
















































































































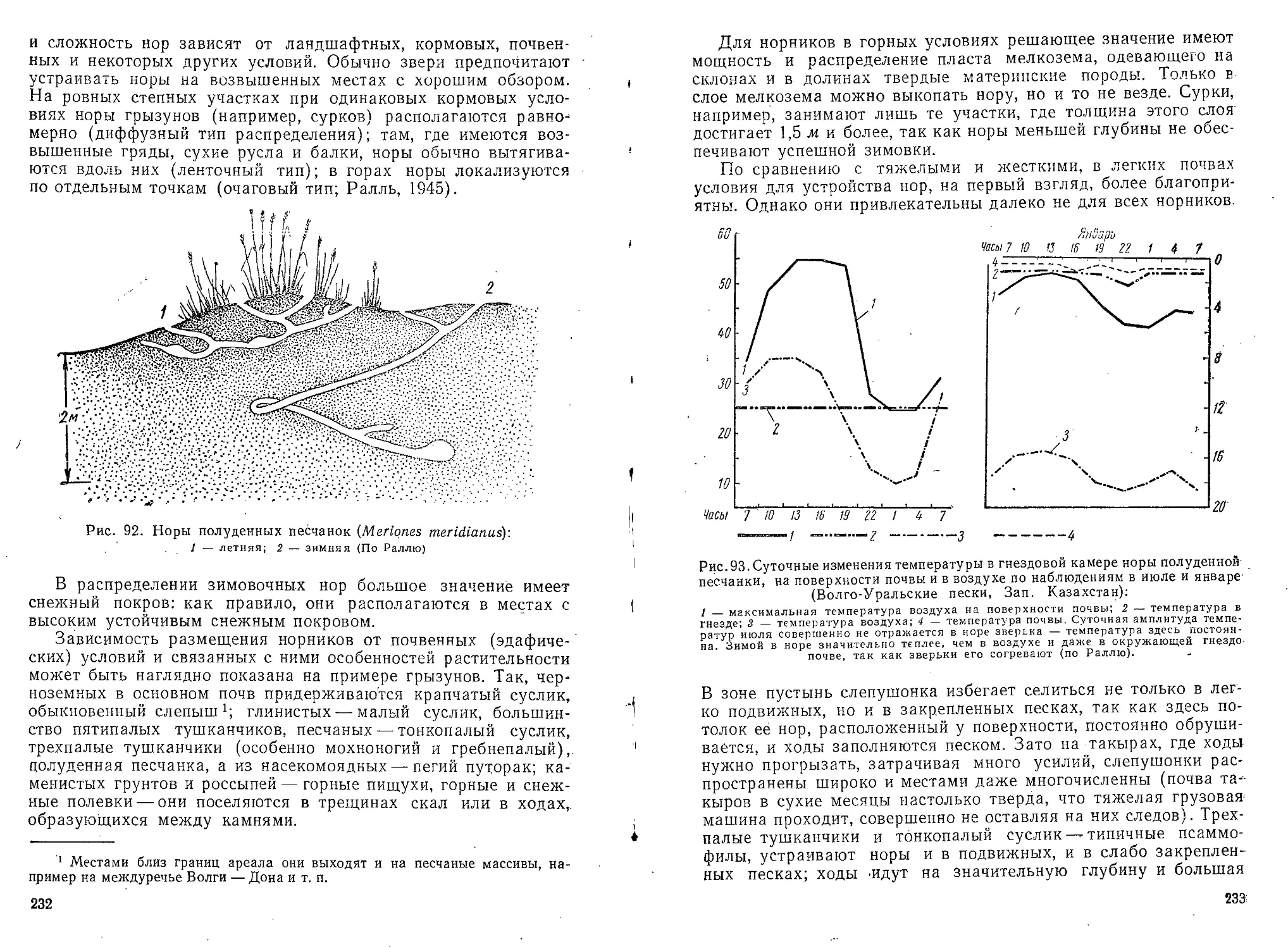



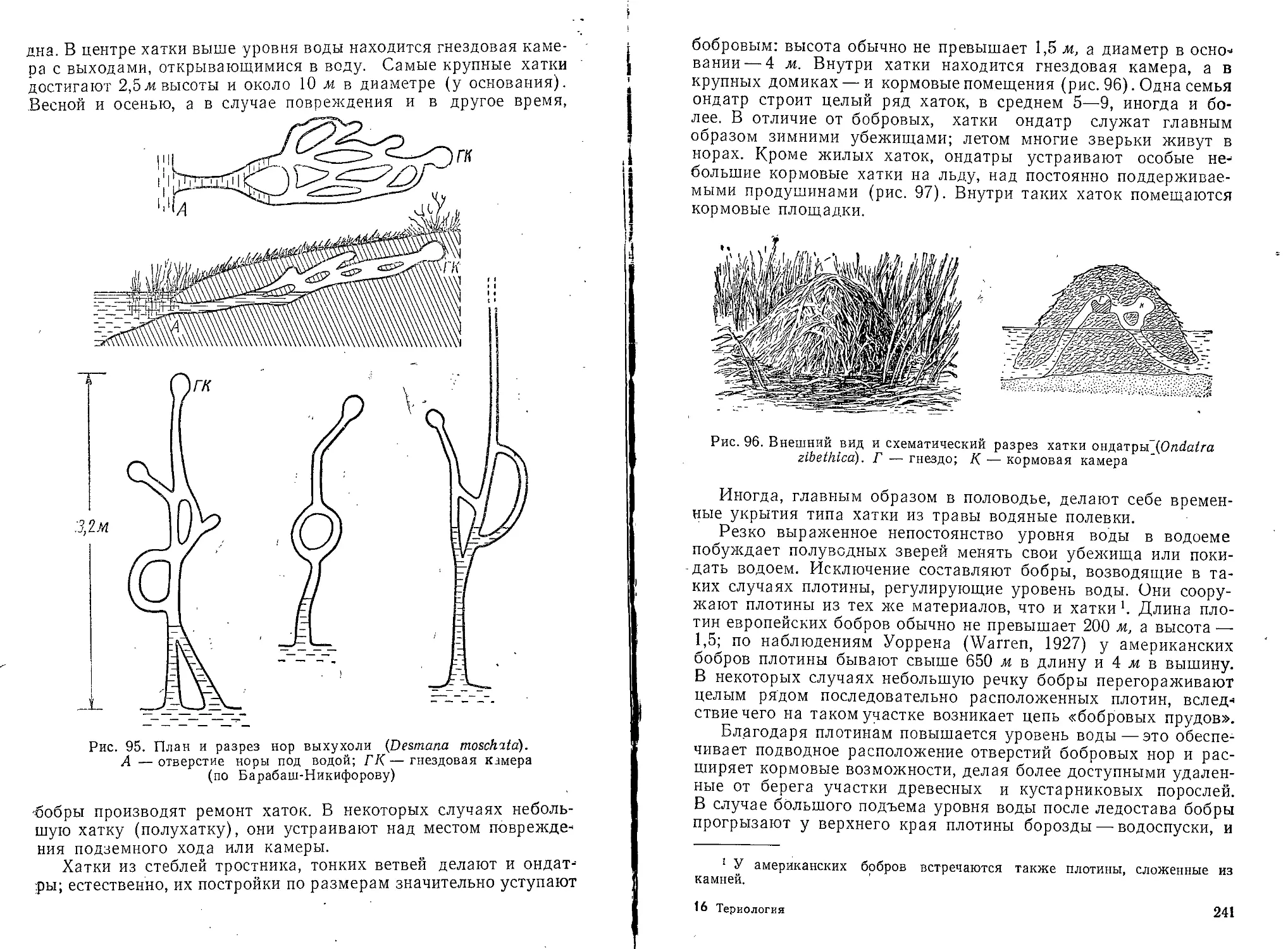






















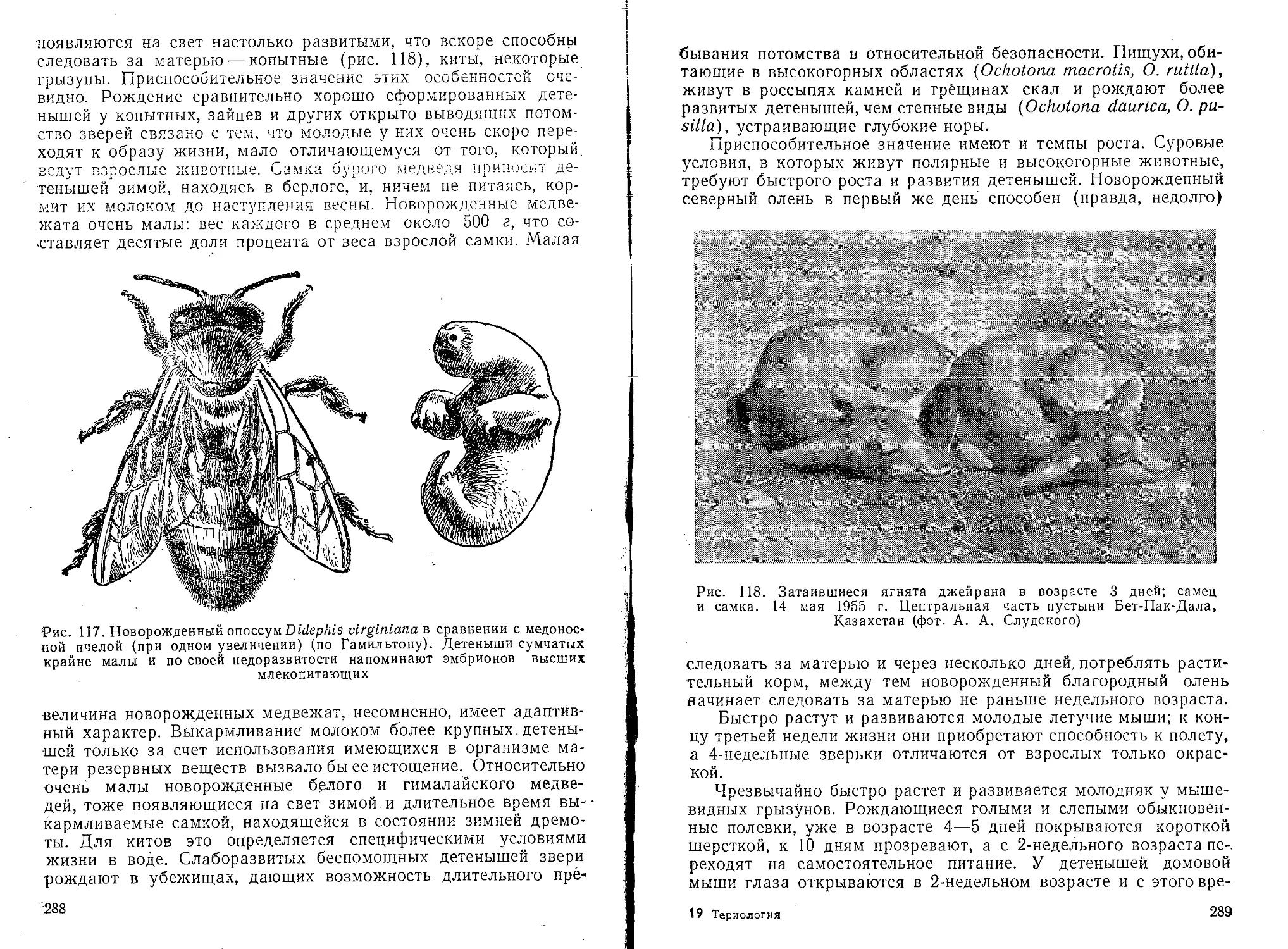
























































Текст
И. И. БАРАБАШ-НИКИФОРОВ, А. Н. ФОРМОЗОВ
ТЕРИОЛОГИЯ
ПОД РЕДАКЦИЕЙ А. Н. ФОРМОЗОВА
Допущено
Министерством высшего и среднего специального
образования СССР
в качестве учебного пособия
для государственных университетов СССР
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «ВЫСШАЯ ШКОЛА»
Москва — 1963
Предисловие
В данном учебном пособии даются в сжатом виде основные
(ведения о строении, классификации, происхождении, биологии
и практическом значении млекопитающих, необходимые при
изучении специального курса териологии и могущие послужить
дополнительным материалом для расширения и углубления зна-
ний, получаемых из общего курса зоологии позвоночных. Класс,
млекопитающих имеет огромное многостороннее значение, и
пособие такого рода так же необходимо, как существующие
аналогичные по другим позвоночным («Ихтиология», «Герпето-
логия», «Орнитология»),
До сих пор мы не имели подобного рода отечественного из-
дания. Часть соответствующих сведений имеется в «Курсе био-
логии позвоночных животных» Д. Н. Кашкарова и В. В. Стан-
чинского (1934). Но эта книга неудобна для пользования из-за
громоздкости, к тому же она в известной мере устарела. Книга
С. И. Огнева «Очерки экологии млекопитающих» (1951), хотя
и содержит весьма ценные материалы, не может, однако, за-
менить учебное пособие.
В «Териологии» излагается переработанный и сокращенный
вариант цикла лекций по одноименному специальному курсу,
читанному авторами на протяжении ряда лет для студентов
старших курсов университетов. Основой при создании курса
послужили крупные сводки и монографии, а также специальные
статьи советских и иностранных авторов, посвященные данной
группе животных, и оригинальные материалы.
Структура книги определяется ее назначением. Наибольшее
место отводится экологии млекопитающих — разделу, в котором
ощущается наибольшая йотребность. Морфологическая часть
предельно сжата, назначение ее — подчеркнуть только общие
характерные особенности организации млекопитающих, чтобы
избежать оторванности биологии от морфологии, функции от
формы. Исключение сделано лишь для некоторых важных раз-
делов морфологии, которые приводятся в расширенном объеме.
Предельно сжато излагаются классификация, происхождение.
Намеченный объем издания не дал возможности подробно
осветить такие важные разделы биологии млекопитающих, как
их болезни (вирусные, бактериальные, протозойные и др.),
3
фауну их экто- и эндопаразитов, средн которых многие имеют
прямое отношение к охране здоровья человека, домашних и цен-
ных диких ЖИВОТНЫХ И 'Г. д,
Заключительная глава посвящена практическим вопросам
народного хозяйства н здравоохранения и намечает дальнейшие
задачи изучения млекопитающих.
Авторы выражают благодарность за ценные указания по от-
дельным разделам проф. В. С. Матвееву и проф. А. И. Акаев-
скому (морфология млекопитающих), проф. В. Г. Гептнеру
(классификация) и доц. |А. Н, Дружинину (происхождение
класса).
Авторы
Введение
(Краткий исторический очерк)
Термин «териология» (от греч. «therion» — зверь и «logos» —
наука) предложен в 1928 г. С. И. Огневым взамен употребляв-
шегося и частично употребляемого в настоящее время неблаго-
звучного термина «маммалогия» или «маммалиология» (от
«mammalia» — млекопитающие). Таким образом, териология —
наука о млекопитающих, или зверях >.
Необходимость выделения этой отрасли зоологии в особую
дисциплину определяется исключительно важным, многосторон-
ним значением млекопитающих. Многие млекопитающие издав-
на приручены человеком и одомашнены с целью получения раз-
ного рода продуктов питания, кожи, шерсти или для использо-
вания в качестве рабочей силы,'осуществления сторожевых,
поисковых и разведывательных функций и пр. Многие дикие
млекопитающие служат объектами промысла и спортивной охо-
ты. Некоторые дикие копытные используются с целью улучше-
ния домашних пород путем гибридизации. Косвенную пользу
человеку приносят звери, истребляющие вредных грызунов и на-
секомых (мелкие хищники, насекомоядные, летучие мыши), а
также составляющие основной источник пищи для ценных пуш-
ных видов (некоторые грызуны). Наконец, важно и большое
эстетическое значение млекопитающих.
Вместе с тем, большое число видов млекопитающих играет
существенную отрицательную роль в жизни людей: они наносят
большой ущерб сельскому и лесному хозяйству, повреждая и
уничтожая ценные дикие и культурные растения, портят про-
дукты и товары, рассеивают вирусы различных опасных болез-
ней, прокармливают и распространяют разнообразных эктопа-
разитов и гельминтов (преимущественно грызуны, в меньшей сте-
пени другие млекопитающие) или угрожают непосредственно
жизни человека и домашних животных (крупные хищники).
Вполне понятен поэтому проявляемый человеком с самых от-
даленных времен особый интерес к млекопитающим. В первую
очередь, конечно, привлекали к себе внимание крупные виды,
служившие объектами охоты. Упоминания о них встречаются
1 А. Кабрера (Cabrera, 1922) предложил для этой науки еще одно назва-
ние— «мастозоология» ’(от греч. «mastos» — сосок млечной железы), однако
распространения оно не получило.
5
уже и npviiiHix лстшин-их, и члгпни'тп, в знаменитом «Поучении»
Влnp.'i Мономаха (/КП пек). Отрывочные данные о про-
мыс,новых :пч‘|И1Х содержатся также в путевых дневниках и до-
несениях первооткрывателей новых земель.
В подобного рода источниках, как и в первых охотничьих
и зоологических сочинениях, затрагивавших в той или иной сте-
1КЧН! м.че|(о1шт:п<иц!1х, точные наблюдения нередко сочетались
с. вымыслом, Соответственно общему уровню знаний того вре-
мени, чти произведения отличались большой элементарностью и
неполнотой. Из них можно назвать интересный английский охот-
ничий трактат, изданный в XV веке («Book of Saint Alban»,
1196), «Историю животных» Конрада Геснера (С. Hesner
«1 listoriae animalium» 1551—1587), «Историю четвероногих жи-
вотных» Э. То-пселла (Е. Topsell «History of Fourefootcd Bcastes»,
1607) и др.1 Все же и эти сочинения сыграли определенную
роль в развитии зоологических (в том числе териологических)
знаний. О большом значении трудов К- Линнея (С. Lin)ie
«Sistema Naturae», 1758), Г. Бюффона (G. Buffon «Histoine na-
turelle», 1749— 1788) и других классиков естествознания XVIII ве-
ка говорить не приходится.
На протяжении XIX и первой половины XX веков в Запад-
ной Европе и Америке появился ряд работ, специально посвя-
щенных млекопитающим или отводящих им соответствующие
большие разделы. Прежде всего нужно указать на известные
работы Ч. Дарвина, использовавшего богатые наблюдения, соб-
ранные за время путешествия на корабле «Бигль», и обширную
литературу для описания адаптаций и процесса эволюции ряда
групп млекопитающих. Значение биологических и палеонтологи-
ческих фактов для решения важнейших теоретических вопросов,
ясно показанное Ч. Дарвиным, было очень важным толчком, на-
правившим усилия натуралистов на изучение тех сторон жизни
зверей, которые раньше почти не привлекали внимания. С этого
времени, кроме множества фаунистических работ, все чаще по-
являются специальные биологические монографии, например,
труд Э. Сетона (Seton, 1929) по биологии охотничьих зверей
Северной Америки. Большое значение для дальнейшего разви-
тия науки о млекопитающих имели труды М. Вебера (М. Weber,
1’927, 1928) по морфологии и систематике млекопитающих,
А. Брема (A. Brehm, 1912—1916), Гамильтона (Hamilton, 1939),
Крумбигеля (Krumbigel, 1930—1931), Бурльера (Bourliere,
1951) и др.— по экологии зверей, множества авторов— по фау-
нам отдельных стран, соответствующие главы зоологического
трактата под редакцией П. Грассе («Traite de zoologie», XVI,
1 Здесь не упоминаются более ранние естественноисторические произве-
дения, в которых, начиная уже с сочинений Аристотеля (IV век до нашей
эры), Плиния Старшего (I век) и некоторых других, уделяется соответст-
вующее место млекопитающим.
6
XVII, v. 1, 2, 1955). Издаются научные журналы, посвященные
млекопитающим и т. д.
В России планомерное изучение млекопитающих, как и
других животных, началось с основания Академии наук
(1725). Особенно важное значение для изучения фауны и от-
части биологии млекопитающих имели работы П. С. Палласа,
И. И. Лепехина, А. Гюльденштедта, С. Гмелина и некоторых
других участников известных «академических экспедиций»
конца XVIII века. Сочинение известного путешественника-
энциклопедиста П. С. Палласа «Novae species Quadrupeduni е
Glirium ordinae» (1778) содержит подробные сведения по систе-
матике, анатомии и географическому распространению многих
наших грызунов. Множество ценных сведений рассеяно и в опи-
саниях его путешествий. Из трудов И. И. Лепехина наибольший
интерес для териологии представляют «Дневные записки...»
(1772—1804) в которых приводятся данные о распространении,
биологии и экономическом значении ряда видов зверей (тар-
пана, сайги, бобра и др.). Лепехину же принадлежит заслуга
обогащения коллекций Академии наук большими сборами мле-
копитающих, позднее обработанных Палласом.
Антон Гюльденштедт в описании своего путешествия по юго-
востоку России (опубликованном посмертно Палласом) приво-
дит ряд ценных териологических материалов, в частности, впер-
вые описывает такие интересные виды млекопитающих, как
джейран, слепыш, перевязка. В сочинении С. Г. Гмелина «Путе-
шествие по России для исследования трех царств естества»
(1771—1785) приведены интересные данные о прежнем распро-
странении тарпана и некоторых других млекопитающих.
Из других работ того времени интересно описание фауны
Берингова моря, составленное Г. В. Стеллером («De bestiis
marinis», 1753); в нем даются первые подробные сведения
о вскоре исчезнувшем с лица земли представителе сиреновых —
морской корове, о морской выдре (калане) и пр.
Накопление сведений о русской териофауне продолжается
и в XIX веке, когда выдвинулись имена таких' видных исследо-
вателей, как Ф. Ф. Брандт, Э. А. Эверсман, А. Ф. Миддендорф,
Н. А. Северцов.
Академику Ф. Ф. Брандту принадлежит большая 'заслуга
создания, на базе старинной петровской кунсткамеры, Зоологи-
ческого музея, в котором видное место заняли млекопитающие.
Брандт — автор описаний многих новых видов зверей. Ему же
принадлежат ценные сводки: «Материалы к познанию млеко-
питающих России» (1855) и «Позвоночные животные Северно-
европейской России» (1856).
1 Полное название этого четырехтомного труда — «Дневные записки пу-
тешествия доктора и Академии наук адъюнкта Ивана Лепехина по разным
провинциям Российского государства».
7
Профессор Казанского уппперситет.ч Л. Эвсрсман посвя-
тил второй том своей ociioinioii работы «Естественная история
Оренбургского края» (1(440 1866) систематике, распростране-
нию и биологии представителей териофауны Поволжья, При-
уралья и Прикаспийской низменности (в том числе ряду инте-
ресных видов).
В классическом сочинении академика А. Ф. Миддендорфа
«Путешествие ча Север и Восток Сибири» (1860—1875) при-
ведено много экологических наблюдений над млекопитающими,
дано монографическое описание бурого медведя, лемминга.
Вопрос о годовом цикле жизни млекопитающих в зависимо-
сти от условий среды получил освещение в замечательном ис-
следовании Н. А. Северцова — «Периодические явления в жиз-
ни зверей, птиц и гад Воронежской губернии» (1855).
Несомненно, положительную роль в развитии отечественной
териологии сыграла обширная сводка Ю. Симашко «Русская
фауна» (1851), второй том которой посвящен млекопитающим.
Большую ценность представляют коллекции и наблюдения по
млекопитающим Уссурийского края и Центральной Азии, соб-
ранные Н.. М. Пржевальским и продолжателем его дела
П. К- Козловым.
На рубеже XIX и XX веков и несколько позднее публикуются
обстоятельные работы по систематике, фаунистике и зоогеогра-
фии млекопитающих нашей страны. Н. Ф. Кащенко дал инте-
ресный «Обзор млекопитающих Западной Сибири и Туркестана»
(1905) и составил «Определитель млекопитающих животных
Томского края» (1900), значение которого далеко не ограничи-
валось пределами названной территории. Млекопитающим Кав-
каза посвящены богатые биологическими данными труды
Н. Я. Динника («Звери Кавказа», 1910—1914) и К- А. Сатунина
(«Млекопитающие Кавказского края», 1915—1920). Н. А. Смир-
нов опубликовал классическое исследование по ластоногим
(«Очерк русских ластоногих», 1908).
Следует отметить также «Материалы по фауне млекопитаю-
щих Симбирской губернии» Б. М. Житкова (1898) и посвящен-
ный млекопитающим отдел его другого труда «Полуостров
Ямал» (1913).-
В развитии исследований по териофауне средней полосы
России видную роль сыграла книга С. И. Огнева «Млекопитаю-
щие Московской губернии» (1913). Были и другие более или
менее значительные териологические исследования.
В развитии прикладной териологии положительную роль сы-
грали труды Л. П. Сабанеева, Н. В. Туркина, А. А. Силантьева,
Г. Г. Доппельмайра, Н. А. Смирнова-, и др.
Однако, оценивая общее состояние русской териологии в
конце XIX и в начале XX веков, приходится констатировать ее
упадок по сравнению с предшествующим периодом. Особенно
8
плачевно обстояло дело с экологией млекопитающих, в разра-
ботке которой царил полный застой.
Положение коренным образом изменилось лишь после Вели-
кой Октябрьской социалистической революции. Как и другие
области науки, териология с этого времени вступила в полосу
мощного подъема. Териологические исследования с этого вре-
мени приобрели государственный, плановый характер; вместо
одиночек-териологов появились коллективы специалистов, под-
готовленных советскими вузами.
Одним из ведущих териологов был профессор Московского
университета С. И. Огнев (1886—1951). Труд его жизни —
семитомник «Звери СССР» (1928—1950) 1 представляет собой
сводку всех сведений о териофауне СССР и сопредельных
стран, в которой использованы также личные наблюдения
автора и собранные им огромные коллекции млекопитающих
(около 15 тысяч экземпляров, позднее поступившие в фонд
Зоологического музея МГУ). Капитальный труд С. И. Огнева,
несомненно, послужил делу дальнейшего развития нашей терио-
логии.
В 20 — 30-е годы достигла высокого уровня деятельность
других крупных зоологов старшего поколения, вступивших на
научное поприще еще в дореволюционный период. Из них осо-
бого упоминания заслуживает профессор Б. М. Житков (1872—
1943)—известный теоретик и практик в области разработки
биологических основ народнохозяйственного использования жи-
вотных (охотничьего промысла, акклиматизации, охраны жи-
вотных й пр.). В частности, Житковым была претворена в
жизнь идея акклиматизации в СССР ценного американского
пушного грызуна ондатры, занявшей вскоре одно из первых:
мест в отечественных пушных заготовках1 2. Благодаря под-
держке Житкова осуществлены завоз нутрии в нашу страну,,
первый опыт перевозки на Мурман для целей акклиматизации
ценнейшего в мире пушного зверя калана (с Командорских ост-
ровов) и пр. Не меньшее значение имеет тот факт, что под его-
руководством в Московском университете сформировалась силь-
ная группа зоологов, энергично взявшихся за изучение эколо-
гии промысловых зверей (С. П. Наумов, Н. П. Лавров,.
А. Ф. Чиркова, Д. М. Вяжлинский и др.). Плодотворно рабо-
тают также Н. М. Кулагин, А. А. Бялыницкий-Бируля, Н. А. Бо-
бринский, Б. С. Виноградов и др. Борис Степанович Виноградов,,
крупный систематик, морфолог и фаунист монографически об-
1 Первые два тома вышли под названием «Звери Восточной Европы и
Северной Азии», последующие — «Звери СССР и сопредельных стран». Де-
вятый том («Китообразные»), написанный учеником Огнева — А. Г. Томи-
линым, опубликован в 1958 г.
2 Идея акклиматизации ондатры была выдвинута проф. Н. А. Смир-
новым.
работал сем. тушканчиковых, нашел и описал ряд новых энде-
мичных родов и видов. Он очень много сделал для изучения
трудной для систематики группы полевок, опубликовал превос-
ходные определители грызунов СССР, вскрыл ряд интересных
•особенностей экологии млекопитающих пустынь Средней Азии
и т. д.
Вместе с тем в советский период выдвигается целая плеяда
териологов младших поколений, которым принадлежит огром-
ное количество более или менее крупных работ, посвященных
млекопитающим !. Небывало широкий размах приобрели в этот
период экологические исследования, причем на первый план вы-
двинулось решение вопросов, связанных с интересами народного
хозяйства и здравоохранения. Продолжаются экспедиционные
работы, создаются специальные научно-исследовательские уч-
реждения, в круг деятельности которых входит изучение млеко-
питающих (вредных грызунов и биологических основ борьбы с
ними, промысловых зверей и вопросов рационального их исполь-
зования). Учреждаются заповедники для сохранения ценных
природных участков, их фауны и флоры. Советские заповедни-
ки—г «лаборатории в природе», стали очень важными опорными
базами для изучения млекопитающих. На материале заповедни-
ков написаны многие ценные монографии охраняемых видов
зверей.
Но несмотря на все указанные достижения в териологии
остается еще много пробелов, обусловленных главным образом
трудностью наблюдения млекопитающих в природе в связи с их
скрытным образом жизни1 2. Многие имеющиеся сведения о зве-
рях уже устарели. Изученность отдельных видов и групп млеко-
питающих крайне неравномерна. -Некоторые даже самые обык-
новенные виды в экологическом отношении нам почти неизве-
стны (землеройки, ряд хищных, грызунов, рукокрылых).
Задача современной зоологии — заполнение имеющихся в
ней пробелов и дальнейшая разработка биологических основ
рационального «управления» животным миром. Решение многих
практических вопросов в приложении к млекопитающим воз-
можно только при наличии соответствующих териологических
знаний.
1 Особо следует отметить предпринятое в самое последнее время изда-
ние трехтомника «Млекопитающие Советского Союза» под редакцией
В. Г. Гептнера и Н. П. Наумова (в 1961 г.' вышел первый том, посвящен-
ный копытным).
2 Этим объясняется странное на первый взгляд явление, что в такой
сравнительно очень немногочисленной группе животных как млекопитающие
возможны еще находки новых, не известных науке видов и родов. Совсем
недавно в девственных лесах Африки были открыты окапи (мелкий пред-
ставитель жирафовых), карликовый бегемот, карликовый слон; в лесах Индо-
китая — гигантский бык; в пустыне Бетпакдала (Казахстан) — своеобразный
грызун селевиния (боялычная соня).
Систематический очерк класса
млекопитающих
Общее, в основе своей правильное определение млекопита-
ющих было дано еще Аристотелем (384—322 гг. до н. э.). В даль-
нейшем это понятие уточнялось многими зоологами. К. Линнеи
(1707—1778) впервые выделил млекопитающих в качестве
высшего класса животных, к которому он отнес и человека.
Однако попытки подразделения класса долгое время оставались
неудачными. В своей системе млекопитающих Линней в один
отряд Bestiae объединял таких далеких друг от друга животных,
как сумчатые, насекомоядные, неполнозубые и свиньи; с прима-
тами он соединял рукокрылых. Подобного же рода недостатки
имеются в системе зверей Ж. Ламарка (1744—1829): в одном
отряде «когтистых» здесь фигурируют, например, насекомояд-
ные, грызуны, сумчатые, рукокрылые и обезьяны. Принятое
этим ученым подразделение класса всего на 4 отряда — копыт-
ных, когтистых, бескопытных и земноводных,— конечно, не мог-
ло обеспечить сколько-нибудь удовлетворительного охвата всего
многообразия форм млекопитающих. Большой искусственно-
стью отличается также система Ж. Кювье (1769—1832), в кото-
рую, правда, входило уже 8 отрядов (киты, жвачные, копытные,
беззубые, грызуны, хищные, четырехрукие и двурукие).
С другой стороны, можно отметить и известный прогресс в
построениях системы млекопитающих. В частности, еще в 1816 г.
французский зоолог А. Бленвиль (1777—1850) предложил де-
ление класса Mammalia на подклассы птицеутробных, двуутроб-
ных и одноутробных, которое под другими наименованиями
сохраняется и в настоящее время (клоачные, сумчатые и пла-
центарные) .
Э. Геккель (1834—1919) установил для млекопитающих уже
30 отрядов, сгруппированных в 9 «легионов».
Линней мог назвать в десятом издании «Systema naturae»
(1758) 1 всего 86 известных ему ныне живущих видов зверей, но
в позднейших системах число их быстро нарастает. В настоящее
время насчитывается около 3,5 тысяч видов млекопитающих1 2.
1 Согласно международным правилам, это издание считается исходным
для зоологической номенклатуры.
2 Число видов млекопитающих обычно определялось в 6—6,5 тысяч, но
в последнее время в связи с «переводом» ряда «видов» в подвиды это число
сократилось вдвое (Гептнер, 1956). Вместе с подвидами в 20-х годах нынеш-
него столетия насчитывалось свыше 12 500 форм млекопитающих; в 30-Х го-
дах — свыше 20 000 форм.
11
Вместе с вымершими они составляют около 30 отрядов, вклю-
чающих около 260 семейств.
Быстрый рост числа выделенных и описанных видов до на-
чала 60-х годов прошлого столетия требовал для ориентировки
в них хотя бы условного подразделения, без учета единства
происхождения членов выделяемых групп. Этим характеризует-
ся вся додарвннская систематика. Последарвинский период
отличается эволюционным подходом к классификации, при кото-
ром широко привлекаются не только морфологические призна-
ки, но и данные палеонтологии. Именно по этому принципу по-
строена система млекопитающих, предложенная Дж. Симпсоном
(1945). Как новейшая и относительно наиболее совершенная,
она положена в основу помещаемого ниже перечня. Лишь в
редких случаях введены коррективы, отражающие взгляды дру-
гих систематиков-териологов, более, на наш взгляд, правильные.
КЛАСС МЛЕКОПИТАЮЩИЕ, ИЛИ ЗВЕРИ (MAMMALIA)1
Подкласс первозвери (Prototkerla)
Отряд клоачные, или однопроходные (Monotremata)
Сем. утконосы (Ornithorhynchidae). Австралия, Тасмания.
Единственный вид — утконос (Ornithorhynchus anatinus).
Сем. ехидны (Echidnidae). Австралия, Тасмания, Новая Гвинея.
Ехидны (Tdchyglossus aculeata, Т. setosa), проехидна
(Zaglossus bruynit).
Подкласс низшие звери (Metatheria)
Отряд сумчатые (Marsupialia)
Подотряд многорезцовые (Polyprotodontia)
Сем. опоссумы (Didelphydae). Южная и отчасти Северная Америка.
Сумчатая крыса опоссум (Didelphys marsupialis), толсто-
хвостый опоссум (Metachirops opossum), карликовая сум-
чатая крыса (Marmosa elegans), плавун (Chironectes mi-
nimus).
Сем. сумчатые куницы (Dasyuridae). Австралия, Тасмания, Новая
Гвинея.
Сумчатая куница (Dasyurus viverrina), сумчатый дьявол
(Sarcophilus harrisii), тафа (Phascogale penicillatd).
Сем. мурашееды (Myrmecobiidae). Австралия.
Единственный вид—мурашеед, или сумчатый муравьед.
(Myrmecobius fasciatus).
1 Подразделение класса доведено до семейств, для которых указываются
некоторые представители. Особо оговариваются случаи, когда семейство
представлено единственным видом.
12
Сем. сумчатые кроты (Notoryctidae). Австралия. .
Сумчатые кроты (Notoryctes typhlops, N. caurinus).
Сем. сумчатые барсуки (Peramelidae). Австралия, Тасмания,
Новая Гвинея.
Сумчатый барсук бандикут (Perameles gunni), длинноухий
сумчатый барсук (Macrotis lagotis)'.
Подотряд ценолестовые (Caenolestoidea)
Сем. ценолестовые (Caenolestidae). Южная Америка (Анды).
Темный ценолест (Caenolestes obscuras), инка (Lestoros
inca), носатый ценолест (Rhyncholestes raphanurus).
Подотряд двурезцовые (Diprotodontia)
Сем. лазающие сумчатые (Phalangeridae). Австралия, Новая
Гвинея, Тасмания, Сулавеси (2 вида кускусов).
Кускус (Phalanger maculatus), лисий кузу (Trichosurus
vulpeculd), сумчатая белка (Acrobates pygmaeus), сумчатая
летяга, или сахарная белка (Petaurus sciureus), пяткоход—
сумчатый медоед (Tarsipes spenserae).
Сем. сумчатые медведи (Phascolarctidae). Австралия, Тасмания,
Новая Гвинея.
Сумчатый медведь коала (Phascolarctos cinereus), вомбат
(Vombatus ursinus), малый вомбат (yombatula gillespiei).
Сем. кенгуровые (Macropodidae). Австралия, Новая Гвинея.
Мускусный кенгуру (Hypsiprymnodon moschatus), кенгу-
ровая крыса потору (Potorous tridactylus), кистехвостый
кенгуру (Bettongia peniciltata), серый, или гигантский
кенгуру (Macropus giganteus), древесный кенгуру (Dend-
rolagus dorianus), горный кенгуру (Petrogale xanthopus).
Подкласс высшие звери, или плацентарные
(Eutheria, или Placentalia)
Когорта когтистые (Unguiculata)
Отряд насекомоядные. (Insectivora)
Сем. щелезубы (Solenodontia). Большие Антильские острова.
Кубинские щелезубы (Solenodon cubanus, S. poeyanus),
гаитянский щелезуб (S. paradoxus).
Сем. тенреки (Tenrecidae). Мадагаскар, Западная Африка.
Обыкновенный тенрек (Tenrec ecaudatus), полосатый тен-
рек (Hemicentetus semispinosus), иглистый тенрек (Setifer
setosus).
Сем. выдровые землеройки (Potamogalidae). Западная Африка.
Выдоовые землеройки (Potamogale velox, Micropotamogale
lamottei).
13
Сем златокроты (Chrysochloridae). Южная Африка.
Капский златокрот (Chrysochloris аигеа), исполинский зла-
токрот (Ch. trevelyani).
Сем. ежи (Erinaceidae). Европа, Азия, Африка.
Гимнура (Gymnura gymnura), малый крысиный еж (Hylo-
mys suilla), обыкновенный еж (Erinaceus europaeus), ушас-
тый еж (Hemiechinus attritus'), длинноиглый еж (Paraechinus
hy pamelas).
Сем. прыгунчики (Macroscelididae). Африка.
Прыгунчики (Elephantulus rozeti, Petrodromus robusius).
Сем. землеройки (Soricidae). Европа, Азия, Африка, Северная
Америка.
Обыкновенная бурозубка (Sorex araneus), кутора (Neotnys
fodiens), обыкновенная белозубка (Crocidura russula), боль-
шая белозубка (С. lasiura).
Сем. кроты (Talpidae). Европа, Азия, Северная Америка.
Землеройкообразные кроты (Uropsilus gracilis, Rhynchonax
andersoni), скалоп (Scalopus aquations), обыкновенный крот
(Talpa europaea), могера (Mogera robusta), звездорыл (Can-
dy lura cristata).
Сем. выхухоли (Desmanidae). Пиренейский полуостров, Восточ-
ная Европа (СССР).
Пиренейская выхухоль (Galetnys pyrenaicus), русская вы-
хухоль (Desmana moschata).
Сем. тупайи (Tupaiidae) \ Индия, Индокитай, Китай, Малайский
за архипелаг, Филиппины.
Малайская тупайя (Tupaia ferruginea), тана (Т. tana).
Отряд шерстокрылы (Dermoptera)
Сем. шерстокрылы (Cynocephalidae). Малайский архипелаг, Ма-
лакка.
Шерстокрыл, кагуан, или летучий маки (Cynocephalus
vo Ians).
Отряд рукокрылые (Chiroptera)
Подотряд плодоядные рукокрылые, или летучие
собаки (Megachiroptera)
Сем. крыланы (Pteropidae). Тропические и субтропические об-
ласти Восточного полушария.
Летучая собака, калонг (Pteropus calaeno), летучая лисица-
(Р. medias), ошейниковый крылан (Rousetus collaris), длин-
1 Некоторые систематики относят тупайи к отряду приматов или выде-
ляют их в самостоятельный отряд.
Н
ноязычные крыланы (Megaloglossus woermanni, Nesonycteris
woodfordi), ночные крыланы (Nyctimene major, N. stimulus)-
крылан гарпия (Harpyionycteris whiiehaidi).
Подотряд насекомоядные рукокрылые,
или летучие мыши (Microchiroptera)
Сем. ланцетоносы (Rhinopomidae). Египет, Южная Азия.
Египетский ланцетонос (Rhinopoma microphy Пит).
Сем. свободнохвостые летучие мыши (Embalonuridae). Тропиче-
ские области Восточного и Западного полушарий.
Эмбалонура (Embalonura peninsularis), голобрюхая летучая
мышь (Taphozous nudiventris).
Сем. никтериды (Nycteridae). Тропическая Африка, Юго-Восточ-
ная Азия.
Nycteris javanica, N. avakubia.
Сем. мегадермы (Megadermidae). Тропическая Африка, Южная
Азия, Австралия.
Megaderma lyra, М. spasma, Macroderma gigas, Lavia frons.
Сем. подковоносы (Rhinolophidae). Тропические и умеренные
области Восточного полушария.
Большой подковонос (Rhinolophus ferrum-equinum), малый
подковонос (Rh. hipposideros).
Сем. ложные подковоносы (Hipposideridae). Тропические и суб-
тропические области Восточного полушария.
Большой ложный подковонос (Hipposideros gigas), персид-
ский ложный подковонос. (Тriaenops persicus).
Сем. зайцеротые летучие мыши (Noctilionidae). Тропическая,
область Америки.
Зайцероты (Noctilio leporinus, N. labialis).
Сем; листоносы (Phyllostomidae). Южная и Центральная Америка.
Копьенос (Phyllostomus hastatus), вампир (Vampirum spe-
ctrum), глоссофага (Glossophaga soricina), большой крово-
сос (Desmodus rotundus), малый кровосос (Diphylla есаи-
data).
Сем. обыкновенные (гладконосые) летучие мыши (Vespertilionidae)^
Все материки, кроме Антарктиды.
Двухцветный кожан (yespertilio tnurinus), широкоушка
(Barbastella barbastellus), большая ночница (Myotis myotis) r
ушан (Plecotus auritus), рыжая вечерница (Nyctalus пос-
tula), серый стрелоух (Otonycteris cinereus), ушастый труб-
конос (Murina aurata), гарпиевый трубконос (Harpioce-
phalus leucogaster), длиннокрылы (Miniopterus schreibersite
M. inflatus).
Сем. новозеландские летучие мыши (Mystacinidae). Новая Зелан-
дия.
15
Единственный вид — Mystacina tuberculata.
Сем. бульдоговые летучие мыши (Mollosidae).
Тропические' и субтропические области Восточного и За-
падного полушарий.
Рыжая бульдоговая мышь (Mo/ossus rufus), широкоухий
складчатогуб (Tadarida teniotis).
Отряд приматы (Primates)
Подотряд полуобезьяны, или лемуры
(Lemuroidea, или Prosimiae)
Сем. лемуры (Lemuridae). Мадагаскар.
Собственно лемуры (Lemur catta, L. variegatus), индри
(Indri indri), руконожка (Chiromis madagascarensis).
Сем. лориевые (Loristidae). Юго-Восточная Азия, Африка.
Лори тонкий (Loris gracilis), лори толстый (L. tardig radus),
ушастый маки (Galago demidoffi).
Подотряд долгопятовые (Тarsioidea)
Сем. долгопяты (Tarsiidae). Филиппинские и Большие Зондские
острова.
Маки-домовой (Tarsius spectrum), темный долгопят (Т.
fuscus).
Подотряд обезьяны (Р ithecoidea, или Simiae) 1
Сем. игрунковые (Hapalidae).'Южная и Центральная Америка.
Игрунки (Hapale jacchus, Н. pygmaeus).
Сем. ревуны (Cebidae) 1 2. Южная и Центральная Америка.
Капуцин (Cebus capucinus), ревун (Alouatta seniculus),
паукообразные обезьяны (Ateles palliata, A. ater).
Сем. мартышковые (Cercopithecidae). Тропические области Аф-
рики и Азии.
Мартышки (Cercopithecus diana, С. pata), макаки (Macacus
maurus, М. fuscata), павиан (Papio papio), гамадрил (P.
hamadrias), мандрилы (Mandrillus sphinx, M. leucophaeus),
тонкотелы (Semnopithecus entellus, S. nemaeus), толстотелы
(Colobus satanas, C. verus), носач (Nasalis barbatus).
Сем. гиббоны (Hylobatidae). Индия, Индокитай, Малайский
архипелаг.
1 Кроме перечисленных ниже семейств этого подотряда, к нему относит-
ся также семейство Hominidae (люди), которое здесь рассматриваться не
может, т. к. является предметом изучения других наук.
2 Семейства игрунковых и ревунов принято объединять в группу широ-
коносых обезьян (Platyrrhini); остальные семейства подотряда — в группу
узконосых обезьян (Catarrhini).
16
Белорукий гиббон, или лар {Hylobates lar), быстрый гиббон
{Н. agilis), сиаманг (Я. syndactylus).
4,ем. человекообразные (Anthropomorphidae). Африка, Суматра,
Калимантан.
Горилла {Gorilla gorilla), орангутан {Simia satyrus), шим-
панзе {Pan troglodytes, P. paniscus).
Отряд неполнозубые (Edentata)
Подотряд беспанцирные (Anicanodonta)
('•ем. ленивцы (Bradipodidae). Южная Америка.
Трехпалый ленивец {Bradipus tridactylus), двупалый ле-
нивец {Choloepus didactylus).
( см. муравьеды (Myrmecophagidae). Южная и Центральная Аме-
рика.
Трехпалый муравьед {Myrmecophaga tridactyla), четырёх-
палый муравьед тамандуа {Tamandua tetradactyla), дву-
палый муравьед {Cyclops didactylus).
Подотряд панцирные (Hicanodonta)
Сем. броненосцы (Dasypodidae). Южная и Центральная Америка.
Девятипоясный броненосец {Dasypus novemcinctus), трех-
поясный броненосец {Talypeutes tricinctus), гигантский
броненосец {Priodontes giganteus), шестипоясный броне-
носец {Euphractes sexcinctus), плащеносные броненосцы
{Chlarrtyphorus truncatus, Burmeisteria setosa).
Отряд ящеры, или панголины (Pholidota)
('.ем.' ящеры (Manidae). Южная Азия, Южная Африка.
Яванский ящер {Manis javanica), гигантский ящер {М.
gigantea).
Когорта грызуны (Glires)
Отряд зайцеобразные (Lagomorpha)
Сем. пищухи, или сеноставки (Ochotonidae). Европа, Азия, Се-
верная Америка.
Северная пищуха {Ochotona alpina), степная пищуха {О.
pusilia), американская пищуха (О. princeps).
Сем. зайцы и кролики (Leporidae). Все материки, кроме Антарк-
тиды (в Австралии — акклиматизированные кролики).
2 Териология 17
Заяц-беляк (Lepus timidus), заяц-русак (L. europaeus),
бразильский заяц (Silvilagus brasiliensis), гималайский
заяц (Caprolagus hispidus), дикий кролик (Oryctolagus
cuniculus), складчатозубые зайцы (Pentalagus furnassi,
Romerolagus nelsoni).
Отряд грызуны (Rodentia)
Сем. беличьи (Sciuridae). Повсеместно, кроме Австралии, Тасма-
нии, Мадагаскара, Антарктиды.
Обыкновенная белка (Sciurus vulgaris), американская серая
белка (S. carolinensis), карликовые белки (Funisciurus
leucostlgma, Dremomys rufigenis), бурундук (Tatnias stria-
tus), крапчатый суслик (Citellus suslica), малый суслик
(C. pygmaeus), полосатый американский суслик (С. tri.de-
cemlineatus), луговая собачка (Cynomys ludovicianus) ,
альпийский сурок (Marmota marmota), степной сурок
(М. bobac), земляные белки (Xerus erythropus, X. inaurus)?
тонкопалый суслик (Spermophilopsis leptodactylus).
Сем. летяги (Pteromyidae). Европа, Азия, Северная Америка.
Обыкновенная евразиатская летяга (Pteromys volans), се-
вероамериканская летяга (Glaucomys volans), индийская
летяга (Petaurista candidulus).
Сем. шипохвостые белки (Anomaluridae). Африка.
Шипохвостые белки-летяги (Anomalurus pelei, Idiurus
macrotis).
Сем. горные бобры (Aplodontidae). Северная Америка.
Горный бобр (Aplodonta rufa).
Сем. сони (Myoxidae). Европа, Азия, Африка.
Евразиатские сони — соня-полчок (Glis glis), лесная соня
(Dyromys nitedula), садовая соня (Eliomys quercinus), аф-
риканские сони (Graphiurus murinus, Gliriscus rupicola).
Сем. селевинии, или боялычные сони ( Seleviniidae). Восточный
Казахстан.
Единственный вид — боялычная соня (Selevinia betpak-
dalensis).
Сем. мешетчатые крысы (Geomyidae). Северная и Центральная
Америка.
Мешетчатые крысы, или гоферы (Geomys bursarius, Tho-
momys perpallidus).
Сем. кенгуровые, или мешетчатые крысы (Heteromyidae). Север-
ная Америка.
Heteromys anomalus, Perognathus hispidus, Dipodomys agi-
lis, D. deserta.
Сем. гунди, или гребнепалые крысы (Ctenodactylidae). Северная
Африка.
Ctenodactylus gundi, Pactinator spekei.
18
( л'м. тушканчиковые (Dipodidae). Европа, Азия, Северная Африка,
Северная Америка.
Лесная мышовка (Sicista betulina), степная мышовка (S.
subtilis), североамериканская мышовка (Zapus hudsonicus),
мохноногий тушканчик (Dipus sagitta), гребнепалый туш-
канчик (Paradipus ctenodactylus), африканский трехпалый
тушканчик (Jaculus jaculus), земляной заяц (Allactaga
jaculus), земляной зайчик (Alactagulus pygmaeus), толсто-
хвостый тушканчик (Pigerethmus platyurus), жирнохвос-
тый и настоящий карликовые тушканчики (Salpingotus-
crassicauda, Cardiocranius paradoxus).
С,ем. хомячьи (Cricetidae). Европа, Азия, Северная и Южная Аме-
рика, Африка, Мадагаскар.
Мышевидные хомячки (Perotnyscus leucopus1, Calotnyscus
bailwardi, Reithrodon caurinus, Neotoma albigula), настоя-
щие хомяки — обыкновенный хомяк (Cricetus cricetus),
серый хомячок (Cricetulus migratorius), джунгарский хо-
мячок (Phodopus songarus)', полевки — обыкновенная по-
левка (Microtus arvalis), водяная полевка (Arvicola ter-
restris), рыжая лесная полевка (Clethrionomys glareolus),
ондатра (Ondatra zibethica), степная пеструшка (Lagurus
lagurus), северная пеструшка, или норвежский лемминг
(Lemmus lemmus), копытный лемминг (Dicrostonyx tor-
quatus), слепушонка (Ellobius talpinus), цокор (Myospalax
myospalax)-, песчанки — большая песчанка (Rhombomys opi-
mus), полуденная песчанка (Meriones meridianus), южная
песчанка (Gerbillus gerbillus).
Сем. мышиные (Muridae)1 2. Жаркий и умеренный пояса Восточного
полушария, кроме Мадагаскара. Еы:е космополиты.
Собственно мыши и крысы — домовая мышь (Mas muscu-
lus), полевая мышь (Apodemus agrarius), мышь-малютка
(Micromys minutus), рыжая крыса-пасюк (Rattus norve-
gicus), черная крыса (Rattus rattus), индийская пластин-
чатозубая крыса (Nesokia indica), азиатская древесная мышь
(Phloemys pallidus), африканские древесные мыши (Den-
drotnys mesomelas, Steatomys pratensis, Prionomys batesi),
австралийские водягыэ крысы (Hydromas chrysogaster,
Mayermys ellermani), ушас"ыэ крысы (Otomys tropicalis,
Parotomys brantsi).
Сем. долгоноги (Pedetidae). Южная Африка.
Единственный вид — кафрский долгоног (Pedetes cafer).
1 «Оленья мышь».
2 Мышиных и хомячьих часто объединяют под общим названием мы-
шсподобные грызуны (Падсем. Muroidea).
2* 19
Сем. бобры (Castoridae). Европа, Азия, Северная Америка.
Евразиатский бобр (Castor fiber), американский бобр (С.
canadensis).
Сем. дикобразы (Hystricidae). Передняя, Средняя и Южная Азия,
Африка, Южная Европа.
Азиатский дикобраз (Hystrix leucura), африканский дико-
браз (Atherurus africanus).
Сем. иглошерсты, или дикобразы Нового Света (Erethizontidae).
Северная и Южная Америка.
Erethizon dorsatum, Coendou prehensilis.
Сем. трионо.мииды, или тростниковые крысы (Thryonomyidae).
Африка.
Thryonomys swinderianus, Choeromys gregorianus.
Сем. горные мыши (Petromyidae). Африка.
Pet готу s typicus.
Сем. пескорои (Bathyergidae). Африка.'
Bathyergus suillus, Heterocephalus glaber.
Сем. слепыши (Spalacidae). Европа, Азия, Африка.
Обыкновенный слепыш (Spalax microphlhalmus), гигант-
ский слепыш (S. giganteus).
Сем. ризомииды (Rhizomyidae). Индо-Малайская область.
Rhizomys sumatrensis, Tachyoryctes cheesmani.
Сем. морские свинки (Caviidae). Южная Америка.
Морская свинка (Cavia porcellus), патагонская свинка
(Dolichotis patagonica).
Сем. водосвинки (Hydrochoeridae). Центр, и Южная Америка.
Единственный вид — капибара (Hydrochoerus hydrochoerus).
Сем. паки (Cunicuiidae). Южная Америка.
Пака (Cuniculus раса), горная пака (Stictomys taczanowskii).
Сем. шиншиллы (Chinchillidae). Южная Америка.
Шиншилла (Chinchilla laniger) ,BucKama(Lagidium peruanum).
Сем. диномииды, или длиннохвостые паки (Dinomyidae). Южная
Америка.
Dinomys brannickii.
Сем. нутриевые (Capromyidae). Южная Америка.
Нутрия (Myocastor coy pus)', ежовая крыса (Caprotnys
pilorides).
Сем. цепкохвостые щетинистые крысы (Echimyidae). Южная и
Центральная Америка.
Echimys armatus, Thrinacodus albicauda.
Сем.’ктеномииды, или тукотуко (Ctenomyidae). Южная Америка.
Ctenomys magellanicus.
Сем. осьмизубые (Octodontidae). Южная Америка.
Octomys mimax, Octodon degas.
Сем. агути (Dasyproctidae). Южная Америка.
Dasyprocta aguti, Myoprocta acouchy.
1 Акклиматизирована в ряде стран Европы и в Северной Америке. ;
Когорта киты (Mutica)
Отряд китообразные (Cetacea)
Подотряд зубатые киты (Odontoceti)
Сем. речные дельфины (Platanistidae). Южная Америка, Южная
Азия.
Сусук (Platanista gangetica), иния (Inta amasonica), лап-
латский дельфин (Stenodelphis blainvillei).
Сем. настоящие дельфины (Delphinidae). Космополиты.
Дельфин-белобочка (Delphinus delphis), афалина (Tursiops
truncatus), морская свинья (Phocaena phocaena), гринда
(Globicephalus melasl), косатка (Orcinus orca), белуха (Del-
phinapterus leucas), нарвал (Monodon monoceros).
Сем. клюворылы (Ziphiidae). Космополиты.
Настоящий клюворыл (Ziphius cavirostris), атлантический
ремнезуб (Mesoplodon bidens), северный плавун (Berardius
baird.it), бутылконос (Hyperoodon ampullatus).
Сем. кашалоты (Physeteridae). Космополиты.
Кашалот (Physeter catodon), карликовый кашалот (Kogia
breviceps).
Подотряд беззубые, или усатые киты (Mystacoceti)
Сем. гладкие киты (Balaenidae). Космополиты.
Гренландский кит (Balaena mysticetus), японский кит
(В. sieboldi), южный кит (В. australis).
Сем. серые киты (Eschrichtiidae). Северная часть Тихого океана.
Единственный вид — серый кит (Eschrichtius gibbosus).
Сем. полосатики (Balaenopteridae). Космополиты.
Синий кит (Balaenoptera musculus), финвал (В. physalus),
сейвал (В. borealis), горбач (Megaptera nodosa).
Когорта хищные и копытные (Ferungulata)
Надотряд хищные (Ferae)
Отряд хищные (Carnivora, или Fissipedia)
Сем. виверровые (Viverridae). Африка, Мадагаскар, Южная Азия,
Малайский архипелаг, Южная Европа.
Циветты (Viverra zibetha, Civettictes civetta, Viverricula
indica), генетта (Genetta genetta), мунго (Mungos mungo),
ихневмон (Herpestes ichneumon) \ суриката (Suricata su-
ricata), пагума (Paguma larvata), бинтуронг (Arctictis
binturong) , фосса (Fossa fossa), выдровая циветта (Супо-
gale bennettii).
1 Мунго и ихневмона часто объединяют под названием мангуст.
21
Сем. гиеновые (Hyaenidae). Африка, Азия, Юго-Восточная Евро-
па (Закавказье).
Пятнистая гиена (Crocuta crocuta), полосатая гиена (Hyaena
hyaena), гривистый волк (Proteles cristatus)1.
Сем. кошачьи (Felidae). Повсеместно, кроме Австралии, Антарк-
тиды и Мадагаскара.
Европейская лесная кошка (Felis silvestris), барс (F. раг-
dus), лев (F. 1ео), тигр (F. tigris), рысь (Lynx lynx), гепард
(Acinonyx jubatus).
Сем. собачьи (Canidae). Повсеместно, за исключением Антарк-
тиды, Новой Зеландии, Полинезии.
Волк (Cants lupus), шакал (С. aureus), лисица (Вulpes
vulpes), фенек (Fennecus zerda), песец (Alopex lagopus),
енотовидная собака (Nyctereutes procyonoides), азиатский
красный волк (Cyan alpinus), американский красный волк
(Cynogale venaticus), гиеновая собака (Lycaon pictus).
Сем. медведи (Ursidae). Европа, Азия, Северная Африка, Северная
и Южная Америка.
Бурый медведь (Ursus arctos), американский черный мед-
ведь-барибал (U. americanus), белый медведь (U. mari-
timus), губач (Melursus ursinus), бамбуковый медведь
(Ailuropus melanoleucus)1 2.
Сем. енотовые (Procyonidae). Северная и Южная Америка, Юж-
ная Азия.
Енот-полоскун (Procyon lotor), носуха (Nasia rufa), цепко-
хвостый медведь, или пото (Potos flavus), панда (Ailurus
fulgens).
Сем. куньи (Mustelidae). Повсеместно, кроме Австралии, Мадага-
скара и Антарктиды.
Горностай (Mustela erminea), лесная куница (Martes
martes), гризон (Grison vittatus), зорилла (Zorilla zorilla),
росомаха (Gulo gulo), медоед (Mellivora capensis), обыкно-
венный барсук (Meles meles), ошейниковый барсук (Arc-
tonyx collaris),. американский барсук (Taxidea taxus),
обыкновенный скунс (Mephitis mephitis), инка (Conepatus
inca), речная выдра (Lutra lutra), калан, морская выдра
(Enhydra lutris), африканская выдра (Aonyx capensis)
Отряд ластоногиг (Pinnipedia)
Сем.’ушастые тюлени (Otariidae). Тихий океан, воды Антарктики.
Морской котик (Callorhinus ursinus), сивуч (Eumetopias.
jubatus), морской лев (Zalophus califomianus).
1 Некоторые зоологи относят к виверровым.
2 Этот вид некоторые зоологи относят к семейству енотов.
22
< ’.ем. моржи (Odobaenidae). Арктические моря.
Единственный вид — морж (Odobaenus rosmarus).
Еем. настоящие тюлени (Phocidae). Космополиты.
Обыкновенный тюлень (Phoca vitulind), гренландский тю-
лень (Pagophoca groenlandica), полосатый тюлень (His-
triophoca fasciatd), морской заяц (Erignathus barbatus),
белобрюхий тюлень (Monachus monachus), тюлень Уэддел-
ла (Leptonichoetes weddelli), тюлень-крабоед (Lobodon car-
cinophagd). хохлач (Cystophora cristata), морской слон
(Macrorhinus leoninus).
Надотряд первичные копытные (Protungulata)
Отряд трубкозубы (Tubulidentata)
Сем. трубкозубы (Orycteropidae). Южная Африка.
Единственный вид — трубкозуб (Orycteropus afer).
Надотряд слонообразные (Paenangalata)
Отряд хоботные (Proboscidea)
Сем. слоны (Elephantidae). Юго-Восточная Азия, Африка, Сула-
веси, Суматра.
Индийский слон (Elephas maximus), африканские слоны
(Loxodonta africana, L. cyclotis).
Отряд даманы, или жиряки (Hyracoidea)
Сем. даманы (Procaviidae). Юго-Западная Азия, Африка.
Абиссинский даман (Procavia. habessinicd), древесный даман
(Dendrohyrax dorsalis).
Отряд сирены (Sirenia)
Сем. ламантины (Manatidae). Прибрежные воды тропического и
субтропического поясов Атлантического океана и соот-
ветствующие реки.
Африканский ламантин (Manatus senegalensis), американ-
ские ламантины (М. manatus, М. inunguis).
Сем. дюгони (Dugongidae). Прибрежные воды тропического и
субтропического поясов Индийского океана, Красное море.
Индийский дюгонь (Dugong dugori), австралийский дюгонь
(D. australe), красноморский дюгонь (D. hemprichi).
23
Надотряд непарнокопытные (Mesaxonia)
Отряд непарнокопытные (Perissodactyla)
Подотряд лошадеобразные (Hippomorpha)
Сем. лошади ( Equidae). Юго-Западная и Центральная Азия,
Африка.
Дикая лошадь Пржевальского (Equus przewalskii), зебры
(Е. zebra, Е. grevyi), кулан, или полуосел (Е. hemionus),
ослы (Е. africanus, Е. somaliensis).
Подотряд тапирообразные (Ceratomorpha)
Сем. тапиры (Tapiridae). Центральная и Южная Америка, Юго-
Восточная Азия.
Американские тапиры (Tapirus terrestris, Т. pinchaque),
индийский, или чепрачный тапир (Tapirus indicus).
Сем. носороги (Rhinocerotidae). Юго-Восточная Азия, Африка.
Азиатские однорогие носороги (Rhinoceros unicornis, R.
sondiacus), азиатский двурогий носорог (Didermoceros su-
matraensis), африканские черный и белый носороги (Diceros
bicornis, D. sitnus).
Надотряд парнокопытные (Paraxonia)
Отряд парнокопытные (Artiodactyla)
Подотряд нежвачные (Suiformes)
Сем. свиные (Suidae). Все материки, кроме Австралии и Антарк-
тиды.
Дикий кабан (Sus scrofa), речная свинья (Potamochoerus
porcus), бабирусса (Babirussa babirussa), бородавочник
(Phacochoerus aethiopicus), пекари (Dicotyles tajacu, D. la-
biatus)1.
Сем. бегемотовые (Hippopotamidae). Африка.
Бегемот (Hippopotamus amphibius), карликовый бегемот
(C hoe гор sis liber iensi s').
Подотряд мозоленогие (Tylopoda)
Сем. верблюдовые (Camelidae). Европа, Азия, Африка, Юж-
ная Америка (в большинстве одомашненные животные).
Одногорбый верблюд (Camelus dromedarius), двугорбый
верблюд (С. bactrianus), гуанако (Lama guanicoe), викунья
(L. vicugna).
1 Некоторые систематики выделяют пекари в особое семейство.
24
Подотряд жвачные ( ghuminantia)
Сем. оленьки (Tragulidae). Южная Азия, Африка.
Яванский оленек (Tragulus javanicus), африканский водя-
ной оленек (Hyemoschus aquaticus).
Сем. олени (Cervidae)1. Европа, Азия, Америка, Северная Африка.
Кабарга (Moschus moschiferus), мунтжаки (Muntjacus
muntjak, М. lacrimans), хохлатый олень (Elaphodus се-
phalophus), водяной олень (Hydropotes inermis), пятнистый
олень (Cervus nippori), настоящий, или благородный олень
(С. elaphus), лань (С. dama), олень Давида (Elaphurus
davidianus), косуля (Capreolus capreolus), лось (Alces alces),
м аз а мы (Mazama americana, М. rufina), пуду (Pudu pud и),
пампасовые олени (Odocoileus virginianus, О. hemionus),.
северный олень (Rangifer tarandus).
Сем. вилорогие антилопы (Antilocapridae). Северная Америка.
Единственный вид — вилорог (Antilocapra americana).
Сем. быки (Bovidae). Европа, Азия, Африка, Северная Америка..
Джейран .(Gazelid subgutturosa), ориби (Ourebia moniana),
болотные козлы (Adenota lechae, Kobus defossa), хохлатые-
антилопы, или дукеры (Cephalophus ogillyi, С. grimi),
лошадиные антилопы (Hippotragus equinus, Н. niger),
сернобыки (Oryx beisa, О. gazella), бубал (Alcelaphus lich-
tensteini), белохвостый гну (Connochaetes gnu), сибирский
горный козел (Capra sibirica), кубанский тур (C. caucasica),
винторогий козел (С. falconeri), полукозел-тар (Hemi-
tragus jemlahicus), снежный баран (Ovis canadensis), серна
(Rupicapra rupicapra), горал (Nemorhaedus caudatus),
мускусный бык, или овцебык (Ovibos moschatus), сайга
(Saiga tatarica), оленебык-канна (Taurotragus oryx), лесные
антилопы (Tragelaphus euryceros, T. angasi), четырехрогая
антилопа (Tetracerus quadricornis), нильгау (Boselaphus-
tragocamelus), африканские буйволы (Syncerus nanus, S.
caffer), азиатский буйвол (Bubalus bubalus), бантенг (Bibos
banteng), американский бизон (Bison bison), зубр (В. bo-
nasus), як (Poephagus mutus).
Сем. жирафы (Giraffidae). Африка.
Обыкновенная жирафа (Giraffa Camelopardalis), сетчатая
жирафа (G. reticulata), окапи (Okapia johnstoni).
1 Оленьков и оленей иногда выделяют в надсемейство плотнорогих.
(Cervoidea) в отличие от вилорогих антилоп и быков, относимых к надсе-
мейству полорогих (Bovoidea).
Происхождение современного состава
класса млекопитающих
Палеонтологические данные и морфологические особенности
современных млекопитающих с полной определенностью говорят
о их происхождении от пресмыкающихся. Предками млекопи-
тающих считают очень древних рептилий (рис. 1), не утратив-
ших еще черт строения амфибий. Иначе, непонятным было бы
.наличие амфибийных признаков у млекопитающих (см. стр. 35).
К таким рептилиям принадлежала существовавшая в перм-
ском периоде и триасе группа, соединяемая в подкласс зверопо-
добных (Theromorpha). Общей особенностью этого подкласса
было наличие в черепе одинарных нижних височных ям — при-
знак группы синапсид, к которой принадлежат .и млекопитаю-
щие. В процессе эволюции у звероподобных развились некото-
рые новые черты, сближающие их с млекопитающими. Особенно
близок к последним отряд зверозубых (Theriodontia), имевший
дифференцированную зубную систему, сильно редуцированную,
квадратную кость, увеличенную зубную кость, сходное с млеко-
питающими деление позвоночника, строение поясов конечностей
и самих конечностей.
Высшие зверозубые были крупными животными. Так, на-
пример, иностранцевия из пермских отложений СССР достигала
3 м длины, циногнат из триасовых отложений Южной Африки
имел в длину около 2 м. Высокая специализация уводит такие
формы более или менее далеко в сторону от родословной линии
млекопитающих.
Мелкие размеры древнейших млекопитающих и особенности
их организации дают основание искать предков зверей среди
-более примитивных и мелких представителей зверозубых. Такие
формы также имелись в перми и триасе. Например, пермоцино-
дон из верхнепермских континентальных отложений СССР пред-
ставлял собой небольшое животное с черепом и зубной систе-
мой, напоминавшими собачьи. Примитивные признаки древней-
ших рептилий сочетались в этом животном с особенностями,
напоминающими млекопитающих. Другой представитель прими-
тивных зверозубых — иктидозавр из верхнего триаса Южной
Африки не превышал по размерам крысу. Череп (особенно вто-
ричное твердое небо), а также остальной скелет этого животного,
насколько можно судить по незначительным сохранившимся
•остаткам, больше напоминают скелет млекопитающих, чем реп-
-.26
Рис. 1. Филогения млекопитающих
тилий (собственно рептильные признаки у иктидозавра ограни-
чиваются только сложностью строения нижней челюсти).
Иктидозавры, как и некоторые другие триасовые териодон-
ты, были очень близки к древним млекопитающим, но кто имен-
но из них был предком последних — пока не ясно. Несомненно,
лишь, что исходной группой для млекопитающих послужили зве-
розубые!.
В связи с изменением условий в течение триасового периода
группа звероподобных пресмыкающихся быстро сокращалась в.
количестве и, дав начало млекопитающим, исчезла к концу это-
го периода.
Находки остатков древнейших млекопитающих относятся к
юрским отложениям. Наиболее примитивными формами были
многобугорчатые (Multituberculata), названные так по нали-
чию у них большого числа бугорков на коренных зубах1 2. Так
как уже при первом своем появлении многобугорчатые обладали
высокой специализацией и были представлены разнообразными
формами, можно полагать, что они возникли еще в триасе.
Особенностями зубной системы многобугорчатые приближа-
лись к грызунам: клыки у них отсутствовали и резцы отделялись
от коренных зубов диастемой. С другой стороны, коренные зубы
этих древнейших млекопитающих приближались по строению к
зачаточным зубам утконосов.
Многобугорчатые были растительноядными, вероятно, яйце-
кладущими, сравнительно мелкими (не крупнее бобра) живот-
ными.
Из представителей этой группы наиболее древний — трити-
лодонт; известны также очень примитивный плагиаулакс из
верхней юры Европы, полимастодон и птилодус из палеоцена
Северной Америки.
Просуществовав длительное время — от триаса до нижнего
эоцена,— многобугорчатые вымерли, не оставив потомков. Не-
смотря на близость этой группы с клоачными, считать ее ис-
ходной для них нет достаточных оснований. Происхождение
крайне примитивного подкласса клоачных (Prototheria, Мопо-
tremata), дошедшего до нашего времени благодаря особо благо-
приятным условиям, сложившимся в Австралии (почти полное
1 Существует точка зрения, согласно которой различные группы млеко-
питающих возникли независимо друг от друга от разных групп рептилий.
Однако большинством палеозоологов эта полифилетическая гипотеза не под-
держивается. Весь палеонтологический материал, а также данные сравни-
тельной анатомии и эмбриологии, склоняют нас к признанию монофилети-
ческого происхождения млекопитающих, т. е. происхождения их от одной
группы рептилий.
2 Примитивность этой группы дала повод Л. Ш. Давиташвили (1949)
причислить ее к зверозубым рептилиям, однако, эта точка зрения большин-
ством палеонтологов не разделяется.
28
отсутствие на этом материке высших млекопитающих), пока еще
не выяснено. Ископаемые остатки этих животных известны толь-
ко из четвертичных отложений, причем имеющиеся формы мало
отличаются от современных.
Возникшие в юрском периоде другие древние ветви млеко-
питающих— трехконусозубые, симметродонты и пантотерии—
были менее долговечны. Они существовали в течение средней
и верхней юры и полностью исчезли в нижнем мелу.
Трехконусозубые (Triconodontia) характеризовались нали-
чием на коренных зубах трех острых конических бугорков по-
чти одинаковой высоты, расположенных в ряд. Это были, по-ви-
димому, хищные животные, размером не более кошки, например
триконодои из верхнеюрских отложений Англии.
Симметродонты (Symmetrodontia) также имели трехконус-
ные зубы, но средний конус у них был значительно выше двух
боковых и лежал не в одной плоскости с ними; вместе конусы
образовывали симметричный треугольник. Симметродонты до-
стигали средних размеров и, вероятно, были хищниками. Типич-
ные представители этой группы — тинодон из верхней юры Се-
верной Америки, спалакотерий из юрских отложений Европы
и др.
Что касается пантотериев, или трехбугорчатых (Pantotheria,
или Trituberculata), то они представляют собой наиболее разно-
образную и интересную группу юрских млекопитающих, открыв-
ших настоящую историю класса. Это были мелкие зверьки, близ-
кие к современным насекомоядным. Коренные зубы их имели
три тупых бугорка, расположенных асимметричным треугольни-
ком, и, в нижней челюсти, низкую «пятку». Такая структура зу-
бов указывает на питание частью животными, частью расте-
ниями. Известно много форм пантотериев, наиболее характерны
архаичный амфитерий из верхней юры Англии и более прогрес-
сивный меланодон из тех же отложений Сев. Америки.
Пантотерии'—наиболее вероятные предки сумчатых и пла-
центарных.
Сумчатые (Metatheria, Marsupialia) появились, по-видимому,
в нижнем мелу; они’были представлены мелкими формами (типа
опоссумов), остатки которых известны из мела Северной Аме-
рики и из нижнетретичных отложений Америки и Евразии. Это
были всеядные или плотоядные зверьки. В миоцене появляются
также более или менее крупные хищные сумчатые, а в плейсто-
цене Австралии — даже гигантские растительноядные формы
как, например, дипротодон, череп которого достигал длины око-
ло 1 м.
Родиной сумчатых были Евразия и Северная Америка, но
под прямым или косвенным давлением заселивших эти конти-
ненты высокоорганизованных плацентарных большинство древ-
них сумчатых вымерло здесь еще до конца третичного периода.
29
Только в Австралии (куда плацентарные не проникли) сумча-
тые получили возможность процветать в той же мере, в какой
процветали плацентарные на других материках.
Высшие звери, или плацентарные (Eutheria, Placentalia)
также возникли в меловом периоде. Центром их развития была
Азия, откуда они расселялись в разных направлениях, особенно
на запад.
Как можно судить на основании ископаемых остатков, мле-
копитающие мелового периода были близки к пантотериям и
представляли собой маленьких, частью насекомоядных, частью
хищных древесных животных. Однако, несмотря на мелкие раз-
меры, по развитию головного мозга древнейшие плацентарные-
стояли выше других современных им животных. С исчезнове-
нием крупных пресмыкающихся (в связи с резким ухудшением
климата в конце мезозоя) перед теплокровными живородя-
щими плацентарными открылась возможность быстрого разви-
тия, приведшего их к господству на земле. Окончательная смена
«века пресмыкающихся» «веком млекопитающих» произошла в.
третичном периоде именно за счет плацентарных, так как мно-
гобугорчатые к этому времени исчезли, а распространение сум-
чатых сократилось — они удержались, за небольшим исключе-
нием, только в Австралии.
Как уже указывалось, древнейшие млекопитающие принад-
лежали к насекомоядным. Таковы вымершие дельтатерии и ца-
ламбдалесты из верхнего мела Монголии и некоторые другие.
В палеоцене появились примитивные ежи, позднее — в олиго-
цене— кроты и землеройки, а в мио-плиоцене и другие совре-
менные насекомоядные (Insectivora). Древние насекомоядные
послужили исходной группой и для других отрядов плацен-
тарных.
Древесные насекомоядные, приспособившиеся к планирующе-
му и активному полету, дали начало шерстокрылам (Dermoptera)
и рукокрылым (Chiroptera). Развившаяся в сторону хищниче-
ского образа жизни ветвь примитивных насекомоядных дала в
палеоцене начало древним примитивным хищным — креодон-
там (Creodonta). Эта группа процветала на протяжении всего
эоцена, нО в конце олигоцена, когда хищнический способ пита-
ния сильно усложнился в связи со сменой медлительных траво-
ядных животных нижнетретичного времени значительно более
подвижными, креодонты были вытеснены новыми прогрессивны-
ми хищными (Carnivora). Из примитивных креодонтов известен
нижнеэоценовый трицентес Северной Америки, похожий на
хорька, но питавшийся в основном насекомыми. Прогрессивны-
ми чертами (главным образом относительно большим мозгом)
отличались куницеподобные миациды из эоцена Северной Аме-
рики и олигоцена Европы. Миациды имели уже настоящие хищ-
ные зубы; они, по-видимому, и дали начало новым хищным.
30
Последние очень рано (на грани между эоценом и олигоценом)
расщепились на две неравные ветви: крупную — наземных хищ-
ных (Fissipedia) и небольшую — водных хищных, или ластоно-
гих (Pinnipedia).
В олигоцене уже существовали предки виверр, куниц, собак,,
кошек, а также своеобразные медведе-собаки (собачьего склада
животные с зубами типа медвежьих). Первые медведи, а также'
гпепы, появились в плиоцене. На протяжении всего мио-плиоце-
нового времени и в плейстоцене были многочисленны саблезу-
бые кошки — махайродусы, с огромными клыками. В плейсто-
цене были также представлены пещерные львы и многие хищные
современного типа.
Часть креодонтов, перейдя на растительное питание, дала е
палеоцене начало первичным копытным — так называемым кон-
дилартрам (Condylarthra).
Из примитивных представителей кондилартр хорошо изве-
стна палеоценовая эупротогония (Euprotogonia) и нижне-
эоценовый фенакод (Phenacodus) Северной Америки и Евро-
пы — довольно крупные пятипалые животные, сохранявшие еще
много черт хищных (большие клыки, когти). Эупротогония, воз-
можно, далекий предок современных непарнокопытных (Perisso-
dactyla).
Представленная в палеоцене своеобразная группа северо-
американских пятипалых копытных — амблипод (Amblipoda)
окончила свое существование в эоцене. Некоторые амблиподы
достигали огромных размеров и были вооружены тремя парами
рогов.
С эоцена в Северной Америке и Евразии развивалась ветвь
гитанотериев (Titanotheria)—гигантских животных с раздвоен-
ным рогом на носовой части черепа. В олигоцене эта ветвь пре-
кратила свое существование.
Собственно носороги того времени были совершенно лишены
рогов. Среди них также имелись гигантские формы (например,
индрикотерий высотой в плечах около 5,5 м). Но в плейстоцене
жил крупный двурогий носорог — эласмотерий, а в конце этой
же эпохи — шерстистый носорог с одним огромным рогом.
Богатую разнообразными формами группу составляли по-
явившиеся в эоцене парнокопытные (Artiodactyla). Из них изве-
стны эоценовые четырехпалые ореодонты, двупалые протило-
пусы, считающиеся предками мозоленогих, олигоценовые антра-
котерии, давшие, как полагают, начало бегемотам, гигантские-
свиньи — энтелодонты и предки современных полорогих и оле-
ней. От оленеобразных предков.в миоцене отделились первые
жирафы.
В эоцене появились первые хоботные (Proboscidea) — предки
слонов: меритерий и мастодонт, а в плейстоцене — мамонт.
зг
Из других копытных большую, но полностью вымершую в
голоцене группу составляли нотунгуляты, или южноамерикан-
ские, копытные (Notungulata).
По-видимому, от древних насекомоядных берет начало и об-
ширная, известная с эоцена группа грызунов (Glires, Rodentia).
Среди примитивных грызунов (Proglires) встречаются формы,
^„Растеризующиеся смешением черт грызунов и насекомоядных.
Большой интерес в этом отношении представляет вымершая
группа тиллодонтов (Tillodontia) из эоцена Северной Америки,
грызунонасекомоядные признаки которой не позволяют с уве-
ренностью указать ей определенное место1. В олигоцене суще-
ствовали уже настоящие белки, а также предки зайцев, дикоб-
разов, мышей; в миоцене — бобры, сони, тушканчики.
От древних же насекомоядных произошли неполнозубые
(Edentata, Xenarthra), трубкозубые (Tubulidentata) и далеко
пошедшие по линии приспособления к водной среде обитания
сирены (Sirenia) и китообразные (Cetacea). Разделение китов
на зубатых и беззубых произошло в мио-плиоценовое время 1 2.
Наконец, от древнейших насекомоядных, по-видимому, еще
•в верхнем мелу, отделилась ветвь примитивных полуобезьян
(Prosimiae). Первые представители этой группы, например пле-
зиадапиды палеоцена Европы, носили еще явственные черты
цзевкомоядных.
В палеоцене же появились предки долгопятов, причисляв-
шихся до последнего времени к. полуобезьянам, но сейчас сбли-
жаемых с обезьянами (Simiae). Примитивные обезьяны — апи-
диумы и парапитеки, а также предки человекообразных обезьян
пропли°питеки> известны в олигоцене. От последних берет нача-
ло и ветвь людей (Hominidae).
Зольшое значение для понимания закономерностей эволю-
ционного процесса представляет изучение эволюционных рядов
-отдельных групп животных, палеонтологические остатки кото-
рых дают достаточный для этого материал. Классическими при-
мерами этого рода служат эволюционные ряды лошадей и
слонов.
Эволюция лошадей шла по пути приспособления к быстрому
„бегу и питанию грубыми травянистыми кормами. Такими при-
1 А. Ш. Ромер (1935) причисляет тиллодонтов к насекомоядным,
д В. Павлова (1929) —к грызунам.
2 Многие авторы считают, что киты произошли от креодонтов. Древние
примитивные киты действительно по некоторым признакам приближаются
•к древним хищным, но, с другой стороны, они обладают также некоторыми
чертами сходства с древними копытными. Поэтому в последнее время пред-
ков китообразных ищут среди очень ранних плацентарных, предшествовав-
ших выделению ветвей хищных и копытных, т. е. среди древних насекомояд-
ных [Слипер (Slijper), 1936].
Существует также гипотеза о дифилетическом происхождении китообраз-
ных. Вопрос этот остается пока дискуссионным.
32
способлениями являлись: сокращение числа пальцев, увеличе-
ние общих размеров (благоприятствующее передвижению в гу-
стой растительности) и повышение коронок коренных зубов (в
противовес быстрому
их стиранию). Эти из-
менения с большой на-
глядностью прослежи-
ваются на ряде лоша-
дей — от нижпеэоценр-
ного эогиппуса, имев-
шего четырехпалые пе-
редние и трехпалые
задние конечности,
рост, не превышавший
рост лисицы, и низкие
(«брахиодонтные») ко-
ренные зубы,— до одно-
палой крупной совре-
менной лошади с ее
очень высокими («гип-
селодонтными») корен-
ными зубами *.
В эволюционном
ряду хоботных можно
проследить постепен-
ное общее увеличение
размеров тела, измене-
ние структуры зубов и
развитие бивней и хобо-
та. Начинается ряд —
небольшим, ростом со
свинью, эоценовым ме-
ритерием, лишенным
еще характерных внеш-
них слоновьих черт и
имеющим многочис-
ленные бугорчатые зу-
бы, и заканчивается —
огромным настоящим
слоном с сокращенным
количеством очень
Рис. 2. Эволюция титанотериев. А — Eoii'a-
nops (нижний эоцен); Б — Manteoceros (верх-
ний эоцен); В — Protitanotherium (верхний
эоцен); Г — Bronthotherium (нижний олиго-
цен) (по Осборну)
крупных складчатых
коренных зубов (до одного в каждой челюсти) и с сильно раз-
витыми бивнями и хоботом.
1 Выдающуюся роль в изучении истории лошадей сыграли исследования
русского палеозоолога. В. О. Ковалевского.
3 Териология
33
Упомянутые выше эволюционные ряды приводятся почти во
всех зоологических учебниках, поэтому подробнее на них можно
здесь не останавливаться. Более или менее хорошо изучены так-
же ряды последовательных форм носорогов, титанотериев и не-
которых других групп.
В эволюционном ряду титанотериев, например, насчитывает-
ся 6 звеньев, древнейшее из которых — нижнеэоценовый безро-
гий эотитанопс величиной с козу. Четвертое звено ряда — верх-,
неэоценовый протитанотерий имел уже крупные размеры и
заметный выступ на носовой части черепа. Наконец, представ-
лявший последнее звено олигоценовый бронтотерий достигал ги-
гантских размеров и был вооружен высоким раздвоенным носо-
вым рогом (рис. 2).
При оценке, с точки зрения эволюционных возможностей,
подклассов млекопитающих обращает на себя внимание общая
примитивность и особенно яйценоскость клоачных. Насиживание
яиц сильно ограничивало подвижность этих животных, а вме-
сте с тем и способность к прогрессивному развитию. Значитель-
но более благоприятным в этом отношении было положение
сумчатых и особенно плацентарных млекопитающих.
Общее направление эволюции млекопитающих выражалось
в повышении жизнедеятельности, выработке теплокровности,
живородности, в приспособлении к различным средам обитания
и различной пище (гетередонтность и пр.). Естественно, что эти
приобретения не могли бы появиться без прогрессивного раз-
вития центральной нервной системы и особенно — головного
мозга.
Очерк морфологии млекопитающих
ОБЩИЕ ОСОБЕННОСТИ ОРГАНИЗАЦИИ МЛЕКОПИТАЮЩИХ
Млекопитающие — наиболее высокоорганизованные позво-
ночные животные. Как высшим позвоночным им свойствен це-
лый ряд прогрессивных черт организации: высоко развитая нерв-
ная система, обусловливающая сложные формы поведения,
живорождение и выкармливание детенышей молоком, большая
интенсивность обмена веществ и совершенная терморегуляция,
обеспечивающая постоянство температуры тела (гомотерм-
ность). Для млекопитающих характерны: наличие млечных
желез *, волосяной покров (имеющийся хотя бы только в за-
родышевый период развития), непарная левая дуга аорты,
разделение грудной и брюшной полостей цельной сухожильно-
мускульной перегородкой — диафрагмой, дифференцированная
зубная система с двумя сменами зубов (молочными и постоян-
ными), сокращение костных элементов нижней челюсти до одной
зубной кости, превращение квадратной и сочленовной костей
в слуховые косточки (наковальню и молоточек), прямое при-
членение нижней челюсти к черепу (кчешуйчатой кости), большое
постоянство числа шейных позвонков, наличие в скелете гор-
тани надгортанника и щитовидного хряща, безъядерность крас-
ных кровяных телец и некоторые другие признаки.
От своих предков — древнейших примитивных амфибиеоб-
разных рептилий, млекопитающие унаследовали такие общие
с амфибиями черты, как сильное развитие кожных желез, рас-
положение сочленений в конечностях (между голенью и заплюс-
ной и между предплечьем и запястьем) и, частично, двойной за-
тылочной мыщелок1 2.
Как животные, освоившие все среды обитания и приспосо-
бившиеся к самым различным способам передвижения, млеко-
питающие обладают многообразными особенностями организа-
1 Эту особенность отражает и научное название класса — Mammalia
(mamma — млечная железа). Выкармливание детенышей молоком представ-
ляет прогрессивное явление, благодаря которому в самых различных усло-
виях среды развивающемуся организму обеспечивается полноценное питание.
2 Двойной затылочный мышелок относят иногда к унаследованным от
амфибий признакам млекопитающих, что, однако, неправильно. Парные заты-
лочные бугры, млекопитающих — вторичное образование, возникшее у неко-
торых древних рептилий путем расщепления первичного простого бугра.
3*
35
цни и внешнего облика. Среди них имеются как наиболее рас-
пространенные— наземные формы, так и подземные (землерои),
водные и воздушные (летающие).
Сильно варьируют также размеры млекопитающих: от самой
мелкой землеройки — белозубки-малютки с длиной тела (без
хвоста), не превышающей 4,5 см, весом 2 г, до самого крупного
кита — синего полосатика, достигающего длины более 30 м и
веса около 150 т
НАРУЖНЫЕ ПОКРОВЫ
Кожа. Кожа млекопитающих характеризуется толщиной и
сильным развитием желез и роговых образований. Она состоит
из двух хорошо обособленных слоев: сравнительно тонкого верх-
него эктодермального — эпидермиса, и более толстого нижнего
мезодермального — кориума (рис. 3).
Эпидермис (epidermis) представляет собой многослойную
эпителиальную ткань, состоящую в глубоких своих частях из ци-
линдрических или призматических живых размножающихся кле-
ток— так называемого мальпигиева слоя. Постепенно переме-
щаясь вверх, к поверхности кожи, эти клетки утрачивают ядра'
и способность делиться, приобретают все более уплощенную
форму и ороговевают, образуя мертвый наружный слой. Эти
клетки слущиваются с поверхности, заменяясь новыми из ниже
лежащих слоев.
Из эпидермиса развиваются волосы и другие роговые обра-
зования, а также кожные железы. Здесь же в основном сосре-
доточен пигмент, обусловливающий окраску кожи.
Роговой слой эпидермиса в значительной степени защищает
организм животного от действия вредных химических веществ,
препятствует проникновению болезнетворных микробов, а также
ограничивает испарение влаги и теплоотдачу.
Кориум, или собственно4 кожа (corium, cutis, derma)
состоит из волокнистой соединительной ткани. Основную толщу
его образует сетчатый слой, состоящий из перекрещивающихся
коллагеновых и эластических волокон, которые придают коже
большую прочность. Верхняя часть кориума — сосочковый слой,
вдается сосочками в мальпигиев слой эпидермиса, снабжая его
кровью (собственных кровеносных сосудов мальпигиев слой не
имеет). Книзу от сетчатого слоя лежит более рыхлый слой —
подкожная клетчатка, между волокнами которой имеются скоп-
ления жировых клеток. Последние образуют иногда мощный
жировой слой, предохраняющий тело от охлаждения и служа-
щий резервом питательного материала на время голодания.
Это соответствует, примерно, весу 30 слонов или 150 быков.
36
В целом кориум служит надежной защитой тела от механиче-.
ских повреждений Ч
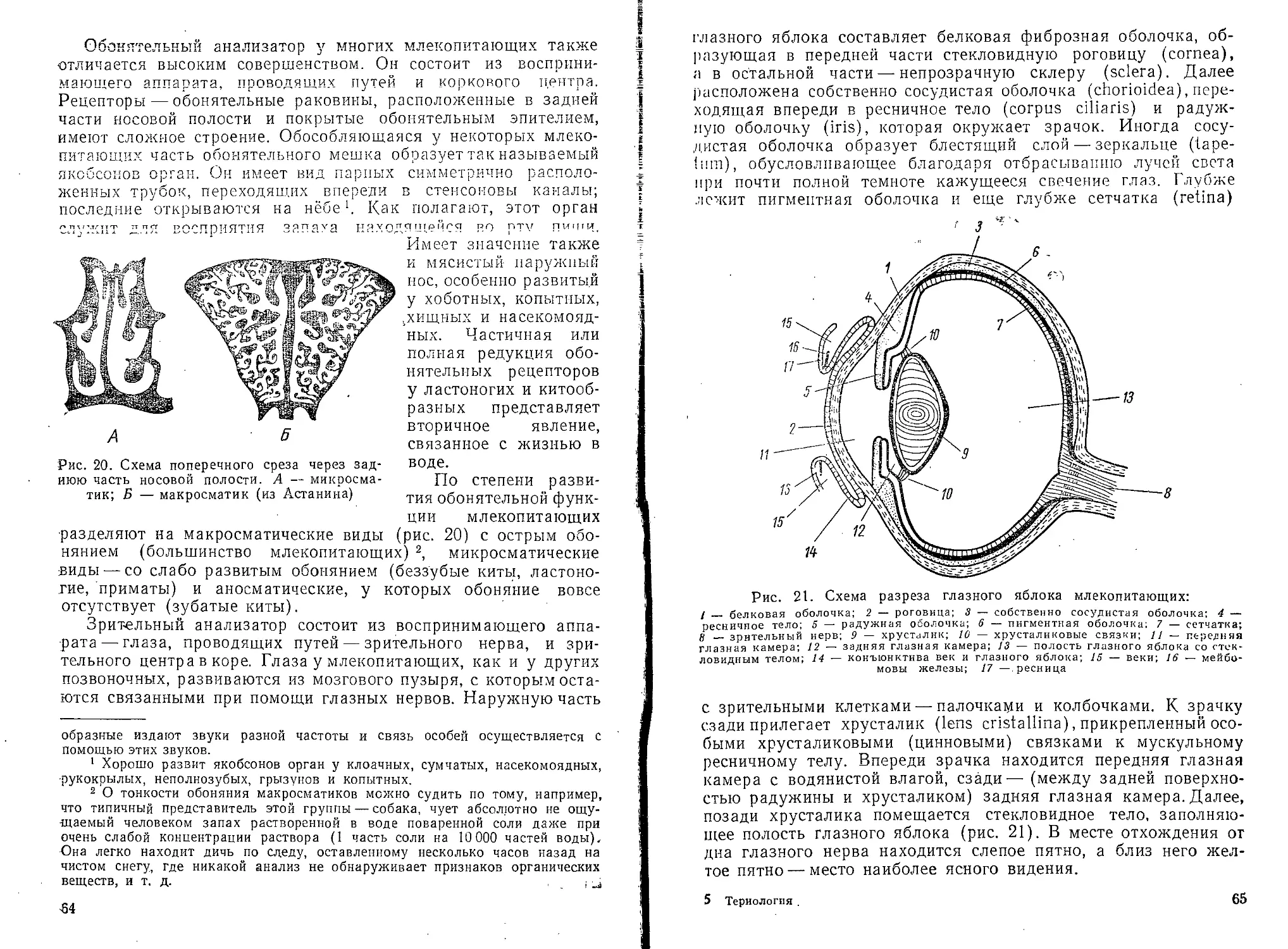
Волосы и их видоизменения. Волосяной покров — наиболее
характерное роговое эпидермическое образование млекопитаю-
щих. Отсутствие его у некоторых видов — вторичное явление.
Закладка волоса происходит путем энергичного деления
отдельных групп кле-
ток эпидермиса, глубоко
внедряющихся в толщу
кориума. Разрастаясь и
дифференцируясь, этот
зачаток дает начало осно-
ванию волоса — волося-
ной луковице, в дно кото-
рой вдается питающий
сосочек кориума с крове-
носными сосудами и нер-
вами— волосяной сосо-
чек. Размножение клеток
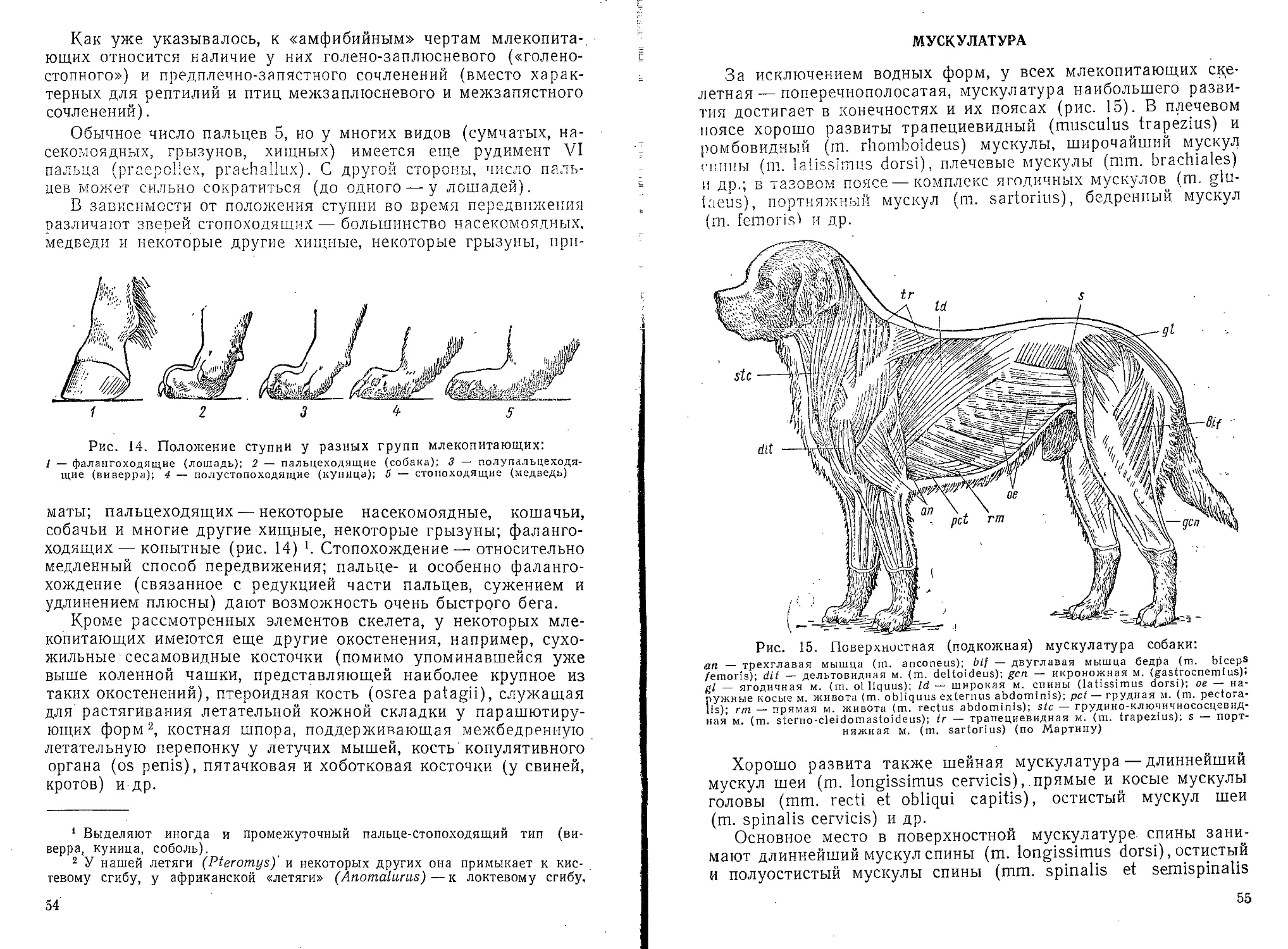
волосяной луковицы обус-
ловливает рост волоса
(рис. 4).
Волос состоит из стер-
жня. выдающегося над
Рис. 3. Вертикальный разрез через кожу
млекопитающего:
SC — роговой слой; SM — мальпигиев слой;
СО — кориум (собственно кожа); SF) потовые
железы; SD' — их выводные протоки; ср — со-
судистые сосочки; пр — нервные сосочки; Н —
волос; D — сальные железы; N — нервы; G —
сосуды; F — жировые клетки (по Видерсгейму)
поверхностью кожи, и
корня, лежащего в воло-
сяной сумке. Внутренняя
поверхность последней
образована волосяным
влагалищем (впячива-
пием мальпигиева слоя
эпидермиса), наружная — волосяным мешком (производным ко-
риума). Между тем и другим находится прозрачная стекловид-
ная оболочка.
1 Интересную особенность имеет кожа зайпев, характеризующаяся от-
носительной непрочностью и слабым развитием кровеносных сосудов. С этим
связана своеобразная автотомия, спасающая иногда жизнь грызуну. Схва-
тивший зайпа.за шкурку хищник легко вырывает из нее кусок, упуская са-
мую добычу. Полученная зайцем рана не кровоточит и быстро затяги-
вается.
Каудальная автотомия (подобная широко распространенной среди яще-
риц) наблюдается у некоторых, видов мышей, сонь и тушканчиков. Кожный
хвостовой футляр у них легко отрывается и соскальзывает с хвостовых по-
звонков, что дает возможность схваченному за хвост зверьку уйти от врага.
Установлено, что фхтляр разрывается непроизвольно в определенных участ-
ках, где кожа особенно тонка. Обнажившийся конец хвоста высыхает и
отпадает: у сонь после этого имеет место ограниченная регенерация, выра-
жающаяся в восстановлении поврежденной части позвонка или межпозво-
37
В волосяную сумку открывается проток сальной железы
(иногда 2—3 желез), выделения которой способствуют сохране-
нию эластичности волоса. От нижней части волосяной сумки
начинаются пучки гладких мышечных волокон, сокращением ко-
торых волос, имеющий обычно несколько наклонное положение,-
может быть выпрямлен. Под влиянием силы тяжести волос и
ворс имеют направление от спины к брюху. Исключение состав-
особенностями древесного
ляют ленивцы, у которых в связи с
Рис. 4. А, Б, В, Г — стадии развития волоса;
со — кориум; ер — эпидермис; f — зачаток влага-
лища волоса; g — зачаток, сальной железы; pl —
зачаток волоса (по Гессе)
образа жизни (посто-
янным пребыванием в
подвешенном состоя-
нии спиной вниз) ворс
имеет направление от
брюха к спине. У ти-
пичных землероев
- (кротов, слепышей и
др.) волосяной покров
очень низкий, бархати-
стый, что дает возмож-
ность зверькам быстро
двигаться в узких под-
земных ходах как впе-
ред, так и назад (не
поворачиваясь) *.
Волос млекопитаю-
щих— сложное много-
клеточное образование,
что отличает его от
«волоса» беспозвоноч-
ных животных, пред-
ставляющего собой кутикулярный вырост одной клетки. Цент-
ральную часть волоса занимает сердцевинный слой, состоящий
из кубических ороговевших клеток, которые разделены воздуш-
ными промежутками (рис. 5). В основании и вершине зрелого
волоса сердцевина вообще отсутствует и на ее месте образуется
канал.
Кнаружи от сердцевины расположен средний, корковый
слой — он состоит из вытянутых по оси волоса спаянных между
собой веретеновидных ороговевших клеток. Этот слой обуслов-
ливает крепость и упругость волоса. В клетках сердцевины и
ночного диска [Томас (Thomas), 1905; Гегль (Gogl), 1930; Гаффер (Gaffer),
1941].
' У других млекопитающих защитным приспособлением служит сильное
утолщение кожи в наиболее уязвимых местах; у кабана — на передней части
туловища, у моржа — на шее.
1 Эта способность доказана экспериментально. Естественно, что ее обус-
ловливают также соответствующее строение конечностей, пропорции тела
и т. д.
38
коркового слоя содержится пигмент, придающий волосам ту или
иную окраску.
Поверхность волоса покрыта тонким чешуйчатым слоем (ко-
жицей, «кутикулой»), образованным плоскими ороговевшими
клетками, которые либо опоясывают стержень волоса сплош-
ными кольцами, либо располагаются по поверхности частично
перекрывающими друг друга
шуйками. В соответ-
ствии с э'тим чешуйча-
тый слой называют
кольцевидным, черепи-
цеобразным или мо-
стовидным.
Чешуйчатый слой Л
защищает волос от раз-
ного рода физико-хи-
мических воздействий.
В зависимости от
или только соприкасающимися че-
гистологического стро-
ения, размеров и фор-
мы волосы млекопи-
тающих могут быть
разделены на две
основные категории:
ость и пух.
Остевые, или по-
кровные, волосы
(шерсть) характери-
зуются длинным тол-
стым, прямым или
слегка изогнутым стер-
жнем с хорошо разви-
той сердцевиной (со-
стоящей из нескольких
рядов клеток) и чере-
пипеобоазным типом
Рис. 5. А —типы строения кутикулярного
слоя волоса:
/—кольцевидный; 2 — черепнцеобразный; 3— мосте-
видный.
Б — схема строения стержня волоса:
/— чешуйчатый (кутикулярный) слой; 2 — корко-
вый слой: 3 — сердцевина с пигментными клет-
ками и воздушными полостями (по Кузнецову)
кожицы. Кроме многочисленных относительно коротких остевых
волос, у многих видов имеются более редкие, но длинные на-
правляющие волосы. Длинные толстые стержни их выдаются над
вершинами волос других категорий.
Пуховые волосы (подшерсток)—это более короткие, по
сравнению с остевыми, тонкие, обычно волнообразно изогнутые
волосы, лишенные сердцевины или со слабо развитой сердцеви-
ной (содержащей один ряд четкообразно расположенных кле-
ток); кожица у них кольцевидного типа.
Различают еще промежуточные волосы, занимающие про-
межуточное положение между остью и пухом.
39
Специальное назначение имеют сильно удлиненные волосы,
образующие хвостовой пучок (обмахивание тела), гриву, баки,
бороду (защита лицевых частей, шеи) и т. п.
Для большинства зверей характерна двуярусность меха, сла-
гающегося из шерсти и подшерстка Г Основное функциональное
значение шерсти — предохранение пуховых волос и кожи от раз-
личных внешних механических воздействий; подшерстка — вы-
полнение терморегуляционных функций.
У небольшой части млекопитающих волосяной покров преи-
мущественно или полностью состоит из волос лишь одной кате-
гории, например у взрослых настоящих тюленей и оленей —
в летнем покрове — из ости, у слепышей, кротов — почти исклю-
чительно из пуховых волос. Отсутствие ярусности (как и полная
безволосость)—результат редукции ранее существовавшего
полного волосяного покрова.
Независимо от принадлежности к той или иной категории
волосы весьма разнообразны как по форме стержня, так и по
характеру изогнутости последнего.
Волосы располагаются на теле либо равномерно поодиночке
(например, у лошади, коровы), либо более или менее сложными
группами (свиньи, хищные и др.). То или иное расположение
строго специфично для каждого вида млекопитающих и служит
хорошим систематическим признаком и показателем филогене-
тических отношений1 2.
В период утробного развития у всех млекопитающих имеется
сплошной эмбриональный пух (lanugo foetalis), который затем
сменяется постоянным волосяным покровом. Последний подвер-
жен у большинства зверей сезонным сменам в процессе так
называемой линьки.
Наибольшего развития волосяной покров достигает у зверей
полярной и бореальной областей; для этих животных его термо-
изолирующие свойства имеют особо важное значение.
Густой мех имеют некоторые тропические формы (обезья-
ны, ленивцы), но у них главное значение приобретает защита от
проливных дождей и укусов насекомых (безволосые тропиче-
ские звери — слоны, носороги, хорошо защищены от насекомых
грубой толстой кожей)3.
1 При наличии промежуточных волос возникает трехъярусность меха.
2 У зародышей многих млекопитающих зачатки волос располагаются по
краям зачатков чешуй. На основании этого полагают, что предшественником
волосяного покрова был чешуйчатый покров.
3 В зависимости от климатических условий, мест обитания и особенностей
образа жизни высота волос и степень опушенности отдельных участков тела
у разных видов зверей не одинакова. У белок и других древолазаюших
млекопитающих наиболее длинные волосы располагаются на спине, крестце
и хвосте, у настоящих норников — сусликов, сурков — на боках, у лисиц —
иа лопатках, у зайцев — на брюхе; у роющих видов длина волос по всему-
40
Видоизменения волос представляют вибриссы, щетины и
иглы. Вибриссы, или осязательные волосы,— очень круп-
ные, слабо изогнутые жесткие волосы с мостовидным типом
ci роения кожицы. Они расположены на голове (вокруг рта, на
щеках, подбородке, над глазами) и на других, наиболее подвер-
женных механическим воздействиям частях тела. Волосяные
мешки вибрисс головы иннервируются ветвями тройничного нер-
па и снабжены поперечнополосатыми мышцами. Окружающие
скрыта под панцирем:
Рис. 6. Броненосец, свернувшийся в шар
незащищенная часть
их венозные лакуны
служат, по-видимому,
амортизаторами сотря-
сений, передаваемых
стержнем (об осяза-
тельной функции виб-
рисс см. стр. 61).
Иглы образуют сво-
его рода колючий пан-
цирь; они характеризу-
ются мощным развити-
ем коркового слоя,
врастающего продоль-
ными перегородками в
сердцевину, прочно-
стью, большой упруго-
стью и очень острой
вершиной '.
Щетина — переход-
ный тип между волоса-
ми и иглами. Она обыч-
на у зверей, обладаю-
щих иглистым панцирем, наибольшего же развития достигает
у диких свиней и тенреков.
Другие роговые образования. Из других роговых образова-
ний млекопитающим свойственны чешуи, когти, рога, небные
ороговения, мозоли и челюстные чехлы.
Чешуи в качестве основного покрова развиты у яще-
ров и броненосцев (рис. 6), причем у последних они подстила-
ются еще костными пластинками. Многие млекопитающие име-
ют чешуйчатый покров на хвосте (бобр, выхухоль, мышевидные
грызуны, африканская колючехвостая белка, многие сумчатые),
лапах (различные землерои) и на других участках тела, подвер-
гающихся механическим раздражениям. Рудименты чешуй име-
телу одинакова. Имеются различия и в распределении плотности (густоты)
волосяного покрова (Церевитинов. 1958).
* Для дикобразов характерна легкость потери части игл хвоста, кото-
рым грызун ударяет нападающего. Это может рассматриваться как своего
рода автотомия.
41
ются также в области спинного плавника некоторых китообраз-
ных (морских свиней, гринд). Обычно чешуйчатый покров соче-
тается у млекопитающих с волосами.
Тщательное изучение кожных покровов дает основание пола-
гать, что чешуевидные образования в коже млекопитающих
представляют собой не остатки чешуйчатого покрова рептиль-
ных предков, а вторичные образования, возникшие из чешуе-
видных эмбриональных зачатков, встречающихся у зародышей
млекопитающих. У большинства видов чешуевидные образова-
ния с возрастом совершенно сглаживаются и лишь у некоторых
продолжают развиваться дальше на отдельных участках тела,
где они играют роль полезных приспособлений (Матвеев, 1949).
Когти и их видоизменения — ногти, копыта, защищают ко-
нечные фаланги пальцев млекопитающих от повреждений, слу-
жат орудиями нападения и защиты, рытья, лазания. Коготь
-состоит из плотной роговой когтевой пластинки, прикрывающей
последнюю фалангу сверху и с боков, и из более рыхлой подош-
венной пластинки, граничащей с мягкой подушечкой пальца *.
У некоторых зверей отдельные когти приобретают специфи-
ческое назначение и изменяют соответственно свой внешний вид.
'Таковы, например, служащие в первую очередь для выжимания
воды из ости, а также для смазывания меха жирным выделе-
нием анальных желез и для вычесывания эктопаразитов из меха,
раздвоенные клещеобразные когти бобров. У кенгуру такие ког-
ти несут только последнюю функцию (рис. 7).
Ноготь приматов отличается уплощением когтевой пластин-
ки, прикрывающей фалангу только сверху, значительной редук-
цией подошвенной пластинки и сильным развитием подушечки
пальца. Копыто, наоборот, имеет мощно развитые когтевую и
подошвенную пластинки и ороговевающую подушечку пальца
(«стрелку»), которые вместе образуют как бы башмак, оде-
вающий со всех сторон концевую фалангу.
Рога характерны для многих копытных.Чисто эпидермаль-
ное образование представляет собой рог носорога (он состоит
как бы из склеенных в сплошную роговую массу волосовидных
1 Более медленное изнашивание когтевой пластинки по сравнению с по-
.дошвенной обусловливает сохранение остроты когтя. Особой остроты дости-
гают втяжные когти кошек (кроме гепарда)—это тонкое приспособление,
«облегчающее захват и удержание очень подвижной добычи. Нарастание
жоггя происходит от его основания за счет размножения клеток эпидермиса.
У многих видов когти подвержены сезонным изменениям. Так, у копыт-
ного лемминга (Dicrostonyx torquatus) летом когти тонки, зимой же вследст-
вие разрастания подошвенной пластинки и ороговения подушечки пальца
-они превращаются в род копытец (приспособление к разрыванию твердого
•снега); весной наросшая часть отпадает. Когти монгольской пищухи
(Ochotona pallasi), песпа и др. зимой длиннее и острее, чем летом. Значи-
тельным сезонным изменениям подвержены также копыта, особенно у се-
верного оленя (летом они менее развиты, чем,зимой, когда служат для рас-
капывания снега).
42
нитей, образующих прочный конус, возвышающийся над носо-
выми костями). У так называемых «полорогих» копытных рога
имеют вид полых роговых чехлов, сидящих на особых костных
стержнях, очень рано срастающихся с лобными костями. Рога
эти- медленно растут в своем основании и сохраняются пожиз-
ненно, за исключением антилопы-вилорога, у которой они перио-
дически сбрасываются. У жираф и оленей на костных стержнях
рогов развивается кожа с волосами. У первых она остается по-
жизненно, у вторых ежегодно отмирает и слущивается. Кост-
ные стержни у большинства видов оленей сильно разрастаются,
и взрослые животные имеют ветвистые рога *. В противополож-
ность полым, такие рога растут в верхней части. Нижняя их
Рис. 7. Задняя йога исполинского кенгуру (Mac-
ropus giganteus} с «чесальными» двойными когтями
на недоразвитых сращенных втором и третьем
пальцах (по Брему)
часть, окостеневая, образует плотный цилиндр — внутри него
находится губчатое костное вещество, пронизанное сосудами, по
которым кровь поступает в вершины рогов. По окончании ро-
ста и отпадания кожи рога представляют собой чисто костные
образования. Такие рога ежегодно сбрасываются при разруше-
нии узкого слоя костной ткани в их основании. Остающийся
костный зачаток покрывается кожей и затем продуцирует новый
рог.
Молодые мягкие, покрытые короткими волосами рога пят-
нистых оленей и маралов носят название пантов (рис. 8). Содер-
жащиеся в них тонизирующие вещества используются в меди-
цине для изготовления лечебного средства — пантокрина.
Многие виды пользуются рогами для защиты и нападения,
у ряда других, они служат «турнирным» оружием самцов (см.
«Размножение»).
1 Возможность определения возраста оленей по количеству отростков на
рогах весьма ограничена, так как абсолютной корреляции между этим пока-
зателем и количеством прожитых животным лет не существует (у старых
животных наблюдается не увеличение, а уменьшение количества отростков).
У многих полорогих возраст довольно хорошо определяется по количе-
ству годичных колец, хорошо выраженных вследствие замедления роста ро-
гов зимой.
43
Небные роговые образования представлены в
виде пластин («китовый ус» беззубых китов) или утолщений
небных валиков (сирены).
роста рогов алтайского
elaphus sibiricus). А, Б,
Рис. 8. Стадии
марала (Cervus
В,— 59, 82 и 105 дней после сбрасы-
вания (рис. Н. Н. Кондакова с фот.
В. И. Цалкина)
в
У некоторых млекопитающих на безволосых участках тела,
подвергающихся частому трению и давлению, развились в про-
' цессе эволюции более или менее значительные утолщения рого-
вого слоя эпидермиса — мозоли. Особенно мощного развития
они достигают на ногах и на груди верблюдов, на запястных
суставах свиней-бородавочников, и на седалищных поверхностях
некоторых обезьян.
<4
Челюстные роговые чехлы («клюв») функционально заме-
няют у клоачных отсутствующие зубы.
Кожные железы. Многоклеточные железы, свойственные
коже млекопитающих, бывают двух основных типов: трубчатые
и альвеолярные.
К первому типу относятся потовые железы, состоящие обыч-
но из штопорообразно изогнутой трубки, конец которой свернут
в клубочек, а выводное отверстие открывается па поверхность
кожи или в волосяную сумку. Секрет потовых желез — пот, со-
держит воду, некоторые соли и мочевину.
Обычно пот жидок и бесцветен, но у лошадей он содержит
много белка, свертывающегося на коже разгоряченного живот-
ного и образующего густую пену; у бегемотов пот слизистый и
окрашен в красный цвет. Красный пигмент содержится также
в секрете потовых желез у самцов исполинских кенгуру. У од-
ного из африканских видов антилоп пот имеет голубую окраску.
Играя частично роль органа выделения, потовые железы вы-
полняют также терморегуляторную функцию. Испаряясь при вы-
сокой температуре с поверхности кожи, пот охлаждает ее, что
способствует поддержанию на определенном уровне темпера-
туры тела.
У большинства млекопитающих потовые железы расположе-
ны равномерно по всему телу, но у некоторых грызунов и куниц
имеются лишь на подошвах ног. У кошек и у собак они развиты
крайне слабо, а у китообразных, сирен, кротов, ящеров и неко-
торых других вовсе отсутствуют (такие млекопитающие обла-
дают другими формами терморегуляции).
Ко второму типу — альвеолярных (гроздевидных) желез,
принадлежат сальные железы, связанные с волосяными сумка-
ми. Жирный секрет этих желез.— продукт распада клеток, слу-
жит смазкой, предохраняющей кожу и волосы от смачивания и
пересыхания, а стало быть и от преждевременного изнаши-
вания х.
Из видоизменений кожных желез наиболее характерны для
млекопитающих млечные железы. Хотя они и имеют гроздевид-
ное строение, но произошли от потовых желез. Это хорошо вид-
но на примере клоачных млекопитающих, млечные железы ко-
торых имеют трубчатое строение. Секрет млечных желез —
молоко, представляет собой, подобно секрету потовых желез,
продукт жизнедеятельности клеток, а не распада последних,
как это свойственно сальным железам. Наибольшего развития
млечные железы достигают в период беременности и выкармли-
вания молодняка.
1 Кроме того, потовые и сальные железы имеют важное биологическое
значение, придавая специфический запах животному и его следам, что об-
легчает общение между Особями одного вида.
45
Выводные протоки млечных желез открываются у клоачных
на брюшном млечном поле, у части сумчатых — на млечных по-
лях, погруженных в особые сосцовые сумки, выворачивающиеся
только во время сосания, у остальных млекопитающих — на по-
стоянных сосках. На последних выводные протоки открываются
Рис. 9. Расположение выводных протоков млечной
железы. А — на железистом поле у клоачных;
Б — на «истинном» соске; В — на «ложном» соске
живородящих млекопитающих
либо отдельными отверстиями («истинные соски» большинства
сумчатых, обезьян, грызунов), либо в общий выводной канал
(«ложные соски» копытных, хищных) (рис. 9).
У плодовитых форм соски располагаются в два ряда вдоль
всей брюшной поверхности тела. При частичной редукции их
числа соски могут занимать одно из следующих положений:
Рис. 10. Мускусные железы хвоста выхухоли (Desmana
moschata)
брюшное (абдоминальные соски многих форм), грудное (пекто-
ральные соски обезьян, сирен, слонов, большинства летучих мы-
шей), паховое (ингвинальные соски копытных, китообразных)..
У части полуобезьян, кроме пары грудных сосков, имеется пара
брюшных. У нутрий соски расположены не на брюшной линии,,
а высоко на боках тела, что принято связывать с выкармлива-
нием детенышей в условиях болотистых мест (самка может кор-
мить, будучи полупогружена в грязь или воду).
46
Количество сосков варьирует у разных видов от двух,
(обезьяны, большинство летучих мышей, многие копытные) до»
25 (тенрек, опоссум), причем у сумчатых расположение их мо-
жет быть непарным, у высших же млекопитающих—всегда,
парное.
Соски имеются как у самок, так и у самцов, но у последних:
они рудиментарны.
Видоизменениями потовых или сальных желез или комбина-
циями тех и других являются различные пахучие железы — спе-
цпфические органы, выделения
которых имеют множественные
функции: облегчают встречу
особей разного пола, использу-
ются' для «мечения» занятой
территории, служат средством
защиты от врагов. Таковы мус-
кусные железы кабарги, овце-
быка, землероек, выхухоли
(рис. 10), ондатры; каудаль-
ные, промежностные и аналь-
ные железы хищных; копытные
и зароговые железы коз, серн и
некоторых других парнокопыт-
ных; предглазничные железы
оленей и антилоп (рис. 11);
комплекс пахучих желез обезь-
ян; спинная железа пекари и
даманов; пекторальная — опос-
сума и пр.'. Исключительно
защитное значение имеют па-
хучие (анальные) железы не-
которых куньих (хорьки, нор-
ки, барсуки, вонючки). У скун-
са выделения этих желез на-
Рис. 11. Пахучие железы. А—пред-
глазничная железа четырехрогой.'
антилопы (Tetraceros quadricornis)'^
Б — межкопытная железа оленя-
СТОЛЬКО едки, что вызывают У мунтжака (Cervus muntjak) (по»,
человека, подвергшегося их Поккоку)
действию, тошноту, а иногда и
обморочное состояние (запах выделений отличается -к тому же-
чрезвычайной стойкостью)1 2.
1 У одних только жвачных насчитывается свыше десятка кожных железе
метатарзальные; тарзальные, карпальные, интердигитальные, копытные, пре-
орбитальные, ингвинальные, ретрокопнальные, окципитальные, каудальные^
препуциальные и некоторые другие. У сайгаков А. Мури (Murie, 1870) нашеда
до 24 различных кожных желез.
2 Биологически активные продукты внешней секреции животных, дейст-
пуюшие на других особей своего вида или иных видов, носят общее название'
телергонов. Вещества, привлекающие других особей своего вида, называются
чпагонами; защитные или отпугивающие вещества — амнионами.
СКЕЛЕТ
Млекопитающие, как и другие позвоночные, имеют внутрен-
ний скелет, выполняющий функции опоры тела, субстрата для
прикрепления мышц и защиты для многих внутренних орга-
нов. Наружный костный скелет в виде органа пассивной защи-
ты— панциря, имеется только у броненосцев (см. рис. 6).
Для скелета млекопитающих характерно слабое развитие
хряща, сохраняющегося у взрослых животных только в местах,
требующих особой гибкости (в позвоночнике, на концах ребер).
Количество костных элементов, составляющих скелет, сильно со-
кращено по сравнению с рептилиями. Кости преимущественно
«замещающие» — образовавшиеся за счет замещения первич-
ного хряща (часто их называют «хрящевыми» костями).
В меньшей степени представлены «покровные» кости — кожного
происхождения.
В черепе млекопитающих (рис. 12), как и у всех позвоноч-
ных, различают два отдела — невральный, вмещающий мозг и
важнейшие органы чувств, и висцеральный, охватывающий пе-
реднюю часть пищеварительного тракта.
Невральный отдел, или просто невральный (мозговой) че-
реп, характеризуется ложной платибазальностью, развившейся
на основе первично тропибазального черепа Ч
Характерно также образование костями черепной коробки
более или менее сложных комплексов, очень поздно срастаю-
щихся между собой. Это обусловливает возможность беспрепят-
ственного длительного роста головного мозга, а в последующем
прочность черепа.
В затылочной области неврального черепа млекопитающих
общие с рептилиями четыре затылочные кости сливаются в одну
затылочную кость (os occipitale), несущую парные затылочные
мыщелки (condyli occipitales), между которыми находится за-
тылочное отверстие (foramen occipitale magnum). Последнее у
высших млекопитающих смещено книзу, к основанию черепа.
В слуховой капсуле ушные кости рано срастаются в мощную
каменистую кость (os petrosum) — вместилище лабиринта внут-
реннего уха и барабанной полости (в последней помещаются
слуховые косточки). С каменистой костью тесно связана свой-
ственная только млекопитающим покровная барабанная кость
(os tympanicum) —производное угловой кости нижней челюсти.
У клоачных, сумчатых и многих насекомоядных барабанная
кость имеет вид кольца, на которое натянута барабанная пере-
1 В связи с сильным разрастанием мозговой коробки межглазничная
перегородка перви’’ного тпопибазального черепа млекопитающих в значи-
тельной степени редуцируется, а глазницы оттесняются в бока,- что и при-
дает черепу платибазальный характер.
48
попка. У грызунов, хищных и других высших зверей она обра-
зует наружную стенку барабанной полости, иногда в виде пузы-
рсобразного вздутия (bulla ossea). С каменистой же костью ча-
сто бываете сращена и другая покровная кость — чешуйчатая
Рис. 12. Схема строения черепа млекопитающих (покровные кости—светлые,
замещающие — темные):
1— решетчатая (обонятельная, ethmoideum) кость; 2 — глазнично-клиновидная
(orbitosphenoideum); <? — крылоклиновидная (alisphenoideum); 4 — передняя клино-
видная (praesphenoideum): 5 — основная клиновидная (basisphenoideum): 6 — ка-
менистая (petrosum, или perioticum); 7 — верхняя затылочная (supraoccipitale); 8 —
боковая затылочная (exoccipitale); 9 — основная затылочная (basioccipitale); 10 —
хрящ носовой перегородки; 11 — носовая (nasale); 12 — слезная (lacrymale);
13 —'лобная (frontale); 14 — теменная (parietale); 15 — межтеменная (interparfetale);
/5 — чешуйчатая (squamosum); 17 — сошник (vomer); 18 — небная (palatinum);
19 — крыловидная (pterygoideum); 20 — межчелюстная (praemaxillare); 21 — верх-
нечелюстная (maxillare); 22 — скуловая (jugale); 23 — зубная (dentale); 24 — стре-
мечко (stapes); 25 — наковальня (incus); 26 — молоточек (malleus); 27 — барабан-
ная (tympanicum); 28 — остатки гиоида (hyoideum); 29 — подъязычный аппарат,
Z— IV— остатки жаберных дуг: 30 — щитовидный хрящ; 31 — черпаловидный хрящ;
32 — перстневидный хрящ; 33 — шиловидный отросток (styloiueus); 34 — трахея
(по Хильцгеймеру)
(os squamosum), к суставной ямке которой причленяется ниж-
няя челюсть. Комплекс сросшихся каменистой, барабанной и
чешуйчатой костей известен под общим названием височной
кости (os temporale).
Под мозговой полостью и в области глазниц непарные основ-
ная и передняя клиновидные кости (ossa basi- и praesphenoide-
lim) срастаются с парными крылоклиновидными и глазоклино-
видными костями (ossa alt- и orbitosphenoideum), образующими
4 Териология
49
их характерные «отростки» В передней части глазницы заклады-
вается слезная кость (os lacrimale). Снаружи и снизу глазница
и височная область черепа ограничиваются скуловой дугой, ос-
новную часть которой образует скуловая кость (os jugale).
В обонятельной области развиваются решетчатая кость (os
ethmoideum) с ее сложными пластинчатыми выростами — обоня-
тельными раковинами (ethmoturbinalia) и срастающиеся с но-
совой и челюстной костями носовая и челюстная раковины
(naso- и maxilloturbinale). Снизу здесь находится непарный
сошник (vomer), принимающий участие в образовании носовой
перегородки.
В крыше черепа теменные кости (ossa parietalia) нередко сра-
стаются между собой. Между ними и затылочной костью разви-
вается свойственная только млекопитающим межтеменная кость
(os inf er parietale) —она служит как бы вставкой, увеличиваю-
щей емкость мозговой коробки. Лобные кости (ossa frontalia)
также могут срастаться между собой. Наконец, обонятельную
область прикрывают парные носовые кости (ossa nasalia).
В верхней части затылочной области у многих млекопитаю-
щих развивается поперечный затылочный гребень (crista occipi-
talis), а по средней линии теменной кости — продольный стре
ловидный гребень (crista sagittalis). Первый из названных греб-
ней служит для прикрепления затылочных, второй — жеватель-
ных мышц.
Висцеральный отдел черепа имеет ряд существенных особен-
ностей. Функцию верхней челюсти выполняют в основном верх-
нечелюстные кости (ossa maxillaria); межчелюстные кости (ossa
intermaxillaria) сильно редуцированы. Небные отростки верхне-
челюстных костей вместе с отростками небных образуют вторич-
ное твердое небо. Небные и крыловидные кости (ossa palatinum и
pterygoideum) занимают вертикальное положение, примыкая с
боков к хоанам. Проксимальная часть небно-квадратного хряща,
соответствующая квадратной кости других позвоночных, у мле-
копитающих сильно редуцируется, переходит в ушную область
и преобразуется во вторую слуховую косточку — наковальню
(incus).
Нижнюю челюсть составляет (с каждой стороны) одна лишь
зубная кость (os dentale). Проксимальная часть первичного
нижнечелюстного меккелева хряща дает начало третьей слухо-
вой косточке — молоточку (os malleus) с его отростком, кото-
рый рассматривается как производное сочленовой кости (os go-
niale). Угловая кость, как уже говорилось выше, превращается в
барабанную.
1 На внутренней поверхности basisphenoideum имеется характерное углуб-
ление— турецкое седло (sella turcica), вмещающее гипофиз.
50
Поскольку квадратная кость утратила функцию подвеска для
нижней челюсти, последняя сочленяется с черепом при помощи
челюстного сустава чешуйчатой кости.
В подъязычной дуге млекопитающих существенных измене-
ний по сравнению с рептилиями не происходит. Верхний эле-
мент дуги (hyomandibulare) дает первую слуховую косточку —
стремя (os stapes), нижний элемент (hyoidetim) образует пе-
Рис. 13. Скелет млекопитающего (собаки):
/ — шейные позвонки (vertebrae cervicales); 2 — грудные позвонки (v. thoracales);
3 — поясничные позв. (v. lumbales); 4 — крестцовые позв. (v. sacrales, или os. sacrum);
Г) — хвостовые позв. (v. caudales); а — атлант (atlas); е — эпистрофей (epistropheus);
tns — рукоятка груднны (manubrium sterni); cs — тело грудины (corpus sterni); со —
ребра (costae); sc ~ лопатка (scapula); fi — плечевая кость (humerus); г — лучевая
кость (radius); и — локтевая кость (ulna); са — запястье (carpus); тс — пястье (me-
tacarpus); d — фаланги пальцев (phalangae digiiorum); i — подвздошная кость (ileum)
р — лобковая кость (pubis); is — седалищная кость (ischium), f — бедренная кость
(femur), ра — коленная чашечка (patella), t — большая берцовая кость (tibia), fi —
малая берцовая кость (fibula), ta — заплюсна (tarsus), mt — плюсна (metatarsus)
редние рожки подъязычного аппарата и шиловидный отросток
височной кости (у орангутана и человека). Из первой жаберной
дуги возникают задние рожки подъязычной кости, а из остатков
копул — тело последней. Остатки остальных жаберных дуг об-
разуют скелет гортани.
Для позвоночника млекопитающих (рис. 13) характерно по-
стоянное число шейных позвонков — 7 (у длинношеей жирафы
4* 51
эти позвонки сильно удлинены, у лишенного шеи кита — упло-
щены и частично сливаются между собой, но и у той и у другого
число их равно 7). Исключение составляют ламантины и дву-
палые ленивцы с 6 шейными позвонками, ящеры — с 8 и трех-
палые ленивцы с 9 шейными позвонками.
Число грудных позвонков, сочленяющихся с ребрами, варьи-
рует от 9 до 25 (максимальное число свойственно ленивцам; v
большинства зверей насчитывается 12—13 грудных позвонков).
Поясничный отдел имеет от 2 до 9 позвонков (чаще 6—7),
крестцовый — от 1 до 10 позвонков (чаще 2—4), в значитель-
ной сражающихся между еибий, хвостовой — щ 3 до 49
(максимальное число у ящеров).
За исключением первых двух шейных позвонков (атласа и
эпистрофея), позвонки лишены свободных сочленений. Они при-
надлежат к платицельному типу (их тела имеют плоские сочле-
новные поверхности). Только у копытных они приближаются к
опистоцельному (выпукло-вогнутому) типу строения. В состав,
позвонка, кроме его тела, входят невральные дуги, образующие
в совокупности спинномозговой канал, верхние остистые и попе-
речные отростки. У некоторых китообразных в хвостовом отделе
имеются также нижние — гемальные, дуги. Между телами по-
звонков заложены соединительнотканные эластические прослой-
ки — мениски (внутри их, в виде студенистых ядер, сохраняются
остатки хорды).
У большинства видов наибольшей подвижностью обладают
шейный и хвостовой отделы позвоночника, менее подвижны по-
ясничный и грудной отделы и совершенно неподвижен крест-
цовый. Гибкость шеи связана с многообразием ее функций,
прежде всего с участием в захватывании пищи. Хвостовой от-
дел играет роль руля, служит для балансирования, опоры,
отпугивания насекомых, у водных форм — для передвижения.
При утрате этих функций хвост в большей или меньшей степени
редуцируется (многие приматы, медведи, зайцы, сеноставки,
слепцы и др.).
Ребра у млекопитающих полного развития достигают в груд-
ной области, где образуют грудную клетку, обеспечивающую
дыхательные движения и служащую защитой для органов груд-
ной полости. Связанные с остальными отделами позвоночника
ребра сильно редуцированы-(в хвостовом отделе они полностью
отсутствуют).
Истинные ребра, путем слияния своих дистальных концов,
дают начало грудной кости, или грудине (os sternum), в кото-
рой различают следующие друг за другом три отдела — руко-
ятку, тело и мечевидный отросток. У клоачных впереди руко-
ятки образуется еще надгрудинник — гомолог соответствующей
кости рептилий. Значительная часть ребер не достигает грудины
и оканчивается свободно (так называемые ложные ребра).
52
В плечевом поясе клоачных, кроме лопаток (scapulae) и
ключиц (claviculae), хорошо развиты как самостоятельные
кости коракоиды и прокоракоиды. У остальных млекопитающих
они редуцированы, причем рудименты коракоидов прирастают
к лопаткам, образуя их коракоидные отростки, а прокоракоиды
исчезают. Ключицы — кости кожного происхождения, также об-
наруживают тенденцию к редукции. Значительного развития они
достигают у форм, передние конечности которых приспособлены
к разнообразным движениям (обезьяны, рукокрылые, кроты), и
полностью редуцированы у форм с однообразными движениями
конечностей (копытные, некоторые хищные, китообразные, си-
рены). На латеральной поверхности лопатки проходит хорошо
развитый продольный гребень, продолжающийся в акромиаль-
ный отросток (acromion).
Тазовый пояс в эмбриональный период состоит из типичных
для этого отдела парных костей — подвздошной (os ileum), се-
далищной (os ischium) и лобковой (os pubis); позднее они сра-
стаются с каждой стороны в одну безымянную кость (innomina-
tum). Внизу правая и левая безымянные кости образуют между
собой сращение (symphysis). Кроме того, в вертлужной впа-
дине развивается небольшая вертлужная кость (os acetabulare).
У клоачных и сумчатых к лобковым костям причленяются так
называемые сумчатые кости (основная функция их — поддержа-
ние брюшной стенки тела).
Скелет конечностей у млекопитающих соответствует общей
схеме строения пятипалой конечности. В отличие от рептилий,
имеющих раздвинутые в стороны конечности, у млекопитающих
они подведены под туловище, что обеспечивает лучшую подвиж-
ность животного1. В передней конечности характерны: сильное
развитие плечевой кости (os humerus), образование проксималь-
ной частью локтевой кости (os ulna) большого выступа (ole-'
cranon), частичная редукция и срастание локтевой кости с луче--
вой (os radius), срастание между собой отдельных элементов
запястья (carpus) например, 4Ц-5 carpalia, centrale -ф radi-
ale, редукция или слияние между собой некоторых пястных
элементов (metacarpus). В задней конечности мощная бедренная
кость (os femur) несет на своем проксимальном конце 2—3 вы-
ступа (trochanteres) для прикрепления мышц; путем'небольшого
местного окостенения сухожилия, проходящего через коленный
сустав, развивается коленная чашка (patella). Малая берцовая
кость (os fibula) у некоторых видов редуцируется, прирастая к
большой берцовой (os tibia). В заплюсне (tarsus) происходит
слияние части элементов с образованием крупной таранной ко-
сти (os astragalus); сильного развития достигает пяточная кость
(os calcaneus).
1 Исключение составляют клоачные, имеющие «рептильную» постановку
конечностей.
53
Как уже указывалось, к «амфибийным» чертам млекопита-.
ющих относится наличие у них голено-заплюсневого («голено-
стопного») и предплечно-запястного сочленений (вместо харак-
терных для рептилий и птиц межзаплюсневого и межзапястного
сочленений).
Обычное число пальцев 5, но у многих видов (сумчатых, на-
секомоядных, грызунов, хищных) имеется еще рудимент VI
пальца (praepoHex, praehallux). С другой стороны, число паль-
цев может сильно сократиться (до одного — у лошадей).
В зависшмости от положения ступни во время передвижения
различают зверей стопоходящих — большинство насекомоядных,
медведи и некоторые другие хищные, некоторые грызуны, прп-
Рис. 14. Положение ступни у разных групп млекопитающих:
/ — фалангоходящие (лошадь); 2 — пальцеходящие (собака); 3 — полупальцеходя-
щие (виверра); 4 — полустопоходящие (куница); 5 — стопоходящие (медведь)
маты; пальцеходящих — некоторые насекомоядные, кошачьи,
собачьи и многие другие хищные, некоторые грызуны; фаланго-
ходящих — копытные (рис. 14) '. Стопохождение — относительно
медленный способ передвижения; пальце- и особенно фаланго-
хождение (связанное с редукцией части пальцев, сужением и
удлинением плюсны) дают возможность очень быстрого бега.
Кроме рассмотренных элементов скелета, у некоторых мле-
копитающих имеются еще другие окостенения, например, сухо-
жильные сесамовидные косточки (помимо упоминавшейся уже
выше коленной чашки, представляющей наиболее крупное из
таких окостенений), птероидная кость (osrea patagii), служащая
для растягивания летательной кожной складки у парашютиру-
ющих форм1 2, костная шпора, поддерживающая межбедренную
летательную перепонку у летучих мышей, кость'копулятивного
органа (os penis), пятачковая и хоботковая косточки (у свиней,
кротов) и др.
1 Выделяют иногда и промежуточный пальце-стопоходящий тип (ви-
верра, куница, соболь).
2 У нашей летяги (Pteromys)' и некоторых других она примыкает к кис-
тевому сгибу, у африканской «летяги» (Anotnalurus)—к локтевому сгибу,
54
МУСКУЛАТУРА
За исключением водных форм, у всех млекопитающих ске-
летная—поперечнополосатая, мускулатура наибольшего разви-
тия достигает в конечностях и их поясах (рис. 15). В плечевом
поясе хорошо развиты трапециевидный (musculus trapezius) и
ромбовидный (m. rhomboideus) мускулы, широчайший мускул
'.шины (т. latissimus dorsi), плечевые мускулы (mm. brachiales)
и др.; в тазовом поясе — комплекс ягодичных мускулов (т. giu-
laeus), портняжный мускул (т. sartorius), бедренный мускул
(т. femorisl и др.
Рис. 15. Поверхностная (подкожная) мускулатура собаки:
<ап — трехглавая мышца (m. anconeus); bif — двуглавая мышца бедра (m. biceps
/emoris); dit — дельтовидная м. (m. deltoideus); gen — икроножная м. (gastrocnemius);
gl — ягодичная м. (m. olliquus); Id — широкая м. спины (latissimus dorsi); oe — на-
ружные косые м. живота (m. obliquus externus abdominis); pct — грудная м. (m. pectora-
iis); rm — прямая м. живота (m. rectus abdominis); sic — грудино-ключнчнососцевнд-
ная м. (m. sterno-cleidomastoideus); tr — трапециевидная м. (m. trapezius); s — порт-
няжная м. (m. sartorius) (по Мартину)
Хорошо развита также шейная мускулатура — длиннейший
мускул шеи (m. longissimus cervicis), прямые и косые мускулы
головы (mm. recti et obliqui capitis), остистый мускул шеи
(т. spinalis cervicis) и др.
Основное место в поверхностной мускулатуре, спины зани-
мают длиннейший мускул спины (m. longissimus dorsi), остистый
и полуостистый мускулы спины (mm. spinalis et semispinalis
55
dorsi), хвостовая мышца (m. coccygeus) и др.; в брюшной и груд-
ной— комплекс брюшных мускулов (mm. rectus, transversus,
obliquus, abdominis), прямой грудной мускул (m. rectus thoracis)
И др.
В области головы хорошо развита жевательная мускулатура.
У растительноядных форм наиболее мощны собственно жева-
тельные мышцы (m. masseter), у плотоядных — «кусательные»—-
ВНСОч 1*1 Ыс МЫШЦЫ сЕШрОГЗпь) . kJiO/aliOn МУСКуЛЗТурОИ ООЛй*'
дают подвижные язык и губы. Характерна для млекопитающих
лицевая подкож’ТсШ мускулэтурй, богато развитая у приматов
(мимические мышцы). К лицевой мускулатуре относятся также
мышцы, приводящие в движение веки, ушные раковины, нос,
хоботок многих насекомоядных, хобот слона.
У млекопитающих развита также подкожная мускула-гура
туловища (особенно подвижна кожа у копытных). Особая коль-
цевая подкожная мускулатура свойственна свертывающимся
формам (ежам, броненосцам).
Специфична для млекопитающих сухожильно-мускульная
грудобрюшная преграда — диафрагма.
НЕРВНАЯ СИСТЕМА
Свойственное млекопитающим совершенство взаимосвязей
составных частей организма и организма с внешней средой обу-
словлено исключительно высоким развитием их нервной. систе-
мы. Как и у других позвоночных, у млекопитающих основной
структурный и функциональный элемент нервной системы
представлен нервной клеткой (нейрон,, или неврон) с отходя-
щими от нее отростками — нервными волокнами. Эти волокна
служат проводниками нервных импульсов от периферии к телу
нейрона (центростремительные, или афферентные волокна) и
от последнего к иннервируемому им органу или к другим ней-
ронам (центробежные, или эфферентные волокна).
Центральная часть нервной системы представлена головным-
и спинным мозгом.
Головной мозг подразделяется на 5 отделов. Для первого из
них — переднего, или большого мозга (telencephalon)—харак-
терны относительно крупные размеры и высокая дифференциа-
ция полушарий, пересеченных у высших зверей бороздами и из-
вилинами (рис. 16, 17) Г Кпереди большой мозг образует обоня-
тельные-луковицы (bulbi olfactorii), наиболее развитые у живот-
ных с хорошим обонянием и сильно редуцированные у чисто
водных форм (китов).
1 Вопреки широко распространенным представлениям, большое количе-
ство мозговых извилин не служит признаком высоких интеллектуальных
способностей животного. Такие крупные млекопитающие как киты и слоны
имеют больше мозговых извилин, чем человек. Наоборот, мозг некоторых
маленьких обезьян (уистити), по своему положению значительно ближе, стоя-
щих к человеку, имеет очень слабо развитые извилины или совсем лишен их.
56
полосатых тел (.corpora
f 6
Рис. 16. Схематизированный сагит-
тальный разрез головного мозга
млекопитающего:
1 — передний мозг; 2 — полосатое тело;
3 — гипофиз; 4 — промежуточный мозг;
5 — средний мозг; 6 — мозжечок; 7 —
варолиев мост; 8 — продолговатый мозг
В поверхностной части переднего мозга расположены тела
нейронов, образующие серое вещество мозга — кору больших,
полушарий. Это — центр высшей (условнорефлекторной) нерв-
ной деятельности, откуда посылаются импульсы к различным
органам и где осуществляется регуляция физиологических про-
цессов соответственно условиям среды. Кора полушарий пред-
ставляет собой в основном вторичный свод, или плащ (neopal-
ijiiiiiy, который iipoi 1са0дит из ззкзткз соответствует';? — ^ ere.?.?
рептилий и, разрастаясь, покрывает весь мозг. Первичный свод
(;ncljipaliiiiin) оттесняется при этом внутрь полушарий, образуя
складку — гиппокамп, или аммонов рог (hippocampus).
Серое вещество .имеется и в подкорковом слое, в виде отдель-
ных скоплении — ядер, напри
striata), связанных с автомати-
ческими движениями (безус-
ловными рефлексами).
<гая составная часть
большого мозга — белое веще-
ство, т. е. скопление нервных
волокон, связывающих между
собой нервные клетки. У выс-
ших зверей эти волокна обра-
зуют спайку полушарий — мо-
золистое тело (corpus callo-
sum). С помощью нервных во-
локон кора больших полуша-
рий связана с остальными от-
делами мозга, а через них и с
органами и тканями тела.
В полушариях имеются две
полости — правый и левый бо-.
ковые желудочки, соединяющиеся при помощи монроевых от-
верстий друг с другом и. с далее лежащим третьим желудочком.
За большим мозгом расположен промежуточный мозг
(diencephalon), главную часть которого составляют зрительные
бугры (thalami optici) — коллектор центростремительных воло-
кон, передающих импульсы от большинства воспринимающих
раздражения концевых образований — рецепторов. Над-
бугорная область промежуточного мозга несет эпифиз (epiphysis-
cerebri)—рудимент теменного органа рептилий, подбугорная —
воронку (infundibulum), составляющую дно третьего желудоч-
ка, и гипофиз (hypophysis cerebri). Последний представляет
важную железу внутренней секреции (что касается эпифиза,
то приписываемая ему секреторная функция, у млекопитающих
животных еще не полностью подтверждена).
За промежуточным мозгом следует средний мозг (mesen-
cephalon) с так называемым четверохолмием (corpora quadrige-
57
mina), передние бугры которого связаны с восприятием свето-
вых, а задние — слуховых раздражений. Желудочек этого от-
дела мозга представлен узким каналом — сильвиевым водо-
проводом.
Четвертый отдел мозга — мозжечок (cerebellum), имеет на
поверхности серое вещество, в которое вдается белое вещество
в виде разветвленного дерева, образуя так называемое «древо
Рис. 17. Головной мозг лошади:
1 — обонятельная луковица; 2 — передний мозг; 3 — промежуточный мозг; 4 —
зрительный нерв; 5 — воронка; 6 — гипофиз: 7 — варолиев мост; 8 — продолговатый
мозг; 9 — диагональная борозда; 10 — предсильвиева борозда; 11 — сильвиева
борозда; 12 — наружная сильвиева борозда: 13 — надсильвиева борозда: 14 —
боковая (латеральная) борозда; 15 — иаружно-боковая борозда; 16—17— базальная
борозда; 18 — мозжечок; 19 — сосудистое сплетение 4-го мозгового желудочка;
III— глазодвигательный нерв; V— тройничный нерв; VI— отводящий нерв; VII—
лицевой нерв; VIII— слуховой нерв (по Сиссон)
жизни» (arbor vitae). Мозжечок состоит из средней доли —
червячка и боковых долей — полушарий, связанных между со-
бой лежащей под ними комиссурой из нервных волокон.—
варолиевым мостом (pons Varolii). Основная функция мозжеч-
ка— координация движений и мышечного тонуса (регуляция
равновесия тела).
Пятый отдел — продолговатый мозг (myelencephalon) со-
держит ряд жизненно важных нервных центров и, в частности,
дыхательный центр. Желудочек продолговатого мозга (четвер-
тый) образует ромбовидную ямку и переходит далее в канал
спинного мозга.
На эволюцию головного мозга зверей в целом большое влия-
ние оказывает внешняя среда и двигательная — пищедобываю-
58
тая, оборонительная деятельность!. У одомашненных животных
-)ти виды деятельности почти отсутствуют, в связи с чем раз-
меры Их мозга уменьшаются (Дарвин, 1868; Мюллер, 1919 и
др.). Более, чем размеры, характеризует прогрессивное разви-
тие головного мозга млекопитающих закономерное и последо-
вательное повышение степени кортикализации больших полуша-
рии (образования коры).
Спинной мозг служит продолжением продолговатого, он
расположен в позвоночном канале и характеризуется у млеко-
питающих значительным укорочением. В поясничной или даже
грудной области позвоночника он уже заканчивается, а пучок
н<-рвов, отходящих от пего, вместе с конечной нитью самого спин-
ного мозга, образует «конский хвост» (catida equina). Как и в про-
долговатом мозгу, серое вещество находится в центре спинного
мозга, а белое — окружает его. В плечевой и поясничной обла-
стях спинной мозг образует утолщения, размеры которых явст-
венно связаны со степенью развития конечностей (например,
у кенгуру, имеющих мощные задние ноги, сильно развито пояс-
ничное утолщение, у летучих мышей, передвигающихся с по-
мощью передних конечностей,— плечевое)/. Функция спинного
мозга — осуществление простых безусловных рефлексов.
Периферическую часть нервной системы составляют «нервы»,
т. е. совокупности нервных волокон, окруженные соединитель-
нотканными оболочками. Различают головные (черепномозго-
вые) и спинномозговые нервы, содержащие и афферентные, и
эфферентные нервные волокна.
Головных нервов у млекопитающих 12 пар:
1) обонятельный нерв (n. olfactorius)—комплекс афферент-
ных волокон, идущих от обонятельного эпителия к обонятель-
ной луковице; 2) зрительный нерв (n. opticus) — афферентные
нервные волокна, отходящие от сетчатки и идущие по стебель-
ку глазного бокала в промежуточный мозг; 3) глазодвигатель-
ный нерв (п. oculomotorius)—отходит от среднего мозга и ин-
нервирует нижние косые, верхние,1 нижние и внутренние
прямые мышцы глаз, а также ресничную мышцу и мышцу раду-
жины; 4) блоковый нерв (n. trochlearis) —берет начало на гра-
нице между средним мозгом и мозжечком и иннервирует верх-
ние косые мышцы глаза; 5) тройничный нерв (n. trigeminus) —
сложный нерв, отходящий от продолговатого мозга и разде-
ляющийся на 3 ветви: глазничную (которая иннервирует кожу
передней части головы), верхнечелюстную и нижнечелюстную;
1 Развитие различных частей головного мозга определяется в основном
способами отыскивания пищи: у собаки, павиана, пользующихся в этом про-
цессе обонянием, больше развита обонятельная область; у кошки,' отыскиваю-
щей пищу с помощью зрения — зрительная; у макаки, пользующейся зрением
и слухом — зрительная и слуховая (Матвеев, , 1951; Л. Шапошников, 1953;
Садыков, 1961).
59
6) отводящий нерв (n. abdtlcens) — отходит от продолговатого
мозга и иннервирует наружную прямую мышцу глаза; 7) лицевой
нерв (п. facialis) —идет от продолговатого мозга и иннервирует
сильно развитую лицевую мускулатуру; 8). слуховой нерв,
(n. acusticus) — представляет собой обособившуюся часть лице-
вого нерва, распадающуюся на ветви во внутреннем ухе;
9) языкоглоточный нерв (n. glossopharyngeus)—отходит от
продолговатого мозга, служит главным вкусовым нервом;
10) блуждающий нерв (n. vagus) — отходит от продолговатого
мозга и иннервирует сердце, пищеварительные и дыхательные
органы; 11) добавочный нерв (n. accessorius)—обособившаяся
(только у млекопитающих) часть блуждающего нерва, иннер-
вирующая трапециевидную и грудино-ключичиососцевидные
мышцы; 12) подъязычный нерв (n. hypoglossus) — берет нача-
ло от задней части продолговатого мозга и иннервирует муску-
латуру языка.
Спинномозговые нервы отходят от боковых сторон спинного
мозга в каждом миомере двумя корешками: дорзальным — с
афферентными и вентральным — с эфферентными нервными во-
локнами. Соединяясь по выходе из позвоночного канала, они дают
с каждой стороны начало смешанному нерву. В организме мле-
копитающего насчитывается свыше 200 нервов и их ответвлений;
Всю нервную систему условно подразделяют на соматиче-
скую, иннервирующую мускулатуру и органы чувств, и вегета-
тивную, которая иннервирует внутренние органы (в состав по-
следней входит так называемая симпатическая нервная систе-
ма). Резкого разграничения между этими системами нет.
АНАЛИЗАТОРЫ
Все раздражения, воспринимаемые организмом, подвергают-
ся тонкому анализу со стороны сложных нервных механиз-
мов— анализаторов. В состав последних входят, во-первых, вос-
принимающие аппараты — рецепторы, во-вторых, проводящие
пути — нервы, в-третьих — корковые центры, расположенные в
коре полушарий. Анализаторы составляют неразрывное целое
с нервной системой: они делятся на зрительные, равновесно-
слуховые, обонятельные, кожные, двигательные. С переходом от
низших форм к высшим изменяется локализация функциональ-
ных центров коры — мозговых концов двигательного, кожного,
зрительного, равновесно-слухового анализаторов. Значительную
часть коры у высших форм занимают центры позднейшего про-
исхождения с более совершенной функцией, расположенные в
лобной и височно-теменной областях (рис. 18).
При общей высокой организации рецепторов (в соответст-
вии с совершенством нервной системы) у разных экологических
групп млекопитающих они имеют свои особенности. Различают
60
экстерорецепторы, расположенные па наружной поверхности ор-
ганизма, интерорецепторы, находящиеся на внутренних поверх-
ностях, и проприорецепторы, локализующиеся в мышцах (кине-
стетические, или двигательные рецепторы). Далее в основном
описаны экстерорецепторы.
Наиболее просто устроен кожный анализатор. Рецепторами
этого анализатора служат I) свободные окончания нервов
в эпителии, которые воспринимают болевые и температурные
ощущения, давление и служат хеморецепторами, 2) осязательные
клетки, оплетенные сетью нервных волокон, и 3) осязательные
i С - 1 и II cl ) U1 р 4-1 О L' С. Г111 Ы ч.' i р j 11 11 ci ivi il Uc л cl 1 Cci 1j 11 L>i s\ PC 1 U 1\, OU ivv 1
ценных в соединительнотканную оболочку (они хорошо развиты
Рис. 18. Локализация мозговых концов анализа-
торов. А —у насекомоядных; Б —у полуобезьян.
Концы анализаторов: 1 — двигательного; 2 — кожного;
3 — зрительного; 4 — слухового. Вторичные ассо-
циационные области: 5 — лобная; 6 — височно-
теменная
на пальцах лазающих млекопитающих, на конце хобота слона,
рыльца крота и т. д.). Но основными рецепторами, восприни-
мающими эти раздражения и отчасти положение тела в прост-
ранстве, у млекопитающих служат волосы, особенно вибриссы.
Вибриссы реагируют не только на прикосновения к окружаю-
щим предметам, но и на колебания воздуха. У норников, имею-
щих широкую поверхность соприкосновения со стенками норы,
вибриссы, кроме головы, разбросаны по всему туловищу; у ла-
зающих форм (например, у белок, лемуров), они расположены
также на брюшной поверхности и на частях конечностей, кон-
тактирующих с субстратом при передвижении по деревьям,
и т. д. -
Восприятия химических раздражений осуществляются
у млекопитающих вкусовыми рецепторами. Раздражение полу-
чают вкусовые почки, расположенные на грибовидных, валько-
ватых и листовидных сосочках языка. У некоторых млекопитаю-
щих вкусовые почки имеются также на небе и надгортаннике.
Наиболее развиты вкусовые рецепторы у млекопитающих,
долго удерживающих при жевании пищу в ротовой полости (на-
пример, у копытных,’ приматов).
61
Равновесно-слуховой анализатор у млекопитающих дости-
гает наивысшего совершенства и представлен слуховыми нер-
вами, внутренним, средним и наружным ухом и центром в ви-
сочной области мозга. Во внутреннем ухе имеется вполне раз-
витая улитка (cochlea), к которой подходит слуховой нерв,
принимающий участие в образовании кортиева органа — слож-
ного слухового аппарата, свойственного только млекопитающим.
Кортнев орган состоит из воспринимающих звуковые раздраже-
ния чувствующих клеток, расположенных над дном улитки
(основная перепонка) и соединенных с нервными волокнами.
Улитка примыкает к круглому мешочку (яяеснЬю), отделен-
ному перехватом от овального мешочка (utriculus). От этого
перехвата к поверхности головы отходит заканчивающийся
слепо эндолимфатический канал. С овальным мешочком соеди-
няются три расположенных во взаимно перпендикулярных
плоскостях полукружных канала, наполненных эндолимфой
с мельчайшими известковыми тельцами. Этот отдел восприни-
мает изменения положения тела в пространстве. Все вместе
составляет перепончатый лабиринт в костном лабиринте каме-
нистой кости. Внутреннее ухо соединяется со средним затянутыми
перепонками овальным и круглым окошечками (рис. 19).
В среднем ухе (барабанной полости), как уже указывалось
выше, к стремечку у млекопитающих присоединяются нако-
вальня и молоточек. Так возникает трехчленная цепочка, пере-
дающая звуковые колебания от барабанной перепонки, отде-
ляющей среднее ухо от наружного, к овальному отверстию внут-
реннего уха и колебательные движения распространяются в
жидкости внутреннего уха. Среднее ухо сообщается с полостью
глотки евстахиевой трубой (рудимент брызгальца).
Для наружного уха млекопитающих характерно значитель-
ное развитие наружного слухового прохода и наружной ушной'
раковины. Последняя обладает большой подвижностью, что
облегчает улавливание звуковых волн. Очень крупных размеров
достигают ушные раковины у летучих мышей, зайцев, тушкан-
чиков, фенеков и других зверей, обладающих острым слухом h
С другой стороны, размеры ушных раковин связаны с их тер-
морегуляционной ролью (теплоотдача). Водные формы (киты,
большинство ластоногих) и настоящие землерои (кроты, сле-
пыши) ушных раковин не имеют. У ряда полуводных зверей
(бобр, калан, ушастые тюлени) ушные раковины способны за-
мыкаться при нырянии.
Слух у зверей достигает такой тонкости, что собака,
например, способна улавливать разницу всего в Vs тона (она
отличает звук высотой в 80 тысяч колебаний от звука в 100 ты-
сяч колебаний; для человеческого слуха эти звуки неощутимы).
1 У ряда видов ушные раковины играют роль резонаторов.
62
У летучих мышей выработался своеобразный механизм
ориентировки с помощью ультразвуковой локации. Сущность
се заключается в улавливании при помощи очень тонкого слу-
ха отраженных предметами звуков высокой частоты, изда-
ваемых голосовым аппаратом зверька; Учащая ультразвуко-
вые импульсы и улавливая их отражения, летучая мышь спо-
собна определять,.не только наличие предмета, но и расстояние
Рис. 19. Схема строения равновесно-слухового анализатора:
1 — улитка; 2 — овальный и 3 — круглый мешочки; 4 — эндолимфатический канал;
<5 — полукружные каналы; 6 — круглое и 7 — овальное окошечки; 8 — стремечко;
!) — наковальня; 10 — молоточек; 11 — евстахиева труба; 12 — барабанная пере-
понка; 13 — часть ушной раковины. Черным обозначен перепончатый лабиринт
до него и т. п. Такая локация почти полностью заменяет слабо
развитое зрение [Галамбос и Гриффин (Galambos a. Griffin),
1942; Кузякин, 1950]х.
Сходного типа устройство имеется и у китообразных, спо-
собных быстро передвигаться в совершенно непрозрачной воде,
не наталкиваясь на препятствия (экспериментальные данные,
полученные в океанариуме) 1 2.
1 В опытных условиях ослепленные летучие мыши свободно летают,
по наталкиваясь на предметы, но зверьки с выключенным слухом или лишен-
ные способности издавать ультразвуки, полностью утрачивают ориентировку.
2 Китообразные имеют очень небольшие слуховые отверстия. Узкий на-
ружный слуховой проход у них частично заполнен водой, барабанная пере-
понка неподатлива, однако по имеющимся наблюдениям, киты могут слы-
шать на далекие расстояния, благодаря способности воспринимать звуки и
ультразвуки, распространяющиеся в воде быстрее, чем в воздухе. Сами кито-
63
Обонятельный анализатор у многих млекопитающих также
•отличается высоким совершенством. Он состоит из восприни-
мающего аппарата, проводящих путей и коркового центра.
Рецепторы — обонятельные раковины, расположенные в задней
части носовой полости и покрытые обонятельным эпителием,
имеют сложное строение. Обособляющаяся у некоторых млеко-
питающих часть обонятельного мешка образует так называемый
якобсонов орган. Он имеет вид парных симметрично располо-
женных трубок, переходящих впереди в стенсоновы каналы;
последние открываются на нёбе'. Как полагают, этот орган
п q пг р м qq ро пт v П w 11 ’ И,
Имеет значение также
и мясистый- наружный
нос, особенно развитый
у хоботных, копытных,
.хищных и насекомояд-
ных. Частичная или
полная редукция обо-
нятельных рецепторов
у ластоногих и китооб-
разных представляет
вторичное явление,
связанное с жизнью в
воде.
По степени разви-
тия обонятельной функ-
ции млекопитающих
(рис. 20) с острым обо-
Рис. 20. Схема поперечного среза через зад-
нюю часть носовой полости. А — микросма-
тик; Б — макросматик (из Астанина)
разделяют на макросматические виды
нянием (большинство млекопитающих) * 1 2, микросматические
виды — со слабо развитым обонянием (беззубые киты, ластоно-
гие, приматы) и аносматические, у которых обоняние вовсе
отсутствует (зубатые киты).
Зрительный анализатор состоит из воспринимающего аппа-
рата— глаза, проводящих путей — зрительного нерва, и зри-
тельного центра в коре. Глаза у млекопитающих, как и у других
позвоночных, развиваются из мозгового пузыря, с которым оста-
ются связанными при помощи глазных нервов. Наружную часть
образные издают звуки разной частоты и связь особей осуществляется с
помощью этих звуков.
1 Хорошо развит якобсонов орган у клоачных, сумчатых, насекомоядных,
рукокрылых, неполнозубых, грызунов и копытных.
2 О тонкости обоняния макросматиков можно судить по тому, например,
что типичный представитель этой группы — собака, чует абсолютно не ощу-
щаемый человеком запах растворенной в воде поваренной соли даже при
очень слабой концентрации раствора (1 часть соли на 10 000 частей воды).
Сна легко находит дичь по сл.еду, оставленному несколько часов назад на
чистом снегу, где никакой анализ не обнаруживает признаков органических
веществ, и т. д. (
54
глазного яблока составляет белковая фиброзная оболочка, об-
разующая в передней части стекловидную роговицу (cornea),
а в остальной части — непрозрачную склеру (sclera). Далее
расположена собственно сосудистая оболочка (chorioidea), пере-
ходящая впереди в ресничное тело (corpus ciliaris) и радуж-
ную оболочку (iris), которая окружает зрачок. Иногда сосу-
дистая оболочка образует блестящий слой — зеркальце (tape-
tum), обусловливающее благодаря отбрасыванию лучей света
при почти полной темноте кажущееся свечение глаз. Глубже
лежит пигментная оболочка и еще глубже сетчатка (retina)
' 3
Рис. 21. Схема разреза глазного яблока млекопитающих:
/ _ белковая оболочка; 2 — роговица; 3 — собственно сосудистая оболочка; 4 —
ресничное тело; 5 — радужная оболочка; 6 — пигментная оболочка; 7 — сетчатка;
8 — зрительный иерв; 9 — хрусталик; 16 — хрусталиковые связки; 11 — передняя
глазная камера; 12 — задняя глазная камера; 13 — полость глазного яблока со стек-
ловидным телом; 14 — конъюнктива век и глазного яблока; 15 — веки; 16 — мейбо-
мовы железы; 17 —.ресница
с зрительными клетками — палочками и колбочками. К зрачку
сзади прилегает хрусталик (lens cristallina), прикрепленный осо-
быми хрусталиковыми (цинновыми) связками к мускульному
ресничному телу. Впереди зрачка находится передняя глазная
камера с водянистой влагой, сзади— (между задней поверхно-
стью радужины и хрусталиком) задняя глазная камера. Далее,
позади хрусталика помещается стекловидное тело, заполняю-
щее полость глазного яблока (рис. 21). В месте отхождения ог
дна глазного нерва находится слепое пятно, а близ него жел-
тое пятно — место наиболее ясного видения.
5 Териология . 65
В качестве защитных образований служат веки, выстланные
слизистой оболочкой — конъюнктивой. Веки снабжены ресни-
цами, назначение которых препятствовать засорению глаза, и
мейбомовыми, или тарсальными железами, выделяющими
глазную смазку. Имеется также рудиментарная мигательная
перепонка (membrana nicticans). У переднего (внутреннего)
угла глаза помещается гардерова железа, выделяющая жир-
ный секрет (ее нет у приматов). В наружном углу глаза —
слезная железа, выделения которой омывают глаз и стекают по
слезному каналу в носовую полость.
Аккомодация глаза осуществляется путем изменения формы
хрусталика. Когда животное смотрит вдаль, ресничная мышца
расслабляется, а связки хрусталика натягиваются — это обус-
ловливает уплощение хрусталика. В том случае, если рассмат-
риваемый предмет находится на близком расстоянии, происхо-
дит сокращение ресничной мышцы, в результате чего хрустали-
ковые связки расслабляются, и хрусталик как эластическое
тело принимает более выпуклую форму. Наибольшей способ-
ностью к аккомодации обладают приматы, наименьшей — ноч-
ные виды.
У форм, ведущих сумеречный или ночной образ жизни,
глаза либо достигают очень крупных размеров (например,
у долгопятов), либо невелики (у летучих мышей) —тогда недо-
статок зрения компенсируется высоко развитым слухом, обоня-
нием, осязанием. У роющих подземных видов (кроты, слепцы,
гоферы, златокроты) глаза в большей или меньшей степени ре-
дуцированы.
Цветовым зрением обладают в основном приматы (у других
млекопитающих оно либо зачаточно, либо полностью отсутству-
ет) х. Им же в наибольшей степени свойственно различение ри-
сунков и форм.
ОРГАНЫ ПИЩЕВАРЕНИЯ
Органы пищеварения осуществляют важнейшие жизненные
функции: захватывание, измельчение, переработку пищи, усвое-
ние ее и выведение наружу непереваренных остатков. К орга-
нам пищеварения относятся рото-глоточная полость, пищевод,
желудок, кишечник и пищеварительные железы.
Пищеварительный тракт млекопитающих начинается рото-
вым отверстием. Оно ограничено свойственными только млеко-
1 Цветовое зрение высших приматов приближается к таковому у чело-
века. У лемуров оно развито слабо. У других млекопитающих, у которых
констатировано цветовое зрение, дифференциация цветов очень ограничена..
Например, зебу различает 6 цветов (серый, красный, оранжевый, желтый,
синий и фиолетовый), но не дифференцирует зеленый от желтого. Котка,
кроме 6 цветов, улавливает до 25 оттенков серого. Лошадь различает только
4 цвета — красный, желтый, зеленый и синий. Белая крыса не дифферен-
цирует зеленый — желтый и синий — зеленый цвета [Виод (Viaud), 1960].
66
I
t
b
Рис. 22. Защечные мешки
хомяка (Cricetus cricetus),
ие заполненные пищей
питающим мягкими подвижными губами, отсутствующими
лишь у клоачных. Слабо развиты и лишены подвижности губы
у китообразных; наоборот, очень развиты и подвижны у млеко-
питающих, захватывающих ими пищу,— жираф, верблюдов,,
лошадей, овец, коз, плодоядных рукокрылых, медведей (осо-
бенно у губача) и некоторых других. Исключительно подвижен
хобот слонов, образовавшийся в результате срастания сильно
удлиненного носа с верхней губой. Между губами и зубами у
млекопитающих образуется так называемое преддверье — пред-
ротовая область, от которой у утконо-
сов, некоторых видов сумчатых, многих
грызунов п обезьян отходят парные за-
щечные мешки, служащие для времен-
ного помещения и переноса корма
(рис. 22). Величина и форма собствен-
но рото-глоточной полости соответству-
ют характеру питания животного.
Огромную пасть имеют киты, особенно
беззубые. Они пропускают через це-
дильный аппарат (из «китового уса»)
большое количество воды, чтобы обес-
печить поступление в желудок доста-
точного количества мелких морских
организмов, которыми они питаются.
Относительно велика также пасть у
летучих мышей, что увеличивает лов-
чую поверхность при ловле насекомых налету. Наоборот, крайне
мала она у муравьеда, ящера, трубкозуба, ехидны, добываю-
щих муравьев и термитов из их узких ходов.
Зубы у млекопитающих имеются только на челюстных
костях — они погружены корнями в отдельные ячейки (теко-
донтный тип).
Зубы представляют собой кожные окостенения, гомологич-
ные плакоидным чешуям. Главную массу зуба составляет осо-
бое органическое вещество — дентцн. В вершинной части—•
коронке, его покрывает эмаль, в корне — костное вещество це-
мент. Внутри зуб имеет полость, заполненную зубной мякотью,
или пульпой (рис. 23), в которую проникают нервы и крове-
носные сосуды.
Зубная система млекопитающих гетеродонтна — представ-
лена несколькими категориями зубов. Передние зубы, или
резцы (incisivi), имеют обычно долотообразную форму и слу-
жат в основном для захватывания пищи. За ними расположена
пара конических крупных клыков (canini), с помощью которых
пища удерживается и разрывается на части. Последующие две
категории зубов — предкоренные (praemolares) и коренные
(molares), отличаются широкой бугорчатой или складчатой по-
5
67
верхностью и служат для перетирания пищи. Особой сложно-
сти достигают коренные зубы копытных.
Если происхождение резцов и клыков может быть легко
выяснено при сопоставлении с зубами рептилий, то вопрос
о возникновении сложных коренных зубов решить значительно
труднее. Согласно существующим взглядам, коренной зуб мог
образоваться либо путем слияния между сойой соседних кони-
Рис. 23. Развитие зуба млекопи-
тающего:
ds —• зубная пластинка; ео — эмалевый
орган; р — мякоть зуба; — зачаток
постоянного зуба (по Гертвигу)
ческих зубов — «теория кон-
креции», либо путем постепен-
ного усложнения простого
конического зуба предков и
превращения его в трсхбугор-
чать’й — «тритуберкулярная
теория», или «теория диффе-
ренциации» Копа — Осборна —
Грегори (рис. 24). Последний
взгляд, несмотря на то, что он
до последнего времени пользо-
вался широким признанием,
оказался ошибочным, так как
его сторонники не уделяли не-
обходимого внимания данным
индивидуального развития жи-
вотных. Выяснено, что в онто-
генезе у представителей совре-
менных отрядов млекопитаю-
щих сохраняются признаки са-
мостоятельного образования
отдельных дентиновых конусов
(формирование сложных ко-
ренных зубов в онтогенезе млекопитающих идет путем сраста-
ния двух рядов отдельных дентиновых конусов), а не диффе-
ренциации отдельного конического зуба (АГатвеев, 1961).
В строении, числе зубов и в развитии их отдельных форм
у разных видов наблюдаются значительные вариации, служа-
щие хорошим систематическим признаком. Для обозначения
структуры зубной системы применяют формулы, в которых от-
мечают зубы одной половины верхней и нижней челюстей. Так,
опоссум имеет зубную формулу
/4” С у рт -3- т-^= 25x2 (для обеих половин) =50,
ИЛИ
волк, лисица
3— ]— 4— °
3—1—4—3
(42;;
68
лошадь • 3—1_3—з 3—1—3-3 (40);
енот • 3—1—4—2 с—1—4—2 (40);
подковонсс, ночница 2—1—3—3 3—1—0—3 (38);
куница 3-1—3—1 3—1—3-2 (34);
OJKHb 0—1—3—3 3—1—3—3 (34);
высшие приматы 2—1—2—3 2—1—2—3 (32);
калан» 2—1—1—3 3—1—2—3 (32);
кошка 3—1—3—1 3—1—2—1 (30);
заяц, кролик 2—Q—3—3 1—0—2—3 (28);
сурок, суслик 1—0—2—3 1—0—1—3 (22);
бобр 1—0—1—3 1—0—1—3 (20);
мышь, пслевка 1—0—0—3 1—0—0—3 (16);
австралийская 1—0—0—2 (12);
водяная крыса 1—и—0—2
слон 1—0—0—1 0—0—0—1 (6)’.
Общее количество зубов у сумчатых достигает 58, у дельфи-
нов, в результате вторичных преобразований — до 250. Сравни-
тельно простые, слабо дифференцированные зубы свойственны
примитивным млекопитающим, например насекомоядным, руко-
крылым, неполнозубым.
Далеко зашедшая специализация-—питание кровью птиц и
млекопитающих, имеет место у американских летучих мышей-
рода десмодовых (Desmodus). Резцы у них выдаются впе-
ред и имеют очень острые режущие края. Благодаря содер-
жащимся в слюне десмодовых анестезирующим и антикоагу-
лирующим веществам спящее животное не чувствует укуса, а
1 Во всех приведенных случаях даны полные ‘зубные формулы. Фаза
развития зубнсй системы и степень изношенности зубов служат критерйем
для определения возраста животного.
вытекающая из ранки кровь длительное время не сверты-
вается К
Вторичное упрощение зубов в связи с заглатыванием нераз-
мельченной пищи произошло у ластоногих. Однако клыки у мно-
гих из них хорошо развиты. У моржей они образуют мощные
бивни (у самцов до 75 см длиной), служащие для выкапывания
беспозвоночных (моллюсков, червей) со дна моря, опорой при
вылезании на лед и как орудие нападения.,
У зубатых китов в результате упрощения зубная система
стала вполне гомодонтной. Интересную особенность имеют зубы
нарвалов. Из нескольких
закладывающихся зубов
только у самцов прорезы-
вается единственный зуб,
который образует огром-
ный направленный вперед
бивень длиной до 3 м
(функция его не ясна; су-
ществует предположение,
что он служит животному
для пробивания льда при
устройстве «продухов»),
У хищных зверей наи-
большего развития дости-
гают клыки, коренные же
зубы в значительной сте-
пени редуцируются (в
связи с почти полным от-
Рис. 24. Происхождение коренных зубов.
А — по теории дифференциации; Б — по
теории слияния:
1 — гаплоидная форма; 2 — триконодонтная
форма; 3 — тритуберкулярная форма; 4 — ту-
беркулосекториальный зуб; 5 — 6— образование
многобугорчатого зуба путем слияния двух
триконодоитных зубов (по Астанину)
сутствием жевательной
функции); наиболее развитые — последний предкоренной зуб
верхней челюсти и лежащий позади него коренной нижней челю-
сти, образуют режущий аппарат. Каждый из них называют хищ-
ническим зубом (dens sectorius). Относительно очень большие
плоские коренные зубы свойственны каланам (морским выдрам),
пищу которых составляют морские ежи, моллюски, ракообраз-
ные— животные с твердыми скелетными образованиями. Очень
массивны зубы у гиен, дробящих трубчатые кости крупных жи-
вотных.
Для грызунов характерно сильное развитие резцов, обла-
дающих, благодаря отсутствию корней, постоянным ростом (сти-
рание резцов на концах непрерывно возмещается нарастанием
у основания). При повреждении одного из резцов противостоя-
щий ему резец, не встречая препятствий, вырастает до такой
1 Название «кровососущие летучие мыши», применяемое к десмодовым,
не соответствует действительности, так как они не сосут кровь, а слизы-
вают капельки ее, выступающие на месте прокусов кожи.
70
Рис. 25. Ненормальный рост верх-
него резца сурка (Marmota bobac),
как следствие повреждения про-
тивостоящего ему и в норме сти-
рающего его нижнего резца (по
Барабаш-Никифорову)
степени, что образует иногда уродливый завиток или кольцо, ме-
шающее питанию животного (рис. 25), что может привести к
гибели. Благодаря тому, что эмаль развивается только на пе-
редней поверхности резцов, снашивание их идет неравномерно
(задняя поверхность стирается скорее передней); этим обеспе-
чивается острота резцов, всегда сохраняющих долотообразную
форму х.
Зайцеобразные (зайцы, пищухи) позади пары нормальных
резцов в верхней челюсти имеют пару недоразвитых, прини-
мающих на себя удары нижних резцов при скусывании твердых
стеблей. У ряда грызунов, напри-
мер, у тушканчиков, слепышей,
слепушонок и др. резцы играют
важную роль при разрыхлении
плотной почвы во время прокла-
дывания ходов нор. С этим свя-
заны крупные размеры резцов -и
их положение: они сильно высту-
пают вперед, а у некоторых видов
(слепцов и др.) находятся вне рта,
так как на губах имеются специ-
альные выступы, которые смыка-
ются позади резцов и ограничи-
вают ротовое отверстие спереди.
Благодаря этой особенности зем-
ля, разрыхляемая резцами, не по-
падает в рот зверька. Сильно вы-
даются вперед резцы и у гренландского зайца-беляка, вынуж-
денного прогрызать при добывании корма корку плотного снега.
Сочленовная головка нижней челюсти грызунов обычно вы-
тянута в продольном направлении и, соответственно, сочленов-
ная ямка имеет корытообразную форму. Грызение происходит
при переднем положении нижней челюсти с нефиксированной
осью челюстного рычага (Б. Виноградов, 1926) и при ее зад-
нем положении, когда ось челюстного рычага фиксируется, упи-
раясь сочленовной головкой в сочленовную поверхность. Раз-
деление функций грызения и перетирания пищи приводит к
образованию дополнительных сочленений нижней челюсти с чере-
пом, способствующих фиксации оси челюстного рычага при гры-
зении. Например, гоферы (Geomyidae) имеют три сочленения
нижней челюсти с черепом (Н. Воронцов, 1962). При сильной
1 У домовой мыши на задней поверхности верхних резцов имеется тон-
кий слой эмали, задерживающий стирание. Поэтому, в отличие от других
мышей (лесной, полевой), на этих резцах у нее образуется характерная
выемка. Это явление рассматривается как приспособление к питанию мел-
кими семенами: в выемке удерживается мелкое семечко, обрабатываемое
«ижними резцами.
71
работе резцов ряды коренных могут совсем не соприкасаться,
что предохраняет их от излишнего снашивания.
Клыки у грызунов отсутствуют, и между резцами и корен-
ными зубами образуется широкое, не занятое пространство—
диастема. Коренные зубы хорошо развиты, хотя количество их
имеет тенденцию к сокращению. У видов, питающихся грубыми
частями растений, коренные не имеют корней и, подобно рез-
цам, непрерывно растут по мере снашивания коронки от пере-
тирания пищи.
В зубной системе копытных первенствующее значение при-
надлежит коренным зубам, резцы же и клыки в значительной
степени редуцируются. Сильное развитие клыков у кабана и
кабарги связано у первого с добыванием корма из земли и обо-
ронительными функциями; у обоих они служат турнирным ору-
жием. Среди коренных зубов копытных различают бунодонтные
(бугорчатые), свойственные всеядным свиньям, лофодонтные (с
поперечными гребнями), характерные для слонов, носорогов и
других видов, питающихся ветвями деревьев и кустов, и селено-
донтные (с лунчатой формой бугорков), имеющиеся у оленей,
лошадей и других животных, которые питаются листьями дре-
весных пород и травой.
Копытные, питающиеся жесткими степными травами (напри-
мер, лошади), имеют высокие, длительно растущие (благодаря
позднему или неполному развитию корней) гипселодонтные
(длиннокоронковые) зубы, которые хорошо противостоят интен-
сивному стиранию. Наоборот, формы, питающиеся мягкой соч-
ной растительностью (например, бегемоты), имеют низкие бра-
хиодонтные (короткокоронковые) зубы с ограниченным
ростом !.
Зубная система хоботных характеризуется превращением
резцов в огромные бивни, служащие для выкапывания корней,
защиты от хищников и пр. Исключительно массивны и сложны
складчатые коренные зубы слонов, легко перетирающие отно-
сительно толстые ветви деревьев. У сирен резцы либо редуци-
руются (ламантин), либо превращаются в бивни (дюгонь), клы-
ки и предкоренные редуцируются, но число коренных зубов воз-
растает.
У млекопитающих, лишенных зубов, последние закладыва-
ются в период зародышевого развития (беззубые киты) или
развиваются в молодости (утконосы), но затем исчезают.
Для зубной системы млекопитающих характерен дифиодон-
тизм, т. е. наличие двух генераций зубов — молочной и постоян-
1 Замена брахиодонтии гипселодонтией в зависимости от твердости пищи
имеет место и у грызунов. Так, например, зайцы, населяющие пустынные
районы и питающиеся жесткими растениями, к которым постоянно приме-
шивается песок, имеют более высокие зубы по сравнению с их сородичами
из других местностей.
72
ной, сменяющих друг друга (рис. 26). Только коренные зубы,
за редким исключением, не имеют смены, чем они в основном
и отличаются от предкоренных Г Слабо выражена смена зубов-
у сумчатых, неполнозубых, грызунов, насекомоядных, полно-
стью отсутствует у рукокрылых, ластоногих, китообразных (мо-
нофиодонтные млекопитающие). Обычно зубы сменяются в вер-
тикальном направлении (дефинитивный зуб по мере роста вы-
талкивает предшествующий
Горизонтальная смена зу-
бов свойственна слонам и
ламантинам. В этом слу-
чае происходит постепен-
ная смена стирающегося
старого зуба новым, за-
кладывающимся позади
него и перемещающимся
в горизонтальном направ-
лении.
Почти всю ротовую
полость у млекопитающих
занимает чрезвычайно
подвижный мускулистый
язык, в функции которого
входит захватывание пи-
щи, перемешивание ее во
рту (для смачивания слю-
ной) , способствование
акту глотания и восприя-
тие вкусовых ощущений.
Огромный лопатообраз-
ный язык имеют беззу-
ему зуб молочной смены).
Рис. 26. Постоянные и молочные (заштри-
хованы) зубы кошки:
/ — резцы (incisivi); с — клыки (canini); р —
предкоренные (praemolares); tn — коренные
(molares)
бые киты, у которых он служит своего рода поршнем для
наполнения ротовой полости водой и выжимания через цедиль-
ный аппарат наружу. У муравьедов и трубкозубов, специализи-
ровавшихся на питании термитами, муравьями, язык имеет чер-
веобразную форму и достигает значительной длины; покрываю-
щая его клейкая слюна облегчает извлечение насекомых из
подземных ходов. Очень длинный язык у летучей мыши Glosso-
phaga (Америка), австралийской сумчатой мыши, пяткохода и
некоторых других форм, питающихся нектаром или мелкими
насекомыми, посещающими нектарники цветков. У Glossonhaga
язык покрыт ворсинками, по которым нектар поднимается сам
в силу закона капиллярности. Важную роль при захватывании
пищи играет очень гибкий язык жвачных; развитые на нем ро-
говые сосочки предохраняют нежный орган от порезов острыми
краями стеблей травы. У хищников покрывающие язык загнутые
! У сумчатых сменяется последний коренной зуб.
73
Рис. 27. Поперечный разрез через го-
лову усатого кита в области средней
части цедильного аппарата
назад роговые сосочки помогают сдирать мясо с костей и чис-
тить мех.
В стенках ротовой полости помещаются крупные парные
слюнные железы: околоушная (glandula parotis) —серозная же-
леза с длинным выводным околоушным протоком, открываю-
щимся у верхних коренных зубов; подъязычная (gl. sublingua-
lis) — слизистая железа, открывающаяся бартолиниевым прото-
ком в месте соединения правой и левой нижних челюстей;
подчелюстная (gl. submaxillaris) — слизисто-серозная железа,
открывающаяся под языком
вартоповым протоком. Кро-
ме того, имеются еще мел-
кие ротовые железы: губ-
ные, щечные, язычные, неб-
ные и др. Секреты всех этих
желез, смешиваясь, образу-
ют слюну, основная функ-
ция которой — смачивание
пищи (поэтому у водных
млекопитающих слюнные
железы развиты слабо или
вовсе отсутствуют)
Свод ротовой полости об-
разует твердое небо, несущее
на своей поверхности поперечные складки — небные валики. Кза-
ди оно переходит в небную занавеску (мягкое небо). У беззубых
(усатых) китов небные валики разрастаются и, ороговевая, об-
разуют свешивающиеся в ротовую полость роговые бахромча-
тые пластины — китовый ус (рис. 27). Эти пластины выполняют
функцию цедильного аппарата, удерживая попадающие вместе
с водой мелкие планктонные организмы — пищу кита. Роговые
утолщения небных валиков сирен служат для перетирания водо-
рослей.
1 Густой клейкий секрет щечных желез летучих мышей, наносимый
зверьками на летательные перепонки, сохраняет их эластичность, защищает
от высыхания. Об особенностях слюны питающихся кровью летучих мышей
говорилось выше.
В последнее время установлено, что слюна североамериканской коротко-
хвостой землеройки (Blarina brevicauda) содержит ядовитые вещества. Вве-
дение небольшой дозы экстракта подчелюстной железы этой землеройки
вызывает смерть мыши менее, чем через 1 минуту после инъекции. Меньшей
силы яд обнаружен в подчелюстной железе некоторых других мелких насе-
комоядных и грызунов (Sorex personatus, Peromyscus leucopus, Microtus
pennsylvanicus).
Ядовит также секрет подчелюстной железы куторы (Neomys fodiens).
В природе яд Blarina и Neomys приводит к потере подвижности мелких
грызунов.
Наличие токсинов в слюне примитивных плацентарных млекопитающих
рассматривается как отражение их филогенетической связи с рептилиями
ТПуцек (Pucek), 1959].
74
За мягким небом находится глотка, в которую открываются
хоаны и евстахиевы трубы.
Непосредственно за глоткой начинается пищевод — длин-
ная, лишенная желез трубка, проводящая пищу в желудок. По-
следний может быть либо простым однокамерным, либо слож-
ным многокамерным.
Наиболее сложен желудок жвачных, состоящий из четырех
отделов. В связи с этим процесс переработки пищи у жвачных
имеет специфические особенности. Слабо пережеванные комки
пищи попадают в первый, наибольший отдел желудка — рубец
(rumen), где кормовая масса подвергается мацерации и биохи-
мическим превращениям в результате действия большого коли-
чества симбиотических микроорганизмов (бактерий, инфузо-
рий) ; населяющих рубец1. Отсюда порции пищи поступают в
•следующий отдел — сетку (reticulum) и отрыгиваются в рот.
После вторичного, более тщательного разжевывания, или жвач-
ки, превращенная в кашицеобразную массу пища снова заглаты-
вается и стекает по особому пищеводному желобку непосред-
ственно в третий отдел — книжку (oriiasus), из которого перехо-
дит в четвертый — сычуг (abomasus) 1 2. Биологическое значение
1 жвачки в том, что благодаря ей животное может наиболее пол-- •
но использовать, ценные компоненты корма, разрушая клетчатку
и древесную паренхиму растительных клеток. Кроме того, жвач-
ка дает возможность сокращать время пастьбы, когда живот-
ное в наибольшей степени подвергается опасности нападения
хищников: наполнив за короткий срок свой огромный рубец, оно
пережевывает и переваривает пищу уже в укрытом месте, в тени,
где не беспокоят кровососущие насекомые, и т. д.
Сложный (многокамерный) желудок имеют также китооб-
разные, сирены, ленивцы и некоторые другие звери, питающие-
ся грубым или трудно переваримым кормом.
Уже упоминавшиеся десмодовые летучие мыши имеют желу-
док, снабженный особым боковым придатком, в котором резер-
вируется заглатываемая кровь.
По особенностям внутреннего строения в желудке можно
различать пищеводную — безжелезистую, область и собственно
желудочную, снабженную пищеварительными железами. По-
следние дифференцируются на кардиальные железы (выделяют
слизистый щелочной секрет), донные, или фундальные (выде-
ляют пепсиноген, прохимозин, липазу и соляную кислоту), и
пилорические железы (секретируют также пепсиноген и прохи-
1 Аналогичное явление наблюдается также в желудке некоторых нежвач-
ных млекопитающих — свиней, грызунов и других, где роль бродильного
агента выполняют развивающиеся в массе дрожжевые грибки.
2 У некоторых жвачных пищевые комки отрыгиваются непосредственно
из рубца и туда же -возвращаются после дополнительного разжевывания
во рту. В следующие отделы проходит уже сильно” разжиженная пища.
75
мозин). По местоположению этих желез получили название и
соответствующие участки собственно желудка (рис. 28). В же-
лудке жвачных только сычуг служит собственно желудком, все
же остальные отделы представляют расширение пищевода1.
За желудком следует кишечник, состоящий из двух отделов:
тонкой и толстой кишок. Передняя часть тонкой кишки носит
название двенадцатиперстной (duodenum). Стенки ее несут мно-
гочисленные ворсинки и, помимо общекишечных желез, содержат
так называемые дуоденальные (бруннеровы) железы, выделяю-
щие секрет, близкий к секрету пилорических желез. В начале
Рис. 28. Схема строения желудка млекопитающих человека:
1 —человек; 2 — собака; 3 — крыса; 4 — хомяк; 5 — лошадь; 6 — жвачное.
Поперечные штрихи — пищевод и пищеводный отдел желудка; косые штрихи — об-
ласть кардиальных желез; пунктир — область донных желез; крестики — область
пилорических желез (по Гессе)
толстого отдела .кишечника у большинства млекопитающих от-
ходит слепая кишка (coecum). Она отсутствует у некоторых на-
секомоядных, неполнозубых, медведей, бегемотов, китов. У не-
жвачных травоядных (некоторые сумчатые, грызуны, лошади),
слепая кишка достигает огромных размеров, выполняя функцию
рубца жвачных, т. е. являясь своего рода бродильным чаном для
грубой пищи. У плотоядных млекопитающих слепая кишка раз-
вита слабо. Заканчивается толстый отдел прямой кишкой (rec-
tum). Стенки толстого отдела кишечника богаты трубчатыми
железами, выделяющими слизь. Наружу прямая кишка откры-
вается анальным отверстием' (лишь у клоачных конечная часть
ее впадает в клоаку).
1 Пищевод и желудок видов, питающихся грубыми кормами (бобра,
некоторых мышевидных грызунов) имеют роговую выстилку — защитное при-
способление против повреждения стенок.
76
Общая длина кишечника сильно варьирует у разных видов
зверей, что зависит от типа питания, строения желудка, разме-
ров тела и пр. Так, длина кишечного канала превышает длину
тела у летучих мышей в 1,5—2, у кошки — в 4, у собаки — в 6,
у лошади — в 12, у свиньи — в 15, у коровы — в 20, у китов —
до 32 раз. Как правило, звери, питающиеся грубой пищей, име-
ют более длинный кишечник (рис. 29). У молодых особей, вслед-
ствие высокой потребности в пище при более интенсивном обме-
не веществ кишечник относительно длиннее, чем у взрослых.
Рис. 29. Кишечный тракт. А — грызуна (морской свинки); Б — хищник
(лисицы):
J — начало двенадцатиперстной кишки; 2 — конец прямой кишки; 3 — слепая кишка;
4 — петля ободочной кишки; 5 — 7— разрезы кровеносных сосудов (по Митчел)
Из крупных пищеварительных желез у млекопитающих хо-
рошо развиты печень и поджелудочная железа.
Печень (hepar) имеет лопастное строение. В ней вырабаты-
вается желчь, которая собирается в желчном пузыре (vesica
fellea), имеющемся у большинства млекопитающих. Проток его,
соединяясь с печеночным, образует общий желчный проток
(ductus choledochus), впадающий в двенадцатиперстную кишку.
Желчного пузыря нет у крыс, лошадей, верблюдов, китообраз-
ных и некоторых других. Желчь способствует расщеплению жи-
ров и активизирует перистальтику кишечника. Печень играет и
важную метаболическую роль, в ней откладывается в качестве
запасного вещества, гликоген (животный крахмал), концентри-
77
руются запасы витаминов, особенно витамина Л1. Печень мле*
копитающих служит не только энергетическим, но и белковым;,
депо. В случае голодания белок печени начинает распадаться.,
служа важным источником питания организма.
Поджелудочная железа (pancreas) имеет компактное или:
диффузное. строение и состоит из собранных в гроздья долек,.,
выводные протоки которых соединяются в главный панкреати-
ческий проток, открывающийся в двенадцатиперстную кишку.
Секрет железы — важнейший пищеварительный панкреатиче-
ский сок, ферменты которого действуют па белки (трипсин),,
углеводы (амилаза) и жиры (липаза). Частично pancreas слу-
жит также железой внутренней секреции (см. стр. 83).
Механическая и химическая переработка пищи начинается,
уже во рту (жевание, смачивание слюной). Под влиянием фер-
ментов слюны (птиалин, мальтаза) крахмал превращается в.
декстрины и сахар. Дальнейший процесс пищеварения проте-
кает в желудке под воздействием ферментов желудочного
сока — пепсина (расщепляющего белки до альбумоз и пепто-
нов), химозина (створаживающего молоко) и липазы (расщеп-
ляющей жиры). Значительно содействует перевариванию соля-
ная кислота желудочного сока. Превратившаяся в химус пища;
поступает в кишечник.
В тонкой кишке действуют выделяемая печенью желчь,..фер-
менты кишечного сока и сока поджелудочной железы,--завер-
шающие пищеварительный процесс. Здесь же в основном проис-
ходит всасывание продуктов пищеварения в кровщ Наконец, в;
толстой кишке происходит, обработка пищевых остатков с по-
мощью бактериального брожения и формирование кала, выде-
ляемого наружу. '
ОРГАНЫ ДЫХАНИЯ
Млекопитающим свойственно легочное и отчасти кожное ды-
хание. В легкие воздух поступает через воздухоносные пути:
носовую полость (область расположения носовых и челюстных:
раковин представляет собой обогревательно-фильтрационный
аппарат), носоглоточные ходы, гортань, трахею и бронхи. Про-
ходя через них, воздух согревается, увлажняется, очищается от
мелких пылевых частиц и обеззараживается (слизь носовой по-
лости обладает бактерицидными свойствами).
Для гортани млекопитающих характерно развитие надгор-
танника и щитовидного хряща, не свойственных другим позво-
ночным, имеются также перстневидный и парные черпаловид-
ные хрящи. Все эти хрящи подвижно соединяются друг с другом:
1 Очень богата этим витамином печень белого медведя, китов. Жир пе-
чени китов используется в лечебных целях, аналогично рыбьему жиру.
78
с помощью связок и снабжены мышцами. Между щитовидным
и черпаловидными хрящами помещаются голосовые связки, при-
водимые в движение особыми мышцами. Вибрация этих связок
производит звуки (у некоторых млекопитающих для этого слу-
жит вибрация других частей гортани). Голоса животных очень
отличаются не только у разных видов, но и у особей одного вида,
но разного пола. В этой же области имеются особые ямки —
так называемые боковые гортанные желудочки; у некоторых ви-
дов они сильно разрастаются и образуют гортанные мешки,
играющие роль резонаторов (у ревунов гортанные мешки поме-
щаются в пузыревидном вздутии подъязычной кости).
Гортань переходит в трахею, продолжением которой служат
парные бронхи, разветвляющиеся при переходе в легкие на
бронхи 2-го, 3-го и 4-го порядков и мельчайшие бронхиоли..
Стенки трахеи и бронхов не спадаются благодаря хрящевым
кольцам.
Легкие в основном состоят из конечных расширений брон-
хиолей — легочных пузырьков, или альвеол, оплетенных густой
сетью кровеносных капилляров —в них происходит газообмен..
Форма и величина легких в значительной степени связаны с об-
разом жизни животных: малоподвижные крупные формы имеют
однодольчатые, относительно небольшие легкие, наоборот, вы-
сокоподвижные мелкие животные характеризуются относительно
большими многодольчатыми легкими.
Дыхательный акт совершается при помощи движений ребер-
и диафрагмы — сужения и расширения грудной клетки. Мелкие-
животные с интенсивным обменом по частоте дыхательных дви-
жений превосходят крупных (у мыши, например, число дыха-
тельных движений в минуту около 200, а у лошади их около 12)..
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Кровеносная система, обеспечивающая обмен веществ между
тканями и наружной средой, достигает у млекопитающих боль-
шого совершенства. Вместе с высокой организацией дыхатель-
ного аппарата это обусловливает постоянство температуры
тела млекопитающих (гомотермность).
Центральный орган кровообращения — сердце (сог) пред-
ставляет собой мускульный четырехкамерный орган, заключен-
ный в околосердечную сумку — перикард (pericardium). Подоб-
но размерам легких, размеры сердца находятся в прямом;
соответствии с положением местообитания над уровнем моря и
с подвижностью животного (отношение веса сердца к весу те-
ла—«сердечный индекс» очень подвижного песца более чем
в два раза превосходит соответствующий индекс сравнительно-
медлительной енотовидной собаки).
7»
От левого желудочка сердца отходит единственная дуга аор-
ты (arcus aortae dorsalis); она заворачивает влево и назад и
разносит артериальную кровь по всему телу. Через стенку ка-
Рис. 30. Схема кровеносной системы
млекопитающих:
1 — внешняя сонная артерия (a, carotis
externa): 2 — внутренняя сонная артерия
(а. с. interna): 3 — подключичная арте-
рия (a. subclaviu); 4 — дуга аорты; 5 —
легочная артерия (a. pulnionulis); 6 —
левое предсердие; 7 — правое предсер-
дие: 8 — левый желудочек; 9 — правый
желудочек; 1С — спинная аорта (ао. dor-
salis): 11— внутренностная артерия (а.
соеИася): 12— почечная артерия (а. гена-
lis); 13 — подвздошная a. (a. iliaca);
14 — яремная вена (v. jugularis); 15 —
подключичная в. (v. sub cl avia); 16 —
левая непарная в. (v. hemiazygos); 17 —
правая непарная в. (v. azygos); 18 —
задняя голая гена (v. cava posterior): 19 —
печеночная вена (v.hepatica): 20 — ворот-
ная вена печени (v. portae henatis); 21 —
печень; 22 — почка; 23 — подвздошная в.
(v. iliaca)
пилляров кровь отдает кисло-
род и питательные вещества
клеткам и тканям, а в нее по-
ступают углекислый газ и про-
дукты обмена. Венозная кровь
собирается в переднюю и зад-
нюю полые вены (vena cava
anterior et v. c. posterior), впа-
дающие в правое предсердие —
так замыкается большой круг
кровообращения.
Из правого желудочка ве-
нозная кровь по легочным ар-
териям (аа. pulmonales). течет
в легкие, откуда, окислившись,
возвращается по легочным ве-
нам (vv. pulmonales) в левое
предсердие, завершая малый,'
или дыхательный, круг крово-
обращения.
Отверстия между предсер-
диями и соответствующими же-
лудочками снабжены створча-
тыми клапанами, препятствую-
щими обратному току крови.
Карманообразные полулунные
клапаны имеются также в на-
чале аорты и легочной артерии.
Деятельность отделов серд-
ца происходит в такой после-
довательности: 1) наполнение
кровью и систола (сокраще-
ние) обоих предсердий, благо-
даря чему кровь поступает в
желудочки, 2) диастола (рас-
слабление) предсердий и си-
стола желудочков — проталки-
вание крови в отходящие от.
сердца сосуды, 3) диастола
желудочков и общая пауза
сердца.
Наиболее сложна, естест-
венно, система сосудов боль-
шого круга кровообращения
80
(рис. 30). От дуги аорты к голове отходят общие сонные арте-
рии, разветвляющиеся на внутренние и наружные сонные (аа. са-
rotides). В передние конечности кровь поступает по подключич-
ным артериям (аа. subclaviae) *. К внутренним органам арте-
риальная кровь приносится в основном чревной (a. coeliaca),
брыжеечной (a. mesenterica) и почечной (a. renalis) артериями.
Наконец, заднюю часть туловища снабжают кровью хвостовая
артерия (a. caudalis) и парная подвздошная артерия (a. iliaca)
с ее разветвлениями.
В состав венозной системы, кроме названных выше вен, вхо-
дят яремная (v. jugularis) и подключичная (v. subclavia) вены,
которые, сливаясь, дают начало передней полой вене, у млеко-
питающих обычно непарной (левая ветвь.ее редуцируется, заме-
няясь безымянной веной). В переднюю полую вену впадают асим-
метрично развитые остатки задних кардинальных вен — непар-
ные правая и левая вены (vv. azygos et hemiazygos), в боль-
шинстве случаев соединяющиеся друг с другом поперечным
анастомозом и несущие кровь из грудной полости.
Из других вен большого развития достигают вены воротной
системы печени: воротная вена (v. portae)—начинается от
кишечника, желудка и селезенки и впадает в печень, обра-
зуя в ней сеть капилляров, и печеночная вена (v. hepatica), —-
по ней кровь поступает из печени в заднюю полую вену. В по-
следнюю впадают также парная почечная вена (v. rena-
lis) 1 2, хвостовая (v. caudalis) и парная подвздошная вена
(v. iliaca).
Кровь состоит из жидкой плазмы и форменных элементов.
Плазма содержит соли, питательные вещества, белки, ферменты
и др. Большое значение 'имеют содержащиеся в плазме анти-
тела— защитные вещества, которые нейтрализуют или разру-
шают инородные тела (и токсины), попадающие в кровь3.
Форменные элементы крови представлены эритроцитами,
лейкоцитами и тромбоцитами. Эритроциты служат транспорте-
рами кислорода, выполняя таким образом дыхательную функ-
цию. Характерная особенность эритроцитов млекопитающих —
их безъядерность и уплощенно-дисковидная форма (только
верблюды и ламы имеют овальные эритроциты) 4 * 6. Лейкоциты
1 У некоторых млекопитающих начальные отделы сонных и подключич-
ных артерий соединяются, образуя безымянные артерии (аа. anonimae).
2 Воротная система почек, свойственная другим позвоночным, у млекЬ-
питающих редуцирована.
3 Кровь некоторых млекопитающих содержит вещества, являющиеся про-
тивоядием змеиного яда. Так, иммунитетом к укусам змей обладают вивер-
ровые, отчасти ежи и некоторые другие.
4 С безъядерностью эритроцитов связано понижение их обмена веществ,
что в свою очередь обусловливает уменьшение потребления кислорода самими
эритроцитами и, как результат, повышение отдачи кислорода тканям. Упло-
щенность имеет некоторые преимущества над шарообразной формой в смысле
более равномерной диффузии газов.
6 Териология
81
выполняют ряд функций: они служат транспортерами питатель- j
ных веществ, поглощают микробов (фагоцитоз), обезвреживают i
бактериальные токсины, образуют в результате переработки
белков антитела. В-отличие от эритроцитов лейкоциты имеют I
ядра и способны к амебовидным движениям. Тромбоциты обус- |
ловливают свертывание крови и, согласно новейшим данным, '
участвуют в выработке иммунитета к действию ядов; они безъ-
ядерны и имеют дискообразную форму.
Кровь образуется в красном костном мозге, лимфатических !
узлах, зобной железе и селезенке. i
Селезенка (lien) представляет собой компактный орган, при- ;
нимающий участие в кровообразовании и в ряде биохимических
процессов. Кроме того, селезенка выполняет защитные функции :
и представляет депо крови — регулирует кровообращение в со- 1
ответствии с изменениями условий среды.
Лимфатическая система. Часть плазмы крови, вы-
ходя через стенки капилляров, образует лимфу. Совершив опре-
деленный путь по специальным лимфатическим сосудам, из ко-
торых наибольшего развития достигает грудной проток (ductus j
thoracicus), она снова поступает в кровяное русло (в веноз-
ную систему). Лимфатические сосуды прерываются лимфатиче- i
скими узлами, которые обогащают лимфу лимфоцитами и, кроме
того, освобождают ее от микроорганизмов и токсинов. Обога- !
щение лимфы клетками обеспечивается также и другими лим-
фоидными органами пищеварительной трубки: одиночными лим-
фатическими узелками и их скоплениями (пейеровыми бляшка-
ми) и миндалинами.
ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
Железы внутренней секреции регулируют взаимоотношения
между различными органами тела, характеризуются отсут-
ствием выводных протоков и выделяют свои инкреты в кровь .
или лимфу.
Важнейшая железа внутренней секреции — щитовидная
(glandula thyreoidea), расположенная возле гортани. Гормон ее 1
тироксин — влияет на процессы обмена веществ, роста и разви-
тия. Вблизи щитовидной железы или непосредственно на ней
(и даже в ней) помещаются околощитовидные железы, или эпи-
телиальные тельца (gl. parathyreoideae)—маленькие округлой ]
формы образования. Они регулируют кальциевый и фосфорный
обмен, влияя тем самым на развитие костей, зубов и т. и.
Зобная, или вилочковая, железа (gl. thymus) представляет
собой дольчатый орган, лежащий в области трахеи и в передней
части грудной полости. Достигая сильного развития у молодых
животных, с возрастом она претерпевает постепенное обратное
развитие. Функции тимуса недостаточно выяснены; установлено
82
влиянйе этой железы на рост и развитие; частично она выпол-
няет функцию кровообразовательного органа. Околопочечникй
(gl. pararenales)—парные органы, топографически связанные
с почками. Они вырабатывают гормон адреналин, влияющий
на кровообращение, углеводный обмен и некоторые другие фи-
зиологические процессы.
Гипофиз, или нижний мозговой придаток, играет исключи-
тельно важную роль в регуляции многих процессов в организме.
Гипофиз состоит из железистой и нервной долей. Железистая
продуцирует большое число гормонов, регулирующих процессы
роста (гормон роста), развитие половых желез (гонадотроп-
ные— фолликулостимулирующий и лютеиноизирующий гормо-
ны), молокообразование (лактогенный гормон), влияющих на
функцию щитовидной железы (тиреотропный гормон), и пр.
Нервная его доля своими гормонами (или одним общим гор-
моном) влияет на кровяное давление, мочеотделение и работу
мускулатуры матки. '
Частично железами внутренней секреции являются подже-
лудочная и половые железы. В поджелудочной железе инкре-
торную роль выполняют так называемые островки Лангерганса;
'выделяемый ими гормон инсулин влияет на углеводный обмен.
Мужские и женские половые железы выделяют в кровь поло-
вые гормоны (группы стеронов), воздействующие нй развитие
наружных половых признаков, цикличность половых процессов,
обмен веществ и особенности поведения животного.'
ОРГАНЫ ВЫДЕЛЕНИЯ
Как и у всех амниот, у млекопитающих процесс выведения
из организма (выделения из крови) конечных продуктов-обме-
на веществ — азотистых продуктов белкового обмена, солей,
воды, осуществляется парной дефинитивной метанефрической
почкой; только у молодых клоачных и сумчатых- некоторое
время функционирует мезонефрос, у высших же млекопитающих
он существует только в эмбриональный период. От него у взрос-
лых животных остаются лишь незначительные части (paradidy*
.mis), расположенные книзу от гонад. .
У большинства млекопитающих почки (renes) имеют глад-
кую поверхность и представляют собой бобовидные компактные
тела. У медведей, китообразных, ластоногих и некоторых дру-
гих почки множественные, т. е. состоят из большого числа долек.
В почке различают наружную корковую (мочеотделитель-
ную) зону, в которой размещены почечные тельца (капсулы
Шумлянского-Боумена с сосудистыми клубочками). и извитые
почечные канальцы с петлями Генле1, и.центральную мозговую
1 В сосудистых клубочках осуществляется фильтрационный процесс, в из-
витых канальцах и петлях Генле — реабсорбция воды; сахара и боли. /
6* 83
(отводящую) зону, содержащую прямые собирательные трубоч-
ки, соединенные в пирамиду (pyramida renalis). Последняя от-
крывается сосочком в воронкообразный резервуар — почечную
лоханку (pelvis renalis), от которой берет начало мочеточник
(ureter), впадающий в мочевой пузырь (vesica urinaria). У кло-
ачных мочеточники впадают в мочеполовой синус, откуда моча
поступает в мочевой пузырь. Наружу моча выводится у самок
через короткое преддверие влагалища, а у самцов через моче-
половой канал, открывающийся на конце копулятивного органа.
Кроме мочеотделения, почки играют существенную роль
в регулировании водного баланса и химического состава крови.
К почкам примыкают околопочечники — железы внутренней
секреции.
ОРГАНЫ РАЗМНОЖЕНИЯ
К органам размножения млекопитающих относятся половые
железы, половые протоки и различные дополнительные органы.
Мужские половые органы. Половыми железами
.самцов служат парные семенники (testes) — компактные эллип-
соидной формы тела, в которых развиваются мужские половые
клетки; .в то же время они играют роль желез внутренней сек-
реции. Семенники (рис. 31) закладываются в брюшной полости,
где у части млекопитающих — так называемых крипторхов, оста-
ются пожизненно (клоачные, некоторые неполнозубые и насеко-
моядные, носороги, слоны, даманы, сирены, китообразные).
У многих млекопитающих семенники в период спаривания опу-
скаются через паховый канал в особый кожный мешок — мо-
шонку (scrotum) и затем снова втягиваются обратно (некото-
рые насекомоядные, рукокрылые, грызуны). У остальных зверей
спустившиеся в раннем возрасте в мошонку семенники, вслед-
ствие зарастания пахового канала, остаются в ней пожизненно.
Мошонка помещается позади копулятивного органа; только у
сумчатых она расположена кпереди от него.
Наружную часть семенника образует соединительнотканная
белочная оболочка, от которой внутрь тела семенника врастают
тяжи, разделяющие семенник на дольки. В последних нахо-
дятся извитые семенные канальцы; стенки их выстланы эпите-
лиальными клетками Сертоли и включают различные стадии1
развития половых клеток. В прослойках между канальцами по-
мещаются крупные интерстициальные клетки Лейдига, выпол-
няющие функции желез внутренней секреции. С семенниками
у некоторых млекопитающих срастаются остатки передних
концов мюллеровых протоков — гидатиды Морганьи (hydatus
Morgagni), в то время как остатки задних концов образуют у
начала мочеполового канала так называемую мужскую матку
(uterus- masculinus; грызуны, хищные, копытные).
84
Семенные канальцы переходят в выносящие канальцы, со-
бирающиеся в большой общий канал. Выносящие канальцы и
общий канал выходят уже за пределы собственно семенника.,
формируя прилежащий к нему придаток (epididymis).
Рис. 31. Схема мочеполового аппарата. А—самца; Б—самки:
h — стебельчатая гидатида; т — мюллеров канал; w — вольфов канал; os — воронка
яйцевода; ер — придаток семенника (epididymis); t — семенник (testis); ov — яичних
(ovarium); mt — почка (metanephros); pd — остаток .первичной -•почки; и —матка (ute-
rus); v — влагалище (vagina); t>s — пузыревидная железа (glandula vesicularis); ит —
мужская матка (uterus masculinus); vu —Умочевой пузырь (vesica urinaria); pr — пред-
стательная железа (gl. prostata); gc — куперова железа (gl. eouperi); cl — клитор (сП
toris); p — копулятивный орган (penis); r — прямая кишка (rectum) (no Студеицову}
В сперматогенезе — развитии мужских половых клеток (спер-
матозоидов) — сменяют друг друга несколько фаз. Началь-
ная фаза сперматогенеза — разрастание семенных канальцев
и размножение в них мелких клеток—сперматогониев (одно-
временно происходит процесс редукции интерстициальной тка-
ни). В результате деления сперматогонии превращаются в бо-
лее многочисленные сперматоциты I порядка. Дальнейший рост
и созревание половых клеток приводят к образованию сперма-
цитов II порядка. В результате нового деления диплоидное
число хромосом превращается в гаплоидное, и такие клетки
получают название' сперматид. В цитоплазме клеток Сертоли
сперматиды снабжаются питательными веществами и превра-
щаются в зрелые половые клетки. Завершение их формирова-
ния происходит в придатке семенника.
В начальных стадиях развития семенные клетки имеют
округлую форму. Зрелые сперматозоиды состоят из головки,
шейки и хвоста; они очень малы (измеряются микронами) и
отличаются* * большой подвижностью. Каналы придатков семен-
ников дают начало парному семепроводу (ductus deferens);
оба эти отдела — производные вольфова канала. На конечной
части семепроводов у некоторых млекопитающих развиваются
пузырьковидные железы (gl. vesicularis) !, студенистые выделе-
ния которых’служат у некоторых насекомоядных, летучих мы-
шей и грызунов для закупоривания половых путей самки после
введения в них спермы. В области мочевого пузыря семяпрово-
ды, соединившись, впадают в непарный мочеполовой канал.
С последним связаны хорошо развитые предстательная железа
(gl. prostata) и ниже расположенные луковичные куперовы же<
лезы (gl. bulbourethrales Couperi). Основная функция всех при-
даточных желез— выделение секретов, разжижающих семя;*
секрет предстательной железы вызывает подвижность спермато-
зоидов. .
Конечная часть мочеполового канала пронизывает копуля-
тивный орган (penis), служащий для внесения семени в поло-
вые пути -самки. Копулятивный орган в основном состоит из
каверноздых (пещеристых) тел и головки2. Переполнение пе-
щеристых тел кровью, перед совокуплением и в момент его
обусловливает общее напряжение пениса — эрекцию — необхо-
димое условие для нормального осуществления этого акта.
Кость копулятивного органа (os penis) служит опорным обра-
зованием'; облегчающий- совокупление3. Головку пениса прикры-
вает ко‘я<на;я; складна—'так называемый препуций, с которым
бывают связаны'особые парные препуциальные железы, выде-
ляющие сильно пахнущие,вещества.
/ У су^ча)тых’ крпулятивныщ орган раздвоен, соответственно.'
двойному влагалищу самки. Вообще форма копулятивного орга-
на у разных групп млекопитающих различна; в последнее
время это'й'-а’рйЙу со "'строением os penis используют в качестве
систематического -признака,"позволяющего различать близкие
Риды ит. п.
Женские половые органы. Половые железы самок
представлены яичниками (ovaria) — небольшими парными те-
;Нередко этим желе шм дают название «семенные пузырьки» («vesiculi-
seminalis»); что неправильно, так как семени они не содержат.
• >.? Головка пениса, рудиментарна . или полностью отсутствует у свиней,
бегемотов,, верблюдов,, китов, У оленей, антилоп, жираф и некоторых других
Пение несеТ на1 конце тонкий отросток, пронизанный каналом мочеточника.
У: грызунЪв : гбловйа ' бывает покрыта твердыми шипиками, усиливающими
в момент копуляции, возбуждение самки.
. 3 Особенно сильного, развития эта кость достигает у моржа (у взрослого
животного’ дйин’а ее может превышать 0,5 м, а ширина 6 см) и некото-
рых других. •..... 1
86
лами, подвешенными на мезентерии к спинной стенке брюшной
полости.
Снаружи яичники покрыты зачатковым эпителием, который
врастает в виде тяжей внутрь соединительнотканной основы —
стромы, с расположенными в ней крупными интерстициальными
клетками. К зачатковому слою примыкает фолликулярная зона,
где в особых клеточные комплексах — фолликулах, развиваются
половые клетки. Здесь же находятся крупные интерстициальные
клетки, продуцирующие половой гормон эстрон. Глубже, лежит
сосудистая зона с кровеносными сосудами.
В овогенезе—развитии женских половых клеток (яиц) сме-
няют друг друга три периода: размножение, рост и созревание.
В первый период происходит деление половых клеток — ово-
гониев, в результате чего возрастает их количество. Во второй —
яйцеклетки растут и превращаются в овоциты I порядка. Эти
этапы яйцеклетка проходит в фолликуле, который по мере
своего' развития постепенно перемещается в поверхностную
часть яичника. Здесь он превращаете^ в так называемый гра-
афов‘пузырек., Лопаясь, граафов пузырек освобождает вместе
с наполняющей его жидкостью и яйцеклетку — этот процесс
носит название овуляции. .На месте лопнувших граафовых пу-
зырьков из оставшихся периферических клеток фолликулярного
эпителия развивается новый, слой'клеток характерного желтого
цвета вследствие находящегося -в ;них пигментно-жирового ве-
щества лютеина. Сформировавшееся таким способом небольшое;
желтоё‘"тело овуляции у самок, оставшихся холостыми, вскоре*
резорбируется, а у. оплрдотворенных разрастается и превра-
щается в желтое тело беременности, временно выполняющее
функции органа внутренней секреции. Выделяемый этим орга-'
ном гормон прогестерон обеспечивает нормальное развитие
зародыша.
Остальные отделы половой системы самки, служащие в ос-
новном для выделения созревших яйцеклеток и вынашивания-
зародышей, представляют собой производные парного яйцево-
да, который соответствует мюллеровым каналам. В начальной
своей части это собственно яйцевод (oviductus), или фаллопиева
труба (tuba Fallopii) — парный трубчатый орган, открываю-
щийся воронками в полость тела близ яичников и впадающий
с другой стороны в матку. Выпавшие из яичника яйцеклетки
попадают через полость тела в .воронку яйцевода и затем в фал-,
лопиеву трубу. После выделения направительных телец они-
превращаются в овоциты II порядка и, наконец, в зрелые яйца
с гаплоидным числом хромосом.
Зрелые яйца млекопитающих имеют шаровидную форму.
У клоачных они содержат большое количество питательных ве-
ществ и достигают относительно-крупных размеров (2,5—4,0 мм;-
в матке они увеличиваются до 15 мм и более). Яйца сумчатых
87'
значительно меньших размеров (0,2—0,4 мм), а у высших мле-
копитающих микроскопически малы (0,05—0,2 мм).
В фаллопиевой трубе происходит оплодотворение яйца,пре-
образующегося таким образом в зиготу1. Последняя тут же на-
чинает дробиться, образуя под яйцевой оболочкой дочерние
Рис. 32. Схема дифференцировки яйцеводов млекопитающих. А —. клоач-
ное; Б — молодая двуутробка; В — взрослое сумчатое; Г — плацентарное
млекопитающее с двойной маткой; Д — с двурогой маткой; Е — с простой
маткой:
cl — клоака; od — яйцеводы; г — прямая кишка; swg — мочеполовой синус; иг — моче*
точник; at — матка; vag — влагалище; vu — мочевой пузырь (по Шмальгаузену)
клетки — бластомеры1 2. В фазе морулы бластомеры дифферен-
цируются на два слоя: наружный — трофобласт (питающий ли-
сток) и внутренний — эмбриобласт (зародышевый листок).
В дальнейшем основная масса клеток эмбриобласта формирует
зародышевую пластинку — зачаток эмбриона, остальные клетки
образуют стенку желточного мешка. Перистальтическим сокра-
щением стенок фаллопиевой трубы и соответствующими дви-
жениями ресничек ее эпителия яйцо продвигается в матку.
Матка (uterus), служащая вместилищем для развивающе-
гося зародыша, у клоачных и сумчатых представлена парными,
полностью раздельными расширениями яйцеводов (рис. 32).
1 Неоплодотворенные яйца погибают и рассасываются. Количество вы-
деляемых яиц варьирует в соответствии с большей или меньшей плодови-
тостью отдельных видов.
2 Яйца млекопитающих проходят полное (голобластическое) дробление.
Исключение составляют яйца клоачных, у которых большее количество пи-
тательного желтка и дробление меробластическое.
88
У высших млекопитающих наблюдаются все переходы от парной
матки к непарной. В зависимости от степени слияния маточных
отделов яйцеводов различают двойную матку (uterus duplex)
с сильно сближенными дистальными концами маточных отде-
лов, но с самостоятельными выводными отверстиями последних
(некоторые рукокрылые, грызуны, трубкозубы, слоны); двураз-
дельную матку (u. bipartitus), образованную частично сливши-
мися маточными отделами с одним общим выводным отвер-
стием (ряд грызунов, хищных, копытных); двурогую матку
(u. bicornis), в значительной степени утратившую парность (на-
секомоядные, ряд рукокрылых, хищных, копытных, китообраз-
ные, полуобезьяны); простую матку (u. simplex), полностью
утратившую парность (некоторые рукокрылые, приматы).
В слизистой оболочке матки имеются железы, секрет кото-
рых— эмбриотроф («маточное молоко»), обеспечивает в пер-
вое время- питание зародыша. Характерно также сильное раз-
витие крупных кровеносных сосудов, доставляющих питатель-
ный материал плоду, и мускулов стенки матки, с помощью
которых выводится наружу созревший плод.
При переходе в матку дробящийся зародыш освобождается
от яйцевой оболочки и функцию наружной оболочки принимает
на себя трофобласт. Клетки трофобласта оказывают биохими-
ческое воздействие на сильно измененную стенку матки, вызы-
вая в месте соприкосновения ответные реакции с ее стороны,
что и обусловливает возможность прикрепления зародыша —
имплантацию.
Вступив в тесный контакт с материнским организмом, заро-
дыш начинает быстро расти, происходит дифференцировка эле-
ментов. Исключительно важное значение при этом имеет форми-
рование плодных оболочек. Ближайшая к зародышу оболочка —
амнион, возникает в результате углубления зародышевой.
пластинки в толщу плодного пузыря и смыкания над ней склад-
ки наружного зародышевого листка. Нижняя часть складки
образует амнион, а верхняя — наружную оболочку — прохорион,
на всей поверхности которого развиваются особые выросты —
ворсины. Выпячивание задней части первичной кишки дает на-
чало третьей свойственной всем амниотам зародышевой обо-
лочке— аллантоису. Разрастаясь, аллантоис, внедряется в про-
странство между амнионом и прохорионом, а его кровеносные
капилляры врастают в ворсины прохориона, превращая тем са-
мым последний в хорион — сосудистую оболочку.
У большинства низших зверей связь зародыша с матерью-
осуществляется только прилеганием его к стенке матки. У не-
которых сумчатых и у всех высших (плацентарных) млекопи-
тающих развивается специальный временный орган связи плода,
с телом матери — плацента — детское место, послед (рис. 33).
Закладывается этот орган путем внедрения ворсин хориона в
8».
«стенку матки, чем обеспечивается тесная связь между кровенос-
ными системами матери и зародыша. -Через плаценту происхо-
дит поступление в зародыш из крови матери питательных ве-
ществ и кислорода и выделение из зародыша продуктов распада
Рис,. 33. Схема строения плаценты. Слева — разрез через матку, справа —
внешний вид. А — дольчатая плацента жвачных; Б — кольцевая плацента
хищников: '
j _ амнион; 2 — аллантоидный пузырь; '3 —тпофобляст (хорион); 4 — котиледоны;
5 — кольцевая плацента; 6 — рудимент желточного мешка (по Портманну)
:й углекислоты. Сама плацента выделяет секреты, действующие
.как на зародыш, так и на организм мл 1 ери.
По способу прикрепления различаются плаценты неотпадаю-
щие и отпадающие. Неотпадающая плацента (placenta adeci-
<iuata) характеризуется слабой связью со стенкой матйи:'ееворк
SO
сины входят в соответствующие углубления слизистой оболочки
матки, не срастаясь с ней, благодаря чему при родах матка не
повреждается1. Отпадающая плацента (pl. deciduata), наоборот,
настолько тесно срастается со стенкой матки, что при родах
часть слизистой оболочки ее отпадает, вызывая более или менее
значительное кровоизлияние1 2. Между этими двумя основными
типами есть переходная форма 3.
После родов в стенке матки на месте имплантации эмбрио-
нов остаются так называемые плацентарные пятна 4.
Конечная часть половых путей! самки, служащая для выве-
дения плода и принятия семени самца при копуляции, носит
название влагалища (vagina). С ней связаны парные бартоли-
ниевы железы (гомологи куперовых желез самцов), секрет ко-
торых увлажняет влагалище. У сумчатых два влагалища, кото-
рые иногда образуют кольцеобразное сращение со слепым, вы-
ростом, отходящим к мочеполовому синусу.. При родах этот
вырост прорывается и тогда возникает так называемое третье
влагалигце (некоторые кенгуру, вомбат).
В конечный (наружный) отдел влагалища открывается мо-
чевой пузырь, далее начинается короткое преддверие — моче-
половой канал, заканчивающийся наружным отверстием — вуль-
вой. С боков вульва ограничена складками — большими губами
(у приматов есть'также более глубоко лежащие малые губы),
а спереди продолговатым выростом —клитором (гомологом
пениса) / Клитор состоит из пещеристых тел и головки и у не-
которых видов (грызунов, медведей, куньих) снабжен особой
косточкой ‘(os clitoridis). У сумчатых, грызунов, копытных и
приматов вход во влагалище суживается кольцевой перепонкой
(девственной плевой), разрываемой при первом совокуплении.
У молодых самок кротов половое отверстие отсутствует, а у
взрослых бывает открыто только в период размножения.
ВТОРИЧНЫЕ ПОЛОВЫЕ ПРИЗНАКИ
Как известно, у многих животных самцы и самки одного
вида отличаются друг от друга не только первичными половыми
признаками, но .и целым рядом вторичных половых признаков,
1 Неотпадающая плацента свойственна некоторым сумчатым, ящерам,
лошадям, свиньям, верблюдам, сиренам, китам, лемурам.
2 В такой плапенте различается материнская, или маточная — отторгаю-
щаяся часть слизистой оболочки матки, и зародышевая часть (ворсины хо-
риона). Отпадающую плаценту имеют насекомоядные, шерстокрылы, руко-
крылые, грызуны, американские неполнозубые, обезьяны;
3 У жвачных.
4 Подсчет плацентарных пятен у добытых самок используют для уста-
новления количества детенышей в помете и числа пометов в сезон размно-
жения. Метод этот еще недостаточно разработан и требует осторожного-
применения Во избежание ошибок.
91.
обусловливающих половой диморфизм. Что касается млекопи-
тающих, то у них половой диморфизм выражен сравнительно
слабо, преимущественно в размерах и общем внешнем облике:
самцы обычно несколько крупнее самок, обладают более креп-
ким телосложением, волосяной покров у них несколько
грубее.
Рис. 34. Половой диморфизм. А — у антилоп гарна (Antilope bezoar-
tied); Б — у кашалотов (Physeter catodon)
Только у небольшого числа видов зверей половой диморфизм
выражен резко (рис. 34). Так, например, самцы кашалотов, уша-
тых тюленей, некоторых обезьян значительно превосходят раз-
мерами самок. У беззубых китов и некоторых других, наоборот,,
самки заметно крупнее самцов. Клыки, бивни, рога, пахучие
железы, грива (у львов, оленей, ушатых тюленей), баки, борода
более развиты у самцов или свойственны только им. У части
видов зверей половой диморфизм проявляется и в окраске.
92
Вторичные половые признаки облегчают взаимные поиски
особей другого пола, способствуют осуществлению полового
отбора, влияя на особенности поведения, содействуют синхрони-
зации протекания половых циклов самцов и самок (см. «Раз-
множение») .
Большинство вторичных половых признаков развивается под
влиянием половых гормонов, что доказывается сглаживанием
различий между особями разных полов, наступающим вслед
.за кастрацией', или, в естественных условиях, в случае патоло-
гических (чаще всего травматических) нарушений деятельности
половых желез.
1 Так, например, кастрированный в раннем возрасте самец оленя остается
всю жизнь безрогим; у кастрированного взрослого оленя развиваются урод-
ливые рога.
Окраска, ее физиологическое
и биологическое значение
Окраску млекопитающих в основном обусловливают пигмен-
ты, сосредоточенные в волосах. Кожа обычно бывает окрашена
лишь при полном отсутствии волос (например, у китов, сирен)
или на отдельных оголенных участках (у летучих мышей„
обезьян),
Как правило, окраска млекопитающих не отличается ярко-
стью и богатством красок; им свойственны лишь две категории
пигментов — зернистые черные и жидкие желтые (рыжие) К
Сочетания этих пигментов и обусловливают общую окраску.
В подавляющем большинстве это бурые, коричневые и серые
тона 1 2.
Волосяной покров млекопитающих бывает одноцветный, по-
лосатый или пятнистый. Согласно существующей гипотезе, ист
ходной окраской в филогенезе следует считать продольнополо-
сатую. Из нее, путем разрыва полос, возникла пятнистая, кото-
рая, в свою очередь, дала начало поперечнополосатой (в ре-
зультате соответствующего слияния пятен) и одноцветной (при:
редукции пятен).
Эта гипотеза основана на анализе изменений окраски, на-
блюдающихся в онтогенезе ряда видов. Детеныши диких сви-
ней, тапиров, пум имеют полосатую окраску, молодые настоя-
щие олени, маленькие львята — пятнистую, для эмбрионов зай-
'цев и жеребят домашних лошадей характерна черная спинная
полоса («ремень»), а взрослые животные всех этих видов, за
редкими исключениями, одноцветны.
В зрительном восприятии окраски волосяного покрова час-
тично играет роль и преломление световых лучей соответствен-
но структуре волос (наиболее ярко иридирующая окраска вы-
ражена у златокротов).
1 Как те, так и другие принадлежат к группе меланинов. Желтые пиг-
менты, ранее относимые к группе липохромов, сейчас рассматриваются как
те же меланины, но неполно окисленные.
2 Отсутствие яркой окраски у подавляющего большинства, млекопитающих
связано с тем, что при отыскании друг друга они пользуются главным об-
разом слухом и обонянием, а не зрением, которое у них развито сравни-
тельно слабо. Исключение в этом отношении составляют обезьяны, в окраске-
которых как раз и встречаются яркие цвета (красный, желтый, зеленый,
голубой).
94
Окраска млекопитающих подвержена индивидуальной, воз-
растной, половой, сезонной и географической изменчивости.
Индивидуальная изменчивость чаще всего вы-
ражена в вариациях тонов (с постепенными переходами между
ними у особей одного вида). В некоторых случаях наблюдается
диморфизм в окраске, например у песцов, представленных,
двумя цветными морфами — «голубой» и белой (те и другие
иногда встречаются в одном и том же выводке).
Рис. 35. Серебристо-черная и белая лисицы как примеры мела-
низма и альбинизма (из Тылинека)
К категории индивидуальных вариаций окраски можно от-
нести также цветовые аберрации (отклонения от нормы): аль-
бинизм, т. е. частичное или полное выпадение пигментов, хро-
мизм (руфинизм)—выпадение черного пигмента при неизмен-
но остающемся желтом, и меланизм — выпадение- или слабое-
развитие желтого пигмента при гипертрофии черногоь
(рис. 35).
Наиболее широко распространены в природе- меланисты,.
образующие у некоторых видов стойкие морфы (черно-серебри-
стые и черно-бурые лисицы, черные бобры, «голубые» песцы).
Довольно часто встречаются также хромисты, но сравнительно-
редкое явление представляют альбиносы, элиминируемые отбо-
1 Все эти аберрации могут передаваться, по наследству, ио чаще они'
представляют случайнее, не наследуемые особенности.
95.
ром в первую очередь вследствие своей заметности. К тому же
альбиносы отличаются общей пониженной жизнеспособностью *.
Как результат одомашнения нередко возникает пегая
окраска.
Из других отклонений наблюдается исчезновение обычной
для большинства млекопитающих зонарности в окраске волос,
появление седины (вследствие депигментации отдельных волос),
пегости (пятнистости) у видов, которым обычно она не свой-
ственна и пр.
Интересна наблюдающаяся у некоторых млекопитающих
коррелятивная связь окраски с другими морфологическими при-
знаками, полом и поведением. Так, рыси-руфинисты обладают
более жестким и низким мехом по сравнению с серыми особя-
ми; белые голубоглазые кошки и белые норки обычно бывают
глухи; трехцветную (бело-черно-рыжую) окраску у кошек име-
ют только самки; все дико живущие альбиносы отличаются
повышенной пугливостью и ,т. д.
Хорошо выражена возрастная изменчивость окрас-
ки, которая может идти не только по пути замены полосатости
и пятнистости детенышей однотонной окраской взрослых (дикие
свиньи, настоящие олени и некоторые другие), о чем говорилось
выше, но и по другим линиям. Например, новорожденные ко-
тики имеют блестяще-черную окраску («черные котики»), заме-
няющуюся у взрослых животных серой или бурой1 2, наоборот,
у настоящих тюленей белые при рождении детеныши («бельки»)
с возрастом приобретают более темную окраску. Черновато-
бурый мех лисенка сменяется у взрослой лисицы рыжим. Мед-
вежата у бурого медведя рождаются почти черными с белым
«ошейником»; с возрастом они светлеют и утрачивают ошейник.
Молодые белухи имеют темную окраску — взрослые становятся
белыми (рис. 36).
У ряда видов окраска подвержена половой изменчи-
вости: у котиков самцы бурого цвета, самки серого, самцы
гренландских тюленей отличаются от самок ярко выраженной
полосатостью окраски, у многих оленей самцы темнее, чем сам-
ки, у самцов мандрилов окраска голых участков кожи ярче,
чем у самок, и т. д.
1 Уже Ч. Дарвин (1895) указывал на то, что белые особи домашних жи-
вотных менее устойчивы к заболеваниям (к отравлению ядовитыми расте-
ниями), чем темноокрашенные. Позднее было установлено пониженное со-
держание антитоксинов в крови светлоокрашенных домашних животных.
Интенсивность обмена веществ у альбиносов ниже, чем у нормально окра-
шенных особей. В качестве примера стойких альбинистических морф можно
привести белых мышей, белых крыс, белых кроликов. Отдельные случай
альбинизма известны у соболей, куниц, каланов, норок, рысей, волков, лисиц,
медведей, кротов, белок, дельфинов и др.
2 Черный цвет поступающего в продажу котикового меха не натуральный.
96
У большинства зверей, обитающих в областях с резко выра-
женной сменой сезонов, существует сезонная изменчи-
вость структуры и окраски меха. Обычно зимний мех бывает
светлее летнего. Многие темноокрашенные в летний период
Рис. 36. Различия в детском и взрослом нарядах. А — косули
(Capreolus capreolus); Б — кабана (Sus scrofa); В — белухи (Del-
phinapterus leucas)
виды зверей северных и умеренных широт на зиму белеют. Тако-
го рода сезонный диморфизм ярко выражен у белого песца, зай-
ца-беляка, некоторых подвидов русака, горностая, ласки, ко-
пытного лемминга, джунгарского хомячка, сайги, некоторых рас
северных оленей и др.
Особого рода сезонные изменения окраски наблюдаются
у оленей. В летний период настоящие олени имеют одноцветный
рыжеватый наряд; к зиме он сменяется многоцветным (появ-
ляется серый тон с темными участками). Пятнистая летняя
окраска пятнистых оленей зимой сменяется одноцветной. Косули
летом имеют рыжую шерсть, зимой — серую.
Сезонные изменения общей окраски могут зависеть также
от роста и изнашивания зонарно окрашенных волос.
7 Териология
97
Наконец, географическая изменчивость окраски
волосяного покрова проявляется в образовании локальных кра-
сочных форм. Особенно хорошо выражены такие формы у ли-
сиц, соболей, белок. С другой стороны, у лесных хорьков, хомя-
ков, бурундуков они почти отсутствуют.
Звери, населяющие холодные области, имеют обычно менее
насыщенную окраску, чем живущие в теплых областях, что до-
статочно хорошо прослеживается на большинстве видов. Белею-
щие на севере своих ареалов звери, на юге ареалов в большей
или меньшей степени сохраняют летнюю окраску и зимой'.Име-
ются, однако, исключения. Например, в крайне континенталь-
ном холодном климате Якутии широко распространены мела-
нистические вариации лисиц, сурков, бурых медведей; соболь,
беляк, суслик представлены здесь темными расами.
Сезонная и географическая изменчивость окраски обнару-
живает более или менее ясно выраженную зависимость от кли-
матических условий. Влияние температуры на пигментацию
развивающихся волос подтверждается экспериментально. Так,
действуя холодом на обритые участки кожи белых кроликов,
можно вызвать появление на них черных волос (Ильин, 1926). На-
оборот, воздействие высокими температурами приводит к раз-
витию светлого меха, что экспериментально показано на неко-
торых куньих (Огнев, 1951). В свете этих данных можно счи-
тать, что темная летняя окраска обусловлена тем, что летний
волос закладывается при сравнительно низких ранневесенних
температурах (в марте — апреле), в то время как более свет-
лый зимний мех закладывается при относительно высоких ран-
неосенних температурах (в августе).
Звери стран с сырым морским климатом имеют более насы-
щенную окраску, чем обитатели стран с сухим континентальным
климатом1 2. Среди многих форм белок, например, наиболее свет-
лым мехом обладает телеутка — обитатель сухих ленточных бо-
ров, белки же сырых лесов лесной зоны заметно темнее. Ту же
закономерность обнаруживает окраска косуль, оленей, некото-
рых кошек, волков и ряда других зверей. Меланистические ва-
риации чаще всего встречаются в местностях с относительно вы-
сокой влажностью.
1 Отмеченные явления послужили в свое время основанием для выдви-
жения так называемой «термической теории окраски» [Калантар (Kalantar,.
1927)], по которой белая (и вообще светлая) окраска уменьшает излучение
тепла и таким образом выгодна животным, обитающим в условиях низких
температур (закон Кирхгофа). Однако произведенные позднее специальные-
исследования не подтвердили эту теорию, показав, что светлая окраска во-
лосяного покрова не оказывает сколько-нибудь заметного влияния на спо-
собность животного сохранять внутреннее тепло [Свила (Svihla), 1956].
2 Эта закономерность соответствует установленному на птицах «клима-
тическому правилу Глогера».
98
Фотопериодизм, действуя через центральную нервную си-
стему, гипофиз, щитовидную железу, контролирует сезонную
смену волос и вместе с тем «перекраску» животных. Сокраще-
ние светового дня (в условиях эксперимента) приводит к потем-
нению меха у лисиц, песцов, соболей.
Наконец, не безразличны для окраски и физиологические
свойства организма животных. Установлена, например, зависи-
мость интенсивности пигментации меха зверей (в эксперимен-
тальных условиях) от состояния их упитанности, подбора кор-
мов и пр.
Все рассмотренные явления указывают на то большое зна-
чение в развитии окраски животных, которое принадлежит фор-
мообразующей роли внешней среды, действующей как непосред-
ственно, так и через изменения физиологических механизмов.
Антидарвинисты отрицают биологический смысл широко
распространенного в.природе соответствия окраски животных
основному фону среды, т. е. явления маскирующей, или криптиче-
ской, окраски (нашедшего свое объяснение в дарвиновской тео-
рии естественного отбора). В качестве серьезных доводов для
такого отрицания указывают на наличие маскирующей окраски
у ряда ночных и подземных форм, образ жизни которых якобы
исключает необходимость маскировки (например, у пустынных
летучих мышей, слепушонки), на существование в одной и той
же полярной обстановке как форм, белеющих зимой (песец, бе-
ляк, горностай) или носящих белый наряд круглый год (белый
медведь), так и форм, остающихся в течение всего года темными
(мускусный бык, росомаха), и на некоторые другие факты.
Однако дарвиновская точка зрения, трактующая происхож-
дение и значение маскирующей окраски как результат естест-
венного отбора, действующего на основе оценки визуального
эффекта окраски (вызывающего обман глаза врага), находит
подтверждение в огромном фактическом материале.
Бурые и серые тона хорошо маскируют млекопитающих на
фоне травяного покрова, светлая или чисто белая окраска де-
лает их незаметными на снегу или на светлом фоне песчаного
ландшафта, пятнистость и полосатость прекрасно гармонирует
с окраской лесной подстилки и солнечными бликами на ней.
Сходство окраски у систематически очень далеких друг дру-
гу животных, например у хищников и их жертв, обусловливает-
ся одинаковой необходимостью маскировки: для первых в целях
нападения, для вторых —в целях защиты. Сезонные изменения
волосяного покрова представляют ярко выраженную адаптацию
к сезонным изменениям температуры и фона внешней среды.
С этой точки зрения понятно, почему в снежных северных ча-
стях своих ареалов ласка полностью белеет, русак сильно свет-
леет, на юге же в условиях незначительного и недолговремен-
ного снежного покрова они сохраняют темную окраску круглый
7* 99
год. Только в областях с длительным снежным покровом (на-
пример, в юго-западной Сибири) становятся белыми зимой
джунгарские хомячки (в Забайкалье или в Монголии с их бес-
снежными зимами они не белеют) и т. д.
В ряде случаев наблюдается поразительная «подогнанность»
окраски животных к цвету субстрата. В пустынях Северной Аме-
рики мелкие грызуны (Peromyscus, Perognathus) представлены
Рис. 37. Расчленяющая
расцветка млекопитающего,
контуры (из Котта)
маскирующая его
на белесоватых песках светлыми (иногда почти белыми) ра-
сами, а на рядом расположенных участках темных извержен-
ных лав — темными или совершенно черными. Так как песчаные
и лавовые участки расположены на одинаковой высоте над
уровнем моря в совершенно одинаковых климатических усло-
виях и прямое воздействие почвы на окраску отсутствует, воз-
никновение соответствующих фону светлых и темных рас гры-
зунов можно объяснить только отбором, действующим через
хищников [Джемсон (Jameson), 1898; Осгуд (Osgood), 1909;
Бенсон (Benson), 1932, 1933; Дайс и Блоссом (Dice a. Blossom),
1937 и др.].
- В соответствующих условиях прекрасно скрывает животных
и пестрый наряд. Пятнистая окраска, например, «растворяется»
на фоне древесной листвы. Понятно поэтому значение интерес-
ного явления, наблюдающегося у японских оленей: в лесах с
опадающей листвой они на зиму сменяют пятнистый наряд на
однотонный (что свойственно также лани и пятнистому оленю),
но в вечнозеленых лесах пятнистость сохраняется у них круглый
год [Котт (Cott), 1940].
л Пятна и полосы служат своего рода камуфляжем — средст-
вом маскировки, оптически расчленяющим и искажающим ре-
альные очертания. животного и тем самым затрудняющим его
Опознавание (рис. 37).
100
Такое значение, по-видимому, имеет пятнистость жирафы,
ягуара, полосатость зебры, тигра ’.
Из частей тела животного наибольшее внимание привлекают
глаза. Даже очень хорошая маскировка в некоторых случаях
Рис. 38. Маскировка .глаза. А —у сернобыка (Oryx gazelid)-,
Б — барсука (Meles meles); В — вискачи (Viscacia maxima)
утрачивает смысл, если она не скрывает резко очерченных глаз-
ных кругов. Поэтому в процессе эволюции у ряда видов выра-
ботались особые элементы окраски, маскирующие глаза, а имен-
но — полосы, проходящие через них или круглые пятна на ли-
цевой поверхности, среди которых теряются пятна глаз. В ка-
1 X. Кирби (Kirby, 1896) о скрадывающем действии окраски жираф го-
ворит следующее: «Таинственность исчезновения (жираф) просто порази-
тельна. Я часто подбирался к ним, и если в этот момент какой-либо дру-
гой объект отвлекал на несколько мгновений мое внимание, то, взглянув
снова, я видел, что они исчезли. Не просто скрылись, отойдя на какое-то
расстояние, но буквально исчезли, пропали, как дымка тумана при восходе
солнца. Сколь это ни кажется странным, немногие животные так легко исче-
зают из вида, если на мгновение отвлечь от них свое внимание». Он же
говорит, что среди растительности в сумерки совершенно незаметными на
самых близких расстояниях остаются зебры.
А. Уоллес (Wallace, 1889) приводит следующие слова известного охот-
ника за тиграми Уолфорда: «Однажды, преследуя раненого тигра, я по край-
ней мере с минуту не мог рассмотреть его в траве под деревом на расстоя-
нии всего 20 ярдов в открытых джунглях, и только туземцы заметили его;
наконец и мне удалось настолько хорошо рассмотреть его, что можно было
стрелять, но и тогда я не мог разобрать, в какую часть тела я стреляю».
По словам наших отечественных тигроловов, в камышах тигр при ярком
освещении совершенно невидим.
101
Рис. 39. Скуис, или вонючка;
пример окраски апосематического
типа
честве хороших примеров камуфлированных глаз можно ука-
зать на глаза перевязки, енота, вискачи, сернобыка, пантеры,
барсука, лесной сони (рис. 38).
Несомненно, защитное значение имеет общая закономер-
ность в распределении окраски верхней и нижней поверхностей
тела: как правило, спинная сторона у млекопитающих, как и у
других животных, темнее нижней. Это приводит к скрадыванию
формы тела, мнимому уплоще-
нию поверхности
Наличие в фауне снежных об-
ластей наряду со светлоокрашен-
ными формами также и темных
обусловлено не одинаковой по-
требностью животных в маски-
ровке. Постоянно темный мускус-
ный бык — стадное животное,
фактически не имеет врагов, от
которых ему нужно было бы
укрываться, пользуясь маскиров-
кой, как это делает заяц-беляк;
не нуждается он как раститель-
ноядный вид и в скрадывании
добычи, к чему часто прибегают
криптически окрашенные хищни-
ки (например, белый медведь,
песец).
Вполне понятно, что маски-
рующая окраска проявляет свои
защитные свойства главным об-
разом при неподвижном состоя-
нии животного (во время отдыха
или затаивания). Однако и в мо-
мент бегства она может очень
помочь отделаться от преследования, особенно в сумерках в гус-
той траве и т. п. Криптическая окраска выражена у самок в
большей степени, чем у самцов, что вполне целесообразно, так
как более сильные и подвижные самцы испытывают в ней мень-
шую нужду.
Ряду млекопитающих свойственна предостерегающая, или
предупреждающая' (апосематическая), окраска, представляю-
щая полную противоположность криптизму. Для нее характер-
1 Согласно оптическому принципу противотени («закон Тайера»), при
однообразной окраске тела , верхняя, наиболее освещенная его поверхность
резко контрастирует с нижней, находящейся в тени, что придает рельефность,
а. стало быть, и заметность телу; наоборот, при наличии тёмноокрашенного
верха и. светлой нижней поверхности действие тени и света уравновешивается,
обусловливая незаметность тела [Тайер (Thayer), 1856—1912].
102
ны бросающиеся в глаза контрастные цветовые пятна, как бы
предупреждающие врага о несъедобности их носителя или о
способности его дать жестокий отпор. Классический пример ок-
раски апосематического типа представляют вонючки, обладаю-
щие яркой (сочетание белых и черных полей) окраской (рис.
39) и крайне едкими выделениями вонючих желез, дикобразы,
способные пучками острых и длинных игл нанести тяжелые
раны даже волку, тигру и др.
Рис. 40. Устрашающая окраска бамбукового медведя (Ailuropus
melanoleucus) — черные круги около глаз, контрастная окраска
частей тела
Представляющую отступление от закона Тайера черную ок-
раску брюха у хомяка и барсука нужно рассматривать тоже
как устрашающую (отпугивающую), поскольку хомяк защища-
ется стоя, а барсук отбивается, лежа на спине. То же значение
придают тем редким случаям, когда, как например, у бамбуко-
вого медведя, глаза не маскируются окраской, а, наоборот, при-
обретают большую заметность благодаря черному окологлаз-
ному кольцу (рис. 40), резко выделяющемуся на светлом фоне
головы (Фаусек, 1913).
Имеется еще один элемент окраски млекопитающих — опо-
знавательные пятна. В наряде многих видов, ориентирующих-
ся с помощью зрения, есть отметины или «сигнальные пятна»,
сильно контрастирующие с общим фоном окраски, но мало или
говеем незаметные во время покоя животных. Стоит им вско-
чить при тревоге с лёжек, и внезапно появившиеся белые «зер-
кала», приподнятые кверху черные султаны хвоста и т. п.
послужат стадным животных сигналом опасности. Наличие
таких пятен, нередко способных сокращаться во время по-
103
коя и увеличиваться при опасности, например, благодаря меня-
ющемуся наклону белых волос у зеркала (вилорогая антило-
па), облегчает стадным животным ориентировку, связь между
особями во время стремительного бега, а молодым — следова-
ние за матерью в темноте и т. д.
Приведенные примеры приспособительной окраски подтвер-
ждают правильность эволюционного учения Дарвина. Критика
этой теории носит односторонний характер, так как антидарви-
нисты недоучитывают того, что адаптивные окраски, как и вся-
кие другие приспособления, имеют относительное значение. Од-
нако общая эффективность их действия не вызывает сомнений
1 Хорошая, хотя и не лишенная некоторых недостатков сводка данных
о приспособительном значении окрасок животных содержится в книге
X. Котта (Cott) «Приспособительная окраска животных», 1940 (русский
перевод, 1950).
Адаптивные типы
млекопитающих
Приспособленность к жизни в определенной среде, опреде-
ленных природно-географических зонах, биотопах находит яр-
кое выражение в общей организации различных групп млеко-
питающих, во внешнем их облике и особенно в строении и функ-
циях органов движения.
По исторически обусловленной связанности с той или иной
средой выделяют группы зверей наземных, подземных (земле-
рои), древесных, воздушных (летающих) и водных (с перехода-
ми между ними).
НАЗЕМНЫЕ ФОРМЫ
В состав этой обширной и разнообразной группы входят как
мало специализированные, широко распространенные виды,,
так и виды определенных ландшафтов — тундровые, лесные,
степные, пустынные, горные. Общая организация наземных мле»
копитающих не представляет существенных отклонений от типа,
поэтому ниже мы рассмотрим главным образом лишь особен-
ности движения отдельных форм данной группы.
Основные типы поступательного движения наземных млеко-
питающих— ходьба и бег; по мере ускорения сменяют друг
друга шаг, рысь, галоп и карьер (рис. 41). Первые два аллюра
характеризуются более или менее быстрым попеременным пере-
мещением ног по диагонали (правая передняя, левая задняя —
левая передняя, правая задняя). Так передвигается большин-
ство копытных, собачьи, кошачьи. У жираф, слонов, верблюдов,
медведей, некоторых лошадей встречается разновидность ры-
си— иноходь, своеобразие которой заключается в попеременной
односторонней перестановке ног (правая передняя, правая зад-
няя— левая передняя, левая задняя). Иноходь способствует до
некоторой степени ускорению бега.
Галопом, т. е. поступательным движением при помощи рит-
мических прыжков, и карьером (предельно быстрым аллюром)
большинство зверей пользуется, убегая от опасности или пре-
следуя добычу.
Способность передвигаться прыжками развита у разных на-
земных зверей далеко не одинаково. Совершенно не прыгают
ежи, дикобразы, барсуки и другие медленно передвигающиеся
виды. Постоянно сочетают разные аллюры с прыжками почти
все хищные (особенно кошачьи), многие грызуны и. копытные.
105
Многие звери, передвигаясь галопом, заносят при каждом
прыжке задние ноги далеко вперед, за линию передних ног, от-
ходящих в этот момент назад. Вследствие этого получается ха-
рактерный след с отпечатками задних конечностей впереди пе-
редних (зайцы, белки и многие др.).
Рис. 41. Бег млекопитающих. А — рикошетирующими прыжками (кенгуру);
£ — прыжками галопом (заяц); В — нормальной рысью (северный олень);
Г — иноходью (слон)
Крайней степени развития способность к прыганию дости-
гает у кенгуру, тушканчиков, кенгуровых крыс (Heteromyidae), -
прыгунчиков (Macroscelididae) и некоторых др. У этих живот-
ных удлинение задних конечностей сопровождается сокращени-
ем количества пальцев (до двух у кенгуру) и прочным соедине-
нием между собой костей заплюсны. В то же время передние
конечности редуцируются настолько, что утрачивают функцию
опоры при быстром беге.
Очень сильное развитие мышц разгибателей спины, увеличи-
вающее мощность толчка вперед, и компактность тела также,
несомненно, связаны со скакательным типом движения '. Типич;
1 Компактность тела обусловлена укорочением шеи (путем частичного
слияния шейных позвонков) и концентрацией мощной мускулатуры конеч-
106
ные прыгуны способны совершать огромные прыжки. Для туш-
канчиков, например, указываются рекордные прыжки длиной
3 м, для кенгуру — 7 и даже 12 м [Тротон (Troughton), 1947].
Характерная особенность тушканчиков и других прыгунов—1
«рикошетирующий бег», осуществляемый прыжками из стороны
в сторону как бы вследствие неровностей почвы. Зигзагообраз-
ное движение и мелькание белой кисточки («знамени») на кон-
це длинного хвоста очень затрудняют преследование зверька
хищником. Длинный хвост у прыгунов играет роль балансира и
руля, позволяющего изменять направление стремительного бега
иногда под прямым углом'. При медленном передвижении и у
сидящего животного хвост служит дополнительной точкой
опоры.
Быстрота передвижения особенно важна в открытых ланд-
шафтах— степях, пустынях, где мало укрытий от врагов и при-
ходится преодолевать большие расстояния в поисках пищи и
убежищ. Лесные наземные звери также могут передвигаться
более или менее быстро, но здесь их бег осложняется необходи-
мостью лавировать между деревьями. В общем лесные формы
уступают в быстроте бега степным и пустынным. Предельную
максимальную скорость бега — около 120 км в час — имеет
житель пустынь гепард [Гильдебранд (Gildebrand), 1959]. Со
скоростью около 100 км в час могут бегать некоторые антило-
пы, львы. Зебры, джейраны имеют скорость бега до 65 км в час,
жирафы, кенгуру — около 50 км. Обитатель степи — бизон раз-
вивает скорость до 40 км и более в час.; лесной дикий бык —
бантенг бегает не быстрее 25 км в час [по А. Хоуэллу (Howell)
и др., 1932]. .
Способность к быстрому бегу обусловлена целым комплек-
сом приспособлений в строении конечностей, мышц, кровенос-
ной системы и органов дыхания (в частности, адаптивное зна-
чение имеет расширение нижнего носового хода у хороших бе-
гунов, что обеспечивает свободный проход воздуха в легкие).
Однопалость лошадей, малая подвижность и сближенность
пальцев у степных парнокопытных — дзеренов, джейранов, по-
зволяют им быстро передвигаться только по твердому грунту в
условиях равнинного рельефа* 1 2. Обитатели песчаных пустынь,
болотистых местностей и многоснежных северных областей, т. е.
звери, передвигающиеся по сыпучему или топкому субстрату,
не увязают в нем благодаря большой опорной площади подошв
ностей в их проксимальной части (в то время, как в дистальной находятся
лишь сухожилия).
1 У некоторых видов тушканчиков хвост лишь немного превышает длину
Тела, но это компенсируется его утолщением. Ампутация хвоста у тушкан-
чика сразу нарушает координацию движений зверька.
2 У степных тушканчиков подобие однопалое™ достигается тесным
сближением пальцев'и когтей трехпалых задних лап.
107
ног относительно веса тела. Так, у живущих в подобных усло-
виях парнокопытных функционирующие III и IV пальцы несут
очень широкие или очень длинные копыта. Кроме того, допол-
нительную опорную площадь у некоторых образуют менее не-
доразвитые, чем у степных, II и V пальцы, имеющие копытца.
То же значение имеют наклонное положение всей стопы, отра-
стание к зиме щеток из жестких волос вокруг копыт и способ-
ность пальцев сильно раздвигаться в стороны (северный олень,
лось, кабан). Острые режущие края роговой кромки копыт у
северных оленей позволяют им свободно бегать по обледенелым
склонам и добывать пищу, раскапывая снег.
У живущих в аналогичных условиях представителей хищ-
ных, грызунов и некоторых других отрядов роль песчаных или
снежных «лыж» играет длина и подвижность пальцев, общая
опушенность лап (рысь, росомаха, соболь, заяц-беляк и др.),
или наличие густых щеток на ногах (мохноногий и гребнепа-
лый тушканчики, тонкопалый суслик и др.) Б
Формообразующая роль снежного покрова сказывается так-
же на высоте ног. Лесные звери, передвигающиеся по глубо-
кому снегу (например рысь, лесной северный олень), обладают
более высокими ногами по сравнению со степными и тундро-
выми формами, имеющими дело с низким снеговым покровом
(степные кошки, тундровый северный олень).
В ином направлении развиваются адаптации к передвиже-
нию в горах. Типично горные копытные (серна, горал, горные
козлы) характеризуются относительно низкими ногами с креп-
кими заостренными спереди копытами, роговая ткань которых
отличается упругостью и эластичностью. Именно эластичность
дает копытам возможность находить точки опоры на ничтожных
неровностях скользких скал. Края копыт, интенсивно сти-
раясь на каменистом грунте, быстро восстанавливаются благо-
даря высокой регенеративной способности ткани, формирующей
роговой чехол. Эти приспособления позволяют горным копыт-
ным подниматься по почти отвесным скалистым стенам ущелий,
не скользить на ледниках, тормозить спуск по гладкой наклон-
ной поверхности (последнему способствуют также копытца бо-
ковых пальцев). Чрезвычайно высоко развита способность этих
животных резко изменять направление бега применительно к
положению точек опоры.
Сильная мускулатура конечностей позволяет горным копыт-
ным совершать огромные прыжки. Даже горал легко перено-
сится через расщелины шириной до 3 м, вскакивает без разбега
на выступы в 2 л высотой (Бромлей, 1956) 1 2.
1 У северных форм опушенность лап имеет, конечно, и термоизоляцион-
ное значение.
2 Несмотря на совершенство приспособлений к лазанию по горам не ис-
ключены случаи падения горных копытных с высоких скал. Старые архары,
108
У грызунов, жизнь которых приурочена к каменистым осы-
пям и скалам (например, у полевок Alticola, Platycranlus и др.),
адаптивное значение имеют цепкие лапы, уплощенный череп и
некоторые другие особенности.
ПОДЗЕМНЫЕ ФОРМЫ
Очень многие млекопитающие по организации и образу жиз-
ни занимают промежуточное положение между наземными и
подземными формами. Приспособления к жизни под землей у
них выражены слабо. Таковы роющие более или менее слож-
ные норы, но добывающие корм на поверхности земли барсук,
кролик, сурки, суслики, полевки и многие другие. Их объ-
единяют под названием «норников» и относят к полуподземным
формам. Уже среди них имеются некоторые виды, обнаружива-
ющие определенные черты строения, связанные с роющей дея-
тельностью. Так, типичные норники кролики отличаются от
близких к ним, но открыто живущих зайцев, заметно укорочен-
ными задними ногами и ушными раковинами, более сильным
развитием предплечий, более прочным и сжатым с боков чере-
пом и некоторыми другими особенностями. Для отличных зем-
лероев барсуков характерны сравнительно низкие конечности,
небольшие ушные раковины, узкая голова с удлиненной лице-
-вой частью, сильная мускулатура и длинные когти передних
конечностей. Мощными передними конечностями и когтями об-
ладают трубкозубы, муравьеды (кроме рытья нор, они служат
им и для раскапывания прочных построек термитов).
Наивысшую степень специализации к подземной жизни об-
наруживают типично подземные формы — настоящие, или по-
стоянные землерои ’, редко выходящие на поверхность земли
(обыкновенные и сумчатые кроты, златокрот, слепыши, цоко-
ры, слепушонки, гоферы и некоторые др.). Общие морфологи-
ческие особенности этой группы зверей: цилиндрическая форма
туловища со слабо выраженной шеей и укороченным хвостом,
сильное развитие грудной мускулатуры и служащих для при-
крепления мышц костных гребней и отростков на лопатках и
плечевых костях, широкие кости передних конечностей, хорошо
развитые ключицы, почти горизонтальное положение узкого
длинного таза и неподвижное соединение его с позвоночником,
большая прочность черепа. В связи со специфическими особен-
например, передвигаясь по узким каменным карнизам, нередко задевают
своими огромными спирально закрученными рогами за выступы скал, сры-
ваются и разбиваются насмерть (существующее представление о том, что
дикие бараны могут падать с большой высоты прямо на свои рога и таким
образом избегать смерти, не имеет основания).
1 В отличие от временных землероев — норников, к постоянным земле-
роям часто применяют неудачный термин — «рытики».
109
ностями подземной жизни у типичных землероев в большей или
меньшей степени редуцируются ушные раковины и глаза (у
слепыша, сумчатого и слепого кротов зачаточные глаза скрыты
под кожей); наружные слуховые проходы у них замыкаются
кожными складками — клапанами1, ноздри смещаются вниз (то
и другое служит защитой от попадания земли).
В связи с выключением или слабым развитием зрения очень
важную роль в жизни этих животных играет осязание. У кротов
основные осязательные функции выполняет сильно вытянутое и
снабженное особой гибкой косточкой (praenasale) рыло — хобо-
ток. У звездорыла (Condylura) конец носа несет звездообразно
расположенные осязательные придатки.
Пушистый густой мех, или, при его отсутствии, хорошо раз-
витый жировой слой, защищают землероев от охлаждения в
сырых норах. Характерно также для типичных землероев отсут-
ствие ворса, о чем уже говорилось в морфологическом очерке.
Большая постоянная работа — рытье и выталкивание земли,—
обусловливает у них относительно крупные размеры и хорошее
развитие сердца.
Строение и функции роющего аппарата обнаруживают зави-
симость от физических свойств почвы. У типичных землероев,
живущих в мягком грунте, функцию роющего орудия выполня-
ют передние конечности. У кротов (разных видов и родов) это—
мощные лопатообразно расширенные, несколько вывернутые на-
ружу лапы с оторочкой из жестких волос и с уплощенными
острыми когтями (поверхность лапы расширена благодаря пре-
образованию praepollex в серповидную кость — falciforme)..
Прокладывая подземный ход, крот вращается вдоль продольной
оси тела, выдвигая вперед то одну, то другую переднюю лапу,
подрезает, а затем отбрасывает назад порции земли. Неподвиж-
ность лопаток усиливает действие лап. Продвигаясь в лесной
подстилке или рыхлом поверхностном слое, зверек просто раз-
двигает частицы, приподнимает их в виде свода над ходом1 2.На
глубине он частично вжимает нарытую землю в стенки хода
при винтообразном движении тела (Депарма, 1951). Основную
массу земли крот выталкивает передней частью туловища че-
рез проделанные вертикальные выводные отнорки.
У цокоров и некоторых других обитателей более плотного-
грунта рытье осуществляется обычного типа передними лапами,
несущими однако большие долотообразные когти. Другие зем-
лерои, живущие в твердом грунте — слепыши, слепушонки и
др.— имеют сравнительно слабо развитые передние лапы. Роют
они в основном с помощью крупных, выдающихся вперед рез-
1 Несмотря на это, замлерои слышат хорошо благодаря высокой звуко-
проводимости почвы.
2 Такой способ — «минирование» широко используют также землеройки;
для продвижения в лесной подстилке. .
Нов, которыми измельчают землю. Срастание губ позади резцов
предохраняет рот от попадания земли. Выбрасывание нарытой
земли производится слепышами главным образом при помощи
плоской лопатообразной головы, отороченной валиком из упру-
гих щетинок; слепушонками — головой и задними лапами.
В заключение следует отметить, что у кротов, некоторых
землероек, ехидны, трубкозуба наблюдается образование рого-
вого вещества — кератина на роговице, что защищает глаза от
повреждения при рытье.
ВОДНЫЕ ФОРМЫ
Водные млекопитающие произошли от наземных форм через;
ряд переходов. Большую группу составляют, виды, жизнь ко-
торых проходит частично на суше, частично в воде.
К полуводным («земноводным», амфибиотическим) млеко-
питающим принадлежат из клоачных утконос, из сумчатых —
плавун (Chlronectes), из насекомоядных — выхухоль, кутора,.
выдровая землеройка (Potamogale), из грызунов — водяная по-
левка, ондатра, бобр, нутрия, водосвинка (капибара), из хищ-
ных— норка, выдра, калан, белый медведь, из копытных — бе-
гемот. к этой же группе примыкают ластоногие — следующая
переходная группа к настоящим водным млекопитающим.
Те млекопитающие, организация которых представляет как
бы первые этапы приспособления к водной жизни, сохраняют
еще общий наземный облик (водяная полевка, норка, белый мед-
ведь). Направление специализации проявляется у них главным
образом в развитии плавательных перепонок или оторочек и»
жестких волос по краям пальцев и ступней, в укорочении шеиг
конечностей и ушных раковин, в некотором уплощении головы..
Для многих амфибионтов характерен плотный волосяной покров-
или значительные жировые отложения. У кормящихся под во-
дой грызунов наблюдается изоляция резцов от ротовой полости-
(путем срастания губ позади резцов).
Многие из приведенных особенностей отмечались и для по-
луподземных форм, что объясняется сходством условий обита-
ния тех и других в плотной среде, а также и тем, что значитель-
ная часть амфибибтических млекопитающих — в то же время и
норники (водяная полевка, ондатра, бобр, выхухоль и др.).
Из других характерных особенностей, выраженных у тех или
иных полуводных зверей, наблюдается уплощение хвоста, отсут^-
ствие scrotum у самцов (лучшая обтекаемость), развитие при-
способлений для замыкания ушей и ноздрей, гибкость позво-
ночника (наклонное положение остистых отростков поясничных
позвонков), параллельное расположение элементов таза, силь-
ное развитие грудной клетки (вместительные легкие), увеличе-
ние удельного веса- тела. Интересная особенность бегемотов —
11 £
Рис. 42. Положение головы бегемота,
отдыхающего в воде (по Гессе)
смещение вверх ноздрей, глаз и ушей, что позволяет животным
оставаться неограниченное время под водой, выставив на по-
верхность лишь верхнюю часть головы (рис. 42) '.
Высокой степени специализации достигают каланы и ласто-
ногие. Задние конечности у калана сильно отодвинуты назад и
веслообразно расширены; у ластоногих они направлены назад
и превратились в настоящие ласты. Ушные раковины у каланов
и ушатых тюленей малы и в воде замыкаются; у настоящих
тюленей и моржей они полностью редуцированы. Ноздри у ка-
лана и у всех ластоногих плотно замыкаются в воде.
Проводящие длительное время на суше каланы и котики
имеют хорошо развитый меховой покров. Его структура так же,
как и у бобров, выдр и мно-
гих других амфибиотических
зверей, благодаря очень гу-
стой подпуши обеспечивает
сохранение воздушной про-
слойки, предохраняющей от
намокания и переохлажде-
ния. У менее связанных с
- землей настоящих тюленей
и моржей волосяной покров
в большей или меньшей сте-
пени редуцирован, и функ-
цию термоизоляции прини-
мает на себя подкожный жировой слой. Уменьшению удельного
веса тела во время отдыха и сна на воде способствуют разви-
тые у некоторых ластоногих (моржей, полосатых тюленей) осо-
бые воздушные мешки, связанные с нижним отделом трахеи
или с пищеводом (Слепцов, 1940).
Плавание в толще воды (при нырянии) у большинства полу-
водных млекопитающих осуществляется или, в основном, с по-
мощью колебательных движений тела (виды с вытянутым те-
лом), или гребных движений задних лап (виды с коротким
телом). Хвост служит также для управления движением.
У калана и ластоногих расширенные задние конечности при
плавании вытягиваются назад и образуют вместе с хвостом ком-
плексный орган, действующий по принципу хвостового плавника
рыб (у каланов он колеблется вертикально, у ластоногих — ла-
терально).
Передние конечности выполняют гребную функцию обычно
лишь при медленных движениях в воде. При быстром плавании
они прижимаются к телу
Движениями передних и
или отчасти служат
задних конечностей
рулями глубины,
амфибиотические
1 То же самое у рептилий
характерно и для ряда
видов крокодилов.
112
звери удерживают тело при желании оставаться на месте на
поверхности воды. Каланы могут неограниченное время лежать
на воде, распластавшись брюхом кверху и не двигая конечно-
стями.
Все полуводные млекопитающие способны нырять. Макси-
мальная продолжительность пребывания под водой утконоса и
калана равна 8—10 мин, ондатры и выхухоли—\2мин\ бобра
и настоящих тюленей — 15, моржа — 161 2[Буррелл (Burell), 1927;
Харниш (Harnisch), 1937, Ирвинг (Irving)’, 1939; Ирвинг и Орр
(Irving а. Огг), 1935; Миллэ (Millais), 1906; Паркер (Parker),
1922; Барабаш-Никифоров, 1933, 1947].
Не способны нырять детеныши бобра, ондатры, калана, тю-
леней и некоторых других амфибионтов, что связано с их не-
большим удельным весом и сильным развитием воздушной
прослойки в мехе. При попытке нырнуть вода тут же выталки-
вает их на поверхность, где животные могут пассивно держаться
подобно поплавкам неопределенно долгое время.
Многие полуводные звери свободно передвигаются по суше,
хотя и уступают в этом отношении наземным формам. Выдра,
калан, а иногда и норка, спускаясь с заснеженного склона бере-
га, обычно скользят на брюхе, почти не прибегая к конечностям.
Но калан испытывает значительные затруднения при передвиже-
нии на суше. Длинное его туловище волочится по земле, движе-
ния требуют больших усилий Сравнительно редко калан пере- ~
двигается по твердому субстрату другим способом: выгнув дуго-
ооразно спину и солизив вследствие этого задние конечности
с передними, он довольно быстро идет на них и делает даже
иногда неуклюжие прыжки, однако такое передвижение его
быстро утомляет (Барабаш-Никифоров, 1947).
Среди ластоногих наибольшей наземной подвижностью обла-
дают ушастые тюлени и моржи. Задние конечности у.них доста-
точно подвижны и могут подводиться под туловище (рис. 43).
Попеременное подгибание и выпрямление их обеспечивает по-
ступательное движение типа галопирования (передняя часть ту-
ловища при этом опирается на хорошо развитые передние лас-
ты); вполне понятно, что такое движение не может, быть продол-
жительным. Более специализированные настоящие тюлени на
суше совершенно беспомощны (основные сегменты обеих пар
1 У выхухоли наблюдается своеобразное приспособление дыхательных
путей, увеличивающее их объем и дающее возможность животному одновре-
менно дышать и глотать на суше и питаться под водой (дыхательное горло
выхухоли способно замыкаться с помощью небной и глоточной мускулатуры).
2 У ластоногих, помимо упоминавшихся выше воздушных мешков, дли-
тельное пребывание под водой облегчается обилием крови, большим количе-
ством миоглобина (мышечного гемоглобина), способностью на длительное
время выключать дыхание и задерживать поступление венозной крови в сердце.
8 Териология
113
конечности скрыты у них в кожно-мышечном мешке, дисталь-
ные, выступающие наружу сегменты очень коротки; задние ко-
нечности направлены назад и
могут) !.
В полном смысле водными
ляются сирены и киты, даже на
водную среду.
подгибаться под туловище не
формами (гидробионтами) яв-
короткое время не покидающие
Рис. 43. Движение на суше. А — настоящего тюленя;
Б — ушастого
По степени специализации к жизни в воде сирены представ-
ляют как бы промежуточную ступень между ластоногими и ки-
тообразными. Для них характерны торпедообразная форма
тела с сильно укороченной шеей (благодаря уплощению и сра-
станию шейных позвонков), листообразные передние конечности,
редукция ушных раковин, почти полная редукция волосяного
покрова и задних конечностей (от последних в толще мышц,
сохраняются лишь рудименты тазовых и бедреных костей) и раз-
витие горизонтального хвостового плавника, служащего основ-
ным локомоторным органом. Однако по способности к нырянию
1 Известны все же единичные случаи далеких передвижений настоящих
тюленей по суше, льду или снегу на расстояние до 30 км от берега мор»
(Врублевский, 1959; А. Вишняков, 1961).
114
сирены не превосходят ластоногих. Так, максимальное время-
пребывания под водой для ламантина равно 16 мин [Ирвинг'
(Irving), 1942].
Наивысшую степень приспособления -к водной среде обита-'
ния обнаруживают киты. Рыбообразная форма тела и редукция-
всех выступающих частей создают у них совершенную обтекаем
мость. Шейный перехват в слабой степени выражен только у
белух; у всех остальных китообразных голова полностью ели-'-
вается с туловищем. Мощный подкожный жировой слой умень-
шает теплоотдачу и умеряет действие давления воды. Пре-:
вращение в плавники передних конечностей и редукция задних-
(от которых остаются дериваты таза и ничтожные остатки ко-
стей самих конечностей), развитие у многих видов спин-
ного жирового плавника и у всех видов — горизонтального двух-
лопастного хвостового плавника, приближают китов в отноше-’
нии плавательной способности к рыбам. Из других особенностей
характерны уплощение очень толстой роговицы глаз, толстая-
склера и шарообразный хрусталик, отсутствие слезных желез и
каналов и сильное развитие гардеровых желез, жирный секрет
которых защищает глаза от вредного действия соленой воды1,
значительное смещение ноздрей в дорзальном направлении,
особое устройство гортани, надежно страхующее дыхательные
пути от попадания в них воды во время принятия пищи, огром-
ная емкость и эластичность легких, имеющих сильно развитую
мускулатуру даже в стенках альвеол. Развитие перечисленных
и некоторых других приспособлений дало возможность китам-'
полностью освоить водную стихию как среду жизни. >
Основным локомоторным органом у китообразных служит хво-;
стовой отдел тела (хвостовой стебель с хвостовым плавником)/
С помощью вертикальных колебаний этого органа даже крупные'
киты могут развивать скорость движения до 40 км в час. Мелкие’
киты — дельфины подолгу следуют за быстроходными судами,-
описывают вокруг них круги и при этом кажутся совершенно'
неподвижными, так как вибрация их хвостовых плавников проис-1
ходит с колоссальной быстротой и не улавливается глазом1 2.
Передние конечности — плавники служат у китов органом управ-
ления и лишь в редких случаях — органом ограниченного,
движения (им присуща и терморегуляторная функция). При,
обычном плавании поступательное движение кита идетповолно-
образной кривой, так как для возобновления запаса воздуха]
1 То, же значение имеет кератинизация поверхности роговицы. (
2 Как показали исследования Крамера (Krammer, I960), вокруг плывут
щего дельфина не образуется вихревое (турбулентное), движение воды, сильной
тормозящее другие тела такой же конфигурации, например подводные лод'-:
ки. «Антитурбулентность» в данном случае, зависит от специфического строе-’
ния кожи дельфина, пронизанной огромным количеством ходов и.трубочек,
заполненных губчатым амортизирующим веществом.
8* 113
в легких животное вынуждено периодически всплывать на по-
верхность. Выдыхаемый с большой силой сжатый воздух обра-
зует специфические для крупных китов «фонтаны» в виде обла-
ка с примесью брызг (последние поднимаются в воздух в том
случае, когда кит начинает выдох прежде, чем его ноздри успе-
ют подняться над водой) L
Киты обладают способностью глубоко нырять (по вертика-
ли). Имеются, например, указания на случаи ныряния финвалов
на глубину до 355 м, кашалотов — до 988 м. Время пребывания
под водой у разных видов различно. Дельфины обычно погру-
жаются не более чем на 20 мин. Большинство китов-полосатиков
может оставаться под водой около 20—30 мин, кашалот до
-75 мин, гренландский кит до 1,5 ч, бутылконосы (Нyperoodon)
. в некоторых случаях до 2 ч. Дельфины нормально выныривают
для дыхания через промежутки не более, чем 5 мин [Скаммон
(Scammon), 1874; Миллз (Millais), 1906; Льювилль (Lionville),
1913; Аллен (Allen), 1916, Грегори (Gregory), 1937; Ирвинг
(Irving), 1939; Слипер (Slijper), 1958; Томилин, 1958 и др.].
Ныряние связано с повышением давления (на глубине 1000 .и
, давление достигает 100 атмосфер!) и длительной задержкой ды-
хания. Естественно, что у китов выработался в связи с этим ряд
приспособлений. Так, в противовес высокому давлению киты
обладают повышенной прочностью эпидермиса и исключительно
большой толщиной жирового слоя; сильного развития у них
достигают хрящевые кольца трахеи и бронхов и запирательная
, мускулатура альвеол1 2. С длительностью пребывания китов под
водой'связаны большая вместимость их легких3, сильное раз-
витие .«чудесной сети» (экономия потребления кислорода),боль-
. ,шое количество миоглобина и. повышенное кровоснабжение моз-
га, слабая чувствительность к накоплению в организме угле-
кислоты и др.. У кашалотов и бутылконосов (наилучших ныряль-
щиков) .один из носовых проходов преобразуется в наполняемый
воздухом .запасный резервуар, что также увеличивает возмож-
ную длительность дыхательной паузы.
1 Облако, по-видимому, состоит не из конденсированных в холодном
воздухе паров, как полагали раньше (оно образуется и в теплом воздухе
тропиков)-, а из большого количества слизи, выбрасываемой из дыхательных
л путей-кита.[Слипер (Slijper), 1958].
. ..4 Давление в легких при нырянии остается близким к атмосферному,
что' обусловливается изоляцией в это время легких от проводящих дыха-
.тельных'отделов (бронхов, трахеи). Довольно длительное время среди зоо-
логов и физиологов велась дискуссия по вопросу о причинах отсутствия
у китов кессонной болезни (поражающей водолазов при быстром подъеме
больших глубин). Однако сама постановка этого вопроса беспочвенна,
\ так как явления, связанные с нырянием, у китов совершенно не соответст-
вуют тем, которые имеют место при погружении на глубину и поднятии
,у водолазов.
3 Вычислено, что у синего кита весом в 122 т емкость легких состав-
ляет 14000 л, а запас кислорода достигает 2800 л [Крог (Krogh), 1934].
116
Благодаря сгоей плавучести киты могут в период покоя по-
долгу лежать без движения, выставив на Поверхность лишь верх-
нюю часть туловища. С другой стороны, сделав соответствую-
щий разгон, даже крупный кит (например, горбач) может вы-
прыгнуть из воды и описать в воздухе довольно большую дугу
или даже сделать сальто (рис. 44).
Рис. 44. Последовательность движений кита-полосатика (Balaeno-
pterd}, плывущего у поверхности воды, и кита-горбача (Megaptera
nodosa}, совершающего сальто (из Слипера)
ДРЕВЕСНЫЕ ФОРМЫ
Среди лесных млекопитающих имеется много видов, жизнь
которых протекает частично на поверхности земли, частично на
деревьях, где они регулярно добывают себе корм, находят убе-
жище на время отдыха, спячки и т. п. (бурый и черный медведи,
лесная куница, лесные кошки, енот, бурундук и др.). Эти полу-
древесные формы уже имеют некоторые адаптивные черты, свя-
занные с лазаньем по деревьям. Так, лесная куница отличается
117
от близких наземных форм (каменной куницы, соболя) укоро-
ченной шеей, относительно более длинными поясничным и
крестцовым отделами позвоночника, сильным развитием муску-
латуры передних конечностей, большей длиной хвоста, более
крупными изогнутыми когтями (Юргенсон, 1950).
К настоящим древесным формам принадлежат белки и сони
(особенно соня-полчок). Благодаря цепким хватательным лапкам
они свободно лазают по гладкой вертикальной поверхности дре-
весных стволов. Длинный, хвост с двурядным расположением
волос служит балансиром и парашютом при больших прыжках
(широко расставив конечности и расправив хвост, белки и полч-
ки совершают прыжки длиной до 4 м и более по наклонной ли-
нии). Помещающиеся на внешних частях конечностей и на боках
вибриссы позволяют зверькам легко лавировать в густых спле-
тениях ветвей, не снижая скорости движения.
Несмотря на такую высокую адаптированность к древесному
образу жизни белки и сони хорошо бегают по земле (первые из
них совершают иногда таким способом настоящие миграции).
Более узко специализированные дендробионты — полуобезья-
ны, большинство обезьян, ленивцы, сумчатый медведь (коала),
кускус, опоссумы и многие др. У одних приспособление к лаза-
нию идет по пути противопоставления одного или двух пальцев
конечностей остальным (первое имеет место у полуобезьян и
обезьян, второе — у коала, кускуса); такого рода хватательные
конечности дают возможность прочно держаться на ветвях.
У долгопята-маки и некоторых других древесных форм на кон-
цах пальцев имеются .присасывательные подушечки, а на по-
дошвах— мозоли; то й другое усиливает хватательную функцию
конечностей и облегчает-лазание по деревьям.
Большинству древолазов свойственны острые искривленные
когти, но особенно сильного развития они достигают у ленив-
цев, находящихся постоянно (как при движении, так и в покое)
в подвешенном состоянии. Стопоходящие обезьяны, у которых
когти превратились.в. широкие тупые ногти (например, мартыш-
ки), ..передвигаются по ветвям (по. верхней их стороне),, опира-
ясь на всю поверхность стопы и кисти.
Длинный хвост помогает обезьянам сохранять равновесие.
Как и у многих других дендробионтов, у обезьян хорошо развита
координация движений, обусловленная функцией соответствую-
щих частей головного мозга.
По особому пут'и’пошла адаптация к древолазанию у гиббо-
1 нов, коати и некоторых других обезьян, передвигающихся по де-
ревьям и перепрыгивающих с дерева на дерево с помощью толь-
-ко передних конечностей (такое передвижение носит название
брахиации). Повиснув на ветке на своих исключительно длин-
ных руках, гиббон раскачивается, отрывается и, пролетев более
'' или менее значительное расстояние по воздуху, повисает на дру-
* Н8
гой ветке (рис. 45). При этом он не делает хватательного дви-
жения рукой, а лишь цепляется за ветку пальцами, согнутыми
крючком. Такого рода цепляние облегчается у гиббонов исчез-
новением I и V пальцев. У человекообразных обезьян, которые
также прибегают к брахиации, более или менее редуцируется
J палец.
Функцию хватательного органа у многих лазающих форм
несет хвост. С его помощью животное фиксирует положение тела
Рис. 45. Переносящийся с дерева на дерево с помощью бра-
хиации гиббон (Hylobates lar) (по Валуа)
на ветке, а иногда и повисает на нем вниз головой (опоссумы,
кускус, древесные формы дикобразов, муравьедов, ящеров, но-
суха, цепкохвостые обезьяны) *.
По земле узко специализированные дендробионты передви-
гаются сравнительно плохо. Гиббон обычно прибегает к двуно-
гому хождению, балансируя поднятыми и расставленными в сто-
роны руками. Горилла, передвигаясь по земле на задних ногах,
поддерживает тело руками, опираясь на тыльную сторону кистей.
Почти полностью лишены способности передвигаться по земле
ленивцы.
1 Цепкий хвост свойствен также некоторым млекопитающим, лазаю-
щим по высоким стеблям травянистых растений, например, мыши-малютке
и др.
119
I
ЛЕТАЮЩИЕ ФОРМЫ
Некоторые древолазающие млекопитающие, как уже указы-
валось, обладают способностью значительно удлинять свои
прыжки путем планирующего полета. У белки, полчка роль пара-
шюта играет хвост. У толстотелых обезьян (Colobus) для этого-
служат очень длинные волосы по бокам тела; развеваясь во
время прыжков, они создают дополнительную площадь опоры о
воздух.
К полувоздушным формам могут быть отнесены звери,, об-
ладающие кожными складками на боках туловища, что дает им
возможность пользоваться планированием не только для спуска
Рис. 46. Млекопитающие, представители разных групп, обладающие
приспособлениями к планирующему полету. А — сумчатая летучая
белка (Ре/aurus); Б — летяга (Pteromysp, В — шипохвостая африкан-
ская белка (Anomalurus)-, Г — шерстокрыл (Cynocephalus) (из Эйзен-
траута)
по наклонной линии, но и для поворотов в воздухе и незначи-
тельных подъемов (рис. 46). К таким формам принадлежат сум-
чатые белки (Petauroides, Petaurus), летяги (Pteromyidae),
Anomaluridae и шерстокрылы (Dermoptera) Летяга, например,
заканчивая свой прыжок-полет, обычно несколько взмывает
кверху, что смягчает толчок о ствол дерева. Еще большей ма-
невренностью отличается полет шерстокрыла, у которого лета-
тельная перепонка достигает наибольшего развития. «Парашют»
позволяет животному переноситься по воздуху на десятки мет-
ров; сахарная белка, например, преодолевает по воздуху рас-
стояние до 30—40 м, шерстокрыл — до 50 и даже 70 м.
К настоящим воздушным формам принадлежат многочис-
ленные представители отряда рукокрылых. Приспособления к
1 Кожная складка поддерживается с каждой стороны у летяг особой
тонкой длинной косточкой, отходящей от запястья назад и в сторону, у аф-
риканских чешуйчатохвостых летяг (Anomalurus)—хрящевым выступом
локтевого сгиба (см. рис. 46, Б, В).
120
активному полету позволяют им длительное время быстро и с
большой маневренностью передвигаться в воздухе, здесь же до-
бывать себе пищу и даже спариваться. Характерная особенность
этой группы млекопитающих — превращение передних конеч-
ностей в длинные гибкие крылья, причем строение последних
не соответствует строению птичьих крыльев (рис. 47). Пальцы
передних конечностей рукокрылых, за исключением свободного
первого, служащего для ползания, сильно удлинены и связа-
ны тонкой летательной перепонкой (patagium), охватывающей
также туловище, задние конечности и хвост. У многих летучих
Рис. 47. А — планирующий полет североамериканской летяги (Glaucotnys
volans) (по Бурльеру); Б — активный полет большой ночницы (Myotis myo-
tis) (по Кузякину)
мышей плечевая кость несет большой бугор, образующий допол-
нительный сустав с лопаткой, что предохраняет крыло от вывиха
при резких его движениях во время полета.
Аналогично птицам у рукокрылых хорошо развиты грудные
(летательные) мышцы, прикрепляющиеся у большинства видов
к небольшому килю грудины. С воздушным образом жизни
связано также раннее срастание костей черепа, быстрое окостене-
ние скелета, слияние некоторых элементов грудной клетки, мощ-
ное развитие ключиц, своеобразный механизм дыхания
1 Дыхательные движения грудной клетки у рукокрылых производятся
во время полета с помощью особых вспомогательных мускулов (а не меж-
реберных мышц, как у других млекопитающих); движения грудной клетки
связаны также с работой крыльев.
121
От формы крыльев зависят быстрота и изворотливость по-
лета: лучшими летунами являются виды, имеющие узкие и длин-
ные крылья; сравнительно медленно летают обладатели широ-
ких и тупых крыльев В качестве тормозного органа служит
межбедренная часть летательной перепонки, поддерживаемая
особой костной или хрящевой шпорой (calcar). Тип полета чаще
всего гребной, осуществляемый последовательными взмахами
крыльев. Возможны также порхающий и планирующий полеты
(но не парящий).
Полет требует большой затраты энергии. С этим связаны
крупные относительные размеры сердца рукокрылых. Сердечный
индекс летучих мышей достигает 10—14,4%, т. ё. мало уступает
сердечному индексу быстро летающих птиц (у ласточек, напри-
мер, величина сердечного индекса равняется 14,7%).
Задние конечности служат рукокрылым главным образом для
ползания и подвешивания (вниз головой) во время покоя. В слу"
чае необходимости летучие мыши могут довольно быстро пере-
двигаться по земле, действуя обеими парами конечностей, лазать
по отвесным стенам и даже плавать; способностью совер-
шать вертикальный взлет с земли они превосходят птиц. Значи-
тельно более крупные рукокрылые — летучие собаки, или кры-
ланы— уступают в легкости полета и подвижности насекомояд-
ным летучим мышам.
* *
Вее, рассмотренные адаптации служат подтверждением того
общего положения, что эволюция организмов совершается не
под действием каких-либо автономных сил нематериального по-
рядка (как это полагают автогенетики), а в неразрывной связи
•с условиями обитания, в результате отбора и закрепления новых
признаков наследственностью.
1 По М. Эйзентрауту (Eusentraut), средняя скорость полета последних
•составляет около 4,3 м/сек. Длиннокрылые виды летают в несколько раз
«быстрее.
Питание и явления, связанные с ним
ОБЩЕЕ ЗНАЧЕНИЕ ПИТАНИЯ
Питание занимает одно из важнейших мест среди других
проявлений жизнедеятельности млекопитающих. Характер пи-
сания определяет отношение данного животного к важнейшему
элементу среды — источнику необходимых пищевых веществ,
т. е. ко многим другим растительным и животным организмам
и, следовательно, в значительной мере обусловливает положе-
ние данного животного в ценозах—естественных группировках
организмов. Выдающийся русский физиолог И. М. Сеченов сто.
лет тому назад (1861) указал, что «организм без внешней
среды, поддерживающей его существование, невозможен, по-
этому в научное определение организма должна входить и
среда, влияющая на него»1. В многочисленных и разнородных
связях животного со средой одной из «существеннейших... яв-
ляется связь через известные химические вещества, которые
должны поступать в состав данного организма, т. е. связь через
пищу» (И. П. Павлов).
Химические связи со средой осуществляются главным обра-
зом чёрез использование готовых органических соединений при
поедании продуктов растительного и животного происхожде-
ния. Кроме этой обычной пищи, организм млекопитающих спо-
собен усваивать ряд необходимых неорганических соединений,
растворенных в питьевой воде (рек, ручьев, минеральных источ-
ников) или находящихся в засоленной почве и т. п.
Пищей млекопитающим служат самые различные растения
от низших — водорослей, лишайников,, грибов до высших — цвет-
ковых, и разнообразнейшие животные, относящиеся к боль-
шинству существующих типов и классов. Так, например, мор-
ская корова Стеллера, водившаяся у Командорских островов, по-
позднее полностью истребленная, питалась крупными бурыми и
красными водорослями, образующими большие заросли по мел-
ководным прибрежным участкам на севере Тихого океана. Не-
которые наземные и древесные лишайники в холодную поло-
вину года — важнейший корм северного оленя, кабарги, ряда
видов полевок; некоторые мхи охотно поедаются леммингами,
плодовые тела высших грибов— лакомая пища многих видов
1 За много лет до Сеченова Эту же мысль высказывал и профессор
К. Ф. Рулье, один из зачинателей отечественной экологии.
12ft
Рис. 48. А — голова цветочной летучей
мыши (Choeronycteris mexicana) из Южной
Америки. Вытянутая лицевая часть головы,
очень длинный язык, на конце несущий
подобие щеточки из удлиненных папилл —
приспособления к питанию пыльцой и
нектаром цветков; Б — молотоголовая,
летучая собака (Hypsignathus (monstrosus),
схватившая плод смоковницы из Эйзент-
раута). По строению головы этот вид
резко отличается от предыдущего
оленей (в том числе лося, косули, северного оленя), ряда видов
полорогих, кабана, белки, зайцев, многих видов полевок. У выс-1
ших растений млекопитающие едят как зеленые ассимилирую-
щие органы (листья, стебли), так и генеративные—завязи
цветков, цветки, семена, мякоть плодов, а также луковицы,
клубни, корни и корневища, у древесных и кустарниковых по-
род— кору и целые ветви.
В тропических странах есть виды и группы млекопитающих,
узко специализированные в отношении питания каким-либо од-
ним видом корма расти-
тельного происхождения,,
например, мелкие летучие
мыши — они сосут нектар
и поедают пыльцу цвет-
ков, раскрывающихся
ночью. Связь между эти-
ми рукокрылыми и их
кормовыми растениями
настолько тесная и древ-
няя, что отразилась на
морфологии венчика,
пыльников и пестиков
цветков: их могут опы-
лять только летучие мы-
ши. Последние, в свою
очередь, отличаются свое-
образным строением язы-
ка, зубов, удлиненностью
лицевой части головы
и т. д. (аналогия с птица-
ми тропических групп —
колибри и нектарницами,
приспособленными к пи-
танию нектаром и опыле-
нию цветков, распускаю-
щихся днем). Представи-
тели другой группы руко-
крылых, по преимущест-
ву наиболее крупных ле-
тучих мышей, питаются
только нежными плодами
(рис. 48). Эти животные служат важными распространителями
семян своих кормовых растений, так как часто совершают боль-
шие перекочевки, отыскивая места с обильным урожаем плодов.
В сухих степях и пустынях Евразии, Африки, Северной и
Южной Америки есть немало видов грызунов, настоящих «кор-
неедов» — все они питаются сочными корневищами, клубнями и
124
луковицами многолетних травянистых растений, служащими
для них источником пищи и необходимой влаги. Даже такой
доступный очень короткое время корм, как сладкий сок деревь-
ев, вытекающий весной из надрезов коры, имеет своих потреби-
телей среди млекопитающих. Из представителей нашей фауны,
надкусывая ветви, пьют сок берез, кленов и др. грызуны древо-
лазы— обыкновенная белка и некоторые виды сонь (интерес-
ная аналогия с дятлами, среди которых есть много видов соко-
сосов как в нашей, так и в североамериканской фауне).
Из беспозвоночных животных земляные черви играют важ-
ную кормовую роль для ряда видов кротов, барсука, диких сви-
ней. Пресноводных моллюсков в массе поедают водяная земле-
ройка кутора, выхухоль, охотно ест их также грызун ондатра; не-
которые донные двустворчатые моллюски — основной корм мор-
жа; крылоногих моллюсков охотно вылавливают беззубые киты
планктофаги, крупных головоногих — зубатые киты, а в их чис-
ле такой ценный вид, как кашалот. Китообразные поедают так-
же голотурий, иногда морских звезд. Морские ежи — основной
корм крупной морской выдры калана и тех речных выдр, кото-
рые живут у побережий океанов. Мелких планктических рако-
образных в огромном количестве вылавливают при помощи со-
вершенного цедильного аппарата огромные беззубые киты, дон-
ных десятиногих раков поедают дельфины, тюлени, выдры,
норки. Из других беспозвоночных большую роль в питании мно-
гочисленных видов и групп млекопитающих (особенно мелких
сумчатых, насекомоядных, рукокрылых, некоторых хищных, не-
полнозубых, части обезьян и грызунов) играют различные на-
секомые.
Велико и количество млекопитающих, охотящихся за ры-
бами; ихтиофаги, часто узко специализированные, есть среди
насекомоядных, хищных, ластоногих, китообразных, грызунов
п даже среди рукокрылых. На амфибий, рептилий и высших по-
звоночных тоже охотятся многие виды. Хищники, специализи-
ровавшиеся в преследовании птиц и млекопитающих, принад-
лежат к относительно немногочисленным, но наиболее высоко-
организованным представителям отряда и класса — таковы
общеизвестные волк, тигр, леопард, гепард, морской леопард/
косатка (рис. 49) и ряд других.
Особую категорию представляют виды некрофагш— пожира-
тели трупов крупных животных (гиены и др.), обладающие це-
лым рядом морфологических черт, облегчающих разгрызание
крепких костей, связок и т. п. Многие группы животноядных
наземных млекопитающих (представителей «белково-липоид-
ного типа питания») уже в третичное время в связи с измене-
нием природных условий стали переходить на потребление ра-
стительной пищи, перегруженной клетчаткой, т. е. к «клетчат-
ковому типу питания». Это стало основным направлением их
125
эволюции. Следовательно, растительноядность — во многих слу-
чаях геологически недавнее приобретение, у ряда групп к тому
же в известной мере обратимое.
Каков бы ни был набор кормов, используемых тем или иным
млекопитающим, «пищедобывательная деятельность» (А. Д. Сло-
ним) и сам акт еды неизбежно занимает в его жизни очень
Рис. 49. Охота косаток (Orca orca) на морских котиков (из
Гамильтона). Эти хищные киты питаются главным образом
крупными теплокровными морскими животными. В желудке
одной особи находили до 34 проглоченных тюленей
большое место. Передвижение в поисках мест, наиболее удоб-
ных для пастьбы или охоты за живой добычей, при определен-
ных условиях (температуры, освещения, ветра, силы течения и
волнения, высоты и структуры снежного покрова, характера
растительности, механического состава и плотности почвы, при
наличии постоянной угрозы со стороны врагов) требует от жи-
вотного четкого восприятия разнородных сигналов, способности
ориентироваться в пространстве и соответствующих целесооб-
разных реакций, осуществляемых через посредство сложных со-
четаний безусловных и условных рефлексов. Скрадывание и
•захват подвижной быстрой жертвы, добывание плодов с тонких
ветвей, выкапывание насекомых и червей из толщи почвы и
другие акты, повторяемые многократно в течение дня, требуют-
126
определенных затрат мышечной и нервной энергии, Подсчи-
тано, например, что северный олень летом использует о коло-
66 % суточного времени на пастьбу и прием корма и только 34%
на отдых. Его двигательная активность при пастьбе измеряется
ежедневно 15—24 тысячами шагов, что соответствует расстоя-
нию примерно в 19—30 км. Зимой при пастьбе на ягельных паст-
бищах длина суточной тропы оленя в 3—5 раз короче, но выка-
пывание корма из-под снега требует большой дополнительной
работы передних ног. В зависимости от высоты и плотности;
снега каждый олень делает за сутки от 2 до 8 тысяч копатель-
ных движений. На лесных пастбищах, отличающихся рыхло-
стью снежного покрова, у оленей более 44% суточного времени
уходит на добывание и поедание корма, около 20% на пере-
ходы, а остальное — на отдых.
За одну кормежку взрослый кавказский кабан съедает
2—3 кг корма; подсчитано, что для этого он должен отыскать,,
подобрать и разжевать около 12 000 опавших на землю буко-
вых орешков или равноценные по весу 1050 желудей, или 37Е>
плодов дикой груши и т. п. Синий кит планктофаг, обладающий
цедильным аппаратом, приспособленным к вылавливанию срав-
нительно крупных (2,4—6,5 см) массовых планктических рач-
ков, должен отыскивать их скопления и процеживать огромное-
количество воды, чтобы наполнить желудок 1,5—2 т корма..
В желудке крупных особей голубых китов антарктических вод
находили до 1,5 млн. экземпляров рачков Euphasia superba.
'Лесная куница, охотясь за полевками и отыскивая спящих
в снегу тетеревиных птиц, оставляет за одну ночь след длиною’
от 5 до 17 км и более. Волки зимою нередко делают суточные
переходы до 30—50 км. Еще длиннее зимние следы охотяще-
гося уссурийского тигра. Между двумя его остановками на от-
дых иногда лежит расстояние более 100 км. С этим «двигатель-
ным поведением животного в процессе добывания пищи»
(А. Д. Слоним) связан и самый акт еды — освобождение пищи;
от несъедобных частей, разгрызание или отрывание кусков,,
пережевывание и процессы собственно пищеварения, включая
двигательную и секреторную деятельность пищеварительного-
тракта. У некоторых видов ярко выражена также деятельность^
направленная на запасание и сохранение корма впрок.
С особенностями пищедобывания и питания, занимающих:
такое важное место среди других проявлений жизнедеятельно-
сти, тесно связан целый комплекс видовых морфофизиологиче-
ских адаптивных черт скелета и мускулатуры, особенно органов:
движения, органов пищеварения, нервной системы и органон
чувств (втяжные острые когти кошки и ее глаза, приспособ?
ленные к дневному и ночному зрению, очень острое обоняние и»
слух лисицы, способность насекомоядных рукокрылых отыски-
вать в темноте и преследовать летящих насекомых с помощью
127
тончайшей ультразвуковой локации и др.). Эти органы корре-
лятивно связаны с другими органами и функциями животного,
не имеющими прямого отношения к питанию, и накладывают
на них свой отпечаток.
Организм животного как единая целостная система всегда
носит и в общем своем облике и в деталях строения следы фор-
мирующей роли среды и в первую очередь кормовой специали-
зации.
Без'изучения пищедобывательной деятельности и питания
нельзя понять ни биологии вида в целом, ни отдельных прояв-
лений жизнедеятельности особей, групп, популяций млекопи-
тающих.
Значение разных видов млекопитающих для жизни и хо-
зяйства человека во многих случаях зависит именно от харак-
тера кормовой специализации животных. Например, вампиоы—
кровопьющие 1 летучие мыши Южной и Центральной Америки,
иногда нападающие ночью на спящих людей, способны зара-
жать их вирусом бешенства, а стадам домашних животных они
нередко приносят огромный ущерб, вызывая массовые эпизо-
отические вспышки этого тяжелого заболевания. Прокормление
одного выводка волков в' черноземной полосе Европейской части
СССР, где они уничтожают рогатый скот, свиней, домашних гу-
сей и ценных охотничьих животных, обходится хозяйству стра-
ны не менее 2500 рублей в год (по заготовительным пенам).
Лисица в одних местах вредна как истребитель дичи, а в дру-
гих— полезна. Например,, в полупустынях междуречья Волги —
Урала, где она истребляет много песчанок и сусликов — храни-
телей опасных инфекций человека, по указанию Министерства
здравоохранения СССР нередко полностью запрещается охота
на этого хищника, помогающего работе санитарно-эпидемиоло-
гических отрядов.
В отличие от класса птиц — группы довольно однородной по
облику входящих в нее видов, класс млекопитающих — яркий
пример огромного разнообразия размеров и формы тела жи-
вотных, разнообразия приемов и органов схватывания и пер-
вичной обработки пищи, а также тех условий, в которых этот
корм добывается. Птицы, отчасти в связи с отсутствием живо-
родности и. необходимостью насиживать яйца, не смогли так
полно освоить пищевые ресурсы океанов," как это сделали мле-
копитающие. В гнездовый период птицы нуждаются в твердом
субстрате для откладывания и насиживания яиц и неизбежно
привязаны к островам или побережьям материков. Китообраз-
ные же и сирены полностью оторвались от суши; твердый суб-
страт не нужен им ни в один из периодов жизни. Именно кито-
1 Название кровососущие для них не подходит, т. к. вампиры не сосут,
а лакают кровь из сделанной ранки.
128
образные смогли освоить пелагиаль океанов в такой степени,
в какой это не удалось птицам. Кормовые богатства глубин
моря тоже лучше освоены млекопитающими, а именно китооб-
разными и ластоногими, способными нырять на значительно
большую глубину, чем лучшие ныряльщики из птиц. Кашалот,
например, питается главным образом головоногими моллю-
сками, отчасти донными рыбами, и ловит их на глубинах 300—
400 м и более (зарегистрировано два случая гибели кашалотов,
запутавшихся в подводном кабеле на глубине 510 и 988 м). На
сотни метров опускается и охотящийся за донными животными
высоколобый бутылконос.
Толща почвы с ее очень богатыми ресурсами растительной
и животной пищи освоена птицами только с самой поверхности;
среди них нет ни одного роющего подземного вида. Очень мало
используют птицы ее глубину и для устройства надежно укры-
тых гнезд, тогда как сложнейшая многоярусная сеть кормовых
ходов и сливающихся с ними жилых нор млекопитающих про-
низывает почву и отчасти подпочву от поверхности до глубины
4—5 м на площадях, измеряемых миллионами квадратных ки-
лометров. Многие виды мелких грызунов, насекомоядных
и хищных млекопитающих хорошо приспособлены к добыванию
корма под высоким слоем снега и проводят длинные холодные
зимы в тех широтах, откуда улетает огромное большинство
птиц. Все ярусы растительного покрова от зарослей злаков до
вершин самых крупных деревьев используются лазящими, мле-
копитающими в качестве источника разнообразных кормов и
места для устройства семейных и одиночных гнезд. Воздушная
среда хорошо освоена представителями отряда рукокрылых,
насчитывающего около 1000 видов. Многие из рукокрылых по
скорости и маневренности полета, способности ловить быстрых
и увертливых насекомых не уступают лучшим воздухореям из
насекомоядных птиц.
Таким образом, из всех позвоночных млекопитающие наибо-
лее широко освоили все экологические подразделения биосферы
с их пищевыми ресурсами. В Арктике они доходят до самого
полюса, в Антарктике вместе с пингвинами населяют воды
у кромки вечных „льдов и выходят на окраины Антарктиды.
Океаны, внутриматериковые воды и суша всех континентов на-
селены млекопитающими за исключением мертвых участков
в поясе вечного снега и льда на вершинах наиболее высоких
гор; за этим исключением все высотные, или вертикальные
пояса суши и первые ступени глубины в морях входят в сферу
жизни современных млекопитающих. На суше • количество ви-
дов и особей растительноядных млекопитающих в большинстве
мест превосходит животноядных; в море соотношение обрат-
ное— растительноядных очень мало, преобладают потребители
беспозвоночных и позвоночных животных.
9 Териология 129
СПЕЦИАЛИЗАЦИЯ В ПИТАНИИ,
ЗНАЧЕНИЕ РАЗНЫХ ГРУПП КОРМОВ
По степени кормовой специализации млекопитающих как и
других животных делят на стенофагов и эурифагов. В усло-
виях умеренных и холодных зон северного полушария типич-
ных стенофагов—животных, использующих ограниченный на-
бор кормов, сравнительно мало, большинство видов наших
млекопитающих — эурифаги, или «всеядные» животные.
К стенофагам можно, например, отнести виды строго насе-
комоядных рукокрылых, некоторых наших дельфинов, питаю-
щихся почти исключительно рыбами, принадлежащими к огра-
Рис. 50. Гигантский муравьед (Myrtnecophaga centralis, из неполнозубых
Ю. Америки) имеет вытянутое трубкообразное рыло и длинный червовидный
язык. Узкоспециализированный термитофаг (из Гамильтона, с изменениями)
ничейному числу родов и видов, некоторых беззубых китов
с очень специализированным цедильным аппаратом и др. Хоро-
ший пример стенофага — гигантский муравьед (рис. 50), питаю-
щийся только термитами и муравьями, крепкие жилища кото-
рых он разрушает большими когтями передних лап. Его гиб-
кий длинный, покрытый липкой слизью язык легко проникает
в ходы и камеры этих насекомых и извлекает приклеившихся
особей. Гигантский муравьед — наземный вид; в Южной Аме-
рике есть более мелкие виды древесных муравьедов, специали-
зировавшихся в поисках термитников, располагающихся на
ветвях деревьев тропических и экваториальных лесов. Лени-
вец — узко специализированный потребитель листвы высоких
деревьев тропических лесов Южной Америки, на ветвях кото-
рых он проводит всю свою жизнь — тоже типичный стенофаг.
130
Рис. 51. Североамериканский хомячок
Onychomys («grasshopper mouse») поедает
в большом количестве кобылок, жуков
и справляется со скорпионами и ящери-
цами (из Гамильтона)
Полную противоположность им представляют такие эури-
фаги, как бурый медведь, енот, барсук, лесная и каменная ку-
ницы, соболь, кабан, серая крыса и ряд др. Беспозвоночные и
позвоночные животные, ягоды, орехи, фрукты, зелень играют
в их питании более или менее постоянную и существенную
роль.
Понятия «плотоядный» (зоофаг) и «растительноядный», или
«травоядный» (фитофаг), при внимательном изучении биоло-
гии видов часто оказываются достаточно условными. Так, на-
пример, из числа типичных хищников («плотоядные») волк,
лесная кошка и даже тигр временами едят ягоды и фрукты,
а растительноядные полев-
ки, хомяки и суслики — на-
секомых (рис. 51). Принад-
лежащие к отряду насеко-
моядных и действительно
насекомоядные по преобла-
дающему характеру пи-
щи— ушастый еж охотно ест
сочные ягоды, а землеройки-
бурозубки — семена ели, со-
сны, березы и др.
Корма, используемые
представителями данного
вида в какой-либо местно-
сти, для удобства их срав-
нения разделяют на основ-
ные, второстепенные,
и с л у ч а й н ы е, в зависи-
мости от большей или мень-
шей частоты случаев их поедания (или встреч в желудках), а
также от среднего числа съеденных особей или, наконец, от
среднего веса или объема в содержимом пищеварительного
тракта вскрытых животных;
Основными кормами считают те, которые поедаются часто
и в большом количестве; в ряде случаев к ним нужно относить
и сравнительно редко поедаемые виды корма, но имеющие
большую биологическую ценность. К случайным кормам отно-
сят используемые редко и по существу мало характерные для
диеты данного вида.
Такое разделение помогает разобраться в пищевых связях
животного, но все же, в известной мере, является формальным,
так как не указывает на причины более частого или редкого
использования тех или иных пищевых объектов. Зависит ли
оно только от обилия данного корма и его доступности или от
особой его привлекательности — вот вопрос, ответить на кото-
рый нередко оказывается довольно трудно. Непосредственные
9*
131
наблюдения за пастьбой и охотой животных в природе (рис. 52),
специальные опыты по кормлению животных, содержимых
в вольерах или в загонах, наконец, тщательные подсчеты на
свежих следах зверя числа съеденных и нетронутых растений
разных видов, раскопанных или оставленных без внимания нор
мелких грызунов и т. п. дают возможность судить об избира-
тельности, о предпочтении, оказываемом определенным кормам.
Рис. 52. Леопард, убивающий оленя (Индия) (из Корбетта, с изменениями).
Хищник удерживает поваленную жертву за шею, перегрызает крупные сосуды
и позвоночник. В этом положении олень не может защищаться копытами —
своим наиболее ' сильным оружием. Сходными приемами пользуется тигр
и др.
По степени их привлекательности корма разделяют на явно
предпочитаемые, поедаемые охотно и вынуж-
денные. Наблюдения показывают, что большинство млеко-
питающих обладает очень тонкой избирательной способностью.
Так, например, на Западном Кавказе многие крупные живот-
ные осенью питаются плодами диких яблонь и груш, причем
кабаны явно предпочитают груши, отличающиеся по сравнению
с яблоками большей сахаристостью и меньшей кислотностью.
Учет по следам показал, что кабаны, медведи, олени, косули и
волки из деревьев, находившихся под наблюдением, посещали
92% груш от общего числа, имевших сладкие плоды, 66%
груш, имевших кислые плоды, и только 28% из приносивших
горькие. Хорошо известно, что виды сельдей, в том числе и кас-
пийских, различать по внешним признакам довольно трудно, но
каспийские тюлени, иногда осматривающие ставные сети рыба-
ков, безошибочно выбирают среди других и съедают прежде
всего запутавшуюся в ячеях сельдь залом, имеющую наиболь-
шую жирность (до 16% жира).
Предпочитаемый корм (это нередко относится даже к от-
дельным частям растений или животных), как правило, биоло-
ги-чески наиболее ценен для потребителя в условиях данного
сезона и географической обстановки. Поедая жирную рыбу,
тюлень быстрее восстанавливает слой подкожного жира в пе-
риод интенсивного питания (нагула). Но потребности организ-
ма меняются по мере протекания сезонных биологических явле-
ний (линьки и отрастания шерстного покрова, сбрасывания и
роста рогов, течки, беременности, лактации, подготовки к спяч-
ке и т. д.), а также в зависимости от пола и возраста животных
и от того, каковы были условия их существования в предшест-
вующем сезоне, году и т. д. Поэтому предпочитаемый корм
может через некоторое время стать поедаемым слабо, неохотно
и наоборот.
Поедаемые охотно корма не всегда составляют основу дие-
ты животного; они не обязательно биолошчески ценны. В неко-
торых случаях они даже вредны для потребителя. Полорогие,
например, охотно поедают черный паслен, лоси — борец и па-
поротник орляк, грызуны — морской лук. В результате может
наступить отравление. С другой стороны, некоторые млекопи-
тающие нечувствительны к растительным и животным ядам.
Бобры, например, поедают без вреда для себя цикуту, смер-
тельную для других животных даже в небольших дозах, туры
потребляют сильно ядовые рододендрон, лютики; зайцы — целый
ряд растений, содержащих такие ядовитые вещества, как си-
нильная кислота, цитизин, эуфорбин, леуцин и др. Ежи охотно
едят ядовитых жуков-нарывников и т. д.
К категории вынужденных относят корма, потребляемые
главным образом при голодовке. Некоторые голодающие звери
иногда поедают собственный кал и кал других животных. Как
обычное явление калоедство, или копрофагия, наблюдается
у гиен, лисиц, песцов, енотовидных собак, свиней, землероек.
Интересные особенности представляет копрофагия у кроликов,
зайцев, сеноставок и бобров. Кролики поедают свой мягкий ноч-
ной кал, богатый протеином и переполненный бактериями. По
Шойнерту и Циммерману (Shounert u. Zimmerman, 1952), он
служит важным источником снабжения организма витаминами
группы В. Лишение кролика возможности поедать ночной кал
приводит к расстройству функций организма и истощению.
Сильно развита копрофагия и у бобров.
Учитывая отличия биологической ценности кормов, их раз-
деляют на продуктивные, поддерживающие и бал-
ластные. • : гЦ
К продуктивным относят те корма, которые полностью обе-
спечивают жизнедеятельность организма, его рост, накопление
им необходимых энергетических резервов и нормальное проте-
кание полового цикла. Поддерживающие корма могут некото-
рое время поддерживать жизнь, но не являются полноценными,
хотя и дают животному возможность уцелеть в трудный по кор-
мовым условиям период или сезон. Таковы, например, ягель-
ные— зимние корма'северного оленя. Ежедневно наедаясь ими
досыта, олень тем не менее непрерывно теряет вес и растрачи-
133
вает накопленные за лето резервы. Его продуктивные корма
летние — зелень травянистых растений, ветви и листья ив, по-
лярной березки. Они содержат много переваримых белков, бо-
гаты витаминами и т. п. На зеленом подножном корме у оле-
ней быстро восстанавливается вес, накапливаются энергетиче-
ские резервы к осени — на период гона и на зиму.
Балластные корма —в то же время и в высшей степени вы-
нужденные: 'животные наполняют ими желудок при полной,
длительной голодовке. Так, например, в многоснежные холод-
ные зимы ветви кустарников, кора деревьев, лишайники служат
вынужденным кормом кабанам, не имеющим доступа к корне-
вищам, орехам, желудям — их продуктивному корму. Иногда
в желудке кабана находят трухлявую древесину гнилых
пней, в желудке волка куски веревок и ремней, тигра — лишай-
ники, песца — тряпки и обрывки старых рыболовных сетей,
подобранных на месте летней стоянки рыбаков. Все это — бал-
ластные корма, они заполняют желудок, но ни в какой степени
не могут утолить голода животных. Путешественником Кэном
описан случай, когда голодные белые медведи съели кофе, па-
руса и американский флаг, а Мак-Клюр в желудке убитого
хищника этого вида нашел табак и липкий пластырь, тоже по-
хищенные из склада полярных исследователей.
Если при вскрытиях встречают много порожних желудков
или наполненных только балластным кормом, можно утверж-
дать, что популяция животных, из которой взята данная проба,
сильно голодает. Но и по количеству поедаемого корма не все-
гда можно с уверенностью судить о его биологической значи-
мости так же, как и по степени наполненности желудков—
о благоприятных кормовых условиях данного сезона и местно--
ста. Нередко объемистые корма, которые при грубом статисти-
ческом методе работы можно бы отнести к основным, в дей-
ствительности— вынужденные, поддерживающие. Так, напри-
мер, у белок Западной Сибири в 1932 г., при хорошем урожае
«орешков»- кедра средний вес содержимого желудка был 5,5 г,
причем ядра кедровых «орехов» встречались в 83% желудков,
семена других хвойных пород — в 1,3%, а грибы — в 15,7%.
В 1935 г. на севере Европейской часта СССР при полном не-
урожае семян хвойных и слабом урожае грибов средний вес
содержимого желудка белок равнялся 25,1 г (в некоторых слу-
чаях доходил до ИЗ а), причем почки ели встречались в 67,6%,
грибы в 27,4% желудков. Если сравнить средний вес пищи, по-
требляемой за кормежку в эти годы (5,5 г и 25,1 а), то может
показаться, что в 1935 г. сложились более благоприятные усло-
вия. В действительности положение было совсем иным. За
сутки белка съедает 15—35 а кедровых «орехов», и это обеспе*
чивает получение 56,2—109 Кал. Калорийность семян ели лишь
немного ниже — 1 а/6720 кал против 1 а/6910 кал для кедра.
134
Калорийность почек ели (в воздушно-сухом состоянии) состав-
ляет 1 а/4040 кал. Тепловой эквивалент в 80 Кал белка может
получить только в том случае, если съест не менее 10 000 почек,
что совершенно непосильно в отношении добывания этого кор-
ма и работы пищеварительного тракта (Данилов, 1944).
В те годы, когда белки питаются почками ели и грибами,
они очень истощаются за зиму, часть органов приобретает пато-
логические необратимые изменения. Многие зверьки погибают,
а уцелевшие, за редкими исключениями, не участвуют в раз-
множении. В результате численность белок резко сокращается.
В годы обилия семян хвойных, наоборот, зверьки хорошо упи-
таны уже с осени, успешно заканчивают зимовку; брачный пе-
риод у них начинается рано, еще в конце зимы. За сезон белки
успевают принести два выводка, и количество зверьков к на-
чалу промысловой охоты резко возрастает. Успешное размно-
жение при питании семенами хвойных — лучший показатель их
высокой биологической ценности для белок. Характерно, что
•грибы и при этой «сытной диете» остаются привлекательными.
.Дело в том, что семена хвойных — жирная, калорийная, но бед-
ная зольными элементами пища. Грибы восполняют этот недо-
статок и, кроме того, дают некоторые ценные витамины. Прене-
брежение к учету биологической ценности кормов может приво-
дить к грубым ошибкам Г Вместе с тем, при изучении питания
диких и домашних млекопитающих следует всегда иметь в виду
авторитетное высказывание И. В. Ларина (1956): «Часто...
охотность поедания растения ближе к действительной кор-
мовой оценке, чем теоретический расчет питательной ценности
по коэффициентам переваримости».
Для многих эурифагов (куница, барсук, медведь, кабан, ли-
сица и др.) характерно наличие нескольких заменяющих друг
друга групп пищи. Это ценное для вида приспособление к жиз-
ни в условиях с большой неустойчивостью кормовой базы. Оно,
как отмечает И. П. Наумов (1955), «составляет сущность эури-
фагии» и у некоторых видов объясняет устойчивость их числен-
ности. Но есть немало видов млекопитающих, использующих
обширный круг (сложный «спектр») кормов, из которых жиз-
ненно важное -значение, имеют очень немногие. Таков, напри-
мер, песец материковых тундр, для которого решающее значе-
ние имеют доминирующие полевки тундры — лемминги Lemmus
lemmus, L. obensis, Dlcrostonyx iorquatus (рис. 53). Только
в годы их высокой численности песцы хорошо упитаны, прино-
1 Так, например, в известный учебник проф. Д. Н. Кашкарова «Основы
экологии животных» (1938, стр. 343) попало ошибочное утверждение
В. Ф. Исакова, что основным кормом белки являются не шишки хвойных,
а грибы, и что корм для белки всегда существует в тайге в достаточном
количестве. Этот вывод обусловлен тем, что срок наблюдений Исакова был
слишком короток и целиком совпал с неурожаем семян хвойных.
135
Тамбей. Темней
щуча» лксарка-ле
| )/ ^^2 ЕЙ 7 ' /
Рис. 53. Зимнее питание песца грызунами в разных подзонах полуострова
Ямал (Зап. Сибирь):
1 — обский лемминг; 2 — копытный лемминг; 3 — виды рода Microtis', 4 — леммчгги
(ближе неопределенные); 5 — водяная крыса. Цифры в центре кружков обозначают го.ьы
и количество исследованных данных; под кружками названия факторий, на которые
были сданы песцы, использованные для изучения их пнтаиия. График показывает, что
повсеместно в тундрах основную роль играет обский лемминг (от 45 до 80 и более % встреч).
В лесотундре иногда (зимой 1937/38 г.) песцы ловят много серых полевок и водяных крыс
(по Дунаевой и Осмоловской)
сят большие жизнеспособные выводки и успешно выкармли-
вают молодых, благодаря чему количество этого пушного зверя
к осени резко возрастает.
Аналогичная закономерность на большем материале уста-
новлена в биологии довольно всеядной лисицы средних и юж-
ных степных районов Союза ССР (Vulpes vulpes cruclgera, V.
v. diluta, V. v. siepensis, V. v. caragan и др.) с тем отличием, что
леммингов заменяют серые полевки (Mtcrotus arvalis, М. socia-
Its) и степная пеструшка (Lagurus lagurus). При низкой чис-
ленности этих грызунов резко увеличивается процент яловых
самок, сокращается величина выводков лисиц, часть производи-
телей в многоснежные месяцы зимы погибает от истощения
(Фоцмозов, 1935; Чиркова, 1941 и др.). Малочисленность или
обилие полевок оказывают решающее влияние на жизнь лисиц
даже в тех районах, где летом много сусликов, ящериц, саран-
чевых и т. п. Видимо, эти летние корма не имеют значения,
свойственного полевкам — предпочитаемому зимнему корму
взрослых лисиц и летнему корму молодых щенят после перехода-
их на животную пищу. Сами серые полевки поедают десятки ви-
дов растений (некоторые виды Mtcrotus — до нескольких сот),,
но и в этих списках ясно выделяется группа основных, наиболее
привлекательных растений, обладающих большой биологиче-
ской ценностью. Для серой полевки (М. arvalis)—это мезо-
фильные широколистные злаки и бобовые.
СЕЗОННЫЕ ИЗМЕНЕНИЯ КОРМОВОГО РЕЖИМА
В природно-географических зонах с резко выраженными се-
зонами' состояние растительного покрова и населения живот-
ных, особенно мелких пойкилотермных и части видов гомотерм-
ных (птицы), так сильно изменяется, например, от весны к зиме,
что кормовая база большинства видов наземных млекопитаю-
щих испытывает ритмические колебания (рис. 54). В условиях
влажных тропиков млекопитающие в течение всего года отно-
сительно равномерно обеспечены самыми различными кормами,
в связи с чем там могут существовать и многие узко специали-
зированные по питанию виды и нет необходимости приспосаб-
ливаться к периодически повторяющемуся отсутствию или пол-
ной недоступности того или иного корма.
В условиях умеренного и холодного поясов нашей страны
даже у видов с очень коротким периодом-активности (напри-
мер, сусликов и сурков, ежегодно проводящих в спячке до 8—
9 месяцев) диета заметно отличается в период, следующий тот-
час за пробуждением, во время выкармливания молодых и, на-
конец, при «нажировывании» в период нагула перед залеганием
в спячку. Весной малых сусликов привлекает свежая сочная
зелень, богатая витаминами; позднее, при огрубении и высыха-
137
нии травы, они в большом количестве выкапывают «луковички»
тонконога и луковицы тюльпанов и птицемлечника, а в период
нагула, кроме зелени, охотно поедают созревшие семена эфеме-
ров, наливающиеся зерновки ковыля, житняка, культурных зла-
ков и т. п.
Еще более велики сезонные отличия питания у млекопитаю-
щих с круглогодичной активностью (рис. 55). На их диете
сказывается влияние сезонных изменений запаса, качества и
доступности кормов, циклические процессы, протекающие в са-
мом организме потребителя, а также перемещение его из одних
Рис. 54. Способы добывания беспозвоночных животных бурым медведем
Кольского полуострова с момента выхода из берлоги по сентябрь (результаты
учета следов деятельности медведей по месяцам в процентах от общего числа
наблюдений за каждый месяц):
/ — раскопка земли и дерна; 2 — перевертывание камней; 3 — разбивание колод
пней; 4 — разорение насыпных ' муравейников^ (из Насимовича и Семенова-Тян-Шаи-
ского). График показывает, что поедание муравьев играет большую роль в деятельности
медведя с конца апреля до половины июня, а добывание животных, скрывающихся под
камнями, — с половины июля по сентябрь
биотопов в другие, если вид принадлежит к числу мигрирую-
щих. Хвоя пихты, сосны, кедра, ели и можжевельника, поеда-
емая лосем зимою в северных лесах, содержит витамин С, а
сосна, кроме того, провитамин А *. Содержание витамина С
в хвое в течение года меняется: бывает наименьшим летом,
постепенно увеличивается в осенне-зимний период и достигает
максимума к апрелю, после чего резко падает. Содержание са-
харов в хвое, обеспечивающее повышение морозостойкости,
1 Провитамин — вещество растительного происхождения, превращающееся
в организме животного в витамин (например, каротин — в витамин А).
138
тоже возрастает к осени. Характерно, что сроки наибольшего
содержания витаминов и сахара в хвое перечисленных пород
совпадают со временем наиболее интенсивного поедания их вет-
вей лосями. Этот корм, всегда доступный и летом, очевидно,
вполне соответствует потребностям лосей, обеспечивая их зимой
теми питательными веществами, которыми в холодную половину
года бедны лиственные породы. Таким образом, в данном слу-
чае сезонность использования обусловлена сезонными отличиями
биологической ценности этой группы кормов для их потреби-
теля.
Рис. 55. Потребление растений разных групп полевкой Брандта Microta
(Lasiopodomys) brandti в Вост. Монголии. После зимнего питания сухими
растениями из заготовленных осенью запасов полевки в марте —• апреле с
жадностью поедают сочную зелень луков, богатую витамином С (по Кучеруку
и Дунаевой)
У лиственных пород — березы, осины, рябины, разных видов
ив, лось в летнее время поедает молодые побеги с листьями или
сдергивает с ветвей одни только листья, предпочитая наиболее
нежные, верхушечные. После листопада в течение поздней
осени, зимы и- ранней весны лось поедает ветви тех же листвен-
ных пород и гложет их не слишком огрубевшую кору. Как из-
вестно, запасные вещества древесных пород откладываются
осенью в камбии — деятельном слое коры и, отчасти, в древе-*
сине. Ценные пищевые вещества содержат также зачатки листь-
ев и соцветий в почках. Весной в период сокодвижения лоси
еще охотно обдирают кору с ив, рябины, вяза, обгладывают
139
ее с осины, но полностью утрачивают к ней интерес, как только’
начнут развертываться листочки этих деревьев; к этому времени
основная масса ценных для растительноядного животного ве-
ществ перемещается в тонкие ветви и листья.
Зайцы, многие виды полевок, лемминги, отчасти бобры так-
же используют веточные корма и кору почти исключительно
в холодный период года (зайцы-беляки в средней полосе
с половины октября до половины или конца апреля, в зависи-
мости от метеорологических условий весны). Это еще один при-
мер того, как сезонность использования корма обусловлена его
ограниченной во времени биологической ценностью. При силь-
ных морозах кора осины становится такой твердой, что не под-
Рис. 56. Схематический план ходов, проложенных в свежем снегу серой по-
левкой за одни сутки при поисках корма на убранном участке картофеля в
окрестностях г. Москвы. Гнездо ее помещалось в куче засохшей ботвы кар-
тофеля. Общая длина ходов на данном участке, бедном зеленым кормом —
45 м (ориг. А. Н. Формозова)
дается рездам лося. Поэтому имеющийся в природе предпочи-
таемый корм становится временно недоступным, и на годовом
графике, показывающем количественное соотношение кормов,
поедаемых лосем, кривая потребления коры имеет резкий
прогиб книзу в части, относящейся к самым морозным меся-
цам зимы.
Временная (полная или частичная) недоступность предпочи-
таемого корма очень часто ограничивает срок его использова-
ния млекопитающими. Так, например, в питании лисицы-кара-
ганки (V. vulpes caragan)— обитательницы сухих степей и
полупустынь Казахстана, большую роль играют суслики,
тушканчики, ушастые ежи, ящерицы, отчасти змеи. Эти живот-
ные проводят длительный период покоя в глубоких норах, от-
куда лисица не может их выкопать; они доступны для нее
только в течение довольно короткого периода. В этом случае
сезонность использования корма хищником полностью зависит
от особенностей годового цикла жизни его жертв. Малый суслик
деятелен около 4 месяцев в году и в течение только этих меся-
140
цев он играет существенную роль как корм лисицы. Основная по
числу видов и особей масса мелких млекопитающих и все реп-
тилии глинистых полупустынь и пустынь Казахстана и Средней
Азии для лисы становятся недоступными в конце лета; отми-
рают и массовые крупные насекомые (саранчевые), улетает
большинство птиц. Поэтому условия добывания корма резко
ухудшаются, что вынуждает лисиц-караганок откочевывать на
зиму в пески, где преобладают по численности грызуны, не впа-
дающие в спячку (песчанки), или уходить в котловины озер,
речные долины, где встречаются полевки. Сезонная недоступ-
ность ряда основных кормовых объектов вызывает сезонные
перекочсвки лисиц и соответственно — изменение набора вылав-
ливаемых животных.
В жизни среднерусских лисиц роль еще более узкосезон-
ного корма играют майские жуки (Melolontha hippocasta.ru).
•Лисицы ловят их по вечерам в момент выхода из земли или
в первые минуты низкого полета, когда жуки с гулом направ-
ляются к лесным опушкам и садам. Массовый лет майских
хрущей в средней полосе Европейской части Союза происходит
в конце мая — июне; в течение этого периода лисицы еже-
дневно, еще в ранних сумерках вечера выходят на лесные
-опушки, прилегающие к полям или большим лесосекам — местам
массового выплода хрущей; каждый хищник вылавливает их
десятками.
В этот же период интенсивно вылавливает хрущей и рыжая
вечерница (Nyctalus noctula), для которой это тоже строго се-
зонный корм. В период массового вылета крупных белых поде-
нок (Polymitarcys Virgo), количество которых в некоторые годы
бывает так велико, что множество вьющихся над рекой насеко-
мых напоминает живую метель (отсюда их .народное название
«метелица»), над Волгой, Окой и другими крупными реками
можно наблюдать концентрацию охотящихся летучих мышей.
Обилие очень доступной добычи привлекает не только мелких
рукокрылых: водяных, прудовых, усатых ночниц (Myotis daubеп-
tonl, М. dasycneme, М. mystacinus), двуцветных кожанов
(Vespertlllo murinus), но и такие крупные в'иды как рыжая и ги-
гантская вечерницы (Nyctalus noctula, N. slculus). Период силь-
ного лёта этой поденки часто бывает короче, чем у хруща, и так
же коротко время, в течение которого она играет существенную
роль в питании рукокрылых.
Концентрация пелагических рыб на местах нереста или на-
гула, подход массы проходных и полупроходных рыб к устьям
рек при нерестовых миграциях и т. п. создают предпосылки для
успешной охоты рыбоядных ластоногих и китообразных. Такая
сезонная концентрация жертв обусловливает четкую сезонность
их вылова млекопитающими ихтиофагами, для которых охота за
единичными особями, распыленными по большой акватории,
невыгодна, а зачастую и просто невозможна по причине их
недоступности (пребывание на больших глубинах, например
лососевых). Изучение питания черноморского дельфина (Delphi-
nus delphis ponticus), выполненное на большом материале
(свыше 10 000 желудков), показало, что зимой и весной основ-
ным, а у южного побережья Кавказа — единственным кормом
этого вида служит хамса, образующая большие скопления. Ле-
том, с отходом хамсы, дельфин переходит на более разнообраз-
ный корм — видимо, ни один отдельно взятый объект его пита-
ния не может целиком заменить хамсу. Пелагические морские
иглы, шпрот, мерланг (черноморская пикша) и значительно
реже другие виды рыб служат второстепенными и случайными
кормами черноморского дельфина летом и осенью. Во все се-
зоны добычу его составляют почти исключительно представи-
тели пелагической фауны и притом виды мелкие по размерам
особей — таков характер пищевой специализации черномор-
ского дельфина (Клейненберг, 1936).
Другой представитель мелких китообразных Черного
моря — морская свинья (Phocaena phocaena) пожирает главным
образом донных рыб, а пелагические (хамса и атеринка) при-
влекают ее лишь тогда, когда находятся в виде больших, густых
косяков. Весной, следуя за мигрирующими скоплениями хамсы,
морская свинья идет из Черного в Азовское море, а осенью —
обратно; в эти сезоны она охотится преимущественно за пела-
гическими рыбами (хамса, атеринка). Зимой и летом в рацион
входят донные рыбы (бычки — главным образом ротан и пе-
сочник)., За год в среднем количество съеденных донных рыб по
весу в два раза превышает пелагических (эти данные получены
в результате исследования 4000 желудков черноморской свиньи;
Цалкин, 1940). Таким образом, сезонность использования неко-
торых кормов бывает обусловлена появлением больших времен-
ных скоплений мелкой добычи. Следуя за идущими на нерест
стадными рыбами большой кормовой ценности (лососями, си-
гами и др.), в эстуариях концентрируются нерпы, белухи, дель-
фины. Часть этих морских зверей, преследуя рыб, проникает
вместе с ними в реки и поднимается на многие десятки кило-
метров вверх по течению Оби, Енисея, Амура.и др.
Изучение сезонных особенностей питания часто дает воз-
можность вскрыть причины перемещений и миграций млекопи-
тающих. Это относится и к водным, и к наземным животным;.
В питании кабанов Западного Кавказа орешки бука, желуди
служат основным кормом в течение 6- -7 месяцев, начиная с
октября, когда плоды осыпаются на землю, и до апреля вклю-
чительно (Донауров и Теплов, 1948). Позднее их остается очень
мало, так как это излюбленный корм многих видов животных —
конкурентов кабана. Уцелевшие и проросшие плоды бука и
дуба поедаются кабанами даже в мае и июне, но уже стано-
143;
вятся случайным кормом. Плоды груши служат кабанам основ-. |
ным кормом только в сентябре, а дикой яблони — в октябре, . I
когда они осыпаются на землю. Но этот корм быстро утрачи- ,
вает свои ценные качества — по мере того как лежащие в сыро- 1
сти плоды загнивают, а содержащиеся в них легко растворимые I
вещества вымываются дождями. В этом случае сезонность ?
использования плодов древесных пород зависит не только от '
сроков их созревания и опадания, но и от длительности сохра- I
нения их кормовой биологической ценности при лежании на |
земле, где они только и доступны для кабанов. Уже с конца
лета распределение стад кабанов по отдельным урочищам зави- !
сит от урожая фруктов, а осенью и весной от наличия основных j
кормов — плодов бука, дуба (рис. 57). Зимой большое значение
имеют глубина и плотность снежного покрова, от которого зави-
сит доступность большинства важнейших кормов. Поэтому ка-
баны— бродячие, непрерывно кочующие животные, несколько
раз тз году меняющие районы обитания в горных лесах не
только Кавказа, но и Средней Азии, Сибири, Дальнего Востока.
ГЕОГРАФИЧЕСКИЕ ИЗМЕНЕНИЯ ПИТАНИЯ
Особенности географических аспектов, а также сезонных • j
изменений пищевого режима широко распространенных всеяд-
ных млекопитающих можно показать на примере медведя.
Обыкновенный бурый медведь (Ursus arctos arctos) в северной
полосе тайги Европейской части СССР проводит в берлоге
около 6 месяцев и пробуждается в первых числах мая. Обычно
к этому времени снежный покров уже осел и по ночам покры-
вается плотной коркой, кормов мало, и они не везде доступны.
Чаще всего медведь раскапывает едва показавшиеся из-под
снега муравейники, добираясь до зимующих в глубине рыжих
лесных муравьев и разбивает трухлявые пни в поисках жирных
личинок жуков ксилофагов. Найдя лосей, упорно гоняет их по
насту и, доведя до изнеможения, убивает иногда по 2—3 из
одного стада, что надолго обеспечивает хищника мясом.
На освободившихся от снега сфагновых болотах собирает пере-
зимовавшую клюкву, на гарях — бруснику, потом выходит пас-
тись на свежей зелени злаков и сочных ростках дудника к луго-
винам по берегам рек, где вегетация начинается раньше, чем
в тенистых местах леса. С началом сокодвижения у молодых
осинок, растущих куртинами, медведь, собирая деревца в
охапку, соскабливает зубами набухшие почки.
Весь июнь и отчасти июль медведи пасутся по разреженным
участкам леса с густым, рослым травостоем и кормятся глав-
ным образом зеленью — медвежьей дудкой (Angelica silvestris),
иван-чаем (Eplloblum angustifolium), молоканом (Mulgedium
siblricum) и др., сдабривая вегетарианскую пищу муравьями,
144
их куколками, личинками шмелей, златок и других насекомых.
По мере огрубения трав и созревания ягод медведи переходят
на чернику и голубику, в заросли малины по зарастающим га-
рям, затем на брусничники и поздней осенью в участки леса
с обилием рябины.
Если среди леса есть участки полей с посевами овса, мед-
веди в конце лета ходят кормиться овсом и сильно выбивают
полосы. Обильные урожаи ягод—важнейшего нажировочного
корма этого медведя, бывают не везде и не ежегодно, поэтому
хищник осенью настойчиво преследует крупных животных и при
случае систематически поедает дичь, попавшую в слопцы и
жердки охотников. Нападения медведей этого подвида на коров
и лошадей, пасущихся близ леса, очень обычны. Описаны слу-
чаи, когда один медведь убивал за лето от 10 до 50 и даже
84 голов крупного рогатого скота из стад 5—6 соседних де-
ревень.
Летом, в период гона, израненный в драке слабый самец не-
редко становится жертвой своего сильного противника и пол-
ностью им съедается. В голодные годы некоторые недостаточно
откормившиеся медведи вынуждены бродить после наступления
зимы. С голода такие «медведи-шатуны» нередко нападают на
особей своего вида, лежащих в берлоге, и пожирают их. Слу-
чаи каннибализма у U. a. arctos отмечаются и летом, и зимой.
Изголодавшиеся «шатуны» или отдельные старые особи, утра-
тившие способность охотиться на крупных животных, иногда
нападают на людей при случайных встречах и даже специально
охотятся на них, подкарауливая или отыскивая по следам..
Однако в Европейской части СССР такие случаи редки; более
часты они на Урале и в особенности в Сибири. Сибирские мед-
веди от Зауралья до Приамурья (здесь выделяют несколько
географических рас) более смелы, злобны и опасны для людей;
очень часто они нападают и на домашних животных.
Сильно отличается биология огромного камчатского медведя
(U. a. berlnglanus), одно из латинских названий которого
U. a. piscator — «медведь-рыболов». Он выходит из берлоги
в середине апреля, когда на многоснежной Камчатке лишь кое-
где намечаются первые проталины. В это время доступные хищ-
нику корма — главным образом прошлогодняя сухая трава,
первая зелень и, при удаче, небольшое количество идущей вес-
ною рыбы (гольца — Salvelinus malma), не могут полностью
его обеспечить. На таком скудном и в основном малопитатель-
ном корме медведю приходится существовать до развития рас-
тительности, расходуя остатки отложений жира, сохранившиеся
после зимнего сна.
В течение конца апреля и части мая медведи используют
разные более или менее случайные корма, но зеленые растения
по мере развития травостоя приобретают все большее значение
10 Териология
145
в их рационе. Из кормов, сохранившихся с осени, медведям
удается в небольшом количестве находить ягоды вороники
(«шикши») и плоды кедрового стланца. Медведи, спустившиеся
на побережье, пользуются «выбросами моря» — моллюсками,
морской капустой, трупами китообразных, ластоногих, птиц и
т. п. Чаще других им достаются выброшенные морем детеныши
нерпы. В июне медведи питаются почти исключительно зеленым
кормом: дудником (Angelica), борщевником (Heraclewn), шело-
майником (Filipendula), вейником (Calamagrostls langsdorfii)
и др. и рассеяны повсюду благодаря обилию зарослей высоко-
травья.
С июля по сентябрь длится летний период жизни медведя,
когда появляются специфические его корма: проходные лососе-
вые рыбы, ягоды и «орехи» кедрового стланца, обеспечивающие
восстановление сил после голодной весны и начало накопления
жира перед зимним сном. Все же основу диеты составляют соч-
ные растения высокотравья, но в начале июля появляются первые
особи горбуши, а рунный ее ход начинается с половины этого
месяца. Далее, с небольшими перерывами идут в верховья рек
на нерестилища кумжа, голец, кижуч, кета, красная. До глубо-
кой осени в рыбных реках встречается рыба нескольких видов.
Появление этого корма вызывает заметную концентрацию зве-
рей около рек, но небольшое количество медведей все же
остается в горах, продолжая питаться только растениями. Все
долины рыбных рек Камчатки отмечены неизгладимыми сле-
дами многих поколений медведей-рыболовов: «торные тропы
зверей тянутся вдоль берегов рек, пересекают их через броды,
выводят из долин по удобным склонам к хребтам...» (Аверин,
1948). В желудке крупного медведя бывает до 20 кг рыбы, но
почти всегда имеется примесь травы, а ближе к осени и ягод.
В конце июля начинают поспевать вороника и голубика;
медведи поедают эти ягоды в огромном количестве. Рыба
остается основным кормом, но ягоды начинают приобретать все
большее значение. В августе медведи часто выходят на' откры-
тые ягодниковые тундры. Голубика и шикша служат важнейшим
кормом до половины сентября, затем первая осыпается, а во-
ронику оттесняют на второй план созревающие к этому времени
рябина и «орешки» кедрового стланца. Зеленая растительность
в сентябре подсыхает, грубеет, поедается редко и в малом ко-
личестве. Осенью медведи часто встречаются по склонам хреб-
тов— в рябинниках и особенно в зарослях кедрового стланца.
За октябрь — ноябрь медведи, питаясь рябиной, «орешками»
стланца, а местами гольцом и кижучем, сильно отъедается пе-
ред залеганием в берлогу. В кормные годы у отдельных особей
к ноябрю слой подкожного жира на спине и бедрах достигает
15 см толщины. Характерно, что у камчатских медведей он
почти до весны сохраняет сильный .запах рыбы — свидетельство-
146
того, что лососевые играют большую роль в нажировочном пе-
риоде этих хищников.
Камчатские медведи, несмотря на свои крупные размеры,—
очень мирные звери; случаи неспровоцированных нападений на
людей совсем неизвестны, нападения на домашних животных
исключительно редки. При большом росте этот медведь имеет
относительно слабые зубы, чем заметно отличается от предста-
вителей подвидов, охотящихся за крупной добычей.
Белокоготный тяньшанский медведь (U. a. pruinosus) в
основном тоже питается растительной пищей. С весны до поло-
вины лета он поедает сочную зелень, корневища, например гре-
чишек (Polygonum viviparum, Р. alplnuin), луковицы, а также
насекомых и их личинок и земляных червей. После созревания
ягод кормится малиной, кислицей, а позднее переходит в ябло-
невые леса, где плоды диких яблонь становятся его основным
кормом. Когда подрастут молодые сурки и начнут выходить на
кормежку, медведь охотится на них, обычно выкапывая зверь-
ков, спрятавшихся при тревоге в короткие временные норы.
Осенью он разрывает даже глубокие .зимовочные норы, уничто-
жая целые семьи ожиревших сурков, находящихся в полной
спячке. Постоянное переворачивание и разбрасывание камней,
, огромная работа по раскапыванию нор наложили отпечаток на
морфологические особенности этого подвида. В частности, когти
передних лап тяньшанского медведя ^относительно длиннее и
более прямые—не столь изогнутые, как у таежных представи-
' телей вида. Случаи нападения этого медведя на домашних жи-
вотных исключительно редки, а на человека совершенно . не-
известны.
Безлесные гористые районы Центральной Азии населяет еще
более мелкий медведь-пищухоед (U. a. lagomyiarius), питаю-
щийся зеленью альпийских растений, корневищами ревеня, соч-
ными плодами эфедры и постоянно раскапывающий норы пи-
щух. Эти мелкие грызуны — важнейший из-животных кормов
пищухоеда; П. К. Козлов нашел 25 пищух в желудке медведя,
еще не закончившего свою охоту. Реже пищухоеды занимаются
выкапыванием сурков из их глубоких нор. Под осень они выхо-
дят к солончаковым понижениям и в массе поедают солоновато-
сладкие сочные ягоды селитрянки (Nitraria). Когти передних
лап у этой формы медведя тоже прямые и сильные, коренные
зубы относительно крупные, может быть в связи с тем, что ему
приходится дробить много крепких костянок селитрянки.
На крупных животных пищухоед никогда не нападает, поэтому
скот спокойно пасется в местах, где постоянно охотится этот
всеядный хищник.
Еще в XVII и XVIII столетиях медведи были обычны в сте-
пях юга нынешней Европейской части СССР. Никаких сведений
о биологии этих степных медведей не сохранилось, но, зная те-
10* 147
перь образ жизни пищухоеда и тяныпанского медведя, можно ’
с уверенностью утверждать, что южнорусские звери выкапывали
сурков и откармливались на-плодах степной вишни и терна,
заросли которых в ту пору занимали обширные площади.
Таким образом, повсюду бурый медведь ведет себя как все-
ядное животное. Зеленые части растений, плоды, беспозвоноч-
ные и позвоночные животные занимают определенное место в
питании всех представителей вида Ursus arctos, но видовой со- !
став поедаемых растений и животных, количественное соотноше-' [
иие отдельных компонентов, размеры жертв хищника сильно
варьируют. Соответственно различаются и размеры вреда, ко-
торый приносят медведи, обитающие в разных областях и при- •
надлежащие к разным географическим расам. I
Кавказская лесная куница (Martes martes lorenzi, очень I
крупный подвид) охотится почти исключительно на земле; на
деревья поднимается только приближаясь к дуплу, где соби-
рается расположиться на отдых. Основная ее добыча зимой —
кустарниковые полевки, мелкие мыши, летом — также и полчки.
Большое место в ее питании занимают ягоды, особенно малина,
ежевика, кизил, рябина, а зимою тисс и омела, и фрукты.
В сомкнутых темнохвойных лесах севера у сравнительно мел-
ких М. martes ruthena и М. martes sabanejevi рыжие лесные
полевки и белка занимают основное' место среди поедаемых
животных, зимой эти куницы ловят также рябчиков и изредка
зайцев. Плоды (и ягоды) играют в питании меньшую роль, чем
у кавказского подвида; из них следует отмстить бруснику, чер-
нику,. морошку, рябину, а на востоке ареала — кедровые
«орешки». В поисках беличьих гнезд эти куницы часто подни-
маются на деревья и перемещаются по ветвям.
Довольно крупная северная куница (М. т. martes), по
наблюдениям, проведенным на Кольском п-ве,— такой же по
преимуществу наземный хищник, как кавказская. В разреженных
хвойных лесах у северной границы ее ареала белки очень мало,
а большие просветы между деревьями лишают куницу возмож-
ности передвигаться поверху, прыгая с ветки на ветку. Основная
ее добыча — мелкие лесные грызуны, из ягод она поедает воро-
нику, чернику и рябину; в зимнее время часто и успешно охо- «
тится на тетеревиных птиц — белых куропаток, глухарей. В це-
лом для лесной куницы, как и ряда других хищников, харак-
терно закономерное возрастание плотоядности с юга на север г
и наоборот — увеличение всеядности, т. е. доли плодов (и ягод)
и насекомых, при движении к югу.
Значительные географические отличия в пищевых связях не-
трудно установить и у других млекопитающих, например, у
волка, лисицы, соболя, горностая и даже многих широко рас-
пространенных фитофагов — лося, зайца-беляка, водяной
крысы. Последней успешно освоены субальпийские луга Кав-
148
каза, тростниковые заросли озер Казахстана, дельты Волги и
Дона, поемные луга Северной Двины, Печоры, Оби и других
рек, берега болот, разного типа ручьев, маленьких лесных речек
и т. д. Уже по одному разнообразию занимаемых биотопов
можно судить о том, что список кормовых растений водяной
крысы достаточно обширен и сильно варьирует в разных при-
родных зонах и вертикальных поясах.
ИЗМЕНЕНИЯ ПИТАНИЯ В ЗАВИСИМОСТИ ОТ ВОЗРАСТА,
ПОЛА, ИНДИВИДУАЛЬНЫХ НАВЫКОВ ОСОБЕЙ
И КОЛЕБАНИЙ УРОЖАЙНОСТИ КОРМОВ
Кормление новорожденных детенышей материнским моло-
ком— характерная для всех представителей класса млекопи-
тающих прогрессивная черта биологии. Молозиво — секрет
млечных желез только что родившей самки, сильно отличается
по составу от молока, которое детеныши будут получать
позднее. Специфические особенности молозива очень важны
именно для новорожденных; например, у северного оленя моло-
зиво очень богато .белками и витаминами. Молозиво перевари-
вается легче, чем молоко, и очень необходимо новорожденному,
так как способствует освобождению желудочно-кишечного
тракта от первородного кала и обладает бактерицидными свой-
ствами. Если новорожденный олененок не пососет молозива, он
через 6—8 часов настолько ослабеет, что может погибнуть
(Друри, 1955).
Переход с молочного питания на пищу взрослых обычно
осуществляется постепенно, соответственно ^ перестройке дея-
тельности всего пищеварительного аппарата, начиная с форми-
рования зубной системы, функционирования слюнных и других
желез, разных отделов желудка (у жвачных) и т. д. Молодняк
растительноядных видов уже в период молочного питания начи-
нает понемногу есть свежую траву, причем роль этой пищи
быстро возрастает.
У большинства травоядных видов деторождение приурочено
к периоду года, когда и мать, и ее молодняк могут получать
полноценный зеленый, легко переваримыи корм (олени, анти-
лопы, горные бараны, зайцы, суслики, сурки и многие др.). Мо-
лочное кормление мелких грызунов и зайцев, приносящих по-
следовательно несколько пометов, непродолжительно, и переход
молодых на растительный корм происходит очень рано. Неко-
торые хищники еще в период лактации начинают добавлять
к молочной пище молодых порции разжеванной и слегка пере-
варенной животной пищи — так называемую отрыжку. В даль-
нейшем переходом на корм взрослых служит период поедания
сравнительно мелких животных, которых доставляют родители,
так как зубы молодых еще не могут справиться с разгрызанием
149
крепких костей и сухожилий крупных жертв. Волки, например,
обычно ловят для своих щенят сусликов, леммингов, полевок,
молодых зайчат, птенцов, тогда как осенью уже подросшие
«прибылые» волки успешно участвуют в семейных охотах на
оленей, косуль, крупный рогатый скот и т. п. Но у молодых
хищников, добывающих корм путем одиночной охоты, отсут-
ствие опыта и соответствующих навыков часто обусловливает
значительные отличия качественного состава их корма от пищи
взрослых. Например, молодые белухи (Delpfiinapterus leucas),
достигшие роста взрослых, но легко отличающиеся своей том-
ной или голубовато-серой окраской, часто и в большом коли-
честве поедают медлительных ракообразных, тогда как взрос-
лые (белые) дельфины успешно охотятся на быстрых и увертли-
вых рыб — более ценных в пищевом отношении.
Как известно, для ряда видов и групп млекопитающих ха-
рактерны хорошо выраженные половые различия в кормодо-
бывающей деятельности. У лесной куницы, соболя, солонгоя,
горностая и, вероятно, у некоторых других крупные и более
сильно вооруженные самцы имеют более обширные охотничьи
участки и чаще, чем относительно мелкие самки, ловят сравни-
тельно крупных животных. Это1 вызывает заметные отличия
в качественном составе кормов, используемых особями разного
пола. Самки лесной куницы, например, в основном питаются
полевками, растительными кормами, мелкими птицами, отчасти
белками и рябчиками; куницы-самцы, кроме мелких грызунов
и белок, ловят рябчиков, белых куропаток, тетеревов, глухарей,
зайцев-беляков. Самцы горностая чаще самок ловят крупных
водяных полевок, нападают на птиц и т. п. В зимнее время они
оставляют длинные следы на поверхности снега, тогда как
самки питаются почти исключительно мелкими зверьками и,
подобно ласке, большую часть зимы проводят под снегом, в хо-
дах полевок.
Экологическое значение возрастных и половых различий пи-
тания очень существенно; оно обеспечивает расширение круга
пищевых связей вида, а следовательно, и более- полное исполь-
зование имеющихся в природе ресурсов, что в какой-то степени J
способствует устойчивости численности населения вида и его
расселению.
Индивидуальные особенности питания особей представляют - ’
большой интерес с точки зрения практической (выборочное
истребление хищников-людоедов, медведей-«стервятников»,
хорьков-куроедов и т. п.) и в аспекте изучения поведения жи- .
вотных. Узко локальная специфика условий существования, на-
копленный своеобразный индивидуальный опыт, разного рода
случайные обстоятельства (легкое ранение, вызвавшее частич-
ную инвалидность, разрушение зубов оТ старости или болезни
ы т. д.) —-все это может стать причиной довольно резких укло-
150
нений в «пищедо'бывательной деятельности» (А. Д. Слоним) и
специализации отдельных особей или даже небольших популя-
ций млекопитающих.
Известно, с какой осторожностью относятся лисицы и даже
шакалы к предметам, побывавшим в руках человека и сохра-
нившим его запах. Но это относится только к особям, обитаю-
щим вдали от поселков, торных дорог и т. п. На юге Дагестана
близ линии Кавказской железной дороги хищники в течение
многих поколений привыкли отыскивать остатки пищи, выбро-
шенные пассажирами проходящих поездов. Очень часто кури-
ные кости, головы копченой рыбы выбрасывают завернутыми в
бумагу; вид такой упаковки настолько прочно ассоциируется у
«придорожных» шакалов и лисиц с возможностью поживиться,
что здесь лучший способ завлечь их в капкан —положить око-
ло него промасленную бумагу. Лисица, выросшая в глубине
тайги, наткнувшись зимою на лыжный след, не решается через
него перепрыгнуть и обходит стороной, хотя бы для этого ей
пришлось сделать несколько лишних километров. Лисицы гу-
стонаселенных районов, где развит лыжный спорт, настолько
привыкают к удобствам передвижения по торным лыжням и тро-
пам, что в период многоснежья зверей очень трудно согнать с
них. Такие лисицы зимою смело подходят к окраинам селений
в поисках падали, а летом нередко становятся самыми дерзки-
ми похитителями кур.
Среди медведей Европейского Севера далеко не все охотно
и успешно нападают на пасущийся в лесу скот. Отдельные осо-
би, видимо случайно получившие ценный для них индивидуаль-
ный опыт, становятся затем специализированными охотниками
на коров и режут по несколько десятков голов за пастбищный
сезон (таких медведей в народе называют «стервятниками», в
отличие от мирных «муравьятников» и т. п.). Очень убедитель-
но описаны Дж. Корбеттом (1959) случаи, показывающие, как
отдельные леопарды и тигры становятся людоедами. Например,
после большой эпидемии, унесшей много жертв среди насе-
ления Гарвала (горно-лесной район в верховьях бассейна
Ганга), один леопард настолько привык питаться человеческим
мясом, что в конце вспышки, за неимением трупов, стал охо-
титься за крестьянами, работающими в лесу, на полях и даже
уносить их ночью из жилищ. За период с 1918 по 1926 гг. этот
«леопард из Рудрапраяга» убил более 175 людей, прежде чем
был застрелен. Одним из самых опасных тигров-людоедов ока-
залась самка, потерявшая правые клыки верхней и нижней
челюсти в результате пулевого ранения. Из-за этого увечья она
не могла добывать привычную дичь и предпочла самую легкую
охоту — на людей. Жертвой ее стали 434 человека в Непале и
Кумаоне, пока не удалась одна из многих попыток ее уничто-
жить.
151
Леопарды Копет-Дага прежде никогда не нападали на до-
машних животных, так как в горах было достаточно диких ба-
ранов и козлов. За последние десятилетия в связи с уменьше-
нием количества диких копытных в этом хребте отдельные лео-
парды охотятбя на лошадей и временами наносят урон хозяйст-
ву колхозов Туркмении в предгорьях (рис. 58).
Рис. 58. Леопард, убитый 16 марта 1950 г. в Бахарденском районе
Туркмении.
Количество многих, в том числе наиболее ценных естествен-
ных кормов, очень непостоянно, в разные годы, что вызывает
сильные колебания обеспеченности пищей целого комплекса ее
потребителей, в частности и млекопитающих (рис. 59). Так, на-
пример, высокие урожаи семян ели, лиственницы, «орешков»
сибирского кедра повторяются с интервалами в 3—5 и более
лет, периодически сменяясь полными неурожаями, которые
охватывают области тайги, измеряемые сотнями тысяч квад-
ратных километров (рис. 60, 61). Столь же велики площади
с одновременными высокими урожаями или неурожаями
грибов и т. п. (Формозов, 1935, 1936; Данилов, 1940, 1943,
1949, 1951, 1952). Лесные полевки, бурундуки, белки, лесные
куницы, соболи, медведи, кабаны, населяющие эти территории,
в одни годы имеют возможность многие месяцы питаться та-
ким полноценным кормом, как кедровые «орешки», а в другие
152
Рис 60 Распределение урожая «орешков» сибирского кедра в 1946 г..
/ - высокий урожай; - средний урожай; 5 -^неУРО-8; J
СВИт6аИйге<О3Га°ураелья «Зап^Среда ’Жри’ Алтая, т. е. на огромной территории, были хорошо обеспечены этим кормом,
в тайге Зауралья, dan. и ър д g BO£oqHofi части ареала кедра был средний урожай
Рис. 61. Урожайность семян ели на территории лесной золы СССР осенью 1938 г.:
условные обозначения 1—4_те же, что на рис. 60; 5 — северная и южная границы ареала ели (по Данилову). Около
половины площади, занятой еловыми лесами, было поражено неурожаем, на остальном пространстве урожай был сред-
ним. Зима 1938/39 гг. была для белки в большинстве мест голодной
вынуждены пользоваться различными заменяющими кормами,,
часто имеющими для потребителей гораздо меньшую биологи-
ческую ценность.
Там, где среди сплошных еловых массивов есть небольшие-
пятна насаждений сосны, белки при неурожае еловых шишек
переходят на питание семенами сосны и уже с половины лета
переселяются в субори и боры — участки с преобладанием этой
лесообразующей породы. Но средняя величина большого уро-
жая семян сосны на единицу площади леса всегда меньше хо-
рошего урожая ели, чешуи сосновых шишек гораздо тверже
еловых — их труднее разгрызать, в разреженных кронах сосен
белки хуже защищены от холодных зимних ветров, нападений
ястреба-тетеревятника и крупных сов. Охота на белку в сосно-
вых лесах легче, чем в ельниках, и промысловики убивают в
них больший процент зверьков.
Следовательно, переход на питание семенами сосны вызыва-
ет в жизни местной популяции белок ряд взаимно связанных,
на первый взгляд незначительных, но в действительности очень
важных изменений, сказывающихся на физиологическом со-
стоянии самих зверьков и условиях их борьбы за существо-
вание.
Нередки случаи совпадения неурожая шишек всех хвой-
ных пород: в такие годы для животных-семеноедов создаются
исключительно трудные кормовые условия.
В зоне тундр, в лесостепи, степи и, в меньшей степени, в
лесной зоне и зоне пустынь, непостоянна численность населения
мелких млекопитающих — предпочитаемого корма многих хищ-
ников.
Резко изменяется по годам количество леммингов, серых по-
левок, степных пеструшек, общественных полевок и некоторых,
других массовых, «ландшафтных» видов мелких грызунов. Со-
ответственно изменяются и условия охоты песцов, лисиц, кор-
саков, ласок, горностаев, их поведение, величина охотничьих
участков, подвижность, упитанность, устойчивость против бо-
лезней (резистентность) и т. п.
При бескормице даже типичные хищники могут пополнять
свой рацион за счет растительных кормов. Горностай, например,
на Кольском полуострове в таких случаях ест плоды можже-
вельника.
Чередование лет, хорошо обеспеченных продуктивными кор-
мами, слабо обеспеченных и совершенно голодных хорошо про-
слеживается при непосредственных наблюдениях в природе и
по анализам содержимого желудков животных, собранных в од-
ной и той же местности за достаточно большой период. Ясно,,
что эти годовые изменения питания оказывают прямое и кос-
венное влияние на различные важные стороны жизнедеятельно-
сти млекопитающих и всегда должны учитываться.
156
СУТОЧНАЯ НОРМА ПИЩИ,
СПОСОБНОСТЬ ПЕРЕНОСИТЬ ГОЛОДАНИЕ
Количество пищи, необходимое в сутки, зависит от ее каче-
ства и переваримости, интенсивности обмена веществ, свой-
ственной данному животному и, в очень большой степени, от
массы его тела. При этом между размерами животных разных
видов и потребной для них суточной нормой пищи нет прямой
пропорциональности. Организм млекопитающих, как гомотерм-
ных и гетеротермных животных, значительную часть получае-
мых с пищей калорий использует на поддержание температу-
ры тела. По закону Рубнера зависимость между количеством
поддерживающего корма и весом животных обусловливается
отношением поверхности тела к его весу. Мелкие зверьки при
малой массе тела имеют относительно большую поверхность,
излучающую тепло, и соответственно находятся в худших усло-
виях терморегуляции, чем крупные. Поэтому их потребность в
пище всегда относительно больше. Эту закономерность удобно
показать, сравнивая потребность в корме у крупнейших из ныне
живущих млекопитающих — беззубых китов и более мелких
китообразных, тоже потребителей животной пищи (табл. 1);
Таблица 1
Средняя суточная норма пищи (в % от веса тела)
взрослых особей китообразных в условиях относительного
равновесия (поддерживающего питания) и в условиях
нагула (накопления резервных веществ)
(по Томилину, 1946)
Виды1
Синий кит ...................
Финвал.......................
Горбач ......................
Серый кит....................
Белуха (охотская) ......
Вес тела, кг Средняя суточная норма пищи
при равно- весии при иагуле
96810 2,0 4,3
59000 2,6 5,2
34810 3,6 7,2
31550 3,9 7 ,8
1100 7,4 И,4
Как видно из таблицы, потребность в пище закономерно
возрастает от более крупных животных к сравнительно мелким.
Этот показатель иногда называют показателем «относительной
прожорливости». Следует обратить внимание и на то обстоя-
тельство, что у всех приведенных в таблице видов. китообраз-
ных суточная норма пищи при нагуле в 1,5—2 раза выше, чем
1 Вес животных вычислен по формулам.
157
в период поддерживающего питания. Значит, и сезонные изме-
нения биологии сказываются на количестве пищи, поедаемой
животными за сутки. У многих видов, например у настоящих
тюленей, с периодом линьки связана длительная голодовка, а
у котиков, оленей, ряда полорогих в период образования гаре-
мов и спаривания потребление пищи самцами резко сокращает-
ся, происходит своего рода «брачный пост», как известно, осо-
бенно ярко выраженный у некоторых рыб. Наконец, существо-
вание периода нагула, или «нажировываиия», характерно для
очень многих млекопитающих, особенно тех видов, которые
впадают в длительную спячку (ежи, селевинии, сони, суслики,
сурки, мышовки, тушканчики), в зимний сои (медведи, ското-
видные собаки, барсуки и др.) или совершают большие сезон-
ные миграции (рукокрылые, китообразные, ластоногие).
Таким образом, показатель «относительной прожорливости»
вида изменяется по сезонам. В некоторые периоды он зависит
также от пола животных. Если взять для завершения ряда,
представленного в табл. 1, самых мелких зоофагов нашей ф.ау*
ны — землероек1, то получатся очень убедительные цифры.
Водяная землеройка, или кутора (Neomys fodtens, вес 18—
22 г), съедает за сутки корма около 75—80% от веса собствен-
ного тела; обыкновенная бурозубка (Sorex araneus, вес 8—
14,7 г, средний 9,8)—142—200%, малая бурозубка (Sorex ml-
nutus, вес 3—5 г)—203—250%, а бурозубка крошечная
(Sorex minutissimus, вес 1,8—2,9 г)—один из самых мелких
зверьков нашей и мировой фауны,— до 420%. Таким образом
показатели относительной прожорливости млекопитающих
укладываются в ряд от 2% (синий кит) до 420% (крошечная
бурозубка).
Интересно ознакомиться с некоторыми абсолютными цифра-
ми. Синий кит в период нагула съедает за сутки до 4111 кг
мелких ракообразных, белуха—до 124 кг рыбы. Лисице для
полного насыщения нужно около 500 г мясной пищи, что равно-
примерно 15—20 экз. серых полевок. Для лесной куницы суточ-
ная норма — 120—150 г, т. е. 5—7 мелких грызунов или всего
половина белки (оставшуюся заднюю часть пойманного зверь-
ка куница обычно съедает на другой день) 1 2. Степному хорю-
нужно 100—125 г в сутки, т. е. 1 суслик или 4—5 полевок. Пе-
гий путорак (Dlplomesodon pulchellum, вес 10 г) летом съедает
за сутки 10—17 г мяса ящериц.
Фитофаги, как. правило, имеют несколько большую суточ-
ную норму, чем зоофаги, пища которых по составу белков бо-
1 Многие из них едят и растительную пищу, но основные корма — жи-
вотные (насекомые, земляные черви и т. п.).
2 Как и большинство хищников этого семейства, куница начинает пожи-
рать жертву с головы — в первую очередь наиболее привлекательный голов-
ный мозг, затем область груди и т. д.
158
лее соответствует потребностям организма животных и мен^е
перегружена плохо перевариваемой клетчаткой и т. п. Белки-
служат тем пластическим материалом, который идет на по-
строение клеток организма. Хищные звери получают животные
белки вместе с пищей (растительный белок ими усваивается
плохо). Растительноядные млекопитающие также нуждаются
в животном белке. Бобры, питаясь летом водноприбрежными
травянистыми растениями, заглатывают вместе с ними и нахо-
дящихся на них брюхоногих моллюсков, насекомых и их клад-
ки и пр. Таким путем они непроизвольно вводят в свой орга-
низм животные белки. В рубце жвачных развивается огромное,
измеряемое килограммами, количество инфузорий. Вместе с пи-
щей они переходят в последующие отделы желудка, где и пе-
ревариваются, служа таким образом источником поступления
в организм жвачных животных белков.
Данные, имеющиеся по питанию лося, показывают, что су-
точная его потребность в корме не постоянна, а изменяет-
ся по сезонам. Взрослые лоси северо-востока Европейской
части СССР (вес 345—§20 кг) зеленые корма (травянистые рас-
тения, листья кустарников и деревьев) в наибольшем количе-
стве (30—40 кг за сутки) поедают в летний период; 15—20 кг
веточного корма необходимо в осенне-зимний период, а весной
(во время линьки — март—апрель) лось довольствуется всего
6—12 кг ветвей, хвои и коры. Показательно, что в полном со-
ответствии с сезонными колебаниями качества и количества по-
едаемого корма находятся и закономерные сезонные изменения
живого веса лося. Эти данные получены путем тщательного
изучения пастьбы прирученных лосей в лесу и говорят о значи-
тельных сезонных изменениях потребностей организма
(Кнорре, 1959).
Вскрытия взрослых лосей, убитых на охоте, дают несколько
иные цифры (в любое время года — 30—50 кг измельченной
пищи, занимающей все отделы желудка), так как каждая пор-
ция съеденного корма находится в желудке и кишечнике зна-
чительно больше суток. У северного оленя переход пищи из
рубца в сычуг начинается спустя 3—4 ч после поедания корма,
но постепенное опорожнение рубца занимает 12—13 суток, при-
чем за первые 4 суток переходит 50—60% пищевой массы.
Дольше всего задерживаются более грубые части корма (Дру-
ри, 1955).
' Число кормежек за день, а следовательно, ритм суточной
активности, продолжительность пребывания в норе, в гнезде
или логове — с одной стороны, и на местах выпаса или охо-
ты--- с другой, зависят от величины потребной суточной нормы
корма, его качества, переваримости и распределения па участ-
ке, обитаемом животным.
159-
У серых полевок, обычно питающихся грубыми зелеными,
недостаточно калорийными кормами («объемистыми», по тер-
минологии животноводов), перерывы между кормежками колеб-
лются от 20 мин до 2 ч как днем, так и ночью (по данным эк-
спериментов в лаборатории). В течение суток они много раз
выходят из норы, каждый раз наполняя освободившийся желу-
док новой порцией пищи. Отчасти поэтому серые полевки со-
ставляют постоянную добычу сов и дневных хищных птиц.
Мыши значительное время проводят в убежищах, но вместе с
тем каждая отлучка мышей из гнезда в среднем продолжитель-
нее, чем у серых полевок (16—30 мин у желтогорлой мыши,
13—17 мин у лесной, 20—47 мин у полевой и 26—53 мин у
мыши-малютки), так как при поисках семян, редко и споради-
чески рассеянных по территории, им приходится обследовать
относительно большие участки; несмотря на это проворные
мыши гораздо реже подвергаются нападениям хищников. Такой
тип пищедобывательной деятельности возможен только для
зверьков с достаточно'быстрым бегом, хорошо развитым обо-
нянием, зрением и слухом (что, действительно, характерно для
мышей), иначе они очень скоро были бы уничтожены хищни-
ками.
Опытами установлено также, что по мере удаления от опти-
мальных частей ареала увеличивается ночная активность
зверьков, так как удлиняется продолжительность каждого вы-
хода и укорачиваются интервалы между ними, увеличивается
число особей, выходящих на поиски корма днем, хотя величина
дневной активности каждой особи остается почти неизменной,,
наконец, учащается посещение убежищ во время кормежки.
Все эти факты свидетельствуют об ухудшении условий сущест-
вования ближе к границам ареала.
У землероек-бурозубок ночная деятельность несколько про-
должительнее дневной, но становится более равномерной в раз-
ные часы суток, если расположить виды в ряд от более круп-
ных к мелким. Число кормежек за день в условиях экспери-
мента у куторы — 52 со средней продолжительностью перерывов
между ними в 27 мин, у обыкновенной бурозубки —54 с длиной
перерывов в 15 мин, у крошечной бурозубки —121 кормежка в
сутки с интервалами в 10 мин (Благосклонов, 1957). Эти зверь-
ки, хотя и отыскивают корм главным образом под опавшими
.листьями, в ходах под валежником, сухой травой и т. п., все
:же при своей почти непрерывной пищедобывательной деятель-
ности часто попадаются на глаза пернатым хищникам и стано-
вятся их добычей (благодаря сильному запаху мускусных же-
лез землеройки мало привлекательны для хищных млекопитаю-
щих— лисиц, куниц, которые, однако, убивают их, хотя едят
только при сильной нужде зимой).
Лось, питающийся грубыми растительными кормами' и с
160
этой стороны напоминающий серых полевок, благодаря боль-
шой величине тела нуждается в относительно меньшей суточ-
ной норме пищи и пасется в течение суток значительно меньшее
число раз. По данным Е. П. Кнорре (1959), в тайге верховьев
р. Печоры поздней осенью и в первую половину зимы’(15/Х—
15/1) в сутки у лося 5 периодов пастьбы, каждый продолжи-
тельностью по 1,5—3 ч и пять периодов покоя по 1,5—5 ч, в те-
чение которых лось лежит на снегу, отдыхая или пережевывая
жвачку. Кормежки занимают 46% суточного времени и ппиуро-
чены в основном к светлым дневным часам, отдых (54%) —
преимущественно к ночным.
В морозный и глубокоснежный период зимы и ранней весны
(15/1—15/IV) у лося 4 периода пастьбы по 1,5—4 ч (42% вре-
мени), приуроченные в основном к вечеру и утру и 3 периода
покоя (58%), приходящихся на темные часы суток. Весной
(15/IV—1/VI) в течение суток 9 периодов пастьбы, распреде-
ляющихся равномерно в течение всего дня, в общей сложности
занимающие 58% времени, и 10 довольно коротких периодов
отдыха (42%). Весна для печорских лосей — период нагула,
они пасутся почти круглые сутки, выбирая среди свежей зеле-
ни наиболее привлекательные и ценные корма. Летом (VII—
VIII) из-за жаркой погоды и главным образом из-за беспокой-
ства, причиняемого слепнями, комарами, оводами, лось пасет*
ся меньше, чем весной. В период VII—VIII у него всего 5 вы-
ходов на пастьбу, приуроченных к прохладным часам ночи
(46% суток) и 6 периодов покоя (54%). Продуктивность лес-
ных пастбищ лося очень велика, и ему нет необходимости де-
лать большие переходы в поисках корма. Лось — одно из очень
малоподвижных животных леса во все сезоны года, кроме пе-
риода гона и миграций.
Полную противоположность представляют крупные хищни-
ки: охота за крупной добычей требует от них большой подвиж-
ности, длительного настойчивого преследования, скрадывания
или подкарауливания и т. п. Так, например, амурский тигр
Felis (Tigris) tigris longipilis зимой, отыскав тропу кабана,
изюбря или другого зверя, идет по следам, пока не доберется
до самых свежих; здесь заходит с подветренной стороны на путь
жертвы и залегает, ожидая ее приближения. Одиночный тигр
(самец или холостая самка), голодавший несколько дней, пой-
мав зверя весом в 100—.150 кг, тотчас наедается доотвала и
остается около добычи 5—10 дней, совершенно не оставляя сле-
дов по сторонам, так как вместо водопоя довольствуется сне-
гом (летом тигр после кормежки ходит на ближайшие водо-
пои). Съев добычу полностью или бросив часть мяса, тигр от-
правляется в путь по своему охотничьему участку, двигаясь
ночью и днем, и первое время не обращая внимания на встре-
чающихся зверей или их следы. Время от времени он останав-
11 Териология . 161
ливается на отдых и лежит на месте по 10—12 ч. В сутки тигр
проходит 20—50 км, иногда до 100, и вся зимняя жизнь одиноч-
ного хищника представляет чередование больших переходов
длительностью по несколько суток, пожирания пойманной до-
бычи и отдыха около нее в течение ряда дней, соответственно
с ее размерами.
По расчетам, тигру весом 150—200 кг нужно в год около
3000 кг мясной пищи, что составляет в среднем всего 8 кг в
сутки (Капланов, 1948). В действительности порция, поедаемая
за один прием, значительно больше, но 5—10-дневные периоды
полного насыщения чередуются с длительными периодами до-
бровольного или вынужденного голодания (тигрица с вывод-
ком тигрят или пара взрослых в период спаривания ведут
себя несколько отлично от описанного). Взрослый тигр, в от-
личие от очень мелких хищников, совершенно не нуждается в
многочисленных ежедневных приемах пищи и способен к про-
должительному полному голоданию.
Важная адаптивная особенность всех крупных хищников,
четвероногих и пернатых — способность к длительному полному
голоданию, выработавшаяся у них как приспособление к неиз-
бежным случайным неудачам охоты за подвижными осторож-
ными жертвами из числа высокоорганизованных позвоночных.
Эта способность обеспечивает успешное выживание хищников
в трудные периоды года. В лабораторных опытах собаки вы-
держивают 60-дневное и даже 98-дневное полное голодание, те-
ряя до 49—65% от первоначального веса тела, и после этого
зходят в норму. Такой же выносливостью обладают белые мед-
веди, леопарды, волки, лисицы, песцы. Последних, например,
встречали зимой на мощном ледниковом щите Гренландии в
300 км от побережья, где хищник не может найти никакого
корма. Переход песца по льдам с Ямала на западный берег Се-
верного острова Новой Земли (более 1000 км по прямой), уста-
новленный методом мечения (Сдобников, 1940), тоже свиде-
тельствует о необычайной приспособленности этого зверька к
голодному существованию и преодолению пространства.
Известны случаи появления северных оленей, носивших мет-
ки ямальских ненцев, на Шпицбергене, для чего животные
должны были совершить огромные переходы по льдам Карско-
го и Баренцева морей, совершенно не имея пищи (способность
северных оленей к большим зимним пробегам без выпаса хоро<
шо известна оленеводам). Следовательно, приспособленность к
длительному голоданию, помимо ее большого значения в борь-
бе за существование, обеспечивает животным также возмож-
ность совершать дальние перекочевки и расселение.
Из мелких млекопитающих впадающие в спячку и зимний
сон формы, способные накоплять в теле энергетические резер-
вы и медленно их расходовать (ежи, летучие мыши, сони, сус-
162
лики и др.), значительно лучше переносят длительное голода-
ние, чем виды с постоянной высокой относительной прожорли-
востью и интенсивным обменом. Так, например, по опытам
И. В. Зильберминц (1950), срок жизни разных видов землероек
без корма тем меньше, чем выше потребление корма относи-
тельно веса тела. Кутора погибает через 25—26 ч голодания,
обыкновенная бурозубка—через 9 ч 30 мин, средняя бурозуб-
ка (Sorex caecutiens) —через 6 ч 40 мин, а малая—-через 5 ч
30 мин.
МИНЕРАЛЬНОЕ ПИТАНИЕ
Опыты по кормлению животных показали, что млекопитаю-
щие в тех случаях, когда органические соединения содержат
мало необходимых минеральных веществ, могут пополнять их
из неорганических соединений. Это относится к неорганическим
соединениям Na, К, Са, Mg, Cl, J, Р, Fe и некоторым другим.
Минеральные вещества регулируют осмотическое давление в
клетках тела животных, влияют на коллоидальное состояние
белковых соединений, участвуют в непрерывно идущем обмене
веществ, играют большую роль как структурный материал при
образовании клеток, тканей, органов. Без участия этих веществ
не может идти ни онтогенетическое развитие, ни размножение
млекопитающих.
Приток минеральных веществ должен осуществляться не-
прерывно; если они отсутствуют в корме, животные погибают,
при их недостатке развиваются заболевания, хорошо известные
животноводам: рахит, остеомаляция, анемия, тетания, остеопа-
рез, послеродовой парез, лизуха и др. Вопрос о нормальной
потребности диких млекопитающих в определенных минераль-
ных веществах почти не изучен, да и в отношении одомашнен-
ных видов он еще далек от окончательного решения. Данные
о потребностях сельскохозяйственных животных в отдельных
элементах золы и о содержании их в кормах имеют важное
значение для организации правильного минерального питания
животных
Хищные и всеядные млекопитающие получают необходимое
количество минеральных веществ вместе с животной пищей, так
как в крови, железистых органах, мышцах и костях их жертв
эти вещества содержатся в гораздо больших концентрациях,
чем в растениях. (Народы со специальной диетой, питаясь гу-
стым наваром из костей, мяса и самим мясом морских зверей,
никогда не испытывали потребности в поваренной соли.)
При растительноядном режиме возникает потребность в соли.
Травоядные млекопитающие получают основную массу необхо-
1 См. проф. М. Ф. Томмэ и др, «Минеральный состав кормов СССР»,
1948. ;
11
163
димых им минеральных веществ из поедаемых растений и отча-
сти из питьевой воды, но в них ряд важных биогенных элемен-
тов и соединений нередко содержится в ничтожном количестве.
Так, например, травянистые корма, особенно в областях с уме-
ренным или избыточным количеством осадков, где почвы силь-
но промыты, крайне бедны солями, в том числе и NaCl. Эта
соль — настоящее лакомство для крупного рогатого скота и
овец; посоленное сено они едят с жадностью; домашние север-
ные олени тоже очень падки на соль, соленую рыбу и т. п. Вхо-
дящий в состав соли С1 служит одним из исходных материалов
при образовании соляной кислоты желудочного сока животных,
a Na необходим для мышечной деятельности.
Таким образом, привлекательность поваренной соли для
травоядных животных обусловлена ее большой биологической
ценностью и крайне .недостаточным содержанием в естествен-
ных кормах. Деревянное топорище забытого в лесу топора,
впитавшее вместе с потом рук небольшое количество соли, вско-
ре обгрызают зайцы-беляки, полевки, белки. Землю, пропитан-
ную солью, случайно рассыпанной на месте, где был стан охот-
. ников или сборщиков кедровых «орешков», полностью выедают
олени, косули, зайцы.- Зимой снег, смоченный мочой человека,
содержащей-небольшое количество солей, без остатка съедают
северные олени и зайцы. ,
Такого рода случайные наблюдения и существование естест-
венных «солонцов» уже давно натолкнули охотников на мысль
устраивать искусственные солонцы — прикормку из поваренной
соли на специально вйбранных местах, чтобы подкарауливать
и стрелять осторожных-оленей, косуль, туров, серн и других ко-
пытных, когда они станут, регулярно посещать приваду. Искус-
ственный солонец не теряет двоей привлекательности для траво-
ядных животных в течейие многих лет, пока, в земле останутся
хоть малейшие следы NaCl. При зимней ловле зайцев-беляков
в Сибири в опадные орудия — «пасти» для приманки обычно
кладут пучки ивовых прутьев, обрызганных соленой водой или
-мочой. В заповедниках и охотничьих-хозяйствах искусственные
солонцы устраивают с целью улучшить минеральное питание
оленей, серн, туров, зайцев, а- также чтобы облегчить наблюде-
ние за ними. Ю. М. Салмин, зоолог Сихотэ-Али некого заповед-
ника, выкладывая соль в небольших деревянных корытцах и
постепенно перемещая эти миниатюрные солонцы все ближе к
избушке — своему наблюдательному пункту, добился того, что
белки, летяги, азиатские лесные мыши пользовались прикорм-
кой прямо на подоконнике. - -
Питьевые воды в зонах тундры и. хвойных лесов относятся к
- гидрокар-бонатному классу с минерализацией всего до 0,3 г/л.
Летние зеленые корма .в тундре как. травянистые, так и кустар-
никовые богаты белками, витаминами, отличаются высокой пе-
164
реваримостью, но бедны зольными элементами. Грибы, очень
многочисленные в южной подзоне тундры, в лесотундре и тай-
ге, так охотно поедаются северным оленем, лосем, маралом,
косулей, крупным и мелким рогатым скотом, зайцами, белкой,
полевками — в первую очередь, возможно, по причине высокого
содержания зольных элементов (6,6—11% от сухого веса, чаще
7—8%). В золе грибов много калия (20—50%), фосфорной
кислоты (0,2—40%), кроме того Na, Са, Fe, S, Mg, Cl, Si.
Рис. 62. Северный олень с трудом добывает ягель из-под толщи снега
в лесотундре
В грибах, особенно в шляпках, много белков (25—45% от сухо-
го веса), 2,5—7,5% жира при 20—40% клетчатки, много вита-
минов, особенно витаминов роста—А и В, а также D и С.
В целом плодовые тела грибов — корм высокой биологиче-
ской ценности, но период, в течение которого ими могут поль-
зоваться обитатели тундры, очень короток— I—1,5 месяца.
Период обилия и легкой доступности зеленых кормов тоже не
велик и продолжается всего 2—3 месяца. 8—9 месяцев в году
северный олень кормится в основном ягелем и некоторыми дру-
гими лишайниками, отличающимися очень низким содержани-
ем белков и зольных элементов (рис. 62). В результате дли-
тельного неполноценного питания и невозможности добыть не-
достающие вещества из других источников, у северного оленя,
особенно в конце зимы, обнаруживаются все признаки азоти-
165
стого и зольного голодания. Не удивительно, что олень ведет
себя весной и летом подобно всеядному животному: он жадно
поедает свежую траву, ловит леммингов, полностью уничтожает
кладки яиц в случайно найденных гнездах, объедает выветрив-
шиеся кости, оленьи рога на старых стойбищах 1 и т. п.
Прежде на Мурмане из-за недостатка сена коров и овец ча-
сто подкармливали ягелем, который с осени заготовляли впрок.
Зная по опыту неполноценность тундровых пастбищных кор-
мов, поморы в большом количестве запасали для рогатого ско-
та сушеные головы трески. Но животным, видимо, и этого было
мало: они нередко собирались около брошенных на берегу обо-
дранных туш тюленей и обгладывали их, насколько могли. Бе-
лые овцы, перемазавшиеся в тюленьей крови,— зрелище, кото-
рое можно увидеть только в тундре с ее своеобразным химиз-
мом пастбищных кормов.
На большей части лесной зоны минерализация питьевой
воды столь же низкая, как и в тундрах, растительные корма в
. среднем отличаются недостаточным содержанием белков. Зна-
чительная часть остающейся на зиму травы, вследствие вымо-
кания осенью и в предзимье, теряет большую часть питатель-
ности. Поэтому ветви кустарников и кора деревьев для многих
видов растительноядных млекопитающих служат зимой основ-
ным кормом; но он беден витаминами, перегружен клетчаткой.
Минеральное голодание' животных, обусловленное в основном
практическим отсутствием солей в питьевых водах и малой
зольностью растений,-—широко распространенное здесь явле-
ние. Сброшенные рога лосей, оленей, косуль, кости и даже зубы
животных без остатка и быстро уничтожаются зайцами-беля-
ками, полевками и другими грызунами всюду в лесной зоне
Евразии и Северной Америки. Во влажных тропиках и эквато-
риальной зоне отмечены сходные явления. .
В Африке охотники за слоновой костью давно убедились,
что совсем нельзя рассчитывать на собирание ее от животных,
погибших естественной смертью: бивни очень быстро уничто-
жаются грызунами. Д. Хантер (1960) застал однажды двух ди-
кобразов, поедавших бивень мертвого слона. От клыка, весив-
шего 90 анг. фунтов, осталось не более 2 фунтов. В Конго ло-
шади и ослы без специальной минеральной подкормки болели
остеомаляцией, так как почвы там бедны Са и Р, а содержание
Са в воде тоже крайне низкое. В условиях минерального голо-
дания разного типа естественные солонцы, встречающиеся в лес-
ной зоне как на равнинах, так и в горах, привлекают многих
растительноядных животных, особенно копытных, и некоторых
всеядных.
1 В оленьих рогах содержится очень много протеина; обгладывание их
приводит к введению в организм не только минеральных веществ, но и белков.
166
О химическом составе естественных солонцов дают пред-
ставление результаты анализа проб, взятых из грунта несколь-
ких солонцов сибирских косуль в Забайкалье (Фетисов, 1953).
Эти данные приведены в таблице 2.
Таблица 2
Химический состав естественных солонцов косули
(в миллиграммах на 100 г вещества)
Номера солонцов н со3 SO, Cl Са Mg FezO; К Р3О5 Сухой остаток
1 164,4 358,3 3,2 35,2 29,2 1,6 138,9 нет 1216,0
2 81,9 198,9 6,0 35,0 8,3 9,6 74,7 нет 379,0
3 80,0 838,0 0,6 214,6 51,3 1,6 91 9 5,0 1411,0
4 66,2 10,0 Н,4 20,8 5,6 3,2 2,9 нет 128,0
Таблица показывает, что почво-грунты солонцов содержат
серу, хлор, кальций, марганец, окись железа, калий, фосфор.
Количество этих веществ на разных солонцах отличается, что,
естественно, сказывается на ценности их для копытных.1
Солонцы, наиболее часто посещаемые животными несколь-
ких видов, за многие годы использования превращаются в
сильно вытоптанные, покрытые ямами участки с целой сетью
хорошо заметных троп, соединяющих пастбища с местами со-
лонцевания. Ямы, вырытые многими поколениями зверей, иногда
бывают настолько глубоки, что подкопанные вековые деревья
валятся на землю.
Иногда солонцы — это лишенные воды пятна с легкими вы-
цветами солей на поверхности почвы, чаще — минеральные
источники разного типа от едва сочащихся до ручьев с доста-
точным стоком, не сходные и по химизму (например, на Кавказе
животные посещают серные, нарзанные, углекисло-щелочные,
соляные, железистые источники). В Баргузинском районе При-
байкалья известны солонцы изюбрей, где эти олени используют
отложения горячих минеральных источников. Большинство есте-
ственных солонцов все же лишено NaCl, и потому животные
охотнее посещают искусственные — с поваренной солью.
По наблюдениям А. А. Насимовича (1939), в западной части
Кавказа период наибольшей посещаемости естественных и ис-
кусственных солонцов — весна, т. е. время смены кормового ре-
1 Результаты химического анализа других солонцов приведены в следую-
щих изданиях: Г. Г. Доппельмайр. Соболиный ппомысел на северо-
восточном побережье Байкала. Материалы экспедиции 1914—15 гг. Верхне-
удинск, 1926; А. А. Насимович. К познанию минерального питания
диких животных Кавказского заповедника. 'Груды Кавказского гос. з-ка,
в. 1, 1938; и др.
167
жима (переход с веточных кормов на свежую зелень), линьки
волосяного покрова и роста молодых рогов у самцов косули и , ;
оленя, у самок копытных-—донашивания плода, отела и после- I
дующей лактации. Большой расход организмом самок мине- '
ральных веществ обусловливает их повышенную потребность в
минеральных кормах и более частое посещение солонцов сам- 1
ками, чем самцами. Второй подъем интенсивности использова-
ния солонцов следует за летним снижением посещаемости и
приходится на вторую половину августа и сентябрь, что совпа-
дает с периодом гона у косули и оленя, а также с засыханием
и огрубением большинства кормовых трав и осенней линькой
шерсти у копытных. Кабан и бурый медведь (последний на I
Кавказе тоже по преимуществу растительноядное животное)
тоже приходят «солонцоваться», но реже и менее постоянно, чем
представители оленей и полорогих. Осенью и весной, когда ка-
бан и медведь питаются однообразными кормами, по прей;
муществу богатыми жирами и белками (плоды бука, лещины,
дуба, каштана и т. п.), они солонцуются чаще, чем летом.
В областях с недостаточным количеством осадков, например
в полупустынях Казахстана, воды рек по О. А. Алекину (1948)
чаще всего относятся к натриевой группе хлоридного и суль-
фатного классов с минерализацией порядка 1 г/л. Вместе с тем. ’
многие растения полупустыни и пустыни отличаются высокой
зольностью (особенно солянки) при резком преобладании (до
40 раз) Са над Р. В депрессиях рельефа здесь встречаются
сильно минерализированные непроточные водоемы и обширные
участки засоленых почв. Поэтому в полупустынях и пустынях
не может быть такого минерального голодания травоядных жи-
вотных, как в областях с избыточным количеством осадков (воз- ;
можно, что они все же ощущают недостаток Р). И действи- !
тельно, случаи, длительного устойчивого солонцевания у диких ।
копытных животных этих, зон неизвестны; многочисленные, ле- ‘|
жащие на земле выветрившиеся кости никогда не имеют следов
использования их грызунами. Но большие песчанки иногда
грызут солонцеватые корочки на краях котловинок и охотнее
селятся близ развалин старых зимовок, где благодаря обилию j
золы растения видимо отличаются более высоким содержанием: 1
зольных элементов.
Замечено (А. Н. Формозов),-что в степях суслики и пролет-
ные гуси весной охотнее кормятся свежей зеленью на местах, где
поздней осенью прошли пожары, уничтожившие старую траву.
Наконец, и в лесной зоне, «гари», зарастающие иван-чаем, мо-
лодыми лиственными деревцами — излюбленные летние па-
стбища зайца, косули, оленя, лося. Растворенные водой золь-
ные элементы пожарища, поступившие в почву, служат удобре-
нием и улучшают кормовые достоинства растений. Овцеводы
США давно подметили, что в стадах, выпасаемых на недавно
168
выжженных участках прерии, плодовитость овец выше, чем на
не выжженных; это служит еще одним доказательством биоло-
гической ценности зольных элементов, содержащихся в расте-
ниях пастбищ.
Не малую роль играет также минеральный состав материн-
ских пород, который всегда нужно учитывать при оценке усло-
вий существования млекопитающих. Установлено, что на изве-
стняках даже в районах с обильным увлажнением размеры ро-
гов у диких оленей больше, а рогатый скот развивается лучше
и бывает крупнее, чем в соседних местностях на лесных участ-
ках с подзолистыми почвами. За последние годы накопляется
все больше данных о большом значении микроэлементов (ред-
ких и рассеянных элементов) в жизни растений, животных и
человека. Трудами отечественных учёных, продолжателей дела
акад. В. И. Вернадского — А. П. Виноградовым и В. В. Коваль-
ским 1 создано учение о «биогеохимических провинциях», вскры-
вающее очень важные зависимости между геологической исто-
рией отдельных областей нашей страны, химизмом их материн-
ских пород, почв, химической средой существования раститель-
ного, животного мира и условиями ведения сельского хозяйства,
охраны здоровья населения. Давно известно, например, суще-
ствование эндемичных очагов заболевания щитовидной железы
человека, домашних и диких млекопитающих в районах преиму-
щественно горных, где материнские породы, почвы и питьевые
воды совсем не содержат йода. Известны районы, где высокое
содержание в растениях селена, олова или некоторых других
элементов оказывает угнетающее воздействие на животных, и т. д.
ПОТРЕБЛЕНИЕ ВОДЫ И ВОДОПОЙ
Вода совершенно необходима для нормального функциони-
рования организма, так как без ее участия не могут осуще-
ствляться разнообразные процессы, протекающие в каждой жи-
вой клетке тела. В среднем клетки млекопитающих (Палладии,
1935) содержат около 80% воды (в частности, скелет — 22,0;
белое вещество мозга — 70,0; мышцы — 76,0; кровь — 79,1; по-
товые выделения — 99,5%). Вода — один из важнейших «строи-
тельных материалов» при образовании и росте клеток; она
растворяет вещества и образует растворы разного типа, играет
важную роль в процессах терморегуляции. Тело в очень малой
степени способно утрачивать воду: при голодании, потеряв весь
жир и половину белков, оно утрачивает только 10% воды.
1 См. например, А. ГТ. Виноградов. Биогеохимические провинции,
Тр. Юбилейной сессии АН СССР, посвящ. столетию со дня рождения
В. В. Докучаева, М., 1949; В. В. Ковальский. Физиологическая роль,
микроэлементов у животных. Сб. «Микроэлементы в жизни растений и жи-
вотных», М., 1952.
169»
Недостаток воды большинство млекопитающих переносит
гораздо труднее, чем недостаток пищи. Лабораторные мыши,
лишенные одной воды, умирают в 10 раз скорее, чем лишенные
всякой пищи, но получающие воду (это в полной мере относится
и к человеку: без пищи летальный исход наступает, при сравни-
тельно благоприятной температуре воздуха, на 25—30-й день,
без воды человек погибает уже на 10-й день; потребность взрос-
лого человека в воде равна 35 г в сутки на 1 кг веса тела; у ре-
бенка она выше в 3—4 раза).
В процессе эволюции у сухопутных млекопитающих вырабо-
талась более высокая терпимость к жажде, чем у полуводных
форм, всегда имеющих возможность удовлетворять ее. В экспе-
риментальных условиях, например, обыкновенные землеройки
выдерживают отсутствие питьевой воды значительно дольше,
чем связанная в своем распространении с водоемами кутора.
Некоторые виды, приспособленные к жизни в крайне засушли-
вом климате пустынь, могут долго жить, питаясь только сухим
кормом; организм их крайне экономно расходует воду.
Животные пьют воду непосредственно из водоемов или полу-
чают ее вместе с пищей, содержащей достаточное количество
влаги, или, наконец, в областях с холодной зимой,— поедая снег.
Последний отличается от питьевой воды тем, что почти не со-
держит минеральных примесей, поэтому снежная вода «без-
вкусна» и при длительном потреблении (например, в тундре, где
продолжительность снежного периода достигает 240—260 и бо-
лее дней в году) вызывает у растительноядных млекопитающих
острое минеральное голодание. У одомашненного северного
оленя, питающегося зимой кормами, бедными зольными элемен-
тами, и около 8 месяцев получающего воду из поедаемого снега,.,
кости к весне становятся крайне хрупкими: недостаток мине-
рального питания организм восполняет за счет мобилизации
своих внутренних ресурсов, до предела расходуя то, что отло-
жено в скелете При ловле оленей арканом и других опера*
днях упавшие животные нередко получают переломы ребер,
костей ног и т. п., что осенью бывает крайне, редко. Замечено
также, что во всех случаях, когда зимой есть доступ к воде
ключей, наледей или незамерзающих порожистых участков рек,
и растительноядные и хищные млекопитающие отдают ей пред-
почтение по сравнению со снегом (А. Н. Формозов).
С другой стороны, сильная минерализация воды, избыток
растворенных в ней солей делает воду океанов, морей и многих
внутренних водоемов засушливых областей совершенно непри-
годной для ряда млекопитающих, происходящих из областей с
достаточным или избыточным увлажнением, где они всегда
1 Длительное употребление снежной воды не проходит бесследно и для
человека, вызывая ухудшение состояния зубов и т. п.
170
пользовались «хорошей питьевой водой». Двугорбый верблюд,
происходящий из аридных областей Центральной Азии, охотно
пьет сильно солоноватую и горько-соленую воду озер пустыни,
ст которой отказываются даже долго не имевшие воды лошади
и коровы. А джейраны, обитающие на восточном побережье
Каспия, ходят на водопой даже к Кара-Богаз-Голу, крайне на-
сыщенному солями (человек может пить океанскую воду в огра-
ниченном количестве в продолжении всего 4—5 дней; в дальней-
шем организм настолько перегружается солями, что почки не
справляются с их выведением, вследствие чего развивается ост-
рое заболевание почек—нефрит). К сожалению, в литературе
еще нет научно обоснованных оценок воды по ее пригодности
для водопоя даже сельскохозяйственных животных; много неяс-
ного и в вопросе о нормах потребления воды разными породами
животных. В отношении диких млекопитающих неясностей еще
больше.
По сравнению с привычной для нас, питьевая вода, получав
мая из колодцев сухих степей и пустынь, оказывается минера-
лизованной выше всех общепринятых гигиенических норм. Ме-
стные породы скота, созданные длительным отбором, например
каракульские овцы и одногорбые верблюды Каракумов Туркме-
нии, охотно пьют эту воду без вреда для здоровья. Что касается
норм потребления воды, то можно привести следующие цифры:
каракульская овца в пустыне при летней пастьбе на участках с
высохшим травостоем выпивает в сутки 10—12 л (редко до 15),
а весной, питаясь сочной свежей травой — только 1—2,5 л воды.
По сравнению с другими породами, каракульские овцы отли-
чаются водным обменом, позволяющим более экономно расхо-
довать полученную влагу.
Верблюду летом при каждом водопое нужны десятки литров,
а весной стада верблюдов могут проводить на пастбищах до
2—2,5 месяцев, не приближаясь к колодцам и получая всю не-
обходимую влагу вместе с поедаемыми растениями. Значение
«метаболической воды», образующейся в организме при исполь-
зовании жировых отложений, значительно меньше того, какое
ему ранее приписывали (Минервин, 1958). Вопреки прежним
представлениям, выпитая верблюдом вода не резервируется в
его желудке, а чрезвычайно быстро всасывается. В дальнейшем
она расходуется очень экономно: верблюд выделяет мало мочи,
кал его необычайно сух, животное почти не потеет и не изме-
няет режима дыхания. Благодаря особенностям кровяной
плазмы даже острый дефицит воды в организме этого живот-
ного не вызывает сгущения крови, т. е. не нарушает нормальной
циркуляции последней.
В опытах Н. -В.' Тупиковой мелкие грызуны, содержавшиеся
в клетках и получавшие в качестве корма овес, овсянку и
свеклу, потребляли каждый в среднем за сутки следующее ко-
171
личество воды: полевая мышь (Apodemus agrarius, из Подмо-
сковья) — 1,5 см3, азиатская лесная мышь (Apodemus speciosusr
из Приморского края) — 1,25 см3, домовая мышь (Mus musculus?
из Подмосковья) — 1,5 см3, мышь-малютка (Micromys minutus,
из Подмосковья) — 1,3 см3, лесная мышь (Apodemus sylvaticus,
из Подмосковья) —0,7 см3, рыжая лесная полевка (Clethriono-
mys glareolus, из Подмосковья) — 1,0 см3. Перечисленные зверь-
ки— обитатели районов с мезофитной растительностью и доста-
точно обильными осадками. Резко отличается от них степная
пеструшка (Lagurus lagurus), представитель степной фауны: из
12 пеструшек, участвовавших в опыте, воду пили только 4; их
суточная норма потребления равнялась всего 0,1 смл, наличия
сочного корма — свеклы было вполне достаточно, чтобы удов-
летворить потребность пеструшек в воде. В природе им нередко
в течение месяцев приходится питаться довольно сухими ли-
стьями степных злаков, полыней и т. п., совсем не имея питьевой
воды.
Таежная влаголюбивая полевка-экономка (Microtus оесопо-
mus) при содержании на сухом овсе потребляла воды на 1 г
живого веса в среднем 0,344, обыкновенная полевка (М. arva-
lis)—0,228, полевая мышь—0,171 г, домовая — 0,152 (Кара-
сева и др., 1957).
Мало нуждаются или совсем не нуждаются в водопоях мно-
гие сотни видов из сумчатых, насекомоядных, неполнозубых,
грызунов, хищников и копытных, живущих в пустынях и сухих
степях всех континентов. Мелкие зверьки, например суслики,
песчанки, тушканчики, ушастые ежи, использующие в качестве
убежищ более или менее постоянные норы, густо населяют об-
ширные участки наших пустынь, на десятки километров удален-
ные от каких-либо водопоев Всю необходимую им влагу они
получают в пище, весной поедая сочную зелень и насекомых, а
когда наступит период летней жары и земля днем начнет рас-
каляться до 60—80°С, выкапывают луковицы, клубни, надку-
сывают мощные стержневые корни многолетних растений и сли-
зывают выступающий сок.
В сухой и морозный период зимы в песках Каракумов гры-
зуны получают необходимую влагу, обгладывая сочную кору
кустов и деревьев — кандымов, саксаула, песчаной акации и др.
Эти растения имеют мощную корневую систему, достигающую'
грунтовой воды. Кроме того, грызуны поедают мелкую расти-
тельную труху, покрытую инеем, который выпадает по ночам.
Хищники, например обычные в наших пустынях перевязка,
корсак, лисица, барханная и пятнистая кошки, в Африке — фе-
1 Однако не следует думать, что эти зверьки вовсе не умеют пить. Когда
весной на такырах скопляется снеговая вода, ее пьют суслики, песчанки,
ушастые ежи и зайцы-толаи, обитающие поблизости. Но это удел ничтож-
иого числа особей.
172
нек и др., получают достаточное количество влаги, поедая свои
жертвы — грызунов, насекомых, и т. п. При случае хищники
тоже охотно пользуются вегетационной, или «растительной во-
дой», поедая, например, плоды дикого арбуза, широко распро-
страненного в пустынях Африки и Передней Азии, а на бахчах
наших степей и пустынь — культивируемые арбузы и дыни.
Волки и лисицы — частые посетители бахчей и местами серьез-
ные враги бахчеводства. В сухих подгорных районах Туркмении
установлено, что шакалы сильнее объедают грозди винограда
на плантациях, когда в арыках нет воды и наоборот. Однако
более высокая численность выводковых нор волков, шакалов,
лисиц, гиен и ряда других видов поблизости от водопоев и явная
спорадичность распределения в безводных участках пустынь
указывают, что питьевая вода в жизни крупных хищников
играет большую роль, чем в жизни мелких представителей сем.
куньих, ежей и т. п.
Исключительно большую роль открытые водопои играют в
жизни летучих мышей — группы, особенно богатой видами и осо-
бями в субтропических районах нашей страны и еще более в
тропиках Азии, Африки, Америки. Покидая вечером свои убе-
жища, летучие мыши прежде всего направляются к водоемам
.(ключам, широким арыкам, прудам, озерам) и пьют налету,
после чего начинают охоту за насекомыми. Охлаждение орга-
низма при напряженной мускульной работе во время полета
осуществляется через испарение воды с поверхности легких,
рта и, возможно, других участков тела летучих мышей
Поэтому, питаясь насекомыми, подобно ушастым ежам, руко-
крылые в отличие от последних не могут обходиться без
водопоя. Все колонии и поселения летучих мышей, находящиеся
в пустынях, степях и горах аридных континентальных районов,
расположены поблизости от водоемов, пригодных для подчер-
пывания воды налету. Это обстоятельство очень четко сказы-
вается на географическом распределении колоний летучих мы-
шей в аридных областях нашей страны; последнее без учета
водопоев не может быть правильно понято (регулярно пьют и
насекомоядные птицы, охотящиеся налету — стрижи, ласточки,
щурки, козодои, тогда как виды с другой биологией, но живу-
щие в тех же районах, могут обходиться без водопоя).
В литературе часто встречается очень обычное утверждение,
что копытные животные пустынь и степей, обладающие способ-
ностью к быстрому бегу, регулярно пользуются водопоями,
иногда расположенными, даже на расстоянии десятков километ-
ров от их пастбищ. В действительности в этом для большинства
видов животных нет особой необходимости, так как они находят
1 Установлено, что мелкие птицы во время сезонных миграций теряют
вчень много воды при беспосадочных перелетах над морем и т. п.
173
зеленые растения, содержащие достаточное количество водьъ
Весной кормовые травы, например наших песчаных пустынь (в
основном эфемеры и эфемероиды) содержат 75—80% влаги, а
некоторые до 90% и даже выше, т. е. влажность их измеряется
отношениями 1:3, 1 :4 и даже 1 :9. При выпадении дождей или
росы это отношение у смоченного травостоя повышается до
1 : 14 и даже 1:19. Многие массовые кормовые растения содер-
жат очень много воды и без наружного их смачивания. В пусты-
нях Средней и Центральной Азии таковы солянки, в пустынях
Африки — молочаи и уже упоминавшийся дикий арбуз, в пусты-
нях Америки — кактусы, которые содержат воды до 91%—94%.
и более.
Поедая очень сочный и водянистый корм на весенних паст-
бищах в пустыне или на летних в лесной зоне, животные одно-
временно получают вполне достаточное, а иногда даже избы-
точное количество воды. Несомненно, избыточное ее количество
получают олени, лоси, косули, серны, пасущиеся на сочных гор-
ных лугах (например, Кавказа, Алтая, Саян), где выпадает
много летних осадков. Замечено, что у животных при такой
пище расстраивается пищеварение и они жадно поедают мелко-
структурные глины и суглинки, отыскивая их в обнажениях
склонов, в конечных моренах и т. п. Такие места нередко назы-
вают солонцами, хотя в поглощаемом зверями материале почти
полностью отсутствуют важные биогенные соли. Ошибочность
этого представления вскрыл Ф. Д. Шапошников (1953), обсле-
довавший ряд «псевдосолонцов» горнотаежной части Алтая..
У животных, поедающих мелкоструктурную глину, пищевая
кашица становится более густой, и дефекация приходит в норму
(поедание глины домашними копытными также чаще отмечают
при выпасе на свежей траве и в дождливую погоду).
Животноводы знают, что излишнее потребление воды домаш-
ними животными приводит к значительному скоплению жид-
кости в тканях, вызывает их вялость и отечность, у молодых жи-
вотных ухудшает пищеварение. Рабочие лошади при чрезмерно»
водянистом корме слабеют и потеют, овцы подвергаются тяже-
лым общим заболеваниям и т. д. С этими фактами следует
сопоставить установленное наблюдениями улучшение биологи-
ческой ценности пастбищ диких животных после подсыхания
части растений и созревания их семян. Избыток воды устра-
няется, а семена, содержащие много ценных жиров, белков,,
углеводов, служат для накопления энергетических резервов —
подкожного и полостного жира, гликогена и т. п., необходимых
как при спячке, так и для активной жизни в сезоны с резко'
ухудшенными кормовыми условиями.
В наших пустынях по мере высыхания весеннего зеленого
Ковра часть видов грызунов впадает в длительную спячку, дру-
гие начинают в большем количестве выкапывать сочные луко-
174
вицы и корневища, поедать побеги поздно вегетирующих кустар-
ников и деревьев, подвижные копытные начинают кочевать в
поисках менее выгоревших пастбищ и приближаются к водо-
поям. Кормящие самки джейранов с ягнятами собираются близ
рек, озер на равнинах или у ключей в предгорьях. Лактирующие:
животные и подрастающие молодые более нуждаются в воде,,
чем взрослые самцы. Последние даже в жаркие месяцы доволь-
ствуются «вегетационной водой»: они ощипывают сочные листья
каперса, верхушки верблюжьей колючки и других долго не вы-
сыхающих растений с мощными корнями. Местами жарким
летом, как, например, по течению р. Сарысу в 1948 г., встре-
чается масса самок джейранов, откочевавших с молодыми
ближе к воде, тогда как самцы остаются в безводных местах
пустыни Бет-Пак-Дала на многие десятки километров южнее
(Формозов). Антилопы и зебры Африки в случае пересыхания
водоемов также совершают кочевки и настоящие миграции или
выкапывают глубокие ямы в песке, добираясь до водоносного
слоя. Ямы-колодцы на обсохших озерах Казахстана делают
волки.
Несомненно, физиологическая роль «растительной» и обыч-
ной питьевой воды не равноценна. Вода сочных растительных
кормов, связанная с коллоидами, лучше усваивается организ-
мом, положительно влияет на интенсивность роста тканей, в том
числе и на формирование половых клеток. У свиней скармли-
вание сочных корнеплодов сильно повышает плодовитость (уве-
личение числа поросят в каждом помете; Н. М. Кулагин)-
Необходимо учитывать, что постоянное потребление млекопитаю-
щими вегетационной воды — явление, очень широко распростра-
ненное. Предлагаемые некоторыми авторами термины «снего-
пой» и «травопой» заслуживают признания наряду с привычным
«водопой», так как с точки зрения экологии'млекопитающих
каждому из них соответствует особое содержание.
ЗАПАСАНИЕ КОРМА
Резкие сезонные изменения качества или доступности корма,,
колебания урожайности семян и «мякотных» плодов, правильное-
чередование обильных пищей и голодных сезонов обусловили
возникновение и развитие у многих млекопитающих важной
адаптивной черты пищедобывательной деятельности — способ-
ности при известных условиях собирать и сохранять запасы
корма впрок. Эта черта биологии, отсутствующая у представи-
телей низших классов позвоночных (рыб, амфибий, рептилий),,
хорошо выражена лишь у немногих, филогенетически молодых
и прогрессивных групп птиц (воробьиные, дятлы, совы) и очень,
характерна для огромного числа видов млекопитающих, осо-
бенно из отряда грызунов, отчасти хищных, насекомоядных
175-
и др. Спячка и дальние миграции млекопитающих также пред-
ставляют собой важные приспособительные черты биологии,
обеспечивающие успешное переживание животными неблаго-
приятных сезонов года. Животные, впадающие в спячку, в своем
организме накопляют необходимые энергетические резервы,
крайне экономно используемые в течение многих месяцев спяч-
ки; широко мигрирующие виды (китообразные, ластоногие,
копытные, рукокрылые) покидают летние участки ареала, не
пригодные для зимовки, направляясь в другие, более благо-
приятные.
Естественно, что виды, способные к длительной полной
спячке или далеким миграциям, не нуждаются в собирании за-
пасов корма; эта адаптивная черта поведения в наиболее ярко
выраженной форме характерна для видов, относительно оседлых
и деятельных в течение всего года. Однако и часть видов, совер-
шающих непериодические миграции и местные перекочевки, а
именно те, у которых нет широкого разрыва между участками
летовки и зимовки, например песцы и белки, при случае соби-
рают запасы даже во время самих кочевок (мигрирующие
белки осенью, находясь в пути, продолжают развешивать на
ветвях грибы, которыми сами они не смогут воспользоваться, но
грибы достанутся белкам, кочующим позднее или оставшимся
зимовать на месте). Таким образом, с точки зрения интересов
сохранения вида и такое запасание корма целесообразно (Фор-
мозов, 1936).
Наконец, некоторые виды, проводящие всю зиму в норе, но
не впадающие в полную спячку (обыкновенный хомяк, бурун-
дук, длиннохвостый суслик) собирают запасы семян, так как
нередко пробуждаются от зимнего сна и кормятся ими. Их кла-
довые располагаются обязательно в норе, рядом с зимовочной
камерой. Весенний выход этих зверьков из нор происходит на-
столько рано, что они не могут найти достаточного количества
полноценных кормов, еще скрытых под плотным снегом. Сохра-
нившиеся с осени запасы кладовых играют большую роль в
жизни бурундука и, видимо, длиннохвостого суслика именно в
этот ранневесенний период, когда у них происходит спаривание.
В тех частях земного шара, где нет резкой смены сезонов,
например, во влажных областях тропической и экваториальной
зоны, лесная растительность которых всегда имеет свежую ли-
ству, а зрелые плоды встречаются в течение всего года, не было
и нет экологических предпосылок для развития способности жи-
вотных создавать большие, долговременные запасы корма. Дей-
ствительно, некоторые обезьяны, собирающие плоды на дере-
вьях, насытившись, прячут небольшое количество пищи за щеки
и уносят ее, чтобы использовать позднее. Однако такие акты
пищедобывательной деятельности трудно считать за подлинное
проявление хорошо развитой способности к собиранию запасов.
176
В тех же тропических областях, где чередуются резко выражен-
ные сухие и дождливые сезоны, например в саваннах Африки,
часть видов млекопитающих собирает большие запасы корма
перед наступлением сухого, зимнего периода.
Таким образом, влияние географической среды на распро-
странение этой приспособительной черты поведения среди мле-
копитающих различных фаун совершенно очевидно. В нашей
стране число видов и особей млекопитающих, которые собирают
запасы, особенно велико в континентальных областях степей и
пустынь с их резкими сменами жаркого лета и длительной очень
холодной зимы, а также в области листопадных и хвойных ле-
сов умеренного и холодного поясов. Так, например, из 40 видов
млекопитающих, зарегистрированных в тайге верхнего течения
р. Печоры (Печоро-Илычский государственный заповедник),
более половины (21 вид) в той или иной форме запасают корм.
В полупустынях Центрального Казахстана таких видов 29.
В зоне тундр виды мелких зверьков, собирающих запасы корма,
предназначенные для длительного использования зимой, немно-
гочисленны, но составляют большой процент в фауне: это по-
левки узкочерепная, Миддеидорфа, экономка, а также северная
пищуха — формы не аборигенные, а пришлые с юга. Характер-
ные и массовые тундровые грызуны — пеструшки, или лемминги
(у нас четыре вида), запасов не собирают и довольствуются в
течение всей продолжительной полярной зимы зеленым и веточ-
ным кормом, который с осени на корню законсервирован хо-
лодом.
Многие хищники при успешной охоте избыток добычи прячут
в укромных местах с тем, чтобы использовать позднее. Лесная
куница, поймав зайца-беляка или глухаря, прячет отдельные
куски добычи под колоды, камни, иногда затаскивает на деревья
и помещает на ветвях у ствола. Леопарды в Африке, как пра-
вило, укрывают часть добычи на деревьях, так как оставленную
на земле быстро уничтожают гиены и шакалы. Росомаха, убив-
шая зимой северного оленя или лося, умело «разделывает» тушу
на части, уносит их под упавшие деревья, в бурелом, закапы-
вает в снег и живет близ такого склада, пока полностью не ис-
пользует добычу. Так же поступает и лисица при удачной охоте
на зайца: в северных районах она закапывает добычу по частям
в снег, тщательно уминая его поверхность над запасами, в пес-
чаных пустынях — зарывает в песок или закапывает в пустующие
норы грызунов. В перечисленных случаях все запасы укры-
ваются так, чтобы не привлекать внимания пернатых нахлебни-
ков— воронов, ворон, сорок, соек; они нередко следят за круп-
ными хищниками, растаскивают часть их добычи и своим кри-
ком привлекают более опасных четвероногих конкурентов.
Бурый медведь крупную добычу не разделывает на куски, а,
оттащив в укромное место, заваливает ветвями, мхом и посте-
12 Терио.'о'ия
177
пенно использует, временами даже охраняя от назойливых на-
хлебников. Некоторые мелкие хищники из сем. куньих заготов-
ляют относительно большие запасы, которые у ласки и горно-
стая иногда содержат до 20—40 полевок и мышей, собранных в
одном месте, у черного хоря — многие десятки лягушек, у евро-
пейской норки — 2—5 кг мелкой рыбы, у американской норки —
до 20 кг и т. д. Эти запасы хорошо укрыты от большинства воз-
можных конкурентов и, кроме того, расположены в полостях
ниже уровня почвы и под защитой высокого слоя снега, где они
не могут промерзнуть и затвердеть так сильно, как оставленные
на поверхности (ласка и горностай, имеющие мелкие зубы, с
трудом грызут промерзшее мясо).
Но у всех хищников наиболее ценный для них корм (если он
сложен про запас не в холодное время года и не у слоя вечной
мерзлоты) не может долго сохраняться. Исключение составляют
только яйца птиц, которые куницы, хорьки и колонки, спрятав
в начале лета в прохладные, затененные укрытия, нередко ис-
пользуют уже зимой, что установлено изучением следов зверь-
ков и анализом содержимого желудков.
Характерные особенности собирания запасов хищниками сле-
дующие: они устраивают их не в специально приготовленных
хранилищах, а в любом месте на большом индивидуальном
охотничьем участке, где удается добыть количество корма
больше суточной потребности. Запасы собирает и использует
обычно одна особь; в целом они играют в ее жизни весьма огра-
ниченную подсобную роль и не имеют решающего значения в
борьбе за существование.
В зачаточной форме эта адаптивная черта поведения свой-
ственна и насекомоядным — группе довольно примитивных мле-
копитающих. Небольшие запасы беспозвоночных собирают севе-
роамериканские короткохвостые землеройки (Blartna brevica-
uda), водных беспозвоночных запасает наша выхухоль. Опытами
И. И. Барабаш-Никифорова (1949) установлен интересный
прием, с помощью которого выхухоль поедает крупных, обла-
дающих очень прочными, плотно замыкающимися раковинами
двустворчатых моллюсков (перловиц и беззубок). Затащив не-
сколько живых моллюсков в надводную часть своей норы, зве-
рек оставляет их здесь до тех пор, пока ослабевшие животные
не приоткроют створки своих раковин. Легко расправившись с
таким моллюском, выхухоль выбрасывает пустые створки на-
ружу (таково происхождение скоплений пустых неповрежденных
створок моллюсков, часто находимых у отверстий выхухолевых
нор). Несомненно, описанный прием возник у выхухолей на
почве запасания кормов.
Установлено, что в некоторых участках ареала кроты (Talpa
еигораеа) запасают впрок земляных червей: по данным Ф. Бурль-
ера (Burlier, 1951), находили до 1000 червей, собранных в один
178
из ходов кротовой норы. По наблюдениям С. Скочена (Skoczen,
1961), сделанным в Польше, основной период собирания запа-
сов кротами — осень, а внешним сигналом служит начало пони-
жения температуры почвы. Чаще встречаются запасы из 100—
300 червей, расположенные на дне ходов вблизи гнездовой
камеры норы. Все черви имеют повреждения от укусов на го-
ловном отделе; черви остаются живыми, но лишаются способ-
ности передвигаться. По материалам Оатса (Oates, 1956), со-
державшиеся в неволе кроты в любые месяцы закапывали изли-
шек червей, тоже предварительно иммобилизовав их укусами.
Группа реакций, связанных с запасанием кормов, в более
совершенной форме проявляется у грызунов, тоже живущих по-
одиночке или парами в течение всего неблагоприятного периода
года, но концентрирующих запасы в ближайших, лишь для них
доступных окрестностях постоянной норы или гнезда и, наконец,
непосредственно в самой норе. Летяга поздней осенью запасает
и складывает в нескольких дуплах по 30—40 а тонких побегов
березы и ольхи с сережками (мужскими соцветиями), служащих
ей основным кормом зимой (А. Н. Формозов). Казалось бы,это
не дает зверьку никаких преимуществ, так как Сережкина дере-
вьях для него вполне доступны всю зиму. В действительности
дело обстоит сложнее. В течение всего холодного периода года
сережки истребляются рябчиками, живущими в тех же биото-
пах, где и летяга; запасание этого корма в дуплах устраняет
их конкуренцию. Кроме того, сильные морозы, ветер, появление
ожеледи на ветвях затрудняют кормежку летяги на деревьях;
наличие кладовых дает ей возможность при неблагоприятных
условиях погоды кормиться запасами в хорошо защищающем
ее дупле.
Белки уже с середины лета, когда начнут созревать семена
хвойных деревьев, орехи и желуди, перемещаются в насажде-
ния, наиболее богатые этим осенне-зимним кормом. Совершен-
но созревшие «орешки» кедра и желуди каждый осевший на ме-
сте зверек во множестве прячет в опавшие листья, лишайники
и мох, небольшие дупла по всему кормовому участку, окру-
жающему гнездо.
При недостаточном урожае этих кормов и обилии грибов
белки в августе — октябре развешивают их плодовые тела по
ветвям на высоте 1,5—5 м от земли. Эти грибы хорошо провя-
ливаются и сохраняются, находясь выше наибольшего уровня
снега, т. е. легко доступны для белки зимой, но мало доступны
для большинства ее конкурентов. В1 темнохвойных лесах тайги
белка запасает от нескольких десятков грибов до 150—300
(А. Н. Формозов) (рис. 63). В ленточных борах Западной Сибири,
где семенной корм дает только ..сосна и кормовые условия
значительно хуже, чем в тайге, белки местной географической
расы — телеутки (Sciurus vulgaris exalbidus) собирают назиму
12!
179
каждая от 1500 до 2000 грибов, среди которых преобладают
масляники {Boletus luteus) и др. На Кольском полуострове за-
пасания грибов, видимо, совсем не бывает (Г. А. Новиков). Это
один из примеров того, как в соответствии с местными геогра-
фическими условиями у животных .одного вида, но живущих в
разных частях ареала, инстинкт запасания корма проявляется
то сильнее, то слабее и может совсем не влиять на характер
пищедобывательной деятельности там, где по природным усло-
виям в этом нет необходимости.
Рис. 63. Часть большого запаса грибов обыкновенной белки. Осенние опен-
ки (Armillaria mellea) развешаны зверьком по ветвям молодых елок у ствола
на высоте, превышающей мощность снежного покрова, в густом насажде-
нии, где ветер не может сбросить грибы на землю. Костромская обл., октябрь
1931 г. (ориг. А. Н: Формозова)
Полевка-экономка, описанная П: С. Палласом по экземпля-
рам из Сибири, была им названа так в связи с тем, что соби-
рает большие запасы корневищ и клубней (до 10—15 кг и более
на одну семью небольших зверьков). Для хранения запасов
экономки еще осенью, пока земля не промерзла, выкапывают
большие камеры — расширения ходов норы по соседству с гнез-
дом. Эти запасы играют существенную роль в жизни экономок
Камчатки и Восточной Сибири, где почва сильно промерзает,
что лишает зверьков возможности выкапывать корневища в те-
чение всей длинной- и очень морозной зимы. По мере движения
180
к западу, соответственно уменьшению континентальности клима-
та и суровости зим, размеры запасов, собираемых полевками-
экономками местных популяций, становятся все меньше. В Ко-
стромской области большинство экономок еще выкапывают
специальные кладовые, а в Московской области и тем более на за-
падной границе ареала, где зима относительно очень мягкая,
только ничтожный процент этих зверьков и не ежегодно соби-
рает небольшие запасы корма (Формозов, 1948).
То же самое наблюдается и у водяной крысы (Arvicola ter-
restris). На западе Европейской части СССР у этого полувод-
ного грызуна запасы корма па зиму встречаются редко и не
всегда; по массе они не превышают нескольких сот граммов, тогда
как на северо-востоке ареала — в Якутии, вес запасов дости-
гает 25—30 кг (Соломонов, 1961).
У серых полевок (Microtus arvalis), населяющих Европей-
скую часть СССР, запасы корма (корневища, клубни, зерно
хлебных злаков) невелики (не более нескольких сот граммов),
встречаются не ежегодно и обычно только в ничтожном процен-
те обитаемых нор. Узкочерепные полевки Microtus (Stenocra-
nius) gregalis, принадлежащие к той же биологической группе,
но населяющие север Азиатского материка, нередко собирают
очень большие запасы (до нескольких килограммов) корневищ,
клубней, зерна хлебных злаков.
Обитатель континентальных степей Северной Монголии и
Забайкалья—-полевка Брандта Microtus (Lasiopodomys) brand-
ti, с августа начинает прочищать старые подземные камеры и
роет новые помещения для запасов. В первых числах сентября
в каждой зимовочной колонии собираются группы из 10—20
зверьков и сообща в течение двух с половиной месяцев запа-
сают корм (рис. 64). Они или срезают кусочки побегов полыни
или целиком выкапывают узколистную осоку, лапчатку, некото-
рые злаки и т. п. Плотно уложенные растения заполняют все
камеры-кладовые; вес запасов достигает 10 кг и более. Крайне
малоснежная, морозная с сильными ветрами зима континен-
тальных высоких степей по своим условиям очень неблагопри-
ятна для жизнедеятельности мелких зверьков. С наступлением
холодов полевки Брандта забивают ходы в нору и в продолже-
ние 2—2,5 месяцев не показываются на поверхности земли, пи-
таясь только собранными запасами. В ясные дни конца января
и в феврале они начинают изредка выходить греться на солн-
це и только с половины марта переходят к весеннему образу
жизни, для которого характерны большая продолжительность
пребывания на поверхности почвы и поиски свежего корма.
В. В. Кучерук и Т. Н. Дунаева (1948) выяснили, что предпо-
читаемый корм этих полевок в период с февраля до появления
свежей зелени —- луковицы двух видов луков (Allium tenuissi-
тит и A. polyrhiziim), которые зверьки усиленно выкапывают
181
и с жадностью поедают, хотя в.кладовых еще имеются запасы
корма. Возникла мысль, что после зимовки полевки остро нуж-
даются в полноценном витаминозном корме. Анализ нескольких
образцов, взятых из запасов, это подтвердил — в сухом корме
полностью отсутствовал витамин С.
Рис. 64. Зимовочная нора полевки Брандта с обширными
камерами для запасов. Вверху — план, внизу — разрез
(по Кучеруку и Дунаевой)
Для полевки Брандта характерна групповая, или обществен-
ная, заготовка и использование зимних запасов, что свойствен-
но ряду других видов, грызунов, зимующих семьями или сезон-
ными группами. Это установлено, например, для больших,
краснохвостых, монгольских, малоазийских и полуденных песча-
нок, курганчиковых мышей, горных полевок, малых, даурских и
монгольских пищух, речных бобров и др. Некоторые из перечис-
ленных грызунов собирают только запасы семян и небольшого
182
количества клубней (мыши, многие песчанки), другие заготов-
ляют сено, складывая его в специальные стожки (даурская пи-
щуха, местами большая песчанка) или укрывая в нишах и есте-
ственных ходах под защитой скал и в специальных камерах нор
(горные сеноставки, снежные полевки и др.).
Заслуживает внимания тот факт, что запасание сена харак-
терно или для обитателей скалистых биотопов, где его запасы
можно легко уберечь от намокания, используя защиту камен-
ных плит, или для видов, свойственных степям и пустыням с
континентальным климатом и большой сухостью воздуха, благо-
приятной для длительного хранения сухого корма. Многие виды
зверьков подсушивают запасаемые растения, раскладывая их на
короткий срок на камнях или оголенной земле, а затем уносят
в кладовые. Из практики сельского хозяйства известно, что дли-
тельная сушка травы на ярком солнце вызывает снижение ко-
личества каротина в заготовленном сене, т. е. ухудшает его
биологическую ценность как корма. Многие грызуны, например,
наши виды полевок рода Alttcola и большие песчанки подсуши-
вают сено в утренние часы еще при слабой инсоляции или вече-
ром и убирают на хранение слегка подсушенным, что дает им
возможность создавать запасы достаточно полноценного корма
(многие образцы сена из запасов горных полевок, пищух и пес-
чанок долго сохраняют зеленоватый цвет и приятный аромат).
Конечно, это обусловлено не сознательно направленными
действиями зверьков. Летом сильная полуденная жара и резкая
инсоляция угнетают деятельность даже дневных зверьков, по-
этому они наиболее активны в утренние и вечерние часы, что,
независимо от их воли, автоматически обеспечивает наилучшие
условия для заготовки сена.
Установлено, что живущие в скалах зверьки прикрывают со-
бранное сено мелкими камешками,— это предохраняет запасы
от раздувания ветром и расхищения сибирскими козерогами,
косулями и др. Кучки мелких камней примерно одного размера
всегда лежат наготове около нор Alttcola, монгольской пищу-
хи и др.
Семена и плоды с плотными оболочками использует для се-
зонных запасов гораздо большее число видов млекопитающих,
чем сено, и в условиях самых различных ландшафтов (исклю-
чение составляет лишь арктическая зона и зона тундр, где се-
менная продукция растительности ничтожно мала). Обыкновен-
ный хомяк, серый, темноцветный, барабинский хомячки, лесная
и желтогорлая мыши, бурундук и некоторые другие виды соби-
рают запасы семян в одиночку или парой и упорно их охра-
няют; полуденная, монгольская, краснохвостая песчанки, кур-
ганчиковая мышь зимуют семьями и, видимо, иногда образуют
группы даже из неродственных особей. Они собирают семейные
или общественные запасы корма. Среди потребителей корневищ
183
и луковиц, собираемых в зимние «погреба», тоже есть виды, жи-
вущие и по-одиночке (слепцы, цокоры, слепушонки) и семьями
(полевки-экономки и др.).
У бобров пары и семьи более постоянны, чем у большин-
ства грызунов, так как их молодые свыше года остаются при
родителях. В областях с длительной и холодной зимой, где лед
рек и глубокий снежный покров крайне затрудняют бобрам до-
бывание веточного корма и поиски укрытий при появлении
опасности, они в течение всей осени достают и транспортируют
зимние запасы (рис. 65). Бобры приступают к заготовке после
Рис. 65. Схематический разрез через зимовочную хатку канадских бобров и
водоем, образовавшийся у их плотины. Зимний запас ветвей расположен в
глубокой части бобрового пруда, подо льдом. Один зверек отгрызает под
водой ветку, второй плывет с кормом к жилой части хатки, третий грызет
кору, сидя в жилой камере. Изолированные от внешнего мира сводом хатки
и льдом бобры могут длительное время жить за счет запасов веточного корма,
собранных осенью (по Гамильтону, с изменениями)
первых ночных похолоданий, которые, видимо, служат внешним,
сигналом, вызывающим эту специфическую реакцию. Они под-
грызают и валят ивы, осины, тополя, реже березы, дубы, лещи-
ны. и некоторые другие породы, разделяют толстые ветви на до-
вольно большие куски и по воде доставляют их к зимовочной
норе или хатке. Здесь ветви погружают в воду близ берега, но
на достаточно глубоком месте и укрепляют, зарывая толстые,
концы в дно. Из травянистых растений в зимние запасы они в
184
небольшом количестве кладут крупные корневища кувшинок,
кубышек, ириса, тростника и т. д. Каждую ночь в течение всей
осени все члены семьи заняты заготовкой, поэтому у больших
семей зимние запасы оказываются соответственно большими,
чем у бездетных пар или одиночек (по количеству и величине
стволов, срубленных за осень бобрами каждого поселения, мож-
но с известной долей вероятности судить о величине семей этих
очень скрытных животных).
Полный запас зимнего корма большой семьи бобров нередко
содержит до 25—30 куб. м ветвей, стволов, корневищ. Верхние
ветви такой кучи вмерзают в лед, нижние лежат на дне. Прого-
лодавшийся бобр ныряет подо льдом к запасам, отгрызает ку-
сок ветви и возвращается с ним в свое жилье. Тонкие побеги
эти грызуны съедают целиком с почками, корой и древесиной,
с толстых ветвей обгладывают кору. В холодной воде веточный
корм несколько месяцев сохраняет свои пищевые достоинства;
бобры, имеющие запасы, проводят время в достаточно теплой
камере хатки или норы и совсем не подвергаются опасностям,
грозящим неуклюжему тяжелому зверю при появлении в мороз
на глубоком рыхлом снегу. Очищенные от коры сучья или вы-
брасываются в воду, или используются для укрепления стенок
и кровли хаток.
Необычно высокие осенние паводки, затопляющие жилища
бобров, мешают заготовке корма и могут сильно ухудшить ус-
ловия зимовки. Так случилось, например, на р. Березине (Бело-
русская ССР) в 1952 г.; лишенные запасов бобры были зимою
сильно истощены, часть их погибла. Сам по себе процесс запа-
сания корма, за которым в течение суток приходится десятки
раз удаляться от норы или гнезда, увеличивает для зверьков
опасность гибели от хищников. Повышение смертности. от на-
падений хищников, главным образом пернатых, среди особей,
занятых поисками и переноской корма, было отмечено у бурун-
дуков, узкочерепных полевок, полевок-экономок, хомячков и
ряда других видов (в погадках сов вместе с костями хомячков
часто встречаются целые порции зерна хлебных злаков, семян
бобовых и т. п. из защечных мешков этих зверьков). Однако
сам период заготовки кормов, как правило, значительно короче,
чем время, в течение которого использование запасов дает жи-
вотным возможность избегать неблагоприятной погоды, голода
и нападений пернатых хищников.
Таким образом, положительное значение этой адаптивной
черты поведения несомненно. Это подтверждается и существо-
ванием у видов, запасающих мелкие семена, специального мор-
фологического приспособления для их переноски — пары объ-
емистых защечных мешков: они имеются у бурундуков, некото-
рых сусликов, многих хомяков, а также у мешетчатых крыс, или
гоферов (Geomyidae), и кенгуровых крыс (Heteromyidae) Се-
185
Geomyidae). Прокладывание
, Рис. 66. Один из видов североамери-
канских роющих грызунов гоферов
(сем.
ходов производится сильными ост-
рыми когтями передних ног и рез-
цами. Щечные мешки служат для
переноса запасов корма в подземные
кладовые (по Гамильтону)
верной Америки1. У одних видов (гоферы, кенгуровые крысы)
их щечные мешки открываются наружу и представляют карма-
нообразные выпячивания кожи, внутри покрытые шерстью
(рис. 66). У других щечные мешки лишены шерсти, открыва-
ются внутрь ротовой полости (бурундуки, суслики, хомяки).
Особенно объемисты щечные мешки у хомяков — задний ко-
нец их тянется за лопатки и прикреплен особой мышцей к обла-
сти крестца (см. рис. 22). Эволюция мешков шла в связи с есте-
ственным отбором особей, лучше приспособленных к собиранию
и доставке в нору запасов семян. Кроме запасов обычной пищи,
у белок установлено затаски-
вание на ветви выветрившихся
костей, т. е. заготовка впрок
минеральной подкормки.
Чтобы оценить биологиче-
ское значение запасания кор-
мов в борьбе за существование
представителей того или дру-
гого вида, нужно прежде все-
го знать, какое количество су-
точных норм пищи («пайков»)
они успевают сосредоточить и
укрыть у своего жилья до на-
чала наступления неблаго-
приятного периода. Подсчеты
показывают, что обыкновен-
ный бурундук, запасающий от
1.5 до 6 кг семян (редко боль-
ше), имеет в норе от 100 до
400 суточных пайков, если зи-
мует в одиночку. Однако в зи-
мовочной норе нередко нахо-
дят пару (самца и самку); в
таком случае продолжитель-
ность периода, обеспеченного
меньшей. В действительности
запасами, окажется в два раза
запасов и паре обычно хватает даже на весну, так как зверьки,
находящиеся в состоянии зимнего сна, пробуждаются и кормят-
ся редко.
Европейская норка иногда хранит в укромных местах до-
5 кг рыбы, что составляет не менее 50 суточных норм. При на-
личии таких запасов и сильном ожирении с осени норка по не-
скольку недель не выходит на поверхность снега и, естественно,
реже подвергается опасности.
1 Птицы, в большом количестве запасающие семена, например кедровки,
тоже имеют специальное вместилище для их переноски — объемистый подъ-
язычный мешок.
186
Нередко запасы даже мелких зверьков бывают столь велики,
что часть их остается на следующее лето. Так, например, од-
нажды в Беловежской пуще тяжестью огромного запаса желу-
дей, собранных желтогорлыми мышами (возможно, за ряд лет),
была выломлена подгнившая стенка дупла и 47 кг желудей вы-
сыпались на землю. Конечно, пара мышей не могла бы уничто-
жить этот запас даже и за более продолжительную зиму, чем в
Белоруссии. Но нередко случается и так, что собранные запасы
расхищают другие, более сильные млекопитающие. Кабаны в
пограничных районах Сибири и Монголии в течение осенних
месяцев тщательно разыскивают кладовые узкочепепных поле-
вок (Stenocranius) и поедают запасы корневищ живородящей
гречишки, медведи осенью и весной разрушают норы бурунду-
ков, добывая собранные ими кедровые «орешки»; стада север-
ных оленей на болотистых тундрах севера Сибири с большой
жадностью поедают сочные ростки осоки, собранные в кладовых
полевкой Миддендорфа. Большие запасы привлекают крупных
расхитителей; мелкие подсобные запасы и остатки пищи, укры-
тые в различных местах охотничьего участка норки, куницы или
лисицы, истребляет целый ряд врагов и конкурентов этих жи-
вотных. Таких примеров много.
В течение столетий у народов аборигенов Восточной Сибири,
Камчатки, Забайкалья и Северной Монголии бытовал обычай
осенней заготовки на еду корневищ и луковиц, выкопанных из
кладовых полевок. Установлено, что плоды лещины, кедра, дуба,
липы в кладовых бурундука, желтогорлых и лесных мышей от-
личаются исключительно высокими качествами — грызуны отби-
рают их. пользуясь своим тонким чутьем. Лесоводы Прибалти-
ки и УССР уже давно используют запасы плодов липы, взятые
из кладовых желтогорлых мышей,— этот семенной материал от-
личается лучшей всхожестью, чем собранный обычным способом,
кроме того, и заготовка обходится дешевле (в одной норе их
иногда находят до 30 кг} '. Растаскивание и укрывание грызу-
нами орехов и доугих плодов приводит к расселению древесных
пород, к обсеменению вырубок и гарей, хотя в этом отношении
деятельность птиц (ореховка, сойка) оказывается гораздо более
эффективной. С другой стороны, выяснено, что ряд видов ам-
барных клещей, очень опасных вредителей зерна и мучных про-
дуктов в наших хранилищах, по своему происхождению — оби-
татели нор грызунов, пользующиеся их запасами семян и т. п.
Таким образом, изучение этой стороны биологии млекопитаю-
щих представляет большой и разносторонний интерес.
1 В >Kvpva.ne «Лесное хозяйство» была недавно опубликована статья,
рекомендующая этот способ заготовки семенного материала. Заготовка лес-
ных семян, изъятых из кладовых грызунов, практикуется и в США.
Суточная активность. Спячка
СУТОЧНАЯ АКТИВНОСТЬ
Млекопитающие по их отношению к свету могут быть раз-
делены на дневных, ночных и нейтральных — активных как
днем, так и ночью. Однако это деление довольно условно — у
одного и того же вида отношение к свету существенно изменяет-
ся под влиянием разных внешних и внутренних причин ( глав-
ным образом голода, степени преследования и особенностей по-
ведения, связанных с брачным периодом). В большинстве слу-
чаев характер активности определяется не прямым влиянием
светового режима, а связанными с последним условиями пи-
тания.
Дневной или ночной образ жизни и закономерное чередова-
ние периодов деятельного состояния и покоя на протяжении со-
ответствующей части суток составляют суточный цикл, или ритм’
активности животного.
Различают монофазную активность (графически ее можно
изобразить одновершинной кривой), свойственную, например,
барсукам, дикобразам, желтогорлым мышам, обезьянам; дву-
фазную (с двувершинной кривой), хорошо выраженную у неко-
торых видов летучих мышей, сусликов, сурков, лесной мыши, и
полифазную (с многовершинной кривой), обычную для земле-
роек, кошек, енотовидных собак, полевок (рис. 67, 68).
Ритмика активности у млекопитающих подвержена геогра-
фическим изменениям. В полярных странах, где день длится на
протяжении почти всего лета, а ночь-— в течение всей зимы,
преобладают нейтральные формы с полифазным типом активно-
сти (ночные виды здесь легко приспособляются к полярному
дню, дневные — к полярной ночи, что до некоторой степени
сглаживает разницу между ними). В остальной части зоны арк-
тических пустынь и тундровой и в умеренном поясе повышается
число дневных форм, а в субтропиках и тропиках преобладают-
ночные формы.
На активность многих млекопитающих оказывает влияние
яркость освещения. В пасмурную погоду, например, независимо
от температуры, утренний выход сурков, сусликов и других днев-
ных норников на поверхность значительно запаздывает, общая
активность их снижается, а вечерний уход в норы ускоряется
по сравнению с солнечными днями. Не меньшее влияние на
188 ' .
ритм активности зверей оказывает температура. Летом, в часы
наиболее высокой температуры наблюдается резкое снижение
деятельности большинства зверей; они укрываются в порах
и т. п. убежищах, что предохраняет их от перегрева. Весной,
осенью и зимой эти же звери могут оставаться деятельными всю
светлую часть суток1. Выпадение осадков, сильные ветры обыч-
но снижают активность зверей. Во всех перечисленных явлениях
ясно выражена зависимость суточной активности от метеороло-
гических условий. i
Связь ритмики активности с типом питания яснее всего вы-
ражена у ночных и круглосуточно активных видов. Многофаз-
ность суточной активности ряда мышевидных грызунов, земле-
Рис. 67. Двуфазная суточная активность лесной мыши
{Apodemus silvaticus) (по Элтону, Форду и Бекеру)
роек и некоторых других млекопитающих обусловлена тем, что
они вынуждены несколько раз наполнять желудок пищей; обес-
печить себе суточную кормовую норму за один прием они не
могут.
Опыты по изучению суточной активности мелких млекопи-
тающих, проведенные в специально оборудованных клетках —
актографах, дали много интересных сведений (табл. 3). Выяс-
нилось, например, что мыши лесная Apodemus sylvaticus, желто-
горлая A. tauricus (flavicollis), азиатская A. speciosus, полевая
А. agra.ri.us деятельны преимущественно ночью и могут в днев-
ное время не выходить из гнезда в течение 2—8 ч. Такие про-
должительные перерывы поиска и поедания корма среди мелких
грызунов свойственны только видам, питающимся высоко-
1 Хорошо выражена эта закономерность у джейрана, в летний период
пасущегося два раза в сутки (утром и вечером), а зимой — на протяжении
целого дня (конечно, с перерывами на отдых).' Кривая суточной ритмики
сурков, сусликов, большой песчанки летом двувершинна, весной и осенью —
одновершинна. Такое же соотношение летней и зимней активности отмечено
для полевки Брандта и ряда других видов.
189
калорийной пищей (семена, орехи, насекомые), которая насы-
щает зверьков на продолжительный срок.
Млекопитающие
Таблица 3
Продолжительность перерывсв в активности мелких млекопитающих?
по даньым эксперимента в актографах
(по Тупиковой и Кулик, 1954)
Мьчпи
Серые полевки . . .
Землеройки-бурозубки
Продолжительность переръ вов акт ивност и
днем НОЧЬЮ
ч С4 мин—4 ч 59 мин 48 мин—1 ч 37 мин
40 мин—2 ч 2J мин—2 ч
30 мин—2 ч 20 мин
Многим млекопитающим свойственна большая пластичность
и лабильность ритма активности, что позволяет им приспосаб-
ливаться к меняющимся условиям среды. Синантропные формы,,
например, легко «подгоняют» свойственную им суточную ритми-
ку к многообразию режимов, существующих в поселениях че-
ловека. При усиленном преследовании охотниками дневные
промысловые звери иногда переходят на ночной образ жизни
Наоборот, отсутствие опасности, например в малолюдных райо-
нах или заповедниках, приводит иногда к восстановлению днев-
ной активности у очень осторожных зверей (бобров, барсуков,
рысей). У лисицы, енотовидной собаки, соболя в зависимости;
от обилия корма и интенсивности преследования преобладает то>
ночной, то дневной тип активности.
Высокая плотность популяции также может вызвать изме-
нение активности. Сумеречные летучие мыши при недостатке'
пищи частично переходят на ночное питание, а ночные — на:
утреннее и дневное (такие зверьки иногда летают даже при
ярком солнечном свете).
Несомненна связь суточной активности с полом, возрастом
и физиологическим состоянием животного. Резко повышается
активность самцов и самок ряда млекопитающих в период течки
и, наоборот; у беременных самок и больных животных она рез-
ко падает. В разгар гона ночные и дневные формы часто ста-
новятся деятельными круглосуточно. У копытных наблюдаются
нарушения ритма суточной активности при массовом появлении
кровососущих двукрылых (гнуса). Как правило, в зонах с пес-
симальными условиями питания суточная активность млекопи-
тающих повышается, в оптимальных — понижается и т. д. Таким
образом, суточный ритм активности представляет собой слож-
1 Так, например, дневные по своей природе дикие кролики во многих
местах, где на них интенсивно охотятся, превратились уже в ночных животных-
190
ную реакцию поведения животных на целый ряд экзогенных и
эндогенных факторов, действующих либо комплексно, либо от-
Рис. 68. Полифазная суточная активность темной
полевки (MicrotUs agrestis) и монофазная — желтогор-
лой мыши (Apodemus flavicollis) (по Дэвису и Кала-
бухову)
цельными своими элементами, в числе которых для млекопитаю-
щих основную роль играют световой, температурный режимы и
пищевой фактор.
СПЯЧКА
Спячка, или состояние пониженной жизнедеятельности, ха-
рактеризующееся неподвижностью животного и рядом специфи-
ческих изменений в его физиологических отправлениях, пред-
ставляет собой явление сезонной цикличности, связанное у мле-
копитающих, как и суточная активность, главным образом с
условиями питания.
Спячка позволяет животному длительное время обходиться
без пищи и таким образом предохраняет его от гибели в небла-
гоприятный в кормовом отношении период года. Это своеобраз-
ное приспособление свойственно только представителям опреде-
ленных групп животных, обладающих соответствующими эколо-
19Г
гическими и физиологическими особенностями: в спячку впа-
дают представители клоачных, сумчатых, насекомоядных, руко-
крылых, грызунов, неполнозубых, трубкозубых, хищных и полу-
обезьян.
По приуроченности к холодному или теплому времени года
различают зимнюю и летнюю спячку.
Зимняя спячка, по-видимому, возникла еще в плейстоцене
как одна из ответных реакций организма на резкое изменение
климата в сторону похолодания. Однако, сложившись историче-
ски под влиянием холода, явление зимней спячки в настоящее
время обнаруживает зависимость не столько от непосредствен-
ного воздействия температуры среды, сколько от недостатка
пищи. У части видов млекопитающих (сурки, суслики, сони, ежи
и некоторые др.) спячка в процессе эволюции превратилась в
наследственно закрепленную потребность организма, проявляю-
щуюся в определенное время даже при наличии обстановки,
вполне благоприятной для активной жизни.
Глубина зимней спячки у разных групп зверей не одинакова.
Можно принять следующую градацию типов:
1. Факультативная спячка, или зимний сон —состояние
сравнительно легкого оцепенения, не сопровождающееся сколь-
ко-нибудь существенным снижением температуры тела и легко
прерываемое. Свойственна медведям, барсукам, скунсам, ено-
там, енотовидным собакам. В районах с благоприятными зимни-
ми климатическими и кормовыми условиями эти животные по-
чти или вовсе не впадают в спячку. Барсуки в Западной Европе
и на Кавказе всю зиму остаются активными. Разбуженные в
берлоге медведи могут на протяжении всей оставшейся части
зимы вести бродячий образ жизни («шатуны»). У белых медве-
дей в спячку, как правило, впадают только самки (преимущест-
венно беременные).
К категории факультативной спячки можно также отнести
зимнюю дремоту, в которую временно впадают в очень мороз-
ные дни белки, летяги, норки, соболи и некоторые другие мле-
копитающие.
2. Настоящая спячка, периодически прерываемая,— состоя-
ние более или менее глубокого оцепенения с понижением тем-
пературы тела, но с сохранением способности пробуждаться и
короткое время бодрствовать среди зимы (главным образом, в
оттепели). Такая спячка свойственна хомякам, бурундукам,
многим видам летучих мышей (особенно ушанам).
3. Настоящая сезонная непрерывная спячка-—глубокое оце-
пенение с сильным понижением температуры тела, длящееся
всю зиму без перерывов. Так спит часть видов летучих мышей,
ежи, сурки, суслики, мышовки, тушканчики, сони.
Готовиться к зимней спячке звери начинают уже с осени.
Одни из них усиленно кормятся, накопляя резервные вещества в
192
организме (сурки, суслики, ежи) >, другие делают запасы кор-
мов на время зимних пробуждений (хомяки, бурундуки).
Очень важное значение имеют выбор и подготовка убежищ
для спячки. Как правило, температура воздуха в убежищах не
должна опускаться ниже нуля. Однако слишком теплые убежи*
ща также не пригодны для спячки (в тепле расходование орга-
низмом резервных материалов идет настолько быстро, что они
иссякают раньше, чем наступит нормальный срок пробуждения
животного). Обязательно постоянство температуры и вообще
микроклимата в убежище.
Большинство животных перед залеганием в спячку тщатель-
но готовит специальные зимовочные убежища, натаскивая в них
мягкие растительные материалы, забивая входные отверстия нор
большими земляными пробками и пр. Ошибка в выборе убежи-
ща для зимовки приводит к гибели животного. В северных и
умеренных широтах нередки, например, случаи замерзания лету-
чих мышей, оставшихся на зимовку в дуплах деревьев. В суро-
вые зимы с необычно глубоким промерзанием почвы погибают
ежи, суслики.
Сроки начала спячки у одного и того же вида довольно
сильно колеблются в зависимости от географического положе-
ния местности, климатических условий, упитанности, возраста и
пола животных. В северных .частях ареалов и в горах млекопи-
тающие погружаются в спячку раньше, чем в южных и равнин-
ных. Суслики на склонах холмов, обращенных к северу, начи-
нают зимовку раньше, чем на склонах южной экспозиции;
взрослые самцы, рано успевающие запастись жиром, залегают
первыми, за ними следуют взрослые самки и позднее всех —
молодняк.
Спят млекопитающие либо поодиночке (большинство ви-
дов), либо группами. Последние могут состоять, из членов одной
семьи (сурки) или из неродственных особей, нередко враждую-
щих друг с другом в период активной жизни (сони) 1 2. Для
уменьшения излучения тепла важное значение имеет свертыва-
ние животного в клубочек, прикрывание тела пушистым хвостом
(сони), окутывание с помощью перепончатых крыльев (летучие
мыши) (рис. 69, 70, 71).
Механизм зимней спячки связан с рядом факторов как экзо-
генного порядка (температура, свет, наличие корма и пр.), так
и эндогенных (упитанность, терморегуляция, гормональная дея-
тельность, состояние нервной системы и пр.). Зависимость от
1 За счет жировых накоплений вес суслика, например, к моменту зале-
гания в спячку почти удваивается по сравнению с летним.
2 Возможно, что такие группы возникают постепенно путем присоеди-
нения к заснувшим уже зверькам вновь прибывших (побудительной причиной
образования подобного рода групп может служить недостаток подходящих
зимних убежищ).
13 Териология
193
низкой температуры доказывается хотя бы тем, что с помощью'
искусственного охлаждения любое способное к зимней спячке
животное может быть среди лета переведено в состояние оцепе-
нения. Падение температуры воздуха ниже 15° С вызывает спяч-
ку у ежей ит. п.1 Определенное значение имеет также измене-
ние светового режима: сокращение светлой части суток стиму-
лирует подготовку к
спячке. Однако основ-
ным ее стимулом, несо-
мненно, служит отсут-
ствие или недоступ-
ность пищи в зимний
период (зеленых частей
растений, насекомых).
Из факторов эндо-
генного порядка важ-
нейшее значение имеет
состояние упитанности
к началу спячки; отло-
жению жира у зверей
способствует снижение
метаболизма, наблю-
дающееся у них осе-
нью 1 2. При погружении
животного в спячку у
него выключается тер-
Рис. 69. Соня-полчок (fills glis) в спячке
морегуляция, вследствие чего температура тела резко снижается
(табл. 4).
У летучих мышей температура тела в некоторых случаях мо-
жет упасть без летального исхода значительно ниже нуля
(рис. 72) 3. Обычно температура тела во время спячки держит-
ся на уровне, лишь немного превышающем температуру окру-
жающего воздуха, в связи с чем впадающих в спячку зверей и
называют гетеротермными животными. Гетеротермность служит
защитным приспособлением к длительному голоданию, гибель-
ному при сохранении обычной температуры. Еж в состоянии
1 В южных частях своего ареала некоторые звери впадают в спячку
не каждый год, а лишь в особенно суровые зимы.
2 Жир, накопляемый млекопитающими перед спячкой, отличается осо-
быми физико-химическими свойствами, например низкой температурой за-
стывания (иначе он не мог бы расходоваться при низкой температуре тела
спящего животного). Можно полагать также, что жир имеет' антисептиче-
ское значение, убивая или ослабляя вредные для спящего ?кивотного мик-
роорганизмы.
3 При постепенном отогревании переохлажденные летучие мыши возвра-
щаются в активное состояние, если в их клетках и тканях не произошла
кристаллизация воды. Оживление полностью промерзшего зверька, вопреки:
представлению сб анабиозе ГТ. И. Бахметьева (1912), невозможно.
194
Таблица 4
Температура тела некоторых млекопитающих в состоянии
бодрствования и спячки
(по Эйзснтрауту (Eisentraut), 1953)
Температура, °C
Млекопитающие
в состоянии
бОДрСТВОВиНиЯ
в состоянии
спячки
Еж европейский ................
Рыжая вечерница................
Сурок .........................
Сони...........................
Хомяк .........................
33,7
37,9
36,0—38,0
36,8—38,1
37,8—39,3
1,8
0,1
4,6—7,6
3,0—3,7
4,0—5,0
спячки остается без пищи в течение около 240 суток, бодрст-
вующий же зверек не переносит и 30 дней голодания. Сурок при
резком снижении температуры тела в период глубокой спячки
теряет в среднем около 0,1% веса тела в сутки, голодовка же
во время бодрствования вызывает у него потерю около 4% веса
в сутки (Скориченко, 1891).
Частота сердцебиений и дыхательных движений во время
спячки резко замедляется. У летучих мышей, например,, количе-
ство сердцебиений падает с 420 до 15—16, а количество дыха-
тельных — с 96 до 5—6 в минуту, у сурка — соответственно —
со 100 до 10 и с 20 дыханий до 3 в минуту. Вместе с тем нару-
шается ритм дыхания; между дыхательными движениями на-
блюдаются паузы длительностью до четверти часа'. Сокращает-
ся также газообмен. У сурка, например, дыхательный коэффи-
циент, т. е. отношение количества потребляемого кислорода
к количеству выделяемого углекислого газа, в период спячки
снижается с 0,7 до 0,4. В печени во время спячки происходит
накопление гликогена (животного крахмала), что может приве-
сти в начале ее к некоторому увеличению веса тела. Прибав-
ление в весе может происходить и от накопления метаболиче-
ской воды в организме. В дальнейшем, конечно, по мере расхо-
дования резервных материалов происходит потеря в весе.
Наблюдаются и другие более или менее глубокие изменения
физиологического состояния погруженного в спичку животного,
например, состава крови (возрастание содержания углеки-
1 У убитого во время спячки сурка сердце продолжает сокращаться
несколько часов, в то время как у убитого бодрствовавшего зверька оно
останавливается через несколько минут. При погружении спящего ежа
в воду он начинает задыхаться только через полчаса, бодрствующий — че-
рез 2—3 минуты.
13
195
слоты, снижение содержания сахара) *, деятельности эндокрин-
ных желез (ослабление их функций вследствие инволюции) 1 2,
биохимических процессов, протекающих в мозгу, в состоянии
нервной системы вообще (торможение функций коры больших
Рис. 70. Подковоносы (Rhinolophus) в состоянии зимней
спячки
полушарий, снижение тонуса симпатической нервной системы
и пр.) и, наконец, в состоянии половой сферы (подавление спер*
матогенеза у ежей и некоторых других спящих зимой живот-
ных) 3.
1 Снижение содержания сахара (глюкозы) в крови и отложение запасов
гликогена в печени указывают на то, что углеводный обмен у находящегося
в спячке животного сведен к минимуму [Кайзер (Kayser), 1953].
2 Ясно выраженная гипофункция щитовидной железы во время спячки
наблюдается, например, у ежей, летучих мышей, сурков (при введении тирео-
идина эти животные пробуждаются). Наоборот, инкреторная функция под-
желудочной железы во время спячки сильно возрастает.
3 У самцов сурков, сусликов, бурундуков и некоторых других активность
половых желез во время спячки полностью не прекращается, и с момента
пробуждения эти зверьки способны участвовать в гоне.
196
Несмотря на резкое общее снижение жизнедеятельности ор-
ганизм животного во время спячки сохраняет все же способ-
ность реагировать на внутренние и внешние раздражения. На-
пример, чрезмерное накопление углекислоты в крови, перепол-
нение мочевого пузыря, повышение температуры в убежище мо-
гут вызвать пробуждение животного. Тот же эффект наблю-
дается при охлаждении животного ниже свойственной ему «ми-:
нимальной температуры спячки», так как в таком случае вклю-
чается в действие терморегуляция, а вместе с тем повышаются
до предела активность и тем-
пература тела [Эйзентраут
(Eisentraut), 1956] \ С другой
стороны, у некоторых видов
зимнее оцепенение достигает
такой глубины, что почти пол-
ностью исключает реагирова-
ние животного даже на силь-
ные внешние воздействия (у
суслика, например, спячка не
прекращается при нанесении
ему серьезных ранений вплоть
до ампутации конечностей).
Общая продолжительность
зимней спячки колеблется от
нескольких недель до 9 меся-
цев (наиболее кратковремен-
ная спячка свойственна земля-
ным зайчикам и некоторым
другим южным формам, наибо-
лее продолжительная — длин-
нохвостым сусликам и камчат-
ским суркам в северных райо-
нах их распространения).
Весеннее пробуждение от
спячки во многих случаях вы-
зывается стойким повышением
температуры среды. Сроки это-
Рис. 71. Ушан (Plecotus auritus)
в состоянии зимней спячки. Огром-
ные уши спрятаны под крылья; вниз
торчат козелки (по Эйзентрауту)
го явления изменяются в зависимости от тех же причин, что и
сроки залегания, но последовательность явлений, конечно, обрат-
ная (на юге пробуждение начинается раньше, чем на севере,
и т. д.).
В момент пробуждения животное производит конвульсивные
движения, температура его тела быстро повышается, находив-
1 Пробуждение может быть кратковременным, при - котором животное
освобождается от накопившихся продуктов обмена веществ, согревается и
затем снова засыпает, Или полным. В ряде случаев оно может иметь гибель-
ные для жизотного последствия,
19
щиеся в угнетенном состоянии физиологические процессы вое- •
станавливаются.
Вследствие хотя и очень замедленного, но не прекращающе-
гося на протяжении всего периода спячки расходования резерв-
ных материалов, животные к весне теряют часть веса. Однако
основные резервы они используют в голодное весеннее время,
у ряда видов совпадающее с брачным периодом. Виды, впадаю-
щие в зимний сон, гораздо менее экономно расходуют резервные
вещества и, как правило, пробуждаются истощенными. Они уси-
Р ис. 72. Изменения мпературы тела лету-
чих мышей (рыжих вечерниц Nyctalus noctula)
при охлаждении ниже 0° С. А — замерзание
без переохлаждения; Б — замерзание после
переохлаждения; В —длительное переохлаж-
дение (по Калабухов'у)
наливая подорванные
длительной голодовкой
силы.
Помимо большого
теоретического интере-
са, явление зимней
спячки имеет и практи-
ческое значение, по-
скольку среди впадаю-*
щих в спячку млекопи* -
тающих имеются про-
мысловые виды, вреди*
тели сельского хозяй» .
ства, переносчики ин-
фекций. Изучение явле-
ний спячки открывает
также некоторые пер*
спективы лечебного
применения искусст-
венно вызванного (ох-
лаждением) оцепене-
ния. Экспериментально установлено, что на протяжении всего
периода спячки у млекопитающих не развиваются инфек-
ционные болезни, даже такие смертоносные как чума; подав-
ляется также рост злокачественных опухолей. Путем охлажде-
ния удавалось полностью освобождать организм некоторых жи-
вотных от болезнетворных простейших (трипанозом, спирохет).
В последнее время холодовое оцепенение начали применять в
медицинской практике (при сложных операциях и т. п.).
Летняя спячка свойственна некоторым пустынным и степным
млекопитающим. Ее вызывает недостаток влаги в пересыхающей
растительности, а также изменение нервного механизма живот-
ных под влиянием неблагоприятных условий среды (Калабухов,
1956; Слоним, 1952). Наиболее полно изучена летняя спячка
у ряда видов сусликов. По мере снижения содержания влаги в
растениях «роющая деятельность» сусликов повышается; они
устраивают себе норы для спячки. В годы с влажным дождли-
.9.8
зым летом переход в спячку происходит значительно позднее,
чем в засушливые годы. Желтые суслики, живущие в пустынях
Средней Азии, залегают в спячку уже в конце мая — начале
июня; в северной части их ареала этот срок отодвигается на
июль — август. Продление периода активности наблюдается
также у зверьков, поселяющихся возле арыков, где свежая ра-
стительность сохраняется значительно дольше (Кашкаров и
Лейн-Соколова, 1927).
Летняя спячка непосредственно (без перерыва) переходит в
зимнюю.
Имеются указания, что в летнюю спячку впадают, кроме сус-
ликов, некоторые виды сурков, орешниковая соня. С исчезнове-
Рис. 73. Изменения темпера-
туры тела малого суслика
(Citellus pygmaeus) в летней
спячке (по Калабухову)
1 — температура тела бодрствую-
щего суслика; 2 — температура
тела спящего суслика; 3 — тем-
пература воздуха
высокой (оис. 73). Вслед-
нием насекомых в жаркие месяцы
связана непродолжительная летняя
спячка у некоторых летучих мышей.
Сенегальские ежи проводят в летней
спячке около 3 месяцев, мадагаскар-
ское насекомоядное тенрек — около
4 месяцев.
Перед впадением в летнюю спяч-
ку в организме животных также на-
капливаются резервные вещества (в
том числе и жир). Существенное от-
личие летней спячки от зимней за-
ключается в том, что она протекает
при температуре воздуха, близкой к
температуре бодрствования. Кроме
того, и температура тела спящего
животного остается относительно
ствие этого обмен веществ в организме происходит более интен-
сивно, чем в зимнюю спячку. Однако некоторые физиологиче-
ские процессы все же . замедляются (например, дыхание).
В общем же летняя спячка менее глубока, чем зимняя.
Как зимняя, так и летняя спячка млекопитающих должна
рассматриваться с позиций учения И. П. Павлова как сложно-
рефлекторная реакция на изменения условий среды, как биоло-
гически целесообразное наследственно закрепленное свойство,
исторически выработавшееся у отдельных видов млекопитаю-
щих, дающее им .возможность противостоять неблагоприятным
сезонным явлениям природы и существовать в областях, где
круглогодичная деятельность для них совершенно невозможна.
Миграции. Линька
МИГРАЦИИ
Сезонные, обратимые передвижения млекопитающих на бо-
лее или менее значительные расстояния — кочевки, или мигра-
ции,— носят приспособительный характер и связаны либо с се-
зонными изменениями метеорологических и кормовых условий,
либо с определенными стадиями жизненного цикла (размноже-
нием, расселением), либо с теми и другими вместе. Нередко ко-
чевки рассматриваются как особый тип передвижений, отличаю-
щийся от собственно миграций своей сравнительно небольшой
протяженностью. Однако провести грань между кочевками и ми-
грациями очень трудно, тем более, что дальность передвижений
у одного и того же вида может в зависимости от обстоятельств
изменяться в самых широких пределах. Изучение миграций зна-
чительно подвинулось вперед с того времени, как начали смете--
матически метить животных '. Применение мечения дало ключ
к пониманию сущности этих явлений, хотя многие детали тре-
буют еще уточнения.
Различают регулярные, периодические и обратимые мигра-
ции и нерегулярные, обычно необратимые. Существуют и другие,
второстепенные типы миграций: горизонтальные и вертикальные,
суточные и сезонные, активные и пассивные2.
1 Мечение млекопитающих чаще всего производится при помощи нуме-
рованных металлических колец, продеваемых в ушные раковины (у боль-
шинства видов зверей) или закрепляемых на конечностях (например, у лету-
чих мышей — на предплечья), на корне хвоста (у выхухолей) и пр. Для
мечения китов служат заостренные полые цилиндры из нержавеющей стали,
которыми стреляют из гарпунных ружей с судов, а в последние годы и
с вертолетов (вонзаясь в толщу подкожного жирового слоя, эти метки
не причиняют вреда животным). Применяется также татуировка (накалывание,
татуировочными щипцами номера на внутренней голой поверхности уха с по-
следующим втиранием туши или краски), а также надрезы или пробивание
специальными щипцами отверстий в ушных раковинах и в перепонках лап
по условному ключу. Мышевидных грызунов метят, отрезая у них отдельные
пальцы (также по особому ключу). В последнее время для мечения. живот-
ных начинают применять радиоактивные изотопы, стойкие красящие веще-
ства и портативные радиостанции, прикрепленные к телу животных.
. 2 Попытки классификации миграций производились многими авторами
[Кан (Кап), 1925; Томсон (Thomson), 1926; Уильямс (Williams), 1930; Хип
(Неаре), 1931]. В последнее время Эркарт (Urquhart, 1958) предложил
следующее подразделение передвижений животных на территории; пассивное
расселение, номадизм и миграции. Миграции подразделяются им на эмигра-
200
Периодические, направленные, исторически сложившиеся
миграции представляют наиболее распространенное явление.
Чаще всего это правильно повторяющиеся сезонные передвиже-
ния по сложившимся веками путям или направлениям. Суточ-
ные передвижения совершаются млекопитающими в основном от
мест их отдыха к местам кормежек, водопоев или к солонцам.
Чем дальше отстоят эти места от мест отдыха, тем большей про-
тяженности достигают перекочевки. Сезонные миграции также
в большинстве случаев связаны с питанием (сезонными изме-
нениями кормовой базы).
Сравнительно небольшие масштабы имеют вертикальные и
отчасти горизонтальные сезонные передвижения типично торных
копытных — снежных баранов, горных козлов, горалов, серн.
С наступлением зимы они спускаются в ниже расположенные ча-
сти склонов с перемежающейся снежностью или переходят в дру-
гие участки той же зоны, где снежный покров менее глубок и,,
стало быть, более доступны корма (рис. 74). Весной происходит
обратное их перемещение. Значительные вертикальные передви-
жения — от субальпийских лугов до лесов подошвы гор, совер-
шают дикие кабаны, косули, благородные олени Кавказа и гор >
Сибири. Некоторые летучие мыши (двуцветный кожан, Кавказ-’
ский кожанок) перелетают осенью с гор в низины, а весной-
обратно.
Регулярные горизонтальные миграции в условиях гор хорошо-
выражены у лосей и косуль, населяющих Уральский хребет:
зимой они переходят с западного, ранее и обильнее покрываю-
щегося снегом склона на восточный малоснежный и весной воз-
вращаются обратно.
В условиях равнин и водных пространств горизонтальные
миграции могут иметь самую различную протяженность. Неболь-
шие, связанные с состоянием кормовой базы суточные и сезон-
ные перемещения из одной стации в другую совершают многие-
ведущие бродячий образ жизни копытные, ластоногие и другие-
«номады». Лоси, ограничивающиеся в малоснежных районах
небольшими местными перекочевками, в многоснежных местах
предпринимают миграции протяженностью в несколько сотен
километров. Дальние сезонные миграции совершают северные
олени. Осенью они сбиваются в большие стада и сосредоточи-
ваются в лесотундре или в северной части тайги, где находят
надежную защиту от снежных буранов, а на лето переходят в
северные подзоны тундры, где их меньше беспокоят кровососу-
щие насекомые и где в это время лучше кормовые условия
(рис. 75, 76). Во время таких передвижений олени покрывают
сотни километров, преодолевая на своем пути крупные озера и
<ии, иммиграции и ремиграции (т. е. миграции с возвращением). Однако-
все эти построения носят довольно искусственный характер.
sot
реки. Джейраны пустынь Средней Азии на зимовку уходят из
северной полосы более многоснежных пустынь к югу или в пред-
горья, где находят лучшие укрытия от непогоды и больше до-
ступных кормов. Миграции африканских копытных — разных
антилоп, газелей, буйволов, зебр — связаны со сменами сухих
и влажных сезонов. Период дождей вызывает временное буйное
:Рис. 74. Распределение оленей Odocoileus hemionus по вертикальным поясам
западного склона хребта Сьерра-Невада (Калифорния) в зависимости от
времени года, температуры воздуха и высоты снежного покрова. В много-
снежную зиму 1948/49 г. вся масса оленей сосредоточилась на небольшой
площади. В теплую половину года животные распределены широко в
.большинстве поясов обоих макросклонов хребта (по Леопольд с соавторами)
1 — высокая, 2_________средняя, 3 — низкая плотность населения оленей
развитие растительности в саваннах и полупустынях, куда и
устремляются многочисленные табуны копытных. Выгорание рас-
тительности по окончании дождливого периода обусловливает
обратное их движение.
Своеобразны сезонные миграции летучих мышей, улетающих
на зиму из полосы тайги, смешанных лесов и даже лесостепи и
202
60 30 100 120 140 160 180
Рис. 75. Основные направления осенних кочевок диких северных' оленей (Rangifer tarandus) в Арктике
и Субарктике к местам зимовок:
1 — основные направления современных кочевок; 2 — основные направления бывших кочевок (прекратившихся после умень-
шения поголовья); 3 — северная граница лесотундры; 4 — северная граница тайги; Б — места зимовок (по Насимовичу)
Рис. 76. Пути осенних миграций наиболее крупных стад тундровых северных
оленей в Канаде (по Бэнфилд)
степи как в Евразии, так и в Сев. Америке. Эти перелеты имеют
много общего с перелетами птиц, по-видимому, связаны с анало-
гичным наследственным инстинктом. У бухарских подковоносов,
например, осенний отлет начинают молодые особи, летящие к
тому же на большом расстоянии друг от друга, что полностью
исключает возможность научения (Кузякин, 1956).
Интересно, что летучие мыши иногда совершают перелеты
в дневные часы, при этом они могут присоединяться к ласточ-
кам, образуя с ними общие смешанные стаи [Штадлер (Stadler),
1922; Финк фон Финкельштейн и Шефер (Fink von Finkelstein
u. Schaefer), 1934; Эйзентраут (Eisentraut), 1937). Расстояния
между летним и зимним ареалом у разных видов и популяций
сильно разнятся. Некоторые виды перелетают на значительные
расстояния. Путем кольцевания удалось установить, что летние
места обитания рыжих вечерниц (Nyctalus noctula), зимующих
в окрестностях Дрездена, расположены в Польше и Литве, т. е.
на расстоянии до 750 км (Эйзентраут, 1957). Нетопырь-карлик
(Vespertilio ptplstrellus), окольцованный в районе г. Днепропет-
ровска в июне 1939 г., был пойман в сентябре того же года в
южной части Болгарии (у Пловдива) на расстоянии по прямой
более 1000 км от места кольцевания. Нетопырь Натузиуса (Ves-
pert'dio nathusil) с кольцом, надетым в Воронежском заповед-
нике в июле 1958 г., был найден в ноябре того же года вблизи
г. Стамбула (Турция). Он преодолел расстояние около 1750 км,
но и это, несомненно, не предел. Мигрирующие летучие мыши
способны перелетать открытые моря, покрывая при этом огром-
ные расстояния. Например, ряд североамериканских видов ре-
гулярно появляется осенью на Бермудских островах, отстоящих
от ближайшего материкового берега на расстоянии более
1000 км, а весной возвращается обратно.
Сезонные миграции морских млекопитающих обнаруживают
связь как с питанием, так и с размножением. Гренландские тю-
лени (Pagophoca groenlandlca), проводящие летний период на-
гула в прибрежных водах Гренландии и Канадского архипе-
лага, у кромки льдов от Шпицбергена до Земли Франца Иосифа
и отчасти в Карском море, образуют три стада, из которых одно
мигрирует в конце зимы к о. Ньюфаундленду, другое — ко. Ян-
майену и третье — в Горло Белого моря, где расположены
места их щенки на морских льдах, отличающихся меньшей по-
движностью от льдов открытого океана (рис. 77). Размножаю-
щиеся в северной части Тихого океана котики (Callorhmus
ursinus) образуют две расы и два стада — азиатское и американ-
ское. Первое из ни^ на зиму мигрирует от Командорских остро-
вов и о. Тюленьего к восточным берегам Кореи и островов Хок-
кайдо и Нипон, второе — от Прибыловых островов вдоль запад--
кого берега Америки в район Южной Калифорнии. После
зимовки в южных морях они снова возвращаются на север к ост-
205
ровам, где у них всегда происходит щенка, выкармливание моло-
дых и спаривание.
Миграции моржей с юга на север весной и обратно осенью
связаны в основном с тем, что мелководья, где они могут добы-
вать донных моллюсков, становятся для них недоступными при
появлении льдов, и отчасти с переходами к местам, удобным для.
размножения.
Рис. 77. Направление осенних миграций трех стад гренланд-
ского тюленя из районов нагула к местам щенки и линьки.
Тюлени беломорского стада трогаются в путь в конце сентября,
в декабре они появляются у берегов Мурманска и в Горле
Белого моря (по Фишеру и др. с добавлениями)
Миграции китов тоже зависят от необходимости менять ме-
ста нагула и переходить в районы, благоприятные для родов и
кормления молодняка. Рыбоядные -киты (белухи, дельфины.)
мигрируют в зависимости от распределения стадных рыб и гид-
рологических условий. Так, например, белухи Карского стада
в летние месяцы держатся в прибрежных водах Карского моря
от Новой Земли до Северной'Земли. Здесь они охотятся за сай-
кой, а в опресненных заливах и устьях рек главным образом за
206
сиговыми рыбами, образующими большие косяки перед ходом на
нерестилища. В конце ноября — начале декабря белухи в массе
уходят из Карского моря на запад через проливы между остро-
вами Новой Земли и проводят зиму у кромки льдов в восточной
и юго-восточной части Баренцева моря. Нередко массовый ход
этих дельфинов через проливы совпадает с миграцией сайки,
следом за которой часто идут и так называемые «юрова»— ста-
да гренландских тюленей.
Основными местами нагула кашалотов служат области с бо-
гатой фауной головоногих моллюсков (животных стенотермных
и стеногалинных). Они рас-:
ПОлОЖСПЫ В ВОДДХ ТрОПИЧС"
ского, субтропического и от-
части умеренного поясов
(холодные воды с низкой
температурой и пониженной
соленостью для этих моллю-
сков неблагоприятны). Сам-
ки кашалота за редкими ис-
ключениями не выходят за
пределы тропической и суб-
тропической зон, где они
растят своих детенышей, а
кочующие крупные самцы в
летнее полугодие в северном
полушарии достигают Ба-
ренцева и Берингова морей,
а в южном — Антарктики.
У беззубых китов планк-
тофагов на севере и юге
хорошо выражены миграции
на лето в холодные воды, к
I п ш а ли ш ядххш-
кромке полярных льдов, где
. Рис. 78. Схема сезонных изменений
но мере их таяния и отступа-
ния в массе развивается зоо-
планктон (рис. 78). Его ос-
новные представители —
рачки Calantis finmarchtcus,
Thysanessa tnermts, Eupha-
sla pactftca и др., а также
биомассы планктона в морях и океанах
на разных широтах. Сплошная линия —
фитопланктон, прерывистая — зооплан-
ктон:
а — арктическая область; б — субарктиче-
ская; в — северная бореальная; г — южная
бореальная; д — субтропическая; е — тро-
пическая (по Богорову)
привлеченные ими рыбы,- служат китам обильным нажиро-
вочным кормом во время пребывания на богатых «полях
питания», где они образуют достаточно плотные скопле-
ния. Здесь и производится наиболее успешный китобойный про-
мысел. К осени зоопланктон исчезает, льды смыкаются, и киты,
разбившись на небольшие группы, уходят зимовать в умерен-
ные и. субтропические воды (рис. 79), где кормовая база
207
Рис. 79. Схема миграции китов:
гренландский кит (Balaena mysticetus)', 2 — серый кит (Eschrich.tius glbbosus)-, 3 — горбатый кит (Megaptera nodosa) (по Келлогу)
беднее. Переход из холодных вод в теплые необходим китам
и для щенки, так как новорожденные при отсутствии под-
кожных жировых отложений могут существовать только в усло-
виях достаточно высокой температуры. Исключение составляют
гренландские киты, белухи и нарвалы, приспособившиеся к раз-
множению у кромки льда, а также финвалы и некоторые другие
киты, щенка которых происходит в северных морях в сфере влия-
ния Гольфстрима [Кларк (Clarke), 1957; Томилин, 1958 и др.]1.
В отличие от регулярных обратимых миграций нерегулярные
необратимые миграции, или эмиграции, представляют собой мас-
совые выселения животных обычно без последующего возвраще-
ния на места постоянного обитания. Вызываются такие выселе-
ния резкими ухудшениями жизненных условий, бескормицей в
связи с высокой плотностью популяции, лесными или степными
пожарами, сильными засухами, наводнениями, чрезмерным мно-
госнежьем и другими причинами.
Классическим примером нерегулярных миграций могут слу-
жить массовые выселения норвежских и обских леммингов и
обыкновенных белок в годы их высокой численности и недо-
статка кормов. Обычно в течение многих дней или даже недель
зверьки движутся поодиночке на некотором расстоянии друг от
друга в одном направлении, которого придерживаются с боль-
шим постоянством. Движение носит стихийный характер. На-
толкнувшись на препятствие, зверьки не обходят его, а идут «на-
пролом», стараясь преодолеть. Переселяющиеся зверьки нередко
появляются в поселках, городах; они очень возбуждены и теряют
обычную осторожность. Встречая на своем пути большие реки,
озера, лемминги или белки пытаются их переплыть, что, однако,
удается немногим. Достигнув берега моря, зверьки бросаются в
волны и в таком случае все погибают. А. Ф. Миддендорф образ-
но назвал такие эмиграции «кочеванием до смерти». Эмиграции
белок часто начинаются с расселения молодых после разделения
выводков. Если данный год на большой лесной площади неуро-
жайный на корма, следом за молодыми или одновременно с ними
начинают откочевывать и взрослые. Вскоре в «поток» эмигран-
тов вливаются тысячи зверьков. Сезонные и многолетние коле-
бания частоты эмиграций белок определяются в основном усло-
виями питания, реже изменениями защитных свойств стаций, вы-
званных лесными пожарами, метеорологическими условиями и
т. п. (Формозов, 1936) 1 2.
У мелких мышевидных грызунов массовые выселения пред-
ставляют более редкое явление, они кратковременны и не захва-
1 В редких случаях небольшая часть видов китов может мигрировать и
через экватор.
2 Во время выселений белки проходят расстояния до 500 км. Длитель-
ность эмиграции может достигать 6 месяцев. Ширина фронта, которым дви-
жется масса эмигрирующих белок, достигает 300 км и более.
14 Териелогия
209
тывают больших пространств. Яркую картину массового пере-
движения общественных полевок в степях Закавказья нарисовал
К- Н. Россиков (1887, 1916) !. Осенью 1940 г., при массовом раз-
множении грызунов в степях Предкавказья наблюдались круп-
ные передвижения домовых мышей (Н. Наумов, 1948). В 1945 г.
при подобных обстоятельствах на территории Центрально-черно-
земной полосы отмечались массовые передвижения обыкновен-
ных и водяных полевок (Барабаш-Никифоров, 1949).
С высокой плотностью популяции и сокращением кормовой
базы связаны, по-видимому, наблюдающиеся время от времени
выселения лосей, северных оленей, косуль, кабанов. В литера-
туре имеется много указаний на вынужденные выселения зайцев
и копытных, связанные с многоснежьем и буранами2. Сильные
засухи и бескормица вызывают откочевку джейранов.
Особую категорию представляют передвижения расселяю-
щихся молодых животных после распада семей. Это обратимые
и полезные для вида передвижения. Направления «путей рассе-
ления» определяются экологическими руслами, которые могут
служить и путями сезонных миграций соответствующих видов.
Сопряженными миграциями называют связанные друг с дру-
гом передвижения двух или нескольких видов. Например, мигра-
ции и эмиграции хищников, связанные с передвижениями их
жертв. В основе своей это кормовые миграции, но стимулирует
их не бескормица, а обилие пищи. Многие хищники, следуя за
мигрирующими животными, охотятся за ними, другие подбирают
остатки трапезы более сильных собратий или поедают трупы
павших животных (гиены, шакалы, песцы). Можно привести
много примеров таких миграций: за горными копытными сле-
дуют барсы, за оленями, косулями — рыси и росомахи, за север-
ными оленями — волки и песцы, за сайгаками — волки, за афри-
канскими пустынными копытными — львы, гепарды, гиены, ша-
калы, за кабанами — тигры и т. д.
1 Краски в этом описании сильно сгущены.
2 В чрезвычайно снежную зиму 1923 г. в Мариупольском районе (Украи-
на) наблюдалось движение огромного количества зайцев-русаков на юг, при-
чем зверьки продолжали следовать и по льду замерзшего Азовского моря.
Они погибали здесь массами (Неженцев, 1924). В феврале 1928 г. в районе
степного хозяйства Аскания-Нова (Украина) разразился сильнейший буран,
длившийся двое суток. В результате тысячи русаков, спасаясь от голодной
смерти, ринулись в поселок Аскания и заполнили его площадь и дворьь
В степи было обнаружено много мертвых зайцев. Большое количество осла-
бевших животных было выбито населением просто палками (Фортунатов,
1929). В феврале 1952 г. на льду и на островах северного побережья Каспия
появились тысячные стада сайгаков. С потеплением и таянием льда они
оказались отрезанными от берега, что привело к массовой гибели животных
от голодовки (Адольф, 1955). В Сибири в обильную снегом зиму 1913/1914 гг.
лоси, маралы и косули вынуждены были оставить места постоянного обита-
ния й искать себе пропитание вблизи человеческих поселений; отдельные
особи при этом заходили даже в деревни (Формозов, 1946).
210
В некоторых случаях особенности передвижения млекопи-
тающих не находят еще полного объяснения, например, пере-
ходы песцов за один зимний сезон на расстояние более тысячи
километров, по-видимому, представляющие крайние варианты
обычных осенних выселений (рис. 80).
j 36’ 42’ 48° 54° 60° .55’ 72’ 78° 84°
Рис. 80. Направления кочевки песцов, помеченных на Ямале (по Сдоб-
никову)
Все рассмотренные примеры относятся к категории активных
миграций. Пассивные миграции также имеют важное значение,
хотя и представляют у млекопитающих сравнительно редкое яв-
ление. К ним можно отнести перемещения моржей на дрейфую-
щих льдинах; нередко на большие расстояния на льдах уносит
белых медведей и песцов. Совершенно случайный характер носят
перемещения мелких млекопитающих на речных льдинах во
время ледохода, на плавучих островках и бревнах в половодье.
Более часты имеющие серьезное практическое значение переме- -
щения с различными видами транспорта.
Оценивая биологическое значение миграций,следует прежде :
всего иметь в виду их роль в удовлетворении жизненно важных
требований организма, связанных с питанием и размножением.
Миграции обеспечивают также более равномерное и полное ис-
пользование видом территории, широкий обмен особями между
разными популяциями (устранение близкородственного спари-
вания, предотвращение перенаселения); они придают необходи-
мую гибкость в распределении особей по угодьям. Даже эмигра-
ции леммингов и белок; сопровождающиеся массовым отходом
зверьков, полезны для вида в целом, так как изреживают достиг-
шие высокой плотности популяции. [Хильцхеймер (Hllzheimer),
1929; Хип (Неаре), 1931; Метыоз (Matthews), 1954; Н. Наумов,
1955 и-др.]. В отдельных случаях выселения приводят и к расши-
рению ареалов. Например, с эмиграцией белок связано заселе-
ние ими в 20-х годах нашего столетия Камчатки, где вид этот
ранее отсутствовал. Принявшее й последнее время широкие мас-
штабы эмиграционное движение лосей и кабанов привело К бы-
строму восстановлению их населения на территориях, где; они
были истреблены 100—200 лет назад. (Слудский, 1953; Бара-4
баш-Нийифоров, 1956).
ЛИНЬКА
Смена шерстного покрова и тесно связанные с ней изменения
кожи млекопитающих представляют собой очень тонкий биоло-
гический процесс, обеспечивающий сохранение целостности по-
кровов тела, как основного защитного образования. Остевые,
направляющие и отчасти пуховые волосы млекопитающих,'.щет-
ки упругих волос на подошвах ног и другие относительно неж-
ные образования, часто, соприкасающиеся с субстратом и окру-
жающими предметами, быстро изнашиваются. Охотники хорошо
знают, как быстро портится мех степных лисиц, скрывающихся
на день в густых тростниковых зарослях; жесткие стебли трост-
ника уже в самом начале зимы настолько вытирают ость на
боках лисиц, что пушные шкурки их в значительной степени
утрачивают свою ценность. Преждевременная сильная потер-
тость меха бывает у соболей, часто скрывающихся в узких хо-
дах между камнями, у многих видов землероев (слепыши, сле-
пушонки, цокоры, кроты и др.). В процессе линьки эти дефекты
устраняются.
Периодическая смена шерстного покрова необходима также
й потому, что в условиях климата с резко выраженной сменой
теплых и холодных сезонов однотипный покров с определенной
теплоизолирующей способностью не может быть пригодным в те-
чение всего года.
212
Например, у ряда арктических животных с хорошо развитой
физической терморегуляцией зимой сохранение постоянного
уровня температуры в самые жестокие морозы обеспечивается
высокими теплоизоляционными свойствами волосяного покрова.
В летнее же время постоянство температуры тела у них дости-
гается в значительной степени за счет увеличения теплопровод-
ности покровов в 3—4 раза по сравнению с зимой, а также бла-
годаря хорошо развитому механизму полипное и теплоотдаче
через конечности [Харт (Hart), 1956].
Ход линьки определяет качество пушных шкурок, добывае-
мых охотниками или получаемых звероводами,— в этом практи-
ческое значение его изучения. В основе механизма линьки
млекопитающих как и других наземных позвоночных лежит
гормональное воздействие гипофиза и щитовидной железы.
Гипофиз воздействует на щитовидную железу, а ее гормон
тиреоидин вызывает линьку покровов защитного и термоизоли-
рующего типа. Но эти процессы не автономны; они проходят
под контролем и влиянием внешней среды.
Температура — основной фактор, влияющий на сезонную
линьку. Однако стимулятором начала этого процесса служит
свет. У некоторых видов свету, действующему через зрительные
восприятия на гипофиз, принадлежит первенствующая роль в
развитии процесса линьки. У беляка, например, линька зависит
от фотопериодизма, а температура служит фактором, ускоряю-
щим или задерживающим смену волос (Афонская, 1949). В экс-
периментальных условиях, сокращая или удлиняя продолжи-
тельность освещения, можно изменять сроки линьки, намного
ускорять «созревание» меха, что в отношении пушных видов
имеет существенное хозяйственное значение
Влияние внешней среды на сроки линьки и даже на харак-
тер сезонного диморфизма волосяного покрова доказывается
также практикой акклиматизации млекопитающих. Например,
у видов, вывезенных из стран северного полушария и выпущен-
ных в Австралии, Новой Зеландии и Южной Америке, посте-
пенно сместились сроки линьки, а также спячки и размножения.
Млекопитающие, выпущенные в районы с относительно более
суровыми условиями, чем на их родине, приобрели более пыш-
ный зимний мех (енотовидная собака в ряде областей СССР).
Наоборот, акклиматизированные виды, попавшие в условия
сравнительно теплого климата (белка-телеутка в Крыму, алтай-
ская белка на Кавказе), утратили характерный для них неж-
ный и высокий мех: он стал более грубым и коротким (Гера-
1 Так, например, сокращением длительности светового дня летом, т. е.
в период наиболее длительного естественного светового дня. можно ускорить
более чем на месяц созревание зимнего меха у норок и лисий. [Биссонет и
Вильсон (Bissonnette a. Wilson), 1939; Биссонет и Левелин (Bissonnette а.
Lovelin), 1947; Беляев,' 1953].
213
симова, 1955; А. Шапошников, 1960; Насимович, 1961). Инте-
ресно, что зайцы-беляки, выловленные в Норвегии и выпущенные
в половине прошлого века на Фарерских островах, в пер-
вый период акклиматизации еще одевали белый зимний наряд,
а теперь и в холодную половину года носят рыжевато-бурый
мех. подобный летнему. В условиях бесснежных зим белый
наряд невыгоден, так как слишком заметен; островная попу-
ляция примерно за столетие утратила эту бесполезную и, быть
может, даже вредную особенность сезонного наряда [А. де Вос
(A. de Vos) и др., 1956].
В то время как у амфибий и рептилий — животных с непо-
стоянной температурой тела, смена покровов одновременно
охватывает все его части, у теплокровных животных — птиц и
млекопитающих, при линьке, как правило, последовательно сме-
няются покровы отдельных участков тела. Эта особенность свя«
зана с усложнением строения и функций покровов у млекопи-
тающих и птиц.
Развитие нового меха начинается с закладки остевых волос,
от сумок которых, как полагают, отпочковываются уже зачатки
пуховых. Процесс замещения волос у разных групп млекопи-
тающих протекает не одинаково. У хищников зачаток нового
волоса закладывается из клеток дна луковицы старого. По мере
роста новый волос выталкивает отделившийся от луковицы, но
довольно долго удерживающийся в волосяной сумке старый.
У грызунов закладка зачатков новых волос происходит совер-
шенно независимо от волосяных сумок старых, выпадающих.
Поэтому, в противоположность хищникам, группировки волос
нового меха у них не соответствуют таковым старого. В зачат-
ках новых волос концентрируются зерна пигмента. Просвечи-
вая через подкожную клетчатку, они придают синеватый цвет
мездре (нижней поверхности кожи). Так как в разных участ-
ках линька обычно протекает не одновременно, на мездре обра-
зуется характерный рисунок, состоящий из так называемых
«пятен линьки». По их расположению и форме можно судить
о наступлении той или иной стадии линьки’. С ростом волос,
выносящих пигмент из кожи, происходит посветление мездры,
идущее в той же последовательности, в какой шло ее потемне-
ние. Полностью «очистившаяся» от пятен мездра — признак
окончания процесса линьки (рис. 81, 82, 83).
С сезонной линькой бывает нередко связано изменение гус-
тоты меха и его окраски, выраженное у некоторых видов очень
резко (например, у белеющих на зиму зверей).
При линьке не только изменяется волосяной покров, но и
кожа. Так, в ходе линьки кориум разрыхляется развивающи*
Естественно, что при развитии белых (не содержащих пигмента) во-
лос пятна линьки на мездре не образуются.
214
мися зачатками новых волос и соответственно этому утолщает-
ся; в межлинечные периоды он уплотняется. Жировой слой,
сильно развитый зимой, летом уменьшается или полностью
исчезает.
В сложном физиологическом процессе, связанном с линькой,
участвует весь организм животного. В период линьки увеличив
Рис. 81. Последовательные стадии изменения цвета
мездры во время осенней линьки белки (по Котову)
вается потребность животных в минеральном питании и в вита-
минах, повышается белковый обмен, усиливается возбудимость,
падает вес тела.
Большинству зверей холодной и умеренной зон свойствен-
ны две линьки в’году—весенняя, при которой густой высо-
215
кии зимний мех сменяется редким и низким летним, и осенняя,
когда происходит обратный процесс.
Перед началом весенней линьки мех тускнеет, волосы утра-
чивают свойственную им эластичность, ость ломается, пуховые
часто свойлачиваются. Далее начинается развитие новых и вы-
падение старых волос (рис. 84). Весенняя линька может быть
более или менее неполной. У кротов, например, после весенней
Рис. 82. Последовательные стадии линьки белки (Sciurus vulgaris).
А .— весенней; Б — осенней (по Огневу)
216
линьки часто остаются участки зимнего меха (Депарма, 1951).
Норки в весеннюю линьку теряют пуховой волос, остевой же
выпадает лишь во время осенней (Попов, 1949). Осенняя
линька отличается от весенней большой растянутостью сроков
и полной сменой волос. Весенняя линька обычно начинается
с головы и спины, распространяясь от-
сюда кзади на бока и брюшко; осенняя
линька протекает в обратной последо-
вательности (см. рис. 82).
У впадающих в зимнюю спячку зве-
рей и у тюленей бывает одна линька
в году, протекающая на протяжении
лета (рис. 85). С другой стороны,
у типичных землероев (кротов, слепу-
шонок), волосяной покров которых
быстро изнашивается вследствие по-
стоянного трения в узких ходах нор,
кроме двух обычных линек наблюдает-
ся еще третья — восстановительная,
или компенсационная. Она прослеже-
на у кротов, слепышей и слепушонок
(Вяжлинский, 1928; Депарма, 1951;
Крыльцов, 1959). Компенсационная
линька в основном приурочена к лет-
нему периоду, но частично (у кротов)
наблюдается и зимой (Вяжлинский,
1928; Григорьев, 1956).
Высказанное многими авторами
мнение о том, что у зайца-беляка,гор-
ностая и песца летний мех во время
осенней линьки не выпадает, а сохра-
няется на всю зиму, отрастая и депиг-
ментируясь, новейшими исследования-
ми не подтвердилось. Оказалось, что
зимний наряд целиком состоит из
вновь развившихся волос, имеющих
иные размеры и форму, чем летние.
Густота и соотношение категорий во-
лос летнего и зимнего меха также не
одинаковы. Так, например, у зайца-
русака юго-востока Европейской ча-
5
l!
j
3
Рис. 83. Рисунок линьки на мездре степной
мышовки (Sicista subtilis). Благодаря разной
интенсивности пигментации луковиц новых
волос точно отражены расположение и шири-
на темных и светлых полос на спинке зверь-
ка (ориг. А. Н. Формозова)
217
сти СССР на 1 кв. см летом среднее число остевых волос — 382,
промежуточных—504, пуховых — 8156 при средней длине по-
следних 18,5 мм. Зимой тот же ряд цифр имеет такой вид: 968,
1250 и 18012, средняя длина волос подпуши — 22,2 мм. Всего на
1 кв. см летом приходится волос 9042, а зимой 20240. Густота
шерстного покрова увеличивается более чем в два раза глав-
ным образом за счет резкого возрастания количества пуховых
волос.
Еще более резки сезонные изменения меха у тонкопалого
суслика (Spermophllopsls leptodactylus), обитателя песчаных
пустынь советской Средней Азии, Афганистана и северного
Р ис. 84. Взрослый песец в летнем наряде, кое-где сохранившем немного
зимних волос. Окрести, бухты Тикси (фот. Н. Немнонова)
Ирана. На зиму этот зверек в спячку не залегает и таким обра-
зом активен как летом, когда песок раскаляется до 60—80° С,
так и зимой при достаточно сильных морозах. Его летние во-
лосы более похожи на короткие плоские иголочки, плотно при-
легающие к телу. На спине число остевых и направляющих
волос на 0,25 кв. см — 217, промежуточных и пуховых — 258,
всего — 475 при длине от 1 до 7,5 — 8,5 мм: То же зимою: осте-
вых, направляющих, промежуточных—132, пуховых—1109,
всего—1241. Длина зимних волос от 9,2 мм до 18,1—20,9 мм;
они мягкие, шелковистые. Нежный зимний мех тонкопалого сус-
лика очень отличается от жесткого и грубого летнего. Столь
сильно выраженный сезонный диморфизм меха у этого вида
218
вполне соответствует большой годовой амплитуде температур
песчаной пустыни.
Нередко смена одного сезонного наряда на другой совершен-
но преображает облик животного. Летний мех соболя темный,
короткий, плотно прилегающий к телу. В этом наряде зверек
выглядит поджарым, тощим, большеухим и довольно длинноно-
гим. После осенней линьки в высоком блестящем и густом меху
почти полностью скрываются уши, одетый длинными волосами
хвост становится пышным, а ноги кажутся короче и толще. Зи-
мой соболь — коренастый, крепко сложенный зверек, настолько
непохожий на летнего, что неопытный человек может принять
Рис. 85. А — продолжительность единственной в году линьки и ношения
сменившегося наряда у взрослых гренландских тюленей беломорского стада.
Б — продолжительность двух (весенней и осенней) линек и ношения зимнего
я летнего наряда у белок северо-востока Костромской области (ориг.
А. Н. Формозова)
«го за другой вид. Еще более разительно меняется облик оде-
тых летним и зимним мехом песца, зайца-беляка, белок неко-
торых подвидов, сайгака и ряда других.
Помимо усиления термоизолирующих свойств, волосяной
покров'у многих видов при осенней линьке приобретает еще
ряд особенностей, необходимых и выгодных именно в условиях
зимы. Например, строение кутикулы остевых и направляющих
Волос зимнего меха росомахи таково, что даже при самых силь-
ных морозах на них не садится иней (Церевитинов, 1961). Это
характерно также для остевых волос хвоста лисицы и песца,
Оба последних вида при отдыхе на снегу свертываются «кала-
чиком» и укрывают голову хвостом (морда покрыта относи-
тельно очень -коротким мехом и, естественно, должна сильнее
страдать от холода).. Если бы иней, образующийся от дыхания,
219
оседал на волосах хвоста, эти животные неизбежно примерзали,
бы головой к хвосту и при пробуждении повреждали бы шер-
стный покров.
Подошвы ног рыси, песца, росомахи, северных рас лисиц,
куниц, белок и некоторых других видов к концу осени густо
зарастают довольно длинными упругими волосами, почти пол-
ностью скрывающими оголенные летом участки. Образовав-
шиеся густые щетки волос не только «утепляют», но и защища-
ют пальцы и ступни от возможных повреждений при раскапы-
вании обындевевшего снега, плотного наста и т.п. Вместе с тем,
эти щетки увеличивают опорную поверхность лап, создавая по-
добие лыж, или «снегоступов» ', что облегчает животным пере-
движение по рыхлому глубокому снегу. Особенно существенно
значение такого густого опушения лап в жизни росомахи, со-
боля, лесной куницы, суточные переходы которых зимой, в пе-
риод многоснежья, бывают очень большими. Волосы щеток вы-
линивают в период сильного снеготаяния весной, как только
становятся ненужными. Показательно, что у подвидов лисиц,
населяющих степи и пустыни с морозной, но малоснежной зимой
эти щетки отсутствуют; мало опушаются на зиму и ступни лап
- южных подвидов русаков, а также зайца-толая. Наоборот, у
русаков, занимающих северную часть ареала, ступни на зиму
обрастают щеткой, почти столь же густой и длинной, как у зай-
ца-беляка, лучше других наших зайцев приспособленного к жиз-
ни в многоснежных областях.
У обыкновенной белки при смене летнего меха на зимний
отрастают довольно длинные и густые волосяные кисточки,
укрывающие дистальный, наиболее зябнущий край уха. Они
достигают полного роста к моменту окончания осенней линьки,
и охотники в первые дни промысла нередко по длине кисточек
определяют, стоит или не стоит стрелять ту или иную белку,
спрятавшуюся на верхушке дерева. Волосы кисточек весной вы-
падают довольно быстро, но отдельные уцелевшие исчезают
только в июне — июле. В летнем наряде уши взрослой белки
покрыты очень короткой шерстью. Крайне медленно сменяются
волосы хвоста; Он у белки выполняет ряд функций и, в частно-
сти, при больших прыжках с дерева на дерево поддерживает
зверька в воздухе, облегчая планирование. Эту роль он играет
в течение всего года, независимо от сезона. Бурная весенняя
линька меха белки, начавшись с головы, дойдя в начале мая
до основания хвоста, резко замедляется. У взрослого зверька,
получившего летний наряд, потертые и выгоревшие зимние во-
лосы хвоста полностью выпадают и сменяются новыми, тоже
зимними, только к сентябрю. Благодаря постепенной линьке в
любые месяцы года хвост, одетый длинными волосами, спосо-
1 Применяются местами вместо лыж в горных областях.
220
бен выполнять роль парашюта; он линяет один раз в году,
тогда как голова, тело, ноги — дважды. Функции волосяного по-
крова разных частей тела не равнозначны, в связи с этим и
линька проходит не по одной схеме, а по нескольким.
Приспособительное значение сезонных изменений окраски,
возникающих при смене шерстного покрова, особенно прояв-
ляется у видов, полностью белеющих на зиму. Средняя продол-
жительность ношения зимнего белого меха, хорошо гармони-
рующего с фоном укрытой снегом земли, довольно точно соот-
ветствует средней продолжительности постоянного снежного
покрова в том или ином районе. Так, например, горностай в
южной полосе Европейской части СССР около 6,5 месяцев в
году носит рыжевато-бурый летний мех под цвет почвы и около
5,5 — белый зимний; в северной полосе — только 4 месяца лет-
ний и около 8 месяцев — зимний. На Кольском полуострове близ
Полярного круга зайца-беляка можно видеть в белом меху при-
мерно с 20 октября по 20 мая; здесь устойчивый снежный по-
кров в лесу лежит в среднем с 31 октября по 21 мая (с 4.Х по
31.Х бывают частые снегопады, но покров неустойчив — време-
нами сходит, снова появляется и т. д.). Сроки весенней линьки
нашего беляка примерно совпадают с периодом интенсивного
снеготаяния и схода снега, а осенней — с «предзимьем» — вре-
менем холодных дождей, сменяющихся все более частыми сне-
гопадами (О. Семенов-Тян-Шанский, 1937).
Близкий к нашему беляку гренландский заяц (Lepus arcticus
groenlandicus) большую часть года носит зимний белый мех,
да и летний у него не бурый, а почти белый, лишь слегка дым-
чатый на спине. С другой стороны, географические расы беля-
ков, проникшие в Северной Америке по горным хребтам к югу,
в малоснежные районы США, не белеют на зиму. Из европей-
ских форм шотландский беляк (L. timidus scoticus) летом бу-
ровато-серый, зимой чисто белый, но с невысоким и не пышным
мехом, а ирландский подвид (L. t. hibernlcus) осенью становит-
ся заметно серее; лишь единичные особи делаются белыми.
В районах с короткой или малоснежной зимой как теплой
(юг Западной Европы, юг Украины, Закавказье, многие районы
Средней Азии), так и морозной (Монголия) зимний мех ласки
•более густой, чем летний, но бурого или рыжевато-серого, а не
белого цвета. Та же закономерность прослеживается и в се-
зонном диморфизме горностая на всем протяжении Голарк-
тики.1.
1 В литературе встречаются указания, что белые, т. е. бесцветные волосы
и перья якобы отличаются от пигментированных меньшей теплопроводностью,
в связи с чем белую зимнюю окраску нужно рассматривать как адаптацию,
возникшую под влиянием холода, а не снежного покрова. Из эксперимен-
тальных исследований и данных экологии следует, что взгляд этот совер-
шенно ошибочен.
221
Хотя ход линьки тесно связан с внешними условиями, все
же такой сложный процесс не может всегда и очень точно сле-
довать за всеми «капризами погоды». Действительно, бывают
годы, когда снежный покров устанавливается позднее обычного
и белый зимний наряд ласки, горностая, зайца-беляка оказы-
вается очень заметным на темном фоне земли, покрытой отмер-
шей травой и опавшими листьями. Беляки в такое время ищут’
более надежные укрытия для дневного отдыха: ложатся под
защитой нижних ветвей елок, под вершинами упавших на землю-
деревьев или в болоте на кочках, поросших густой осокой.
Ласка большую часть времени проводит в норах полевок, кро-
тов и появляется на поверхности земли сравнительно редко и
на короткий срок.
Иногда при ранней весне и ускоренном снеготаянии перечис-
ленные зверьки «опаздывают» сменить зимний наряд на летний
и недели две, а случается и больше, живут при невыгодном для
них отсутствии маскирующей окраски меха. Заяц-беляк, как
более заметный и имеющий много врагов, сильнее, чем ласка
и горностай, реагирует на такое стечение обстоятельств. Он вы-
ходит кормиться только- в темноте, днем нередко укрывается
на последних сугробах снега, где заметить его очень трудно-.
Конечно, в такие годы популяции зверьков некоторое время
несут большие, чем обычно, потери от нападений хищников.
Однако в среднем за большой ряд лет значение преимуществ
в борьбе за существование, которые дает сезонная смена крип-
тических окрасок обладающим . ими видам, не вызывает со-
мнений.
Как правило у видов, обитающих в условиях резких смен
холодного и теплого сезонов (большинство зверей умеренного
и холодного поясов), процесс линьки протекает бурно, в опре-
деленные сжатые сроки. Но у зверей, не испытывающих резкой
смены сезонных условий (обитателей тропических стран, полу-
водных форм), сезонных отличий волосяного покрова нет или
они незначительны, линька протекает незаметно, часто в виде
растянутого на протяжении всего года выпадания старых и по-
явления новых волос.
Ондатра — один из видов, хорошо приспособленных к полу-
водной жизни. Отсутствие достаточно существенных сезонных от-
личий меха обусловлено основной особенностью ее биологии —
очень частым и длительным пребыванием в воде при поисках
корма, постройке хаток, расселении, преследовании конкурентов-
и т. п. Температура воды во все сезоны значительно ниже тем-
пературы тела ондатры и ослабление защитной роли волося-
ного покрова могло бы вызвать неблагоприятные для зверька
последствия. Соотношение числа волос разных категорий (на-
правляющих, остевых, промежуточных и пуховых), приходя-
щихся на единицу площади кожи, почти одинаково в течение?
222
года и не зависит от сезонов. Линька взрослых особей длится
почти круглый год. Только в течение короткого отрезка времени
(в апреле или мае у ондатр северной половины Европейской
части СССР), приходящегося на конец зимы, шкурки не имеют
следов линьки. Но уже в мае мездра начинает утолщаться, а
затем на ней появляется синева — просвечивают скопления пиг-
мента в луковичках, закладывающих новый волос. Растянуто-
стью, замедленным течением линьки обусловливается хорошее
состояние меха ондатры во все месяцы года. Только на спин-
ной стороне тела, которая реже соприкасается с водой, густота
меха несколько изменяется по сезонам: в июле она примерно
в два раза меньше, чем в конце зимы. С августа густота меха
снова увеличивается. Молодые ондатры ранних выводков в те-
чение осенне-летнего периода имеют две возрастные линьки,
а зверьки из запоздалых выводков — одну, которая к тому же
проходит у них более быстро.
Замедленная, растянутая линька характерна и для других
полуводных млекопитающих нашей страны: калана, речной выд-
ры, выхухоли и в меньшей степени — европейской норки.
Замедленно и постепенно сменяются производные кожи, но-
сящие ярко выраженные оборонительные функции. Например,
иглы дикобразов и ежей выпадают всего по нескольку штук
за сутки. У ушастого ежа в день выпадает 5—20 иголок, благо-
даря чему животное все время сохраняет свой колючий панцирь
пригодным для обороны (А. Н. Формозов). По одному выпадают
и сменяются осязательные волосы (вибриссы), жесткие щетин-
ки оторочек на лапах полуводных зверей и т. п.
Бурная линька с выпадением волос пучками и с одновремен-
ным сбрасыванием эпидермиса большими лоскутами характерна
для северных тюленей родов Phoca, Erlgnathiis, PagophoccK
Эти ластоногие в период линьки лежат на льду или берегу и не
кормятся в течение длительного времени. Из наземных зверей
такая же интенсивная линька наблюдается у забайкальского
сурка-тарбагана (Marmota sibtrica) и боялычной сони (Selevlnia
betpakdalensls) (Дубинин, 1945; Зверев, 1940).
Линька свойственна почти всем роговым образованиям мле-
копитающих: периодически происходит смена когтей, слущива-
ние ороговевших клеток поверхностного слоя эпидермиса, еже-
годное сбрасывание рогов у большинства оленьих и т. п.
(рис. 86, 87).
Кроме сезонных смен волосяного покрова, существует еще
возрастная линька, при которой ювенильный наряд (наряды)
сменяется дефинитивным взрослым. У некоторых видов по-
следний появляется после нескольких возрастных линек (у кро-
ликов, например, их может быть до четырех). Возрастные
линьки у ряда настоящих тюленей связаны со сменой ювениль-
ного (утробного) наряда белька (белый высокий мех с осте-
223
еыми и густыми пуховыми волосами, непригодный для ныря-
ния; у детенышей бывает около 20 дней) на наряд серки' из
грубых коротких волос; серка уже ловит пищу в море. При
последующих ежегодных линьках, одновременно являющихся
и сезонными и возрастными, окраска животного через 2—3 года
приближается к свойственной половозрелым особям.
У грызунов, приносящих за год несколько выводков, моло-
дые при первой ювенильной линьке получают различные наряды
в зависимости от сезона. Например, молодые белки, родившиеся
летом, получают летний взрослый наряд, а появившиеся в конце
Рис. 86. Передняя лапка копытного лемминга. Когти III и IV пальцев имеют
зимой большие размеры и вильчатую форму, так как разрастается не только
сам коготь, но и ороговевающая подушечка пальцев. Весной большая часть
вильчатого когтя отпадает — он приобретает обычные размеры и острый
конец (рисунок по экземпляру с Ямала, добытому 5 апреля 1939 г.)
зимы, еще не достигнув полного роста — пышный зимний мех
и густые кисточки на ушах. Молодые копытные-лемминги, ро-
дившиеся в подснежных гнездах, при первой же линьке полу-
чают густой белый наряд, похожий на зимний взрослых лем-
мингов.
Так как сроки линьки изменяются в зависимости от пола и
возраста1, а также и физиологического состояния животных1 2,
кормовых и погодных условий, бывает довольно трудно точно
охарактеризовать состояние мехового покрова той или иной
1 У кротов, например, самцы линяют значительно позже самок, у нето-
пырей-карликов, наоборот, начинают линьку самцы (Депарма, 1951; Кузя-
кин, 1950).
2 Упитанные животные линяют раньше истощенных. У беременных самок
и больных особей лииька длительное время задерживается на какой-либо
стадии; заметное влияние на ход. линьки оказывает и сильная зараженность
гельминтами.
224
популяции зверей и в частности, установить надежно обосно-
ванные сроки начала охоты в данном году. Прежде неодно-
кратно сталкивались с фактом низкого качества миллионов
беличьих шкурок, заготовленных в годы, когда при созревании
свежего урожая корма белки приносили обильные сентябрь-
ские выводки. Молодые зверьки таких поздних выводков, со-
Рис. 87. Схема смены рогов у оленей:
/ — начальная”стадия (одновершинный рог, покрытый кожей); 2 — рог обнажен,
в основании его образуется полость (результат разрушения костного вещества); 3 — 4 —
рог сброшен, кожа затягивает вершину оставшегося пенька; 5 — двувершинный рог,
покрытый-кожей; 6 — обнажившийся рог. Е — эпидермис; С — собственно кожа;
О — кость (по Нитше)
ставляя значительную часть популяции, не имели еще полного
зимнего меха к традиционной дате начала промысловой охоты
(14 октября). Их шкурки принимали 2-м и 3-м сортом, в связи
с чем и охотники, и хозяйство несли серьезные материальные
потери. В настоящее время, чтобы определить целесообразный
срок начала охоты, введены в практику пробные отстрелы. Их
осуществляют специально выделенные лица или бригады.
Убежища. Участки обитания
УБЕЖИЩА РАЗНОГО ТИПА
Млекопитающих можно разделить на биологические группы
ПО ИХ ОТНОШЕНИЮ К у !И!! щ , Г]пц ^ТОМ ВЗЖПО V’JT*ТЫВ3ть двя
независимых ряда признаков: характер и постоянство связи
особей данного вида с убежищем и тип используемых убежищ.
По характеру и постоянству связи с убежищем выделяются три
группы видов: 1) не нуждающиеся в убежищах в течение всей
жизни — китообразные, многие ластоногие, сирены, копытные,
отчасти зайцеобразные; 2) использующие убежища лишь в -оп-
ределенные биологические периоды, например во время рожде-
ния и выкармливания молодых, для спячки и т. п.— крупные и
среднего размера хищники, кабан, ежи и др.; 3) тесно связан-
ные с убежищами и не способные существовать без них на про-
тяжении всей жизни. К третьей группе относится подавляющее
большинство наземных и полуводных мелких и среднего раз-
мера видов — многие сумчатые, неполнозубые, насекомоядные,
рукокрылые, мелкие и среднего размера хищники, почти все
грызуны, часть видов зайцеобразных и др. Эти животные поль-
зуются убежищами разного типа при отдыхе, для защиты от
неблагоприятных воздействий среды, резкой инсоляции, очень
низких и высоких температур, сильного ветра, осадков, напа-
дения хищников и для выращивания потомства в относительно
устойчивых благоприятных и безопасных условиях (Кучерук,
1960).
В качестве убежищ млекопитающие используют естествен-
ные пустоты, норы и наземные гнезда. К первым относятся
пещеры, трещины в скалах и почве, ходы между камнями, на-
висы подмытого берега, трещины в коре и древесине, дупла,
пустоты в сооружениях человека. Например, полости в снегу
под береговыми нависами и стволами упавших деревьев, пусто-
ты в каменистых россыпях и в разветвленной корневой системе
широко используются многими грызунами и мелкими хищни-
ками. В дуплах деревьев укрываются и выводят потомство бел-
ки, сони, некоторые мышевидные грызуны, летучие мыши, ку-
ницы, черные медведи (последние в них же проводят зимний
сон). Пещеры служат жилищем для крупных кошек, гиен и не-
которых других хищников, а также летучих мышей, которые
занимают их как летом, так и зимой (в период спячки).
226
Некоторые виды рукокрылых, мышевидных грызунов и зем-
лероек тесно связаны в своем обитании с постройками человека
(синантропные виды). Мышевидные грызуны используют также
в качестве убажищ и мест размножения стога и копны. В стогах
часто укрываются енотовидные собаки, реже барсуки, медведи,
кабаны. В старых, брошенных стогах иногда мечут детенышей
волчицы.
Из млекопитающих, использующих убежища лишь в опре-
деленные биологические периоды, можно выделить группу «ло-
говиков», или «логовников», являющуюся как бы переходной
к следующей категории — «норников». Детеныши логовиков
рождаются хорошо развитыми, способными иногда в первые
же дни следовать за взрослыми. Поэтому убежища логовинам
нужны в большинстве случаев на короткий срок и, естественно,
они не отличаются сложностью. Простейшее логово представ-
ляет собой небольшое естественное или вырытое животным
(в грунте или в снегу) углубление под кустом, деревом, скалой,
в густом травостое и пр. Такое логово («лежка») служит для
отдыха, сна и реже для укрытия детенышей, например, у зай-
цев (рис. 88). Некоторые звери «благоустраивают» свои логова.
Кабаны, например, нагромождают в избранном под логово
месте большую кучу сухой травы, мха, веток и листьев, кото-
рые служат им как постелью, так и крышей. Самка с новорож-
денными поросятами занимает отдельное логово. Зимой иногда
образуются большие групповые логова, в которых собирается
целое стадо из 10—15 голов.
Ежи прикрывают вырытое в земле углубление сухой лист-
вой (получается своего рода шалаш). В южных областях такие
логова служат и зимним убежищем. Погребенные под снегом,
они превращаются в укрытия, где зверьки проводят спячку (на
севере ежи зимуют в норах).
Сходно с описанным происхождение медвежьей берлоги.
Первоначально — это логово (иногда с постелью из мха, сухих
листьев), устроенное где-либо в буреломе, под корневым выво-
ротом и пр. Занесенное снегом вместе с лежащим в нем зверем,
логово превращается в берлогу, т. е. большую камеру, в кото-
рой медведь проводит зимний сон и где у самки появляется
потомство. Белый медведь устраивает глубокую снежную нору,
отыскивая места, где имеется большая толща снежного покро-
ва, наметенного ветрами (рис. 89). В областях с очень морозной
малоснежной зимой бурые медведи зимуют не в логовах,
а в неглубоких земляных норах (Средняя и Восточная Си-
бирь). Тюлени, связанные в своем распространении с замер-
зающими водоемами, устраивают для новорожденных логова
в сугробах снега возле постоянно поддерживаемых отверстий
во льду — «продушин», или «лазок».
15*
227
В некоторых случаях логовики настолько углубляют свою
лежку, что превращают ее в полунору, а иногда и в настоящую
нору. Так поступают, например, зайцы в местах, где их посто-
янно тревожат пернатые хищники или требуется укрытие от
сильного мороза (снежные норы) и реже от палящих лучей
солнца (летние норы в сыром песке).
Норы звери выкапывают или выгрызают в почве, древесине,
снегу, слежавшейся соломе или другом плотном субстрате.
Рис. 88. Логова зайца-русака (Lepus europaeus)
в разрезе:
1 — логово на твердом грунте нли на плотном снегу;
2 — летнее логово в песчаных дюнах с шелюгой (пески
Подонья); 3 — снежная нора (Присивашская степь) (ориг.
И. И. Барабаш-Никифорова)
Норы — наиболее совершенный тип убежищ млекопитающих.
Они служат надежным укрытием как от неблагоприятных абио-
тических воздействий, так и от хищников (кроме тех немногих
видов, которые приспособились к преследованию добычи в уз-
ких ходах). Благоприятный режим тепла и отчасти влажности
воздуха поддерживается с помощью теплоизолирующих свойств
гнезда, прогревания его телом зверька и благодаря расположе-
нию гнездовой камеры на той глубине, где в данный период
температура почвы наиболее соответствует потребностям вида.
228
По назначению и особенностям строения норы млекопи-
тающих могут быть подразделены на две основные категории;
временные, или защитные (кормовые) и постоянные (выводко-
вые, гнездовые).
Временные обеспечивают укрытие животным, застйгнутым
опасностью или непогодой вдали от основного жилища (чаще
всего на местах кормежки); они примитивны и, как правило,
лишены гнезда. При переходе молодняка к самостоятельной
жизни (в период расселения) такие норы часто занимают мо-
лодые животные и могут переделывать их в постоянные.
Рис. 89. Берлога самки белого медведя; слева — вертикальный раз-
рез, справа — в плане (по Педерсену)
Некоторые млекопитающие, например бобры, сурки, хомяки
и некоторые др., на протяжении всего теплого периода года
живут в простых летних норах, расположенных в наиболее
кормных участках, а на зиму переходят в зимовочные (рис. 90).
Наконец, звери, обитающие в речных поймах, .устраивают на
время половодья простые «норы переживания» на незатопляе-
мых возвышенных участках берега.
Постоянные норы служат для родов и выкармливания дете-
нышей, имеют гнездовую камеру и отличаются более или менее
сложным строением (рис. 91). Кроме гнездовой камеры и основ-
ного хода, в сложной постоянной норе имеются боковые ответ-
вления ходов и различные отнорки, которые открываются на
поверхность или заканчиваются слепо (рис. 92). Сложная си-
стема ходов позволяет норнику успешно маневрировать в слу--
229
чае нападения врага. Слепые отнорки (тупики) могут служить
кладовыми или использоваться в качестве уборных.
Многие виды очень чувствительны к изменениям температу--
ры, влажности воздуха в норах и т. п. (рис. 93) '. Повреждения
ходов быстро исправляются хозяином норы (к этому его побуж-
дает и инстинкт самосохранения). Залегающие в спячку нор-
ники запивают входное отверстие норы плотной земляной проб-
кой, что также способствует устойчивости температуры и влаж-
ности в подземном жилище и увеличивает общие защитные
свойства норы'. Некоторые виды закрывают отверстия нор и на
время обычного отдыха (тушканчики).
Рис. 90. Зимняя нора хомяка (Cricetus cricetus):
t — камера для спячки; 2 — кладовые с запасами. Заштрихована
часть ходов, забитая землей (по Эйзентрауту)
С другой стороны, во многих случаях норы имеют так на-
зываемые вентиляционные отнорки — узкие ходы, открываю-
щиеся на поверхность небольшими отверстиями — отдушинами.
Благодаря им устраняется чрезмерное накопление углекислоты
в норе, снижается влажность, регулируется температура (зимой
вентиляционные ходы прокладываются и через толщу снега) 1 2.
В полунорах и норах часто выводят потомство волки (осо-
бенно в степных районах, где мало естественных убежищ).
В условиях морозных, но малоснежных областей роют себе
зимние полуноры и норы бурые медведи.
У полуводных млекопитающих, как правило, вход в нору
расположен под водой; у наземных он помещается в корнях де-
ревьев, кустов, под плотными дерновинами злаков или камня-
ми. Этим достигается незаметность норы и меньшая доступность
ее для большинства хищников. Однако во многих случаях, от-
верстия нор не только не маскируются, но, напротив, привлёка-
ют к себе внимание кучами выброшенной на поверхность земли,
1 Нормально микроклимат нор отличается большой устойчивостью.
2 В легких почвах и при наличии естественных «воздухопроводов» (в виде
различных скважин или прикорневых полостей) отдушины в норах отсут-
ствуют.
230
наружными уборными, тропами и т. п. В этих случаях у обита-
телей нор имеются глубокие ходы или сложный подземный ла-
биринт.
Одна и та же нора может служить зверю на протяжении
ряда лет. Такая нора из года в год «разрабатывается», допол-
няется новыми ходами и отнорками и превращается в агрегат
нор — «городок». Иногда агрегат занимают несколько семей,
живущих рядом, т. е. образуется настоящая колония.
Рис. 91. Схематический план части поселения
больших песчанок (Rhombotnys opimus). Заштри-
хованы камеры, заполненные сеном (по Камбу-
лину)
Выедание кормов вблизи норы, сильное загрязнение гнездо-
вых камер или размножение в них паразитов заставляют обла-
дателей норы переселяться в новые убежища Одну нору мо-
гут последовательно занимать разные особи или семьи того же
вида, а иногда и других видов.
Самцы и самки многих норников (например, хомяков, сус-
ликов и др.) живут в обособленных норах; иногда самец сам не
роет нору, пользуясь жилищем, приготовленным самкой. Моло-
дые животные имеют менее сложные норы, чем старые. Выбор
места для норения, характер и плотность размещения, глубина
1 Иногда гнездовую, камеру, сильно заселенную блохами, зверьки пол-
ностью изолируют, забивая ее ход плотной земляной пробкой (монгольские
тарбаганы в Забайкалье), а взамен рядом устраивают новую.
231
и сложность нор зависят от ландшафтных, кормовых, почвен-
ных и некоторых других условий. Обычно звери предпочитают
устраивать норы на возвышенных местах с хорошим обзором.
На ровных степных участках при одинаковых кормовых усло-
виях норы грызунов (например, сурков) располагаются равно-*
мерно (диффузный тип распределения); там, где имеются воз-
вышенные гряды, сухие русла и балки, норы обычно вытягива-
ются вдоль них (ленточный тип); в горах норы локализуются
по отдельным точкам (очаговый тип; Ралль, 1945).
Рис. 92. Норы полуденных песчанок (Merion.es meridianus):
. 1 — летняя; 2 — зимняя (По Раллю)
В распределении зимовочных нор большое значение имеет
снежный покров: как правило, они располагаются в местах с
высоким устойчивым снежным покровом.
Зависимость размещения норников от почвенных (эдафиче-
ских) условий и связанных с ними особенностей растительности
может быть наглядно показана на примере грызунов. Так, чер-
ноземных в основном почв придерживаются крапчатый суслик,
обыкновенный слепыш1; глинистых — малый суслик, большин-
ство пятипалых тушканчиков, песчаных — тонкопалый суслик,
трехпалые тушканчики (особенно мохноногий и гребнепалый),,
полуденная песчанка, а из насекомоядных — пегий пудорак; ка-
менистых грунтов и россыпей — горные пищухи, горные и снеж-
ные полевки — они поселяются в трещинах скал или в ходах,
образующихся между камнями.
1 Местами близ границ ареала они выходят и на песчаные массивы, на-
пример на междуречье Волги — Дона и т. п.
232
Для норников в горных условиях решающее значение имеют
мощность и распределение пласта мелкозема, одевающего на
склонах и в долинах твердые материнские породы. Только в
слое мелкозема можно выкопать нору, но и то не везде. Сурки,
например, занимают лишь те участки, где толщина этого слоя
достигает 1,5 м и более, так как норы меньшей глубины не обес-
печивают успешной зимовки.
По сравнению с тяжелыми и жесткими, в легких почвах
условия для устройства нор, на первый взгляд, более благопри-
ятны. Однако они привлекательны далеко не для всех норников.
Рис.93. Суточные изменения температуры в гнездовой камере норы полуденной-
песчанки, на поверхности почвы и в воздухе по наблюдениям в июле и январе
(Волго-Уральские пески, Зап. Казахстан):
1 — максимальная температура воздуха на поверхности почвы; 2 — температура в
гнезде; 3 — температура воздуха; 4 — температура почвы. Суточная амплитуда темпе-
ратур июля совершенно не отражается в норе зверька — температура здесь постоян-
на. Зимой в норе значительно теплее, чем в воздухе и даже в окружающей гнездо-
почве, так как зверьки его согревают (по Раллю).
В зоне пустынь слепушонка избегает селиться не только в лег-
ко подвижных, но и в закрепленных песках, так как здесь по-
толок ее нор, расположенный у поверхности, постоянно обруши-
вается, и ходы заполняются песком. Зато на такырах, где ходы
нужно прогрызать, затрачивая много усилий, слепушонки рас-
пространены широко и местами даже многочисленны (почва та-
кыров в сухие месяцы настолько тверда, что тяжелая грузовая^
машина проходит, совершенно не оставляя на них следов). Трех-
палые тушканчики и тонкопалый суслик —- типичные псаммо-
филы, устраивают норы и в подвижных, и в слабо закреплен-
ных песках; ходы 'Идут на значительную глубину и большая
233;
часть галерей расположена во влажном, не осыпающемся слое
песка. Тушканчики часто выкапывают временные защитные
норы, в которых проводят день, и вскоре их бросают, чтобы сде-
лать новые. В какой-то степени это связано с легкостью прокла-
дывания ходов, однако и на плотных солонцах, где копать труд-
нее, другие виды тушканчиков также часто сменяют свои защит-
ные норки.
Для зверей нередко имеет большее значение не легкость или
трудность прокладывания ходов, а те особенности почвы, кото-
рые обеспечивают условия, соответствующие требованиям ее хо-
зяина к hoop, как жилью. Малый суслик ппеппочитает селиться
на солонцах, так как здесь дождевая и талая снежная вода впи-
тывается только у самой поверхности, а на уровне гнездовой
камеры, где зверек проводит в спячке 6—8 месяцев, грунт всег-
да остается сухим.
В оврагах барсуки и лисы очень охотно роют норы на скло-
нах, где выходит супесчаный слой, выше и ниже которого ле-
жит слой глины; верхний из них не пропускает атмосферных
осадков, нижний — защищает гнездовую камеру от затопления
грунтовыми водами, а в супеси легко копать. Кроме того ходы,
расположенные в супесчаном слое, теплее и суше, чем окружен-
ные глиной. Такие условия — удобные для постройки постоян-
ных нор, встречаются не часто, поэтому однажды возникший го-
родок служит многим поколениям животных и даже разруше-
ние части ходов браконьерами не может отпугнуть зверей.
В охотничьих хозяйствах такие места, полностью удовлетворяю-
щие всем требованиям промысловых животных к условиям но-
рения, должны находиться на особом учете и бережно охра-
няться.
Уровень воды в почвенно-грунтовом слое имеет большое зна-
чение и для других норников. Например, на заболоченных участ-
ках низин с близким к поверхности почвы уровнем воды серым
и водяным полевкам, экономкам и др. приходится рыть норы в
высоких кочках — там, где они имеются, в противном случае —
делать шаровидные гнезда в траве. На возвышенных участках
они роют довольно сложные норы с гнездами, расположенными
на глубине 20—80 см.
Очень своеобразны условия норения в районах вечной мерз-
лоты. Глубина сезонного оттаивания зависит от состава почво-
грунтов, экспозиции склонов, характера растительного покрова
и т. п. Так, например, сплошной мохово-лишайнйковый ковер
мешает летнему прогреванию почвы, и оттаивание в таких усло-
виях происходит на незначительную глубину. Слабо заросшие
песчаные или мелкощебнистые холмы, склоны долин, обращен-
ные на юг или юго-запад, хорошо прогреваются и глубоко от-
таивают. Именно в таких местах чаще всего устраивают свои
норы песцы, лисицы, волки, живущие в тундре и лесотундре.
234
Наличие системы ходов вызывает промерзание грунта зимой,
но ускоряет его оттаивание с наступлением теплых дней.
Грызуны, впадающие в спячку (длиннохвостый суслик, чер-
ношапочный сурок), вынуждены все же зимовать в камерах, ок-
руженных мерзлотой. Зимовку им облегчает очень пышная под-
стилка из сухой травы, в которую зверьки полностью закапы-
ваются; сурки при этом, как и в других областях, собираются
семьями, это тоже помогает сохранять в камере положительную
температуру.
Мелкие зверьки (мышевидные грызуны, землеройки) обычно
устраивают гнездовые камеры в поверхностных слоях почвы
(на глубине 10—80 см). Норы относительно крупных млекопи-
тающих (сусликов, сурков, дикобразов, барсуков) расположены
в нижних горизонтах почвы и в подпочвенном слое (на глубине
до 2—5 м). Глубина расположения гнездовой камеры даже у
животных одного вида колеблется в зависимости от условий се-
зона и географического положения местности. Так, например,
домовые мыши Нижнего Поволжья или Предкавказья, остав-
шиеся зимовать в степи *, по мере понижения ночных темпера-
тур углубляют свою нору и переносят гнездо в более глубокие
слои. Если в октябре оно находится у большинства зверьков на
расстоянии 20—30 см от поверхности почвы, то к январю ока-
зывается уже на глубине 60—80 см. Зимовочные гнезда полуден-
ных песчанок в Волго-Уральских песках располагаются на глу-
бине 1,2—1,5 м. Весной, когда поверхностный слой песка уже
прогреется, в глубине еще держится низкая температура, так
как зимние условия в толще почвы наступают и кончаются с
большим опозданием. В этот период песчанки бросают свои
зимние гнезда и устраивают новые — летние, на значительно
меньшей глубине, где господствуют умеренные температуры
(см. рис. 92).
Географическая изменчивость строения нор, а также гнезд
млекопитающих, еще недостаточно изучена, но, по имеющимся
данным, свойственна большинству широко распространенных
видов. Желтогорлые мыши средней полосы Зап. Европы нередко
зимуют в дуплах и в них же устраивают запасы корма на зиму.
На той же'широте в Среднем Поволжье и Предуралье — у во-
сточной границы своего ареала, эти мыши зимуют только в глу-
боких норах и кладовые устраивают под землей. Здесь, в усло-
виях континентального климата зимние морозы настолько силь-
ны и продолжительны, что зимовка в дуплах для этих мышей
была бы гибельной. В процессе естественного отбора выработа-
лись у представителей восточной популяции вида особенности
поведения, более выгодные в местных условиях.
1 Значительная часть домовых мышей при наступлении первых холодных
ночей (в октябре — ноябре) мигрирует из полей и степи и поселяется на
зиму в скирдах соломы, хозяйственных и жилых постройках и т. п.
235
Довольно просты самостоятельно вырытые лисьи норы. У мо-
лодых животных они неглубоки и состоят из одного короткого-
прямого хода, заканчивающегося расширением. Занимаемые
много лет подряд выводковые норы имеют несколько ходов, ту-
пиков и камер. Выброшенная из выводковой норы земля утап-
тывается начинающими выходить на поверхность лисятами. Во-
круг образовавшейся площадки, а частью и на ней, скопляются
остатки пищи и экскременты, издающие специфический запах..
Песцовая нора, вырытая в рыхлом грунте тундры, нередко пред-
ставляет сложный лабиринт, занимающий площадь свыше
150 кв. м. В каменистом грунте норы значительно проще (в го-
рах песцы часто занимают естественные убежища). Кроме по-
стоянной (выводковой) норы, песцы роют многочисленные не-
глубокие временные (защитные) норы.
Большой сложности достигают барсучьи норы — они пред-
ставляют собой нередко целую систему ходов, расположенных в
несколько этажей и уходящих в глубину до 5 м, где помещается
гнездовая камера. Последняя имеет мягкую подстилку из су-
хого, регулярно обновляемого растительного материала. Общая
площадь, занятая «барсучим городком», может достигать мно-
гих тысяч квадратных метров. Выброшенный из подземного ла-
биринта грунт образует на поверхности группу холмов — они
пронизаны траншеями и тропами, идущими от многочисленных
входных отверстий (до двух десятков и более). Постоянно ис-
пользуются немногие лазы, от которых отходят хорошо прото-
ренные тропы, некоторые протяженностью до 2—3 км. В проти-
воположность лисьим и песцовым, барсучьи норы содержатся в
большой чистоте! В них имеются специальные отнорки, служа-
щие уборными, где животные засыпают экскременты землей.
Имеются также вентиляционные отнорки.
Иногда барсуки устраивают себе летние неглубокие норы (с
одним выходом), на местах, богатых кормом, из которых пере-
бираются осенью в постоянные норы, приспособленные для зи-
мовки.
Со сложными барсучьими норами сходны постоянные норы
сурков. Но как дополнение к ним, сурки устраивают себе на ме-
стах кормежки целый пояс защитных нор, которые могут слу-
жить временным укрытием. Уборные у сурков помещаются ле--
том вне норы, на выбросе земли — сурчине, а осенью в специ-
альных отнорках. На время спячки сурки забивают ходы изнут-
ри земляной пробкой и щебнем, цементируя все это каловыми
массами. Длина пробки достигает 1,5—2 м. Подстилка гнезда,
как и у барсуков, регулярно сменяется (при этом сурки либо
выбрасывают старый материал наружу, либо замуровывают всю
гнездовую камеру, устраивая вместо нее новую).
Глубокие норы (обособленные у самцов и самок) роют хомя-
ки. Нора самца несложна; у самки она представляет сильно раз-
236
ветвленную систему ходов, что облегчает бегство матери ю мно-
гочисленными детенышами в случае опасности. Характерны для
нор хомяков вместительные кладовые (до 5 в одной норе), со-
общающиеся с гнездовой камерой. Нарытую землю зверьки вы-
брасывают через наклонный ход, с которого начинают рытье
норы. Кроме него имеется вертикальный ход, отрываемый изнут-
ри и потому не имеющий кучки земли у наружного отверстия.
Интересные особенности имеют норы сусликов и тушканчи-
ков. Начиная строить нору, суслик, подобно хомяку, роет длин-
ный наклонный ход, заканчивающийся гнездовой камерой — из
ное о.сеш-.ю ведет почт!! до поверхности вертикальный ход, а
нарытой при этом землей забивает наклонный. В закрытой та-
ким образом со всех сторон норе суслик проводит зимнюю спяч-
ку. Весной, после пробуждения зверек доводит до поверхности
вертикальный ход, через который и выбирается наружу. Естест-
венно, что кучка нарытой земли (бутан, или сусликовина) отме-
чает лишь отверстие наклонного хода и отсутствует у выхода
вертикального. В последующие годы близ первичной норы появ-
ляются новые наклонные ходы, что приводит к образованию це-
лого агрегата нор (рис. 94). Он возникает благодаря тому, что
образовавшийся холмик — сусликовина привлекает зверьков;
даже с небольшого возвышения лучше скатывается вода, не
проникая в нору. Близ постоянных сусличьих нор тоже имеются
защитные неглубокие норки. В постоянных норах у длиннохво-
стых сусликов (Citellus undulatus) устроены иногда так назы-
ваемые «спасательные камеры» — высокб расположенные рас-
ширенные тупики, в которых грызуны отсиживаются при затоп-
лении ходов талыми водами. Каждую нору занимает только
одно животное (кроме, конечно, периода вывода потомства).
Тушканчики имеют летние (постоянные и временные) и зи-
мовочные норы. Наиболее сложна летняя постоянная нора
большого тушканчика. В начальной стадии ее сооружения зве-
рек копает главный ход и устраивает гнездовую камеру. Нары-
тую землю он сперва выбрасывает наружу, но по мере углубле-
ния норы все более заполняет главный ход и, наконец, полно-
стью закрывает его (в дальнейшем зверек им не пользуется).
От гнездовой камеры к поверхности тушканчик прорывает вто-
рой ход, прикрываемый изнутри (во время пребывания зверька
в норе) короткой земляной пробкой. Кроме того, изнутри же он
выкапывает доходящий почти до поверхности отнорок, служа-
щий запасным выходом. При разрушении пробки хищником
тушканчик выскакивает через запасный выход, легко проламы-
вая тонкий земляной свод;в других случаях он выходит, раз-
брасывая землю пробки. Летние временные, а также зимовоч-
ные норы тушканчиков, значительно проще только что описан-
ной.
237
Сложную картину представляют поселения серых полевок
(Microtus arvalis, М. socialis) и степных пеструшек (Lagurus
lagurus). Нора, принадлежащая отдельной семье, состоит из
гнездовой камеры с 5—6 ходами, ведущими на поверхность, и
нескольких камер, служащих кладовыми или «столовыми». Все
они связаны между собой системой ходов. Как бы продолже-
Рис. 94. Схема последовательных этапов расширения сети нор
малого суслика (Citellus pygmaeus) (по Бируле).
нием последних на поверхности служит густая сеть хорошо очи-
щенных тропинок и траншей, проложенных в траве. Они соеди-
няют основную нору с расположенными вокруг защитными
норами и «кормовыми столиками» (площадками, на которых по-
едается пища). Зимой полевки прокладывают ходы в толще
снега и здесь же устраивают гнезда. По мере роста семьи и об-
разования новых нор на периферии («хуторков», или «высел-
ков»), что связано с уничтожением корма близ материнского
238
гнезда, площадь, занимаемая поселением, увеличивается, дости-
гая до 2—3 тысяч квадратных метров и более (Формозов, 1939,
Н. Наумов, 1948).
Очень сложны также норы больших песчанок.
По сравнению с норами полевок и песчанок поселения мы-
шей отличаются большой простотой. Защитные норы в них от-
сутствуют, так как проворные зверьки в случае опасности успе-
вают скрыться в жилую пору. Ходы мышиных пор разработаны
слабо, поверхностные дорожки отсутствуют.
Очень сложны норы типичных землероев, так как ходы жи-
лой части постройки непосредственно сливаются с очень слож-
ной сетью, кормовых ходов, непрерывно расширяющейся и ус-
ложняющейся. Таковы норы кротов, слепышей, слепушонок, по-
коров, гоферов и ряда других землероев. Их норы не имеют на
поверхности определенных входных отверстий; нарытую землю
зверьки выбрасывают по мере накопления через специально от-
рываемые короткие отиорки, отверстия которых скрыты под куч-
ками земли. Эти кучки в известной мере обозначают располо-
жение кормовых ходов. Над жилой частью норы земляные вы-
бросы обычно имеют больший объем. Когда нужно выйти на
поверхность, землерои прорывают или проламывают земляной
потолок в любой части кормовых ходов.
Как уже указывалось, для нор большинства полуводных
зверей характерно подводное расположение входов (рис. 95).
Норы бобров, ондатр, выхухолей состоят из переплетающейся
системы галерей, из которых нижние (примыкающие к входным
отверстиям) заполнены водой. По мере опускания уровня воды
в водоеме, животные прорывают новые более глубокие ходы,
сохраняя подводное расположение входных отверстий. Таким об-
разом возникает многоярусность ходов, хорошо заметная на кру-
тых береговых склонах. Семейные норы имеют неглубоко рас-
положенные гнездовые камеры и большое количество отнорков,
в том числе и вентиляционных. Кроме построек, в которых вы-
ращиваются детеныши, имеются также особые летние (у бобров)
и защитные норы на местах кормежки.
Передвигаясь по одному и тому же пути из норы в нору, к
местам кормежек и обратно, выхухоли, а частично и другие по-
луводные виды непроизвольно прочищают на дне водоема канав-
ки— траншеи. Бобры прокладывают также наземные дорожки,
связывающие урез берега с местами расположения кормовой
растительности. В заболоченных участках такие дорожки посте-
пенно углубляются, заполняются водой и образуют каналы,
очень облегчающие грызунам транспортировку «сплавом» отрез*
ков ветвей и стволов к жилищу.
На низменных берегах бобры строят так называемые хатки —
конусообразные или куполообразные сооружения из сучьев и дру-
гих древесных материалов, скрепленных илом, доставленным со
239
дна. В центре хатки выше уровня воды находится гнездовая каме-
ра с выходами, открывающимися в воду. Самые крупные хатки
достигают 2,5ж высоты и около 10 ж в диаметре (у основания).
Весной и осенью, а в случае повреждения и в другое время,
Рис. 95. План и разрез нор выхухоли (Desmana moschata).
А — отверстие норы под водой; ГК — гнездовая камера
(по Барабаш-Никифорову)
•бобры производят ремонт хаток. В некоторых случаях неболь-
шую хатку (полухатку), они устраивают над местом поврежде-
ния подземного хода или камеры.
Хатки из стеблей тростника, тонких ветвей делают и ондат-
ры; естественно, их постройки по размерам значительно уступают
бобровым: высота обычно не превышает 1,5 м, а диаметр в осно-
вании— 4 м. Внутри хатки находится гнездовая камера, а в
крупных домиках — и кормовые помещения (рис. 96). Одна семья
ондатр строит целый ряд хаток, в среднем 5—9, иногда и бо-
лее. В отличие от бобровых, хатки ондатр служат главным
образом зимними убежищами; летом многие зверьки живут в
норах. Кроме жилых хаток, ондатры устраивают особые не-
большие кормовые хатки на льду, над постоянно поддерживае-
мыми продушинами (рис. 97). Внутри таких хаток помещаются
кормовые площадки.
Рис. 96. Внешний вид и схематический разрез хатки опдатры_(О/г^а/га
zibethica). Г — гнездо; К. — кормовая камера
Иногда, главным образом в половодье, делают себе времен-
ные укрытия типа хатки из травы водяные полевки.
Резко выраженное непостоянство уровня воды в водоеме
побуждает полуводных зверей менять свои убежища или поки-
дать водоем. Исключение составляют бобры, возводящие в та-
ких случаях плотины, регулирующие уровень воды. Они соору-
жают плотины из тех же материалов, что и хатки'. Длина пло-
тин европейских бобров обычно не превышает 200 м, а высота —
1,5; по наблюдениям Уоррена (Warren, 1927) у американских
бобров плотины бывают свыше 650 м в длину и 4 л в вышину.
В некоторых случаях небольшую речку бобры перегораживают
целым рядом последовательно расположенных плотин, вслед-
ствие чего на таком участке возникает цепь «бобровых прудов».
Благодаря плотинам повышается уровень воды—это обеспе-
чивает подводное расположение отверстий бобровых нор и рас-
ширяет кормовые возможности, делая более доступными удален-
ные от берега участки древесных и кустарниковых порослей.
В случае большого подъема уровня воды после ледостава бобры
прогрызают у верхнего края плотины борозды — водоспуски, и
1 У американских бобров встречаются также плотины, сложенные из
камней.
16 Териология
241
уровень пруда стабилизируется на определенной отметке. При
разрушении плотины бобры незамедлительно восстанавливают
ее, сооружая иногда для уменьшения напора воды вспомогатель-
ные «аварийные» плотины.
Своеобразные строительные работы производят обитающие
в каменистых биотопах Алтая, Монголии и некоторых других
горных областей плоскочерепные полевки (Alttcola strelzovt).
Эти грызуны огораживают естественные пустоты между камня-
ми, где помещаются их гнезда или склады запасов, стеной из.
щебня и разных случайных материалов, скрепляя их собствен-
Рис. 97. Внешний вид и схематический разрез кормовой^хатки^ондатры
на льду (по Околовичу и Корсакову)
ными экскрементами. Такая стена в некоторых случаях дости-
гает длины 10 л и высоты до 1 м. Обладая большой прочно-
стью, она служит хорошей защитой от непогоды и хищников.
Самое гнездо представляет собой небольшое углубление, сделан-
ное в затвердевшем слое помета и выстланное перьями, шерстью
и т. п. На зиму зверьки переселяются в подснежные гнезда (Обо-
ленский, 1948; Шубин, 1-956). Кучки мелких камней для защиты
запасов сена, сложенного в трещинах скал, собирают и гобий-
ские пищухи (Ochotona pricei).
Как общее правило, в гнездовой камере нор млекопитающих
имеется подстилка или гнездо из мягкого, теплого материала.
Некоторые норники устраивают также гнезда на поверхности
земли. К наземным гнездам можно отнести сделанные как на
поверхности почвы (полевки, ежи, орешниковая соня), на пла-
вучих кучах отмерших стеблей растений (водяная полевка, он-
датра— хатки и др.), так и подвешенные к травянистым рас-
242
тениям (мышь-малютка, американский хомячок Peithrodontomys
tnegalotis) или устроенные на деревьях (красная древесная по-
левка Phenacomys longtcaudatus Северной Америки, черная
крыса, сони, многие белки и др.) (рис. 98). Гнезда предохраняю^
животных от неблагоприятного воздействия абиотических факто-
ров, но не могут служить защитой при нападении большинства
хищников. Все обитатели подобных гнезд при тревоге покидают
их, ища спасения в траве, на деревьях и т. п. Обитатели же нор,
как правило, при опасности
отсиживаются в глубине сво-
их убежищ или поспешно
роют в них новые отнорки,
забивая их землей с той сто-
роны, откуда грозит опас-
ность.
Искусные летние шаро-
образные гнезда из расти-
тельного материала подве-
шивают на высоких стеблях
травы мыши-малютки (рис.
99). Более крупные и грубые
шарообразные гнезда уст-
раивают на древесных вет-
вях сони, еще более круп-
ные — белки. Основу бе-
личьего гнезда, или «тайна»,
составляет шарообразный
каркас, сплетенный из мел-
ких ветвей. В нем находится
тоже толстостенное гнездо,
изготовленное из сухой тра-
вы, прядей лишайников и
размочаленного луба.
Внутрь гнезда ведет один,
реже два хода, прикрывае-
мые пучком лишайников или
луба. Даже в зимнее время
Рис. 98. Зимние гнезда сони-полчка
(Glis glis):
1 — в трухе дупла; 2 — под корнями дерева,
14 в земле (по Спангенбергу)
после ухода белки на кормежку в ее гнезде сохраняется тепло
до возвращения зверька на отдых. Особенно массивные, утеплен-
ные внутри перьями, пухом и шерстью, гнезда делают белки в
областях с морозной зимой (рис. 100). Кроме жилых гайн, белки
летом иногда строят так называемые теневые гайна (без мягкой
внутренней выстулки), служащие, по-видимому, для защиты от
солнечных лучей и хищных птиц (Гольцмайер, 1935).
Теплые гнезда используются млекопитающими в качестве по-
стоянного убежища, гнезда легкой постройки — только летом
(на зиму их обладатели переходят в лучше защищенные убежи-
16*
243
ща—норы). Оригинально укрытие, устраиваемое тропической
летучей мышью Artibeus из раскроенного листа пальмы (рис. 101).
Грубые примитивные гнезда в виде помостов из сучьев и над-
ломленных ветвей сооружают на
Рис. 99. Гнездо мыши-малютки (Л4!с-
romys minutus)
деревьях человекообразные
обезьяны (рис. 102) '. Такие
помосты служат для ночле-
га (нередко только на одну
ночь) или для дневки семьи 1 2.
Некоторые млекопитаю-
щие охотно занимают гото-
вые ч-ужие норы и гнезда.
Лисицы поселяются в бар-
сучьих и сурочьих норах,
степные хорьки — в сусличь-
их, сони и лесные полевки
часто занимают птичьи гнез-
да, куницы и летяги — 6ej
личьи гайна и т. п.
В то время как многие
виды зверей занимают лю-
бое более или менее подходя-
щее укрытие, другие — узко
специализированы и поль-
зуются только убежищами
определенного типа. Послед-
нее особенно ясно выражено
у разных видов летучих мы-
шей: подковоносы и длинно-
крылы, например, занимают
только пещеры, вечерницы
почти исключительно посе-
ляются в дуплах с округ-
лым летком, нетопырь На-
тузиуса предпочитает дупла с щелевидным отверстием, поздний
кожан и нетопырь-карлик — человеческие постройки. Самки пру-
1 Горилла строит гнездо и на земле. Оно представляет собой простое
углубление, в лесной подстилке, окруженное небольшим валом из листьев
папоротника. Гнездо, помещающееся на дереве (на высоте 1—1,5 л), имеет
вид платформы из веток, обрамленной кольцом из отломанных или согнутых
веток. В этом очень прочном гнезде горилла, видимо, лежит на спине, подло-
жив руки под голову и свесив ноги через край.
Гнездо шимпанзе проще и более рыхло (в нем нет переплетения мате-
риала); располагается оно на высоте 5—30 м. Самка строит такое гнездо
для себя и детеныша всего примерно за 3 минуты [Болвиг (Bolwig), 1959].
2 Гнездами («беседками») часто называют нагромождения сучьев,
оставляемые в кронах деревьев гималайскими медведями. На самом деле
это. не гнезда, а случайные скопления ветвей, обломанных зверем при добы-
вании плодов (Бромлей, 1956).
2Ф1
довой ночницы тяготеют к строениям, в то время как самцы это-
го вида поселяются в убежищах любого типа. Непритязатель-
ностью к убежищам отличаются ушан, усатая ночница, двух-
цветный и северный кожаны (Кузякин, 1950).
Для каждой природно-географической зоны характерен свой
набор и преобладание определенного типа убежищ.
В степях, полупустынях, прериях, кампос серрадос наблю-
дается резкое преобладание обитателей нор. Огромное количест-
во ходов, подземных галерей и туннелей, гнездовых камер про-
Рис. 100. Схематический разрез зимнего гнезда обыкно-
венной белки (Костромская обл.):
1 — крыша, боковые стенки и основание гнезда из мелких
веток; 2 — собственно гнездо, сделанное из мягкого материала;
<3 — гнездовая камера (ориг. А. Н. Формозова)
низывает здесь почву и даже толщу подпочвы. В норах и у нор
сосредоточена почти вся жизнь аридных областей, так как не
только звери, но и птицы, рептилии, насекомые, пауки, клещи и
др. охотно пользуются норами млекопитающих.
В северных хвойных лесах млекопитающие используют убе-
жища всех типов; из них на первом месте стоят естественные
пустоты как в пнях, так и деревьях, на втором — норы. В тро-
пических лесах относительное количество нор еще меньше, так
как обилие муравьев и термитов делает жизнь мелких и средне-
го размера млекопитающих в земле и на уровне земли совершен-
ие
но невозможной. Большинство гнезд или убежищ в естественных
пустотах они занимают высоко над землей, на уровне крон круп-
ных деревьев. Так, например, в ветвях на высоте 30—50 м над
землей проходит вся жизнь ленивцев и многих обезьян, где они
всегда обеспечены пищей и необходимыми укрытиями. Некото-
рые из обезьян, живущих в кронах высоких деревьев,— храните-
ли возбудителя желтой лихорадки, а их эктопаразиты — комары,
специфические переносчики этой инфекции, живут тоже на уров-
не крон, выплаживаются в воде небольших «чаш», имеющихся
в пучках эпифитов, густо одевающих ветви деревьев влажного
Рис. 101. Убежище летучей мыши Artibeus watsoni (тропиче-
ская Америка), устроенное из соответствующим образом под-
готовленного листа пальмы Geonoma (вид сверху и сбоку)
тропического леса. Неоднократно отмечалось отсутствие заболе-
вания людей желтой лихорадкой в ее очагах, пока не начиналась
рубка леса. Только вылетая с крон поваленных на землю дере-
вьев, комары получали возможность нападать на людей и через
укусы заражали их желтой лихорадкой.
В наших" степях, полупустынях и пустынях сильная инсоля-
ция, резкие скачки температур, крайняя сухость приземного слоя
воздуха очень неблагоприятны для жизни мелких членистоногих
животных на поверхности почвы. Особенно чувствительны к
этим условиям мокрицы, скорпионы, многие жуки, тараканы,
сверчки, некоторые пауки, а из паразитических форм москиты,
блохи, иксодовые и гамазовые клещи и др. Те виды членисто-
ногих, которые не способны устраивать собственные норки, укры-
ваются в норах грызунов и мелких хищников. Микроклимат этих
убежищ вполне удовлетворяет требования к абиотической среде
246
Рис. 102. Орангутан (Simla satyrus) на гнезде
этих членистоногих; многие из них находят в норах и необхо-
димую пищу. Ряд видов эктопаразитов млекопитающих размно-
жается только в их норах и в них же проходит весь жизненный
цикл.
Эволюция представителей такого отряда насекомых как бло-
хи непосредственно связана с их длительным существованием
в норах грызунов (Беклемишев, 1951, 1954). Каждый из зверей-
норников имеет целый набор этих эктопаразитов, в том числе
нередко и свои специфические виды. Паразитологи разделяют
блох на две естественные группы •— блох гнезда и блох шерсти,
> 247
отличающихся рядом экологических особенностей. Примечатель-
но, что на большинстве логовников (копытных, зайцах и некото-
рых других) блохи не паразитируют. Микроклимат логовищ,
мало отличающийся от микроклимата поверхности почвы, слу-
чайность и кратковременность использования их хозяевами дела-
ет логова непригодными для размножения и существования блох.
С очень сложным и богатым миром эктопаразитов, населяю-
щих норы, связано существование обширных природных очагов
болезней домашних животных и человека: чесотки, чумы, туля-
ремии, клещевого возвратного и сыпного тифов, лихорадки Ку,
кожного лейшманиоза и мн. др. Все это — зоонозы, т. е. болезни
диких животных, которыми люди заражаются при посещении
очагов через укусы инфицированных блох, клещей, москитов
или иным путем.
Оздоровление природных очагов таких болезней специальные
санитарно-эпидемиологические отряды осуществляют путем ист-
ребления грызунов носителей инфекций и их эктопаразитов.
Использование отравленных приманок, ядовитых дустов и газов,
которые обычно вводят в норы, бывает успешным только тогда,
когда хорошо известно строение нор, длина ходов, глубина зале-
гания гнездовой камеры и другие особенности жилищ зверьков.
При подготовке истребительных работ проводят предваритель-
ный учет нор, определяют процент жилых и пустующих. Это не-
обходимо для расчета потребных ядохимикатов, специального
снаряжения, рабочей силы и т. д. При планировании отлова и
отстрела многих промысловых зверей, при определении продук-
тивности тех или иных охотничьих угодий тоже приходится учи-
тывать норы и убежища млекопитающих, для чего необходимо
уметь различать их видовую принадлежность. Таким образом из-
учение их имеет не только теоретическое, но и практическое зна-
чение.
УЧАСТКИ ОБИТАНИЯ
Каждое животное, занимающее нору или построенное логово,
использует прилегающий участок для поисков корма, воды и
встреч с себе подобными. Постоянное достаточно длительное об-
следование участка дает возможность выявить все удобные пути
передвижения, укрытые места, где имеется тот или другой корм,
и т. п. Постепенно участок становится освоенным и в какой-то
степени обжитым. С этой обжитостью возникает привязанность
хозяина норы или гнезда не только к последним, но и к приле-
гающему участку.
Размеры таких участков обитания зависят от видовой при-
надлежности, биологических особенностей, возраста, пола живот-
ного и, конечно, от кормовых условий. Богатые кормами участки
имеют обычно меньшую площадь по сравнению с малокормными,
248
участки самцов, как правило, бывают крупнее участков самок и
молодых животных.
Виды, питающиеся грубыми растительными кормами, имею-
щимися в изобилии, занимают меньшие участки, чем потребите-
ли семян, луковиц и т. п., вынужденные при поисках корма об-
следовать значительно большие площади. Естественно, что хищ-
ники, охотящиеся на зверьков только что упомянутых групп,
обследуют еще более обширные участки. Поэтому хищники, как
правило, населяют местность с плотностью в сотни раз меньшей,
чем грызуны или копытные, непосредственно трофически связан-
Рис. 103. Гнездово-кормовые участки рыжих полевок (Clethrionomys
glareolus). Заштрихованы налегающие друг на друга отрезки участ-
ков разных зверьков (по Н. Наумову)
ные с растениями-продуцентами, составляющими основу каждого
биоценоза. Ленивцы, питающиеся преимущественно листьями и
отчасти плодами — крайне малоподвижные животные, способные
в течение многих дней довольствоваться кормами, имеющимися
на ветвях нескольких крупных деревьев с хорошо развитой кро-
ной. Несмотря на их размеры, превышающие размеры многих
видов мелких обезьян, ленивцы имеют очень ограниченные участ-
ки обитания, тогда как обезьяны — потребители плодов и мелких
животных, очень подвижны и обследуют за короткий срок сотни
гектаров леса.
Крайне мал участок обитания кустарниковой полевки (грызу-
на, питающегося корневищами, зеленью и ведущего преимуще-
ственно подземный образ жизни) — он равен нескольким квад-
ратным метрам; у рыжей лесной полевки (рис. 103) величина уча-
стка колеблется от 500 кв. м (самки) до 2 500 кв. м. (самцы);
249>
участок нутрии занимает площадь около 10 000 кв. м (1 га); у
соболя — около 10 кв. км\ у тигра (самки) на севере ареала —
свыше 4000 кв. км.
Наблюдаются изменения величины участка по сезонам, свя-
занные в основном с изменением кормовых условий в разные
годы. У зверей, пользующихся временным жилищем (лисица,
белка и др.), обитаемая площадь подразделяется на гнездовый
участок (на нем находится жилище) и кормовой. В период вы-
вода детенышей такие звери обнаруживают большую привязан-
ность к гнездовому участку, но по мере приобретения молодыми
самостоятельности постепенно первенствующее значение в жиз-
ни семьи приобретает кормовой участок.
Млекопитающие, пользующиеся постоянными жилищами
(барсук, сурок), не меняют занимаемой территории, вследствие
чего их участки не имеют указанного выше подразделения.
У некоторых видов зверей обладатель участка ревниво охра-
няет его от вторжения других особей своего вида. Если это слу-
чится, он энергично преследует пришельца. Борьба на этой почве
может принимать ожесточенный характер. Так, например, схват-
ки бобров с появляющимися на занятом ими участке другими
бобрами (чаще всего расселяющимися молодыми особями) не-
редко кончаются гибелью пришельца от укусов. Вражда из-за
индивидуальных участков иногда имеет место и у соболей. Хо-
рошо выраженные изолированные участки имеются у белок, ко-
торые весьма активно охраняют их, вступая в драки со всеми
пришлыми особями. Наоборот, у мышевидных грызунов участки
обитания могут широко перекрываться.
Для млекопитающих, образующих пары (моногамов), харак-
терна сильно выраженная привязанность к участку обитания; у
полигамов это явление выражено слабее, однако и у них также
можно заметить стремление держаться на определенной терри-
тории, не допуская на нее «чужих» особей (некоторые предста-
вители оленей, ластоногих). Интересно, что у бегемотов ожесто-
ченные драки происходят и из-за имеющих для них большое
значение мест отдыха — отмелей [Ленькова (Lenkowa), 1956].
Остроту конкуренции из-за территории до некоторой степени
предотвращает маркировка занятого участка, выполненная его
«хозяином». Явление это широко распространено среди млекопи-
тающих и осуществляется путем оставления на видных местах
•своих следов; «меток» в виде выделений пахучих желез, экскре-
ментов, затесов или царапин на коре деревьев (рис. 104), частой
подачей голоса при выходе на охоту и т. п. Олени и некоторые
.антилопы «метят» занятую ими территорию обильно выделяе-
мым пахучим секретом предглазничных желез, для чего трутся
мордой о ветки и стволы. Косули, серны, снежные козы в пе-
риод гона бодают кусты, оставляя на них пахучие выделения
зароговой железы. Мускусный пекари прокладывает пахучую
250
трассу, стирая на своем пути о свисающие ветви секрет спиннои
мускусной железы. Медведь также иногда оставляет пахучий
след, поднимаясь на задние лапы у стволов деревьев и потира-
ясь о них мордой и спиной, чаще же он сдирает кору когтями,
т. е. прибегает к визуальной маркировке.
Конкурентные межвидовые и внутривидовые отношения воз-
никают не только из-за жилых участков, но и из-за самих убе-
жищ. С этим связано наличие у многих видов норников спе-
циальных желез, постоянно оставляющих пахучие следы на стен-
Рис. 104. Зубр (Bison bonasus), «маркирующий» дерево
ках обитаемой норы. Наблюдаются, например, иногда драки за
обладание дуплом между белками или из-за нор у сусликов.
Шершни и осы, захватывая дупла, вытесняют из них белок и ле-
тучих мышей. Часто имеет место чередование в пользовании
одним и тем же дуплом; сменяют, например, друг друга лес-
ная соня, мухоловка-пеструшка и осы. Из-за обладания убежи-
щем конкурируют между собой лисица и енотовидная собака,
желтогорлая и лесная мыши и пр.
Нередко захват чужой норы сопровождается агрессивными
действиями. Горностай, приспосабливая для себя нору водяных
полевок, уничтожает хозяев норы. Так же поступает степной хо-
рек, завладевая норой суслика.
Достаточно сильные животные оказывают сопротивление
претендентам на их норы, например, барсук выгоняет из своих
ходов енотовидную собаку и даже уничтожает лисят, найдя вы-
водок в своей летней норе. Но если он уже залег в спячку, ено*
251
товидная собака располагается в одном из заброшенных ходов.
Пробуждаясь раньше барсука, она избегает столкновения с хо-
зяином норы и весной.
С другой стороны, нередкое Явление представляет сожитель-
ство разных видов животных в одной норе. В некоторых случаях
это сожительство может быть безразличным для обеих сторон.
На равных основаниях, например, обитают в одном дупле лету-
чие мыши и сони (первые занимают верхнюю часть дупла, вто-
рые — нижнюю).
Несравненно чаще имеет место использование жилищ с одно-
сторонней пользой (квартирантство). Не только многие беспоз-
воночные, о чем говорилось выше, но и некоторые позвоночные
(жабы, ящерицы), постоянно живут или временно укрываются в
норах зверей, привлекаемые благоприятными микроклиматиче-
скими и защитными условиями. С хозяевами нор они почти не
соприкасаются.
В Южной Америке наблюдается совместное обитание в норах
грызуна вискачи трех видов птиц, мелкого представителя куньих
и лисицы [Хадсон (Hudson, 1896]. В норах пищух поселяются
земляные вьюрьки, в норах сусликов — каменки. Совместно оби-
тают в норах корсак и пеганка.
Лисицы в случае преследования или в непогоду укрываются
в периферийных частях системы ходов жилых сурочьих и барсу-
чьих нор и здесь же иногда выводят потомство. В качестве вре-
менных убежищ и скрытых путей передвижения мелкие зверьки
(ласки, полевки, сони, землеройки) используют подземные ходы
кротов, слепышей, водяных полевок и др. Обширные норы боб-
ров дают временный приют самым разнообразным животным, в
том числе и другим млекопитающим — выхухолям, водяным по-
левкам, куторам, горностаям, норкам. Широко используются
также многими животными норы барсуков, сурков. Летяга, жел-
тогорлая мышь, рыжая полевка занимают иногда запасные
гнезда белок.
По существу квартирантство, если оно и не было сопряжено
с насилием, вполне безразличным для хозяина быть не может.
Даже кратковременный, а тем более продолжительный, контакт
на почве использования одних и тех же убежищ всегда может
вызвать обмен паразитами. Водяные полевки, например, посе-
щая норы бобров, роняют в них своих клещей, которые частично
переходят на хозяев норы. Блох сусликов разносят забегающие
в их норы домовые мыши, песчанки и т. д. Существование в не-
которых районах стойких природных очагов чесотки лисиц, ку-
ниц-белодушек и др. связано с постоянным посещением нор, из-
давна зараженных чесоточными клещами.
К квартирантству приближается по своему значению явление
сопряженности нор. Очень часто бывают связаны между собой
норы бобров, выхухолей и водяных полевок, норы сурков и сус-
252
ликов, сурков и пищух, хомячков и полевок, хомячков и тушкан-
чиков и т. д.
На почве совместного пользования жилищами возникают и
взаимополезные отношения. Так, обоюдную пользу приносят
друг другу бобры и выхухоли, живущие в одном водоеме. Выху-
холь очень часто использует бобровые норы для устройства соб-
ственных нор: преодолеть плотный барьер береговой раститель-
ности для самостоятельного рытья норы ей бывает не под силу,
кроме того, забирающиеся в просторные подводные «устья»
бобровых нор рыбы, водные насекомые, моллюски служат кор-
мом выхухоли. Для бобров сожительство с выхухолями полезно
тем, что последние истребляют брюхоногих моллюсков, являю*
щихся промежуточными хозяевами опасных для бобров гель*
минтов Stichorchis subtriquetrus (Барабаш-Никифоров, 1950).
Бобровые и ондатровые хатки в период половодья служат
островками спасения для водяных полевок и многих других
зверьков.
Обоюдно полезно норовое сожительство млекопитающих с
различного рода сапро- и 'копрофагами, способствующими под-
держанию нормальных санитарных условий в норе, а также с
хищными членистоногими, поедающими мелких паразитов (кле-
шей, блох).
Размножение и явления, связанные с ним
ОБЩАЯ ХАРАКТЕРИСТИКА РАЗМНОЖЕНИЯ
Размножение — важнейший биологический процесс, обеспе-
чивающий поддержание и увеличение численности вида, воз-
можность его расселения и, в конечном итоге, успех борьбы за
существование. С интенсивностью размножения вредного вида
связаны трудности его подавления при защите посевов. Про-
мысловый вид с высокой плодовитостью способен выдерживать
большую «промысловую нагрузку» (процент ежегодного изъя-
тия особей из популяции), что выгодно отличает его от менее
плодовитых видов, дающих продукцию той же ценности. Все это
говорит о большом практическом значении изучения размноже-
ния в целях овладения методами воздействия на него.
У млекопитающих процесс размножения имеет как некото-
рые общие для высших позвоночных черты (внутреннее опло-
дотворение, забота о потомстве), так и специфические особен-
ности (живорождение, выкармливание детенышей молоком).
По способу размножения звери могут быть подразделены на
три биологические группы, соответствующие основным система-
тическим подразделениям класса.
Наименее типичную для млекопитающих группу, обладаю-
щую унаследованным от рептильных предков примитивным спо-
собом размножения — откладкой яиц, составляют клоачные
(Monotremata). Отложенное оплодотворенное яйцо клоачные
либо высиживают в гнезде, либо вынашивают во временной
выводковой сумке, образующейся на брюхе самки.
Более прогрессивна живородящая, но еще сравнительно-
низко организованная группа зверей — сумчатые (Marsupialia).
Связь зародыша с материнским организмом у этой группы бо-
лее тесная (у некоторых появляется даже зачаточная плацента),,
однако срок внутриутробного развития кратковременный; дете-
ныши появляются на свет сильно недоразвитыми и длительно-
донашиваются в выводковой сумке матери.
Высшие млекопитающие (Placentalia)—наиболее совер-
шенная по типу размножения группа зверей. После более или
менее продолжительной беременности, во время которой плод,
находится в теснейшем контакте с матерью при помощи особого
временного органа — плаценты, высшие звери рождают относи-
тельно хорошо развитых детенышей.
254
Размножение становится возможным лишь по достижении
животным половой зрелости. Сроки наступления ее у разных
видов млекопитающих сильно отличаются, но, как правило, у
мелких форм с небольшой продолжительностью жизни способ-
ность к воспроизводству наступает раньше. Некоторые мелкие
грызуны способны приносить потомство уже в месячном воз-
расте, еще не достигнув размеров взрослого животного. Зайцы
достигают половой зрелости в возрасте около года, тигры —
5 лет, носороги — 12—15 лет, слоны — 15—20 лет.
Самцы и самки многих видов зверей становятся половозре-
лыми не одновременно (у оленей, котиков, моржей расхождение
в этом отношении достигает 1—3 лет).
Воспроизводство потомства связано с половой цикликой,т. е.
с периодическим повторением комплекса специфических для
размножения явлений. В течение полового цикла в организме
животного происходит ряд изменений. Наибольшей глубины эти
изменения в период размножения достигают у самки. Половой
цикл должен закончиться оплодотворением, если же этого не
произошло, самка остается холостой. Каждый половой цикл
состоит из стадий, которые в свою очередь можно разделить
на фазы:
Стадии Фазы
I. Уравновешивания, или ’ покоя 1. Предтечкбвого покоя
II. Возбуждения, или фолли- кулярная 2. Подготовки, к течке (пред- течки) 3. Течки
III. Торможения, или лютеино- вая (желтого тела) 4. Послетечкового восстанов- ления
5. Послетечкового покоя
В случае оплодотворения наступают еще стадии:
IV. Беременности и V. Лактации.
Половые циклы повторяются до тех пор, пока самка способ-
на к размножению. По достижении определенного возраста эта.
способность утрачивается, наступает так называемый климакте-
рический период (climax).
По количеству половых циклов различают виды полицикли-
ческие, у которых половые циклы (у самки) при отсутствии бе-
ременности повторяются много раз на протяжении года (часть
видов грызунов и копытных, обезьяны, лев, гиена и некоторые-
др.), и моноциклические, имеющие один цикл в году, приурочен-
ный к определенному сезону (клоачные, часть сумчатых, бобр,,
сурок, волк, лисица, хорек, крот и др.).
Половая циклика у самцов выражена слабее и заключается
в основном в изменениях интенсивности сперматогенеза (и вместе-
255-
с тем размера семенников) и в изменении поведения животных.
Половая система млекопитающих высоко чувствительна к
изменениям внешней среды. Как показали исследования Биссо-
нетта (Bissonnette, 1932), Бекера и Ренсона (Baker a. Ranson,
1932), Рерига и Кнохе (Rorig u. Knoche, 1916), Калабухова
(1935), Н. Наумова (1936), Стрельникова (1940) и др., на ритм
половой циклики влияют главным образом пища, температура,
свет.
Весь круг явлений, связанный с процессом размножения у
зверей, складывается из нескольких этапов: 1) подготовка к
размножению, 2) соединение полов и совокупление, 3) беремен-
ность, 4) роды, 5) подсосный период и воспитание потомства,
6) распад семей и расселение молодых.
ПОДГОТОВКА К РАЗМНОЖЕНИЮ,
ТЕЧКА (ГОН), СПАРИВАНИЕ
Первая стадия полового цикла—стадия уравновешивания
характеризуется уравновешенностью общего состояния орга-
низма, необходимой для накопления сил и подготовки к раз-
множению. Применяемый для этого состояния термин — «стадия
предтечкового покоя» (anoestrum) условен, так как покой, в
котором находится в это время организм, относителен: половые
железы продолжают исподволь продуцировать половые клет-
ки Ч Лишь внешние проявления взаимоотношений обоих полов
действительно отвечают такому определению.
По мере приближения срока размножения стадия уравнове-
шивания сменяется стадией возбуждения. Эта стадия характе-
ризуется рядом изменений в половом аппарате самки и общим
возбужденным ее состоянием. Под влиянием усиленного проду-
цирования передней долей гипофиза гонадотропного гормона
в яичниках происходит интенсивное развитие фолликулов. Со-<
зревающие фолликулы выделяют в кровь самки фолликулярный
гормон, который вызывает (через центральную нервную систему)
гиперемию половых органов, увеличение размеров яичников и
более или менее ярко выраженное влечение к самцу.
Начальные, слабые еще проявления этих процессов состав-
ляют фазу предтечки (prooestrum), которая вскоре переходит в
фазу течки (oestrus). Яичники в этой последней фазе достигают
наибольших размеров. У зайцев-беляков, например, вес яичню
ков в стадии уравновешивания составляет около 180—200 мг,
с наступлением пика возбуждения—1000—2140 мг, а у отдельных
особей до 5350 мг (С. Наумов, 1947). Из половых путей самки
1 Глубокую ошибку совершали животноводы и звероводы, резко ослаб-
лявшие уход за животными в стадии «покоя». Уход, полноценное кормление
в стадии уравновешивания — залог нормального размножения и высокой
производительности животных.
?56
обильно выделяется слизь — начинается в собственном смысле
течка
Процесс выведения и миграции яйцеклеток — овуляция, у
большинства видов млекопитающих совпадает с течкой, у не-
многих отделен от нее более или менее значительным промежут-
ком времени. У сумчатой куницы, например, овуляция наступает
через неделю после окончания течки, у многих летучих мы-
шей— через 6—7 месяцев. Овуляция происходит либо спонтанно
(самопроизвольно), либо только после или во время полового
акта как реакция на него. Первый тип овуляции широко рас-
пространен, второй свойственен немногим видам (кроликам,
хорькам, некоторым видам кошек, сусликов, землероек). По-
скольку у большинства млекопитающих течка (охота) и овуля-
ция более или менее совпадают во времени, их обычно объеди-
няют под общим названием течки.
Течка у моноциклических видов, как сказано выше, приурочена
к определенному сезону, но сроки в пределах сезона под влия-
нием разных причин могут сильно варьировать. Молодые жи-
вотные обычно приходят в течку позднее взрослых, что часто
обусловливает растянутость этого явления. Так, у молодых вол-
чиц «переярков» течка на 2—3 недели запаздывает по сравне-
нию со старыми особями; такая же примерно запоздалая течка
наблюдается у молодых енотовидных собак и у ряда других зве-
рей. Важное значение имеют также условия внешней среды, ре-
гулирующие общий ход размножения.
Продолжительность течки у разных видов сильно отличается:
у коровы 1 —1,5 дня, у овцы, козы 1—2 дня, у лошади 5—6 дней,
у собаки 4—9 дней. Обычно течковый период сопровождается
набуханием гениталий, что особенно резко выражено у пави-
анов 1 2.
Одновременно с изменениями, происходящими в период теч-
ки в половых органах самок, соответствующим образом изме-
няется и половая сфера самцов. Подготовка к размножению у
них заключается в основном в интенсивном созревании и форми-
ровании мужских половых клеток — сперматозоидов. Подобно
яичникам, семенники в период подготовки к размножению также
сильно увеличиваются в размерах (у зайцев-беляков вес их с
1 Фаза течки у отдельных видов млекопитающих проявляется по-раз-
ному. У значительного числа видов зверей (грызунов, хищников и др.) она
характеризуется не обильным выделением слизи из половых путей, а, наобо-
рот, повышенной сухостью слизистой оболочки.
2 Внешне с течкой очень сходно явление менструации, свойственное при-
матам, Менструация — это периодическое более или менее обильное кровоте-
чение из половых органов самки, вызванное отторжением части слизистой
оболочки матки. Таким образом, хотя менструации и связаны с половым
циклом, отождествлять их с течкой не следует (Киршенблат, 1951). У многих
видов обезьян у одной и той же самки существуют как течка, так и' менст-
руации.
17 Териология
257
3 г в период «покоя» возрастает в период размножения до 36 г
и более; у соболей семенники, не превышающие в обычное время
размеров конопляного семени, ко времени размножения раз-
бухают до размеров лесного ореха). Увеличиваются также при-
датки семенников и пузыревидные железы — «семенные пу-
зыри».
Важное значение для активации мужских половых желез
имеет их температура. В особых условиях в этом отношении
находятся крипторхи, у которых внутреннее расположение об-
условливает постоянно высокую температуру семенников.
У крипторхов продуцирование сперматозоидов начинается
только при соответствующем охлаждении семенников (бобры,
например, начали размножаться в неволе только после того, как
им была предоставлена возможность охлаждать брюшную по-
верхность тела, лежа на снегу или на холодном цементном
полу) !.
Недостаток влаги в пище вызывает торможение как сперма-
тогенеза, так и овогенеза (в экспериментальных условиях у
обыкновенной полевки даже при частичном недостатке воды в-
корме в первые же дни прекращается половой цикл).
Наличием овогенеза и течки у самок и активного сперматоге-
неза у самцов определяется готовность особей обоих полов к
совершению полового акта.
Важную роль в встрече особей разного пола играют разного
рода сигналы: звуки, запахи и т. п. Так, рев оленей быков ока-
зывает сильное стимулирующее воздействие на половую сферу
самок — это обеспечивает синхронизацию полового созревания.
У оленей в брачный период ревут только самцы. У лисиц, ко-
шек голос подают как самцы, так и самки. У лосей первой сиг-
нализирует храпом о месте своего нахождения самка — на него
откликается самец.
По запаху отыскивают друг друга многие звери. Эту задачу
облегчает усиливающаяся в период гона деятельность пахучих
кожных желез (например, у баранов, козлов), особенно же мус-
кусных желез (у ондатры, выхухоли, кабарги, мускусного овце-
быка) 1 2. У кабанов-самцов препуциальные железы начинают
обильно выделять густую жидкость, оставляющую на следу рез-
кий запах. У бобров задерживающаяся в особых мешках в об-
ласти гениталий моча в результате соответствующих биохимиче-
ских процессов приобретает темный цвет и специфический очень
стойкий запах. Оставляемые как самцами, так и самками на
1 В природе с приближением периода размножения бобры все чаще-
выходят на снег. Охлаждают семенники перед размножением слоны, носо-
роги, бегемоты и другие крипторхи.
2 Реакция на мускус у млекопитающих в 'брачный период положитель-
ная, в другое время она отрицательна, что способствует равномерному рас-
пределению особей на территории.
258
возвышенных участках берега порции этой так называемой
«бобровой струи» служат одним из средств, облегчающих
встречи самцов и самок. Возбуждающе действующий на самцов
запах появляется в моче пришедших в охоту самок у большин-
ства зверей.
Значительно меньшую роль в соединении полов играют зри-
тельные сигналы; восприятие внешнего вида партнера служит
главным образом для ориентировки действий при сближении.
Брачный наряд у самцов млекопитающих не развивается, но
такие вторичные половые признаки, как грива, борода, баки,
рога и пр. в известной мере играют роль раздражителя самки.
Рис. 105. Гоньба у косуль (по Бурльеру)
Присутствие самца, его вид, голос, запах действуют стиму-
лирующе на процесс овуляции у самки. У изолированной самки
образование фолликулов происходит замедленно; такая самка
нередко после полового акта остается бесплодной (Машковцев,
1940).
Вся совокупность воспринимаемых от другого пола ощуще-
ний возбуждающе действует на партнеров. У многих видов зве-
рей вошедшие в охоту самки начинают искать сближения с сам-
цами, заигрывают с ними, производят ряд характерных движе-
ний, учащают мочеиспускание, перед самым совокуплением при-
нимают позу, облегчающую этот акт. У других видов самки
менее активны и внешне остаются спокойными. Самцы, движи-
мые половым инстинктом, начинают настойчиво преследовать
самок, гоняются за ними (рис. 105). Отсюда другое название
этой фазы — «гон» (гоньба), употребляемое наравне с терми-
ном «течка».
Кроме собственно погони за самкой, самцам свойственны в
брачный период и другие действия, составляющие вместе с от-
ветными действиями самки «брачные игры». Самец привлекает
внимание самки характерными телодвижениями, звуками (кри-
ками, храпом, щелканием зубами, ударами ног о землю) и пр.
(рис. 106). При наличии возле самки нескольких самцов между
ними обычно происходят драки.
Хорошо изучено протекание гона у оленей. Брачный период
у них начинается осенью с рева самцов, который вначале слы-
17*
259
шится только по ночам (или на рассвете), но затем все более
учащается и в разгар гона звучит на протяжении почти суток.
Ревущее животное имеет крайне возбужденный вид; шея
набухает, глаза наливаются кровью, тело издает резкий запах.
Олень бьет землю копытами, топчется на месте, выбивая харак-
терные площадки — «точки», бьет и ломает рогами ветки кустов.
Рис. 106. Брачные игры у лисиц (по Темброку)
На рев начинают собираться самки, но вместе с тем он привле-
кает и других самцов — это приводит к дракам, заканчиваю-
щимся преследованием более слабого, а в некоторых случаях
гибелью одного из соперников1 (рис. 107). Победитель — взрос-
лый сильный самец — удерживает возле себя «гарем» — достав-
шихся ему самок, до тех пор, пока не покроет их всех.
Характерные особенности имеет брачный период у другого
гаремного вида — морского котика. Весной приваливающие пер-
выми к островам северной части Тихого океана (из вод умерен-
ной зоны) самцы этого вида, выходят на отдельные участки бе-
рега, образуя лежбища. Каждый старый самец — «секач» рас-
полагается на некотором расстоянии от соседнего и захватывает
1 Ожесточенные драки происходят также между самцами косуль. В 1938 г.
в Ильменском государственном заповеднике найден погибший козел, голова
и грудь которого были в сильнейших ссадинах и кровоподтеках, одна из
передних ног сломана, лицевая часть черепа и нижняя челюсть раздроблены,
а мозговая коробка продавлена. Кроме того, один рог был сломан в верх-
ней трети и висел. В другом случае в том же заповеднике был найден труп
козла 4-летнего возраста. Левый рог его был вырван вместе с костью черепа,
мозг обнажен. Рядом с трупом на пространстве 3—5 м копытами животных
была выбита вся трава (Ушков, 1938; Аверин, 1949).
260
отдельный участок лежбища. По
участках, секачи объединяют их
создавая таким образом гаремы
мере появления самок на этих
в группы под своей защитой,
численностью до 30—50 и бо-
Рис. 107. Олень-самец, павший в драке с соперником. Воро-
нежский заповедник (фот. П. А. Мертц)
лее самок в каждом. Каждый секач стремится завладеть как
можно большим числом самок, и между самцами часто проис-
ходят жестокие схватки (рис. 108). Раны, наносимые разъярен-
ными секачами друг другу при помощи мощных клыков, не-
261
редко бывают смертельными. Формирование гаремов продол-
жается и после, того, как на лежбище появятся новорожденные
котики (самки выходят на .берег за 1—2 дня перед родами).
В сутолоке драк и погони за самками секачи давят часть мо-
лодняка. Иногда к смертельному для самки исходу приводит
одновременное нападение на нее двух секачей, каждый из ко-
торых стремится присоединить ее к своему гарему. Отмечались
случаи похищения самок секачами из соседних гаремов. Охрана
гаремов секачами (рис. 109), как и у оленей, длится до окон-
чания оплодотворения самок.
Рис. 108. Дерущиеся самцы котики (Callorhinus ursinus) (рис. Н. Н. Конда-
кова по фот. И. И. Барабаш-Никифорова)
В течение брачного периода, около 3 месяцев, секачи нахо-
дятся на берегу и почти не кормятся. К концу периода гаремной
жизни и длительного «брачного поста» (в последних днях
июля — начале августа) самцы оказываются крайне истощен-
ными, начинают уходить в море на кормежку и вскоре совсем
покидают лежбища, мигрируя к югу на места зимовок.
У сивуча — другого представителя семейства ушастых тюле-
ней, тоже обитающего в северной части Тихого океана, гаремы
меньше, чем у котика: обычно они не превышают 15—20 самок.
Сильное истощение самцов в период гона и брачного поста
оказывает влияние на смертность1 полигамов. Так, например,
самцы сайгаки, захватывающие группы от 2—3 до 20—25 и в
редких случаях 50 самок, пасут эти «семейные стада», или га-
232
Рис. 109. Котик секач (взрослый самец) и самки — часть его гарема. О-в
Тюлений. Сентябрь 1946 г. (фот. Г. А. Белянкина)
ремы, в течение 15—20 дней декабря, постоянно затевая стычки
с соперниками. Все это время они почти не кормятся, но часто
едят снег. Ослабевшие от брачного поста самцы значительно-
чаще, чем самки, становятся добычей волков, а в случае небла-
гоприятной зимовки в массе погибают от истощения. Это отме-
чено и в отношении самцов джейранов, диких свиней и др.
Если учесть, что некоторое количество самцов сайгаков, ка-
банов умирает от тяжелых ранений, полученных в драках с со-
перниками во время гона, то станет ясным, как своеобразно
идет элиминация половозрелых особей. Доля взрослых самцов
сайгаков в популяции составляет 18,3% в декабре — январе и
снижается до 11,9% к началу лета (Банников с соавторами,
1961). Иначе протекает брачный период у зверей, не собираю-
щих гаремов. У лисиц гон начинается с особого «подлаивания»,
издаваемого пришедшими в охоту самками. На этот звук сбе-
гается несколько самцов; между ними завязываются драки.
Самка не стоит на месте, и самцы следуют за ней вереницей;
то же наблюдается у волков — это получило название лисьих и
волчьих «свадеб». У белок также одну самку упорно преследует
несколько самцов, причем эта погоня происходит частью на
земле, частью на деревьях. У зайцев гоняющуюся группу состав-
ляет несколько самцов и самок, при этом самцы издают осо-
бого рода крики, наскакивая друг на друга на бегу и -нанося
сильные удары задними ногами.
Типичные проявления гона отмечены и у китов. Самцы дель-
фины, например, гоняясь с большой скоростью за самкой, хва-
тают друг друга зубами за плавники. Самцы горбатых китов
во время гона обнаруживают возбуждение, хлопая друг друга
огромными грудными плавниками, перекатываясь с бока на бок,
выпрыгивая из воды. Особенно бурно, с драками, протекает гон
у полигамного кашалота.
Не всегда гон идет по описанной определенной схеме. У оле-
ней, например, типичные для гона драки между самцами иногда
предшествуют началу рева, а не следуют за ним. Нередко опе-
режает гон соединение в пары у белок и некоторых других зве-
рей. У лисиц наблюдается оставление самцами покрытых уже
самок и вторичное соперничание из-за беременных самок.
Своеобразная черта гона у ондатр — очень сходный характер
поведения особей обоего пола. Находящиеся в охоте самки сами
активно разыскивают самцов, вступая в драки между собой.
В практике животноводства и звероводства известны случаи,
когда самка, нарушая обычно свойственное ей подчинение силь-
нейшему самцу, оказывает одному из самцов явное предпочте-
ние перед другими, яростно ею отгоняемыми.
В связи с драками, происходящими в брачный период между
самцами за обладание самкой, у оленей развилось особое тур-
нирное оружие в виде рогов, обычно не употребляемых для за-
264
щиты от хищников. То, что рога имеются только у самцов, раз-
виваются к сезону размножения и сбрасываются по окончании
его, говорит об их истинном назначении1. Ветвистость рогов
снижает их смертоносность, что, конечно, в интересах вида
Рис. 110. Погибшие белохвостые олени, «намертво»
сцепившиеся рогами (по Гамильтону)
(вместо ударов острыми концами, действие рогов при таком их.
строении чаще ограничивается лишь отталкиванием рогов со-
перника). Но иногда эта ветвистость служит причиной гибели
обоих соперников, когда при ожесточенном бое, сцепившись
етростками рогов, они не в состоянии разойтись. Описаны слу-
чаи гибели сцепившихся рогами косуль, лосей, благородных
оленей, маралов и др. (рис. ПО). Турнирное применение имеют
рога и у значительной части полорогих (загнутые назад или
внутрь рога антилоп, козлов, баранов и некоторых быков отно-
1 Особняком в этом отношении стоят северные олени, самки которых
тоже имеют рога, причем у самок они сбрасываются значительно позднее-
(к весне, в то время как взрослые самцы сбрасывают их в начале зимы).
Эта особенность связана с питанием: разгребая передними ногами снег и-
склоняясь над вырытой в нем ямой, самка непроизвольно прикрывает по-
следнюю своими ветвистыми рогами, закрывая доступ к найденному корму
другим оленям, хотя бы и более сильным.
265
сительно безобидны). У кабарги турнирное значение имеют
длинные, острые, сильно выдающиеся вниз клыки.
Опасность применяемого в брачных драках оружия не тур-
нирного характера несколько снижается особенностями спосо-
бов борьбы и развитием защитных приспособлений, чаще всего
в виде длинных волос на наиболее уязвимых местах. У кабана-
секача к периоду гона на передней части туловища под кожей
образуется своеобразный панцирь-«калкан» — очень плотная
соединительнотканная прослойка, хорошо защищающая от уда-
ров соперника. Сходное приспособление имеется у самцов мор-
жей. Шея и грудь самцов котиков защищена густой длинной
шерстью1.
Таким образом, гон (течка) представляет собой сложный
комплекс действий, направленных на соединение разнополых
особей, т. е. на спаривание, в результате которого совершается
акт совокупления (часто эти два понятия отождествляют, что
не совсем правильно).
Характер брачных отношений у млекопитающих, принадле-
жащих к полигамам и моногамам, имеет существенные отличия.
Число полигамов, т. е. видов, у которых один самец за период
размножения оплодотворяет несколько самок, значительно
больше, чем представителей другой биологической группы.
Самцы основной массы полигамных видов после спаривания
не удерживают самок возле себя (многие грызуны, хищные, на-
секомоядные); таким образом у них не образуются постоянные
лары или существование их ограничивается немногими днями.
В некоторых случаях оплодотворенная самка, начиная строить
семейное гнездо или расширять нору, где она принесет детены-
шей, сама настойчиво и решительно отгоняет самца. Он ищет
другое пристанище, используя следы, и обычно находит другую
неоплодотворенную самку. Во многих случаях это не может по-
мешать ему через некоторое время вернуться к первой самке,
успевшей выкормить детенышей и готовой к новому спариванию
(полевки, мыши, хомяки и др.).
Значительно меньшую группу млекопитающих составляют
моногамы, характеризующиеся образованием более или менее
стойких супружеских пар. У некоторых видов существование
пары ограничивается только временем совокупления, после ко-
торого самец оставляет самку (настоящие тюлени). Для ряда
видов характерно сохранение пары на протяжении одного се-
зона размножения (волк, шакал, лисица, песец, ондатра и неко-
1 Однако указанные приспособления далеко не полностью обеспечивают
защиту. Так, например, борющиеся самцы кабаны стараются ударить про-
тивника по незащищенному калканом брюху и более слабые нередко поги-
бают от тяжелых повреждений кишечника или нагноения ран. У самцов
котиков драки часто приводят к повреждениям глаз, серьезным нарушениям
целости покровов и другим увечьям.
260
торые др.). У хищников сильный самец удерживает иногда возле
•себя одну и ту же самку в течение нескольких лет. Наконец,
редкое явление составляют постоянные пары, не распадающиеся
на протяжении всей жизни животных (некоторые обезьяны).
Существует ряд переходных форм. Так, у причисляемых
обычно к моногамам лосей пары очень неустойчивы, самцы не-
редко меняют самок или удерживают возле себя несколько са-
мок. Бобры в разных географических областях ведут себя либо
как полигамы, либо как моногамы. Моногамные в основном ли-
сицы проявляют иногда явно полигамные наклонности. При
фермовом же разведении один самец кроет до десяти самок за
сезон, превращаясь таким образом в настоящего полигама.
Сходные явления наблюдаются и у других видов зверей.
Кроме моногамии и полигамии, как редкое явление, у мле-
копитающих наблюдается полиандрия, или многомужество, при
котором с одной самкой спаривается несколько самцов. Это
свойственно летучим мышам и некоторым обезьянам (например,
плащеносным ревунам Alouatta palliata).
Наиболее ожесточенные драки, как уже было сказано, про-
исходят между самцами полигамных видов. В отношении к сам-
кам они часто применяют принуждение, удары рогами и т. п.,
иногда наносят травмы. Самцы моногамы если и дерутся, то
менее ожесточенно, в их отношениях с самкой чаще проявля-
ются элементы «ухаживания». При полиандрии самцы вовсе не
враждуют между собой. В связи с этим и условия, в которых
происходит акт совокупления, или копуляции (coitus), весьма
различны.
Осуществление совокупления обеспечивается рядом безус-
ловных и условных рефлексов, проявляющихся в определенной
последовательности.
Как говорилось выше, у многих видов самец во время гона
приобретает очень сильный специфический запах (олени, анти-
лопы, козы, кабаны). При спаривании он сообщает «самцовый
запах» и самке, чем предупреждает покрытие другими самцами.
При совокуплении может не произойти оплодотворение —
слияние мужской и женской половых клеток. Это приводит к
прохолостанию самок — яловости, которая широко распростра-
нена среди млекопитающих.
Слияние половых клеток осуществляется в фаллопиевой
трубе (воронке яйцевода) С В яйцевую клетку может проникать
огромное количество сперматозоидов, но оплодотворяет ее один
из них (исключение составляют клоачные; им присуща поли-
спермия). У летучих мышей совокупление происходит осенью и
весной. Осенью сперма, полученная от нескольких самцов, пере-
полняет матку и влагалище самки (вытеканию ее препятствует
У тенреков (Centetes) оплодотворение яйца происходит в яичнике.
26?
образующаяся «вагинальная пробка»). Овуляции в это время
не происходит, но сперматозоиды сохраняются, не теряя жизне-
способности, вплоть до пробуждения самки из спячки (в течение-
5—7 месяцев). Весной происходит овуляция, и мигрирующая
яйцеклетка оплодотворяется перезимовавшими сперматозои-
дами. При весенней копуляции оплодотворение происходит
обычным путем.
По окончании брачного периода возбуждение самцов и са-
мок спадает, в половом цикле самок наступает стадия тормо-
жения (лютеиновая стадия, или стадия желтого тела), характе-
ризующаяся угасанием полового тонуса, уменьшением яични-
ков, ослаблением гиперемии гениталий. Для первой фазы стадии
торможения — фазы послетечкового восстановления (metaoest-
rum) — характерны из упоминавшихся выше процессов: прекра-
щение секреторной деятельности, постепенное возвращение по-
ловых органов к состоянию покоя. Во второй фазе — послетеч-
кового покоя (dioestrum) происходит общее восстановление-
сил, израсходованных за время течки.
У самцов после гона также наступает общее физиологиче-
ское равновесие, а у полигамов в результате длительного голо-
дания и непрерывной половой деятельности — временный общий
упадок сил. Вследствие затухания сперматогенеза увеличивается
интерстициальная ткань семенников и резко уменьшается диа-
метр семенных канальцев, отчего происходит уменьшение семен-
ников. Сильно сокращаются и придаточные железы.
Гон и связанные с ним явления сильно истощают самцов,
при большой затрате энергии длительное время почти или
вовсе не принимающих пищи («брачный пост» секачей, котиков,
оленей и др.). Отсюда значительный процент гибели самцов,
принимавших участие в размножении, что наблюдается, на-
пример, у оленей и сайгаков в зимние месяцы. Большая связан-
ная с гоном подвижность, утрата обычной осторожности вызы-
вают повышенную смертность самцов от хищников, что наблю-
дается у полевок, мышей и др.
МЕХАНИЗМ РЕГУЛЯЦИИ ПОЛОВЫХ ПРОЦЕССОВ САМКИ
В настоящее время можно считать вполне установленным,
что ритм полового цикла самки определяется как внешними
факторами (фотопериодизмом, освещенностью, температурой),
так и внутренними (взаимодействием нервной и гуморальной
систем). Чрезвычайно важное'значение имеет участие гормонов:
фолликулина, гонадотропного гормона, выделяемого гипофизом
и распадающегося на две фракции (пролай А и продан Б), и
лютеостерона, вырабатываемого желтым телом.
В центре механизма регуляции находится гипофиз — он ис-
пытывает со стороны других органов целый ряд воздействий,
268
е
У а, Н ;
Воспой/1 ° „
^„няте/вд (?. " if
и 11
CU
ft
П«^оРь'ого
реЛ"ь,'°
fiUt^nip^L
Рог матки
Ж®
Фолликул
Желтое
тело
/ Эндокринная
1р др. системы
V организма
V*
%
^головлого
мозга
Г ипосриз
Плод
и
плацента.
7ечка
Фолликулин
плаценты
Влагалище
Рис. 111. Схема регуляции половых процессов организма самки
(по Студенцову)
вызывающих соответствующие его реакции; общее стимули-
рующее влияние на гипофиз оказывают попадающие в орга-
низм с пищей и образующиеся в тканях витамины и стероны1.
С другой стороны, регулирующее действие оказывает на гипо-
физ кора головного мозга, в которой, в результате восприятий
рецепторами внешних сексуальных раздражений и передачи их
по нервам в соответствующие центры коры, возникают соответ-
ствующие импульсы. Воздействуют также на гипофиз гормоны,
выделяемые фолликулами, яйцом и плацентой (рис. 111).
Кора головного мозга побуждает гипофиз к выделению про-
дана А, обусловливающего созревание фолликулов и овуляцию,
а фолликулин вместе с названными выше гормонами — выделе-
ние пролана Б, стимулирующего развитие желтого тела.
БЕРЕМЕННОСТЬ, ЗАРОДЫШЕВОЕ РАЗВИТИЕ
С момента оплодотворения начинается стадия беремен-
ности — физиологическое состояние организма самки, обеспечи-
вающее вынашивание плода.
Как уже указывалось в морфологическом очерке, у беремен-
ной самки разрастающееся желтое тело овуляции превращается
в крупное желтое тело беременности. Оно занимает значитель-
ную часть яичника и сохраняется на протяжении всей беремен-
ности, а у некоторых видов — и периода молочного кормления
детенышей. Желтое тело замещает полость разорвавшегося фол-
ликула и выделяет в кровь свой сложный инкрет, тормозящий
дальнейшее развитие яйцевых фолликулов1 2, прекращающий
овуляцию и поддерживающий пролиферативные (связанные с
воспроизводством потомства) процессы в матке. Слизистая обо-
лочка матки еще более гиперемируется и гипертрофируется.
Матка сильно увеличивается—^увеличиваются и млечные же-
лезы.
В начале беременности образовавшаяся в результате слия-
ния мужской и женской гамет зигота свободно плавает в жид-
кости половых путей самки; интимной связи с материнским ор-
ганизмом она еще не имеет и питается за счет вырабатываемого
маткой так называемого маточного молока (эмбриотрофа).
В дальнейшем зигота имплантируется в стенку матки, и с этого-
времени получает питание через кровеносную систему.
Промежуток времени между оплодотворением яйцеклетки и
имплантацией зиготы носит название латентной (скрытой) фа-
зы беременности, или диапаузы. У подавляющего большинства
зверей эта фаза занимает не более нескольких дней, у части же
видов (у ряда куньих, котика, косули, некоторых тропических
1 Стероны — группа органических веществ, основу которых составляет
углеводород.
2 Как правило, число желтых тел соответствует количеству эмбрионов.
270
форм летучих мышей) она достигает нескольких месяцев, удли-
няя соответственно и общую продолжительность беременности.
Это явление, несомненно, имеет приспособительное значение,
«подгоняя» рождение и воспитание молодняка к наиболее бла-
гоприятному по метеорологическим и кормовым условиям вре-
мени года. Так, у куницы и соболя течка и оплодотворение про-
исходят летом, развитие зародыша укладывается в месячный
срок, но затем, благодаря 8—9-месячному латентному периоду
роды сдвигаются на весну следующего года. У барсука спари-
вание бывает либо в середине лета (в июле), либо осенью (в ок-
тябре). В первом случае беременность с продолжительной ла-
тентной стадией длится около 8 месяцев, во втором, благодаря
сокращению диапаузы,— 5-месяцев. В обоих случаях роды на--
ступают в весеннее время (в конце марта).
Латентный период представляет собой задержку развития,,
но не полное его прекращение.
У названных выше представителей куньих имплантация и
начало развития зародыша после латентной стадии приходятся
на конец зимы. К этому времени (вернее, к ранней весне) бы-
вает приурочено новое возбуждение животных. Самцы опять
гоняются . за самками, делают попытки покрыть их, дерутся
между собой, однако в результате такого «лжегона» оплодотво-
рения не происходит. Некоторые авторы (Топорков, 1941; Раев-
ский, 1947) отрицают сексуальную основу такого весеннего
оживления, связывая его с перераспределением угодий, случай-
но совпадающим с моментом имплантации зародыша у самок.
Возможно, что «ложный гон» определяется как половым воз-
буждением, вызванным глубокими изменениями, происходящи-
ми в половой системе беременной самки, так и конкуренцией
из-за жилого участка. Однако у сибирской косули, спариваю-
щейся, как правило, ранней осенью, весенний лжегон в виде
исключения приводит к оплодотворению отдельных самок,
оставшихся яловыми. Ягнята у таких самок рождаются осенью и к
наступлению зимы не успевают достаточно вырасти и окрепнуть,
что приводит к их гибели в морозный и малокормный период..
Млекопитающим свойственна как одноплодная, так и мно-
гоплодная беременность — одновременное развитие в матке од-
ного или нескольких плодов, каждый из которых образуется из
отдельной зиготы. Интересно многоплодие у броненосцев, пред-
ставляющее результат расщепления зиготы (у короткохвостого-
броненосца таким путем одна зигота может дать до 12 однопо-
лых зародышей). Это явление носит название полиэмбрионии1.
Полиэмбриония может иметь место и как ненормальное явле-
ние: в случае неполного расщепления бластомеров зиготы воз-
1 Возможно, она имеет место и у некоторых других зверей, например
кротов, тарбаганов, у которых число эмбрионов обычно превышает количе-
ство желтых тел.
271-
никают сращенные двойники. Из других отклонений от нормы
в период плодоношения у млекопитающих изредка наблюдает-
ся внематочная беременность, обусловленная тем, что яйцеклет-
ка в отдельных случаях может быть оплодотворена и начать
развиваться, находясь еще в яичнике, в брюшной полости или
в фаллопиевой трубе. У домашних млекопитающих эта анома-
лия хорошо известна; отмечалась она и у диких зверей. Как
правило, плод при внематочной беременности погибает и под-
вергается рассасыванию или мумификации.
Не превышающее известного предела рассасывание (резорб-
ция) эмбрионов представляет у млекопитающих обычное явле-
ние. У свиньи, например, резорбции подвергается каждый вто-
рой эмбрион. На ранних стадиях беременности у нее заклады-
вается до 30 эмбрионов, но далее происходит гибель половины
из них (часто при вскрытии матки наблюдается чередование в
ней — через одного — живых и мертвых эмбрионов). У 70% ко-
ров, забиваемых на бойнях, обнаруживается по 2 эмбриона,
между тем рождается у этих животных только 1 теленок. Когда
.-существовал промысел каланов, у добытых самок этого вида не-
редко находили по 2 зародыша, но новорожденный детеныш при
матери всегда бывает один.
Превышающий норму процент резорбирующихся эмбрионов
у многих самок служит показателем депрессивного состояния
организма животных (и популяции в целом). Особенно ярко это
-явление бывает выражено у грызунов в неблагоприятные для
них годы. Нередко при этом наблюдается эмбриональная смерт-
ность преимущественно одного пола (чаще самцов). Высокая
эмбриональная смертность может быть причиной малоплодия.
С истощением организма, тяжелой физической перегрузкой
на поздних стадиях беременности, с авитаминозом и заболева-
ниями связаны различные абортивные явления (выкидыши).
Например, в многоснежные зимы, сильно затрудняющие пере-
движение, массовое абортирование отмечено у сайгаков, джей-
ранов, лошадей при табунном их выпасе и др.
В некоторых случаях в матке имеются эмбрионы разной ве-
личины. Это может зависеть либо от неодинаковых темпов их
-развития (и начавшейся резорбции части из них), либо от раз-
новозрастное™ эмбрионов (у некоторых млекопитающих, на-
-пример зайцев, норок, самки могут подвергаться новому опло-
дотворению в период беременности; в результате наряду с круп-
-ными, хорошо развитыми эмбрионами в матке оказываются и
находящиеся на начальных этапах развития).
Продолжительность беременности варьирует в больших пре-
делах (табл. 6). .
Хотя время спаривания у разных видов млекопитающих с
длительной беременностью весьма различно (у волков, лисиц —
в конце зимы, у хорьков, норок, зайцев — в начале весны, у со-
Таблица 6
Продолжительность жизни, беременности и плодовитость
разных видов млекопитающих
(сводные данные из различных источников)
Виды Продолжи- тельность жизни Сроки наступ- ления половой зрелости Средняя продолжи- тельность беремен- ности в ДНЯХ; Количество детенышей в помете количество пометов в год
Опоссум 12 8—16 1
Гигантский кенгуру 39 1 1
Еж 42 3—6 (до 8) 1
Крот 37 3—9 1
Выхухоль .... 45 3—5 1
Рыжая вечерница 70 2 1
Водяная ночница . до 20,5 лет2 54 1 1
Норка европейская 10—11 мес. 43 4—7 1
Степной хорь . . Лесной (черный) 10 мес. 36 8—11 1
хорь » 43 4—8 1
Горностай .... 4—7 лет 3 мес. 300-320* 3—12 1
Лесная куница . . до 15—17 лет 2—3 года 260* 3—6 1
Соболь ...... » 265* 2—4 (до 6) 1
Выдра до 11 лет 65 2—4 1
Барсук 10—12 лет 2 года 230* 3—5 1
Енотовидная со- -
бака до 10 лет ок. 1 года 59 5—7 (до 14) 1
Лисица до 11—12 лет » 51—52 4—6 (до 13) 1
Волк до 15—20 лет ок. 2 лет 61 3—8 (до 14) 1
Лесной кот . . . 10—12 лет 63 3—6 1
Рысь до 20 лет ок. 3 лет 65 2—3 1
Тигр до 40—50 лет после 5 лет 104 2—3 (до 5) 1 (через 2—3 го-
Бурый медведь . . 30—50 лет 4 года 200 2—3 (до 5) да) 1 (через . год) ‘
Морж 3—6 лет (самки—3 го- да самцы— 5—блет) 300 1
1 (через год)
Котик ок. 15 лет 3—4 года (са мки — —2—Згода самцы —4 года) 355* 1
1
1 Звездочками отмечены сроки беременности с латентным периодом.
2 Холл, Клутьер и Гриффин (Hall, Cloutier a. Griffin), 1957.
18 Териология
Продолжение
Виды Продолжи- тельность жизни Сроки наступ- ления половой зрелости Средняя Продо Л/1..И- тельность берем ности в днях Количество детенышей в помете Количество пометов в год
Гренландский тю- лень 3—4 года 350 1 1
Заяц-русак .... до 10 лет (самки — 2—3 года самцы —4 года) ок. 1 года 46 3-5 (до 7) 2-4
Заяц-беляк . . . » » 50 3—5 до 9) 2—3
Кролик дикий . . 5—7 лет ок. мес. 30 4-5 2—3
Сурок-байбак . . ок. 15 лет 3 года 40 4—5 (до 7) 1
Белка до 10 лет 35 3—8 (до 1 | 1—2
Крапчатый суслик ок. 10 лет 21 3-6 (до 11) 1
Б обр до 50 лет 2- года 106 2—3 до 5) 1
Соня-полчок . . . до 3 лет 10—11 мес. 22—25 3-8 (до 10) 1 '
Крыса-пасюк . . . 2—3 года 2—3 мес. 22 8—10 (до 22) 4—8 2
Домовая мышь . . ок. 1 года 2—3 мес. 22 до 10
Хомяк ок. 2,5 лет 4 мес. 30 4—10 (до 18) 6—7 (до 0) 2—3
Ондатра до 4 лет 3—5 мес. 26 2—4
Обыкновенная по- левка до 2 лет 47—6 0 дн. 20 4—7 (до 10) 3—8
Рыжая лесная по- левка » 60—70 дн. 18 4—5 (до 9) 3-4
Нутрия до 10 лет 4—5 мес. 130 4-6 2—3
Косуля до 15 лет ок. 2 лет 270* 1—2 1
Марал до 25 лет 3—4 года 250 1 (редко 2 1
Лось 15—20 лет 2,5 года 245 1—2 1
Овцы, козы . . . до 15 лет 8 мес. 230 1—3 1
Зубр до 50 лет 5—6 лет 265 1 1 (через
Верблюд до 40 лет 2—5 лет 365 1 2—3 го- да) 1 (через год) 1
Дикий кабан . . . до 20 лет 2—3 года 125 3- 5 (до 12)
Лошадь до 30 лет 2—3 года 340 1 1
Слон индийский . 70—80 лет 15— (лет 630 1 1 (ч. год)
Гренландский кит до 20 лет 2—3 года ок. 36 0 1 1 (через
Орангутанг . . . 15—25 лет 272 1 2—3 го- да) 1
Шимпанзе .... » 236 1 1
Низшие, обезьяны до 20—30 лет 165 1-2 1
болей, куниц, медведей — среди лета, у оленей — осенью). Роды
у них одинаково приурочены к концу весны — началу лета,
исключение составляют медведи, многие ластоногие, у которых
детеныши появляются зимой. Однако в таком случае обычно
имеются особые приспособления, защищающие детенышей От
вредных воздействий среды. Мелкие зверьки с высокой плодо-
витостью и малой продолжительностью беременности (полевки,
мыши, землеройки) способны приносить и выкармливать моло-
дых в разные сезоны.
Сроки утробного развития связаны со степенью организации
(сумчатые и плацентарные млекопитающие), размерами жи-
вотных, а также степенью защищенности детенышей от небла-
гоприятных климатических условий и хищников. Так, менее про-
должительна беременность у видов, имеющих возможность вос-
питывать беспомощный молодняк в норах, гнездах и других убе-
жищах, более продолжительна — у видов, выращивающих потом-
ство открыто. У типичного норника — кролика — беременность
длится около 30 дней, у открытоживущего зайца—около 50 дней.
Самки одного и того же вида имеют продолжительность бе-
ременности в большей или меньшей степени различную в зави-
симости от сезона спаривания, физиологического состояния ор-
ганизма, возраста животного, пола плода, количества зароды-
шей и пр.1
Нормальное завершение беременности — родовой акт, заклю-
чается в выведении из половых путей самки доношенного жиз-
неспособного плода с оболочками и плацентой. Роды происхо-
дят в связи со сложным нервно-гуморальным процессом, регу-
лируемым в основном корой головного мозга.
С приближением родов самка становится беспокойной, пере-
стает кормиться, ищет удобное укромное место: у многих видов
она подготавливает гнездо или логово для себя и будущего по-
томства. У обезьян и некоторых других зверей эти действия сти-
мулируются предродовыми болями. У части видов копытных, ла-
стоногих, китов, летучих мышей с приближением родов самки
обособляются от самцов и образуют особые однополые стада.
Обычно у самок открытоживущих видов роды протекают
ночью. Днем беспокойная обстановка и непосредственное влия-
ние света угнетают родовую деятельность. Напротив, ночью, не
отвлекаемая возбуждениями извне кора головного мозга актив-
но реагирует на раздражения, идущие от половой сферы, обус-
ловливая нормальное течение родового процесса.
Родовой акт начинается с раскрытия шейки матки и приня-
тия плодом положения, обеспечивающего выход наружу. Далее
следует выведение плода. У большинства зверей плод продви-
1 Есть указания на то, что длительность плодоношения у молодых самок
сельскохозяйственных животных больше, чем у старых (Мышкин, 1937), за-
родыши мужского пола .вынашиваются несколько дольше, чем женского и т.н.
18*
275
гается по родовым путям головой вперед; хвостовой частью
вперед появляются детеныши у китов, части видов ластоногих,
рукокрылых, грызунов и хищных. Вслед за рождением детены-
ша выходит и послед (плацента).
Поведение самки в родовой период у разных видов зверей
имеет свои особенности. Обычно самка у наземных зверей ро-
жает в лежачем положении, но с появлением на свет детеныша
вскакивает на ноги или делает другие резкие движения, в ре-
зультате которых пупочный канатик обрывается, а детеныш ос-
вобождается от плодных оболочек. Самки многих видов пере-
грызают пуповину. Слоны, жирафы и, в отдельных случаях, би-
зоны, олени и некоторые другие млекопитающие рожают стоя.
Обычное явление у млекопитающих, особенно плотоядных,
представляет поедание родившей самкой плаценты и плодных
оболочек. Обезьяны съедают также большой комок слизи, вы-
деляющийся из влагалища во время родов. Объяснить эти вы-
работавшиеся в процессе эволюции действия можно следующим
образом.
Поскольку в период беременности зародыш получает пита-
ние и выделяет продукты обмена через организм матери, самка
к моменту родов сильно истощается, а во время родов она те-
ряет много крови. Поедание обладающей высокой питательно-
стью плаценты служит одним из средств восстановления сил на
первое время, пока она не может отлучиться от новорожден-
ного; вместе с тем плацента и влагалищные выделения играют
роль стимуляторов послеродовой инволюции (обратного разви-
тия) матки, а также процесса молокоотделения. Некоторое по-
слабление кишечника, вызываемое поеданием плаценты, также
полезно ввиду обычного застоя каловых масс у самок в предро-
довой период1.
Своеобразен родовой акт у китообразных. Самки гренлад-
ского кита, белухи, дельфинов в момент родов принимают вер-
тикальное положение, выставляя из воды хвостовую часть. Та-
ким образом, детеныш появляется в воздухе и может осущест-
вить свой первый вдох. Однако в последнее время установлено,
что детеныши китов могут рождаться и под водой. Очень корот-
кий и непрочный пупочный канатик легко обрывается при пер-
вом движении матери, а освободившийся детеныш тотчас же
всплывает на поверхность и может дышать.
Летучие'мыши в момент родов принимают горизонтальное
или вертикальное (головой вверх) положение, подвешиваясь
Ц Поедание плаценты у диких млекопитающих имеет и другое значение —
оно способствует защите детенышей от хищников (предотвращение запаха
разложения, который выдавал бы местонахождение выводка). У домашних
копытных й' результате поедания плаценты иногда возникают тяжелые рас-
стройства функций пищеварительных органов, но это вторичное явление,
возникшее в связи с доместикацией.
276
когтями больших пальцев крыльев. Появившийся детеныш по-
падает в подогнутую к брюху межбедренную перепонку или в
полость, образуемую брюшной поверхностью тела и сложенны-
ми крыльями. Самка кенгуру рожает, лежа на спине.
ПЛОДОВИТОСТЬ
Величина выводка и частота размножения у разных видов
зверей сильно варьируют: от 1 до 22 детенышей в помете, от
1 помета в 2—3 года до 10 пометов в год (см. табл. 6). В зави-
симости от этого диапазон плодовитости очень велик.
Для каждого вида характерны определенные размеры пло-
довитости, но она может сильно меняться под влиянием воздей-
ствий пищи, условий погоды, географического положения и пр.
Величина выводка связана с количеством млечных желез у
самки: как правило, на пару млечных желез приходится один
детеныш. У крупных, с относительно большой продолжительно-
стью жизни видов зверей количество детенышей в помете и чис-
ло пометов меньше. Слон, живущий 70—80 лет, не каждый год
рождает по одному детенышу, в то время как у мелких грызу-
нов, потенциальная продолжительность жизни которых не пре-
вышает трех лет, в году бывает до 10 пометов с числом детены-
шей в каждом до 10—20 и даже 22. Относительно более высо-
кая плодовитость характерна для северных млекопитающих,
которые выращивают потомство в суровых климатических усло-
виях (северный олень, песец, лемминги). Лесные мелкие гры-
зуны, лучше защищенные от хищников и резких колебаний ме-
теорологических условий, менее плодовиты, чем степные.
Несомненна зависимость плодовитости от возраста самки.
У молодых особей величина помета обычно меньше, чем у
взрослых. Молодая волчица, например, мечет 3—4, взрослая
6—8 волчат, молодая лосиха — одного, взрослая двух лосят, у
молодых белок в помете бывает 3—6 бельчат, у взрослых 5—
8 и т. д. Старые самки имеют меньшее количество детенышей,
чем взрослые; то же наблюдается у часто рожавших, истощен-
ных многочисленными беременностями особей. У-'самцов уча-
стие в размножении доступно разным возрастным категориям
не в одинаковой степени. Отсюда ясно значение возрастного со-
става популяции для интенсивности размножения в целом.
Совершенно очевидно также влияние на интенсивность раз-
множения популяции относительной численности полов. Среди
эмбрионов и новорожденных обычно преобладают самцы, реже
самки (белки, дикие свиньи). Однако у взрослых животных со-
отношение полов часто становится равным 1:1. Исключение со-
ставляют некоторые полигамы, у которых в популяции преоб-
ладают самки (ушатые тюлени, дикие кабаны, некоторые ан*
277
тилопы, большинство оленей). В благоприятные годы наблюда-
ется преобладание самцов в пометах, в неблагоприятные (бес-
кормные) резко возрастает количество самок. Таким образом, в
годы с неудовлетворительными условиями существование попу-
ляции поддерживается повышенной размножаемостью за счет
роста числа самок '.
При прочих равных условиях относительно высокой плодо-
витостью всегда отличаются более здоровые, крепкие, жизне-
способные самки. Поставленные в лучшие условия домашние
животные, даже при отсутствии направленной селекции, значи-
тельно более плодовиты, чем их дикие сородичи: у дикого кро-
лика, например, в помете бывает до 5 крольчат, у домашнего —
до 10; дикая кошка приносит один помет в году, домашняя —
несколько.
Неблагоприятные условия могут на короткий или длитель-
ный срок подавлять размножение. Например, в условиях дель-
ты Волги многочисленные там водяные полевки приступают к
размножению в зависимости от погоды в конце февраля .или в
марте. В начале июня наступает паводок; на время его зверьки
лишаются не только своих убежищ, но и самых ценных кормов,
так как они скрываются глубоко под водой. В течение месяца
или более, иногда до начала второй декады июля включитель-
но, в дельте трудно найти размножающихся водяных полевок.
Новый период размножения начинается после спада воды и про-
должается до ноября (изредка бывает размножение немногих
особей и в декабре; рис. 112). Таким образом, в условиях ланд-
шафта дельты Волги из года в год период размножения водя-
ных полевок распадается на два отрезка — предпаводковый и
послепаводковый, чего нет в условиях других ландшафтов
(Кондрашкин, 1955).
Хорошо известна общая зависимость интенсивности размно-
жения от физиологического состояния животных, связанного с
кормовыми условиями.
У кабанов Беловежской пущи в сезоны размножения после
.богатого урожая желудей среднее число поросят в пометах уве-
личивается на 12,5% (Северцов и Саблина, 1953). По наблю-
дениям В. Ф. Гаврина, там же, следом за полным неурожаем
ценных кормов среднее число поросят в выводке равнялось 2,9,
а послеурожаев средних или выше среднего—4,1—6,6 (Слудский,
1'956). По данным последнего автора, летом 1945 г. в низовьях
1 В голодные годы наблюдается повышенная смертность эмбрионов и
новорожденных мужского пола, что в свою очередь приводит к преобладанию
числа самок. Среди взрослых животных в такие годы также в большей сте-
пени гибнут самны, сильно истощающиеся за время гона. На соотношение
полов среди взрослых промысловых животных может оказывать влияние и
избирательность промысла, охватывающего в основном более активных
самцов.
278
р. Или (Казахстан) было много перелетной саранчи, и кормив-
шиеся этим биологически ценным кормом дикие свиньи к осени
были отлично упитаны. Кроме того, и зима 1945/46 г. оказалась
многоснежной, но теплой, что дало кабанам возможность рыться
в талой почве, добывая корневища и т. п. Весной 1946 г. убитые
свиньи имели в среднем по 6,2 эмбриона (при полном отсут-
ствии резобирующихся)—это на 15,5% выше среднего числа
поросят в выводке, равного 5,37 (по многолетним данным).
Месяцы I U Ш IS V Л Л К I 3
Рис. 112. Изменение процента беременных взрослых водяных полевок (во-
дяных крыс) дельты Волги в течение года как показатель хода размножения.
В июне и первой декаде июля процент беременных самок падает до нуля в
связи с паводком:
1 — 1944 г.; 2 — 1945 г.; 3 — 1946 г. (по Кондрашкнну)
Особенно чувствительны к недостатку пищи воспроизводи-
тельные функции видов, потребляющих концентрированные кор-
ма (семена, плоды, беспозвоночных и позвоночных животных).
Грызуны, делающие большие зимние запасы (лесные мыши, на-
пример), приступают к размножению значительно раньше, чем
не запасающие корма. Упитанные самки, как правило, отлича-
ются повышенной плодовитостью по сравнению с тощими (но
ожиревшие часто остаются бесплодными). В годы (или в мест-
ностях) с плохими кормовыми условиями повышаются яловость
самок, эмбриональная и детская смертность. У самцов плохое
питание вызывает нарушение сперматогенеза, подавление сек-
реторной функции семенников, снижение интенсивности гона.
279
Влияние кормов на половую функцию определяется не столь-
ко их количеством, сколько качеством. Так, содержание в расте-
ниях определенного количества так называемых эстрогенов—
веществ, действующих на воспроизводительную способность жи-
вотных, обеспечивает нормальное размножение растительнояд-
ных видов (как недостаток, так и избыток эстрогенов отрица-
тельно действует на половую сферу копытных). Особенно резко
на функциях размножения сказывается авитаминозность и не-
достаточная калорийность кормов, вызывающие нарушение хо-
да овуляции, течки, сперматогенеза и в результате — бесплодие.
Содержание в пище антистерильного витамина Е очень важно
для стимуляции размножения и нормальной лактации. Из дру-
гих витаминов большое влияние на половую сферу оказывают
витамины А и В. Отсутствие или недостаток их в диете вызы-
вают серьезные нарушения деятельности мужских и женских
половых желез, нарушение родового акта у самок, половую
инактивацию самцов и пониженную жизнеспособность потом-
ства.
Плотоядные звери при успешной охоте получают в составе
пищи необходимые для размножения вещества в достаточном
количестве. Растительноядные иногда нуждаются в пополнении
их обычного рациона животными кормами. При фермбвом их
разведении подкормка кроликов, ондатр, нутрий куколкой туто-
вого шелкопряда повышает плодовитость и молочность произ-
водителей, благотворно влияет на выживаемость, рост и разви-
тие молодняка.
Свет — один из важнейших стимуляторов половой активно-
сти, действующий через центральную нервную систему. Свето-
вой раздражитель активизирует деятельность гипофиза, влияю-
щего через гонадотропные гормоны на половые железы. Усилен-
ное выделение последними половых гормонов приводит в доми-
нантное состояние половые центры головного мозга, что и сти-
мулирует размножение (Ухтомский, 1927). Принято считать, что
свет воздействует на гипофиз через органы зрения. Однако он
обнаруживает тот же эффект и у ослепленных животных, у ви-
дов землероев и у норников, деятельных только ночью. -Благо-
даря специальным исследованиям Перри (Раггу, 1938) и др.
выяснено, что свет может воздействовать на половую систему
не только путем зрительного анализатора, но и через пищу —
удавалось стимулировать развитие гонад с помощью дополни-
тельного к дневному облучения корма кварцевой лампой.
Искусственное удлинение светового дня осенью и зимой в
экспериментальных условиях приводит к ускорению полового
созревания, сокращению беременности и повышению плодовито-
сти у соболей, куниц, норок, лисиц, белок и некоторых других
пушных зверей [Биссонет и Чех (Bissonnette a. Csech), 1939;
А. Кузнецов, 1953 и др.]. Темные полевки (Mtcrotus agrestis),
280
подвергавшиеся ежедневно освещению в течение 15 ч, прино-
сили в среднем в 3,5 раза больше выводков и рождали в 6 раз
больше детенышей, чем зверьки, освещавшиеся ежедневно толь-
ко по 9 часов в сутки (Калабухов, 1951).
Кроме «длиннодневных» форм животных, половые функции
которых стимулируются увеличением светового дня, имеются
также формы «короткодневные», на которых аналогичное воз-
действие оказывает сокращение продолжительности освещения
(козы, овцы и некоторые другие парнокопытные). Такое несо-
ответствие объясняется тем, что у первых течка приурочена к
весенне-летнему (полнодневному) периоду, у вторых — к осен-
нему (короткодневному) 1.
Ясно выражена также связь размножения с температурой.
У некоторых млекопитающих (например, у ежей) в эксперимен-
тальных условиях одно повышение температуры среды (без до-
бавочного освещения) способно вызывать половую активность.
Пищухи, сурки, суслики, обитающие на южных, прогреваемых
весной в первую очередь склонах гор, начинают размножаться
на 10—15 и более дней раньше живущих на северных склонах,
где холоднее и поздно появляется свежий зеленый корм. Хо-
лодная затяжная весна задерживает размножение большинства
зверей. Она нарушает сперматогенез и овогенез, вызывает по-
вышение эмбриональной смертности. Раннее осеннее похолода-
ние тормозит наступление гона у некоторых диких копытных и
иногда обусловливает высокий процент яловых самок. Но у ка-
банов на Кавказе в годы с ранним наступлением зимы появле-
ние течки и гона ускоряется: начинается 15—18 ноября и кон-
чается 1—5 января. Запоздалая теплая зима удлиняет период
гона до' 2. мес.; он начинается 25—28 ноября и заканчивается
20—25 января (Логинов, 1936). Смещение сроков течки в теп-
лые зимы объясняют тем, что в такие сезоны свиньи медленнее
теряют упитанность и дольше сохраняют высокий тонус орга-
низма. В некоторых областях ареала, например, в Казахстане,
задержка наступления гона у кабана — явление даже благопри-
ятное для Вида (Слудский, 1956), так как ранние выводки, по-
являющиеся в марте, могут погибнуть от больших холодов и от
нападений волков, еще не имеющих в это время достаточного
количества других кормов (сусликов, гнездящихся птиц и т. п.)..
Поздние выводки попадают в значительно лучшие условия.
В суровую длительную зиму у многих копытных увеличи-
вается количество абортов, гибель молодняка. Как общее пра-
вило, после тяжелых зим снижается плодовитость у всех не
впадающих в спячку зверей. Так, например, у одомашненных се-
верных оленей в годы с малоснежной мягкой зимой и ранней:
1 Свет оказывает тормозящее влияние и на размножение морских свинок.
Кролики, ежи и некоторое другие млекопитающие не обнаруживают зависи-
мости размножения от условий освещения.
281
весной, т. е. при наиболее благоприятных условиях зимнего паст-
бищного кормления самок (важенок), телята рождаются более
крупные, крепкие, лучше развитые, легче переносящие весенние
бураны и т. п. Мелкие телята обычно слабее развиты, менее
жизнеспособны и часто погибают весной. Многочисленные на-
блюдения показали, что вес мертворожденных телят или поги-
бающих в первые дни после рождения редко бывает выше 4—
4,5 кг. Важенки весом 50—60 кг приносят телят со средним ве-
сом 5,5 кг, важенки весом 70—80 кг — 6,1 кг, а весящие 90 кг и
более — телят по 7,0 кг (Друри, 1955). Однако для судьбы при-
плода северных оленей большое значение имеют не только усло-
вия зимнего периода, но и предшествующего лета. После зи-
мовки северные олени всегда оказываются сильно истощенными
и больше всего от условий летнего выпаса зависит, насколь-
ко упитанными и восстановившими силы они станут к осени.
Замечено, что в жаркое лето, когда комары, слепни и овода не
дают животным спокойно пастись, олени встречают зиму плохо
отъевшимися, слабыми. Если следом за таким летом наступит
многоснежная зима, телята родятся мелкие, мало жизнеспособ-
ные. В прохладное, облачное лето с ветрами со стороны Ледо-
витого океана олени пасутся спокойно и хорошо накапливают
силы перед гоном и зимовкой.
Засуха и чрезмерно высокая температура также отрица-
тельно влияют на размножение млекопитающих, резко ухудша-
ют качество кормов, вызывают депрессию функций половой си-
стемы, тормозят развитие молодняка. Количество выпадающих
осадков особенно отражается на интенсивности размножения
степных и пустынных грызунов.
Несомненно, более или менее сильное влияние на размноже-
ние оказывают и другие факторы среды. Обилие хищников, а
также особей своего вида, снижает плодовитость. Сильная глист-
ная инвазия может быть причиной бесплодия, высокого процента
резорбирующихся эмбрионов, отставания в развитии молод-
няка. К снижению плодовитости или полной стерильности при-
водят в некоторых случаях резкие изменения условий сущест-
вования при акклиматизации, доместикации, переводе на кле-
точное содержание’ и т. п.
ЛАКТАЦИЯ
В период беременности происходит постепенное увеличение
размеров млечных желез (рис. 113), обусловленное разраста-
нием млечных альвеол и каналов. Ко времени родов млечные
железы достигают своего максимального развития и становятся
1 Хорошо размножаются в неволе медведи, волки, гиены, львы, многие
представители семейства оленей, большинство грызунов. Плохо размножаются
в неволе обезьяны, слоны.
282
способными выполнять функцию образования и выделения мо-
лока. Таким образом начинается последняя стадия полового
цикла самки — стадия лактации (лат. lacto — кормлю молоком).
Выделение молока стимулируется в основном действием соот-
ветствующих гормонов и актом сосания — одним из первых
врожденных (безусловных) рефлексов детенышей. При сосании
раздражаются нервные окончания млечной железы, что рефлек-
торно передается гипофизу, вызывая выделение им лактогенных
гормонов, способствую-
щих молочной секреции.
В первые дни лакта-
ции молочные железы вы-
деляют молозиво—гус-
тую, вязкую жидкость, ха-
рактеризующуюся повы-
шенным по сравнению с
молоком содержанием со-
лей, белков, витаминов и
некоторых других ве-
ществ. Молозиво облада-
ет также бактерицидными
свойствами. В дальней-
шем, когда новорожден-
ные несколько окрепнут,
113. Вымя кормящей самки сибир-
ского козла (Capra sibirica)
(рис. А. Н. Формозова)
овой железы. У клоачных моло-
молозиво заменяется мо-
локом, химический состав Рис
которого сильно варьиру-
ет у разных групп зверей.
Молочная железа — гомолог пс
ко еще мало отличается по составу от пота, у вышестоящих
млекопитающих молоко приобретает свои характерные особен-
ности (табл. 7).
Состав молока имеет важное значение для роста и развития
детенышей. Установлена, например, прямая зависимость скоро-
сти роста молодняка от содержания белков в молоке.
Продолжительность лактационного периода зависит от спо-
собности детенышей разных видов в различном возрасте перехо-
дить к другим видам питания. Так, чрезвычайной длительностью
характеризуется лактационный период моржей, так как свойст-
венное этому виду питание донными организмами становится
возможным лишь при наличии достаточно длинных клыков (они
развиваются лишь на втором году жизни). У волков, лисиц, со-
болей, рано начинающих подкармливать свое потомство отрыж-
кой собственной пищи, срок лактации сравнительно короток. Се-
верный олень имеет исключительно питательное молоко (табл. 7),
и лактация у этого вида длится 4—5 месяцев; у коровы, лоша-
ди— 9—10 месяцев.
283
Таблица 7
Состав молока у некоторых видов млекопитающих (в %)
(сводные данные из различных источников)
Млекопитающие Состав молока Калорийность молока (Кал.)
Вода Жир Велок Сахар Минераль- ные ьеще- ства
Корова 88,0 3,4 3,3 4,7 0,7 713
Коза 86,2 4,5 4,0 4,6 0,8 760
Лошадь 90,0 2,0 2,5 6,5 0,3 500
Северный олень . . . 67,7 17,1 10,9 2,8 1,5 2500
Свинья 84,0 4,6 7,2 3,1 1,1 872
Кролик 69,4 10,5 15,5 2,0 2,6 1708
Собака 77,0 9,3 9,7 3,1 0,9 1411
Кошка 82,2 3,3 9,1 4,9 0,5 915
Гренландский тюлень . 43,8 42,8 12,0 — 0,9 —
Дельфин 48,8 43,7 — — 0,5 —
Кит-полосатик .... 54,1 30,6 13,1 — 1,4 (2600)
В большинстве случаев матери кормят новорожденных лежа,,
реже сидя (зайцы) или стоя (большинство копытных, иногда
волки). У не имеющих сосков клоачных детеныши слизывают
молоко с поверхности железистого поля на брюхе матери. Не-
доразвитые, неспособные самостоятельно сосать новорожденные
кенгуру и некоторых других сумчатых как бы прирастают к со-
ску, сильно разбухающий конец которого заполняет всю рото-
вую полость. Молоко впрыскивается в рот такого детеныша при
помощи сокращения специального мускула, сжимающего млеч-
ную железу матери. У китов детеныш захватывает концом рта
сосок, выдвигающийся в подсосный период из особой кармано-
образной складки брюха матери, и молоко, как у сумчатых,
струей впрыскивается ему в рот сокращением особых мус-
кулов. .
В случае гибели сосунков остающееся в молочных железах
молоко утрачивает свои свойства — «перегорает». При нормаль-
ном окончании лактации молочные железы постепенно переста-
ют функционировать, происходит их обратное развитие — инво-
люция.
Лактация вызывает у самок части видов зверей длительную
задержку течки и овуляции.
У других видов эти процессы нормально продолжаются, и
оплодотворение в это время может привести к новой беремен*
ности, которая протекает однако тем медленнее, чем больше со*,
сунков у матери.
284
РОСТ И РАЗВИТИЕ МОЛОДНЯКА
В развитии млекопитающих сменяют друг друга три этапа:
дозародышевый (овогенез и сперматогенез), зародышевый (эм-
бриональный), уже рассмотренные выше, и послезародышевый
(постнатальный).
Молодое животное растет значительно медленнее, чем эм-
брион. Рост молодого животного графически может быть пред-
ставлен в виде логистической кривой, I. е. симметричной S-об-
разной кривой, вначале быстро поднимающейся, а затем, с се-
редины снова снижающейся (рис. 114, Д).
Рис. 114. А—логистическая кривая роста (N) и ее
производная — кривая прироста D; х — место пере-
гиба на половине роста; Б —схема изменений интен-
сивности роста млекопитающих в разных направ-
лениях:
а — рост в длину; б — в высоту; в — в ширину и глубину;
г — рождение травоядных; д — рождение свиней, грызу-
нов и плотоядных (по Пшеничному)
Наибольшая интенсивность роста наблюдается в лактацион-
ный период. При переходе молодняка на самостоятельное пита-
ние и в период полового созревания рост задерживается.
Вследствие того, что детеныши у разных видов млекопитаю-
щих рождаются не одинаково развитыми, интенсивность роста
тела и его частей у них также существенно различается.
Таким образом, определяются типы роста, характерные для
отдельных видов. Так, у видов, рождающих после того, как рост
эмбриона в высоту начинает спадать, конечности успевают
настолько вырасти, .что детеныш появляется на свет относитель-
285
но высоконогим (копытные). В противоположность этому, у ви-
дов, потомство которых рождается на более ранних стадиях, ко-
гда кривая роста в высоту находится еще на середине подъема,
рост конечностей происходит в основном уже в постэмбриональ-
ный период, и новорожденный детеныш характеризуется отно-
сительно низкими ногами (рис. 114,Б).
На разных этапах жизни млекопитающие, как и все вообще
организмы, требуют различных условий внешней среды: усло-
вий питания, газообмена и пр. Эти условия резко отличаются в
утробный и послеутробный периоды развития.
Рис. 115. Слева — зародыш ехидны с яйцевым зубом; справа—
новорожденный кенгуру (обратить внимание на сильное
развитие передних конечностей)
В ходе утробного развития различают этапы яйцеводный, за-
родышевый, предплодный и плодный (Матвеев, 1956). Постна-
тальный период развития также распадается на ряд этапов, от-
личающихся интенсивностью относительного роста, изменением
пропорций тела. Стадия молозивного питания сменяется ста-'
дней молочного и затем — самостоятельного питания. Голые но-
ворожденные некоторое время пойкилотермны — температура их
тела и интенсивность обмена веществ зависят от температуры
окружающей среды. По мере развития они превращаются в го-
мотермных животных с установившейся постоянной температу-
рой тела.
'286
Клоачные (Monotremata) размножаются путем откладки яиц,
которые или высиживают в гнезде (утконос), или вынашивают
в кожистой сумке (ехидна). Сумчатые (Marsupialia) рождают
недоразвитых, очень мелких детенышей (рис. 115). Они дажене
могут сосать мать и держатся на соске благодаря срастанию с
ним краев рта. Плацентарные (Placentalia) рождают более или
менее развитых детенышей, способных сосать молоко. Степень
развития и величина новорожденного у этой группы зверей связа-
ны с высокой организацией этих животных, а также с продол-
жительностью беременности и образом жизни. Чем дольше
плод находится в матке (не считая латентной фазы), тем круп-
Рис. 116. Однодневные детеныши: кролика (слева) и зайца-беляка (справа)>
(по Мантейфелю)
нее и более развитым появляется он на свет (рис. 116). У китов,
беременность которых длится около года, новорожденные до-
стигают трети или даже половины длины тела матери и хорошо .
сформированы. Небольшой грызун морская свинка после 68—
70 суток беременности рождает относительно крупных, зрячих,
покрытых шерстью детенышей. С другой стороны, кенгуру ве-
личиной с козу — после 38—40 дней беременности рождает-
эмбрионоподобного детеныша величиной с наперсток. Опос-
сум— величиной с кошку после 12—13-дневной беременности
производит на свет детенышей, не превышающих по размеру
пчелу (рис. 117).
Большинство зверей рождает слабо развитых, слепых, не спо-
собных к передвижению детенышей (большинство насекомояд-
ных, хищных, грызунов); у небольшой части видов детеныши
287
появляются на свет настолько развитыми, что вскоре способны
следовать за матерью — копытные (рис. 118), киты, некоторые
грызуны. Приспособительное значение этих особенностей оче-
видно. Рождение сравнительно хорошо сформированных дете-
нышей у копытных, зайцев и других открыто выводящих потом-
ство зверей связано с тем, что молодые у них очень скоро пере-
ходят к образу жизни, мало отличающемуся от того, который,
ведут взрослые животные. Самка бурого медведя приносит де-
тенышей зимой, находясь в берлоге, и, ничем не питаясь, кор-
мит их молоком до наступления весны. Новорожденные медве-
жата очень малы: вес каждого в среднем около 500 г, что со-
ставляет десятые доли процента от веса взрослой самки. Малая
Рис. 117. Новорожденный опоссум Didephis virginiana в сравнении с медонос-
ной пчелой (при одном увеличении) (по Гамильтону). Детеныши сумчатых
крайне малы и по своей недоразвитости напоминают эмбрионов высших
млекопитающих
величина новорожденных медвежат, несомненно, имеет адаптив-
ный характер. Выкармливание молоком более крупных, детены-
шей только за счет использования имеющихся в организме ма-
тери резервных веществ вызвало бы ее истощение. Относительно
очень малы новорожденные белого и гималайского медве-
дей, тоже появляющиеся на свет зимой, и длительное время вы-
кармливаемые самкой, находящейся в состоянии зимней дремо-
ты. Для китов это определяется специфическими условиями
жизни в воде. Слаборазвитых беспомощных детенышей звери
рождают в убежищах, дающих возможность длительного прё-
288
бывания потомства в относительной безопасности. Пищухи, оби-
тающие в высокогорных областях (Ochotona macrotis, О. rutila),
живут в россыпях камней и трещинах скал и рождают более
развитых детенышей, чем степные виды (Ochotona daurlca, О. pu-
silia), устраивающие глубокие норы.
Приспособительное значение имеют и темпы роста. Суровые
условия, в которых живут полярные и высокогорные животные,
требуют быстрого роста и развития детенышей. Новорожденный
северный олень в первый же день способен (правда, недолго)
Рис. 118. Затаившиеся ягнята джейрана в возрасте 3 дней; самец
и самка. 14 мая 1955 г. Центральная часть пустыни Бет-Пак-Дала,
Казахстан (фот. А. А. Слудского)
следовать за матерью и через несколько дней, потреблять расти-
тельный корм, между тем новорожденный благородный олень
начинает следовать за матерью не раньше недельного возраста.
Быстро растут и развиваются молодые летучие мыши; к кон-
цу третьей недели жизни они приобретают способность к полету,
а 4-недельные зверьки отличаются от взрослых только окрас-
кой.
Чрезвычайно быстро растет и развивается молодняк у мыше-
видных грызунов. Рождающиеся голыми и слепыми обыкновен-
ные полевки, уже в возрасте 4—5 дней покрываются короткой
шерсткой, к 10 дням прозревают, а с 2-недельного возраста пе-.
реходят на самостоятельное питание. У детенышей домовой
мыши глаза открываются в 2-недельном возрасте и с этого вре-
19 Териология 289
Рис. 119. Молочные зубы ново-
рожденного детеныша большой
ночницы. С их помощью дете-
ныши крепко держатся за сос-
цы матери во время ее полета
мени молодые начинают уже выбегать из гнезда и самостоя-
тельно кормиться; в 30—45 дней они переходят к самостоятель-
ной жизни.
Значительные различия в темпах роста на разных стадиях
развития установлены у домашних животных. Так, например,
резкий подъем относительной скорости роста у свиней происхо-
дит в первый месяц после рождения, а у крупного рогатого ско-
та— в конце внутриутробного развития. Для ягнят среднеази-
атской каракульской породы овец характерен чрезвычайно бы-
стрый рост в первый месяц жизни. Только благодаря этому они
успевают нагулять достаточное количество резервных питатель-
ных веществ в курдюке и выдер-
жать голодный зимний период
(Кашкаров и Коровин, 1936).
Из опыта звероферм хорошо из-
вестны существенные различия в хо-
де развития молодняка у американ-
ских норок и серебристых лисиц.
Норки рождаются слепыми, с очень
редким волосяным покровом, беспо-
мощными. Развиваются они доволь-
но медленно: на 20—25-й день у них
прорезываются зубы, открывается
слуховой проход; на 30—35-й день —
открываются глаза. Новорожденные
лисята имеют хорошо развитый во-
лосяной покров, прозревают они уже
на 14—16-й день; в это же время у
них прорезаются зубы и открывает-
ся слуховой проход. Однако в даль-
нейшем росте и развитии молодые
норки несколько обгоняют лисят. Первоначальный суточный при-
вес у них составляет 8—10%, тогда каку лисят — 6—7%. С мо-
мента перехода от чисто молочной к диете с добавлением других
кормов среднесуточный привес у тех и других, резко снижается
(у норок до 3 %, у лисят до 2 %). В дальнейшем темпы роста де-
тенышей обоих видов ускоряются и достигают наибольшей вели-
чины к концу лактационного периода. Прекращение лактации вы-
зывает новое снижение прироста (до 1 %), и лишь по мере полного
приспособления молодых животных к новым условиям питания
прирост у них снова повышается. По достижении норками 3- и
лисятами 4-месячного возраста начинается не прекращающееся
уже снижение среднесуточного привеса, сходящего на нет с
окончанием роста и развития животных1.
1 По материалам наблюдений в Сомовском зверосовхозе Воронежской
области (Барабаш-Никифоров, 1956).
290
Основным фактором среды, определяющим рост и развита^
детеныша в ранний, наиболее критический период жизни, слу-
жит его мать и связанное с ее физиологическим состоянием мо-
лочное питание. Фактор питания сохраняет свое первостепенное
значение и в дальнейшем. Установлено решающее влияние на
рост и созревание молодняка количества и качества молока ма-
тери, витаминности кормов и пр. Кит достигает половой зрело-
сти в возрасте 2—3 лет, слон — в возрасте 15—20 лет. Быстрые
темпы развития морского гиганта обусловлены чрезвычайно вы-
сокой питательностью китового молока и несравненно большим
богатством и полноценностью кормов в океанических водах по
сравнению с сушей.
Некоторые отклонения в темпах роста и развитии детенышей
связаны с их полом. У норок, лисиц и некоторых других зверей
наблюдается начинающееся уже в утробный период отставание
в росте самок от самцов. В дальнейшем эта разница обычно вы-
равнивается. Исключение составляют виды с половым димор-
физмом, у которых отличия сохраняются и могут достигать
большой величины.
Отклонения в темпах роста могут быть связаны и с сезонами
года. Так, например, у молодых полевок, мышей, землероек, до-
стигших самостоятельности осенью в условиях умеренной и
холодной зоны, рост в зимнее время прекращается. Он возобнов-
ляется только с наступлением'весны. Ясно, что судить о возра-
сте мелких зверьков по их весу нужно с большой осторожно-
стью, так как разница в возрасте особей с одинаковой массой
тела, встреченных в начале лета, может достигать 5—7-ме-
сяцев,
С наступлением половой зрелости рост и развитие молодых
замедляется, а затем прекращается, но не у всех. Опоссумы,
мелкие грызуны (полевки, мыши, хомячки), например, продол-
жают расти и после наступления половой зрелости, уподобля-
ясь, таким образом, неотеническим формам. То же наблюдается
у копытных, у китообразных.
Большой интерес представляет зависимость развития детены-
шей от величины выводка. Уже при рождении детеныши в
крупных выводках обычно бывают менее развитыми, чем в не-
больших. Дальнейшее отставание их развития может быть обус-
ловлено несоответствием числа детенышей числу сосков матери
или вообще недостатком молока у нее.
Рост частей тела молодых животных идет неравномерно. На
ранних этапах развития у всех млекопитающих голова непро-
порционально велика, но поскольку рост ее отстает от роста
туловища (в связи с ранним прекращением роста мозга), то с
возрастом пропорции выравниваются.
Большинство молодых млекопитающих обладает относитель-
но более крупными размерами сердца, печени, почек, что связа-
19* - 291
но с интенсивным обменом, необходимым при росте и развитии.
В дальнейшем у большинства млекопитающих происходит
уменьшение относительных размеров этих органов.
Киты в эмбриональном состоянии имеют пропорции тела,
почти не отличающиеся от взрослых животных. Это, по-види-
мому, имеет адаптивное значение, поскольку в водной среде
молодой должен вести тот же образ жизни, что и взрослые
(Земский, 1953).
У новорожденных летучих мышей непропорционально малы
крылья, что позволяет им избегать гибельного воздействия из-
менений температуры среды, поскольку перепончатые крылья
представляют поверхность тела, наиболее интенсивно излучаю-
щую тепло.
Небольшие размеры крыльев, вероятно, связаны также с ус-
ловиям^ родового акта и с тем, что крылья вначале не функцио-
нируют.
У некоторых видов новорожденные имеют специальные
временные приспособления, связанные с особенностями их
жизни.
Заканчивающие яйцевой период своего развития клоачные
имеют подобно птенцам птиц «яйцевой зуб», с помощью кото-
рого проламывают скорлупу для выхода наружу (см. рис. 115).
Новорожденные кенгуру обладают относительно крупными и
сильными передними конечностями, с помощью которых они,
цепляясь за шерсть матери, совершают путь от полового отвер-
стия до сумки. У детенышей сумчатых, кроме того, имеются осо-
бые приротовые органы, которые дают возможность прочно при-
крепиться к соскам; маленьким летучим мышам для той же цели
служат изогнутые назад двух- или трехвершинные молочные
зубы (рис. 119).
Хорошо развитые когти имеет новорожденный ленивец; сей-
час же после рождения он вцепляется ими в шерсть матери.
Детенышу обезьян также свойственен этот врожденный реф-
лекс— появившись на свет, он прицепляется когтями к телу ма-
тери. Эти приспособления развились в связи с жизнью ленив-
цев и обезьян на ветвях, в кронах леса.
Таким образом, среда, в которой проходит развитие млеко-
питающих, существенно изменяется на разных этапах онтоге-
неза. В предзародышевый период средой для половых клеток
служат половая железа и половые пути, в зародышевый —
матка матери, в послезародышевый — факторы, действующие
Вне материнского организма. Оказывая воздействие на среду, в
соответствии с проходимым этапом развития, человек может
влиять на темпы роста, дифференцировку частей тела и органов
развивающегося организма, а также повышать плодовитость,
превращать моноциклических животных в полициклических
.и т. д.
192
ЗАБОТА О ПОТОМСТВЕ (РОДИТЕЛЬСКИЕ ИНСТИНКТЫ)
Ко времени деторождения у самок млекопитающих начинает
проявляться забота о потомстве — родительские инстинкты, вы-
ражающиеся прежде всего в устройстве логовищ, нор и других
убежищ для будущего потомства. Самки, рождающие детены-
шей открыто на поверхности (антилопы, бараны, олени, зайцы
и др.), ко времени родов уходят в малодоступные или густо за-
росшие растительностью места; у стадных оленей самки на
время уединяются; обитающие в северных морях киты мигри-
руют в теплые южные воды, где температурные условия и шти-
левые погоды благоприятны для молодняка. У ластоногих раз-
множение происходит на особых летных залежках: беременные и
щенные самки каланов обосновываются в спокойных, хорошо
защищенных от штормов бухтах, образуя в них однополые груп-
пы. Самки некоторых млекопитающих запасают корм на первые
дни после родов, что. позволяет им в этот трудный период оста-
ваться неотлучно при выводке (хомяк, тушканчики, соболь, ку-
ницы и некоторые др.).
Материнский инстинкт особенно сильно начинает проявляться
с момента появления на свет детенышей. Заботы о потомстве
могут принимать различные формы. Самка ехидны вынашивает
снесенное яйио в образовавшейся на брюхе сумке. Утконос вы-
сиживает 1—2 яйца в норе, где устраивает для этого гнездо. Сам-
ка кенгуру, лежа в момент родов на спине, вылизывает дорожку
между половым отверстием и сумкой (рис. 120), облегчая тем
самым преодоление этого пространства своему эмбрионоподоб-
ному детенышу (в некоторых случаях она сама помещает дете-
ныша в сумку, схватывая его губами); в дальнейшем мать дли-
тельное время (до 8 месяцев) донашивает детеныша в сумке.
Подросший и начавший уже самостоятельно кормиться молодой
долго еще пользуется ею как временным убежищем.
В флоридском океанариуме наблюдали, как самка дельфина-
афалины (Tursiops truncatus) поддерживала своего новорож-
денного в плавучем положении, на поверхности во время первых
его дыхательных движений (в этом ей помогали и другие нахо-
дившиеся тут же самки). Известен случай, когда шимпанзе-мать
до тех пор трясла, подбрасывала и тормошила своего не пода-
вавшего никаких признаков жизни новорожденного, пока он не
начал двигаться и дышать. Обезьяны нередко поддерживают
или подсаживают детенышей при лазании, образуют своими те-
лами «мост», по которому детеныши переправляются с дерева
на дерево и пр.
Благоустройство гнезда', поддержание в нем чистоты, охра-
на выводка также представляют собой яркое выражение роди-
1 Самка кролика утепляет гнездо выщипанным с брюха пухом, другие
звери устраивают подстилку из мягких растительных материалов.
293
тельских инстинктов. Поедание матерью мертвых плодов, кала
детенышей, перевод их из загрязненного убежища в другое, сме-
на подстилки — все это имеет важное гигиеническое значение и
до некоторой степени способствует сокрытию места нахождения
выводка от врагов (устранение сильного запаха). Правда, за-
бота о чистоте проявляется не у всех видов: нора лисицы, логово
волка после подрастания молодых нередко издают зловоние от
вылизывающая дорожку на брюхе — на пути
передвижения новорожденного (X.) из полового
отверстия (XX) в сумку (XXX) (по Метьюзу)
скопляющихся остатков мясной пищи; крайне нечистоплотны
также лесные сони и некоторые другие звери.
Что касается поддержания чистоты тела детенышей, то этот
инстинкт, по-видимому, свойствен всем без исключения млеко-
питающим. ААать часто вылизывает шерсть детенышей, выиски-
вает у них блох. Самки енотовидных собак и барсуки часто вы-
носят маленьких щенков из нор «на воздух» и через некоторое
время снова бережно возвращают в гнездо. У многих зверей
294
самка массирует языком брюшко и промежность неспособных
еще к самостоятельной дефекации новорожденных, что вызы-
вает у них выделение кала (самка тут же заглатывает его).
Оставляя логово или гнездо, родители прикрывают детены-
шей материалом подстилки (дикая свинья, ондатра, белка) или
закупоривают входное отверстие норы (бобр, тушканчики). Воз-
вращаясь к выводку, родители обычно некоторое время задер-
живаются в отдалении, обходят логово кругом, проверяя отсут-
ствие опасности (волк, лисица). В период вывода потомства ма-
терые волки, как правило, не нападают на домашний скот,
пасущийся вблизи логова; если это «правило» нарушается, то
обычно не взрослыми животными, а задержавшимися недалеко
от логова переярками.
Непослушных детенышей родители «наказывают», приводя
этим в повиновение. Наблюдая, например, лисий выводок у
норы, можно сделаться свидетелем того, как один из родителей,
схватив замешкавшегося на поверхности после тревожного сиг-
нала детеныша, сильно встряхивает его несколько раз и утаски-
вает в нору. Обезьяны пользуются такими «воспитательными»
приемами по отношению к непослушным детенышам, как шлеп-
ки, покусывание, подталкивание, потягивание за руку и пр.
К концу лактации родители начинают постепенно переводить
детенышей на корм взрослых, выводя их на места кормежки или
поднося пищу к гнезду. При этом хищники часто доставляют
своему выводку полузадушенную живую еще добычу. Расправа
с ней вырабатывает у маленьких хищников приемы и охотничьи
навыки, необходимые для их. будущей самостоятельной жизни.
В случае частого беспокойства, затопления логова талой или
полой водой, накопления паразитов в гнезде или истощения за-
пасов корма в данном районе -родители переносят или перево-
дят своих детенышей в другое место. Чаще это делает самка,
держа детеныша в зубах за складку кожи на загривке, на боку
или на животе. Обезьяна переносит детеныша с помощью пе-
редних конечностей, если детеныш не держится на ней сам
(рис. 121). Медведица забирает в рот всю голову перетаскивае-
мого новорожденного медвежонка. Губами за загривок держат
детенышей многие грызуны и насекомоядные. У ондатры дете-
ныши так прочно присасываются к соскам, что, спасаясь от
врага и ныряя в воду, мать сразу уносит на себе весь выводок'.
У некоторых млекопитающих самки более или менее посто-
янно носят детенышей на себе.-Держась на спине матери, пере-
мещаются молодые опоссумы, коала, бегемоты, муравьеды, яще-
ры; детеныши ленивцев, каланов, обезьян помещаются на груди
1 Самки белобрюхой белозубки и некоторых других землероек переводят
своих подросших детенышей «караваном»: молодые зверьки последовательно
держатся за шерсть впереди идущего, а первый — за шерсть матери [Бурльер
(Bourliere), 1951; Циппелиус (Zippelius), 1958 и др.].
295'
I
।
I
I
»
Рис. 121. Способы переноски или длительного ношения детенышей:
I — лисица; 2 — кенгуру; 3 — опоссум; 4 - ленивец; 5 — ящер; 6 -летучая мышь:
7—калан; 8—гиббон
296
или на брюхе (обезьяны носят их и на спине). Молодые лету-
чие мыши удерживаются на матери во время ее полета, прице-
пившись зубами к соску или к особому придатку паховой обла-
сти, а когтями к телу.
Особенно ярко проявляются родительские инстинкты при не-
посредственной опасности, грозящей гибелью потомству. У неко-
торых видов мать в таких случаях пытается отвлечь внимание
врага. Так, например, поступают соболи, лисы, хорььи. ь иногда
и зайцы, олени, лоси, детеныши которых в момент опасности за-
таиваются.
Самки моржей и китов даже тогда, когда им самим грозит
смерть, не покидают детенышей. Стадо копытных при нападении
хищников образует вокруг молодых защитное кольцо (рис. 122).
Рис. 122. Защитное кольцо взрослых овцебыков, готовых отразить нападение,-
волков; в центре кольца молодняк
В крайнем случае мать может самоотверженно защищать потом-
ство, что свойственно многим хищникам, а также моржам, ки-
там, диким свиньям и даже зайцам и белкам. Самка ящера при
нападении врага помещает своего детеныша на брюшную по-
верхность и отбивается длинным хвостом, хорошо защищенным,
острыми чешуями.
Самки разных видов иногда проявляют заботу о чужих де-
тенышах. Чаще это бывает в случае потери собственного потом-
ства в период лактации, когда молоко начинает «перегорать».
Но иногда и вне периода лактации у самок проявляется мате-
ринский инстинкт по отношению к чужим находящимся побли-
зости осиротевшим детенышам (в условиях содержания в неволе
это наблюдалось у обезьян, лисиц и некоторых других зверей,
а в естественных условиях у джейранов, сайгаков, косуль и
др.1). У ластоногих, образующих плотные детные залежки, и у
1 Интересный случай выкармливания собакой маленькой обезьяны описан
М. Р. Сани (1959). В одной из деревень Шаньских гор потерявшая свою
297
некоторых других видов, выводки которых свободно перемеши-
ваются между собой, самки могут кормить как своих, так и чу-
жих детенышей *. Отмечались случаи выкрадывания песцами
детенышей из соседних выводков или совместного воспитания
двумя-тремя самками объединенного в одной норе сборного вы-
водка. Чаще, однако, самки проявляют враждебность по отно-
шению к чужим детенышам 2.
Материнский инстинкт не всегда проявляется сразу в полную
силу. Молодые самки обычно меньше заботятся о своем вывод-
ке, чем старые. Самка бурого медведя, имсюшая новорожден-
ных медвежат, в случае опасности легко покидает их, но не-
сколько позже, когда медвежата начинают следовать за ней,
самоотверженно защищает их от врага.
Доля участия обоих родителей в заботах о потомстве не
одинакова. Самцы большинства моногамных видов в той или
.иной мере участвуют в воспитании детенышей, выкармливании
их, охране гнезда и гнездового участка. У полигамов, как пра-
вило, заботится о выводке только самка.
Самцы некоторых видов (например, тигров, соболей) не
только не заботятся о своих детенышах, но даже пожирают их,
если самка не успеет вовремя укрыть выводок или отогнать
-самца.
Редкое явление представляют самки, которые не проявляют
заботы о потомстве (если не считать лактации), легко покидают
детенышей в случае опасности и не всегда к ним возвращаются.
Наоборот, часто молодые самки проявляют излишнюю заботу,
затаскивая детенышей до смерти.
Общая особенность млекопитающих — более или менее рев-
ностная охрана самкой или обоими родителями места нахожде-
ния выводка, а в некоторых случаях и всего используемого семь-
ей участка. Однако и эта особенность сильно варьирует даже в
пределах одного вида. Пары песцов обычно проявляют крайнюю
нетерпимость ко всем другим песцам, появляющимся в их гнез-
довом районе, но в то же время у них наблюдается и объедине-
ние нескольких семей в одной норе (значительную роль при
этом играет состояние кормовой базы).
мать обезьянка направилась к собаке, кормившей щенят, и принялась вместе
с ними сосать молоко. Войдя в новую семью, обезьянка, как более агрессив-
ная по природе, получала свою порцию в первую очередь. Прицепившись к
брюху собаки, она не разлучалась с ней даже во время ее отлучек от вывод-
ка в поисках корма. Собака относилась к ней так же заботливо, как и к соб-
ственным своим детенышам.
1 Но у котиков, несмотря на то, что новорожденные сбиваются вместе
в очень плотные большие группы, каждая самка кормит только своего де-
теныша, погибающего в случае ее смерти.
2 Самки многих зверей отличают своих детенышей от чужих только по
запаху. Сообщив детенышу запах соответствующей самки, можно добиться
принятия его этой самкой.
298
Таким образом, забота о потомстве — это цепь последова-
тельных рефлексов, выработавшихся в процессе эволюции, обес-
печивающих сохранение вида. Выпадение хотя бы одного из
звеньев этой цепи вызывает исчезновение заботы о потомстве, а
иногда и резкие извращения материнского инстинкта.
В практике животноводства и звероводства хорошо извест-
ны случаи поедания матерью своих живых детенышей, что
обычно обусловлено либо патологическими нарушениями обмена
веществ на почве неправильного кормления, либо обостренной
нервной реакцией на какие-либо внешние раздражения (поеда-
ние приплода особенно часто наблюдается у свиней, кошек, ли-
сиц, норок, кроликов). Аналогичные случаи иногда имеют место
и у .диких зверей в природных условиях.
У хищников больные или отставшие в развитии детеныши
(«заморыши») обычно подвергаются жестокой расправе, в кото-
рой участвуют как родители, так и другие члены выводка.
Как нормальное явление, ослабление и исчезновение роди-
тельских инстинктов у взрослых происходит по мере приобрете-
ния молодыми животными способности к самостоятельной
жизни.
РАСПАД СЕМЕЙ
По мере того как молодые становятся все более самостоя-
тельными, родительская привязанность к ним постепенно осла-
бевает. Наконец, родители начинают отгонять от себя детены-
шей, ускоряя тем самым их переход к самостоятельной жизни,
который у разных видов наступает в разные сроки. Молодые ди-
кие свиньи остаются при матери до достижения ими половой
зрелости, молодые тигры — вплоть до следующей течки матери,
что происходит только через 2—3 года. Долго держатся с ро-
дителями волчьи выводки. Самка бурого медведя ходит с мед-
вежатами на протяжении всего лета и вместе с ними залегает
осенью в берлогу. Семьями зимуют сурки. До следующей весны
остаются при матери детеныши рыси, выдры и многих других
относительно крупных зверей.
Распад семей мелких видов происходит уже к осени или еще
раньше (у мышевидных грызунов через 1—2 месяца после рож-
дения детенышей). Сроки распада семей в значительной степени
зависят от обеспеченности кормом. По мере истощения количе-
ства пищи в районе норы, не опекаемые больше родителями
молодые песцы, лисицы все более удлиняют свои охотничьи вы-
лазки, подолгу задерживаются вдали от норы и, наконец, со-
всем расстаются с ней. В начальный период самостоятельной
жизни молодые зверьки очень неосторожны, и значительная
часть их становится добычей хищников или гибнет от случайных
причин. '
299
Обилие кормов отдаляет распад семей. У тундровых песцов
ранний распад семей в связи с недостатком кормов — обычное
явление. Молодые зверьки, лишившиеся заботы родителей и
мало еще приспособленные к самостоятельному существованию,
часто голодают, плохо развиваются и в большом числе поги-
бают. В соответствии с большими требованиями к кормовой
базе крупные семьи, как правило, распадаются быстрее мелких.
У многих видов млекопитающих распад семьи начинается ухо-
дом самки, вслед за тем уже разбредается и выводок.
Таким образом, молодые животные переходят к самостоя-
тельной жизни в разном возрасте:
рыжая лесная полевка
малый суслик
белка, заяц-русак
степной хорь
лисица
волк
сурок-байбак
тигр, дикий кабан
в возрасте около 3 нед.,
« « 1 мес.,
« «2 мес.,
« «3 мес.,
« « 4—5 мес.,
« « 10 — 11 мес
« « 1 года,
« « 2—3 лет
.Дальнейший этап жизни молодняка — расселение.
Начавшись с беспорядочных передвижений, расселение по-
степенно может приобретать направленность и превращаться в
миграции.
РАССЕЛЕНИЕ МОЛОДНЯКА
У малых сусликов в глинистой полупустыне Заволжья весь
период расселения (по наблюдениям за мечеными особями в
1951 г.) от момента распадения выводков и до поселения моло-
дых зверьков в собственных постоянных норах занял около
трех недель (23—30/V—21—22/VI) (Ходашова и Солдатова,
1955). В открытой полупустыне суслики, как грызуны, не обла-
дающие способностью к быстрому бегу, не могут выселяться
на большое расстояние от гнездовой норы. Преобладающая
часть молодых (74%) осела всего на расстоянии 20—90 м, по-
селившись в пределах своего выводкового участка или в непо-
средственной близости от него. Два зверька поселились на рас-
стоянии 104 м от выводковой норы, один — в 126 м и один —
в 278 м. Часть молодых особей совсем не выселялась: три вы-
рыли норы на холмиках земли материнских нор, а один всего-
в 12 м от нее.
В конце июня, когда был проведен заключительный отлов,
на опытном участке, оказалось 18 вселившихся молодых (неме-
ченых) сусликов и всего 11 меченых (27% от первоначального-
их числа). Остальные меченые зверьки, видимо, погибли, так
как в противном случае они бы попали в учет на прилегающих
участках. Новые суслики вселились в пограничную часть терри-
тории, окружающую опытный участок. Следовательно, расселе-.
300
яие молодых зверьков в равнинной местности с однообразными
на всем протяжении условиями идет во всех возможных на-
правлениях.
В разных частях ареала сроки и продолжительность рассе-
ления молодых сусликов могут сильно отличаться, так как за-
висят от обеспеченности кормом и скорости выгорания расти-
тельности. В южных ковыльных степях Ростовской обл., где
растительность значительно богаче, чем в глинистой полупу-
стыне Заволжья, период расселения длится 45—50 дней и за-
канчивается в начале июля. Кроме того, продолжительность
времени расселения сокращается в засушливые годы и растя-
гивается во влажные.
У грызунов, приносящих несколько пометов в течение сезона
размножения, расселение молодых идет как бы волнами, сле-
дующими одна за другой. Массовое мечение рыжих полевок и
многочисленные повторные их отловы в Михневском р-не Мо-
сковской обл. показали, что наиболее интенсивно передвигаю-
щейся группой в течение всего сезона наблюдений были моло-
дые созревающие зверьки. В июне это были самцы и самки
(соответственно 50 и 29%) первого выводка (в это время коче-
вало всего 10% взрослых перезимовавших полевок). В июле
особи первого выводка все еще продолжали перемещаться в по-
исках мест, где можно осесть, и началось расселение молодых
второго выводка. В августе и сентябре старые полевки совсем
не кочевали, и основную массу перемещающихся зверьков со-
ставляли молодые второго и третьего выводков.
|0 К осени стало заметным общее сокращение числа мигран-
тов, что свидетельствовало об оседании полевок на постоянных
зимних участках и о начале подготовки к зиме — собирание за-
пасов, постройка теплых гнезд (Н. Наумов, 1951). Большинство
молодых рыжих полевок оседает на новых местах, расположен-
ных в немногих десятках и в редких случаях — сотнях метров
от выводкового гнезда.
Молодые особи крупных видов полевок при распаде семей
расселяются значительно дальше. Из 17 меченых молодых он-
датр на озерах северо-запада штата Айова только три высели-
лись за 3,2—5,2 км, остальные 14 осели в пределах 90—855 м
от родительской хатки [Эррингтон (Errington), 1944].
Молодые меченые ондатры в дельте р. Или, по наблюдениям
А. А. Слудского (1948), при частичном осеннем расселении
оседали в 200—360 м от родительской норы или хатки. У этого
вида полный распад семьи происходит только на следующую
весну, так как совместная жизнь самки и ее 8—10 молодых
в одной хатке дает целый ряд преимуществ. Весеннее расселе-
ние созревших молодых ондатр, совпадающее по времени с на-
чалом гона, происходит в дельте р. Или в марте. В это время
встречали расселяющихся ондатр даже в несвойственной им
30)
обстановке — в пустыне на расстоянии 4—6 км от ближайших
водоемов. Вниз по течению они уплывают значительно дальше-,
так как средняя скорость расселения основной массы ондатр
за год даже в таежной зоне СССР равна 9—10 км. Отдельные
особи за один сезон уходят по течению реки на 100—150 км
и более. Именно благодаря этой биологической особенности и„
конечно, высокой плодовитости акклиматизированная у нас
ондатра очень быстро заселила огромную площадь.
Большая подвижность популяций многих видов, обусловлен-
ная расселением молодых, постоянное перемешивание особей,,
происходящих из разных выводков, имеет существенное биоло-
гическое значение. Прежде всего, этим предотвращается близ-
кородственное скрещивание (инбридинг). Расселение дает
возможность наиболее полно осваивать все пригодные для жиз-
ни вида участки и обеспечивает относительно равномерное
распределение особей и пар в сходных по условиям биотопах.
Вместе с тем беспорядочное перемещение зверьков усиливает
внутривидовые и межвидовые контакты, что содействует быст-
рому распространению возбудителей болезней, широкому обме-
ну эктопаразитами и т. п. Так, например, молодые полевки-эко-
номки, расселяющиеся из травяных болот, способны выносить
лептоспирозную инфекцию в другие стации, в частности на па-
хотные угодья. Это увеличивает возможность заражения людей
(Лаврова и Андреева, 1960). Молодые зверьки нередко отли-
чаются большей восприимчивостью и меньшей резистентностью
в отношении многих инфекций, поэтому с моментом массового
расселения совпадает начало эпизоотий (например, чумы у ма-
лых сусликов и т. п.).
ГЕОГРАФИЧЕСКАЯ, СЕЗОННАЯ И МНОГОЛЕТНЯЯ
ИЗМЕНЧИВОСТЬ ХОДА РАЗМНОЖЕНИЯ
В свете приведенных выше данных о влиянии условий среды
на физиологию размножения, понятны причины изменчивости
хода размножения млекопитающих в разных географиче-
ских районах. Даже в соседних биоте пах, отличающихся микро-
климатом, кормовыми и другими условиями, в размножении,
особей одного и того же вида могут быть существенные откло-
нения. Особенно ярко это выражено у мелких грызунов (от
мышей и.мелких полевок до ондатры включительно). Ондатры,
населяющие малокормные водоемы, приносят в течение года
не более одного помета, тогда как в богатых кормами водоемах
' у них бывает два помета. В местообитаниях с плохими защит-
ными условиями рост и половое созревание зверьков идут-
быстрее.
Естественно, что в разных географических районах отклоне-
ния интенсивности размножения соответствующих популяций.
02
вида, оказываются еще более существенными. Так, напримещ
общественные полевки в полынно-злаковой степи предгорий
Азербайджана (высота 600 м над уровнем моря) размножают-
ся круглый год, но преимущественно с февраля по декабрь,
причем наибольшее количество беременных самок (от 50 до
90%) у них бывает в период с мая по сентябрь. Зима здесь от-
носительно мягкая, осадки благодаря значительной высоте над
уровнем моря выпадают в большем количестве, чем на равшше.
В той же республике, но на низменности с полынно-солянко-
вой полупустыней, уже в феврале бывает более 20% беремен-
ных самок, а в марте наступает весенний максимум (90% самок
с эмбрионами). Однако к июню сильная жара, засуха и выго-
рание растительности полностью подавляют половую актив-
ность полевок. Новая волна размножения наступает в сентяб-
ре, вслед за окончанием жары и осенней вегетацией. Причем
в октябре число самок с эмбрионами превышает 80%, а к де-
кабрю снижается до 70%, но держится еще на достаточно вы-
соком уровне в январе. В этой зоне избыток тепла и недоста-
ток влаги летом представляют основные особенности среды,
ограничивающие плодовитость общественной полевки. В неко-
торые годы сильное понижение температуры может привести
к некоторому сокращению доли участвующих в размножении
зверьков и среди зимы.
Период интенсивной жизнедеятельности общественной по-
левки в зоне полупустыни восточной части Закавказья совпа-
дает с ранневесенним периодом вегетации эфемеретума и разно-
травья.
В расположенной значительно севернее глинистой полупу-
стыне Заволжья (Западный Казахстан) у популяции общест-
венной полевки свои особенности годового цикла жизни —
в полном соответствии с местными условиями существования.
Семилетние наблюдения К. С. Ходашовой (1960) показали, что-
размножение этого грызуна начинается здесь в апреле и идет
наиболее интенсивно по конец мая. Затем наступает перерыв,
продолжительностью около трех месяцев, обусловленный жар-
кой погодой, засухой, выгоранием трав. Осенний сезон размно-
жения продолжается с начала сентября по конец октября, пока
не наступят холода. Однако при выпадении достаточно глубо-
кого снега (правда, это бывает далеко не ежегодно), предостав-
ляющего укрытие от холода, размножение возобновляется, но-
вее же не принимает массового характера. Наибольшее коли-
чество беременных самок здесь бывает весной. Именно от ин-
тенсивности размножения в апреле—мае, которая связана с ис-
пользованием свежего урожая трав, появляющегося за счет
увлажнения почвы зимними осадками, зависит уровень числен-
ности общественных полевок в этой зоне.
Зима здесь суровее и продолжительнее, чем в Закавказье,
лето засушливо, как и во всех наших полупустынях. Поэтому
большую роль в ограничении периода размножения заволжской
популяции общественной полевки играет не только летний
избыток тепла и нехватка влаги, но и длительный зимний не-
достаток тепла.
В процессе исторического развития у млекопитающих выра-
боталось целесообразное приспособление к поддержанию чис-
ленности вида: плодовитость меняется в соответствии с усло-
виями сохранения потомства. Плодовитость особей одного и
того же вида или близких форм в большинстве случаев выше
в северных и вообще неблагоприятных условиях и ниже в зна-
чительно более мягких и благоприятных.
В отличиях плодовитости многих полевок и мышей (видов
разной специализации) легко обнаружить параллелизм и связь
с зональными географическими условиями.
Лесные мыши и рыжие полевки имеют почти вдвое мень-
шую плодовитость по сравнению с видами — обитателями лугов
и степей. Главным образом это происходит за счет удлинения
промежутков между выводками до 50—60 дней (вместо 25—30).
Лесные серые полевки (экономки и темные) по этому показа-
телю занимают промежуточное положение. Можно считать, что
отмеченная закономерность связана с лучшей защищенностью
лесных видов от неблагоприятных воздействий погоды и хищ-
ников, т. е. с пониженной смертностью.
Слепушонка, населяющая степи и полупустыни — открытые
биотопы, защищена тем не менее от многих неблагоприятных
воздействий тем, что живет в сложной системе подземных хо-
дов, редко появляясь на поверхности. Плодовитость этого земле-
роя невысока и тоже за счет больших перерывов между появ-
лением выводков (Н. Наумов, 1948).
На Крайнем Севере среднее число молодых в помете у раз-
ных видов полевок значительно выше, чем у особей тех же ви-
дов из популяций, обитающих в более южных зонах. Среднее
число эмбрионов пашенной (темной) полевки в Субарктике — 8,1,
экономки—около 8, красной полевки — 9,8, а полярного под-
вида узкочерепной полевки (Microtus gregalis major) — 9,9
(Шварц, 1959).' Защитные условия мест обитания этих зверьков
в лесотундре и тундре значительно хуже, чем в лесной зоне,
а неблагоприятные типы погоды повторяются чаще.
Плодовитость типичного арктического хищника — песца почти
в 2 раза выше, чем у лисицы, распространенной преимущест-
венно в зонах южнее тундры. Взрослая самка лося — обитателя
тайги севера Евразии, как правило, приносит два теленка,
тогда как самка самого мелкого из наших подвидов — чеечвий-
ского лося (Alces alces pjlzenmayerl) жителя Уссурийского
края и Приамурья — только одного.
304
Географическое положение местности обусловливает наступ-
ление сроков и продолжительность генеративного периода. У зай-
цев, белок, ондатр и некоторых других зверей количество по-
метов по направлению с юга на север уменьшается, величина
же выводка увеличивается, в результате чего при сокращении
периода размножения на севере разница в общей плодовитости
несколько сглаживается.
Хорошо известна сезонная изменчивость хода размножения,
связанная с изменениями метеорологических условий в разные
сезоны года. В известной мере эти изменения зависят от дли-
тельности светового дня, воздействующей на половые органы
через центральную нервную систему. Свет сигнализирует о на-
ступлении в природе сезонных изменений раньше, чем темпера-
тура или кормовая база, которые в* момент начала изменений
половой сферы могут еще оставаться неизменными.
Худшими по метеорологическим условиям и обеспеченности
кормами в северной части Евразии и Америки обычно бывают
ранневесенний, позднеосенний и зимний сезоны, лучшими — позд-
невесенний и летний. Связанную с этим сезонную изменчивость
плодовитости можно показать на примере зайцев. Ранневесен-
ние и осенние выводки у них, как правило, уступают в величи-
не поздневесенним и летним. По Ц. Олдоусу (Aldous, 1937), у
американского беляка на одну беременную самку весной прихо-
дится в среднем 2 эмбриона, летом 3 и осенью 2,8. У наших
беляков в весенне-летних пометах принимают участие все самки,
в осеннем — лишь около половины. Соотношение полов среди
эмбрионов тоже подвержено у беляка сезонным изменениям:
в лучший'—летний сезон у самок беляка преобладают эмбри-
оны мужского пола, в худший — ранневесенний — основную
часть эмбрионов составляют самки. То же наблюдается и у
многих других грызунов. Не выражена эта закономерность (или
проявляется в значительно меньшей степени) только у синант-
ропных грызунов, живущих в относительно однородных усло-
виях круглый год.
Изменения сочетаний метеорологических и других условий
отдельных лет, влияя на половую деятельность животных, тем
самым определяют многолетнюю изменчивость хода их раз-
множения.
РОДСТВЕННОЕ СПАРИВАНИЕ, МЕЖВИДОВЫЕ
И МЕЖРОДОВЫЕ ПОМЕСИ
В результате распада семей и расселения молодняка спари-
вание обычно происходит между особями, происходящими из
разных семей. При 'Таком неродственном или отдаленно-родст-
20 Териология
305
венном спаривании обеспечивается наибольшая жизненность1
потомства.
В тех случаях, когда расселение почему-либо затруднено
или вовсе невозможно, например, для лесных животных—в не-
больших изолированных островках леса, или для полуводных
зверей — в изолированных водоемах, в спаривание вовлекаются
родственные особи вплоть до членов одной семьи. О значении
родственного спаривания у диких зверей в природных условиях
мнения зоологов расходятся. Для окончательного решения это-
го вопроса требуются специальные исследования.
В природе инбридинг в значительной степени устраняется
некоторыми особенностями биологии животных. Так, неодновре-
менное половое созревание самцов и самок одного выводка
исключает у многих млекопитающих возможность оплодотворе-
ния самок своими братьями; молодые самцы раньше покидают
выводок и ко времени полового созревания своих сестер успе-
вают уже удалиться на более или менее значительное расстоя-
ние от них.
С давних времен установлено появление ценных качеств
у гибридов — помесей, полученных в результате скрещивания
особей не только неродственных, но даже принадлежащих
к разным породам и систематическим категориям. Успешно осу-
ществляются межвидовая и межродовая гибридизация в усло-
виях звероводческих хозяйств. Получены, например, помеси
между соболями и куницами, между лисицами и песцами
(к лисо-песцовым гибридам принадлежит платиновая лисица).
Среди диких млекопитающих очень обычно скрещивание
между особями разных подвидов одного вида, наблюдаемое, как
правило, в местах перекрытия ареалов. Не представляет боль-
шой редкости скрещивание между особями близких видов.
Гибриды соболя и лесной куницы, например, существуют не
только на зверофермах, но и в природе (так называемые «ки-
дасы», или «кидусы»). Довольно часто встречаются помеси
между лесными и степными хорьками, между хорьками и нор-
ками, между крапчатыми и малыми сусликами, зайцами-
русаками и беляками («тумаки») и пр.2; описаны помеси между
1 В агробиологии применяется понятие «жизненность», под которой под-
разумевается высокая ассимиляционная способность, крепкое телосложение,
устойчивость против болезней и высокая плодовитость животных. Необхо-
димое условие повышения жизненности нового поколения — достаточная
степень различия объединяющихся при оплодотворении гамет (как наслед-
ственной основы). Естественно, что у родительских особей, находящих-ся
в близком родстве, такие различия очень незначительны, что и приводит
к снижению жизненности в ряде поколений (Милованов. 1953).
2 Существование значительных различий в строении полового аппарата
русаков и беляков ставило под сомнение возможность появления их помесей
в природе. Однако специальные опыты, проведенные в последнее время в Ка-
занском зоопарке, полностью подтвердили такую возможность.
волками и шакалами, волками и собаками. Имеются указания
на встречи помесей между лесными и каменными куницами
С другой стороны, некоторые близкие виды не скрещивают-
ся между собой. Неизвестны, например, помеси между русским
и канадским бобрами (при их совместном фермовом содержа-
нии), между близкими видами мышей, крыс и полевок.
По сравнению с межвидовыми помесями значительно более
редкое явление представляют межродовые помеси. Специально
поставленные опыты показали невозможность получения поме-
сей даже между представителями таких близких родов, как
кролики и зайцы, овцы и козы, лисицы и собаки.
Для большинства гибридов характерно увеличение разме-
ров по сравнению с исходными формами (гетерозис) и пони-
женная плодовитость или даже полная стерильность. Однако
путем обратного скрещивания малоплодовитых гибридов с ис-
ходными родительскими видами животноводам удается восста-
новить плодовитость вновь созданных или улучшенных в ре-
зультате гибридизации пород. Вместе с тем имеются гибриды,
которым свойственна вполне нормальная плодовитость. Таковы
гибриды зебу и крупного рогатого скота, одногорбого и дву-
горбого верблюдов, муфлона и овцы и др.
1 До последнего времени считалось, что эти виды не дают помесей.
20*
Движение численности населения зверей
УСЛОВИЯ, ВЛИЯЮЩИЕ НА ДВИЖЕНИЕ
ЧИСЛЕННОСТИ
Вопрос о численности животных какого-либо вида в пределах
всего ареала или в отдельных участках, районах и областях и
о ее изменениях во времени и пространстве — один из важней-
ших в теоретическом и практическом отношении. Идет ли речь
о ценных промысловых видах или о важных вредителях сель-
ского хозяйства и переносчиках опасных инфекций, первое, что
нужно о них знать, это 1) каковы область распространения и
плотность населения, 2) как животные распределены по биото-
пам, 3) насколько постоянна их численность — не изменяется ли
она в разные сезоны и годы.
Планирование заготовок пушнины, организация промысловой
и спортивной охоты, также как истребление вредных видов и
защита от них урожая, не могут быть успешными без постоян-
ного учета распределения и численности животных.
Каждый вид млекопитающих населяет пространство ареала
не сплошь; особи, пары, семьи, стада живут более или менее
разобщенно, они распределены отдельными пятнами, полосами,
занимая лишь пригодные биотопы. От специфики требований
вида к среде, от места, занимаемого в «цепях питания», от его
эвритопности или стенотопности и, естественно, от природно-гео-
графических особенностей территории зависит степень равномер.;
ности или дисперсности, спорадичности распределения населе-
ния животных.
Сравнительное изучение показывает, что в различных биото-
пах плотность населения животных далеко не одинакова. Нали-
чие ряда градаций заселенности (если они установлены не по
случайным данным) свидетельствует о разной степени пригод-
ности биотопов для жизни вида и дает возможность судить о его
требованиях к среде. Благодаря длительным наблюдениям с
применением количественного учета выяснено, что численность
млекопитающих, особенно мелких грызунов, насекомоядных и
хищников, населяющих отдельные угодья и целые области, да-
леко не стабильна и колеблется по сезонам и годам в очень ши-
роких пределах.
У видов, отличающихся высокой плодовитостью и малой про-
должительностью жизни особей (землеройки, полевки, мыши,
308
некоторые песчанки) сезонные изменения численности выражены
особенно резко. Максимальная численность их обычно прихо-
дится на осень (сентябрь — октябрь)—время непосредственно
после периода размножения, когда, кроме производителей,
имеется много подросших молодых, а вылов зверьков хищни-
ками, гибель от неблагоприятных условий погоды и т. п. еще
не вызвали заметного снижения численности популяций.
Деятельная часть населения малых сусликов, приносящих
только один помет в течение весенне-летнего периода, достигает
наиболее высокого уровня численности в мае, в период выхода
молодых зверьков из выводковых нор Г Взрослые суслики вскоре
залегают в спячку, а молодые продолжают расселяться, усилен-
но откармливаются и по причине недостаточной осторожности
часто становятся добычей хищников. Поэтому количество их ко
времени залегания в спячку (июль — август) оказывается уже
сократившимся наполовину и даже более.
Для серых полевок, обитающих в степных и луговых стациях,
большое значение имеют метеорологические условия осеннего
периода. В теплую и не слишком сухую осень удлиняется период
вегетации растений, идет массовое прорастание семян урожая
данного года. Степь, поля, луга уходят под снег с обилием кор-
ма. Это удлиняет сезон размножения взрослых полевок на всю
зиму и, ускоряя созревание, дает возможность начать размноже-
ние молодым зверькам, составляющим до 80—90 °/о осеннего
населения (в обычные годы эти молодые зверьки зимуют непо-
ловозрелыми) . В результате наступает бурный рост численности,
и популяции достигают максимума к концу зимы (Пидопличко,
1930; Мигулин, 1925, 1927; Калабухов и Раевский, 1935; Н. Нау-
мов, 1948 и др.).
Дальнейшее часто зависит от характера весенней погоды,
условий снеготаяния и особенностей биотопов. При бурном тая-
нии снега, сильных дождях, чередующихся с похолоданиями,
когда вода затопляет норы и большие участки лугов и полей,
значительная часть зверьков погибает в ледяной воде или
делается жертвой различных хищников. Обычно с периодом сне-
готаяния совпадает массовый пролет специализированных хищ-
ников-мышеедов — луней, болотных сов, канюков-зимняков и
канюков обыкновенных, пустельг и др., особенно успешно пресле-
дующих грызунов, лишенных убежищ. Грачи, вороны, сороки,
чайки, цапли в этот период тоже занимаются охотой на полевок,
в другие сезоны для них менее или совсем недоступных. В по-
добных случаях следом за максимумом численности наступает
глубокая депрессия. Но в местах с увалистым или холмистым
рельефом, на участках, изобилующих буграми, зарослями ку-
1 Беспомощные молодые, находящиеся в норах и гнездах, составляют
особую часть населения и в данном'случае в расчет не принимаются.
309
старников или густых бурьянов, зверьки легче находят убежища
от снеговой воды и нападений хищников. В этих условиях гибель
полевок не столь катастрофична, как на открытых ровных уча-
стках.
Таким образом, от условий весенней погоды зависит успеш-
ность охоты хищников, а от особенностей рельефа и раститель-
ного покрова—вероятность сохранения части полевок в крити-
ческий период снеготаяния. В природе факторы, влияющие на
движение численности населения животных, никогда не дей-
ствуют изолированно, они тесно переплетаются между собой и
обычно образуют сложные комплексы.
Чаще же всего на весну приходится минимум численности
землероек, мышей, хомячков, полевок, некоторых видов песча-
нок, так как осенью размножение прекращается, а гибель зверь-
ков идет непрерывно. Однако имеется и немало исключений из
этого правила. Например, в широколиственных лесах юго-запад-
ной части СССР и на Кавказе после опадения на землю богатого
урожая желудей, семян граба, орехов бука и лещины создаются
особенно благоприятные условия для зимнего размножения се-
меноядных грызунов — лесных и желтогорлых мышей, местами
и рыжих полевок. Подъем их численности отмечается на сле-
дующий год после большого урожая корма (Першаков, 1934;
Жарков, 1938; Н. Наумов, 1948 и др.).
В некоторые годы происходит интенсивное зимнее размноже-
ние обских и копытных леммингов, степных пеструшек под за-
щитой снежного покрова, защищающего зверьков от морозов и
преследования большинства хищников.
При благоприятном сочетании условий бурное размножение
полевок и мышей приводит иногда к такому увеличению их чис-
ленности, что зверьки буквально наводняют поля, огороды, сады,
постройки сельских населенных пунктов и там, где не была свое-
временно проведена борьба, причиняют большой ущерб урожаю,
распространяют заразные болезни, опасные для человека и до-
машних животных. Это явление в народе образно называют «мы-
шиной напастью» — оно представляет собой один из частных
случаев непрерывных изменений численности мелких зверьков.
Гибель хлебов в годы «мышиной напасти» неоднократно упоми-
нается в древнейших русских летописях; еще более ранние све-
дения такого характера имеются в документах античного вре-
мени и хрониках европейского средневековья.
Уже много столетий назад охотники знали, что в любой мест-
ности количество лисиц, песцов, горностаев, зайцев, белок не
постоянно. Тверская летопись в 1408 г. отмечает неожиданную
гибель белок из-за голода, вызванного сильным повреждением
лесов какими-то гусеницами. Академик И. И. Лепехин (1770)
указывал, что в Сибири белка «в известные годы бывает в ве-
ликом множестве»... В охотничье-промысловых районах издавна
310
бытуют выражения «урожай» и «неурожай» песца, белки, зай-
ца-беляка и т. п.
Иногда говорят также «набег песца», «набег зайца»,
«белка ушла», считая, что все изменения численности обуслов-
лены миграциями, притоком животных со стороны или их отко-
чевкой, Местную вспышку «мышиной напасти» лица, не имею-
щие специальных зоологических 'знаний, тоже нередко объяс-
няют массовой иммиграцией зверьков, причем всегда находятся
Рис, 123. Схема пути выселявшейся обыкновенной полевки (общая длина
около 350 л), блуждавшей при плотном снеге на зяби и частично на суходоле
с кустами (пунктир — граница между полем и суходолом).. Такие вынужден-
ные переселения полевск часто бывают зимой из-за недостатка или недоступ-
ности корма и нередко кончаются гибелью зверьков. 8 декабря 1946 г. Пуш-
кинский район Московской обл. (по Формозову)
люди, свидетельствующие, что они, якобы, сами видели, как це-
лые «полчища мышей» переправлялись через реки, полотно же-
лезной дороги и т. п. Угасание местной популяции куниц, собо-
лей, косуль под влиянием неограниченного отстрела охотники
склонны объяснять откочевкой животных. Эти ошибочные пред-
ставления дезориентируют практических работников, уводят в
сторону от понимания подлинных причин, вызывающих подъемы
и падения численности промысловых животных и вредных гры-
зунов.
Вселения и выселения действительно играют немалую роль в
перераспределении части популяций, особенно таких подвижных
животных как песец, белка, колонок, местами заяц-беляк, но
всегда бывают следствием успешного их размножения, роста
численности на какой-то территории. Так, например, через каж-
дые три-четыре года значительная часть песцов, расплодившихся
311
на п-ове Ямал, откочевывает осенью на запад, благодаря чему
хорошая охота на пришлых зверьков в течение одного сезона
бывает в низовьях Печоры, Большеземельской тундре и т. д. Но
мелкие мышевидные грызуны не откочевывают на большие рас-
стояния, и кажущаяся внезапность появления огромного количе-
ства полевок или мышей всегда объясняется недостатком наблю- ,
дений за жизнью популяций этих зверьков в начальный период
«мышиной напасти» — массового размножения. Их перекочевки
носят узко местный характер (из одних угодий в другие) и
обычно не превышают сотен метров или немногих километров
(рис. 123).
Таким образом, выселения и изменения численности млеко-
питающих— взаимно связанные явления, но первое из них имеет
подчиненное значение, не может вызывать таких резких увели-
чений населенности, как успешное размножение местной попу- |
ляции. Каждый из этих процессов изучают отдельно, всегда имея
в виду их взаимную связь и, конечно, не преувеличивая роль
массовых вселений, если не удается вскрыть истинные причины
интенсивного роста местных популяций.
Движение состава населения животных отражает два проти-
воположных процесса, протекающих в любой популяции: размно-
жение и отмирание особей — это итог борьбы за существование
(в ее широком дарвиновском понимании). Изменения численно-
сти 1 отражают количественную сторону этой борьбы. |
ПРИЧИНЫ СМЕРТНОСТИ
I
Гибель особей в естественных условиях зависит от многих
причин, далеко не равноценных по эффективности их воздейст-
вия на численность населения животных в целом. Эти причины
можно разбить на следующие группы: 1) достижение предель-
ного возраста (смертность от старости), 2) несчастные случаи,.
3) недостаток корма (голодание), 4) болезни (вирусные, бакте-
риальные, вызываемые простейшими, гельминтами, членистоно-
гими), 5) деятельность хищников и конкурентов, 6) стихийные
бедствия, неблагоприятные метеорологические условия (крайне
суровые морозы, наводнения, глубокое зимнее промерзание поч- I
вы, завальные снега, гололедицы, лесные пожары и т. п.), 7) хо-
зяйственная деятельность людей (непосредственное истребление
животных или радикальное изменение условий их существова-
ния, вызывающее гибель от голода, уничтожения лучших мест ।
обитания и т. д.). ।
1 Вместо точных и понятных выражений «ход численности», «движение
численности населения» употребляют крайне неудачные и не отражающие- (
сущности процессов выражения «динамика населения», «динамика числен-
ности». В. Н. Беклемишев (Зоологический журн.,№ 2,1961) убедительно по-
казал, что слово «динамика» в описаниях, касающихся изменений популяции
животных, употреблять не следует.
312
Чрезвычайно важно знать все причины, вызывающие болезни
и смертность тех или иных видов. Целенаправленное устранение
одних или снижение интенсивности воздействия других факто-
ров — важный метод, содействующий охране и увеличению по-
головья ценных видов. Искусственное усиление действия усло-
вий, вызывающих гибель вредных видов, должно занимать по-
стоянное место в системе мероприятий по защите урожаев и
борьбе с вредителями. Например, на полях, на участках посева
и посадки лесных пород нельзя оставлять стога и ометы, необ-
ходимо уничтожать кучи хвороста и пни — укрытия мышей и
полевок, нужно охранять и привлекать хищных птиц-мышеедов,
проводить раннюю, тщательную послеуборочную обработку по-
лей, разрушающую норки грызунов, лишающую их семенного
корма, укрытий от хищников и т. д.
Достижение предельного возраста
Огромное большинство млекопитающих в природных усло-
виях не доживает до предельного возраста, которого они могут
достигнуть при лабораторном содержании, в зоопарках и т. п.
Состарившееся животное, утратившее необходимую выносли-
вость, быстроту и т. п., на воле неизбежно погибает от голода
или становится жертвой хищника. Только там, где крупные хищ-
ники полностью уничтожены, отдельные олени, косули, горные
козлы и др. доживают до предельного возраста, снашивают зубы
и умирают от старости.
Специальные исследования показали, что у многих видов до
50—60% молодых погибает уже в первые месяцы жизни (сусли-
ки, белки, косули, кабаны), а мелкие грызуны и землеройки
имеют фактическую среднюю продолжительность жизни, колеб-
лющуюся в пределах 2—5 мес., максимально едва доживая до
9—15 мес.; их популяции полностью обновляются в течение од-
ного года.
Несчастные случаи
В результате несчастных случаев разного рода погибает раз-
личное число жертв, за редкими исключениями независимо от
пола и возраста животных..Например, зимой мелкие зверьки с
несовершенной терморегуляцией (землеройки, полевки, мыши),
уйдя далеко от норы, замерзают на поверхности снега, покры-
того коркой плотного наста, гололеда, препятствующей им за-
рыться в глубину снежного покрова. Летом, наоборот, отмечены
случаи гибели от теплового удара не только мелких грызунов, но
и таких крупных как речной бобр. Многие наземные виды тонут
в период высоких паводков или при переправе через реки и озера
во время миграций (лемминги, полевки, землеройки, белки, пес-
315
цы). Отмечены случаи гибели при переправах через водоемы;
покрытые слабым «молодым» льдом даже таких могучих живот-
ных как бурый медведь, лось и др. Тонкий лед ломается, не вы-
держивая их тяжести, и в то же время препятствует передвиже-
нию вплавь. Животные, вынужденные плыть и ломать грудью
лед. быстро теряют силы и замерзают в ледяной воде.
В Печорском заповеднике лоси наиболее часто тонут при
осенней миграции (ноябрь) в годы с затяжным ледоставом, когда
между появлением шуги и ледоставом проходит более 20 дней.
Из 27 найденных погибших лосей (за период 1937—1946 гг.)
14 утонули, провалившись под тонкий лед, одномесячный теле-
нок умер от истощения, отстав от самки из-за вывиха ноги, и
12 были убиты хищниками (росомахой и медведем). Таким об-
разом, несчастные случаи вызвали гибель 56% лосей, найден-
ных мертвыми, а хищники — 44% (Теплов, 1948).
Морские млекопитающие (киты, дельфины, ламантины), в от-
личие от тонущих наземных, гибнут-от «обсыхания», когда, пре-
следуемые хищниками или гоняясь за рыбой и т. п., налетают
на прибрежные мели и обсыхают на них во время отлива. Ткн
лени, особенно'бельки у видов, щенящихся на льду (тюленей-
«пагофилов»), нередко гибнут задавленные льдами при сильных
подвижках торосов. В горах целые стада туров, серн, оленей
погибают при падении снежных лавин; сходную роль играют
сильные камнепады, реже — селевые потоки. На больших ледни-
ках крупные животные нередко проваливаются в глубокие, за-
маскированные слоем снега трещины и уже не могут из них
выбраться. На равнинах в топких местах торфяников, соленых
грязей степных озер и т. п. увязают и гибнут как крупные, так
и мелкие млекопитающие. Естественный «палеонтологический
музей» — богатое захоронение скелетов разнообразных млекопи-
тающих, а также рептилий, птиц, погибших в вязкой массе би-
тума, находится в Бинагадах (Азербайджан) и. представляет
хороший пример опасной ловушки такого типа, действовавшей
в течение многих столетий.
Молодняк стадных копытных (северные олени, некоторые ан-
тилопы) нередко гибнет под копытами взрослых при переполо-
хах, вызванных внезапным . появлением опасных хищников.
У морских котиков большое количество новорожденных давят
огромные самцы-секачи во время постоянных драк с обладате-
лями соседних «гаремов».
Натуралистами и охотниками зарегистрировано немало слу-
чаев гибели лосей и оленей разных видов, запутавшихся ветви-
стыми рогами в сучьях деревьев, и пар самцов, намертво сце-
пившихся рогами во время схваток в период гона. Лишенные
возможности освободиться, животные умирают медленной голод-
ной смертью. Наконец, для самцов кабанов, оленей, некоторых
полорогих, морских котиков и др. отмечены нередкие случаи
314
гибели от тяжелых травматических повреждений, нанесенных
соперниками во время гона. Таким образом, некоторый процент
гибели от несчастных случаев носит избирательный характер,
вызывая элиминацию самцов у полигамных форм, отличающихся
резко выраженным половым диморфизмом. Примеры другого
порядка — несчастные случаи, вызывающие гибель только са-
мок, малочисленны. Например, случаи неудачных родов со смер-
тельным исходом отмечены у некоторых видов оленей, тюленей
и др. форм, приносящих относительно крупных детенышей.
Несмотря на большое разнообразие условий, вызывающих
гибель от несчастных случаев, значение последних в движении
численности животных ничтожно; в лучшем случае они вызы-
вают изменения соотношения полов, что для полигамов имеет
даже положительное значение. Болезни, особенно эпизоотии, но-
сящие повальный характер, и глистные инвазии, а также голод,
стихийные бедствия и в меньшей степени деятельность хищников
способны оказывать решающее влияние на движение численно-
сти млекопитающих.
Недостаток корма
Значение неурожаев корма или его недоступности из-за глу-
бокого и плотного снега и т. п. довольно полно изучено, так как
во многих случаях они вызывают гибель ценных промысловых
животных. Особенно большое значение это имеет в жизни попу-
ляций видов, питающихся концентрированными калорийными
кормами, которые в случае нужды не могут быть заменены на
грубые объемистые, так как узко специализированные органы
пищеварения не справляются с их переработкой и усвоением.
.Для кабанов Беловежской пущи желуди дуба — основной
источник белковой пиши, и отсутствие их сказывается на всей
биологии популяции, обитающей в неурожайном массиве, если
из него нельзя откочевать. По данным Т. Б. Саблиной (1955), с
августа 1948 по август 1949, кабаны пущи были лишены этого
корма. В первых числах апреля 1949 г. по ночам отмечались за-
морозки и тогда же на исходе зимы среди истощенных живот-
ных шел падеж. Из найденных трех павших животных два были
подсвинки (рождения 1948 г.) и один трехлеток, все самцы. Ко-
нечно, это только небольшая часть животных, погибших от го-
лодного истощения, большинство их теряется в лесу, поедается
хнгцниками и т. д.
. 'Обильный урожай желудей дает.кабанам возможность встре-
тить период гона и зиму, имея хорошую упитанность. Упитан-
ность сказывается на сроках гона и на количестве поросят, появ-
ляющихся в следующем году, а именно на количестве самок, дав-
шем приплод, и на среднем числе молодых в помете.
Так, например, в 1947 г. после хорошего урожая желудей
315
начало гона было 15 сентября, первые молодые появились в
следующем году 12 февраля, среднее число их в выводке было
5,8, участвовало в размножении 100% половозрелых самок.
В 1948 г., когда урожая не было, гон начался только 15 декаб-
ря, первые молодые появились 22 апреля, среднее число их в
выводке было 3,1, в гоне участвовали только старые крупные
животные (Саблина, 1955). Особенно тяжело переносят ка-
баны отсутствие желудей в суровые многоснежные зимы; то-
гда надеж их особенно велик и возрастает гибель от хищ-
ников.
Это хорошо видно также из ряда примеров, тщательно рас-
смотренных в монографии Д. Лэка (1957). Один из них ка-
сается вида, питающегося более грубыми и массовыми кормами,
чем кабан — чернохвостого оленя (Odocoileus hemionus). В се-
верной Юте к 1917 г. их оставалось около 250 голов. К 1939 г.
в результате охраны их количество возросло до 6000. Истоще-
ние пастбищ (в основном зимних, т. е. веточного корма) было
замечено уже в начале 30-х годов, а зимой 1935/36 г. при очень
глубоком снежном покрове много животных погибло. Как здесь,
так и в других местах особенно пострадал молодняк. Много
оленей погибло от голода; среди повторно отловленных мече-
ных животных относительная потеря веса была выше у моло-
дых, а не у взрослых. Были отмечены и заболевания, но их рас-
сматривали как следствие недостатка пищи [Доман и Расмус-
сен (Doman a. Rasmussen), 1944].
Во всех этих случаях хищники, способные уничтожить осла-
бевших животных, почти не играли роли, так как в районах
разведения оленей они давно истреблены. Лэк в заключение
указывает, что у виргинского оленя недостаток пищи регулиру-
ет численность вида, изменяя как плодовитость, так и смерт-
ность (причем влияние на гибель имеет, вероятно, большее
значение).
Более сложный механизм воздействия выяснился при изуче-
нии зимнего падежа европейских, или благородных оленей, у
нас в Воронежском заповеднике. В период с февраля по ап-
рель 1951 г. здесь, впервые за многие годы охраны и разведе-
ния этого животного, произошла гибель значительного числа
оленей. Всего был найден 41 труп, из них 29 взрослых самцов
и 12 телят рождения 1950 г. Детальное изучение причин паде-
жа дало довольно неожиданные и поучительные результаты.
Наибольшее число животных погибло в конце марта и на-
чале апреля. Павшие олени были очень истощенными, у них
полностью отсутствовали жировые отложения, потеря веса до-
ходила до 30% по сравнению с осенним периодом, когда они-
имеют наилучшую упитанность. При патолого-анатомическом’
вскрытии трупов и бактериологических исследованиях не обна-
ружено признаков какой-либо инфекции или глистной инвазии.
316
Выяснилось, что основная масса (22 взрослых самца и 11 те-
лят) пали на площади участка в 4 тыс. га, где зимой обитало
не более 120 оленей, тогда как за его пределами было 380 го-
лов, из которых погибло всего 8 (7 самцов и 1 теленок). Запа-
сы веточного корма на участке падежа были примерно такие
же, как и в других частях массива, кроме того, здесь имелись
3 постоянных места подкормки и более 10 стогов сена. Желудки
павших животных были наполнены пищевой массой в количест-
ве, обычном для нормального питания в зимний период.
Причина гибели связана с тем, что олени этого участка,,
имевшего в 1950 г. удовлетворительный урожай желудей, ока-
зались лишенными возможности использовать свой единствен-
ный концентрированный осенний корм, так как с середины сен-
тября по конец октября здесь работало около 100 сборщиков
желудей,' заготовлявших семенной материал для лесопосадок.
В период гона, осенью, многие половозрелые олени-самцы из-за
повышенной возбудимости мало едят и быстро худеют, утрачи-
вая весь подкожный жировой слой. Но после гона они могут,
используя хорошие корма, в данном случае желуди, восстано-
вить упитанность и в удовлетворительном состоянии встретить
зиму.
С брачным периодом совпадает осенняя линька оленей; к
зимним холодам волосяной покров удлиняется, полностью об-
разуется густая подпушь. Однако у слабо упитанных животных
линька задерживается, ость растет медленно, а подпушь почти
не образуется. Исследованные погибшие самцы в основном
были среднего и выше среднего возраста (7—12 лет), все они
имели следы бурно проведенного брачного периода: у 26 — по-
вреждены в драках отростки рогов, у 7 — имелись срастающие-
ся переломы ребер, разрывы кожи на шее, брюхе и т. п. Вме-
сте с тем все они, в отличие от самок и телят, имели очень
слабо развитую подпушь и редкую ость.
Зима 1950/51 г. оказалась исключительно тяжелой: за
снежный период отмечено 58 дней с температурой от —15° до
—35°С; в первой декаде февраля высота снежного покрова в
лесу достигала 45 см, а в середине марта — 70 см. Длительные
январские и февральские холода вызвали у худых и плохо вы-
.линявших самцов чрезмерную теплоотдачу. Веточный корм и
по временам доступное им сено не. могли компенсировать рас-
ходы энергии, и наиболее истощенные олени начали гибнуть
после первых же сильных морозов в феврале.
Телята, достаточно упитанные с осени и хорошо вылиняв-
шие, удовлетворительно перенесли январские и февральские
морозы, но наступившее в марте многоснежье вызвало их боль-
тпую физическую перегрузку из-за трудностей передвижения по
глубокому снегу. Вынужденные ходить по следам, проложен-
„ным самками, телята поедали только оставшийся после них
317
грубый, малокалорийный веточный корм. Это вызвало повы-
шенный расход энергетических резервов организма и быстрое
истощение. Падеж телят начался только в третьей декаде мар-
та, когда уже стало теплее и снег начал сходить. За пределами
участка, где была изъята подавляющая часть урожая желудей,
погиб только один слабый, поздно родившийся теленок.
Таким образом П. А. Мертцу (1953) удалось расшифровать
все детали гибели оленей, случайно вызванной изъятием их
естественного продуктивного корма.
Болезни
Изучение особенностей эпизоотий диких животных представ-
ляет очень большой теоретический и практический интерес.
В природе массовая гибель животных от заразных болезней
распространена чрезвычайно -широко. Падежи «являются не
случайным, но закономерно повторяющимся и широко распро-
страненным явлением борьбы за существование’, о котором мы,
до недавнего времени, не имели правильного представления»—
писал С. А. Северцов (1941).
За последние десятилетия широкая сеть специальных меди-
цинских и отчасти ветеринарных учреждений нашей страны до-
стигла больших успехов в своевременном обнаружении эпизоо-
тий, в организации разрыва контактов между инфицированными
дикими животными и людьми, в подавлении эпизоотий и пол-
ной ликвидации природных очагов некоторых зоонозов (болез-
ней животных, передающихся людям). Особенно широкое раз-
витие, благодаря работам школы акад. Е. Н. Павловского,
получило учение о природной очаговости таких болезней, пере-
дающихся людям, как чума, туляремия, эпидемические энцефа-
литы, клещевые тифы, лихорадка Ку, лептоспирозы, кожный
лейшманиоз и др. Все это болезни диких животных, преимуще-
ственно грызунов; вспышки эпизоотий, вызванных возбудителя-
ми перечисленных инфекций, нередко наносят большой урон
популяциям млекопитающих. Очень широкое распространение
эпизоотий среди млекопитающих освещает сводка Ч. Элтона
(Elton, 1925), теперь уже значительно устаревшая; в моногра-
фии А. Ольта и А. Штрёзе (Olt und Strose, 1914) описаны ин-
фекционные заболевания, существовавшие, как оказывается, у
всех видов охотничьих животных Германии, и предложены не-
которые меры борьбы с ними. Нашими учеными спонтанная
(естественная) зараженность туляремией доказана бактериоло-
гически для 36 видов грызунов, 5 видов насекомоядных и 6. ви-
дов хищных1. Список этот в дальнейшем, несомненно, будет.
1 Кроме того, естественная инфипивованность установлена и у 8 видов
домашних млекопитающих (Дунаева, 1960).
318
еще расширен. Более чем у 30 видов зверей нашей страны
констатированы спонтанные заболевания чумой (Фенюк, 1939).
Несмотря на высокую патогенность микроба туляремии, да-
леко не все млекопитающие нашей фауны одинаково' легко за-
ражаются туляремией и переносят это заболевание. В связи с
выяснившейся различной степенью восприимчивости и инфек-
ционной чувствительности животных к заболеванию туляреми-
ей млекопитающие подразделяются на 3 группы.
К первой группе относятся виды, высоковосприимчивые и
высокочувствительные к туляремии; они заражаются легко при
подкожном введении минимальных доз возбудителя, т. е. 1 —
10 бактерий. Заболевание протекает в острой форме и заканчи-
вается гибелью зверьков на 3—16 сутки, в зависимости от дозы
заражения. В эту группу входят зайцы беляк и русак, ондатра,,
водяная крыса, многие виды серых и рыжих полевок, степная,
пеструшка, норвежский лемминг, слепушонка, домовая, лесная
и желтогорлая мыши, мышь-малютка, хомяки и хомячки, неко-
торые песчанки, лесная мышовка, орешниковая соня, алтайский
цокор, слепыш, а из насекомоядных — крот и землеройки-буро-
зубки— всего 42 вида (Дунаева, 1960).
Вторую группу составляют животные, столь же восприимчи-
вые, как и виды предыдущей группы, но с гораздо более низ-
кой инфекционной чувствительностью; они погибают только при
заражении массивными дозами, порядка 100 млн.— 1 млрд,
микробных клеток. Принадлежность к этой группе эксперимен-
тально доказана пока для 17 видов грызунов и 5 видов насеко-
моядных, а именно для всех изученных представителей сем. бе-
личьих (суслики, бурундук, белка), некоторых видов мышиных,
(крысы, полевая мышь, хомячок Эверсмана), соневых, а так-
же речного бобра и нутрии, из насекомоядных выхухоли, куто-
ры, малой белозубки и обоих видов ежей — обыкновенного и
ушастого. У животных этой группы алиментарное (через пищу)
заражение туляремией обычно не ведет к гибели, так как при-
живающееся количество микробов не достигает величины смер-
тельной дозы. Полевые мыши, серые крысы и ежи, поедавшие
павших от туляремии полевок, в большинстве выживали, при-
обретая к ней иммунитет. Отсюда понятны факты сохранения
- полевых мышей в скирдах и стогах с массой трупов погибших
от туляремии обыкновенных полевок и мышей-малюток.
Третья группа включает виды мало восцриимчивые и прак-
тически не чувствительные к туляремии, в основном хищников,,
постоянно питающихся грызунами: ласка, горностай, оба вида
хорьков, лисица, барсук, енотовидная собака, а из домашних
животных'—собака и кошка1. Эти виды не способны служить,.
1 Колонок, солонгой и некоторые другие виды не подвергались еще ис-
следованию.
319?
источниками инфекции в природе, хотя при некоторых условиях
могут подвергаться заражению и некоторое время хранить, в
организме возбудителя туляремии.
Большая сложность комплекса животных, непрерывно под-
держивающих существование возбудителя этой болезни в при-
родных очагах, связана с тем, что помимо млекопитающих зна-
чительное количество членистоногих (иксодовые, аргасовые и
гамазовые клещи, вши грызунов и в меньшей степени их блохи,
слепни и комары) служат активными переносчиками туляре-.
мии, а некоторые клещи и постоянными хранителями ее микро-
бов. Иксодовые клещи легко инфицируются, кормясь на туля-
ремийных животных, возбудитель сохраняется в их организме
в течение всего метаморфоза от личинки до половозрелой особи
и легко передается восприимчивым животным при сосании кле-
щами крови. Кроме того, в организме иксодовых клещей туля-
ремийные бактерии размножаются, и количество их за время
развития от личинки до упитанной половозрелой самки может
увеличиться в 1 —10 тыс. раз, достигнуть 10 млрд, микробных
клеток в 1 клеще (Петров и Олсуфьев, 1953). Возбудитель мо-
жет сохраняться в иксодовом клеще пожизненно; в опыте до-
казано сохранение микробов туляремии половозрелыми клеща-
ми Dermacentor marginatus в течение 710 дней (Петров, 1959),
причем вирулентность возбудителя совершенно не изменилась.
Именно клещи служат основными переносчиками инфекции
и ее наиболее важными хранителями в период между эпизоо-
тиями, когда численность грызунов слишком низка (например, в
луго-полевых очагах средней полосы страны эту роль выполня-
ет Dermacentor pictus и, возможно, некоторые гамазовые
клещи).
В пределах очаговых территорий имеются более мелкие
участки — микроочаги (элементарные очаги), в. которых инфек-
ция удерживается относительно постоянно даже и в межэпизоо-
тические периоды. В годы высокой численности грызунов ин-
фекция из микроочагов распространяется на обширную терри-
торию и может оказаться занесенной на сельскохозяйственные
угодья и в населенные пункты *.
Среди водяных крыс интенсивные туляремийные эпизоотии
чаще бывают весной и летом, так как передача возбудителя
идет через зараженную воду и через укусы кровососущих дву-
крылых.
У обыкновенных полевок интенсивные эпизоотии обычно
протекают в зимнее время после массового летнего размно-
жения зверьков и достижения очень высокой численности позд-
ней пг-римо.
1 Советскими учеными Б. Я. Эльбертом, М. А. Гайским и М. М. Фабичем
блестяще решен вопрос о вакцинопрофилактике туляремии. Теперь эта ин-
фекция не представляет для людей такой опасности, как прежде.
320
Массовое размножение обыкновенных полевок нередко
охватывает большие территории, в связи с чем локальные или
широко разлитые эпизоотии туляремии почти одновременно
возникают во многих районах с высокой плотностью населения
зверьков. Например, зимой 1937/38 г. интенсивная эпизоотия
туляремии среди полевок была отмечена в Московской, Туль-
ской, Рязанской и Орловской областях, зимой 1945/46 г.— в
Тамбовской, Рязанской, Московской, Воронежской и др. (Ол-
суфьев и Дунаева, 1960).
Разлитые эпизоотии у обыкновенных полевок в средней по-
лосе Европейской части СССР бывают при плотности населе-
ния не ниже 50—100 особей на объединенный гектар1. При
обычном для этого вида осеннем вселении зверьков в скирды
мигранты приносят в них инфекцию из микроочагов, в резуль-
тате чего возникают интенсивные эпизоотии.
Соотношение числа трупов и живых полевок в скирде зави-
сит от длительности эпизоотии. К концу ее соотношение- может
быть 3: 1 и даже 1 : 0,1 (т. е. на 10 погибших 1 живая полевка).
В отдельных густо населенных зверьками скирдах число трупов
может достигать нескольких сотен (Олсуфьев и Дунаева, I960).
Одновременно с туляремией гибель’ полевок могут вызывать
листериоз и другие инфекции, но они обычно оказываются ме-
нее губительными, чем первая.
В степных районах такие же интенсивные осенне-зимние
эпизоотии туляремии возникают среди сильно размножившихся
домовых мышей, особенно при большой скученности их в скир-
дах, стогах и постройках.
Другие виды млекопитающих, восприимчивых к этой инфек-
ции, обычно вовлекаются в эпизоотии, уже протекающие на
перечисленных выше массовых видах мелких грызунов.
Однако зайцы, на которых в большом количестве нападают
иксодовые клещи, заражаются от них и в межэпизоотические
периоды.
Эта краткая характеристика туляремии — одного из зооно-
зов, наиболее детально у нас изученных, говорит о том, что
интенсивные эпизоотии возникают, как правило, при высокой
численности животных, одновременно с усилением контактов
между особями и особенно часто в больших скоплениях зверь-
ков. Некоторое значение, видимо, имеют и условия погоды
(дожди, холода поздней осени, мокрые снегопады), снижаю-
щие сопротивляемость организма зверьков против инфекции.
При изучении любых эпизоотий, особенно в целях борьбы с
ними, важно получить ответ на три основных вопроса: как и в
1 Средняя плотность для определенной местности, подсчитанная по плот-
ностям, установленным для отдельных стаций, с учетом и в соответствии
с их относительной площадью.
21 Териология
321
каких формах сохраняется инфекция в межэпизоотические пе-
риоды, как она рассеивается по территории и каковы причины-,
вызывающие развитие и затухание острой эпизоотии, сопро-
вождающейся падежом (Кучерук, 1948). Многое зависит от
того, способен ли вид или группа видов, восприимчивых к дан-
ной инфекции, к интенсивному размножению и быстрому уве-
личению численности, или популяция и плотность населения
вида остается относительно стабильной. Существенное значение
имеет также характер годового цикла, в частности ход рассе-
ления молодняка, наличие зимней спячки, укрупнение групп
зверьков в период совместной заготовки запасов корма и под-
готовки к зимовке и т. п.
Чумные эпизоотии изучены с большой детальностью у мало-
го суслика. Ранней весной в очагах этой инфекции при пробуж-
дении сусликов от спячки отмечается наличие только латент-
ной (растянутой, вяло протекающей) хронической формы чумы,
которая затем сменяется отдельными спорадическими генера-
лизованными формами. При генерализации процесса кровь боль-
ного зверька переполняется бактериями чумы, и блохи в норах,
которые он посещает, становятся при сосании крови инфици-
рованными. Так создаются'условия для накопления инфекцион-
ности в очаге в период, который можно назвать предэпизооти-
ческим. Во время расселения молодых сусликов при посещении
ими многочисленных нор, в том числе и тех, где чумные зверь-
ки погибли от заражения, блохи нападают на каждого нового-
посетителя. Так складывается комплекс условий, вызывающий
быстрое возникновение интенсивной эпизоотии. Но она длится
всего 30—40 дней. По окончании ее снова появляются хрониче-
ские формы, которые при впадении сусликов в спячку становят-
ся латентными.
Многочисленными исследованиями давно установлена высо-
кая сопротивляемость зверьков, находящихся в спячке, против
воздействия инфекций и токсинов. Даже столь острая инфек-
ция, как чума, у спящих сусликов и сурков принимает латент-
ную, хроническую форму. Но в ветеринарной практике хорошо
известны обратные случаи перехода латентных форм инфекций
и даже бессимптомного бациллоносительства в острые формы
заболевания под влиянием ухудшившегося питания, резкого по-
холодания, сильного физического утомления, ослабляющего со-
противляемость организма (Гутира и Марек, 1938). Таким об-
разом и воздействие эпизоотий на численность животных осу-
ществляется не изолированно, но в тесной связи с другими
экологическими условиями, а также особенностями жизни кон-
кретных популяций (в частности зависит от их возрастного со-
става, плотности населения, интенсивности внутривидовых и
межвидовых контактов, индекса зараженности эктопаразитами.
И Др.).
3?2
Деятельность хищников и конкурентов
Чем меньше размеры зверька и ограниченнее его возможно-
сти активной и пассивной защиты от нападений, тем большее
количество хищных животных пользуется им как добычей.
Летучие мыши умеренных широт — животные мелкие, но
обладающие сильным и маневренным полетом. Они деятельны
ночью, день проводят в хорошо укрытых, часто трудно досягае-
мых для хищников убежищах. Таким образом, эти зверьки до-
ступны лишь для ограниченного круга врагов. Кроме того, вы-
деления особых желез придают летучим мышам сильный специ-
фический запах, благодаря которому они явно не привлека-
тельны для хищных млекопитающих. В итоге врагов у них
мало: изредка ловят совы, деятельные ночью, и сокол-чеглок,
заканчивающий свой охотничий день в поздних сумерках, еще
реже — кошки. Следовательно, они достаточно хорошо защище-
ны и несут очень небольшие потери от хищников.
Совсем иное положение с мелкими насекомоядными и гры-
зунами, особенно с медлительными землеройками, полевками,
хомячками и др., обитающими в сравнительно неглубоких но-
рах. На леммингов, полевок; землероек, кротов и белок при их
переправах через реки и озера успешно нападают даже неко-
торые рыбы (таймень, щука, сом, хариус и др.). Молодые он-
датры в старицах сибирских рек нередко становятся добычей
крупных щук; часть ондатр, схваченных ими, но упущенных,
вскоре погибает от нагноения ран, оставленных острыми щучь-
ими зубами. /
В южных зонах нашей страны на мелких зверьков постоян-
но охотятся-змеи; в Средней Азии эту роль иногда играет так-
же крупная ящерица — серый варан. Если в сухих степях от-
носительно часто охотятся на грызунов только степная гадюка
и щитомордник, то в южных полупустынях и пустынях количе-
ство видов и, что важнее, особей рептилий, питающихся мел-
кими зверьками, возрастает во много раз.
Еще большую роль как враги млекопитающих рептилии
играют в тропиках. Змеи проникают в норы и наземные гнезда
зверьков й целиком уничтожают выводки беспомощных моло-
дых; взрослых зверьков эти хищники подкарауливают у выхода
из нор или на местах кормежки.
Очень велико количество видов птиц, питающихся мелкими
млекопитающими, причем нередко охотой на них занимаются
представители групп, весьма далеких от настоящих хищников.
Тац, например, молодых узкочерепных полевок находили в же-
лудке лиловых дроздов (Myophonus temminki) на Тянь-Шане
(Формозов, 1940); фазаны в Уссурийском крае охотно поедают
детенышей мыши-мдлютки, разоряя ее гнезда, расположенные
в траве невысоко над землей (Слепцов, 1947). Полевок, обыч-
21* «823
них в степи в «мышиные годы», хватают дрофы, серые жу-
равли, белые аисты, серые и белые цапли, серебристые чайки
и др.
Но, конечно, наибольший урон поголовью зверьков наносят
дневные хищные птицы и совы, а и,з воробьиных — врановые и
сорокопуты. Некоторые хищные виды узко специализированы
в охоте преимущественно на мелких млекопитающих, главным
образом мышевидных грызунов, поэтому на языке многих наро-
дов эти птицы называются «мышеедами», или «мышатниками».
В действительности их роль в биоценозах гораздо сложнее, так
как, кроме грызунов, они нередко поедают и специализированных
истребителей грызунов — ласок, горностаев, степных хорей,
степных гадюк и др.
Хищные птицы не обладают развитым чувством обоняния,
поэтому многие виды одинаково охотно поедают как полевок,
мышей, хомячков, так и землероек, кротов, издающих сильный
мускусный запах. Последний в какой-то степени защищает
мелких насекомоядных от некоторых четвероногих хищников —
обладателей тонкого обоняния, например обыкновенной лисицы,
песца., корсака. Они никогда не охотятся за землеройками так
длительно и настойчиво, как за грызунами. Однако даже сытая
лисица при случайных встречах убивает землероек и кротов,
но бросает их на месте поимки. В голодные месяцы зимы она
поедает и только что пойманного крота, и полуистлевшие трупы
кротов, сохранившиеся местами под снегом еще с лета. Случаи
поедания лисицей ласок отмечены тоже только в трудные для ее
охоты зимние месяцы бедного полевками года. Секрет желез,
свойственных всем куньим, видимо, крайне неприятен для ли-
сицы. От съеденной ласки у нее иногда начинается рвота, после
чего голодный хищник снова проглатывает разжеванного
зверька, обладателя отталкивающего запаха. Соболь, горностай,
ласка менее чувствительны к запахам, чем лисица, и поедают
землероек-бурозубок регулярно во все сезоны. Таким образом,
железы, выделяющие секрет с неприятным запахом, как сред-
ство пассивной защиты некоторых млекопитающих оказыва-
ются полезными далеко не во всех случаях.
Также относительно и значение иглистого панциря ежа.
Орел-могильник, филин, лисица легко справляются со взрослым
ежом, свернувшимся в клубок; иглы защищают его лишь от
более мелких хищников. Но молодого ежа ростом в половину
взрослого убивает и расклевывает даже серая ворона. Прежде
чем начать его есть’, она выщипывает иглы (Формозов, 1925).
Независимо от имеющихся защитных образований и особен-
ностей поведения, большинство мелких млекопитающих пресле-
дуется многими видами хищников. На обыкновенную полевку
(Microtus arvalls), например, охотится более 40 видов позво-
ночных животных. У крупных млекопитающих количество вра-
324
гов значительно меньше.' На лося охотятся только медведь, тигр,
волк, росомаха и рысь, причем последние два вида предпочи-
тают молодых, а на взрослых лосей нападают чаще в обста-
новке, обеспечивающей явное преимущество преследователю,
например при необычно высоком снежном покрове или появле-
нии наста, крайне затрудняющих передвижение жертвы.
На значительной части ареала наибольшие потери стаду лосей
наносят волки, как правило, действующие группой в 5—8 особей,
что очень облегчает им охоту на сильного и смело обороняюще-
гося зверя. Бурый медведь — наиболее опасный враг лося там,
где нет волков. Он всегда преследует лося в одиночку, причем
особенно успешно весной по насту. Для огромных усатых китов
единственный опасный враг — косатка, охотящаяся стаями, по-
добно волкам.
При преследовании крупных стад подвижных животных хищ-
ники вообще вынуждены объединяться в большие группы, так
как охота в одиночку крайне утомительна и, как правило, кон-
чается неудачей. Групповые охоты с помощью загонов или
окружения («охота котлом») характерны для хищников, пре-
следующих копытных в открытых ландшафтах тундр, степей,
саванн, пустынь. Копытные этих зон в большинстве очень
быстры, обладают дальним зрением, особой системой сигнали-
зации об опасности и обычно пасутся так, что враги лишены
возможности подкрасться к ним незамеченными. В тундре зи-
мой около больших стад северных оленей длительное время
кормятся группы, включающие до 20 волков, состоящих из пар-
тий в 3—5 штук (Романов, 1941). Для охоты на сайгаков, соби-
рающихся в огромные стада на местах зимних кочевок, волки
объединяются в группы до 20—30 и даже 42 особей (Слудский,
1962). В Центральной и Южной Африке гиеновые собаки, пита-
ющиеся главным образом антилопами (пала, водяной козел,
бушбок и др.), охотятся стаями по 20—30 и даже 100 голов.
При охоте стаей крупная добыча, как правило, используется
полнее и «экономнее».
Хищникам, охотящимся мелкими группами, например львам,
обычно не удается уберечь недоеденную часть жертвы отнекро-
фагов (шакалов, гиен, грифов), которые ее полностью уничто-
жают, едва только львы отправятся на водопой или уйдут на
отдых. По имеющимся подсчетам в Восточной Африке половину
добычи львов съедают некрофаги, что заставляет хищников
убивать в два раза больше зебр и антилоп, чем им нужно было
бы без подкармливания «нахлебников» [Райт (Wraht), 1960].
С другой стороны, в период особенно успешной охоты мно-
гие хищники убивают гораздо больше, чем им необходимо для
питания в ближайшие дни, например черный хорь. Волки зимой
нередко съедают у загнанных ими сайгаков только жир с
крупа, а всю тушу бросают. Птицы миофаги в год, обильный
325
грызунами, съедают иногда только головы жертв, а тушки бро-
сают. Ласка в скирде, плотно населенной полевками, убивает
десятки зверьков и складывает их про запас. Однако такими
запасами ей чаще всего не приходится пользоваться, так же как
и воробьиному сычику (Glaucldium passerinum), в осень с бога-
тым уловом лесных полевок и землероек.
Из приведенных фактов как будто можно сделать вывод об
односторонней зависимости жертв от хищников. В действитель-
ности их взаимоотношения, сформировавшиеся в процессе дли-
тельной сопряженной эволюции, гораздо сложнее и полны про-
тиворечий.
Прежде всего численность и распределение в пространстве
специализированных хищников полностью зависят от численно-
сти и распределения их жертв (например, белых сов — от коли-
чества леммингов, болотных сов и луней — от распределения
степных пеструшек и обыкновенных полевок и т. д.). При рез-
ком сокращении численности мелких грызунов на обширной
площади, вызванном стихийными бедствиями или эпизоотиями,
вскоре резко и на целый ряд лет уменьшается количество всех
узкоспециализированных мышеедов (от степной гадюки, ласки,
болотной совы и полевого луня до лисицы и корсака). Движение
численности этих хищников во времени и пространстве в из-
вестной мере повторяет (с некоторым отставанием во времени
и с меньшей амплитудой из-за меньшей плодовитости) все ко-
лебания изменений населения их жертв.
Не только существование жертв зависит от деятельности
хищников, но и размножение, жизнь и смертность хищников
в значительной степени связаны с благополучием популяций
животных, на которых они охотятся. Хищники и их жертвы
исторически так давно и так тесно взаимно связаны, что во мно-
гих отношениях приспособились друг к другу.
Хищные птицы-миофаги, несмотря на их высокую специали-
зацию, большую подвижность, прожорливость, способность при
обилии добычи концентрироваться в определенных районах,
увеличивать число яиц в кладках и число кладок за сезон (бо-
лотная сова) даже вместе с хищными млекопитающими все же
часто не способны играть роль решающего фактора в движении
численности мелких, очень плодовитых зверьков. Их значение
становится явным только при определенных ситуациях, напри-
мер, когда хищников относительно много, а условия жизни
жертв неблагоприятны. Тогда хищники могут сдерживать на-
растание численности ряда вылавливаемых видов и вызывать
относительно длительную стабилизацию их количества на низ-
ком уровне. Если при высоком уровне своей численности хищ-
ные птицы найдут в период послегнездовых кочевок ограничен-
ные участки территории, где произошли местные вспышки раз-
множения полевок, последние будут почти полностью ими
326
уничтожены (уцелеет лишь часть особей, поселившихся в скир-
дах. под защитой густых бурьянов и т. п.').
В гнездовой период пернатые хищники вылавливают преиму-
щественно наиболее деятельных взрослых леммингов, полевок,
мышей и др.— чаще самцов, занятых поисками самок, и в огра-
ниченном количестве мало активных молодых зверьков. По-
этому к осени в популяциях грызунов резко изменяется соотно-
шение числа половозрелых самцов и самок в пользу последних.
Рис. 124. Взрослый изюбрь самец, загрызенный волками. Сихотэ-Алинский
заповедник. 1941 г. (фот. Л. Г. Капланова)
Это воздействие миофагов на половой и возрастной состав
населения очень плодовитых животных (прослежено в зоне
тундры, тайги, лесостепи, степи) —убедительное доказательство
глубокого влияния пернатых хищников на жизнь их жертв.
У дикого северного оленя самцы гибнут от нападений волков
в 16 раз чаще, чем самки .(Семенов-Тян-Шанский, 1948), а у
сайги — в 17 раз (Слудский, 1962). По наблюдениям Г. Ф. Бром-
лея, в Судзухинском заповеднике волки уничтожают ежегодно
от 5 до 30% поголовья пятнистых оленей, в зависимости от
условий охоты и состояния жертв, но преимущественно тоже
самцов. В Южном Казахстане, где много джейранов и сайга-
ков, один волк в течение года уничтожает несколько десятков
этих антилоп, а все хищники ежегодно истребляют сотни тысяч
голов. Волки режут не только больных и ослабленных живот-
ных, но и вполне здоровых (рис. 124). Но даже и столь значи-
тельное истребление хищниками особей этих медленно размно-
327
жающихся видов не влияет сколько-нибудь заметно на движе-
ние численности антилоп. Преимущественное истребление зимой
старых самцов, ослабленных гоном, по имеющимся взглядам,
даже полезно для популяций копытных, так как сохраняет
жизнь беременных самок, сокращает конкуренцию животных
из-за корма на зимних пастбищах, обычно обладающих низкой
емкостью и т. п. (Слудский, 1962).
Больные животные часто утрачивают обычную подвижность
и осторожность, поэтому чаще, чем здоровые, становятся добы-
чей хищников. Из остатков 178 грызунов, найденных в гнездах
степных орлов и канюков Забайкалья и исследованных бакте-
риологически, чума установлена у трех, или в 1,7% случаев,
тогда как из 21 929 зверьков, пойманных капканами в том же
районе, не оказалось ни одного зараженного чумой (Тарасов,
1944; Безрукова и Липнин, 1944). На оз. Неро (Ярославская
обл.) в очаге безжелтушного лептоспироза учет полевок-эко-
номок, принесенных в гнезда полевыми лунями, показал, что
в их уловах явно преобладали особи, больные лептоспирозом
(Карасева, Герман,. Коренберг, 1955). То же отмечали в отно-
шении избирательного вылова пернатыми хищниками мышей и
полевок, пораженных гельминтозами, на Западном Кавказе и
в Беловежской пуще. Таким образом, деятельность хищников
может иногда содействовать оздоровлению популяции жертв
путем постоянного отбора больных особей. Однако в тех слу-
чаях, когда хищник — окончательный хозяин гельминта, а жерт-
ва— промежуточный или при рассеивании пожирателями погиб-
ших от антракса животных, спор бактерий сибирской язвы,
роль хищников в отношении санитарного состояния популяции
поедаемых животных становится резко отрицательной.
Деятельность хищников можно и должно использовать как
необходимое звено в системе мероприятий, направленных на
регулирование численности вредных животных. Управление
численностью ценных промысловых видов должно быть по воз-
можности полностью в руках человека, насколько знание био-
ценологии и экологии видов это позволяет.
Значение как внутри-, так и межвидовой конкуренции в дви-
жении численности млекопитающих менее бросается в глаза и
менее изучено, чем воздействие хищников. При высокой плот-
ности населения соперничество между особями одного вида
возникает из-за удобных убежищ, пищи, площади участков,
обладания самкой и других причин. Часто более слабые особи
вытесняются в менее благоприятные местообитания, где мало-
корма, хороших укрытий, в связи с чем они скоро становятся
жертвой хищников. Недостаток убежищ ведет иногда к сниже-
нию процента размножающихся особей.
Поиски молодыми половозрелыми особями свободных участ-
ков нередко проходят в условиях ожесточенной конкуренции,
328
что у видов, расселяющихся по узким ленточным биотопам,
например у бобров и ондатр, нередко вызывает даже гибель
мигрантов. В Воронежском заповеднике, где бобров много, ча-
сто находят трупы двухлеток, вынужденных при поисках участ-
ка для поселения проплывать через плесы реки, уже давно-
занятые и тщательно охраняемые осевшими тут бобровыми
семьями. Изгоняя пришельца, бобры наносят ему множество-
укусов; израненный выселяющийся бобр умирает от гноящихся
ран. Жестокие ранения иногда получают от конкурентов и рас-
селяющиеся молодые ондатры. Вероятно, именно трудности про-
движения по водной поверхности, уже распределенной между
семьями, вынуждают часть бобров и ондатр двигаться сушей,
где эти полуводные грызуны несут большой урон от собак, ли-
сиц и других хищников. Серьезного значения для движения чис-
ленности популяций эти потери не имеют.
Межвидовая конкуренция приводит к различным результа-
там в зависимости от численности населения и особенностей
экологии соревнующихся форм. Так как пищедобывательная
деятельность играет огромную роль в жизни млекопитающих,
именно конкуренция из-за корма часто имеет наибольшее зна-
чение.
А. А. Шорыгин (1952) предложил различать следующие по-
нятия: объем пищевой конкуренции, т. е. степень сходства пи-
щевых спектров конкурентных видов, и напряжение конку-
ренции— отношение величины потребностей конкурентов к на-
личному количеству пищи, из-за которой идет борьба. Про-
изведение объема конкуренции на напряженность состав-
ляет силу конкуренции, от которой и зависит ход борьбы.
В уже упоминавшихся (из Тверской летописи за 1408 г.) слу-
чаях гибели белки в лесах, где гусеницы объели хвою деревь-
ев, конкуренция множества насекомых лишила лесного грызуна
и пищи и укрытий. В последние десятилетия столь же большую
силу конкуренции лесные млекопитающие встречают со сторо--
ны кедрового шелкопряда, уничтожающего значительные масси-
вы кедровников и отчасти лиственничников в Средней, Восточ-
ной Сибири и Прибайкалье. Иногда гусеницы бабочек, уничто-
жающие семена в шишках ели, резко сокращают запас этого
наиболее ценного продуктивного корма белки. Грандиозные
налеты странствующей саранчи-шистоцерки, перелетной азиат-
ской и марокканской саранчи вызывали голодание мелких
зверьков фитофагов из числа норников.
Гораздо труднее установить значение постоянной конкурен-
ции между близкими видами, занимающими сходные биотопы.
Иногда только случай дает возможность оценить эффективность,
подобных отношений. Так, например, малоснежной, но морозной
зимой 1938/39 г. весьма значительно упала численность рыжих
полевок (Clethrionomys glareolus) в Приветлужье Костромской
32»
обл. Более выносливый к морозам сибирский вид — красная-
полевка (С/. rutilus) перезимовал успешно и на следующую
осень составлял в уловах полевок этого рода 72,7%. Ранее крас-
ная полевка заселяла всего 25—45% обследуемых участков, но
в результате депрессии популяции рыжей полевки в 1939 г. за-
няла уже 54,2% участков, хотя абсолютная ее численность
почти не увеличилась. Очевидно, конкуренция со стороны
Cl. glareolus лишала ее в обычные годы возможности существо-
вать на многих пригодных для нее участках (Формозов, 1948).
Нередко межвидовая конкуренция переходит в отношения
хищника и жертвы. Домовая .мышь всегда малочисленна в по-
стройках, где обычны серые крысы. Они не только лишают ее
корма, но и активно преследуют, пожирают. Черный хорь —
конкурент горностая и одновременно его враг. Соболь изго-
няет из обитаемых им участков и горностая и колонка, причем
иногда охотится на них и пойманных съедает. Лет 30—40 назад,
когда соболь был сильно истреблен, колонок стал многочислен-
ным в горной тайге Алтая и равнинной тайге Западной Сибири.
За последние 20—25 лет в результате охрана численность со-
боля восстановилась, и он стал быстро вытеснять колонка из
временно захваченных им районов. Таким образом, внутриви-
довая и межвидовая конкуренция вносит ряд особых штрихов
в основные процессы, определяющие ход изменений численности.
Стихийные бедствия, неблагоприятные
метеорологические условия
Стихийные бедствия, наносящие заметный урон поголовью
млекопитающих, обычно связаны с крайне неблагоприятными
условиями погоды: длительными сильными морозами, глубоким
промерзанием почвы, с выпадением снежного покрова, резко
превышающего по высоте обычную для той или иной местности
норму (рис. 125), с гололедицами, наводнениями, лесными по-
жарами, возникающими в засушливые годы, и т. п.
Влияние подобных условий удобно рассмотреть на примере
сайгака — типичной степной антилопы, питающейся в основном
травянистыми растениями. Неоднократно были отмечены слу-
чаи массового падежа сайгаков при зимней бескормице, вызван-
ной появлением сплошного снежного покрова высотой 25—40 см
и более, сохраняющегося свыше двух недель. Плотный наст или
корки гололедицы, иногда появляющиеся на поверхности сне-
га, увеличивают трудности добывания подножного корма, так
как сайгаки, подобно большинству степных антилоп — обитателей
районов с малоснежными зимами, не умеют раскапывать снег
копытами, как это делают северный олень, косуля, лошади (при
330
табунном содержании) и др. Если многоснежная зима наступит
в области, населенной сайгаками и джейранами, после засуш-
ливого малокормного лета, происходит массовая гибель антилоп
от истощения, холода и усиливающегося в таких случаях хищ-
ничества волков. (Домашний скот, при пастбищной' кочевой
форме животноводства, в такие годы тоже погибал десятками
и сотнями тысяч голов. Бедствие это у казахов носит название
«джут», у монголов «зут»), Джучы повторяются примерно че-
рез 10—12 лет. О гибели сайгаков от джутов писали С. Гмелин
(1806), Н. Ребров (1849), К- Глич (1886) и многие др. позднее.
Рис. 125. Взрослый изюбрь самец в беспомощном состоянии в многоснеж-
ную зиму 1940/1941 г. Бассейн р. Чукем, притока р. Хора,. Хабаров-
ский край (фот. Мельникова)
По сведениям, собранным А. А. Слудским (1955), джуты в
Казахстане за последние сто лет были в зимы 1855/56, 1866/67,
1879/80, 1891/92, 1903/04, 1917/18, 1927/28, 1948/49 и 1950/51 гг.
Отход-бывает очень большой. Например, после джута зимы
1949/50 г. численность сайгаков в Западном Казахстане сокра-
тилась примерно в два раза. Высота снежного покрова в эту
зиму достигала 40—60 см., а куда снег наносило ветром и боль-
ше. Снежные бураны сопровождались морозами до —40° С.
Зимой 1950/51 г. в Центральном Казахстане основной при-
чиной гибели сайгаков был не столько снежный покров, сколько
331
ледяная корка толщиной до 2 см, совершенно закрывшая до-
ступ к корму. (Раков, 1956). После тяжелой (буранной, холод-
ной и многоснежной) зимы 1953/54 г. в западной части Прикас-
пийской низменности (западнее Волги), несмотря на принятые-
меры помощи, погибло несколько десятков тысяч сайгаков (Бан-
ников с соавторами, 1961).
При джутах в первую очередь гибнут взрослые сайгаки-сам-
цы (рис. 126), истощенные в период гона, затем молодые сам-
цы в возрасте до года, взрослые и молодые самки. Среди сай-
гаков, павших во время джута зимой 1949/50 и 1953/54 гг. в Ка-
захстане самиы составляли около 70—80% (Слудскпй, 1953,
1955, Раков, 1956 и др.).
!
Рис. 126/Истощенный ослабевший сайгак самец, загнанный буранами
в камыши дельты р. Терека зимой 1953/54 г. (фот. А. Т. Текучева)
Иное соотношение погибших животных по полу и возрасту
бывает при эпизоотиях. Например, весной 1957 г. в Калмыцкой
АССР много сайгаков, по свидетельству ветеринарных работни-
ков, погибло во время эпизоотии ящура. О количестве погиб-
ших можно судить по тому, что местами один труп взрослого
сайгака приходился в среднем на 18 га площади. Из осмотрен-
ных 182 павших животных 124 оказались самками и только 58:
(35,2%) самцами. В ту весну больше всего сайгаков погибло
в период окота (Клименко и Фомушкин, 1959). Возможно, ав-
торы осматривали только участок, где преобладали самки, не-
332
редко образующие особые временные скопления на местах, удоб-
ных для родов и первых дней жизни новорожденных сайгачат.
Только этим можно объяснить такое выборочное поражение
ящуром самок. Важно подчеркнуть, что разгар эпизоотии при-
шелся на период, когда животные уже имели свежий зеленый
корм. Многие остро заразные и высоковирулентные болезни
способны наносить урон поголовью животных даже в периоды
хорошей обеспеченности их кормами (чума сусликов, песчанок,
сибирская язва лосей, оленей и др.), но недостаток полноцен-
ных кормов, как правило, усиливает любую эпизоотию.
Даже такой сильный и всеядный зверь как бурый медведь
в некоторые годы не может найти в тайге достаточного коли-
чества пищи, необходимой, чтобы откормиться перед длитель-
ным зимним сном. Истощенные животные бродят зимой по глу-
бокому снегу и нередко гибнут от голода; в такие годы среди
них учащаются и случаи каннибализма.
Самые различные проявления разрушительных сил природы
(извержения . вулканов, землетрясения и вызываемые ими
страшной силы волны — цунами, мощные ураганы) тоже вызы-
вают гибель значительного количества млекопитающих. Но есть
стихийные бедствия, повторяющиеся с гораздо большим посто-
янством, чем землетрясения или сильные ураганы, и охватываю-
щие более обширные территории. Таковы, например, засухи или
длительные, обильные все затопляющие дожди, исключительно
сильные и устойчивые морозы и др. ~~
В тех случаях, когда период сильных морозов начинается до
выпадения снежного покрова или когда слой его слишком тонок,
почва промерзает на большую глубину, чем обычно. В пред-
горьях Урала при полном промерзании тонкого слоя почвы, ле-
жащего на каменистой материнской породе, почти повсеместно
полностью вымирает обыкновенный крот. Такие бедствия повто-
ряются периодически через 8—9 лет (Кириков, 1935, 1952).
После малоснежных морозных зим с глубоким промерзанием
почвы в 10—30 и более раз сокращается численность кротов, вы-
мирают землеройки (рис. 127) в лесной полосе Европейской час-
ти СССР (Московская, Горьковская, Костромская области, Та-
тарская АССР), но механизм этого явления еще далеко не ясен.
Видимо основную роль играет вызываемое промерзанием уплот-
нение почвы, не поддающейся когтям крота и, как следствие,
недоступность корма, а также гибель части компонентов почвен-
ной фауны, используемых этим видом. Так, например, в 1937 г.
в некоторых лесных массивах Татарии почва промерзала на глу-
бину до 90—140 см, т. е. значительно глубже, чем обычно роется
крот (Попов, 1960). При глубоком промерзании почвы только
те кроты переживают зиму, которым удалось найти доста-
точно просторный участок, свободный от сезонной мерзлоты
благодаря согреванию его выходящими к поверхности грунтовы-
333
ми водами (берега родникового ключа, болотистый участок
грунтового питания и т. п. (Формозов, 1961).
Еще большее значение имеет засушливая погода лета или
ряда лет. Земляные черви при высыхании почвы в конце глубо-
ких вертикальных ходов устраивают камеры и, свернувшись
плотным клубочком, впадают в оцепенение. В этих условиях кро-
ты лишаются возможности добывать достаточное количество!
своего основного корма, так как насекомые почти повсюду иг-
рают в их питании меньшую роль. Сильные засухи, как извест-
но, нередко охватывают большие территории, и на соответствую-
щей площади происходит резкое сокращение численности кротов.
Рис. 127. Изменения численности землероек-бурозубок в лесах
б. Шарьинского района Костромской области по данным учета ло-
вушками на постоянных линиях за 1931—1940 гг.:
/ —среднее число землероек на 100 ловушко-суток осенью, 2—максимальная
высота снежного покрова в предшествующую зиму
Значение крайне суровых морозных зим в движении числен-
ности животных нельзя преуменьшать, так как изучением исто-
рии наземной фауны Европы за ряд столетий установлены слу-
чаи полного вымирания черного хоря, зайца-русака и некоторых
других видов в северной части их ареалов в циклы лет с часто
повторяющимися холодными зимами и, наоборот, их обратное-
расселение на север при потеплении. Такое расселение идет и в
текущем столетии, начиная с 20-х годов, когда начало проявлять-
ся заметное потепление климата в северном полушарии. Благо-
приятное для жизни одних видов, оно оказывает отрицательное
воздействие на другие. В Западной Сибири за последние десяти-
летия отступила к северу южная граница «норения» арктического1
вида — песца, т. е. сократилась территория, где он размножается,
и соответственно расширилась на север область норения лисицы.
(Скробов, сообщение на Первом всесоюзном совещании по мле-
копитающим). ‘
334
Речной бобр тесно связан с водной средой, но для отдыха,,
сна и т. п. нуждается в суше, поэтому жизнь его часто зависит
от характера, длительности паводков и условий погоды во время
подъема уровня воды в обитаемых им реках. Ежегодно повто-
ряющиеся и потому привычные весенние паводки бобры относи-
тельно легко переживают во временных убежищах и укрытиях, а
довольно редкие зимние паводки в некоторых районах становят-
ся для этих грызунов стихийным бедствием, так же как и для
ондатры.
Зима 1954/55 г. в бассейне р. Воронежа была необычайно
теплой, и температура ряда декад декабря, января и февраля
оказалась выше многолетних средних на 7—8 и даже 10 граду-
сов. Количество осадков, выпавших в зимние месяцы, сильно
превысило норму, причем осадки чаще были в форме дождя, а
не снега. Продолжительные оттепели, перемежавшиеся с силь-
ными похолоданиями, вызвали значительный подъем уровни
воды в реках. Бобры были вынуждены покинуть затопленные'
хатки, норы и оказались на льду без защиты от холода, ледя-
ного ветра, а местами лишились также и доступа к запасам
корма. В эту зиму в районе Воронежского заповедника много
бобров погибло от истощения, реже от травм и нападений хищ-
ников (Жарков, 1956).
Так, например, один взрослый бобр был найдён вмерзшим
между двумя слоями льда, где он застрял при попытке выбрать-
ся на поверхность; другой бобр-двухлеток — придавлен крышей
домика, обрушившейся при оседании ледового покрова и т. п.
В соответствии с особенностями этих паводков, их продол-
жительностью и высотой подъема воды больший урон понесли
бобры в пойме р. Воронеж и в приустьевых участках его прито-
ков. Подсчет показал, что в пойме количество бобров сократи-
лось на 40—50% от поголовья, учтенного осенью. В верхнем и
среднем течении малых рек и по ключам число найденных трупов-
бобров не превышало 0,7% осеннего поголовья. Таким образом,
это стихийное бедствие, приуроченное только к речным долинам,,
действует в известной мере выборочно в разных участках бас-
сейна каждой реки.
Примером того, что описанные здесь уклонения зимней погоды
от нормы, способны вызвать в мире млекопитающих стихийное
бедствие, «широкого разлитого характера», захватывающее об-
ширную площадь, может служить картограмма, помещенная на
рис. 128. После дождей, выпавших на снежный покров, и после-
дующих похолоданий уже в январе 1955 г. образовалась сплош-
ная притертая к земле ледяная корка на пространстве несколь-
ких областей, расположенных в северной половине черноземной
полосы Европейской части СССР. К февралю местами тол-
щина ледяной корки достигала 15 см, и озимые поля к этому вре-
мени, например, в Курской области, находились под ледяным
335-.
Рис. 128, Распространение ледяной корки, притертой к земле, и корки, взве-
шенной в толще снежного покрова, в Европейской частиСССР в третью декаду
февраля 1955 г. (по данным Центрального института прогнозов)
панцирем более двух месяцев. (Корки сильно влияют на состоя-
ние озимых полей, поэтому агроклиматологи внимательно изу-
чают их возникновение, распространение и исчезновение).
Сведения о массовой гибели обыкновенных полевок и отчасти
домовых и полевых мышей в эту зиму были получены из многих
районов Орловской, Курской и Липецкой областей. Грызуны по-
гибают от затопления их норок водой, от недоступности корма,
вмерзшего в лед, от истощения и холода на поверхности снега
при попытках выселиться из участков, охваченных бедствием.
Местами до 60% площади полей было затоплено снеговой водой,
вскоре превратившейся в лед. Вымирание полевок было настоль-
ко полным, что численность их не могла восстановиться за лето
и оставалась очень низкой даже осенью 1955 г.
Ледяную корку как причину гибели мелких грызунов на об-
ширных участках неоднократно отмечали в зонах тундр, степей
и полупустынь нашей страны (Горбунов, 1929; Браунер, 1928;
Семенов, 1955; Хрусцелевский, 1954; Формозов, 1961 и многие
др.) Часть зверьков, благодаря особенностям некоторых биото-
пов, сохраняется от гибели и служит тем исходным поголовьем,
от размножения которого через короткое время восстанавливает-
ся популяция и постепенно начинает вновь заселять участки,
подвергшиеся опустошению.
В лесной зоне наличие защитного полога крон деревьев, кус-
тарников, обилие пней, бурелома и т. п. делает невозможным
появление сплошной притертой ледяной корки, и уже одно это
обусловливает понижение уровня смертности мелких наземных
зверьков .(землероек, полевок, мышей) при отклонениях зимней
погоды от нормы, подобных только что описанным.
Хозяйственная деятельность людей
' Роль хозяйственной деятельности общества во многих слу-
чаях оказывается очень важной. Охотники ежегодно добывают
некоторую, иногда очень значительную часть поголовья промыс-
ловых зверей, нередко избирательно выбивая преимущественно
молодых или только взрослых животных, самцов или самок.
В результате сельскохозяйственных работ на обширных масси-
вах при использовании современной мощной техники зверьки
то разом лишаются запасов корма, нор и хороших укрытий на
, многих тысячах гектаров, то затопляются водой их норы при
' поливе полей, уничтожаются их кормовые сорные растения с по-
мощью гербицидов и т. п. С другой стороны, истребление кустар-
ников и густых бурьянов лишает хищников-мышеедов (луней и
. болотных сов) удобных мест для гнездования. Количество этих
пернатых врагов мелких грызунов в районах интенсивного сель-
| скохозяйственного производства- резко сокращается, отчего раз-
множение и зимовка зверьков идут без сдерживающего воздей-
ствия со стороны хищников.
22 Териология
337
Превращение естественных ландшафтов в культурные, насаж-
дение лесных массивов и полезащитных лесных полос, обводне-
ние степей, регулирование стока рек доказывает, что непрерывно
увеличивается власть общества над природой, в том числе и над
миром млекопитающих.
ТИПЫ ДВИЖЕНИЯ ЧИСЛЕННОСТИ
Движение численности населения млекопитающих в извест-
ной мерс специфично у каждого вида, но некоторые его особен-
ности позволяют различать формы, принадлежащие к одному се-
мейству или отряду. С. А. Северное (1937, 1941, 1942) отметил
основные биологические свойства, характеризующие отдельные
«типы динамики» у представителей разных классов, в том числе
и у млекопитающих; это — плодовитость и продолжительность
жизни.
Плодовитость исторически связана со стойкостью против не-
благоприятных воздействий и средней продолжительностью жиз-
ни особей. Появление адаптаций, уменьшающих гибель и увеличи-
вающих среднюю продолжительность жизни особей, сопровож-
дается соответственным снижением плодовитости. Следуя за
Ч,- Дарвином, С. А. Северцов рассматривает уровень плодови-
тости как приспособление, сложившееся в ходе естественного от-
бора и компенсирующее смертность. Таким образом известная
сбалансированность размножения и гибели, обеспечивающая от-
носительную стабильность популяции при относительно устой-
чивых условиях существования, имеет глубокие исторические
корни. Взятые С. А. Северцовым показатели удачны потому, что
они определяют основные черты движения численности населе-
ния— скорость его оборота- и связанную с ней изменчивость
численности: период колебаний — короткий или длинный, обу-
словливаемый долговечностью особей, и амплитуду колебаний,
зависящую в основном от плодовитости.
С. А. Северцов различал у млекопитающих пять «типов ди-
намики населения». Позднее Н. П. Наумов (1948) внес в эту
схему некоторые изменения и уточнения. Для краткости можно
свести движение населения к трем типам: 1) крайне неустойчи-
вого (скачкообразного) движения населения, характерного для
«эфемеров» — мелких грызунов и землероек с очень быстрым
.оборотом населения; 2) неустойчивого движения населения с до-
вольно частыми и относительно резкими изменениями численно-
сти (белка, песец, лисица и др.); 3) устойчивого движения на-
селения с медленными, растянутыми во времени и плавными
подъемами и сокращениями численности.
Основные черты движения населения одного вида или груп-
пы родственных видов определяются их главными биологически-
ми свойствами, а именно плодовитостью, стойкостью против
338
неблагоприятных воздействий и связанных с последней продол-
жительностью жизни и возрастным составом популяции. Внеш-
ним условием, стимулирующим рост численности, для одних
групп служит обилие корма, для других — благоприятная по-
года.
Общая черта движения населения у разных видов — харак-
тер нарастания численности после ее депрессии. Нарастание
идет по S-образной или логистической кривой. Быстро нарастая
вначале, она на каком-то определенном этапе поднимается за-
медленно, затем прогрессивно падает1. Примерно так идет рост
численности мелких грызунов после резких депрессий, очень
характерных для движения населения этой группы (Виноградов,
1934; Свириденко, 1934; Фенюк, 1934, 1941, 1951; Формозов,
1937; Башенина, 1947; Н. Наумов, 1948 и др.).
Другая важная и общая черта движения численности — за-
кономерно развивающаяся по мере нарастания плотности насе-
ления смена условий его существования и, как следствие, смена
ведущих факторов движения численности. Различают фазу
подъема численности населения (размножение резко превышает
смертность), пик численности (высокая плотность населения
тормозит размножение и выживание молодых), фазу ,падения
численности (сложившиеся условия вызывают большую смерт-
ность от голода, неблагоприятной погоды или заболеваний) и
фазу депрессии — снижение численности прекращается, начина-
ют восстанавливаться условия для ее нового подъема (Формо-
зов, 1935, 1937; Н. Наумов, 1958).
Тип крайне неустойчивого (скачкообразного)
движения численности
Мелкие грызуны (мыши, полевки, лемминги, в меньшей мере
хомячки) отличаются наиболее высокой плодовитостью среди
всех млекопитающих и очень большими ее колебаниями в раз-
ные сезоны и годы. Биологическая целесообразность столь ши-
рокой лабильности плодовитости не подлежит сомнению. Раз-
множение сокращается при неблагоприятных условиях, когда
выживание многочисленного потомства мало вероятно, и макси-
мально быстро используется возникающая благоприятная об-
становка — обильный урожай корма, теплая затяжная осень,
мягкая зима и т. п. При интенсивном подъеме численности мо-
лодняк в короткий срок занимает потерянные при вымирании
участки, и распределение.популяций грызунов из пятнистого,
спорадичного, становится сплошным или почти сплошным. Tas
1 По такой же логистической кривой идет.и размножение акклиматизи-
руемого вида, встретившёго на новой родине благоприятные условия-жизни.
22* '339
ким образом, в фазу пика численности территория и ее ресурсы
используются ими наиболее полно, резко увеличивается коли-
чество заселенных биотопов. Кроме того, часть животных мигри-
рует в разных направлениях, благодаря чему ареал временно
Чоирен Баишинту-Сумз -L
Рис. 1291 Ареал полевки Брандта в Восточной Монголии.
Вверху — весной 1941 г. в год низкой численности, внизу-—
осенью 1943 г. в год пика численности. Площадь, густо засе-
ленная полевкой, резко увеличилась, граница ареала при пике
Численности продвинулась к северу (по Кучеруку и Дунаевой)
расширяется, как это бывает, например, у полевки Брандта
(рис. 129), норвежского лемминга {Lemmas lemmas), появ-
ляющегося на Скандинавском полуострове, в Финляндии и у нас
на Кольском полуострове на десятки километров южнее обыч-
340
ной границы распространения (здесь возникают временные ко-
лонии иммигрантов).
При интенсивном размножении численность полевок, пестру-
шек и мышей за короткий период увеличивается в сотни раз,
так как молодые зверьки становятся половозрелыми, еще не до-
стигнув полного роста, и в свою очередь приносят один выводок
за другим. Благодаря крайнему обилию господствующих по
численности зверьков двух-трех видов, на которых сосредоточи-
вается охота хищников, получают возможность почти беспре-
пятственно размножаться и такие зверьки, которые в другие
годы интенсивно вылавливаются совами, лунями и пр., например
серые хомячки (Cricetulus tnigratorius), землеройки-белозубки
(Crocidura suaveolens) и др.
Некоторые исследователи считают, что в южных районах
страны, с большой продолжительностью вегетационного пе-
риода, массовое размножение полевок и мышей вследствие спо-
собности этих зверьков при благоприятных условиях размно-
жаться и в течение зимы, т. е. круглогодично, может произойти
даже за один год.
В период так называемого массового размножения количе-
ство отверстий нор полевок достигает в степной зоне 20000—
35 000 на 1 га; зверьки десятками в течение всего дня то и
дело попадаются на глаза, в скирдах беспрестанно слышен шо-
рох и писк грызунов, множество дорожек, кучек нарытой земли,
поврежденных растений говорит об их, интенсивной деятельно-
сти. Известны случаи, когда размножившиеся степные пеструш-
ки в Средней Сибири и полевки Брандта в Забайкалье и Мон-
голии полностью уничтожали растительный покров пастбищ и
сенокосов, крайне затрудняя выпас домашних стад. Большие
повреждения осоковых участков вызывает и зимняя деятель-
ность обских леммингов в тундрах Сибири. От сильно размно-
жившихся полевок-экономок на лесных луговинах и покосах
таежной зоны может страдать разнотравье.
Сотого времени, когда деятельность сильно размножившихся
зверьков начинает оставлять на заселенных участках многочис-
ленные следы повреждений, вступают в действие эндогенные
силы (связанные с природой самой популяции, достигшей высо-
кого уровня численности), которые сдерживают дальнейшее раз-
множение. Так например, при изучении домовых мышей, насе-
ляющих клади необмолоченного хлеба в Англии, было установ-
лено, что среднее количество зверьков на 1 м3 изменялось от
0,5 экз. в скирдах, простоявших 4 месяца, до 2—6 и более мы-
шей в скирдах, простоявших 7*/2—8'/2 месяцев. При этом выяс-
нилось, что в скирдах с высокой плотностью населения количе-
ство размножающихся самок уменьшается до 85%, а самцов до
92%; процент беременных самок от 58,5% в скирдах с низкой
плотностью падает до 43,4% в имеющих наивысшую плотность..
341
Средняя величина выводка была 6,2—5,1; при этом последняя
цифра тоже относилась к мышам из скирд с высокой плотно-
стью населения. Наконец, количество самок с резорбирующи-
мися эмбрионами — 27,4% оказалось больше тоже в скирдах
с высокой плотностью населения и меньше (14%) при низкой
заселенности.
Таким образом, все показатели согласно говорят о том, что
с увеличением плотности населения мышей их размножение на-
чинает затормаживаться [Саусуик (Southwick), 1958]. Сходные
выводы получены и при изучении размножения мелких грызунов
в открытых биотопах (поле, луг, степь) и в лесу.
Уже давно было замечено, что в период массовых размноже-
ний, когда численность полевок и мышей достигает наивысшего
уровня, плодовитость их резко снижается (Кучерук и Рюмин,'
1'938; Мигулин, 1938). В Тульских засеках в 1938 г. Н. П. Нау-
мов (1948) установил резкое падение интенсивности размноже-
ния лесных грызунов, которое нельзя было объяснить ни усло-
виями погоды, ни состоянием запасов корма. В этом году из
630 исследованных самок рыжей полевки (Clethrlonomys gla-
reolus) беременных было всего 2,8%, а из 267 самок желтогор-
лых мышей (Apodemus flavicollis) только 1,8%, тогда как в
1939 г. при низкой численности зверьков соответствующий по-
казатель равнялся 53,1 % и 20,0%. Низкая интенсивность раз-
множения лесных грызунов в 1938 г. при исключительно высокой
их численности была следствием исключительной «тесноты» в
лесу (Н. Наумов, 1948), Большая часть зверьков ютилась в
убежищах, непригодных для размножения? что, видимо, и было
Основной причиной холостания подавляющего количества самок.
Немалое значение имеет и фактор беспокойства, связанный с
высокой плотностью населения. Постоянная беготня множества
зверьков создает неблагоприятную обстановку при спариваний,
родах и выкармливании новорожденных, что доказано экспери-
ментами. Снижение плодовитости лисиц при сильной перена-
селенности Тульских засек отметил Г. Н. Лихачев (1958).
Другая вероятная причина, сдерживающая размножение
грызунов в годы высокой численности,— глистные инвазии, рас-
пространение которых (экстенсивность и интенсивность инвазит
рованности зверьков) увеличивается при высокой плотности их
населения. Известно, что продукты жизнедеятельности гельмин-
тов способны оказывать токсическое действие на организм хо-
зяина и угнетать его половую продуктивность вплоть до появ-
ления полной стерильности (Тереза и Шалимов, 1937).
Кроме того, мышевидные грызуны, особенно полевки, как
зверьки с несовершенной терморегуляцией, очень чувствщ
тельны к воздействию погоды; она влияет на их размножение,
перемещения, распределение по стациям. Резкие уклонения ее
342
от нормы вызывают массовую гибель взрослых и молодых зверь-
ков. Очень велико и косвенное влияние погоды через состав и
состояние растительности (высота, сомкнутость травостоя, нали-
чие или отсутствие витаминов, урожайность плодов и т. д.). В ли-
тературе отмечены многочисленные случаи массовой гибели по-
левок и пеструшек от длительных холодных ливней, при зимних
оттепелях, затопляющих норки снежной водой с последующим
образованием притертой к земле ледяной корки, от сильных мо-
розов, наступающих до образования достаточно высокого снеж-
ного покрова х, при бурном таянии снега в дождливую весну
и т. д. Такие губительные условия погоды оказывают на зверь-
ков не только непосредственное влияние, вызывая большую их
смертность от намокания, переохлаждения, обострения болез-
ней, но и служат причиной вынужденных массовых перемеще-
ний зверьков, облегчающих распространение инфекций и дея-
тельность многочисленных хищников.
В конце массового размножения осенью 1932 г., начиная с
октября — ноября, среди домовых мышей и серых полевок, раз-
множившихся на территории быв. Северо-Кавказского края, на-
чались интенсивные эпизоотии, в результате которых погибла
значительная часть грызунов, а местами они совершенно исчез-
ли. Эпидемиологами было исследовано более 6000 грызунов
(главным образом Mus musculus и единичные Microtus arvalis)
из западной части края. Авторы отчета (Миллер, Квашнина,
Гржебина, 1935) указывают, что в 1932 г. спонтанные эпизоотии
мышей в большинстве случаев были вызваны гемолитическим
стрептококком, отчасти микробами-из групп, вызывающих ге-
моррагическую септицемию, паратифозных и др,
Отличия мехового покрова, особенности физиологии и пове-
дения, характерные для разных видов полевок и мышей, обус-
ловливают то, что при резких уклонениях погоды от нормы
популяции этих зверьков сокращаются далеко не сходно. В дуб-
равах Тульской обл. зимой 1938/39 г., отличавшейся при бес-
снежье в декабре сильными морозами, сменившимися затем
сильными снегопадами, оттепелями и гололедицами, мыши ока-
зались гораздо более стойкими, чем полевки. Численность се-
рой полевки сократилась в 100 раз, рыжей лесной полевки в 50,
а лесной и желтогорлой мышей — всего в 4—5 раз. (Н. Наумов,
1948). Сходные наблюдения сделал С. И. Оболенский (1926) в
Каменной степи в неблагоприятную для мелких грызунов зиму
1921/22 г. Большую стойкость полевой мыши (Apodemus agra-
rius) к морозам установили при разборке скирд Кучерук,
А. Кротов, А. Рюмин (1935). Эта мышь устойчива и к туляре-
мийной инфекции.
1 Защитой от потери тепла служит слой рыхлого снега, обычно не менее
20 см высотой.
343
Таким образом, падение численности населения мелких гры-
зунов после вспышки размножения обычно вызывается сочета-
нием целого ряда факторов как эндогенного, так и экзогенного
происхождения.
При свойственной мышам и полевкам высокой плодовитости
их население может достигнуть высокого уровня численности за
короткий ряд лет. При малой стойкости против условий, вызы-
вающих смертность, и, следовательно, малой продолжительно-
сти жизни столь же быстро происходит депрессия численности.
По многолетним наблюдениям Тульской санитарно-эпидемиоло-
гической станции (Мясников, 1955), подъемы численности серой
полевки, составляющей в среднем 50—95% всего населения гры-
зунов в полях этой области, возникают через 2, 3, 4 года и бы-
вают даже в течение двух лет подояд (1941 и 1942 гг.). В годы
пика численности серой полевки обычно развиваются туляремий-
ные и иные эпизоотии. С такой же частотой повторяются подъ-
емы численности этой полевки под Москвой; северные пеструш-*
ки (лемминги) в массе появляются каждые 3—4 года, подъемы
численности мелких лесных грызунов обычно бывают через
2—5 лет (Н. Наумов, 1948).
Помимо этих частых, местных подъемов численности
(рис. 130) немногих видов грызунов на небольших площадях
(так называемые «малые волны») в движении населения степ-
ных и полевых грызунов нашей страны различают массовые
размножения, происходящие обычно один раз в десятилетие и
охватывающие огромные территории («большие волны»). В эти
грандиозные массовые размножения вовлекаются многие виды
грызунов1. Б. С. Виноградов (1934) выделил эти массовые раз-
множения как подъемы численности, имеющие много своеобраз-
ных черт, использовав большие материалы за длинный ряд лет.
Позднее детали их были проанализированы рядом авторов (Фе-
нюк, 1934; Формозов, 1937; Н. Наумов, 1948 и др.).
Изучению движения численности мышевидных грызунов по-
священа большая литература. В настоящее время разработаны
и апробируются различные приемы построения прогнозов чис-
ленности грызунов для своевременного предупреждения потерь
урожая и рассеивания инфекций. При прогнозировании прини-
мают во внимание численность исходного поголовья, его возра-
стной состав и жизненность, условия погоды (температура воз-
1 Среди млекопитающих наблюдается взаимосвязь в движении числен-
ности нескольких видов. «Сопряженность вспышек массового размножения»
недавно установлена и у насекомых. Сопряженность вспышек бывает пря-
мой и обратной. Первая объясняется наличием у некоторых видов общих
благоприятных и неблагоприятных условий. Обратная сопряженность наблю-
дается в том случае, когда условия массового размножения одного вида
исключают возможность массового размножения другого вида. Некоторые
зоологи (П. А. Свириденко, И. Я. Поляков) отрипают существование «боль-
ших волн» размножения и их относительно регулярную повторяемость.
344
духа и количество осадков) в решающие периоды года — позд-
ней осенью, весной, летом, состояние снежного покрова зимой,
качество уборки урожая, состояние полей (количество сорняков,
бурьянов), количество хищников-мышеедов и т. п.
Вследствие широкого расселения грызунов в период пика их
численности и увеличения количества биотопов, занятых ими в
этот период, истребление вредных зверьков требует затраты
большого труда, огромного количества ядохимикатов, приманок
и т. д. Поэтому в каждом случае следует организовать борьбу
так, чтобы не допускать подъема численности грызунов до угро-
жающего уровня, приурочивая ее к тому моменту, когда площа-
ди очагов размножения сравнительно невелики и урожай еще
не пострадал (Формозов, 1937).
После очередного сокращения численности полевки и мыши
сохраняются обычно в немногих изолированных участках-«ре-
зервациях», или «стациях переживания». Именно в них-то и
выгоднее всего истреблять весной сохранившееся маточное по-
головье, способное в ближайший благоприятный сезон дать
вспышку размножения и расселения. Проведенные опыты ранне-
весенних затравок грызунов в резервациях показали преимуще-
ства этого приема (Н. Наумов, 1948).
Тип неустойчивого движения численности
Еще Ч. Дарвин («Происхождение видов», 1859) писал: «Мно-
гочисленные факты ясно указывают, как особенно чувствительна
воспроизводительная система даже к самым слабым изменениям
в окружающих условиях». Из этих условий важнейшими являт
ются кормовые, трофические. Уже среди зверьков «эфемеров»
есть целый ряд узкоспециализированных семеноедов, в движе-
нии численности которых урожай плодов (и семян) играет роль
важнейшего внешнего толчка к массовому размножению
(рис. 131). Типичные представители таких грызунов — лесная,
желтогорлая и домовая мыши, в некоторых лесных районах так-
же рыжая лесная (Clethrionomys glareolus) и красная (С. rutt-
lus) полевки. Длительные наблюдения В. А. Попова (1960) в ле-
сах левобережья Волги у г. Казани показали, что наибольшее
значение из факторов, определяющих численность рыжих поле-
вок, имеет обеспеченность продуктивными кормами; на втором
месте стоят условия зимовки, и, в частности, высота снежного
покрова, и на третьем — условия погоды в период размножения.
Сходные выводы на 11-летнем материале были получены и при
изучении рыжих полевок хвойных лесов северной части Кост-
ромской области (Формозов, 1948).
Даже среди видов, питающихся грубыми растительными
кормами, способных усваивать не только свежую зелень, но и за-
сохшую, выгоревшую траву, сухие побеги и т. п., установлены
346
случаи значительных изменений численности в связи с недостат-
ком или обилием пищи.
Большая песчанка (Rhombomys opimus)— грызун, свойст-
венный полупустыням и пустыням Азии, отличается крайней не-
прихотливостью в отношении выбора кормов. Она поедает боль-
шинство видов растений, имеющихся по соседству с ее норой, в
том числе формы, совершенно непривлекательные для других
«фитофагов — жгучие, эфироносные, колючие, ядовитые. Так, на-
пример, она без видимого вреда для себя питается гармалой,
или адраспаном (Peganum harmala), ядовитым для скота рас-
тением, содержащим несколько алкалоидов. Гармала часто и
Рис. 131. Изменение осенней численности курганчиковых мышей (Mas mus-
culus hortulanus) на юге Украины за годы 1934—1938 в сопоставлении с
величиной осадков за август — сентябрь. Осенние осадки обусловливают
увеличение урожая семян сорняков, за счет которых обеспечивается кормом
поголовье половозрелых мышей и их успешное размножение (по Н. Наумову)
в большом количестве селится на выбитых скотом пастбищах,
так как домашние животные в период вегетации ее не трогают,
а конкурентов поедают и угнетают. Казалось бы, что столь не-
требовательный грызун, как большая песчанка, всегда может
найти достаточное количество корма, Выяснилось, однако, что
в условиях засушливого климата полупустынь и пустынь не-
редки годы, крайне бедные осадками, когда растительность бы-
вает угнетена и дает ничтожно малую продукцию. Большая пес-
чанка •— типичный норник; она кормится не далее 40—60 м от
норы, куда готова скрыться при первом признаке опасности.
В засушливые годы участок диаметром 100—120 м не может
полностью удовлетворить потребности семьи песчанок в корме.
При более далеких рейсах зверьки чаще становятся добычей
хищников. Состояние растительности в пустынях, особенно наи-
более ценных в кормовом отношении эфемеров, эфемероидов, и
347
части кустарничковых многолетников (полыней, солянок) зави-
сит от запасов почвенной влаги, непосредственно обусловливае*-
мых величиной осадков, выпадающих в холодную половину года
(летом осадков в части наших пустынь или совсем не бывает,,
или, там, где выпадают, они не успевают впитываться в почву
и полностью испаряются).
Длительные наблюдения зоологов противочумных станций:
показали, что численность больших песчанок зависит в основном
от состояния растительности в период их размножения и, в ко-
рне. 132. Изменение численности песчанок
в зависимости от количества осадков:
J —количество осадков за октябрь — май 1944—1952 гг. в Панфиловском районе
Южного Казахстана; 2 — среднее число песчанок на гектар (по Бурделову и Касаткину>
нечном счете, от количества осадков за предшествующую зиму
(Варшавский и Шилов, 1955, Бурделов и Касаткин, 1958; Шеха-
нов, 1959 и др.).
Установлено также, что сходное движение численности до-
статочно постоянно повторяется у популяций больших песчанок,,
занимающих целые природно-географические области. Это лег-
ко понять, зн'ая, что условия погоды обычно бывают одинако-
выми на больших пространствах равнин Южного Казахстана к
Средней Азии. Рис. 132 показывает, как следом за сезонами с
хорошим зимне-весенним увлажнением поднималась числен-
ность больших песчанок в Панфиловском районе Казахской
ССР (идеального совпадения кривых нет, так как на амплитуде-
изменений численности этих грызунов сказывается и величина'
поголовья производителей, сохранившихся до весны — момента
348
начала размножения). Хорошая осенняя вегетация солянок при
теплой погоде обусловливает появление новой, третьей волны
размножения и смещение пика численности на позднюю осень.
Неблагоприятный ход весенних температур, а именно дефицит
тепла, поздние заморозки могут задержать размножение и ока-
зать отрицательное воздействие и на второй генеративный цикл
>(Бурделов и Касаткин, 1958).
В целом наиболее важны обеспеченность кормом, с которой
связан и ход смертности производителей, и интенсивность их
размножения. Поэтому при прогнозах численности больших пес-
чанок, занимающих отдельные природно-географические обла-
сти, основное внимание уделяют характеристике погоды холод-
ного периода и состояния растительного покрова в весенние ме-
сяцы и осенью. Большая песчанка играет первостепенную роль
в эпизоотии чумы в пустынях Средней Азии и Казахстана. Про-
гнозирование движения численности этого грызуна облегчает
организацию борьбы с эпизоотиями чумы и помогает предупре-
ждать возможность заболевания людей.
У обыкновенной белки, находящейся в такой же зависимо-
сти от урожаев семян лесных пород, как, например, желтогор-
лая мышь, все же вследствие ее меньшей плодовитости не бы-
вает столь быстрых и резких подъемов и падений численности.
Малая плодовитость (по сравнению с мышиными) обусловлена
ее большей стойкостью и относительно высокой продолжитель-i
ностью жизни: в природных условиях небольшая часть белок
доживает даже до 6—7 лет. Для мелких мышей в урожайный
на семена год характерно длительное осенне-зимнее размноже-
ние, начинающееся обычно после опадения на землю желудей
дуба, орешков бука, липы и др. и продолжающееся до начала
лета следующего года включительно. Белки же при высоком
урожае, например семян ели, успевают за счет высокоценного
свежего корма принести лишь один выводок в конце лета того
же года (конец августа — сентябрь) *. В основном же использу-
ют его в теплую половину следующего года, поэтому и подъемы
их численности всегда отстают на один год по сравнению с уро-
жайными годами лесных пород.
Список кормов, используемых белкой, довольно велик: в него
входят семена и орехи лесных пород, ягоды, почки ели, березы
и других деревьев, грибы шляпочные и почвенные (трюфели),
нежная кора и сок побегов сосны, ели, пихты, березы, лишай-
ники, трава и листья, личинки мух, куколки и взрослые
.муравьи, изредка яйца и птенчики птиц. Несмотря на такое раз-
нообразие кормов, давно установлено, что белка и по морфоло-
1 Они начинают использовать семена нового урожая при их молочной
спелости, в лесах северной половины нашей страны — с конца июня или на-
чала июля.
349
гическим (строение зубов, желудка, кишечника), и по биологи-
ческим особенностям — узкоспециализированный потребитель
концентрированных кормов, а именно семян лесных пород.
На преобладающей части ареала важнейшую роль в питаний
белки играют семена ели (рис. 133, 134), лиственницы и кедров
(сибирского, корейского, кедрового стланика), в меньшей сте-
пени — сосны, дубов и лещин. Семена хвойных пород бывают
в изобилии не всегда, а только в годы высоких урожаев, повто-
ряющихся раз в несколько лет. Перерывы между урожаями за-
висят от биологии данной древесной породы, а также от ши-
роты местности и степени континентальности климата в разных
частях ее ареала. В условиях равнинной тайги нередко про-
странства в сотни тысяч квадратных километров еловых или
кедровых насаждений одновременно приносят высокий урожай
или поражаются полным неурожаем (Формозов, 1935, 1936; Да-
нилов, 1940, 1952; Кирис, 1944 и др.).
Условия добывания корма белкой зависят не только от вели-
чины урожая, но отчасти и от деятельности многочисленных кон-
курентов: насекомых, повреждающих шишки, клестов, дятлов,
кедровок, в массе уничтожающих семена. Если корм с осени
осыпается на землю (шишки кедра, желуди дуба), большую
роль играет также структура снежного покрова в данную зиму:
появление наста, корок гололеда очень затрудняет отыскивание
и выкапывание белкой пищи из-под снега. Вызываемое недо-
статком семян изменение диеты белки (использование вынуж-
денных и балластных кормов) обусловливает истощение орга-
низма животного, заболевания, иногда, кроме того, откочевки
и в конечном итоге —• массовую гибель. Особенно много исто-
щенных белок погибает в морозные месяцы зимы и при поздне-
осенних миграциях, что сильно сокращает количество произво-
дителей перед началом гона. Длительное использование биоло-
гически неполноценного корма сильно сказывается на половой
функции белки, вызывая резкое снижение плодовитости и даже,
полное прекращение размножения.
В нормальных условиях большинство самок приносит в те-
чение года два помета (первый весной, второй — летом); пара
взрослых в среднем приносит 9 бельчат. Осенью, перед началом:
промысла молодые белки составляют в популяции около 8'2%
(большинство из весеннего помета). Соотношение полов в по-
пуляции 1 : 1. В годы, несколько худшие по кормовым условиям,
число пометов остается обычным, но среднее число бельчат на-
пару взрослых к концу сезона размножения не превышает
4,9—5,8 (большинство из летнего помета), молодые составляют
в популяции около 70% (Кирис, 1944).
После бескормной зимы размножения совсем не бывает, реже
часть белок приносит один весенний или один летний помет
(процент яловых очень велик). Количество молодых к осенйв
350
Рис. 133. Урожай семян хвойных пород осенью 1935 г. по глазомерной оценке в баллах. (Картограмма составлена
во Всесоюзном институте охотничьего' $озяйства_на основе использования сообщений корреспондентов)
ний охотников-корреспондентов)
составляет значительно меньше 50%; число самок становится
ниже нормы. В итоге после неурожайного года, а особенно по-
сле двух последовательных неурожайных лет численность белки
резко сокращается; из обычного массового грызуна она вре-
менно превращается в редкое и малозаметное животное леса.
Условия погоды непосредственно в гораздо меньшей степени
влияют на движение численности этого грызуна, чем на нестой-
ких гетеротермных полевок.
В зависимости от чередования семенных и неурожайных лет
основной кормовой породы «урожаи» белки повторяются с про-
межутками в 3, 4, 5, 6 лет. Даже в пределах одной области или
республики (например, Якутской) разные ее районы могут суще-
ственно отличаться в частоте «урожаев» белки, в их величине
и т. д. Это обусловливается особенностями лесных площадей,
средними размерами урожая семян, продолжительностью перио-
да размножения белки и т. п.
Еще 140 лет тому назад путешественник Ф. Врангель (1841)
со слов якутских промышленников записал, что «урожаи пес-
цов» на северо-востоке Сибири повторяются с известной пра-
вильностью через каждые 3—4 года. В наше время специальные
исследования, проведенные в той же Якутии, показали, что этот
3—4-летний ритм колебаний численности остался неизменным.
Для севера Канады имеются цифры за большой ряд лет, пока-
зывающие, как такие же изменения количества песцов отража-
ются на заготовке его шкурок. Так, например, по X. Элтону (Elton,
1924), в одном из участков северной Канады численность песца
была наибольшей в годы: 1856, 1861, 1864, 1869, 1873, 1878, 1884,
1888, 1892, 1896, 1899, 1903, 1907 и 1911. Следовательно, «уро-
жайные» годы повторялись здесь через промежутки в 4, 3, 3, 5,
3, 3, 4, 4, 4, 4, 4, 4, 4, 5, 4 лет, в среднем — 3,9 года. Сходная
цикличность подъемов и падений численности песцов установ-
лена наблюдениями зоологов и промышленников на Европей-
ском Севере СССР, включая Новую Землю, и в тундрах Сибири,
причем годы урожая песца в Якутии не совпадают с таковыми
на Ямале и т. п.х.
Следовательно, одновременным урожаем или неурожаем;
охватывается только треть или четвертая часть огромного про-
тяжения наших тундр. Повсеместно высокая численность песцов
обусловливается массовым размножением пеструшек (леммин-
гов)— на западе — норвежского лемминга, а восточнее Белого
моря — главным образом обского и отчасти копытного. ч
Наблюдениями на'Ямале (Дунаева и Осмоловская, 1948)
установлено, что в зимы, когда идет подснежное размножение
пеструшек, в желудке песцов часто находят целые выводКи,' вы-
1 Тем не менее некоторые авторы (например, В. Н. Сдобников) отри-
цают существование ритмичных колебаний численности песцов. .
23 Териология
353
копанные из гнезд. После такой зимы обычно и летняя числен-
ность пеструшек оказывается высокой, что обеспечивает вы-
кармливание больших пометов песца Ч
Плодовитость песца выше, чем у лисицы: в выводке нередко
бывает 8—10 и более подросших молодых. Наибольшее извест-
ное число эмбрионов — 16 (Житков, 1913), а количество после-
родовых пигментных пятен в матке до 26 (Тюлин, 1938). Хорошо
обеспеченные продуктивным кормом осенью и в течение всей
зимы песцы не рассеиваются по тундре, лесотундре и льдам
моря, а держатся близ нор и рано приступают к. гону; овуляция
и беременность самок протекает нормально, они легко вынаши-
вают и выкармливают многочисленных щенков. В такие годы
заселенность нор бывает высокой. Так, например, по данным
В. Д. Скробова (1958), в Карском и Пайхойском очагах размно-
жения песца в 1955 г. было заселено 100% осмотренных нор, но
в 1956 г. только 20%, в 1957 — 20% и 10% (в Пайхойском), а в
1958 снова 80% и 100%. Среднее число выкормленных щенков
в выводке, по подсчетам Скробова, в августе было 8 (1955 г.,
обилие пеструшек), 4 (1953 и 1956 гг., пеструшек крайне мало
или нет) и даже 3 (1957 г., лемминги отсутствовали).
В годы низкой численности пеструшек и узкочерепных поле-
вок Microtus (Stenocranius) gregalis, которые тоже служат цен-
ным кормом, но играют в жизни песца второстепенную роль,
многие самки этого хищника остаются бесплодными или прино-
сят слабых щенят, погибающих уже осенью. При остром недо-
статке пищи отдельные пары бросают свои выводки. В течение
Одного-двух бескормных лет население песцов резко уменьшает-
ся, так как при предельно сократившейся плодовитости одно-
временно идет интенсивное вымирание производителей.
Осенью в год высокой численности песцов, часто совпадаю-
щий с вымиранием пеструшек, начинаются массовые перекочев-
ки хищников, среди них распространяются широко разлитые
эпизоотии, наносящие огромный урон поголовью. Преобладают
случаи поражения нейротропным вирусом.— болезнь, известная
на севере как «дикованье». «Дикующий» песец крайне возбуж-
ден, беспокоен, подвижен, утрачивает всякую осторожность —
смело бросается на упряжки ездовых собак, нередко заскаки-
вает в освещенные окна промысловых избушек и т. д. Ездовые
-собаки, лисицы и волки тоже вовлекаются в эти эпизоотии.
Среди охотников существует мнение, что падеж ездовых собак
и песцов вызывается бешенством.
1 Авторы этого исследования предложили остроумный способ прогноза
численности песпов на следующий промысловый сезон, основанный на учете
встреч молодых леммингов в желудках песцов, добытых в течение предшест-
вующего сезона. Сбор тушек для вскрытия легко организовать на факториях
и заготпунктах.
354
«Дикованье», действительно, напоминает его некоторыми
симптомами.
В тундрах большинство песцов умирает в первую же зиму
после начала эпизоотии. Количество трупов, вытаивающих вес-
ной с сохранившимся, хорошим мехом, нередко бывает огром-
ным (рис. 135). Практикуется даже специальный промысел и
заготовка шкурок павших песцов. На упряжках оленей промыш-
ленники обследуют проталины тундры и отыскивают белых, из-
далека заметных зверьков.
Так, например, в районе одной только речки западносибир-
ской тундры однажды нашли 130 павших песцов, большинство
которых было в зимнем меху и отличалось высокой упитанно-
Рис. 135. Труп песца в полном зимнем меху, вытаявший из снега. Март 1950 г.
Нижне-Колымский р-н Якутии (фот. Н. Н. Немнонова)
стью (Цецевинский, 1940). На протяжении примерно 20 км пути
А. Ф. Чиркова (1951) насчитала остатки 16 трупов песцов.
В Большеземельской и Малоземельской тундрах после сильных
вспышек заболевания весной находили иногда до двух трупов
на 1 км2 (Скробов, 1958).
Годы появления большого числа «песцовых мышей», как
нередко называют на севере пеструшек, в Большеземельской и
Малоземельской тундрах были за последний период следующие:
1926, 1929, 1932, 1935, 1940, 1943, 1946, 1949, 1952, 1958. Таким
образом, урожай пеструшек в наших тундрах повторяется через
два года на третий. Это дает возможность составлять многолет-
ние прогнозы урожаев леммингов и песцов (Скробов, 1958),
23*
355
основываясь на длительном изучении экологии песца северо-
востока Европейской части СССР.
Таким образом, движение населения песца с его очень ла-
бильной, резко колеблющейся по годам плодовитостью, в благо-
приятные годы почти в два раза превышающей плодовитость
лисицы, отличается частыми подъемами численности и быстрыми
ее сокращениями. Пик численности длится всего один сезон;
цикличность изменений копирует свойственную массовым гры-
зунам тундры — пеструшкам — важнейшему продуктивному
корму песцов.
Движение численности лисицы в основных чертах сходно с
описанным для песца, но имеет и существенные отличия. Лисица
распространена необычайно широко и населяет все природно-
географические зоны СССР, отличающиеся и по климатическим
условиям и по характеру фауны наземных животных, многие из
которых служат кормом этого хищника. Популяции лисиц, за-
нимающие разные природно-географические зоны, характеризу-
ются, каждая своими особыми вариантами движения численно-
сти, черты которых обусловлены как естественными биотиче-
скими и абиотическими факторами, так и влиянием хозяйствен-
ной деятельности человека.
В средней и степной полосе страны, где в населении мелких
грызунов господствуют обыкновенные (Microtus arvalis) и об-
щественные (М. socialis) полевки и степные пеструшки (Lagurus
lagurus) подъемы и падения их численности, повторяющиеся
обычно с промежутками в 2—3 года, вызывают соответствую-
щие, но более сглаженные, изменения количества лисиц. Кар-
тина осложняется проявлением влияния «больших волн» массо-
вого размножения грызунов, повторяющихся с интервалами в
9—10 лет.
Исключительное, но кратковременное обилие мелких зверьков,
на огромных пространствах степной, лесостепной и отчасти лес-
ной полос нашей страны, не может не вызывать соответствующих
сдвигов в плодовитости и смертности лисиц. Наличие обуслов-
ленных этими «большими волнами» и почти синхронных с ними
размножений лисиц служит убедительным доказательством ре-
альности существования вспышек размножения грызунов с
10-летней цикличностью, в чем некоторые авторы необоснован-
но сомневались
При всем разнообразии используемых кормов полевки и пе-
струшки играют для лисицы роль самого важного продуктив-
ного корма, особенно осенью и зимой. В годы обилия полевок
хищники хорошо упитаны всю зиму и весной успешно размно-
жаются, В 1921—24 гг. и через 10 лет —в 1932—1934 гг. мас-
совым размножением грызунов были охвачены средняя и южная
части Европейской территории Союза. На 1—2 года позднее
(время, необходимое для использования хищниками благопри-
356
ятной для размножения ситуации) следовали большие подъе-
мы численности лисиц, хорошо отраженные в изменениях заго-
товки шкурок этого вида на Кавказе, Украине, в Нижнем и
Среднем Поволжье, черноземной полосе и областях Промыш-
ленного Центра за 1924—1929 и 1933—1935 гг. (следующая
грандиозная вспышка размножения пришлась на 1943—1945 гг.
и была далеко не полностью использована охотниками).
Судя по количеству добываемых лисиц, численность их в
годы, благоприятные для размножения, увеличивается не менее,
чем в 5—7 раз. Это подтверждается и непосредственными на-
блюдениями. В период предзимья, в год большого «урожая»
лисиц с одного места можно одновременно увидеть в Ставро-
полье и Нижнем Поволжье до 10—12 мышкующих на полях ли-
сиц, а в Подмосковье — до 4—5, тогда как в неурожайные—•
не более 1—2.
Ухудшение условий добывания корма в осенне-зимний сезон,
предшествующий периоду гона, вызывает сильное снижение
плодовитости лисиц. Прежде всего возникает несоответствие в
сроках готовности к спариванию у самцов и самок. Например,
в малокормном 1939 г. в Ставрополье максимум готовых к гону
самцов приходился на январь, большинство же самок было в
состоянии течки во второй половине февраля, а у некоторых она
наступила только в марте. В этих условиях самцы, рано закан-
чивающие гон, не могут покрыть всех самок, и процент прохоло-
ставших лисиц резко возрастает. Кроме того, в связи с длитель-
ным полуголодным существованием истощенный организм уже
оплодотворенной самки во многих случаях оказывается не спо-
собным обеспечить нормальное развитие эмбрионов. В упомяну-
том 1939 г. из шести добытых в Ставрополье самок у пяти
резорбировались полностью все зародыши и только у одной
была частичная резорбция эмбрионов (из 8 погибли 2) (Чир-
кова, 1947).
Все это приводит к тому, что после голодной зимы , количе-
ство пустующих -выводковых лисьих нор увеличивается соответ-
ственно большому проценту прохолоставших или не ощенив-
шихся лисиц. В 1939 г. в Ставрополье к концу весны таких са-
мок было 67%. Но среднее число лисят в выводках и в этих ус-
ловиях оставалось близким к обычному. В 1937 г., благополуч-
ном по обеспеченности кормом, средняя величина выводка была
6, в 1938 г.— 5,4. Таким образом, уменьшение плодовитости ли-
сиц в неблагоприятные годы идет, видимо, прежде всего за счет
сокращения числа выводков, увеличения процента холостых осо-
бей, а не за счет уменьшения числа щенков в помете В Кра-
1 Однако по наблюдениям Г. Н. Лихачева .(1958) в Тульских засеках
в 1938 г. при хороших кормовых условиях среднее число лисят в выводке
было 5,5, а в следующем малокормном 1939 г.— всего 2 щенка.
357
пивенском районе Тульских засек на учетной площади леса в
4465 га, пересеченной множеством оврагов и имевшей много
нор, количество выводков лисиц за десятилетие (1936—1945 гг.)
изменялось следующим образом (см. табл. 8).
Таблица 8
Количество выводков лисицы по годам
Годы 1936 1937 1938 1939 194 0 194 1 1942 1943 1944 1945
Количество . . Численность по- 5 6 7 — 6 9 9 8 2 4
левок .... + + + + — + + + + + + + 4—И
Примечание: годы высокой численности полевок отмечены знаком
средней +, бедные кормом —.
Поскольку одновременное снижение численности полевок не- 61
редко охватывает обширные территории (например, все степные
и лесостепные области юга СССР от Украины до Западного Ка-
захстана и от Предкавказья до Нижнего Поволжья), количе-
ство лисиц сокращается на таком же пространстве.
Помимо снижения плодовитости длительный недостаток про-
дуктивного корма вызывает увеличение смертности взрослых
лисиц: часть их погибает от голода, часть от болезней, усили-
вающихся в связи с истощенностью животных, их большей по-
движностью и т. д:
Количество лисиц становится особенно большим вскоре по-
сле массового размножения грызунов, которое, как уже отме-
чено выше, обычно заканчивается быстрым вымиранием. При
этом сильно размножившиеся лисицы в течение двух-трех лет
оказываются в крайне неблагоприятных кормовых условиях. Та-
кое положение возникло в средних областях Европейской части
СССР в 1945—1946 гг. По причине полного прекращения охоты
в годы Великой Отечественной войны маточное поголовье лисиц
было особенно большим, а после массового появления полевок
в 1945 г. и успешного размножения количество хищников не-
обычайно увеличилось. «Начиная с осени 1945 г. и весь 1946 г.
к нам поступали донесения от- охотников и лесников о наличии
среди лисиц какой-то эпизоотии,— писал Г. Н..Лихачев (1958)
о популяции Крапивенского района Тульской области.— Лисицы
не уходили от гончих собак, иногда падали мертвыми во время
гона; некоторые... лежали на земле, не имея сил подняться при
приближении людей. Часто попадались лисьи трупы...» В эти_.
годы в Калужской, Московской и других областях среди лисиц
8 58
были отмечены отдельные случаи бешенства и более многочис-
ленные— другой болезни, видимо, энцефалита. В западных
областях резко увеличилось количество лисиц, пораженных
чесоткой.
В зоне тайги и лесотундры лисица зимой испытывает силь-
ные затруднения в добывании мышевидных грызунов из-за боль-
шой мощности снежного покрова. Как. показали наблюдения
В. П. Теплова (1950), проведенные в тайге верхнего течения
р. Печоры, основу зимнего питания местной лисицы составляют
заяц-беляк и тетеревиные птицы; по частоте встреч — 59% —бе-
ляк стоит на первом месте среди других кормов. Лисица систе-
матически пользуется тропами беляка при передвижении в мно-
госнежный период и на них же подкарауливает и скрадывает
зайцев. Учеты лисиц и данные заготовок показали наличие зна-
чительных колебаний их численности в Печорской тайге: мини-
мальные и максимальные показатели разнятся в 9—12 раз.
После зим, обильных кормом (беляком или тетеревиными), ко-
личество лисиц в районе остается стабильным, а в течение наи-
более голодных зим их погибает около 75%.
Судя по имеющимся данным, основная масса лисиц уми-
рает от крайнего истощения и небольшую часть ослабевших ло-
вят росомахи. Зимой 1940/41 г. у погибших от голода лисиц от-
ношение веса тушки (в граммах) к длине (в сантиметрах) рав-
нялось 30, тогда как этот показатель упитанности у особей,
добытых в обильную кормами зиму 1937/1938 г., равнялся 54.
Глубокий и рыхлый снежный покров тайги крайне ограни-
чивает набор кормов, доступных лисице, и ставит хищника в
тесную зависимость от зайца-беляка, подверженного очень рез-
ким колебаниям численности. Это в основном и обусловливает
движение населения таежных лисиц по годам, что давно подме-
чено при сопоставлении кривых заготовок лисьих и заячьих шку-
рок в тайге Сибири и Северной Америки. Здесь высокая чис-
ленность лисиц отмечается с промежутками в 8—10 лет. Очень
отчетливо по данным Ц. Хьюитта (Hewitt, 1921) цикличность
колебаний количества лисиц выражена в северной Канаде.
- В песчаных пустынях Средней Азии мелкие лисицы местной
географической расы (Volpes vulpes flavescens) связаны по пи-
танию более всего с песчанками, отчасти с сусликами, тушканчи-
ками. Зимние условия кормодобывания здесь мало отличаются
от летних, так как снежного покрова почти не бывает. Несо-
мненно, что детали динамики численности лисиц здесь иные, чем
на севере, но они еще недостаточно выяснены.
Изучение в течение пяти смежных лет кормового режима и
плодовитости соболя Зауралья (Залекер и Полузадов, 1959) 1
1 Учитывалась урожайность кормов в тайге, изучено содержимое 778 же-
лудков соболей, добытых ,в промысловый сезон, и произведен анализ органов
размножения 397 самок.
359
показало, что хорошее размножение этого хищника обус- 1
ловливается обилием рыжих лесных полевок (Clethrionomys), |
серых полевок (Mtcrotus) и лесных леммингов (Myopus). Прочие !
корма (птицы, белки, зайцы, землеройки, насекомые, орехи, яго-
ды и др.), каждый в отдельности, имеют для соболей второсте- ,
пенное значение, но в совокупности играют роль, в количествен-
ном отношении близкую к таковой мышевидных грызунов. i
В годы обилия рыжих, серых полевок и лесных леммингов,— ;
продуктивного корма соболей, 90—100% соболюшек участвуют i
в гоне и оказываются оплодотворенными. Число эмбрионов, судя
по подсчету желтых тел беременности, колеблется от 2 до 6, 1
а в среднем равняется 4—4,5. Число соболей сеголетков в пробе
из популяции составляет около 50%, соотношение полов равно-
мерное. В год с ухудшением запасов корма (уменьшение коли- J
чества полевок, неурожай «орешков» кедра) число оплодотво- i
ренных самок равнялось 70% от общего числа исследованных,
среднее количество эмбрионов было 3,2 (при колебаниях 2—4).
Число сеголетков в популяции составляло 32%. Среди добытых
зверьков резко повысилось (до 48% от общего числа) количе-
ство взрослых самцов.
Для близкого вида — лесной куницы Д. И. Асписов (1959)
считает наличие 50% прибылых животных в добыче охотников
показателем хорошего состояния популяции, от 50 до 30%— '
-удовлетворительного и ниже 30% —плохого, т. е. отличавшегося
низкой плодовитостью в предшествующий сезон размножения.
Колебания плодовитости несомненно оказывают влияние на
изменение численности соболей, но, при свойственной виду зна- J
чительно?! стойкости, падения и подъемы кривой, показывающие 1
движение их населения, гораздо более плавны, и не столь резки, ’I
как у грызунов или узкоспециализированных хищников мыше- fl
едов, отличающихся относительно высокой плодовитостью. fl
По данным С. С. Фолитарека (1947), урожайные и неуро- Я
жайные на соболя годы повторяются с интервалами в 11 лет. fl
Американские зоологи Е. Сетон (Seton, 1912) и Ц. Хьюитт (Не-
witt, 1921), располагавшие статистическими сведениями о заго- fl
товке шкурок близкой к соболю американской куницы (Martes Я
americana) за длинный ряд лет, установили, что подъемы чис- Д
ленности этого вида повторяются в среднем через 9.5—10 лет.. Л
К группе' видов с неустойчивой численностью населения при-
надлежат и зайцы, в частности наши русак и беляк. Зайцы при-
•надлежат к своеобразной группе Lagomorpha, приближающих- ||
ся по некоторым чертам биологии, в частности характеру пище-
варения, к копытным. fl
Беляк (Lepus timidus) — вид с огромным ареалом, в разных Я
частях которого движение населения имеет некоторые местные
отличия. Он приспособлен, к жизни в различных природно-гео- Я
графических зонах от тундры, лесотундры, тайги и смешанных Я
360
лесов до лесостепи и даже степи, если находит в ней осиновые'
колки и участки, поросшие ивами, таволгой и т. п. Как уже ука-
зывалось, обычно он не устраивает постоянных, нор и даже дете-
нышей приносит в простых, почти ничем не укрытых логовах.
Несмотря на это зайчата, быстро приобретающие самостоятель-
ность, относительно мало страдают от нападений хищников и
непогоды (целиком погибают только ранние выводки в случае
возврата холодов). По своей неприхотливости в отношении пи-
тания беляк напоминает лесных оленей, питающихся большую
часть года веточным кормом, и некоторых серых полевок. Как
и для этих грызунов, состояние и доступность корма в движе-
нии численности популяции беляка играют второстепенную роль.
Беляки почти оседлы, массовые перекочевки бывают у них ис-
ключительно редко.
В годы высокой численности за зимний период питания ве-
точным кормом и корой они наносят очень большие и хорошо
заметные повреждения кустарникам и молодым деревцам (ивам,
осине, рябине, можжевельнику, дубу, лещине и многим др.).
Особенно серьезно страдают от них лесные питомники и моло-
дые плодовые сады. Однако год высокой численности вскоре
сменяется периодом депрессии, и беляк становится иногда на-
столько редким зверьком, что за целое лето работы в лесу его
совсем не удается встретить. В течение нескольких лет лесная
растительность полностью оправляется от повреждений, а из-
любленные биотопы беляка очищаются от рассеянных зайцами
бесчисленных яичек гельминтов.
Сильные «заячьи моры» известны издавна в Евразии и на -
материке Северной Америки. В северной полосе нашей страны
весной после «мора» в мелколесьях и на зарастающих вырубках
иногда почти под каждой кучей хвороста находят трупы зайцев
в зимнем меху. В канадской и американской литературе есть
много описаний опустошительных эпизоотий, вызывавших рез-
кое падение численности американского беляка (Lepus ameri-
canus) 1 на пространстве от северной Аляски до южных штатов
(рис. 136) Е. Сетон (Seton, 1910) пишет, что в 1887 г. из окна ва-
гона Канадско-Тихоокеанской железной дороги на протяжении
50 миль можно было видеть «трупы зайцев, которые лежали сот-
нями, если не тысячами»;
В СССР в таежной полосе численность беляка колеблется в
4—6-кратных пределах, в лесостепной в 3—4-кратных. У северо-
восточной границы ареала, в отдельных районах Якутии по не-
давним исследованиям количество беляков всего за 5—10 лет
изменяется в сотни и даже в 2500 раз (С. Наумов, 1956, 1961).
Якутия давно известна крайней неустойчивостью популяций
зайца-беляка; здесь продолжительность циклов (от одного пика
До другого) составляет около 12 лет. В Западной Сибири она
1 Очень близок к 'нашему Lepus timidus и, возможно, идентичен с ним.
361
равна 8—10 годам, на Европейском Севере СССР 9—11, в
Ленинградской области — 5—7, в Московской — 4—5 и т. д.
Следовательно, в областях с более суровыми природными усло-
виями нужно большее число лет, чтобы сохранившиеся произво-
дители могли обеспечить достаточный рост популяции.
Продолжительность жизни зайца равна 6—7 годам, плодо-
витость его довольна велика. Наш беляк чаще приносит в год
только 2 помета; в первом, ранневесением гоне принимает уча-
стие до 83—100% самок, во втором, весеннем — практически все
половозрелые самки. В третьем, летнем гоне в средних и южных
т 1885 1890 1895 1300 1905 1910 1915 1920 1925
Рис. 136 Изменения количества рыси в Канаде по данным заготовки
шкур факториями Кампании Гудзонова залива за период 1822—
1930 гг. Подъемы численности этого хищника следуют за годами
обилия американского беляка (по Элтону)
областях СССР участвуют не более 30—40% самок беляка, а в
северных— 10—15%. В полосе тундр больше двух выводков во-
обще не бывает по причине малой продолжительности теплого
периода года (зайчата уже в 9—10-дневном возрасте начинают
есть траву; таким образом их благополучие зависит от наличия
свежей зелени, а длительность всего периода размножения бе-
ляка— от продолжительности вегетационного периода). Числа
молодых в помете колеблется от 1 до 9, в среднем 3,5—5,2 зай-
чат. Наибольший приплод дает второй — летний помет.
Выяснено, что плодовитость беляков меняется по годам,,
причем это обусловливается изменением величины выводка, то-
гда как процент самок, участвующих в размножении, остается
постоянным (несомненное влияние относительно равномерной
обеспеченности кормом в разные годы). Основная причина ко-
лебаний плодовитости — интоксикация организма самок, возни-
кающая на почве глистных и протозойных инвазий. Она умень-
362
шает вероятность имплантации оплодотворенных яиц, вызывает
резорбцию эмбрионов. Но изменение плодовитости по годам —
не единственная причина колебаний численности беляка; оно-
только усиливает несоответствие между рождаемостью и смерт-
ностью. Последняя в фазу падения численности велика и в-
основном обусловлена гибелью зайцев от болезней и деятельно-
стью хищников, уничтожающих много ослабевших зайцев.
Беляки бывают сильно инвазированы нематодами Protostron-
gylus (до 100% зараженных при среднем количестве глистов-
на 1 зайца от 25 до 180 экз.) и Trichostrongylus (до 100% зара-
женности при среднем числе на 1 зайца в 4—570 экз.), а также-
заражены кокцидиозом. Токсичность легочных нематод выше,,
чем кишечных. Наиболее широкое распространение болезни по-
лучают в холодные дождливые годы при достаточно высокой1
численности беляков. Своеобразная особенность пищеварения
зайцев заключается в том, что дефекация у них происходит йе-
на местах отдыха, а в основном во время кормежки; поэтому
участки с зарослями привлекательных кормовых растений всегда
особенно густо усеяны их «орешками». Это приводит к сильной
зараженности таких участков яичками гельминтов, что естест-
венно облегчает обмен эндопаразитами между зайцами и усили-
вает интенсивность инвазий. На постоянно влажной почве и лес-
ной подстилке дольше сохраняют жизнеспособность яички и ли-
чинки гельминтов, ооцисты кокцидий, и, следовательно, легче
происходит заражение. Засушливые годы задерживают распро-
странение инвазий даже при большой численности беляков.
С этими особенностями этиологии гельминтозов и кокцидиоза:
связан и тот факт, что сильные заячьи моры более обычны в рав-
нинных заболоченных местностях с подзолистыми почвами..
В щебнистых и песчаных районах с хорошим дренажем эпизо-
отии случаются реже и уносят меньше жертв.
Таким образом, на движение численности беляка условия
погоды воздействуют не прямо, а через посредство паразитиче-
ских организмов. Они вызывают у беляков снижение плодови-
тости и тяжелые заболевания, нередко кончающиеся смертью-
наиболее инвазированных особей; В то время как для мелких
грызунов большое значение имеет погода осенью, зимой и вес-
ной, в жизни беляка эту роль играют метеорологические усло-
вия лета, когда происходит развитие во внешней среде его
паразитов или их промежуточных хозяев.
Тип устойчивого движения численности
Большое число видов млекопитающих отличается достаточ-
ной стойкостью против неблагоприятных внешних влияний и зна-
чительной продолжительностью жизни особей. Численность их
населения изменяется .медленно, без резких катастрофических
36$
подъемов и депрессий, так как плодовитость невелика, мало из-
меняется по годам и процент смертности тоже держится на
относительно постоянном низком уровне. К этой группе отно-
сится большинство крупных млекопитающих — копытных, хищ-
ных, китообразных, ластоногих и др., а из мелких — рукокрылые.
Летучие мыши отличаются очень низкой плодовитостью —
самка приносит в год только одного, а у некоторых видов двух
детенышей, по продолжительность жизни этих зверьков много
•больше, чем у мелких насекомоядных и грызунов. Так, например,
Myotis lucifugus доживает до 2О'/2 лет, Myotic keenl — до
1872 лет [Холл, Клутьер, Гриффин (Hall, Cloutier, Griffin), 1957];
давно известно, что тропические плодоядные летучие мыши, пой-
манные уже взрослыми, жили в зоопарках по 15—20 лет. Дли-
тельное наблюдение за колониями летучих мышей в пещерах,
дуплах, старых зданиях показывает, что численность особей из-
меняется медленно.. Картина осложняется только сменой убе-
жищ, при которой значительная часть особей может внезапно
исчезнуть из участка, находящегося под наблюдением.
Лось в естественных условиях доживает до 13—15 лет, может
быть немного более. Показатель плодовитости популяции в тайге
верховьев Печоры, установленный на большом материале со-
трудниками Печор о-Илычског-о заповедника, равен 1,32—
иными словами, ежегодный приплод в 132 теленка на каждые
100 половозрелых лосих. Смертность наиболее высока среди
телят в возрасте до 6—7 месяцев; взрослые лоси от болезней,
стихийных бедствий и хищников страдают относительно мало и
погибают в небольшом количестве. Возрастной состав стада,
установленный по картине стирания коренных зубов у осмотрен-
ных. 523 лосей из числа мигрирующих через заповедник, пока-
зан в табл. 9.
Продолжительность жизни аляскинского морского котика
•^Callorhinus alascanus) равна 14—15 годам; как у нашего
•командорского котика (Callorhinus ursinus), взрослая самка
приносит за год только одного детеныша (двойни очень редки).
Гибель котиков велика только в первые месяцы жизни, так как
364
много молодых давят тяжелые секачи при драках на лежбищах:
(до 6% новорожденных), часть (до 3%) умирает от сильной
инвазированности нематодами Uncinaria или от голода в случае
гибели матери и т. п. Всего в первые 3 месяца жизни погибает-
не менее 10% молодых, «черных» котиков. Позднее в море не-
которое число неокрепших животных уничтожают косатки и т. п.
Показатели естественной убыли аляскинских котиков по возраст-
ным-группам приведены в табл. 10. Эти цифры получены путем
многочисленных повторных подсчетов меченых животных на всех,
лежбищах островов, где в летний период концентрируется пол-
ностью вся популяция котиков.
Таблица 10
Смертность котиков на островах Прибылова по возрастным
группам
(в % от числа животных в группе)
Возраст в годах 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Убыль В % ... 35'15 10 10 10 20 20^33 40 50 60 70 80 100
У видов с таким ходом размножения и смертности заметные-
подъемы и депрессии численности возможны только при воз-
действии каких-то длительно действующих факторов, например
постоянного интенсивного промысла, больших многолетних сдви-
гов гидрологических условий (изменение направления и напря-
женности морских течений и т. п.). Так, например, в русской,
охотничьей литературе за последнее столетие есть указания на:
период значительного увеличения численности лосей, но этот-
процесс не был в свое время достаточно полно изучен. Тип устой-
чивого движения населения вообще не привлекал к себе такого
внимания исследователей, как массовые размножения грызунов
или вспышки эпизоотий среди промысловых животных с неустой-
чивой численностью.
Пути направленного воздействия на млекопитающих
в интересах человека
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК
Уже в эпоху палеолита, когда люди перешли от собирания
съедобных растений и мелких животных к добыванию более
крупных, т. е. к охоте, млекопитающие имели в жизни людей
большое значение. Крупные хищники (пещерный медведь, лев
и др.) были опасными врагами человека. Но крупные звери слу-
жили и наиболее ценной добычей, так как от них охотники полу-
чали много мяса, жира, костного мозга (лакомства наших да-
леких предков), шкуры для изготовления одежды и крепкие
жилы для сшивания этих шкур. Многочисленные кости оленей,
быков, диких лошадей, мамонтов и других зверей, переполняю-
щие культурный слой палеолитических стоянок — яркое свиде-.
тельство успешных охот того времени.
После приручения собаки, рогатого скота, овцы, козы, осла
зарождается новая отрасль хозяйства — животноводство, кото-
рое в течение многих тысячелетий развивалось одновременно
с охотой, во многих случаях дополняя, облегчая ее и делая более
успешной (использование верховой лошади, оленя как транс-
портных животных и для преследования и загона диких копыт-
ных, использование собак при поисках, удерживании, захвате
животных и т. п.).
Вознйкновение земледелия, появление первых обработанных
участков, засеянных'хлебными злаками, корнеплодами, сразу же
поставили земледельцев перед необходимостью защищать свои
посевы от различных вредителей, среди которых первое место
зачастую занимали тоже млекопитающие. В Африке это много-
численные обезьяны, слоны, носороги,' бегемоты, и в наши дни
уничтожающие там посевы кукурузы, маниока и др., в северной
Евразии — кабаны, олени, суслики, полевки, мыши, зайцы и др.
Со временем число видов прирученных животных и их общее
количество увеличивалось, развивались и совершенствовались
приемы ведения сельского хозяйства, все большее число видов
растений вводилось в культуру, расширялись пахотные площади,
появлялись новые орудия обработки земли и т. Д., но задача
охраны урожая, домашних стад, плодовых деревьев, лесных по-
садок и даже построек от вредителей, как и прежде, оставалась
актуальной.
366
В нашу эпоху человечество осваивает природные ресурсы
всех материков и океанов. Огромное количество видов живот-
ных мировой фауны, в том числе и млекопитающих, так или
иначе включено в сферу хозяйственных, культурных или науч-
ных интересов людей.
ОХРАНА ЦЕННЫХ ДИКИХ МЛЕКОПИТАЮЩИХ,
РЕГУЛИРОВАНИЕ ЧИСЛЕННОСТИ
Столетия стихийного, ничем не регулируемого использования
животного мира Земли уже оставили много печальных следов.
Только в историческое время исчезло (точнее, уничтожено)
105 видов крупных 'млекопитающих; из них 33 вида примерно
до 1700 г., 36 видов в XVIII—XIX вв. и около 40 — за последние
50 лет1. Каждый уничтоженный вид — это невосстановимая
потеря для хозяйства, науки, культуры.
В 1741 г. участники Второй Камчатской экспедиции при кру-
шении корабля у неизвестного в то время и необитаемого о. Бе-
ринга открыли невиданное европейцами животное, позднее на-
званное морской коровой Стеллера. Близ Командорских остро-
вов эти звери паслись целыми стадами, поедая массу морской
капусты.
Промышленники, плававшие на Алеутские острова в погоне
за ценной пушниной, запасались на Командорских островах мя-
сом только морских коров, отличавшимся прекрасными качест-
вами. (От взрослой сирены получали до 3,5 т мяса.) Это живот-
ное было одним из самых беззащитных и доверчивых предста-
вителей морских млекопитающих. К пасущейся сирене можно
было спокойно подъехать на лодке и вонзить в ее тело железный
крюк, привязанный к прочному канату, закрепленному на бе-
регу. При отливе пойманная таким простым способом морская
корова «обсыхала» и промышленникам оставалось только раз-
делать тушу на части. Примерно через 30 лет после открытия
Командорских островов этот вид был полностью истреблен
{Греков, 1958). Как правильно отметил еще А. Ф. Миддендорф,
морскую корову Стеллера, видимо, легко можно было приру-
чить, что дало бы в руки обитателей островов доверчивое и не-
прихотливое прирученное животное, питающееся только водо-
рослями и производящее много отличного мяса. Но такая воз-
можность утрачена навсегда. Угроза исчезновения ряда других
ценных видов выдвинула необходимость организовать их охрану.
Полное истребление даже крупных и опасных хищников те-
перь, когда накоплен большой опыт борьбы с ними и стала хо-
рошо известна их роль в биоценозах, является нежелательным.
Так, например, во многих частях Африки применение кап-
1 В основном после 1756 г. уничтожено также 44 видов и подвидов птиц.
367
канов, ядов, собак и ружей для истребления леопардов, на-
падавших на домашний скот, привело за несколько десятиле-
тий к почти полному уничтожению этого хищника. Последствия
оказались довольно неожиданными: всюду, где исчезли лео-'
парды, чрезвычайно размножились бабуины, и сельское хо-
зяйство стало нести огромные потери от постоянных набегов
на плантации хорошо организованных и очень подвижных стай
этих обезьян. Раньше леопарды ежегодно убивали тысячи ба-
буинов; популяции последних, освобожденные от «пресса» хищ-
ников, стали быстро размножаться. В настоящее время лео-
парды в Африке взяты под охрану (Хантер, 1960), а местами,
где они совсем исчезли, осуществляют их реакклиматизацию.
Многолетний опыт широких кампаний по истреблению ма-
лого суслика (Citellus pygmaeus) на землях Предкавказья, Кал-
мыкии, Астраханской области и др. показал, что при имеющейся
сейчас технике полное уничтожение этого вредного грызуна на
миллионах гектаров степей и полупустынь потребует затраты
огромных средств. Вместе с тем выяснилось, что организованное
истребление сусликов с сокращением плотности их населения
до 2—3 особей на гектар, способно радикально оздоровлять
природные очаги чумы, существование которых поддерживают
малые суслики и паразитирующие на них блохи. Таким образом,
и в данном случае вопрос о полном истреблении вида отпадает
или откладывается на долгий срок, и практически, более ценным
является регулирование его численности.
Охрана одних видов и научно обоснованное регулирование
численности других — важнейшие пути направленного воздей-
ствия на популяции диких млекопитающих.
Охрана природы, как цель широкого общественного движе-
ния, вскоре ставшая также одной из задач государственных
учреждений во многих странах мира, возникла во второй
половине прошлого века1. Особенно больших успехов она до-
стигла в первые десятилетия нашего века. С 1949 г. Междуна-
родный Союз охраны природы и природных ресурсов изучает
вопросы сохранения видов млекопитающих, находящихся под
угрозой истребления или вымирания. В этом списке 23 вида:
сумчатый тасманийский волк, азиатский лев, 3 вида азиатских
носорогов, дикий двугорбый верблюд, зубр, карликовый бело-
хвостный олень (Odocoileus virginianus clavium), 2 вида анти-
лоп, кулан, лошадь Пржевальского, шиншиллы, 3 вида тюленей
и др. Полный запрет охоты и преследования вместе с надлежа-
щим надзором за выполнением таких постановлений играют
большую роль в охране видов, которым грозит уничтожение.
1 У многих народов еще ранее обычаи, установленные на основе веко-
вого опыта, были направлены на ограничение количества добываемых охот-
никами животных и охрану отдельных участков территории. ’
368
В отношении морских млекопитающих, совершающих мигра-
ции вдали от территориальных вод (котики, киты), существуют
международные договоры, направленные на предотвращение
чрезмерного уничтожения этих животных заинтересованными
организациями и гражданами разных стран.
В угрожаемом положении находится сейчас амурский тигр.
Количество особей Fells tigris longlpilis, имеющихся на терри-
тории советского Дальнего Востока, не превышает 200. Вред,
приносимый тиграми диким животным, невелик; нападения на
домашних животных совсем прекратились, а значение самого
хищника как объекта зооэкспорта и редкого представителя
фауны уссурийской тайги трудно переоценить. Поэтому охоту на
тигра запретили; в редких случаях, только по специальным ли-
цензиям разрешается отлов тигрят для зоопарков.
Под полной охраной находятся зубр, кулан, горал, дикий пят-
нистый олень, а в республиках Прибалтики — бурый медведь.
Длительный запрет охоты на лося, сайгу, соболя уже дал поло-
жительные результаты — количество этих животных настолько
увеличилось, что разрешен плановый отстрел, основанный на
подсчетах имеющегося поголовья в республиках, областях и
районах.
В октябре 1960 г. в РСФСР принят «Закон об охране при-
роды», имеющий большое значение для организации рациональ-
ного использования и сохранения природных богатств, в том
числе фауны Российской Федерации. Усилилась охрана природы
и в союзных республиках.
Учитывая богатый накопленный опыт, специалисты экологи
всего мира считают сейчас очень важным обеспечивать охрану
не только самих животных, но и благоприятной для них жиз*
ненной среды, т. е. естественной обстановки, достаточно обшир-
ных природных участков и целых ландшафтов. Разработаны
различные приемы «оздоровления» сильно измененных, испор-
ченных неумелым хозяйствованием ландшафтов одновременно
с охраной ценных животных.
ДЕЯТЕЛЬНОСТЬ ЗАПОВЕДНИКОВ, ЗАКАЗНИКОВ
И ОХОТНИЧЬИХ хозяйств
Крайне важную роль в сохранении ценных по красоте, науч-
ному и историческому значению районов играют национальные
парки и, особенно, заповедники. Охрана животного мира и, в
первую очередь, ценных млекопитающих, особенно сильно по-
страдавших от истребления в предшествующие столетия, а так-
же богатой естественной растительности и вод — одна из основ-
ных задач их деятельности.
24 Териология
369
Обширная сеть заповедников — участков, изъятых из хозяй-,
ственного использования, имеется в Африке, Северной Америке,
многих странах Евразии. В некоторых африканских заповедни-
ках туристы прямо из машины могут любоваться на близком
расстоянии большими пасущимися стадами различных антилоп,
зебр, жирафами, слонами, львами.
В СССР государственные заповедники возникли после Вели-
кой Октябрьской социалистической революции; первые из них
были организованы по указанию В. И. Ленина и играют сейчас
важную роль в деле охраны и изучения наших ценнейших мле-
копитающих. В сохранении многих видов — амурского тигра, со-
боля, изюбря, лося, кабарги, косули, черного и бурого медведей
и др. большую роль сыграл Сихоте-Алинский государственный
заповедник, в Кавказском заповеднике вместе с другими видами
находятся под охраной большие стада западнокавказских ту-
ров, серн, кавказских оленей, кабанов, лесные и каменные ку-
ницы; в нем идут работы по восстановлению зубра. В Воронеж-
ском заповеднике много бобров, европейских оленей, куниц,
в Печоро-Илычском — соболей и лесных куниц, лосей, выдр, ди-
ких северных оленей и т. д.
Заповедники представляют собой не только отличные убе-
жища для млекопитающих, где имеются почти ненарушенные
хозяйственной деятельностью условия существования, но и за-
мечательные исследовательские лаборатории в природе; многие
десятки лучших трудов по экологии промысловых зверей опуб-
ликованы зоологами заповедников. Здесь научные сотрудники
проводят большую опытную работу, намечая и проверяя пути
направленного воздействия на популяции млекопитающих.
На зиму для копытных заготовляют сено и используют его для
подкормки в многоснежный период года. Устраивают искусствен-
ные солонцы, что дает возможность устранять чрезмерную кон-
центрацию животных около естественных солонцов и умень-
шать опасность заражения гельминтозами и инфекционными
болезнями. Для летучих мышей вывешивают искусственные
«гнездовья» — дуплянки и т. д. Болезни охраняемых животных
привлекают особое внимание персонала заповедников; они ис-
следуют их этиологию, ставят опыты по предупреждению забо-
леваний и их лечению.
Сходные работы осуществляются специалистами в охот-
ничьих хозяйствах промыслового и спортивного типа. Постоян-
ное наблюдение за санитарным состоянием бобров, соболей,
стад оленей, косуль, кабанов, отстрел больных, слабых и подо-
зрительных в отношении заболеваний особей, улучшение усло-
вий существования дают возможность активно вмешиваться в
жизнь целых популяций.
В наших государственных ондатровых хозяйствах осущест-
вляют целую систему биотехнических мероприятий, чтобы улуч-
370
шить кормовые и защитные условия водоемов, населенных он-
датрой. Для увеличения «гнездопригодной площади» создают
десятки помостов, на которых зверьки могут строить хатки.
С целью улучшения условий добывания корма специальными
косилками делают прокосы в густых зарослях тростника; тем
самым «омолаживают» пастбища ондатры, вызывают появление
молодых, более нежных и питательных стеблей. Одновременно
персонал охотничьих хозяйств ведет истребление наиболее опас-
ных врагов ондатры — камышовых луней, выпей, крупных ца-
пель, шакалов и др.
Улучшая кормовые условия для белки, во многих местах уже
начали акклиматизировать лиственницу, кедр, кедровый стла-
ник. При увеличении числа лесообразующих хвойных пород,
несомненно, урожаи будут более частыми — это придаст неко-
торую устойчивость кормовой базе белки. В островных песцовых
хозяйствах удалось пойти дальше: там для подкормки взрослых
и молодых животных используют кормушку-ловушку; через нее
практически проходит все поголовье песцов, при этом худших
производителей отлавливают, а оставляют наиболее ценных.
Это действенный прием улучшения племенных качеств песцов
. путем селекции.
Соболь, один из наиболее ценных пушных зверьков тайгй
Евразии, одно время был близок к'полному уничтожению. Дли-
тельный запрет охоты, организация специальных заповедников
и заказников 1 обеспечили сохранение и успешное размножение
этого хищника. За последние годы почти полностью восстано-
вился былой ареал соболя, а численность его в Сибири превы-
шает ту, которая была там 200—250 лет назад. Восстановление
ареала шло быстро благодаря тому, что многие десятки партий
специально отловленных соболей были перевезены на самолетах,
,, автомашинах, вьючных оленях для выпуска в те районы тайги,
> куда, при естественном расселении зверьки могли бы проникнуть
только через десятилетия. Искусственное расселение дало воз-
можность подойти к решению и еще одной важной задачи —
улучшить племенные качества (качество меха) некоторых red-
графических-рас и «кряжей» соболя. Лучшие по цвету и нежно-
сти меха темноокрашенные баргузинские соболи (Martes zibellinb
I princeps) были выпущены большими партиями в те районы, где
местные соболи имели светлую окраску. (Например, в Западной
Сибири распространен светлый М. z. zibeUina). Позднейший
наблюдения показали, что местами «прослойка» темноокрашен-
' ных соболей в районах, окружающих места выпуска, увели-
чилась.
1 В отличие от заповедников, учреждаемых на неограниченно длитель-
ный срок, заказники—участки, где охота на определенные виды животных
или всякая охота запрещается на несколько сезонов.
24* 371
Многие, промысловые охотники подкармливают летом лисьи,
песцовые выводки и соболей — это помогает успешно сохранять
приплод животных в малокормные сезоны и одновременно об-
легчает охоту. В ряде мест приморской тундры, где летом про-
изводится лов рыбы и охота на тюленей, промысловики закла-
дывают в ямы большие запасы корма для песцов. Наличие при-
кормки предотвращает откочевку песцов, создает концентрацию
зверьков в заранее известных местах и обеспечивает успех
рхоты. Таким образом, даже такое стихийное явление, как мас-
совая миграция песцов, в какой-то мере поддается регулиро-
ванию.
Эти примеры показывают, что в современных условиях пра-
вильно ориентированное, научно обоснованное вмешательство
людей в жизнь популяций промысловых млекопитающих посте-
пенно стирает границы между дикими и домашними животными
.и, в конечном счете, увеличивает власть человека над природой.
ОБОГАЩЕНИЕ ФАУНЫ ПУТЕМ АККЛИМАТИЗАЦИИ
МЛЕКОПИТАЮЩИХ
Уже в отдаленные эпохи люди,- переселяясь, уводили с собой
прирученных животных и тем обогащали фауну новых мест.
Искусственное расселение млекопитающих приняло особенно
широкие размеры в последнее столетие.
В недавно опубликованном обзоре А. А. Насимович (1961)
приводит . сведения (по-видимому, еще неисчерпывающие) о
.150 видах млекопитающих, которые были объектами акклимати-
заций— преднамеренной или случайной, удачной и неудачной
в разных странах мира. В этом числе парнокопытных — 49 ви-
дов, грызунов— 27, хищных — 34, сумчатых—11, зайпеобраз-
ных— 9, приматов и непарнокопытных — по 6, неполнозубых — 4,
однокопытных — 3 и рукокрылых—1. Кроме, того, более десяти
.видов домашних и полудомашних млекопитающих были заве-
зены в различные области, выпущены на волю и, одичав, обра-
зовали, большие популяции.
Особенно . насыщенной иноземными видами стала фауна
-Новой Зеландии. По данным монографии К. Водзицкого (Wodzic-
ki, 1950), там были успешно акклиматизированы 19 видов млеко-
питающих из Европы, 5 —из Азии, 4 — из Северной Америки,
6 — из Австралии и Полинезии. Наиболее высокую способность
к акклиматизации в новозеландских условиях проявили виды,
..происходящие из Европы и, в известной мере, из Палеарктики
вообще'. Многие палеарктические виды зверей успешно прижи-
лись также в Северной и Южной Америке, в Австралии и на
ряде островов мирового океана.
Наоборот, виды, происходящие из Африки и Южной Аме-
рики,1 животный мир которых сохраняет многие архаические
372
черты, труднее приспособляются к новым условиям; попытки
акклиматизировать их чаще кончаются неудачей.
Известно немало примеров, когда выпуск небольшого числа
животных приводил к быстрому увеличению их количества и
расселению вида на обширном пространстве — к существенному
экономическому эффекту. Так, например, на о-в Ньюфаундленд
в 1878 г. была выпущена пара американских лосей, а в 1904 г.
еще две. В настоящее время их насчитывают 30—40 тыс. и еже-
годно добывают более 4500 голов [Пимлот и Карберри, (Pimlott
a. Carberry), 1958].
Добиваясь обогащения фауны млекопитающих какой-либо
территории, очень важно хорошо знать ее жизненные условия,'
емкость, наличие свободных экологических ниш, все детали эко-
логии вселяемого вида и его возможных конкурентов из числа’
аборигенных животных. Просчеты могут причинять большие'
бедствия. ;
Так произошло, например, с акклиматизацией в Австралии'
дикого (европейского) кролика, начатой в 1859 г. Быстро раз-'
множившиеся кролики сильно снизили продуктивность пастбищ,
и овцеводству — основе хозяйства многих областей Австралии—'
грозил крах. Для борьбы с кроликами стали применять различ-
ные ловушки и капканы, собак, яды, разрушение нор, выкури-’
ванне ,и.т. п. Была произведена акклиматизация некоторых хищт
ников, истребляющих кроликов. Но все эти меры оказались,
мало эффективными. Численность кроликов - менее чем, через,-
100 лет после начала акклиматизации, уже в период напряжен-'
ной борьбы с ними, достигла в Австралии сотен миллионов го-,
лов. Лишь в последние годы в результате применения вируса»
миксомы, вызвавшего эпизоотию огромного масштаба, числен-
ность кроликов удалось резко сократить (Герман, 1953). Аккли-.
матизация на о. Маврикия макаки и мангусты имела катастро-.
фические последствия для местных птиц, так как первые стали
их злейшими врагами [Бенсон (Vinson), 1956] и т. д. <
В нашей стране в дореволюционный период отдельные по-:
пытки акклиматизации животных носили кустарный характер;
к ним относятся расселение небольшого количества ланей
(Dama dama) в некоторых поместьях Европейской России, рас-
селение бобра, благородного оленя и т. п.
После Великой Октябрьской социалистической революции
учеными совместно с практическими работниками был разрабо-’
тан план акклиматизации целого ряда экзотических пушных
млекопитающих и расселения некоторых ценных отечественных
грызунов и копытных. При этом принимали во внимание нали:
чие более или менее значительных территорий, не заселенных
в силу ряда исторических причин, ценными видами животных^
но имеющих для этого большие возможности.
373
Наибольший хозяйственный эффект дала акклиматизация
полуводного североамериканского грызуна — мускусной крысы,
или ондатры, занявшей теперь в СССР огромный ареал. Пло-
щадь водоемов, покрытая зарослями прибрежных и плавающих
гидрофитов, в нашей стране очень велика. Эти запасы корма,
почти не используемые аборигенными животными, дали возмож-
ность поселить множество ондатр, т. е. занять пустовавшую
«экологическую нишу». Во многих озерных и речных районах
охота на ондатру дает теперь отличной пушнины на суммы зна-
чительно большие, чем все аборигенные виды зверей. Ондатра
с 1926 г. прочно вошла в нашу фауну; ежегодная заготовка ее
шкурок достигает нескольких миллионов штук *.
Успешно идет восстановление численности и расселение реч-
ного бобра, который в период перед Великой Октябрьской со-
циалистической революцией сохранился в России в очень немно-
гих местах в количестве лишь сотен особей. Хорошо размно-
жается и занимает все новые районы на востоке страны аккли-
матизированная у нас крупная и ценная американская норка.
Нутрия, или болотный бобр (Myopotamus coypus) —крупный
теплолюбивый полуводный грызун, вывезенный из Южной Аме-
рики, был поселен у нас на зарастающих водоемах южной части
Союза — в Закавказье, Средней Азии. Нутрия обладает отлич-
ной пушной шкуркой. К тому же этот зверь истребляет массу
жесткой растительности, которая ухудшает условия жизни кар-
пов в рыбоводных прудах и затрудняет использование ороси-
тельных каналов1 2. Поэтому нутрия вдвойне полезна. Однако
даже на юге во время редких морозных зим нутрии у нас стра-
дают и гибнут от холода, замерзания водоемов и делаются лег-
кой добычей хищников (шакала, лисицы и др.). При таком по-
ложении продвижение нутрии в более северные части страны
казалось неосуществимым. Выход был найден в так называемом
полувольном разведении, широко применяемом теперь в сред-
ней части страны. На лето имеющихся производителей выпу-
скают в богатые прибрежной водной растительностью водоемы,
охраняемые от хищников, осенью по возможности всех нутрий
вылавливают, часть забивают на мех, а часть содержат в волье-
рах до весны.
1 Почти не используются аборигенными животными неисчислимые за-
пасы морских ежей, моллюсков и ракообразных у берегов Восточного Мур-
мана. Поэтому выдвинута задача: заселить мурманское побережье каланом
(Enhydra lutris) — основным потребителем этих кормоЪых объектов на се-
верных островах Тихого океана. Первые шаги, сделанные в этом направле-
нии, дали обнадеживающие результаты.
2 В отличие от ондатры, нутрия не роет нор и длинных ходов в стенках
каналов, земляных плотинах и т. п. Роющая деятельность ондатры местами
сильно вредит гидротехническим сооружениям, тогда как нутрия охраняет
земляные каналы и искусственные водоемы от зарастания и быстрого заили-
вания.
374
Из других американских животных у нас акклиматизирован
енот и были осуществлены опыты расселения скунса, но сколько-
нибудь существенного хозяйственного эффекта они пока не дали.
Из отечественных животных успешно были расселены заяц-русак
в Западной Сибири, белка в степных борах Казахстана, горных
лесах Тянь-Шаня и Кавказа, пятнистый олень в Прибайкалье и
Европейской части СССР, марал на Южном Урале, осущест-
влены небольшие переброски выхухоли и поселение енотовидной
собаки во многих местах юга Сибири и всей Европейской ча-
сти СССР.
Енотовидная собака дает хорошего качества пушную шкур-
ку, хорошо размножается и местами уже превышает числен-
ность местных лисиц. Но, как хищник-полифаг, енотовидная со-
бака оказалась в некоторых районах (дельта р. Волги, цент-
ральная полоса Европейской России) серьезным вредителем
охотничьего хозяйства: уничтожает много гнезд уток, гусей, фа-
занов и тетеревиных птиц, что значительно снижает хозяйствен-
ную ценность акклиматизации этого вида.
В некоторых случаях акклиматизацию млекопитающих осу-
ществляют с целью расселения животных, способных истреблять
или даже подавлять местных и завозных вредителей сельского
хозяйства. Так, например, индийская мангуста, или мунго
(Herpestes mungo), еще в 70-х годах прошлого столетия была
поселена на о. Ямайке и приносит там большую пользу истреб-
лением крыс, повреждающих сахарный тростник на плантациях.
Мангусты поселены также и на некоторых островах Тихого
океана, где случайно завезенные крысы стали бичом хозяйства.
Многообразие видов мировой фауны млекопитающих, эколо-
гическая пластичность многих из них обеспечивает любой стране
ценный материал для обогащения своей фауны. В этом отноше-
нии имеется простор и большие неиспользованные возможности
для новых опытов. Но, как было отмечено выше, к этому делу
нужно подходить с большой осторожностью, чтобы не вызвать
непредвиденных отрицательных последствий.
ОДОМАШНЕНИЕ МЛЕКОПИТАЮЩИХ
Одомашнение важнейших млекопитающих было начато еще
в далеком прошлом. Это была важная ступень в развитии че-
ловечества, новый смелый шаг в овладении естественными про-
изводительными силами природы. Благодаря животноводству
вскоре обогатились пищевые ресурсы ряда племен и народов,
стало более устойчивым их хозяйство, увеличились возможности
преодоления пространства, переброски грузов и т. д. Тысячелет-
ний кропотливый труд многих поколений по выведению пород
домашних животных, несмотря на то, что он долгое время шел
как бы наощупь, не опираясь на точные научные знания, дал
375
изумительные результаты. Развились сила, выносливость и быст-
рота ездовых животных, хорошая мясная, шерстная, молочная
и мясо-молочная продуктивность, плодовитость, скороспелость и
другие ценные качества выводимых пород. Таким образом, еще
до возникновения научной генетики, селекции и зоотехнии че-
ловек в известной мере уже владел основами искусства изме-
нять природу животных. В наше время, применяя селекцию,
гибридизацию, искусственное осеменение, специальные режимы
кормления, содержания молодых и взрослых животных, социа-
листическое сельское хозяйство, опираясь па передовую мате-
риалистическую биологию, может творить в области животно-
водства подлинные чудеса. Достаточно указать на рекорды
удойности коров, на замечательные настриги шерсти тонкорун-
ных овец, высокое качество шкурок каракульских овец и многие
другие достижения. Помимо практической значимости, эти до-
стижения имеют и огромное теоретическое значение. В форми-
ровании новых ценных наследственных особенностей домашних
животных наиболее ярко и продуктивно осуществляется направ-
ленное воздействие людей на млекопитающих в интересах об-
щества. Уже Ч. Дарвин неоднократно указывал, как вредна''
недооценивать факты, касающиеся эволюционного значения по-
родообразования, как важно для зоологов хорошо разбираться
в вопросах происхождения домашних животных, в животновод-
стве. Опыт столетия, истекшего со времени появления выдаю-
щегося труда Дарвина, полностью подтвердил эту точку зрения.
Первые успешные попытки приручения млекопитающих были
сделаны по одним взглядам уже в палеолите, по другим — в ме-
золите; объектами их были виды рода собак (Cants). Несомнен-
ные остатки этих животных, например на территории СССР,
найдены при раскопках пещерных мезолитических стоянок
Крыма (14—12 тыс. лет до нашей эры) и, по-видимому, в па-
леолите Афонтовой горы в Сибири. Для неолита северной и
средней полосы Европейской части СССР (Ладожское озеро,
Владимирская, Ярославская области за 3—2 тыс. лет до на-
чала нашей эры) характерно наличие собак уже двух пород при
отсутствии других домашних животных. В это время на юге
Украины, в Молдавии, Средней Азии, а также в неолите Швей-
царии, Египта и др. уже были, кроме собак, одомашненные
овцы, козы, свиньи и "крупный рогатый скот.
Считают, что дикие предки домашней собаки — шакалы и
волки часто появлялись около древних стоянок бродячих охот-
ников и, возможно, селились поблизости, привлекаемые постоян-
ным наличием корма: костей, внутренностей животных и тому
подобных кухонных отбросов. И в наше время песцы, лисицы,
шакалы, волки, енотовидные собаки охотно берут на себя роль
«санитаров» на рыбных промыслах, местах разделки туш тюле-
ней, дельфинов, забоя стад домашних оленей и. т. и.
376
Близкое соседство таких хищников было выгодно обитателям
мезолитических стоянок. Благодаря чуткому слуху, острому
обонянию эти животные невольно заранее предупреждали людей
о появлении крупных и опасных зверей — пещерного медведя,,
льва — и, возможно, помогали их отгонять, преследуя как конку-
рентов в использовании пищи. Можно предполагать, что первым
толчком к приручению видов рода Cants было именно желание
приобрести сторожевых животных. Таким образом, первым эта-
пом одомашнения собаки, как и других видов млекопитающих,,
было приручение. При дальнейшем одомашнении их стали ис-
пользовать на охоте, после чего были выведены породы типа
современных лаек (в неолите), затем борзой и др. В настоящее
время, как всем известно, существует огромное разнообразие-
(более 350) пород домашних собак. Используя изменчивость
признаков, применяя племенной подбор, меняя условия кормле-
ния, содержания и т. п., люди создали все это разнообразие
пород с самыми различными целями. Существуют очень миниа-
тюрные комнатные- собачки, своего рода забавные живые иг-
рушки, мелкие низкорослые, но крепкие и сильные таксы и‘
фокстерьеры, в одиночку справляющиеся с барсуком или лисой
в их норе, проворные, похожие на белого песца оленегонные
лайки севера, огромные пастушьи овчарки, способные защищать
стадо от волков, выносливые и неприхотливые ездовые собаки —
незаменимые транспортные животные там, где невыгодно или
невозможно использовать лошадей й оленей, и многие др. При
наличии большого числа культурных пород, сильно отличаю-
щихся от своих предков, сохранились и примитивные, по всем7
признакам напоминающие шакала, волка и др. 1
Домашние в точном смысле слова, или культурные животные,
имеют многочисленные отличия, возникшие под влиянием созна-
тельного отбора и содержания в условиях, сильно отличаю-
щихся от условий жизни их предков. Доказано, что уже в антич-
ное время греки и римляне умели правильно вести селекцию
домашних животных; сохранились их достаточно детально и.
точно изложенные наставления для животноводов и владельцев,
стад. К- Маркс подчеркивал, что домашние животные — это-
животные, «измененные посредством труда, выращенные чело-
веком». В литературе нередко употребляют термин «полуодо-
машненные животные» по отношению к видам, занимающим,
промежуточное положение между прирученными и полностью-
одомашненными. Проф. К. Ф. Рулье, более 100 лет назад с боль-
шим интересом изучавший вопросы акклиматизации и домести-
1 В ряде мест еще и сейчас, чтобы улучшить выносливость, силу ездовых-
собак, производят их скрещивание с волками. Эволюция домашних пород со-
бак продолжается, так же как и использование ценных особенностей исход-
ных форм.
37Z-
кации животных, употреблял термин «порабощение», противо-
поставляя его «приручению», и подчеркивал этим полную
зависимость одомашненных форм от воли человека. В действи-
тельности провести четкую границу между только прирученной
и уже одомашненной формой удается не всегда.
Среди разводимых северных оленей довольно часто встре-
чаются чисто белые, пегие и пестрые животные, чрезвычайно
редкие среди диких оленей. Это один из показателей довольно
длительной доместикации. Нередко встречаются так называемые
«авки» — телята, почему-либо брошенные важенкой или поте-
рявшие мать. Их выкармливают искусственно, держа около
чума; подросшие авки — это совершенно ручные олени, не рас-
стающиеся с обитателями чума, в особенности с детьми, которые
обычно больше взрослых хлопочут около брошенных оленят-пы-
жиков. Наконец, существует так называемое «избяное оленевод-
ство»— разведение северных оленей около населенных пунктов
при более или менее постоянном скармливании им заранее за-
готовленных кормов. Но основная масса — миллионы разводи-
мых северных оленей по внешнему виду, биологии почти не от-
личается от своих диких родичей. Очень часто отдельные особи,
группы и даже целые стада, потерянные пастухами при сильном
тумане или угнанные волками, оказываются вдали от владель-
цев, продолжают пастись и кочевать без надзора, пока не воль-
ются в стада диких оленей и не одичают совсем. Дело в том, что
к прирученному северному оленю преобразующий труд людей
был приложен в меньшей степени, с меньшей интенсивностью,
мем к другим сельскохозяйственным копытным животным. На
первый взгляд это может показаться странным, если учесть, что
оленеводство представляет достаточно древнюю отрасль хозяй-
ства. Исторические, археологические и этнографические мате-
риалы доказывают, что олень в Северной Евразии был приру-
чен раньше, чем там появились лошади и овцы *. (По одним ав-
торам начало оленеводства датируется поздним неолитом, по
другим — началом железного века.)
Основная ценная биологическая особенность оленя — его
способность круглый год существовать на подножном корме, не
требуя от владельца других забот, кроме правильного перегона
с одних сезонных пастбищ на другие и охраны от хищников.
При постоянных кочевках и частой гибридизации с дикими оле-
нями проводить селекционную работу довольно трудно, да и
вести ее начали сравнительно недавно. Еще важнее то обстоя^
тельство, что условия жизни, в частности режим питания до-
1 Здесь оленеводство формировалось совершенно независимо в ряде цент-
ров с разницей в сотни лет. Население Северной Америки совершенно не
знало оленеводства. Начало ему было положено в конце прошлого и в два
первых десятилетия текущего века путем завоза одомашненных оленей на
Аляску с востока Сибири и севера Скандинавского полуострова.
378
машних и диких стад, не имеют существенных отличий; кочев-
ники в силу социально-бытовых условий своей жизни не могли
•создать для этого другую, новую среду и переделать природу
вида в своих интересах. Естественно, что у разводимых север-
ных оленей нет таких пород, как среди овец или крупного рога-
того скота. Отличаются они одна от другой не столь резко,
поэтому Н. Друри (1955) считает даже, что их еще нельзя призна-
вать за отдельные продуктивные породы или породные группы.
Все же никто не смешает рослых и тяжелых верховых (рис. 137)
и вьючных оленей горных районов Южной Сибири (их разводят
тофалары, тувинцы и др.) с относительно мелкими и быстрыми
упряжными тундровыми оленями Крайнего Севера Евразии (их
разводят саамы, коми, ненцы, чукчи, якуты, коряки и др.).
Хотя у некоторых народов северный олень используется
одновременно как транспортное, мясное и молочное животное
(например, у эвенков, тувинцев, якутов), молочная продуктив-
ность домашней важенки очень невелика — в среднем за сутки
около 400 мл и лишь в редких случаях более 1 л. За всю лакта-
цию (с мая по сентябрь) удой составляет всего 60—70 л.
(Друри, 1955). Между тем, по вкусовым и пищевым качествам
оленье молоко — ценнейший продукт, отличающийся от молока
других сельскохозяйственных животных высоким содержанием
жира и легко усвояемых белков. По калорийности оленье мо-
локо почти в 4 раза превосходит коровье. Оно настолько жирно
(17,1% против 3,9% в среднем у коровы), что для получения
масла его можно сбивать без предварительного сепарирования.
Но меры, необходимые для увеличения надоя молока (обильное
кормление и раздой важенок, племенной отбор в направлении
повышения молочности, правильное воспитание телят), начали
применять только в последние годы; видимо, дело улучшения
породы в этом отношении у народов Сибири не двигалось с ме-
ста в течение многих столетий. Таким образом, породообразо-
вание северного оленя — дело будущего, так как в прошлом
ч работа по разведению не была нацелена на изменение продук-
тивных качеств оленей.
О -происхождении оленеводства можно строить догадки и
судить из сопоставлений с данными этнографических наблюде-
ний. Бродячие охотники далекого прошлого, видимо, получали
иногда возможность захватывать живыми отдельных оленей
или небольшие группы их и содержать около своих временных
жилищ. Эти животные облегчали существование людей, как «са-
модвиж-ущийся» запас пищи. Возможно, что, используя инстинкт
стадности диких оленей, люди стали применять прирученных,
животных в качестве «манщиков» на охоте. Наконец, приспо-
собить отдельных пойманных и обжившихся животных к пере--
возке скудного кочевого имущества было уже не трудно (Дру-
ри, 1955). J
379
Далеко не все попытки одомашнения диких млекопитающих
были вполне успешными и дали большой экономический эффект.
Известно, например, что в древнем Египте имелись приручен-
ные антилопы, более 10 видов, но ни один из них не вошел прочно
в хозяйство и не был широко расселен. Африканские слоны
когда-то служили транспортными животными и с успехом ис-
пользовались в наступательных действиях на войне; позднее
интерес к ним почему-то исчез, и сейчас они не играют той роли,
которую в течение веков сохраняет индийский слон. Гепард, как
легко приручаемое исключительно ценное ловчее животное,
в старину широко применялся для охоты на степных и пустын-
ных копытных в Африке, Передней Азии. Индии и даже на на-
шем степном юге (во времена Киевской Руси). Однако настоя-
щей охотничьей кошки из него не получилось, и этот вид, как
объект разведения, видимо, навсегда утратил свое значение.
В подавляющем большинстве случаев особенно успешным
оказалось одомашнение животных, принадлежащих к ролам,
отличающихся обилием видов, или к видам, имеющим обилие
географических рас, т. е. к биологически' процветающим, про-
грессивным формам. Нередко виды стадные приручаются
успешнее, чем ведущие одиночную или семейную жизнь. Точно
подсчитать общее число видов, ставших одомашненными, до-
вольно трудно, так как в образовании некоторых пород участво-
вали по 2—3 диких предка. Всего насчитывается около 25 пол-
ностью одомашненных и более 16 видов разводимых, которые
доже могут стать одомашненными.
Еще в неолите разводимые животные были мелкими, отлича-
лись наличием инфантильных признаков, а в эпоху бронзы они
стали значительно крупнее, продуктивнее. На имеющих давность
5 тысяч лет изображениях овец, коз, крупного рогатого скота,
буйволов, ослов, верблюдов, собак Египта и Месопотамии уже
можно найти признаки их длительного одомашнения, черты фор-
мирования пород. Следовательно, работа над ними была начата
значительно раньше. Десятки тысяч лет прошли прежде, чем
люди достигли эпохи земледелия и животноводства, а затем
менее чем за 10 тыс. лет по всему земному шару они рас^
пространили лучших домашних животных, уже резко отличаю-
щихся от своих дикйх предков. За последние 400—450 лет ими
заселены материки Северной и Южной Америки, Австралия и
множество океанических островов.
Трудно привести точное число уже созданных человеком
пород; только крупного рогатого скота (с учетом местных отро-
дий) насчитывается примерно около 400. v овен — более 250, ло-
шадей до 150 и т. д. (Боголюбский, 1940). В то время как особи
примитивных пород рогатого скота весят всего 80—100 кг, вес
культурных пород достигает 1500 кг. Вес овец колеблется от
25 кг (у малопородйстых) до 200 кг, свиней от 30 кг до 500 кг
381
и т. д. При этом ряд пород отличается резко увеличенной про-
дуктивностью. Так, например, убойный вес от 50% повышается
до 70% и выше (следовательно, соответственно меньше отхо-
дов) ; годовой настриг шерсти у овец примитивных пород равен
1 кг; а у культурных — до 16 кг и более, причем качество шер-
сти значительно выше, так как ее составляют только волосы
пуха или пух и ость в очень выгодных для промышленной об-
работки соотношениях. Низкопородные, неумело содержимые
коровы дают около 700 л молока в год (его едва хватает для
выпаивания теленка), тогда как высокопородные — до 16 тыс. л
и более в год.
Некоторые домашние животные обладают исключительно
ценными биологическими особенностями. Такова, например, спо-
собность верблюдов перевозить грузы и людей через пустыни
с очень скудными и несъедобными для лошадей и быков тра-
вами, отсутствием хороших водопоев и т. д. Горбатый индийский
скот зебу устойчив против заражения пироплазмозом; значитель-
ные области Южной Америки, где обычный рогатый скот совер-
шенно не выживает от повальных заболеваний, заселены зебу
и его гибридами. Азиатские буйволы прекрасно уживаются
в заболоченных местах; грандиозная долина р. Амазонки, непри-
годная для разведения других видов и пород крупного рогатого
скота, с населением по этой причине лишенным свежих молоч-
ных продуктов, оказалась вполне доступной для заселения буй-
волами.
По сравнению с числом диких млекопитающих количество
одомашненных все еще очень невелико. Уже это указывает на
возможность расширения их списка путем приручения и одо-
машнения новых видов, использования диких форм для гибри-
дизации с домашними и т. д. Но нужно ли это? Ведь даже от
имеющегося сейчас разнообразия видов и пород домашних
животных путем их дальнейшего совершенствования можно по-
лучить гораздо больше продукции, чем они дают сейчас. Прак-
тика, хозяйственный опыт показывают, что процесс одомаш-
нения новых видов идет непрерывно.
В прошлом столетии и начале текущего большой спрос на
молодые рога оленей — панты, используемые в Китае для из-
готовления лекарства, вызвал сильное истребление пятнистого-
оленя на Дальнем Востоке и марала в южной Сибири. Охо-
титься за «пантачами» (самцами оленей в период отрастания
рогов) с каждым годом становилось труднее. Необходимость,
заставила некоторых охотников перейти от добывания пант
путем отстрела оленей к содержанию пойманных животных в не-
воле. Маралов и пятнистых оленей содержали в особых загонах
и летом, в период «полной спелости пантов» их спиливали,,
фиксируя животных во время операции в особых станках. Позд-
нее, кроме самцов, в маральниках стали содержать и самок,.
382
получая от них приплод и выращивая его в неволе. Так, на
Алтае, в Забайкалье и Приморье возникла новая отрасль жи-
вотноводства «пантовое оленеводство». В последние десятилетия
экстракт из специально обработанных пантов, так называемый
«пантокрин», широко применяется в советской медицине. Это»
очень ценный препарат — он поднимает тонус организма, уско-
ряет выздоровление людей, ослабленных тяжелой болезнью
и т. п. Таким образом, развилась новая область животновод-
ства — пантовое оленеводство. Продукция его используется сей-
час не только для экспорта пантов, но и для удовлетворения
потребностей внутри нашей страны.
За последние 25 лет в заповедниках СССР успешно осущест-
влялось изучение биологии лося, проводились опыты по его
одомашнению. Ценные результаты получены в Печоро-Илыч-
ском государственном заповеднике, где уже имеется значитель-
ное стадо хорошо прирученных и объезженных лосей. Очень
рослый, неприхотливый в отношении корма лось служит отлич-
ным вьючным и упряжным животным в самых глухих районах
тайги, где лошадей и оленей невозможно обеспечить кормом*
а дороги очень плохи или совсем отсутствуют. Широкое исполь-
зование лосей как ездовых и скороспелых мясных животных
в хозяйстве таежных колхозов и совхозов,— дело будущего, но
уже сейчас эти опыты (работы Е. П. Кнорре и др.) дали воз-
можность изучить экологию лося так детально и глубоко, как
никогда раньше Этот пример еще раз показал, насколько прав,
был Ч. Даовин, говоривший о необходимости изучения опыта
животноводства.
Непрерывно растущий спрос на пушнину, резкое сокращение
численности некоторых особенно упорно преследуемых видов и
географических форм пушных животных вызвали необходимость
разведения их в условиях неволи. Первые опыты (1887 г., в Ка-
наде) были осуществлены с серебристо-черными лисицами —
они были очень редки в природе и когда-то ценились необы-
чайно высоко. Вскоре стали разводить в неволе и голубых пес-
цов, американских норок, скунсов, а затем некоторых других
представителей отряда хищных и наиболее ценных грызунов..
Преодолев- большой ряд неудач и ошибок, энтузиасты новой-
отрасли животноводства добились больших успехов.
Сейчас во многих странах мира, в том числе и в СССР,
существует множество благоустроенных звероводных хозяйств^
разного типа. Миллионы отличных шкурок от зверьков, выра-
щенных в клетках, поступают на пушные аукционы, служат
предметом экспорта или широко используются в странах, со-
здавших хорошо поставленное звероводство. Селекционеры и
1 См. Труды Печоро-Илычского гос. заповедника, Коми. Книжное изда-
тельство, Сыктывкар, 195'9.
383
здесь сыграли большую роль — вывели ряд исключительно кра-
сивых новых и оригинальных цветовых форм животных, напри-
мер, «платиновую лису», «голубую» и «черную» норку, скунса,
лишенного характерных белых полос — с одноцветной темно-бу-
рой блестящей шкуркой и т. п. Приручение, содержание в не-
воле, контроль за подбором пар производителей и в этом случае
.дали возможность переделывать природу животных, повышая
качество получаемой пушнины в соответствии с интересами
покупателей (считаясь с быстро меняющимися требованиями
рынка).
Приручение и одомашнение новых видов млекопитающих, не-
сомненно, даст человечеству еще много новых источников цен-
ных продуктов и одновременно будет стимулом и средством для
углубленного изучения этих животных.
ИСТРЕБЛЕНИЕ И ПОДАВЛЕНИЕ ВРЕДНЫХ МЛЕКОПИТАЮЩИХ
Наряду с охраной и осуществлением мер, направленных на
расселение и размножение полезных видов млекопитающих,
приходится вести борьбу с вредными. Они истребляют ценных
диких животных и нападают на домашних, повреждают различ-
ные культурные растения, пастбища или распространяют болез-
ни, поражающие человека и домашних животных.
Количество вредных млекопитающих очень велико. Доста-
точно сказать, что огромное большинство видов грызунов, со-
ставляющих около половины числа видов млекопитающих СССР,
приносят вред полеводству, садоводству, лесоводству и служат
носителями различных инфекций. Очень вредны для хозяйства
летучие мыши — крыланы — они кочуют большими стаями и
уничтожают урожай фруктов в Австралии, на островах Тихого
океана, на юго-востоке матеоика Азии и т. д. Наппимер. на Са-
моа до 90% плодов манго уничтожают летучие собаки. Из кито-
образных значительный вред приносит косатка (Orca orca), ист-
ребляющая котиков, тюленей, дельфинов, в то воемя как сама
не имеет большой промысловой ценности. В США используют
самолеты, вооруженные пулеметами, для истребления косаток,
нападающих на аляскинских котиков во время их сезонных ми-
граций.
Очень вредны для животноводства и охотничьего хозяйства
многие представители семейств Canidae, Felidae и отчасти Mus-
telidae (например, росомаха). Из них в нашей стране наиболь-
ший вред приносит волк (Cants lupus)—убытки от волков еже-
годно измеряются десятками миллионов рублей. Помимо истреб-
ления домашних животных, волк вреден как опасный впаг пуш-
ных животных, дичи и носитель вируса бешенства. Борьба с
волками ведется в течение столетий; территории ряда стран За-
падной Европы уже полностью освобождены от него. В СССР
384
за последние годы тоже достигнуты большие успехи в сокраще-
нии численности волков, благодаря применению самолетов, аэро-
саней и быстроходных автомобилей. Поиски и уничтожение вы-
водков, истребление волков с помощью отстрела благодаря при-
менению новых средств транспорта стали значительно более
эффективны ми.
В деле борьбы с вредными грызунами, помимо улучшения
системы землепользования, повышения уровня агротехники, все
большую роль начинают играть массовые истребительные меро-
iipioiши с широким применением приманок с ядами, пропылива-
ния нор ядовитыми дустами, паро- и газообразными веществами
удушающего действия, заражения болезнетворными бактериями
и вирусами. /Механизация обработки участков, заселенных гры-
зунами, рассеивание отравленных приманок с самолетов, авто-
мобилей и т. п. дает возможность быстро и с минимальной за-
тратой труда уничтожать зверьков там, где возникает опасность
для урожая или начавшиеся на грызунах эпизоотии могут рас-
пространиться на людей.
Пример успешного применения вируса против кроликов,
чрезмерно размножившихся в Австралии, говорит о том, какими
могущественными средствами для борьбы с вредителями воору-
жен сейчас человек. Успех истребительных мероприятий обус-
ловливается не только достижениями химии токсических ве-
ществ, бактериологии и техники, но и детальным знанием эко-
логии вредителей, их требований к среде, а также «слабых
мест» в их поведении, органах чувств и т. п. Очень часто именно
териологические исследования прокладывают путь для лучшего
применения химических веществ и новой техники против вреди-
телей, обитающих в полях, участках лесных культур и т. п.
Проблему большого народнохозяйственного и санитарно-
•эпидемиологического значения представляет борьба с синантроп-
ными грызунами — группой видов, приспособившихся к обита-'
нию в различных жилых и хозяйственных постройках, в том чи-
сле в зернохранилищах, мясокомбинатах, холодильниках, про-
дуктовых магазинах, столовых, городских вокзалах, рынках, а
в сельских районах — на животноводческих фермах и т. д.
Как известно, развитие торгового мореплавания привело к
расселению домовой мыши, серой и черной крыс почти по всем
материкам и огромному большинству островов земного шара
(за исключением арктических и антарктических районов). Про-
никновение на корабли происходит во время стоянки и погрузки,
особенно в примитивных, неблагоустроенных портах, густо на-
селенных грызунами. Питаясь имеющимися на судах продук-
тами, грызуны нередко интенсивно размножаются во время
пути. При представившей,ся возможности они выселяются на бе-
рег, пользуясь трапами, причальными канатами и т. п., находят
на новом месте удовлетворяющие их биотопы и становятся чле-
25 Териология
385
нами местных биоценозов, принеся с собой специфических экто-
и эндопаразитов. Расселение синантропных грызунов идет также
с речным транспортом, по железным и проселочным дорогам.
Значение этого все еще продолжающегося процесса нельзя не-
дооценивать, так как синантропные грызуны причиняют огром-
ный вред.
Они истребляют и портят культурные растения на корню,
различные продукты и товары от дынь, помидоров, орехов, зер-
на. масла, мяса до мыла, тканей, книг и т. п. Серые крысы на
фермах нападают на поросят, кроликов, цыплят и наносят им
смертельные раны, в массе расхищают яйца, иногда нападают
даже на крупных откармливаемых свиней. В трущобах неблаго-
устроенных городов и селений капиталистических стран неодно-
кратно отмечались случаи сильного поранения крысами спящих
детей, оставленных без надзора. При укусах и загрязнении про-
дуктов и питьевой воды выделениями синантропные грызуны
рассеивают болезнетворные начала, вирулентные для человека и
домашних животных. Эктопаразиты серых и черных крыс, домо-
вых мышей, например блохи, клещи, в свою очередь служат хра-
нителями и переносчиками ряда вирусов и бактерий, опасных
для человека. Так, например, «влажнотропические очаги кры-
синой чумы Юго-Восточной Азии», особенно Индии, Цейлона,
Вьетнама, Таиланда, своим существованием в значительной сте-
пени обязаны разнообразию видов и обилию крыс, в том числе
и синантропных (Ралль, 1958). Из стран Южной Азии корабли
вместе с крысами неоднократно завозили чуму в портовые го-
рода Европы, Северной и Южной Америки. Например, послед-
няя «портовая вспышка» чумы в Одессе (1911 г.) была крыси-
ного происхождения.
Прогрызая ходы в деревянных, глинобитных и каменных по-
стройках, крысы и мыши портят стены, полы, потолки, изоля-
цию электропроводов и т. п. Описаны случаи полного уничтоже-
ния домовыми мышами легких (например, камышитовых) по-
строек в Казахстане за одну зиму (зверьки вселялись в дома на
зимовку в год массового размножения).
В кратком обзоре невозможно сколько-нибудь полно осве-
тить огромную и многообразную вредоносную деятельность си-
нантропных грызунов. Считают, что в одних только США, для
которых серая крыса — относительно недавний иммигрант, этот
грызун ежегодно наносит потери, оцениваемые в 200 млн. дол-
ларов (Гамильтон, 1939). По другим подсчетам, в которых при-
няты во внимание и различные косвенные неблагоприятные по-
следствия, причиняемые деятельностью крыс, стоимость потерь
по США достигает 2,5 млрд, долларов в год (Голсендорф и
Кларк, 1937). Несомненно, что серая крыса — наиболее вредный
вид не только из числа синантропных грызунов, но и вообще из
всех млекопитающих земного шара.
386
Борьба с этой группой грызунов ведется давно и хотя ока-
залась далеко нелегкой, в последние десятилетия становится все
более успешной. Для предупреждения дальнейшего расселения
крыс и мышей — возможности распространения ими чумной ин-
фекции на морских судах, в портах и на железнодорожном
транспорте ведется непрерывная борьба с грызунами; погрузка
и разгрузка судов идет под надзором санитарно-эпидемиологи-
ческих станций. Периодически большие группы отловленных
зверьков исследуются специалистами — паразитологами и бак-
териологами. Таким образом, ни одна эпизоотия не может прой-
ти незамеченной и обнаружение зараженных зверьков служит
сигналом к усилению дератизационной (крысоистребительной)
работы, которая служит одним из основных методов защиты на-
селения от заражения зоонозами. (Сходным образом организо-
вано наблюдение за природными очагами чумы в степях, полупу-
стынях, пустынях и горных районах СССР. Разница только в
том, что здесь под постоянным надзором находятся популяции
песчанок, сусликов, сурков, а отряды в составе врачей, зоологов
и паразитологов обеспечены лучшими транспортными сред-
ствами, так как ежегодно обследуют большие площади).
Вред от крыс и мышей в городах настолько велик, что во мно-
гих странах в архитектуру жилых и торговых помещений вне-
сены усовершенствования, придающие зданиям «крысо- и мыше-
непроницаемость». Это, естественно, сводит к минимуму количе-
ство мест, где синантропные грызуны могут поселяться, если они
проникают в черту города. С другой стороны, огромную роль сы-
грала рационализация очистки дворов и улиц от пищевых отхо-
дов и мусора. Исследования показали, что именно там, где по-
стоянно и в доступных для зверьков условиях накопляются
пищевые отходы, возникают городские очаги размножения си-
нантропных грызунов. Устранение этой «кормовой базы» очи-
щает целые кварталы от назойливых и крайне вредных «спутни-
ков» человека.
Естественным дополнением к этим предупредительным мерам
(крысонепроницаемость зданий, быстрая и полная очистка насе-
ленных пунктов от пищевых отходов) служит дератизация.
Очень важная для санитарного благополучия населения горо-
дов непрерывная работа дератизаторов, уничтожающих зверь-
ков ловушками и отравленными приманками, обеспечивает конт-
ролирование численности, доводя ее до минимума, при котором
грызуны уже не могут нанести вред хозяйству и здоровью на-
селения.
В населенных пунктах сельского типа борьба с синантроп-
ными грызунами значительно сложнее, чем в городе. При весен-
ней миграции мыши и крысы выселяются из домов на посевы,
луга, берега речек и т. п. В течение всего лета они мало доступ-
25*
387
ны для наблюдений и борьбы с ними. Осенью размножившиеся
зверьки в массе вселяются в жилые и другие постройки и в ко-
роткий срок могут принести большой вред. Кроме подлинных
синантропов в постройках появляются на время и другие виды
грызунов (серые и рыжие полевки, лесные мыши, серые хомяч-
ки, а в пустынях даже песчанки), несущие на себе эктопарази-
тов, нередко прямо из природных очагов опасных инфекций
человека. Между домовыми мышами, крысами и прикочевав-
шими обитателями степи и леса происходит интенсивный обмен
эктопаразитами, что еще более осложняет обстановку.
Там, где всегда имеется большое количество легко доступного
корма (например, на птицефермах, в свинарниках), возникают
оседлые популяции серых крыс. Обычного типа хозяйственные
постройки вполне проницаемы для этих грызунов, истребление
их отравленными приманками очень затруднено (много корма,
есть опасность отравить домашних животных и т. п.), а вред
крыс в' этих условиях велик. Он не ограничивается одним
расхищением корма, яиц, цыплят и т. д.— от крыс свиньи зара-
жаются трихинозом, а рогатый скот через загрязненные водо-
пои — лептоспирозом. Лептоспироз грызунов может переда-
ваться и человеку.
г В сельских местностях нужна целая система мероприятий,
хорошо приспособленных к конкретным природным и хозяйст-
венным условиям каждого колхоза и совхоза и направленных
на создание наиболее неблагоприятных условий для жизни гры-
зунов, а также периодически повторяемое их истребление с по-
мощью отравленных приманок. Конструкция и размещение по-
строек, способы хранения, раздачи животным корма и т. п. дол-
жны быть такими, чтобы синантропным грызунам не было до-
ступа к пище и хорошо защищенным убежищам.
ИСПОЛЬЗОВАНИЕ МЛЕКОПИТАЮЩИХ
В КАЧЕСТВЕ ЛАБОРАТОРНЫХ ЖИВОТНЫХ
Целый ряд видов издавна служит лабораторными живот-
ными, необходимыми для физиологических, генетических, био-
химических, гистологических и медицинских исследований. Бе-
лых мышей и крыс, хомячков, морских свинок, кроликов, собак,
хорьков, ежей, обезьян содержат в тысячах лабораторий, в ко-
торых и разводят как подопытных животных для различных
экспериментов. Так, например, лишь благодаря использованию
обезьян паразитологам удалось расшифровать до конца цикл
развития малярийных плазмодиев в теле теплокровного хозяи-
на и экспериментально проверить на этих животных — «моде-
лях» действие новейших медикаментов. Борьба с малярией —
одним из жесточайших бичей человечества, стоит теперь на вер<
ном пути, и заболеваемость в странах с достаточно развитой эко-
388
номикой и санитарной культурой может быть быстро и полно-
стью уничтожена.
Для изучения разных форм вируса гриппа в качестве лабо-
раторных животных используют светлых хорей, заражая их че-
рез нос. Появление раковых опухолей у лабораторных мышей и
крыс легко вызывают в экспериментах, что дает возможность
изучать рост злокачественных новообразований и испытывать
различные лечебные средства. Перед наукой возникают новые
задачи и соответственно непрерывно расширяется число видов
л а бор а тор н ых млекопитающих.
ЭСТЕТИЧЕСКОЕ ЗНАЧЕНИЕ МЛЕКОПИТАЮЩИХ
Велико и эстетическое значение представителей этой группы
позвоночных. Многие дикие звери так красивы и так гармони-
руют с окружающей их естественной обстановкой, что наблю-
дать за ними доставляет большое наслаждение каждому, кто
не лишен эстетического чувства. Проворные изящные белки
оживляют и украшают любой парк; недаром в общественных
садах многих стран Европы, Азии, Африки и Америки их бе-
режно охраняют, подкармливают, и доверчивые зверьки охотно
идут за «угощением» к рукам посетителей, обследуют их кар-
маны в поисках орехов и т. п. В тех же парках и пригородных
лесных массивах нередко разводят оленей, ланей, косуль.
Туристам, карабкающимся по извилистым тропам через ска-
лы и альпийские лужайки к вершинам гор, большую радость
доставляет наблюдение за сернами, горными козлами, сурками
и т. п. Без этих животных своеобразный мир высокогорий утра-
тит значительную часть своего очарования. С ростом культуры
все большее число людей стремится проводить дни отдыха в
районах с хорошо сохранившейся природой и достаточно бога-
тым животным миром.
*
Таким образом, млекопитающие в практическом, научном и
культурном отношении — одна из наиболее важных групп жи-
вотного мира. Большое разнообразие их биологических типов,
высокая организация, широкое географическое распространение,
экологическая пластичность, свойственные им сложные формы
нервной деятельности, богатая палеонтологическая история де-
лают их исключительно интересным объектом ^ля теоретических
исследований. В Советском Союзе в связи с разнообразием его
природных условий, обширностью недостаточно освоенных тер-
риторий и широкими задачами строительства коммунизма изу-
889
чение млекопитающих имеет гораздо большее значение, чем в
ряде других стран.
Достижения советской териологии велики и бесспорны, од-
нако, остается еще очень много нерешенных вопросов как в об-
ласти теоретических проблем морфологии, систематики, зооге-
ографии, истории и экологии млекопитающих нашей фауны, так
и в области практических задач направленного воздействия на
млекопитающих в интересах нашего общества.
Перед териологами—обширное поле для плодотворной дея-
тельности. Организация в Москве Первого всесоюзного совеща-
ния по млекопитающим (25—31 января 1962 г.)—свидетель-
ство того, что в нашей стране исследования этой группы живот-
ных вступают в новый этап с большей согласованностью, сла-
женностью и целенаправленностью коллективных усилий армии
териологов. Несомненно, что по мере развития науки и техники
будут найдены новые, более совершенные пути пользования
ресурсами териофауны.
ЛИТЕРАТУРА1
Астанин Л. А. Органы тела млекопитающих и их работа, М., 1958.
Банников А. Г., Жирнов Л. В., ЛебедевЛ.С., Фандеев
А. А. Биология сайгака, М., 1961.
Б а р а б а ш-Н икифоровИ. И. Бобр и выхухоль, как компоненты
водноберегового комплекса, Воронеж, 1950.
Б а р а б а ш-Н икифоровИ. И. Калан (Enhydra lutris L.) его биоло-
гия и вопросы хозяйства, М., 1947.
Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Опреде-
литель млекопитающих СССР, М., 1944.
Браунер А. А. Сельскохозяйственная зоология, Харьков, 1923.
Б рем А. Э. Жизнь животных. Млекопитающие, т. X, Спб., 1913.
Б р е м А. Э. Жизнь животных. Млекопитающие, т. V, М., 1941.
Виноградов Б. С. иГромовИ. М. Грызуны фауны СССР, М.—Л.,
1952.
Виноградов Б. С. иОболенскийС. И. Вредные и полезные в
сельском хозяйстве млекопитающие, М., 1932.
ГептнерВ. Г., Морозова-Турова Л. Г., Ц а л к и н В. И-
Вредные и полезные звери районов полезащитных насаждений, М., 1959
ГептнерВ. Г., НасимовичА. А., Банников А. Г. Млеко
питающие Советского Союза, т. I, Парнокопытные и непарнокопытные,
М., 1961.
Гольцмайер Г. К. Белка-телеутка, М., 1935.
Давиташвили Л. Ш. Курс палеонтологии, изд. 2., 1949.
Доппел ьмайрГ. Г., Мальчевский А. С., Новиков Г. А.,
Фалькенштейн Б. Ю., Биология лесных зверей и птиц, М.—Л.,
1951.
Д р у р и Н. В. Дикий северный олень Советской Арктики и Субарктики,
Л.—М., 1949.
Ж и т к о в Б. М. Биология лесных зверей и птиц, М,,“1928.
Ж и т к о в Б. М. Акклиматизация животных, М., 1934.
КалабуховН. И. Спячка животных (изд. 1—1936; изд. 2—1946),
Харьков, 1956.
Капланов Л. Г. Тигр, изюбрь, лось, М., 1918.
КашкаровД. Н. Основы экологии животных, изд. 2, Л., 1945.
1 Приводятся только крупные работы типа монографий (вышедшие от-
ельными изданиями).
391
КашкаровД. Н. и Ста н чи нс кий В. В. Курс биологии позво-
ночных, М.—Л., 1934.
КлимовА. Ф. Анатомия домашних животных, т. I, М., 1955.
КлимовА. Ф. иАкаевский А. И. Анатомия домашних животных,
т. II, М., 1955.
КолосовА. М. иБакеевН. Н. Биология зайца-русака. Мат. к
позн. фауны и флоры СССР, Нов. серия, вып. 9, М., 1947.
Колосов А. М., Лавров Н. П., Н а у м о в С. П. Биология про-
мысловых зверей СССР, М., 1961.
Котт X. Приспособительная окраска животных. М., 1950.
К р я ж е в В. Я- Высшая нервная деятельность животных в условиях
общения, М., 1955.
Кузнецов Б. А. Основы товароведения пушномехового сырья, М., 1941,
Кузякин А. П. Летучие мыши, М., 1950.
Куле ш'о в П. Н. Влияние питания на формы живого тела и на характер
продуктивности, Изв. Петровск. землед. и лесн. акад., вып. 3, 1879
Л а в р о в Н. П. Акклиматизация и реакклиматизация пушных зверей в
СССР, М„ 1946.
Лэк Д. Численность животных и ее регуляция в природе, М., 1957.
Мак Ко л л юм и Саймондс. Новое в учении о питании и кормлении.
М—Л., 1934.
НасимовичА. А. Роль режима снежного покрова в жизни копытны •
животных на территории СССР, М., 1955.
НаумовН. П. Очерки сравнительной экологии мышевидных грызунов,!
М.— Л., 1948.
НаумовН. П. Экология животных (изд. 1 —1955), М., 1962.
НаумовС. П. Экология зайца-беляка, М., 1947.
Новиков Г. А. Полевыеисследования по экологии наземных позвоноч-
ных, М., 1953.
Новиков Г. А. Хищные млекопитающие фауны СССР, М.—Л., 1956.
Новиков Г. А. Экология зверей и птиц лесостепных дубрав, Л., 1959-
О г н е в С. И. Звери Восточной Европы и Северной Азии, т. I—1928’’
т. II—1931, М,—Л.
О г н е в С. И. Звери СССР и прилежащих стран, т. III—1935, т. IV—1940
т. V—1947, т. VI—1948, т. VII —1950, М.—Л.
О г и е в С. И. Очерки экологии'млекопитающих, М., 1951.
Павлова М. В. Палеозоология, ч. 2, М.—Л., 1929.
П о п о в В. А. Материалы по экологии норки (Mustela vison Вг.) и резуль-
таты акклиматизации ее в Татарской АССР, Казань, 1949.
Раевский В. В. Жизнь Кондо-Сосвинского соболя, М., 1947.
Р о м е р А. Ш. Палеонтология позвоночных, М.—Л., 1939.
СвириденкоП. А. Запасание корма животными, Киев, 1957.
Сев ер цо в Н. А. Периодические явления в жизни зверей, птиц и гад
Воронежской губернии, 1855, (изд. 2—1950).
Северцов С. А. Проблемы экологии животных, т. I, М., 1951.
С ка л о н В. Н. Речные бобры Сев; Азии. Мат. к позн. фауны и флоры
СССР, Нов. сер., вып. 25, М., 1961.
С л о н и м А. Д. Животная теплота И ее регуляция в организме млекопи-
тающих, М.—Л., 1952.
С л о н и м А. Д. Основы общей экологической физиологии млекопитающих.
М,—Л., 1961.
Слудский А. А. Ондатра и акклиматизация ее в Казахстане, Алма-Ата,
1948.
С л у д с к и й А. А. Кабан (морфология, экология, хозяйств, и эпизоотолог.
значение, промысел), Алма-Ата, 1956.
Тимофеев В. В. и Н а д еев В. Н. Соболь, М., 1955.
Томилин А. Г. Звери СССР и прилежащих стран. Т. IX, Китообраз-
ные, М., 1957.
Федюшин А. В. Речной бобр, его история, жизнь и опыты по размно-
жению, М., 1935.
Ф л е р о в К- К. Кабарги и олени. Фауна СССР, Млекопитающие. М,— Л.*
1952.
Формозов А. Н. Колебания численности промысловых животных,
М.— Л., 1935.
ФормозовА. Н. Снежный покров в жизни млекопитающих и птиц
СССР, М., 1946.
ФридериксК. Экологические основы прикладной зоологии и энтомоло-
гии, Л.—М., 1932.
Ц а л к и н В. И. Горные бараны Европы и Азии. Материалы к позн. фауны
и флоры СССР, Нов. сер., отд. зоолог., вып. 27 (42), М., 1951.
Allen G. М. Bats. Cambridge, Massachusetts, Harvard University Press, 1939.
BeddardF. E. Mammalia, London, 1902.
Bourliere F. Vie et moeurs des mammiferes, Paris, 1951.
Bourl iere F. Mammals of the World, London, 1955.
Bourliere F. The Natural History of Mammals, London, 1955.
Brehms. Tierleben. Die Saugetiere, neubearb. von M. Hilzheimer und L.
Heck, В. IV, 1920.
Brink, van den, F. H. Die Saugetiere Europas, T. Haltenroten, Berlin,
1956.
Cabrera A. Manual de mastozoologia, Madrid, 1922.
Cabrera A., and Y e p e s J. Marriiferos sudamericanos... Tucuman and
Buenos Aires, Argentina. Compania Argentina de Editores, 1940.
Darlington Ph. J. Zoogeography. The geographical distribution of ani-
mals, New York, 1957.
Eisentraut M., Die deutschen Fl edermause. 1937.
Eisentraut M. Der Winterschlaf mit seinen okologischen und physiolo-
gischen Begleiterscheinungen, Jena, 1956.
Eisentraut M. Aus dem Leben der Fledermause und Flughunde, Jena,
1957.
Ellenberger — Baum. Handbuch £der Vergleichenden Anatomie der
Haustiere 18te Auflage, bearbeitet ton Zimmermann O., Ackerknecht E.,
Gran H,, Berlin, 1943.
Eller m a n I. R., Morrison — Scott T. C. Checklist ofPalaearctic and
Indian mammals, L, 1951.
393
Е 11 e r m a n I. R., Morr iso n-S с о 11 T. C. and Hayman, K. W. Sauthern
African Mammals, London, 1953.
Hamilton W. J. American Mammals, their Lives, Habits and Economic
Relations, New—Jork, 1939.
H a r p e r F. Extinct and vanishing Mammals of the Old World; Americ. Com-
mittee for Internal. Wild Life Protection,Spec. Publ. N 12, New York, 1945.
Heape W. Migration, Emigration and Nomadism. Cambr., 1931.
Henderson J., Craig E. L-, Economic Mammology, C. Thomas edif.,
Springfield Jll., Baltimore Marylend, 1932.
Howell B. Aquatic Mammals. Their adaptations to life in the water, Bal-
timore, 1930.
Krumbiegel 1. Biologie der Tiere Deutschlands (Mammalia), Berlin,
1930—31.
Krumbiegel I. Biologie der Sangetiere, В. I, II, 1954—55.
Matthews L. H. British Mammals, London, 1952.
Matthews L. H. The Migration of Mammals. Ann. Report of the Board
of Regents of the Smithsonian Instit, Waschington, 1954.
M I 1 n e-E d w a r d s A. Recherches pour servir a 1’historie naturelle des
mammiferes, Paris, 1868—1874.
N e 1 s о n E. W. Wild Animals of Northern America, Washington, 1918.
О 1 t A. a. und S t r 6 s e A. Die Wildkrankheiten und ihre Bekampfung.
J, Neumann in Neudqmm Verlag, 1914.
P о cock R. J. The Fauna of British India including Ceylon and Burma
v. I. Mammalia, London, 1939.
Seton E. T. Life histories of the northern animals, Loud. v. 1, 1909, v. 2,
1910.
Simpson G. G. The principles of classification and a classification of Mam
mals. Bull, of the Amer. Mus. of nat History, v. 85, New York, 1945.
Tembrock G. Tierpsychologie. Wittenberg Lutherstadt, 1956.
T r a i t e de zoologie (anatomie, systematique, biologie), Dr. P. Grasse. Tome
XVI, XVII, Mammiferes fasc. 1, 2, 1955.
long I. Z. The Life of Mammals, Oxford, 1S57.
Weber M. Die Saugetiere, Jena. Bd. 1, 2.
ОГЛАВЛЕНИЕ
Предисловие ..................................................... 3
Введение.......................................................... 5
Систематический очерк класса млекопитающих ...................... 11
Происхождение современного состава класса млекопитающих.......... 26
Очерк морфологии млекопитающих................................... 35
Общие особенности организации млекопитающих .................. 35
Наружные покровы.............................................. 36
Скелет....................................................... 48
Мускулатура................................................. 55
Нервная система............................................. 56
Анализаторы................................................... 60
Органы пищеварения.......................................... 66
Органы дыхания ............................................... 78
Сердечно-сосудистая система ................................ 79
Железы внутренней секреции................................... 82
Органы выделения.............................................. 83
Органы размножения........................................... 84
Вторичные половые признаки................................... 91
Окраска, ее физиологическое и биологическое значение............. 94
Адаптивные типы млекопитающих .................................. 105
Наземные формы............................................... 105
Подземные формы •............................................ 109
Водные формы................................................. 111
Древесные формы.............................................. 117
Летающие формы............................................... 120
Питание и явления, связанные с ним ............................. 123
Общее значение питания................................... . 123
Специализация в питании, значение разных групп кормов........ 130
Сезонные изменения кормового режима.......................... 137
Географические изменения питания............................ 144
Изменения питания в зависимости от возраста, пола, индивидуальных
навыков особей и колебаний урожайности кормов............. 149
Суточная норма пищи, способность переносить голодание........ 157
Минеральное питание........................;................. 163
Потребление воды и водопой................................... 169
Запасание корма.............................................. 175
Суточная активность. Спячка..................................... 188
Суточная активность ........................................ 188
Спячка....................................................... 191
Миграции. Линька................................................ 200
Миграции.................................................... 200
Линька ..................................................... 212
Убежища. Участки обитания ...................................... 226
Убежища разного типа........................................ 226
Участки обитания............................................ 248
Размножение и явления, связанные с ним ............................. 254
Общая характеристика размножения................................. 254
Подготовка к размножению, течка (гон), спаривание............... 256
Механизм регуляции половых процессов самки.................. 268
Беременность, зародышевое развитие............................... 271
Плодовитость..................................................... 277
Лактация.................................'....................... 282
Рост и развитие молодняка........................................ 2t5
Забота о потомстве (родительские инстинкты) ..................... 293
Распад семей .................................................... 299
Расселение молодняка............................................. 300
Географическая, сезонная и многолетняя изменчивость хода размно-
жения ........................................................ 302
Родственное спаривание, межвидовые и межродовые помеси........... 305
Движение численности населения зверей .............................. 308
Условия, влияющие на движение численности ....................... 308
Причины смертности............................................... 312
Достижение предельного возраста . .......................... 313
Несчастные случаи ............................................ 313
Недостаток корма.............................................. 315
Болезни....................................................... 318
Деятельность хищников и конкурентов .......................... 323
Стихийные бедствия, неблагоприятные метеорологические усло-
вия ....................................................... 330
Хозяйственная деятельность людей ............................. 337
Типы движения численности........................................ 338
Тип крайне неустойчивого скачкообразного движения численности 339
Тип неустойчивого движения численности........................ 346
Тип устойчивого движения численности.......................... 363
Пути направленного воздействия на млекопитающих в интересах че-
ловека ....................................................... 366
Краткий исторический очерк..................................... 366
Охрана ценных диких млекопитающих, регулирование численности. 367
Деятельность заповедников, заказников и охотничьих хозяйств . . . 369
Обогащение фауны путем акклиматизации млекопитающих.............. 372
Одомашнение млекопитающих........................................ 375
Истребление и подавление вредных млекопитающих............. . . 384
Использование, млекопитающих в качестве лабораторных животных. 388
Эстетическое значение млекопитающих ............................. 389
Литература......................................................... 391
Илья Ильич Барабаш-Никифоров
Александр Николаевич Формозов
ТЕРИОЛОГИЯ
Редактор издательства Л. П. Петровская
Технический редактор Л. А. Григорчук
Корректор Г. П. Меркулова
Сдано в набор 14/VIII-62 г. Подписано к печати 23/1-63 г.
Бумага 60 X SO1/, 24,75 печ. л..24,74 уч.-изд. л.
Тираж 3500. Т-0260». Изд. № Е,.'36-60 г. Цена 89 коп.
Государственное издательство «Высшая школа»,
Москва К-62, Подсосенский пер., 20
Набрано в.первой Образцовой типографии имени А. А. Жданова
Мо/коТзуого городского совнархоза. Москва, Ж-54, Валовая, 28
Отпач£5н0 в тип. изд-ва „Высшая школа”, Москва, Неглинная, 29/14. 3. 640
Замеченные опечатки
Страница Строка Напечатано Следует читать
12 15 снизу Didelphydae Didelphiidae
18 20 снизу Aplodonta Aplodontia
24 16 сверху sondiacus sondaicus
54 6 снизу osrea patagii os pea patagii
206 Подпись к у берегов Мурман- у берегов Мурмана
рис. 77 ' ска
.353 22 снизу X. Элтону Ч. Элтону
;С>7 5 сверху за 1924—1929 за 1924—1926
,35‘.> И снизу Ц, Хыоитта Г. Хьюитта
14 снизу Ц. Хьюитт Г. Хьюитт
Внкнз № (ИО И. И. Барабаш «Териология».