/


































Текст
-4 -
ФУНКЦИИ ПОЧЕК В ОРГАНИЗМЕ ЧЕЛОВЕКА
В живом организме всегда образуются или попадают в него из внеш-
ней среды вещества, которые необходимо удалить. Физиологический про-
цесс, направленный на выведение из внутренней среды организма конеч-
ных продуктов метаболизма, чужеродных, токсических веществ, избытка
ионов и воды называется выделением или экскрецией. Исключительную
роль в процессе выделения играют почки, где кровь очищается 350 раз за
сутки и без которых жизнь невозможна. В состав выделительной системы
организма входят также легкие, желудочно-кишечный тракт, железы (слюн-
ные, потовые, сальные, печень).
Основная функция почек - гомеостатическая. Во внутренней среде
организма почками поддерживается pH, ионный состав крови (цзоиония),
осмотическое давление (изоосмия), объем жидкости (изоволюмия). От их
работы зависит системное гидростатическое давление крови.
Почки выполняют и другие функции:
- синтезируют и секретируют в кровь гуморальные факторы регуля-
ции кровообращения (ренин, простагландины, кинины), эритропоэза (эри-
тропоэтины), свертывания крови (урокиназа, тромбопластин, тромбоксан),
кальций-фосфорного обмена (кальшприол);
- участвуют в глюконеогенезе: во время длительного голодания поч-
ки синтезируют глюкозу из аминокисло i;
- выводят из организма чужеродные и токсические вещества, избы-
ток азота.
СТРОЕНИЕ НЕФРОНА
Отличительной особенностью почек млекопитающих является четкое
деление на две зоны: наружную корковое вещество и внутреннюю -
мозговое вещество. Корковое вещество темное, имеет гранулированность.
Мозговое вещество более светлое, делится на наружную и внутреннюю зо-
ны. Все эти отличия отражают расположение различных компонентов неф-
рона - структурного элемента почки на микроскопическом уровне. В каж-
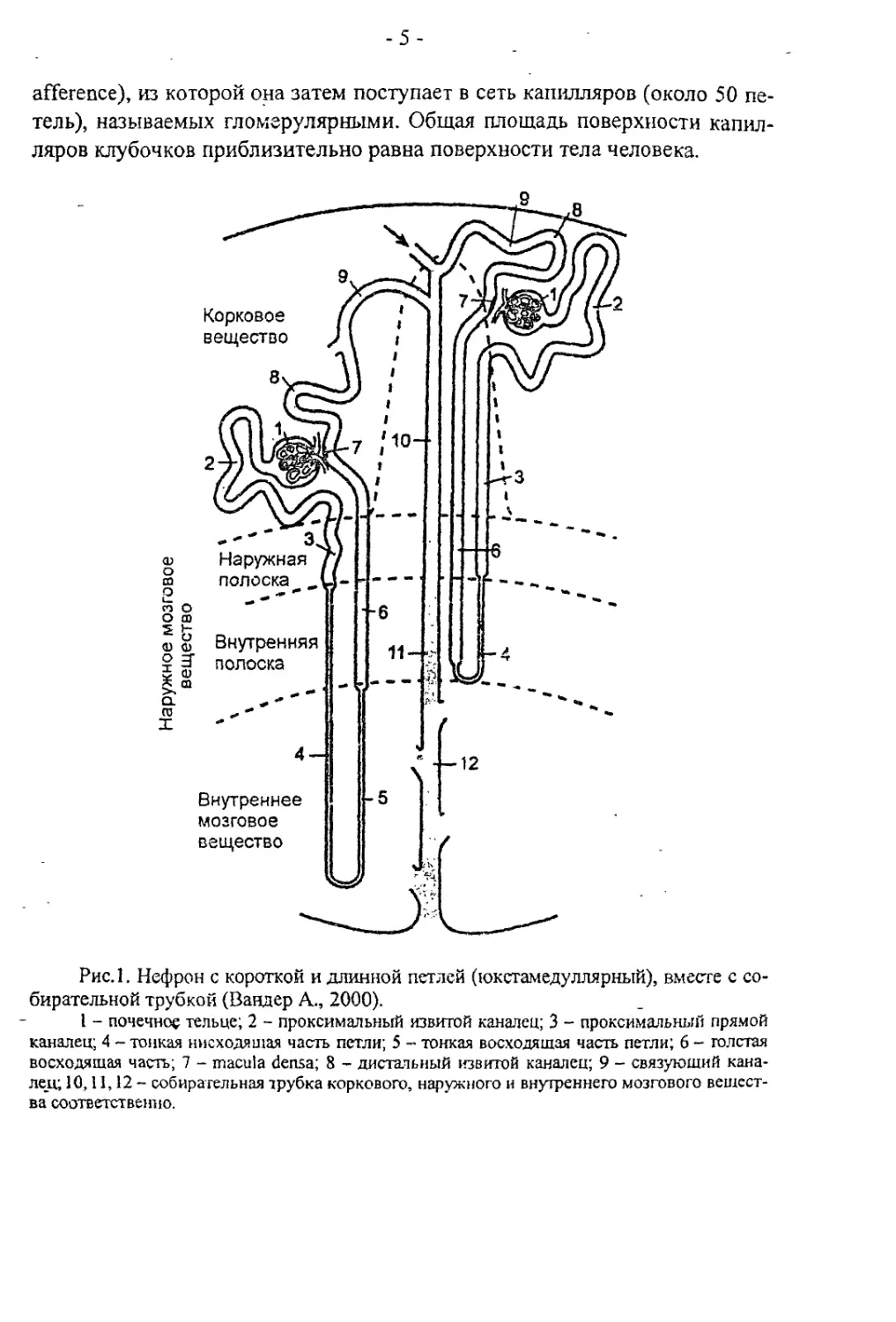
дой почке более миллиона нефронов (рис. 1).
Нефрон состоит из почечного тельца и канальцев. Почечное тельце
расположено в корковом веществе. Оно представлено капсулой Боумена-
Шумлянекого диаметром 0,2 мм, в которую включен сосудистый клу-
бочек. К почечному тельцу кровь приходит по приносящей артериоле (vas
-5-
afference), из которой она затем поступает в сеть капилляров (около 50 пе-
тель), называемых гломерулярными. Общая площадь поверхности капил-
ляров клубочков приблизительно равна поверхности тела человека.
4-
Внутреннее
мозговое
вещество
Рис.1. Нефрон с короткой и длинной петлей (юкстамедуллярный), вместе с со-
бирательной трубкой (Вандер А., 2000).
I - почечное тельце; 2 - проксимальный извитой каналец; 3 - проксимальный прямой
каналец; 4 - тонкая нисходящая часть петли; 5 - тонкая восходящая часть петли; 6 - толстая
восходящая часть; 7 - macula densa; 8 - дистальный извитой каналец; 9 - связующий кана-
лец; 10,11,12 - собирательная трубка коркового, наружного и внутреннего мозгового вещест-
ва соответственно.
- 6 -
Капсула образована двумя листками: париетальным и висцеральным
между которыми есть просвет. Часть плазмы крови переходит в просвет
капсулы, остальная кровь выходит из почечного тельца по выносящей ар-
териоле (vas efference). Между капиллярами клубочка расположены мезан-
гиальные клетки, представляющие собой видоизмененные м иоэ питал иаль-
ные клетки. При их сокращении уменьшается площадь клубочка, что при-
водит к уменьшению объема жидкости, попадающей в просвет капсулы.
Полость капсулы переходит в просвет канальца, который имеет дли-
ну 35-55 мм и делится на отделы: проксимальный сегмент, петлю Генле
и дистальный сегмент.
Проксимальный сегмент, куда поступает жидкость из капсулы, состо-
ит из извитого и прямого канальца, который переходит в петлю Генле, со-
стоящую из нисходящей и восходящей частей.
Тонкая нисходящая часть петли опускается из коркового вглубь моз-
гового вещества, изгибается на 180° и переходит в тонкую восходящую
часть петли. Тонкий каналец этой части затем переходит в толстый, кото-
рый поднимается до уровня клубочка своего же нефрона в корковом ве-
ществе почки, где начинается дистальный извитой каналец.
Конечный участок восходящего колена петли Генле подходит к клу-
бочку и соприкасается с его полюсом между приносящей и выносящей
артериолами. Клетки эпителия извитого канальца, имеющие гиперхромные
ядра, здесь тесно контактируют с гранулированными миоэпителиальными
клетками артериолы. Поэтому этот участок выглядит темным, и его называ-
ют плотным пятном (macula densa) юкстагломерулярного аппарата почки.
Дистальный сегмент состоит из прямого (толстая часть петли), изви-
того и связующего канальцев. Связующий каналец впадает в собиратель-
ную трубку. Собирательная трубка не относится к нефрону, но функцио-
нально и топографически связана с ним.
На всем протяжении каналец и собирательная трубка образованы
одним слоем клеток однорядного эпителия, покоящихся на базальной мем-
бране. Структурная и цитохимическая характеристика клеток канальца и со-
бирательной трубки варьирует от сегмента к сегменту, но одной обшей
чертой является наличие плотного контакта между смежными клетками.
Плотные контакты делят мембрану на две части: апикальную (люминаль-
ную) и базолатеральную мембраны (рис.2).
Апикальная (люминальная) мембрана контактирует с просветом ка-
нальца, а базолатеральная - с интерстициальным пространством. Мембра-
- 7-
ны отличаются строением, химическими характеристиками, мембранным
потенциалом и функциональной нагрузкой.
Базальная
мембрана
Базолатеральная
мембрана
Интерстициальная
жидкость
Плотное
соединение
Люминальная
мембрана
Рис.2. Схематическое изображение канальцевого эпителия (Вандер А.,2000).
КЛАССИФИКАЦИЯ НЕФРОНОВ
Выделяют три типа нефронов:
- суперфициальные - 20-30%;
- корковые - 60-70%;
- юкстамедуллярные - 10—15%
(соотношение в процентах приводится для нефронов почки человека).
Нефроны отличаются локализацией почечного тельца, длиной петли
Генле, кровоснабжением и функцией.
Название нефрона определяет локализация его почечного тельца: у су-
перфициальногб нефрона оно расположено на глубине 1 мм от поверхнос-
ти почки, у коркового - в середине коркового вещества, у юкстамедулляр-
ных - ближе к границе с мозговым веществом (рис.З).
Петля Генле у юкстамедуллярных нефронов намного больше, чем
у корковых, и спускается глубоко, почти до вершинки сосочка пирамиды.
Юкстамедуллярные нефроны играют ведущую роль в процессах концен-
трирования и разведения мочи.
Зыноояцдая
Корковый
Рис.З. Принципиальна* схема строения и кровоснабжения юкст<%мед.уляярнО’
го и коркового нефронов (Guyton А.С., Half Y.E., 2001).
ОСОБЕННОСТИ КРОВОСНАБЖЕНИЯ ПОЧКИ
Кровь поступает в почку через почечную артерию, которая затем раз-
ветвляе-тся на see более мелкие ветви: междолевую, дуговую к радиальную
(междольковую) артерии. Каждая радиальная артерия делится на прино-
сящие (афферентю?,е) артериолы, но которым кровь поступает в капилляр
ры почечного клубочка. Из гломерулярных капилляроц почечного тельца
кровь поступает в выносящую артериолу, которая образует вторичную ка-
пиллярную систему, окружающую .чана»ьцы. Эти капилляры затем объеди-
няются, и образуетет радиальная, дуговая, мр-жда.певаи и почечная вена, но
которой кровь в конечном тоге покидает почку. Наличие ?»?ух чипов ^рте-
-9-
'риол двух типов капилляров является особенностью сосудистой системы
почки (см. рис.З).
У коркового нефрона приносящая артериола шире выносящей. У юкс-
^медуллярного нефрона диаметр приносящей артериолы меньше или равен
диаметру выносящей артериолы. Выносящая артериола разветвляется на
вторичные капиллярные сети ©округ- канальцев - перитубулярные (около»
канальцевые) капилляры. У корковых нефронов выносящая артериола об-
разует сеть капилляров, оплетающих каналец. У юкстамедуллярных нефро-
нов выносящая артериола переходит в прямые сосуды (vasa recta), стенка
которых состоит из одного слоя эндотелиальных клеток. Эти капилляры
идут параллельно петле Генле.
Почечная артерия широкий, короткий сосуд, отходящий от брюшной
аорты. Поэтому гидростатическое давление крови в гломерулярных ка-
пиллярах высокое (60-70 мм рт.ст.), от него зависит переход порядка 20%
плазмы из гломерулярных капилляров в просвет капсулы. В перитубуляр-
ных капиллярах гидростатическое давление крови низкое (20 мм рт.ст.),
что необходимо для перехода веществ из интерстициального пространства
почки в кровь.
В почку поступает 20-25% от сердечного выброса крови, то есть 400 мл
крови на 100 г ткани почки за 1 минуту. Это очень большая величина орган-
ного кровотока. В почке кровь не только очищается, но и удовлетворяет
метаболические потребности клегок тканей почки.
90% крови притекает в корковое вещество, где кровоток может оста-
ваться неизменным при колебаниях системного артериального давления
в диапазоне 90-190 мм рт.ст., что имеет большое значение прежде всего
для процессов клубочковой фильтрации.
Постоянство коркового кровотока обеспечивается следующими ме-
ханизмами.
а Миогенешй, Повышение гидростатического давления крови в при-
носящей артериоле вызывает ее растяжение. В ответ на растяжение про-
исходит констрикция артериолы, что предотвращает дальнейшее увеличе-
ние объема притекающей по ней крови. При понижении давления тонус
артериолы падает, и в нее поступает прежнее количество крови.
© Каншш$ева~клубочковая обратная связь. При повышении почеч-
ного кровотока в юкстагломерулярном аппарате выделяется аденозин, ко-
торый избирательно суживает приносящую артериолу, что приводит к сни-
жению притока крови. При снижении почечного кровотока в юкстаглома-
рулярном аппарате почки выделяется ренин. Выделение ренина ведет к об-
оазованито внеиотеныжв-П, являющегося мощным ва'юконстрнк!-згхлт
сосудов почки. Помимо этого, ангиогешин-Н стимулирует выработку г.те-
и АДГ, которые тоже оказывают на большинство сосудс-?= пазо
консгрикторкый эффект. Альдостерон и АДГ увеличивают реабсорбцию
ноды в гючкйх. В результате артериальное давление повышается и приток
крови к почке возрастает.
Почечный кровоток б-, большей степени зависит от гуморальных фак-
торов. чем нервных. Простагландины и кинины, вырабатываемые в поч-
ках, вызывают вазодилатацию ее сосудов. Катехоламины при действии
на ^--адренорецепторы суживают сосуды почки, а реакция сосудов на до-
фамин зависит от его дозы: в малых дозах дофамин вызывает конструк-
цию, а в больших - дилатацию.
Симпатические нервы через агадренорецепторьг вызывают вазоксн-
сфикцию. Скшпатическне холинергические нервы вызывают вазодилата-
цию сосудов мозгового вещества почки.
ПРОЦЕССЫ, ЛЕЖАЩИЕ В ОСНОВЕ МОЧЕО’БРАЗС’Н^ГЯЙЛ
Моча образуется в почках из плазмы крови. Очищение крови в почках
связано с тремя процессами мочеобразо^аикя; реабсорбци-
ей и секрецией (рис.4).
Фильтрация •- это переход части крови из гломерулярных, успипля-
ров в просвет капсулы Боумена-Шумлянскогс. Жидкость.. попавшую п про-
свет капсулы, называют перюнной мочой, а. из-за ее кшзкомслекуляркого
состава - ул&трафилыпратом.
Из крови fj просвет капсулы поступает большое количество полезных
веществ, которые необходимо вернуть в кроль. Ультрафильтрат ш капсулы
попадает в каналец, клетки которого переносят нужные вещества :? кровь.
Перенос веществ из канальца в крс-щ- называетсяреабсорбцией.
Клетки канал i-na также могут переносить вещества из кренш в кз на ли-
цевую жидкость. Перенос веществ из крови в канальцевую жидкость назы-
вается секрецией. Реабсорбция п секреция идут в канальце и собиратель-
ной трубке.
По величине филырации, реабсорбции и секреции судят о функш-ю-
нальвом состоянии почек. В основе методов оценки функционального со-
стояний почки лежит сравнение количества вещества в первичной и ковеч-
ной моче Еспи вещество фщулин, креатинин) только те его
количество в первичной моче (улюрафильграте) и в конечной моче гщи.чл-
- и -
ковое. Если вещество фильтруется и реабсорбируется (глюкоза), ио не се-
кретируется, то его количество в первичной моче больше, чем в конечной
на величину реабсорбции. Если вещество фильтруется и секретируется
(парааминогиппурозая кислота), но не реабсорбируется, то его количество
в конечной моче больше, чем в первичной на величину секреции.
Артерия Афферентная Гломерулярный
артериола
капилляр
Эфферентная
артериола
Выделение
мочи
Рис.4. Основные процессы мочеобразования (Вандер А.,2000).
I - гломерулярная фильтрация; 2 - канальцевая секреция; 3 - канальцевая реабсорбция.
КЛУБОЧКОВАЯ ФИЛЬТРАЦИЯ
И ФАКТОРЫ ЕЕ ОПРЕДЕЛЯЮЩИЕ
Образование мочи начинается с фильтрации плазмы крови, от кото-
рой фильтрат по химическому составу отличается незначительно. Основное,
отличие состоит в содержании основных белков крови, которых в фильт-
рате практически нет. Первичная моча (или ультрафильтрат) по составу ами-
нокислот, глюкозы, мочевины и других низкомолекулярных компонентов
идентична плазме крови. Однако в ней мало белковых анионов, но на 5%
больше анионов хлора и бикарбонатов и меньше катионов натрия, калия.
Преградой для фильтрации форменных элементов крови и крупных
белковых молекул является фильтрационный барьер, который не пропуска-
ет веихестеа с размером молекулы бшн.а’е 6-5 нм. Электрический жд-:
яйдяется вторым по значимости фактором^ определяющим фильтруемое-).-
молекул- Отрицательно заряженные макромолекулы фильтруются а ме:н
шей степени, а аг.егпроположг^тельные - в большей степени, чем электро^
нейтральнее.
Фильтрационный барьер состоит из трех слоев, эндотелия гломеру-
лярных капилляров., базальной мембраны и слоя, эпителиальных клеток
висцерального листка капсулы Боумена-Шум лежкого (рис.5).
i
Рис.5. Фйльгрш ровный барьер включает- внутренний слой фенестрнрояянко-
го эндотелия, базальную мембрану к наружный слой отростков ножек пслошгю^
между которыми проходит фильтруемая жидкость (Шейман Дж.А., 200;}.
Первый слой представлен эндотелием фенестрирозаниьа капилляров,
30% поверхности ме.мбракы этих клеток приходится из поры размером
50—100 нм. Базальная мембрана - это гелепод-...-бп-ч-., исюклеточнсс, ячеис-
тое образование с порами 3--6 нм. В ее еостаь нхолят оилианиогш., пропят
ствующие фильтрации низкомолекулярных - >piдян-тееккх отрицательно
заряженных веществ. Из-за отрицательного заряда, например, адьбумж-ьч
с размером молеку.ты 5 им, практг-тх-ки не поидпгкуг в ультрафильтра
Эпителиальные клетки висцерального листка капсулы называют по.доожа-
ми. Эти клетки жжго множество папьпеБВДЕЫх отростков, упирающихся
- 13 -
з базальную мембрану. Пространства между отростками смежных подо-
цитов перекрываются диафрагмами, имеющими поры размером 5-12 им.
Поры покрыты гликокаликсом, оставляющим отверстия диаметром 6 нм.
Пальцевидные отростки покрыты слоем внеклеточного материала - сна-
лигликопротеина, имеющего отрицательный заряд. Подоциты имеют ак-
томиозиновые миофибриллы. Они могут сокращаться и расслабляться,
действуя как микронасосы, перекачивающие фильтрат в полость капсулы.
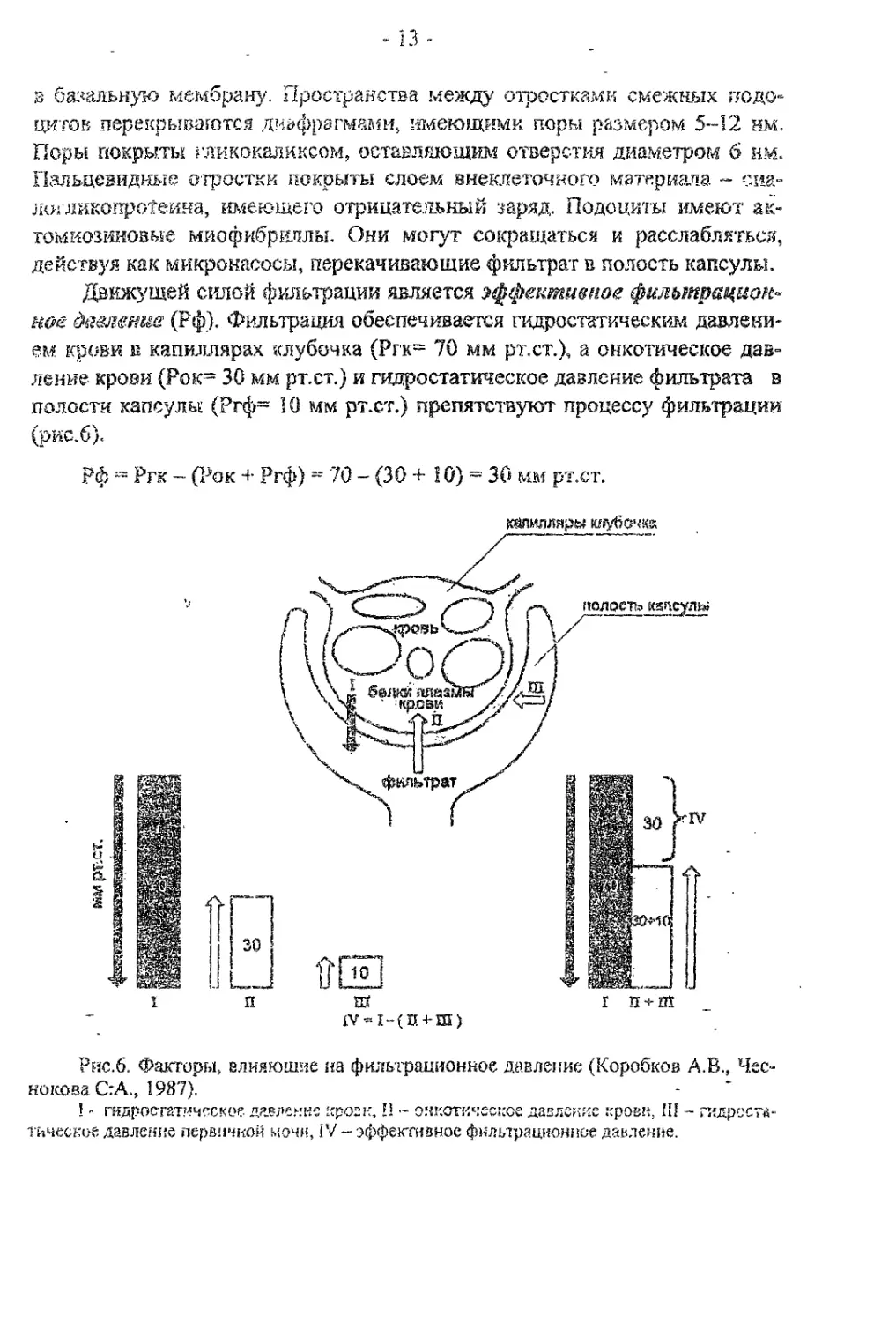
Движущей силой фильтрации является эффективное фильтрациом»
к&е давление (Рф). Фильтрация обеспечивается гидростатическим давлени-
ем крови в капиллярах клубочка (Ргк= 70 мм рт.ст.), а онкотическое дав-
ление крови (Рок~ 30 мм рт.ст.) и гидростатическое давление фильтрата в
полости капсулы (Ргф= 10 мм рт.ст.) препятствуют процессу фильтрации
(рис.б).
Рф = Ргк ~ (Рок 4- Ргф) = 70 - (30 +Ю) = 30 мм рт.ст.
I п + ш
Рис.6. Факторы, влияющие на фильтрационное давление (Коробков А.В., Чес-
нокова С:А., 1987).
1 - гидростатическое давле.чие кроен, Н - онкотическое давление крови. Ш - гидроста-
тическое давление первичной мочи, IV ~ эффективное фильтрационное давление.
’ 14 -
Фильтрация протекает по всей длине гломерулярного капилляра.
Однако силы, способствующие фильтрации, в начальных участках плпия-
ляра выше, чем в конечных. так как по мере прохождения крови во кнтип-
ляру гидростатическое давление уменьшается, а онкотическое - увеличи-
вается.
О фильтрационной способности почки судят по скорости клубочко-
вой фильтрации (СКФ), то есть объему ультрафильтрата, образованному
за единицу времени. Для определения СКФ в кровь вводится и-чулях
инертное вещество, которое хорошо фильтруется, но не реабсорбируй гс?
и не секретируется. Поэтому концентрация инулина в фильтрате, такая хе,
как и в плазме крови (Рин). Количество профильтровавшегося инулина
(произведение его концентрации и плазме крови Рин на СКФ), и количссу-
зо кнулина в конечной моче (концентрация инулина в моче Син,
женкзя на объем мочи V), равны, иначе говоря Рин х СКФ ~ Uhe к V.
Это позволяет определить величину СКФ:
скфЛ^ЛХ.
Рин
Инулин в организме отсутствует, поэтому при определении СКФ е-го
;необходимо капельно вводить я кровоток в течение всего времени измере-
ния, чтобы концентрация инулина в плазме (Рин) оставалась досфгщфй.
В связи с этим чаще определяют СКФ z:-o креатинину (проба Реберн.
Крёмтшыя - эндогенное вещество, продукт метаболизма мышечных
клеток, концентрация которого а нла?ме постоянна. Недогг.ъток пробы по
креатинину связан со способностью ночек незначительно реабсорбировать
и секретировать это вещество,
Вместо термина СКФ чаще нспользуется термин юшренс ккужж
(Син) или креатинина (Скг). Клиренс характеризует способность почки
очищать кровь от какого-либо вещества. Клиренс эешсстна (Сх) щ>каз?»1вч-
ет какой объем плазмы очищен от даниогс вещества .ы йремя.нц.
Его величина равна количеству Рсщееттй в конечной мане й)ромз₽е?;.сннс
концентрации ьегцества в моче (Ux) на объе.м. м-тчя V), деленному на кон-
центрацию зтого вещества в плазме кропи (Рх):
Норма СКФ для женщин - 110 млКццт для му^чцг. - 125 шг/мии.
15 -
сутки в почке человека образуется примерно 180 л ультрафильтра-
та» который имеет такое же осмотическое давление и такую же концен-
трацию пшкомолекулярных веществ, как и плазма крови.
РЕАБСОРБЦИЯ И ЕЕ ВИДЫ
После поступления ультрафильтрата в каналец начинается реабсорб-
цйя полезных веществ в кровь. Вещества могут реабсорбироваться через
мембрану клетки канальца (транхшдаюляпкый путь) и через межклеточ-
кые пространства (парацеллюлярный путь). Реабсорбция бывает облигат-
ной (или обязательной} и факультативной.
Облигатная реабсорбция протекает & проксимальном канальце, со-
стоящем «гз одного слоя клеток кубического эпителия с щеточной каемкой
на аитжальмой мембране. Здесь реабсорбируется из фильтрата за сутки
120 л воды, -50-70% Na.\ 90% К\ глюкоза, витамины, низкомолекулярные
белиц и другие компоненты крови (рис.7).
За реабсорб1фуем»жи веществами по осмотическом)' градиенту обя-
зательно выходит вода, поэтому в петлю Генле поступает жидкость, изо-
тс-ничная плазме крови. Клетки пронсямального канальца хорошо пронима-
емы №$ вода/ благодаря п авмкалъной мембране водных каналов.
образованных особыми белковыми молекулами акваноринами i-ro тип??.
реабсорбция протекает в дистальном отделе и^ф-
ропа и собирательной трубке, Объем факультативной реабсорбции може*
вменяться к зависимости от потребности организма в ионах и заде.
КЛАССИФИКАЦИЯ М^ХАНШМО^ ТРАНСПОРТА ВЕЩЕСТВ
& Диффузия', переход веществ без затраш энергии АТФ по электро-
химическому, осмотическому', концентрационном)' градиенту. Например,
иода переносится по осмотическому гр8.дие-ц}у (из раствора с меньшей
кохщенгрйцин в раствар с большей концентрацией).
® Оил£гчё$ж№я диффузш етлр^аетсх от простой тем, что вещество
гранспортнру^гся.по градиенту' конценто.ццц при участии специфическо-
го белка-переносчика. Например., такны способом через базолатерзльиую
^?мбртггу переносятся гюОлЧ^-^а. а.мннокисл.-гпя, жирные кислоты, витами-
i55k Для каждой группы вещестэ <-.с’;ъ слои переносчики.
« S7i:p3f.fri^a перенос вещества протия граду-
е-гча концентрации с обязательным цспс^ьзонншсм энергии АТФ. Извест
ны следующие системы первично активного транспорта: Ж+ДС-АТФаза;
Са2+-АТФаза; 1-Г-АТФаза; ЬГ,К+~АТФаза. Например, через базолатср&ль-
ф/ю мембрану Na+ переносится из клетки в околокональцевую жидкость,
а К+ наоборот, _в клетку за счет энергии Ка\К\АТФазы.
Глюкоза
Гидрокарбонат
Натрий
Калий
Магний
Кальций
Фосфат
Сульфат
Аминокислоты
Витамины
Белок
Мочевина
Хлор
Натрий
Вода
нсо;
Na
Вода
Вода ф
5>
*
1Й1
ы
Ген
Гш
Н‘
NH
- Холин
Na* '
Мд2*
Iе
ПАГ . ®
Диодр.
-• '° Я
Пенициллин;!
СГ
Na*“asi
сг
Вода
^Мочевина
Na
NH
и
§
а
в
К
_ Рис.7. Локализация реабсорбции и секреции веществ в почечных канальцах.
(Покровский В.М., Коротысо Г.Ф., 1998).
Направление стрелок указывает на фильтрацию, реабсорбцию и оькрсцщо веществ.
-17-
& вторично м&швныи ^ракспорт-. перенос вещества против гра-
диента кошдсь'тралщи. когда для переноса одного вещества используется
энергия; освобождаемая при одновременном транспорте по градиенту кон-
центрации другого вещества. Б этом случае два вещества связываются на
мембране клетки со специфическим переносчиком и оба транспортируют-
ся через мембрану ютетки. Движение обоих веществ в одном направлении
называется квтрамекертвм, а движение каждого в противоположном на-
правлении - аМНШ&0рПШ£.
Транспорт глюкозы в клетку канальца щт.ет против концентрационно-
го градиента следующим образом. На апикальной мембране клетки есть
специфический переносчик глюкозы, к которому присоединяется ион Na*..
Этот комплекс поступает' в клетку за счет энергии, выделяемой при пере-
носе натрия по градиенту концентрации. Апикальная мембрана дня пере-
чйедеияых элементов проницаема только в одном направлении. В клетке
комплекс распадается. Переносчик для глюкозы возвращается ж апикаль-
ную /Мембрану. Из клетки через базолатеральную мембрану Na+ перено-
сится первично активным транспортом, а глюкоза - облегченной диффу-
зией. Если в комплекс© с игприем переносится органическое вещество,
какой котранспорт называют Подобным способом реабсор-
бируютоя аминокислоты,, жирные кислоты, витамины.
Пшюцытог.. перенос вещества в клетку с образованием вакуоли.
Нзпрщтр, белок или пептид подходит к апикальной мембране клетки, зэа-
н-модсйствует со специфическим рецеггтором, мембрана впячивается и об-
зояакивает малекулу, образуется пиноцитозная вакуоль, которая движется
сторону базолатеральной мембраны и сливается с лизосомами. Под влия-
нием лизосомных ферментов белки или пептида расщепляются до амино-
кислот, которые переносятся облегченной диффузией через базолатераль-
ную мембрану в околоканальцевую жидкость. Пш-юцитоз протекает с за-
тратой энергии АТФ. - -
Пептида могут переноситься и другим, пугем. На апикальной мш-
брюзе есто аминопептидазы, расщепляющие пептиды до аминокислот, ко-
торые переносятся вторично активным транспортом через апикальную
мембртлу клетки и облегченной диффузией через базолатеральную мем-
брану. Рисунок 8 иллюс'грирует различные виды фане порта.
Механизмы реабсорбции поной Ж+.
Джрфузш через апикальную мембран); если она пропинаема ,ц;£Я Ма'\.
через базолатеральиую мембрану,
через апикальную мембрану'.
18
ПРОСВЕТ
канальца
- ИНТЕРСТИЦ/i АЛЫ iC> Е
ПРОСТРАНСТВО
Толстая.
восходящая часть
петли Генле
Дистапьчнй
извитой кэналец
Собирательная
трубка
Р'^с.8. Наиболее важные механизмы реабсорбции натрия и хлора в разных сег-
-тйх канальца (Вандер A.,2Q00).
Ионные насосы /первично активный транспорт) показан черными кружочками и сплоят-
'и стрелками. Перенос с участием переносчиков кпужочк&мо н сплошными серелкамн.
',ри;1С(.ОрТ _ цщэихоаы-чи стрелками.
- i9 -
Через стенку проксимального канальца вслед за пере-
косммыми веществами пассивно во осмотическому градиенту следует вода..
Начальные участки проксимальных канальцев практически не прошщаемы
д ля хлора, концентрация которого возрастает из-за реабсорбции воды. Меж-
клеточные контакты конечных участков проксимапьчых канальцев хорошо
проницаемы для иона хлора, который по градиенту концентрации уходит
из канальца в интерстидий. За отрицательно заряженным ионом СГ по элек-
трохимическому п>адиенту выходит положительно заряженный ион NaT.
А-ШШ-тярт. Через апикальную мембрану в клетку по градиенту кон-
центрации поступает ион натрия, а ига клетки секретируется ион водорода
или ион гГ. Из-за конкурентных отношений ионов калия и водорода уве-
личение концентрации одного из них влияет на секрецию другого. Поэтому
гиперкалиемия ведет к задержке ионов водорода, и к ацидозу, а гипокалие-
мии - к алколозу.
Для оценки реабсорбщ-юнной способности почки определяют вели-
чину максимального транспорта {Тт«) вещества, которое хорошо фильт-
руется, не секретируется и имеет порог реабсорбции. Порогом реабсорбции
называют максимальную концентрацию вещества в плазме крови, при ко-
торой зто вещество полностью реабсорбируется. К таким веществам опю-
сйтся глюкоза, торог реабсорбции которой равен 1.0 ммсл.ь/л.
При увеличении концентрации глюкозы в плазме кропи сьыше
10 ммсль/л глюкоза появляется в конечной моче. Это обусловлено тем, что
р люминальной мембране клеток проксимального ка.нальо.а находится огра-
ниченное количество переносчиков глюкозы. Когда они полностью насы-
щаются глюкозой, достигается ее максимальная реабсорбция, а глюкоза
сверх порога реабсорбций удаляется с конечной мочой. Определение ве-
личины максимальной реабсорбции глюкозы имеет значение для функ-
циональной оценки реабсорбционной способности клеток проксимальных
канальцев почки.
Для определения величины максимального транспорта глюкозы
(Ттзхгл) необходимо создать условия полного насыщения системы ее ка-
нальцевого транспорта, когда концентрация глюкозы в крови (Ргл) превы-
сит порог ее реабсорбций. С этой цевью в кровь вводят глюкозу, повышая
ее концентрацию в клубочковом фильтрате до тех .пор, пока не будет дос-
тигнут порог реабсорбций и глюкоза не начнет в значительном количестве
выделяться с мочой. Вместе с глкиюзой в кровь вводят инулин, чтобы мож-
но было nr. его клиренсу (Син) определить объем клубочкового фильтрата.
- 20 -
Величину максимального транспорта рассчитывают по разнице меж
ду количеством профильтровавшейся глюкозы, ранным произнедснию кон-
центрации глюкозы в плазме крови (Ргл) на объем ютубочковоп? фильтра-
та (Син), и количеством глюкозы з конечной моче, равным произведению
концентрации гликозы в моче (игл) на объем выделившейся мочи V:
Тт8Хгл ~ Ргл х Син - игл х V.
Ттв!4гл, характеризующий полную загрузку системы транспорта глю-
козы, у женщин ЗОЗ мг/мин, у мужчин 375 мг/мин, или 1,8-2 ммояь/мин.
СЕКРЕЦИЯ И ЕЕ ВИДЫ
Термин канальцевая секреция показывает только направление транс-
порта в отличие от канальцевой реабсорбции. Специфические механизмы
мембранного транспорта э случае секреции остаются такими же, как и в сл;/-
ча« реабсорбции.
Хан&Фьцемй секрецией называют транспорт в мочу продуктов обме
на й чужеродных веществ, содержащихся в крови иди образушых в самих
кяетках канальца. Секреция осуществляется, как правило, против когъцен-
трашючйого «ли эдекгеохимического градиента с затратами А ГФ. Спо-
собностью к сеьреции обладают клетки эпителия как прокс^альиого, так
и диетадьного отде.:Еок? поэтому секреция бывает проксимальной: и ден-
тальной.
В проксимальном канальце секретируются слабые органические кис-
лоты и основания как эндогенной {жирные кмслопш, уркты, соли ж^лч-
них кислот, адреналин^ гшшк, крешшкш*), так и экзо
генной природы, среди которых много лекарственных препаратов: фурасе-
Mudf пенициллин, люрф^н, хинин. 1) це-
большом количестве секретируются ионы водорода и аммиак. В /дист-аль-
ном канадьце и собирательной трубке преимущественно секретируютоэ
ионы колия^ и который образуется клетками канальца.
Для определения секреторной работы почки вместе с и^ливом
» кровь вводят шсложу (ПАГК 1>к как .ПАГ фильт-
руется, не реабсорбируется, но секретируется, ее количество в конечной
моче больше, чем в первичной, на аелгяину секреции. Количество ПАЕ
вЬ1дел!-штаейСЯ с мочой, равно произведению концентрации ПАГ й моч&
Шпаг) на объем мочи V, а количество профипыроваешейся ПАГ равно
произведению ее концентрации в плазме крови (Рпаг) на объем клубочксь
-21 -
вого фильтрата (Син). По максимальному транспорту/ (Ттах) ПАГ опреде-
ляют величину секреции:
Ттахпаг = Unar х V - Рпаг х Син.
Величина Ттах ПАГ равна 90-80 мг/мин, или 0,5-0,6 ммоль/мин.
Поскольку путем фильтрации и секреции плазма крови в почке очи-
щается от ПАГ' полностью, ее клиренс (Спаг) соответствует величине плаз-
мотока, то есть объему плазмы, протекающему в сосудах почки за едини-
цу времени. Зная показатель гематокрита (Ht), можно рассчитать величину
почечного кровотока (Q):
_ Спаг
В норме Спаг= 550-650 мл/мин, Ht= 0,4-0,45.
О способности вещества в почках реабсорбироваться или секретиро-
ваться можно судить по концентрационному показателю (Ux/Px), то есть
отношению концентрации вещества в конечной моче (Ux) к концентрации
в крови (Рх). Такое отношение концентраций по инулину (Uhh/Phh) ука-
зывает, во сколько раз уменьшился объем фильтрата при прохождении но
канальцам. Если концентрационный показатель для какого-либо вещества
больше показателя по инулину, то это вещество в почках секретируется,
а если меньше - то реабсорбируется. Отношение концентрационного по-
казателя вещества к концентрационному показателю по инулину называ-
ется экскретируемой фракцией (EF).
СЕКРЕЦИЯ ИОНОВ ВОДОРОДА
Секреция ионов водорода происходит в проксимальном канальце, толс-
той восходящей части петли Генле, в системе собирательной трубки и игра-
ет важную роль в регуляции pH крови. От секреции иона водорода зависит
реабсорбция HCOj. Реабсорбция в кровь одного иона НСО3 оказывает
такой же эффект на pH крови, как и удаление иона водорода из нее.
В процессе реабсорбции бикарбонатов важную роль играет обмен
части реабсорбируемых ионов Na+ на внутриклеточные ионы водорода,
которые механизмом антинерта переходят в просвет канальцев.
- В канальцевой жидкости ионы Н+, соединяясь с ионами НСО3, -обра-
зуют угольную кислсзу, которая при содействии карбоангидразы щеточной
- 22 -
каймы клеток канальпев расщепляется на СО2 и Н2О. Растворенный С(Ъ.
легко проникает путем диффузш? в клетки эпителия канальцев, где взаимо-
действует с водой. В результате этой реакции образуется снова угольная
кислота, которая внутри клеток канальцев при участии карбоангзддты
диссонирует на ионы И+ и НССх. Водород активно секретируется s яро*
свет канальца, а НСО/ реабсорбируется через базолатеральную мембрану
в интерстициальную жидкость путем облегченной диффузии и поступает ’
в кровь перитубулярных капилляров.
Посредством секреции ионов J-Г а проксимальном канальце реабсор-
бируется примерно 80% профильтровавшегося бикарбоната. В толстой вос-
ходящей части петли Генле реабсорбируется 10-15% бикарбоната, а почти
весь оставшийся бикарбонат - в дистальном извитом канальце и системе
собирательной трубки.
Кроме того, секретируемый нон водорода в просвете канальца сведи-
няется с иебикарбонаткым буфером, в основном с фосфатом (НРОф ). об-
разуется Н2РОд. Таким путем ионы водорода, связанные с фосфатным бу-
фером, выводятся с мочой.
Длй перемещения через мембрану как иона водорода, так и бикарбо-
ната, необходимы специальные переносчики. Активный перенос иона недо-
рода через апикальную мембрану и» клетки в просвет канальца осу шесто-*
льется "ф?лмя различными переносчиками.
I. Первично активная ГГ-АТФаза присутствует во всех секретирую--
щих ион водорода сегментах канальца и собирательной трубке.
2. В проксш&едьном канальце и толстой восходящей части петли
Генле имеется большое количество Na+»H+ переносчиков - антипортеров,
поэтому секреция ионов Н* в этих сегментах, в основном, осуществляется
вторично активным транспортом и отшлифуется реабсорбцией иона Ыа+.
3, В собирательной трубке имеются вставочные клетки типа А. На
нх мембране локализованы К+>Н+-АТФазы, которые одновременно пере-
носят ионы водорода в просвет канальца, я иоиы К* в клетку.
Еще один механизм регулирования кислотио-щелечкого баланса ре-
ализуется почечной глутаминазам. Под ее влиянием осуществляется гид-
ролиз на гяутамат & бммиак. Аммиак секретируется в £фто
свет канальца, где соединяется с ионом ЬГ и выводится с конечной мо°ой
в виде ионов аммония,
’ 23 -
ПССТУШ1ЕНМЕ ЖИДКОСТИ
В П^И’УБУЛЯРНЫЕ КАПИЛЛЯРЫ
Перенос вещест» из интерстициального пространства в кровь периту-
булярных капилляров (рис,9) обусловлен тем, что силы, определяющие ре-
абсорбцию (Рр) тканевой жидкости в кровь, больше сил, способствующих
фильтрации (Рф) из крови в интерстиций. Реабсорбцию обеспечивает сум-
ма гидростатического давления жидкости ивтерстиция (Рои== б мм рт.ст.)
и онкотического давление крови (Ро№ 33 мм рт.ст.), а фильтрацию- сум-
ма гидростатического давления крови (Ргк= 20 «мм рт.ст.) и онкотического
давления в интерстиции (Рои=6 мм рт.ст.)':
Рр ~ Рги Jr Рок ~ 36 мм рт.ст.
Рф = Ргк + Рои 26 мм рт.ст.
Рис.9. Транспорт ионов и молекул из канальцевой жидкости в кровь перитубу-
лярных капилляров.
Стрелками показаны силы, способствующие (4-) или препятствующие (Т) транспорту
’iciuc ,;ь из iiiirepc'oiujtfl s кровь пертубулярнм* капилляров.
- 24 •
МЕХАНИЗМ СОЗДАНИЯ
КОНЦЕНТРАЦИОННЫХ ГРАДИЕНТОВ В ПОЧКЕ
.Конечная моча может быть как гипотонической, так и гипертоничес-
кой особенно при повышении осмотического давления крови, когда необ-
ходимо экономить воду и удалять из внутренней среды организма осмоти-
чески активные вещества. Способностью выделять гипертоническую мочу
обладают только почки млекопитающих и птиц. Механизм концентриро-
вания мочи обусловлен деятельностью поворотно-противоточной системы.
Доеоротно-пропшвоточкая система представлена петлей Генле, со-
бирательной трубкой, прямыми кровеносными сосудами (vasa recta) и ин~
терстицием мозгогового вещества почки.
Главный элемент системы - петля Генле состоит из нисходящего тон-
кого колена, которое у вершины почечного сосочка поворачивается под
углом 180° и переходит в восходящее колено, имеющее вначале тонкий,
а затем толстый отдел, который еще называют прямой частью дистального
канальца.
В нисходящее колено поступает за сутки около 60 л изотоничной жид-
кости из проксимального извитого канальца. Стенка нисходящего колена
хорошо проницаема для воды и почти не пропускает ионы СГ и Na+. Стен-
ка восходящего колена не проницаема для воды, но через нее активно транс-
портируются ионы С1 и Na+ в окружающий интерстиций, повышая его
осмотическое давление (рис. 10). По осмотическому градиенту вода выхо-
тит из нисходящего колена петли э интерстиций. Жидкость в нисходящем
юлене становится гипертонической, причем ее осмотическое давление рав-
ным давлению в интерстиции на данном уровне мозгового вещества почки.
Несмотря на поступление воды из нисходящего колена, осмотическое
явление в интерстиции не снижается из-за продолжающегося активного
ранспорта ионов С1 и Na+ из восходящего колена. Эти процессы совер-.
даются на всём пути движения жидкости по петле, поэтому происходит
□степенное увеличение осмотичности в нисходящем колене и в интер-
.иции по мере удаления от корковой зоны вглубь мозгового вещества.
Йо время движения жидкости по восходящему колену ее осмотич-
?сть понижается из-за активного транспорта ионов, и она становится ги-
)т°нической. Возникает поперечный осмотический градиент. В корковую
113 исходящего колена притекает около 20 л гипотоничной жидкости -
'утки. -
-25-
Рис. 10 Противоточно-множительная канальцевая система (Ван дер а.,2000).
Цифрами обозначены величины осмотического давления жидкости в ннтерспниш. ка-
нальцах и собирательной трубочке при образовании концентрированной мочи.
Дальнейшее концентрирование мочи идет в связующем '•> со*
бврателькой трубке, стенки которых могут пропускать воду в ирису гствии
АДГ. к вода по осмотическому градиенту выходит в интерсгищдй. При
этом объем конечной мочи уменьшается, а ее осмотическое давление уве-
личивается. В норме из организма выделяется 1,5-2 л гипертонической
мочи в сутки. У человека осмотичность конечной мочи может достигать
1450 Рлосмоль, а у пустынных животных еще выше.
Ионы Na+ и С1 активно реабсорбируются только в толстой части пет-
ли Генле. В глубине мозгового вещества, где расположена тонкая часть
петли, активный транспорт Na+ и CI не происходит. Повышение осмоти-
ческого давления здесь в большей степени определяет не активный транс-
порт ионов Na+ и О, а пассивный транспорт мочевины из нижнего сег-
мента собирательной трубки.
АДУ делает этот сегмент проницаемым не только для воды, ио и для
мочевины, концентрация которой возрастает из-за реабсорбции воды. Из
- 26 -
собирательной трубки мочевина выходит в интерстиций, попадает в кровь
И в жидкость, текущую по петле Генле. Осмотечность названных сред во>
растает, что способствует увеличению реабсорбции воды. Из петли Генле
мочевина попадает в собирательную трубку, из нее в интерстиций и т.д.,
наблюдается кругооборот мочевины.
При движении крови по vasa recta в глубь мозгового вещества в нее
поступают ионы Na+, Cf и пассивно выходит вода. При движении крови
из мозгового вещества в корковое в нее пассивно поступает вода и выхо-
дят ионы Na+, CL Таким образом, vasa recta функционируют как противо-
точный обменник, который препятствует вымыванию из интерс твдия осмо-
тически активных ионов. Повышение кровотока в прямых сосудах ведет
к вымыванию осмотически активных веществ и уменьшению реабсорбции
воды. Какая-то часть воды с растворенными в нем веществами из интер-
стация попадает в кровь перитубулярных капилляров и уносится из почки.
Благодаря поворотно-противоточной системе, реабсорбируется около 60 л
воды в сутки, почти весь натрий, не успевший реабсорбироваться в про-
ксимальных канальцах. Механизм большинства диуретиков базируется на
их способности уменьшать реабсорбцию ионов, прежде всего Na+, что ве-
дет к снижению реабсорбции воды и увеличению диуреза.
РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОНОВАЯ СИСТЕМА
Большое значение в регуляции артериального давления, почечного кро-
вотока и работы почки имеет ренин-ангиотензин-альдостероновая система
(РААС). В месте соприкосновения дистального извитого канальца с сосу-
дами: vas afference и vas efference располагается юкстагломерулярный ап-
парат почки (ЮГА), имеющий форму треугольника (рис.11).
Основание этого треугольника образовано гиперхромными клетками
канальца - плотным пятном (macula densa). Боковые стороны образованы
гранулированными эпителиальными - юкстагломерулярными клетками, ле -
жащими в артериолах, особенно их много a vas afference. Основная функ-
ция этих клеток — секрециярвниня.
Клетки macula densa играют роль своеобразных рецепторов, реагирую-
^™еНеНИе К0Нце^пРагЩи ионов Na' и Cl в жидкости, протекаю -
щей в дистальном канальце.
При увеличении почечил™
и“счного кровотока растет ооъем ультрафильтрата
и скорость его движения по о 7 1 г к
канальцах не успевает пеаХДГ В Связи С в пРоксималь™х
Р «сорбироваться должная часть профильтровав-
шихся ионов Na* и СД . Поэтому увеличивается загрузка ими дистальных
отделов канальцев, а также реабсорбция их клетками macula densa.
Рис.11. Структура юкстагломерулярного аппарата (Guyton А.С., Hall YE.,
2001).
При увеличении концентрации ионов Na* и С1 в просвете канальца,
Na , К+, С1 переносчик на апикальной мембране клеток плотного пятна
способствует повышению содержания этих ионов и в- цитоплазме эпите-
лиальных клеток. В результате открытия катионных каналов плазмолеммы
Происходит поступления Сат* в цитоплазму. Прирост Cav* в цитоплазм
стимулируем секреции; из клеток плотного пятна паракринных агента
в виде аденозина.
О Гй
-28-
Гладкомышечные клетки приносящей артериолы имеют рецепторы
к аденозину. Воздействие аденозина на эти рецепторы приводит к поступ-
лению в цитоплазму мышечных клеток ионов Са++, а вызванная ими кон-
струкция артериолы снижает СКФ.
При уменьшении общего объема жидкости в организме снижается по-
чечный кровоток и скорость клубочковой фильтрации, и поэтому в дис-
тальный каналец притекает меньше жидкости с ионами натрия и хлора.
Снижение концентрации в дистальных канальцах ионов Na+ и CI улавли-
вается клетками macula densa, последние посылают сигнал к юкстагломе-
рулярным клеткам, что стимулирует секрецию ренина (рис. 12).
Такая система сигнал - эффектор позволяет почкам регулировать ре-
абсорбцию ионов Na+ и осуществлять контроль над АД, почечным крово-
током, электролитным составом крови.
Рекин является ферментом, который действует на белок крови ангио-
тензиноген (а-глобулин), который вырабатывается в печени и циркул!фу-
ет в крови. Ангиотензиноген в присутствии ренина превращается в ангио-
тензин I, который под влиянием ангиотензинпревращающего фермента пе-
реходит в ангиотензин II.
Ангиотензин II вызывает разнообразные биологические эффекты: со-
кращение гладких мышц кровеносных сосудов и мезангиоцитов, стимули-
рует жажду и секрецию альдостерона, но тормозит секрецию ренина, явля-
ясь одним из звеньев отрицательной обратной связи в регуляции секреции
ренина.
Растяжение гранулярных клеток при увеличении почечного кровото-
ка и давления в приносящей артериоле угнетает секрецию ренина. Боль-
шинство этих эффектов ведет к поддержанию уровня кровяного давления.
В клинике часто для снижения артериального давления используются пре-
параты, ингибирующие знгиотензинпревращающий фермент.
Секреция ренина усиливается через ргадренорецепторы симпатичес-
кой нервной системой, а также под влиянием адреналина, АДГ, при повы-
шении концентрации К4' и снижении Na? в крови. Секреция ренина снижа-
ется под влиянием предсердного натрийуретического гормона, ангиотен-
зина II, аденозина.
Ангиотензин К стимулирует секрецию альдостерона корой надпочеч-
таов. Альдостерон проходит через базолатеральную мембрану ?сиеток по-
чечных канальцев и соединяется с рецепторами в цитоплазме. Гормонрецен-
торный комплекс проникает в ядро клетки и влияет на синтез РНК, что
^едет к образованию, как минимум, двух белков. Один из них активирует
s ,1 NaCi / внеклеточного объема жидкости
i 1 артериального давления
Помогает корректировать
I Печень
Легкие
Кора
надпочечников
Почки
Задержка
натрия
Задгркка ] J
зодьг К"
Почки
Pe;-.v<^
——а—;—ч
Конаертир.
фермент
ангиотензиноген —'*> ангиотензин J
ангиотёнзин В
альдостерон
Н2О реабсорби-
руется осмоти-
чески вторично
за реабсорбци-
ей натрия
Кровоток
I Вазопреосин |
Жажда J
Сужение
артериол
Т реабсорб-
ции натрия
в почечных
канальцах
ГГреабсорбции
| воды
в канальцах
t потребления
жидкости
Рис.12. Ренин-ангиотензин-альдостероновая система (Sherwood L., 1999).
-30-
Па+,К’-АТФазу на базолатеральной мембране, другой увеличивает прони-
цаемость апикальной мембраны для иона Na+.
НЕЙРОГУМОР А ЛЬНЫЕ МЕХАНИЗМЫ
РЕГУЛЯЦИИ РАБОТЫ ПОЧКИ
К почкам подходит большое количество симпатических норадренер-
гических нервов, иннервирующих гладкую мускулатуру сосудов почки,
юкстагломерулярный аппарат и многие участки канальца.
Через «1 -адренорецепторы симпатические нервы вызывают сосудо-
суживающий эффект в почке. В покое влияние симпатических нервов на
сосуды почки мало и они не оказывают существенного влияния на почеч-
ную гемодинамику.
Повышение тонуса симпатических нервов вызывает спазм артериол
в почке и снижение почечного кровотока. Однако сужение артериол не
обязательно вызовет снижение скорости клубочковой фильтрации (СКФ),
поскольку констрикция обеих артериол ведет к незначительному повыше-
нию фильтрационного давления. Кроме того, уменьшение почечного кро-
вотока стимулирует секрецию простагландина Е в почке. Простагландин
расширяет сосуды почки, кровоток несколько возрастает, а в результате
СКФ практически остается неизменной.
Сосудистые эффекты симпатической регуляции связаны также с умень-
шением кровотока в сосудах мозгового вещества почки, от которого зави-
сит эффективность противоточной системы и концентрирования мочи.
Симпатические нервы через |3-адренорецепторы стимулируют выде-
ление ренина юкстагломерулярными клетками. Активация ренин-ангио-
тензин-альдостероновой системы ведет к повышению артериального дав-
ления и увеличению реабсорбции натрия. Перерезка симпатических нер-
вов, иннервирующих почки, ведет к повышению выведения ионов Na+ с ко-
нечной мочой (натрийурезу).
Симпатические нервы оказывают трофические влияния на канальцы,
повышая их реабсорбционную способность.
Влияние парасимпатических нервов на почку выражено слабо. Стиму-
ляция парасимпатических нервов снижает реабсорбцию натрия и увеличи-
вает реабсорбцию глюкозы.
Гуморальные влияния на почку выражены сильнее, чем нервные.
Большое значение в регуляции почечного кровотока и работы почки имеет
ренин-ангиотензин-альдостероновая система.
-31 -
Работа почки существенно зависит от предсердного натрийурети-
ческого фактора (ПНФ) - гормона пептидной природы, секретируемого
клетками предсердий. Этот гормон действует на дистальные канальцы и со-
бирательные трубки во внутренней зоне мозгового вещества почки. Он
угнетает реабсорбцию Na\ уменьшая проницаемость апикальной мембра-
ны для этого иона. Кроме того, он косвенно тормозит реабсорбцию Na+,
угнетая секрецию ренина. Главным стимулом секреции ПНФ является рас-
тяжение предсердий при увеличении венозного возврата крови к сердцу.
На работу почки большое влияние оказывает антидиуретический
гормон (АДГ). Основная функция АДГ заключается в увеличении реаб-
сорбции воды. Он увеличивает проницаемость стенки собирательной труб-
ки для воды двумя путями. На базолатеральной мембране собирательной
трубки есть У2-рецепторы, чувствительные к АДГ. Образование гормон-
рецепторного комплекса активирует систему вторичных посредников:
аденилатциклаза-цАМФ. Активация цАМФ ведет к встраиванию в апикаль-
ную мембрану белков - водных каналов (аквапоринов - 2), через которые
вода поступает в клетку. Вода по микротубулярной системе клеток пере-
мещается к базолатеральной мембране, где постоянно встроены вазопрес-
синнезависимые белки аквапорины 3-го и 4-го типов. Через них вода выхо-
дит в интерстициальную жидкость по осмотическому градиенту.
Согласно другой теории, АДГ активирует гиалуронидазу, которая рас-
щепляет гиалуроновую кислоту - мукополисахарид, "цемент", склеиваю-
щий клетки собирательной трубки. В связи с этим через межклеточные
контакты вода может поступать из собирательной трубки в нтерстиций по
осмотическому градиенту.
АДГ стимулирует реабсорбцию NaT в толстой части петли Генле
и в корковой зоне собирательной трубки. Реабсорбция иона Na+ сопровож-
дается увеличением реабсорбции воды.
АДГ увеличивает проницаемость стенки собирательной трубки в зо-
не внутреннего мозгового вещества для мочевины. По градиенту концен-
трации мочевина выходит из собирательной трубки. Это ведет к повыше-
нию осмотичности интерстиция мозгового вещества и способствует уве-
личению реабсорбции воды.
Сокращение мезангиоцитов под влиянием АДГ реализуется через Vr
рецепторы, что ведет к уменьшению площади гломерулярных капилля-
ров, и поэтому скорость клубочковой фильтрации уменьшается.
Vj-рецепторы есть в гладкомышечных клетках сосудов. Их возбуж-
дение активирует систему вторичных посредников: фосфолипаза С - ино-
-32 -
зитол-трифосфат. При этом увеличивается концентрация Са4-4 в цитоплаз-
ме мышечных клеток, повышается тонус артериол и артериальное давление.
В почках этот системный эффект проявляется в ускорении кровотока в пря-
мых капиллярах мозгового вещества, что облегчает удаление из интерсти-
ция в кровь воды, поступающей в него в процессе факультативной реаб- *
сорбции под воздействием АДГ на Уг-рецепторы клеток собирательной
трубки.
В гипоталамусе вырабатывается натрийуретический гормон, кото-
рый усиливает экскрецию ионов Na+ и задерживает ионы К+ в организме.
Этот гормон угнетает активность №+,К+-АТФазы на базолатеральной мем-
бране и поэтому называют его гипоталамическим калийсберегающим
гормоном.
Нейроны гипоталамуса секретируют кроме АДГ гормон - окситоцин,
который оказывает слабое влияние на почку. Этот гормон усиливает реаб-
сорбцию иона К+ и тормозит реабсорбцию иона Na+. Натрийуретический
эффект окситоцина проявляется в присутствии АДГ.
Паратгормон увеличивает реабсорбцию кальция и уменьшает реаб-
сорбцию фосфатов. Эффект гормона реализуется через систему вторичных
посредников (аденилатциклаза - цАМФ). 60-70 % профильтровавшегося
кальция реабсорбируются в проксимальных канальцах. Реабсорбция Са44"
в проксимальном канальце сопряжена с реабсорбцией Na+, которую парат-
гормон угнетает, что в свою очередь ведет к уменьшению реабсорбции
Са44. Оставшийся в канальцевой жидкости Са++ реабсорбируется в дис-
тальном отделе нефрона. Паратгормон уменьшает реабсорбцию фосфатов
в проксимальном канальце.
Кальцитонин - гормон щитовидной железы уменьшает реабсорбцию
и кальция, и фосфатов.
И паратгормон, и кальцитонин вызывают фосфатурию, но паратгор-
мон за счет угнетения проксимальной, а кальцитонин - дистальной реаб-
сорбции аниона.
Синтез калъцитриола (активной формы витамина D3) завершается
в почках. Этот гормон увеличивает реабсорбцию кальция и фосфатов.
Катехоламины - гормоны мозгового вещества надпочечников в низ-
ких дозах увеличивают фильтрацию и повышают диурез, а в высоких -
снижают. При действии на Ргадренорецепторы катехоламины увеличи-
вают секрецию ренина, а действуя на агадренорецепторы, повышают то-
нус сосудов почки.
-’33 -
Глюкокортикоиды и адреналин в ситуации стресса приводят к уве-
личению концентрации глюкозы в крови. Если содержание глюкозы ста-
новится выше пороговой величины, наблюдается увеличение диуреза.
Глюкокортикоиды в больших дозах оказывают на почку такое же влияние,
как альдостерон.
Простагландины Е2 и 12 являются вазодилататорами. В норме они
образуются в малых количествах. В случае повышения тонуса почечных
нервов, возрастания концентрации адреналина и ангиотензина в крови про-
исходит резкое сужение сосудов почки. Одновременно эти же факторы
стимулируют секрецию простагландинов, которые локально расширяют
артериолы. В конечном счете, многочисленные сосудосуживающие сти-
мулы нейтрализуются сосудорасширяющим действием простагландинов
Е2 и 12, и сосудистое сопротивление кровотоку в почке изменяется сущест-
венно меньше. Значимость таких приспособительных эффектов определя-
ется, с одной стороны, необходимостью увеличения АД за счет повыше-
ния ОПСС, а с другой стороны, вероятностью возникновения поврежде-
ния почки при сильно выраженной констрикции ее сосудов.
НЕЙРОГУМОР АЛЬНЫЕ МЕХАНИЗМЫ
РЕГУЛЯЦИИ ОБЪЕМА И ОСМОТИЧЕСКОГО ДАВЛЕНИЯ
ВНУТРЕННЕЙ СРЕДЫ ОРГАНИЗМА
Объем жидкости в организме улавливается барорецепторами, которые
есть в сердце и сосудах. Барорецепторы в области низкого давления (в по-
лых венах, в предсердиях, в легочных венах) называют волюморецептора-
ми. Повышение объема жидкости ведет к увеличению артериального дав-
ления и активности барорецепторов. Рефлекторно снижается активность
симпатических нервов, контролирующих выделение решена. Торможение
секреции ренина ведет через РААС к снижению реабсорбции воды.
Растяжение предсердий большим объемом жидкости стимулирует се-
крецию предсердного натрийуретического фактора (ПНФ), который уси-
ливает экскрецию натрия и воды почками несколькими способами. Он по-
вышает скорость клубочковой фильтрации, избирательно снижая сопротив-
ление приносящей артериолы, угнетает секрецию ренина, снижает реабсорб-
цию Na+ и воды в почечных канальцах. Увеличение общего объема жид-
кости сопряжено с ростом венозного возврата. Этот параметр улавливается
волюморецепторами и рефлекторно тормозит секрецию АДГ. Увеличение
общего объема жидкости ведет к увеличению почечного кровотока и растя-
-34-
жению приносящей артериолы. Растяжение приносящей артериолы тор-
мозит секрецию ренина.
Печень и почки имеют собственные волюморецепторы. Эти органы
покрыты капсулой. При повышении в них интерстициального гидростати-
ческого давления происходит рефлекторное торможение секреции АДГ
и увеличение выделения воды из организма. Гибель нейронов гипоталаму-
са, продуцирующих АДГ, приводит к увеличению диуреза и заболеванию,
которое называется несахарным диабетом.
Осмотическое давление во внутренней среде организма улавливается
периферическими и центральными осморецепторами. Периферические ре-
цепторы есть в сосудах, селезенке, в печени и других тканях. Ведущую
роль в регуляции осмотического давления играют центральные осморецеп-
торы, локализованные в гипоталамусе.
При повышении осмотического давления клетки гипоталамуса, сами
обладающие осморецепторной функцией, а также получающие сигналы от
периферических осморецепторов, увеличивают секрецию АДГ, в связи
с чем реабсорбция воды возрастает и осмотическое давление снижается.
Своеобразными осморецепторами являются натриевые рецепторы ги-
поталамуса. Введение гипертонических растворов NaCl в область третьего
желудочка мозга вызывает увеличение секреции АДГ. Однако введение
других гипертонических растворов без натрия не влечет за собой такого
же воздействия на секрецию АДГ.
Регуляция объема и осмотического давления во внутренней среде
организма взаимосвязаны. Если объем жидкости уменьшился менее, чем
на 10%, то регулируется прежде всего осмотическое давление. При умень-
шении объема жидкости более, чем на 10%, регуляция направлена прежде
всего на поддержание объема в ущерб осмотическому давлению.
-35 -
Приложение
Показатели функционального состояния почки:
- суточный диурез 0,8-2,0 л/сутки;
- скорость клубочковой фильтрации 110-125 мл/мин (клиренс креатинина);
- процент реабсорбции воды - 98-99,5%;
- максимальная реабсорбция глюкозы - 1,8-2 ммоль/мин;
- максимальная секреция ПАГ - 0,5-0,6 ммоль/мин;
концентрация осмотически активных веществ в сыворотке крови - 280-295
мосмоль/л.
Концентрация в ультрафильтрате (первичной моче):
Na+- 135-150 ммоль/л;
К+ - 3,5-5,0 ммоль/л;
Са2+ - 2,3-2,8 ммоль/л; .
Mg2+ - 0,8-1,0 ммоль/л;
С1 - 101-105 ммоль/л;
НСО3 - 21-26 ммоль/л;
фосфата - 0,8-1,5 ммоль/л;
NH/ - 0,01-0,005 ммоль/л;
аминокислот - 2—4 ммоль/л;
креатинина - 0,05-0,1 ммоль/л;
мочевой кислоты - 0,3-0,4 ммоль/л;
мочевины - 3-6 ммоль/л;
белка - 10 мг/л.
Выделение почками за сутки:
жидкости - 1-1,5 л;
Na" - 1,4-4,6 г;
К+- 1,2-3,9 г;
Са2+- 0,1-0,25 г;
Mg2+- 0,1-0,2 г;
NH/ - 0.5-0,9 г;
Cf - 4,3-9,5 г;
НСО3" - 0,1-3,0 г;
фосфата - 0,6-1.3 г;
сульфата -2 г;
мочевой кислоты - 0,5-1,0 г;
мочевины - 12-36 г;
креатинина: муж. - 1,0-2,0 г, жен. - 0,8-1,8 г;
белка-125 мг.
Осмотическое давление - 50-1450 мосмоль/л.