприятные периоды года
массово развиваются однолетники из числа «ложных» экспле-
рентов. Р.Уиттекер, изучавший фитоценозы пустыни Сонора,
пишет, что для разных вариантов благоприятных периодов
имеются свои наборы видов. Поэтому для выявления всего по-
тенциально возможного видового состава пустынных фитоцено-
зов нужно не менее 10 лет.
Фитоциклические флюктуации связаны с особенностями
биологических ритмов растений, входящих в состав фитоценоза.
Дуб (Quercus robur) обильно плодоносит в среднем один раз в
четыре года, и на следующий после этого год в фитоценозе бу-
дет много его всходов. Клевер (p.Trifolium) обильно цветет один
раз в четыре-пять лет (в так называемые клеверные годы). В
клеверные годы, кстати, можно наблюдать формируемую клеве-
ром горным (Т. montanum) фитоэнвайроментальную мозаику, о
которой говорилось п. 12.2.2.
Зоогенные флюктуации обусловлены массовым развитием
какого-либо вида животных (этот вид может появиться в сооб-
ществе из соседней или даже весьма удаленной экосистемы, на-
пример, прилет саранчи). Так, в широколиственных лесах в го-
ды массового развития непарный шелкопряд уничтожает прак-
тически всю листву на деревьях, экскременты гусениц обильно
удобряют почву. Это дополнительное питание и улучшение ре-
жима освещения повышает продуктивность травяного яруса в
1,5-2 раза.
Интересные зоогенные флюктуации фитоценозов наблюда-
ются в степях Монголии. В отдельные годы там массово разви-
вается полевка Брандта, которая почти полностью выедает дер-
нины ковылей и разрыхляет поверхность почвы. Это приводит к
тому, что из почвенного «банка корневищ» массово развивается
колосняк хангайский (Leymus chinensis), и степь изменяется до
неузнаваемости. Однако через 4-5 лет ковыли восстанавливают-
ся, а колосняк снова становится второстепенным видом, хотя с
обильным «банком» вегетативных зачатков в почве.
Антропические флюктуации связаны с изменениями антро-
погенного влияния на фитоценозы. Например, при разном ре-
жиме пастьбы может изменяться состав травостоя пастбищ.
При периодическом внесении минеральных удобрений изменя-
ется состав естественного луга. У фитоценозов, расположенных
в сфере влияния промышленных предприятий, состав может
изменяться под действием однократных или периодических
эмиссий веществ, загрязняющих атмосферу и почву.
218
12.3. Динамика растительности
В последние десятилетия роль НОР и фитоценологии как ее
части возросла. Это связано с принципиальной возможностью
по состоянию растительности осуществлять мониторинг за из-
менениями природы под усиливающимся воздействием челове-
ка. Поэтому изучение динамики фитоценозов, как под влияни-
ем человека, так и связанной с естественными процессами, се-
годня является одним из самых важных разделов НОР.
12.3.1. Классификация форм динамики растительности
Рассмотрим векторизованные (направленные) изменения
фитоценозов, при которых изменяется видовой состав или ко-
личественное соотношение видов, т.е. структура фитоценоза.
Этим векторизованные изменения отличаются от «структуры
во времени» - сезонных и разногодичных колебаний состава и
структуры фитоценозов, при которых видовой состав сохраня-
ется постоянным, а изменения выражаются только в колебаниях
обилия доминантных видов.
Различают три основных класса динамики фитоценозов
(рис. 44).
Нарушения. При нарушениях (пожар, горный сель, вырубка
леса, разлив большого количества нефти и т.д.) за короткое
время уничтожается весь фитоценоз или его часть. В последнем
случае нарушения имеют локальный характер, например вывал
дерева в лесу под влиянием ветра, когда при выворачивании де-
рева с корнем образуется холмик земли.
219
Сукцессии. Эти постепенные изменения фитоценозов вызы-
ваются внутренними (взаимоотношения растений, отношения
растений и условий среды) или внешними по отношению к фи-
тоценозам причинами. Первые сукцессии называются автоген-
ными, вторые - аллогенными.
Эволюция. Эти изменения подобны сукцессиям, так как со-
став фитоценозов изменяется постепенно. Отличие заключается
в том, что в результате сукцессии новые фитоценозы не возни-
кают, и формируются сочетания видов, уже существовавшие в
данном районе, а в ходе эволюции возникают новые фитоцено-
зы. Эволюция - это не обязательно прогресс и формирование
более богатых видами фитоценозов, чем существующие.
По предложению В.Н.Сукачева (1954) различают по два ти-
па автогенных и аллогенных сукцессий.
Автогенные сукцессии разделяются на сингенез и эндоэко-
генез. При сингенезе изменения происходят под влиянием
взаимоотношений между растениями. Этот быстрый процесс
можно наблюдать на богатом субстрате, где под влиянием био-
логических особенностей растений (наличие или отсутствие
банка семян в почве, способ доставки семян к месту сукцессии,
длительность жизни) за несколько лет происходит формиро-
вание фитоценоза вначале из видов R-стратегии, на смену кото-
рым приходят виды со стратегиями RS, CR и CRS. При этом
растения-однолетники заменяются многолетниками. При эндо-
экогенезе в результате жизнедеятельности растений изменя-
ются условия среды (на скалах формируется почва, зарастает и
мелеет озеро и т.д.).
Оба типа сукцессий тесно связаны. При сингенезе также
происходит изменение условий среды, но роль этих изменений
по сравнению с ролью взаимоотношений растений незначитель-
на, так как почва не успевает существенно и, тем более, необ-
ратимо преобразоваться. При эндоэкогенезе взаимоотношения
между растениями также определенным образом влияют на из-
менения в фитоценозе, но слабее, чем преобразование фитоце-
нозом собственной среды обитания.
Аллогенные сукцессии разделяются на гейтогенез - ло-
кальные изменения конкретных фитоценозов и гологенез - из-
менение фитоценозов в пределах целого ландшафта (например,
в процессе развития форм рельефа, под влиянием строительства
водохранилищ и др.).
Сукцессии, кроме того, классифицируются по следующим
критериям:
220
по масштабу времени - на быстрые, протекающие в мас-
штабе десятилетий; средние, длящиеся столетиями; медленные,
продолжающиеся тысячелетиями, и очень медленные (десятки
тысяч лет);
по обратимости - на обратимые и необратимые;
по степени постоянства процесса - на постоянные и пре-
рывающиеся;
по происхождению - на первичные, т. е. те, которые начи-
наются на субстратах, где никогда не было растительности, и
вторичные, происходящие в местообитаниях, где растительность
была, но уничножена;
по характеру изменения структуры и видового состава -
на прогрессивные (в результате сукцессии происходит увеличе-
ние видового богатства, повышение продуктивности) и регрес-
сивные (в результате сукцессии уменьшается видовое богатст-
во, снижается продуктивность). Между видовым богатством
фитоценоза и его продуктивностью нет жесткой положительной
зависимости, и поэтому иногда при обеднении видового состава
продуктивность повышается. В таких случаях по тенденциям
изменения видового состава и продуктивности отдельно разли-
чают прогрессивные и регрессивные сукцессии;
по антропогенности - на антропогенные (вызванные влия-
нием человека) и природные (происходящие по естественным
причинам).
12.32. Автогенные сукцессии
В современной растительности автогенные сукцессии очень
разнообразны, они могут быть первичными и вторичными, про-
грессивными и регрессивными, быстрыми и медленными. Как
правило, автогенные сукцессии необратимы и протекают посто-
янно. Несмотря на спонтанность этих сукцессий, их может вы-
звать человек, нарушающий растительность. На месте наруше-
ний начинаются восстановительные автогенные сукцессии.
Все разнообразие автогенных сукцессионных процессов
сводится к четырем принципиальным моделям сукцессии, пред-
ложенным Дж.Кэнелем (J.Connel) и П.Слейтером (P.Slatyer).
Модель благоприятствования. В ходе сукцессии смена видов
связана с постепенным улучшением условий среды, и поэтому
сукцессия протекает как прогрессивная. Такую сукцессию при
зарастании скал изучал крупнейший американский фитоценолог
XX в. Ф.Клементс. Для процесса зарастания скал он выделял
стадии накипных и кустистых лишайников, а также мхов, трав,
221
кустарников, деревьев. Растения (и, разумеется, связанные с
ними гетеротрофные организмы) в ходе этой сукцессии преоб-
разуют горную породу и создают почву. По мере улучшения
условий среды возрастает продуктивность фитоценоза, услож-
няется его структура (появляется ярусность) и повышается
видовое богатство.
Этой же модели соответствует естественный процесс зарас-
тания отвалов пустой породы от горных разработок, если поро-
да пригодна как субстрат для жизни растений. Если порода не
пригодна для растений, человек может помочь началу сукцес-
сии, покрывая поверхность породы слоем гумусированной поч-
вы или торфа. Ускорить сукцессию можно высевом травосмеси
или внесением удобрений (это практиковали американские эко-
логи при восстановлении растительности после нефтяных раз-
работок на Аляске).
Модель толерантности. Сукцессии, протекающие по этой
модели, распространены в природе не менее часто, чем сукцес-
сии, соответствующие модели благоприятствования. Как прави-
ло, эти сингенетические сукцессии наблюдаются в тех случаях,
если растения, заселяя местообитания с исходно благоприят-
ными условиями среды, постепенно расходуют ресурсы, при
этом усиливается конкуренция. В результате видовой состав
меняется в направлении усиления роли видов с выраженным
свойством патиентности.
Примером проявления модели толерантности может слу-
жить сукцессия на вырубке. Если вырубка используется в ка-
честве сенокоса или пастбища, то лес не восстанавливается, а
формируются сенокосные и пастбищные травостои, типичные
для данной зоны. В лесной зоне на смену крупнотравью, кото-
рое развивается вследствие минерализации лесной подстилки и
корней вырубленных деревьев (иван-чай Hatnerion angusti-
folium, лабазник вязолистный - Filipendula ulmaria), приходят
злаково-разнотравные фитоценозы с участием вейников (р. Са-
lamagrostis), пырея ползучего (Elytrigia repens), овсяницы луго-
вой (Festuca pratensis), ежи сборной (Dactylis glomerata). По ме-
ре того, как с урожаем из почвы выносятся питательные эле-
менты, продуктивность травостоя постепенно снижается и
крупные злаки замещаются мелкими - полевицей тонкой (Ag-
rostis tenuis), трясункой средней (Briza media), душистым колос-
ком (Anthoxathum odoratum).
Если интенсивное сенокошение продолжается, то сукцессия
приводит к формированию травостоя из белоуса (Nardus stricta).
Если вырубка используется как пастбище, то преобладающими
222
становятся виды из рода манжетка (Alchemilla). Описанные сук-
цессии, протекающие по модели толерантности, нежелательны.
Применяя удобрения и правильно используя вырубки, можно
воспрепятствовать этому и задержать сукцессию на стадии цен-
ных в кормовом отношении и продуктивных злаков.
Сукцессии по модели толерантности протекают и при само-
восстановлении елового леса, когда вначале появляются расте-
ния- «няни» (ольха, березы, ивы), а затем под их пологом - ус-
тойчивая к затенению и, следовательно, с более выраженным
свойством патиентности ель. Под пологом ели развиваются еще
большие патиенты - мхи и теневыносливые кустарнички и травы.
Модель ингибирования. Эта модель соответствует регрес-
сивным сукцессиям, когда процесс приостанавливается в ре-
зультате появления в фитоценозе видов, которые создают усло-
вия, непригодные для поселения новых видов. Такие сукцессии
наблюдались в Англии на гарях, где развивается густой покров
мха кукушкин лен (Polytrichum commune). Семена деревьев не
достигают поверхности почвы, и фитоценозы десятки лет со-
храняются в виде зеленомошной гари.
Аналогичный процесс происходит в пустынях Средней
Азии, где при отсутствии выпаса поверхность почвы покрывает-
ся плотной коркой из мхов и водорослей (ее называют карахар-
санг). В результате нарушается семенное возобновление пус-
тынных деревьев и кустарников и происходит деградация всей
экосистемы.
Характерно, что и на гари, и в пустыне сукцессия по моде-
ли ингибирования обусловлена тем, что из экосистем исключа-
ются животные. Если моховую гарь посещают олени, то они
копытами нарушают моховой покров и создают регенерацион-
ные ниши для возобновления деревьев. Подобным же образом
пасущиеся в пустыне верблюды или овцы создают регенераци-
онные ниши для поселения растений.
Модель нейтральности. Этой модели соответствуют сукцес-
сии, при которых изменения фитоценозов протекают как попу-
ляционный процесс, а роль взаимодействия популяций незначи-
тельна. Такие сукцессии крайне редки, причем если видовой со-
став конкретной стадии сукцессии определяется популяцион-
ными механизмами (биологическими особенностями видов), то
количественные соотношения видов (т.е. структура) в большей
степени зависят от взаимоотношений популяций.
В ходе сукцессий может протекать смена моделей (на ран-
них стадиях сукцессия соответствует модели благоприятство-
вания, а на более поздних - модели толерантности). Примером
223
сукцессии со сменой модели является процесс зарастания озера
и последующего развития на месте озера верхового болота.
Вначале, когда чаша озера заполняется отмирающими расте-
ниями и повышается уровень дна, условия для роста растений
могут улучшаться и на смену плавающим макрофитам (рде-
стам - р. Potamogeton, телорезу - Stratiotes aloides, ряскам
р. Letnna) приходят высокие полупогруженные растения - тро-
стник (Phragmites australis), рогоз (Thypa latifolia), хвощ болот-
ный (Equisetum palustre).
Постепенно озеро окончательно зарастает и превращается в
так называемое низинное, или эвтрофное (богатое), болото: на
нем начинается процесс накопления торфа и поселяются осоки
(р. Carex) и ольха черная (Alnus glutinosa). Год от года мощ-
ность слоя торфа увеличивается, и растения, корни которых
уже не достигают почвы, переходят на питание за счет торфа.
Минерализация торфа происходит медленно и не до конца, и
каждому новому поколению растений достается все меньше пи-
тательных элементов.
На смену растениям, требовательным к условиям питания,
приходят виды, приспособленные к обитанию в условиях огра-
ниченных ресурсов минеральных элементов: береза пушистая
(Betula pubescens), вахта трехлистная (Menyanthes trifoliata),
белокрыльник болотный (Calla palustris). При этом формируется
переходное болото. При дальнейшем нарастании торфяного
слоя условия питания еще больше ухудшаются, и все болото
покрывается сфагновым мхом.
Низинное болото превращается в верховое. На нем появля-
ется толерантная форма сосны (Pinus sylvestris f. pumila), а так-
же клюква (Oxycoccus palustris) и другие растения из семейства
вересковых - багульник болотный (Ledum palustre), болотный
мирт (Chamaedaphne calyculata), подбел (Andromeda polifolia).
12.3.3. Аллогенные сукцессии
Аллогенные сукцессии обусловлены влиянием внешнего фак-
тора, который действует на фитоценоз. В его отсутствие алло-
генная сукцессия сменяется автогенной восстановительной.
В большинстве случаев аллогенные сукцессии соответствуют
модели толерантности: по мере ухудшения условий среды проис-
ходит смена видов в направлении их более высокой патиентности
и приспособленности к фактору, вызвавшему сукцессию. Реже
аллогенные сукцессии протекают по модели благоприятствова-
ния, например, при удобрении лугов или эвтрофикации (повы-
224
шении содержания в воде питательных элементов) водоемов. Но в
этом случае уменьшается видовое богатство фитоценозов.
При гологенезе меняется растительность целого ландшафта
или его существенной части. Такие сукцессии могут быть при-
родными и антропогенными. Примером природной гологенети-
ческой сукцессии являются изменения растительности при раз-
витии речной долины (рис. 45). Каждый год река углубляет
русло (базис эрозии), и за счет наилка паводковых вод повыша-
ется поверхность поймы. Кроме того, при постепенном измене-
нии русла реки (меандрировании) разрушается часть высокого
берега и наращивается песчаный пляж. В результате на вновь
намываемых местообитаниях формируются пионерные (первич-
ные) сообщества, которые по мере удаления от них реки и по-
вышения уровня изменяются в ходе сукцессии (формируются
ивняки, затем тополевники, вязовники и липняки).
Рис. 45. Зоны речной поймы средней полосы европейской части России:
А—Б-поперечный профиль; И - ива; Т - тополь; В - вяз; Л -
липа; О - ольха черная
Примером антропогенного гологенеза является изменение
растительности под влиянием строительства гидротехнических
сооружений. Водохранилище изменяет климат, который стано-
вится более мягким. Выше плотины происходит подтопление
почв, в результате леса и травяная растительность сменяются
фитоценозами, устойчивыми к высокому уровню грунтовых вод
(вместо липово-дубовых лесов развиваются ольшаники, а вместо
225
сухих лугов и степей - осоковые болота). Ниже плотины имеет
место иссушение поймы, а в южных районах - еще и засоление
почв.
Оба рассмотренных примера показывают основную особен-
ность гологенеза: под влиянием внешних факторов изменяется
совокупность растительности обширных территорий, причем
изменения состава разных фитоценозов взаимосвязаны.
Гейтогенетические сукцессии - это изменения отдельных
фитоценозов, на которые действует внешний фактор. Приведем
некоторые примеры.
Сукцессии под влиянием радиации. Эти сукцессии изучали
Г.Вудвелл (G.Woodwell) и Р.Уиттекер (R.Whittaker) на экспери-
ментальном полигоне на острове Лонг-Айленд (США). С по-
вышением радиационной нагрузки, задаваемой гамма-излуча-
телем, изменения растительности шли в обратной последова-
тельности к сукцессии по модели благоприятствования при за-
растании скал. Вначале исчезали деревья, потом кустарники, за-
тем травы, мхи и лишайнику.
В окрестностях Чернобыльской АЭС после катастрофы
также произошло усыхание деревьев (эту зону назвали «бу-
рым лесом»), но кустарники и травы сохранились, т.е. сук-
цессия дошла до первой стадии.
Сукцессии под влиянием выпаса. Это самый распространен-
ный вариант сукцессии. Сегодня связанные с избыточным пого-
ловьем скота перевыпас и обеднение фитоценозов — одно из са-
мых тяжелых последствий хозяйственной деятельности человека.
Выпас действует на травянистые фитоценозы прямо из-
бирательно скусываются растения и косвенно - через уплотне-
ние почвы и изменение ее водно-воздушного и солевого режи-
ма. В результате высокотравные и малоустойчивые к выпасу
растения сменяются пастбищным низкотравьем: подорожником
средним (Plantago media), клевером ползучим (Trifolium repens),
одуванчиком лекарственным (Taraxacum officinale), лапчаткой
гусиной (Potentilla anserina) и др. При этом уменьшается видо-
вое богатство фитоценоза, упрощается структура, снижается
общее проективное покрытие. Сукцессия при сильном выпасе
(перевыпасе) называется пастбищной дигрессией.
В степных сообществах (рис. 46а) под влиянием выпаса вна-
чале исчезают ковыли и усиливается типчак (рис. 466). При
усилении вый&са типчак замещается полынью австрийской (Ar-
temisia austriaca, рис. 46в).
В результате пастбищной дигрессии продуктивность траво-
стоев может снижаться в 10 и более раз. Воспрепятствовать
226
столь нежелательным последствиям можно внедрением пастби-
щеоборота, при котором нагрузка скота на пастбище нормиру-
ется и разным участкам периодически предоставляется отдых.
Рекреационные сукцессии. Эти сукцессии протекают в рас-
тительности под влиянием отдыха людей. Основной фактор -
вытаптывание, и потому рекреационная сукцессия отличается
от пастбищной только тем, что здесь отбор идет на устойчивые
к вытаптыванию виды без учета их кормовых качеств. В лесах
рекреационная сукцессия вначале захватывает травяной ярус, в
котором лесные виды трав замещаются более устойчивыми к
вытаптыванию луговыми растениями. При этом видовое богат-
ство фитоценозов повышается.
б)
)
Рис. 46. Пример пастбищной дигрессии степного фитоценоза
При усилении вытаптывания преобладание переходит к рас-
тениям розеточных форм, видовое разнообразие падает, сук-
цессия захватывает подлесок и даже древесный ярус, так как
уплотнение почвы препятствует возобновлению деревьев, ухуд-
шает их состояние и может привести к усыханию древостоя.
Правильной организацией территории (обозначением пешеход-
ных троп, отведением специальных площадок для игр, обуст-
ройством мест отдыха и т.д.) можно удержать сукцессию на
ранних стадиях, на которых при достаточно интенсивном отды-
хе формируется разреженный парковый лес с травяным ярусом
лугового типа.
Человек управляет аллогенными сукцессиями экологиче-
ским нормированием, т.е. регулированием нагрузки на фитоце-
ноз того фактора, который вызывает сукцессию. Считается, что
если в фитоценозе сохранилась хотя бы половина видов, то по-
сле прекращения действия фактора, вызвавшего аллогенную
сукцессию, возможно самовосстановление фитоценоза в ре-
зультате процесса обратного изменения, т.е. вторичной автоген-
ной сукцессии, которая называется восстановительной.
12.3.4. Концепция климакса в растительности
Концепция климакса (устойчивого состояния, к которому
«стремится» сообщество в ходе автогенной сукцессии) разраба-
тывалась американскими экологами в конце прошлого и начале
нашего столетия. Наиболее полно она была развита Ф.Кле-
ментсом, представления которого получили название монокли-
макса в растительности.
Суть взглядов Клементса сводилась к следующему. В пре-
делах одной климатической зоны все фитоценозы в ходе сук-
цессии должны конвергировать к одному климаксовому фито-
ценозу. Например, в зоне тайги болота при достижении опреде-
ленной мощности торфяной залежи должны зарасти лесом, на
скалах должна образоваться почва и вырасти такой же лес и
т.д. В каждой климатической зоне на любых типах субстратов в
результате сукцессии все фитоценозы сходятся в один тип. Для
тайги это будет еловый лес, для степной зоны - ковыльная
степь, для тундр - сообщества мелких ив, осок и лишайников.
Клементс считал, что климаксовый фитоценоз формируется
очень долго - тысячи и десятки тысяч лет, причем он допускал
возможность различных отклонений от основного климакса, ко-
торый назывался климатическим. Так, если был фактор, кото-
рый препятствовал формированию климакса (например, выпас),
то возникающие фитоценозы назывались дисклимаксобыми, а
если формированию климакса мешали местные экологические
факторы (например, паводки в пойме), то образованные фито-
ценозы получали название субклимакса и т.д. К концу жизни
(Клементс публиковал свои работы в течение почти пятидесяти
лет) он выделил около 30 различных вариантов климакса. Кли-
максовые устойчивые, фитоценозы Клементс противопоставлял
серийным, т.е. сукцессионным. Смена серийных фитоценозов
приводит к климаксу.
Представления Ф.Клементса о моноклимаксе стали предме-
том критики. В результате была сформулирована концепция по-
ликлимакса'. в одной климатической зоне фитоценозы разных
местообитаний изменяются в ходе сукцессии, но в один тип не
конвергируют, а на песках, скалах, болотистых местообитаниях
и т.д. формируют свои климаксы.
Р.Уиттекер (R.Whittaker) предложил третий вариант кон-
цепции климакса - климакс-континуум. Он считал, что между
климаксовыми фитоценозами существуют переходы, и поэтому
количество климаксов в поликлимаксе стремится к бесконечно-
сти. В каждой точке, утверждал Уиттекер, свой индивидуальный
климакс.
228
В современной фитоценологии концепция климакса играет
важную роль, но климакс не абсолютизируется, а понимается
как тенденция формирования фитоценозов зонального типа.
12.3.5. Эволюция фитоценозов
Фитоценоз не представляет собой целостности («организма
второго порядка») с жестко связанными популяциями растений.
По этой причине эволюция растительности представляет собой
сеткообразный процесс: виды эволюционируют независимо друг
от друга, а в одном фитоценозе они совмещаются за счет
дифференциации экологических ниш. При этом устойчивые свя-
зи между видами (кроме случаев «хозяин-паразит») не возни-
кают. Каждый вид по-своему приходит в фитоценоз и по-
своему уходит из него. При этом важную роль играет диверси-
фикация: эъомоцпя не «к», а «от», т.е. повышение несходства
видов. Несходство позволяет видам занимать разные экологиче-
ские ниши в одном фитоценозе и, следовательно, снижать кон-
куренцию. Различающиеся по биологии и экологии виды легче
сочетаются в фитоценозах, чем виды со сходными потребностя-
ми в ресурсах.
Эволюция растительности как природный процесс протека-
ет в геологических масштабах времени. В последние два столе-
тия он полностью подавлен быстрым процессом эволюции
под влиянием человека - антропогенной эволюцией.
Антропогенная эволюция может быть целенаправленной,
когда человек ставит задачу создать новые типы фитоценозов,
соответствующие его потребностям, и стихийной, когда эволю-
ция вне зависимости от намерений человека сопровождает его
хозяйственную деятельность.
Стихийная антропогенная эволюция играет сегодня несрав-
ненно большую роль.
Рассмотрим основные варианты того и другого процесса.
Целенаправленная эволюция. Этот процесс широко распро-
странен практически во всех странах и проявляется в следующем:
замене естественной растительности на искусственные фи-
тоценозы;
интродукции видов из других районов и включении их в со-
став естественных или искусственных фитоценозов;
изменении состава естественных фитоценозов за счет вве-
дения новых видов и культиваров;
конструировании новых типов искусственных фитоцено-
зов (полевых, лесных, парковых, рекультивационных на тех-
ногенных субстратах и т.д.).
229
Для каждого варианта такой антропогенной эволюции не-
трудно подобрать примеры, так как ее результатом являются и
садово-парковые комплексы с множеством экзотов, и лесные
посадки с интродуцированными видами древесных растений
(вплоть до американской секвойи в Крыму). И все же, роль це-
ленаправленной эволюции сравнительно невелика.
Стихийная эволюция под влиянием человека. Этот процесс
проявляется в следующем:
занос видов из других районов. Во флоре любого района ми-
ра значительная часть видов - заносные. Из 6000 видов, состав-
ляющих флору Италии, около 1000 заносных. Впрочем, в боль-
шинстве районов России на долю заносных видов приходится 10-
20% флоры. Поразительным примером эволюции такого рода яв-
ляется процесс «африканизации» американских саванн. Под дей-
ствием высоких пастбищных нагрузок, вызванных большим пого-
ловьем скота, разведенного заселившими Америку европейцами,
произошла деградация злаковников. В них распространились ру-
деральные виды африканского происхождения, которые были за-
везены в Америку вместе с черными рабами (Panicum maximum,
Brachiaria mutica, Hypperrhenia ruta и др.);
уничтожение части видов и снижение генотипического разно-
образия сохранившихся видов. Сегодня каждый десятый вид рас-
тений является угрожаемым, и Красные книги постоянно попол-
няются новыми видами. Происходит обеднение фитоценозов. На-
пример, в Башкортостане как угрожаемые на страницы Красной
книги попали все виды ковылей, кроме ковыля-волосатика. А в
недалеком прошлом Башкортостан был краем, где пастбища были
представлены богатыми ковыльными степями;
смещение границ растительности в результате хозяйствен-
ной деятельности человека. Сегодня граница степной зоны по-
всеместно смещена к северу под влиянием интенсивного ис-
пользования леса. В Африке по той же причине саванна насту-
пает на сухие тропические леса;
возникновение новых устойчивых к влиянию человека вто-
ричных типов фитоценозов. Пример этого варианта эволюции -
развитие бедных видами пастбищных сообществ, которые от-
сутствовали до того, как распашкой земель была сокращена
площадь естественных кормовых угодий, а поголовье скота уве-
личилось в связи с ростом народонаселения и повышением
спроса на продукты животноводства;
формирование новых фитоценозов на антропогенных суб-
стратах вследствие их самозарастания. На отвалах горных пород
возникают сочетания видов, которые отсутствовали в природе.
230
Антропогенная эволюция растительности протекает в горо-
дах, в которых формируются особые сообщества из видов,
адаптированных к высоким антропогенным нагрузкам. Среди
этих видов много заносных, поэтому растительные сообщества
городов, достаточно удаленных друг от друга, сходны. Возник
особый раздел фитоценологии - урбанофитоценояогия (от лат.
урбанус - городской), которая изучает процесс антропогенной
эволюции растительности в городах.
Для обозначения такой стихийной антропогенной эволюции
используется термин синантропизация растительности.
Задача человечества - поставить под контроль стихийные
формы эволюции, разрушающие естественную растительность.
12.4. Классификация растительности
Несмотря на то, что растительность представляет контину-
ум и в ней нет четко отделенных друг от друга фитоценозов,
важным разделом фитоценологии является классификация. Без
классификации невозможно инвентаризировать растительность,
составлять карты, оценивать продуктивность, запасы раститель-
ных ресурсов и другие хозяйственно полезные качества. Без
классификации невозможно решать и вопросы охраны расти-
тельности.
Существуют различные подходы к классификации раститель-
ности, но наиболее распространенными на сегодняшний день яв-
ляются два: эколого-флористический и физиономический.
12.4.1. Эколого-флористический подход
При обсуждении факторов организации фитоценозов и
множественности моделей их организации мы подчеркивали,
что главным фактором всегда является внешняя среда. Именно
внешняя среда, т.е. разнообразие наборов экологических усло-
вий экотопов, определяет, какие типы фитоценозов могут
сформироваться в данном районе. Все прочие факторы (конку-
ренция, влияние животных, нарушения) действуют на фоне этих
различных условий внешней среды. Поэтому для систематиза-
ции фитоценозов экологические критерии - самые важные.
Учет экологических критериев возможен и при рассматривае-
мом далее физиономическом подходе, однако наиболее полно
эту задачу удается решить при использовании эколого-флорис-
тического подхода.
Принципы и метод эколого-флористической классификации
были разработаны в 20-е годы минувшего столетия крупнейшим
европейским фитоценологом Ж.Браун-Бланке (J.Braun-Blanqu-
et). Этот метод достаточно быстро «завоевал» не только все ев-
231
ропейские страны, но и Японию, и Африку, а с 80-х годов стал
распространяться в США и бывшем СССР. Единицы, выделяе-
мые этим методом, называются синтаксонами, а их система -
синтаксономией.
По аналогии с таксономией, в синтаксономии используется
хорошо продуманная система классификационных рангов с со-
ответствующими окончаниями, по которым ранг легко опозна-
ется. Кроме того, если в таксономии при наименовании семей-
ства, рода или вида указывается автор, то в синтаксономии при
наименовании синтаксона, кроме автора, указывается еще и да-
та его установления. Последнее связано с большей условно-
стью синтаксонов по сравнению с таксонами, недостаточной
четкостью критериев их различения и потому возможностью
появления синонимов (разных названий для одного и того же
типа фитоценоза) и омонимов (одного названия для разных ти-
пов фитоценозов).
Таблица 20
Номенклатура синтаксонов
эколого-флористической классификации
Синтаксо- номиче- ский ранг Окончание Пример
сегетальная расти- тельность луговая раститель- ность
Класс -etea Secalietea Br.-Bl. 1951 Molinio-Arrhena- theretea R.Tx. 1937
Порядок -etalia Secalietalia Br.-Bl. 1931 ern. J. et R.Tx. 1960 Galietalia veri Mir- kin et Naumova 1986
Союз -ion Galeopsion bifidae Abramova in Mirkin et al. 1985 Trifolion montani Naumova 1986
Подсоюз -enion Caricenion prae- cocis Mirkin et Nau- mova 1986
Ассоциа- ция -etum Galeopsetum bifi- dae Abramova in Mirkin et al. 1985 Polygono sibirici- Puccinellietum tenui- florae Mirkin et al. 1985
Субассо- циация -etosum Galeopsetum bifi- dae fumaretosum Ab- ramova in Mirkin et al. 1985 Polygono sibirici- Puccinellietum tenui- florae alopecuretosum Mirkin et al. 1985
Вариант Typicum Typicum
Фация -osum Chenopodiosum al- bae Glaudosum mariti- mae
232
В табл. 20 приведены примеры названий синтаксонов для
двух типов растительности - сегетальной (сообществ полевых
сорняков) и луговой.
Конструирование названий, их видоизменение и отмена ре-
гулируются специальным «Кодексом фитосоциологической но-
менклатуры».
Синтаксоны эколого-флористической классификации уста-
навливаются на основе диагностических видов, среди которых
могут различаться характерные, дифференцирующие и кон-
стантные (рис. 47). Характерные виды встречаются только в од-
ном синтаксоне или встречаются в этом синтаксоне чаще, чем в
других (т.е. центрированны в нем). Дифференцирующие виды
Рис. 47. Схема, показывающая основные варианты диагностических ви-
дов, используемых в системе Браун-Бланке: А, Б, В - ассоциации; а, б,
в - субассоциации; 1, 2 - характерные виды ассоциации Б; 3, 4 - ха-
рактерные виды ассоциаций А; 5 - характерный вид ассоциации В; 4,
6,7,8 - дифференциальные виды субассоциаций в ассоциации Б; 9, 10,
11 - константные виды, которые могут быть характерными для клас-
сификационных единиц более высокого ранга
233
диагностируют границей своего ареала и входят в состав не-
скольких синтаксонов. Константные виды встречаются с высо-
ким постоянством и часто с высоким обилием, но проходят
через этот синтаксон «транзитом». Виды, константные для ас-
социации или союза, могут быть дифференцирующими или ха-
рактерными для порядка или класса.
Впрочем, различия характерных и дифференцирующих ви-
дов весьма условны, и потому очень часто их рассматривают
как единую группу диагностических видов (но в этом случае к
ней не относят константные виды).
Основной высшей единицей в синтаксономии является
класс. Этот синтаксон по своему объему приближается к об-
щепринятым географическим единицам растительности, таким
как тайга, широколиственные леса, луга, степи, ольховые боло-
та и т.д. Иметь представление об этих классах студент должен
так же, как и об основных семействах цветковых растений.
Список основных классов растительности средней полосы
европейской части России с указанием их диагностических ви-
дов приводится в Приложении 2.
12.4.2. Физиономический подход
Физиономические классификации (по составу доминантов)
могут использоваться для предварительной ориентации в мате-
риале. При этом выделяются доминантные типы (например, ли-
пово-дубовый лес со снытью, пырейно-кострецовый луг, разно-
травно-перистоковыльная степь и т.д.).
Фундаментальное значение для науки имеет использование
физиономических критериев для установления высших еди-
ниц биогеографической классификации Земли - биомов. В тра-
диции отечественной географии биому соответствует природная
зона (тундровая, хвойных лесов, широколиственных лесов, ле-
состепная, степная, полупустынная и т.д.). Существует не-
сколько систем биомов (Х.Вальтера, Ю.Одума, Р.Уиттекера).
Самой простой является система Ю.Одума. Приведем ее с крат-
кими комментариями.
Тундры — безлесные пространства, расположенные севернее
лесного биома.
Тайга - лесные территории с доминированием хвойных де-
ревьев (ели - р. Picea, сосны - р. Pinus, пихты - р. Abies, лист-
венницы - р. Larix).
Широколиственные листопадные леса - южная часть лес-
ной зоны с более теплым, чем в биоме тайги, климатом. Основ-
ные доминанты лесов - дуб (р. Quercus), липа (р. Tilia), ильм
(р. Ulmus), береза (р. Betula), осина (р. Populus).
234
Степи - безлесные пространства с черноземными почвами,
растительный покров которых сформирован главным образом
злаковыми сообществами с преобладанием ковылей (р. Stipa),
типчака (Festuca valesiaca) и других засухоустойчивых растений.
Между биомами широколиственных лесов и степей расположен
экотон - лесостепь, где чередуются участки леса и степи, в ос-
новном благодаря влиянию хозяйственной деятельности человека.
Саванны - травяные сообщества с разреженным покровом
деревьев в теплых и достаточно увлажненных (осадков 1000-
1500 мм/год) условиях.
Чапараль и жестколистные леса - биом засушливого кли-
мата с обильными зимними дождями, распространен в Среди-
земноморье, Северной Америке и Австралии.
Пустыни — пространства в условиях, где количество осад-
ков меньше 250 мм/год. Растительность представлена кустарни-
ками, кустарничками, реже деревьями (например, саксаулом -
р. Haloxylon).
Полувечнозеленые сезонные тропические леса распрост-
ранены в тропических районах с периодами засухи летом или
зимой.
Тропические дождевые леса - тропические районы, где ко-
личество осадков превышает 2000 мм/год при отсутствии дли-
тельных периодов без дождей.
Одум предложил также классификацию биомов пресно-
водных и морских экосистем, которую мы не рассматриваем.
Глава 13
ИЗУЧЕНИЕ ФЛОР
13.1. Региональная и конкретная флора
Флора совокупность видов растений на определенной
территории объект флористики (раздела ботанической гео-
графии).
В современной флористике наиболее распространен тради-
ционный подход - изучение региональных флор, т.е. выявление
списка видов растений территорий, границы которых достаточ-
но произвольны. Обычно это границы административной еди-
ницы (может быть изучена флора России, Республики Башкор-
тостан, Башкирского Предуралья, одного административного
района, одного города или села и т.д.).
При другом подходе - изучении конкретных флор - флора
представляет не произвольно выделенный регион, а террито-
рию с однородным географическим ландшафтом и с площадью,
которая достаточна, чтобы эта флора могла быть выявлена. Од-
235
нако метод конкретных флор менее удобен, так как накладыва-
ет на выбираемый объект жесткие ограничения. Например, тер-
ритория населенного пункта оказывается недостаточной для ус-
тановления списка видов, соответствующего своим содержани-
ем конкретной флоре, площадь выявления которой составляет
сотни квадратных километров. По этой причине чаще изучают
региональные флоры.
13.2. Анализ флор
После выявления региональной флоры (т.е. списка видов
конкретной территории), анализируется ее структура - состав-
ляются спектры по различным признакам:
систематическому составу,
жизненным формам,
географической структуре,
происхождению,
фитосоциологическому составу (доле представленности ви-
дов разных классов),
ресурсному значению.
При анализе систематического состава флор особо значимы
первые 10 семейств, представленность которых уже отражает и
комплекс почвенно-климатических факторов, и историю, и со-
временное состояние флоры, испытывающей влияние человека
(как пример см. табл. 21).
Зачастую изучают также степень синантропизации флоры,
т.е. участия в ней заносных видов, называемых адвентивными.
Таблица 21
Количественный состав ведущих семейств
флоры Башкортостана
Семейство Число родов Число видов
Asteraceae 60 204
Роасеае 42 149
Rosaceae 20 100
Сурегасеае 11 98
Brassicaceae 42 97
Fabaceae 23 97
Caryophyllaceae 23 73
Lamiaceae 21 57
Scrophulariaceae 14 55
Apiaceae 41 54
236
13.3. Анализ адвентивного компонента флоры
Изучение процесса пополнения флоры заносными (адвен-
тивными) растениями представляет собой один из наиболее
информативных вариантов биомониторинга за состоянием ок-
ружающей среды. Доля заносных растений в большинстве со-
временных флор (степень их синантропизации) колеблется от
10 до 30%, что связано с интенсивностью трансформации расти-
тельности человеком.
Адвентивные виды различаются по времени заноса, спосо-
бу заноса, а также по степени натурализации, т.е. уровню
адаптированности к новым географическим условиям.
По времени заноса различают археофиты, занесенные до
XVI века, и кенофиты (неофиты), появившиеся в более позднее
время.
По способу заноса выделяют: ксенофиты занесенные
случайно; эргазиофиты - занесенные преднамеренно; ксеноэрга-
зиофиты - переходная группа с неясным сценарием появления
в новом районе.
По степени натурализации различают: эфемерофиты - флюк-
туирующие виды, которые то появляются, то исчезают в локаль-
ных местообитаниях (например, Oenothera depressa, Padus virginia-
na в Удмуртии); колонофиты - виды, прочно закрепившиеся на
новых местообитаниях, но не распространяющиеся из них; эпеко-
фиты - виды-пришельцы, которые расселяются по нарушенным
местообитаниям (виды рудеральных сообществ, подобные предста-
вителям рода Ambrosia); агриофиты — заносные растения, которые
внедряются в естественные сообщества (например, зверобой в
США, опунция в Австралии и др.).
Приложение 1
СПИСОК НАИБОЛЕЕ РАСПРОСТРАНЕННЫХ
КУЛЬТУРНЫХ РАСТЕНИЙ, НЕ ВХОДЯЩИХ
В РАССМОТРЕННЫЕ СЕМЕЙСТВА ЦВЕТКОВЫХ
Русское название растения Латинское название вида Семейство
Ананас Ananas comosus Bromeliaceae
Апельсин Citrus sinensis Rutaceae
Бамия Hibiscus esculentus Malvaceae
Банан Musa paradisiaca Musaceae
Батат Ipomoea batatas Convolvulaceae
Виноград Vitis vinifera Vitaceae
Гвоздичное дерево Syzygium aromaticum Myrtaceae
Гранат Punica granatum Punicaceae
Грейпфрут Citrus paradis Rutaceae
Грецкий орех Juglans regia Juglandaceae
Дерево какао Theobroma cacao Sterculiaceae
Имбирь Zingiber officinale Zingibiraceae
Инжир Ficus carica Moraceae
Кардамон Elettaria cardamomum Zingibiraceae
Киви Actinidia chinensis Actinidiaceae
Клюква Oxycoccus palustris Vacciniaceae
Кола Cola nitida Sterculiaceae
Кофейное дерево Coffea arabica Rubiaceae
Крыжовник Grossularia reclinata Grossulariaceae
Кунжут Sesamum orientate Pedaliaceae
Лен культурный Linum usitatissimum Linaceae
Лимон Citrus limon Rutaceae
Манго Mangifera indica Anacardiaceae
Мандарин Citrus unshiu Rutaceae
Маслина Olea europaea Oleaceae
Мускатное дерево Myristica fragrans Myristicaceae
Облепиха Hippophae rhamnoi- des Elaeagnaceae
Папайя Carica papaya Caricaceae
238
Русское название растения Латинское название вида Семейство
Перец черный Piper nigrum Piperaceae
Смородина черная Kibes nigrum Grossular iaceae
Унаби Ziziphus jujuba Rhamnaceae
Фейхоа Feijoa sellowiana Myrtaceae
Фисташка Pistacia vera Anacardiaceae
Хлопчатник Gossypium hirsutum Malvaceae
Хурма восточная Diospyros kaki Ebenaceae
Чайный куст Thea sinensis Theaceae
Шелковица Morus alba Moraceae
Ямс Dioscorea alata Dioscoreaceae
Приложение 2
СПИСОК ОСНОВНЫХ КЛАССОВ
РАСТИТЕЛЬНОСТИ СРЕДНЕЙ ПОЛОСЫ
ЕВРОПЕЙСКОЙ ЧАСТИ РОССИИ
Синантропная растительность
(поля, сообщества пустырей, опушек, сильно сбитых пастбищ)
CHENOPODIETEA Вг.-В1. 1952 em. Lohm., J. et R.Tx. 1961 ex
Matusz. 1962 - сообщества однолетников, представляющие начальные
стадии восстановительных сукцессий после нарушений.
Флористическая комбинация: Amaranthus blitoides, A. retroflexus,
Anisanthe tectorum, Arenaria serpyllifolia, Atriplex nitens, A. patula,
A. tatarica, Axyris amaranthoides, Brassica campestris, Chenopodium
album, Ch. glaucum, Descurainia spphia, Echinochloa crusgalli, Erigeron
canadensis, Erodium cicutarium, Erysimum cheiranthoides, Galeopsis bifida,
G. speciosa, G. ladanum, Lactuca serriola, L. tatarica, Lappula squarrosa,
Malva pusilia, Matricaria perforata, Oberna behen, Panicum miliaceum
Potentilla norvegica, Salsola collina, Senecio vulgaris, Setaria viridis,
S. glauca, Sinapis arvensis, Sisymbrium loeselii, S. officinale, Solanum nig-
rum, Thlaspi arvense, Tussilago farfara, Urtica urens, Xanthium struma-
rium.
SECALIETEA Br.-Bl. 1951 - сегетальные (полевые) сообщества.
Флористическая комбинация: Avena fatua, Buglossoides arvensis,
Cannabis ruderalis, Capsella bursa-pastoris, Cirsium setosum, Consolida
regalis, Convolvulus arvensis, Eqiusetum arvense, Fallopia convolvulus,
Lycopsis arvensis, Neslia paniculata, Silene noctiflora, Sonchus arvensis,
Spergula arvensis, Stachys annua, Stellaria media, Rumex acetosella.
ARTEMIS1ETEA VULGARIS Lohm., Prsg. et Tx. in Tx. 1950 - ру-
деральные сообщества высокорослых дву- и многолетних видов.
Флористическая комбинация: Arctium lappa, A. minus, A. nemo-
rosum, A. tomentosum; Artemisia absinthium, A. vulgaris, Berteroa incana,
Carduus acanthoides, C. crispus, C. nutans, Cichorium inthybus, Conium
maculatum, Hyoscyamus niger, Leonurus quinquelobatus, Linaria vulgaris,
Melandrium album, Nepeta cataria.
GALIO-URTICETEA Passarge 1967 - естественные и антропоген-
ные нитрофильные сообщества при ограниченном освещении в лесо-
парках, скверах, в поймах рек и ручьев.
Флористическая комбинация: Anthriscus sylvestris, Calystegia
sepium, Carduus crispus, Chaerophyllum prescottii, Chelidonium majus,
Cucubalus baccifer, Cuscuta europaea, Galium aparine, Geum rivale,
G. urbanum, Glechoma hederacea, Heracleum sibiricum, Impatiens noli-
tangere, Lamium album, L. maculatum, Myosoton aquaticum, Rumex
obtusifolius, Saponaria officinalis, Solanum dulcamara, Stellaria nemorum,
Urtica dioica.
AGROPYRETEA REPENTIS Oberd., Th. Muller et Gors in Oberd. et
al. 1967 - рудеральные сообщества с преобладанием многолетних злаков,
представляющие продвинутую стадию восстановительных сукцессий.
240
Флористическая комбинация: Convolvulus arvensis, Elytrigia repens,
Falcaria vulgaris.
PLANTAGINETEA MAJORIS Tx. et Prsg. in Tx. 1950 - сообщества
низкорослых, устойчивых к вытаптыванию и выпасу мезофитов и гиг-
рофитов на пастбищах, спортивных площадках, во дворах, вдоль дорог.
Флористическая комбинация: Agrostis stolonifera, Chamomilla
suaveolens, Chenopodium glaucum, Inula britannica, Junciis compressus,
Lepidium ruderale, Odontites vulgaris, Plantago major, Poa annua,
Polygonum aviculare, Potentilla anserina, Ranunculus repens, Rorippa syl-
vestris, Rumex crispus, Taraxacum officinale, Trifolium repens.
EPILOBIETEA ANGUSTIFOLII Tx. et Prsg. in Tx. 1950 - сообще-
ства вырубок и гарей.
Флористическая комбинация: Calamagrostis epigeios, Chamerion
angustifolium, Fragaria vesca, Hypericum hirsutum, Omalotheca
sylvatica, Rubus idaeus.
BIDENTETEA TRIPARTITAE Tx., Lohm et Prsg. in Tx. 1950 -
синантропные сообщества переувлажненных мест.
Флористическая комбинация: Bidens cernua, В. tripartita, Cheno-
podium polyspermum, Polygonum hydropiper, P. lapathifolium, P. mite,
Ranunculus sceleratus, Rorippa palustris.
Водная, прибрежно-водная
и болотная растительность
LEMNETEA Тх. 1955 сообщества свободноплавающих на по-
верхности и в толще воды неукореняющихся растений.
Флористическая комбинация: Hydrocharis morsus-ranae, Lemna
minor, L. trisulca, L. gibba, Salvinia natans, Spirodela polyrhiza, Stratiotes
aloides, Utricularia vulgaris, U. minor.
POTAMETEA Klika in Klika et Novak 1941 - сообщества гидатофи-
тов - прикрепленных к дну растений с плавающими на поверхности
или погруженными в толщу воды листьями.
Флористическая комбинация: Batrachium circinatum, В. tricho-
phyllum, Callitriche cophocarpa, C. hermaphroditica Elodea canadensis,
Myriophyllum spicatum, M. verticillatum, Nuphar lutea, N. pumila,
Nymphaea alba, N. Candida, Potamogeton compressus, P. crispus, P. lu-
cens, P. natans, P. pectinatus, P. perfoliatus. P. pusillus, Polygonum
amphibium.
PHRAGMITI-MAGNOCARICETEA Klika in Klika et Novak 1941
сообщества гелофитов - прикрепленных к дну возвышающихся над
водой растений.
Флористическая комбинация: Alisma plantago-aquatica, Butomus
umbellatus, Cladium mariscus, Equisetum fluviatile, Eleocharis palustris,
Glyceria maxima, Hippuris vulgaris, Lycopus europaeus, Lythrum
salicaria, Oenanthe aquatica, Phragmites australis, Polygonum amphibium,
Rorippa amphibia, Rumex hydrolapathum, Sagittaria sagittifolia, Scirpus
lacustris, Sium latifolium, Sparganium emersum, S. erectum, Stachys
palustris, Typha angustifolia, T. latifolia.
241
ALNETEA GLUTINOSAE Br.-Bl. et Tx. ex Westhoff et al. 1946 -
эутрофные лесные и кустарниковые болота на торфянистой почве.
Флористическая комбинация: Alnus glutinosa, Calamagrostis cane-
scens, Calla palustris, Carex caespitosa, C. elongata, C. juncella, C. pseu-
docyperus, Cicuta virosa, Comarum palustre, Dryopteris carthusiana,
D. cristata, Frangula alnus, Galium palustre, G. uliginosum, Molinia
caerulea, Naumburgia thyrsiflora, Ribes nigrum, Salix aurita, S. cinerea,
S. pentandra.
SCHEUCHZERIO-CARICETEA NIGRAE Tx. 1937 - мелкоосоковые
моховые мезотрофные болота, обычно с пологом Be tula pubescens.
Флористическая комбинация: Betula pubescens, Carex canescens,
C. dioica, C. flava, C. limosa, Carex nigra, Drosera anglica, D. rotundifolia,
Eleocharis pauciflora, Epilobium palustre, Equisetum palustre, Eriophorum
latifolium, E. polystachyon, Juncus alpino-articulatus, Menyanthes trifoliata,
Pamassia palustris, Pinguicula vulgaris, Rhynchospora alba, Scheuchzeria
palustris, Triglochin palustre.
OXYCOCCO-SPHAGNETEA Br.-Bl. et Tx. ex Westhoff et al. 1946 -
верховые олиготрофные моховые болота голарктических областей.
Флористическая комбинация: Andromeda polifolia, Aulacomnium
palustre, Baeothryon cespitosum ssp. cespitosum, Cephalozia connivens,
Chamaedaphne calyculata, Drosera rotundifolia, Eriophorum vaginatum,
Milia anomala, Oxycoccus palustris, Pohlia nutans var. sphagnetorum,
Sphagnum angustifolium, S. magellanicum, S. nemoreum, S. papillosum,
S. rubellum.
Растительность засоленных почв
THERO-SALICORNIETEA Tx. in Tx. et Oberd. 1958 - сообщества
однолетних суккулентных галофитов на переувлажненных местооби-
таниях.
Флористическая комбинация: Salicomia europaea, Suaeda
comiculata, S. prostrata.
ASTERETEA TRIPOLIUM Westhoff et Beeftink in Beeftink 1962
сообщества многолетних луговых растений несуккулентного облика
на засоленных почвах в различных условиях увлажнения (от сырых до
опустыненных).
Флористическая комбинация: Alopecurus arundinaceus, Cirsium
esculentum, Glaux maritime, Hordeum brevisubulatum, Juncus gerardii,
Triglochin tnaritimum, Plantago cornuti, P. maxima, P. salsa, Puccinellia
distans, Rumex maritimus, Scorzonera parviflora Taraxacum bessarabicum,
Tripolium vulgare.
Луга на незасоленных почвах и степи
MOLINIO-ARRHENATHERETEA Тх. 1937 ет. Тх. 1970 - луга Ев-
разии, чаще послелесные.
Флористическая комбинация: Achillea millefolium, Agrostis gigantea,
A. tenuis, Alopecurus pratensis, Bromopsis inermis, Carum carvi, Centaurea
242
jacea, Dactylis glomerata, Festuca pratensis, F. rubra, Geranium pratense,
Leucanthemum vulgare, Lotus corniculatus, Phleum pratense, Pimpinella
saxifraga, Poa pratensis, Ranunculus acris, Sanguisorba officinalis, Stellaria
graminea, Thalictrum simplex, Trifolium pratense, Vicia cracca.
TRIFOLIO-GERANIETEA SANGUINEI Th.Muller 1961 - луговые
сообщества лесных опушек и редколесий.
Флористическая комбинация: Agrimonia eupatoria, Clinopodium
vulgare, Coronilla varia, Geranium sanguineum, Hypericum perforatum,
Inula aspera, I. hirta, I. salicina, Lathyrus sylvestris, L. tuberosus, Melam-
pyrum cristatum, Origanum vulgare, Peucedanum alsaticum, Seseli liba-
notis, Silene nutans, Trifolium medium, Veronica chamaedrys, Vicia
tenuifolia.
FESTUCO-BROMETEA Br.-Bl. et Tx. 1943 - ксеротермные и полу-
ксеротермные травяные сообщества.
Флористическая комбинация: Achillea setacea, Alyssum tortuosum,
Amygdalus папа, Artemisia latifolia, Artemisia sericea, Astragalus
austriacus, A. helmii, A. onobrychis, Campanula sibirica, Caragana frutex,
Centaurea ruthenica, C. scabiosa, Cerasus fruticosa, Festuca rupicola,
F. valesiaca, Galium ruthenicum, Goniolimon speciosum, Helictotrichon
desertorum, Hieracium virosum, Koeleria cristata, Medicago falcata,
M. romanica, Onosma simplicissima, Orostachys spinosa, Oxytropis pilosa,
Phleum phleoides, Phlomis tuberosa, Plantago urvillei, Poa stepposa,
Potentilla humifusa, Pulsatilla flavescens, Sedum hybridum, Scabiosa
ochroleuca, Stipa capillata, S. pennata, S. zalesskii, Salvia nutans,
S. stepposa, Spiraea crenata, S. hypericifolia, Thymus marschallianus.
Аесная растительность
QUERCO-FAGETEA Br.-Bl. et Vlieger in Vlieger 1937 - сообщества
мезофитных и мезоксерофитных широколиственных листопадных лесов
на богатых почвах.
Флористическая комбинация:
деревья и кустарники - Acer platanoides, Alnus incana, Corylus
avellana, Euonymus verrucosa, Lonicera xylosteum, Padus avium, Picea
abies, Quercus robur, Tilia cordata, Ulmus laevis, Viburnum opulus;
травы Aconitum septentrionale, Actaea spicata, Aegopodium
podagraria, Anemonoides ranunculoides, Asarum europaeum, Bromopsis
benekenii, Cacalia hastata, Campanula latifolia, Chrysosplenium alternifol-
ium, Cicerbita uralensis, Circaea lutetiana, Convallaria majalis, Dryopteris
filix-mas, Elymus caninus, Epipactis helleborine, Equisetum pratense,
E. sylvaticum, Festuca gigantea, Ficaria verna, Galium odoratum, Lathyrus
vernus, Lilium martagon, Matteuccia struthiopteris, Milium effusum, Paris
quadrifolia, Polygonatum multiflorum, Primula macrocalyx, Pulmonaria
obscura, Stachys sylvatica, Stellaria bungeana, S. holostea, Viburnum opu-
lus, Viola mirabilis.
VACCINIO-PICEETEA Br.-Bl. in Br.-Bl., Siss. et Vlieger 1939 -
хвойные леса с развитым моховым покровом.
Флористическая комбинация:
243
деревья и кустарники Lonicera altaica, Picea abies, P. obovata,
Pinus sylvestris;
кустарнички - Arctostaphylos uva-ursi, Empetrum hermaphroditum,
Linnaea borealis, Vaccinium myrtillus, V. uliginosum, V. vitis-idaea;
травы Dryopteris austriaca, Goodyera repens, Gymnocarpiutn
dryopteris, Huperzia selago, Listera cordata, Luzula pilosa, Lycopodium
annotinum, Maianthemum bifolium, Moneses uniflora, Oxalis acetosella,
Pyrola minor, P. rotundifolia, Stellaria longifolia, Trientalis europaea;
мхи и лишайники Barbilophozia floerkei, Blechnum spicant,
Cetraria islandica, Cladonia arbuscula, C. rangiferina, C. cenotea, C. chlo-
rophaea, C. coccifera, C. cornuta, C. crispata, C. gracilis, C. bellidiflora,
C. squamosa, C. uncialis, Corallorhiza trifida, Dicranum majus, D. scopa-
rium, D. undulatum, Hylocomnium splendens, H. umbratum, Nephroma
arcticum, Peltigera aphthosa, Pleurozium schreberi, Ptilidium ciliare,
Ptilium crista-castrensis, Plagiochila major, Plagiothecium undulatum,
Rhytiadelphus calvescens, Rh. loreus, Rh. triquetrus, Sphagnum girgen-
sohnii, Webera nutans.
SALICETEA PURPUREAE Moor 1958 - сообщества пойменных
ивняков и ивово-тополевых лесов.
Флористическая комбинация: Humulus lupulus, Lysimachia vulgaris,
Роа trivialis, Populus nigra, Rubus caesius, Salix alba, S. triandra, S. vi-
minalis, Stachys palustris.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Ботаника. Систематика растений /Н.А.Комарницкий,
Л.В.Кудряшов, А.А.Уранов. - М.: Просвещение, 1975.
Вальтер Г. Общая геоботаника: Пер. с нем. - М.: Мир, 1982.
Горышина Т.К. Экология растений. М.: Высшая школа,
1979.
Губанов И.А. Энциклопедия природы России. Пищевые рас-
тения: Справочное издание. - М., 1996.
Еленевский А.Г., Соловьева М.П., Тихомиров В.Н. Бота-
ника высших, или наземных растений. - М.: Центр Академия,
2000
Жизнь растений: В 6 т. - М.: Просвещение.- 1978- 1982.
Миркин Б.М. Что такое растительные сообщества. - М.:
Наука, 1986.
Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная
наука о растительности. Учебник. - М.: Логос, 2000.
Миркин Б.М., Розенберг Г.С., Наумова А.Г. Словарь поня-
тий и терминов современной фитоценологии. - М.: Наука, 1989.
Рейвн П., Эверт Р., Айкхорн С. Современная ботаника:
В 2 т. Т.1. - М.: Мир, 1990.
Тахтаджян А.Л. Система магнолиофитов. - Л.: Наука, 1987.
Хржановский В.Г. Курс общей ботаники (систематики рас-
тений): Учебник для сельхозвузов. - М.: Высшая школа, 1982.
Работнов ТА. Фитоценология. 3-е изд. - М.: Изд-во МГУ,
1992.
Уиттекер Р. Сообщества и экосистемы. - М.: Прогресс,
1980.
Указатель латинских названий растений
Abies 62 Angelica archangelica 122
Abies sibirica 63 Angelica sylvestris 122
Acacia dealbata 120 Angiopteris 42
Achillea 135, 136 Angiopteris evecta 42
Aconitum 82, 83, 84 Anthriscus 121
Adianthum 42 Antirrhinum 129, 130
Adonis 82, 83 Apium 121
Adonis vernalis 84 Apium graveolens 123
Aegopodiutn 121 Aquilegia 82, 84
Aegopodium podagraria 122 Arachis 116
Aesculus hippocastanum 95 Arachis hypogaea 118
Agathis 61, 62 Aralia 123
Agathis australis 62 Araucaria 62
Agropyron 153 Araucaria araucana 62
Agrostis 150 Arctium 135, 137
Ajuga reptans 132 Areca catechu 162
Alchemilla 113 Arenga 158
Aleurites 111, 112 Arenga pinnata 161
Aleurites moluccana 113 Armeniaca 113
Allium 140 Armeniaca vulgaris 114
Allium сера 140 Armoracia rusticana 105
Allium fistulosum 141 Aronia melanocarpa 115
Allium obliquum 141 Artemisia 135
Allium polyrrhizum 142 Artemisia absinthium 137
Allium porrum 141 Artemisia vulgare 137
Allium sativum 140 Asparagus officinalis 141
Allium schoenoprasum 141 Asplenium 42
Allium victorialis 141 Asplenium septentrionale 42
Alnus 96 Aster 135, 137
Alnus glutinosa 98 Astragalus 116
Alnus incana 98 Athyrium 42
Alopecurus 150 Atriplex 88, 90
Alopecurus pratensis 157 Atriplex сапа 90
Alyssum 103 Atriplex nitens 89
Amaranthus 91 Atropa 123
Ambrosia 135 Atropa belladonna 124, 125,
Amygdalus 113 126
Amygdalus communis 115 Avena 150
Anemone 82 Avena sativa 151, 155
Anemone sylvestris 84 Anemonoides 82 Azolla 45
Anemonoides altaica 84 Barbarea 103
Anemonoides nemorosa 84 Barbarea vulgaris 105
Anemonoides ranunculoides 84 Bassia 90
Anethum 121 Bellis 135, 137
Anethum graveolens 123 Berberis 84
246
Berteroa 103
Beta 91
Beta vulgaris 88, 89
Beta vulgaris subsp. maritime 91
Betula 96
Betula nana 96
Betula pendula 97, 98
Betula pubescens 98
Bidens 135, 136, 137
Bistorta 92
Bistorta vivipara 93
Boehmeria 109
Boehmeria nivea 110
Bolboschoenus 147
Bolboschoenus maritimus 149
Borago Y2.7
Borago officinalis 128
Botrychium 39
Botrychium simplex 41
Brassica 103, 105
Brassica juncea 105
Brassica napus var. napus 105
Brassica nigra 105
Brassica oleracea 104
Brassica oleracea var. botrytis 105
Brassica oleracea var. capitata 105
Brassica oleracea var. gem-
mifera 105
Brassica oleracea var. gongyloi-
des 105
Brassica rapa* 105
Bromopsis 150
Bromopsis inermis 152, 157
Bryonia alba 102
Bunias 103, 104
Bunias orientalis 105
Cakile maritima 105, 106
Calamagrostis 150
Catamites 34
Calamus 158
Calendula 137
Calligonum 93
Caltha 82
Calymmatotheca 54
Calypso bulbosa 146
Calystegia 128
Camelina 104
Cannabis 106
Cannabis ruderalis 108
Cannabis sativa 106, 107
Capsella bursa-pastoris 104, 105
Capsicum 123
Capsicum annuum 124
Caragana 116
Caragana arborescens 120
Cardamine 105
Cardamine pratense 106
Carduus 135, 137
Carex 147, 148
Carex acuta 147
Carex cespitosa 147
Carex duriuscula 147
Carex muricata 147
Carex pediformis 147
Carex pilosa 147
Carex praecox 147
Carex rhizina 147
Carex supina 147
Carex sylvatica 147
Carex vesicaria 147, 148
Carpinus 96
Carpinus betulus 99
Carum 121, 122
Carum carvi 122, 123
Caryota urens 160
Castanea 93, 95
Castanea dentata 95
Castanea sativa 95
Cedrus 64
Cedrus libani 64
Celosia argentea 91
Centaurea 135, 136, 137
Cephalanthera 142
Cerapadus 115
Ceraprunus 115
Cerastium 86
Cerasus 113
Cerasus avium 115
Cerasus vulgaris 114
Ceratonia siliqua 120
Chamaerops humilis 161
Cheilanthes kuhnii 43
Chelidonium majus 85
Chenopodium 88, 90
Chenopodium foliosum 90
Chenopodium album 89, 90
Chosenia 99
247
Chrisanthemum 135, 137
Cicer 116
Cicer arietinum 118
Cicuta virosa 122
Cinnamomum 79
Cirsium 135, 136
Cirsium arvense 137
Citrullus 101
Citrullus lanatus 102
Cladium 147
Cleistogenes 151, 153
Clematis 82, 84
Cocos 158
Cocos nucifera 159, 160
Colobanthus quitensis 88
Conium 121, 122
Conium maculatum 122
Consolida 84
Convallaria 141
Convallaria majalis 141
Convolvulus arvensis 128
Coriandrum 122
Coriandrum sativum 123
Coronaria 86
Coronilla 116
Corylus 96, 97
Corylus avellana 97, 98
Corylus maxima 98
Corylus pontica 98
Corypha elata 159
Cosmos 137
Crambe 103, 105
Crambe maritime 104
Crataegus 113
Cucumis 101
Cucumis sativus 102
Cucurbita 101
Cucurbita pepo 102
Cupressus 65
Cupressus lusitanica 65
Cupressus sempervirens var.
sempervirens 65
Cupressus sempervivus var.
gorisontalis 65
Cuscuta 128
Cycadeoidea 58
Cycas 56
Cycas circinalis 55
Cycas revoluta 55
248
Cydonia 113
Cydonia oblonga 114
Cynara 135
Cynoglossum 127
Cyperus 147, 148
Cyperus alternifolius 149
Cyperus esculentus 147, 149
Cyperus fuscus 149
Cyperus papyrus 149
Cypripedium 142, 143
Cypripedium calceolus 146
Cystopteris 42
Dactylis glomerata 157
Dactylorhiza 142, 143
Dahlia 135
Datura 123, 124
Datura stramonium 125
Daucus 121
Daucus carota 122, 123
Degeneria 70
Delphinium 82, 83, 84
Deschampsia 150
Deschampsia cespitosa 152
Descurainia sophia 105
Dianthus 86, 87
Dianthus barbatus 88
Dianthus chinensis 88
Dianthus caryophyllus 88
Digitalis 129, 130
Drimys 70
Dryopteris 42
Dryopteris filix-mas 38, 40, 43
Ecballium elaterium 103
Echinocystis lobata 102
Echium 127
Elaeis guineensis 160
Eleocharis 147, 149
Eleocharis dulcis 147, 149
Eleutherococcus senticosus 8,
123
Elytrigia 150
Elytrigia repens 151, 152
Encephalartos inopinus 56
Entada scandens 120
Ephedra 67
Epipactis 142, 143, 146
Equisetum 34
Equisetum arvense 34, 35
Equisetum fluviatile 35
Equisetum hyemale 35
Equisetum myriochaetum 35
Equisetum sylvaticum 35
Eriobotrya japonica 115
Eriophorum 147, 149
Eryngium 121
Eryngium planum 121, 122
Erythronium 139
Euphorbia 111, 112
Euphorbia virgata 112
Euphrasia 129
Euryale ferox 81
Fagopyrum 92
Fagopyrum esculentum 93
Fagus 93, 95
Fagus orientalis 95
Fagus sylvatica 95
Festuca 150, 151, 153
Festuca pratensis 151, 157
Festuca pseudovina 153
Festuca rupicola 153
Festuca valesiaca 152, 153
Filipendula 113
Foeniculum 121
Foeniculum vulgare 123
Fragaria 113
Fragaria ananassa 115
Fragaria magna 115
Fragaria vesca 114, 115
Fragaria virginiana 115
Fragaria chiloensis 115
Fritillaria 139, 140
Fritillaria ruthenica 140
Gagea 139, 140
Galanthus 84
Galega 116, 118
Galeobdolon 131
Galeopsis bifida 134
Galeopsis ladanum 134
Genista 116
Gerardinia 110
Geum 113
Geum rivale 114
Ginkgo biloba 59
Gladiolus 141
Glechoma 131
Glechoma hederacea 132
Glycine 116
Glycine max 119
Gnetum 67
Goodyera 142
Gymnadenia 142
Gypsophila aretioides 88
Gypsophila paniculata 87
Haloxylon 88, 89
Hedera 123
Helianthus 135
Helianthus annuus 136, 137
Helianthus tuberosus 137
Heracleum 121
Heracleum sibiricum 122
Hesperis matronalis 105
Hevea 111
Hevea brasiliensis 111
Hordeum 150
Hordeum vulgare 155
Humulopsis 106
Humulus 106
Humulus lupulus 106, 107
Hyacinthus 141
Hyenia 34
Hyoscyamus 123, 124
Hyoscyamus niger 125
Indig ofera 116
Ipomoea purpurea 128
Iris 141
Isatis 105
Isoetes 32
Isoetes lacustris 30
Juniperus 65, 66
Juniperus communis 66
Juniperus excelsa 66
Juniperus oxycedrus 66
Juniperus sabina 66
Juniperus sibirica 66
Juniperus pseudosabina 66
Kalidium 90
Kobresia 147, 149
Koeleria 150, 153
249
Lactuca 137
Lactuca tatarica 137
Lamium 131
Lamium album 132
Laportea 110
Lappula 127, 128
Larix 62, 63
Larix gmelinii 63
Larix sibirica 63
Lathraea squamaria 130
Lathyrus 116, 118
Laurus nobilis 79
Lavandula 131
Lavandula angustifolia 134
Lavandula latiFolia 134
Ledum palustre 23
Leersia 151
Lemna minor 70
Leonurus 131
Lepidium 103
Lepidium ruderale 106
Leucaena leucocephala 120
Leucanthemum 135, 137
Lilium 139
Lilium martagon 140
Linaria 129
Linaria vulgaris 130
Liriodendron 77
Liriodendron tulipifera 79
Listera 142, 146
Livistona 161
Lodoicea maldivica 159, 161
Luffa 101
Luffa cylindrica 102
Lupinus 116, 117, 120
Lychnis 86
Lycopersicon 123
Lycopersicon esculentum 124
Lycopodium 30
Lycopus 131
Macrozamia 58
Magnolia 77, 78
Magnolia grandiflora 78
Magnolia hypoleuca 78
Maianthemum 141
Majorana hortensis 134
Malus 113, 114
Malus domestica 115
250
Manihot 111
Manihot esculenta 112
Marattia 42
Marchantia polymorpha 18, 21
Marsilea 43
Marsilea drummondii 44
Marsilea quadrifolia 40, 43
Matricaria 137
Matteuccia 42
Matteuccia struthiopteris 42
Matthiola annua 105
Medicago 116, 117, 118
Medullosa 54
Melampyrum 129, 130
Melandrium 86
Melilotus 116, 117, 118
Melissa officinalis 132, 134
Melo 101
Melo sativa 102
Mentha 131
Mentha arvensis 132
Mentha piperita 134
Mercurialis 111
Metasequoia 64
Metroxylon sagu 159
Microbiota decussata 66
Mimosa pudica 120
Moehringia 87
Mora oleifera 120
Morus 106
Muscari 141
Myosotis 127, 128
Nardosmia 137
Nelumbo 81
Nelumbo nucifera 81
Neottia 146
Nepeta 134
Nephrolepis 42
Neslia paniculata 105
Nicotiana 123, 124
Nicotiana rustica 124
Nicotiana tabacum 124
Nonea 127
Nothofagus 93
Nuphar 79, 80
Nuphar lutea 81
Nuphar pumila 81
Nymphaea 79, 80
Nymphaea alba 80, 81 Nymphaea Candida 81 Nymphaea tetragona 81 Physalis 123, 124 Physalis franchettii 125 Physalis peruviana 124 Picea 62, 63
Ocimum 131 Ocimum basilicum 134 Odontites 130 Oncidium 144 Onobrychis 116, 117, 118 Onopordum 137 Ophioglossum 39, 40 Ophioglossum vulgatum 39 Ophris 145 Ophris insectifera 143, 145 Oplopanax elatus 8, 123 Orchis 142 Origanum 131 Origanum vulgare 134 Ornithogalum 141 Oryza 150 Oryza saliva 156 Oxycoccus palustris 23 Oxytropis 116 Picea abies 63 Picea canadensis 63 Picea engelmannii 63 Picea obovata 63 Picea pungens 63 Pilea 109, 110 Pimpinella 121 Pimpinella anisum 123 Pimpinella saxifraga 122 Pinus 63 Pinus koraiensis 63 Pinus longaeva 64 Pinus palustris 61 Pinus pinea 64 Pinus sibirica 51, 63 Pinus sylvestris 63 Piper betle 162 Pisum 116, 117 Pisum sativum 119 Platanthera 142
Padus 113 Panax ginseng 8, 123 Panicum 150 Panicum miliaceum 156 Papaver somniferum 84 Pastinaca 121 Pastinaca sativa 118, 123 Pedicularis 129, 130 Persea americana 79 Persica 113 Persica vulgaris 115 Petroselinum 121 Petroselinum crispum 123 Peucedanum 121 Phalaroides arundinacea 153 Phaseolus 116, 118 Phaseolus vulgaris 118, 119 Phleum pratense 151, 157 Phlomis 131 Phlomoides tuberosa 132 Phoenix 158 Phoenix dactylifera 159, 160 Phragmites 150 Phragmites australis 153 Phyllitis scolopendrium 37 Pleurospermum uralense 122 Poa 150, 151 Poa annua 151 Podocarpus 61, 66 Polygonatum 141 Polygonum 92 Polygonum bistorta 93 Polygonum arenastrum 93 Polypodium 42 Polypodium vulgare 42 Polystichum 42 Poly trichum commune 18, 22 Populus 99 Populus alba 100 Populus nigra 100 Populus tremula 100 Potentilla 113 Prunus 113, 114 Prunus cerasifera 114 Prunus domestica 114 Pteridium 42 Pteridium aquilinum 38, 43 Puccinellia 150 Pulmonaria 127 Pulsatilla 82, 83
251
Pulsatilla patens 84
Pyrrosia lingua 43
Pyrus 113, 114
Pyrus communis 115
Quercus 93, 94
Quercus ilex 95
Quercus robur 94
Quercus suber 95
Ranunculus 82, 83
Raphanus 103
Raphanus raphanistrum 104,
105
Rhapis 161
Rheum 92, 93
Rhinanthus 129, 130
Rhynchospora 147
Ricinus 111
Ricinus communis 111
Robinia 116
Rorippa 103
Rosa 113, 114
Rosa damascena 115
Rubus 113
Rubus idaeus 114, 115
Rudbeckia 135
Rumex 93, 93
Rumex obtusifolius 92
Rumex thyrsiflorus 93
Sabal 161
Saccharum 150
Saccharum officinarum 157
Salicomia 88, 89, 90
Salix 99
Salix acutifolia 101
Salix alba 100
Salix caprea 100
Salix cinerea 100
Salix pentandra 100
Salix triandra 100
Salix viminalis 100
Salsola 88
Salvia 131, 132
Salvia sclarea 134
Salvinia 45
Salvinia auriculata 45
Salvinia natans 40, 45
252
Sanguisorba 113
Saponaria 86
Sasa 152
Schisandra 70
Schistostega pennata 26
Scirpus 147, 148
Scirpus lacustris 148
Scorzonera 135
Scrophularia 129, 130
Scutellaria 131
Secale 150
Secale cereale 155
Selaginella 30
Selaginella sanguinolenta 33
Selaginella selaginoides 33
Senecio 135
Sequoia 64
Sequoiadendron 64
Serratula 137
Seseli 121
Seseli libanotis 122
Setaria 150
Silene 86
Silene nutans 87
Sinapis 103
Sisymbrium 103
Sisymbrium loeselii 105
Sisymbrium officinale 105
Sium 121
Sium latifolium 122
Solanum 123
Solanum melongena 124
Solanum tuberosum 124
Solidago 135, 136
Sonchus 135
Sonchus oleraceus 137
Sorbus 113
Sorghum 150, 152
Spergula 87
Sphaerocephalum 135
Sphagnum 18, 22
Sphenopyllum 34
Spinacia oleracea 88, 91
Spiraea 113, 114
Splachnum 26
Stachys 131
Stachys annua 134
Stachys palustris 132
Stachys sylvatica 132
Stellaria 86
Stellaria decumbens 88
Stellaria nemorum 87
Steris 86
Stipa 150, 152, 153
Suaeda 88, 90
Symphytum 127
Tagetes 135, 137
Tamarindus indica 120
Tanacetum 135, 137
Taraxacum 135, 136, 137
Taxus 67
Taxus baccata 67
Taxus cuspidata 67
Thalictrum 82, 83
Thalictrum minus 84
Theophrasta 9
Thlaspi 103
Thlaspi arvense 104, 105, 106
Thuja occidentalis 65
Thuja orientalis 66
Thymus 131, 132
Thymus vulgaris 134
Tortula laevipila 26
Trachycarpus fortunei 161
Trifolium 116, 117, 118
Triticum 150
Triticum aestivum 151, 155
Triticum durum 155
Trochodendron 70
Trollius 83
Trollius asiaticus 84
Trollius europaeus 85
Tulipa 139, 140
Tussilago 135, 136
Typha 149
Ulmus 106
Urtica 109
Urtica cannabina 109
Urtica dioica 109
Urtica urens 109
Vanilla planifolia 146
Verbascum 129
Verbascum nigrum 130
Vanilla 142
Vanilla planifolia 146
Veronica 129
Veronica chamaedrys 130
Vicia 116, 117, 118
Vicia faba 119
Victoria 79, 80
Vigna 118
Welwitschia 67
Welwitschia mirabilis 67
Williamsonia 58
Williamsoniella 58
Woodsia 42
Xanthium 136, 137
Zea 150, 152
Zea majus 151, 156
Zelkova 106
Zostera marina 70
Указатель русских названий растений
Абрикос 114
Авокадо 79
Агатис 61, 62
Адонис весенний 84
Азолла 45
Айва обыкновенная 114
Акация серебристая 120
Аконит 84, 85
Алыча культурная 114
Амарант 91
Амброзия 135
Ангиоптерис 42
Ангиоптерис вознесенный 42
Анис обыкновенный 123
Аралия 123
Араукария 61, 62
Араукария чилийская 62
Арахис 118, 120
Арбуз 102
Багульник 23
Базилик 134
Баклажан 124, 125
Барбарис 84
Бассия 90
Бедренец камнеломка 122
Белена черная 125
Белладонна 125, 126
Береза карликовая 96
Береза повислая 98
Береза пушистая 98
Бешеный огурец 103
Бобы кормовые 119
Бодяк полевой 137
Болиголов крапчатый 122
Болотница 149
Болотница сладкая 149
Борщевик сибирский 122
Будра плющевидная 132
Бук 95
Бук восточный 95
Бук европейский 95
Бурачник лекарственный 128
Вайда 105
Ваниль 146
Василек 137
Василистник малый 84
Вельвичия 67, 68
Вельвичия удивительная 67
Венерин башмачок 143
Верба 101
Вероника 129
Ветреница лесная 84
Ветреничка 85
Ветреничка алтайская 84
Ветреничка дубравная 84
Ветреничка лютиковидная 84
Вех ядовитый 122
Вигна 118
Вика 118
Виктория 80
Вильямсониелла 58
Вильямсония 58
Вишня садовая 114
Водосбор 84
Вьюнок 128
Вьюнок полевой 128
Вяз 106
Гвоздика 88
Гевея бразильская 111
Гиацинт 141
Гиения 34
Гинкго 59
Гинкго двулопастный 59
Гладиолус 141
Гнездовка 146
Гнетум 67
Горец 93
Горец живородящий 93
Горох 118
Горох посевной 119
Горчица сарептская 105
Горчица черная 105
Граб европейский 99
Гречиха 93
Гроздовник 39, 41
Гроздовник простой 41
Груша домашняя 115
Гулявник Лезеля 105
254
Гулявник лекарственный 105
Гусиный лук 10
Дельфиниум 84
Дескурайния Софьи 105
Джузгун 93
Дзельква 106
Донник 118, 120
Дремлик 146
Дуб 9, 94, 95
Дуб пробковый 95
Дуб черешчатый 94, 95
Дудник лесной 122
Дурман вонючий 125
Дурнишник 137
Душица 134
Дыня 102
Дягиль 122
Ежа сборная 157
Ель 60, 62, 63
Ель канадская 63
Ель обыкновенная 63
Ель сибирская 63
Жабрица порезниковая 122
Женьшень 8, 41, 123
Жерардиния ПО
Живокость 84
Живучка 133
Живучка ползучая 132
Житняк 153
Жминда 90
Заманиха 8, 123
Заразиха 138
Звездчатка 86
Земляника лесная 115
Земляника садовая 115
Змеевка 153
Зопник 134
Зопник клубненосный 132
Ива 99, 100
Ива белая 100
Ива козья 100
Ива корзиночная 100
Ива пепельная 100
Ива пятитычинковая 100
Ива трехтычинковая 100
Ипомея пурпурная 128
Ирис 9, 141
Кабачок 102
Каламит 34
Каламус 158
Калимматотека 54
Камыш озерный 148
Капуста 105
Капуста брюссельская 105
Капуста кольраби 105
Капуста кочанная 105
Капуста огородная 105
Капуста цветная 105
Карагана древовидная 120
Картофель 124, 125
Катран 105
Каури 62
Качим аретиевидный 88
Качим метельчатый 87
Каштан 95
Каштан зубчатый 95
Каштан настоящий 95
Кедр 63, 64
Кедр ливанский 64
Кипарис 51, 65
Кипарис вечнозеленый 65
Кипарис мексиканский 65
Клевер 117, 118
Клещевина 111
Клинолист 34
Клоповник сорный 106
Клубнекамыш морской 149
Клубника 115
Клюква 23
Кобрезия 149
Ковыль 152, 153
Козлятник 118
Кокпек 90
Колобантус кито 88
Кольраби 105
Конопля 106, 108
Конский каштан 95
Консолида 84
Кориандр посевной 123
Корифа высокая 159
Коричник 79
Коровяк 129
255
Кострец безостый 152, 157
Котовник 134
Краекучник Куна 43
Крапива 109
Крапива двудомная 109, ПО
Крапива жгучая 109
Крапива коноплевидная 109
Красавка обыкновенная 125
Кубышка 81
Кувшинка 81
Кукуруза 152,156
Кукушкин лен 10, 22
Купальница 83
Купальница азиатская 84
Купальница европейская 85
Купена 141
Лаванда 132,134
Лавр 79
Лавр благородный 79
Ландыш 141
Ландыш майский 141
Лапортея ПО
Лапчатка 113
Латук 137
Лебеда 90
Левкой однолетний 105
Лещина 96, 98, 130
Лещина обыкновенная 98
Ливистона 161
Лилия саранка 140
Липучка 128
Лисохвост луговой 157
Лиственница 60, 62, 63
Лиственница Гмелина 63
Лиственница сибирская 63
Листовик 37
Ломонос 84
Лопух 137
Лотос 81
Лотос орехоносный 81
Лук 140, 141
Лук косой 141
Лук многокорневой 142
Лук репчатый 140
Лук-батун 141
Лук-порей 140
Львиный зев 129, 131
Льнянка 129
Люпин 120
Лютик 83
Люффа 102
Люцерна 118
Магнолия 78
Магнолия крупноцветковая 78
Магнолия обратнояйцевидная 78
Майник 141
Майоран 134
Мак 85
Мак снотворный 84
Макрозамия 58
Малина обыкновенная 115
Мамонтово дерево 65
Мангольд 91
Манжетка 113
Маниок съедобный 112
Мараттия 42
Марсилея 43, 50
Марсилея четырехлистная 43
Маршанция многообразная 21
Марь 90
Марь белая 90
Марьянник 130
Медуллоза 54
Медуница 127
Мелисса 134
Мерингия 87
Метасеквойя 64
Микробиота перекрестнопарная
66
Мимоза стыдливая 120
Миндаль обыкновенный 115
Можжевельник 61, 65, 66
Молокан татарский 137
Молочай 111, 112
Молочай лозный 112
Молуккское дерево 113
Мора маслоносная 120
Морковь 122
Морская горчица 105, 106
Мускари 141
Мушмула японская 115
Мытник 130, 131
Мышиный гиацинт 141
Мята 134
Мята полевая 132
256
Наперстянка 129
Нардосмия 137
Незабудка 128
Неслия метельчатая 105
Нивяник 137
Нимфея 80
Ночная фиалка 105
Нут 118
Овес посевной 155
Овсяница 153
Овсяница луговая 157
Огурец 102
Огуречная трава 128
Одуванчик 137
Ольха 96, 98, 130
Ольха серая 98
Ольха черная 98
Онцидиум 144
Орляк 38, 43
Осина 100
Осока 148
Осока дернистая 147
Осока острая 147
Осока приземистая 147
Осока пузырчатая 147
Осока ранняя 147
Осока стоповидная 147
Осока твердоватая 147
Осокорь 100
Осот огородный 137
Офрис 145
Офрис насекомоносная 145
Очанка 130
Пальма бетелевая 162
Пальма винная 160
Пальма кокосовая 159, 160
Пальма масличная 160
Пальма саговая 159
Пальма сахарная 161
Пальма сейшельская 159, 161
Пальма финиковая 159, 160
Папирус 149
Пастернак посевной 123
Пастушья сумка обыкновенная
105
Переступень белый 102
Персик 115
Петров крест 130
Петрушка кудрявая 123
Петуший гребень 91
Пижма 137
Пикульник двунадрезанный 134
Пикульник ладанниковый 134
Пилея ПО
Пиния 64
Пиррозия язычковая 43
Пихта 60, 62, 63
Пихта сибирская 63
Плаун 30
Плющ 123
Повилика 128
Повой 128, 131
Погремок 130
Подокарп 61, 66
Подснежник 84
Подсолнечник 135, 137, 138
Полушник 32
Полынь 135, 137
Полынь горькая 137
Полынь обыкновенная 137
Помидор 124
Поручейник широколистный 122
Поташник 90
Примула 9
Пролесник 111
Просо обыкновенное 156
Прострел раскрытый 84
Птицемлечник 141
Пузыреплодник лопастнолистный
102
Пушица 149
Пшеница мягкая 155
Пшеница твердая 155
Пырей ползучий 152
Рами ПО
Рапис 161
Рапс 105
Реброплодник уральский 122
Ревень 93
Редька дикая 105
Рис 158
Рис посевной 156
Рогоз 9, 149
Рожковое дерево 120
257
Рожь посевная 155
Роза 115
Роза дамасская 115
Розмарин 132
Ромашка 137
Рубус 113
Рябина черноплодная 115
Рябчик 140
Ряска малая 70
Сабаль 161
Саговник 56
Саговник завитой 55
Саговник поникающий 55, 57
Саза 152
Саксаул 89, 90, 91
Сальвиния 45
Сальвиния плавающая 45
Сальвиния ушковидная 45
Сахарный тростник 157
Сведа 90
Свекла 91
Свекла морская 91
Свербига восточная 105
Секвойя 64, 65
Секвойядендрон 64, 65
Селагинелла 33, 50
Сельдерей пахучий 123
Сердечник 105
Сердечник луговой 106
Серпуха 137
Синеголовник 121
Синяк 127
Сладкий перец 124
Слива домашняя 115
Смолевка 86
Смолка 86
Сныть обыкновенная 122
Солерос 90
Солерос обыкновенный 90
Сон-трава 83, 85
Сорго 152
Сосна 51, 52, 60, 62, 63
Сосна болотная 61
Сосна долговечная 64
Сосна корейская 63
Сосна обыкновенная 63
Сосна сибирская 8, 51, 63
Соя 118, 119
Спаржа 141
Сплахн 26
Спорыш 93
Страусиное перо 42
Сурепка обыкновенная 105
Сфагн 22
Схистостега перистая 26
Сыть бурая 149
Сыть очереднолистная 149
Сыть съедобная 149
Табак 124
Тайник 146
Тамаринд 120
Татарник 137
Тимофеевка луговая 157
Тимьян 132, 134
Типчак 152, 153
Тис 67
Тис остроконечный 67
Тис ягодный 67
Тмин обыкновенный 122, 123
Тонконог 153
Топинамбур 137
Тописолнечник 137
Тополь 99, 100, 101
Тополь белый 100
Тополь дрожащий 100
Тополь черный 100
Трахикарпус 161
Тростник 153
Тунг 112
Турнепс 105
Туя 9
Туя восточная 66
Туя западная 65
Тыква 102
Тюльпан 140, 141
Тюльпанное дерево 79
Ужовник 39
Укроп огородный 123
Фасоль 118
Фасоль обыкновенная 118,119
Фенхель обыкновенный 123
Физалис овощной 124
Физалис Франше 125
Фундук 98
258
Хамеропс 161
Хвощ 33, 34
Хвощ зимующий 35
Хвощ лесной 35
Хвощ многощитинковый 35
Хвощ полевой 35
Хвощ приречный 35
Хмель 106, 108
Хрен дикий 105
Хризантема 138
Хумулопсис 106
Цикадеоидея 58
Череда 137
Черемша 141
Черешня 115
Чертополох 137
Чеснок 140
Чина 118
Чистец болотный 132
Чистец лесной 132
Чистец однолетний 134
Чистотел большой 85
Чозения 99
Чуфа 149
Шалфей 132, 133, 134
Шалфей мускатный 134
Шароголовник 135
Шелковица 106
Шильник 32, 33, 48, 55, 76
Шиповник 113
Шнитт-лук 141
Шпинат 91
Щавель 93
Щавель пирамидальный 93
Щирица 91
Щитовник 38
Щитовник мужской 43
Щучка дернистая 152
Эвкалипт 58, 70
Эвриала устрашающая 81
Элеутерококк 123
Элеутерококк колючий 8
Энтада лазящая 120
Эспарцет 118
Эфедра 67
Эхиноцистис 102
Яблоня домашняя 115
Ярутка полевая 105, 106
Ясколка 86
Яснотка 133
Яснотка белая 132
Ячмень культурный 155
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ................................................ 3
ЧАСТЬ 1. ВВЕДЕНИЕ В СИСТЕМАТИКУ ВЫСШИХ РАСТЕНИЙ 5
ГЛАВА1. ДЛЯ ЧЕГО НУЖНА СИСТЕМАТИКА ВООБЩЕ И
ВЫСШИХ РАСТЕНИЙ В ЧАСТНОСТИ............................. 5
1.1. Понятие о таксономии.............................. 5
1.2. Задачи, для решения которых нужна систематика
РАСТЕНИЙ............................................ 6
1.2.1. Определение областей экстраполяции данных, получа-
емых другими науками............................. 6
1.2.2. Изучение флор................................. 7
1.2.3. Прогностическая роль систематики растений. 8
1.3. История систематики растений...................... 8
1.4. Особенности современного этапа развития.......... 10
Глава 2. ОБЩАЯ ХАРАКТЕРИСТИКА ВЫСШИХ РАСТЕНИЙ.. 11
2.1. Отличия высших растений от низших............. 11
2.2. Приспособления к наземному образу жизни....... 12
2.3. Жизненный цикл................................ 13
2.4. Основные отделы.................................. 14
2.5. Заключение: сравнение высших и низших растений .. 15
Часть 2. СПОРОВЫЕ И ГОЛОСЕМЕННЫЕ.......................... 17
Глава 3. МОХОВИДНЫЕ (BRYOPHYTA)........................... 17
3.1. Общая характеристика............................. 17
3.1.1. Гаметофит.................................... 17
3.1.2. Оплодотворение и образование спорофита.... 19
3.2. Обзор основных таксонов.......................... 20
3.2.1. Класс Hepaticopsida (печеночники) 20
3.2.2. Класс Bryopsida (настоящие мхи).............. 21
3.3. Экологическое разнообразие и механизмы устойчивости
моховидных......................................... 24
3.4. Значение моховидных.............................. 26
3.5. Заключение: сравнение основных таксонов....... 26
Глава 4. СОСУДИСТЫЕ СПОРОВЫЕ РАСТЕНИЯ..................... 28
4.1. Общая характеристика............................. 28
4.1.1. Гаметофиты................................... 28
4.1.2. Строение спорофита........................... 29
4.2. Отдел Lycopodiophyta (плауновидные).............. 29
4.2.1. Порядок Lycopodiales (плауновые)............. 30
4.2.2. Порядок Isoetales (полушниковые)............. 32
4.2.3. Порядок Selaginellales (селагинелловые) 33
4.3. Отдел Equisetophyta (хвощевидные)................ 33
4.4. Отдел Polypodiophyta (папоротниковидные) 36
4.4.1. Общая характеристика......................... 36
4.4.2. Класс Ophioglossopsida (ужовниковые) 39
4.4.3. Класс Marattiopsida (мараттиевые)............ 41
4.4.4. Класс Polypodiopsida......................... 42
4.5. Экологическое разнообразие и механизмы устойчивости
сосудистых споровых растений....................... 46
260
4.6. ЗАКЛЮЧЕНИЕ: СРАВНЕНИЕ ОТДЕЛОВ.................. 47
ГЛАВА 5. ОТДЕЛ PINOPHYTA (ГОЛОСЕМЕННЫЕ)................. 48
5.1. Общая характеристика........................... 48
5.1.1. Особенности жизненного цикла.............. 50
5.1.2. Репродуктивные органы..................... 51
5.1.3. Основные таксоны.......................... 53
5.2. Класс Pteridospermae (семенные папоротники).. 54
5.3. Класс Cycadopsida (саговниковые)............... 54
5.3.1. Общая характеристика...................... 54
5.3.2. Жизненная форма........................... 55
5.3.3. Генеративные органы....................... 56
5.3.4. Значение саговниковых..................... 57
5.4. Класс Bennettitopsida (беннеттиты)........... 5 8
5.5. Класс Ginkgoopsida (гинкговые)................. 59
5.6. Класс Pinopsida, подкласс pinidae (хвойные).. 60
5.6.1. Общая характеристика...................... 60
5.6.2. Порядок Araucariales (араукариевые) 62
5.6.3. Порядок Pinales........................... 62
5.6.4. Порядок Cupressales (кипарисовые)......... 64
5.6.5. Порядок Podocarpales (подокарповые)....... 66
5.6.6. Порядок Taxales (тисовые)................. 67
5.7. Класс Gnetopsida (гнетовые)................... 67-
5.8. Заключение: сравнение саговниковых и хвойных. 68
Часть 3. ЦВЕТКОВЫЕ...................................... 69
ГЛАВА 6. ОБЩАЯ ХАРАКТЕРИСТИКА ЦВЕТКОВЫХ РАСТЕНИЙ
И ОБЗОР НЕКОТОРЫХ ГИПОТЕЗ ИХ ПРОИСХОЖДЕ-
НИЯ.................................................... 70
6.1. Общая характеристика цветковых................. 70
6.2. Гипотезы происхождения цветковых............... 72
6.3. Однодольные и двудольные....................... 75
6.4. Заключение: причины успеха цветковых........... 76
Глава 7. Класс Magnoliopsida - Двудольные............... 77
7.1. Семейство Magnoliaceae - Магнолиевые........... 77
7.2. Семейство Nymphaeaceae - Нимфейные............. 79
7.3. Семейство Ranunculaceae - Лютиковые............ 82
7.4. Семейство Caryophyllaceae - Гвоздичные......... 86
7.5. Семейство Chenopodiaceae - Маревые............. 88
7.6. Семейство Polygonaceae - Гречишные............. 92
7.7. Семейство Fagaceae - Буковые................... 93
7.8. Семейство Betulaceae - Березовые............... 96
7.9. Семейство Salicaceae - Ивовые.................. 99
7.10. Семейство Cucurbitaceae - Тыквенные.......... 101
7.11. Семейство Brassicaceae (Cruciferae) - Капустные
(Крестоцветные)..................................... ЮЗ
7.12. Семейство Cannabaceae - Коноплевые........... 106
7.13. Семейство Urticaceae - Крапивные.............. Ю9
7.14. Семейство Euphorbiaceae - Молочайные......... 111
7.15. Семейство Rosaceae - Розовые.................. ИЗ
261
7.16. Семейство Fabaceae - Бобовые................. 116
7.17. Семейство Apiaceae (Umbelliferae) - Сельдерейные
(Зонтичные)...................................... 121
7.18. Семейство Solanaceae - Пасленовые............ 123
7.19. Семейство Boraginaceae - Бурачниковые........ 127
7.20. Семейство Scrophulariaceae - Норичниковые.... 129
7.21. Семейство Lamiaceae (Labiatae) - Яснотковые
(Губоцветные).................................... 131
7.22. Семейство Asteraceae (Compositae) - Астровые
(Сложноцветные).................................. 135
ГЛАВА 8. КЛАСС LILIOPSIDA - ОДНОДОЛЬНЫЕ................ 139
8.1. Семейство Liliaceae - Лилейные................ 139
8.2. семейство Orchidaceae - Орхидные.............. 142
8.3. Семейство Cyperaceae- Осоковые................ 147
8.4. Семейство Poaceae (Gramineae) - Мятликовые (Злаки) 150
8.5. Семейство Arecaceae (Palmaceae) - Арековые
(Пальмовые)...................................... 158
часть 4. ОСНОВЫ НАУКИ О РАСТИТЕЛЬНОСТИ................. 163
ГЛАВА 9. ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ......................... 164
9.1. Классификация экологических факторов.......... 164
9.2. Комплексные градиенты......................... 165
9.3. Ведущие экологические факторы................. 165
9.4. Биотические факторы........................... 166
9.5. Антропогенные факторы......................... 166
ГЛАВА 10. ЭКОЛОГИЯ ВИДОВ............................... 166
10.1. Основные подходы при изучении экологии видов. 166
10.2. Жизненные формы.............................. 167
10.3. География видов.............................. 169
10.4. Экологические группы видов................... 171
10.5. Стратегии видов растений..................... 172
10.5.1. Конкуренция............................. 172
10.5.2. Экологическая ниша...................... 174
10.5.3. Типы стратегий растений Маклиода - Пианки.... 177
10.5.4. Типы стратегий растений Раменского - Грайма.. 177
10.5.5. Стратегии культурных растений........... 183
ГЛАВА 11. ПОПУЛЯЦИИ РАСТЕНИЙ........................... 184
11.1. Локальные и менделевские популяции........... 184
11.2. Сравнение популяций растений и животных...... 184
11.3. Плотность популяции.......................... 185
11.4. Масса популяции.............................. 187
11.5. Распределение популяций в пространстве....... 188
11.6. Гетерогенность популяций..................... 189
11.7. Особенности популяций споровых растений...... 192
ГЛАВА 12. РАСТИТЕЛЬНЫЕ СООБЩЕСТВА (ФИТОЦЕНОЗЫ).. 193
12.1. Факторы и модели организации фитоценозов..... 193
12.1.1. Экотоп.................................. 193
12.1.2. Взаимоотношения растений................ 194
12.1.3. Роль гетеротрофных организмов........... 199
262
12.1.4. Роль режима нарушений...................... 202
12.1.5. Множественность моделей организации фитоце-
нозов....................................... 203
12.2. Основные признаки фитоценозов................... 205
12.2.1. Видовое богатство.......................... 205
12.2.2. Структура фитоценоза....................... 209
12.3. Динамика растительности......................... 219
12.3.1. Классификация форм динамики растительности. 219
12.3.2. Автогенные сукцессии....................... 221
12.3.3. Аллогенные сукцессии....................... 224
12.3.4. Концепция климакса в растительности........ 228
12.3.5. Эволюция фитоценозов....................... 229
12.4. Классификация растительности.................... 231
12.4.1. Эколого-флористический подход.............. 231
12.4.2. Физиономический подход..................... 234
ГЛАВА 13. ИЗУЧЕНИЕ ФЛОР.................................. 235
13.1. Региональная и конкретная флора. 235
13.2. Анализ флор..................................... 236
13.3. Анализ адвентивного компонента флоры............ 237
Приложение 1. СПИСОК НАИБОЛЕЕ РАСПРОСТРАНЕННЫХ
КУЛЬТУРНЫХ РАСТЕНИЙ, НЕ ВХОДЯЩИХ В РАССМОТРЕН-
НЫЕ СЕМЕЙСТВА ЦВЕТКОВЫХ............................... 238
Приложение 2. СПИСОК ОСНОВНЫХ КЛАССОВ РАСТИТЕЛЬ-
НОСТИ СРЕДНЕЙ ПОЛОСЫ ЕВРОПЕЙСКОЙ ЧАСТИ
РОССИИ................................................ 240
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ.......................... 245
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ РАСТЕНИЙ.................... 246
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ РАСТЕНИЙ...................... 254
Учебное издание
Миркин Борис Михайлович
Наумова Лениза Гумеровна
Мулдашев Альберт Акрамович
Высшие растения: краткий курс
систематики с основами науки о растительности
Учебник
Редактор Э.М. Горелик
Переплет Е. Молчанова, С. Носова
Компьютерная верстка Т.Н. Лебедевой
Корректор М.Д. Шунина
Изд. лиц. ИД № 01670 от 24.04.2000
Налоговая льгота - общероссийский классификатор
продукции ОК-005-93, том 2: 953000
Подписано в печать 29.12.2000. Формат 60x90/16.
Бумага офсетная № 1. Печать офсетная. Гарнитура MyslCTT.
Печ. л. 16,5. Тираж 3000 экз. Заказ № 684
Издательско-книготорговый дом «Логос»
105318, Москва, Измайловское ш., 4
Отпечатано Марийским полиграфическо-издательским комбинатом
424000, Йошкар-Ола, ул. Комсомольская, 112
По вопросам приобретения литературы
обращаться по адресу:
105318, Москва, Измайловское ш., 4.
Тел./факс: (095) 369-5819,369-5668, 369-7727
Электронная почта: universitas@mail.ru
Интернет-магазин «Университетская книга»:
L http://www.chat.ru/~universltas/ .